
9. - Ministério do Meio Ambiente · significativamente entre as estações do ano. ... Lagoa do...
12
144 9. Macroinvertebrados bentônicos Cecilia Volkmer-Ribeiro, Rosária De Rosa-Barbosa, Demétrio Luis Guadagnin, Carolina Coimbra Mostardeiro & Ana Paula da Silva Pedroso
Transcript of 9. - Ministério do Meio Ambiente · significativamente entre as estações do ano. ... Lagoa do...

144
9.
Macroinvertebrados
bentônicos
Cecilia Volkmer-Ribeiro,Rosária De Rosa-Barbosa,Demétrio Luis Guadagnin,
Carolina CoimbraMostardeiro & Ana Paula
da Silva Pedroso

145
Material e métodos
A proposta visava inicialmente seguir a metodologia de
dragagem descrita no Aquarap (Takeda et al., 2000), utilizando
draga de Petersen modificada (0,0345m2
). No entanto, constatou-
se que os barcos disponíveis não teriam condições (espaço e
calagem reduzidos) de operá-la, devido à necessidade de
instalação de molinete para a suspensão da draga fechada, devido
ao peso do sedimento capturado. Os demais procedimentos para
fixação das amostras seguiram Takeda et al. (2000).
Em razão do exposto, optou-se por amostrar o sedimento/
bentos com draga de Ekman, correspondendo a uma captação de
0.0225m2
de superfície de sedimentos (abertura da draga),
coletando-se 4 amostras em cada estação georreferenciada, para
análise de macrofauna bêntica e mais uma amostra para análise
granulométrica e de matéria orgânica dos sedimentos, totalizando
portanto 5 dragadas em cada estação. Nove estações de coleta (seis
na Lagoa do Casamento e ecossistemas associados e três no Butiazal
de Tapes e ecossistemas associados) foram selecionadas para
representar a diversidade de habitats em cada ambiente e duas
campanhas de amostragem representaram a estacionalidade do
clima – de 5 a 9 de maio (outono) e de 18 a 20 de novembro de
2003 (primavera) na região da Lagoa do Casamento e de 3 a 5 de
junho (outono) e de 2 a 3 de dezembro (primavera) de 2003 na
região do Butiazal de Tapes. A tabela I apresenta as coordenadas
geográficas/características para os ambientes selecionados na
região da Lagoa do Casamento e na região dos Butiazais de Tapes.
Nos locais de coleta, foram registradas as seguintes
variáveis físicas e químicas dos ambientes: transparência e
profundidade com disco de Secchi; pH, temperatura da água e
temperatura do ar com medidor de pH EC 10 Portable Modelo
50050; oxigênio dissolvido e porcentagem de saturação de
oxigênio dissolvido com medidor de oxigênio dissolvido DO 175
Dissolved Oxigen Meter Modelo 50175; a condutividade,
salinidade e sólidos totais dissolvidos, registrados com medidor de
condutividade CO 150 Conductivity Meter - Modelo 50150.
O sedimento amostrado para detecção do zoobentos foi
acondicionado em sacos plásticos, conservados em formol, e
levado ao laboratório para lavagem e posterior triagem dos
componentes do bentos sob microscópio estereoscópico. O
material para análise sedimentilógica e de matéria orgânica não
sofreu qualquer fixação.
Dado ao grande volume de sedimento para exame de
zoobentos, optou-se por triar três amostras de cada estação. Do
total de 54 amostras (36 amostras da região da Lagoa do
Casamento e 18 da região do Butiazal de Tapes), 22 foram
totalmente examinadas (19 da região da Lagoa do Casamento e
três da região do Butiazal de Tapes). O restante das amostras,
devido à grande quantidade de areia nas mesmas, sofreu
processamento diferenciado. Cada uma delas foi dividida em oito
porções iguais, após serem espalhadas homogeneamente em placa
de Petry com a área compatível com o tamanho da amostra.
Introdução
Levantamentos quantitativos de comunidades de macro-invertebrados bênticos em lagoas costeiras do Brasil foram
realizados de modo mais intenso nas faixas costeiras dos Estadosdo Rio Grande do Sul e do Rio de Janeiro.
Nessimian (1995 a, b) registrou Crustacea, Oligochaeta e Insectacomo sendo os grupos mais abundantes e os maiores
contribuintes em termos de biomassa, em um brejo entre dunasna restinga da Barra de Maricá, Rio de Janeiro.
Callisto et al. (1998) inventariaram os macroinvertebradosbentônicos em três lagoas do Município de Macaé, Rio de
Janeiro - Imboassica, Cabiúnas e Comprida, sendo apenas aúltima dulcícola, as outras duas sofrendo influência marinha. Na
lagoa Imboassica predominaram Polychaeta e Bivalvia,principalmente o gastrópode Heleobia australis; nas lagoas
Cabiúnas e Comprida foi observado o predomínio de larvas deinsetos aquáticos, especialmente Chironomidae e Chaoboridae,
além do tricóptero Oxyethira hyalina Gonçalves Jr. et al. (1998),demonstraram forte influência do tipo de sedimento na estrutura
das comunidades de macroinvertebrados bentônicos nas mesmaslagoas estudadas por Callisto et al. (1998).
Würdig et al. (1998) constataram que a variação nascomunidades de macroinvertebrados de duas lagunas e duaslagoas costeiras do RS (Osório e Tramandaí) deveu-se mais a
mudanças espaciais do que temporais, sendo que a presença demacrófitas, quantidade de matéria orgânica e o tamanho médio
das partículas do sedimento foram os fatores mais significativos.Wiedenbrug et al. (1997), em estudo da comunidade bentônicada lagoa Emboaba (também em Osório, RS) listaram 87 táxons
(Insecta – Chironomidae mais abundante) e baixa densidade (640ind.m-2 no inverno e 921 ind.m-2 no verão), se comparado a outrosestudos. Nessimian (1995 a, b), registra uma densidade média de
20905 ind.m-2, a densidade média sendo maior no verão, porémnem a densidade nem a composição do bentos diferiram
significativamente entre as estações do ano. Rodrigues & Hartz(2001) detectaram 45 táxons na lagoa Caconde (Osório, RS),
sendo Diptera o grupo mais comum, seguido por Crustacea,Annelida, insetos (exceto Diptera) e Mollusca.
A importância do estudo agora concluído reside não apenas nacomparação com os levantamentos já feitos em outros locais, mas,
sobretudo, na oportunidade de compararem-se comunidades deinvertebrados bênticos de lagoas de água doce de duas áreas
costeiras situadas em contexto diferenciado em relação ao oceanoe a Laguna dos Patos pois, enquanto os corpos lênticos da regiãodo Butiazal de Tapes encontram-se na margem interna da laguna,
e portanto mais afastados do litoral, os da região da Lagoa doCasamento situam-se na borda externa da laguna e mais próximos
da linha de costa. Os primeiros de idade mais antiga do que osúltimos, no que toca à formação dessa planície costeira (Tomazelli
et al., 2000).
Foram escrutinadas sempre 3/8 de cada amostra, conforme
determinado por suficiência amostral. Os valores obtidos nestas
três porções foram somados e feitas as médias para cada táxon
detectado, sendo esse valor multiplicado por oito.
O material triado foi encaminhado aos especialistas do MCN
para identificação. No caso da espongofauna considerou-se como
indivíduo cada gêmula encontrada na amostra (Tavares et al., no
prelo) uma vez que cada uma tem o potencial de formar um novo
indivíduo e constitui o elemento imprescindível para chegar-se à
identificação específica.
Os padrões espaciais e temporais na composição e
abundância dos macroinvertebrados foram descritos através de
análises exploratórias de ordenação indireta e direta, empregando
o programa Multiv 2.0 (Pillar, 2000). A abundância nas três
réplicas foi somada e os dados log-tranformados foram
submetidos à Análise de Coordenadas Principais. A relação entre a
composição e abundância de macroinvertebrados e as
características físico-químicas do sedimento foi realizada através
de Análise de Redundância, utilizando apenas os dados da coleta
de inverno, previamente centralizados e normalizados. Em ambas
análises foi empregada a distância euclidiana como medida de
dissimilaridade.
Resultados
Os resultados da análise sedimentológica, de matéria
orgânica e os parâmetros físicos e químicos para os ambientes da
região da Lagoa do Casamento e dos Butiazais de Tapes são
apresentados nas tabelas II e III. A tabela IV relaciona os táxons
específicos amostrados nas duas regiões. As tabelas V e VI
apresentam a quantificação dos táxons específicos amostrados na
região da Lagoa do Casamento respectivamente na primeira
(outono) e na segunda (primavera) expedições. As tabelas VII e
VIII apresentam igualmente a quantificação dos táxons específicos
amostrados na região dos Butiazais de Tapes respectivamente na
primeira (outono) e na segunda (primavera) expedições. Os
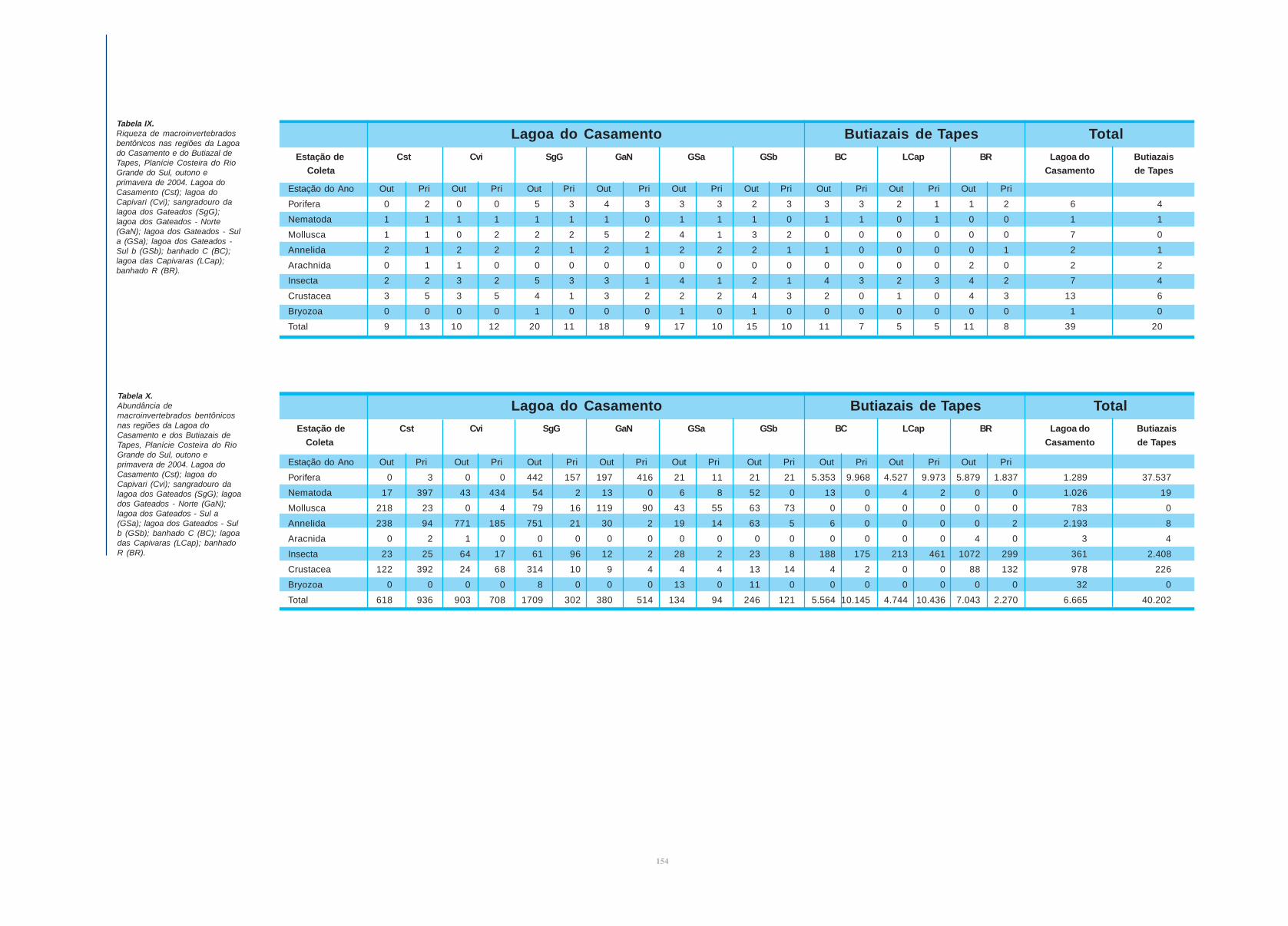
resultados da avaliação da riqueza e da abundância,
respectivamente, nas tabelas IX e X.
Foram registrados 46867 indivíduos pertencentes a 45
táxons. Os ambientes amostrados na Lagoa do Casamento e
ecossistemas associados apresentaram maior riqueza (39 espécies)
e menor abundância (6665 indivíduos) que os amostrados no
Butiazal de Tapes e ecossistemas associados (20 espécies e 40202
indivíduos). Ao todo, 14 morfoespécies foram compartilhadas
entre os ambientes, 25 foram encontradas exclusivamente na
região da Lagoa do Casamento e seis na dos Butiazais de Tapes.
Porifera, Mollusca, Insecta e Crustacea foram os grupos
taxonômicos com maior riqueza e abundância em ambos
ambientes. Na região do Butiazal de Tapes chama a atenção a
ausência de Nematoda e Anellida, bem representados na área da
Lagoa do Casamento, bem como pela grande abundância de
Porifera, Mollusca e Aracnida. Cabe lembrar que o número de
146
pontos de amostragem (seis na região da Lagoa do Casamento e
três na dos Butiazais de Tapes) foi proporcional ao tamanho das
áreas indicadas para inventário, de forma que são esperadas,
apenas por este fator, maior riqueza e abundância na Lagoa do
Casamento. De um modo geral a riqueza de espécies foi maior no
outono que na primavera.
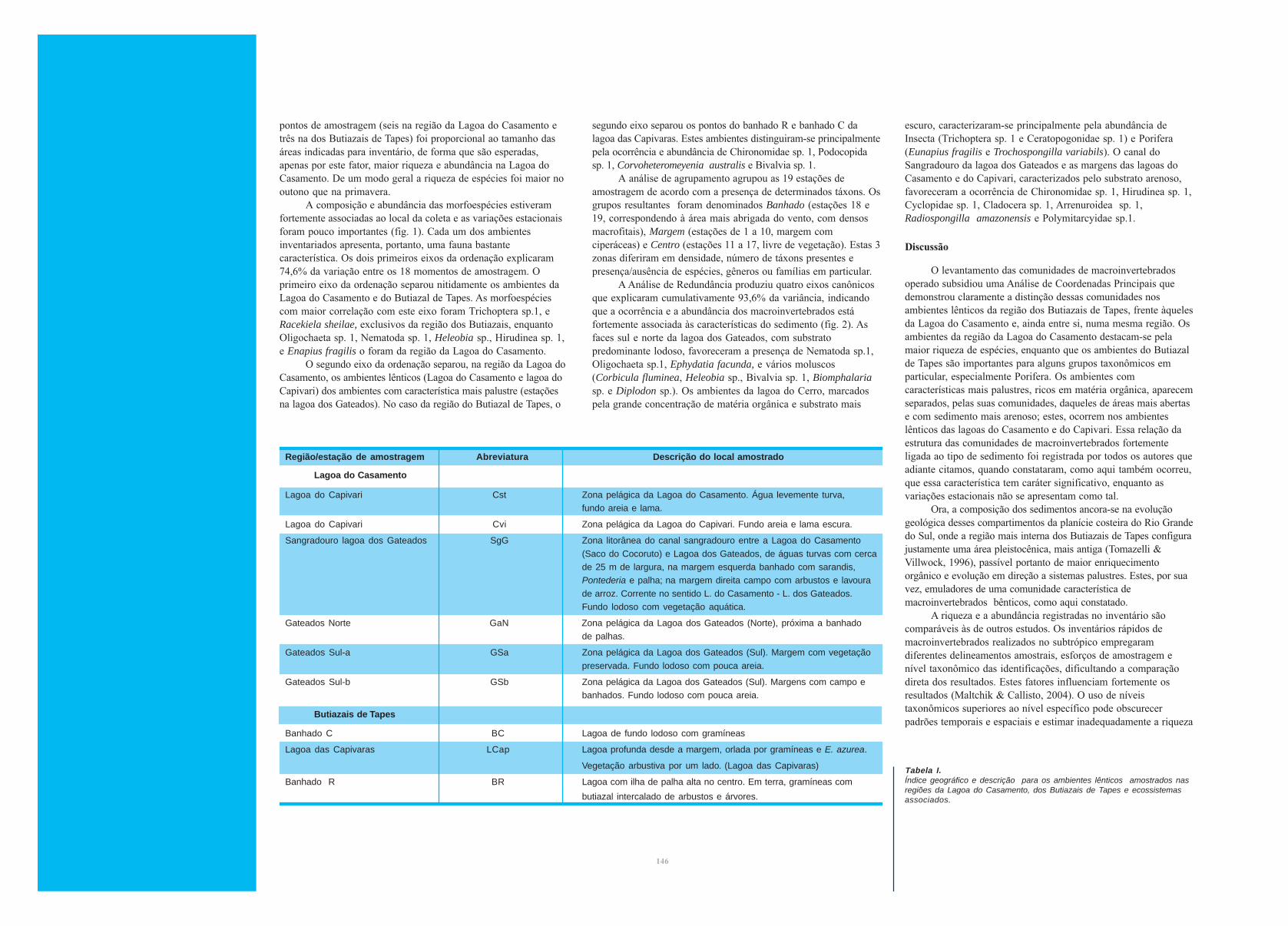
A composição e abundância das morfoespécies estiveram
fortemente associadas ao local da coleta e as variações estacionais
foram pouco importantes (fig. 1). Cada um dos ambientes
inventariados apresenta, portanto, uma fauna bastante
característica. Os dois primeiros eixos da ordenação explicaram
74,6% da variação entre os 18 momentos de amostragem. O
primeiro eixo da ordenação separou nitidamente os ambientes da
Lagoa do Casamento e do Butiazal de Tapes. As morfoespécies
com maior correlação com este eixo foram Trichoptera sp.1, e
Racekiela sheilae, exclusivos da região dos Butiazais, enquanto
Oligochaeta sp. 1, Nematoda sp. 1, Heleobia sp., Hirudinea sp. 1,
e Enapius fragilis o foram da região da Lagoa do Casamento.
O segundo eixo da ordenação separou, na região da Lagoa do
Casamento, os ambientes lênticos (Lagoa do Casamento e lagoa do
Capivari) dos ambientes com característica mais palustre (estações
na lagoa dos Gateados). No caso da região do Butiazal de Tapes, o
segundo eixo separou os pontos do banhado R e banhado C da
lagoa das Capivaras. Estes ambientes distinguiram-se principalmente
pela ocorrência e abundância de Chironomidae sp. 1, Podocopida
sp. 1, Corvoheteromeyenia australis e Bivalvia sp. 1.
A análise de agrupamento agrupou as 19 estações de
amostragem de acordo com a presença de determinados táxons. Os
grupos resultantes foram denominados Banhado (estações 18 e
19, correspondendo à área mais abrigada do vento, com densos
macrofitais), Margem (estações de 1 a 10, margem com
ciperáceas) e Centro (estações 11 a 17, livre de vegetação). Estas 3
zonas diferiram em densidade, número de táxons presentes e
presença/ausência de espécies, gêneros ou famílias em particular.
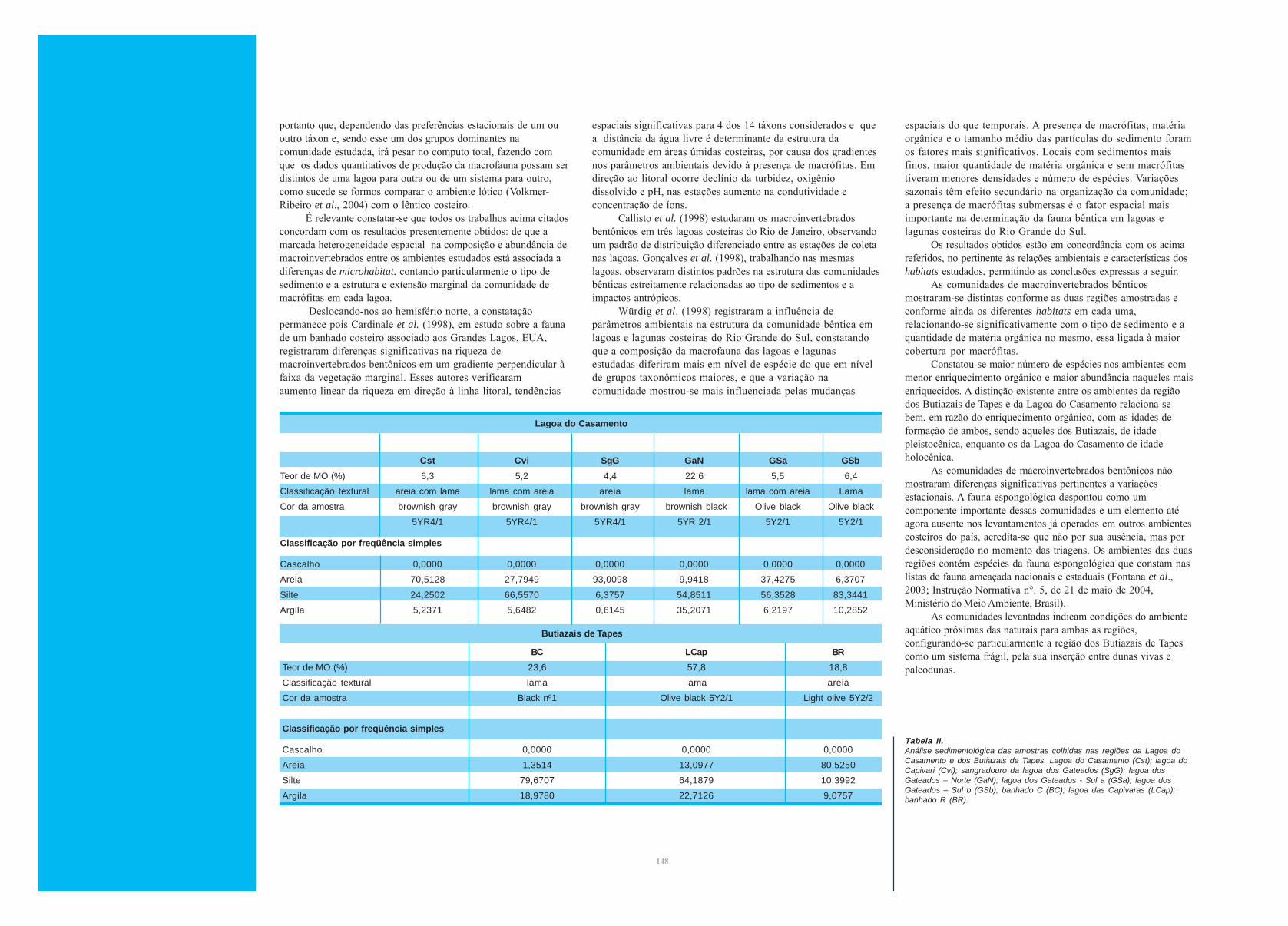
A Análise de Redundância produziu quatro eixos canônicos
que explicaram cumulativamente 93,6% da variância, indicando
que a ocorrência e a abundância dos macroinvertebrados está
fortemente associada às características do sedimento (fig. 2). As
faces sul e norte da lagoa dos Gateados, com substrato
predominante lodoso, favoreceram a presença de Nematoda sp.1,
Oligochaeta sp.1, Ephydatia facunda, e vários moluscos
(Corbicula fluminea, Heleobia sp., Bivalvia sp. 1, Biomphalariasp. e Diplodon sp.). Os ambientes da lagoa do Cerro, marcados
pela grande concentração de matéria orgânica e substrato mais
escuro, caracterizaram-se principalmente pela abundância de
Insecta (Trichoptera sp. 1 e Ceratopogonidae sp. 1) e Porifera
(Eunapius fragilis e Trochospongilla variabils). O canal do
Sangradouro da lagoa dos Gateados e as margens das lagoas do
Casamento e do Capivari, caracterizados pelo substrato arenoso,
favoreceram a ocorrência de Chironomidae sp. 1, Hirudinea sp. 1,
Cyclopidae sp. 1, Cladocera sp. 1, Arrenuroidea sp. 1,
Radiospongilla amazonensis e Polymitarcyidae sp.1.
Discussão
O levantamento das comunidades de macroinvertebrados
operado subsidiou uma Análise de Coordenadas Principais que
demonstrou claramente a distinção dessas comunidades nos
ambientes lênticos da região dos Butiazais de Tapes, frente àqueles
da Lagoa do Casamento e, ainda entre si, numa mesma região. Os
ambientes da região da Lagoa do Casamento destacam-se pela
maior riqueza de espécies, enquanto que os ambientes do Butiazal
de Tapes são importantes para alguns grupos taxonômicos em
particular, especialmente Porifera. Os ambientes com
características mais palustres, ricos em matéria orgânica, aparecem
separados, pelas suas comunidades, daqueles de áreas mais abertas
e com sedimento mais arenoso; estes, ocorrem nos ambientes
lênticos das lagoas do Casamento e do Capivari. Essa relação da
estrutura das comunidades de macroinvertebrados fortemente
ligada ao tipo de sedimento foi registrada por todos os autores que
adiante citamos, quando constataram, como aqui também ocorreu,
que essa característica tem caráter significativo, enquanto as
variações estacionais não se apresentam como tal.
Ora, a composição dos sedimentos ancora-se na evolução
geológica desses compartimentos da planície costeira do Rio Grande
do Sul, onde a região mais interna dos Butiazais de Tapes configura
justamente uma área pleistocênica, mais antiga (Tomazelli &
Villwock, 1996), passível portanto de maior enriquecimento
orgânico e evolução em direção a sistemas palustres. Estes, por sua
vez, emuladores de uma comunidade característica de
macroinvertebrados bênticos, como aqui constatado.
A riqueza e a abundância registradas no inventário são
comparáveis às de outros estudos. Os inventários rápidos de
macroinvertebrados realizados no subtrópico empregaram
diferentes delineamentos amostrais, esforços de amostragem e
nível taxonômico das identificações, dificultando a comparação
direta dos resultados. Estes fatores influenciam fortemente os
resultados (Maltchik & Callisto, 2004). O uso de níveis
taxonômicos superiores ao nível específico pode obscurecer
padrões temporais e espaciais e estimar inadequadamente a riqueza
Tabela I.Índice geográfico e descrição para os ambientes lênticos amostrados nasregiões da Lagoa do Casamento, dos Butiazais de Tapes e ecossistemasassociados.
Região/estação de amostragem Abreviatura Descrição do local amostrado
Lagoa do Casamento
Lagoa do Capivari Cst Zona pelágica da Lagoa do Casamento. Água levemente turva,fundo areia e lama.
Lagoa do Capivari Cvi Zona pelágica da Lagoa do Capivari. Fundo areia e lama escura.
Sangradouro lagoa dos Gateados SgG Zona litorânea do canal sangradouro entre a Lagoa do Casamento
(Saco do Cocoruto) e Lagoa dos Gateados, de águas turvas com cercade 25 m de largura, na margem esquerda banhado com sarandis,Pontederia e palha; na margem direita campo com arbustos e lavoura
de arroz. Corrente no sentido L. do Casamento - L. dos Gateados.Fundo lodoso com vegetação aquática.
Gateados Norte GaN Zona pelágica da Lagoa dos Gateados (Norte), próxima a banhadode palhas.
Gateados Sul-a GSa Zona pelágica da Lagoa dos Gateados (Sul). Margem com vegetação
preservada. Fundo lodoso com pouca areia.
Gateados Sul-b GSb Zona pelágica da Lagoa dos Gateados (Sul). Margens com campo ebanhados. Fundo lodoso com pouca areia.
Butiazais de Tapes
Banhado C BC Lagoa de fundo lodoso com gramíneas
Lagoa das Capivaras LCap Lagoa profunda desde a margem, orlada por gramíneas e E. azurea.
Vegetação arbustiva por um lado. (Lagoa das Capivaras)
Banhado R BR Lagoa com ilha de palha alta no centro. Em terra, gramíneas com
butiazal intercalado de arbustos e árvores.

147
específica (Wellborn et al., 1996) mas podem facilitar
comparações entre áreas e estudos, especialmente em grupos
taxonômicos pouco conhecidos.
Wiedenbrug et al. (1997), em estudo sobre a composição e
distribuição espacial da comunidade bentônica da lagoa Emboaba,
ambiente dulcícola na zona costeira do RS, registraram 87 táxons
(Insecta – Chironomidae, mais abundante) e baixa densidade (640
ind.m-2
no inverno e 921 ind.m-2
no verão), se comparado a outros
estudos (Nessimian, 1995b, densidade média 20.905 ind.m-2
).
Densidade média foi maior no verão, porém nem a densidade nem
a composição do bentos diferiram significativamente entre as
estações do ano.
A comparação feita com outros ambientes mostrou que os
insetos (Chironomidae, Trichoptera, Chaoboridae) são os que mais
caracterizaram lagoas costeiras dulcícolas (Gonçalves et al., 1998;
Callisto et al.,1998; Wiedenbrug et al., 1997). Quando se avalia
lagoas costeiras mixohalinas, despontam como grupos
característicos moluscos gastrópodes, particularmente Heleobiasp., moluscos bivalves e anelídeos poliquetos (Gonçalves et al.,1998; Callisto, 1998; Chomenko & Schäfer, 1984).
Os resultados presentemente obtidos confirmam os grupos
indicadores acima listados, já que os moluscos em geral e os
anelídeos ocorreram em maior abundância na região da Lagoa do
Casamento, portanto mais relacionada, dada sua contigüidade e
abertura, com a Laguna dos Patos. Nos ambientes dulcícolas da
região dos Butiazais de Tapes, os tricópteros dominaram,
registrando-se ainda a presença de Chaoboridae (Diptera).
No entanto, os resultados presentes destacam-se, de todos os
anteriormente citados, pela constatação da presença significativa
de gêmulas de esponjas nos sedimentos das duas regiões
abordadas, particularmente naqueles ambientes mais ricos em
macrófitas, indicando a ocorrência significativa de espécimes
nesses locais. Resultado semelhante já havia sido constatado na
área lagunar de Mundaú/Manguaba no litoral da Paraíba
(Volkmer-Ribeiro & Tavares, 1990; Pinheiro et al., 2004) e em
ambientes dulcícolas no Rio Grande do Sul (Volkmer-Ribeiro etal., neste volume). Acredita-se que a falta de referência deva-se,
não à ausência das esponjas nesses ambientes, mas à ausência de
familiaridade com essa fauna.
No pertinente às variações estacionais apresentadas pelas
comunidades bênticas estudadas, constatou-se de um modo geral
que a riqueza de espécies foi maior no outono que na primavera, em
ambas as regiões amostradas. Os resultados indicaram, no entanto,
reduzida heterogeneidade temporal na composição das comunidades
bênticas, a qual acreditamos deva-se, em parte, a que não houve
amostragem de inverno, mas somente de outono e primavera, duas
estações que até agora vem aportando riquezas e abundâncias
maiores em estudo de comunidades bênticas nessa região do RS.
Volkmer-Ribeiro et al. (1984, 2004), em amostragens
abarcando as quatro estações do ano no curso inferior do rio Caí,
próximos ao delta do Jacuí (portanto na planície costeira),
constataram diferenças significativas das comunidades bênticas
somente entre o inverno e a primavera, os menores valores
ocorrendo sempre no inverno. Os mesmo autores registram
heterogeneidade e relações faunísticas mais significativas devidas
aos ambientes amostrados do que às variações sazonais.
Wiedenbrug et al. (1997), em estudo sobre a composição e
distribuição espacial da comunidade bentônica da lagoa
Emboaba (Osório, planície costeira do RS), com amostragem
realizada no verão (fevereiro) e inverno (agosto), registraram
densidade média maior de táxons no verão (fevereiro), porém
nem a densidade nem a composição do bentos diferiram
significativamente entre essas duas estações do ano. Rodrigues &
Hartz (2001), investigando a lagoa Caconde, próxima à lagoa
Emboaba, registram que os maiores números de
macroinvertebrados bentônicos foram encontrados no outono
(43.800 ind.m-2
) e no inverno (46.414), seguidos pela primavera
(23.453) e verão (11.626).
Ozorio (1993) verificou que a lagoa das Custódias
(Tramandaí, RS), com características lagunares mesoalinas,
apresentou uma variação sazonal que incidiu sobre o aumento da
densidade total no inverno, mantendo-se porém a composição
faunística do macrobentos quase a mesma.
Nessimian (1995 a, b) estudou a abundância e biomassa de
macroinvertebrados em um brejo entre dunas no litoral do Estado
do Rio de Janeiro. O brejo estudado apresentou flutuações
sazonais no nível da água, com 4 períodos distintos: período seco
de verão (exposição do solo, decomposição acelerada por
microrganismos aeróbicos); ascendente de outono (aumento da
condutividade elétrica, devido à grande disponibilidade de íons);
cheia de inverno (maior diluição, diminui a condutividade) e
descendente de primavera (diminui o volume de água, aumenta a
condutividade elétrica). Foram realizadas 15 amostras mensais (de
janeiro de 1987 a fevereiro de 1988). O autor registrou a maior
produção de macrobentos nos período descendente de primavera e
no início do verão.
Todas as pesquisas citadas para a faixa costeira do Rio
Grande do Sul incidem na conclusão de que a sazonalidade não é
o fator que detém importância significativa na estrutura das
comunidades bênticas desses ambientes dulcícolas lênticos
costeiros, a maioria indicando o outono como a época, no
mínimo, de maior abundância. Já na altura do Rio de Janeiro,
Nessimian (1995 a, b) relaciona a maior abundância não tanto à
temperatura, mas aos níveis de água no sistema.
Por outro lado, existem disparidades estacionais nos picos de
abundância dos distintos grupos faunísticos que compõem essas
comunidades na planície costeira do Rio Grande do Sul
(Rodrigues & Hartz, 2001; Nessimian, 1995 a, b) admitindo-se
Figura 1.Ordenação (Análise de Coordenadas Principais) do inventário rápido dacomposição e abundância de macroinvertebrados bentônicos realizadono outono (figuras cheias) e primavera (figuras vazadas) nas regiões daLagoa do Casamento (losangos) e dos Butiazais de Tapes (quadrados).Lagoa do Casamento (Cst); lagoa do Capivari (Cvi); sangradouro dalagoa dos Gateados (SgG); lagoa dos Gateados – Norte (GaN); lagoa dosGateados - Sul a (GSa); lagoa dos Gateados – Sul b (GSb); banhado C(BC); lagoa das Capivaras (LCAP); banhado R (BR).
Figura 2.Biplot da Análise de Redundância da composição e abundância demacroinvertebrados bentônicos (losangos) de acordo com ascaracterísticas do sedimento (setas = correlação com os eixos canônicos)nas regiões da Lagoa do Casamento (losangos) e dos Butiazais de Tapes(quadrados). Lagoa do Casamento (Cst); lagoa do Capivari (Cvi);sangradouro da lagoa dos Gateados (SgG); lagoa dos Gateados – Norte(GaN); lagoa dos Gateados - Sul a (GSa); lagoa dos Gateados – Sul b(GSb); banhado C (BC); lagoa das Capivaras (LCAP); banhado R (BR).
148
portanto que, dependendo das preferências estacionais de um ou
outro táxon e, sendo esse um dos grupos dominantes na
comunidade estudada, irá pesar no computo total, fazendo com
que os dados quantitativos de produção da macrofauna possam ser
distintos de uma lagoa para outra ou de um sistema para outro,
como sucede se formos comparar o ambiente lótico (Volkmer-
Ribeiro et al., 2004) com o lêntico costeiro.
É relevante constatar-se que todos os trabalhos acima citados
concordam com os resultados presentemente obtidos: de que a
marcada heterogeneidade espacial na composição e abundância de
macroinvertebrados entre os ambientes estudados está associada a
diferenças de microhabitat, contando particularmente o tipo de
sedimento e a estrutura e extensão marginal da comunidade de
macrófitas em cada lagoa.
Deslocando-nos ao hemisfério norte, a constatação
permanece pois Cardinale et al. (1998), em estudo sobre a fauna
de um banhado costeiro associado aos Grandes Lagos, EUA,
registraram diferenças significativas na riqueza de
macroinvertebrados bentônicos em um gradiente perpendicular à
faixa da vegetação marginal. Esses autores verificaram
aumento linear da riqueza em direção à linha litoral, tendências
espaciais significativas para 4 dos 14 táxons considerados e que
a distância da água livre é determinante da estrutura da
comunidade em áreas úmidas costeiras, por causa dos gradientes
nos parâmetros ambientais devido à presença de macrófitas. Em
direção ao litoral ocorre declínio da turbidez, oxigênio
dissolvido e pH, nas estações aumento na condutividade e
concentração de íons.
Callisto et al. (1998) estudaram os macroinvertebrados
bentônicos em três lagoas costeiras do Rio de Janeiro, observando
um padrão de distribuição diferenciado entre as estações de coleta
nas lagoas. Gonçalves et al. (1998), trabalhando nas mesmas
lagoas, observaram distintos padrões na estrutura das comunidades
bênticas estreitamente relacionadas ao tipo de sedimentos e a
impactos antrópicos.
Würdig et al. (1998) registraram a influência de
parâmetros ambientais na estrutura da comunidade bêntica em
lagoas e lagunas costeiras do Rio Grande do Sul, constatando
que a composição da macrofauna das lagoas e lagunas
estudadas diferiram mais em nível de espécie do que em nível
de grupos taxonômicos maiores, e que a variação na
comunidade mostrou-se mais influenciada pelas mudanças
espaciais do que temporais. A presença de macrófitas, matéria
orgânica e o tamanho médio das partículas do sedimento foram
os fatores mais significativos. Locais com sedimentos mais
finos, maior quantidade de matéria orgânica e sem macrófitas
tiveram menores densidades e número de espécies. Variações
sazonais têm efeito secundário na organização da comunidade;
a presença de macrófitas submersas é o fator espacial mais
importante na determinação da fauna bêntica em lagoas e
lagunas costeiras do Rio Grande do Sul.
Os resultados obtidos estão em concordância com os acima
referidos, no pertinente às relações ambientais e características dos
habitats estudados, permitindo as conclusões expressas a seguir.
As comunidades de macroinvertebrados bênticos
mostraram-se distintas conforme as duas regiões amostradas e
conforme ainda os diferentes habitats em cada uma,
relacionando-se significativamente com o tipo de sedimento e a
quantidade de matéria orgânica no mesmo, essa ligada à maior
cobertura por macrófitas.
Constatou-se maior número de espécies nos ambientes com
menor enriquecimento orgânico e maior abundância naqueles mais
enriquecidos. A distinção existente entre os ambientes da região
dos Butiazais de Tapes e da Lagoa do Casamento relaciona-se
bem, em razão do enriquecimento orgânico, com as idades de
formação de ambos, sendo aqueles dos Butiazais, de idade
pleistocênica, enquanto os da Lagoa do Casamento de idade
holocênica.
As comunidades de macroinvertebrados bentônicos não
mostraram diferenças significativas pertinentes a variações
estacionais. A fauna espongológica despontou como um
componente importante dessas comunidades e um elemento até
agora ausente nos levantamentos já operados em outros ambientes
costeiros do país, acredita-se que não por sua ausência, mas por
desconsideração no momento das triagens. Os ambientes das duas
regiões contém espécies da fauna espongológica que constam nas
listas de fauna ameaçada nacionais e estaduais (Fontana et al.,2003; Instrução Normativa n°. 5, de 21 de maio de 2004,
Ministério do Meio Ambiente, Brasil).
As comunidades levantadas indicam condições do ambiente
aquático próximas das naturais para ambas as regiões,
configurando-se particularmente a região dos Butiazais de Tapes
como um sistema frágil, pela sua inserção entre dunas vivas e
paleodunas.
Tabela II.Análise sedimentológica das amostras colhidas nas regiões da Lagoa doCasamento e dos Butiazais de Tapes. Lagoa do Casamento (Cst); lagoa doCapivari (Cvi); sangradouro da lagoa dos Gateados (SgG); lagoa dosGateados – Norte (GaN); lagoa dos Gateados - Sul a (GSa); lagoa dosGateados – Sul b (GSb); banhado C (BC); lagoa das Capivaras (LCap);banhado R (BR).
Lagoa do Casamento
Butiazais de Tapes
BC LCap BR
Teor de MO (%) 23,6 57,8 18,8
Classificação textural lama lama areia
Cor da amostra Black nº1 Olive black 5Y2/1 Light olive 5Y2/2
Classificação por freqüência simples
Cascalho 0,0000 0,0000 0,0000
Areia 1,3514 13,0977 80,5250
Silte 79,6707 64,1879 10,3992
Argila 18,9780 22,7126 9,0757
Cst Cvi SgG GaN GSa GSb
Teor de MO (%) 6,3 5,2 4,4 22,6 5,5 6,4
Classificação textural areia com lama lama com areia areia lama lama com areia Lama
Cor da amostra brownish gray brownish gray brownish gray brownish black Olive black Olive black
5YR4/1 5YR4/1 5YR4/1 5YR 2/1 5Y2/1 5Y2/1
Classificação por freqüência simples
Cascalho 0,0000 0,0000 0,0000 0,0000 0,0000 0,0000
Areia 70,5128 27,7949 93,0098 9,9418 37,4275 6,3707
Silte 24,2502 66,5570 6,3757 54,8511 56,3528 83,3441
Argila 5,2371 5,6482 0,6145 35,2071 6,2197 10,2852
149
Agradecimentos
Os autores agradecem aos Mestres Hilda Alice de Oliveira
Gastal, Marcelo Pereira de Barros e Sílvia Drügg Hahn, pela
identificação, respectivamente, da entomofauna, carcinofauna e
malacofauna detectadas.
Referências bibliográficas
Callisto, M.; Gonçalves Jr., J. F.; Leal, J. J. F. & Petrucio, M. M. 1998.
Macroinvertebrados bentônicos nas lagoas Imboassica, Cabiúnas e
Comprida. In: Esteves, F. A. ed. Ecologia das Lagoas Costeiras do
Parque Nacional da Restinga de Jurubatiba e do Município de
Macaé (RJ). Rio de Janeiro, NUEPM-UFRJ. p. 283-297.
Cardinale, B. J.; Brady, V. J. & Burton, T. M. 1998. Changes in
the diversity of a coastal wetland fauna from the open water/
macrophyte edge towards shore. Wetlands Ecology and
Management, 6:59-68.
Chomenko, L. & Schäfer, A. 1984. Interpretação biogeográfica da
distribuição do gênero Littoridina (Hydrobiidae) nas lagoas
costeiras do Rio Grande do Sul, Brasil. Amazoniana, 9(1):127-146.
Fontana, C. S.; Bencke, G. A. & Reis, R. E. eds. 2003. Livro
Vermelho da Fauna Ameaçada de Extinção no Rio Grande do
Sul. Porto Alegre, Edipucrs. 632 p.
Gonçalves Jr., J. F.; Callisto, M. & Leal, J. J. F. 1998. Relações
entre a composição granulométrica do sedimento e as
comunidades de macroinvertebrados bentônicos nas lagoas
Imboassica, Cabiúnas e Comprida. In: Esteves, F. A. ed.
Ecologia das Lagoas Costeiras do Parque Nacional da Restinga
de Jurubatiba e do Município de Macaé (RJ). p. 299-310.
Maltchik, L. & Callisto, M. 2004. The use of rapid assessment
approach to discuss ecological theories in wetland systems,
southern Brazil. Interciencia, 29(4):219-222.
Nessimian, J. L. 1995a. Abundância e biomassa de
macroinvertebrados bentônicos em um brejo entre dunas no
litoral do Estado do Rio de Janeiro. Rev. Bras. de Biol.,
55(4):661-683.
Nessimian, J. L. 1995b. Composição da fauna de invertebrados de
um brejo entre dunas no litoral do Estado do Rio de Janeiro.
Acta Limnol. Brasil., 7:41-59.
Ozorio, C. P. 1993. Estrutura espacial e sazonal da macrofauna
bentônica da Lagoa das Custódias, Tramandaí (RS), Brasil :
situações de verão e inverno. Dissertação de Mestrado, PPG
Ecologia, UFRGS, Porto Alegre. 167 p.
Pillar, V. D. 2000. Multiv - Multivariate exploratory analysis,
randomization testing and bootstrap resampling: User’s Guide.
2000. Version 2.0. Porto Alegre, UFRGS.
Pinheiro, U. S.; Hajdu, E. & Correa, M. D. 2004. First description
of gemmules of Ephydatia facunda Weltner, 1895 (Porifera,
Haplosclerida, Spongillidae) by scanning electron microscopy,
with underwater observations of a large population from north-
eastern Brazil. J. Nat. Hist., 38:1071-1080.
Rodrigues, G. & Hartz, S. M. 2001. Food dynamics of fish and
the interaction with macroinvertebrates from a shallow lake in
Southern Brazil. Verh. Internat. Verein Limnol., 27:3309 -
3314.
Takeda, A. M.; Callisto, M. & Barbosa, F. A. R. 2000.
Zoobenthos survey of the Pantanal, Mato Grosso do Sul,
Brazil. In: Willink. P. W.; Chernoff, B.; Alonso, L. E.;
Montambault, J. R. & Lourival, R. orgs. RAP Bulletin of
Biological Assessment, 18:49-55.
Tavares, M. C. M.; Volkmer-Ribeiro, C. & Hermany, G. (no prelo).
Seasonal variation of abundance in a freshwater sponge
assembly at a Southern Neotropical inner Delta. J. Coastal Res..
Tomazelli, J. L.; Dillenburg, S. R. & Villwock, J. A. 2000. Late
quaternary geological history of Rio Grande do Sul Coastal
plain, Southern Brazil. Rev. Bras. de Geol., 30(3):470 - 472.
Tabela III.Variáveis físicas e químicas registradas nosambientes das regiões da Lagoa do Casamento edos Butiazais de Tapes. Estações deamostragem: Lagoa do Casamento (Cst); lagoado Capivari (Cvi); sangradouro na lagoa dosGateados (SgG); lagoa dos Gateados – Norte(GaN); lagoa dos Gateados – Sul a (GSa); lagoados Gateados – Sul b (GSb). Variáveis: tempera-tura do ar (Tar); temperatura da água (T_água);sólidos totais dissolvidos (STD); salinidade (sal);potencial de hidrogênio (pH); condutividade(Cond); transparência da água (Transp); oxigêniodissolvido (O2); porcentagem de saturação deoxigênio dissolvido (O2 %); profundidade (Z).Outono (out); primavera (pri).
Casamento out pri out pri out pri out pri out pri out pri out pri out pri out pri out pri
Cst 20,4 22,4- 45 0 7,6 6,9 94,5 80 45 30 7,8 87 200 200
22,6
Cvi 20,5 20,6 22,3- 52 0,1 6,7 6,9 107 119 35 25 5,9 65,8 200 200
22,6
SgG 15,5 16,2 21,7- 0 6,2 6,1 13,7 55 50 5,9 60,2 130 130
21,9
GaN 16,6 20,6- 93 0,1 6,8 7,2 195 230 25 12,5 7,2 73,6 100 100
20,0
GSa 19-20 17 20,3 92 0,1 6,8 187 116,5 30 25 8,5 86,6 250 250
GSb 17,2 20,4 86 0,1 6,8 6,9 187 12,6 25 25 8,1 84 275 275
Lagoa do Tar T_água STD Sal pH Cond Transp O2 O2 Z
Tomazelli, L. J. & Villwock, J. A. 1996. Quaternary Geological
Evolution of Rio Grande do Sul Coastal Plain, Southern Brazil.
An. Acad. Bras. Ci., 68(3):373-382.
Volkmer-Ribeiro C.; Guadagnin, D. L.; De Rosa-Barbosa, R.;
Silva, M. M.; Drügg-Hahn, S.; Lopes-Pitoni, V.; Gastal, H. A.
O.; Barros, M. P. & Demaman, L. V. 2004. A
polyethylenetherephthalate (pet) device for sampling freshwater
benthic macroinvertebrates. Braz. J. of Biol., 64(3A):531-541.
Volkmer-Ribeiro, C. & Tavares, M. C. M. 1990. Esponjas de água
doce do complexo lagunar Mundaú-Manguaba e dos seus rios
formadores, Alagoas, Brasil. Iheringia, Sér. Zool., (70):171-
172.
Volkmer-Ribeiro, C.; Mothes de Moraes, B.; De Rosa-Barbosa,
R.; Mansur, M. C. D. & Veitenheimer-Mendes, I. L. 1984.
Um estudo do bentos em raízes de Eichhornia azurea (Sw.)
Kunth, do curso inferior de um rio subtropical sul-americano.
Rev. Bras. Biol., 44(2):125 - 132.
Wellborn, G. A.; Skelly, D. K. & Werner, E. E. 1996.
Mechanisms creating community structure across a freshwater
habitat gradient. Annu. Rev. Ecol. and Syst., 27:337-363.
Wiedenbrug, S.; Nolte, U. & Würdig, N. L. 1997.
Macrozoobentos of a coastal lake in Southern Brazil. Arch.
Hydrobiol., 140 (4):533 - 548.
Würdig, N. L.; Albertoni, E. F.; Ozorio, C. P.; Wiedenbrug, S. &
Rodrigues, G. G. 1998. The influence of environmental
parameters in the structure of the benthic community in
coastal lakes and lagoons of Rio Grande do Sul, Brazil. Verh.
Internat. Verein Limnol, 26:1514-1517.
150
Filo Subfilo Classe Subclasse Ordem Superfamília Família Gênero/espécie LC BTmorfotipo
Tabela IV.Zoobentos das regiões da Lagoa doCasamento, dos Butiazais de Tapese ecossistemas associados (planíciecosteira do Rio Grande do Sul).Táxon não determinado (ni). Porifera Demospongiae Haplosclerida Spongillidae Eunapius fragilis X
Radiospongilla amazonensis X
Ephydatia facunda X X
Racekiela sheilae X
Trochospongilla lanzamirandai X
Trochospongilla variabilis X X
Corvoheteromeyenia australis X X
Nematoda sp. 1 X X
Mollusca Gastropoda Mesogastropoda Hydrobiidae Heleobia sp. X
Basommatophora Planorbidae Biomphalaria sp. X
Bivalvia sp. 1 X
Veneroida Sphaeriidae Pisidium sp. X
Corbiculidae Corbicula fluminea X
Unionoida Hyriidae Diplodon sp. X
Mycepodidae Anodontites sp. X
Annelida Hirudinea sp. 1 X
Oligochaeta sp. 1 X X
Arthropoda Arachnida Acari Arrenuridae sp. 1 X
sp. 2 X
ni sp. 1 X
Hygrobatoidea ni sp. 1 X
Insecta Ephemeroptera Polymitarcyidae sp. 1 X
Diptera Chironomidae sp. 1 X X
Ceratopogonidae sp. 1 X X
Trichoptera ni sp. 1 X X
ni sp. 2 X
Ephemeroptera Leptophlebiidae sp. 1 X
Coleoptera Gyrinidae sp. 1 X
Odonata Gomphidae sp. 1 X
Crustacea Maxillopoda ni ni sp. 1 X
Ostracoda Podocopida ni sp. 1 X X
ni sp. 2 X
ni sp. 3 X
ni sp. 4 X X
Cyprididae sp. 1 X X
sp. 2 X
Cytheridae sp. 1 X
Copepoda Cyclopoida ni sp. 1 X
ni sp. 2 X
ni sp. 3 X
Ciclopidae sp. 1 X X
sp. 2 X
Malacostraca Amphipoda Hyalellidae Hyalella sp. X
Branchiopoda Cladocera ni sp. 1 X
Bryozoa sp. 1 X
Chordata Actinopterygii Synbranchiformes Synbranchidae Synbanchus marmoratus X

151
Filo Subfilo Classe Ordem Super- Família Espécie/ Cst Cvi SgG GaN GSa GSbfamília morfotipo
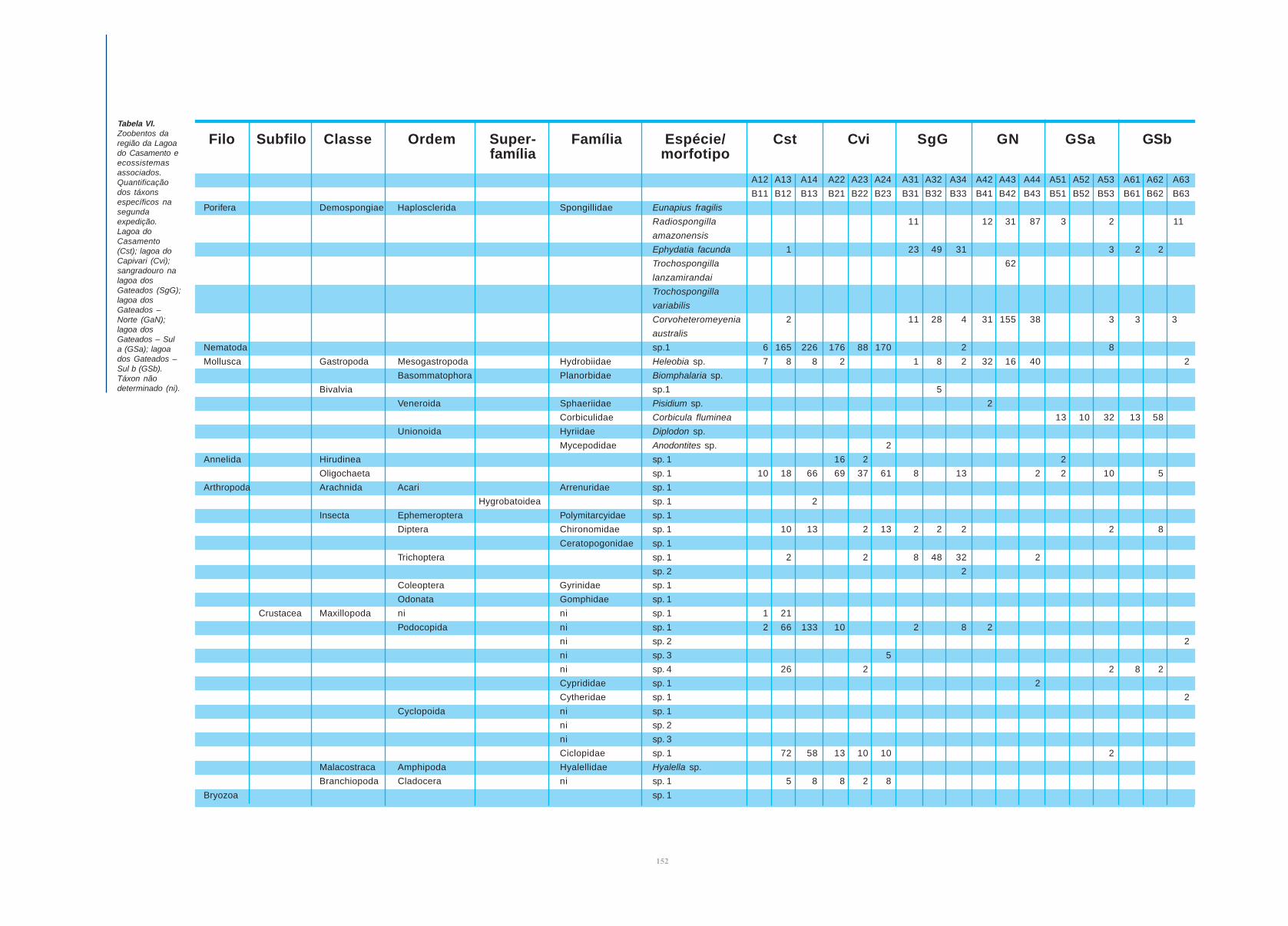
Tabela V.Zoobentos daregião daLagoa doCasamento eecossistemasassociados.Quantificaçãodos táxonsespecíficos naprimeiraexpedição.Lagoa doCasamento(Cst); lagoa doCapivari (Cvi);sangradouro nalagoa dosGateados(SgG); lagoados Gateados –Norte (GaN);lagoa dosGateados – Sula (GSa); lagoados Gateados –Sul b (GSb).Táxon nãodeterminado (ni).
A12 A13 A14 A22 A23 A24 A31 A32 A34 A42 A43 A44 A51 A52 A53 A61 A62 A63
Porifera Demospongiae Haplosclerida Spongillidae Eunapius fragilis 33
Radiospongilla 80 5
amazonensis
Ephydatia 19 107 65 10 5 3 4
facunda
Trochospongilla 7
lanzamirandai
Trochospongilla 33 5
variabilis
Corvoheteromeyenia 7 98 58 119 3 3 4 11 2
australis
Nematoda sp. 1 12 5 4 23 16 12 1 41 3 9 1 6 8 5 39
Mollusca Gastropoda Mesogastropoda Hydrobiidae Heleobia sp. 25 23 170 5 73 20 38 45 3 7 6 4 5
Basommatophora Planorbidae Biomphalaria sp. 1 1
Bivalvia sp. 1 1 1
Veneroida Sphaeriidae Pisidium sp. 1
Corbiculidae Corbicula fluminea 12 1 16 6 9 24 13 10
Unionoida Hyriidae Diplodon sp. 1 1
Mycepodidae Anodontites sp.
Annelida Hirudinea sp. 1 5 2 7 1 10 1 3 1 1 1 2
Oligochaeta sp. 1 154 10 69 223 269 269 214 201 326 5 16 5 8 5 5 23 12 24
Arthropoda Arachnida Acari Arrenuridae sp. 1 1
Hygrobatoidea sp. 1
Insecta Ephemeroptera Polymitarcyidae sp. 1 1 8 8 9 17 1 3 1
Diptera Chironomidae sp. 1 13 8 13 15 18 4 2 15 4 4 5 14 6 10 5 4
Ceratopogonidae sp. 1 3 2 1
Trichoptera sp. 1 2 1 5 3 1 2 1
sp. 2
Coleoptera Gyrinidae sp. 1 1
Odonata Gomphidae sp. 1 1
Crustacea Maxillopoda ni ni sp. 1 0
Podocopida ni sp. 1 44 4 17 2 3 2 1 3 1 1 5 2 5 2
ni sp. 2 1 5 18
ni sp. 3 51 4
ni sp. 4 3 1
Cyprididae sp. 1
Cytheridae sp. 1
Cyclopoida ni sp. 1 2 14 4 11 2 1
ni sp. 2 1
ni sp. 3 3
Ciclopidae sp. 1
Malacostraca Amphipoda Hyalellidae Hyalella sp. 2
Branchiopoda Cladocera ni sp. 1 270 1
Bryozoa sp. 1 0 5 3 11 2 6 5
152
A12 A13 A14 A22 A23 A24 A31 A32 A34 A42 A43 A44 A51 A52 A53 A61 A62 A63
B11 B12 B13 B21 B22 B23 B31 B32 B33 B41 B42 B43 B51 B52 B53 B61 B62 B63
Porifera Demospongiae Haplosclerida Spongillidae Eunapius fragilis
Radiospongilla 11 12 31 87 3 2 11
amazonensis
Ephydatia facunda 1 23 49 31 3 2 2
Trochospongilla 62
lanzamirandai
Trochospongilla
variabilis
Corvoheteromeyenia 2 11 28 4 31 155 38 3 3 3
australis
Nematoda sp.1 6 165 226 176 88 170 2 8
Mollusca Gastropoda Mesogastropoda Hydrobiidae Heleobia sp. 7 8 8 2 1 8 2 32 16 40 2
Basommatophora Planorbidae Biomphalaria sp.
Bivalvia sp.1 5
Veneroida Sphaeriidae Pisidium sp. 2
Corbiculidae Corbicula fluminea 13 10 32 13 58
Unionoida Hyriidae Diplodon sp.
Mycepodidae Anodontites sp. 2
Annelida Hirudinea sp. 1 16 2 2
Oligochaeta sp. 1 10 18 66 69 37 61 8 13 2 2 10 5
Arthropoda Arachnida Acari Arrenuridae sp. 1
Hygrobatoidea sp. 1 2
Insecta Ephemeroptera Polymitarcyidae sp. 1
Diptera Chironomidae sp. 1 10 13 2 13 2 2 2 2 8
Ceratopogonidae sp. 1
Trichoptera sp. 1 2 2 8 48 32 2
sp. 2 2
Coleoptera Gyrinidae sp. 1
Odonata Gomphidae sp. 1
Crustacea Maxillopoda ni ni sp. 1 1 21
Podocopida ni sp. 1 2 66 133 10 2 8 2
ni sp. 2 2
ni sp. 3 5
ni sp. 4 26 2 2 8 2
Cyprididae sp. 1 2
Cytheridae sp. 1 2
Cyclopoida ni sp. 1
ni sp. 2
ni sp. 3
Ciclopidae sp. 1 72 58 13 10 10 2
Malacostraca Amphipoda Hyalellidae Hyalella sp.
Branchiopoda Cladocera ni sp. 1 5 8 8 2 8
Bryozoa sp. 1
Tabela VI.Zoobentos daregião da Lagoado Casamento eecossistemasassociados.Quantificaçãodos táxonsespecíficos nasegundaexpedição.Lagoa doCasamento(Cst); lagoa doCapivari (Cvi);sangradouro nalagoa dosGateados (SgG);lagoa dosGateados –Norte (GaN);lagoa dosGateados – Sula (GSa); lagoados Gateados –Sul b (GSb).Táxon nãodeterminado (ni).
Filo Subfilo Classe Ordem Super- Família Espécie/ Cst Cvi SgG GN GSa GSbfamília morfotipo
153
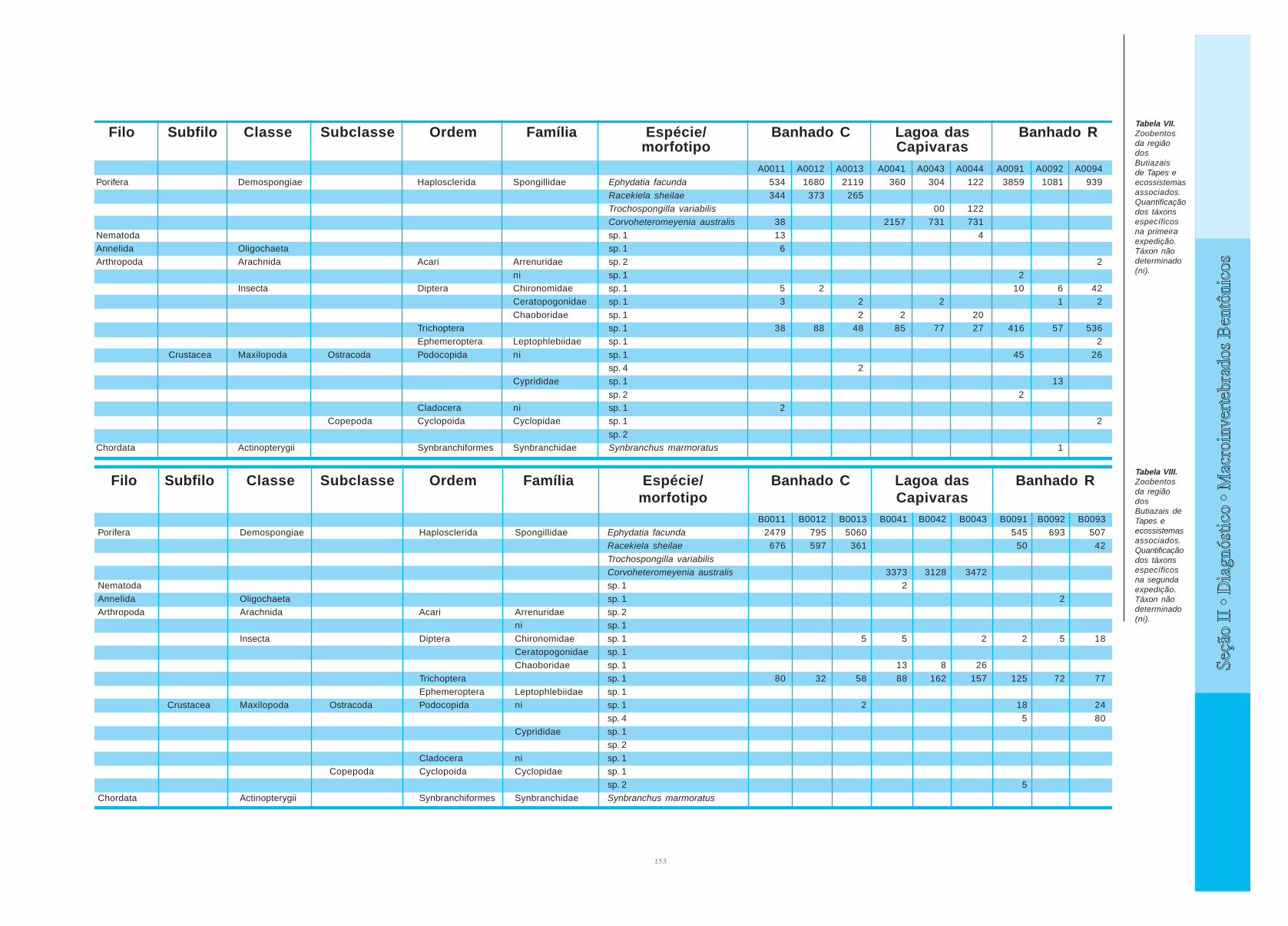
Filo Subfilo Classe Subclasse Ordem Família Espécie/ Banhado C Lagoa das Banhado Rmorfotipo Capivaras
Tabela VIII.Zoobentosda regiãodosButiazais deTapes eecossistemasassociados.Quantificaçãodos táxonsespecíficosna segundaexpedição.Táxon nãodeterminado(ni).
B0011 B0012 B0013 B0041 B0042 B0043 B0091 B0092 B0093Porifera Demospongiae Haplosclerida Spongillidae Ephydatia facunda 2479 795 5060 545 693 507
Racekiela sheilae 676 597 361 50 42
Trochospongilla variabilisCorvoheteromeyenia australis 3373 3128 3472
Nematoda sp. 1 2
Annelida Oligochaeta sp. 1 2Arthropoda Arachnida Acari Arrenuridae sp. 2
ni sp. 1
Insecta Diptera Chironomidae sp. 1 5 5 2 2 5 18Ceratopogonidae sp. 1Chaoboridae sp. 1 13 8 26
Trichoptera sp. 1 80 32 58 88 162 157 125 72 77Ephemeroptera Leptophlebiidae sp. 1
Crustacea Maxilopoda Ostracoda Podocopida ni sp. 1 2 18 24
sp. 4 5 80Cyprididae sp. 1
sp. 2
Cladocera ni sp. 1Copepoda Cyclopoida Cyclopidae sp. 1
sp. 2 5
Chordata Actinopterygii Synbranchiformes Synbranchidae Synbranchus marmoratus
Filo Subfilo Classe Subclasse Ordem Família Espécie/ Banhado C Lagoa das Banhado Rmorfotipo Capivaras
A0011 A0012 A0013 A0041 A0043 A0044 A0091 A0092 A0094
Porifera Demospongiae Haplosclerida Spongillidae Ephydatia facunda 534 1680 2119 360 304 122 3859 1081 939
Racekiela sheilae 344 373 265Trochospongilla variabilis 00 122Corvoheteromeyenia australis 38 2157 731 731
Nematoda sp. 1 13 4Annelida Oligochaeta sp. 1 6Arthropoda Arachnida Acari Arrenuridae sp. 2 2
ni sp. 1 2Insecta Diptera Chironomidae sp. 1 5 2 10 6 42
Ceratopogonidae sp. 1 3 2 2 1 2
Chaoboridae sp. 1 2 2 20Trichoptera sp. 1 38 88 48 85 77 27 416 57 536Ephemeroptera Leptophlebiidae sp. 1 2
Crustacea Maxilopoda Ostracoda Podocopida ni sp. 1 45 26sp. 4 2
Cyprididae sp. 1 13
sp. 2 2Cladocera ni sp. 1 2
Copepoda Cyclopoida Cyclopidae sp. 1 2
sp. 2Chordata Actinopterygii Synbranchiformes Synbranchidae Synbranchus marmoratus 1
Tabela VII.Zoobentosda regiãodosButiazaisde Tapes eecossistemasassociados.Quantificaçãodos táxonsespecíficosna primeiraexpedição.Táxon nãodeterminado(ni).
154
Estação de Cst Cvi SgG GaN GSa GSb BC LCap BR Lagoa do Butiazais
Coleta Casamento de Tapes
Lagoa do Casamento Butiazais de Tapes TotalTabela IX.Riqueza de macroinvertebradosbentônicos nas regiões da Lagoado Casamento e do Butiazal deTapes, Planície Costeira do RioGrande do Sul, outono eprimavera de 2004. Lagoa doCasamento (Cst); lagoa doCapivari (Cvi); sangradouro dalagoa dos Gateados (SgG);lagoa dos Gateados - Norte(GaN); lagoa dos Gateados - Sula (GSa); lagoa dos Gateados -Sul b (GSb); banhado C (BC);lagoa das Capivaras (LCap);banhado R (BR).
Tabela X.Abundância demacroinvertebrados bentônicosnas regiões da Lagoa doCasamento e dos Butiazais deTapes, Planície Costeira do RioGrande do Sul, outono eprimavera de 2004. Lagoa doCasamento (Cst); lagoa doCapivari (Cvi); sangradouro dalagoa dos Gateados (SgG); lagoados Gateados - Norte (GaN);lagoa dos Gateados - Sul a(GSa); lagoa dos Gateados - Sulb (GSb); banhado C (BC); lagoadas Capivaras (LCap); banhadoR (BR).
Estação de Cst Cvi SgG GaN GSa GSb BC LCap BR Lagoa do Butiazais
Coleta Casamento de Tapes
Lagoa do Casamento Butiazais de Tapes Total
Estação do Ano Out Pri Out Pri Out Pri Out Pri Out Pri Out Pri Out Pri Out Pri Out Pri
Porifera 0 3 0 0 442 157 197 416 21 11 21 21 5.353 9.968 4.527 9.973 5.879 1.837 1.289 37.537
Nematoda 17 397 43 434 54 2 13 0 6 8 52 0 13 0 4 2 0 0 1.026 19
Mollusca 218 23 0 4 79 16 119 90 43 55 63 73 0 0 0 0 0 0 783 0
Annelida 238 94 771 185 751 21 30 2 19 14 63 5 6 0 0 0 0 2 2.193 8
Aracnida 0 2 1 0 0 0 0 0 0 0 0 0 0 0 0 0 4 0 3 4
Insecta 23 25 64 17 61 96 12 2 28 2 23 8 188 175 213 461 1072 299 361 2.408
Crustacea 122 392 24 68 314 10 9 4 4 4 13 14 4 2 0 0 88 132 978 226
Bryozoa 0 0 0 0 8 0 0 0 13 0 11 0 0 0 0 0 0 0 32 0
Total 618 936 903 708 1709 302 380 514 134 94 246 121 5.564 10.145 4.744 10.436 7.043 2.270 6.665 40.202
Estação do Ano Out Pri Out Pri Out Pri Out Pri Out Pri Out Pri Out Pri Out Pri Out Pri
Porifera 0 2 0 0 5 3 4 3 3 3 2 3 3 3 2 1 1 2 6 4
Nematoda 1 1 1 1 1 1 1 0 1 1 1 0 1 1 0 1 0 0 1 1
Mollusca 1 1 0 2 2 2 5 2 4 1 3 2 0 0 0 0 0 0 7 0
Annelida 2 1 2 2 2 1 2 1 2 2 2 1 1 0 0 0 0 1 2 1
Arachnida 0 1 1 0 0 0 0 0 0 0 0 0 0 0 0 0 2 0 2 2
Insecta 2 2 3 2 5 3 3 1 4 1 2 1 4 3 2 3 4 2 7 4
Crustacea 3 5 3 5 4 1 3 2 2 2 4 3 2 0 1 0 4 3 13 6
Bryozoa 0 0 0 0 1 0 0 0 1 0 1 0 0 0 0 0 0 0 1 0
Total 9 13 10 12 20 11 18 9 17 10 15 10 11 7 5 5 11 8 39 20

155


















