AVALIAÇÃO DOS EFEITOS GENOTÓXICOS E … · peixes expostos a FSG na concentração de 5% durante...
50
6 ESTHER DO LAGO E PRETTI AVALIAÇÃO DOS EFEITOS GENOTÓXICOS E NEUROTÓXICOS DA FRAÇÃO SOLÚVEL DA GASOLINA PARA Prochilodus lineatus Londrina – Paraná 2007
Transcript of AVALIAÇÃO DOS EFEITOS GENOTÓXICOS E … · peixes expostos a FSG na concentração de 5% durante...

6
ESTHER DO LAGO E PRETTI
AVALIAÇÃO DOS EFEITOS GENOTÓXICOS E NEUROTÓXICOS DA FRAÇÃO SOLÚVEL DA
GASOLINA PARA Prochilodus lineatus
Londrina – Paraná
2007

7
ESTHER DO LAGO E PRETTI
AVALIAÇÃO DOS EFEITOS GENOTÓXICOS E NEUROTÓXICOS DA FRAÇÃO SOLÚVEL DA
GASOLINA PARA Prochilodus lineatus
Supervisor de Estágio Cláudia Bueno Dos Reis Martinez
Londrina – Paraná 2007
Monografia apresentada ao Curso de Graduação em Ciências Biológicas da
Universidade Estadual de Londrina como um dos requisitos à obtenção do
título de Bacharel em Ciências Biológicas.

8
BANCA EXAMINADORA
Profa. Dra. Cláudia Bueno dos Reis Martinez
Profa. Dra. Silvia Helena Sofia
Ms. Juliana Delatim Simonato
Londrina, 23 de novembro de 2007

9
AGRADECIMENTOS
Agradeço à minha orientadora Profa. Dra. Cláudia Bueno dos Reis Martinez pela
oportunidade, confiança, paciência e verdadeira orientação num turbulento último ano
de graduação.
Agradeço à amiga Dalita Cavalcante pelos ensinamentos e paciente cooperação a
cada experimento realizado.
Agradeço aos meus pais Humberto e Esther por todos os anos de suporte e apoio,
mesmo que à distância. Aos meus irmãos, Giselle, Gerson e Humberto por fazerem
de mim, tudo o que sou hoje.
Agradeço aos meus grandes amigos pelos anos de estudos desesperados, favores
de última hora, jantares experimentais, reflexões científicas e filosóficas e
divertidíssima companhia: Júnior, Kenia, Jeanne Marie e especialmente à Cibele, por
me surpreender e acompanhar todos os dias.
Agradeço a Universidade Estadual de Londrina pelos grandes professores e
ensinamentos adquiridos dentro e fora de aula.

10
SUMÁRIO
PÁG.
1. RESUMO 01
2. INTRODUÇÃO 02
3. REVISÃO BIBLIOGRÁFICA 04
3.1 ENSAIO DO COMETA EM MEIO ALCALINO 08
3.2 TESTE DO MICRONÚCLEO 09
3.3 ANÁLISE DA AÇÃO DA ACETILCOLINESTERASE 11
4. ARTIGO 14
RESUMO 15
INTRODUÇÃO 16
MATERIAL E MÉTODOS 18
ACLIMATAÇÃO 18
PREPARO DA FSG 19
AMOSTRAGEM – COLETA DE SANGUE, CÉREBRO E MÚSCULO 19
ENSAIO DO COMETA 20
TESTE DO MICRONÚCLEO 21
ANÁLISE DA ATIVIDADE DA ACETILCOLINESTERASE 22
ANÁLISE ESTATÍSTICA 23
5. RESULTADOS 23
5. DISCUSSÃO 29
7. CONCLUSÃO 33
REFERÊNCIAS BIBLIOGRÁFICAS 34

1
1. RESUMO
Os possíveis efeitos genótoxicos, mutagênicos e neurotóxicos da gasolina
foram avaliados para espécimes jovens de Prochilodus lineatus (Characiformes,
Prochilodontidae), usando-se o ensaio do cometa, teste do micronúcleo, (ambos
utilizando sangue periférico) e análise da atividade da enzima acetilcolinesterase
(AchE) em cérebro e musculatura axial. Outras anormalidades eritrocíticas nucleares
(AENs) também foram analisadas além do micronúcleo, como indicativo de dano
genético. Como controle positivo foi utilizado ciclofosfamida na concentração de
0,04mg/g de animal. A FSG foi obtida simulando-se um vazamento desse poluente
em águas neotropicais durante 24 horas (6 horas de exposição solar direta) e os
peixes expostos a FSG na concentração de 5% durante os tempos experimentais de
6, 24 e 96 horas (exposição aguda). Os resultados mostraram um efeito mutagênico
desse poluente sobre os animais em todos os tempos experimentais como mostrado
pelos escores significativamente maiores obtidos com o ensaio do cometa nos
peixes expostos a FSG comparados aos animais expostos somente à água limpa
(controle negativo). Nossos resultados também indicam aumento significativo de
AENs, como núcleo lobulado, segmentado e em forma de rim em todos os tempos
experimentais em relação ao controle negativo, indicando também, dano genotóxico.
A atividade da Acetilcolinesterase (AchE) não foi significativamente diferente nos
peixes expostos a FSG comparados aos respectivos controles negativos em nenhum
dos tempos experimentais.

2
2. INTRODUÇÃO
O petróleo, seus produtos refinados e seus hidrocarbonetos
aromáticos policíclicos (PAHs – do inglês: polycyclic aromatic hydrocarbons) são
encontrados como poluentes acidentais em rios, mares e águas subterrâneas com
ocorrências detectadas nos componentes vivos e não vivos do ecossistema
(ALBERS, 2002). O petróleo pode ser refinado em diversos produtos, como óleo
diesel, gasolina, gás natural e durante esse processo de destilação e transporte,
freqüentemente há vazamentos e derrames, contaminando solos e água. A
toxicidade do petróleo é, em sua maioria, atribuída à sua parte solúvel (ALMEIDA-
VAL et al., 2002) e seus hidrocarbonetos aromáticos policíclicos, encontrados em
efluentes de refinarias de petróleo, são descritos como os principais componentes
capazes de causar danos no DNA (HAMOUTENE et al. 2002).
A escolha pelo peixe Neotropical Prochilodus lineatus se deu
principalmente por ser um animal de biologia e fisiologia conhecidas, além de ser
sensível aos efeitos de poluentes como a gasolina e pelo fato dos peixes serem
capazes de bio-acumulação devido à exposição direta a produtos químicos e
ingestão de alimento contaminado (RAND & PETROCELLI, 1984). Além de esses
animais incorporarem os poluentes presentes na água pela respiração, o fato de
serem detritívoros também permite que eles incorporem os mesmos poluentes pela
ingestão de sedimentos.
Nesse trabalho serão avaliadas as respostas à exposição aguda de
P. lineatus à fração solúvel da gasolina (FSG) através dos seguintes parâmetros
biológicos: i) avaliação da ação da acetilcolinesterase em cérebro e músculo e ii)
dano no DNA através dos testes do cometa e micronúcleo e presença de
anormalidades eritrocíticas nucleares. Esse estudo foi delineado para detectar as

3
possíveis mudanças bioquímicas e fisiológicas em um peixe de água doce exposto a
uma concentração subletal da FSG, como forma de se ter um alerta precoce em
casos de verdadeiros derrames antes que comunidades inteiras e até todo o
ecossistema seja comprometido permanentemente.
O uso de exemplares de espécies nativas de peixes para a
avaliação da mutagenicidade de xenobióticos é relevante, visto que há uma
necessidade de preservar tais espécies, além do que, através da alimentação,
muitos deles são agentes de transferência de contaminantes ao homem (AL-SABTI
& METCALFE, 1995).
O objetivo desse trabalho é avaliar o potencial genotóxico de um
derivado de petróleo, a gasolina, através de testes como o ensaio do cometa e do
micronúcleo, além de quantificação da atividade da acetilcolinesterase em cérebro e
músculo usando como instrumento o peixe neotropical Prochilodus lineatus.
Esse estudo vem então de encontro a uma necessidade de
monitoramento através da utilização de biomarcadores, da contaminação por
hidrocarbonetos, no caso, da gasolina, evidenciando necessidades de maior
fiscalização e monitoramento de estações de abastecimento e distribuição de
derivados de petróleo em nosso país.

4
3. REVISÃO BIBLIOGRÁFICA
O ambiente aquático é um habitat para grande número de plantas,
animais e espécies microbianas e sua composição influencia diretamente estes
organismos (ALLOWAY & AYRES, 1997). Poluentes orgânicos como os pesticidas,
compostos químicos industriais (como é o caso das bifenilas policloradas - BPCs),
solventes, detergentes e os derivados de petróleo são prejudiciais para o
ecossistema aquático (WRIGHT & NEBEL, 2000).
A liberação proposital ou acidental de compostos químicos danosos
para o ambiente tem o potencial de perturbar a estrutura e o funcionamento dos
ecossistemas naturais. Conforme a tecnologia industrial avança, as características
das fontes de dejetos industriais se tornam um grande problema e a toxicidade da
água residual de uso industrial também pode se tornar mais perigosa. Uma vez que
o ambiente aquático é o destinatário final dos poluentes produzidos por fontes
naturais ou antropogênicas, o acúmulo e persistência de efluentes industriais no
ambiente aquático constituem-se em uma ameaça à vida biológica (FLEEGER et al.,
2003).
O processamento do petróleo pelas indústrias necessita de grande
quantidade de água e os resíduos gerados e descartados pela indústria de petróleo
tipicamente contém uma vasta gama de poluentes orgânicos e metálicos, incluindo
fenóis, óleos e graxas, sulfetos, nitrato de amônia e hidrocarbonetos policíclicos
aromáticos (BROWN & DONELLY, 1986; DASGUPTA & ZDUNEK, 1992).
O petróleo pode ser destilado em diversos produtos, como óleo
diesel, gasolina, óleo cru, gás natural dentre outros. A toxicidade do petróleo é, em
sua maioria, atribuída à sua parte solúvel (ALMEIDA-VAL et al., 2002). Os
hidrocarbonetos aromáticos policíclicos presentes em efluentes de refinarias de

5
petróleo são descritos como os principais componentes capazes de causar danos no
DNA (HAMOUTENE et al., 2002), além de conhecidamente afetarem a
neurotransmissão e a locomoção nos animais.
Após um derrame de petróleo ou de seus derivados em águas
naturais, ocorre uma variedade de efeitos físicos, químicos e biológicos. Processos
de intemperismo iniciam-se com a evaporação, seguida de fotodegradação, a qual
favorece o processo de degradação biológica (NICODEM et al., 1998).
Os peixes são geralmente usados como organismos sentinela por
que eles têm uma série de papéis na rede trófica, acumulando substâncias tóxicas e
respondendo a baixas concentrações de mutagênicos (ÇAVAS & ERGENE-
GOZUKARA, 2005). Assim, o uso de peixes como indício de efeitos da poluição
através da biomarcação é de grande importância e pode permitir a detecção de
problemas aquáticos ambientais no início dos mesmos (LÓPZ-BAREA, 1996; VAN
DER OOST et al., 2003).
O peixe de água doce neotropical Prochilodus lineatus
(VALENCIENNES, 1847) é uma escolha adequada para testes de toxicidade, por ser
sensível a agentes tóxicos (MAZON & FERNANDES, 1999; MARTINEZ & SOUZA,
2002; DA SILVA et al., 2004; MARTINEZ et al., 2004; ALMEIDA et al., 2005) e,
portanto, ser considerado um potencial vertebrado para monitoramento ambiental
(CERQUEIRA & FERNANDES, 2002; CAMARGO & MARTINEZ, 2006).
Biomarcadores são quaisquer alterações biológicas relacionadas à
presença de um composto químico no ambiente no nível sub-individual, medida
dentro do organismo ou em seus produtos (urina, fezes, pêlos e penas) que indica
um desvio do estado normal e que não pode ser detectada no organismo intacto
(RIBEIRO et al., 2003).

6
Os biomarcadores em peixes são excelentes ferramentas para
monitorar a saúde do ecossistema aquático e têm sido incluídos em vários
programas de monitoramento ambiental modernos (WALKER et al., 1996). A
principal razão para a sua utilização é o fato de fornecerem informações sobre os
efeitos biológicos dos poluentes, bem como pistas sobre os mecanismos de ação
dos contaminantes, e não correspondem apenas à quantificação desses poluentes
no ambiente. Assim, os biomarcadores podem ser usados para identificar sinais
iniciais de danos aos peixes, sugerir as relações de causa-efeito entre exposição aos
contaminantes e os efeitos observados e documentar os efeitos integrados do
estresse químico nos animais. Considerando-se ainda que muitos biomarcadores
são indicadores rápidos de efeitos adversos crônicos, eles podem permitir uma
intervenção antes que efeitos prejudiciais irreversíveis tornem-se inevitáveis
(RIBEIRO et al., 2003).
A maioria dos xenobióticos que entra no organismo é lipofílica, uma
propriedade que permite que esses compostos exógenos penetrem as membranas
lipídicas e sejam transportados por lipoproteínas nos fluidos corpóreos (HODGSON
& GOLDSTEIN, 2001). Embora uma seqüência de processos de detoxificação ocorra
após a entrada do xenobiótico, em alguns casos os produtos intermediários ou finais
podem ser ainda mais tóxicos que os compostos originais, e a seqüência, nesse
caso, pode ser denominada de bioativação. Os efeitos tóxicos de um xenobiótico ou
seus metabólitos podem se manifestar quando o composto se liga a macromoléculas
celulares, levando à ruptura da membrana, a danos celulares ou efeitos genotóxicos,
que por sua vez, podem levar ao desenvolvimento ou progressão de doenças (VAN
DER OOST et al., 2003).

7
A biotransformação de xenobióticos é uma importante fonte de
espécies reativas de oxigênio (EROs), que correspondem aos produtos da redução
do oxigênio molecular, como o ânion radical superóxido (O2-), o radical hidroxil (OH˙)
e o peróxido de hidrogênio (H2O2) (STEGEMAN et al., 1992) Essas espécies
reativas de oxigênio são oxidantes extremamente potentes, capazes de reagir com
macromoléculas celulares críticas, causando diferentes tipos de danos biológicos
que podem levar à morte celular (MARTINEZ, 2004).
Os organismos aeróbicos apresentam diferentes mecanismos,
enzimáticos e não enzimáticos, de defesa antioxidante que podem prevenir a
formação das EROs, reagir com esses intermediários reativos, bem como reparar os
danos causados pelos mesmos (STOREY, 1996). Caso o sistema de defesa
antioxidante esteja comprometido, pode ser estabelecido um desequilíbrio pró-
oxidante celular, denominado estresse oxidativo, que, por sua vez, está associado a
vários processos patológicos e pode comprometer a sobrevivência dos peixes.
Muitos poluentes ou seus metabólitos podem exercer sua toxicidade gerando
estresse oxidativo (MARTINEZ, 2004). No decorrer da vida, o DNA sofre alterações
denominadas mutações, que podem ser causadas por erros durante a sua
duplicação ou durante a divisão celular (RIBEIRO & MARQUES, 2003). Danos na
desoxirribose, que usualmente resultam em quebra nas fitas, também pode ocorrer
em conseqüência da oxidação do DNA. A detecção de quebras nas cadeias de DNA
representa uma técnica valiosa da ecotoxicologia, uma vez que tem revelado
sensibilidade maior do que outros métodos comumente empregados na avaliação de
danos no material genético (LEE & STEINERT, 2003).
O ensaio do cometa é extremamente sensível e bastante útil para a
detecção de danos no DNA (SINGH et al., 1998) e tem sido amplamente utilizado

8
para avaliação de possíveis danos mediados por EROs causados por contaminantes
ambientais (MARTINEZ & CÓLUS, 2002).
3.1 Ensaio do cometa em meio alcalino
O teste do cometa não é utilizado para detectar mutações, mas sim
lesões genômicas, que, após serem processadas, podem resultar em mutação.
Diferente das mutações, as lesões detectadas pelo teste do cometa são passíveis de
correção. Uma vez que danos no DNA são freqüentemente célula e tecido
específicos, uma metodologia como o teste do cometa que permite a detecção de
danos e seu reparo em uma única célula, e conseqüentemente, em determinada
subpopulação celular, é de extrema relevância para a avaliação de compostos
genotóxicos (TICE, 1995).
O ensaio do cometa tem sido proposto para estudos de
toxicogenética devido às suas vantagens e peculiaridades quando comparado a
outros testes que detectam substâncias genotóxicas, sendo uma ferramenta
fundamental de investigação em estudos de reparo de DNA, biomonitoramento
ambiental e teste de genotoxicidade (ROSS et al., 1995). As vantagens de se
empregar o ensaio do cometa para avaliar danos no DNA incluem: mensuração de
danos em células individuais; o baixo número de células necessário para a
realização do teste; possibilidade de ser realizado em qualquer tipo de célula
eucariótica nucleada e ser um método muito sensível para detectar danos no DNA
(LEE & STEINERT, 2003).
Deve-se ter em mente que não existe célula sem dano no DNA, visto
que o próprio metabolismo celular pode gerar em torno de 1000 lesões diárias no
DNA/célula. O que se faz, rotineiramente é modular as condições técnicas (tempo de

9
relaxamento e eletroforese) para que um mínimo de DNA migre da cabeça para a
cauda do cometa nos controles negativos (TICE, 1995). O ensaio do cometa tem
sido amplamente aplicado em eritrócitos e linfócitos do sangue periférico porque
estes tipos celulares podem ser facilmente amostrados e a dissociação celular não é
necessária (BELPAEME et al., 1996).
O ensaio do Cometa, por suas diversas vantagens já citadas e por
ser rápido, relativamente econômico, preciso e reproduzível, torna-se um sistema-
teste adequado para o biomonitoramento ambiental e na genética toxicológica
(TICE, 1995).
3.2 Teste do micronúcleo
O Teste do micronúcleo serve como o primeiro passo no estudo de
qualquer substância mutagênica. É um teste rápido e sensível, tanto para detectar
alterações cromossômicas estruturais como numéricas (HEDDLE et al., 1983).
Também é um ensaio citogenético comumente usado em vários sistemas biológicos
para o monitoramento de genotoxicidade ambiental (MERSCH & BEAUVAIS, 1997).
O teste do micronúcleo é amplamente utilizado para estimar o dano
citogenético induzido por agentes químicos ou físicos. Embora a maior parte dos
estudos com empregando o teste do micronúcleo tenha sido realizada com
mamíferos (especialmente roedores), este teste tem também se mostrado uma
ferramenta muito útil também em estudos com outros animais. Particularmente, em
ambientes em aquáticos, ele permite por meio de estudos da biota aquática, detectar
as propriedades genotóxicas de compostos presentes no ambiente (UDROIU, 2006).
Como peixes aparentemente respondem a xenobióticos da mesma maneira que os

10
mamíferos, eles podem ser usados para testar as possíveis propriedades
genotóxicas de agentes químicos e físicos (UDROIU, 2006).
Micronúcleos são massas de cromatina citoplasmática com a
aparência de pequenos núcleos que emergem de fragmentos cromossômicos ou são
cromossomos inteiros deixados para trás na anáfase, durante a divisão celular. Sua
presença na célula é um reflexo de aberração cromossômica estrutural ou numérica
durante a mitose (FENECH et al., 1999).
Micronúcleos aparecem quando todo um cromossomo ou um
fragmento dele falha durante migração juntamente com um dos dois novos núcleos
formados durante a mitose. O primeiro caso (perda do cromossomo) se deve a um
evento aneugênico, enquanto o segundo caso se deve a quebra de um
cromossomo. Essas inclusões podem ocorrer em todo tipo de célula, tanto somática
quanto germinativa. Assim, o teste do micronúcleo pode ser aplicado em qualquer
tecido do animal (UDROIU, 2006).
Segundo BRUNETTI et al. (1988), a freqüência de micronúcleos na
avaliação da poluição ambiental é um marcador sensível da presença de
contaminantes genotóxicos. Monitorar efeitos clastogênicos de poluentes é um dos
primeiros interesses da mutagênese ambiental aquática para determinar a relação
da poluição com o estresse em organismos vivos (KIM & HYUM, in press).
A aplicação desse teste em eritrócitos é muito popular, em particular
com amostras sanguíneas onde há centenas de células disponíveis para o escore
(UDROIU, 2006). Enquanto eritrócitos micronucleados de órgãos hematopoiéticos
refletem um dano que ocorreu durante um tempo equivalente ao ciclo celular,
aqueles da circulação periférica refletem eventos que ocorreram num tempo
equivalente à vida do eritrócito na circulação (SCHLEGEL & McGREGOR, 1982).

11
Pesquisas com micronúcleos em peixes têm sido conduzidas principalmente
analisando eritrócitos de sangue periférico, mas a sensibilidade e facilidade de obter
células epiteliais de brânquias como alternativa têm sido mostrado em vários
estudos (TAKAI et al., 2004; ÇAVAS & ERGENE-GÖZÜKARA, 2005).
Recentemente, vários estudos descreveram a presença de
anormalidades eritrocíticas nucleares (AENs) além do micronúcleo, em células de
peixe expostas a substâncias genotóxicas (AYLLON & GARCIA-VAZQUEZ, 2000;
ÇAVAS & ERGENE-GOZUKARA, 2005). Em geral essa anormalidades são
consideradas indicadoras de dano genotóxico e, portanto, podem complementar o
escore de micronúcleo em pesquisas de genotoxicidade (ÇAVAS & ERGENE-
GOZUKARA, 2005). As anormalidades eritrocíticas nucleares (AENs) podem ser
classificadas da seguinte forma: núcleo lobado (L), segmentado (S) e em forma de
rim (K).
Nos últimos anos, a expressão simultânea de anormalidades
nucleares juntamente com micronúcleos recebeu atenção considerável. Embora os
mecanismos responsáveis pela formação das anormalidades nucleares não tenham
sido completamente esclarecida, muitos estudos indicam que ENAs são induzidas
em resposta à exposição a agentes genotóxicos (TOLBERT et al., 1992; SERRANO-
GARCIA & MONTERO-MONTOYA, 2001).
3.3 Atividade da acetilcolinesterase
A acetilcolina é um neurotransmissor encontrado no cérebro e nas
junções neuromusculares, compondo parte do sistema nervoso parassimpático.
Seus efeitos incluem a contração dos músculos lisos, dilatação dos vasos
sanguíneos e regulação da taxa de batimentos cardíacos; no cérebro está envolvido

12
nas sinapses associadas ao controle motor, memória e cognição. Sua atividade e
permanência na fenda sináptica são reguladas por hidrólise catalisada pela
acetilcolinesterase (AChE), que regenera a colina, seu precursor (VIEGAS JUNIOR
et al., 2004).
A acetilcolina é o principal neurotransmissor nos sistemas sensorial
e neuromuscular em peixes; seus níveis são regulados pela acetilcolinesterase
(AChE) a partir de hidrólises da acetilcolina em ácido acético e seu produto ativo, a
colina. Esses são reabsorvidos e usados como substrato para a produção contínua
de acetilcolina. A síntese initerrupta ou a inabilidade de hidrolisar a acetilcolina é
responsável pela estimulação contínua ou excessiva das fibras musculares ou
nervosas (KIRBY, 2000).
A atividade da acetilcolinesterase (AChE) é um biomarcador muito
usado em estudos de ecotoxicologia aquática (KIRBY, 2000), visto que é uma
enzima bastante sensível a baixas concentrações de poluentes (ALMEIDA, 2005).
Inseticidas, como o metil paration inibem a atividade da AChE em peixes, resultando
num bloqueio do sistema neuronal. Nesse caso, o sistema muscular pode continuar
se movimentando sem controle até a paralisia, convulsão e morte (AGUIAR et al.,
2004). A maioria dos estudos com AChE são feitos a partir de cérebros de peixes,
pois os efeitos mais evidentes são observados em tecidos nervosos (ALMEIDA et
al., 2005).
Num estudo realizado por AGUIAR (2002), o peixe conhecido
popularmente como matrinxã (Brycon cephalus) foi exposto a concentrações de 2
ppm de metil paration por 24 horas. O cérebro do matrinxã mostrou a maior taxa de
inibição de AChE, como esperado. A inibição no cérebro foi seguida pelos músculos,

13
brânquias e fígado. A inibição da AChE está muito relacionada com o nível de
enervação de tecidos, portanto pode ser dito que quanto maior a concentração de
AchE, maior a susceptibilidade a inibição (ALMEIDA et al., 2005).

14
4. ARTIGO
AVALIAÇÃO DOS EFEITOS GENOTÓXICOS E NEUROTÓXICOS DA FRAÇÃO SOLÚVEL DA GASOLINA PARA Prochilodus lineatus
Esther L. Pretti e Cláudia B.R.Martinez

15
RESUMO
Os possíveis efeitos genótoxicos, mutagênicos e neurotóxicos da gasolina
foram avaliados para espécimes jovens de Prochilodus lineatus (Characiformes,
Prochilodontidae), usando-se o ensaio do cometa, teste do micronúcleo, (ambos
utilizando sangue periférico) e análise da atividade da enzima acetilcolinesterase
(AchE) em cérebro e musculatura axial. Outras anormalidades eritrocíticas nucleares
(AENs) também foram analisadas além do micronúcleo, como indicativo de dano
genético. Como controle positivo foi utilizado ciclofosfamida na concentração de
0,04mg/g de animal. A FSG foi obtida simulando-se um vazamento desse poluente
em águas neotropicais durante 24 horas (6 horas de exposição solar direta) e os
peixes expostos a FSG na concentração de 5% durante os tempos experimentais de
6, 24 e 96 horas (exposição aguda). Os resultados mostraram um efeito mutagênico
desse poluente sobre os animais em todos os tempos experimentais como mostrado
pelos escores significativamente maiores obtidos com o ensaio do cometa nos
peixes expostos a FSG comparados aos animais expostos somente à água limpa
(controle negativo). Nossos resultados também indicam aumento significativo de
AENs, como núcleo lobulado, segmentado e em forma de rim em todos os tempos
experimentais em relação ao controle negativo, indicando também, dano genotóxico.
A atividade da Acetilcolinesterase (AchE) não foi significativamente diferente nos
peixes expostos a FSG comparados aos respectivos controles negativos em nenhum
dos tempos experimentais.

16
INTRODUÇÃO
Dentre os diferentes tipos de poluentes, os derivados de petróleo
são dos mais relevantes para a toxicologia aquática (PACHECO e SANTOS, 2001).
O petróleo pode ser destilado em diversos produtos, como óleo
diesel, gasolina, gás natural e durante esse processo de destilação e transporte,
freqüentemente há vazamentos e derrames, contaminando solos e água sendo a
toxicidade do petróleo, em sua maioria, atribuída à sua parte solúvel (ALMEIDA-VAL
et al., 2002) que contém compostos de baixo peso molecular, como hidrocarbonetos
monoaromáticos (benzeno, tolueno e xileno – BTX) e hidrocarbonetos aromáticos
policíclicos (PAHs) (PACHECO e SANTOS, 2001), encontrados em efluentes de
refinarias de petróleo, que são descritos como os principais componentes capazes
de causar danos no DNA (HAMOUTENE et al., 2002).
Embora grandes vazamentos de óleo sejam mais conhecidos,
acredita-se que a principal fonte de contaminação de águas continentais por petróleo
e seus derivados se deva a pequenos e contínuos vazamentos de tanques de
armazenagem de combustível subterrâneos, atingindo, portanto, corpos de água
subterrânea e finalmente, rios (TIBURTIUS et al., 2005). A exposição ao óleo cru e
seus derivados pode induzir uma variedade de sintomas devido à toxicidade em
animais experimentais. Os hidrocarbonetos do petróleo podem atuar como
mediadores na geração de radicais livres em peixes (ACHUBA e OSAKWE, 2003).
Apesar de alguns experimentos prévios com peixes sobre os
efeitos e respostas toxicológicas de derivados de petróleo, algumas respostas
toxicológicas permanecem pouco esclarecidas revelando a falta de informações
sobre mecanismos de estresse bem como biotransformação e respostas
genotóxicas (PACHECO e SANTOS, 2001), mostrando assim a grande necessidade

17
por informação relacionada à contaminação por derivados de petróleo em peixes
neotropicais.
Os peixes são geralmente usados como organismos sentinela por
que eles têm uma série de papéis na rede trófica, acumulando substâncias tóxicas e
respondendo a baixas concentrações de mutagênicos (ÇAVAS e ERGENE-
GOZUKARA, 2005). Assim, o uso de peixes como indício de efeitos da poluição
através da biomarcação é de crescente importância e pode permitir a detecção de
problemas aquáticos ambientais no início dos mesmos (LÓPZ-BAREA, 1996; VAN
DER OOST et al.2003).
O peixe de água doce Prochilodus lineatus (VALENCIENNES, 1847)
é uma escolha adequada para testes de toxicidade, por ser um animal de biologia
bastante conhecida, sensível a agentes tóxicos (MAZON e FERNANDES, 1999;
MARTINEZ e SOUZA, 2002; DA SILVA et al., 2004; MARTINEZ et al., 2004;
ALMEIDA et al., 2005) e detritívoro, portanto, sendo considerado um potencial
vertebrado para monitoramento ambiental (CERQUEIRA & FERNANDES, 2002;
CAMARGO & MARTINEZ, 2006).
Os organismos aeróbicos apresentam diferentes mecanismos,
enzimáticos e não enzimáticos, de defesa antioxidante que podem prevenir a
formação das espécies reativas de oxigênio (EROs), reagir com esses intermediários
reativos, bem como reparar os danos causados pelos mesmos (STOREY, 1996). Em
peixes, caso o sistema de defesa antioxidante esteja comprometido, pode ser
estabelecido um desequilíbrio pró-oxidante celular, denominado estresse oxidativo,
que, por sua vez, está associado a vários processos patológicos e pode
comprometer a sobrevivência desses animais. Muitos poluentes ou seus metabólitos
podem exercer sua toxicidade gerando estresse oxidativo (MARTINEZ, 2004).

18
No decorrer da vida, o DNA sofre alterações denominadas
mutações, que podem ser causadas por erros durante a sua duplicação ou durante a
divisão celular (RIBEIRO & MARQUES, 2003). Danos na desoxirribose, que
usualmente resultam em quebra nas fitas, também pode ocorrer em conseqüência
da oxidação do DNA. A detecção de quebras nas cadeias de DNA representa uma
técnica valiosa de ecotoxicologia, uma vez que tem revelado sensibilidade maior do
que outros métodos comumente empregados na avaliação de danos no material
genético (LEE & STEINERT, 2003).
Assim, tendo em vistao grande número de acidentes e vazamentos
de derivados de petróleo, como a gasolina, em águas neotropicais, torna-se evidente
a necessidade de levantamento de informações a respeito de seus efeitos na cadeia
trófica. Assim, o objetivo deste trabalho foi avaliar o potencial genotóxico,
mutagênico e neurotóxico de um derivado de petróleo, a gasolina, através do ensaio
do cometa e teste do micronúcleo, além de avaliação da atividade da enzima
acetilcolinesterase em cérebro e músculo usando como parâmetro o peixe
neotropical Prochilodus lineatus.
MATERIAL E MÉTODOS
Aclimatação
Espécimes jovens do peixe neotropical Prochilodus lineatus
(Characifomes, Prochilodontidae) conhecido popularmente como curimba, pesando
aproximadamente 10 g (8 - 12 g), foram fornecidos pela estação de piscicultura da
Universidade Estadual de Londrina (EPUEL) e aclimatados durante
aproximadamente sete dias em tanques de 300L, contendo água desclorada no

19
laboratório de Bioensaios do Laboratório de Ecofisiologia Animal (LEFA-UEL).
Durante esse período os animais foram alimentados com ração comercial a cada 48
horas. Buscando melhorar a condição de vida dos peixes durante a aclimatação e
assim aumentar a confiabilidade dos experimentos realizados, foi utilizada uma
metodologia para o combate a patógenos aplicando-se no tanque 100mg de gesso
para cada litro de água e 100mg de sal por litro de água. Essa metodologia se
mostrou efetiva uma vez que a mortalidade e acometimento por patógenos foram
sensivelmente reduzidos.
Preparo da FSG
Para obter a fração solúvel da gasolina (FSG) a gasolina foi diluída
em água na proporção do 1:4 em um aquário de vidro e exposta a luz solar durante
o fotoperíodo de 6 horas com o intuito de simular um derrame verdadeiro de gasolina
em ambiente tropical, havendo assim fotodegradação e evaporação. Foi coletada
então somente a fração solúvel da gasolina e descartada a gasolina sobrenadante.
Essa fração solúvel foi então diluída para obtenção de concentração final de 5% da
FSG na água.
Amostragem
Os peixes foram submetidos a testes de toxicidade agudos (6, 24 e
96h) em aquários de 100L, cada um contendo oito peixes. Um grupo controle,
também contendo oito animais, exposto somente à água desclorada também foi
amostrado a cada experimento dos grupos expostos a FSG. Foram feitas réplicas
para cada tempo experimental bem como grupos controle negativos. Durante o
período de experimentação (6, 24 ou 96 horas) os valores de pH, oxigênio

20
dissolvido, temperatura e condutividade da água. foram monitorados diariamente
Imediatamente após serem retirados da água, os animais foram
anestesiados com benzocaína (0,1g/L) e amostras de sangue retiradas através da
veia caudal por seringas plásticas de 1mL previamente heparinizadas. A seguir os
peixes foram sacrificados por secção medular e amostras de cérebro e músculo
imediatamente removidos e congelados a – 80ºC para análises posteriores.
Ensaio do Cometa
O ensaio do cometa empregado no presente trabalho teve como
base a metodologia descrita por Tice (1990). Lâminas foram preparadas com
agarose a ponto de fusão normal a 1,5% cinco dias antes da amostragem. Após a
amostragem 10µL de sangue foram adicionado a 1mL de solução salina (17mM
NaCl, 4.8mM KCl, 1.5mM CaCl2, 1.2mM NaHCO3, 4.5mM Na2HPO4, 2.9mM
NaH2PO4). Em uma sala escura 50µL da mistura de sangue e solução salina foram
adicionados a 120 µL de agarose a baixo ponto de fusão. Essa mistura foi então
depositada sobre a lâmina com cobertura de agarose 1,5% e a mesma coberta com
lamínula e as lâminas mantidas em temperatura de 4-8ºC por cerca de trinta
minutos. Após esse período a lamínula foi retirada e as lâminas acondicionadas em
uma cuba com solução de uso de lise (2,5 M NaCl, 100 mM EDTA, 10mM Tris, 10%
DMSO e 1 mL Triton X-100), pH 10,0 em escuro em temperatura de 4-8ºC por pelo
menos uma hora. As lâminas então passaram pelo processo de desnaturação com
solução de eletroforese por 30 minutos e então por eletroforese a 25V, 300mA por
20 minutos.
As lâminas foram então neutralizadas com solução de neutralização
(0,4 M Tris, pH 7,5) por cinco minutos. Esse procedimento foi realizado três vezes e

21
após as lâminas foram fixadas em álcool 100% por 10 minutos. A coloração foi feita
em sala escura com brometo de etídeo (agente intercalante) e as lâminas foram
analisadas em microscópio de fluorescência. Foram contados 100 nucleóides por
animal e classificados quando ao dano apresentado (classes 0, 1, 2 e 3). O cálculo
do escore foi feito multiplicando-se o número de nucleóides de uma dada classe pelo
número correspondente a essa classe, sendo: (no. nucleóides classe 0 X 0) + (no.
céls classe 1 X 1) + (no. céls classe 2 X 2) + (no. céls classe 3 X 3) (Figura 1).
Figura1. Classificação dos nucleóides de sangue de peixes submetidos ao ensaio do
cometa.
Teste do Micronúcleo
O procedimento para o teste do micronúcleo foi seguindo a
metodologia de AL-SABTI; METCALFE, 1995. Do sangue extraído 5µL foram
usados para fazer um esfregaço sobre uma lâmina limpa. Após secarem por 24

22
horas em temperatura ambiente as lâminas foram fixadas em metanol 100% por 10
minutos. A coloração foi feita com corante Giemsa a 5% em tampão fosfato 0,1M em
pH 6,8 por 20 minutos. O preparo das lâminas permanentes foi feito com Entellan e
a análise feita em microscópio de luz comum. Foram contadas 3000 células por
animal e analisadas quanto à presença de micronúcleos e de anormalidades
eritrocíticas nucleares sendo elas núcleo Lobulado (L) núcleo segmentado (S) e
núcleo em forma de rim (K) (CARRASCO et al., 1990).
Como controle positivo foi utilizado ciclofosfamida na concentração
de 0,04mg/g (concentração padronizada) de animal injetada intraperitonialmente em
cada animal. Essa substância é conhecidamente mutagênica, servindo assim de
comparação para os resultados obtidos com os animais expostos a FSG.
Atividade da acetilcolinesterase
A Atividade da Acetilcolinesterase (AchE) foi quantificada de acordo
com o ensaio de Ellmann em leitora de microplaca e o procedimento para
quantificação da atividade da enzima e dosagem de proteínas foi o mesmo para o
cérebro e para o músculo.
Amostras do tecido muscular e cerebral (cerca de 50mg) foram
homogeneizadas em tampão fosfato 0,1M pH 7,5 na proporção de 1:10. As amostras
foram centrifugadas a 10000 rpm a 4ºC durante 20 minutos. Após a centrifugação o
sobrenadante foi retirado. Numa microplaca limpa foram colocados nas primeiras
duas cavidades da placa somente 300µL de tampão fosfato. Nas demais foram
aplicados, em réplicas, 50 µL de sobrenadante (amostra) 200µL de DTNB e 50µL de
ATC (tiocolina) com a ATC sendo aplicada imediatamente antes da leitura. A
microplaca foi então submetida à leitura na leitora de microplaca a 415 nm no tempo

23
zero (adição de ATC) e após três e seis minutos. Para a dosagem da proteína foi
empregado o método de dosagem de Lowry et al. (1951), usando se as seguintes
concentrações de BSA (proteína bovina) para a confecção da curva de calibração:
10, 20, 40, 80 e 100 µL. Ao final do procedimento do método de Lowry et al. (1951)
foi feita a leitura em espectrofotômetro a 700 nm.
Análise estatística
Diferenças entre os grupos controle negativos e animais
expostos a FSG quanto ao escore do ensaio do cometa, teste do micronúcleo, AENs
e atividade da enzima AChE, para cada tempo experimental foram determinadas
usando-se o teste estatístico paramétrico t de Student. Para se comparar esses
resultados ao grupo controle positivo foi utilizado o teste estatístico não paramétrico
de Tukey. Foram considerados significativos valores de P ≤ 0,05.
RESULTADOS
Os resultados obtidos usando o ensaio do cometa em eritrócitos de
Prochilodus lineatus expostos a FSG e seus respectivos controle negativo e controle
positivo estão apresentados na Tabela 1.
Os resultados obtidos mostraram diferença significativa nos escores
de peixes expostos à FSG nos tempos: 6h (P≤ 0,019), 24h (P≤0,001) e 96h
(P≤0,001) em relação aos seus respectivos controles negativos (CTR). Animais
expostos a FSG nos tempos experimentais de 24 (P ≤ ,001) e 96 (P≤0,001) horas
mostraram um aumento significativo no número de nucleóides danificados em
relação ao seu respectivo controle negativo (Tabela 2).
24
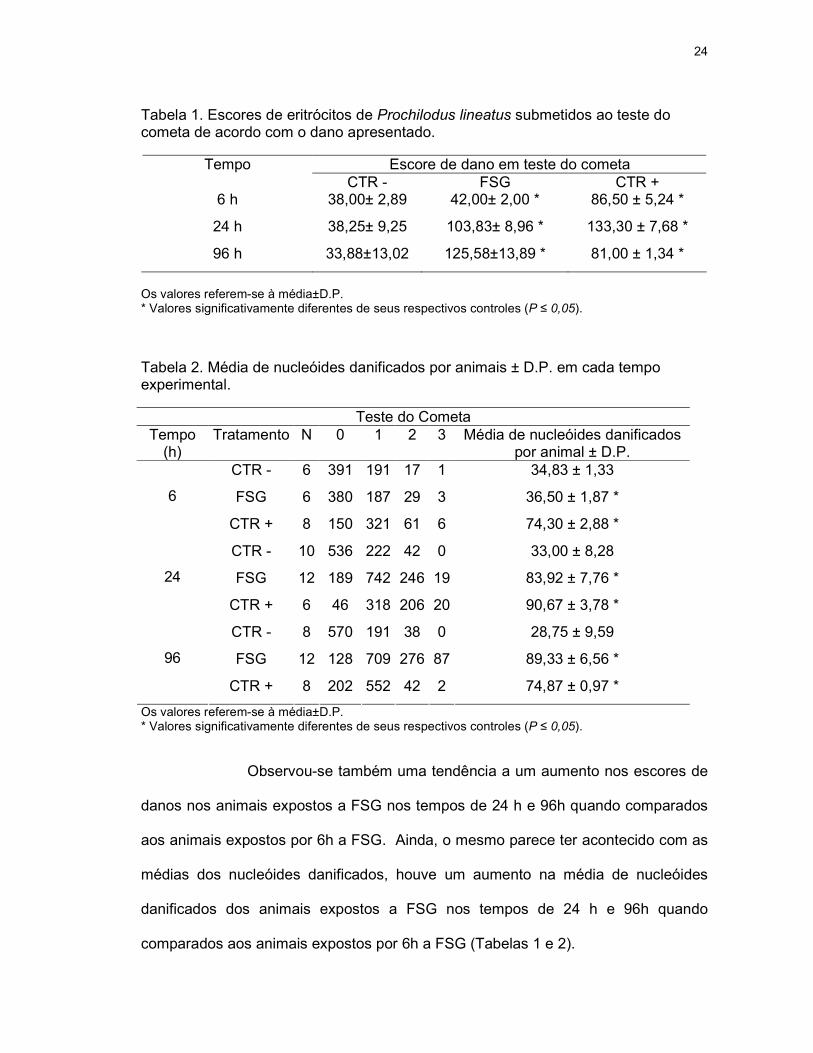
Tabela 1. Escores de eritrócitos de Prochilodus lineatus submetidos ao teste do cometa de acordo com o dano apresentado.
Escore de dano em teste do cometa Tempo CTR - FSG CTR +
6 h 38,00± 2,89 42,00± 2,00 * 86,50 ± 5,24 *
24 h 38,25± 9,25 103,83± 8,96 * 133,30 ± 7,68 *
96 h 33,88±13,02 125,58±13,89 * 81,00 ± 1,34 *
Os valores referem-se à média±D.P. * Valores significativamente diferentes de seus respectivos controles (P ≤ 0,05). Tabela 2. Média de nucleóides danificados por animais ± D.P. em cada tempo experimental.
Teste do Cometa Tempo (h)
Tratamento N 0 1 2 3 Média de nucleóides danificados por animal ± D.P.
CTR - 6 391 191 17 1 34,83 ± 1,33
FSG 6 380 187 29 3 36,50 ± 1,87 *
6
CTR + 8 150 321 61 6 74,30 ± 2,88 *
CTR - 10 536 222 42 0 33,00 ± 8,28
FSG 12 189 742 246 19 83,92 ± 7,76 *
24
CTR + 6 46 318 206 20 90,67 ± 3,78 *
CTR - 8 570 191 38 0 28,75 ± 9,59
FSG 12 128 709 276 87 89,33 ± 6,56 *
96
CTR + 8 202 552 42 2 74,87 ± 0,97 *
Os valores referem-se à média±D.P. * Valores significativamente diferentes de seus respectivos controles (P ≤ 0,05).
Observou-se também uma tendência a um aumento nos escores de
danos nos animais expostos a FSG nos tempos de 24 h e 96h quando comparados
aos animais expostos por 6h a FSG. Ainda, o mesmo parece ter acontecido com as
médias dos nucleóides danificados, houve um aumento na média de nucleóides
danificados dos animais expostos a FSG nos tempos de 24 h e 96h quando
comparados aos animais expostos por 6h a FSG (Tabelas 1 e 2).
25
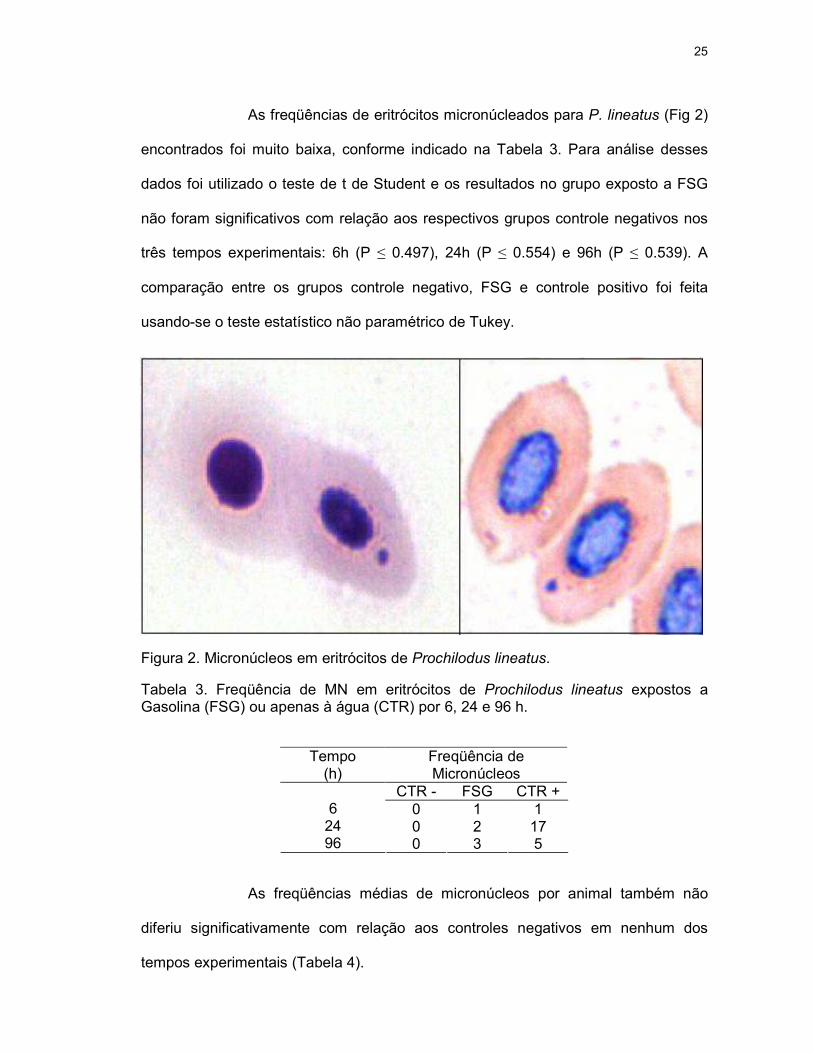
As freqüências de eritrócitos micronúcleados para P. lineatus (Fig 2)
encontrados foi muito baixa, conforme indicado na Tabela 3. Para análise desses
dados foi utilizado o teste de t de Student e os resultados no grupo exposto a FSG
não foram significativos com relação aos respectivos grupos controle negativos nos
três tempos experimentais: 6h (P ≤ 0.497), 24h (P ≤ 0.554) e 96h (P ≤ 0.539). A
comparação entre os grupos controle negativo, FSG e controle positivo foi feita
usando-se o teste estatístico não paramétrico de Tukey.
Figura 2. Micronúcleos em eritrócitos de Prochilodus lineatus.
Tabela 3. Freqüência de MN em eritrócitos de Prochilodus lineatus expostos a Gasolina (FSG) ou apenas à água (CTR) por 6, 24 e 96 h.
Tempo (h)
Freqüência de Micronúcleos
CTR - FSG CTR + 0 1 1 0 2 17
6 24 96 0 3 5
As freqüências médias de micronúcleos por animal também não
diferiu significativamente com relação aos controles negativos em nenhum dos
tempos experimentais (Tabela 4).

26
Tabela 4. Freqüência média de Micronúcleos por animal ± D.P. em cada tempo experimental.
Tempo (h)
Tratamento N Total de Micronúcleos
Freqüência média de Micronúcleos ± D.P.
6
CTR – FSG CTR +
8 10 8
0 1 1
0 ± 0 0,111± 0,333 0,125± 0,354
24
CTR – FSG CTR +
16 16 8
0 2 17
0 ± 0 0.125± 0.342 2.125 ± 1.88 *
96
CTR - FSG CTR +
9 12 8
0 3 5
0 ± 0 0.25± 0.62 0.625 ± 0.744
A presença de anormalidades eritrocíticas nucleares foi marcante
em todos os grupos experimentais, sendo significativa a diferença entre os grupos
experimentais de 6h (P≤0,023), 24h (P≤0,001) e 96h (P≤0,039) e seus respectivos
grupos controle negativo (Tabela 5). Os resultados para a detecção de AENs foi
dado como uma média (‰) das somas das anormalidades: núcleo lobulado (L)
(Figura 3), núcleo segmentado (S) (Figura 4) e núcleo em forma de rim (K) (Figura
5).
Os mecanismos responsáveis pela formação das AENs ainda não
foram completamente elucidados, porém elas são consideradas indicadoras de dano
genotóxico e servem como um complemento ao teste do micronúcleo.
Abaixo, estão representadas, algumas das lesões encontradas,
fotografadas em aumento 1000x.

27
Figura 3. Núcleos lobulados de eritrócitos de Prochilodus lineatus. Tabela 5. Ocorrência de AENs em eritrócitos de Prochilodus lineatus dados como uma média (‰) das somas (L+S+K).
Tempo(h) N Ocorrência de AENs
CTR - FSG CTR +
6 8 3.12± 1,81 5,70 ± 2,41 * 6.85 ± 2.79 *
24 8 2.53± 2.44 7.43± 3.20* 7.00 ± 2.92 *
96 8 1,33±1,00 3,58±2,90 * 12,00 ± 1,00 *
Os valores referem-se à média±D.P. * Valores significativamente diferentes de seus respectivos controles (P ≤ 0,05). Figura 4. Núcleos segmentados de eritrócitos de Prochilodus lineatus.
28
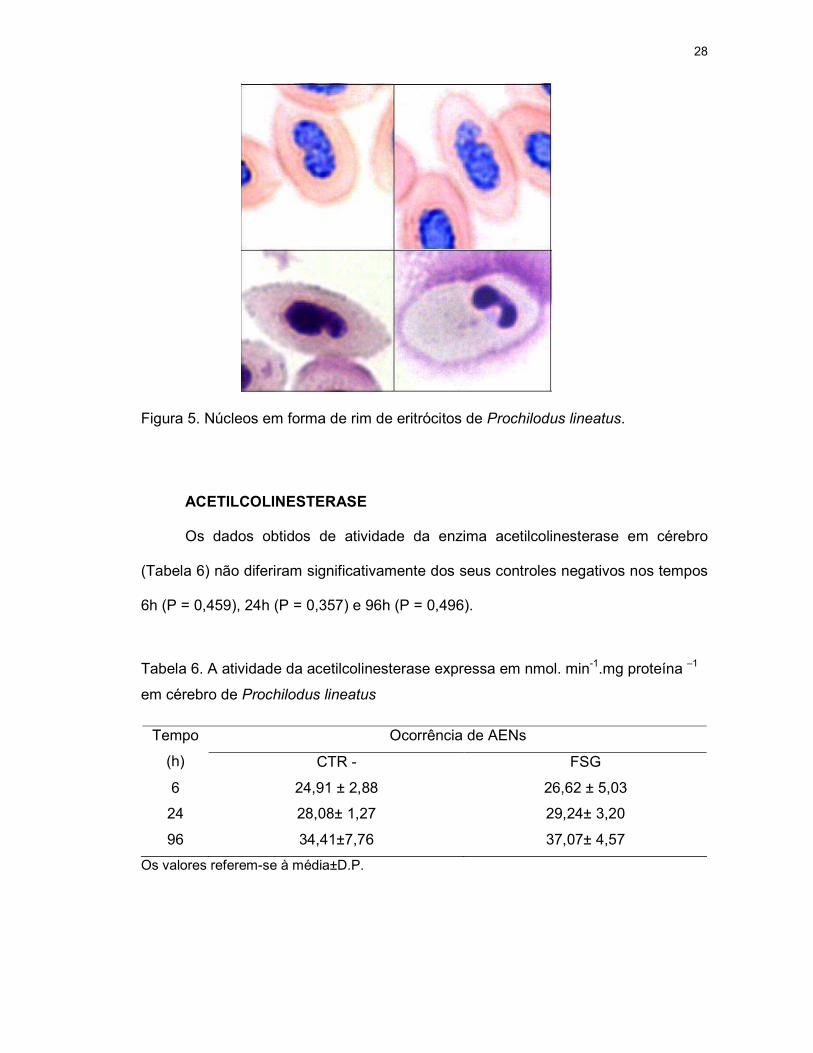
Figura 5. Núcleos em forma de rim de eritrócitos de Prochilodus lineatus.
ACETILCOLINESTERASE
Os dados obtidos de atividade da enzima acetilcolinesterase em cérebro
(Tabela 6) não diferiram significativamente dos seus controles negativos nos tempos
6h (P = 0,459), 24h (P = 0,357) e 96h (P = 0,496).
Tabela 6. A atividade da acetilcolinesterase expressa em nmol. min-1.mg proteína –1
em cérebro de Prochilodus lineatus
Ocorrência de AENs Tempo
(h) CTR - FSG
6 24,91 ± 2,88 26,62 ± 5,03
24 28,08± 1,27 29,24± 3,20
96 34,41±7,76 37,07± 4,57
Os valores referem-se à média±D.P.

29
Assim como em cérebro a atividade da acetilcolinesterase em
músculo não diferiu significativamente dos seus controles negativos nos tempos
experimentais de 6h (P = 0,051), 24h (P = 0,673) e 96h (P = 0,868) (Tabela 7).
Tabela 7. A atividade da acetilcolinesterase expressa em nmol. min-1.mg proteína –1
em músculo de Prochilodus lineatus.
Ocorrência de AENs Tempo (h)
CTR - FSG
6 64,64 ± 19,41 85,77 ± 18,57
24 87,29 ± 21,50 91,87± 17,92
96 57,91 ± 15,19 59,15 ± 5,06
Os valores referem-se à média±D.P. DISCUSSÃO
O presente estudo aponta dano genotóxico em eritrócitos do peixe
neotropical dulcícola Prochilodus lineatus expostos a FSG. No ensaio do cometa, o
alto número de nucleóides danificados (Tabela 2) em peixes expostos a gasolina,
mostram um alto nível de genotoxicidade da FSG, similar aquela vista em peixes
expostos a ciclofosfamida por 96h (controle positivo). Da mesma maneira, o escore
de todos os tempos experimentais foi significativamente superior nos animais
expostos a FSG em relação aos respectivos controles negativos, comprovando a
atividade genotóxica da gasolina. Observou-se também uma tendência a um
aumento nos escores de danos nos animais expostos a FSG nos tempos de 24 h e
96h quando comparados aos animais expostos por 6h a FSG. Ainda, o mesmo
parece ter acontecido com as médias dos nucleóides danificados, havendo um
aumento na média de nucleóides danificados dos animais expostos a FSG nos
tempos de 24 h e 96h quando comparados aos animais expostos por 6h a FSG

30
(Tabelas 1 e 2). Esse aumento é um reflexo principalmente de maior quantidade de
nucleóides classificados em classe 2 e 3 (danos mais severos).
Como se sabe, a gasolina contem compostos tóxicos e
hidrocarbonetos de dois grupos relevantes, BTX (benzeno, tolueno, xileno) e
hidrocarbonetos poliaromáticos (PAHs) de baixo peso molecular, como naftaleno e
fluoreno (PACHECO & SANTOS, 2001). A toxicidade do petróleo é, em sua maioria,
atribuída à sua parte solúvel (ALMEIDA-VAL et al., 2002). Os hidrocarbonetos
aromáticos policíclicos presentes em efluentes de refinarias de petróleo são
descritos como os principais componentes capazes de causar danos no DNA
(HAMOUTENE et al., 2002), sendo comumente detectados pelo ensaio do cometa.
O ensaio do cometa tem sido proposto para estudos de toxicogenética devido às
suas vantagens e peculiaridades quando comparado a outros testes que detectam
substâncias genotóxicas, sendo uma ferramenta fundamental de investigação em
estudos de reparo de DNA, biomonitoramento ambiental e teste de genotoxicidade
(ROSS et al., 1995). As vantagens de se empregar o ensaio do cometa para avaliar
danos no DNA incluem: mensuração de danos em células individuais; o baixo
número de células necessário para a realização do teste; possibilidade de ser
realizado em qualquer tipo de célula eucariótica nucleada e ser um método muito
sensível para detectar danos no DNA (LEE & STEINERT, 2003).
Os efeitos que os peixes expostos a FSG sofreram pela presença
desse poluente podem ser ampliados pelo fato dessa espécie possuir grande valor
comercial e representar um importante item alimentar para consumo humano (ORTÍ
et al. 2001), portanto, ela pode atuar como vetor desse contaminante do ambiente
para humanos e outros vertebrados.

31
O processo de biotransformação de PAHs em peixes, ao contrário
do que acontece com a maioria dos compostos químicos, muitas vezes converte o
xenobiótico em substancias intermediárias reativas e de alta toxicidade (MARIA et al.
2002) que podem causar dano oxidativo no DNA. Os efeitos genéticos, como
quebras na fita de DNA e presença de micronúcleos podem causar mutagenicidade,
carcinogenicidade, teratogenicidade e alterações populacionais subseqüentes
(TUVIKENE et al. 1999, VAN DER OOST et al. 2003). O dano celular detectado pelo
ensaio do cometa em P. lineatus expostos a FSG no presente trabalho pode ter
resultado desse processo oxidativo no qual os hidrocarbonetos aromáticos
policiclicos (PAHs) presentes na em associação com a molécula de DNA causaram
as diversas lesões. Dano semelhante foi observado por VANZELLA (2007) para a
mesma espécie de peixe expostos a fração solúvel do diesel (FSD). Além disso,
FRENZELLI (2004) descreve dano significativo no DNA de Zoarces viviparus
expostos a óleo bruto no estuário de Göteborg, Suécia através do ensaio do cometa.
Dentre as técnicas para detecção de efeitos genotóxicos em
peixes, o teste do micronúcleo é muito usado e relativamente fácil de adaptar à
várias espécies (ÇAVAS & ERGENE-GOZUKARA, 2005). A frequência de
micronúcleos encontrados foi muito baixa em todos os tempos experimentais (6, 24
e 96h), não sendo significativamente superior àquela encontrada nos respectivos
controles negativos, conforme indicado pela Tabela 2. Os valores das médias do
número de micronúcleos encontrados por animal também não diferiram
significativamente daquele encontrado nos controles negativos em nenhum dos
tempos experimentais, não havendo assim, nessa concentração, atividade
mutagênica da gasolina para P. lineatus.

32
Recentemente, vários estudos descreveram a presença de
anormalidades nucleares (AENs) além do micronúcleo, em células de peixe
expostas a substâncias genotóxicas (AYLLON e GARCIA-VAZQUEZ, 2000; ÇAVAS
e ERGENE-GOZUKARA, 2003). Em geral essa anormalidades são consideradas
indicadoras de dano genotóxico e, portanto, podem complementar o escore de
micronúcleo em pesquisas de genotoxicidade (ÇAVAS & ERGENE-GOZUKARA,
2005).
Neste estudo a diferença entre a freqüência de todas as
anormalidades eritrocíticas somadas (L+S+K) foi significativamente superior àquela
encontrada em seus respectivos controles negativos em todos os tempos
experimentais, 6h (P=0,015), 24h (P<0.001) e 96h (P = 0,017) como indicado pela
Tabela 5. Sendo assim, fica demonstrado que a gasolina causou danos
permanentes nos núcleos dos eritrócitos de P. lineatus.
A acetilcolina é um importante neurotransmissor presente nos
neurônios colinérgicos. Ela encontra-se armazenada dentro de vesículas nas
terminações das células nervosas, sendo secretada por exocitose na fenda sináptica
ou nas junções neuromusculares. Ao ser liberada, a acetilcolina liga-se a moléculas
específicas da membrana pós-sináptica de um neurônio, ou da placa motora,
abrindo os canais iônicos de sódio e potássio. O fluxo de íons através desses canais
causa uma mudança no potencial de membrana da célula pós-sináptica,
desencadeando o potencial de ação (RANDALL et al., 1994). A enzima
acetilcolinesterase (AChE), é responsável pela hidrólise da acetilcolina em colina e
ácido acético, limitando o tempo de ação desse transmissor na condução do impulso
nervoso (HUGGET et al., 1992; RANDALL et al., 1994).

33
A medida da atividade da enzima acetilcolinesterase em cérebro de
animais expostos a FSG não apresentou diferenças significativas (P>0,05) quanto
aos grupos controles negativos em nenhum dos tempos experimentais 6h (P =
0,459), 24h (P = 0,357) e 96h (P = 0,496). O mesmo padrão ocorreu nas medidas
referentes a atividade da enzima em músculo do peixe. No tempo experimental de
6h P = 0,051, em 24h P = 0,673 e 96h P = 0,868 não havendo diferença significativa
entre o grupo de animais expostos a FSG e grupos controle negativo (Tabelas 6 e
7). Portanto, nessa concentração o poluente gasolina não atuou como agente
anticolinesterásico no peixe neotropical dulcícola Prochilodus lineatus.
CONCLUSÃO
Os dados obtidos corroboram a hipótese de que a gasolina, em
concentração subletal, enquanto contaminante de águas tropicais atua como agente
genotóxico, no peixe neotropical Prochilodus lineatus.
Esses resultados enfatizam a necessidade de monitoramento
através da utilização de biomarcadores, de vazamentos de hidrocarbonetos, no
caso, da gasolina, evidenciando a urgência por efetiva fiscalização e monitoramento
de estações de abastecimento e distribuição de derivados de petróleo em toda
região neotropical.

34
REFERÊNCIAS BIBLIOGRÁFICAS
ACHUBA, F.I., OSAKWE, S.A. Petroleum-induced free radical toxicity in African cat
fish (Clarias gariepinus). Fish Physiology Biochemistry, v. 29, p. 97–103, 2003.
ADAMS, S.M., SHUGART, L.R., SOUTHWORTH, G.R., HINTON, D.E. Application of
bioindicators in accessing the health of fish populations experiencing contaminant
stress. In: McCARTHY, J.F., & SHUGART, L.R. (eds). Biomarkers of
environmental contamination, Lewis Publishers, p. 333-353, 1990.
AGUIAR, L.H. Efeitos do inseticida organofosforado metil paration (Folidol
600®) sobre o teleósteo de água doce, matrinxã, Brycon cephalus (Gunther,
1869): aspectos do metabolismo intermediário. Doutorado em Ecologia e
Recursos Naturais – Curso de Pós-graduação em Ecologia e Recursos Naturais,
Universidade Federal de São Carlos, 2002.
AGUIAR, L.H., MORAES, G.,AVILEZ, I.M., ALTRAN, A.E., CORRÊA, C.F.
Metabolical effects of Folidol 600® on the neotropical freshwater fish matrinxã,
Brycon cephalus. Environmental Research, v.95, p.224-230, 2004.
ALBERS, P.H. Petroleum and individual polycyclic aromatic hydrocarbons. In:
HOFFMAN, D. J., RATTNER, B. A., BURTON JR, G., CAIRNS JR, A.J. Handbook
of ecotoxicology, p. 341-371, 2002.
ALLOWAY, B.J.; AYRES, D.C. Chemical Principles of Environmental Pollution
2.ed. London: Chapman and Hall, p. 17-62, 1997.

35
ALMEIDA, L.C., AGUIAR, L.H., MORAES, G. Efeito do metil paration na atividade da
acetilcolinesterase muscular e cerebral de matrinxã (Brycon cephalus). Ciência
Rural, v. 35 N.6 , 2005.
ALMEIDA, V. M. F., VAL, W. P. DUNCAN, A. L. Crude oil effects on fish of the
Amazon: Current Status. In: Tropical Fish: News and Reviews. Internation
Congress on The Biology of Fish, p. 49 – 60, 2002.
AL-SABTI, K., METCALFE, C.D., Fish micronuclei for assessing genotoxicity in
water. Mutatation Research, v. 343, p. 121-135, 1995.
AYLLON, F.; GARCIA-VAZQUEZ, E. Induction of micronuclei and other nuclear
abnormalities in European minnow Phoxinus phoxinus and mollie Poecilia latipinna:
an assessment of the fish micronucleus test. Mutation Research, v. 467, p. 177-186,
2000.
BASU, H.S., WRIGHT, W.D., DEEN, D.F., ROTI ROTI, J. MARTOON, L.J. Treatment
with a polyamine analog alters DNA-matrix association in HeLa cell nuclei: a nucleoid
halo assay. Biochemistry, v. 32, p. 4073-4076, 1993.
BELPAEME, K., DELBEKE, L., ZHU, L., KIRSCH-VOLDERS, M. Cytogenetic
studies of PCB77 on brown trout (Salmo trutta fario) using the micronucleus test and
the alkaline comet assay. Mutagenesis, v. 11 p. 485-492, 1996.
BROWN, K.W., DONELLY, K.C. Mutagenic activity of runoff and leachate water from
hazardous waste land treatment. Environmental Pollution, v. 35, p. 229-246, 1984.

36
BRUNETTI,R., MAJONE, F., GOLA, I. BELTRAME, C. The Micronuclous Test:
Examples of Application to marine Ecology. Marine Ecology Progress Series. v. 44,
p. 65-68, 1988.
CAMARGO, M.M.P., & MARTINEZ, C.B.R. Biochemical and phisiological biomarkers
in Prochilodus lineatus submitted to in situ tests ins na urban stream in southern
Brazil. Environmental Toxicology Pharmacology, v. 21, p. 61-69, 2006.
CARRASCO, K. R.; TILBURY, K. L.; MYERS, M. S. Assessment of the piscine
micronucleus test as an in situ biological indicator of chemical contaminant effects.
Canadian Journal of Fisheries and Aquatic Sciences, v. 47, p. 2123–2136, 1990.
ÇAVAS, T.; ERGENE-GÖZÜKARA, S. Micronuclei, nuclear lesions and interphase
silver-stained nuclolar organizer regions as cyto-genetic indicator in Oreochromis
niloticusexposed to textile mill effluent. Mutation Research, v. 534, p. 93-99, 2003.
ÇAVAS, T.; ERGENE-GÖZÜKARA, S. Induction of micronuclei and nuclear
abnormalities in Oreochromis niloticus following exposure to petroleum refinery and
chromium processing plants effluents. Aquatic Toxicology, v. 74, p. 264-271, 2005.
CERQUEIRA, C.C.C., FERNANDES, M.N. Gilltissue recovery after copper exposure
and blood parameter responses in the tropical fish Prochilodus scrofa.
Ecotoxicology and Environmental Safety, v. 52, p. 83-91, 2002.
COOK, P.R., BRAZELL, I.A. Conformations constraints in nuclear DNA. Journal of
Cell Science, v. 22, p. 287-302, 1976.

37
COOK, P.R., BRAZELL, I.A. Spectrofluorometric measurement of the binding of
ethidium to superhelical DNA from cell nuclei. European Journal of Biochemistry,
v. 18, p. 465-477, 1978.
DA SILVA, M.E.F., SILVA, J.A., MARANGONI, S., NOVELLO, J.C., MEIRELLES,
N.C. A new method to purify hepatic CYP1A of Prochilodus scrofa, a Brazilian
freshwater fish. Comparative Biochemistry and Physiology Part C: Toxicology
and Pharmacology, v. 138, p. 67-74, 2004.
DASGUPTA, A., ZDUNEK, T., In vitro lipid peroxidation of human serum catalysed by
cupric ion:antioxidant rather than peroxidant role of ascorbate. Life Sciences, v. 50,
p. 875-882, 1992.
FENECH, M., HOLLAND, N., CHANG, W.P., ZEIGER, E., BONASSI, S. The human
micronucleous project-an international collaborative study on the use of
micronucleous technique for measuring DNA damage in humans. Mutation
Research, v. 428, p. 271-283, 1999.
FLEEGER, J.W.; CARMAN, K.R.; NISBET, R.M. Indirect effects of contaminants in
aquatic ecosystems. The Science of the Total Environment, v. 317, p. 207-233,
2003.
FRENZILLI, G., SCARELLI, V., DEL BARGA, I., NIGRO, M., FÖRLIN, L.,
BOLOGNESI, C., STURVE, J. DNA in eelpout (Zoarces viviparous) from Göteborg
harbour, Mutation Research, v. 552, p. 187–195, 2004.

38
HAMOUTENE, D., PAYNE, J. F., RAHIMTULA, A., LEE, K.. Use of the Comet assay
to assess DNA damage in hemocytes and digestive gland cells of mussels and clams
exposed to water contaminated with petroleum hydrocarbons. Marine
Environmental Research, v. 54 p. 471-474, 2002.
HEDDLE, J.A., HITE, M., JRKHART, B., MACGREGOR, J.T., SALAMONE, M.F. The
Indutio of Micronuclei as a Measure of Genotoxicity. Mutation Research, v. 123, p.
61-118, 1983.
HEDDLE, J. A.,; CIMINO, M.C., HAYASHI, M.,; ROMAGNA, F., SHELBY, M.D,
TUCKER, J.D., VANPARYS, P.H., McGREGOR, J.T. Micronuclei as an Index of
Citogenetic Damage: Past, Present, Future. Environmental and Molecular
Mutagenesis, v. 18, p. 277-291, 1991.
HODGSON, E. & GOLDSTEIN, J.A. Metabolism of toxicants: phase I reactions and
pharmacogenetics. In: HODGSON, E. & SMART, R.C. Introduction to biochemical
toxicology. Wiley-Interscience, 3º ed, p. 67-113, 2001.
HUGGETT, R.J.; KIMERLE, R.A.; MEHRLE JR, P.M. Biomarkers. Biochemical,
physiological, and histological markers of anthropogenic stress. BERGMAN HL
(ed). Boca Raton, FL, USA. 1992.
KIM, I.Y. & HYUM, C-K. Comparative evaluation of the alkaline comet assay with the
micronucleous test for genotoxicity monitoring using aquatic organisms.
Ecotoxicology and Environmental Safety. Article in press, acesso em 12 de
Outubro de 2007.

39
KIRBY, M.F. The use of cholinesterase activity in flounder (Platichthys flesus)
muscle tissue as a biomarker of neurotoxic contamination in UK estuaries. Marine
Pollution Bulletin, v.40, n.9, p.780-791, 2000.
LACKNER, R. “Oxidative stress” in fish by environmental pollutants. apud
BRAUNBECK, T., HINTON, D.E., STREIT, B. Fish Ecotoxicology, p. 203-224,
1998.
LEE, R. F.; STEINERT, S. Use of the single cell gel electrophoresis/comet assay for
detecting DNA damage in aquatic (marine and freshwater) animals. Mutation
Research, v. 544, p. 43-64, 2003.
LIVINGSTONE, D.R. The fate of organic xenobiotics in aquatic
ecossystems:quantitative and qualitative differences in biotransformation by
invertebrates and fish. Comparative Biochemistry and Physiology Part A:
Molecular & Integrative Physiology, v. 120, p. 43-49, 1998.
LÓPEZ-BAREA, J., Biomarkers to detect environmental pollution. Toxicology
Letters, v. 88, p. 79, 1996.
LOWRY, O.H., ROSENBROUGH, N.J., FARR, A.L., RANDALL, R.J. Protein
measurements with the Folin phenol reagent. Journal of Biological Chemistry. v.
193, p. 265–275, 1951.
MARIA V.L CORREIA A.C. SANTOS M.A. Anguilla anguilla L. Biochemical and
Genotoxic Responses to Benzo[a]pyrene, Ecotoxicology and Environmental
Safety v. 53, p. 86-92, 2002.

40
MARTINEZ, C.B.R. Parâmetros bioquímicos de peixes para avaliação da qualidade
da água. In: RANZANI-PAIVA, M.J.T., TAKEMOTO, R.M., LIMA, L.M.A.P. Sanidade
de organismos aquáticos no Brasil. Ed. Varella. P. 43-62, 2004.
MARTINEZ, C.B.R., CÓLUS, I.M.S. Biomarcadores em peixes neotropicais para o
monitoramento da poluição aquática na bacia do rio Tibagi. In: MEDRI, M.E.,
BIANCHIN, E., SHIBATTA, O.E., PIMENTA, J.A. A bacia do rio Tibagi, cap. 29, p.
551-557, 2002.
MARTINEZ, C.B.R., NAGAE, M.Y., ZAIA, C.T.B.V., ZAIA, D.A.M. Acute
morphological and physiological effects of lead in theneotropical fish Prochilodus
lineatus. Brazilian Journal of Biology, v. 64, p. 797–807, 2004.
MARTINEZ, C.B.R., SOUZA, M.M., Acute effects of nitrite on íon regulation in two
neotropical fish species. Comparative Biochemistry and Physiology – Part A:
Molecular & IntegrativePhysiology, v. 133, p. 151-160, 2002.
MAZON, A.F., FERNANDES, M.N. Toxicity and differential tissue accumulation of
copper in the tropical freshwater fish, Prochilodus scrofa. Bulletin of Environmental
Contamination and Toxicology, v. 63, p. 797–804, 1999.
MERSCH, J., BEAUVAIS, M-N. The micronucleous assay in the zebra mussel,
Deissena polymorpha, to in situ monitor genotoxicity in freshwater environments.
Mutation Research, v. 383, p. 141-149, 1997.
NICODEM, D.E., GUEDES, C.L.B., CORREA, R.J. Photochemistry of petroleum. I
Systematic study of a Brazilian intermediate crude oil. Marine Chemistry, v. 63, p.
93–104, 1998.

41
OLIVE, P.L., BANÁTH, J.P., DURAND, R.E. Heterogeneity in radiation-induced DNA
damage and repair in tumor and normal cells measured using the ‘comet’ assay.
Radiation Research, v. 122, p. 69-72, 1990.
ORTÍ, G. ARJUN, A. BERMINGHAN, E. Population structure and biogeography of
migratoryfreshwater fishes (Prochilodus: Characiformes) in major South American
rivers. Molecular Ecology, v. 10, p. 407-417, 2001.
ÖSTLING, O., JOHANSON, K.J. Microeletrophoretic study of radiation-induced DNA
damages in individual mammalian cells. Biochemical and Biophysical Research
Communications, v. 136, p. 291-298, 1984.
PACHECO, M., SANTOS, M.A. Biotransformation, endocrine, and genetic responses
of Anguilla anguilla L. to petroleum distillate products and environmentally
contaminated waters. Ecotoxicology of Environmental Safety, v. 49, p. 64–75,
2001.
RANDALL W.R.; RIMER M.; GOUGH, N.R. Cloning and analysis of chicken
acetylcholinesterase transcripts from muscle and brain. Biochimica et Biophysica
Acta v. 218, p. 453–456, 1994.
RAND, G.M.; PETROCELLI, S.M. Fundamentals of aquatic toxicology methods
and applications. McGraw-Hill, p. 666, 1984.
RIBEIRO, L.R., MARQUES, E.K. A importância da mutagênese ambiental na
carcinogênese humana. In: RIBEIRO, L.R., SALVADORI, D.M.F., MARQUES, E.K.
(Orgs.). Mutagênese ambiental, Canoas: Ulbra, p. 21-28, 2003.

42
ROSS, G.M., McMILLAN, T.J., WILCOX, P., COLLINS, A.R. The single cell gel
electrophoresis assay (comet assay): Technical Aspects and applications. Report on
the 5th LH Gray Trust Workshop, Institute of Cancer Research. Mutation Research,
v. 337, p. 57-60, 1995.
ROTI ROTI, J.L., WRIGHT, W.D. Visualization of DNA loops in nucleoids from HeLa
cells:assays for DNA damage and repair. Cytometry, v. 8, p. 461-467, 1987.
SCHLEGEL & MACGREGOR, The persistence of micronuclei in peripheral blood
erythrocytes: detection of chronic chromosome breakage in mice, 1982 In: GRAWE,
J; ZETTERBERG, G; AMNEUS, H Effects of extended low-dose-rate exposure to
super(137)Cs detected by flow-cytometric enumeration of micronucleated
erythrocytes in mouse peripheral blood. International Journal of Radiation
Biology, v. 63, p. 339-347, 1993.
SERRANO-GARCIA L, MONTERO-MONTOYA R. Micronuclei and chromatid buds
are the result of related genotoxic events. Environmental and Molecular
Mutagenesis, v. 38(1), p. 38–45, 2001.
SIMONATO, J.D., ALBINATI, A.C.L., MARTINEZ, C.B.R. Effects of the water soluble
fraction of diesel fuel oil on some functional parameters of the neotropical freshwater
fish Prochilodus lineatus Valenciennes. Bulletin of Environmental Contamination
and Toxicology, v. 76, p. 505–511, 2006.
SINGH, N.P.; MCCOY, M.T.; TICE, R.R.; SCHNEIDER, E.L. A Single Tecnique for
Quantification of Low Levels of DNA Damage in Individual Cells. Experimental Cell
Research, v. 175, p. 184-191, 1988.

43
STAHL, R.G. The genetic toxicity of organic compounds in natural waters and waste
waters. Ecotoxicology and Environmental Safety, v. 22, p.94-125, 1991.
STEGEMAN, J.J., BROUWER, M., DI GIULIO, R.T., FÖRLIN, L., FOWLER, B.A.,
SANDERS, B.M., VAN VELD, P.A. Molecular responses to environmental
contamination: enzyme and protein systems as indicators os chemical exposure and
effect. apud HUGGET, R.J., KIMERLE, R.A., MEHRLEJR, P.M., BERGMAN, H.L.
Biomarkers. Biochemical, physiological, and histological markers of
antropogenic stress. Lewis Publishers, p. 235-335, 1992.
STOREY, K. B. Metabolic adaptations supporting anoxia tolerance in reptiles: recent
advances. Comparative Biochemistry and Physiology – Part B: Biochemistry &
Molecular Biology, v. 113, p. 23-35, 1996.
TAKAI, A., KAGAWA, N. FUJIKAWA, K. Suceptibility of male and female medaka
(Oryzias latipes) fish to spontaneous and X-ray induced micronucleous formation in
gill cells. Mutation Research, v. 558, p. 131-136, 2004.
TIBURTIUS, E.R.L., PERALTA-ZAMORA, P., EMMET, A., LEAL, E.S., Degradacão
de BTXs via processos oxidativos avançados. Química Nova v. 28, p. 61–64, 2005.
TICE, R. The Single Cell/ Comet Assay: A Microgel Eletroforetic Technique for the
Detection of DNA Damage and Repair in Individual Cell. In: PHILLIPS, D. H.;
VENNIT, S. (Eds.). Environmental mutagenesis. Bios Scientific Publishers Ltd.
Oxford, UK; p. 315-339, 1995.

44
TOLBERT, P.E., SHY, C.M., ALLEN, J.W. Micronuclei and other nuclear anormalies
and buccal smears: methods development. Mutation Research. v. 271, p. 69-77,
1992.
TUCKER, J.D.; VANPARYS, P.H.; McGREGOR, J.T. Micronuclei as an index of
citogenético damage: past, present, future. Environmental and Molecular
Mutagenesis, v. 18, p.277-291, 1991.
TUVIKENE, A.; HUUSKONEN, S.; KOPONEN, K..; RITOLA, O.; MAUER,
LINDSTRÖM-SEPPÄ P, U. Oil shale processing as a source of aquatic pollution:
monitoring of the biologic effects in caged and feral freshwater fish. Environmental
Health Perspectives, v. 107, p. 745-752, 1999.
UDROIU, I. Environmental and molecular mutagenesis. Wiley-Liss, Inc., A Wiley
Company , 2006.
VAN DER OOST, R.; BEYER, J.; VERMELEN, N.P.E. Fish bioaccumulation and
biomarkers in risk assessment: a review. Environmental Toxicology and
Pharmacology, v.13, p. 57-149, 2003.
VANZELLA, T.P., MARTINEZ, C.B.R., CÓLUS, I.M.S. Genotoxic and mutagenic
effects of diesel oil water soluble fraction on a neotropical fish species. Mutation
Research, v 631, p. 36-43, 2007.
VIEGAS JUNIOR, C. BOLZANI, V.S., FURLAN, M., FRAGA, C.A.M, BARREIRO,
E.J. Natural products as candidates for useful drugs in the treatment of Alzheimer's
disease. Química Nova, v. 27, n. 4, 2004.

45
VOGELSTEIN, B., PARDOLL, D.M., COFFEY, D.S. Supercoiled loops and
eukaryotic DNA replication. Cell, v. 22, p. 79-85, 1980.
WALKER, C.H., HOPKIN, S.P., SILBY, R.M., PEAKALL, D.B.Principles of
ecotoxicology, Taylor & Francis, p. 321, 1996.
WRIGHT, R. T.; NEBEL, B.T. In: Environmental Science: The Way World Works. 7.
ed., cap. 18, p 402-420, 2000.