
Andreia Maria da Anunciação Gomes - LETC/UFRJletc.biof.ufrj.br/sites/default/files/publicacoes/D...
186
Andreia Maria da Anunciação Gomes ESTUDO ECOFISIOLÓGICO DE CIANOBACTÉRIAS FORMADORAS DE FLORAÇÕES NA LAGOA DE JACAREPAGUÁ TESE DE DOUTORADO SUBMETIDA À UNIVERSIDADE FEDERAL DO RIO DE JANEIRO VISANDO A OBTENÇÃO DO GRAU DE DOUTOR EM CIÊNCIAS BIOLÓGICAS (BIOFÍSICA) Universidade Federal do Rio de Janeiro Centro de Ciências da Saúde Instituto de Biofísica Carlos Chagas Filho 2011
Transcript of Andreia Maria da Anunciação Gomes - LETC/UFRJletc.biof.ufrj.br/sites/default/files/publicacoes/D...

Andreia Maria da Anunciação Gomes
ESTUDO ECOFISIOLÓGICO DE CIANOBACTÉRIAS FORMADORAS DE FLORAÇÕES NA LAGOA DE
JACAREPAGUÁ
TESE DE DOUTORADO SUBMETIDA À
UNIVERSIDADE FEDERAL DO RIO DE JANEIRO VISANDO A
OBTENÇÃO DO GRAU DE DOUTOR EM CIÊNCIAS BIOLÓGICAS
(BIOFÍSICA)
Universidade Federal do Rio de Janeiro Centro de Ciências da Saúde Instituto de Biofísica Carlos Chagas Filho 2011

ii
Andreia Maria da Anunciação Gomes
ESTUDO ECOFISIOLÓGICO DE CIANOBACTÉRIAS FORMADORAS DE FLORAÇÕES NA LAGOA DE
JACAREPAGUÁ
Tese de Doutorado submetida à Universidade Federal do Rio de Janeiro visando a obtenção do grau
de doutor em Ciências Biológicas (Biofísica).
Orientadora: Dra. Sandra Maria Feliciano de Oliveira e Azevedo Universidade Federal do Rio de Janeiro
Centro de Ciências da Saúde Instituto de Biofísica Carlos Chagas Filho
2011

iii
Gomes, Andreia Maria da Anunciação
Estudo Ecofisiológico de Cianobactérias Formadoras de Florações na Lagoa de
Jacarepaguá / Andreia Maria da Anunciação Gomes. Rio de Janeiro, 2011.
xxv, 143 f.: il.
Tese de Doutorado – Universidade Federal do Rio de Janeiro, Instituto de Biofísica
Carlos Chagas Filho / Programa de Pós-graduação em Ciências Biológicas (Biofísica),
2011.
Orientador: Sandra Maria Feliciano de Oliveira e Azevedo.
1. Eutrofização. 2. Fitoplâncton. 3. Cianobactérias. 4. Alelopatia. 5. Competição. 6.
Temperatura. I. Azevedo, Sandra Maria Feliciano de Oliveira e. II. Universidade Federal
do Rio de Janeiro, Instituto de Biofísica Carlos Chagas Filho, Programa de
Pós-graduação em Ciências Biológicas (Biofísica). III. Título.

iv
Andreia Maria da Anunciação Gomes
Estudo Ecofisiológico de Cianobactérias Formadoras de Florações na Lagoa de Jacarepaguá
Rio de Janeiro, de de 2011
Aprovada por:
_______________________________________ PROF.
_______________________________________ PROF.
_______________________________________ PROF.
_______________________________________ PROF.
_______________________________________ PROF.
_______________________________________ PROF.
_______________________________________ Dra. Sandra Maria Feliciano de Oliveira e Azevedo / IBCCFo
(Orientadora)

v
Ao meu pai, pelo seu incansável
apoio, incentivo
e dedicação.

vi
“A Ciência, como um todo, não é nada mais que um refinamento do pensar diário”.
Albert Einstein

vii
AGRADECIMENTOS
Os anos de doutoramento foi um período de grande aprendizagem para mim, mas não apenas no sentido acadêmico. Aprendi bastante sobre amizade, companheirismo, solidariedade, paciência e muito sobre mim mesma. Esses foram anos de grandes transformações na minha vida. Transformações que me deixaram mais forte, segura... e ainda me fizeram mostrar quem realmente sou. Antigas amizades foram consolidadas, novas amizades foram feitas. Descobri muitas pessoas especiais e por isso devo muitos agradecimentos....
Primeiro, agradeço a CAPES pela concessão da minha bolsa de doutorado, que permitiu o desenvolvimento desta tese e ao CNPq pela concessão da bolsa sanduíche no exterior.
Agradeço a minha orientadora Sandra Azevedo que vem me apoiando desde a minha iniciação científica sendo incisiva quando tinha que ser e solidária e encorajadora nos momentos mais difíceis dessa carreira que escolhi.
Agradeço ao Prof. Miquel Lürling pela confiança depositada em mim e pela oportunidade de trabalhar com um grupo tão competente que é o do laboratório de Ecologia Aquática da Universidade de Wageningen.
Agradeço enormemente a revisora Professora Bia pela dedicação, cuidado e preciosas contribuições feitas a esse trabalho.
Agradeço aos meus amigos e companheiros de jornada. Antigos e Novos Companheiros do LETC: Valéria, Raquel, Simone, Ana Cláudia, Ana Lúcia, Isabel, Ronaldo, Rodrigo, Betinha, Bárbara, Pedro, Carol, Laís, Elisângela, Luana, Priscila, Roberta, Ramon, Iamê, Rafael, Adriana, Tais, Fernanda, Gustavo, César, Daniel, Roberta e Ricardinho.
Agradeço especialmente as professoras Valéria e Raquel pelos seus incentivos, além da sincera e grande amizade.
Ao professor MMM, Marcelo Manzi Marinho, agradeço pela grande ajuda no desenvolvimento dessa tese, pela confiança e incentivo.
Agradeço a queridíssima mineirinha, paulista Ana Lúcia pela amizade e todo o mimo. Obrigada por sempre emitir muita força na piruca, rsrsrs...
Agradeço meu amigo, mais que querido, Pedro Sampaio pelas ilustríssimas discussões, que muitas vezes viraram brigas, mas que no dia seguinte voltavam as admirações. Agradeço também pela ajuda nos experimentos e ainda pela oportunidade de me sentir orientando um pouquinho um aluno de iniciação científica já iniciado, comprometido e interessado.

viii
Agradeço ao meu anjo da guarda Carolzita, que só dá para agradecer abraçando, porque não consigo transformar em palavras a gratidão, a importância que tens na minha vida, amiga querida...é pra sempre!!!
Tenho que agradecer muito, muito a minha amiga Betinha por toda paciência que sempre teve comigo. Pela grande e eterna amizade estabelecida!! Obrigada pela força que me deste em todos os âmbitos da minha vida.
Agradeço a querida Belzinha por estar sempre disposta a me escutar e a ajudar quando necessário. Uma bela pessoa e amiga!!!
Muito obrigada Bêzinha!!!! Obrigada por TUDO!!! Em um momento impar da minha vida te conheci… Não acredito em coincidências. Você é muito especial para ser meramente coincidência. A partir de então usaremos pra sempre meias vermelhas... muita sorte pra nós!!! Obrigada minha mais nova e querida irmã!!
Maszar!!! Thank you a lot!!! Obrigada por transformar as noites frias e os dias nublados de Wageningen em fantásticos e calorosos momentos de sol. Você é uma das pessoas mais especiais que apareceram na minha vida. Você é meu anjo e estaremos conectados para sempre, mesmo que estejamos hoje em continentes diferentes... Conexões Brasil-Paquistão….estaremos conectados pelo nosso puro sentimento de amizade.
Agradeço também aos demais amigos paquistaneses: Príncipe Hamid, Maszar, Mobarik e Luck pelos momentos mais encantadores que experimentei na vida. Vocês são as pessoas mais bonitas que já encontrei. Obrigada pela oportunidade de conhecer um outro lado do mundo de vocês e pelos inúmeros jantares especialíssimos e cheios de significados!!!
Aos queridos brasileiros que juntos em uma cidade pequenina no meio da Holanda vão transformando os dias frios da região temperada no calor humano do nosso país tropical... são eles: Maurício, Gustavo, Cimille, Camila, Marcela, Rafa, Fernanda, Júlio, Felipe, Priscila, Charles, Aninha e a linda Bezinha. Agradeço muito a vocês!!
Agradeço a querida Irenita pela calorosa recepção espanhola em Wageningen, pela ótima convivência, pela amizade e apoio...sem nunca esquecer que "rapadura é doce, mas também não é mole não!!!"
Queridíssimo Dieguito, muito obrigada por todo seu carinho e amizade, além das riquíssimas discussões ecológicas e estatísticas. Tem pessoas que sem muito tempo ou esforço se transformam nas mais especiais e queridas... com você foi assim... Obrigada também por todos os mais divertidos momentos que tivemos juntos naquele país sem sol. Essa amizade está selada...
Agradeço aos meus amigos do laboratório ao lado, que de longa data vem me acompanhando. Os queridos Sérgio, Cláudio, Márcio, Antônio, Rodrigo e Petrus.

ix
Agradeço a minha mais nova "room-mate", querida Camila, pelo companheirismo, amizade, incentivo, ajuda e paciência.
Agradeço a minha irmã Adriana e as minhas sobrinhas lindas, maravilhosas, fantásticas, perfeitas Mayara e Marcelle por todo carinho de vocês.
Obrigada Tereza pelo seu carinho e pela harmonia do ambiente familiar que você me oferece.
Agradeço ao meu pai pela eterna dedicação e apoio em todas as minhas escolhas.
Agradeço ao meu querido R pela sutileza e despretensão com que chegou... sem sentir me deu a mão e me encorajou a finalizar esta tese...

x
RESUMO
GOMES, Andreia Maria da Anunciação. Estudo Ecofisiológico de Cianobactérias Formadoras de Florações na Lagoa de Jacarepaguá. Rio de Janeiro, 2011. Tese (Doutorado em Ciência Biológicas/ Biofísica) – Instituto de Biofísica Carlos Chagas Filho, Universidade Federal do Rio de Janeiro, Rio de Janeiro, 2011.
A lagoa de Jacarepaguá está localizada na costa sul do estado do Rio de
Janeiro, na zona metropolitana da cidade do Rio de Janeiro, Brasil. É uma lagoa rasa,
oligohalina que vem sofrendo um intenso processo de eutrofização nas últimas
décadas, com ocorrências de florações de cianobactérias reportadas desde a década
de 90. Esporadicamente ocorre quebra da dominância das cianobactérias
acompanhada de aumento da diversidade fitoplanctônica. Este estudo teve como
objetivo avaliar, através de estudo de campo e laboratório, que fatores influenciam a
dinâmica fitoplanctônica deste ambiente. Durante um ciclo sazonal (2007-2008), foram
monitorados a comunidade fitoplanctônica, e fatores físicos e químicos da água. No
período de floração de cianobactérias as espécies de maior contribuição foram
Microcystis aeruginosa, Aphanizomenon sp. e Planktothrix sp. Apesar de continuarem
dominantes no sistema, uma significativa redução da sua biomassa foi observada no
inverno, quando a temperatura da água passou de 30oC (verão) para 18oC. O colapso
da floração acarretou na liberação de grande quantidade de nitrogênio, permitindo que
outros grupos ocupassem o nicho liberado pelas cianobactérias: entre as diatomáceas
Cyclotella sp. e entre as clorofíceas Desmodesmus quadricauda. Em vista destas
observações de campo, verificou-se, em laboratório, o efeito da redução de nutrientes
sobre a comunidade fitoplanctônica, assim como o efeito da temperatura na competição
direta e indireta entre as principais espécies da lagoa de Jacarepaguá. Embora a
redução da concentração de nutrientes tenha reduzido a biomassa de cianobactérias,
não foi capaz de reduzir sua dominância. Ainda assim, o sistema mais empobrecido
favoreceu outros grupos, especialmente diatomáceas. A redução do aporte de fosfato
na lagoa de Jacarepaguá poderia recuperar esse sistema. A temperatura pode
influenciar no crescimento e substituição de espécies fitoplactônicas. Em laboratório,
estabelecemos as melhores temperaturas para o crescimento de P. agardhii (27oC), C.

xi
meneghiniana (18 e 21oC), D. quadricauda (18 e 30oC) e M. aeruginosa, (todas as
testadas entre 18 e 30oC). Nos experimentos de competição direta em cultivos mistos,
M. aeruginosa foi melhor competidora do que as demais em ambas as temperaturas
testadas (18 e 30oC). Nos cultivos com exudatos observamos diferentes respostas
alelopáticas (inibição ou estímulo do crescimento) de acordo com a temperatura.
Observando a dinâmica da comunidade na lagoa, M. aeruginosa é favorecida nos
períodos de temperaturas mais elevadas. Nos cultivos, os exudatos das suas
competidoras estimularam seu crescimento a 30oC, mas não a 18oC. A adaptação de
uma cepa a uma certa temperatura determina não só sua vulnerabilidade a compostos
alelopáticos, como sua capacidade de expressá-los. Por fim, é difícil apontar um fator
determinante para a ocorrência de florações de cianobactérias na lagoa de
Jacarepaguá. Um sinergismo de fatores ambientais e biológicos aliados a própria
fisiologia das espécies presentes irão determinar a dinâmica da comunidade
fitoplanctônica neste sistema.

xii
ABSTRACT
GOMES, Andreia Maria da Anunciação. Estudo Ecofisiológico de Cianobactérias Formadoras de Florações na Lagoa de Jacarepaguá. Rio de Janeiro, 2011. Tese (Doutorado em Ciência Biológicas/ Biofísica) – Instituto de Biofísica Carlos Chagas Filho, Universidade Federal do Rio de Janeiro, Rio de Janeiro, 2011.
Jacarepaguá Lagoon is located in the South coast of Rio de Janeiro State, in the
metropolitan zone of Rio de Janeiro City, Brazil. It is an oligohaline, shallow lagoon that
has undergone intense eutrophication in the past decades, with cyanobacterial blooms
reported since the 90'. Sporadically, a break of cyanobacteria dominance is followed by
an increase of phytoplankton diversity. The aim of this study was to evaluate, through
field and laboratory studies, which factors influence phytoplankton succession in this
environment. During a seasonal cycle (2007-2008) we analyzed phytoplankton
community and water chemical and physical parameters. During cyanobaterial blooms,
the most prominent species were Microcystis aeruginosa, Aphanizomenon sp. and
Planktothrix sp. Although still dominant, a significant reduction in biomass was observed
during winter, when water temperature decreased from about 30oC (summer) to 18oC.
Bloom collapse resulted in the release of a large quantity of DIN, allowing other species
to occupy the niche released by cyanobacteria: Cyclotella sp. among diatoms and
Desmodesmus quadricauda. among green algae. Considering these field observations,
we tested, in laboratory, the effect of nutrient reduction on phytoplankton, as well as the
effect of temperature on direct and indirect competition among the main species of
Jacarepaguá lagoon. The reduction in nutrient concentration decreased cyanobacterial
biomass but did not reduce their dominance. Even so, the low nutrient condition favored
other phytoplanktonic groups, especially diatoms. The reduction of phosphate input in
Jacarepaguá lagoon could recover this system. Temperature can influence growth and
phytoplanktonic species replacement. We established, under laboratory conditions,
optimum growth temperatures for P. agardhii (27oC), C. meneghiniana (18 and 21oC), D.
quadricauda (18 and 30oC) and M. aeruginosa, (all those tested between 18 and 30oC).
In competition tests with mixed cultivation M. aeruginosa was more competitive than
others in both temperatures (18 and 30oC). In cultivation with exudates, different
allelopathic responses (growth inhibition or stimulation) were attained in different

xiii
temperatures. Observing the community dynamics in the lagoon, M. aeruginosa is
favored during periods of higher temperatures. Exudates of its competitors stimulated its
growth at 30oC, but not at 18oC. Adaptation of one strain to a specific temperature will
determine not only its vulnerability to allelopathic compounds, but also its ability to
express the allelochemical potential. Finally, it is difficult to point one factor that dictates
the occurrence of cyanobacterial blooms in the Jacarepaguá lagoon. A synergy of
environmental and biological factors combined with the physiology of the species will
determine the dynamics of phytoplankton in this system.

xiv
LISTA DE ILUSTRAÇÕES
ÁREA DE ESTUDO
Figura 1: Complexo Lagunar de Jacarepaguá................................................................14
CAPÍTULO I: AVALIAÇÃO DA DINÂMICA E DOS POSSÍVEIS FATORES REGULADORES DA
DOMINÂNCIA DE ESPÉCIES DE CIANOBACTÉRIAS NA LAGOA DE JACAREPAGUÁ
Figura 1: Lagoa de Jacarepaguá com a marcação das duas estações de
coleta...............................................................................................................................22
Figura 2: Variação dos valores de profundidade de disco de Secchi (Zsec),
profundidade máxima (Zmax) e profundidade da zona eufótica (Zeu) da lagoa de
Jacarepaguá....................................................................................................................26
Figura 3: Isolínhas dos valores de temperatura (oC), oxigênio dissolvido (mg.L-1),
condutividade elétrica (mS.cm-1) e salinidade (ppt) da água da lagoa de
Jacarepaguá....................................................................................................................29
Figura 4: Variação dos valores de pH da água na estação 1 (a) e estação 2 (b) da lagoa
de Jacarepaguá...............................................................................................................31
Figura 5: Variaçãol dos valores de fósforo solúvel reativo (PSR) e fósforo total (P-total)
da água na estação 1 (a, c) e estação 2 (b, d) da lagoa de
Jacarepaguá....................................................................................................................33
Figura 6: Variação dos valores de nitrogênio inorgânico dissolvido (NID) da água na
estação 1 (a) e estação 2 (b) da lagoa de Jacarepaguá.................................................36

xv
Figura 7: Variação da biomassa total e da contribuição relativa dos principais grupos
fitoplanctônicos na estação 1 (a, c) e na estação 2 (b, d) da lagoa de
Jacarepaguá....................................................................................................................35
Figura 8: Variação da biomassa total e da contribuição relativa das principais espécies
de cianobactérias, da Cyclotella e outras algas na estação 1 (a, c) e na estação 2 (b, d)
da lagoa de Jacarepaguá................................................................................................37
Figura 9: Diagrama de Ordenação da Análise de Componentes Principais (ACP) das
variáveis limnológicas das estações de coleta 1 e 2 da lagoa de Jacarepaguá. Zmax:
profundidade máxima, Zsecchi: profundidade de disco de Secchi, Temp: temperatura da
água, cond: condutividade elétrica, sal: salinidade da água, NID: nitrogênio inorgânico
dissolvido, PSR: fósforo solúvel reativo, P-total: fósforo total, clo-a: clorofila-a, Cyan:
Cianobactérias, Chlor: Chlorophyta, Bacil: Bacillariophyta, Cryp: Cryptophyta, Outras:
outras algas.....................................................................................................................38
CAPÍTULO II: EFEITO DA REDUÇÃO DA CONCENTRAÇÃO DE NUTRIENTES NA COMUNIDADE
FITOPLANCTÔNICA DA LAGOA DE JACAREPAGUÁ
Figura 1: Variação dos valores de temperatura da água (a), intensidade luminosa (b),
condutividade elétrica (c), salinidade (d), oxigênio dissolvido (e) e pH (f) da água, ao
longo do experimento sem sedimento.............................................................................57
Figura 2: Variação dos valores de temperatura da água (a), intensidade luminosa (b),
condutividade elétrica (c), salinidade (d), oxigênio dissolvido (e) e pH (f) da água, ao
longo do experimento com sedimento.............................................................................58
Figura 3: Variação das concentrações de PSR e NID, ao longo dos experimentos sem
sedimento (a) e (b) e com sedimento (c) e (d)................................................................59

xvi
Figura 4: Variação da concentração de clorofila-a ao longo dos experimentos sem
sedimento (a) e com sedimento (b).................................................................................60
Figura 5: Variação da biomassa total e da contribuição relativa dos principais grupos
fitoplanctônicos do controle (a) e (b), tratamento A (c) e (d) e tratamento B (e) e (f), ao
longo do experimento sem sedimento.............................................................................62
Figura 6: Variação da biomassa total e da contribuição relativa dos principais grupos
fitoplanctônicos do controle (a) e (b), tratamento A (c) e (d) e tratamento B (e) e (f), ao
longo do experimento com sedimento.............................................................................64
Figura 7: Variação da biomassa total e da contribuição relativa dos principais espécies
de cianobactérias do controle (a) e (b), tratamento A (c) e (d) e tratamento B (e) e (f), ao
longo do experimento sem sedimento.............................................................................67
Figura 8: Variação da biomassa total e da contribuição relativa dos principais espécies
de cianobactérias do controle (a) e (b), tratamento A (c) e (d) e tratamento B (e) e (f), ao
longo do experimento com sedimento.............................................................................68
CAPÍTULO III: EFEITO DA TEMPERATURA NO CRESCIMENTO E COMPETIÇÃO ENTRE ESPÉCIES
FITOPLANCTÔNICAS
Figura 1: Taxas de Crescimento baseadas na concentração de clorofila-a de M.
aeruginosa (MIJAC-01) em cinco diferentes temperaturas. As letras correspondem a
diferenças significativas (p<0.01)....................................................................................83
Figura 2: Variação diária da concentração de clorofila-a (µg L-1) (a) e dos valores de
rendimento do fotossistema II (b) da cepa de M. aeruginosa (MIJAC-01) em cinco
diferentes temperaturas...................................................................................................83

xvii
Figura 3: Taxas de Crescimento baseadas na concentração de clorofila-a de P. agardhii
(PAT-1) em cinco diferentes temperaturas. As letras correspondem a diferenças
significativas (p<0.01)......................................................................................................85
Figura 4: Variação diária do logaritmo natural da concentração de clorofila-a (µg L-1) (a)
e dos valores de rendimento do fotossistema II (b) da cepa de P. agardhii (PAT-1) em
cinco diferentes temperaturas.........................................................................................85
Figura 5: Taxas de Crescimento baseadas na concentração de clorofila-a de C.
meneghiniana (CCAP1070/5) em cinco diferentes temperaturas. As letras
correspondem a diferenças significativas (p<0.01).........................................................87
Figura 6: Variação diária do logaritmo natural da concentração de clorofila-a (µg L-1) (a)
e dos valores de rendimento do fotossistema II (b) da cepa de C. meneghiniana (CCAP)
em cinco diferentes temperaturas...................................................................................87
Figura 7: Taxas de Crescimento baseadas na concentração de clorofila-a de D.
quadricauda (DEJAC-02) em cinco diferentes temperaturas. As letras correspondem a
diferenças significativas (p<0.01)....................................................................................88
Figura 8: Variação diária do logaritmo natural da concentração de clorofila-a (µg L-1) (a)
e dos valores de rendimento do fotossistema II (b) da cepa de D. quadricauda (DEJAC-
02) em cinco diferentes temperaturas.............................................................................88
Figura 9: Taxas de crescimento das monoculturas e das culturas mistas de M.
aeruginosa (MIJAC) e P. agardhii (PAT) em duas diferentes temperaturas: (a) 18oC e
(b) 30oC. As letras correspondem a diferenças significativas
(p<0.01)...........................................................................................................................91
Figura 10: Taxas de crescimento das monoculturas e das culturas mistas de M.
aeruginosa (MIJAC) e C. meneghiniana (CCAP) em duas diferentes temperaturas: (a)

xviii
18oC e (b) 30oC. As letras correspondem a diferenças significativas
(p<0.01)...........................................................................................................................91
Figura 11: Taxas de crescimento das monoculturas e das culturas mistas de P. agardhii
(PAT) e C. meneghiniana (CCAP) em duas diferentes temperaturas: (a) 18oC e (b)
30oC. As letras correspondem a diferenças significativas
(p<0.01)...........................................................................................................................92
Figura 12: Taxas de crescimento das monoculturas e das culturas mistas de M.
aeruginosa (MIJAC), P. agardhii (PAT) e C. meneghiniana (CCAP) em duas diferentes
temperaturas: (a) 18oC e (b) 30oC. As letras correspondem a diferenças significativas
(p<0.01)...........................................................................................................................93
CAPÍTULO IV: EFEITO DA TEMPERATURA NA INTERAÇÃO ALELOPÁTICA ENTRE TRÊS
DIFERENTES ESPÉCIES FITOPLANCTÔNICAS
Figura 1: Esquema simplificado do desenho experimental para verificar o efeito da
temperatura no potencial alelopático das espécies M. aeruginosa (MIJAC), P. aghardii
(PAT) e C. meneginiana (CCAP) em monoculturas e cultivos mistos. O controle de cada
cepa foi cultivado em meio WC modificado (Lürling & Beekman,
1999).............................................................................................................................109
Figura 2: Taxas de Crescimento baseadas na concentração de clorofila-a de M.
aeruginosa (MIJAC) cultivada nos exudatos de M. aeruginosa (Ma exud), P. agardhii
(Pa exud), C. meneghiniana (Cm exud) e do cultivo misto dessas três espécies (Ma +
Pa + Cm exud) em duas temperaturas: 18oC (a) e 30oC (b). O controle foi crescido em
meio WC........................................................................................................................113
Figura 3: Variação dos valores de rendimento do fotossistema II da cepa de M.
aeruginosa (MIJAC) cultivada nos exudatos de M. aeruginosa (Ma exudato), P. agardhii
(Pa exudato), C. meneghiniana (Cm exudato) e do cultivo misto dessas três espécies

xix
(Ma + Pa + Cm exudato) em duas temperaturas: 18oC (a) e 30oC (b). O controle foi
crescido em meio WC...................................................................................................113
Figura 4: Variação do logaritmo natural da concentração de clorofila-a (µg L-1) da cepa
de M. aeruginosa (MIJAC) cultivada nos exudatos de M. aeruginosa (Ma exudato), P.
agardhii (Pa exudato), C. meneghiniana (Cm exudato) e do cultivo misto dessas três
espécies (Ma + Pa + Cm exudato) em duas temperaturas: 18oC (a) e 30oC (b). O
controle foi crescido em meio WC.................................................................................114
Figura 5: Variação dos valores de pH do cultivo da cepa de M. aeruginosa (MIJAC)
cultivada nos exudatos de M. aeruginosa (Ma exudato), P. agardhii (Pa exudato), C.
meneghiniana (Cm exudato) e do cultivo misto dessas três espécies (Ma + Pa + Cm
exudato) em duas temperaturas: 18oC (a) e 30oC (b). O controle foi crescido em meio
WC.................................................................................................................................115
Figura 6: Taxas de Crescimento baseadas na concentração de clorofila-a de P. agardhii
(PAT) cultivada nos exudatos de M. aeruginosa (Ma exud), P. agardhii (Pa exud), C.
meneghiniana (Cm exud) e do cultivo misto dessas três espécies (Ma + Pa + Cm exud)
em duas temperaturas: 18oC (a) e 30oC (b). O controle foi crescido em meio
WC.................................................................................................................................116
Figura 7: Variação dos valores de rendimento do fotossistema II da cepa de P. agardhii
(PAT) cultivada nos exudatos de M. aeruginosa (Ma exudato), P. agardhii (Pa exudato),
C. meneghiniana (Cm exudato) e do cultivo misto dessas três espécies (Ma + Pa + Cm
exudato) em duas temperaturas: 18oC (a) e 30oC (b). O controle foi crescido em meio
WC.................................................................................................................................117
Figura 8: Variação do logaritmo natural da concentração de clorofila-a (µg L-1) da cepa
de P. agardhii (PAT) cultivada nos exudatos de M. aeruginosa (Ma exudato), P. agardhii
(Pa exudato), C. meneghiniana (Cm exudato) e do cultivo misto dessas três espécies

xx
(Ma + Pa + Cm exudato) em duas temperaturas: 18oC (a) e 30oC (b). O controle foi
crescido em meio WC...................................................................................................117
Figura 9: Variação dos valores de pH do cultivo da cepa de P. agardhii (PAT) cultivada
nos exudatos de M. aeruginosa (Ma exudato), P. agardhii (Pa exudato), C.
meneghiniana (Cm exudato) e do cultivo misto dessas três espécies (Ma + Pa + Cm
exudato) em duas temperaturas: 18oC (a) e 30oC (b). O controle foi crescido em meio
WC.................................................................................................................................118
Figura 10: Taxas de Crescimento baseadas na concentração de clorofila-a de C.
meneghiniana (CCAP) cultivada nos exudatos de M. aeruginosa (Ma exud), P. agardhii
(Pa exud), C. meneghiniana (Cm exud) e do cultivo misto dessas três espécies (Ma +
Pa + Cm exud) em duas temperaturas: 18oC (a) e 30oC (b). O controle foi crescido em
meio WC........................................................................................................................119
Figura 11: Variação dos valores de rendimento do fotossistema II da cepa de C.
meneghiniana (CCAP) cultivada nos exudatos de M. aeruginosa (Ma exudato), P.
agardhii (Pa exudato), C. meneghiniana (Cm exudato) e do cultivo misto dessas três
espécies (Ma + Pa + Cm exudato) em duas temperaturas: 18oC (a) e 30oC (b). O
controle foi crescido em meio WC.................................................................................120
Figura 12: Variação do logaritmo natural da concentração de clorofila-a (µg L-1) da cepa
de C. meneghiniana (CCAP) cultivada nos exudatos de M. aeruginosa (Ma exudato), P.
agardhii (Pa exudato), C. meneghiniana (Cm exudato) e do cultivo misto dessas três
espécies (Ma + Pa + Cm exudato) em duas temperaturas: 18oC (a) e 30oC (b). O
controle foi crescido em meio WC.................................................................................120
Figura 13: Variação dos valores de pH do cultivo da cepa de C. meneghiniana (CCAP)
cultivada nos exudatos de M. aeruginosa (Ma exudato), P. agardhii (Pa exudato), C.
meneghiniana (Cm exudato) e do cultivo misto dessas três espécies (Ma + Pa + Cm

xxi
exudato) em duas temperaturas: 18oC (a) e 30oC (b). O controle foi crescido em meio
WC.................................................................................................................................121

xxii
LISTA DE TABELAS
ÁREA DE ESTUDO
Tabela 1: Dados morfométricos das lagoas do complexo lagunar de
Jacarepaguá....................................................................................................................17
CAPÍTULO I: AVALIAÇÃO DA DINÂMICA E DOS POSSÍVEIS FATORES REGULADORES DA
DOMINÂNCIA DE ESPÉCIES DE CIANOBACTÉRIAS NA LAGOA DE JACAREPAGUÁ
Tabela 1: Valores das medianas, mínimos e máximos das variáveis limnológicas e
concentração de clorofila-a na sub-superfície nas Estações 1 e 2 nos períodos I (PI), II
(PII) e III (PIII) da lagoa de Jacarepaguá. Temp. água, temperatura da água; OD,
oxigênio dissolvido; Cond., condutividade elétrica; Sal., salinidade da água; Zsecchi,
profundidade do disco de Secchi; Zmax., profundidade máxima; DIN, Nitrogênio
inorgânico dissolvido; PSR, Fósforo solúvel reativo; P-total, Fósforo total.....................28
Tabela 2: Autovalores e Porcentagem da variância total e cumulativa resultantes da
Análise de Componentes Principais (ACP) das variáveis ambientais da lagoa de
Jacarepaguá....................................................................................................................39
Tabela 3: Correlação das variáveis ambientais com os eixos 1 e 2 resultantes da
Análise de Componentes Principais (ACP) das variáveis ambientais da lagoa de
Jacarepaguá....................................................................................................................39
CAPÍTULO II: EFEITO DA REDUÇÃO DA CONCENTRAÇÃO DE NUTRIENTES NA COMUNIDADE
FITOPLANCTÔNICA DA LAGOA DE JACAREPAGUÁ
Tabela 1: Soma das contribuições relativas (%) da biomassa de Microcystis aeruginosa
e Aphanizomenon sp. para biomassa total de cianobactérias, ao longo dos
experimentos...................................................................................................................66

xxiii
CAPÍTULO III: EFEITO DA TEMPERATURA NO CRESCIMENTO E COMPETIÇÃO ENTRE ESPÉCIES
FITOPLANCTÔNICAS
Tabela 1: Valores de Rendimento máximo - R (µg L-1) baseado na concentração de
clorofila-a das quatro cepas cultivadas em cinco temperaturas
diferentes........................................................................................................................89
Tabela 2: Efeito das competições entre as cepas de M. aeruginosa (MIJAC), P. agardhii
(PAT), e C. meneghiniana (CCAP) em cultivos mistos em duas temperaturas (18 e 30 oC)...................................................................................................................................90
CAPÍTULO IV: EFEITO DA TEMPERATURA NA INTERAÇÃO ALELOPÁTICA ENTRE TRÊS
DIFERENTES ESPÉCIES FITOPLANCTÔNICAS
Tabela 1: Densidade celular (no. céls mL-1) de cada espécie do cultivo misto das cepas
M. aeruginosa (MIJAC), P. agardhii (PAT) e C. meneghiniana (CCAP) após 10 dias de
cultivo para obtenção do exudato.................................................................................108
Tabela 2: Concentração de nutrientes dos exudatos livres de células das cepas de M.
aeruginosa (MIJAC), P. agardhii (PAT), C. meneghiniana (CCAP) e da Tricultura
dessas cepas (MIJAC+PAT+CCAP) cultivadas a 18 e 30oC, antes e depois da adição
das soluções de nutrientes do meio de cultura WC.....................................................110
Tabela 3: Efeito dos exudatos das monoculturas de M. aeruginosa (MIJAC), P. agardhii
(PAT), C. meneghiniana (CCAP) e do cultivo misto das três cepas (MIJAC+PAT+CCAP)
no crescimento das mesmas cepas cultivadas em duas temperaturas (18 e
30oC).............................................................................................................................110

xxiv
SUMÁRIO
APRESENTAÇÃO............................................................................................................1
1 .INTRODUÇÃO..............................................................................................................4
1.1. LAGOAS COSTEIRAS......................................................................................................4
1.2. CIANOBACTÉRIAS…………………….............................................................................5
1.3. CIANOBACTÉRIAS X INTERAÇÕES COM O AMBIENTE........................................................6
1.4. CIANOBACTÉRIAS X INTERAÇÕES COM OUTROS ORGANISMOS..............…………............9
1.5. COMPOSTOS BIOATIVOS DAS CIANOBACTÉRIAS.............................................................10
1.6.EUTROFIZAÇÃO NO COMPLEXO LAGUNAR DE JACAREPAGUÁ...........................................11
2. ÁREA DE ESTUDO....................................................................................................14
2.1. LOCALIZAÇÃO.............................................................................................................14
2.2.CLIMA.........................................................................................................................15
2.3.GEOMORFOLOGIA........................................................................................................15
2.4.HIDROGRAFIA .............................................................................................................16
3. OBJETIVO GERAL.....................................................................................................18
4. OBJETIVOS ESPECÍFICOS.......................................................................................18
CAPÍTULO I: AVALIAÇÃO DA DINÂMICA E DOS POSSÍVEIS FATORES REGULADORES DA
DOMINÂNCIA DE ESPÉCIES DE CIANOBACTÉRIAS NA LAGOA DE JACAREPAGUÁ.......................20
CAPÍTULO II: EFEITO DA REDUÇÃO DA CONCENTRAÇÃO DE NUTRIENTES NA COMUNIDADE
FITOPLANCTÔNICA DA LAGOA DE JACAREPAGUÁ..................................................................48
CAPÍTULO III: EFEITO DA TEMPERATURA NO CRESCIMENTO E COMPETIÇÃO ENTRE ESPÉCIES
FITOPLANCTÔNICAS...........................................................................................................76
CAPÍTULO IV: EFEITO DA TEMPERATURA NA INTERAÇÃO ALELOPÁTICA ENTRE TRÊS
DIFERENTES ESPÉCIES FITOPLANCTÔNICAS.......................................................................103

xxv
5. DISCUSSÃO GERAL................................................................................................130
6. CONCLUSÕES.........................................................................................................135
7. REFERÊNCIAS BIBLIOGRÁFICAS ........................................................................137
APÊNDICE....................................................................................................................144

1
APRESENTAÇÃO:
A lagoa de Jacarepaguá, área de estudo escolhida para o desenvolvimento
desta tese, é um ambiente hipereutrófico com freqüentes florações de cianobactérias
e que já vem sendo estudada pela equipe do laboratório de Ecofisiologia e
Toxicologia de Cianobactérias da UFRJ desde 1996. Embora muitos trabalhos já
tenham reportado vários fatores como sendo determinantes na dominância das
cianobactérias em ambientes aquáticos, pouco se sabe sobre as condições que
levam uma espécie de cianobactéria dominar sobre outras espécies desse mesmo
grupo ou sobre outras espécies de grupos fitoplanctônicos distintos. O presente
trabalho buscou respostas para compreender o padrão de variação de espécies
fitoplanctônicas na lagoa de Jacarepaguá, além de identificar os possíveis fatores
reguladores da ocorrência e/ou dominância de cianobactérias neste ambiente.
Os resultados obtidos nesta tese são apresentados na forma de capítulos. No
capítulo I foram reunidos dados do monitoramento dos fatores físicos e químicos e
da comunidade fitoplanctônica durante um ciclo sazonal (2007/2008) na lagoa de
Jacarepaguá, para tentar compreender a dinâmica de substituições de espécies
fitoplanctônicas neste mesmo ambiente. O capítulo II teve o objetivo de avaliar o
efeito do empobrecimento nutricional da água da lagoa de Jacarepaguá (através de
experimentos em laboratório) na comunidade fitoplanctônica.
Um dos principais resultados obtidos durante o trabalho de campo foi a
observação da comunidade fitoplanctônica dominada pelas cianobactérias
Microcystis aeruginosa, Aphanizomenon e Planktothrix sp. na maior parte do ano, no
entanto, com a chegada do período de inverno, quando a temperatura da água
passou de aproximadamente 30oC para até 18oC, houve um aumento da

2
contribuição das espécies dos grupos das clorofíceas e diatomáceas. Dentre as
espécies ocorridas durante esse período, Cyclotella meneghiniana foi uma das mais
representativas entre as diatomáceas e Desmodesmus quadricauda entre as
clorofíceas. Diante deste cenário, surgiu uma segunda etapa desta tese de
doutorado, que foi buscar entender as relações entre as espécies fitoplanctônicas
dominantes e a relação com a variação da temperatura. Concomitantemente, surgiu
a oportunidade de um estágio no exterior, onde esta segunda etapa do trabalho
(capítulos III e IV) foi desenvolvida em colaboração com o Professor Dr. Miquel
Lürling da Universidade de Wageningen, na Holanda e para a viabilidade desta
etapa contou-se também com a concessão pelo CNPq da bolsa sanduíche no
exterior. O capítulo III teve como objetivo verificar o efeito da temperatura no
crescimento e na competição direta (através de cultivos mistos) das espécies
fitoplanctônicas Microcystis aeruginosa, Planktothrix agardhii, Cyclotella
meneghiniana e Desmodesmus quadricauda. O capítulo IV buscou avaliar o efeito
da temperatura nas relações alelopáticas entre as três espécies: Microcystis
aeruginosa, Planktothrix agardhii e Cyclotella meneghiniana.
Por fim, os resultados das experimentações em laboratório das interações
intra e interespecíficas das espécies estudadas foram comparados com as
observações de ocorrência das espécies fitoplanctônicas dominantes na lagoa de
Jacarepaguá em uma discussão geral de todos os capítulos, com a intenção de
auxiliar no entendimento da dinâmica fitoplanctônica deste ambiente e compreender
os fatores que estão contribuindo para ocorrência de florações de cianobactérias em
ambientes eutrofizados.
Além disso, no anexo I estão apresentados os dados de uma revisão
organizada pela presente autora dos compartimentos bióticos e abióticos da lagoa

3
de Jacarepaguá, gerados desde 1996 em colaboração com vários grupos de
pesquisa em ecologia de microalgas e cianobactérias do Município do Rio de
Janeiro, que foi publicada na revista Oecologia Brasiliensis durante o período de
desenvolvimento desta tese.

4
1. INTRODUÇÃO GERAL
1.1. LAGOAS COSTEIRAS
As lagoas costeiras são ecossistemas de grande importância econômica
devido a sua alta produção pesqueira, intenso uso para aquicultura, recreação e
depósito de resíduos alóctones (Spaulding, 1994). São frequentemente apontadas
como um dos sistemas mais produtivos do mundo por apresentarem altas taxas de
produção primária e secundária (Knoppers, 1994). Ocorrem em praticamente todo
litoral brasileiro e são consideradas como um dos principais sistemas lênticos do
país (Esteves, 1988). Geralmente são caracterizadas como sistemas rasos,
separados do oceano por uma barreira terrestre e que podem ser conectados ao
mesmo, pelo menos temporariamente, por uma ou mais entradas restritas (Kjerfve,
1994). Sofrem uma forte influência dos ventos, o que leva a uma maior
homogeneização da coluna d’ água. Devido a sua proximidade com o mar, as lagoas
costeiras apresentam uma grande variação da salinidade. O tamanho e a
morfometria desses ambientes também influenciam muitas de suas características
físicas, químicas e biológicas (Panosso et al., 1998). Os diversos usos desses
sistemas aquáticos pelo homem vêm promovendo profundas alterações em suas
condições naturais. Como consequência, é comum verificar acelerados processos
de eutrofização – enriquecimento artificial dos ecossistemas aquáticos por nutrientes
a partir, sobretudo, de descargas de esgotos domésticos e industriais dos centros
urbanos e das regiões agricultáveis. Uma das principais consequências da
eutrofização é a diminuição da diversidade fitoplanctônica devido à dominância e
aumento da abundância das cianobactérias.

5
1.2. CIANOBACTÉRIAS
A origem das cianobactérias, inicialmente conhecidas como cianofíceas ou
algas azuis, foi estimada ser de cerca de 3,5 bilhões de anos. Acredita-se que as
cianobactérias tenham contribuído para oxigenação inicial da atmosfera terrestre e
esse tenha sido o seu principal papel na evolução das condições no nosso planeta
(Carmichael, 1994).
As cianobactérias são amplamente distribuídas em ecossistemas de água
doce, marinho e estuarino, pois apresentam características estruturais e metabólicas
específicas que lhes conferem grande plasticidade adaptativa. Várias espécies
também são capazes de viver em solos e rochas onde desempenham um importante
papel nos processos funcionais do ecossistema e na ciclagem de nutrientes
(Azevedo, 1998). Esses microrganismos procariotos usam a fotossíntese como o
seu principal modo de obtenção de energia para o metabolismo. Seus processos
vitais requerem somente água, dióxido de carbono, substâncias inorgânicas e luz.
Possuem clorofila-a, carotenóides e ainda outros pigmentos acessórios como as
ficobilinas: ficocianina, ficoeritrina e aloficocianina (Kromkamp, 1987). As
cianobactérias apresentam uma série de adaptações que as auxiliam a sobreviver
em ambientes em que outros microrganismos fotossintetizantes não são capazes de
sobreviver. Uma importante adaptação é a sua capacidade de fixar nitrogênio
molecular (N2). Em algumas cianobactérias filamentosas, a fixação de nitrogênio
ocorre nos heterocitos, que são células especializadas (Yoo et al., 1995). Outra
adaptação importante são os acinetos, células de resistência que permitem que as
cianobactérias sobrevivam durante condições desfavoráveis, mesmo na ausência de
luz, e depois regenerem em melhores condições. A presença de vacúolos gasosos é
uma terceira característica adaptativa importante para cianobactérias formadoras de

6
florações, como Microcystis, Anabaena e Aphanizomenon. Os vacúolos permitem
que essas cianobactérias flutuem na zona fótica e explorem de forma otimizada a
coluna d’ água (Padisák, 2004).
As células de cianobactérias são microscópicas, frequentemente menores
que 10 µm de comprimento ou diâmetro. Muitos gêneros de cianobactérias formam
colônias ou agregados filamentosos. Uma colônia pode conter centenas e até
milhares de células, atingindo milímetros de diâmetro. Florações de cianobactérias
são formadas quando milhões dessas colônias ou agregados são produzidos em um
corpo d’água sob condições favoráveis ao seu crescimento. Nem todas as
cianobactérias formam florações superficiais. Algumas cianobactérias filamentosas
são bastante sensíveis a altas intensidades luminosas e por isso migram
verticalmente na coluna d’ água. O gênero Planktothrix, por exemplo, pode existir
tanto disperso no epilímnio, quanto na zona metalimnética de um ambiente aquático
termicamente estratificado, especialmente se possuir um grande conteúdo de
pigmento vermelho (ficoeritrina) (Padisak, 2004; Reynolds, 2006).
1.3. CIANOBACTÉRIAS X INTERAÇÕES COM O AMBIENTE
As diferentes características fisiológicas das cianobactérias têm permitido a
esse grupo explorar diferentes habitats do ecossistema aquático. A sua dominância
em determinados ambientes aquáticos vem sendo associada a alguns fatores
ambientais característicos, tais como: regime de mistura com estratificação
duradoura da coluna d’água (Reynolds, 1987) ou diária (constância ambiental)
(Ganf, 1974); baixa disponibilidade luminosa (Zevenboom & Mur, 1980; Smith,
1986); reduzida razão Zona eufótica/Zona de mistura (Jensen et al., 1994); elevadas
temperaturas (Shapiro, 1990); pH elevado com baixa disponibilidade de CO2 (King,

7
1970; Shapiro, 1990; Caraco & Miller, 1998); alta concentração de P-total (McQueen
& Lean, 1987; Trimbee & Prepas, 1987; Seip, 1988; Watson et al., 1997); baixos
teores de N-total (Smith, 1983) e de nitrogênio inorgânico dissolvido (Blomqvist et
al., 1994) e baixa razão N/P (Smith, 1983). Além disso, as cianobactérias
apresentam vantagens adaptativas para estocar fósforo, capacidade de fixar
nitrogênio atmosférico, habilidade de minimizar herbivoria e de controlar sua
flutuabilidade (Pettersson et al., 1993; Haney, 1987; Reynolds, 1987, 2006, Paerl,
2006). Embora esses estudos tenham abordado somente algumas espécies de
cianobactérias, especialmente as formadoras de florações, seus resultados acabam
gerando conclusões para todo o grupo. Entretanto, as cianobactérias constituem um
grupo bastante antigo e diverso de espécies, que apresentam estratégias
ecofisiológicas muito diferentes e algumas vezes até contrastantes. Estas
generalizações nem sempre podem ser extrapoladas para todo o grupo, e, enquanto
não forem adequadamente testadas, a aplicação para todas as cianobactérias deve
ser necessariamente tratada com cautela (Marinho, 2006).
Embora florações possam ocorrer em condições favoráveis ao crescimento
fitoplanctônico, a dominância de uma única espécie em detrimento de outras irá
depender das interações entre os organismos e o seu ambiente (Oliver & Ganf,
2000). Muitas vezes, a dominância das cianobactérias é correlacionada a fatores
como condições luminosas, temperatura e disponibilidade de nutrientes (Reynolds,
1987). No entanto, para uma espécie ser capaz de formar florações ela deve ser
favorecida pelo efeito somatório desses fatores.
A temperatura é um dos fatores ambientais que mais afetam o crescimento
das microalgas e cianobactérias, uma vez que influencia diretamente processos
metabólicos relacionados à fotossíntese e outras vias biossintéticas (Robarts &

8
Zohary, 1987; Davidson, 1991; Cole & Jones, 2000). Cada espécie possui uma
temperatura ótima para seu crescimento (Raven & Geider, 1988). Além disso,
mudanças de temperatura também podem afetar as características morfológicas das
espécies, uma vez que tamanho celular e formação de colônias parecem ser
inversamente relacionados ao aumento da temperatura (Morgan & Kalff, 1979;
Lürling & Van Donk, 1999; Montagnes & Franklin, 2001; Soares, 2008). Embora a
variação anual da temperatura nos trópicos não seja tão grande quanto na região
temperada, a ocorrência de cianobactérias em muitos sistemas brasileiros tem sido
relacionada a períodos de temperaturas mais elevadas (Branco & Senna, 1994;
Bouvy et al., 2000; Huszar et al., 2000; Marinho & Huszar, 2002).
Quanto aos nutrientes, nitrogênio e fósforo são os principais elementos que
determinam a produção primária, a periodicidade e o tipo de comunidade
fitoplanctônica presente em um dado ambiente (Henry & Tundisi, 1982; Elser et al.,
1990; Elser & Goldman, 1991; González & Ortaz, 1998; González, 2000). Diversos
estudos mostram uma correlação positiva entre a concentração de clorofila-a e a
concentração de P-total e N-total em corpos d’ água (Vollenweider & Kerekes, 1980;
Bachmann, 2001). Embora tenha sido sugerido que cianobactérias possuam maior
habilidade para estocar fósforo do que outros microrganismos fotossintetizantes
(Sommer, 1985), nenhuma diferença genética entre cianobactérias e microalgas foi
encontrada para sustentar essa hipótese. No entanto, a capacidade de flutuar na
coluna d`água pela presença de aerótopos, confere às cianobactérias a vantagem
de explorar nichos com concentrações específicas de fósforo com mais eficiência.
A capacidade de fixar nitrogênio atmosférico pelas cianobactérias de água
doce da ordem Nostocales também é uma grande vantagem competitiva. Logo, as

9
cianobactérias são favorecidas em condições de deficiência de nitrogênio no sistema
(Blomqvist et al., 1994).
Portanto, identificar fatores que favoreçam o crescimento de uma espécie de
cianobactéria sobre outros organismos fitoplanctônicos torna-se bastante relevante,
principalmente no sentido de prever a ocorrência de florações desse grupo.
1.4. CIANOBACTÉRIAS X INTERAÇÕES COM OUTROS ORGANISMOS
A abundância e composição fitoplanctônicas não estão apenas relacionadas
com o ambiente físico e/ou químico, mas também com a presença de outras
espécies algais. Estas interações podem ser indiretas, envolvendo a exploração de
recursos comuns, ou diretas, quando um competidor causa danos diretamente ao
outro (Lampert & Sommer, 1997).
Alelopatia deve ser incluída como um importante fator de competição direta
entre espécies, uma vez que os efeitos das substâncias alelopáticas podem conferir
vantagens competitivas sobre outras espécies (Legrand et al., 2003). Embora
relativamente pouco estudado em ambientes aquáticos, o papel da alelopatia pode
ser importante nas mudanças na comunidade planctônica, competição e formação
de florações (Keating, 1977; Figueredo et al., 2007). Alguns estudos com
organismos fitoplanctônicos já registraram atividade alelopática em primnesiofíceas
(Fistarol et al., 2003; Granéli & Johansson, 2003), dinoflagelados (Fistarol et al.
2004; Wu et al., 1998), clorofíceas (Chiang et al., 2004) e cianobactérias (Suikkanen
et al., 2004; 2005; Fistarol et al., 2003 Leão et al., 2009). Outros trabalhos também
vêm demonstrando uma função alelopática para as toxinas de cianobactérias
(Sedmak & Kosi, 1998; Kearns & Hunter, 2000; Hu et al., 2004; Sedmak & Elersek,
2006; Bar-Yosef et al,. 2010; Leão et al., 2010;)

10
1.5. COMPOSTOS BIOATIVOS DAS CIANOBACTÉRIAS
O crescimento excessivo das cianobactérias (florações) não é indesejável
apenas por causar problemas estéticos (formação de nata verde e odores fétidos
nas margens de rios, lagos e lagoas), mas também por gerar efeitos danosos para a
biota aquática, além de sérios riscos para população humana, uma vez que vários
gêneros e espécies de cianobactérias que formam florações podem produzir
compostos tóxicos.
As cianobactérias produzem um grande número de metabólitos secundários,
que são substâncias não essenciais para o metabolismo primário. Alguns
metabólitos secundários inibem o crescimento e desenvolvimento de outros
organismos. Apesar das cianobactérias serem organismos aquáticos, muitas das
suas toxinas são mais nocivas para mamíferos do que para outros organismos
aquáticos, podendo ser letalmente tóxicas tanto para animais selvagens e
domésticos quanto para os seres humanos (Carmichael 1997; Carmichael, 2001a;
Cood et al., 2005). Esses metabólitos podem ser incluídos em três grandes grupos,
de acordo com suas estruturas químicas: os peptídeos cíclicos, os alcalóides e os
lipopolissacarídeos. Entretanto, por sua ação farmacológica, as duas principais
classes de cianotoxinas até agora caracterizadas são as neurotoxinas (anatoxina-a,
anatoxina-a(s) e saxitoxinas) e hepatotoxinas (microcistinas, nodularinas e
cilindrospermopsinas). Além desses, diversos outros grupos de compostos bioativos
podem ser produzidos por cianobactérias.
Existem vários relatos de intoxicação humana relacionados a florações de
cianobactérias na Austrália, Inglaterra, China, África do Sul (Falconer, 1994; Chorus
& Bartram, 1999). No Brasil já foram relatados dois casos: em 1988 foi constatada
uma correlação epidemiológica entre a floração de cianobactérias no Reservatório

11
de Itaparica, Bahia e a morte de 88 pessoas, entre 2000 intoxicadas (Teixeira et al.,
1993). Em outro caso, um pouco mais recente, em Caruaru, Pernambuco, 130
pessoas que faziam tratamento de hemodiálise passaram a desenvolver os sintomas
típicos de hepatotoxicoses, o que levou ao óbito de mais de 60 pessoas, sendo
detectada a presença de microcistinas (média de 223 ng/g) no sangue e nos fígados
das pessoas afetadas, assim como no filtro de carvão ativado, na areia e nas resinas
de troca iônica (valores entre 0,5 - 2,1 µg/g) da clínica de hemodiálise. Também
foram encontrados valores de até 19,7 µg/g de cilindrospermopsinas no carvão
ativado desta mesma clínica. Este foi o primeiro caso confirmado de morte humana
por intoxicação com cianotoxinas (Jochimsen et al., 1998; Carmichael et al., 2001b;
Azevedo et al., 2002).
Problemas associados a cianobactérias e suas toxinas vem aumentando
significativamente no Brasil, especialmente em regiões onde ocorre crescimento
populacional desordenado e tratamento de água e esgoto inadequados.
1.6. EUTROFIZAÇÃO NO COMPLEXO LAGUNAR DE JACAREPAGUÁ
No Complexo Lagunar de Jacarepaguá o impacto antropogênico é
evidenciado pela ocupação urbana desordenada, que resulta no aporte de efluentes
sem nenhum tipo de tratamento, além dos assoreamentos e aterros acelerados das
lagoas, causando o estrangulamento dos canais de ligação com o mar. Tal fato tem
acelerado modificações significativas na composição das águas lagunares,
aumentando excessivamente a proporção de água doce. Como consequência há
dificuldades de troca de água entre as lagoas e o mar, o que tem provocado,
periodicamente, grandes taxas de mortandade de peixes.

12
Em 2002, um estudo da Secretaria Municipal de Meio Ambiente (SMAC,
2002), responsável pelo monitoramento das águas das lagoas do município,
caracterizou o Arroio Pavuna e o Arroio Fundo como as principais fontes de poluição
do sistema lagunar da região. Os canais se localizam na lagoa do Camorim,
poluindo principalmente as lagoas adjacentes, sendo elas, Jacarepaguá e Tijuca. No
caso da lagoa de Jacarepaguá há que se destacar a intensa atividade industrial
exercida na região, cujos resíduos vão desaguar na lagoa (Feema, 1982; Feema,
1984). Cabe destacar que as lagunas desse complexo se intercomunicam,
distribuindo de forma mais ou menos homogênea as descargas que recebem
(Domingos, 2001).
O processo de eutrofização observado nesse sistema lagunar vem sendo
apontado há mais de 20 anos (Saieg-Filho, 1986), muito embora na década anterior,
Semeraro e Costa (1972) já relatassem a primeira ocorrência de florações de
cianobactérias, além de destacar a frequente ocorrência de florações de
dinoflagelados (Gymnodinium spp.), que conferiam coloração castanha às águas da
lagoa de Jacarepaguá.
Uma das principais consequências do processo de eutrofização é o aumento
relativo de cianobactérias na comunidade fitoplanctônica, resultante, dentre outros
fatores, do acúmulo de nutrientes inorgânicos dissolvidos. No complexo lagunar
Jacarepaguá-Camorim-Tijuca, florações frequentes de cianobactérias vêm sendo
registradas desde 1990 (Fernandes, 1993), inclusive sendo registrada a ocorrência
de cepas produtoras de cianotoxinas (Magalhães et al., 2001). Essa ocorrência se
tornou constante a partir de meados da década de 90 e tem sido objeto de estudo do
grupo de pesquisadores do Laboratório de Ecofisiologia e Toxicologia de
Cianobactérias/UFRJ, evidenciando o predomínio de cianobactérias nesta lagoa

13
(Domingos, 2001; Ferrão-Filho et al., 2002a, 2002b) com a presença de cepas de
Microcystis aeruginosa produtoras de microcistinas (Magalhães et al., 2001).
Entretanto, se desconhecem estudos que contemplem a proposição de limites
máximos recomendáveis de nutrientes na coluna d’água deste sistema para limitar o
crescimento de cianobactérias e sua consequente dominância no fitoplâncton.
A presença de cianobactérias tóxicas nessas lagoas também acarreta
problemas de saúde pública na região costeira onde o sistema lagunar comunica-se
com o mar. Devido à hidrodinâmica deste sistema, suas águas são lançadas em
uma praia de grande uso recreativo. No verão de 2007 foram observados valores
superiores a 0,3 µg L-1 de microcistinas dissolvidas na água da praia da Barra da
Tijuca, o que levou as autoridades estaduais de meio ambiente e saúde a interditar
um trecho desta praia.
Em face dos conhecimentos expostos acima e da escassez de dados
recentes sobre o processo de eutrofização da lagoa de Jacarepaguá, faz-se
necessário, portanto, estudar a dinâmica da ocorrência das florações de
cianobactérias, assim como identificar fatores que contribuam para o seu
estabelecimento nesse ambiente. Além disso, esses estudos propiciarão o avanço
do conhecimento sobre processos ecofisilógicos e ecotoxicológicos dos principais
grupos de cianobactérias que comumente apresentam-se como dominantes em
vários ecossistemas costeiros brasileiros.

14
2. ÁREA DE ESTUDO
2.1. LOCALIZAÇÃO
O Complexo Lagunar de Jacarepaguá localiza-se na Planície Costeira de
Jacarepaguá, no litoral sul do Estado do Rio de Janeiro e na região sudeste do
município de mesmo nome, entre os pontos de coordenadas 43º 17’ e 43º 30’ W e
22º 55’ e 23º 00’ S (Fig. 1). Esse complexo lagunar possui aproximadamente 13 km2
de extensão e é formado por três lagoas principais: Jacarepaguá, Tijuca e
Marapendi, que são bastante extensas e alongadas, e pela lagoa de Camorim –
trecho entre as lagoas de Jacarepaguá e da Tijuca (Domingos, 2001).
Figura 1: Complexo Lagunar de Jacarepaguá

15
2.2. CLIMA
A baixada de Jacarepaguá possui um microclima típico de região litorânea
tropical, influenciada pela sua localização geográfica (latitude e longitude),
proximidade com o mar, topografia local, natureza da cobertura vegetal e ação da
circulação local, como as brisas marítimas e terrestres. O clima dessa região é
tropical chuvoso megatérmico, caracterizando-se por um verão úmido e um inverno
seco (Zee, 1994). As mais altas temperaturas e os valores mais altos de
pluviosidade ocorrem no período de verão/outono (dezembro a abril) e os menores
valores ocorrem no inverno (maio a agosto).
2.3. GEOMORFOLOGIA
A Planície Costeira de Jacarepaguá tem uma extensão de 140 Km e está
encaixada entre os relevos montanhosos, a oeste, pelo Maciço da Pedra Branca e a
leste, pelo Maciço da Tijuca; tendo ainda como referência o Oceano Atlântico ao sul.
Essas montanhas são formadas por rochas do embasamento cristalino mostrando
vertentes abruptas, podendo atingir altitudes de até 1025 m. O relevo dessa região
ainda é formado pelo clinoplano periférico, que pode ser visto como a zona de
transição entre o relevo de montanhas e as planícies paludiais. O clinoplano
periférico é uma superfície com inclinação convergente para o centro da baixada,
envolvendo-a pelos flancos oeste, norte e leste. As duas planícies paludiais
existentes nessa região têm topografia plana. A planície paludial mais extensa
apresenta-se delimitada pelo clinoplano periférico e pela barreira alongada interna
(restinga interna ou restinga de Jacarepaguá) e nela se localizam as lagoas da
Tijuca, Camorim e Jacarepaguá. A planície paludial de menor área está contida
entre as barreiras alongadas interna e externas (restingas de Jacarepaguá e da

16
praia do Recreio dos Bandeirantes), e nela se encontra a lagoa de Marapendi
(Fernandes, 1993). Hoje, as regiões de restingas estão quase todas ocupadas por
construções de grandes condomínios de casas e apartamentos.
O aporte sedimentar proveniente de outras áreas é pouco significativo, uma
vez que as projeções rochosas formam barreiras ao trânsito litorâneo. As duas
zonas de progradação foram, portanto, construídas principalmente com material
exposto da plataforma continental, através de regressões e mobilizações pelas
ondas (Strag,1965).
2.4. HIDROGRAFIA
A bacia hidrográfica de Jacarepaguá é formada pelos rios que descem das
vertentes dos Maciços da Tijuca e da Pedra Branca e do escudo rochoso situado ao
norte da baixada, e pelas lagoas da Tijuca, Camorim, Jacarepaguá e Marapendi (Fig
1). A drenagem tem como destino as lagoas, em primeira instância, e em seguida o
mar. A área total da bacia hidrográfica de Jacarepaguá é de aproximadamente 402
Km2, sendo que cerca de 176 Km2 referem-se às superfícies drenadas pelos rios e
226 Km2 pelas lagoas. A lagoa de Jacarepaguá é a que recebe maior contribuição
de rios que drenam para sua bacia. Os principais rios afluentes da lagoa de
Jacarepaguá são Arroio Pavuna (5,0 Km), rio Pavuninha (4,0 Km), rio Passarinho
(2,5 Km), rio Caçambé (3,2 Km), rio Camorim (6,5 Km) e rio Marinho (3,5 Km)
(SEMADS, 2001).
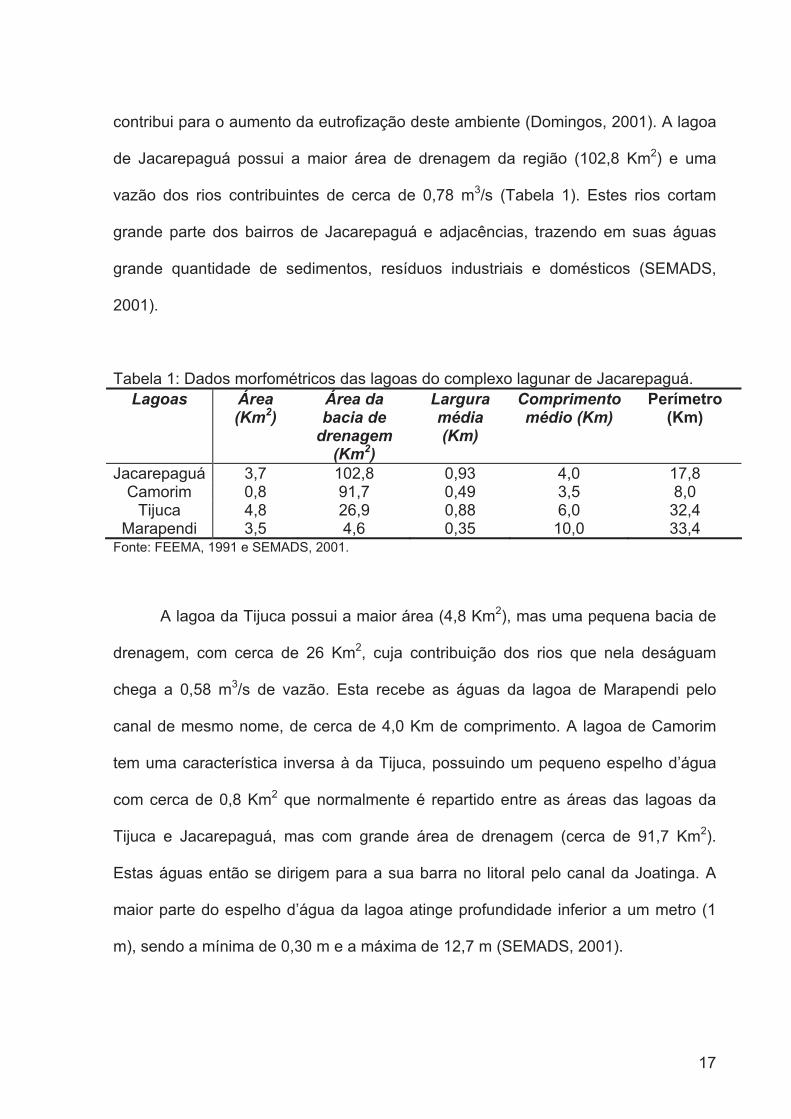
A lagoa de Jacarepaguá apresenta uma área de 3,7 Km2, profundidade média
de 3,3 m e comunica-se com o mar através das lagoas do Camorim, da Tijuca e do
canal artificial da Joatinga. Essa lagoa é a mais continental do complexo, com pouca
troca de água com o mar e apresenta maior tempo de retenção da água. Isso
17
contribui para o aumento da eutrofização deste ambiente (Domingos, 2001). A lagoa
de Jacarepaguá possui a maior área de drenagem da região (102,8 Km2) e uma
vazão dos rios contribuintes de cerca de 0,78 m3/s (Tabela 1). Estes rios cortam
grande parte dos bairros de Jacarepaguá e adjacências, trazendo em suas águas
grande quantidade de sedimentos, resíduos industriais e domésticos (SEMADS,
2001).
Tabela 1: Dados morfométricos das lagoas do complexo lagunar de Jacarepaguá. Lagoas Área
(Km2)Área da bacia de
drenagem(Km2)
Larguramédia(Km)
Comprimento médio (Km)
Perímetro(Km)
Jacarepaguá 3,7 102,8 0,93 4,0 17,8 Camorim 0,8 91,7 0,49 3,5 8,0
Tijuca 4,8 26,9 0,88 6,0 32,4 Marapendi 3,5 4,6 0,35 10,0 33,4
Fonte: FEEMA, 1991 e SEMADS, 2001.
A lagoa da Tijuca possui a maior área (4,8 Km2), mas uma pequena bacia de
drenagem, com cerca de 26 Km2, cuja contribuição dos rios que nela deságuam
chega a 0,58 m3/s de vazão. Esta recebe as águas da lagoa de Marapendi pelo
canal de mesmo nome, de cerca de 4,0 Km de comprimento. A lagoa de Camorim
tem uma característica inversa à da Tijuca, possuindo um pequeno espelho d’água
com cerca de 0,8 Km2 que normalmente é repartido entre as áreas das lagoas da
Tijuca e Jacarepaguá, mas com grande área de drenagem (cerca de 91,7 Km2).
Estas águas então se dirigem para a sua barra no litoral pelo canal da Joatinga. A
maior parte do espelho d’água da lagoa atinge profundidade inferior a um metro (1
m), sendo a mínima de 0,30 m e a máxima de 12,7 m (SEMADS, 2001).

18
3. OBJETIVO GERAL
Através de estudo de campo e de laboratório, conhecer a dinâmica sazonal
da comunidade fitoplanctônica na lagoa de Jacarepaguá, investigando os fatores
bióticos ou abióticos que influenciam na substituição de espécies e dominância de
cianobactérias neste sistema.
4. OBJETIVOS ESPECÍFICOS
CAPÍTULO I:
� Conhecer a dinâmica sazonal das espécies de cianobactérias formadoras de
florações na lagoa de Jacarepaguá;
� Identificar os possíveis fatores reguladores da ocorrência e dominância de
cianobactérias neste ambiente;
CAPÍTULO II:
� Avaliar os efeitos da redução de nutrientes na água da lagoa de Jacarepaguá
sobre a comunidade fitoplanctônica;
CAPÍTULO III:
� Através de experimentos em laboratório, avaliar a influência da temperatura
no crescimento e na competição entre quatro espécies fitoplanctônicas que
ocorrem na lagoa de Jacarepaguá;

19
CAPÍTULO IV:
� Verificar o efeito da temperatura no potencial alelopático do exudato de cepas
de cianobactérias e uma diatomácea que ocorrem com abundâncias
alternadas na lagoa de Jacarepaguá.

20
CAPÍTULO I:AVALIAÇÃO DA DINÂMICA E DOS POSSÍVEIS FATORES REGULADORES DA DOMINÂNCIA DE
ESPÉCIES DE CIANOBACTÉRIAS NA LAGOA DE JACAREPAGUÁ
INTRODUÇÃO
A dominância das cianobactérias, assim como a sua capacidade de formar
florações, está entre as principais consequências do acelerado processo de
eutrofização de lagos e reservatórios (Moss et al., 1997). Estas florações têm se
tornado cada vez mais frequentes em corpos d'água do mundo inteiro. A alta
densidade celular de cianobactérias não produz apenas problemas estéticos, devido
à densa camada de células na superfície dos corpos d'água, mas também afeta o
sabor e o odor da água potável e pode causar a morte da biota aquática devido às
condições associadas a sua senescência (p.e., redução de oxigênio dissolvido e alta
concentração de amônia) e a produção de toxinas (Paerl et al., 2001).
O entendimento das causas da dominância das cianobactérias nos ambientes
aquáticos tem sido foco de interesse de muitas pesquisas limnológicas. Determinar
fatores ambientais específicos que promovem florações de cianobactérias é uma
questão crucial para muitos pesquisadores, mas parece óbvio que um único fator
não irá servir como preditor. No entanto, o papel de alguns fatores tem sido
discutido. Schindler (1977) reportou que altas concentrações de fósforo são
favoráveis à formação de florações de cianobactérias. Smith (1983) propôs que sob
baixas razões NT:PT, as cianobactérias se tornam melhores competidoras por
nitrogênio em ambiente com escassez deste elemento. Outros fatores como regime
de mistura com estratificação duradoura da coluna d’água (Reynolds, 1987) ou diária
(constância ambiental) (Ganf, 1974); baixa disponibilidade luminosa (Zevenboom &
Mur, 1980; Smith, 1986); reduzida razão Zona eufótica/Zona de mistura (Jensen et

21
al., 1994); elevadas temperaturas (Shapiro, 1990); pH elevado com baixa
disponibilidade de CO2 (King, 1970; Shapiro, 1990; Caraco & Miller, 1998); baixos
teores de N-total (Smith, 1983) e de nitrogênio inorgânico dissolvido (Blomqvist et
al., 1994), também têm sido apontados como favoráveis à dominância de
cianobactérias. Além disso, o fato de apresentarem vantagens adaptativas para
estocar fósforo, capacidade de fixar nitrogênio atmosférico, habilidade para
minimizar herbivoria e para controlar sua flutuabilidade (Pettersson et al., 1993;
Haney, 1987; Reynolds, 1987, 2006, Paerl, 2006) e mudanças climáticas globais,
como aquecimento global e o fenômeno El Nino (Bouvy et al., 1999; Paerl &
Huisman, 2009) também fazem parte desta lista de fatores favoráveis.
A lagoa de Jacarepaguá vem sofrendo um processo de eutrofização artificial
há mais de 20 anos (Saieg-Filho, 1986) e florações frequentes de cianobactérias
vêm sendo registradas desde 1990 (Fernandes, 1993; Gomes et al., 2009), inclusive
sendo registrada a ocorrência de cepas produtoras de cianotoxinas (Magalhães et
al., 2001). Neste ambiente, as cianobactérias ocorrem sob condição de grande
estabilidade, elevadas temperaturas (Gomes et al, 2009) e elevadas concentrações
de nutrientes (Domingos, 2001). Entretanto, picos de maior diversidade
fitoplanctônica foram observados nos meses de inverno (Gomes et al., 2009). Logo,
surge a pergunta: Que fatores seriam responsáveis pela quebra dessa estabilidade e
supressão da vantagem competitiva das cianobactérias que permitiriam que
espécies de outros grupos taxonômicos ocorressem neste período? A hipótese
levantada é que, em um sistema rico em nutrientes, outros fatores estariam
controlando a dominância das cianobactérias. Portanto, o objetivo deste capítulo foi
avaliar os fatores que controlam a floração de cianobactérias e a composição
taxonômica na comunidade fitoplanctônica da lagoa de Jacarepaguá.

22
MATERIAL E MÉTODOS
ÁREA DE ESTUDO
A lagoa de Jacarepaguá, área de estudo do presente capítulo, está
devidamente descrita no item 2 desta tese.
PERIODICIDADE E ESTAÇÕES DE COLETA
Foram realizadas coletas quinzenais, durante um ano (fev/2007 a fev/2008),
em duas estações de coleta na lagoa de Jacarepaguá, em um total de 48
amostragens. As estações de amostragem foram definidas com base em
amostragem prévia e dados pré-existentes (Gomes et al., 2009).
Figura 1: Lagoa de Jacarepaguá com a marcação das duas estações de coleta.

23
ESTUDOS DE CAMPO
Medidas de Campo – O perfil térmico da coluna d’água foi obtido, através de um
termístor digital, medindo-se a temperatura da água a cada 0,2 m até o fundo da
lagoa. Valores de oxigênio dissolvido (OD), condutividade elétrica, salinidade e pH
foram obtidos com o uso de eletrodos específicos. Também foram realizados perfis
de OD, condutividade e salinidade. A transparência d’água foi estimada pela
extinção do disco de Secchi. A intensidade da luz que chega na coluna d’água foi
medida com auxílio de um quantameter.
Clorofila-a – As coletas de água foram realizadas com garrafa de Van – Dorn. Parte
da amostra coletada foi filtrada em campo, em filtros de fibra de vidro. Os filtros
foram preservados congelados para posterior análise de clorofila-a. A concentração
de clorofila-a foi obtida por espectrofotometria, seguindo a metodologia descrita em
APHA (1998). Esta técnica foi realizada a partir da extração de clorofila-a das células
retidas nos filtros com acetona 90%, durante 20-24h. Realizou-se em seguida a
centrifugação da amostra a 1000 g durante 10 minutos, sendo a absorbância do
sobrenadante determinada em espectrofotômetro a 665 e 750nm, antes e após
acidificação com solução de ácido clorídrico (HCl) 0,3 M.
Nutrientes – As amostras filtradas foram destinadas à análise de nutrientes
dissolvidos. Os nutrientes analisados foram nitrogênio amoniacal, nitrito, nitrato e
fosfato solúvel reativo. Todos os nutrientes dissolvidos foram analisados através da
metodologia de separação e análise por cromatografia de íons, utilizando o
cromatógrafo Dionex ICS-1000. A análise do fósforo total foi realizada a partir da
amostra de água não filtrada, seguida de uma digestão do seu conteúdo orgânico

24
feita em autoclave com persulfato de potássio. A metodologia foi descrita por Murphy
& Riley (1962) e consistiu na formação de um ácido complexo (ácido
fosfomolíbdico), a partir da reação entre fósforo e molibdato de amônio em meio
ácido. Este complexo é reduzido a azul de molibdênio na presença de ácido
ascórbico. A absorbância dessa solução azul foi determinada em espectrofotômetro,
em comprimento de onda de 885 nm.
Fitoplâncton – As amostras de fitoplâncton foram coletadas na sub-superfície (0,1
m) de cada estação de coleta e fixadas com solução de lugol. As populações foram
identificadas sempre que possível em nível de espécie. A densidade fitoplanctônica
(ind./ml) foi estimada segundo Utermöhl (1958). Os indivíduos (células, colônias,
cenóbios, filamentos) foram enumerados em campos aleatórios (Uhelinger, 1964),
em número suficiente para alcançar 100 indivíduos da espécie mais freqüente,
sendo o erro inferior a 20%, a um coeficiente de confiança de 95%. O biovolume
(mm3.L-1) foi estimado multiplicando-se as densidades de cada espécie pelo volume
médio de suas células, sempre que possível considerando as dimensões médias de
cerca de 25 indivíduos. O volume de cada célula foi calculado a partir de modelos
geométricos aproximados à forma dos indivíduos. A biomassa específica foi
expressa em mg L-1 (peso fresco) calculada pelo produto da densidade populacional
vezes o volume médio de cada unidade, assumindo a densidade das células
fitoplanctônicas de 1 g.cm-3.
Análise de Dados – Considerando os dados de temperatura da água, biomassa dos
principais grupos fitoplanctônicos e de nitrogênio inorgânico dissolvido, o ano de
estudo foi dividido em três períodos: período I (PI) de fev/07 a abr/07, período II (PII)

25
de mai/07 a jul/07 e o terceiro período chamado de PIII, foi de ago/07 a fev/08. Os
valores de profundidade da zona eufótica foram definidos de acordo com Cole
(1994). Os dados de nitrogênio inorgânico dissolvido (NID) foram obtidos a partir da
soma das concentrações de N-NO3- e N-NH4
+. O N-NO2- não foi utilizado na soma do
NID devido a problemas análíticos como a sobreposição do seu pico com o pico de
cloreto. Este último íon é bastante abundante na água da lagoa devido ao fato da
mesma ser uma lagoa costeira (alta salinidade). A análise de componentes
principais foi realizada com objetivo de indentificar padrões de distribuição espacial e
temporal. Apenas os dados de biomassa populacional dos principais grupos
taxonômicos da comunidade fitoplanctônica foram transformados em log (x+1). Esta
análise foi realizada utilizando o programa de Análise Multivariada para dados
Ecológicos PC-ord 5.0 (MjM Software Desing).
RESULTADOS E DISCUSSÃO
Segundo a definição de Scheffer (2004), que caracteriza uma lagoa com
profundidade média menor que 3 m como lagoa rasa, a lagoa de Jacarepaguá pode
ser descrita como tal, uma vez que sua profundidade máxima foi de 2,4 m, atingida
no mês de dezembro na estação de coleta 1 (fig.2), e a média anual da profundidade
foi de 1,5 m.
Flutuações nos valores de penetração de luz na coluna d'água da lagoa foram
observadas entre os períodos, embora os valores indiquem que este seja um
ambiente túrbido. O PII teve o maior valor de profundidade de disco de Secchi (Zsec)
(máximo: 0,7 m estação1 (E1) e 0,8 m estação2 (E2)) e consequentemente maior
zona eufótica (Zeu) (máximo: 2,2 m E1 e 1,9 m E2), porém atingindo o fundo da
lagoa nos meses de março, junho, dezembro e janeiro na E1 e maio, junho e
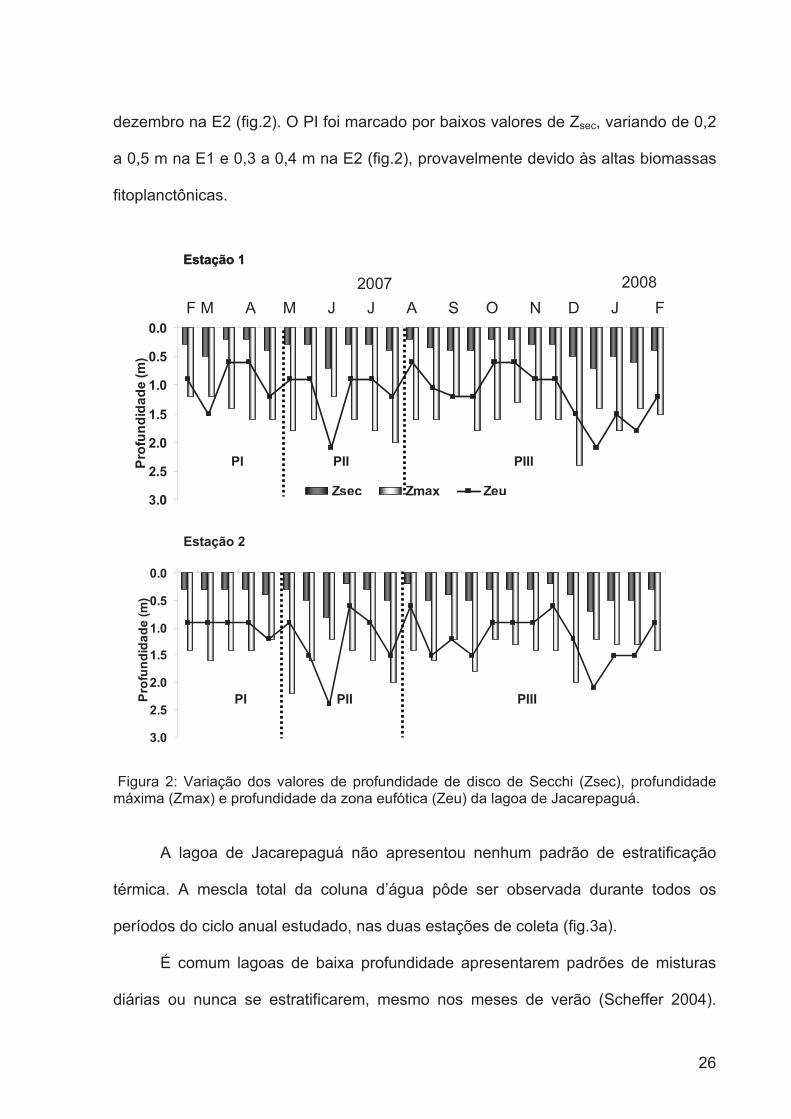
26
dezembro na E2 (fig.2). O PI foi marcado por baixos valores de Zsec, variando de 0,2
a 0,5 m na E1 e 0,3 a 0,4 m na E2 (fig.2), provavelmente devido às altas biomassas
fitoplanctônicas.
Figura 2: Variação dos valores de profundidade de disco de Secchi (Zsec), profundidade máxima (Zmax) e profundidade da zona eufótica (Zeu) da lagoa de Jacarepaguá.
A lagoa de Jacarepaguá não apresentou nenhum padrão de estratificação
térmica. A mescla total da coluna d’água pôde ser observada durante todos os
períodos do ciclo anual estudado, nas duas estações de coleta (fig.3a).
É comum lagoas de baixa profundidade apresentarem padrões de misturas
diárias ou nunca se estratificarem, mesmo nos meses de verão (Scheffer 2004).
0.0
0.5
1.0
1.5
2.0
2.5
3.0
08/02
/07
14/03
/07
30/03
/07
11/04
/07
25/04
/07
11/05
/07
30/05
/07
13/06
/07
29/06
/07
16/07
/07
31/07
/07
15/08
/07
29/08
/07
12/09
/07
26/09
/07
10/10
/07
1/11/2
007
14/11
/2007
30/11
/2007
14/12
/2007
27/12
/2007
11/1/
2008
25/1/
2008
8/2/20
08
Prof
undi
dade
(m)
Zsec Zmax Zeu
Estação 1
Estação 2
0.0
0.5
1.0
1.5
2.0
2.5
3.0
08/02
/07
14/03
/07
30/03
/07
11/04
/07
25/04
/07
11/05
/07
30/05
/07
13/06
/07
29/06
/07
16/07
/07
31/07
/07
15/08
/07
29/08
/07
12/09
/07
26/09
/07
10/10
/07
1/11/2
007
14/11
/2007
30/11
/2007
14/12
/2007
27/12
/2007
11/1/
2008
25/1/
2008
8/2/20
08
Prof
undi
dade
(m)
Zsec Zmax Zeu
PI PIIIPII
PI PIIIPII
F M A M J J A S O N D J F2007 2008
0.0
0.5
1.0
1.5
2.0
2.5
3.0
08/02
/07
14/03
/07
30/03
/07
11/04
/07
25/04
/07
11/05
/07
30/05
/07
13/06
/07
29/06
/07
16/07
/07
31/07
/07
15/08
/07
29/08
/07
12/09
/07
26/09
/07
10/10
/07
1/11/2
007
14/11
/2007
30/11
/2007
14/12
/2007
27/12
/2007
11/1/
2008
25/1/
2008
8/2/20
08
Prof
undi
dade
(m)
Zsec Zmax Zeu
Estação 1
Estação 2
0.0
0.5
1.0
1.5
2.0
2.5
3.0
08/02
/07
14/03
/07
30/03
/07
11/04
/07
25/04
/07
11/05
/07
30/05
/07
13/06
/07
29/06
/07
16/07
/07
31/07
/07
15/08
/07
29/08
/07
12/09
/07
26/09
/07
10/10
/07
1/11/2
007
14/11
/2007
30/11
/2007
14/12
/2007
27/12
/2007
11/1/
2008
25/1/
2008
8/2/20
08
Prof
undi
dade
(m)
Zsec Zmax Zeu
PI PIIIPII
PI PIIIPII
F M A M J J A S O N D J F2007 2008

27
Embora nenhuma estratificação térmica tenha sido observada, houve uma
estratificação química na lagoa de Jacarepaguá, haja visto os dados de perfil de
condutividade, salinidade e oxigênio dissolvido (OD) na água (fig.3). Em alguns
períodos do ano (PI e PIII) as concentrações de OD foram mais elevadas nas
primeiras profundidades da coluna d’água, em consequência da elevada produção
primária. Altas biomassas fitoplanctônicas foram a causa dos elevados valores de
OD e da menor penetração de luz na coluna d'água. Na maioria das amostragens, a
hipóxia ou anóxia foi registrada já a partir dos primeiros 0,50 m da coluna d’água
(Fig.3b).
Um dos fatores que apresentou maior variabilidade temporal na lagoa de
Jacarepaguá foi a temperatura da água. Esta variação esteve de acordo com a
variação climática (verão/ inverno) da região. A temperatura da água variou de 31,4
oC (valor máximo) no PI (fev – abr/07) para 18,5 oC (mínimo) no PII (mai – jul/07), na
E1. Já na E2, esta variação foi de 30,6 a 19,1 oC (fig 3; tab.1). A variação de um
pouco mais de 10 oC de temperatura da água entre os meses de inverno e verão
observada na lagoa de Jacarepaguá parece ter sido um fator que contribuiu de
forma importante para a mudança na composição e biomassa fitoplanctônica neste
ambiente. Embora alguns pesquisadores defendam que a variação anual da
temperatura nos trópicos não seja tão grande quanto na região temperada, a
dinâmica das principais espécies fitoplanctônicas em muitos sistemas brasileiros tem
sido relacionada a períodos de grandes variações de temperatura (Branco & Senna,
1994; Bouvy et al., 2000; Huszar et al., 2000; Marinho & Huszar, 2002).
A condutividade elétrica é uma medida indireta da concentração de íons na
água (Esteves, 1988). Muitos destes íons podem ser nutrientes para o fitoplâncton,
embora no caso da lagoa de Jacarepaguá, a variação da condutividade tenha

28
estado bem intimamente relacionada com a salinidade da água. Para as duas
estações de coleta, os maiores valores foram encontrados no PII (medianas:12.3 mS
cm-1, E1 e 12,9 mS cm-1, E2).
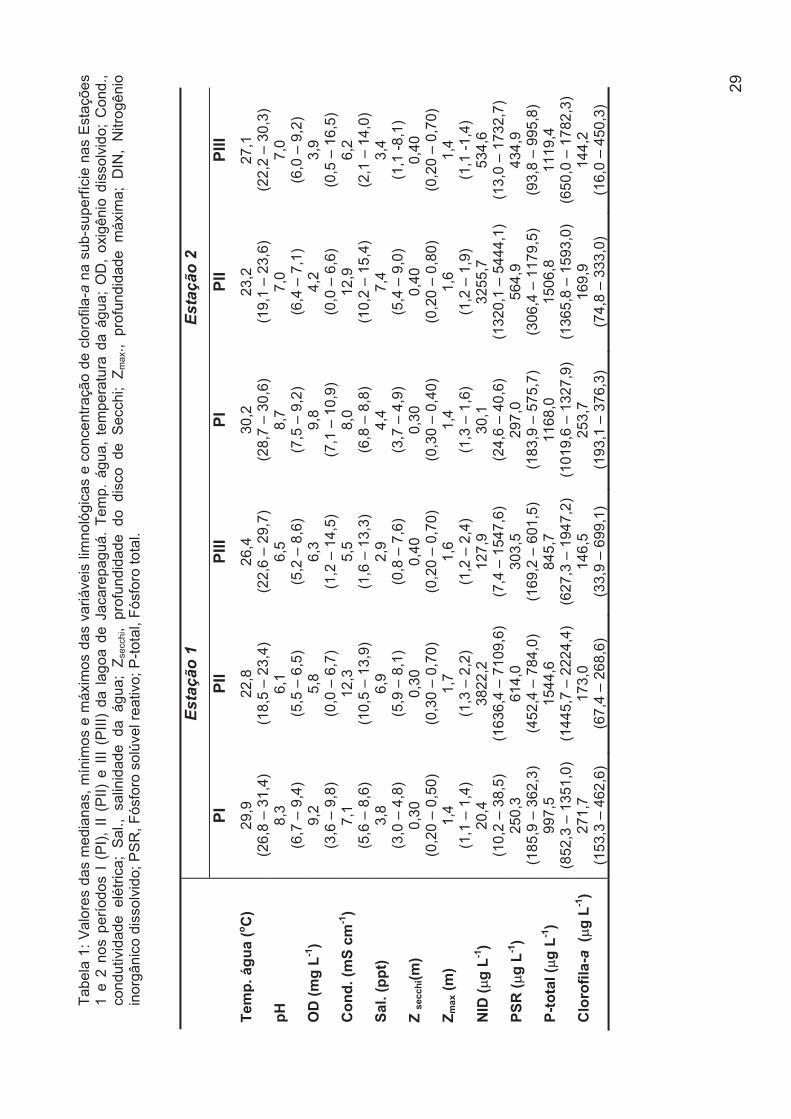
29
Tabe
la 1
: Val
ores
das
med
iana
s, m
ínim
os e
máx
imos
das
var
iáve
is li
mno
lógi
cas
e co
ncen
traçã
o de
clo
rofil
a-a
na s
ub-s
uper
fície
nas
Est
açõe
s 1
e 2
nos
perío
dos
I (P
I), I
I (P
II) e
III
(PIII
) da
lago
a de
Jac
arep
aguá
. Te
mp.
águ
a, t
empe
ratu
ra d
a ág
ua;
OD
, ox
igên
io d
isso
lvid
o; C
ond.
, co
ndut
ivid
ade
elét
rica;
Sal
., sa
linid
ade
da á
gua;
Zse
cchi,
prof
undi
dade
do
disc
o de
Sec
chi;
Z max
., pr
ofun
dida
de m
áxim
a; D
IN,
Nitr
ogên
io
inor
gâni
co d
isso
lvid
o; P
SR
, Fós
foro
sol
úvel
reat
ivo;
P-to
tal,
Fósf
oro
tota
l.
Esta
ção
1 Es
taçã
o 2
PI
PII
PIII
PI
PII
PIII
Tem
p. á
gua
(o C)
29,9
(26,
8 –
31,4
) 22
,8(1
8,5
– 23
,4)
26,4
(22,
6 –
29,7
) 30
,2(2
8,7
– 30
,6)
23,2
(19,
1 –
23,6
) 27
,1(2
2,2
– 30
,3)
pH8,
3(6
,7 –
9,4
) 6,
1(5
,5 –
6,5
) 6,
5(5
,2 –
8,6
) 8,
7(7
,5 –
9,2
) 7,
0(6
,4 –
7,1
) 7,
0(6
,0 –
9,2
) O
D (m
g L-1
)9,
2(3
,6 –
9,8
) 5,
8(0
,0 –
6,7
) 6,
3(1
,2 –
14,
5)
9,8
(7,1
– 1
0,9)
4,
2(0
,0 –
6,6
) 3,
9(0
,5 –
16,
5)
Con
d. (m
S cm
-1)
7,1
(5,6
– 8
,6)
12,3
(10,
5 –
13,9
) 5,
5(1
,6 –
13,
3)
8,0
(6,8
– 8
,8)
12,9
(10,
2 –
15,4
) 6,
2(2
,1 –
14,
0)
Sal.
(ppt
) 3,
8(3
,0 –
4,8
) 6,
9(5
,9 –
8,1
) 2,
9(0
,8 –
7,6
) 4,
4(3
,7 –
4,9
) 7,
4(5
,4 –
9,0
) 3,
4(1
,1 -8
,1)
Zse
cchi
(m)
0,30
(0,2
0 –
0,50
) 0,
30(0
,30
– 0,
70)
0,40
(0,2
0 –
0,70
) 0,
30(0
,30
– 0,
40)
0,40
(0,2
0 –
0,80
) 0,
40(0
,20
– 0,
70)
Z max
(m)
1,4
(1,1
– 1
,4)
1,7
(1,3
– 2
,2)
1,6
(1,2
– 2
,4)
1,4
(1,3
– 1
,6)
1,6
(1,2
– 1
,9)
1,4
(1,1
-1,4
) N
ID (�
g L-1
)20
,4(1
0,2
– 38
,5)
3822
,2(1
636,
4 –
7109
,6)
127,
9(7
,4 –
154
7,6)
30
,1(2
4,6
– 40
,6)
3255
,7(1
320,
1 –
5444
,1)
534,
6(1
3,0
– 17
32,7
) PS
R (�
g L-1
)25
0,3
(185
,9 –
362
,3)
614,
0(4
52,4
– 7
84,0
) 30
3,5
(169
,2 –
601
,5)
297,
0(1
83,9
– 5
75,7
) 56
4,9
(306
,4 –
117
9,5)
43
4,9
(93,
8 –
995,
8)
P-to
tal (�g
L-1
)99
7,5
(852
,3 –
135
1,0)
15
44,6
(144
5,7
– 22
24,4
) 84
5,7
(627
,3 –
194
7,2)
11
68,0
(101
9,6
– 13
27,9
) 15
06,8
(136
5,8
– 15
93,0
) 11
19,4
(650
,0 –
178
2,3)
C
loro
fila-
a (�
g L-1
)27
1,7
(153
,3 –
462
,6)
173,
0(6
7,4
– 26
8,6)
14
6,5
(33,
9 –
699,
1)
253,
7(1
93,1
– 3
76,3
) 16
9,9
(74,
8 –
333,
0)
144,
2(1
6,0
– 45
0,3)
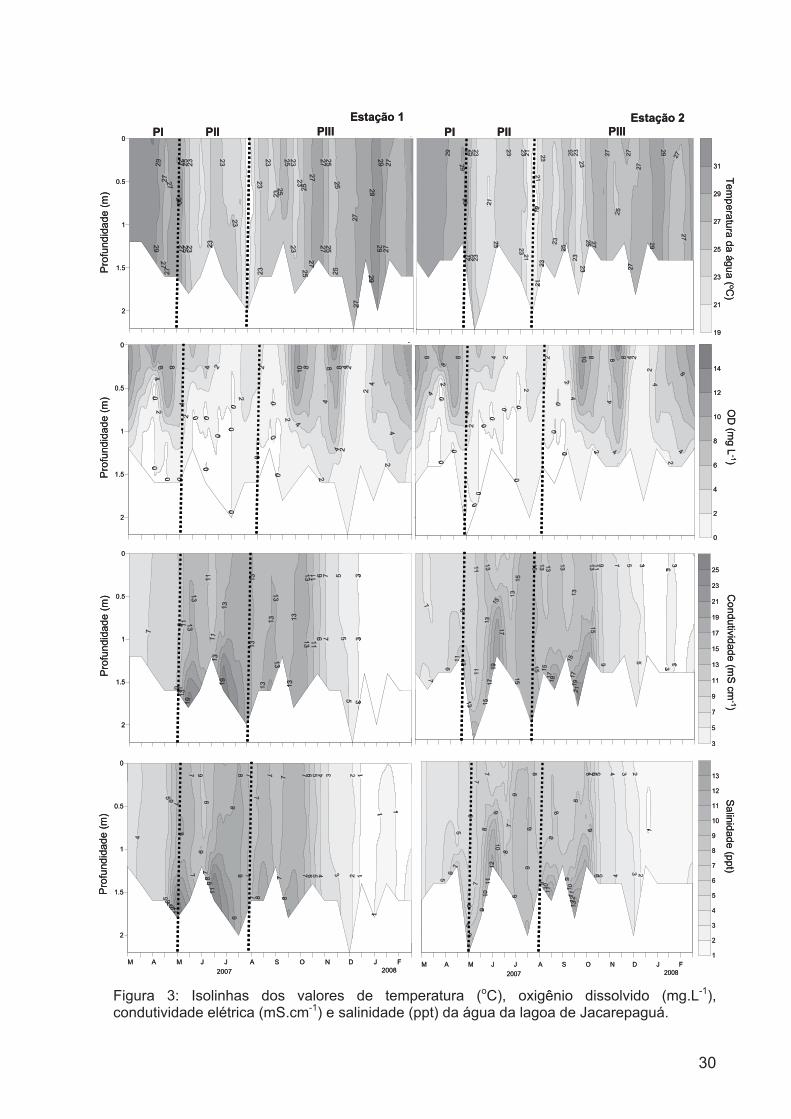
30
Figura 3: Isolinhas dos valores de temperatura (oC), oxigênio dissolvido (mg.L-1),condutividade elétrica (mS.cm-1) e salinidade (ppt) da água da lagoa de Jacarepaguá.
2
1.5
1
0.5
0
19
21
23
25
27
29
31
2
1.5
1
0.5
0
Pro
fund
idad
e (m
)
Temperatura da água ( oC
)
2
1.5
1
0.5
0
0
2
4
6
8
10
12
14
2
1.5
1
0.5
0
Pro
fund
idad
e (m
)
OD
(mg L
-1)
2
1.5
1
0.5
0
3
5
7
9
11
13
15
17
19
21
23
25
2
1.5
1
0.5
0
Pro
fund
idad
e (m
) Condutividade (m
Scm
-1)
2
1.5
1
0.5
0
M A M J J A S O N D J F2007 2008
1
2
3
4
5
6
7
8
9
10
11
12
13
2
1.5
1
0.5
0
M A M J J A S O N D J F2007 2008
Pro
fund
idad
e (m
)
Salinidade (ppt)
Estação 1 Estação 2PI PII PIII PI PII PIII
2
1.5
1
0.5
0
19
21
23
25
27
29
31
2
1.5
1
0.5
0
Pro
fund
idad
e (m
)
Temperatura da água ( oC
)
2
1.5
1
0.5
0
0
2
4
6
8
10
12
14
2
1.5
1
0.5
0
Pro
fund
idad
e (m
)
OD
(mg L
-1)
2
1.5
1
0.5
0
3
5
7
9
11
13
15
17
19
21
23
25
2
1.5
1
0.5
0
Pro
fund
idad
e (m
) Condutividade (m
Scm
-1)
2
1.5
1
0.5
0
M A M J J A S O N D J F2007 2008
1
2
3
4
5
6
7
8
9
10
11
12
13
2
1.5
1
0.5
0
M A M J J A S O N D J F2007 2008
Pro
fund
idad
e (m
)
Salinidade (ppt)
Estação 1 Estação 2PI PII PIII PI PII PIII

31
A salinidade da água da lagoa de Jacarepaguá variou desde valores próximos
a zero ppt (PIII, E1 e E2) até 9,0 ppt (PII, E2) (Fig.3; tab. 1). Em PIII, ocorreram
salinidades mais baixas nos meses de verão (período chuvoso), se comparadas aos
meses de seca do inverno, sugerindo uma possível diluição da água da lagoa pela
água da chuva. Estes valores de salinidade não se comparam com aqueles
observados nas lagoas localizadas no norte do estado do Rio de Janeiro, que
chegam a 53,4 ppt (Enrich-Prast et al. 2004), possivelmente devido à escassa
circulação da água do mar provocada pelo estrangulamento e fechamento dos
canais de comunicação da lagoa com a região costeira.
Os valores de pH nas estações 1 e 2 variaram de neutro a levemente alcalino
no PI (medianas: 8,3 - 8,7; máximo: 9,3) (Fig. 4, tab. 1). Uma considerável redução
nos valores de pH foi registrada nos meses de inverno (medianas: 6,1 - 7,0; máximo:
7,1) (PII), coincidindo com os menores valores de biomassa fitoplanctônica total e
redução da dominância de cianobactérias, observada no período anterior, e aumento
da contribuição de espécies dos grupos das clorofíceas e diatomáceas na
composição fitoplanctônica. É importante ressaltar que, embora as cianobactérias
apresentem vantagens em colonizar ambientes com pH mais elevado (Caraco &
Miller 1998), a dominância desse grupo na comunidade fitoplanctônica aqui
estudada não se restringiu a épocas em que as águas estiveram alcalinas. Na E1
durante PIII, mesmo com o retorno da elevada biomassa de cianobactérias, o
valores de pH permaneceram neutros na maioria dos meses (mediana: 6,5) (Fig. 4,
tab. 1).
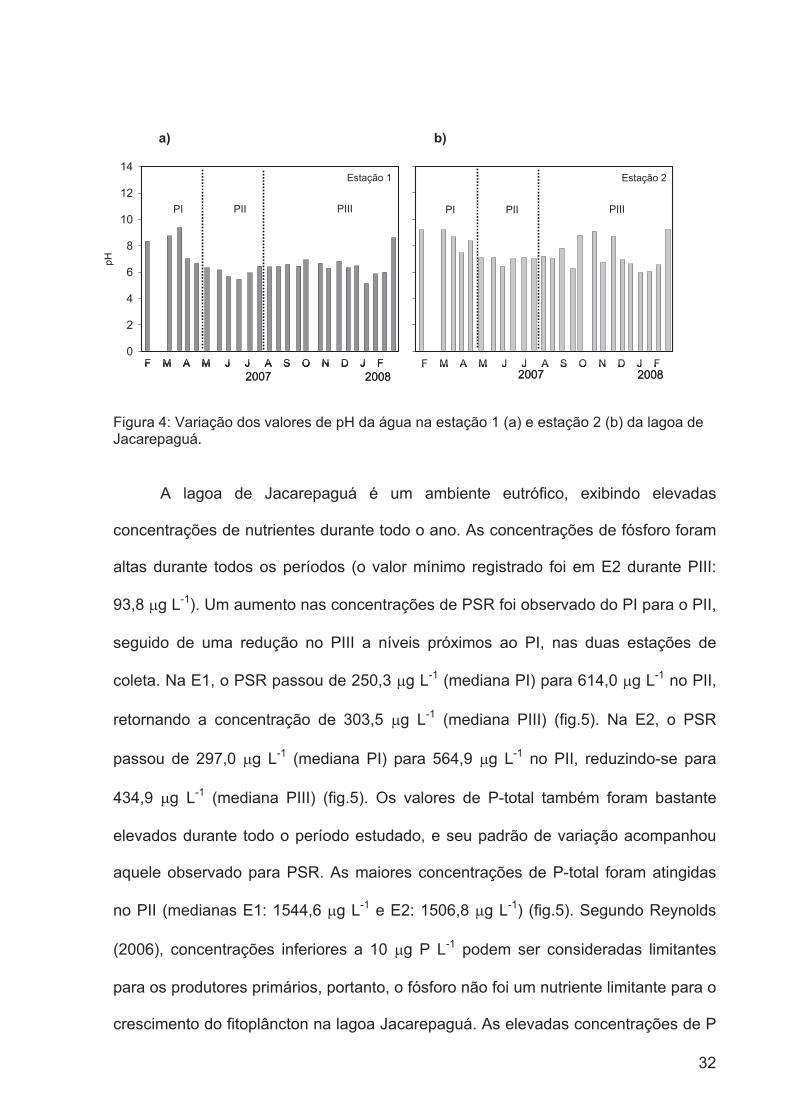
32
Figura 4: Variação dos valores de pH da água na estação 1 (a) e estação 2 (b) da lagoa de Jacarepaguá.
A lagoa de Jacarepaguá é um ambiente eutrófico, exibindo elevadas
concentrações de nutrientes durante todo o ano. As concentrações de fósforo foram
altas durante todos os períodos (o valor mínimo registrado foi em E2 durante PIII:
93,8 �g L-1). Um aumento nas concentrações de PSR foi observado do PI para o PII,
seguido de uma redução no PIII a níveis próximos ao PI, nas duas estações de
coleta. Na E1, o PSR passou de 250,3 �g L-1 (mediana PI) para 614,0 �g L-1 no PII,
retornando a concentração de 303,5 �g L-1 (mediana PIII) (fig.5). Na E2, o PSR
passou de 297,0 �g L-1 (mediana PI) para 564,9 �g L-1 no PII, reduzindo-se para
434,9 �g L-1 (mediana PIII) (fig.5). Os valores de P-total também foram bastante
elevados durante todo o período estudado, e seu padrão de variação acompanhou
aquele observado para PSR. As maiores concentrações de P-total foram atingidas
no PII (medianas E1: 1544,6 �g L-1 e E2: 1506,8 �g L-1) (fig.5). Segundo Reynolds
(2006), concentrações inferiores a 10 �g P L-1 podem ser consideradas limitantes
para os produtores primários, portanto, o fósforo não foi um nutriente limitante para o
crescimento do fitoplâncton na lagoa Jacarepaguá. As elevadas concentrações de P
a)
Estação 1
pH
0
2
4
6
8
10
12
14
F M A M J J A S O N D J F
Estação 2
F M A M J J A S O N D J F
PI PIIIPII PI PIIIPII
2007 2008 2007 2008
Estação 1
pH
0
2
4
6
8
10
12
14
F M A M J J A S O N D J F
Estação 2
F M A M J J A S O N D J F
PI PIIIPII PI PIIIPII
Estação 1
pH
0
2
4
6
8
10
12
14
F M A M J J A S O N D J F
Estação 2
F M A M J J A S O N D J F
PI PIIIPII PI PIIIPII
2007 2008 2007 2008
b)

33
observadas neste sistema são resultado do grande aporte deste elemento devido a
forte ocupação urbana e industrial em seu entorno (Silva, 2006). Também é sabido
que o P acumula-se facilmente no sedimento em ambientes eutrofizados (Jensen et
al., 1992) e que em lagoas rasas há uma forte interação entre o sedimento e a
coluna d' água (Kosten et al., 2009). A anoxia da região de interface entre a coluna
dá água e o sedimento favorece a liberação de P do sedimento para água. Este fato,
aliado à pequena profundidade do sistema e constante circulação da água, também
pode explicar as elevadas concentrações de P na lagoa durante todo período de
estudo.
As concentrações de NID nos meses de verão do ano de 2007 (PI) não
ultrapassaram 40 �g L-1 nas duas estações de coleta (fig 6; tab.1), ou seja,
estiveram sempre abaixo de 100 �g L-1, que é a concentração considerada por
Reynolds (2006) como limitante para o crescimento do fitoplâncton. Possivelmente,
este foi um período em que as elevadas biomassas fitoplanctônicas (mediana: 271,7
�g L-1 E1 e 253,7 �g L-1 E2) (fig. 7) estavam consumindo a maior parte do nitrogênio
disponível. Quando houve o colapso da comunidade fitoplanctônica, o nitrogênio
incorporado foi liberado para o sistema. Este período (PII) foi marcado pelo drástico
aumento da concentração de NID (mediana: 3822,2 �g N L-1 E1 e 3255,7 �g N L-1
E2) (fig.8). Além da lise celular, a decomposição da matéria orgânica também
normalmente permite a liberação do nitrogênio na forma de amônia (Scheffer, 2004)
e mais de 90% da concentração de NID era composta por esta molécula. A amônia
é a forma preferida de NID do fitoplâncton devido as suas vantagens energéticas
(McCarthy et al., 1982; Miyazaki et al., 1989; Mallet et al., 1998), uma vez que o
nitrato tem que ser reduzido a amônia para ser utilizado metabolicamente pelas
células (Wetzel 1983; Reynolds, 1984). Uma vez liberado no sistema, a
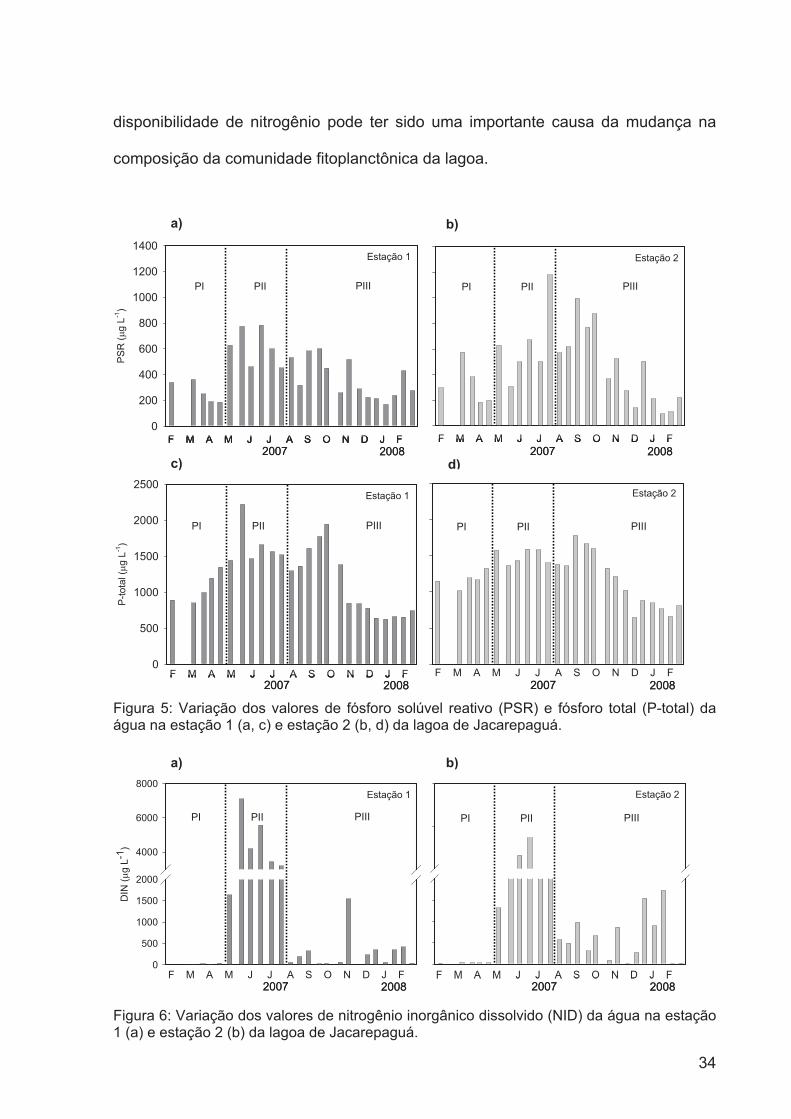
34
disponibilidade de nitrogênio pode ter sido uma importante causa da mudança na
composição da comunidade fitoplanctônica da lagoa.
Figura 5: Variação dos valores de fósforo solúvel reativo (PSR) e fósforo total (P-total) da água na estação 1 (a, c) e estação 2 (b, d) da lagoa de Jacarepaguá.
Figura 6: Variação dos valores de nitrogênio inorgânico dissolvido (NID) da água na estação 1 (a) e estação 2 (b) da lagoa de Jacarepaguá.
d)c)
b)a)
b)a)
Estação 1
PSR
(�g
L-1)
0
200
400
600
800
1000
1200
1400
F M A M J J A S O N D J F
Estação 2
F M A M J J A S O N D J F
PI PIIIPII PI PIIIPII
20082007 20082007
Estação 1
PSR
(�g
L-1)
0
200
400
600
800
1000
1200
1400
F M A M J J A S O N D J F
Estação 2
F M A M J J A S O N D J F
PI PIIIPII PI PIIIPII
Estação 1
PSR
(�g
L-1)
0
200
400
600
800
1000
1200
1400
F M A M J J A S O N D J F
Estação 2
F M A M J J A S O N D J F
PI PIIIPII PI PIIIPII
20082007 20082007
Estação 1
P-to
tal (�g
L-1
)
0
500
1000
1500
2000
2500
F M A M J J A S O N D J F
Estação 2
F M A M J J A S O N D J F
PI PIIIPII PI PIIIPII
20082007 20082007
Estação 1
P-to
tal (�g
L-1
)
0
500
1000
1500
2000
2500
F M A M J J A S O N D J F
Estação 2
F M A M J J A S O N D J F
PI PIIIPII PI PIIIPII
Estação 1
P-to
tal (�g
L-1
)
0
500
1000
1500
2000
2500
F M A M J J A S O N D J F
Estação 2
F M A M J J A S O N D J F
PI PIIIPII PI PIIIPII
20082007 20082007
Estação 1
DIN
(�g
L-1)
0
500
1000
1500
2000
4000
6000
8000
F M A M J J A S O N D J F
Estação 2
F M A M J J A S O N D J F
PI PIIIPII PI PIIIPII
2008200720082007
Estação 1
DIN
(�g
L-1)
0
500
1000
1500
2000
4000
6000
8000
F M A M J J A S O N D J F
Estação 2
F M A M J J A S O N D J F
PI PIIIPII PI PIIIPII
Estação 1
DIN
(�g
L-1)
0
500
1000
1500
2000
4000
6000
8000
F M A M J J A S O N D J F
Estação 2
F M A M J J A S O N D J F
PI PIIIPII PI PIIIPII
2008200720082007

35
A comunidade fitoplanctônica foi dominada por espécies do grupo das
cianobactérias em todos os períodos, com uma única exceção em agosto/07 onde
uma espécie de diatomácea, Thalassiosira sp., apresentou uma contribuição relativa
maior que 50% (fig.7). O PII foi caracterizado como um período de menores
biomassas e de maior contribuição de espécies de outros grupos taxonômicos para
a composição do fitoplâncton. O grupo das bacilariofíceas foi o segundo grupo de
maior contribuição (fig.7). A mudança na proporção de espécies fitoplanctônicas foi
coincidente com uma expressiva variação da temperatura da água (fig.7). Embora
dominantes quase todo o tempo, a elevada temperatura, aliada à alta disponibilidade
de nutrientes, parece ter favorecido ainda mais o crescimento de espécies de
cianobactérias. Alguns autores vêm reportando a importância da temperatura como
fator determinante para a dominância de cianobactérias, mesmo em ambientes
tropicais (Bouvy et al., 2000; Huszar et al., 2000; Marinho & Huszar, 2002; Soares et
al., 2009). Quando a temperatura passou de aproximadamente 30 oC para em torno
de 18 oC no PII, a população de cianobactérias entrou em colapso, disponibilizando
nitrogênio, principalmente na forma de amônia, para o sistema e deu oportunidade
para o crescimento de espécies dos demais grupos. No PIII, a temperatura voltou a
aumentar e as cianobactérias voltaram a dominar quase completamente a
comunidade fitoplanctônica.
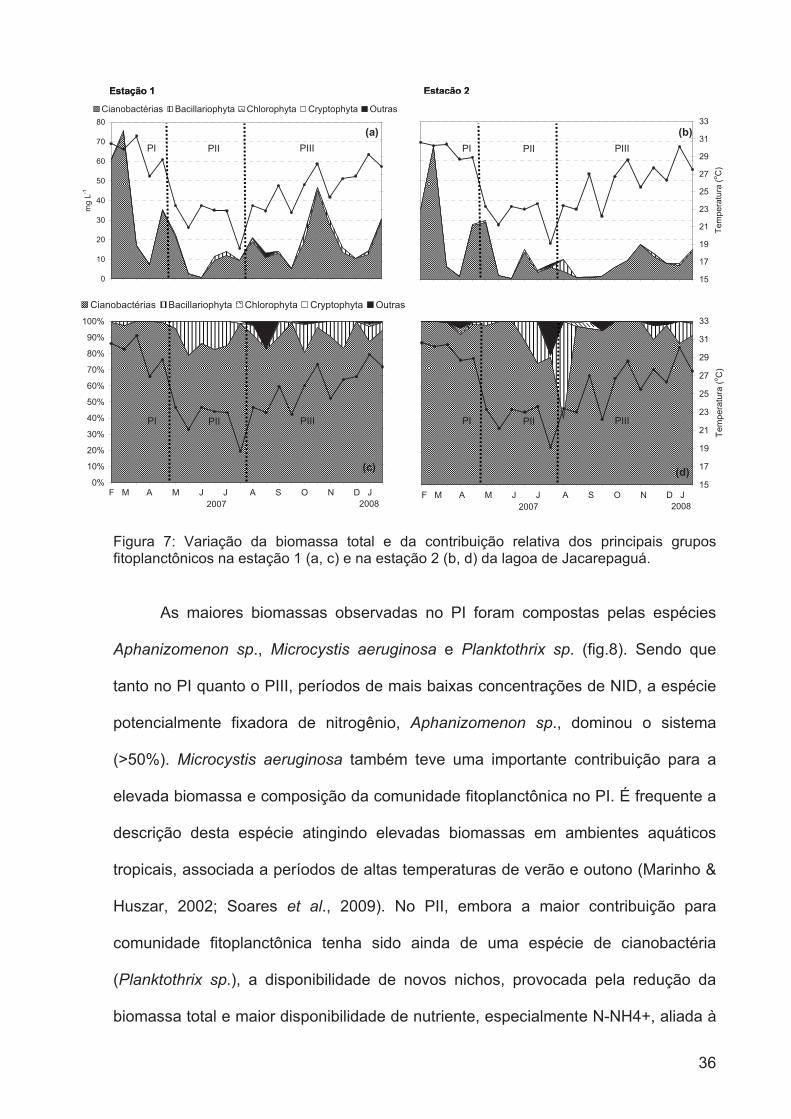
36
Figura 7: Variação da biomassa total e da contribuição relativa dos principais grupos fitoplanctônicos na estação 1 (a, c) e na estação 2 (b, d) da lagoa de Jacarepaguá.
As maiores biomassas observadas no PI foram compostas pelas espécies
Aphanizomenon sp., Microcystis aeruginosa e Planktothrix sp. (fig.8). Sendo que
tanto no PI quanto o PIII, períodos de mais baixas concentrações de NID, a espécie
potencialmente fixadora de nitrogênio, Aphanizomenon sp., dominou o sistema
(>50%). Microcystis aeruginosa também teve uma importante contribuição para a
elevada biomassa e composição da comunidade fitoplanctônica no PI. É frequente a
descrição desta espécie atingindo elevadas biomassas em ambientes aquáticos
tropicais, associada a períodos de altas temperaturas de verão e outono (Marinho &
Huszar, 2002; Soares et al., 2009). No PII, embora a maior contribuição para
comunidade fitoplanctônica tenha sido ainda de uma espécie de cianobactéria
(Planktothrix sp.), a disponibilidade de novos nichos, provocada pela redução da
biomassa total e maior disponibilidade de nutriente, especialmente N-NH4+, aliada à
Estação 1 Estação 2
07
Mar./07
Mar./07
Abr./07
Abr./07
Mai./07
Mai./07
Jun./
07
Jun./
07
Jul./0
7
Jul./0
7
Ago./0
7
Ago./0
7
Set./07
Set./07
Out./07
Out./07
Nov./0
7
Nov./0
7
Dez./0
7
Jan./
08
Jan./
0815
17
19
21
23
25
27
29
31
33
Tem
pera
tura
(o C)
nobactérias Bacillariophyta Chlorophyta Cryptophyta Outros
0
10
20
30
40
50
60
70
80
Fev./0
7
Mar./07
Mar./07
Abr./07
Abr./07
Mai./07
Mai./07
Jun./
07
Jun./
07
Jul./0
7
Jul./0
7
Ago./0
7
Ago./0
7
Set./07
Set./07
Out./07
Out./07
Nov./0
7
Nov./0
7
Dez./0
7
Jan./
08
Jan./
08
mg
L-1
15
17
19
21
23
25
27
29
31
33
Tem
pera
tura
(o C)
Cianobactérias Bacillariophyta Chlorophyta Cryptophyta Outros
0
10
20
30
40
50
60
70
80
Fev./0
7
Mar./07
Mar./07
Abr./07
Abr./07
Mai./07
Mai./07
Jun./
07
Jun./
07
Jul./0
7
Jul./0
7
Ago./0
7
Ago./0
7
Set./07
Set./07
Out./07
Out./07
Nov./0
7
Nov./0
7
Dez./0
7
Jan./
08
Jan./
08
mg
L-1
15
17
19
21
23
25
27
29
31
33
Tem
pera
tura
(o C)
Cianobactérias Bacillariophyta Chlorophyta Cryptophyta Outras
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
Fev./0
7
Mar./07
Mar./07
Abr./07
Abr./07
Mai./07
Mai./07
Jun./
07
Jun./
07
Jul./0
7
Jul./0
7
Ago./0
7
Ago./0
7
Set./07
Set./07
Out./07
Out./07
Nov./0
7
Nov./0
7
Dez./0
7
Jan./
08
Jan./
0815
17
19
21
23
25
27
29
31
33
Tem
pera
tura
(oC
)
Cianobactérias Bacillariophyta Chlorophyta Cryptophyta Outras
(a) (b)
(c) (d)
PI PII PIII PI PII PIII
PI PII PIII PI PII PIII
F M A M J J A S O N D J2007 2008
F M A M J J A S O N D J2007 2008
Estação 1 Estação 2
07
Mar./07
Mar./07
Abr./07
Abr./07
Mai./07
Mai./07
Jun./
07
Jun./
07
Jul./0
7
Jul./0
7
Ago./0
7
Ago./0
7
Set./07
Set./07
Out./07
Out./07
Nov./0
7
Nov./0
7
Dez./0
7
Jan./
08
Jan./
0815
17
19
21
23
25
27
29
31
33
Tem
pera
tura
(o C)
nobactérias Bacillariophyta Chlorophyta Cryptophyta Outros
0
10
20
30
40
50
60
70
80
Fev./0
7
Mar./07
Mar./07
Abr./07
Abr./07
Mai./07
Mai./07
Jun./
07
Jun./
07
Jul./0
7
Jul./0
7
Ago./0
7
Ago./0
7
Set./07
Set./07
Out./07
Out./07
Nov./0
7
Nov./0
7
Dez./0
7
Jan./
08
Jan./
08
mg
L-1
15
17
19
21
23
25
27
29
31
33
Tem
pera
tura
(o C)
Cianobactérias Bacillariophyta Chlorophyta Cryptophyta Outros
0
10
20
30
40
50
60
70
80
Fev./0
7
Mar./07
Mar./07
Abr./07
Abr./07
Mai./07
Mai./07
Jun./
07
Jun./
07
Jul./0
7
Jul./0
7
Ago./0
7
Ago./0
7
Set./07
Set./07
Out./07
Out./07
Nov./0
7
Nov./0
7
Dez./0
7
Jan./
08
Jan./
08
mg
L-1
15
17
19
21
23
25
27
29
31
33
Tem
pera
tura
(o C)
Cianobactérias Bacillariophyta Chlorophyta Cryptophyta Outras
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
Fev./0
7
Mar./07
Mar./07
Abr./07
Abr./07
Mai./07
Mai./07
Jun./
07
Jun./
07
Jul./0
7
Jul./0
7
Ago./0
7
Ago./0
7
Set./07
Set./07
Out./07
Out./07
Nov./0
7
Nov./0
7
Dez./0
7
Jan./
08
Jan./
0815
17
19
21
23
25
27
29
31
33
Tem
pera
tura
(oC
)
Cianobactérias Bacillariophyta Chlorophyta Cryptophyta Outras
07
Mar./07
Mar./07
Abr./07
Abr./07
Mai./07
Mai./07
Jun./
07
Jun./
07
Jul./0
7
Jul./0
7
Ago./0
7
Ago./0
7
Set./07
Set./07
Out./07
Out./07
Nov./0
7
Nov./0
7
Dez./0
7
Jan./
08
Jan./
0815
17
19
21
23
25
27
29
31
33
Tem
pera
tura
(o C)
nobactérias Bacillariophyta Chlorophyta Cryptophyta Outros
0
10
20
30
40
50
60
70
80
Fev./0
7
Mar./07
Mar./07
Abr./07
Abr./07
Mai./07
Mai./07
Jun./
07
Jun./
07
Jul./0
7
Jul./0
7
Ago./0
7
Ago./0
7
Set./07
Set./07
Out./07
Out./07
Nov./0
7
Nov./0
7
Dez./0
7
Jan./
08
Jan./
08
mg
L-1
15
17
19
21
23
25
27
29
31
33
Tem
pera
tura
(o C)
Cianobactérias Bacillariophyta Chlorophyta Cryptophyta Outros
0
10
20
30
40
50
60
70
80
Fev./0
7
Mar./07
Mar./07
Abr./07
Abr./07
Mai./07
Mai./07
Jun./
07
Jun./
07
Jul./0
7
Jul./0
7
Ago./0
7
Ago./0
7
Set./07
Set./07
Out./07
Out./07
Nov./0
7
Nov./0
7
Dez./0
7
Jan./
08
Jan./
08
mg
L-1
15
17
19
21
23
25
27
29
31
33
Tem
pera
tura
(o C)
Cianobactérias Bacillariophyta Chlorophyta Cryptophyta Outras
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
Fev./0
7
Mar./07
Mar./07
Abr./07
Abr./07
Mai./07
Mai./07
Jun./
07
Jun./
07
Jul./0
7
Jul./0
7
Ago./0
7
Ago./0
7
Set./07
Set./07
Out./07
Out./07
Nov./0
7
Nov./0
7
Dez./0
7
Jan./
08
Jan./
0815
17
19
21
23
25
27
29
31
33
Tem
pera
tura
(oC
)
Cianobactérias Bacillariophyta Chlorophyta Cryptophyta Outras
(a) (b)
(c) (d)
PI PII PIII PI PII PIII
PI PII PIII PI PII PIII
F M A M J J A S O N D J2007 2008
F M A M J J A S O N D J2007 2008

37
diminuição da temperatura, permitiu o crescimento da espécie Cyclotella sp. (figs. 8b
e 8d). Zhang & Prepas (1996) associaram a dominância de diatomáceas na lagoa
Alberta (Canadá) à baixa estabilidade da coluna d'água e a baixas temperaturas da
água. Embora a circulação da água tenha sido constante ao longo do ano, no PII, a
temperatura da água na lagoa de Jacarepaguá variou de 18,5 a 23,5 oC na E1 e de
19,1 a 23,6 oC na E2, as menores registradas neste sistema. A alta concentração de
NID no PII, também permitiu o aumento da biomassa da espécie de cianobactéria
não fixadora de nitrogênio Planktothrix sp. no final deste período.
No terceiro período (PIII), as biomassas totais voltaram a aumentar,
especialmente na E1, atingindo valores próximos a 50 mg L-1. Durante os primeiros
meses desse período, a espécie Synecocystis aquatilis dominou a comunidade
fitoplanctônica (fig. 8). Segundo Reynolds (1988, 1997, 2006), as espécies C
estrategistas, grupo no qual a espécie S. aquatilis pode ser inserida, têm uma alta
razão superfície/volume e consequentemente altas taxas de crescimento, além de
mais alto requerimento por nutrientes. Portanto, as elevadas concentrações de
nitrogênio no PII parecem ter favorecido o crescimento dessa espécie no período
seguinte. Possivelmente, o crescimento de S. aquatilis estava sendo limitado por
nitrogênio nos períodos anteriores (especialmente no PI), quando outras espécies de
cianobactérias S estrategistas (tolerantes ao estresse), que suportam altas
intensidades luminosas e têm um menor requerimento por nutrientes, dominaram o
sistema em temperaturas elevadas.
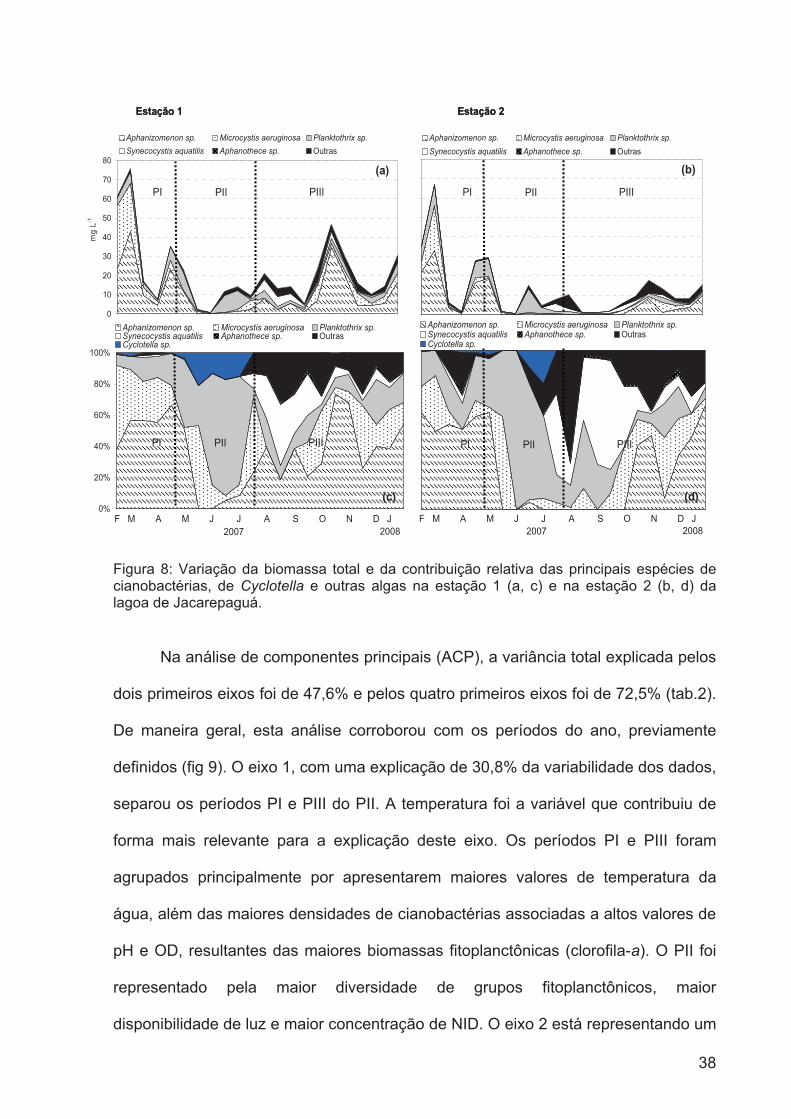
38
Figura 8: Variação da biomassa total e da contribuição relativa das principais espécies de cianobactérias, de Cyclotella e outras algas na estação 1 (a, c) e na estação 2 (b, d) da lagoa de Jacarepaguá.
Na análise de componentes principais (ACP), a variância total explicada pelos
dois primeiros eixos foi de 47,6% e pelos quatro primeiros eixos foi de 72,5% (tab.2).
De maneira geral, esta análise corroborou com os períodos do ano, previamente
definidos (fig 9). O eixo 1, com uma explicação de 30,8% da variabilidade dos dados,
separou os períodos PI e PIII do PII. A temperatura foi a variável que contribuiu de
forma mais relevante para a explicação deste eixo. Os períodos PI e PIII foram
agrupados principalmente por apresentarem maiores valores de temperatura da
água, além das maiores densidades de cianobactérias associadas a altos valores de
pH e OD, resultantes das maiores biomassas fitoplanctônicas (clorofila-a). O PII foi
representado pela maior diversidade de grupos fitoplanctônicos, maior
disponibilidade de luz e maior concentração de NID. O eixo 2 está representando um
0
10
20
30
40
50
60
70
80
Fev./0
7
Mar./07
Mar./07
Abr./07
Abr./07
Mai./07
Mai./07
Jun./
07
Jun./
07Ju
l./07
Jul./0
7
Ago./0
7
Ago./0
7
Set./07
Set./07
Out./07
Out./07
Nov./0
7
Nov./0
7
Dez./0
7
Jan./
08
Jan./
08
mg
L-1
Aphanizomenon sp. Microcystis aeruginosa Planktothrix sp.Synecocystis aquatilis Aphanothece sp. Outras
0
10
20
30
40
50
60
70
80
Fev./0
7
Mar./07
Mar./07
Abr./07
Abr./07
Mai./07
Mai./07
Jun./
07
Jun./
07Ju
l./07
Jul./0
7
Ago./0
7
Ago./0
7
Set./07
Set./07
Out./07
Out./07
Nov./0
7
Nov./0
7
Dez./0
7
Jan./
08
Jan./
08
mg
L-1
Aphanizomenon sp. Microcystis aeruginosa Planktothrix sp.Synecocystis aquatilis Aphanothece sp. Outras
0%
20%
40%
60%
80%
100%
Fev./0
7
Mar./07
Mar./07
Abr./07
Abr./07
Mai./07
Mai./07
Jun./
07
Jun./
07Ju
l./07
Jul./0
7
Ago./0
7
Ago./0
7
Set./07
Set./07
Out./07
Out./07
Nov./07
Nov./0
7
Dez./0
7
Jan./
08
Jan./
08
Aphanizomenon sp. Microcystis aeruginosa Planktothrix sp.Synecocystis aquatilis Aphanothece sp. OutrasCyclotella sp.
0%
20%
40%
60%
80%
100%
Fev./0
7
Mar./07
Mar./07
Abr./07
Abr./07
Mai./07
Mai./07
Jun./
07
Jun./
07Ju
l./07
Jul./0
7
Ago./0
7
Ago./0
7
Set./07
Set./07
Out./07
Out./07
Nov./0
7
Nov./0
7
Dez./07
Jan./
08
Jan./
08
Aphanizomenon sp. Microcystis aeruginosa Planktothrix sp.Synecocystis aquatilis Aphanothece sp. OutrasCyclotella sp.
PI PII PIII
F M A M J J A S O N D J2007 2008
F M A M J J A S O N D J2007 2008
PI PII PIII
PI PII PIII PI PII PIII
Estação 1 Estação 2
(a) (b)
(c) (d)
0
10
20
30
40
50
60
70
80
Fev./0
7
Mar./07
Mar./07
Abr./07
Abr./07
Mai./07
Mai./07
Jun./
07
Jun./
07Ju
l./07
Jul./0
7
Ago./0
7
Ago./0
7
Set./07
Set./07
Out./07
Out./07
Nov./0
7
Nov./0
7
Dez./0
7
Jan./
08
Jan./
08
mg
L-1
Aphanizomenon sp. Microcystis aeruginosa Planktothrix sp.Synecocystis aquatilis Aphanothece sp. Outras
0
10
20
30
40
50
60
70
80
Fev./0
7
Mar./07
Mar./07
Abr./07
Abr./07
Mai./07
Mai./07
Jun./
07
Jun./
07Ju
l./07
Jul./0
7
Ago./0
7
Ago./0
7
Set./07
Set./07
Out./07
Out./07
Nov./0
7
Nov./0
7
Dez./0
7
Jan./
08
Jan./
08
mg
L-1
Aphanizomenon sp. Microcystis aeruginosa Planktothrix sp.Synecocystis aquatilis Aphanothece sp. Outras
0%
20%
40%
60%
80%
100%
Fev./0
7
Mar./07
Mar./07
Abr./07
Abr./07
Mai./07
Mai./07
Jun./
07
Jun./
07Ju
l./07
Jul./0
7
Ago./0
7
Ago./0
7
Set./07
Set./07
Out./07
Out./07
Nov./07
Nov./0
7
Dez./0
7
Jan./
08
Jan./
08
Aphanizomenon sp. Microcystis aeruginosa Planktothrix sp.Synecocystis aquatilis Aphanothece sp. OutrasCyclotella sp.
0%
20%
40%
60%
80%
100%
Fev./0
7
Mar./07
Mar./07
Abr./07
Abr./07
Mai./07
Mai./07
Jun./
07
Jun./
07Ju
l./07
Jul./0
7
Ago./0
7
Ago./0
7
Set./07
Set./07
Out./07
Out./07
Nov./0
7
Nov./0
7
Dez./07
Jan./
08
Jan./
08
Aphanizomenon sp. Microcystis aeruginosa Planktothrix sp.Synecocystis aquatilis Aphanothece sp. OutrasCyclotella sp.
PI PII PIII
F M A M J J A S O N D J2007 2008
F M A M J J A S O N D J2007 2008
PI PII PIII
PI PII PIII PI PII PIII
Estação 1 Estação 2
(a) (b)
(c) (d)

39
gradiente de trofia do sistema. Ele está positivamente ligado às maiores
profundidade de Secchi, maior diversidade fitoplanctônica e negativamente ligado às
elevadas concentrações de clorofila-a, PSR e P-total, além das medidas mais
indiretas de eutrofização, que são condutividade elétrica e salinidade. Cabe ressaltar
a posição do eixo NID, quase diametralmente oposta às cianobactérias,
evidenciando mais uma vez que o colapso da sua população, possivelmente,
promoveu o aumento da concentração de nitrogênio. Nos períodos de maior
densidade de cianobactérias, as concentrações de NID foram baixas, devido ao seu
consumo.
Figura 9: Análise de Componentes Principais (ACP) das variáveis limnológicas das estações de coleta 1 e 2 da lagoa de Jacarepaguá. Zmax: profundidade máxima, Zsecchi: profundidade de disco de Secchi, Temp: temperatura da água, cond: condutividade elétrica, sal: salinidade da água, NID: nitrogênio inorgânico dissolvido, PSR: fósforo solúvel reativo, P-total: fósforo total, clo-a: clorofila-a, Cyan: Cianobactérias, Chlor: Chlorophyta, Bacil: Bacillariophyta, Cryp: Cryptophyta, Outras: outras algas.
Gra
u de
Tro
fia
PI e PIIIPII
Gra
u de
Tro
fia
PI e PIIIPII
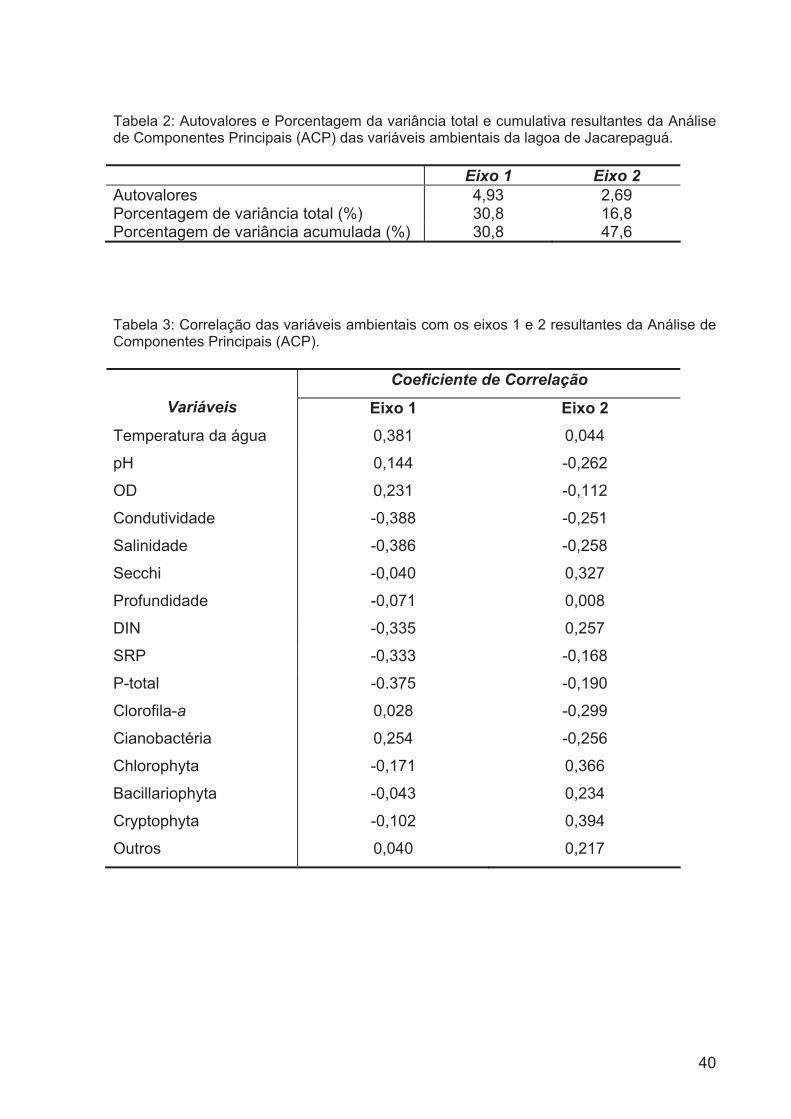
40
Tabela 2: Autovalores e Porcentagem da variância total e cumulativa resultantes da Análise de Componentes Principais (ACP) das variáveis ambientais da lagoa de Jacarepaguá.
Eixo 1 Eixo 2 Autovalores 4,93 2,69 Porcentagem de variância total (%) 30,8 16,8 Porcentagem de variância acumulada (%) 30,8 47,6
Tabela 3: Correlação das variáveis ambientais com os eixos 1 e 2 resultantes da Análise de Componentes Principais (ACP).
Coeficiente de Correlação Variáveis Eixo 1 Eixo 2
Temperatura da água 0,381 0,044
pH 0,144 -0,262
OD 0,231 -0,112
Condutividade -0,388 -0,251
Salinidade -0,386 -0,258
Secchi -0,040 0,327
Profundidade -0,071 0,008
DIN -0,335 0,257
SRP -0,333 -0,168
P-total -0.375 -0,190
Clorofila-a 0,028 -0,299
Cianobactéria 0,254 -0,256
Chlorophyta -0,171 0,366
Bacillariophyta -0,043 0,234
Cryptophyta -0,102 0,394
Outros 0,040 0,217

41
CONSIDERAÇÕES FINAIS:
A dominância constante de cianobactérias na lagoa de Jacarepaguá está
estreitamente relacionada com o intenso processo de eutrofização deste sistema. A
elevada biomassa fitoplanctônica, nos períodos I e III, causou diminuição na
disponibilidade de luz por auto-sombreamento e aumento nos valores de pH. As
elevadas concentrações de OD também foram associadas às intensas atividades
fotossintéticas nas camadas superficiais da lagoa.
Embora as concentrações de P-total e clorofila-a permitam a classificação da
lagoa de Jacarepaguá como hipereutrófica, além da alta disponibilidade de PSR
durante todo o ano e elevadas biomassas terem sido observadas, o crescimento
fitoplanctônico parece ser limitado por nitrogênio em alguns meses.
A temperatura foi o fator de maior variação temporal. Nos meses de verão a
temperatura da água atingiu valores máximos de até 30 oC, mas sofreu uma redução
atingindo valores mínimos de 18 oC nos meses de inverno. Os períodos de menor
temperatura da água coincidiram com o período de colapso da floração de
cianobactérias, disponibilizando uma alta concentração de nitrogênio, principalmente
na forma de amônia. Cabe ressaltar que este foi o único período (PII) de entrada de
uma cunha salina na lagoa, o que também pode ajudar a explicar a drástica redução
da biomassa fitoplanctônica observada.
Segundo Nunes-Flores (2000), quando há uma mudança de estado trófico
para condições hipereutróficas, as variáveis físicas se tornam mais importantes para
promover o desenvolvimento de diferentes espécies com diversas estratégias de
adaptação. No entanto, a interpretação da relação entre variáveis físicas e químicas
e a biomassa algal pode ser problemática, uma vez que esta relação pode ser causa
ou consequência da atividade fitoplanctônica. Contudo, a variação da temperatura e

42
o aumento da disponibilidade de nitrogênio e luz, além de promoverem variação na
biomassa, também provocaram mudança na composição do fitoplâncton.
A lagoa de Jacarepaguá apresentou uma grande variabilidade sazonal, e
suas características limnológicas levaram à identificação de três períodos distintos.
O período I se caracterizou por elevadas temperaturas (mediana da E1: 29,9 oC e
E2: 30,2 oC), reduzida concentração de NID (mediana da E1: 20,4��g L-1 e E2: 30,4
�g L-1) e alta biomassa de cianobactérias (mediana da E1: 21,9 mg L-1 e E2: 27,9 mg
L-1) e consequentemente, baixa penetração de luz (mediana Zsecchi da E1 e E2: 0,3
m). Neste período houve uma co-dominância entre as espécies Microcystis
aeruginosa e Aphanizomenon sp., também com uma considerável contribuição de
Planktothrix sp. Possivelmente as diferentes estratégias adapatativas para captação
de N entre estas espécies permitiram a co-dominância. M. aeruginosa, assim como
Planktothrix sp. são espécies de cianobactérias não fixadoras de nitrogênio
atmosférico e, provavelmente, estavam utilizando todo o NID para a produção de
sua biomassa. No entanto, Aphanizomenon sp. pode dividir o nicho com estas
espécies por poder utilizar nitrogênio atmosférico para o seu crescimento. Marinho &
Huszar (2002) também observaram a dominância de duas cianobactérias fixadoras
de nitrogênio, Anabaena spiroides e Cylindrospermopsis raciborskii, associada a
períodos de baixa disponibilidade de N no reservatório de Juturnaíba, RJ.
O período II foi marcado principalmente pela reduzida biomassa
fitoplanctônica (mediana da E1: 12,89 mg L-1 e E2: 6,21 mg L-1). Neste período, a
temperatura da água chegou a atingir o mínimo de aproximadamente 18 oC e uma
mediana de 22,8 oC na E1 e 23,2 oC na E3, o que coincidiu com o colapso da
floração de cianobactérias observada no período anterior. Esse colapso acabou
disponibilizando uma alta concentração de NID (mediana da E1: 3822,2 �g L-1 e E2:

43
3255,7 �g L-1), o que permitiu que a Planktothrix sp., embora em baixa biomassa,
dominasse a comunidade. A composição fitoplactônica também ganhou uma maior
contribuição das diatomáceas, principalmente representadas pela espécie Cyclotella
sp., o que foi associado à reduzida temperatura da água desses meses de inverno.
A mais alta disponibilidade de luz, promovida pelo colapso da floração neste
período, também pode ter favorecido as diatomáceas.
As diferentes estratégias adaptativas do fitoplâncton (estratégias C-S-R,
descritas por Reynolds 1997, 2006) também poderiam explicar as substituições de
espécies no final do período II e início do período III. Synecocystis sp., por ser uma
espécie competidora (C-estrategista), pode aproveitar o NID disponibilizado pela
redução da biomassa de Planktothrix sp e investir rapidamente no seu crescimento.
No entanto, no retorno dos meses mais quentes de verão do final do ano de 2007 e
início de 2008 (final do PIII), a lagoa de Jacarepaguá retornou a condições
semelhantes a do período I. Houve um aumento de biomassa (máx da E1: 46,9 mg
L-1 e E2: 17,8 mg L-1), baixas concentrações de NID (min da E1:7,4 mg L-1 e
E2:13,0mg L-1) e co-dominância das espécies Microcystis aeruginosa,
Aphanizomenon sp. e Planktothrix sp. No entanto, neste período, houve uma maior
contribuição de outros grupos taxonômicos do que no período I.
Em síntese, a lagoa de Jacarepaguá apresentou uma grande dinâmica
sazonal, tendo como principais fatores reguladores a temperatura da água e a
concentração de nutrientes, especialmente nitrogênio. Desta forma, os capítulos
seguintes tiveram como objetivo avaliar a redução das concentrações de nutrientes,
assim como a variação da temperatura no crescimento e competição entre espécies
fitoplanctônicas de maior representatividade na lagoa de Jacarepaguá.

44
REFERÊNCIAS
APHA. 1998. Standard Methods for Examination of Waste and Wastewater – American Public Health Association, New York.
BLOMQVIST, P.; PETTERSSON A. & HYENSTRAND, P. 1994. Ammonium-nitrogen: a key regulatory factor causing dominance of non-nitrogen-fixing Cianobacteria in aquatic systems. Arch. Hydrobiol. 132: 141-164.
BOUVY, M., FALCÃO, D., MARINHO, M., PAGANO, M., MOURA, A. 2000. Occurrence of Cylindrospermopsis (Cyanobacteria) in 39 Brazilian tropical reservoirs during the 1998 drought. Aquat Microb Ecol. 23: 13-27.
BOUVY, M., MOLICA, R., OLIVEIRA, S., MARINHO, M., BEKER, B. 1999. Dynamics of toxic cyanobacterial Bloom (Cylindrospermopsis raciborskii) in a shallow reservoir in the semi arid region of northeast Brazil. Aquat Microbiol Ecol. 20: 285-297.
BRANCO, C.W.C.; SENNA, P.A.C. 1994. Factors influencing the development ofCylindrospermopsis raciborskii and Microscystis aeruginosa in the Paranoá Reservoir, Algological Studies, v. 75, p. 85-96.
CARACO, N. & MILLER, R. 1998. Direct and indirect effects of CO2 on competition between a cyanobacteria and eukaryotic phytoplankton. Can. J. Fish. Aquat. Sci. 55:54-62.
COLE, G.A. 1994. Texbook of limnology. Illinois: Waveland Press Inc. 412p.
DOMINGOS, P. 2001. Dinâmica de Cianobactérias produtoras de microcistinas na Lagoa de Jacarepaguá (RJ). Centro de Ciências da Saúde, UFRJ. 107p. (Tese de Doutorado em Biotecnologia Vegetal).
ENRICH-PRAST, A.; BOZELLI, R.; ESTEVES, F.A. & MEIRELLES, F. 2004. Lagoas Costeiras da Restinga de Jurubatiba: Descrição de suas variáveis limnológicas. Pp. 245-254. In: C.F.D. Rocha, F.A. Esteves & F.R. Scarano. (eds). Pesquisas de Longa duração na Restinga de Jurubatiba Ecologia, História Natural e Conservação. RiMa, São Carlos. 37.
ESTEVES, F. A. 1998. Fundamentos de Limnologia. 2 ed. Rio de Janeiro, Interciência. 602p.
FERNANDES, V. O. 1993. Estudos sucessionais da comunidade perifítica em folhas de Typha dominguensis Pers, relacionado às variáveis limnológicas na Lagoa de Jacarepaguá, RJ. Tese de Mestrado, UFSCar, SP. 131p.
GANF, G.G. 1974. Diurnal mixing and the vertical distribution of phytoplankton in a shallow equatorial lake (Lake George) Uganda. J. Ecol. 62: 611-629.
GOMES, A. M. A., SAMPAIO, P. L., FERRÃO-FILHO, A.S., MAGALHÃES, V. F., MARINHO, M.M. OLIVEIRA, A. C. P., SANTOS, V. B., DOMINGOS, P., AZEVEDO, S.M.F.O. (2009). Florações de Cianobactérias Tóxicas em uma Lagoa Costeira

45
Hipereutrófica do Rio de Janeiro/Rj (Brasil) e suas Consequências para Saúde Humana. Oecologia Brasiliensis. 13 (2), 329-345.
HANEY, J. F. 1987. Field studies on zooplankton-Cyanobacteria interactions. N.Z.J. Mar. freshwat. Res. 21: 467-475.
HUSZAR,V.L., SILVA, L.H.S., MARINHO, M.M., DOMINGOS, P., SANT'ANNA, C. 2000. Cyanoprocaryote assembleges in eigth productive tropical brazilian Waters. Hydrobiologia. 424: 67-77.
JENSEN, J.P.; JEPPESEN, E.; OLRIK, K. & KRISTENSEN, P. 1994 Impact of nutrientes and physical factors on the shift from cyanobacterial to chlorophyte. Can.J. Fish. Aquati Sci., 51: 1692-1699.
JENSEN, J.P.; ANDRESEN, F.O. 1992. Importance of temperature, nitrate, and pH for phosphate release from aerobic sediments of 4 shallow, eutrophic lakes. Limnology and Oceanography, 37: 577-589.
KING, D.L. 1970. The role of carbon in eutrophication. J. Wat. Pollut.42: 2035-2051.
KOSTEN, S., HUSZAR, V.L., MAZZEO, N., SCHEFFER, M., STERNBERG, L.S.L., JEPPESEN, E. 2009. Lake and watershed characteristics rather than climate influence nutrient limitation in shallow lakes. Ecological applications. 19(7): 1791-1804.
MAGALHÃES, V.F.; SOARES, R.M. & AZEVEDO, S.M.F.O. 2001. Microcystin contamination in fish from the Jacarepaguá (RJ, Brazil): ecological implication and human health risk. Toxicon, 39: 1077-1085.
MALLET, C., CHARPIN, M. F. & DEVAUX, J. 1998. Nitrate reductase activity of plankton population in eutrophic Lake Aydat and meso-oligotrophic Lake Pavin: a comparison. Hydrobiol., 373/374: 135-148.
MARINHO, M.M., HUSZAR, V.L. 2002. Nutrient availability and physical condictions as controlling factors of phytoplankton composition and biomass in tropical reservoir (South-eastern Brazil). Arch. Hydrobiol. 153: 443-468.
McCARTHY, J. J., WYNNE, D. & BERMAN, T. 1982. The uptake of dissolved nitrogenous nutrients by Lake Kinnerest (Israel) microplankton. Limnol. Oceanogr. 24(2): 243-252.
MIYAZAKI, T., WATASE, M. & MIYAKE, K. 1989. Daily changes of uptake of inorganic carbon and nitrogen, and their relation to phytoplankton blooms in late spring-early summer in Lake Nakanuma, Japan. Hydrobiol., 185: 223-231.
MOSS, B., MADGWICK, J., PHILLIPS, G. 1997. A guide to the restoration of nutrient-enriched shallow lakes. W.W. Hawes, UK.
MURPHY, J., RILEY, J.P. 1962. A modified single solution method for the determination of phosphate in natural waters. Anal. Chim. Acta. 27: 31-36.

46
PAERL, H. & HUISMAN, J. (2009). Climate change: a catalyst for global expansion of harmful of cyanobacterial blooms. Environmental Microbiology Reports, 1 (1), 27-37.
PAERL, H.W. 2006. Nutrient and other environmental controls of harmful cyanobacterial blooms along the freshwater-marine continuum. In: H.K. Hudnell (ed): Proceedings of the Interagency, International Symposium on Cyanobacterial Harmful Algal Blooms. Advances in Experimental Medicine & Biology.
PAERL, H. W., FULTON, R. S., MOISANDER, P. H. & DYBLE, J. 2001. Harmful freshwater algal blooms, with an emphasis on cyanobacteria. The Scientific World, 1: 76-113.
PETTERSSON, K., HERLITZ, E. & ISTVANOVICS, V. (1993). The role of Gloeotrichia echinulata in the transfer of phosphorus from sediments to water in lake Erken. Hydrobiologia, 25: 123-129.
REYNOLDS, C.S. 2006. Ecology of Phytoplankton. Cambridge. 535p.
REYNOLDS, C.S. 1997. Vegetation Processes in the Pelagic: A Model for Ecosystem Theory. Ecology Institute, Oldendorf/Luhe.371p.
REYNOLDS, C.S. 1988. Functional morphology and the adaptive strategies of freshwater phytoplankton. p. 388-433 In C. D. Sandgren [ed.] Growth and reproductive strategies of freshwater phytoplankton. Cambridge University Press. Cambridge.
REYNOLDS, C.S. 1987. Cyanobacterial water-blooms. In: J. Callow (ed). Advances in botanical research. Academic Press., London. PP:67-143 (vol 13).
REYNOLDS, C.S. 1984. The Ecology of Freshwater Phytoplankton. Cambridge University Press, Cambridge.
SAIEG-FILHO, E. 1986. Ecologia do Fitoplâncton Marginal das Lagunas da Baixada de Jacarepaguá, Rio de Janeiro- RJ. Instituto de Biologia, UERJ. 150p. (Monografia de Bacharelado).
SCHEFFER, M. 2004. Ecology of shallow lakes.Kluwer Academic Publishers, The Netherlands.357p.
SCHINDLER, D.W. 1977. Evolution of phosphorus limitation in lakes. Science. 195: 260-262.
SHAPIRO, J. 1990. Currents beliefs regarding dominance by blue-greens: the case of the importance of CO2 and pH. Verh. Int. ver. Limnol. 24: 38-54.
SILVA, G. 2006. Processo de Ocupação Urbana da Barra da Tijuca (RJ): Problemas Ambientais, Conflitos Sócio-Ambientais, Impactos Ambientais Urbanos. Pesquisa em arquitetura e Construção. 28p.

47
SMITH, V. 1983. Low nitrogen to phosphorous rations favor dominance by blue-green algae in lake phytoplankton. Science 221: 669-671.
SMITH, V. 1986. Light and nutrient effects on the relative biomass of blue-green algae in lake phytoplankton. Can. J. Fish. Aquat. 43: 148-153.
SOARES, M.C.S., ROCHA, M.I., MARINHO, M.M., AZEVEDO, S.M.F.O., BRANCO, C.W.C., HUSZAR, V.L. 2009. Changes in species composition during annual cyanobacterial dominance in tropical reservoir: physical factors, nutrients and grazing effects. Aquat. Microb Ecol. 57, 137-149.
UHELINGER, V. (1964) Étude statisque des méthodes de dénobrement planctonique. – Arch. Sci. 17: 121-123.
UTERMÖHL, H. 1958. Zur Vervollkommung der quantitativen Phytoplankton Methodik. Mitt. Int. Verein. Limno., 9: 1-39.
WETZEL, R. G. 1983. Limnology. 2ed. Saunders College Publishing. New York. 767p.
ZHANG, Y., PREPAS, E.E. 1996. Regulation of dominance of planktonic diatoms and cyanobacteria in four eutrophic hardwater lakes by nutrients, water column stability, and temperature. Can. J. Fish. Aquatic Sci. 53:621-633.
ZEVENBOOM, W. & MUR, L.R. 1980. N2-fixing cyanobacteria: Why they do not become dominat in Dutch, hypertrophic lakes.Develop. Hydrobiol. 2: 123-130.

48
CAPÍTULO II:EFEITO DA REDUÇÃO DA CONCENTRAÇÃO DE NUTRIENTES NA COMUNIDADE
FITOPLANCTÔNICA DA LAGOA DE JACAREPAGUÁ
INTRODUÇÃO
Um dos principais impactos da atividade humana nos ambientes aquáticos é o
aporte de elevadas concentrações de nutrientes, através do lançamento de esgotos
domésticos e industriais (Tundisi & Matsumura-Tundisi, 1992). Uma das
consequências desse enriquecimento por nutrientes (eutrofização) é o acúmulo de
elevada biomassa de cianobactérias, chamada de floração. Conhecer a estrutura e a
dinâmica da comunidade fitoplanctônica é necessário para se entender o impacto
deste processo de eutrofização. Já é sabido que o nitrogênio (N) e fósforo (P) são os
mais importantes elementos neste processo, porque influenciam diretamente no
crescimento do fitoplâncton (Vollenweider & Kerekes, 1980; Paerl, 2006) e
consequentemente podem determinar a sua dinâmica, composição e biomassa em
um dado ambiente (Paerl, 2006; Reynolds 2006).
Um esforço intenso tem sido feito para produzir um modelo de classificação
que vise prever os efeitos de impactos antropogênicos no ambiente. Estes modelos
se tornam úteis na medida em que agrupam espécies de acordo com suas respostas
específicas a mudanças ambientais (Lavorel et al., 1997). A capacidade de prever
mudanças ambientais decorrentes de impactos pode ser uma poderosa ferramenta
no manejo dos ecossistemas. No entanto, é relativamente limitado o conhecimento
sobre as mudanças no fitoplâncton durante um processo de restauração de um lago,
como por exemplo, durante a redução do aporte de P (Padisák & Reynolds, 1998),
principalmente em ecossistemas tropicais e subtropicais.

49
A redução de nutrientes tem sido a principal técnica estudada e aplicada para
diminuir os efeitos nocivos da eutrofização artificial. Como parte desse processo,
experimentos têm sido feitos visando conhecer a dinâmica de nutrientes e seu efeito
nas comunidades de um lago (Paerl & Bowles, 1987; Ortega-Mayagoitia et al., 2002,
2003). Entretanto, ainda há escassez de informação sobre os efeitos do processo de
oligotrofização na comunidade fitoplanctônica.
Como já discutido, a lagoa de Jacarepaguá é um ambiente bastante
impactado, com elevado aporte de nutrientes e constantes florações de
cianobactérias (Gomes et al., 2009; capítulo I). Entretanto, permanecem as
seguintes perguntas: (i) a redução de nutrientes nesse ambiente seria capaz de
diminuir estas recorrentes florações? (ii) qual seria a concentração de nutrientes que
limitaria o crescimento exacerbado das cianobactérias neste ambiente?
O presente trabalho teve como objetivo, através de estudo com microcosmos
em laboratório, simular um processo de redução de nutrientes (especialmente P) na
tentativa de verificar seu efeito na dinâmica, estrutura e composição do fitoplâncton,
dominado por cianobactérias, da lagoa de Jacarepaguá.
MATERIAL E MÉTODOS
Visando inferir valores críticos de concentração de macro-nutrientes que
limitem o crescimento e a dominância de cianobactérias no fitoplâncton na lagoa de
Jacarepaguá, foram realizados experimentos em laboratório com água desse
sistema (Experimento de Oligotrofização). O processo de oligotrofização foi simulado
através da diluição da água da lagoa de Jacarepaguá, considerando um cenário de
interrupção de lançamentos de poluentes nos rios contribuintes. Para fazer tais
diluições foi utilizada água do rio Camorim. Esse sistema foi escolhido por estar

50
localizado na mesma bacia hidrográfica da lagoa de Jacarepaguá e por ser um
ambiente preservado e com baixas concentrações de nutrientes.
Montagem dos Experimentos: Amostras de água da lagoa de Jacarepaguá e do
rio Camorim foram coletadas em 11/09/2008 e trazidas para experimentação em
laboratório. Essas amostras foram filtradas em rede de 68 µm para eliminar a
comunidade zooplanctônica. Foram também coletadas amostras do sedimento da
lagoa de Jacarepaguá com a finalidade de melhor simular as condições da
qualidade da água no ambiente, onde certamente os nutrientes estocados no
sedimento estão em permanente troca com a coluna d'água. A quantidade de
sedimento utilizada em cada microcosmo foi calculada para manter, nas mesmas
proporções de 1m3, a que seria encontrada no valor médio de profundidade de
coluna d'água da lagoa. Os experimentos foram conduzidos em balões de 3 litros
sob intensidade luminosa de 140 µmol fótons.m2.s
-1, temperatura de 24 + 2
oC,
aeração contínua, fotoperíodo de 12 horas, em condições não axênicas. O
experimento teve uma duração 32 dias, durante os quais foram monitoradas
algumas variáveis físicas e químicas (temperatura da água, pH, OD, salinidade,
condutividade elétrica e intensidade luminosa) e coletadas amostras para avaliação
da comunidade fitoplanctônica, análises de nutrientes e clorofila-a, a cada 3 dias até
o 12º dia e após essa data, a cada 7 dias até o 32º dia. As metodologias
empregadas nessas análises estão descritas abaixo.
O experimento incluiu 6 grupos diferentes, cada um consistindo de um tipo de
tratamento em triplicata:
� Controle - 3 balões com água da lagoa de Jacarepaguá, com concentração
de P-PO4- = 516,54 ug/L;

51
� Tratamento A - 3 balões com água da lagoa de Jacarepaguá diluída com
água do rio Camorim, chegando à concentração de P-PO4- = 189,14 ug/L;
� Tratamento B - 3 balões com água da lagoa de Jacarepaguá diluída com
água do rio Camorim, chegando à concentração de P-PO4- = 46,63 ug/L;
� Controle c/ sedimento - 3 balões com água da lagoa de Jacarepaguá, com
concentração de P-PO4- = 516,54 ug/L e com sedimento da lagoa de
Jacarepaguá;
� Tratamento A c/ sedimento - 3 balões com água da lagoa de Jacarepaguá
diluída com água do rio Camorim, chegando à concentração de P-PO4-=
189,14 ug/L e com sedimento da lagoa de Jacarepaguá;
� Tratamento B c/ sedimento - 3 balões com água da lagoa de Jacarepaguá
diluída com água do rio Camorim, chegando à concentração de P-PO4- =
46,63 ug/L e com sedimento da lagoa de Jacarepaguá.
Clorofila-a: As amostragens de água de cada frasco experimental foram realizadas
por sucção com pipeta e cada amostra foi filtrada em filtros de fibra de vidro. Os
filtros foram preservados congelados para posterior análise de clorofila-a. A
concentração de clorofila-a foi obtida por espectrofotometria, seguindo a
metodologia descrita em APHA (1998). Esta técnica foi realizada a partir da extração
de clorofila-a das células retidas nos filtros com acetona 90%, durante 20-24h.
Realizou-se em seguida a centrifugação da amostra a 1000 g durante 10 minutos,
sendo a absorbância do sobrenadante determinada em espectrofotômetro a 665 e
750nm, antes e após acidificação com solução de ácido clorídrico (HCl) 0,3 M.

52
Nutrientes: O volume filtrado das amostras foi destinado à análise de nutrientes
dissolvidos. Os nutrientes analisados foram nitrogênio amoniacal, nitrito, nitrato e
fosfato solúvel reativo. Todos os nutrientes dissolvidos foram analisados através da
metodologia de separação e análise por croamtografia iônica, utilizando o
cromatógrafo Dionex ICS-1000.
Fitoplâncton: As amostras de fitoplâncton foram coletadas por sucção de água por
pipeta e fixadas com solução de lugol. As populações foram identificadas sempre
que possível em nível de espécie. A densidade fitoplanctônica (ind./ml) foi estimada
segundo Utermöhl (1958). Os indivíduos (células, colônias, cenóbios, filamentos)
foram enumerados em campos aleatórios (Uhelinger, 1964), em número suficiente
para alcançar 100 indivíduos da espécie mais frequente, sendo o erro inferior a 20%,
a um coeficiente de confiança de 95%. O biovolume (mm3.l-1) foi estimado
multiplicando-se a densidade de cada espécie pelo volume médio de suas células,
sempre que possível considerando a dimensão média de cerca de 25 indivíduos. O
volume de cada célula foi calculado a partir de modelos geométricos aproximados à
forma dos indivíduos. A biomassa específica foi expressa em mg (peso fresco) L-1
calculada pelo produto da densidade populacional vezes o volume médio de cada
unidade, assumindo a densidade das células fitoplanctônicas de 1 g.cm-3.
Análise Estatítica: Para comparar a diferença entre as variáveis ambientais dos
diferentes tratamentos foi realizada uma de análise de variância com medidas
repetidas (Repeated-measures ANOVA). As variações que apresentaram diferenças
significativas foram submetidas ao teste de comparação múltipla de Tukey. Estas
análises foram feitas utilizando o programa estatístico SPSS® 17.0.

53
RESULTADOS
VARIÁVEIS ABIÓTICAS
Embora tenha se tentado manter a temperatura controlada ao longo dos
experimentos, ocorreu uma variação significativa (p<0,05) de aproximadamente 4oC,
do primeiro para o sexto dia, no experimento sem sedimento (fig. 1a). Já no
experimento com sedimento essa variação foi um pouco menos pronunciada (3oC)
(fig. 2a). No entanto, as três condições experimentais (controle, tratamento A,
tratamento B) variaram de modo equivalente durante todo experimento, não
prejudicando a comparação, tanto para os experimentos com sedimento quanto para
os sem sedimento. Apesar de a temperatura ser um importante fator determinante
da composição fitoplanctônica de um corpo d'água, a variação da temperatura
observada ao longo dos experimentos parece ter sido resultado da sua variação na
sala de experimentação e não de uma alteração do metabolismo dentro dos
microcosmos. Portanto, podemos afirmar que a temperatura não foi um fator
importante para as mudanças observadas na dinâmica da comunidade
fitoplanctônica entre os tratamentos, durante os experimentos, já que sua variação
foi igual para todas as condições.
Os maiores valores de intensidade luminosa foram observados nos
tratamentos com maiores diluições da água da lagoa de Jacarepaguá (tratamento B
e tratamento B com sedimento) e os menores valores nas condições controle e
controle com sedimento (figs.1b e 2b). A variação da intensidade luminosa, portanto,
parece ter sido resultado da diluição feita com a água do rio Camorim. Quanto maior
a diluição realizada, menor foi a biomassa fitoplanctônica observada e a variação da
intensidade luminosa foi resultado desse fenômeno. As condições controle
apresentaram valores iniciais de intensidade luminosa de 56,5 µmol fóton m-2 cm-1, o

54
tratamento A de 109,6 µmol fóton m-2 cm-1 e o tratamento B de 143,9 µmol fóton m-2
cm-1. No experimento com sedimento esses valores foram um pouco menores
(controle com sed. = 45,9 µmol fóton m-2 cm-1, tratamento A com sed. = 81,9 µmol
fóton m-2 cm-1 e tratamento B com sed. = 119,0 µmol fóton m-2 cm-1). Estas
diferenças permaneceram relativamente constantes ao longo dos experimentos e as
variações foram inversamente relacionadas às biomassas fitoplanctônicas.
A condutividade elétrica e a salinidade da água também variaram de acordo
com as diluições realizadas com a água do rio Camorim. Os maiores valores de
condutividade elétrica (mediana: 8780,8 µS cm-1 p/ controle e 8889,8 µS cm-1 p/
controle com sedimento) e salinidade (mediana: 4,88 ppt p/ controle e 4,96 ppt p/
controle com sedimento) foram encontrados na água da lagoa de Jacarepaguá.
Consequentemente, as condições com maiores proporções de água do rio Camorim
(o tratamento B e tratamento B com sedimento) apresentaram os menores valores
de salinidade (mediana: 0,77 ppt p/ tratamento B e 0,33 ppt p/ tratamento B com
sedimento) e condutividade elétrica (mediana: 156,3 µS cm-1 p/ tratamento B e 691,3
µS cm-1 p/ tratamento B com sedimento) (figs. 1c e 1d; 2c e 2d). É comum observar
que quanto maior o estado trófico de um sistema, maior será a concentração de
íons, fruto principalmente das elevadas concentrações de nutrientes. Os maiores
valores de salinidade encontrados nos controles foram resultado da ausência de
diluição da água da lagoa de Jacarepaguá, sistema esse caracterizado como
oligoalino. Cabe ainda ressaltar que em nenhuma condição experimental houve uma
variação significativa (p>0,05) destas variáveis ao longo do tempo de
experimentação (exceção para o valor de salinidade do 20º dia da condição
controle).

55
Os valores de oxigênio dissolvido sofreram flutuações entre 11 e 7 mg L-1 no
experimento sem sedimento, para os tratamentos A e B. Na condição controle a
redução do OD do primeiro para o segundo dia foi de um pouco mais que 4 mg L-1
(fig. 1e). No experimento com sedimento, os valores de oxigênio dissolvido da água
foram um pouco mais elevados (p<0,05) no primeiro dia (12,4 -15,6 mg L-1) e se
mantiveram com menores flutuações (em torno de 10 mg L-1) no resto do período
experimental (fig. 2e). Todos os tratamentos estavam em condições saturantes de
oxigênio dissolvido na água. Isso se deve ao fato de que foi usada aeração contínua
em todos os balões experimentais. A escolha pela aeração dos balões foi para
melhor simular as condições da lagoa, uma vez que a coluna d'água desse ambiente
esteve misturada durante todo o período de estudo em 2007 (capítulo I). Os maiores
valores de OD medidos no primeiro dia, possivelmente foram resultados das
manipulações no momento de montagem do experimento.
Para os experimentos sem sedimento, o valor de pH da água no tratamento B
no início do experimento foi levemente ácido (6,1), mas se manteve na faixa de pH
neutro durante o resto do período experimental (fig. 1f). Já a água do tratamento A e
controle apresentaram valores de pH mais alcalinos (mediana: 8,51 p/ tratamento A
e 8,66 p/ controle). Essas últimas condições apresentaram maiores biomassas
fitoplanctônicas, sendo as cianobactérias o grupo mais representativo. Cabe
ressaltar que as cianobactérias são um dos grupos fitoplanctônicos capazes de
explorar ambientes alcalinos. No experimento com sedimento, as três condições
apresentaram valores de pH variando na faixa alcalina desde o início do
experimento (mediana: 8,82 p/ controle, 9,12 p/ tratamento A e 8,34 p/ tratamento B)
(fig. 2f). Altos valores de pH (maiores 8,9) podem indicar ambientes limitados por C,
também influenciando na estrutura e composição das comunidades fitoplanctônicas.

56
As concentrações de PSR foram elevadas nos 13 primeiros dias de
experimento no controle (fig. 3a) e durante todo o experimento no controle com
sedimento (fig. 3c). No experimento sem sedimento, as condições mais eutróficas
foram observadas no início e as altas concentrações de PSR foram sendo
consumidas ao longo do tempo, tornando-se limitante ao crescimento fitoplanctônico
(menor que 10 µg L-1, segundo Reynolds 2006) a partir do 13º dia nos tratamentos A
e A com sedimento e já a partir do 3º dia nos tratamentos B e B com sedimento. Em
todos os microcosmos, as maiores concentrações de NID também foram registradas
no início do experimento (figs. 3b e 3b). Entretanto, ao contrário da dinâmica do
PSR, os maiores valores de NID foram observados nos tratamentos A e B com e
sem sedimento. As águas do rio Camorim, contribuíram com os mais altos valores
de nitrogênio (Máx: 347,19 µg L-1 p/ tratamento A com sedimento e 479,10 µg L-1 p/
tratamento B com sedimento), especialmente na forma de nitrato. Esses valores
passaram a ser limitantes para o crescimento fitoplanctônico (valores menores que
100 µg L-1, segundo Reynolds 2006) a partir do sexto dia para os experimentos sem
sedimento e desde o primeiro dia para a condição controle com sedimento.
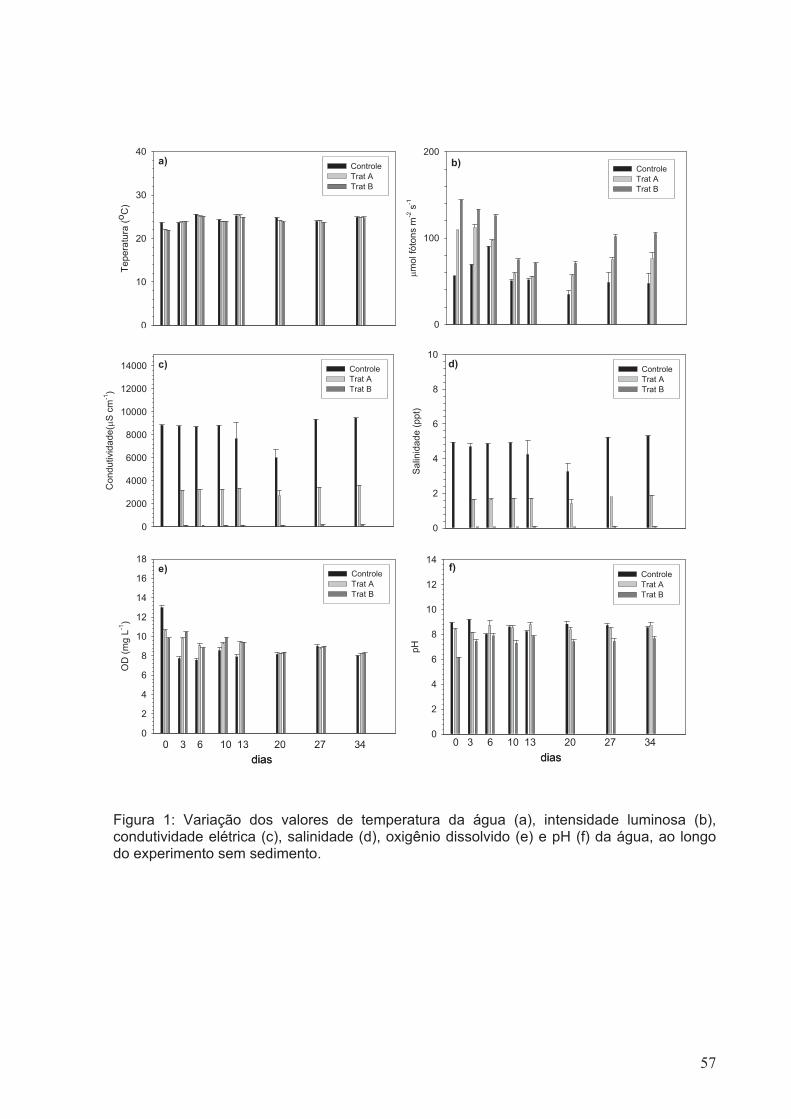
57
Figura 1: Variação dos valores de temperatura da água (a), intensidade luminosa (b), condutividade elétrica (c), salinidade (d), oxigênio dissolvido (e) e pH (f) da água, ao longo do experimento sem sedimento.
Tepe
ratu
ra (o C
)
0
10
20
30
40Controle Trat A Trat B
� mol
fóto
ns m
-2 s
-1
0
100
200
Controle Trat A Trat B
Con
dutiv
idad
e(�S
cm
-1)
0
2000
4000
6000
8000
10000
12000
14000 Controle Trat A Trat B
Salin
idad
e (p
pt)
0
2
4
6
8
10Controle Trat A Trat B
OD
(mg
L-1)
0
2
4
6
8
10
12
14
16
18Controle Trat A Trat B
pH
0
2
4
6
8
10
12
14Controle Trat A Trat B
0 3 6 10 13 20 27 34 dias
0 3 6 10 13 20 27 34 dias
a) b)
c) d)
e) f)
Tepe
ratu
ra (o C
)
0
10
20
30
40Controle Trat A Trat B
� mol
fóto
ns m
-2 s
-1
0
100
200
Controle Trat A Trat B
Con
dutiv
idad
e(�S
cm
-1)
0
2000
4000
6000
8000
10000
12000
14000 Controle Trat A Trat B
Salin
idad
e (p
pt)
0
2
4
6
8
10Controle Trat A Trat B
OD
(mg
L-1)
0
2
4
6
8
10
12
14
16
18Controle Trat A Trat B
pH
0
2
4
6
8
10
12
14Controle Trat A Trat B
0 3 6 10 13 20 27 34 dias
0 3 6 10 13 20 27 34 dias
a) b)
c) d)
e) f)
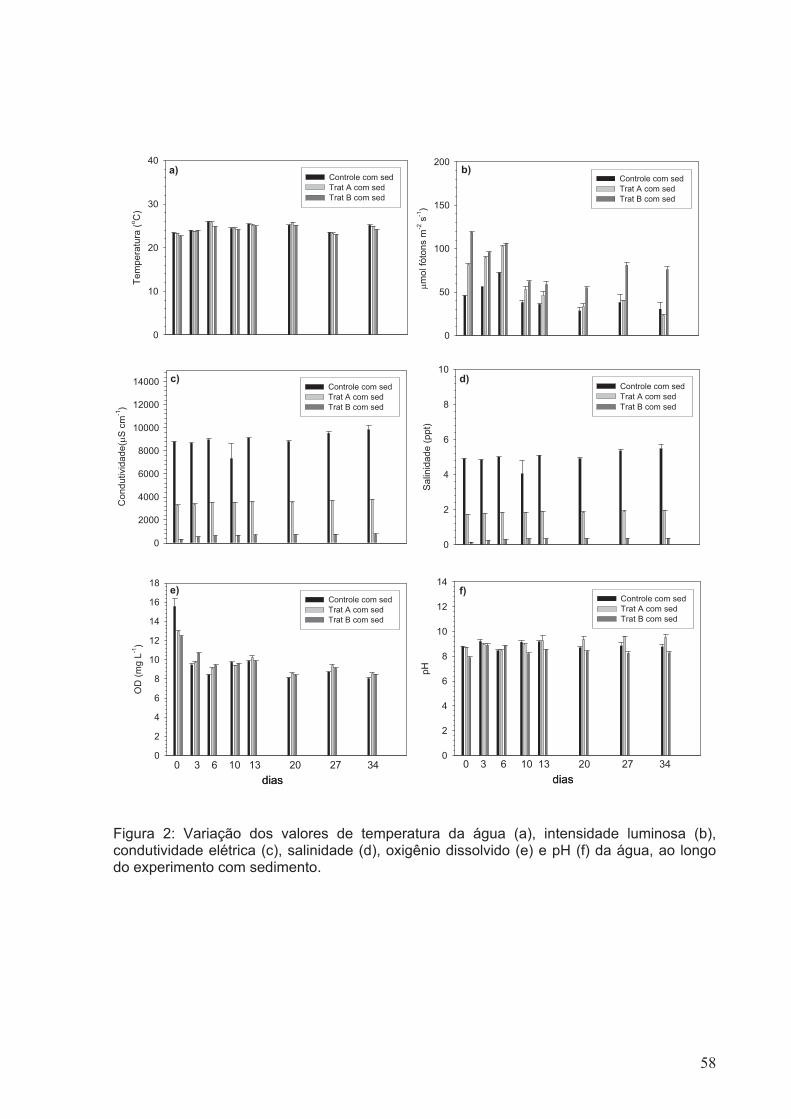
58
Figura 2: Variação dos valores de temperatura da água (a), intensidade luminosa (b), condutividade elétrica (c), salinidade (d), oxigênio dissolvido (e) e pH (f) da água, ao longo do experimento com sedimento.
Con
dutiv
idad
e(� S
cm
-1)
0
2000
4000
6000
8000
10000
12000
14000 Controle com sed Trat A com sed Trat B com sed
Sal
inid
ade
(ppt
)
0
2
4
6
8
10
Controle com sed Trat A com sed Trat B com sed
OD
(mg
L-1)
0
2
4
6
8
10
12
14
16
18
Controle com sed Trat A com sed Trat B com sed
pH
0
2
4
6
8
10
12
14
Controle com sed Trat A com sed Trat B com sed
Tem
pera
tura
(o C)
0
10
20
30
40
Controle com sed Trat A com sed Trat B com sed
�mol
fóto
ns m
-2 s
-1)
0
50
100
150
200
Controle com sed Trat A com sed Trat B com sed
0 3 6 10 13 20 27 34 dias
0 3 6 10 13 20 27 34 dias
a) b)
c) d)
e) f)
Con
dutiv
idad
e(� S
cm
-1)
0
2000
4000
6000
8000
10000
12000
14000 Controle com sed Trat A com sed Trat B com sed
Sal
inid
ade
(ppt
)
0
2
4
6
8
10
Controle com sed Trat A com sed Trat B com sed
OD
(mg
L-1)
0
2
4
6
8
10
12
14
16
18
Controle com sed Trat A com sed Trat B com sed
pH
0
2
4
6
8
10
12
14
Controle com sed Trat A com sed Trat B com sed
Tem
pera
tura
(o C)
0
10
20
30
40
Controle com sed Trat A com sed Trat B com sed
�mol
fóto
ns m
-2 s
-1)
0
50
100
150
200
Controle com sed Trat A com sed Trat B com sed
0 3 6 10 13 20 27 34 dias
0 3 6 10 13 20 27 34 dias
a) b)
c) d)
e) f)
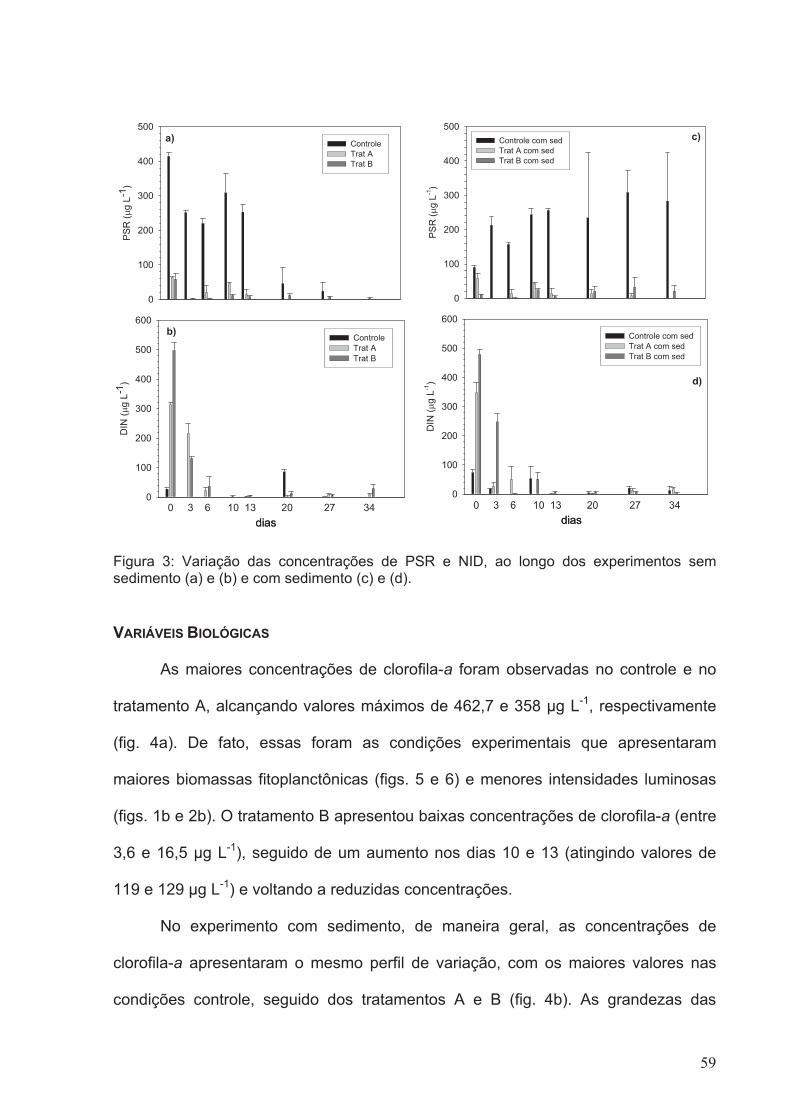
59
Figura 3: Variação das concentrações de PSR e NID, ao longo dos experimentos sem sedimento (a) e (b) e com sedimento (c) e (d).
VARIÁVEIS BIOLÓGICAS
As maiores concentrações de clorofila-a foram observadas no controle e no
tratamento A, alcançando valores máximos de 462,7 e 358 µg L-1, respectivamente
(fig. 4a). De fato, essas foram as condições experimentais que apresentaram
maiores biomassas fitoplanctônicas (figs. 5 e 6) e menores intensidades luminosas
(figs. 1b e 2b). O tratamento B apresentou baixas concentrações de clorofila-a (entre
3,6 e 16,5 µg L-1), seguido de um aumento nos dias 10 e 13 (atingindo valores de
119 e 129 µg L-1) e voltando a reduzidas concentrações.
No experimento com sedimento, de maneira geral, as concentrações de
clorofila-a apresentaram o mesmo perfil de variação, com os maiores valores nas
condições controle, seguido dos tratamentos A e B (fig. 4b). As grandezas das
DIN
(�g
L-1)
0
100
200
300
400
500
600
Controle com sed Trat A com sed Trat B com sed
PSR
(�g
L-1)
0
100
200
300
400
500Controle com sed Trat A com sed Trat B com sed
DIN
(�g
L-1)
0
100
200
300
400
500
600
Controle Trat A Trat B
PSR
(�g
L-1)
0
100
200
300
400
500
Controle Trat A Trat B
0 3 6 10 13 20 27 34 dias
0 3 6 10 13 20 27 34 dias
a)
b)
c)
d)
DIN
(�g
L-1)
0
100
200
300
400
500
600
Controle com sed Trat A com sed Trat B com sed
PSR
(�g
L-1)
0
100
200
300
400
500Controle com sed Trat A com sed Trat B com sed
DIN
(�g
L-1)
0
100
200
300
400
500
600
Controle Trat A Trat B
PSR
(�g
L-1)
0
100
200
300
400
500
Controle Trat A Trat B
0 3 6 10 13 20 27 34 dias
0 3 6 10 13 20 27 34 dias
a)
b)
c)
d)
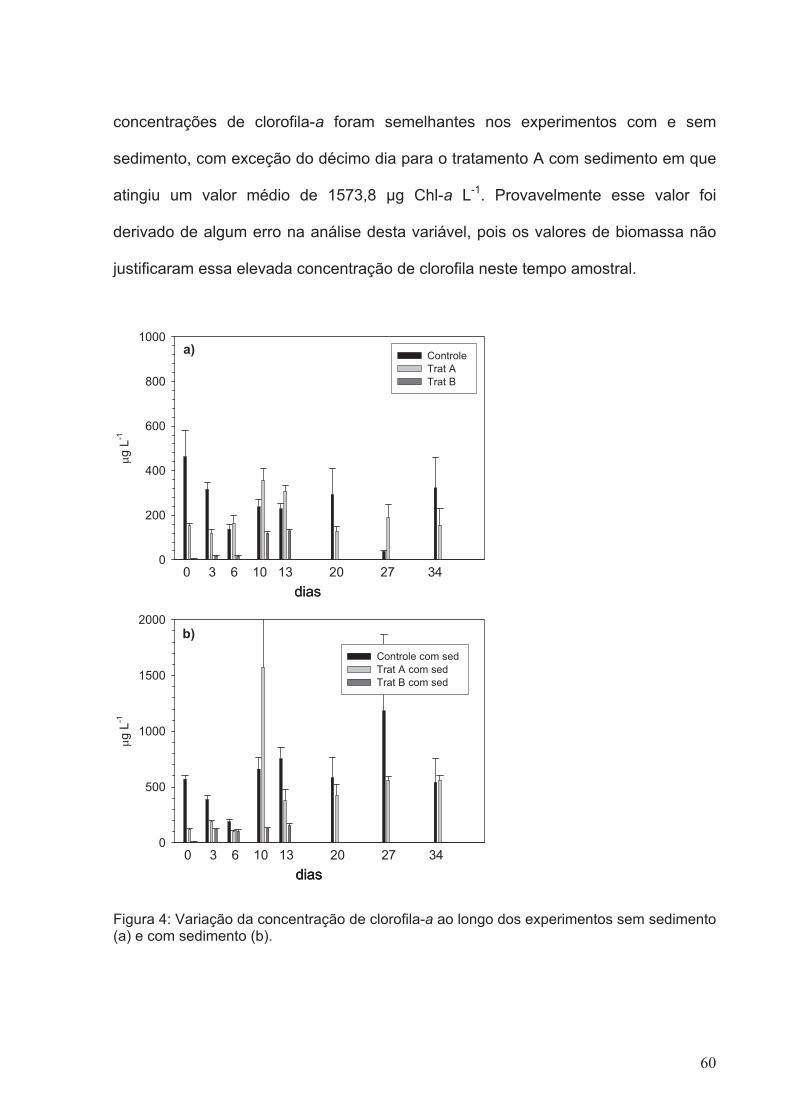
60
concentrações de clorofila-a foram semelhantes nos experimentos com e sem
sedimento, com exceção do décimo dia para o tratamento A com sedimento em que
atingiu um valor médio de 1573,8 µg Chl-a L-1. Provavelmente esse valor foi
derivado de algum erro na análise desta variável, pois os valores de biomassa não
justificaram essa elevada concentração de clorofila neste tempo amostral.
Figura 4: Variação da concentração de clorofila-a ao longo dos experimentos sem sedimento (a) e com sedimento (b).
�g L
-1
0
500
1000
1500
2000
Controle com sed Trat A com sed Trat B com sed
0 3 6 10 13 20 27 34 dias
�g L
-1
0
200
400
600
800
1000Controle Trat A Trat B
0 3 6 10 13 20 27 34 dias
a)
b)
�g L
-1
0
500
1000
1500
2000
Controle com sed Trat A com sed Trat B com sed
0 3 6 10 13 20 27 34 dias
�g L
-1
0
500
1000
1500
2000
Controle com sed Trat A com sed Trat B com sed
0 3 6 10 13 20 27 34 dias
�g L
-1
0
200
400
600
800
1000Controle Trat A Trat B
0 3 6 10 13 20 27 34 dias
�g L
-1
0
200
400
600
800
1000Controle Trat A Trat B
0 3 6 10 13 20 27 34 dias
a)
b)

61
Principais Grupos Fitoplanctônicos
Bacillariophyta, Cryptophyta, Chlorophyta e Cyanobacteria foram os principais
grupos fitoplanctônicos observados em todos os tratamentos com e sem sedimento.
No entanto, as cianobactérias e as diatomáceas foram os grupos mais
representativos. Na condição controle sem sedimento, as cianobactérias e as
diatomáceas estiveram em proporções iniciais bastante semelhantes. No terceiro dia
experimental, houve uma redução da biomassa das diatomáceas, que se manteve
até o décimo terceiro dia, sofrendo uma nova redução a partir do vigésimo dia (figs.
5a e 5b). Já as cianobactérias, embora tenham aumentado sua biomassa no terceiro
dia, sofreram uma pronunciada redução de biomassa no sexto dia experimental,
voltando a crescer exponencialmente a partir no décimo terceiro dia até o final do
experimento, atingindo valores de até 190 mg L-1.
No tratamento A, o crescimento exponencial das cianobactérias pode ser
observado já a partir do primeiro dia amostral. As diatomáceas sofreram pequenas
oscilações, não alcançando uma biomassa superior a 18 mg L-1, sendo
pronunciadamente reduzidas ao final do experimento (figs 5c e 5d).
Embora no tratamento B as cianobactérias tenham sido as espécies
dominantes também na maioria dos tempos amostrais, uma maior diversidade de
espécies foi observada (figs. 5e e 5f). Esses dados podem ser observados com o
aumento da biomassa de outras espécies fitoplanctônicas ao longo do experimento
nesse tratamento. Entretanto, as biomassas alcançadas nesta condição não
ultrapassaram 16 mg L-1.
No controle e tratamento A com sedimento, as diatomáceas apresentaram
maiores biomassas no primeiro dia de experimento. Em seguida, sua biomassa foi
reduzida e as cianobactérias dominaram a comunidade fitoplanctônica até o final do
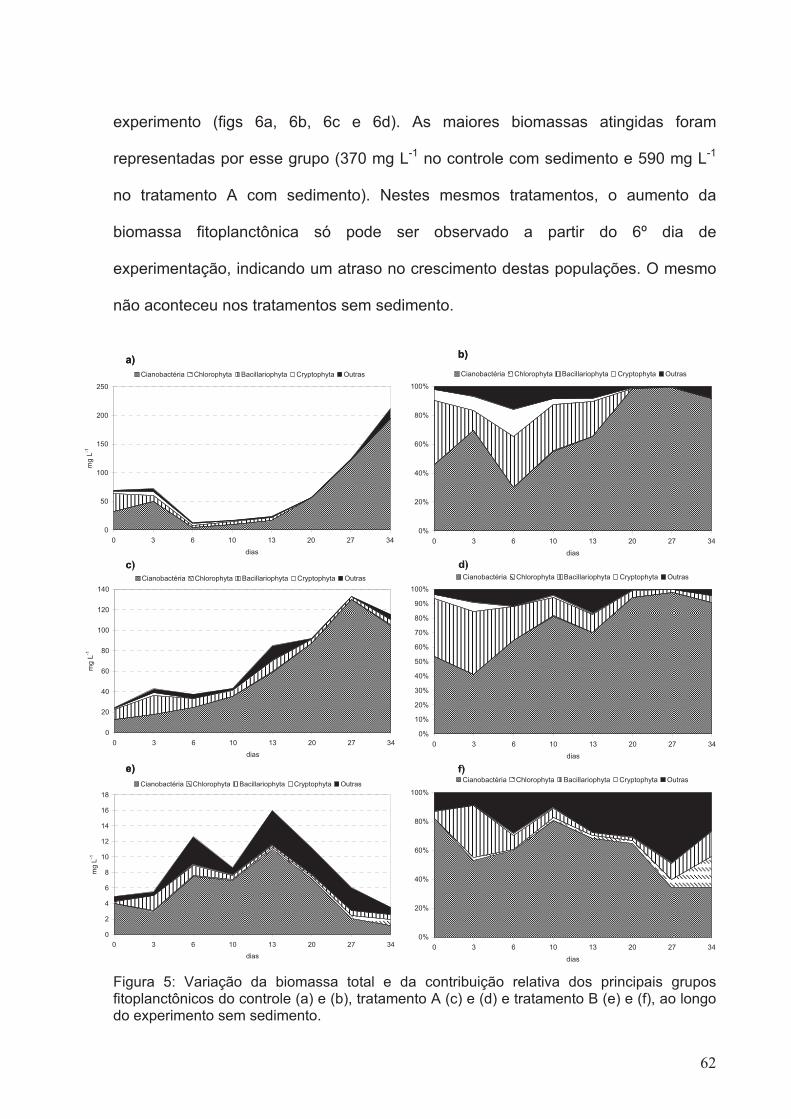
62
experimento (figs 6a, 6b, 6c e 6d). As maiores biomassas atingidas foram
representadas por esse grupo (370 mg L-1 no controle com sedimento e 590 mg L-1
no tratamento A com sedimento). Nestes mesmos tratamentos, o aumento da
biomassa fitoplanctônica só pode ser observado a partir do 6º dia de
experimentação, indicando um atraso no crescimento destas populações. O mesmo
não aconteceu nos tratamentos sem sedimento.
Figura 5: Variação da biomassa total e da contribuição relativa dos principais grupos fitoplanctônicos do controle (a) e (b), tratamento A (c) e (d) e tratamento B (e) e (f), ao longo do experimento sem sedimento.
0
50
100
150
200
250
0 3 6 10 13 20 27 34dias
mg
L-1
Cianobactéria Chlorophyta Bacillariophyta Cryptophyta Outras
0%
20%
40%
60%
80%
100%
0 3 6 10 13 20 27 34dias
Cianobactéria Chlorophyta Bacillariophyta Cryptophyta Outras
0
20
40
60
80
100
120
140
0 3 6 10 13 20 27 34dias
mg
L-1
Cianobactéria Chlorophyta Bacillariophyta Cryptophyta Outras
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
0 3 6 10 13 20 27 34dias
Cianobactéria Chlorophyta Bacillariophyta Cryptophyta Outras
0
2
4
6
8
10
12
14
16
18
0 3 6 10 13 20 27 34dias
mg
L-1
Cianobactéria Chlorophyta Bacillariophyta Cryptophyta Outras
0%
20%
40%
60%
80%
100%
0 3 6 10 13 20 27 34dias
Cianobactéria Chlorophyta Bacillariophyta Cryptophyta Outras
a) b)
c) d)
e) f)
0
50
100
150
200
250
0 3 6 10 13 20 27 34dias
mg
L-1
Cianobactéria Chlorophyta Bacillariophyta Cryptophyta Outras
0%
20%
40%
60%
80%
100%
0 3 6 10 13 20 27 34dias
Cianobactéria Chlorophyta Bacillariophyta Cryptophyta Outras
0
20
40
60
80
100
120
140
0 3 6 10 13 20 27 34dias
mg
L-1
Cianobactéria Chlorophyta Bacillariophyta Cryptophyta Outras
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
0 3 6 10 13 20 27 34dias
Cianobactéria Chlorophyta Bacillariophyta Cryptophyta Outras
0
2
4
6
8
10
12
14
16
18
0 3 6 10 13 20 27 34dias
mg
L-1
Cianobactéria Chlorophyta Bacillariophyta Cryptophyta Outras
0%
20%
40%
60%
80%
100%
0 3 6 10 13 20 27 34dias
Cianobactéria Chlorophyta Bacillariophyta Cryptophyta Outras
a) b)
c) d)
e) f)

63
Uma dinâmica diferente foi observada no tratamento B com sedimento, onde
os maiores valores de biomassa fitoplanctônica foram representados pelo grupo das
bacilariofíceas (fig. 6e). Inicialmente, essas diatomáceas atingiram valores de 19 mg
L-1, seguido de uma drástica redução da sua biomassa, chegando até a serem
excluídas no final do experimento (figs. 6e e 6f). Nesse tratamento, as clorofíceas
também apresentaram maiores valores de biomassa, nos períodos iniciais, do que
nos demais tratamentos. No entanto, assim como as diatomáceas, as clorofíceas
também foram praticamente excluídas após 34 dias de experimento. Embora as
cianobactérias tenham conseguido permanecer até o final do experimento, nos três
últimos dias houve um conspícuo decréscimo da biomassa fitoplanctônica total (fig.
6e).
Principais espécies de Cianobactérias
Uma grande representatividade em biomassa de Microcystis aeruginosa e
Aphanizomenon sp. foi observada, dentre as cianobactérias, nos controles e
tratamentos A, o que pode ser confirmado pela soma das contribuições relativas das
espécies desse grupo ao longo do experimento (tab. 1). As duas espécies chegaram
a atingir valores de mais de 95% de contribuição relativa no controle com sedimento
e tratamento A com sedimento, deixando clara a sua dominância diante das outras
espécies no grupo. Enquanto Microcystis aeruginosa teve maior contribuição no
controle com sedimento e tratamento A com sedimento (fig. 8, tab 1),
Aphanizomenon sp. superou esta, em representatividade de biomassa, no controle
sem sedimento e suas contribuições foram bastante semelhantes no tratamento A
sem sedimento (fig. 7, tab. 1). Em ambos os tratamentos B, a representatividade das
duas espécies, quando somadas, se mostrou menor do que em todos outros (figs. 7f
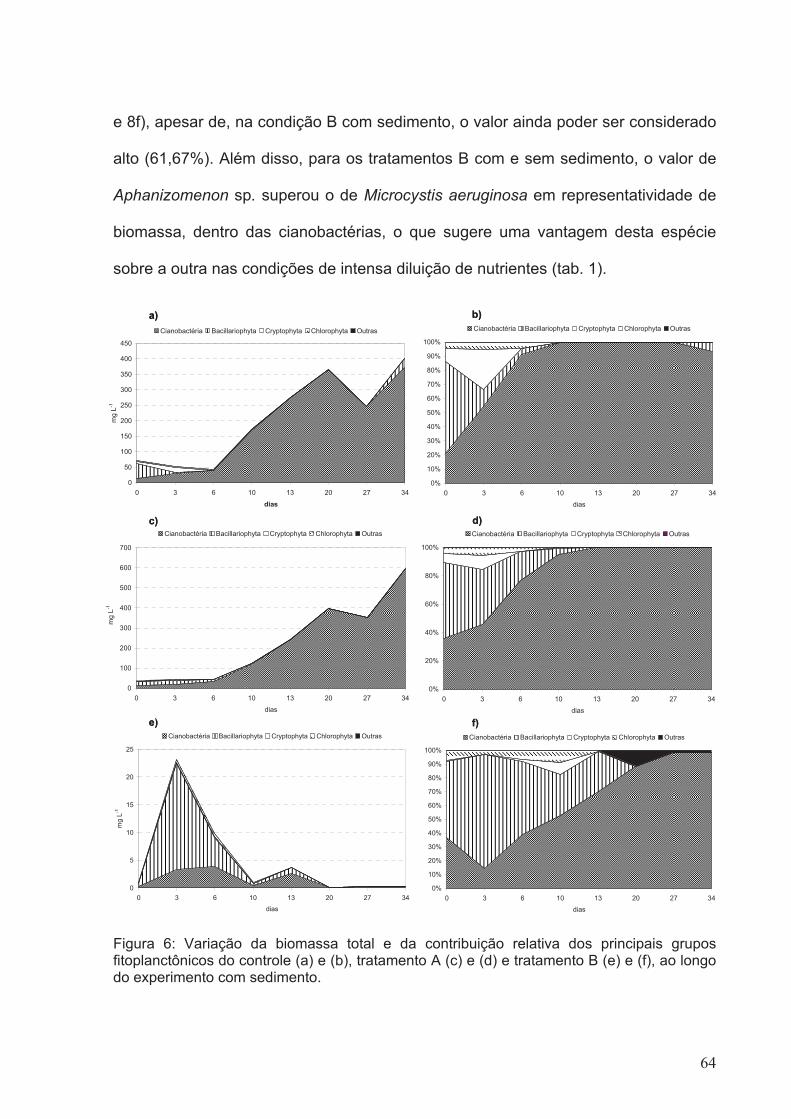
64
e 8f), apesar de, na condição B com sedimento, o valor ainda poder ser considerado
alto (61,67%). Além disso, para os tratamentos B com e sem sedimento, o valor de
Aphanizomenon sp. superou o de Microcystis aeruginosa em representatividade de
biomassa, dentro das cianobactérias, o que sugere uma vantagem desta espécie
sobre a outra nas condições de intensa diluição de nutrientes (tab. 1).
Figura 6: Variação da biomassa total e da contribuição relativa dos principais grupos fitoplanctônicos do controle (a) e (b), tratamento A (c) e (d) e tratamento B (e) e (f), ao longo do experimento com sedimento.
0
50
100
150
200
250
300
350
400
450
0 3 6 10 13 20 27 34
dias
mg
L-1
Cianobactéria Bacillariophyta Cryptophyta Chlorophyta Outras
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
0 3 6 10 13 20 27 34dias
Cianobactéria Bacillariophyta Cryptophyta Chlorophyta Outras
a) b)
0
100
200
300
400
500
600
700
0 3 6 10 13 20 27 34
dias
mg
L-1
Cianobactéria Bacillariophyta Cryptophyta Chlorophyta Outras
0%
20%
40%
60%
80%
100%
0 3 6 10 13 20 27 34
dias
Cianobactéria Bacillariophyta Cryptophyta Chlorophyta Outras
0
5
10
15
20
25
0 3 6 10 13 20 27 34dias
mg
L-1
Cianobactéria Bacillariophyta Cryptophyta Chlorophyta Outras
c) d)
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
0 3 6 10 13 20 27 34dias
Cianobactéria Bacillariophyta Cryptophyta Chlorophyta Outras
e) f)
0
50
100
150
200
250
300
350
400
450
0 3 6 10 13 20 27 34
dias
mg
L-1
Cianobactéria Bacillariophyta Cryptophyta Chlorophyta Outras
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
0 3 6 10 13 20 27 34dias
Cianobactéria Bacillariophyta Cryptophyta Chlorophyta Outras
a) b)
0
100
200
300
400
500
600
700
0 3 6 10 13 20 27 34
dias
mg
L-1
Cianobactéria Bacillariophyta Cryptophyta Chlorophyta Outras
0%
20%
40%
60%
80%
100%
0 3 6 10 13 20 27 34
dias
Cianobactéria Bacillariophyta Cryptophyta Chlorophyta Outras
0
5
10
15
20
25
0 3 6 10 13 20 27 34dias
mg
L-1
Cianobactéria Bacillariophyta Cryptophyta Chlorophyta Outras
c) d)
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
0 3 6 10 13 20 27 34dias
Cianobactéria Bacillariophyta Cryptophyta Chlorophyta Outras
e) f)

65
Apesar da dominância dessas espécies ao final do experimento ter
contribuído de forma marcante para o incremento da biomassa no sistema, os
períodos iniciais apresentaram uma maior diversidade, com espécies de menor
volume. Dentre estas, se destacam Aphanotece sp., Aphanocapsa sp. e
Geitlerinema sp.
A substituição dessa diversidade pela dominância observada ao final do
experimento se deu de modo mais gradual no controle e tratamento A com
sedimento (figs. 8b e 8d), enquanto as condições sem sedimento apresentaram uma
substituição rápida, a partir do sexto dia (figs. 7b e 7d). Nos tratamentos B, essa
diversidade foi sustentada até o fim do experimento, quando, na última semana,
observou-se uma queda acentuada de todas as espécies, o que ocorreu antes na
condição com sedimento (20º dia) (figs. 7e e 8e). Aphanizomenon sp. e Microcystis
aeruginosa foram as principais espécies de cianobactérias responsáveis pelos
aumentos de biomassa no tratamento B com sedimento, embora Aphanotece sp. e
Planktothrix agardhii também tenham contribuído em alguns períodos (fig. 8f). No
entanto, o tratamento B (sem sedimento) teve sua biomassa total muito influenciada
pelas espécies filamentosas Planktothrix agardhii e Aphanizomenon sp. (fig. 7f).
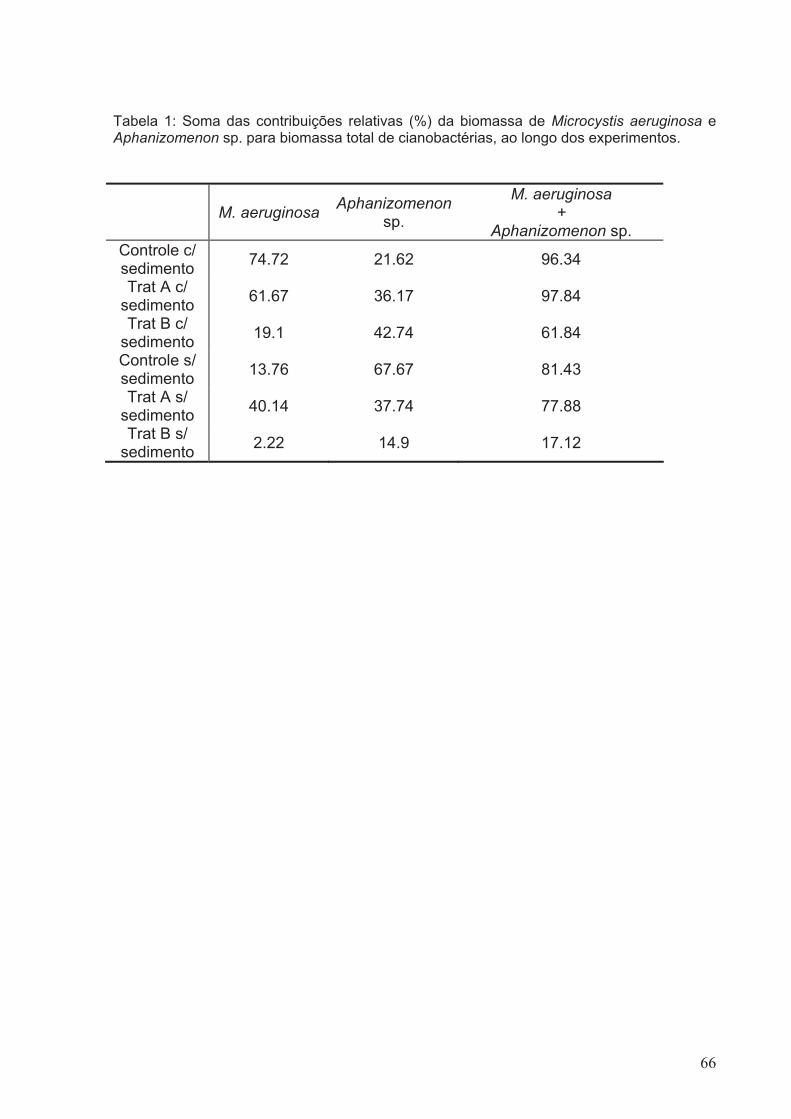
66
Tabela 1: Soma das contribuições relativas (%) da biomassa de Microcystis aeruginosa e Aphanizomenon sp. para biomassa total de cianobactérias, ao longo dos experimentos.
M. aeruginosa Aphanizomenon sp.
M. aeruginosa +
Aphanizomenon sp.Controle c/ sedimento 74.72 21.62 96.34
Trat A c/ sedimento 61.67 36.17 97.84
Trat B c/ sedimento 19.1 42.74 61.84
Controle s/ sedimento 13.76 67.67 81.43
Trat A s/ sedimento 40.14 37.74 77.88
Trat B s/ sedimento 2.22 14.9 17.12
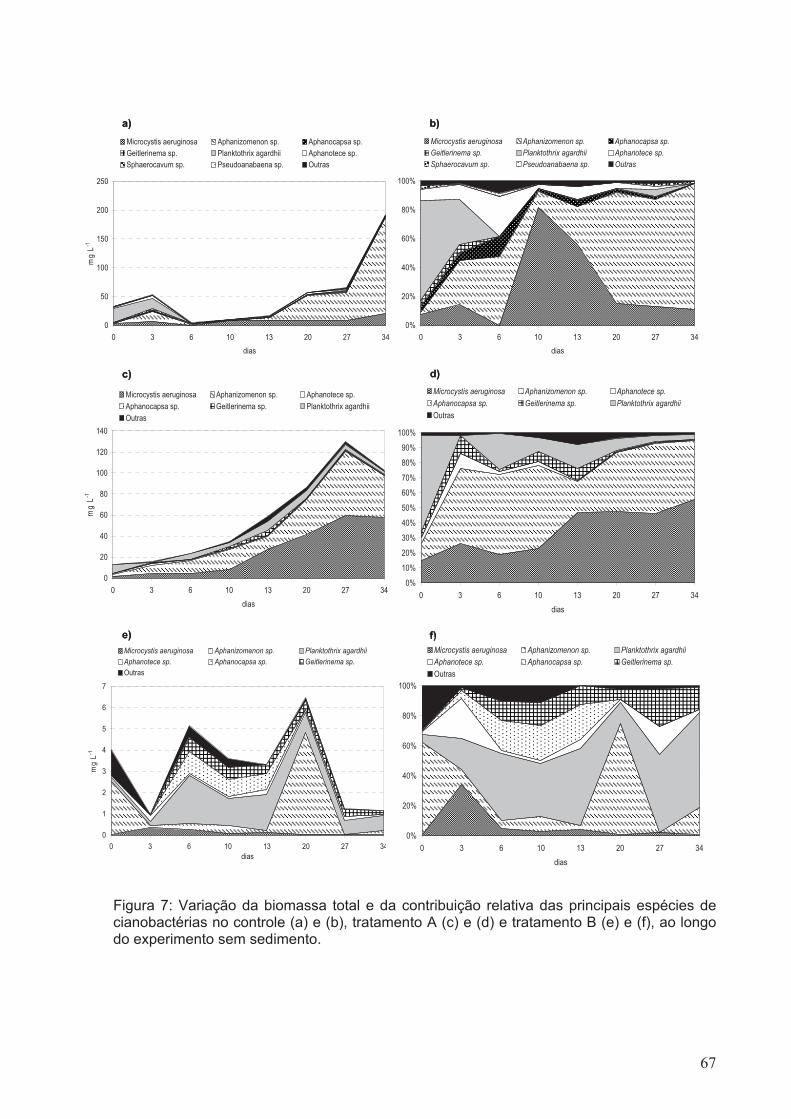
67
Figura 7: Variação da biomassa total e da contribuição relativa das principais espécies de cianobactérias no controle (a) e (b), tratamento A (c) e (d) e tratamento B (e) e (f), ao longo do experimento sem sedimento.
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
0 3 6 10 13 20 27 34dias
Microcystis aeruginosa Aphanizomenon sp. Aphanotece sp.Aphanocapsa sp. Geitlerinema sp. Planktothrix agardhiiOutras
0%
20%
40%
60%
80%
100%
0 3 6 10 13 20 27 34dias
Microcystis aeruginosa Aphanizomenon sp. Aphanocapsa sp.Geitlerinema sp. Planktothrix agardhii Aphanotece sp.Sphaerocavum sp. Pseudoanabaena sp. Outras
0
20
40
60
80
100
120
140
0 3 6 10 13 20 27 34dias
mg
L-1
Microcystis aeruginosa Aphanizomenon sp. Aphanotece sp.Aphanocapsa sp. Geitlerinema sp. Planktothrix agardhiiOutras
0
1
2
3
4
5
6
7
0 3 6 10 13 20 27 34dias
mg
L-1
Microcystis aeruginosa Aphanizomenon sp. Planktothrix agardhiiAphanotece sp. Aphanocapsa sp. Geitlerinema sp.Outras
0%
20%
40%
60%
80%
100%
0 3 6 10 13 20 27 34
dias
Microcystis aeruginosa Aphanizomenon sp. Planktothrix agardhiiAphanotece sp. Aphanocapsa sp. Geitlerinema sp.Outras
0
50
100
150
200
250
0 3 6 10 13 20 27 34dias
mg
L-1
Microcystis aeruginosa Aphanizomenon sp. Aphanocapsa sp.Geitlerinema sp. Planktothrix agardhii Aphanotece sp.Sphaerocavum sp. Pseudoanabaena sp. Outras
a) b)
c) d)
e) f)
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
0 3 6 10 13 20 27 34dias
Microcystis aeruginosa Aphanizomenon sp. Aphanotece sp.Aphanocapsa sp. Geitlerinema sp. Planktothrix agardhiiOutras
0%
20%
40%
60%
80%
100%
0 3 6 10 13 20 27 34dias
Microcystis aeruginosa Aphanizomenon sp. Aphanocapsa sp.Geitlerinema sp. Planktothrix agardhii Aphanotece sp.Sphaerocavum sp. Pseudoanabaena sp. Outras
0
20
40
60
80
100
120
140
0 3 6 10 13 20 27 34dias
mg
L-1
Microcystis aeruginosa Aphanizomenon sp. Aphanotece sp.Aphanocapsa sp. Geitlerinema sp. Planktothrix agardhiiOutras
0
1
2
3
4
5
6
7
0 3 6 10 13 20 27 34dias
mg
L-1
Microcystis aeruginosa Aphanizomenon sp. Planktothrix agardhiiAphanotece sp. Aphanocapsa sp. Geitlerinema sp.Outras
0%
20%
40%
60%
80%
100%
0 3 6 10 13 20 27 34
dias
Microcystis aeruginosa Aphanizomenon sp. Planktothrix agardhiiAphanotece sp. Aphanocapsa sp. Geitlerinema sp.Outras
0
50
100
150
200
250
0 3 6 10 13 20 27 34dias
mg
L-1
Microcystis aeruginosa Aphanizomenon sp. Aphanocapsa sp.Geitlerinema sp. Planktothrix agardhii Aphanotece sp.Sphaerocavum sp. Pseudoanabaena sp. Outras
a) b)
c) d)
e) f)
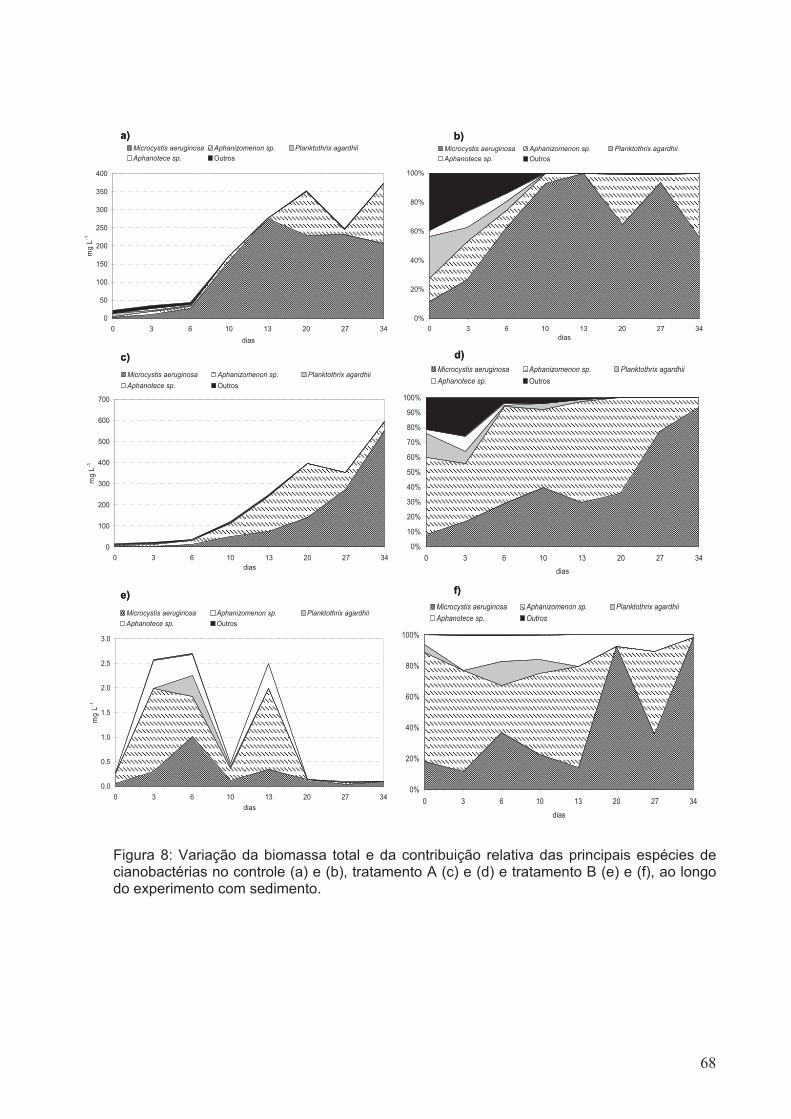
68
Figura 8: Variação da biomassa total e da contribuição relativa das principais espécies de cianobactérias no controle (a) e (b), tratamento A (c) e (d) e tratamento B (e) e (f), ao longo do experimento com sedimento.
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
0 3 6 10 13 20 27 34dias
Microcystis aeruginosa Aphanizomenon sp. Planktothrix agardhiiAphanotece sp. Outros
0%
20%
40%
60%
80%
100%
0 3 6 10 13 20 27 34
dias
Microcystis aeruginosa Aphanizomenon sp. Planktothrix agardhiiAphanotece sp. Outros
0
50
100
150
200
250
300
350
400
0 3 6 10 13 20 27 34dias
mg
L-1
Microcystis aeruginosa Aphanizomenon sp. Planktothrix agardhiiAphanotece sp. Outros
0%
20%
40%
60%
80%
100%
0 3 6 10 13 20 27 34dias
Microcystis aeruginosa Aphanizomenon sp. Planktothrix agardhiiAphanotece sp. Outros
0
100
200
300
400
500
600
700
0 3 6 10 13 20 27 34dias
mg
L-1
Microcystis aeruginosa Aphanizomenon sp. Planktothrix agardhiiAphanotece sp. Outros
0.0
0.5
1.0
1.5
2.0
2.5
3.0
0 3 6 10 13 20 27 34dias
mg
L-1
Microcystis aeruginosa Aphanizomenon sp. Planktothrix agardhiiAphanotece sp. Outros
a) b)
c) d)
e) f)
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
0 3 6 10 13 20 27 34dias
Microcystis aeruginosa Aphanizomenon sp. Planktothrix agardhiiAphanotece sp. Outros
0%
20%
40%
60%
80%
100%
0 3 6 10 13 20 27 34
dias
Microcystis aeruginosa Aphanizomenon sp. Planktothrix agardhiiAphanotece sp. Outros
0
50
100
150
200
250
300
350
400
0 3 6 10 13 20 27 34dias
mg
L-1
Microcystis aeruginosa Aphanizomenon sp. Planktothrix agardhiiAphanotece sp. Outros
0%
20%
40%
60%
80%
100%
0 3 6 10 13 20 27 34dias
Microcystis aeruginosa Aphanizomenon sp. Planktothrix agardhiiAphanotece sp. Outros
0
100
200
300
400
500
600
700
0 3 6 10 13 20 27 34dias
mg
L-1
Microcystis aeruginosa Aphanizomenon sp. Planktothrix agardhiiAphanotece sp. Outros
0.0
0.5
1.0
1.5
2.0
2.5
3.0
0 3 6 10 13 20 27 34dias
mg
L-1
Microcystis aeruginosa Aphanizomenon sp. Planktothrix agardhiiAphanotece sp. Outros
a) b)
c) d)
e) f)

69
DISCUSSÃO
As diferenças nos valores de condutividade elétrica, salinidade e intensidade
luminosa entre os tratamentos foram consequência do processo de diluição da água
da lagoa e, portanto permaneceram estáveis ao longo do experimento.
Embora a água de Lagoa de Jacarepaguá apresentasse 516,5 µg L-1 PSR e a
água proveniente do Rio Camorim 43,7 µg L-1 PSR no momento da coleta, as
concentrações de P observadas já no primeiro dia de experimentação foram
diferentes dos valores teóricos esperados. Entretanto, as diluições realizadas não
atingiram valores iniciais limitantes de nutrientes, embora este processo tenha
permitido uma redução de biomassa da comunidade fitoplanctônica do tratamento
controle para o tratamento B (de maior diluição).
O esgotamento de nutrientes dissolvidos pôde ser observado rapidamente em
todos os tratamentos, com exceção do PSR da condição controle com sedimento. O
sedimento da lagoa de Jacarepaguá parece ser um forte contribuinte para o aporte
de P para a coluna d'água. A água do rio tributário Camorim contribuiu com maior
aporte de N, principalmente na forma de nitrato, que a lagoa de Jacarepaguá. Como
visto no capítulo anterior, esta lagoa tem uma forte limitação por nitrogênio em
alguns períodos do ano.
O empobrecimento por nutrientes permitiu uma redução da biomassa da
comunidade fitoplanctônica, mas a dominância de cianobactérias foi observada em
todos os tratamentos do experimento sem sedimento e nos controles e tratamento A
com sedimento. Os tratamentos B apresentaram as menores biomassas
fitoplanctônicas.
Ao contrário do observado em outros trabalhos com redução de nutrientes
(Sommer et al. 1993; Ruggiu et al. 1998; Crosseti & Bicudo 2005), o confinamento

70
da comunidade fitoplanctônica promoveu um aumento de sua biomassa ao longo do
experimento. A justificativa dos já referidos trabalhos foi que a redução da biomassa
tenha sido provocada pelo esgotamento de nutrientes livres na cultura. No presente
estudo, não foi possível observar a redução de biomassa nos controles e
tratamentos A com e sem sedimento, devido ao seu estado hipereutrófico, muito
embora os nutrientes tenham sido consumidos logo nos primeiros dias de
experimento.
A composição da comunidade fitoplanctônica foi similar em todos os
tratamentos e sofreu alterações ao longo dos 34 dias de experimento. Segundo
Gaedke & Schweizer (1993) alterações na composição florística são comuns durante
o processo oligotrofização, e geralmente precedem a redução da biomassa.
A dominância da comunidade fitoplanctônica pelas cianobactérias ocorreu em
todos os experimentos, comprovando que não basta apenas reduzir a carga de
nutrientes na água para que deixe de haver dominância por parte das mesmas.
Entretanto, vale ressaltar a redução da biomassa total fitoplanctônica no tratamento
de maior redução de nutrientes e a quebra da dominância das cianobactérias pode
ser dependente da variação temporal. O experimento teve uma duração de 34 dias.
Esse tempo não foi suficiente para observarmos uma quebra da dominância das
cianobactérias, mas os resultados de biomassa sugerem que a redução de P pode
ser uma metodologia eficiente para remediação deste ecossistema.
De acordo com Huszar et al. (2000) a dominância de cianobactérias em um
corpo d'água pode estar diretamente relacionada com a temperatura e pH, e
inversamente relacionada com a luz, reforçando os resultados do experimento, onde
podemos observar a redução da biomassa de cianobactérias nos tratamentos A e B.
Nestas condições, o pH e a temperatura foram inferiores, embora não significativos,

71
e a intensidade luminosa superior, aos valores apresentados no controle,
influenciando para a diminuição da população de cianobactérias em ambos os
tratamentos, que corresponde à principal causa da redução da biomassa total. Os
grupos restantes variaram sua biomassa, mas não contribuíram diretamente para a
variação da biomassa total, com exceção das diatomáceas.
O grupo das bacilariofíceas foi o segundo mais representativo na
comunidade, foi possível observar que os valores mais elevados de biomassa de
diatomáceas estiveram concentrados nos dias com menores valores de biomassa de
cianobactérias, indicando uma substituição entre as espécies destes dois grupos. De
acordo com Reynolds (1984) o crescimento das diatomáceas está relacionado com
dois principais fatores, a disponibilidade de luz e a relação entre sílica e fósforo. A
concentração de sílica não foi mensurada no presente trabalho, mas as maiores
biomassas de diatomáceas coincidiram com os maiores valores de PSR no inicio do
experimento. Além disso, o tratamento B foi o que apresentou maior contribuição
relativa deste grupo.
Dentre as principais espécies de cianobactérias, Microcystis aeruginosa e
Aphanizomenon sp. foram as que apresentaram maiores contribuições relativas para
biomassa fitoplanctônica total, principalmente nas condições controles e tratamentos
A. Nos tratamentos B com e sem sedimento, o valor de Aphanizomenon sp. superou
o de Microcystis aeruginosa em representatividade de biomassa, dentro das
cianobactérias, o que sugere uma vantagem desta espécie sobre a outra em
condições de intensa diluição de nutrientes. Cabe ressaltar que a espécie
Aphanizomenon sp. pode apresentar heterocitos, o que lhe permite a fixação de
nitrogênio atmosférico e uma vantagem em ambiente empobrecidos. Microcystis
aeruginosa apresentou maiores biomassas nos experimentos com sedimento do que

72
nos sem sedimento. É possível que o sedimento da lagoa de Jacarepaguá
apresentasse células desta espécie, que quando colocadas sob as condições de
experimentação foram ressuspensas e crescidas, aumentando sua
representatividade em tais tratamentos.
Apesar da dominância das espécies Microcystis aeruginosa e
Aphanizomenon sp. ao final do experimento, contribuindo de forma marcante para o
incremento da biomassa no sistema, os períodos iniciais apresentaram uma maior
diversidade, com espécies de menor volume contribuindo para a densidade total.
Dentre essas, se destacam Aphanotece sp., Aphanocapsa sp., Geitlerinema sp. e
Phormidium sp (tratamentos B com e sem sedimento). Esse fato é comumente
observado nos estágios iniciais de sucessão em ambientes pouco eutrofizados
(Reynolds 2006).
Em síntese, a redução inicial da concentração de P não foi suficiente para
acabar com a dominância das cianobactérias, embora tenha havido um esgotamento
dos nutrientes ao longo do experimento. A dinâmica do N e P foi importante para
determinar a composição da comunidade fitoplanctônica, mas não promoveu a
redução da sua biomassa fitoplanctônica nos tratamentos com condições iniciais
mais eutróficas (controles e tratamentos A). A ciclagem desses nutrientes permitiu a
substituição das espécies. Assim como visto no capítulo anterior, as espécies M.
aeruginosa, Aphanizomenon sp. e P. agardhii apresentaram grande
representatividade na comunidade fitoplanctônica e a dinâmica dessas espécies
provavelmente esteve associada principalmente à dinâmica do N, uma vez que o
requerimento por P para o fitoplâncton é menor (Reynolds, 2006) e este elemento
esteve presente durante boa parte do período de experimentação. Portanto, o
melhoramento da qualidade da água da lagoa de Jacarepaguá só irá ocorrer quando

73
o aporte de P neste sistema for cessado ou reduzido para concentrações ainda
menores que as testadas nestes experimentos (menor que 46 µg L-1), uma vez que
o sedimento da lagoa contribui com a entrada de P no sistema, assim como já
classicamente proposto para outros sistemas por Vollenweider & Kerekes (1980) e
Reynolds (1992), entre outros.

74
REFERÊNCIAS
APHA. 1998. Standard Methods for Examination of Waste and Wastewater – American Public Health Association, New York.
CROSSETI, L.O., BICUDO, C.E.M. 2005. Sructural and Functional phytoplankton responses to nutrient impoverishment in mesocosms placed in a shallow eutrophic reservoir (Graças Pond), São Paulo, Brazil. Hydrobiologia. 541: 71-85.
DOS SANTOS, A.C.A, CALIJURI, M.C. 1998. Survival strategies of some species of phytoplankton community in Barra Bonita reservoir (São Paulo, Brazil). Hydrobiologia. 367: 139-152.
GAEDKE, U., SCHWEIDER, A. 1993. The first decade of oligotrophication in Lake Constance, I: The response of phytoplankton biomass and cell size. Oecologia. 93: 268-275.
GOMES, A. M. A., SAMPAIO, P. L., FERRÃO-FILHO, A.S., MAGALHÃES, V. F., MARINHO, M.M. OLIVEIRA, A. C. P., SANTOS, V. B., DOMINGOS, P., AZEVEDO, S.M.F.O. (2009). Florações de Cianobactérias Tóxicas em uma Lagoa Costeira Hipereutrófica do Rio de Janeiro/Rj (Brasil) e suas Consequências para Saúde Humana. Oecologia Brasiliensis. 13 (2), 329-345.
HUSZAR, V.L., SILVA, L.H.S., MARINHO, M.M., DOMINGOS, P., SANT'ANNA, C. 2000. Cyanoprocaryote assembleges in eigth productive tropical Brazilian Waters. Hydrobiologia. 424: 67-77.
LAVOREL, S., MCINTYRE, S., LANDSBERG, J., FORBES, T.D.A. 1997. Plant functional classifications: from general groups to specific group base on response to disturbance. Trends in Ecology and Evolution. 12: 474-478.
ORTEGA-MAYAGOITA, E., RODRIGO, M.A., ROJO, C. 2002. Factors masking the trophic cascade in shallow eutrophic wetlands: evidence from a mesocosm study. Archv für Hydrobiologie. 155: 43-63.
ORTEGA-MAYAGOITA, E., RODRIGO, M.A., ROJO, C. 2003. Controlling factors of phytoplankton assemblages in wetlands: an experimental approach. Hydrobiologia. 502: 177-186.
PADISAK, J., REYNOLDS, C.S. 1998. Selection of phytoplankton associations in Lake Balaton, Hungary, in response to eutrophication and restoration measures, with special reference to the cyanoprokariotes. Hydrobiologia. 384: 41-53.
PAERL, H.W. 2006. Nutrient and other environmental controls of harmful cyanobacterial blooms along the freshwater-marine continuum. In: H.K. Hudnell (ed): Proceedings of the Interagency, International Symposium on Cyanobacterial Harmful Algal Blooms. Advances in Experimental Medicine & Biology.

75
PAERL, H.W, BOWLES, N.D. 1987. Dilution bioassays: their application to assessments of nutrient limitation in hypereutrophic waters. Hydrobiologia. 146: 265-273.
REYNOLDS, C.S. 2006. Ecology of Phytoplankton. Cambridge University Press. Cambridge. 535p.
REYNOLDS, C.S. 1992. Eutrophication and management of planktonic algae: what Vollenweider couldn't tell us. In: Eutrophication: Research and application to water supply. Ed: David W Sutcliffe and J. Gwynfryn Jones.
REYNOLDS, C.S. 1988. Functional morphology and the adaptive strategies of freshwater phytoplankton. p. 388-433 In C. D. Sandgren [ed.] Growth and reproductive strategies of freshwater phytoplankton. Cambridge University Press. Cambridge.
REYNOLDS, C.S. 1984. The ecology of freshwater phytoplankton. Cambridge University Press. Cambridge.
RUGGIU, D., MORABITO, G., PANZANI, P., PUGNETTI, A. 1998. Trends and relations among basic phytoplankton characteristics in the course of the long term oligotrophication of Lake Maggiore (Italy). Hydrobiologia. 369-370: 243-257.
SOMMER, U., GAEDKE, U., SCHWEIZER, A. 1993. The first decade of oligotrophication in Lake Constance, 2: the response of phytoplankton taxonomic composition. Oecologia. 93: 276-284.
TUNDISI, J. G. & MATSUMURA-TUNDISI, T. 1992. Eutrophication of lakes and reservoirs: a comparative analysis, case studies, perspectives. In: Cordeiro Marino et al., editors. Algae and Environment – A general approach. Sociedade Brasileira de Ficologia. P 1-33.
UHELINGER, V. 1964. Étude statisque des méthodes de dénobrement planctonique. Arch. Sci. 17: 121-123.
UTERMÖHL, H. 1958. Zur Vervollkommung der quantitativen Phytoplankton Methodik. Mitt. Int. Verein. Limno., 9: 1-39.
VOLLENWEIDER, R., KEREKES, J. 1980. The loading concept as basis for controlling eutrophication philosophy and preliminary results of the OECD programme on eutrophication. Prog. Wat. Tech. 12: 5-38.

76
CAPÍTULO III: EFEITO DA TEMPERATURA NO CRESCIMENTO E COMPETIÇÃO ENTRE ESPÉCIES
FITOPLANCTÔNICAS
INTRODUÇÃO
O tema “Mudanças Globais” tem despertado interesse no mundo acadêmico,
principalmente pelo fato de os modelos preditivos sugerirem um aumento da
temperatura global de 1,4 a 5,8 oC até o ano de 2100 (Houghton et al., 2001). Esse
aumento da temperatura pode promover efeitos nas interações bióticas de vários
ambientes, incluindo os aquáticos (Houghton et al., 2001, Winder & Schindler, 2004).
Nos ambientes aquáticos, é sabido que a temperatura pode exercer uma forte
influência na sucessão entre espécies fitoplanctônicas (Goldman & Ryther, 1976;
Rhee & Gotham, 1981; Descamps-Julien & Gonzalez, 2005). Há algumas décadas
vem sendo discutido o efeito da temperatura na fisiologia da utilização dos recursos
pelo fitoplâncton, como por exemplo, na absorção de nutrientes (Goldman, 1977;
Rhee & Gotham, 1981), assim como na composição química das células desses
organismos (Goldman & Ryther, 1976; Thompson et al., 1992). Entretanto, cada
espécie responde à variação de temperatura de maneira diferente. Algumas
espécies do grupo das diatomáceas são associadas a ambientes de baixas
temperaturas (Zhang & Prepas, 1996), enquanto que temperaturas mais altas
parecem favorecer o crescimento das cianobactérias (Elliott et al., 2006; Jöhnk et al.,
2008; Kosten, 2010).
A importância da temperatura na dinâmica da comunidade fitoplanctônica em
ambientes aquáticos das regiões temperadas vem sendo frequentemente discutida
(Zhang & Pepas, 1996; Jöhnk et al., 2008). Um antigo consenso entre
pesquisadores da limnologia sugerem que variações de temperatura no ambiente

77
não seriam suficientes para influenciar a sucessão de espécies fitoplanctônicas em
regiões tropicais. As temperaturas seriam sempre altas e favoráveis às
cianobactérias. Entretanto, trabalhos mais recentes vêm mostrando a importância da
variação da temperatura sobre a dinâmica do fitoplâncton em reservatórios
brasileiros (Marinho & Huszar, 2002; Soares et al., 2009).
Como relatado no capítulo anterior, as cianobactérias dominaram a
comunidade fitoplanctônica da lagoa de Jacarepaguá durante todo o ciclo sazonal
de 2007. No entanto, no período de inverno, quando a temperatura da água reduziu-
se de 30 oC para até 18 oC, houve um aumento da contribuição relativa das espécies
dos grupos das clorofíceas e diatomáceas. Dentre as espécies presentes durante
esse período, Cyclotella sp. foi uma das mais representativas entre as diatomáceas
e Desmodesmus quadricauda entre as clorofíceas. No período de dominância das
cianobacatérias, Microcystis aeruginosa e Planktothrix sp. foram espécies co-
dominantes, além da Aphanizomenon sp.
Microcystis aeruginosa e Planktothrix sp. são cianobactérias conhecidas como
formadoras de florações e potencialmente produtoras de toxinas (Krishnamurthy et
al., 1986; Meriloto et al., 1989; Sivonen et al., 1990; Cood et al., 1999; Akcaalan et
al., 2006). Essas espécies são cosmopolitas e estão frequentemente associadas a
ambientes eutrofizados (Reynolds et al., 1981; Reynolds, 1987; Barco et al., 2004 ).
Seu sucesso na formação de florações também pode depender da temperatura
(Jöhnk et al., 2008; Pearl & Huisman 2008, 2009). A temperatura ótima para o
crescimento de Microcystis aeruginosa fica entre 25 oC e 30 oC, embora ela também
seja capaz de se tornar dominante em temperaturas de até 13 oC (Watanabe &
Oishi, 1985). Planktothrix apresenta maiores taxas de crescimento entre 25 oC e 30
oC (Paerl & Huisman, 2009).

78
Por outro lado, as diatomáceas tornam-se competidoras mais fracas quando
submetidas a altas temperaturas (Elliott et al 2006; Paerl & Huisman, 2009).
Experimentos de laboratório e estudos de campo mostraram uma significante
redução da biomassa das diatomáceas quando submetidas a temperaturas
superiores a 15 oC (Tilman et al., 1986; Zhang & Prepas, 1996).
Segundo a revisão realizada por Padisak (2004), as melhores temperaturas
para o crescimento das espécies do grupo das clorofíceas variavam entre 20 oC e 35
oC. A espécie Desmodesmus quadricauda (anteriormente descrita como
Scenedesmus quadricauda) apresentou temperatura ótima de 32 oC.
Uma questão relevante, porém não discutida nesses trabalhos, é a origem
das cepas testadas. É possível que cepas de uma mesma espécie, mas originadas
da região temperada, possam ter uma tolerância menor a temperaturas mais
elevadas, quando comparadas com cepas da região tropical.
Portanto, é preciso refinar o conhecimento acerca de fatores classicamente
considerados como determinantes na ecologia e fisiologia do fitoplâncton, assim
como na sua sucessão na comunidade aquática. Uma vez que mudanças na
comunidade fitoplanctônica na lagoa de Jacarepaguá foram acompanhadas de uma
significante variação de temperatura, o presente trabalho teve como objetivos
avaliar, em laboratório, o efeito da temperatura sobre o crescimento de duas
espécies de cianobactérias, uma de clorofícea e uma de diatomácea, que ocorrem
em abundâncias alternadas neste ambiente. Além disso, foi avaliado o efeito da
temperatura em experimentos de competição em cultivos mistos envolvendo as
cianobactérias e a diatomácea.

79
MATERIAL E MÉTODOS
CONDIÇÕES DE CULTIVO DO FITOPLÂNCTON
Para realização dos experimentos foram utilizadas cepas de Microcystis
aeruginosa (MIJAC-01) e Desmodesmus quadricauda (DEJAC-02/ anteriormente
descrita como Scenedesmus quadricauda), isoladas da lagoa de Jacarepaguá
durante o período de trabalho de campo (2007). Porém, a cepa de Planktothrix
isolada da lagoa de Jacarepaguá não sobreviveu em cultivo monoespecífico e a
cepa de diatomácea isolada (gênero Cyclotella) foi contaminada com outro
microrganismo fotossintetizante. Portanto, embora a intenção inicial fosse estudar
cepas isoladas da lagoa de Jacarepaguá, essas cepas precisaram ser substituídas e
utilizou-se uma cepa de P. agardhii cedida pelo Laboratório “Aquatic Ecology and
Water Quality/AEW” da Universidade de Wageningen e uma cepa de Cyclotella
meneghiniana adquirida do “Culture Colection of Algae and Protozoa /CCAP”, ambas
isoladas de lagos da região temperada. Estas cepas foram mantidas em cultivos
estanques monoespecíficos com meio WC modificado (Lürling & Beekman, 1999),
sob condições controladas de intensidade luminosa, temperatura, pH e fotoperíodo,
como descrito abaixo.
EXPERIMENTO I: EFEITO DA TEMPERATURA NO CRESCIMENTO DE QUATRO ESPÉCIES
FITOPLANCTÔNICAS
Para verificar o efeito de diferentes temperaturas nos crescimentos das cepas
de M. aeruginosa, P. agardhii, D. quadricauda e C. meneghiniana, as mesmas foram
cultivadas em meio WC modificado (Lürling e Beekman, 1999), sob intensidade
luminosa de 60 �mol fóton m-2 s-1, fotoperíodo de 18:6h (luz/escuro), agitação de 60
rpm. As temperaturas testadas foram de 18, 21, 24, 27 e 30 o C. A escolha destas

80
temperaturas baseou-se na variação da temperatura da água na lagoa de
Jacarepaguá, possivelmente um fator que influencia as substituições dessas
espécies. Os cultivos foram realizados em Erlenmeyer de 200 mL, com um volume
de cultura de 100 mL, dentro de incubadoras aclimatadas. O experimento teve
duração de 10 dias, sendo retiradas amostras diárias para as análises de clorofila-a
e da eficiência do fotossistema II. Essas análises foram realizadas in vivo com
auxílio de um fluorímetro - PHYTO-PAM (Heins Walz GmgH, Effeltrich, Germany).
EXPERIMENTO II: EFEITO DA TEMPERATURA NA COMPETIÇÃO ENTRE TRÊS ESPÉCIES
FITOPLANCTÔNICAS
Para verificar o efeito da temperatura na competição entre espécies, foram
realizados experimentos de cultivos mistos com M. aeruginosa (MIJAC-01), P.
agardhii (PAT-3) e C. meneghiniana (CCAP 1070/5) em duas temperaturas distintas
(18 e 30 oC). Os inóculos das culturas mistos foram realizados utilizando o mesmo
biovolume inicial para cada espécie (107 �m3 mL-1). As combinações das culturas
mistas foram: (a) MIJAC + PAT; (b) PAT + CCAP; (c) MIJAC + CCAP e (d) MIJAC +
PAT + CCAP. Os cultivos monoespecíficos (controle), além dos cultivos mistos
dessas cepas foram realizados em triplicata, durante 10 dias. A intensidade
luminosa (60 �mol fótons m-2 s-1), o fotoperíodo (18:6h) e a agitação (60 rpm) foram
controlados. Uma alíquota amostral foi retirada diariamente para análises de
clorofila-a e eficiência do fotossistema II (PHYTO-PAM - Heins Walz GmgH,
Effeltrich, Germany). A contagem do número de células de cada espécie dentro de
uma mesma amostra do cultivo misto foi realizada através do uso de um
hemocitômetro de Fuchs-Rosenthal e um microscópio óptico.

81
ANÁLISE DE DADOS
As taxas de crescimento foram calculadas a partir da equação:
�= ln (Nt-N0)/�t,
onde, � é a taxa de crescimento, N0 e Nt são as concentrações de clorofila no início
e final da fase exponencial e �t é o intervalo de tempo, em dias, da fase
exponencial. O �t foi definido pelo intervalo de tempo no qual a concentração de
clorofila apresentou um aumento exponencial, verificado através do valor de
coeficiente de correlação (r2) acima de 95%.
O rendimento máximo (R) das culturas nas diferentes condições foi calculado
a partir da razão entre a concentração máxima de clorofila atingida em cada cultura
e a concentração de clorofila inicial.
Para comparar a diferença das taxas de crescimento entre as espécies e as
temperaturas foi realizada uma de análise de variância (ANOVA). As variações que
apresentaram diferenças significativas foram submetidas ao teste de comparação
múltipla de Tukey. Para os dados de biomassa, de clorofila e rendimento
fotossintético foi utilizado o modelo de análise de variância com medidas repetidas
(Repeated-measures ANOVA). Estas análises foram feitas utilizando o programa
estatístico SPSS® 17.0.
RESULTADOS
EXPERIMENTO I: EFEITO DA TEMPERATURA NO CRESCIMENTO DE QUATRO ESPÉCIES
FITOPLANCTÔNICAS
No presente trabalho, assumimos a medida da concentração de clorofila-a
para avaliar a biomassa total e utilizamos o logaritmo dessa concentração para
medir o crescimento das espécies sob cinco diferentes temperaturas. Essa

82
metodologia foi adotada com base no fato de que a concentração de clorofila por
célula não variou ao longo do tempo (dados não mostrados).
Na maioria das temperaturas testadas, a taxa de crescimento da cepa de M.
aeruginosa (MIJAC-01) não apresentou nenhuma diferença significativa (fig.1). A
exceção foi a temperatura de 21 oC, na qual houve uma pequena redução da sua
taxa de crescimento (F=10,02; p=0,002) (fig. 1). Entretanto, os maiores valores de
rendimento máximo foram observados nos cultivos de M. aeruginosa a 18 oC e 30 oC
(tab. 1). Nenhuma diferença significativa na eficiência do fotossistema II foi
observada para esta cepa nas diferentes temperaturas (fig. 2b).
M. aeruginosa apresentou menor biomassa (medida a partir das
concentrações de clorofila-a) na temperatura de 18 oC (p<0,001) ao longo de todo o
período experimental, enquanto diferenças menos pronunciadas foram observadas
entre as temperaturas de 21 a 30 oC (fig. 2a). No entanto, ao final de 10 dias de
experimento, esta cepa alcançou a mesma biomassa em todas as temperaturas. A
18 oC M. aeruginosa parecia estar ainda na fase de crescimento exponencial até o
10º dia de cultivo, enquanto nas demais temperaturas, neste período, esta cepa já
estava ou na fase estacionária ou na senescência.
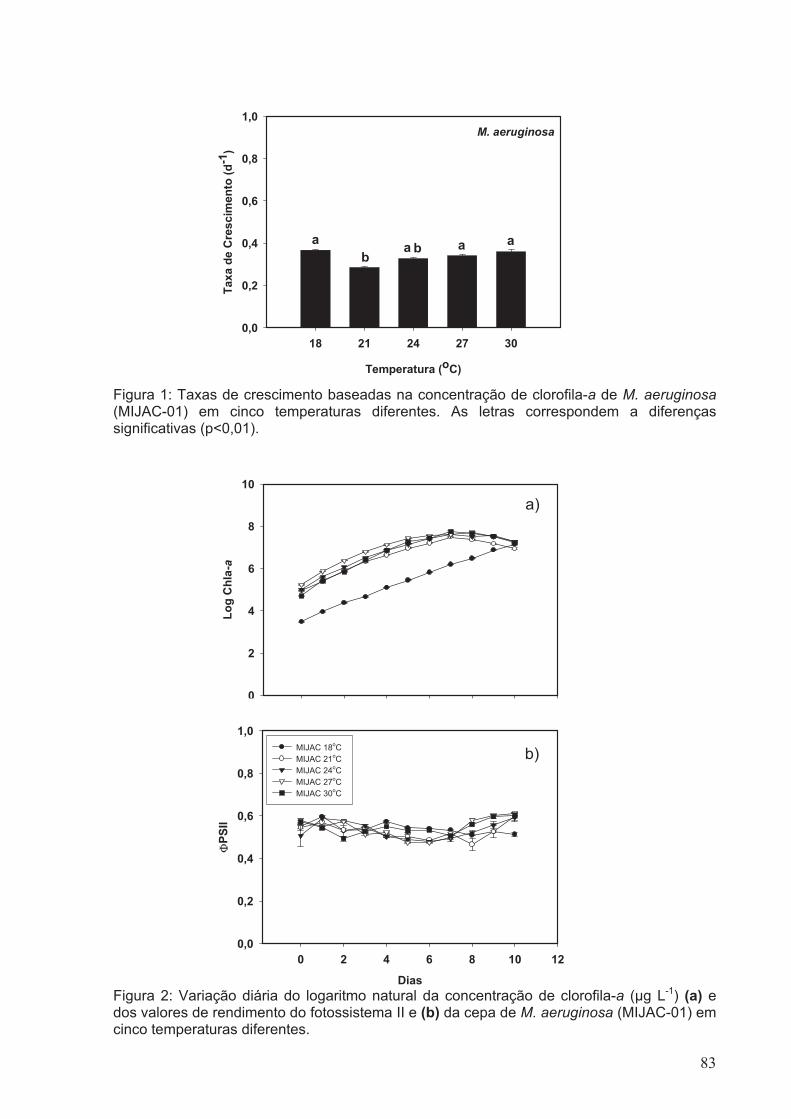
83
Figura 1: Taxas de crescimento baseadas na concentração de clorofila-a de M. aeruginosa (MIJAC-01) em cinco temperaturas diferentes. As letras correspondem a diferenças significativas (p<0,01).
Figura 2: Variação diária do logaritmo natural da concentração de clorofila-a (µg L-1) (a) e dos valores de rendimento do fotossistema II e (b) da cepa de M. aeruginosa (MIJAC-01) em cinco temperaturas diferentes.
M. aeruginosa
Temperatura (oC)
18 21 24 27 30
Taxa
de
Cre
scim
ento
(d-1
)
0,0
0,2
0,4
0,6
0,8
1,0
a a abb
a
M. aeruginosa
Temperatura (oC)
18 21 24 27 30
Taxa
de
Cre
scim
ento
(d-1
)
0,0
0,2
0,4
0,6
0,8
1,0
a a abb
aLo
g C
hla-
a
0
2
4
6
8
10
Dias
0 2 4 6 8 10 12
�PS
II
0,0
0,2
0,4
0,6
0,8
1,0MIJAC 18oCMIJAC 21oCMIJAC 24oCMIJAC 27oCMIJAC 30oC
a)
b)
Log
Chl
a-a
0
2
4
6
8
10
Dias
0 2 4 6 8 10 12
�PS
II
0,0
0,2
0,4
0,6
0,8
1,0MIJAC 18oCMIJAC 21oCMIJAC 24oCMIJAC 27oCMIJAC 30oC
a)
b)

84
P. agardhii (PAT-3) apresentou uma maior taxa de crescimento a 27 oC
(p<0,001), seguido das temperaturas de 30, 21 e 24 oC. Na menor temperatura (18
oC), sua taxa de crescimento foi reduzida (p<0,001) (fig. 3). Resultados semelhantes
foram observados para os valores de biomassa, medidos a partir das concentrações
de clorofila. A partir do terceiro dia, também a biomassa total dessa cepa, cultivada a
27 oC, foi superior aos demais tratamentos (fig. 4a).
O maior rendimento máximo atingido pela cepa de P. aghardii foi à
temperatura de 21 oC (124,01��g L-1) e o menor a 18 oC (30,68��g L-1 ) (tab. 1).
Diferenças significativas entre os valores de rendimento fotossintético foram
observadas ao longo do tempo em uma mesma temperatura (F=223,62; p<0,001) e
em um mesmo tempo em diferentes temperaturas (F=167,42; p<0,001). As maiores
variações de rendimento fotossintético foram observadas nos dois primeiros dias (fig.
4b). Porém, a partir do terceiro dia, essas diferenças foram bastante reduzidas. Isso
sugere que, apesar da temperatura influenciar na eficiência fotossintética, P.
agardhii consegue recuperar o seu fotossistema posteriormente.
A temperatura também influenciou o crescimento da diatomácea C.
meneghiniana (CCAP 1070/5). As maiores taxas de crescimento observadas foram
nos cultivos nas temperaturas de 21 e 18 oC (p<0,001) (fig. 5).
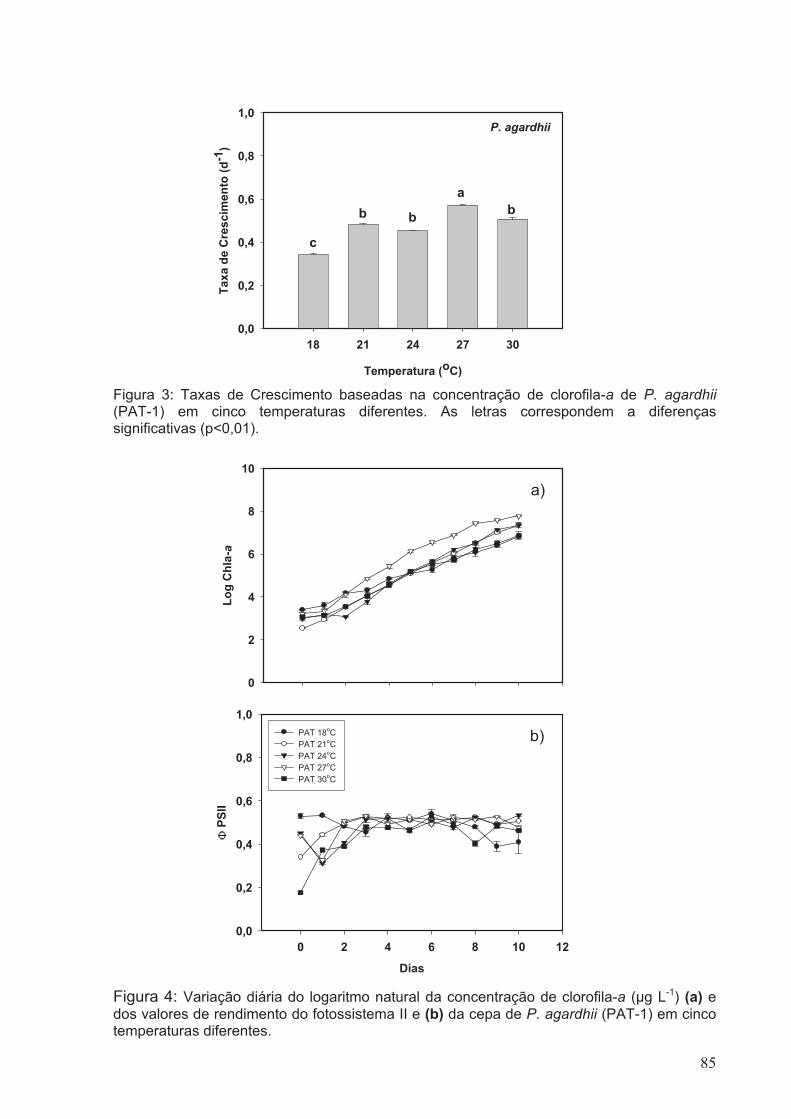
85
Figura 3: Taxas de Crescimento baseadas na concentração de clorofila-a de P. agardhii(PAT-1) em cinco temperaturas diferentes. As letras correspondem a diferenças significativas (p<0,01).
Figura 4: Variação diária do logaritmo natural da concentração de clorofila-a (µg L-1) (a) e dos valores de rendimento do fotossistema II e (b) da cepa de P. agardhii (PAT-1) em cinco temperaturas diferentes.
Dias
0 2 4 6 8 10 12
� P
SII
0,0
0,2
0,4
0,6
0,8
1,0PAT 18oCPAT 21oCPAT 24oCPAT 27oCPAT 30oC
Log
Chl
a-a
0
2
4
6
8
10
a)
b)
Dias
0 2 4 6 8 10 12
� P
SII
0,0
0,2
0,4
0,6
0,8
1,0PAT 18oCPAT 21oCPAT 24oCPAT 27oCPAT 30oC
Log
Chl
a-a
0
2
4
6
8
10
a)
b)
P. agardhii
Temperatura (oC)
18 21 24 27 30
Taxa
de
Cre
scim
ento
(d-1
)
0,0
0,2
0,4
0,6
0,8
1,0
ab b
c
b
P. agardhii
Temperatura (oC)
18 21 24 27 30
Taxa
de
Cre
scim
ento
(d-1
)
0,0
0,2
0,4
0,6
0,8
1,0
ab b
c
b

86
Diferenças significativas foram observadas tanto ao longo do tempo em uma
mesma temperatura (F=3661,71; p<0,001) quanto em um mesmo tempo em
diferentes temperaturas (F=309,39; p<0,001), seja nos valores de biomassa total
(fig.6a), ou de rendimento fotossintético (fig.6b) (tempo F=223,62; p<0,001; tempo x
temperatura F=16742; p<0,001). Além disso, essa cepa apresentou biomassas
bastante reduzidas quando cultivada a 30 oC (p<0,001) (fig. 6a).
Além da alta taxa de crescimento, o maior valor de rendimento máximo
observado para C. meneghiniana também foi a 18 oC, chegando a 194,26 �g L-1. A
24 oC, esta cepa ficou extremamente comprometida, apresentando a menor taxa de
crescimento e o menor valor de rendimento máximo (7,44��g L-1) (fig. 5; tab 1).
Os menores rendimentos fotossintéticos foram observados em C.
meneghiniana cultivada a 27 e 30 oC (fig. 6b). A 27 oC, esse rendimento foi
rapidamente recuperado (2 dias), no entanto, na temperatura de 30 oC, C.
meneghiniana só apresentou rendimento fotossintético semelhante às demais
temperaturas a partir do sétimo dia de cultivo (fig. 6b).
A clorofícea D. quadricauda (DEJAC-02) teve maiores taxas de crescimento
nas temperaturas extremas, de 18 e 30 oC, que não apresentaram diferenças
significativas (p=0,998). Já nas temperaturas intermediárias, o crescimento foi
significativamente reduzido (p<0,001) (fig. 7). De acordo com os dados de taxa de
crescimento, os maiores valores de rendimento máximo também foram observados
nos cultivos nas temperaturas de 18 oC e 30 oC, respectivamente (tab. 1).
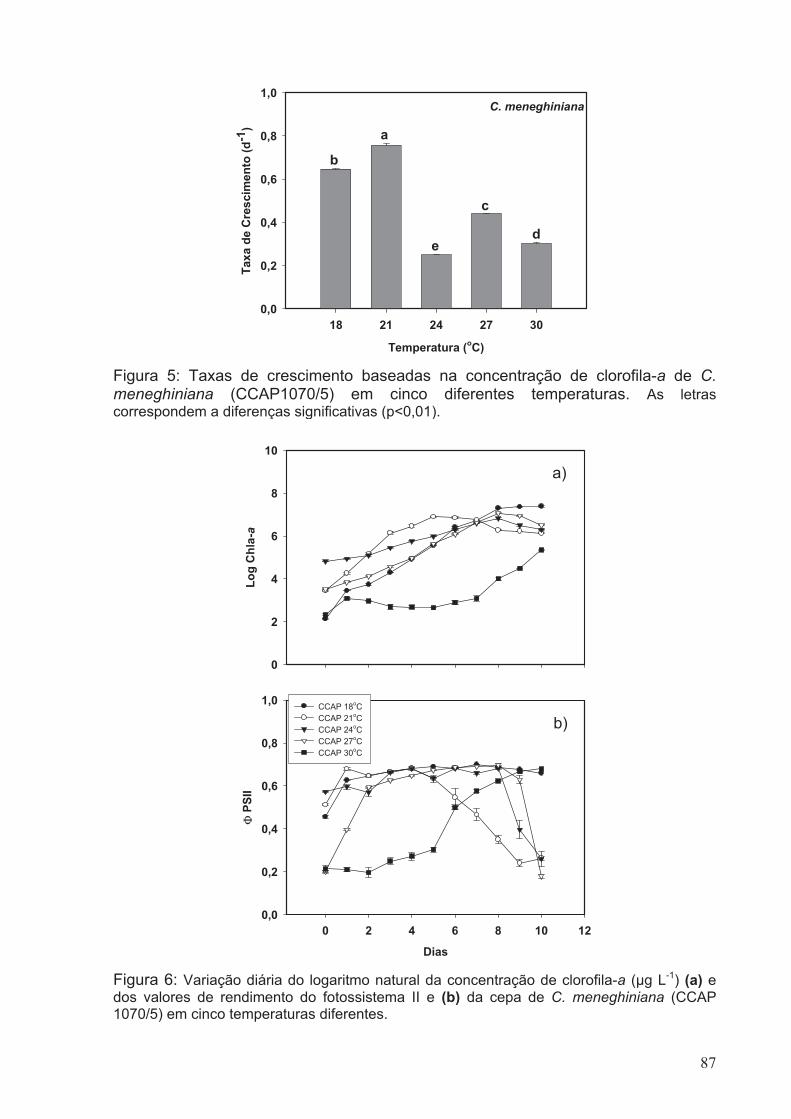
87
Figura 5: Taxas de crescimento baseadas na concentração de clorofila-a de C. meneghiniana (CCAP1070/5) em cinco diferentes temperaturas. As letras correspondem a diferenças significativas (p<0,01).
Figura 6: Variação diária do logaritmo natural da concentração de clorofila-a (µg L-1) (a) e dos valores de rendimento do fotossistema II e (b) da cepa de C. meneghiniana (CCAP 1070/5) em cinco temperaturas diferentes.
Dias
0 2 4 6 8 10 12
��P
SII
0,0
0,2
0,4
0,6
0,8
1,0 CCAP 18oCCCAP 21oCCCAP 24oCCCAP 27oCCCAP 30oC
b)
Log
Chl
a-a
0
2
4
6
8
10
a)
Dias
0 2 4 6 8 10 12
��P
SII
0,0
0,2
0,4
0,6
0,8
1,0 CCAP 18oCCCAP 21oCCCAP 24oCCCAP 27oCCCAP 30oC
b)
Log
Chl
a-a
0
2
4
6
8
10
a)
C. meneghiniana
Temperatura (oC)
18 21 24 27 30
Taxa
de
Cre
scim
ento
(d-1
)
0,0
0,2
0,4
0,6
0,8
1,0
a
b
c
de
C. meneghiniana
Temperatura (oC)
18 21 24 27 30
Taxa
de
Cre
scim
ento
(d-1
)
0,0
0,2
0,4
0,6
0,8
1,0
a
b
c
de
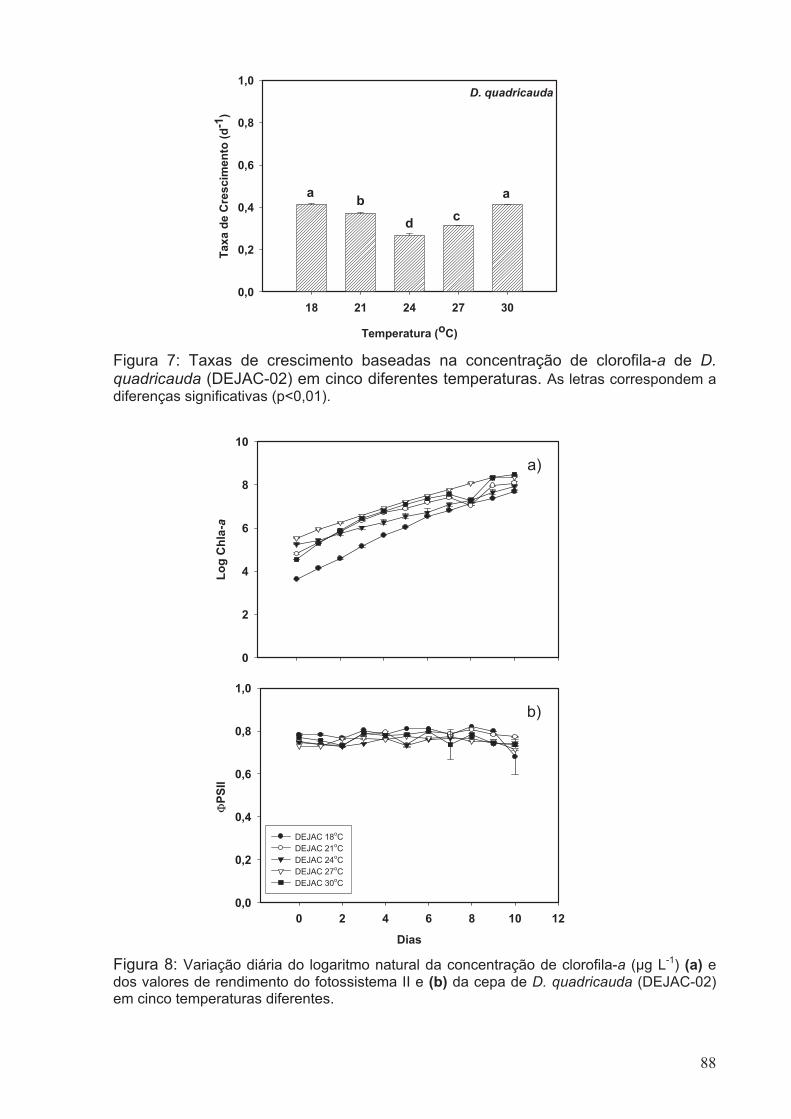
88
Figura 7: Taxas de crescimento baseadas na concentração de clorofila-a de D. quadricauda (DEJAC-02) em cinco diferentes temperaturas. As letras correspondem a diferenças significativas (p<0,01).
Figura 8: Variação diária do logaritmo natural da concentração de clorofila-a (µg L-1) (a) e dos valores de rendimento do fotossistema II e (b) da cepa de D. quadricauda (DEJAC-02) em cinco temperaturas diferentes.
Dias
0 2 4 6 8 10 12
�PS
II
0,0
0,2
0,4
0,6
0,8
1,0
DEJAC 18oCDEJAC 21oCDEJAC 24oCDEJAC 27oCDEJAC 30oC
b)
Log
Chl
a-a
0
2
4
6
8
10
a)
Dias
0 2 4 6 8 10 12
�PS
II
0,0
0,2
0,4
0,6
0,8
1,0
DEJAC 18oCDEJAC 21oCDEJAC 24oCDEJAC 27oCDEJAC 30oC
b)
Log
Chl
a-a
0
2
4
6
8
10
a)
D. quadricauda
Temperatura (oC)
18 21 24 27 30
Taxa
de
Cre
scim
ento
(d-1
)
0,0
0,2
0,4
0,6
0,8
1,0
a abd c
D. quadricauda
Temperatura (oC)
18 21 24 27 30
Taxa
de
Cre
scim
ento
(d-1
)
0,0
0,2
0,4
0,6
0,8
1,0
a abd c
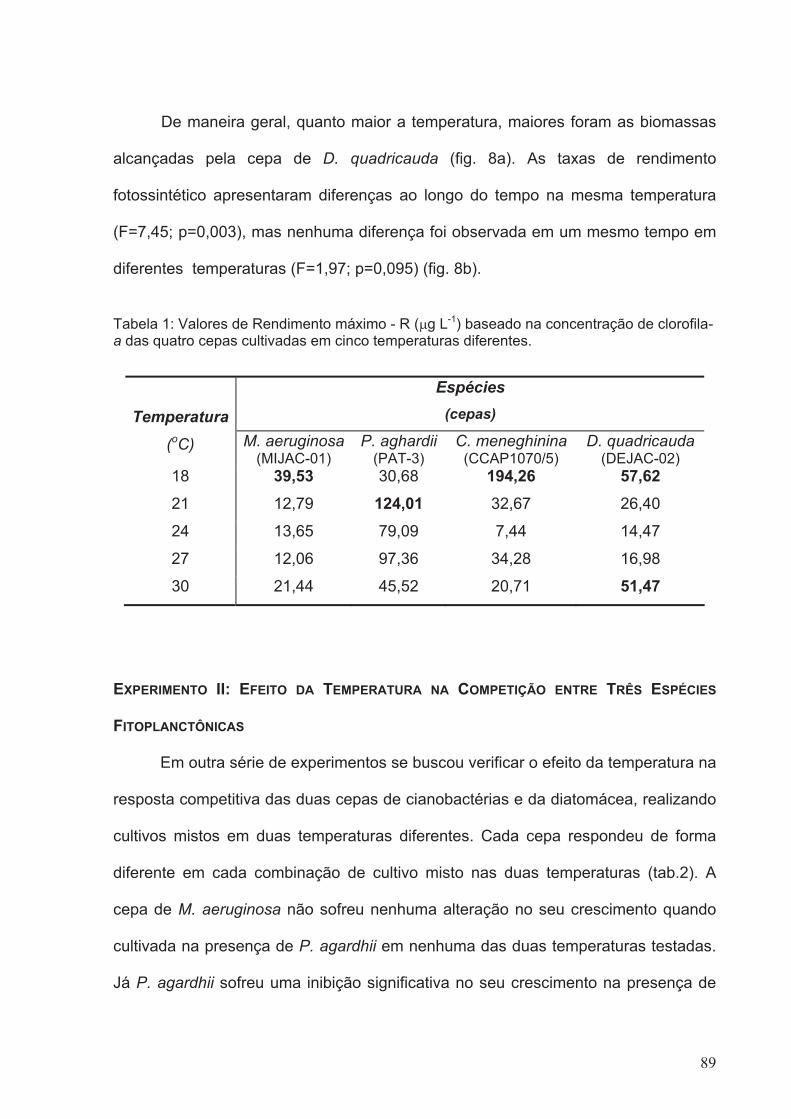
89
De maneira geral, quanto maior a temperatura, maiores foram as biomassas
alcançadas pela cepa de D. quadricauda (fig. 8a). As taxas de rendimento
fotossintético apresentaram diferenças ao longo do tempo na mesma temperatura
(F=7,45; p=0,003), mas nenhuma diferença foi observada em um mesmo tempo em
diferentes temperaturas (F=1,97; p=0,095) (fig. 8b).
Tabela 1: Valores de Rendimento máximo - R (�g L-1) baseado na concentração de clorofila-a das quatro cepas cultivadas em cinco temperaturas diferentes.
EXPERIMENTO II: EFEITO DA TEMPERATURA NA COMPETIÇÃO ENTRE TRÊS ESPÉCIES
FITOPLANCTÔNICAS
Em outra série de experimentos se buscou verificar o efeito da temperatura na
resposta competitiva das duas cepas de cianobactérias e da diatomácea, realizando
cultivos mistos em duas temperaturas diferentes. Cada cepa respondeu de forma
diferente em cada combinação de cultivo misto nas duas temperaturas (tab.2). A
cepa de M. aeruginosa não sofreu nenhuma alteração no seu crescimento quando
cultivada na presença de P. agardhii em nenhuma das duas temperaturas testadas.
Já P. agardhii sofreu uma inibição significativa no seu crescimento na presença de
Espécies(cepas)Temperatura
(oC) M. aeruginosa(MIJAC-01)
P. aghardii (PAT-3)
C. meneghinina (CCAP1070/5)
D. quadricauda (DEJAC-02)
18 39,53 30,68 194,26 57,62 21 12,79 124,01 32,67 26,40
24 13,65 79,09 7,44 14,47
27 12,06 97,36 34,28 16,98
30 21,44 45,52 20,71 51,47

90
M. aeruginosa e essa inibição foi mais pronunciada (p<0,05) a 30 oC do que a 18 oC,
quando sua taxa de crescimento foi reduzida em 6,5 vezes (figs.9a e 9b; tab.2).
A presença da diatomácea C. meneghiniana inibiu o crescimento de M.
aeruginosa à temperatura de 18 oC (p<0,001; figura 10a), mas nenhum efeito foi
observado a 30 oC (p=0,465; figura 10b). Para C. meneghiniana, a presença da M.
aeruginosa afetou significativamente o crescimento em ambas as temperaturas
(p<0,001; figura 10a e 10b; tab. 2), no entanto, na temperatura elevada o efeito foi
ainda mais acentuado (p<0,001; fig. 10b). Enquanto a presença da M. aeruginosa
reduziu 5,9 vezes o crescimento da C. meneghiniana a 18 oC, quando cultivadas a
30 oC, esta redução foi de 7,1 vezes.
Tabela 2: Efeito das competições entre as cepas de M. aeruginosa (MIJAC), P. agardhii(PAT), e C. meneghiniana (CCAP) em cultivos mistos em duas temperaturas (18 e 30 oC).
COMPETIÇÃO* TEMP. EFEITO
MIJAC+PAT PAT+CCAP MIJAC+CCAP MIJAC+PAT+CCAPMIJAC 0 + - 0
PAT - - 18 oC CCAP - - -
MIJAC+PAT PAT+CCAP MIJAC+CCAP MIJAC+PAT+CCAPMIJAC 0 0 0 0
PAT - - 30 oC CCAP + - -
* +, estímulo do crescimento, -, inibição do crescimento, 0, nenhum efeito no crescimento da espécie alvo. Estes efeitos foram determinados a partir das taxas de crescimento, que foram significativamente diferentes do controle (p<0,01), como determinado pela Análise de Variância (1-way ANOVA).
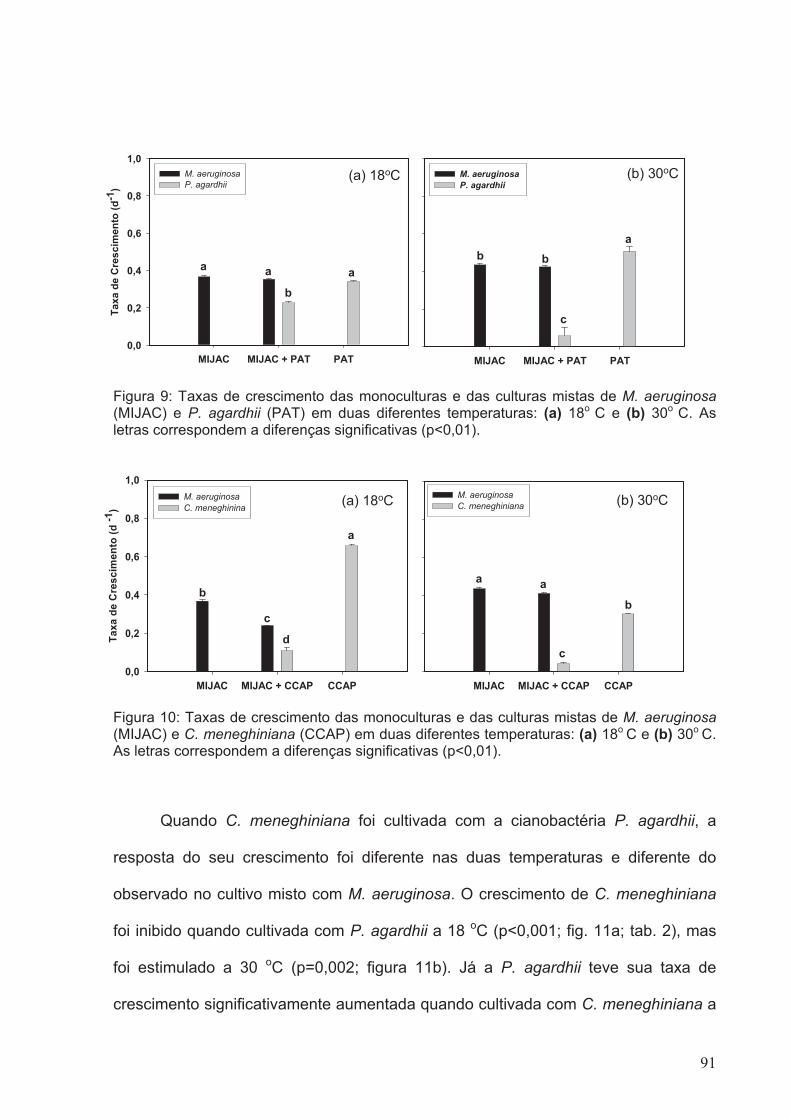
91
Figura 9: Taxas de crescimento das monoculturas e das culturas mistas de M. aeruginosa(MIJAC) e P. agardhii (PAT) em duas diferentes temperaturas: (a) 18o C e (b) 30o C. As letras correspondem a diferenças significativas (p<0,01).
Figura 10: Taxas de crescimento das monoculturas e das culturas mistas de M. aeruginosa(MIJAC) e C. meneghiniana (CCAP) em duas diferentes temperaturas: (a) 18o C e (b) 30o C.As letras correspondem a diferenças significativas (p<0,01).
Quando C. meneghiniana foi cultivada com a cianobactéria P. agardhii, a
resposta do seu crescimento foi diferente nas duas temperaturas e diferente do
observado no cultivo misto com M. aeruginosa. O crescimento de C. meneghiniana
foi inibido quando cultivada com P. agardhii a 18 oC (p<0,001; fig. 11a; tab. 2), mas
foi estimulado a 30 oC (p=0,002; figura 11b). Já a P. agardhii teve sua taxa de
crescimento significativamente aumentada quando cultivada com C. meneghiniana a
MIJAC MIJAC + CCAP CCAP
M. aeruginosaC. meneghiniana
MIJAC MIJAC + CCAP CCAP
Taxa
de
Cre
scim
ento
(d -1
)
0,0
0,2
0,4
0,6
0,8
1,0M. aeruginosa C. meneghinina (a) 18oC (b) 30oC
b
a
c
d
b
a
c
a
MIJAC MIJAC + CCAP CCAP
M. aeruginosaC. meneghiniana
MIJAC MIJAC + CCAP CCAP
Taxa
de
Cre
scim
ento
(d -1
)
0,0
0,2
0,4
0,6
0,8
1,0M. aeruginosa C. meneghinina (a) 18oC (b) 30oC
b
a
c
d
b
a
c
a
MIJAC MIJAC + PAT PAT
Taxa
de
Cre
scim
ento
(d-1
)
0,0
0,2
0,4
0,6
0,8
1,0M. aeruginosaP. agardhii
(a) 18oC
b
a a a
a
MIJAC MIJAC + PAT PAT
M. aeruginosa P. agardhii
abb
c
(b) 30oC
MIJAC MIJAC + PAT PAT
Taxa
de
Cre
scim
ento
(d-1
)
0,0
0,2
0,4
0,6
0,8
1,0M. aeruginosaP. agardhii
(a) 18oC
b
a a a
a
MIJAC MIJAC + PAT PAT
M. aeruginosa P. agardhii
abb
c
(b) 30oC
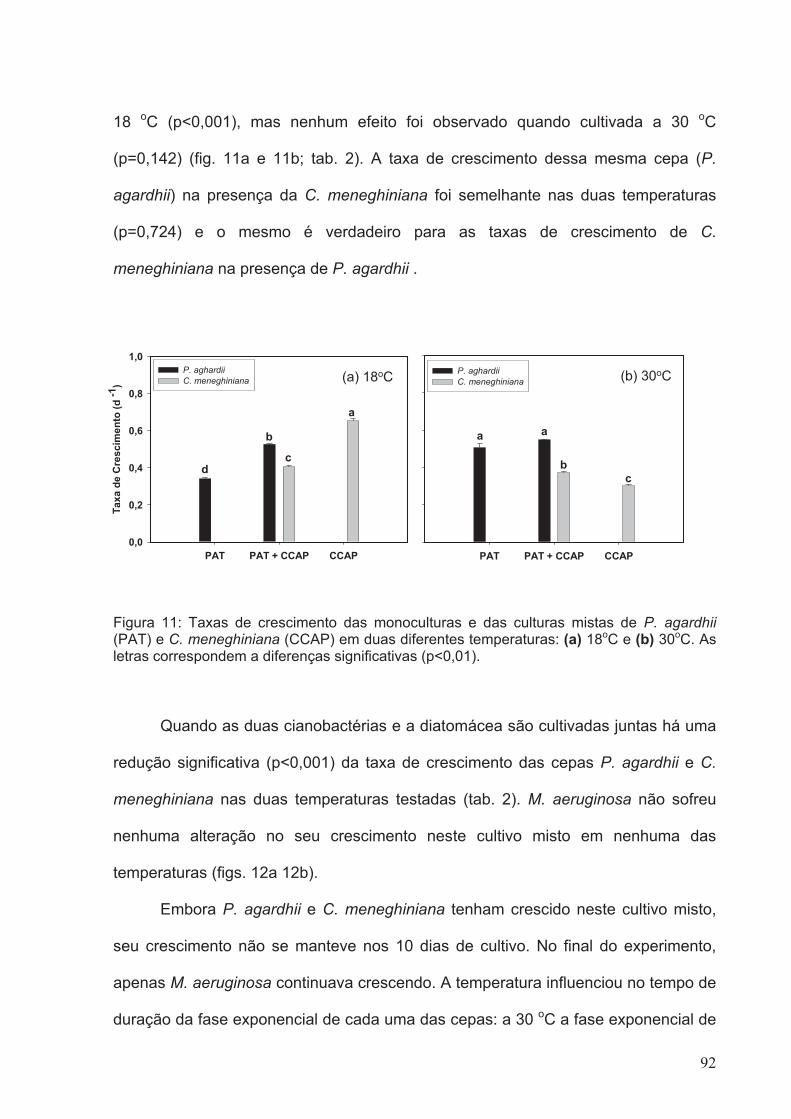
92
18 oC (p<0,001), mas nenhum efeito foi observado quando cultivada a 30 oC
(p=0,142) (fig. 11a e 11b; tab. 2). A taxa de crescimento dessa mesma cepa (P.
agardhii) na presença da C. meneghiniana foi semelhante nas duas temperaturas
(p=0,724) e o mesmo é verdadeiro para as taxas de crescimento de C.
meneghiniana na presença de P. agardhii .
Figura 11: Taxas de crescimento das monoculturas e das culturas mistas de P. agardhii(PAT) e C. meneghiniana (CCAP) em duas diferentes temperaturas: (a) 18oC e (b) 30oC. As letras correspondem a diferenças significativas (p<0,01).
Quando as duas cianobactérias e a diatomácea são cultivadas juntas há uma
redução significativa (p<0,001) da taxa de crescimento das cepas P. agardhii e C.
meneghiniana nas duas temperaturas testadas (tab. 2). M. aeruginosa não sofreu
nenhuma alteração no seu crescimento neste cultivo misto em nenhuma das
temperaturas (figs. 12a 12b).
Embora P. agardhii e C. meneghiniana tenham crescido neste cultivo misto,
seu crescimento não se manteve nos 10 dias de cultivo. No final do experimento,
apenas M. aeruginosa continuava crescendo. A temperatura influenciou no tempo de
duração da fase exponencial de cada uma das cepas: a 30 oC a fase exponencial de
PAT PAT + CCAP CCAP
Taxa
de
Cre
scim
ento
(d -1
)
0,0
0,2
0,4
0,6
0,8
1,0P. aghardiiC. meneghiniana (a) 18oC
a
b
cd
PAT PAT + CCAP CCAP
P. aghardiiC. meneghiniana (b) 30oC
aa
bc
PAT PAT + CCAP CCAP
Taxa
de
Cre
scim
ento
(d -1
)
0,0
0,2
0,4
0,6
0,8
1,0P. aghardiiC. meneghiniana (a) 18oC
a
b
cd
PAT PAT + CCAP CCAP
P. aghardiiC. meneghiniana (b) 30oC
aa
bc
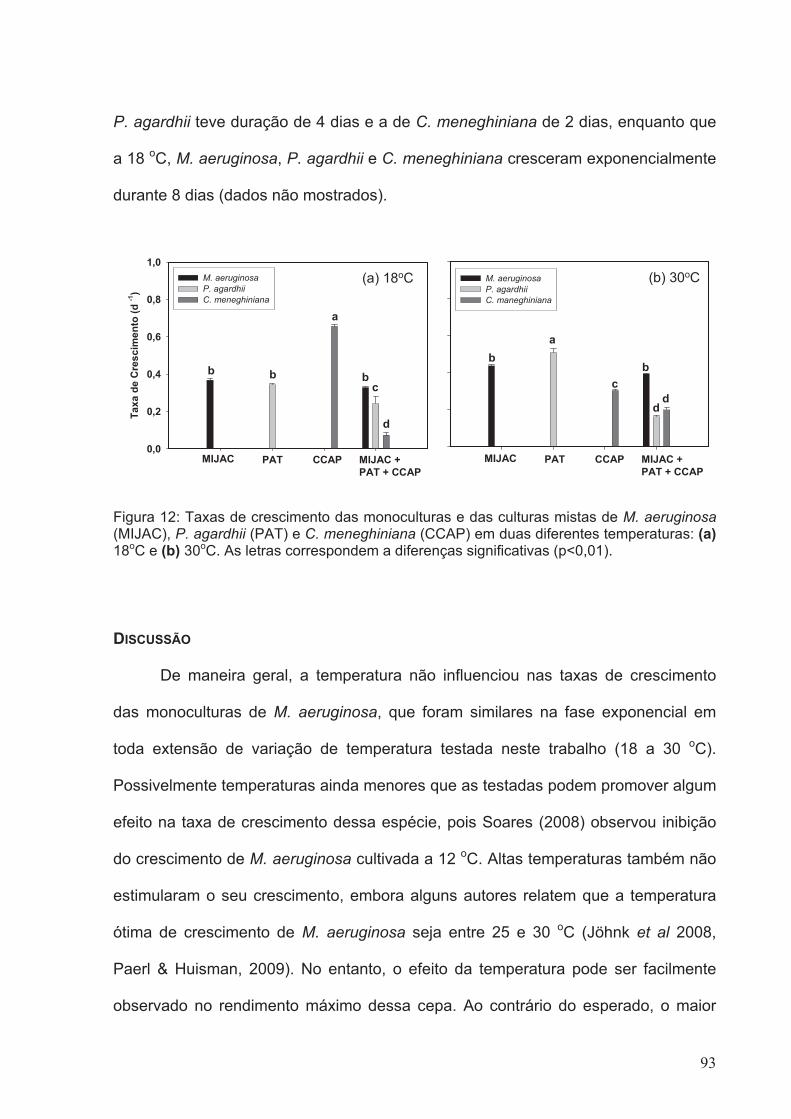
93
P. agardhii teve duração de 4 dias e a de C. meneghiniana de 2 dias, enquanto que
a 18 oC, M. aeruginosa, P. agardhii e C. meneghiniana cresceram exponencialmente
durante 8 dias (dados não mostrados).
Figura 12: Taxas de crescimento das monoculturas e das culturas mistas de M. aeruginosa(MIJAC), P. agardhii (PAT) e C. meneghiniana (CCAP) em duas diferentes temperaturas: (a)18oC e (b) 30oC. As letras correspondem a diferenças significativas (p<0,01).
DISCUSSÃO
De maneira geral, a temperatura não influenciou nas taxas de crescimento
das monoculturas de M. aeruginosa, que foram similares na fase exponencial em
toda extensão de variação de temperatura testada neste trabalho (18 a 30 oC).
Possivelmente temperaturas ainda menores que as testadas podem promover algum
efeito na taxa de crescimento dessa espécie, pois Soares (2008) observou inibição
do crescimento de M. aeruginosa cultivada a 12 oC. Altas temperaturas também não
estimularam o seu crescimento, embora alguns autores relatem que a temperatura
ótima de crescimento de M. aeruginosa seja entre 25 e 30 oC (Jöhnk et al 2008,
Paerl & Huisman, 2009). No entanto, o efeito da temperatura pode ser facilmente
observado no rendimento máximo dessa cepa. Ao contrário do esperado, o maior
MIJAC PAT CCAPMIJAC + PAT + CCAP
M. aeruginosaP. agardhiiC. maneghiniana
MIJAC PAT CCAPMIJAC + PAT + CCAP
Taxa
de
Cre
scim
ento
(d -1
)
0,0
0,2
0,4
0,6
0,8
1,0M. aeruginosaP. agardhiiC. meneghiniana
(a) 18oC (b) 30oC
b
a
cbb
d
b
d
c
a
b
d
MIJAC PAT CCAP MIJAC + PAT + CCAP
MIJAC PAT CCAP MIJAC + PAT + CCAP
MIJAC PAT CCAPMIJAC + PAT + CCAP
M. aeruginosaP. agardhiiC. maneghiniana
MIJAC PAT CCAPMIJAC + PAT + CCAP
Taxa
de
Cre
scim
ento
(d -1
)
0,0
0,2
0,4
0,6
0,8
1,0M. aeruginosaP. agardhiiC. meneghiniana
(a) 18oC (b) 30oC
b
a
cbb
d
b
d
c
a
b
d
MIJAC PAT CCAP MIJAC + PAT + CCAP
MIJAC PAT CCAP MIJAC + PAT + CCAP

94
rendimento foi observado na menor temperatura. A 18 oC, M. aeruginosa se dividiu
na mesma velocidade que as demais temperaturas testadas, no entanto, por ter
ficado mais tempo na fase exponencial de crescimento (10 dias) o seu rendimento
máximo foi maior. Estes resultados não concordam com aqueles da literatura, que
descrevem aumento de biomassa fitoplanctônica dominada por cianobactérias
durante os períodos mais quentes do ano (Zhang & Prepas, 1996; Soares 2008;
Gomes et al., 2009), incluindo trabalhos com simulações preditivas (Elliott et al.,
2006; Jöhnk et al., 2008; Mehnert et al., 2010).
P. agardhii apresentou maior taxa de crescimento a 27 oC. Essa é a
temperatura ótima descrita para outra espécie de cianobactéria, Cylindrospermopsis
rasciborskii (Padisak, 1997), que coincidentemente apresenta morfologia
semelhante. Tal fato corrobora a hipótese de Reynolds de que a morfologia pode
influenciar na sensibilidade térmica da taxa de crescimento de uma microalga
(Reynolds, 2006). Isso porque os processos metabólicos celulares são determinados
pela capacidade de absorção das células, que por sua vez depende da razão
superfície volume, parâmetro este influenciado pela temperatura. Portanto, cabe
discutir se a influência da temperatura no crescimento do fitoplâncton seria
específica para espécie ou para tipo morfológico. Outros autores também vêm
dando importância para as características morfológicas das espécies, inclusive para
classificação de distintos grupos funcionais do fitoplâncton (Kruk et al., 2010).
Assim como visto para M. aeruginosa, a clorofícea D. quadricauda também
teve os maiores valores de rendimento máximo e de taxas de crescimento nas
temperaturas extremas (18 e 30 oC). Além disso, seu rendimento fotossintético
também não sofreu influência da temperatura. Esses resultados sugerem que a
extensão da variação de temperatura experimentada neste trabalho não

95
comprometeu o processo fotossintético dessas duas espécies. Cabe ressaltar, que
ambas são originárias da lagoa de Jacarepaguá e por isso estão sujeitas a variações
de temperatura da água menores, enquanto que as demais (P. agardhii e C.
meneghiniana) têm origem em sistemas aquáticos temperados, onde os lagos são
congelados no inverno. Esta pode ser a principal justificativa das diferenças
observadas nas respostas à temperatura neste trabalho.
As variações das taxas de crescimento máximo de C. meneghiniana em
resposta à temperatura estão de acordo com as observações encontradas para
espécies do grupo das diatomáceas. O seu crescimento foi favorecido em
temperaturas mais baixas. Quando observamos sua distribuição no ambiente
natural, as diatomáceas dominam a comunidade fitoplanctônica nos períodos de
temperatura mais baixa da água (Zhang & Prepas, 1996; Gomes et al., 2009).
A mais elevada temperatura testada (30 oC) prejudicou o sistema
fotossintético da C. meneghiniana resultando em um longo período de ausência de
crescimento. Durante 8 dias, C. meneghiniana esteve em um provável período de
adaptação, também chamado de prolongamento da fase lag. Em seguida, seu
rendimento fotossintético foi recuperado e ela começou a crescer. Para algumas
espécies de microalgas, altas temperaturas podem reduzir a assimilação de
carbono, consequentemente, comprometer a fotossíntese e o seu crescimento
(Goldman, 1977; Reynolds, 2006). Mesmo que tardio, o crescimento de C.
meneghiniana pôde ser observado em monocultura, no entanto, em um sistema
natural existem outros organismos que podem levar vantagens nessa condição de
temperatura e ganhar a competição por recursos, substituindo essa espécie na
comunidade.

96
A temperatura pode ter influenciado no “trade off” entre taxa de crescimento e
rendimento máximo para as cepas de P. agardhii e C. meneghiniana. Apesar da
maior taxa de crescimento para P. agardhii ter sido observada a 27 oC, seu maior
rendimento máximo foi a 21 oC, acompanhado de uma fase exponencial mais longa
(10 dias a 21 oC; 8 dias a 27 oC). A espécie C. meneghiniana apresentou maior taxa
de crescimento a 21 oC e permaneceu em exponencial por apenas 4 dias, enquanto
que a 18 oC seu rendimento final foi maior e a duração da sua fase exponencial foi
de 8 dias. Tais resultados sugerem que a temperatura pode ter influenciado na
aquisição e utilização de recursos, como nutrientes. Na temperatura mais elevada
(21 oC para C. meneghiniana e 27 oC para P. agardhii) estas cepas investiram em
crescimento, explorando ao máximo os recursos externos. Nas menores
temperaturas (18 oC para C. meneghiniana e 21 oC para P. agardhii), o investimento
pode ter sido em estocar nutrientes, continuar crescendo com menor taxa por um
período maior e ter um maior rendimento final. Apesar deste trabalho não ter
analisado as concentrações de nutrientes nos cultivos, essa pode ser uma
explicação plausível, uma vez que sabemos que a temperatura influencia
diretamente nas taxas metabólicas do fitoplâncton (Darley, 1982).
Nos cultivos mistos, a cepa M. aeruginosa levou vantagem na competição
com todas as cepas estudadas e em ambas as temperaturas. Embora a 18 oC M.
aeruginosa tenha sofrido redução da taxa de crescimento no cultivo misto com C.
meneghiniana em relação ao controle (cultivo em monocultura), ainda assim ela
ganhou a competição. As baixas temperaturas favoreceram o crescimento de C.
meneghiniana, tanto em monocultura quanto em cultivo misto, ainda que neste com
uma taxa de crescimento menor que a de M. aeruginosa. Possivelmente, o melhor
crescimento de C. meneghiniana a 18 oC inibiu o crescimento de M. aeruginosa. No

97
entanto, tanto a 18 quanto a 30 oC M. aeruginosa leva vantagem na competição com
C. meneghiniana. Essa desvantagem da C. meneghiniana pode ser devida a sua
maior sensibilidade à mudança de temperatura, uma vez que essa cepa sofreu a
maior variação na taxa de crescimento nas diferentes temperaturas.
O sucesso competitivo de M. aeruginosa também pode estar relacionado à
sua maior habilidade na aquisição de recursos, como por exemplo, na absorção de
nutrientes. Espécies do gênero Microcystis apresentam médias de valores de
constantes de semi-saturação para absorção de fósforo (Ks) igual a 0,5 �g P L-1,
enquanto o Ks(P) para C. meneghiniana é de 8��g P L-1 (Padisak, 2004). Isso
significa que M. aeruginosa, além de ter uma maior taxa de crescimento que as
demais cepas, tem um menor requerimento por P, o que faz desta espécie uma
excelente competidora.
Neste mesmo sentido, pode-se discutir a competição entre C. meneghiniana
e P. agardhii. Esta última ganhou a competição em ambas as temperaturas. Sua
taxa de crescimento foi maior e seu requerimento por P é menor (1 �g P L-1) do que
os de C. meneghiniana. Além disso, o gênero de Planktothrix também é conhecido
pelo seu baixo requerimento por luz, devido ao fato de possuir alto conteúdo do
pigmento ficoeritrina (Padisak, 2004; Reynolds, 2006). A presença de C.
meneghiniana ainda estimulou o crescimento da P. agardhii. Algumas diatomáceas
são capazes de estimular o crescimento de indivíduos de outros grupos
fitoplanctônicos, incluindo cianobactérias, porque podem excretar vitaminas para o
meio (Carlucci & Bowes 1970 appud in Fogg & Thake, 1987), o que pode ter
acontecido no cultivo misto com C. meneghiniana e P. agardhii. No entanto,
nenhuma destas características favoreceu P. agardhii na competição com M.
aeruginosa. Tanto na bicultura com M. aeruginosa, quanto na tricultura com M.

98
aeruginosa e C. meneghiniana, especialmente a 30 oC, P. agardhii foi bastante
comprometida. O tamanho de seus filamentos se reduziu consideravelmente
(observação em microscópio, dados não mostrados) e a duração da sua fase de
crescimento também foi significativamente diminuída. Na tricultura, após 4 dias de
cultivo, P. agardhii começou a entrar na senescência e a partir de 7 dias apresentou
taxas de crescimento negativas, enquanto que na monocultura sua fase de
crescimento durou até o 8º. dia.
Observou-se que a temperatura influenciou o crescimento das microalgas e
cianobactérias em monocultura, os valores mais baixos favoreram o crescimento da
diatomácea, enquanto as cianobactérias cresceram melhor em temperaturas
elevadas e a clorofícea foi favorecida tanto na mais alta quanto na mais baixa
temperatura. Porém, em cultivo misto as cianobactérias sempre ganharam a
competição com a diatomácea. Portanto, podemos sugerir que não só a
temperatura, mas a sua combinação com outros fatores ambientais como luz,
concentração de nutrientes (ver capítulo I), pode ter sido a responsável pela
variação na abundância de espécies na lagoa de Jacarepaguá.
Apesar disso, diante do cenário de aquecimento global, a preocupação se
justifica já que o aumento de alguns graus na temperatura da água poder promover
a ocorrência e dominância de cianobactérias em ambientes eutrofizados
(Christoffersen et al., 2006; Elliott et al., 2006). Um problema adicional é que muitos
gêneros de cianobactérias são produtores de toxinas e segundo Davis et al (2009),
além de favorecer o crescimento de cianobactérias, altas temperaturas adicionadas
a elevadas concentrações de nutrientes podem também selecionar as espécies
tóxicas, aumentando as implicações para saúde pública.

99
Por fim, estes resultados podem contribuir para conhecer o efeito da
temperatura sobre o crescimento de microalgas e cianobactérias e sobre a dinâmica
do fitoplâncton, o que permitirá discutir de forma preditiva o efeito do aquecimento
global nos ambientes aquáticos.

100
REFERÊNCIAS
AKCAALAN, R.A., YOUNG, F.M., METCALF, J.S., MORRISON, L.F., ALBAY, M., CODD, G.A. 2006. Microcystin analysis in single filaments of Planktothrix spp. in laboratory cultures and environmental blooms. Water Reasearch. 40, 1583-1590.
BARCO, M., FLORES, C., RIVERA, J., CAIXACH, J. 2004. Determination of microcystin variants and related peptides present in water Bloom of Planktothrix (Oscillatoria) rubencens in a Spanish drinking water reservoir by LC/ESI-MS. Toxicon. 44, 881-886.
CHISTOFFERSEN, K.; ANDERSEN, N.; SONDERGAARD, M.; LIBORIUSSEN, L.; JEPPERSEN, E. 2006. Implications of climate enforced temperature on freshwater pico and nanoplankton populations studied in artificial ponds during 16 months. Hydrobiologia. 560, 259-266.
CODD, G.A., BELL, S.G., KAYA, K., BEATTIE, K.A. METCALF, J.S. 1999. Cyanobacterial toxins, exposure routes and human health. European Journal of Phycology. 34, 405-415.
DARLEY, W. M. 1982. Algal Biology: a physiological approach. 1 ed. London. Blackwell Scientific Publications. 168p.
DAVIS, T.W., BERRY, D.L. BOYER, G.L. GOBLER, C.J. 2009. The effects of temperature and nutrients on growth and dynamics of toxic and non-toxic strains of Microcystis during cyanobacteria blooms. Harmful Algae. 8, 715-725
DESCAMPS-JULIEN, B., GONZALEZ, A. 2005. Stable coexistence in fluctuating environmental an experimental demonstration. Ecology. 86(10), 2815-2824.
ELLIOTT, J.A., JONES, I.D., THACKERAY, S.J. 2006. Testing the sensitivity of phytoplankton communities to changes in water temperature and nutrient load, in a temperate lake. Hydrobiologia. 559, 401-411.
FOGG, G.E., THAKE, B. 1987. Algal cultures and Phytoplankton Ecology. The University of Wisconsin Press, Wisconsin. 269p.
GOLDMAN, J.C., RYTHER, J.H. 1976. Temperature influenced species competition in mass cultures of marine phytoplankton. Biotechnol. Bioeng. 18, 1125-1144.
GOLDMAN, J.C. 1977. Temperature effects on phytoplankton growth in continuous culture. Limnology and Oceanography, 22 (5), 932-936
GOMES, A. M. A., SAMPAIO, P. L., FERRÃO-FILHO, A.S., MAGALHÃES, V. F., MARINHO, M.M. OLIVEIRA, A. C. P., SANTOS, V. B., DOMINGOS, P., AZEVEDO, S.M.F.O. 2009. Florações de Cianobactérias Tóxicas em uma Lagoa Costeira Hipereutrófica do Rio de Janeiro/RJ (Brasil) e suas Consequências para Saúde Humana. Oecologia Brasiliensis. 13 (2), 329-345.

101
KOSTEN, S. 2010. Aquatic ecosystems in hot water. Effects of climate on the functioning of shallow lakes. Thesis Wageningen University, Wageningen, NL. 160p.
KRISHNAMURTHY, T.; CARMICHAEL, W.W.; SARVER, E.W. 1986. Toxic peptides from freshwater cyanobacteria blue-green algae. Isolation, purification and characterization of peptides from Microcystis aeruginosa and Anabaena flos-aquae.Toxicon.24(9),865-873.
KRUK, C. 2010. Morphology Captures Function in Phytoplankton. A Large scale analysis of phytoplankton community in relation to their environmental. Thesis Wageningen University, Wageningen, NL. 160p.
HOUGHTON, J., DING, Y., GRIGGS, D.J., NOGUER, M., VAN DER LINDEN, P.J., XIAOSU, D. 2001. Climate change 2001: The scientific basis contribution of working group I to the third assessment report of the Intergovernamental Panel on Climate Change (IPCC). Cambridge University Press, Cambridge, UK.
JÖHNK, D.K., HUISMAN, J., SHARPLES, J.M., SOMMEIJER, B., VISSER, P., STROOM, J. 2008. Summer heatwaves promote blooms of harmful cyanobacteria. Global Change Biology. 14, 495-512.
LÜRLING, M.; BEEKMAN, W. 1999. Grazer induced defenses in Scenedesmus (Chlorococcales; Chlorophyceae): coenobium and spine formation. Phycologia. 38, 368-376.
MARINHO, M. M.; HUSZAR, V. L. M. 2002. Nitrogen availability and physical conditions as controlling factors of phytoplankton composition and biomass in a tropical reservoir (Southern Brasil). Archiv für Hydrobiologie, v. 153, p. 443-468.
MEHNERT, G., LEUNERT, F., CIRÉS, S. JÖHNK, D.K., RÜCKER, J. NIXDORF, B., WIEDNER, C. 2010. Competitiveness of invasive and native cyanobacteria from temperate freshwater under various light and temperature conditions. Journal of Plankton Research. 32(7), 1009-1021.
MERILOTO, J.A.O.,SANDSTROM, J.E., ERIKSSON, G., REMAUD, G., CRAIG, A.G., CHATTOPADHYAYA, J. 1989. Toxicon (27) 1021-1034.
PADISÁK, J. 2004. Phytoplankton. In O'Sullivan, P. E. & Reynolds, C. S. (editors) The Lakes Handbook 1. Limnology and Limnetic Ecology: 251-308. Blackwell Science Ltd. Oxford.
PADISÁK, J. 1997. Cylindrospermopsis raciborskii (Woloszynska) Seenaya et Subba Raju, an expanding, highly adaptative cyanobacterium: worldwide distribution and review of its ecology. Archiv für Hydrobiologie. 107: 563-593.
PAERL, H.W. & HUISMAN, J. 2008. Blooms like it hot. Science. 320: 57-58.
PAERL, H. & HUISMAN, J. 2009. Climate change: a catalyst for global expansion of harmful of cyanobacterial blooms. Environmental Microbiology Reports, 1 (1), 27-37.

102
REYNOLDS, C.S., JAWORSKI, G.M.H., CMIECH, H.A., LEEDALE, GF. 1981. On the annual cycle of the blue green alga Microcystis aeruginosa Kütz. Emend. Elenkin. Phlilosophical Transactions of the Royal Society London, Series B. 293, 419-177.
REYNOLDS, C.S. 1987. Cyanobacterial water-blooms. In: J. Callow (ed). Advances in botanical research. Academic Press., London. 13, 67-143.
REYNOLDS, C.S. 2006. Ecology of Phytoplankton. Cambridge University Press, Cambridge. 535p.
RHEE, G.Y. & GOTHAM, I.J. 1981. The effect of environmental factors on phytoplankton growth: Temperature and the interactions of temperature with nutrient limitation. Limnology and Oceanography. 26 (4), 635-648.
SIVONEN, K.; HIMBERG, K.; LUUKKAINEN, R.; NIEMELA, S.; POON, G.K.; CODD, G.A. 1989. Preliminary characterization of neurotoxic cyanobacterial blooms and strains from Finland. Toxicity Assessment, 4: 339-352.
SOARES, M.C. 2008. Cianobactérias Tóxicas no Reservatório do Funil: Uma Batalha Pelágica.Tese de Doutorado em Botânica. Universidade Federal do rio de Janeiro.419p.
SOARES, M.C., ROCHA, M.I.A., MARINHO, M.M., AZEVEDO, S.M.F.O., BRANCO, C.C., HUSZAR, V.L. 2009. Changes in species composition during annual cyanobacterial dominance in tropical reservoir: physical factors, nutrients and grazing effects. Aquatic Microbial Ecology. 57, 137-149
THOMPSON, P.A., GUO, M., HARRISON, P.J. 1992. Effets of variation in temperature. I. On the biochemical composition of eight species of marine phytoplankton. Journal of Phycology. 28, 481-488.
TILMAN, D., KIESLING, R., STERNER, R., KILHAM, S.S., JOHNSON, F.A. 1986. Green, blue-green and diatom algae: taxonomic differences in competitive ability for phosphorus, silicon and nitrogen. Arch. Hydrobiol. 106, 473-485.
WATANABE, M.F., OISHI, S. 1985. Effects of Environmental Factors on Toxicity of a Cyanobacterium (Microcystis aeruginosa) under Culture Conditions. Applied Environmental Microbiology. 1342-1344.
WINDER, M., SCHINDLER, D.E. 2004. Climate Change uncouples trophic interactions in an aquatic ecosystem. Ecology. 85(8), 2100-2106.
ZHANG, Y. & PREPAS, E.E. 1996. Regulation of dominance of planktonic diatoms and cyanobacteria in four eutrophic hardwater lakes by nutrients, water column stability, and temperature. Canadian Journal of Fisheries Aquatic Science. 53, 621-633.

103
CAPÍTULO IV: EFEITO DA TEMPERATURA NA INTERAÇÃO ALELOPÁTICA ENTRE TRÊS DIFERENTES
ESPÉCIES FITOPLANCTÔNICAS
INTRODUÇÃO
O termo “alelopatia” (do grego “allelon” que significa mútuo e “pathos” que
significa afetar) foi introduzido por Molisch (1937) para descrever o processo em que
uma planta influenciava outra por meio de interações químicas. Posteriormente, Rice
(1984) incluiu os microrganismos nessa definição e considerou tanto os efeitos
positivos quanto negativos no organismo alvo. Mais recentemente, a Sociedade
Internacional de Alelopatia definiu esse termo como sendo qualquer processo
envolvendo metabólitos secundários produzidos por plantas, algas, bactérias e
fungos que influenciam no crescimento e desenvolvimento dos sistemas agrícolas e
biológicos (International Allelopathy Society, 1996).
Entretanto, ainda hoje, o conceito de alelopatia é discutido devido às
diferentes definições adotadas pelos autores e pela dificuldade de se evidenciar um
processo alelopático no ambiente, especialmente em sistemas aquáticos. Leflaive &
Tem-Hage (2007) ainda discutem na sua revisão, a inclusão das interações entre
predadores e presas no conceito de alelopatia. Neste capítulo foram apenas
consideradas como interações alelopáticas os efeitos positivos e negativos entre
microrganismos fotoautotróficos, ou seja, no conceito de competição e não de
predação.
Os organismos que exibem aleleopatia liberam compostos químicos que
estimulam ou inibem diretamente o crescimento do organismo competidor ou
indiretamente, afetando a disponibilidade dos recursos em comum (Legrand et al.,
2003). O mecanismo de ação dos compostos alelopáticos vai depender da natureza

104
da interação entre os organismos produtores e os organismos alvos. Os principais
mecanismos de ação descritos para o fitoplâncton são inibição da fotossíntese,
inibição enzimática, paralisia celular, inibição da síntese de ácidos nucléicos e
formação de espécies reativas de oxigênio (Leflaive & Tem-Hage, 2007). Quando o
efeito do aleloquímico é estimular o crescimento de outro organismo, os
mecanismos sugeridos são liberação de nutrientes ou vitaminas previamente
estocados pelo organismo produtor, produção de metabólitos secundários bioativos
ou a produção de compostos antibacterianos ou antifúngicos que irão beneficiar
outros organismos fitoplanctônicos (Gross, 2003; Legrand et al., 2003; Carey &
Rengefors, 2010).
Alguns fatores ambientais, tais como concentração de nutrientes, intensidade
luminosa, pH e temperatura, podem influenciar na produção de compostos
alelopáticos ou na sensibilidade dos organismos alvo a esses compostos (Von Elert
& Juttner, 1996; Issa, 1999; Ray & Bagchi, 2001; Reigosa et al., 1999; Granéli &
Hansen, 2006). Além disso, a alelopatia vem sendo considerada muitas vezes mais
efetiva em situações de estresse ambiental, como por exemplo, sob limitação de
nutrientes (Von-Elert & Jüttner, 1997; Reigosa et al., 1999). Granéli & Johansson
(2003) observaram um aumento na produção de substâncias alelopáticas pelas
células de Prymnesium parvum crescidas sob condições de deficiência de
nutrientes. Outro fator importante pode ser a temperatura. Issa (1999) observou que
a produção de antibiótico pelas cianobactérias Oscillatoria angustissima e Calothrix
parietina foi dependente da temperatura, mas não de sua biomassa. A intensidade
luminosa também pode influenciar na eficiência da estratégia alelopática em
cianobactérias (De Nobel et al., 1998). No entanto, a intensidade da interação
alelopática não é somente influenciada por fatores abióticos, mas também vai

105
depender de fatores bióticos, como a fase de crescimento, ou da concentração de
células, tanto do organismo produtor quanto do alvo. Contudo, a integração de
fatores que influenciam a produção de compostos aleloquímicos é essencial para o
entendimento do papel ecológico da alelopatia para o fitoplâncton.
As cianobactérias e microalgas são conhecidas como produtoras de uma
grande variedade de metabólitos secundários que têm várias atividades biológicas
distintas (Leflaive & Tem-Hage, 2007). Muitos desses compostos podem ser
liberados para o meio durante o crescimento ou pela lise celular e
consequentemente ter efeito alelopático e afetar a estrutura do ecossistema. Vários
organismos fitoplanctônicos já foram descritos como produtores de compostos
alelopáticos. Entre os gêneros de cianobactérias já citados estão: Anabaena,
Anabaenopsis, Arthrospira, Calothrix, Gomphosphaeria, Aphanizomenon,
Hapolosiphon, Fischerella, Microcystis, Nodularia, Nostoc, Oscillatoria, Phormidium,
Scytonema, Trichormus, Cylindrospermopsis, Leptolyngbya, Chroococcus,
Synechococcus (Gross, 2003; Legrand et al., 2003; Van der Grinten et al., 2005;
Figueiredo et al., 2007; Suikkanen, 2008; Leão et al., 2009; Bar-Yosef et al. 2010;
Leão et al., 2010). Além das cianobactérias, outros grupos de microalgas como
diatomáceas, dinoflagelados, clorofíceas, rafidofíceas, haptofíceas também foram
relatados como produtores de aleloquímicos (Leflaive & Tem-Hage, 2007; Legrand
et al., 2003).
Atualmente, as interações alelopáticas têm sido consideradas importantes nos
processos de dominância ou mudanças na estrutura da comunidade aquática, que
aparentemente não podem ser explicados pelas interações tróficas ou por
competição por recursos (Legrand et al., 2003; Gross, 2003). Além disso, seu papel
na formação, estabelecimento e finalização das florações também vem sendo

106
discutido (Keating, 1977; 1978; Suikkanen et al., 2005; Bar-Yosef et al. 2010; Carey
& Rengefors, 2010).
Na lagoa de Jacarepaguá (RJ), a comunidade fitoplactônica foi dominada
pelas cianobactérias Microcystis aeruginosa e Planktothrix sp., entre outras, na
maior parte do ano de 2007. No entanto, quando a temperatura variou de 30 para 18
oC, essa a composição fitoplanctônica sofreu uma alteração, aumentando a
contribuição relativa das diatomáceas, especialmente da espécie Cyclotella sp. (ver
capítulo I). Diante de tal cenário, surgem as perguntas: Quais fatores controlam a
dinâmica da comunidade fitoplanctônica, a formação e o colapso de uma floração?
Além de fatores abióticos, também interações bióticas podem estar envolvidas? A
combinação entre eles pode explicar a dinâmica da comunidade fitoplanctônica? A
temperatura pode afetar estas interações biológicas? Para tentar esclarecer algumas
dessas perguntas e entender a sucessão de espécies fitoplanctônicas na lagoa de
Jacarepaguá, o presente trabalho teve como objetivo avaliar o efeito de duas
temperaturas (18 e 30 oC) na interação alelopática entre três espécies
fitoplanctônicas (Microcystis aeruginosa, Planktothrix agardhii e Cyclotella
meneghiniana) que foram escolhidas para representar as espécies que ocorrem em
maior prevalência e alternância nessa lagoa.

107
MATERIAL E MÉTODOS
DESENHO EXPERIMENTAL:
Para a obtenção dos exudatos, monoculturas das cepas de M. aeruginosa
(MIJAC-01), P. agardhii (PAT-3) e C. meneghiniana (CCAP 1070/5), além do cultivo
misto destas três espécies (MIJAC-01 + PAT-3 + CCAP1070/5) foram crescidos em
sistema de cultivo tipo “batch”, sem limitação de nutrientes, sob condições luminosas
controladas (60 �mol fótons m-2 s-1), fotoperíodo (18:6h), agitação (60 rpm) e
temperaturas de 18 oC ou 30 oC. O biovolume inicial de cada monocultivo e de cada
cepa que compõe a cultura mista foi 5,0 x 107 µm3.mL-1. No cultivo misto, a
densidade de células final (após 10 dias) de cada espécie estão descritas na tabela
1.
Após os 10 dias crescimento, essas culturas foram cuidadosamente filtradas
em filtros de 0,2 µm de porosidade (Schleicher & Schell Microscience, Germany) em
sistemas de filtração esterilizados. Foram adicionados nutrientes aos filtrados nas
concentrações encontradas no meio WC modificado (Lürling & Beekman, 1999) para
evitar o efeito da limitação por nutrientes. As concentrações de nitrogênio (nitrito,
nitrato e amônia) e fósforo dissolvidos foram analisadas nos exudatos de cada
espécie (antes após reconstituição do meio) antes do inóculo da cepa alvo, seguindo
a metodologia descrita em APHA (1998). As concentrações de carbono orgânico e
inorgânico dissolvidos (COD e CID) também foram determinadas nesses exudatos e
estão discriminadas na tabela 2.
As séries experimentais foram realizadas da seguinte forma: (a) crescimento
monoespecífico de MIJAC em exudato de PAT, (b) crescimento monoespecífico de
PAT em exudato de MIJAC, (c) crescimento monoespecífico da CCAP em exudato
de MIJAC, (d) crescimento monoespecífico da CCAP em exudato de PAT, (e)

108
crescimento monoespecífico de MIJAC em exudato de CCAP, (f) crescimento
monoespecífico de PAT em exudato de CCAP (g) crescimento monoespecífico das
três cepas nos seus próprios exudatos e (h) crescimento monoespecífico de PAT,
MIJAC e CCAP em exudato de cultivo misto dessas três espécies (esquema do
desenho experimental na fig 1). O controle de cada cepa foi cultivado em meio WC
modificado (Lürling & Beekman, 1999). Todas as culturas tiveram biomassas iniciais
iguais (107 µm3.mL-1). As séries de experimentação foram cultivadas sob as mesmas
condições já descritas para os cultivos para obtenção dos exudatos, ocorrendo sob
as temperaturas de 18 oC ou 30 oC. Os experimentos foram realizados em triplicata
e ao longo do período experimental (12 dias) foram retiradas sete amostras, em
intervalos de 2 dias, para analisar as concentrações de clorofila-a e a eficiência do
fotossistema II, utilizando o aparelho PHYTO-PAM (Heins Walz GmgH, Effeltrich,
Germany). Os valores de pH também foram monitorados em cada frasco durante
todo o período experimental.
Tabela 1: Densidade celular (no. céls mL-1) de cada espécie no cultivo misto de M. aeruginosa (MIJAC), P. agardhii (PAT) e C. meneghiniana (CCAP) após 10 dias de cultivo para obtenção do exudato.
Densidade Celular (no. céls mL-1)Temperaturado cultivo MIJAC PAT CCAP
18 oC 2,6 x 106 8,6 x 10 4 2,4 x 104
30 oC 3,2 x 105 1,7 x 105 2,0 x 104
ANÁLISE DE DADOS:
Análises de variância com medidas repetidas (Repeated-measures ANOVA)
foram usadas para testar as diferenças entre os parâmetros: rendimento
fotossintético, biomassa clorofiliana e valores de pH, entre as culturas alvo tratadas

109
com os exudatos e o controle, ao longo do período experimental. O teste post-hoc
de Tukey foi usado para verificar se essas diferenças foram significativas. As taxas
de crescimento foram calculadas como já descritas no capítulo anterior e a
existência de diferenças entre os tratamentos e controle foi testada por análise de
variância (ANOVA) com teste de significância de Tukey. Estas análises foram feitas
utilizando o programa estatístico SPSS® 17.0.
Figura 1: Esquema do desenho experimental realizado para verificar o efeito da temperatura no potencial alelopático das espécies M. aeruginosa (MIJAC), P. aghardii (PAT) e C.meneginiana (CCAP) em monoculturas e cultivos mistos. O controle de cada cepa foi cultivado em meio WC modificado (Lürling & Beekman, 1999).
MEIO WC MIJAC exudato PAT exudato CCAP exudato MIJAC + PAT + CCAP exudato
MEIO WC MIJAC PAT CCAPMIJAC +
PAT + CCAP
Adição de Nutrientes
MIJAC
PAT
CCAP
10 d
ias
cultu
ra
inóc
ulo
18oC e 30oC
MIJAC
PAT
CCAP
MIJAC
PAT
CCAP
MIJAC
PAT
CCAP
MIJAC
PAT
CCAP
MEIO WC MIJAC exudato PAT exudato CCAP exudato MIJAC + PAT + CCAP exudato
MEIO WC MIJAC PAT CCAPMIJAC +
PAT + CCAP
Adição de Nutrientes
MIJAC
PAT
CCAP
10 d
ias
cultu
ra
inóc
ulo
18oC e 30oC
MIJAC
PAT
CCAP
MIJAC
PAT
CCAP
MIJAC
PAT
CCAP
MIJAC
PAT
CCAP
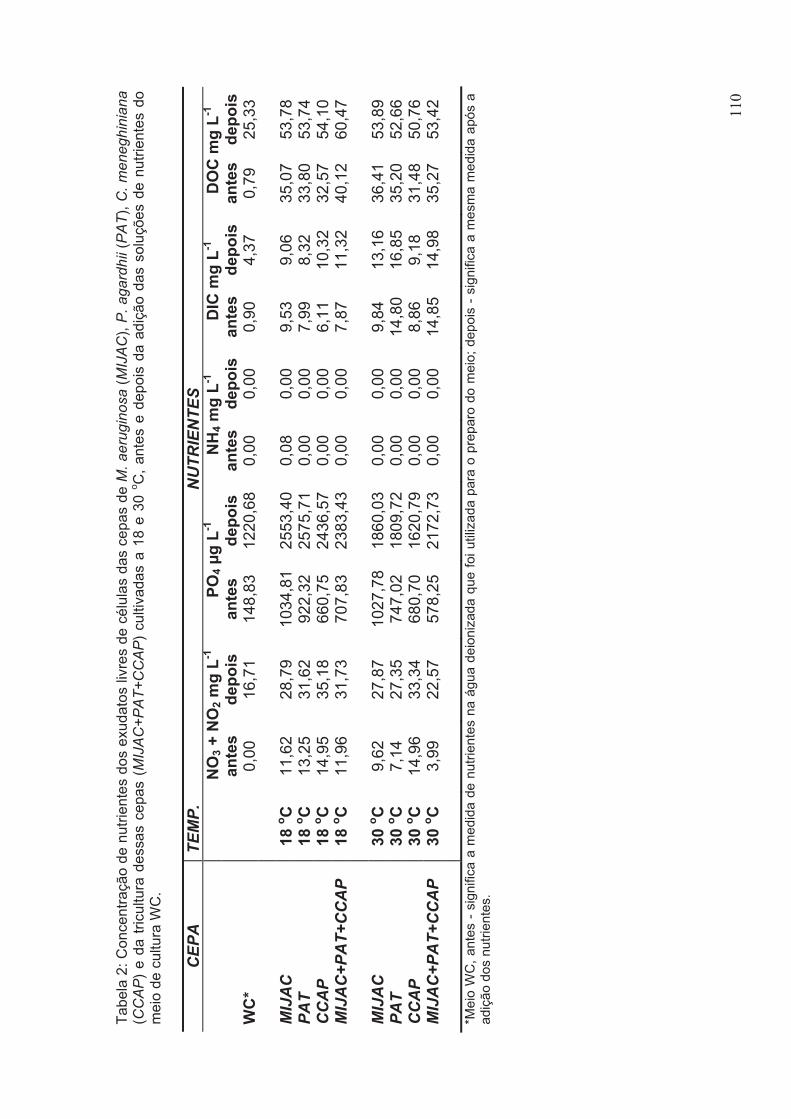
110
Tabe
la 2
: Con
cent
raçã
o de
nut
rient
es d
os e
xuda
tos
livre
s de
cél
ulas
das
cep
as d
e M
. aer
ugin
osa
(MIJ
AC
), P
. aga
rdhi
i (P
AT)
,C. m
eneg
hini
ana
(CC
AP
) e
da tr
icul
tura
des
sas
cepa
s (M
IJA
C+P
AT+
CC
AP
) cu
ltiva
das
a 18
e 3
0 o C
, ant
es e
dep
ois
da a
diçã
o da
s so
luçõ
es d
e nu
trien
tes
do
mei
o de
cul
tura
WC
.
CEP
A
TEM
P.
NU
TRIE
NTE
SN
O3 +
NO
2 mg
L-1
PO4 μ
g L-1
N
H4 m
g L-1
D
IC m
g L-1
D
OC
mg
L-1
ante
s de
pois
an
tes
depo
is
ante
s de
pois
an
tes
depo
is
ante
s de
pois
W
C*
0,
00
16,7
1 14
8,83
12
20,6
8 0,
00
0,00
0,
90
4,37
0,
79
25,3
3
MIJ
AC
18
o C11
,62
28,7
9 10
34,8
1 25
53,4
0 0,
08
0,00
9,
53
9,06
35
,07
53,7
8 PA
T18
o C13
,25
31,6
2 92
2,32
25
75,7
1 0,
00
0,00
7,
99
8,32
33
,80
53,7
4 C
CA
P18
o C14
,95
35,1
8 66
0,75
24
36,5
7 0,
00
0,00
6,
11
10,3
2 32
,57
54,1
0 M
IJA
C+P
AT+
CC
AP
18o C
11,9
6 31
,73
707,
83
2383
,43
0,00
0,
00
7,87
11
,32
40,1
2 60
,47
MIJ
AC
30
o C9,
62
27,8
7 10
27,7
8 18
60,0
3 0,
00
0,00
9,
84
13,1
6 36
,41
53,8
9 PA
T30
o C7,
14
27,3
5 74
7,02
18
09,7
2 0,
00
0,00
14
,80
16,8
5 35
,20
52,6
6 C
CA
P30
o C14
,96
33,3
4 68
0,70
16
20,7
9 0,
00
0,00
8,
86
9,18
31
,48
50,7
6 M
IJA
C+P
AT+
CC
AP
30o C
3,99
22
,57
578,
25
2172
,73
0,00
0,
00
14,8
5 14
,98
35,2
7 53
,42
*Mei
o W
C, a
ntes
- si
gnifi
ca a
med
ida
de n
utrie
ntes
na
água
dei
oniz
ada
que
foi u
tiliz
ada
para
o p
repa
ro d
o m
eio;
dep
ois
- sig
nific
a a
mes
ma
med
ida
após
a
adiç
ão d
os n
utrie
ntes
.

111
RESULTADOS
Como produtoras de alelopáticos, as cepas testadas nesse estudo
apresentaram três tipos de efeito: estimularam ou inibiram o crescimento de sua
competidora, ou seu próprio crescimento, ou não promoveram nenhum efeito. O tipo
de efeito dependeu tanto da combinação de cepa produtota-cepa alvo testada
quanto da temperatura do cultivo (tab. 3).
Tabela 3: Efeito dos exudatos das monoculturas de M. aeruginosa (MIJAC), P. agardhii(PAT), e C. meneghiniana (CCAP) e do cultivo misto das três cepas (MIJAC+PAT+CCAP) sobre o crescimento das mesmas cepas cultivadas em duas temperaturas (18 e 30 oC).
ATIVIDADE* EXUDATO
18 oCTEMP. Sp. ALVO
MIJAC PAT CCAP MIJAC+PAT+CCAPMIJAC + 0 0 0 PAT - + - - 18 oC
CCAP + + 0 +30 oC
MIJAC PAT CCAP MIJAC+PAT+CCAPMIJAC + + + + PAT - - 0 0 30 oC
CCAP - + + + * +, estímulo do crescimento, -, inibição do crescimento, 0, nenhum efeito no crescimento da espécie alvo. Estes efeitos foram determinados a partir das taxas de crescimento, que foram significativamente diferentes do controle (p<0,01), como determinado pela Análise de Variância (1-way ANOVA).
Considerando o efeito dos exudatos sobre o crescimento de M. aeruginosa a
18 ºC, quando cultivada em seu próprio exudato seu crescimento aumentou em
comparação com o controle (meio WC) (p=0,032) (fig. 2a). No entanto, nenhuma
diferença significativa (p=0,996) foi encontrada entre os valores de rendimento
fotossintético destas condições. Os cultivos em exudato de outras cepas, não
surtiram efeito sobre o crescimento de M. aeruginosa (fig. 2a, tab. 3). Quanto ao
rendimento fotossintético, em exudato de P. agardhii, foram observados os menores
valores deste parâmetro para M. aeruginosa a 18 oC (fig. 3a). Os valores de clorofila

112
da cepa de M. aeruginosa (18 oC) apresentaram diferenças significativas entre os
tempos em uma mesma condição (F= 3702,37; p<0,001) e entre um mesmo tempo
em diversos na interação tempo e tratamentos (F =101,22; p<0,001). O controle
entrou na fase de senescência dois dias antes (8º dia) dos demais tratamentos (10º
dia) (fig. 4a), possivelmente devido ao esgotamento de nutrientes, uma vez que as
concentrações de nitrogênio e fósforo são em média pelo menos duas vezes
maiores no meio WC, do que nos exudatos reconstituídos (tab. 2). Os valores de pH
seguiram o mesmo padrão de variação da concentração de clorofila, com diferenças
significativas entre os tempos (F=1059,13; p<0,001) e entre tempo e tratamento
(F=31,71; p<0,001) (fig. 5a). Esses resultados sugerem que a variação de pH
acompanhou o crescimento da cepa. Houve aumento do pH conforme a biomassa
aumentava e não em função dos diferentes tratamentos, e uma redução desses
valores quando as células entraram em senescência.
Quando M. aeruginosa foi cultivada a 30 oC, sua taxa de crescimento foi
significativamente aumentada em todos os exudatos, comparados com o controle
(p<0,001 para todos) (tab. 1), sendo os efeitos mais pronunciados observados em
seu próprio exudato e no de P. agardhii (fig. 2b). O seu rendimento fotossintético não
apresentou grandes variações entre os tratamentos, embora diferenças significativas
ao longo do tempo (F=18,85); p<0,001) tenham sido observadas (fig. 3b).
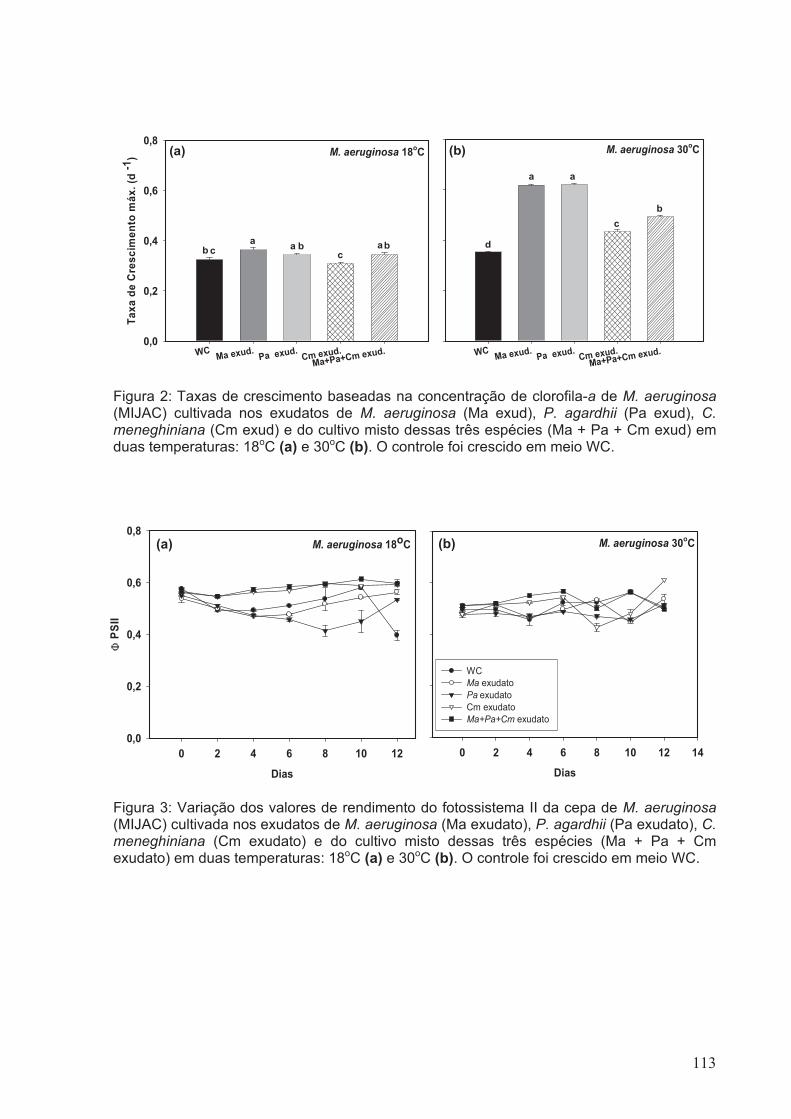
113
Figura 2: Taxas de crescimento baseadas na concentração de clorofila-a de M. aeruginosa(MIJAC) cultivada nos exudatos de M. aeruginosa (Ma exud), P. agardhii (Pa exud), C. meneghiniana (Cm exud) e do cultivo misto dessas três espécies (Ma + Pa + Cm exud) em duas temperaturas: 18oC (a) e 30oC (b). O controle foi crescido em meio WC.
Figura 3: Variação dos valores de rendimento do fotossistema II da cepa de M. aeruginosa(MIJAC) cultivada nos exudatos de M. aeruginosa (Ma exudato), P. agardhii (Pa exudato), C. meneghiniana (Cm exudato) e do cultivo misto dessas três espécies (Ma + Pa + Cm exudato) em duas temperaturas: 18oC (a) e 30oC (b). O controle foi crescido em meio WC.
M. aeruginosa 30oC
WC Ma exud. Pa exud. Cm exud.Ma+Pa+Cm exud.
M. aeruginosa 18oC
WC Ma exud. Pa exud.Cm exud.
Ma+Pa+Cm exud.
Taxa
de
Cre
scim
ento
máx
. (d
-1)
0,0
0,2
0,4
0,6
0,8
a a ab bb c c
a a
cb
d
(a) (b) M. aeruginosa 30oC
WC Ma exud. Pa exud. Cm exud.Ma+Pa+Cm exud.
M. aeruginosa 18oC
WC Ma exud. Pa exud.Cm exud.
Ma+Pa+Cm exud.
Taxa
de
Cre
scim
ento
máx
. (d
-1)
0,0
0,2
0,4
0,6
0,8
a a ab bb c c
a a
cb
d
M. aeruginosa 30oC
WC Ma exud. Pa exud. Cm exud.Ma+Pa+Cm exud.
M. aeruginosa 18oC
WC Ma exud. Pa exud.Cm exud.
Ma+Pa+Cm exud.
Taxa
de
Cre
scim
ento
máx
. (d
-1)
0,0
0,2
0,4
0,6
0,8M. aeruginosa 30oC
WC Ma exud. Pa exud. Cm exud.Ma+Pa+Cm exud.
M. aeruginosa 18oC
WC Ma exud. Pa exud.Cm exud.
Ma+Pa+Cm exud.
Taxa
de
Cre
scim
ento
máx
. (d
-1)
0,0
0,2
0,4
0,6
0,8
a a ab bb c c
a a
cb
d
(a) (b)
M. aeruginosa 30oC
Dias
0 2 4 6 8 10 12 14
WCMa exudatoPa exudatoCm exudatoMa+Pa+Cm exudato
Dias
0 2 4 6 8 10 12
� P
SII
0,0
0,2
0,4
0,6
0,8M. aeruginosa 18oC(a) (b) M. aeruginosa 30oC
Dias
0 2 4 6 8 10 12 14
WCMa exudatoPa exudatoCm exudatoMa+Pa+Cm exudato
Dias
0 2 4 6 8 10 12
� P
SII
0,0
0,2
0,4
0,6
0,8M. aeruginosa 18oC M. aeruginosa 30oC
Dias
0 2 4 6 8 10 12 14
WCMa exudatoPa exudatoCm exudatoMa+Pa+Cm exudato
Dias
0 2 4 6 8 10 12
� P
SII
0,0
0,2
0,4
0,6
0,8M. aeruginosa 18oC(a) (b)
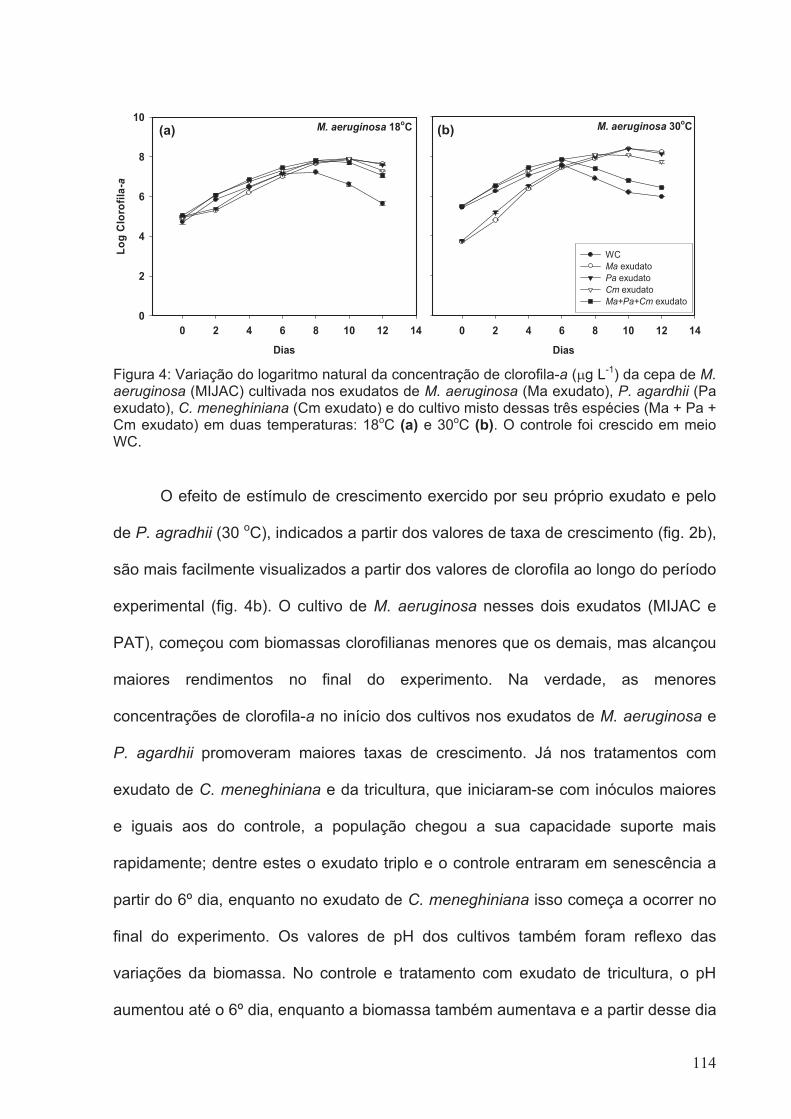
114
Figura 4: Variação do logaritmo natural da concentração de clorofila-a (�g L-1) da cepa de M. aeruginosa (MIJAC) cultivada nos exudatos de M. aeruginosa (Ma exudato), P. agardhii (Pa exudato), C. meneghiniana (Cm exudato) e do cultivo misto dessas três espécies (Ma + Pa + Cm exudato) em duas temperaturas: 18oC (a) e 30oC (b). O controle foi crescido em meio WC.
O efeito de estímulo de crescimento exercido por seu próprio exudato e pelo
de P. agradhii (30 oC), indicados a partir dos valores de taxa de crescimento (fig. 2b),
são mais facilmente visualizados a partir dos valores de clorofila ao longo do período
experimental (fig. 4b). O cultivo de M. aeruginosa nesses dois exudatos (MIJAC e
PAT), começou com biomassas clorofilianas menores que os demais, mas alcançou
maiores rendimentos no final do experimento. Na verdade, as menores
concentrações de clorofila-a no início dos cultivos nos exudatos de M. aeruginosa e
P. agardhii promoveram maiores taxas de crescimento. Já nos tratamentos com
exudato de C. meneghiniana e da tricultura, que iniciaram-se com inóculos maiores
e iguais aos do controle, a população chegou a sua capacidade suporte mais
rapidamente; dentre estes o exudato triplo e o controle entraram em senescência a
partir do 6º dia, enquanto no exudato de C. meneghiniana isso começa a ocorrer no
final do experimento. Os valores de pH dos cultivos também foram reflexo das
variações da biomassa. No controle e tratamento com exudato de tricultura, o pH
aumentou até o 6º dia, enquanto a biomassa também aumentava e a partir desse dia
M. aeruginosa 30oC
Dias
0 2 4 6 8 10 12 14
WCMa exudatoPa exudatoCm exudatoMa+Pa+Cm exudato
M. aeruginosa 18oC
Dias
0 2 4 6 8 10 12 14
Log
Clo
rofil
a-a
0
2
4
6
8
10(a) (b) M. aeruginosa 30oC
Dias
0 2 4 6 8 10 12 14
WCMa exudatoPa exudatoCm exudatoMa+Pa+Cm exudato
M. aeruginosa 18oC
Dias
0 2 4 6 8 10 12 14
Log
Clo
rofil
a-a
0
2
4
6
8
10M. aeruginosa 30oC
Dias
0 2 4 6 8 10 12 14
WCMa exudatoPa exudatoCm exudatoMa+Pa+Cm exudato
M. aeruginosa 18oC
Dias
0 2 4 6 8 10 12 14
Log
Clo
rofil
a-a
0
2
4
6
8
10(a) (b)
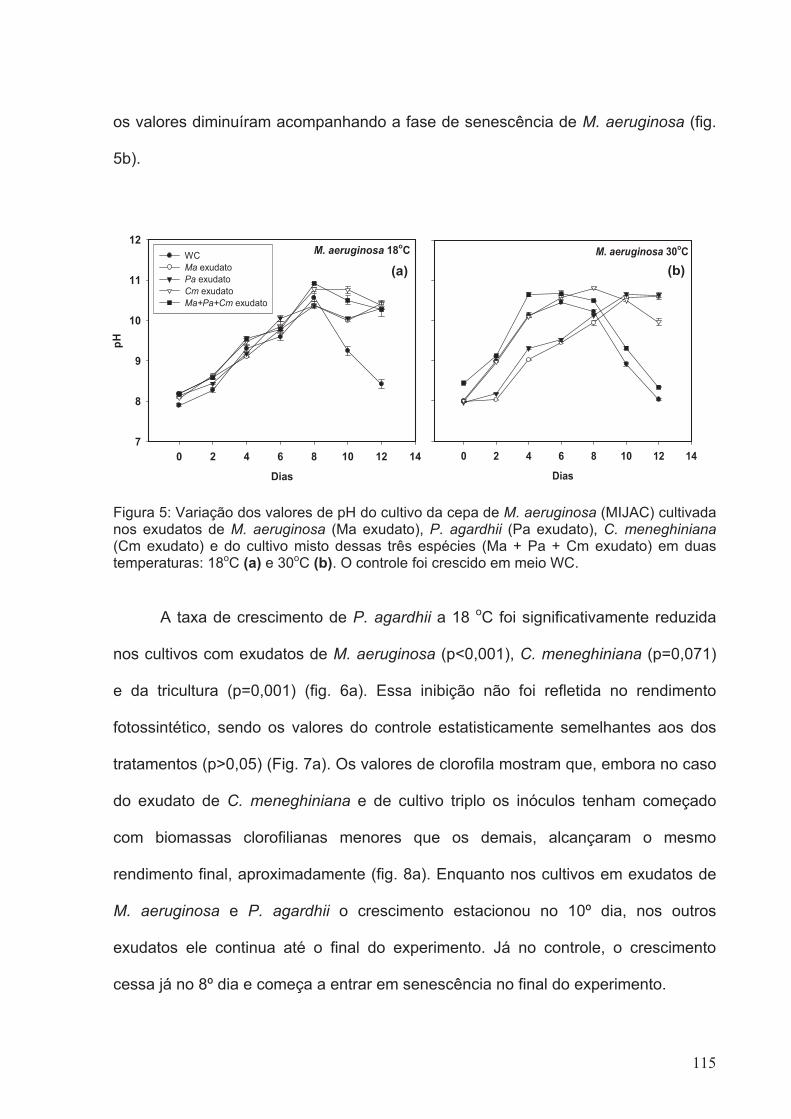
115
os valores diminuíram acompanhando a fase de senescência de M. aeruginosa (fig.
5b).
Figura 5: Variação dos valores de pH do cultivo da cepa de M. aeruginosa (MIJAC) cultivada nos exudatos de M. aeruginosa (Ma exudato), P. agardhii (Pa exudato), C. meneghiniana(Cm exudato) e do cultivo misto dessas três espécies (Ma + Pa + Cm exudato) em duas temperaturas: 18oC (a) e 30oC (b). O controle foi crescido em meio WC.
A taxa de crescimento de P. agardhii a 18 oC foi significativamente reduzida
nos cultivos com exudatos de M. aeruginosa (p<0,001), C. meneghiniana (p=0,071)
e da tricultura (p=0,001) (fig. 6a). Essa inibição não foi refletida no rendimento
fotossintético, sendo os valores do controle estatisticamente semelhantes aos dos
tratamentos (p>0,05) (Fig. 7a). Os valores de clorofila mostram que, embora no caso
do exudato de C. meneghiniana e de cultivo triplo os inóculos tenham começado
com biomassas clorofilianas menores que os demais, alcançaram o mesmo
rendimento final, aproximadamente (fig. 8a). Enquanto nos cultivos em exudatos de
M. aeruginosa e P. agardhii o crescimento estacionou no 10º dia, nos outros
exudatos ele continua até o final do experimento. Já no controle, o crescimento
cessa já no 8º dia e começa a entrar em senescência no final do experimento.
M. aeruginosa 30oC
Dias
0 2 4 6 8 10 12 14
M. aeruginosa 18oC
Dias
0 2 4 6 8 10 12 14
pH
7
8
9
10
11
12WC Ma exudatoPa exudatoCm exudatoMa+Pa+Cm exudato
(a) (b)M. aeruginosa 30oC
Dias
0 2 4 6 8 10 12 14
M. aeruginosa 18oC
Dias
0 2 4 6 8 10 12 14
pH
7
8
9
10
11
12WC Ma exudatoPa exudatoCm exudatoMa+Pa+Cm exudato
M. aeruginosa 30oC
Dias
0 2 4 6 8 10 12 14
M. aeruginosa 18oC
Dias
0 2 4 6 8 10 12 14
pH
7
8
9
10
11
12WC Ma exudatoPa exudatoCm exudatoMa+Pa+Cm exudato
(a) (b)
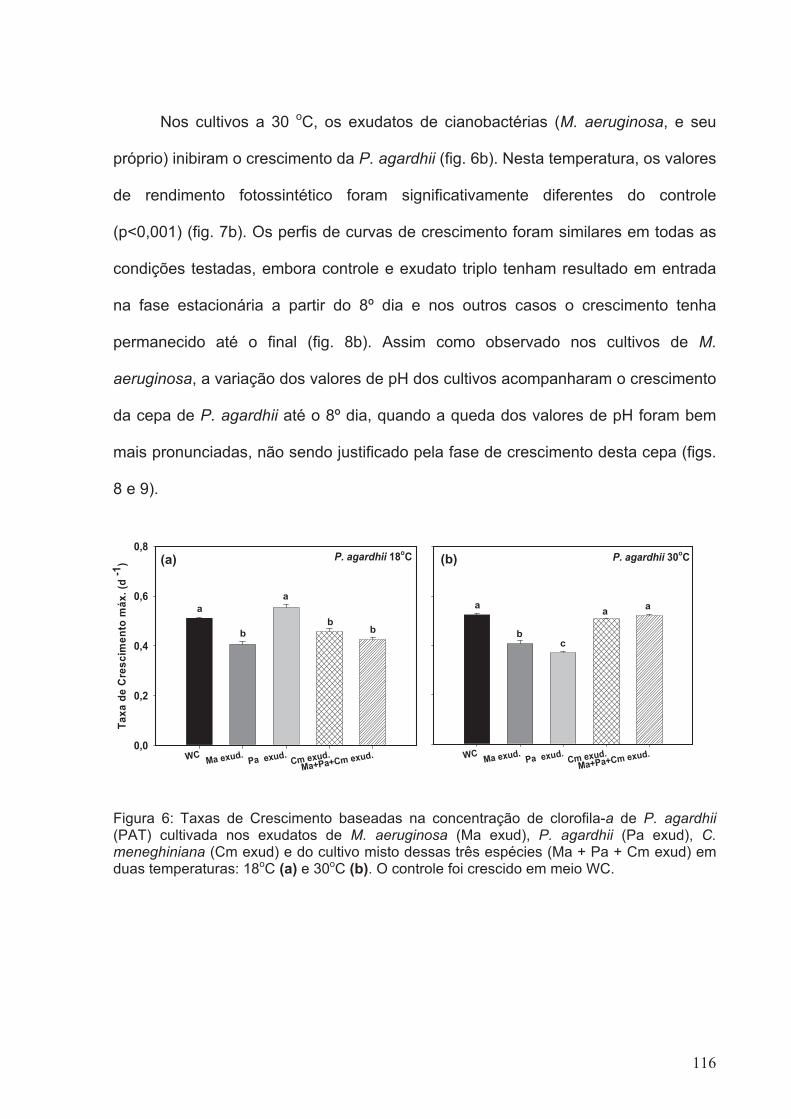
116
Nos cultivos a 30 oC, os exudatos de cianobactérias (M. aeruginosa, e seu
próprio) inibiram o crescimento da P. agardhii (fig. 6b). Nesta temperatura, os valores
de rendimento fotossintético foram significativamente diferentes do controle
(p<0,001) (fig. 7b). Os perfis de curvas de crescimento foram similares em todas as
condições testadas, embora controle e exudato triplo tenham resultado em entrada
na fase estacionária a partir do 8º dia e nos outros casos o crescimento tenha
permanecido até o final (fig. 8b). Assim como observado nos cultivos de M.
aeruginosa, a variação dos valores de pH dos cultivos acompanharam o crescimento
da cepa de P. agardhii até o 8º dia, quando a queda dos valores de pH foram bem
mais pronunciadas, não sendo justificado pela fase de crescimento desta cepa (figs.
8 e 9).
Figura 6: Taxas de Crescimento baseadas na concentração de clorofila-a de P. agardhii (PAT) cultivada nos exudatos de M. aeruginosa (Ma exud), P. agardhii (Pa exud), C. meneghiniana (Cm exud) e do cultivo misto dessas três espécies (Ma + Pa + Cm exud) em duas temperaturas: 18oC (a) e 30oC (b). O controle foi crescido em meio WC.
P. agardhii 30oC
WC Ma exud. Pa exud. Cm exud.Ma+Pa+Cm exud.
P. agardhii 18oC
WC Ma exud. Pa exud.Cm exud.
Ma+Pa+Cm exud.
Taxa
de
Cre
scim
ento
máx
. (d
-1)
0,0
0,2
0,4
0,6
0,8
aa
bb
b b
a a a
c
(a) (b) P. agardhii 30oC
WC Ma exud. Pa exud. Cm exud.Ma+Pa+Cm exud.
P. agardhii 18oC
WC Ma exud. Pa exud.Cm exud.
Ma+Pa+Cm exud.
Taxa
de
Cre
scim
ento
máx
. (d
-1)
0,0
0,2
0,4
0,6
0,8
aa
bb
b b
a a a
c
P. agardhii 30oC
WC Ma exud. Pa exud. Cm exud.Ma+Pa+Cm exud.
P. agardhii 18oC
WC Ma exud. Pa exud.Cm exud.
Ma+Pa+Cm exud.
Taxa
de
Cre
scim
ento
máx
. (d
-1)
0,0
0,2
0,4
0,6
0,8P. agardhii 30oC
WC Ma exud. Pa exud. Cm exud.Ma+Pa+Cm exud.
P. agardhii 18oC
WC Ma exud. Pa exud.Cm exud.
Ma+Pa+Cm exud.
Taxa
de
Cre
scim
ento
máx
. (d
-1)
0,0
0,2
0,4
0,6
0,8
aa
bb
b b
a a a
c
(a) (b)
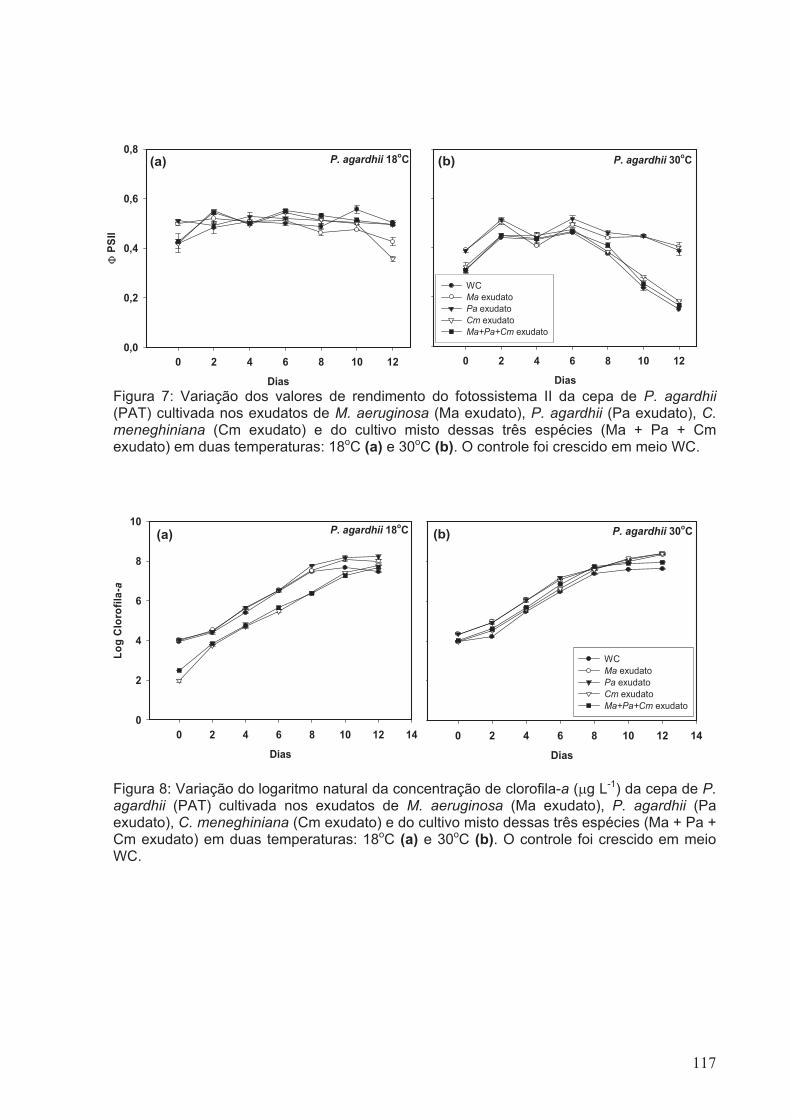
117
Figura 7: Variação dos valores de rendimento do fotossistema II da cepa de P. agardhii(PAT) cultivada nos exudatos de M. aeruginosa (Ma exudato), P. agardhii (Pa exudato), C.meneghiniana (Cm exudato) e do cultivo misto dessas três espécies (Ma + Pa + Cm exudato) em duas temperaturas: 18oC (a) e 30oC (b). O controle foi crescido em meio WC.
Figura 8: Variação do logaritmo natural da concentração de clorofila-a (�g L-1) da cepa de P. agardhii (PAT) cultivada nos exudatos de M. aeruginosa (Ma exudato), P. agardhii (Pa exudato), C. meneghiniana (Cm exudato) e do cultivo misto dessas três espécies (Ma + Pa + Cm exudato) em duas temperaturas: 18oC (a) e 30oC (b). O controle foi crescido em meio WC.
P. agardhii 30oC
Dias
0 2 4 6 8 10 12
WCMa exudato Pa exudato Cm exudato Ma+Pa+Cm exudato
P. agardhii 18oC
Dias
0 2 4 6 8 10 12
� P
SII
0,0
0,2
0,4
0,6
0,8(a) (b) P. agardhii 30oC
Dias
0 2 4 6 8 10 12
WCMa exudato Pa exudato Cm exudato Ma+Pa+Cm exudato
P. agardhii 18oC
Dias
0 2 4 6 8 10 12
� P
SII
0,0
0,2
0,4
0,6
0,8P. agardhii 30oC
Dias
0 2 4 6 8 10 12
WCMa exudato Pa exudato Cm exudato Ma+Pa+Cm exudato
P. agardhii 18oC
Dias
0 2 4 6 8 10 12
� P
SII
0,0
0,2
0,4
0,6
0,8(a) (b)
P. agardhii 30oC
Dias
0 2 4 6 8 10 12 14
WCMa exudatoPa exudatoCm exudatoMa+Pa+Cm exudato
P. agardhii 18oC
Dias
0 2 4 6 8 10 12 14
Log
Clo
rofil
a-a
0
2
4
6
8
10(a) (b) P. agardhii 30oC
Dias
0 2 4 6 8 10 12 14
WCMa exudatoPa exudatoCm exudatoMa+Pa+Cm exudato
P. agardhii 18oC
Dias
0 2 4 6 8 10 12 14
Log
Clo
rofil
a-a
0
2
4
6
8
10P. agardhii 30oC
Dias
0 2 4 6 8 10 12 14
WCMa exudatoPa exudatoCm exudatoMa+Pa+Cm exudato
P. agardhii 18oC
Dias
0 2 4 6 8 10 12 14
Log
Clo
rofil
a-a
0
2
4
6
8
10(a) (b)
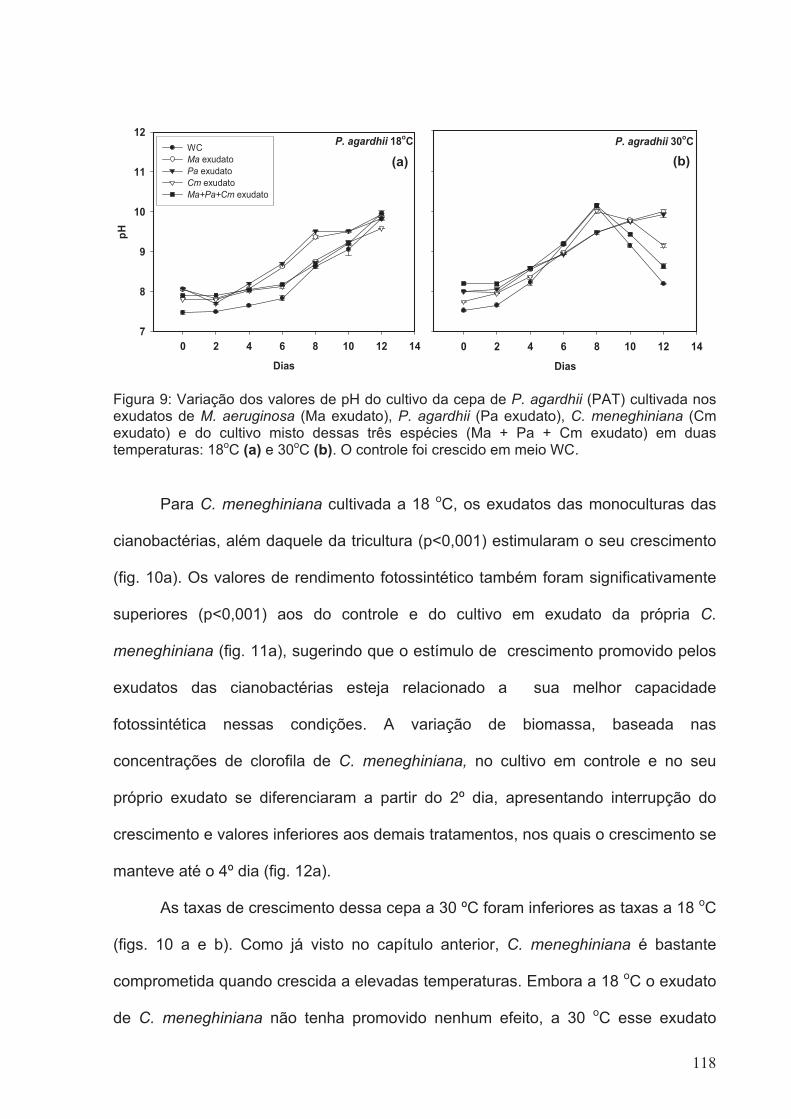
118
Figura 9: Variação dos valores de pH do cultivo da cepa de P. agardhii (PAT) cultivada nos exudatos de M. aeruginosa (Ma exudato), P. agardhii (Pa exudato), C. meneghiniana (Cm exudato) e do cultivo misto dessas três espécies (Ma + Pa + Cm exudato) em duas temperaturas: 18oC (a) e 30oC (b). O controle foi crescido em meio WC.
Para C. meneghiniana cultivada a 18 oC, os exudatos das monoculturas das
cianobactérias, além daquele da tricultura (p<0,001) estimularam o seu crescimento
(fig. 10a). Os valores de rendimento fotossintético também foram significativamente
superiores (p<0,001) aos do controle e do cultivo em exudato da própria C.
meneghiniana (fig. 11a), sugerindo que o estímulo de crescimento promovido pelos
exudatos das cianobactérias esteja relacionado a sua melhor capacidade
fotossintética nessas condições. A variação de biomassa, baseada nas
concentrações de clorofila de C. meneghiniana, no cultivo em controle e no seu
próprio exudato se diferenciaram a partir do 2º dia, apresentando interrupção do
crescimento e valores inferiores aos demais tratamentos, nos quais o crescimento se
manteve até o 4º dia (fig. 12a).
As taxas de crescimento dessa cepa a 30 ºC foram inferiores as taxas a 18 oC
(figs. 10 a e b). Como já visto no capítulo anterior, C. meneghiniana é bastante
comprometida quando crescida a elevadas temperaturas. Embora a 18 oC o exudato
de C. meneghiniana não tenha promovido nenhum efeito, a 30 oC esse exudato
P. agradhii 30oC
Dias
0 2 4 6 8 10 12 14
P. agardhii 18oC
Dias
0 2 4 6 8 10 12 14
pH
7
8
9
10
11
12WC Ma exudatoPa exudatoCm exudatoMa+Pa+Cm exudato
(a) (b)P. agradhii 30oC
Dias
0 2 4 6 8 10 12 14
P. agardhii 18oC
Dias
0 2 4 6 8 10 12 14
pH
7
8
9
10
11
12WC Ma exudatoPa exudatoCm exudatoMa+Pa+Cm exudato
P. agradhii 30oC
Dias
0 2 4 6 8 10 12 14
P. agardhii 18oC
Dias
0 2 4 6 8 10 12 14
pH
7
8
9
10
11
12WC Ma exudatoPa exudatoCm exudatoMa+Pa+Cm exudato
(a) (b)
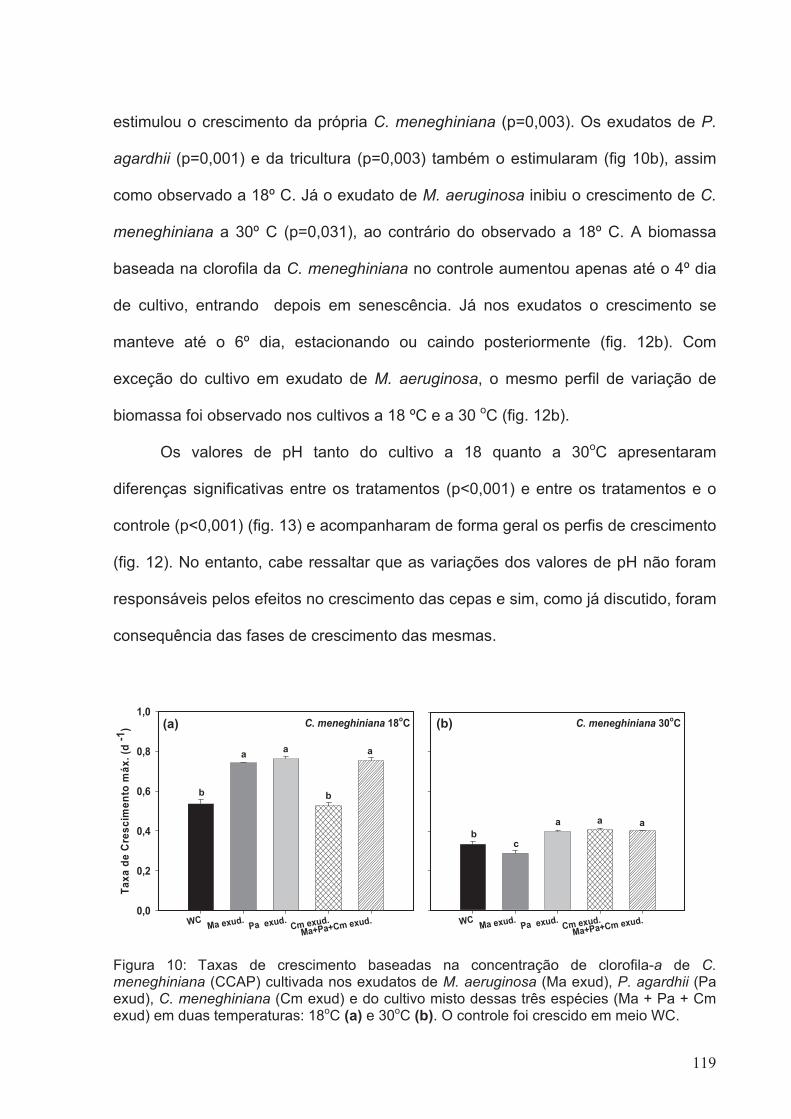
119
estimulou o crescimento da própria C. meneghiniana (p=0,003). Os exudatos de P.
agardhii (p=0,001) e da tricultura (p=0,003) também o estimularam (fig 10b), assim
como observado a 18º C. Já o exudato de M. aeruginosa inibiu o crescimento de C.
meneghiniana a 30º C (p=0,031), ao contrário do observado a 18º C. A biomassa
baseada na clorofila da C. meneghiniana no controle aumentou apenas até o 4º dia
de cultivo, entrando depois em senescência. Já nos exudatos o crescimento se
manteve até o 6º dia, estacionando ou caindo posteriormente (fig. 12b). Com
exceção do cultivo em exudato de M. aeruginosa, o mesmo perfil de variação de
biomassa foi observado nos cultivos a 18 ºC e a 30 oC (fig. 12b).
Os valores de pH tanto do cultivo a 18 quanto a 30oC apresentaram
diferenças significativas entre os tratamentos (p<0,001) e entre os tratamentos e o
controle (p<0,001) (fig. 13) e acompanharam de forma geral os perfis de crescimento
(fig. 12). No entanto, cabe ressaltar que as variações dos valores de pH não foram
responsáveis pelos efeitos no crescimento das cepas e sim, como já discutido, foram
consequência das fases de crescimento das mesmas.
Figura 10: Taxas de crescimento baseadas na concentração de clorofila-a de C.meneghiniana (CCAP) cultivada nos exudatos de M. aeruginosa (Ma exud), P. agardhii (Pa exud), C. meneghiniana (Cm exud) e do cultivo misto dessas três espécies (Ma + Pa + Cm exud) em duas temperaturas: 18oC (a) e 30oC (b). O controle foi crescido em meio WC.
C. meneghiniana 30oC
WC Ma exud. Pa exud. Cm exud.Ma+Pa+Cm exud.
C. meneghiniana 18oC
WC Ma exud. Pa exud. Cm exud.Ma+Pa+Cm exud.
Taxa
de
Cre
scim
ento
máx
. (d
-1)
0,0
0,2
0,4
0,6
0,8
1,0
a a a
bb
ba a a
c
(a) (b) C. meneghiniana 30oC
WC Ma exud. Pa exud. Cm exud.Ma+Pa+Cm exud.
C. meneghiniana 18oC
WC Ma exud. Pa exud. Cm exud.Ma+Pa+Cm exud.
Taxa
de
Cre
scim
ento
máx
. (d
-1)
0,0
0,2
0,4
0,6
0,8
1,0
a a a
bb
ba a a
c
C. meneghiniana 30oC
WC Ma exud. Pa exud. Cm exud.Ma+Pa+Cm exud.
C. meneghiniana 18oC
WC Ma exud. Pa exud. Cm exud.Ma+Pa+Cm exud.
Taxa
de
Cre
scim
ento
máx
. (d
-1)
0,0
0,2
0,4
0,6
0,8
1,0
a a a
bb
ba a a
c
(a) (b)
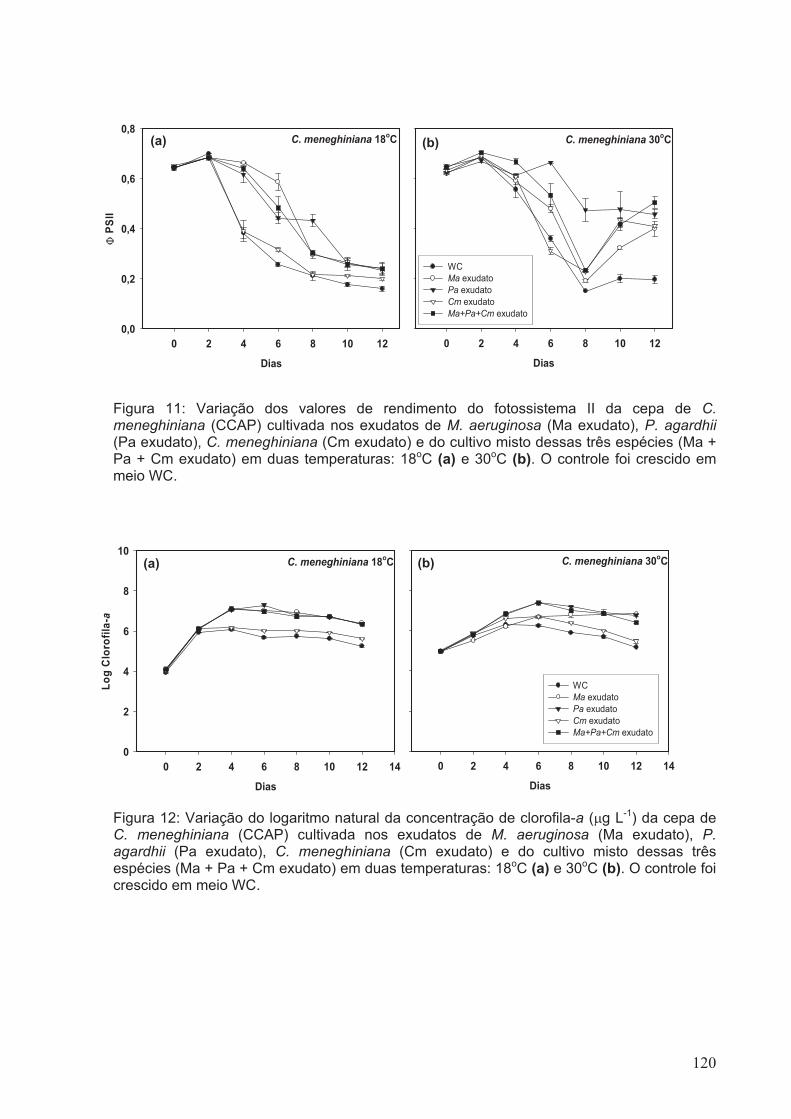
120
Figura 11: Variação dos valores de rendimento do fotossistema II da cepa de C.meneghiniana (CCAP) cultivada nos exudatos de M. aeruginosa (Ma exudato), P. agardhii(Pa exudato), C. meneghiniana (Cm exudato) e do cultivo misto dessas três espécies (Ma + Pa + Cm exudato) em duas temperaturas: 18oC (a) e 30oC (b). O controle foi crescido em meio WC.
Figura 12: Variação do logaritmo natural da concentração de clorofila-a (�g L-1) da cepa de C. meneghiniana (CCAP) cultivada nos exudatos de M. aeruginosa (Ma exudato), P. agardhii (Pa exudato), C. meneghiniana (Cm exudato) e do cultivo misto dessas três espécies (Ma + Pa + Cm exudato) em duas temperaturas: 18oC (a) e 30oC (b). O controle foi crescido em meio WC.
C. meneghiniana 30oC
Dias
0 2 4 6 8 10 12
WCMa exudato Pa exudatoCm exudato Ma+Pa+Cm exudato
C. meneghiniana 18oC
Dias
0 2 4 6 8 10 12
� P
SII
0,0
0,2
0,4
0,6
0,8(a) (b) C. meneghiniana 30oC
Dias
0 2 4 6 8 10 12
WCMa exudato Pa exudatoCm exudato Ma+Pa+Cm exudato
C. meneghiniana 18oC
Dias
0 2 4 6 8 10 12
� P
SII
0,0
0,2
0,4
0,6
0,8C. meneghiniana 30oC
Dias
0 2 4 6 8 10 12
WCMa exudato Pa exudatoCm exudato Ma+Pa+Cm exudato
C. meneghiniana 18oC
Dias
0 2 4 6 8 10 12
� P
SII
0,0
0,2
0,4
0,6
0,8(a) (b)
C. meneghiniana 30oC
Dias
0 2 4 6 8 10 12 14
WCMa exudatoPa exudatoCm exudatoMa+Pa+Cm exudato
C. meneghiniana 18oC
Dias
0 2 4 6 8 10 12 14
Log
Clo
rofil
a-a
0
2
4
6
8
10(a) (b) C. meneghiniana 30oC
Dias
0 2 4 6 8 10 12 14
WCMa exudatoPa exudatoCm exudatoMa+Pa+Cm exudato
C. meneghiniana 18oC
Dias
0 2 4 6 8 10 12 14
Log
Clo
rofil
a-a
0
2
4
6
8
10C. meneghiniana 30oC
Dias
0 2 4 6 8 10 12 14
WCMa exudatoPa exudatoCm exudatoMa+Pa+Cm exudato
C. meneghiniana 18oC
Dias
0 2 4 6 8 10 12 14
Log
Clo
rofil
a-a
0
2
4
6
8
10(a) (b)

121
Figura 13: Variação dos valores de pH do cultivo da cepa de C. meneghiniana (CCAP) cultivada nos exudatos de M. aeruginosa (Ma exudato), P. agardhii (Pa exudato), C. meneghiniana (Cm exudato) e do cultivo misto dessas três espécies (Ma + Pa + Cm exudato) em duas temperaturas: 18oC (a) e 30oC (b). O controle foi crescido em meio WC.
DISCUSSÃO
A alelopatia vem sendo apontada como um importante fator que regula a
sucessão de espécies fitoplanctônicas, em adição aos tradicionais fatores abióticos
e bióticos (Keating, 1977). No presente trabalho, avaliamos se interações
alelopáticas, assim como a influência da temperatura nestas interações, poderiam
ajudar a entender o padrão de substituição de espécies que ocorreu na lagoa de
Jacarepaguá (RJ).
Nos experimentos de alelopatia a 18 oC, o exudato de M. aeruginosa
estimulou o crescimento de C. meneghiniana e inibiu o crescimento de P. agardhii
(tab. 3). Na lagoa de Jacarepaguá, no ano de 2007, quando a temperatura da água
atingiu 18 oC, houve uma drástica redução da densidade de células de M.
aeruginosa e uma das espécies que aumentou sua biomassa e contribuição relativa
no sistema foi uma diatomácea do gênero Cyclotella. A lise celular de M. aeruginosa
e consequente liberação do seu conteúdo celular pode ter favorecido o crescimento
da espécie sucessora. P. agardhii foi sempre inibida pelo exudato de suas
C. meneghiniana 30oC
Dias
0 2 4 6 8 10 12 14
C.meneghiniana 18oC
Dias
0 2 4 6 8 10 12 14
pH
7
8
9
10
11
12WC Ma exudatoPa exudatoCm exudatoMa+Pa+Cm exudato
(a) (b)
C. meneghiniana 30oC
Dias
0 2 4 6 8 10 12 14
C.meneghiniana 18oC
Dias
0 2 4 6 8 10 12 14
pH
7
8
9
10
11
12WC Ma exudatoPa exudatoCm exudatoMa+Pa+Cm exudato
C. meneghiniana 30oC
Dias
0 2 4 6 8 10 12 14
C.meneghiniana 18oC
Dias
0 2 4 6 8 10 12 14
pH
7
8
9
10
11
12WC Ma exudatoPa exudatoCm exudatoMa+Pa+Cm exudato
(a) (b)

122
competidoras a 18 oC, no entanto, nas observações de estudo de campo, essa
espécie aumentou a sua biomassa no período de mais baixas temperaturas.
Nas temperaturas elevadas (PI e PIII), embora M. aeruginosa e Planktothrix
sp coexistissem na lagoa de Jacarepaguá, suas biomassas se alternavam. A
alelopatia pode ajudar a justificar esta dinâmica, uma vez que o exudato de M.
aeruginosa estimulou o seu próprio crescimento e inibiu o crescimento de P. agardhii
a 30 oC e esta última estimulou o crescimento da primeira nesta mesma
temperatura.
As diferentes respostas nas interações alelopáticas nas temperaturas de 18 e
30 oC podem ajudar a entender a substituição das espécies M. aeruginosa e
Cyclotella sp. quando há uma variação de temperatura da água. A espécie
antecessora é capaz de estimular o crescimento de sua sucessora quando esta
última estiver em sua temperatura ótima de crescimento. Quando a temperatura for
favorável à antecessora, a mesma inibe o crescimento da seguinte. Por exemplo, 30
oC favorece M. aeruginosa e seu exudato inibe o crescimento de C. meneghiniana,
que tem crescimento ótimo a 18 oC (ver capítulo III). A 30 oC, C. meneghiniana é
comprometida e seu exudato estimula o crescimento de M. aeruginosa (o que pode
ser somado ao fato de nesta temperatura de M. aeruginosa estimular o seu próprio
crescimento). A 18º C, ocorre o quadro inverso, o exudato de M. aeruginosa estimula
o crescimento de C. meneghiniana.
Outros casos do impacto diferencial de cianobactérias sobre seus
antecessores e sucessores na comunidade fitoplanctônica, especialmente espécies
do grupo das diatomáceas, também foram observados por Keating (1977) e Keating
(1978). Este autor sugeriu o papel direto da alelopatia, combinando três anos de
observações em campo com experimentos em laboratório, para justificar a sucessão

123
de espécies fitoplanctônicas no lago Linsley, nos Estados Unidos (Keating
1977,1978).
É sabido que sob condições de estresse ambiental, os efeitos alelopáticos
podem ser aumentados, enquanto a espécie alvo se torna mais sensível (von Elert &
Jüttner, 1997; Granéli & Johansson, 2003; Rengefors & Legrand, 2001; Fistarol et
al., 2005). A temperatura pode ser um estresse ambiental que regula as interações
alelopáticas entre as espécies fitoplactônicas na lagoa de Jacarepaguá. Este fato
pode justificar as diferentes respostas observadas para C. meneghiniana sob efeito
do exudato nas duas temperaturas. Em sua temperatura ótima de crescimento, 18
oC, seu crescimento foi estimulado pelo exudato de M. aeruginosa e a 30 ºC, quando
esta cepa foi significativamente comprometida e estaria sob estresse, este exudato a
inibiu. Também, em seu ótimo de temperatura, 30 oC, M. aeruginosa foi estimulada a
crescer por todos os exudatos, o que não ocorreu na temperatura desfavorável de
18 oC. Fato parecido se observou com o crescimento de P. agardhii , que ainda que
tenha sido quase sempre inibido, sofreu menor efeito na condição favorável de 30º C
do que na temperatura adversa de 18 oC (tab. 1).
Outra idéia que também pode justificar as diferenças nas respostas
alelopáticas de acordo com a temperatura, seria o efeito desta variável não só atuar
sobre a espécie alvo, como dito anteriormente, mas também sobre a espécie
produtora, que pode ser capaz de sintetizar compostos com bioatividades opostas
ou com biodisponibilidades diferentes de forma dependente deste fator ambiental.
Por exemplo, o exudato de M. aeruginosa estimulou o crescimento de C.
meneghiniana a 18 oC, mas o inibiu 30 oC. O mesmo foi observado para o
crescimento de P. agardhii cultivada em seu próprio exudato. Leão e colaboradores
(2009) também encontraram padrões de respostas opostas ao longo do tempo,

124
quando cultivaram Ankistrodesmus falcatus em exudato de cianobactérias. A
atividade de dois ou mais compostos bioativos não depende apenas da variação
abiótica, como encontrado nesse trabalho, mas também de variações biológicas,
como por exemplo, densidade celular. Atualmente, vem sendo discutido que a
interação alelopática entre cianobactérias deve ser mediada pelo sinergismo de uma
série de compostos bioativos e não apenas de um único aleloquímico (Leão et al.,
2010).
Embora pareça óbvio que organismos fotoautotróficos produzam compostos
bioativos que irão comprometer os processos dependentes de luz do seu
competidor, nem sempre os efeitos de inibição e estímulo do crescimento
observados nas cepas testadas nesse trabalho foram acompanhados de efeitos no
fotossistema II. Em poucos casos isto ocorreu: C. meneghiniana aumentou seu
rendimento fotossintético nos cultivos em exudato a 30 oC que estimularam seu
crescimento (todos exceto M. aeruginosa) e o diminuiu naquele que inibiu seu
crescimento, M. aeruginosa (figs. 3b e 11b). Em outros casos não houve esta
correlação. Existem também outros efeitos que podem reduzir o crescimento do
competidor que não apenas danos no sistema fotossintético, como inibição da
síntese protéica, modificação ou ativação de alguma função metabólica (Legrand et
al., 2003). No presente trabalho, essas atividades não foram testadas.
Um composto excretado pelo fitoplâncton pode também estimular o
crescimento de seu competidor por disponibilizar mais nutrientes no meio, ou como
no caso das diatomáceas, excretarem vitaminas, ou ainda por liberar compostos
bioativos ou antibacterianos e antifúngicos que irão favorecer outro organismo
(Carey & Rengefors, 2010). No entanto, a hipótese de que a maior concentração de
macro-nutrientes no material excretado presente nos exudatos estimularia o

125
crescimento não se sustenta pois no caso de C. meneghinina a 18 oC (fig. 10a), ao
contrário de todos os outros exudatos testados, o seu próprio exudato não foi capaz
de estimular seu crescimento, ainda que todos tivessem concentrações de nutrientes
iniciais semelhantes (tab. 2). Tal fato sugere a existência de outro(s) composto(s)
que favoreça o crescimento e que não está presente no meio WC (controle) nem no
seu próprio exudato.
As espécies de cianobactérias M. aeruginosa (MIJAC) e P. agardhii (PAT) são
produtoras das toxinas chamadas microcistinas. Essas cianotoxinas são conhecidas
pelos seus efeitos letais em organismos da comunidade aquática, animais
domésticos e até nos seres humanos (Carmichael, 1994; Welker & Von Dohren,
2006). Existem várias especulações sobre a função dessas toxinas, com evidências
do seu papel anti-predação, comunicação celular, interação alelopática (DeMott et
al., 1991; Carmichael, 1994; Sedmak & Kosi, 1998). No entanto, ainda não está claro
o papel desses compostos no processo alelopático. Primeiro, pelo fato das cepas
não tóxicas destas mesmas espécies também promoverem efeitos, assim como no
caso da diatomácea C. meneghiniana entre outros grupos descritos na literatura,
segundo porque quando é testada a toxina purificada a resposta muitas vezes é
inexistente ou reduzida (Suikkanen et al., 2004; 2006; Sedmak et al., 2006).
O processo alelopático no ambiente aquático é muitas vezes questionado
pelo fato do aleloquímico, quando liberado para água, sofrer imediatamente um
processo de diluição. Entretanto, Jonsson e colaboradores (2009) mostraram que as
concentrações efetivas que alcançam a célula competidora são mínimas e reais.
Além disso, alelopatia também pode ser eficaz em baixas densidades celulares
(Leão et al., 2009).

126
A intrigante questão sobre a coexistência e a substituição de espécies no
fitoplâncton, que muitas vezes é atribuída apenas às variações de fatores abióticos
como temperatura, concentração de nutrientes ou à hidrodinâmica do sistema, pode
também ser atribuída às interações alelopáticas e à influência desses fatores nessas
interações. Além disso, a alelopatia pode oferecer uma explicação para a
característica mais ofensiva do acelerado processo de eutrofização que é a
ocorrência de florações de cianobactérias. Isso pode explicar os contrastes das
respostas fitoplanctônicas, e ser aplicado em um ambiente com constante excesso
de nutrientes, como o caso da lagoa de Jacarepaguá, em que as florações variam
em composição de espécies e temporalmente.
Em um ambiente aquático, nunca vai existir um único fator a ser apontado
como determinante para a dinâmica da comunidade fitoplanctônica, mas sim um
sinergismo entre as condições abióticas, os processos biológicos e a hidrodinâmica
do sistema.

127
REFERÊNCIAS
APHA. 1998. Standard Methods for Examination of Waste and Wastewater. American Public Health Association, New York. 894p.
BAR-YOSEF, Y; SUKENIK, A.; HADAS, O.; VINER-MOZZINI, Y.; KAPLAN, A. 2010. Enslavement in the Water Body by Toxic Aphanizomenon ovalisporum, Inducing alkaline Phosphatase in phytoplanktons. Current Biology 20:1557–1561.
CAREY, C.C., RENGEFORS, K. 2010. The cyanobaterium Gloetrichia echinulatastimulates the growth of other phytoplankton. Journal of Plankton Research. 32 (9), 1349-1354.
CARMICHAEl, W. W. 1994. The toxins of cyanobacteria. Scientific American. 270(1): 78-86
DEMOTT, W.R., ZHANG, Q.X., CARMICHAEL, W.W. 1991. Effects of toxic cyanobacteria and purified toxins on the survival and feeding of a copepod and three species of Daphnia. Limnology and Oceanography. 36, 1346-1357.
DE NOBEL, W.T., MATTHIJS,H., VON ELERT, E., MUR, L.R. 1998. Comparison of light-limited growth of the nitrogen-fixing cyanobacteria Anabaena and Aphanizomenon. New Phytol. 138 (4), 579-587.
FIGUEIREDO, C.C., GIANI, A., BIRD, D. 2007. Does allelopathy contribute to Cylindrospermopsis raciborskii (Cyanobacteria) Bloom occurrence and geographic expansion? Journal of Phycology. 43, 256-265.
FISTAROL, G.O., LEGRAND, C., GRANÉLI, E. 2005. Alleopathic effect on a nutrient limited phytoplankton species. Aquatic Microbiol Ecology. 41, 153-161.
GRANÉLI, E., JOHANSSON, N. 2003. Increase in production of allelopathic substances by Prymnesium parvum cells grown under N or P deficient conditions. Harmful Algae. 2, 135-145.
GRANÉLI, E., HANSEN, P.J. 2006. Allelopathy in Harmful Agae: A mechanism to compete for resources? Pp. 189-201. In: E. Granéli, J. T. Turner (Eds). Ecology of Harmful Algae. Springer-Verlag Berlin Heidelberg, Germany.
GROSS, E.M. 2003. Alleopathy of aquatic autotrophs. Critical Reviews in Plant Science. 22, 313-339.
INTERNATIONAL ALLELOPATHY SOCIETY, 1996. Constitution. http://www-ias.uca.es/bylaws.htm#CONSTI.
ISSA, A.A.1999. Antibiotic production by the cyanobacteria Oscillatoria angustissima and Calothrix parietina. Environmental Toxicology and Pharmacology. 8, 33-37.
JONSSON, P.R., PAIVA, H., TOTH, G. 2009. Formation of harmful algal blooms cannot de explained by allelopathic interactions. PNAS. 106 (27), 11177-11182.

128
KEATING, K.1978. Blue-green algal inhibition of diatom growth: Transition from mesotrophic to eutrophic community structure. Science. 199, 971-973.
KEATING, K.1977. Allelopathic influence on blue green bloom sequence in eutrophic lake. Science. 196, 885-887.
LEÃO, P.N., PEREIRA, A.R., LIU, W.T., NG, J., PEVZNER, P.V., DORRESTEIN, P., KÖNIG, G.M., VASCONCELOS, V.M., GERWICK, W. 2010. Synergistic allelochemicals from a freshwater cyanobacterium. PNAS. 107 (25), 11183-11188.
LEÃO, P.N., VASCONCELOS, M.T.S.D., VASCONCELOS, V.M. 2009. Alleopathic activity of cyanobacteria on Green microalgae at low densities. Eur. J. Phycology. 44 (3), 347-355.
LEÃO, P.N., VASCONCELOS, M.T.S.D., VASCONCELOS, V.M. 2009. Allelopathy in freshwater cyanobacteria. Critical Reviews in Microbiology. 35 (4), 271-182.
LEFLAIVE, J., TEN-HAGE, L. 2007. Algal and cyanobacterial secondary metabolites in freshwaters: a comparison of allelopathic compounds and toxins. Freshwater Biology. 52, 199-214.
LEGRAND, K, RENGEFORS, K., FISTAROL, G.O., GRANÉLI, E. 2003. Allelopathy in phytoplankton – biochemical, ecological and evolutionary aspects. Phycologia. 42 (4), 406-419.
LÜRLING, M.; BEEKMAN, W. 1999. Grazer induced defenses in Scenedesmus (Chlorococcales; Chlorophyceae): coenobium and spine formation. Phycologia. 38, 368-376.
MOLISCH, H. 1937. Der Einfluss einer Pflanze auf die andere: Allelopathie. Ficher Verlag, Jena.
RAY, S., BAGCHI, S.N. 2001. Nutrients and pH regulate algicide accumulation in culture of the cyanobacterium Oscillatoria laetevirens. News Phytologist. 149, 455-460.
RICE, E.L. 1984. Alellopathy. Academics Press, New York.
REIGOSA, M.J., SANCHEZ-MOREIRAS, A., GONZALES, L. 1999. Ecophysiological approach in allelopathy. Critical Reviews in Plant Sciences. 18, 577-698.
RENGEFORS, K., LEGRAND, C. 2001. Toxicity in Peridinium aciculiferum – an adaptive strategy to outcompete other winter phytoplankton? Limnology and Oceanography. 46 (8), 1990-1997.
SEDMAK, B., KOSI, G. 1998. The role of microcystins in heavy cianobacterial bloom formation. Journal of Plankton Research. 20, 691-708.

129
SEDMAK, B, ELERSEK T. 2006. Microcystins induce morphological and physiological changes in selected representative phytoplanktons. Microbiol Ecol 51:508-515.
SUIKKANEN, S. 2008. Allelopathic effects of filamentous cyanobacteria on phytoplankton in the Baltic sea. Academic dissertation of the Faculty of Biosciences of the University of Helsinki, Finland. 41p.
SUIKKANEN, S., ENGSTRÖM-OST, J., JOKELA, J., SIVONEN, K., VIITASALO, M. 2006. Allelopathy of Baltic Sea cyanobacteria: no evidence for the role of nodularin. Journal of Plankton Research. 28 (6), 543-550.
SUIKKANEN, S., FISTAROL, G.O., GRANÉLI, E. 2005. Effects of cyanobacterial allelochemicals on a natural plankton community. Marine Ecology Progress Series. 287, 1-9.
SUIKKANEN, S., FISTAROL, G.O., GRANÉLI, E. 2004. Allelopathic effects of Baltic cyanobacteria Nodularia spumigena, Aphanizomenon flos-aquae and Anabaena lemmermannii on algal monocultures. Journal of Experimental marine Biology and Ecology. 308, 85-101.
VAN DER GRINTEN, E., JANSSEN, A., DE MUTSERT, K., BARRANGUET, C., ADMIRAAL, W. 2005. Temperature and light dependent performance of the cyanobacterium Leptolyngbya foveolarum and the diatom Nitzchia perminuta in mixed biofilms. Hydrobiologia. 548, 267-278.
VON-ELERT, E, JÜTTNER, F. 1997. Phosphorus limitation and not light controls the extracellular release of allelopathic compounds by Trichormus doliolum(Cyanobacteria). Limnology and Oceanography. 42 (8), 1796-1802.
VON-ELERT, E, JÜTTNER, F. 1996. Factors influencing the allelopathic activity of planktonic cyanobacterium Trichormus doliolum. Phycology. 35,68-73.
WELKER, M., VON DOHREN, H. 2006 Cyanobacterial peptides - Nature’s own combinatorial biosynthesis. FEMS Microbiol Rev 30:530–563.

130
5. DISCUSSÃO GERAL
A lagoa de Jacarepaguá vem sofrendo um acelerado processo de
eutrofização artificial a mais de duas décadas (Saieg-Filho, 1986). Estudos
anteriores mostraram mudanças na comunidade fitoplanctônica com perda da
dominância das clorofíceas e surgimento de florações de cianobactérias desde 1990
(Fernandes, 1993, Gomes et al., 2009), possivelmente, provocadas pela elevada
concentração de nutrientes e forte estabilidade da coluna d'água (Domingos, 2001).
Entretanto, nos meses de inverno, ocorre a redução da dominância das
cianobactérias, juntamente com o surgimento de um pico de maior diversidade da
comunidade (Gomes et al, 2009).
No período estudado, as cianobactérias dominaram a comunidade de
fitoplâncton na maior parte do tempo. Elevadas biomassas foram observadas
principalmente no período I (fev-abr/07), mas também no período III (ago/07 –
jan/08), sendo Aphanizomenon sp., M. aeruginosa e Planktothrix sp. as principais
espécies. Estes períodos foram caracterizados por altas concentrações de PSR,
baixas concentrações de NID e elevadas temperaturas. Os elevados valores de pH,
OD e menores Zsecchi estiveram muito mais associados às conseqüências dessas
florações do que às causas. No período II, o colapso da comunidade fitoplanctônica
promoveu uma maior disponibilidade de NID, especialmente na forma de amônia, o
que permitiu que outras espécies pudessem ocupar esse nicho. Cyclotella sp foi a
espécie que aumentou sua contribuição relativa nesse período, no momento de mais
baixas temperaturas. Segundo alguns autores, as diatomáceas são melhores
competidoras em reduzidas temperaturas (Zhang & Prepas, 1996; Elliott et al., 2006;
Paerl & Husman, 2009). Além disso, o fato delas terem uma taxa de absorção
máxima de nutrientes mais elevada do que outros grupos, como por exemplo, das

131
clorofíceas, torna-as especialistas mais efetivas na utilização dos pulsos de
nutrientes (Litchman et al., 2006). Quando a temperatura voltou a aumentar, no
início do período seguinte (PIII), a espécie S. aquatilis dominou o sistema,
possivelmente favorecida pela sua alta taxa de crescimento em ambientes
enriquecidos (Reynolds 1997, 2006).
Uma vez que as concentrações de PSR foram elevadas durante todo o ano,
as variações temporais das concentrações de NID e da temperatura da água
pareceram ter sido os fatores mais relevantes para justificar a dinâmica do
fitoplâncton, separando os períodos de maiores biomassas e a substituição das
espécies de maior representatividade na comunidade.
Quando a redução da concentração de nutrientes na comunidade
fitoplanctônica da lagoa de Jacarepaguá foi testada, através de experimentos em
laboratório, observou-se que, mesmo nas condições mais empobrecidas, a
dominância das cianobactérias permaneceu. Embora a biomassa total tenha
diminuído em cultivos em que a água da lagoa de Jacarepaguá foi diluída com a do
rio Camorim, a composição da comunidade fitoplanctônica foi similar em todos os
tratamentos. O sedimento da lagoa de Jacarepaguá pode atuar como uma fonte de
P para o sistema, contribuindo para a contínua ocorrência de elevadas biomassas
de cianobactérias. De fato este grupo é conhecido por levar vantagem em ambientes
com altas concentrações de P (Schindler, 1977; Trimbee & Prepas, 1987; Seip,
1988; Watson et al., 1997).
Tanto a observação do estudo de campo quanto a experimentação em
laboratório mostraram que o NID é facilmente incorporado pelo fitoplâncton e o seu
esgotamento favorece a dominância da espécie fixadora de N atmosférico,
Aphanizomenon sp. Mesmo quando a composição fitoplanctônica está dividida entre

132
diatomáceas e cianobactérias, espécies deste último grupo, como M. aeruginosa, P.
agardhii e Aphanizomenon sp. mostram vantagens adaptativas e acabam
dominando a comunidade. Estas espécies têm recebido particular atenção,
justamente por dominar o plâncton de ambientes aquáticos eutrofizados (Reynolds
1984; Smith, 1986; Trimbee & Prepas, 1987; Scheffer et al., 1997). M. aeruginosa
está entre os gêneros que ocorrem mais comumente nos sistemas aquáticos
brasileiros (Huszar & Silva, 1999) e se destaca pela sua capacidade de controlar sua
flutuabilidade na coluna d'água (Pettersson et al., 1993; Reynolds 1987, 2006; Paerl,
2006) e evitar herbivoria, pela produção de toxinas (Ferrão-Filho et al., 2002).
Planktothrix, além de ser um gênero produtor de cianotoxinas, assim como
Aphanizomenon (Carmichael, 2001; Codd et al., 2005), também leva vantagem em
ambientes com baixa penetração de luz (Scheffer et al., 1997; Padisak, 2004;
Legnani et al., 2005). O gênero Aphanizomenon, por formar células especializadas
em fixar N atmosférico (heterocitos), é citado como dominante em ambientes com
baixa disponibilidade de nitrogênio (Jensen et al., 2004).
Compreender como os fatores ambientais estariam influenciando na
substituição das espécies fitoplanctônicas da lagoa de Jacarepaguá foi um dos
principais objetivos desta tese. Uma vez que a redução dos nutrientes não foi
suficiente para diminuir a dominância das cianobactérias, o efeito da temperatura no
crescimento e competição entre as espécies de maior contribuição relativa foi
testado. M. aeruginosa não sofreu nenhum efeito da temperatura no seu
crescimento, embora a literatura descreva que seu crescimento ótimo esteja entre
25 e 30 oC (Jöhnk et al., 2008; Paerl & Husman, 2009). Esta espécie também foi a
melhor competidora nos cultivos mistos com as demais espécies testadas (P.
agardhii e C. meneghiniana), tanto a 18 oC quanto a 30 oC. Esse sucesso pode ser

133
atribuído ao seu menor requerimento por nutrientes (Padisak, 2004). Em
competição, M. aeruginosa apresentou uma taxa de crescimento maior a 30 oC do
que a 18 oC. Segundo Fujimoto e colaboradores (1997) esta espécie tem menor
quota celular de N e P em temperaturas mais elevadas, permitindo melhor
aproveitamento desses nutrientes para transformá-los em biomassa.
Os processos alelopáticos também podem ajudar a justificar os resultados de
competição, principalmente entre as cepas de M. aeruginosa e P. agardhii. Tanto a
18 quanto a 30 oC, P. agardhii foi inibida pela presença de M. aeruginosa e o mesmo
efeito foi observado quando a primeira foi crescida em exudato da última.
P. agardhii teve maior taxa de crescimento a 27 oC e foi uma competidora
superior a C. meneghiniana em todas as temperaturas. A temperatura ótima para o
crescimento desta última foi 21 oC. Quando observamos sua distribuição no
ambiente natural, as diatomáceas dominam a comunidade fitoplanctônica nos
períodos de mais baixos valores de temperatura da água (Zhang & Prepas, 1996;
Gomes et al., 2009). Isso pode ajudar a explicar o aumento da sua biomassa quando
a temperatura da água da lagoa de Jacarepaguá diminuiu. Planktothrix sp. também
aumentou consideravelmente sua biomassa, dominando a comunidade por alguns
meses de inverno (jun-jul/07).
As interações alelopáticas aliadas à influência da temperatura na eficiência
destas interações também ajudaram a entender o padrão de substituição de
espécies que ocorreu na lagoa de Jacarepaguá. A espécie antecessora foi capaz de
estimular o crescimento de sua sucessora quando esta última esteve em sua
temperatura ótima de crescimento. Porém, quando espécie antecessora estava em
sua temperatura favorável, inibiu o crescimento da sua sucessora. A temperatura de
30 oC favoreceu M. aeruginosa e seu exudato inibiu o crescimento de C.

134
meneghiniana, cujo crescimento ótimo ocorre a 18 oC. O exudato de C.
meneghiniana a 18 oC não foi capaz de promover nenhum efeito no crescimento da
M. aeruginosa, mas a 30 oC o estimulou. Nas temperaturas elevadas (PI e PIII),
embora as espécies M. aeruginosa e Planktothrix sp. coexistissem na lagoa de
Jacarepaguá, suas biomassas se alternavam. Nos experimentos de alelopatia a 30
oC, o exudato de M. aeruginosa inibiu o crescimento de P. agardhii enquanto o desta
última estimulou o crescimento de M. aeruginosa nesta mesma temperatura. Porém
os resultados destes experimentos não podem ser diretamente transpostos para a
realidade do ambiente natural. P. agardhii foi sempre inibida pelo exudato de suas
competidoras a 18 oC, no entanto, nas observações de estudo de campo, essa
espécie aumentou a sua biomassa no período de mais baixas temperaturas. Por fim,
as interações alelopáticas, aliadas aos tradicionais fatores bióticos e abióticos,
ajudaram a interpretar o padrão de substituição das espécies fitoplanctônicas da
lagoa.
Em síntese, é difícil apontar um único fator que esteja determinando a
ocorrência de cianobactérias formadoras de florações na lagoa de Jacarepaguá. É o
sinergismo entre fatores ambientais e biológicos, aliado à própria fisiologia das
espécies presentes, que irão direcionar a dinâmica da comunidade fitoplanctônica de
um ecossistema.

135
6. CONCLUSÕES
1. A Lagoa de Jacarepaguá, apesar de ser hipereutrófica e apresentar
constantes florações de cianobactérias, ainda é um sistema muito dinâmico,
com elevada variabilidade temporal;
2. O regime temporal da disponibilidade de N na coluna d’água parece ter um
efeito significativo na dinâmica das espécies dominantes e nas tendências
das competições na lagoa de Jacarepaguá;
3. Embora a dinâmica dos nutrientes, assim como a disponibilidade de luz e as
elevadas temperaturas possam ter favorecido a dominância de
cianobactérias, as temperaturas mais baixas nos meses de inverno parecem
ter reduzido a biomassa das principais espécies;
4. A redução das concentrações de nutrientes na água da lagoa parece reduzir
a biomassa fitoplanctônica, embora a dominância das cianobactérias tenha se
mantido;
5. A temperatura influenciou na fisiologia de crescimento das espécies
fitoplanctônicas e também afetou os resultados de competição indireta
(utilização de recursos) nos experimentos em laboratório. M. aeruginosa foi
uma competidora superior às demais espécies testadas, tanto em alta quanto
em baixa temperatura;

136
6. A temperatura também pode influenciar nas interações competitivas diretas
(compostos alelopáticos), devido a sua capacidade de alterar a
vulnerabilidade de uma espécie a um composto alelopático ou a sua
capacidade de expressá-lo;
7. É difícil apontar um fator que esteja determinando a ocorrência de
cianobactérias formadoras de florações na Lagoa de Jacarepaguá. Um
sinergismo entre fatores ambientais (físicos e químicos) e biológicos
(competição direta e indireta), aliado à própria fisiologia das espécies
presentes, direciona a dinâmica da comunidade fitoplanctônica nesse
sistema.

137
7. REFERENCIAS BIBLIOGRÁFICAS
AZEVEDO, S.M.F.O. 1998. Toxinas de Cianobactérias: causas e conseqüências para saúde pública. Medicina on line 1(3): 1-22.
AZEVEDO, S.M.F.O., CARMICHAEL, W.W., JOCHINSEN, E.M., RINEHART, K.L., LAU, S., SHAW, G.R., EAGLESHAM, G.K. 2002. Human intoxication by microcystins during renal dialysis treatment in Caruaru- Brazil. Toxicology (181-182): 441-446.
BACHMANN, R. W. 2001. The Limiting Factor Concept . What Stops Growth? Spring Lakeline 26-28 p.
BRANCO, C.W.C.; SENNA, P.A.C. 1994. Factors influencing the development of Cylindrospermopsis raciborskii and Microscystis aeruginosa in the Paranoá Reservoir, Brasília, Brazil. Algological Studies, v. 75, p. 85-96.
BLOMQVIST, P.; PETTERSSON A. & HYENSTRAND, P. 1994. Ammonium-nitrogen: a key regulatory factor causing dominance of non-nitrogen-fixing Cianobacteria in aquatic systems. Arch. Hydrobiol. 132: 141-164.
BOUVY, M. et al. 2000. Occurrence of Cylindrospermopsis (Cyanobacteria) in 39 Brazilian tropical reservoirs during the 1998 drought. Aquatic Microbial Ecology, v. 23, p. 13-27.
CARACO, N. & MILLER, R. 1998. Direct and indirect effects of CO2 on competition between a cyanobacteria and eukaryotic phytoplankton. Can. J. Fish. Aquat. Sci. 55:54-62.
CARMICHAEL, W.W.; MAHMOOD, N.A.; HYDE, E.G. 1990. Natural Toxins from Cyanobacteria (blue-green algae). In Marine Toxins: Origin, Structure, and Molecular Pharmacology, eds. Hall, S. � Strichartz, G.. Washington, DC: American Chemical Society. 87-106.
CARMICHAEl, W. W. 1994. The toxins of cyanobacteria. Scientific America.270(1): 78-86.
CARMICHAEL, W.W. 2001a. Health Effects of Toxin-Producing Cyanobacteria: The CyanoHABs. Human and Ecological Risk Assessment, 7(5): 1393-1407.
CARMICHAEL, W.W.; AZEVEDO, S.M.F.O.; AN, J.S.; MOLICA, R.J.R.; JOCHIMSEN, E.M.; LAU, S.; RINEHART, K.I.; SHAW, G.R.; EAGLESHAM, G.K. (2001b). Human fatalities from cyanobacteria: chemical and biological evidence for cyanotoxins. Journal of Environmental Science and Health Part A - Environmental, v. 109, n. 7, p. 663-668.
CHIANG, I.Z.; WUANG, W.-Y.; WU, J.-T. 2004. Allelochemicals of Botryococcus braunii (Chlorophyceae). Journal of Phycology, v. 40, p. 474-480.

138
CHORUS I., BARTRAM J. 1999. Toxic Cyanobacteria in water: A guide to the Public Health Consequences, Monitoring and Management. E and FN Spon, London. p.416.
COOD, G.A.; MORRISON, L. & METCALF, J. 2005. Cyanobacterial toxins: risk management for health protection. Toxicology and Applied Pharmacology (203): 264-272.
COLE, J. F.; JONES, C. 2000. Effect of temperature on photosynthesis-light response and growth of four phytoplankton species isolated from a tidal freshwater river. Journal of Phycology, v. 36, p. 7-16.
DAVIDSON, I. R. 1991. Environmental effects on algal photosynthesis: Temperature. Journal of Phycology, v. 27, p. 2-8.
DOMINGOS, P. 2001. Dinâmica de Cianobactérias produtoras de microcistinas na Lagoa de Jacarepaguá (RJ). Centro de Ciências da Saúde, UFRJ. 107p. (Tese de Doutorado em Biotecnologia Vegetal).
ELSER, J. J. & GOLDMAN, C.R. 1991. Zooplankton effects on phytoplankton in lakes of contrasting trophic status. Limnol. Oceanog., 36: 64-90.
ELSER, J. J., MARZOLF, E. R. & GOLDMAN, C. R. 1990. Phosphorus and nitrogen limitation of phytoplankton growth in the freshwaters of North America: a review and critique of experimental enrichments. Can. J. Fish. Aquat. Sci., 47: 1468-1477.
ESTEVES, F.A. 1988. Fundamentos de limnologia. Interciência/ FINEP. Rio de Janeiro. 575p.
FALCONER, I.R. 1994. Health implications of Cyanobacterial (blue-green algae) toxins. In: Toxic Cyanobacteria current status of research and management. Eds. STEFFENSEN, D.A. & Nicholson, B.C. Proceedings of an International Workshop. Adelaide. Australia.
FEEMA – Fundação Estadual de Engenharia de Meio Ambiente. 1982. Estudo da Barra da Tijuca – Fase I: Lagoa de Jacarepaguá. 37p.
FEEMA – Fundação Estadual de Engenharia de Meio Ambiente. 1984. Quadro da situação ambiental da Baixada de Jacarepaguá. Rio de Janeiro. DIPLAN. 107p.
FEEMA – Fundação Estadual de Engenharia de Meio Ambiente. 1991. Qualidade das águas do Estado do Rio de Janeiro. Período 1987-1989. Rio de Janeiro.
FERNANDES, V. O. 1993. Estudos sucessionais da comunidade perifítica em folhas de Typha dominguensis Pers, relacionado às variáveis limnológicas na Lagoa de Jacarepaguá, RJ. Tese de Mestrado, UFSCar, SP. 131p.

139
FERRÃO-FILHO, A.D., KOZLOWSKY-SUZUKI, B., & AZEVEDO, S.M.F.O. 2002a. Accumulation of microcystins by a tropical zooplankton community. Aquatic Toxicology, 59, 201-208.
FERRÃO-FILHO, A.S., DOMINGOS, P., & AZEVEDO, S.M.F.O. 2002b. Influences of a Microcystis aeruginosa Kutzing bloom on zooplankton populations in Jacarepagua Lagoon (Rio de Janeiro, Brazil). Limnologica, 32, 295-308.
FIGUEREDO, C. C.; GIANI, A.; BIRD, D. 2007. Does allelopathy contribute to Cylindrospermopsis raciborskii bloom occurrence and geographic expansion? Jounal of Phycology, v. 43, p. 256-265.
FISTAROL, G.O. et al. 2004. Allelopathy in Alexandrium spp.: effect on a natural plankton community and on algal monocultures. Aquatic Microbial Ecology, v. 35, p. 45-56.
FISTAROL, G.O.; LEGRAND, C.; GRANÉLI, E. 2003. Allelopathic effect of Prymnesium parvum on a natural plankton community. Marine Ecology Progress Series, v. 255, p.115-125.
GANF, G.G. 1974. Diurnal mixing and the vertical distribution of phytoplankton in a shallow equatorial lake (Lake George) Uganda. J. Ecol. 62: 611-629.
GONZÁLEZ, E, J. 2000. Nutrient enrichment and zooplankton effects on phytoplankton community in microcosns from El Andino reservoir (Venezuela). Hydrobiol., 434: 81-96.
GONZÁLEZ, E, J. & Ortaz, M. 1998. Efectos del enriquecimiento con N y P sobre La comunidad del fitoplancton en microcosmos de un embalse tropical (La Mariposa, Venezuela). Verh. Int. Ver. Limnol., 46: 27-34.
GRANÉLI, E.; JOHANSSON, N. 2003. Increase in the production of allelopathic substances by Prymnesium parvum cells grown under N- or P-deficient conditions. Harmful Algae, v. 2, p. 145.
KJERFVE, B. 1994. Coastal lagoons. Pp. 1-8. In: B. Kjerfve, (ed). Coastal lagoon processes. Elsevier Oceanography Series, Amsterdam. 599p.
KNOPPERS, B. 1994. Aquatic primary production in coastal lagoons. Pp. 243-286. In: B. Kjerfve. (ed). Coastal lagoon processes. Elsevier Oceanography Series, Amsterdam. 599p.
HANEY, J. F. 1987. Field studies on zooplankton-Cyanobacteria interactions. N.Z.J. Mar. freshwat. Res. 21: 467-475.
HENRY, R. & TUNDISI, J.G. 1982a. Efeitos de Enriquecimento artificial por nitrato e fosfato no crescimento da comunidade fitoplanctônica da Represa do Lobo (“Broa”, Brotas – Itirapina, SP). Ciência e Cultura, 34(5): 518-524.

140
HU, Z., LIU, Y., LI, D. 2004. Physiological and Biochemical Analyses of Microcystin-RR Toxicity to the Cyanobacterium Synechococcus elongates. Wiley Periodicals, Inc. 571-577.
HUSZAR, V. L. M, SILVA, L.H.S., MARINHO, M., DOMINGOS, P., SANT'ANNA, C.L. 2000. Cyanoprokaryote assemblages in eight productive tropical Brazilian waters. Hydrobiologia, v. 424, p. 67:77.
JENSEN, J.P.; JEPPESEN, E.; OLRIK, K. & KRISTENSEN, P. 1994. Impact of nutrientes and physical factors on the shift from cyanobacterial to chlorophyte.Can. J. Fish. Aquati Sci., 51: 1692-1699.
JOCHIMSEN, E.M.; CARMICHAEL, W.W.; AN, J.; CARDO, D.; COOKSON, S.T.; HOLMES, C.E.M.; ANTUNES, M.B.C.; MELO FILHO, D.A.; LYRA, T.M.; BARRETO, V.; AZEVEDO, S.M.F.O. & JARVIS, W.R. 1998. Liver failure and death following exposure to microcystin toxins at a hemodialysis center in Brazil. The New England Journal of Medicine. 36: 373-378.
KEARNS, K. D. & HUNTER, M. 2000. Green algal extracellular products regulate antialgal toxin production in a cyanobacterium. Environmental Microbiology. 2(3): 291-297.
KEATING, K.I. 1978. Blue green algal inhibition of diatom growth: Transition from mesotrophic to eutrophic community structure. Science. (199) 971-973.
KING, D.L. 1970. The role of carbon in eutrophication. J. Wat. Pollut.42: 2035-2051.
KROMKAMP, K. 1987. Formation and functional significance of storage products in cyanobacteria. New Zeal. J. Mar. and Fresh. Res 21:457-465.
LAMPERT, W.; SOMMER, U. 1997. Limnoecology: The Ecology of Lakes and Streams.Oxford: Oxford University Press,. 400 p.
LEGRAND, K, RENGEFORS, K., FISTAROL, G.O., GRANÉLI, E. 2003. Allelopathy in phytoplankton – biochemical, ecological and evolutionary aspects. Phycologia, v. 42, p. 406-419.
LÜRLING, M.; VAN DONK, E. 1999. Grazer-induced colony formation in Scenedesmus acutus (Chlorophyceae): ecomorph expression at different temperatatures. Journal of Phycology, v. 35, p. 1120-1126.
MAGALHÃES, V.F.; SOARES, R.M. & AZEVEDO, S.M.F.O. 2001. Microcystin contamination in fish from the Jacarepaguá (RJ, Brazil): ecological implication and human health risk. Toxicon, 39: 1077-1085.
MARINHO, M. M.; HUSZAR, V. L. M. 2002. Nitrogen availability and physical conditions as controlling factors of phytoplankton composition and biomass in a tropical reservoir (Southern Brasil). Archiv für Hydrobiologie, v. 153, p. 443-468.

141
Mc QUEEN, D.J. & LEAN, D.R.S. 1987. Influence of water temperature and nitrogen to phosphorus rations on the dominance of blue-green algae in lake St. George, Ontario. Can. J. aquat. Sci. 44: 598-604.
MONTAGNES, D. J. S.; FRANKLIN, D. J. 2001. Effect of temperature on diatom volume, growth rate, and carbon and nitrogen content: Reconsidering some paradigms. Limnology and Oceanography, v. 46, p. 2008-18.
OLIVER, R. L.; GANF, G. G. 2000. Freshwater Blooms. In: B. A., WHITTON; M. POTTS (eds.). The ecology of Cyanobacteria. The diversity in time and space. Dordrecht: Kluwer, pp. 149-194.
PADISÁK 2004. Phytoplankton In: O’Sullivan, P.E. & Reynolds,C.S. (ed). Limnology and Ecology. Blackwell., PP:251-308 (vol 1)
PAERL, H.W. 2006. Nutrient and other environmental controls of harmful cyanobacterial blooms along the freshwater-marine continuum. In: H.K. Hudnell (ed): Proceedings of the Interagency, International Symposium on Cyanobacterial Harmful Algal Blooms. Advances in Experimental Medicine & Biology.
PANOSSO, R.F.; ATTAYDE, J.L.; MUEHE, D. 1998. Morfometria das lagoas Imboassica, Cabiúnas, Comprida e Carapebus: Implicações para seu funcionamento e manejo. Pp. 91-108 In: F.A Esteves, (Ed.) Ecologia das Lagos Costeiras do Parque Nacional da Restinga de Jurubatiba e do Município de Macaé (RJ). Universidade Federal do rio de janeiro. 464p.
PETTERSSON, K., HERLITZ, E. & ISTVANOVICS, V. 1993. The role of Gloeotrichia echinulata in the transfer of phosphorus from sediments to water in lake Erken. Hydrobiologia, 25: 123-129.
RAVEN, J. A.; GEIDER, R. J. 1988. Temperature and algal growth. New Phytologist, v. 110,p. 441-61.
REYNOLDS, C.S. 2006. Ecology of Phytoplankton. Cambridge. 535p.
REYNOLDS, C.S. 1987. Cyanobacterial water-blooms. In: J. Callow (ed). Advances in botanical research. Academic Press., London. PP:67-143 (vol 13).
ROBARTS, R. D.; ZOHARY, T. 1987 Temperature effects on photosynthetic capacity, respiration, and growth rates of bloom-forming cyanobacteria. New Zealand Journal of Marine and Freshwater Research, v. 21, p. 391-399
SAIEG-FILHO, E. 1986. Ecologia do Fitoplâncton Marginal das Lagunas da Baixada de Jacarepaguá, Rio de Janeiro- RJ. Instituto de Biologia, UERJ. 150p. (Monografia de Bacharelado).
SEDMAK, B. & KOSI, G. 1998. The role of microcystins in heavy cyanobacterial bloom formation. Journal of Plankton Research. 20 (4):691-708.

142
SEIP, K. L. 1988. A system for identifying lakes response to reductions in phosphorus loading. Part II. Ocurrence of bluegreen algae (cyanobacteria) (Etregelsystem for a identifisere innsjoers rspons pa redusjoner fosforbelastning. Del II. Forekormst av blagronnlger(cianobacteria)).Limnos.,3:8-12.
SEMADS. Secretaria Estadual de Meio Ambiente e Desenvolvimento Sustentável. Ambiente das águas do estado do Rio de Janeiro. Rio de Janeiro: Sholna Reproduções gráficas, 203p.
SEMERARO, J.; COSTA, A.F. 1972. O Plâncton e a Poluição nas lagoas da Tijuca, Camorim e Jacarepaguá. XII Congresso Interamericano de Engenharia Sanitária, Assunção, Paraguai. Tema 4, Publ. no.3:31p.
SHAPIRO, J. 1990. Currents beliefs regarding dominance by blue-greens: the case of the importance of CO2 and pH. Verh. Int. ver. Limnol. 24: 38-54.
SILVA, G. 2006. Processo de Ocupação Urbana da Barra da Tijuca (RJ): Problemas Ambientais, Conflitos Sócio-Ambientais, Impactos Ambientais Urbanos. Pesquisa em arquitetura e Construção. 28p.
SMITH, V. 1983. Low nitrogen to phosphorous rations favor dominance by blue-green algae in lake phytoplankton. Science 221: 669-671.
SMITH, V. 1986. Light and nutrient effects on the relative biomass of blue-green algae in lake phytoplankton. Can. J. Fish. Aquat. 43: 148-153.
SMAC - Secretaria Municipal de Meio Ambiente. (2002) Qualidade Ambiental – primavera, Verão, Outono. Rio de Janeiro.
SOARES, M.C. 2008. Cianobactérias Tóxicas no Reservatório do Funil: Uma Batalha Pelágica.Tese de Doutorado em Botânica. Universidade Federal do rio de Janeiro.419p.
SOMMER, U. 1985. Seasonal succession of phytoplankton in Lake Constance. BioScience,v.5, p. 351-357,
SPAULDING, E. 1994. Modeling of circulation and dispersion in coastal lagoons. Pp 69-101 In: B. Kjerfve, (Ed). Coastal lagoon processes. Elsevier Oceanography Series, Amsterdam. 577p.
STRANG, H.E.; CASTELLANOS, A.; CHAVES, C.M.; ATALA, F.; MARTINS, H.F.; COIMBRA-F, A.F.; SOARES, I.L.; CAMPOS, T.T. 1965. A restinga de Jacarepaguá. Bol. Geogr. 187.
SUIKKANEN, S., FISTAROL, G.O., GRANÉLI, E. 2005. Effects of cyanobacterial allelochemicals on a natural plankton community. Marine Ecology Progress Series. 287, 1-9.

143
SUIKKANEN, S.; FISTAROL, G. O.; GRANÉLI, E. 2004. Allelopathic effects of the Baltic cyanobacteria Nodularia spumigena, Aphanizomenon flos-aquae andAnabaena lemmermannii on algal monocultures. Journal of Experimental Marine Biology and Ecology, v. 308, p. 85-101.
TEIXEIRA, M.G.L.C.; Costa, M.C.N.; Carvalho, V.L.P.; Pereira, M.S.; Hage,;. 1993. Epidemia de gastroenterite na área da barragem de Itaparica, Bahia. Bol. of Sanit. Panam. 114 (6): 502-511.
TRIMBEE, A.M. & PREPAS, E.E. 1987. Evaluation of total phosphorus as a predictor of the relative biomass of blue-green algae with emphasis on Alberta lakes. Can. Fish. Aquat. Sci.44: 1337-1342.
VOLLENWEIDER, R & KEREKES, J. 1980. The loading concept as basis for controlling eutrophication philosophy and preliminary results of the OECD programme on eutrophication. Prog. Wat. Tech. 12: 5-38.
WATSON, S.B.; MC CAULEY, E. & DOWNING, J.A. 1997. Patterns in phytoplankton taxonomic composition across temperate lakes of differing nutrient status. Limnol. Oceanogr. 42: 487-495.
YOO, R. S.; CARMICHAEL, W.W.; HOEHN, R.C.; HRUDEY, S.C. 1995. Cyanobacterial (blue-green algal). Toxins: A Resource Guide. AWWA Research Foudation and American Water Works Association, USA: 1-229.
ZEE 1994. Estudo ambiental: Poluição dos Recursos hídricos da baixada de Jacarepaguá. Relatório Técnico. Departamento de Oceanografia, UERJ, 83p.
ZEVENBOOM, W. & MUR, L.R. 1980. N2-fixing cyanobacteria: Why they do not become dominat in Dutch, hypertrophic lakes.Develop. Hydrobiol. 2: 123-130.

APÊNDICE

329FLORAÇÕES DE CIANOBACTÉRIAS TÓXICAS EM UMA LAGOA COSTEIRA HIPEREUTRÓFICA
Oecol. Bras., 13(2): 329-345, 2009
FLORAÇÕES DE CIANOBACTÉRIAS TÓXICAS EM UMA LAGOA COSTEIRA HIPEREUTRÓFICA DO RIO DE JANEIRO/RJ (BRASIL) E SUAS CONSEQUÊNCIAS
PARA SAÚDE HUMANA
Andreia Maria da Anunciação Gomes1*, Pedro Lima Sampaio1, Aloysio da Silva Ferrão-Filho2, Valeria de Freitas Magalhães1, Marcelo Manzi Marinho3, Ana Cláudia Pimentel de Oliveira4, Vitor Barbosa dos Santos1,4, Patrícia Domingos4 & Sandra Maria Feliciano de Oliveira e Azevedo1
1 Laboratório de Ecofisiologia e Toxicologia de Cianobactérias, Instituto de Biofísica Carlos Chagas Filho, Centro de Ciências da Saúde, Universidade Federal do Rio de Janeiro, Ilha do Fundão. CEP 21949-900. Rio de Janeiro, RJ, Brasil.2 Laboratório de Avaliação e Promoção da Saúde Ambiental, Departamento de Biologia – Instituto Oswaldo Cruz, FIOCRUZ, Av. Brasil 4365, Manguin-hos. CEP 21045-900. Rio de Janeiro, RJ, Brasil. 3 Laboratório de Taxonomia e Ecologia de Algas, Departamento de Biologia Vegetal – Instituto de Biologia Roberto Alcântara Gomes, Universidade Estadual do Rio de Janeiro, Rua São Francisco Xavier N°524 – PHLC Sala 511, Maracanã. CEP 20550-900. Rio de Janeiro, RJ, Brasil. 4 Fundação Estadual de Engenharia do Meio Ambiente, Divisão de Laboratórios, Laboratório de Biologia, Avenida Salvador Allende, 5500, Recreio dos Bandeirantes. CEP: 22780-160. Rio de Janeiro, RJ, Brasil.*Email: [email protected]
RESUMOA baixada de Jacarepaguá (RJ/Brasil) vem sofrendo há pelo menos quatro décadas um processo desordenado
de ocupação urbana e industrial, o que tem se refletido em mudança na qualidade da água e na incidência de florações de cianobactérias no Complexo Lagunar de Jacarepaguá (CLJ). O presente estudo reuniu uma série de dados temporais objetivando avaliar a qualidade hídrica da Lagoa de Jacarepaguá (LJ) através do monitoramento físico (pH, transparência da água e temperatura), químico (salinidade, O.D. e clorofila a) e da comunidade planctônica. Foi também analisado a concentração de microcistinas no pescado comercializado na região. Os dados foram divididos em três períodos de acordo com os diferentes ciclos sazonais estudados: PI (ago/96 a set/97), PII (jan/99 a nov/99) e PIII (fev/07 a jan/08). Dominância de cianobactérias ocorreu na maior parte do tempo, intercalada por pulsos de maior diversidade fitoplanctônica, observados a partir de 1999 (PII e III). Esta diversidade foi representada pelas clorofíceas, diatomáceas, prasinoficeas e criptoficeas. Foi interessante notar que a dinâmica observada em 1999 tenha se repetido quase 10 anos depois, o que sugere uma tendência sazonal de estruturação da comunidade no CLJ. O desaparecimento da floração coincidiu com épocas de menores temperaturas, sugerindo ser esse um importante fator regulador da dinâmica observada. A LJ apresentou uma elevada abundância zooplanctônica, com baixa diversidade de espécies. Durante alguns meses, as concentrações de microcistinas encontradas nos músculos dos peixes estiveram acima dos valores recomendados pela OMS para consumo humano. Palavras-chave: Lagoa costeira, eutrofização, cianobactérias, microcistinas.
ABSTRACTTOXIC CYANOBACTERIAL BLOOMS IN AN EUTROPHICATED COASTAL LAGOON IN RIO
DE JANEIRO, BRAZIL: EFFECTS ON HUMAN HEALTH. The Jacarepaguá Basin, Rio de Janeiro, Brazil has over the last four decades undergone a disordered process of urban and industrial growth with direct consequences on water quality and incidence of cyanobacterial blooms in the Jacarepaguá Lagoon Complex (JLC). The present study combines data aiming to evaluate water quality in Jacarepaguá Lagoon (JL) from monitoring of physical variables (pH, water transparency, temperature), chemical variables (salinity, dissolved oxygen, chlorophyll), the planktonic community, and the concentration of microcystins in fish from the region. The data herein presented were divided into three periods according to seasonal cycles: PI (aug/96 to sep/97), PII (jan/99 to nov/99), and PIII (feb/07 to jan/08). Dominance by cyanobacteria characterized most of the study period, interspersed with sudden peaks (e.g. during PII and PIII) of high diversity of phytoplankton. Phytoplanktonic peaks were represented by Chlorophyta, diatoms, Prasinophyta and Criptophyta. Interestingly, the events observed in 1999 repeated almost 10 years later, suggesting roughly seasonal community structure

330 GOMES, A.M.A. et al.
Oecol. Bras., 13(2): 329-345, 2009
dynamism in the JLC. Disappearance of the bloom coincided with the lowest recorded water temperatures, suggesting that temperature is an important factor in the observed dynamics. The JL had a high abundance of zooplankton, yet species diversity was relatively low. Concentrations of microcystins found in muscles of fish from JL often contained values above the levels recommended by OMS for human consumption.Keywords: Coastal lagoon, eutrophication, cyanobacteria, mycrocistins.
RESUMENFLORACIONES DE CIANOBACTERIAS TOXICAS EN UNA LAGUNA COSTERA
HIPEREUTROFICA DE RIO DE JANEIRO/RJ (BRASIL) Y SUS CONSECUENCIAS PARA LA SALUD HUMANA. La Cuenca de Jacarepaguá (RJ/Brasil) ha sufrido, en las últimas cuatro décadas, un proceso desordenado de ocupación urbana e industrial, que se refleja en un cambio en la calidad del agua y en la incidencia de floraciones de las cianobacterias en el Complejo Lagunar Jacarepaguá (CLJ). El presente estudio reúne una serie de datos con el objeto de evaluar la calidad del Lago Jacarepaguá (LJ), a través del monitoreo físico (pH, transparencia del agua y temperatura), químico (salinidad, O.D. y clorofila a) y de la comunidad planctónica. También fue analizada la concentración de microcistinas en pescados comercializados en la región. Los datos fueron divididos en tres períodos de acuerdo con los diferentes ciclos estacionales estudiados: PI (ago/96 a sept/07), PII (ene/99 a nov/99) y PIII (feb/07 a ene/08). La dominancia de cianobacterias ocurre en la mayor parte del período de estudio, intercalada con pulsos de mayor diversidad fitoplanctónica, observada desde 1999 (PII y PIII). Dicha diversidad estuvo representada por las clorofíceas, diatomeas, prasinoficeas y criptoficeas. Cabe destacar que la dinámica observada en 1999 se repitió casi 10 años después, lo que sugiere una tendencia estacional para la estructura de la comunidad del CLJ. La desaparición de la floración coincide con épocas de bajas temperaturas sugiriendo que ésta es un factor regulador importante de la dinámica observada. El LJ presentó una elevada abundancia zooplanctónica, con baja diversidad de especies. Durante algunos meses, las concentraciones de microcistinas encontradas en los músculos de los peces estuvieron por encima de los valores recomendados por la OMS para el consumo humano.Palabras claves: Laguna costera, eutrofización, cianobacterias, microcistinas.
INTRODUÇÃO
As lagoas costeiras são ecossistemas de grande importância econômica devido a sua alta produção pesqueira, intenso uso para aqüicultura, recreação e depósito de resíduos alóctones (Spaulding 1994). São freqüentemente apontadas como um dos sistemas mais produtivos do mundo por apresentarem altas taxas de produção primária e secundária (Knoppers 1994). Ocorrem em praticamente todo litoral brasileiro e são consideradas como um dos principais sistemas lênticos do país (Esteves 1988). Geralmente são caracterizadas por serem sistemas rasos, separados do oceano por uma barreira terrestre e podem ser conectados ao mesmo, pelo menos temporariamente, por uma ou mais entradas restritas (Kjerfve 1994). Sofrem uma forte influência dos ventos, o que leva a uma maior homogeneização da coluna d’ água. Devido a sua proximidade com o mar, as lagoas costeiras apresentam uma grande variação da salinidade. O tamanho e a morfometria desses ambientes também
influenciam muitas de suas características físicas, químicas e biológicas (Panosso et al. 1998).Os diversos usos desses sistemas aquáticos pelo homem vêm promovendo profundas alterações em suas condições naturais. Como conseqüência, é comum a verificação de acelerados processos de eutrofização – enriquecimento artificial dos ecossistemas por nutrientes a partir, sobretudo, de descargas de esgotos domésticos e industriais dos centros urbanos e das regiões agricultáveis.
No Complexo Lagunar de Jacarepaguá (CLJ) o impacto antropogênico é evidenciado pela ocupação urbana desordenada, que resulta no aporte de efluentes sem nenhum tipo de tratamento, além dos assoreamentos e aterros acelerados das lagoas, causando o estrangulamento dos canais de ligação com o mar (Silva 2006). Tal fato tem modificado a composição de suas águas, aumentando excessiva-mente a proporção de água doce em relação à água do mar. Ainda como conseqüência das dificuldades de troca de água entre as lagoas e o mar, ocorre

331FLORAÇÕES DE CIANOBACTÉRIAS TÓXICAS EM UMA LAGOA COSTEIRA HIPEREUTRÓFICA
Oecol. Bras., 13(2): 329-345, 2009
estagnação e desoxigenação da coluna d’água, o que tem provocado periodicamente mortandade maciças de peixes.
Um estudo da Secretaria Municipal de Meio Ambiente (SMAC 2002), responsável pelo monitoramento das águas das lagoas do município do Rio de Janeiro, caracterizou o Arroio Pavuna e o Arroio Fundo como as principais fontes de poluição do sistema lagunar da região. Esses canais se localizam na lagoa do Camorim, poluindo principalmente as lagoas adjacentes Jacarepaguá e Tijuca. No caso da lagoa de Jacarepaguá há que se destacar a intensa atividade industrial exercida na região, cujos resíduos vão desaguar na lagoa (Feema 1982, 1984). Cabe destacar ainda que as lagoas do CLJ se intercomunicam, distribuindo de forma mais ou menos homogênea as descargas que recebem (Domingos 2001).O processo de eutrofização observado nesse sistema lagunar vem sendo apontado há cerca de 20 anos (Saieg-Filho 1986), muito embora na década anterior, Semeraro & Costa (1972) já relatavam a primeira ocorrência de florações de cianobactérias, além de destacarem a freqüente ocorrência de florações de dinoflagelados (Gymnodinium spp.) que conferiam coloração castanha às águas da lagoa de Jacarepaguá.
Uma das principais conseqüências do processo de eutrofização é o aumento relativo de cianobac-térias na comunidade fitoplanctônica, resultante, dentre outros fatores, do acúmulo de nutrientes inorgânicos dissolvidos. No complexo lagunar Jacarepaguá-Camorim-Tijuca, florações freqüentes de cianobactérias vêm sendo registradas desde a década de 90 (Fernandes 1993, Ferrão-Filho et al. 2002a), inclusive sendo registradas a ocorrência de cepas produtoras de cianotoxinas e o acúmulo dessas toxinas pelo zooplâncton e pelo pescado (Ferrão-Filho et al. 2002b, Magalhães et al. 2001).Uma das toxinas mais comuns produzidas por ciano-bactérias são as microcistinas, as quais são classifi-cadas como heptapeptídeos cíclicos hepatotóxicos. São extremamente tóxicas para animais e já foram envolvidas em acidentes de intoxicação em humanos levando-os a morte e também em casos de envene-namento em animais (Jochimsen et al. 1998, Duy et al. 2000). A presença de cianobactérias tóxicas no CLJ também pode acarretar problemas de saúde pública na região costeira marinha, uma vez que o
sistema lagunar comunica-se com o mar e, devido a sua hidrodinâmica, as águas são lançadas em uma praia de grande uso recreativo.
O presente trabalho teve como objetivo avaliar a qualidade hídrica da Lagoa de Jacarepaguá através de monitoramento físico, químico e da comunidade planctônica; além disso avaliou-se a qualidade do pescado comercializado na região, quanto à contaminação por microcistinas.
ÁREA DE ESTUDO
O Complexo Lagunar de Jacarepaguá localiza-se na Planície Costeira de Jacarepaguá, no litoral sul do Estado do Rio de Janeiro (43º17’ e 43º30’W; 22º55’ e 23º00’S, Figura 1). Esse complexo possui 130km2 de extensão e é formado por três lagoas principais alongadas: Jacarepaguá, Tijuca e Marapendi, e pela lagoa de Camorim, trecho também alongado que conecta as lagoas de Jacarepaguá e Tijuca (Domingos 2001). A bacia hidrográfica de Jacarepaguá é formada também pelos rios que descem das vertentes dos Maciços da Tijuca e da Pedra Branca e do escudo rochoso situado ao norte da baixada. A drenagem tem as lagoas como primeiro destino e em seguida, o mar. A área total da bacia hidrográfica de Jacarepaguá é de aproximadamente 226km2, sendo que cerca de 176km2 se referem às superfícies drenadas pelos rios. A lagoa de Jacarepaguá é a que recebe maior contribuição de rios que drenam para sua bacia. Os principais afluentes da lagoa de Jacarepaguá são o Arroio Pavuna (5,0km), rio Pavuninha (4,0km), rio Passarinho (2,5km), rio Caçambé (3,2km), rio Camorim (6,5km) e rio Marinho (3,5km) (SEMADS 2001).
A lagoa de Jacarepaguá apresenta uma área de 3,7km2, profundidade média de 3,3m e comunica-se com o mar através das lagoas do Camorim, da Tijuca e do canal artificial da Joatinga. Essa lagoa é a mais continental do CLJ, com escassa troca de água com o mar e com o maior tempo de retenção da água, o que contribui para o aumento da eutrofização (Domingos 2001). A lagoa de Jacarepaguá possui a maior área de drenagem da região (102,8km2) e uma vazão dos rios contribuintes de cerca de 0,78m3/s. Estes rios cortam grande parte dos bairros de Jacarepaguá e adjacências, trazendo em suas águas grande quantidade de sedimentos, resíduos industriais e domésticos (SEMADS 2001).

332 GOMES, A.M.A. et al.
Oecol. Bras., 13(2): 329-345, 2009
MÉTODOSPERIODICIDADE E ESTAÇÕES DE COLETA
Foram realizadas amostragens a cada quinze dias, durante os meses de agosto a dezembro de 1996, janeiro a setembro de 1997, janeiro a novembro de 1999 e fevereiro de 2007 a fevereiro de 2008 em duas estações de coleta na lagoa de Jacarepaguá. As estações de amostragem foram definidas com base em amostragens prévias e dados pré-existentes e suas localizações estão assinaladas na Figura 1. As coletas de zooplâncton e peixes para análises de microcistinas foram realizadas mensalmente, entre os períodos de agosto de 1996 a novembro de 1999 e de março de 2006 a abril de 2007.
MEDIDAS DE CAMPO
A temperatura da coluna d’água foi obtida através de um termístor digital. Valores de oxigênio dissolvido, condutividade elétrica, salinidade e pH
foram medidos utilizando eletrodos específicos para essas variáveis. A transparência d’água foi estimada pela extinção do disco de Secchi, no lado sombreado do barco (Lund & Talling 1957).
CLOROFILA-A
As coletas de água foram realizadas com garrafa de Van Dorn. Parte da amostra foi filtrada em campo, em filtros de fibra de vidro (Whatman GF/C). Os filtros foram preservados congelados para posterior análise de clorofila-a. As concentrações de clorofila-a foram estimadas por espectrofotometria, segundo metodologia descrita em APHA (1998), apartir de extração com acetona 90%, durante 20-24h.
FITOPLÂNCTON
As amostras de fitoplâncton foram coletadas na sub-superfície (0,1m) das estações de coleta e fixadas com solução de Lugol. As populações
Figura 1. Mapa e localização do complexo lagunar de Jacarepaguá, assinaladas as estações de coleta.Figure 1. Regional map and location of the Jacarepaguá lagoon complex. Sampling sites are indicated with numbers.

333FLORAÇÕES DE CIANOBACTÉRIAS TÓXICAS EM UMA LAGOA COSTEIRA HIPEREUTRÓFICA
Oecol. Bras., 13(2): 329-345, 2009
foram identificadas sempre que possível em nível de espécie. As densidades das populações (células.mL-1) foram estimadas segundo a técnica da sedimentação (Utermöhl 1958). Os indivíduos (células, colônias, filamentos) foram enumerados em campos aleatórios (Uhelinger 1964), em número suficiente para alcançar 100 indivíduos da espécie mais freqüente, sendo o erro inferior a 20%, a um intervalo de confiança de 95% (Lund et al. 1958).
ZOOPLÂNCTON
Para análise da densidade do zooplâncton foram coletados 9,0L de água da sub-superfície da lagoa e concentrados em rede de plâncton com abertura de malha de 68μm. O concentrado foi acondicionado em frascos de vidro, contendo solução de formol a 4%. Para identificação e quantificação das populações, foi utilizado microscópio óptico, sendo os animais identificados em nível de espécie, sempre que possível. Para a quantificação foi utilizada uma câmara de Sedgwick-Rafter reticulada com capacidade para 1mL. Para melhor confiabilidade dos resultados, essas contagens foram feitas três vezes por amostra.
CIANOTOXINAS
Microcistinas foram analisadas a partir de amostras de séston da lagoa retida nos filtros de borosilicato. Cerca de 2,0L de água da lagoa foram filtrados em membranas de borosilicato (Sartorius AG 37070, Goettingen, Germany), sendo os mesmos congelados até a análise. A extração de toxina dos filtros foi feita segundo método descrito em Krishnamurthy et al. (1986), utilizando-se uma solução butanol-metanol-água (5:20:75 vol/vol), seguida de centrifugação do extrato e evaporação do sobrenadante a 1/3 do volume. Em seguida, os extratos foram passados em cartucho C18 (Varian) para uma primeira purificação da fração contendo microcistinas. Este cartucho foi então eluído com 20mL de água deionizada, 20mL de metanol 20% e finalmente com 20mL de metanol 100%. Esta fração foi então evaporada até a secagem completa, ressuspensa em 1,0mL de água deionizada e congelada para posterior análise.
A verificação da presença de microcistinas foi feita através de análise por Cromatografia Líquida de Alta Eficiência (HPLC), em cromatógrafo Shimadzu
com detetor UV/Vis. SPD-M10A. As análises foram realizadas em condições isocráticas, sendo utilizada uma coluna de fase reversa Supercosil LC-18 (5μm, 25cm x 10mm), fase móvel de acetonitrila e acetato de amônio 20mM (28:72 v/v), fluxo de 1,0mL.min-1 e absorbância a 238nm.
O espectro de cada um dos picos obtidos nos cromatogramas foi analisado entre 200-300nm, com o auxílio de um fotodetector de diodo (SPD-M10A - Diode Array-PDA) e comparados a um espectro padrão de microcistina-LR, no sentido de confirmar a presença da toxina. A quantificação da toxina foi feita através da comparação de área do pico da amostra em relação a uma área obtida num cromatograma feito com um padrão de microcistina-LR, no qual a concentração de microcistinas é conhecida.
Para verificação da presença e/ou acumulação de microcistinas no tecido muscular, os peixes foram dissecados com bisturi e as amostras do tecido muscular foram extraídas com metanol 100%, seguida de uma partição com hexano para a retirada do excesso de lipídios e pré-purificada em cartucho de octadecilsilano (C18) seguindo a metodologia descrita por Magalhães et al. (2001).
Os peixes coletados a partir de 2006 foram extraídos com metanol:TFA (0,1%) (Chorus & Bartran 1999) e pré purificada em cartucho de sílica HP20. O material extraído e seco foi analisado por técnicas de imunoensaio do tipo ELISA (kit de placas Beacon®), desenvolvido por Chu et al. (1990). Neste caso, é importante ressaltar que toda a quantidade de microcistinas extraída dos tecidos se refere a microcistinas livres, pois no momento em que essas toxinas se ligam as proteínas fosfatases não podem ser extraídas, por ser esta uma ligação covalente irreversível.
RESULTADOS E DISCUSSÃOVARIÁVEIS FÍSICAS, QUÍMICAS E BIOLÓGICAS
A lagoa de Jacarepaguá tem sido caracterizada como um ambiente raso, túrbido, hipereutrófico, e com altas densidades fitoplanctônicas (Fernandes 1993, Domingos 2001, Ferrão-Filho et al. 2002a).
Os perfis de temperatura observados durante o período de amostragem do ano de 2007 sugerem uma constante circulação de toda a coluna d’água ao longo de todo ciclo sazonal (dados não mostrados).

334 GOMES, A.M.A. et al.
Oecol. Bras., 13(2): 329-345, 2009
Para melhor interpretação dos dados, os mesmos foram divididos em três períodos de acordo com os ciclos sazonais estudados, são eles: Período I (PI) de agosto/96 a setembro/97, Período II (PII) de janeiro/99 a novembro/99 e Período III (PIII) de fevereiro/07 a janeiro/08. Em PI a temperatura variou de 20,0 e 21,0oC (agosto/96) à 31,2 e 32,6oC (janeiro/97) nos pontos 1 e 2, respectivamente (Figura 2). A variação da temperatura em PII foi de 19oC em agosto/99 (nos dois pontos de coleta) à 29,6oC (ponto 1) e 30,5oC (ponto 2). Já em PIII, os menores valores de temperatura foram observados no mês de julho/07 (18, 5oC, ponto 1; 19,1oC, ponto 2) e os maiores em março/07 (31,4oC; ponto 1) e em fevereiro/07 (30,6oC; ponto 2). A variação de um pouco mais de 10oC de temperatura da água entre os meses de inverno e verão observados na lagoa de Jacarepaguá parece ter sido um fator importante que contribuiu para a mudança na composição e densidade fitoplanctônica nesse ambiente.
Os valores de pH no ponto 1 variaram de neutro à levemente alcalino na maioria dos meses em PI e PII (Figura 2). Um considerável aumento nos valores de pH foi registrado nos meses de verão (pH 10; novembro/99), coincidindo com o maior valor de densidade fitoplanctônica total (1,2 x 107cél.mL-1) e dominância de cianobactérias observado em todo período de estudo. Em PIII (ponto 1), apenas os meses de fevereiro e março/07 apresentaram altos valores de pH, nos demais meses desse período, o pH da água variou de neutro a ácido. No ponto 2, os valores de pH também variaram de neutro a alcalino na maioria dos meses, com valores ligeiramente ácidos nos meses de verão em PIII. É importante ressaltar que embora as cianobactérias apresentem vantagens em colonizar ambientes com pH mais elevado (Caraco & Miller 1998), a dominância desse grupo sobre comunidade fitoplanctônica não se restringiu a épocas em que as águas estiveram alcalinas.
A salinidade da água da lagoa de Jacarepaguá variou desde valores próximos a zero ppt (fevereiro e março/97; janeiro e fevereiro/99) a 10ppt (novembro/99; Figura 2). Em PIII, ocorreram salinidades mais baixas nos meses do verão (período chuvoso), se comparadas aos meses de seca do inverno, sugerindo uma possível diluição da água da lagoa pela água da chuva.
A salinidade não parece ser um fator tão importante quanto nas lagoas localizadas no norte do estado do Rio de Janeiro que atingem valores de até 53,4ppt (Enrich-Prast et al. 2004), possivelmente devido à escassa circulação da água do mar provocada pelo estrangulamento e fechamento dos canais de comunicação da lagoa com a região costeira.
Ao longo dos três períodos estudados, foi registrada uma grande variabilidade das concentrações de oxigênio dissolvido (OD, Figura 2). As maiores concentrações de OD (máx = 16,40mg.L-1 em agosto/97) encontradas na superfície na coluna d’água são resultado da elevada atividade fotossintética fitoplanctônica. Em PIII, maiores concentrações de OD foram observadas na superfície e anoxia, no fundo da coluna d’água (dados não mostrados). Na maioria das amostragens, a anoxia foi registrada já a partir dos primeiros 50cm da coluna d’água.
A transparência da água na lagoa de Jacarepaguá foi reduzida ao longo de todos os três períodos estudados, com a maioria dos valores entre 10 e 50cm de coluna d’água (Figura 2). A baixa transparência da água pode ser atribuída à alta densidade fitoplanctônica.
Durante os três períodos estudados, as concentrações de clorofila-a foram bastante elevadas (Figura 3). Em PI, alcançaram os valores máximos de 207 e 304,3μg.L-1 (pontos 1 e 2, respectivamente). As maiores concentrações dessa variável foram observadas em PII, atingindo valores de 9770,4μg.L-1 (ponto 2; outubro/99). As elevadas concentrações de clorofila-a estão relacionadas com a alta disponibilidade de nutrientes neste sistema lagunar (Domingos 2001).
A reduzida transparência da água, associada às elevadas concentrações de clorofila-a, caracterizam a lagoa de Jacarepaguá como um ambiente hipereutrófico, de acordo com qualquer índice de estado trófico utilizado (Nürnberg 1996, Vollenweider & Kerekes 1980, Salas & Martino 1991). A ocupação urbana desordenada na região da Barra da Tijuca (RJ) pode ser apontada como uma das principais causas do acelerado processo de eutrofização dessa lagoa, que vem sendo intensificada desde a década de 70 (Semeraro & Costa 1972). As lagoas costeiras do Estado do Rio de Janeiro que apresentam uma intensa ocupação urbana e/ou industrial em seu entorno são freqüentemente reportadas como ambientes de alta
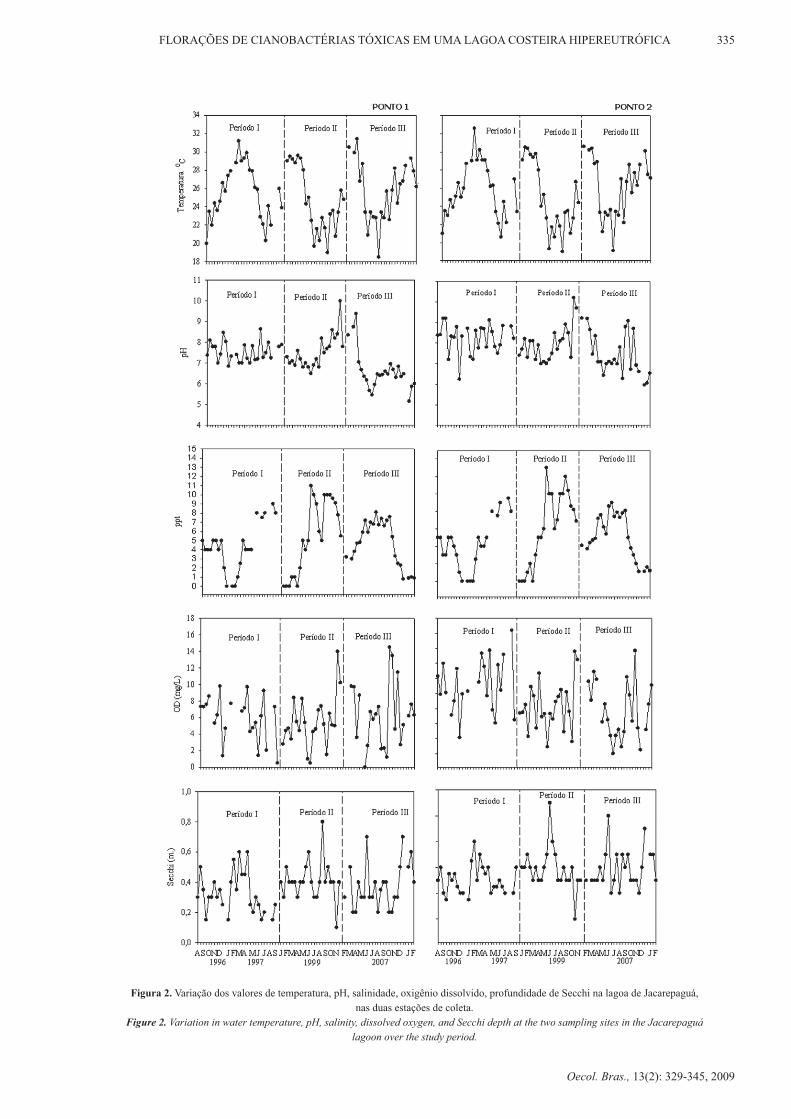
335FLORAÇÕES DE CIANOBACTÉRIAS TÓXICAS EM UMA LAGOA COSTEIRA HIPEREUTRÓFICA
Oecol. Bras., 13(2): 329-345, 2009
Figura 2. Variação dos valores de temperatura, pH, salinidade, oxigênio dissolvido, profundidade de Secchi na lagoa de Jacarepaguá, nas duas estações de coleta.
Figure 2. Variation in water temperature, pH, salinity, dissolved oxygen, and Secchi depth at the two sampling sites in the Jacarepaguá lagoon over the study period.
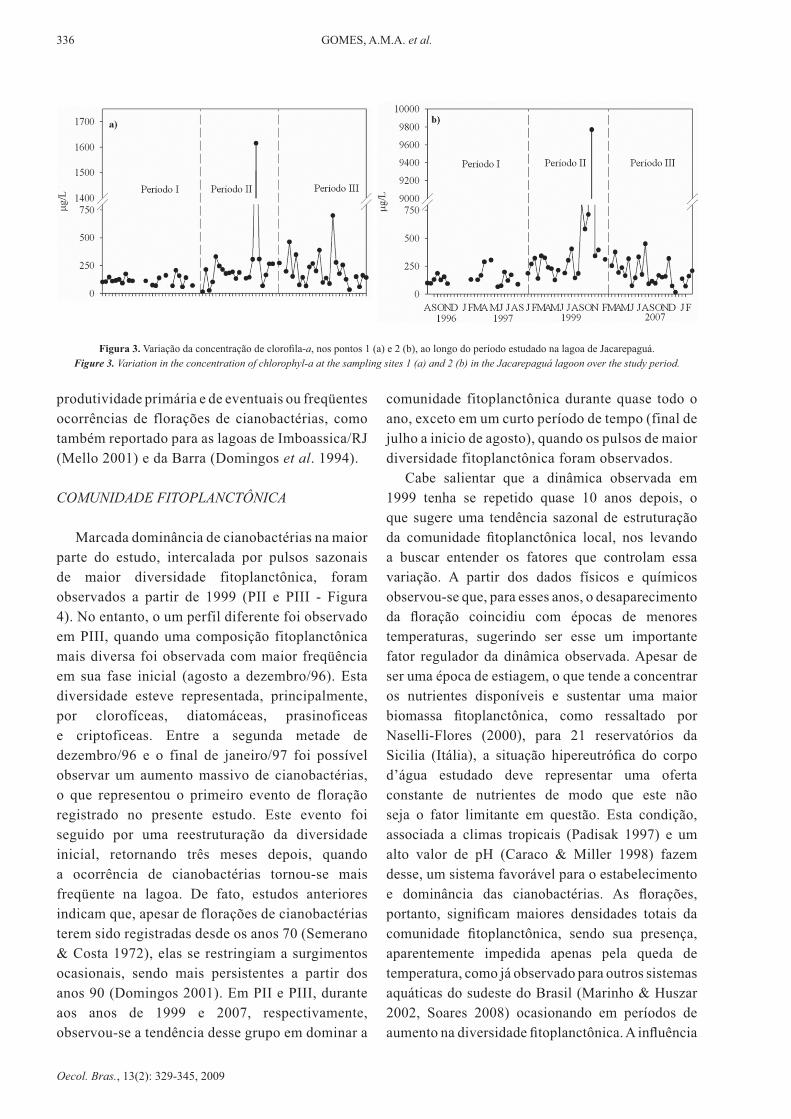
336 GOMES, A.M.A. et al.
Oecol. Bras., 13(2): 329-345, 2009
produtividade primária e de eventuais ou freqüentes ocorrências de florações de cianobactérias, como também reportado para as lagoas de Imboassica/RJ (Mello 2001) e da Barra (Domingos et al. 1994).
COMUNIDADE FITOPLANCTÔNICA
Marcada dominância de cianobactérias na maior parte do estudo, intercalada por pulsos sazonais de maior diversidade fitoplanctônica, foram observados a partir de 1999 (PII e PIII - Figura 4). No entanto, o um perfil diferente foi observado em PIII, quando uma composição fitoplanctônica mais diversa foi observada com maior freqüência em sua fase inicial (agosto a dezembro/96). Esta diversidade esteve representada, principalmente, por clorofíceas, diatomáceas, prasinoficeas e criptoficeas. Entre a segunda metade de dezembro/96 e o final de janeiro/97 foi possível observar um aumento massivo de cianobactérias, o que representou o primeiro evento de floração registrado no presente estudo. Este evento foi seguido por uma reestruturação da diversidade inicial, retornando três meses depois, quando a ocorrência de cianobactérias tornou-se mais freqüente na lagoa. De fato, estudos anteriores indicam que, apesar de florações de cianobactérias terem sido registradas desde os anos 70 (Semerano & Costa 1972), elas se restringiam a surgimentos ocasionais, sendo mais persistentes a partir dos anos 90 (Domingos 2001). Em PII e PIII, durante aos anos de 1999 e 2007, respectivamente, observou-se a tendência desse grupo em dominar a
comunidade fitoplanctônica durante quase todo o ano, exceto em um curto período de tempo (final de julho a inicio de agosto), quando os pulsos de maior diversidade fitoplanctônica foram observados.
Cabe salientar que a dinâmica observada em 1999 tenha se repetido quase 10 anos depois, o que sugere uma tendência sazonal de estruturação da comunidade fitoplanctônica local, nos levando a buscar entender os fatores que controlam essa variação. A partir dos dados físicos e químicos observou-se que, para esses anos, o desaparecimento da floração coincidiu com épocas de menores temperaturas, sugerindo ser esse um importante fator regulador da dinâmica observada. Apesar de ser uma época de estiagem, o que tende a concentrar os nutrientes disponíveis e sustentar uma maior biomassa fitoplanctônica, como ressaltado por Naselli-Flores (2000), para 21 reservatórios da Sicilia (Itália), a situação hipereutrófica do corpo d’água estudado deve representar uma oferta constante de nutrientes de modo que este não seja o fator limitante em questão. Esta condição, associada a climas tropicais (Padisak 1997) e um alto valor de pH (Caraco & Miller 1998) fazem desse, um sistema favorável para o estabelecimento e dominância das cianobactérias. As florações, portanto, significam maiores densidades totais da comunidade fitoplanctônica, sendo sua presença, aparentemente impedida apenas pela queda de temperatura, como já observado para outros sistemas aquáticas do sudeste do Brasil (Marinho & Huszar 2002, Soares 2008) ocasionando em períodos de aumento na diversidade fitoplanctônica. A influência
Figura 3. Variação da concentração de clorofila-a, nos pontos 1 (a) e 2 (b), ao longo do período estudado na lagoa de Jacarepaguá.Figure 3. Variation in the concentration of chlorophyl-a at the sampling sites 1 (a) and 2 (b) in the Jacarepaguá lagoon over the study period.
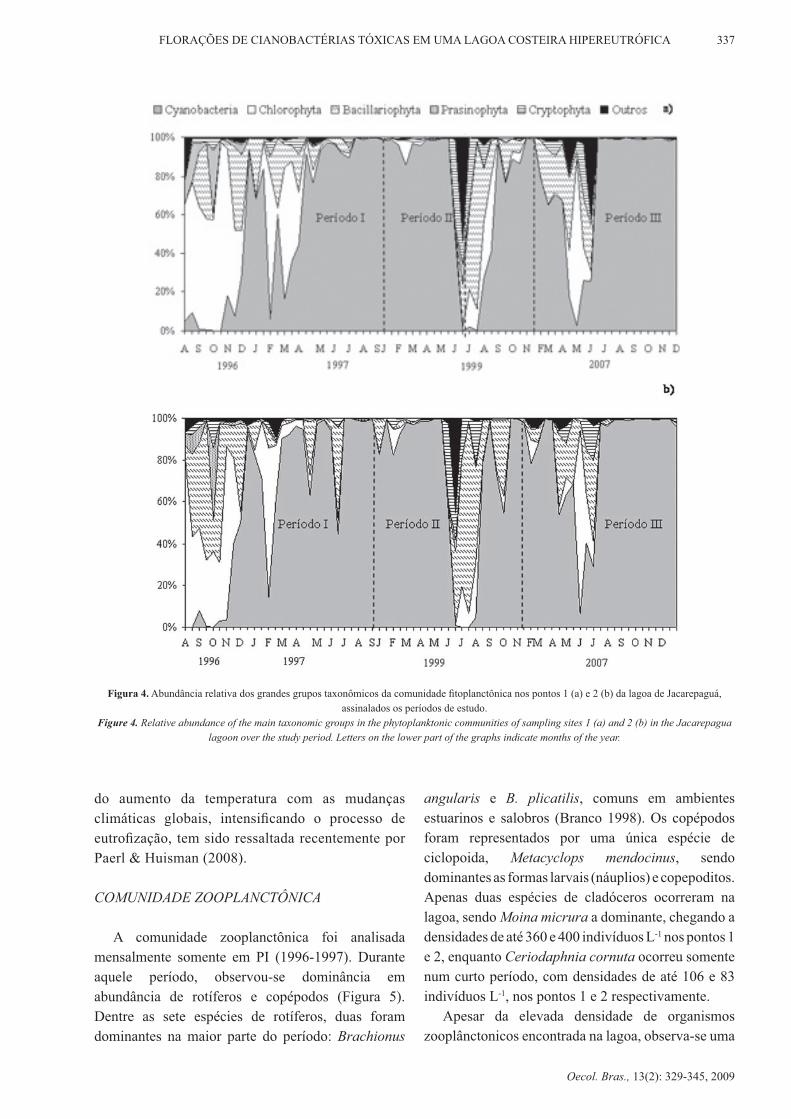
337FLORAÇÕES DE CIANOBACTÉRIAS TÓXICAS EM UMA LAGOA COSTEIRA HIPEREUTRÓFICA
Oecol. Bras., 13(2): 329-345, 2009
do aumento da temperatura com as mudanças climáticas globais, intensificando o processo de eutrofização, tem sido ressaltada recentemente por Paerl & Huisman (2008).
COMUNIDADE ZOOPLANCTÔNICA
A comunidade zooplanctônica foi analisada mensalmente somente em PI (1996-1997). Durante aquele período, observou-se dominância em abundância de rotíferos e copépodos (Figura 5). Dentre as sete espécies de rotíferos, duas foram dominantes na maior parte do período: Brachionus
angularis e B. plicatilis, comuns em ambientes estuarinos e salobros (Branco 1998). Os copépodos foram representados por uma única espécie de ciclopoida, Metacyclops mendocinus, sendo dominantes as formas larvais (náuplios) e copepoditos. Apenas duas espécies de cladóceros ocorreram na lagoa, sendo Moina micrura a dominante, chegando a densidades de até 360 e 400 indivíduos L-1 nos pontos 1 e 2, enquanto Ceriodaphnia cornuta ocorreu somente num curto período, com densidades de até 106 e 83 indivíduos L-1, nos pontos 1 e 2 respectivamente.
Apesar da elevada densidade de organismos zooplânctonicos encontrada na lagoa, observa-se uma
Figura 4. Abundância relativa dos grandes grupos taxonômicos da comunidade fitoplanctônica nos pontos 1 (a) e 2 (b) da lagoa de Jacarepaguá, assinalados os períodos de estudo.
Figure 4. Relative abundance of the main taxonomic groups in the phytoplanktonic communities of sampling sites 1 (a) and 2 (b) in the Jacarepagua lagoon over the study period. Letters on the lower part of the graphs indicate months of the year.
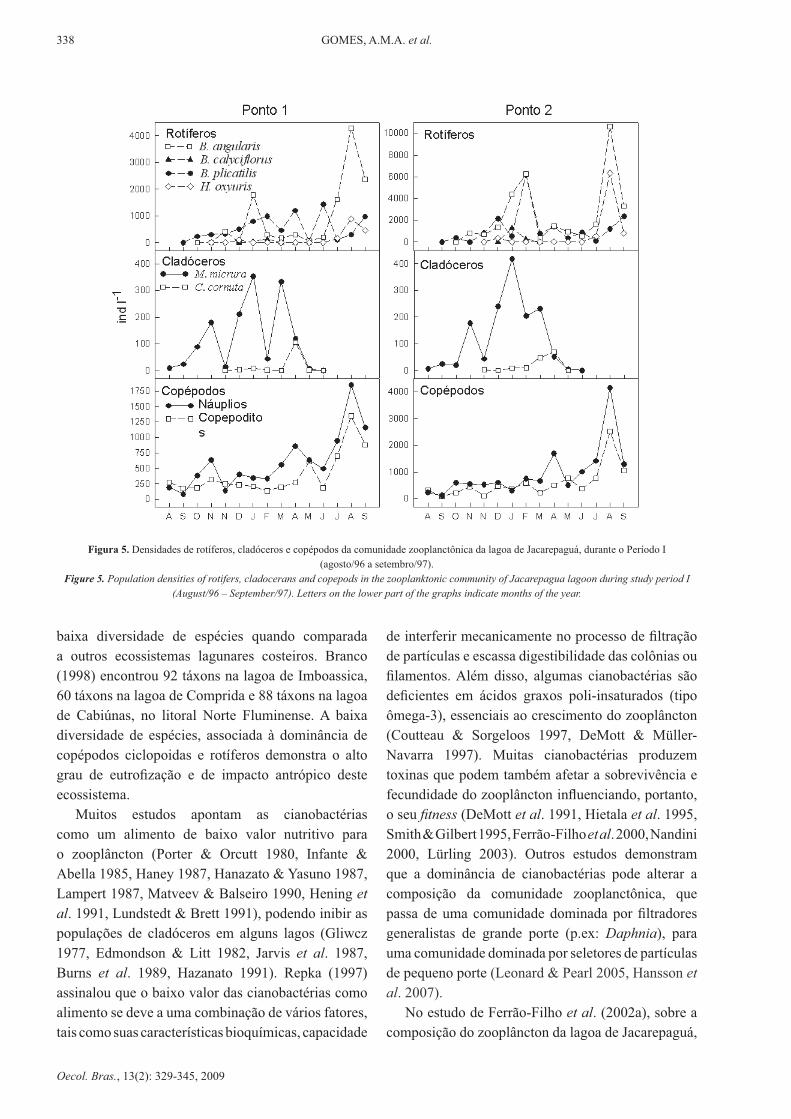
338 GOMES, A.M.A. et al.
Oecol. Bras., 13(2): 329-345, 2009
baixa diversidade de espécies quando comparada a outros ecossistemas lagunares costeiros. Branco (1998) encontrou 92 táxons na lagoa de Imboassica, 60 táxons na lagoa de Comprida e 88 táxons na lagoa de Cabiúnas, no litoral Norte Fluminense. A baixa diversidade de espécies, associada à dominância de copépodos ciclopoidas e rotíferos demonstra o alto grau de eutrofização e de impacto antrópico deste ecossistema.
Muitos estudos apontam as cianobactérias como um alimento de baixo valor nutritivo para o zooplâncton (Porter & Orcutt 1980, Infante & Abella 1985, Haney 1987, Hanazato & Yasuno 1987, Lampert 1987, Matveev & Balseiro 1990, Hening et al. 1991, Lundstedt & Brett 1991), podendo inibir as populações de cladóceros em alguns lagos (Gliwcz 1977, Edmondson & Litt 1982, Jarvis et al. 1987, Burns et al. 1989, Hazanato 1991). Repka (1997) assinalou que o baixo valor das cianobactérias como alimento se deve a uma combinação de vários fatores, tais como suas características bioquímicas, capacidade
de interferir mecanicamente no processo de filtração de partículas e escassa digestibilidade das colônias ou filamentos. Além disso, algumas cianobactérias são deficientes em ácidos graxos poli-insaturados (tipo ômega-3), essenciais ao crescimento do zooplâncton (Coutteau & Sorgeloos 1997, DeMott & Müller-Navarra 1997). Muitas cianobactérias produzem toxinas que podem também afetar a sobrevivência e fecundidade do zooplâncton influenciando, portanto, o seu fitness (DeMott et al. 1991, Hietala et al. 1995, Smith & Gilbert 1995, Ferrão-Filho et al. 2000, Nandini 2000, Lürling 2003). Outros estudos demonstram que a dominância de cianobactérias pode alterar a composição da comunidade zooplanctônica, que passa de uma comunidade dominada por filtradores generalistas de grande porte (p.ex: Daphnia), para uma comunidade dominada por seletores de partículas de pequeno porte (Leonard & Pearl 2005, Hansson et al. 2007).
No estudo de Ferrão-Filho et al. (2002a), sobre a composição do zooplâncton da lagoa de Jacarepaguá,
Figura 5. Densidades de rotíferos, cladóceros e copépodos da comunidade zooplanctônica da lagoa de Jacarepaguá, durante o Período I (agosto/96 a setembro/97).
Figure 5. Population densities of rotifers, cladocerans and copepods in the zooplanktonic community of Jacarepagua lagoon during study period I (August/96 – September/97). Letters on the lower part of the graphs indicate months of the year.
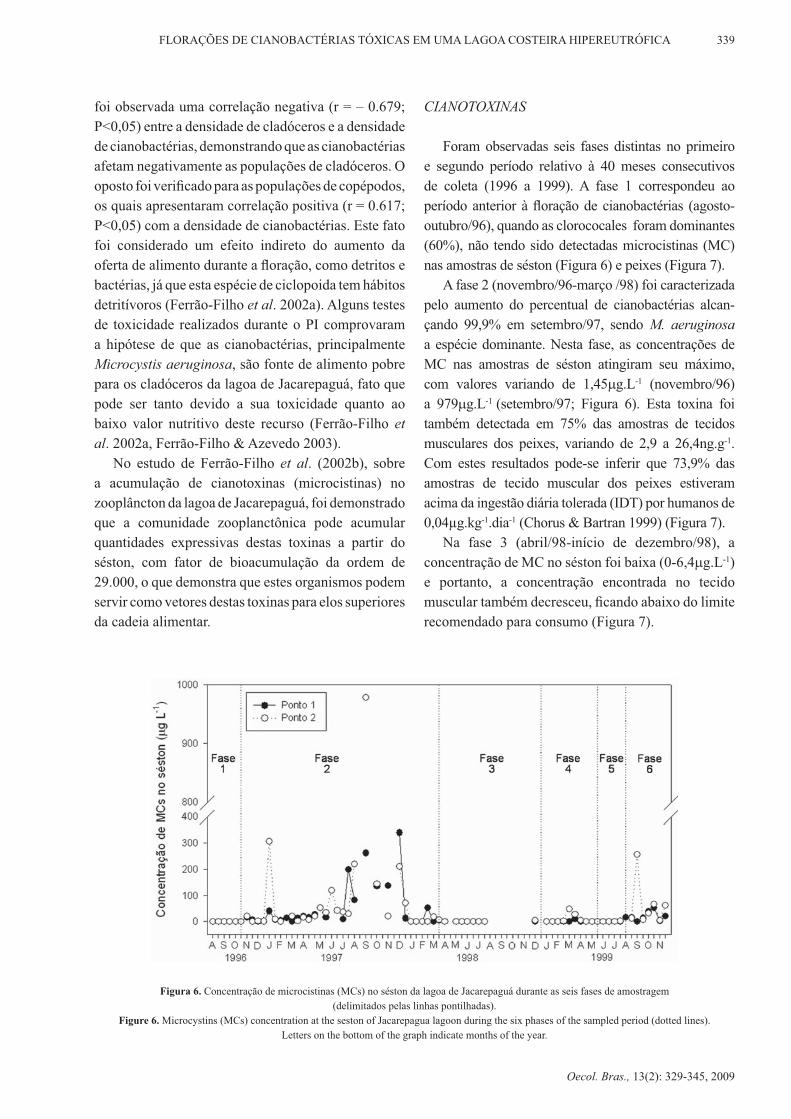
339FLORAÇÕES DE CIANOBACTÉRIAS TÓXICAS EM UMA LAGOA COSTEIRA HIPEREUTRÓFICA
Oecol. Bras., 13(2): 329-345, 2009
foi observada uma correlação negativa (r = – 0.679; P<0,05) entre a densidade de cladóceros e a densidade de cianobactérias, demonstrando que as cianobactérias afetam negativamente as populações de cladóceros. O oposto foi verificado para as populações de copépodos, os quais apresentaram correlação positiva (r = 0.617; P<0,05) com a densidade de cianobactérias. Este fato foi considerado um efeito indireto do aumento da oferta de alimento durante a floração, como detritos e bactérias, já que esta espécie de ciclopoida tem hábitos detritívoros (Ferrão-Filho et al. 2002a). Alguns testes de toxicidade realizados durante o PI comprovaram a hipótese de que as cianobactérias, principalmente Microcystis aeruginosa, são fonte de alimento pobre para os cladóceros da lagoa de Jacarepaguá, fato que pode ser tanto devido a sua toxicidade quanto ao baixo valor nutritivo deste recurso (Ferrão-Filho et al. 2002a, Ferrão-Filho & Azevedo 2003).
No estudo de Ferrão-Filho et al. (2002b), sobre a acumulação de cianotoxinas (microcistinas) no zooplâncton da lagoa de Jacarepaguá, foi demonstrado que a comunidade zooplanctônica pode acumular quantidades expressivas destas toxinas a partir do séston, com fator de bioacumulação da ordem de 29.000, o que demonstra que estes organismos podem servir como vetores destas toxinas para elos superiores da cadeia alimentar.
CIANOTOXINAS
Foram observadas seis fases distintas no primeiro e segundo período relativo à 40 meses consecutivos de coleta (1996 a 1999). A fase 1 correspondeu ao período anterior à floração de cianobactérias (agosto-outubro/96), quando as clorococales foram dominantes (60%), não tendo sido detectadas microcistinas (MC) nas amostras de séston (Figura 6) e peixes (Figura 7).
A fase 2 (novembro/96-março /98) foi caracterizada pelo aumento do percentual de cianobactérias alcan-çando 99,9% em setembro/97, sendo M. aeruginosa a espécie dominante. Nesta fase, as concentrações de MC nas amostras de séston atingiram seu máximo, com valores variando de 1,45μg.L-1 (novembro/96) a 979μg.L-1 (setembro/97; Figura 6). Esta toxina foi também detectada em 75% das amostras de tecidos musculares dos peixes, variando de 2,9 a 26,4ng.g-1. Com estes resultados pode-se inferir que 73,9% das amostras de tecido muscular dos peixes estiveram acima da ingestão diária tolerada (IDT) por humanos de 0,04μg.kg-1.dia-1 (Chorus & Bartran 1999) (Figura 7).
Na fase 3 (abril/98-início de dezembro/98), a concentração de MC no séston foi baixa (0-6,4μg.L-1) e portanto, a concentração encontrada no tecido muscular também decresceu, ficando abaixo do limite recomendado para consumo (Figura 7).
Figura 6. Concentração de microcistinas (MCs) no séston da lagoa de Jacarepaguá durante as seis fases de amostragem (delimitados pelas linhas pontilhadas).
Figure 6. Microcystins (MCs) concentration at the seston of Jacarepagua lagoon during the six phases of the sampled period (dotted lines). Letters on the bottom of the graph indicate months of the year.
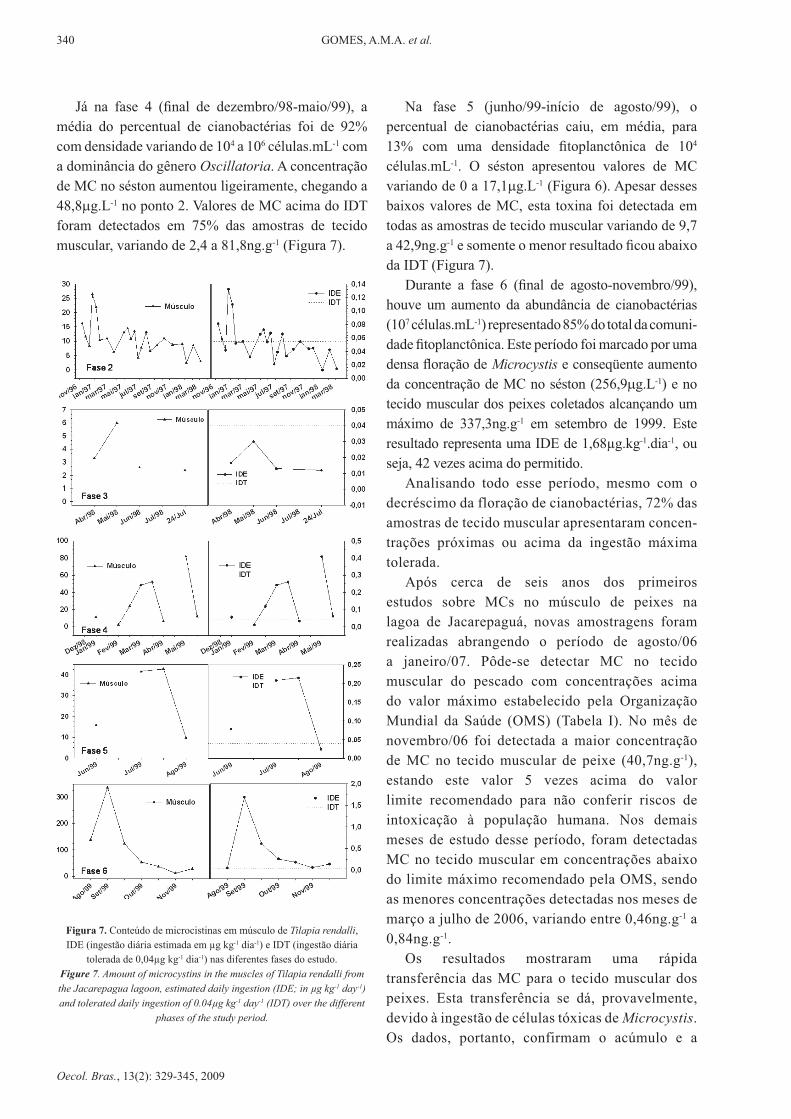
340 GOMES, A.M.A. et al.
Oecol. Bras., 13(2): 329-345, 2009
Já na fase 4 (final de dezembro/98-maio/99), a média do percentual de cianobactérias foi de 92% com densidade variando de 104 a 106 células.mL-1 com a dominância do gênero Oscillatoria. A concentração de MC no séston aumentou ligeiramente, chegando a 48,8μg.L-1 no ponto 2. Valores de MC acima do IDT foram detectados em 75% das amostras de tecido muscular, variando de 2,4 a 81,8ng.g-1 (Figura 7).
Figura 7. Conteúdo de microcistinas em músculo de Tilapia rendalli, IDE (ingestão diária estimada em μg kg-1 dia-1) e IDT (ingestão diária
tolerada de 0,04μg kg-1 dia-1) nas diferentes fases do estudo.Figure 7. Amount of microcystins in the muscles of Tilapia rendalli from the Jacarepagua lagoon, estimated daily ingestion (IDE; in μg kg-1 day-1) and tolerated daily ingestion of 0.04μg kg-1 day-1 (IDT) over the different
phases of the study period.
Na fase 5 (junho/99-início de agosto/99), o percentual de cianobactérias caiu, em média, para 13% com uma densidade fitoplanctônica de 104 células.mL-1. O séston apresentou valores de MC variando de 0 a 17,1μg.L-1 (Figura 6). Apesar desses baixos valores de MC, esta toxina foi detectada em todas as amostras de tecido muscular variando de 9,7 a 42,9ng.g-1 e somente o menor resultado ficou abaixo da IDT (Figura 7).
Durante a fase 6 (final de agosto-novembro/99), houve um aumento da abundância de cianobactérias (107 células.mL-1) representado 85% do total da comuni-dade fitoplanctônica. Este período foi marcado por uma densa floração de Microcystis e conseqüente aumento da concentração de MC no séston (256,9μg.L-1) e no tecido muscular dos peixes coletados alcançando um máximo de 337,3ng.g-1 em setembro de 1999. Este resultado representa uma IDE de 1,68μg.kg-1.dia-1, ou seja, 42 vezes acima do permitido.
Analisando todo esse período, mesmo com o decréscimo da floração de cianobactérias, 72% das amostras de tecido muscular apresentaram concen-trações próximas ou acima da ingestão máxima tolerada.
Após cerca de seis anos dos primeiros estudos sobre MCs no músculo de peixes na lagoa de Jacarepaguá, novas amostragens foram realizadas abrangendo o período de agosto/06 a janeiro/07. Pôde-se detectar MC no tecido muscular do pescado com concentrações acima do valor máximo estabelecido pela Organização Mundial da Saúde (OMS) (Tabela I). No mês de novembro/06 foi detectada a maior concentração de MC no tecido muscular de peixe (40,7ng.g-1), estando este valor 5 vezes acima do valor limite recomendado para não conferir riscos de intoxicação à população humana. Nos demais meses de estudo desse período, foram detectadas MC no tecido muscular em concentrações abaixo do limite máximo recomendado pela OMS, sendo as menores concentrações detectadas nos meses de março a julho de 2006, variando entre 0,46ng.g-1 a 0,84ng.g-1.
Os resultados mostraram uma rápida transferência das MC para o tecido muscular dos peixes. Esta transferência se dá, provavelmente, devido à ingestão de células tóxicas de Microcystis. Os dados, portanto, confirmam o acúmulo e a
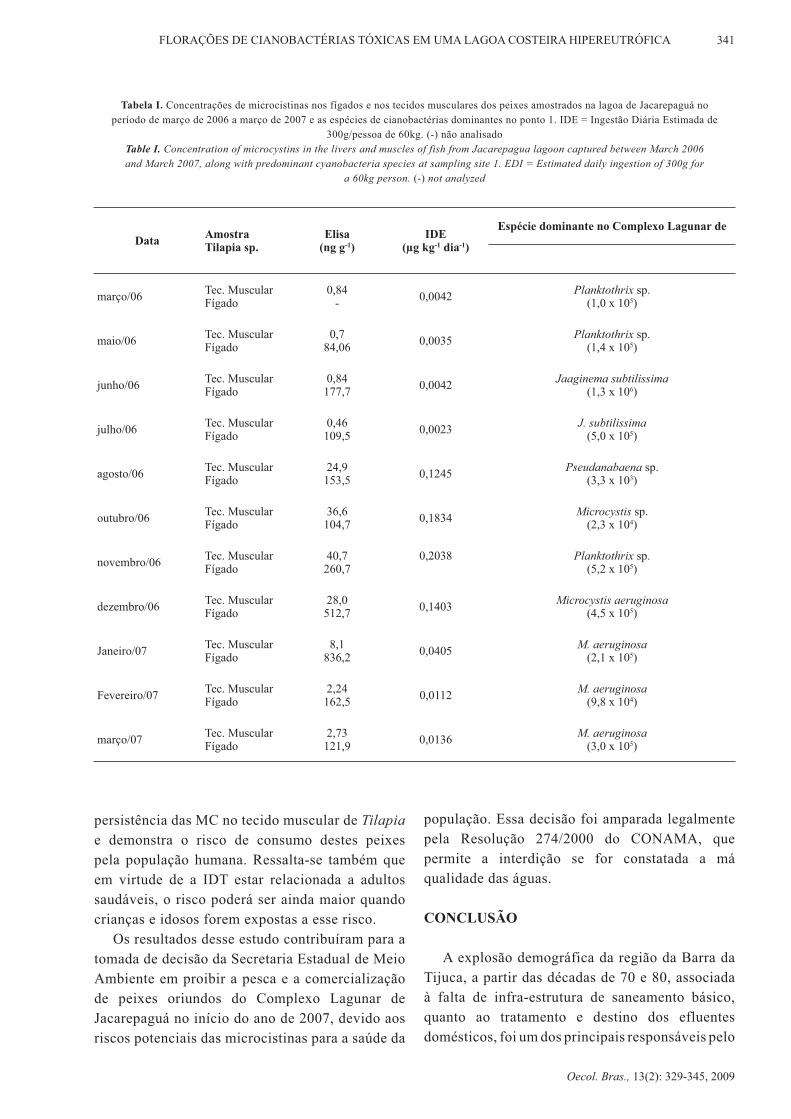
341FLORAÇÕES DE CIANOBACTÉRIAS TÓXICAS EM UMA LAGOA COSTEIRA HIPEREUTRÓFICA
Oecol. Bras., 13(2): 329-345, 2009
persistência das MC no tecido muscular de Tilapia e demonstra o risco de consumo destes peixes pela população humana. Ressalta-se também que em virtude de a IDT estar relacionada a adultos saudáveis, o risco poderá ser ainda maior quando crianças e idosos forem expostas a esse risco.
Os resultados desse estudo contribuíram para a tomada de decisão da Secretaria Estadual de Meio Ambiente em proibir a pesca e a comercialização de peixes oriundos do Complexo Lagunar de Jacarepaguá no início do ano de 2007, devido aos riscos potenciais das microcistinas para a saúde da
Tabela I. Concentrações de microcistinas nos fígados e nos tecidos musculares dos peixes amostrados na lagoa de Jacarepaguá no período de março de 2006 a março de 2007 e as espécies de cianobactérias dominantes no ponto 1. IDE = Ingestão Diária Estimada de
300g/pessoa de 60kg. (-) não analisado Table I. Concentration of microcystins in the livers and muscles of fish from Jacarepagua lagoon captured between March 2006 and March 2007, along with predominant cyanobacteria species at sampling site 1. EDI = Estimated daily ingestion of 300g for
a 60kg person. (-) not analyzed
Data AmostraTilapia sp.
Elisa(ng g-1)
IDE(µg kg-1 dia-1)
Espécie dominante no Complexo Lagunar de
março/06 Tec. MuscularFígado
0,84- 0,0042 Planktothrix sp.
(1,0 x 105)
maio/06 Tec. MuscularFígado
0,784,06 0,0035 Planktothrix sp.
(1,4 x 105)
junho/06 Tec. MuscularFígado
0,84177,7 0,0042 Jaaginema subtilissima
(1,3 x 106)
julho/06 Tec. MuscularFígado
0,46109,5 0,0023 J. subtilissima
(5,0 x 105)
agosto/06 Tec. MuscularFígado
24,9153,5 0,1245 Pseudanabaena sp.
(3,3 x 103)
outubro/06 Tec. MuscularFígado
36,6104,7 0,1834 Microcystis sp.
(2,3 x 104)
novembro/06 Tec. MuscularFígado
40,7260,7
0,2038 Planktothrix sp.(5,2 x 105)
dezembro/06 Tec. MuscularFígado
28,0512,7 0,1403 Microcystis aeruginosa
(4,5 x 105)
Janeiro/07 Tec. MuscularFígado
8,1836,2 0,0405 M. aeruginosa
(2,1 x 105)
Fevereiro/07 Tec. MuscularFígado
2,24162,5 0,0112 M. aeruginosa
(9,8 x 104)
março/07 Tec. MuscularFígado
2,73121,9 0,0136 M. aeruginosa
(3,0 x 105)
população. Essa decisão foi amparada legalmente pela Resolução 274/2000 do CONAMA, que permite a interdição se for constatada a má qualidade das águas.
CONCLUSÃO
A explosão demográfica da região da Barra da Tijuca, a partir das décadas de 70 e 80, associada à falta de infra-estrutura de saneamento básico, quanto ao tratamento e destino dos efluentes domésticos, foi um dos principais responsáveis pelo

342 GOMES, A.M.A. et al.
Oecol. Bras., 13(2): 329-345, 2009
acelerado e intensificado processo de eutrofização das águas da lagoa de Jacarepaguá. Uma das conseqüências da eutrofização nesse ambiente tem sido a freqüente ocorrência de florações de cianobactérias. O grupo das cianobactérias vem dominando a comunidade fitoplanctônica desse sistema há vários anos, salvo alguns curtos pulsos de redução da representatividade desse grupo e aumento da diversidade, fato este que parece estar também associado a uma brusca redução da temperatura nos meses de inverno. A ocorrência de espécies de cianobactérias produtoras de microcistinas assim como altas concentrações destas toxinas foram reportadas na lagoa de Jacarepaguá. Essas toxinas são acumuladas nos peixes dessa lagoa e estes são consumidos e comercializados pela população humana local, principalmente pela comunidade que vive em torno dessa lagoa. Durante 60% do meses de estudo, as concentrações de microcistinas encontradas no tecido muscular do pescado estiveram acima dos valores recomendados pela OMS para consumo humano. Em decorrência disso, no início do ano de 2007, a pesca na lagoa de Jacarepaguá foi proibida. Nessa mesma época, um trecho da praia da Barra da Tijuca ficou interditado para uso recreativo devido à presença de toxinas de cianobactérias na água do mar. De fato, a freqüente ocorrência de florações de cianobactérias tóxicas na lagoa de Jacarepaguá, leva-nos a concluir que é de grande importância o constante estudo de monitoramento tanto da qualidade da água quanto do pescado, principalmente porque essa lagoa ainda é utilizada para recreação e produção pesqueira.
AGRADECIMENTOS: A Fundação Estadual de Engenharia do Meio Ambiente (Feema), a Divisão de Laboratório (Dilab) e em especial as biólogas Gisele Gomara e Gláucia F. Sampaio.
REFERÊNCIAS
APHA. 1998. Standard Methods for Examination of Waste and Wastewater. American Public Health Association, New York.
894p.
BRANCO, C.W.C. 1998. Comunidades zooplanctônicas e aspectos limnológicos de três lagoas costeiras da região Norte Fluminense. (Macaé, RJ). Tese de Doutorado. Universidade
Federal do Rio de Janeiro. 277p.
BURNS, C.W.; FORSYTH, D.J.; HANEY, J.F.; JAMES,
M.R.; LAMPERT, W. & PRIDMORE, R.D. 1989. Coexistence
and exclusion of zooplankton by Anabaena minutissima
var. attenuata in Lake Rotongaio. New Zelaand. Archive für Hydrobioloy, 32: 63-82.
CARACO, N. & MILLER, R. 1998. Direct and indirect effects
of CO2 on competition between a cyanobacteria and eukaryotic
phytoplankton. Canadian Journal of Fisheries and Aquaic. Scence, 55: 54-62.
CHORUS, I & BARTRAM, J. 1999. Toxic cyanobacteria in water. A guide to public health consequences, monitoring and management. Published by E & FN Spon on behalf of WHO. 416p.
CHU, F.S.; HUANG, X. & WEI, R.D. 1990. Enzime linked
imunosorbent assay for microcystins in blue-green algae
blooms. Journal-Association of Official Analytical Chemists. 73: 451-456.
COUTTEAU, P. & SORGELOOS, P. 1997. Manipulation of
dietary lipids, fatty acids and vitamins in zooplankton cultures.
Freshwater Biology, 38: 501-512.
DEMOTT, W.R. & MÜLER-NAVARRA, D.C. 1997. The impor-
tance of highly unsaturated fatty acids in zooplâncton nutrition:
evidence from experiments with Daphnia, a cyanobacterium and
lipid emulsions. Freshwater Biology, 38: 649-664.
DEMOTT, W.R.; ZHANG, Q.X.; CARMICHAEL, W.W.
1991. Effects of toxic cyanobcteria and purified toxins on the
survival and feeding of a copepod and three species of Daphnia.
Limnology and Oceanography, 36: 1346-1357.
DOMINGOS, P. 2001. Dinâmica de Cianobactérias produtoras de microcistinas na Lagoa de Jacarepaguá (RJ). Tese de
Doutorado em Biotecnologia Vegetal. Universidade Federal do
Rio de Janeiro, Rio de Janeiro, Brasil. 111p.
DOMINGOS, P., HUSZAR, V. L., CARMOUZE, J. 1994.
Composition et biomasse de phytoplancton d´une lagune
tropicale (Brèzil) marquée par une mortalité de poissons. Revue D´Hydrobiologie Tropicale, 27 (3): 235-250.
DUY, T.N.; LAM, P.K.S.; SHAW, G.R. & CONNELL, D.W. 2000.
Toxicology and risk assessment of freshwater cyanobacterial
(blue green algal) toxins in water. Reviews of Environmental
Contamination and Toxicology. 163: 115-136.
EDMONDSON, W.T. & LITT, A.H. 1982. Daphnia in Lake
Washington. Limnology and. Oceanography. 27: 272-293.
ENRICH-PRAST, A.; BOZELLI, R.; ESTEVES, F.A. &
MEIRELLES, F. 2004. Lagoas Costeiras da Restinga de
Jurubatiba: Descrição de suas variáveis limnológicas. Pp.

343FLORAÇÕES DE CIANOBACTÉRIAS TÓXICAS EM UMA LAGOA COSTEIRA HIPEREUTRÓFICA
Oecol. Bras., 13(2): 329-345, 2009
245-254. In: C.F.D. Rocha, F.A. Esteves & F.R. Scarano. (eds).
Pesquisas de Longa duração na Restinga de Jurubatiba Ecologia,
História Natural e Conservação. RiMa, São Carlos. 376p.
ESTEVES, F.A. 1988. Fundamentos de limnologia. Interciência/
FINEP. Rio de Janeiro. 575p.
FEEMA (Fundação Estadual de Engenharia de Meio
Ambiente). 1982. Estudo da Barra da Tijuca – Fase I: Lagoa
de Jacarepaguá. 37p.
FEEMA (Fundação Estadual de Engenharia de Meio Ambiente).
1984. Quadro da situação ambiental da Baixada de Jacarepaguá.
Rio de Janeiro. DIPLAN. 107p.
FERNANDES, V.O. 1993. Estudos sucessionais da comunidade perifítica em folhas de Typha dominguensis Pers, relacionado às variáveis limnológicas na Lagoa de Jacarepaguá, RJ. Disser-
tação de Mestrado. Universidade Federal de São Carlos, São
Paulo, Brasil. 131p.
FERRÃO-FILHO, A.S. & AZEVEDO S.M.F.O. 2003. Effects of
unicellular and colonial forms of toxic Microcystis aeruginosa
from laboratory cultures and natural populations on tropical
cladocerans. Aquatic Ecology, 37: 23-35.
FERRÃO-FILHO, A.S.; AZEVEDO, S.M.F.O. & DEMOTT,
W.R. 2000. Effects of toxic and nontoxic cyanobacteria on the
life history of tropical and temperate cladocerans. Freshwater Biology, 43: 1-19.
FERRÃO-FILHO A.S.; DOMINGOS P. & AZEVEDO S.M.F.O.
2002a. Influences of a Microcystis aeruginosa Kützing bloom on
zooplâncton populations in Jacarepaguá Lagoon (Rio de Janeiro,
Brazil). Limnologica, 32: 295-308.
FERRÃO-FILHO, A.S; SUZUKI, B.K. & AZEVEDO, S.M.O.
2002b. Accummulation of microcystins by a tropical zooplâncton
community. Aquatic Toxicology, 59: 201-208.
GLIWICZ, Z.M. 1977. Food size selection and seasonal
succession of filter feeding zooplankton in an eutrophic lake.
Ekologia Polska. 25: 179-225.
HANAZATO, T. 1991. Interrelations between Microcystis and Clado-
cera in the higlhy eutrophic Lake Kasumigaura, Japan. Internationale Revue der gesamten Hydrobiolgie und Hydrographie, 76: 21-36.
HAZANATO, T. & YASUNO, M. 1987. Evaluation of Microcystis
as food for zooplankton in an eutrophic lake. Hydrobiologia,
144: 251-259.
HANEY, J.F. 1987. Field studies on zooplankton-cyanobacteria
interactions. New Zealand Journal of Marine and Freshwater Research, 21: 467-475.
HANSSON, L.; GUSTAFSSON, S.; RENGEFORS, K. &
BOMARK, L. 2007. Cyanobacterial chemical warfare affects
zooplankton community composition. Freshwater Biology, 52:
1290-1301.
HENING, M.; HERTEL, H.; WALL, H. & KOHL, J.G.
1991. Strain-specific influence of Microcystis aeruginosa
on food ingestion and assimilation of some cladocerans and
copepods. Internationale Revue der gesamten Hydrobiolgie und Hydrographie, 76: 37-45.
HIETALA, J.; REINIKAINEN, M. & WALLS, M. 1995.
Variation in life history responses of Daphnia to toxic Microcystis aeruginosa. Journal of Plankton Research, 17: 2307-2318.
INFANTE, A.; ABELLA, S.E.B. 1985. Inhibition of Daphnia by
Oscilatoria in Lake Washington. Limnology and Oceanography,
30: 1046-1052.
JARVIS, A.C.; HART, R.C. & COMBRINK, S. 1987.
Zooplankton feeding on size fractionated Microcystis colonies
and Chlorella in a hypereutrophic lake (Hartbeespoort Dam,
South Africa): Implications to resource utilization and
zooplankton succession. Journal of Plankton Research. 9:
1231-1249.
JOCHIMSEN, E.M.; CARMICHAEL, W.W.; AN, J.; CARDO,
D.; COOKSON, S.T.; HOLMES, C.E.M.; ANTUNES, M.B.C.;
MELO FILHO, D.A.; LYRA, T.M.; BARRETO, V.; AZEVEDO,
S.M.F.O. & JARVIS, W.R. 1998. Liver failure and death following
exposure to microcystin toxins at a hemodialysis center in Brazil.
The New England Journal of Medicine, 36: 373-378.
KJERFVE, B. 1994. Coastal lagoons. Pp. 1-8. In: B. Kjerfve,
(ed). Coastal lagoon processes. Elsevier Oceanography Series,
Amsterdam. 599p.
KNOPPERS, B. 1994. Aquatic primary production in coastal
lagoons. Pp. 243-286. In: B. Kjerfve. (ed). Coastal lagoon
processes. Elsevier Oceanography Series, Amsterdam. 599p.
KRISHNAMURTHY, T.; CARMICHAEL, W.W.; SARVER,
E.W. 1986. Toxic peptides from freshwater cyanobacteria
blue-green algae. Isolation, purification and characterization of
peptides from Microcystis aeruginosa and Anabaena flos-aquae. Toxicon, 24 (9): 865-873.
LAMPERT, W. 1987. Laboratory studies on zooplankton-
cyanobacteria interactions. New Zealand Journal of Marine and Freshwater Research, 21: 483-490.
LEONARD, J.A. & PEARL, H.W. 2005. Zooplâncton
community structure, micro-zooplâncton grazing impact, and
seston energy content in the St. Johns river system, Florida as

344 GOMES, A.M.A. et al.
Oecol. Bras., 13(2): 329-345, 2009
influenced by the toxic cyanobacterium Cylindrospermopsis raciborskii. Hydrobiologia, 537: 89-97.
LUND, J.W.G. & TALLING, J.F. 1957. Botanical limnological
methods with special reference to the algae. Botanical Review,
23: 485-583.
LUNDSTED, L. & BRETT, M.T. 1991. Differential growth rates
of three cladoceran species in response to mono- and mixed-algal
cultures. Limnology and Oceanography, 36: 159-165.
LÜRLING, M. 2003. Daphnia growth on microcystin-producing
and microcystin-free Microcystis aeruginosa in different mixtures
with the green alga Scenedesmus obliquus. Limnology and Oceanography, 48: 2214–2220.
MAGALHÃES, V.F.; SOARES, R.M. & AZEVEDO, S.M.F.O.
2001. Microcystin contamination in fish from the Jacarepaguá
(RJ, Brazil): ecological implication and human health risk.
Toxicon. 39: 1077-1085.
MATVEEV, V.F. & BALSEIRO, E.G. 1990. Contrasting responses
of two cladocerans in the nutritional value of nanoplânkton.
Freshwater Biology, 23: 197-204.
MELLO, S. 2001. Fitoplâncton da Lagoa de Imbossica (Macaé, RJ – Brasil): Flora, Estrutura de Comunidade e Variações espaciais e temporais. Tese de Doutorado em Ecologia. Universidade
Federal do Rio de Janeiro, Rio de Janeiro, Brasil.154p.
MARINHO, M.M. & HUSZAR, V.L.M. 2002 Nutrient availability
and physical conditions as controlling factors of phytoplankton
composition and biomass in a tropical reservoir (Southeastern
Brazil). Archive für Hydrobiologie. 153: 443-468.
NANDINI, S. 2000. Responses of rotifers and cladocerans to
Microcystis aeruginosa (Cyanophyceae): a demographic study.
Aquatic Ecology, 34: 227-242.
NASELLI-FLORES, L. 2000. Phytoplankton assemblages in
twenty one Sicilian reservoirs: relatioships between species compo-
sition and environmental factors. Hydrobiologia, 424: 1-11.
NÜRNBERG, G. 1996. Trophic state of clear and colored, soft-
and hardwater lakes with special consideration of nutrients,
anoxia, phytoplankton and fish. Lake and Reservoir Management, 12: 432-447.
PADISÁK, J. 1997. Cylindrospermopsis raciborskii
(Woloszynska) Seenaya et Subba Raju, an expanding, highly
adaptative cyanobacterium: worldwide distribution and review of
its ecology. Archiv für Hydrobiologie, 107: 563-593.
PANOSSO, R.F.; ATTAYDE, J.L.; MUEHE, D. 1998.
Morfometria das lagoas Imboassica, Cabiúnas, Comprida e
Carapebus: Implicações para seu funcionamento e manejo. Pp.
91-108 In: F.A Esteves, (Ed.) Ecologia das Lagos Costeiras do
Parque Nacional da Restinga de Jurubatiba e do Município de
Macaé (RJ). Universidade Federal do rio de janeiro. 464p.
PAERL, H.W. & HUISMAN, J. 2008. Blooms like it hot. Science,
320: 57-58.
PORTER, K.G. & ORCUTT, J.D. 1980. Nutritional adequacy,
manageability and toxicity as factors that determine the food
quality of green and blue-green algae for Daphnia. Pp. 268-281.
In: W.C. Kerfoot (ed.). Evolution and ecology of zooplankton
communities. Hanover University Press of New England.793p.
REPKA, S. 1997. Effects of food type on the life history of
Daphnia clones from lakes differing in trophic state. I. Daphnia galeata feeding on Scenedesmus and Oscillatoria. Freshwater Biology, 37: 675-683.
SAIEG-FILHO, E. 1986. Ecologia do Fitoplâncton Marginal das Lagunas da Baixada de Jacarepaguá, Rio de Janeiro-RJ.
Monografia de Bacharelado em biologia. Universidade Federal
do Rio de Janeiro, Rio de Janeiro, Brasil. 150p.
SALAS, H.J. & MARTINO, P. 1991. A simplified phosphorus
trophic state model for warm-water tropical lakes. Water Research, 25: 341-350.
SEMADS (Secretaria Estadual de Meio Ambiente e
Desenvolvimento Sustentável). 2001. Ambiente das águas do estado do Rio de Janeiro. Rio de Janeiro: Sholna Reproduções
gráficas. 203p.
SEMERARO, J. & COSTA, A.F. 1972. O Plâncton e a Poluição
nas lagoas da Tijuca, Camorim e Jacarepaguá. XII Congresso
Interamericano de Engenharia Sanitária, Assunção, Paraguai.
Tema 4, Publ. no.3: 31p.
SILVA, G. 2006. Processo de Ocupação Urbana da Barra da Tijuca (RJ): Problemas Ambientais, Conflitos Sócio-Ambientais, Impactos Ambientais Urbanos. Pesquisa em arquitetura e
Construção. 28p.
SMAC (Secretaria Municipal de Meio Ambiente) 2002. Qualidade
Ambiental – primavera, Verão, Outono. Rio de Janeiro.172p.
SMITH, A.D. & GILBERT, J.J. 1995. Relative susceptibilities of
rotifers and cladocerans to Microcystis aeruginosa. Archive für Hydrobiologie, 132: 309-336.
SOARES, M. C. S. 2008. Cianobactérias Tóxicas no Reservatório do Funil: Uma batalha Pelágica. Tese de
Doutorado em Botânica. Universidade Federal do Rio de
Janeiro, Rio de Janeiro, Brasil. 419p.

345FLORAÇÕES DE CIANOBACTÉRIAS TÓXICAS EM UMA LAGOA COSTEIRA HIPEREUTRÓFICA
Oecol. Bras., 13(2): 329-345, 2009
SPAULDING, E. 1994. Modeling of circulation and dispersion in
coastal lagoons. Pp 69-101 In: B. Kjerfve, (Ed). Coastal lagoon
processes. Elsevier Oceanography Series, Amsterdam. 577p.
UHELINGER, V. 1964. Étude statisque des méthodes de
dénobrement planctonique. Archives of Sciences, 17: 121-123.
UTERMÖHL, H. 1958. Zur Vervollkommung der quantitativen
Phytoplankton Methodik. Mitteilungen der Internationalen Vereinigung für Limnologie, 9: 1-39.
VOLLENWEIDER, R. & KEREKES, J. 1980. The loading
concept as basis for controlling eutrophication philosophy and
preliminary results of the OECD programme on eutrophication.
Program Water Technoloy, 12: 5-38.
Submetido em 09/12/2008.Aceito em 30/01/2009.


















