
AÇÃO DO ÁCIDO KÓJICO SOBRE NEUTRÓFILOS HUMANOS … · PROGRAMA DE PÓS-GRADUAÇÃO EM ... Dr....
97
UNIVERSIDADE FEDERAL DO PARÁ INSTITUTO DE CIÊNCIAS BIOLÓGICAS PROGRAMA DE PÓS-GRADUAÇÃO EM BIOLOGIA DE AGENTES INFECCIOSOS E PARASITÁRIOS AÇÃO DO ÁCIDO KÓJICO SOBRE NEUTRÓFILOS HUMANOS E DURANTE INFECÇÃO COM Leishmania (Leishmania) amazonensis PAULA CRISTINA RODRIGUES FRADE Belém–Pará 2015
-
Upload
nguyennguyet -
Category
Documents
-
view
217 -
download
0
Transcript of AÇÃO DO ÁCIDO KÓJICO SOBRE NEUTRÓFILOS HUMANOS … · PROGRAMA DE PÓS-GRADUAÇÃO EM ... Dr....

UNIVERSIDADE FEDERAL DO PARÁ
INSTITUTO DE CIÊNCIAS BIOLÓGICAS
PROGRAMA DE PÓS-GRADUAÇÃO EM
BIOLOGIA DE AGENTES INFECCIOSOS E PARASITÁRIOS
AÇÃO DO ÁCIDO KÓJICO SOBRE NEUTRÓFILOS HUMANOS E
DURANTE INFECÇÃO COM Leishmania (Leishmania) amazonensis
PAULA CRISTINA RODRIGUES FRADE
Belém–Pará
2015


PAULA CRISTINA RODRIGUES FRADE
AÇÃO DO ÁCIDO KÓJICO SOBRE NEUTRÓFILOS HUMANOS E
DURANTE INFECÇÃO COM Leishmania (Leishmania) amazonensis
Dissertação apresentada ao Programa de Pós-
Graduação em Biologia de Agentes Infecciosos e
Parasitários do Instituto de Ciências Biológicas da
Universidade Federal do Pará como requisito para a
obtenção do grau de Mestre em Biologia de Agentes
Infecciosos e Parasitários.
Orientadora: Profª. Drª. Edilene Oliveira da Silva
BELÉM – PARÁ - BRASIL
2015


1
PAULA CRISTINA RODRIGUES FRADE
AÇÃO DO ÁCIDO KÓJICO SOBRE NEUTRÓFILOS HUMANOS E DURANTE
INFECÇÃO COM Leishmania (Leishmania) amazonensis
Dissertação apresentada ao Programa de Pós-Graduação em Biologia de Agentes Infecciosos
e Parasitários, do Instituto de Ciências Biológicas da Universidade Federal do Pará, como
requisito para a obtenção do grau de Mestre em Biologia de Agentes Infecciosos e
Parasitários.
Orientadora: Profª. Drª. Edilene Oliveira da Silva
Laboratório de Parasitologia, ICB-UFPA
Banca Examinadora:
Membro: Dr. Adriano Penha Furtado
Professor do Instituto de Ciências Biológicas – UFPA
Membro: Dra. Barbarella Matos Macchi
Professora do Instituto de Ciências Biológicas – UFPA
Membro: Dr. Chubert Bernardo Castro de Sena
Professor do Instituto de Ciências Biológicas – UFPA
Suplente: Dr. José Luiz Martins do Nascimento
Professor do Instituto de Ciências Biológicas – UFPA
Belém, 22 de Janeiro de 2015.

2
“...Vida - É o amor existencial.
Razão - É o amor que pondera.
Estudo - É o amor que analisa.
Ciência - É o amor que investiga...
Tudo é amor...”
André Luiz

3
Dedico aos alicerces da minha vida!
À Deus pelo Seu Amor Incondicional que conduz a minha vida ...
À minha família por serem o meu porto seguro...
Aos meus amigos por todo o afeto e estímulo que me proporcionam.

4
AGRADECIMENTOS
Mais um passo foi dado e, cada vez mais, tenho a certeza de que sozinhos nada somos.
Primeiramente, agradeço a Deus pela vida e pelas infinitas oportunidades colocadas em meu
caminho.
Este trabalho é fruto de uma somatória de esforços. Agradeço a minha linda família,
pelo carinho e pela oportunidade de crescimento entre vocês! Em especial, aos meus pais
Paulo e Silvia e meu irmão Pedro, que me cercam de amparo, amor, cuidado e auxílio ao
longo desta minha trajetória de vida.
Serei eternamente grata a minha orientadora Dra. Edilene pela oportunidade de
aprender, crescer e vivenciar neste maravilhoso universo da pesquisa. Obrigada professora por
me orientar desde a iniciação científica, por me ensinar desde as mínimas coisas como
apresentar um seminário e montar um desenho experimental até a dar uma aula e escrever um
artigo.
Como também, agradeço a todos os meus companheiros do Laboratório de
Protozoologia. As sugestões, as ideias, o compartilhamento de técnicas e de conhecimentos
foram fundamentais para a realização desse trabalho. Meu muito obrigada a todos vocês!
À Neide, minha “mãe científica”, pela amizade, pelo carinho e por todo o apoio.
Foram tantas trocas de favores, tantas madrugadas na UFPA fazendo experimentos, tantas
palavras de incentivo que não teria como agradecer tanto carinho em poucas palavras. És um
exemplo para mim, te amo amiga!
Ao Bruno, minha dupla de iniciação científica, graduação e mestrado. Obrigada por
dividir comigo um pouquinho do seu gigantesco conhecimento, pela paciência com meus
inúmeros pedidos de ajuda em e-mails “hiper mega super urgentes” e por sempre estar tão
disposto a colaborar!
Ao Rodrigo, meu bandido favorito (hahahaha), pelos momentos de descontração,
alegria e diversão. Sempre com um pensamento positivo, com uma piadinha, ou com um
simples “vai dar certo, deixa de drama” melhoraste os meus dias. Obrigada também por
sempre atender meus pedidos desesperados “mandaaaa leishmaniaaa pra miiim”. Ah, sei que
me amas muito (hahaha).
À Ana Paula, praticamente minha co-orientadora, pelas discussões tão enriquecedoras,
pelas ideias, pelas técnicas ensinadas, pelas discussões tão enriquecedoras... Enfim, por toda a
orientação que recebi ao longo desses anos. Obrigada principalmente pela paciência para
responder minhas intermináveis dúvidas e pelas longas horas de análises de microscopia.

5
Ao Luis pelas muitas vezes em que me ajudou, dentre elas na resolução de cálculos,
construção de gráficos e análises no citômetro. Além de sempre ter um tempinho para uma
análise rápida no microscópio confocal (hahaha).
À duplinha querida Amanda e Raquel por me ensinarem as primeiras técnicas, pelas
sugestões, pelos momentos de descontração e por tantas vezes ficarem me esperando apenas
para irmos embora juntas.
Ao Davi por toda ajuda recebida para conseguir passar no mestrado. Obrigada por ter
se disponibilizado a assistir os seminários dos artigos, a discutir, devo meu ingresso no
mestrado ao seu auxílio!
À Carol, minha parceira de western blotting, obrigada principalmente pela companhia
durante o estágio na UFRJ.
Aos alunos mais novos do lab, Evelen e Sandro, obrigada pelos momentos de alegria.
À Evelen pelos abraços tão cheios de energia e pela amizade construída. Ao Sandro por tirar
meu título de mais furona (hahaha).
À Lienne e ao Jorge pelos momentos de atenção e de descontração.
Além deles, devo um agradecimento especial à Fernanda, técnica do Laboratório de
Biologia Estrutural, e à Neidiane, técnica do Laboratório de Neuroquímica, por serem tão
atenciosas e por terem me ajudado tantas vezes.
Ao Profº. Dr José Luiz do Nascimento, pelas valiosas contribuições e por permitir a
utilização dos reagentes e dos equipamentos do Laboratório de Neuroquímica.
Ao Profº. Dr. Chubert por ceder reagentes e pelo auxílio no manuseio dos
equipamentos do LBE.
À Profª Dra Debora Foguel por me permitir estagiar em seu laboratório e por ter me
recebido tão bem. E a sua aluna Estefânia sempre disposta a ensinar todas as técnicas
possíveis e a colaborar para o andamento dos trabalhos.
As instituições que colaboraram: HEMOPA por ter cedido às bolsas de sangue,
Laboratório de Investigação Sistemática em Biotecnologia e Biodiversidade Molecular da
Universidade Federal do Pará por fornecer o metabólito utilizado neste estudo e ao Instituto
Evandro Chagas pelo fornecimento das cepas de Leishmania e por ter cedido os microscópios
para a aquisição das imagens.
À CAPES e ao INBEB pelo importante suporte financeiro.
Aos Membros da banca Drª. Barbarela, Dr. Adriano e Dr. Chubert por terem aceitado
participar e pelas contribuições com esse trabalho.
Ao Programa de Pós Graduação em Biologia dos Agentes Infecciosos e Parasitários.

6
Aos meus amigos tão amáveis Vanessa Amaral, Vanessa Figueiredo, Taysa, Daniela,
Lorena, Adriano, Stéphanie, e André por aturarem me ouvir falar tantas vezes sobre minha
pesquisa e pelos desabafos ouvidos tanto carinho e atenção. A nossa amizade levarei para
sempre!
Enfim, esse trabalho foi construído por muitas pessoas e não conseguiria colocar o
nome de todos em um agradecimento de poucas folhas. Assim, agradeço cada palavra de
apoio, cada ensinamento, cada sugestão e cada conselho... Meu muitíssimo obrigado a todos
que contribuíram!

7
SUMÁRIO
LISTA DE FIGURAS ............................................................................................................. 10
LISTA DE SÍMBOLOS E ABREVIATURAS ..................................................................... 12
RESUMO.................................................................................................................................14
ABSTRACT ............................................................................................................................ 15
1 INTRODUÇÃO ........................................................................................................... 16
1.1 ASPECTOS GERAIS .................................................................................................... 16
1.2 AS LEISHMANIOSES ................................................................................................. 17
1.2.1 Leishmaniose Tegumentar Americana (LTA) ............................................................ 17
1.2.2 Aspectos Epidemiológicos, Clínicos e Imunológicos ................................................... 17
1.3 O GÊNERO Leishmania .................................................................................................... 20
1.3.1 Classificação ................................................................................................................... 20
1.3.2 Morfologia ...................................................................................................................... 21
1.3.3 Ciclo Biológico ............................................................................................................... 22
1.4 NEUTRÓFILOS: CARACTERÍSTICAS GERAIS, MIGRAÇÃO E ATIVAÇÃO .......... 24
1.5 NEUTRÓFILOS E MECANISMOS MICROBICIDAS .................................................... 26
1.6 INTERAÇÃO NEUTRÓFILO E Leishmania .................................................................... 29
1.7 TRATAMENTO DAS LEISHMANIOSES ....................................................................... 32
1.8 ÁCIDO KÓJICO (AK) ....................................................................................................... 33
1.9 OBJETIVOS ....................................................................................................................... 36
1.9.1 Objetivo geral: ............................................................................................................... 36
1.9.2 Objetivos específicos: .................................................................................................... 36
2 MATERIAL E MÉTODOS ................................................................................................ 37
2.1 FLUXOGRAMA ................................................................................................................ 37
2.2 CULTIVO E MANUTENÇÃO DO PARASITO .............................................................. 37
2.3 OBTENÇÃO E CULTIVO DE NEUTRÓFILOS DO SANGUE PERIFÉRICO
HUMANO ................................................................................................................................ 38
2.4 OBTENÇÃO E DILUIÇÃO DO METABÓLITO AK ...................................................... 38
2.5 TRATAMENTO DAS CÉLULAS ..................................................................................... 39
2.6 AVALIAÇÃO DA VIABILIDADE DAS CÉLULAS TRADADAS COM AK ............... 39
2.6.1 Método thiazolyl blue (MTT) ....................................................................................... 39
2.6.2 Detecção de Potencial da Membrana Mitocondrial (JC-1) ....................................... 40

8
2.6.3 Iodeto de Propídio (IP) .................................................................................................. 40
2.7 ANÁLISE MORFOLÓGICA DE NEUTRÓFILOS TRATADOS COM AK ................... 41
2.7.1 Microscopia Óptica (MO) e Análise de área celular (Morfometria) ........................ 41
2.7.2 Microscopia Eletrônica de Varredura (MEV) ............................................................ 41
2.7.3 Microscopia Eletrônica de Transmissão (MET) ......................................................... 42
2.7.4 Detecção de filamentos de actina por Microscopia Óptica de Fluorescência...........42
2.8 RESPOSTA MICROBICIDA ............................................................................................ 43
2.8.1 Produção de Radicais Superóxidos pelo teste do Nitro Blue Tetrazolium (NBT) ... 43
2.8.2 Produção de Espécies Radicais de Oxigênio (ERO) pela marcação com CellROX. 43
2.8.3 Avaliação da atividade da Mieloperoxidase (MPO) ................................................... 44
2.8.4 Detecção da produção de Óxido Nítrico (NO) pela marcação com DAF-MT ......... 45
2.9 INTERAÇÃO DE Leishmania (Leishmania) amazonensis COM NEUTRÓFILOS
TRATADOS COM AK ............................................................................................................ 45
2.9.1 Índice Endocítico ........................................................................................................... 45
2.9.2 Índice de Infectividade .................................................................................................. 46
2.9.3 Produção de Radicais Superóxidos pelo teste do Nitro Blue Tetrazolium (NBT) ... 46
2.10 ANÁLISE ESTATÍSTICA ............................................................................................... 46
3 RESULTADOS .................................................................................................................... 48
3.1 AVALIAÇÃO DA VIABILIDADE DE NEUTRÓFILOS TRADADOS COM AK ........ 48
3.1.1 Método thiazolyl blue (MTT) ....................................................................................... 48
3.1.2 Detecção de Potencial da Membrana Mitocondrial (JC-1) ....................................... 49
3.1.3 Iodeto de Propídio (IP) .................................................................................................. 49
3.2 ANÁLISE MORFOLÓGICA DE NEUTRÓFILOS TRATADOS COM AK ................... 51
3.2.1 Microscopia Óptica e Morfometria ............................................................................. 51
3.2.2 Microscopia Eletrônica de Varredura ......................................................................... 51
3.2.3 Microscopia Eletrônica de Transmissão...................................................................... 51
3.2.4 Detecção de filamentos de actina por Microscopia Óptica de Fluorescência...........55
3. 3 AVALIAÇÃO DA RESPOSTA MICROBICIDA ........................................................ 57
3.3.1 Produção de Radicais Superóxidos em neutrófilos tratados com AK ...................... 57
3.3.2 Produção de Espécies Reativas de Oxigênio ............................................................... 58
3.3.3 Avaliação da atividade da Mieloperoxidase (MPO) ................................................... 59
3.3.4 Produção de óxido nítrico em neutrófilos tratados com AK ..................................... 62

9
3.4 Avaliação da INTERAÇÃO DE Leishmania (leishmania) amazonensis COM
NEUTRÓFILOS TRATADOS COM AK ................................................................................ 63
3.4.1 Determinação do Índice Endocítico ............................................................................. 63
3.4.2 Determinação do Índice de Infectividade .................................................................... 64
3.4.3 Produção de Radicais Superóxidos .............................................................................. 65
6 REFERÊNCIAS BIBLIOGRÁFICAS .............................................................................. 72
7 MISSÃO DE ESTUDOS ..................................................................................................... 87
8 PARTICIPAÇÃO EM EVENTOS .................................................................................... 87
9 PREMIAÇÃO ...................................................................................................................... 87
ANEXO 1 ................................................................................................................................. 88
ANEXO 2 ................................................................................................................................. 89
ANEXO 3 ................................................................................................................................. 90
ANEXO 4 ................................................................................................................................. 91
ANEXO 5 ................................................................................................................................. 92
ANEXO 6 ................................................................................................................................. 93

10
LISTA DE FIGURAS
Figura 1 – Formas clínicas da Leishmaniose Tegumentar Americana ...................................18
Figura 2 – Taxonomia do protozoário Leishmania .................................................................21
Figura 3 – Formas evolutivas do parasita Leishmania ............................................................22
Figura 4 – Ciclo biológico do parasita Leishmania sp ............................................................23
Figura 5 – Morfologia de neutrófilos humanos ......................................................................24
Figura 6 – Degranulação e produção de ERO no interior do neutrófilo .................................27
Figura 7 – Mecanismo oxidativo dos neutrófilos ...................................................................28
Figura 8 – Estrutura química do ácido kójico ........................................................................ 34
Figura 9 – Fluxograma da metodologia utilizada ...................................................................37
Figura 10 – Viabilidade celular pelo método MTT de neutrófilos tratados com AK
.................................................................................................................................................. 48
Figura 11 – Potencial de membrana mitocondrial avaliado por citometria de fluxo após
incubação com JC-1 de neutrófilos tratados com AK ............................................................. 49
Figura 12 – Ensaio de apoptose por citometria de fluxo utilizando Iodeto de Propídio como
marcador de neutrófilos tratados com AK .............................................................................. 50
Figura 13 – Análise morfológica e morfométrica de neutrófilos tratados com AK e corados
com Giemsa ............................................................................................................................. 52
Figura 14 – Análise ultraestrutural por MEV de neutrófilos tratados com AK ..................... 53
Figura 15 – Análise ultraestrutural por MET de neutrófilos tratados com AK ......................54
Figura 16 – Detecção dos filamentos de actina do citoesqueleto de neutrófilos tratados com
AK ........................................................................................................................................... 56
Figura 17 – Detecção da produção de radicais superóxidos em neutrófilos tratados com 50
µg/mL de AK por 1 hora através da reação citoquímica com NBT ........................................57
Figura 18 – Detecção da produção de Espécies Reativas de Oxigênio em neutrófilos tratados
com 50 µg/mL de AK por 1 hora pela marcação com CellROX® ..........................................58
Figura 19 – Avaliação qualitativa da atividade da Mieloperoxidase em neutrófilos tratados
com AK por microscopia de fluorescência...............................................................................60
Figura 20 – Avaliação quantitativa por citometria de fluxo da atividade da Mieloperoxidase
em neutrófilos tratados com 50 µg/mL de AK por 1 hora .......................................................61
Figura 21 - Detecção da produção de óxido nítrico em neutrófilos tratados com por citometria
de fluxo pela marcação com DAF-MT ................................................................................... 62

11
Figura 22 - Índice endocítico de neutrófilos tratados com 50 µg/mL de AK por 1 hora e
submetidos a interação com Leishmaia (Leishmania) amazonensis por 3 horas .....................63
Figura 23 – Índice de infectividade de neutrófilos tratados com 50 µg/mL de AK por 1 hora,
nos tempos de 0, 3 e 6 horas após infecção com Leishmania amazonensis ............................64
Figura 24 – Detecção da produção de radicais superóxidos em neutrófilos tratados com 50
µg/mL de AK por 1 hora e em interação com Leishmaia (Leishmania) amazonensis através da
reação citoqímica com NBT ....................................................................................................65

12
LISTA DE SÍMBOLOS E ABREVIATURAS
ºC – Graus Celsius
µg/mL – Microgramas por mililitros
µL – Microlitros
µm – Micrometros
AK – Ácido kójico
ANOVA – Análise de Variância
Cl- - íon cloreto
DAF-MT – 4 amino–5–metilamino-2’,7-diclorofluoresceína
DAPI – Diaminofenilindole
DMEM – Duleccos’ Modified Eagle’s Medium
DMSO - Dimetilsufóxido
DNA – Ácido desoxirribonucleico
EDTA – Sigla inglesa paraácido etilenodiamino tetra-acético
ERK 1/2 – Proteína quinase reguladora de sinais extracelulares
ERN – Espécie reativa de nitrogênio
ERO – Espécie reativa de oxigênio
GM-CSF – Fator estimulador de colônias de granulócitos-macrófagos
HMP – 5-hidroxi-2-hidroximetil-γ-pirona
H2O2–Peróxido de hidrogênio
HOCl – Ácido hipocloroso
IL-1 – Interleucina1
IL-4 – Interleucina 4
IL-5 – Interleucina 5
IL-8 – Interleucina 8
IL-10 – Interleucina 10
IL-12 – Interleucina 12
INF-γ – Interferon - γ
IP – Iodeto de Propídio
JC-1 – Potencial de Membrana Mitocondrial
LCD – Leishmaniose Cutânea Difusa
LCF – fator quimiotático
LCL – Leishmaniose Cutânea Localizada

13
LPS – Lipopolissacarídeo
LT – Leishmaniose Tegumentar
LTA – Leishmaniose Tegumentar Americana
M – Molar
M-CFS - Fator Estimulador de Colônia de Macrófago
MET – Microscopia Eletrônica de Transmissão
MEV – Microscopia Eletrônica de Varredura
mg/mL – Miligramas por mililitros
MPO –Mieloperoxidase
MTT Thiazolyl Blue Tetrazolium Bromide
NAHCO3 – Bicarbonato de sódio
NADPH – nicotinamida adenina dinucleotídeofosfatasehidrogenase
NBT – Nitrobluetetrazolium
NE – Elastase neutrofílica
NET –Sigla inglesa para rede extracelular de neutrófilos
NH4Cl – Cloreto de amônio
nm – Nanômetros
NNN – Neal, Novy e Nicolle
NO – Óxido nítrico
O2 – Oxigênio molecular
O2- - Íon superóxido
1O2 – Oxigênio Singlate
OH- – Radical hidroxila
ONOO- - Peroxinitrito
PAF – Fator de ativação de plaquetas
PBS – Sigla inglesa para tampão fosfato salino
PGE2 – Prostaglandina E2
PMA – 13-acetato de forbol éster 12-miristato
PMN – Polimorfonuclear
Sb5+ - Antimonial pentavalente
SBF – Soro bovino fetal
SOD – Superóxido dismutase
TGF-β – Sigla inglesa para fator modulador de crescimento
TNF – Sigla inglesa para fator de necrose tumoral

14
RESUMO
Neutrófilos são fagócitos envolvidos na resposta imune primária para eliminar patógenos.
Leishmania é um protozoário responsável por uma das doenças infecciosas mais importantes
do mundo. A quimioterapia é o único tratamento efetivo, mas além do custo elevado, os
fármacos são tóxicos e requerem um longo período de tratamento. Por isso a busca por novos
quimioterápicos é de grande interesse. Neste trabalho, foi utilizado Ácido Kójico (AK), um
metabólito secundário sintetizado por algumas espécies de fungos dos gêneros Aspergillus,
Penicillium e Acetobacter. AK tem várias aplicações, sendo usado como aditivo alimentar,
cosméticos, agente antitumoral, ativador de macrófagos e também como agente
leishmanicida. Assim, este estudo foi realizado para determinar os efeitos do AK como
indutor da ativação de neutrófilos humanos e durante a interação com Leishmania
(Leishmania) amazonensis. Neutrófilos foram tratados previamente por 1 hora com 50 μg/ml
de AK e três ensaios foram utilizados para avaliar a viabilidade celular. Os resultados
mostraram que AK não possui efeito citotóxico em neutrófilos. Análises morfológicas
mostraram grande habilidade de espraiamento, formação de pseudópodes, rearranjo dos
filamentos de actina, intensa vacuolização e aumento do volume celular, características que
são frequentemente observadas em células ativadas. O aumento da área citoplasmática foi
confirmado por análise morfométrica. Posteriormente, foi avaliado o “burst” oxidativo em
neutrófilos induzido por AK (espécies reativas de oxigênio - ERO, atividade da
mieloperoxidase e óxido nítrico - NO), que são importantes mediadores leishmanicidas.
Células tratadas com AK demonstraram atividade microbicida que ocorreu por meio da
produção de ROS, aumento da atividade da mieloperoxidase (MPO), porém não ocorreu
produção de NO. Neutrófilos tratados apresentaram maior capacidade de fagocitar parasitas
Leishmania e o número de parasitas intracelulares diminuiu após 6 horas. Essa diminuição foi
associada com a produção de radicais superóxidos. Em conclusão, esses resultados mostram
que AK pode estar envolvido na ativação de neutrófilos por meio da produção de ROS,
sugerindo um possível mecanismo de controle em neutrófilos infectados por Leishmania.
Palavras-chave: ativação de neutrófilos humanos; Leishmania (L.) amazonensis e Ácido
Kójico.

15
ABSTRACT
Neutrophils are phagocytes involved in the primary immune responses to eliminate
pathogens. Leishmania is a protozoan responsible for the one of the most important infectious
diseases in the world. Chemotherapy is the only effective treatment but besides the high cost,
these drugs are toxic and require a long period of treatment. Therefore the search for new
chemotherapeutics is of great interest. In this work, we have used Kojic Acid (KA), a
secondary metabolite synthesized by some species of fungi from Aspergillus, Penicillium and
Acetobacter genus. KA has several applications, being used as a food additive, cosmetics,
antitumor agent, macrophage activator and also as anti-leishmanial agent. Thus, this study
was designed to determine the effects of KA as inductor of human neutrophils activation and
during interaction with Leishmania (Leishmania) amazonensis. Neutrophils were treated
previously for 1 hour with 50 μg/mL of KA and three tests were used to assess cell viability.
These results showed that KA has no citotoxicity effect on neutrophils. Morphological
analysis showed great ability of spreading, pseudopodia formation, rearrangement of actin
filaments, intense vacuolization and increase in cell volume, features that are often observed
in activated cells. The increased cytoplasmic area was confirmed by morphometric analyzes.
Subsequently, was assessed KA induced neutrophil oxidative burst (reactive oxygen species –
ROS, myeloperoxidase activity and nitric oxide – NO), which are important leishmanicidal
mediators. The treatment of cells with KA demonstrated microbicide activity that occurred
through ROS production, increase myeloperoxidase activity, but not NO production. The
treatment of neutrophils had a higher capacity to phagocytose Leishmania parasites and
decrease the number of intracellular parasites after 6 hours. That effect was associated with
the production of superoxide radicals. In conclusion, these results show that KA is involved in
neutrophil activation might be do pathway of ROS production, suggesting a possible control
mechanism for Leishmania infected neutrophils.
Keywords: human neutrophil activation; Leishmania (L.) amazonensis and Kojic Acid.

16
1 INTRODUÇÃO
1.1 ASPECTOS GERAIS
As leishmanioses representam um grave problema de saúde pública e acometem mais
de 12 milhões de pessoas. Estima-se que dois milhões de pessoas sejam infectadas pelo parasito
por ano e são responsáveis por 20.000 a 40.000 mortes por ano (Okwor & Uzonna, 2009;
WHO, 2010).
Além disso, as leishmanioses afetam cerca de 98 países, dentre os quais 72 são
considerados países em desenvolvimento. E por estarem relacionadas à pobreza, fazem parte de
um grupo de doenças negligenciadas que não despertam o interesse de indústrias farmacêuticas.
Esse complexo de doenças está entre as seis doenças tropicais de maior relevância no mundo e
o Brasil está entre os cinco países com maior número de casos (Brasil, 2010, WHO, 2010,
Alvar et al., 2012).
Essa infecção tem caráter antropozoonótico e é causada por 20 espécies de protozoários
flagelados do gênero Leishmania. Apresenta três principais manifestações clínicas: cutânea,
mucocutânea e visceral (WHO, 2010). A manifestação clínica depende da espécie do agente
etiológico, e do tipo de resposta imune gerada pelo hospedeiro (David & Craft, 2009). Todavia,
apesar de se conhecer o agente etiológico da leishmaniose há mais de um século, ainda não está
totalmente elucidada a ação da resposta imune do hospedeiro sobre o protozoário.
No hospedeiro vertebrado, a infecção inicia-se através do repasto sanguíneo realizado
pelo inseto vetor. Rapidamente, ocorre o recrutamento de neutrófilos para o local onde os
parasitas são inoculados, iniciando assim, uma resposta inflamatória (Malafaia & Rezende,
2009).
Os neutrófilos são considerados a primeira linha de defesa do organismo, pois são as
primeiras células a serem recrutadas da corrente sanguínea para os sítios de inflamação (Luster
et al., 2005; Nathan, 2006). Estudos mostram que durante o repasto sanguíneo do flebotomíneo
no homem, a saliva do vetor induz uma reação inflamatória que atrai neutrófilos para o local da
infecção (Teixeira et al., 2005; Silva et al., 2005).
Nesse contexto, os neutrófilos podem internalizar as leishmanias e destruí-las ou
favorecer sua sobrevivência (Laufs et al., 2002). Neutrófilos podem fagocitar protozoários,
destruí-los por meio de seus mecanismos microbicidas e com isso possibilitar o controle inicial
da infecção (Tacchini-Cottier et al., 2000; Chen et al., 2005). De outro modo, podem fagocitar
os parasitos, sem entretanto destruí-los e sofrer apoptose. Em seguida, ao serem reconhecidos e

17
fagocitados por macrófagos, com suas respostas microbicidas silenciadas, facilitam a
sobrevivência e a multiplicação desses parasitos, auxiliando assim o processo de infecção
(Peters et al., 2008).
Portanto, estudos que possam identificar os principais fatores envolvidos na resposta
imune inata e sua interferência no desenvolvimento da infecção são muito úteis para adoção de
medidas terapêuticas precoces e eficazes. Outro aspecto que valoriza estudos dessa natureza é
que apesar de existir uma variedade de tratamentos atualmente disponíveis, ainda não existe
uma terapia eficaz para o combate à leishmaniose cutânea (Bailey et al., 2007).
Neste sentido, bioprodutos apresentam-se como alvo terapêutico potencial para o
tratamento das leishmanioses. Metabólitos como o ácido kójico capaz de ativar células do
sistema imunológico (Niwa & Akamatsu, 1991, Rodrigues et al., 2011, Costa 2012) e, ainda,
destruir o parasito Leishmania (Rodrigues et al., 2014), mostram-se um promissor agente
leishmanicida.
1.2 AS LEISHMANIOSES
As Leishmanioses apresentam um conjunto de manifestações clínicas diferentes de
acordo com a espécie causadora. Sendo que essas manifestações podem ser englobadas em dois
conjuntos de formas clínicas: Leishmaniose Tegumentar Americana e Leishmaniose Visceral
(Kaye & Scott, 2011).
1.2.1 Leishmaniose Tegumentar Americana (LTA)
Há relatos da leishmaniose tegumentar (LT) desde a antiguidade, século I d.C., contudo
no Brasil seu primeiro relato foi em 1827 (Lainson, 2010). É considerada uma doença não
contagiosa causada por protozoários intracelulares obrigatórios, de evolução crônica, que se
caracteriza por atingir principalmente as estruturas da pele de forma localizada ou difusa
(Desjeux, 2004; David & Craft, 2009).
1.2.2 Aspectos Epidemiológicos, Clínicos e Imunológicos
A LTA é uma das principais doenças transmitidas por vetores, cerca de 60.000 casos
são registrados por ano na América do Sul e Central. Brasil, Peru e Colômbia concentram
aproximadamente 80% dos mesmos (Who, 2010).
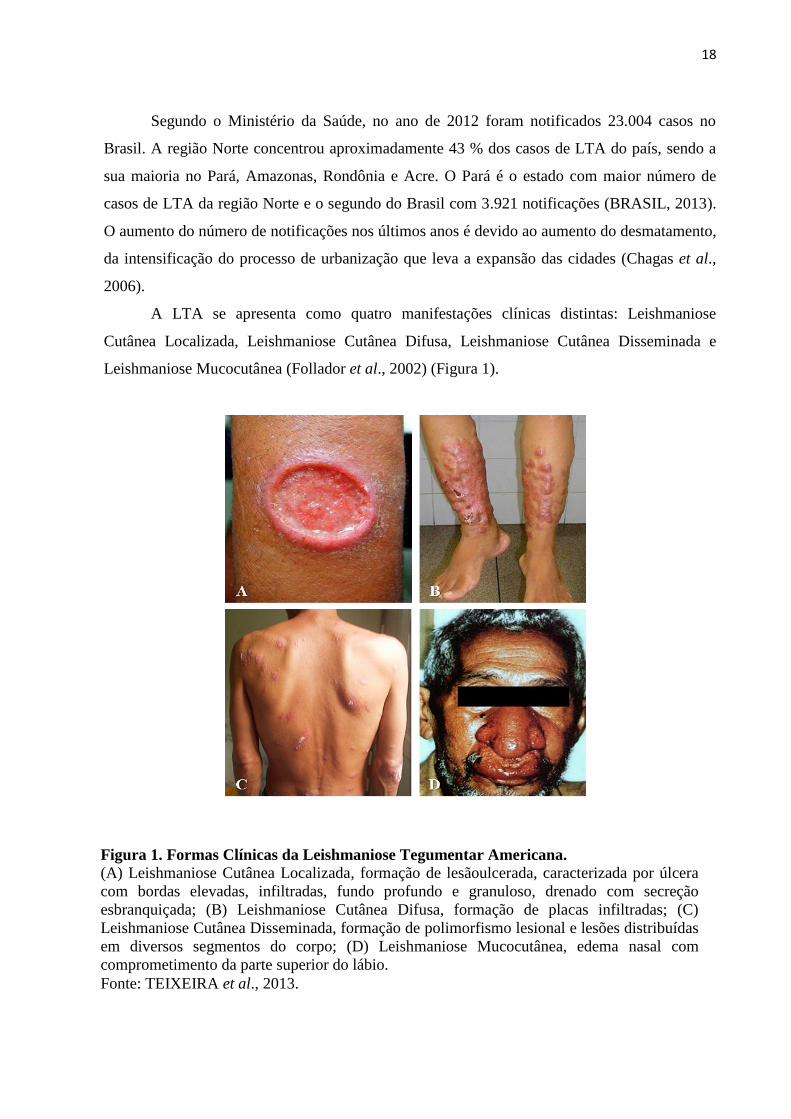
18
Segundo o Ministério da Saúde, no ano de 2012 foram notificados 23.004 casos no
Brasil. A região Norte concentrou aproximadamente 43 % dos casos de LTA do país, sendo a
sua maioria no Pará, Amazonas, Rondônia e Acre. O Pará é o estado com maior número de
casos de LTA da região Norte e o segundo do Brasil com 3.921 notificações (BRASIL, 2013).
O aumento do número de notificações nos últimos anos é devido ao aumento do desmatamento,
da intensificação do processo de urbanização que leva a expansão das cidades (Chagas et al.,
2006).
A LTA se apresenta como quatro manifestações clínicas distintas: Leishmaniose
Cutânea Localizada, Leishmaniose Cutânea Difusa, Leishmaniose Cutânea Disseminada e
Leishmaniose Mucocutânea (Follador et al., 2002) (Figura 1).
Figura 1. Formas Clínicas da Leishmaniose Tegumentar Americana.
(A) Leishmaniose Cutânea Localizada, formação de lesãoulcerada, caracterizada por úlcera
com bordas elevadas, infiltradas, fundo profundo e granuloso, drenado com secreção
esbranquiçada; (B) Leishmaniose Cutânea Difusa, formação de placas infiltradas; (C)
Leishmaniose Cutânea Disseminada, formação de polimorfismo lesional e lesões distribuídas
em diversos segmentos do corpo; (D) Leishmaniose Mucocutânea, edema nasal com
comprometimento da parte superior do lábio.
Fonte: TEIXEIRA et al., 2013.

19
A leishmaniose cutânea localizada caracteriza-se pela formação de uma lesão indolor,
de formato arredondado, ulcerada ou não e de tamanho variado no local da picada do inseto. A
leishmaniose cutânea difusa representa uma forma clínica rara, contudo crônica e muito grave.
Ocorre em pacientes com resposta imune debilitada, que apresentam múltiplas lesões não
ulceradas por grandes extensões do corpo. A leishmaniose cutânea disseminada apresenta-se
com múltiplas lesões papulares por várias partes do corpo. E a leishmaniose mucocutânea em
que os parasitas se alojam nas mucosas da boca, nariz, palato, faringe e laringe e levam a
formação de lesões desfigurantes (Teixeira et al., 2013).
As diferentes manifestações clínicas estão relacionadas à espécie do parasita e ao tipo
de resposta imunológica que será desencadeada a partir dessa relação “parasito-hospedeiro”
(Pires et al., 2012). Devido à alta complexidade da infecção por Leishmania, a resposta imune
nos seres humanos ainda não está muito bem descrita. Todavia já é consenso que a resistência
ou a susceptibilidade a esse tipo de infecção está relacionada com a expansão de células T
auxiliares (Th) dos tipos 1 e 2 (Reis et al., 2006).
A L. amazonensis é causadora das manifestações clínicas: leishmaniose cutânea
localizada (LCL) em que pacientes desenvolvem predominantemente ativação de linfócitos
Th1. Como também pode causar a leishmaniose cutânea difusa (LCD) em que pacientes
desenvolvem ativação quase que exclusiva de linfócitos Th2 (Reis et al., 2006).
Pacientes com LCL apresentam uma lesão característica com um elevado infiltrado de
macrófagos intensamente vacuolizados e repletos de amastigotas, além da produção de
citocinas que auxiliam a ativação de macrófagos na eliminação do parasita, tais como:
interleucina 12 (IL-12) e interferon γ (INF-γ), que atuam no recrutamento de células de defesa
ativadas. Enquanto que pacientes com LCD apresentam resposta imune deficiente, com elevado
níveis de citocinas que inibem a ativação do macrófago, como: interleucina 10 (IL-10),
interleucina 4 (IL-4) e interleucina 5 (IL-5) e, consequentemente, favorecem a sobrevivência do
parasita (Pires et al., 2012; Mougneau et al., 2012). Por isso, mesmo após o tratamento, são
frequentes as recaídas (Soong et al., 2012).
As leishmanias, ao serem fagocitadas por macrófagos, são capazes de manipular a
resposta microbicida dessas células através da redução de espécies reativas de oxigênio (ERO),
óxido nítrico (NO) e outros metabólitos tóxicos. Além da redução de produção de citocinas,
entre outras estratégias que esse parasita utiliza para garantir sua sobrevivência (Cunningham,
2002; Campanelli et al., 2010).

20
1.3 O GÊNERO Leishmania
1.3.1 Classificação
Os protozoários do gênero Leishmania pertencem ao reino Protista, sub-reino Protozoa,
filo Sarcomastigophora, subfilo Mastigophora, classe Zoomastigophora, ordem Kinetoplastida,
subordem Trypanosomatida, família Trypanosomatidae, gênero Leishmania (Shaw, 2006;
Bañuls et al., 2007).
O gênero Leishmania ainda pode ser subdividido em dois subgêneros: Viannia e o
Leishmania. Essa subdivisão é baseada principalmente na diferença quanto à localização do
parasito no intestino do vetor. As promastigotas do subgênero Leishmania colonizam o
intestino anterior, médio e posterior do vetor, já o subgênero Viannia colonizam apenas o
intestino anterior e médio (Shaw, 2006; Bañuls et al., 2007).
Desde a descrição desse gênero em 1903 até o início da década de 1960 a classificação
das leishmanias se baseava na distribuição geográfica, nas espécies de vetores que transmitem o
parasito, no reservatório, no tropismo e nas manifestações clínicas. Contudo com o advento da
biologia molecular e aprimoramento de testes bioquímicos e imunológicos passou-se a
considerar esses testes na classificação atual desse parasito (Bañuls et al., 2007).
Levando-se em consideração todas essas características, foi criado um novo esquema de
classificação taxonômica. Atualmente mais de 30 espécies desse gênero já foram descritas e
sabe-se que 20 são consideradas patogênicas para o homem (Bañuls et al., 2007; Lainson,
2010) A espécie utilizada neste estudo foi a Leishmania amazonensis, classificada no
subgênero Leishmania e está agrupada no complexo L. (L) mexicana (Figura 2).

21
1.3.2 Morfologia
O ciclo biológico da Leishmania é digenético, pois se desenvolve em dois hospedeiros
distintos e possui dois estágios de desenvolvimento: a forma promastigota, presente no trato
digestório do hospedeiro invertebrado e a forma amastigota no interior da célula hospedeira dos
vertebrados (Shaw, 2006).
As formas promastigotas são móveis, possuem corpo celular fusiforme e alongado com
flagelo externalizado na porção anterior e medem cerca de 6 a 8 μm de comprimento.
Apresentam organelas típicas dos eucariotos como mitocôndria, retículo endoplasmático e
complexo de Golgi, como também, organelas exclusivas da Família Trypanosomatidae tais
como cinetoplasto e acidocalcissomo (Shaw, 2006; Kate & Scott, 2011) (Figura 3 A-B).
As formas amastigotas são arredondadas, com flagelo reduzido e internalizado, medem
aproximadamente 3 μm de largura e 6 μm de comprimento (Bates, 1994). Essa forma é
intracelular obrigatória de células do sistema fagocítico mononuclear de mamíferos. É
resistente e capaz de desenvolver mecanismos de escape no interior do vacúolo parasitóforo
dessas células para garantir sua sobrevivência (Thi et al., 2012) (Figura 3 C-D).
Figura 2. Taxonomia do protozoário Leishmania.
Fonte: Bañuls et al., 2007, com modificações.

22
As duas formas evolutivas possuem as mesmas organelas e estruturas com exceção da
estrutura paraflagelar, exclusiva de promastigotas e os megassomos, exclusivos de amastigotas
(Thi et al., 2012).
1.3.3 Ciclo Biológico
O parasita Leishmania apresenta um ciclo heteroxênico (Figura 4). A transmissão da
Leishmania sp inicia quando a fêmea do flebotomíneo infectada inocula, durante o repasto
sanguíneo, a promastigota metacíclica (Kaye & Scott, 2011).
Promastigotas metacíclicas penetram na pele do hospedeiro e são capazes de induzir o
recrutamento de neutrófilos para o local da lesão (Muller et al., 2001; Laufs et al., 2002; Van
Zandbergen et al., 2002; Peters et al., 2008). Esses neutrófilos podem fagocitar essas formas
promastigotas e as destruírem por meio de suas respostas microbicidas ou podem entrar em
Figura 3. Formas biológicas do parasita Leishmania sp.
Forma Promastigota: A - Desenho esquemático. B – Ultraestrutura de promastigotas L.
amazonensis. Observar corpo alongado e flagelo externalizado na porção anterior. Forma
Amastigota: C – Desenho esquemático. D – Ultraestrutura de amastigota de L. amazonensis.
Observar corpo arredondado com flagelo reduzido e internalizado.
Fonte: Teixeira et al., 2013.

23
processo de apoptose. Os neutrófilos ao sinalizarem que estão em apoptose são fagocitados por
macrófagos que passam a albergar esses parasitas. As leishmanias, no interior dos neutrófilos,
não se transformam em amastigotas e não se dividem. Somente no interior do vacúolo
parasitóforo do macrófago, sua principal célula hospedeira, que as formas promastigotas
transformam-se em amastigotas por influência de alterações de pH, enzimas e temperatura
(Stuart et al., 2008; Alcolea et al., 2010).
As amastigotas são resistentes e capazes de burlar a resposta microbicida do macrófago
para se multiplicar até ocupar grande parte do citoplasma, romper a membrana dessa célula e
invadir outros macrófagos e outras células do sistema imune. Como também, podem infectar
uma nova fêmea de flebotomíneo durante o repasto sanguíneo (Kaye & Scott, 2011).
O fleblotomíneo ao picar um hospedeiro infectado, ingere células sanguíneas e outras
células, principalmente macrófagos contendo amastigotas em seu interior. Em seu trato
digestório, as células infectadas são destruídas por enzimas digestivas e as amastigotas são
liberadas. No intestino transformam-se em promastigotas procíclicas e passam a se multiplicar.
Após sucessivas multiplicações, migram para a região anterior do intestino e passam por um
processo denominado metaciclogênese, em que se tornam altamente infectantes. Ao realizar um
novo repasto sanguíneo em um hospedeiro vertebrado, inicia-se novamente o ciclo (Wilson et
al., 2010).
Figura 4. Ciclo evolutivo do parasita Leishmania sp.
Fonte: National Institute of Allergy and Infectous Diseases, 2012 (com modificações).

24
1.4 NEUTRÓFILOS: CARACTERÍSTICAS GERAIS, MIGRAÇÃO E ATIVAÇÃO
Os neutrófilos são considerados a primeira linha de defesa do organismo, pois são as
primeiras células a serem recrutadas da corrente sanguínea para os sítios de inflamação (Luster
et al., 2005; Nathan, 2006). São os leucócitos mais numerosos na corrente sanguínea (50 a
70%) (Lorenzi, 2005), um adulto humano produz cerca de 1011 neutrófilos diariamente e este
número pode aumentar para 1012 com uma inflamação aguda, representando 70% dos
leucócitos circulantes (Amulic et al., 2011). Apresentam vida média na circulação sanguínea de
6 a10 horas e nos tecidos podem viver de 1 a 2 dias. No entanto, o tempo de vida dessas células
pode ser prolongado ou encurtado dependendo dos sinais do micro-ambiente em que se
encontra (Kobayashi et al., 2003; Maianski et al., 2004 a).
Os neutrófilos (Figura 5) presentes nos vasos sanguíneos possuem um aspecto
arredondado, com cerca de 10 a 15 µm de diâmetro, núcleo segmentado (polimorfonuclear) de
dois a cinco lóbulos (multilobulado) conectados entre si por finas pontes de cromatina, nucléolo
ausente e diversos tipos de grânulos presentes no citoplasma (Borregaard & Cowle,1997;
Zychlinsky et al., 2003; Lorenzi, 2005).
Essas células permanecem em sua forma característica até serem ativadas por algum
sinal quimiotático, como citocinas, quimiocinas, produtos de microorganismos invasores,
dentre outros. Ao receber esse sinal, as células saem da circulação e migram em direção ao
gradiente quimiotático (Kobayashi et al., 2003).
Figura 5. Morfologia de neutrófilos humanos. Observar núcleo segmentado conectados
entre si por finas pontes de cromatina e diversos tipos de grânulos presentes no citoplasma
Fonte: Kennedy & DeLeo, 2009.
Neutrófilos
Eosinófilo

25
Esse mecanismo de ativação desencadeia uma série de eventos celulares: atração,
rolamento sobre o endotélio, adesão, diapedese e movimentação da célula no tecido (Wagner &
Roth, 2000; Theilgaard-Monch et al, 2006). Esses eventos promovem alterações morfológicas
graduais na célula, caracterizando três estágios morfológicos distintos: repouso ou quiescente,
estimulado e ativado (Swain et al., 2002; Sheppard et al., 2005).
Os neutrófilos quiescentes são aqueles que circulam livremente na corrente sanguínea,
não aderem ao endotélio e possuem morfologia arredondada (Swain et al., 2002; Sheppard et
al., 2005). Ao receber um sinal quimiotático, ocorrem alterações na membrana celular, com o
aumento na expressão de moléculas de adesão, sendo esta expressão a responsável por
promover adesão das células ao endotélio. As interações transitórias entre as glicoproteínas
transmembranas de adesão chamadas selectinas, presentes no endotélio e no neutrófilo
promovem o rolamento dessas células pela parede do vaso sanguíneo adjacente ao sítio de
inflamação (Kuijpers, 2001; Borregaard, 2010).
As células endoteliais expressam a selectina P e a selectina E. A selectina P é
armazenada em grânulos do citoplasma e é rapidamente redistribuída à superfície em resposta a
produtos microbianos e citocinas. A selectina E, expressa na membrana celular, é sintetizada
em resposta a interleucina1 (IL-1) e ao fator de necrose tumoral (TNF), que são citocinas
responsáveis pela ativação do endotélio, como também em resposta a produtos microbianos
(Kennedy & DeLeo, 2009; Zhang et al., 2009).
Os neutrófilos expressam a selectina L e os ligantes dos carboidratos para selectinas P e
E, facilitando as interações com as células endoteliais. Como essas interações são transitórias,
por serem de baixa afinidade, os neutrófilos podem se ligar e se destacar repetidas vezes,
permitindo a rolagem ao longo da superfície endotelial (Kennedy & DeLeo, 2009; Zhang et al.,
2009).
Os neutrófilos também expressam uma família de moléculas de adesão, chamadas
integrinas. A ativação dessas moléculas permite a adesão das células ao endotélio, o qual
também aumenta a expressão dos ligantes das integrinas, propiciando a fixação dessas células
ao endotélio, através da reorganização do citoesqueleto e espalhamento sobre a superfície
endotelial (Kennedy & DeLeo, 2009).
Nessa primeira etapa do processo inflamatório, os neutrófilos encontram-se no estágio
estimulado, onde realizam o rolamento sobre o endotélio, diapedese e migração. Esse fenótipo
é marcado pela mudança de morfologia relacionada à aderência, tornando-se aderentes. Em
seguida ocorre a diapedese, que é a migração dos neutrófilos entre as células endoteliais e a
movimentação do neutrófilo no tecido. Esta migração é orientada pela concentração de

26
quimiocinas, as quais estimulam os neutrófilos aderentes a migrar através dos espaços entre as
células endoteliais para o local de infecção (Dallegri & Ottonelo, 1997; Burg & Pillinger, 2001;
Theilgaard-Monch et al., 2006).
No sítio inflamatório, onde há maior concentração de ativadores e patógenos, os
neutrófilos encontram-se ativados. Características típicas desse fenótipo são: aumento do
volume celular, emissão de pseudópodes e atividade microbicida. Nesse estágio, neutrófilos
reconhecem e entram em contato com a partícula ou microorganismo invasor, em seguida, os
neutrófilos emitem pseudópodes que se fundem em torno do patógeno, formando o fagossomo.
Este mecanismo de fagocitose ocorre através da interação entre receptores específicos na
membrana dos neutrófilos com ligantes presentes na superfície do agente invasor (Theilgaard-
Monch et al., 2006; Borregaard, 2010).
Subsequentemente à fagocitose, ocorre à fusão do fagossoma com grânulos
citoplasmáticos, seguida pela degranulação, formando o fagolisossoma. Dentro deste, o
microorganismo é degradado por peptídeos antimicrobianos e espécies reativas de oxigênio
(ERO) ou via espécies reativas de nitrogênio (ERN) (Mayer-Scholl et al., 2004; Zychlinsky et
al., 2003; Theilgaard-Moch, 2006).
A quantidade e a duração da reação inflamatória estão relacionadas ao tipo de estímulo
recebido pelas células. Essas diferenças são percebidas na adesão celular, nas diferentes
expressões de receptores, nos tipos de leucócitos envolvidos, na liberação dos grânulos
citoplasmáticos, na realização ou não de fagocitose e na regulação da apoptose (Kuijpers,
2001).
1.5 NEUTRÓFILOS E MECANISMOS MICROBICIDAS
A atividade desses fagócitos profissionais é importante para atuação da resposta imune
celular e apresentam como principais funções a fagocitose, produção de citocinas e
quimiocinas, liberação de grânulos sobre o patógeno e liberação de redes extracelulares
(Brinkman et al., 2004; Nathan 2006).
Após fagocitose de microrganismos por neutrófilos, dois processos microbicidas são
ativados concomitantemente para criar um ambiente altamente tóxico dentro do fagossomo: a
degranulação, que corresponde à liberação do conteúdo dos grânulos dentro do fagossomo,
seguida pela secreção de enzimas presentes nesses grânulos para o meio extracelular e a
produção de O2- dependente do complexo enzimático NADPH-oxidase que é um precursor de
outras ERO (Figura 6) (Nauseef, 2007).

27
Além desses dois mecanismos microbicidas clássicos, um estudo realizado por
Brinkman e colaboradores (2004) descreveu um novo mecanismo microbicida associado à
morte de neutrófilos, chamado de NETose. Nesse mecanismo, a morte acontece após ativação
do neutrófilo levando a liberação de redes extracelulares (Neutrophil Extracellular Traps -
NET) compostas por DNA, proteínas nucleares e dos grânulos. Essas NETs são capazes de
prender bactérias, fungos e protozoários, além de proporcionar alta concentração de moléculas
antimicrobianas (Brinkmann et al., 2004; Guimarães-Costa et al., 2009; Guimarães-Costa et
al., 2011).
A degranulação é um mecanismo de defesa independente de oxigênio e baseia-se na
liberação do conteúdo dos grânulos (azurófilos, específicos e terciários). Os neutrófilos
apresentam três tipos de grânulos em seu citoplasma compostos de proteínas distintas. Os
grânulos primários ou azurófilos, que são os primeiros a serem formados e os que se
apresentam em maior quantidade, possuem enzimas conversoras dos reativos intermediários do
Figura 6. Degranulação e produção de ERO no interior do neutrófilo.
Mecanismos microbicidas que ocorrem simultaneamente para eliminação do patógeno.
Abreviaturas: O2 – Oxigênio Molecular; O2- – Íon superóxido; H2O2 –Peróxido de hidrogênio;
Cl- – Íon Cloreto; MPO – Mieloperoxidase; HOCl – Ácido hipocloroso.
Fonte: Kennedy & De Leo, 2009.

28
oxigênio como a mieloperoxidase, α-defensinas, proteínas ligadas à membrana e proteases; os
grânulos secundários contêm enzimas proteolíticas antibacterianas (lisozima e lactoferrina) e os
grânulos terciários contêm catepsinas e gelatinases (Faurshou & Borregaard 2003; Nathan,
2006; Borregaard et al., 2007).
Os constituintes dos diferentes tipos de grânulos dos neutrófilos são responsáveis pelos
mecanismos de destruição dos patógenos. A fusão dos grânulos nos fagossomas é seguida pela
liberação de substâncias microbicidas dentro dos mesmos, levando a morte do patógeno. Tais
substâncias microbicidas são, principalmente, proteínas antimicrobianas como a defensina e a
lisozima (que funcionam rompendo a superfície aniônica bacteriana) e as proteases (que
degradam proteínas bacterianas) (Mayer-Schol et al., 2004; Pham, 2006).
Além da degranulação, pode ocorrer a geração de ERO. As mitocôndrias e o complexo
NADPH são as maiores fontes de ERO. A produção de ERO é importante para a fisiologia
normal dos neutrófilos, pois funcionam como moléculas de sinalização e regulam funções
celulares (Fialkow et al., 2007; Finkel, 2011)
O complexo NADPH oxidase se agrupa na membrana plasmática do fagossomo e inicia
a cascata de ERO a partir da conversão do oxigênio molecular (O2), sendo denominado de
“burst” oxidativo (Figura 7). As ERO são moléculas altamente reativas, por isso são potentes
agentes oxidantes, que atuam em conjunto com os constituintes dos grânulos e são capazes de
destruir os agentes fagocitados (Zychlinsky et al., 2003; Nauseef, 2008).
Figura 7. Mecanismo oxidativo de neutrófilos.
O oxigênio molecular pode reagir com óxido nítrico (NO) e formar o peroxinitrito (ONOO-).
Ou pode ativar a via de ERO e a partir da ação do complexo NADPH-oxidase produzir radicais
superóxidos (O2-). Por sua vez, pela ação da SOD pode dar origem ao peróxido de hidrogênio
(H2O2). E, pode ainda pela ação da MPO produzir o ácido hipocloroso (HOCl).

29
Sabe-se que o mecanismo microbicida oxidativo se inicia com a atuação do complexo
enzimático nicotinamida adenina dinucleotídeo fosfato oxidase (NADPH oxidase). Esse
complexo enzimático é responsável pela produção do radical ânion superóxido (O2-) a partir da
redução do oxigênio molecular. Esse radical superóxido é a principal espécie reativa de
oxigênio (Klebanoff, 2005).
O aumento da atividade da NADPH oxidase deve ocorrer restritamente ao interior do
fagossomo e de forma rápida, pois a produção excessiva de ERO pode acarretar danos aos
tecidos. Um dos tipos de regulação da atividade desse complexo enzimático é a sua separação
em subunidades em diferentes compartimentos subcelulares (citosol e membrana plasmática)
durante o estado de repouso dessas células (Babior, 1999). A polimerização dos filamentos de
actina também possui um papel importante na regulação da montagem e estabilização dessa
enzima (Bengtsson et al., 2006).
O O2- produzido pela NADPH oxidase ainda pode ser dismutado pela enzima
superóxido dismutase (SOD) produzindo peróxido de hidrogênio (H2O2). Esse metabólito pode
ser utilizado como substrato na produção de outras ERO e/ou outras ERN tais como HOCL e
ONOO (Babior, 2000; Brigagão et al., 2000; Nathan, 2006; Segal, 2007).
Os radicais de nitrogênio também podem ser formados por sua reação com radicais
superóxidos. Como por exemplo o peroxinitrito (ONOO-) que é um radical altamente reativo,
produzido a partir da reação do NO com o O2- (Granfeldt et al., 2002; Segal, 2007).
A reação do H2O2 com o íon Fe2+ forma outro composto reativo, o radical HO. E
também o H2O2 pode dar origem a outras moléculas altamente reativas (Klebanoff, 2005). Um
importante composto é o ácido hipocloroso (HOCl), que é formado pela enzima
mieloperoxidase (MPO) na presença de íons Cl-, que é mais reativo que o superóxido
(Winterbourn et al., 2000).
Neste contexto, a enzima MPO apresenta-se como de vital importância na destruição de
patógenos, sendo armazenada nos grânulos azurofílicos e liberada apenas após o neutrófilo ser
estimulado. O HOCl é considerado o mais danoso na destruição da maioria dos patógenos
invasores (Lacy, 2005; Dale et al., 2008).
1.6 INTERAÇÃO NEUTRÓFILO E Leishmania
Os neutrófilos são rapidamente recrutados para o local da infecção após a inoculação
das promastigotas metacíclicas na pele, seja por meio do seu vetor natural, o flebotomíneo ou
por injeção de agulha (Peters et al., 2008; Ribeiro-Gomes et al., 2012). Peters et al., (2008)

30
mostraram por microscopia intravital, através da captação de imagens em tempo real, a
migração e o acúmulo dessas células no local da picada do mosquito Phlebotomus duboscqi,
infectado com Leishmania major, em camundongos C57BL/6. Os autores mostraram ainda que
inicialmente os parasitas são fagocitados por neutrófilos e não por monócitos/macrófagos ou
células dendríticas.
O recrutamento dessas células em murinos após infecção com L. major foi associado
aos altos níveis de produção das quimiocinas MIP-2 e KC (equivalente a interleucina 8 – IL-8)
pelas células residentes estimuladas pelos parasitas (Muller et al., 2001). E em culturas in vitro
com neutrófilos humanos também foi observada alta produção de IL-8 (Laufs et al., 2001).
Outra pesquisa demonstrou que as formas promastigotas são capazes de induzir a
migração de neutrófilos através da liberação de um fator quimiotático (LCF). Como esse fator
atua somente no recrutamento desse tipo de fagócito, foi sugerido que ele seja o principal
responsável pela indução dessa migração para o sítio de infecção e que a IL-8 seja apenas um
aliado amplificador desse sinal (Van Zandbergen et al., 2002).
Após o contato dos neutrófilos e as promastigotas de Leishmania podem ocorrer vários
tipos de interação. Promastigotas de L. major podem sobreviver dentro de neutrófilos e isso
pode favorecer o estabelecimento da doença. Na interação de L. major e neutrófilos humanos
foi observado que esse parasita impede a fusão do vacúolo com os grânulos primário e
terciário, que contêm espécies reativas de oxigênio (ERO) e outras substâncias tóxicas para o
mesmo, isso garante a sobrevivência do parasita (Ribeiro-Gomes et al., 2012).
Pode ainda ocorrer, das leishmanias conseguirem sobreviver dentro de neutrófilos
apoptóticos por um determinado período de tempo, essa hipótese foi chamada de “Cavalo de
Tróia”. Foi baseada em estudos in vitro em que neutrófilos humanos entraram em processo de
apoptose após interação com L. major e liberaram CCL4, responsável pelo recrutamento de
monócitos/macrófagos. Assim, essas células ao serem fagocitadas por macrófagos, facilitam a
entrada dos parasitos na sua principal célula hospedeira de forma silenciosa (Van Zandbergen
et al., 2004).
Van Zandbergen et al., (2004) relataram ainda que neutrófilos humanos infectados com
L. major entraram em apoptose cerca de 42 horas após a infecção, sugerindo que esse parasita
seja capaz de retardar o processo de apoptose. O parasita é capaz de ativar uma quinase
reguladora de sinais extracelulares 1/2 (ERK 1/2), levando ao aumento das moléculas anti-
apoptóticas Bfl-1 e Bcl-2, que por sua vez, inibem a liberação do citocromo c e,
consequentemente, bloqueiam a ativação das vias das caspase-3 e caspase-6 (Sarkar et al.,
2013).

31
Neste sentido, neutrófilos apoptóticos parecem contribuir para o estabelecimento da
infecção no modelo experimental com a espécie L. major. Com relação a infecção por L.
amazonensis foi demonstrado que a ingestão de neutrófilos humanos apoptóticos por
macrófagos infectados aumentou a carga parasitária através de um mecanismo dependente de
fator modulador de crescimento - beta 1 (TGF-β1) e prostaglandina E2 (PGE2). Ao contrário
do que foi observado com neutrófilos necróticos em que ocorreu diminuição da carga
parasitária por meio da produção de fator de necrose tumoral (TNF-α) e elastase neutrofílica
(NE) (Afonso et al., 2008).
Uma pesquisa recente demonstrou ainda que neutrófilos em contato com promastigotas
de L. amazonensis podem sofrer um tipo de morte celular especializada, denominada NETose.
E que esse tipo celular, por meio da formação de redes extracelulares, é capaz de aprisionar e
degradar o parasita (Guimarães-Costa et al., 2009), os neutrófilos restantes morrem por
apoptose na sua maioria (Xin et al., 2010).
A destruição do parasita também pode ser ocasionada através de outros meios, como a
produção de ERO após a fagocitose, como também pela produção de NO, importantes
mediadores leishmanicidas (Chang, 1981; Segal, 2005).
Para avaliar a influência dos neutrófilos na infecção o uso de anticorpos para depleção
desse tipo celular em camundongos vem sendo bastante utilizado. Todavia, apesar de muitos
trabalhos realizados, não existe um consenso em relação a real função desses
polimorfonucleares para a progressão ou não da doença (Tacchini-Cottier et al., 2000; Chen et
al., 2005).
Apesar de serem reportados papeis contraditórios para neutrófilos, essas células são de
grande importância para o controle dessa doença e seu papel não se restringe apenas ao início
da infecção. Os neutrófilos foram observados nas lesões ulceradas dos doentes cronicamente
infectados com L. major, e em alguns pacientes, essas células foram o tipo celular
predominante no local da infecção (Daboul, 2010) e nas lesões de até 36 meses de pacientes
infectados com L.tropica (Dabiri et al., 1998).
Um estudo recente mostrou ainda que há diferença entre as respostas geradas por
neutrófilos na infecção com Leishmania amazonensis. Foi realizada a infecção in vitro com
promastigotas e amastigotas provenientes de culturas axênicas e de lesão e ambas as formas
foram fagocitadas por neutrófilos. Contudo, as promastigotas desencadearam a liberação de
TNF-α, enquanto que as amastigotas estimularam mais a secreção de IL - 10 por essas células.
Além disso, a maioria das promastigotas foi morta por esses polimorfonucleares, já as

32
amastigotas tanto as axênicas como as de lesão apresentaram-se altamente resistentes à resposta
microbicida dessas células (Carlsen et al., 2013).
Há ainda pesquisas feitas em modelos de co-cultura de neutrófilos e macrófagos em
interação com L. amazonensis in vitro. Um estudo revelou que em modelos de co-culturas com
macrófagos infectados e neutrófilos, os parasitas intracelulares foram mais eficientemente
destruídos. Essa atividade leishmanicida foi associada com a produção de TNF-α, NO, ERO,
NE e fator de ativação de plaquetas (PAF) (Carmo et al., 2010).
Neste contexto, neutrófilos desempenham um papel fundamental durante infecção por
Leishmania, assim o desenvolvimento de drogas que atuem na ativação da resposta microbicida
dessas células pode levar a um controle inicial da infecção.
1.7 TRATAMENTO DAS LEISHMANIOSES
Segundo a Organização Mundial de Saúde (OMS), as drogas de primeira escolha são os
antimoniais pentavalentes (Sb+5), apesar de serem utilizados nos últimos sessenta anos (Frézard
& Demicheli, 2010). Esse medicamento inibe a atividade das vias glicolítica e oxidativa dos
ácidos graxos na forma amastigota (Croft et al., 2006). É administrado pelas vias parental ou
oral e é indicado para o tratamento de todos os tipos de leishmanioses (Frézard & Demicheli,
2010). Entretanto, devido ao tratamento com o uso diário e elevado desse antimonial, algumas
espécies de Leishmania apresentam resistência à droga (Palumbo, 2010). Além disso, a alta
toxidade, o elevado custo e os vários efeitos colaterais dificultam o tratamento (Ashutosh et al.,
2007; Chakravarty & Sundar, 2010).
Em caso de resistência aos antimoniais ou tratamento insatisfatório, as drogas de
segunda escolha são as anfotericinas e as pentamidinas (Pavli & Maltezou, 2010). A
anfotericina B possui ação no ergosterol da membrana da superfície do parasita, tanto na forma
promastigota como na amastigota, causando um desequilíbrio de membrana que leva ao
aumento da permeabilidade e entrada de íons (Ellis et al., 2009).
Por outro lado, as pentamidinas atuam na síntese de DNA, que leva a alteração da
morfologia do cinetoplasto e fragmentação da membrana mitocondrial do parasita. Não há
muitos estudos com relação à utilização dessa droga nas Américas, todavia as pentamidinas
apresentam eficácia no tratamento contra algumas espécies, por exemplo, L. guyanensis e L.
panamensis (Goto & Lindoso, 2010).
Em meados de 1980 foi descoberta a atividade leishmanicida da miltefosina, que foi
desenvolvida inicialmente para o tratamento do câncer. Essa droga atua na membrana celular

33
do protozoário, promovendo sua morte (Costa Filho et al., 2008). Vários ensaios clínicos
foram realizados desde então em alguns países, como Índia, Colômbia e Paquistão (Soto et al.,
2004; Mitropoulos et al., 2010). Todas essas pesquisas clínicas possibilitaram a aprovação
recente desse medicamento pelo órgão americano Food and Drug Administration (FDA) para
tratamento da leishmaniose dentro deste país.
Na ausência de uma vacina eficaz e o uso dos mesmos medicamentos para tratar todas
as formas clínicas da leishmaniose, muitos estudos estão sendo realizados na procura de uma
nova terapia para essa doença como as drogas Paromomicina, Sitamaquina e Itraconazol
(Sundar et al., 2007; El-Sayed & Anwar, 2010). Além disso, vem sendo realizados testes com
imunomoduladores como o GM-CSF, que é um fator estimulador de colônias maduras de
granulócitos e/ou macrófagos. Os resultados obtidos mostraram aumento das funções
microbicidas de monócitos e macrófagos. Além disso, ensaios clínicos revelaram que o GM-
CSF associado ao antimonial foi capaz de reduzir o tempo de cura em pacientes tratados
quando comparados aos tratados somente com o antimonial e ao placebo na leishmaniose
cutânea. Além de acelerar o processo de cicatrização, sem efeitos colaterais e sem recidiva da
doença durante um ano de acompanhamento (Almeida et al., 1999; Santos et al., 2004)
Outros compostos que também vem sendo testados são os metabólitos e as substâncias
provenientes de fungos. Por exemplo, o extrato produzido do fungo Agaricus blazei foi eficaz
na redução de formas promastigotas de L. amazonensis, L. chagasi e L. major, e ainda
estimulou a produção NO (Valadares et al., 2011).
Neste contexto, a busca por novos quimioterápicos com propriedades leishmanicidas de
baixo custo, de baixa toxidade e ainda com capacidade de ativar células do sistema
imunológico, apresentam-se como uma alternativa terapêutica importante para o tratamento das
Leishmanioses.
1.8 ÁCIDO KÓJICO (AK)
O Ácido Kójico (AK), 5-hidroxi-2-hidroximetil--pirona (HMP) (Figura 8), é um
metabólito secundário sintetizado por vários microorganismos, em especial fungos dos gêneros
Aspergillus, Acetobacter e Penicillium (Burdock et al., 2001; Nohynek, 2004; Parvez et al.,
2006; Rho et al., 2007; Rodrigues et al., 2011). É produzido a partir de um processo de
fermentação, envolvendo vários tipos de substratos que atuam como fonte de carbono (Rho et
al., 2007; Mohamad et al., 2010). Foi isolado pela primeira vez a partir de fungos do gênero

34
Aspergillus em 1907 e é uma substância cristalina altamente solúvel em água, etanol e acetona
(Gomes et al., 2001).
Figura 8: Estrutura química do ácido kójico.
Fonte: Rodrigues et al., 2011.
O AK possui uma ampla variedade de aplicações. Em relação à aplicabilidade
biológica, ele é comercializado no Japão há muitos anos como aditivo alimentar para vegetais e
frutos do mar (Burdock et al., 2001; Blumenthal, 2004; Nohynek, 2004; Bentley, 2006;
Chusiriet al., 2011). Também é encontrado em baixos níveis em comidas tradicionais japonesas
como molho de soja ou sakê (Nohynek, 2004). É utilizado nos alimentos como antioxidante e
intensificador de sabores (Burdock et al., 2001; Mohamad et al., 2010). Burdock e cols (2001)
mostraram que o AK é uma substância inofensiva para a saúde humana na quantidade
consumida como aditivo alimentar.
Em relação à aplicação terapêutica, o AK é amplamente utilizado em preparações
cosméticas como agente clareador (Lin & Huang, 2007), por possuir uma atividade inibitória
na formação da melanina e por sua ação protetora contra raios ultravioleta. Sendo bastante
utilizado também como componente fundamental em cremes para o tratamento de sardas e
manchas de idade e até mesmo produtos de clareamento dentário (Mohamad et al., 2010).
O AK é um inibidor seletivo da tirosinase (Gupta et al., 2006; Sato et al., 2007;
Mohamad et al., 2010), que desempenha papel chave na cascata melanogênica (Cabanes et al.,
1994. Por atuar como inibidor desta enzima, o AK auxilia no tratamento de melasma, onde
ocorre uma intensa produção da melanina (Lim, 1999; Parvez et al., 2006; MiHa et al., 2007),
devido à intensificação da ação da tirosinase (Gupta et al., 2006; Smit et al., 2009).

35
Para avaliar a segurança (toxicidade e genotoxicidade) na utilização do AK, Nohynek e
colaboradores (2004), mostraram que seu uso pela via tópica não oferece riscos ao consumidor.
Possui, ainda, ação antioxidante contra espécies reativas de oxigênio liberados no meio
extracelular (Niwa & Akamatsu, 1991; Gomes et al., 2001).
Além das funções já descritas, o AK possui outras aplicações, dentre elas: industrial
(Mohamad et al., 2010), antitumoral (Moto et al., 2006; Tamura et al., 2006; Higa et al., 2007;
Chusiri et al., 2011), agente radioprotetor (Emami et al., 2007), ação microbicida contra fungos
das espécies Candida albicans, Cryptococcus neoformanse, Trichophyton rubrum, que são
considerados importantes patógenos humanos (Chee & Lee , 2003) e ação cicatrizante em
feridas após tratamento tópico em modelo animal (Mohammadpour et al., 2012).
Em relação à ação antiparasitária, AK foi capaz de reduzir o número de ovos do parasito
Schistosoma mansoni (Fitzpatrick et al., 2007). E, recentemente, revelou-se um promissor
agente leishmanicida, como tratamento tópico contra a leishmaniose cutânea (Rodrigues et al.,
2014).
Com relação à ativação de células de defesa, é capaz de ativar macrófagos peritoneais
de murinos (Rodrigues et al., 2011) e induzir diferenciação de monócitos humanos em
macrófagos (Costa, 2012). E em relação à ação do AK sobre neutrófilos humanos, foi mostrado
que o metabólito, nas concentrações de 0,05, 0,5 e 20 µg/ml, potencializou a produção
intrínseca de radicais de oxigênio, aumentou a atividade fagocítica e a produção de Ca++
intracelular, importante íon relacionado com a ativação celular. (Niwa&Akamatsu, 1991).
Até o presente momento não existem trabalhos na literatura relacionados à ação do
metabólito em neutrófilos humanos durante interação com parasitos do gênero Leishmania.

36
1.9 OBJETIVOS
1.9.1 Objetivo geral:
Avaliar o papel do metabólito Ácido Kójico sobre neutrófilos humanos e durante
infecção com Leishmania (Leishmania) amazonensis.
1.9.2 Objetivos específicos:
1. Analisar a viabilidade de neutrófilos humanos após o tratamento com AK;
2. Analisar alterações estruturais e ultraestruturais de neutrófilos humanos tratados
com AK;
3. Verificar a resposta microbicida de neutrófilos após tratamento com AK, através
da produção de Espécies Reativas de Oxigênio via ativação do complexo
NADPH-oxidase, de Óxido Nítrico, a atividade da enzima mieloperoxidase e a
produção de radicais superóxidos durante interação com Leishmania
(Leishmania) amazonensis;
4. Determinar o índice endocítico e o índice de infectividade de neutrófilos tratados
com AK após interação com Leishmania (Leishmania) amazonensis;
.

37
2 MATERIAL E MÉTODOS
2.1 FLUXOGRAMA
Figura 9: Fluxograma da metodologia utilizada.
Metodologia utilizada no presente estudo, inicialmente foi avaliada a ação do AK sobre
neutrófilos humanos. Primeiramente foram realizados testes de viabilidade, seguidos das
análises morfológicas e, por fim a verificação da resposta microbicida. Posteriormente, foram
realizados testes com neutrófilos tratados com AK e infectados com Leishmania (L.)
amazonensis para avaliar o comportamento dessas células após tratamento em interação com o
parasita.
2.2 CULTIVO E MANUTENÇÃO DO PARASITO
As formas promastigotas de Leishmania (Leishmania) amazonensis
(MHOM/BR/2009/M26361) foram obtidas em meio NNN provenientes do Programa de
Leishmanioses do Instituto Evandro Chagas, Belém – Pará. Essas formas foram cultivadas em meio
RPMI 1640, suplementado com Soro Bovino Fetal (SBF) a 10% e GPPS a 1% (Glutamina 0,2M;
Ácido Pirúvico 0,0125 M; Penicilina/Estreptomicina 1%) e mantidas em estufa B.O.D (Biological
Oxygen Demand) à 27ºC.

38
2.3 OBTENÇÃO E CULTIVO DE NEUTRÓFILOS DO SANGUE PERIFÉRICO HUMANO
A purificação das células polimorfonucleares foi realizada conforme descrito por
Guimarães-Costa et al (2009). Esse método consiste na mistura do sangue com compostos que
agregam as hemácias (polímeros de polissacarídeos) e aumentam a taxa de sedimentação dos
leucócitos (Peres & Curi, 2005).
Os neutrófilos foram obtidos a partir de bolsas com concentrado de leucócitos (buffy
coats) provenientes da Fundação Centro de Hemoterapia e Hematologia do Pará (HEMOPA).
Esse concentrado de leucócitos foi adicionado a tubos de centrífuga contendo Histopaque®-
1077 (Sigma ChemCo, St Louis, MO, U.S.A) na proporção de 4,0 ml de Histopaque/8 ml de
concentrado de leucócitos.
Posteriormente, essas amostras foram centrifugadas por 40 minutos a 1900 rpm em
temperatura de 23°C. Após isso, ocorreu à formação de um halo contendo células
polimorfonucleares, acima da camada de hemácias. As células foram aspiradas com auxílio de
uma pipeta Pasteur estéril. Em seguida, foram incubadas com solução de hemólise (150
MmNH4Cl, 10 mM NaHCO3, 0,1 mM EDTA, pH 7,4) por 6 minutos e depois foram
centrifugadas a 1000 rpm por 10 minutos a temperatura de 23ºC. O sobrenadante, contendo as
hemácias hemolisadas, foi desprezado, os neutrófilos foram lavados com PBS e,
posteriormente, ressuspendidos em 1,0 ml de meio de cultura DMEM sem soro.
A viabilidade celular foi avaliada pelo teste de coloração com azul de Tripan. Foram
utilizadas amostras com mais de 80% de viabilidade. Os polimorfonucleares foram mantidos
em suspensão em tubo estéril contendo meio de cultivo DMEM. A contagem das células foi
feita em Câmara de Neubauer e a concentração ajustada de acordo com o número de células
utilizadas em cada experimento.
2.4 OBTENÇÃO E DILUIÇÃO DO METABÓLITO AK
O AK é um metabólito secundário obtido a partir de fungos do gênero Aspergillus por
um processo biotecnológico de acordo com Ferreira et al., 2010. Essa substância foi fornecida
pelo grupo do Laboratório de Investigação Sistemática em Biotecnologia e Biodiversidade
Molecular da Universidade Federal do Pará.
A solução estoque foi preparada na concentração de 1mg/ml de AK diluída em meio
DMEM. A concentração final utilizada para cada experimento foi obtida a partir dessa solução.

39
2.5 TRATAMENTO DAS CÉLULAS
Os neutrófilos foram tratados na concentração de 50 μg/mL de AK por 1 hora. Essa
concentração foi escolhida por apresentar efeito leishmanicida, induzir a ativação de
macrófagos peritoneais e diferenciação de monócitos em macrófagos em estudos realizados
pelo nosso grupo (Rodrigues et al., 2011; Costa, 2012; Rodrigues et al., 2014). Contudo para o
teste de viabilidade Método thiazolyl blue foram utilizadas as concentrações de 10, 25, 50, 100
e 200 μg/mL de AK no tratamento por 1 hora e o tramento das células com 50 μg/mL de AK
por 4, 6, 12 e 24 horas. As células foram mantidas em estufa de 5% de CO2 a temperatura de
37°C. Neutrófilos não tratados foram utilizados como controles em todos os experimentos. Para
controle positivo, neutrófilos foram ativados com 100 nM de 13- acetato de forbol éster 12-
miristato (PMA) por 1 hora ou ativados com 15 μg/mL de lipopolissacarídeo (LPS) por 4 horas.
2.6 AVALIAÇÃO DA VIABILIDADE DAS CÉLULAS TRADADAS COM AK
Para análise do efeito do AK sobre a viabilidade de neutrófilos humanos, foram
utilizados três testes: Método thiazolyl blue (MTT), detecção do Potencial da Membrana
Mitocondrial- ΔΨM (JC1) e Iodeto de Propídio (IP).
2.6.1 Método thiazolyl blue (MTT)
O MTT, (3-(4,5-dimetiltiazol-2yl)-2,5-difenil brometo de tetrazolina), é um sal
tetrazolium solúvel em água. Ele é convertido pelas desidrogenases mitocondriais em cristais
azuis de formazan insolúveis em água. O produto formazan é impermeável às membranas
celulares, acumulando-se em células viáveis, sendo posteriormente diluído em DMSO para
solubilização desses cristais. O produto final da reação é quantificado por espectrofotometria
(Fotakis &Timbrell, 2006).
Neutrófilos foram obtidos e cultivados como descrito no item 2.3 em placas de 96 poços
e submetidos a tratamento com o AK nas concentrações de 10, 25, 50, 100 e 200 μg/mL por 1
hora. Em seguida, neutrófilos foram tratados com a concentração de 50 μg/mL, escolhida para
os demais ensaios, por 4, 6, 12 e 24 horas para verificar se o metabólito seria tóxico para a
célula em um período de tratamento maior.
O sobrenadante foi retirado e as células foram lavadas com PBS. Logo após a lavagem,
foi adicionado 0,5 mg/mL MTT diluído em PBS e incubados à 37ºC/5% de CO2 por 3 horas.

40
Em seguida, novamente as células foram lavadas com PBS e foi adicionado 200 μL de DMSO
em cada poço para solubilização dos cristais de formazan seguido da incubação em agitação
por 10 minutos. Posteriormente, a solução resultante foi lida em leitor de ELISA (BIO-RAD
Model 450 Microplate Reader) em um comprimento de onda de 570 nm. Como controle da
reação, células foram mortas com solução de 10% de formol em PBS (Fotakis &Timbrell,
2006).
2.6.2 Detecção de Potencial da Membrana Mitocondrial (JC-1)
O potencial de membrana mitocondrial (ΔΨ) é um importante parâmetro da função
mitocondrial utilizado como indicador de viabilidade celular. O JC-1 é um marcador
fluorescente que mensura o potencial da membrana mitocondrial das células. A perda de
potencial da membrana mitocondrial é utilizada como indicador de apoptose. O JC-1 possui
formas conhecidas como “J-agregados” que marcam células não-apoptóticas com fluorescência
vermelha intensa. Por outro lado, células apoptóticas com baixo ΔΨ, permanecem na forma
monomérica, apresentando apenas fluorescência verde (Maianski et al., 2004 b).
Para realização do teste, neutrófilos foram cultivados em tubos de centrífugas e
submetidos ao tratamento como descrito no item 2.5. Após o tratamento, essas células foram
centrifugadas a 1000 rpm por 10 minutos e ressuspendidas em solução de PBS pH 7,4. Em
seguida, as células foram novamente centrifugadas a 1000 rpm por 10 minutos e incubadas por
30 min com 10 mM de JC-1 a 37°C. Por fim, foram novamente centrifugadas, ressuspendidas
em PBS e a fluorescência das células foi imediatamente determinada por citometria de fluxo.
As células foram analisadas utilizando citômetro de fluxo BD FACS Canto II TM em
comprimento de onda de excitação de 488 nm, sendo que os monômeros de JC-1 emitem a 529
nm e os agregados a 590 nm. Os dados foram obtidos utilizando o software BD FACSDiva.
Um total de 10.000 eventos foi coletado para cada amostra e analisados com auxílio do
programa WinMDI 2.9. Foram realizados três experimentos e os resultados foram expressos em
porcentagens.
2.6.3 Iodeto de Propídio (IP)
A detecção da apoptose de neutrófilos foi realizada utilizando a marcação com Iodeto
de Propídio. O IP é impermeável à membrana plasmática íntegra, assim as células negativas
para a marcação com IP são consideradas viáveis. Já nas células mortas, o IP tem acesso ao

41
DNA celular devido à perda da integridade das membranas plasmática e nuclear dessas células
(Koopman et al., 1994).
Para realização deste teste, neutrófilos foram cultivados em tubos de centrífuga e
submetidos ao tratamento conforme item 2.5. Após o tratamento, células foram centrifugadas,
lavadas com PBS e novamente centrifugadas. Em seguida, as células foram marcadas com 10
μL de IP por 30 minutos. Para finalizar, as células foram lavadas com PBS e a leitura feita
imediatamente por citometria de fluxo. Um total de 10.000 eventos foi coletado para cada
amostra e analisados conforme descrito no item 2.6.2. Foram realizados três experimentos e os
resultados foram expressos em porcentagens.
2.7 ANÁLISE MORFOLÓGICA DE NEUTRÓFILOS TRATADOS COM AK
2.7.1 Microscopia Óptica (MO) e Análise de área celular (Morfometria)
Neutrófilos foram cultivados e tratados com AK na concentração de 50 μg/mL, fixados
com uma solução de tampão PHEM 0,1 M e 4% de paraformaldeído durante 30 minutos e
posteriormente mantidos em tampão PHEM 0,1 M até ser realizada a coloração. A coloração
foi feita com o corante Giemsa, diluído a 10 % em água destilada, durante 30 minutos à
temperatura ambiente. Após esse período, as células foram desidratadas em acetona 100% e
passadas em misturas crescentes de acetona–xilol, até duas passagens finais em xilol puro. As
lamínulas foram montadas em lâminas de vidro com Entellan®. As células foram analisadas
em Microscópio Axioscope A1 Zeiss. Células ativadas com 13-acetato de forbol éster 12-
miristato (PMA) na concentração de 100 nM por 1 hora foram utilizadas como controle.
Para avaliar se o tratamento com AK em neutrófilos estaria promovendo o aumento da
área citoplasmática, foi utilizada a morfometria para medir a área total da célula. A partir de
fotos de MO, foram feitos o cálculo da área celular e a média de 50 células de cada amostra
(grupo controle sem tratamento, grupo tratado com PMA, grupo tratado com AK). A área total
do citoplasma das células foi medida com recurso do programa Image J.
2.7.2 Microscopia Eletrônica de Varredura (MEV)
Após tratamento com AK como descrito no item 2.5, as células foram desidratadas em
uma série de etanol em água a 30, 50, 70, 90% (10 minutos cada etapa) e 100% (3 vezes

42
durante 10 minutos). As amostras foram secas pelo método do ponto crítico (Modelo K 850 -
Marca EMITECH) usando CO2. Em seguida, foram fixadas em suporte apropriado e
metalizadas com uma película de ouro, usando o aparelho Emitech K550-England. As células
foram analisadas no microscópio eletrônico de varredura LEO 1450VP (Instituto Evandro
Chagas). Células ativadas com 100 nM de PMA foram utilizadas como controle positivo.
2.7.3 Microscopia Eletrônica de Transmissão (MET)
Neutrófilos foram cultivados e tratados como descrito no item 2.5. E células ativadas
como 100 nM de PMA foram utilizadas como controle. Posteriormente, foram fixados em uma
solução contendo 2,5% de glutaraldeido a 25%, 4% de paraformaldeído, 2,5% de sacarose, em
tampão cacodilato de sódio 0,1 M, pH 7.2. Após a fixação, as células foram lavadas 3 vezes em
Tampão cacodilato 0,1 M seguido de pós-fixação com: 1% tetróxido de ósmio e ferrocianeto de
potássio 0,8% por 1 hora à temperatura ambiente. As células foram lavadas três vezes em
tampão cacodilato 0,1 M e então desidratadas em série crescente de acetona durante 10 minutos
à temperatura ambiente. Após desidratação as células foram lentamente impregnadas com
resina Epon nas seguintes concentrações: 2:1, 1:1 e 1:2 (acetona 100%: Epon - 12 horas em
cada etapa). Em seguida o material foi colocado em Epon puro por 6 horas e depois no suporte
para polimerização a 60o C por 48 horas. Os blocos polimerizados foram cortados em
ultramicrótomo (Leica EM UC6) e os cortes obtidos foram contrastados com acetato de uranila
5% por 20 minutos, e posteriormente com citrato de chumbo por 5 minutos e observados em
MET LEO 906 E (UFPA).
2.7.4 Detecção de filamentos de actina por Microscopia Óptica de Fluorescência
Neutrófilos foram cultivados e tratados como descrito no item 2.5. As células foram
fixadas em paraformaldeído 4% em tampão PHEM 0,1 M por 1 hora. Posteriormente, foram
realizadas lavagens com PBS pH 8,0, seguido do processo de permeabilização com Triton-X a
0,3%. Em seguida, as células foram novamente lavadas em PBS e os sítios de ligação foram
bloqueados com solução de NH4Cl 50mM.
Após o bloqueio, as células foram lavadas com PBS suplementado com 1 e 3% de BSA.
Para assim, serem incubadas com Faloidina (Molecular ProbesInvitrogen®), para detecção de
filamentos de actina, diluído 1:198 em PBS e DAPI, para marcação do núcleo, diluído 1:10 por
40 minutos.

43
Por fim, as células foram lavadas em PBS e em água. Após isso, as lamínulas foram
montadas em lâmina contendo o Pro Long Gold antifade reagente (Molecular Probes
Invitrogen®). As células foram analisadas em Microscópio Axioscope A1 Zeiss. As células
marcadas com Faloidina foram analisadas em filtro de 594 nm. A marcação com DAPI foi
analisada em filtro de 365 nm. Células ativadas como 100 nM de PMA foram utilizadas como
controle.
2.8 RESPOSTA MICROBICIDA
2.8.1 Produção de Radicais Superóxidos pelo teste do Nitro Blue Tetrazolium (NBT)
Para verificar se AK induz a produção de radicais superóxidos foi realizado o teste do
NBT. O NBT possui coloração amarela e na presença de oxigênio reativo (O2-) é oxidado,
levando a formação de partículas de coloração azul no meio intracelular (formazan NBT) (Choi
et al., 2006).
Após obtenção, as células foram incubadas com meio contendo 1 mg/ml de NBT e
tratadas com 50 μg/mL do AK ou 100nM de PMA. E para confirmar a via de produção de
radicais através da ativação do complexo NADPH foi utilizado como inibidor da NADPH
oxidase, a apocinina, na concentração de 1mM por 1hora e 20 minutos (Ximenes et al, 2007;
Stefanska & Pawliczak, 2008).
Após 1 h a 37 ºC e em atmosfera de 5% de CO2, as células foram lavadas com PBS e
fixadas com metanol. Para solubilização da membrana celular, foi adicionado hidróxido de
potássio (KOH) 2M. Em seguida, para dissolver o formazan NBT foi acrescentado DMSO. Por
fim, as amostras foram transferidas para uma microplaca de 96 poços e a leitura foi realizada
em leitor ELISA (BIO-RAD Model 450 Microplate Reader).
2.8.2 Produção de Espécies Radicais de Oxigênio (ERO) pela marcação com CellROX®
O teste do CellRox® foi realizado conforme descrito por Hyo-Yang et al. (2012), com
algumas modificações. Neutrófilos foram cultivados em tubos de centrífugas e submetidos ao
tratamento com AK. Após o tratamento, essas células foram centrifugadas a 1000 rpm por 10
minutos para remoção do meio e ressuspendidas em solução de PBS pH 7,4. Em seguida, as

44
células foram incubadas por 30 min com 5 µM de CellROX® em estufa a 37°C. Esta etapa do
experimento foi realizada ao abrigo da luz, pois o marcador é fotossensível.
Por fim, as células foram novamente centrifugadas e transferidas para placas específicas
para leitura por fluorimetria. A leitura da fluorescência foi realizada em fluorímetro (VICTOR
X Multilabel Plate Readers) com comprimento de onda de 490 nm. Neutrófilos ativados como
100 nM de PMA foram utilizadas como controle.
2.8.3 Avaliação da atividade da Mieloperoxidase (MPO)
Para avaliação da atividade da MPO, neutrófilos foram tratados com AK por 1 hora e
foi feito um controle com células ativadas com PMA na concentração de 100nM. Foram
realizadas imunomarcação e análise por microscopia óptica de fluorescência para a aquisição
de imagens e a citometria para a quantificação de células marcadas e intensidade de
fluorescência.
Neutrófilos foram cultivados, tratados e fixados em paraformaldeído 4% em tampão
PHEM 0,1 M por 1 hora. Posteriormente, as células foram lavadas em PBS e os sítios de
ligação foram bloqueados com solução de NH4Cl 50mM por 45 minutos. Foram realizadas
lavagens com PBS suplementado com 1 e 3% de BSA. Para marcação da MPO, as células
foram incubadas com anticorpo primário anti-mieloperoxidase neutrofílica (Sigma-Aldrich®)
diluído 1:500 em PBS com 1% de BSA durante 1 hora, seguido de lavagens sucessivas em PBS
para total remoção de anticorpos não ligantes. Após isso, as células foram incubadas com
anticorpo secundário goat anti-rabbit alexa 488 (Molecular Probes Invitrogen®), com diluição
de 1:500 por 1 hora, seguido de sucessivas lavagens em PBS. Além disso, foi realizada a
marcação com DAPI para identificação do núcleo, diluído 1:10 por 10 minutos. Por fim, as
células foram lavadas em PBS e em água e as lamínulas foram montadas em lâmina contendo o
Pro Long Gold antifade reagente (Molecular Probes Invitrogen®). As células foram analisadas
em Microscópio Axioscope A1 Zeiss. As células marcadas com Mieloperoxidase foram
analisadas em filtro de 488 nm A marcação com DAPI foi analisada em filtro de 365 nm.
Células ativadas como 100 nM de PMA foram utilizadas como controle.
Para citometria, células foram cultivadas e tratadas em tubos de centrífuga de 15 mL.
Em seguida, as células foram centrifugadas a 1500 rpm por 10 minutos para lavagem com PBS
pH 8, centrifugadas novamente e permeabilizadas com Triton-X a 0,3%. Novamente foram
lavadas com PBS e centrifugadas. A solução de NH4Cl 50mM foi utilizada como bloqueio e
seguida essas células passaram por uma nova centrifugação seguida de lavagem com PBS.

45
Para assim, serem incubadas com anticorpo primário anti-mieloperoxidase neutrofílica (Sigma-
Aldrich®) diluído 1:499 em PBS por 1 hora e anticorpo secundário goat anti-rabbit alexa 488
(Molecular Probes Invitrogen®) 1:499 também diluído em PBS por 1 hora. Por fim, uma nova
centrifugação foi realizada e as células ressuspendidas em PBS pH 8.0. Um total de 10.000
eventos foram coletados para cada amostra e analisados conforme descrito no item 2.6.2. O
experimento foi realizado três vezes e células ativadas com PMA foram utilizadas como
controle positivo para atividade da MPO.
2.8.4 Detecção da produção de Óxido Nítrico (NO) pela marcação com DAF-MT
O 4-amino-5-metillamino-2`,7-diclorofluoresceína (DAF-FM) é um reagente que
permite a determinação de NO intracelular. Não possui fluorescência até reagir com NO para
formar um benzotriazol fluorescente. A medida da fluorescência foi feita pela leitura da
amostra no fluorímetro e é proporcional à concentração de NO nas células (Kojima et al., 1998;
Sheng et al., 2005).
Para realização do teste, neutrófilos foram cultivados em tubos de centrífugas e
submetidos ao tratamento com AK. Como controle negativo, foram utilizadas células sem
tratamento e como controle positivo, células incubadas com 15 µg/mL de LPS. Após o
tratamento, essas células foram centrifugadas a 1000 rpm por 10 minutos para remoção do
meio e ressuspendidas em solução de PBS pH 7,4. Em seguida, as células foram incubadas por
30 min com 5 µM de DAF-FM (Molecular Probes Invitrogen®) em estufa a 37°C. Essa etapa
do experimento foi realizada ao abrigo da luz, pois o marcador é fotossensível.
Por fim, foram novamente centrifugadas e as células foram transferidas para placas para
leitura pelo fluorímetro (VICTOR X Multilabel Plate Readers) com comprimento de onda de
488 nm.
2.9 INTERAÇÃO DE Leishmania (Leishmania) amazonensis COM NEUTRÓFILOS
TRATADOS COM AK
2.9.1 Índice Endocítico
Para verificar se neutrófilos tratados com AK eram capazes de fagocitar o protozoário
Leishmania (Leishmania) amazonensis foi determinado o índice endocítico. Células tratadas
foram infectadas na proporção de (1:5) durante 3 horas a 37 ºC em atmosfera de 5% de CO2.

46
Em seguida, as células foram fixadas com paraformaldeído 4% e tampão PHEM 0,1 M
e coradas com Giemsa. As lamínulas foram montadas em lâminas de vidro, tendo Entellan
como meio de montagem. Foram contados 200 neutrófilos por lamínula e o índice foi obtido
calculando-se a porcentagem de células que endocitaram e a média de parasitos por células. As
células foram contadas em microscópio óptico Olympus BX41.
2.9.2 Índice de Infectividade
Para verificar se neutrófilos tratados com AK eram capazes de destruir os parasitas
fagocitados, foi determinado o índice de infectividade. Células tratadas foram infectadas com
Leishmania (Leishmania) amazonensis, na proporção de (1:5) durante 3 horas a 37 ºC em
atmosfera de 5% de CO2. Após esse período, o sobrenadante foi desprezado e os neutrófilos
lavados com PBS, pH 7.2, por três vezes, para retirar as leishmanias não internalizadas. Foram
avaliados três períodos de tempo após a infecção: 0, 3 e 6 horas.
Em seguida, as células foram fixadas com paraformaldeído 4% e tampão PHEM 0,1 M
e coradas com Giemsa. As lamínulas foram montadas em lâminas de vidro, tendo Entellan
como meio de montagem. Foram contados 200 neutrófilos por lamínula e o índice foi obtido
calculando-se a porcentagem de células que endocitaram e a média de parasitos por células. As
células foram contadas em microscópio óptico Olympus BX41.
2.9.3 Produção de Radicais Superóxidos pelo teste do Nitro Blue Tetrazolium (NBT)
Para verificar se AK induz a produção de radicais superóxidos em neutrófilos mesmo
durante interação com Leishmania (L.) amazonensis foi realizado o teste do NBT. Para
realização deste teste, neutrófilos foram cultivados em placa de cultura de 24 poços, infectados
com Leishmania (Leishmania) amazonensis na proporção de (1:5), tratados com 50 μg/mL de
AK e incubados com 1 mg/ml de NBT por 1 hora. Foi realizado o teste como descrito no item
2.8.1.
2.10 ANÁLISE ESTATÍSTICA
Todos os experimentos foram realizados em triplicada e todos os dados são expressos
com média e desvio padrão. Os dados obtidos foram analisados utilizando o GraphPedPrism
Versão 5.0. A análise estatística das diferenças entre os valores médios de dois grupos

47
experimentais foi realizada utilizando o Teste T Student, e para comparações múltiplas de
grupos usamos ANOVA seguida do Teste de Tukey. Os resultados foram considerados
estatisticamente significativos para valores de p inferior a 0,05.
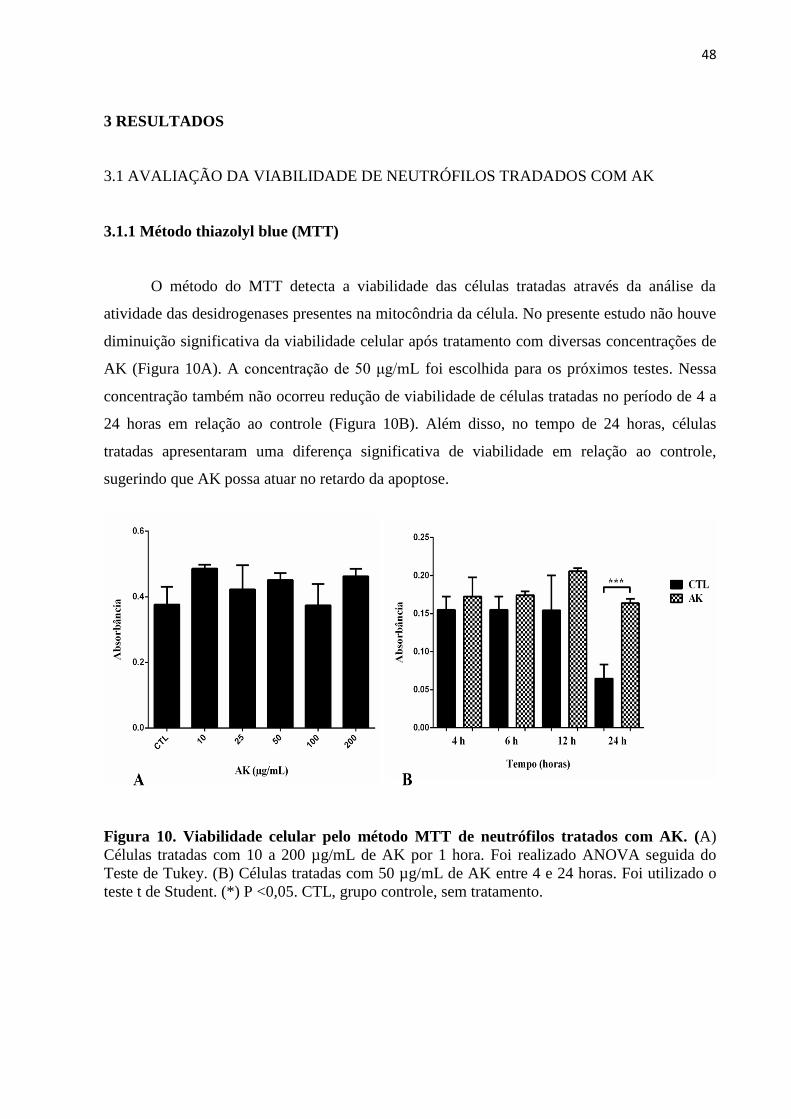
48
3 RESULTADOS
3.1 AVALIAÇÃO DA VIABILIDADE DE NEUTRÓFILOS TRADADOS COM AK
3.1.1 Método thiazolyl blue (MTT)
O método do MTT detecta a viabilidade das células tratadas através da análise da
atividade das desidrogenases presentes na mitocôndria da célula. No presente estudo não houve
diminuição significativa da viabilidade celular após tratamento com diversas concentrações de
AK (Figura 10A). A concentração de 50 μg/mL foi escolhida para os próximos testes. Nessa
concentração também não ocorreu redução de viabilidade de células tratadas no período de 4 a
24 horas em relação ao controle (Figura 10B). Além disso, no tempo de 24 horas, células
tratadas apresentaram uma diferença significativa de viabilidade em relação ao controle,
sugerindo que AK possa atuar no retardo da apoptose.
Figura 10. Viabilidade celular pelo método MTT de neutrófilos tratados com AK. (A)
Células tratadas com 10 a 200 µg/mL de AK por 1 hora. Foi realizado ANOVA seguida do
Teste de Tukey. (B) Células tratadas com 50 µg/mL de AK entre 4 e 24 horas. Foi utilizado o
teste t de Student. (*) P <0,05. CTL, grupo controle, sem tratamento.
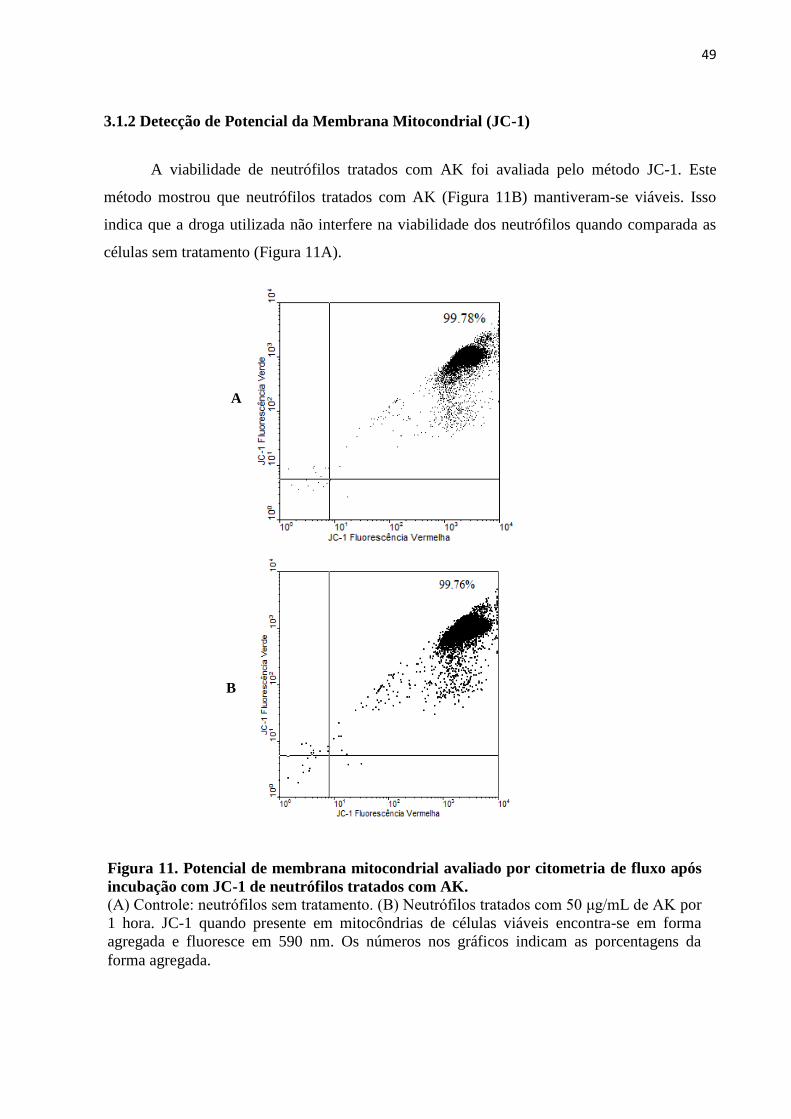
49
3.1.2 Detecção de Potencial da Membrana Mitocondrial (JC-1)
A viabilidade de neutrófilos tratados com AK foi avaliada pelo método JC-1. Este
método mostrou que neutrófilos tratados com AK (Figura 11B) mantiveram-se viáveis. Isso
indica que a droga utilizada não interfere na viabilidade dos neutrófilos quando comparada as
células sem tratamento (Figura 11A).
Figura 11. Potencial de membrana mitocondrial avaliado por citometria de fluxo após
incubação com JC-1 de neutrófilos tratados com AK. (A) Controle: neutrófilos sem tratamento. (B) Neutrófilos tratados com 50 μg/mL de AK por
1 hora. JC-1 quando presente em mitocôndrias de células viáveis encontra-se em forma
agregada e fluoresce em 590 nm. Os números nos gráficos indicam as porcentagens da
forma agregada.
B
A
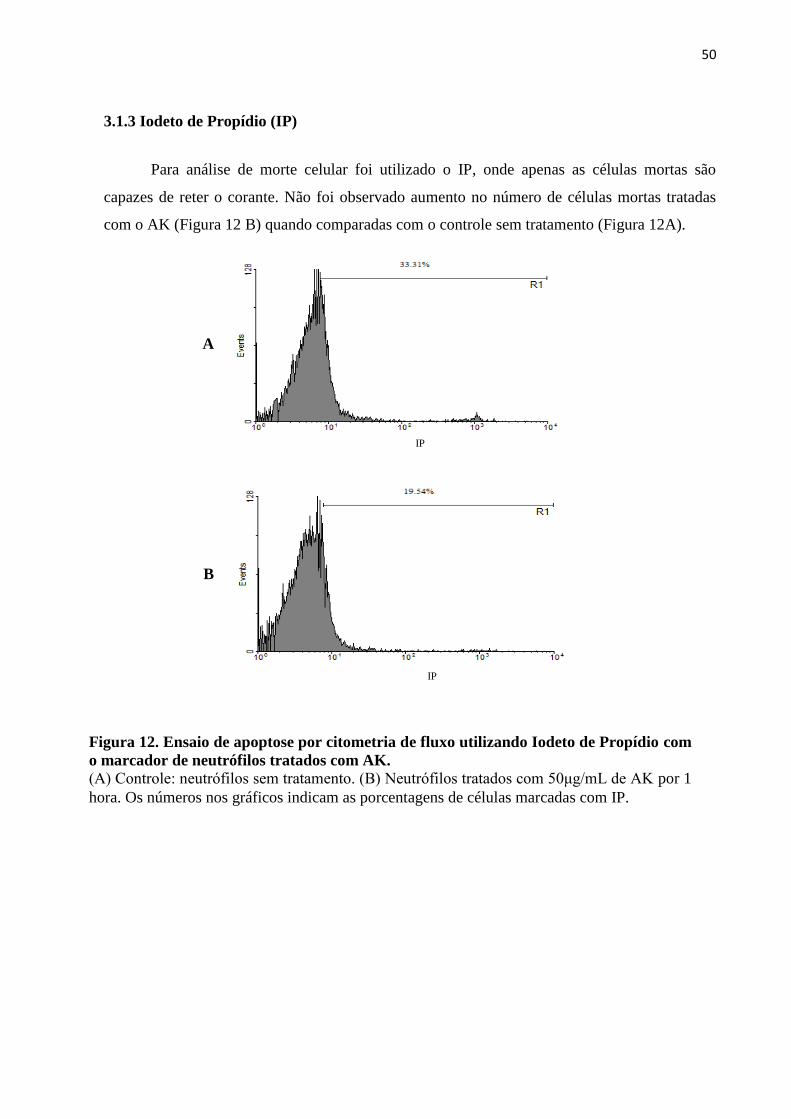
50
3.1.3 Iodeto de Propídio (IP)
Para análise de morte celular foi utilizado o IP, onde apenas as células mortas são
capazes de reter o corante. Não foi observado aumento no número de células mortas tratadas
com o AK (Figura 12 B) quando comparadas com o controle sem tratamento (Figura 12A).
Figura 12. Ensaio de apoptose por citometria de fluxo utilizando Iodeto de Propídio com
o marcador de neutrófilos tratados com AK. (A) Controle: neutrófilos sem tratamento. (B) Neutrófilos tratados com 50μg/mL de AK por 1
hora. Os números nos gráficos indicam as porcentagens de células marcadas com IP.
IP
IP
B
A

51
3.2 ANÁLISE MORFOLÓGICA DE NEUTRÓFILOS TRATADOS COM AK
3.2.1 Microscopia Óptica e Morfometria
Na análise morfológica de neutrófilos tratados por microscopia óptica, foi observado
aumento na quantidade de vacúolos, presença de projeções citoplasmáticas e aumento no
volume de neutrófilos tratados (Figura 13C) em relação ao controle sem tratamento (Figura 13
A). E para comparação morfológica com neutrófilos ativados, as células foram tratadas com
PMA (Figura 13B).
A medida da área celular confirmou o aumento da área citoplasmática. Neutrófilos
tratados com AK por 1 hora apresentaram aumento significativo da área celular em relação ao
grupo controle. Esse aumento foi semelhante ao observado com células ativadas com PMA
(Figura 13D).
3.2.2 Microscopia Eletrônica de Varredura
Para confirmação dos resultados observados em MO, foi realizada a MEV para análise
mais detalhada das alterações morfológicas na superfície celular. Neutrófilos tratados com 50
μg/mL de AK (Figura 14C) apresentaram maior adesão, espraiamento celular e projeções
citoplasmáticas, semelhantes as alterações induzidas pelo PMA (Figura 14B). Já as células sem
tratamento (Figura 14A) permaneceram com morfologia característica de neutrófilos no estágio
de repouso.
3.2.3 Microscopia Eletrônica de Transmissão
Para obtenção de maiores detalhes acerca das alterações intracelulares induzidas pelo AK nos
neutrófilos foi realizada a MET. Neutrófilos tratados com 50 µg/mL de AK apresentaram maior
concentração de grânulos e vacúolos, além de não apresentarem características apoptóticas
(Figura 15C). Como controles, foram utilizadas células sem tratamento (Figura 15A) e células
ativadas com PMA (Figura 15B).

52
Figura 13. Análise morfológica e morfométrica de neutrófilos tratados com AK e
corados com Giemsa.
(A) Controle sem tratamento. Observar o tamanho da célula e ausência de projeções
citoplasmáticas. (B) Neutrófilos ativados com PMA. Observar a presença de projeções
citoplasmáticas e vacúolos. (C) Neutrófilos tratados com 50 µg/mL de AK por 1 hora.
Observar nas células tratadas o aumento do tamanho celular, a presença de vacúolos e
projeções citoplasmáticas. Barra 10μm. (D) Medida da área citoplasmática de neutrófilos
tratados com AK. Foi utilizado ANOVA seguido do Teste de Tukey. (***) p< 0,001.
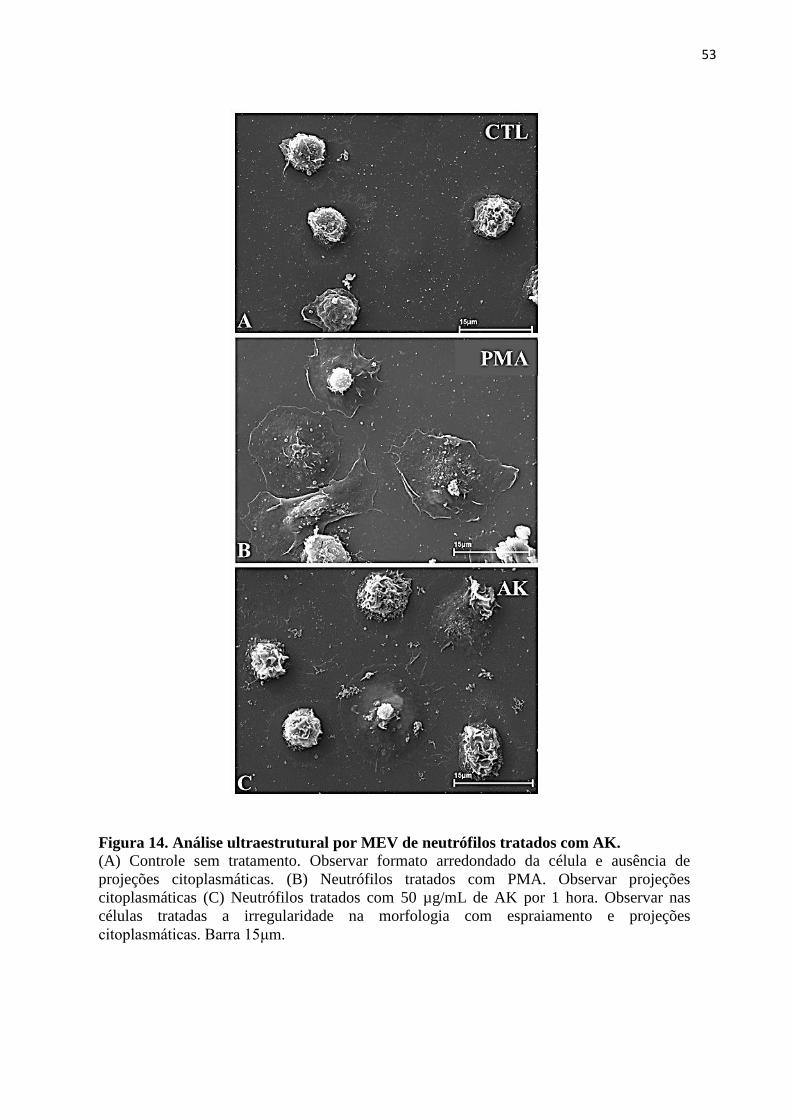
53
Figura 14. Análise ultraestrutural por MEV de neutrófilos tratados com AK.
(A) Controle sem tratamento. Observar formato arredondado da célula e ausência de
projeções citoplasmáticas. (B) Neutrófilos tratados com PMA. Observar projeções
citoplasmáticas (C) Neutrófilos tratados com 50 µg/mL de AK por 1 hora. Observar nas
células tratadas a irregularidade na morfologia com espraiamento e projeções
citoplasmáticas. Barra 15μm.
CTL
A
AK
B
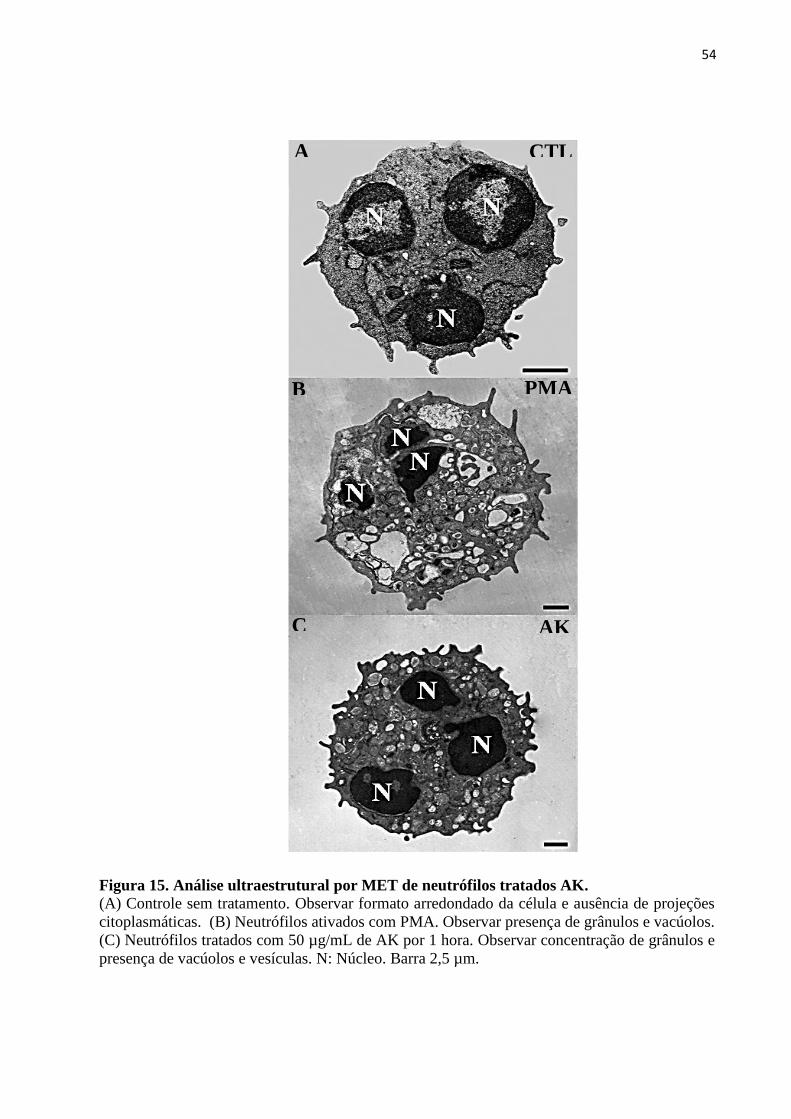
54
Figura 15. Análise ultraestrutural por MET de neutrófilos tratados AK.
(A) Controle sem tratamento. Observar formato arredondado da célula e ausência de projeções
citoplasmáticas. (B) Neutrófilos ativados com PMA. Observar presença de grânulos e vacúolos.
(C) Neutrófilos tratados com 50 µg/mL de AK por 1 hora. Observar concentração de grânulos e
presença de vacúolos e vesículas. N: Núcleo. Barra 2,5 µm.
C AK
B PMA
CTL A

55
3.2.4 Detecção de filamentos de actina por Microscopia Óptica de Fluorescência
Os filamentos de actina do citoesqueleto de neutrófilos tratados e não tratados foram
analisados utilizando a microscopia de fluorescência através da marcação com Faloidina. O
DAPI foi utilizado para a identificação e análise de alterações nucleares. Células não tratadas
demonstraram características semelhantes a neutrófilos quiescentes (Figura 16 A, B, C).
Células tratadas com PMA (Figura 16 D, E, F) apresentaram características de células ativadas.
Neutrófilos tratados com 50 μg/mL de AK (Figura 16 G, H, I) apresentaram polimerização dos
filamentos de actina e a sua redistribuição no citoplasma possibilitando o espraiamento celular
e a formação de filopódios.
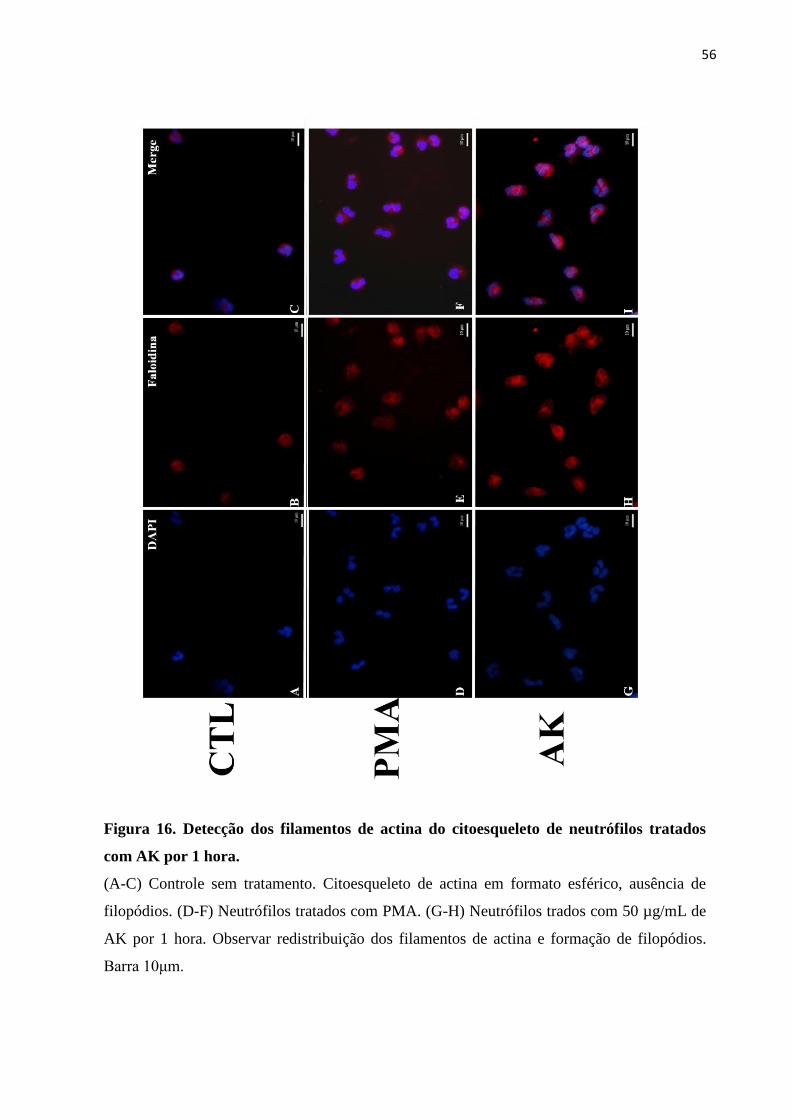
56
Figura 16. Detecção dos filamentos de actina do citoesqueleto de neutrófilos tratados
com AK por 1 hora.
(A-C) Controle sem tratamento. Citoesqueleto de actina em formato esférico, ausência de
filopódios. (D-F) Neutrófilos tratados com PMA. (G-H) Neutrófilos trados com 50 µg/mL de
AK por 1 hora. Observar redistribuição dos filamentos de actina e formação de filopódios.
Barra 10μm.
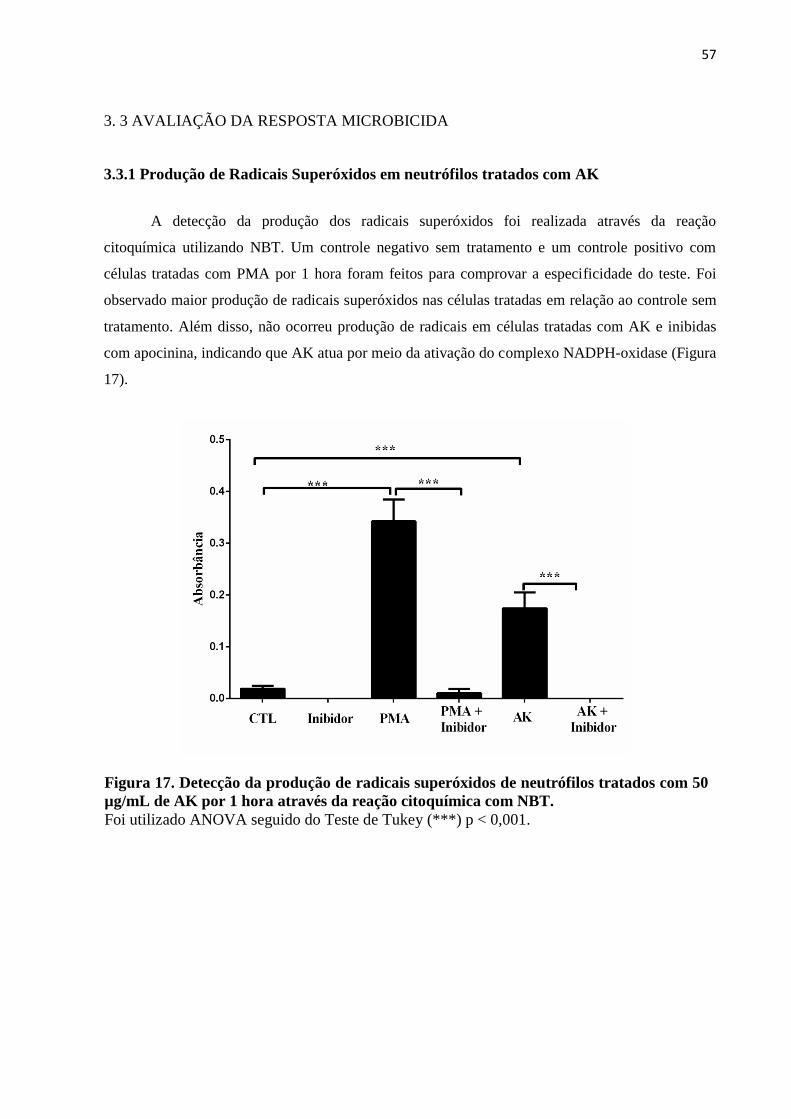
57
3. 3 AVALIAÇÃO DA RESPOSTA MICROBICIDA
3.3.1 Produção de Radicais Superóxidos em neutrófilos tratados com AK
A detecção da produção dos radicais superóxidos foi realizada através da reação
citoquímica utilizando NBT. Um controle negativo sem tratamento e um controle positivo com
células tratadas com PMA por 1 hora foram feitos para comprovar a especificidade do teste. Foi
observado maior produção de radicais superóxidos nas células tratadas em relação ao controle sem
tratamento. Além disso, não ocorreu produção de radicais em células tratadas com AK e inibidas
com apocinina, indicando que AK atua por meio da ativação do complexo NADPH-oxidase (Figura
17).
Figura 17. Detecção da produção de radicais superóxidos de neutrófilos tratados com 50
µg/mL de AK por 1 hora através da reação citoquímica com NBT.
Foi utilizado ANOVA seguido do Teste de Tukey (***) p < 0,001.
Merge

58
3.3.2 Produção de Espécies Reativas de Oxigênio
Para confirmar o resultado do NBT e avaliar a produção total de ERO em culturas de
neutrófilos tratados com o metabólito, foi realizada a marcação com CellROX. Ocorreu
significativa produção de ERO em células tratadas por 1 hora com AK quando comparadas
com o controle sem tratamento (Figura 18). Como controle positivo, as células foram ativadas
com PMA.
Figura 18. Detecção da produção Espécies Reativas de Oxigênio de neutrófilos
tratados com 50 µg/mL de AK por 1 hora pela marcação com CellROX®.
Foi utilizado ANOVA, Teste de Tukey. (**) p< 0,01; (***) p < 0,001.

59
3.3.3 Avaliação da atividade da Mieloperoxidase (MPO)
A avaliação da atividade da MPO foi feita qualitativamente por microscopia óptica de
fluorescência (Figura 19) e quantitativamente (Figura 20) por meio da citometria de fluxo. Foi
observada maior atividade dessa enzima em células tratadas quando comparado ao controle
sem tratamento. Além disso, ocorreu um maior número de células marcadas após o tratamento
com AK quando comparado ao controle sem tratamento e ao controle positivo, estimulado com
PMA.
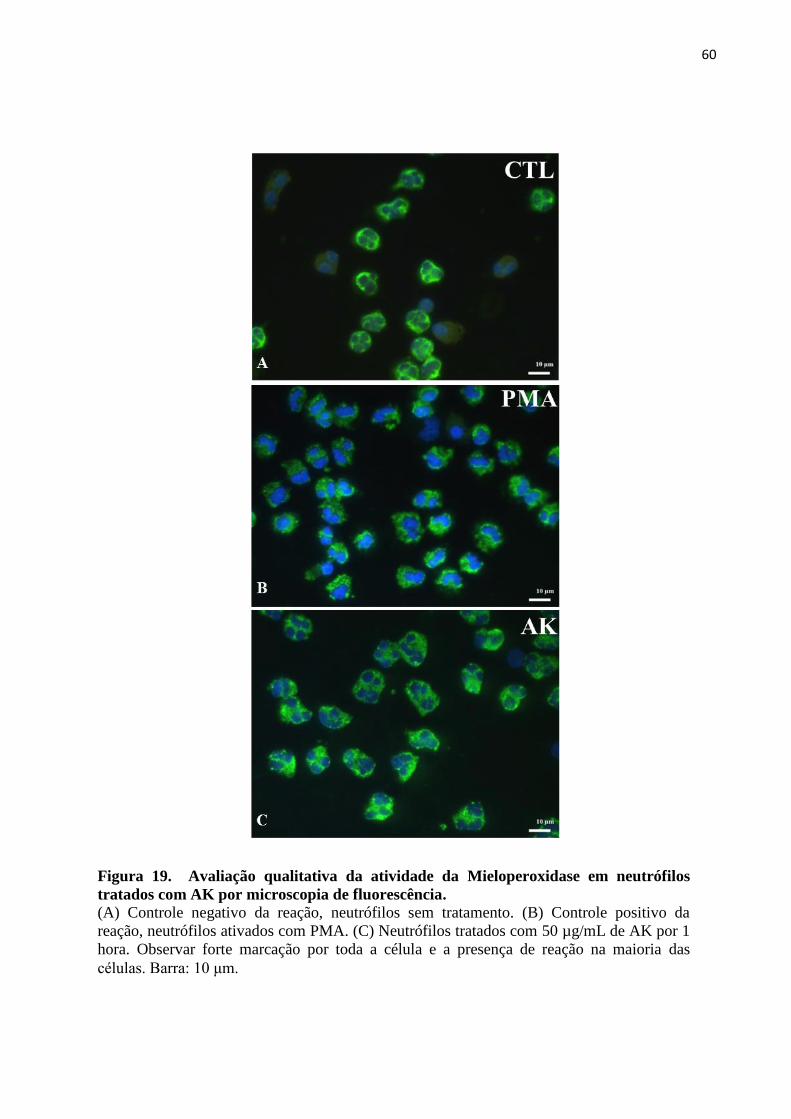
60
Figura 19. Avaliação qualitativa da atividade da Mieloperoxidase em neutrófilos
tratados com AK por microscopia de fluorescência. (A) Controle negativo da reação, neutrófilos sem tratamento. (B) Controle positivo da
reação, neutrófilos ativados com PMA. (C) Neutrófilos tratados com 50 µg/mL de AK por 1
hora. Observar forte marcação por toda a célula e a presença de reação na maioria das
células. Barra: 10 μm.

61
Figura 20. Avaliação quantitativa por citometria de fluxo da atividade da
Mieloperoxidase em neutrófilos tratados com 50 µg/mL de AK por 1 hora.
(A) Intensidade de fluorescência da Mieloperoxidase. (B) Percentagem de células marcadas.
Para análise estatística, foi utilizado ANOVA seguido do teste Tukey. (*) p < 0,05, (**) p<
0,01, (***) p< 0,001.
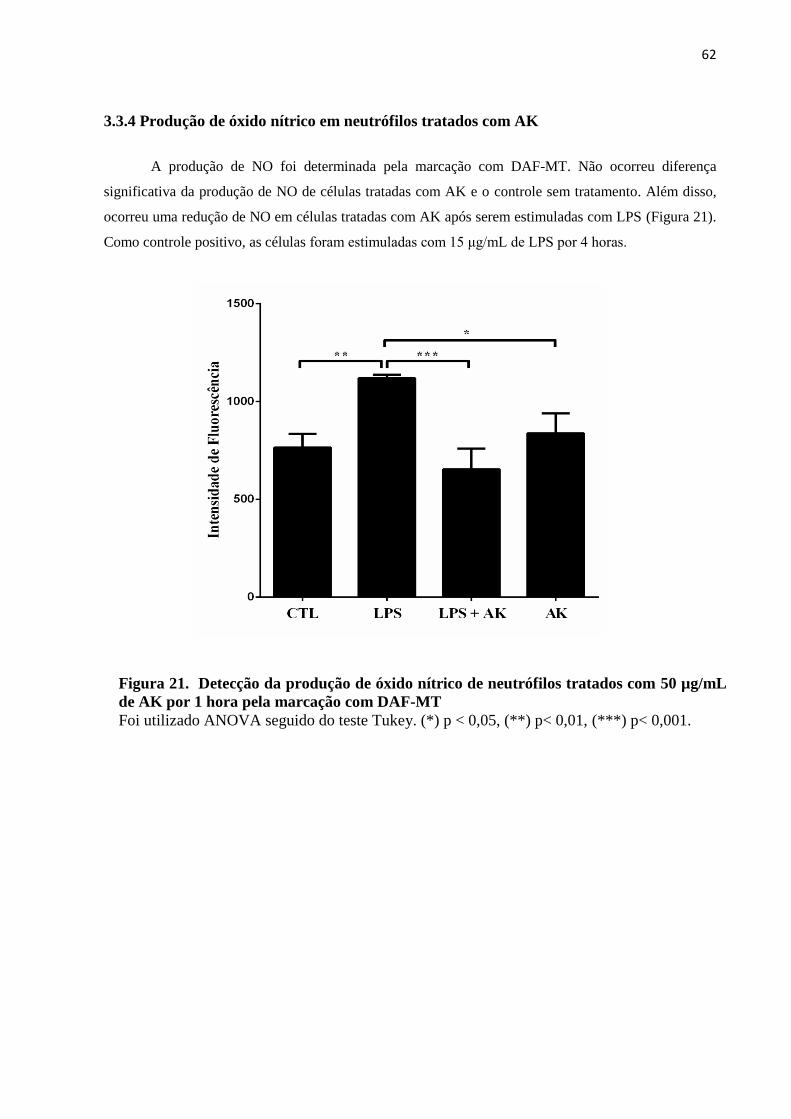
62
3.3.4 Produção de óxido nítrico em neutrófilos tratados com AK
A produção de NO foi determinada pela marcação com DAF-MT. Não ocorreu diferença
significativa da produção de NO de células tratadas com AK e o controle sem tratamento. Além disso,
ocorreu uma redução de NO em células tratadas com AK após serem estimuladas com LPS (Figura 21).
Como controle positivo, as células foram estimuladas com 15 μg/mL de LPS por 4 horas.
Figura 21. Detecção da produção de óxido nítrico de neutrófilos tratados com 50 µg/mL
de AK por 1 hora pela marcação com DAF-MT
Foi utilizado ANOVA seguido do teste Tukey. (*) p < 0,05, (**) p< 0,01, (***) p< 0,001.
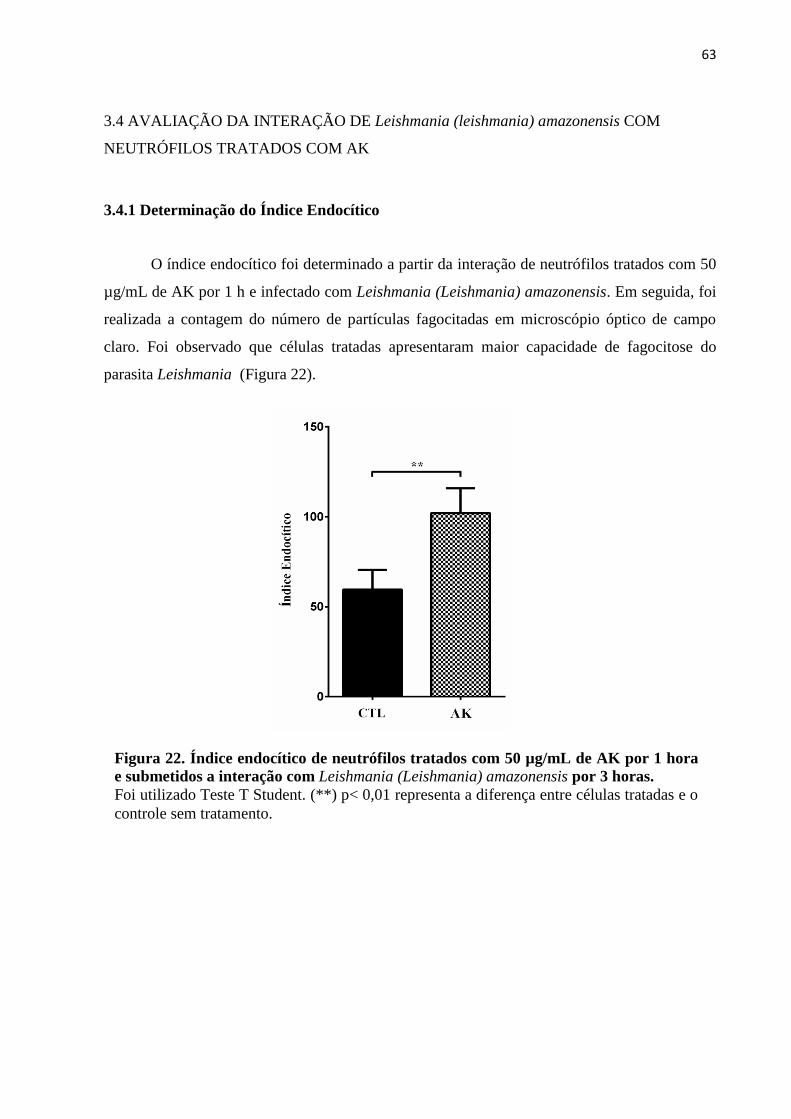
63
3.4 AVALIAÇÃO DA INTERAÇÃO DE Leishmania (leishmania) amazonensis COM
NEUTRÓFILOS TRATADOS COM AK
3.4.1 Determinação do Índice Endocítico
O índice endocítico foi determinado a partir da interação de neutrófilos tratados com 50
µg/mL de AK por 1 h e infectado com Leishmania (Leishmania) amazonensis. Em seguida, foi
realizada a contagem do número de partículas fagocitadas em microscópio óptico de campo
claro. Foi observado que células tratadas apresentaram maior capacidade de fagocitose do
parasita Leishmania (Figura 22).
Figura 22. Índice endocítico de neutrófilos tratados com 50 µg/mL de AK por 1 hora
e submetidos a interação com Leishmania (Leishmania) amazonensis por 3 horas.
Foi utilizado Teste T Student. (**) p< 0,01 representa a diferença entre células tratadas e o
controle sem tratamento.
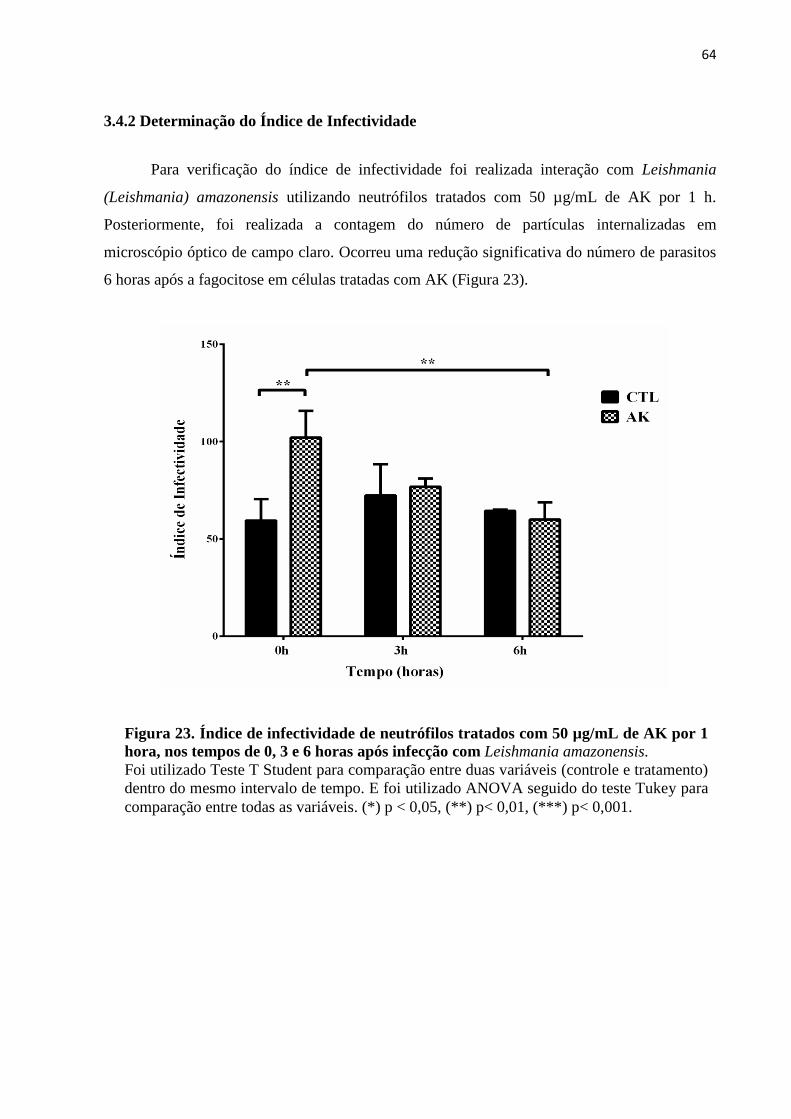
64
3.4.2 Determinação do Índice de Infectividade
Para verificação do índice de infectividade foi realizada interação com Leishmania
(Leishmania) amazonensis utilizando neutrófilos tratados com 50 µg/mL de AK por 1 h.
Posteriormente, foi realizada a contagem do número de partículas internalizadas em
microscópio óptico de campo claro. Ocorreu uma redução significativa do número de parasitos
6 horas após a fagocitose em células tratadas com AK (Figura 23).
Figura 23. Índice de infectividade de neutrófilos tratados com 50 µg/mL de AK por 1
hora, nos tempos de 0, 3 e 6 horas após infecção com Leishmania amazonensis.
Foi utilizado Teste T Student para comparação entre duas variáveis (controle e tratamento)
dentro do mesmo intervalo de tempo. E foi utilizado ANOVA seguido do teste Tukey para
comparação entre todas as variáveis. (*) p < 0,05, (**) p< 0,01, (***) p< 0,001.

65
3.4.3 Produção de Radicais Superóxidos
Através da reação citoquímica utilizando NBT foi determinada a produção de radicais
superóxidos em neutrófilos durante interação com Leishmania (Leishmania) amazonensis.
Neutrófilos tratados com AK produziram maior quantidade de radicais superóxidos quando
comparado ao controle sem tratamento (Figura 24).
Figura 24. Detecção da produção de radicais superóxidos em neutrófilos tratados com
50 µg/mL de AK e em interação com Leishmania (L.) amazonensis através da reação
citoquímica com NBT. Foi utilizado ANOVA seguido do Teste de Tukey (***) p < 0,001.

66
4 DISCUSSÃO
Neutrófilos são os leucócitos mais abundantes nos seres humanos e o primeiro tipo
celular a ser recrutado para locais de infecção. Os neutrófilos participam da defesa primária
contra infecções bacterianas e fúngicas, além de serem capazes de fagocitar patógenos e
destruí-los pela combinação de espécies reativas de oxigênio, nitrogênio e de componentes
citotóxicos (Faurschou & Borregaard, 2003; Klebanolff, 2005; Kobayashi et al., 2005; Quinn et
al., 2006; Nauseef, 2007).
O desenvolvimento de bioprodutos que atuem na ativação de células fagocíticas e
consequentemente no sistema imune inato é de suma importância no combate a invasão do
organismo por microrganismos. Muitas substâncias de origem natural vêm sendo estudadas
com a finalidade de buscar agentes ativadores dessas células (Lopes, et al., 2006; Maity, et al.,
2009; Fuzissaki, 2009). Contudo, existem poucos estudos relacionados à ação de metabólitos
sobre neutrófilos humanos, por isso a necessidade da realização de trabalhos nesta área.
No presente trabalho avaliou-se a ação de AK sobre neutrófilos humanos e durante
infecção com L. amazonensis. O bioproduto AK é um metabólito secundário proveniente de
fungos do gênero Aspergillus e apresenta uma grande variedade de funções, dentre elas, atua
como agente leishmanicida (Rodrigues et al., 2014), ativador de macrófagos peritoneais
(Rodrigues et al., 2011) e antioxidante em neutrófilos humanos (Niwa & Akamatsu, 1991).
Inicialmente foram realizados três testes de viabilidade para verificar se o AK
apresentava algum efeito citotóxico em neutrófilos humanos. O MTT que avalia a atividade das
desidrogenases mitocondriais, o JC-1 que avalia o potencial mitocondrial e o IP que avalia a
integridade da membrana plasmática. Em todos os testes utilizados não houve redução
significativa do número de células viáveis após tratamento quando comparadas ao controle. AK
também não apresentou efeito citotóxico para macrófagos peritoneais provenientes de
camundongos após tratamento por 1 hora (Rodrigues et al., 2011) e monócitos humanos, após
tratamento por 24, 48, 72 e 96 horas (Costa et al., 2014).
Além disso, AK mostrou retardar a apoptose de neutrófilos até 24 horas em culturas in
vitro. Agentes ativadores de neutrófilos como o GM-CSF são capazes de retardar a apoptose de
neutrófilos in vitro (Larbi et al., 2005). In vivo, neutrófilos ativados no sítio inflamatório,
sofrem ação de citocinas como Interleucina-1 e Interleucina-8 que estendem sua vida útil para
poderem contribuir na eliminação de patógenos (Turina et al., 2005). A apoptose tardia

67
promove o aumento da produção de mediadores inflamatórios importantes, como quimiocinas e
citocinas (Mantovani et al., 2011).
Características apoptóticas também não foram observadas na análise morfológica por
MO e por MET. Neutrófilos apresentaram morfologia típica, com núcleo com vários lóbulos
interligados por finas pontes de cromatina, já em neutrófilos apoptóticos essa estrutura é
rapidamente perdida e ocorre condensação da cromatina e formação do núcleo picnótico
(Derouet et al., 2006; Kennedy & Deleo, 2009).
A mudança na morfologia celular, com emissão de filopódios e lamelipódios e o
aumento da área citoplasmática podem ser induzidos por agentes ativadores. Neutrófilos
tratados apresentaram maior espraiamento, aumento do número de pseudópodes, aumento de
volume celular e intensa vacuolização na análise por MO. Essas características são típicas de
neutrófilos ativados (Swain et al., 2002; Sheppard, 2005; Amulic et al., 2012). O aumento
celular aparente foi confirmado pela análise morfométrica, em que neutrófilos tratados com AK
sofreram aumento significativo quando comparados ao controle sem tratamento.
Para uma análise morfológica mais detalhada das possíveis alterações na superfície
celular induzidas por AK foi feita a MEV. Os resultados obtidos confirmaram os resultados
encontrados na MO, em que neutrófilos tratados com AK possuíram maior capacidade de
aderência e emissão de pseudópodes. Características semelhantes foram apresentadas por
neutrófilos estimulados com fMLP, que possui propriedade quimioatraente, levando a formação
de pseudópodes bem definidos (Xu et al., 2003).
A análise por MET revelou maiores detalhes sobre as alterações intracelulares induzidas
por AK. Polimorfonucleares tratados com AK apresentaram maior concentração de grânulos e
vesículas, que também são características de células ativadas. Agentes ativadores induzem ao
processo de vacuolização, concentração de grânulos, fusão entre grânulos e membrana
plasmática (secreção) e fusão intracelular entre grânulos e vesículas (Kobayashi et al., 1998;
Karlsson et al., 2000).
Essas alterações morfológicas são acompanhadas pela reorganização do citoesqueleto
de actina. O citoesqueleto possui várias funções, dentre elas, é responsável pela migração,
adesão, espraiamento e fagocitose (Stramer et al., 2010). AK promoveu a polimerização e a
redistribuição dos filamentos de actina em células tratadas, semelhante à atuação de alguns
ativadores já conhecidos, como o PMA, que induz reorganização da actina (Downey et al.,
1992). A polimerização dos filamentos de actina é uma das primeiras e mais sensíveis respostas
funcionais dos neutrófilos. Esses eventos do citoesqueleto próximos à membrana da célula
permitem adesão e emissão de projeções citoplasmáticas que estão relacionados à fagocitose,

68
trânsito de grânulos, regulação e montagem do complexo NADPH-oxidase e geração de
superóxido (Bengtsonn et al., 2006; Groves et al., 2008).
O radical ânion superóxido é o primeiro ERO formado durante o “burst” oxidativo, que
pode ser gerado no espaço extracelular ou fagossomal, por meio da ativação do complexo
NADPH-oxidase (Sheppard et al., 2005; El-Benna et al., 2008). Espécies reativas de oxigênio
apresentam importante papel na atividade microbicida dessas células, tornando-se um fator
fundamental na defesa do organismo (Siddiqui et al., 1995; Granfeldt, 2002).
Através do ensaio de NBT, foi observado que neutrófilos tratados com 50 μg/mL de AK
apresentaram acúmulo significativo de radicais superóxidos em relação ao controle sem
tratamento. A marcação com CellROX, confirmou o resultado do NBT e mostrou que AK
induz não só a produção de radicais superóxidos, como também de outras ERO.
Muitos produtos de origem natural são antioxidantes, inibindo a produção de ERO, e
protegendo os tecidos de lesões (Liz et al., 2012; Suzuki et al., 2012). Contudo, um estudo
realizado por Niwa e Akamatsu (1991) mostrou que em neutrófilos humanos tratados com 0.05,
0.5, 5 e 20 µg/mL de AK a produção de ERO foi restrita ao espaço fagossomal, impedindo a
lesão tecidual.
A NADPH-oxidase é responsável pela produção de radicais superóxidos, ânion
precursor da produção de outras ERO mais potentes. Por possuir um papel importante nessa
via, sua ativação e sua inibição vêm sendo estudada inclusive como estratégia terapêutica
(Jaquet et al., 2009). A adição de apocinina, inibidor do complexo NADPH-oxidase, em
culturas de neutrófilos inibiu completamente a produção de radicais superóxidos induzidos por
AK. Esse resultado sugere que o acúmulo de ERO necessita dessa via em funcionamento.
Dentre as ERO formadas, está o peróxido de hidrogênio, que é convertido pela enzima
MPO em ácido hipocloroso, que é considerado o composto mais tóxico para patógenos. MPO
também está envolvida em inúmeros processos inflamatórios promovendo a regulação de
proteínas e mediadores inflamatórios secretados por neutrófilos (Winterbourn et al., 2000; Van
den Veen et al., 2009).
AK induziu o aumento da atividade dessa enzima, importante integrante de uma
resposta microbicida efetiva. A atividade dessa enzima é fundamental no combate a agentes
infecciosos. Camundongos knockout para MPO mostraram-se mais susceptíveis a infecções
(Aratani et al, 2000) e foi observado que pacientes completamente deficientes para MPO
apresentaram complicações infecciosas ou inflamatórias graves (Kutter, 1998).
A ativação de neutrófilos desencadeia não apenas a produção de ERO, mas também de
óxido nítrico. NO, por sua vez, pode reagir com radicais superóxidos e formar o peroxinitrito

69
(Bentz et al., 2002; Ren et al, 2007). A presença excessiva de espécies reativas no meio
extracelular estabelece um estresse oxidativo, que é o desequilíbrio celular entre a quantidade
de oxidantes e antioxidantes nesse meio, característico em inúmeras doenças (Valko, 2007;
Filippin et al., 2008).
A avaliação de produção de NO foi realizada através da marcação com DAF-MT.
Neutrófilos tratados com AK não produziram NO e neutrófilos estimulados com LPS
reduziram a produção de NO após tratamento com o metabólito. Resultados semelhantes foram
encontrados com macrófagos tratados com 50 μg/mL de AK, onde não ocorreu produção de
NO (Rodrigues et al., 2011). Neste sentido, sugere-se que AK possa estar interferindo na
produção de NO. Maiores estudos serão necessários para evidenciar se AK está inibindo a
enzima iNOS ou através do sequestro de moléculas de NO formadas.
A geração de NO e de ERO está relacionada com a capacidade fagocítica de neutrófilos,
como também, com a sua capacidade de matar patógenos (Charmoy et al, 2010; Horta et al.,
2012). Uma resposta microbicida eficiente dessas células pode auxiliar no combate ao parasito.
Ribeiro-Gomes e colaboradores (2004) mostraram em um modelo in vivo que a depleção de
neutrófilos ocasionou o aumento da carga parasitária em camundongos C57BL/6 infectados
com L. major.
Diversos estudos apontam os neutrófilos como as primeiras células a fagocitar L.
major ou L. donovani (Peters et al., 2008; Thalhofer et al., 2011; Ribeiro-Gomes et al., 2012).
Neste trabalho, foi possível observar que neutrófilos tratados com AK durante interação com
Leishmania amazonensis aumentaram sua capacidade fagocítica. Em um estudo realizado por
Niwa & Akamatsu (1991) também foi observado que neutrófilos tratados com 0,05, 0,5, 5 e 20
μg/mL de AK possuíram maior capacidade de fagocitose em um ensaio com gotículas de óleo
de parafina. Esses resultados permitem inferir que AK pode induzir o processo de fagocitose
em neutrófilos humanos.
Além do aumento da fagocitose, foi observado que em neutrófilos tratados com AK
ocorreu uma redução do número de parasitos internalizados. Seis horas após a interação, havia
uma diminuição significativa em relação à hora zero. Um estudo in vitro recente com
neutrófilos e Leishmania amazonensis mostrou que essas células são capazes de fagocitar as
formas promastigotas e seu arsenal microbicida é eficiente na eliminação do mesmo (Carlsen et
al., 2013).
Neutrófilos de camundongos BALB/c foram capazes de induzir a morte do parasito.
Esse mecanismo auxiliou no estabelecimento de uma resposta protetora na infecção por L.
donovani ou L. infantum através da produção de NO e de ERO (Rousseau et al., 2001;

70
McFarlane et al., 2008). A produção de ERO também foi avaliada em um estudo com co-
cultura de neutrófilos murinos vivos e macrófagos infectados com L. braziliensis. Onde ocorreu
a morte do parasita concomitante a produção de radicais superóxidos (Novais et al., 2009).
Neste estudo também foi avaliada a produção de radicais superóxidos por neutrófilos
tratados com AK durante interação com Leishmania amazonensis. Como resultado, células
tratadas produziram mais radicais superóxidos quando comparado ao controle. Resultados
semelhantes foram encontrados em macrófagos tratados com 50 μg/mL de AK por 1 hora.
Através do ensaio NBT foi observado que macrófagos infectados com L. amazonensis tiveram
uma maior produção de radicais superóxidos quando comparados ao controle sem tratamento
(Rodrigues et al., 2014). Esses resultados sugerem que AK pode atuar no combate a L.
amazonensis mediante via de geração de ERO, indicado pela grande produção de radicais
superóxidos durante a interação.
Assim, pode-se inferir que o AK induziu a ativação de neutrófilos humanos através de
alterações morfológicas, remodelamento do citoesqueleto de actina, produção de ERO,
aumento da atividade da enzima mieloperoxidase, sem causar efeitos citotóxicos. Além disso,
durante interação com o parasito Leishmania amazonensis, AK atuou no aumento de
fagocitose, na diminuição de parasitos internalizados e na produção de radicais superóxidos.
Todavia, mais estudos devem ser realizados, inclusive em modelos in vivo, para evidenciar
como AK atua em neutrófilos e como pode auxiliar na ativação dessas células e no combate do
parasito.

71
5 CONCLUSÃO
Os resultados apresentados e discutidos permitem concluir que o metabólito AK:
1. Não possui efeito citotóxico para neutrófilos humanos. Além disso, sugere-se que o AK
possa atuar no retardo da apoptose dessas células.
2. Promoveu alterações morfológicas em neutrófilos, uma vez que houve maior espraiamento,
aumento da área celular, intensa vacuolização, emissão de pseudópodes e reorganização dos
filamentos de actina.
3. Induziu a produção de espécies reativas de oxigênio pela ativação do complexo NADPH-
oxidase.
4. Não induziu a produção de óxido nítrico em neutrófilos humanos e sugere-se que AK possa
interferir na produção de óxido nítrico.
5. Promoveu aumento da atividade da mieloperoxidase em neutrófilos.
6. Induziu aumento da capacidade fagocítica em neutrófilos.
7. Promoveu diminuição de Leishmania amazonensis internalizadas pelos neutrófilos seis horas
após a infecção.
8. Aumentou a produção de radicais superóxidos em neutrófilos durante interação com
Leishmania amazonensis.

72
6 REFERÊNCIAS BIBLIOGRÁFICAS
AFONSO, L., BORGES, V. M., CRUZ, H., RIBEIRO-GOMES, F. L., DOSREIS, G. A, DUTRA,
A. N., CLARÊNCIO, J. Interactions with apoptotic but not with necrotic neutrophils increase
parasite burden in human macrophages infected with Leishmania amazonensis. Journal of
leukocyte biology, 2008. 84(2), 389–96.
ALCOLEA, P.J.; ALONSO, A.; GÓMEZ, M. J.; SÁNCHEZ-GOROSTIAGA, A.; MORENO-PAZ,
M.; SONZÁLEZ-PASTOR, E.; TORANO, A.; LARRAGA, V. Temperature increase prevails
over acidification in gene expression modulation of amastigote differentiation in Leishmania
infantum. BMC Genomics, v. 14, p.11:31, 2010.
ALMEIDA, R.; D’OLIVEIRA, A.; MACHADO, P; BACELLAR, O.; KO, A. I.; DE JESUS, A. R.;
MOBASHERY, N.; BRITO SANTOS, J.; CARVALHO, E. M. Randomized, double-blind
study of stibogluconate plus human granulocyte macrophage colony-stimulating fator versus
stibogluconate alone in the treatment of cutaneous Leishmaniasis. Journal Infect Disease,
180(5):1735-7, 1999.
ALVAR, J.; VÉLEZ, I.D.; BERN, C.; HERRERO, M.; DESJEUX, P.; CANO, J.; JANNIN, J.;
DEN BOER, M. Leishmaniasis worldwide and global estimates of its incidence. PLoS One
7:e35671, 2012.
AMULIC, B.; CAZALET, C.; HAYES, G. L.; METZLER, K. D.; ZYCHLINSKY, A. Neutrophil
function: From mechanisms to disease. Annuals review of Immunology. 30:459-89, 2012.
ARATANI ,Y.; KURA, F.; WATANABE, H.; AKAGAWA, H.; TAKANO; Y.; SUZUKI, K.;
MAEDA, N.; KOYAMA, H. Differential host susceptibility to pulmonary infections with
bacteria and fungi in mice deficient in myeloperoxidase. J Infect Dis;182(4):1276-1279, 2000.
ASHUTOSH; SUNDAR, S.; GOYAL, N. Molecular Mechanisms of Antimony Resistance In
Leishmania. J Med Microbiol, v. 56 (PT 2), p. 143-1453, 2007.
BABIOR, B. M. NADPH oxidase: an update. Blood, New York, v. 93, n. 5, p. 1464-1475, 1999.
BABIOR, B. M. Phagocytes and oxidative stress. American Journal of Medice, Marion, v. 109,
n.1, p. 33-44, 2000.
BAÑULS, A.L.; HIDE, M.; PRUGNOLLE, F. Leishmania and the leishmaniases: a parasite genetic
update and advances in taxonomy, epidemiology and pathogenicity in humans. Adv Parasitol,
v. 64, p. 1-109, 2007.
BATES, P. A., Complete developmental cycle of Leishmania Mexicana in axenic culture.
Parasitology 108; 1-9, 1994.

73
BAILEY, M.S.; LOCKWOOD, D.N.J. Cutaneous leishmaniasis. Clinics in Dermatology, v. 25 p.
203 – 221, 2007.
BENGTSSON, T.; ORSELIUS, K.; WEITTERO, J. Role of the cytoskeleton during respiratory
burs in chemoattractant-stimulated neutrophils. Cell Biology International, London, v. 30, n.
2, p. 154-163, 2006.
BENTLEY, R. From miso, sake and shoyuto cosmetics: a century of science for kojic acid. Nat
Prod Rep23:1046-62, 2006.
BENTZ, B. G., CHANDRA, R., HAINES, G. K., ROBINSON, A. M., SHAP, P., RADOSEVICH,
J. A. Nitric oxide and apoptosis during human head and neck squamous cell carcinoma
development. Am J Otolaryngol, 23 (1), 4-11, 2002.
BLUMENTHAL, C. Z. Production of toxic metabolites in Aspergillusniger, Aspergillusoryzae, and
Trichodermareesei: justification of mycotoxin testing in food grade enzyme preparations
derived from the three fungi. RegulToxicolPharmacol 39:214-28, 2004.
BORREGAARD, N.; COWLAND, J. B. Granules of the Human Neutrophilic Polymorphonuclear
Leukocyte. Blood, New York, v. 89, n. 10, p. 3503-3521, 1997.
BORREGAARD, N. SORENSEN, O. E.; THEILGAARD-MONCH, K. Neutrophil granules: a
library of innate immunity proteins. Trends in Immunology, vol. 28, nº 8, 340-345, 2007.
BORREGAARD, N. Neutrophils, from Marrow to Microbes. Immunity Review, n. 33, p. 657-670,
2010.
BRASIL. MINISTERIO DA SAUDE, DEPARTAMENTO DE VIGILANCIA DA SAUDE.
Sistema de Informação de Agravos de Notificação. Serviço de Vigilância em Saúde. 2010
BRASIL. Ministério da Saúde. DATASUS/Indicadores e dados básicos (IDB). Indicadores de
Morbidade - Leishmaniose Tegumentar Americana. Serviço de Vigilância em Saúde, 2013.
BRIGAGÃO, M. R. P. L.; BARROSO, A. S.; COLEPICOLO, P. Flutuation of reactive oxygen
species released by inflammatory cells. The Redox State and Circadian Rhythms. P. 177-
191, 2000.
BRINKMANN, V.; REICHARD, U.; GOOSMANN, C.; FAULER, B.; UHLEMANN, Y.; WEISS
D. S.; WEINRAUCH, Y.; ZYCHLINSKY, A. Neutrophil extracellular traps kill bacteria.
Science. 2004. 303, 1532-5.
BURDOCK, G.A. SONI, M.G. CARABIN.I.G. Evaluation of Health Aspects of Kojic Acid in
Food.Regulatory Toxicology and Pharmacology, v.33, p.80–101, 2001.
BURG, N. D. & PILLINGER, M. H. The neutrophil: function and regulation in innate and humoral
immunity. Clinical Immunology, Orlando, v. 99, n.1,p.7-17, 2001.

74
CABANES, J. CHAZARRA, S. CARMONA, F.G. Kojic acid, a cosmetic skin whitening agent, is a
slow-binding inhibitor of catecholase activity of tyrosinase. J Pharm Pharmacol, v.46, n.12,
p.982-985, 1994.
CAMPANELLI, A.P.; B, C. I.; OLIVEIRA, V. S. B.; SILVA, C.; ROSELINO, A. M.; COSTA, J.;
SALDANHA, A. C.; FREITAS, L. A. R.; OLIVEIRA, C. I.; BARRAL NETO, M.; SILVA, J.
S. BARRAL, A. M. P. Chemokines and chemokine receptors coordinate the inflammatory
immune response in human cutaneous leishmaniasis. Human immunology, v. 71, n. 12, p.
1220-7, 2010.
CARLSEN, E. D.; HAY, C.; HENARD, C. A.; POPOV, V.; GARG, N. J.; B, SOONGB, L.
Leishmania amazonensis Amastigotes Trigger Neutrophil Activation but Resist Neutrophil
Microbicidal Mechanisms. Infection and Immunity, 2013.
CARMO, E. V. S.; KATZ, S.; BARBIERI, C. L. Neutrophils reduce the parasite burden in
Leishmania (Leishmania) amazonensis-infected macrophages. PLoS One, 2010. 5:e13815.
CHAGAS, A.C.; PESSOA, F. A. C.; MEDEIROS, J. F.; PY-DANIEL, V.; MESQUITA, E C;
BALESTRASSI, D. A. Leishmaniose Tegumentar Americana (LTA) em uma vila de
exploração de minérios - Pitinga, municipio de Presidente Figueiredo, Amazonas, Brasil.
Brazilian journal of epidemiology, v. 9, n. 2, p. 186-192, 2006.
CHANG K P. Leishmanicidal mechanisms of human polymorphonuclear phagocytes. Am. J. Trop.
Med. Hyg. 1981.30:322–333.
CHAKRAVARTY, J.; SUNDAR, S. Drug resistance in leishmaniasis. J Glob Infect Dis, v. 2, n. 2,
p.167-176, 2010.
CHARMOY, M., BRUNNER-AGTEN, S., AEBISCHER, D., AUDERSET, F., LAUNOIS, P.,
MILON, G., PROUDFOOT, A. E. I. Neutrophil-derived CCL3 is essential for the rapid
recruitment of dendritic cells to the site of Leishmania major inoculation in resistant mice.
PLoS pathogens, 2010 6(2), 1-12.
CHEE, H.Y. &LEE, E.H. Fungistatic Activity of Kojic Acid Against Human Pathogenic Fungi
and Inhibition of Melanin-production in Cryptococcus neoformans.Mycobiology 31(4): 248-
250, 2003.
CHEN L, ZHANG ZH, WATANABE T, YAMASHITA T, KOBAYAKAWA T, KANEKO A,
FUJIWARA H, SENDO F. The involvement of neutrophils in the resistance to Leishmania
major infection in susceptible but not in resistant mice. Parasitol. Int. 54:109–118, 2005.
CHOI, H. S.; KIM, J. W.; CHA, Y.; KIM, C. A quantitative nitroblue tetrazolium assay for
determining intracelular superoxide anion production in phagocytic cells. Journal of
Immunoassay and Immunochemistry, v 27, p.31-44, 2006.

75
CHUSIRI, Y.; WONGPOOMCHAI, R.; KAKEHASHI, A.;WEI, M.; WANIBUCHI, H.;
VINITKETKUMNUAN, U.; FUKUSHIMA, S. Non-genotoxic mode of action and possible
threshold for hepatocarcinogenicity of Kojic acid in F344 rats. FoodandChemicalToxicology
49: 471–476, 2011.
COSTA, J. P. Ação do Metabólito Secundário 5-hidroxi-2-hidroximetil gama-pirona isolado de
fungos do gênero Aspergillussobre monócitos humanos in vitro. Dissertação de Mestrado.
Belém. Universidade Federal do Pará, 2012. 78p.
COSTA FILHO, A.V.; LUCAS, I.C.; SAMPAIO, R.N. Comparative study between oral miltefosine
and parenteral N-metil glucamine antimoniate for the treatment of experimental leishmaniasis
caused Leishmania (Leishmania) amazonensis. Rev Soc Bras Med Trop, v. 41,
n. 4, p. 424-427, 2008.
CROFT, S. L.; SUNDAR, S.; FAIRLAMB, A. H. Drug Resistance in Leishmaniasis. Clin
Microbiol Rev, v. 19, n. 1, p. 111-126, 2006.
CUNNINGHAM, A. C. Parasitic Adaptive Mechanisms in Infection by Leishmania. Exp Mol
Pathol, v. 72, p. 132-141. 2002.
DABIRI S, HAYES MM, MEYMANDI SS, BASIRI M, SOLEIMANI F, MOUSAVI M R.
Cytologic features of “dry-type” cutaneous leishmaniasis. Diagn. Cytopathol, 1998. 19:182–
185.
DABOUL, M. W. Role of neutrophils in cutaneous leishmaniasis. East Mediterr. Health J. 2010.
16:1055–1058.
DALE, D. C.; BOXER, L.; LILES, W. C. The phagocytes: Neutrophils and monocytes. Blood,
New York, v. 112, n. 4, 2008.
DALLEGRI, F.; OTTONELO, L. Tissue injury in neutrophilic inflammation. Inflamation
Research, Basel, v.46, n.10, p.382-391, 1997.
DAVID, C.V.; CRAFT, N. Cutaneous and mucocutaneous leishmaniasis. Dermatol Ther, v. 22(6),
p. 491-502, 2009.
DEROUET, M.; THOMAS, L.; MOULDING, D. A.; AKGUL, C.; CROSS, A.; MOOTS, R J.;
EDWARDS, S W. Sodium salicylate promotes neutrophil apoptosis by stimulating caspase-
dependent turnover of Mcl-1. J Immunol, v. 176, n. 2, Jan 15, p. 957-65. 2006
DESJEUX, P. Leishmaniasis. Nature Reviews Microbiology 2:692-693, 2004.
DOWNEY, G. P.; CHAN, C. K.; LEA, P.; TAKAI, A.; GRINSTEIN, S. Phorbol ester-induced
actin assembly in neutrophils: role of protein kinase C. J Cell Biol, 116(3): p. 695-706, 1992.
ELLIS, M.; BERNSEN, R.; ALI-ZADEH, H.; KRISTENSEN, J.; HEDSTROM, U.; POUGHIAS,
L. A safety and feasibility study comparing an intermittent high dose with a daily standard dose

76
of liposomal amphotericin B for persistent neutrophenic fever. J Med Microbiol, 2009. 58 (Pt
11): 474-85.
EL-BENNA, J.; DANG, P. M. C.; GOUGEROUT-POCIDALO, M. A. Primins of the neutrophil
NADPH oxidase activation: role of p4phox phosphorylation and NOX2 mobilization to the
plasma membrande. Semin Immunopathol, Berlim, v. 30, p. 279-289, 2008
EL-SAYED, M. & ANWAR, A.E. Intralesional sodium stibogluconate alone or its combination
with either intramuscular sodium stibogluconate or oral ketoconazole in the treatment of
localized cutaneous leishmaniasis: a comparative study. J Eur Acad Dermatol Venereol, v.
24, n. 3, p. 335-340, 2010.
EMAMI, S.; HOSSEINIMEHR, S. J.; TAGHDISI, S. M.; AKHLAGHPOOR, S. Kojic acid and its
manganese and zinc complexes as potential radio protective agents. Bioorg Med ChemLett
17:45-48, 2007.
FAURSCHOU, M. & BORREGAARD. N. Neutrophil granules and secretory vesicles in
inflammation. Microbes Infect, v. 5, n. 14, p. 1317-17. 2003.
FERREIRA, N. R.; SARQUIS, M.I.M; ALVES, C. N.; SANTOS, A. S. Biotransformation of
sucrose into 5-hydroxy-2-hydroxymethil-γ-pirone by Aspergillus flavus. Annals of the
Brazilian Academy of Sciences. 2010; 82(3):569-76.
FIALKOW, L.; WANG, Y.; DOWNEY, G.P. Reactive oxygen and nitrogen species assignaling
molecules regulating neutrophil function. Free Radic. Biol. Med. 42,153–164, 2007.
FILIPPIN, L. I.; VERCELINO, R.; MARRONI, N. P.; XAVIER, R. M. Redox signaling and the
inflammatory response in rheumatoid arthritis. Clinical and Experimental Immunology,
Oxford, v.152, p. 415-422, 2008.
FINKEL, T. Signal transduction by reactive oxygen species. J. Cell Biol. 194, 7–15, 2011.
FITZPATRICK, J.M; HIRAI, Y.; HIRAI, H.; HOFFMANN, K. F. Schistosome egg production is
dependent upon the activities of two developmentally regulated tyrosinases. FASEB J.
21(3):823-35, 2007.
FOLLADOR, I.; ARAÚJO C.; BACCELAR, O.; ARAUJO C. B.; CARVALHO L. P.; ALMEIDA,
R. P.; CARVALHO, E. M. Epidemiologic and immunologic findings for the subclinical form
of Leishmania braziliensis infection. Clin Infect Dis 34:E54-8, 2002.
FOTAKIS, G. & TIMBRELL, J. A. in vitro cytotoxicity assays: comparison of LDH, vermelho
neutro, MTT and protein assay in hepatoma cell lines following exposure to cadmium chloride.
Toxicol Letters, 160:171–177, 2006.
FRÉZARD, F.; DEMICHELI, C. New delivery strategies for the old pentavalent antimonial drugs.
Expert Opin Drug Deliv, v. 7 (12), p.1343-1358, 2010.

77
FUZISSAKI, C. N. Estudo da modulação de funções efetoras de neutrófilos humanos por
erivados cumarínicos: avaliação do efeito biológico sobre a produção de espécies reativas
de oxigênio e a degranulação. Riobeirão Preto. Universidade de São Paulo. 2009
GOMES, A.J.; LUNARD, C. N.; GONZALEZ, S.; TEDESCO, A. C. The antioxidant action of
Polypodium leucotomos extract and kojic acid: reactions with reactive oxygen species. Braz J
Med Biol Res, v.34, p.1487-1494, 2001.
GOTO, H.; LINDOSO, J.A. Current diagnosis and treatment of cutaneous and mucocutaneous
leishmaniasis. Expert Rev Anti Infect Ther, v. 8 (4), p. 419-33, 2010.
GRANFELDT, D.; SAMUELSSON, M.; KARLSSON, A. Capacitative Ca2+ influx and activation
of the neutrophil respiratory burst. Different regulation of plasma membrane- and granule-
localized NADPH-oxidase. J Leukoc Biol. 2002 Apr; 71(4):611-7.
GROVES, E.; DART, A. E.; COVARELLI, V.; CARON, E. Molecular mechanisms of phagocytic
in mammalian cells. Cell Mol Life Sci, v. 65, p. 1957-1976, 2008.
GUIMARÃES-COSTA, A. B.; NASCIMENTO, M. T. C.; FROMENT, G. S.; SOARES, R. P. P.;
MORGADO, F. N.; CONCEIÇÃO-SILVA, F.; SARAIVA, E. M.
Leishmaniaamazonensispromastigotesinduceand are killedbyneutrophilextracellulartraps.
Proceedings of the National Academy of Sciences of the United States of America,
106(16):6748-53, 2009
GUIMARÃES-COSTA, A. B.; NASCIMENTO, M. T. C.; WRDINI, A. B.; PINTO-DA-SILVA,
L.; SARAIVA, E. M. ETosos: A MicrobicidalMechanismbeyoundCellDeath. Journal of
Parasitology Research. 2011. V. 2012, article ID 929743.
GUPTA, A.K.; GOVER, M.D; NOURI, K.; TAYLOR, S.He treatment of melasma: a review of
clinical trials. J Am AcadDermatol 55 (6) :1048-65, 2006.
HIGA, Y.; KAWABE, M.; NABAE, K.; TODA, Y.; KITAMOTO, S., HARA,; TANAKA, N.;
KARIYA, K.; TAKAHASHI, M. Kojic Acid – Absence of tumor-initianing activity in rat liver,
and of carcinogenic an photo-genotoxicpotencial in mouse skin. J ToxicolSci 32(2):143-159,
2007.
HORTA, M. F., MENDES, B. P.; ERIC, H. R.; NORONHA, F. S. M.; MACEDO, J. P.;
OLIVEIRA, L. S.; DUARTE, M. M.; VIEIRA, L. Q. Reactive Oxygen Species and Nitric
Oxide in Cutaneous Leishmaniasis. Journal of Parasitology Research, 2012.
HYO-YANG, A., KATHRYN, E., SMITH, F. Two-photon fluorescence microscopy imaging of
cellular oxidative stress using profluorescent nitroxides. Journal of the American Chemical
Society 134 (10): 4721-4730, 2012.

78
JAQUET, V; SCAPOZZA, L.; ROBERT, A.; KARL-HEINZ KRAUSE, C.; LAMBETH, J. D.
Small-Molecule NOX Inhibitors: ROS-Generating NADPH Oxidases as Therapeutic Targets.
Antioxidants & Redox Signaling, v. 11, n. 10, 2009.
KARLSSON, A.; NIXON, J. B.; MCPHAIL, L. C. Phorbol myristate acetate induces neutrophil
NADPH-oxidase activity by two separate signal transduction pathways: dependent or
independent of phosphatidylinositol 3-kinase. J. Leukoc. Biol. 67, 396–404, 2000.
KAYE, P.; SCOTT, P. Leishmaniasis: complexity at the host-pathogen interface. Nature reviews.
Microbiology, v. 9, n. 8, p. 604-15, 2011.
KENNEDY, A. D.& DELEO, F. R. Neutrophil apoptosis and the resolution of infection. Immunol
Res, 43:25-61, 2009.
KLEBANOLFF, V. S. J. Myeloperoxidase: friend and foe. Journal of Leucocyte Biology,
2005;77:598–625.
KOBAYASHI, T.; ROBINSON, J. M.; SEGUCHI, H. Identification of intracellular sites of
superoxide production in stimulated neutrophils. Journal of Cell Science, 111, p. 81-91, 1998.
KOBAYASHI, S. D.; VOYICH, J. M.; DELEO, F. R. Regulation of the neutrophil-medisted
inflammatory response to infection. Microbes and Infection, Paris, v.5, n.14, p. 1337-1344,
2003.
KOBAYASHI, S. D.; JOVANKA, M. V.; BURLAK, C.; DELEO, F. R. Neutrophils in the innate
imune response. Arch ImmunolTherExp, 2005, 53, 505-517.
KOJIMA, H.; SAKURAI, K.; KIKUCHI, K.; KAWAHARA, S.; KIRINO, Y.; NAGOSHI, H.;
HIRATA, Y.; NAGANO, T. Development of a fluorescent indicator for nitric oxide based on
the fluorescein chromophore. Chemical & Pharmaceutical Bulletin, v. 46, n. 2, p. 373-375,
1998.
KOOPMAN G, REUTELINGSPERGER CP, KUIJTEN GA, KEEHNEN RM, PALS ST, VAN
OERS MH. Annexin V for flow cytometric detection of phosphatidylserine expression on B
cells undergoing apoptosis. Blood. Sep 1.84(5):1415-20, 1994.
KUIJPERS, T. W. Neutrophils. Encyclopedia of life Sciences/Nature Publishing Group. 2001.
KUTTER, D. Prevalence of myeloperoxidase deficiency: population studies using Bayer-Technicon
automated hematology. J Mol Med 1998;76(10):669-675.
LACY, Paige. The Role of Rho GTPases And SNAREs In mediator release from granulocytes.
Pharmacology & therapeutics,v.107,n.3,p.358 - 376,set.2005.
LAINSON, R. The Neotropical Leishmania species: a brief historical review of their discovery,
ecology and taxonomy. Revista pan-Amazônica de saúde, v. 1, n. 2, p. 13-32, 2010.

79
LARBI, A; DOUZIECH, N; FORTIN, C; LINTEAU, A.; DUPUIS, G.; FULOP, T. The role of the
MAPK pathway alterations in GM-CSF modulated human neutrophil apoptosis with aging.
Immunity & Ageing, 2:6, p 1-16, 2005.
LAUFS, H.; MULLER, K.; FLEISCHER, J.; REILING, N.; JAHNKE, N.; JENSENIUS, J. C.
Intracellular survival of Leishmania major in neutrophil granulocytes after uptake in absence of
heat-labile serum factors. Infect Immun, Feb; 70(2):826-35, 2002.
LIM, J.T. E. Treatment of melasma using kojic acid in a gel containing hydroquinone and glycolic
acid. DermatolSurg 25: 282-84, 1999.
LIN, C.; WU, H.; HUANG, Y. Combining high-performance liquid chromatography with on-line
microdialysis sampling for the simultaneous determination of ascorbylglucoside, kojic acid,
and niacinamide in bleaching cosmetics. Anal Chim Acta 581:102-07, 2007.
LIZ, R., HORST, H., PIZZOLATTI, M.G., FRODE, T.S., GIRARD, D. Activation of human
neutrophils by the anti-inflammatory mediator Esenbeckia leiocarpa leads to atypical
apoptosis. Mediators Inflamm 20: 1-10, 2012.
LOPES, L.; GODOY, L.M.F.; DE OLIVEIRA, C.C.; GABARDO, J.; SCHADECK, R.J.G.;
BUCHI, D.F. Phagocytosis, endosomal/lysosomal system and other cellular aspects of
macrophage activation by Canova medication. Micron; 37:277-87, 2006.
LORENZI, T. F. Manual de Hematologia: propedêutica e clínica. 3. Ed. Rio de Janeiro: Medsi,
2005.
LUSTER A. D., ALON R., VON ANDRIAN U.H. Immune cell migration in inflammation: present
and future therapeutic targets. Nature immunology, New York, v.6, n 12, p. 1182-1190, 2005
MAIANSKI, N. A.; MAIANSKI, A. N.; KUIJPERS, T. W.; ROOS, D. Apoptosis of neutrophils.
Acta Haematol 2004;111:56–66.
MAIANSKI, N. A.; GEISSLER, J.; SRINIVASULA S. M.; ALNEMRI, E. S.; ROOS, D.;
KUJIPERS, T. W. Functional characterization of mitochondria in neutrophils: a role restricted
to apoptosis. Cell Death and Differentiation 2004 11, 143-153.
MAITY, P.C.; BHATTACHARIEE, S.; MAIUMDAR, S.; SIL, A.K. Potentiation by cigarette
smoke of macrophage function against Leishmania donovani infection. Inflamm Res 2009;
58:22-9.
MALAFAIA, G.; REZENDE, S. A. O papel dúbio dos neutrófilos na infecção por parasitos do
gênero Leishmania: uma breve discussão. Rev. Saúde e Biol., v. 4, n. 1, p. 38-44, 2009.

80
MANTOVANI, A.; CASSATELLA, M. A; COSTANTINI, C; JAILLON, S Neutrophils in the
activation and regulation of innate and adaptative immunity. Nature Reviews Immunology 11,
519-531, 2011.
MAYER-SCHOLL, A.; AVERHOFF, P.; ZYCHLINSKY, A. How do neutrophils and pathogens
interact? Current Opinion in Microbiology, London, v. 7, n. 1, p. 62-66, 2004.
MCFARLANE, E., PEREZ, C., CHARMOY, M., ALLENBACH, C., CARTER, K. C.,
ALEXANDER, J., & TACCHINI-COTTIER, F. Neutrophils contribute to development of a
protective immune response during onset of infection with Leishmania donovani. Infection
and immunity, 76(2), 532–41, 2008.
MI HA,Y.; CHUNG,S. W.; SONG, S.; LEE,H.;SUH,H.; CHUNG, H. Y. 4-(6-Hydroxy-2-
naphthyl)-1,3-bezendiol: A Potent, New Tyrosinase Inhibitor. Biol Pharm Bull30(9):1711-15,
2007.
MITROPOULOS, P.; KONIDAS, P.; DURKIN-KONIDAS, M.; New World cutaneous
leishmaniasis: updated review of current and future diagnosis and treatment. J Am Acad
Dermatol. 2010. 63(2):309-22.
MOHAMAD, R.; MOHAMED, M.S.; SUHAILI, N.; SALLEH, M.M.; ARIFF, A.B. Kojic acid:
Applications and development of fermentation process for production. Biotechnol. Mol. Biol.
Rev. 5(2): 24-37, 2010.
MOHAMMADPOUR, M.; BEHJATI, M.; SADEGUI, A.; FASSIHI, A. Wound healing by topical
application of antioxidant iron chelators: kojic acid and deferiprone. International Wound
Journal 2012.
MOTO, M.; MORI, T., OKAMURA, M.; KASHIDA, Y.; MITSUMORI, K. Absence of liver
tumor-initiating activity of kojic acid in mice.Arch Toxicol, 80: 299-304, 2006.
MOUGNEAU, E.; BIHL, F.; GLAICHENHAUS, N. Cell biology and immunology of Leishmania.
Immunol Rev, v. 240(1), p. 286-96, 2011.
MULLER, K.; VAN ZANDBERGEN, G.; HANSEN, B.; et al. Chemokines, natural killer cells and
granulocytes in the early course of Leishmania major infection in mice. Medical Microbiology
and Immunology, Berlin, v. 190, n. 1/2, p. 73-76, 2001.
NATHAN, C. Neutrophils and immunity: challenges and opportunites. Nature, USA, v. 6. 2006.
National Institute of Allergy and Infectious Diseases (NIAID). (2012). (Disponível em:
http://www.niaid.nih.gov/topics/leishmaniasis/Pages/lifecycle.aspx; acesso em 04/05/2014).
NAUSEEF, W. M. How human neutrophils kill and degrade microbes: an integrated review.
Immunol Rev. 2007;219:88–102.

81
NAUSEEF, W. M. Biological roles for the NOX family NADPH oxidases, J. Biol. Chem. 283,
16961–16965, 2008
NIWA, Y. & AKAMATSU, H. Kojic acid scavenges free radicals while potentiating leukocyte
functions including free radical generation. Inflammation 15 (4):4, 1991.
NOHYNEK, G.J.; KIRKLAND, D.; MARZIN, D.; TOUTAIN, H.; LECLERC-RIBAUD, C.;
JINNAI, H. An assessment of the genotoxicity and human health risk of topical use of kojic
acid [5-hydroxy-2-(hydroxymethyl)-4H-pyran-4-one]. Food and Chemical Toxicology, v.42,
p.93-105, 2004.
NOVAIS, F. O., SANTIAGO, R. C., BÁFICA, A., KHOURI, R., AFONSO, L., BORGES, V. M.,
BRODSKYN, C. Neutrophils and macrophages cooperate in host resistance against Leishmania
braziliensis infection. Journal of immunology, 183(12), 8088–98, 2009.
OKWOR, I.; UZONNA, J. Vaccines and vaccination strategies against human cutaneous
leishmaniasis. Hum Vaccin, v. 5 (5), p. 291-301, 2009.
PALUMBO, E. Treatment strategies for mucocutaneous leishmaniasis. J Glob Infect Dis, v. 2 (2),
p. 147-50, 2010.
PARVEZ, S.; KANG, M.; CHUNG, H.S.; CHO, C.; HONG, M.C; SHIN, MK , BAE H. Survey and
mechanism of skin depigmenting and lightening agents. Phytother. Res. 20:921–934, 2006.
PAVLI. A., MALTEZOU, H. C. Leishmaniasis, An Emerging Infection In Travelers. Int. J. Infect
Dis. 14(12): 1032-1039, 2010.
PERES, C. M., CURI, R.; Como cultivar células. Ed. Guanabara Koogan, 283p; 2005.
PETERS, N. C.; EGEN, J. G.; SECUNDINO, N.; DEBRABANT, A.; KIMBLIN, N.; KAMHAWI,
S.; LAWYER, P.; FAY, M. P. In vivo imaging reveals an essential role for neutrophils in
leishmaniasis transmitted by sand flies. Science 321:970-974, 2008.
PHAM, C. T. N. Neutrophil serine proteases: specific regulator of inflammation. Nature reviews
immunology, London, v. 6, n. 7, p. 541-550, 2006.
PIRES, A.; COSTA, G. C.; GONÇALVES, E. G. R.; ALVIM, A. C.; NASCIMENTO, F. R. F.
Aspectos imunológicos e clínicos da Leishmaniose Tegumentar Americana: Uma revisão.
Revista de ciências da saúde, v. 14, n. 1, p. 30-39, 2012.
QUINN, M. T.; AMMONS, M. C.; DELEO, F. R. The expanding role of NADPH oxidases in
health and disease: no longer just agents of death and destruction. ClinSci (Lond). 2006;111:1–
20.
REIS, L. C.; BRITO, M. E. F.; SOUZA, M. A.; PEREIRA, V. R. A. Mecanismos imunológicos na
resposta celular e humoral na leishmaniose tegumentar americana. Rev Patol Trop 2006;
35(2): 103-115.

82
REN, J., CHUNG, S. H. Anti-inflammatory effect of alpha-linolenic acid and its mode of action
through the inhibition of nitric oxide production and inducible nitric oxide synthase gene
expression via NF-kappaB and mitogen-activated protein kinase pathways. J Agric Food
Chem, 55, 5073–5080, 2007.
RHO, H.S.; BAEK, H.S.;YOU, J.W.; KIM, S.; LEE, J.Y.; KIM, D.H.; CHANG, I.S. New 5-
Hydroxy-2-(hydroxymethyl)-4H-pyran-4-one Derivative Has Both Tyrosinase Inhibitory and
Antioxidant Properties. Bull. Korean Chem. Soc.28( 3): 471-473, 2007.
RIBEIRO-GOMES, F. L.; OTERO, A. C.; GOMES, N. A.; et al. A. Macrophage interactions with
neutrophils regulate Leishmania major infection. The Journal of Immunology, Bethesda, v.
172, n. 7, p. 4454–4462, 2004.
RIBEIRO-GOMES, F.L.; PETERS, N.C.; DEBRABANT, A., SACKS, D. L. Efficient capture of
infected neutrophils by dendritic cells in the skin inhibits the early anti-Leishmania response.
PLoS Pathog. 2012. 8:e1002536.
RODRIGUES, A.P.; CARVALHO, A.S.; SANTOS, A.S.; ALVES, C.N.; DO NASCIMENTO
S.N.R.J.L.; SILVA, E.O. Kojic acid, a secondary metabolite from Aspergillus sp., acts as na
inducer of macrophage activation. Cell Biol. Int. 35: 335–343, 2011.
RODRIGUES, A.P.; FARIAS, L. H.; CARVALHO, A.S.; SANTOS, A.S.; ALVES, C.N.; DO
NASCIMENTO, J.L.; SILVA, E.O. A novel function for Kojic Acid, a secundar metabolite
from Aspergillus fungi, as antileishmanial agent. Plos One, 2014.
ROUSSEAU, D., DEMARTINO, S., FERRUA, B., FRANÇOIS, J., ANJUÈRE, F., FRAGAKI, K.,
FICHOUX, Y. LE. In vivo involvement of polymorphonuclear neutrophils in Leishmania
infantum infection. BMC Microbiology, 2001.
SANTOS, J. B.; DEJESUS, A. R.; MACHADO, P. R.; MAGALHAES, A.; SALGADO, K.;
CARVALHO, E. M. Antimony plus recombinant human granulocyte-macrophage colony-
stimulating fator applied topically in low doses enhances healing os cutaneous Leishmaniasis
ulcers: a randomized, double-blind, placebo-controlled study. J Infect Dis. 2004 15; 190(10):
1793-6.
SARKAR A., AGA E., BUSSMEYER U., BHATTACHARYYA A., MÖLLER S., HELLBERG
L., BEHNEN M., SOLBACH W., & LASKAY T. Infection of neutrophil granulocytes with
Leishmania major activates ERK 1/2 and modulates multiple apoptotic pathways to inhibit
apoptosis. Med. Microbiol. Immunol. 202, 25-35, 2013.
SATO, M. E.O.; PONTAROLO, F. G. R..; ANDREAZZA, I.F.; ZARONI, M. Permeação cutânea
in vitro do ácido kójico. Brazilian Journal of Pharmaceutical Sciences 43 (2): 195-203,
2007.

83
SEGAL, A. W. How neutrophils kill microbe. Ann Rev Immunol 2005;23:197–223.
SEGAL, A. W. How Neutrophils Kill Microbes. Europe PMC Funders Group. 2007. 2(2).
SHAW, J. J. Further thoughts on the use of the name Leishmania (leishmania) infantum chagasi for
the aetiological agent of visceral leismanisis. Memórias do Instituto Oswaldo Cruz, 101:
577-579, 2006.
SHENG, J.; WANG, D.; BRAUN, A. P. DAF-FM (4-Amino-5-methylamino-2_,7_-
difluorofluorescein) Diacetate Detects Impairment of Agonist-Stimulated Nitric Oxide
Synthesis by Elevated Glucose in Human Vascular Endothelial Cells: Reversal by Vitamin C
and L-Sepiapterin. The Journal Of Pharmacology And Experimental Therapeutics, v. 315,
n. 2, p. 931-940, 2005.
SHEPPARD, F. R.; KELHER, M. R.. Structural organization of the neutrophil NADPH oxidase:
phosphorylation and translocation during priming and activation. Journal of Leucocyte
Biology, v. 78, n. 5, Nov, p. 1025-42. 2005.
SIDDIQUI, R. A.; ENGLISH, D., et al. Phorbol ester-induced priming of superoxide generation by
phosphatidic acid-stmulated neutrophils and granule-free neutrophil cytoplasts.Journal of
Leucocyte Biology, v. 58, n.2, Aug, p. 189-95. 1995.
SILVA, F.; GOMES, R.; PRATES, D.; MIRANDA, J. C., ANDRADE, B., BARRAL-NETTO, M.;
BARRAL, A. Inflammatory cell infiltration and high antibody production in BALB/c mice
caused by natural exposure to Lutzomia longipalpis bites. Am J Trop Med Hyg, 2005.
Jan;72(1)?94-8
SMIT, N.; VICANOVA, J.; PAVEL, S. The Hunt for Natural Skin Whitening Agents. Int. J. Mol.
Sci.10: 5326-5349, 2009.
SOONG, L., CALVIN, A., HENARD., MELBY, P.C. Immunopathogenesis of non-healing
American cutaneous leishmaniasis and progressive visceral leishmaniasis. Semin
Immunopathol 34: 735–751, 2012.
SOTO, J.; ARANA, B. A.; TOLEDO, J.; RIZZO, N.; VEJA, J. C.; DIAZ, A. Miltefosine for new
world cutaneous leishmaniasis. Clin Infect Dis, 2004. 1;38(9): 1266-72.
STEFANSKA J, PAWLICZAK R. Apocynin: molecular aptitudes. Mediat Inflamm 2008:106507
STRAMER, B.; MOREIRA, S.; MILLARD, T.; EVANS, I.; HUANG, C. Y.; SABET, O.;
MILNER, M.; DUNN, G.; MARTIN, P.; WOOD, W. Clasp- mediated microtubule budling
regulates persistent motility and contact repulsion in Drosophila macrophages in vivo. J Cell
Biol, v. 189, n, 4, p. 681-689, 2010.

84
STUART, K.; VRUN, R.; CROFT, S.; FAIRLAMB, A.; GUTLER, R. E., MCKERROW, J.;
REED, S.; TARLETON, R. Kinetoplastids: related protozoan pathogens, different diseases.
Journal of Clinical Investigation 118: 1301-1310, 2008.
SUNDAR, S. et al. Injectable paromomycin for Visceral leishmaniasis in India. N Engl J Med v.
21, 356(25), p. 2571-2581, 2007.
SUZUKI, K., OHNO, S., SUZUKI, Y., OHNO. Y., OKUYAMA, R., ARUGA, A., YAMAMOTO,
M., ISHIHARA, K.O., NOZAKI, T., MIURA, S., YOSHIOKA, H., MORI, Y. Effect of green
tea extract on reactive oxygen species produced by neutrophils from cancer patients.
Anticancer Res. 32 (6): 2369-2375, 2012.
SWAIN, S. D.; R. T. T.; QUINN, M. T. Neutrophil priming in host defense: role of oxidants as
priming agents. Antioxidants & Redox Signaling, v.4, n.1, p.69-83. 2002.
TACCHINI-COTTIER F, ZWEIFEL C, BELKAID Y, MUKANKUNDIYE C, VASEI M,
LAUNOIS P, MILON G, LOUIS JA. An immunomodulatory function for neutrophils during
the induction of a CD4_ Th2 response in BALB/c mice infected with Leishmania major. J.
Immunol, 2000. 165:2628–2636.
TAMURA, T.; MITSUMORI, K.; TOTSURA, Y.; WAKABAYASHI, K.; KIDO, R.; KASAI, H.;
NASU, M.; HIROSE, M. Absence of in vivo genotoxic potential and tumor initiation activity
of kojic acid in the rat thyroid. Toxicology 222:213-224, 2006.
TEIXEIRA, C. R.; TEIXEIRA M. J.; GOMES, R. B.; SANTOS C. S.; ANDRADE B. B.;
RAFAELE-NETO, I. Saliva from Lutzomyia loongipalpis induces CC chemokine ligand
2/monocyte chemoattractant protein-1 expression and macrophage recruitment. J Imunol, 2005
dec 15; 175 (12):8346-53.
TEIXEIRA, D. E.; BENCHIMOL, M.; RODRIGUES, J. C. F.; CREPALDI, P. H.; PIMENTA, P.
F. P.; SOUZA, W. Atlas didático: Ciclo de vida da Leishmania. 1º Ed. Rio de Janeiro.
Fundação CECIERJ, Consórcio CEDERJ, 2013.
THEILGAAR-MONCH, K.; PORSE, B. T.; BORREGAARD, N. Systems biology of neutrophil
differentiation and immune response. Curr Opin Immunol. 2006; 18(1):54-60.
THI, E.P.; LAMBERTZ, U., REINER, N.E. Sleeping with the enemy: how intracellular pathogens
cope with a macrophage lifestyle. Plos pathogens. v. 8(3): e1002551, 2012.
TURINA, M.; MILLER, F. N.; MCHUGH, P. P.; CHEADLE, W. G.; POLK, H. C. Endotoxin
inhibits but induces primary necrosis in neutrophils. Inflammation, v. 29, n.1, Feb, p. 55-63.
2005.
VALADARES, D.G. et al. Leishmanicidal activity of the Agaricus blazei Murill in different
Leishmania species. Parasitology international, v. 60, n. 4, p. 357-63, 2011.

85
VALKO, M., LEIBFRITZ, D., MONCOL, J., CRONIN, M. T., MAZUR, M., TELSER, J. Free
radicals and antioxidants in normal physiological functions and human disease. Int J Biochem
Cell Biol, 39, 44 – 84, 2007.
VAN ZANDBERGEN, G.; HERMANN, N.; LAUFS, H.; et al. Leishmania promastigotes release a
granulocyte chemotactic factor and induce interleukin-8 release but inhibit gamma interferon-
inducible protein 10 production by neutrophil granulocytes. Infection and Immunity,
Washington, v. 70, n. 8, p. 4177- 4184, 2002.
VAN DEN VEEN, B. S.; WINTER, M. P. J.; HEERINGA, P. Myeloperoxidase: Molecular
mechanisms action and relevance to human health and disease. Antioxidant Redox Signal,
Larchmont, v.11, p. 2899-2937, 2009.
VAN ZANDBERGEN G, KLINGER M, MUELLER A, DANNENBERG S, GEBERT A,
SOLBACH W, LASKAY T. Neutrophil granulocyte serves as a vector for Leishmania entry
into macrophages. J. Immunol. 2004. 173:6521–6525.
WAGNER, J. G. & ROTH R. A. Neutrophil migration mechanisms, with an emplhasis on the
pulmonary vasculature. Pharmacol Rev, v.52, n.3, Sep, p.349-74. 2000.
WILSON, R. et al. Stage-specific adhesion of Leishmania promastigotes to sand fly midgets
assessed using an improved comparative binding assay. PLoS Negl Trop Dis, v.7;4(9), pii:
e816, 2010.
WINTERBOURN, C. C.; KETTLE, A. Biomarkers of myeloperoxidase-derived hypochlorous acid.
Free Radical Biology and Medicine, New York, v. 29, n.5, p. 403-409, 2000.
WHO EXPERT COMMITTEE. Control of the leishmaniases. WHO technical report series; no.
949. Report of a meeting of the WHO Expert Committee on the Control of Leishmaniases,
Geneva, 22–26 March 2010.
XIMENES, V. F.; KANEGAE, M. P.; RISSATO, S. R. The oxidation of apocynin catalyzed by
myeloperoxidase: proposal for NADPH oxidase inhibition. Arch Biochem Biophys 457:134–
141, 2007.
XIN L, VARGAS-INCHAUSTEGUI DA, RAIMER SS, KELLY BC, HU J, ZHU L, SUN J,
SOONG L. Type I IFN receptor regulates neutrophil functions and innate immunity to
Leishmania parasites. J. Immunol, 2010. 184:7047–7056.
XU, J.; WANG, F.; VAN KEYMEULEN, A; HERZMARK, P.; STRAIGHT, A.; Divergent signals
and cytoskeletal assemblies regulate self-organizing polarity in neutrophils. Cell 114:201–14,
2003.

86
ZHANG, X., MAJLESSI, L., DERIAUD, E., LECLERC, C., & LO-MAN, R. Coactivation of Syk
kinase and MyD88 adaptor protein pathways by bacteria promotes regulatory properties of
neutrophils. Immunity, 31(5), 761–71, 2009.
ZYCHLINSKY, A.; WEINRAUCH, Y.; WEISS, J. Forum in immunology. Introduction: Forum in
immunology on neutrophils. Microbes and Infection, Paris, v. 5, n. 14, p. 1289-1291, 2003.

87
7 MISSÃO DE ESTUDOS
Estágio realizado no Laboratório de Agregação de Proteínas e Amiloidoses da
Universidade Federal do Rio de Janeiro no período de 15 de Julho a 16 de Agosto de 2013
(Anexo 1).
8 PARTICIPAÇÃO EM EVENTOS
IV Encontro Anual Instituto Nacional de Ciência e Tecnologia de Biologia Estrutural e
Bioimagem (INBEB) – 08 a 10 de maio de 2013
Trabalho apresentado (oral e pôster): Effects of 5-hydroxy-2-hydroxymethyl-gamma-pyrone
(HMP), a bioproduct obtained from Aspergillus fungi, on human neutrophils in vitro (Anexo
2).
V Encontro Anual Instituto Nacional de Ciência e Tecnologia de Biologia Estrutural e
Bioimagem (INBEB) – 07 a 09 de maio de 2014
Trabalho apresentado (oral e pôster): Effects of kojic acid on human neutrophils and during
interaction with Leishmania (Leishmania) amazonensis in vitro (Anexo 3).
18th International Microscopy Congress (IMC) – 07-12 de setembro de 2014
Trabalho apresentado (Oral e pôster): Human neutrophil activation by Kojic Acid (Anexo 4).
XXX Annual Meeting of the Society of Protozoology / XLI Annual Meeting on Basic
Research on Chagas’ disease – 03 a 05 de novembro de 2014.
Trabalho apresentado (pôster): Kojic Acid induces oxidative mechanism and increase
phagocytosis of Leishmania (Leishmania) amazonensis by human neutrophils (Anexo 5).
9 PREMIAÇÃO
Melhor pôster, na categoria mestrado, pelo trabalho “Effects of kojic acid on human
neutrophils and during interaction with Leishmania (Leishmania) amazonensis in vitro” no V
Encontro Anual Instituto Nacional de Ciência e Tecnologia de Biologia Estrutural e
Bioimagem (INBEB) (Anexo 6).

88
ANEXO 1

89
ANEXO 2

90
ANEXO 3

91
ANEXO 4

92
ANEXO 5

93
ANEXO 6


















