ANÁLISE BIOSTRATIGRÁFICA DE UMA SONDAGEM DO CANAL DE ... · Foram estudados os foraminíferos...
7
ANÁLISE BIOSTRATIGRÁFICA DE UMA SONDAGEM DO CANAL DE MOÇAMBIQUE, , , BASEADA NO ESTUDO DOS FORAMINIFEROS PLANCTONICOS por MARIA DE LOURDES UBALDO * RESUMO Foram estudados os foraminíferos planctónicos de três amostras de testemunho colhido no Canal de Moçambique, durante um cruzeiro efectuado em 1965 (AL3/65). Estão presentes, no conjunto, três associações de foraminiferos planctónicos, com as seguintes idades: Miocénico médio, Miocénico superior ou Pliocénico inferior e Actual. Pensa-se que a idade do testemunho é Miocénico médio e que as faunas mais modernas podem ser explicadas por deslizamentos . '. . ... _.' .. _.. . ." . . ... Discutem-se os resultados comparando-os com os obtidos em cruzeiro anterior (AL2/64) que, para a mesma estação, tinha indicado idade eocénica, e admite-se que os respectivos testemunhos correspondem a camadas diferentes de um mesmo corte estratigráfico. ABSTRACT Planktonic foraminifera were studied in 3 samples from a core of a drilled hole in the Mozambique Channel, during an ocea- nographic cruise carried out 1965 (AL3/65). On the whole, three planktonic foraminiferal assemblages are present, with the. following ages: Middle Miocene, Late Miocene or Early Pliocene and Recent. It is thought that the core age is Middle Miocene and that the more recent fauna can be explained by local sliding. , . Results are discussed comparing them with those obtained during a former oceanographic cruise (AL2/64) which had indi- cated for the same site the eocenic age. It is admitted that such cores correspond to düferent layers of the same stratigraphic section. I - INTRODUÇÃO O estudo que efectuámos, há alguns anos, sobre os foraminiferos de sedimentos do Canal de Moçam- bique (UBALDO, 1973) baseou-se em material colhido durante o cruzeiro oceanográfico AL2/64, organizado pelo Instituto Hidrográfico e realizado pelo N. H. «Almirante Lacerda». Nas sondagens para a colheita dos sedimentos foi utilizado um pequeno colhedor «Phleger», de 60 cm de comprimento. Os testemunhos, de dimen- sões reduzidas, apresentavam, como seria de esperar, associações faunisticas recentes. No entanto, em duas estações (N. os 5 e 44) os sedimentos eram cons- tituidos por microfaunas fósseis ainda que mistu- radas com formas actuais. Nas amostras da estação N. o 5, ao sul do Canal, estava representado o Eocénico médio e nas da estação N. o 44, ao norte, fauna fóssil mais recente, do Miocénico superior ou Pliocénico inferior. Dado o óptimo estado de conservação destas microfaunas, formulámos então a hipótese de que se encontravam «in situ» e tinham sido postas a des- coberto pela erosão. Observando a topografia do Canal (UBALDO, 1973, figs. 23 e 24), verificámos também que os tes- munhos :referentes àquelas estações estavam situa- dos em locais de pendor muito acentuado, o que dificultava a deposição de sedimentos actuais. Con- siderámos então que as amostras fósseis poderiam corresponder a camadas de verdadeiros cortes estra- tigráficos. Em 1965, o Instituto Hidrográfico organizou novo cruzeiro oceanográfico (AL3/65), como comple- mento dos cruzeiros anteriores, que foi realizado também pelo N. H. «Almirante Lacerda». Abran- gendo área mais limitada, a zona sudoeste do Canal de Moçambique, este cruzeiro teve como objectivo o estudo das águas e' da geologia dos sedimentos do fundo. As estações oceanográficas coincidiram, em parte, com as executadas nos cruzeiros anteriores, tendo-se realizado trabalhos semelhantes e utilizado o mesmo tipo de colhedor nas sondagens. Os testemunhos dos sedimentos marinhos colhi- dos no cruzeiro AL3/65 foram entregues ao LEPPU para estudos geológicos, tal como os do cruzeiro anterior (AL2/64). • Laboratório de Estudos Petrológicos e· Paleontológicos do Ultramar, Alameda D. Afonso Henriques, 41-4. 0 D, 1000 Lisboa. 377
Transcript of ANÁLISE BIOSTRATIGRÁFICA DE UMA SONDAGEM DO CANAL DE ... · Foram estudados os foraminíferos...

ANÁLISE BIOSTRATIGRÁFICA DE UMA SONDAGEM DO CANAL DE MOÇAMBIQUE, , , BASEADA NO ESTUDO DOS FORAMINIFEROS PLANCTONICOS
por
MARIA DE LOURDES UBALDO *
RESUMO
Foram estudados os foraminíferos planctónicos de três amostras de testemunho colhido no Canal de Moçambique, durante um cruzeiro efectuado em 1965 (AL3/65). Estão presentes, no conjunto, três associações de foraminiferos planctónicos, com as seguintes idades: Miocénico médio, Miocénico superior ou Pliocénico inferior e Actual. Pensa-se que a idade do testemunho é Miocénico médio e que as faunas mais modernas podem ser explicadas por deslizamentos locai~._.. . '. . ... _.' .. _.. . ." . . ...
Discutem-se os resultados comparando-os com os obtidos em cruzeiro anterior (AL2/64) que, para a mesma estação, tinha indicado idade eocénica, e admite-se que os respectivos testemunhos correspondem a camadas diferentes de um mesmo corte estratigráfico.
ABSTRACT
Planktonic foraminifera were studied in 3 samples from a core of a drilled hole in the Mozambique Channel, during an oceanographic cruise carried out 1965 (AL3/65). On the whole, three planktonic foraminiferal assemblages are present, with the. following ages: Middle Miocene, Late Miocene or Early Pliocene and Recent. It is thought that the core age is Middle Miocene and that the more recent fauna can be explained by local sliding. , .
Results are discussed comparing them with those obtained during a former oceanographic cruise (AL2/64) which had indicated for the same site the eocenic age. It is admitted that such cores correspond to düferent layers of the same stratigraphic section.
I - INTRODUÇÃO
O estudo que efectuámos, há alguns anos, sobre os foraminiferos de sedimentos do Canal de Moçambique (UBALDO, 1973) baseou-se em material colhido durante o cruzeiro oceanográfico AL2/64, organizado pelo Instituto Hidrográfico e realizado pelo N. H. «Almirante Lacerda».
Nas sondagens para a colheita dos sedimentos foi utilizado um pequeno colhedor «Phleger», de 60 cm de comprimento. Os testemunhos, de dimensões reduzidas, apresentavam, como seria de esperar, associações faunisticas recentes. No entanto, em duas estações (N. os 5 e 44) os sedimentos eram constituidos por microfaunas fósseis ainda que misturadas com formas actuais.
Nas amostras da estação N. o 5, ao sul do Canal, estava representado o Eocénico médio e nas da estação N. o 44, ao norte, fauna fóssil mais recente, do Miocénico superior ou Pliocénico inferior.
Dado o óptimo estado de conservação destas microfaunas, formulámos então a hipótese de que se encontravam «in situ» e tinham sido postas a descoberto pela erosão.
Observando a topografia do Canal (UBALDO, 1973, figs. 23 e 24), verificámos também que os tes-
munhos :referentes àquelas estações estavam situados em locais de pendor muito acentuado, o que dificultava a deposição de sedimentos actuais. Considerámos então que as amostras fósseis poderiam corresponder a camadas de verdadeiros cortes estratigráficos.
Em 1965, o Instituto Hidrográfico organizou novo cruzeiro oceanográfico (AL3/65), como complemento dos cruzeiros anteriores, que foi realizado também pelo N. H. «Almirante Lacerda». Abrangendo área mais limitada, a zona sudoeste do Canal de Moçambique, este cruzeiro teve como objectivo o estudo das águas e' da geologia dos sedimentos do fundo.
As estações oceanográficas coincidiram, em parte, com as executadas nos cruzeiros anteriores, tendo-se realizado trabalhos semelhantes e utilizado o mesmo tipo de colhedor nas sondagens.
Os testemunhos dos sedimentos marinhos colhidos no cruzeiro AL3/65 foram entregues ao LEPPU para estudos geológicos, tal como os do cruzeiro anterior (AL2/64).
• Laboratório de Estudos Petrológicos e· Paleontológicos do Ultramar, Alameda D. Afonso Henriques, 41-4.0 D, 1000 Lisboa.
377
SGP
Referência bibliográfica
Boletim da Sociedade Geológica de Portugal, Vol. XXII, 1980-1981.

Dado que, entre as estações repetidas, se enconI trava a N. ° 5 que, como acabámos de descrever, , tinha levantado, do ponto de vista estratigráfico, ; alguns problemas interessantes, resolvemos iniciar
o estudo micropaleontológico pelas amostras provenientes desta estação.
O nosso objectivo foi o de comparar os resultados obtidos neste trabalho com os determinados em 1973 e analisar a hipótese que então expusemos ao tentar explicar a presença de faunas fósseis, em sedimentos marinhos quase superficiais que, continuamos a pensar, se encontram «in situ».
, . \; ~, ..
OCEANO rI
I'NDICO
4S"
16·'&-------~r_~----~~~~~------------~
~.,&-------~r_------------~------------~~ o
H,~--------~~----------~~--------_+~~ , .. . . 120 mKm
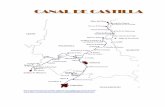
Fig. 1 - Localização da estação n.O 5 nos cruzeiros AL2/64 e AL3/65.
A localização da estação N. ° 5 está indicada no mapa da Fig. 1 e os dados referentes à profundidade a que se efectuou a sondagem e ao comprimento do testemunho colhido, bem como as coordenadas geográficas, vêm expressos no Quadro I, comparativamente, para os dois cruzeiros (AL2/64 e AI3/65).
O método de estudo utilizado no tratamento das amostras foi o descrito no nosso trabalho de 1973 1.
QUADRO I
Localização da estação N. ° 5 nos cruzeiros AL2/64 e AL3/65 e profundidades dos tes-
temunhos colhidos
Compri-
Cruzeiros Estação N.O tes- Latitude Longi- Profun- mento N.O temunho tude didade testemu-
(m) nho (m) ------ ---AL2/63 5 C5 25° 12'S 36° 42'E 2380 0,21
------ --- --- ---AL3/65 5 C34 25° I1'S 36° 33'E 2060 0,56
378
II - CONSIDERAÇÕES GERAIS SOBRE A MICROFAUNA DE FORAMINÍFEROS PLANCTÓNICOS
O testemunho C34, colhido na estação N. ° 5, é constituído por pelito argiloso com zonas de pelito arenoso, irregularmente distribuídas. Para facilidade de estudo, tomaram-se 3 amostras, uma na parte superior do' testemunho (C34/1) outra na média (C34/2) e outra na inferior (C34/3).
Do ponto de vista da microfauna (Quadro II), os foraminiferos são muito abundantes, principalmente na amostra da parte superior, onde o número de foraminíferos por grama de sedimento seco é de cerca de 9 800. Este valor corresponde unicamente à fauna planctónica, pois os foraminíferos bentónicos, em todo o testemunho, são raríssimos.
As associações de foraminíferos planctónicos, nas três amostras, podem considerar-se anómalas, pois contêm formas que, normalmente, não ocorrem juntas. A par de fauna actual, estão presentes abundantes formas fósseis, tornando bastante difícil o estudo biostratigráfico deste testemunho.
Verifica-se que, na amostra da parte superior, as maiores percentagens dizem respeito a formas actuais, embora estejam também presentes, mas em quantidades relativamente pequenas, as espécies fósseis que caracterizam as duas amostras inferiores. Por outro lado, uma parte da fauna actual está também representada nestas duas amostras, mas sempre em fracas percentagens.
Em resumo,' a associação de foraminiferos planctónicos apresenta mistura de faunas, diminuindo as formas actuais para zonas mais baixas do testemunho e observando-se o contrário no que se refere às formas fósseis. Estas, do ponto de vista estratigráfico, não se podem definir claramente pois, por sua vez, também se encontram misturadas, englobando formas do Miocénico médio, Miocénico supe'; Iior e/ou PliQcénico inferior.
Assim, a microfauna do testemunho C34 apresenta, no conjunto, pelo menos, três associações de foraminiferos planctónicos: uma actual e duas (ou mais) fósseis.
Entre as espécies recentes mais comuns assinalam-se: Hastigerina siphonifera, Globorotalia cultrata menardii, Turborotalia inflata, Globigerinita glutinata, Globigerinoide$ trilo bus, G. ruber e Globoquadrina dutertrei.
Em percentagens menores identificaram-se, entre outras, Globorotalia truncatulinoides, Turborotalia crassaformis, T. scitula, Globigerina calida, G. falconensis, G. rubescens, Globigerinoides conglobatus, G. quadrilobatus (s. 1.), Orbulina universa e Pulleniatina obliquiloculata.
A fauna fóssil mais· antiga, característica do Miocénico médio, tem, como espécies bem representadas em número de indivíduos, Turborotalia peripheroacuta e T. mayeri, com percentagens máximas de 8 % e 19 %, respectivamente, na amostra mais profunda. Segundo W. BLOW (1969) estas duas espécies têm como limites estratigráficos as biozonas N 10 e N 13. Com o mesmo intervalo, identificaram-se, também, ainda que em percentagens muito reduzidas, Globigerinopsis aguasayensis e G. martinkayei.
1 Agradecemos ao sr. Helder Borges a valiosa ajuda na execução das fotografias.

Dada a mistura da fauna fóssil, é difícil discernir, entre as espécies acompanhantes com largo _ intervalo estratigráfico, quais as que fazem parte desta associação e quais .as que pertencema. associações fósseis mais modernas. Assim, Globoquadrina altispira (s. 1.), G. dehiscens (s. 1.), G. venezuelana, Sphaeroidinel/opsis seminulina, & subdehiscens, Globigerinoides obliquus, Turborotalia obesa e Globorotalia miozea conoidea são, de maneira geral, formas miocénicas, estendendo-se algumas ao Pliocénico.
Além destas espécies, está ainda presente um pequeno grupo constituído por Globorotalia tumida plesiotumida, Turborotalia acostaensis acostaensis, T. acostaensis humerosa, Globigerina nepenthes e Globigerinoides obliquus extremUs. A ·maior parte destas espécies apareceram no Miocénico superior e extinguiram-se no Pliocénico. O intervalo estratigráfico mais reduzido é o correspondente a Globorotalia tumida plesiotumida que, segundo W. BLow (1969) abrange apenas as biozonas N 17 - N 18. Com base nesta espécie, a idade desta associação pode considerar-se do Miocénico superior ou Pliocénico inferior pois, para W. BLow, o Pliocénico começa na parte média da zona N 18.
A presença de outras espécies, ainda que em percentagens muito reduzidas, tais como Globorotalia hirsuta praehirsuta, Globigerinoides quadriolobatus fistulosus e Pul/eniatina obliquiloculata praecursor que, segundo W. Blow, aparecem na zona N 18, levam a pensar no Pliocénico inferior como idade mais provável. No entanto, dada a mistura de fauna observada em todas as amostras, não é possível a datação exacta dos sedimentos. Por agora, pode apenas dizer-se que estão presentes, pelo menos, três associações de foraminíferos planctónicos, sendo a mais moderna constituída por fauna actual, a mais antiga percentente ao Miocénico médio (biozonas de Blow entre N 10 e N 13) e uma intermédia considerada miocénica superior ou pliocénica inferior (biozonas N 17 ou N 18).
III - DISCUSSÃO DOS RESULTADOS COMPARANDO-OS COM OS OBTIDOS NO CRUZEIRO AL2/64
Algumas das estações oceanográficas executadas no cruzeiro AL3/65 foram, como atrás referimos, a repetição de estações realizadas em cruzeiros anteriores. É evidente que as coordenadas geográficas tiveram variações de alguns minutos, pelo que as sondagens efectuadas na estação N. o 5 durante os cruzeiros AL2/64 (testemunho C5) e AL3/65 (testemunho C34) sofreram desvios. Assim, a profundidade a que os sedimentos foram colhidos teve uma variação de cerca de 300 m (2 380 m no cruzeiro AL2/64 e 2060 m no AL3/65). O testemunho C34 foi pois colhido a profundidade menor do que o C5 o que poderá explicar a diferença de fauna registada entre eles.
Como vimos (UBALDO, 1973), os foraminíferos planctónicos identififlcados no testemunho C5 indícaram uma idade eocénica média enquanto, no pre.sente trabalho, o estudo da microfauna planctónica do testemunho C34 apontou para o Miocénico médio e o Miocénico superior ou Pliocénico inferior.
No trabalho de 1973, a propósito da fauna eocénica considerámos que ela se encontrava «in situ» e que teria sido posta a descoberto pela erosão. O perfil topográfico (UBALDO, 1973, fig. 23) levou-nos a admitir a existência de um corte estratigráfico em que -estaria representado o Eocénico médio. Com os resultados do presente trabalho, parece-nos possível admitir que, no referido corte, o Miocénico se encontra a cerca de 300 m acima do nível eocénico.
A mistura de faunas fósseis comprovada no testemunho C34 deve ter a sua origem em pequenos deslizamentos locais, pois a microfauna não apresenta sinais de transporte. Além disso, como assinalámos em 1973, a topografia local, de pendor bastante acentuado, poderia explicar estes escorregamentos, assim como a presença de fauna actual (se bem que em pequena percentagem) misturada com a fóssil.
Com base nestas considerações, pensamos que a idade dos sedimentos colhidos na sondagem C34 deve corresponder ao Miocénico médio e que a fauna conjunta mais moderna pode ser explicada por deslizamentos - locais.
A corroborar esta hipótese, está o facto de termos assinalado (UBALDO, 1973), aquando da descrição da microfaqna do testemunho C5, a presença muito rara, na amostra superior, de três espécies pertencentes ao Miocénico: Turborotalia peripheroronda, Globoquadrina altispira e Sphaeroidinel/opsis seminulina.
Em conclusão, admite-se pois a hipótese de que as sondagens efectuadas na estação oceanográfica N. o 5, durante os cruzeiros AL2/64 e AL3/65 foram realizadas em locais de pendor acentuado, correspondendo os respectivos testemunhos a diferentes camadas de um mesmo corte estratigráfico.
BIBLIOGRAFIA
BLOW, W. H. (1969) - Late Middle Eocene to Rect'nt planktonic foraminiferal biostratigraphy. Proc. 1st. Inter. Conf. Plankt. Microfossils, Geneve, 1967, 1, pp. 199-421. Leiden.
ELLIS, B. F. & MESSINA, A. (1940 & seg.) - Catalogue of Foraminifera. Amer Mus. Nat. Hist., Spec. Pub/., New York.
INSTITUTO HIDROGRÁFICO (1973) - Cooperação na Expedição Internacional ao Oceano índico: Resultados das observações oceanográficas no Canal de Mocambique. Cruzeiro AL3/65: Abril-Maio 1965. Publ. Inst. Hidrogr. Lisboa, n.0 7.
LoEBUCH, A. R. & TAPPAN, H. (1964) - Protista 2. Sarcodina, chiefiy <<Thecamoebians» and Foraminiferida. ln R. C. MOORE: Treatise on Invertebrate Paleontology, Part C, 2 voIs. Geol. Soe. America & Univ. -Kansas Press, Lawrence.
UBALDO, M. L. (1973) - Étude des Foraminiferes de sondages du Canal de Mozambique (Océan Indien). Mem. J. l. U., Lisboa, 2." série, D. o 62.
379

QUADRO II
Distribuição das espécies
Testemunho C34
Amostras C34/1 C34/2 C34/3 - .
Distância do topo, em cm 2,5-3,5 27-28 55-56
N.O de foraminíferos planctónicos/grama de sedimento seco 9800 4500 6000
Foraminíferos planctónicos:
HANTKENINIDAE
Hastigerina siphonifera (D'ORBIGNY) 7 X
GLOBOROTALIIDAE
G/oborotalia cu/trata menardii (PARKER, JONES & BRAOY) 4 X » hirsuta praehirsuta BLOW X » miozea conoidea WALTERS 1 X 1 » truncatulinoides (O'ORBIGNY) 2 » tumida p/esiotumida BLOW & BANNER 3 7 4 » » tumida (BRAOY) 1
Turborotalia acostaensis acostaensis (BLOW) 1 1 X » » humerosa (TAKAYANAGI & SAlTO) 2 » crassaformis (GALLOWAY & WISSLER) 2 X » inflata (O'ORBIGNY) . 4 X » mayeri (CUSHMAN & ELLISOR) 1 12 19 » obesa (BOLLI) X » oscitans (Tooo) 2 » peripheroacuta BLOW & BANNER 1 3 8 » scitu/a (BRAOY) 3 X X
GLOBIGERINIDAE
Candeina nitida O'ORBIGNY X G/obigerina cf. angiporoides HORNIBROOK X X X
» bradyi WIESNER X » calida P ARKER 3 X » cf. decoraperta TAKAYANAGI & SAlTO 1 » falconensis BLOW 2 X » nepenthes Tooo 2 X 2 » pachyderma (EHRENBERG) X X » praebulloides BLOW X » rubescens HOFKER 2
G/obigerinita glutinata (EGGER) 8 2 4 Globigerinoides cf. bollii BLQW X 5 X
» conglobatus (BRAOY) 2 » elongatus (O'ORBIGNY) 3 X » obliquus extremus BOLLI & BERMÚDEZ X 1 1 » » obliquus BOLLI 2 12 6 » quadrilobatus jistulosus (SCHUBERT) X » » immaturus (LE Roy) 3 7 8 » » irregularis (LE Roy) X » » quadrilobatus BANNER & BLOW X 2 5 » » sacculifer (BRAOY) 3 6 5 » » tri/obus (REuss) 4 1 2 » ruber (O'ORBIGNY) 4 X X » tenellus P ARKER X » indeterminados X 1 X
Globigerinopsis aguasayensis BOLLI X X » martinkayei BOLLI X X X
380

Globoquadrina altispira altispira (CUSHMAN & JARVIS) X X » » globosa BOLLI 4 17 11 » dehiscens advena BERMÚDEZ X X X » » dehiscens (CHAPMAN, PARR & COLLINS) X X » dutertrei f. C (ZOBEL) 2 » » f. eggeri (RHUMBLER) 9 » venezuelana (HEDBERG) X X X
Globorotaloides hexagona (NATLAND) X Orbulina universa D'ORBIGNY 2 3 1
» » f. bilobata (D'ORBIGNY) X X
» » f. suturalis BRONNIMANN X 4 4 Pulleniatina obliquiloculata obliquiloculata (PARKER & JONES) 2
» » praecursor BANNER & BLOW X » primalis BANNER & BLOW X
Sphaeroidinellopsis seminulina (SCHWAGER) X 2 3 » subdehiscens (BLOW) X X X
Indeterminados 4 5 7
Os números representam percentagens, com arredondamento. O sinal X indica percentagem inferior a 1 %.
381

Figs. 1-3 - Globorotalia tumida plesiotumida BLOW & BANNER, X 50. Figs. 4-6 - Globorotalia miozea conoidea WALTERS, X 80 e x 60' Fig. 7 - Globorotalia hirslIta praehirsuta BLOW, X 50. Figs. 8-9 - Turborotalia peripheroacllta BLOW & BANNER, X 90. Figs. 10-11 - Globoqlladrina delziscens advena BERMÚDEZ, X 70. Figs. 12-13 - Globoquadrina dehiscens dehiscens (CHAPMAN, PARR & COLLINS), X 70.
ESTAMPA I

Figs. 1-2 - Globoquadrina venezuelana (HEDBERG), x 60. Figs. 3-4 - Turborotalia mayeri (CUSHMAN & ELLISOR), X 70. Figs. 5-6 - Sphaeroidinellopsis seminulina (SCHWAGER), x 50. Fig. 7 - Globigerinoides quadrilobatus fistulosus (SCHUBERT), x 50. Figs. 8-9 - Globigerina nepenthes TODO, x 90. Figs. 10-11 - Globigerinoides obliquus extremus BOLLI, x 90. Figs. 12-13 - Globigerinopsis martinkayei BOLLI, x 90· Figs. 14-15 - Globigerinopsis aguasayensis BOLLI, x 80.
ESTAMPA II


















