
ATIVIDADE REDOX-PROTETORA DA Passiflora cincinnata Mast ... · lipoperoxidação plasmática...
96
UNIVERSIDADE FEDERAL DE SERGIPE PRÓ-REITORIA DE PÓS-GRADUAÇÃO E PESQUISA MESTRADO EM CIÊNCIAS DA SAÚDE CLÉSIO ANDRADE LIMA ATIVIDADE REDOX-PROTETORA DA Passiflora cincinnata Mast SOBRE O ESTRESSE OXIDATIVO INDUZIDO PELO EXERCÍCIO FÍSICO ARACAJU 2011
Transcript of ATIVIDADE REDOX-PROTETORA DA Passiflora cincinnata Mast ... · lipoperoxidação plasmática...

UNIVERSIDADE FEDERAL DE SERGIPE
PRÓ-REITORIA DE PÓS-GRADUAÇÃO E PESQUISA
MESTRADO EM CIÊNCIAS DA SAÚDE
CLÉSIO ANDRADE LIMA
ATIVIDADE REDOX-PROTETORA DA Passiflora
cincinnata Mast SOBRE O ESTRESSE OXIDATIVO
INDUZIDO PELO EXERCÍCIO FÍSICO
ARACAJU
2011

CLÉSIO ANDRADE LIMA
ATIVIDADE REDOX-PROTETORA DA Passiflora
cincinnata Mast SOBRE O ESTRESSE OXIDATIVO
INDUZIDO PELO EXERCÍCIO FÍSICO
ARACAJU
2011
Dissertação apresentada ao Núcleo de Pós-Graduação em Medicina da Universidade Federal de Sergipe como requisito parcial à obtenção do grau de Mestre em Ciências da Saúde. Orientador: Prof. Dr. Charles dos Santos Estevam

CLÉSIO ANDRADE LIMA
ATIVIDADE REDOX-PROTETORA DA Passiflora
cincinnata Mast SOBRE O ESTRESSE OXIDATIVO
INDUZIDO PELO EXERCÍCIO FÍSICO
Aprovada em ____/___/____
__________________________________________________________
Orientador: Prof. Dr. Charles dos Santos Estevam
Universidade Federal de Sergipe – Orientador
_________________________________________________________
1º Examinador: Prof. Dr. Amaro Mendonça Cavalcante
Universidade Federal de Alagoas – UFAL
__________________________________________________________
2º Examinador: Profª. Dr. Brancilene Silva Araujo
Universidade Federal de Sergipe – DFS/UFS
Dissertação apresentada ao Núcleo de Pós-Graduação em Medicina da Universidade Federal de Sergipe como requisito parcial à obtenção do grau de Mestre em Ciências da Saúde.

Aos meus avós paterno, Manoel Lima e Venina
Nunes Lima, e avô materno, Marcolino Pereira de
Andrade, grandes avós cujos exemplos de vida,
dignidade, honradez, amor e carinho sempre me
inspiraram e iluminaram meu caminho (in
memoriam).
Dedico

"A mente que se abre a uma nova idéia jamais voltará ao seu tamanho original."
Albert Einstein

AGRADECIMENTOS
À Deus e aos meus pais pela vida e dignidade.
Ao prof. Dr. Charles dos Santos Estevam pelo ensinamento, compreensão, incentivo
e paciência durante toda a orientação.
A todos os professores do núcleo de pós-graduação em medicina da UFS que
contribuíram para essa nova etapa de minha vida.
A profª Dr. Brancilene pela enorme contribuição com está dissertação (co-
orientação). Kkkkk
A todos os estudantes que compõem o laboratório de bioquímica e química de
produtos naturais pela enorme força e ajuda prestada, direta ou indiretamente, na
construção desta obra.
Aos amigos (Vitor, André e Jymmys) pela enorme contribuição com a construção
desta dissertação.
A Iara Samir, que sem entender como, fez enorme diferença para elaboração desta
obra.
A todos, o meu muito obrigado!!!

RESUMO
Nos últimos anos, têm aumentado o interesse em avaliar o efeito de constituintes
fitoquímicos presentes em alimentos e chás de ervas na atenuação do estresse
oxidativo associado ao exercício físico de alta intensidade. Nesse contexto, o chá das
folhas de plantas do gênero Passiflora pode inibir o dano celular causado pelo
estresse oxidativo devido à presença de componentes fenólicos. No presente estudo,
o perfil fitoquímico do extrato etanólico das folhas de P. cincinnata Mast foi
investigado através de reações colorimétricas ou de precipitação, enquanto o teor de
fenóis totais foi quantificado usando o método de Folin-Ciocalteu. Para estudar o
efeito agudo e crônico do extrato sobre o estresse oxidativo, ratos Wistar (250-300g)
foram divididos aleatoriamente em grupos (n=8 e n=6, respectivamente) que foram
tratados com o extrato (200 mg.kg-1) e submetidos ou não a exercícios físicos de alta
intensidade. Os níveis de marcadores do estresse oxidativo foram analisados. Devido
ao elevador teor fenólico (430,63 ± 46,71 µg GAEq.g-1 extrato), foi constatada
atividade seqüestradora dos radicais DPPH● e ABTS●+ acima de 97%. O extrato
inibiu, também, valores elevados da lipoperoxidação in vitro induzida por AAPH
(acima 95 %), FeSO4 (até 40,82 %) e H2O2 (até 62,50 %), sugerindo o ação ativo
contra radicais peroxil e hidroxil. A administração aguda do extrato protegeu ratos da
lipoperoxidação plasmática (70,41%), hepática (64,1%) e cardíaca (20,0%), além de
proteger a oxidação protéica (94,08%) hepática. Os níveis de sulfidrila plasmático
(264,32%), hepático (261,86%) e cardíaco (478,8%) foram elevados. Constatamos,
também, que o treinamento físico utilizado foi capaz de aumentar à oxidação lipídica
no aparelho urinário (201 %), tecido sanguíneo (123 %) e hepática (161 %), além de
oxidação a proteínas cardíaca (226 %) os animais exercitados (GT) em comparação
ao controle sedentário (GC) . Em contrapartida, o tratamento com o extrato, após 8
semanas de tratamento, foi capaz de prevenir a lipoperoxidação de estruturas do
aparelho urinário (50,96%), sanguínea (39,91%), hepático (83,64%) e cardíaca
(15,82%), bem como, lesões oxidativas a proteínas hepáticas (9,62%) induzidas pelo
exercício físico. Desta forma, os resultados sugerem que o consumo diário do extrato
etanólico da P. cincinnata pode trazer benefícios à saúde.
Palavras-Chave: Passiflora cincinnata Mast, atividade antioxidante, atividade
redox-protetora, exercício físico.

ABSTRACT
The interest in evaluating the effect of the phytochemical constituents in food and
herb teas in attenuating the oxidative stress associated with high intensity physical
exercise has increased in the last years. In this context, leaf tea of plants of the
Passiflora genus can inhibit the cell damage because of the oxidative stress due to
the presence of phenolic compounds. In the present study, the phytochemical profile
was investigated by colorimetric or precipitation methods, while the total phenol
content was quantified by the Folin-Ciocalteu method. To study the acute and
chronic effect of the extract on the oxidative stress, Wistar rats (250-300 g) were
divided randomly into groups (n=8 and n=6, respectively) that were treated with the
extract and were submitted or not to high intensity physical exercises. The oxidative
stress marker levels were analyzed. Because of the high content of phenolic
constituents (430.63 ± 46.71 µg EAG.g-1 extract), it was verified a scavenger activity
of the free radicals DPPH● and ABTS●+ up of 97%. The extract inhibited also levels
of lipid peroxidation induced by AAPH (above 95 %),
FeSO4 (up 40.82%) and H2O2 (up62.50%), suggesting the the extract is active against
peroxil and hydroxyl radical. The acute administration of the extract protected rats
from lipoperoxidation with reductions in levels in plasmatic (70.41%), hepatic
(64.1%) and cardiac (20.0 %) samples, and protected liver against protein oxidation
(94.08%). Sulfhydryl levels in plasmatic (264.32%), hepatic (261.86%) and cardiac
(478.8%) tissues were high. The research showed that physical
training used was able to increase lipid oxidation in the urinary tract (201%), blood
tissue (123%) and liver (161%), and the protein oxidation rate (226%)
rats exercised compared to sedentary control group.
In contrast, treatment with the extract , after 8 weeks of treating the rats submitted to
high intensity physical exercises with the extract, there was a chronic prevention of
the lipoperoxidation in structures of the urinary (50.96%), plasmatic (39.91%),
hepatic (83.64%) and cardiac (15.82%) tissues, as well as, oxidative lesion of hepatic
proteins (9.62%). Therefore, these results suggest benefits of P. cincinnata leaf
ethanol extract to health.
Keywords: Passiflora cincinnata Mast, antioxidant activity, redox-protective
activity, physical exercises.

LISTA DE FIGURAS
Figura 1: Doenças associadas as EROs ..................................................................... 8
Figura 2: Representação metabólica da geração de EROs no organismo.................. 9
Figura 3: Conceito de estress oxidativo baseado no desequilíbrio entre as ações
pró-oxidante e antioxidante.................... .................................................................... 14
Figura 4: Reações de oxidação da 2’-desoxiguanosina (dGuo) com radical hidroxila
(•OH), elétron livre (-e-) e oxigênio singlete (1O2), na geração de 8-oxo-7,8-diidro-
2’-deoxiguanosina (8-oxodGuo).................... ............................................................ 18
Figura 5: Determinação quantitativa do teor de fenóis totais do extrato etanólico e
frações da P. cincinnata Mast .................................................................................... 45
Figura 6: Efeito do extrato de P. cincinnata Mast na formação de TBARs induzido
por AAPH.................... .............................................................................................. 48
Figura 7: Efeito do extrato de P. cincinnata Mast na formação de TBARs induzido
por FeSO4.................... .............................................................................................. 48
Figura 8: Efeito do extrato de P. cincinnata Mast na formação de TBARs induzido
por H2O2.................... ................................................................................................ 49
Figura 9: Efeito redox agudo do extrato de P. cincinnata Mast na redução da
lipoperoxidação em ratos.................... ....................................................................... 50
Figura 10: Efeito redox agudo do extrato de P. cincinnata Mast na redução da
oxidação de proteínas em ratos .................................................................................. 50

Figura 11: Efeito redox agudo do extrato de P. cincinnata Mast no conteúdo de
grupamentos sulfidrilas totas em amostra de ratos .................................................... 51
Figura 12: Efeito do extrato de P. cincinnata Mast sobre os níveis de TBARs em
amostras sanguínea, urinária, hepática e cardíaca de ratos ........................................ 52
Figura 13: Efeito do extrato de P. cincinnata Mast sobre os níveis de derivados
carbonil em amostras hepática e cardíaca de ratos..................................................... 53
Figura 14: Efeito do extrato de P. cincinnata Mast sobre os níveis de derivados
sulfidrila em amostras hepática e cardíaca de ratos ................................................... 54

LISTA DE TABELAS
Tabela 1: Espécies reativas de oxigênio e sua meia-vida .......................................... 5
Tabela 2: Detecção colorimétrica de antocianidinas, antocianinas e flavonóides ... 36
Tabela 3: Detecção colorimétrica de leucoantocianinas,catequinas e flavononas ... 36
Tabela 4: Protocolo de exercício físico para indução de estresse oxidativo ............ 42
Tabela 5: Atividade antioxidante do extrato etanólico e fração acetato de etíla de P.
cincinnata Mast determinada através da redução do radical livre DPPH• ................. 46
Tabela 6: Atividade antioxidante do extrato etanólico de P. cincinnata Mast
determinada através da redução do radical livre ABTS•+ .......................................... 47

LISTA DE FORMULAS, ABREVIATURA E SIGLAS
AA – Atividade antioxidante
AAPH - dihidrocloreto de 2,2’azobis-2-amidopropano
ABTS – Radical 2,2'-azinobis-3-etilbenzotiazolino-6-sulfonato
AG – Ácido gálico
AMP – Monofosfato de adenosina
ATP – Trifosfato de adenosina
BHT – Hidroxitolueno butilado
Ca2+ - íon cálcio
CAT – enzima catalase
CE50 – Concentração eficiente em 50%
DNA – Adenina dinucleotídeo
DNPH – Ácido dinitrofenilhidrazina
DPPH - Radical 2,2-difenil-1-picrilhidrazina
DTNB – Ácido 2-nitrobenzóico 5,5-ditio-bis
EA - Exercício aeróbio
EAG – equivalente de ácido gálico
EO – Estresse Oxidativo
EROs – espécies reativas de oxigênio
ExEt – Extrato etanólico bruto de P. cincinnata Mast
FActE – Fração acetate de etila
FADH - Flavina adenina dinucleotídeo (forma reduzida)
FCl – Fração clorofórmica
FeCl3 – Cloreto férrico
FeSO4 – Sulfato ferroso
FHMoH – Fração hidrometanólica
FT – Fenóis totais
GPx – Glutationa peroxidase
Gr – glutationa redutase
GSH – Glutationa reduzida
GSSG – Glutationa oxidada

H- - íon hidrogênio
H2O2 – Peróxido de hidrogênio
H2SO4 – Ácido sulfúrico
HCl – Ácido clorídrico
HOCl – Ácido hipocloroso
IAA – Índice de atividade antioxidante
KCl – Cloreto de potássio
L• – Ácido graxo livre
LOO• – Radical peroxil
MDA - Malonaldeído
MeOH – Metanol
NADPH - Nicotinamida adenina dinucleotídeo fosfato (Forma reduzida)
NaOH – Hidróxido de sódio
NO• – Radical oxido nítrico
O2 – Oxigênio molecular 1O2 – Oxigênio singlet
O2• – Radical superóxido
OH• – Radical hidroxila
ONOO• – Radical peróxido nítrico
OOH• – Radical hidroperóxido
PMSF – Floreto de Fenilmetilsulfonila
RL – Radial livre
RO• – Radial alcoxila
ROO• – Radical peroxila
SDS – Sulfato dodecil de sódio
SH – Grupos sulfidrilas
SOD – Enzima superóxido dismutase
TBA – Ácido tiobarbitúrico
TBARs – Espécies reativas ao ácido tiobarbitúrico
TCA – Ácido tricloracético
VO2máx – Volume máximo de oxigênio

SUMÁRIO
1. INTRODUÇÃO ....................................................................................................... 3 2. REVISÃO DE LITERATURA ................................................................................ 5
2.1 Radicais Livres ................................................................................................... 5 2.1.1 Espécies reativas de oxigênio...................................................................... 7 2.1.2 Mecanismos de geração de EROs no organismo ........................................ 8 2.1.3 Produção de EROs no exercício físico ...................................................... 10
2.2 O Sistema de Defesa Antioxidante .................................................................. 11 2.2.1 Defesa antioxidante no exercício agudo ................................................... 12 2.2.2 Adaptações antioxidantes ao treinamento físico ....................................... 13
2.3 O Estresse Oxidativo ........................................................................................ 13 2.3.1 Lipoperoxidação ........................................................................................ 15 2.3.2 Oxidação de proteínas ............................................................................... 16 2.3.3 Oxidação de ácidos nucléicos ................................................................... 17
2.4 Exercício Físico e Estresse Oxidativo .............................................................. 18 2.4.1. Estresse oxidativo no músculo esquelética .............................................. 19 2.4.2. Estresse oxidativo no coração .................................................................. 21 2.4.3. Estresse oxidativo no fígado .................................................................... 22 2.4.4. Estresse oxidativo no sangue ................................................................... 23 2.4.4.1. Estresse oxidativo e eritrócitos ....................................................... 23 2.4.4.2. Estresse oxidativo e sistema imune ................................................. 24 2.4.4.3. Estresse oxidativo e plasma ............................................................ 25 2.4.4.4. Estresse oxidativo e lipoproteínas .................................................. 26 2.4.5. Estresse Oxidativo e DNA ....................................................................... 28
2.5 Exercício Físico, Estresse Oxidativo e Suplementação Antioxidante ............. 28 2.6 Passiflora cincinnata Mast................................................................................ 30
3. OBJETIVOS .......................................................................................................... 33 3.1 Geral ..................................................................................................................... 33 3.2 Específicos ........................................................................................................... 33 4. METODOLOGIA .................................................................................................. 34
4.1 Coleta e Identificação do Material Vegetal ...................................................... 34 4.2 Preparação e Fracionamento do Extrato Etanólico Bruto ................................ 34 4.3 Ensaios Químicos ............................................................................................. 35
4.3.1 Triagem fitoquímica .................................................................................. 35 4.3.1.1 Teste para fenóis e taninos ............................................................... 35 4.3.1.2 Teste para antocianinas, antocianidinas e flavonóides ................... 35 4.3.1.4 Teste para flavonóis, flavononas, flavononóis e xantonas ............... 36 4.3.1.5 Teste para esteróides e triterpenóides (Liebermann-Buchard) ....... 36 4.3.1.6 Teste para saponinas ....................................................................... 37 4.3.1.7 Teste para alcalóides ....................................................................... 37 4.3.2 Quantificação de fenóis totais (FT) ........................................................... 38 4.3.3 Avaliação da atividade antioxidante (AA) ................................................ 38 4.3.3.1 Capacidade seqüestradora do radical DPPH• ................................ 38 4.3.3.2 Capacidade seqüestradora do radical ABTS•+ ................................ 40 4.3.4 Avaliação da atividade redox (in vitro) ..................................................... 40
4.4 Ensaios Biológicos ........................................................................................... 41 4.4.1 Animais ..................................................................................................... 41

4.4.2 Delineamento experimental ...................................................................... 41 4.4.3 Preparo do material biológico ................................................................... 42 4.4.4 Determinação de lipoperoxidação ............................................................. 43 4.4.5 Determinação de proteína carbonilada ...................................................... 43 4.4.6 Determinação de sulfidrilas totais (tióis) .................................................. 44
4.5 Análise Estatística ............................................................................................ 44 5. RESULTADOS ...................................................................................................... 45
5.1 Triagem Fitoquímica ........................................................................................ 45 5.2 Fenóis Totais .................................................................................................... 45 5.3 Atividade Antioxidante (in vitro) ..................................................................... 46
5.3.1 Atividade sequestradora do radical DPPH• ............................................... 46 5.3.1 Atividade sequestradora do radical ABTS•+ ............................................. 47
5.4 Atividade Redox (in vitro) ............................................................................... 47 5.5 Atividade Redox-Protetora Após Tratamento Agudo (in vivo) ....................... 49 5.6 Atividade Redox-Protetora Após Tratamento Crônica (in vivo) ..................... 51
7. DISCUSSÃO ......................................................................................................... 54 8. CONCLUSÕES ..................................................................................................... 59 REFERENCIAS ......................................................................................................... 61

3
1. INTRODUÇÃO
É consenso na literatura que a prática regular de exercício físico é uma importante
ferramenta na manutenção da saúde e prevenção de diversas doenças como as
cardiovasculares, diabetes e hipertensão. Por conta disso, nas últimas décadas tornou-se
crescente o número de adesões a programas de exercício físico pela população em geral.
Paralelamente, novas evidências no campo da atividade física e desporto vem
surgindo, principalmente, quanto ao exercício de alta intensidade, o qual está associado à
elevada produção de radicais livres (RL) e consequentes danos celulares. Isso porque, durante
o exercício, há um aumento no consumo de oxigênio pelo organismo, bem como, um aumento
na captação desse oxigênio pelo tecido muscular, o qual favorece o aumento de espécies
reativas de oxigênio (EROs) mitocondrial, danos oxidativos e alterações da função celular.
Além disso, foi evidenciado, também, que a deficiência de alguns nutrientes associado ao
esforço físico de alta intensidade contribui para a depleção do sistema de defesa antioxidante
e redução da capacidade do organismo em remover EROs e prevenir lesões.
Por conta disso, novas estratégias têm alterado o foco de diversos estudos no campo
do exercício físico e da saúde, uma delas é a utilização de constituintes fitoquímicos
específicos, presentes em alimentos e chás de ervas, na atenuação do estresse oxidativo
associado ao esforço físico, principalmente, os constituintes polifenólicos. Neste sentido,
estudos têm demonstrado que o consumo de chás rico em polifenóis protege rins e fígado de
ratos contra o estresse oxidativo induzido pelo exercício. Não obstante, foi observado em
seres humanos, que a ingestão aguda ou regular de alimentos ricos em polifenóis pode reduzir
os danos oxidativos em resposta ao esforço. Desta forma, o consumo de produtos naturais
ricos nestes constituintes pode representar uma potencial ferramenta na prevenção de
possíveis danos celulares provocado pelo aumento de EROs e estresse oxidativo (EO) em
resposta ao esforço físico.
Dentre os diversos gêneros de plantas ricas nestes compostos, podemos destacar o
gênero Passiflora, o qual apresenta propriedade farmacológica e medicinal comprovada,
principalmente, em relação às substâncias fenólicas, as quais estão relacionadas à prevenção e
a terapia de diversas doenças associadas ao aumento de RL e espécies oxi-redutoras. As
catequínas, por exemplo, grupos de polifenóis presentes em espécies do gênero passiflora,
consistem em eficientes antioxidantes naturais que diminuem a lipoperóxidação renal em

4
ratos sedentários após sessão aguda de exercício aeróbico. No entanto, não há, ainda, relatos
na literatura quanto aos efeitos redox protetores das diversas espécies do gênero passiflora,
como por exemplo, a P. cincinnata Mast, a qual é bastante consumida por diferentes
comunidades do semi-árido nordestino do Brasil.
A Passiflora cincinnta Mast (Passifloraceae), conhecida popularmente como
“maracujá-do-mato” e típica de regiões semi-áridas, é bastante encontrada no nordeste
brasileiro, principalmente, nos Estados da Bahia, Alagoas, Pernambuco e Sergipe, a qual é
comumente utilizada como planta ornamental e medicinal. O chá de suas folhas é
rotineiramente utilizado na medicina popular como ansiolítico, hipoglicemiante, hipotensor e
antioxidante.
Foi comprovado recentemente, em ensaios in vitro, que esta espécie possui potencial
antioxidante e baixa toxidade biológica. No entanto, investigações in vivo são necessárias para
justificar o uso popular e conduzir novas pesquisas na descoberta do(s) constituinte(s)
ativo(s), o qual, num futuro bem próximo, poderá ser utilizado como suplemento nutricional
por atletas e praticantes de atividades físicas amadores ou profissionais, objetivando
minimizar os efeitos redox de agentes radicalares ao organismo, durante e após o exercício,
bem como, obter maior usufruto dos benefícios desta prática à saúde.
Assim sendo, a investigação do potencial antioxidante e redox protetor das diversas
espécies do gênero passiflora, em especial a P. cincinnata Mast, é de suma importância, tanto
do ponto de vista cientifico, por ser capaz de prevenir lesões oxidativas em órgãos e tecidos
induzido por EROs, bem como econômico, tornando-se uma nova opção de renda para os
pequenos agricultores do semi-árido nordestino, uma vez que se trata de uma espécie
adaptada às condições locais de cultivo e nativa da região. A qual poderá ser comercializada
para chás, na forma de saquês, na fabricação de doces e geléias, bem como, servir de matéria
prima à indústria farmacêutica para formulação de medicamentos e suplementos nutricionais.

5
2. REVISÃO DE LITERATURA
2.1 Radicais Livres
O termo radical livre é definido como qualquer átomo, grupo de átomo ou
molécula que possui em sua estrutura elétrons desemparelhado, e, são capazes de existir de
maneira independente (livre) (SIGNORINI & SIGNORINI, 1995). Eles possuem tempo de
vida muito curto, na ordem de milésimos de segundo, o qual está associado a sua reatividade
(Tabela 1) (BACURAU & ROSA, 2004).
Tabela 1: Epécies reativas de oxigênio e sua meia-vida.
Espécies Reativas de Oxigênio e Nitrogênio Meia-Vida (segundos)
OH•
OOH•
RO•
ROO•
ONOO•
H2O2
O2•
1O2
NO•
HOCl
Radical hidroxila
Radical hidroperóxido
Radical alcoxila
Radical peroxila
Radical peroxinitrico
Peróxido de hidrogênio
Radical superóxido
Oxigênio singleto
Radical oxido nítrico
Ácido hipocloroso
10-9
10-8
10-6
7
0,05 - 1
Variável
Variável
10-5
1 - 10
Variável
obs: R é um radical lipídico, por exemplo, o linoleato
Fonte: VANNUCCHI et al. (1998).
Conforme Koury & Donangelo (2003) os RL, elementos químicos instáveis,
tendem a ligar seus elétrons desemparelhados a outros elementos e estruturas celulares
próximas, podendo, com isso, ceder um elétron (radical redutor) ou captar um elétron (radical
oxidante). Como consequência dessa ação redox, a homeostase celular pode ser alterada e,
com isso, o funcionamento celular e tecidual normal ser modificado (SEN, 2001; VANCINI
et al., 2005).
Para Signorini & Signorini (1995), Ferreira & Matsubara (1997), Olszewer
(2001), Lancha Jr., (2004), o radical OH• é o mais reativo que existe no organismo. Porém, se
tratando de importância biológica, tanto ele quanto o O2• são importantes, uma vez que os

6
mesmos podem ser formados tanto no metabolismo normal, quanto no processo exacerbado
de redução do O2 molecular no interior das mitocôndrias, ou mesmo, através da
metabolização de bases purínicas pelo ciclo de Lowenstein, e ainda, através da redução do
peróxido de hidrogênio (H2O2) pelo ânion O2•, pelo Fe2+ e Cu+ através de reções de Fenton e
Haber-Weiss, como mostrado abaixo (LOWENSTEIN, 1990; BENZI, 1993; YU, 1994).
Fe+2 + H2O2 → Fe+3 + OH• + OH- → Reação de Fenton
O2•- + H2O2 → O2 + H2O + OH•
→ Reação de Haber-Weiss
Cu+ + H2O2 → Cu+2 + •OH + OH–
O O2• é formado no organismo através da redução monovalente do O2 molecular, o
qual adquire um único elétron (RODRIGUES, 2005). Conforme o mesmo autor, o principal
local de formação desse radical no organismo é a cadeia transportadora de elétrons, cuja
produção pode ser aumentada à medida que se aumenta a concentração do O2 no mesmo. Não
obstante, o mesmo autor referencia ainda que, mesmo sendo menos reativo que outros
radicais, há muitos alvos sensíveis a ele que a outras espécies radiculares. Além disso, esta
espécie radicalar é uma das principais fontes geradores de hidroxila radicalar (OH•), a qual é
responsável pela maioria dos danos oxidativos induzidos por RL no organismo, podendo
atacar todos os tipos de biomoléculas, originando reações em cadeia e induzindo, assim,
alterações redox ao sistema biológico, como peroxidação lipídica e oxidação de proteínas
(POLLACK & LEEUWNBURGH, 1999; CLARKSON & THOMPSON, 2000; EVANS,
2000).
Entretanto, o termo RL é muito amplo para designar todas as espécies radicalares
produzidas no organismo, haja vista que, há espécies que não apresentam elétrons
desemparelhados, porém, mesmo assim, apresenta potencial reatividade biológica como, por
exemplo, o oxigênio singlet (1O2), o peróxido de hidrogênio (H2O2) e o ácido hipocloroso
(HOCl), os quais são estáveis (FERREIRA & MATSUBARA, 1997). Neste sentido, passa a
ser apropriado designar essas espécies químicas reativas de acordo com seu elemento químico
de formação: espécies reativas de oxigênio ou EROs (oriunda do metabolismo do oxigênio) e
espécies reativas de nitrogênio ou ERNs (oriunda do metabolismo do nitrogênio), sendo que
esta ultima não será abordada neste trabalho (LI Jr., 1999; BACURAU & ROSA, 2004).

7
2.1.1 Espécies reativas de oxigênio
Por definição, podemos conceituar que as EROs são RL que, mesmo não
apresentando elétrons desemparelhados, são oriundos do metabolismo do oxigênio e
apresentam potencial reatividade biológica (RODRIGUES, 2005).
Apesar do O2 ser imprescindível à vida, ele pode apresentar uma elevada toxidade
para o organismo (YU & CHUNG, 2006). Por exemplo, a cadeia transportadora de elétrons é
responsável por mais de 90% do consumo de O2 total do organismo. Nela o oxigênio é o
aceptor final de elétrons do NADH e do FADH2 que são derivados da oxidação de fontes
energéticas (carboidratos e lipídios, por exemplo). Nesta via, o O2 é fundamental para geração
de energia (ATP). Porém, neste processo de redução do oxigênio e geração de energia,
oxidases dão origem a EROs, tais O2•, H2O2 e/ou OH• (RODRIGUES, 2005).
Neste sentido, à medida que o organismo necessita de maior aporte energético
e/ou maior concentração de O2, como no exercício físico, por exemplo, há inevitavelmente
uma maior produção de EROs pelo mesmo, uma vez que cerca de 2 a 5% do O2 consumido
pode resultar na geração dessas espécies reativas (RODRIGUES, 2005).
Todavia, apesar da reatividade das EROs e da capacidade das mesmas de poderem
desencadear ações deletérias ao reagir com proteínas, lipídios, carboidratos e ácidos nucléicos
(DNA e RNA), as mesmas, em baixas concentrações exercem importantes papeis fisiológicos
em reações redox essenciais à vida, como aumento da permeabilidade ao Ca2+, aumento da
força de contratilidade muscular, regulação da expressão gênica, regulação metabólica,
regulação do fluxo sanguíneo, destruição de microorganismos por fagócitos, síntese de
mediadores inflamatórios e detoxificação tecidual. Contudo, em excesso, se tornam
prejudiciais, podendo ocasionar ações deletérias ao mesmo, como redução da força de
contratilidade muscular, aceleração dos processos de fadiga e contribuição no aumento das
lesões musculares devido a oxidações das biomoléculas (lipídios, proteínas, DNA e RNA),
cujo resultado altera o metabolismo e o funcionamento normal da célula (CHOW, 1979;
FERREIRA & MATSUBARA, 1997, HALLIWELL & GUTTERIDGE, 1999; LEITE &
SARNI, 2003; SILVEIRA, 2004).
Conforme Ferreira & Matsubara (1997), Leite & Sarni (2003) e Schneider &
Oliveira (2004), já são mais de 50 patologias associadas às ações deletérias das EROs (Figura
1), tais como enfisema pulmonar, displasia bronco-pulmonar, pneumoconiose, asma,
síndrome de angústia respiratória do adulto (SARA), artrite reumatóide, lesão de retina,
mutações, mal de Alzheimer e Parkinson, envelhecimento, aterosclerose e câncer, sendo as

8
duas ultimas, as principais causas de óbito da sociedade moderna. Desta forma, conhecer o
mecanismo de como essas substâncias são geradas é de fundamental importância.
Figura 1: Doenças associadas as EROs
2.1.2 Mecanismos de geração de EROs no organismo
Conforme Olszewer (2001), quando o oxigênio ingressa no organismo, este tenderá a
se transformar numa ERO. De acordo com Vancini et al. (2005); Souza, Fernandes & Cyrino
(2006); Gandra, Macedo & Alves (2006), há dois fatores primordiais para geração de EROs
no organismo: um é endógeno e outro exógeno. Contudo, ambos podem atuar de três
maneiras distintas: a) através da clivagem homolítica de uma ligação covalente entre duas
moléculas; b) perda de um elétron; ou c) adição de um elétron à molécula (CHEESEMAN &
SLATER, 1996).
É descrito na literatura que as EROs podem ser geradas endogenamente em seis
locais metabólicos distinto (Yu, 1994; Morrisey & O’ Brien, 1998; Silveira, 2004). a) na
cadeia transportadora de elétrons mitocondrial, devido ao extravasamento de elétrons (íons H-
), principalmente, no complexo I e a coenzima Q, promovendo a redução univalente do
oxigênio molecular; b) nos peroxissomos que contém acetil-coA oxidase, dopamina β-

9
hidroxilase e urato oxidase; c) no sistema enzimático citocromo P-450 durante a defesa contra
xenobióticos; d) nas células do sistema imunológico como monócitos e fagócitos; e) Pela
enzima xantina oxidase, durante um episódio isquemico, cuja molécula de AMP (monofosfato
de adenosina) é continuamente degradada a hipoxantina, podendo, em seguida, ser convertida
para xantina e ácido úrico pela xantina oxidase, que usa o oxigênio como aceptor final de
elétrons, podendo, dessa maneira, formar o radical O2•; f) através da peroxidação lipídica,
onde há a formação de radical de ácido graxo livre (L•), o qual reage facilmente com O2
formando o radical peroxil (LOO•), que age oxidando outras moléculas numa reação em
cadeia até que o mesmo venha a reagir com outra EROs ou com um agente antioxidante, o
qual possa o estabilizar (Figura 2).
Quanto ao fator exógeno, este contribui para o aumento de EROs através da elevada
ingestão de metais de transição como ferro e cobre na alimentação (reações de Fenton e
Haber-Weiss), bem como de gorduras saturadas, além da exposição à radiação, ao fumo, a
antibióticos, ao álcool, a poluição e a prática de exercício físico de alta intensidade (YU,
1994; OLSZEWER, 2001; CHIHUAILAF, CONTRERAS & WITTWER, 2002).
Figura 2: Representação metabólica da geração de EROs no organismo. Fonte: GASTELL & ALEJO (2000).

10
2.1.3 Produção de EROs no exercício físico
Durante o exercício há um aumento acentuado na produção de EROs, principalmente
durante os exercício aeróbicos de alta intensidade, afinal a ventilação pulmonar triplica nestes
exercício, ocorrendo, paralelamente, um aumento no consumo de oxigênio pelo mesmo de 10
a 20 vezes comparado ao repouso, bem como, aumento na captação desse oxigênio de 100 a
200 vezes pelo tecido muscular, o qual favorece o aumento de EROs mitocôndrial durante o
processo de produção de energia (ATP), danos oxidativos e alterações da função celular
(BACURAU & ROSA, 2004).
Paralelamente, além do aumento de O2, outros processos também contribuem para o
aumento de EROs durante o exercício, tais como aumento de catecolaminas, produção de
ácido láctico, elevação da taxa de auto-oxidação de hemoglobina e aumento da temperatura
corporal (BACURAU & ROSA, 2004).
Não obstante, durante e, principalmente, após o exercício, um outro fenômeno
denominado de isquemia-reperfusão contribui significativamente para o aumento de EROs no
organismo, devido à hipóxia e à reoxigenação temporária que ocorrem no músculo exercitado,
em função das contrações e relaxamentos estabelecidos ciclicamente. (SCHNEIDER &
OLIVEIRA, 2004; FINAUD et al., 2006; CRUZAT et al., 2007). Durante a contração, a
compressão vascular estabelece um quadro de isquemia e, portanto, de hipóxia; no
relaxamento ocorre a reperfusão e, consequentemente, a reoxigenação (SCHNEIDER &
OLIVEIRA, 2004). Uma vez que durante o exercício a circulação sanguínea é desviada para
os músculos em atividade, outros tecidos também podem sofrer hipóxia temporária
(CRUZAT et al., 2007). Como consequência, esses tecidos recebem uma grande
quantidade de oxigênio após o exercício, favorecendo a geração de EROs (SCHNEIDER &
OLIVEIRA, 2004; FINAUD et al., 2006; CRUZAT et al., 2007). No processo de
isquemia/reperfusão, o aumento EROs durante e após o exercício é favorecido pelo
catabolismo das purinas. Tendo em vista que, a conversão da enzima xantina-desidrogenase à
sua forma oxidada (xantina-oxidase), mediante proteases intracelulares ativadas por Ca2+,
favorece o aumento de EROs. Durante a hipóxia, o ATP é degradado até hipoxantina, que se
acumula nos tecidos. Como resultado, há falência da homeostase celular, permitindo o influxo
de Ca2+ para as células, o que ativa proteases intracelulares a converterem a enzima xantina
desidrogenase em xantina oxidase. Na reperfusão a xantina oxidase utiliza O2 para promover
a conversão de hipoxantina em xantina e posteriormente em ácido úrico, ocorrendo redução
univalente do oxigênio molecular e consequente geração de H2O2 e HO• (JI, 1999; CAMPOS

11
& YOSHIDA, 2004; FINAUD et al., 2006; CRUZAT et al., 2007; FERREIRA et al.,
2007).
Em contrapartida, com o intuito de manter a integridade celular contra os ataques
redox das diversas espécies radicalares, os organismos aeróbicos (a exemplo do ser humano)
no transcurso de sua evolução desenvolveram um sofisticado e eficiente sistema de defesa,
denominado antioxidante (PÔRTO, 2001).
2.2 O Sistema de Defesa Antioxidante
Conforme Signorini & Signorini (1995) o sistema antioxidante é a principal defesa
orgânica contra a ação das EROs, o qual pode ser enzimático ou não enzimático. O primeiro
inclui as enzimas catalase (CAT), superóxido dismutase (SOD), glutationa peroxidase (GPx),
Glutationa redutase (GR) e Glutationa reduzida (GSH), que são produzidas pelo próprio
organismo (agentes endógenos) e que estão presentes em maior quantidade no fígado, cérebro
e músculos, principalmente, nos músculos com predomínio oxidativo, ou seja, aqueles que
apresentam uma maior concentração de fibras do tipo I.
O segundo tipo inclui compostos não enzimáticos produzidos pelo próprio organismo
(endógeno) como bilirrubina, ceruloplasmina, hormônios sexuais, melantonina, coenzima Q,
ácido úrico, além de compostos exógenos, que são ingeridos através da dieta ou
suplementação alimentar, como ácido ascórbico (vitamina C), α-tocoferol (vitamina E), β-
caroteno (o precursor de vitamina A) e grupos fenólicos de plantas tais como flavonóides
(SIGNORINI & SIGNORINI, 1995; SCHNEIDER & OLIVEIRA, 2004).
Conforme Ferreira & Matsubara (1997), a ação dos antioxidantes no organismo pode
ocorrer de duas maneiras distintas: a primeira como detoxificadora, atuando antes que ocorra
lesão às estruturas biológicas, sendo constituída pela GSH, SOD, GPx e vitamina E. A
segunda linha tem a função de reparar a lesão ocorrida, e é constituída pela vitamina C, GR e
GPx, entre outras. Entretanto, apenas a vitamina E é considerada um antioxidante estrutural
de membrana, não estando presente no meio intracelular.
Para Yu (1994), os antioxidantes enzimáticos não agem separadamente, elas agem
em conjunto e são responsáveis pela eliminação do O2•-, H2O2, OH• e outros hidroperóxidos
que podem oxidar as estruturas celulares no meio intra e extracelulares. A SOD, por exemplo,
age transformando dois ânions radicais superóxidos em um peróxido de hidrogênio (reação
1). A CAT possui a capacidade de transformar o peróxido de hidrogênio em água e oxigênio

12
(reação 2), porém, sua localização está nos peroxissomas, tendo, por isto, sua ação diminuída
em órgãos como coração, pulmão e cérebro, os quais possuem pouco peroxissomas. Nestes
órgãos a ação antioxidante ocorre, prioritariamente, pela GPx (reação 3), localizada no citosol
e na matriz mitocondrial. Sua ação ocorre através da redução do peróxido de hidrogênio ou
hidropeptídeos orgânicos através da utilização da GSH, substrato da GPx, transferindo dois
hidrogênios dos grupamentos sulfidrílas para os peróxidos, convertendo-os em álcool e/ou
água, resultando em glutationa dissulfeto (GSSG).
SOD + O2• + 2H+
→ SOD + H2O2 (1)
2H2O2 + CAT → O2 + 2H2O (2)
H2O2 ou RO2H + GPx + GSH → GSSG + H2O ou ROH (3)
Desta forma, o sistema antioxidante é responsável pela manutenção do equilíbrio
redox celular, uma vez que são capazes de reduzir a formação e os efeitos indesejáveis das
EROs (HALLIWELL & GUTTERIDGE, 1999).
2.2.1 Defesa antioxidante no exercício agudo
Segundo Mcardle et al. (1998), não há possibilidade de se parar a produção de EROs
no organismo, principalmente, durante a prática de atividade física. Cuja é possível identificar
um aumento acentuado na produção de EROs e, consequentemente, maior requerimento dos
mecanismos de defesa antioxidante.
Durante o exercício agudo foi comprovado haver aumento na atividade enzimática,
principalmente, naquelas que constituem o escalão de defesa primária (CAT, SOD e GPx),
tanto em animais, quanto em humanos (ISIDÓRIO, 2007). Em contrapartida, até o momento,
não foi comprovado que estas mesmas enzimas mantêm-se capazes de sustentar o estado
redox até o fim do exercício, devido, principalmente, o aumento exacerbado na síntese de
EROs pelo mesmo, bem como, o tempo é reduzido para promover adaptações a nível
quantitativo durante o exercício (COSTA, 2010).
Di Meo et al. (1996), descreve que a capacidade antioxidante dos diversos tecidos e
órgãos mostram-se diferentes durante o exercício. Sendo o fígado, órgão de elevado teor
metabólico, aquele que exibe maior capacidade antioxidante, seguido por sangue, coração e
músculo. No entanto, mesmo sendo comprovado que o treinamento físico melhora a

13
capacidade antioxidante dos diversos tecidos supracitados, o músculo esquelético é aquele
que adquire maior adaptação a produção de espécies radicalares impostas pelo exercício,
principalmente, a nível de enzima antioxidante (AGUILAR-SILVA et al., 2002).
2.2.2 Adaptações antioxidantes ao treinamento físico
Quando o exercício promove uma depleção dos estoques antioxidantes não-
enzimáticos, tais como as vitaminas A, E, C e glutationa, e, caso a ingestão destes não esteja
em equilíbrio com a demanda, como conseqüência biológica preventiva, o organismo, a longo
prazo, cria mecanismos de ativação adaptativos e aumenta a taxa de síntese das enzimas
antioxidantes para compensar o estresse oxidativo intenso (ISIDÓRIO, 2007).
Cada enzima apresenta uma forma diferente de adaptação ao estresse oxidativo,
dependendo do modelo específico de síntese de cada enzima e das taxas de EROs produzidas.
A SOD, por exemplo, apresenta alta atividade em todos os tecidos e tipos de fibras
musculares, onde a remoção de superóxido não é fator limitante. Enquanto, a GPx, que
degrada os produtos finais das EROs (H2O2 e peróxidos orgânicos), tem atividade baixa e
apresenta grande adaptação ao treinamento, em comparação a SOD e CAT.
As adaptações antioxidantes são fortemente influenciadas por uma série de fatores
fisiológicos e ambientais, tais como sexo, idade, dieta e medicação. Conforme Oliveira
(2010), em estudo como animais, ratas têm demonstrado menor grau de lesão oxidativa
muscular induzida pelo exercício e adaptações de enzimas antioxidantes ao treinamento que
ratos, possivelmente devido ao efeito antioxidante do estrógeno. Bem como, animais e seres
humanos idosos, geralmente apresentam menor grau de adaptação de enzimas antioxidantes
que indivíduos jovens submetidos à mesma carga de trabalho. A razão para esta diferença é
complexa e pode está relacionada a alterações intrínsecas no sistema de defesa antioxidante,
assim como na transdução de sinais, a qual em animais velhos tornam-se falhas.
2.3 O Estresse Oxidativo
Há situações em que o sistema de defesa não consegue proteger as estruturas
biológicas da ação das EROs (Figura 3). Quando o organismo excede o nível de esforço físico
ao qual está adaptado, há um maior na produção de EROs e maior depleção do sistema de

14
defesa antioxidante, o qual não consegue manter o balanço redox adequado dessas espécies
reativas no organismo (SIGNORINI & SIGNORINI, 1995). Quando isso ocorre, o organismo
fica em condições de excesso de agentes oxidantes e deficiência de antioxidante, esse
desequilíbrio redox implica em perda de funções celulares vitais e instauração de um quadro
ou condição orgânica prejudicial para o organismo denominado de estresse oxidativo (EO)
(FERREIRA & MATSUBARA, 1997).
Figura 3: Conceito de estress oxidativo baseado no desequilíbrio entre as ações pró-oxidante e antioxidante. Fonte: Ferreira, Ferreira & Duarte (2007)
Hábitos de vida inapropriados, tais como o consumo de álcool, fumo, dieta
inadequada e prática de exercício de alta intensidade, assim como, condições ambientais
impróprias como a exposição à radiação UV, a poluição, a umidade e temperatura elevada,
bem como, estado psicológico que provoque estresse emocional e o envelhecimento, são
fatores que estão diretamente relacionados com o EO (LIU et al.,2000; MCARDLE et al.,
2001; VANCINI et al., 2005).
Urso & Clarkson (2003) e Mota et al., (2004) relataram que uma situação de EO
representa uma incapacidade do organismo de impedir os efeitos deletérios das EROs as
diversas biomoléculas, a qual pode ocorrer através da lipoperoxidação, oxidação de proteínas
e oxidação de ácidos nucléicos (NOHL et al., 2005).

15
2.3.1 Lipoperoxidação
De acordo com Sen (2001) e Rodrigues (2005), a lipoperoxidação é um fenômeno
que ocorre quando uma ERO reage com um ácido graxo (ácido linoléico, por exemplo) dando
origem a outras espécies radicalares. Embora os ácidos graxos saturados possam reagir com o
oxigênio, à susceptibilidade de lipídeos à oxidação é maior em ácidos graxos com
insaturações. Ácidos graxos contendo uma ou mais duplas ligações são especialmente
sensíveis, pois o ataque do oxigênio ocorre principalmente nas posições adjacentes às duplas
ligações (NAWAR, 1998). Este mecanismo pode ser descrito em três etapas distintas:
iniciação, propagação e terminação (SEN, 2001; RODRIGUES, 2005).
Na Iniciação, radicais alquil, alcoxil, hidroxil, peróxidos e hidroperóxido (X•), por
exemplo, reagem com um átomo de hidrogênio de um grupo metileno (-CH2-) da cadeia
lateral de um ácido graxo poliinsaturado, deixando um radical centrado no carbono, uma vez
que os hidrogênios desse grupo metileno são bastante reativos, podendo ser removidos seja na
forma de íons H+ ou na forma de hidrogênio radical (H•) devido à proximidade com as duplas
ligações (reação 1).
X• + L-CH → L-C• + H2O (1)
O radical carbono centrado (L-C•), recém formado, reage muito rapidamente com o
oxigênio molecular formando radical peróxil (L-C-O-O•). Esses radicais são altamente
reativos e capazes de remover átomos de hidrogênio de outros ácidos graxos insaturados,
formando hidroperóxido e outros radicais lipídicos. Os novos radicais reagem com o oxigênio
e ocorre uma propagação desta reação (reação 2).
L-C• + O2 → L-C-O-O• (2)
Devido à ressonância na estabilização de radicais, pode haver, também, uma
mudança na posição de duplas ligações resultando na formação de hidroperóxidos isômeros
contendo dienos conjugados. Hidroperóxidos são produtos primários de autoxidação
relativamente instáveis, que participam de numerosas reações de degradação e interação
resultando em um grande número de compostos de diferentes pesos moleculares, limiares de
percepção de flavor e importância biológica (NAWAR, 1996).

16
O término da lipoperóxidação (reação 3) ocorre quando os radicais reagem entre si
formando um produto não radicalar. As reações também podem terminar quando um dos
radicais reage com um antioxidante, removendo um hidrogênio e formando um radical inerte
do antioxidante (RODRIGUES, 2005).
L-C• + L-C• → L-C-C-L (3)
Segundo Sen (2001), quando EROs são produzidas, elas podem ser transportadas
pelo sangue para alvos distantes, podendo iniciar diversas lesões oxidativas. Caso não haja
interrupção, as reações em cadeia podem gerar danos extensivo à lipídios de membrana.
Como consequência, as membranas sofrerão alterações em sua permeabilidade,
comprometendo, com isso, toda a integridade e funcionalidade celular (SELMAN et al.,
2002). Para Chang et al. (2006), o principal antioxidante responsável pela prevenção da
lipoperoxidação é a vitamina E, que devido sua liposolubilidade, consegue neutralizar
diretamente os radicais ROO•.
Conforme Halliwell & Gutteridge (1999) há vários métodos para se determinar a
lipoperóxidação. Contudo, o teste de substâncias reativa ao ácido tiobarbitúrico (TBARs) é o
mais amplamente utilizado como biomarcador da lipoperoxidação em sistemas biológicos:
sangue, urina e órgãos (YAGI, 1976; OHKAWA et al., 1979; LAPENNA et al., 2001).
Durante a lipoperóxidação hidrocarbonetos voláteis (etano e pentano), aldeídos saturados e
insaturados, cetonas, entre outros produtos são produzidos, dentre estes o malondialdeído
(MDA), por exemplo, o qual é um aldeído saturado que reage prontamente com o ácido
tiobarbitúrico (TBA), formando um complexo colorido que pode ser quantificado
espectrofotometricamente (YAGI, 1976; OHKAWA et al., 1979; HALLIWELL &
GUTTERIDGE, 1999; LAPENNA et al., 2001).
2.3.2 Oxidação de proteínas
Devido sua complexa estrutura e aos grupos funcionais oxidáveis dos aminoácidos,
as proteínas também são alvo fáceis das EROs (FRIGUET, 2006). Assim, durante a oxidação
protéica pode acontecer uma série de modificações químicas nas proteínas, que podem
produzir alterações das estruturas das mesmas, inativando-as e provocando danos nas
atividades biológicas a elas associadas (DALLE-DONNE et al., 2003). Dentre as principais

17
modificações químicas ocasionadas pelas EROs podemos destacar a hidroxilação de
aminoácidos aromáticos, oxidação de aminoácidos que possuem enxofre (cisteína),
peroxidação de aminoácidos alifáticos e formação de grupos carbonilas. Todas estas
alterações estão associadas à perda ou mudanças nas funções biológicas das mesmas
(STADMAN & LEVINE, 2000).
Conforme Linton et al. (2001), os mecanismos para a lesão oxidativa das proteínas
podem ocorre através da formação de EROs em local específico através da ativação redox de
metais de transição que não estão ligados fortemente à proteína ou através de EROs não
dependentes de metais. Neste caso, o dano pode ocorrer diretamente nos aminoácidos por
excisão ou por quebra da ligação peptídica.
Quando uma proteína ou aminoácido é oxidado, subprodutos carbonílicos são
gerados, os quais servem como indicadores de lesão oxidativa e podem ser quantificados
espectrofotometricamente através de reações específicas com o reagente 2,4-dinitrofenil
hidrazina (DNPH) (REZNICK & PACKER, 1994).
2.3.3 Oxidação de ácidos nucléicos
Um terceiro alvo do EO são os ácidos nucléicos (DNA e RNA). Segundo Remmen et
al. (2003), uma situação de EO pode induzir uma lesão oxidativa significativa ao DNA
através de modificações de suas bases purínicas e formação de produtos finais deletérios,
sendo o HO• o principal responsável por essas lesões. As quais são favorecidas pela intensa
força negativa dos grupamentos fosfato do DNA, capazes de ligar-se com metais de carga
positiva. Entre estes, estão o ferro e o cobre, desencadeadores de reações de Fenton e Haber-
Weiss.
São identificadas atualmente mais de cem alterações químicas EROs-dependente
sobre ácidos nucléicos, as quais são capazes de comprometer todo o seu desempenho
fisiológico (RIEGEL, 2005). Ainda, conforme o autor, a maioria destas lesões afeta,
principalmente, o DNA mitocondrial, uma vez que é na mitocôndria que mais se forma EROs.
E, entre todas as alterações, a mais comum é a conversão de 2’-deoxoguanosina em 8-oxi-2’-
deoxoguanosina (oxo8dG) (Figura 4), a qual é utilizado comumente como marcador de lesão
oxidativa de DNA, podendo ser quantificado espectrofotometricamente ou por cromatografia
de massa (CADENAS et al., 1997; HERRERO & BARJA, 1998).
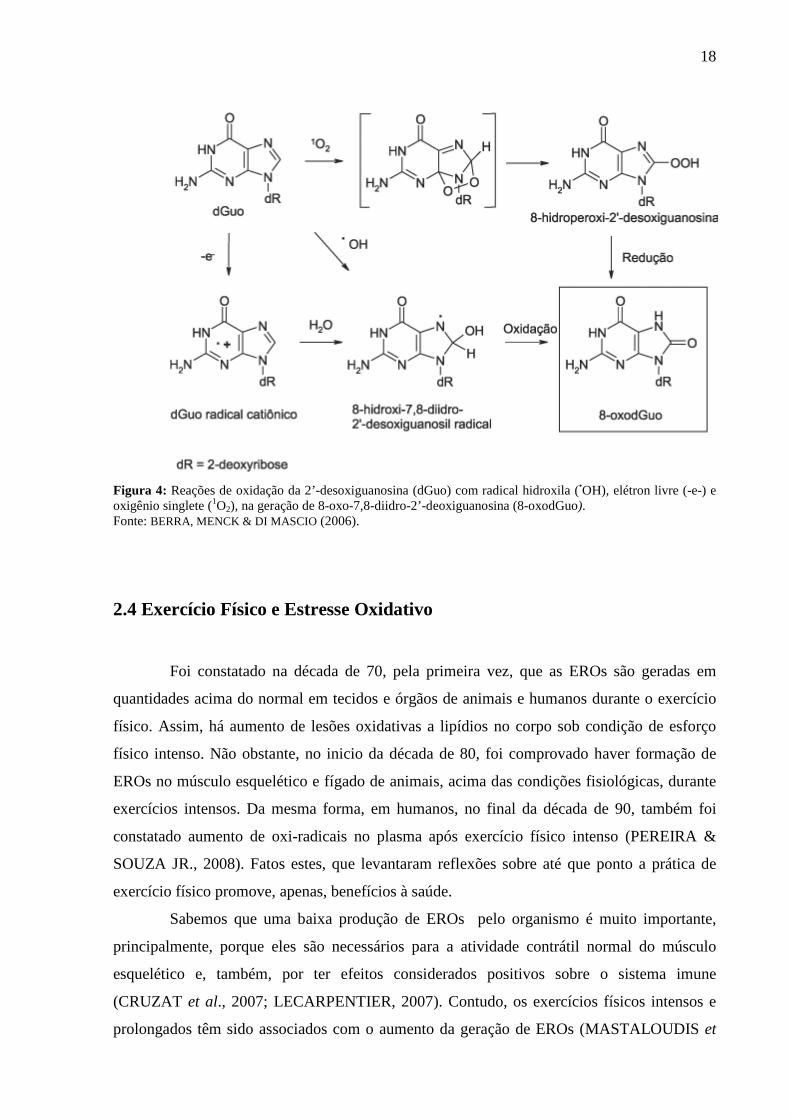
18
Figura 4: Reações de oxidação da 2’-desoxiguanosina (dGuo) com radical hidroxila (•OH), elétron livre (-e-) e oxigênio singlete (1O2), na geração de 8-oxo-7,8-diidro-2’-deoxiguanosina (8-oxodGuo). Fonte: BERRA, MENCK & DI MASCIO (2006).
2.4 Exercício Físico e Estresse Oxidativo
Foi constatado na década de 70, pela primeira vez, que as EROs são geradas em
quantidades acima do normal em tecidos e órgãos de animais e humanos durante o exercício
físico. Assim, há aumento de lesões oxidativas a lipídios no corpo sob condição de esforço
físico intenso. Não obstante, no inicio da década de 80, foi comprovado haver formação de
EROs no músculo esquelético e fígado de animais, acima das condições fisiológicas, durante
exercícios intensos. Da mesma forma, em humanos, no final da década de 90, também foi
constatado aumento de oxi-radicais no plasma após exercício físico intenso (PEREIRA &
SOUZA JR., 2008). Fatos estes, que levantaram reflexões sobre até que ponto a prática de
exercício físico promove, apenas, benefícios à saúde.
Sabemos que uma baixa produção de EROs pelo organismo é muito importante,
principalmente, porque eles são necessários para a atividade contrátil normal do músculo
esquelético e, também, por ter efeitos considerados positivos sobre o sistema imune
(CRUZAT et al., 2007; LECARPENTIER, 2007). Contudo, os exercícios físicos intensos e
prolongados têm sido associados com o aumento da geração de EROs (MASTALOUDIS et

19
al., 2004; TRABER, 2006). E este aumento pode causar dano oxidativo ao DNA, na
musculatura esquelética e nos tecidos, o que pode desencadear envelhecimento prematuro e
graves lesões oxidativa com consequente processo inflamatório, fatos que implicam em
prejuízo no desempenho físico e à saúde do atleta (JI & LEICHTWEIS, 1997; SUREDA et
al., 2005; FINAUD et al., 2006; TRABER, 2006; CRUZAT et al., 2007; FERREIRA et al.,
2007; LECARPENTIER, 2007; RADAK et al., 2007; VALKO et al., 2007). Não obstante,
o treinamento físico, exercitado em níveis moderados, é um bom indutor de adaptações do
sistema de defesa antioxidante enzimático (JI & LEICHTWEIS, 1997; VILLA-
CABALLERO et al., 2000; BELVIRANLI & GÖKBEL, 2006; CRUZAT et al., 2007;
FERREIRA et al., 2007; JUDGE & LEEUWENBURGH, 2007; RADAK et al., 2007; JI et
al., 2008). Haja vista que, foi demonstrado um aumento na capacidade antioxidante
enzimática no tecido cardíaco, hepático e pulmonar de ratos após período de treinamento fisco
moderado (FERREIRA et al., 2007).
Embora o treinamento físico induza adaptações benéficas, a realização de exercícios
intensos e/ou prolongados, acima da intensidade habitual de esforço ou com frequência de
treinamento muito elevada, ele suplanta a capacidade do sistema antioxidante endógeno e
geralmente resulta em lesão oxidativa. Esta sobrecarga oxidativa, apesar de se fazer sentir
mais intensamente nos músculos esqueléticos, tem sido relatada, também, em muitos outros
órgãos e sistemas corporais responsáveis pela regulação e manutenção da homeostase
(CRUZAT et al.,2007; FERREIRA et al., 2007; KIM et al., 2007; LECARPENTIER, 2007),
incluindo coração (FERREIRA et al.,2007), fígado (NAGEL et al., 1990; DE PAZ et al.,
1995; FERREIRA et al., 2007; KIM et al., 2007), rins e pulmões (FERREIRA et al., 2007),
eritrócitos (SCHMIDT et al., 1988; SUREDA et al., 2005; YUSOF et al., 2007), sistemas
imune (JI, 1999; MOOREN et al., 2002; SUREDA et al., 2005; BELVIRANLI & GÖKBEL,
2006; CRUZAT et al., 2007; KIM et al., 2007) e ósteo-articular (FERREIRA et al., 2007;
KIM et al., 2007). De fato, as acentuadas alterações hormonais, assim como, as alterações
térmicas, associadas ao exercício intenso e exaustivo far-se-ão sentir, de forma mais ou menos
intensa, na maior parte das células corporais (FERREIRA et al., 2007).
2.4.1. Estresse oxidativo no músculo esquelética
No músculo esquelético, apesar da mitocôndria constituir a principal fonte e,
simultaneamente, o principal alvo de EROs, a atividade das enzimas xantina oxidase e

20
fosfolipase A2, a desaminação das catecolaminas e a infiltração de leucócitos pós-exercício,
juntamente com a oxidação da mioglobina por RL ou por sua auto-oxidação, contribuem
fortemente como fontes adicionais de EROs, consequentemente, lesões oxidativas e
estresse oxidativo (FINAUD et al., 2006; FERREIRA et al., 2007).
Lesões no tecido muscular induzidas pelo exercício podem ser decorrentes de vários
fatores: ruptura de tecidos conectivos ligados a miofibrilas adjacentes, da própria célula
muscular, da lâmina basal adjacente à membrana plasmática, da membrana plasmática da
célula muscular, do sarcômero, do retículo sarcoplasmático, ou ainda de uma combinação
desses componentes (CRUZAT et al., 2007).
Para comprovação de lesões musculares induzidas pelo exercício várias técnicas
podem ser utilizadas, desde a avaliação direta, através de técnicas histológicas ou microscopia
eletrônica, às avaliações indiretamente, através da determinação do efluxo de enzimas
citosólicas específicas para a circulação sanguínea (CRUZAT et al., 2007; FOSCHINI et al.,
2007), podendo variar desde uma lesão ultra-estrutural de fibras musculares até traumas
envolvendo a completa ruptura do músculo. As características morfológicas e ultra-
estruturais da lesão induzida pelo exercício estão bem documentadas em modelos animais e
humanos (ARMSTRONG et al., 1983; GIBALA et al., 1995; MORGAN & ALLEN, 1999;
PROSKE & MORGAN, 2001; STUPKA et al., 2001; MAGAUDDA et al., 2004).
Em humanos, por razões éticas e logísticas, as evidências de estresse e lesão
oxidativa induzidos pelo exercício têm sido essencialmente estudadas em nível sanguíneo,
quer no plasma, quer nas células circulantes (FERREIRA et al., 2007). Neste contexto, o
aumento da concentração de proteínas citosólicas na circulação após o exercício reflete a
lesão muscular. As proteínas avaliadas frequentemente são a creatina quinase (CPK), a lactato
desidrogenase (LDH), a transaminase glutâmico-oxalacética (TGO, também chamada de
aspartato aminotransferase ou AST) e a mioglobina, que, normalmente, são incapazes de
atravessar a membrana plasmática. A presença dessas proteínas na circulação sanguínea
reflete significativa alteração na estrutura e permeabilidade da membrana miofibrilar
(CRUZAT et al., 2007; APPLE et al., 1988; AGARWAL & ANKRA- BADU, 1999;
BARBOSA et al., 2003; LAC & MASO, 2004; GASPER & GILCHRIST 2005;
FOSCHINI et al., 2007). Dentre elas, a CPK é frequentemente descrita como principal
marcador indireto de dano muscular, sobretudo após o exercício de força ou outros exercícios
que exijam ações predominantemente excêntricas (BARBOSA et al., 2003; FOSCHINI et al.,
2007).

21
2.4.2. Estresse oxidativo no coração
Semelhante ao que foi descrito para o músculo esquelético, hoje é unanimemente
aceito que o exercício, sobretudo de alta intensidade, promove, também, elevação do estresse
oxidativo no coração, induzindo um aumento em marcadores de lesão oxidativa (FERREIRA
et al., 2007). O miocárdio parece ser particularmente susceptível a todas as situações que
promovam uma elevação do seu metabolismo, nas quais se incluem o exercício físico, uma
vez que a sua capacidade metabólica oxidativa é muito elevada devido à abundância de
mitocôndrias (FERREIRA et al., 2007; JUDGE & LEEUWENBURGH, 2007). O
estresse oxidativo aumentado neste órgão durante o exercício é resultado de aumentos
dramático no consumo de oxigênio (JUDGE & LEEUWENBURGH, 2007). Nestas
condições, a reação da CPK é importante para a rápida ressíntese de ATP a partir de creatina
fosfato e ADP, uma vez que o coração aumenta seu trabalho (PUTNEY et al., 1984;
NASCIMBEN et al., 1996; FOSCHINI et al., 2007). Considerando que CPK é uma enzima
citosólica, níveis séricos aumentados dessa enzima após exercício podem ser também
indicativos de lesão oxidativa no miocárdio.
Várias evidências sugerem que o estresse oxidativo desempenha um papel
importante na patogênese de doenças cardiovasculares (WATTANAPITAYAKUL &
BAUER, 2001; MOLAVI & MEHTA, 2004; ABRAMSON et al., 2005; BELARDINELLI,
2007; KASAP et al., 2007). A presença de níveis plasmáticos elevados de alguns marcadores
de estado inflamatório, como a proteína C-reativa (PCR) é um fator preditivo de risco de
síndrome coronariana aguda, infarto do miocárdio, doença arterial periférica oclusiva e morte
cardíaca súbita, tanto em indivíduos sadios, como em pacientes com doença aterosclerótica
estabelecida (MITKA, 2004; DUMMER et al., 2007). Níveis séricos aumentados da
enzima TGO foram usados na diagnose de infarto agudo do miocárdio desde 1954. Essa
transaminase é encontrada no citoplasma e nas mitocôndrias de muitas células, principalmente
do fígado, coração, músculos esqueléticos, rins, pâncreas e hemácias (DEWAR et al., 1958).
Apesar dos marcadores citados não serem específicos para verificar lesões no miocárdio,
aumentos nos níveis plasmáticos de PCR e TGO após exercício também podem ser
indicativos de lesão oxidativa no miocárdio.

22
2.4.3. Estresse oxidativo no fígado
É descrito na literatura que exercício físico, igualmente ao que ocorre em outros
órgãos, induz a formação de EROs e danos oxidativos no fígado, tais como a lipoperoxidação
e oxidação a proteínas (NAGEL et al., 1990; WITT et al., 1992; DE PAZ et al., 1995;
FERREIRA et al., 2007; KIM et al., 2007). A quantidade de dano depende da intensidade
do exercício e do estado de treinamento (WITT et al., 1992). O fígado apresenta elevada
taxa metabólica, a qual está naturalmente associada com o alto fluxo de oxigênio. Entretanto,
este fluxo diminui significativamente durante o exercício, devendo ser similar ao fenômeno
de isquemia/reperfusão (RADAK et al., 2008). Diferentemente do músculo esquelético, o
fígado contém altos níveis de xantina desidrogenase, a qual, durante o exercício, é convertida
em xantina oxidase, que por sua vez contribui como fonte adicional de ERO e,
consequentemente, com danos oxidativos (FERREIRA et al., 2007; RADAK et al., 2008).
Enquanto um único turno de exercício intenso estimula adaptações no sistema antioxidante do
músculo esquelético (JI & FU, 1992; ALESSIO et al., 2002; CRUZAT et al. 2007;
FERREIRA et al., 2007; RADAK et al. 2007), o mesmo não ocorre com o fígado, que fica
oxidativamente estressado (JI & FU, 1992; RADAK et al. 2007). Todavia, semelhantemente
ao músculo esquelético, tem sido demonstrado em animais que o treinamento físico pode
promove melhorias na capacidade antioxidante do fígado (WATARU et al., 2003;
FERREIRA et al., 2007). Em ratos, como resultado de adaptação induzida pelo treinamento
físico, foi evidenciado aumento no conteúdo de algumas enzimas antioxidantes com reduzida
produção de EROs (WATARU et al., 2003; RADAK et al., 2008).
A lesão hepática pode ser evidenciada pelo efluxo de algumas enzimas citosólicas
para a circulação sanguínea, particularmente a TGO e a TGP (transaminase glutâmico-
pirúvica), também chamada de ALT (alanina aminotransferase); esta última é encontrada em
altas concentrações apenas no fígado (BRUCE et al., 1958; DEWAR et al., 1958;
ALMERSJÖ et al., 1968; CHALIFOUX & LAGACÉ, 1969). Consequentemente, níveis
séricos aumentados dessas enzimas após exercício servem como indicativos de lesão
oxidativa neste órgão.

23
2.4.4. Estresse oxidativo no sangue
2.4.4.1. Estresse oxidativo e eritrócitos
Tem sido sugerido que exercícios intensos de longa duração e treinamentos
exaustivos podem, também, comprometer nossa capacidade de detoxificar EROs dentro das
células sanguíneas e os eritrócitos parecem ser bastante vulneráveis aos danos oxidativos
(PETIBOIS & DÉLÉRIS, 2005; SUREDA et al., 2005). Isto decorre devido sua capacidade
biossintética ser muito limitada (ausência de núcleo), mecanismo de reparo pobre (SANTOS-
SILVA et al., 2001; CAZZOLA et al., 2003; SUREDA et al., 2005), presença de grandes
quantidades de tióis (R-SH) e ácidos graxos poliinsaturados (AGPI) na sua membrana,
alta concentração interna de oxigênio e hemoglobina, fontes potenciais de processos
oxidativos (FERREIRA & MATSUBARA, 1997; CAZZOLA et al., 2003; FEKSA et al.,
2008).
Apesar de possuírem um elaborado sistema de defesa antioxidante que inclui as
enzimas CAT, SOD e GPx, se a eficiência deste sistema for superada pela magnitude dos
processos oxidativos, ocorrerá EO, o qual resultará em hemólise (FERREIRA &
MATSUBARA, 1997; FEKSA et al., 2008). Neste caso, a hemoglobina extracelular se torna
tóxica devido à natureza oxidativa dos íons ferro contidos no grupo heme, os quais participam
da reação de Fenton para produzir EROs, que causam injúria celular (STERN, 1985;
JOSEPHY et al., 1997).
Durante o EO as alterações mais comuns são a peroxidação dos lipídeos e das
proteínas de membrana, o que pode desestabilizar o citoesqueleto e comprometer a
sobrevivência da célula (PETIBOIS & DÉLÉRIS, 2005; SUREDA et al., 2005; YUSOF et
al., 2007). Adicionalmente, os eritrócitos também estão altamente expostos ao estresse
mecânico, assim como a mudanças de pH citosólico e extracelular (PETIBOIS & DÉLÉRIS,
2005).
As EROs podem alterar as propriedades químicas e físicas da membrana eritrocitária
por modificar a composição, o empacotamento e a distribuição de seus lipídeos, o que conduz
a uma alteração estrutural com redução de sua fluidez. Isto pode modificar a atividade de
diversas proteínas de membrana e acelerar a senescência do eritrócito, ou mesmo, provocar
sua remoção prematura da circulação (CAZZOLA et al., 2003; PETIBOIS & DÉLÉRIS,
2005). Os agentes oxidantes podem converter os grupamentos tióis em componentes

24
dissulfeto (GSSG), levando à desnaturação das proteínas da membrana. Neste processo, pode
ocorrer lesão intracelular com oxidação da hemoglobina (Hb) e ocorrendo precipitação da
metahemoglobina (meta-Hb), com isso, formação de corpos de inclusão eritrocitários
denominados corpúsculos de Heinz (FERREIRA & MATSUBARA, 1997; FEKSA et al.,
2008). No corpo humano, 3% do total de hemoglobina (cerca de 750 g) são transformados por
auto-oxidação. Essa reação, que produz meta-Hb e O2•, pode aumentar com o exercício
(BELVIRANLI & GÖKBEL, 2006; FINAUD et al., 2006). Os produtos da peroxidação dos
componentes lipídicos da membrana eritrocitária, também, podem induzir o EO intracelular.
A associação dos fenômenos de oxidação dos grupamentos SH, formação de corpúsculos de
Heinz e lipoperoxidação poderão promover a lesão da membrana do glóbulo vermelho
(FERREIRA & MATSUBARA, 1997; FEKSA et al., 2008).
Durante exercícios ou treinamento físico exaustivos pode ocorrer a destruição de
eritrócitos, não só pelos motivos anteriormente mencionados, mas também em decorrência da
redução do volume do plasma. O deslocamento de água para fora do espaço vascular pode
atuar como estresse químico, uma vez que a perda de água pelo sangue conduz à desidratação
do eritrócito, geralmente como consequência da perda de potássio. A desidratação dos
eritrócitos também é influenciada pelo fenômeno de isquemia/reperfusão. No entanto, em
resposta ao estresse químico, o organismo libera renina, aldosterona e vasopressina (hormônio
antidiurético ou ADH) e o resultado final é um aumento no volume plasmático (CARLSON &
MAWDSLEY, 1986; EICHNER, 2002; PETIBOIS & DÉLÉRIS, 2005). A consequência
imediata é a então chamada “anemia do esporte”, caracterizada por valores reduzidos de
eritrócitos, hemoglobina e hematócrito e alteração do volume celular médio (VCM)
(CARLSON & MAWDSLEY, 1986; EICHNER, 1992; EICHNER, 1996; EICHNER, 1998;
FALLON et al., 1999; VILARDI et al., 2001; EICHNER, 2002).
2.4.4.2. Estresse oxidativo e sistema imune
Além de aumentar o consumo de oxigênio e induzir o EO como resultado da
produção aumentada de EROs, a atividade física exaustiva pode iniciar reações que se
assemelham à fase aguda da resposta imune à infecção, induzindo liberação de proteínas de
fase aguda e mudanças na contagem das células imunes (SANTOS-SILVA et al., 2001;
MOOREN et al., 2002; CAZZOLA et al. 2003; PETIBOIS & DÉLÉRIS, 2004; PETIBOIS
& DÉLÉRIS, 2005; SUREDA et al., 2005; YUSOF et al., 2007). Enquanto os neutrófilos

25
aumentam após exercício exaustivo, a contagem de linfócitos diminui rapidamente, podendo
esta alteração permanecer por diversas horas após o exercício. O treinamento moderado
aumenta a função imune, mas o exercício exaustivo causar perturbações no sistema imune e
depressão imunológica, aumentando o risco de infecções nas vias respiratórias superiores em
decorrência da redução da função antiviral dos macrófagos alveolares nos pulmões (WOODS
et al., 2000; PEIJIE et al., 2003; MOOREN et al., 2002; SUREDA et al., 2005).
Após o exercício exaustivo, inicialmente os neutrófilos e posteriormente os
monócitos e os linfócitos são recrutados para o local de inflamação, onde produzem EROs e
enzimas proteolíticas.
A infiltração de neutrófilos é estimulada por fatores quimiotáticos, incluindo
prostaglandinas, fator de necrose tumoral (TNF)-Į, interleucinas IL-1 e IL-6. Essas duas
últimas citocinas são conhecidas por aumentar em resposta ao exercício. A IL-6 atua como
mediador primário da reação de fase aguda, estimulando a produção hepática de proteínas de
fase aguda, como PCR e inibidores de proteases. Também restringe a extensão da resposta
inflamatória por ativar a síntese de citocinas antiinflamatórias e estimular a glândula hipófise
a liberar o hormônio adrenocorticotrófico (ACTH), que promove o aumento da liberação do
hormônio cortisol pelo córtex adrenal (CRUZAT et al., 2007).
As alterações hormonais que acompanham o exercício e a geração de EROs podem
inibir a proliferação de linfócitos, enquanto o dano oxidativo pode ativar processos
apoptóticos nestas células (SUREDA et al., 2005). Diferentes estudos sugerem que os
processos apoptóticos ativados nos linfócitos após exercício intenso contribuem para a
regulação da resposta imune (MOOREN et al., 2002; SUREDA et al., 2005) e parecem
estar relacionados ao status do treinamento e à intensidade do exercício (PETERS et al.,
2006).
2.4.4.3. Estresse oxidativo e plasma
O aumento do metabolismo imposto pelo exercício promove acréscimo na produção
de EROs, o que pode induzir dano oxidativo nos eritrócitos e no plasma, frações mais
suscetíveis à peroxidação lipídica (SUREDA et al., 2005).
Em humanos, a maioria dos estudos realizados para investigar a peroxidação lipídica
plasmática tem examinado a presença de peróxidos lipídicos ou de bioprodutos da
peroxidação lipídica, tais como dieno conjugados, hidrocarbonetos lipídicos (hidrocarbonetos

26
alifáticos e hidrocarbonetos policíclicos aromáticos–HPAs). Destes, o mais comum é a
quantificação de dieno conjulgados como, por exemplo, o malonaldeído ou MDA, através do
teste de TBARs, cujos resultados são expressos geralmente em milimols (mmol) ou nanomol
(nmol) de equivalentes de MDA (CLARKSON & THOMPSON, 2000). Alterações nas
concentrações plasmáticas das vitaminas C e E e de glutationa oxidada (GSSG) têm sido
também usadas para indicar aumento das reações oxidativas. Pensa-se que essas vitaminas
antioxidantes podem ser mobilizadas dos tecidos para combater o EO em outra parte do
corpo. O efluxo de GSSH para o plasma e a redução de grupamentos sulfídrilas totais é
considerado, atualmente, indicativos de EO, uma vez que a glutationa reduzida (GSH) é
oxidada a GSSG nas células em resposta ao aumento de EROs (CLARKSON &
THOMPSON, 2000).
Após o exercício exaustivo, também ocorrem alterações nas concentrações séricas de
ferro e ferritina, na capacidade total de ligação do ferro e na saturação de transferrina. Apesar
de tais alterações serem similares àquelas encontradas na anemia crônica, apenas refletem a
resposta de fase aguda (FALLON et al., 1999).
Durante o exercício intenso, grande quantidade de ácido lático pode ser produzido
pelo músculo esquelético e liberada na circulação sanguínea para posterior remoção pelos
tecidos periféricos. Os exercícios lactacidêmicos, ou seja, com intensidade de esforço acima
da qual o lactato passa a se acumular na corrente sanguínea (limiar anaeróbio - LAN), têm
também uma influência direta no nível da lipoperoxição plasmática, por inibir o sistema de
defesa antioxidante dos eritrócitos (RIBEIRO et al., 2004; PETIBOIS & DÉLÉRIS, 2005;
MANCHADO et al., 2006).
2.4.4.4. Estresse oxidativo e lipoproteínas
Existem evidências significativas do envolvimento de EROs na patogênese da
aterosclerose. A modificação oxidativa de lipoproteínas de baixa densidade (LDL)
desempenha um papel fundamental na amplificação da resposta inflamatória, mediando uma
variedade de processos imunes pró-inflamatórios que determinam a progressão da
aterosclerose (MAGNUSSON et al., 1994; STEINBERG, 1997; SANTOS-SILVA et al.,
2001; LIU et al., 2004; LAUFS et al., 2005; KIECHL et al., 2007). As EROs contribuem
para o início e a progressão de lesões ateroscleróticas, ao favorecerem a infiltração e o
acúmulo de lipídios no espaço subendotelial. EROs produzidas por células musculares lisas,

27
endoteliais ou macrófagos podem modificar as LDLs que, quando oxidadas, promovem o
recrutamento de monócitos circulantes, o acúmulo de macrófagos residentes, a fagocitose
acelerada e finalmente a formação de “células espumosas” (constituídas por macrófagos ricos
em lipídeos) e estrias gordurosas, característicos das lesões iniciais (STEINBERG, 1997;
SANTOS-SILVA et al., 2001). Adicionalmente, estudos epidemiológicos sugerem que o
aumento da viscosidade do sangue também está relacionado à aterogênese e ao risco
cardiovascular (SANTOS-SILVA et al., 2001).
Neste contexto, exercícios físicos exaustivos e de alta intensidade podem favorecer a
aterosclerose e o risco cardiovascular (SCHNEIDER & OLIVEIRA, 2004), uma vez que têm
sido associados com o aumento do EO no endotélio vascular (LAUFS et al., 2005; PETIBOIS
& DÉLÉRIS, 2005) e com o aumento da viscosidade do sangue (SANTOS-SILVA et al.,
2001). Apesar de exercícios prolongados estarem associados com uma pequena redução
plasmática de triglicerídeos, que pode refletir o uso de LDL (lipoproteína rica em
triglicerídeos) como combustível ou ser atribuída à redução da secreção hepática dessas
lipoproteínas (Hardman, 1998), a inibição do sistema de defesa antioxidante dos eritrócitos
pelo estresse químico induzido por exercícios de intensidade elevada (lactacidêmico) pode
favorecer a oxidação de LDL e, consequentemente, a aterosclerose (SANTOS-SILVA et al.,
2001; PETIBOIS & DÉLÉRIS, 2005).
Por outro lado, evidências epidemiológicas indicam que a atividade física está
associada com risco reduzido de doença coronariana e mortalidade decorrente de problemas
cardiovasculares na meia idade (WANNAMETHEE & SHAPER, 2001). A atividade física
moderada melhora a função endotelial, a capacidade de exercício e a colateralização da
vascularização em pacientes com doença coronariana e previne a progressão da aterosclerose
da carótida. Entre outros efeitos benéficos, o treinamento físico favorece a redução do peso
(massa) corporal, melhora o humor, a pressão sanguínea e a sensibilidade à insulina
(WANNAMETHEE & SHAPER, 2001; LAUFS et al., 2005). A longo prazo, exercícios
moderados também favorecem o aumento da expressão (upregulation) das enzimas
antioxidantes com consequente redução do EO (LAUFS et al., 2005).

28
2.4.5. Estresse Oxidativo e DNA
Uma vez que o metabolismo celular normal está bem estabelecido como fonte
endógena de EROs e que estas espécies se relacionam com níveis basais de dano no DNA
detectados em tecidos normais (COOKE et al., 2003), o exercício exaustivo pode levar ao
EO, o qual pode ser evidenciado pela detecção de produtos da peroxidação lipídica, oxidação
de proteínas e de bases danificadas do DNA (MOLLER et al., 2001; MASTALOUDIS et al.,
2004). De fato, tem sido mostrado que o exercício exaustivo induz dano no DNA de
leucócitos circulantes (NIES et al., 1996; MASTALOUDIS et al., 2004; DEMIRBAG et al.,
2006) e que pode induzir apoptose através de mecanismos diferentes tais como redução
intracelular dos níveis de glutationa, alteração oxidativas à proteínas mitocondriais ou
danificando diretamente o DNA (MOOREN et al., 2002). Por outro lado, tem sido observado
que a extensão dos danos ao DNA de indivíduos treinados é pequena se comparada à de
indivíduos não-treinados, sugerindo que a adaptação ao treinamento de resistência aeróbica
pode reduzir os efeitos do estresse oxidativo, como danos ao DNA (NIES et al., 1996;
MASTALOUDIS et al., 2004; SCHNEIDER & OLIVEIRA, 2004). Neste contexto, além da
técnica de quantificação de oxo8dG para avaliar danos oxidativos indiretos a ácidos nucléicos,
o teste do cometa passou a ser bastante utilizado para determinar danos oxidativos a DNA
induzidos pelo exercício de forma direta e mais precisa (MASTALOUDIS et al., 2004), uma
vez que detecta quebras de fita simples e de fita dupla, sitios alcali-lábeis, sítios incompletos
de reparo, ligação cruzada de DNA-DNA e de DNA-proteína em células individuais
(MASTALOUDIS et al., 2004; BRENDLER-SCHWAAB et al., 2005).
Em contrapartida, frente a tudo que foi exposto, novas pesquisas vêm propondo,
como forma alternativa, a suplementação de antioxidante na atenuação do estresse oxidativo
induzido pelo exercício (ANDRADE et al., 1998; MCBRIDE et al., 1998; MCBRIDE &
KRAEMER, 1999; REID et al., 1994).
2.5 Exercício Físico, Estresse Oxidativo e Suplementação Antioxidante
Segundo Alessio (1993), Smolka et al. (2000) e Ji (2002), a ação das EROs sobre o
sistema enzimático antioxidante tem relação direta com seu nível de produção. Estudos
mostraram que indivíduos treinados podem apresentar níveis mais elevados de enzimas
antioxidantes, e por isso, demonstrarem uma maior resistência ao EO gerado pelo exercício

29
(SEN, 1995; RADÁK et al., 1999; SMOLKA et al., 2000; JI, 2002). Em contrapartida, outros
autores mostraram que sob situação de exercício de alta intensidade há um efeito inibitório na
atividade de diversas enzimas antioxidantes e, como consequência, aumento de ataque
oxidativo celular (DAVIES et al., 1982; SJÖDIN et al., 1990; JI, FU & MITCHELL, 1992;
SMOLKA et al., 2000).
Paralelamente, alguns autores sugerem que a suplementação ou o consumo diário de
antioxidantes naturais como vitamina E (α-tocoferol), vitamina C, e composto polifenólicos
podem atenuar o estresse oxidativo induzido pelo exercício (SIGNORINI & SIGNORINI,
1995; BACURAU, 2000; SCHNEIDER & OLIVEIRA, 2004).
Conforme Goldfarb et al. (1994) um consumo médio diário de 200 mg de vitamina E
pode ser suficiente para prevenir os danos oxidativos causados pelo exercício e, com isso,
aumentar os benefícios proporcionados pelo exercício físico. Em estudos, os mesmos autores
observaram menores concentrações de hidroperóxidos lipídicos e TBARS no plasma e fibras
musculares de ratos submetidos a exercício intenso suplementados por cinco semanas com
vitamina E (250 UI Vit.E/kg). Metin et al. (2002) também verificaram a redução na
concentração de TBARs em ratos suplementados com vitamina E (30 mg/kg/ dia) e treinados
com natação. Rokitzki et al.(1994) relataram que ciclistas ingerindo 300 mg de vitamina E/dia
durante 20 semanas apresentaram menor concentração de MDA após exercício extenuante,
quando comparados ao grupo controle.
Em estudo mais recente, Goldfarb et al. (2005) ministraram doses de 500 ou 1.000
mg de vitamina C/dia a voluntários durante duas semanas e, ao final do período, foi realizada
uma corrida de 30 minutos a 75% do VO2máx. Após o exercício, o grupo suplementado
apresentou menor quantidade de proteínas carboniladas em relação ao grupo controle;
entretanto, não houve influência da suplementação nas concentrações de TBARS. A vitamina
C é hidrossolúvel e está presente em maior concentração no meio aquoso, por isso torna-se
menos eficiente em neutralizar radicais lipofílicos gerados na membrana lipídica (CRUZAT et
al., 2007).
Em mesma linha de estudo, pesquisas recentes sugerem que o consumo de
constituintes fitoquímicos, a exemplo dos compostos fenólicos, presentes em alimentos e chás
de ervas pode atenuar o estresse oxidativo induzido pelo exercício físico e reduzir os riscos de
desenvolvimento de patologias associadas ao EO como o câncer e as doenças
cardiovasculares (HERTOG et al., 1993; ALESSIO et al., 2002; McKAY et al., 2002;
MANACH et al., 2004; McANULTY et al., 2004; MORILLAS-RUIZ et al., 2005; PANZA
et al., 2008).

30
Estudos in vitro têm demonstrado que os flavonóides são potentes neutralizadores de
diversas espécies reativas de oxigênio e de nitrogênio (VAN ACKER, 1996; BIXBY et al.,
2005; ODONTUYA et al., 2005). Além disso, os flavonóides podem interferir na ação
oxidante de metais de transição (VAN ACKER, 1996). A inibição da peroxidação de lipídeos
promovida por flavonóides parece estar relacionada à doação de hidrogênio as espécies
radicalares durante a fase de propagação, bem como, a quelação de metais de transição e ao
efeito preservativo da vitamina E e do β-caroteno no organismo (SALAH et al., 1995; RICE-
EVANS et al., 1996; LOTITO & FRAGA, 2000).
Erba et al. (2005) sugerem que o consumo de chás de produtos naturais ricos em
polifenóis melhora o estado antioxidante geral do organismo e protege contra danos
oxidativos em seres humanos. Neste sentido, a Passiflora cincinnata Mast, conhecida
popularmente como maracujá-do-mato, poderá ser uma excelente ferramenta no controle e
atenuação do estresse oxidativo induzido pelo exercício físico, haja vista que é consenso na
literatura que espécies do gênero Passiflora apresentam atividade antioxidante in vitro e in
vivo por serem rica em compostos fenólicos (DHAWAN et al., 2004; RUDNICKI et al.,
2007).
2.6 Passiflora cincinnata Mast
A Passiflora cincinnata Mast pertence à família Passifloraceae, cujo gênero é
originário da America do Sul, conhecido popularmente como maracujá, possuindo mais de
650 espécies e 16 gêneros distribuídos nas regiões tropicais e semitropicais da América,
África tropical e, principalmente, centro-norte do Brasil (VANDERPLANK, 1996; POZZI,
2007). As espécies mais comuns são a P. incarnata, P. alata e P. edulis, as quais apresentam
efeitos ansiolíticos e sedativos confirmados na literatura, além de valor comercial elevado
(DHAWAN et al., 2004).
O nome Passiflora provém do latim passio, passiones: flor da paixão. Já o termo
maracujá é de origem tupi guarani (muruku’ia: comida feita em cuia, uma alusão ao fruto
édulos do vegetal). As suas flores são vistas como as mais belas da flora mundial. É citado na
literatura seu uso contra ansiedade, enxaqueca, insônia e inflamações (CAPASSO & PINTO,
1995; DHAWAN, KUMAR & SHARMA, 2004).
No Brasil, há aproximadamente 130 espécies de plantas nativas do gênero Passiflora,
das quais a P. cincinnata Mast. faz parte (KILLIP, 1938; OLIVEIRA & RUGGIERO, 2005).

31
No entanto, apenas duas delas são largamente comercializadas: P. edulis, usada comumente
para fazer suco, e P. alata, conhecida como “maracujá doce”, sendo esta última a espécie
oficial da Farmacopéia brasileira (I, II e III), a qual possui efeito ansiolítico e depressor do
sistema nervoso central (OGA et al., 1984; PETRY et al., 2001; DE PARIS et al., 2002;
PEREIRA & VILEGAS, 2000).
As folhas do maracujá passaram a ser utilizadas pela medicina clássica em 1867,
sendo empregadas por sua ação calmante em casos de insônia e irritabilidade (FREITAS,
1985). No entanto, para o gênero Passiflora também é reportada ação hipotensora,
antiinflamatória, antiespasmódica, antimicrobiana, antidiabética e antioxidante (MULLER et
al., 2005; EDWIN et al., 2007).
Embora diversos experimentos in vivo tenham sido realizados a fim de precisar o
constituinte farmacológico ativo da Passiflora, não foi possível atribuir todos os seus efeitos a
uma única classe de compostos ou espécie, mas sim a provável ação conjunta de alcalóides e
flavonóides (POZZI, 2007). Conforme Miller et al. (2005), as folhas do gênero Passiflora são
ricas em compostos fenólicos como taninos e flavonóides, principalmente, os flavonóides C-
glicosílados derivados de apigenina e luteolina (vitexina, isovitexina, orientina, isoorientina).
Além de alcalóides, glicosídeos e terpenos (PEREIRA & VILLEGAS, 2000; DHAWAN et
al., 2004).
Quanto a P. cincinnata Mast., esta é nativa de regiões semi-áridas, e é bastante
utilizada pela medicina popular, principalmente, no nordeste brasileiro, nos Estados da
Alagoas, Bahia, Pernambuco e Sergipe (OLIVEIRA & RUGGIERO, 2005), a qual, também,
está passando a ganhar destaque comercial através da utilização de seu fruto no preparo de
geléias, doces e sucos (FEITOZA et al., 2006). Contudo, há uma deficiência muito grande de
estudos para esta espécie, principalmente, quanto as suas propriedades farmacológicas e
medicinais. Apenas David et al., (2007) verificaram o efeito antioxidante da mesma, a qual
apresentou boa atividade seqüestradora para o radical livre DPPH (CE50 22,3 ± 3,5 µg.mL-1
DPPH) em relação a outras espécies do gênero como a P. coerulea L. (245,84 µg.mL-1
DPPH) e P. edulis (113,41 µg mL-1 DPPH) (VICENTINO & MENESES, 2007; JORGE,
2009).
Neste sentido, apesar das propriedades medicinais do maracujá serem reconhecidas
mundialmente, ainda são precárias as informações científicas sobre algumas espécies. Pouco
se sabe, ainda, a respeito da composição química, princípios ativos e efeitos sobre a saúde de
várias espécies do gênero usadas como medicinais, como exemplo, a P.cincinnata.

32
Não obstante, sabemos que a progressiva destruição da flora brasileira representa
grande ameaça a estas espécies nativas, que se quer são conhecidas ou avaliadas em todo seu
potencial biológico, medicinal e químico. Por disso, torna-se urgente o estudo das plantas
conhecidas popularmente como medicinais e utilizadas empiricamente na medicina popular.
Haja vista que, a comprovação científica do efeito benéfico à saúde do consumo destes
produtos, abrirá um grande leque para novos estudos quanto à descoberta de novos
fitoquímicos e medicamentos para prevenção e terapia das diversas doenças associadas às
ações oxi-redotoras dos RL. Bem como, poderá servir como uma nova opção de renda para
pequenos agricultores do semi-árido nordestino, tanto na comercialização de chás, na forma
de saquês, quanto na fabricação de doces e geléias, bem como, na comercialização do vegetal
como matéria prima à indústria farmacêutica para formulação de medicamento. Sendo assim,
tanto a população em geral será beneficiada, direta e indiretamente, quanto à natureza, a qual
terá seu espaço verde aumentando, e a comunidade científica que estará justificando sua
função social, econômica, ambiental e científica perante a sociedade.

33
3. OBJETIVOS
3.1 Geral
Avaliar a atividade antioxidante e redox-protetora do extrato e frações das folhas de
P. cincinnata Mast. sobre o estresse oxidativo induzido pelo exercício físico em ratos.
3.2 Específicos
� Identificar os grupos fitoquímicos presentes no extrato bruto das folhas P. cincinnata
Mast.;
� Determinar o conteúdo fenólico no extrato e frações das folhas P. cincinnata Mast.;
� Avaliar a atividade seqüestradora dos radicais 2,2-difenil-1-picrilhidrazina (DPPH•) e
2,2'-azinobis-3-etilbenzotiazolino-6-sulfonato (ABTS●+) do extrato e frações das
folhas P. cincinnata Mast.;
� Avaliar a atividade redox-protetora sobre a lipoperoxidação induzida por dihidrocloreto
de 2,2’azobis-2-amidopropano (AAPH); sulfato ferroso (FeSO4) e peróxido de
hidrogênio (H2O2);
� Avaliar a atividade redox-protetora em órgãos e tecidos de ratos após administração
aguda de P. cincinnat Mast;
� Avaliar a atividade redox-protetora da P. cincinnat Mast em órgãos e tecidos de ratos
após dois meses de treinamento físico de alta intensidade;

34
4. METODOLOGIA
4.1 Coleta e Identificação do Material Vegetal
Folhas de P. cincinnata Mast foram coletadas pela manhã, em março de 2008, na
cidade de Moita Bonita, agreste sergipano, Brasil, e levadas para a Universidade Federal de
Sergipe (UFS). Um voucher da espécie foi depositado no Herbário da instituição sob número
ASE 11.112. A identificação Botânica foi realizada pela Drª. Ana Paula do Nascimento Prata
do departamento de Biologia da UFS. As amostras foram submetidas à secagem em estufa
(MARCONI, modelo 037/18) com renovação e circulação de ar a temperatura constante de 40
°C até completa desidratação.
4.2 Preparação e Fracionamento do Extrato Etanólico Bruto
As folhas secas foram reduzidas a pó (293,3 g) utilizando moinho de facas, e em
seguida, submetidas à extração com etanol a 95% durante 5 dias, por maceração exaustiva.
Após este período, o material foi filtrado e concentrado em rota-evaporador sob pressão
reduzida (BUCHI, modelo/461) a uma temperatura de 55 ºC, obtendo-se 103,3 g de extrato
etanólico das folhas de “maracujá-do-mato” (ExEt), rendimento de 35,22%. Parte do ExEt (50
g) foi diluído em uma solução metanol/água (2:3) e submetido à extração líquido-líquido com
clorofórmio e acetato de etila. Obtendo, assim, as frações clorofórmica (FCl), acetato de etila
(FActE) e hidrometanólica (HMoH).
Para melhor delineamento experimental do nosso estudo, conduzimos os
experimentos em duas etapas. A primeira consistindo em ensaios químicos, que buscaram
identificar os constituintes fitoquímicos presentes no extrato e quantificar aqueles que,
segundo a literatura, apresentaram potencial antioxidante. O extrato e/ou frações que
apresentaram maiores quantidades desses constituintes foram testados em ensaios biológicos.

35
4.3 Ensaios Químicos
4.3.1 Triagem fitoquímica
O extrato etanólico foi submetido à prospecção fitoquímica, seguindo metodologia
descrita por Matos (1998). O objetivo foi detectar a ocorrência de diversas classes de
constituintes e, principalmente, as que apresentam atividade antioxidante, conforme a
literatura. Para tanto, sete porções de 3-4 mL do extrato foi dissolvido em etanol e foram
colocados em tubos de ensaio enumerados de 1 a 7. Todos os testes foram realizados em
triplicata.
4.3.1.1 Teste para fenóis e taninos
No tubo de número 1, foram adicionadas três gotas de solução alcoólica de FeCl3 1
mol.L-1 em HCl 60 mmol.L-1. Após agitação, foram observadas quaisquer variações de cor
e/ou formação de precipitado escuro abundante. O resultado foi comparado com um branco,
usando-se água e FeCl3.
A variação da coloração entre azul e vermelho foi indicativa de fenóis. A formação
de um precipitado azul escuro indicou à presença de taninos pirogálicos (taninos
hidrolisáveis) e de cor verde a presença de taninos flobabênicos (taninos condensados ou
catéquicos).
4.3.1.2 Teste para antocianinas, antocianidinas e flavonóides
A porção do extrato nos tubos de número 2 foram acidificadas a pH 3 com HCl 3
mol L-1, enquanto as porções nos tubos 3 e 4 foram alcalinizadas a pH 8,5 e 11 com NaOH 1
mol L-1. As mudanças de coloração foram interpretadas conforme descrito na tabela 2.
4.3.1.3 Teste para leucoantocianidinas, catequinas e flavononas
A porção do extrato no tubos de número 5 foram acidificadas a pH 3 por adição de
HCl 3 mol.L-1, enquanto o material no tubos 6 foram alcalinizadas a pH 11 com NaOH 1

36
mol.L-1. Os tubos foram aquecidos cuidadosamente, sendo observada qualquer modificação
na coloração por comparação com os tubos correspondentes usados no teste anterior. A
interpretação dos resultados foi feita como mostrado tabela 3.
Tabela 2. Detecção colorimétrica de antocianinas, antocianidinas e flavonóides.
Constituintes Cor
Ácido Ph=3 Alcalino pH=8,5 Alcalino pH=11
Antocianinas e antocianidinas Vermelha Lilás Azul-púrpura
Flavonas, flavonóis e xantonas - - Amarela
Chalconas e auronas Vermelha - Vermelho Púrpuro
Flavononóis - - Vermelho Laranja
Tabela 3. Detecção colorimétrica de leucoantocianidinas, catequinas e flavononas.
Constituintes
Cor
Meio Ácido Meio Alcalino
Leucoantocianidinas Vermelha -
Catequinas (taninos catéquicos) Pardo-amarelada -
Flavononas - Vermelho Laranja
4.3.1.4 Teste para flavonóis, flavononas, flavononóis e xantonas
Nos tubos de número 7 foram adicionados 10 mg de magnésio granulado e 0,5 mL
de HCl concentrado. O término da reação foi indicado pelo fim da efervescência. Os
resultados foram observados por comparação a mudança na cor da mistura da reação nos
tubos 5 e 7. O aparecimento ou intensificação da cor vermelha foi indicativo da presença de
flavonóis, flavanonas, flavanonóis e/ou xantonas, livres ou seus heterosídios.
4.3.1.5 Teste para esteróides e triterpenóides (Liebermann-Buchard)
10 mL do extrato foi adicionado a um becker e deixado secar em banho-maria.
O resíduo seco de cada becker foi extraído três vezes com porções de 1-2 mL de CHCl3.

37
Sendo os extratos colocados em tubos diferentes e colocaram-se algumas gotas de CHCl3. A
solução clorofórmica foi filtrada em um pequeno funil fechado com algodão, coberta com
Na2SO4 anidro, para um tubo de ensaio bem seco. Adicionou-se 1 mL de anidrido acético e
agitou-se suavemente. Adicionaram-se cuidadosamente três gotas de H2SO4 concentrado.
Agitou-se suavemente e observou-se o rápido desenvolvimento de cores. A coloração azul
seguida da verde permanente é um indicativo da presença de esteróides livres. Coloração
parda até vermelha indica triterpenóides pentacíclicos livres.
4.3.1.6 Teste para saponinas
Os resíduos insolúveis em clorofórmio, separados no teste anterior, foram
solubilizou-se em água destilada e filtrado para um tubo de ensaio. O tubo com a solução foi
então fortemente agitado por dois a três minutos e observou a formação da espuma. O
aparecimento de espuma persistente e abundante indica a presença de saponinas.
Para confirmar a presença de saponinas, 2 mL de HCl concentrado foram
adicionados ao conteúdo do tubo de ensaio, que foi deixando por uma hora imerso em banho-
maria. Após período de imersão, o material no tubo foi neutralizando, resfriado e novamente
agitado. A presença de precipitado e a não formação de espuma confirmou a presença de
saponina.
4.3.1.7 Teste para alcalóides
O extrato após ser dissolvido em etanol foi filtrado e posto em um erlermayer. Ao
filtrado foi adicionado uma solução de NH4OH 6 N até atingir pH 11. Posteriormente, para a
extração de bases orgânicas do filtrado foram adicionados três porções sucessivas de 30, 20 e
10 mL de clorofórmio. Utilizando um funil de separação com Na2SO4 anidro a fase aquosa foi
separada para o teste de bases quaternárias, obtendo assim duas soluções, uma delas foi posta
para secar sendo utilizada para o teste em cromatoplaca e a outra adicionada três porções
sucessivas de HCl diluído a 0,1 N para extração das bases orgânicas, a qual foi colocada em
três tubos de ensaios. Nestes foram adicionados, distintamente, três gotas dos reagentes de
precipitação de alcalóides, “Hager”, ”Mayer” e “Dragendorff”. A presença de precipitado
floculoso, pesado em pelo menos dois tubos sugeriu a presença de alcalóides.

38
4.3.2 Quantificação de fenóis totais (FT)
A quantificação espectrofotométrica de compostos fenólicos foi realizada através do
método de Folin-Ciocalteau (SOUSA at al., 2007). ExEt e FActE (10 mg) foram dissolvidos
em metanol, sendo transferidas quantitativamente para um balão volumétrico de 10 mL e o
volume completado com metanol para dar uma solução de concentração final 1 mg.mL-1.
Alíquotas de 0,1 mL desta solução foram transferidas para tubos Falcon e agitadas em
presença de 0,5 mL do reagente de Folin-Ciocalteau e 6 mL de água destilada por 1 min.
Passado este tempo, 2 mL de Na2CO3 a 20% foram adicionados à mistura e foi então agitada
por 30 s. Por fim, a solução foi acertada com água destilada para um volume final de 10 mL e
deixada por 2 h em ambiente livre de luz. Após este período de repouso, as absorbâncias das
amostras foram medidas a 750 nm utilizando-se cubetas de vidro, tendo como “branco” o
metanol e todos os reagentes, menos o extrato ou frações. O teor de FT foi determinado por
interpolação da média de absorbância de três leituras da amostra contra uma curva de
calibração construída com padrões de ácido gálico (10 a 350 µg.mL-1) e expresso como mg de
equivalentes de ácido gálico (EAG) por g de extrato. A equação da curva de calibração do
ácido gálico foi C = 809,0200A + 5,0827, onde C é a concentração do ácido gálico, A é a
absorbância a 750 nm e o coeficiente de correlação R = 0,999. Todas as análises foram
realizadas em triplicata.
4.3.3 Avaliação da atividade antioxidante (AA)
A avaliação da AA do extrato e fração da P. cincinnata Mast que apresentaram maior
conteúdo fenólico foram realizadas através da capacidade sequetradora dos radicais DPPH• e
ABTS•+.
4.3.3.1 Capacidade seqüestradora do radical DPPH•
A capacidade de sequestrar o radical DPPH• foi determinada conforme metodologia
descrita por Soler-Rivas et al. (2000) e Argolo et al. (2004). Uma alíquota de 50 mL de
solução estoque de DPPH• em metanol na concentração de 40 µg.mL-1 foi preparada e
mantida sob refrigeração e protegida da luz. Em seguida, diluições para dar as concentrações

39
finais de 30, 25, 20, 15, 10, 5 µg.mL-1 da solução estoque de DPPH• foram preparadas. A
curva de calibração foi construída a partir dos valores da absorbância a 515 nm de todas as
soluções medidas em cubetas de vidro com percurso óptico de 1 cm, tendo como “branco” o
metanol. As medidas de absorbância foram efetuadas em triplicata e nos tempo de 1, 5, 10 20,
30, 40, 50 e 60 min. A Equação da curva de calibração do DPPH• foi C = 35,846A – 0,230,
onde C corresponde à concentração de DPPH• no meio, A é a absorbância medida no
comprimento de onda a 515 nm e o coeficiente de correlação R = 0,9997.
Igualmente, soluções do extrato e da fração de passiflora (500 µg.mL-1), bem como
do controle positivo (ácido gálico) em metanol foram diluídas para dar concentrações finais
de 30, 25, 20, 15, 10 e 5 µg.mL-1. As medidas das absorbâncias das misturas reacionais (0,3
mL da amostra ou do controle positivo e 2,7 mL de DPPH•) foram realizadas a 515 nm nos
mesmos tempos da curva de calibração do DPPH•. Misturas de metanol (2,7 mL) e solução
metanólica do extrato (0,3 mL) foram utilizadas como branco.
A partir da equação da curva de calibração e dos valores de absorbância para cada
concentração testada, foram determinados os percentuais de DPPH• remanescentes
(%DPPH•REM), conforme a equação:
%DPPH•REM = [DPPH•]T=t ÷ [DPPH•]T=0 x 100
Onde, [DPPH•]T=t corresponde à concentração de DPPH• no meio, após a reação com
o extrato e [DPPH•]T=0 é a concentração inicial de DPPH•.
Os valores de absorbância em todas as concentrações testadas foram convertidos em
porcentagem de inibição (IP).
A concentração eficiente, quantidade de antioxidante necessária para decrescer a
concentração inicial de DPPH• em 50% (CE50), foi determinada usando o programa Microcal
Origin 7.5, a partir de uma curva exponencial de primeira ordem, obtida plotando no eixos das
abscissa as concentrações da amostra (µg.mL-1) ou do controle positivo e no eixo das
ordenadas, a porcentagem de DPPH• remanescente (% DPPH•REM).
AA foi determinada através do índice de atividade antioxidante (IAA) conforme
Scherer e Godoy (2009) e calculada através da equação abaixo.
IAA = %DPPH• REM (µg.mL-1) ÷ CE50 (µg.mL-1)

40
A atividade antioxidante foi considerada insatisfatória quando o valor do IAA for
inferior a 0,5, moderada quando o IAA estiver entre 0,5 e 1,0, forte quando o IAA estiver
entre 1,0 e 2,0, e muito forte quando o valor do IAA for superior a 2,0.
4.3.3.2 Capacidade seqüestradora do radical ABTS•+
A capacidade de sequestrar o radical ABTS•+ foi avaliada seguindo metodologia
descrita Re et al. (1999). O radical livre ABTS•+ se obtém mediante a reação de ABTS com
persulfeto de potássio.
Soluções estoque de ABTS (7 mM) com persulfeto de potássio (2,45 mM) foram
utilizadas para gerar o radical ABTS•+ por misturas em partes iguais, sendo a mesma
armazenada a temperatura de ±20 ºC num prazo de 12 h ao abrigo da luz. Formado o radical
ABTS•+, este foi diluído misturando 1 mL da solução ABTS•+ com metanol (~35 mL) até a
obtenção de um valor de absorbância de 0,700 (±0,03) unidades a 734 nm, usando o
espectrofotômetro.
Soluções do extrato e fração de Passiflora (500 µg/mL) e do controle positivo (ácido
gálico e hidroxitolueno butilato - BHT) em metanol foram diluídas nas concentrações de 30,
25, 20, 15, 10 e 5 µg.mL-1 . As medidas das absorbâncias das misturas reacionais (0,3 mL das
amostras ou controles positivo e 2,7 mL de ABTS•+) foram realizadas a 734 nm (±25 ºC) no
tempo de 5 minutos. Misturas de metanol (2,7 mL) e solução metanólica do extrato ou fração
(0,3 mL) foram utilizadas como branco.
Os resultados da AA estarão expressos em IAA, calculados como descrito no item
4.3.3.1.
4.3.4 Avaliação da atividade redox (in vitro)
A capacidade do extrato para inibir a lipoperoxidação foi determinada através do
monitoramento da produção de substâncias reativas ao ácido tiobarbitúrico (TBARs) em meio
rico em lipídio conforme protocolo descrito por Silva et al. (2007). Para a quantificação de
TBARs foi realizada protocolo de Lapenna et al. (2001).
Em síntese, 1,0 mL de homogenato de gema de ovo (1% m/v) em tampão fosfato 20
mM (pH 7,4), foi sonicado (10 s na potência 4) e, em seguida, homogeneizado com 0,1 mL do
ExEt e controles positivos em diferentes concentrações (5, 50 e 100 µg.mL-1) preparadas

41
imediatamente antes do experimento com metanol. A peroxidação lipídica foi induzida pela
adição de 0,1 mL de solução de AAPH (0,17 M), FeSO4 (0,17 M) e H2O2 (0,4 M) em
momentos distintos. Trolox e ácido gálico foram usados como moléculas antioxidantes de
referência (controle positivo); o controle negativo foi apenas extrato e veículo (água ou
metanol). As reações foram realizadas por 30 min a 37 ºC. Após o resfriamento, as amostras
(0,5 mL) foram centrifugadas com 0,5 mL de ácido tricloroacético (15%) a 1200 g por 10
min. Uma alíquota de 0,5 mL do sobrenadante foi misturada com 0,5 mL de TBA (0,67%) e
aquecida a 95 °C por 60 min. Após o resfriamento, as absorvâncias foram medidas usando
espectrofotômetro a 532 nm. O coeficiente de extinção molar de 1,54 x 10-5 M-1.cm-1 foi
usado no presente estudo e o resultado foi expresso em equivalentes de malondialdeído (Eq
MDA).mL -1 de substrato.
.
4.4 Ensaios Biológicos
4.4.1 Animais
Foram utilizados 40 ratos machos da linhagem Wistar com 3 meses de idade (250-
300 g) obtidos do biotério central da UFS. Os mesmos foram aleatoriamente alojados em
gaiolas apropriadas sob temperatura controlada (22 ± 3 ºC) com ciclo claro escuro de 12 horas
(luzes acesas, 06h00 – 18h00), com livre acesso à alimentação específica para roedores e água
de torneira. Todos os procedimentos descritos no presente trabalho foram aprovados pelo
Comitê de Ética em Pesquisa Animal da UFS (protocolo 58/09).
4.4.2 Delineamento experimental
Os ensaios biológicos foram conduzidos em duas etapas. A primeira teve por
objetivo avaliar a capacidade redox-protetora do ExEt das folhas da P. cincinnata Mast em
condições de tratamento agudo, após uma única administração de 200 mg.Kg-1 do ExEt
(previamente selecionado, após estudo piloto realizado em nosso laboratório, quanto a
redução da lipoperóxidação sanguínea de ratos que ingeriram distintamente 50, 100 e 200
mg.kg-1 do ExEt), sobre parâmetros de estresse oxidativo em ratos sob condições fisiológicas

42
basais. Para tanto, os animais foram divididos aleatoriamente em dois grupos (n=8), GC
(salina, vo) e GP (Passiflora 200 mg.Kg-1, vo), após uma hora à administração do extrato, os
animais foram anestesiados com tiopental sódico (45 mg.Kg-1, ip), e o sangue (5 mL) coletado
por punção cardíaca com seringas heparinizadas. Paralelamente, os órgãos também foram
removidos (fígado e coração) para posteriores análises dos marcadores de estresse oxidativo.
Na segunda etapa dos experimentos, foi avaliada a capacidade redox-protetora do
ExEt das folhas de P. cincinnata Mast em condições de tratamento crônico, consumo diário
de 200 mg.Kg-1 do extrato ao final de 8 semanas, quanto aos parâmetros de estresse oxidativo
induzido por protocolo de exercício físico de alta intensidade. Para tanto, quatro grupos foram
divididos aleatoriamente (n=6): GC (Salina, vo), GP (Extrato, vo), GT (Exercício aeróbico) e
GPT (Extrato (vo) + Exercício aeróbico). Em seguida, foram tratados com 200 mg.kg-1 do
ExEt das folhas de P. cincinnata (GP e GPT) durante 8 semanas, uma vez por dia, 5 vezes por
semana. Paralelamente, os animais (GT e GPT) realizaram exercício físico como descrito por
Smolka et al. (2000) na tabela 4. Ao final do período de treinamento, um dia após a ultima
sessão para evitar o efeito agudo do exercício, os animais foram anestesiados com tiopental
sódico (45 mg.kg-1 de peso, ip) e o sangue coletado (5 mL) por punção cardíaca com seringas
heparinizadas. Paralelamente, urina e órgãos (fígado e coração), também, foram removidos
para análise dos marcadores de estresse oxidativo.
Tabela 4: Protocolo de exercício físico para indução de estresse oxidativo.
Semana Velocidade (m.min-1) Duração (min)
1ª 15 20
2ª 20 30
3ª 22,5 45
4ª 25 60
5ª – 8ª 25 60 *Os animais exercitaram 5 dias por semana.
4.4.3 Preparo do material biológico
Os procedimentos para o preparo do material biológico foram realizados segundo
metodologia descrita por Sant´Anna (2005). O sangue, após a coleta, foi imediatamente
centrifugado a 800 × g por 15 minutos a ± 4º C e o sobrenadante armazenado a ± -70º C.
Paralelamente, os órgãos removidos (fígado e coração) foram lavados por 3x em solução de

43
cloreto de potássio (KCl) 1,15%, secos e pesados. Em seguida, homogeneizados onde cada
grama de tecido foi misturada com 5 mL de KCl + 10 µL de Fluoreto de Fenilmetilsulfonila
(PMSF - 100 m.mol-1) + 15 µL de solução Triton a 10%, centrifugada 3000 x g por 10
minutos a ± 4°C e o sobrenadante armazenando a ± -70º C para análises posterior dos
marcadores de estresse oxidativo.
A urina dos animais, coletada após o período de treinamento, recebeu 100 µL de
EDTA e foi armazenada a ± -70º C para análise posterior dos marcadores de estresse
oxidativo.
4.4.4 Determinação de lipoperoxidação
A oxidação de lipídios foi determinada pela medida de substâncias reativas ao ácido
tiobarbitúrico (TBARS) de acordo com método descrito por Lapenna et al. (2001). Alíquotas
de 200 µL das amostras (sangue, urina e órgãos) foram adicionadas a uma mistura formada
por partes iguais de ácido tricloroacético (TCA) 15%, HCl 0,25 N e TBA 0,375%, mais 2,5
mM de hidroxitolueno butilado (BHT), e 40 µL de dodecil de sódio (SDS) a 8,1%, sendo
aquecida por 30 mim à 95ºC em estufa. O pH da mistura foi ajustado para 0,9 com HCl. O
BHT foi usado para prevenir a peroxidação lipídica durante o aquecimento. Em seguida, após
resfriamento à temperatura ambiente e adição de 4 mL de n-butanol, o material foi
centrifugado a 800xg por 15 minutos a ± 4º C e a absorbância do sobrenadante foi medida a
532 nm. O coeficiente de extinção molar utilizado foi 1,54 x 105 M-1 cm-1 e o resultado de
TBARS expresso em nmol Eq MDA.mL-1 de plasma para as amostras de sangue, em nmol Eq
MDA.mL-1 de urina para as amostras de urina ou em nmol Eq MDA.mL-1 soro para os
órgãos.
4.4.5 Determinação de proteína carbonilada
A oxidação de proteínas foi avaliada através da determinação de grupos carbonil de
acordo com a metodologia descrita por Faure & Lafond (1995). Alíquotas de 200 µL das
amostras (fígado e coração) foram misturadas a 800 µL de DNPH (dinitrofenilhidrazina) e
incubadas por uma hora a ± 37 ºC, no escuro, sendo agitado a cada 15 minutos. Em seguida,
1000 µL de TCA 15% foram adicionados às amostras, as quais foram centrifugadas a 1000 x
g por 10 minutos a ± 4º C. Após esse tempo, acrescentou-se ao precipitado 800 µL de TCA

44
15% e agitou-se. O precipitado foi lavado 3x com 800 µL de etanol/acetato de etila (1:1) para
remover o DNPH livre e lipídios contaminantes. O precipitado final foi dissolvido em 400 µL
de Guanidina 6M e incubado por 10 minutos a ± 37°C. O material insolúvel foi removido por
centrifugação adicional a 1000 x g por 5 minutos a ± 4 ºC. O sobrenadante foi recolhido e a
absorbância foi determinada espectrofotometricamente a 380 nm. O coeficiente de extinção
molar utilizado foi de 22 mM-1 cm-1 e o resultado expresso em nmol.mg-1 de tecido.
4.4.6 Determinação de sulfidrilas totais (tióis)
Os grupos sulfidrilas (SH) são estruturas associadas a proteínas e, portanto,
susceptíveis a danos oxidativos. Sua quantificação revela o nível antioxidante do tecido. A
determinação dos grupos sulfidrilas foi realizada conforme metodologia descrita por Faure &
Lafond (1995). Alíquotas de 50 µL de amostra de fígado e/ou coração foram misturadas em 1
mL de tampão Tris-EDTA, pH 8,2. Em seguida, foi realizada a primeira leitura (A) no
espectrofotômetro a 412 nm. Após a leitura, as amostras foram transferidas para tubos de
ensaio e misturadas a 20 µL de DTNB 10 mM diluído em metanol (4 mg.mL-1), ficando em
repouso no escuro. Ao final de 15 minutos, a segunda leitura de absorbância (A2) foi
realizada. A concentração de SH foi calculada conforme equação: (A2 - A1) – B x 1,57 mM x
1000. O resultado expresso em nmol.mg-1 tecido.
4.5 Análise Estatística
Os dados obtidos foram expressos em média ± EPM. Para observar a normalidade
dos dados foi utilizado o teste Shapiro-Wilk através do pacote estatístico XLSTAT-Pro. Para
avaliar a significância das diferenças entre as médias foram utilizados os testes t de Student e
a análise de variância de uma via ou duas vias seguido do pós-teste de Bonferroni ou Tukey,
quando apropriado. Os valores foram considerados estatisticamente significativos quando p <
0,05. Para todos estes procedimentos foi utilizado o programa estatístico GraphPad Prism
versão 5.0 (GraphPad Software, San Diego,CA, E.U.A.). Todos os experimentos avaliados
foram realizados em triplicata.
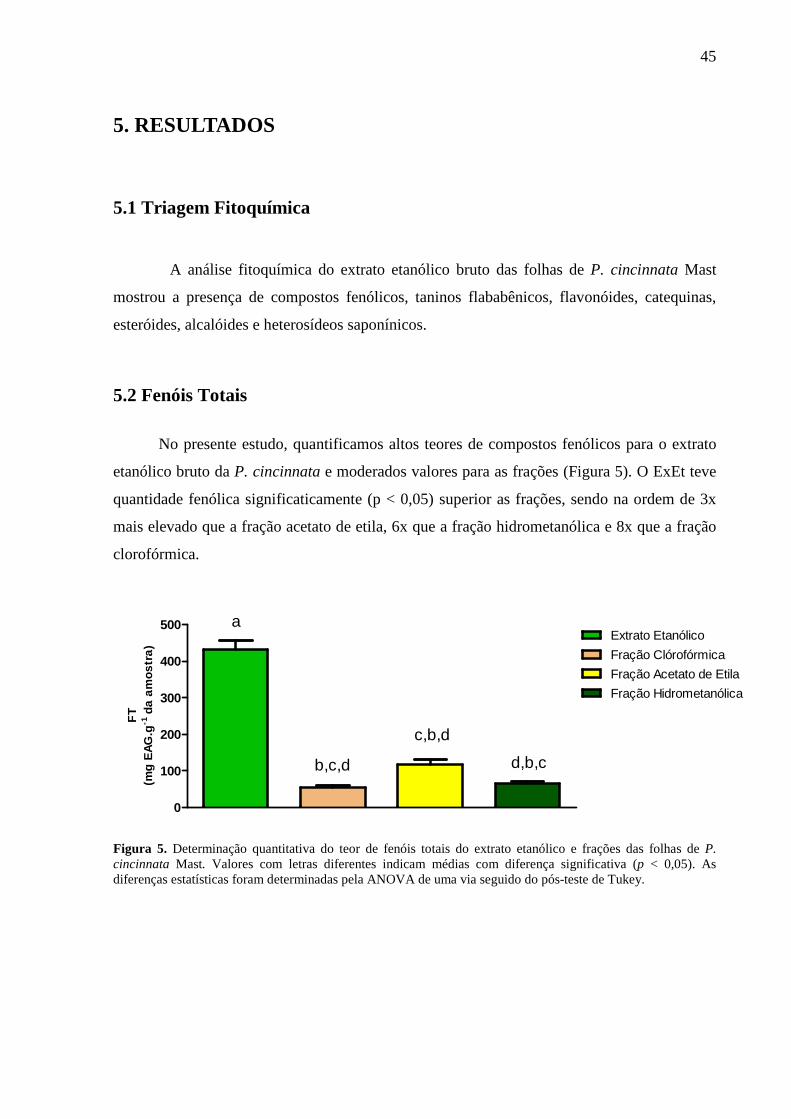
45
5. RESULTADOS
5.1 Triagem Fitoquímica
A análise fitoquímica do extrato etanólico bruto das folhas de P. cincinnata Mast
mostrou a presença de compostos fenólicos, taninos flababênicos, flavonóides, catequinas,
esteróides, alcalóides e heterosídeos saponínicos.
5.2 Fenóis Totais
No presente estudo, quantificamos altos teores de compostos fenólicos para o extrato
etanólico bruto da P. cincinnata e moderados valores para as frações (Figura 5). O ExEt teve
quantidade fenólica significaticamente (p < 0,05) superior as frações, sendo na ordem de 3x
mais elevado que a fração acetato de etila, 6x que a fração hidrometanólica e 8x que a fração
clorofórmica.
0
100
200
300
400
500Extrato Etanólico
Fração Clórofórmica
Fração Acetato de Etila
Fração Hidrometanólica
a
b,c,d
c,b,d
d,b,c
FT
(mg
EAG
.g-1
da
am
ostr
a)
Figura 5. Determinação quantitativa do teor de fenóis totais do extrato etanólico e frações das folhas de P. cincinnata Mast. Valores com letras diferentes indicam médias com diferença significativa (p < 0,05). As diferenças estatísticas foram determinadas pela ANOVA de uma via seguido do pós-teste de Tukey.

46
5.3 Atividade Antioxidante (in vitro)
5.3.1 Atividade sequestradora do radical DPPH•
As amostras que apresentaram os maiores teores de FT (ExEt e FActE) foram
utilizadas para a quantificação da atividade seqüestradora do radical DPPH•. Foi constatado
maior percentual de inibição (IP) ao radical DPPH• pelo ExEt em comparação a FActE (p <
0,05), sendo seu valor similar aos controles BHT e AG (p > 0,05). Da mesma forma, quando
foi constatado o consumo do ExtE para reduzir o radical em 50% (CE50), ele foi menor e,
portanto, melhor que a FActE e o controle BHT (p < 0,05), apresentando resposta similar ao
controle AG ( p > 0,05) como podemos observar na tabela 5.
Quanto a determinado do potencial antioxidante através do índice de atividade
antioxidante (IAA), foi observado que o ExEt apresentou maior IAA na redução do radical
DPPH• quanto a FActE e BHT (p > 0,05), e valor similar aquele observado para AG (p > 0,05,
tabela 5), mostrando com isso características semelhantes do ExEt com o controle AG na
interação antioxidante ao radical DPPH•.
Tabela 5. Atividade antioxidante do extrato etanólico e fração Acetato de etila de P. cincinnata Mast determinado através da redução do radical livre DPPH•
IP
(% ± EPM)
CE50
(µg.mL-1 ± EPM) IAA
ExEt
97,01 ± 0,72a
7,26 ± 1,85 a
4,13 ± 0,16 a
FActE 23,69 ± 0,80 b 21,59 ± 0,80 b 0,41 ± 0,01 b
AG 96,24 ± 0,12 c,a 9,33 ± 0,22 c,a 3,21 ± 0,13 c,a
BHT 96,91 ± 0,25 d,a,c 12,33 ± 3,25 d,c 2,43 ± 0,16 d,c
Extrato etanólico bruto (ExEt) e fração acetato de etila (FActE) de folhas de P. cincinnata Mast. Valores com letras diferentes indicam médias com diferença significativa (p < 0,05). Percentual de inibição (IP) das amostras (30 µg/mL) foram calculados no tempo de 60 min. A concentração eficiente em 50% (CE50)
das amostras foram calculadas no tempo de 60 min. O índice de atividade antioxidante (IAA) foi calculado dividindo o valor da concentração final da amostra pelo CE50 no tempo de 60 min e classificado como fraco quando IAA < 0,5, moderado quando IAA > 0,5 < 1, forte quando IAA > 1 < 2 e muito forte quando IAA > 2. As diferenças estatísticas foram determinadas pela ANOVA de uma via seguido do pós-teste de Tukey.

47
5.3.1 Atividade sequestradora do radical ABTS•+
O ExEt apresentou excelente resposta na redução do radical catiônico ABTS•+,
reduzindo o radical de forma quantitativamente superior os controles AG e BHT (p < 0,05).
Porém, o extrato foi menor que os controles devido o mesmo apresentar maior valor de CE50
(p < 0,05). Em contrapartida, o extrato teve forte índice de atividade antioxidante, assim
como, os controles positivos (Tabela 6).
Tabela 6. Atividade antioxidante do extrato etanólico de P. cincinnata Mast determinado através da redução do radical livre ABTS•+
IP
(% ± EPM)
CE50
(µg mL-1 ± EPM) IAA
ExEt
97,05 ± 0,21 a
7,91 ± 0,38 a
3,79 ± 0,78 a
AG 95,89 ± 0,01 b 1,31 ± 0,01 b 11,53 ± 0,03 b
BHT 92,28 ± 0,30 c 2,60 ± 0,06 c 22,90 ± 0,05 c
Extrato etanólico bruto (ExEt) e fração acetato de etila (FActE) de folhas de P. cincinnata Mast. Valores com letras diferentes indicam médias com diferença significativa (p < 0,05). Percentual de inibição (IP) das amostras (30 µg/mL) foram calculados no tempo de 60 min. A concentração eficiente em 50% (CE50)
das amostras foram calculadas no tempo de 60 min. O índice de atividade antioxidante (IAA) foi calculado dividindo o valor da concentração final da amostra pelo CE50 no tempo de 60 min e classificado como fraco quando IAA < 0,5, moderado quando IAA > 0,5 < 1, forte quando IAA > 1 < 2 e muito forte quando IAA > 2.. As diferenças estatísticas foram determinadas pela ANOVA de uma via seguido do pós-teste de Tukey.
5.4 Atividade Redox (in vitro)
A atividade redox in vitro do ExEt foi determinada através da capacidade de inibir a
lipoperoxidação induzida por três agentes químicos AAPH, FeSO4 e H2O2 e quantificada pela
dosagem de TBARs. As diferentes concentrações testadas do ExEt das folhas de P. cincinnata
Mast reduziram os níveis de TBARs nos três indutores testado (p < 0,05) (Figuras 6, 7 e 8). O
ExEt apresentou característica redox semelhante ao controle ácido gálico e trolox em todas as
concentrações testadas quanto à prevenção da lipoperoxidação induzida pelo AAPH (p >
0,05), com valores acima de 95% em todas as concentrações avaliadas (Figura 6).
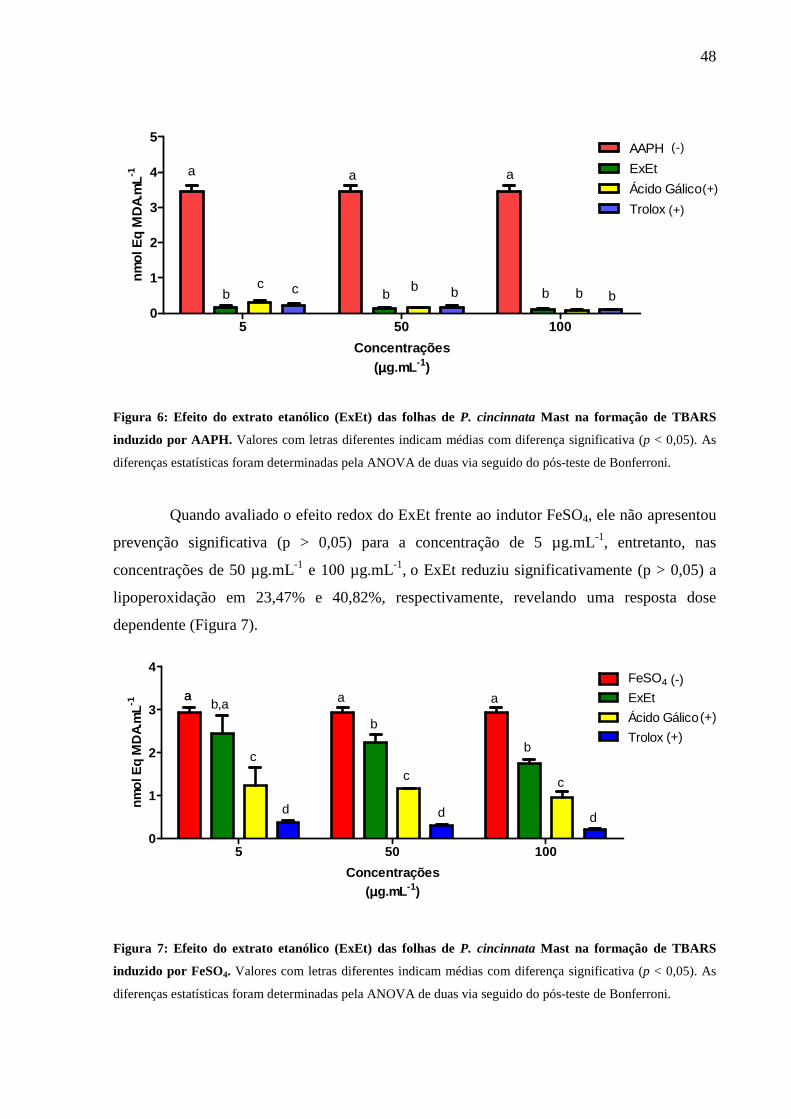
48
5 50 1000
1
2
3
4
5AAPH
ExEt
Ácido Gálico
Trolox
a a a
b b bc b bc b b
(-)
(+)
(+)
Concentrações(µµµµg.mL -1)
nmol
Eq
MD
A.m
L-1
Figura 6: Efeito do extrato etanólico (ExEt) das folhas de P. cincinnata Mast na formação de TBARS
induzido por AAPH. Valores com letras diferentes indicam médias com diferença significativa (p < 0,05). As
diferenças estatísticas foram determinadas pela ANOVA de duas via seguido do pós-teste de Bonferroni.
Quando avaliado o efeito redox do ExEt frente ao indutor FeSO4, ele não apresentou
prevenção significativa (p > 0,05) para a concentração de 5 µg.mL-1, entretanto, nas
concentrações de 50 µg.mL-1 e 100 µg.mL-1, o ExEt reduziu significativamente (p > 0,05) a
lipoperoxidação em 23,47% e 40,82%, respectivamente, revelando uma resposta dose
dependente (Figura 7).
5 50 1000
1
2
3
4FeSO4
ExEt
Ácido Gálico
Trolox
a a aa
b
b
b,a
c c
c
d dd
Concentrações(µµµµg.mL -1)
nmol
Eq
MD
A.m
L-1
(-)
(+)
(+)
Figura 7: Efeito do extrato etanólico (ExEt) das folhas de P. cincinnata Mast na formação de TBARS
induzido por FeSO4. Valores com letras diferentes indicam médias com diferença significativa (p < 0,05). As
diferenças estatísticas foram determinadas pela ANOVA de duas via seguido do pós-teste de Bonferroni.

49
Observando o comportamento do ExEt mediante a lipoperoxidação induzida pelo
peróxido de hidrogênio (Figura 8), identificamos que o ExEt apresentou efeito redox
preventivo semelhante aos controles ácido gálico e trolox (p > 0,05), com valores de
(45,19%, 61,54% e 62,50%) para as concentrações de 5, 50 e 100 µg.mL-1 ,respectivamente.
5 50 1000.0
0.5
1.0
1.5H2O2
ExEt
Ácido Gálico
Trolox
a aa
bb b
b
bb
c bb
Concentrações(µµµµg.mL -1)
nmol
Eq
MD
A.m
L-1
(-)
(+)
(+)
Figura 8: Efeito do extrato etanólico (ExEt) das folhas de P. cincinnata Mast na formação de TBARS
induzido por H2O2. Valores com letras diferentes indicam médias com diferença significativa (p < 0,05). As
diferenças estatísticas foram determinadas pela ANOVA de duas via seguido do pós-teste de Bonferroni.
5.5 Atividade Redox-Protetora Após Tratamento Agudo (in vivo)
O efeito redox-protetor do consumo agudo do ExEt de P. cincinnata Mast no
organismo de ratos em condições fisiológicas basais mostrou uma redução significativa (p <
0,05) na lipoperoxidação hepática e sanguínea dos animais tratados com a Passiflora. Bem
como, uma leve tendência, mas não significativo (p > 0,05), na redução da lipoperoxidação no
tecido cardíaco dos mesmos animais (Figura 9).
Da mesma forma, verificamos que o consumo agudo do ExEt das folhas de P.
cincinnata, também, foi capaz de prevenir significativamente (p < 0,05) a oxidação de
proteínas hepáticas. Mas, em contrapartida, não foi capaz de prevenir a oxidação a proteínas
sanguíneas e hepáticas, (Figura 10).
Por outro lado, constatamos que o consumo do ExEt possibilitou o aumento do
conteúdo de tióis sanguíneo, hepático e cardíaco de forma significativa (p < 0,05), sugerindo
um aumento no estado antioxidante para os tecidos avaliados (Figura 11).
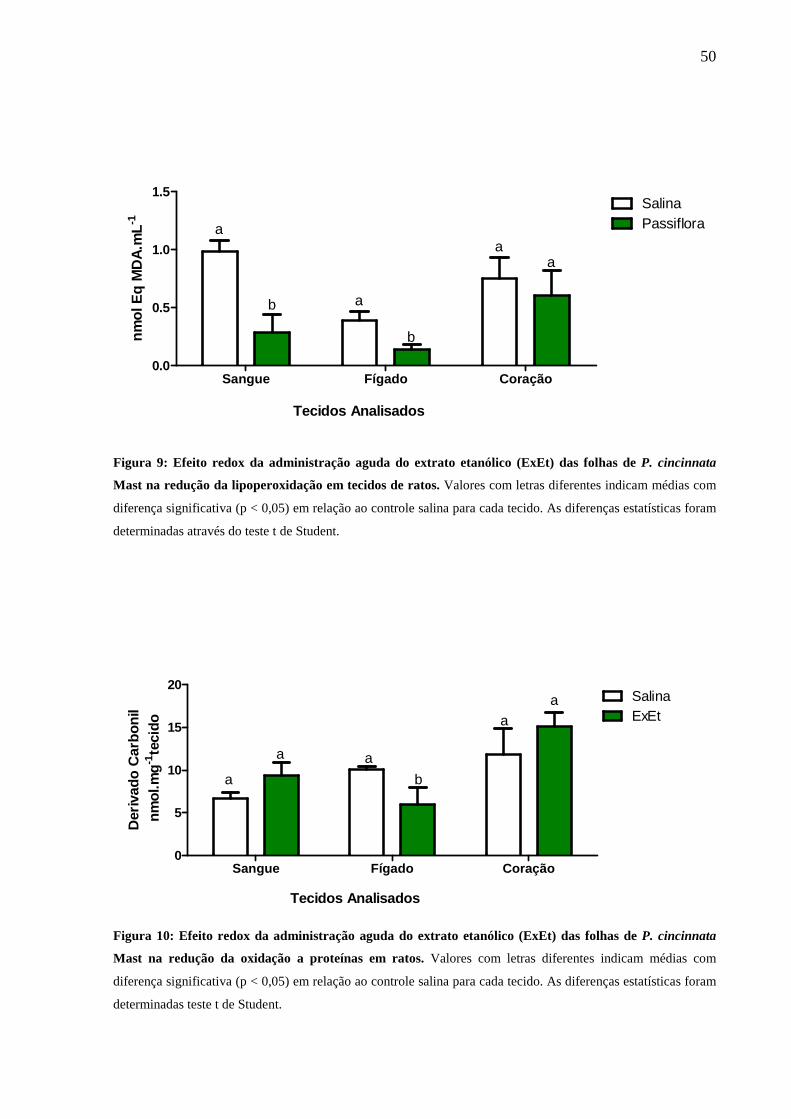
50
Sangue Fígado Coração0.0
0.5
1.0
1.5SalinaPassiflora
b
b
Tecidos Analisados
a
a
aa
nmol
Eq
MD
A.m
L-1
Figura 9: Efeito redox da administração aguda do extrato etanólico (ExEt) das folhas de P. cincinnata
Mast na redução da lipoperoxidação em tecidos de ratos. Valores com letras diferentes indicam médias com
diferença significativa (p < 0,05) em relação ao controle salina para cada tecido. As diferenças estatísticas foram
determinadas através do teste t de Student.
Sangue Fígado Coração0
5
10
15
20SalinaExEt
b
Tecidos Analisados
aa
a
aa
Der
ivad
o C
arbo
nil
nm
ol.m
g-1
teci
do
Figura 10: Efeito redox da administração aguda do extrato etanólico (ExEt) das folhas de P. cincinnata
Mast na redução da oxidação a proteínas em ratos. Valores com letras diferentes indicam médias com
diferença significativa (p < 0,05) em relação ao controle salina para cada tecido. As diferenças estatísticas foram
determinadas teste t de Student.
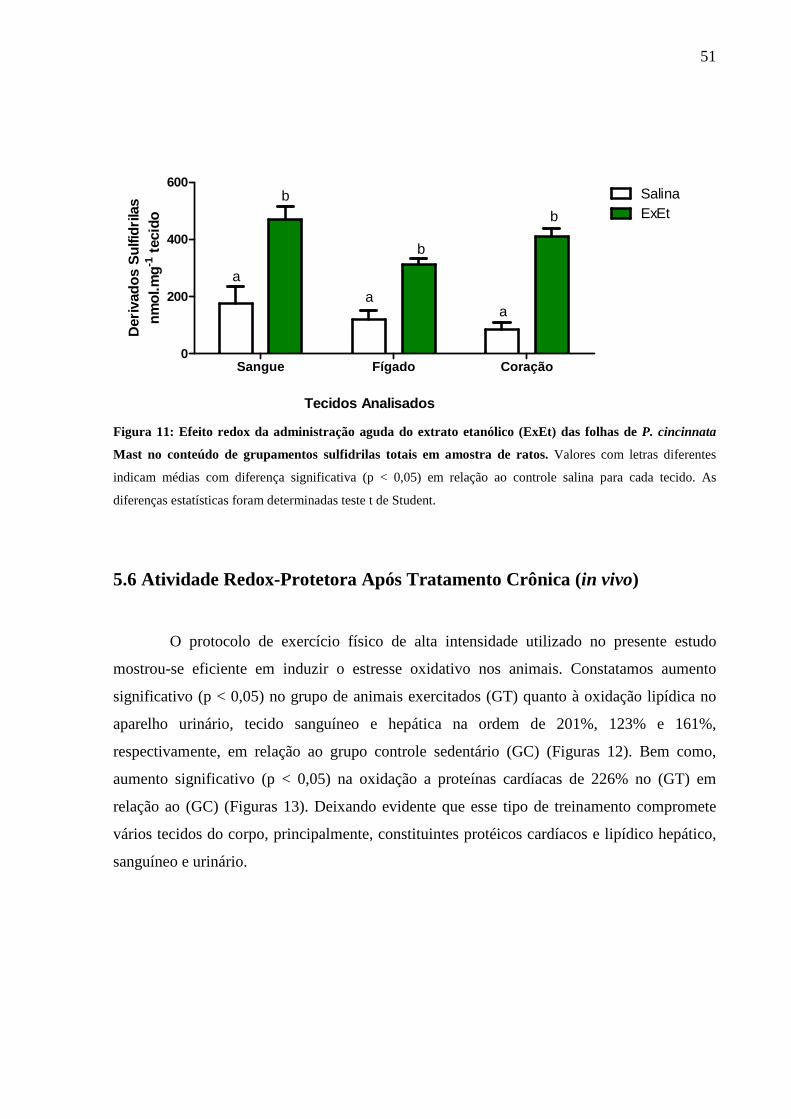
51
Sangue Fígado Coração0
200
400
600SalinaExEt
b
aa
Tecidos Analisados
b
a
b
Der
ivad
os S
ulfid
rilas
nmol
.mg
-1 t
ecid
o
Figura 11: Efeito redox da administração aguda do extrato etanólico (ExEt) das folhas de P. cincinnata
Mast no conteúdo de grupamentos sulfidrilas totais em amostra de ratos. Valores com letras diferentes
indicam médias com diferença significativa (p < 0,05) em relação ao controle salina para cada tecido. As
diferenças estatísticas foram determinadas teste t de Student.
5.6 Atividade Redox-Protetora Após Tratamento Crônica (in vivo)
O protocolo de exercício físico de alta intensidade utilizado no presente estudo
mostrou-se eficiente em induzir o estresse oxidativo nos animais. Constatamos aumento
significativo (p < 0,05) no grupo de animais exercitados (GT) quanto à oxidação lipídica no
aparelho urinário, tecido sanguíneo e hepática na ordem de 201%, 123% e 161%,
respectivamente, em relação ao grupo controle sedentário (GC) (Figuras 12). Bem como,
aumento significativo (p < 0,05) na oxidação a proteínas cardíacas de 226% no (GT) em
relação ao (GC) (Figuras 13). Deixando evidente que esse tipo de treinamento compromete
vários tecidos do corpo, principalmente, constituintes protéicos cardíacos e lipídico hepático,
sanguíneo e urinário.
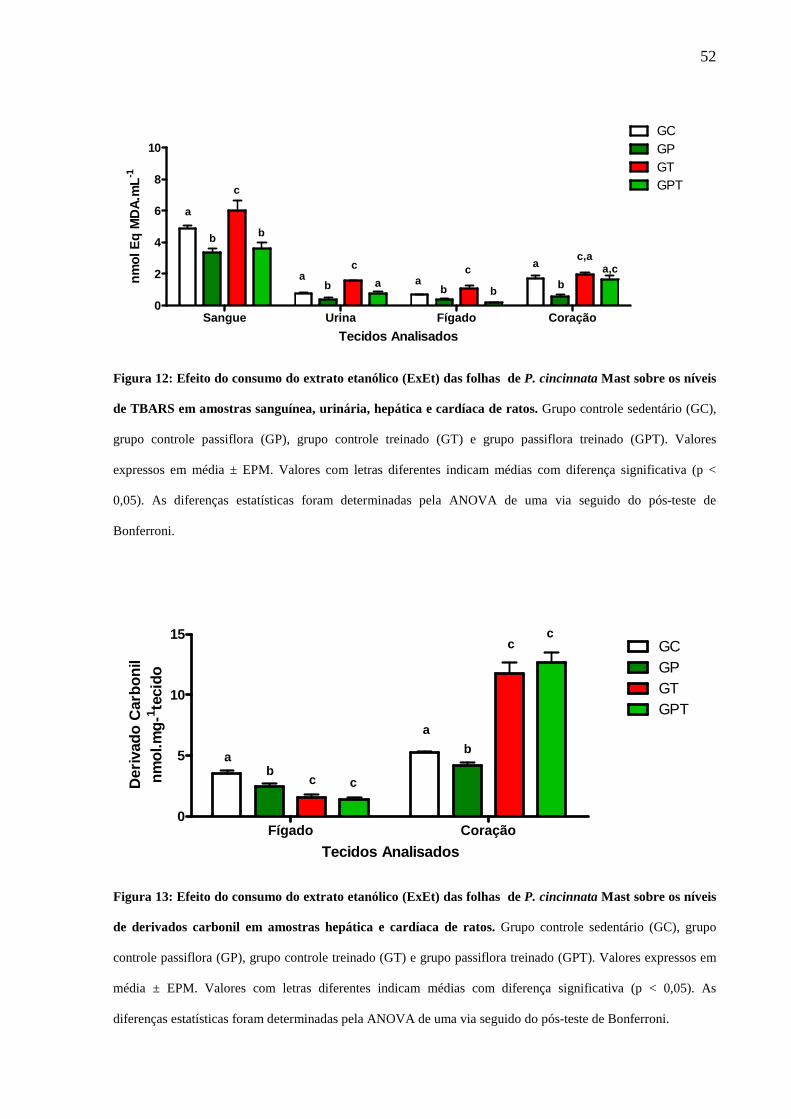
52
Sangue Urina Fígado Coração0
2
4
6
8
10GCGPGTGPT
a
b
a
ba
b ab
c
c cc,a
b
ab
a,c
Tecidos Analisados
nmol
Eq
MD
A.m
L-1
Figura 12: Efeito do consumo do extrato etanólico (ExEt) das folhas de P. cincinnata Mast sobre os níveis
de TBARS em amostras sanguínea, urinária, hepática e cardíaca de ratos. Grupo controle sedentário (GC),
grupo controle passiflora (GP), grupo controle treinado (GT) e grupo passiflora treinado (GPT). Valores
expressos em média ± EPM. Valores com letras diferentes indicam médias com diferença significativa (p <
0,05). As diferenças estatísticas foram determinadas pela ANOVA de uma via seguido do pós-teste de
Bonferroni.
Fígado Coração0
5
10
15GCGPGTGPT
ab
c c
a
b
cc
Tecidos Analisados
Der
ivad
o C
arbo
nil
nmol
.mg-
1 teci
do
Figura 13: Efeito do consumo do extrato etanólico (ExEt) das folhas de P. cincinnata Mast sobre os níveis
de derivados carbonil em amostras hepática e cardíaca de ratos. Grupo controle sedentário (GC), grupo
controle passiflora (GP), grupo controle treinado (GT) e grupo passiflora treinado (GPT). Valores expressos em
média ± EPM. Valores com letras diferentes indicam médias com diferença significativa (p < 0,05). As
diferenças estatísticas foram determinadas pela ANOVA de uma via seguido do pós-teste de Bonferroni.

53
O tratamento crônico com o ExEt das folhas de Passiflora (200 mg.kg-1), preveniu a
lipoperoxidação no tecido sanguíneo, cardíaco e hepático, bem como, no aparelho urinário
dos animais do grupo GP em relação ao GC (p < 0,05). Além disso, constatamos também
redução significativa (p < 0,05) nos níveis de MDA no aparelho urinário, tecido sanguíneo e
hepático dos animais do grupo GPT em relação ao GT (p < 0,05). Quanto à prevenção
oxidativa no músculo cardíaco dos animais do grupo GPT em relação ao grupo GT, este
apresentou leve tendência redutiva, porém não significativa (p > 0,05).
Paralelamente, foi observada uma redução significativa (p < 0,05) na oxidação ao
conteúdo protéico hepático e cardíaco dos animais do GP em relação ao GC. Semelhança não
evidenciada nos demais grupos (figura 13).
O conteúdo de grupamento sulfídrila avaliado no presente estudo indica,
indiretamente, o recrutamento do sistema de defesa antioxidante das glutationas para o tecido
testado. Neste sentido, podemos verificar que o consumo diário de Passiflora não alterou de
forma significativa (p > 0,05) o conteúdo de tióis nos animais do GP em relação ao GC, tanto
no tecido hepático quanto no cardíaco. Em contrapartida, este sistema foi bastante requisitado
nos animais em exercício, sofrendo redução significativa (p < 0,05) no tecido hepático dos
animais do GPT em relação ao GT. Bem como, uma redução não significativa (p > 0,05) de
78% no tecido cardíaco dos animais do GPT em relação ao GT (Figura 14).
Fígado Coração0
10
20
30GCGPGTGPTa
a,c
b
c aa,c
b
c
Tecidos Analisados
Der
ivad
os S
ulfid
rila
nmol
.mg
-1te
sido
Figura 14: Efeito do consumo do extrato etanólico (ExEt) das folhas de P. cincinnata Mast sobre os níveis
de derivados sulfidrilas em amostras hepática e cardíaca de ratos. Grupo controle sedentário (GC), grupo
controle passiflora (GP), grupo controle treinado (GT) e grupo passiflora treinado (GPT). Valores expressos em
média ± EPM. Valores com letras diferentes indicam médias com diferença significativa (p < 0,05). As
diferenças estatísticas foram determinadas pela ANOVA de uma via seguido do pós-teste de Bonferroni.

54
7. DISCUSSÃO
No presente estudo verificamos que as classes dos constituintes químicos
encontrados no extrato da P. cincinnata Mast são condizentes com o previamente encontrado
para o gênero Passiflora (PEREIRA & VILLEGAS, 2000; DHAWAN et al., 2004; MULLER
et al., 2005). Estes resultados sugerem que os diferentes usos medicinais dessa espécie podem
ser validados, principalmente, devido à presença dos constituintes fenólicos, cujo consenso na
literatura é de que eles estão associados a diversas propriedades medicinais, dentre elas, as
propriedades antioxidantes (RUDNICKI et al., 2007), prevenindo doenças associadas ao
envelhecimento, além de cardiopatias e câncer (VAN DER SLUIS et al., 1997; RICE-
EVANS, MILLER & PAGANGA, 1997).
Há evidências na literatura sobre a existência de uma associação direta e significativa
entre o teor de compostos fenólicos em extratos e frações e a atividade antioxidante dos
mesmos (HARBONE & WILLIAMS, 2001; MENSOR et al., 2001; RUDNICKI et al., 2007;
OLIVEIRA, 2008). Diante disso, foi constatado que o teor fenólico do ExET está acima
daqueles já encontrados para outras espécies do gênero Passiflora, a exemplo, Passiflora sp
(20,0 ± 2,6 mg Eq ácido gálico.g-1) (KUSKOSKI et al., 2006), a P. edulis (92,5 ± 2,2 mg Eq
ácido tanico.mg-1 extrato) (RUDNICKI et al., 2007) e a P. alata (103,10 ± 10,4 mg Eq àcido
gálico.100g-1) (OLIVEIRA, 2008), as quais tiveram atividade antioxidante abaixo dos valores
encontrados em nosso estudo.
A concentração eficiente de um produto natural (extrato ou fração) em reduzir um
radical livre em 50% (CE50) é uma característica muito importante quanto à capacidade
antioxidante do mesmo. Souza et al. (2007) avaliaram a atividade antioxidante de algumas
plantas medicinais e encontraram valores de CE50 3,8 e 15,3 vezes maior no extrato da casca
de Terminalia brasiliensis e extrato da raiz de Copernicia prunifera, respectivamente, em
relação a P. cincinnata Mast. Da mesma forma, Boscolo et al. (2007) avaliaram o potencial
antioxidante de plantas medicinais da restinga, encontrando valores de CE50 115 vezes maior
que a P. cincinnata Mast. Estes dados corroboram com a hipótese de que a P. cincinnata Mast
tem grande potencial antioxidante em relação a diversas outras espécies de plantas com
propriedade medicinais comprovadas na literatura.
Contudo, segundo Scherer & Godoy (2009), o índice de atividade antioxidante ou
IAA pode representar, com maior exatidão, o potencial antioxidante de um composto sintético
ou produto natural frente a um radical livre que simplesmente o CE50. Haja vista que o IAA
anula os erros na comparação de resultados decorrente da concentração final do radical livre

55
utilizado, uma vez que a formula para o IAA depende da concentração final do radical
utilizado e o CE50 encontrado para no experimento (SCHERER & GODOY, 2009). Assim,
observamos que o ExEt das folhas de P. cincinnata Mast apresentou IAA muito forte
(superior a 3 para o radical ABTS●+ e a 4 para o radical DPPH), o que nos sugere elevado
potencial antioxidante.
Argolo et al. (2004) encontraram valores muito forte IAA ao radical DPPH pelos
extratos etanólico (2,0 ± 0,02) e aquoso (2,15 ± 0,05), fração acetato de etila (2,32 ± 0,01), e
clorofómica (2,43 ± 0,02) em folhas de B. monandra, espécie bastante utilizada no Brasil
como antidiabete. Da mesma forma, Sousa et al. (2007), avaliaram a atividade antioxidantes
de diversas plantas de uso medicinal frente ao radical DPPH e encontraram valores muito
forte de IAA nos extratos etanólicos de T. brasiliensis (3,62 ± 0,08 casca e 2,25 ± 0,19
folhas) e T. fagifolia (2,37 ± 0,07 - folhas), porém, forte IAA nos extratos etanólicos das C.
macrophyllum (1,27 ± 0,05 - folhas) e Q. grandiflora (1,99 ± 0,08 - folhas), além de
moderada atividade para o extrato etanólico da C. prunifera (0,89 ± 0,03 - raiz).
Segundo Oliveira (2008), além do teor fenólico do produto natural, a estrutura
química do componente ativo do mesmo (extrato ou fração) são fatores importantes que
influenciam na eficácia do antioxidante natural frente a diversos RLs. A posição e o número
de hidroxilas presentes na molécula dos polifenóis, principalmente, na posição orto é um fator
relevante para esta atividade. Neste sentido, Masteikova et al. (2008) verificaram que o
extrato etanólico de P. incarnata possui 2,5 e 2,3 mais vezes atividade de seqüestro de radical
DPPH• e ABTS•+, respectivamente, que o extrato aquoso. Isso deve ter ocorrido por causa da
maior presença de compostos fenólicos com orto-dihidroxilação no extrato etanólico em
comparação ao extrato aquoso, bem como, ao maior conteúdo fenólico presente no extrato
etanólico (OLIVEIRA, 2008).
Segundo Lima, Cunha & Estevam (2009), o ExEt da P. cincinnata apresenta uma
cinética de reação muito rápida com estabilização entre 5 a 10 minutos. Essa característica é
especialmente importante quando consideramos que, quanto mais rápida a reação de uma
molécula ou substância antiradicalar a um radical, melhor será sua efetividade antioxidante
em organismos vivos, haja vista que a meia vida de um RL no organismo é muito curta (10-6
segundos) (LIMA, 2008). Por este motivo, e, pelo elevado teor fenólico do ExEt, foi
constatado no presente estudo que uma única dose de 200 mg.kg-1 do ExEt das folhas da P.
cincinnta Mast foi capaz de reduzir significativamente a peroxidação lipídica de tecido
sanguíneo e hepático, bem como, a oxidação de proteínas do tecido hepático de ratos.

56
Fazendo-nos crê, com maior veemência, no potencial efeito redox positivo do ExEt sobre a
prevenção de lesões oxidativas por EROs à estruturas celulares.
Rodrigues et al. (2005), Lima et al. (2005) e Wang et al. (2008) encontraram níveis
de peróxidação lipídica reduzidos em tecido hepático de ratos tratados com amostras de vinho
tinto, infusão aquosa de sálvia (Salvia officinalis) e soja fermentada que apresentam como
principal característica o fato de serem ricos em polifenóis. Lima (2008) descreve que altas
doses de extratos ricos em polifenóis são efetivos na redução da lipoperóxidação e oxidação
de diversos componentes celulares (LIMA, 2008). Porém, o mecanismo pelo qual isso ocorre,
ainda, não está totalmente elucidado, entretanto, especula-se que polifenóis possam atuar
como sequestradores de radicais ou atuarem como moduladores na sinalização da expressão
gênica de diversas enzimas antioxidantes (Middleton et al., 2000).
Neste sentido, o ExEt das folhas da P. cincinnata Mast, em nosso estudo, pode ter
agido como seqüestrador direto de radicais, uma vez que, o conteúdo de tióis, representado
por grupamento sulfidríla, foi aumentado no tecido hepático e cardíaco dos ratos que
ingeriram em caráter agudo 200 mg.kg-1 do mesmo (Figura 7), indicando um menor
recrutamento do sistema de defesa antioxidante dependente de glutationa como GPx e GSH,
uma vez que, acreditamos que não houve tempo hábil para aumento na expressão gênica
destas enzimas durante o experimento (VANNUCCHI et al., 1998; ROVER JR et al., 2001).
Rudnicki et al. (2007) investigaram o efeito da administração oral do extrato
hidroetanólico de P. alata contra danos oxidativos em órgãos de ratos e mostraram que o
extrato reduziu os níveis de lipoperoxidação hepática e cardíaca dos animais tratados. Além
disso, foi constatado aumento na atividade de algumas enzimas antioxidantes como catalase
(CAT) e superóxido dismutase (SOD), mostrando com isso que, os polifenóis do extrato da
Passiflora preservam o conteúdo enzimático antioxidante através de sua ação antioxidante
(MIDDLETON et al., 2000). Dados estes que tornam nossos achados condizentes com a
literatura.
Nossos resultados sobre o consumo crônico do ExEt das folhas de P. cincinnata
Mast, avaliado após 8 semanas, reforça a hipótese de que a Passiflora em estudo, protege o
tecido sanguíneo, aparelho urinário, além do fígado e coração de ratos (GP) contra lesões
oxidativa a estruturas lipídica celulares, bem como, oxidações à estruturas protéicas do fígado
e coração dos mesmos animais. A justificativa é a mesma apresentada anteriormente, após o
consumo de uma única dose do ExEt, cujos animais apresentaram proteção redox nos tecidos
avaliados através do seqüestro direto de RLs, uma vez que, o conteúdo de tióis, grupamento

57
sulfidríla, não sofreu alterações significativas (p > 0,05) no tecido hepático e cardíaco (Figura
10).
O exercício físico é uma fonte potencial para aumentar a produção de Eros no
organismo (ALESSIO et al., 2002). Contudo, é a intensidade da atividade física o fator
determinante para o aumento destas espécies radicalares durante o exercício (MUSARO et al.,
2010). Souza, Oleveira & Pereira (2005) encontraram valores elevados de MDA e proteínas
carboniladas em jogadores de futebol, maratonistas e praticantes de outras modalidades
esportivas após uma sessão de exercício de alta intensidade.
Neste sentido, verificamos em nossos resultados que o protocolo de exercício foi
eficiente em aumento a produção de EROs e danos oxidativos aos tecidos avaliados, como já
relatado anteriormente. Em contrapartida, o ExEt das folhas de Passiflora, quando consumida
antes do exercício físico, foi capaz de prevenir significativamente (p < 0,05) lesões oxidativas
a estruturas lipídicas celulares (lipoperoxidação) no tecido sanguíneo, hepático e aparelho
urinário dos animais (GPT). Porém, não foi capaz de prevenir de forma significativa a
lipoperoxidação cárdica, nem a oxidação de proteínas hepática e cardíaca.
Para tanto, há duas explicações, a primeira é a comprovação da elevada produção de
EROs com o protocolo de exercício utilizado, uma vez que, mesmo com o aumento de
constituintes fenólicos no organismo através do consumo do ExEt, verificamos uma depleção
no teor de tióis totais dos tecidos (Figura 14), mostrando, com isso, um maior nível de
estresse oxidativo e recrutamento do sistema de defesa antioxidante das glutationas.
A segunda explicação está na dosagem do ExEt, a qual pode não ter sido suficiente
para prevenir lesões oxidativas em todos os tecidos de maneira igualitária. Haja vista que os
compostos polifenólicos quando ingeridos, antes de serem secretados na circulação sanguínea,
sofrem metabolização no fígado (MANACH & DONAVAN, 2004). Portanto, esse órgão
pode servir como um depósito de compostos polifenólicos, combatendo os radicais livres de
maneira mais rápida e eficiente que outros órgãos como o coração que receberá quantidade
inferior de constituinte fenólico.
Rudnicki et al. (2007) ainda em seu estudo, verificaram que a dosagem utilizada do
extrato da P. alata não foi capaz de reduzir significativamente os níveis de lipoperóxidativos
e oxidação as proteínas hepática e cardíaca de maneira igualitária. Porém, foi possível
verificar, uma tendência na redução deste parâmetro oxidativo com aumento da dosagem. Isso
sugere que devido ao elevado consumo dos polifenóis no fígado, há uma redução na
biodisponibilidade destes compostos a outros órgãos como coração, mostrando não ser

58
possível a prevenção oxidativa a lipídios e proteínas nos diversos órgãos de maneira
igualitária. Dados que tornam nossos achados condizentes com a literatura.
Buscando entender, em parte, a propriedade redox dos constituintes presentes no
ExEt frente a oxidação lipídica induzida por três geradores químicos de radicais livres, o
mesmo apresenta uma maior afinidade em neutralizar a lipoperoxidação induzida por AAPH e
H2O2 que FeSO4. O AAPH é um indutor químico que apresenta como característica a geração
de radical carbono centrado, o qual reage prontamente com oxigênio para formar o radical
peroxil. H2O2 é uma fonte geradora de radical hidroxila, em quanto o FeSO4 é o promotor das
reações de Fenton e gerador de radical alcoxil, todos responsáveis potencialmente pela cascata
lipoperoxidativa (RUBERTO, 2000).
Segundo Budni et al. (2007), compostos fenólicos presentes em extratos vegetais
apresentam uma relação direta entre o seu o poder redox e a quantidade de grupos hidroxilas
ligadas a ele. Van Acker et al. (1996) demonstraram haver diferença no potencial redox de
compostos fenólicos a depender de sua estrutura química, pois sua propriedade depende do
rearranjo dos grupos funcionais em torno da estrutura nuclear. Fato que justifica a interação
entre os compostos fenólicos e grupos radicalares na prevenção da lipoperoxidação.
Apesar dos achados em nosso estudo serem com extrato hidroetanólico, é possível
fazer equivalências dos mesmos com o consumo de chás de folhas da Passiflora pela
população, haja vista que achados cromatográficos mostraram que há relação entre o conteúdo
de fenóis totais do material seco do extrato aquoso e hidroetanólico que são semelhantes,
sendo, o segundo com maior quantidade de constituintes (DE-PARIS et al., 2002; BIRK et
al., 2005).

59
8. CONCLUSÕES
No presente estudo, a quantidade de fenóis totais, atividade antioxidante e
propriedade redox-protetora da P. cincinnata mast foi investigada. Os achados encontrados
sugerem as seguintes conclusões:
• O ExEt e a FActE apresentaram alto teores de fenóis totais;
• A FActE apresentou fraca capacidade seqüestradora para radical DPPH•;
• O ExEt apresentou forte capacidade seqüestradora para os radicais DPPH• e ABTS●+;
• O ExEt apresentou excelente capacidade redox-protetora para inibir a lipoperoxidação
induzida por AAPH e H2O2 e moderada atividade para o indutor FeSO4;
• O tratamento agudo dos animais com 200 mg.kg-1 de ExEt de P. cincinnata Mast foi
capaz de proteger estruturas do aparelho sanguíneo e hepático contra danos oxidativos
produzidos pelo organismo;
• O tratamento agudo dos animais com 200 mg.kg-1 de ExEt de P. cincinnata Mast foi
capaz de aumentar o conteúdo de grupamento sulfidríla totais (tióis) do tecido
sanguíneo, hepático e cardíaco dos mesmos.
Em adição, os resultados encontrados após o consumo diário de 200 mg.kg-1 do
ExEt, durante dois meses, no modelo de dano oxidativo induzido pelo exercício físico de alta
intensidade sugerem as seguintes conclusões:
• O consumo diário, 200 mg.kg-1 de ExEt de P. cincinnata Mast, durante 2 meses, foi
capaz de prevenir a oxidação lipídica e protéica das estruturas celulares do aparelho
urinário, sanguíneo, bem como, hepática e cardíaca em ratos sobre condições de
estresse fisiológico;
• O consumo diário, 200 mg.kg-1 de ExEt de P. cincinnata Mast, durante 2 meses,
preservou o conteúdo de grupamento sulfidrila no tecido hepática e cardíaco de ratos
sob condições de estresse fisiológico;
• O consumo diário, 200 mg.kg-1 de ExEt de P. cincinnata Mast, antes da prática de
exercício físico de alta intensidade, durante 2 meses, foi capaz de prevenir lesões
oxidativas a componentes lipídicos do tecido sanguíneo e hepático, bem como do

60
aparelho urinário de ratos sob condições de estresse oxidativo induzido pelo exercício
físico;
• O consumo diário, 200 mg.kg-1 de ExEt de P. cincinnata Mast, antes da prática de
exercício físico de alta intensidade, durante 2 meses, não foi capaz de prevenir lesões
oxidativas de componentes lipídico cardíaco, bem como de proteínas do tecido
hepático e cardíaco dos ratos sob condições de estresse oxidativo induzido pelo
exercício físico;
• O consumo diário, 200 mg.kg-1 de ExEt de P. cincinnata Mast, antes da prática de
exercício físico de alta intensidade, durante 2 meses, não foi suficiente para preservar
o conteúdo de tióis totais hepático e cardíaco dos animais tratados sob condições de
estresse oxidativo induzido pelo exercício físico.

61
REFERENCIAS
ABRAMSON, J. L.; HOOPER, W. C.; JONES, D. P.; ASHFAQ, S.; RHODES, S. D.;
WEINTRAUB, W. S.; HARRISON, D. G.; QUYYUMI, A. A.; VACCARINO, V. (2005).
Association between novel oxidative stress markers and C-reactive protein among adults
without clinical coronary heart disease. Atherosclerosis 178: 115-121.
AGARWAL, M. M.; ANKRA-BADU, G. A. (1999). Serum creatine kinase: a marker
for muscle damage in sickle cell painful crisis. Ann. Saudi Med. 19(3): 264-266.
AGUILAR-SILVA, R.H.; CINTRA, B.B.; MILANI, S.; MORAES, T.P.; TSUJI, H. (2002).
Estado antioxidante do sangue como indicador da eficiência do treinamento em nadadores.
Rev. Bras. Ciên. Mov. 10(3):7-11.
ALBUQUERQUE, M.M. (2006). Implicações do estresse oxidativo sobre os eritrócitos e
desenvolvimento neuropsicomotor em adultos com síndrome de Down. Trabalho de
Conclusão de Curso. (Graduação em Fisioterapia) - Universidade do Sul de Santa Catarina.
ALESSIO, H. M. (1993). Exercise-induced oxidative stress. Medicine and Science in Sports
and Exercise. 25(2):218-224.
ALESSIO, H. M., HAGERMAN, A. E.; ROMANELLO, M.; CARANDO, S.;
THRELKELD, M. S.; ROGERS, J.; DIMITROVA, Y.; MUHAMMED, S.; WILEY, R.
L. (2002). Consumption of green tea protects rats from exercise-induced oxidative
stress in kidney and liver. Nutr. Res. 22(10): 1177–1188.
ALMERSJÖ, O.; BENGMARK, S.; ENGEVIK, L.; HAFSTRÖM, L. O.;
LOUGHRIDGE, B. P.; NILSSON, L. A. (1968). Serum enzyme changes after hepatic
dearterialization in man. Ann. Surg. 167(1): 9-17.
ANDRADE, J.A.B.; GOMES T.A.T.; NETO, U.F. (1998). Letalidade em lactantes com
diarréia persistente: fatores de risco associados ao óbito. Arq Gastroenterol. 35(1): 62-8.

62
APONTE, Y.; JÁUREGUI, D. (2004). Algunos aspectos de la biología floral de Passiflora
cincinnata Mast. Revista de la Facultad de Agronomia. 21(3):211- 219.
APPLE, F. S.; HELISTEN, Y.; CLARKSON, P. M. (1988). Early detection of skeletal muscle
injury by assay of creatine kinase MM isoforms in serum after acute exercise. Clin. Chem.
34(6):1102-1104.
ARGOLO, A. C. C.; SANTANA, A. E. G.; PLETSCH, M.; COELHO, L. C. B. B. (2004).
Antioxidant activity of leaf extracts from Bauhinia monandra. Bioresouce Technology. 95:
229 233.
ARMSTRONG, R. B.; OGILVIE, R. W.; SCHWANE, J. A. (1983). Eccentric exercise-
induced injury to rat skeletal muscle. J. Appl. Physiol. 54: 80-93.
ASCENSÃO, A.; MAGALHÃES, J.; SOARES, J.; OLIVEIRA, J.; DUARTE, J. (2003).
Exercício e Estresse Oxidativo Cardíaco. Rev. Port. Card, 22(5):651-678.
BACURAU, R. F. (2000). Nutrição e suplementação esportiva. São Paulo: Phorte. 292p.
BACURAU, R.F.P. & ROSA, L.F.B.P.C. (2004). Produção de espécies reativas de oxigênio
durante a atividade física. In: LANCHA JR, A.H. Nutrição e Metabolismo Aplicado à
Atividade Motora. São Paulo: Atheneu, p.151-153.
BARBOSA, T. M.; MAGALHÃES, P. M.; LOPES, V. P.; NEUPARTH, M.; DUARTE, J. A.
(2003). Comparação da variação da actividade neuromuscular, da creatina quinase e da força
isométrica máxima voluntária entre dois protocolos exaustivos e inabituais. Rev. Port.
Cien. Desp. 3(1):7-15.
BELARDINELLI, R. (2007). Exercise Training in Chronic Heart Failure - How to
Harmonize Oxidative Stress, Sympathetic Outflow, and Angiotensin II. Circulation 115:
3042-3044.
BELVIRANLI, M.; GÖKBEL, H. (2006). Acute exercise induced oxidative stress and
antioxidant changes. Eur. J. Gen. Med. 3(3): 126-131.

63
BENZI, G. Aerobic performance and oxygen free radicals. The J. Sports Med. Physical
Fitness. 33:205-222, 1993.
BERNACCI, L.C.; VITTA, F.A.; BAKKER, Y.V . (2003). Passiflora L. In: Wanderley MGL,
Shepherd GJ, Giulietti AM, Melhem TS (org.) Flora Fanerogâmica do Estado de São Paulo
v. 3. São Paulo: RiMa/FAPESP, p. 248-274.
BERRA, C.M.; MENCK, C.F.M.; DI MASCIO, P. (2006). Estresse oxidativo, lesões no
genoma e processos de sinalização no controle do ciclo celular. Quím. Nova. 29(6):1340-
1344.
BIXBY, M. et al. (2005). Ilex paraguariensis extracts are potent inhibitors of nitrosative
stress: a comparative study with green tea and wines using a protein nitration model and
mammalian cell cytotoxity. Life Sci. 77(3):345-358.
BRENDLER-SCHWAAB, S.; HARTMANN, A.; PFUHLER, S.; SPEIT, G. (2005). The in
vivo comet assay: use and status in genotoxicity testing. Mutagenesis 20(4): 245-254.
BRUCE, R.; TODD, J. K.; LE DUNE, L. (1958). Serum Transaminase: Its Clinical Use in
Diagnosis and Prognosis. Br. Med. J. 2(5105):1125-1128.
CADENAS, S.; BARJA, G.; POULSEN, H.E.; LOFT, S. (1997). Oxidative DNA damage
estimated by oxo8dG in the liver of guinea-pigs supplemented with graded dietary doses of
ascorbic acid and a-tocopherol. Carcinogenesis. 18(12):2373–2377.
CAMPOS, E. B. P.; YOSHIDA, W. B. (2004). O papel dos radicais livres na fisiopatologia
da isquemia e reperfusão em retalhos cutâneos: modelos experimentais e estratégias de
tratamento. J. Vasc. Br. 3(4): 357-366.
CAPASSO, A.; PINTO, A. (1995). Experimental investigations of the synergistic-sedative
effect of passiflora and kava. Acta Therapeutica. 21:127-140.
CARLSON, D. L.; MAWDSLEY, R. H. (1986). Sports anemia: A review of the literature.
Am. J. Sports Med. 14(2): 109-112.

64
CASTELL, P.P.; ALEJO P.J. (2000). Métodos para medir el daño oxidativo. Rev Cubana
Med Milit. 29(3): 192-198.
CAZZOLA R, RUSSO-VOLPE S, CERVATO G, CESTARO B. (2003). Biochemical
assessments of oxidative stress, erythrocyte membrane fluidity and antioxidant status in
professional soccer players and sedentary controls. Eur. J. Clin. Invest. 33: 924-930.
CHALIFOUX, A.; LAGACÉ, A. (1969). Enzymes Sériques pour le Diagnostic de la
Nécrose Hépatique Aigüe Expérimentale. Can. J. Comp. Med. 33(3): 178-186.
CHANG, C.K.; HUANG, H.Y.; TSENG, H.F.; HSUUW, Y.D.; TSO, T.K. (2006). Interação
da vitamina E e treinamento físico sobre o estresse oxidativo e atividade das enzimas
antioxidantes em músculos esqueléticos de ratos. J Nutr Biochem., 39:18-45.
CHEESEMAN, K.H.; SLATER, T.F. (1996). Radicais livres em medicina. Rio de Janeiro:
Interlivros, 251 p.
CHIHUAILAF, R.H.; CONTRERAS, P.A.; WITTWER, F.G. (2002). Patogénesis del estrés
oxidativo: consecuencias y evoluación em salud animal. Vet. Méx., 33(3), 265-283.
CHOW, C. K. (1979). Nutritional influence on cellular antioxidant defense systems. Am J.
clin Nutr., 32:1066-1081.
CLARKSON, P. M.; THOMPSON, H. S. (2000). Antioxidants: what role do they play
in physical activity and health? Am. J. Clin. Nutr. 72(suppl): 637S–46S.
COOKE, M. S.; EVANS, M. D.; DIZDAROGLU, M.; LUNEC, J. (2003). Oxidative
DNA damage: mechanisms, mutation and disease. FASEB J. 17: 1195-1214.
CRUZAT, V.F; ROGERO, M.M; BORGES, M.C; TIRAPEGUI, J. (2007). Aspectos atuais
sobre estresse oxidativo, exercícios físicos e suplementação. Rev. Bras. Med. Esp. 13(5):
336-342.

65
DALLE-DONNE, I.; ROSSI, R.; GIUSTARINI, D.; MILZANI, A.; COLOMBO, R. (2003).
Protein carbonyl groups as biomarkers of oxidative stress. Clinica Chimica Acta. 329:23-38.
DAVID, J.P,; MEIRA, M.; DAVID, J.M.; BRANDÃO, H.N.; BRANCO, A.; AGRA, M.F.;
BARBOSA, M.R.V.; QUEIROZ, L.P.; GIULIETTI, A.M. (2007). Radical scavenging,
antioxidant and cytotoxic activity of Brazilian Caatinga plants. Fitoterapia 78:215-218.
DAVIES, K.J.A.; QUINTANILHA, A.T.; BROOKS, G.A.; PACKER, L. (1982). Free
radicals and tissue damage produced by exercise. Biochemical and Biophysical Research
Communications. 107(04)1198-1205.
DE PAZ, J. A.; VILLA, J. G.; LOPEZ, P.; GONZALEZ-GALLEGO, J. (1995). Effects
of long-distance running on serum bilirubin. Med. Sci. Sports Exerc. 27(12): 1590-1594.
DE PARIS, F.; PETRY, R.D.; REGINATTO, F.H.; GOSMANN, G.; QUEVEDO, J.;
SALGUEIRO, J.B.; KAPCZINSKI, F.; ORTEGA, G.; SCHENKEL, E.P. (2002).
Pharmacochemical Study of Aqueous Extracts of Passiflora alata Dryander and Passiflora
edulis Sims. Acta Farmacéutica Bonarense. 21(1):5-8.
DEMIRBAG, R.; YILMAZ, R.; GÜZEL, S.; ÇELIK, H.; KOÇYIGIT, A.; ÖZCAN, E.
(2006). Effects of treadmill exercise test on oxidative/antioxidative parameters and DNA
damage. Anadolu Kardiyol. Derg. 6: 135-40.
DEWAR, H. A.; ROWELL, N. R.; SMITH, A. J. (1958). Serum glutamic oxalacetic
transaminase in acute myocardial infarction. Brit Med 2(5105):1121–1125.
DHAWAN, K.; KUMAR, S.; SHARMA, A. (2004). Passiflora: a review update. Journal of
Ethnopharmacology. 94(1):1-23.
DI MEO, S., VENDITT, P. e De LEO, T. (1996). Tissue protection against oxidative stress.
Experientia.15:52(8), 786-94.

66
DILLARD, C.J.; LITOR, R.E.; SAVIN, W.M.; DUMELIN, E.E.; TAPPEL, A.L. (1978).
effects of exercise vitamin E, and ozone on pulmonary function and lipid peroxiodation.
Journal of Applied Physiology. 45: 927-932.
DUMMER, C. D. THOMÉ, F. S.; VERONESE, F. V. (2007). Doença renal crônica,
inflamação e aterosclerose: novos conceitos de um velho problema. Rev. Assoc. Med. Bras.
53(5): 446-50.
EDWIN, E.; CEJA, E.; DHANBAL, S.P.; ZURRES, B. (2007). Antihiperglicemic Activity of
Pasiflora mollissima Bailey. Indian jornal of Pharmaceutical Sciences. 69 (4): 570-571.
EICHNER, E. R. (1996). “Anemia do Esportista”: terminologia inadequada para um
Fenômeno Real. Sports Science Exchange. Gatorade Sports Science Institute (GSSI), Sports
Science Exchange 08-Novembro/Dezembro-1996. Disponível em:
<http://www.gssi.com.br/publicacoes/sse/pdf/gatoradesse8.pdf> Acesso em 10/01/2009.
EICHNER, E. R. (1992). Sports anemia, iron supplementation, and blood doping. Med. Sci.
Sports Exerc. 24(9): S315-S318.
EICHNER, E. R. (1998). Sports Anemia. In: Safran, M. R.; McKeag, D.; Van Camp, S. P.
(Eds). Manual of Sports Medicine. Lippincott Williams & Wilkins. p 255-259.
EICHNER, E. R. (2002). Anemia e formação de sangue (hematopoiése). Gatorade Sports
Science Institute (GSSI), Sports Science Exchange 32- Janeiro/Fevereiro/Março. Disponível
em <http://www.gssi.com.br/scripts/publicacoes/sse/sse_artigo.asp?IDTipo=1&IDPublicac
ao=3 7&DscArquivo=gatoradesse32.pdf&DscArquivoHtm=/SSE/Html/32.htm> Acesso em
10/01/2009.
ERBA, D. et al. (2005). Effectiveness of moderate green tea consumption on antioxidative
status and plasma lipid profile in humans. J. Nutr. Biochem. 6(3):144-149.
EVANS, W.J. (2000). Vitamin E, vitamin C, and exercise. Am J Clin Nutr ,
72(suppl):647S–52S.

67
FALLON, K E; SIVYER, G; SIVYER, K; DARE, A. (1999). Changes in haematological
parameters and iron metabolism associated with a 1600 kilometre Ultramarathon. Br. J.
Sports Med.33:27–32.
FEITOZA, E.A.; MONTEIRO, S.P.; LEMO, I.B.; KIILL, L.H.P.; MELO, N.F.; ARAÚJO,
F.P. (2006). Fenologia de passiflora cincinnata mast. (passifloraceae), na região do vale do
são francisco, Petrolina-Pe. In: Reunião Nordestina de Botânica, 29, Mossoró, diversidade,
conservação e uso sustentável da flora nordestina: resumos, Mossoró: UERN, CD-ROM.
FEKSA, L. R.; LUCHESE, M. D.; FREZZA, R. B.; OLIVEIRA, E.; BISI, S. L.;
MOREIRA, M. C.; BENETTI, E.; TROMBINI, T.; BERLEZE, D. B. (2008). Exercício
intenso e intoxicação pelo chumbo podem gerar estresse oxidativo e prejuízo à saúde humana.
Lecturas: EF & Deportes. Revista Digital - Buenos Aires - Ano 12 - N° 117.
Disponivel em http://www.efdeportes.com/efd117/ejercicio-intenso-e-intoxicacao-pelo-
chumbo.htm. Acesso em 08/10/2010.
FERREIRA F.; FERREIRA, R.; DUARTE, J. A. (2007). Stress oxidativo e dano oxidativo
muscular esquelético: influência do exercício agudo inabitual e do treino físico. Rev. Port.
Cien. Desp. 7(2) 257–275.
FERREIRA, A.L.A.; MATSUBARA, L.S. (1997). Radicais livres: conceitos, doenças
relacionadas, sistema de defesa e estresse oxidativo. Rev. Assoc. Med. Bras. 43(1):61-68.
FERREIRA, F.; FERREIRA, R.; DUARTE, J. A. (2007).Stress oxidativo e dano oxidativo
muscular esquelético: influência do exercício agudo inabitual e do treino físico. Rev. Port.
Cien. Desp. 7(2):257-275.
FIAMONCINI, R. L. (2002). Análise do Estresse Oxidativo em Jogadores Juniores de
Futebol: Comparação Entre Pré e Pós-Exercício Aeróbio e Anaeróbio. Dissertação de
Mestrado. Florianópolis: Universidade Federal de Santa Catarina.
FINAUD, J.; LAC, G.; FILAIRE, E. (2006). Oxidative Stress: Relationship with Exercise
and Training. Sports Med. 36(4): 327-358.

68
FLORINDO, A.; LATORRE, M.; TANAKA, T.; JAIME, P.; ZERBINI, C.(2001). Fatores
associados à prática de exercícios físicos em homens voluntários adultos e idosos residentes
na Grande São Paulo, Brasil. Rev Bras Epid, 4(2):105-13.
FOSCHINI, D.; PRESTES, J.; CHARRO, M. A. (2007). Relação entre exercício físico,
dano muscular e dor muscular de início tardio. Rev Bras. Cineantropom Desempenho
Hum. 9(1):101-106.
FREITAS, P.C.D. (1985). Estudo Farmacognóstico Comparativo de Espacies Brasileiras
do gênero Pasiflora L. Dissertação (Mestrado) – Faculdade de ciências Farmacêuticas –
Ubniversidade de São Paulo: São Paulo, 133p.
FRIGUET, B. (2006). Oxidized protein degradation and repair in ageng and oxidative stress;
FEBS Lett. 580 2910–2916.
GANDRA, P. G.; MACEDO, D. V.; ALVES, A. A. (2006). Fontes de espécies reativas de
oxigênio na musculatura esquelética durante o exercício. Revista Brasileira de Ensino de
Bioquímica e Biologia Molecular. (02):01-11.
GASPER, M. C.; GILCHRIST, J. M. (2005). Creatine kinase: a review of its use in the
diagnosis of muscle disease. Med. Health R. I. 88(11): 398- 404.
GIBALA, M. J.; MACDOUGALL, J. D.; TARNOPOLSKY, M. A.; STAUBER, W. T.;
ELORRIAGA, A. (1995). Changes in human skeletal muscle ultrastructure and force
production after acute resistance exercise. J Appl Physiol, 78: 702–708.
GOLDFARB, A.H.; BLOOMER, R.J.; McKENZIE, M.J. (2005). Combined antioxidant
treatment effects on blood oxidative stress after eccentric exercise. Med. Sci. Sports Exerc.
37(2):234-239.
GOLDFARB, A.H.; MCINTOSH, M.K.; BOYER, B.T.; FATOUROS, J. (1994). Vitamin E
effects on indexes of lipid peroxidation in muscle from DHEA-treated and exercised rats. J
Appl Physiol. 76:1630-5.

69
HALLIWELL, B.; GUTTERIDGE, J.M.C. (1999). Free radicals in biology and medicine.
3ª ed., Oxford: Claredon Press.
HARDMAN, A. E. (1998). The influence of exercise on postprandial triacylglycerol
metabolism. Atherosclerosis 141(1): S93-S100.
HERRERO, A.; BARJA, G. (1998). Heart and Brain OXO8dG in the Genomic DNA of Rats
and Pigeons and Maximum Longevity. Annals of the New York Academy of Sciences.
854: 518.
HERTOG, M.G.L.; E.J.M. FESKENS, P.C.H.; HOLLMAN, M.B.; KATAN, A.N.D.;
KROMHOUT, D. (1993). Content of potentially anticarcinogenic flavonoids of tea infusions,
wines and fruit juices. J. Agric. Food Chem., 41: 1242-1246.
KUSKOSKI, E. M.; ASUERO, A.G.; MORALES, M.T.; FETT, R. (2006). Frutos tropicais
silvestres e polpas de frutas congeladas: atividade antioxidante, polifenóis e antocianinas.
Ciência Rural. 36(4): 1283-1287.
ISIDÓRIO, M. S. (2007). Exercício e estresse oxidativo. Revista Mineira de Educação
Física. 15(1):70-86.
JI, L. L.; FU, R. (1992). Responses of glutathione system and antioxidant enzymes to
exhaustive exercise and hydroperoxide. J. Appl. Physiol. 72: 549-554.
JI, L.L.; FU, R.; MITCHELL, E.W. (1992). Glutathione and antioxidant enzymes in skeletal
muscle: effects of fiber type and exercise intensity. Journal of Applied Physiology, 73:1854-
1859.
JI, L. L.; LEICHTWEIS, S. (1997). Exercise and oxidative stress: sources of free radicals and
their impact on antioxidant systems. Age. 20: 91-106.
JI, L. L. (1999). Antioxidants and oxidative stress in exercise. Proc. Soc. Exp. Biol. Med.
222: 283-292.

70
JI, L.L. (2002). Increasing healthy life span: conventional measures and slowing the innate
aging process. Ann. N. Y. Acad. Sci. 959: 82-92.
JI, L. L.; RADAK, Z.; GOTO, S. (2008) Hormesis and Exercise: How the Cell Copes with
Oxidative Stress. Am. J. Pharmacol. Toxicol. 3(1): 41-55.
JOSEPHY, P. D., MANNERVIK, B., MONTELLANO, P. D. (1997). Molecular
Toxicology, In: Oxidative stress in the erythrocytes. Oxford Universitary Press, pp. 105-
123.
JUDGE, S.; LEEUWENBURGH, C. (2007). Cardiac mitochondrial bioenergetics, oxidative
stress, and aging. Am. J. Physiol. Cell Physiol. 292:1983-1992.
KASAP, S.; GÖNENÇ, A.; ùENER, D. E.; HISAR, I. (2007). Serum Cardiac Markers in
Patients with Acute Myocardial Infarction: Oxidative Stress, C-Reactive Protein and N-
Terminal Probrain Natriuretic Peptide. J. Clin. Biochem. Nutr. 41: 50–57.
KIECHL, S.; WILLEIT, J.; MAYR, M.; VIEHWEIDER, B.; OBERHOLLENZER, M.;
KRONENBERG, F.; WIEDERMANN, C. J.; OBERTHALER, S.; XU, Q.; WITZTUM,
J. L.; TSIMIKAS, S. (2007). Oxidized Phospholipids, Lipoprotein(a), Lipoprotein-
Associated Phospholipase A2 Activity, and 10-Year Cardiovascular Outcomes. Thromb.
Vasc. Biol. 27: 1788-1795.
KILLIP, E. P. (1938). The American species of Passifloraceae. Publ. Field Mus. Nat. Hist.,
Bot. Ser. 19(1-2): 1-613.
KIM, H. J.; LEE, Y. H.; KIM, C. K. (2007). Biomarkers of muscle and cartilage
damage and inflammation during a 200 km run. Eur. J. Appl. Physiol. 99: 443–447.
KOURY, J.C.; DONANGELO, C.M. (2003). Zinco, estresse oxidativo e atividade física.
Rev. Nutr. 16(4):433-441.
LAC, G.; MASO, F. (2004). Biological markers for the follow-up of athletes throughout the
training season. Pathol. Biol. 52(1): 43-9.

71
LANCHA JR, A.H. (2004). Nutrição e Metabolismo Aplicado à Atividade Motora. São
Paulo: Atheneu, p. 151-153.
LAPENNA, D.; CIOFANI, G.; PIERDOMENICO, S.D.; GIAMBERARDINO, M.A.;
CUCCURULLO, F. (2001). Reaction conditions affecting the relationship between
thiobarbituric acid reactivity and lipid peroxides in human plasma. Free Rad Biol Med 31:
331-335.
LAUFS, U.; WASSMANN, S.; CZECH, T.; MÜNZEL, T.; EISENHAUER, M.; BÖHM, M.;
NICKENIG, G. (2005). Physical Inactivity Increases Oxidative Stress, Endothelial
Dysfunction, and Atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 25: 809-814.
LECARPENTIER Y. (2007). Physiological role of free radicals in skeletal muscles. J. Appl.
Physiol. 103:1917-1918.
LEITE, H.P.; SARNI, R.S. (2003). Radicais livres, anti-oxidantes e nutrição. Rev. Brás.
Nutr. Clin. , 18(2): 87-94.
LIMA, C.A,; CUNHA, G.C.; ESTREVAN, C.S. (2009). Quantificação de fenóis totais e
avaliação da atividade antioxidante in vitro e in vivo da Passiflora cincinnata Mast. In: 32ª
Reunião Anual da Sociedade Brasileira de Química, 32, resumos, Fortaleza-Ce: SBQ, CD-
ROM.
LINTON, S.; DAVIES, M.J.; DEAN, R.T. (2001). Protein oxidation and ageing. Exp
Gerontol, 36:1503-1518.
LIU, J.; YEO, H.C.; OVERVIK-DOUKI, E.; STEPHANIE, J.T.; DANIEL, D.W.; GEORGE,
C.; BROOKS, A.; AMES, B.N. (2000). Chronically and acutely exercised rats: biomarkers of
oxidative stress and endogenous antioxidants. Journal of Applied Physiology, 89: 21-28.
LIU, M. L.; YLITALO, K.; SALONEN, R.; SALONEN, J. T.; TASKINEN, M. R. (2004).
Circulating Oxidized Low-Density Lipoprotein and Its Association With Carotid Intima-
Media Thickness in Asymptomatic Members of Familial Combined Hyperlipidemia
Families. Arterioscler. Thromb. Vasc. Biol. 24: 1492-1497.

72
LOTITO, S.; FRAGA, C. G. (2000). Catechins delay lipid oxidation and α-tocopherol and β-
carotene depletion following ascorbate depletion in human plasma. Proc. Soc. Exp. Biol.
Med. 225(1):32-38.
LOWENSTEIN, J.M. (1990). The purine nucleotide cycle revised. J. Sports Med. 11:S37-
S46.
LUKASKI, H.C. (2004). Vitamin and mineral status: effects on physical performance.
Nutrition . 20(7-8) : 632-44.
MAGAUDDA, L.; DI MAURO, D.; TRIMARCHI, F.; ANASTASI, G. (2004). Effects
of Physical Exercise on Skeletal Muscle Fiber: Ultrastructural and Molecular Aspects.
Basic Appl. Myol.14(1): 17-21.
MAGNUSSON, M. K.; SIGFUSSON, N.; SIGVALDASON, H.; JOHANNESSON,
G. M.; MAGNUSSON, S.; THORGEIRSSON, G. (1994). Low iron-binding capacity as a
risk factor for myocardial infarction. Circulation . 89: 102-108.
MANACH, C.; DONAVAN, J. (2004). Pharmacokineties and metabolism of dietary
flavonoids in humans. Free Rad Res. 38: 771-785.
MANCHADO, F. B; CLAUDIO GOBATTO, A.; CONTARTEZE, R. V. L.; PAPOTI, M.;
MELLO, M. A. R. (2006). Máxima fase estável de lactato é ergômetro-dependente em
modelo experimental utilizando ratos. Rev. Bras. Med. Esporte 12(5): 259-262.
MASTALOUDIS, A.; YU, T.W.; O’DONNELL, R. P.; FREI, B.; DASHWOOD, R. H.;
TRABER, M. G. (2004). Endurance exercise results in DNA damage as detected by the comet
assay. Free Rad. Biol. Med. 36(8): 966 – 975.
MATOS, F. J. A. (1998). Introdução à fitoquímica experimental. Fortaleza: UFC.
MCANULTY, S.R.; MCANULTY, L.S.; NIEMAN, D.C.; DUMKE, C.L.; MORROW, J.D.;
UTTER, A.C.; HENSON, D.A.; PROULX, W.R.; GEORGE, G.L. (2004). Consumption of

73
blueberry polyphenols reduces exercise-induced oxidative stress compared to vitamin C.
Nutr. Res. 24:209-221.
MCARDLE, A.; PATTWELL, A.; VASILAKI, A.; GRIFFITHS, R.D.; JACKSON, M.J.
(2001). Contractile activity-induced oxidative stress: cellular origin and adaptive responses.
Am J Physiol Cell Physiol. 280:C621-7.
MCARDLE, W.D.; KATCH, F.I.; KATCH, V.L. (1998). Fisiologia do exercício: energia,
nutrição e desempenho humano. 4ª ed. Editora Guanabara Koogan. p.695.
McBRIDE , J .M. ; KRAEMER, W. J . (1999). Free radicals, exercise and antioxidants.
J. Strength. Cond. Res. 13:175-83.
McBRIDE, J.M.; KRAEMER, W.J.; TRIPLETT-MCBRIDE, T.; SEBASTIANELLI,
MCKAY, D.L.; BLUMBERG, J. B. (2002). The role of tea in human health: an update. J.
Am. Coll. Nutr. , 21(1):1-13.
MENSOR, L. L.; MENEZER, F. S. LEITÃO, G. G.; REIS. A. S. SANTOS, T. C.; COUBE,
C. S.; LEITÃO, S. G. (2001). Screening of Brazilian plant extracts for antioxidant activity by
the use of DPPH free radical method. Phytotherapy Research. 15:127-130.
METIN, G.; ATUKEREN, P.; GÜMÜSTAS, M.K.; BELCE, A.; KAYSERILIOGLU, A.
(2002). The effect of vitamin E treatment on oxidative stress generated in trained rats.
Tohoku J Exp Med. 198:47-53.
MITKA, M. (2004). Biomarkers for Coronary Heart Disease: Predictive Value or Background
Noise?JAMA 292(23): 2824-2825.
MOLAVI, B.; MEHTA, J. L. (2004). Oxidative stress in cardiovascular disease: molecular
basis of its deleterious effects, its detection, and therapeutic considerations. Curr. Opin.
Cardiol. 19:488-493.

74
MOLLER, P.; LOFT, S.; LUNDBY, C.; OLSEN, N. V. (2001). Acute hypoxia and hypoxic
exercise induce DNA strand breaks and oxidative DNA damage in humans. The FASEB
Journal. 15:1181-1186.
MOOREN, F. C.; BLÖMING, D.; LECHTERMANN, A.; LERCH, M. M.; VÖLKER,
K. (2002). Lymphocyte apoptosis after exhaustive and moderate exercise. J. Appl. Physiol.
93: 147- 153.
MORGAN, D. L.; ALLEN, D. G. (1999). Early events in stretch-induced muscle damage. J.
Appl. Physiol. 87:2007-2015.
MORILLAS-RUIZ, J.M.; VILLEGAS, G.J.A.; LÓPEZ, F.J.; VIDAL-GUEVARA, M.L.;
ZAFRILLA, P. (2006). Effects of polyphenolic antioxidants on exercise-induced oxidative
stress. Clinical Nutrition 25: 444–453.
MORRISEY, P.A.; O’ BRIEN, N.M. (1998). Dietary antioxidants in health and disease. Crit
Rev Toxical, 8:463-472.
MOTA, M.P.; FIGUEIREDO, P.; DUARTE, J.A. (2004). Teorias biológicas do
envelhecimento. Rev. Port. Cienc. Desp. 4:81-110.
MULLER, S.D.; VASCONCELOS, S.B.; COELHO, M.; BIAVATTI, M.W. (2005). LC and
UV determination of flavonoids from Passiflora alata medicinal extracts and leaves. Jornal of
pharmaceutical and Biomedical Análisis. 37:399-403.
MUSARO, A.; FULLE, S.; FANO, G. (2010). Oxidative stress and muscle homeostasis.
Current Opinion in Clinical Nutrition and Metabolic Care.13:236-242.
NAGEL D.; SEILER D.; FRANZ H. JUNG K. (1990). Ultra-long-distance running and the
liver. Int. J. Sports Med. 11(6): 441-445.
NASCIMBEN, L.; INGWALL, J. S.; PAULETTO, P.; FRIEDRICH, J.; GWATHMEY, J. K.;
SAKS, V.; PESSINA, A. C.. ALLEN, P.D. (1996). Creatine Kinase System in Failing and
Nonfailing Human Myocardium. Circulation . 94: 1894-1901.

75
NAWAR, W.W. (1996). Lipids. In: FENNEMA, O.R. Food Chemistry. 3 ed. New York:
Marcel Dekker. 1069p.
NAWAR, W.W. (1998). Biochemical processes: lipid instability. In: TAUB, I.A.; SINGH,
R.P. Food Storage Stability. Washington: CRC Press. 539p.
NIES, A. M.; HARTMANN, A.; GRUNERT-FUCHS, M.; POCH, B.; SPEIT, G. (1996).
DNA damage after exhaustive treadmill running in trained and untrained men. Int. J.
Sports Med. 17:397-403.
NOHL, H.; GILLE, L.; STANIEK, K. (2005). Intracellular generation of reactive oxygen
species by mitochondria. Biochem. Pharmacol. 69:719 – 723.
ODONTUYA, G.; HOULT, J.R.S.; HOUGHTON, P.J. (2005). Structure-activity relationship
for antiinflamatory effect of luteolin and derived glycosides. Phytother. Res. 19:782-786.
OGA, S.; DE FREITAS, P.C.; SILVA, A.C.G.; HANADA, S. (1984). Pharmacological trials
of crude extract of Passiflora alata. Planta Medica. 27:303-306.
OHKAWA, H.; OHISHI, Ν.; YAGI, Κ. (1979). Assay for lipid peroxides in animal tissues
by thiobarbituric acid reaction; Anal. Biochem. 95:351-358.
OLIVEIRA, J.C.; RUGGIERO, C. (1998). Aspectos sobre o melhoramento do maracujazeiro
amarelo. Simpósio Brasileiro sobre a Cultura do Maracujazeiro. Jaboticabal, Brasil.
Oliveira, A.C (2008). Capacidade antioxidante de farinhas de resíduos de frutas
tropicais. Dissertação (Mestrado em Ciências da Saúde) – Maceió: Universidade Federal de
Alagoas, 88p.
OLSZEWER, E. (2001). Microscopia óptica como método de medida de radicais livres:
análise celular “in vitro”- HBL. 2ª ed. São Paulo: Ícone.
PACKER, L. (1997). Oxidants, antioxidant nutrients and the athlete. J Sports Sci. 15:353-
363.

76
PANZA, V.S.; WAZLAWIK, E.; SCHÜTZ, R.G.; COMIN, L.; HECHT, K.C.; SILVA, E.L.
(2008). Consumption of green tea favorably affects oxidative stress markers in weight-trained
men. Nutrition . 24(5):433-42.
PEIJIE, C.; HONGWU, L.; FENGPENG, X.; JIE, R.; JIE, Z. (2003). Heavy load exercise
induced dysfunction of immunity and neuroendocrine responses in rats. Life Sci. 72: 2255-
2262.
PEREIRA, B.; SOUZA Jr., T.P. (2008). Exercício Físico Intenso como Pró-oxidante:
mecanismos de indução de estresse oxidativo e conseqüências. Revista Brasileira de Ciência
e Movimento. 16(3):2-26.
PEREIRA, C.A.M.,;VILEGAS, J.H.Y. (2000). Constituintes químicos e farmacólogicos do
genêro Passiflora com ênfase a P. alata Dryander, P. Edulis Sims e P. Incarnata. Revista
Brasileira de Plantas Medicinais. 3(1):1-12.
PETERS, E.; EDEN, M.; TYLER, N.; RAMAUTAR, A.; CHUTURGOON, A. (2006).
Prolonged exercise does not cause lymphocyte DNA damage or increased apoptosis in well-
trained endurance athletes. Eur. J. Appl. Physiol. 98(2): 124-131.
PETIBOIS C.; DÉLÉRIS, G. (2005). Erythrocyte Adaptation to Oxidative Stress in
Endurance Training. Arch. Med. Res. 36: 524-531.
PETIBOIS C.; DÉLÉRIS, G. (2004). Oxidative stress effects on erythrocytes determined by
FT-IR Spectrometry. Analyst. 129: 912 - 916.
PETRY, R.D.; REGINATTO, F.; PARIS, G.; GOSMANN, J.; SALQUEIRO, J.; QUEVEDO,
F. KAPEZINSKI, G.; ORTEGA, G.G.; SCHENKEl., E.P. (2001). Comparative
pharmacological study on hydroethanol extracts of Passiflora alata and Passiflora edulis
leaves. Pytotherapy Research. 15:162-164.

77
POLIDORI, M.; MECOCCI, P.; CHERUBINI, A.; SENIN, U. (2001). Physical activity and
oxidative stress During aging. International Journol of Sport s Medicine. 21:154-157.
POLLACK, M.; LEEUWENBURGH, C. (1999). “Molecular mechanisms of oxidative stress
in aging: Free radicals, aging, antioxidants and disease,” in C. K. Sen, L. Packer, and O.
Hanninen (eds.), Handbook of Oxidants and Antioxidants in Exercise. Amsterdam:
Elsevier Science B.V., pp. 881–923.
PÔRTO, W.G. (2001). Radicais livres e neurodegeneração. Entendimento fisiológico: base
para nova terapia. Rev. Neurociências, 9(2):70-76.
POZZI, A. C. S. (2007). Desenvolvimento de métodos de análise Espectrofotométrica de
flavonóides do “maracujá”. Dissertação de Mestrado. São Paulo: Universidade de São
Paulo.
PROSKE U., MORGAN D. L. (2001). Muscle damage from eccentric exercise:
mechanism, mechanical signs, adaptation and clinical applications. J Physiol. 537:333-45.
PUTNEY, S.; HERLIHY, W.; ROYAL, N.; PANG, H.; APOSHIAN, H. V.;
PICKERING, L.; BELAGAJE, R.; BIEMANN, K.; PAGE, D.; KUBY, S.;
SCHIMMEL, P. (1984). Rabbit Muscle Creatine Phosphokinase. J. Biol. Chem. 259(23):
14317-14320.
RADAK, Z.; CHUNG, H. Y.; GOTO, S. (2008). Systemic adaptation to oxidative challenge
induced by regular exercise. Free Rad. Biol. Med. 44: 153-159.
RADAK, Z.; KUMAGAI, S.; NAKAMOTO, H; GOTO, S. (2007). 8-Oxoguanosine and
uracil repair of nuclear and mitochondrial DNA in red and white skeletal muscle of
exercise-trained old rats. J. Appl. Physiol. 102:1696-1701.
RADÀK, Z.; TAHARA, S.; NAKAMOTO, H.; OHNO, H.; SASVÁRI, M.; NYAKAS, C.;
GOTO, S. (1999). The effect of exercise training on oxidative damage of lipids, proteins and
DNA in rat skeletal muscle: evidence for beneficial outcomes. Free Radical Biology &
Medicine. 27:69-74.

78
RAHMAN, I. & MACNEE, W. (2000) Regulation of redox Glutathion levels and gene
trasncription in lung inflammation: therapeutic approaches. Free Radic Biol Med, 28:1405-
20.
RE, R.; PELLEGRINI, N.; PROTEGGENTE, A.; PANNALA, A.; YANG, M.; RICE-
EVANS, C. (1999). Antioxidant activity applying an improved ABTS radical cation
decolorization assay. Free Radical Biol. Med. 26:1231-1237.
REID, M. B; DOBRIVOJE, S; STOKIC, S. M, FADI, A; LEIS, A. A. (1994). N-
acetylcysteine inhibits muscle fatigue in humans. J Clin Invest. 94:2468-2474.
REMMEN, HV.; IKENO, Y.; HAMILTON, M. (2003). Life-long reduction in MnSOD
activity results in increased DNA damage and higher incidence of cancer but does not
accelerate aging. Physiol. Genomics. 16, 29–37.
RESNICK, AZ.; PACKER, K. (1994). Oxidative damage to proteins: spectrophotometric
method for carbonyl assay. Methods Enzymol. 233: 263-357.
RIBEIRO, L. F. P.; BALDISSERA. V.; BALAKIAN, P.; SOARES, A. R. (2004). Limiar
anaeróbio em natação: comparação entre diferentes protocolos. Rev. bras. Educ. Fís. Esp.
18(2): 201-212.
RICE-EVANS, C.A.; MILLER, N.J.; PAGANGA, G. (1996). Structure-antioxidant activity
relationships of flavonoids and phenolic acids. Free Radic. Biol. Med. 20(7): 933-956.
RIEGEL, R. E. (2005). Bioquímica Nutricional do Exercício Físico. Ed. UNISINOS, São
Leopoldo, 1°ed.
RODRIGUES, É. L. (2005). O efeito da suplementação de vitaminas antioxidantes no
equilíbrio pró e antioxidante de ratos submetidos ao treinamento de natação a 80% da
carga máxima. Dissertação (Mestrado em Ciências Biológicas) – São José dos Campos:
UniVap, 92p.

79
RODRÍGUEZ, M. El ejercicio fisico moderado como redutor de la Formación de
Radicales libres consecuencia del estrés. 2005. (Tese) Universidad de Cádiz.
ROKITZKI, L.; LOGEMANN, E.; HUBER, G.; KECK, E.; KEUL, J. (1994). Alpha-
tocopherol supplementation in racing cyclists during extreme endurance training. Int J Sport
Nutr. 4:253-64.
ROVER JÚNIOR, L.; HÖEHR, N.F.; VELLASCO, A.P.; KUBOTA , L.T. (2001). Sistema
antioxidante envolvendo o ciclo metabólico da glutationa associado a métodos eletroanalíticos
na avaliação do estresse oxidativo. Quím. Nova. 24(1): 112-119.
RUDNICKI, M.; OLIVEIRA, M.R.; PEREIRA, T.V.; REGINATTO, F.H.; DAL-PIZZOL,
F.; MOREIRA, J.C.F. (2007). Antioxidant and antiglycation properties of Passiflora alata and
Passiflora edulis extracts. Food Chem. 100: 719-724.
SALAH, N. et al. (1995). Polyphenolic flavanols as scavengers of aqueous phase radicals and
as chainbreaking antioxidants. Arch. Biochem. Biophys., 322(2):339-346.
SANTOS-SILVA A, REBELO MI, CASTRO EMB, BELO L, GUERRA A, REGO C,
QUINTANILHA A. (2001). Leukocyte activation, erythrocyte damage, lipid profile and
oxidative stress imposed by high competition physical exercise in adolescents. Clin. Chim.
Acta 306:119-126.
SCHERER, R.; GODOY, H.T. (2009). Antioxidant activity index (AAI) by the 2,2-diphenyl-
1-picrylhydrazyl method. Food Chem. 112: 654-658.
SCHMIDT, W.; MAASSEN, N.; TROST, F.; BÖNING, D. (1988). Training induced effects
on blood volume, erythrocyte turnover and haemoglobin oxygen binding properties. Eur
J. Appl. Physiol. 57:490-498.
SCHNEIDER, C. D.; OLIVEIRA, A. R. (2004). Radicais livres de oxigênio e exercício:
mecanismos de formação e adaptação ao treinamento físico. Rev. Bras. Med. Esporte 10(4):
314-318.

80
SELMAN, C.; McLAREN, J.C; COLLINS, A.L.I.; DUTHIE, G.G.; SPEAKMAN, J.R.
(2002). Antioxidant enzymes activities, lipid peroxidation and DNA oxidative damage:
effects of short-term voluntary where running. Arch. Biochem. Biophys. 401:255-261.
SEN, C. K. (1995). Oxidants and antioxidants in exercise. Journal of Applied Physiology.
79(3):675-686.
SEN, C. K. (2001). Antioxidants in Exercise Nutrition. Sports Medicine. 31(13):891-908.
SIGNORINI, J.L.; SIGNORINI, S.L. (1995). Atividade física e radicais livres: aspectos
biológicos, químicos, fisiopatológicos e preventivos. São Paulo: Ícone. 192 p.
SILVEIRA, L.R. (2004). Considerações Críticas e Metodológicas na Determinação de
Espécies Reativas de Oxigênio e Nitrogênio em Células Musculares Durante Contrações. Arq
Bras Endocrinol Metab. 48(6):812-822.
SJÖDIN, B.; WESLING, H.; APPLE, S. (1990). Biochemical mechanisms for oxygen free
radical formation during exercise. Sports Medicine. 10:236-254.
SMOLKA, M.B.; ZOPPI, C.C.; ALVES, A.A.; SILVEIRA, L.R.; MARANGONI, S.;
PEREIRA-DA-SILVA, L.; NOVELLO, J.C.; MACEDO, D.V. (2000). HSP72 as a
complementary protection against oxidative stress induced by exercise in the soleus muscle of
rats. American Journal of Physiology. 279: R1539-R1545.
SOLER-RIVAS, C.; ESPÍN, J. C.; WICHERS, H. J. (2000). An easy and fast test to compare
total free radical scavenger capacity of foodstuffs. Phytochem. Anal. 11(1):330-338.
SOUZA, C. M. M.; SILVA, H. R.; VIEIRA-JR., G. M.; AYRES, M. C. C.; DA COSTA, C.
L. S.; ARAÚJO, D. S.; CAVALCANTE, L. C. D.; BARROS, E. D. S.; ARAÚJO, P. B. M.;
BRANDÃO, M. S.; CHAVES, M. H. (2007). Fenóis totais e atividade antioxidante de cinco
plantas medicinais. Quim. Nova. 30(2):351-355.

81
SOUZA, C.F.; FERNANDES, L.C.; CYRINO, E.S. (2006). Produção de espécies reativas de
oxigênio durante o exercício aeróbio e anaeróbio. Revista Brasileira de Cineantropometria
& Desempenho Humano. 8(2):102-109.
STADTMAN, E.R.; LEVINE, R.L. (2000). Protein oxidation. Ann. N. Y. Acad. Sci.
899:191-208.
STEINBERG, D. (1997). Low Density Lipoprotein Oxidation and Its Pathobiological
Significance. J. Biol. Chem. 272(34): 20963-20966.
STERN, A. (1985). Red cell oxidative damage. In: H. Sies (ed), Oxidative stress. Academic
Press, London. pp. 331-349.
STUPKA, N.; TARNOPOLSKY, M. A.; YARDLEY, N. J.; PHILLIPS, S. M. (2001).
Cellular adaptation to repeated eccentric exercise-induced muscle damage. J. Appl.
Physiol. 91:1669-1678.
SUREDA, A.; TAULER, P.; AGUILÓ, A.; CASES, N.; FUENTESPINA, E.; CÓRDOVA,
A.; TUR, J. A.; PONS, A. (2005). Relation between oxidative stress markers and
antioxidant endogenous defences during exhaustive exercise. Free Rad. Res. 39(12): 317-
1324.
TRABER, M. G. (2006). Relationship of vitamin E metabolism and oxidation in exercising
human subjects. Br. J. Nutr. 96(S1): S34-S37.
URSO, M.L.; CLARKSON, P.M. (2003). Oxidative stress, exercise, and antioxidant
supplement. Toxicology. 189: 41-54.
VALKO, M.; LEIBFRITZ, D.; MONCOL, J.; CRONIN, M. T. D.; MAZUR, M.; TELSER, J.
(2007). Free radicals and antioxidants in normal physiological functions and human disease.
Int. J. Biochem. Cell Biol. 39:44-84.
VAN ACKER, S.A.B.E. (1996). Structural aspects of antioxidant activity of flavonoids. Free
Radic. Biol. Med. 20(3):331-342.

82
VANCINI, R.L.; LIRA, C.A.B.; ABOULAFIA, J.; NOUAILHETAS, V.L.A. (2005).
Radicais Livres, estresse oxidativo e exercício. São Paulo: UNIFESP. Disponível em
<www.centrodeestudos.org.br/pdfs/oxidativo.pdf>, acesso em 23/11/2009.
VANDERPLANK, J. (1996). Passion Flowers. Cambridge: MIT Press. 224p.
VANNUCCHI, H. et al. (1998). Papel dos nutrientes na peroxidação lipídica e no sistema de
defesa antioxidante. Medicina. 31:31-34.
VILARDI, T. C. C.; RIBEIRO, B. G.; SOARES, E. A. (2001). Distúrbios nutricionais em
atletas femininas e suas inter-relações. Rev. Nutr. 14(1): 61-69.
VILLA-CABALLERO, L.; NAVA-OCAMPO, A. A. ; FRATI-MUNARI, A.; PONCE-
MONTER, H. (2000). Oxidative stress, acute and regular exercise: are they really harmful in
the diabetic patient? Med. Hypotheses. 55(1):43-46.
W. Effect of resistance exercise on free radical production. Med. Sci. Sports Exerc.
WANNAMETHEE, S. G.; SHAPER, A. G. (2001). Physical Activity in the
Prevention of Cardiovascular Disease. Sports Med. 31(2): 101-114.
WATARU, A.; YUJI, N; TOSHIKAZU, Y. (2003). Chronic Exercise and Protection of
Oxidative Damage Induced by Ischemia-Reperfusion. Descente Sports Sci. 24:139-144.
WATTANAPITAYAKUL, S. K.; BAUER, J. A. (2001). Oxidative pathways in
cardiovascular disease Roles, mechanisms, and therapeutic implications. Pharmacol. Ther.
89: 187- 206.
WITT, E. H.; REZNICK, A. Z.; VIGUIE, C. A.; STARKE-REED, P.; PACKER, L. (1992).
Exercise, oxidative damage and effects of antioxidant manipulation. J. Nutr. 122(S3): 766-
773.
WOODS, J. A.; LU, Q.; LOWDER, T. (2000). Exercise-induced modulation of macrophage
function. Immunol. Cell Biol. 78: 545-553.

83
YAGI, K. A. (1976). fluorometric ensaio simples para lipoperóxidos no plasma sanguíneo.
Bioquímico Medicina. 15:216 -212.
YU, B.; CHUNG, H. (2006). Mecanismos de adaptação ao estresse oxidativo durante o
envelhecimento. Mechanisms of Ageing and Development. 127:436–443.
YU, B.P. (1994). Cellular defenses against damage from reactive oxygen species.
Physiological Reviews. 74(1):139-162.
YUSOF, A; LEITHAUSER, R. M.; ROTH, H. J.; FINKERNAGEL, H.; WILSON, M. T.;
BENEKE, R. (2007). Exercise-induced hemolysis is caused by protein modification and
most evident during the early phase of an ultraendurance race. J. Appl. Physiol. 102: 582-
586.
ZANUSSO, M. C. (2010). Exercício Físico e Estresse Oxidativo: Evidências de Risco à
Saúde. Artigo. (Especialização em atividade física adaptada e saúde) - Universidade Gama
Filho, UGF, Brasil, 28p.


















