
Nem tudo que parece ser, é lesão: aspectos anatômicos, não ...
Upload
nguyentrucCategory
view
217download
0
UNIVERSIDADE FEDERAL DE GOIÁS
ESCOLA DE VETERINÁRIA E ZOOTECNIA
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIA ANIMAL
ASPECTOS ANATÔMICOS DA INERVAÇÃO DO MEMBRO PÉLVICO DO
TAMANDUÁ-BANDEIRA (Myrmecophaga tridactyla, Linnaeus, 1758)
Viviane Souza Cruz
Orientador: Prof. Dr. Eugênio Gonçalves de Araújo
GOIÂNIA 2013

ii

iii
VIVIANE SOUZA CRUZ
ASPECTOS ANATÔMICOS DA INERVAÇÃO DO MEMBRO PÉLVICO DO
TAMANDUÁ-BANDEIRA (Myrmecophaga tridactyla, Linnaeus, 1758)
Tese apresentada para obtenção do
grau de Doutor em Ciência Animal
junto à Escola de Veterinária e
Zootecnia da Universidade Federal de
Goiás
Área de Concentração:
Patologia, Clínica e Cirurgia Animal
Orientador:
Prof. Dr. Eugênio Gonçalves de Araújo - EVZ - UFG
Comitê de Orientação:
Prof. Dr. Júlio Roquete Cardoso - ICB - UFG
Profa. Dra. Luciana Batalha de Miranda Araújo - EVZ - UFG
GOIÂNIA 2013

iv

v

vi
Aos meus queridos filhos Lauro Cruz e
Souza e Vitória Cruz Silvano.
Ao José Itamarcos Rodrigues Silvano,
companheiro fiel e amado. Pessoas
indispensáveis nesta trajetória.
Aos animais, em especial à aqueles que serviram
ao conhecimento científico.
Dedico

vii
AGRADECIMENTOS
Agradeço a Deus pela vida e por me dar coragem para seguir em
frente, apesar das dificuldades.
Ao meu querido companheiro José Itamarcos Rodrigues Silvano, pelo
amor, incentivo, compreensão e paciência. Com certeza você teve uma grande
participação nesta conquista.
Aos meus queridos filhos Lauro Cruz e Souza e Vitória Cruz Silvano,
pela compreensão pelas minhas ausências e até mesmo pela falta de paciência
para com eles.
Aos meus colegas de Doutorado pela amizade e ajuda em vários
momentos desta trajetória.
Ao meu orientador Prof. Dr. Eugênio Gonçalves de Araújo, pelo aceite,
apoio e confiança na execução deste doutorado.
Ao meu co-orientador Prof. Dr. Júlio Roquete Cardoso, antes de tudo
pela amizade e exemplo de dedicação, e ainda pelo acompanhamento em todas
as etapas do doutorado, em especial, na execução deste trabalho.
À minha querida co-orientadora Profa. Dra. Luciana Batalha de Miranda
Araújo e ao chefe do Departamento de Morfologia Prof. Dr. Paulo Cesar Moreira,
pela ajuda, apoio e dedicação na aquisição dos tamanduás-bandeira junto ao
IBAMA.
À Profa. Dra. Naida Cristina Borges, pela amizade, apoio e
disponibilidade no auxílio aos exames radiográficos, assim como as acadêmicas
de Medicina Veterinária Aaline Anarelli Araújo e Nadine Bou Fares.
Aos professores do Departamento de Morfologia, do Instituto de
Ciências Biológicas da Universidade Federal de Goiás, no apoio quando da
solicitação de afastamento para cursar o doutorado.
Aos professores da Escola de Veterinária e Zootecnia da Universidade
Federal de Goiás, pelos ensinamentos e por me receberem com amor, carinho e
amizade em mais esta jornada.

viii
Aos funcionários da Escola de Veterinária e Zootecnia e do
Departamento de Morfologia, do Instituto de Ciências Biológicas ambos da
Universidade Federal de Goiás, pelo carinho e ajuda durante todas as etapas, em
especial ao Sr. Darci José dos Santos “in memorian”.
Ao CETAS/ IBAMA, pela liberação dos exemplares utilizados neste
projeto.
À Escola de Veterinária e Zootecnia e ao Departamento de Morfologia,
do Instituto de Ciências Biológicas ambos da Universidade Federal de Goiás, por
fornecer a estrutura e recursos materiais, de suma importância para a conclusão
de mais uma etapa de minha carreira docente.
Finalmente agradeço aos animais que mesmo após a morte
contribuíram para a ciência e estudo se tornando peças indispensáveis para a
realização e conclusão deste trabalho.

ix
Tamanduá-bandeira
Tua figura não se classifica de beleza. Aparentas ser até bicho desagradável Mas és um ser como eu, com certeza.
Animal que sente, ama. És afável! No habitat, atacas apenas uma zona: Depredador de cupins e formigueiro,
Usando patas com garras e línguona, Alimentas teu corpo por inteiro.
Bonito é o teu papel na natureza.
Ao lado do tatu e do bicho preguiça, No reino dos Xenarthra, és realeza.
De insetos vives, mas se fogo te incendeia, Esturricado ficas, sem chance de defesa. Investir em ti, tirar-te dessa intricada teia,
Recuperar tua espécie será grande proeza, A consagrar político, cientista e até princesa.
Sandra Fayad

x
SUMÁRIO
CAPÍTULO 1 – CONSIDERAÇÕES INICIAIS 1
1 Introdução..................................................................................................... 1
2 Superordem Xenarthra.................................................................................. 3
3 Myrmecophaga tridactyla.............................................................................. 6
4 Aspectos morfofuncionais do sistema nervoso periférico............................. 11
4.1 Formação e distribuição dos nervos do plexo lombossacral..................... 13
4.1.1 Plexo lombar........................................................................................... 15
4.1.2 Plexo sacral............................................................................................. 16
5 Referências................................................................................................... 21
CAPÍTULO 2 – ASPECTOS ANATÔMICOS DO PLEXO LOMBOSSACRAL
DE Myrmecophaga tridactyla (Linnaeus, 1758)...............................................
29
RESUMO......................................................................................................... 30
INTRODUÇÃO................................................................................................. 31
MATERIAL E MÉTODOS................................................................................. 32
RESULTADOS E DISCUSSÃO....................................................................... 33
AGRADECIMENTOS....................................................................................... 44
ABSTRACT...................................................................................................... 44
REFERÊNCIAS................................................................................................ 45
CAPÍTULO 3 – ASPECTOS ANATÔMICOS DOS NERVOS DA COXA DO
TAMANDUÁ-BANDEIRA (Myrmecophaga tridactyla, Linnaeus, 1758)...........
48
RESUMO......................................................................................................... 49
INTRODUÇÃO................................................................................................. 50
MATERIAL E MÉTODOS................................................................................. 50
RESULTADOS ................................................................................................ 52
DISCUSSÃO.................................................................................................... 57
CONCLUSÕES................................................................................................ 60
AGRADECIMENTOS....................................................................................... 61
ABSTRACT...................................................................................................... 61
REFERÊNCIAS................................................................................................ 62
CAPÍTULO 4 – ANATOMICAL ASPECTS OF THE NERVES OF THE LEG

xi
AND FOOT OF THE GIANT ANTEATER (Myrmecophaga tridactyla,
Linnaeus, 1758)...............................................................................................
65
ABSTRACT...................................................................................................... 66
RESUMO......................................................................................................... 67
INTRODUCTION.............................................................................................. 67
MATERIAL AND METHODS............................................................................ 68
RESULTS ........................................................................................................ 69
DISCUSSION................................................................................................... 74
ACKNOWLEDGEMENTS................................................................................ 77
REFERENCES................................................................................................. 78
CAPÍTULO 5 – CONSIDERAÇÕES FINAIS.................................................... 80
ANEXOS.......................................................................................................... 82

xii
LISTA DE FIGURAS
CAPÍTULO 1
Figura 1 Espécime empalhado de Tamanduá-bandeira (Myrmecophaga
tridactyla)......................................................................................
2
Figura 2 A - Classificação morfológica clássica evidenciando quatro
grandes clados de mamíferos placentários (Xenarthra,
Ungulata, Archonta e Anagalida) (árvore de SHONANI &
MCKENNA (1998);. B - Classificação molecular com quatro
outros clados de mamíferos placentários. (Afrotheria,
Xenarthra, Laurasiatheria e Euarchontoglires) (árvore de
Murphy, et al., 2001). As cores distinguem os quatro clados
placentários basais da árvore molecular......................................
4
Figura 3 Espécime de Tamanduá-bandeira (Myrmecophaga tridactyla).
com sua classificação científica..................................................
6
Figura 4 Distribuição do tamanduá-bandeira (Myrmecophaga tridactyla).
Azul - presente, vermelho - possivelmente extinto (RJ e
RS).....................................................................................
7
Figura 5 Fotografias de peças anatômicas (Vista lateral esquerda: A e
B) e imagens radiográficas (Projeção látero-lateral esquerda: A’
e B’) do segmento lombossacral da coluna vertebral e cintura
pélvica de esqueleto de Tamanduá-bandeira (Myrmecophaga
tridactyla) da coleção do Museu Arlindo Coelho de
Souza/ICB/UFG. Indivíduo com 15 vértebras torácicas (T),
duas lombares (L) e cinco sacrais (S)..........................................
8
Figura 6 Fotografia de peça antômica (Vista ventral: A) e imagens
radiográficas (Projeção ventro-dorsal: A’ e ventro-dorsal
oblíqua: B) da coluna vertebral e cintura pélvica de esqueleto
de Tamanduá-bandeira (Myrmecophaga tridactyla) da coleção
do Museu Arlindo Coelho de Souza/ICB/UFG.............................
9
Figura 7 Representação esquemática da estrutura de um nervo.............. 11

xiii
Figura 8 Estrutura da medula espinhal e formação dos nervos
espinhais......................................................................................
12
Figura 9 Esquema da formação do plexo lombossacral no cão................ 17
CAPÍTULO 2
Figura 1 A, B, C, D, E – Imagens radiográficas em projeção ventro-
dorsal da coluna vertebral torácica caudal, lombar, sacral e
coccígea cranial de tamanduás-bandeira adultos. Figuras A e D
correspondentes a um exemplar com 15 vértebras torácicas,
duas lombares e cinco sacrais. Em B e E, exemplar com 15
vértebras torácicas, três lombares e quatro sacrais e C,
exemplar com 16 vértebras lombares, duas lombares e cinco
sacrais. F - Radiografia em projeção látero-lateral direita da
coluna vertebral lombar caudal, sacral e coccígea cranial de
tamanduá-bandeira adulto............................................................
35
Figura 2 Vista ventral do plexo lombossacral de M. tridactyla adultos,
sendo A, indivíduo com 16 vértebras torácicas e duas lombares
e B, indivíduo com 15 vértebras torácicas e três
lombares.......................................................................................
38
Figura 3 Vista ventral do plexo lombossacral de M. tridactyla adulta, com
15 vértebras torácicas, duas lombares e cinco sacrais...............
40
CAPÍTULO 3
Figura 1 Vista ventral do plexo lombossacral de M. tridactyla adulta,
com 15 vértebras torácicas, duas lombares e cinco sacrais.....
52
Figura 2 A- Vista medial da coxa direita de M. tridactyla adulto e
B- Vista lateral da coxa esquerda de M. tridactyla adulto ..........
53
CAPÍTULO 4
Figure 1 A- Medial view of the thigh and leg of an adult M. tridactyla ,
B and C- Dorsal view of the right foot of an adult M. tridactyla ..
70
Figure 2 Lateral view of the distal extremity of thigh and proximal
extremity of the right leg of M. tridactyla…………………………..
72
Figure 3 Plantar surface of the left foot of M. tridactyla............................ 74

xiv
LISTA DE QUADROS
CAPÍTULO 1
Quadro 1 Formação do plexo lombossacral em animais domésticos........ 14
Quadro 2 Formação do plexo lombossacral em algumas espécies de
animais selvagens......................................................................
14
Quadro 3 Composição dos nervos do plexo lombossacral nos animais
domésticos.................................................................................
20
CAPÍTULO 3
Quadro 1 Nervos da coxa do M. tridactyla e seus respectivos territórios
de inervação...............................................................................
54

xv
LISTA DE TABELAS
CAPÍTULO 2
Tabela 1 Segmentos nervosos que compõe o plexo lombossacral do
tamanduá-bandeira e de outros mamíferos...............................
36
Tabela 2 Segmentos nervosos que compõe os nervos do plexo lombar
em Myrmecophaga tridactyla.....................................................
37
Tabela 3 Segmentos nervosos que compõe os nervos do plexo sacral
em Myrmecophaga tridactyla.....................................................
41
CAPÍTULO 4
Table 1 Digital nerves of the foot of M. tridactyla………………………… 71

xvi
LISTA DE ABREVIATURAS
C Vértebra cervical
T Vértebra torácica
L Vértebra lombar
S Vértebra sacral
Cc Vértebra coccígea
NE Nervos Espinhais
N. Nervo
Nn. Nervos
M. Músculo
Mm. Músculos
A. Artéria
V. Veia
IUCN International Union for Conservation of Nature
CETAS Centro de Triagem de Animais Silvestres
IBAMA Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais
Renováveis
CEUA Comissão de Ética no Uso de Animais
UFG Universidade Federal de Goiás
EVZ Escola de Medicina Veterinária e Zootecnia
ICB Instituto de Ciências Biológicas
DMORF Departamento de Morfologia

xvii
RESUMO
Os tamanduás são acometidos por vários tipos de desordens clínicas, porém tanto em seu habitat, como em cativeiro, os traumas estão entre as causas mais comuns de afecções nesta espécie. Logo, há uma crescente demanda de tratamentos clínicos e cirúrgicos para esses animais, procedimentos estes que dependem do conhecimento de sua anatomia. Sendo assim, o objetivo do trabalho foi descrever a origem e distribuição dos nervos do membro pélvico do tamanduá-bandeira. Para tanto, foram utilizados seis cadáveres adultos de Myrmecophaga tridactyla. Os cadáveres foram cedidos pelo IBAMA-GO (licença 99/2011, Protocolo CEUA-UFG 015/11). Os animais foram descongelados em temperatura ambiente durante 48h, fixados e preservados em soluções de formaldeído a 10% e 4%, respectivamente. A dissecação teve início após um período mínimo de 72h a partir da fixação. A presente tese gerou a produção de três artigos. O primeiro intitulado “ASPECTOS ANATÔMICOS DO PLEXO LOMBOSSACRAL DE Myrmecophaga tridactyla (Linnaeus, 1758)”. Observou-se que o plexo lombossacral originou-se em: T16, L1, L2, S1-S5, Cc1 (33,3%), T15, L1-L3, S1-S5 (16,6%), T15, L1, L2, S1-S5, Cc1 (16,6%), L1-L3, S1-S5 (16,6%), T15, L1-L3, S1-S4, Cc1 (8,3%) e L1-L3, S1-S4 (8,3%). Os nervos derivados do plexo, com suas formações mais frequentes foram: genitofemoral (T15, L1, L2 e T16, L1), cutâneo femoral lateral (T15, L1, L2 e T16, L1), femoral (T16, L1, L2), obturador (L1-L3), isquiático (L3, S1-S3), glúteo cranial (L2, S1), glúteo caudal (L2, S1-S3 e L3, S1, S2), pudendo (S4-S5) e retal caudal (S5 e S4, S5, Cc1). O segundo artigo intitulado “ASPECTOS ANATÔMICOS DOS NERVOS DA COXA DO TAMANDUÁ-BANDEIRA (Myrmecophaga tridactyla, Linnaeus, 1758)” observou-se que os nervos responsáveis pela inervação da coxa do M. tridactyla foram o genitofemoral, cutâneo femoral lateral, femoral, safeno, obturador, glúteo cranial, isquiático e cutâneo femoral caudal. O terceiro artigo intitulado “ASPECTOS ANATÔMICOS DOS NERVOS DA PERNA E PÉ DO TAMANDUÁ-BANDEIRA (Myrmecophaga tridactyla, Linnaeus, 1758)”, observou-se que os nervos responsáveis pela inervação da perna e pé foram o safeno, fibular comum, tibial, cutâneo lateral da sura e cutâneo caudal da sura. O plexo lombossacral do M. tridactyla apresenta como particularidade o envolvimento de ramos torácicos e coccígeos na composição de seus nervos e uma maior variação anatômica em sua formação, bem como de seus nervos, que pode ser atribuída principalmente à variação numérica de vértebras torácicas, lombares e sacrais nesta espécie. Os nervos responsáveis pela inervação da coxa, perna e pé são os mesmos relatados em animais domésticos e selvagens, porém com algumas diferenças, como uma maior inervação da face medial da coxa, a divisão mais distal do nervo isquiático e do fibular comum, ausência de ramos metatarsianos dorsais do N. fibular profundo e uma maior participação do nervo safeno na inervação digital, contribuindo com ramos inclusive para os dedos III e IV, além dos dedos I e II.
Palavras-chave: Anatomia, animais selvagens, Myrmecophagidae, nervos espinhais, plexo lombossacral

xviii
ABSTRACT
The anteaters are affected by several kinds of clinical disorders, but both in their habitat as in captivity, the injuries are among the most common causes of disorders in this species. Therefore, there is an increasing demand for medical and surgical treatments for these animals, which depend on knowledge of their anatomy. Thus, the aim of the study was to describe the origin and distribution of the nerves of the pelvic limb of the giant anteater. For this purpose, six adult cadavers of M. tridactyla provided by IBAMA-GO (license 99/2011, UFG-CEUA Protocol 015/11) were used. The animals were thawed at room temperature for 48h, fixed and preserved in formaldehyde solutions at 10% and 4%, respectively. The dissection was initiated after a minimum period of 72 hours from the fixation procedure. This thesis rendered possible the production of three articles. The first one is entitled “ANATOMICAL ASPECTS OF THE LUMBOSSACRAL PLEXUS OF THE Myrmecophaga tridactyla (Linnaeus, 1758)”. It was observed that the lumbosacral plexus of M. tridactyla comprised T16, L1, L2, S1-S5, Cc1 (33,3%), T15, L1-L3, S1-S5 (16,6%), T15, L1, L2, S1-S5, Cc1 (16,6%), L1-L3, S1-S5 (16,6%), T15, L1-L3, S1-S4, Cc1(8,3%) and L1-L3, S1-S4 (8,3%). The nerves derived from the plexus with their most common arrangement were as follow: genitofemoral (T15, L1, L2 and T16, L1), lateral femoral cutaneous nerve (T15, L1, L2 and T16, L1), femoral (T16, L1, L2), obturator (L1-L3), sciatic (L3, S1-S3), cranial gluteal (L2, S1, S2), caudal gluteal (L2, S1-S3 and L3, S1, S2), pudendal (S4, S5) and rectal caudal (S5 and S4, S5, Cc1). The secund article is entitled “ANATOMICAL ASPECTS OF THE NERVES OF THE THIGH OF THE GIANT ANTEATER (Myrmecophaga tridactyla, Linnaeus, 1758)”, observed that the nerves responsible for the innervation the thigh of the M. tridactyla were the genitofemoral, lateral cutaneous femoral, femoral, saphenous, obturator, cranial gluteal, sciatic and caudal cutaneous femoral nerves. The nerves of the leg and foot were the saphenous nerve, fibular, tibial, lateral sural cutaneous nerve and caudal sural cutaneous nerve. The lumbosacral plexus of the M. tridactyla shows as peculiarities the involvement of thoracic and coccygeal rami in its nerve composition and a greater anatomical variation in its formation as well as in their nerves, which can be mainly related to the numerical variation of thoracic, lumbar and sacral vertebrae in this species. The third article is entitled “ANATOMICAL ASPECTS OF THE NERVES OF THE LEG AND FOOT OF THE GIANT ANTEATER (Myrmecophaga tridactyla, Linnaeus,1758)” observed that the nerves responsible for the thigh, leg and foot innervation were the same as those reported in domestic and wild animals, but with some differences, such as the greater innervation of the medial surface of the thigh, the more distal division of the sciatic and common fibular nerve, absence of dorsal metatarsal branches of the deep fibular nerve and a greater involvement of the saphenous nerve in the digital innervation with branches to the digits III and IV, in addition to digits I and II.
Keywords: Anatomy, animals, lumbosacral plexus, Myrmecophagidae, spinal nerves

CAPÍTULO 1 – CONSIDERAÇÃOES INICIAIS
1 Introdução
O estudo do comportamento, reprodução e doenças de animais
selvagens tem sido objeto de muitas investigações e ganhado importância em
pesquisas científicas. Neste aspecto, se destaca o crescente número de artigos
sobre o tamanduá.
A anatomia animal é um ramo da Medicina Veterinária que serve como
embasamento para várias áreas, em especial para clínica e cirurgia, e, apesar do
bem estabelecido conhecimento que se tem da anatomia topográfica dos animais
domésticos, o mesmo não ocorre com os animais selvagens, dos quais pouco se
conhece e muito se tem a pesquisar.
Muitas espécies de animais selvagens tem sofrido redução em suas
populações de forma que algumas espécies já estão classificadas como em risco
ou ameaçadas de extinção em determinadas regiões.
No caso do Myrmecophaga tridactyla, espécie pertencente à
Superordem Xenarthra, as causas da queda na população desta espécie estão
ligadas à caça e a destruição ou degradação de seu habitat, acarretando provável
modificação e redução de comunidades de formigas e cupins, que compreendem
sua principal fonte de alimento (SILVEIRA et al., 1999; MEDRI & MOURÃO, 2005;
MIRANDA & COSTA, 2007). É relatado que estes animais voltam a utilizar áreas
queimadas pouco tempo após a passagem do fogo (BRAGA et al., 2010),
aumentando o risco de sua predação devido à drástica redução na cobertura
vegetal (BRAGA & SANTOS, 2009). Não obstante a interferência humana, as
taxas reprodutivas do tamanduá-bandeira são baixas. Estes fatores tornam o
tamanduá mais vulnerável à pressão antrópica, intensificando sua vulnerabilidade
à extinção (EISENBERG & REDFORD, 1999).
Em decorrência destes fatores vem ocorrendo uma mudança no perfil
de atendimentos em hospitais e clínicas veterinárias e hoje os atendimentos
clínico-cirúrgicos em animais selvagens está se tornando uma rotina. Com a
escassez de conhecimento anatômico nessas espécies, vem surgindo à

2
necessidade cada vez maior de estudos e pesquisas nesta área, a fim de
contribuir na qualidade e especificidade nos atendimentos, procedimentos e
tratamentos necessários, possibilitando assim um melhor prognóstico a estes
animais.
Até o momento, os dados sobre o membro pélvico do tamanduá-
bandeira se restringem à descrição dos ossos (OLIVEIRA, 2001) e dos músculos
intrínsecos do pé (GAMBARYAN et al., 2009), o que decisivamente limita a ação
do médico-veterinário. Ademais, quando se trata do membro pélvico em especial,
a demanda por um tratamento eficiente é importante para o tamanduá-bandeira,
pois estes apêndices além de importantes na deambulação, também são
fundamentais para a defesa, uma vez que apoiam-se nestes membros e na base
da cauda, formando um tripé que o possibilita elevar o tronco adquirindo uma
posição bípede durante o ataque, ou mesmo durante a alimentação, momento
este que requer a liberação dos membros torácicos para o ato de cavar (Figura 1).
Logo, qualquer deficiência neste membro pode dificultar o retorno do animal ao
seu habitat.
FIGURA 1. Espécime empalhado de Tamanduá-bandeira (Myrmecophaga tridactyla). Fonte:http://2.bp.blogspot.com/_kc89sKVUMWM/TLnbA_cryfI/AAAAAAAAG1k/W9SQ8FcpA3s/s400/69+DSC03758.jpg

3
Sendo assim, este trabalho objetivou-se fornecer dados anatômicos
acerca da inervação do membro pélvico do M. tridactyla, de forma a contribuir
com estudos em anatomia comparada e ainda nas condutas clínicas, cirúrgicas e
anestésicas nesta espécie.
2 Superordem Xenarthra
A Superordem Xenarthra (xeno = estranho e arthron = articulação) é
formada pelos tatus (ordem Cingulata), tamanduás e preguiças (ordem Pilosa)
(VIZCAÍNO & LOUGHRY, 2008).
Os processos históricos responsáveis pelas distribuições geográficas
dos táxons podem ser inferidos pelo registro dos fósseis, além da inferência
filogenética. Dos quatro principais grupos de mamíferos placentários identificados
pelos taxonomistas moleculares: Afrotherios, Xenarthras, Laurasiatérios e
Euarchontoglires; os dois ramos mais antigos são os Afrotérios e os Xenarthras
(DAWKINS, 2009).
Os Xenarthras foram um dos poucos grupos que se diversificaram em
um espaço geográfico restrito (ENGELMANN, 1985). Muitos estudos procuram
desvendar a origem deste grupo e o início do seu isolamento, sugerindo que a
sua origem, provavelmente, foi antes da separação da Gondwana. O isolamento
geográfico entre os grupos ocorreu durante o Mesozóico, principalmente durante
o Cretáceo, com a deriva continental (HEDGES et al., 1996). A divergência entre
Xenarthra e os outros mamíferos placentários provavelmente ocorreu no
Cretáceo, há aproximadamente 106 milhões de anos (DELSUC et al., 2001).
Depois migraram para a América do Sul durante o Plioceno, a cerca de 85
milhões de anos. Pelo istmo do Panamá, os xenartros provavelmente migraram
em direção à América do Norte entre o final do Plioceno e o início do Pleistoceno
(ENGELMMAN, 1985).
Embora dentro da classificação geral dos mamíferos haja controvérsias
com relação ao parentesco dos Xenarthra com outras ordens, não há dúvidas
sobre o seu monofiletismo. As classificações morfológicas colocam Xenarthra
como sendo o táxon de mamíferos placentários mais basais com outros três

4
clados bem definidos (Ungulata, Archonta e Anagalida) (Figura 2A) (SHOSHANI &
MCKENNA, 1998), enquanto as mais recentes classificações moleculares
reconhecem quatro outros diferentes clados (Afrotheria, Xenarthra, Laurasiatheria
e Euarchontoglires) (Figura 2B) (SPRINGER et al., 2004).
FIGURA 2. A - Classificação morfológica clássica evidenciando quatro grandes clados de mamíferos placentários (Xenarthra, Ungulata, Archonta e Anagalida) (árvore de SHONANI & MCKENNA (1998); B - Classificação molecular com quatro outros clados de mamíferos placentários. (Afrotheria, Xenarthra, Laurasiatheria e Euarchontoglires) (árvore de Murphy, et al., 2001) As cores distinguem os quatro clados placentários basais da árvore molecular. Fonte: Modificado de Springer et al. (2004).
As Subordens de Eutheria são representadas por quatro clados, dois
com origem no hemisfério sul (Gondwana) – Afrotheria (que inclui seis ordens,
Tubulidentata, Macroscelidae, Afrosoricida, Sirenia, Hyracoidea e Proboscidae) e
Xenarthra (com duas ordens, Pilosa e Cingulata) – e dois clados do Norte
(Laurasia) – Euarchontoglires (constutuído pelas Ordens Dermoptera, Scadentia,
Primata, Rodentia e Lagomorpha) e Laurasiatheria (inclui as Ordens Eulipotyphia,

5
Carnivora, Pholidota, Cetartiodactyla, Perissodactyla e Chiroptera) (SPRINGER et
al., 2004).
LUCKETT (1993) agrupou as preguiças, tamanduás e tatus baseado
na análise cladística das membranas fetais e características crânio-esqueleto.
Entretanto, a comparação entre os resultados dos dados morfológicos e
moleculares mostram que dentro dos mamíferos existem vários casos de
correspondência (SHOSHANI & MCKENNA, 1998).
A partir de 1960, os estudos da Superordem Xenarthra têm se
apropriado de algumas técnicas bioquímicas e moleculares. Os marcadores
moleculares, como o DNA mitocondrial e o DNA nuclear, são de grande
importância nos estudos evolutivos, como marcadores de diversidade genética.
O Xenarthra é um clado de mamíferos Sul Americanos com uma
mistura intrigante de características primitivas e derivadas, sugerindo que eles
podem ser o membro mais primitivo dos mamíferos placentários e um clado irmão
de todos os outros eutérias (ENGELMANN, 1985).
A Superordem Xenarthra é considerada monofilética por diferentes
métodos de análises (GAUDIN, 2003). MESS & CARTER (2007) afirmaram que
existe uma dicotomia entre os tatus e as preguiças e tamanduás. MURPHY et al.
(2001) juntaram preguiças (Cloloepus spp.) e tamanduás (Myrmecophaga,
Tamandua) na subordem Pilosa, tornando-se um grupo irmão dos tatus. Para
definir as relações filogenéticas entre preguiça (Bradypus variegatus) e tamanduá
(Tamandua mexicana e Myrmecophaga tridactyla), De JONG et al. (1985)
isolaram e seqüenciaram os aminoácidos da proteína a- cristalina, encontrando
uma origem monofilética.
Sendo assim, os xenartros apresentam características comuns ao
grupo, entre estas, se destaca a presença de articulações extras nas vértebras
lombares, a redução ou ausência de dentes, a fusão do ísquio com a primeira
vértebra caudal e a presença de placas dorsais de origem dérmica, no entanto,
esta última característica se perdeu em algumas preguiças e tamanduás
(ENGELMMAN, 1985).
No cerrado brasileiro são encontrados 10 espécies de Xenarthra
terrestre (Myrmecophagidae e Dasypodidae) além de um representante da família
Bradypodidae (EISENBERG & REDFORD, 1999).

6
3 Myrmecophaga tridactyla
O tamanduá-bandeira (Myrmecophaga tridactyla) é um mamífero
pertencente ao Reino Animalia, Filo Chordata, Classe Mammalia, Subclasse
Theria, Infraclasse Eutheria, Superordem Xenarthra, Ordem Pilosa, Subordem
Vermilingua, Família Myrmecophagidae, Gênero Myrmecophaga e Espécie
Myrmecophaga tridactyla, sendo o maior tamanduá existente no mundo (Figura 3)
(SMITH, 2009).
FIGURA 3. Espécime de Tamanduá-bandeira (Myrmecophaga tridactyla) com sua classificação científica. Fonte:http://www.infoescola.com/wp-content/uploads/2009/08/full-12-f373f1a5a7 .jpg
Sua distribuição geográfica é ampla, sendo encontrado desde o sul de
Belize e Guatemala, oeste dos Andes, a noroeste do Equador, Colômbia, sul da
Venezuela, sudeste da Bolívia, oeste do Paraguai, noroeste da Argentina, Brasil e
leste do Uruguai, sendo que neste último já esteja extinto (Figura 4) (EISENBERG
& REDFORD, 1999; SAMPAIO et al., 2006; COLLEVATTI, et al., 2007;
GREGORINI et al., 2007; MOURÃO & MEDRI, 2007).
É encontrado nos mais variados habitats, desde florestas, cerrados,
campos abertos e pastagens. No Brasil, ocorrem em todos os biomas, desde a
Amazônia até os campos sulinos, no entanto, as maiores concentrações estão

7
nos cerrados e campos. A sua densidade populacional é relativamente baixa,
variando de 1,3 a dois animais/km2 (NOWAK, 1999; SMITH, 2009). A área de vida
da espécie é de 12 a 25 km2, no entanto, pesquisa realizada no Parque Nacional
de Brasília relatou ser em média 3,7 km2 para fêmeas adultas e 2,7 km2, para
machos adultos (SHAW et al., 1987).
FIGURA 4. Distribuição do tamanduá-bandeira (Myrmecophaga tridactyla). Azul – presente, vermelho – possivelmente extinto (RJ e RS). Fonte:http://upload.wikimedia.org/wikipedia/commons/thumb/7/75/Giant_Anteater_area.png/200px-Giant_Anteater_area.png
Seus parentes mais próximos e integrantes da mesma família são o
Tamandua tetradactyla (tamanduá-mirim) e o Cyclopes didactylus (tamanduaí ou
tamanduá-anão ou tamanduá-seda), ambos arborícolas e de menor porte. O M.
tridactyla possui o maior porte, medindo entre 1,00 a 1,20 m de comprimento, sem
incluir a cauda, que mede entre 65 e 90 cm (SHAW et al., 1987; NAPLES, 1999).
Pesa entre 18 e 39 kg, havendo o registro de um exemplar mantido no Zoológico
de São Paulo, que pesou 51,7 kg (BERESCA & CASSARO, 2001).
A coluna vertebral apresenta uma articulação adicional nas vértebras
pós-diafragmáticas (torácicas caudais e lombares), denominada de “xenarthrales”,

8
formada pelos processos xenarthros, localizados ventralmente aos processos
mamilares, cuja função é evitar a superextensão da coluna vertebral, e ainda a
presença de fusão entre a primeira vértebra caudal e o ísquio (Figuras 5 e 6)
(GAUDIN & BRANHAM, 1998; ENDO et al., 2009; NYAKATURA & FISHER,
2010). Apresentam ainda outros aspectos anatômicos bastante peculiares, como
a presença de dupla veia cava caudal e uma rete mirable em cada extremidade
dos apêndices locomotores e cauda, com a finalidade de atuar em um mecanismo
de contracorrente para a troca de calor (GILLESPIE, 1993), reduzindo a sua
perda para os membros, e assim minimizando o gasto metabólico necessário à
manutenção da temperatura corpórea (WITHERS, 1992).
FIGURA 5. Fotografias de peças anatômicas (Vista lateral esquerda: A e B) e imagens radiográficas (Projeção látero-lateral esquerda: A’ e B’) do segmento lombossacral da coluna vertebral e cintura pélvica de esqueleto de Tamanduá-bandeira (Myrmecophaga tridactyla) da coleção do Museu Arlindo Coelho de Souza/ICB/UFG. Indivíduo com 15 vértebras torácicas (T), duas lombares (L) e cinco sacrais (S). T, L, S, Cc - Processos espinhosos dos segmentos vertebrais torácicos, lombares, sacrais e coccígeos, respectivamente; I - Ílio; Is - Ísquio; F - Fêmur; Cost - Costela; PX - Processo xenarthro; PM - Processo mamilar; Setas - Articulação do último segmento sacral e primeiro coccígeo com o ísquio. Fonte: Arquivo pessoal
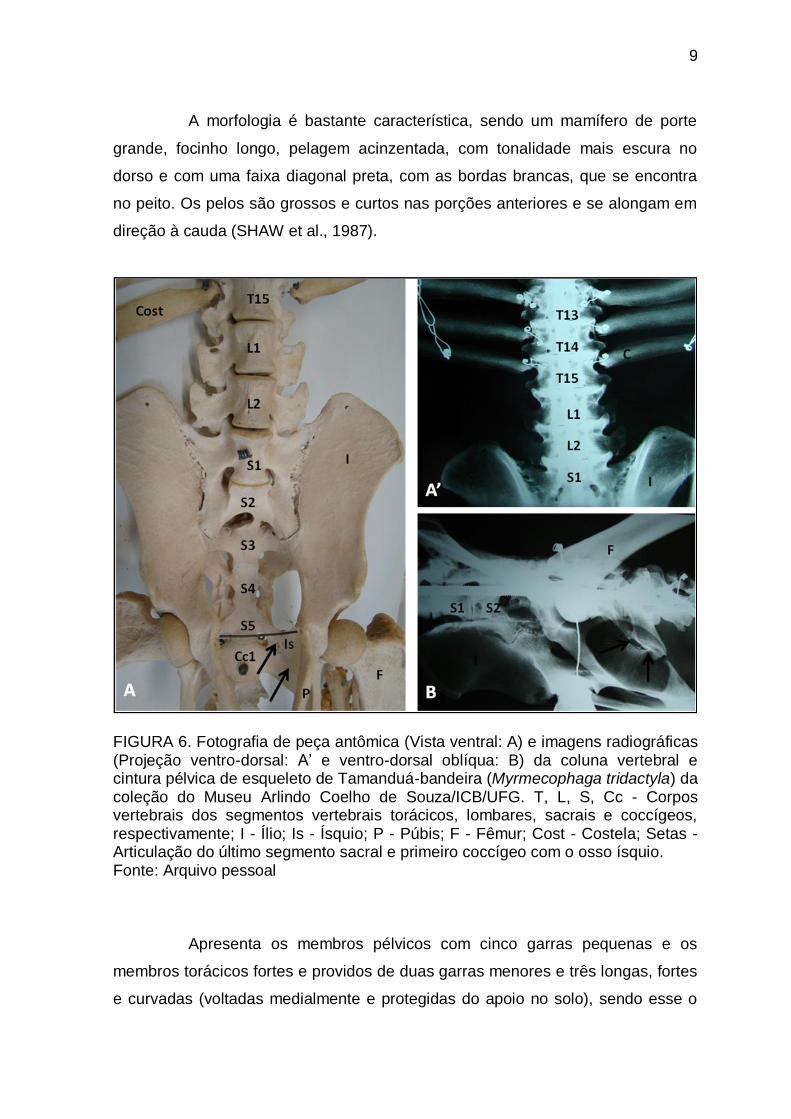
9
A morfologia é bastante característica, sendo um mamífero de porte
grande, focinho longo, pelagem acinzentada, com tonalidade mais escura no
dorso e com uma faixa diagonal preta, com as bordas brancas, que se encontra
no peito. Os pelos são grossos e curtos nas porções anteriores e se alongam em
direção à cauda (SHAW et al., 1987).
FIGURA 6. Fotografia de peça antômica (Vista ventral: A) e imagens radiográficas (Projeção ventro-dorsal: A’ e ventro-dorsal oblíqua: B) da coluna vertebral e cintura pélvica de esqueleto de Tamanduá-bandeira (Myrmecophaga tridactyla) da coleção do Museu Arlindo Coelho de Souza/ICB/UFG. T, L, S, Cc - Corpos vertebrais dos segmentos vertebrais torácicos, lombares, sacrais e coccígeos, respectivamente; I - Ílio; Is - Ísquio; P - Púbis; F - Fêmur; Cost - Costela; Setas - Articulação do último segmento sacral e primeiro coccígeo com o osso ísquio. Fonte: Arquivo pessoal
Apresenta os membros pélvicos com cinco garras pequenas e os
membros torácicos fortes e providos de duas garras menores e três longas, fortes
e curvadas (voltadas medialmente e protegidas do apoio no solo), sendo esse o

10
único meio de defesa e importante ferramenta alimentar (SHAW et al., 1987;
RODRIGUES et al., 2009). Seu modo de locomoção é plantígrado podendo
adquirir a posição bípede ao escavar para obtenção de alimento ou durante a
defesa (GAMBARYAN et al., 2009).
Sua característica fisiológica marcante é a baixa temperatura corporal,
variando de 32 a 34oC (DICKMAN, 2001), ou de 30,7 a 37oC (ROSA, 2007;
SMITH, 2009), associada a sua baixa taxa metabólica que pode variar entre 33 e
60% daquelas esperadas para o seu peso, quando comparada à de outros
mamíferos. A sua alimentação é de baixo teor calórico, composta de formigas
(família Formicidae) e térmitas (famílias Kalotermitidae, Rhinotermitidae,
Serritermitidae e Termitidae (DICKMAN, 2001).
Têm hábitos de vida solitários, exceto no período de acasalamento, ou
quando se trata da mãe e seu filhote (MIRANDA & COSTA, 2007). O padrão de
atividade varia entre uma região e outra, tendendo os animais a se tornarem mais
noturnos como resposta à influência humana (FONSECA et al., 1994), sendo
assim, parece ser mais ativo durante o dia em áreas livres de habitações
humanas e durante à noite em áreas muito povoadas, embora este ritmo de
atividade possa ter influência drástica da temperatura ambiente, regime de
chuvas, ou mesmo em regime de cativeiro (EISENBERG & REDFORD, 1999;
NOWAK, 1999; MEDRI & MOURÃO, 2005; CAMILO-ALVES & MOURÃO, 2006;
ROSA, 2007).
Não possuem dimorfismo sexual, sendo a sexagem possível somente
por meio de exame detalhado (POCOCK, 1924; SHAW & CARTER, 1980; DINIZ
et al., 1995). Os machos são criptorquidas, com saco urogenital similar ao da
fêmea, e não é possível a exposição manual do pênis do sistema urogenital
(NOWAK, 1999). As fêmeas são menos corpulentas e apresentam peso menor do
que os machos (MIRANDA, 2004). A gestação é de 190 dias e parem apenas um
filhote, que depende exclusivamente da mãe para se alimentar até os dois anos
de idade (NOWAK, 1999; CHEBEZ & CIRIGNOLI, 2008).

11
4 Aspectos morfofuncionais do sistema nervoso periférico
O sistema nervoso periférico é constituído por nervos, terminações
nervosas e gânglios. Os nervos são cordões esbranquiçados constituídos por
feixes de fibras nervosas reforçadas por tecido conjuntivo (endoneuro, perineuro e
epineuro), que unem o sistema nervoso central aos órgãos periféricos. Os nervos
podem ser espinhais ou cranianos, conforme sua conexão com o sistema nervoso
central (TRUMBLE, 2000; MACHADO, 2006).
O endoneuro é a camada mais interna e delicada de tecido conjuntivo,
distensível, vascularizada e com função protetora contra alterações pressóricas
(GAMBLE & EARNES, 1964; SALGADO, 2003). O perineuro é uma membrana
fina mais densa, bastante vascularizada, rígida e forte, de tecido conjuntivo que
envolve cada fascículo (HAMILTON, 1982; SALGADO, 2003; BUTLER, 2009). Os
fascículos são mantidos unidos devido ao epineuro, o qual é frouxo e permite
certa mobilidade ao nervo (Figura 7) (HAMILTON, 1982; TRUMBLE, 2000).
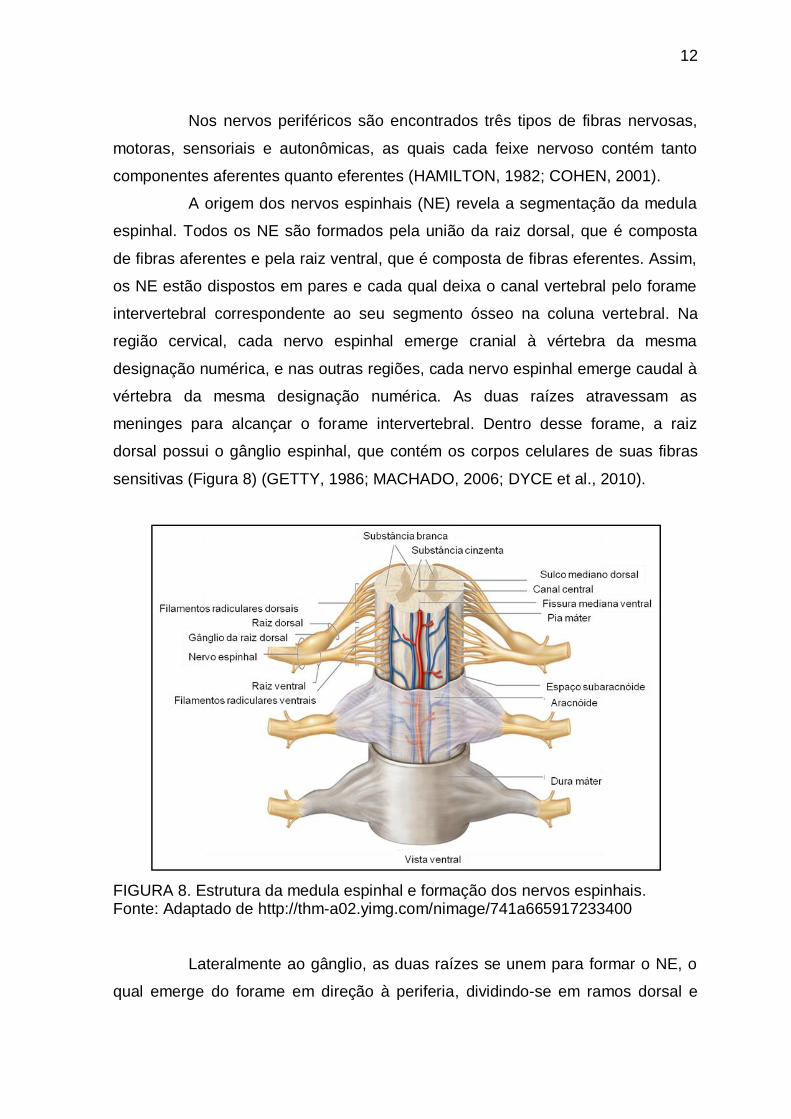
FIGURA 7. Representação esquemática da estrutura de um nervo. Fonte: http://healthguide.howstuffworks.cm/myelin-and-nerve-structure-picture-htm
12
Nos nervos periféricos são encontrados três tipos de fibras nervosas,
motoras, sensoriais e autonômicas, as quais cada feixe nervoso contém tanto
componentes aferentes quanto eferentes (HAMILTON, 1982; COHEN, 2001).
A origem dos nervos espinhais (NE) revela a segmentação da medula
espinhal. Todos os NE são formados pela união da raiz dorsal, que é composta
de fibras aferentes e pela raiz ventral, que é composta de fibras eferentes. Assim,
os NE estão dispostos em pares e cada qual deixa o canal vertebral pelo forame
intervertebral correspondente ao seu segmento ósseo na coluna vertebral. Na
região cervical, cada nervo espinhal emerge cranial à vértebra da mesma
designação numérica, e nas outras regiões, cada nervo espinhal emerge caudal à
vértebra da mesma designação numérica. As duas raízes atravessam as
meninges para alcançar o forame intervertebral. Dentro desse forame, a raiz
dorsal possui o gânglio espinhal, que contém os corpos celulares de suas fibras
sensitivas (Figura 8) (GETTY, 1986; MACHADO, 2006; DYCE et al., 2010).
FIGURA 8. Estrutura da medula espinhal e formação dos nervos espinhais. Fonte: Adaptado de http://thm-a02.yimg.com/nimage/741a665917233400
Lateralmente ao gânglio, as duas raízes se unem para formar o NE, o
qual emerge do forame em direção à periferia, dividindo-se em ramos dorsal e

13
ventral. Os ramos dorsais, à exceção das regiões cervical e coccígea, são
menores que os ventrais. Normalmente se dividem em ramos medial e lateral,
indo suprir os músculos e a pele da parte dorsal do pescoço e tronco. Os ramos
ventrais são, em geral, maiores que os dorsais e suprem as partes ventral e
lateral do tronco e todas as partes dos membros. Na região torácica, permanecem
independentes um do outro, porém nas regiões cervical caudal, lombar e sacral,
os ramos ventrais permutam fibras para formar plexos dos quais emergem os
nervos para os membros.
Além dos ramos ventral e dorsal, o NE possui um ramo comunicante.
Este último é de dois tipos: branco e cinzento. Ambos são formados por fibras
conectadas com o tronco simpático, sendo formado por uma cadeia de gânglios
ao lado da coluna vertebral. O ramo comunicante branco surge a partir das
vértebras torácicas até as duas primeiras vértebras lombares e é formado por
fibras nervosas simpáticas pré-ganglionares. O ramo comunicante cinzento liga o
tronco simpático a todos os nervos espinhais, sendo composto por fibras
simpáticas pós-ganglionares. Por isso, a lesão dos nervos espinhais pode
culminar também com déficits autônomos (GETTY, 1986; MACHADO, 2006;
DYCE et al., 2010).
O número de nervos espinhais varia entre as espécies de acordo com
o número de vértebras (FLOWER, 1885; GODINHO et al., 1985; GETTY, 1986).
4.1 Formação e distribuição dos nervos do plexo lombossacral
Segundo LACERDA et al. (2006) há discordância entre os autores de
tratados de anatomia quanto aos nervos que originam-se do plexo lombossacral.
GETTY (1986) o separam em plexo lombar e sacral, não considerando sua
formação em conjunto. HABEL et al. (1986) consideraram como constituintes do
plexo lombossacral em animais domésticos os nervos femoral, obturatório
(obturador) e isquiático. FRANDSON et al. (2011), por outro lado, descreve os
nervos glúteo cranial, glúteo caudal, femoral, obturatório, isquiático, fibular e tibial,
como integrantes desse plexo.
14
O plexo lombossacral é formado pelas raízes nervosas dos segmentos
lombares (plexo lombar) associadas às raízes nervosas dos segmentos sacrais
(plexo sacral), e apresenta variações na sua formação entre as espécies de
animais domésticos (GETTY, 1986; FRANDSON et al., 2011). A formação do
plexo lombossacral nos animais domésticos está sumarizada no Quadro 1.
QUADRO 1 - Formação do plexo lombossacral em animais domésticos Espécie Ramos do plexo Referências
Bovideo L2-L6 e S1
GETTY (1986); FRANDSON et al. (2011)
Equideo L2-L6 e S1 L1-L6 e S1
GETTY (1986); FRANDSON et al. (2011)
Suideo L2-L6 e S1
GETTY (1986); FRANDSON et al. (2011)
Canideo L3-L7 e S1-S3 L6-L7 e S1-S2
MILLER (1964) GETTY (1986)
Assim, como observado nos animais domésticos, há também uma
grande variação dos segmentos formadores dos nervos do plexo lombossacral
entre os animais selvagens (Quadro 2). Essas variações podem ter implicações
diretas em condutas clínicas, cirúrgicas e anestésicas. Há uma grande carência
de informações dessa natureza, sendo poucas as espécies estudadas.
QUADRO 2 - Formação do plexo lombossacral em algumas espécies de animais selvagens
Espécie Nervos espinhais Referências Gorila, Gibbon, Chimpanzé, Orangotango
L6-L7 e S1-S2 HEPBURN (1892)
Cutias-douradas (Dasprocta aguti) (L3-L5)* L6-L7 e S1 NUNES & SILVINO (1996)
Macaco-prego (Cebus apella) L2-L5 (L3-L4)* (P. lombar) L4-S4 (P. sacral)
BARROS et al. (2003)
Mocó (Kerondon rupestres) L5-L7, S1-S3 (50%) LACERDA et al. (2006) Porquinho-da-índia (Galea spixii spixii)
L5-L7, S1-S3 (65%) OLIVEIRA et al. (2008)
Porco-espinho (Hystrix cristata) T15, L1-L4, S1-S2 AYDIN (2009) Lobo-marinho (Arctocephalus australis)
L1-S3 CASTRO et al. (2009)
Esquilo-vermelho (Sciurus vulgaris) L6, L7, S1, S2 AYDIN (2010) Raposa (Dusicyon vetulus) L4-L7, S1-S3 SILVA et al. (2010) Tamanduá-mirim (Tamandua tetradactyla)
T17, T18, L1-L3, S1-S4 BENETTI et al. (2012)
Jaguatirica (Leopardus pardalis) L4-L5, S1 LOPES et al. (2012)
*Segmentos inconstantes estão entre parênteses

15
4.1.1 Plexo lombar
Este plexo é formado pelos nervos (Nn.) ilioipogástrico cranial e caudal,
ilioinguinal, genitofemoral, cutâneo lateral da coxa, femoral e obturador (KÖNING
& LIEBICH, 2004).
Os Nn. ilioipogástricos cranial e caudal são os ramos ventrais de L1 e
L2, respectivamente. Ambos penetram no músculo (M.) quadrado lombar e
seguem pela região aponeurótica do M. transverso do abdome até se distribuir em
sua musculatura (GETTY, 1986).
Seu terceiro nervo (N.) denomina-se ilioinguinal, sendo formado pelo
ramo ventral de L3, podendo ter uma contribuição de L2 (GETTY, 1986) ou L4
(MILLER, 1964). Emite um ramo para o M. psoas maior e segue se dividindo em
dois ramos, um lateral e outro medial. O ramo lateral atravessa o M. oblíquo
externo do abdome e segue ventralmente na superfície cranial da coxa e lateral
do joelho, onde emite um ramo cutâneo lateral. O ramo medial corre junto ao N.
ilioipogástrico e emite ramos para os músculos abdominais (GETTY, 1986).
Seguindo caudalmente, têm-se o N. genitofemoral, sendo formado pelo
ramo ventral de L3, podendo ter contribuições inconstantes de L2 e L4 (GETTY,
1986, SCHALLER, 1999). Segue caudalmente junto à musculatura lombar
profunda onde emite um ramo para o M. psoas. Divide-se em dois ramos, um
muscular que emerge cranialmente aos vasos circunflexos profundo do ílio e
segue para os músculos (Mm) cremáster (externo) e oblíquo interno do abdome.
O outro, o ramo genital emerge caudalmente aos vasos anteriormente citados, e
segue lateral e paralelamente à A. ilíaca externa até o canal inguinal. Emerge do
anel inguinal superficial junto à A. pudenda externa e se ramifica nos órgãos
genitais externos e na pele da região inguinal (GETTY, 1986). Nos machos, inerva
o escroto e o prepúcio e nas fêmeas, as mamas inguinais. Além disso, o N.
genitofemoral contém fibras autonômicas, por meio das quais o fluxo de leite é
regulado durante a amamentação (KÖNING & LIEBICH, 2004).
O N. cutâneo femoral lateral recebe suas fibras principalmente de L4,
podendo, no entanto receber fibras de L3 e L5 (SCHALLER, 1999). Inerva a
musculatura lombar profunda e segue com seu ramo caudal pela parede

16
abdominal junto à A. circunflexa ilíaca profunda, inervando a pele lateral e distal
da coxa e em torno da articulação do joelho (KÖNING & LIEBICH, 2004).
Emergindo de L4-L6, encontra-se o nervo femoral, sendo que, no seu
segmento proximal, emite ramos para os Mm. ilíaco e o psoas maior. Na face
medial proximal da coxa, emite o N. safeno, que penetra no canal femoral. A
porção distal do N. femoral inerva o M. quadríceps femoral e, ao nível da
tuberosidade pré-púbica, sua lesão pode ocasionar paralisia desse grupo
muscular (KÖNING & LIEBICH, 2004; DYCE et al., 2010). O N. safeno emite
ramos na sua origem, para os Mm. sartório, pectíneo e grácil. Passa pelo canal
femoral cranialmente à artéria (A) femoral, com um trajeto superficial. Ao nível da
articulação do joelho, emite ramos que acompanham a A. descendente do joelho,
continuando paralelo à A. safena e à veia (V) safena medial, suprindo a região
medial da coxa e da perna, até a articulação tibiotársica (KÖNIG & LIEBICH,
2004; DYCE et al., 2010).
Tendo praticamente a mesma origem (L4-L6) do nervo femoral, surge o
N. obturador. Este, transita medialmente ao corpo do ílio em direção ao forame
obturador, inervando os músculos que atuam na adução do membro, ou seja, os
Mm. pectíneo, grácil, obturador externo e o grupo adutor (VAUGHAN, 1964;
HABEL, 1968; COX et al., 1975; SCHWARZE & SCHRODER, 1979; GODINHO et
al., 1985; GETTY, 1986; KÖNING & LIEBICH, 2004; DYCE et al., 2010).
SCHALLER (1999), sem se referir à espécie, e MIRANDA et al. (2007), em
bovinos, citaram também a participação de S1 na formação do N. obturador,
sendo que o último autor não observou a participação de L4 na sua formação. No
asinino, o N. obturador é constituído por L3 e L4, podendo ser também L3 a L5 e
inclusive L2 a L4 (BRUNI & ZIMMERL, 1977). As origens medulares dos nervos
do plexo lombar no cão estão esquematizadas na Figura 9.
4.1.2 Plexo sacral
Seus nervos incluem o glúteo cranial e caudal, isquiático, cutâneo
femoral caudal, pudendo e o retal caudal (SCHALLER, 1999).

17
FIGURA 9 – Esquema da formação do plexo lombossacral no cão. Fonte: Adaptado de CORDEIRO (1996)
Originando dos ramos ventrais de L6-S1, encontra-se o N. glúteo
cranial (GETTY, 1986; CHRISMAN et al., 2005) e transita juntamente com a
artéria e a veia glúteas craniais, dorsalmente à incisura isquiática maior. Seus
ramos inervam os Mm. glúteo médio e profundo, piriforme e tensor da fáscia lata
(KÖNING & LIEBICH, 2004), e, em algumas espécies, parte do M. glúteo
superficial (DYCE et al., 2010).
Ramificando-se caudalmente ao longo da superfície lateral do
ligamento sacrotuberal largo, suprindo o M. bíceps femoral ou glúteo bíceps
(ruminantes e suínos), têm-se o N. glúteo caudal. Dependendo da espécie,
também inerva os Mm. glúteo superficial, abdutor crural caudal, semitendinoso e
semimembranoso (KÖNING & LIEBICH, 2004; DYCE et al., 2010). Esse nervo
tem a mesma origem do N. glúteo cranial, sendo L6-S1 (CHRISMAN et al., 2005),
ou S1-S2 (GETTY, 1986) .
O N. isquiático é o maior nervo do corpo, sendo a continuação
extrapélvica do tronco lombossacral até a extremidade distal do membro pélvico
(SEIFERLE, 1975; SCHWARZE & SCHRODER, 1979; ELLENBERGER & BAUM,
1985; KÖNING & LIEBICH, 2004). Emerge da cavidade pélvica pelo forame
isquiático maior como um cordão amplo, plano e acinzentado, que se dirige
caudal e distalmente na face lateral do ligamento sacrotuberal largo (GETTY,
1986), e, próximo à sua emergência da cavidade pélvica, fornece ramos

18
musculares para os Mm. gluteobíceps, semimembranoso e semitendinoso
(GETTY, 1986). É formado principalmente pelas raízes L6 e S1, recebendo
também uma frequente contribuição de L5 e outra do S2 (SCHWARZE &
SCHRODER, 1970; COX et al., 1975; GETTY, 1986; DYCE et al., 2010). No
canino, o mesmo deriva de L6, L7 e S1 e, às vezes, de S2 (GETTY, 1986).
VAUGHAN (1964) ressalta ainda a usual contribuição do ramo ventral de L5 em
sua formação.
Antes de atingir o M. gastrocnêmio, o N. isquiático se divide em Nn.
tibial e fibular comum, os quais compartilham a responsabilidade pela inervação
de todas as estruturas distais ao joelho, exceto a pele medial (DYCE et al., 2010).
O N. isquiático inerva os músculos extensores do quadril, os flexores do joelho e a
maior parte dos músculos distais do membro (DYCE et al., 2010).
O N. fibular comum se distribui nos músculos flexores do tarso e
extensores dos dedos. Superficializa-se e fica exposto a lesões ao cruzar o
côndilo lateral da fíbula. A condição é comumente observada em todas as
espécies animais de grande porte, sendo comum em vacas leiteiras que
permanecem em decúbito lateral no pós-parto, como resultado de hipocalcemia
(DYCE et al., 2010). Após passar entre os Mm. flexor longo do dedo I e fibular
longo, divide-se em ramos superficial e profundo (GETTY, 1986; KÖNING &
LIEBICH, 2004).
Após a divisão, o N. fibular superficial se projeta distalmente ao longo
da margem lateral do M. extensor digital longo, suprindo a região cranial da perna.
Na face dorsal do tarso, se ramifica em ramos medial e lateral, os quais se
dividem dorsalmente, constituindo os nervos digitais (GETTY, 1986; KÖNING &
LIEBICH, 2004).
Profundamente na perna junto com a A. tibial cranial, segue o N. fibular
profundo. No terço proximal deste segmento, emite ramos para os Mm. tibial
cranial, fibular longo, fibular terceiro, fibular curto, assim como para os extensor
digital longo, extensor digital lateral e extensor digital curto. Assim como ocorre
com o N. fibular superficial, divide-se na face dorsal do tarso, em ramos lateral e
medial. O ramo lateral emite fibras ao M. extensor digital curto e supre a pele da
face lateral do metatarso e boleto e o medial emite fibras ao músculo extensor
digital curto, fáscia e pele da face medial do boleto e fornece ramificações

19
cutâneas para a cápsula articular da referida articulação (GETTY, 1986; KÖNING
& LIEBICH, 2004).
Na coxa, o N. tibial é o principal ramo do N. isquiático. Passa entre as
duas cabeças do M. gastrocnêmio, inervando os músculos flexores dos dedos e
extensores do tarso, e continua com um predomínio de fibras aferentes, com um
pequeno ramo motor para os músculos intrínsecos do pé. Segue cruzando o tarso
e se ramifica para inervar as estruturas plantares (GETTY, 1986; DYCE et al.,
2010).
Apresentando número variável de acordo com as espécies, têm-se os
Nn. sacrais. O N. cutâneo femoral caudal (cutâneo caudal do fêmur) origina-se de
S1-S3 (SCHALLER, 1999). Projeta-se em direção à tuberosidade isquiática e
emite ramos para o M. semitendinoso e se superficializa. Seus ramos cutâneos
suprem a pele ao redor da tuberosidade isquiática e caudal da coxa e se
conectam com o N. pudendo. Nos ruminantes este nervo é delgado e as regiões
supridas pelos ramos cutâneos são inervadas pelo N. pudendo (KÖNING &
LIEBICH, 2004).
O N. pudendo mostra-se variável em sua formação, sendo S1-S3 no
cão, S2-S4 em ruminantes e S3-S4 no equino, podendo ter contribuição de S2.
Segue caudoventralmente, medial ao ligamento sacrotuberal largo. Por meio de
ramos comunicantes, se une com o N. cutâneo femoral caudal. Inerva os órgãos
copuladores, assim como os músculos das regiões anal e perineal. Com fibras
sensoriais e motoras, supre os Mm. isquiocavernoso, bulboesponjoso, retrator do
pênis, uretral, constritor da vulva, coccígeo, o elevador do ânus e o esfícnter anal
externo. Origina os Nn. perineais profundos e superficiais, além dos vários ramos
cutâneos e continua, nos machos, como o N. dorsal do pênis até a glande e
prepúcio, onde se ramifica. Corpúsculos sensoriais estão localizados nas
extremidades dos seus ramos mais finos. Nas fêmeas, correspondentemente,
termina no clitóris (KÖNING et al., 2004; DYCE et al., 2010).
As fibras parassimpáticas do N. pudendo, que se originam da medula
espinhal sacral, separam-se desse no seu segmento inicial e correm
superficialmente ao peritônio, como Nn. pélvicos, para o plexo pélvico, onde se
unem com fibras simpáticas dos Nn. hipogástricos e Nn. esplâncnicos sacrais,
alcançando os órgãos da cavidade pélvica. Nesse local, formam os plexos de
20
acordo com os respectivos órgãos. As sinapses dos neurônios pré-ganglionares
com os pós-ganglionares parassimpáticos situam-se em pequenos gânglios de
posição intramural (KÖNING & LIEBICH, 2004).
Emergindo dos ramos ventrais de S2-S4, encontra-se o N. retal caudal.
É motor para os músculos estriados da parte dorsal do períneo e sensorial para o
reto, parede do canal anal e pele adjacente. Os ramos ventrais dos nervos
caudais suprem os músculos ventrais ou depressores da cauda (DYCE, et al.,
2010). Portanto, corre caudalmente, sendo coberto pelos músculos sacrocaudais
ventrais, e termina no M. esfíncter anal externo e no M. levantador do ânus
(GETTY, 1986). A composição dos nervos do plexo lombossacral dos animais
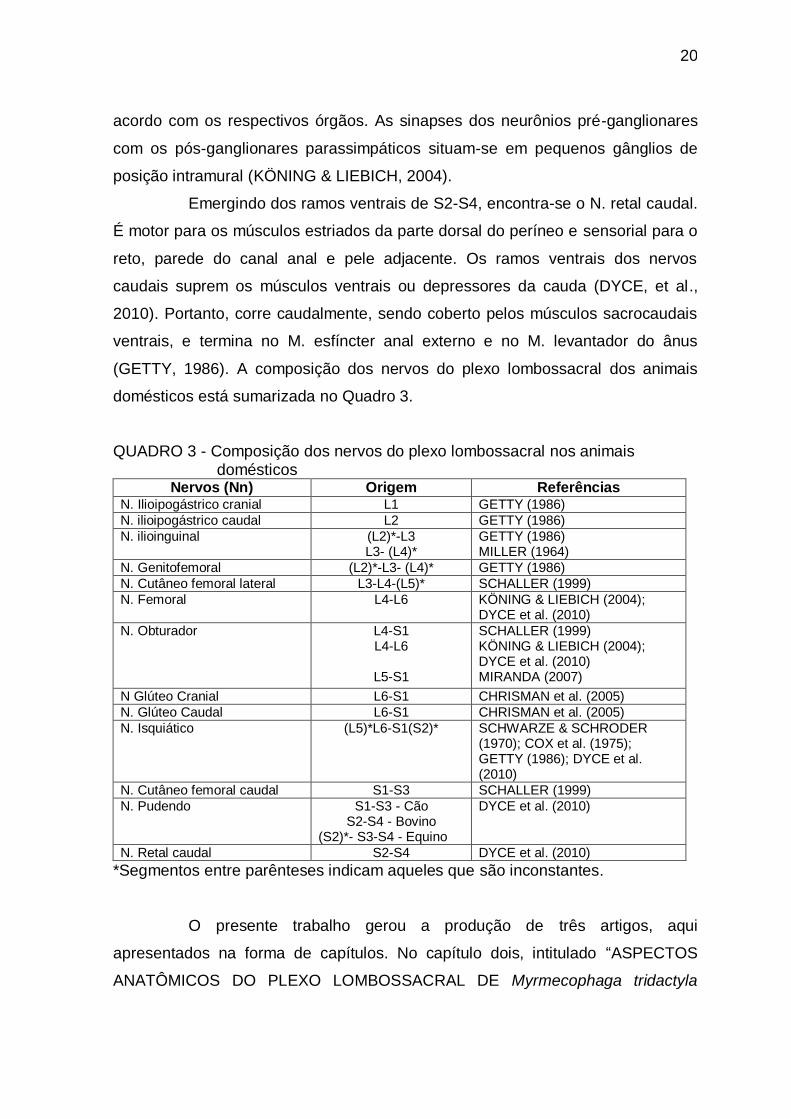
domésticos está sumarizada no Quadro 3.
QUADRO 3 - Composição dos nervos do plexo lombossacral nos animais domésticos
Nervos (Nn) Origem Referências N. Ilioipogástrico cranial L1 GETTY (1986)
N. ilioipogástrico caudal L2 GETTY (1986)
N. ilioinguinal (L2)*-L3 L3- (L4)*
GETTY (1986) MILLER (1964)
N. Genitofemoral (L2)*-L3- (L4)* GETTY (1986)
N. Cutâneo femoral lateral L3-L4-(L5)* SCHALLER (1999)
N. Femoral L4-L6 KÖNING & LIEBICH (2004); DYCE et al. (2010)
N. Obturador L4-S1 L4-L6
L5-S1
SCHALLER (1999) KÖNING & LIEBICH (2004); DYCE et al. (2010) MIRANDA (2007)
N Glúteo Cranial L6-S1 CHRISMAN et al. (2005)
N. Glúteo Caudal L6-S1 CHRISMAN et al. (2005)
N. Isquiático (L5)*L6-S1(S2)* SCHWARZE & SCHRODER (1970); COX et al. (1975); GETTY (1986); DYCE et al. (2010)
N. Cutâneo femoral caudal S1-S3 SCHALLER (1999)
N. Pudendo S1-S3 - Cão S2-S4 - Bovino
(S2)*- S3-S4 - Equino
DYCE et al. (2010)
N. Retal caudal S2-S4 DYCE et al. (2010)
*Segmentos entre parênteses indicam aqueles que são inconstantes.
O presente trabalho gerou a produção de três artigos, aqui
apresentados na forma de capítulos. No capítulo dois, intitulado “ASPECTOS
ANATÔMICOS DO PLEXO LOMBOSSACRAL DE Myrmecophaga tridactyla

21
(Linnaeus, 1758)” propôs-se descrever o plexo lombossacral do M. tridactyla no
que se refere à sua origem e composição de seus nervos.
No capítulo três intitulado “ASPECTOS ANATÔMICOS DOS NERVOS
DA COXA DO TAMANDUÁ-BANDEIRA (Myrmecophaga tridactyla, Linnaeus,
1758)” objetivou-se descrever os nervos da coxa do M. tridactyla, abordando sua
topografia, ramificação e território de inervação.
No capítulo quatro intitulado “ASPECTOS ANATÔMICOS DOS
NERVOS DA PERNA E PÉ DO TAMANDUÁ-BANDEIRA (Myrmecophaga
tridactyla, Linnaeus, 1758)” propôs-se descrever os nervos da perna e pé do M.
tridactyla, com ênfase na sua ramificação, distribuição e território de inervação.
5 Referências
1. AYDIN, A. The dissemination of the pelvic limb nerves originating from the lumbosacral plexus in the porcupine (Hystrix cristata). Veterinární Medicína,
Praha, v. 54, n. 7, p. 333-339, 2009.
2. AYDIN, A. The spinal nerves that constitute the plexus lumbosacrales of the red squirrel (Sciurus vulgaris). Veterinární Medicína, Praha, v. 55, n. 4, p. 183-186,
2010.
3. BARROS, R. A. C.; PRADA, I. L. S.; SILVA, Z.; RIBEIRO, A. R.; SILVA, D. C. O. Constituição dos plexos lombar em macaco Cebus apella. Brazilian journal of Veterinary Research and Animal Science, São Paulo, v. 40, n. 5, p. 373-381, 2003.
4. BENETTI, E. J.; CARDOSO, J. R.; CRUZ, V. S.; SOUZA, P. R.; SILVA, M. S. B;
MOREIRA, P. C. Anatomical Study of the Lumbosacral plexus of the Tamandua tetradactyla. In: INTERNATIONAL SYMPOSIUM ON MORPHOLOGICAL SCIENCES, 22., 2012, São Paulo. Resumos... São Paulo: International Symposium on Morphological Sciences, 2012.
5. BERESCA, A. M.; CASSARO, K. Biology and captive management of
armadillos and anteaters. In: FLOWER, M. E.; CUBAS, Z. S. Biology, medicine and surgery of South American Wild Animals. Ames: Iowa:
University Press, 2001. p. 238-245.
6. BRAGA, F. G.; SANTOS, R. E. F. Relações entre a fauna e o fogo. In: SOARES, R. V.; BATISTA, A. C.; NUNES, J. R. S. (Org.). Incêndios florestais no Brasil: o estado da arte. 1.ed. Curitiba: Ronaldo Viana Soares e Antonio Carlos Batista editores independentes, 2009. 251 p.

22
7. BRAGA F. G.; SANTOS, R. E. F.; BATISTA, A. C. Marking behaviour of the
giant anteater Myrmecophaga tridactyla (Mammalia: Myrmecophagidae) in Southern Brazil. Zoologia, Curitiba, v. 27, n. 1, p. 7-12, 2010.
8. BRUNI, A. C.; ZIMMERL, U. Anatomia degli animalii domestici. 2.ed. Milano:
Casa Editrici Dr. Francisco Vallardi, 1977. p. 557-559.
9. BUTLER, D. S. Mobilização do sistema nervoso. 1.ed. São Paulo: Manole,
2009. 270 p.
10. CAMILO-ALVES, C. S. P.; MOURÃO, G. Responses of a specialized insectivorous mammal (Myrmecophaga tridactyla) to variation in ambient. Biotropica, Washington, v. 38, n. 1, p. 52- 56, 2006.
11. CASTRO, T. F.; SOUZA, D. A. S.; SILVA-FILHO, R. P.; PEREIRA, M. A. M.
Sistematização e distribuição da inervação lombar e sacral em Arctocephalus australis. Brazilian Journal of Veterinary Research and Animal Science, São Paulo, v. 46, n. 5, p. 404-411, 2009.
12. CHEBEZ, J. C.; CIRIGNOLI, S. In: CHEBEZ, J.C. (Ed.). Los que se van:
Fauna argentina amenazada – Tomo 3 (Mamíferos). Albatros. Buenos Aires, Argentina, 2008. p. 31-40.
13. CHRISMAN, C; MARIANI, C; PLATT, S; CLEMMONS, R. Neurologia para o
Clinico de Pequenos Animais. 1.ed. São Paulo: Roca, 2005. 306 p.
14. COHEN, H. Neurociência para fisioterapeutas: incluindo correlações clínicas. 2.ed. São Paulo: Manole, 2001. 494 p.
15. COLLEVATTI, R. G.; LEITE, K. C. E.; MIRANDA, G. H. B.; RODRIGUES, F. H.
G. Evidence of high inbreeding in a population of the endangered giant anteater, Myrmecophaga tridactyla (Myrmecophagidae), from Emas National Park, Brazil. Genetics and Molecular Biology, Ribeirão Preto, v. 30, p. 112-120, 2007.
16. CORDEIRO, J. M. C. Exame Neurológico de Pequenos Animais. Pelotas:
EDUCAT, 1996. 270 p.
17. COX, N. S.; BREAZILE, J. E.; HOOVER, T. R. Surgical and anatomic study of calving paralysis. American Journal of Veterinary Research, Missouri, v. 36,
p. 427-430, 1975.
18. DAWKINS, R. Xenarthros. In: R. Dawkins (Ed.). A Grande História da Evolução.São Paulo: Companhia das Letras, 2009. 792 p.
19. DELSUC, F.; VIZCAINO, S. F.; DOUZERY, E. J. P. Influence of Tertiary paleoenvironmental changes on the diversification of South American

23
mammals: a relaxed molecular clock study within Xenarthrans. BMC Evolutionary Biology, London, v. 4, n. 1, p. 11, 2004.
20. DICKMAN, C. R. Anteaters. In: MACDONALD, D. (Ed.). The new
encyclopedia of mammals. Oxford: Oxford University Press, 2001, p. 786-
787.
21. DINIZ, L. S. M.; COSTA, E. O.; OLIVEIRA, P. M. A. Clinical disorders observed in anteaters (Myrmecophagidae Edentata) in captivity. Veterinary Research Communication, New York, v. 19, n. 5, p. 409-415, 1995.
22. DYCE, K. M.; SACK, W. O.; WENSING, C. J. G. Tratado de anatomia
veterinária, 4.ed., Rio de Janeiro: Elsevier, 2010. 834 p.
23. EISENBERG, J. F.; REDFORD, K.H. Mammals of the Neotropics: The
Central Neotropics (Ecuador, Peru, Bolivia, Brazil), Chicago: Univer University of Chicago Press, 1999. 609 p. v. 3.
24. ELLENBERGER, W. BAUM, H. Handbuch der vergleichenden anatomie der
haustiere, 18.ed., Berlin: Springer, 1985. 1070 p.
25. ENDO, H.; KOMIYA, T.; KAWADA, S.; HAYASHIDA, A.; KIMURA, J.; ITOU, T.; KOIE, H.; SAKAI, T. Three-dimensional reconstruction of the xenarthrous process of the thoracic and lumbar vertebrae in the giant anteater. Mammal Study, Tokyo, v. 34, p. 1-6, 2009.
26. ENGELMANN, G. F. The Phylogeny of the Xenarthra. In: MONTGOMERY, G.
G.(Org.). The evolution and ecology of armadillos, sloths, and vermilinguas.Washington: Smithsonian Institution, 1985. p. 51-64.
27. FLOWER, W. H. An introduction to the osteology of the mammalian. 3.ed.
London: Macmillan and Co., 1885. 382 p.
28. FONSECA, G. A. B.; RYLANDS, A. B.; COSTA, C. M. R.; MACHADO, R. B.; LEITE, Y. L. R. Livro vermelho dos mamíferos brasileiros ameaçados de extinção. Biodiversitas: Belo Horizonte, 1994. 460 p.
29. FRANDSON, R. D; WILKE, W. L.; FAILS, A. D. Anatomia e fisiologia dos
animais de fazenda. 7.ed. Rio de Janeiro: Guanabara Koogan, 2011. 413 p.
30. GAMBARYAN, P. P.; ZHEREBTSOVA, O. V.; PEREPELOVA, A. A.;
PLATONOV, V. V. Pes muscles and their action in giant anteater Myrmecophaga tridactyla (Myrmecophagidae, Pilosa) compared with other plantigrade mammals. Russian Journal of Theriology. Rússia, v. 8, n. 1, p. 1-15, 2009.
31. GAMBLE, H. J.; EARNES, R. A. An electron microscopic observations on the
connective tissues of human peripheral nerve. Journal of Anatomy, London, v. 98, n. 5, p. 17-25, 1964.

24
32. GAUDIN, T. J. Phylogeny of the Xenarthra. Senckenbergiana Biologica, Germany, v. 83, n. 1, p. 27-40, 2003.
33. GAUDIN, T. J.; BRANHAM, D. G. The Phylogeny of the Myrmecophagidae
(Mammalia, Xenarthra, Vermilingua) and the Relationship of Eurotamandua to the Vermilingua. Journal of Mammalian Evolution, New York, v. 5, n. 3. p.
237-265, 1998.
34. GETTY, R.; SISSON/ GROSSMAN: Anatomia dos animais domésticos. 5.ed. Rio de Janeiro: Guanabara Koogan, 1986. 2000 p. v. 1 e 2.
35. GILLESPIE, D. S. Edentata: diseases. In: FOWLER, M. E. Zoo and wild
animal medicine. 3.ed. Philadelphia: W.B. Saunders, 1993, p. 304-309.
36. GODINHO, H. P.; NASCIMENTO, J. F.; CARDOSO, F. M. Anatomia dos ruminantes domésticos. Belo Horizonte: Instituto de Ciências Biológicas da
UFMG, p. 89-126, 1985.
37. GREGORINI, M. Z.; RODOLFO, A. M.; CÂNDIDO Jr.; J. F.; TÔRRES, N. M. Modelagem de distribuição geográfica do tamanduá-bandeira (Myrmecophaga tridactyla) e sua ocorrência em unidades de conservação no estado do paraná. In: CONGRESSO DE ECOLOGIA DO BRASIL, 8., 2007, Caxambu. Anais...
Congresso de Ecologia do Brasil, Caxambu, 2007. p. 1-3.
38. HABEL, R. E. Manual de disección de los ruminantes domésticos. Zaragoza: Acribia, 1968. p. 71-73.
39. HABEL, R. E.; DE LAHUNTA, A.; ROBERTY. W. B. Appliedy veterinaria
anatomy. 1.ed. Washington: Saunders Company, 1986. 330 p.
40. HAMILTON, W. J. Tratado de anatomia humana. 2.ed. Rio de Janeiro, Interamericana, 1982. 789 p.
41. HEPBURN, D. The comparative anatomy of the muscles and of the superior
and inferior extremities of the anthropoid apes Part II. Journal Anatomy of Physiology, London, p. 324-356, 1892.
42. JONG, W. W.; ZWEERS, A.; JOYSEY, K. A.; GLEAVES, J. T.; BOULTER, D. Protein Sequence Analysis Applied to Xenarthran and Pholidote Phylogeny. In: G. G. Montgomery (Ed.). The evolution and ecology of armadillos, sloths, and vermilinguas. Washington and London: Smithsonian Institution Press, 1985, p.65-76.
43. KÖNIG, H. E.; LIEBICH, H. G. Anatomia dos animais domésticos: texto e
atlas colorido. Porto Alegre: Artmed, 2004. 399 p. v. 2.

25
44. LACERDA, P. M. O.; MOURA, C. E. B.; MIGLINO, M. A.; OLIVEIRA, M. F.; ALBUQUERQUE, J. F. G. Origem do plexo lombossacral de mocó (Kerondorupestris). Brazilian Journal of Veterinary Research and Animal Science, São Paulo, v. 43, n. 5, p. 620-628, 2006.
45. LOPES, J. A.; FÉ, L. C. M.; LIMA, A. R.; PEREIRA, L. C.; BRANCO, E.
Morfologia do plexo lombossacral da jaguatirica (Leopardus pardalis). Biotemas, Florianópolis, v. 25, n. 4, p. 216-220, 2012.
46. MACHADO, A. B. M. Neuroanatomia funcional. 2.ed. São Paulo: Atheneu,
2006. 366p.
47. MEDRI, I. M.; MOURÃO, G. Home range of giant anteaters (Myrmecophaga tridactyla) in the Pantanal wetland, Brazil. Journal of Zoology, London, v. 266, n. 4, p. 365-375, 2005.
48. MESS, A.; CARTER, A. M. Evolution of the placenta during the early radiation
of placental mammals. Comparative Biochemistry and Physiology, Philadelphia, v. Part A 148, p. 769-779, 2007.
49. MILLER, M. E. Anatomy of the dog. Philadelphia: W. B. Saunders, 1964.
941p.
50. MIRANDA, G. H. B. Ecologia e conservação do tamanduá-bandeira (Myrmecophaga tridactyla, Linnaeus 1758) no Parque Nacional das Emas.
2004. 81f. Tese (Doutorado em Ecologia) - Universidade de Brasília, Brasília.
51. MIRANDA, F.; COSTA, A. M. Xenarthra (Tamanduá, Tatu, Preguiça). In:__Tratado de Animais Selvagens – Medicina Veterinária. 1.ed. Roca,
2007. cap. 26, p. 402-414.
52. MIRANDA, R. L.; CARNEIRO E SILVA, F. O.; SEVERINO, R. S.; DRUMMOND, S. S.; GONÇALVEZ, R. C. Origens e distribuições dos nervos obturatórios em fetos fêmeas de bovinos azebuados. Bioscience Journal, Uberlândia, v. 23, n. 4, p. 120-127, 2007.
53. MOURÃO, G.; MEDRI, M. Activity of a specialized insectivorous mammal (Myrmecophaga tridactyla) in the Pantanal of Brazil. Journal of Zoology, London, v. 271, p. 187-192, 2007.
54. MURPHY, W. J.; EIZIRIK, E.; JOHNSON, W. E.; ZHANG, Y. P.; RYDER, O. A.; O'BRIEN, S. J. Molecular phylogenetics and the origins of placental mammals. Nature, Philadelphia, v. 409, n. 6820, p. 614-618, 2001.
55. NAPLES, V. L. Morphology, evolution and function of feeding in the giant
anteater (Myrmecophaga tridactyla). Journal of Zoology, London, v. 249, p.
19-41, 1999.

26
56. NOWAK, R. M. “Anteaters”. Walker’s Mammals of the World. 6.ed.
Baltimore and London: The John Hopkins University Press, 1999. 836 p. v. 1.
57. NUNES, L. C.; SILVINO, M. J. Formação do plexo lombossacral em cutia-dourada (Dasprocta aguti). In: CONGRESSO BRASILEIRO DE ANATOMIA, 17. 1996, Fortaleza. Anais..., CONGRESSO BRASILEIRO DE ANATOMIA, 1996, p.139.
58. NYAKATURA, J. A.; FISCHER, M. Functional morphology and three-
dimensional kinematics of thoraco-lumbar region of the spine of the two-toed sloth. The Journal of Experimental Biology, London, v. 213, p. 4278-4290,
2010.
59. OLIVEIRA, M. F. Morfologia funcional e desenho corporal da cintura pélvica e membros posteriores dos tamanduás (MAMMALIA: XENARTHRA: MYRMECOPHAGIDAE). 2001. 80f. Tese (Mestrado em Ciências) - Centro de Ciências Biológicas, Universidade Federal do Pará e do Museo Paraense Emílio Goeldi, Belém.
60. OLIVEIRA, G. B.; ALBUQUERQUE, J. F. G.; OLIVEIRA, M. F.; RODRIGUES,
M. N.; SOUSA, R. S.; SOUSA, E. S.; MIGLINO, M. A. Source and distribution of the lumbosacral plexus in spix’s yellow-toothed cavy (Galea spixii spixii). Brazilian Journal Morphology Science, São Paulo, v. 25, n. 1-4, p. 1-34,
2008.
61. POCOCK, R. L. The external characters of the South American Edentates. Proceedings of the Zoological Society of London, London, v. 65, p. 983-
1031, 1924.
62. RODRIGUES, M. C.; QUESSADA, A. M.; DANTAS, D. A. S. B.; ALMEIDA, H. M.; COELHO, M. C. O. C. Amputação do membro pélvico de tamanduá-mirim (Tamandua tetradactyla): relato de caso. Ciência Animal Brasileira, Goiânia, v. 10, n. 1, p. 330-334, 2009.
63. ROSA, A. L. M. Efeito da temperatura ambiental sobre atividade, uso de
habitat e temperatura corporal do tamanduá-bandeira (Myrmecophaga tridactyla) na fazenda Nhumirim, Pantanal. 2007. 32f. Dissertação (Mestrado
em Ecologia e Conservação), Universidade Federal do Mato Grosso do Sul, Corumbá.
64. SALGADO, A. F. Mobilização neural. Cascavel: CESUMAR, Curso de terapia
manual e postural internacional, 2003.
65. SAMPAIO, C.; ALVES, P. C.; MOURÃO, G. M. Responses of a specialized insectivorous mammal (Myrmecophaga tridactyla) to variation in ambient temperature. Biotropica, Washington, v. 38, n. 1, p. 52-56, 2006.
66. SCHALLER, O. Nomenclatura anatômica veterinária ilustrada. 1.ed. São
Paulo: Manole, 1999. 494 p.

27
67. SCHWARZE, H.; SCHRODER, L. Compêndio de anatomia veterinária.
Zaragoza: Acríbia, 1979. 206 p. v. 4.
68. SEIFERLE, E. Nervensystem sinnesorgane endokrine drüsen. Berlim: Paul
Parey, 1975. 240 p.
69. SHAW, J. H.; CARTER, T. S. Giant anteaters. Natural History, New York, v. 89, p. 62-67, 1980.
70. SHAW, J. H.; MACHADO-NETO, J.; CARTER, T. S. Behavior of free-living
Giant Anteaters (Myrmecophaga tridactyla). Biotropica, Washington, v. 19, p. 255-259, 1987.
71. SILVA, M. S. B.; MOREIRA, P. C.; SOUZA, P. R.; CRUZ, V. S.; MENDONÇA,
A. C.; BENETTI, E. J.; CARDOSO, J. R. Formação dos nervos do plexo lombossacral de raposa Dusicyon vetulus (Lund, 1842). In: CONGRESSO DE PESQUISA, ENSINO E EXTENSÃO (CONPEEX), 7., 2010, Goiânia. Anais..., Goiânia: 2010. p. 8036-8039 (Resumo expandido).
72. SILVEIRA, L.; RODRIGUES, F. H. G.; JACOMO, A. T. D.; DINIZ, J. H. F.
Impact of wildfires on the megafauna of Emas National Park, Central Brazil. Oryx, Cambridge, v. 33, n. 2, p. 108-114, 1999.
73. SMITH, P. Giant anteater. Myrmecophaga tridactyla, Linnaeus, 1758. Fauna
Paraguay-Handbook of the Mammals of Paraguay, [on line], n. 2, p. 1-12, 2009. Disponível em: http://www.faunaparaguay.com/mammhb2.html. Acesso 27 de setembro, 2012.
74. SHOSHANI, J. & MCKENNA, M.C. 1998. Higher taxonomic relationships among extant mammals based on morphology, with selected comparisons of results from molecular data. Molecular Phylogenetics and Evolution,
Philadelphia, v.9, p. 572–584, 1998.
75. SPRINGER, M. S.; STANHOPE, M. J.; MADSEN, O. & DE JONG, W. W. Molecules consolidate the placental mammal tree. Trends in Ecology and Evolution.Philadelphia, v.19, n.8, p. 430-438, 2004.
76. TRUMBLE, T. E. Peripheral nerve injury: pathophysiology and repair. In: FELICIANO, D. V.; MOORE, E. E.; MATTOX, K. L. (eds). Trauma. 4.ed. New
York: Mmcgraw-Hill, 2000, p. 1047-1055.
77. VAUGHAN, L. C. Peripheral nerve injuries: an experimental study in cattle. The Veterinary Record, London, v. 72, n. 46, p. 1293-1301, 1964.
78. VIZCAÍNO, S. F.; LOUGHRY, W. J. The Biology of the Xenarthra. 1.ed.
Gainesville: University Press of Florida, 2008. 370p.

28
79. WITHERS, P. C. Temperature. In:__ Comparative animal physiology. Fort
Worth: Saunders College, 1992, p. 122-191.

29
CAPÍTULO 2 - ASPECTOS ANATÔMICOS DO PLEXO LOMBOSSACRAL DE
Myrmecophaga tridactyla (Linnaeus, 1758)
ANATOMICAL ASPECTS OF THE LUMBOSSACRAL PLEXUS OF THE
Myrmecophaga tridactyla (Linnaeus, 1758)
Viviane Souza CRUZ1, Júlio Roquete CARDOSO2, Luciana Batalha de
Miranda ARAÚJO3, Paulo Roberto de SOUZA1, Naida Cristina BORGES3,
Eugênio Gonçalves de ARAÚJO3
1.Doutorandos no Programa de Pós-Graduação em Ciência Animal, Escola de Veterinária e Zootecnia, Universidade Federal de Goiás, UFG, Goiânia, GO; Professores mestres no Departamento de Morfologia, Instituto de Ciências Biológicas, Universidade Federal de Goiás, UFG, Goiânia, GO. [email protected]; [email protected], Caixa Postal 131 – Campus Samambaia (Campus II), CEP: 74001-970 – Goiânia – Goiás 2.Professor doutor, Departamento de Morfologia, Instituto de Ciências Biológicas, Universidade Federal de Goiás, UFG. Goiânia, GO. [email protected], Caixa Postal 131 – Campus Samambaia (Campus II), CEP: 74001-970 – Goiânia – Goiás 3.Professores doutores, Departamento de Medicina Veterinária, Escola de Veterinária e Zootecnia, Universidade Federal de Goiás, UFG, Goiânia, GO. [email protected], [email protected], [email protected], Caixa Postal 131 – Campus Samambaia (Campus II), CEP: 74001-970 – Goiânia – Goiás
Submetido na Revista Bioscience Journal em 24 de setembro de 2012
Situação: Em avaliação (última alteração em 20 de novembro de 2012)

30
ASPECTOS ANATÔMICOS DO PLEXO LOMBOSSACRAL DE Myrmecophaga
tridactyla (Linnaeus, 1758)
ANATOMICAL ASPECTS OF THE LUMBOSSACRAL PLEXUS OF THE
Myrmecophaga tridactyla (Linnaeus, 1758)
RESUMO: Apesar de sua importância para os biomas de muitos países, pouco se
conhece acerca da morfologia do M. tridactyla. Sendo assim, objetivou-se descrever nesta espécie a origem do plexo lombossacral, bem como a composição de seus nervos. Para tanto, foram analisados 12 plexos de seis cadáveres adultos e de ambos os sexos, fornecidos pelo IBAMA-GO (licença 99/2011, CEUA-UFG protocolo nº 015/11), por meio de dissecação da face ventral das regiões torácica caudal, lombar e sacral para a evidenciação dos ramos ventrais dos nervos espinhais, suas comunicações e os nervos derivados do plexo. Os animais foram descongelados em temperatura ambiente durante 48h, fixados e conservados em solução de formaldeído a 10% e 4%, respectivamente e, posteriormente, dissecados após um período mínimo de 72h a partir da fixação. Para constatação do número de vértebras, foi realizado exame radiográfico da região torácica, lombar e sacral da coluna vertebral. Às radiografias verificaram-se 15 ou 16 vértebras torácicas, duas ou três lombares e quatro ou cinco sacrais. O plexo lombossacral originou-se em: T16, L1, L2, S1-S5, Cc1 (33,3%), T15, L1-L3, S1-S5 (16,6%), T15, L1, L2, S1-S5, Cc1 (16,6%), L1-L3, S1-S5 (16,6%), T15, L1-L3, S1-S4, Cc1 (8,3%) e L1-L3, S1-S4 (8,3%). Os nervos derivados do plexo com suas formações mais frequentes foram: genitofemoral (T15, L1, L2 e T16, L1), cutâneo femoral lateral (T15, L1, L2 e T16, L1), femoral (T16, L1, L2), obturador (L1-L3), isquiático (L3, S1-S3), glúteo cranial (L2, S1), glúteo caudal (L2, S1-S3 e L3, S1, S2), pudendo (S4-S5) e retal caudal (S5 e S4, S5, Cc1). O nervo cutâneo femoral caudal originou-se do nervo isquiático. O plexo lombossacral do M. tridactyla assemelha-se em geral ao dos demais mamíferos, porém apresenta como particularidade o envolvimento de ramos torácicos e coccígeos na composição de seus nervos e uma maior variação anatômica em sua formação, que pode ser atribuída principalmente à variação numérica de vértebras torácicas, lombares e sacrais nesta espécie.
PALAVRAS-CHAVE: Anatomia, Inervação. Sistema nervoso. Xenartros.

31
INTRODUÇÃO
Os xenartros são considerados como a Superordem mais variada de
mamíferos, que agrupa animais de morfologia, comportamento e ambientes
completamente diferentes (LARRÁZABAL, 2004). Estes animais são divididos nas
ordens Cingulata e Pilosa. Os tamanduás, pertencentes à ordem Pilosa,
representam a família Myrmecophagidae e são divididos em três gêneros, o
Myrmecophaga (tamanduá-bandeira), o Tamandua (tamanduá-mirim) e o
Cyclopes (pigmeu), com características anatomofisiológicas e comportamentais
similares (MIRANDA; COSTA, 2007).
O Myrmecophaga tridactyla é o maior tamanduá existente no mundo, e
sua distribuição geográfica é ampla em campos e cerrados do sul de Belize e
Guatemala até o norte da Argentina, incluindo todo o território do Brasil
(MOURÃO; MEDRI, 2007). Além disso é classificado como uma espécie
“vulnerável” (VU), na List of the International Union for Conservation of Nature
(IUCN, 2012).
Os estudos envolvendo a neuroanatomia de animais selvagens são
escassos, principalmente no que se refere ao estudo da formação dos plexos
nervosos. Sendo assim, a formação do plexo lombossacral é bem estabelecida
nos animais domésticos, no entanto, poucos são os estudos da formação deste
plexo nos demais mamíferos não primatas, tendo sido realizado até o momento
em jumento (ERDEN, 1993), mocó (LACERDA et al., 2006), porquinho-da-índia
(OLIVEIRA et al., 2008), lobo-marinho (CASTRO et al., 2009), porco-espinho
(AYDIN, 2009), esquilo-vermelho (AYDIN, 2010) e chinchila (MARTINEZ-
PEREIRA; RICKES, 2011).
Lesões no plexo lombossacral ou em seus nervos associados podem
acarretar entre outros, a alteração de reflexos espinhais e de propriocepção
consciente no membro pélvico, atrofia, paralisia, perda de flexão ou extensão
voluntária de músculos e incapacidade de sustentar o peso (LORENZ;
KORNEGAY, 2006). Sendo assim, o conhecimento da formação deste plexo,
além de contribuir para o estudo da anatomia comparada, pode ter implicações
diretas em condutas clínicas, cirúrgicas e anestésicas.

32
Nesta perspectiva, o objetivo deste trabalho foi descrever o plexo
lombossacral do M. tridactyla no que se refere à sua origem e composição de
seus nervos.
MATERIAL E MÉTODOS
Foram utilizados seis exemplares adultos de M. tridactyla, sendo três
fêmeas, dois machos e outro de sexo indefinido devido à retirada dos órgãos
genitais em procedimento de necropsia. Os dois antímeros foram dissecados,
resultando na análise de 12 plexos lombossacrais.
Os cadáveres dos animais foram cedidos pelo Centro de Triagem de
Animais Silvestres (CETAS), órgão vinculado ao Instituto Brasileiro do Meio
Ambiente e dos Recursos Naturais Renováveis (IBAMA), mediante licença
número 99/2011. O estudo foi aprovado pela Comissão de Ética no Uso de
Animais da Universidade Federal de Goiás (CEUA-UFG, 015/11). O preparo,
acondicionamento e dissecação foram realizados no Laboratório de Anatomia
Animal do Instituto de Ciências Biológicas da UFG.
No Laboratório, os animais foram descongelados a temperatura
ambiente por um período de 48h, higienizados em água corrente e submetidos à
fixação por meio de injeções com solução de formol a 10%, num volume
correspondente a 10% do peso corporal (RODRIGUES, 2010). Posteriormente
foram imersos em tanques com solução conservadora de formol a 4% e após um
período mínimo de 72h de fixação, iniciou-se a dissecação.
O acesso à cavidade abdominal e pélvica se deu a partir de uma
incisão longitudinal na linha alba, remoção da sínfise pélvica e de parte dos púbis
e ísquios. O cólon e reto, os órgãos geniturinários e os vasos sanguíneos
lombares e sacrais foram afastados. O peritônio parietal, as fáscias pélvicas e
parte da musculatura sublombar foram removidos, permitindo assim, a
visualização dos ramos ventrais dos nervos espinhais e, por conseguinte, o plexo
lombossacral, suas comunicações e os nervos originados. As estruturas nervosas
foram dissecadas e os achados foram documentados por meio de esquemas e

33
fotografias, obtidas com uma câmera digital (DSC-H50, Sony, Brasil). A
nomenclatura empregada para a descrição dos resultados está de acordo com o
proposto pelo International Committee on Veterinary Gross Anatomical
Nomenclature (2012)
Para confirmação do número de vértebras torácicas, lombares e
sacrais, cada animal foi submetido a exame radiográfico em projeções ventro-
dorsal e látero-lateral. As radiografias foram efetuadas em um aparelho de
radiografia fixo da marca TUR, modelo T-350, com capacidade de 600mA e grade
antidifusora Potter-Bucky. Foram utilizadas películas radiográficas (T-MAT,
Kodak, Brasil) montadas em chassis rígidos de tamanho 30cm x 40cm, equipados
com par de écrans intensificadores. A distância foco-filme foi mantida em 90cm e
o cálculo da kilovoltagem e miliamperagem da técnica radiográfica utilizada foi
realizado de acordo com a espessura, em centímetros, de cada segmento
vertebral do animal, de acordo com Lavin (2003). As películas radiográficas foram
identificadas e reveladas em processadora automática da linha Vision Line (LX-2,
Lotus, Brasil). Os exames foram realizados no Setor de Radiologia do Hospital
Veterinário da Escola de Veterinária e Zootecnia da UFG.
A estatística descritiva foi utilizada na análise dos dados, por meio de
frequência percentual, conforme os estudos de Lacerda et al. (2006); Oliveira et
al. (2008) e Oliveira et al. (2011).
RESULTADOS E DISCUSSÃO
Com base na literatura consultada, pode-se afirmar que este é o único
estudo sobre o plexo lombossacral em membros da Superordem Xenarthra.
Portanto, os resultados deste estudo foram compilados com relatos em outras
espécies de animais mamíferos, incluindo os animais domésticos.
A composição do plexo lombossacral está diretamente relacionada ao
número de vértebras lombares e sacrais e suas variações. Mediante esta
constatação, optou-se pelo exame radiográfico para identificação do número de
vértebras torácicas, lombares e sacrais na espécie estudada.

34
Na coluna vertebral verificou-se a presença de 15 ou 16 vértebras
torácicas e duas ou três lombares, coincidindo com o descrito por Flower (1885),
Sánchez-Villagra et al. (2007) e Endo et al. (2009). Flower (1885) apontou ainda,
a correlação de 15 vértebras torácicas para três lombares ou 16 torácicas para
duas lombares, o que é sustentado pelos resultados do presente estudo, haja
vista que esta correlação esteve presente na maioria dos animais (83,3%). A
exceção foi na relação de 15 vértebras torácicas para duas lombares em um
indivíduo. Em relação às vértebras sacrais, Flower (1885) descreveu a presença
de três ou cinco, no entanto, foi observada neste estudo a presença de quatro
(16,6%) ou cinco (83,3%) vértebras (Figura 1).
O plexo lombossacral do M. tridactyla foi formado predominantemente
pelos ramos ventrais dos nervos espinhais T16, L1, L2, S1-S5, Cc1, o que o difere
do plexo da maioria dos mamíferos no que se refere à participação de todos os
segmentos lombares e a contribuição de segmentos torácicos e coccígeos na sua
composição (Tabela 1). Esta diferença se estende também aos animais
domésticos, onde o plexo lombossacral é constituído pelos ramos ventrais de L2-
L6, S1 em equinos, bovinos e suínos (GETTY, 1986), podendo ter participação de
L1 em equinos (GETTY, 1986) e por L3-L7 e S1-S3 em cães (MILLER, 1964).
Em 11 plexos (91,6%), o plexo lombar e sacral estava unido,
constituindo um plexo lombossacral verdadeiro, sendo o último segmento lombar
(L2 ou L3), o elo entre eles, pois sendo bifurcado, este ramo contribuiu tanto com
a formação de nervos do plexo lombar, quanto do sacral. Em um plexo (8,3%), o
último segmento lombar (L2) não contribuiu para a formação do plexo sacral.
Todavia, para melhor apresentação dos resultados, os plexos lombar e sacral
serão discutidos separadamente.
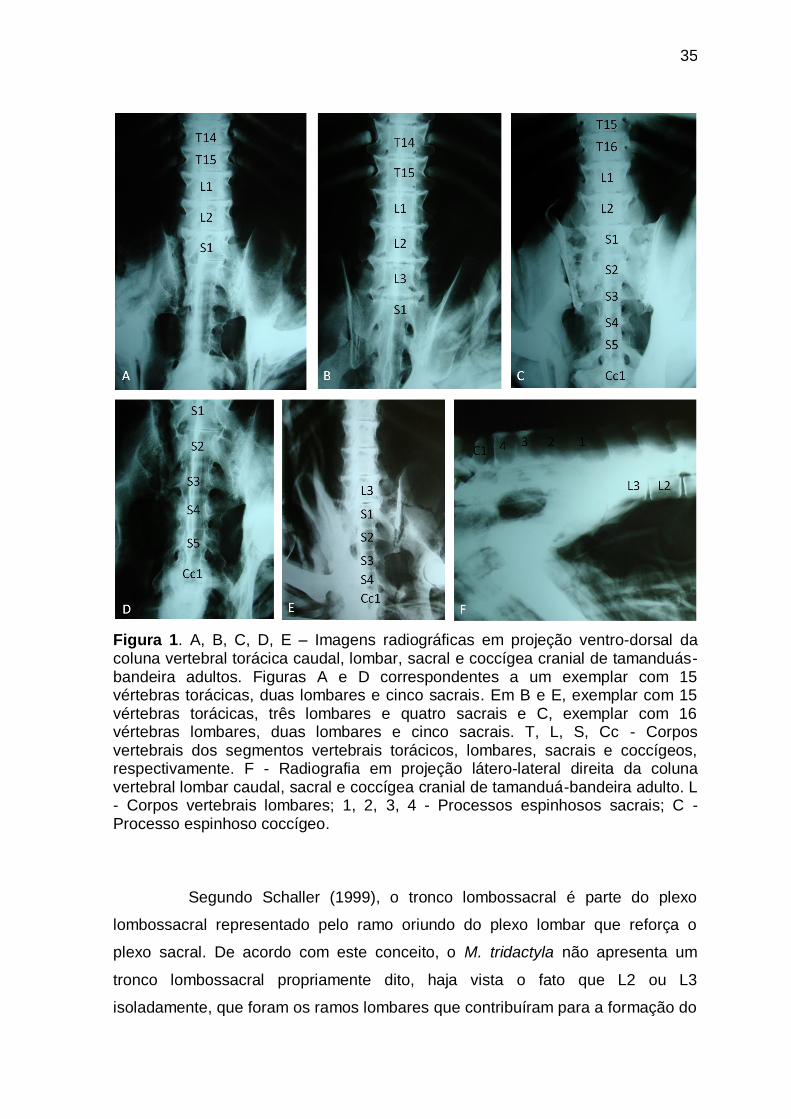
35
Figura 1. A, B, C, D, E – Imagens radiográficas em projeção ventro-dorsal da coluna vertebral torácica caudal, lombar, sacral e coccígea cranial de tamanduás-bandeira adultos. Figuras A e D correspondentes a um exemplar com 15 vértebras torácicas, duas lombares e cinco sacrais. Em B e E, exemplar com 15 vértebras torácicas, três lombares e quatro sacrais e C, exemplar com 16 vértebras lombares, duas lombares e cinco sacrais. T, L, S, Cc - Corpos vertebrais dos segmentos vertebrais torácicos, lombares, sacrais e coccígeos, respectivamente. F - Radiografia em projeção látero-lateral direita da coluna vertebral lombar caudal, sacral e coccígea cranial de tamanduá-bandeira adulto. L - Corpos vertebrais lombares; 1, 2, 3, 4 - Processos espinhosos sacrais; C - Processo espinhoso coccígeo.
Segundo Schaller (1999), o tronco lombossacral é parte do plexo
lombossacral representado pelo ramo oriundo do plexo lombar que reforça o
plexo sacral. De acordo com este conceito, o M. tridactyla não apresenta um
tronco lombossacral propriamente dito, haja vista o fato que L2 ou L3
isoladamente, que foram os ramos lombares que contribuíram para a formação do
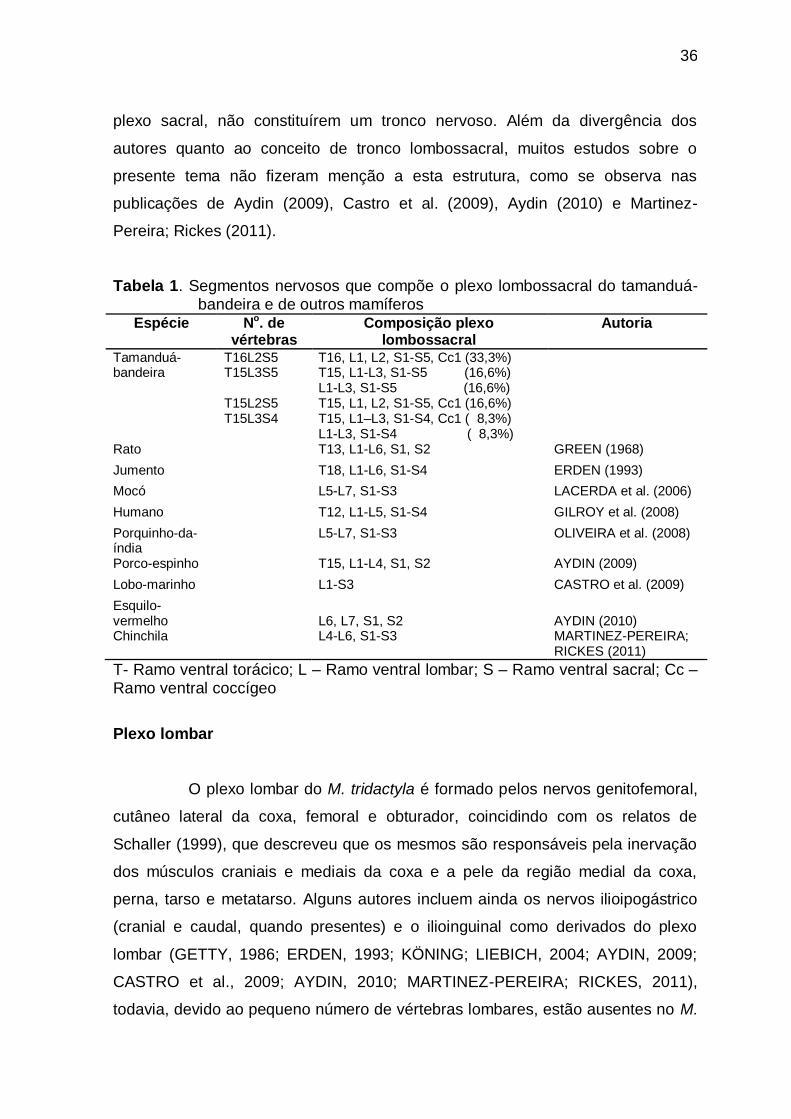
36
plexo sacral, não constituírem um tronco nervoso. Além da divergência dos
autores quanto ao conceito de tronco lombossacral, muitos estudos sobre o
presente tema não fizeram menção a esta estrutura, como se observa nas
publicações de Aydin (2009), Castro et al. (2009), Aydin (2010) e Martinez-
Pereira; Rickes (2011).
Tabela 1. Segmentos nervosos que compõe o plexo lombossacral do tamanduá-bandeira e de outros mamíferos
Espécie No. de vértebras
Composição plexo lombossacral
Autoria
Tamanduá-bandeira
T16L2S5 T15L3S5 T15L2S5 T15L3S4
T16, L1, L2, S1-S5, Cc1 (33,3%) T15, L1-L3, S1-S5 (16,6%) L1-L3, S1-S5 (16,6%) T15, L1, L2, S1-S5, Cc1 (16,6%) T15, L1–L3, S1-S4, Cc1 ( 8,3%) L1-L3, S1-S4 ( 8,3%)
Rato T13, L1-L6, S1, S2 GREEN (1968)
Jumento T18, L1-L6, S1-S4 ERDEN (1993)
Mocó L5-L7, S1-S3 LACERDA et al. (2006)
Humano T12, L1-L5, S1-S4 GILROY et al. (2008)
Porquinho-da-índia
L5-L7, S1-S3 OLIVEIRA et al. (2008)
Porco-espinho T15, L1-L4, S1, S2 AYDIN (2009)
Lobo-marinho L1-S3 CASTRO et al. (2009)
Esquilo-vermelho
L6, L7, S1, S2
AYDIN (2010)
Chinchila L4-L6, S1-S3 MARTINEZ-PEREIRA; RICKES (2011)
T- Ramo ventral torácico; L – Ramo ventral lombar; S – Ramo ventral sacral; Cc – Ramo ventral coccígeo
Plexo lombar
O plexo lombar do M. tridactyla é formado pelos nervos genitofemoral,
cutâneo lateral da coxa, femoral e obturador, coincidindo com os relatos de
Schaller (1999), que descreveu que os mesmos são responsáveis pela inervação
dos músculos craniais e mediais da coxa e a pele da região medial da coxa,
perna, tarso e metatarso. Alguns autores incluem ainda os nervos ilioipogástrico
(cranial e caudal, quando presentes) e o ilioinguinal como derivados do plexo
lombar (GETTY, 1986; ERDEN, 1993; KÖNING; LIEBICH, 2004; AYDIN, 2009;
CASTRO et al., 2009; AYDIN, 2010; MARTINEZ-PEREIRA; RICKES, 2011),
todavia, devido ao pequeno número de vértebras lombares, estão ausentes no M.

37
tridactyla, sendo o nervo genitofemoral o mais cranial dos nervos lombares. O
território de inervação dos referidos nervos ausentes nesta espécie é suprido pelo
nervo costo-abdominal, que corresponde ao ramo ventral do último nervo torácico,
o que também é relatado em suínos, no caso de ausência do nervo ilioipogástrico
cranial, quando estão presentes seis vértebras lombares ao invés de sete
(GETTY, 1986). A composição dos nervos do plexo lombar está demonstrada na
Tabela 2.
Tabela 2. Segmentos nervosos que compõe os nervos do plexo lombar em
Myrmecophaga tridactyla Nervo Composição Frequência
Genitofemoral T15, L1, L2 33,3%
T16, L1 33,3%
T15, L1 16,6%
L1, L2 16,6%
Cutâneo femoral lateral T15, L1, L2 25,0%
T16, L1 25,0%
L1, L2 25,0%
T15, L1 16,6%
T16, L1 ,L2 8,3%
Femoral T16, L1, L2 33,3%
T15, L1, L2, L3 25,0%
L1, L2, L3 25,0%
T15, L1, L2 16,6%
Obturador L1, L2, L3 25,0%
T16, L1, L2, S1 16,6%
T16, L1, L2 16,6%
T15, L1, L2, S1 16,6%
T15, L1, L2, L3, S1 16,6%
T15, L1, L2, L3 8,3%
T- Ramo ventral torácico; L – Ramo ventral lombar; S – Ramo ventral sacral
A participação de nervos torácicos na formação do nervo
genitofemoral, como observado em 83,3% dos casos deste estudo (Tabela 2,
Figura 2) e em porco-espinho (AYDIN, 2009) é incomum. Esta diferença se deve
provavelmente à menor quantidade de vértebras lombares nestas espécies,
sendo duas ou três no M. tridactyla e quatro no porco-espinho. Os resultados aqui
apresentados diferem-se dos demais mamíferos, como lobo-marinho (CASTRO et
al., 2009) e animais domésticos, onde este nervo é formado exclusivamente por
38
ramos lombares, sendo L3 e L4 (MILLER, 1964; GETTY, 1986; SCHALLER,
1999), podendo ter participação de L2 em ruminantes, equinos (GETTY, 1986;
SCHALLER, 1999) e suínos (GETTY, 1986). Pode originar-se ainda de uma única
raiz lombar, representada por L2 em jumentos (ERDEN, 1993), L3 em chinchilas
(MARTINEZ-PEREIRA; RICKES, 2011) e L4 no esquilo-vermelho (AYDIN, 2010).
Figura 2. Vista ventral do plexo lombossacral de M. tridactyla adultos, sendo A, indivíduo com 16 vértebras torácicas e duas lombares e B, indivíduo com 15 vértebras torácicas e três lombares. T, L e S, ramos ventrais dos nervos torácicos, lombares e sacrais, respectivamente; C, N. costoabdominal; Gf, N. genitofemoral; CL, N. cutâneo femoral lateral; F, N. femoral; O, N. obturador; I, N. isquiático; Pd e P, Tronco comum dos nervos pudendo e pélvico; GCa, N. glúteo caudal; GCr, N. glúteo cranial; TS, tronco simpático; *, Ramo de S1 para o N. obturador; 1, Músculo psoas menor.

39
O nervo cutâneo femoral lateral (n. cutâneo lateral da coxa) do M.
tridactyla (Figura 2, Tabela 2) recebeu na maioria dos casos (75%), a contribuição
do último nervo torácico, o que corrobora os achados de Aydin (2009) em porco-
espinho. Nas demais espécies, é formado exclusivamente por segmentos
lombares, sendo L3-L5 nos animais domésticos (GETTY, 1986; SCHALLER,
1999), podendo ter contribuição de L2 no equino (GETTY, 1986). Pode originar-se
ainda de uma única raiz lombar, representada por L3 no lobo-marinho (CASTRO
et al., 2009) e L4 em chinchilas (MARTINEZ-PEREIRA; RICKES, 2011).
Para contribuir com a formação dos nervos genitofemoral e cutâneo
femoral lateral, o último nervo torácico se bifurca, sendo que o ramo cranial desce
paralelo à última costela constituindo com o ramo caudal do penúltimo nervo
intercostal, o denominado nervo costoabdominal, enquanto o ramo caudal une-se
aos segmentos lombares para integrar os nervos do plexo lombar. Esta
característica é padrão para todos os demais nervos torácicos nesta espécie,
onde cada nervo espinhal torácico bifurca-se, fornecendo um ramo que contribui
com a formação do nervo intercostal correspondente e outro que contribui com o
nervo intercostal seguinte. De acordo com os achados atuais, não há relatos
desta característica em qualquer outra espécie estudada (Figura 3).
Do plexo lombar, o nervo femoral foi o mais espesso, o que confirma a
descrição de Schaller (1999), referindo-se aos animais domésticos e também
apresenta composição similar ao do porco-espinho (AYDIN, 2009) no que se
refere à participação de segmentos torácicos em sua composição (Tabela 2,
Figura 2). Por outro lado, diferencia-se substancialmente dos animais com maior
número de segmentos lombares, sendo formado por L3-L6 no equino, suíno e em
parte dos cães (GETTY, 1986), L4-L6 em bovinos (GETTY, 1986), L2-L4 no
jumento (ERDEN, 1993), L3-L4 no lobo-marinho (CASTRO et al., 2009), L4-L5 no
esquilo-vermelho (AYDIN, 2010) e chinchila (MARTINEZ-PEREIRA; RICKES,
2011), L5-L6 no porquinho-da-índia (OLIVEIRA et al., 2008) e mocós (OLIVEIRA
et al., 2011).

40
Figura 3. Vista ventral do plexo lombossacral de M. tridactyla adulta, com 15 vértebras torácicas, duas lombares e cinco sacrais, onde observa-se a formação dos nervos intercostais. T, L - Ramos ventrais torácicos e lombares, Gf - N. Genitofemoral, CL - N. Cutâneo femoral lateral, F - N. Femoral, O - N. Obturador, C - N. Costoabdominal e * - Ramos caudais dos nervos espinhais torácicos T13 e T14, contribuindo para a formação dos nervos intercostais 14 e 15, respectivamente.
Caudalmente, o nervo obturador (Figura 2) teve a mesma constituição
do nervo femoral em 50,0% dos casos (Tabela 2), coincidindo com o descrito em
animais domésticos por König; Liebich (2004), Lorenz; Kornegay (2006) e Dyce et
al. (2010). A composição do nervo obturador se diferencia da descrita em outras
espécies, não somente pela contribuição torácica, mas como também pela sacral,
que apenas é descrito por Miranda et al. (2007) em fêmeas de bovinos azebuados
e Schaller (1999), apesar de não se referir à espécie. Assim, segundo Getty
(1986), em cães e bovinos o mesmo é formado pelos segmentos L4-L6, podendo
ter a contribuição de L3 em equinos e suínos, por L3, L4 no lobo-marinho
(CASTRO et al., 2009), L4, L5 no esquilo-vermelho (AYDIN, 2010) e chinchila
(MARTINEZ-PEREIRA; RICKES, 2011) e por L5, L6 nos mocós (LACERDA et al.,
2006) e porquinho-da-índia (OLIVEIRA et al., 2008).

41
Plexo sacral
Emergindo do plexo sacral do M. tridactyla observou-se os nervos
glúteos cranial e caudal, isquiático, cutâneo caudal do fêmur, pudendo e o retal
caudal. Os segmentos nervosos que os compõem estão descritas na Tabela 3.
O nervo glúteo cranial (Figura 2) deixa dorsalmente o tronco do plexo
sacral, sendo formado predominantemente por L2, S1, S2 (Tabela 3). Sua
composição difere-se da demonstrada nas outras espécies, sendo formado por L3
ou L3 e L4 no porco-espinho (AYDIN, 2009), L7 no porquinho-da-índia (OLIVEIRA
et al., 2008) e mocós (LACERDA et al., 2006), L4-S1 no jumento (ERDEN, 1993)
e L6, S1 em bovinos (GETTY, 1986). Em cães e equinos (GETTY, 1986), lobo-
marinho (CASTRO et al., 2009) e chinchila (MARTINEZ-PEREIRA; RICKES,
2011), este nervo é formado pelos dois últimos segmentos lombares, junto com
S1.
Tabela 3. Segmentos nervosos que compõe os nervos do plexo sacral em
Myrmecophaga tridactyla Nervo Composição Frequência
Glúteo cranial L2, S1, S2 41,6%
L3, S1 33,3%
L2, L3, S1 16,6%
S1, S2 8,3%
Glúteo caudal L2, S1, S2, S3 25,0%
L3, S1, S2 25,0%
L3, S1, S2, S3 25,0%
L2, S1, S2, S3, S4 16,6%
S1, S2, S3 8,3%
Isquiático L3, S1, S2, S3 50,0%
L2, S1, S2, S3, S4 41,6%
S1, S2, S3, S4 8,3%
Cutâneo femoral caudal Nervo isquiático 100,0%
Pudendo S4, S5 75,0%
S3, S4 16,6%
S3, S4, S5 8,3%
Retal caudal S5 33,3%
S4, S5, Cc1 33,3%
S4, Cc1 16,6%
S5, Cc1 16,6%
L – Ramo ventral lombar; S – Ramo ventral sacral; Cc – Ramo ventral coccígeo

42
Na maioria dos casos (58,3%), o nervo glúteo caudal originou-se do
plexo sacral, cranialmente ao último segmento de formação do nervo isquiático
(Figura 2), coincidindo com os relatos de Castro et al. (2009) em lobo-marinho e
Lacerda et al. (2006) em mocós. Segundo a literatura destes animais, o nervo
glúteo caudal teve a mesma origem do glúteo cranial, assim como ocorre no suíno
(GETTY, 1986), padrão este, não observado no M. tridactyla. Nos demais cinco
antímeros (41,6%), foi ramo do nervo isquiático, como ocorre nos ruminantes,
suínos e gatos (GETTY, 1986), macaco Cebus apella (BARROS et al., 2003) e
chinchila (MARTINEZ-PEREIRA; RICKES, 2011).
Derivando predominantemente de L3, S1-S3 (Tabela 3) têm-se o nervo
isquiático (Figura 2). Apesar de grupos de animais compartilharem do mesmo
padrão de formação deste nervo, como se nota a seguir, o do M. tridactyla não se
enquadra em nenhum deles. Desta forma, o nervo isquiático é formado pelo
último nervo lombar e dois primeiros sacrais em ruminantes (GETTY, 1986; LIMA
et al., 2008), equino (GETTY, 1986), jumento (ERDEN, 1993) e mocós (SANTOS
et al., 2006), pelos dois últimos nervos lombares e os dois primeiros sacrais no
cão (GETTY, 1986) e no esquilo-vermelho (AYDIN, 2010), pelos dois últimos
nervos lombares e primeiro sacral no porquinho-da-índia (OLIVEIRA et al., 2008),
no lobo-marinho (CASTRO et al., 2009), no porco-espinho (AYDIN, 2009), no preá
(OLIVEIRA et al., 2010), na chinchila (MARTINEZ-PEREIRA; RICKES, 2011) e no
mão-pelada (PEREIRA et al., 2011).
Nota-se ainda, que na maioria das espécies, a contribuição nervosa
para o nervo isquiático termina mais cranialmente do que ocorre no M. tridactyla,
sendo S2 o último segmento participante, mas, a contribuição de S3 é reportada
em humanos (GILROY et al., 2008) e javali (IGLESIAS et al., 2011), podendo
estar presente em bovinos azebuados (CAMPOS et al., 2003; FERRAZ et al.,
2006) e caprinos da raça Saanen (LIMA et al., 2008). Até onde se pode constatar,
a participação de S4, como observado neste estudo, ainda não fora reportada em
outra espécie. Esta diferença pode ser atribuída mais uma vez ao menor número
de vértebras lombares em relação às sacrais no tamanduá-bandeira.
O nervo cutâneo femoral caudal originou-se do nervo isquiático em
100% dos plexos, discordando com o demonstrado em outras espécies. Ele
deriva da borda ventral do nervo glúteo caudal em equinos ou da borda dorsal do

43
nervo isquiático em ruminantes, enquanto no gato (GETTY, 1986) e no jumento
(ERDEN, 1993), deriva de S2-S3, podendo receber contribuição de S1 em suínos
(GETTY, 1986). Em cães, deriva de S1, S2 (GETTY, 1986). No lobo-marinho
(CASTRO et al., 2009) e na chinchila (MARTINEZ-PEREIRA; RICKES, 2011)
provém do mesmo tronco nervoso do plexo isquiático, que emite os nervos glúteo
cranial, glúteo caudal e isquiático.
Predominantemente por S4-S5 (Tabela 3), o nervo pudendo foi
formado. Estes ramos reúnem-se num tronco comum (Figura 2), de onde também
deriva o nervo pélvico, que segue cranialmente para formar o plexo pélvico. O
nervo pudendo dos mamíferos apresenta uma grande variação na sua formação,
mas geralmente, é mais caudal no M. tridactyla do que nos demais animais. É
formado por S2 no porco-espinho (AYDIN, 2009), S1, S2, podendo ter
contribuição de S3 em chinchila (MARTINEZ-PEREIRA; RICKES, 2011), mais
frequentemente por S2, S3 em suínos, cães e gatos (GETTY, 1986; SCHALLER,
1999), no mocó (LACERDA et al., 2006), no porquinho-da-índia (OLIVEIRA et al.,
2008) e no lobo-marinho (CASTRO et al., 2009) e por S3, podendo ter
participação de S2 e S4 em ruminantes e equinos (GETTY, 1986; SCHALLER,
1999).
Com a participação principalmente de S5 (Tabela 3), encontra-se o
nervo retal caudal. O padrão de formação monossegmentar também é reportado
em equinos, bovinos e suínos (GETTY, 1986), sendo S4 o segmento envolvido,
ou S3 na chinchila (MARTINEZ-PEREIRA; RICKES, 2011) e S2 no porco-espinho
(AYDIN, 2009), diferente do observado em cães (GETTY, 1986) e lobo-marinho
(CASTRO et al., 2009), cuja formação deriva de dois segmentos medulares, S2 e
S3. Em cães, o nervo retal caudal pode ainda ser ramo do nervo pudendo
(MILLER, 1964; SCHALLER, 1999), o que não foi observado no presente estudo.
No jumento, deriva de S3-S5 (ERDEN, 1993). A participação de segmento
coccígeo foi relatada apenas em suínos, nos quais o nervo retal caudal se origina
de S2-S4, sendo que na maioria dos casos, o ramo ventral de S4 se relacionou
com o ramo ventral de Cc1 (CHAGAS et al., 2010).
Assim, pode-se concluir que a composição do plexo lombossacral e de
seus nervos no M. tridactyla apresenta similaridades aos dos demais mamíferos,
em especial daqueles que também possuem um menor número de vértebras

44
lombares. Contudo, apresenta algumas peculiaridades na sua composição, como
a participação de ramos torácicos e coccígeos e uma maior variação anatômica,
que por sua vez está associada à variação do número de vértebras torácicas,
lombares e sacrais.
AGRADECIMENTOS
Ao Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais
Renováveis do estado de Goiás (IBAMA-GO) pela doação dos cadáveres de M.
tridactyla utilizados neste estudo.
ABSTRACT: Despite its significance to the biomes of many countries, little is
known about the morphology of the M. tridactyla. Therefore, this study aimed to describe in M. tridactyla the origin of the lumbosacral plexus and the composition of its nerves. For this purpose, 12 plexuses from six adult cadavers of both sexes donated by IBAMA-GO (license 99/2011 UFG-CEUA Protocol 015/11) were used. The animals were thawed at room temperature for 48h, fixed and preserved in formaldehyde solution at 10% and 4%, respectively, and subsequently dissected after a minimum period of 72 hours from the fixation procedure. In order to verify the number of vertebrae, radiographic examination of the thoracic, lumbar and sacral segments of the vertebral column was performed. The ventral rami of spinal nerves, their communications and the nerves arising from the plexus were verified after the dissection of the ventral face of the caudal thoracic, lumbar and sacral regions. The radiographs showed 15 or 16 thoracic vertebrae, two or three lumbar and four or five sacral one. The lumbosacral plexus of M. tridactyla originated in T16, L1, L2, S1-S5, Cc1 (33,3%), T15, L1-L3, S1-S5 (16,6%), T15, L1, L2, S1-S5, Cc1 (16,6%), L1-L3, S1-S5 (16,6%), T15, L1-L3, S1-S4, Cc1 (8,3%) and L1-L3, S1-S4 (8,3%). The nerves derived from the plexus with their most common arrangement were as follow: genitofemoral (T15, L1, L2 and T16, L1), lateral femoral cutaneous nerve (T15, L1, L2 and T16, L1), femoral (T16, L1, L2), obturator (L1-L3), sciatic (L3, S1-S3), cranial gluteal (L2, S1, S2), caudal gluteal (L2, S1-S3 and L3, S1, S2), pudendal (S4, S5) and rectal caudal (S5 and S4, S5, Cc1). The caudal femoral cutaneous nerve originated from the sciatic nerve. In general, the lumbosacral plexus of the M. tridactyla resembles that of other mammals, but it shows some peculiarities, such as the involvement of thoracic and coccygeal rami in its nerve composition and a greater anatomical variation in its formation, which can be mainly related to the numerical variation of thoracic, lumbar and sacral vertebrae in this species.
KEYWORDS: Anatomy. Innervation. Nervous system. Xenarthra.

45
REFERÊNCIAS
AYDIN, A. The dissemination of the pelvic limb nerves originating from the lumbosacral plexus in the porcupine (Hystrix cristata). Veterinární Medicína,
Praha, v. 54, n. 7, p. 333-339, 2009. AYDIN, A. The spinal nerves that constitute the plexus lumbosacrales of the red squirrel (Sciurus vulgaris). Veterinární Medicína, Praha, v. 55, n. 4, p. 183-186,
2010. BARROS, R. A. C.; PRADA, I. L. S.; SILVA, Z.; RIBEIRO, A. R.; SILVA, D. C. O. Constituição do plexo lombar em macaco Cebus apella. Brazilian Journal of Veterinary Research and Animal, São Paulo, v. 40, n. 5, p. 373-381, 2003. CAMPOS, D. B.; SILVA, F. O. C.; SEVERINO, R. S.; DRUMMOND, S. S.; LIMA, E. M. M.; BOMBONATO, P. P.; SANTANA, M. I. S. Origem e distribuição dos nervos isquiáticos em fetos de bovinos azebuados. Ars Veterinaria, Jaboticabal, v. 19, n. 3, p. 219-223, 2003. CASTRO, T. F.; SOUZA, D. A. S.; SILVA FILHO, R. P.; PEREIRA, M. A. M. Sistematização e distribuição da inervação lombar e sacral em Arctocephalus australis. Brazilian Journal of Veterinary Research and Animal, São Paulo, v.
46, n. 5, p. 404-411, 2009. CHAGAS, L. G. S.; DRUMMOND, S. S.; SILVA, F. O. C.; CHAGAS R. G. Origem e distribuição do nervo retal caudal em suínos (Sus scrofa domesticus- LINNAEUS, 1758) da linhagem Per Ar Lan. Horizonte científico, Uberlândia, v. 4, n. 2, p. 1-11, 2010. DYCE, K. M.; SACK, W. O.; WENSING, C. J. G. Tratado de anatomia veterinária, 4. ed. Rio de Janeiro: Elsevier, 2010, 834 p. ENDO, H.; KOMIYA, T.; KAWADA, S.; HAYASHIDA, A.; KIMURA, J.; ITOU, T.; KOIE, H.; SAKAI, T. Three-dimensional reconstruction of the xenarthrous process of the thoracic and lumbar vertebrae in the giant anteater. Mammal Study, Tokyo, v. 34, p. 1-6, 2009. ERDEN, H. Merkebin (equus asinus l.) plexus lumbosacralis'I üzerinde makroanatomik ve subgros çalişmalar. Uludağ Üniversitese Veteriner Fakültesi Dergisi, Konya, v. 9, n. 1, p. 64-71, 1993.
FERRAZ, R. H. S. et al. Estudo anatômico da porção intrapélvica do nervo isquiático em fetos de bovinos azebuados. Brazilian Journal of Veterinary Research and Animal, São Paulo, v. 43, n. 3, p. 302-308, 2006.

46
FLOWER, W. H. An introduction to the osteology of the mammalian. 3. ed.
London: Macmillan and Co., 1885, 382 p. GETTY, R. Anatomia dos animais domésticos. 5. ed. Rio de Janeiro: Guanabara Koogan, 1986. 2000p. v. 1 e 2. GILROY, A. M.; MacPHERSON, B. R.; ROSS, L. M. Atlas de Anatomia. 1. ed.,
Rio de Janeiro: Guanabara Koogan, 2008, 656p. GREEN, C. E. Anatomy of the Rat. In:__The nervous system. Hafner Publishing Company, New York and London, 1968. cap. 26, p. 115–175. IGLESIAS, L. P.; SILVA, F. O. C; BRITO, T. R. Origem e distribuição do nervo isquiático em fetos de javalis (Sus scrofa scrofa). Biotemas, Florianópolis, v. 24, n. 4, p. 141-145, 2011. INTERNATIONAL COMMITTEE ON VETERINARY GROSS ANATOMICAL NOMENCLATURE. Nomina anatomica veterinaria. 5. ed. Ithaca: Cornell University, 2012. 160 p. IUCN 2012. IUCN Red List of Threatened Species. Version 2012.1.
<www.iucnredlist.org>. Acessado em 26 agosto 2012. KÖNIG, H. E.; LIEBICH, H. J. Anatomia dos animais domésticos. 1. ed. Porto Alegre: Artmed, 2004. 399 p. v. 2. LACERDA, P. M. O.; MOURA, C. E. B.; MIGLINO, M. A.; OLIVEIRA, M. F.; ALBUQUERQUE, J. F. G. Origem do plexo lombossacral de mocó (Kerondo rupestris). Brazilian Journal of Veterinary Research and Animal, São Paulo, v.
43, n. 5, p. 620-628, 2006. LARRAZÁBAL, L. B. Crianza en cautiverio de perezoso de dos dedos (Choloepus didactylus). Edentata, v. 6, p. 30-36, 2004.
LAVIN, L. M. Radiography in veterinary technology. 4. Ed. Philadelphia:
Elsevier, 2006. 400p. LIMA, E. M. M.; SILVA, F. O. C.; SEVERINO, R. S.; DRUMMOND, S. S.; CAMPOS, D. B.; SANTANA, M. I. S.; MORAES, D. D. A. Origem e distribuição dos nervos isquiáticos em caprinos da raça Saanen. Ciência Rural, Santa Maria, v. 38, n. 2, p. 372-377, 2008. LORENZ, M. D.; KORNEGAY, J. N. Neurologia Veterinária. 2. ed. Barueri:
Manole, 2006. 467 p. MARTINEZ-PEREIRA, M. A.; RICKES, E. M. The spinal nerves that constitute the lumbosacral plexus and their distribution in the chinchilla. Journal of the South African Veterinary Association, Pretoria, v. 82, n. 3, p. 150-154, 2011.

47
MILLER, M. E. Anatomy of the dog. Philadelphia: W. B. Saunders, 1964, 941 p.
MIRANDA, F.; COSTA, A. M. Xenarthra (Tamanduá, Tatu, Preguiça). In: CUBAS, Z. S.; SILVA, J. C. R.; CATÃO-DIAS, J. L. Tratado de animais selvagens – medicina veterinária. 1. ed. Roca: São Paulo, 2007, p. 402-414.
MIRANDA, R. L.; CARNEIRO E SILVA, F. O.; SEVERINO, R. S.; DRUMMOND, S. S.; GONÇALVES, R. C. Origens e distribuições dos nervos obturatórios em fetos fêmeas de bovinos azebuados. Bioscience Journal, Uberlândia, v. 23, n. 4, p.
120-127, 2007. MOURÃO, G.; MEDRI, M. Activity of a specialized insectivorous mammal (Myrmecophaga tridactyla) in the Pantanal of Brazil. Journal of Zoology, London,
p. 271: 187-192, 2007. OLIVEIRA, G. B.; ALBUQUERQUE, J. F. G.; OLIVEIRA, M. F.; RODRIGUES, M. N.; SOUSA, R. S.; SOUSA, E. S.; MIGLINO, M. A. Source and distribution of the lumbosacral plexus in spix’s yellow-toothed cavy (Galea spixii spixii). Brazilian Journal Morphology Science, São Paulo, v. 25, n. 1-4, p. 1-34, 2008.
OLIVEIRA, G. B.; RODRIGUES, M. N.; SOUSA, E. S.; PAIVA, A. L. C.; MOURA, C. E. B.; MIGLINO, M. A.; OLIVEIRA, M. F. Origem e distribuição dos nervos isquiáticos do preá. Ciência Rural, Santa Maria, v. 40, n. 8, p. 1741-1745, 2010.
OLIVEIRA, G. B.; ALBUQUERQUE, J. F. G.; RODRIGUES, M. N.; PAIVA, A. L. C.; MOURA, C. E. B.; MIGLINO, M. A.; OLIVEIRA, M. F. Origem e distribuição do nervo femoral em mocó, Kerodon rupestris (Cavidae). Pesquisa Veterinária Brasileira, Rio de Janeiro, v. 31, n. 1, p. 84-88, 2011. PEREIRA, K. F.; PARANAIBA, J. F. F. S.; HELRIGLE, C.; ARAÚJO, E. G. Origem e distribuição anatômica do nervo isquiático de mão-pelada. Pesquisa Veterinária Brasileira, Rio de Janeiro, v. 31, n. 1, p. 74-78, 2011. RODRIGUES, H. Técnicas anatômicas. 4. ed. Vitória: GM. Gráfica e Editora, 2010, 270p. SÁNCHEZ-VILLAGRA, M. R.; NARITA, Y.; KURATANI, S. Thoracolumbar Vertebral Number: First Skeletal Synapomorphy For Afroterian Mammals. Systematics and Biodiversity, Cambridge, v. 5, n. 1, p. 1-7, 2007.
SANTOS, R. C.; ALBUQUERQUE, J. F. G.; SILVA, M. C. V.; MOURA, C. E. B, CHAGAS, R. S. N; BARBOSA, R. R.; MIGLINO, M. A. Anatomia do nervo isquiático em mocós (Kerodon rupestris WIED, 1820) aplicada à clínica de animais silvestres. Brazilian Journal of Veterinary Research and Animal, São Paulo, v. 43, n. 43, p. 647-653, 2006. SCHALLER, O. Nomenclatura anatômica veterinária ilustrada. 1. ed. São
Paulo: Manole, 1999. 494 p.

48
CAPÍTULO 3 - ASPECTOS ANATÔMICOS DOS NERVOS DA COXA DO
TAMANDUÁ-BANDEIRA (Myrmecophaga tridactyla, Linnaeus, 1758)
ANATOMICAL ASPECTS OF THE NERVES OF THE THIGH OF THE GIANT
ANTEATER (Myrmecophaga tridactyla, Linnaeus, 1758)
Viviane Souza CRUZ1, Júlio Roquete CARDOSO2, Luciana Batalha de
Miranda ARAÚJO3, Paulo Roberto de SOUZA1, Eugênio Gonçalves de
ARAÚJO3
1.Doutorandos no Programa de Pós-Graduação em Ciência Animal, Escola de Veterinária e Zootecnia, Universidade Federal de Goiás, UFG, Goiânia, GO; Professores mestres no Departamento de Morfologia, Instituto de Ciências Biológicas, Universidade Federal de Goiás, UFG, Goiânia, GO. [email protected]; [email protected], Caixa Postal 131 – Campus Samambaia (Campus II), CEP: 74001-970 – Goiânia – Goiás 2.Professor doutor, Departamento de Morfologia, Instituto de Ciências Biológicas, Universidade Federal de Goiás, UFG. Goiânia, GO. [email protected], Caixa Postal 131 – Campus Samambaia (Campus II), CEP: 74001-970 – Goiânia – Goiás 3.Professores doutores, Departamento de Medicina Veterinária, Escola de Veterinária e Zootecnia, Universidade Federal de Goiás, UFG, Goiânia, GO. [email protected], [email protected], Caixa Postal 131 – Campus Samambaia (Campus II), CEP: 74001-970 – Goiânia – Goiás
Submetido na Revista Bioscience Journal em 05 de março de 2013
Situação: Em avaliação (última alteração em 18 de março de 2013)

49
ASPECTOS ANATÔMICOS DOS NERVOS DA COXA DO TAMANDUÁ-
BANDEIRA (Myrmecophaga tridactyla, Linnaeus, 1758)
ANATOMICAL ASPECTS OF THE NERVES OF THE THIGH OF THE GIANT
ANTEATER (Myrmecophaga tridactyla, Linnaeus, 1758)
RESUMO: A carência de conhecimento básico acerca das espécies silvestres, em especial de anatomia topográfica, tem constituído um obstáculo à prática de procedimentos clínico-cirúrgicos em animais selvagens. Portanto, o objetivo deste trabalho foi descrever os nervos da coxa do M. tridactyla, abordando sua topografia, ramificação e território de inervação. Para tanto, foram utilizados seis cadáveres adultos, fornecidos pelo IBAMA-GO (licença 99/2011, CEUA-UFG protocolo nº 015/11), fixados e preservados em soluções de formaldeído a 10% e 4%, respectivamente. A dissecação teve início após um período mínimo de 72h a partir da fixação. Os nervos responsáveis pela inervação da coxa do M. tridactyla foram o genitofemoral, o cutâneo femoral lateral, o femoral e seu principal ramo, o nervo safeno, o obturador, o glúteo cranial, o isquiático e o cutâneo femoral caudal. O ramo femoral do nervo genitofemoral inerva a região cutânea medial da coxa e linfonodo inguinal superficial. O nervo cutâneo femoral lateral distribui na região cutânea craniomedial e craniolateral da coxa. O nervo Femoral se ramifica nos músculos ilíaco, psoas maior e menor, iliopsoas, pectíneo, sartório, quadríceps e a pele da região medial da coxa. O nervo safeno inerva a pele da face medial da coxa e joelho. O nervo obturador envia ramos aos músculos obturador externo, inclusive sua parte intrapélvica, adutores magno, longo e curto e o grácil. O nervo glúteo cranial inerva o músculo tensor da fáscia lata. O nervo isquiático distribui nos músculos gêmeos, quadrado femoral, semimembranoso, semitendinoso, bíceps femoral longo e curto e na região cutânea lateral da coxa e joelho. O nervo cutâneo femoral caudal inerva o músculo semitendinoso e a região da tuberosidade isquiática além da face caudolateral da coxa. Há consideráveis diferenças entre o território de inervação dos nervos da coxa do M. tridactyla em relação ao dos animais domésticos, o que na prática inviabiliza a extrapolação de técnicas de procedimentos clínico-cirúrgicos e protocolos anestésicos adotados em animais domésticos para a espécie em questão.
PALAVRAS-CHAVE: Inervação, Plexo lombossacral. Sistema nervoso periférico.
Tamanduás

50
INTRODUÇÃO
O tamanduá-bandeira (Myrmecophaga tridactyla) é um mamífero
pertencente à Superordem Xenarthra, ordem Pilosa e família Myrmecophagidae,
sendo esta formada por três gêneros: Myrmecophaga (tamanduá-bandeira),
Tamandua (tamanduá-mirim) e Cyclopes (tamanduá-pigmeu). O M. tridactyla está
classificado como “vulnerável” (VU) na lista da União Internacional para a
Conservação da Natureza e dos Recursos Naturais (IUCN, 2012), sendo que esta
queda na população é decorrente de vários fatores, dentre eles a caça, o ataque
por cães, a degradação de seu habitat pelas queimadas, o avanço da pecuária e
agricultura, e atropelamentos (MIRANDA; COSTA, 2007; MOURÃO; MEDRI,
2007).
Dentre as consequências dos traumatismos, estão as neuropatias, que
podem ser resultantes de lesões mecânicas, fraturas, compressão, estiramento,
laceração e infusão de medicamentos no interior ou nos arredores dos nervos,
podendo acarretar a lesão de um único ou de um grupo de nervos (FORTERRE et
al., 2007). Assim, clínicos e cirurgiões devem ter o conhecimento da
neuroanatomia para um atendimento adequado e que favoreça a sobrevida e
preservação desta espécie. No entanto, o estudo do sistema nervoso ainda é uma
área pouco explorada em animais selvagens e no que se refere à Superordem
Xenarthra, não há até o momento estudos acerca da distribuição nervosa nos
membros pélvicos de qualquer uma de suas espécies.
Nesta perspectiva, o objetivo deste trabalho foi descrever os nervos da
coxa do M. tridactyla, abordando sua topografia, ramificação e território de
inervação.
MATERIAL E MÉTODOS
Foram utilizados seis exemplares de tamanduá-bandeira, adultos,
sendo três fêmeas, dois machos e outro indefinido devido à retirada dos órgãos
genitais em procedimento de necropsia. Todos os espécimes tiveram os dois
antímeros dissecados, portanto os resultados deste estudo são produto da análise

51
de 12 plexos lombossacrais com seus respectivos membros pélvicos. Os animais
foram cedidos pelo Centro de Triagem de Animais Silvestres (CETAS) – IBAMA-
GO (licença 99/2011), logo, nenhum animal foi submetido ao procedimento de
eutanásia. Toda conduta foi previamente aprovada pela Comissão de Ética no
Uso de Animais – CEUA (Processo nº 015/11). O preparo, acondicionamento e
dissecação foram realizados no Laboratório de Anatomia Animal do Instituto de
Ciências Biológicas (ICB), da Universidade Federal de Goiás (UFG).
Ao dar entrada no Laboratório, os animais foram descongelados a
temperatura ambiente por um período de 48 horas, higienizados em água
corrente e posteriormente submetidos à fixação por meio de injeções musculares
e na artéria carótida, com solução de formol a 10%, num volume correspondente
a 10% do peso corporal e posteriormente imersos em tanques com solução
conservadora de formol a 4% e após um período mínimo de 72h de fixação,
iniciou-se a dissecação (RODRIGUES, 2010). O acesso à cavidade abdominal e
pélvica se deu a partir de uma incisão longitudinal na linha alba, remoção da
sínfise pélvica e de parte dos púbis e ísquios.
Para a visualização dos ramos ventrais dos segmentos lombares e
sacrais junto aos forames intervertebrais, foram afastados o intestino, os órgãos
genitourinários e os vasos sanguíneos lombares e sacrais e ainda, a remoção do
peritônio parietal. Os nervos e suas ramificações foram dissecados até a
extremidade distal da coxa. Os resultados foram documentados por meio de
esquemas e fotografias, obtidas com uma câmera digital (DSC-H50, Sony, Brasil).
A nomenclatura utilizada para a descrição dos resultados está de acordo com o
proposto pelo International Committee on Veterinary Gross Anatomical
Nomenclature (2012).
A estatística descritiva foi utilizada na análise dos dados, por meio de
frequência percentual, conforme os estudos de Campos et al. (2003); Santos et al.
(2006); Miranda et al. (2007); Lizardo et al. (2009); Iglesias et al. (2011); Oliveira
et al. (2011) e Pereira et al. (2011).
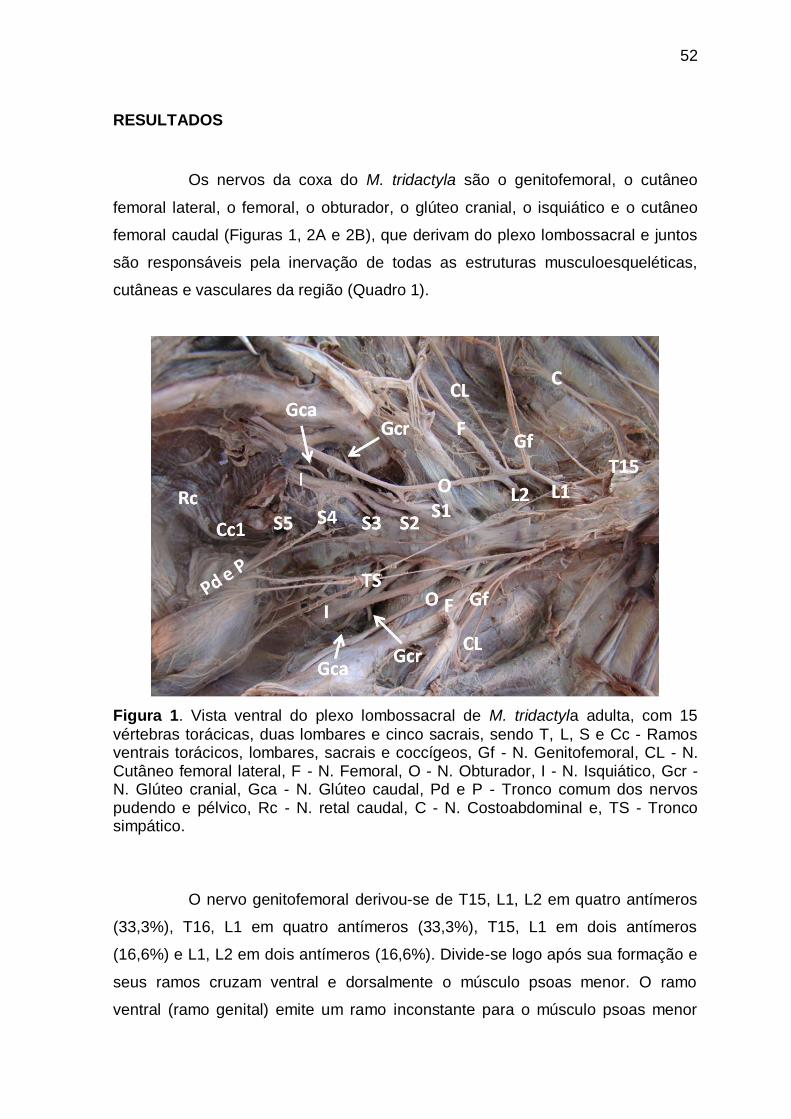
52
RESULTADOS
Os nervos da coxa do M. tridactyla são o genitofemoral, o cutâneo
femoral lateral, o femoral, o obturador, o glúteo cranial, o isquiático e o cutâneo
femoral caudal (Figuras 1, 2A e 2B), que derivam do plexo lombossacral e juntos
são responsáveis pela inervação de todas as estruturas musculoesqueléticas,
cutâneas e vasculares da região (Quadro 1).
Figura 1. Vista ventral do plexo lombossacral de M. tridactyla adulta, com 15
vértebras torácicas, duas lombares e cinco sacrais, sendo T, L, S e Cc - Ramos ventrais torácicos, lombares, sacrais e coccígeos, Gf - N. Genitofemoral, CL - N. Cutâneo femoral lateral, F - N. Femoral, O - N. Obturador, I - N. Isquiático, Gcr - N. Glúteo cranial, Gca - N. Glúteo caudal, Pd e P - Tronco comum dos nervos pudendo e pélvico, Rc - N. retal caudal, C - N. Costoabdominal e, TS - Tronco simpático.
O nervo genitofemoral derivou-se de T15, L1, L2 em quatro antímeros
(33,3%), T16, L1 em quatro antímeros (33,3%), T15, L1 em dois antímeros
(16,6%) e L1, L2 em dois antímeros (16,6%). Divide-se logo após sua formação e
seus ramos cruzam ventral e dorsalmente o músculo psoas menor. O ramo
ventral (ramo genital) emite um ramo inconstante para o músculo psoas menor

53
(16,6%), segue em direção à parede abdominal para suprir o músculo oblíquo
abdominal interno e termina distribuindo-se caudalmente na face interna da região
púbica. O ramo dorsal (ramo femoral) do nervo genitofemoral pode ser único ou
duplo. Destes, um segue distalmente, atravessa o canal inguinal em direção à
região cutânea medial da coxa, distribuindo-se no linfonodo inguinal superficial e
pele local. O outro é inconstante e se une a um ramo do nervo femoral em
posição interna ou externa à cavidade abdominal, atravessa a parede abdominal
junto aos vasos femorais e continua distribuindo-se na região medial da coxa e
joelho (Figura 2A).
Figura 2. A- Vista medial da coxa direita de M. tridactyla adulto, mostrando: L e S - Ramos ventrais dos nervos lombares e sacrais, respectivamente. Gf - N. genitofemoral; CL - N. cutâneo femoral lateral; F - N. femoral; O - N. obturador; f e g - Ramos femoral e genital do N. genitofemoral, respectivamente; S – N. safeno; rcS - Ramo cutâneo do N. safeno; 1 - M. grácil; 2 - M. pectíneo; 3 - M. semimembranoso; 4 - M. adutor; 5 - M. sartório; 6 - M. vasto medial; 7 - M. reto femoral; 8 - Fáscia superficial medial; a - Ramo do N. femoral para o M. sartório. B- Vista lateral da coxa esquerda de M. tridactyla adulto, mostrando: I - N.
isquiático, Gcr - N. glúteo cranial, Gca - N. glúteo caudal, Cfc - N. cutâneo femoral caudal, rm - Ramo muscular do nervo isquiático, Cls - N. cutâneo lateral da sura, T e F - N. tibial e fibular comum, respectivamente (ramos do nervo isquiático), 1 - M. glúteo profundo, 2 - M. piriforme, 3 - M. Glúteo médio, 4 - M. Glúteo superficial, 5 - M. Glúteo femoral, 6 - M. bíceps femoral longo, 7 - M. semitendinoso, 8 - M. semimembranoso, 9 - M. quadrado femoral, 10 - M. quadríceps, 11 - M. tensor da fáscia lata.
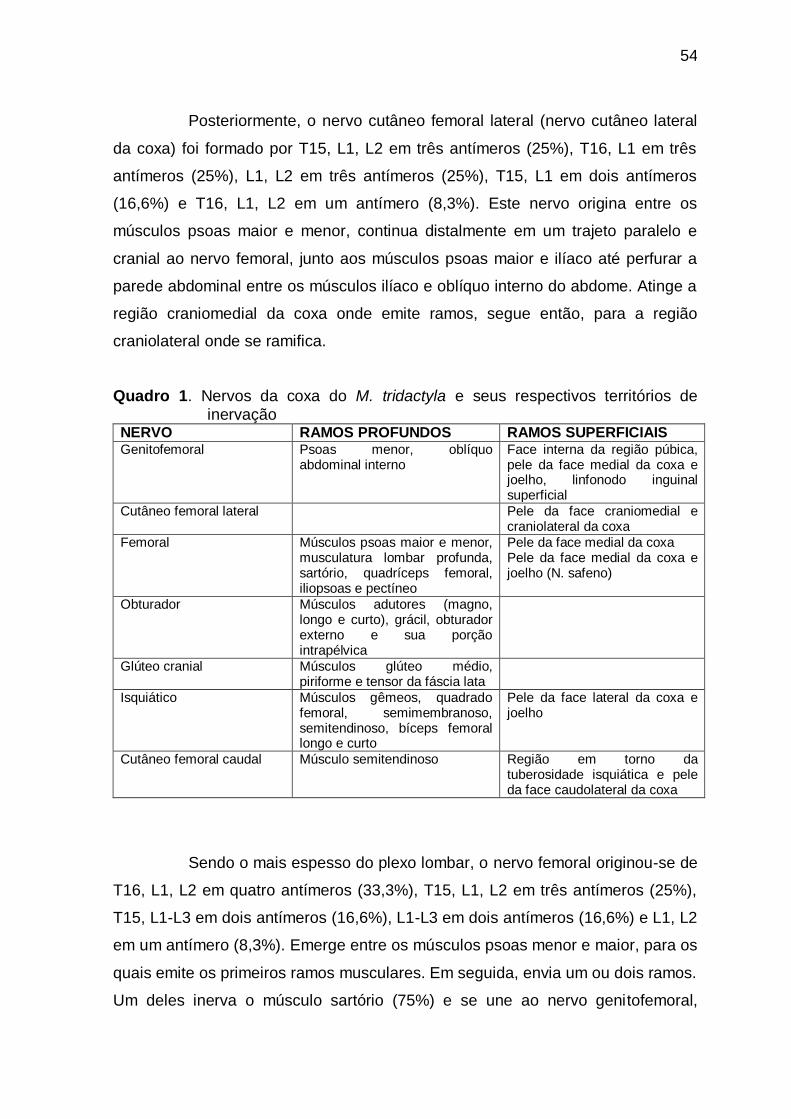
54
Posteriormente, o nervo cutâneo femoral lateral (nervo cutâneo lateral
da coxa) foi formado por T15, L1, L2 em três antímeros (25%), T16, L1 em três
antímeros (25%), L1, L2 em três antímeros (25%), T15, L1 em dois antímeros
(16,6%) e T16, L1, L2 em um antímero (8,3%). Este nervo origina entre os
músculos psoas maior e menor, continua distalmente em um trajeto paralelo e
cranial ao nervo femoral, junto aos músculos psoas maior e ilíaco até perfurar a
parede abdominal entre os músculos ilíaco e oblíquo interno do abdome. Atinge a
região craniomedial da coxa onde emite ramos, segue então, para a região
craniolateral onde se ramifica.
Quadro 1. Nervos da coxa do M. tridactyla e seus respectivos territórios de inervação
NERVO RAMOS PROFUNDOS RAMOS SUPERFICIAIS Genitofemoral Psoas menor, oblíquo
abdominal interno Face interna da região púbica, pele da face medial da coxa e joelho, linfonodo inguinal superficial
Cutâneo femoral lateral Pele da face craniomedial e craniolateral da coxa
Femoral Músculos psoas maior e menor, musculatura lombar profunda, sartório, quadríceps femoral, iliopsoas e pectíneo
Pele da face medial da coxa Pele da face medial da coxa e joelho (N. safeno)
Obturador Músculos adutores (magno, longo e curto), grácil, obturador externo e sua porção intrapélvica
Glúteo cranial Músculos glúteo médio, piriforme e tensor da fáscia lata
Isquiático Músculos gêmeos, quadrado femoral, semimembranoso, semitendinoso, bíceps femoral longo e curto
Pele da face lateral da coxa e joelho
Cutâneo femoral caudal Músculo semitendinoso Região em torno da tuberosidade isquiática e pele da face caudolateral da coxa
Sendo o mais espesso do plexo lombar, o nervo femoral originou-se de
T16, L1, L2 em quatro antímeros (33,3%), T15, L1, L2 em três antímeros (25%),
T15, L1-L3 em dois antímeros (16,6%), L1-L3 em dois antímeros (16,6%) e L1, L2
em um antímero (8,3%). Emerge entre os músculos psoas menor e maior, para os
quais emite os primeiros ramos musculares. Em seguida, envia um ou dois ramos.
Um deles inerva o músculo sartório (75%) e se une ao nervo genitofemoral,

55
seguindo para a região cutânea medial da coxa. O segundo ramo, quando
presente, acompanha o ramo anterior. Em 25% dos antímeros, a inervação do
músculo sartório surgiu de outro ramo oriundo do nervo femoral em seu trajeto no
trígono femoral.
Seguindo distalmente, o nervo femoral atravessa o canal femoral e
segue no trígono femoral, emitindo ramos para o músculo iliopsoas e o nervo
safeno. Posteriormente, segue profundamente aos vasos femorais penetrando
entre o músculo reto femoral e vasto medial. Termina emitindo ramos para todas
as porções do músculo quadríceps femoral.
Sendo bastante espesso no M. trydactyla, o nervo safeno origina-se do
nervo femoral em seu trajeto na face craniomedial do terço distal do músculo
iliopsoas. Em seu trajeto ao longo da coxa, emite um ramo para a inervação
cutânea medial da coxa e joelho. Na extremidade distal da coxa, cruza a
articulação do joelho, próximo à inserção do músculo vasto medial, atingindo a
face medial da perna, onde emite ramos cutâneos craniais e mediais para a
inervação das superfícies cranial e medial da perna, respectivamente.
Dos nervos do plexo lombar, o nervo obturador é o mais caudal, sendo
formado pelos ramos ventrais de L1-L3 em três antímeros (25%), T16, L1, L2, S1
em dois antímeros (16,6%), T16, L1, L2 em dois antímeros (16,6%), T15, L1, L2,
S1 em dois antímeros (16,6%), T15, L1-L3, S1 em dois antímeros (16,6%) e T15,
L1-L3 em um antímero (8,3%). Logo após sua origem, percorre a face medial do
ílio até atingir o forame obturado, porém, antes de atravessá-lo, divide-se em dois
ramos, um cranial e outro caudal.
Seu ramo cranial é o mais espesso e ao atravessar o forame obturado,
emite um pequeno ramo para o músculo adutor curto e penetra entre o músculo
adutor longo e adutor magno, para os quais emite três ou quatro ramos. O
primeiro segue distalmente junto à face medial do músculo adutor magno até o
terço médio da coxa, onde termina penetrando neste músculo; o segundo,
penetra no terço proximal do músculo adutor longo. O terceiro ramo, mais
profundo, penetra entre as fibras do músculo adutor longo, para emergir
distalmente entre este e o músculo grácil. Segue distalmente junto à face medial
do músculo grácil até seu terço médio, onde se aprofunda. O quarto ramo segue

56
cranialmente à origem do músculo adutor longo até atingir a face medial do
músculo grácil, onde termina se ramificando.
Sendo mais delgado, o ramo caudal atravessa o forame obturado e
termina penetrando junto à união das porções do músculo obturador externo.
O nervo glúteo cranial é formado por L2, S1, S2 em cinco antímeros
(41,6%), L3, S1 em quatro antímeros (33,6%), L2, L3, S1 em dois antímeros
(16,6%) e S1, S2 em um antímero (8,3%). Deixa dorsalmente o tronco do plexo
sacral, atravessa junto à incisura isquiática e logo se curva em sentido cranial.
Segue entre o músculo glúteo médio e profundo onde emite dois ramos. Um
destes se distribui no músculo glúteo médio e piriforme e o outro segue em
contato com a face dorsal do músculo glúteo profundo, o inervando e termina
penetrando no músculo tensor da fáscia lata.
O nervo isquiático é o maior do plexo sacral e originou-se de L3, S1-S3
em seis antímeros (50%), L2, S1-S4 em cinco antímeros (41,6%), S1-S4 em um
antímero (8,3%). Foi observada assim a presença de um ramo ventral lombar em
onze antímeros (91,6%), e ausência deste em um antímero (8,3%).
O nervo isquiático se aprofunda para atravessar o forame isquiático.
Segue profundamente ao músculo piriforme e caudal ao glúteo profundo junto ao
ísquio, onde emite o nervo glúteo caudal (41,6%) dos casos, e o nervo cutâneo
femoral caudal (100%) dos casos e ramos para o músculo gêmeos. Continua em
direção à coxa, cruzando caudalmente a articulação coxofemoral e
superficialmente ao tendão de inserção do músculo gêmeos, continuando entre o
músculo gluteofemoral, e quadrado femoral. Neste trajeto emite ramos
musculares bastante desenvolvidos que penetram no terço proximal do músculo
quadrado femoral, músculo este bastante desenvolvido no tamanduá, no terço
proximal do músculo semimembranoso, e no terço médio do músculo
semitendinoso e bíceps femoral longo. Segue distal, cruza profundamente ao
músculo bíceps curto onde emite alguns ramos para este músculo. Torna-se mais
superficial, onde emite ramos cutâneos para a região lateral da coxa e joelho e
musculares para o terço distal do músculo bíceps longo.
Este nervo, termina se dividindo no terço distal da coxa nos nervos
cutâneo lateral da sura, fibular comum e nervo tibial. O nervo cutâneo lateral da
sura após se originar do nervo isquiático segue na fáscia craniolateral da coxa,

57
entre o músculo bíceps femoral longo e o fêmur, se distribuindo em vários ramos
que seguem para a região cutânea craniolateral do joelho e perna. Os demais
ramos seguem distalmente para inervar a perna e pé.
Originando-se diretamente do nervo isquiático caudalmente a origem
do nervo glúteo caudal, observou-se o nervo cutâneo femoral caudal. Segue
caudalmente, e se divide em três ramos. Um deles segue para o músculo
semitendinoso, o segundo ramo segue profundamente aos músculos
glúteofemoral e semitendinoso no sentido da tuberosidade isquiática se
ramificando nos ramos glúteos, suprindo ao redor da tuberosidade isquiática e
região caudal da coxa. O terceiro ramo segue distalmente na fáscia laterocaudal
da coxa cranialmente ao músculo semitendinoso, contribuindo para a inervação
cutânea lateral da coxa.
DISCUSSÃO
Os nervos da coxa do M. tridactyla são o genitofemoral, o cutâneo
femoral lateral, o femoral, o obturador, o glúteo cranial, o isquiático com seu ramo
cutâneo lateral da sura e o cutâneo femoral caudal. Juntos são responsáveis pela
inervação de todas as estruturas musculoesqueléticas, cutâneas e vasculares da
região craniomedial e caudolateral da coxa, coincidindo com os relatos em
animais domésticos (GETTY, 1986) e selvagens (ERDEN, 1993; AYDIN, 2009;
CASTRO et al., 2009).
A divisão do nervo genitofemoral no M. tridactyla em ramos femoral e
genital é descrita também nos animais domésticos (GETTY, 1986; SCHALLER,
1999; KÖNIG; LIEBICH, 2004), mas não há relatos desta característica em
animais selvagens. Não há ramos do nervo genitofemoral destinados a estruturas
do aparelho reprodutor ou glândula mamária no M. tridactyla, uma vez que esta
espécie não apresenta músculo cremáster, escroto e prepúcio, além do fato de
apresentarem somente o par de glândula mamárias torácicas.
O nervo cutâneo femoral lateral (nervo cutâneo lateral da coxa) do M.
tridactyla atravessa a parede abdominal separadamente da artéria e veia
circunflexa profunda do ílio, diferentemente do que ocorre nos animais domésticos

58
(GETTY, 1986; KÖNIG; LIEBICH, 2004). Erden (1993) estudando jumentos,
relatou ainda a inervação dos músculos psoas menor, ilíaco e oblíquo abdominal
interno, o que não se observou neste estudo.
No M. tridactyla, o nervo femoral inervou os músculos psoas maior e
menor, sartório, quadríceps femoral, iliopsoas e pectíneo. A inervação do músculo
quadríceps femoral sucede como nas demais espécies (GETTY, 1986; KÖNING;
LIEBICH, 2004; MARTINEZ-PEREIRA; RICKES, 2011; OLIVEIRA et al., 2011). O
músculo sartório também é suprido por este nervo nos animais domésticos
(SCHALLER, 1999; LORENZ; KORNEGAY, 2006; LIZARDO et al., 2009; SILVA
et al., 2011), chinchilas (MARTINEZ-PEREIRA; RICKES, 2011), mocós
(OLIVEIRA et al., 2011) e humanos (GILROY et al., 2008).
A contribuição do nervo femoral com a inervação do músculo iliopsoas
coincide com as descrições em cães (MILLER, 1964; KÖNING; LIEBICH, 2004),
equinos (GETTY, 1986) e humanos (GILROY et al., 2008), enquanto que a
contribuição com o músculo ilíaco é relatada por Getty (1986) e Lizardo et al.
(2009) no bovino, Silva et al. (2011) em ovinos e Oliveira et al. (2011) em mocó e
para o músculo psoas maior por Getty (1986) e Lizardo et al. (2009) em bovinos,
Silva et al. (2011) em ovinos e Martinez-Pereira; Rickes (2011) em chinchila. O
músculo psoas menor também é inervado pelo N. femoral no lobo-marinho
(CASTRO et al., 2009) e na chinchila (MARTINEZ-PEREIRA; RICKES, 2011).
A inervação do músculo pectíneo pelo nervo femoral também é
relatada por Schaller (1999), Lizardo et al. (2009), Oliveira et al. (2011) e Silva et
al. (2011), no entanto, é comumente inervado pelo nervo obturador (GETTY,
1986; ERDEN, 1993; MIRANDA et al., 2007; AYDIN, 2009; DYCE et al., 2010;
MARTINEZ-PEREIRA; RICKES, 2011) ou mesmo pelos dois nervos (GILROY et
al., 2008).
Além da ampla inervação motora, o nervo femoral contribui com ramos
cutâneos para a face medial da coxa no M. tridactyla, assim como em animais
domésticos (GETTY, 1986; SCHALLER, 1999). Não há, entretanto, descrição
deste fato para outras espécies de animais selvagens.
Uma característica curiosa no M. tridactyla foi a presença de um maior
número de nervos para a face medial da coxa. Em adição, estes nervos são mais
espessos do que usualmente se observa em outras espécies. Enquanto em

59
outros animais, como o porco-espinho (AYDIN, 2009), esta região recebe
inervação somente dos nervos genitofemoral e cutâneo femoral lateral, e no
jumento (ERDEN, 1993), cães (MILLER, 1964; LORENZ; KORNEGAY, 2006),
pelos nervos genitofemoral e safeno, no M. tridactyla, recebe ramos espessos
oriundos dos nervos genitofemoral, cutâneo femoral lateral e femoral. Isto talvez
possa ser justificado pela presença de uma extensa fáscia na face medial da coxa
para a inserção do músculo cutâneo do tronco, que nesta espécie se estende até
o joelho.
O nervo safeno é espesso no M. tridactyla, como descrito por Castro et
al. (2009) no lobo-marinho, todavia não emite ramos musculares, como ocorre em
cães (MILLER, 1964), bovinos (GETTY, 1986; LIZARDO et al., 2009), equinos
(GETTY, 1986) e chinchilas (MARTINEZ-PEREIRA; RICKES, 2011).
A divisão do nervo obturador em ramos cranial está em concordância
com os relatos de Schaller (1999), que descreveu que o nervo emite ramos para
os músculos adutores curto, magno e longo, grácil, obturador externo e sua parte
intrapélvica (músculo obturador interno). Não contribui, entretanto, com ramos
para o músculo pectíneo, como observado em outras espécies (GETTY, 1986;
ERDEN, 1993; MIRANDA et al., 2007; AYDIN, 2009; DYCE et al., 2010;
MARTINEZ-PEREIRA; RICKES, 2011).
No M. tridactyla, o nervo glúteo cranial apresentou a mesma
distribuição muscular do que a comumente descrita em outras espécies (MILLER,
1964; GETTY, 1986; LORENZ; KORNEGAY, 2006; DYCE et al., 2010;
MARTINEZ-PEREIRA; RICKES, 2011).
O nervo cutâneo femoral caudal no M. tridactyla inervou o músculo
semitendinoso, como relatado por König; Liebich (2004) em animais domésticos,
Aydin (2009) no porco-espinho, Castro et al. (2009) no lobo-marinho e Martinez-
Pereira; Rickes (2011) na chinchila. O território de inervação cutânea envolveu a
região em torno da tuberosidade isquiática, como descrito por König; Liebich
(2004), e a face caudolateral da coxa, diferindo do descrito em porco-espinho
(AYDIN, 2009), lobo-marinho (CASTRO et al., 2009) e chinchila (MARTINEZ-
PEREIRA; RICKES, 2011), onde não há ramos para face lateral da coxa, ficando
restrita sua distribuição à sua face caudal.

60
O nervo isquiático inerva, no M. tridactyla, os músculos gêmeos,
quadrado femoral, semimembranoso, semitendinoso, bíceps femoral longo e
curto. Além destes, supre os músculos obturador interno no cão, equino (GETTY,
1986; SCHALLER, 1999; DYCE et al., 2010), lobo-marinho (CASTRO et al., 2009)
e chinchila (MARTINEZ-PEREIRA; RICKES, 2011), adutor no suíno (GETTY,
1986), porco-espinho (AYDIN, 2009), javali (IGLESIAS et al., 2011), mão-pelada
(PEREIRA et al., 2011), pectíneo no suíno (GETTY, 1986) e o tensor da fáscia
lata em javali (IGLESIAS et al., 2011).
Embora a musculatura glútea receba inervação direta do nervo
isquiático em várias espécies (CAMPOS et al., 2003; SANTOS et al., 2006; LIMA
et al., 2008; IGLESIAS et al., 2011; PEREIRA et al., 2011), no M. tridactyla, os
músculos deste grupo foram inervados exclusivamente pelos nervos glúteos
cranial e caudal.
O nervo isquiático do M. tridactyla termina se dividindo no terço distal
da coxa nos nervos cutâneo lateral da sura, fibular comum e tibial. Esta divisão
mais distal difere daquela relatada por Getty (1896), Castro et al. (2009) e Pereira
et al. (2011), que ocorre logo após o nervo atingir a coxa ou até o terço médio da
mesma. Os resultados deste estudo sustentam os de Schaller (1999), König;
Liebich (2004), Aydin (2009) e Dyce et al. (2010) no que se refere à origem do
nervo cutâneo caudal da sura, como ramo do nervo tibial, porém, contrastam com
os relatos dos mesmos autores quanto à origem do nervo cutâneo lateral da sura,
que neste estudo derivou diretamente do nervo isquiático e não do fibular comum.
CONCLUSÕES
Apesar dos nervos da coxa do M. tridactyla serem os mesmos
descritos em animais domésticos, foram identificadas diferenças importantes,
como a maior variação anatômica na formação dos nervos, maior inervação da
face medial da coxa, a divisão mais distal do nervo isquiático e um padrão de
distribuição do nervo genitofemoral peculiar, devido às características do aparelho
reprodutor desta espécie. Na prática, estas diferenças podem inviabilizar a

61
extrapolação de técnicas de procedimentos clínico-cirúrgicos e protocolos
anestésicos adotados em animais domésticos para a espécie em questão.
AGRADECIMENTOS
Ao Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais
Renováveis do estado de Goiás (IBAMA-GO) pela doação dos cadáveres de M.
tridactyla utilizados neste estudo.
ABSTRACT: The lack of basic data about wild species, especially concerning the
topographic anatomy has been a barrier to the practice of clinical and surgical procedures in wild animals. Therefore, the aim of this study was to describe the nerves of the thigh of M. tridactyla, approaching its topography, ramification and territory of innervation. For this purpose, six adult cadavers provided by IBAMA-GO (license 99/2011, UFG-CEUA Protocol 015/11), fixed and preserved in formaldehyde solutions at 10% and 4%, respectively were used. The dissection was initiated after a minimum period of 72 hours from the fixation procedure. The nerves responsible for the innervation the thigh of the M. tridactyla were the genitofemoral, lateral cutaneous femoral, the femoral and its main branch, the saphenous nerve, the obturator, the cranial gluteal, the sciatic and caudal cutaneous femoral nerves. The femoral branch of the genitofemoral nerve supplies the medial cutaneous region of the thigh and superficial inguinal lymph nodes. The lateral cutaneous femoral nerve distributes in the craniomedial and craniolateral cutaneous region of the thigh. The femoral nerve branches the iliacus, psoas major and minor, iliopsoas, pectineus, sartorius and quadriceps muscles and the skin of the medial surface of the thigh. The saphenous nerve innervates the medial aspect of the thigh and stifle joint. The obturator nerve sends branches to external obturator muscle, including its intrapelvic portion, gracilis and adductor magnus, long and short muscles. The cranial gluteal nerve innervates the tensor fascia lata muscle. The sciatic nerve distributes in gemelli, quadratus femoris, semimembranosus, semitendinosus, long and short biceps femoris muscles and in skin of the lateral surface of the thigh and stifle joint. The caudal cutaneous femoral nerve innervates the semitendinosus muscle, the region of the ischial tuberosity and caudolateral surface and the thigh. There are considerable differences between the territory of innervation of the nerves of the thigh of the M. tridactyla compared to domestic animals, which in turn makes unfeasible the extrapolation of clinical-surgical procedures and anesthetic protocols used in domestic animals to the giant anteater.
KEYWORDS: Innervation, Lumbosacral plexus. Peripheral nervous system.
Xenarthra.

62
REFERÊNCIAS
AYDIN, A. The dissemination of the pelvic limb nerves originating from the lumbosacral plexus in the porcupine (Hystrix cristata). Veterinární Medicína,
Praha, v. 54, n. 7, p. 333-339, 2009. CAMPOS, D. B.; SILVA, F. O. C.; SEVERINO, R. S.; DRUMMOND, S. S.; LIMA, E. M. M.; BOMBONATO, P. P.; SANTANA, M. I. S. Origem e distribuição dos nervos isquiáticos em fetos de bovinos azebuados. Ars Veterinaria, Jaboticabal, v. 19, n. 3, p. 219-223, 2003. CASTRO, T. F.; SOUZA, D. A. S.; SILVA FILHO, R. P.; PEREIRA, M. A. M. Sistematização e distribuição da inervação lombar e sacral em Arctocephalus australis. Brazilian Journal of Veterinary Research and Animal, São Paulo, v.
46, n. 5, p. 404-411, 2009. DYCE, K. M.; SACK, W. O.; WENSING, C. J. G. Tratado de anatomia veterinária, 4. ed. Rio de Janeiro: Elsevier, 2010, 834p.
ERDEN, H. Merkebin (equus asinus l.) plexus lumbosacralis'I üzerinde makroanatomik ve subgros çalişmalar. Uludağ Üniversitese Veteriner Fakültesi Dergisi, Konya, v. 9, n. 1, p. 64-71, 1993.
FORTERRE, F.; TOMEK, A.; RYTZ, U.; BRUNNBERG, L.; JAGGY, A.; SPRENG, D. Iatrogenic sciatic nerve injury in eighteen dogs and nine cats (1997-2006). Veterinary Surgery, v. 36, p. 464-471, 2007.
GETTY, R. Anatomia dos animais domésticos. 5. ed. Rio de Janeiro:
Guanabara Koogan, 1986. 2000p. v. 1 e 2. GILROY, A. M.; MacPHERSON, B. R.; ROSS, L. M. Vascularização e inervação. In: ___Atlas de Anatomia. 1. ed. Rio de Janeiro: Guanabara Koogan, 2008. p.
420-448. IGLESIAS, L. P.; SILVA, F. O. C; BRITO, T. R. Origem e distribuição do nervo isquiático em fetos de javalis (Sus scrofa scrofa). Biotemas, Florianópolis, v. 24,
n. 4, p.141-145, 2011. INTERNATIONAL COMMITTEE ON VETERINARY GROSS ANATOMICAL NOMENCLATURE. Nomina anatomica veterinaria. 5. ed. Ithaca: Cornell
University, 2012, 160p. IUCN 2012. IUCN Red List of Threatened Species. Version 2012.1. <www.iucnredlist.org>. Acessado em 26 agosto 2012. KÖNIG, H. E.; LIEBICH, H. J. Sistema nervoso. In: ___Anatomia dos animais domésticos. 1. ed. Porto Alegre: Artmed, 2004. cap.14, p. 203-275. v.2.

63
LIMA, E. M. M.; SILVA, F. O. C.; SEVERINO, R. S.; DRUMMOND, S. S.; CAMPOS, D. B.; SANTANA, M. I. S.; MORAES, D. D. A. Origem e distribuição dos nervos isquiáticos em caprinos da raça Saanen. Ciência Rural, Santa Maria,
v. 38, n. 2, p. 372-377, 2008. LIZARDO, F. B.; SILVA, F. O. C.; SEVERINO, R. S.; GUIMARÃES, E. C.; SANTOS, L. A.; EULÁLIO, F. H. F.; SOUSA, G. C.; FACURY NETO, M. A.; BERNARDINO JÚNIOR, R.; CABRAL, L. G. Origin and distribution of the femoral nerve in fetuses of zebu-crossed bovines. Brazilian Journal of Morphological Sciences, São Paulo, v. 26, n. 2, p. 91-96, 2009. LORENZ, M. D.; KORNEGAY, J. N. Localização das lesões no sistema nervoso. In: ___Neurologia Veterinária. 2. ed. Barueri: Manole, 2006. cap. 2, p. 45-66.
MARTINEZ-PEREIRA, M. A.; RICKES, E. M. The spinal nerves that constitute the lumbosacral plexus and their distribution in the chinchilla. Journal of the South African Veterinary Association, Pretoria, v. 82, n. 3, p. 150-154, 2011.
MILLER, M. E. Anatomy of the dog. Philadelphia: W. B. Saunders, 1964, 941p.
MIRANDA, F.; COSTA, A. M. Xenarthra (Tamanduá, Tatu, Preguiça). In: CUBAS, Z. S.; SILVA, J. C. R.; CATÃO-DIAS, J. L. Tratado de animais selvagens – medicina veterinária. 1. ed. Roca: São Paulo, 2007. cap. 26, p. 402- 414.
MIRANDA, R. L.; CARNEIRO E SILVA, F. O.; SEVERINO, R. S.; DRUMMOND, S. S.; GONÇALVES, R. C. Origens e distribuições dos nervos obturatórios em fetos fêmeas de bovinos azebuados. Bioscience Journal, Uberlândia, v. 23, n. 4, p.
120-127, 2007. MOURÃO, G.; MEDRI, M. Activity of a specialized insectivorous mammal (Myrmecophaga tridactyla) in the Pantanal of Brazil. Journal of Zoology, London,
v. 271, p. 187–192, 2007. OLIVEIRA, G. B.; ALBUQUERQUE, J. F. G.; RODRIGUES, M. N.; PAIVA, A. L. C.; MOURA, C. E. B.; MIGLINO, M. A.; OLIVEIRA, M. F. Origem e distribuição do nervo femoral em mocó, Kerodon rupestris (Cavidae). Pesquisa Veterinária Brasileira, Rio de Janeiro, v. 31, n. 1, p. 84-88, 2011.
PEREIRA, K. F.; PARANAIBA, J. F. F. S.; HELRIGLE, C.; ARAÚJO, E. G. Origem e distribuição anatômica do nervo isquiático de mão-pelada. Pesquisa Veterinária Brasileira, Rio de Janeiro, v. 31, n. 1, p. 74-78, 2011.
RODRIGUES, H. Técnicas anatômicas. 4. ed. Vitória: GM. Gráfica e Editora,
2010, 270p. SANTOS, R. C.; ALBUQUERQUE, J. F. G.; SILVA, M. C. V.; MOURA, C. E. B, CHAGAS, R. S. N; BARBOSA, R. R.; MIGLINO, M. A. Anatomia do nervo isquiático em mocós (Kerodon rupestris WIED, 1820) aplicada à clínica de

64
animais silvestres. Brazilian Journal of Veterinary Research and Animal, São
Paulo, v. 43, n. 43, p. 647-653, 2006. SCHALLER, O. Sistema nervoso. In: ___Nomenclatura anatômica veterinária ilustrada. 1. ed. São Paulo: Manole, 1999. p. 414-502.
SILVA, F. O. C.; BRITO, T. R.; VASCONCELOS, B. G.; CANABRAVA, H. A. N.; PEREIRA, C. C. H.; NOLASCO, R. M.; HONORATO, A. G. O. Origens e distribuições dos nervos femorais em ovinos sem raça definida. Bioscience Journal, Uberlândia, v. 27, n. 6, p. 978-981, 2011.

65
CAPÍTULO 4 - ANATOMICAL ASPECTS OF THE NERVES OF THE LEG AND
FOOT OF THE GIANT ANTEATER (Myrmecophaga tridactyla, Linnaeus, 1758)
ASPECTOS ANATÔMICOS DOS NERVOS DA PERNA E PÉ DO TAMANDUÁ-
BANDEIRA (Myrmecophaga tridactyla, Linnaeus, 1758)
V.S. CruzI*; J.R. CardosoII; L.B.M. AraújoIII; P.R. SouzaII ; M.S.B. SilvaII; E.G.
AraújoIII
I*Departamento de Morfologia - Instituto de Ciências Biológicas - Universidade Federal de Goiás – Goiânia, GO e Doutoranda no Programa de Pós-Graduação em Ciência Animal, Escola de Veterinária e Zootecnia - Universidade Federal de Goiás - Goiânia, GO. *E-mail: [email protected] IIDepartamento de Morfologia - Instituto de Ciências Biológicas - Universidade Federal de Goiás - Goiânia, GO. E-mail: [email protected], [email protected], [email protected]. IIIDepartamento de Medicina Veterinária - Escola de Veterinária e Zootecnia - Universidade Federal de Goiás - Goiânia, GO. E-mail: [email protected], [email protected].
Submetido na Revista Arquivo Brasileiro de Medicina Veterinária e Zootecnia
(ABMVZ) em 06 de fevereiro de 2013
Situação: Aguardando avaliação do Comitê Editorial

66
ANATOMICAL ASPECTS OF THE NERVES OF THE LEG AND FOOT OF THE
GIANT ANTEATER (Myrmecophaga tridactyla, Linnaeus, 1758)
ASPECTOS ANATÔMICOS DOS NERVOS DA PERNA E PÉ DO TAMANDUÁ-
BANDEIRA (Myrmecophaga tridactyla, Linnaeus, 1758)
ABSTRACT
Although nerve distribution distal to the stifle joint has been well established in domestic animals, this approach is rare in wild animals. Therefore, the aim of this study was to describe the nerves of the leg and foot of M. tridactyla with emphasis on its ramification, distribution, topography and territory of innervation. For this purpose, six adult cadavers provided by IBAMA-GO (license 99/2011, UFG-CEUA Protocol 015/11) were used. They were fixed and preserved in formaldehyde solution at 10% and 4%, respectively. The nerves of the leg and
foot of the M. tridactyla were saphenous nerve (branch of the femoral nerve), fibular and tibial nerves and lateral sural cutaneous nerve (branches of the sciatic nerve) and caudal sural cutaneous nerve (branch of the tibial nerve). The saphenous nerve sends branches to the skin, craniomedial surface of the leg, medial surface of the tarsal and metatarsal regions and dorsomedial surface of the foot and dorso of the digits I and II (100%), III (50%) and IV (25%). The lateral sural cutaneous nerve innervates the skin of the craniolateral region of the knee and leg. The fibular nerve innervates the flexor and extensor muscles of the tarsal region of the digits and skin of the craniolateral surface of the leg and dorsolateral surface of the foot. The tibial nerve innervates the extensor muscles of the tarsal joint and flexor, adductor and abductor muscles of the digits and the skin of the plantar surface. The caudal sural cutaneous nerve innervates the skin of the caudal surface of the leg. It can be concluded that the nerves responsible for the leg and foot innervation were the same as reported in domestic and wild animals, but with some differences, such as the more distal division of the common fibular nerve, absence of dorsal metatarsal branches of the deep fibular nerve and a greater involvement of the saphenous nerve in the digital innervation with branches to the digits III and IV, in addition to digits I and II.
Keywords: Extremities, foot, peripheral nervous system, Xenarthra

67
RESUMO
Apesar de bem estabelecida nos animais domésticos, a abordagem da distribuição nervosa distal ao joelho é rara em animais selvagens. Portanto, o objetivo deste estudo foi descrever os nervos da perna e pé do M. tridactyla, com ênfase na sua ramificação, distribuição, topografia e território de inervação. Para tanto, foram utilizados seis cadáveres adultos fornecidos pelo IBAMA-GO (licença 99/2011, CEUA-UFG processo nº 015/11). Eles foram, fixados e conservados em solução de formaldeído a 10% e 4%, respectivamente. A dissecação envolveu desde a formação dos nervos femoral e isquiático pelos ramos ventrais dos nervos espinhais lombares e sacrais até sua distribuição nos territórios propostos. Os nervos responsáveis pela inervação da perna e pé do M. tridactyla foram o N. safeno (ramo do N. femoral), os nervos fibular comum e tibial e o N. cutâneo lateral da sura (derivados do N. isquiático) e o N. cutâneo caudal da sura (ramo do N. tibial). O nervo safeno emite ramos cutâneos para a superfície craniomedial da perna, medial do tarso e metatarso, dorsomedial do pé e dorso dos dedos I e II (100%), III (50%) e IV (25%). O nervo cutâneo lateral da sura inerva a região cutânea craniolateral do joelho e perna. O nervo fibular inerva os músculos flexores do tarso e extensores dos dedos e a região cutânea craniolateral da perna e dorsolateral do pé. O nervo tibial inerva os músculos extensores do tarso e flexores, adutores e abdutores dos dedos e região cutânea plantar. O nervo cutâneo caudal da sura inerva a pele da face caudal da perna. Pode-se concluir que os nervos responsáveis pela inervação da perna e pé foram os mesmos relatados em animais domésticos e selvagens, porém com algumas diferenças, como a divisão mais distal do nervo fibular comum, ausência de ramos metatarsianos dorsais do N. fibular profundo e uma maior participação do nervo safeno na inervação digital, contribuindo com ramos inclusive para os dedos III e IV, além dos dedos I e II.
Palavras-chave: membros, pé, sistema nervoso periférico, xenartros
INTRODUCTION
The Myrmecophaga tridactyla is the largest mammal in the Xenarthra
Superorder, which has recently been subdivided into the Cingulata (represented
by the armadillo) and Pilosa (represented by the anteater and the sloth) orders
(Vizcaíno and Loughry, 2008). Its closest relatives who belong to the same family
are the Tamandua tetradactyla (lesser anteater) and the Cyclopes didactylus (silky
anteater), both arboreal and smaller (Shaw et al., 1987). They inhabit the fields
and savannas of the south of Belize and Guatemala up to the north of Argentina

68
including all of the Brazilian territory (Mourão and Medri, 2007). They have laterally
flattened bodies, fairly short limbs and plantigrade locomotion. The hands have
long nails, which bend inwards whilst it is in motion (Shaw et al., 1987).
The number of anteaters that have been run over has increased due to
the growing number of roads running through its habitat and their necessity to
move around searching for food (Braga, 2009). Additionally, the giant anteater is
the large mammal most affected by bushfires (Silveira et al., 1999). For these and
other reasons this species is described as “vulnerable” (VU) in the International
Union for Conservation of the Nature and Resources (IUCN, 2012) list.
The femoral, saphenous, sciatic, common fibular, tibial and lateral
cutaneous and caudal cutaneous sural nerves are responsible for the sensitive
and motor innervation of the leg and foot of domestic (Getty, 1986; Schaller, 1999;
Dyce et al., 2010) and wild animals (Aydin, 2009; Castro et al., 2009). Injuries to
these nerves cause a variety of deficiencies, such as loss of sensitivity or
muscular movement, or both, in a restricted area supplied by the nerve or nerves
involved (Lorenz and Kornegay, 2006).
There is a lack of data related to distal innervation of the locomotive
limbs in wild animals and to date there are no records of studies of this nature in
the pelvic limbs of any species in the Xenarthra superorder. Thus, this study
presents unpublished and important information in many areas, especially in
comparative anatomy, clinical practice, anaesthesiology and surgery of wild
animals.
Therefore, the objective of this study was to present a description of the
nerves of the leg and foot of the M. tridactyla, with emphasis on its ramification,
distribution, topography and territory of innervation.
MATERIAL AND METHODS
Twelve pelvic limbs of six adult giant anteaters (three females, two
males and one undefined due to removal of genital organs in necropsy) were
studied. The cadavers were provided by IBAMA - Brazilian Institute of
Environment and Renewable Natural Resources) (license 99/2011) and the
experimental protocol has been approved by the Ethics Committee for Animal Use

69
of the Federal University of Goiás - Brazil (Process CEUA-UFG no. 015/11). The
preparation, lodgement and dissection of the specimens occurred in the
Laboratory of Animal Anatomy at the Institute of Biological Sciences (ICB) at the
Federal University of Goiás (UFG).
After thawing, the specimens were fixed and preserved in 10% aqueous
solution of formaldehyde (Rodrigues, 2010). The access to the abdominal cavity
was performed by an incision trough the linea alba and removal of part of the
pubis and pelvic symphysis. In order to dissect the ventral face of the lumbar and
sacral regions, the colon, rectum, genitourinary organs, lumbar and sacral blood
vessels, parietal peritoneum and pelvic fascia were removed. Posteriorly, the
ventral rami of the spinal nerves which form the sciatic and femoral nerves were
dissected, and the pelvic limb skin was removed to allow exposure up to the
branches that are responsible for the innervation of the leg and foot regions. All
dissection stages were documented by drawings and digital images taken with a
Canon digital camera (Sony DSC-H50)® and the terms were used in agreement
with the Nomina Anatomica Veterinaria (International Committee on Veterinary
Gross Anatomical Nomenclature, 2012).
Descriptive statistics was used for data analysis by means of frequency
percentage as studies Campos et al. (2003); Santos et al. (2006); Lizardo et al.
(2009); Iglesias et al. (2011); Oliveira et al. (2011) e Pereira et al. (2011).
RESULTS
The radiographic exams showed that the M. tridactyla specimens used
in this study had 15 (66.6%) or 16 (33.3%) thoracic vertebras, two (50%) or three
(50%) lumbar and four (16.6%) or five (83.3%) sacral vertebras. The difference in
the number of vertebras consequently leads to a variation in the nerve composition
of the leg and foot.
The nerves which supply the leg and foot of the M. tridactyla were the
saphenous (S) (branch of the femoral nerve), common fibular (F), tibial (T) and
lateral sural cutaneous nerves (LSC) (derived from the sciatic nerve) and the
caudal sural cutaneous nerve (CSC) (branch of the tibial nerve).
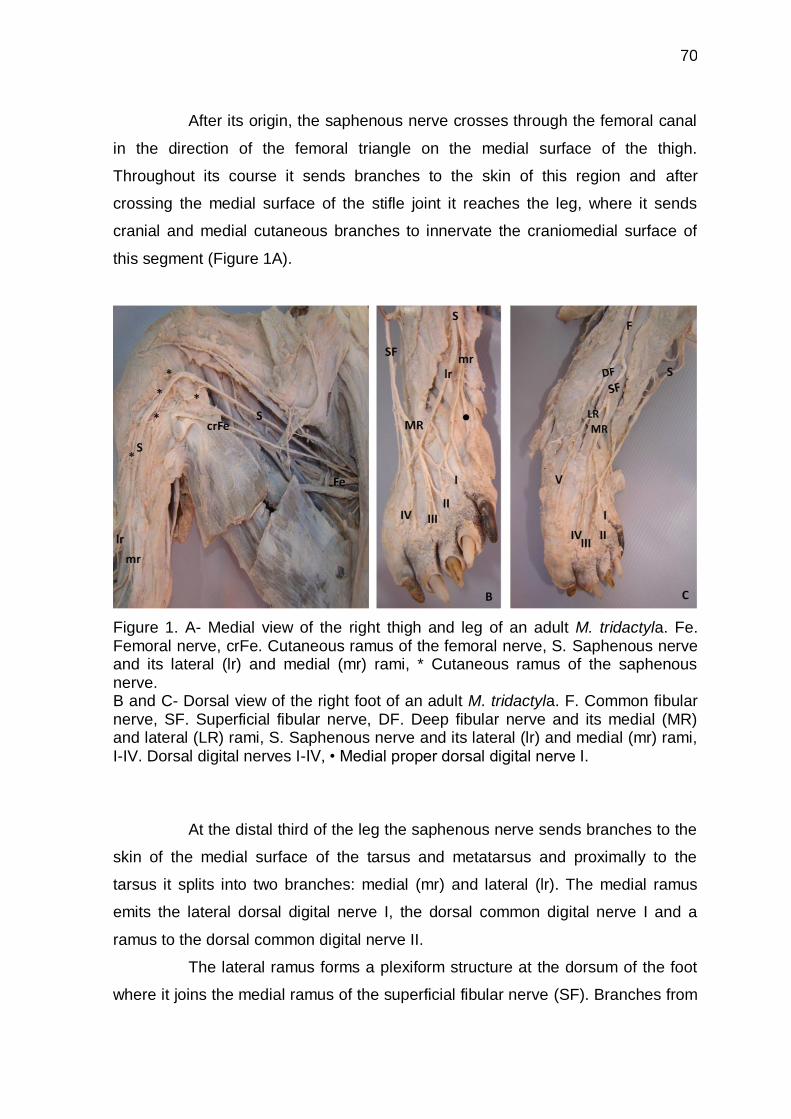
70
After its origin, the saphenous nerve crosses through the femoral canal
in the direction of the femoral triangle on the medial surface of the thigh.
Throughout its course it sends branches to the skin of this region and after
crossing the medial surface of the stifle joint it reaches the leg, where it sends
cranial and medial cutaneous branches to innervate the craniomedial surface of
this segment (Figure 1A).
Figure 1. A- Medial view of the right thigh and leg of an adult M. tridactyla. Fe. Femoral nerve, crFe. Cutaneous ramus of the femoral nerve, S. Saphenous nerve and its lateral (lr) and medial (mr) rami, * Cutaneous ramus of the saphenous nerve. B and C- Dorsal view of the right foot of an adult M. tridactyla. F. Common fibular nerve, SF. Superficial fibular nerve, DF. Deep fibular nerve and its medial (MR) and lateral (LR) rami, S. Saphenous nerve and its lateral (lr) and medial (mr) rami, I-IV. Dorsal digital nerves I-IV, • Medial proper dorsal digital nerve I.
At the distal third of the leg the saphenous nerve sends branches to the
skin of the medial surface of the tarsus and metatarsus and proximally to the
tarsus it splits into two branches: medial (mr) and lateral (lr). The medial ramus
emits the lateral dorsal digital nerve I, the dorsal common digital nerve I and a
ramus to the dorsal common digital nerve II.
The lateral ramus forms a plexiform structure at the dorsum of the foot
where it joins the medial ramus of the superficial fibular nerve (SF). Branches from

71
this plexus reinforce the common dorsal digital nerves I and II in both antimeres.
However, in six antimeres (50%) it sends a ramus to the dorsal common digital
nerve III (ramus of the superficial fibular nerve) and in three antimeres (25%) it
sends a ramus to the dorsal common digital nerve IV (Figure 1B and Table 1).
Table 1. Digital nerves of the foot of M. tridactyla
Digit Surface
Common fibular n. Tibial n. Saphenous n.
SF PF MP LP MR LR
MR LR
I Dorsal X X Plantar X
II Dorsal X X X Plantar X
III Dorsal X X (50%) Plantar X
IV Dorsal X X X (25%) Plantar X
V Dorsal X Plantar X
SF. Superficial fibular nerve, DF. Deep fibular nerve, MP. Medial plantar nerve, LP. Lateral plantar nerve, MR. Medial ramus, LR. Lateral ramus.
The sciatic nerve (I) is the largest nerve in the sacral plexus of the M.
tridactyla and is formed by the last lumbar segment and the three or four first
sacral segments. After its origin, it crosses the greater sciatic foramen in distal
direction and ends up splitting into the lateral sural cutaneous, common fibular and
tibial nerves at the distal third of the thigh (Figure 2).
After arising from the sciatic nerve the lateral sural cutaneous nerve
follows the craniolateral fascia of the thigh between the femoral biceps and the
femur and splits into many rami which follow up to the craniolateral cutaneous
region of the stifle joint and leg (Figure 2).
Upon leaving the sciatic nerve at the distal third of the thigh, the
common fibular nerve runs downward across the craniolateral surface of the thigh
where it becomes related to the lateral sural cutaneous nerve. Next, it crosses
between the distal extremity of the femoral biceps and the proximal extremity of
the lateral head of the gastrocnemius, emerging at the proximal third of the lateral
surface of the leg, distally to the condyle of the fibula. It then dips between the long

72
digital extensor and long fibular muscles. Throughout its course, it supplies the
long and short fibular muscles, the cranial tibial and the long digital extensor
muscles.
Figure 2. Lateral view of the distal extremity of thigh and proximal extremity of the right leg of M. tridactyla. T. Tibial nerve, F. Common fibular nerve, LSC. Lateral sural cutaneous nerve, CSC. Caudal sural cutaneous nerve, 1. M. biceps femoris longus, 2. M. biceps femoris brevis.
The common fibular nerve splits into the superficial (SF) and deep
fibular nerves (DF) at the level of the middle third of the leg (Figure 1C). The
superficial fibular nerve runs distally in contact with the lateral surface of the long
digital extensor muscle and in 50% of the cases it splits into medial (MR) and
lateral rami (LR) at the craniolateral surface of the tibiotarsal articulation. The other
50% split more distally at the proximal region of the metatarsus, where two rami
are emitted.
These rami establish connections in various ways to innervate the
dorsal surface of the foot, making it difficult to establish a standard for the species.
In general, the lateral ramus gives off the lateral proper dorsal digital nerve V and

73
occasionally the dorsal common digital nerve IV. The medial ramus gives off
cutaneous rami to the dorsum of the tarsus and metatarsus and also branches
which join with rami of the saphenous nerve to form the dorsal common digital
nerves II, III and IV (Figure 1C and Table 1).
The deep fibular nerve runs cranially up to the tibia and the tibiotarsal
joint continuing deeply to the belly of the extensor digiti I longus, to which it sends
branches and ends up dividing in the short digital extensor muscles.
The tibial nerve is the main continuation of the sciatic nerve in its course
through the distal third of the thigh. In the deep surface of the biceps femoris
brevis muscle, it gives off the caudal sural cutaneous nerve (Figure 2), responsible
for the cutaneous innervation of the caudal surface of the leg.
After sending the caudal sural cutaneous nerve, the tibial nerve extends
distally between the medial and lateral heads of the gastrocnemius muscle
reaching the caudomedial region of the leg. On its course, it sends branches to the
gastrocnemius, plantar, soleus, long digital flexor, caudal tibial, flexor digiti I longus
and popliteus muscles.
After innervating the caudal musculature of the leg, the tibial nerve
travels distally between the medial head of the gastrocnemius and the soleus
muscles to the middle third of the leg, where it then deviates medially and
superficially when it crosses the distal surface of the muscular belly of the medial
head of the gastrocnemius. In this route, it runs alongside with the rete mirabile of
the medial plantar artery and at the middle third of the leg it sends two branches to
the skin of the caudal surface of the leg.
At the distal third of the leg it splits into medial and lateral plantar
nerves, which are responsible for the innervation of the flexor digitorum brevis,
quadratus plantae, flexor digiti V, abductor digiti V, abductor brevis digiti I,
abductor metatarsal V, adductor brevis digiti I, adductor digiti II, flexor brevis digiti
I, interossei and lumbricales muscles.
On the plantar surface of the tarsometatarsal joint, the medial plantar
nerve splits into three branches, which give off muscular branches and continue as
common digital plantar nerves of the digits I, II and III (Figure 3 and Table 1).

74
Figure 3. Plantar surface of the left foot of M. tridactyla. T. Tibial nerve, LP. Lateral plantar nerve, MP. Medial plantar nerve (Branches of the tibial nerve), I-IV. Common plantar digital nerves, V. lateral proper dorsal digital nerve V, 1. M. flexor brevis digiti, a. Tendon of the M. flexor pollicis brevis, b, c and d. Tendons of the M. flexor brevis digitorum, e. Tendon of the M. flexor digiti quinti brevis.
The lateral plantar nerve divides at the same level as the medial plantar
nerve into the common digital plantar nerve IV and the proper plantar nerve V
(Figure 3 and Table 1). Each digital nerve splits into the proper digital nerves at
the level of the distal interphalangeal articulation.
DISCUSSION
There are many studies related to the origin and distribution of the
sciatic and femoral nerves (Aydin, 2009; Castro et al., 2009; Lizardo et al., 2009;
Martinez-Pereira and Rickes, 2011; Pereira et al., 2011) however, the anatomy of
the rami which branch from these nerves and that reach the leg remain unknown
in most of these species, since the authors rarely address the distribution of
nerves distally to the knee. Possibly due to the difficulty in the dissection process,
the cutaneous rami of the plantar and dorsal nerves of the foot are even less

75
addressed, having been described in full only in the porcupine (Aydin, 2009) and
partially in the sea lion (Castro et al., 2009) and the chinchilla (Martinez-Pereira
and Rickes, 2011).
The nerves of the lumbosacral plexus responsible for the innervation of
the foot and leg of the M. tridactyla were the saphenous, common fibular, tibial,
caudal cutaneous sural and lateral cutaneous sural nerves, which coincides with
that reported in domestic animals (Getty, 1986; Schaller, 1999; Köning and Liebich
2004; Lorenz and Kornegay, 2006; Dyce et al., 2010) and wild animals (Erden,
1993; Aydin, 2009; Castro et al., 2009).
The distribution of the saphenous nerve on the cranial and medial
surface of the leg of the M. tridactyla was similar to that described in domestic
animals (Getty, 1986; Lizardo et al., 2009; Dyce et al., 2010). Among the wild
animals, its distribution was addressed in the sea lion (Castro et al., 2009), in
which the territory of innervation was restricted to the sural caudolateral region,
which differs from that observed in this study.
A plexiform structure was formed by the saphenous and fibular nerves
in the foot of the M. tridactyla, which is a feature also described in dogs and cats
by Getty (1986). However, the author does not specify which digits the rami of this
plexus lead to. In pigs, the same author reports that the saphenous nerve splits
into lateral and medial rami at the distal third of the leg as also presented in this
study. The lateral ramus joins the medial ramus of the superficial fibular nerve to
form the dorsal common digital nerve II, while the medial ramus descends as the
dorsal medial digital nerve II. In the M. tridactyla, apart from the digit II, the digits III
and IV also receive branches from the saphenous nerve, which is not reported as
a feature in any other species.
The sciatic nerve in the M. tridactyla ends by dividing into the lateral
sural cutaneous nerve, common fibular and tibial nerves. This division is more
distal than that observed in domestic animals (Getty, 1986), in the sea lion (Castro
et al., 2009) and in the crab-eating raccoon (Pereira et al., 2011), in which the
ramification usually occurs after the sciatic nerve reaches the thigh or even in its
middle third.
The origin of the caudal sural cutaneous nerve varies among different
species. In the M. tridactyla, as in the porcupine (Aydin, 2009) and in the chinchilla

76
(Martinez-Pereira and Rickes, 2011), it arises from the tibial nerve. In dogs and
bovines (Getty, 1986), sea lions (Castro et al., 2009) and in the chinchilla
(Martinez-Pereira and Rickes, 2011), it arises directly from the sciatic nerve.
The same occurs with the lateral sural cutaneous nerves origin. It
branches directly from the sciatic nerve in dogs (Getty, 1986) and in the chinchilla
(Martinez-Pereira and Rickes, 2011), as also observed in this study. On the other
hand, Aydin (2009) in the porcupine, Schaller (1999), König and Liebich (2004)
and Dyce et al. (2010) in domestic animals describe it as a branch of the common
fibular nerve.
The division of the common fibular nerve into superficial and deep
fibular nerves occurs as soon as the common fibular nerve emerges in the leg
(Getty, 1986; Schaller, 1999; König and Liebich, 2004; Castro et al., 2009),
however, in the M. tridactyla this division takes place at the level of the middle third
of the leg. As a consequence, the craniolateral muscles of the leg of the M.
tridactyla are innervated by branches directly from the common fibular nerve, not
by branches from the superficial and deep fibular nerves as in other species.
The muscular rami that branch directly off the common fibular nerve in
the M. tridactyla were addressed to the long fibular, short fibular, cranial tibial and
long digital extensor muscles, which are all innervated by the deep fibular nerve in
domestic animals (Getty, 1986). Therefore, the deep fibular nerve in the M.
tridactyla only innervates two muscles: the extensor hallucis longus and the
extensor digitorum brevis.
The lateral digital extensor muscle and the fifth fibular muscle are
mentioned by Schaller (1999) in domestic animals and Castro et al. (2009) in sea
lion, respectively as innervated by the superficial fibular nerve, but they are absent
in the M. tridactyla.
Apart from the motor innervation of the tarsal flexor and digital extensor
muscles, Miller (1964) and Dyce et al. (2010) also mention the sensitive
innervation of the foot structures, which is intermediated by the union of dorsal
metatarsal structures derived from the deep fibular nerve with dorsal digital rami,
which in turn are derived from the superficial fibular nerve. This does not occur in
the M. tridactyla, since the deep fibular nerve does not send dorsal metatarsal
branches.

77
In the M. tridactyla, the tibial nerve innervated the gastrocnemius,
plantar, soleus, long flexor of the digits, caudal tibial, flexor hallucis longus and
popliteus muscles, which is in accordance with that described in humans (Gilroy et
al. 2008). This is possibly due to the muscular similarity between these two
species. The division of the tibial nerve into medial and lateral plantar nerves
corroborates with the standard described in the other species (Miller, 1964; Getty,
1986; König and Liebich, 2004; Gilroy et al., 2008; Dyce et al., 2010). However,
the formation of the digital nerves is directly correlated to the number of digits in
such a manner that the distal distribution of this nerve was closer to that described
in animals with a larger number of digits, such as carnivores. Thus, the
ramification of the medial and the lateral plantar nerves for the formation of the
common digital nerves was in accordance to what Schaller (1999) described in
swine. Miller (1964) and Schaller (1999), though, described the plantar digital
nerves in dogs as derived from the lateral plantar nerve since the two join to form
the common digital nerve. This was not observed in the M. tridactyla.
It can be concluded that the nerves responsible for the innervation of
the leg and foot were the same as those reported in domestic and wild animals but
with a few differences, such as the more distal division of the common fibular
nerve, which results in significant differences in the innervation of the leg muscles
in relation to the standard described in domestic animals. Also, the absence of
dorsal metatarsal branches of the deep fibular nerve to innervate the dorsum of
the foot and a more effective participation of the saphenous nerve in the digital
innervation, contributing also with branches to the digits III and IV, apart from the
digits I and II.
ACKNOWLEDGEMENTS
The authors would like to thank IBAMA - Brazilian Institute of
Environment and Renewable Natural Resources for donating the M. tridactyla
specimens used in this study.

78
REFERENCES
AYDIN, A. The dissemination of the pelvic limb nerves originating from the lumbosacral plexus in the porcupine (Hystrix cristata). Vet. Med-Cezech, v.54, n.7, p.333-339, 2009. BRAGA, F.G. Plano de conservação para tamanduá-bandeira (Myrmecophaga tridactyla). In: Instituto Ambiental do Paraná, Planos de ação para Espécies de mamíferos ameaçados. IAP: Curitiba, p.14-30, 2009. CASTRO, T.F.; SOUZA, D.A.S.; SILVA FILHO, R.P.; PEREIRA, M.A.M. Sistematização e distribuição da inervação lombar e sacral em Arctocephalus australis. Braz. J. Vet. Res. Anim. Sci., v.46, n.5, p.404-411, 2009. DYCE, K.M.; SACK, W.O.; WENSING, C.J.G. Tratado de anatomia veterinária, 4a
ed. Rio de Janeiro: Elsevier, 2010. 834p. ERDEN, H. Merkebin (equus asinus l.) plexus lumbosacralis'I üzerinde makroanatomik ve subgros çalişmalar. S. Ü. Vet. Fak. Derg., v.9, n.1, p.64-71, 1993. GETTY, R. Anatomia dos animais domésticos. 5a ed. Rio de Janeiro: Guanabara Koogan, 1986. 2000p. v. 1 e 2. GILROY, A.M.; MACPHERSON, B.R.; ROSS L.M. Atlas de Anatomia. 1a ed. Rio de Janeiro: Guanabara Koogan, 2008. 656p. INTERNATIONAL COMMITTEE ON VETERINARY GROSS ANATOMICAL NOMENCLATURE. Nomina anatomica veterinaria. 5a ed. Ithaca: Cornell University, 2012. 160p. IUCN, Red List of Threatened Species. Version 2012.1. Disponível em: <http//www.iucnredlist.org>. Acessado em: 26 Ago. 2012. KÖNIG, H.E.; LIEBICH, H.J. Anatomia dos animais domésticos. 1a ed. Porto Alegre: Artmed, 2004. 399 p. v. 2. LIZARDO, F.B.; SILVA, F.O.C.; SEVERINO, R.S. et al. Origin and distribution of the femoral nerve in fetuses of zebu-crossed bovines. Braz. J. Morphol. Sci., v.26, n.2, p.91-96, 2009. LORENZ, M.D.; KORNEGAY, J.N. Neurologia Veterinária. 2a ed. Barueri: Manole, 2006. 467p. MARTINEZ-PEREIRA, M.A.; RICKES, E.M. The spinal nerves that constitute the lumbosacral plexus and their distribution in the chinchilla. J. S. Afri. Vet. Ass., v.82, n.3, p.150–154, 2011.

79
MILLER, M.E. Anatomy of the dog. Philadelphia: W. B. Saunders, 1964. 941p. MOURÃO, G.; MEDRI, M. Activity of a specialized insectivorous mammal (Myrmecophaga tridactyla) in the Pantanal of Brazil. J. Zool., v.271, p.187-192, 2007. PEREIRA, K.F.; PARANAIBA, J.F.F.S.; HELRIGLE, C.; ARAÚJO, E.G. Origem e distribuição anatômica do nervo isquiático de mão-pelada. Pesq. Vet. Bras., v.31, n.1, p.74-78, 2011. RODRIGUES, H. Técnicas anatômicas. 4a ed. Vitória: GM. Gráfica e Editora, 2010, 270p. SCHALLER, O. Nomenclatura anatômica veterinária ilustrada. 1a ed. São Paulo: Manole, 1999. 494p. SHAW, J.H.; MACHADO-NETO, J.; CARTER, T.S. Behavior of free-living giant anteaters (Myrmecophaga tridactyla). Biotropica, v.19, p.255-259, 1987.
SILVEIRA, L.; RODRIGUES, F.H.G.; JACOMO, A.T.D.; DINIZ, J.H.F. Impact of wildfires on the megafauna of Emas National Park, Central Brazil. Oryx, v.33, n.2, p.108-114, 1999. VIZCAÍNO, S.F.; LOUGHRY, W.J. The Biology of the Xenarthra. 1a ed. Gainesville: University Press of Florida, 2008. 370p.

80
CAPÍTULO 5 - CONSIDERAÇÕES FINAIS
Apesar de o Brasil ser detentor de uma das faunas mais ricas do
planeta, o cenário da pesquisa em animais selvagens ainda é insipiente em nosso
país. Tomando como exemplo os tamanduás, apesar de serem animais típicos do
Brasil e de alguns outros países da América Latina, a maioria e os principais
estudos acerca de sua morfologia, inclusive a dos demais xenartros, foram
realizados em países que nem mesmo possuem tamanduás em seus biomas.
Diferente das espécies selvagens carnívoras ou roedoras, por exemplo,
que compartilham características morfológicas muito próximas de seus
representantes domesticados, os tamanduás não têm um representante que
possa servir de referencial em atendimentos médicos de qualquer natureza. Os
xenartros, de forma geral, são indivíduos que apresentam características
morfológicas e fisiológicas que transcendem as superficiais comparações
naturalmente estabelecidas com outros animais que tem sua morfologia
conhecida, por isso demandam especial e devida atenção científica.
O M. tridactyla não é apenas mais um mamífero do cerrado, mas é
uma espécie que requer atenção por parte das autoridades e comunidades
acadêmicas, pois pelos vários motivos já expostos no presente estudo, se não
houver uma ação mais efetiva no sentido de preservá-la, sua classificação pela
IUCN (International Union for Conservation of Nature) tende a piorar nos próximos
anos. Nesse sentido, consideramos esta pesquisa como parte de um esforço na
tentativa de contribuir com sua preservação.
O estudo apresentado trouxe à luz do conhecimento informações
completas acerca de um tópico importante em anatomia, que é a inervação de
determinada parte do corpo. No caso do M. tridactyla, são as primeiras
informações que se tem relacionadas a inervação de seus membros pélvicos. Na
verdade, este é, até o momento, o estudo mais completo da inervação do membro
pélvico de uma espécie do cerrado brasileiro e uma das raras abordagens da
inervação do pé em espécies selvagens não primatas. Portanto, espera-se que os
resultados deste trabalho possam ser úteis aos estudos em anatomia
comparativa, taxonomia e nas mais diversas áreas da medicina veterinária de

81
animais selvagens, e que a linha de pesquisa iniciada se consolide e se fortaleça
com a inclusão de outras espécies da Superordem Xenarthra.

82
ANEXOS

83

84

85

86

87

88

89

90

91





![[Anatomia] Termos Anatômicos: Movimentos](https://static.fdocumentos.com/doc/165x107/587baf621a28ab81758b7a3f/anatomia-termos-anatomicos-movimentos.jpg)











