
2012 1º Vol Morfologia e função
125
Carlos Aguiar Botânica para Ciências Agrárias e do Ambiente Volume I • Morfologia e função Instituto Politécnico de Bragança 2012
-
Upload
alysson-vitor -
Category
Documents
-
view
290 -
download
17
Transcript of 2012 1º Vol Morfologia e função

Carlos Aguiar
Botânica para Ciências Agrárias e do Ambiente
Volume I • Morfologia e função
Instituto Politécnico de Bragança
2012

Publicado pelo Instituto Politécnico de Bragança
Imagem da capa: Antirrhinum lopesianum Rothm. (Plantaginaceae).
Versão de 18-IV-2012
© Carlos Aguiar
ISBN 978-972-745-123-4

1 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
Crescem as flores no seu dever
biológico,
e as cores que patenteiam, por sua
natureza,
só podem ser aquelas, e não outras.
Vermelhas, amarelas, cor de fogo,
lilazes, carmesins, azuis, violetas,
assim, e só assim,
tudo conforme a sua natureza.
Ásperas são as folhas, macias,
recortadas
ou não, tudo conforme;
e o aprumo como tal,
ou rasteiras, ou leves, ou pesadas,
tudo no seu dever,
por sua natureza.
[…]
«Poema da Minha Natureza» in Novos
Poemas Póstumos, António Gedeão

2 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente

3 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
Índice
Prólogo aos quatro volumes ......................................................................................... 5
1. Conceito de planta .............................................................................................. 11
2. O corpo das plantas-com-semente .................................................................... 14
3. Sistema vegetativo ............................................................................................. 23
3.1. Aspectos morfológicos comuns a todo o corpo vegetativo ......................... 23
3.2. Raiz ............................................................................................................... 27
3.2.1. Funções da raiz ..................................................................................... 27
3.2.2. Alongamento e ramificação da raiz ...................................................... 28
3.2.3. Aspectos gerais da morfologia externa da raiz ..................................... 29
3.2.4. Metamorfoses da raíz ........................................................................... 31
3.2.5. Modificações causadas por microrganismos ........................................ 33
3.2.6. As raízes das árvores ............................................................................. 35
3.3. Caule ............................................................................................................. 35
3.3.1. Funções do caule .................................................................................. 36
3.3.2. Alongamento, ramificação e cladoptose .............................................. 36
3.3.3. Aspectos gerais da morfologia externa do caule .................................. 45
3.3.4. Metamorfoses do caule ........................................................................ 48
3.3.5. Lenho de reacção .................................................................................. 49
3.3.6. Produção de látex ................................................................................. 49
3.4. Folha ............................................................................................................. 49
3.4.1. Funções da folha ................................................................................... 50
3.4.2. Os filomas .............................................................................................. 50
3.4.3. Aspectos gerais da morfologia externa da folha .................................. 52
3.4.4. Metamorfoses da folha ......................................................................... 67
3.5. O corpo das gramíneas ................................................................................. 70
4. Sistema reprodutivo ........................................................................................... 71
4.1. Gimnospérmicas ........................................................................................... 71
4.1.1. Estruturas reprodutivas ........................................................................ 71
4.1.2. Frutificações e sementes ...................................................................... 73

4 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
4.2. Angiospérmicas ............................................................................................ 74
4.2.1. Inflorescência ........................................................................................ 74
4.2.1. Flor ........................................................................................................ 80
4.2.1.1. Definição de flor. Iniciação floral. Ciclo floral ................................. 80
4.2.1.2. Constituição da flor e expressão sexual ......................................... 81
4.2.1.3. Filotaxia e simetria da flor .............................................................. 82
4.2.1.4. Receptáculo .................................................................................... 83
4.2.1.5. Perianto ........................................................................................... 83
4.2.1.6. Hipanto............................................................................................ 89
4.2.1.7. Androceu ......................................................................................... 89
4.2.1.8. Pólen ............................................................................................... 91
4.2.1.9. Gineceu ........................................................................................... 93
4.2.1.10. Nectários florais e osmóforos ....................................................... 97
4.2.1.11. Primórdios seminais ...................................................................... 97
4.2.1.12. Fórmulas florais............................................................................. 98
4.2.2. Fruto .................................................................................................... 100
4.2.3. Semente .............................................................................................. 109
4.2.4. Estruturas reprodutivas das gramíneas .............................................. 111
5. Fisionomia e fenologia das plantas-com-semente ........................................... 114
5.1. Tipos fisionómicos ...................................................................................... 114
5.2. Posição das inflorescências nos ramos de plantas lenhosas ..................... 116
5.3. Ciclos fenológicos ....................................................................................... 117
6. Referências ....................................................................................................... 120

5 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
Prólogo aos quatro volumes
Este documento não é um livro texto de referência e muito menos um tratado. Trata-se de uma
revisão bibliográfica actualizada em torno de temas chave de organografia vegetal1, de botânica
sistemática de plantas-com-semente e de botânica económica, complementada pela experiência de 20
anos de ensino e investigação do seu autor. Tem por destinatários os estudantes do ensino superior que
necessitam, num curto período de tempo, de apreender os conceitos e os vocábulos fundamentais de
organografia vegetal, percepcionar a diversidade morfológicas e taxonómica das plantas-com-semente e
de dominar os grupos mais importantes de plantas com interesse económico. Por isso, a organização do
texto, a linguagem, e, muitas vezes, a estrutura das frases nem sempre será fácil de perseguir. De início
a sua leitura será trabalhosa, quando não cansativa, porém, a persistência e a perseverança abrirão as
portas a um conjunto de conhecimentos essenciais para todos aqueles que têm a plantas como objecto
de trabalho.
Certamente persistem alguns erros, omissões e gralhas: contamos com todos os utilizadores para os
corrigir. Para tal está à disposição dos utilizadores deste texto o seguinte endereço de e-mail:
Objecto
A forma externa, a biologia da reprodução e a organização sistemática, como produtos de um
processo evolutivo, e o uso dos taxa de plantas-com-semente são os principais objectos deste livro. O
seu estudo pode ter diferentes pendores. Por exemplo, pode ser dirigido aos mecanismos evolutivos
que subjazem a génese dos taxa, ter uma abordagem descritiva-formal, uma perspectiva histórico-
evolutiva ou insistir em aspectos metodológicos. Pretendeu-se acima de tudo conferir competências a
futuros profissionais de “biologia aplicada”. Com este objectivo em mente foi dada uma ênfase especial
aos seguintes temas:
a) Descrição dos caracteres morfológicos internos e externos de maior interesse taxonómico;
b) Relações morfologia-função;
c) Biologia da reprodução das plantas-com-semente;
d) Sistemática e filogenética dos grandes grupos de plantas-com-semente;
e) Tipologia, origem e evolução das plantas cultivadas;
f) Taxonomia, características morfológicas diagnóstico e distribuição das plantas úteis de maior
interesse económico.
A segunda parte do II volume, depois de uma pequena introdução à biologia da evolução, inclui uma
história evolutiva das plantas-terrestres. Pode parecer estranho que um tema tão especializado e volátil
como este seja desenvolvido num livro de botânica que se pretende aplicada. As plantas, ao longo da
sua evolução, foram, umas vezes agentes de mudança, outras sujeitos passivos nas alterações climáticas
e na dinâmica da composição química da atmosfera terrestre. Sem noções de evolução das plantas é
impossível entender estes dois temas chave das ciências do ambiente (vd. Beerling, 2007). Depois, como
escrevia em 1973 o evolucionista norte-americano de origem ucraniana Theodosius Dobzhansky, "Nada
em biologia faz sentido excepto à luz da evolução”.
A sistemática molecular ganhou, nas últimas décadas, uma importância acrescida. Os caracteres
moleculares são tão ou mais valorizados do que os caracteres morfológicos no estabelecimento de
filogenias, na exploração de padrões filogeográficos ou na caracterização de taxa. Porém, tendo em
atenção o perfil e as necessidades de informação botânica dos destinatários deste documento, não
1 A organografia vegetal ou fitomorfologia tem por objecto a morfologia externa das plantas (= estrutura externa). A anatomia vegetal dedica-se à morfologia interna (= estrutura interna). Neste texto consideram-se redundantes os conceitos de estrutura, morfologia e forma.

6 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
serão abordados no texto. De qualquer modo importa ter presente que o fenótipo é um produto directo
da expressão génica. Por outro lado, a exploração das relações genoma-fenótipo exige uma perfeita
compreensão da estrutura das plantas e o uso de termos precisos. Deste modo, o estudo da morfologia
externa e interna e da biologia da reprodução das plantas antecede, necessariamente, a “descida” à
fisiologia e ao gene. A discussão da indispensabilidade do estudo da morfologia externa na botânica
moderna é retomada no Volume III, aquando da apresentação dos sistemas de classificação cladística.
Breve reflexão epistemológica
Na organização dos seres vivos reconhecem-se vários níveis de complexidade (vd. figura). As células,
as unidades elementares da vida, organizam-se em tecidos, os tecidos em órgãos e sistemas, e estes,
por sua vez, integram os organismos. Os indivíduos ocupam nichos ecológicos e trocam informação
genética entre si no âmbito de uma população. A componente viva dos ecossistemas, a biocenose,
compreende indivíduos de diferentes espécies. Finalmente, os ecossistemas organizam-se em sistemas
ecológicos de complexidade variável (e.g. comunidade vegetal, série de vegetação, geossérie e bioma).
Os diferentes níveis de complexidade interactuam entre si, de forma tanto mais intensa quanto mais
próximos estiverem na escala de complexidade, e ajustam-se às flutuações e variações direccionais do
ambiente abiótico e biótico, através de rearranjos na sua estrutura (e.g. composição florística de uma
comunidade vegetal ou plasticidade fenotípica dos indivíduos) e pela acção da selecção natural à escala
do indivíduo. A biologia e a ecologia, à semelhança de outras ciências fundamentais, procuram explicar
e prever a estrutura e função de cada nível de complexidade, em função dos imediatamente anteriores.
Constata-se, porém, que “em cada salto de complexidade” este esforço esbarra na emergência de novas
propriedades, não previstas nos níveis de complexidade inferiores. Por exemplo, a estrutura do genoma
é insuficiente para uma compreensão total do funcionamento celular, ou a autoecologia das espécies é
insuficiente para explicar e prever o funcionamento de um ecossistema. Os epistemólogos – os
especialistas em filosofia da ciência – repartem-se em dois grupos para explicar a emergência de novas
propriedades. Muitos são de opinião que a escassez de conhecimento está na origem das limitações do
reducionismo2 e, por conseguinte, as propriedades ditas emergentes são um artefacto das limitações
epistémicas da mente humana. Os holistas, pelo contrário, admitem que a emergência de propriedades
é uma característica constitutiva dos sistemas complexos.
Qualquer uma destas hipóteses não impede que actividades humanas tão complexas como a
agronomia, a silvicultura ou a restauração ecológica, todas elas parte de uma grande disciplina que
poderíamos denominar por biologia aplicada, se possam aproveitar das abordagens reducionistas-
mecanicistas próprias da ciência moderna. Sob esta perspectiva, uma botânica dirigida à sistematização
das formas, dos sistemas de reprodução e da diversidade do mundo vegetal, e ao uso humano das
plantas, conforme se segue neste texto, oferece informação indispensável para a prática da biologia
aplicada, impossível de obter noutros domínios da biologia. Isto é, o acervo de informação e métodos da
botânica sistemática, da biologia de reprodução de plantas e da botânica económica são insubstituíveis
2 O reducionismo é uma doutrina filosófica que sustenta que a fragmentação em partes da realidade é necessária, e suficiente, para explicar o todo. Portanto, para os reducionistas, sistemas complexos como os seres vivos ou os ecossistemas não são mais do que a soma das suas partes. A ciência é intrinsecamente reducionista. Na prática da ciência a realidade é decomposta em níveis de complexidade. Cada um destes níveis, por seu lado, é explorado per se, procurando-se estabelecer as conexões causais entre diferentes níveis de complexidade.
Níveis de complexidade do vivo

7 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
mas não suficientes para o agrónomo, o silvicultor e ao especialista em restauração ecológica exercerem
a sua actividade de forma eficaz e eficiente.
O conhecimento científico resulta da observação meticulosa da realidade através da concorrência de
instrumentos conceptuais e observacionais dos mais diversos tipos. Os instrumentos conceptuais (e.g.
conceitos, teorias, hipóteses e modelos), formalizados matematicamente ou não, categorizam e
organizam a realidade, i.e. atribuem-lhe uma estrutura por nós percepcionável. Os instrumentos
observacionais (e.g. microscópio e sondas de DNA) permitem ultrapassar as limitações físicas dos nossos
sentidos. Todo o conhecimento científico é, por definição, não dogmático porque está sujeito a um
escrutínio permanente por parte da comunidade científica e dos utilizadores da ciência (e.g.
agricultores, silvicultores e conservacionistas). Os mecanismos de revisão e de aperfeiçoamento do
conhecimento atribuem à ciência uma enorme capacidade de explicar, de prever o funcionamento e de
actuar na realidade. Por outras palavras, o conhecimento científico está particularmente adequado para
a solução de problemas. No entanto, o conhecimento científico deve ser aceite sempre de forma
provisória porque não pode ser comprovado. Não existem verdades absolutas em ciência, existem sim
conceitos mais ou menos consistentes, e teorias e hipóteses mais ou menos corroboradas. O
conhecimento científico não é definitivo e não deve (pode) ser tomado como tal.
As relações forma-função nas plantas são um bom exemplo da humildade que deve caracterizar a
atitude científica. A função de muitas formas, internas e externas, das plantas é autoevidente: as
gavinhas servem para ancorar as plantas aos suportes e o néctar das flores é uma recompensa para os
polinizadores. No entanto, muitas formas actuais foram evolutivamente adquiridas num passado
remoto, e no presente não desempenham qualquer função ou têm uma função distinta da sua função
inicial. As formas podem, ainda, nunca ter desempenhado uma função e a sua retenção ser uma obra do
acaso. Os raciocínios de tipo adaptativo, que relacionam a forma com o desempenho de uma dada
função, exigem uma ampla base indutiva observacional ou, de preferência, corroboração experimental.
As proposições teleológicas do género “as plantas desenvolveram espinhos para evitar a herbivoria”
simplesmente devem ser evitadas. A especulação em torno das relações forma-função envolve, por
conseguinte, elevados níveis de incerteza. Propõe-se, por isso, que o utilizador deste texto tenha uma
atitude crítica em relação aos temas mais adiante discutidos.
A observação empírica da forma das plantas faculta a detecção de padrões, por exemplo, na posição
relativa dos órgãos vegetais, na forma das folhas ou na estrutura da flor e do fruto. A conceptualização e
a inventariação das regularidades na forma das plantas são uma das tarefas mais antigas e importantes
da botânica. Os conceitos são representações mentais, que no âmbito da organografia vegetal resumem
as propriedades de um objecto natural: o corpo das plantas. No capítulo dedicado à botânica
sistemática os objectos naturais conceptualizados serão os taxa.
Uma correspondência biunívoca inequívoca entre os conceitos, representados por vocábulos ou
símbolos (e.g. fórmulas florais), e os objectos ou ideias conceptualizados melhora a qualidade e acelera
as trocas da informação, por exemplo, entre professores e alunos, ou entre os praticantes de uma
ciência. Quanto maior a precisão e o detalhe de um corpo conceptual, maior o seu valor heurístico, i.e.
maior a sua utilidade para gerar hipóteses e mais longe se pode chegar na compreensão do objecto de
estudo. Logo, no estudo científico da forma das plantas, da biologia da reprodução ou na sistemática de
plantas, a observação e a construção de hipóteses – sustentadas na grande teoria unificadora da
biologia que é a teoria da evolução – devem caminhar lado a lado com a construção de um corpo
consistente de conceitos.
No que à organografia vegetal diz respeito, pese embora uma história de quase três séculos de
observação e descrição atenta da forma das plantas, falta ainda percorrer um longo caminho em busca
da universalidade e consistência terminológico-conceptual. Como referem Voght et al. (2009) a
descrição da morfologia dos entes viventes continua dificultada pela falta: (i) de uma terminologia

8 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
estandardizada de uso comum; (ii) de um método comum estandardizado de descrição morfológica; (iii)
e um conjunto de princípios a aplicar na delimitação de caracteres morfológicos.
A botânica sistemática desenvolve-se na confluência de um conjunto alargado ciências fundamentais
– evolução, histologia, ecologia, etc. Esta disciplina oferece um conjunto de princípios, métodos e
informação descritiva que permite apreender a diversidade vegetal que nos rodeia de uma forma rápida
e estruturada. Uma vez que a partilha de caracteres entre as plantas, excluindo os casos de
convergência evolutiva, se deve à partilha de antepassados comuns, a sistemática vegetal, por seu
turno, permite que os seus utilizadores aperfeiçoem as suas capacidades inatas de antecipar a
organização do mundo vivo à escala da sua percepção visual. A botânica sistemática é uma ciência
moderna e de vanguarda. No passado reduzia-se à prática da classificação biológica das plantas. Hoje,
por exemplo, é fundamental em ecologia – em ecologia as biocenoses são geralmente segmentadas ao
nível da espécie ou da família –, em paleoclimatologia e na testagem de hipóteses de biogeografia e de
biologia da evolução. A botânica económica tem por objectivo principal a sistematização dos usos das
plantas. A botânica conforme é abordada neste texto, é uma disciplina de charneira entre a evolução, a
biogeografia, a etnobiologia e a agronomia.
Fontes de informação
Este documento divide-se em quatro volumes que correspondem a quatro grandes objectos da
botânica: morfologia externa, biologia reprodutiva, sistemática e botânica económica. Num capítulo
menor do segundo volume foi feita uma pequena introdução à biologia da evolução das plantas.
Morfologia e relações morfologia-função de plantas-com-semente
As Noções de Morfologia Externa de Plantas Superiores do Prof. João de Carvalho e Vasconcellos
(1968) fixaram a terminologia botânica de uso corrente em Portugal, e não se antevê, para breve, a sua
substituição por outras publicações mais actualizadas. Ainda assim, no estudo da morfologia externa das
plantas são também indispensáveis o Diccionario de Botánica de Pio Font Quer (1985) e o Glossário de
Termos Botânicos de Prof. Rosette Battarda Fernandes (1972). Na preparação deste documento foram
também de grande importância os livros de Pérez Morales (1999), Judd et al. (2002), Vozzo (2002),
Keller (2004), Ingrouille & Eddie (2006), Bell (2008) e de Ronse de Craene (2010), e um grupo alargado
de artigos citados nas referências bibliográficas. Estes últimos trabalhos foram essenciais para explorar
as relações morfologia-função.
Biologia da reprodução de plantas-com-semente
O ensino da biologia da reprodução tem sido descurado no ensino superior agrário. No entanto, é
um tema indispensável para compreender a fisiologia da produtividade e no melhoramento de plantas.
Com mais de 100 anos de edições sucessivas, o Strasburger: Tratado de Botánica (Sitte et al., 2003)
continua a ser uma das fontes mais valiosas de informação sobre a biologia da reprodução de plantas. O
livro de texto de Díaz et al. (2004) contém uma descrição cuidadosa e precisa dos ciclos de vida das
plantas.
Estrutura das descrições das famílias e taxa superiores
Desde a publicação do Genera Plantarum de Antoine de Jussieu, no final do séc. XVIII, que a família é
a categoria taxonómica superior ao género mais utilizada na organização do mundo vegetal. A
generalização do uso de designações ao nível da família deveu-se, por um lado, à necessidade de reunir
a enorme diversidade das plantas num número mais pequeno de entidades taxonómicas (taxa) de fácil
memorização e, por outro, ao facto de muitas famílias serem de fácil reconhecimento com
conhecimentos rudimentares de morfologia externa de plantas. Apesar da atribuição da categoria
familiar a um determinado grupo taxonómico ser eminentemente arbitrária, o conhecimento da
diversidade das plantas organiza-se em torno da categoria da família (Stevens, 2001+).

9 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
A organização taxonómica das famílias e grandes grupos taxonómicos seguida neste livro funda-se
no APG III (Angiosperm Phylogeny Group, 2009). A descrição das famílias botânicas foi, em grande parte,
adaptada do Guia de Árvores e Arbustos de Portugal Continental de Bingre et al. (2007), que por sua vez
foi beber grande parte da informação a Heywood (1993), Stevens (2001+), Spichiger et al. (2004) e Judd
et al. (2007). Para facilitar a percepção da morfologia das famílias, de uma forma muito concisa, é
oferecida informação sobre o número de espécies à escala global ou presentes em Portugal continental
e um apontamento sobre as plantas de maior interesse económico. Os exemplos incluídos nos dois
primeiros volumes e nas descrições das famílias, salvo indicação em contrário, referem-se à flora de
Portugal (inc. arquipélagos atlânticos) ou a plantas cultivadas.
Para saber mais sobre as famílias das plantas-com-semente recomendam-se dois livros de
referência: W. Judd et al. (Plant Systematics. A Phylogenetic Approach. Sinauer. 2002) e V. H. Heywood
(Flowering Plants of the World. Oxford University Press. 1993). O site
http://www.mobot.org/MOBOT/research/APweb/welcome.html é indispensável para quem se quiser
manter actualizado com as descobertas taxonómicas mais recentes.
Botânica económica
Uma vez que os alunos de ciências agrárias e do ambiente são o público-alvo deste documento foi
atribuída uma grande importância ao uso das plantas. A bibliografia usada no capítulo de botânica
económica envolveu livros-texto de proveniências várias e um alargado conjunto de papers da área da
arquelogia e da história da agricultura.
Abreviaturas, siglas e expressões latinas
Az – arquipélago dos Açores
ca. – circa, aproximadamente
cf. – confer, conferir, ver
cv. – cultivar
e.g. – exempli gratia, por exemplo
fam. – família
gén. – género
i.e. – isto é
ing. – na lígua inglesa
ICNAFP – Código Internacional de
Nomenclatura Botânica (desde 1 de Janeiro de
2012 está em vigor o código de Melbourne)
ICNCP – Código Internacional de
Nomenclatura das Plantas Cultivadas (Brickell
et al., 2004)
inc. – inclui
Lu – Portugal continental
Ma – arquipélago da Madeira (inc. ilhas da
Madeira, Porto Santo e Desertas)
M.a. – milhões de anos antes do presente
n.b. – nota bene, prestar atenção
o.m.q. – o mesmo que
s.l. – sensu lato, num sentido alargado do
termo
s.str. – sensu stricto, num sentido estrito do
termo
sin. – sinónimo
sing. – singular
sp. – espécie não determinada
sp.pl. – várias espécies
subsp. – subespécie
vd. – vide, ver
Convenções
Nomes latinos específicos e genéticos – escritos em itálico, e.g. Celtis australis e Magnoliophyta;
refira-se que na literatura mais actual existe a tendência de italicizar apenas os nomes genéricos,
específicos e infraespecíficos.
Vernaculização de nomes latinos – à excepção dos nomes genéricos, específicos e infraespecíficos
admite-se que todos os nomes latinos podem ser vernaculizados. Os sufixos previstos pelo ICNAFP

10 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
foram aportuguesados no feminino e no plural: -phyta em “-fitas” (divisão ou filo), -phytina em “-fitinas”
(subdivisão ou subfilo), -opsida em “-ópsidas” (classe), -idae em “-idas” (subclasse), -ales em “-ales”
(ordem), -ineae em “-íneas” (subordem), -aceae em “-áceas” (família), -oideae em “-óideas”
(subfamília), -eae em “-eas” (tribo) e -inae em “-inas” (subtribo). Exemplos: equisetópsidas, rosales e
magnoliáceas. Nomes vulgares de uso corrente foram retidos depois de clarificado o seu significado; e.g.
fetos, musgos. Os nomes vernáculos de taxa foram tomados como substantivos comuns e escritos em
minúsculas; e.g. angiospérmicas, coníferas e asteráceas.
Tradução dos clados – nas publicações de filogenética vegetal em língua inglesa os clados
geralmente terminam em “ids”. Na sua tradução para português optou-se por substituir “ids” por “idas”;
e.g. rosids em rosidas e lamiids em lamiidas. O sufixo “ideas” usado por alguns autores de língua
portuguesa, e.g. rosidas ou lamiidas, não nos parecede adequado.
Sufixo “fita” – em harmonia com a literatura mais recente em inglês o sufixo “fita” foi usado de
forma indiscriminada para designar grandes grupos taxonómicos sem uma categoria taxonómica
definida; e.g. briófitas, cormófitas, traqueófitas pteridófitas e licófitas.
Redundâncias fonéticas – para evitar redundâncias fonéticas os nomes específicos, foram, por vezes,
abreviados; e.g. Q. robur (= Quercus robur).
Nomes vulgares – em minúsculas, hifenizados e entre aspas; e.g. «lódão-bastardo».
Nomes de cultivares – em acordo com o ICNCP, primeira letra maiúscula, hifenizados e entre aspas
simples ou, sem aspas, e então precedidos da abreviatura “cv.”; e.g. alface ‘Orelha-de-mula’ e macieira
cv. Starking.
Clados (grupos monofiléticos) – em minúsculas. As relações filogenéticas entre os grupos que
constituem os clados foram pontualmente expressas através de parêntesis rectos; e.g. clado fixador de
azoto das fabidas = Fabales [Rosales [Cucurbitales + Fagales]].
Grados (grupos parafiléticos) – em minúsculas e entre aspas simples; e.g. ‘carófitas’, ‘briófitas’ e
‘angiospérmicas basais’.
Imagens
As representações esquemáticas sem autoria são originais. As fotografias sem identificação autoral
provêm do acervo de fotografia botânica do autor.

11 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
1. Conceito de planta
Aristóteles (séc. IV a.C.) reconheceu dois grandes grupos de seres vivos, depois de Carlos Lineu (séc.
XVIII) categorizados ao nível do reino: reino Plantae e reino Metazoa (= Animalia). Durante a maior
parte do século XX os livros-texto de botânica, além das plantas-terrestres (= embriófitas), incluíram no
reino das plantas as algas, os fungos e, em alguns casos, as bactérias e outros procariotas. Os
procariotas assim como os protozoários, as algas e fungos haviam já sido segregados num reino
autónomo – reino Protista – pelo zoólogo alemão Ernst H. Haeckel, em 1866. Robert Whittaker (1969)
propôs, com êxito, um sistema alternativo de cinco reinos de seres vivos: Monera, Protista, Animalia,
Fungi e Plantae. Este conhecido sistema tem uma índole funcional e ecológica – R. Whittaker era um
ecólogo de vegetação – porque é independente de qualquer tipo de relação evolutiva, e se baseia no
nível de organização (procariotas e eucariotas, unicelulares ou multicelulares), no modo de nutrição
(autotrofia, ingestão e absorção) e no papel desempenhado pelos seres vivos nos ecossistemas naturais
(produtores, consumidores e decompositores). Os conhecimentos acumulados nas últimas décadas de
bioquímica, fisiologia, genética e biologia evolutiva, a par da progressiva aceitação do princípio da
monofilia – os taxa têm de incluir todos, e apenas, os descendentes de um ancestral comum –
implicaram uma restrição significativa do conceito de planta e a substituição do sistema de Whittaker
por sistemas de classificação filogenéticos.
As plantas são eucariotas (domínio Eukaryota), um dos três domínios (= super-reinos) da vida celular
– por definição excluindo vírus e priões – definidos por Carl Woese et al. (1990). Os eucariotas são
actualmente organizados em 5 ou 6 grandes grupos, consoante os autores. Parfrei et al. (2006), por
exemplo, reconhecem seis grandes grupos. Grosso modo quatro deles resultam da pulverização dos
antigos protistas, os Opistokonta reúnem fungos e animais e as plantas aglomeram algas-verdes e
vermelhas, glaucófitas (um pequeno grupo de algas de água doce) e plantas-terrestres. As relações
filogenéticas entre os eucariotas não estão, por enquanto, resolvidas e, por essa razão, na bibliografia, o
nome Plantae é usado para designar diferentes combinações de organismos fotossintéticos, algo que
dificulta seriamente o seu estudo a nível não graduado.
Os autores mais modernos reduzem o conceito de planta, concretizado no reino Plantae (=
Archaeplastida), a um agregado constituído pelas plantas-terrestres e por um número variável de
grupos de algas. Um ponto da situação actualizado deste conceito, e da sistemática dos grandes grupos
de eucariotas, pode ser apreciado na “Tree of Life”3. A designação Plantae é aplicada, por estes autores,
a um grande grupo monofilético constituído pelas algas-vermelhas (Rodophyta) + plantas-verdes (sub-
reino Chlorobionta4). As plantas-verdes, por sua vez, englobam ‘algas-verdes’ (Chlorophyta s.l.5) e
plantas-terrestres (Equisetopsida6). As ‘algas-verdes’ incluem um conjunto muito diverso de linhagens,
que não cabe aqui explorar, uma delas, as Charophyta7, é evolutivamente muito próxima das actuais
plantas-terrestres (vd. Volume II), constituindo a reunião de ambas um grupo monofilético, as
Streptophyta (= Charophyta s.l.8). A classe Equisetopsida (plantas-terrestres ou embriófitas) abrange as
‘briófitas’ (= hepáticas + antóceros + musgos), as ‘pteridófitas’ (= licopódios + fetos) e as plantas-com-
semente (= angiospérmicas + gimnospérmicas). As hepáticas são basais relativamente às restantes
plantas-terrestres, portanto, foram o primeiro grupo, entre as plantas-terrestres actuais, a diferenciar-se
e a colonizar a terra emersa, algures durante o período Ordovícico.
No quadro que se segue faz-se um enquadramento taxonómico dos principais grupos de plantas-
terrestres para facilitar a percepção das suas relações evolutivas e diversidade. Embora se tenha optado
3 http://tolweb.org/tree/ 4 Viridaeplantae, ou Viridiplantae, para outros autores. 5 As Chlorophyta s. str. contêm todas as algas-verdes menos as Charophyta. 6 Sub-reino Embryobionta ou classe Embryophyta (= Metaphyta) noutros sistemas de classificação. 7 Ou Charophyceae, classe das charofíceas. 8 As Charophyta s.str. não incluem todos os decendentes de um ancestral comum, são um grupo parafilético.

12 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
por uma categorização formal dos grandes grupos de seres vivos, e de plantas, é importante referir que
as categorias entre o domínio e a classe, inclusive, são, na prática, supérfluas, por duas razões:
a) Os grupos de seres vivos ao nível do domínio e do reino estão marcados por numerosos eventos
de endossimbiose (e.g. formação das mitocôndrias e dos cloroplastos, vd. Volume II) e,
implicitamente, por trocas genéticas em rede (evolução reticulada). Os sistemas hierárquicos de
classificação, como o regulado pelo CNB, são inadequados para lidar com casos de evolução
reticulada;
b) Os grandes grupos estão aninhados (ing. “nested”) num cladograma com um grande número de
ramos longos sucessivos, difícil de plasmar numa nomenclatura hierarquizada com um escasso
número de categorias (vd. Volumes II e III).
Por conseguinte, na bibliografia é frequente o uso indiscriminado do sufixo de divisão “phyta”. A
taxonomia das plantas vasculares é retomada no III Volume.
Categoria taxonómica Taxa Nome vulgar
Domínio Eukaryota Eucariotas
Reino Plantae Plantas
Subreino Chlorobionta Plantas-verdes
Divisão Streptophyta
Classe Equisetopsida Plantas-terrestres, embriófitos
Subclasse Marchantiidae Hepáticas
Subclasse Anthocerotidae Antóceros
Subclasse Bryidae Musgos
Subclasse Lycopodiidae Lycopodiidas, licófitas, licopódios
Subclasse Ophioglossidae Ophioglossidas, fetos-ophioglossidos
Subclasse Equisetidae Equisetidas, fetos-equisetidos, equisetófitas, equisetos
Subclasse Marattiidae Marattiidas, marattiófitas, fetos-marattiidos
Subclasse Polypodiidae Polypodiidos, polipodiófitas, fetos-verdadeiros, fetos-leptoesporangiados
Subclasse Ginkgoidae Ginkgoidas, ginkgófitas, ginkgos
Subclasse Cycadidae Cycadidas, cicadófitas, cicas
Subclasse Pinidae Pinidas, pinófitas, coníferas
Subclasse Gnetidae Gnetidas, gnetófitas
Subclasse Magnoliidae Angiospérmicas, magnoliidas, plantas-com-flor, magnoliófitas
Uma vez que este texto versa as plantas-com-semente, e as plantas-terrestres cormofíticas serão
ciclicamente recordadas, impõe-se, desde já, uma importante questão: o que é uma planta terrestre? As
plantas-terrestres (Equisetopsida) são um taxon monofilético, cujos elementos partilham um conjunto
alargados de características, maioritariamente devidas a uma ancestralidade comum. Estas
Quadro 1. Enquadramento taxonómico da classe Equisetopsida «plantas-terrestres» (baseado em Chase &
Reveal 2009, Christenhusz et al. 2011a, Christenhusz et al. 2011b e na “Tree of Life”)

13 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
características serão desenvolvidas nos capítulos que se seguem. Ingrouille & Eddie (2006) resumem-nas
do seguinte modo:
a) Eucariotas – organismos com um núcleo e outras estruturas celulares complexas encerradas por
membranas;
b) Multicelulares – indivíduos com mais de uma célula, com diferentes funções e interdependentes
entre si;
c) Sésseis – organismos fixos a um substrato durante grande parte do seu ciclo biológico, com fases
de dispersão breves, normalmente sob a forma de esporos ou de sementes;
d) Foto-autotróficos – produzem compostos orgânicos complexos e ricos em energia a partir de
moléculas inorgânicas simples (e.g. H2O, CO2, K+, e NO-3), e da energia (sob a forma de ATP) e do
poder redutor (sob a forma de NAPH2) gerados pelas moléculas de clorofila excitadas pela luz do
sol; secundariamente9 sem clorofila e heterotróficas (plantas parasitas e plantas saprófitas);
e) Que concentram compostos inorgânicos e carbono a partir de soluções gasosas (ar) ou aquosas
(regra geral água do solo) muito diluídas;
f) Com uma elevada relação superfície/ volume dos seus órgãos;
g) Com uma estrutura modular – vd. Estrutura modular das plantas;
h) Elevada plasticidade fenotípica – vd. Variação morfológica intraespecífica;
i) Crescimento indeterminado – vd. Organização do corpo das plantas-com-semente.
As plantas caracterizam-se ainda por possuírem:
a) Paredes celulares celulósicas;
b) Cutícula – camada cerosa de protecção das folhas e outros órgãos aéreos não atempados (sem
crescimento secundário) produzida por células epidérmicas;
c) Rizóides (nas ‘briófitas’ e no protalo das ‘pteridófitas’) ou sistema radicular;
d) Sistemas de ramos com folhas s.l., i.e. folhas dos musgos, micrófilos e megáfilos (vd. Teoria
telomática de W. Zimmermann);
e) Estomas – pequenas aberturas na superfície das folhas por onde se processam as trocas
gasosas;
f) Ciclo de vida haplodiplonte heteromórfico, i.e. com meiose desfasada da fecundação e
alternância de duas gerações – fases haplóide (gametófito) e diplóide (esporófito) – de distinta
morfologia;
g) Anterídeos e arquegónios – órgãos onde se diferenciam, respectivamente, gâmetas ♂ e ♀;
h) Esporângios – órgãos onde se diferenciam esporos;
i) Esporopolenina nas paredes dos esporos – biopolímero complexo, de composição química
pouco conhecida, muito resistente à agressão química;
j) Embrião multicelular geralmente protegido por um tecido multicelular haplóide (nas ‘briófitas’,
‘pteridófitas’ e gimnospérmicas) ou triplóide (nas angiospérmicas).
9 Secundariamente porque os ancestrais destas plantas eram foto-autotróficos.

14 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
2. O corpo das plantas-com-semente
Organização do corpo das plantas-com-semente
O esporófito (fase diplóide) das plantas-com-semente é constituído por três órgãos fundamentais:
raiz, caule e folhas. O seu conjunto constitui o cormo (= corpo da planta). As raízes e os caules
patenteiam a nível anatómico uma simetria radial (vd. Simetria) e, a maioria, demonstra um
crescimento indeterminado. As folhas, pelo contrário, são, salvo raríssimas excepções10, determinadas e
de simetria bilateral (vd. Simetria). Nos órgãos de crescimento determinado, como sejam as folhas e
alguns tipos de caules (e.g. esporões), o crescimento e a diferenciação estão sujeitos a um estreito
controlo genético, pouco sensível a factores ambientais. As folhas representam um caso extremo de
determinação porque, concluída a sua diferenciação, mantêm a mesma forma e estrutura interna até à
senescência (= morte).
Tipos de conceito
Conceito Definição
Organografia vegetal
Concrescência (= coerência)
Partes semelhantes unidas entre si, desenvolvendo-se e crescendo em conjunto; e.g. em muitas espécies as pétalas estão soldadas entre si (i.e. concrescentes), formando um tubo, dizendo-se a corola simpétala
Adnação (= aderência)
Partes distintas unidas entre si, desenvolvendo-se e crescendo em conjunto; e.g. no clado das asteridas os estames estão soldados (= aderentes ou adnados) ao tubo da corola dizendo-se, por isso, epipétalos
Conivência Partes, embora não concrescentes, indistintas de tão encostadas.
Livre Termo usado para designar partes não concrescentes, nem aderentes; e.g. carpelos livres de um gineceu apocárpico
Proximal (= anterior ou adaxial)
Diz-se da parte de um órgão que está mais próxima do eixo ou do ponto onde se insere; e.g. o pecíolo corresponde à parte proximal da folha
Distal (= posterior ou abaxial)
O inverso de proximal; e.g. o ápice da folha corresponde à parte distal da folha e o estigma tem uma posição distal no pistilo
Segmento Partes em que um todo se divide; e.g. segmento de folha secta
Adventício Termo usado para coadjuvar a designação de órgãos situados posições atípicas; e.g. raízes de origem caulinar (= raízes adventícias)
Deiscência Processo ou mecanismo natural mediante o qual um fruto, uma antera ou um esporângio, entre outros órgãos, abrem espontaneamente e libertam para o exterior o respectivo conteúdo
Biologia da evolução
Adaptação Carácter morfológico ou funcional, produzido por selecção natural, que incrementa a probabilidade de sucesso reprodutivo dos indivíduos portadores no seu ambiente natural; por consequência, um indivíduo diz-se adaptado quando a sua forma, fisiologia e comportamento (nos animais) lhe conferem uma elevada probabilidade de sobrevivência e reprodução em condições naturais
Metamorfose Conjunto das modificações morfológicas ocorridas numa planta, no decurso da sua história evolutiva, à escala do corpo (e.g. corpo das plantas aquáticas flutuantes do género Lemna [Araceae]) ou do órgão (e.g. folhas de protecção dos gomos). As metamorfoses adaptativas conferem vantagens reprodutivas aos indivíduos; algumas metamorfoses representam adaptações a ambientes do passado e não evidenciam uma função clara no presente; outras metamorfoses são exaptações, desempenham actualmente uma função distinta daquela sob cuja influência evoluíram.
10 Estão descritas algumas excepções. As folhas têm crescimento indeterminado, por exemplo, na Welwitschia mirabilis (Welwitschiaceae, Pinidae) e vários membros da família das Gesneriaceae (Magnoliidae).
Quadro 2. Alguns conceitos fundamentais de organografia e biologia da evolução usados neste capítulo

15 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
As folhas inserem-se, num padrão regular,
obliquamente nos nós (= verticilos caulinares). A
porção de caule entre dois nós sucessivos chama-
se entrenó. Nas Ephedraceae «éfedras» e nas
Casuarinaceae «casuarinas» os entrenós
destacam-se com facilidade, são articulados. Na
axila de cada folha encontra-se, geralmente, pelo
menos uma gema, i.e. um aglomerado de células
indiferenciadas com capacidade meristemática (=
capacidade de divisão celular), envolvido por
esboços de folhas, por vezes, com um
revestimento externo de folhas de protecção
escamiformes (catáfilos, vd. Tipos de filomas).
Estes aglomerados de células estaminais são,
genericamente, designados nas plantas por
meristemas. Na extremidade distal dos ramos (vd.
quadro 2) situa-se uma gema apical; as gemas
axilares localizam-se, como se depreende do
termo, na axila das folhas. A queda das folhas
deixa uma cicatriz foliar no nó, cuja forma tem
valor diagnóstico em algumas famílias de plantas-
com-flor (e.g. Moraceae).
O cormo das plantas-com-semente apresenta
outros padrões que interessa explorar. As raízes
inserem-se, geralmente, no colo (região de
encontro do caule com o sistema radicular) ou
noutras raízes. As raízes adventícias, por definição, surgem em qualquer ponto da parte aérea,
sobretudo nos nós dos caules, imediatamente abaixo da inserção das folhas. As raízes jamais possuem
folhas embora, por vezes, possam diferenciar gemas adventícias que mais tarde dão origem a novos
caules (designados, respectivamente, nas plantas lenhosas por pôlas radiculares e nas plantas herbáceas
vivazes por rebentos de raiz). Nas raízes inserem-se outras raízes; nos caules, folhas com uma ou mais
gemas axilares que, entretanto, podem evoluir em novos caules.
A morfologia externa e a posição espacial no corpo da planta facilitam a identificação de órgãos
evolutivamente muito modificados. Os espinhos situados na axila de uma folha, ou com pequenas folhas
dispersas, têm, certamente, uma origem caulinar. Um par de espinhos inseridos na proximidade do
pecíolo de uma folha provavelmente corresponde a um par de estípulas modificadas (vd. Apêndices
foliares). Nos casos extremos em que a morfologia externa e a posição espacial sejam inconclusivos, as
raízes, as folhas e os caules podem ser diferenciados através de cortes histológicos e de estudos
ontológicos (de desenvolvimento). A adnação de órgãos, por exemplo de caules com folhas ou de
estames com o cálice, dificulta este tipo de interpretação. As emergências (vd. Emergências) não são
interpretáveis como caules, folhas, raízes ou gemas modificadas porque têm origem na epiderme ou em
tecidos subepidérmicos caulinares ou foliares (e.g. acúleos de uma roseira e espinhos foliares).
Nas plantas-com-semente a formação do cormo inicia-se com a germinação da semente e a retoma
do crescimento do embrião. Este consta geralmente de:
a) Radícula (= raiz embrionária) – esboço de raiz;
b) Cotilédones – folhas embrionárias normalmente ricas em reservas;
c) Plúmula – esboço de caule com folhas embrionárias a envolverem um meristema apical.
Estrutura do embrião e do cormo. A) Embrião jovem: Co – cotilédone. B) Embrião maduro (de uma semente): Hy – hipocótilo; Ra – radícula. C) Plântula: Pw – raiz primária. D) Estrutura do cormo de uma angiospérmica: Gk – gema apical; Sw – raízes laterais; w – raízes adventícias (Sitte et al., 2003)

16 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
Dois entrenós caulinares – o hipocótilo e o epicótilo – conectam, respectivamente, os cotilédones
das eudicotiledóneas s.l. com a radícula e a plúmula. Nas Poaceae (= gramíneas) o primeiro entrenó do
caule acima do cotilédone designa-se por mesocótilo. O colo corresponde à zona de transição entre a
raiz e o caule. A polaridade embrionária, concretizada pelos ápices radicular e caulinar, mantém-se ao
longo de todo o ciclo de vida da planta, desde a germinação até à senescência.
O termo desenvolvimento (= ontogénese) refere-se à história das transformações estruturais vividas
por um indivíduo, ou por uma parte de um indivíduo, desde o embrião, ou de um meristema, até à
senescência. Diz-se, por exemplo, que os espinhos foliares e caulinares têm uma ontogénese distinta
porque nos primeiros são folhas modificadas e os segundos caules modificados (vd. Espinhos). O
desenvolvimento das plantas envolve dois processos distintos: crescimento e diferenciação. O
crescimento é um acréscimo da massa de células vivas originado pela multiplicação e alongamento das
células. O número de tipos celulares, tecidos, órgãos e, em último caso, a fisionomia das plantas, são
uma consequência da diferenciação11 celular.
Nas plantas, a capacidade de produzir novas células está restringida a tecidos particulares de células
estaminais, os meristemas. As células de tecidos diferenciados, ocasionalmente, podem dividir-se, por
regra sem grande significado na estrutura do corpo das plantas. As plantas, ao contrário dos animais,
não “crescem de forma difusa”. Nas plantas com crescimento secundário, tanto mono como
dicotiledóneas s.l., o crescimento processa-se em dois tempos governado por meristemas distintos: as
raízes e os caules primeiro alongam-se pela extremidade e só posteriormente aumentam de diâmetro
com a produção do corpo secundário. O crescimento dos caules e das raízes nas plantas com corpo
secundário decompõe-se, assim, em duas componentes principais:
a) Alongamento – o alongamento terminal é liderado pelo meristema apical; nas
monocotiledóneas o meristema apical é coadjuvado por um meristema de engrossamento
primário; entre as monocotiledóneas, sobretudo nas gramíneas resistentes à herbivoria,
formam-se meristemas intercalares na base dos entrenós sendo, por isso, possível um
alongamento intercalar dos entrenós;
b) Aumento de diâmetro (engrossamento) – consequência directa do crescimento secundário
(formação do corpo secundário); nas dicotiledóneas s.l. depende da presença de um meristema
especializado, do câmbio vascular (= câmbio libero-lenhoso); nas raras monocotiledóneas com
crescimento secundário o engrossamento é conduzido pelo meristema de engrossamento
secundário (Rudal, 2007).
As plantas sem crescimento secundário têm apenas corpo primário porque não dispõem de câmbio
vascular ou de um meristema de engrossamento secundário.
Volume e superfície nas plantas
As plantas, como todos os seres vivos que povoam o planeta Terra, são máquinas biológicas sujeitas
à segunda lei da termodinâmica: para crescerem, reproduzirem-se ou, simplesmente, para
permanecerem vivas, consomem e dissipam energia. A interrupção do consumo de energia implica,
inevitavelmente, a desorganização das células e a morte dos indivíduos. A energia consumida pelas
plantas provém directamente da luz visível emitida pelo sol; as plantas são seres autotróficos. A
radiação solar é pouco energética e as plantas são pouco eficientes na sua conversão em energia
química (ATP) e poder redutor (NADPH) através da fotossíntese. Por conseguinte, ao longo da sua
história evolutiva a parte aérea das plantas foi pressionada no sentido de um aumento da superfície em
detrimento do volume. O sistema radicular foi sujeito a uma pressão semelhante porque a solução do
solo é muito diluída. A captura de nutrientes, nestas condições, depende da exploração de um grande
volume de solo através de uma fina e extensa rede de raízes. O aumento da relação área/volume nas
11 Alguns autores identificam diferenciação com desenvolvimento.

17 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
plantas fez-se à custa de folhas laminares e de raízes e caules de pequeno diâmetro. Hallé (2002) estima
que a superfície externa da parte aérea de uma árvore com 40 m de altura pode ultrapassar 1 ha. A
superfície das raízes é ainda maior. A relação superfície da parte área/superfície da parte subterrânea é
muito variável. Hallé (2002) refere um valor meramente indicativo de 1:130. Portanto, a superfície
externa das raízes de uma planta com 40 m de altura pode atingir os 130 ha!
Estrutura modular das plantas
A compreensão das semelhanças, e das dissemelhanças, no modo como são construídos os corpos
dos animais unitários e das plantas facilita a percepção da configuração e da autoecologia das plantas,
assim como a sua identificação.
A maioria dos animais exibem um crescimento e estrutura determinados, i.e. são seres unitários (=
não modulares). A estrutura externa e interna dos animais unitários – e.g. o número de membros e a
posição espacial do aparelho digestivo ou dos pulmões de um mamífero ou de uma ave – mantêm-se
praticamente inalterados durante todo o crescimento pós-embrionário. O volume e a massa do corpo
estabilizam atingido o estado adulto. Os insectos de metamorfoses completas (= insectos
holometábolos) embora sofram modificações estruturais muito profundas durante o seu
desenvolvimento, transitam de forma determinada entre os estádios de ovo, larva, crisálida e adulto, e
os adultos são muito semelhantes entre si. A dimensão e as estruturas interna e externa dos seres
unitários encontram-se sob um controlo apertado do genoma e são, por isso, pouco sensíveis aos
factores ambientais. Consequentemente, o seu corpo só em parte pode ser ajustado à disponibilidade
de recursos. Este ajustamento ocorre ao nível da população através de variações da densidade
populacional: se o alimento abunda multiplicam-se com celeridade; na falta de alimente morrem de
fome em massa.
As partes em que se divide o corpo dos animais unitários estão organizadas hierarquicamente, são
interdependentes e têm, aproximadamente, a mesma idade, ainda que coexistam células mais velhas e
mais jovens. Hierarquizados porque são constituídos por um elevado número de tipos celulares
especializados, organizados em números tipos de tecidos, de órgãos e, finalmente, de sistemas, com
funções definidas e permanentes. A interdependência é tal que os indivíduos necessitam de todas, ou
de quase todas, as suas partes para se manterem funcionais, e é impossível isolar os órgãos (e.g. numa
cirurgia, durante grandes períodos de tempo). A organização hierárquica e a interdependência funcional
das partes resultaram, em muitas linhagens evolutivas de animais unitários, numa acentuada
complexificação e numa estreita integração funcional do corpo. Em contrapartida, a totipotência
celular12 reduziu-se, o que dificulta ou impede a reposição de partes perdidas por efeito da idade,
doença, acidente ou predação (e.g. senescência células cerebrais, tecido cardíaco necrosado, perda de
membros ou órgãos).
As plantas, por oposição aos animais unitários, são organismos:
a) Modulares – são constituídos pela repetição de unidades multicelulares discretas designadas
por módulos (= metâmeros), de grande autonomia funcional (semi-autónomas);
b) De crescimento indefinido – as plantas crescem permanentemente até à senescência (= morte),
com períodos de quiescência (= suspensão do desenvolvimento) mais ou menos alargados (e.g.
repouso vegetativo invernal das plantas temperadas).
A estrutura modular das plantas só se verifica na parte aérea do corpo vegetativo. As raízes não se
decompõem em módulos, ou pelo menos em módulos exteriormente evidentes, e têm um crescimento
oportunístico, dirigido pelos gradientes de oxigénio, água e nutrientes no solo. Os ramos, pelo contrário,
são construídos através do “encaixe” sucessivo, como numa construção de lego, de um módulo
12 A totipotência celular é a capacidade de uma célula retomar a capacidade de se multiplicar (= capacidade meristemática) e dar origem a todos os tipos de células diferenciadas do organismo.

18 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
(metâmero) elementar – o fitómero – constituído por um entrenó, um nó e uma ou mais folhas com os
respectivos meristemas axilares. Este módulo elementar, por sua vez, organiza-se em módulos de
complexidade crescente, e.g. sistemas de ramos. O crescimento da parte aérea é menos oportunístico
do que o das raízes, caso contrário, as copas das árvores seriam fortemente assimétricas e, nas latitudes
mais elevadas do hemisfério norte, tombariam com a idade todas para sul. As flores são o módulo
reprodutivo das angiospérmicas.
As células vegetais são, genericamente, totipotentes. Esta propriedade, conjugada com a estrutura
modular, permite que as plantas possam, com alguma facilidade, repor ou compensar a perda de partes.
Quando uma árvore perde uma fracção significativa da sua copa activam-se gomos dormentes, ou
diferenciam-se meristemas adventícios, que iniciam a reconstrução da região danificada da copa. Em
casos extremos a copa de uma árvore pode ser rolada (= totalmente eliminada) e novamente
restaurada. Rolam-se as copas das árvores de Castanea sativa (Fagaceae) «castanheiro» infectadas com
doença da tinta (Phythophtora cinnamomi, Chromalveolata) para estimular a emissão de raízes sãs e
conter o avanço da doença no sistema radicular; poucos anos depois as árvores têm uma nova copa e
um sistema radicular parcialmente renovado. Uma argumentação análoga é utilizada para explicar a
facilidade com que se propagam vegetativamente as plantas, com fragmentos de caules, de raízes ou
folhas, com gomos isolados ou, ainda, com pequenos aglomerados de células nas técnicas de
micropropagação laboratorial.
Nas plantas, em alternativa ou em complemento à reposição, as partes perdidas podem ser
compensadas por um crescimento mais vigoroso, mais ou menos descentralizado, de outras. Os frutos
das árvores-de-fruto comerciais têm maior calibre, e mais sementes, se um número significativo de
flores for eliminado com uma poda em verde ou por uma geada tardia. Algumas espécies arbustivas e as
plantas herbáceas com intensa propagação vegetativa (e.g. por rizomas ou bolbos) são virtualmente
imortais porque as partes que, por qualquer razão, colapsam são continuamente substituídas por outras
novas. A resistência à herbivoria das plantas pratenses resulta, também, da sua estrutura modular: os
animais herbívoros consomem biomassa aérea que posteriormente é restituída por meristemas
intercalares e por meristemas axilares, localizados na proximidade da superfície do solo (vd. Corpo
vegetativo das gramíneas).
As plantas têm um corpo flexível em massa, volume e forma porque ajustam o número, a disposição
espacial e, como se refere mais adiante, a forma e a natureza dos módulos às condições ambientais (e.g.
temperatura) e à disponibilidade de recursos (e.g. luz e nutrientes). Diz-se, por isso, que as plantas têm
uma grande plasticidade fenotípica. Quando os recursos são abundantes os meristemas caulinares
activos são mais numeroso e produzem mais módulos, geralmente de maior dimensão. Por
consequência, os caules são mais ramificados e mais longos, e as inflorescências são mais numerosas e
têm mais flores. Em condições de escassez são construídos menos módulos, por vezes mais curtos, e em
casos extremos é reduzido o número de partes, e.g. pela abcisão de ramos (= cladoptose) e/ou folhas
por efeito do ensombramento ou da escassez de água. O crescimento por módulos possibilita que as
plantas ultrapassem, parcialmente, a limitações impostas à captura de recursos pela sua natureza séssil
(imobilidade). Pela mesma razão, os animais sésseis geralmente também têm uma estrutura modular
(e.g. corais). A totipotência e a semi-autonomia dos módulos que compõem as plantas permitem que o
crescimento, ao nível do indivíduo, seja matematicamente modelado de forma análoga a um
comunidade de organismos similares e independentes, correspondendo, neste caso, cada “organismo” a
um módulo individual.
Entre as plantas, indivíduos com a mesma idade podem ter um tamanho e fisionomia muito
distintas. Por outro lado, as células, os tecidos e os órgãos das plantas, ao contrário do ocorrido nos
animais unitários, não têm a mesma idade. Num tronco as células diminuem de idade de dentro para
fora e num ramo as folhas proximais são mais velhas do que as folhas distais. As plantas-com-flor
(angiospérmicas) são mais plásticas que as restantes plantas-vasculares (fetos e gimnospérmicas), facto

19 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
que, aparentemente, ajuda a explicar o seu sucesso evolutivo. A modularidade tem ainda outra
vantagem importante: permite que os módulos possam evoluir de forma quase independente sem
alterar significativamente o funcionamento de outras partes. Por exemplo, as flores podem estar
sujeitas a uma grande pressão de selecção pelos polinizadores enquanto o corpo vegetativo se mantém
inalterado (i.e. em estase evolutiva). Nos seres unitários as mutações somáticas (nas células não
reprodutivas) não são, geralmente, transmitidas à descendência. Nos seres modulares nada impede que
uma mutação somática ocorrida num determinado ponto da copa não possa ser transmitida, por via
assexual ou sexual, à descendência.
O crescimento dos organismos modulares, além no número de módulos e da forma como estão
espacialmente organizados, envolve duas outras componentes: a forma e a natureza dos módulos. Por
exemplo, num mesmo indivíduo, ao nível do módulo elementar caulinar, os entrenós podem ser longos
ou curtos e as folhas apresentarem modificações mais ou menos acentuadas: os ramos mais expostos ao
sol têm, frequentemente, folhas mais pequenas; os ramos estiolados pela falta de luz exibem entrenós
mais longos e folhas maiores e mais delgadas; as plantas pratenses sujeitas a uma herbivoria intensa
têm folhas mais pequenas, entrenós mais curtos e um hábito prostrado, etc. Todos estes casos são
exemplos de plasticidade fenotípica porque as alterações na forma são controladas por factores
ambientais. Pelo contrário, a diferenciação de flores, a formação de tubérculos ou a formação de
rosetas foliares implicam mudanças radicais na natureza dos módulos e têm um controlo genético
directo.
As plantas, ao contrário dos animais unitários, estão “condenadas” a crescer continuamente porque
a imobilidade confere-lhes uma grande susceptibilidade à herbivoria e à competição pela luz. A
herbivoria só pode ser compensada pela reposição de partes perdidas e o acesso à luz depende da
capacidade de expandir folhas acima da canópia dos competidores mais directos. Nas plantas perenes a
degradação dos sistemas fotossintético e vascular com o tempo é, também, resolvida pela continuidade
do crescimento porque as plantas têm capacidades limitadas de reparação daquelas funções. Ao nível
do tecido vegetal e do órgão a substituição de partes é essencial parar assegurar a perenidade no
funcionamento do corpo das plantas.
Os mecanismos de degradação dos sistemas vascular e fotossintético estão bem esclarecidos. A água
que preenche os vasos e traqueídos do xilema encontra-se sob tensão. Consequentemente, os gases
têm tendência a segregar-se em bolhas microscópicas que podem coalescer, formar folhas maiores e
obstruir os elementos condutores do xilema. Designa-se este fenómeno por embolia. As plantas têm
mecanismos para se defenderem da embolia que não evitam, porém, uma lenta perda de eficiência do
sistema condutor (i.e. da condutância hidráulica). A capacidade de conversão da energia da luz em
cadeias carbonadas de alta energia vai decaindo à medida que as folhas vão envelhecendo. Por três
razões:
a) Fotodegradação do sistema fotossintético – a incidência da luz nas folhas acarreta a formação
de radicais livres que danificam as membranas cloroplásticas e os fotopigmentos;
b) Degradação mecânica, herbivoria e parasitismo das folhas – as folhas estão sujeitas a abrasão
(e.g. danos causados por grãos de areia ou sais transportados a grande velocidade pelo vento) e
rasgam-se, sendo consumidas ou parasitadas com alguma facilidade;
c) Ensombramento das folhas por efeito do crescimento – uma vez que os ramos, enquanto
funcionais, se alongam continuamente, as folhas mais velhas vão ficando relegadas para as
camadas mais profundas da copa, afastadas da exposição directa da luz, até que os seus
consumos respiratórios de energia ultrapassam os ganhos fotossintéticos.
As estruturas reprodutivas das plantas, ao invés do corpo vegetativo, têm uma organização
hierárquica e uma estrutura determinada. Na flor completa as pétalas sucedem-se às sépalas, os
estames às pétalas e o gineceu ao androceu. Esta sequência tem um controlo genético preciso. Com a
formação da flor extingue-se o meristema que lhe deu origem. A dimensão e a estrutura das partes dos

20 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
órgãos reprodutivos das plantas são, por regra, intraespecificamente mais estáveis do que o corpo
vegetativo. Este facto, somado com a diversidade morfológica e funcional das estruturas reprodutivas e
a sua estabilidade evolutiva, explica a sua importância na identificação e classificação do mundo vegetal.
A natureza modular das plantas torna a sua identificação
bastante mais complexa que a dos animais unitários.
Muitos destes animais podem ser positivamente
identificados com base na silhueta, no tamanho, na cor,
na postura corporal ou até no movimento. Por isso as
aves ou os mamíferos são normalmente representados
em corpo inteiro nos guias de campo. A fisionomia das
plantas, por exemplo de uma árvore ou de um arbusto, é
francamente menos informativa. A identificação das
plantas obriga a uma observação visual de proximidade,
por vezes com recurso a lupas de bolso, da inserção e
forma das folhas, dos pêlos das folhas, da estrutura da
flor, entre outros aspectos. Ainda assim, sobretudo em
ambientes tropicais, a forma das árvores e arbustos pode
ser de grande utilidade para determinar a identidade de
indivíduos, ou de populações de indivíduos de uma
mesma espécie. Quando se observam povoamentos
arbóreos em fotografia aérea ou, a contraluz, é fácil
distinguir a silhueta de uma Castanea sativa (Fagaceae)
«castanheiro», de uma Tilia (Malvaceae, Tilioideae)
«tília» ou de um Pinus pinea (Pinaceae) «pinheiro-
manso».
Simetria
A simetria, por definição, é a repetição regular,
geneticamente determinada, de elementos estruturais
iguais ou similares. Reconhecem-se três formas
fundamentais de simetria:
a) Simetria por metameria – repetição de elementos estruturais ao longo de um eixo; e.g.
repetição do módulo elementar caulinar – o fitómero – constituído por um entrenó, um nó e
uma ou mais folhas com os respectivos meristemas axilares;
b) Simetria radial – repetição de um número variável de elementos estruturais, com um mesmo
ângulo, em torno de um eixo; e.g. pétalas em redor do eixo floral (= receptáculo) ou a disposição
radial dos tecidos no caule e na raiz;
c) Simetria bilateral – repetição de elementos estruturais nos dois lados de um plano de simetria;
simetria predominante no reino animal; e.g. flores zigomórficas (com um plano de simetria) e
folhas dorsiventrais.
Nas plantas, as simetrias por metameria e radial emergem, directamente, da sua natureza modular.
A simetria bilateral é comum nas flores polinizadas por insectos.
As regras de simetria por metameria e radial controlam a disposição espacial dos módulos e, por
essa via, a forma dos indivíduos. A programação total da forma exigiria imensa informação genética –
muito mais do que a requerida pelo controlo genético da metameria, da simetria radial e da estrutura
dos fitómeros – necessariamente rígida do ponto de vista evolutivo (como acontece nos animais). A
simetria, conjugada com a estrutura modular, é, então, uma solução evolutiva parcimoniosa (simples e
que exige pouca informação) para gerar, em resposta ao ambiente, formas complexas e plásticas
a)
b)
Simetria. a) Cistus ladanifer (Cistaceae): simetria radial: corola e androceu. Simetria por metameria: unidade entrenó e nó com duas folhas opostas. b) Viola riviniana (Violaceae): simetria bilateral: corola (n.b. existe apenas um plano de simetria porque as pétalas são distintas entre si, uma delas formando um esporão na base) (de Bonnier & Douin, 1912-1935).

21 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
(plasticidade fenotípica), mas, ao mesmo tempo, evolutivamente flexíveis, i.e. sensíveis a pressões de
selecção de índole diversa. A prontidão para mudar é uma característica fundamental das plantas.
Homologia e analogia
Na determinação das relações de parentesco entre taxa – basilar nos sistemas de classificação
cladísticos – é essencial a distinção entre analogia e homologia. As analogias são semelhanças
geneticamente determinadas, morfológicas, fisiológicas ou moleculares, que resultam da convergência
evolutiva de linhagens independentes. As analogias são uma consequência de pressões de selecção
semelhantes que arrastam consigo adaptações, a nível morfológico ou fisiológico, também semelhantes.
A forma cactiforme (semelhante a cactos) de várias Euphorbia africanas e canarinas é um exemplo
clássico de convergência evolutiva. A homologia implica a partilha de ancestrais comuns nos quais
tiveram origem os caracteres responsáveis pelas semelhanças actuais.
Variação morfológica intraespecífica
A forma e fisiologia das plantas variam intraespecificamente (entre os indivíduos da mesma espécie)
por três causas fundamentais:
a) Variação ontogénica (= plasticidade ontogénica) – reside nas diferenças entre os indivíduos, ou
partes de indivíduos, juvenis e adultos; a transição para a fase adulta está associada à
capacidade de produzir flores (só as plantas ou as partes adultas de uma planta produzem
flores); as alterações morfológicas e fisiológicas da transição da fase juvenil para a adulta são
geneticamente determinadas; nas plantas lenhosas estas alterações frequentemente ocorrem
na extremidade da copa, permanecendo a base da planta juvenil; nas plantas herbáceas esta
transição é acompanhada por uma acentuada redução das taxas de crescimento; as folhas
juvenis, muitas vezes, são maiores (e.g. Eucalyptus), mais dentadas ou espinhosas (e.g. Quercus
rotundifolia «azinheira»); a presença de espinhos é também frequente nos estádios juvenis (vd.
espinhos);
b) Variação ambiental (= plasticidade fenotípica) – a plasticidade fenotípica, por definição, é a
capacidade demonstrada pelas plantas de modificar a sua morfologia e fisiologia em resposta a
a) b) c)
Homologia e analogia. a) Carlina vulgaris (Asteraceae). b) Eryngium campestre (Apiaceae). c) Daucus carota (Apiaceae) (imagens extraídas de Bonnier & Douin, 1911-1935). O E. campestre é mais parecido com a C. vulgaris mas filogeneticamente mais próximo do D. carota (pertencem à mesma família). A semelhança entre C. vulgaris e E. campestre resulta de convergência evolutiva: o hábito espinhoso é uma homologia.

22 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
alterações ambientais; esta capacidade tem um controlo genético porque nem todas as plantas
têm a mesma plasticidade fenotípica, o produto dessa capacidade goza, porém, de um controlo
ambiental directo; a plasticidade fenotípica é, então, um produto da interacção ambiente-
genoma. Como se referiu anteriormente, a plasticidade fenotípica é facilitada pela natureza
modular das plantas; esta plasticidade tem um grande valor adaptativo porque as plantas são
sésseis e habitam um mundo com uma distribuição dos recursos espacial e temporalmente
heterogénea;
c) Variação genética (= plasticidade genética) – as características genéticas de cada indivíduo são
fixadas durante a fecundação, consequentemente, não são influenciadas pelo ambiente
(embora a sua expressão o possa ser); portanto nem todas as diferenças observáveis na forma e
na função entre os indivíduos de uma mesma espécie têm um controlo ambiental; a
variabilidade genética intraespecífica é a matéria-prima da evolução sobre a qual actua a
selecção natural (vd. Volume II).
As relações entre os mecanismos de regulação génica, os mecanismos fisiológicos e a variação
morfológica intraespecífica (=plasticidade intraespecífica) estão para além dos objectivos deste
documento.
Adaptação e aclimatação
Uma adaptação é uma característica morfológica ou fisiológica, geneticamente transmissível,
favorecida por selecção natural. De acordo com o modelo de evolução por selecção natural
originalmente postulado por Darwin, as características adaptativas primeiro surgem por acaso e só
posteriormente a sua frequência é incrementada por selecção (vd. Volume II). O corpo vegetativo de um
cacto xerófito (adaptado a climas de grande secura) envolveu a acumulação e a concatenação de um
grande número de “acasos felizes”, i.e. de adaptações. Os raciocínios adaptativos simplistas do tipo “as
plantas têm espinhos para se defenderem dos animais herbívoros” envolvem grandes riscos porque se
sustentam em evidências circunstanciais potencialmente mal interpretadas (vd. Prólogo). A identificação
de adaptações, i.e. a demonstração das vantagens adaptativas de caracteres, é complexa e morosa.
O conceito de aclimatação é aplicado às modificações verificadas no corpo dos seres vivos em
resposta a alterações do habitat, geralmente envolvendo variáveis climáticas ou de solo. A adaptação é
um processo genericamente lento, dirigido pela selecção natural, acompanhado de alterações genéticas
qualitativas e quantitativas sexualmente transmissíveis. A aclimatação, pelo contrário, não acarreta
alterações genéticas mas tão-somente modificações fenotípicas, na morfologia e/ou na fisiologia da
planta: é uma consequência directa da plasticidade fenotípica. A colocação ao ar livre de plantas
propagadas em estufa para melhor suportarem condições climáticas mais extremas é um exemplo
prático de aclimatação. Recentemente, foi provado que alguns tipos de aclimatação são, pelo menos em
parte, sexualmente transmissíveis. Os descendentes de plantas aclimatadas a determinadas condições
ambientais – e.g. solos secos ou pobre de nutrientes – frequentemente crescem mais, e reproduzem-se
com mais sucesso, nestas condições do que os descendentes de indivíduos não aclimatados. A
transmissão de caracteres adquiridos por aclimatação está relacionada com modificações ao nível da
repressão e expressão génica (e.g. metilação de genes), ainda não totalmente compreendidas. O estudo
da transmissão de caracteres sem alterações do código genético – a epigenética – é um área recente e
de ponta da biologia evolutiva em franco progresso (vd. volume II).

23 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
3. Sistema vegetativo
3.1. Aspectos morfológicos comuns a todo o corpo vegetativo
Espinhos
No corpo vegetativo das plantas é frequente a presença de vários tipos de estruturas aguçadas,
geralmente, com a função de dissuadir a herbivoria e/ou de fincar as plantas a suportes. Reserva-se o
termo espinho para as estruturas pontiagudas, rijas e difíceis de destacar, providas de feixes vasculares,
resultantes da modificação total ou parcial de ramos, folhas, estípulas ou gomos. Os acúleos, como mais
adiante de esclarece, são emergências.
A posição dos espinhos no cormo permite, muitas vezes, identificar o órgão que lhes deu origem (vd.
figura):
a) Espinhos de origem caulinar – situam-se na axila de uma folha ou da sua cicatriz; a superfície
destes espinhos podem apresentar folhas mais ou menos modificadas ou as suas cicatrizes; e.g.
espinhos de Echinospartum ibericum (Fabaceae) «caldoneira», Ulex (Fabaceae) «tojos» e Olea
europaea var. sylvestris (Oleaceae) «zambujeiro»;
b) Espinhos de origem foliar – regra geral axilam uma gema ou um caule com origem numa gema
axilar; e.g. espinhos de Cactaceae «cactos»;
a) b) c)
Espinhos e acúleos. a) Tipos de espinhos e acúleos (adaptado de van Wyk & van Wyk, 2006). b) Ulex airensis (Fabaceae), um endemismo do CW de Portugal Continental. c) Ramo com acúleos (adaptado de Coutinho, 1898).

24 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
c) Espinhos de origem estipular – dispostos aos pares, geralmente um de cada lado de uma folha,
de uma gema ou de caule desenvolvido a partir de uma gema axilar; e.g. Robinia pseudoacacia
(Fabaceae) «robínia».
Emergências (acúleos e indumento)
As emergências são estruturas constituídas por células de origem epidérmica e subepidérmica não
sendo, por isso, identificáveis com raízes, caules, gomos ou folhas modificadas (Bell, 2008). Como não
possuem conexões vasculares destacam-se com alguma facilidade do órgão onde se inserem.
As emergências espinhosas, +/- lenhosas, são muito frequentes nas plantas-com-flor. Designam-se
genericamente por acúleos, embora na bibliografia a sua nomenclatura seja inconsistente. Os acúleos
são particularmente abundantes nas Rosaceae, tanto nos caules (acúleos caulinares), como no pecíolo
(acúleos peciolares), como ainda nas nervuras das folhas (acúleos foliares); e.g. Rubus ulmifolius
(Rosaceae) «silvas» e Rosa (Rosaceae) «roseiras». As emergências espinhosas localizadas na margem
das folhas (e.g. em Ilex aquifolium [Aquifoliaceae] «azevinho») são, inapropriadamente, designadas por
espinhos foliares. As folhas com emergências espinhosas nas margens dizem-se espinescentes.
Genericamente, designa-se por indumento o revestimento de origem epidérmica constituído por pó
de natureza cerosa ou salina, placas cerosas, papilas, escamas, tricomas (= pêlos) ou glândulas, que
recobre as superfícies das folhas, caules herbáceos (sem crescimento secundário) ou peças da flor
(sobretudo o cálice). As plantas podem combinar mais de um tipo de indumento. Por exemplo, as
Lamiaceae «labiadas» possuem um ou mais tipos de glândulas e de pêlos glandulosos, combinados com
pêlos não glandulosos, simples ou ramificados. Os órgãos desprovidos de indumento dizem-se glabros;
glabrescentes se este for raro e esparso. O indumento é particularmente importante na identificação
das plantas ao nível da espécie. O início do crescimento secundário implica a eliminação do indumento
dos caules.
As papilas são pequenas projecções epidérmicas unicelulares em forma de mamilo. Nas escamas
uma pequena projecção aplanada conecta-se à epiderme por um pequeno “pé”. Os tricomas têm uma
forma muito variável. Reconhecem-se dois tipos principais:
a) Simples (não ramificados) – podem ser unicelulares ou pluricelulares, glandulares ou não
glandulares, peltados, gancheados, urticantes, etc.;
b) Ramificados – podem ser bifurcados (em forma de T ou de Y), estrelados (em forma de estrela,
sésseis ou pediculados), dendríticos, etc.
As glândulas13 possuem capacidade secretória. A sua morfologia é muito diversa assim como os
critérios usados na sua classificação; e.g. glândulas unicelulares ou pluricelulares e glândulas sésseis ou
providas de um pequeno pé (pêlos glandulosos). As substâncias segregadas incluem enzimas
proteolíticas, sais, resinas, óleos essenciais e substâncias urticantes. As glândulas produtoras de enzimas
proteolíticas das plantas carnívoras participam na digestão externa de insectos e de outros organismos.
As glândulas salinas desenvolvem-se, com abundância, nas plantas adaptadas a solos salinos ricos em
cloreto de sódio, como acontece nas Amaranthaceae dos sapais holárticos (e.g. gén. Atriplex) e em
algumas plantas dos mangais tropicais (e.g. Avicennia africana [Acanthaceae] «mangue-branco»). As
glândulas que segregam óleos essenciais e/ou resinas têm, muitas vezes, uma forma globosa, quase
microscópica, e cor amarela ou vermelha brilhante. Rompem-se e libertam o seu conteúdo quando
perturbadas (e.g. passagem de um insecto). A abundância deste tipo de glândulas nas plantas-com-flor
indicia a sua importância na protecção dos órgãos herbáceos contra a herbivoria, sobretudo por
insectos. Nas Urtica (Urticaceae) «urtigas» os pêlos glandulosos são ocos e preenchidos com um líquido
venenoso e urticante. A extremidade do pêlo é arredondada e parte-se com facilidade em contacto com
13 Este termo é geralmente aplicado às glândulas epidérmicas. No entanto, as glândulas podem situar-se em camadas celulares mais profundas e nesse caso não cabem no conceito de emergência.

25 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
um corpo estranho. A parte remanescente toma a forma de uma agulha e penetra a pele dos mamíferos
libertando o seu conteúdo no interior do corpo do animal.
A terminologia associada ao indumento é muito especializada e diversa porque uma correcta
descrição das características da superfície dos órgãos aéreos das plantas é essencial em taxonomia. Os
termos e conceitos mais utilizados na bibliografia de referência são os seguintes:
a) Indumento não constituído por pêlos:
i) Pruinoso – superfície com placas de cera; e.g. turiões de Rubus ulmifolius (Rosaceae)
«silva-comum»;
ii) Farinoso – superfície coberta com um pó de natureza cerosa;
iii) Papiloso – com pequenas papilas.
b) Indumento de pêlos:
i) Indumento de pêlos compridos a intermédios:
Acetinado (ceríceo) – de pêlos aplicados e densos que atribuem um brilho de
cetim; e.g. página inferior do Salix alba (Salicaceae) «salgueiro-branco»;
Ciliado (= celheado) – pêlos concentrados na margem das folhas; e.g. Erica
tetralix (Ericaceae);
Flocoso – de pêlos que se destacam em flocos irregulares; e.g. Verbascum
pulverulentum (Scrophulariaceae);
Hirsuto – de pêlos densos, um pouco rígidos mas flexíveis, patentes ou quase;
Híspido – de pêlos muito rígidos, quase picantes, patentes e não muito densos;
muitas Boraginaceae;
Lanoso – de pêlos crespos (ondulados) e macios, como a lã;
Piloso – de pêlos macios erguidos e não ondulados;
Setígero – com sedas, i.e. pêlos ásperos mais ou menos rígidos e fortes;
Tearâneo (= aracnóide) – com pêlos finos e macios, tenuemente entrelaçados,
como uma teia de aranha;
Tomentoso – com pêlos moles enleados formando um enfeltrado denso; e.g.
página inferior do Salix atrocinerea (Salicaceae) «borrazeira-preta»;
Urticante – de pêlos urticantes; e.g. Urtica (Urticaceae) «urtigas»;
Viloso – de pêlos longos, macios, direitos ou sinuosos, não muito densos,
patentes ou subpatentes e não entrecruzados.
ii) Indumento de pêlos curtos
Aveludado – de pêlos finos, densos e erguidos, de toque e aspecto semelhante
ao do veludo;
Lanuginoso – de pêlos crespos e macios;
Puberulento – de pêlos muito curtos e esparsos;
Pubescente – de pêlos fracos e pouco densos.
Além do indumento de pêlos a superfície dos órgãos herbáceos pode ser:
a) Viscida – superfície viscosa, e.g. folhas e caules de Cistus ladanifer (Cistaceae) «esteva»;
b) Alveolada – com pequenas depressões separadas por pequenas arestas;
c) Perfuradas – com pequenas perfurações.

26 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
O indumento desempenha uma, ou mais, das seguintes funções (mecanismo identificado após o
travessão):
a) Protecção contra o excesso de radiação – ensombramento da superfície foliar através da
reflexão ou absorção da radiação solar;
b) Incremento da captura de luz para a fotossíntese – reflexão da radiação solar em direcção às
células fotossintéticas do mesofilo foliar;
c) Redução das perdas de água – aumento da resistência à difusão de vapor de água conseguida
com um aumento da espessura da camada limite e da formação de uma camada gasosa estável,
rica em água, entre os pêlos e a superfície foliar;
d) Isolamento térmico – formação de uma camada gasosa estável entre os pêlos e a superfície
foliar e aumento da espessura da camada limite;
e) Retenção de nutrientes – redução das perdas de iões por lixiviação da superfície foliar;
f) Tolerância à salinidade – movimento activo de sais do interior do mesofilo foliar para o exterior;
g) Defesa contra a herbivoria – e.g. repulsão de insectos ou vertebrados com pêlos glandulosos;
h) Defesa contra microrganismos patogénicos – e.g. afastamento do inóculo (e.g. esporos de
fungos) da superfície foliar;
i) Protecção dos estomas – e.g. redução dos riscos de bloqueio dos estomas com água ou
partículas sólidas;
j) Digestão de insectos e outras “presas” – produção de enzimas proteolíticas.
Corpos nutritivos, hidátodos e nectários extraflorais
Os corpos nutritivos são secreções sólidas de
substâncias nutritivas, usualmente proteicas, com
função de recompensa em relações mutualistas
planta-insecto, geralmente com formigas
(Hymenoptera, Formicidae). Podem localizar-se nas
folhas, na base do pecíolo, nos caules ou mesmo na
flor. Por exemplo, as formigas sul-americanas do
género Pseudomyrmex constroem formigueiros em
espinhos ocos e consomem corpos nutritivos situados
no ápice dos folíolos de algumas Acacia (e.g. Acacia
cornigera, Fabaceae), em contrapartida defendem a
árvore do ataque de insectos e mamíferos herbívoros,
eliminam folhas e caules de outras espécies de plantas
que contactem com a árvore colonizada e suprimem
as plantas que germinem na sua vizinhança.
Os hidátodos são cavidades com um poro por onde é excretada solução xilémica; este fenómeno
designa-se por gutação e ocorre com mais frequência quando o ar está saturado de água. Os hidátodos
são particularmente frequentes nas plantas das florestas tropicais húmidas. Os nectários extraflorais, ao
contrário dos hidátodos excretam soluções açucaradas – as meladas – que actuam como recompensa
em relações mutualistas com insectos. Enquanto os nectários florais oferecem recompensas a
polinizadores, os nectários extraflorais servem recompensas a insectos mutualistas que defendem a
planta contra a herbivoria (e.g. formigas que atacam larvas herbívoras de borboletas e outros insectos
herbívoros). Os nectários extraflorais geralmente localizam-se no limbo ou no pecíolo das folhas, por
vezes, em bolsas especializadas (domácias); e.g. nectários extraflorais dos pecíolos das Passifloraceae
Nectários extraflorais. Base do limbo de Prunus avium (Rosaceae) «cerejeira»

27 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
«maracujazeiros» e da Prunus avium «cerejeira». Os apicultores conhecem bem o mel de melada, que
tem um valor de mercado inferior ao mel de néctar.
Galhas
As galhas são crescimentos anormais causados por insectos, ácaros, fungos, bactérias ou feridas.
Frequentemente, mostram formas características que permitem identificar, de forma indirecta, o
agente causal. As galhas são muito comuns em Quercus (Fagaceae) «carvalhos», e.g. galhas de Andricus
quercustozae (Hymenoptera, Cynipidae) em Q. pyrenaica ou Q. faginea subsp. faginea, vulgarmente
conhecidas por bugalhos, ou as galhas de Dryomyia lichtensteini (Diptera, Cecydomidae) em Q. suber e
Q. rotundifolia. O nome vulgar da Pistacia terebinthus (Anacardiaceae) – «cornalheira» – deve-se às
galhas folheares corniformes produzidas pela Baizongia pistaciae (Homoptera, Pemphigidae). Nos ramos
das oliveiras (Olea europaea, Oleaceae) varejadas com violência são comuns galhas de origem
bacteriana – «tuberculose» – causadas por uma bactéria, a Pseudomonas savastanoi.
3.2. Raiz
A raiz é um dos três órgãos fundamentais das plantas. Ao invés dos caules e das folhas, o sistema
radicular tem um geotropismo positivo e permanece, na maioria dos casos, oculto no solo durante todo
o ciclo de vida das plantas.
3.2.1. Funções da raiz
As funções mais comuns das raízes são:
a) Absorção de água e nutrientes – dos 27 elementos essenciais que constituem o corpo das
plantas-terrestres apenas o carbono e o oxigénio não são total, ou maioritariamente, absorvidos
pela raiz a partir da solução do solo (água do solo + nutrientes em solução); nas plantas epífitas
os nutrientes são extraídos de resíduos orgânicos (e.g. folhas mortas), de partículas inorgânicas
arrastadas pelo vento (e.g. argilas) e da água da chuva ou dos nevoeiros; nas plantas parasitas a
nutrição e o abastecimento em água são garantidos, total ou parcialmente, pelos seus
hospedeiros;
Galhas. Bugalho de Andricus quercustozae (Hymenoptera, Cynipidae) em Quercus faginea subsp. faginea (ca. 3,5 cm) (duas primeiras fotos). Galha de Neuroterus quercusbaccarum (Hymenoptera, Cynipidae) em Q. faginea subsp. faginea (ca. 6 mm) (terceira foto). Galhas de Baizongia pistaciae (Homoptera, Pemphigidae) em Pistacia terebinthus (última foto à direita)

28 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
b) Síntese de compostos orgânicos – e.g. síntese de hormonas vegetais, em particular de
citoquininas;
c) Ancoragem ao substrato – ao solo e a outros tipos de suporte (e.g. outras plantas, afloramentos
rochosos, etc.).
As raízes podem ainda desempenhar as seguintes funções:
a) Reserva – função particularmente importantes nas raízes tuberosas;
b) Trocas gasosas – directa ou indirectamente todas as células vegetais efectuam trocas gasosas
com a atmosfera ou com a atmosfera do solo; algumas espécies possuem raízes especializadas
nessa função (pneumatóforos);
c) Assimilação – em raras orquídeas tropicais; e.g. na Dendrophylax lindenii «orquídea-fantasma»
os caules e as folhas estão ausentes ou muito reduzidos e a função fotossintética é
desempenhada pelas raízes.
A absorção de nutrientes é uma das funções mais importantes das raízes. Macronutrientes como o
azoto, o fósforo, o potássio, o cálcio e o magnésio são, na prática, exclusivamente absorvidos pelas
raízes. A morfologia e a configuração espacial (arquitectura) das raízes têm uma grande influência na
eficiência da absorção de nutrientes, sobretudo dos nutrientes de menor mobilidade no solo, como o
fósforo. Características que aumentem o volume de solo explorado pelas raízes, com reduzidos custos
metabólicos, são muito vantajosas em solos pobres em nutrientes de baixa mobilidade. A absorção do
fósforo, por exemplo, é favorecida pela presença de aerênquima, de raízes pequeno diâmetro, de raízes
superficiais muito ramificadas (porque as formas biodisponíveis deste nutriente estão concentradas nas
camadas superficiais do solo), e pela produção abundante de pêlos radiculares de grande comprimento.
3.2.2. Alongamento e ramificação da raiz
Alongamento
O alongamento da raiz é conduzido pelo meristema apical radicular. O meristema apical radicular
está permanentemente encapsulado no interior do ápice radicular pela calíptra (= coifa), uma estrutura
em forma de capuz que protege as células meristemáticas do contacto directo com as partículas do solo
e lubrifica o avanço das raízes. À medida que as raízes se alongam, a calíptra liberta para o exterior uma
mucilagem lubrificante – o mucigel – e as suas camadas celulares externas desagregam-se, sendo
substituídas por outras provenientes do meristema apical.
As raízes recém-formadas são delgadas e frágeis. O diâmetro do ápice radicular ronda os 0,2 mm.
Imediatamente atrás do ápice radicular situa-se uma curta zona de alongamento e logo a seguir a zona
pilosa. As células da raiz multiplicam-se no meristema, e expandem-se e diferenciam-se nos vários
tecidos que compõem a raiz na zona de alongamento. As células recém-formadas alongam-se mais de
dez vezes empurram para diante a raiz através do solo. A zona pilosa está revestida de pêlos radiculares
de origem epidérmica, que aumentam a superfície de contacto da raiz com as partículas e a solução do
solo. A maior parte da água e dos solutos consumidos pelas plantas são absorvidos nesta região da raiz;
os pêlos radiculares desempenham um importante papel na absorção de nutrientes presentes em
baixas concentrações na solução solo, sobretudo do fósforo. Os pêlos radiculares degradam-se
rapidamente e a zona pilosa dá lugar à zona de ramificação.
Ramificação
A ramificação lateral da raiz, o tipo de ramificação dominante entre as plantas-vasculares, tem uma
origem endógena, nas células do periciclo ou em células adjacentes à endoderme. Antes de entrar em
contacto com o solo, as raízes laterais recém-diferenciadas têm de irromper pelo córtex radicular e pela

29 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
epiderme da raiz onde se inserem. As raízes laterais, derivadas da raiz primária, designam-se por raízes
secundárias; as raízes terciárias inserem-se nas secundárias, e assim sucessivamente.
Nas dicotiledóneas s.l. partir do momento em que se inicia a formação do corpo secundário da raiz,
através da actividade dos câmbios libero-lenhoso (= cambio vascular) e subero-felodérmico (= felogene),
as raízes perdem a capacidade de absorver água e nutrientes, deixam de se ramificar, aumentam de
diâmetro e na sua superfície diferencia-se uma periderme (casca), em substituição da epiderme (o
tecido que recobre e protege o corpo primário das plantas). No entanto, ao mesmo tempo que a parte
mais velha do sistema radicular aumenta de diâmetro são produzidas novas raízes na sua extremidade
distal.
3.2.3. Aspectos gerais da morfologia externa da raiz
Tipos de radicação
Entende-se por sistema radicular o conjunto de todas as raízes de uma planta. A raiz dita primária ou
principal tem origem na radícula (= raiz embrionária) e funde-se com o caule no colo. Em algumas
espécies, sobretudo entre as monocotiledóneas, a raiz primária atrofia-se rapidamente e é substituída
por raízes adventícias diferenciadas no caule (raízes caulógenas), geralmente nos nós a partir de células
localizadas na região do periciclo. Distinguem-se assim dois tipos de radicação:
a) Sistema radicular aprumado (= sistema radicular magnolióide) – uma raiz principal de origem
embrionária lateralmente ramificada; grande número de raízes espessas e, em contrapartida,
poucas raízes finas e pêlos radiculares; baixa relação entre o volume de raízes com capacidade
de absorção e o volume total do sistema radicular, compensada pelas associações micorrízicas;
sistema característico das gimnospérmicas e da grande maioria das dicotiledóneas s.l.;
b) Sistema radicular fasciculado (= sistema radicular graminóide) – sistema radicular homogéneo,
sem uma raiz principal, constituído por raízes adventícias semelhantes entre si, finas, delicadas,
com abundantes pêlos radiculares; primórdios radiculares adventícios diferenciados nos caules
(regra geral em nós subterrâneos ou aéreos e próximos da superfície do solo) ou, mais
raramente, na base de folhas; elevada relação entre o volume de raízes com capacidade de
absorção e o volume total do sistema radicular; admite-se que é mais eficiente do que o sistema
radicular aprumado (e por isso competitivamente vantajoso) na captura de água e nutrientes
das camadas superficiais do solo, contudo é menos adequado para ancorar plantas de grande
dimensão ao solo; característico das monocotiledóneas e de algumas dicotiledóneas s.l. (e.g.
Ranunculus [Ranunculaceae] «ranúnculos»).
Em algumas árvores (e.g. Salix [Salicaceae] «salgueiros») a raiz principal é tão rudimentar que a
arquitectura do sistema radicular se aproxima do modelo fasciculado. As plantas obtidas por estaca, ou
por outros métodos de propagação vegetativa que impliquem a formação de raízes adventícias,
apresentam um sistema radicular de morfologia intermédia. As raízes adventícias são normalmente
caulógenas mas podem ter origem no pecíolo, no limbo das folhas ou diferenciarem-se a partir de
calos14.
Direcção do sistema radicular
A raiz principal de um sistema radicular aprumado possui um geotropismo positivo: diz-se que é
profundante. O geotropismo característico da raiz principal não se mantém, todavia, em todas as raízes
laterais de um sistema radicular aprumado. Quanto à direcção, as raízes secundárias, e de ordem
superior, podem ser profundantes, pouco profundantes ou plagiotrópicas (próximas da
14 Os calos são proliferações celulares que se produzem em feridas e cortes a partir de células parenquimatosas que mantiveram a capacidade de se multiplicar em tecidos diferenciados (não meristemáticos).

30 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
horizontalidade). A plagiotropia facilita a disseminação tridimensional das raízes pelo solo e maximiza o
volume de solo explorado. As raízes de plantas lenhosas que progridem na horizontal próximo da
superfície do solo têm direito a uma designação especial: raízes pastadeiras. Estas raízes são
fundamentais na nutrição das plantas porque exploram as camadas superiores do solo (horizonte A),
mais ricas em nutrientes – resultantes da deposição de resíduos orgânicos ou da aplicação de
fertilizantes – e de maior actividade biológica. Por essa razão, as oliveiras e outras fruteiras crescem
mais, e são mais produtivas, nos pomares não mobilizados do que nos pomares ciclicamente
mobilizados (lavrados ou escarificados).
Nas monocotiledóneas as raízes secundárias, pelo menos as de maior dimensão, são geralmente
profundantes. No arroz (Oryza sativa, Poaceae), a planta modelo dos geneticistas de cereais, foram
identificados quatro tipos de raízes. A raiz embrionária emerge da semente aquando da germinação. Em
seguida formam-se raízes adventícias, primeiro no nó do coleóptilo, e depois noutros nós da base do
caule. Tanto a raiz embrionária como as raízes adventícias apresentam um geotropismo positivo. A
ramificação destas raízes dá origem a mais dois tipos de raízes: raízes laterais de crescimento
indeterminado profundantes, e raízes laterais determinadas, curtas e de geotropismo indefinido.
Situação
Quanto à situação as raízes podem ser:
a) Subterrâneas – se imersas no solo; tipo mais frequente;
b) Aquáticas – próprias de plantas aquáticas, i.e. de plantas que vivem submersas ou na superfície
de massas de água livre;
c) Aéreas – muito frequentes em lianas e em plantas epífitas, i.e. plantas não enraizadas no solo,
suportadas por outras plantas. Nas raízes aéreas de muitas epífitas de clima tropical húmido,
sobretudo de Orchidaceae, forma-se um velâmen, i.e. uma camada de revestimento exterior de
células mortas, muito espessa, esbranquiçada quando seca, que se mantém inchada e húmida
por muito tempo quando para de chover.
Consistência
Sob o critério consistência admitem-se três tipos de raízes:
a) Herbáceas – raízes tenras, delgadas e flexíveis;
b) Lenhosas – raízes lenhificadas e rijas;
a) b)
Tipos de sistema radicular. a) Morfologia das raízes jovens de uma angiospérmica (adaptado de Hickey & King, 2000). b) Sistema radicular aprumado de Malva (Malvaceae) «malvas» e sistema radicular fasciculado de uma Poaceae (adaptado de Coutinho, 1898).
Tipos de sistema radicular. a) Morfologia das raízes jovens de uma angiospérmica (adaptado de Hickey & King, 2000). b) Sistema radicular aprumado de Malva (Malvaceae) «malvas» e sistema radicular fasciculado de uma Poaceae (adaptado de Coutinho, 1898).

31 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
c) Carnudas (= tuberosas) – de grande volume e ricas em água e substâncias de reserva.
3.2.4. Metamorfoses da raíz
As raízes apresentam vários tipos de adaptações com funções especializadas, por vezes associáveis a
habitats particulares (e.g. zonas húmidas, florestas tropicais, etc.). As metamorfoses da raiz mais
frequentes são:
a) Raízes trepadoras – raízes adventícias, aéreas, geralmente adesivas, que auxiliam as plantas
trepadeiras a aderirem aos seus suportes; e.g. fáceis de observar em Hedera sp.pl. (Araliaceae)
«heras» e em várias espécies de Piper (Piperaceae);
b) Raízes tuberosas – raízes de grande volume de forma cónica a fusiforme. Entende-se por
tuberização das raízes o processo de engrossamento causado pela acumulação de reservas;
consoante as espécies, e por vezes entre variedades da mesma espécie, a tuberização das raízes
pode prolongar-se até ao colo ou ao caule; e.g. prolonga-se até ao caule na Brassica rapa
(Brassicaceae) «nabo», concretamente até ao hipocótilo, e a mais de um entrenó caulinar em
algumas variedades de Beta vulgaris subsp. vulgaris (Amaranthaceae) «beterraba-sacarina». As
substâncias de reserva mais frequentes são o amido (raízes amiláceas), a inulina ou os açúcares;
acumulam amido as raízes de Manihot esculenta (Euphorbiaceae) «mandioca» e de Ipomoea
patatas (Convolvulaceae) «batata-doce»; inulina, as raízes de Cichorium intybus var. sativum
(Asteraceae) «chicória», de Helianthus tuberosus (Asteraceae) «tupinambo» e dos inhames do
género Dioscorea (Dioscoriaceae); sacarose, a raiz de beterraba-sacarina. As raízes tuberosas
são, geralmente, uma adaptação a habitats onde, chegada a estação favorável, é
particularmente vantajoso iniciar no cedo o crescimento vegetativo (e.g. comunidades
herbáceas de regiões com uma estação seca pronunciada) ou a produção de flores (e.g. prados e
florestas caducifólias). Dois tipos de raízes tuberosas:
i) Tuberoso-aprumadas (= tuberculoso-aprumadas) – derivam de um sistema radicular
aprumado; e.g. Daucus carota (Apiaceae) «cenoura»;
ii) Tuberoso-fasciculadas (= tuberculoso-fasciculadas) – derivam de um sistema radicular
fasciculado; e.g. Asphodelus (Asphodelaceae) «asfódelos»; vários géneros de orquídeas
terrestres – e.g. Dactylorhiza, Ophrys e Orchis – possuem, durante o período de
floração, duas raízes tuberosas, uma delas, formada no ano anterior e de aspecto
engelhado, suportou o arranque do crescimento do caule actual, a outra, mais lisa, com
poucos meses de crescimento, sustentará o crescimento inicial do caule no ano
seguinte;
c) Haustórios (= raízes sugadoras) – raízes próprias de plantas parasitas especializadas na
penetração e extracção de água e nutrientes dos seus hospedeiros; podem ser aéreas ou
subterrâneas (vd. figura);
d) Raízes contrácteis – produzidas periodicamente; inicialmente alongam-se em profundidade,
depois contraem-se, aumentando de diâmetro, e puxam os órgãos caulinares para o solo;
frequentes em plantas bulbosas (e.g. Lilium [Liliaceae]), nas gramíneas e nas Arecaceae
«palmeiras»; distinguem-se, morfologicamente, das restantes raízes pelo maior diâmetro e pela
superfície enrugada;
e) Pneumatóforos (= raízes respiratórias) – raízes aéreas emitidas por espécies adaptadas a zonas
húmidas tropicais, para facilitar as trocas gasosas entre o sistema radicular e a atmosfera;
frequentes nas plantas de mangal, e.g. Rizophora mangle (Rizophoraceae) «mangue-vermelho»
e Avicennia africana (Acanthaceae) «mangue-branco»;

32 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
f) Raízes estranguladoras – raízes aéreas emitidas por algumas espécies tropicais de Ficus
(Moraceae) «figueiras» de germinação epífita (na parte aérea do hospedeiro); após a
germinação da semente, as raízes descem pelo troco ou suspendem-se da copa dos hospedeiros
até atingirem o solo; finda esta fase, as plantas aceleram o seu crescimento, competem com o
hospedeiro pelos recursos ambientais e envolvem-no com uma rede raízes que acabam por
coalescer e estrangulá-lo; este processo termina quando os Ficus estranguladores ocupam o
espaço anteriormente preenchido pelo hospedeiro;
g) Raízes tabulares – raízes plagiotrópicas, com função de suporte, muito engrossadas na face
oposta ao solo, geralmente emergentes à superfície nas árvores mais velhas; frequentes nas
grandes árvores tropicais, e.g. Ceiba pentandra (Bombacoideae, Malvaceae) «sumaumeira»;
também nas árvores mais velhas e maior dimensão de Ficus carica (Moraceae) «figueira»;
a) b) c)
a) b) C)
Tipos de raiz. a) Raízes tuberoso-aprumada de Daucus carota (Apiaceae) «cenoura» e tuberoso-fasciculada Asphodelus (Asphodelaceae) «asfódelos». b) Duas plantas parasitas sem clorofila – Orobanche (Orobanchaceae) «orobanca» à esquerda e Cuscuta (Convolvulaceae) «cuscuta» à direita – a parasitar por intermédio de haustórios (não visíveis na figura) os respectivos hospedeiros. c) Raízes aéreas trepadoras de um ramo juvenil de Hedera helix (Araliaceae) «hera» (Coutinho, 1898).
Tipos de raiz. a) Dois nós com raízes escora em Panicum miliaceum (Poaceae). b) Raiz tabular e raízes escora (canto esquerdo e lado direiro da imagem) de Ficus macrophylla (Moraceae). c) Pneumatóforos (raízes que emergem da água em torno da árvore) em Avicennia africana (Acanthaceae) «mangue-branco» (foto da autoria dos alunos dos 12º ano da Escola Portuguesa de Luanda, 2007)

33 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
h) Raízes escora – raízes com função de suporte emitidas da base do tronco ou das pernadas; e.g.
Pandanus (Pandanaceae) «pandanos» e Rizophora mangle (Rizophoraceae) «mangue-
vermelho»; a Zea mays (Poaceae) «milho-graúdo» e o Panicum milliaceum «milho-miúdo»
emitem raízes adventícias anormalmente espessas a partir dos nós inferiores do caule, visíveis
acima do solo, interpretáveis como raízes escora.
3.2.5. Modificações causadas por microrganismos
As micorrizas são associações simbióticas radiculares entre fungos e plantas. As plantas recebem do
fungo água, nutrientes, sobretudo fósforo, e protecção contra infecções radiculares fúngicas e toxinas
presentes no solo (e.g. metais pesados). Os fungos, em troca, têm acesso aos produtos da fotossíntese
da planta. 80% a 90% das plantas-com-semente têm micorrizas. As micorrizas são mais frequentes em
solos pobres em nutrientes ou tóxicos. A dependência das plantas-vasculares das associações
simbióticas micorrízicas é variável: as Proteaceae não têm micorrizas, as Betula (Betulaceae) «bidoeiros»
desenvolvem-se adequadamente sem estas associações, o crescimento dos Quercus (Fagaceae)
«carvalhos» e dos Pinus (Pinaceae) «pinheiros» é muito deprimido sem micorrizas, as sementes de
Orchidaceae «orquídeas» dependem de associações com fungos para germinar.
Existem dois tipos fundamentais de micorrizas :
a) Vesículo-arbusculares (= endomicorrizas, micorrizas endotróficas) – o fungo invade as células
corticais das plantas; a maior parte da massa fúngica situa-se no interior das raízes; limitam-se a
complementar o papel das raízes na absorção de nutrientes do solo; as raízes infectadas são
semelhantes às raízes normais; dominantes nas florestas tropicais; consomem 1-15% do
carbono fixado pelos hospedeiros; baixa especificidade fungo-planta hospedeira; mais
frequentes em solos com matéria orgânica bem humificada; fungos Glomeromycota (=
Zygomycetes p.p.);
b) Formadoras de manto (= ectomicorrizas, micorrizas ectotróficas) – o fungo invade com
haustórios os espaços intercelulares do córtex radicular e forma uma fina camada de micélio,
conhecida por manto micorrízico, que cobre a raízes finas; as raízes infectadas perdem os pelos
radiculares e param de crescer; o fungo emite longos sistemas ramificados de hifas através do
solo que substituem, quase por completo, as raízes infectadas na absorção de nutrientes;
presentes em 90% das árvores temperadas e boreais; consomem 15% ou mais do carbono
fixado pelas plantas hospedeiras; grande especificidade fungo-planta hospedeira; mais
a) b)
Nódulos e micorrizas. a) Nódulos indeterminados em Trifolium (Fabaceae). b) Micorrizas formadoras de manto: corte histológico (à direita); aspecto das raízes normais e raízes micorrizadas (à esquerda) http://www.mhhe.com/biosci/pae/botany/vrl/images/0147.jpg

34 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
frequentes em solos de matéria orgânica ácida e pouco humificada; fungos das divisões
Basidiomycota, Ascomycota e Glomeromycota.
As micorrizas de Ericaceae (e famílias evolutivamente próximas) e de Orchidaceae enquadram-se em
dois tipos especiais de micorrizas não desenvolvidas no texto.
O azoto é o macronutriente mais escasso e que mais condiciona a produtividade vegetal nos
agroecossistemas e nos ecossistemas naturais. Para fazer face a esta limitação alguns grupos de plantas-
vasculares conseguiram estabelecer relações simbióticas com bactérias fixadoras de azoto (= bactérias
diazotróficas), macromorfologicamente expressas pela formação de nódulos radiculares15. As plantas-
com-flor com simbiotes fixadores de azoto formadores de nódulos estão acantonadas no clado das
rosidas I. As Gunneraceae, uma família basal das eudicotiledóneas superiores (ing. “core eudicots”, vd.
vol. III), são a única excepção. As simbioses mais frequentes na Natureza desenvolvem-se entre plantas
não leguminosas e actinomicetas do género Frankia. Os seus nódulos observam-se com alguma
facilidade, por exemplo, em Alnus glutinosa (Betulaceae) «amieiro», Myrica faya (Myricaceae)
«samouco» e Casuarina (Casuarinaceae) «casuarinas». As simbioses com alfa-proteobactérias são
exclusivas da família das Fabaceae, porém, são as mais conhecidas, mais estudadas e de maior
importância económica.
As plantas da família das Fabaceae estabelecem simbioses com pelo menos 12 géneros de alfa-
proteobactérias fixadoras de azoto (e.g. Rhizobium e Bradyrhizobium). Num primeiro estádio da
infecção bacteriana das raízes, os rizóbios16 penetram nas plantas através dos pêlos radiculares. Uma
vez estabelecidos estimulam a actividade meristemática e a proliferação de células do córtex ou do
periciclo. Os nódulos têm uma forma esférica ou mais ou menos alongada e, quando funcionais, após
esmagamento, exibem uma cor rosada causada por uma substância quimicamente próxima da
hemoglobina, a leghemoglobina. Os simbiontes bacterianos localizam-se na zona mais central dos
nódulos radiculares.
Reconhecem-se dois tipos de nódulos entre as Fabaceae:
a) Nódulos determinados – forma esférica; alguns dias a pouca semanas de duração;
b) Nódulos indeterminados – forma alongada; vários meses de duração; alongam-se à custa de um
meristema apical.
As plantas-terrestres estabeleceram relações simbióticas com fungos Glomeromycota – uma das sete
divisões actualmente reconhecidas no reino Fungi – num momento muito recuado da sua evolução. As
endomicorrizas são a condição ancestral da micorrizia; provavelmente auxiliaram as primeiras plantas
terrestres a conquistar a terra emersa (Smith & Read, 2008). Este grupo, certamente muito antigo, de
fungos forma micorrizas arbusculares ancoradas nas células corticais das plantas. As ectomicorrizas e as
endossimbioses com bactérias fixadoras de azoto são termos evolutivos mais tardias. As endossimbioses
radiculares com bactérias desenvolveram-se no clado das rosidas I (angiospérmicas), no Mesozóico.
15 Nem todas as realções simbióticas com bactérias diazotróficas formam nódulos. Por exemplo, bactérias fixadoras de azoto do género Burkholderia foram detectadas em Zea mays «milho-graúdo» e de Saccharum officinarum «cana-de-açúcar», duas plantas cultivadas da família das Poaceae «gramíneas» (Perin et al., 2006). 16 O termo rizóbio, num sentido estrito, refere-se apenas às bactérias do género Rhizobium. Na bibliografia tem geralmente um sentido mais lato sendo aplicado às espécies capazes de nodular as raizes das leguminosas e fixar azoto atmosférico do género Rhizobium, ou de géneros aparentados. Recentemente, foram isoladas em leguminosas bactérias não pertecentes aos grupos tradicionais de alfa-proteobacterias fixadoras de azoto (Willems, 2006).

35 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
3.2.6. As raízes das árvores
Tradicionalmente, admite-se que a estrutura do sistema radicular das árvores reflecte, de algum
modo, a estrutura da copa: as raízes pouco ultrapassam a projecção vertical da copa e ocupam
homogeneamente o solo em profundidade. Na realidade, as raízes das árvores acumulam-se nas
camadas superficiais do solo e estendem-se muito para lá da projecção da copa (4 a 7 vezes o raio da
copa). Nos solos mais espessos as árvores emitem raízes profundantes (em maior número em solos
pouco compactos) até encontrarem algum imperme, água ou atmosferas do solo demasiado pobres em
oxigénio. As raízes profundantes atingem 1-2 m de profundidade; nos solos com características físicas
mais favoráveis (menos compactos e mais arejados) atingem os 3-5 m de profundidade (Thomas, 2000).
Para facilitar a ancoragem ao solo, a maioria das árvores desenvolve uma rede complexa de raízes
laterais mais ou menos horizontais, rígidas e de grande espessura na proximidade do colo, que a poucos
metros da projecção da copa se assemelham a cordas. Estas raízes ramificam-se, sobrepõem-se,
enxertam-se umas nas outras, envolvem grandes pedras e penetram fissuras formando uma estrutura
sólida de grande resistência à tracção. A enxertia radicular é um fenómeno frequente entre indivíduos
vizinhos da mesma espécie.
3.3. Caule
Nas plantas-com-semente a formação do caule inicia-se com a germinação da semente. À medida
que a planta cresce e se desenvolve, novos caules são gerados a partir de outros de ordem inferior.
Pontualmente, os caules têm origem em raízes (pôlas radiculares e rebentos de raiz). Caules vigorosos
diferenciados de raízes são utilizados na propagação de algumas plantas lenhosas (e.g. Salix [Salicaceae]
«salgueiros»). A diferenciação de caules a partir de folhas é muito rara na natureza.
Os caules são entendidos como eixos quando nele se inserem, de forma hierarquizada, outros
elementos; e.g. o tronco (eixo primário das árvores) relativamente aos ramos, estes relativamente às
folhas, o pedúnculo de uma inflorescência relativamente aos pedicelos das flores, etc. De acordo com a
a) b)
As raízes das árvores. a) Modelo tradicional (em cima) e estrutura real dos sistema radicular (em baixo) das árvores (Thomas, 2000). b) Rede de raízes laterais, mais ou menos horizontais, com a função de ancorar a árvore ao solo (Thomas, 2000)

36 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
ordem de inserção reconhecem-se eixos principais (= primários), secundários, etc. O mesmo raciocínio
foi já anteriormente aplicado às raízes.
3.3.1. Funções do caule
O caule desempenha as seguintes funções:
a) Expor as folhas à luz;
b) Expor as flores aos agentes polinizadores;
c) Transportar água e substâncias nutritivas entre a raiz e as folhas (em ambos os sentidos);
d) Armazenar substâncias de reserva;
e) Realizar a fotossíntese nos caules não lenhosos com clorofila.
A presença de folhas e, concomitantemente, de nós são as características macromorfológicas mais
marcantes dos caules, que permitem a sua diferenciação imediata das raízes.
3.3.2. Alongamento, ramificação e cladoptose
O crescimento dos caules nas plantas com corpo secundário tem duas componentes – o
alongamento e o engrossamento – respectivamente conduzidas pelo meristema apical e pelo câmbio
vascular, nas dicotiledóneas s.l., ou pelo meristema de engrossamento secundário, nas
monocotiledóneas com crescimento secundário (vd. Organização do corpo das plantas-com-semente). A
ramificação é governada por meristemas axilares ou adventícios. Uma vez activados, estes meristemas
adquirem a função e as características dos meristemas apicais.
Gemas
Os meristemas não contactam directamente com o exterior: enquanto funcionais permanecem
envolvidos por várias camadas de folhas. Estas podem ser simples esboços foliares (= folhas em início de
desenvolvimento) ou folhas modificadas, geralmente escamiformes, rijas e sem clorofila, designadas por
catáfilos (vd. Tipos de filomas). A estrutura constituída pelos esboços foliares, pelos catáfilos (nem
sempre presentes) e pelas células caulinares recém-diferenciadas, mais o meristema que lhes deu
origem designa-se por gema. A gema pode achar-se activa, a diferenciar caule e folhas, ou quiescente (=
inactiva), apresentando-se, ou não, envolvida por folhas de protecção especializadas.
a) b) c)
Importância taxonómica dos gomos. A forma das escamas dos gomos e a presença de resinas permite distinguir alguns Pinus (Pinaceae): a) Pinus pinaster «pinheiro-bravo» e b) P. pinea «pinheiro-manso». c) Sorbus domestica (Rosaceae) «sorveira-comum» tem gomos viscosos; embora morfologicamente semelhante o S. aucuparia «sorveira-dos-passarinhos» tem gomos secos.

37 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
Os meristemas, sobretudo enquanto activos, são muito frágeis: quebram ou esmagam-se com
facilidade e necrosam em condições ambientais extremas. O funcionamento dos meristemas é
interrompido no início do período desfavorável ao crescimento vegetal nos territórios com uma estação
demasiado fria, ou demasiado seca, para evitar danos irreparáveis nas células meristemáticas e nos
tecidos jovens. Até ao final da estação desfavorável os meristemas permanecem protegidos em gemas
apicais, ou axilares. Na maioria das plantas perenes das regiões com uma estação fria a quiescência dos
gomos, i.e. a interrupção da sua actividade, tem um controlo genético, por via hormonal. Por isso, a
exposição a condições ambientais favoráveis não é suficiente para uma retoma da actividade
meristemática. Este tipo de quiescência, conhecido por dormência, assegura a sincronização do período
de crescimento com a estação favorável.
A prefolheação e a vernação, são, respectivamente, a forma como os esboços foliares se arrumam
individualmente ou em grupo no gomo (vd. Ptixia e vernação). O número de folhas diferenciadas retidas
no interior dos gomos varia de espécie para espécie. Nos trópicos as folhas expandem-se pouco depois
da sua diferenciação: verifica-se uma neo-formação de folhas sempre que os gomos estejam activos.
Nos gomos hibernantes da maioria das árvores de clima temperado (e.g. maioria das Rosaceae de frutos
edíveis) existe uma pré-formação das folhas: os gomos encerram os esboços de todas as folhas a serem
expandidas, por alargamento celular, na Primavera. A forma dos esboços foliares pré-formados
assemelha-se à das folhas adultas e cada esboço axila já um pequeno meristema axilar. Exemplos
intermédios entre a neo e a pré-formação de folhas são também frequentes.
Na quebra da dormência, além de um relógio interno das próprias plantas – a dormência esbate-se
com o tempo –, participam diversos sinais ambientais, e.g. exposição a temperaturas baixas, exposição a
temperaturas elevadas e comprimento do dia. A combinação e a intensidade dos sinais necessários para
a quebra da dormência nas plantas cultivadas varia de espécie para espécie, e ao nível da cultivar. No
início da estação favorável os gomos abrolham: a actividade meristemática é reiniciada, os gomos
incham, o alongamento do caule afasta as folhas de protecção e o meristema encapsulado por folhas
imaturas emerge do interior do gomo. Os catáfilos acabam por se destacar do caule e tombar no solo,
deixando uma cicatriz. As cicatrizes dos catáfilos apresentam-se agrupadas num anel (anel cicatricial),
mais ou menos marcado (por vezes ligeiramente deprimido), muito útil para monitorizar o crescimento
dos ramos do ano. Estes anéis permitem, em muitas espécies (e.g. Rosaceae lenhosas temperadas),
identificar a idade dos ramos não podados – o número de anéis coincide com número de anos – e
determinar a idade das plantas lenhosas (sobretudo nas árvores) mais jovens.
Nos territórios de clima mais benigno as plantas crescem permanentemente, com pequenas
interrupções irregulares nem sempre correlacionadas com variáveis ambientais. No caso de existir uma
interrupção mais ou menos longa do crescimento induzida por condições ambientais desfavoráveis, os
meristemas regra geral não estão protegidos por catáfilos. Os meristemas dos caules aquáticos também
raramente têm catáfilos.
Reconhecem-se dois tipos de gemas:
a) Gomos – gemas protegidas por catáfilos; e.g. olhos da batata;
b) Olhos (= gomos nus) – gemas não ou escassamente protegidas por catáfilos.
A classificação das gemas tem uma grande importância prática, por exemplo, é indispensável em
propagação e na poda de plantas ornamentais ou de árvores fruteiras. Os gomos (e os olhos) são
classificados de acordo com seis critérios:
a) Quanto à situação:
i) Aéreos – formados em caules aéreos (inc. superficiais – se formados na proximidade da
superfície do solo, e.g. no colo de uma árvore);
ii) Subterrâneos – localizados abaixo da superfície do solo;

38 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
iii) Aquáticos – submersos na água;
b) Quanto à natureza:
i) Folheares – dão origem a caules estéreis (sem flores), curtos (braquiblastos estéreis) ou
longos (macroblastos estéreis); e.g. frequentes nas pomóideas e prunóideas;
ii) Florais (= botões) – produzem um caule curto (braquiblasto florífero) com uma flor ou
uma inflorescência, provido ou não de folhas especializadas na função de assimilação (=
nomófilos); e.g. frequentes nas pomóideas e prunóideas;
iii) Mistos – produzem caules, mais ou menos longos (macroblastos floríferos), com flores
solitárias ou inflorescências, estando estas previamente diferenciadas na axila de
esboços foliares contidos no gomo; e.g. videira-europeia;
c) Quanto à posição:
i) Terminais (= apicais) – localizados na extremidade dos eixos caulinares; abrigam, no seu
interior, meristemas apicais;
ii) Axilares (= laterais) – formados na axila das folhas, consequentemente localizados
abaixo de um gomo terminal; acolhem, no seu interior, um ou mais meristemas axilares
(= meristemas laterais);
iii) Adventícios – formados, a posteriori, numa posição atípica, sem relação com a
extremidade dos eixos caulinares ou a axila das folhas; contêm meristemas adventícios;
d) Quanto à inserção:
i) Alternos – posicionados na axila de folhas alternas. Podem ainda ser: espiralados –
dispostos em espiral ao longo dos caules; dísticos (= disticados) – dispostos no mesmo
plano;
ii) Opostos – diferenciados na axila de folhas opostas; quando os gomos de dois nós
sucessivos se encontram em dois planos ortogonais (perfazendo um ângulo de 90°)
dizem-se oposto-cruzados (= decussados);
iii) Verticilados – formados na axila de folhas verticiladas;
e) Quanto à disposição:
i) Colaterais – dois ou mais gomos, uns ao lado dos outros; e.g. pessegueiro (3 gomos) e
videira-europeia (2 ou 3 gomos);
ii) Sobrepostos – dois ou mais gomos, na direcção do eixo caulinar (longitudinalmente
justapostos); e.g. nogueira-europeia;
f) Quanto à evolução:
i) Prontos – evoluem na mesma estação de crescimento em que são formados; e.g. gomo
lateral da videira-europeia;
ii) Hibernantes – abrolham no ano seguinte à sua formação; e.g. a maioria dos gomos das
árvores fruteiras de climas temperados;
iii) Dormentes – permanecem num estado de vida latente durante dois ou mais anos,
podendo nunca abrolhar; condição frequente nas plantas lenhosas;
g) Quanto ao número de meristemas:
i) Simples – com um meristema; condição mais frequente nas plantas-com-flor;
ii) Compostos – com dois ou mais meristemas; e.g. gomos compostos hibernantes da
videira-europeia.

39 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
A “inserção dos gomos” (e dos olhos) depende da disposição
espacial das folhas (= filotaxia) e tem um efeito muito marcado na
estrutura das copas das árvores (vd. Filotaxia). O critério “disposição
dos gomos” reflecte o facto de na axila das folhas poderem co-existir
vários gomos.
Os gomos são particularmente difíceis de classificar na videira-
europeia. Os nós dos sarmentos (caule das vitáceas) do ano de
videira-europeia apresentam um gomo lateral pronto, mais um ou,
raramente, dois gomos compostos hibernantes (Vasconcelos et al.,
2009). O gomo lateral pronto dá origem às netas (ramos laterais); se
não abrolhar acaba por senescer (morre) no Outono ou no Inverno
deixando para trás uma cicatriz. Os gomos compostos hibernantes
são folheares ou mistos; na Primavera seguinte à sua formação dão
origem a um sarmento que poderá apresentar, ou não, cachos de
flores na axila das folhas basais. O gomo lateral é axilado pela folha do
sarmento; os gomos hibernantes diferenciam-se na primeira, ou na
primeira e segunda folhas (prófilos muito modificados) do gomo
lateral. Cada gomo composto hibernante, por sua vez, comporta 3
gemas, uma principal, e duas secundárias, formadas na axila dos dois
primeiros esboços (prófilos) foliares da gema primária; a gema
primária, consoante as variedades, produz 10 a 12 esboços foliares
antes de entrar em dormência, destinados a expandirem-se na
estação de crescimento seguinte.
Número
As plantas podem ter um a muitos caules. Em algumas famílias,
sobretudo entre as Poaceae «gramíneas», Cyperaceae e Juncaceae
«juncos», muitas espécies afilham com facilidade, i.e. diferenciam um
grande número de novos caules a partir dos meristemas axilares das
folhas basais. De cada axila emerge um único caule; os caules filhos,
por sua vez, podem originar novos caules, e assim por diante. Nas
plantas ditas cespitosas estes caules brotam compactados em grande
número numa pequena toiça, próximo da superfície do solo. Nas plantas lenhosas o conceito de número
só é aplicado aos arbustos. A formação de novos caules nas plantas vivazes – plantas perenes que
renovam anualmente a parte aérea –, nos arbustos e nas árvores, salvo raras excepções, deve-se à
diferenciação e activação de gomos apicais ou axilares.
Alongamento rameal
Consoante o comportamento do meristema apical de um eixo caulinar assim se reconhecem dois
tipos de alongamento rameal: monopodial e simpodial.
Nos ramos de alongamento monopodial (ing. “monopodial growth”), i.e. nos monopódios, o
alongamento faz-se pela justaposição de unidades monopodiais formadas durante os períodos de
actividade do meristema apical. Este sistema de alongamento é dominante nas gimnospérmicas. O Ilex
aquifolium (Aquifoliaceae) «azevinho» é uma conhecida angiospérmica de alongamento monopodial.
No alongamento de tipo simpodial (ing. “sympodial growth”), o meristema apical do eixo caulinar no
final, ou ainda durante o período de crescimento aborta ou diferencia-se numa flor, numa gavinha ou
num espinho. No período de crescimento seguinte o alongamento dos ramos é continuado por um dos
meristemas axilares mais próximos do ápice caulinar, necessariamente situado na axila de uma folha ou
de uma cicatriz foliar. Um simpódio é constituído por unidades simpodiais (= caulómeros). O
Gomos sobrepostos. Juglans regia (Juglandaceae) «nogueira» (em sima). Gomos colaterais. Prunus persica (Rosaceae) «pessegueiro»: os dois gomos laterais são florais e o central folhear (em baixo).

40 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
alongamento simpodial é dominante nas angiospérmicas, sobretudo nas monocotiledóneas. A
identificação de um simpódio nem sempre é fácil porque o gomo apical pode estar condensado na
extremidade distal do ramo com um grande número de gomos axilares, como acontece Quercus
(Fagaceae). O alongamento simpodial é fácil de reconhecer em Vitis vinifera (Vitaceae) «videira-
europeia», Corylus avellana (Betulaceae) «aveleira», Aesculus (Sapindaceae) «castanheiros-da-índia» e
em numerosas plantas anuais, e.g. Ranunculus (Ranunculaceae) «ranúnculos» e Solanum lycopersicum
(Solanaceae) «tomateiro». Na videira e nas solanáceas cultivadas, entre outros exemplos, o
alongamento simpodial coloca os cachos numa posição oposta a uma folha (vd. figura).
Os sistemas de alongamento monopodial e simpodial repetem-se nos rizomas e nas inflorescências.
Assim, existem rizomas monopodiais e simpodiais. As inflorescências monopodiais dizem-se indefinidas
e as simpodiais definidas (vd. Inflorescência, Tipos fundamentais).
Intensidade do alongamento
Como se referiu anteriormente, os caules alongam-se pela extremidade. O comprimento atingido
por um caule durante uma estação de crescimento depende do comprimento dos entrenós e do número
de entrenós produzidos pelo meristema apical. O comprimento dos entrenós – i.e. a intensidade do
alongamento – varia de espécie para espécie, e entre os caules de um mesmo indivíduo. Distinguem-se
dois tipos fundamentais de caules quanto à intensidade do alongamento:
a) b)
a) b)
Ramos simpodiais e monopodiais. a) Ramo simpodial de Aesculus hippocastanum (Sapindaceae) «castanheiro-da-índia»; n.b. que o alongamento se fez com um gomo axilar e, por esse motivo, se identifica uma ondulação no contacto entre da madeira do 2º ano e o ramo do ano (imagem da esquerda). b) Ramo monopodial de Malus domestica (Rosaceae) «macieira»; n.b. lentículas (pequenas pontuações suberosas dispersas no ramo do ano).
Simpódio. a) Formação de um simpódio no qual o meristema apical se diferencia numa flor, ou numa inflorescência, que acaba por tomar uma posição lateral; n.b. ausência de folha axilante na flor ou inflorescência (adaptado de van Wyk & van Wyk, 2006). b) No simpódio de videira o meristema apical diferencia-se numa gavinha caulinar, o alongamento do caule é continuado por meristemas situados na axila da folha (Hickey & King, 2000)

41 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
a) Macroblastos – caules geralmente compridos e com entrenós longos; e.g. sarmento, turião e
ramos vegetativos da maioria das árvores e arbustos;
b) Braquiblastos – caules de entrenós curtos, por vezes indistintos ou quase a olho nu e, quando
presentes, com folhas dispostas em roseta (i.e. que parecem sair todas do mesmo ponto,
fazendo fascículos); os géneros Cedrus «cedros», Larix «larícios» ou Pinus «pinheiros», da família
das Pinaceae, produzem todas ou a maioria das folhas em braquiblastos; também muito
frequentes nas angiospérmicas lenhosas (e.g. Rosaceae).
Muitas das adaptações demonstradas pelas plantas ao nível dos órgãos vegetativos envolveram o
alongamento ou o encurtamento dos entrenós. Estas adaptações são recorrentes em diversas linhagens
de plantas-com-flor, e evolutivamente reversíveis, talvez porque exigem pouca informação génica
(reduzido número de genes envolvidos). Os escapos e os estolhos – dois tipos particulares de caule mais
adiante descritos – têm os entrenós desmesuradamente longos. As rosetas de folhas, os bolbos e a flor,
a maior de todas as inovações evolutivas das angiospérmicas, implicaram um encurtamento dos
entrenós. Nos Populus (Salicaceae) «choupos», nos Fraxinus (Oleaceae) «freixos», nos Acer
(Sapindaceae) «bordos» e em muitas árvores com frutos de interesse económico – e.g. Juglans regia
(Juglandaceae) «nogueira» ou entre as Rosaceae, Prunus dulcis «amendoeira», Prunus persica
«pessegueiro», Malus «macieiras», Pyrus «pereiras» e Sorbus «sorveiras» – coexistem macroblastos e
braquiblastos, respectivamente especializados na produção de gomos folheares e florais.
Os braquiblastos rugosos e muito curtos são designados por esporões. A rugosidade é devida às
cicatrizes resultantes da inserção de folhas e pedúnculos florais. A maioria dos gomos dos esporões é de
tipo floral, sendo o alongamento operado por um ou mais gomos folheares. Podem ser rectos (de
grande comprimento em Prunus avium «cerejeira») ou tortuosos (e.g. Pyrus «pereiras») consoante se
alonguem por gomos folheares apicais ou axilares; raramente são ramificados. Nos esporões de Pyrus
«pereiras», Malus «macieiras» e na Magnolia x soulangeana (Magnoliaceae) «magnólia-de-soulange»
formam-se um ou mais entrenós engrossados, com função de reserva, designados por bolsas. Os
esporões jovens com um gomo terminal folhear aguçado são conhecidos em pomicultura17 por dardos.
Os esporões são muito frequentes e longevos entre as Rosaceae.
Nas plantas lenhosas, além dos esporões, distinguem-se outros tipos peculiares de ramos em função
das características do alongamento. Os ramos epicórmicos18 são ramos estéreis (sem flores), de
entrenós longos, geralmente de comprimento variável, provenientes de gomos dormentes ou
adventícios situados em qualquer ponto da copa, na proximidade do colo – pôlas – ou, menos
frequentemente, nas raízes – pôlas radiculares19. Se muito possantes retiram vigor às partes mais velhas
da copa sendo, neste caso, designados por ramos ladrões ou chupões.
Prolepsia e silepsia
O alongamento e a ramificação nas plantas perenes pode ser imediata (= silepsia) ou protelada no
tempo (= prolepsia). Nas regiões de clima temperado ou mediterrânico a ramificação rameal, regra
geral, ocorre através de gomos hibernantes, por definição, situados nos ramos do ano anterior.
Dominam, portanto, os ramos prolépticos. A ramificação imediata, implicitamente baseada em gomos
prontos, é mais frequente em famílias de plantas de origem tropical, e.g. Lauraceae. Os ramos
prolépticos normalmente têm cicatrizes na base resultantes da inserção dos catáfilos; o mesmo não
acontece nos ramos silépticos. Este pormenor tem grande importância prática na interpretação do
crescimento e do vigor das árvores fruteiras.
17 Cultivo de espécies de pomóideas, como as macieiras e as pereiras, entre outras. 18 Correctamente o termo ramo epicórmico dever-se-ia restringir aos lançamentos provenientes de gomos dormentes, por definição de origem caulinar. 19 Nas plantas herbáceas vivazes, as estruturas equivalentes às pôlas e às pôlas radiculares das plantas lenhosas podem ser, respectivamente, designadas por rebentos do colo e rebentos de raiz.

42 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
Tipo, grau e disposição da ramificação
Após a diferenciação do caule persistem na axila das folhas pequenos agregados de células com
capacidade meristemática designados por meristemas axilares ou laterais (vd. Organização do corpo das
plantas-com-semente). A ramificação lateral (= ramificação axilar), o tipo de ramificação mais frequente
entre as plantas-com-semente, depende destes meristemas. Todas as folhas de angiospérmicas axilam
um ou mais meristemas axilares; nas gimnospérmicas a presença de meristemas axilares não é
constante. Na maioria das ‘pteridófitas’ os meristemas laterais situam-se obliquamente, por debaixo da
inserção das folhas, consequentemente, neste grupo de plantas, é pouco apropriado utilizar as
designações meristema e ramificação axilar. Os Lycopodiidae «licófitos» e as Psilotidae «psilotófitos»
ramificam-se de forma dicotómica: o meristema apical fende-se e dá origem a dois ramos inicialmente
semelhantes entre si. Pontualmente, as Cycadaceae «cicas», uma família de gimnospérmicas, as
Arecaceae «palmeiras» e os Pandanus (Pandanaceae) «pandanos», dois grupos de plantas-com-flor,
dividem-se dicotomicamente.
A ausência de câmbio vascular nas monocotiledóneas dificulta e limita a sua ramificação aérea. Nas
mocotiledóneas, em particular nas lenhosas, os primeiros nós das ramificações laterais são curtos e de
diâmetro progressivamente maior. Este facto cria uma zona de fragilidade mecânica na proximidade das
ramificações (óbvias em Dracaena draco «dragoeiro»), aproveitadas por algumas espécies para se
propagarem vegetativamente.
A fisionomia (= aspecto) dos ramos e também o aspecto geral das plantas, i.e. o hábito (= porte),
depende ainda de vários outros aspectos da ramificação:
a) Grau de ramificação – a partir de um eixo principal (e.g. um troco de uma árvores) formam-se
ramos de primeira ordem ou primários; os ramos de segunda ordem ou secundários partem dos
ramos de primeira ordem, e assim sucessivamente; nas plantas lenhosas reserva-se o termo
raminho para os ramos de última ordem formados no próprio ano ou no ano anterior, enquanto
não se reinicia o crescimento vegetativo; os raminhos não ou escassamente atempados, i.e. de
cor ainda verde ou em processo de lenhificação, são genericamente conhecidos por rebentos ou
renovos; o termo lançamento é indiferentemente usado para raminhos ou para caules vigorosos
emitidos em qualquer local da copa, caule ou mesmo raízes;
b) Disposição – depende, em grande medida, da posição dos gomos nos caules; a disposição dos
caules pode ser oposta, alterna ou verticilada.
Direcção e orientação
O caule, ao invés das raízes, tem geralmente um geotropismo negativo: alonga-se em direcções
opostas ao solo. Consoante a direcção, ou direcções, tomadas em relação ao plano representado pelo
solo, os caules são classificados em oito tipos (termos indistintamente aplicados a caules herbáceos,
lenhosos, pouco ou muito ramificados e ramos):
a) Ascendente – inicialmente prostrados curvando, de seguida, para uma posição quase vertical;
quando dobram em joelho num nó dizem-se geniculados;
b) Erecto – verticais ou quase na vertical;
c) Suberecto – quase verticais;
d) Subprostrado – quase aderentes ao solo;
e) Prostrado – aderentes ao solo;
f) Difuso – caule muito ramificado com várias direcções;
g) Decumbente – inicialmente erecto ou suberecto vergando distalmente em direcção ao solo;
h) Trepador ou escandente – direcção em função dos suportes (e.g. muros, ramos, taludes, etc.);
tipo próprio das lianas (= plantas trepadeiras ou plantas escandentes).

43 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
São várias as soluções evolutivas seguidas pelas plantas trepadoras para se suspenderem nos seus
tutores: caules volúveis (vd. Metamorfoses do caule), pecíolos volúveis (que se enrolam em torno do
tutor, e.g. Clematis sp.pl. [Ranunculaceae]), raízes trepadoras (vd. Metamorfoses da raiz), acúleos (vd.
Emergências), espinhos (vd. Espinhos), gavinhas caulinares (vd. Metamorfoses do caule) e gavinhas
foliares (vd. Metamorfoses da folha). As Bougainvillea (Nyctaginaceae) «bougainvílleas», entre outras
plantas, suspendem-se emitindo caules por entre os ramos ou anfractuosidades do tutor.
A orientação do crescimento dos ramos laterais frente ao eixo principal pode ser: ortotrópico – na
vertical – ou plagiotrópico – oblíquo ou na horizontal. Consoante o ângulo de inserção no eixo principal
os eixos plagiotrópicos podem ser:
a) Fastigiados – ângulo de inserção muito agudo;
b) Erecto-patentes – ângulo com cerca de 45°;
c) Patentes – ângulo próximo de 90°;
d) Divaricados – ângulo muito aberto > 90°.
Frequentemente, numa mesma planta, os eixos plagiotrópicos e ortotrópicos têm uma morfologia
distinta, e.g. na forma das folhas e potencial para produzir flores. Na oliveira os ramos ortotrópicos não
produzem fruto sendo activamente eliminados pela poda. Em fruticultura existe a preocupação de
colher garfos em ramos plagiotrópicos porque induzem entradas mais precoces em frutificação.
Dominância e controlo apicais
A activação dos meristemas laterais de um caule é condicionada, em muitas plantas, pela sua
proximidade ao meristema apical (ou a um meristema lateral que o substitua, vd. Alongamento rameal).
Este fenómeno, conhecido por dominância apical, pode ser forte, e suprimir o desenvolvimento de um
grande número de gemas, ou fraco, e apenas uma pequena proporção das gemas se manter dormente.
Nas plantas perenes a dominância apical pode exercer-se no ramo do ano ou propagar-se, com variável
intensidade, aos ramos com mais de um ano.
As plantas lenhosas produzem mais meristemas axilares do que os necessários. Nem todos os
meristemas axilares originam novos caules, caso contrário os ramos ensombrar-se-iam uns aos outros,
num emaranhado caótico e energeticamente ineficiente de caules e folhas. Consequentemente, a maior
parte dos meristemas laterais acaba por nunca ser activado, aborta ou permanece dormente no interior
da copa, com grande probabilidade, durante todo o ciclo de vida da planta. A supressão dos meristemas
laterais nas partes velhas e profundas da copa já não cabe no conceito de dominância apical.
Os lançamentos dos meristemas laterais podem ser, ou não, mais curtos e débeis do que o
lançamento do meristema apical, falando-se neste caso de controlo apical. Os conceitos de acrotonia,
de mesotonia e de basitonia20 são definidos em função do vigor (controlo apical) dos ramos
diferenciados a partir de gomos dormentes, desde a parte proximal à parte distal, numa determinada
unidade de crescimento de uma planta lenhosa. Nos caules acrótonos os lançamentos laterais distais
(mais próximos do ápice) alongam-se mais do que os lançamento próximos da base. Na condição
basítona os crescimentos proximais alongam-se mais do que os lançamentos próximos do ápice. A
mesotonia corresponde a uma condição intermédia. Os efeitos de qualquer uma destas condições
propagam-se no tempo, sendo legíveis na arquitectura das plantas adultas.
A interacção da dominância apical com o controlo apical tem um efeito muito marcado na
configuração espacial (arquitectura) das plantas lenhosas. Na fase de plântula a dominância e o controlo
apical são exercidos por um único meristema apical. Há medida que as estações de crescimento se
20 A terminologia em torno da ramificação e alongamento das plantas é bastante confusa. Alguns autores com o mesmo sentido de basitonia e acrotonia preferem os conceitos de ramificação monopodial e ramificação simpodial.

44 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
sucedem os ramos laterais, assumem, mais ou menos rapidamente, um papel semelhante ao meristema
apical original.
Muitos arbustos, como sejam as giestas (Cytiseae, Fabaceae) e as urzes (Erica, Ericaceae), misturam
uma forte dominância apical com basitonia. Os ramos têm tendência a ramificar-se com pouca
intensidade, particularmente na extremidade distal, e as gemas distais, por seu turno, abortam com
frequência ou diferenciam flores. As ramificações mais próximas da base da planta alongam-se com
mais vigor. Os ramos mais longos são ciclicamente renovados a partir da base em consequência do fogo
ou corte. Em algumas espécies a toiça a alarga-se desmesuradamente formando-se então um xilopódio
com abundantes reservas (particularmente evidente nas plantas sujeitas fogos cíclicos).
Nas árvores a dominância apical é também intensa mas o desenvolvimento dos ramos é
tendencialmente acrótono. Os crescimentos ocorrem predominantemente na extremidade dos ramos
(zonas periféricas da copa). Com o tempo acaba por se formar um tronco bem definido e um hábito
arbóreo. Nas gimnospérmicas arbóreas a dominância dos meristemas apicais mantém-se quase
inalterado durante todo o ciclo de vida, por conseguinte o número de meristemas laterais activados (e
de ramificações laterais) mantêm-se escasso. As árvores crescem permanentemente em altura e
mantêm um eixo primário bem definido do solo até à flecha (parte distal da copa). As angiospérmicas
arbóreas, regra geral, seguem um sistema intermédio – acrotonia dominante. À medida que as árvores
envelhecem, a dominância e o controlo apical esbatem-se: os raminhos do ano (cada vez mais curtos)
ramificam-se abundantemente e o padrão de crescimento dos ramos situados na orla da copa aproxima-
se do modelo basítono. A forma e a altura da copa acabam por se estabilizar nas árvores adultas (o eixo
primário desvanece-se), ao mesmo tempo que se intensifica a desrama natural (queda natural de ramos,
vd. Cladoptose). Ao contrário do que acontece nas gimnospérmicas, o interior da copa fica pouco denso
em ramos enquanto a orla permanece densamente revestida de raminhos. No início de cada estação de
crescimento, regra geral, novos caules são emitidos pelos meristemas apicais e axilares dos caules
formados no ano imediatamente anterior, porém, sobretudo nas árvores velhas, ou com alterações
profundas e recentes na conformação da copa (e.g. por poda, parasitismo ou acidente), surgem ramos
epicórmicos, por vezes muito possantes e eficientes na competição pelos nutrientes radiculares com os
ramos da copa.
Cladoptose
Nas primeiras páginas deste livro defendeu-se que as plantas estão condenadas a crescer (vd.
Estrutura modular das plantas): a produção de ramos e folhas e a expansão da copa para capturar a luz
é uma inevitabilidade nestes organismos. Nas plantas lenhosas a acumulação de ramos em grande
número, ainda que moderado pela dominância apical, aumenta os gastos energéticos (as células vivas
consomem energia), a resistência à deslocação dos fluidos floémicos e xilémicos, o risco de
ensombramento e de lesões mecânicas nas folhas, e o risco de ruptura de ramos e pernadas por efeito
do peso ou da acção mecânica do vento. Por conseguinte, a rejeição dos ramos em excesso na copa e a
aquisição evolutiva de mecanismos para este efeito são potencialmente vantajosos.
A abcisão de ramos chama-se cladoptose. Esta redução activa da massa rameal, à semelhança da
abcisão de folhas, flores abortadas, frutos maduros ou sementes, envolve a formação de camadas de
tecidos especializados que acabam por cortar as conexões vasculares e provocar a morte dos ramos.
Num processo distinto da cladoptose, o peso e o vento forçam também a queda passiva dos ramos em
excesso, seleccionando, preferencialmente, ramos ensombrados, doentes ou mal inseridos (os ramos
cruzados e sobrepostos são mais resistentes à força do vento e, por isso, passíveis de serem arrastados
pelos filetes de ar). As árvores servem-se do vento para limpar as suas copas. As ventanias e os
temporais afinal podem ter um papel importante na saúde das árvores.
Buck-Sorlin & Bell (1998, cit. Bell 2008) recolheram do solo, durante um ano, sob a copa de uma
árvore de Quercus robur, cerca de 37.000 fragmentos (excluindo folhas). As árvores não são eficientes

45 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
por igual a libertarem-se dos ramos em excesso. O Pinus pinaster (Pinaceae) «pinheiro-bravo» desrama
naturalmente, sobretudo em povoamentos densos. Os Cupressus (Cupressaceae) «ciprestes», não. À
medida que os troncos de Cupressus engrossam incorporam no lenho a base dos ramos mais velhos,
estejam eles vivos ou mortos. Para se obterem boas madeiras, por exemplo de C. lusitanica «cipreste-
do-buçaco», é necessário desramar ciclicamente as árvores.
3.3.3. Aspectos gerais da morfologia externa do caule
Situação
Quanto à situação os caules podem ser:
a) Aéreos – tipo dominante;
b) Aquáticos – e.g. caules de Ranunculus subgén. Batrachium (Ranunculaceae) «ranúnculos-
aquáticos»;
c) Subterrâneos – e.g. tubérculos de Solanum tuberosum (Solanaceae) «batateira».
Consistência e superfície
Quanto à consistência reconhecem-se três tipos de caule:
a) Herbáceo – caule tenro normalmente verde; consoante a estrutura da medula pode ser:
i) Fistuloso – oco, i.e. com uma cavidade tubulosa; e.g. caule da maioria das Poaceae
«gramíneas»;
ii) Meduloso – preenchido por um tecido esponjoso; e.g. Sambucus nigra (Adoxaceae)
«sabugueiro»;
iii) Maciço (= sólido) – medula não esponjosa; condição mais frequente;
b) Lenhoso – não verde e com a consistência da madeira; todos os caules lenhosos são inicialmente
herbáceos, a sua consistência é uma consequência do crescimento secundário;
c) Suculento ou carnudo – caules engrossados ricos em água e substâncias de reserva.
Os caules herbáceos normalmente são verdes, lisos, de secção mais ou menos circular, glabros (sem
pêlos) ou revestidos de indumento. Menos vezes são estriados, sulcados ou angulosos. À semelhança do
que acontece nas folhas, as células vivas dos caules lenhosos realizam as trocas gasosas necessárias à
respiração celular através de estomas dispersos pela epiderme. A formação do corpo secundário pelos
câmbios líbero-lenhoso e subero-felodérmico implica a lenhificação dos caules herbáceos. Numa fase
inicial do crescimento secundário os estomas são circundados por tecido suberoso, que se destaca no
fundo verde da epiderme pela sua cor acastanhada ou avermelhada. Nos ramos lenhosos os estomas
acabam por se transformar em pequenas saliências, com um poro no centro, designadas por lentículas.
As lentículas são difíceis de identificar nos caules lenhosos de periderme (casca) espessa e escura.
A superfície dos caules lenhosos é particularmente variável. A periderme pode ser castanha, cinzenta
(e.g. Celtis australis [Cannabaceae] «lódão-bastardo»), creme ou mesmo branca (e.g. Betula
[Betulaceae] «bidoeiro»). Depois de extraída a cortiça no Quercus suber «sobreiro» a periderme é
inicialmente rosa tomando, pouco depois, uma cor vermelho-ocre e mais tarde castanha. A periderme
pode ainda ser brilhante (e.g. Pinus nigra [Pinaceae] «pinheiro-negro») ou baça (Quercus [Fagaceae]),
espessa (e.g. Q. suber) ou delgada, mole (e.g. Sequoia sempervirens [Cupressaceae] «sequoia») ou dura,
lisa (e.g. Celtis australis), sulcada ou destacar-se em placas (e.g. Platanus orientalis [Platanaceae]
«plátano»), em fitas verticais (e.g. Eucalyptus [Myrtaceae] «eucaliptos») ou em fitas horizontais (e.g.
Prunus avium [Rosaceae] «cerejeira»). No Populus nigra (Salicaceae) «choupo-negro», entre outras

46 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
espécies, formam-se bolsas com rebentos epicórmicos no tronco, algo que não acontece nos choupos-
híbridos (e.g. P. x canadensis).
Tipos de caules
As plantas sem caules aparentes dizem-se acaules. Os tipos de caule mais frequentes são os
seguintes:
a) Rizoma – caule subterrâneo, vertical (Elytrigia juncea, Poaceae), horizontal (e.g. Ammophila
arenaria «estorno», Poaceae) ou oblíquo (e.g. Chrysanthemum «crisântemos», Asteraceae),
ramificado ou não, com abundantes raízes adventícias caulógenas, geralmente inseridas nos
nós, revestido de folhas escamiformes (catáfilos) que axilam gemas ou os caules aéreos gerados
pelas gemas; com frequência engrossado (rizoma tuberoso) e com funções de reserva, e.g. Iris
«lírios» (Iridaceae) e Zingiber officinale «gengibre» (Zingiberaceae); as partes mais velhas dos
rizomas vão morrendo, sendo substituídas por outras de forma monopodial (rizomas
monopodiais) ou simpodial (rizomas simpodiais, tipo mais comum);
b) Estolho – caule aéreo, prostrado e com raízes adventícias caulógenas emitidas nos nós; o
Cynodon dactylon «grama» e o Stenotaphrum secundatum «gramão, grama-de-jardim», entre
muitas outras Poaceae, emitem caules radicantes nos nós, os quais, consoante mergulham ou
emergem do solo, são classificados como rizomas ou estolhos;
c) Tubérculo – caule engrossado, com abundantes reservas, folhas reduzidas a pequenas escamas
com uma gema axilar e sem raízes adventícias; geralmente formam-se na extremidade de caules
aéreos ou de rizomas; em condições apropriadas as gemas axilares dão origem a novos caules;
os tubérculos podem ser:
i) Subterrâneos – ocultos no solo; e.g. Solanum tuberosum (Solanaceae) «batateira»;
ii) Aéreos – formados acima da superfície do solo; e.g. Dioscorea alata «inhame-de-são-
tomé»;
d) Prato ou disco dos bolbos – caule curto e de entrenós curtos, geralmente subterrâneo ou
localizado à superfície do solo, de crescimento vertical e com um grande número de raízes
adventícias na base, revestido por uma ou mais folhas carnudas de reserva e, nos bolbos
entunicados, por uma túnica externa constituída por uma ou mais folhas membranosas de
protecção (catáfilos); o conjunto prato do bolbo + folhas recebe o nome de bolbo (vd. Bolbos e
bolbilhos );
a) b)
Rizoma e tubérculo. a) Rizoma de Cynodon dactylon (Poaceae) «grama» e tubérculo de Solanum tuberosum (Solanaceae) «batateira» (Coutinho, 1898). b) Estolhos de Fragaria vesca (Rosaceae) «morangueiro-bravo» (Coutinho, 1898).

47 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
e) Cormos (= bolbos sólidos) – resultam do engrossamento, na
vertical, de um segmento de caule com um ou mais entrenós,
revestido por uma ou mais folhas de protecção e com raízes
adventícias na base; as inflorescências inserem-se no ápice
dos cormos; a renovação dos cormos faz-se pela produção de
um ou mais cormos-filho pela gema apical ou por gemas
laterais; a subordinação do cormo ao conceito de bolbo (vd.
Bolbos e bolbilhos) é imprópria porque os cormos são
morfológica e funcionalmente mais próximos do rizoma (os
rizomas distinguem-se dos cormos por não se apresentarem
revestidos por folhas membranosas); os cormos muito
pequenos caem no conceito de bolbilho (vd. Bolbos e
bolbilhos); e.g. Poa bulbosa (Poaceae), Crocus sativus
(Iridaceae) «açafrão», Gladiolus (Iridaceae) «gladíolos»,
Freesia (Iridaceae) «frésia», Sinningia speciosa (Gesneriaceae)
«gloxinia» e Begonia (Begoniaceae) «begónias» tuberosas; os
pseudobolbos de muitas orquídeas tropicais são, na
realidade, cormos revestidos de folhas espessas;
f) Escapo – caule mais ou menos longo, geralmente sem folhas (áfilo), por vezes com brácteas (vd.
Brácteas) ou escamas, que termina numa flor ou numa inflorescência, com ou sem uma roseta
de folhas na base, as quais, como ele, se originam num bolbo, num rizoma ou em raízes
tuberosas; e.g. Bellis (Asteraceae) «margaridas» e Hyacinthus (Hyacinthaceae) «jacintos»;
g) Colmo – caules, geralmente herbáceos, de nós bem
marcados, frequentemente ocos (fistulosos),
revestidos pelas bainhas das folhas; caule característico
da família das Poaceae;
h) Espique – caule não ramificado, revestido por restos de
folhas, geralmente cilíndrico e esguio, culminado por
uma roseta de grandes folhas, com feixes líbero-
lenhosos fechados em grande número e dispostos
irregularmente; caule das Arecaceae «palmeiras»;
i) Tronco – caule lenhoso, próprio das árvores
gimnospérmicas e da maioria das angiospérmicas de
hábito arbóreo, desprovido de ramos na base,
geralmente cónico e engrossando com a idade; o
tronco (= fuste, na terminologia silvícola) suporta a
copa, a parte aérea ramificada e com folhas das
árvores; as primeiras ramificações do tronco são
designadas por pernadas; a seguir inserem-se as braças
e depois os ramos e os ramos do ano (raminhos);
j) Sarmento – caule lenhoso, muito longo, delgado e flexível que, apoiado em outras plantas, se
pode elevar; os sarmentos jovens, não atempados (i.e. de cor ainda verde), recebem o nome de
pâmpanos; e.g. Vitis vinifera (Vitaceae) «videira-europeia»;
k) Turião – rebentos, frequentemente vigorosos, de origem subterrânea (e.g. Asparagus
(Asparagaceae) «espargos») ou aérea (e.g. Rosa (Rosaceae) «roseiras» e Rubus (Rosaceae)
«silvas»), com folhas (por vezes rudimentares) e sem flores, emitidos na estação de crescimento
por plantas vivazes ou perenes; em Rubus e Rosa as flores formam-se nos ramos do ano laçados
pelos turiões nascidos no ano anterior;
Cormo. Cormos de Arrhenatherum bulbosum subsp. baeticum (Poaceae) «balanquinho»
Designações correntes das ramificações das árvores. a - tronco, b - pernada, c - braça, d - ramo, e - raminho e f - rebento (adaptado de Vasconcellos, 1968)

48 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
l) Filocládio21 (= cladódio) – caule achatado, mais ou menos laminar, que desempenha a função
clorofilina, no qual por vezes se inserem ramos, folhas reduzidas ou flores; na maior parte dos
casos são adaptações à secura edáfica; os caules de Ruscus aculetus (Ruscaceae) «gilbardeira» e
de Opuntia (Cactaceae) são dois exemplos clássicos de filocládio;
m) Caules suculentos – caules volumosos, ricos em água, de diâmetro variável consoante a
disponibilidade de água no solo; folhas frequentemente ausentes ou reduzidas a espinhos; e.g.
Cactaceae «cactos» e muitas Euphorbiaceae africanas e macaronésicas.
3.3.4. Metamorfoses do caule
O caule é evolutivamente mais versátil do que a raiz. Entre os casos descritos de adaptações
caulinares referem-se:
a) Caules de reserva – mais frequentes em climas sazonais quando são necessárias reservas de fácil
mobilização para iniciar o crescimento no início da estação favorável; e.g. tubérculos, rizomas e
bolbos (vd. Tipos de caules);
b) Suculência – vd. Tipos de caules;
c) Caules com funções foliares – vd. Tipos de caules;
d) Caules com função de defesa – vd. Emergências e Espinhos;
e) Caules com função de suporte – além dos caules espinhosos ou aculeados, entre as lianas
observam-se as seguintes soluções adaptativas com função de suporte:
i) Caules volúveis – caules que se adaptam à superfície ou enrolam em torno do tutor;
e.g. Aristolochia sp.pl. (Aristolochiaceae), Ipomoea sp.pl. (Convolvulaceae) e Jasminum
sp.pl. (Oleaceae) «jasmineiros»; consoante as espécies os caules volúveis enrolam-se
tanto para a direita – caules dextrorsos, e.g. Phaseolus (Fabaceae) «feijoeiros» – como
para a esquerda – caules sinistrorsos, condição mais rara, e.g. Humulus lupulus
(Cannabaceae) «lúpulo»;
21 Alguns autores restringem o termo filocládio aos braquiblastos análogos a folhas (e.g. Ruscus) e apelidam de platiclado os macroblastos laminares (e.g. Opuntia).
a) b)
Filocládio. a) Ruscus aculeatus (Ruscaceae): filocládios (ca. 4 cm) com flor (em cima) e fruto tipo baga (em baixo). b) Opuntia elongata (Cactaceae) «figueira-da-índia»

49 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
ii) Gavinhas caulinares – extremidade delgadas de caules flexíveis, ramificadas ou não,
desprovidas de nomófilos, adaptadas a envolver ramos ou outros tipos de suportes;
e.g. gavinhas de Vitis vinifera (Vitaceae) «videira» e Passiflora (Passifloraceae)
«maracujazeiros»; as gavinhas podem ter ainda origem foliar (vd. Outras metamorfoses
da folha); em ambos os casos, depois de contactarem com os tutores, o enrolamento
da gavinha é forçado por um crescimento mais lento dos tecidos mais próximos do
tutor do que os tecidos externos.
3.3.5. Lenho de reacção
O crescimento e a consequente acumulação de massa suscitam, nas plantas lenhosas, fortes tensões
na proximidade do ponto de inserção dos ramos próximos da horizontal (patentes). Os tecidos lenhosos
estão sujeitos a forças de tracção e de compressão consoante se situem na parte superior, ou inferior,
da base dos ramos patentes. O stress imposto pela gravidade incrementa a velocidade da divisão celular
no câmbio vascular e uma sobreprodução de xilema. Este excesso de xilema é designado por lenho de
reacção. Nas angiospérmicas o lenho de reacção acumula-se na parte superior dos ramos – lenho de
tracção – e nas gimnospérmicas na parte inferior – lenho de compressão. O lenho de reacção diminui a
qualidade tecnológica das madeiras e, geralmente, é perceptível através da observação macroscópica
dos ramos, particularmente nas árvores de casca lisa.
3.3.6. Produção de látex
Algumas angiospérmicas exsudam, naturalmente ou por feridas, líquidos de diferente cor e
viscosidade. Os exsudados mais ou menos viscosos, não translúcidos, de cor branca, amarela, laranja,
vermelha ou negra são designados por látex. O látex é uma emulsão complexa de proteínas, açúcares,
resinas, gomas e alcalóides, por vezes de grande toxicidade, que coagula e seca quando exposta ao ar.
Podem existir canais lacticíferos nos caules, raízes, folhas e frutos, porém são sempre mais abundantes
no caule. O látex desempenha várias funções:
a) Protecção contra fungos e bactérias;
b) Protecção contra a herbivoria;
c) Eliminação de subprodutos tóxicos do metabolismo.
Mais de 12.000 espécies de plantas-com-flor produzem látex. A sua presença, cor e abundância têm
grande interesse taxonómico, sobretudo entre a flora arbórea e lianóide tropical. As espécies lacticíferas
mais frequentes na flora Portuguesa pertencem à subfamília Chicorioideae das Asteraceae e ao género
Euphorbia (Euphorbiaceae) «eufórbias». Pertence, igualmente, à família Euphorbiaceae a Hevea
brasiliensis «árvore-da-borracha» cujo látex é utilizado no fabrico da borracha-natural. Muitas outras
famílias compreendem espécies produtoras de látex: Moraceae (e.g. géneros Ficus «figueiras» e Morus
«amoreiras»), Caricaceae (e.g. Carica papaya «mamão») e Anacardiaceae (e.g. Rhus coriaria
«sumagre»). O látex é também muito frequente nas plantas da família Papaveraceae: o látex de Papaver
«papoilas» é rico em alcalóides psicotrópicos (e.g. morfina, um derivado da fenilalanina) e o látex
amarelo de Chelidonium majus «celidónia ou erva-das-verrugas» serve para cauterizar cravos e
verrugas.
3.4. Folha
A folha é um órgão lateral inserido nos caules, usualmente de forma laminar e estrutura dorsiventral
(= bifacial), de crescimento rápido e por regra finito (crescimento determinado). A dorsiventralidade

50 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
implica que as superfícies (páginas) superior e inferior sejam claramente distintas e que, geralmente,
exista um só plano de simetria. A página superior está especializada na intercepção e no processamento
bioquímico da sua luz, a página inferior está optimizada para absorver CO2. As folhas têm origem nas
folhas primordiais do embrião ou nos esboços foliares produzidos pelos meristemas caulinares. As
plantas-vasculares só secundariamente22 não têm folhas ou estas estão reduzidas a bainhas ou
pequenas escamas, dizendo-se então áfilas.
3.4.1. Funções da folha
As folhas desempenham várias funções. As mais relevantes são:
a) Assimilação (fotossíntese) – a produção de fotoassimilados, na maior parte das plantas, ocorre
ao nível das folhas;
b) Transpiração – por definição a perda de água sob a forma de vapor pela cutícula ou pelos
estomas; fundamental no arrefecimento das plantas e na génese das forças de sucção
responsáveis pela circulação de água e nutrientes nas plantas; a turgidez excessiva reduz o
crescimento, a transpiração ao reduzir o teor de água das células, dentro de determinados
limites, incrementa a taxa de crescimento das plantas;
c) Protecção – os meristemas estão sempre protegidos por esboços foliares (= folhas recém-
diferenciadas ainda imaturas) ou por folhas “modificadas” para o efeito (catáfilos);
d) Reserva – as folhas que revestem os bolbos são o exemplo mais conhecido de folhas de reserva;
e.g. Allium cepa (Alliaceae) «cebola»;
e) Reprodução – as peças florais são folhas muito modificadas (e.g. sépalas e pétalas); raramente,
por via assexuada, obtêm-se novas plantas a partir de pequenas gemas diferenciadas na
margem das folhas ou de folhas com capacidade de emitir raízes adventícias peciolares, e.g.
Begonia (Begoniaceae) «begónias»;
f) Absorção de nutrientes – 90% da matéria seca em peso das plantas é constituída, em partes
aproximadamente iguais, por carbono e oxigénio; as folhas capturam do ar o carbono e a
maioria do oxigénio; nos ecossistemas naturais as folhas desempenham um papel significativo
na absorção do azoto (sob a forma de amoníaco gasoso e dióxido de azoto) e do fósforo
(sobretudo nos ecossistemas tropicais);
g) Suporte – através de folhas reduzidas a gavinhas (vd. Outras metamorfoses da folha).
3.4.2. Os filomas
22 Secundariamente porque os ancestrais das plantas áfilas dispunham de folhas.
Eventos elementares ocorridos ao nível dos telomas. Formação dos megáfilos a partir de um sistema de telomas envolvendo uma sequência de três processos elementares: A – culminação (ing. “overtopping”), B – planação (ing. “planation”) e C – concrescência (ing. “webbing”) (Bold et al., 1987).

51 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
Teoria telomática de W. Zimmermann
A teoria telomática de W. Zimmermann tem
servido, desde os anos 30 do século passado, para
explicar as inovações evolutivas verificadas ao nível
do caule e da folha, nas primeiras fases da evolução
das plantas-vasculares. De acordo com esta teoria,
a condição ancestral do cormo das plantas-
vasculares resumir-se-ia a um sistema
tridimensional de caules áfilos (sem folhas), com
um único feixe vascular, dicotomicamente
ramificados, representado pelos fósseis de Rhynia
(Rhyniophyta). A evolução da fisionomia e da
anatomia das traqueófitas ancestrais poderia ser
explanada com base num conjunto reduzido de
processos elementares ocorridos ao nível dos
segmentos terminais estéreis dos caules (telomas), entre os quais (Beerling, 2005):
a) Culminação – diferenciação de ramos laterais determinados (telomas) dominados por caules
(eixos) de crescimento indeterminado;
b) Planação – torção dos sistemas de ramos laterais (telomas) de modo a ocuparem um mesmo
plano;
c) Concrescência – união de telomas;
d) Redução – simplificação e redução dos telomas a estruturas de pequena dimensão.
As folhas, num sentido lato, evoluíram pelo menos por duas vezes nas plantas-vasculares23:
a) Micrófilos – folhas pequenas, em forma de agulha (aciculares), com uma única nervura, sem
lacunas foliares; os micrófilos crescem de forma difusa, de forma mais intensa na base, pela
acção de meristemas intercalares; presentes em Psilotidae «ophioglossidas» e Lycopodiidae
«licófitas»; os micrófilos dos fetos-psilotópsidos são secundários, i.e. resultam de uma
simplificação de megáfilos;
b) Os megáfilos (= eufilos ou folhas verdadeiras) – folhas laminares, secundariamente aciculares,
de sistema vascular complexo e lacunas foliares diversificadas; os megáfilos diferenciam-se em
meristemas de posição apical ou lateral; característicos da maioria dos Euphyllophyta (=
cormófitas): Equisetidae «equisetos», Polypodiidae «fetos-verdadeiros», gimnospérmicas e
Magnoliidae «plantas-com-flor»).
De acordo com W. Zimmermann os micrófilos são telomas reduzidos. Os megáfilos teriam evoluído a
partir de sistemas tridimensionais de telomas, envolvendo uma série hipotética de três transformações:
diferenciação de telomas dominados (culminação); planação dos sistemas de telomas dominados;
diferenciação de limbos foliares através da conexão dos telomas aplanados por expansões laterais de
mesofilo clorofilino (concrescência). Actualmente, consideram-se mais plausíveis duas hipóteses
alternativas à teoria telomática:
a) Micrófilos – evoluíram a partir de excrescências (enações) espiniformes (em forma de espinho)
do caule;
b) Megáfilos – são, originalmente, um caule determinado com expansões laterais concrescentes.
23 Os filódios, nome atribuído às estruturas foliares das brófitas, desenvolvem-se nos gametófitos enquanto os micrófilos e os megáfilos das traqueófitas são característicos da geração esporofítica: têm uma origem evolutiva, não são homólogos.
Traços e lacunas foliares e rameais (adaptado de Raven et al., 2005). Os traços são constituídos pelos feixes vasculares que vão abastecer folhas ou ramos. Nas cormófitas os traços deixam hiatos nos feixes vasculares do eixo caulinar acima da inserção das folhas ou ramos.

52 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
Ainda no âmbito da teoria telomática, os caules providos de sistemas vasculares complexos, que
caracterizam as traqueófitas mais evoluídos, terão resultado da concrescência de dois ou mais telomas.
Tipos de filomas
Os múltiplos tipos de megáfilos das plantas vasculares são genericamente designados por filomas. A
construção do corpo das espermatófitas tem início na germinação de uma semente. Desde o estádio de
plântula (planta recentemente germinada) até à senescência (morte) sucedem-se e coexistem, em
maior ou menor número, no corpo das plantas, vários tipos de filomas:
a) Cotilédones – folhas embrionárias, frequentemente ricas em reservas; interpretáveis como
prófilos de um caudículo embrionário (caule embrionário);
b) Folhas primordiais – folhas de transição, próprias das plântulas recém-germinadas, localizadas
entre os cotilédones e os nomófilos;
c) Nomófilos – filomas especializados na função de respiração e assimilação;
d) Prófilos – filoma(s) do primeiro nó de um caule lateral ou de um caule emitido por um gomo
apical; 1 nas monocotiledóneas e dicotiledóneas basais, geralmente 2 nas dicotiledóneas; regra
geral mais pequenos do que os nomófilos; escamiformes, adpressos ao caule e em posição
adaxial nas monocotiledóneas; de posição lateral nas dicotiledóneas;
e) Catáfilos – folhas com função de protecção, geralmente em forma de escama (escamiformes),
sem clorofila, rígidas e sem meristemas na sua axila; frequentes a envolver bolbos, cormos,
rizomas e gomos;
f) Hipsófilos (= brácteas) – folhas, geralmente modificadas, situadas nas inflorescências (vd.
Brácteas);
g) Antófilos – folhas profundamente modificadas que constituem a flor; dois tipos:
i) Estéreis – antófilos especializados na protecção da flor ou na atracção de polinizadores,
e.g. tépalas, sépalas e pétalas;
ii) Férteis – antófilos com funções reprodutivas, e.g. estames e carpelos.
3.4.3. Aspectos gerais da morfologia externa da folha
Situação
Quanto à situação as folhas podem ser:
a) Aéreas – tipo mais frequente;
b) Aquáticas – flutuantes ou submersas na água livre;
c) Subterrâneas – ocultas no solo; normalmente do tipo catáfilo.
Diferenciação
Nas folhas ditas completas reconhecem-se as seguintes partes:
a) Bainha – parte proximal da folha, alargada e que envolve parcialmente o caule, com a função de
proteger o meristema axilar;
b) Pecíolo – parte da folha, normalmente cilíndrica, situada entre o limbo e a bainha ou que
conecta, directamente, o limbo ao caule;
c) Limbo – parte distal da folha, de ordinário laminar onde, geralmente, se concentra a função
fotossintética da folha.

53 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
A bainha é muito comum nas monocotiledóneas e rara nas dicotiledóneas. Na fase vegetativa das
Poaceae herbáceas e nas Musa (Musaceae) «bananeiras» as bainhas das folhas estão sobrepostas e
comprimidas num pseudocaule. Nas Arecaceae «palmeiras» as bainhas, frequentemente, desfazem-se
em fibras ou dispõem de espinhos. Os pecíolos podem estar reduzidos a espinhos (vd. Adaptações). Nas
folhas compostas observa-se, muitas vezes, uma pequena articulação no pecíolo (vd. Divisão ou
composição). A morfologia do limbo é descrita em pormenor mais adiante (vd. Forma do limbo).
No movimento reversível de folhas ou folíolos (e.g. exposição do limbo ao sol ao longo do dia) estão
envolvidos pequenos engrossamentos circunscritos à base, ou ao ápice do pecíolo, conhecidos por
pulvinos. Nas Fabaceae os pulvinos comandam o movimento dos folíolos ao toque na Mimosa pudica
«mimosa-púdica» e a sobreposição dos folíolos em Trifolium em consequência da falta de água, ou, na
Samanea saman «árvore-da-chuva», em resposta aos dias nebulados. O movimento dos folíolos em
Trifolium diminui a exposição sol e as perdas por evaporação; na Samanea reduz os potenciais efeitos
negativos das chuvas e dos ventos violentos tropicais. Os nós salientes das Poaceae da subfamília
Pooideae também cabem no conceito de pulvino.
Dois tipos frequentes de folhas incompletas:
a) Folhas pecioladas – folhas sem bainha;
b) Folhas sésseis – folhas sem pecíolo.
A folha completa não é um carácter primitivo a partir do qual teriam emergido as folhas pecioladas
ou sésseis. O conceito de folha completa é um artifício pedagógico. Os pecíolos das dicotiledóneas s.l. e
das monocotiledóneas têm uma ontogénese distinta, não são homólogos. Por esse motivo, a constrição
localizada entre a bainha e o limbo nos Bambus (Poaceae, Bambusoideae) e noutras monocotiledóneas
deveria ser designada por pseudopecíolo.
Posição
Quanto à posição as folhas são:
a) Folhas dorsiventrais (= bifaciais) – quando a página superior (= ventral ou adaxial) e a página
inferior (= dorsal ou abaxial) diferem na anatomia e na morfologia externa, por exemplo pela
presença de parênquima em paliçada adaxial e de parênquima lacunoso abaxial, na saliência das
nervuras, no número de estomas e no tipo e disposição do indumento; a maior parte das
angiospérmicas têm folhas dorsiventrais;
b) Folhas equifaciais – faces da folha de morfologia e anatomia semelhante; e.g. folhas adultas de
Eucalyptus globulus (Myrtaceae) «eucalípto-comum»;
Diferenciação da folha. 1. Folha completa de Arum (Araceae). 2. Folha peciolada. 3. Folha séssil de Poaceae. 4. Folha séssil. 5. Folhas reduzidas à bainha de Juncus (Juncaceae) «juncos». 6. Filódios e folhas recompostas de Acacia (Fabaceae) «acácias» (Coutinho, 1898)

54 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
c) Folhas unifaciais – nestas folhas ocorre um sobredesenvolvimento da parte central da página
superior (conduzido pelo meristema adaxial da folha) em detrimento da página inferior e da
expansão lateral da folha; as folhas unifaciais podem ser comprimidas lateralmente (folhas
ensiformes) (e.g. tipo comum nas iridáceas) ou cilíndricas, maciças (e.g. folhas gordas de Sedum
[Crassulaceae]) ou ocas (e.g. Allium [Alliaceae] «alhos»).
Nervação
As nervuras foliares surgem, a olho nu, como espessamentos lineares, muito delgados, normalmente
em alto ou baixo-relevo e de cor mais clara do que o parênquima foliar. Cortes histológicos transversais
mostram que as nervuras são constituídos por feixes vasculares – xilema na face superior da folha e
floema na inferior – frequentemente reforçados por tecidos mecânicos (esclerênquima e/ou
colênquima). As folhas das angiospérmicas, e da maioria das gimnospérmicas, evidenciam uma nervação
hierárquica: subsistem uma ou mais nervuras principais (= nervuras de primeira ordem) que se
desmultiplicam em nervuras secundárias, terciárias, e assim sucessivamente (= nervuras de segunda,
terceira ... ordem). Os feixes vasculares das nervuras principais são mais espessos e reforçado com
tecidos mecânicos do que os restantes.
As nervuras das folhas desempenham duas funções:
a) Transporte de substâncias – importação de água, nutrientes e hormonas via xilema; exportação
dos produtos da fotossíntese via floema;
b) Estabilização mecânica da folha – através do xilema e tecidos mecânicos associados aos feixes
vasculares; a cutícula, a epiderme e a pressão de turgescência das células parenquimatosas
também é importante na estabilização mecânica da folha.
A nervação das folhas é um tema muito especializado e de grande importância em filogenética das
plantas-vasculares. A categorização da nervação das folhas inicia-se com o reconhecimento de dois
modos de ramificação (Roth-Nebelsick et al., 2001):
a) Nervação aberta – nervuras organizadas em árvore e que terminam, de forma livre, no interior
do mesofilo foliar, ou na sua margem; tipo mais primitivo de nervação; e.g. Ginkgo biloba
(Ginkgoaceae) «ginkgo», comum em fetos, raríssima e secundária nas angiospérmicas;
b) Nervação fechada (= nervação reticulada ou nervação em rede) – na nervação fechada as
nervuras apresentam-se anastomosadas e organizadas de forma hierárquica. As nervuras dizem-
se anastosomadas quando se ramificam numa rede onde as nervuras mais finas circundam
pequenas células fechadas (= areólas foliares); a anastomosação facilita a difusão de líquidos na
folha, a nutrição de zonas danificadas da folha e a sobrevivência em climas áridos. A natureza
hierárquica da nervação fechada evidencia-se pela presença uma, ou mais, nervuras principais
(nervuras de 1ª ordem), nas quais se inserem nervuras de 2ª ordem, por sua vez subdivididas
em nervuras de ordem superior. A nervação fechada evoluiu mais que uma vez a partir de tipos
primitivos de nervação aberta; dominante em fetos, gimnospérmicas e angiospérmicas.
Reconhecem-se três tipos maiores de nervação fechada:
a) Peninérvea (= penada) – uma nervura principal (nervura média) que herda os feixes vasculares
principais do pecíolo; nervuras secundárias inseridas ao longo da primária, como as barbas na
ráquis numa pena de ave que terminam, frequentemente, num hidátodo na margem da folha;
quando se identifica apenas a nervura média as folhas dizem-se uninérveas; na maioria das
plantas-vasculares as nervuras secundárias atingem a margem, terminando ou não num dente,
anastosomam-se sem atingir a margem ou arqueiam-se em direcção ao ápice sem o alcançar;
nervação característica de eudicotiledóneas e magnoliidas; rara entre as monocotiledóneas (e.g.
Zingiberales); algumas nervuras das folhas peninérveas merecem designações especiais, e.g.
nervura marginal – percorre todo o limbo próximo da margem da folha;

55 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
b) Palminérvea (= palmatinérvea) – três ou mais nervuras principais inseridas na base (partindo do
ápice do pecíolo), ou próximo da base do limbo, de onde divergem radialmente, como os dedos
numa mão; consoante as espécies as nervuras secundárias são rectas e divergentes (e.g. Malva
[Malvaceae] «malvas») ou curvilíneas e convergentes em direcção ao ápice (e.g. Tamus
communis [Dioscoriaceae] «norça-preta»), neste caso impropriamente cunhadas de curvilíneo-
paralelinérveas; tipo de nervação frequente nas mono e eudicotiledóneas;
c) Paralelinérvea – duas ou mais nervuras principais, normalmente de espessura e dimensão
semelhante, paralelas ao longo da folha e convergentes no ápice; nervuras principais conectadas
por nervuras de ordem superior geralmente inseridas perpendicularmente às nervuras de
ordem inferior; nervação característica das monocotiledóneas, rara nas eudicotiledóneas (e.g.
Plantago lanceolata, Plantaginaceae).
A tipologia enunciada não abarca todos os tipos de nervação fechada conhecidos nas plantas-com-
flor, sobretudo nos trópicos ou no registo fóssil. Para obviar esta dificuldade é cada mais utilizada a
classificação de Hickey (1979) (versão simplificada):
a) Craspedódroma – folhas peninérveas com as nervuras secundárias atingindo a margem da folha;
e.g. Quercus suber (Fagaceae) «sobreiro»;
b) Camptódroma – folhas peninérveas com as nervuras secundárias não atingindo a margem; e.g.
Q. rotundifolia (Fagaceae) «azinheira»;
c) Hifódroma (= uninérvea) – nervuras secundarias ausentes, rudimentares ou ocultas num
mesofilo carnudo ou coriáceo; e.g. muitas Apocynaceae;
d) Actinódroma (= palminérvea s.str.) – três ou mais nervuras primárias inseridas na base, ou
próximo da base, de onde divergem radialmente; e.g. Tilia (Tilioideae, Malvaceae) «tílias»;
e) Acródroma (= curvinérvea) – nervuras primárias e/ou secundárias formando arcos convergentes
em direcção ao ápice da folha; e.g. Celtis australis (Cannabaceae) «lódão-bastardo»;
f) Campilódroma (= curvilíneo-paralelinérvea) – de arcos mais acentuados do que na nervação
acródroma; e.g. Smilax aspera (Smilacaceae) «salsaparilha-bastarda»;
g) Paralelódroma (= paralelinérvea) – duas ou mais nervuras primárias paralelas entre si até ao
ápice, onde convergem; e.g. Poaceae;
a) b)
Nervação da folha. a) 1. Folha peninérvea. 2. Folha palminérvea. 3. Folha paralelinérvea. 4. Folha palminérvea (curvilíneo-paralelinérvea) (adaptado de Coutinho, 1898). b) Tipos de nervação de Hickey (1973)

56 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
h) Palinactinódroma (= pedada) – nervuras primárias divergindo numa série de ramificações
dicotómicas; e.g. Platanus orientalis (Platanaceae) «plátano».
Forma do limbo e recorte
Na descrição da forma do limbo das
folhas há que considerar os seguintes
aspectos: forma geral do limbo, ângulo
da base, ângulo do ápice, forma da base,
forma do ápice, recorte e simetria.
A classificação da forma do limbo
baseia-se na posição do eixo de maior
largura e na relação
comprimento/largura. Em função do
primeiro critério definem-se quatro
formas fundamentais de folhas planas e
simétricas a reter: elíptica, obovada,
ovada e oblonga.
Forma do limbo em folhas planas e simétricas. E.g. as folhas semelhantes ao modelo D3 são ovadas e as semelhantes ao modelo E2 estreitamente-obovadas. N.b. nem todos valores da relação comprimento/lagura e nem todas as posições do eixo de maior largura estão considerados na figura; para as condições intermédias usam-se designações compósitas tipo “folha oblongo-elíptica” ou “linear-oblonga” (adaptado de Systematics Association Committee for Descriptive Biological Terminology, 1962)
Formas do limbo mais frequentes. Consoante a posição do eixo de maior largura das folhas assim se definem os tipos: A) elíptico, B) obovado, C) ovado e D) oblongo (adaptado de Leaf Architecture Working Group, 1999)

57 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
Cruzando os critérios posição do eixo de maior largura, angulosidade da folha e a relação
comprimento/largura obtem-se uma terminologia mais complexa, resumida na figura que se segue.
Alguns exemplos: folhas elípticas – Diospyros kaki (Ebenaceae) «diospireiro»; folhas orbiculares –
Eichornia crassipes (Pontederiaceae); folhas oblongas – Pittosporum undulatum (Pittosporaceae)
«incenso»; folhas ovadas – Citrus sinensis (Rutaceae) «laranjeira-doce»; folhas obovadas – Salix
atrocinerea (Salicaceae) «borrazeira-preta»; folhas lineares – gramíneas; folhas ovado-angulosas –
Solanum nigrum (Solanaceae) «erva-moira»; folhas triangulares – Atriplex prostrata (Amaranthaceae).
Existem muitas outras formas de uso corrente entre as quais:
a) Acicular – em forma de agulha; e.g. folhas clorofiladas de Pinus (Pinaceae) «pinheiros»;
b) Cilíndrica – em forma de cilindro, oco ou maciço; e.g. Allium cepa (Alliaceae) «cebola»;
c) Cordiforme – maior largura próximo da base do limbo e duas aurículas na base, em forma de
coração estilizado; e.g. Tamus communis (Dioscoriaceae) e Ipomoea purpurea (Convolvulaceae)
«batata-doce»;
d) Deltóide (= triangular) – de contorno triangular; e.g. Populus nigra (Salicaceae) «choupo-negro»
e Tetragonia tetragonoides (Aizoaceae);
e) Escamiforme – em forma de escama; e.g. folhas de Cupressus (Cupressaceae) «ciprestes»;
f) Espatulada – em forma de espátula; e.g. Pittosporum tobira (Pittosporaceae);
g) Folha ensiforme – folha unifacial lateralmente comprimida; e.g. Iridaceae;
h) Falciforme – em forma de foice; e.g. folhas adultas de Eucalyptus globulus (Myrtaceae);
i) Flabelada – em forma de leque; e.g. Ginkgo biloba (Ginkgoaceae) «ginkgo»;
j) Hastada – com a folha ponteaguda com lóbulos da base muito divergentes, com a forma da
folha das alabardas; e.g. Rumex angiocarpus (Polygonaceae);
k) Peltada – de mais ou menos contorno circular com o pecíolo inserido no centro ou na sua
proximidade; e.g. Tropaeolum majus (Tropaeolaceae) «chagas» e Ricinus communis
(Euphorbiaceae) «rícino»;
l) Reniforme – em forma de rim; e.g. Calystegia soldanella (Convolvulaceae);
m) Sagitada – de contorno triangular agudo prolongando-se, na base, em duas aurículas ou lóbulos
agudos, dirigidos mais ou menos divergentes; Zantedeschia aetyopica (Araceae) «jarro».
A designação “folha lanceolada” (em forma de lança) é dúbia e deve, por isso, ser evitada embora
seja recorrente na bibliografia.

58 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
O Leaf Architecture Working Group (1999) propõe que o ângulo da base seja medido definindo três
pontos: um vértice situado na inserção do pecíolo no limbo e outros dois pontos resultantes da
intercepção da margem com uma linha perpendicular à linha média, situada a ¼ do comprimento da
folha (a partir da base). A avaliação do ângulo do ápice segue uma lógica similar. Os ângulos do ápice e
da base são agudos ou obtusos consoante os respectivos ângulos sejam inferiores ou superiores a 90°.
Quanto à forma a base das folhas pode ser:
a) Acunheada – em forma de cunha, i.e. triangular;
b) Decurrente – base de margens côncavas que se prolongam pelo pecíolo;
e ainda:
c) Convexa – base de margens convexas;
d) Arredondada – base quase circular;
e) Truncada – base abruptamente interrompida numa margem perpendicular ao eixo de maior
comprimento;
f) Auriculada – base com lóbulos (= aurículas); a forma auriculada pode ser subdividida nos
seguintes tipos:
i) Hastada – auriculada de lóbulos divergentes;
ii) Sagitada – auriculada de lóbulos ligeiramente divergentes a mais ou menos
convergentes, definindo um seio peciolar;
iii) Cordiforme (= cordada na base) – com aurículas pequenas e arredondadas;
iv) Amplexicaule – aurículas envolvendo parcialmente o caule onde se inserem; termo
ainda aplicado a brácteas e estípulas;
v) Assimétrica – aurículas direita e esquerda de distinta forma ou dimensão.
O ápice foliar, quanto forma, pode ser:
a) Acuminado – ápice agudo e de margens côncavas;
b) Atenuado – ápice agudo e de margens pouco côncavas ou rectas;
e ainda:
a) b)
Forma do limbo, do ápice e da base das folhas. a) Outras formas do limbo das folhas: 1 espatulada, 2 cordiforme, 3 hastada, 4 sagitada, 5 obcordiforme, 6 peltada, 7 falcada (adaptado de Diaz et al., 2004). b) Ângulos da base e do ápice. Ângulo da base: A agudo e B obtudo. Ângulo do ápice: C agudo, e D obtuso (adaptado de Leaf Architecture Working Group, 1999)

59 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
c) Arredondado – convexo em semi-círculo;
d) Assovelado (= subulado) – terminando num ponta aguda, mais ou menos longa, semelhante a
uma sovela;
e) Truncado – ápice abruptamente interrompido numa margem perpendicular ao comprimento;
termo ainda aplicado a brácteas, sépalas e pétalas;
f) Retuso – com um entalhe superficial;
g) Chanfrado (= emarginado, = escotado) – com um entalhe mais ou menos profundo; e.g. folíolos
de Ceratonia siliqua «alfarrobeira»;
h) Mucronado – ápice agudo ou obtuso provido de um mucrão, i.e. de uma ponta curta mais ou
menos rígida.
As folhas não recortadas dizem-se inteiras. Dois tipos fundamentais de recorte: marginal e profundo,
cada um dos quais com vários subtipos:
a) Recorte marginal:
i) Crenado – com crenos, i.e. com recortes arredondados;
Crenulado – com crenos muito pequenos;
ii) Dentado – com dentes, i.e. com recortes triangulares, não inclinados e mais ou menos
pontiagudos;
Denticulado – com dentes não inclinados muito pequenos;
iii) Serrado – com dentes inclinados, como os de uma serra, dirigidos para o ápice;
Serrilhado – com dentes inclinados muito pequenos;
b) Recorte profundo:
i) Lobadas – recorte não alcança a ½ da aba da folha;
ii) Fendidas – recortes até ca. ½ da aba da folha;
iii) Partidas – recorte ultrapassa ½ da aba da folha;
iv) Sectas – recorte prolongado até à nervura.
Os recortes das folhas lobadas ou fendidas designam-se por lobos; nas folhas partidas a sectas
reconhecem-se segmentos. Consoante o tipo de nervação os tipos de recorte profundo anteriormente
enunciados são combinados com os prefixos “palmati” e “penati”; e.g. palmatilobado, palmatissecto,
penatifendido e penatipartido. As folhas podem combinar recorte marginal com recorte profundo; e.g. a
Malva sylvestris (Malvaceae) têm folhas palmatifendidas com lobos dentados.
Regra geral as folhas são simétricas, i.e. a aba esquerda e a aba direita são a imagem no espelho uma
da outra. Dois exemplos clássicos de folhas assimétricas: folhas adultas falciformes de Eucalyptus
globulus (Myrtaceae) «eucalipto-comum» e folhas assimétricas na base de Ulmus (Ulmaceae)
«ulmeiros».

60 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
Divisão ou composição
As folhas simples têm apenas um limbo, não articulado. Nas folhas compostas reconhecem-se um ou
mais limbos independentes – os folíolos – geralmente semelhantes entre si e providos de uma pequena
articulação no peciólulo. A articulação identifica-se pela presença de uma constrição anelar,
acompanhada ou não por um pequeno engrossamento. A abcisão (=queda) das folhas compostas dá-se,
geralmente, pela articulação. As folhas compostas não devem ser confundidas com folhas sectas, nem
com ramos de folhas oposto-disticadas. Os segmentos das folhas sectas, ao contrário dos folíolos, são
distintos entre si, frequentemente assimétricos e nunca possuem uma articulação na base. As folhas
compostas têm crescimento determinado e não dispõem de meristemas na axila dos folíolos.
Em função do número de folíolos as folhas compostas são:
a) Unifolioladas (= 1-folioladas) – com 1 folíolo; e.g. Citrus (Rutaceae) «citrinos»; em algumas
espécies de citrinos (e.g. Citrus aurantium «laranjeira-amarga») o pecíolo é alado sendo, por
isso, a articulação do folíolo particularmente evidente;
b) Bifolioladas (= 2-folioladas) – com 2 folíolos; e.g. Colophospermum mopane (Fabaceae), um dos
arbustos mais frequentes do Sul de Angola e de Moçambique;
c) Trifolioladas (= 3-folioladas) – com 3 folíolos; e.g. Trifolium (Fabaceae) «trevos»;
d) Multifolioladas – com 4 ou mais folíolos; e.g. Pistacia lentiscus (Anacardiaceae) «aroeira».
Dois tipos fundamentais de folhas compostas multifolioladas:
a) Palmaticompostas (= digitadas) – todos os folíolos inseridos no ápice do pecíolo, como os dedos
de uma mão; e.g. Aesculus hippocastanum (Sapindaceae) «castanheiro-da-índia»;
b) Penaticompostas (= pinadas24) – folíolos inseridos em duas fiadas opostas, ao longo de um eixo
(= ráquis) disposto no prolongamento do pecíolo; consoante o número de folíolos seja par ou
ímpar (com um folíolo na ponta) as folhas penaticompostas são imparifolioladas (e.g. Ailanthus
altissima (Simaroubaceae) «ailanto») ou parifolioladas (e.g. Ceratonia siliqua (Fabaceae)
«alfarrobeira»).
24 A nomenclatura das folhas compostas é inconsistente. Na bibliografia as folhas pinadas são também impropriamente designadas por pinuladas. Com a mesma raiz resistem os termos bipinulada, 2-pinulada, etc. A folha penada tem uma nervação peninérvea, não confundir com o conceito de folha pinada.
Recorte da folha. Recorte marginal: 1. folha crenada; 2. folha dentada; 3. folha serrada. Recorte profundo: 4. folha penatilobada; 5. folha palmatilobada; 6. folha penatifendida; 7. folha penatissecta. Simetria: 8. Folha assimétrica de Ulmus minor (Ulmaceae) «ulmeiro» (adaptado de Coutinho, 1898)

61 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
As folhas duas ou mais vezes compostas denominam-se recompostas. Nas folhas bipinadas (=2-
penaticompostas ou 2-pinadas), o tipo mais frequente de folhas recompostas, os folíolos estão inseridos
em duas fiadas opostas ao longo de eixos secundários, constituindo pínulas. As pínulas, por sua vez,
inserem-se em duas fiadas opostas na ráquis da folha; e.g. Albizia julibrissin (Fabaceae) «albízia-de-
constantinopla». As folhas bipinadas são paripinuladas ou imparipinuladas consoante disponham um
número par ou ímpar de pínulas. As folhas recompostas podem ainda ser 3, 4 ou mesmo 5 vezes
compostas.
Chave dicotómica 1. Tipos de folhas compostas
1. Folhas com 1 limbo não articulado ................................................................................................................ folhas simples
- Folhas com 1 limbo articulado ou com 2 ou mais limbos semelhantes entre si .................................................................... 2
2. Folhas uma vez compostas ................................................................................................................................................. 3
- Folhas recompostas (= 2 ou mais vezes compostas) ............................................................................................................. 6
3. Folhas com 1, 2 ou 3 limbos ...................................... Respectivamente, folhas unifolioladas, bifolioladas ou trifolioladas
- Folhas com 4 ou mais limbos .......................................................................................................... [folhas multifolioladas] 4
4. Folíolos inseridos no mesmo ponto, como os dedos numa mão ................................................. folhas palmaticompostas
- Folíolos inseridos em 2 fiadas ao longo de um eixo ..................................................................... [folhas penaticompostas] 5
5. Número par de folíolos ........................................................................................... folhas penaticompostas parifolioladas
- Número ímpar de folíolos ..................................................................................... folhas penaticompostas imparifolioladas
6. Pínulas com 3 folíolos inseridos no mesmo ponto, como os dedos numa mão .................................................................. 7
- Pínulas com 4 ou mais folíolos inseridos em 2 fiadas opostas ao longo de um eixo ............................................................. 8
7. Folhas 2 vezes compostas .........................................................................................................................folhas biternadas
- Folhas 3 vezes compostas .......................................................................................................................... folhas triternadas
8. Folhas 2 vezes compostas .......................................................................................................................... folhas 2-pinadas
- Folhas 3 vezes compostas ............................................................................................................................ folhas 3-pinadas
As folhas da bananeira e de muitas palmeiras embora se assemelhem a folhas composta são, na
realidade, folhas sectas cujo recorte resulta da rotura de um único limbo através de linhas de
fragilidade. Alguns autores atribuem-lhes a designação de folhas pseudocompostas.
Não é claro se as folhas compostas são homologáveis a caules de crescimento determinado ou a
folhas simples. No primeiro caso os folíolos e as folhas simples seriam homólogos; no segundo, os
folíolos corresponderiam a subdivisões de uma folha simples e a ráquis coincidiria com a nervura média
(Champagne & Sinha, 2004).
A produção de folhas grandes é, teoricamente, vantajosa quando as plantas competem ferozmente
pela luz. No entanto, as folhas de grande dimensão, por estarem envolvidas por uma espessa camada
limite, acabam por ter dificuldade em absorver o dióxido de carbono e estão submetidas a tensões
mecânicas que as podem partir ou rasgar. As folhas compostas, à semelhança das folhas profundamente
recortadas, sobrevieram, de forma independente, em várias linhagens de plantas-com-flor por serem
uma solução evolutiva eficiente para este trade-off.

62 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
Apêndices foliares
As estípulas e as lígulas são os apêndices foliares mais frequentes e de maior interesse taxonómico.
As estípulas inserem-se no pecíolo das folhas ou nos nós. Geralmente, possuem uma forma laminar,
chegando em alguns grupos de plantas a tomar forma e a dimensão de folhas (estípulas foliáceas) ou
folíolos. As estípulas são raras nas monocotiledóneas e frequentes nas dicotiledóneas. As
dicotiledóneas, geralmente, apresentam duas estípulas, uma de cada lado de um pecíolo ou de um nó.
Primordialmente, têm a função de proteger os primórdios foliares ou os meristemas axilares, após a
expansão da folha. As estípulas podem apresentar-se transformadas em espinhos (vd. Espinhos) ou,
raramente, em gavinhas (vd. Metamorfoses da folha). As estípulas são classificadas em função do local
onde se inserem e da sua concrescência (vd. Figura Tipos de estípulas). O tipo de estípulas é muito
importante na identificação de algumas famílias de óptimo tropical; e.g. Rubiaceae. Estas estruturas
surgiram de forma independente em muitas linhagens de angiospérmicas; e.g. as estípulas de
Rubiaceae, Fabaceae, ou de Brassicaceae não homólogas (Bell, 2008).
As lígulas são apêndices membranosos de origem epidérmica, por vezes transformados numa fiada
de pêlos, situados no encontro da bainha com o limbo de muitas monocotiledóneas. São
particularmente frequentes na família das Poaceae. Também, por exemplo, e Hedychium gardnerianum
(Zingiberaceae) «conteira». Admite-se que se destinam a proteger a bainha da água da chuva ou de
insectos.
a) b)
Composição da folha. a) Composição das folhas: 1. folha parifoliolada; 2. folha imparifoliolada; 3. folha digitada; 4. folha 2-pinada paripinulada (adaptado de Coutinho, 1898). b) Hipóteses explicativas da origem das folhas compostas. Hipótese A: a folha composta é homologável a um caule de crescimento determinado. Hipótese B: a folha composta é homologável de uma folha (Champagne & Sinha, 2004)
Tipos de estípulas. 1 estípulas livres, 2 estípulas aderentes ao pecíolo, 3 ócrea (estrutura estipular tubulosa), 4 estípula inserida por debaixo do pecíolo. 5 estípula intrapeciolar, 6 estípula interpeciolar (adaptado de http://www.anbg.gov.au/glossary/webpubl/glossik.htm )

63 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
Superfície e epifilia
A superfície das folhas e demais órgãos aéreos herbáceos – caules herbáceos e peças da flor – é
muito diversa e, por isso, rica em informação taxonómica (vd. Indumento). Designa-se por epifilia o
desenvolvimento de estruturas na superfície das folhas. Entre outras estruturas surgem na superfície
das folhas:
a) Gemas adventícias – muito raras na
natureza; e.g. em Begonia (Begoniaceae)
«begónias» e Bryophyllum (Crassulaceae)
formam-se pequenas gemas na margem
das folhas que dão origem as novos
caules, usados na multiplicação
vegetativa comercial destas espécies;
b) Raízes adventícias – também infrequente;
e.g. Saintpaulia ionantha (Gesneriaceae)
«saintpaulia»;
c) Espinhos – as folhas podem apresentar
espinhos tanto na superfície como na
margem da folha (vd. Espinhos);
d) Indumento – revestimento de origem
epidérmica (vd. Indumento).
Consistência e cor
Quanto à consistência as folhas podem ser:
a) Herbáceas – folhas tenras; e.g. maioria das gramíneas;
b) Coriáceas – consistência semelhante à do couro; e.g. Quercus suber (Fagaceae) «sobreiro»;
c) Carnudas – espessas e ricas em água; e.g. Sedum sp.pl. (Crassulaceae).
As folhas são geralmente verdes podendo, consoante a concentração relativa dos pigmentos,
mostrar-se amarelas, purpúreas, etc. Entre as plantas ornamentais são frequentes cultivares, ou clones,
de folhas variegadas ou listadas de diversas cores; e.g. Agave americana cv. Variegata (Agavaceae)
«piteira». As folhas (e outros órgãos) com tons azulados ou verde-mar dizem-se glaucas. Pouco antes da
abcisão (queda) das folhas, muitos constituintes foliares são desmantelados e deslocados para centros
de crescimento activo (e.g. gemas) ou armazenamento, e as folhas mudam de cor. Nas plantas
caducifólias a desmobilização dos nutrientes das folhas é despoletada e sincronizada por factores
ambientais (e.g. comprimento do dia) o que explica a mudança, quase simultânea, da cor da copa das
árvores dos bosques temperados de folha caduca no Outono.
Duração
As folhas são órgãos de duração muito limitada. Nas plantas anuais e bienais as folhas basais, muitas
vezes organizadas em rosetas, entram em senescência à medida que os caules se alongam e as
ensombram. Findo o ciclo de vida as folhas sobreviventes morrem quase em simultâneo. Nas plantas
perenes as folhas mais velhas têm que ser, de algum modo, substituídas por outras novas e
fotossinteticamente mais eficientes (vd. Estrutura modular das plantas). As plantas perenes seguem
duas estratégias na renovação das folhas:
a) Caducifolia – renovação da copa no início da estação favorável caindo as folhas, quase em
simultâneo, no início da estação desfavorável; as plantas de folha caduca podem ser caducifólias
Gemas adventícias. Gemas adventícias foliares em Bryophyllum (Kalanchoe) daigremontiana (Crassulaceae), n.b. folhas e raízes adventícias suspensas nas gemas adventícias

64 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
de inverno ou de estação seca25; a Adansonia digitata (Bombacoideae, Malvaceae)
«embondeiro», uma importante árvore das zonas áridas africanas, e a Euphorbia pedroi
(Euphorbiaceae), um endemismo da Serra da Arrábida, estão despidas de folhas na estação
seca; uma boa parte da flora lenhosa portuguesa é caducifólia de Inverno, e.g. Quercus
pyrenaica (Fagaceae) «carvalho-negral», Betulaceae «bidoeiros, amieiros e aveleiras»,
Salicaceae «salgueiros», Ulmaceae «ulmeiros» e ainda as Rosaceae arbustivas e arbóreas
indígenas de Portugal (excepto Prunus azorica, P. hixa e P. lusitanica);
b) Perenifolia – copa revestida de folhas durante todo ano; nas plantas de folha perene (= folha
persistente) raramente permanecem folhas funcionais (i.e. verdes) com mais de 3 anos; nas
regiões com uma estação desfavorável bem marcada a queda das folhas nas plantas perenes
normalmente é concentrada no tempo e antecede ou é simultânea com a produção de novas
folhas.
As plantas semi-caducifólias retêm parte das folhas durante a estação desfavorável, permanecendo
muitas delas funcionais. O Quercus faginea subsp. broteroi (Fagaceae) «carvalho-cerquinho» é o
exemplo mais conhecido da flora portuguesa. Curiosamente, os indivíduos indígenas de Q. robur
«carvalho-roble», apesar de pertencerem ao subgén. Quercus, são também semi-caducifólios. A
retenção de órgãos secos – e.g. folhas, sépalas, pétalas, frutos e frutificações – designa-se por
marcescência. Os indivíduos jovens de Q. pyrenaica «carvalho-negral» têm folhas marcescentes,
carácter que se esbate e desaparece nos indivíduos adultos. No Pinus halepensis (Pinaceae) são
marcescentes as pinhas e em muitas Lamiaceae os cálices.
Filotaxia
Num sentido lato entende-se por filotaxia a descrição abreviada dos padrões de disposição espacial
de estruturas similares nas plantas, e.g. caules, folhas, brácteas, flores e peças da flor. A regularidade da
disposição dos ramos de ordem superior nos ramos de ordem inferior, das folhas nos caules ou das
brácteas e das flores nas inflorescências, são uma consequência directa da regularidade temporal e
espacial da diferenciação de estruturas na periferia dos meristemas apicais. Estes padrões têm um
apertado controlo genético que se expressa ao nível dos meristemas.
25 Nos territórios mediterrânicos a estação mais quente é a mais seca; nas áreas tropicais as temperaturas médias mais baixas verificam-se na estação seca.
a) b)
Perenifolia e caducifolia. Marcescência. a) Euphorbia pedroi (Euphorbiaceae) uma espécie caducifólia de Verão. b) Pinus halepensis (Pinaceae), uma árvore de folhas persistentes e pinhas marcescentes

65 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
Ao nível da folha reconhecem-se três tipos de fundamentais de filotaxia:
a) Alterna – uma folha por nó; tipo mais frequente de filotaxia; as folhas arrosetadas – folhas de
caules com entrenós muito curtos e, por isso, colapsadas umas sobre as outras – têm, regra
geral, filotaxia alterna. As folhas alternas dispõem-se em hélice em torno do caule; o ângulo
estabelecido entre folhas alternas sucessivas (ângulo de divergência φ) permite diferenciar os
seguintes subtipos de folhas alternas:
i) Dísticas (= disticadas) – folhas de dispostas num mesmo plano (φ = 90°); e.g. Fagus
sylvatica (Fagaceae) «faia» e Ulmus (Ulmaceae) «ulmeiros»;
ii) Trísticas – folhas dispostas em três planos (φ = 120°); e.g. Carex (Cyperaceae) «cárices»
iii) Helicoidais (= espiraladas) – termo genericamente usado para designar os restantes
tipos de divergência; tipo mais frequente; e.g. Quercus (Fagaceae) «carvalhos». As
folhas alternas helicoidais podem ocupar um mesmo plano por torção do pecíolo (e.g.
Abies alba [Pinaceae] «abeto-europeu») ou do caule (e.g. comum no género Asclepias
[Apocynaceae]), dizendo-se então pseudodísticas;
b) Oposta – duas folhas por nó; as folhas opostas, sésseis, soldadas na base parecendo perfuradas
pelo caule dizem-se adunadas (e.g. Lonicera implexa [Caprifoliaceae] «madresilva-entrelaçada»);
consoante a disposição relativa das folhas de dois nós sucessivos distinguem-se:
i) Folhas oposto-disticadas – folhas de dois nós sucessivos no mesmo plano;
ii) Oposto-cruzadas (= decussadas) – folhas de dois nós sucessivos em planos
perpendiculares; e.g. Olea europaea (Oleaceae) «oliveira»;
c) Verticilada – três ou mais folhas por nó; e.g. Nerium oleander (Apocynaceae) «loendro» e
Catalpa bignonioides (Bignoniaceae) «catalpa».
Numa mesma planta pode co-existir mais de um tipo de filotaxia. Por exemplo, os ramos juvenis de
Eucalyptus globulus (Myrtaceae) têm folhas opostas e os adultos folhas alternas. Em outras plantas as
folhas dos ramos verticais (ramos ortotrópicos, vd. Orientação) são alternas helicoidais e as dos ramos
próximos da horizontalidade (ramos plagiotrópicos) alternas disticadas.
A regularidade da disposição das folhas nas plantas tem sido explorada pelos botânicos desde o
século XVII. A disposição espacial das folhas é, normalmente, representada por um rácio em que o
numerador corresponde ao número de espirais necessárias até à sobreposição de duas folhas (quando
observadas do topo do caule) e o denominador ao número de folhas que medeiam duas folhas
a) b)
Inserção das folhas. a) 1. Folhas alternas; 2. Folhas opostas; folhas verticiladas (adaptado de Coutinho, 1898). b) Plantago coronopus (Plantaginaceae): roseta de folhas

66 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
sobrepostas. Por exemplo, o rácio 2/5, característico de plantas como os Quercus (Fagaceae)
«carvalhos» ou o Prunus avium (Rosaceae) «cerejeira», significa que são necessárias duas espirais para
que duas folhas se sobreponham e, entre estas (contando apenas uma das folhas dos extremos da
espiral), contam-se 5 folhas. Na Natureza observam-se os rácios (espirais de Fibonacci) 1/2, 1/3, 2/5,
3/8, 5/13, 8/21, etc. Constata-se que o denominador de cada rácio corresponde à soma dos dois
denominadores imediatamente anteriores – e.g. 13=8+5 ou 21= 13+8 – perfazendo a sequência
numérica principal de Fibonacci. A divisão entre denominadores sucessivos – e.g. 8/5, 13/8, 21/13, etc. –
converge no número de Fibonacci ou proporção áurea, 0.618034, usada em arquitectura desde a
antiguidade e recorrente na organização de muitos seres vivos (e.g. conchas de Nautilus «nautilus»,
Mollusca, Nautilidae). A sequência de ângulos 360°*1/2, 360°*1/3, 360°*2/5, etc., por sua vez, converge
no chamado ângulo de Fibonacci.
A diferenciação de primórdios no ápice do meristema apical é controlada por processos de repulsão
ou inibição entre primórdios, de fisiologia não esclarecida, que compelem o primórdio em diferenciação
a formar-se na posição o mais distante possível dos primórdios anteriores. Tendo em consideração esta
constatação empírica, conhecida por regra de Hofmeister (Kirchoff, 2003), foi demonstrado que as
espirais de Fibonacci anteriormente enunciadas, emergem espontaneamente se:
a) O ápice do meristema for longitudinalmente simétrico;
b) A velocidade a que se diferenciam os primórdios foliares, i.e. o plastocrono, for constante;
c) A velocidade da migração radial dos primórdios foliares no ápice caulinar for constante;
d) Não houver uma reorganização espacial dos primórdios foliares após a sua diferenciação;
Por conseguinte, a sequência numérica de Fibonacci não está inscrita no genoma: tem um controlo
genético indirecto. Não existe uma numerologia do vivo!
Ptixia e vernação
Entende-se por ptixia a forma como os esboços foliares se dobram, individualmente, nos gomos. A
vernação (= prefolheação) é um conceito distinto de prefolheação porque se refere ao arranjo das
folhas, umas em relação às outras, no gomo folhear. O estudo da ptixia e da vernação envolve o corte
transversal de gemas dormentes ou recém-abrolhadas, e a sua observação à lupa. Estes dois conceitos
são considerados sinónimos na bibliografia mais antiga. A terminologia da ptixia da folha e da vernação
são discutidas mais adiante, visto serem comuns com a flor (vd. Ptixia e estivação da flor).
Heterofilia
As plantas heterofilas possuem dois ou mais tipos morfológicos de folhas. Já anteriormente se aludiu
às folhas juvenis e adultas de Eucalyptus ou de Quercus rotundifolia «azinheira» (vd. Variação
morfológica intraespecífica). O mesmo fenómeno ocorre na Hedera helix (Araliaceae) «hera». Um
dimorfismo foliar igualmente evidente ocorre em muitas plantas aquáticas entre as folhas submersas e
flutuantes; e.g. Ranunculus sp.pl. (Ranunculaceae) e Callitriche sp.pl. (Plantaginaceae). Na Pyrancantha
coccinea (Rosaceae) «piracanta» a forma e a dimensão dos nomófilos varia, de forma contínua, à escala
do indivíduo e entre indivíduos. Nos Quercus caducifólios as folhas de sombra e de sol, e a folhas de
Posição dos prófilos nos ramos laterais nas mono e dicotiledóneas s.l. N.b. a folha tetriz insere-se no eixo primário e axila uma ramificação lateral

67 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
Primavera e de Verão, são, por vezes, tão distintas que é difícil identificar a espécie em alguns
exemplares de herbário.
A heterofilia é patente ao longo do eixo das plântulas ou dos ramos laterais provenientes do
abrolhamento de gomos dormentes. Estas sequências temporais de folhas, designadas por séries
heteroblásticas, nos eixos das plântulas incluem 1 ou 2 cotilédones, folhas primordiais e nomófilos. Nos
lançamentos caulinares das plantas lenhosas das regiões com uma estação desfavorável verifica-se,
geralmente, a seguinte sequência temporal de filomas: 1 (nas monocotiledóneas e dicotiledóneas
basais) ou 2 (nas dicotiledóneas s.l.) prófilos, um número variável de nomófilos e, no final da estação de
crescimento, a formação de catáfilos a envolver o meristema apical. Esta sequência é modificada com a
diferenciação de flores ou inflorescências: aos nomófilos sucedem-se então hipsófilos e, nas
inflorescências determinadas, os antófilos que compõem a flor.
Os nómofilos das angiospérmicas primitivas
Admite-se que as primeiras plantas com flor tinham nomófilos simples, inteiros, de nervação
fechada, peninérveos e alternos. De facto, este tipo de nomófilo é o mais comum nos grupos mais
antigos de plantas com flor (angiospérmicas basais e magnoliidas).
3.4.4. Metamorfoses da folha
A folha é dos três órgãos fundamentais das plantas o órgão evolutivamente mais versátil na forma e
na função. A bibliografia é pródiga em exemplos de adaptações especializadas ao nível da folha.
Bolbos e bolbilhos
Os bolbos são constituídos por um caule curto (prato ou disco do bolbo, vd. Tipos de caules),
geralmente vertical, de entrenós curtos e com um grande número de raízes adventícias muitas vezes
contrácteis na base, revestido por folhas carnudas de reserva e com uma gema apical muito
desenvolvida. Reconhecem-se dois tipos de bolbos:
a) Entunicados – com um grande número de folhas carnudas de reserva perfeitamente
sobrepostas e exteriormente envolvidas por uma ou mais folhas membranosas (túnica); as
folhas de reserva podem estar reduzidas a bainhas (e.g. Allium cepa [Alliaceae] «cebola»),
possuírem uma base alargada de reserva e prologarem-se num limbo mais ou menos longo
a) b)
Heterofilia, dimorfismo foliar. Hedera helix s.l. (Araliaceae) «hera». a) Ramos juvenis (ramos estéreis): monopodiais, prostrados, fincados no tutor através de raízes adventícias adesivas e com folhas lobadas de inserção dística. b) Ramos adultos (ramos férteis): simpodiais, erectos, sem raízes adventícias e com folhas inteiras de inserção espiralada

68 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
(falsos bolbos) (e.g. Amaryllis [Amaryllidaceae]) ou incluírem folhas escamiformes e folhas com
limbo (e.g. Narcissus [Amaryllidaceae] «narcisos»);
b) Escamosos – com escamas imbricadas, i.e. imperfeitamente sobrepostas, como as telhas de um
telhado; e.g. Lilium candidum (Liliaceae) «açucena».
A cabeça-de-alho (Allium sativum) corresponde a um terceiro tipo de bolbo que poderíamos apelidar
de bolbo de bolbilhos. Na cabeça-de-alho os bolbilhos (dentes de alho) não se sobrepõem
completamente, como no bolbo escamoso, sendo revestidos por uma túnica membranosa.
A renovação dos bolbos nas plantas perenes geralmente faz-se pela diferenciação de um bolbo-filho
na extremidade do bolbo-mãe (bolbos monopodiais) ou pela diferenciação de novos bolbos a partir de
gemas situadas na axila de folhas carnudas ou membranosas (bolbos simpodiais); os bolbos-filhos
geralmente ficam encapsulados pelos catáfilos do bolbo-mãe (e.g. cabeças-de-alho). Nas bulbosas
bienais (e.g. Allium cepa «cebola») o bolbo forma-se no final do primeiro ano de crescimento; no final
do segundo ano as plantas renovam-se por semente. Os bolbos diferenciam inflorescências terminais a
partir da gema localizada no centro do bolbo, ou inflorescências laterais inseridas na axila de uma folha
carnuda. Tanto os bolbos como os bolbilhos são mais frequentes nas monocotiledóneas.
Os bolbilhos são bolbos ou cormos de pequenas dimensões diferenciados numa inflorescência (e.g.
Allium vineale), na axila de uma folha, tipo catáfilo ou não, de um bolbo-mãe (e.g. dentes de Allium
sativum [Alliaceae] «alho»), ou ainda pelo engrossamento de um ou mais entrenós, geralmente na base
do caule (e.g. Poa bulbosa [Poaceae]). No alho os bolbilhos (dentes de alho) são constituídos por uma
folha externa membranosa adaxial (corresponde a um prófilo) que reveste uma espessa folha carnuda
tubulosa, no interior da qual se dispõem várias folhas, não carnudas e de limbo reduzido, em torno de
uma gema.
Os bolbos e os bolbilhos desempenham duas funções: reprodução e reserva de energia, água e
nutrientes. A separação dos bolbos-filho e dos bolbilhos da planta-mãe é uma forma de reprodução
assexuada. As reservas têm um grande valor adaptativo em climas com uma estação seca muito longa
(e.g. clima mediterrânico ou tropical com estação seca) ou em habitats onde seja vantajoso produzir
flores e folhas muito cedo (e.g. antes do abrolhamento das árvores nos bosques caducifólios ou antes
do encanamento das gramíneas nos prados). Nestes ambientes as reservas dos bolbos antecipam a
acumulação de biomassa e o crescimento no arranque da estação favorável ao crescimento das plantas.
As espécies bulbosas podem então cumprir o seu ciclo fenológico antes das plantas mais competitivas.
Os rizomas, os cormos e as raízes tuberosas desempenham funções análogas aos bolbos. A posição
horizontal próxima da superfície do solo de muitos rizomas facilita a disseminação vegetativa das
Bolbo e cormo. 1. bolbo entunicado (corte longitudinal); n.b. bolbo envolvido por folhas escamosas e raízes adventícias fasciculadas inseridas no prato do bolbo. 2. bolbo entunicado (cortes longitudinal e transversal); n.b. prato do bolbo e folhas carnudas. 3. bolbo escamoso. 4. cormo (bolbo sólido) (adaptado de Coutinho, 1898)

69 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
plantas. Não surpreende, por isso, que os rizomas sejam muito frequentes em habitats ciclicamente
perturbados que promovam a fragmentação e transporte dos rizomas; e.g. margens de cursos de água e
sistemas dunares.
Outras metamorfoses
Entre as metamorfoses mais frequentes da folha contam-se:
a) Filódios – folhas reduzidas a um pecíolo (e.g. Asparagus [Asparagaceae] «espargos») ou a um
ráquis (e.g. Acacia [Fabaceae] «acácias») dilatado e achatado, semelhante a um limbo foliar; a
presença de meristemas axilares comprova a natureza foliar dos filódios; em muitas Acacia –
e.g. A. melanoxylon «acácia-austrália» e A. longifolia «acácia-de-espigas» – observam-se ramos
com filódios, folhas recompostas e folhas de morfologia intermédia, com um filódio encimado
por uma ou mais pínulas (vd. Divisão ou composição); geralmente os filódios são uma adaptação
à secura;
b) Folhas gordas – folhas espessas, ricas em água, frequentemente de contorno arredondado;
representam, geralmente, adaptações à secura; e.g. crassuláceas do género Sedum;
c) Espinhos – as folhas podem estar reduzidas a espinhos – espinhos foliares – de origem peciolar
ou estipular (vd. Espinhos); os espinhos têm uma ontogénese distinta das emergências
espinhosas dispersas na margem ou no limbo das folhas epinescentes (vd. Emergências);
d) Gavinhas foliares – folhas modificadas adaptadas a envolver ramos ou outros tipos de suporte;
presentes em muitas lianas; as gavinhas de origem foliar podem resultar da modificação do
limbo de uma folha simples, de um ou mais folíolos (e.g. Vicia [Fabaceae] «ervilhacas) de uma
folha composta, ou muito raramente, de estípulas (e.g. gavinhas estipulares de Smilax
[Smilacaceae]);
e) Armadilhas de origem foliar – a carnivoria é entendida como uma adaptação a habitats pobres
em nutrientes, e.g. turfeiras; as plantas ditas carnívoras servem-se de vários tipos de armadilhas
de origem foliar para capturar insectos ou pequenos organismos multicelulares:
i) Armadilhas pegajosas ou adesivas – os géneros indígenas de Portugal Drosophyllum
(Drosophyllaceae) «erva-pinheira-orvalhada» e Drosera (Droseraceae) têm pêlos
gradulosos pegajosos; em Pinguicula (Lentibulariaceae) todas a superfície das folhas é
pegajosa;
ii) Armadilhas articuladas – e.g. Dionaea muscipula (Droseraceae), uma carnívora norte-
americana;
iii) Armadilhas escorregadias – e.g. os géneros extra-europeus Sarracenia (Sarraceniaceae)
e Nepenthes (Nepenthaceae);
iv) Armadilhas por sucção – e.g. Utricularia (Utriculariaceae), um género cosmopolita
presente em Portugal continental.

70 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
Em pontos anteriores foram descritos mais exemplos de adaptações: catáfilos e hipsófilos. Os
antófilos são a mais surpreendente metamorfose foliar, a ser discutida, mais adiante nos pontos
referentes à flor.
3.5. O corpo das gramíneas
As gramíneas têm um sistema radicular fasciculado. A raiz primária nesta família de
monocotiledóneas senesce num estádio precoce do desenvolvimento. Nos cereais de Outono-Inverno a
raiz primária permanece funcional até à entrada do Inverno, quando tem início o afilhamento.
As gramíneas possuem 5 tipos de meristemas: apicais, axilares, folheares, intercalares do caule e
intercalares da raiz. Os meristemas apicais e axilares, no início do ciclo vegetativo, produzem fitómeros
encaixados de forma linear e, mais tarde, inflorescências. A velocidade a que se formam novos
fitómeros depende, sobretudo, das características genéticas das plantas e da temperatura. À medida
que os meristemas caulinares apicais progridem deixam para trás agregados de células com capacidade
meristemática com a função de alongar as folhas (meristemas folheares) ou de alongar os entrenós do
caule (meristemas intercalares do caule). Ao contrário do meristema apical, estes dois meristemas estão
encravados entre tecidos definitivos. O mesmo acontece com os meristemas intercalares da raiz.
Num momento muito precoce da diferenciação da folha o meristema folhear fracciona-se em duas
partes na região de contacto entre a bainha e o limbo. O alongamento foliar realiza-se, então, pela
extremidade distal da bainha e na base (parte proximal) do limbo. Por esta razão, muitas espécies
gramíneas são capazes de repor, parcialmente, a perda por herbivoria da extremidade do limbo na
última fase do crescimento da folha, quando esta emerge da bainha da folha imediatamente anterior.
Os meristemas intercalares do caule desempenham um papel importante no alongamento dos entrenós
na fase reprodutiva, e na emergência da inflorescência.
a) b)
Adaptações ao nível da folha. a) Folhas gordas, equifaciais e cilíndricas, de Sedum caespitosum (Crassulaceae); n.b. fruto múltiplo de folículos semelhante a uma estrela. b) Nepenthes x ventrata (Nepenthaceae); n.b. parte distal da folha metamorfizada num armadilha escorregadia em forma de jarra (ascídeo) com uma tampa que evita a entrada da água da chuva e a diluição do conteúdo

71 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
Nas gramíneas temperadas e mediterrânicas os
metâmeros caulinares (fitómeros) formados no Outono e
no Inverno são muito curtos. Nesta altura do ano as
folhas apresentam um limbo pequeno e as bainhas
sobrepostas e comprimidas num pseudocaule. Pouco
depois da emergência – nos cereais a partir do estádio
fenológico de 3-folhas –, ainda no Outono nas regiões de
climas não tropical, em consequência da activação de
meristemas axilares, formam-se novos caules na axila das
folhas recém-diferenciadas, ou na axila das folhas de
estolhos ou rizomas que persistiram no solo durante a
estação desfavorável ao crescimento. Iniciada formação
dos novos caules, estes meristemas passam a ser
identificados como meristemas apicais. Em cada axila
foliar insere-se apenas um novo caule: um colmo, um
estolho ou um rizoma. A origem axilar obriga os novos
estolhos ou rizomas a perfurar uma ou mais bainhas
foliares da planta-mãe, antes despontarem no interior ou
à superfície do solo. Os filhos – i.e. os colmos pós-
embrionários, inseridos noutros colmos mais velhos –
alongam-se entre a bainha e o colmo da planta-mãe.
Nas Poaceae anuais a formação de novos caules
axilares pelos meristemas alojados na axila das folhas da
base designa-se afilhamento. Nos cereais de Outono-
Inverno (e.g. trigo, centeio, cevada e aveia) este processo
ocorre durante o Inverno e depende factores internos
(características genéticas das plantas) ou externos (e.g.
temperatura, precipitação, nutrientes e radiação solar). A
radiação solar tem um efeito muito marcado no
afilhamento: quanto maior a quantidade de luz recebida
pelas plantas, por exemplo, em resultado de baixas
densidades de sementeiras, mais intenso é o
afilhamento. Nas Poaceae vivazes, nas Cyperaceae e em
outras famílias de monocotiledóneas, os filhos designam-
se por inovações. Salvo raras excepções, as inovações
produzem flores ou inflorescências no próprio ano.
4. Sistema reprodutivo
4.1. Gimnospérmicas
4.1.1. Estruturas reprodutivas
As gimnospérmicas não têm flores: todos os fenómenos relacionados com reprodução sexuada
ocorrem em estruturas reprodutivas unissexuais. Nas espécies ditas monóicas (vd. Expressão sexual)
a)
b)
a) Corpo vegetativo das gramíneas. N.b. folhas alternas disticadas». b) Pseudocaule e filhos. Lolium rigidum (Poaceae) «erva-febra»

72 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
cada indivíduo possui, em diferentes pontos da copa, estruturas reprodutivas ♂ e ♀; e.g. Pinus
(Pinaceae) «pinheiros» e Abies (Pinaceae) «abetos». Nas espécies dióicas, numa mesma população
coexistem indivíduos ♂ e ♀; e.g. Ginkgo biloba (Ginkgoaceae) «ginkgo» e Taxus baccata (Taxaceae)
«teixo».
As estruturas reprodutivas ♂ das gimnospérmicas, também designadas por estróbilos ♂, são
compostas por um número variável de escamas polínicas (= microsporófilos), com sacos polínicos (=
microsporângios), regra geral 2, inseridos na face dorsal (vd. Volume II).
Neste grupo de plantas os primórdios seminais contactam directamente com o exterior: não estão
encerrados num pistilo como nas angiospérmicas. Consoante as espécies os primórdios seminais
apresentam-se:
a) Solitários; e.g. em Taxaceae;
b) Aos pares (raramente solitários ou em grupos de três) na extremidade de um pequeno caule
(pedúnculo) em Gingkoaceae;
c) Na margem de megasporófilos solitários em forma de leque (flabeliformes) em Cycadaceae;
d) Na superfície ou na margem de megasporófilos organizados em estróbilos ♀; tipo de dominante
nas gimnospérmicas, característicos das famílias Araucariaceae, Cupressaceae, Ephedracea,
Podocarpaceae, Pinaceae e Zamiaceae.
Os estróbilos ♀ são as estruturas reprodutivas ♀ dominantes nas gimnospérmicas. O estróbilo ♀ das
coníferas (Pinidae) é interpretado como um sistema de ramos muito modificados, reduzidos a escamas
de tamanho variável (megasporófilos), que sustêm na sua superfície os primórdios seminais. Estas
escamas, designada por escamas ovulíferas ou escamas férteis, são axiladas por uma escama estéril (=
escama tectriz), nem sempre observável.
a) b)
Estruturas reprodutivas das Gimnospérmicas. a) Estróbilo ♂ e B) estróbilo ♀ de Chamaecyparis lawsoniana (Cupressaceae) (muito ampliados); n.b. em a) sacos polínicos, por abrir, inseridos na face dorsal de microsporófilos; em b) primórdios seminais livres, com uma gota de polinização exsudada pelo micrópilo, inseridos na axila de megasporófilos que mais tarde, na maturação, darão origem às escamas dos gálbulos (vd. imagem mais adiante)

73 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
Os primórdios seminais (=
megasporângio + 1 tegumento) nas
gimnospérmicas têm apenas um tegumento
a envolver a nucela (tecido maternal
diplóide de 2n cromossomas). A nucela, por
sua vez, cinge um saco embrionário
haplóide (n cromossomas). À semelhança
das angiospérmicas, a nucela é homologada
a um megasporângio carnudo (vd. Volume
II). O saco embrionário (= gametófito ♀)
tem muito mais células do que nas
angiospérmicas. Numa das extremidades do
primórdio seminal existe uma abertura para
o exterior (micrópilo). Em muitas espécies
imediatamente abaixo do micrópilo situa-se
uma câmara micropilar.
4.1.2. Frutificações e sementes
Frutificações
O fruto, num sentido estrito, resulta do desenvolvimento de um ovário, regra geral após a
fecundação: o fruto é exclusivo das angiospérmicas. Quando presentes, as estruturas protectoras das
sementes nas gimnospérmicas resultam do amadurecimento dos estróbilos ♀ e designam-se por
frutificações. As frutificações são, portanto, análogas aos frutos das plantas-com-flor. As frutificações
ocorrem nas Cycadidae – família Zamiaceae –, Gnetidae – família Ephedraceae – e Pinidae – famílias
Araucariaceae, Cupressaceae, Podocarpaceae e Pinaceae. As sementes das Taxaceae, Cycadaceae e
Gingkoaceae não estão protegidas por qualquer tipo de estrutura.
a) b)
Estruturas reprodutivas femininas de Ginkgo biloba. N.b. folhas em forma de lequen (flabeliformes) e pequenos caules (megaesporófilos) com dois primórdios seminais na extremidade.
Frutificações das Gimnospérmicas. a) Gálbulos de Chamaecyparis lawsoniana; n.b. no canto superior direito um estróbilo ♀ imaturo e as sementes aladas na axila de escamas férteis peltadas (em forma de guarda chuva). b) Pinha de Pseudosuga menziesii, n.b. escamas estéreis salientes (= excertas) na axila das quais se inserem escamas férteis arredondadas, cada uma com duas sementes apensas (não visíveis na foto)

74 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
Entre as coníferas (Pinidae), o grupo de gimnospérmicas mais diverso das gimnospérmicas e de
maior interesse económico, reconhecem-se dois tipos de frutificações:
a) Gálbulos (= gálbulas) – estróbilo de forma globosa com escamas, normalmente, peltadas (em
forma de guarda-chuva) inseridas mais ou menos no mesmo ponto; e.g. frutificações dos
Cupressus (Cupressaceae) «ciprestes»;
i) Gálbulos baciformes – gálbulos de escamas carnudas, semelhantes a uma drupa; e.g.
frutificações características de Juniperus (Cupressaceae) «zimbros» e de
Podocarpaceae;
b) Pinhas – estróbilo de forma alongada com as escamas inseridas num eixo evidente; e.g. pinhas
de Pinus (Pinaceae) «pinheiros».
Sementes
As reservas nutritivas das sementes de gimnospérmicas resumem-se a um endosperma primário
haplóide, formado antes da fecundação. Por conseguinte, o endosperma primário é um tecido
gametofítico de origem maternal. O número de cotilédones é muito variável. Em Taxus (Taxaceae)
«teixo» as sementes surgem envolvidas por uma estrutura carnuda (arilo), comestível, que actua como
recompensa para a dispersão endozoocórica. As sementes de Ginkgo biloba (Ginkgoaceae) «ginkgo»
têm uma sarcostesta (vd. Episperma) espessa de odor desagradável.
4.2. Angiospérmicas
4.2.1. Inflorescência
Definição e constituição
Por definição a inflorescência é um sistema de caules com flores. Numa inflorescência reconhecem-
se:
a) Flores – vd. Flor;
b) Pedúnculos – porção de caule que sustenta uma flor solitária ou uma inflorescência; numa
inflorescência grupada reserva-se o termo pedicelo para o pequeno caule que suporta cada uma
das flores; as inflorescências sésseis não têm pedúnculo;
c) Ráquis (= eixo primário da inflorescência) – troço de caule onde se inserem as flores ou ramos
da inflorescência;
d) Brácteas – folhas, geralmente modificadas, situadas na proximidade das flores ou dispersas
numa inflorescência.
Brácteas e bractéolas
O termo bráctea é aplicado a três situações distintas:
a) Brácteas (designação usada neste caso sem adjectivação) – folhas modificadas (na cor, forma,
dimensão, consistência, etc.) de cuja axila desponta uma inflorescência ou um ramo da
inflorescência; frequentes na maior parte das inflorescências grupadas (vd. Tipos fundamentais
de inflorescências);
b) Brácteas florais – folhas, modificadas ou não, em cuja axila se insere uma flor; tipo mais
frequente de brácteas;

75 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
c) Brácteas involucrais – folhas modificadas, verticiladas, que envolvem uma ou mais flores, ou,
uma ou mais ramificações da inflorescência, geralmente sem as axilarem de forma evidente; e.g.
valvas de um ouriço de Castanea e brácteas que revestem exteriormente os capítulos das
Asteraceae.
As bractéolas são folhas modificadas inseridas nos pedicelos acima das brácteas, na proximidade das
flores sem, no entanto, as axilarem.
Vários tipos de brácteas ou de estruturas bracteolares merecem uma designação especial:
a) Invólucro – o conjunto de brácteas involucrais que nas asteráceas revestem exteriormente um
capítulo; nas umbelíferas o conjunto de brácteas situado na base de uma umbela composta;
b) Cúpula – estrutura bracteolar característica da família Fagaceae; em Castanea e Fagus um
ouriço com 4 valvas; em Quercus com a forma de taça, rija e coberta por numerosas brácteas
imbricadas e inferiormente concrescentes (vd. Volume III, fam. Fagaceae);
c) Epicálice – pequeno grupo de bractéolas, livres ou concrescentes, localizado na base do cálice e
aparentando a um segundo cálice; frequente em muitas Malvaceae;
d) Espata – bráctea de grandes dimensões, frequentemente colorida e vistosa, que envolve certas
inflorescências; surgem solitárias, e.g. Araceae «família do jarro» e Arecaceae «palmeiras», ou
aos pares, e.g. gén. Allium (Alliaceae) «alhos».
As inflorescências podem ou não ter brácteas dizendo-se, respectivamente, bracteadas ou
ebracteadas. A morfologia das brácteas varia de espécie para espécie, de indivíduo para indivíduo, ou
mesmo longo das inflorescências. As brácteas podem ser semelhantes aos nomófilos, ou reduzidas a
pequenas escamas ou espinhos, sendo possíveis todo o tipo de morfologias intermédias. Nas
inflorescências folhosas as brácteas assemelham-se aos nomófilos e, por isso, são frequentemente
confundidas com caules floríferos. As glumas e glumelas que compõem as espiguetas das gramíneas são
também brácteas.
As brácteas escamiformes serão, muitas vezes, caracteres não funcionais, i.e. resíduos evolucionários
sem uma função clara. Outras protegem os meristemas que dão origem às flores ou às ramificações da
inflorescência; para tal podem estar transformadas em espinhos. Na Euphorbia pulcherrima
(Euphorbiaceae) «poinsétia» e nas Bougainvillea (Nyctaginaceae) brácteas de grande dimensão exibem
cores berrantes e atraem os polinizadores. Na frutificação as brácteas podem envolver e proteger os
a) b) c)
Brácteas. a) Capítulo de Taraxacum gr. officinale (Asteraceae) «dente-de-leão». N.b. algumas das brácteas involucrais deflexas (incurvadas para baixo). b) Arum italicum (Araceae). N.b. espata a envolver a inflorescência carnuda amarela (espata). c) Bougainvillea glabra (Nyctaginaceae). N.b. duas flores envolvidas por três brácteas vermelhas.

76 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
frutos (e.g. Fagaceae e Corylus avellana [Betulaceae] «aveleira») ou facilitar a sua dispersão (e.g.
brácteas com ganchos de Arctium minus [Asteraceae]).
Nas inflorescências bracteadas das eudicotiledóneas cada flor tem uma bráctea abaxial muitas vezes
distalmente acompanhada por duas bractéolas (dois prófilos) em posição lateral. Nas monocotiledóneas
ocorrem duas brácteas, uma abaxial sucedida por outra adaxial (que corresponde a um prófilo) em
substituição das duas bractéolas (vd. Tipos de filomas).
Tipos fundamentais
Dois tipos fundamentais de inflorescências:
a) Inflorescências solitárias – constituídas por uma única flor;
b) Inflorescências grupadas – constituídas por duas ou mais flores; as inflorescências grupadas
podem ser:
i) Inflorescências simples – flores, sésseis ou pediceladas, inseridas directamente no eixo
principal da inflorescência;
ii) Inflorescências compostas26 – eixo principal ramificado em inflorescências parciais; a
inflorescência, no seu todo, é designada por sinflorescência; as inflorescências parciais
podem ser do mesmo tipo da inflorescência de primeira ordem (e.g. umbela de
umbelas, cachos de cachos e dicásio de dicásios [= dicásio composto]) ou não (e.g.
cacho de espigas e corimbo de capítulos); nas inflorescências compostas os eixos de
ordem superior são, tendencialmente, alternos ou opostos, respectivamente, nas
plantas de filotaxia alterna e oposta;
Consoante a sua posição nos caules as inflorescências podem ser:
a) Inflorescências axilares – situadas numa posição lateral, na axila de uma folha;
b) Inflorescências terminais – situadas na extremidade de um caule.
As inflorescências podem apresentar diferentes combinações de flores hermafroditas, unissexuais
e/ou estéreis, por sua vez agrupadas na base, no centro ou no topo da inflorescência. Existe uma vasta
nomenclatura para designar cada uma das combinações possíveis que não cabe aqui desenvolver. Os
amentos de Castanea sativa (Fagaceae) «castanheiro» ilustram bem esta complexidade. Pouco depois
do abrolhamento formam-se amentos unissexuais ♂ a partir de gomos prontos localizados na axila de
algumas das folhas recém-diferenciadas. 10 a 15 dias depois, numa zona mais jovem do ramo do ano em
alongamento e, portanto, numa região mais exterior da copa, surgem amentos androgínicos com
numerosas flores ♂ acompanhadas por 1 a 6, raramente mais, glomérulos de flores ♀ na base (parte
proximal do amento).
Salvo alguns casos especiais, as inflorescências podem ser organizadas em dois grupos fundamentais
em função do comportamento, durante o crescimento, do meristema apical do eixo ou eixos da
inflorescência:
a) Inflorescências indefinidas (= monopodiais, centrípetas ou racemosas) – o meristema localizado
no ápice do eixo principal da inflorescência origina periodicamente flores em posição lateral,
regra geral na axila de uma bráctea; findo o período de crescimento e meristema apical aborta
(inflorescências indefinidas abertas) ou origina uma flor (inflorescências indefinidas fechadas); o
eixo principal da inflorescência tem assim um crescimento indeterminado (inflorescências
indefinidas abertas) ou determinado (inflorescências indefinidas abertas), e a inflorescência
segue um sistema de alongamento monopodial;
26 Na obra de referência de Webberling (1989) é seguido um conceito mais lato de inflorescência simples. No entanto, uma vez que não há consenso em torno da terminologia e interpretação das inflorescências, para facilitar a apreensão do tema, optou-se por considerar todas as cimeiras com mais de um nó como inflorescências compostas.

77 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
b) Inflorescências definidas (= simpodiais, centrífugas ou cimosas) – o meristema localizado no
ápice do eixo primário da inflorescência rapidamente se diferencia numa flor; na axila da(s)
bráctea(s) localizadas imediatamente abaixo da primeira flor emergem novos eixos que por sua
vez diferenciam novas flores, repetindo-se este processo duas ou mais vezes nas inflorescências
definidas compostas; os eixos das inflorescências definidas têm um crescimento determinado e
a inflorescência alonga-se e ramifica-se de forma simpodial.
Inflorescências grupadas simples indefinidas
Tipos fundamentais:
a) Cacho – flores pediceladas inseridas ao longo de um eixo; um tipo de grande importância com
designação especial:
i) Amento (= amentilho) – cacho pêndulo constituído por flores unissexuais nuas (e.g.
Salix [Salicaceae] «salgueiros») ou de perianto sepalóide (e.g. Quercus [Fagaceae]);
b) Espiga – flores sésseis inseridas ao longo de um eixo; e.g. Brassica oleracea (Brassicaceae)
«couve»; dois subtipos com designação especial:
i) Espádice – espiga de eixo carnudo, geralmente revestida por flores pequenas e pouco
vistosas e envolvida por uma espata; e.g. Zantedeschia aetyopica (Araceae) «jarro»;
ii) Espigueta – inflorescência característica das Poaceae;
c) Corimbo – tipo particular de cacho com as flores mais ou menos dispostas no mesmo plano
embora os pedicelos partam de pontos diversos do eixo da inflorescência;
d) Capítulo – inflorescência achatada, côncava ou convexa, raramente mais ou menos globosa, com
flores geralmente sésseis, inseridas num receptáculo capitular que corresponde ao eixo primário
da inflorescência; receptáculo capitular revestido exteriormente por um número variável de
brácteas (brácteas involucrais; vd. Brácteas);
e) Umbela – pedicelos das flores inseridos num mesmo ponto, frequentemente algo dilatado e
com um verticilo de brácteas (invólucro).
Inflorescências simples definidas
As inflorescências simples definidas podem ser:
a) Uníparas (= monocásios) – apenas uma flor (de segunda ordem) inserida sob a flor terminal (de
primeira ordem);
b) Bíparas (= dicásios) – duas flores opostas sob a flor terminal;
c) Multíparas (= pleiocásios) – mais de duas flores verticiladas sob a flor terminal.
Inflorescências compostas
Tipos mais frequentes de inflorescências compostas:
a) Cacho composto – cacho de cachos;
i) Panícula – tipo particular de cacho de cachos, de forma piramidal, mais ramificado na
base do que no topo, no qual o eixo principal e os eixos das inflorescências parciais
terminam numa flor, i.e. são fechados; muito frequente nas poáceas;
b) Tirso – cacho de inflorescências cimosas tipo monocásio ou dicásio, simples ou compostos; e.g.
cachos de dicásios de videira-europeia;
c) Cacho de espigas – inflorescências parciais tipo espiga organizadas num cacho;

78 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
d) Umbela composta – sinflorescência e inflorescências parciais tipo umbela; característica da
família das umbelíferas; a grama (Cynodon dactylon, Poaceae) apresenta uma umbela de 3-4
espigas;
e) Espiga de espigas – sinflorescência e inflorescências parciais tipo espiga; e.g. inflorescência
masculina do milho-graúdo;
f) Dicásio composto – sinflorescência e inflorescências parciais tipo dicásio; frequente na família
das cariofiláceas;
g) Monocásio composto – sinflorescência e inflorescências parciais tipo monocásio; dois tipos
maiores cada um dos quais com dois subtipos:
i) Cimeiras helicóides – cimeiras uníparas compostas com eixos consecutivos inseridos
sempre na mesma posição (relativamente ao eixo anterior):
Bóstrix – eixos consecutivos inseridos sempre para a direita ou sempre para a
esquerda, formando uma espiral em torno de um eixo imaginário da
inflorescência; e.g. inflorescências parciais de Hypericum perforatum
(Hypericaceae) «milfurada»;
Drepânio (= cimeira falciforme) – eixos consecutivos dispostos num mesmo
plano; brácteas, quando presentes, todas no mesmo lado da inflorescência;
e.g. Gladiolus (Iridaceae) «gladíolos» e Juncus bufonius (Juncaceae);
ii) Cimeiras escorpióides – cimeiras uníparas compostas com eixos consecutivos de
inserção alterna; brácteas, se presentes, alternas:
Cincino – eixos consecutivos inseridos, alternadamente, para a esquerda e
para a direita num zig-zag tridimensional; e.g. característico de muitas
Boraginaceae, e.g. Myosotis «miosotis» e inflorescências parciais de Echium
«soagens»;
Ripídio (= cimeira flabeliforme) – eixos consecutivos inseridos,
alternadamente, para a esquerda e para a direita no mesmo plano: e.g. Canna
(Cannaceae) «canas» e Iris sp.pl. (Iridaceae) «lírios».
Tipos especializados de inflorescências definidas compostas mais frequentes:
a) Ciato – inflorescência semelhante a uma flor hermafrodita, constituída por um invólucro em
forma de taça, normalmente culminado por 1-5 glândulas nectaríferas, no interior do qual se
encontra uma flor ♀ nua, na extremidade de um pedicelo, e 5 grupos de flores ♂ nuas com 1
estame, dispostos em torno da flor ♀; característica de Euphorbia (Euphorbiaceae) «eufórbias»
e de Chamaesyce (Euphorbiaceae);
b) Glomérulos – inflorescências cimosas, multifloras, muito contraídas (com os eixos da
inflorescência e os pedicelos muito pequenos), frequentemente globosa ou subglobosa;
frequente em algumas famílias de flores muito pequenas e inconspícuas (e.g. Amaranthaceae);
c) Verticilastros – inflorescências cimosas, multifloras, mais ou menos contraídas, axiladas por um
par de brácteas opostas, com um aspecto verticilado (assemelham-se a um anel de flores em
torno de um eixo); os verticilastros são frequentes entre as Lamiaceae, normalmente
organizados em sinflorescências do tipo cacho (cacho de verticilastros).
Pseudantos
Os pseudantos são inflorescências que se assemelham a flores. Geralmente resultam da agregação
de flores pequenas e reduzidas em inflorescências compactas que tomam, então, a forma de uma flor. O
capítulo das asteráceas é o exemplo mais óbvio de pseudanto. Em Leontopodium (Asteraceae)
«edelweiss» forma-se um pseundanto de capítulos.

79 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
a) b)
Inflorescência. a) Tipos de inflorescências simples e compostas. Inflorescências definidas ou cimeiras: 1. Monocásio simples, 2. Cimeira helicóide tipo bóstrix (projecção num plano vertical em cima e projecção num plano horizontal em baixo), 3. Cimeira helicóide tipo drepânio; 4. Cimeira escorpióide tipo cincino; 5. Cimeira escorpióide tipo ripídio; 6. Dicásio. Inflorescências indefinidas ou racemosas: 7. Cacho; 8. Espiga; 9. Espigueta (g – glumas, l – lema, p – pálea), n.b. duas lodículas imediatamente abaixo dos estames; 10. Espádice; 11. Corimbo; 12. Umbela; 13. Capítulo (i - brácteas involucrais, p - brácteas interflorais). b) Tipos de inflorescências compostas. 1. Cachos de cachos; 2. Umbela de umbelas; 3. Espiga de espiguetas; 4. Capítulo de capítulos; 5. Corimbo de corimbos; 6. Dicásio composto; 7. Tirso; 8. Corimbo de capítulos; 9. Cacho de espiguetas; 10. Antela; 11. Capítulo; 12. Amento; 13. Verticilastro; 14. Ciáto (g – glândulas nectaríferas, b – receptáculo, m – flores ♂ com um estame). Adaptado de Diaz et al. (2004)

80 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
4.2.1. Flor
4.2.1.1. Definição de flor. Iniciação floral. Ciclo floral
Definição de flor
A flor é ramo curto de crescimento determinado (braquiblasto), com entrenós muito curtos e folhas
profundamente modificadas, onde se consuma a reprodução sexuada nas angiospérmicas. Como se
refere no Volume II, a interacção com os agentes polinização e os mecanismos de promoção da
alogamia tiveram um efeito determinante na evolução e na forma da flor. Neste sentido, a flor, no seu
todo, pode ser entendida como uma adaptação à polinização.
Iniciação floral
A vida pós-embriónica das plantas-com-flor pode ser dividida em duas fases: fase vegetativa e fase
reprodutiva. Durante a fase vegetativa os meristemas produzem caule e folhas, possuindo, cada folha,
um meristema axilar. A capacidade de produzir flores – indução floral – é induzida por um conjunto
complexo de sinais endógenos (e.g. activação endógena dos genes envolvidos na iniciação floral) e
exógenos (e.g. exposição temporária ao frio [vernalização], comprimento do dia e exposição à secura).
Por exemplo, nas variedades mais produtivas de Triticum aestivum (Poaceae) «trigo-mole» a iniciação
floral depende de uma exposição a um prolongado período de frio. Na Mangifera indica
(Anacardiaceae) «mangueira» a iniciação floral é impulsionada pela escassez de água no solo. A indução
floral não tem uma expressão morfológica imediata a nível meristemático, é um fenómeno fisiológico. A
transição da fase vegetativa para a fase reprodutiva, i.e. a conversão de meristemas vegetativos em
meristemas reprodutivos, designa-se por iniciação floral27.
Os meristemas reprodutivos começam por diferenciar a inflorescência. Em seguida, acomodam-se
meristemas florais na axila das folhas diferenciadas na inflorescência, i.e. das brácteas28. Cada um destes
meristemas, por sua vez, produz uma flor. A diferenciação das peças florais (sépalas, pétalas, estames e
carpelos) segue, geralmente, a regra de Hofmeister enunciada aquando da discussão da filotaxia dos
caules vegetativos (vd. Filotaxia): os novos primórdios, de qualquer dos órgãos da flor (cálice, corola,
androceu e gineceu), formam-se nos espaços mais amplos disponíveis entre os primórdios mais
próximos já diferenciados. Por esta razão, por regra, a primeira sépala emerge no espaço mais distante
da bráctea que axila a flor, e nas flores cíclicas as pétalas alternam com as sépalas e os estames
alternam com as pétalas.
Ciclo floral
As flores cumprem um conjunto de fases ao longo da sua existência que, genericamente, constituem
o ciclo floral:
a) Indução floral – etapa hormonalmente regulada, ainda sem evidências anatómicas, durante a
qual os meristemas vegetativos, que produzem caule e folhas, se tornam competentes para
produzir flores;
b) Iniciação floral (= diferenciação floral) – conversão anatómica (observável ao microscópio) dos
meristemas vegetativos em meristemas florais;
c) Botão floral – fase que antecede a ântese durante a qual são visíveis flores por abrir (botões
florais), organizados, ou não, em inflorescências;
27 Os termos indução floral e iniciação floral são, muitas vezes, usados como sinónimos. 28 Por perda evoluitva nem sempre presentes.

81 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
d) Ântese (= floração) – concluída a diferenciação da flor verifica-se a ântese, i.e. a abertura da flor
ao exterior, geralmente através da deflexão de sépalas e pétalas; durante a ântese ocorrem a
deiscências das anteras, a polinização, a fecundação e o início da formação do fruto e da
semente; a deiscência das anteras pode anteceder, ser simultânea, ou suceder a polinização; a
ordem das restantes etapas da ântese é constante;
e) Senescência da flor – finda a ântese a flor senesce, i.e. o perianto e os estames escurecem e
perdem turgidez e morrem; geralmente, ambas as estruturas acabam por tombar no solo;
f) Formação do fruto e da semente – os ovários começam a aumentar de volume e as sementes,
no seu interior, a diferenciar-se e a acumular reservas (excepcionalmente as sementes podem
não acumular reservas, e.g. orquídeas);
g) Maturação do fruto e da semente – o fruto para de crescer e adquire o fenótipo (cor, forma,
composição, etc.) que lhe é característico; nesta fase a maior parte das sementes (sementes
ortodoxas) perde água, adquire resistência à secura e entra em quiescência (= suspensão do
desenvolvimento);
h) Dispersão – consoante as espécies os frutos libertam as sementes (frutos deiscentes) ou
dispersam-se em conjunto com estas (frutos indeiscentes).
4.2.1.2. Constituição da flor e expressão sexual
Constituição
A flor dita completa inclui:
a) Receptáculo (= eixo floral) – eixo caulinar, frequentemente alargado, onde se inserem as peças
florais;
b) Perianto duplo – com sépalas e pétalas; característico das flores heteroclamídeas;
c) Androceu – parte ♂ da flor formada pelo conjunto dos estames;
d) Gineceu – parte ♀ da flor constituídas pelos carpelos.
O receptáculo situa-se na parte proximal da flor e o gineceu, invariavelmente, no seu extremo distal,
por cima dos estames29. As flores completas têm os dois sexos funcionais são, portanto, hermafroditas
(= bissexuais). A partir do modelo de flor completa30 definem-se vários tipos de flor incompleta:
a) Flor nua – sem perianto;
b) Flor estéril – não funcional, pela ausência de antófilos ou pelo facto destes não serem
funcionais;
c) Flor apétala – sem pétalas;
d) Flor unissexual – apenas um dos sexos funcional e peças do sexo não funcional ausentes ou
morfologicamente muito modificadas; as flores unissexuais podem ser:
i) Flor pistilada – flor ♀;
ii) Flor estaminada – flor ♂.
As flores solitárias são sustentadas por uma pequena porção de caule designada por pedúnculo. Nas
flores integradas em inflorescências grupadas este caule toma o nome de pedicelo. A flor séssil não
possui pedúnculo (ou pedicelo), i.e. insere-se directamente num nó.
29 Nas estruturas reprodutivas da família Hydatellaceae os estames situam-se acima dos carpelos porém não se sabe se estas estruturas são verdadeiras flores, ou inflorescências condensadas semelhantes a flores (pseudantos). 30 O conceito de flor completa, à semelhança do conceito de folha completa, não envolve inferências evolutivas.

82 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
Expressão sexual (= sistemas sexuais)
A expressão sexual é discutida em profundidade na secção dedicada à polinização. Nesta fase, ainda
inicial, da exploração da flor apresentam-se três sistemas sexuais: homoicia, monoicia e dioicia. As
plantas de flores hermafroditas dizem-se homóicas. As plantas monóicas possuem flores exclusivamente
unissexuadas, estando ambos os sexos presentes num mesmo indivíduo; e.g. Fagaceae e Betulaceae. As
plantas dióicas têm também flores unissexuadas, porém as flores ♂ e as ♀ ocorrem em indivíduos
diferentes; e.g. Salix (Salicaceae) «salgueiros», Populus (Salicaceae) «choupos» e Ilex (Aquifoliaceae)
«azevinhos».
Nas flores funcionalmente unissexuais – funcionalmente ♂ ou ♀ – estão presentes estames e
carpelos aparentemente funcionais, i.e. de morfologia “normal”, porém, por mecanismos vários apenas
um dos sexos é funcional. Do mesmo modo também se podem utilizar as designações: planta
funcionalmente monóica e planta funcionalmente dióica.
4.2.1.3. Filotaxia e simetria da flor
Filotaxia
Quanto à filotaxia as flores podem ser:
a) Acíclicas – peças florais alternas (uma por nó) e helicoidais (vd. Filotaxia); e.g. Nymphaea alba
(Nymphaeaceae) «nenúfar-branco»;
b) Hemicíclicas (= filotaxia intermédia) – parte das peças florais disposta em verticilos (duas ou
mais peças por nó), a restante alterna (uma por verticilo) e helicoidal (vd. Filotaxia); e.g.
Clematis (Ranunculaceae) «clematides»; esta condição é bastante frequente porque o cálice
geralmente segue a filotaxia das folhas caulinares (nomófilos), assim, se as folhas forem alternas
o cálice muitas vezes apresenta o mesmo tipo de inserção enquanto a corola é verticilada;
c) Cíclicas (= filotaxia verticilada) – peças florais organizadas em verticilos (duas ou mais por nó);
condição mais frequente nas angiospérmicas.
As plantas de flores acíclicas estão concentradas nas angiospérmicas basais, i.e. nos grupos mais
próximos da base (mais antigos) da grande árvore filogenética das plantas com flor. As angiospérmicas
basais e nas eudicotiledóneas basais apresentam uma filotaxia variável, desde flores acíclicas a flores
cíclcias. As flores hemicíclicas e cíclicas derivam de flores acíclicas (condição ancestral). As
angiospérmicas mais avançadas são genericamente hemicíclicas ou cíclicas. Alguns grupos recentes de
angiorpérmicas flores são secundariamente acíclicos; e.g. Theaceae e Paeoniaceae. O desenvolvimento
de verticilos estáveis nas flores constituiu uma enorme aquisição evolutiva porque possibilitou a
concrescência de peças, a aderência de órgãos e alterações na simetria da flor, com todas as vantagens
que daí advieram (e.g. interacção com insectos polinizadores e protecção do ovário).
Merismo
O merismo refere-se ao número de peças por verticilo floral. As flores dímeras, trímeras, tetrâmeras,
pentâmeras ou de merismo indefinido são os tipos mais frequentes. As flores acíclicas apresentam um
merismo indefinido. Como se referiu no ponto anterior, o merismo indefinido primário caracteriza
alguns grupos de angiospérmicas basais.Nas angiospérmicas basais, nas eudicotiledóneas basais e nas
monocotiledóneas ocorrem com frequência plantas dímeras (e.g. Lauraceae e Buxaceae) e trímeras (e.g.
mocotiledóneas). As flores das eudicotiledóneas são, genericamente, tetrâmeras (e.g. Brassicaceae) ou
pentâmeras (e.g. Fabaceae).

83 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
Simetria
A simetria é um dos caracteres florais de maior importância taxonómica nas plantas-com-flor.
Quanto à simetria as flores podem ser:
a) Actinomórficas31 (= flor regular) – flor de simetria radial, i.e. qualquer plano divide a flor em
duas partes iguais; nas flores actinomórficas as peças de cada verticilo são iguais entre si; e.g.
Rosaceae; condição ancestral;
b) Zigomórficas (= flor monossimétrica) – flor de simetria bilateral, i.e. com um único plano de
simetria; e.g. grande parte das Lamiaceae;
c) Bissimétricas – flor com dois planos de simetria; e.g. Brassicaceae pelo facto de ter estames
didinâmicos e Papaver rhoeas (Papaveraceae) «papoila-comum» porque as pétalas externas são
maiores do que as internas;
d) Assimétricas – flor sem planos de simetria; e.g. Canna «canas» (Cannaceae) e Lonicera
«madressilvas» (Caprifoliaceae).
Estes termos podem ser aplicados, isoladamente, ao cálice e à corola.
Como se refere do Volume II, a simetria da flor está relacionada com o sistema de polinização e o
tipo vetor polínico. As flores actinomórficas entomófilas são pouco seletivas quanto aos insectos
polinizadores. As espécies que as polinizam acedem ao pólen vindos de qualquer direção. Os
himenópteros são os polinizadores mais frequentes das flores zigomórficas. As flores assimétricas
rareiam na natureza, facto que estará relacionado com a preferência dos insectos polinizadores por
formas simétricas.
4.2.1.4. Receptáculo
O receptáculo32 (= eixo floral ou tálamo) é um braquiblasto, ou seja um caule muito curto, de
entrenós geralmente colapsados, no qual se inserem as peças que constituem a flor. Na maioria das
angiospérmicas cada peça da flor é abastecida por um feixe vascular proveniente do receptáculo. A
ramificação deste feixe varia consoante o órgão em causa, sendo mais pronunciada nas peças do
perianto.
Os entrenós do receptáculo podem alargar-se de forma diferenciada e dar origem a diferentes
estruturas. Por exemplo, nas Caryophyllaceae o entrenó que separa a corola do androceu surge com
frequência alargado constituindo um antóforo. Se o alargamento ocorre entre o androceu e o gineceu
forma-se um ginóforo, como acontece em Capparis spinosa (Capparaceae) «alcaparra». A partir do
receptáculo podem ainda diferenciar-se gibas (pequenas bolsas) e esporões (estruturas mais longas do
que as gibas). Os esporões da corolinos (vd. corola personada) ou calicinais desempenham uma função
análoga aos esporões receptaculares: a oferta de recompensas em néctar.
4.2.1.5. Perianto
Definição de perianto. Tipos fundamentais
Entende-se por perianto o verticilo mais externo da flor, composto por antófilos estéreis. Na grande
maioria das angiospérmicas as peças do perianto evoluíram a partir de brácteas (Ronse de Craene,
2010). As pétalas derivadas de estames são um carácter raro e secundário.
31 Alguns autores alargam o conceito de flor actinomórfica às flores bissimétricas. 32 O termo receptáculo também é utilizado para designar alguns tipos particulares de eixos de inflorescência, geralmente em forma de disco, taça ou copo, como acontece nas famílias Euphorbiaceae, Moraceae e Asteraceae.

84 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
As flores sem perianto dizem-se nuas; e.g. Fraxinus angustifolia (Oleaceae) «freixo-de-folhas-
estreitas» e todas as Poaceae. Quando presente o perianto pode ser:
a) Simples – perianto indiferenciado (= perigónio), i.e. com peças todas iguais entre si;
b) Duplo – perianto diferenciado, i.e. com dois tipos de peças, as sépalas e as pétalas.
Consoante o número de verticilos do perianto e a sua diferenciação, as flores são classificadas do
seguinte modo:
a) Aclamídeas – flores nuas; e.g. Fraxinus angustifolia (Oleaceae) «freixo»;
b) Haploclamídeas – de perianto simples, i.e. com um único verticilo; e.g. Ulmus (Ulmaceae)
«ulmeiros) e Santalaceae «família do sândalo»;
c) Diploclamídeas – de perianto duplo. Dois tipos:
i) Homoclamídeas – de peças iguais; condição comum nas monocotiledóneas (e.g. Tulipa
«tulipas» [Liliaceae]) e nas magnoliidas (e.g. Lauraceae).
ii) Heteroclamídeas (= diclamídeas) – com cálice e corola; e.g. Brassicaceae, Rosaceae e
muitas outras eudicotiledóneas.
O perianto simples pode ser primário, ou resultar da perda evolutiva de sépalas ou pétalas (perianto
simples secundário). A distinção destas três condições pode ser difícil na prática. O perianto simples
primário tem uma grande expressão nas angiospérmicas basais, nas magnoliidas e nas
monocotiledóneas, estando geralmente associado à filotaxia helicoidal ou a flores trímeras (Ronse de
Craene, 2010). Este carácter surge ainda em algumas famílias de eudicotiledóneas basais (e.g.
Ranunculaceae).
As pétalas e, implicitamente, o perianto duplo, evoluíram de forma independente em várias
linhagens de angiospérmicas, a partir de flores haploclamídeas ou homoclamídeas. A transição entre
brácteas e sépalas nas flores heteroclamídeas nem sempre é clara (e.g. Camelia [Cameliaceae]
«camélias). O mesmo pode acontecer entre as sépalas e as pétalas. Algumas famílias de plantas cíclicas
ou hemicíclicas apresentam mais de um verticilo corolino (e.g. Berberidaceae).
A concrescência das peças dos verticilos periantais – i.e. a união de tépalas, sépalas e pétalas,
desenvolvendo-se e crescendo em conjunto – é um dos caracteres de maior interesse taxonómico na
morfologia da flor. A concrescência das peças do perianto dificulta o acesso de parasitas e animais ao
néctar e a sua perda por evaporação. Portanto, a concrescência nos verticilos periantais aumenta a
duração do néctar, reduz os riscos do seu consumo por não polinizadores e, possivelmente, incrementa
a probabilidade de se virem a estabelecer relações planta-polinizador mais vantajosas por co-evolução.
Este conjunto tão significativo de vantagens explica por que razão a concrescência das peças do perianto
surgiu de forma independente em tantos grupos de plantas com flor.
Tipos de perianto. 1. e 2. Flor heteroclamídea de Erysimum cheirii (Brassicaceae) «goivo». 3. Flor haploclamídea de Ulmus minor (Ulmaceae) «ulmeiro». 4. Flor hermafrodita nua de Fraxinus angustifolia (Oleaceae) «freixo-de-folhas-estreitas». 5 Flor ♂ nua de Salix (Salicaceae) «salgueiro» na axila de uma bráctea. 6. Flor ♀ nua de Salix na axila de uma bráctea (adaptado de Coutinho, 1898)

85 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
Perigónio
Designa-se por perigónio o conjunto das tépalas, i.e. dos antófilos estéreis das flores de perianto
simples primário, tenham elas um ou mais de um verticilos de peças. As tépalas podem ser sepalóides
ou petalóideas. A restante terminologia relativa ao perigónio é análoga à aplicada às flores com perianto
descrita em seguida. Existem perigónios dialitépalos e sintépalos, unha e limbo nas tépalas dos
perigónios dialipétalos, e assim por diante. Como se referiu anteriormente, algumas flores
haploclamídeas perderam o verticilo das pétalas ou das sépalas no decurso da sua história evolutiva. O
conceito de tépala não deve ser aplicado a estes casos; e.g. flores das Amaranthaceae e flores tubulosas
e liguladas de Anthemis (Asteraceae).
Cálice
O Cálice é o conjunto das sépalas, as peças florais do verticilo mais externo de um perianto duplo. As
sépalas, normalmente, segue a mesma filotaxia dos nomófilos, têm uma consistência herbácea, cor
esverdeadas e desempenham a função fotossintética. Com frequência encontram-se tricomas, glândulas
e estomas a revestir a superfície exterior das sépalas. O cálice desempenha duas importantes funções:
proteger os verticilos mais interiores da flor no botão floral e produzir fotoassimilados para serem
consumidos pelas da peças flor. O cálice diz-se petalóide quando as sépalas se assemelham a pétalas.
Por vezes o cálice apresenta-se reduzido a uma coroa de escamas, sedas (pêlos rígidos e fortes) ou de
pêlos, simples (não ramificados) ou plumosos (se ramificados). O cálice de pêlos das Asteraceae,
Dipsacaceae e Valerianaceae leva o nome de papilho.
A concrescência do cálice é um carácter de enorme interesse taxonómico. Reconhecem-se dois
tipos:
a) Cálice dialissépalo – com sépalas livres; e.g. magnoliidas;
b) Cálice sinsépalo (= gamossépalo) – com sépalas concrescentes, i.e. soldadas entre si; fácil de
observar em qualquer família de asteridas.
Outros caracteres taxonómicos muito valorizados no cálice são a consistência, a forma e a duração.
Quanto à consistência o cálice pode ser herbáceo, escarioso, membranoso, etc. Quanto à forma:
campanulado, tubuloso, bilabiado, etc. Quanto à duração: caduco ou persistente (visível ainda no fruto).
Na Physalis peruviana (Solanaceae) «physalis» e na família tropical Dipterocarpaceae), entre outras
angiospérmicas, o cálice diz-se acrescente porque continua a crescer após a fecundação.
Corola
O conjunto das pétalas designa-se por corola. Na flor completa a corola situa-se entre os verticilos do
cálice e do androceu. As pétalas geralmente alternam com as sépalas. Nas plantas polinizadas por
insectos as pétalas são, normalmente, maiores e mais delicadas e coloridas do que as sépalas porque
têm a função de atrair animais polinizadores. A cor destas peças periantais depende da concentração e
tipo de pigmentos – e.g. antocianinas, carotenóides, betalaínas e flavonóides – que se acumulam nas
células da epiderme ou do mesofilo. Normalmente, a superfície das pétalas não tem estomas e
encontra-se coberta de papilas. A superfície pode ainda apresentar-se ornada com guias de néctar e/ou
de pólen – e.g. linhas e manchas de cor ultravioletas e linhas de tricomas – a indicar a localização de
recompensas aos insectos polinizadores. Nas plantas polinizadas pelo vento (anemófilas) as pétalas são,
por regra, diminutas ou estão ausentes.
Quanto à concrescência reconhecem-se dois tipos de corola:
a) Dialipétala – de pétalas livres (não soldadas entre si); as corolas dialipétalas podem ter algumas
pétalas soldadas entre si (e.g. corola papilionácea, com 2 pétalas parcialmente unidas formando
uma quilha) porém nunca formam um tubo na base; a corola dialipétala das Malvaceae e das
Theaceae, entre outras famílias, confunde-se facilmente com uma corola simpétala porque as
pétalas estão adnadas, na base, a um tubo formado pela concrescência dos filetes;

86 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
b) Simpétala (= gamopétala) – com todas as pétalas concrescentes formando um tubo, mais ou
menos longo, a partir da base; e.g. famílias do clado das Asteridas.
As pétalas livres das corolas dialipétalas são constituídas por:
a) Unha – parte inferior, mais estreita e por vezes descorada, por onde se faz a inserção da pétala
no receptáculo; de grande dimensão nas Brassicaceae e as Caryophyllaceae;
b) Limbo – parte terminal, geralmente laminar.
Nas corolas simpétalas a porção livre de uma pétala é designada por segmento (vd. Alguns conceitos
fundamentais de organografia e biologia da evolução), e a entrada do tubo por fauce (fauce da corola ou
fauce do tubo da corola). Os lóbulos e os dentes são, respectivamente, segmentos da corola
arredondados ou triangulares e mais ou menos pontiagudos. Estes termos podem ser aplicados aos
cálices tubulosos.
Na corola diferencia-se um sem número de estruturas, por regra envolvidas na atracção (e.g. coroa)
ou selecção de polinizadores (e.g. palato), ou no fornecimento de recompensas nectaríferas (e.g. gibas e
esporões):
a) Coroa – apêndice circular, inteiro a fimbriado (dividido em muitos segmentos finos), resultante
da concrescência total ou parcial de expansões da corola (e.g. Passiflora, Passifloraceae) ou do
perigóneo (e.g. Narcissus, Amaryllidaceae), ou da conivência (encosto) de expansões de pétalas
(e.g. conivência de escamas das pétalas em algumas espécies de Silene, Caryophyllaceae) ou
tépalas livres;
b) Palato – saliência na corola que fecha a fauce (= entrada do tubo da corola) ao exterior, comum
nas corolas personadas; e.g. Linaria e Antirrhinum, Plantaginaceae;
c) Gibas – pequenas bolsas localizadas na base do tubo da corola ou do cálice, geralmente providas
de uma recompensa nectarífera;
d) Esporões – estrutura cónica, cilíndrica ou saciforme (em forma de saco), fechada no ápice, mais
longa do que as gibas, situada na base de uma pétala (e.g. Aquilegia, Ranunculaceae) ou
resultante da concrescência de mais de uma pétala nas corolas simpétalas (e.g. Linaria,
Plantaginaceae), geralmente contendo uma recompensa em néctar para os insectos
polinizadores; o esporão pode ter origem no cálice (esporões calicinos).
A forma da corola é muito variável. Alguns tipos merecem designações particulares:
e) Corola dialipétala:
i) Crucífera – corola actinomórfica, de 4 pétalas com unha mais ou menos longa e limbos
dispostos em cruz; característica da fam. Brassicaceae;
ii) Papilionácea – corola zigomórfica, de 5 pétalas; a superior (estandarte) geralmente
levantada, de maior dimensão e envolvendo as restantes 4 no botão; as 2 pétalas
laterais (asas) por vezes ligeiramente soldadas à quilha (e.g. em Vicia); as 2 pétalas
inferiores concrescentes numa peça com a forma da quilha de um barco (quilha); corola
característica da subfam. Faboideae (Fabaceae);
iii) Rosácea – corola actinomórfica, de 5 pétalas com unha curta e limbo largo.
f) Corola simpétala
i) Bilabiada – corola zigomórfica, de fauce aberta e tubo mais ou menos longo, com as
pétalas concrescentes em dois lábios (3 num lábio inferior e 2 num lábio superior);
característica da fam. Lamiaceae, embora em alguns géneros desta família (e.g.
Teucrium e Ajuga) a corola tenha apenas um lábio superior (corola unilabiada) ou seja
tubulosa (e.g. Mentha);

87 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
ii) Tubulosa – corola actinomórfica de tubo comprido, mais ou menos cilíndrico, e
segmentos pequenos; frequente, entre outras famílias, nas Asteraceae;
iii) Ligulada – corola zigomórfica, de tubo curto, com um lábio alongado, em forma de
língua e dentado na extremidade (cada dente correspondendo a uma pétala);
frequente na família Asteraceae.
Outros tipos, menos relevantes, de corola:
a) Assalveada (= corola hipocrateriforme) – corola simpétala actinomórfica, de tubo longo e
estreito, e região distal de concrescência variável e mais ou menos patente (perpendicular ao
tubo); e.g. Pistorinia hispanica (Crassulaceae);
b) Afunilada – corola simpétala actinomórfica em forma de funil, com as pétalas concrescidas a
todo o comprimento; e.g. Convolvulus (Convolvulaceae) «corriolas»;
c) Campanulada – corola simpétala actinomórfica, de tubo mais ou menos longo, rapidamente
alargado na base na forma de um sino; e.g. Campanula (Campanulaceae) «campânulas»;
d) Personada – corola simpétala zigomórfica, bilabiada, de fauce fechada por uma saliência do
lábio inferior (palato), frequentemente provida de gibas ou de esporões; e.g. Antirrhinum
(Plantaginaceae) «bocas-de-lobo»;
e) Rodada – corola simpétala actinomórfica, de tubo curto, e região distal de concrescência
variável, mais ou menos longa e patente (perpendicular ao tubo); e.g. Solanum tuberosum
(Solanaceae) «batateira»;
Tipos de corola. 1. Corola crucífera. 2. Pétala de corola crucífera (n.b. unha comprida). 3. Corola rosácea. 4. Pétala bilobada de uma corola rosácea. 5. Corola papaverácea (não discutida no texto, n.b. cálice duas sépalas caducas aquando da abertura da flor). 6. Corola papilionácea. 7. Corola urceolada. 8. Corola campanulada. 9. Corola afunilada. 10. Corola cravinosa (não discutida no texto). 11. Pétala de corola cravinosa. 12. Corola personada (n.b. fauce fechada por um palato, retirado na segunda figura). 13. Corola ligulada. 14. Corola tubulosa (n.b. estigmas a emergirem acima de um tubo formado pela concrescência das anteras [estames sinantéricos]). 15. Corola rodada. 16. Corola bilabiada (n.b. giba na base da corola e lábio inferior com três lóbulos correspondentes a três pétalas) (adaptado de Coutinho, 1898)

88 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
f) Unilabiada – corola zigomórfica, de fauce aberta e tubo mais ou menos longo, com um único
lábio; nas Orchidaceae e em certas Fabaceae tropicais (e.g. género Clitoria) o ovário sofre uma
rotação de 180°, correspondendo o lábio à pétala superior (designada por labelo entre as
Orquidáceas); as flores que evidenciam uma torção do ovário dizem-se resupinadas;
g) Urceolada (= corola gomilosa) – corola actinomórfica, bruscamente alargada num tubo bojudo,
estreitado na fauce, e com segmentos muito curtos; e.g. Arbutus unedo (Ericaceae)
«medronheiro».
A corola é ainda classificada quanto à coloração, odor e duração (caduca ou persistente).
Ptixia, estivação e posição espacial das peças da flor
A ptixia33 refere-se à forma como os esboços foliares e os antófilos estéreis (tépalas, sépalas ou
pétalas) se dobram, individualmente, nas gemas (vd. Ptixia e vernação). Ao nível da flor normalmente só
se aplica às pétalas ou tépalas. Os tipos mais frequentes de ptixia são:
a) Conduplicada – folhas ou peças periantais dobradas pela nervura média com as duas abas
encostadas; tipo mais frequente nas plantas-com-flor; e.g. folhas de Quercus (Fagaceae)
«carvalhos» e de Prunus avium (Rosaceae) «cerejeira»;
b) Convoluta (= convolutosa) – folhas ou peças periantais enroladas longitudinalmente sobre si
mesmas e de corte transversal em espiral; geralmente encapsulam no seu interior as folha que
se lhes seguem; e.g. folhas de Tulipa (Liliaceae) «tulipas, tipo dominante nas folhas de Poaceae;
c) Circinada – folhas ou peças periantais enroladas longitudinalmente em espiral em direcção ao
ápex; e.g. folhas dos pteridófitos;
d) Involuta (= involutiva) – abas das folhas ou peças periantais enroladas para a página superior;
e.g. Populus (Salicaceae) «choupos» e Viola (Violaceae) «violetas»;
e) Revoluta (= revolutiva) – abas das folhas ou peças periantais enroladas para a página inferior;
e.g. folhas de Rosmarinus officinalis (Lamiaceae) «alecrim»;
f) Plicada – abas das folhas ou peças periantais dobradas ao longo de várias pregas longitudinais,
de modo que o seu corte transversal seja em zig-zag; e.g. folhas das Arecaceae «palmeiras».
A estivação (= prefloração) versa o estudo da disposição dos antófilos estéreis – tépalas, sépalas ou
pétalas – uns em relação aos outros (no mesmo verticilo) nas gemas. Num perianto duplo o cálice e a
corola não partilham, obrigatoriamente, o mesmo tipo de estivação. No entanto, geralmente só se
estuda a estivação da corola. É um conceito análogo ao de vernação (a terminologia é comum, embora
mais vasta), com um grande interesse taxonómico porque plantas filogeneticamente próximas têm
tendência a partilhar o mesmo tipo de estivação. Reconhecem-se quatro tipos fundamentais:
a) Aberta – as folhas ou as peças periantais não se alcançam;
b) Valvar – as folhas ou as peças periantais tocam-se pelas margens, sem se sobreporem; e.g.
corola de Vitis (Vitaceae) «videira»;
c) Imbricativa – as folhas ou as peças periantais de margens mais ou menos sobrepostas;
d) Enrugada – as folhas ou as peças periantais amarfanhadas por pregas irregulares; e.g. corola de
Papaver (Papaveraceae) «papoilas».
A estivação imbricativa é muito diversa. Tipos mais frequentes:
a) Controcida (= contorta) – cada folha ou peça periantal cobre a margem da peça seguinte, sendo
a outra margem coberta pela da peça anterior, encontrando-se o conjunto enrolado
helicoidalmente, para a esquerda ou para a direita; e.g. corolas de Oxalis pes-caprae
(Oxalidaceae) «erva-pata», de Apocynaceae e de Convolvulaceae;
33 Muitos autores não distinguem ptixia de estivação.

89 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
b) Quincuncial – perianto pentâmero com duas das peças completamente externas e outras duas
completamente internas, a quinta cobre uma interior com uma das suas margens e tem a outra
coberta pela margem de uma das peças exteriores; e.g. corolas de Digitalis (Scrophulariaceae)
«dedaleiras» e Dianthus (Caryophyllaceae) «cravinas»;
c) Coclear – perianto pentâmero com uma peça totalmente externa, outra totalmente interna, e as
três restantes com uma margem interna e outra externa; e.g. corola das Fabaceae:
i) Vexilar (= coclear descendente) – própria das fabóideas (Fabaceae, Faboideae) onde a
peça de maior dimensão – o estandarte – cobre as restantes peças;
ii) Carenal (= coclear ascendente) – pré-floração de algumas Fabaceae Caesalpinioideae
em que a quilha (= carena) cobre as asas e o estandarte.
A maioria das eudicotiledóneas pentâmeras exibe uma sépala inferior, duas laterais e uma superior
(Ronse de Craene, 2010). Uma vez que as pétalas alternam com sépalas, uma pétala é inferior (anterior
ou abaxial), duas laterais e as duas restantes superiores (posteriores ou adaxiais). Nas flores
zigomórficas com lábio inferior este pode resultar da modificação da pétala inferior, ou da
concrescência da pétala inferior com as duas pétalas laterais (e.g. Lamiaceae). As leguminosas
Faboideae escapam a este padrão: têm duas pétalas inferiores (unidas numa quilha), duas laterais (asas)
e uma superior (estandarte). As flores tetrâmeras geralmente apresentam duas sépalas alinhadas com o
eixo onde se inserem (posição mediana) e duas sépalas transversais. As pétalas, ao alternarem com as
sépalas, tomam uma posição oblíqua. A orientação das peças do perianto nas monocotiledóneas é mais
variável do que nas eudicotiledóneas (Ronse de Craene, 2010).
4.2.1.6. Hipanto
O hipanto (= tubo floral) é uma estrutura contínua, em forma de disco, taça, cálice ou tubo, que
rodeia e envolve o gineceu nas flores perigínicas de ovário súpero, ou que emerge da sua extremidade
nas flores perigínicas de ovário ínfero. Nas flores com hipanto – flores perigínicas – as peças do perianto
e os estames inserem-se, geralmente, no bordo deste (vd. imagens a respeito da posição do ovário). O
hipanto pode assemelhar-se pela cor com o cálice ou a corola dificultando a sua identificação (e.g.
Grossulariaceae e Onagraceae).
A presença de hipanto é muito frequente na natureza e tem um grande interesse taxonómico. Em
muitas espécies aloja nectários na sua superfície interior, e funciona como uma “cisterna” de néctar
(e.g. Prunus, Rosaceae). O hipanto serve ainda para afastar potenciais predadores dos primórdios
seminais e separar estames dos estígmas
O hipanto apareceu de forma independente em muitos grupos de plantas-com-semente, tendo
origem numa expansão do receptáculo (receptáculo alargado, hipanto s.str., e.g. Rosaceae) ou na
adnação, pela base, das peças do perianto e dos filetes (e.g. Thymelaeaceae). Alguns autores preferem
designar esta última estrutura por pseudo-hipanto. A distinção do hipanto s.str. e do pseudo-hipanto
não é fácil. Para complicar ainda mais o tema, nas flores simpétalas de estames epipétalos o tubo da
corola inclui tecidos dos estames, o que o aproxima do conceito de hipanto. Para evitar mais
inconsistências é conveniente, pelo menos, não sinonimizar receptáculo e hipanto.
4.2.1.7. Androceu
Estrutura e função dos estames
Os estames são os órgãos masculinos das plantas. Nas flores completas o androceu, i.e. o conjunto
dos estames, situa-se entre a corola e o gineceu. Os estames produzem pólen, secundariamente podem

90 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
atrair polinizadores, produzir néctar ou serem consumidos pelos polinizadores (e.g. coleópteros),
servindo de recompensa.
Os estames são, geralmente, constituídos por:
a) Filete – parte estéril dos estames, normalmente filamentosa, que sustem a antera;
b) Antera – parte dos estames onde se forma e está contido o pólen.
O filete normalmente insere-se no dorso (anteras dorsifixas) ou na extremidade (anteras basifixas)
da antera. O basculamento da antera na inserção do filete facilita a libertação e a dispersão do pólen.
Nas anteras geralmente reconhecem-se duas tecas aglutinadas pelo conectivo, um prolongamento do
filete percorrido por um feixe líbero-lenhoso. Algumas famílias dispõem de uma única teca fértil (e.g.
Cannaceae), noutras o conectivo prolonga-se para além das tecas (e.g. Violaceae). Por norma compõem
cada teca dois sacos polínicos, separados por um septo, perfazendo 4 sacos polínicos por antera. Os
estaminódios são estames estéreis, geralmente com um papel importante na atracção de polinizadores
(e.g. pela cor, pela produção de néctar, de odores, etc.). Em termos evolutivos os sacos polínicos podem
ser identificados com microsporângios (= esporângios ♂) e os estames com microsporófilos (=
esporófilos ♂).
A parede das anteras é constituída por várias camadas celulares. O endotécio (= camada mecânica) e
o tapetum sãos as camadas mais importantes. O endotécio situa-se por debaixo da epiderme. Próximo
da ântese (libertação do pólen) as suas paredes celulares engrossam e rompem-se de modo a facilitar a
deiscência do pólen. O interior dos sacos polínicos está revestido pelo tapetum, um tecido constituído
por células secretórias, metabolicamente muito activas que alimentam os microsporos (= grãos de pólen
uninucleados) e auxiliam a formação da esporoderme (vd. Pólen) durante a diferenciação em grãos de
pólen.
Número e forma
Quanto ao número os estames podem ser:
a) Definidos – estames em número igual ou inferior ao dobro do número de pétalas; e.g. asteridas;
b) Indefinidos – estames em número superior ao dobro do número de pétalas; as flores com
estames indefinidos dizem-se poliândricas; e.g. muitas magnoliidas.
Os filetes, quanto à forma, podem ser alados (com asas), capilares (quando muito delgados), etc. As
anteras, por sua vez, podem ser globosas, lineares, etc. Os estames dizem-se petalóides quando
semelhantes a pétalas.
Inserção e posição em relação às pétalas
A inserção dos filetes no receptáculo pode ser verticilada ou helicoidal. Se verticilada, os estames
apresentam-se organizados num ou dois verticilos, raramente mais (e.g. algumas monocotiledóneas;
Ronse de Craene, 2010). Nos grupos mais “primitivos” de angiospérmicas (e.g. clado das magnoliidas) os
estames dispõem-se helicoidalmente e o filete pouco se distingue da antera.
Os estames de uma flor, quando comparados entre si, podem ser:
a) Iguais – na forma e no tamanho;
b) Desiguais – distintos na forma e no tamanho; dois casos de especial interesse:
i) Didinâmicos – 4 estames, 2 compridos e 2 mais curtos; tipo frequente na fam.
Lamiaceae;
ii) Tetradinâmicos – 6 estames, 4 compridos e 2 mais curtos; tipo frequente na fam.
Brassicaceae.
Nas flores de perianto duplo, caso o androceu seja constituído por um verticilo de estames em
número igual às pétalas, estes podem alternar com as pétalas (flores haplostémonas) ou, com menos

91 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
frequência, opor-se às pétalas (flores ob-haplostémonas), qualificando-se os estames, respectivamente,
de alternipétalos (= antisépalos) e de oposipétalos (= antipétalos). Quando ocorrem dois verticilos de
estames em número igual às pétalas, os estames do verticilo externo (mais próximo da corola) alternam
com as pétalas das flores diplostémonas e opõem-se às pétalas nas flores obdiploestémonas.
Concrescência e aderência
Reconhecem-se três grandes tipos de concrescência dos filetes (= adelfia):
a) Monadelfos – estames unidos pelo filete num único grupo; e.g. frequente em muitas Fabaceae,
Malvaceae e em numerosas famílias tropicais, e.g. Meliaceae, Menispermaceae, Canellaceae e
Salvadoraceae;
b) Diadelfos – estames unidos pelo filete em dois grupos; e.g. frequente em Fabaceae, como sejam
as Vicia «ervilhacas»;
c) Poliadelfos – estames unidos pelo filete em mais de dois grupos; e.g. Hypericum (Hypericaceae)
«hipericões».
Os estames dizem-se sinantéricos quando concrescentes pelas anteras, permanecendo os filetes
livres. Todas as Asteraceae possuem estames sinantéricos. Algumas famílias de angiospérmicas, em
particular no clado das asteridas, têm estames epipétalos (e.g. Asteraceae, Oleaceae, Rubiaceae). Nesta
condição os estames inserem-se directamente na corola, por vezes bem perto da base, porque os filetes
estão total ou parcialmente adnados às pétalas. Nas Orchidaceae, e num pequeno número de famílias
de dicotiledóneas s.l., os estames e gineceu formam uma pequena coluna, o ginostemo.
Deiscência
A desidratação das anteras maduras força a sua abertura por zonas de deiscência e a libertação do
pólen, i.e. a sua deiscência.
No que às anteras diz respeito a deiscência pode ser:
a) Longitudinal – cada teca rompe-se longitudinalmente; e.g. Nicotiana e Datura, entre outras
solanáceas;
b) Transversal – pólen libertado por aberturas transversais; e.g. Verbascum (Scrophulariaceae);
c) Valvar – pólen libertado através de pequenas valvas (aberturas em forma de janela); e.g. Laurus
nobilis (Lauraceae) «loureiro»;
d) Poricida – pólen libertado através pequenos orifícios (poros); e.g. Solanum (Solanaceae).
Morfologia do androceu. 1. Estames livres; n.b. presença de hipanto. 2. Estames didinâmicos de um brassicácea. 3. Estames monadelfos de uma malvácea. 4. Estames sinantéricos de uma composta. 5. Deiscência longitudinal. 6. Deiscência poricida. 7. Deiscência valvar, n.b. dois estaminódios na base da antera (adaptado de Coutinho, 1898)

92 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
Reconhecem-se ainda dois tipos de direcção da deiscência:
a) Introrsa – pólen libertado para o interior da flor; e.g. Nerium e Vinca, entre outras apocináceas;
b) Extrorsa – pólen libertado em direcção ao exterior da flor; e.g. Annonaceae e
Potamogetonaceae.
4.2.1.8. Pólen
O pólen é exclusivo das espermatófitas, por conseguinte, é comum às gimnospérmicas e
angiospérmicas. Guarda no seu interior o microgametófito, servindo de veículo de transporte da
informação genética masculina. A parede do pólen (= esporoderme) tem por função proteger as células
que constituem o microgametófito, da dessecação e da radiação solar, durante o percurso desde a
antera até ao estigma.
O invólucro do gametófito ♂, i.e. a esporoderme, tem duas camadas:
a) Intina – camada celulósica;
b) Exina – camada externa composta por esporopolenina, um biopolímero complexo, de
composição química pouco conhecida, muito resistente à degradação por enzimas e agentes
químicos reactivos.
Na exina, por sua vez, reconhecem-se uma camada interna (endexina, ing. endexine) e outra externa
(ectexina, ing. ectexine). A endexina é laminada (compostas por camadas sobrepostas) nas
gimnospérmicas e homogénea nas angiospérmicas. Neste último grupo de plantas a ectexina está
diferenciada, de dentro para fora, numa camada inferior (ing. foot layer), numa camada columelar (ing.
columellar layer) e num tecto (ing. tectum) onde se inserem elementos esculturais. A camada columelar
é constituída por pequenas colunas (columelas) que conectam a camada inferior com o tecto. Outros
termos de uso comum em palinologia estão explicitados na figura que se segue.
Na superfície da exina observam-se pequenas aberturas circulares, os poros, ou em forma de fenda,
os colpos. Os grãos de pólen são primariamente classificados em função do tipo de abertura. Assim, o
pólen pode ser colpado, porado, colporado (com aberturas que combinam um colpo e um pequeno
poro) ou zonados (aberturas aneladas ou com bandas). Estes termos podem ser precisados com base no
número de aberturas; e.g. pólen monocolpado (com um colpo), tricolpado (com três colpos),
monoporado, policolporado, etc. Os pólenes monocolpado e tricolpado são, respectivamente,
característicos das magnoliidas e das eudicotiledóneas. As ornamentações da superfície da exina
permitem, em muitos casos, a identificação dos grãos de pólen ao nível do género ou mesmo da
espécie.
a) b)
Estrutura do pólen. a) Estrutura da esporoderme (Punt et al., 2007). b) Fotografias de microscópio de varrimento de grãos de pólen, n.b. presença de colpos e a variabilidade dos elementos esculturais (Edlund et al., 2004)

93 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
4.2.1.9. Gineceu
Estrutura e função dos carpelos. Conceito de pistilo
A parte feminina da flor, o gineceu, situa-se no ápice do receptáculo, na região mais interna da flor,
imediatamente acima do androceu. Os órgãos femininos da flor, os carpelos, são folhas muito
modificadas. Admite-se que num estádio evolutivo ainda inicial, eram abertos, dispondo-se os
primórdios seminais livremente na sua margem. Em algum momento, entre os ancestrais mais próximos
das angiospérmicas, os carpelos dobraram-se e soldaram-se pela margem (com a página inferior virada
para o exterior), ficando os primórdios seminais retidos no interior de uma nova estrutura designada por
pistilo. Dadas as dificuldades em estabelecer homologias (relacionar evolutivamente) entre as estruturas
reprodutivas das gimnospérmicas e das gimnospérmicas, o termo carpelo só se aplica às plantas-com-
flor. Uma vez que a megasporogénese ocorre no interior dos primórdios seminais, estes são
interpretados como megasporângios envolvidos por um ou dois tegumentos, e os carpelos como
megasporófilos (vd. Volume II).
O pistilo é constituído por uma ou mais folhas carpelares. Os primórdios seminais inserem-se na face
interna dos pistilos, por regra na sutura carpelar. Os pistilos podem ser estéreis – pistilódios – e, como
os estaminódios, desempenhar outras funções (e.g. atracção de polinizadores). Geralmente
reconhecem-se três partes num pistilo:
a) Ovário – parte basal e dilatada do pistilo que encerra os primórdios seminais;
b) Estilete – parte do pistilo, nem sempre presente, geralmente delgada, intercalada entre o ovário
e o estigma;
c) Estigma – parte terminal do pistilo especializada na captura de grãos de pólen.
Inserção
Os pistilos inserem-se no receptáculo em espiral ou em um ou mais verticilos. As flores com um
pistilo (condição mais frequente) têm, necessariamente, um único verticilo.
Número e concrescência dos carpelos
Compõem o gineceu das plantas-com-flor um ou mais carpelos:
a) Monocarpelar – 1 carpelo;
a) Bicarpelar – 2 carpelos;
b) Pluricarpelar – 3 ou mais carpelos.
Os carpelos, por sua vez, podem ser livres ou concrescentes. No estudo da concrescência carpelar
dos gineceus pluricarpelares aplicam-se os seguintes adjectivos:
a) Apocárpico (= corocárpico) – gineceu com mais de um carpelo livre;
Evolução do carpelo. O carpelo ancestral era aberto dispondo-se os primórdios seminais (megaesporângios) na sua margem. A evolução do carpelo fechado encerrou os primórdios seminais no seu interior. Os pistilos pluricarpelares são uma aquisição evolutiva posterior (Purves et al., 2001)

94 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
b) Sincárpico s.l. (= cenocárpico) – gineceu de carpelos concrescentes.
Em função da morfologia do interior do ovário o gineceu sincárpico s.l. tipifica-se do seguinte modo:
a) Sincárpico s.str. – gineceu de carpelos concrescentes pelas faces externas dos carpelos,
definindo mais de um compartimento no interior do ovário;
b) Paracárpico – gineceu de carpelos concrescentes pelas margens, com um único compartimento
ovarial.
Mais de 80% das angiospérmicas são sincárpicas s.l.; ca. de 10% são apocárpicas e os outros 10%
monocarpeladas (Endress, 1994). Tanto o gineceu34 monocarpelar, como os gineceus pluricarpelares
sincárpicos são constituídos por um único pistilo. O gineceu de tipo apocárpico implica a presença de
dois ou mais pistilos, todos eles compostos por um único carpelo. Os compartimentos ovariais são
designados por lóculos. No gineceu sincárpico s.str. os lóculos são separados por um ou mais septos. Na
família das brassicáceas o ovário embora seja originalmente paracárpico apresenta dois lóculos. No
pistilo das plantas desta família forma-se um falso septo com tecidos da placenta, ficando os primórdios
seminais retidos nas duas suturas carpelares. As Linaceae e as Lamiaceae, entre outras famílias, também
têm falsos septos. Num gineceu sincárpico s.str. de dois carpelos sem falsos septos, os primórdios,
quando inseridos na sutura carpelar (placentação axilar), concentram-se no centro do ovário (e.g.
Solanum lycopersicum [Solanaceae] «tomateiro»).
Cada carpelo, à semelhança dos restantes órgãos da flor, geralmente é servido por um único feixe
vascular. Ainda assim, em muitas espécies, cada carpelo é abastecido por dois feixes vasculares: um
feixe ventral dirige-se aos primórdios seminais, e outro, o dorsal, encaminha-se pelo estilete até ao
estigma. Nos gineceus sincárpicos, geralmente cada carpelo tem conexões vasculares autónomas.
O gineceu apocárpico abunda nos grupos mais primitivos das plantas com flor; e.g. angiospérmicas
basais e magnoliidas. Geralmente não possuem estiletes e estigmas bem diferenciados. A evolução da
sincarpia é um dos aspectos mais estudados da evolução da morfologia das flores. A sincarpia, frente à
apocarpia, aumenta a competição (e selecção) entre os grãos de pólen e permite que os tubos de pólen
atravessem os carpelos e fecundem primórdios que de outra forma abortariam. Consequentemente, a
sincarpia aumenta a quantidade a qualidade da descendência por semente. As vantagens evolutivas da
sincarpia explicam a sua emergência de forma independente em 17 linhagens de angiospérmicas
(Armbruster et al., 2002).
34 Na bibliografia os termos apocárpico, sincárpico e paracárpico são indiferentemente usados para qualificar o gineceu, o pistilo ou o ovário. A bem da consistência conceptual da morfologia botânica era conveniente restringir a sua aplicação ao gineceu.
a) b) c) d) e)
C oncrescência e placentação. a) gineceu monocarpelar de placentação marginal [r – receptáculo, pr – primórdios seminais, pl – placenta, h – feixes vasculares], b) gineceu tricarpelar apocárpico, c) gineceu bicarpelar sincárpico de placentação axilar, d) gineceu tetracarpelar sincárpico de placentação axilar, e) gineceu tricarpelar de placentação central livre (adaptado de Diaz et al., 2004)

95 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
Ovário
A posição do ovário, a inserção das peças do perianto relativamente ao ovário e o tipo de
placentação são os caracteres morfológicos de maior interesse taxonómico sediados no ovário.
Existem três tipos de inserção das peças do perianto relativamente ao ovário:
a) Flor hipogínica – flores sem hipanto; peças do perianto e estames inseridas por baixo do
gineceu; e.g. Brassicaceae «crucíferas»;
b) For perigínica35 – flores com hipanto; ovário súpero ou ínfero; e.g. flor perigínica de ovário
súpero de Prunus avium (Rosaceae) «cerejeira» e flor perigínica de ovário ínfero de
Chaenomeles (Rosaceae);
35 Conceito inconsistente na bibliografia. Alguns autores reservam o termo perigínico para as plantas com hipanto e
a) b)
c)
D)
Inserção das peças do perianto relativamente ao ovário. A) Flor hipogínica de ovário súpero de Brassica napus (Brassicaceae) «nabo»; n.b. visíveis 2 das 4 pétalas e 2 das 4 sépalas, 6 estames didinâmicos (4 maiores e 2 mais pequenos). B) Flor perigínica de ovário ínfero de Chaenomeles x superba (Rosaceae); n.b. estames indefinidos, inserção dos estames na superfície interna do hipanto. C) Flor perigínica de ovário súpero de Prunus avium (Rosaceae) «cerejeira»; n.b. estames indefinidos. D) A. Flor hipogínica de ovário súpero, B. Flor epigínica de ovário ínfero, C. flor perigínica de ovário súpero, D. flor perigínica de ovário ínfero (adaptado de Smith, 2005).

96 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
c) Flor epigínica – hipanto aderente ao ovário (ovário ínfero) não se estendendo para além deste;
peças do perianto e estames inseridos no bordo do hipanto embora pareçam estar implantados
na extremidade do ovário; e.g. Asteraceae «compostas».
O tecido que conecta os funículos dos primórdios seminais com a parede do ovário designa-se por
placenta. Reconhecem-se 5 tipos fundamentais de placentação, i.e. de localização das placentas no
interior do ovário:
a) Marginal – primórdios localizados na proximidade da sutura carpelar num ovário monocarpelar;
b) Axilar (= axial) – ovário di a pluricarpelar sincárpico com primórdios inseridos no ângulo interno
definido pelos septos do ovário;
c) Central livre – ovário unilocular com primórdios seminais inseridos numa coluna central;
d) Parietal – primórdios dispersos na parede do ovário;
e) Basal – primórdios inseridos na base do ovário.
Estilete
Os estiletes, quando presentes, são usualmente classificados de acordo com os seguintes critérios:
a) Concrescência e número – em muitos grupos de plantas identificam-se vários estiletes livres
(e.g. Passifloraceae e Hypericaceae) ou parcialmente concrescentes (e.g. muitas Malvaceae);
noutras plantas os estiletes são concrescentes em todo o comprimento e culminam num único
estigma;
b) Quanto à inserção:
i) Terminal – estilete inserido no topo (extremidade distal) do ovário; condição mais
frequente;
ii) Lateral – lateralmente inserido;
iii) Ginobásico – estilete inserido na base (extremidade proximal) do ovário; característico
das Lamiaceae, das Boraginaceae e das Chrysobalanaceae, uma família tropical com
grande expressão na floresta atlântica brasileira.
O comprimento dos estiletes tem interesse taxonómico e/ou desempenha um importante papel na
biologia da polinização em alguns géneros (e.g. Ficus [Moraceae] e Primula [Primulaceae]). O interior
dos estiletes pode ser fistuloso ou sólido. Os estiletes fistulosos abundam entre as monocotiledóneas. A
cavidade destes estiletes está, geralmente, preenchida com mucilagens. Os estiletes sólidos têm no seu
ovário súpero, outros ainda para as flores de ovário semi-ínfero.
Placentação. A. Gineceu paracárpico tricarpelar de placentação parietal. B. Gineceu sincárpico s.str. tricarpelar de placentação axilar. C. Gineceu paracárpico tricarpelar de placentação central livre. D. Gineceu paracárpico tricarpelar de placentação basal (adaptado de Diaz et al., 2004)

97 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
interior um tecido especial – tecido de transmissão – que desempenha um importante papel na
fecundação.
Estigma
Os grãos de pólen aderem e germinam na superfície da epiderme do estigma. Os estigmas podem
ser “secos” ou glandulosos, caso exsudem secreções viscosas ao tacto. O estigma é séssil quando
assenta directamente sobre o ovário, i.e. quando não existem estiletes. As angiospérmicas basais (e.g.
Nymphaeaceae) e algumas magnoliidas não têm estilete e exibem uma margem estigmática extensa ao
longo de toda, ou de parte, da sutura carpelar. Um estilete pode terminar em um ou mais estigmas. Se
mais do que um então, por regra, em número igual ao número de carpelos. Quanto à forma, os estigmas
podem ser globosos, aclavados (em forma de clava), plumosos (cobertos de pêlos e tomando a forma de
uma pluma), etc.
4.2.1.10. Nectários florais e osmóforos
Os nectários florais secretam para o exterior ou interior da flor ou, por vezes, para estruturas em
forma de bolsa, fluidos ricos em açúcar (néctar). Os nectários florais têm uma localização muito variada
na flor: no receptáculo (e.g. disco glanduloso de Rutaceae, Sapindaceae e Ericaceae, e tubérculo
glanduloso de Brassicaceae), epicálice (e.g. Gossypium [Malvaceae] «algodoeiros»), face interior do
hipanto (e.g. Prunus e outras Rosaceae), sépalas (muitas Malvaceae e Malpighiaceae), pétalas (e.g.
alguns Ranunculus [Ranunculaceae]), esporões com origem no, receptáculo, no cálice ou na corola,
estaminódios (e.g. muitas Lauraceae) e gineceu (e.g. nectários septais de muitas monocotiledóneas,
circunscritos à sutura carpelar).
Para atrair insectos polinizadores muitas flores libertam odores de glândulas apropriadas – os
osmóforos – sediados nos mais diversos órgãos florais, com mais frequência no perianto (e.g. asas ou
estandarte em muitas leguminosas e coroa dos Narcissus).
4.2.1.11. Primórdios seminais
Como se referiu a respeito do gineceu, o primórdio seminal36 é um megasporângio envolvido por um
ou dois tegumentos. Num primórdio seminal identificam-se as seguintes partes:
a) Funículo – cordão delgado e curto que conecta o primórdio seminal à placenta;
b) Tegumento – constituído por uma (primórdios unitegumentados ou unitégmicos) ou duas
(primórdios bitegumentados ou bitégmicos) camadas com função de protecção; os tegumentos
arrancam da base da nucela e prologam-se até ao ápice do primórdio, deixando uma pequena
abertura – o micrópilo – por onde penetrará o tubo polínico durante a fecundação;
c) Nucela – tecido maternal diplóide, com muitas (primórdios crassinucelados) a escassas camadas
de células (primórdios tenuinucelados), situado entre o tegumento e o saco embrionário;
homologado ao megasporângio;
d) Saco embrionário – gametófito ♀ (vd. Volume II).
36 Também designado por óvulo, sobretudo na literatura inglesa. Este termo deve ser evitado porque pode ser confundido com o óvulo animal.
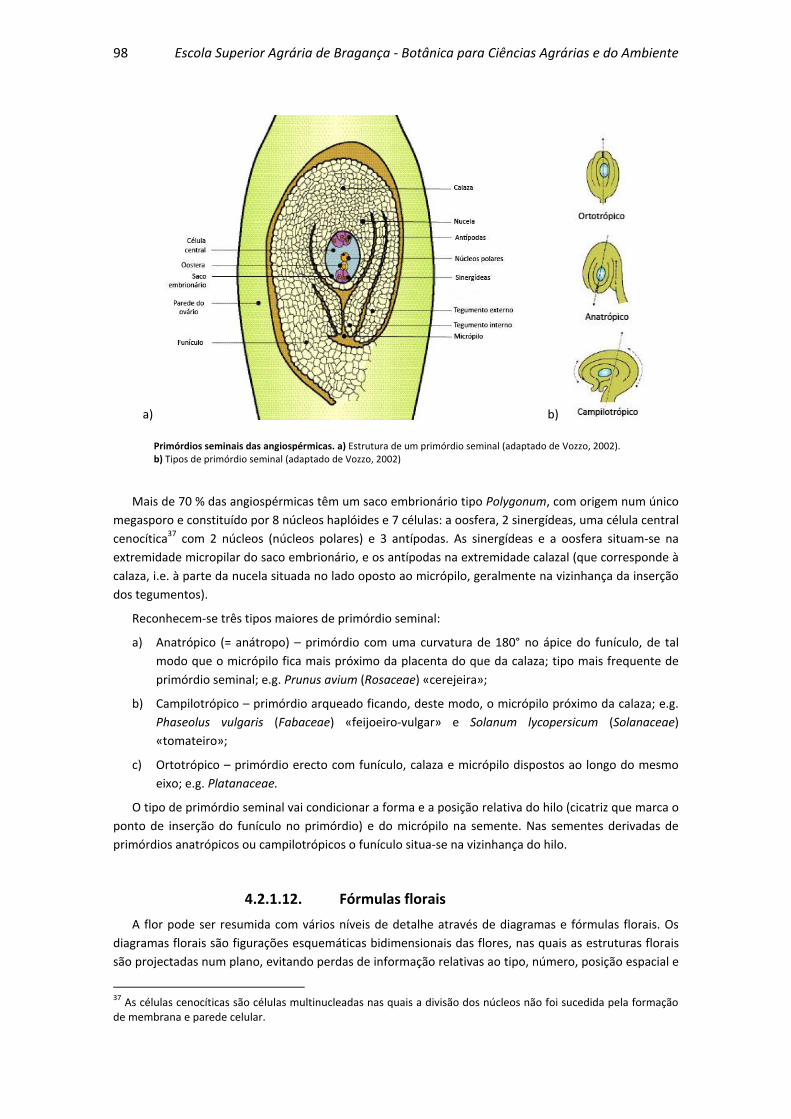
98 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
Mais de 70 % das angiospérmicas têm um saco embrionário tipo Polygonum, com origem num único
megasporo e constituído por 8 núcleos haplóides e 7 células: a oosfera, 2 sinergídeas, uma célula central
cenocítica37 com 2 núcleos (núcleos polares) e 3 antípodas. As sinergídeas e a oosfera situam-se na
extremidade micropilar do saco embrionário, e os antípodas na extremidade calazal (que corresponde à
calaza, i.e. à parte da nucela situada no lado oposto ao micrópilo, geralmente na vizinhança da inserção
dos tegumentos).
Reconhecem-se três tipos maiores de primórdio seminal:
a) Anatrópico (= anátropo) – primórdio com uma curvatura de 180° no ápice do funículo, de tal
modo que o micrópilo fica mais próximo da placenta do que da calaza; tipo mais frequente de
primórdio seminal; e.g. Prunus avium (Rosaceae) «cerejeira»;
b) Campilotrópico – primórdio arqueado ficando, deste modo, o micrópilo próximo da calaza; e.g.
Phaseolus vulgaris (Fabaceae) «feijoeiro-vulgar» e Solanum lycopersicum (Solanaceae)
«tomateiro»;
c) Ortotrópico – primórdio erecto com funículo, calaza e micrópilo dispostos ao longo do mesmo
eixo; e.g. Platanaceae.
O tipo de primórdio seminal vai condicionar a forma e a posição relativa do hilo (cicatriz que marca o
ponto de inserção do funículo no primórdio) e do micrópilo na semente. Nas sementes derivadas de
primórdios anatrópicos ou campilotrópicos o funículo situa-se na vizinhança do hilo.
4.2.1.12. Fórmulas florais
A flor pode ser resumida com vários níveis de detalhe através de diagramas e fórmulas florais. Os
diagramas florais são figurações esquemáticas bidimensionais das flores, nas quais as estruturas florais
são projectadas num plano, evitando perdas de informação relativas ao tipo, número, posição espacial e
37 As células cenocíticas são células multinucleadas nas quais a divisão dos núcleos não foi sucedida pela formação de membrana e parede celular.
a) b)
Primórdios seminais das angiospérmicas. a) Estrutura de um primórdio seminal (adaptado de Vozzo, 2002). b) Tipos de primórdio seminal (adaptado de Vozzo, 2002)

99 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
dimensão relativa dos órgãos e peças da flor. Os diagramas florais realçam, ainda, alguns aspectos da
estrutura da flor, e.g. estrutura do ovário, tipo de primórdios seminais, e forma, concrescência, adnação
e simetria das peças da flor. Os diagramas florais não são desenvolvidos neste texto.
As fórmulas florais veiculam menos informação do que os diagramas florais. Nestas fórmulas a
representação da flor reduz-se a um conjunto abreviado de símbolos referentes ao tipo, número,
simetria, concrescência e adnação das peças da flor. As fórmulas florais são comummente usadas para
representar taxa de categoria igual ou inferior à família, e.g. família, subfamília, género ou espécie. A
simbologia varia de autor para autor. No quadro que se segue descrevem-se, com pequenas alterações,
os símbolos propostos por Ronson de Craene (2010) de uso mais frequente.
Simetria da flor
* flor actinomórfica
↑ flor zigomórfica
↔ flor com dois planos de simetria (bissimétrica)
Órgãos florais
P perigónio
K cálice
C corola
A androceu
G gineceu (G súpero, -G- semi-ínfero, Ğ ínfero)
A˚ estaminódios
G˚ pistilódios
[…] fusão de peças de diferentes órgãos
(…) fusão de peças pertencentes ao mesmo órgão
+ usado para evidenciar o número de verticilos
/ “ou”
, “e”
: quando se verificam diferenças morfológicas assinaláveis entre peças do mesmo órgão
∞ número indefinido de peças do órgão
- utilizado para evidenciar uma variação do número de peças
Alguns exemplos:
a) Orchidaceae «família das orquídeas»: ↑ P3+2:1 [A1+2˚ Ğ(3)] – perigónio “P” de dois verticilos
“… 3+2:1 …” de três tépalas, uma das tépalas do verticilo interno “… :1 …” distinta das restantes;
androceu e gineceu concrescentes “… […]…”; androceu de três estames “…A1+2˚…”, dois deles
estéreis “…2˚…”; gineceu ínfero “…Ğ…” sincárpico e tricarpelar “… (3) …”;
b) Triticum «trigos» (Poaceae): ↔ P2 A3 G(1,2˚) – flores bissimétricas (“↔…”, com dois planos de
simetria porque ocorrem duas lodículas); perigónio reduzido a 2 lodículas “…P2…”; androceu
com 3 estames; gineceu súpero, sincárpico, com 1 carpelo fértil e 2 estéreis “…(G1,2˚)”;
Quadro 3. Simbolos a usar nas fórmulas florais (adaptado de Ronson de Craene, 2010)

100 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
c) Brassica oleracea «couves» (Brassicaceae): ↔ K4 C4 A2+4 G(2) – flores bissimétricas; cálice de 4
sépalas livres; corola dialipétala de 4 pétalas; androceu com 6 estames, 2 no verticilo externo e 4
no interno “… [A2+4] …”; gineceu súpero, sincárpico de 2 carpelos;
d) Solanum tuberosum «batateira» (Solanaceae): * K(5) [C(5) A5] G(2) – flores actinomórficas;
cálice sinsépalo de 5 sépalas; 5 pétalas soldadas num tubo; 5 estames epipétalos “…[…]…”;
gineceu bicarpelar, sincárpico e súpero;
e) Salvia officinalis «salvia» (Lamiaceae): ↑ K(3:2) [C(3:2) A2] G(2) – flores zigomórficas; cálice
pentâmero sinsépalo, com 2 grupos de sépalas, o inferior com 3 sépalas e o superior com 2 “…
(3:2) …”; 5 pétalas soldadas num tubo com dois grupos de pétalas, o inferior com 3 pétalas e o
superior com 2 “… (3:2) …”; androceu de 2 estames epipétalos; gineceu sincárpico, súpero, de 2
carpelos.
4.2.2. Fruto
Definição e função do fruto
O fruto s.str. (num sentido estrito),
resulta do desenvolvimento do(s)
ovário(s), regra geral após, e em
consequência, da fecundação. Num
sentido lato – fruto s.l. – o fruto pode
incluir tecidos não provenientes do
ovário com origem no cálice, no
receptáculo (pseudofrutos) ou nos
caules da inflorescência
(infrutescências). O conceito de
infrutescência é geralmente usado
num sentido lato: a infrutescência
pode ser qualquer sistema de caulinar
com frutos; e.g. uma espiga de milho-
graúdo (Zea mays), um ouriço com
castanhas (Castanea sativa), uma
umbela de cerejas (Prunus avium) um
figo (Ficus carica) ou um ananás
(Ananas comosus).
O fruto tem um papel fundamental no sucesso da germinação das sementes e no estabelecimento
das plântulas. Para tal desempenha uma ou mais das seguintes funções:
a) Protecção da semente contra predadores e parasitas;
b) Promoção da dispersão da semente;
c) Melhoria das condições nutritivas do solo onde germina a semente através da libertação, por
mineralização microbiana, dos nutrientes contidos nos seus tecidos;
d) Facilitação do enterramento das sementes.
Estrutura do fruto
Durante a formação do fruto a parede do ovário modifica-se e dá origem ao pericarpo. De fora para
dentro do fruto distinguem-se três camadas no pericarpo:
Fruto s.str. vrs. pseudofruto. Flor hipogínica de ovário súpero e fruto com sépalas persistentes. B. Flor epigínica de ovário ínfero e pseudofruto com sépalas persistentes. N.b. posição das sépalas num fruto (A) e num pseudofruto (B) (adaptado de van Wyk & van Wyk, 2006)

101 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
a) Epicarpo – corresponde à epiderme exterior (da página inferior) da(s) folha(s) carpelar(es) que
compõem o pistilo;
b) Mesocarpo – tecido formado a partir do parênquima e dos feixes vasculares das folhas
carpelares;
c) Endocarpo – corresponde à epiderme interior (da página superior) das folha(s) carpelar(es).
Os mesmos termos são aplicados aos pseudofrutos, porém, neste caso, não são verdadeiras as
correspondências com a anatomia do carpelo estabelecidas para os frutos verdadeiros. Em linguagem
corrente apelida-se de pedúnculo o “pé” dos frutos mesmo que este tenha origem num eixo da
inflorescência.
Critérios de classificação dos frutos s.l.
Os critérios mais importantes na sistemática dos frutos são a origem, a consistência, a deiscência, o
número de carpelos e o número de sementes. Com base nestes critérios definem-se os seguintes
termos:
a) Quanto à origem:
i) Frutos s. str. – procedentes de uma só flor de ovário súpero; de acordo com a
concrescência do ovário subdivididos em:
Frutos simples – gineceu unipistilado, qualquer que seja o número de carpelos;
e.g. cereja e pêssego;
Frutos múltiplos (= frutos agregados) – gineceu multipistilado, i.e. apocárpico;
cada pistilo dá origem a um frutículo; o fruto múltiplo é constituído pelo
conjunto dos frutículos; e.g. drupéolas na amora (Rubus) e aquenióides no
morango;
ii) Pseudofrutos (= frutos complexos ou falsos frutos) – provenientes de uma só flor de
gineceu ínfero; incorporam tecidos do hipanto, e.g. maçã, pêra e marmelo;
iii) Infrutescências38 – incorporam tecidos carnudos ou secos com origem nos eixos da
inflorescência, nos pedicelos das flores e, por vezes, das brácteas; e.g. ananás e figo;
b) Quanto à consistência:
i) Frutos s.l. secos – pericarpo delgado e mesocarpo com um baixo teor em água; e.g.
cápsula da papoila;
ii) Frutos s.l. carnudos – pericarpo normalmente espesso, e mesocarpo rico em água e de
consistência carnuda; o endocarpo pode ser brando, coriáceo ou apresentar-se
endurecido, i.e. lenhoso; nos pseudofrutos carnudos os tecidos carnudos têm
geralmente origem no hipanto e não nos tecidos carpelares. Exemplos: de endocarpo
brando – bago de uva; de endocarpo coriáceo – maçã; endocarpo lenhoso – cereja;
c) Quanto à deiscência:
i) Frutos s.l. deiscentes – abrem espontaneamente libertando as sementes; a unidade de
dispersão (diásporo) é a semente; os frutos carnudos muito raramente são deiscentes;
e.g. vagem de feijão;
ii) Frutos s.l. indeiscentes – as sementes dispersam-se inclusas no fruto; unidade de
dispersão (diásporo) é o fruto; e.g. fruto do girassol;
38 Alguns autores designam as infrutescências por frutos múltiplos.

102 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
Esquizocarpo – tipo particular de fruto indeiscente que se fragmenta na
maturação em mericarpos; diásporo constituído por um fragmento de fruto
com uma ou mais sementes inclusas; e.g. fruto da malva;
d) Quanto ao número de carpelos:
i) Unicarpelar – com origem num ovário com um carpelo; e.g. cereja;
ii) Bicarpelar – com origem num ovário de 2 carpelos; e.g. fruto da couve;
iii) Pluricarpelar – com origem num ovário com 3 ou mais carpelos; maçã;
e) Quanto ao número sementes:
i) Monospérmico – uma semente; e.g. cereja;
ii) Polispérmico – mais de uma semente; e.g. pimento;
Na classificação dos frutos s.l. é ainda tomada em consideração a presença de asas, a aderência do
pericarpo à semente, a consistência do endocarpo e a organização das brácteas que envolvem os frutos.
Tipos de frutos s.l.
A terminologia carpológica é muito variada e inconsistente. Antes de usar uma Flora ou monografias
taxonómicas é importante consultar, a este respeito, as introduções metodológicas ou os glossários
anexos. O sistema de classificação de frutos seguido neste texto inspira-se em Font Quer (1985), com
actualizações. Em função dos critérios expostos anteriormente admitem-se seis grandes grupos de
frutos s.l. com numerosos tipos (apenas se referem os tipos mais relevantes para a flora portuguesa ou
com valor económico):
a) Frutos múltiplos:
i) Múltiplo de aquénios – frutículos tipo aquénio (aquenióides); e.g. Ranunculus
(Ranunculaceae) e Clematis (Ranunculaceae); na Fragaria x ananassa (Rosaceae)
«morangueiro» pequenos aquenióides estão dispersos na superfície de um hipanto
carnudo;
Cinorrodo – fruto múltiplo de aquénios no qual os aquenióides estão inclusos
(= encerrados) num hipanto em forma de saco, carnudo e de cor vermelha ou
amarelada quando maduro (= úrnula); e.g. característico das Rosa (Rosaceae)
«roseiras»;
ii) Múltiplo de drupas – frutículos tipo drupa (drupéolas); e.g. amoras de Rubus
(Rosaceae) «silvas»;
iii) Outros frutos múltiplos – plurissâmara (e.g. Ailanthus altissima [Simaroubaceae]
«ailanto»), fruto múltiplo de folículos (e.g. Spiraea [Rosaceae] «grinaldas-de-noiva») e
bifolículo (e.g. Nerium oleander [Apocynaceae] «loendro»);
b) Frutos simples esquizocárpicos:
i) Clusa – seco, bicarpelar, ovário profundamente 4-partido, com quatro sementes (2 por
lóculo); fragmenta-se em 4 mericarpos cada um com uma semente inclusa;
característico das Lamiaceae e das Boraginaceae;
ii) Regma – seco, pluricarpelar, com estiletes muito longos, rígidos e persistentes no fruto;
divide-se em 5 mericarpos aristados higroscópicos; característico das Geraniaceae;
iii) Dissâmara (= samarídeo) – seco, bicarpelar, na maturação dividido em 2 mericarpos
alados; característico do género Acer (Sapindaceae) «aceres ou bordos»;
iv) Lomento – seco, mono ou bicarpelar e polispérmico; dividido na maturação num
número variável de mericarpos; dois subtipos bastante frequentes:

103 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
Vagem lomentácea; característica de alguns géneros de Fabaceae, e.g.
Ornithopus «serradelas» e Scorpiurus «cornilhões»;
Silíqua lomentácea; tipo particular de silíqua característico de algumas
Brassicaceae; e.g. Raphanus raphanistrum (Brassicaceae) «saramago»;
c) Frutos simples secos:
i) Cariópse – indeiscente, monospérmico, com semente aderente ao fruto; característico
das Poaceae;
ii) Aquénio – indeiscente, uni a pluricarpelar, monospérmico, com uma semente mais ou
menos livre (não aderente ao fruto, excepto no ponto de encontro do funículo com a
parede interna do fruto); fruto muito frequente na Natureza; e.g. Urtica (Urticaceae)
«urtigas», Fumaria (Papaveraceae) e algumas Fabaceae como os Trifolium «trevos»;
alguns tipos de aquénio merecem uma designação particular:
Sâmara - aquénio alado; e.g. Ulmus (Ulmaceae) «ulmeiros»;
Noz – aquénio de grande dimensão de pericarpo endurecido; e.g. Corylus
avellana (Betulaceae) «aveleira»;
Utrículo – aquénio envolvido por duas brácteas concrescentes; característico
das Cyperaceae;
iii) Cápsula – deiscente, pluricarpelar, polispérmico, uni ou plurilocular; consoante o tipo
de deiscência a cápsula pode ser poricida (por poros, e.g. Papaver [Papaveraceae]
«papoilas»), loculicida (pela nervura média do carpelo, e.g. Lilium [Liliaceae]), septicida
(pelas suturas carpelares, e.g. Aristolochia [Aristolochiaceae]), septifraga (pela nervura
média e pelas suturas carpelares, e.g. Datura stramonium [Solanaceae] «figueira-do-
inferno») ou transversal;
Pixídeo – cápsula de deiscência transversal; frequente nas Amaranthaceae;
iv) Vagem – geralmente deiscente pela sutura carpelar e pela nervura média do carpelo
(duas linhas de deiscência), monocarpelar, polispérmico, com as sementes dispostas
numa única série (fiada) na proximidade da sutura carpelar; característica da grande
maioria das Fabaceae; algumas fabáceas (e.g. Medicago «luzernas») e a maioria das
Caesalpinioideae (e.g. Ceratonia siliqua «alfarrobeira») produzem vagens indeiscentes;
v) Silíqua – deiscente pelas suturas carpelares (duas linhas de deiscência), bicarpelar,
polispérmico, sementes dispostas em duas séries na vizinhança de cada uma das
suturas carpelares (vd. Número e concrescência dos carpelos); característico da maioria
das Brassicaceae;
vi) Folículo – deiscente pela sutura carpelar (uma linha de deiscência), monocarpelar,
polispérmico; e.g. Grevillea (Proteaceae) e Hakea (Proteaceae);
d) Frutos simples carnudos:
i) Baga – indeiscente, polispérmico, de epicarpo delgado, mesocarpo carnudo e
endocarpo membranáceo; e.g. Solanum lycopersicum «tomateiro» e Capsicum annuum
«pimenteiros» na família das solanáceas, e Vitis vinifera (Vitaceae) «videira-europeia»;
ii) Hesperídeo – semelhante a uma baga, proveniente de um gineceu pluricarpelar,
plurilocular, de epicarpo rico em glândulas com óleos essenciais, mesocarpo esponjoso
(geralmente branco) e endocarpo membranoso (película dos gomos), no qual se
inserem pêlos suculentos de grande tamanho; e.g. fruto dos citrinos (várias espécies e
híbridos de Citrus, Rutaceae);

104 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
iii) Drupa – indeiscente, mono a pluricarpelar, endocarpo lenhoso (caroço), com uma
(raramente mais) semente incluída num único caroço; e.g. Olea europaea (Oleaceae)
«oliveira»; característico da subfam. Prunoideae (Rosaceae);
iv) Nuculânio (= pirenário) – drupa derivada de um ovário bi ou pluricarpelar com 2 ou
mais caroços (núculas) individualizados; e.g. Frangula (Rhamnaceae) «sanguinho-de-
água». Alguns Crataegus (Rosaceae) «pilriteiros» produzem um pomo com dois ou mais
caroços, com pouca propriedade também designado por nuculânio;
e) Pseudofrutos:
i) Glande – seco, indeiscente, monospérmico, de pericarpo coriáceo e envolvido por uma
cúpula; característico das Fagaceae;
ii) Balaústia – pericarpo coriáceo, interior dividido em cavidades por lamelas delgadas e
repleto de sementes de episperma carnudo (= sarcotesta); e.g. Punica granatum
(Lythraceae) «romãzeira»;
iii) Pomo – carnudo, polispérmico, pluricarpelar (geralmente 5 carpelos), endocarpo não
lenhoso; muito frequente na subfam. Maloideae (Rosaceae); e.g. Pyrus communis
(Rosaceae) «pereira»;
iv) Pseudoaquénio – pseudofruto análogo a um aquénio mas proveniente de um gineceu
ínfero;
Cipsela – pseudoaquénio rematado por um papilho, i.e. por uma coroa de
escamas, sedas (pêlos rígidos e fortes) ou de pêlos, simples (não ramificados)
ou plumosos (se ramificados); característico da família Asteraceae; frequente
em Dipsacaceae e Valerianaceae;
v) Pseudobaga – semelhante a uma baga mas com origem num ovário ínfero; e.g. Musa
(Musaceae) «bananeiras»;
Pepónio – pseudobaga frequentemente de grande tamanho, derivada de um
gineceu pluricarpelar, polispérmico, de epicarpo rijo, mesocarpo carnudo,
endocarpo muito ténue e placentas muito desenvolvidas; e.g. característico
das Cucurbitaceae;
vi) Pseudocápsula – semelhante a uma cápsula mas com origem num ovário ínfero; e.g.
Eucalyptus (Myrtaceae) «eucaliptos»;
vii) Pseudodrupa (trima) – semelhante a uma drupa mas com origem num ovário ínfero;
e.g. Juglans regia (Juglandaceae) «nogueira»;
viii) Cremocarpo – esquizocárpico, seco, indeiscente e bicarpelar; separa-se na maturação
em dois mericarpos; característico das Apiaceae (= Umbelliferae);
f) Infrutescências:
i) Sorose – derivado de uma inflorescência tipo espádice; são carnudos e participam na
infrutescência o eixo da inflorescência, pedicelos, ovários e brácteas; e.g. Ananas
commosum (Bromeliaceae) «ananás» e Morus (Moraceae) «amoreiras»;
ii) Sícono – resulta de uma inflorescência de pedúnculo piriforme, carnudo na maturação,
que envolve por completo as flores, tendo uma abertura para o exterior pelo qual
penetram os insectos polinizadores (Hymenoptera, Agaonidae) (vd. Volume II); as flores
♀ dão origem a frutos verdadeiros carnudos; característico dos Ficus (Moraceae)
«figueiras»;
iii) Ouriço – com brácteas espinhosas que envolvem 2-3 pseudoaquénios (castanhas);
característico das fagáceas dos géneros Castanea «castanheiros» e Fagus «faias».

105 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
Chave dicotómica 2. Tipos mais frequentes de frutos s.l.
1. Estrutura complexa, derivada de uma inflorescência e em cuja constituição estão envolvidos tecidos com origem caulinar e/ou antófilos estéreis ........................................................................................................ [Infrutescência] Grupo 6
- Estrutura constituída por tecidos do ovário ou do ovário e de um hipanto, ainda que por vezes com sépalas e pétalas, carnudas ou secas, aderentes ................................................................................................................................ [Fruto s.l.] 2
2. Gineceu pluricarpelar apocárpico ................................................................................................. [Fruto múltiplo] Grupo 1
- Gineceu monocarpelar ou pluricarpelar sincárpico .............................................................................................................. 3
3. Ovário ínfero ..................................................................................................................................... [Pseudofruto] Grupo 5
- Ovário súpero .................................................................................................................................................. [Fruto s.str.] 4
4. Fruto fragmentando-se na maturação, dispersando-se 1 ou mais sementes inclusas numa porção de ovário .................................................................................................................................. [Fruto simples esquizocárpico] Grupo 2
- Fruto não se fragmentando na maturação; fruto com sementes inclusas ou sementes per se como unidades de dispersão 5
5. Fruto carnudo ................................................................................................................................ [Fruto carnudo] Grupo 4
- Fruto seco .............................................................................................................................................. [Fruto seco] Grupo 3
Grupo 1. Frutos múltiplos
1. Frutículos alados ............................................................................................................................................... Plurisâmara
- Frutículos não alados ............................................................................................................................................................ 2
2. Frutículos carnudos de endocarpo lenhoso (com caroço) ..................................................................... Múltiplo de drupas
- Frutículos não carnudos ........................................................................................................................................................ 3
3. Frutículos com uma semente .............................................................................................................................................. 4
- Frutículos com duas ou mais sementes ................................................................................................................................ 5
4. Com uma úrnula carnuda envolvendo os frutículos ............................................................................................. Cinorrodo
- Sem úrnula .................................................................................................................................. Múltiplo de aquénios s.str.
5. Dois frutículos ....................................................................................................................................................... Bifolículo
- Mais de dois frutículos .......................................................................................................................... Múltiplo de folículos
Grupo 2. Frutos simples esquizocárpicos
1. Dois mericarpos com asa membranosa ............................................................................................................... Dissâmara
- Mericarpos não alados.......................................................................................................................................................... 2
2. Gineceu monocarpelar .......................................................................................................................... Vagem lomentácea
- Gineceu di ou pluricarpelar ................................................................................................................................................... 3
3. Gineceu bicarpelar .............................................................................................................................................................. 4
- Gineceu pluricarpelar, com estiletes persistentes organizados numa coluna rígida .................................................... Regma
4. Semelhante a uma silíqua, fragmentado dois ou mais mericarpos ........................................................ Silíqua lomentácea
- Derivado de um ovário profundamente 2-partido que se divide em 4 mericarpos monospérmicos na maturação ...... Clusa
Grupo 3. Frutos simples secos
1. Sementes inclusas no fruto na maturação (fruto como unidade de dispersão) ................................... [Fruto indeiscente] 2
- Sementes libertas do fruto na maturação (semente como unidade de dispersão) .................................. [Fruto deiscente] 6
2. Semente intimamente soldada às paredes do ovário ............................................................................................ Cariópse
- Semente livre, conectada à parede do ovário por um funículo ...................................................................... [aquénio s.l.] 3
3. Frutos alados .............................................................................................................................................................Sâmara
- Frutos não alados.................................................................................................................................................................. 4
4. Fruto de grande dimensão de pericarpo lenhoso e endurecido .....................................................................................Noz
- Frutos de pequena dimensão ............................................................................................................................................... 5
5. Frutos envolvidos por duas brácteas concrescentes ................................................................................................ Utrículo
- Frutos não envolvidos por brácteas ................................................................................................................. Aquénio s.str.
6. Gineceu monocarpelar ........................................................................................................................................................ 7
- Gineceu pluricarpelar ........................................................................................................................................................... 8
7. Deiscente por uma fenda longitudinal ..................................................................................................................... Folículo

106 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
- Deiscente por duas fendas longitudinais .................................................................................................................... Vagem
8. Gineceu bicarpelar com dois lóculos separados por um falso septo .......................................................................... Silíqua
- Gineceu pluricarpelar ................................................................................................................................................ Cápsula
Grupo 4. Frutos simples carnudos
1. Endocarpo lenhoso, fruto monospérmico .......................................................................................................................... 2
- Endocarpo membranáceo ..................................................................................................................................................... 3
2. Um caroço ................................................................................................................................................................... Drupa
- Dois ou mais caroços ............................................................................................................................................. Nuculânio
3. Interior do endocarpo revestido de pelos glandulosos ...................................................................................... Hesperídeo
- Interior do endocarpo não glanduloso ............................................................................................................................ Baga
Grupo 5. Pseudofrutos
1. Pseudofruto seco ................................................................................................................................................................ 2
- Pseudofruto carnudo ............................................................................................................................................................ 7
2. Pseudofruto monospérmico ............................................................................................................................................... 3
- Pseudofruto polispérmico ..................................................................................................................................................... 5
3. Pseudofruto envolvido por uma cúpula .................................................................................................................... Glande
- Pseudofruto não envolvido por uma cúpula ......................................................................................................................... 4
4. Pseudofruto alado ......................................................................................................................................... Pseudosâmara
- Pseudofruto não alado, rematado por um papilho (cálice reduzido a um conjunto de pelos) ................................... Cipsela
5. Pseudofruto esquizocárpico com dois mericarpos ............................................................................................ Cremocarpo
- Pseudofruto não esquizocárpico e polispérmico .................................................................................................................. 6
6. Sementes de pericarpo carnudo ............................................................................................................................ Balaústia
- Sementes de pericarpo não carnudo .............................................................................................................. Pseudocápsula
7. Pseudofruto monospérmico de endocarpo lenhoso (com um caroço) ........................................................... Pseudodrupa
- Gineceu polispérmico ........................................................................................................................................................... 8
8. Placenta muito desenvolvida .................................................................................................................................. Pepónio
- Placenta pouco desenvolvida ............................................................................................................................................... 9
9. Endocarpo membranáceo ........................................................................................................................................... Pomo
- Endocarpo não membranáceo .................................................................................................................. Pseudobaga s.str.
Grupo 6. Infrutescências
1. Infrutescências secas com 2-3 pseudoaquénios envolvidos por brácteas espinhosas ............................................... Ouriço
- Infrutescências carnudas ...................................................................................................................................................... 2
2. Flores encerradas numa estrutura carnuda de origem caulinar ................................................................................ Sícono
- Flores livres ao longo do eixo (ráquis) da inflorescência .............................................................................................. Sorose
Na documentação taxonómica é corrente não diferenciar aquénio de pseudoaquénio, cápsula de
pseudocápsula, baga de pseudobaga e drupa de pseudodrupa. Para evitar inconsistências
terminológicas na concretização a natureza do fruto usam-se os adjectivos aqueniforme, capsular,
baciforme e drupáceo.

107 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
Frutos, frutificações e sementes (Coutinho, 1898): 1. Ramo de Prunus dulcis (Rosaceae) «amendoeira»; 2. Cálice sinsépalo de Hyoscyamus sp. (Solanaceae) «meimendro»; 3. Vagem de Phaseolus vulgaris (Fabaceae) «feijoeiro-vulgar»; 4. Drupa de Prunus persica (Rosaceae) «pessegueiro»; 5. Endocarpo com semente inclusa (caroço) de P. persica; 6. Endocarpo e semente de P. persica; 7. Drupa de P. dulcis; 8. Caroço de P. dulcis; 9. Cipsela de Asteraceae; 10. Sâmara de Fraxinus angustifolia (Oleaceae) «freixo»; 11. Tegumento a destacar-se da semente de Ph. vulgaris; 12. Semente revestida de pêlos compridos em Salix sp. (Salicaceae) «salgueiros»; 13. Semente alada de Pinus pinaster (Pinaceae) «pinheiro-bravo»; 14. Cipsela sem papilho de Asteraceae; 15. Cariópse de Triticum aestivum (Poaceae) «trigo-mole»; 16. Sâmara de Ulmus sp. (Ulmaceae) «ulmeiro»; 17. Fruto múltiplo de folículos de Delphinium sp. (Ranunculaceae) «espora»; 18. Vagem de Cytisus scoparius (Fabaceae) «giesta-das-vassouras»; 19. Cápsula de Viola (Violaceae) «violeta»; 20. Cápsula de Papaver rhoeas (Papaveraceae) «papoila-comum»; 21. Cápsula de Anagallis sp. (Myrsinaceae) «morrião»; 22. Drupas de Prunus cerasus (Rosaceae) «gingeira»; 23. Baga de Atropa belladonna (Solanaceae) «beladona»; 24. Baga de Vitis vinifera (Vitaceae) «videira-europeia»; 25. Fruto múltiplo de aquénios de Ranunculus sp. (Ranunculaceae) «ranúnculo»; 26. Fruto múltiplo de folículos Consolida ajacis (Ranunculaceae) «esporas-de-jardim»; 27. Dissâmara de Acer pseudoplatanus (Sapindaceae) «plátano-bastardo»; 28. Fruto múltiplo de drupas de Rubus sp. (Rosaceae) «amora»; 29. Fruto múltiplo de aquénios de Fragaria x ananassa (Rosaceae) «morangueiro». Infrutescências: 30. Sorose de Morus nigra (Moraceae) «amoreira-negra»; 31. Sorose de Ananas commosum (Bromeliaceae) «ananás»; 32. Sícono de Ficus carica (Moraceae) «figueira». Frutificações: 33. Pinha de P. pinaster; 34. Escama fértil com duas sementes de P. pinaster

108 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
Partes edíveis nos frutos s.l.
As partes edíveis num fruto s.l. variam de espécie para espécie. Em diferentes combinações, podem
envolver:
a) Tecidos do ovário – em grande parte dos frutos s.l. temperados consome-se todo o pericarpo
(e.g. uva) ou o mesocarpo (e.g. pêssego e demais prunóideas). No tomate, além do pericarpo,
são muito importantes as placentas e os septos;
b) Tecidos de um hipanto (com origem no receptáculo ou no cálice) – nas pomóideas (Maloideae,
Rosaceae) os tecidos carpelares são pouco relevantes no volume do fruto porque grande parte
da polpa dos pomos tem origem nos tecidos do hipanto;
c) Tecidos do receptáculo – no morango o receptáculo é vermelho, carnudo e na sua superfície
encontram-se simetricamente distribuídos minúsculos aquenióides;
d) Placenta – importante nas cucurbitáceas; as placentas são comestíveis no Citrullus lanatus
«melancia» e no Cucumis sativus «pepino», o mesmo não acontecendo no Cucumis melo
«melão» ou na Cucurbita pepo «abóbora-menina»;
e) Pêlos glandulosos – cada carpelo (gomo) do hesperídeo característico dos citrinos está
preenchido com pêlos glandulosos ricos em água;
f) Pedunculo do fruto – no Anarcardium occidentale (Anacardiaceae) «cajuzeiro» o fruto é seco, o
pedúnculo, em contrapartida, é carnudo e doce; a castanha-de-caju é uma semente que se
consome torrada;
g) Eixo primário da inflorescência (= ráquis) – no figo (Ficus carica, Moraceae) consomem-se o
pedúnculo da infrutescência e o pericarpo dos frutos produzidos pelas flores femininas
encerradas no interior do sícono; a parte carnuda do ananás compreende tecidos do ráquis da
infrutescência, dos pedicelos e das flores;
h) Toda ou partes da semente – na Juglans regia (Juglandaceae) «nogueira» consome-se a
semente que tem que ser previamente extraída de um fruto carnudo drupáceo (pseudodrupa);
algo semelhante acontece com a Prunus dulcis (Rosaceae) «amendoeira»; na romã o pericarpo é
seco e as sementes têm uma sarcotesta muito apreciada; na Garcinia mangostana (Clusiaceae)
«mangostão» o pericarpo é adstringente mas os arilos das sementes, pelo contrário, são
deliciosos; em várias plantas cultivadas são edíveis os frutos e as sementes, como acontece com
as vagens e as sementes imaturas do Phaseolus vulgaris (Fabaceae) «feijoeiro» e da Pisum
sativum (Fabaceae) «ervilheira».
Em número, fazem parte da dieta humana mais tipos de frutos carnudos do que de frutos secos. No
entanto, a maior parte das calorias das dietas actuais provêm das cariópses de gramíneas, como se
referiu anteriormente, um fruto seco intimamente soldado com a semente, à qual cabe a maior parte da
biomassa.

109 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
4.2.3. Semente
Constituição da semente
A semente é um primórdio seminal maduro, por regra, fecundado.
A semente madura é constituída por:
a) Episperma (= tegumento ou casca da semente) – cobertura da semente, de origem maternal,
resultante do desenvolvimento dos tegumentos do primórdio seminal;
b) Tecidos especializados de reserva (nem sempre presentes):
i) Endosperma – tecido de reserva triplóide, geralmente constituído por células vivas
(mortas nas gramíneas), com origem total ou parcial em células do saco embrionário;
ii) Perisperma – tecido esporofítico (diplóide) de origem maternal (células da nucela),
pouco frequente nas angiospérmicas;
c) Embrião – rudimento do esporófito, com origem no zigoto, i.e. na célula resultante da
conjugação da oosfera com um núcleo espermático; os cotilédones, com ou sem função de
reserva, são parte integrante do embrião.
Episperma
O episperma tem uma ou duas camadas consoante o primórdio seminal possua um ou dois
tegumentos. A inserção do funículo no primórdio seminal deixa uma pequena cicatriz na sua superfície
designada por hilo. Nas sementes de Vigna unguiculata (Fabaceae) «feijão-frade», e de muitas outras
Partes edíveis nos frutos s.l. Adaptado, com modificações, de http://www.fao.org/DOCREP/005/Y2515E/y2515e04.htm#P0_0

110 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
leguminosas, o hilo é muito fácil de identificar porque está assinalado por uma mancha mais ou menos
circular. O micrópilo por vezes evidencia-se através de um pequeno orifício na superfície do episperma.
Nas sementes derivadas de primórdios com dois tegumentos a camada externa do episperma
designa-se por testa (ou primina) e interna por tégmen (ou secundina). A testa é mais espessa e,
frequentemente, colorida e/ou rígida; o tégmen é membranoso. Portanto, o termo testa da semente só
deve ser confundido com episperma nas sementes derivadas de primórdios seminais com um único
tegumento.
Normalmente, a forma e a posição relativa do hilo frente ao micrópilo na semente está
correlacionada com o tipo de primórdio seminal. Os primórdios ortotrópicos originam sementes ovais
ou arredondadas com o hilo e o micrópilo em pólos opostos, respectivamente, nas extremidades
proximal e distal da semente. As sementes derivadas de primórdios campilotrópicos ou anatrópicos têm
uma forma reniformes e o micrópilo localiza-se na vizinhança do hilo (e.g. feijão). Geralmente, num dos
lados destas sementes, observa-se um rebordo que termina no hilo, conhecido por rafe, que resulta da
fusão lateral do funículo com o primórdio seminal.
O episperma é geralmente seco. Se seco e muito rígido denomina-se sclerotesta. Se pelo contrário é
carnudo e mole toma o nome de sarcotesta; e.g. nas sementes de Punica granatum (Lythraceae)
«romãzeira». O tegumento das sementes pode ser liso ou apresentar ornamentos – e.g. espinhos, pêlos,
papilas e tubérculos –, de grande interesse taxonómico em alguns grupos de plantas-com-flor (e.g.
Linaria [Plantaginaceae] e Montia [Portulacaceae]). A expansão laminar foliácea ou membranosa,
presente em espécies de dispersão anemocórica designa-se por asa; e.g. sementes de Acer
[Sapindaceae] «bordos». Na superfície das sementes podem desenvolver-se vários tipos de
excrescências nutritivas que actuam como recompensa para os vectores de dispersão endozoocórica,
sendo a mais relevante o arilo. O arilo é uma excrescência de dimensão (pode cobrir por completo a
semente) e localização variável na superfície do episperma, frequentemente de consistência carnuda
(Font Quer, 1985). Dois tipos de arilo merecem uma designação especial:
a) Carúncula – excrescência de pequena dimensão situada na proximidade do micrópilo; e.g.
sementes de Euphorbia (Euphorbiaceae) «eufórbias»;
b) Estrofíolo – excrescência, geralmente em forma de asa, que se forma a partir da rafe na
proximidade do hilo; e.g. sementes de Chelidonium majus (Papaveraceae) «celidónia» e de
Pterospartum tridentatum (Fabaceae) «carqueja».
Reservas nutritivas da semente
As reservas nutritivas destinam-se a alimentar o embrião em crescimento, durante a germinação da
semente. Nas sementes angiospérmicas ocorrem três tipos de reservas nutritivas:
a) Cotilédones – órgãos de reserva das sementes cotiledonares, e.g. leguminosas para grão e
Quercus (Fagaceae); as monocotiledóneas nunca têm sementes cotiledonares, somente as
eudicotiledóneas s.l.;
b) Endosperma – tecido de reserva das sementes albuminosas (= sementes endospérmicas), e.g.
cereais; mais frequente nas monocotiledóneas do que nas dicotiledóneas s.l.; as sementes sem
endosperma dizem-se, genericamente, exalbuminosas; O endosperma das sementes
albuminosas pode ser:
i) Quanto à composição:
Oleaginoso – reservas ricas em gordura; e.g. Cocus nucifera «coqueiro» e
Brassica rapa «colza»;
Amiláceo – reservas ricas em amido; e.g. fruto das gramíneas (cariópse);
ii) Quanto à textura:

111 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
Córneo – albúmen muito duro, como se fora marfim; e.g. algumas palmeiras
(fam. Arecaceae);
Gelatinoso – com a consistência da gelatina;
Líquido – e.g. leite-de-coco Cocus nucifera «coqueiro» (só parte do
endosperma do coco é líquido, o “coco ralado” é obtido da parte sólida).
c) Perisperma – característico das sementes perispérmicas, e.g. sementes de Beta vulgaris
(Amaranthaceae) «beterraba» e da fam. Caryophyllaceae; ao contrário das sementes
cotiledonares e albuminosas, nas sementes de perisperma abundante a nucela não se esgota
com a diferenciação do saco embrionário e a formação do embrião.
Algumas sementes têm, simultaneamente, albúmen e perisperma; e.g. Piper nigrum (Piperaceae)
«pimenteira». Raramente, o hipocótilo pode servir como o órgão de reserva.
As reservas das sementes podem ser constituídas por lípidos, hidratos de carbono (sobretudo amido)
e/ou proteínas. O teor em lípidos atinge os 50% nas sementes de Zea mays (Poaceae) «milho-graúdo» e
de Arachis hypogaea (Fabaceae) «amendoim». As sementes de cereais (excepto milho) contêm 70-80 %
de amido e as de leguminosas cerca de 50%. Quase todas as sementes contêm proteínas sob a forma de
pequenos grânulos proteicos genericamente designados por grânulos de aleurona.
Embrião
As principais estruturas do embrião são a radícula, o(s) cotilédone(s) e a plúmula. O hipocótilo é um
entrenó caulinar que conecta a radícula aos cotilédones. O epicótilo localiza-se entre os cotilédones e a
plúmula. Na semente a radícula está situada na proximidade do micrópilo e a plúmula na zona da calaza.
Normalmente, as monocotiledóneas têm um cotilédone e as dicotiledóneas s.l. dois, embora existam
bastantes excepções entre as dicotiledóneas s.l. (e.g. as Nymphaeaceae têm apenas um cotilédone).
4.2.4. Estruturas reprodutivas das gramíneas
Com a aproximação da fase reprodutiva o meristema apical das gramíneas ganha uma forma
alongada. Em dado momento, dá-se a iniciação floral: o meristema vegetativo converte-se num
meristema reprodutivo, deixa de produzir metâmeros vegetativos e inicia a diferenciação da
inflorescência.
Muitas gramíneas temperado-mediterrânicas, para diferenciarem flores, necessitam de frio invernal,
de dias em crescimento (e.g. algumas variedades de trigo-mole) ou de uma combinação, sequencial, de
ambos os factores ambientes (e.g. Secale cereale «centeio» e algumas variedades de trigo-mole). Nestas
plantas a transição do meristema vegetativo para meristema reprodutivo acontece ainda durante o
afilhamento, pelo menos nos caules mais velhos dos cereais de Outono-Inverno. A activação dos
meristemas intercalares caulinares nos entrenós distais e o início do alongamento do colmo, i.e. do
encanamento, são um pouco mais tardios. O alongamento dos entrenós caulinares distais “empurra” a
inflorescência para o exterior. A velocidade do “encanamento” depende sobretudo da temperatura do
ar: as Primaveras precoces provocam uma antecipação da floração das gramíneas. O período final da
“subida” do meristema pelo interior da bainha da última folha – a bandeira – é exteriormente visível por
inchaço. Nos cereais esta fase no ciclo fenológico é conhecida por emborrachamento.
As flores das Poaceae estão organizadas em inflorescências parciais especializadas designadas por
espiguetas. No meristema reprodutivo as espiguetas formam-se a partir de gemas cujas folhas axilantes
desapareceram numa fase recuada da evolução das gramíneas. O conhecimento da estrutura das
espiguetas é crucial na identificação das gramíneas cultivadas e silvestres.
As espiguetas podem ser sésseis ou posicionarem-se na extremidade de um caule filiforme,
incorrectamente apelidado por pedicelo. Em geral, cada espigueta é delimitada inferiormente por duas

112 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
glumas – gluma inferior e gluma superior. Secundariamente, pode existir uma única gluma; e.g. Lolium
«azevéns». As glumas têm uma origem bracteolar e uma disposição alterna disticada (uma por nó e
dispostas no mesmo plano). Geralmente são escariosas, i.e. secas, membranáceas, um tanto firmes, de
cores mortiças e algo translúcidas. As glumas, e as restantes peças da espigueta, inserem-se num
pequeno eixo caulinar: a ráquila. O número de flores por espigueta é muito variável; e.g. 1 em Agrostis,
2 em Holcus e muitas em Bromus. As flores apresentam, também, uma disposição alterna disticada na
espigueta. Consequentemente, nas espiguetas mutifloras a primeira flor localiza-se acima da gluma
inferior, a segunda flor da gluma superior, a terceira flor, por sua vez, situa-se imediatamente acima da
primeira flor, e assim por diante. Muitos géneros de gramíneas têm 1 (e.g. Panicum) ou mais (e.g. 2 em
Anthoxanthum) flores estéreis. Noutros géneros coexistem espiguetas estéreis e férteis (e.g. Cynosurus),
as primeiras geralmente com uma função de protecção. Em Setaria e Pennisetum as espiguetas estão
rodeadas por pedicelos estéreis (sem espiguetas) com uma função de protecção.
Cada flor de Poaceae está envolvida por duas glumelas. As glumelas são peças bracteolares39,
escariosas, de inserção alterna disticada, dispostas no mesmo plano das glumas. A glumela inferior – a
lema – é normalmente maior do que a superior – a pálea. Na maior parte das gramíneas a lema envolve
a pálea, formando-se uma cavidade fechada que encerra a flor. Na extremidade ou no dorso, quer das
glumas quer das glumelas, observa-se frequentemente uma estrutura delgada ou setiforme, mais ou
menos longa e rígida, conhecida por arista (= pragana ou saruga). A arista tem um importante papel no
enterramento da semente no solo. Nas espécies com aristas geniculadas (com um pequeno “cotovelo”)
– e.g. Avena «aveias» – as espiguetas ou as flores, com as sementes inclusas, retidas em pequenas
irregularidades dos solos, são empurradas para o interior do solo por movimentos higroscópicos40 da
arista. A vibração induzida nas aristas das glumelas pelo vento também auxilia o enterramento das
sementes. O calo da semente – um tufo de pêlos localizado na base das glumelas – funciona de forma
análoga a um anzol dificultando o arranque da semente do solo. A arista, consoante as espécies, poderá
ainda precaver a herbivoria (e.g. as aristas dificultam o corte, o arranque e a deglutição das plantas),
auxiliar a dispersão (e.g. sementes suspensas no pêlo dos mamíferos – dispersão ectozoocórica) e, já no
solo, facilitar a queda das glumelas e a germinação da semente.
As Poaceae têm flores nuas: o perianto está reduzido a 2 (3) escamas muito pequenas (lodículas),
situadas entre as glumelas e o verticilo dos estames, que incham e forçam a abertura das espiguetas na
ântese. Acima das lodículas encontram-se primeiro 3 estames (6 estames em Oryza e 6 ou mais nos
bambus) e depois um ovário tricarpelar, sincárpico, com um primórdio seminal e dois estigmas sésseis e
plumosos. As gramíneas são autogâmicas (e.g. Secale cereale «centeio») ou, mais frequentemente,
alogâmicas de polinização estritamente anemófila. Nestas espécies o vento faz vibrar as anteras e
transporta o pólen. Os colmos e as inflorescências impulsionados pelas mesmas forças vibram e oscilam
facilitando a suspensão do pólen no ar ou a sua captura. Os indivíduos conespecíficos (da mesma
espécie), com frequência, florescem subitamente e em simultâneo. Admite-se que desta forma a
polinização seja facilitada e os riscos de herbivoria diminuídos.
Nas gramíneas não domesticadas as espiguetas desarticulam-se acima ou abaixo das glumas na
maturação. Caso as espiguetas se desarticulem acima das glumas, estas permanecem na planta
suspensas na extremidade dos pedicelos. Nas espécies com espiguetas multifloras desarticuladas acima
das glumas, as sementes tombam no solo protegidas pelas glumelas, individualizadas ou agrupadas.
39 A glumela inferior (= lema) é uma bráctea porque axila o pequeno eixo onde se insere a flor. A glumela superior (= pálea) corresponde à primeira folha (= prófilo) muito modificada desse eixo sendo, por isso, interpretada como uma bractéola (Bell, 2008). 40 Movimentos higroscópicos – movimentos originados por ciclos de embebição e secagem.

113 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
As espiguetas agrupam-se em inflorescências de ordem superior de estrutura muito variada, por
norma de posição terminal, i.e. situadas no ápice dos colmos (as espigas femininas do milho-graúdo são
uma excepção). A panícula de espiguetas (e.g. Avena «aveias»), a espiga de espiguetas (e.g. Hordeum
vulgare «cevada», Secale cereale «centeio», Triticum «trigos» e Zea mays «milho-graúdo») e o cacho de
espigas de espiguetas (e.g. várias espécies dos géneros pratenses tropicais Panicum e Brachiaria)
contam-se entre as sinflorescências mais frequentes na família das gramíneas. As ramificações das
inflorescências das gramíneas são sempre alternas. Nas panículas de Avena, e de muitos outros géneros,
entrenós longos alternam com entrenós curtos formando-se pseudoverticilos de ramificações, por
torção dos entrenós por vezes orientadas para o mesmo lado. Os eixos das panículas, i.e. o ráquis (eixo
principal) e os ramos (eixos secundários) da sinflorescência, são, geralmente, filiformes. Em Z. mays as
cariópses surgem embebidas num eixo engrossado: o carolo.
A) B)
Estruturas reprodutivas das gramíneas. A) Panícula, espigueta e flor de Avena sp.; n.b. na espigueta de Avena não são visíveis as páleas porque estão ocultas por dentro das lemas; n.b. na flor de Avena foi retirada a lema para facilitar a observação das lodículas, estames e ovário; (adaptado de http://appliedweeds.coafes.umn.edu/app/herbarium/images/Hrb-AVEFA-014.jpg). B) Espigueta (em baixo) e flor (em cima) de Avena barbata subsp. lusitanica; n.b. pêlos abundantes e arista inseridos no dorso da lema

114 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
5. Fisionomia e fenologia das plantas-com-semente
5.1. Tipos fisionómicos
A sistematização das plantas em grupos deveu-se, originalmente, à necessidade de tornar mais
compreensível a sua diversidade para fins utilitários (vd. Volume III). A classificação biológica é um
modo, entre muitos outros, de arrumar as plantas em grupos internamente consistentes. Embora
reflicta relações de parentesco e possua um elevado valor preditivo, a classificação biológica das plantas
tem uma utilidade prática limitada em alguns contextos. O agrupamento das plantas em função da
forma exterior é uma alternativa vantajosa, por exemplo, no estudo da vegetação em territórios de flora
mal conhecida, ou na exploração de síndromes de adaptação ao fogo ou à secura edáfica à escala do
ecossistema. Uma vez que a evolução produziu uma imensa variedade de formas nas plantas, existem
múltiplas soluções para as organizar em tipos fisionómicos (ing. “growth form”)41, i.e. em grupos cujas
plantas partilham uma morfologia externa similar.
A classificação das plantas em ervas, arbustos, árvores e lianas é, talvez, o mais simples sistema de
classificação fisionómico. As ervas são plantas anuais ou perenes, de consistência herbácea ou sub-
herbácea. Se ligeiramente lenhosas na base dizem-se sufruticosas. As plantas perenes ou vivazes têm
um ciclo de vida de duração igual ou superior a três anos. Existe a tendência para aplicar o termo vivaz
às plantas de parte aérea herbácea, que se renova anualmente a partir de rizomas, tubérculos, bolbos,
etc. Por definição as árvores têm um tronco indiviso ramificado a uma distância variável do solo. Os
arbustos, pelo contrário, ramificam-se desde a base e raramente têm mais de 5 m de altura. As plantas
herbáceas, anuais, bienais ou perenes, e plantas as lenhosas, dizem-se monocárpicas se florescem uma
única vez, senescendo logo em seguida. Tal é o caso das Agave (Agavaceae): para permanecerem vivas é
necessária eliminar as inflorescências nascentes. As plantas pluricárpicas passam por mais de um
período reprodutivo.
O sistema de tipos fisionómicos mais utilizado à escala global foi proposto em 1934, pelo botânico
dinamarquês Christen C. Raunkjær:
a) Terófitos (= plantas anuais) – concluem o ciclo de vida num único ano atravessando o período
desfavorável na forma de semente;
b) Criptófitos – plantas vivazes com gemas de renovação ocultas no solo ou na água:
i) Geófitos – gemas ocultas no solo em rizomas, cormos, tubérculos, bolbos ou raízes; e.g.
Iris (Iridaceae) «lírios»;
ii) Hidrófitos (= plantas aquáticas) – plantas com gemas de renovação sob ou à superfície
da água, que colapsam ou mudam radicalmente de forma caso a água livre que as
sustenta se extinga; Ranunculus sect. Batrachium (Ranunculaceae) «ranúnculos-de-flor-
branca» ou Lemna (Araceae) «lentilhas-de-água»;
41 Existe alguma confusão na literatura entre tipo biológico (“plant life form”) e tipo fisionómico (“plant growth form”), dois conceitos sinonimizados por muitos autores. Para Barkman (1988) pertencem ao mesmo tipo biológico (ou biótipo) as plantas que partilham as mesmas adaptações morfológicas e/ou fisiológicas a um dado factor ecológico. Os tipos fisionómicos são grupos de plantas de morfologia externa similar, cujas semelhanças foram identificadas sem referências a hipotéticas vantagens adaptativas. Na bibliografia verifica-se uma certa tendência para reservar o conceito de “life form”, e por conseguinte de “tipo biológico” (menos vezes o de “tipo fisionómico”), para a classificação de Raunkjaer (1934), ou para outros sistemas de classificação das plantas baseados nas adaptações morfológicas aos periodos desfavoráveis do ano.

115 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
iii) Helófitos (= plantas anfíbias) – plantas adaptadas a zonas húmidas, capazes de suportar
pequenos períodos de dessecação do solo, com gemas de renovação imersas na água
ou em solos saturados de água; e.g. Typha latifolia (Typhaceae) «tabúa-larga»;
c) Hemicriptófitos – plantas vivazes ou bienais com gemas de renovo localizadas à superfície do
solo; os hemicriptófitos regra geral são estolhosos (e.g. Fragaria vesca [Rosaceae]
«morangueiro-bravo»), cespitosos (e.g. numerosas gramíneas vivazes) ou arrosetados; neste
último caso dispõem uma roseta de folhas à superfície do solo durante a estação desfavorável
que protege as gemas de renovo (e.g. Echium vulgare [Boraginaceae] «soagem»);
d) Caméfitos – plantas com gemas de renovo a menos de 25 cm da superfície do solo; o conceito
de caméfito abarca pequenos arbustos (e.g. Thymus zygis [Lamiaceae] «sal-da-terra»)
(caméfitos fruticosos), por vezes decumbentes ou em forma de almofada, plantas rizomatosas
ou estolhosas (caméfitos reptantes) (e.g. Vinca [Apocynaceae] «pervinca»), plantas cespitosas
(e.g. Dactylis glomerata [Poaceae] «panasco») e algumas plantas gordas (e.g. Sedum
[Crassulaceae]);
e) Fanerófitos – plantas perenes com gemas de renovo a mais de 25 cm da superfície do solo:
i) Fanerófitos escandentes – lianas (= trepadeiras) lenhosas; elevam-se acima do solo
apoiadas em outras plantas ou em suportes (e.g. Bougainvillea glabra
[Nyctaginaceae]), por meio de gavinhas, de raízes aéreas (e.g. Hedera helix
[Araliaceae]), espinhos, acúleos (e.g. Rosa e Rubus [Rosaceae]), ou por enrolamento
(caules volúveis) (e.g. Humulus lupulus (Cannabaceae);
ii) Nanofanerófitos – fanerófitos com gemas de renovo 25 cm a 2 m acima do solo; e.g.
Cistus ladanifer (Cistaceae) «esteva»;
iii) Microfanerófitos – fanerófito com gemas de renovo 2 a 8 m acima do solo; pequenas
árvores e arbustos altos; e.g. Cytisus scoparius (Fabaceae) «giesta-das-vassouras»;
iv) Mesofanerófitos – árvores médias a grandes com gemas de renovo 8 a 30 m acima do
solo; maioria das árvores da flora portuguesa; e.g. Quercus subgén. Quercus (Fagaceae)
«carvalhos»;
v) Megafanerófitos – fanerófito com gemas de renovo a mais de 30 m altura da superfície
do solo; não existem megafanerófitos na flora lenhosa de Portugal; muitas das árvores
das florestas tropicais húmidas são megafanerófitos (e.g. Ceiba pentandra (Malvaceae,
Bombacoideae] «sumaumeira».
O sistema de Raunkjær é difícil de aplicar no mundo tropical húmido. Existem outros sistemas
alternativos na bibliografia como o sistema de Du Rietz ou o sistema de Raunkjær alargado por Mueller-
Dombois & Elemberg (1974).

116 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
5.2. Posição das inflorescências nos ramos de plantas lenhosas
As inflorescências, solitárias (= flores solitárias) ou grupadas, ocorrem em cinco posições
fundamentais nas plantas lenhosas dicotiledóneas s.l.:
a) Macroblastos mistos – o ciclo de crescimento inicia-se com a formação de 2 prófilos (vd. Tipos
de filomas), segue-se um número variável de nomófilos; no final do ciclo vegetativo o meristema
apical diferencia-se numa flor, ou numa inflorescência com as respectivas brácteas; nas plantas
temperadas e mediterrânicas o ramo e respectivas flores têm origem em gomos mistos
hibernantes; de fácil observação em Aesculus (Sapindaceae) «castanheiros-da-índia», Acer
pseudoplatanus «bordo-comum», Catalpa bignonioides (Bignoniaceae) «catalpa», etc.;
b) Braquiblastos floríferos – lançamentos curtos com prófilos e nomófilos total, ou parcialmente,
suprimidos, geralmente reduzidos a brácteas; origem numa gema floral pronta ou hibernante,
quase sempre de posição lateral; consoante as espécies estes braquiblastos floríferos (e os
gomos que lhes deram origem) inserem-se:
i) Em ramos do ano – neste caso os braquiblastos floríferos provêm de gomos prontos
formados, numa posição lateral, nos ramos do próprio ano; as plantas que seguem este
modelo produzem, tendencialmente, flores e frutos no tarde; e.g. Diospyros kaki
(Ebenaceae) «diospireiro», Castanea sativa (Fagaceae) «castanheiro», Tilia (Tilioideae,
Malvaceae) «tílias» e Vitis vinifera (Vitaceae) «videira-europeia»;
ii) Em ramos curtos de dois anos – estes ramos são parte integrante de esporões ou
inserem-se directamente em macroblastos de três anos; quase sempre de inserção
lateral; tipo comum entre as Rosaceae arbóreas, em Juglans regia (Juglandaceae)
«nogueira» e Fraxinus (Oleaceae) «freixos»;
Tipos fisionómicos de Raunkjær. A) caméfito estolhoso. B) caméfito fruticoso. C) fanerófito. D) hemicriptófito arrosetado. E) hemicriptófito estolhoso e de raiz tuberosa. F) hemicriptófito rizomatoso. G) geófito rizomatoso. H) geófito bulboso. I) terófito.

117 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
iii) Em ramos longos de dois anos42 – caso mais frequente; e.g. Cytisus (Fabaceae)
«giestas», Olea europaea var. europaea (Oleaceae) «oliveira», Prunus persica
(Rosaceae) «pessegueiro» e Salix (Salicaceae) «salgueiros»;
i) Em madeira com mais de dois anos (caulifloria) – diferenciação de braquiblastos
floríferos a partir de gomos dormentes localizados no tronco e partes velhas da copa;
frequente em muitas espécies tropicais, e.g. Theobroma cacao (Sterculioideae,
Malvaceae) «cacaueiro», Artocarpus (Moraceae) «fruta-pão» e Myrciaria cauliflora
(Myrtaceae) «jabuticaba»; também na mediterrânica Cercis siliquastrum (Fabaceae)
«olaia».
Os quatro primeiros tipos são difíceis de discriminar em plantas lenhosas de climas sem uma estação
de repouso bem marcada (e.g. plantas de regiões tropicais húmidas). Muitas espécies combinam a
floração na extremidade de macroblastos mistos com a floração em braquiblastos floríferos inseridos
em ramos curtos. Mais rara é a produção, numa única estação de crescimento, de flores ou
inflorescências, em ramos de dois anos e em ramos do ano. Assim acontece nas variedades de figueira
com figos lampos (figos de Primavera – Maio-Junho – inseridos nos ramos longos do ano anterior) e
figos vindimos (figos diferenciados solitários ou aos pares na axila das folhas dos ramos do ano).
5.3. Ciclos fenológicos
A fenologia refere-se ao estudo da fenologia das plantas, e da sua variabilidade em função das
condições ambientais. O ciclo fenológico é entendido como uma sequência de estádios fenológicos (=
estádios de desenvolvimento fenológico, ing. “development stages”). Nas plantas anuais, estende-se da
germinação da semente até à colheita ou senescência da planta. Nas plantas perenes o estudo dos ciclos
fenológicos pode incluir todo o ciclo de vida, um ciclo de crescimento, ou parte deste, por exemplo do
início do ciclo de crescimento (por vezes restringido ao abrolhamento dos gomos florais) até à colheita.
42 Implicitamente formados no ciclo de crescimento ocorrido no ano anterior. Na contagem da idade dos ramos, o ano 1 inicia-se com o abrolhamento do novo ramo e termina com o abrolhamento das gemas inseridas no ramo entretanto diferenciado. O mesmo raciocínio aplica-se aos ramos de maior idade.
a) b)
Localização das inflorescências nas plantas lenhosas. a) Braquiblastos em Prunus avium (Rosaceae) «cerejeira»; n.b. gomos florais que produziram na Primavera braquiblastos floríferos. b) Caulifloria em Theobroma cacao (Sterculioideae, Malvaceae) «cacaueiro»

118 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
Os ciclos fenológicos das plantas têm um forte controlo genético. Embora sejam distintos de espécie
para espécie, e possam variar a nível infraespecífico (e.g. entre variedades de videira ou de macieira), os
ciclos fenológicos anuais respondem a diversos factores ambientais, sobretudo de ordem climática. Os
factores ambientais que exercem um controlo mais significativo nos ciclos fenológicos anuais são o
número de horas de sol, a precipitação e o integral de temperatura (soma das temperaturas a partir de
um determinado limiar e de uma determinada data).
Os estudos fenológicos têm uma grande utilidade em vários domínios da biologia e da agronomia:
a) Prática agrícola – permitem estabelecer momentos mais precisos para fertilizações, plantações,
podas, tratamentos fitossanitários e polinização artificial; facilitam a aplicação de seguros
agrícolas;
b) Melhoramento de plantas – são instrumento indispensável nos programas de selecção de
variedades precoces ou tardias, e na selecção de variedades com ciclos fenológicos desfasados
dos ciclos biológicos de parasitas ou de pragas chave;
c) Ecologia e biologia – fornecem bases metodológicas muito úteis para o estudo de fenómenos
tão importantes como aquecimento global e as flutuações climáticas interanuais.
No estudo da fenologia das plantas são usados protocolos estandardizados com códigos dos estados
fenológicos acompanhados por desenhos demonstrativos. Estes protocolos, e os estádios fenológicos
neles reconhecidos, variam consoante a espécie e os objectivos dos estudos (e.g. variações climáticas,
avisos de tratamentos fitossanitários).
A escala fenológica BBCH
A escala fenológica BBCH (acrónimo de “Biologische Bundesanstalt, Bundessortenamt and CHemical
Industry”) é talvez a tentativa mais bem sucedida de normalização dos estádios fenológicos das plantas-
com-semente (mono e dicotiledóneas s.l.). Foi desenvolvida para um alargado leque de espécies
cultivadas a partir da conhecida escala fenológica dos cereais de Zadoks. Pode ser generalizada a
espécies sem escalas publicadas, cultivadas ou não. A BBCH serve-se de um sistema decimal de
codificação dos estádios fenológicos principais – numerados de 0 a 10 – por sua vez subdividido em
estádios fenológicos secundários, novamente numerados de 0 a 10. Em algumas plantas é conveniente
subdividir os estádios fenológicos uma terceira vez (e.g. cucurbitáceas).
Na aplicação da escala fenológica BBCH é fundamental ter em consideração o seguinte:
a) Nas plantas anuais a semente não semeada é codificada com o número “00”;
b) O número “0” codifica a germinação nas plantas anuais e o abrolhamento nas plantas lenhosas,
dois fenómenos biológicos distintos;
c) Um código mais avançado significa que a planta se encontra num estádio fenológico mais tardio;
d) A duração de um estádio fenológico pode ser calculada a partir das datas de dois estádios
fenológicos consecutivos;
e) Às plantas após colheita (ou à disseminação das sementes) aplica-se o código “99”;
f) O estádio fenológico é avaliado no caule principal nas espécies com mais de um caule (e.g.
gramíneas);
g) À escala da parcela cultivada (ou da população no caso das plantas não cultivadas), o estádio
fenológico é avaliado nos indivíduos mais representativos;
h) Um estádio fenológico só é codificado quando é atingido por mais de 50% das plantas presentes
na parcela cultivada;
i) Pode ser anotada a ocorrência simultânea de dois estádios fenológicos (e.g. 16/22).

119 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
Código Descrição
0 Germinação/abrolhamento
(“Germination / sprouting / bud development”)
1 Desenvolvimento das folhas (caule pricinpal)
(“Leaf development (main shoot)”)
2 Formação de caules laterais/afilhamento
(“Formation of side shoots / tillering”)
3 Alongamento dos caules ou crescimento das rosetas/desenvolvimento do caule (caule principal)
(“Stem elongation or rosette growth / shoot development (main shoot)”)
4 Desenvolvimento das partes vegetativas de utilidade económica ou de órgãos vegetativamente propagados/emborrachamento (nas gramíneas)
(“Development of harvestable vegetative plant parts or vegetatively propagated organs / booting (main shoot)”)
5 Emergência das inflorescências (no caule principal)
(“Inflorescence emergence (main shoot) / heading”)
6 Floração (no caule principal)
(“Flowering (main shoot)”)
7 Desenvolvimento do fruto
(“Development of fruit”)
8 Maturação do fruto e da semente
(“Ripening or maturity of fruit and seed”)
9 Senescência, início da dormência (nas plantas lenhosas)
(“Senescence, beginning of dormancy”)
Estádios fenológicos. Cereais e outras gramíneas anuais (Meier, 2001)
Quadro 4. Estádios fenológicos principais da escala fenológica BBHC (Meier, 2001)

120 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
6. Referências
Angiosperm Phylogeny Group (2009) An update of the Angiosperm Phylogeny Group classification for
the orders and families of flowering plants: APG III. Bot. J. Linn. Soc. 161: 105-121.
Armbruster, W.S, E.M. Debevec & M.F. Willso (2002) Evolution of syncarpy in angiosperms: theoretical
and phylogenetic analyses of the effects of carpel fusion on offspring quantity and quality.
Journal of Evolutionary Biology 15: 657-672.
Barkman, J. J. (1988) New system of plants growth forms and phenological plant types. In Werger, M. J.
A., P. J. M. van der Aart, H. J. During & J. T. A. Verhoeven (eds.) Plant Form and Vegetation
Structure. The Hague: 9-14.
Beerling, D.J. (2005) Leaf Evolution: Gases, Genes and Geochemistry. Annals of Botany 96: 345–352.
Beerling, D. (2007) The Emerald Planet: How Plants Changed Earth's History. Oxford University Press.
Bell, A.D. (2008) Plant Form. An Ilustrated Guide to Flowering Plant Morphology. Timber Press. Portland.
Bingre, P., C. Aguiar, D. Espírito-Santo, P. Arsénio & T. Henriques (coord. cient.) (2007) Guia de Árvores e
Arbustos de Portugal Continental. In J. Sande Silva (coord.) Árvores e Florestas de Portugal. Vol.
IX. LPN.
Bold, H. C., C. J. Alexopoulos & T. Delevoryas (1987) Morphology of Plants and Fungi. 5ª ed. Harper
Collins Publishers, Nova York.
a) b)
Estádios fenológicos. a) Vicia faba «faveira». b) Malus domestica «macieira» (Meier, 2001).

121 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
Bonnier, G. & R.C.V. Douin (1912-1935) Flore complète illustrée en couleurs de France, Suisse et Belgique
(comprenant la plupart des espèces d'Europe). Paris, Neuchâtel, Bruxelles.
Brickell, C. D. et al. (eds.) (2004) International Code of Nomenclature for Cultivated Plants (ICNCP). Acta
Horticulturae: 1-123.
Champagne, C. & N. Sinha (2004) Compound leaves: equal to the sum of their parts? Development 131:
4401-4412.
Christenhusz, J. M. M., J. L. Reveal, F. G. Martin, R. M. Robert & W. M. Chase (2011a) A new classification
and linear sequence of extant gymnosperms. Phytotaxa 19: 55–70.
Christenhusz, J. M. M., X.-C. Zhang & H. Schneider (2011b) A linear sequence of extant families and
genera of lycophytes and ferns. Phytotaxa 19: 7–54.
Coutinho, A.X.P. (1898) Atlas de Botânica: para uso dos lyceus (I, II, III e IV classes). Companhia Nacional
Editora. Lisboa.
Díaz Gonzalez, T.E., M.C. Fernandez-Carvajal Alvarez & J.A. Fernandez Prieto (2004) Curso de Botánica.
Ediciones Treaa-Ciencia. Gijón.
Edlund, A. F., R. Swanson & D. Preuss (2004) Pollen and Stigma Structure and Function: The Role of
Diversity in Pollination. The Plant Cell 16: S84–S97.
Endress, P. (1994) Diversity and Evolutionary Biology of Tropical Flowers. Cambridge University Press.
Endress, P. K. & J.A: Doyle (2009) Reconstructing the ancestral angiosperm flower and its initial
spetializations. American Journal of Botany 96: 22–66.
Fernandes, R. B. (1972) Glossário de termos botânicos. Anuário da Sociedade Broteriana 38: 181-292.
Actualizado por Fátima Sales http://www.uc.pt/herbario_digital/glossario/
Font Quer, P. (1985) Diccionario de Botânica. 9a ed. Editorial Labor. Barcelona.
Hallé, F. (2002) In Praise of Plants. Timber Press.
Heywood, V. H. (1993) Flowering Plants of the World. Oxford University Press.
Hickey, M. & C. King (2000) The Cambridge Illustrated Glossary of Botanical Terms. Cambridge University
Press.
Hickey, L. J. (1973) Classification of the architecture of dicotyledonous leaves. American Journal of
Botany 60: 17-33.
Hickey, L.J. (1979) A revised classifi cation of the architecture of dicotyledonous leaves. In C. Metcalfe &
L. Chalk (eds.) Anatomy of the Dicotyledons. 2nd edition. Vol. 1. Clarendon Press. Oxford: 25-39.
Ingrouille, M. & B. Eddie (2006) Plants: Diversity and Evolution. Cambridge University Press.
Judd, W., C. Campbell, E. Kellogg, P. Stevens & M. Donoghue (2007) Plant Systematics. A Phylogenetic
Approach. Sinauer. 2ª ed.
Keller, R. (2004) Identification of Tropical Woody Plants in the Absence of Flowers. A Field Guide. 2nd
Edition. Birkhäuser Verlag. Basel.
Kirchoff, B.K. (2003) Shape matters: Hofmeister's rule, primordium shape and flower orientation. Int. J.
Plant Sci. 164: 505-517.
Leaf Architecture Working Group (1999) Manual of Leaf Architecture: Morphological Description of
Dicotyledonous and Net-Veined Monocotyledonous Angiosperms. Smithsonian Institution.
Washington, D.C.

122 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
Meier, U. (2001) (ed.) Growth Stages of Mono-and Dicotyledonous Plants. BBCH Monograph. Federal
Biological Research Centre for Agriulture and Forestry. Berlin.
Müeller-Dombois, D. & H. Ellenberg (1974) Aims and methods of Vegetation Ecology. Willey & Sons.
Parfrey, LW, E. Barbero, E. Lasser, M. Dunthorn, D. Bhattacharya, D. J. Patterson & L. A. Katz (2006)
Evaluating support for the current classification of eukaryotic diversity. PLoS Genet 2: 2062-2073.
Perin, L., L. Martínez-Aguilar, R. Castro-González, P. Estrada-de los Santos, T. Cabellos-Avelar, H. V.
Guedes, V. M. Reis & J. Caballero-Mellado (2006) Diazotrophic Burkholderia Species Associated
with Field-Grown Maize and Sugarcane. Appl Environ Microbiol. 72(5): 3103–3110.
Perez-Morales, C. (1999) Morfología de Espermatófitos. Ed. Celarayn. León.
Punt, W., P.P. Hoena, S. Blackmore, S. Nilsson & A. Le Thomas (2007) Glossary of pollen and spore
terminology. Review of Palaeobotany and Palynology 143: 1–81.
Purves, W.K., D. Sadava, G.H. Orians, H.C. Heller (2001) Life: the Science of Biology. 6th edition. Sinauer
Associates, Inc.
Raunkiær, Ch. (1934) The Life Forms of Plants and Statistical Plant Geography. Oxford University Press.
Raven, P.H., R. F. Evert & S.E. Eichhorn (2005) Biology of Plants. W.H.Freeman & Co Ltd.
Ronson de Craene, L.P. (2010) Floral Diagrams. An Aid to Understanding the Flower Morphology and
Evolution. Cambridge University Press. Cambridge.
Roth-Nebelsick, A., D. Uhl, V. Mosbrugger & H. Kerp (2001) Evolution and function of leaf venation
architecture: A review. Annals of Botany 87: 553-566.
Rudall, P. (2007) Anatomy of Flowering Plants. An Introduction to Structure and Development. 3rd
edition. Cambridge University Press.
Sitte, P., E. A. Weiler, A. Bresinsky, J. W. Kadereit & Ch. Korner (2003) Strasburger: Tratado de Botánica.
Ed. Omega. Barcelona.
Smith, S.E. & D.J. Read (2008) Mycorrhizal Symbiosis, 3ª Ed. Academic Press, Londres.
Spichiger, R-E, V. V. Savolainen, M. Figeat & D. Jeanmonod (2004) Systematic Botany of Flowering
Plants: A New Phytogenetic Approach of the Angiosperms of the Temperate and Tropical Regions.
Science Publishers.
Stevens, P. F. (2001+) Angiosperm Phylogeny Website. Version 9, June 2008 [and more or less
continuously updated since]. http://www.mobot.org/MOBOT/research/APweb/
Systematics Association Committee for Descriptive Biological Terminology (1962) II. Terminology of
Simple Symmetrical Plane Shapes (Chart 1). Taxon 11: 145-156.
Thomas, P. (2000) Trees: Their Natural History. Cambridge University Press.
van Wyk, B. & P. van Wyk (2006) Field Guide to Trees of Southern Africa. Struik Publishers.
Vasconcelos, J.C. (1969) Noções sobre a Morfologia Externa das Plantas Superiores. Série Estudos e
Informação Técnica nº 25. Ministério da Economia, Direcção Geral dos Serviços Agrícolas. Lisboa.
Vasconcelos, M. C., M. Greven, C. S. Winefield, M. C.T. Trought & V. Raw (2009) The Flowering Process
of Vitis vinifera: A Review. Am. J. Enol. Vitic. 60: 411-434.
Vogt, L., Th. Bartolomaeus & G. Giribet (2009) The linguistic problem of morphology: structure versus
homology and the standardization of morphological data. Cladistics 26: 301 – 325.

123 Escola Superior Agrária de Bragança - Botânica para Ciências Agrárias e do Ambiente
Vozzo, J.A. (ed.) (2002) Tropical Tree Seed Manual. U.S. Department of Agriculture, Forest Service,
Southern Research Station. Washington, D.C.
Weberling, F. (1992) Morphology of Flowers and Inflorescences. Cambridge University Press.
Whittaker, R.H. (1969) New concepts of kingdoms of organisms. Science 163: 150–160.
Willems, A. (2006) The taxonomy of rhizobia: an overview. Plant Soil 287: 3-14.
Woese, C., O. Kandler & M. Wheelis (1990) Towards a natural system of organisms: proposal for the
domains Archaea, Bacteria, and Eucarya. Proc Natl Acad Sci USA 87 (12): 4576-4579.

![2013 1º Vol Morfologia e função Fasc II[1]](https://static.fdocumentos.com/doc/165x107/55cf9d36550346d033acb1f9/2013-1o-vol-morfologia-e-funcao-fasc-ii1.jpg)
















