APOSTILA tecidos vegetais 01 (1)
40
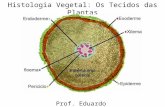
Histologia Vegetal – prof Arlindo Costa – UDESC - 2001 1 1 1. CÉLULA VEGETAL CONSIDERAÇÕES FINAIS Um dos grande avanços na biologia no século XIX foi a descrição de uma unidade estrutural, que forma todos os seres vivos, sejam animais ou vegetais. Esta unidade é a célula, nome que tem se mantido até nossos dias, apesar de todas as modificações que ocorreram nos conceitos de sua estrutura e função. As células são universalmente consideradas as unidades de vida pois podem viver por si só, como no caso de muitas algas e protozoários ou artificial- mente em culturas de laboratório. A cinética da célula vegetal (Fig. 1) é semelhante é da célula animal, ou seja, vários processos metabólicos são comuns a elas. Existem, entretanto, carac- terísticas que são peculiares à célula dos vegetais, principalmente no que se refere á parede celular que envolve o protoplasto, isto é, todo o conteúdo celular. No pro- toplasto encontramos como características da célula vegetal vacúolo(s), plastos e substâncias ergásticas. Figura 1. Diagrama de uma célula vegetal. CG - complexo de Golgi; Cl - cloroplasto; CP - campo primário de pontuação; Mi - microtúbulos; Mi - mitocôndrias; N - núcleo; Nu - nucléolo; PC - parede celular; Pd - plasmodesmo; Pl - plasmalema; RE - retículo endoplasmático. PAREDE CELULAR A parede celular é um componente típico da célula vegetal. Devido à presença da parede celular, a distensão do protoplasto fica restrita e, as- sim, o tamanho e a forma da célula tornam-se fixos na maturidade. O principal composto da parede celular é a celulose, um polissaca- rídeo de fórmula empírica (C6H10O5) cujas moléculas são cadelas lineares de glicose. A celulose se apresenta em forma de estruturas filamentosas chamadas de microfibrilas (com 4 a 20nm de diâmetro), que se reúnem em feixes maiores constituindo a armação fundamental da parede celular. A sín- tese das microfibrilas de celulose é realizada por enzimas situadas na plas- malema e que tem a forma de rosetas ou de rosetas e glóbulos. Nas microfi- brilas há partes em que as moléculas de celulose mostram um arranjo orde- nado; esse arranjo é que dá é celulose propriedade cristalina, apresentando dupla refração (birrefringência), o que a torna brilhante em condições de luz polarizada. Na parede celular a celulose está associada com outros polissa- carídeos, principalmente hemiceluloses e compostos pécticos. Muitas outras substâncias, orgânicas e inorgânicas, ocorrem nas paredes celulares em quantidades variáveis, dependendo do tipo de célula. Entre as orgânicas destacam-se as de natureza protéica e as de natureza lipídica com cutina, suberina e ceras. Estas últimas se encontram nos tecidos protetores superfi- ciais da planta. Em muitos tipos de células a parede é incrustada pela lignina, com a qual adquire maior rigidez. Na parede de muitas células pode ser vista uma certa lamelação, que é conseqüência do modo e do grau de cres-
-
Upload
shemuellsm -
Category
Documents
-
view
251 -
download
0
Transcript of APOSTILA tecidos vegetais 01 (1)

Histologia Vegetal – prof Arlindo Costa – UDESC - 2001
1
1
1. CÉLULA VEGETAL CONSIDERAÇÕES FINAIS
Um dos grande avanços na biologia no século XIX foi a descrição de uma unidade estrutural, que forma todos os seres vivos, sejam animais ou vegetais. Esta unidade é a célula, nome que tem se mantido até nossos dias, apesar de todas as modificações que ocorreram nos conceitos de sua estrutura e função.
As células são universalmente consideradas as unidades de vida pois podem viver por si só, como no caso de muitas algas e protozoários ou artificial-mente em culturas de laboratório.
A cinética da célula vegetal (Fig. 1) é semelhante é da célula animal, ou seja, vários processos metabólicos são comuns a elas. Existem, entretanto, carac-terísticas que são peculiares à célula dos vegetais, principalmente no que se refere á parede celular que envolve o protoplasto, isto é, todo o conteúdo celular. No pro-toplasto encontramos como características da célula vegetal vacúolo(s), plastos e substâncias ergásticas.
Figura 1. Diagrama de uma célula vegetal. CG - complexo de Golgi; Cl - cloroplasto; CP - campo primário de pontuação; Mi - microtúbulos; Mi - mitocôndrias; N - núcleo; Nu - nucléolo; PC - parede celular; Pd - plasmodesmo; Pl - plasmalema; RE - retículo endoplasmático.
PAREDE CELULAR A parede celular é um componente típico da célula vegetal. Devido
à presença da parede celular, a distensão do protoplasto fica restrita e, as-sim, o tamanho e a forma da célula tornam-se fixos na maturidade.
O principal composto da parede celular é a celulose, um polissaca-rídeo de fórmula empírica (C6H10O5) cujas moléculas são cadelas lineares de glicose. A celulose se apresenta em forma de estruturas filamentosas chamadas de microfibrilas (com 4 a 20nm de diâmetro), que se reúnem em feixes maiores constituindo a armação fundamental da parede celular. A sín-tese das microfibrilas de celulose é realizada por enzimas situadas na plas-malema e que tem a forma de rosetas ou de rosetas e glóbulos. Nas microfi-brilas há partes em que as moléculas de celulose mostram um arranjo orde-nado; esse arranjo é que dá é celulose propriedade cristalina, apresentando dupla refração (birrefringência), o que a torna brilhante em condições de luz polarizada. Na parede celular a celulose está associada com outros polissa-carídeos, principalmente hemiceluloses e compostos pécticos. Muitas outras substâncias, orgânicas e inorgânicas, ocorrem nas paredes celulares em quantidades variáveis, dependendo do tipo de célula. Entre as orgânicas destacam-se as de natureza protéica e as de natureza lipídica com cutina,
suberina e ceras. Estas últimas se encontram nos tecidos protetores superfi-ciais da planta. Em muitos tipos de células a parede é incrustada pela lignina, com a qual adquire maior rigidez.
Na parede de muitas células pode ser vista uma certa lamelação, que é conseqüência do modo e do grau de cres-

Histologia Vegetal – prof Arlindo Costa – UDESC - 2001
2
2
cimento dessa parede e do arranjo apresentado pelas microfibrilas nos sucessivos acréscimos de material constituinte. As camadas que se formam primeiro constitu-em a parede primária. As microfibrilas apresentam uma disposição intercalar e o crescimento ocorre por intussuscepção. Em muitas células são depositadas ainda, internamente à parede primária, camadas adicionais, que constituem a parede se-cundária. Tais camadas são designadas respectivamente por S1, S2 e S3. O arran-jo das microfibrilas varia nas diferentes camadas (Fig. 2), sendo que a última cama-da (S3) pode estar ausente. A deposição das microfibrilas se dá por aposição. A parede secundária ocupa assim uma posição mais interna. A linha de união entre as paredes primárias de duas células contíguas é denominada lamela média ou lamela intercelular ou substância intercelular e tem natureza péctica. A parede pri-mária é geralmente mais fina nas células que possuem paredes secundárias. O tipo de parede celular vai determinar a textura do tecido vegetal. Figura 2. Diagrama da estrutura em camadas da parede de uma fibra. O ângulo de deposição das microfibrilas é mostrado pelas linhas oblíquas na parede primária e camadas S1 (externa), S2 (central), S3 (interna) na parede secundária. LM - lamela média; PP - parede primária; PS - parede secundária. A formação da parede celular ocorre na telófase com a formação da placa celular. O desenvolvimento da placa celular é associado com o fragmoplasto constituído por microtúbulos e pequenas vesículas (originadas dos dictiossomos e do retículo en-doplasmático), que surge, inicialmente, no centro do plano equatorial do fuso. A placa celular forma-se por coalescência daquelas vesículas e cresce radialmente (para fora) até fusão com a parede da célula-mãe. Conforme a placa cresce, os microtúbulos e as vesículas são encontrados apenas mais externamente, sugerindo que os microtúbulos atuam no direcionamento das vesículas.
Na placa celular há deposição de material da parede. Admite-se que a placa celular é composta de substâncias pécticas e torna-se a lamela média, que se situa entre as células filhas que estão se formando. Concomitantemente há deposição de material celular sobre a antiga parede da célula-mãe, visto que as células-filhas estão se alongando. Dessa forma, cada célula filha
forma a sua parede primária completa. A parede secundária quando formada aparece mais inter-namente á parede primária. Durante a formação da lamela média e parede primária elementos do retículo endoplasmático ficam retidos entre as vesículas que estão se fundindo, origi-nando os futuros plasmodesmas. Estes são continuidades protoplasmáticas entre uma célula e outra, localizando-se em pequenas depressões da parede primária. Essas depressões da parede celular primária são conhecidas como campos primários de pontuação ou pontuações primárias. Posteriormente, durante a deposição da parede secundária, nenhum material de parede se deposita sobre o campo na pontuação primário formando diferentes tipos de pontuações (Fig. 3A, 3B). Essas surgem em conseqüência da deposição diferencial da parede secundária sobre a primária. Há vários tipos de pontua-

Histologia Vegetal – prof Arlindo Costa – UDESC - 2001
3
3
ções sendo que dois tipos são mais comuns: pontuação simples e pontuação areo-lada. Figura 3. Diagrama de pares de pontuações em vista tridimensional. A. Pontuação simples, B. Pontuação areolada. AbPo - abertura de pontuação; CaPo - Canal de pontuação; LM - lamela média; MePo - membrana de pontuação; PL - plasmodesmos; PP - parede primária; PS - parede secundária Na pontuação simples (Fig. 3A) ocorre apenas uma interrupção da parede secundá-ria. O espaço em que a parede primária não é recoberta pela secundária constitui a chamada cavidade na pontuação. Entre as paredes de duas células podem existir e pontuações que se correspondem e constituem um par de pontuações. Neste caso, além das cavidades de pontuação, existe a membrana de pontuação. Esta mem-brana é formada pelas paredes primárias de ambas as células do par mais a lamela média entre elas. Note-se se que é a única situação em que, referindo-se à parede celular, fala-se em membrana (Fig. 3A, 3B). A pontuação areolada (Fig. 3B) recebe este nome porque em vista frontal se mostra como uma aréola, ou seja, apresenta uma saliência de contorno circular e no centro desta encontra-se uma abertura, também circular. Nesta pontuação é a parede secundária que forma a aréola e a interrupção desta corresponde a abertura da anota. Como a parede secundária apresenta-se bem separada da parede primária, delimita-se internamente a chamada câmara de pontuação. O par de pontuações areoladas apresenta também a membrana de pontuação pontuações areoladas, com características descritas acima, são encontradas em células do sistema condu-tor (xilema), isto é, nos elementos de vaso e traqueídeos. Numa célula que possui pontuação areolada, quando a parede secundária continua a se espessar depois de estar formada a câmara de pontuação e a abertura, distingue-se, além da abertura original, uma segunda abertura. Esta segunda abertura constitui a abertura interna
da pontuação, pois está voltada para o lume da célula e a primeira constitui a abertura externa da pontuação, pois está mais próxima da parede primária. Entre as aberturas, há o canal da pontuação que pode ser cilíndrico quando se mantém como uma projeção da antiga abertura que é circular como se vê na pontuação simples dos esclereídeos. Em outros casos, tem a forma de um funil achatado e a abertura interna pode ultrapassar os limites da aréola como se vê nos fibrotraqueídeos que ocorrem no xilema. As aberturas das duas pontuações que constituem uma par formam um ângulo de 45° entre si. Nas paredes dos traqueídeos, um tipo de célula condutora do xilema, de co-níferas e algumas angiospermas primitivas, ocorre, na membrana da pontua-ção areolada, um espessamento especial chamado toro. O restante da mem-brana em volta do toro é denominado margo. As células podem possuir mais de um tipo de pontuação e com tamanhos diferentes (Fig. 4), variações estas decorrentes das outras células circunvizinhas. Por exemplo, um elemento de vaso pode apresentar par de pontuações areoladas (Fig. 3B) quando estiver contíguo a outros elementos de vaso ou par de pontuações semi-areoladas quando estiver contíguo a outro tipo de célula, como por exemplo uma célula do parênquima.
Figura 4. Diagrama de elementos de vaso, mostrando as paredes com diferentes tipos de pontuação 3. CONTEÚDO

Histologia Vegetal – prof Arlindo Costa – UDESC - 2001
4
4
CELULAR Do conteúdo de uma célula vegetal merecem especial destaque o vacúolo, os plas-tos e as substâncias ergásticas. O vacúolo (Fig. 1), limitado por uma membrana unitária denominada tonoplasto, contém água e uma cena variedade de substâncias inorgânicas e orgânicas, muitas das quais estão dissolvidas. Entre as substâncias existentes no vacúolo podem ser encontrados açúcares, ácidos orgânicos, proteínas, etc. Um exemplo conhecido ocorre no endosperma da semente de mamona (Ricinus communis) que contém proteínas nos microvacúolos. Encontram-se ainda oxalato de cálcio na forma de cristais (prismáticos, drusas, estilóides, rafídeos ou ráfides, etc), substâncias fenóli-cas em forma de inclusões e pigmentos como betalaínas e antocianinas. Em células diferenciadas é comum a ocorrência de um só vacúolo, o qual ocupa um considerá-vel volume celular, em células parenquimáticas, por exemplo, o vacúolo chega a ocupar 90% do espaço celular. O vacúolo tem participação ativa em processos metabólicos celulares, tais como: senescência (envelhecimento), diferenciação e mobilização de reservas. Também dentro dos vacúolos, especialmente nos bem desenvolvidos e de posição central, ocorrem enzimas envolvidas tom a digestão de constituintes celulares Nesse pro-cesso, em alguns locais dos limites do vacúolo, o tonoplasto sofre invaginações, e tais invaginações "carregam" material citoplasmático com constituintes celulares: mitocôndrias, plastídios, ribossomos e outros. Cada invaginação se destaca do tonoplasto e forma uma vesícula que fica suspensa no vacúolo; numa fase final ocorre a lise dos materiais trazidos para dentro do vacúolo. A ação hidrolítica do vacúolo faz com que muitos pesquisadores o considerem como parte relevante do sistema lisossômico da célula vegetal. Os plastos ou plastídios (Figs. 5 e 6) são organelas que possuem um envelope formado por duas membranas unitárias e que, internamente, possui uma matriz (ou
estroma). Na matriz se situa um sistema de membranas, que consiste de formações saculiformes achatadas e que são chamados tilacóides. O grau de expressão atingido pelo sistema de tilacóides depende do tipo de plastídio. O plastídio contém ácido desoxirribonucleico e ribossomos e poderia, portanto, ser geneticamente autônomo. Os plastos são organelas que se apresentam com formas e tamanhos dife-rentes e são classificados de acordo com a ausência ou presença de pigmen-to ou com o tipo deste último. Há três grandes grupos de plastos: cloroplasto, cromoplasto e leucoplasto Os plastos podem passar de um tipo para o outro.

Histologia Vegetal – prof Arlindo Costa – UDESC - 2001
5
5
Figura 5. Diagrama de cloroplasto (corte mediano e detalhe). Observe os envoltó-rios e o estroma contendo os tilacóides. Veja a interconexão dos tilacóides e os plastoglóbulos. E - envoltório; Es -estroma; G - grano; Plg - plastoglóbulo; TG - tilacóide do grano; TEs - tilacóide do estroma. Os cloroplastos (Fig. 5) contém clorofila e estio associados à fotossíntese, ocorren-do em todas as partes verdes da planta, mas sendo mais numerosos e mais dife-renciados em folhas. No sistema de tilacóides do cloroplasto de plantas superiores distinguem-se pilhas de tilacóides em forma de discos chamados de granos (singu-lar:grano) e os tilacóides de estroma, que conectam os granos entre si. A clorofila encontra-se no sistema de tilacóides; tal sistema é, portanto, a sede das reações fotoquímicas responsáveis pela captação e transformação da energia luminosa em energia química. A matriz ou estroma é o local de ocorrência das reações envolvi-das na fixação do gás carbônico para produção de carboidratos, além de outros derivados tais como: aminoácidos, ácidos orgânicos e ácidos graxos. Em certas condições, como por exemplo numa longa exposição à luz, o cloroplasto forma e acumula amido (amido de assimilação). Nos cloroplastos podem estar presentes também os lipídios, estes últimos em forma de glóbulos, os chamados plastoglóbu-los. Os cromoplastos são portadores de pigmentos carotenóides (amarelos, vermelhos, alaranjados, etc.), sendo encontrados em pétalas e outras partes coloridas de flo-res, em frutos e em algumas raízes. Os cromoplastos surgem, em grande parte dos casos, a partir de transformações dos cloroplastos, que sofrem modificações diver-sas levando ao rompimento de tilacóides. O cromoplasto tem a capacidade de sin-tetizar e acumular pigmentos. Na cenoura (Daucus carota) acumula caroteno, no tomate (Lycopersium escutelum) licopeno. O cromoplasto que se desenvolveu a
partir de cloroplasto pode retornar à forma original e em tal caso o cromoplas-to perde o caroteno e desenvolve mais o sistema de tilacóides e clorofila.
Figura 6. Amiloplastos da batata inglesa. A e B - grãos de amido compostos; C - grão de amido simples; D - grão de amido semi-composto. Quanto aos leucoplastos, (Fig.6A, B, C e D) estes não possuem pigmentos e podem armazenar várias substâncias. Os leucoplastídios armazenadores de amido são chamados de amiloplastos, ocorrendo por exemplo em tubérculos de batatinha inglesa (Solanum tuberosum). Os que reservam proteínas são chamados de proteinoplastos. Elaioplastos são os plastídios que reservam lipídios e ocorrem por exemplo em abacate(Parsea americana). Nos amilo-plastos pode ocorrer um armazenamento de um a vários grãos de amido, pode ser visto também que o sistema de tilacóides é pobre. Leucoplastos de tecidos que se tornam expostos à luz podem desenvolver se em cloroplastos como no caso de batatinha inglesa. Todos os tipos de plastos derivam dos protoplastos ou prtoplastídios, que são organelas muito pequenas; na presença de luz originam todo o sistema lame-lar. Ocorre na oosfera e em células meristemáticas. O proplasto desenvolvido

Histologia Vegetal – prof Arlindo Costa – UDESC - 2001
6
6
na ausência de luz, apresenta um sistema especial derivado da membrana interna, formando tubos que se fundem e formam o corpo prolamelar, este plasto recebe o nome de estioplasto. Tanto os proplastos como os plastos podem multiplicar-se por divisão. As substâncias ergásticas são produtos do metabolismo celular e o nome vem do grego, "erg" que significa trabalho. Muitos são materiais de reserva e/ou produtos descartados pelo metabolismo. São substâncias encontradas na parede celular e nos vacúolos, podendo também estar associadas a outros componentes protoplas-máticos. Entre as substâncias ergásticas mais conhecidas destacam-se a celulose, amido, corpos de proteína, lipídios e substâncias relacionadas, além de matéria mineral em forma de cristais de oxalato de cálcio (ráfides, drusas, estilóldes, etc.), de carbonato de cálcio (cistólito) e de sílica (estruturas retangulares, cônicas etc.). Também são ergásticas muitas outras substâncias orgânicas, tais como: substân-cias fenólicas, resinas, gomas, borracha, alcalóides. Muitas vezes, as células que contém as substâncias ergásticas são diferentes morto e fisiologicamente das de-mais células do tecido e, neste caso, recebem o nome de idioblastos.
2. MERISTEMAS 1. ORGANIZAÇÃO HISTOLÓGICA Nas raízes e caules altamente organizados, cada célula passa por uma série orde-nada de fases de crescimento. A célula cúbica produzida na região meristemática ou meristema de uma planta em desenvolvimento te muitos vacúolos. A medida dos vacúolos aumenta e estas finalmente se fundem num só vacúolo central grande. O resto da célula acompanha o aumento de tamanho pela produção de mais material da parede celular, do citoplasma e dos vários tipos de organelas celulares.
A diferenciação ocorre, geralmente, junto com o alongamento, mas algumas vezes segue-se a este. As células externas da raiz, por exemplo, adotam uma de duas formas finais: ou se achatam e funcionam como células epidér-mica com uma extensão bastante longa que absorve prontamente água e sais minerais. Durante a rápida fase de crescimento dessas células, o núcleo quase sempre se situa na extremidade do pêlo e parece ser o centro de grande atividade metabólica. Os pêlos absorventes têm vida curta, mas são produzidos em grande quantidade á medida que a ponta da raiz se aprofunda no solo. Os pêlos absorventes aumentam bastante a área da raiz em contac-to com o solo . Os tecidos centrais da raiz diferenciam-se nos elementos vasculares, uma vez que este órgão, caracteristicamente, não possui medula. Isso também ocorre nos tecidos em cultura. Bem no interior das massas de crescimento rápido de uma cultura de tecido não-diferenciada, encontramos pequenos grupos de traquéides. Por isso supomos que há alguma coisa no interior da massa de células ou falta de contacto com o solo ou outro meio externo.
Envolvendo as células xilemáticas da raiz há três estruturas: 1. Feixes de floema; 2. periciclo meristemático, que dá origem às ramificações de raiz, 3. Uma endoderme que circunda todo o cilindro vascular central.
A endoderme tem uma curiosa espessada, a estria de Caspary, que repre-senta um espaçamento em, forma de faixa nas paredes de suas células. Al-guns botânicos imaginam que a estria de Caspry, impermeável á água, fun-cione como uma espécie de barreira que impeça a difusão de água a longo da parede e force o movimento de todos os materiais através das membra-nas diferencialmente permeáveis das células endodérmicas. Esta teoria é ainda incerta.

Histologia Vegetal – prof Arlindo Costa – UDESC - 2001
7
7
Entre o cilindro vascular interno e a epiderme há um grupo de células não-difertenciadas, dispostas frouxamente, que constitui o córtex. Estas células são grandes, de paredes finas, nucleadas e possuem grandes vacúolos centrais. Sua função, provavelmente, é de armazenar materiais de reserva na raiz . Como entre o xilema e o floema do cilindro central se desenvolve um câmbio e co-mo as raízes se espessam graças às divisões radiais de suas células, o córtex se torna cada vez menor, fendendo-se e descamando-se para fora da raiz. Finalmente, numa raiz mais velha, e epiderme e o córtex perdem-se completamente. A nova camada externa é composta por células suberificadas que constituem a periderme. Estas células suberificadas desenvolvem-se a partir de um meristema secundário, o câmbio suberógeno (felogênio). Esse padrão de crescimento e desenvolvimento é particularmente aparente em caules que se espessam com a idade. A extremidade caulinar, como a radicular, tem uma zona meristemática, cujas células se dividem rapidamente e atrás da qual existe uma região de células que se alongam com rapidez. Uma extremidade cauli-nar é mais complicada do que radicular, pois, além de formar os tecidos do caule, deve produzir gemas e folhas. As gemas são visíveis primeiro como pequeninas projeções de tecido, que se de-senvolvem como gemas vegetativas ou floríferas (Fig.4.8). Em muitas plantas, a natureza da gema é controlada pelas condições ambientais, como temperatura e luz. Por trás da região de alongamento, está a zona de diferenciação. Aqui, também, pode-se ver perfeitamente o desenvolvimento dos tecidos epidérmicos, de um cilin-dro vascular central e de células corticais entre os dois. Provavelmente, a diferença anatômica principal entre caules e raízes é que os primeiros têm, geralmente, me-dula central.
O xilema situa-se ao redor da medula e o floema envolve o xilema. Caules que crescem à luz geralmente não tem uma endoderme, ao contrário dos desenvolvidos no escuro. Nos caules, como nas raízes, a camada cambial desenvolve-se entre o xilema e o floema. Por divisões rápidas, para dentro e para fora, o câmbio dá origem a células que se diferenciam no xilema (para dentro) e no floema (para fora). Eventualmente, são originadas grandes pressões devidas a esse crescimento interno que causam rompimento e descamação das camadas externas no caule. À medida que isso ocorre, a planta produz novas células protetoras sob as áreas que descamam. Aqui de novo, é um câmbio suberógeno que surge e as células produzidas por ele são de paredes impermeáveis à água e típicas dos tecidos suberosos contidos na casa das árvores e arbustos. Os anéis anuais dos caules das árvores resultam de condições climáticas diferentes nos diversos períodos do ano. Na primavera, quando existe água em abundância e as outras condições são favoráveis, os câmbio produz célu-las de paredes delgadas que contêm uma cavidade central grande. Especi-almente no outono, quando as condições tendem a ser menos favoráveis, as traqueídes formadas têm paredes mais espessas, com cavidades menores. Esta alternância regular de lenho primaveril e de fim do verão e começo do outono produz um anel anual. A transição do lenho primaveril para o estival é geralmente gradual, mas a parada abrupta no fim da estação de crescimento é perfeitamente distinta do lenho primaveril do ano seguinte A regularidade dos anéis anuais nos permite datar as árvores e, portanto, as civilizações em que foram encontrados os restos desses vegetais. Por exem-plo, sabemos que certos ciclos climáticos ocorreram em várias regiões. Se um ano for particularmente favorável ao crescimento será produzido um anel, anual bastante espesso. Nos anos secos aparecem anéis anuais muito pe-

Histologia Vegetal – prof Arlindo Costa – UDESC - 2001
8
8
quenos. A seqüência de anéis anuais grandes e pequenos, formando um padrão que reflete as mudanças climáticas do passado, tende a ser constante nas árvores que habitam determinada região. Assim, um pedaço de madeira usado como esco-ra numa casa de uma civilização extinta poderia ser comparado com outros materi-ais de idade conhecida e a civilização poderia ser datada por essa técnica. Embora extremamente útil, o método nem sempre é seguro, porque, às vezes, as árvores produzem diversos anéis de crescimento num ano e os anéis anuais de anos su-cessivos nem sempre estão perfeitamente separados. A velocidade do crescimento de uma planta depende de sua constituição genética (genótipo) e do ambiente. Suponhamos que temos um pequeno grupo de plantas que foram autopolinizadas ou cruzadas somente dentro de seu grupo limitado por diversas gerações. Os descendentes dessa plantas são chamados consangüíneos. Se, por outro lado as plantas são polinizadas por plantas de grupos (''linhagens'') diferentes, os descendentes são ditos híbridos. Os geneticistas de há muito sabem que os cruzamentos sucessivos entre consangüíneos diminuem o vigor de uma linhagem, mas os cruzamentos de diferentes linhagens consangüíneas enfraqueci-das frequentemente produzem híbridos muito vigorosos. A isso se chama vigor híbrido ou heterose. As causas de heterose ainda não estão perfeitamente esclare-cidas. Sabemos, todavia, que os híbridos usam os elementos de seu meio com mais eficiência para o crescimento do que os consangüíneos. Os elementos do ambiente exercem enorme controle sobre uma planta. Por exem-plo, pouca água retarda o crescimento. Quantidades reduzidas de nitrogênio, po-tássio, fósforo ou de qualquer elemento essencial retardam o crescimento ou ma-tam a planta. A intensidade luminosa que atinge uma planta fotossintetizante tam-bém determina sua taxa de crescimento e a possibilidade de sua sobrevivência. Sem luz e dióxido de carbono suficientes, uma planta fotossintetizante não pode
armazenar bastante energia para suas necessidades de crescimento e de-senvolvimento. A temperatura ambiente pode ser também extremamente importante na de-terminação da natureza e da taxa de crescimento do vegetal. Na maioria dos processos químicos, a velocidade da reação aumenta constantemente com um aumento de temperatura. Em geral, a velocidade de uma reação química é duplicada para cada aumento de 10ºC na temperatura. Por causas que não compreendemos, plantas diferentes têm temperaturas (ótimos de temperatu-ra) bastante diversas, nas quais crescem melhor. Isso indica que algum pro-cesso bioquímico fundamental é afetado adversamente por temperatura alta. Assim, se elevarmos constantemente a temperatura, atingiremos, eventual-mente, um valor no qual os efeitos negativos e positivos se combinem para torná-la melhor para o crescimento. Se continuarmos a elevar a temperatura acima desse ponto a taxa de crescimento cairá, às vezes muito intensamen-te, em virtude dos efeitos delétricos sobrepujarem os benéficos. Para maioria das plantas, o ótimo de temperatura situa-se na faixa de 28-32ºC. Não sabemos por que as plantas são injuriadas por temperaturas ao redor de 35ºC. Tanto quanto se sabe, enzimas obtidas de plantas não são alteradas por essa temperatura. Uma suposição é que certas substâncias químicas, essenciais ao crescimento, produzidas pela planta, podem ser destruídas ou impedidas de se formarem em quantidade adequada, em temperaturas ele-vadas. Por exemplo, o bolor vermelho do pão, Neurospora, possui genes ''sensíveis'' à temperatura. O gene responsável pela produção da vitamina B2
em uma das linhagens de Neurospora, funciona muito bem quando o orga-nismo é cultivado em temperaturas baixas. Não funciona bem quando o or-ganismo está crescendo em temperaturas mais elevadas. A 35ºC, o organis-mo requer uma fonte externa de B2, mas a 25ºC ele produz essa substância

Histologia Vegetal – prof Arlindo Costa – UDESC - 2001
9
9
em quantidade suficiente. Provavelmente, uma situação geral idêntica deve ocorrer nas plantas superiores. Se soubermos por que uma temperatura alta retarda a taxa de crescimento, poderemos melhorá-lo grandemente, suprindo a planta com o ma-terial de que necessita. PLANTAS EM CRESCIMENTO Introdução: O desenvolvimento de uma semente até uma planta adulta é um processo extraor-dinário. Envolve crescimento por divisão e extensão celulares, diferenciação de novos órgãos como raízes, caules, folhas e flores e uma série complexa de altera-ções químicas. A forma final da planta é uma mistura de sua ''matriz'' genética com os efeitos modificadores do ambiente (fig.41). Uma semente contém um embrião circundado e protegido por um tegumento e nu-trido por uma fonte de alimento armazenado, o endosperma. A planta embrionária contém um ponto de crescimento radicular, outro caulinar e os cotilédones. Estes podem ser longos, delgados e foliáceos. Se não assim, servem inicialmente para digerir o alimento armazenado no tecido do endosperma e que será utilizado pelos pontos de crescimento do embrião; depois disso, passam a funcionar como órgãos fotossintetizadores. Em outros casos, os cotilédones são órgãos armazenadores suculentos, que saem do solo ou permanecem aí mergulhados e absorvem o mate-rial do endosperma antes que a semente amadureça. Tais cotilédones raramente se tornam foliáceos ou fotossintetizadores (fig.43). FIG. PAG 54. Quando a semente começa a germinar, absorve grande quantidade de água e os pontos de crescimento iniciam a divisão celular. Por motivos ainda não esclareci-dos, a raiz quase sempre começa a desenvolver-se antes do caule. Nas termina-
ções da raiz e do caule do embrião, são formadas novas células pelas zonas meristemáticas dos pontos de crescimento. Divisão, alongamento e diferenci-ação celulares ocorrem na raiz em regiões que se sobrepõem. Como a raiz penetra pelo solo, seu delicado ponto de crescimento precisa ser protegido contra o atrito. Essa proteção é dada por um grupo de célula que formam a coifa. Este revestimento protetor está continuamente se descamando e sendo refeito. Uma diferença marcante entre plantas e animais é que o crescimento nas plantas ocorre, quase inteiramente, próximo às áreas meristemáticas; os animais tendem a possuir zonas de crescimento espalhadas por todo o orga-nismo. Podemos observar o crescimento restrito nas plantas, marcando a superfície da raiz ou do caule com traços eqüidistantes. Depois de alguns dias verá que a área atrás da extremidade é a região onde ocorreu o cresci-mento mais rápido. Esta é a região do alongamento celular. A divisão celular em si não contribui muito para aumentar o tamanho do organismo vegetal; colabora com a formação de novas células que aumentam de tamanho algum tempo depois. (figs.42 e 47). FIGU. DA PAG 55
CINÉTICA DE CRESCIMENTO Se medirmos o tamanho de uma planta a vários intervalos de tempo, come-çando pelo início de sua germinação, e traçarmos um gráfico dessas medida em função do tempo (expresso em dias), obteremos uma curva muito inte-ressante, como a mostrada na Fig. 4.4. Esta curva em forma de S ou sigmói-de é típica do crescimento de todos os órgãos, plantas, populações de plan-tas ou de animais e mesmo de civilizações humanas. Tem, pelo menos, qua-tro secções distintas:

Histologia Vegetal – prof Arlindo Costa – UDESC - 2001
10
10
4. Um período de indução (lag period = Intervalo de tempo decorrido entre o estímulo e a resposta) inicial, durante o qual alterações internas preparam o organismo para o crescimento;
5. Uma fase caracterizada por uma taxa constante de crescimento; 6. Uma fase na qual a taxa de crescimento diminui gradualmente; 7. Um ponto onde o organismo atinge a maturidade e o crescimento pára.
Se a curva for prolongada por um tempo maior, chegará o momento em que ocorre-rão a velhice e a morte dos organismos, dando origem a uma ou duas secções a mais na curva de crescimento (fig.4.4) A senescência e a morte que atingem a todos os animais, como parte de seu ciclo de desenvolvimento, parecem não ser uma parte necessária do ciclo das plantas. Sabemos, por exemplo, que alguns pinheiros e sequóias do oeste dos Estados Unidos atingem idades bem superiores a 3.000 anos. Imaginamos que um dia mor-rerão, provavelmente devido a uma infecção ou ao enfraquecimento da base mecâ-nica onde se assentam. Se houvesse um meio de evitar tais problemas, as árvores continuariam a crescer indefinidamente. Tecidos vegetais cultivados em soluções artificiais demonstram a imortalidade po-tencial das células vegetais. Em 1937, um pesquisador, na França, retirou partes de raízes de cenoura e colocou-as em substâncias químicas nutritivas. O resultado foi um bloco indiferenciado de tecido de calo que cresceu rapidamente. Em seguida, este tecido foi subdividido e transferido para frascos novos em intervalos freqüen-tes. O crescimento continuou numa taxa constante que não diminuiu mesmo após 30 anos. A planta de cenoura, de onde foi tirado o tecido original, certamente deve Ter morri-do muitos anos antes. A parada normal do crescimento de uma planta deve, então, depender de algum efeito inibidor. Se conhecermos qual seja pudermos removê-lo
ou impedi-lo de agir, estaremos em condições de produzir uma planta poten-cialmente imortal. As curvas de crescimento fornecem indícios de vários tipos de controles fisio-lógicos do crescimento. Examinando a extensão dos períodos de indução, por exemplo, teremos indicações sobre as mudanças que devem ocorrer antes que o crescimento se inicie. Em muitas sementes, o período de indu-ção tem a duração de algumas horas, embora em outras possa atingir dias, semanas e até meses. Sementes com longos períodos de indução provavel-mente têm substâncias inibidoras que atrasam o crescimento que só se inicia quando estas forem removidas ou destruídas. A taxa de crescimento durante a fase rápida é freqüentemente determinada por substâncias hormonais que serão discutidas ainda neste capítulo. A inclinação da curva sigmóide pode também fornecer indícios sobre o patri-mônio genético do potencial de crescimento da planta e o ambiente onde ela está se desenvolvendo. A altura total da planta e o momento do início da fase de equilíbrio dinâmico são também controlados geneticamente, mas suscetí-veis da ação do ambiente. Finalmente, a senescência e a morte do organis-mo não são determinadas inteiramente pelas suas características genéticas, mas estão sob o controle do pesquisador. 1. CONSIDERAÇÕES GERAIS As células meristemáticas, embora não especializadas contém, os elementos essenciais para a edificação da estrutura das células diferenciadas. Caracte-rizam-se pelo tamanho reduzido, considerável compactação, parede apenas primária e plastídios não diferenciados (proplastídios). O núcleo pode ser proeminente (como nos meristemas apicais), ou não (como nos meristemas

Histologia Vegetal – prof Arlindo Costa – UDESC - 2001
11
11
laterais); o citoplasma pode ser denso, conseqüência de vacúolos minúsculos (co-mo nos meristemas apicais) ou não (como nos meristemas laterais). Os meristemas são os tecidos responsáveis pelo crescimento e pela cicatrização de injúrias nos vegetais. 2. Meristemas apicais A planta fanerógama adulta sempre mantém a capacidade de adicionar novos in-crementos ao seu corpo, em parte através da atividade dos meristemas apicais presentes nos sistemas caulinar e radicular.
Figura 1. Meristema apical caulinar em corte longitudinal (Coleus) Numa planta em desenvolvimento, meristema apical típico ou promeristema é en-contrado na extremidade do eixo principal e dos ramos do sistema caulinar (meris-
temas apicais caulinares), assim como na extremidade do eixo principal e das ramificações do sistema radicular (meristemas apicais radiculares). Através de divisões, as células desse tecido (células meristemáticas) produzem os precursores dos tecidos primários do caule e da raiz. Portanto, os meristemas apicais formam o corpo primário da planta. O meristema apical pode ser ve-getativo quando origina os tecidos e os órgãos vegetativos ou reprodutivo quando origina tecidos e os órgãos reprodutivos.
Figura 2. Meristema apical radicular (Allium) Num meristema apical, certas células sofrem divisões numa forma tal que um dos produtos de uma divisão torna-se uma nova célula acrescentada ao cor-po da planta, e o outro, permanece como célula meristemática. As células que permanecem no meristema são as iniciais, e as que são acrescentadas ao corpo da planta, derivadas. Portanto, certas células nos meristemas atuam como iniciais, principalmente, porque ocupam uma posição apropriada para tal atividade. No grupo das criptógamas, especialmente, briófitas e pteridófi-tas, ocorre o crescimento através de uma célula apical inicial. Em ápices cau-

Histologia Vegetal – prof Arlindo Costa – UDESC - 2001
12
12
linares e em ápices radiculares de gimnospermas e de angiospermas ocorrem gru-pos de iniciais. O meristema apical é um conjunto complexo de células, que abrange as células iniciais e as células derivadas mais recentes. As derivadas também se dividem e produzem uma ou mais gerações de células. Geralmente, a capacidade de divisão existe até o momento em que se instalam na célula modificações marcantes que fazem parte do processo de diferenciação o qual fornece à célula características próprias. Assim, o crescimento, no sentido de divisão celular, não está limitado às regiões mais apicais do caule ou da raiz, mas estende-se, na realidade, a níveis que estão relativamente longe da região do meristema apical. Na atividade meristemática o processo de divisão celular está combinado com o de aumento em dimensões dos produtos das divisões. Na região do caule ou da raiz, em que as células não mais se dividem, mas estão se diferenciando, as mesmas células podem estar ainda crescendo em suas dimensões. Desse modo, na mesma célula, ocorrem, ao mesmo tempo, os fenômenos de crescimento e de diferencia-ção; além disso, num mesmo nível de um caule ou de uma raiz, diferentes regiões podem estar em diferentes estágios de crescimento e de diferenciação. O termo meristema não é restrito ao topo do ápice caulinar ou radicular, porque as modificações que ocorrem, são graduais entre as regiões dos meristemas apicais até aquelas onde estão situados os tecidos primários adultos do caule ou da raiz; e também porque os fenômenos de divisão celular, aumento celular em dimensões e diferenciação celular, aparecem em gradação, seja no caule, ou na raiz. Pode-se usar os termos meristemas apicais e tecidos meristemáticos primários, que estão abaixo destes últimos, para que se possa fazer uma distinção entre o meristema apical propriamente dito e os níveis subjacentes. Os termos ponta de raiz (ápice radicular) e ponta de caule (ápice caulinar) são usados num sentido amplo, para incluir o meristema apical e os tecidos meristemáticos primários. Esses últimos são:
protoderma que origina todo o revestimento da planta, isto é, a epiderme, procâmbio que origina os tecidos vasculares e parte do câmbio, se a planta cresce em espessura, e, meristema fundamental que origina todos os tecidos de preenchimento. 3. Ápice radicular Considerando-se o ápice radicular como um todo, pode-se visualizar o meris-tema apical propriamente dito e os tecidos meristemáticos primários, que estão em processo inicial de diferenciação. O meristema apical propriamente dito é constituído por uma região central, de células com atividades mitóticas muita baixa ou centro quiescente, o qual é parcialmente envolvido por algu-mas camadas de células, com atividade mitótica mais acentuada. A coifa cobre este meristema e em alguns casos, é formada por um meristema inde-pendente chamado caliptrogênio. Devido à presença da coifa, pode-se fazer referência ao meristema radicular, como sub-apical e o caliptrogênio, como apical. Os tecidos meristemáticos primários, derivados do promeristema e que estão em diferenciação são a protoderme, o meristema fundamental e o procâmbio. A protoderme diferencia-se em epiderme, o meristema fundamen-tal em córtex e o procâmbio em cilindro vascular. 4. Ápice caulinar O caule com seus nós e internós, folhas, gemas axilares e ramos resultantes do desenvolvimento das gemas e, mais tarde, as estruturas reprodutivas, resultam, basicamente, da atividade dos meristemas apicais. Como já foi mencionado, no meristema apical caulinar vegetativo de gimnospermas e de angiospermas ocorrem grupos de iniciais. Entre as teorias que descrevem o ápice meristemático caulinar, a de Schmidt, proposta em 1924, apesar de falha, é a mais simples e aplicável em grande parte dos casos das angios-permas. Esta teoria admite o conceito de túnica-corpo, portanto o meristema

Histologia Vegetal – prof Arlindo Costa – UDESC - 2001
13
13
apical consiste da túnica, abrangendo uma ou mais camadas periféricas de células que se dividem em planos perpendiculares é superfície do meristema (divisões anti-clinais); e do corpo, agrupamento situado abaixo da túnica, e no qual as células dividem-se em vários planos. As divisões que ocorrem no corpo permitem que o meristema apical aumente em volume, enquanto que na túnica, permitem um cres-cimento em superfície. Tanto a túnica quanto o corpo sempre formam novas célu-las, sendo que as mais velhas vão se incorporando às regiões do caule abaixo do meristema apical. O corpo, assim como cada uma das camadas da túnica possuem suas próprias iniciais. As iniciais da túnica contribuem para a formação da parte superficial do caule; as células produzidas pelo corpo são adicionadas ao centro do eixo, isto é ao meristema da medula, e, comumente, também é parte de região peri-férica do caule. Assim como no caso da raiz, quando se observa o ápice caulinar como um todo pode-se ver também os tecidos ainda meristemáticos mas que estão em processo inicial de diferenciação, (a protoderme, o meristema fundamental e o procâmbio). A protoderme diferencia-se em epiderme, o meristema fundamental em córtex e me-dula e o procâmbio em tecido vascular. Ainda são observados no ápice caulinar os primórdios de folhas e as gemas axilares. Quando o meristema apical caulinar passa para o estágio reprodutivo ele sofre mo-dificações relativamente conspícuas. Cessa o crescimento indeterminado do está-gio vegetativo de gema que está se transformando em floral e entre as outras modi-ficações que ocorrem, o meristema pára de produzir folhas vegetativas. A partir do meristema floral forma-se uma flor ou uma inflorescência, os aspectos histológicos mais comuns do meristema floral são: profundidade pequena e certa expansão (alargamento) do tecido meristemático propriamente dito. Observa-se que o ápice alargado é ocupado por um manto de células meristemáticas pequenas,
com citoplasma denso, as quais cobrem o miolo com células meristemáticas maiores e vacuolizadas. A organização túnica-corpo é modificada durante a transição e pode não ser discernível nos meristemas florais. A floração, geralmente é precedida por alongamento dos internós e desenvolvimento precoce das gemas axilares. 5. Meristemas laterais O crescimento primário, conforme foi mencionado, depende dos meristemas apicais, e caracteriza o corpo primário da planta. Em muitas espécies, o caule e a raiz crescem em espessura, adicionando tecidos vasculares ao corpo primário, pela atividade do câmbio vascular e do felogênio. Tal crescimento em espessura, é denominado secundário. O câmbio vascular e o felogênio são conhecidos como meristemas laterais, devido à posição que ocupam, isto é, uma posição paralela aos lados do caule e da raiz, portanto, câmbio e fe-logênio formam o corpo secundário da planta. A maneira pela qual o câmbio vascular e o felogênio se instalam no caule e na raiz será estudada quando tais órgãos forem abordados em textos posteriores. Apenas os aspectos bá-sicos mais característicos desses meristemas serão mencionados a seguir. 6. Câmbio vascular o câmbio vascular instala se entre os tecidos vasculares primários e produz os tecidos vasculares secundários. As células cambiais diferem daquelas dos meristemas apicais, que se caracterizam por possuir citoplasma denso, nú-cleos grandes e forma aproximadamente isodiamétrica. Assim, as células cambiais quando em atividade são altamente vacuoladas, com núcleo pouco conspícuo e formas características.

Histologia Vegetal – prof Arlindo Costa – UDESC - 2001
14
14
Figura 3. Ricinus. Corte transversal do hipocótilo em início de estrutura secundária. O câmbio mostra-se como faixas estratificadas, constituídas pelas células iniciais e pelas células derivadas; as iniciais correspondem a apenas uma fileira de células. Cones longitudinais radiais (1) de um caule ou de uma raiz em crescimento secun-dário, mostram que o câmbio vascular que está entre o xilema secundário e o floe-ma secundário, possui dois tipos de células: as iniciais fusiformes e as iniciais radi-ais. A célula inicial fusiforme é geralmente bem alongada; e a inicial radial é quase isodiamétrica. As iniciais fusiformes e suas derivadas originarão o uniam axial (2), e as iniciais radiais e derivadas, o sistema radial (3). Quando as iniciais cambiais pro-duzem células do xilema secundário e do floema secundário, elas dividem-se peri-clinalmente (4). A mesma inicial dá origem à células derivadas em direção ao xile-
ma, e em direção ao floema, embora não necessariamente em alternância. Dessa maneira, cada inicial produz uma fileira radial e células para dentro e outra para fora. Em fase de intensa atividade, em que muitas células são produzidas, forma-se a zona cambial de vários estratos; após a inicial ter se dividido periclinalmente, uma das células resultantes permanece como inicial e a outra é adicionada ao xilema ou ao floema em diferenciação. É difícil dis-tinguir as iniciais das derivadas recentes, pois essas derivadas dividem-se periclinalmente uma ou mais vezes antes que comecem a se diferenciar em células do xilema ou do floema. A inicial de uma dada fileira de células da zona cambial não tem, necessariamente, um perfeito alinhamento com as iniciais das fileiras vizinhas, portanto, em uma fileira radial a inicial pode estar mais próxima ao xilema ou ao floema, do que na outra fileira. As células iniciais também podem sofrer divisões do tipo anticlinal (5); desse modo ocorre um aumento na circunferência do câmbio. A formação de iniciais radiais a partir de iniciais fusiformes é um fenômeno relativamente comum. Um outro aspecto a ser considerado, diz respeito às divisões anticlinais que ocorrem em iniciais, levando á formação dos raios bisseriados ou multisseri-ados. 7. Felogênio Em caule e raiz de plantas que apresentam crescimento secundário em es-pessura, a epiderme (tecido primário) é substituída por outro tipo de tecido protetor denominado periderme (tecido secundário). Bons exemplos de for-mação de periderme são vistos em plantas lenhosas dos grupos das dicotile-dôneas e gimnospermas. A periderme também ocorre em dicotiledôneas herbáceas, principalmente nas regiões mais velhas do caule e da raiz. Entre

Histologia Vegetal – prof Arlindo Costa – UDESC - 2001
15
15
as monocotiledôneas, algumas formam periderme, outras formam um tipo diferente de tecido protetor secundário. A maneira coma a periderme se instala no caule e na raiz, e suas características serão explanadas posteriormente. Serão mencionados a seguir os seus aspectos histológicos e citológicos principais. o felogênio é o meristema lateral que origina a periderme. Ele produz o felema (ou súber) para fora, e a feloderme para dentro, no órgão em que ele se instalou. O felogênio é simples, quando comparado ao câmbio vascular, pois tem um só tipo de células iniciais. Em cones transversais ele aparece, assim como no caso do câmbio vascular, dentro de uma faixa estratificada, mais ou menos contínua na circunferên-cia do órgão em estudo. Tal faixa é formada por fiteiras radiais de células, onde cada fileira possui a célula inicial do felogênio e as suas derivadas para dentro (fa-zendo parte da feloderme) e para fora (fazendo pane do súber ou felema). As célu-las iniciais que constituem o felogênio são retangulares. radialmente achatadas em cortes transversais; apresentam contornos retangulares ou poligonais, ou às vezes irregulares, em cortes longitudinais. Em estágio maduro, as células da feloderme são comumente vivas e parecem-se com células parenquimáticas. As células do súber, geralmente não são vivas na maturidade, mas podem, em cena casos, conter materiais fluidos ou sólidos, sendo alguns deles incolores, e outros, pigmentados. As células do súber contém suberina (substância graxa) em suas paredes. Em certos tipos de súber em que as células têm paredes muito espessadas, ocorre ainda lignina nessas paredes, além de sube-rina. Geralmente o tecido do súber é compacto, sem espaços intercelulares. Uma estrutura comum em periderme, e que é formada por felogênio é a lenticela, que geralmente se instala na região do estômato. Pode-se ter periderme, sem lenti-cela como se pode ter lenticela sem que haja periderme. Em geral, periderme e lenticelas estão presentes conjuntamente. Nesse caso, os felogênios são contínuos.
De qualquer maneira, o felogênio da lenticela tem uma atividade diferente do felogênio que forma a periderme, originando muito mais células para o exteri-or, o que resulta no seu formato encurvado para dentro. O tecido produzido para o exterior é relativamente frouxo, com numerosos espaços intercelula-res. A lenticela permite a entrada de ar através da periderme. O tecido frouxo formado pelo felogênio, para fora, é chamado tecido complementar ou de preenchimento; o tecido formado para dentro é a feloderma. (1) O plano em que é feito o corte longitudinal do tipo radial coincide ou é paralelo ao plano no raio. (2) Sistema axial é constituído pelos elementos do xilema e floema secundá-rios que se situam paralelos ao eixo do órgão. (3) Sistema radial é constituído pelos elementos do xilema e floema secundá-rios que se situam perpendiculares ao eixo do órgão (geralmente representa-dos pelos raios parenquimáticos). (4) O plano da divisão celular denominada periclinal é paralelo à superfície do órgão em que a célula em questão se encontra. (5) O plano da divisão celular denominada anticlinal é perpendicular (ou qua-se) à superfície do órgão em que a célula em questão se encontra Detalhe frontal da epiderme abaxial de Bromelia balansae, evidenciando tri-comas em forma de escamas. 211,63x.

Histologia Vegetal – prof Arlindo Costa – UDESC - 2001
16
16
3. SISTEMAS DE REVESTIMENTO 1. Considerações Gerais A epiderme origina-se da camada mais externa dos meristemas apicais (proto-derme) e reveste a superfície do corpo vegetal em estágio primário. Nos órgãos que não apresentam crescimento secundário, a epiderme persiste por toda a vida da planta. Em tais casos, as células continuam a dividir-se no sentido anticlinal e a alongar-se no sentido tangencial. A duração da epiderme nos órgãos com cresci-mento secundário (caules e raízes) é variável, geralmente ela é substituída pela periderme, tecido de proteção que aparece após considerável atividade do câmbio vascular. 2. Epiderme
(A) Olea - Oliveira. Tricoma peltado em vista frontal e em corte transversal; (B) Pla-
tanus - Plátano. Tricoma dendróide; (C) Mesembryanthenum - Onze horas. Vesícula de água; (D) Lycopersicum - Tomateiro. Tricoma uniseriado; (E) Lycopersicum -
Tomateiro. tricoma glandular; (F) Semelhante em Urtiga. tricoma glandular com célula basal. A epiderme acha-se constituída por células epidérmicas comuns e células especializadas, dispersas entre as primeiras, tais como estômatos e tricomas, entre outras. As células epidérmicas variam na forma, tamanho e arranjo, têm quase sempre formato tabular e estão sempre intimamente unidas, de modo a for-mar uma camada compacta, desprovida de espaços intercelulares. Tais célu-las, geralmente são vivas, altamente vacuoladas, podendo armazenar vários produtos de metabolismo. As paredes das células epidérmicas diferem em espessura e, freqüente-mente, apresentam campos de pontuações primários e plasmodesmas, loca-lizados especialmente nas paredes radial e tangencial interna. A característi-ca mais importante da parede das células epidérmicas das partes aéreas da planta é a presença da cutina, substância graxa encontrada dentro da pare-de, isto é, nos espaços interfibrilares e intermicelares da celulose (processo de cutinização), e posteriormente, sendo depositada externamente, formando a cutícula (a cuticularização). A espessura da cutícula é variável, sendo mais espessa nas plantas de ambiente seco, em grande parte dos casos. Ela se forma durante os estágios iniciais de crescimento dos órgãos. Admite-se que a cutina migre, do interior para o exterior das células epidérmicas, através dos poros existentes na parede celular. Embora mais raramente, a lignina possa também ser encontrada na parede das células epidérmicas; quando isso ocorre, ela pode ser observada em todas as paredes ou só na externa. Como exemplo, pode-se citar a parede externa das células epidér-micas das folhas das Coníferas.

Histologia Vegetal – prof Arlindo Costa – UDESC - 2001
17
17
Células epidérmicas evidenciando estômatos e células de sílica presentes em folhas de gramí-neas A epiderme acha-se formada, geralmente, por uma só camada de células. Ocorre, porém, casos de epiderme pluriestratificada como, por exem-plo, na folha da falsa-seringueira (Ficus ellastica). Nesse caso, ela resulta de divisões periclinais da protoderme. O velame presente nas raízes aé-
reas de orquídeas (Epidendrum sp) é também um exemplo de epiderme múltipla. Depósitos de diversos tipos. tais como: ceras, óleos, resinas, sais sob forma cristalina, podem ocorrer sobre a cutícula das partes aéreas da planta. Pigmentos, como por exemplo, antocianinas podem também estar presentes nos vacúolos das células epidérmicas. A continuidade das células epidérmicas é interrompida por aberturas ou poros, que são espaços intercelulares limitados por duas células, denominadas células guarda as quais constituem o estômato. Estas células-guarda mediante mudanças de formato, ocasionam a abertura e o fechamento do poro estomático. Em muitas plantas podem, ainda, estar presentes duas ou mais células, em associação com as células-guarda: são as células subsidiárias ou anexas, as quais podem ser morfolo-gicamente semelhantes às demais células epidérmicas ou então diferirem daquelas pela sua morfologia e conteúdo. O estômato, juntamente com as células subsidiá-rias, formam o aparelho estomático. Abaixo desse aparelho observa-se, geralmente a câmara subestomática, que se conecta com os espaços intercelulares do mesofi-lo, no caso da folha. As células-guarda da maioria das plantas têm o formato de um rim em vista fron-tal. A parede dessas células é espessada de modo desigual; a mais fina é aquela
em contato com as células subsidiarias. Em corte transversal do estômato, pode-se ver ainda que a parede pode apresentar saliências, no lado superior ou no superior e inferior, delimitando nesses casos as cavidades externa e interna. A cutícula não só recobre as células-guarda mas, também, pode estender-se até a câmara subestomática. Cada célula-guarda tem um núcleo proeminente e cloroplastos que realizam fotossíntese como os demais. Os estômatos variam quanto à sua posição na epiderme; podem estar situados acima, abaixo ou no mesmo nível das demais células epidérmicas, em criptas ou protuberâncias. Com base na relação entre as células subsidiárias, quatro tipos de com-
plexos estomáticos podem ser reconhe-cidos: anomocítico, paracítico, anisocíti-co e diacítico. As células-guarda das Poaceae e Cyperaceae apresentam forma diferente da descrita; elas têm a forma de haltere. As extremidades dessas células são alargadas e têm paredes finas; a porção mediana é mais estreita e apresenta paredes espessas. O lume celular é es-
treito e o núcleo alongado. Os estômatos são, usualmente, encontrados nas partes aéreas da planta, em especial nas folhas e caules jovens. Nas folhas, são observados na face inferior ou em ambas, superior e inferior. Nas folhas flutuantes de plantas aquáticas, os estômatos são encontrados na face supe-rior, exposta ao ar. Nas folhas com venação reticulada, os estômatos dis-põem-se ao acaso e naquelas com venação paralela, acham-se arranjados em fileiras.

Histologia Vegetal – prof Arlindo Costa – UDESC - 2001
18
18
Além dos estômatos, inúmeras outras células especializadas ocorrem na epi-derme das panes aéreas mencionadas. Dentre estas, destacam-se os tricomas, que englobam diferentes tipos de apêndices epidérmicos. Os pêlos podem ser classifi-cados em tectores (ou de cobertura) e glandulares (ou secretores) e, tanto um tipo como outro pode ser uni ou pluricelular. Há casos, ainda, de pêlos glandulares a-presentarem uma região secretora (cabeça) recoberta pela cutícula. A secreção pode acumular-se, então, entre a célula ou células da cabeça e a cutícula e esta, mais tarde, pode romper-se, liberando a secreção. Devem ser lembrados ainda, os pêlos radiculares que são projeções da própria célula epidérmica que se formam inicialmente, como pequenas papilas na zona de absorção da epiderme das raízes jovens de muitas plantas. Os pêlos radiculares são vacuolados, apresentam parede finas e acham-se recobertos por cutícula. 3. Periderme A periderme é um tecido secundário protetor, que substitui a epiderme nas raí-zes e caules com crescimento secundário continuo. A substituição, portanto, ocorre quando o órgão já apresenta crescimento em espessura, decorrente da atividade cambial, o que provoca a compressão das camadas periféricas. A periderme ocorre em plantas herbáceas e lenhosas, geralmente nas panes mais velhas das raízes e caules. Ocorre, também, em superfícies após a abscisão de parte da plante (como folhas e ramos) e ainda se desenvolve como proteção a vários tipos de injúrias. A periderme se forma à partir da diferenciação de um ou mais felogênios. Por-tanto, ela é constituída por este meristema lateral e os seus produtos: feloderme, situada para dentro e súber, situado para fora. À medida que aumenta o número de camadas da periderme ocorre compressão da epiderme e/ou outros tecidos externos à periderme, os quais terminam rompen-
do-se, sendo eliminados gradativamente. A periderme nos caules instala-se, na maioria dos casos, superficialmente, ou seja, o felogênio diferencia-se próximo à epiderme; na raiz o felogênio diferencia-se, geralmente, do perici-clo, portanto, além da epiderme, o córtex é também eliminado. As monocotiledôneas, em geral, apresentam apenas corpo primário, por-tanto, mantém a epiderme todo o tempo de vida. Há, no entanto, algumas que formam tecidos secundários, como é o caso das palmeiras, das Dracae-na, dos Aloe, entre outras. Nestas plantas pode aparecer uma periderme semelhante a das dicotiledôneas, como ocorre no caso do gênero Dracaena com felogênio dando origem aos tecidos secundários, ou então, pode ocorrer a formação de súber sem a presença de felogênio, como acontece no gênero Cordyline. Neste caso, qualquer célula do córtex pode se dividir formando fileiras de células que se suberificam. O conjunto constitui o chamado súber estratificado. Excepcionalmente a periderme pode ocorrer em órgãos que só apresentam tecidos primários, como em raiz de banana-de-macaco (Monste-
ra deliciosa), por exemplo. À medida que uma árvore envelhece, outras peridermes podem se origi-nar em profundidades cada vez maiores, o que ocasiona um acúmulo de tecidos mortos na superfície do caule e raiz. Esta parte morta composta de tecidos isolados pelas peridermes (especialmente floema externo) e de ca-madas de periderme cujo crescimento já cessou, é chamada de ritidoma. Assim, o ritidoma constitui a casca externa e é bem desenvolvido em caules e raízes mais velhos. Um tipo especial de periderme, denominado poliderme, ocorre em raízes e caules subterrâneos de Rosaceae, Myrtaceae, Onagraceae e Hypericaceae. O tecido consiste de camadas de células suberizadas e células não suberi-

Histologia Vegetal – prof Arlindo Costa – UDESC - 2001
19
19
zadas, que se alternam; como na e periderme, apenas as camadas externas são mortas.
4. SISTEMA DE SUSTENTAÇÃO
COLÊNQUIMA 1. Considerações Gerais A célula colenquimática notabiliza-se pela plasticidade e espessamento da paredes, além da capacidade da divisão. É relevante a razão de crescimento, considerada a mais rápida entre os outros tipos de células vegetais. No Triticum sp. (trigo), por ocasião da ântese, os filetes das anteras podem se estender numa razão de 2-3 mm/minuto. Tal crescimento é resultante de mudanças rápidas na forma das célu-las, acompanhadas de alongamento das paredes; ainda relacionadas com a ener-gia requerida neste processo, ocorrem inúmeras mitocôndrias.
2. Características
O colênquima origina-se do meristema funda-mental, é o tecido de sustentação constituído por células vivas que apresentam a parede primária bem espessada. O colênquima, um tecido de sustentação, também é originado do meristema fundamental e constituído de células vivas, ge-ralmente alongadas, com cloroplastos; parede primária desigualmente espessa e formada por
grande quantidade de substâncias pécticas, hemicelulose além da celulose e de
alto teor de água. As células colenquimatosas (colócitos) dão suporte mecâ-nico e também elástico às folhas e caules. São incomuns em raízes. As célu-las colenquimatosas podem aumentar em comprimento e então compensar o crescimento em extensão.
A característica mais marcante deste tecido refere se ao espessamento desi-gual das paredes celulares, áreas bem espessadas. A composição da parede é típica, contendo grande quantidade de substâncias pécticas e água (60% do peso é água), além da celulose. As paredes celulares apresentam várias camadas, chegando até 20 em aipo (Apium graveolens). As camadas mais externas mostram que as microfibrilas estão arranjadas predominantemente no sentido longitudinal, direção paralela ao eixo do alongamento. O depósito das áreas espessadas ocorre antes da célula estar completamente alongada. No caso do pecíolo de aipo as células são curtas, com 20 a 40 um, mas se alongam à medida que o órgão cresce, até atingir 500 a 2000 υm de compri-mento. A plasticidade da parede celular do colênquima possibilita o cresci-mento do órgão ou tecido até atingir a maturidade. As células do colênquima se assemelham ao parênquima por possuírem protoplasto vivo, campos de pontuações primários e também por serem capazes de retomar a atividade meristemática. Suas células podem ainda conter cloroplastos e realizar fotos-síntese. De um modo geral, a forma é variável, sendo curtas, longas ou isodi-amétricas. À medida que as células envelhecem. o padrão de espessamento pode ser e de um modo geral o lume celular aparece redondo, por deposição de camadas adicionais nas paredes celulares. O termo colenquimatoso refe-re-se ao tecido que apresenta certas características de colênquima. Figura 1. Colênquima. A - angular; B - lamelar 3. Ocorrência

Histologia Vegetal – prof Arlindo Costa – UDESC - 2001
20
20
O colênquima é o tecido mecânico das regiões de crescimento e assim ocorre em órgãos jovens. Nos caules é usualmente periférico, localizando-se abaixo, ou pou-cas camadas abaixo, da epiderme (Ex.. caule de Coleus). Pode ocorrer como um cilindro contínuo como no caule de Sambucus (sabugueiro) ou em cordões individuais como em Cucurbita (aboboreira). Nas folhas ocorre no pecíolo, na nervura central ou na borda do limbo. A polpa de frutos quando são maciços e comestíveis geralmente são colenquimatosas. Raízes terrestres rara-mente contém colênquima: uma exceção ocorre em Vitis vinifera (videira). 4. Tipos de colênquima a) Colênquima angular Nesse tipo de tecido, as paredes mostram maior espessamento nos ângulos; e-xemplos desse tipo de espessamento ocorrem no pecíolo de Begonia (begônia), caule de Ficus (figueira) e de Cucurbita (aboboreira). b) Colênquima lamelar As células mostram um espessamento nas paredes tangenciais interna e externa , como no caule de Sambucus. c) Colênquima lacunar O colênquima pode ter espaços intercelulares e quando os espessamentos ocorrem nas paredes próximas ao espaço é chamado lacunar, como no pecíolo de Salvia (sálvia), raiz de Monstera, caule de Asclepias (erva-de-rato) e de Lactuca (alface). 5. Relação forma-função O colênquima, devido a plasticidade e capacidade de alongar-se, adapta-se à sus-tentação das folhas e caules em crescimento. O grau de espessamento parece estar relacionado com as necessidades da planta pois em espécies sujeitas a ação de agentes externos, como por exemplo, ventos fortes, o espessamento das pare-
des celulares inicia-se precocemente e torna-se mais acentuado do que o espessamento de espécies sob condições amenas. Por outro lado, as regiões jovens, geralmente são fotossintetizantes, necessi-tando, portanto, de tecidos que permitam a entrada da luz, como e o caso do colênquima. Ainda estas regiões, geralmente são tenras e portanto mais fa-cilmente atacadas por herbívoros, além de microorganismos, havendo ne-cessidade de cicatrização e regeneração celular; estes fenômenos são con-seguidos devido a capacidade do colênquima de reassumir a atividade meris-temática. Em regiões mais velhas da planta, o colênquima pode se transfor-mar em esclerênquima, pela deposição de paredes secundárias lignificadas. ESCLERÊNQUIMA 1. Considerações gerais Durante o crescimento da planta. a plasticidade da parede celular (do colên-quima) é muito importante pois as células sofrem alongamento, porém quan-do atinge a maturidade, a célula deve assumir uma forma definida e, neste caso, a elasticidade da parede (do esclerênquima) é mais relevante que a plasticidade. Uma parede elástica pode ser deformada por tensão ou pressão, mas reas-sume sua forma e tamanho originais quando essas forças desaparecem. Se um órgão maduro fosse constituído unicamente de tecidos plásticos, as de-formações causadas pelos mais variados agentes como vento, passagem de animais e outros, seriam permanentes. Por outro lado, a planta deve oferecer resistência às peças bucais, unhas e ovopositores de animais. A presença de esclerênquima como uma camada protetora ao redor do caule, sementes e frutos imaturos evita que os animais e insetos se alimentem deles, porque a lignina não é digerida, sendo um mecanismo de defesa da planta.

Histologia Vegetal – prof Arlindo Costa – UDESC - 2001
21
21
2. Características O esclerênquima é também um tecido de sustentação que, como o colênquima, tem origem primária, diferindo deste último porque geralmente suas células não mantém seus protoplastos vivos na maturidade e apresentam parede secundária lignificada, cujo espessamento é uniforme. As paredes secundárias são depositadas após as células terem atingido o seu tamanho final, ou a deposição pode ocorrer enquanto a célula está se alongando. A composição da parede secundária é de celulose, hemi-celulose, substâncias pécticas e de 18 a 35% de lignina. A lignina é uma substância amorfa, encontrada nas plantas vasculares; é formada pela polimerização de vários álcoois como o p-coumaril, coniferil e sinaptil. A lignificação inicia-se na lamela mé-dia e na parede primária e, depois atinge a parede secundária (que contém menos lignina). Os álcoois precursores são sintetizados no protoplasto, atravessam o plasmalema e permeiam a parede celular onde encontram enzimas (lacases). Ocor-re desidrogenação dos álcoois que ficam com os radicais livres e se polimerizam ao acaso, formando uma rede tridimensional que circunda e envolve as microfibrilas de celulose (parece que esta ligação é covalente). A rede formada ao redor das micro-fibrilas dá maior força e rigidez à parede. Sendo a lignina muito inerte, fornece um revestimento estável, evitando o ataque químico, físico e biológico. Enquanto a água e a maioria das substâncias dissolvidas nela passam facilmente pela parede primária, numa parede secundária a passagem é extremamente lenta. A forma das células do esclerênquima pode variar muito e esse caráter é utilizado na classifica-ção do tecido. 3. Ocorrência As células do tecido esclerenquimático ocorrem em faixas ou calotas ao redor dos tecidos vasculares, fornecendo uma proteção e sustentação. Podem também ocor-rer como grupos celulares muito grandes nas cascas de frutos secos ou endocarpos
de drupas, bem como nos envoltórios de sementes duras. Ocorrem também nos tecidos parenquimáticos, como por exemplo na medula e córtex de cau-les e pecíolo de Hoya, mesofilo de Nymphaea, raízes de Monstera deliciosa, etc. 4. Tipos de Esclerênquima a) Esclereídeos

Histologia Vegetal – prof Arlindo Costa – UDESC - 2001
22
22
As células são muito curtas, com paredes secundárias muito espessadas e presen-ça de numerosas pontuações (simples e ramificadas). O tecido formado é muito rígido. Existem diferentes tipos de esclereídeos, classificadas de acordo com sua forma: braquiesclereídeos ou células pétreas, que são isodiamétricas, aparecendo por exemplo no fruto da pera (Pyrus); astrosclereídeos quando mostram muitos
braços geralmente longos, como ocorre em folhas de Nymphaea e
Trochodendron; osteoesclereídeos quando tem a forma de osso, como observam-se em folhas de Haia; macroesclereídeos quando são alongados e têm uma forma colunar, como ocorrem nos envoltórios das sementes de ervilhas e feijões ("células em ampulheta"); os tricoesclereídeos quando apresentam uma forma semelhante a tricomas, como ocorre nas raízes de Monstera, entre outros. Figura 2. Esclereídeos. A, B -
Pyrus (célula pétrea); C, D - Hoya (caule); E, F - Malus (endocarpo); G - Hakea (colunar do mesofilo); H, I - Camelia (pecíolo); J - Trochondendron (as-troesclereíde do caule K - Allium (epiderme); L, M - Olea (folha); O, P - Phaseolus (células em ampulheta)
b) Fibras As células esclerificadas que são longas e apresentam as extremidades afi-ladas, lume reduzido e paredes secundárias espessas recebem o nome de fibras. Servem como elemento de sustentação nas partes vegetais que não mais se alongam. Podem ser classificadas artificialmente como fibras xilemá-ticas e extraxilemáticas. O grupo de fibras extraxilemáticas, são assim deno-minadas porque ocorrem em outros tecidos que não o xilema. As paredes secundárias são muito espessas, freqüentemente preenchendo o lume celular e as pontuações são simples. As paredes celulares são mais lignificadas nas monocotiledôneas do que nas dicotiledôneas. Nas dicotiledô-neas, estas fibras são floemáticas e são chamadas de fibras macias. Muitas destas fibras são usadas no comércio, como é o caso do Cannabis sativa (cânhamo), Linum usitatissimum (linho) e Boehmeria nivea (rami), pois têm pouca lignina. No cânhamo, as células têm cerca de 6 cm de comprimento, enquanto que no rami chega a ter 55 cm. No caso das monocotiledôneas, como Sansivieria zeylanica (espada-de-são-jorge), Phormium tenax (linho-da-nova-zelândia), Agave sisalana (sisal), as fibras são de origem pericíclica, muito lignificadas e são chamadas de fibras duras. O grupo das fibras xilemá-ticas será estudado juntamente com o xilema. 5. Ontogênese As fibras podem ter uma grande variedade de origens: o procâmbio produz células que podem dar origem a fibras do xilema primário e do floema; o câmbio vascular origina as fibras do xilema e do floema secundários. Células do meristema fundamental ou células do parênquima cortical, ou mesofilo e mesmo a epiderme podem desenvolver fibras. Muitos esclereídeos como as que originam os braquiesclereídeos são inicialmente células não

Histologia Vegetal – prof Arlindo Costa – UDESC - 2001
23
23
diferenciadas do parênquima, até que se inicie a deposição da parede secundária. Os outros tipos de esclereídeos são reconhecidos desde o início quando forma longos braços ou terminações lobadas. A formação das esclereídeos pode ocorrer em qualquer período da ontogênese do órgão; em Camellia por exemplo aparece na folha bem jovem. De um modo geral admite-se que. em ambas, esclereídeos e fibras, após completo desenvolvimento de suas paredes secundárias, o protoplasto não mais funcional, seja eliminado. A presença de numerosas pontuações indica que estas células podem manter um protoplasto vivo enquanto necessário. Em mui-tos casos, entretanto, não há necessidade de se manter um protoplasto vivo, como ocorre no esclerênquima das sementes, onde apenas o embrião e o endosperma precisam ter células vivas.
Figura 3. A -Localização do colênquima e esclerênquima em caules B- Pecíolo; C, D- Caule; E- Contíguo ao floema; F -circundando o feixe vascular
5. PARÊNQUIMAS 1. Considerações Gerais As células parenquimáticas, devido as suas características, parecem ter sido as células que constituíram os eucariotos mais primitivos. A origem do tecido parenquimático verdadeiro, ou seja, com ligação entre as células vizinhas por meio de plasmodesmas, parece ter surgido primeiramen-te nas algas Charophyceae. Fósseis de plantas terrestres primitivas mostram que estas eram parenquimáticas e, possivelmente, com as características do parênquima de musgos e hepáticas. Nestes grupos a maior parte do parên-quima está envolvido na fotossíntese. Considera-se que durante a evolução das plantas, para atender as especializações que foram surgindo, o tecido parenquimático teve que sofrer modificações, originando os diversos tipos de tecidos que constituem o corpo do vegetal. A medida que as plantas evoluí-ram, foi se tornando necessária a divisão de trabalho e o tecido parenquimá-tico foi se especializando. No caso das gimnospermas as células acumulam muitas substâncias fenólicas e realizam secreção; nas angiospermas ocor-rem, além destas, células contendo mucilagens, pigmentos, etc. 2. Características As células parenquimáticas, de um modo geral, apresentam paredes primá-rias relativamente delgadas, cujos principais constituintes são celulose, hemi-celulose e substâncias pécticas Nas paredes encontram-se os campos de pontuações primários com plasmodesmas, mostrando que estas células têm protoplasma vivo e que se comunicam entre si. Essas células possuem vacú-olos bem desenvolvidos. Os espaços intercelulares esquizógenos (isto é, que se formam pela divisão e afastamento das células) são comuns e seus tama-nhos variam conforme a função do tecido. O conteúdo celular apresenta-se

Histologia Vegetal – prof Arlindo Costa – UDESC - 2001
24
24
especializado segundo as atividades funcionais e/ou metabólicas Assim as células parenquimáticas podem conter numerosos cloroplastos amiloplastos substâncias fenólicas, entre outras. Essas células são geralmente descritas como isodiamétricas, entretanto, sua forma pode variar muito. Quando isoladas são mais ou menos esféricas, mas adquirem uma forma definida por ação das várias forças quando se agrupam para formar um tecido. As células do parênquima são capazes de retomar a atividade meristemática e de diferenciarem-se, novamente. em outros tipos de células. Em certos casos, as células parenquimáticas podem desenvolver paredes secundárias lignificadas. 3. Ocorrência O tecido parenquimático é encontrado em todos os órgãos da planta, formando tecido contínuo, como ocorre no córtex e medula do caule, córtex da raiz, tecido fundamental do pecíolo, mesofilo da folha, entre outros. Pode, ainda, fazer pane dos tecidos condutores da planta (xilema e floema). As células do parênquima po-dem apresentar características especiais para facilitar, por exemplo, suas funções de síntese, de reserva, de transporte. 4. Tipos de parênquima a) Parênquima de preenchimento As células são aproximadamente isodiamétricas, com espaços intercelulares pe-quenos (meatos). É o tecido encontrado no córtex e medula dos caules e córtex da raiz .
Figura 1. Tipos de células parenquimáticas. b) Parênquima clorofiliano ou clorênquima A função deste tecido é converter energia luminosa em energia química e armazenar a energia em forma de carboidratos, portanto a morfologia das células é adequada à tais funções. Assim, a forma cilíndrica das células favo-rece uma grande superfície de contato, facilitando a absorção do gás carbô-nico. O vacúolo grande empurra os numerosos cloroplastos que formam uma camada uniforme junto à parede, colocando essas organelas no sítio apropri-ado para a absorção do gás carbônico; onde não há problema de perda de água, os espaços intercelulares são grandes. Esse tipo de tecido é encontra-do no mesofilo foliar, constituindo o parênquima paliçádico e esponjoso. O-corre também em caules jovens ou outros órgãos que realizam fotossíntese. Nas plantas que estão em ambiente seco, as células do clorênquima estão bem unidos para minimizar a exposição das superfícies, evitando a perda d'água.

Histologia Vegetal – prof Arlindo Costa – UDESC - 2001
25
25
Figura 2 . Syringa vulgaris - Corte transversal mostrando parênquima paliçádico e lacunoso c) Parênquima de reserva ou armazenador Muitas partes da planta que são usadas para alimentação como é o caso de se-mentes, frutas, rizomas e outros, são constituídas primariamente de parênquima armazenador. Nesse tipo de tecido, as células geralmente acumulam amido (amilo-plastos), proteínas (proteoplastos) ou lipídios (elaioplastos ou gotículas). Um dos exemplos mais conhecidos é o tubérculo de batata (Solanum tuberosum) onde as células acumulam amido. As células desse tipo de tecido são freqüentemente pre-enchidas com o material de acúmulo, levando em muitos casos ao desaparecimen-to do vacúolo e outras organelas. Nestas células a relação núcleo-citoplasma é relativamente grande, porque durante o processo de acúmulo e saída do produto, o metabolismo é ativo e está sob controle nuclear. As plantas suculentas, como cer-tas bromeliáceas, cactáceas e euforbiáceas, têm células que acumulam água (pa-rênquima aqüífero). Neste caso as células são relativamente grandes e apresentam um grande vacúolo contendo água que é envolvido por uma fina camada de cito-plasma. A relação núcleo-citoplasma é menor.
Encontra se parênquima aqüífero, como por exemplo, em folhas de Bromeli-aceae. Em alguns casos pode haver acúmulo de hemicelulose nas paredes celulares que se tornam espessas, porém durante o desenvolvimento, essas paredes tornam-se finas, por digestão dos açúcares que são consumidos pela plântula. Este mecanismo ocorre em sementes de Coffea arabica (café)
e Diospyros kaki (caqui). Figura 3. Solanum tuberosum - Corte transversal mostrando parênquima de re-serva. d) Aerênquima Em muitos casos, as células em si não são tão importantes quanto os espaços interce-lulares ao redor delas. Como a difusão gasosa é mais rápida no meio gasoso que no meio líquido, órgãos volumosos desen-
volvem tecidos nos quais os espaços intercelulares são grandes e interconec-tados, dando origem a uma fase gasosa contínua, que se ramifica por entre as células do tecido. Esse tecido é denominado aerênquima e ocorre princi-palmente no mesofilo, nas raízes, nos caules e nos pecíolos das angiosper-mas aquáticas. Pode ser encontrado em folha de Nvmphaea (nenúfar), e em estruturas es-peciais, geralmente de natureza radicular, que também funcionam como ór-gão e flutuadores, como no caso de Jussiaea. O aerênquima produz um teci-do que é e relativamente forte, apesar de seu peso leve. As suas células podem ser isodiamétricas, retangulares ou braciformes com lobos ou braços. Se o espaço intercelular for muito acentuado formam-se câmaras, como em

Histologia Vegetal – prof Arlindo Costa – UDESC - 2001
26
26
folhas ou caules de aguapé (Eichornia sp.) e de taboa (Tvnha sp.) ou diafragmas, coma em raízes e caules de sempre vivas (Svngonanthus spp.).
Figura 4. Nymphea - Corte transversal da folha. Figura 5. Salsoda - Corte transversal da folha evidenciando o parênquima aquífero e) Parênquima de transporte Em muitas partes da planta, o material é transfe-rido rapidamente em grande quantidade a uma curta distância. Isso se dá por tipos especiais de células parenquimáticas, denominadas células de transferência, as quais apresentam modificações na sua parede formando inúmeras protrusões voltadas para a face interna. A plasmalema a-companha a parede, e com isso aumenta em
área, facilitando o transporte. 5. Ontogênese
As células parenquimáticas se formam a partir do crescimento e diferencia-ção das células do meristema fundamental (que ocorre no ápice do caule e da raiz). Ainda nos tecidos condutores (floema e xilema) ocorrem células parenquimáticas provenientes do corpo primário ou do secundário (parên-quima axial e radial). No caso dos órgãos que apresentam crescimento se-cundário, o parênquima se origina a partir do câmbio (parênquima axial e radial do xilema e do floema) ou do felogênio (feloderma).
6. FLOEMA
SISTEMA VASCULAR Figura 1. Caule mostrando um anel de feixes vascula-res, com xilema e floema. 1. Considerações gerais A conquista do ambiente terrestre pelas plantas só foi possível porque esses orga-nismos desenvolveram um sistema de distribuição inter-na de água e nutrientes,
concomitante com o desenvolvimento de um sistema de absorção da água do solo e de um sistema de revestimento para evitar a perda excessiva de água. A distribuição de água e nutrientes na planta é feita através do sistema vas-cular, que é constituído por dois tipos de tecidos: o xilema responsável pela condução de água e solutos orgânicos e inorgânicos ("seiva bruta") e o floe-

Histologia Vegetal – prof Arlindo Costa – UDESC - 2001
27
27
ma, responsável pela condução de material orgânico em solução ("seiva elabora-da"). Efetivamente diferenciou-se no corpo da planta duas porções diferentes uma área fotossintetizante e autotrófica e uma interna, não fotossintetizante e heterotrófica, capaz de sobreviver devido aos açucares supridos pela porção fotos-
sintetizante. Figura 2. Raiz mostrando xilema e floema O tecido responsável pela translocação de nutrientes orgânicos, especialmente açu-cares produzidos pela fotossín-tese é o floema. O aparecimen-to do floema teve conseqüên-cias importantes, pois além de transportar as substâncias da
parte autotrófica para a heterotrófica, passou a controlar qual pane da planta deve-ria crescer mais rapidamente, não só em função da época do ano, mas também do tipo de estrutura a receber nutrientes: meristema, primórdio foliar, tecido de reserva, etc. O floema pode translocar uma grande quantidade de material muito rapidamen-te. A seiva elaborada contém entre 50 a 300 mg/ml de matéria seca, sendo que de 80% a 90% dana rnatéria é açúcar. Aminoácidos ocorrem em concentração de 20 a 80 mg/ml e outros componentes são álcoois e fosfatos ligados a açúcares, regula-dores de crescimento, ácidos nucleicos, vitaminas e substâncias inorgânicas. A velocidade de translocação varia entre 10 a 100 cm/h, podendo chegar a 300 cm/h. A evolução do floema é menos conhecida que á do xilema. Acredita-se que os elementos de tubo crivado tenham evoluído a partir de células do parênquima e, devido à fisiologia das suas células, devem ter ocorrido profundas mudanças no
sistema enzimático intracelular. Os estudos do floema e da seiva elaborada estão muito associados aos afídeos. O estudo do fluxo da seiva elaborada é feita com esses insetos pois com seu aparelho bucal conseguem atingir os elementos de tubo crivado. Se o seu corpo for retirado, mantendo-se apenas o aparelho bucal, a seiva elaborada é exudada por um período relativamente longo. A seiva pode ser coletada com auxílio de uma micropipeta e sua com-posição estudada. Afídeos têm também sido usados nos estudos dos movi-mentos dos hormônios ou do efeito dos hormônios na translocação dos assi-milados. Algumas viroses patogênicas são restritas ao floema e são transpor-tadas de uma planta para outra pelos afídeos. 2. Classificação Os tecidos vasculares como floema originam-se por diferenciação do pro-câmbio no corpo primário da planta. O floema que se forma do procâmbio é o floema primário. Quando ocorre crescimento secundário em espessura, o floema se forma a partir do câmbio e daí ser chamado de floema secundário. 3. Ocorrência O floema aparece em caules, raízes, folhas, partes florais, etc. Comumente o floema ocupa uma posição relativamente externa ao eixo caulinar e dorsal (inferior ou abaxial) nas folhas e nos órgãos de natureza foliar. Na raiz em crescimento primário, alterna-se com o xilema. Na raiz em crescimento se-cundário e no eixo caulinar, na maioria dos casos, o floema localiza-se exter-namente ao xilema, porém em muitas dicotiledôneas ocorre o floema interno ao xilema denominado floema incluso, como no caso de representantes da família Apocynaceae, Asclepiadaceae, Asteraceae, Myrtaceae, Solanaceae, etc. Devido a sua posição próxima à periferia do órgão, torna-se modificado com o aumento em espessura do órgão. Eventualmente, o floema não fun-cional, é fragmentado pela formação de peridermes sucessivas.

Histologia Vegetal – prof Arlindo Costa – UDESC - 2001
28
28
4. Composição O floema é um tecido formado basicamente por elementos crivados, células paren-quimáticas especializadas (células albuminosas ou células companheiras), células parênquimáticas e esclerênquima (fibras principalmente).
a) Elementos crivados Figura 3. Pormenor de uma placa crivada - A, D em vista frontal; B, C, E e F em corte longitudunal A, B, D e E, em estágio funcional, C e F, em estágio não funcional Os elementos crivados são os elementos condu-tores do floema e podem
ser de dois tipos: células crivadas e elementos de tubo crivado. A expressão tubo crivado designa uma série longitudinal de elementos de tubo crivado. As paredes celulares dos elementos crivados são em geral mais espessas do que as das célu-las parenquimáticas. Quando observadas ao microscópio óptico, em seções de material fresco, mostram um brilho aperolado recebendo a denominação de pare-des nacaradas. Sua composição é de celulose e compostos pécticos e pelo arranjo observado acredita-se que esta seja primária nas células crivadas e nos elementos de tubo crivado, na maioria dos casos. Essas células apresentam as chamadas áreas crivadas, isto é, áreas que apresentam poros através dos quais interconec-tam-se os protoplastos de elementos crivados contíguos, tanto no sentido vertical como lateral.
No floema funcional, cada poro é revestido por calose, polímero de b1,3 gli-cose. O desenvolvimento da área crivada se dá da seguinte maneira: a) depósito de plaquetas de calose na região do poro; b) as plaquetas de calose se espessam e revestem toda a região do poro formando cilindros de calose; c) a seguir, os cilindros de calose se espessam mais e a calose se deposita também na parede da célula entre os cilindros. Outra característica importante dessas células é a ausência, em geral, de núcleo na maturidade, embora sejam mantidos os leucoplastídios (de amido e/ou de proteína), retículo endoplasmático e as mitocôndrias. O tonoplasto em geral desaparece, constituindo o mictoplasma, mas a plasmalema per-manece. É muito comum, em dicotiledôneas, a presença de uma substância proteiná-cea, a chamada proteína-E, que ocorre como filamentos no citoplasma resi-dual desta célula. Esta, em ultra-estrutura, mostra que é constituída de túbu-los ou fibrilas, ou ambos, que se arranjam helicoidalmente. A variação infra-estrutural da proteína-P está associada a uma variação macromolecular du-rante a diferenciação do elemento crivado. A proteína-P pode ter característi-cas semelhantes ás proteínas contrácteis do tipo actomiosina, favorecendo a aceleração e regulação dos assimilados, na oclusão das áreas crivadas e ainda no reconhecimento celular. Quando o elemento crivado está envelhe-cendo ou se preparando para a dormência, mais calose vai se acumulando e esta pode até mesmo obliterar o poro. Quando o floema é reativado, a calose torna-se reduzida, possibilitando o restabelecimento da continuidade proto-plasmática entre as células contíguas. Pode ainda ocorrer um acúmulo de calose em resposta a ferimentos.

Histologia Vegetal – prof Arlindo Costa – UDESC - 2001
29
29
Aparentemente enzimas localizadas na plasmalema estio envolvidas no controle de síntese ou da degradação da calose. As células crivadas, presentes nas gimnos-permas são, alongadas (cerca de 3,5 mm de comprimento) e apresentam áreas crivadas em suas paredes laterais como também nas terminais. Os poros das áreas crivadas são relativamente estreitos (0,05-2,0 um de diâmetro) e freqüentemente uniformes em tamanho. Os elementos de tubo crivado são curtos (50-150 um de comprimento) e ocorrem na maioria das angiospermas. Nas paredes laterais des-sas células são freqüentes as áreas crivadas com poros relativamente pequenos. Nas paredes terminais e, ocasionalmente, nas paredes laterais ocorrem áreas mais especializadas, as placas crivadas, portadoras de poros maiores (10-14 um de diâ-metro). A interconexão entre um elemento de tubo crivado e outro, ao nível da placa crivada, é mais completa do que entre células crivadas. Os elementos de tubo cri-vado mostram um grau mais avançado de especialização do que aquele visto nas células crivadas. As placas crivadas encontradas nas paredes terminais podem apresentar-se em diferentes graus de inclinação, até totalmente transversais. Os elementos de tubo crivado longos, dotados de paredes terminais muito inclinadas podem ter placas crivadas compostas, ou seja, placas crivadas com várias áreas crivadas. Nessas placas crivadas, os poros são relativamente estreitos, características que indicam primitividade. No curso da especialização parece ter ocorrido uma diminuição da inclinação das paredes terminais, aumento de tamanho do poros nas placas criva-das e distinção entre áreas crivadas nas paredes laterais e placas crivadas nas paredes terminais. Assim, o elemento de tubo crivado mais especializado é aquele que possui paredes terminais transversais, dotadas de placas crivadas simples, com poros largos e áreas crivadas pouco especializadas nas paredes laterais. b) Células albuminosas e companheiras
O movimento de materiais orgânicos no floema depende da interação fisioló-gica entre os elementos crivados e as células parenquimáticas especializa-das contíguas. Assim intimamente associadas aos elementos crivados, exis-tem as chamadas células albuminosas nas gimnospermas e células compa-nheiras nas angiospermas. As células albuminosas, análogas às células companheiras, estão associadas às células crivadas por numerosos plasmo-desmas. Estas células não estão relacionadas ontogeneticamente, isto é, não têm uma precursora comum. As células companheiras são células parenquimáticas especializadas que estão ligadas aos elementos de tubo crivado por meio de plasmodesmas. Essa continuidade entre esses dois tipos de células é tão grande, que fre-qüentemente, em material dissociado, o elemento crivado e sua(s) célula(s) companheiras permanecem juntos. As células companheiras possuem nú-cleos e nucléolos relativamente grandes, numerosas mitocôndrias bem de-senvolvidas, plastídios (freqüentemente cloroplastídios) e muitos ribossomos. As células companheiras e o elemento de tubo crivado podem ou não serem relacionados ontogeneticamente, isto é, serem células irmãs. Nas venações menores da folha, os elementos de tubo crivado são menores e as células parenquimáticas são de modo que estas células e as compa-nheiras são chamadas de células intermediárias porque medeiam o acúmulo e o descarregamento de material. Em algumas plantas essas células têm paredes finas, enquanto que em outras mostram protuberâncias na parede interna aumentando a área da plasmalema, facilitando o transporte a curta distância. Neste caso, recebem o nome de células de tranferência, como já mencionado no capítulo de parênquima. c) Células parenquimáticas

Histologia Vegetal – prof Arlindo Costa – UDESC - 2001
30
30
Células parenquimáticas não especializadas são os componentes normais do floe-ma e podem conter substâncias ergásticas, como amido, cristais, substâncias fenó-licas, etc. No floema secundário temos as células parenquimáticas radiais e as axi-ais. d) Esclerênquima No floema primário estão presentes fibras na parte externa desse tecido, sendo que no floema secundário apresentam uma distribuição variada entre as outras células do sistema axial. Podem apresentar-se septadas como em Vitis ou não; podem ainda ser vivas ou mortas na maturidade. Em muitas espécies as fibras têm valor econômico, como no caso do linho (Linum usitatissimum), cânhamo (Cannabis sati-
va), rami (Bohemeria nivea), etc. Esclereídeos são comuns no floema, podendo aparecer tanto no sistema axial quanto no radial do floema secundário (Ex. Robini-a).
Figura 4. Diagrama do floema secundário e câmbio vascular de uma gimnos-perma (Cupressaceae) e uma dicotileônea (Magnoliaceae) 5. Ontogênese Os tecidos vasculares e, consequentemente o floema, formam-se por dife-renciação do procâmbio no corpo primário da planta. O floema que assim se originou e chamado floema primário (proto e metafloema). Quando ocorre

Histologia Vegetal – prof Arlindo Costa – UDESC - 2001
31
31
crescimento secundário em espessura (gimnospermas e a maioria das dicotiledô-neas) o câmbio forma o floema que é denominado floema secundário. O tecido vascular secundário se organiza num sistema axial e outro radial. Os raios formados são contínuos através do câmbio vascular, indo desde o xilema secundá-rio até o floema secundário. 6. Características do floema secundário O floema secundário é constituído pelo proto e metafloema. O protofloema ocorre nas regiões que ainda estão em crescimento por alongamento e, assim, seus ele-mentos crivados sofrem estiramento e logo param de funcionar (1 ou 2 dias); even-tualmente podem até ficar obliterados. O protofloema é constituído por elementos crivados geralmente estreitos e não conspícuos, podendo ou não ter células companheiras. Podem estar agrupados ou isolados entre as células parenquimáticas. Em muitas dicotiledôneas ocorrem "pri-mórdios" de fibras que crescem e desenvolvem paredes secundárias, dando origem às fibras do protofloema, quando os elementos crivados param de funcionar. O metafloema diferencia-se mais tarde e, nas plantas desprovidas de crescimento secundário, constitui o único floema funcional nas partes adultas da planta. Os ele-mentos crivados são em geral mais largos e numerosos quando comparados ao protofloema. As fibras estão, em geral, ausentes. O floema secundário é proveniente do câmbio. A quantidade de floema produzida pelo câmbio vascular é usualmente menor que a de xilema. No caso das coníferas, o sistema axial contém células cribadas, células albuminosas, células parenquimáti-cas, fibras e esclereídeos. O sistema radial é constituído por raios unisseriados com células albuminosas e células parenquimáticas. Eles estão ausentes em Pinus po-rém ocorrem em outras espécies de gimnospermas como em Taxus, Taxodium, Cupressus, etc.
Nas dicotiledôneas, o sistema axial é formado pelos tubos crivados, células companheiras e células parenquimáticas. As fibras podem estar ausentes ou presentes, neste caso, formam uma faixa contínua ao redor de toda a circun-ferência do órgão (caule e raiz) ou faixas isoladas O sistema radial, formado pelos raios uni ou plurisseriados compõe se de células parenquimáticas, po-de, ainda, conter esclereídeos ou células parenquimáticas esclerificadas com cristais. Os raios podem aparecer dilatados, como conseqüência de divisões anticlinais radiais das células em resposta ao aumento de circunferência do eixo.
7. XILEMA SISTEMA VASCULAR 1. Considerações gerais O xilema juntamente com o floema constituem o sistema vascular, sendo o xilema o principal tecido condutor de água. Devido às paredes lignificadas de suas células, o xilema apresenta as seguintes características: é mais conspí-cuo que o floema, pode ser estudado mais facilmente e é melhor conservado nos fósseis. É um tecido muito utilizado na identificação de plantas, especi-almente as que têm hábito arbóreo. Estruturalmente, o xilema é complexo por constituir-se de diferentes tipos de células vivas e mortas. Os componentes mais característicos são os elemen-tos traqueais, que conduzem água e solventes orgânicos e inorgânicos. Al-guns dos elementos traqueais combinam as funções de condução com a de sustentação. O xilema também possui elementos de sustentação (as fibras) e células parenquimáticas vivas, relacionadas com atividades vitais variadas. As fibras também podem reter seus protoplastos no xilema funcional e então,

Histologia Vegetal – prof Arlindo Costa – UDESC - 2001
32
32
combinar a função vital de acúmulo de amido com a função mecânica de sustenta-ção. Evidências fósseis demonstram que o aumento de parede secundária, lignificada, permitiu que os vegetais pudessem explorar diferentes ambientes, culminando com o que se vê hoje, com plantas crescendo nos mais variados ambientes, desde o mais árido até o mais encharcado, desde o mais frio até o mais quente. 2. Classificação Os tecidos vasculares e consequentemente, o xilema, formem-se por diferenciação do procâmbio no corpo primário da planta, isto é, em estágios pós-embrionários, até mesmo no embrião e em extremidades apicais dos órgãos (regiões jovens). O xile-ma que se forma do procâmbio denomina-se xilema primário. Se houver crescimen-to secundário em espessura o xilema forma-se a partir do câmbio, daí ser denomi-nado xilema secundário. Composição O xilema é um tecido formado basicamente por três tipos de células: os elementos traqueais ou condutores, as fibras e as células parenquimáticas. a) Elementos traqueais O termo "elemento traqueal" é derivado de "traquéia", nome originalmente aplicado para certos elementos do xilema primário, semelhante às traquéias dos insetos. Dois tipos fundamentais de elementos traqueais ocorrem no xilema: traqueídeos e elementos de vaso. As traqueídeos são células imperfuradas, que combinam as funções de condução e de sustentação. O fluxo de água se faz principalmente, no sentido longitudinal, através de séries longitudinais de traqueídeos, podendo ocor-rer, também, fluxo lateral entre elementos contíguos. São células relativamente alongadas e com a parede primária presente em toda a sua extensão. A deposição da parede secundária em um traqueídeo, e sua lignificação pode ocor-rer sob a forma de anéis, de espirais frouxas ou mais densas (helicoidal), de escada
(escalariforme, ou então, forrando completamente a parede primária forne-cendo, no final, um aspecto reticulado ou com pontuações areoladas. Estas pontuações areoladas podem ser do tipo escalariforme ou circulares. Figura 1. Elementos traqueais primários associados à células parenquimáti-cas em caule de Aristolochia. Superfície transversal (A) e longitudinal (B).Em ambas a porção inicial do xilema encontra-se à direita. Observe os diferentes tipos de espessamento da parede secundária, desde o anelar, com alguns anéis, até o pontuado , com a parede secundária em toda extensão do elemento traqueal, exceto nas regi-ões de pontuações. A água passa de um traqueíde para a outra através das suas paredes primá-rias, como por exemplo em traqueídeos anelados, ou através das membra-nas de pontuação nos traqueídeos com pontuações areoladas. Como já foi visto anteriormente, a membrana da pontuação é um conjunto formado pelas paredes primárias de duas células contíguas, mais a lamela média. Os limites da membrana de pontuação são os limites da chamada de pontuação. Os
elementos de vaso são células perfuradas, isto é, células cujas comunica-ções com outras células da mesma série longitu-dinal, que constituem o vaso, se faz através de regiões desprovidas de paredes primárias e se-cundárias. A perfuração dos elementos de vaso

Histologia Vegetal – prof Arlindo Costa – UDESC - 2001
33
33
comumente ocorre nas paredes terminais, mas pode estar presente nas paredes laterais também. A parede que contêm uma perfuração é chamada placa perfurada. Uma placa perfurada pode apresentar uma única perfuração, constituindo a placa de perfuração simples, ou pode conter várias perfurações, formando aplaca de per-furação múltipla. Um vaso origina-se, ontogeneticamente, a partir de séries longitu-dinais de células meristemáticas. Estas são as células procambiais no corpo primá-rio da planta ou células cambiais na estrutura secundária. Internamente à parede primária, forma-se parede secundária em todas as células da série. O elemento traqueal primordial pode ou não alongar-se antes que se desenvolva a parede se-cundária, mas usualmente, expande-se lateralmente. Após completar esses cresci-mento, a parede secundária e depositada. A porção da parede primária que poste-riormente será perfurada não é recoberta por parede secundária. Um vaso é forma-do por um número limitado de elementos de vaso. Cada elemento de vaso conecta-se com o seguinte na série longitudinal através da placa de perfuração. Os elemen-tos terminais da série apresentam placa perfurada na extremidade que está em
contato com outros elementos da mesma série. No lado oposto, são imperfurados. Figura 2. Elemento traqueal em vista longitudinal A comunicação entre um vaso e outro, numa série vertical se faz, como nos traqueídeos, através de paredes imperfu-radas. Sabendo-se que apenas água e solutos passam através das membranas de pontuação, pode-se forçar a penetra-
ção de gás de iluminação nas extremidades cortadas de ramos e, através de tenta-tivas de acender a outra extremidade, também cortada, pode-se conseguir medir
vasos de algumas espécies de dicotiledôneas. Verificou-se por exemplo, que em Acer, eles têm aproximadamente 60 cm e em Faxinus até 3 m de com-primento. No entanto, em espécies com vasos formados por elementos lar-gos, eles parecem se estender através de toda a altura da árvore. O exato processo da remoção da parede celular durante o processo da perfuração não é conhecido. Uma das hipóteses é que tanto os componentes celulósicos como os não celulósicos são removidos pela ação do protoplasma da célula. Outra hipótese é que unicamente compostos não celulósicos são removidos enquanto que as microfibrilas celulósicas seriam apenas empurradas de suas posições originais para as margens da perfuração. É ainda controvertida a hipótese que as células apresentariam celulase necessária para degradar a celulose dessas paredes. Após a formação da placa perfurada, o protoplasto não funcional, é elimina-do. As paredes secundárias dos elementos de vaso, como nos traqueídeos, desenvolvem-se com uma grande variação de padrões. Geralmente, na pri-meira porção formada do xilema primário (protoxilema) uma área mais limita-da da parede primária é recoberta por camadas de paredes secundárias do que no xilema primário formado posteriormente (metaxilema) e do que no xilema secundário. Começando com o xilema primário inicialmente formado, os espessamentos secundários são depositados como anéis, hélices conti-nuas, depois como retículos. Tais espessamentos são chamados respecti-vamente, anelar, espiral ou helicoidal e reticulado. Há elementos com espessamentos apenas escalariformes quando derivam de um espessamento helicoidal no qual houve uma maior aproximação dos segmentos, diminuindo-se os espaços sem parede secundária. Nos elemen-tos de vaso com uma parede secundária mais extensa ocupando maior área, têm-se o espessamento pontuado, caracterizado pelas pontuações areola-

Histologia Vegetal – prof Arlindo Costa – UDESC - 2001
34
34
das. Elementos com pontuações areoladas são característicos do xilema primário formado posteriormente, isto é, o metaxilema e do xilema secundário. Como já foi dito, o xilema é o tecido vegetal que mais se presta a estudos filogenéticos. As evidências fósseis demonstram que o elementos traqueais mais primitivo é o traqueídeo. Por ser fina e longa, com paredes bem espessadas, o traqueídeo acu-mula as funções de condução e sustentação. Na evolução, houve uma transposição dessas funções: de um lado, para os elementos de vasos, mais especializados para a condução, e de outro, para as fibras, mais especializadas para a sustentação. Assim sendo, a traqueídeo evoluiu em duas direções: uma, deu origem as fibras e na outra, deu origem aos elementos de vaso. b) Fibras As fibras são células longas, com paredes secundárias, comumente lignificadas, que usualmente variam em espessura. Suas pontuações podem ser simples, sendo então denominadas libriformes, ou podem ser areoladas, recebendo a denominação de fibrotraqueídeos As fibras podem desenvolver paredes transversais finas depois que as paredes secundárias já estão formadas. Denominam-se, então, fibras sep-tadas. Usualmente elas retêm o protoplasma, possuindo assim, função semelhante a do parênquima. Aliás, a distinção entre fibra e célula de parênquima lenhoso é particu-larmente difícil quando as células do parênquima são alongadas. Se ocorrem fibras vivas no lenho, o parênquima é muito escasso ou ausente. Outra variação das fi-bras do lenho é constituída pelas chamadas fibras gelatinosas. Possuem paredes pouco lignificadas e aparecem em lenho que se desenvolve sob pressões (lenho de reação). As fibras assim como os elementos traqueais e o parênquima axial for-mam-se a partir das derivadas das iniciais fusiformes do câmbio. Parênquima As células do parênquima radial são derivadas de células iniciais radiais do câmbio. As células do parênquima axial são derivadas de células cambiais fusiformes. Se a
célula derivada não sofre divisões transversais, forma-se uma célula de pa-rênquima fusiforme. De um modo geral, no entanto ocorrem divisões transversais formando-se um parênquima com células enfileiradas no sentido longitudinal. No xilema se-cundário percebe-se claramente o parênquima axial e o radial. Ambos são fundamentalmente semelhantes quanto a estrutura e ao conteúdo. O parên-quima reserva amido, óleos, e muitas outras substâncias de função desco-nhecida. Componentes fenólicos e cristais também são comuns. 3. Características
Figura . Esquema mostrando a evolução dos elementos de vaso (A-F): Dimi-nuição no comprimento, redução da inclinação das paredes terminais, passa-gem de placa de perfuração escalariforme para simples. G - Terminação com

Histologia Vegetal – prof Arlindo Costa – UDESC - 2001
35
35
de dois elementos de vaso evidenciando a união dos elementos em uma série. O xilema primário, isto é, o xilema que aparece no corpo primário de uma planta contém os seguintes tipos de células: elementos traqueais (traqueídeos e elemen-tos de vaso), fibras e células de parênquima. Nos caules, folhas e flores, o xilema primário e o floema primário associados, comumente ocorrem em cordões, forman-do os feixes vasculares. O xilema primário usualmente consiste de uma parte que se forma primeiro, o pro-toxilema e uma pane que se forma posteriormente, o metaxilema. Embora os dois tipos apresentem algumas peculiaridades, eles são tão integrados que a delimita-ção entre ambos pode ser feita apenas aproximadamente. De um modo geral ele-mentos anelados e espiralados pertencem ao protoxilema, escalariformes e com
pontuações areoladas, ao metaxile-ma. O protoxilema diferencia-se em par-tes da planta que ainda não comple-taram seu crescimento e a sua dife-renciação. Desta maneira, está sujei-ta à pressões contínuas. Seus ele-mentos traqueais são distendidos e muitas vezes, obstruídos (nas raízes eles persistem mais tempo porque só se tornam adultos acima da região de distensão). Figura . Feixe vascular do caule de
Zea O metaxilema comumente atinge a sua maturidade depois que a distensão já se completou, por isso, é menos afetado pelos tecidos a sua volta. Quando os elemen-
tos de protoxilema são distendidos, eles podem se tornar completamente obliterados pelas células do parênquima a sua volta. Em muitas e monocoti-ledôneas, o elemento não funcional pressionado torna-se parcialmente co-lapsado mas não obliterado. Em vez disso, aparecem canais, as chamadas "lacunas do protoxilema", rodeadas por células de parênquima. As paredes secundárias do elemento não funcional, geralmente em forma de anéis podem ser observadas ao longo da margem da lacuna. O metaxilema é um pouco mais complexo que o protoxilema e pode conter fibras e além dos elementos traqueais e células do parênquima. Em plantas que não apresen-tam crescimento secundário, o metaxilema permanece funcional durante todo o tempo. O xilema secundário é formado pelo câmbio. Dicotiledôneas e gimnospermas apresentam crescimento secundário, por conseguinte, formam xilema secundário. De um modo geral, monocotiledô-neas não apresentam crescimento secundário, portanto, não formam xilema secundário. Da mesma maneira que no xilema primário, encontram-se, no xilema secundário, os seguintes tipos de células: elementos traqueais, fibras e parênquima. Distinguem-se, ainda, dois sistemas de células: axial (longitu-dinal ou vertical) e radial (transversal ou horizontal). O sistema axial é deriva-do das iniciais fusiformes do câmbio. É formado por células (elementos tra-queais, fibras e parênquima) com o seu eixo maior orientado verticalmente no caule ou na raiz, paralelamente ao eixo do órgão. O sistema radial é compos-to de fileiras de células orientadas horizontalmente, perpendiculares ao eixo do órgão sendo constituído basicamente de células parenquimáticas. Todo o sistema radial deriva das iniciais radiais do câmbio. A delimitação entre xilema primário e xilema secundário é observada, pela presença dos raios parenquimáticos (melhor observados em corte transversal), ou seja, onde os mesmos tiveram início, o xilema secundário estará também inician-

Histologia Vegetal – prof Arlindo Costa – UDESC - 2001
36
36
do-se. O xilema secundário (ou lenho secundário) do caule de espécies arbóreas recebe a denominação de madeira e tem grande aproveitamento econômico, sendo usada em construção civil, aplicações externas, mobiliário, embalagens. etc. Anéis de crescimento As madeiras, de um modo geral, apresentam-se divididas em zonas que se repetem sucessivamente, observáveis especialmente em cortes transversais. Estas zonas constituem os anéis de crescimento, ou camadas de crescimento, bem evidentes especialmente em madeiras de regiões temperadas, com estações bem marcadas. Comumente, estas camadas de crescimento apre-sentam-se formadas por dois tipos de lenho: inicial (ou primaveril) e tardio (ou esti-val). O lenho inicial é menos denso que o tardio, porque nele predominam células com paredes mais finas. O lenho tardio, por sua vez, apresenta células com paredes mais espessas e. por conseguinte, o lume menor. Pode-se, muitas vezes, saber a idade relativa de uma árvore de região temperada pelo número de anéis de crescimento. Cada anel de crescimento corresponde a um anel de lenho estival mais o anel de lenho primaveril (daí serem chamados também, de anéis anuais) Cerne e Alburno O xilema secundário gradualmente perde as funções de condução e de reserva de alimentos em conseqüência das células vivas do lenho morrerem. Este estado é precedido por numerosas mudanças, muitas das quais, puramente químicas. Com o tempo, a madeira perde água e substâncias de reserva, tornando-se infiltrada com vários compostos orgânicos, tais como: óleos gomas, resinas, taninos, materiais corantes e aromáticos. Algumas dessas substâncias impregnam as paredes, outras apenas penetram no lume da célula. A região do lenho que passa por estas modifi-cações denomina-se cerne. A parte do lenho que é funcional denomina-se alburno.
Em muitas madeiras desenvolvem-se tilos, isto é, protuberâncias das células do parênquima lenhoso (axial ou radial) que penetram no interior de um ele-mento traqueal quando este se torna inativo ou quando o tecido xilemático é injuriado. Tal crescimento ocorre através de um par de pontuações que faz conexão entre dois elementos. Estas mudanças que ocorrem no cerne não afetam a eficácia da madeira, mas torna-a mais durável que o alburno, menos suscetí-vel à ataques por microorganismos e menos penetrável por líquidos (inclusi-
ve, líquidos preservativos). Figura . Seção longitudinal de um vaso, evidenciando a formação de tilos a partir de células parenquimáticas vizinhas. Em algumas espécies, o alburno logo se converte em cerne, em outras necessita de mais tempo. Variações de estrutura do lenho Coníferas - Apresentam tenho muito homogêneo, no qual faltam elementos de vasos. São formadas exclusivamente de
traqueídeos. O lenho inicial é formado por traqueídeos de paredes finas e lume relativa-mente grande e o lenho tardio, por traqueídeos de paredes espessadas e lume relativamente pequeno. Pode ou não haver parênquima axial. Em al-guns casos, o parênquima axial se reduz ao que está associado aos canais resiníferos presentes neste tipo de lenho. Os raios parenquimáticos nas coníferas normalmente são unisseriados. E-ventualmente tornam-se plurisseriados. pela presença de um canal resinífe-ros. Os raios são constituídos de células de parênquima e neles podem ocor-

Histologia Vegetal – prof Arlindo Costa – UDESC - 2001
37
37
rer também as chamadas traqueídeos radiais. Dicotiledôneas - Apresentam lenho bastante variado em aspecto. De um modo geral, elas apresentam vasos e eventualmente traqueídeos, além de fibras. parênquima axial e radial de mais de um tipo O lenho é muito heterogêneo, mas muito característico para cada espécie, ou cada gênero, a tal ponto que ape-nas com a análise da madeira se pode, muitas vezes, chegar à espécie do qual pertence uma determinada árvore. Os elementos de vaso quando observados transversalmente também podem ser denominados de vasos. De um modo geral, em climas tropicais o lenho das árvores apresenta-se com os vasos difusos. Em regiões temperadas, no entanto, freqüentemente os vasos apresentam uma perfeita ordenação, constituindo o que se costuma chamar de lenho de porosidade em anel. O parênquima axial pode se apresentar, nas dicotiledôneas, de duas ma-neiras: ou está associado aos vasos, constituindo o parênquima paratraqueal, ou apresenta-se independente dos vasos, constituindo parênquima apotraqueal. O parênquima apotraqueal pode ser escasso, difuso, ou formando linhas ou faixas. Pode haver madeira que não possui parênquima axial.

Histologia Vegetal – prof Arlindo Costa – UDESC - 2001
38
38
RESUMO - TECIDOS VEGETAIS Meristemas: São tecidos compostos por células praticamente indiferenciadas e que apresentam uma alta taxa de divisões mitóticas, conduzindo assim o cresci-mento da planta. Apresentam-se, em geral, nas pontas de ramos e raízes. Suas células apresentam apenas paredes primárias bem delgadas. Epiderme: É o tecido que reveste o corpo da planta. Suas células apresentam-se achatadas e perfeitamente justapostas. É o tecido que fica em contato direto com o ambiente, protegendo os tecidos internos. Parênquima: É um tecido que apresenta células principalmente com paredes primá-rias e com conteúdos celulares e funções variáveis. É um dos tecido mais importan-tes do corpo vegetal estando a ele associadas várias funções, a fotossíntese e o acúmulo de reservas nutricionais, por exemplo. É encontrado nas mais diversas partes do corpo vegetal. Colênquima e Esclerênquima: São os tecidos de sustentação do corpo vegetal. O colênquima é formado por células com paredes primárias espessadas, e o escle-rênquima caracteristicamente representado por células com paredes secundárias lignificadas. O colênquima é encontrado principalmente na periferia dos caules jovens e pecío-los, logo abaixo da epiderme. O esclerênquima por sua vez apresenta uma distribu-ição mais variável. Xilema: o xilema é o principal tecido condutor de água das plantas vasculares. As células relacionadas com esta condução, as mais especializadas do xilema, são os elementos traqueais. São mais ou menos alongadas e morrem na maturidade. Têm paredes lignificadas com espessamento secundário e variedade de pontuações. Os dois tipos de células traqueais, os traqueídeos e os elementos de vaso, diferem entre si pelo fato do traqueídeos ser uma célula não perfurada, ao passo que os elementos de vaso apresentam uma ou mais perfurações em cada extremidade e
às vezes também na parede lateral. Nos elementos de vaso a água movi-menta-se livremente de célula para célula, através de perfurações existentes na parede. A parte perfurada da parede de um elemento de vaso é chamada placa de perfuração que pode ser simples, com apenas uma perfuração ou multiperfu-rada; esta é escalariforme se as pontuações formam um padrão semelhante a uma rede. a) Protoxilema: consiste de uma parede precoce do desenvolvimento do
xilema primário. O protoxilema se diferencia nas partes primárias do corpo da planta que não completou seu crescimento e diferenciação. O protoxilema contém normalmente só elementos traqueais incluídos no pa-rênquima que é considerado como parte do protoxilema. b) Metaxilema: consiste de uma parte tardia do desenvolvimento do xilema
primário. O metaxilema inicia-se geralmente no corpo primário da planta ainda em crescimento, mas amadurece só muito depois do alongamento haver sido concluído. O metaxilema é mais complexo que o protoxilema e pode conter fibras além dos elementos traqueais e as células parenquimáticas. Floema: É o tecido condutor de alimentos das plantas vasculares e está as-sociado no sistema vascular ao xilema. O floema é menos esclereficado e persistente que o xilema. Devido à sua posição próxima da periferia do caule e da raiz, o floema sofre maiores modificações quando ocorre aumento de circunferência do eixo.

Histologia Vegetal – prof Arlindo Costa – UDESC - 2001
39
39
a) Elemento de tubo crivado: são as células mais especializadas do floema. As suas principais características morfológicas são as áreas crivadas em suas pa-redes e a ausência de núcleos nos protoplastos adultos.
Nos elementos de tubo crivado as áreas mais diferenciadas são as placas crivadas, quase sempre situadas na extremidade das células. Os elementos de tubos criva-dos formam séries verticais, os tubos crivados, ligados entre si pelas placas criva-das. Os elementos crivados possuem células onde o núcleo está desintegrado, desapa-rece a delimitação entre o vacúolo e o citoplasma parietal, o metabolismo é bem reduzido. b) Célula companheira: os elementos de tubos crivados estão associados intima-
mente a células muito especializadas, chamadas células companheiras. Existe uma interdependência entre os dois tipos de células, tanto que o término de função de um elemento crivado é acompanhado pela morte das células compa-nheiras associadas a ele. As paredes entre os dois tipos de células são muito delgadas ou densamente pontuadas.
As células companheiras diferentemente dos elementos de tubo crivado apresen-tam núcleo e atividade metabólica. Estruturas Secretoras: Secreção é o processo que inclui a síntese, a segregação e a liberação de materiais que, ou são funcionalmente especializados, ou são desti-nados ao armazenamento, ou à excreção. Estes materiais podem ficar dentro da própria célula, em espaços internos especiais ou serem eliminados. As cavidades ou ductos são cavidades alongadas que contém substâncias de natu-reza diversa . Os espaços intercelulares podem ter origem esquizógena, quando as células se afastam, ou lisígena, quando as células se dissolvem, ou ainda de ambas. São en-
volvidos por células que constituem o epitélio, que circunda o lume, onde se encontra o material secretado. Em Pinus elliotti são freqüentes ductos esquizógenos contendo resina, na madeira. Em Mangifema indica (manga) ocorrem ductos gomosos-resiníferos de origem lisígena, nos caules e frutos. Nos frutos CICTRUS (laranja e limão) ocorrem cavidades de óleo de origem lisígena. De modo geral, as resinas têm um papel relevante na defesa da planta contra agentes patogênicos e herbívoros. Células isoladas, tricomas, canais e cavidades podem conter mucilagem. Esta é um polissacarídeo que pode ser encontrada em qualquer parte da planta, principalmente em tecidos com reserva de água, como nas cactáceas. Em plantas insetívoras os tricomas secretam mucilagem, auxiliando na captu-ra do inseto. As células do ápice radicular secretam mucilagem, permitindo “lubrificar” a coifa e facilitar seu crescimento entre as partículas do solo. Os laticíferos são cavidades em geral alongadas que secretam látex. Este pode conter carboidratos, ácidos, alcalóides, taninos, mucilagem, cristais, borracha, proteínas, grãos de amido, etc. O látex geralmente é branco, inco-lor, amarelo ou alaranjado. Folha: As folhas são originadas dos caules e é o órgão responsável pela fo-tossíntese e pela respiração. Em plantas sem folhas, o caule assume estas funções. Os estômatos, pequenos poros na epiderme, podem ocorrer apenas na face superior – folha epiestomática, apenas na face inferior – folha hipoestomáti-ca, e em ambas as faces – folha anfiestomática. O mesófilo, a parte interna da folha, é formado geralmente por parênquima clorofiliano. Este pode ser de dois tipos: paliçádico com células alongadas, dispostas lado a lado, resultando poucos espaços intercelulares; e esponjoso

Histologia Vegetal – prof Arlindo Costa – UDESC - 2001
40
40
com células de vários formatos que permitem a formação de muitos espaços inter-celulares. As folhas em que ocorre parênquima paliçádico em ambas as faces são chamadas de isolateral ou unifacial e as folhas em que este tecido ocorre somente na face superior são chamadas de dorsiventral ou bifacial. O sistema vascular é formado por feixes vasculares que formam as nervuras. Os feixes vasculares são colaterais, com xilema voltado para a face superior e floema para a face inferior.


![Aula 4 e 5-TECIDOS VEGETAIS [Modo de Compatibilidade].pdf](https://static.fdocumentos.com/doc/165x107/5695d3451a28ab9b029d57dd/aula-4-e-5-tecidos-vegetais-modo-de-compatibilidadepdf.jpg)