
1-Digestão e absorção dos nutrientes da refeição. 3...
15
1 Integração dos metabolismos dos carbohidratos, gorduras e proteínas ao longo do dia e no jejum prolongado Departamento de Bioquímica da Faculdade de Medicina do Porto [email protected] O período pós-prandial é caracterizado 1- Digestão e absorção dos nutrientes da refeição. 2- Armazenamento de glicogénio no fígado e músculos e de armazenamento de gordura no tecido adiposo 3- Aumento (recuperação) da massa de proteínas endógenas. 4- Aumento da oxidação dos hidratos de carbono e dos aminoácidos da dieta e diminuição marcada da oxidação das gorduras ⇒ ⇒ ⇒ Quociente Respiratório > 0,95. 5- Razão [insulina]/[glicagina] aumentada período pós-prandial Em geral as 4-6 horas que se seguem à refeição 2 3 Na digestão dos hidratos de carbono participam amílases (salivar e pancreática) e dissacarídases da bordadura em escova dos enterócitos: amílases Dextrinas limite + maltose maltase e isomaltase glicose + galactose lactase glicose sacarase glicose + frutose A absorção dos monossacarídeos ocorre via ação de transportadores membranares: Glut2 Sangue Lúmen intestinal Glut5 SGLT1 glicose e galactose glicose + galactose frutose frutose Enterócito Que modificações ocorrem no metabolismo quando se ingere uma refeição contendo glicose (ou outros hidratos de carbono que geram glicose)? A quantidade de glicose livre no organismo (plasma e líquido extracelular) de um adulto em jejum é de cerca de 10-12 g. No entanto, a ingestão de 6 vezes essa quantidade só faz subir a glicemia apenas para o dobro. Porquê? Estudo na FMUP em Out/Nov 2009. 1 g/Kg (19 voluntários); 0,5 g/kg (68 voluntários). O aumento da glicemia tem efeitos homeostáticos que tendem a atenuar e a corrigir esse aumento.
Transcript of 1-Digestão e absorção dos nutrientes da refeição. 3...

1
Integração dos metabolismos dos carbohidratos, gorduras
e proteínas ao longo do dia e no jejum
prolongado
Departamento de Bioquímica da Faculdade de Medicina do Porto
O período pós-prandial é caracterizado 1- Digestão e absorção dos nutrientes da refeição.2- Armazenamento de glicogénio no fígado e músculos e de armazenamento de gordura no tecido adiposo 3- Aumento (recuperação) da massa de proteínas endógenas.4- Aumento da oxidação dos hidratos de carbono e dos aminoácidos da dieta ediminuição marcada da oxidação das gorduras ⇒⇒⇒⇒ Quociente Respiratório > 0,95.
5- Razão [insulina]/[glicagina] aumentada
período pós-prandial
Em geral as 4-6 horas que se seguem à refeição
2
3
Na digestão dos hidratos de carbono participam amílases (salivar e pancreática) e dissacarídases da bordadura em escova dos enterócitos:
amílasesDextrinas limite +maltose
maltase e
isomaltase
glicose + galactose
lactase
glicose
sacarase glicose + frutose
A absorção dos monossacarídeos ocorre via ação de transportadores membranares:
Glut2
SangueLúmen intestinal
Glut5
SGLT1glicose e galactose
glicose + galactose
frutose frutose
Enterócito
Que modificações ocorrem no metabolismo quando se ingere uma refeição contendo glicose (ou outros hidratos de carbono que geram glicose)?
A quantidade de glicose livre no organismo (plasma e líquido extracelular) de um adulto em jejum é de cerca de 10-12 g. No entanto, a ingestão de 6 vezes essa quantidade só faz subir a
glicemia apenas para o dobro. Porquê?
Estudo na FMUP em Out/Nov 2009. 1 g/Kg (19 voluntários); 0,5 g/kg (68 voluntários).
O aumento da glicemia tem efeitos homeostáticos que tendem a atenuar e a corrigir esse aumento.

5
0
5
10
15
g/h
Consumo de glicose no estado pós-prandial
0
5
g/h
Consumo de glicose no estado pós-absortivo
Relativamente ao período pós-absortivo (antes do pequeno almoço) o consumo de glicose pode aumentar mais de 4 vezes.
O cérebro que, no período pós-absortivo, era responsável por ½ da glicose consumida no organismo, mantém o consumo em cerca de 4 g/h mas deixa de ser o maior consumidor de glicose.No período pós-prandial, o fígado e os músculos consomem mais de metade do total.
Gerich (2010) Diabetic Medicine 27:136-142 6
No período pós-prandial, se o indivíduo estiver em repouso, a massa de glicose consumida pelo organismo no seu todo e os diferentes destinos desta glicose dependem do indivíduo, da composição da refeição e dos níveis de glicogénio previamente acumulados. São valores possíveis: 42 g/h de glicose consumidos pelo organismo no seu todo e uma repartição semelhante entre oxidação, armazenamento e formação de diversos derivados (lactato em diversos tecidos, glicerol-3-fosfato no tecido adiposo, palmitato no fígado e tecido adiposo e alanina nos músculos).
Cerca de 14 g/h = armazenamento de glicogénio (sobretudo fígado e músculos).
Cerca de 16 g/h de glicose oxidada a CO2; dos quais 4 g é glicose oxidada pelo cérebro.
Cerca de 12 g/h: formação de lactato (nos eritrócitos, intestino, fígado, músculos, etc.), glicerol-3-P (tecido adiposo), palmitato (fígado e tecido adiposo) e alanina (músculos).
Armazenamento
Oxidação
Glicose consumida no organismo no período pós-
prandial
(g/hora)
42 g/h
diversos derivados (principalmente lactato)
Glicose
A glicose ingerida entra para o sangue no intestino e aumenta a glicemia. Esta subida da glicemia é atenuada porque a glicose (direta e indiretamente, via insulina) inibe a sua produção endógena e estimula o seu armazenamento e a sua degradação.
3- ↑↑↑↑ da degradação da glicose:↑ produção de lactato e ↑ da oxidação da glicose (sobretudo nos músculos).
Os mecanismos envolvidos estão em grande parte dependentes do ↑ da insulina (e da ↓ da glicagina), mas a glicose também tem ações diretas (sobretudo no fígado).
STOP
1- ↓↓↓↓ da produção endógena de glicose
2- Armazenamento no fígado e no músculo (↑ da síntese de glicogénio no fígado e músculos)
Lactato e CO2
Insulina
7 8
PPi
ATP
ADP
Fosfo-glicomútase
Pirofosforílase do UDP-glicose
Hexocínase IV
Síntase do glicogénioUDP
No caso do músculo é igual exceto que estão envolvidos produtos de genes distintos nos casos do (1) GLUT, (2) da hexocínase e da (3) síntase do glicogénio.
Uma parte do glicogénio que se acumula no fígado, forma-se através de uma via designada de glicogénese direta.
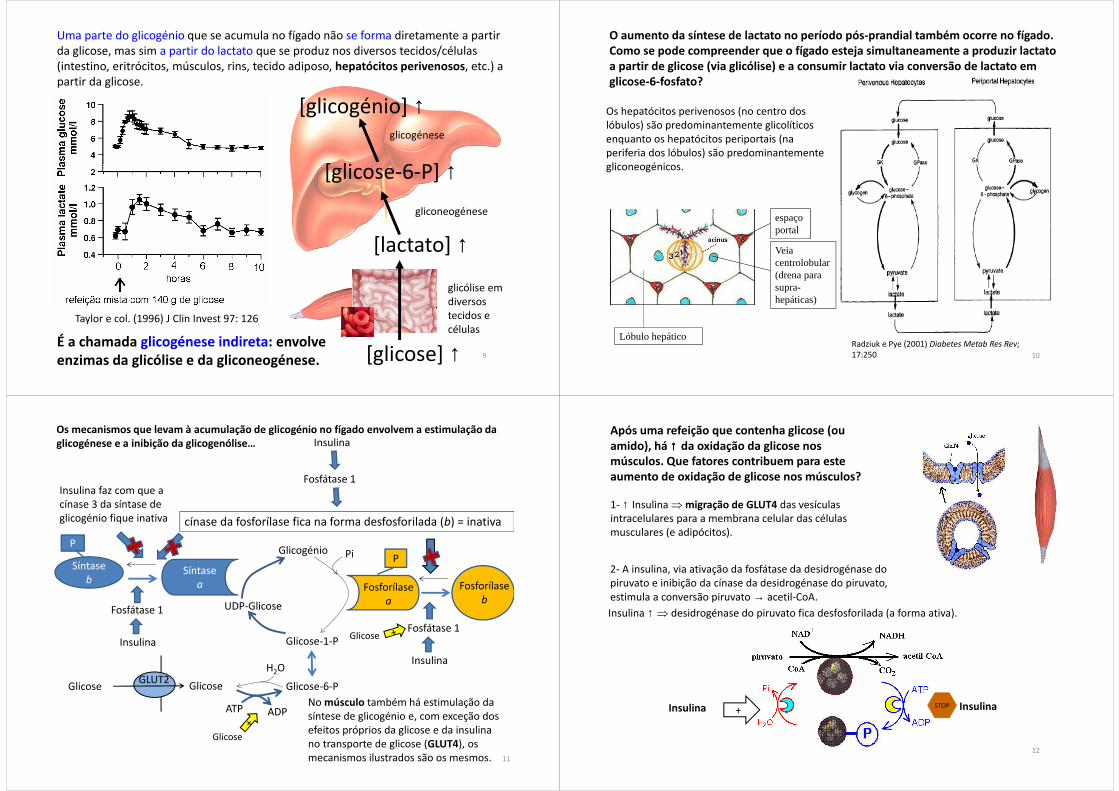
9
Taylor e col. (1996) J Clin Invest 97: 126
Uma parte do glicogénio que se acumula no fígado não se forma diretamente a partir da glicose, mas sim a partir do lactato que se produz nos diversos tecidos/células (intestino, eritrócitos, músculos, rins, tecido adiposo, hepatócitos perivenosos, etc.) a partir da glicose.
[glicose] ↑
[lactato] ↑
glicólise em diversos tecidos e células
gliconeogénese
[glicose-6-P] ↑
[glicogénio] ↑glicogénese
É a chamada glicogénese indireta: envolve enzimas da glicólise e da gliconeogénese.
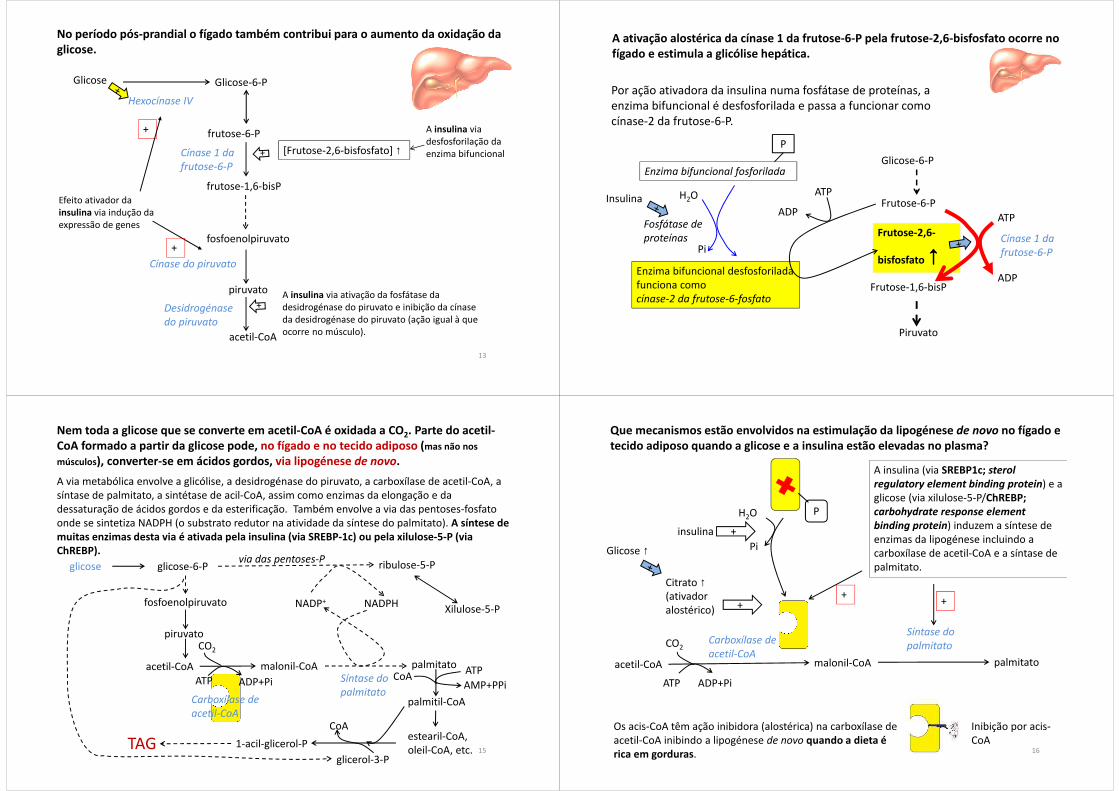
O aumento da síntese de lactato no período pós-prandial também ocorre no fígado. Como se pode compreender que o fígado esteja simultaneamente a produzir lactato a partir de glicose (via glicólise) e a consumir lactato via conversão de lactato em glicose-6-fosfato?
Os hepatócitos perivenosos (no centro dos lóbulos) são predominantemente glicolíticos enquanto os hepatócitos periportais (na periferia dos lóbulos) são predominantemente gliconeogénicos.
Radziuk e Pye (2001) Diabetes Metab Res Rev; 17:250 10
espaço portal
Veia centrolobular (drena para supra-hepáticas)
Lóbulo hepático
Glicogénio
Glicose-1-P
UDP-Glicose
Síntase a
P
Fosforílase a
Glicose-6-PGlicose
Pi
H2O
ATP ADP
GLUT2Glicose
Insulina
Insulina
Síntase b
P
Fosfátase 1
Os mecanismos que levam à acumulação de glicogénio no fígado envolvem a estimulação da glicogénese e a inibição da glicogenólise… Insulina
Fosfátase 1Insulina faz com que a cínase 3 da síntase de glicogénio fique inativa
Glicose
Glicose
No músculo também há estimulação da síntese de glicogénio e, com exceção dos efeitos próprios da glicose e da insulina no transporte de glicose (GLUT4), os mecanismos ilustrados são os mesmos.
cínase da fosforílase fica na forma desfosforilada (b) = inativa
11
Fosforílase b
Fosfátase 1
Após uma refeição que contenha glicose (ou amido), há ↑↑↑↑ da oxidação da glicose nos músculos. Que fatores contribuem para este aumento de oxidação de glicose nos músculos?
1- ↑ Insulina ⇒ migração de GLUT4 das vesículas intracelulares para a membrana celular das células musculares (e adipócitos).
Insulina ↑ ⇒ desidrogénase do piruvato fica desfosforilada (a forma ativa).
Insulina + InsulinaSTOP
2- A insulina, via ativação da fosfátase da desidrogénase do piruvato e inibição da cínase da desidrogénase do piruvato, estimula a conversão piruvato → acetil-CoA.
12
Glicose-6-P
frutose-1,6-bisP
fosfoenolpiruvato
+
[Frutose-2,6-bisfosfato] ↑
No período pós-prandial o fígado também contribui para o aumento da oxidação da glicose.
piruvato
acetil-CoA
A insulina via desfosforilação da enzima bifuncional
A insulina via ativação da fosfátase da desidrogénase do piruvato e inibição da cínase da desidrogénase do piruvato (ação igual à que ocorre no músculo).
frutose-6-P
Glicose
Efeito ativador da insulina via indução da expressão de genes
+
13
Hexocínase IV
Cínase do piruvato
Cínase 1 da
frutose-6-P
Desidrogénase
do piruvato
Frutose-2,6-
bisfosfato ↑↑↑↑
Por ação ativadora da insulina numa fosfátase de proteínas, a enzima bifuncional é desfosforilada e passa a funcionar como cínase-2 da frutose-6-P.
Frutose-6-P
Frutose-1,6-bisP
Piruvato
H2O
Pi
Glicose-6-P
A ativação alostérica da cínase 1 da frutose-6-P pela frutose-2,6-bisfosfato ocorre no fígado e estimula a glicólise hepática.
ATP
ADP
Enzima bifuncional fosforilada
Enzima bifuncional desfosforilada funciona como cínase-2 da frutose-6-fosfato
InsulinaATP
ADP
P
Fosfátase de
proteínas Cínase 1 da
frutose-6-P
Nem toda a glicose que se converte em acetil-CoA é oxidada a CO2. Parte do acetil-CoA formado a partir da glicose pode, no fígado e no tecido adiposo (mas não nos
músculos), converter-se em ácidos gordos, via lipogénese de novo.
A via metabólica envolve a glicólise, a desidrogénase do piruvato, a carboxílase de acetil-CoA, a síntase de palmitato, a sintétase de acil-CoA, assim como enzimas da elongação e da dessaturação de ácidos gordos e da esterificação. Também envolve a via das pentoses-fosfato onde se sintetiza NADPH (o substrato redutor na atividade da síntese do palmitato). A síntese de muitas enzimas desta via é ativada pela insulina (via SREBP-1c) ou pela xilulose-5-P (via ChREBP).
glicose glicose-6-P ribulose-5-P
NADP+ NADPH Xilulose-5-Pfosfoenolpiruvato
piruvato
acetil-CoA malonil-CoA palmitatoATP
AMP+PPi
CO2
palmitil-CoA
CoA
estearil-CoA, oleil-CoA, etc.
glicerol-3-P
ATP ADP+Pi
CoA
1-acil-glicerol-PTAG 15
via das pentoses-P
Carboxílase de
acetil-CoA
Síntase do
palmitato
Que mecanismos estão envolvidos na estimulação da lipogénese de novo no fígado e tecido adiposo quando a glicose e a insulina estão elevadas no plasma?
acetil-CoA malonil-CoA
ATP ADP+Pi
CO2
P
insulina +
Citrato ↑(ativador alostérico) +
A insulina (via SREBP1c; sterol
regulatory element binding protein) e a glicose (via xilulose-5-P/ChREBP; carbohydrate response element
binding protein) induzem a síntese de enzimas da lipogénese incluindo a carboxílase de acetil-CoA e a síntase de palmitato.
H2O
Pi
16
Glicose ↑
++
palmitato
Carboxílase de
acetil-CoA
Síntase do
palmitato
Os acis-CoA têm ação inibidora (alostérica) na carboxílase de acetil-CoA inibindo a lipogénese de novo quando a dieta é rica em gorduras.
Inibição por acis-CoA
2- A dieta mais comum na civilização ocidental contém, relativamente ao caso dos países africanos e da maioria dos asiáticos, um alto teor em gorduras. Os ácidos gordos geram acis-CoA que são inibidores da carboxílase de acetil-CoA impedindo a sua ativação plena.
hidratos de carbono 300 1200 52%gorduras 80 720 31%proteínas 100 400 17%
2320
A lipogénese de novo permite converter glicose em TAG mas, usando uma dieta de tipo ocidental, é incomum que os ácidos gordos de síntese endógena sejam uma parte significativa dos ácidos gordos dos TAG armazenados. Porquê?
acetil-CoA malonil-CoA palmitato
CO2
ATP ADP+Pi
1- Para além dos mecanismos de fosforilação/desfosforilação e alostéricos, a ativação das enzimas da lipogénese envolve mecanismos de indução da síntese de enzimas. Este processo é lento e só se instala plenamente após vários dias com uma dieta rica em carbohidratos (altos níveis de glicogénio). Este padrão alimentar é incomum na Europa e EUA.
Carboxílase de
acetil-CoA Síntase do palmitato
Insulina ↑+ +
acis-CoA
-
17
Após uma refeição que contenha glicose aumenta a oxidação da glicose, mas diminui reciprocamente e marcadamente a oxidação de ácidos gordos nos músculos e no fígado. Porquê?
1 - Insulina ↑ faz com que a perilipina e a lípase hormono-sensível fiquem no estado desfosforilado (inativo) ⇒
↓ da libertação de NEFA (non esterified
fatty acids = FFA = ácidos gordos
livres) para o plasma.
[Frayn et al. (1993) Metabolism 42:504]
3- Diminuição dos NEFA plasmáticos (NEFAcaiem de 0,5 mM para 0,05 mM) e
consequente diminuição da oferta de ácidos gordos aos tecidos.
insulina
fosfodiestérase
AMP
2- A insulina ↑ também ↑ esterificação de ácidos gordos no citoplasma dos adipócitos
18
cAMP
glicerol
NEFA = FFA = ácidos gordos livres
Que mecanismos intracelulares contribuem, nos músculos em repouso, para a diminuição da oxidação em β no período pós-prandial?
[malonil-CoA] ↑…que inibe a CPT1
Acetil-CoA + CO2
2- a concentração de acis-CoA está ↓↓↓↓ e, por isso, a ação inibidora dos acis-CoA na carboxílase de acetil-CoA não se exerce
3- glicose está a ser oxidada ⇒[citrato] ↑↑↑↑
1- os acis-CoA são substratos para a CPT1, mas…[acis-CoA]↓↓↓↓
O passo regulador da oxidação em β é o transporte de ácidos gordos para a mitocôndria que depende da atividade da carnitina-palmitil transférase 1 (CPT1).
No fígado, no estado pós-prandial, também há inibição da oxidação em β. Aqui, para além destes mecanismos, também são relevantes a ativação da carboxílase de acetil-CoA por desfosforilação ativada pela insulina e a indução da transcrição do seu gene (via SREBP-1c e CHREBP). 19
[acis-CoA] ↓↓↓↓
ácidos gordos ↓
CPT1
20
Uma refeição normal, para além de carbohidratos e gordura, contém proteínas que, no lúmen intestinal, são convertidas em AAs e em di- e tripeptídeos que são absorvidos para o interior do enterócito por transporte ativo secundário.
Proteínas da dieta
di- e tripeptídeos
PEPT1
H2O
Na+
H+
Vários simporters AA/Na+
catalisam a absorção de AAs
Os di- e tripeptídeos
terminam a sua hidrólise no
interior dos enterócitos
AAsAAs
AAsno sangue
Síntese proteica(reposição)
Oxidação (com perda do azoto que gera ureia)
Derivados de Aas (quantitativamente pouco relevante)
Conversão noutros AAs
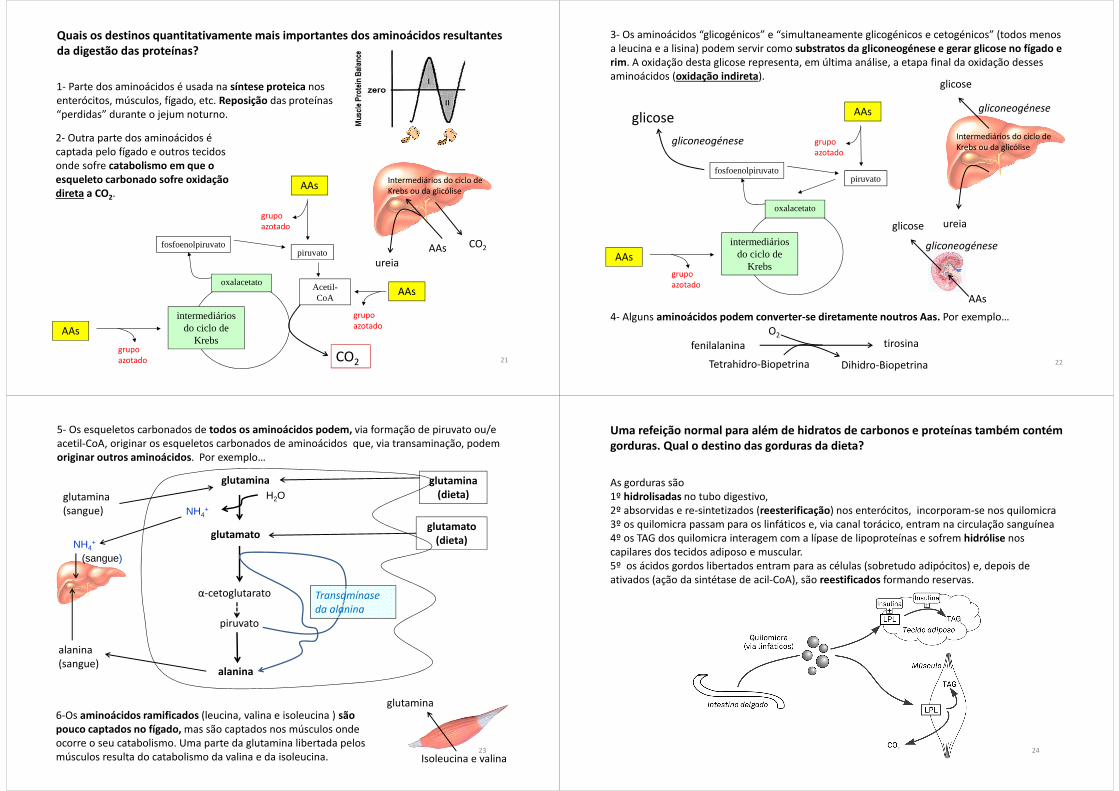
Quais os destinos quantitativamente mais importantes dos aminoácidos resultantes da digestão das proteínas?
AAs
Intermediários do ciclo de Krebs ou da glicólise
1- Parte dos aminoácidos é usada na síntese proteica nos enterócitos, músculos, fígado, etc. Reposição das proteínas “perdidas” durante o jejum noturno.
ureia
2- Outra parte dos aminoácidos é captada pelo fígado e outros tecidos onde sofre catabolismo em que o esqueleto carbonado sofre oxidação direta a CO2.
CO2
AAs
oxalacetato
piruvatofosfoenolpiruvato
AAs
intermediários do ciclo de
Krebs
Acetil-CoA
CO2
AAs
grupoazotado
grupo azotado
grupo azotado
21 22
3- Os aminoácidos “glicogénicos” e “simultaneamente glicogénicos e cetogénicos” (todos menos a leucina e a lisina) podem servir como substratos da gliconeogénese e gerar glicose no fígado e rim. A oxidação desta glicose representa, em última análise, a etapa final da oxidação desses aminoácidos (oxidação indireta).
Intermediários do ciclo de Krebs ou da glicólise
ureia
AAs
oxalacetato
piruvatofosfoenolpiruvato
AAs
intermediários do ciclo de
Krebsgrupo azotado
grupo azotado
glicose
glicose
gliconeogénese
gliconeogénese
glicose
gliconeogénese
AAs
4- Alguns aminoácidos podem converter-se diretamente noutros Aas. Por exemplo…
fenilalanina tirosinaO2
Tetrahidro-Biopetrina Dihidro-Biopetrina
23
6-Os aminoácidos ramificados (leucina, valina e isoleucina ) são pouco captados no fígado, mas são captados nos músculos onde ocorre o seu catabolismo. Uma parte da glutamina libertada pelos músculos resulta do catabolismo da valina e da isoleucina. Isoleucina e valina
glutamina
5- Os esqueletos carbonados de todos os aminoácidos podem, via formação de piruvato ou/e acetil-CoA, originar os esqueletos carbonados de aminoácidos que, via transaminação, podem originar outros aminoácidos. Por exemplo…
NH4+
H2O
alanina
glutamato
glutamina
glutamina (sangue)
glutamato (dieta)
α-cetoglutarato
piruvato
alanina (sangue)
glutamina (dieta)
Transamínase
da alanina
NH4+
(sangue)
Uma refeição normal para além de hidratos de carbonos e proteínas também contém gorduras. Qual o destino das gorduras da dieta?
As gorduras são 1º hidrolisadas no tubo digestivo, 2º absorvidas e re-sintetizados (reesterificação) nos enterócitos, incorporam-se nos quilomicra3º os quilomicra passam para os linfáticos e, via canal torácico, entram na circulação sanguínea 4º os TAG dos quilomicra interagem com a lípase de lipoproteínas e sofrem hidrólise nos capilares dos tecidos adiposo e muscular.5º os ácidos gordos libertados entram para as células (sobretudo adipócitos) e, depois de ativados (ação da sintétase de acil-CoA), são reestificados formando reservas.
24

No tubo digestivo, as lípases gástrica e pancreática catalisam a hidrólise de triacilgliceróis da dieta.
Os produtos da digestão são maioritariamente ácidos gordos e 2-monoacilglicerol que são absorvidos.
Que acontece aos TAG da dieta no tubo digestivo?
Dentro do enterócito ocorre a re-síntese de triacilgliceróis.O processo envolve a ação da sintétase de acil-CoA e de transférases de acilo presentes na face citoplasmática do RE.
Ao mesmo tempo também se formam fosfolipídeos, colesterídeos e apolipoproteínas B48 e A que vão ser importantes constituintes dos quilomicra.
25
Dentro dos enterócitos formam-se lipoproteínas ricas em triacilgliceróis (86% da massa) que se designam por quilomicra. Os quilomicra são vertidos no polo basal dos enterócitos por exocitose e que, via linfáticos, chegam ao sangue.
quilomicra
linfáticos
Veia central (sub-clávia esq.)
base
Apo B48
Apo A
(86%)
(3%)
(2%)
(8%)
(1-2%)
O processo é lento de tal forma que os quilomicra só começam a aparecer no plasma 1 hora após a refeição e só atingem a concentração máxima cerca de 3-4 h depois. 26
No sangue os quilomicra nascentes recebem das HDL apo C e apo E originando os quilomicra maduros que já são capazes de se interagir com a lípase de lipoproteínas.
Apo C e E
HDL
A lípase de lipoproteínas é uma ecto-hidrólase das células endoteliais dos capilares dos tecidos adiposo e muscular.
A hidrólise dos quilomicra ocorre no plasma sanguíneo, maioritariamente nos capilares dos tecidos adiposo, e os ácidos gordos são maioritariamente captados pelos adipócitos. Só uma pequena fração dos ácidos se mantém no sangue (ligação à albumina).
O glicerol não é metabolizado no tecido adiposo nem no músculo: todo o glicerol libertado vai para o fígado e rim.
ALBUMINA
lípase de lipoproteínas
H2Océlu
la e
nd
ote
lial
27
Que acontece à atividade lípase de lipoproteínas no período pós-prandial?
1- A insulina estimula a síntese da lípase de lipoproteínas do tecido adiposo. Tal como a absorção das gorduras e o aparecimento de quilomicra no sangue também este processo é lento: a atividade da lípase de lipoproteínas do tecido adiposo só atinge o máximo 4 h após a refeição.
(A lípase de lipoproteínas dos músculos é inibida pela insulina e estimulada pelo exercício físico.)
2- A lípase de lipoproteínas atua, quer nos TAG das VLDL, quer nos TAG dos quilomicra, mas tem preferência pelos quilomicra ⇒
2.1 Mesmo quando a refeição é rica em gorduras, os quilomicra (que têm semi-vida de apenas alguns
minutos) mantêm concentrações baixas no plasma.
2.2 Devido à competição dos TAG dos quilomicra, a concentração de TAG das VLDL aumenta no plasma após as refeições.
28
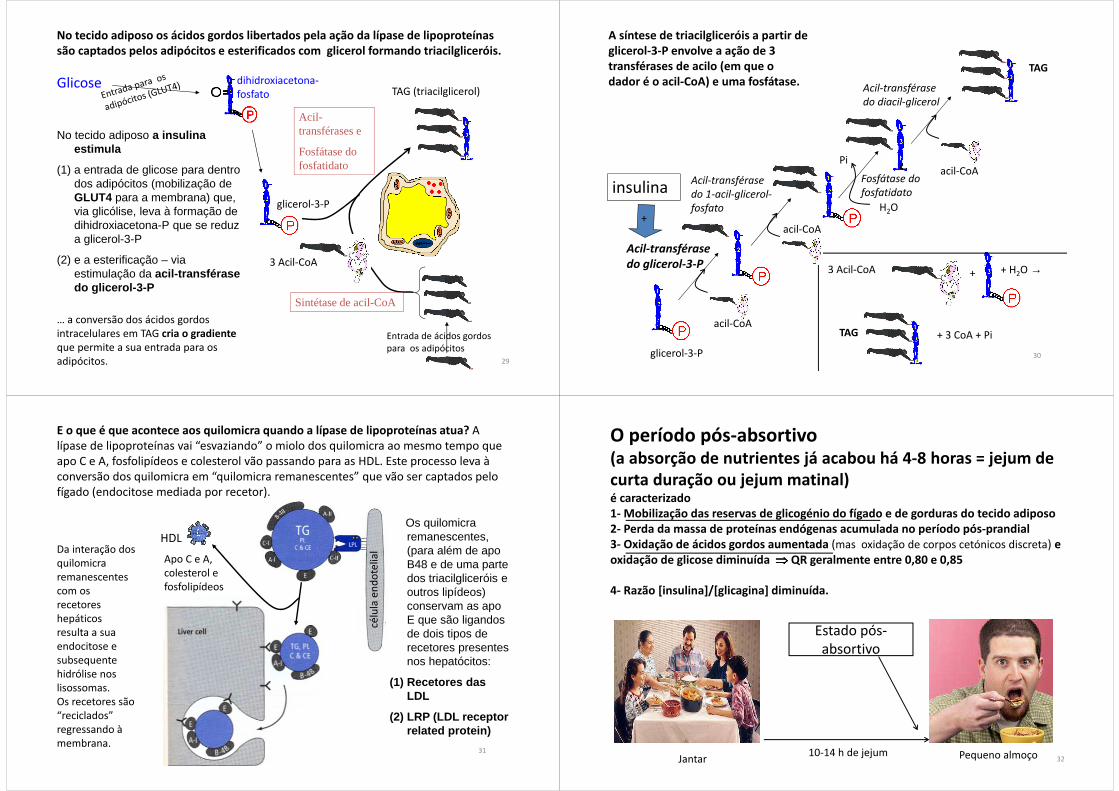
No tecido adiposo os ácidos gordos libertados pela ação da lípase de lipoproteínas são captados pelos adipócitos e esterificados com glicerol formando triacilgliceróis.
3 Acil-CoA
glicerol-3-P
TAG (triacilglicerol)
Sintétase de acil-CoA
Acil-transférases e
Fosfátase do fosfatidato
No tecido adiposo a insulina estimula
(1) a entrada de glicose para dentro dos adipócitos (mobilização de GLUT4 para a membrana) que, via glicólise, leva à formação de dihidroxiacetona-P que se reduz a glicerol-3-P
(2) e a esterificação – via estimulação da acil-transférase do glicerol-3-P
Glicose
Entrada de ácidos gordos para os adipócitos
dihidroxiacetona-fosfato
29
… a conversão dos ácidos gordos intracelulares em TAG cria o gradienteque permite a sua entrada para os adipócitos.
3 Acil-CoA
glicerol-3-P
+ + H2O →
+ 3 CoA + Pi
Acil-transférase
do glicerol-3-P
Acil-transférase
do 1-acil-glicerol-
fosfato
Fosfátase do
fosfatidato
Acil-transférase
do diacil-glicerol
A síntese de triacilgliceróis a partir de glicerol-3-P envolve a ação de 3 transférases de acilo (em que o dador é o acil-CoA) e uma fosfátase.
insulina
+
30
acil-CoA
acil-CoA
acil-CoAPi
TAG
TAG
H2O
E o que é que acontece aos quilomicra quando a lípase de lipoproteínas atua? A lípase de lipoproteínas vai “esvaziando” o miolo dos quilomicra ao mesmo tempo que apo C e A, fosfolipídeos e colesterol vão passando para as HDL. Este processo leva à conversão dos quilomicra em “quilomicra remanescentes” que vão ser captados pelo fígado (endocitose mediada por recetor).
HDL
Apo C e A, colesterol e fosfolipídeos
Os quilomicra remanescentes, (para além de apo B48 e de uma parte dos triacilgliceróis e outros lipídeos) conservam as apo E que são ligandos de dois tipos de recetores presentes nos hepatócitos:
(1) Recetores das LDL
(2) LRP (LDL receptorrelated protein )
Da interação dos quilomicra remanescentes com os recetores hepáticos resulta a sua endocitose e subsequente hidrólise nos lisossomas. Os recetores são “reciclados” regressando à membrana.
célu
la e
nd
ote
lial
31 Pequeno almoço
Estado pós-absortivo
Jantar
O período pós-absortivo (a absorção de nutrientes já acabou há 4-8 horas = jejum de curta duração ou jejum matinal)é caracterizado1- Mobilização das reservas de glicogénio do fígado e de gorduras do tecido adiposo 2- Perda da massa de proteínas endógenas acumulada no período pós-prandial3- Oxidação de ácidos gordos aumentada (mas oxidação de corpos cetónicos discreta) e oxidação de glicose diminuída ⇒⇒⇒⇒ QR geralmente entre 0,80 e 0,85
4- Razão [insulina]/[glicagina] diminuída.
3210-14 h de jejum
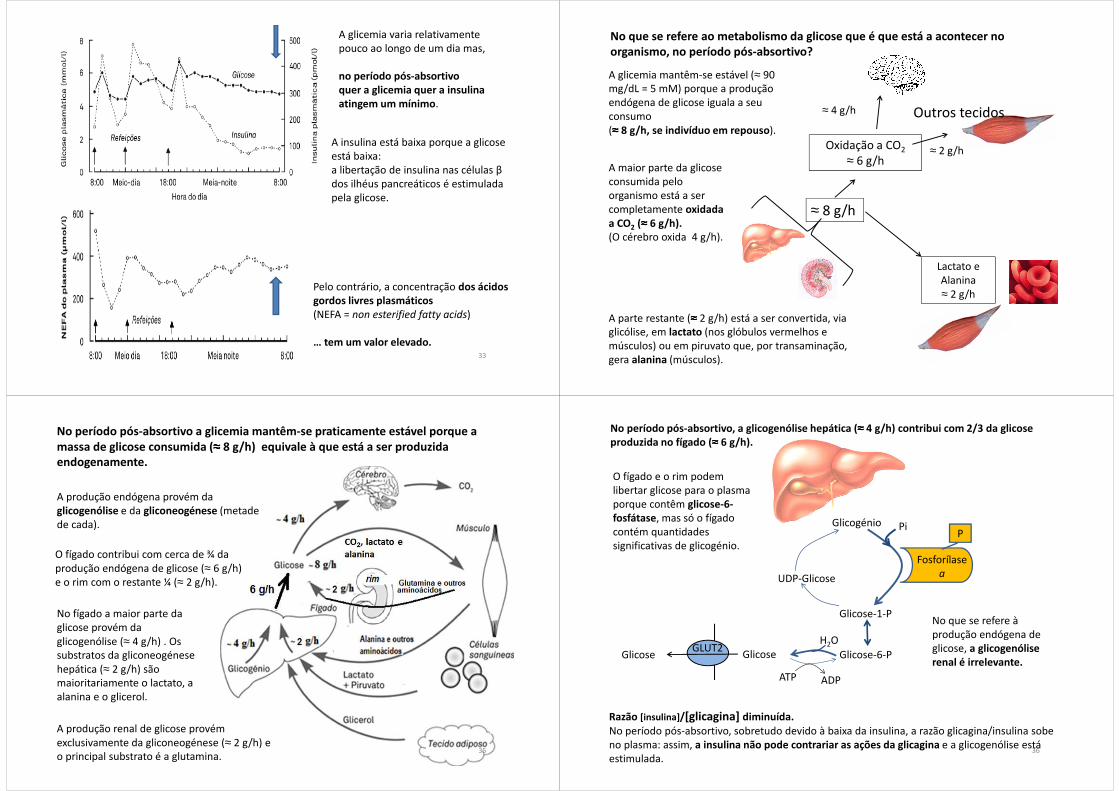
A glicemia varia relativamente pouco ao longo de um dia mas,
no período pós-absortivo quer a glicemia quer a insulina atingem um mínimo.
A insulina está baixa porque a glicose está baixa: a libertação de insulina nas células βdos ilhéus pancreáticos é estimulada pela glicose.
33
Pelo contrário, a concentração dos ácidos gordos livres plasmáticos(NEFA = non esterified fatty acids)
… tem um valor elevado.
No que se refere ao metabolismo da glicose que é que está a acontecer no organismo, no período pós-absortivo?
A maior parte da glicose consumida pelo organismo está a ser completamente oxidada a CO2 (≈≈≈≈ 6 g/h). (O cérebro oxida 4 g/h).
A glicemia mantêm-se estável (≈ 90 mg/dL = 5 mM) porque a produção endógena de glicose iguala a seu consumo (≈≈≈≈ 8 g/h, se indivíduo em repouso).
A parte restante (≈≈≈≈ 2 g/h) está a ser convertida, via glicólise, em lactato (nos glóbulos vermelhos e músculos) ou em piruvato que, por transaminação, gera alanina (músculos).
≈ 8 g/h
Outros tecidos
Oxidação a CO2
≈ 6 g/h
Lactato e Alanina≈ 2 g/h
≈ 4 g/h
≈ 2 g/h
No período pós-absortivo a glicemia mantêm-se praticamente estável porque a massa de glicose consumida (≈≈≈≈ 8 g/h) equivale à que está a ser produzida endogenamente.
35
No fígado a maior parte da glicose provém da glicogenólise (≈ 4 g/h) . Os substratos da gliconeogénese hepática (≈ 2 g/h) são maioritariamente o lactato, a alanina e o glicerol.
A produção endógena provém da glicogenólise e da gliconeogénese (metade de cada).
O fígado contribui com cerca de ¾ da produção endógena de glicose (≈ 6 g/h) e o rim com o restante ¼ (≈ 2 g/h).
A produção renal de glicose provém exclusivamente da gliconeogénese (≈ 2 g/h) e o principal substrato é a glutamina.
36
Glicogénio
Glicose-1-P
UDP-Glicose
Fosforílase a
Pi
H2O
Glicose-6-PGlicose
ATP ADP
GLUT2Glicose
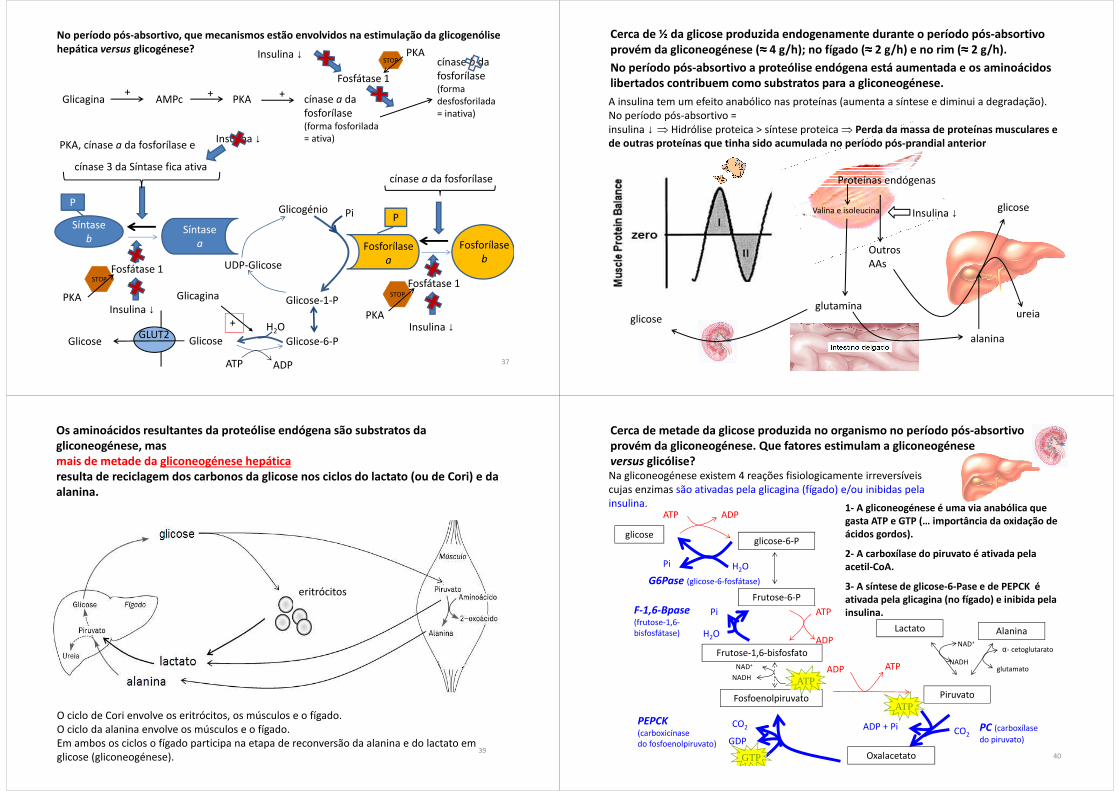
No período pós-absortivo, a glicogenólise hepática (≈≈≈≈ 4 g/h) contribui com 2/3 da glicose produzida no fígado (≈≈≈≈ 6 g/h).
O fígado e o rim podem libertar glicose para o plasma porque contêm glicose-6-fosfátase, mas só o fígado contém quantidades significativas de glicogénio.
No que se refere à produção endógena de glicose, a glicogenólise renal é irrelevante.
Razão [insulina]/[glicagina] diminuída.No período pós-absortivo, sobretudo devido à baixa da insulina, a razão glicagina/insulina sobe no plasma: assim, a insulina não pode contrariar as ações da glicagina e a glicogenólise está estimulada.
P
37
Glicogénio
Glicose-1-P
UDP-Glicose
Síntase a
P
Fosforílase a
Pi
H2O
Insulina ↓
Síntase b
P
Fosfátase 1
Fosforílase b
Glicose-6-PGlicose
ATP ADP
GLUT2Glicose
No período pós-absortivo, que mecanismos estão envolvidos na estimulação da glicogenólise hepática versus glicogénese?
Glicagina AMPc PKA
PKA, cínase a da fosforílase e
cínase a da fosforílase
cínase a da fosforílase(forma fosforilada = ativa)
+++
Insulina ↓
cínase 3 da Síntase fica ativa
Fosfátase 1
Insulina ↓+
GlicaginaPKA
PKA
STOP
STOP
cínase b da fosforílase(forma desfosforilada = inativa)
Fosfátase 1
Insulina ↓ PKASTOP
Cerca de ½ da glicose produzida endogenamente durante o período pós-absortivo provém da gliconeogénese (≈≈≈≈ 4 g/h); no fígado (≈≈≈≈ 2 g/h) e no rim (≈≈≈≈ 2 g/h).
No período pós-absortivo a proteólise endógena está aumentada e os aminoácidos libertados contribuem como substratos para a gliconeogénese.
A insulina tem um efeito anabólico nas proteínas (aumenta a síntese e diminui a degradação).No período pós-absortivo = insulina ↓ ⇒ Hidrólise proteica > síntese proteica ⇒ Perda da massa de proteínas musculares e de outras proteínas que tinha sido acumulada no período pós-prandial anterior
glicose
Outros AAs
Proteínas endógenas
ureia
Insulina ↓
glutamina
Valina e isoleucina
alanina
glicose
39
Os aminoácidos resultantes da proteólise endógena são substratos da gliconeogénese, mas mais de metade da gliconeogénese hepáticaresulta de reciclagem dos carbonos da glicose nos ciclos do lactato (ou de Cori) e da alanina.
O ciclo de Cori envolve os eritrócitos, os músculos e o fígado.O ciclo da alanina envolve os músculos e o fígado.Em ambos os ciclos o fígado participa na etapa de reconversão da alanina e do lactato em glicose (gliconeogénese).
eritrócitos
Cerca de metade da glicose produzida no organismo no período pós-absortivo provém da gliconeogénese. Que fatores estimulam a gliconeogénese versus glicólise?
40
PiruvatoFosfoenolpiruvato
CO2
Oxalacetato
GDP
CO2 ADP + Pi
Frutose-1,6-bisfosfato
Frutose-6-P
ATP
ADPAlaninaLactato
ATP
α- cetoglutarato
glutamatoNADH
NAD+
ADP
H2O
Pi
glicose-6-Pglicose
ATP ADP
Pi H2O
1- A gliconeogénese é uma via anabólica que gasta ATP e GTP (… importância da oxidação de ácidos gordos).
2- A carboxílase do piruvato é ativada pela acetil-CoA.
3- A síntese de glicose-6-Pase e de PEPCK é ativada pela glicagina (no fígado) e inibida pela insulina.
PC (carboxílase
do piruvato)
PEPCK (carboxicínase do fosfoenolpiruvato)
F-1,6-Bpase (frutose-1,6-bisfosfátase)
G6Pase (glicose-6-fosfátase)
NADH
NAD+
ATP
ATP
Na gliconeogénese existem 4 reações fisiologicamente irreversíveis cujas enzimas são ativadas pela glicagina (fígado) e/ou inibidas pela insulina.
GTP
Frutose-6-P
Frutose-1,6-bisfosfato
H2O
Pi
Frutose-2,6-
bisfosfato ↓ ↓ ↓ ↓STOP
Glicose-6-P
Lactato, alanina, glicerol…
glicose
Frutose-2,6-
bisfosfato↓↓↓↓
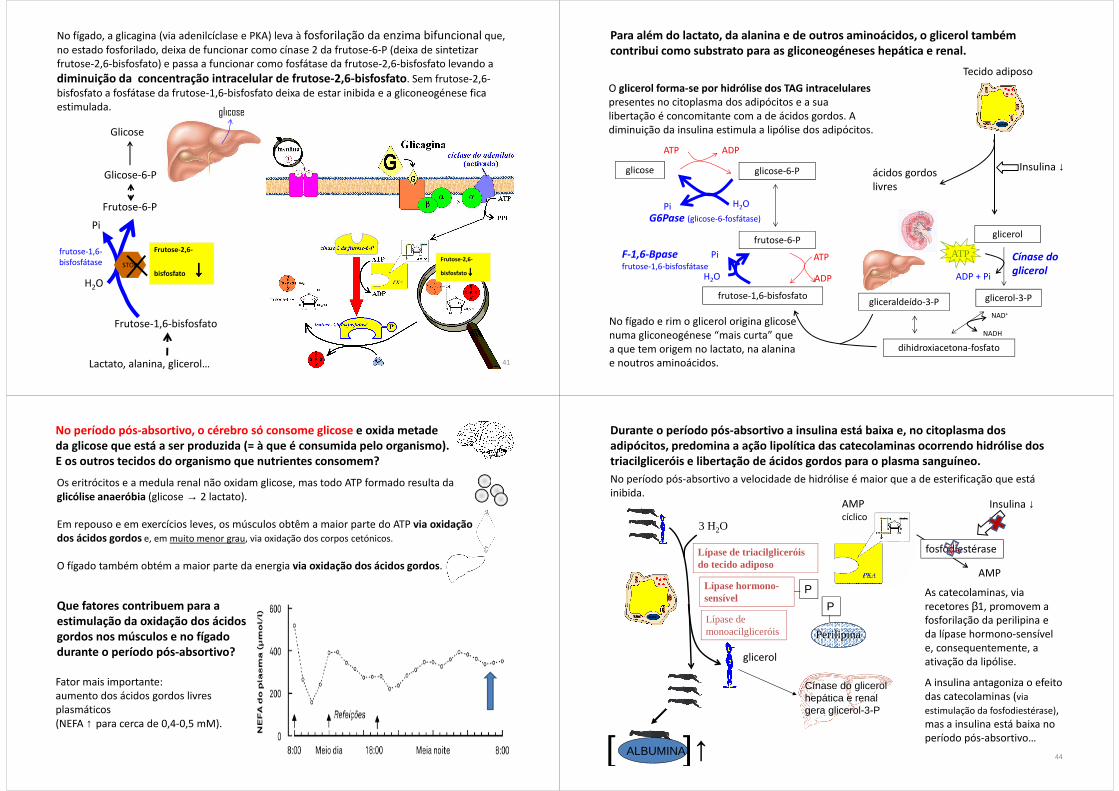
No fígado, a glicagina (via adenilcíclase e PKA) leva à fosforilação da enzima bifuncional que, no estado fosforilado, deixa de funcionar como cínase 2 da frutose-6-P (deixa de sintetizar frutose-2,6-bisfosfato) e passa a funcionar como fosfátase da frutose-2,6-bisfosfato levando a
diminuição da concentração intracelular de frutose-2,6-bisfosfato. Sem frutose-2,6-bisfosfato a fosfátase da frutose-1,6-bisfosfato deixa de estar inibida e a gliconeogénese fica estimulada.
Glicose
frutose-1,6-bisfosfátase
41
Para além do lactato, da alanina e de outros aminoácidos, o glicerol também contribui como substrato para as gliconeogéneses hepática e renal.
O glicerol forma-se por hidrólise dos TAG intracelulares presentes no citoplasma dos adipócitos e a sua libertação é concomitante com a de ácidos gordos. A diminuição da insulina estimula a lipólise dos adipócitos.
frutose-1,6-bisfosfato
frutose-6-P
H2O
Pi
glicose-6-Pglicose
ATP ADP
Pi H2O
F-1,6-Bpase frutose-1,6-bisfosfátase
G6Pase (glicose-6-fosfátase)
glicerol
dihidroxiacetona-fosfato
glicerol-3-Pgliceraldeído-3-P
ADP + Pi
ATP Cínase do
glicerol
No fígado e rim o glicerol origina glicose numa gliconeogénese “mais curta” que a que tem origem no lactato, na alanina e noutros aminoácidos.
ATP
ADP
ácidos gordos livres
Tecido adiposo
Insulina ↓
NADH
NAD+
No período pós-absortivo, o cérebro só consome glicose e oxida metade da glicose que está a ser produzida (= à que é consumida pelo organismo). E os outros tecidos do organismo que nutrientes consomem?
Os eritrócitos e a medula renal não oxidam glicose, mas todo ATP formado resulta da glicólise anaeróbia (glicose → 2 lactato).
Em repouso e em exercícios leves, os músculos obtêm a maior parte do ATP via oxidação dos ácidos gordos e, em muito menor grau, via oxidação dos corpos cetónicos.
O fígado também obtém a maior parte da energia via oxidação dos ácidos gordos.
Que fatores contribuem para a estimulação da oxidação dos ácidos gordos nos músculos e no fígado durante o período pós-absortivo?
Fator mais importante:aumento dos ácidos gordos livres plasmáticos (NEFA ↑ para cerca de 0,4-0,5 mM).
43
Durante o período pós-absortivo a insulina está baixa e, no citoplasma dos adipócitos, predomina a ação lipolítica das catecolaminas ocorrendo hidrólise dos triacilgliceróis e libertação de ácidos gordos para o plasma sanguíneo.
As catecolaminas, via recetores β1, promovem a fosforilação da perilipina e da lípase hormono-sensível e, consequentemente, a ativação da lipólise.
A insulina antagoniza o efeito das catecolaminas (via
estimulação da fosfodiestérase), mas a insulina está baixa no período pós-absortivo…
No período pós-absortivo a velocidade de hidrólise é maior que a de esterificação que está inibida.
44
3 H2O
Cínase do glicerol hepática e renal gera glicerol-3-P
P
Lípase hormono-sensível
Perilipina
ALBUMINA[ ]↑
Lípase de triacilgliceróis do tecido adiposo
Lípase de monoacilgliceróis
P
AMP cíclico
glicerol
Insulina ↓
fosfodiestérase
AMP
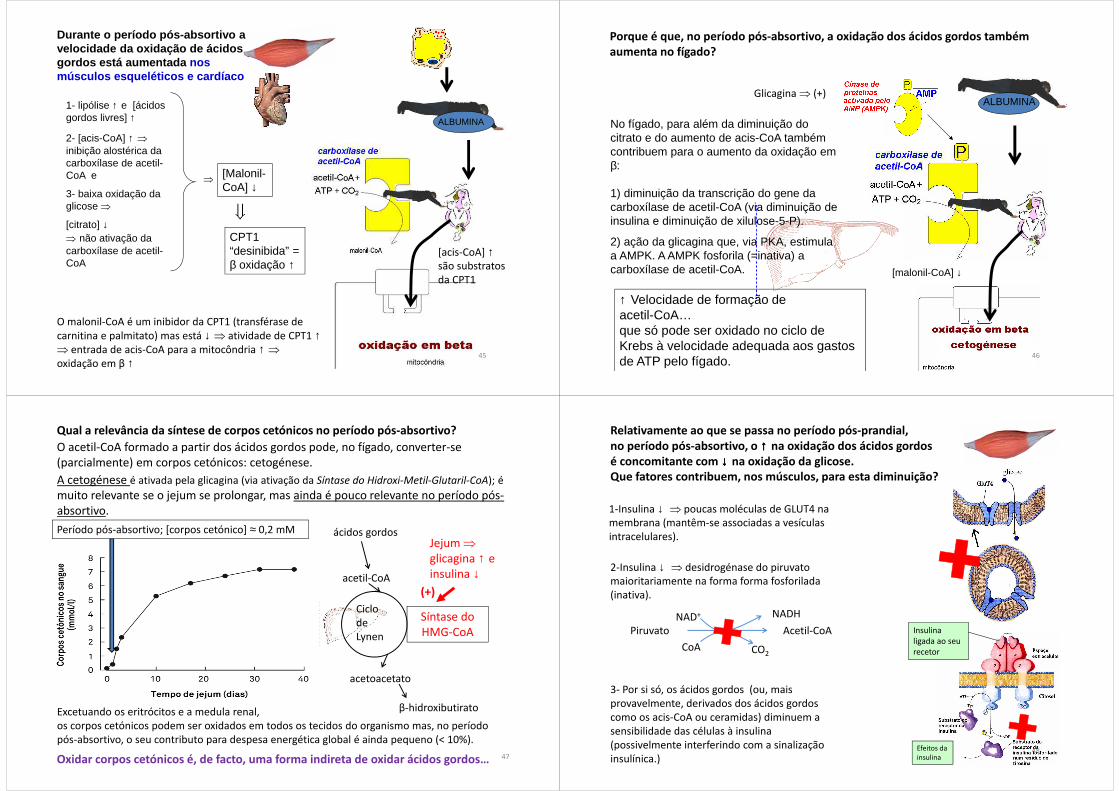
Durante o período pós-absortivo a velocidade da oxidação de ácidos gordos está aumentada nos músculos esqueléticos e cardíaco
ALBUMINA
1- lipólise ↑ e [ácidos gordos livres] ↑
2- [acis-CoA] ↑ ⇒
inibição alostérica da carboxílase de acetil-CoA e
3- baixa oxidação da glicose ⇒
[citrato] ↓⇒ não ativação da carboxílase de acetil-CoA
[Malonil-CoA] ↓
CPT1 “desinibida” = β oxidação ↑
⇒
O malonil-CoA é um inibidor da CPT1 (transférase de carnitina e palmitato) mas está ↓ ⇒ atividade de CPT1 ↑⇒ entrada de acis-CoA para a mitocôndria ↑ ⇒
oxidação em β ↑
[acis-CoA] ↑são substratos da CPT1
45
ALBUMINA
↑ Velocidade de formação de acetil-CoA…que só pode ser oxidado no ciclo de Krebs à velocidade adequada aos gastos de ATP pelo fígado.
No fígado, para além da diminuição do citrato e do aumento de acis-CoA também contribuem para o aumento da oxidação em β:
1) diminuição da transcrição do gene da carboxílase de acetil-CoA (via diminuição de insulina e diminuição de xilulose-5-P).
2) ação da glicagina que, via PKA, estimula a AMPK. A AMPK fosforila (=inativa) a carboxílase de acetil-CoA. [malonil-CoA] ↓
Glicagina ⇒ (+)
Porque é que, no período pós-absortivo, a oxidação dos ácidos gordos também aumenta no fígado?
46
Qual a relevância da síntese de corpos cetónicos no período pós-absortivo?
O acetil-CoA formado a partir dos ácidos gordos pode, no fígado, converter-se (parcialmente) em corpos cetónicos: cetogénese.
Síntase do HMG-CoA
Jejum ⇒glicagina ↑ e insulina ↓
(+)
A cetogénese é ativada pela glicagina (via ativação da Síntase do Hidroxi-Metil-Glutaril-CoA); é
muito relevante se o jejum se prolongar, mas ainda é pouco relevante no período pós-absortivo.
Excetuando os eritrócitos e a medula renal, os corpos cetónicos podem ser oxidados em todos os tecidos do organismo mas, no período pós-absortivo, o seu contributo para despesa energética global é ainda pequeno (< 10%).
47
Período pós-absortivo; [corpos cetónico] ≈ 0,2 mM ácidos gordos
acetil-CoA
Ciclo de Lynen
acetoacetato
β-hidroxibutirato
Oxidar corpos cetónicos é, de facto, uma forma indireta de oxidar ácidos gordos…
Relativamente ao que se passa no período pós-prandial, no período pós-absortivo, o ↑↑↑↑ na oxidação dos ácidos gordos é concomitante com ↓↓↓↓ na oxidação da glicose. Que fatores contribuem, nos músculos, para esta diminuição?
1-Insulina ↓ ⇒ poucas moléculas de GLUT4 na membrana (mantêm-se associadas a vesículas intracelulares).
2-Insulina ↓ ⇒ desidrogénase do piruvato maioritariamente na forma forma fosforilada (inativa).
3- Por si só, os ácidos gordos (ou, mais provavelmente, derivados dos ácidos gordos como os acis-CoA ou ceramidas) diminuem a sensibilidade das células à insulina (possivelmente interferindo com a sinalização insulínica.)
Efeitos da insulina
Insulina ligada ao seu recetor
Piruvato Acetil-CoANAD+ NADH
CoA CO2
48

49
Glicose-6-P
frutose-1,6-bisP
fosfoenolpiruvato
+[Frutose-2,6-bisfosfato] ↓
No fígado, no período pós-absortivo, o balanço glicólise/gliconeogénese favorece a gliconeogénese porque a glicólise se anula. Que fatores contribuem para que a oxidação de glicose seja, no fígado, praticamente nula.
piruvato
acetil-CoA
A glicagina via fosforilação da enzima bifuncional faz com que esta enzima passe a funcionar como frutose-2,6-bisfosfatáse.
O efeito ativador da insulina na desidrogénase do piruvato deixa de existir.
frutose-6-P
Glicose
O efeito ativador da insulina via indução da expressão de genes deixa de existir
+
Hexocínase IV
Cínase do piruvato
Cínase 1 da
frutose-6-P
Desidrogénase
do piruvato
A glicagina via fosforilação pela PKA da cínase do piruvato hepática inativa-a.
STOPP
O efeito ativador da glicose na hexocínase IV deixa de existir
No período pós-absortivo, o cérebro, que não é sensível nem à insulina nem à glicagina, continua a oxidar glicose a uma velocidade semelhante à do período pós-prandial ( ≈≈≈≈ 4 g/h). 50
O Quociente Respiratório varia com o tipo de nutriente que está a ser oxidado.
QR = CO2 / O2
O QR é 1 quando se oxidam glicídeos e 0,7 quando se oxidam lipídeos. O QR das proteínas tem, em média , um valor intermédio ≈0,83.
O Quociente Respiratório (QR) ou Respiratory Exchange Ratio = Razão moles ou volume CO 2 excretado
moles ou volume de O 2 consumido.
glicose (C6H12O6) + 6 O2 →→→→6 CO2 + 6 H2O
6/6 = 1
palmitato (C16H32O2) + 23 O2 →→→→16 CO2 + 16 H2O
16/23= 0,7
Proteína “padrão“( C100H159N26O32S0,7) + 104 O2 →→→→86,6 CO2 + 50,6 H2O + outros produros
86,6/104= 0,83
O QR é 1 se, num dado momento, o único nutriente a ser oxidado é a glicose (ou/e glicogénio).
O QR seria 0,7 se, num dado momento, os únicos nutrientes a serem oxidados fossem lipídeos.
Durante o período pós-prandial a insulina está alta . (1) oxidação da glicose ↑ (pode ser >85% da despesa energética total)(2) oxidação dos ácidos gordos ↓↓↓↓ ou nula .
⇒⇒⇒⇒ Quociente Respiratório geralmente entre 1 e 0,95
⇒⇒⇒⇒ QR ≈≈≈≈ 0,80 a 0,85
Se for assim: % kcalProteínas 10% 10%
Ácidos gordos 60% 45%
Carbohidratos 30% 45%
Quociente Resp = 0,80 0,85
Se for assim: % kcalProteínas 15%
Ácidos gordos 0%
Carbohidratos 85%
Quociente Resp = 0,97
No período pós-absortivoa insulina está baixa. (1) oxidação da glicose ↓; (2) oxidação dos ácidos gordos ↑.
Quer no período pós-absortivo, quer após as refeições o combustível do cérebro continua a ser a glicose mas, noutros órgãos, passam a ser os ácidos gordos (direta e indiretamente, via oxidação de corpos cetónicos) ⇒⇒⇒⇒o QR é mais baixo no período pós-absortivo que no período pós-prandial. 52
No jejum prolongado (mais de 3 dias) o metabolismo distingue-se
qualitativamente do metabolismo no período pós-absortivo em 2 aspetos:
1- O glicogénio hepático esgota-se mas o cérebro continua a usar glicose como combustível.2- Cerca de 1/3 da energia consumida pelo cérebro provém da glicose e cerca de 2/3 passa a depender dos corpos cetónicos.
De onde provém a glicose oxidada pelo cérebro a CO2 (cerca de 40 g/ dia; 1,7 g/h)?
1- Os aminoácidos que resultam da degradação das proteínas endógenas e o glicerol que resulta da lipólise são convertidos pelo fígado e rim em glicose via gliconeogénese de novo.
Glicose 40 g/dia (1/3 da energia consumida pelo cérebro)
CO2
Os ciclos de Cori e da alanina continuam a contribuir para mais de metade da gliconeogénese total que continua a ser ≈ 4g/h. No entanto, os ciclos de Cori e da alanina não podem ser considerados contribuintes para a glicose oxidada no cérebro porque representam apenas reciclagem da glicose.
AAs
Glicogénio esgotado
Proteínas endógenas
2/3 da energia = corpos cetónicos
glicerol
53
2- No jejum prolongado há uma perda contínua de massa muscular, mas a velocidade da proteólise líquida vai diminuindo à medida que o jejum se prolonga ⇒ Excreção diminuída de azoto na urina
Uma das causas dessa diminuição é a diminuição da secreção de TSH (hormona estimulante da tiroide) ⇒diminuição de secreção de T3 (hormona tiroideia) ⇒diminuição do turnover proteico e diminuição da velocidade de perda de proteínas endógenas.
No jejum prolongado a única fonte de glicose é a gliconeogénese. O glicerol dos triacilgliceróis e os aminoácidos das proteínas endógenas fornecem substrato para a síntese de glicose.
1 g de TAG (glicerol)→ ≈ 0,1 g de glicose; 1 g de aminoácidos → ≈ 0,6 g de glicose.
100 g de TAG x 0,1 + 50 g de proteínas x 0,6 = 10 g do glicerol + 30 g dos Aas = 40 g/dia de glicose produzida via gliconeogénese de novo (≈≈≈≈ 40% da gliconeogénese total) 54
Insulina↓ ⇒
Lipólise ↑ ⇒
concentração de ácidos gordos livres plasmáticos ↑⇒ ↑oxidação hepática de ácidos gordos (mas
incompleta; só oxidação em β sem oxidação do acetil-CoA formado a CO2)
No jejum prolongado só o cérebro continua a oxidar glicose a CO2, mas esta glicose deriva de Aas e de glicerol: é uma forma indireta de oxidar Aas e glicerol.
Os músculos passam a oxidar apenas ácidos gordos e corpos cetónicos.… mas oxidar corpos cetónicos é uma forma indireta de oxidar ácidos gordos. 54
Os corpos cetónicos passam a representar cerca de 2/3 da energia gasta pelo cérebro. De onde provêm estes corpos cetónicos? [NEFA ou FFA] ≈ 1,5 mM
(2-3 xs superiores aos valores do período pós-absortivo).
[corpos cetónicos] ≈ 7 mM (30xs superiores aos valores do período pós-absortivo).
55
Lúmen tubular renal Célula tubular renal Plasma sanguíneo
glutaminaglutamina
glutamato
α−cetoglutarato
NH4+
NH3
H+
Na+ Na+
H+
NH3
NH4+
C5H10O3N2 + 4,5 O2 + 2H+ → 2 NH4+ + 5 CO2 + 2 H2O (predomina na acidose)
C5H10O3N2 + 4,5 O2 → CON2H4 + 4 CO2 + 3 H2O (normalmente)
Em situações de acidose (como o jejum prolongado) há aumentoda atividade da glutamínase renal. A glutamina é captada doplasma e leva à formação de amónio (que é excretado) e a α-cetoglutarato que, maioritariamente se converte em glicose (queacaba por ser oxidada).
NAD+
NADH
glicose
H2O
glicose
O catabolismo da glutamina com formação de amónio contribui para o consumo de protões do meio interno. O amónio formado e excretado contém o azoto que fazia parte da glutamina (e do glutamato formado a partir da glutamina), mas também contém protões.
56
Se se está em jejum prolongado o glicogénio hepático e muscular
desapareceu ao fim de poucos dias e oxidamos as nossas gorduras e proteínas e o Quociente Respiratório atinge valores próximos de 0,7.
Admitamos que no dia 10-20 de jejum
∆ gordura =
100 g / dia
∆ proteína =
50 g / dia
BMR = 100 x 9,3 + 50 x 4,2 = 1165 kcal/ dia
Qual o QR?Nutriente
(g)CO2
(L)O2
(L) QR
Proteínas 50 43 52
Gordura 100 143 203
Total 186 255 0,73
Energia consumida pelo organismo no jejum prolongado: ≈ 20% proteínas endógenas e≈ 80% gordura

Baynes JW, Dominiczak MH.: Medical Biochemistry. 3rd. Ed. Elsevier Mosby. New York. 2009.Berg J.M., Tymoczko J.L. and Stryer L.: Biochemistry. 7th. Ed. International Edition. W.H. Freeman and Company. New York. 2012.Frayn K.N.: Regulação Metabólica. Uma Perspetiva Focada no Organismo Humano. U.Porto editorial. Porto. 2012.Gerich, J. E. (2010) Role of the kidney in normal glucose homeostasis and in the hyperglycaemia of diabetes mellitus: therapeutic implications, Diabet Med. 27, 136-42.Kiens, B. (2006) Skeletal muscle lipid metabolism in exercise and insulin resistance, Physiol Rev. 86, 205-43.McGilvery, R. W. & Goldstein, G. W. (1983) Biochemistry, a functional approach, W. B. Saunders Company, Philadelphia.Murray R.K., Granner D.K., Mayes P.A. and Rodwell V.W.: Harpers Illustrated Biochemistry 29th Edition. McGraw-Hill Medical. London. 2012.Nelson D.L., Cox M.M.: Lehningher Principles of Biochemistry. 5th Ed. W.H. Freeman and Company, New York. 2008.Newsholme, E. A. & Leech, T. (2009) Functional Biochemistry in Health and disease, Wiley-Blackwell, Oxford.Nuttall, F. Q., Ngo, A. & Gannon, M. C. (2008) Regulation of hepatic glucose production and the role of gluconeogenesis in humans: is the rate of gluconeogenesis constant?, Diabetes Metab Res Rev. 24, 438-58.Ooiwa, T., Goto, H., Tsukamoto, Y., Hayakawa, T., Sugiyama, S., Fujitsuka, N. & Shimomura, Y. (1995) Regulation of valinecatabolism in canine tissues: tissue distributions of branched-chain aminotransferase and 2-oxo acid dehydrogenase complex, methacrylyl-CoA hydratase and 3-hydroxyisobutyryl-CoA hydrolase, Biochim Biophys Acta. 1243, 216-20.Radziuk, J. & Pye, S. (2001) Hepatic glucose uptake, gluconeogenesis and the regulation of glycogen synthesis, Diabetes Metab Res Rev. 17, 250-72.Stipanuk M.H.: Biochemical, Physiological & Molecular Aspects of Human Nutrition. 3rd Ed. Elsevier Science. St. Louis. 2013.Taylor, R., Magnusson, I., Rothman, D. L., Cline, G. W., Caumo, A., Cobelli, C. & Shulman, G. I. (1996) Direct assessment ofliver glycogen storage by 13C nuclear magnetic resonance spectroscopy and regulation of glucose homeostasis after a mixed meal in normal subjects, J Clin Invest. 97, 126-32.Wallis, G. A., Hulston, C. J., Mann, C. H., Roper, H. P., Tipton, K. D. & Jeukendrup, A. E. (2008) Postexercise muscleglycogen synthesis with combined glucose and fructose ingestion, Med Sci Sports Exerc. 40, 1789-94.
Bibliografia
57


















