
Avaliação experimental da capacidade competitiva de...
77
Universidade Federal de Minas Gerais Instituto de Ciências Biológicas Programa de Pós-graduação em Ecologia, Conservação e Manejo de Vida Silvestre Avaliação experimental da capacidade competitiva de Oocystis lacustris (Chlorophyceae), Microcystis viridis (Cyanobacteria) e Cryptomonas erosa (Cryptophyceae) em diferentes razões Nitrogênio:Fósforo, com ênfase nas suas inter-relações em um reservatório urbano eutrófico. João de Magalhães Lopes Belo Horizonte 2003
-
Upload
truonghanh -
Category
Documents
-
view
216 -
download
0
Transcript of Avaliação experimental da capacidade competitiva de...

Universidade Federal de Minas Gerais Instituto de Ciências Biológicas
Programa de Pós-graduação em Ecologia, Conservação e Manejo de Vida Silvestre
Avaliação experimental da capacidade competitiva
de Oocystis lacustris (Chlorophyceae), Microcystis
viridis (Cyanobacteria) e Cryptomonas erosa
(Cryptophyceae) em diferentes razões
Nitrogênio:Fósforo, com ênfase nas suas inter-relações
em um reservatório urbano eutrófico.
João de Magalhães Lopes
Belo Horizonte 2003

2
2
Avaliação experimental da capacidade competitiva de
Oocystis lacustris (Chlorophyceae), Microcystis viridis
(Cyanobacteria) e Cryptomonas erosa (Cryptophyceae) em
diferentes razões Nitrogênio:Fósforo, com ênfase nas suas inter-
relações em um reservatório urbano eutrófico.
João de Magalhães Lopes
Dissertação apresentada ao programa de pós graduação em Ecologia, Conservação e Manejo de Vida Silvestre, da Universidade Federal de Minas Gerais, como requisito parcial à obtenção do título de mestre.
Orientadora: Dra. Alessandra Giani
Belo Horizonte, 2003.

3
3
À memória de Hugo de Lucena Lopes, um ser
humano excepcional a quem tive o privilégio de
chamar de pai.

4
4
``Suspeito que estejamos vivendo um momento de metamorfose da nossa condição
humana. Até agora temos sido habitantes do mundo da vida. Nosso hábitat é constituído por
florestas, animais, rios e mares. Somos seres biológicos, corpos. Mas agora estamos mudando
de casa. Estamos trocando nossa casa biológica por uma outra casa, eletrônica (...). Nossas
inteligências estão cada vez mais ligadas aos vídeos e computadores e cada vez mais distantes
da natureza. Há crianças que nunca viram uma galinha de verdade, nunca sentiram o cheiro
de um pinheiro, nunca ouviram o canto do pintassilgo e não têm o prazer em brincar com
terra. Pensam que terra é sujeira. Não sabem que terra é vida.´´
Rubem Alves

5
5
AGRADECIMENTOS
Agradeço à minha família pelo apoio, em especial a minha mãe, Vera Lúcia de Magalhães Lopes, que tem
me ensinado, através de sua conduta nestes últimos meses, os verdadeiros significados das palavras força e
superação.
À minha orientadora, Dra. Alessandra Giani, que com seu vasto conhecimento de limnologia e ecologia
sempre direcionou este trabalho, evitando que me perdesse ao longo do caminho.
Ao Professor Ricardo Motta Pinto-Coelho, por me permitir utilizar o seu laboratório para a realização de
análises químicas.
Às Professoras Nádja e Maria Rita, do laboratório de Interação Microrganismos-Plantas, que também me
cederam gentilmente alguns aparelhos para a realização de análises químicas.
Aos meus colegas de laboratório pela inestimável ajuda que prestaram para a realização deste trabalho. Ao
Cléber pelos conselhos e atencioso interesse com o qual acompanhou o projeto. À Luciana pela alegria e sensatez
que tanto me ajudaram a manter a lucidez em momentos difíceis. À Gabriela, companheira de graduação e pós-
graduação, pelas idéias que muito contribuíram para a melhora deste trabalho e pelas análises realizadas com total
desprendimento nos períodos nos quais não pude estar presente em Belo Horizonte. À Elenice que nem imagina o
quanto foi importante para a conclusão do trabalho e para quem é impossível expressar minha gratidão em palavras.
À Sandra, Sávio, Patrícia, Gustavo e Marcos pela saudável convivência e aos agradáveis momentos que me
proporcionaram dentro e fora do ambiente de trabalho.
Aos meus colegas de pós-graduação com os quais compartilhei esta experiência única. Me permito plagiar
um deles que ao final da apresentação de sua dissertação, tão bem explicou o motivo deste agradecimento: “
Agradeço aos meus colegas de pós-graduação por me ajudarem a manter a sanidade mental durante todo este
tempo”.
Aos meus colegas de graduação pela amizade e exemplo que sempre foram para mim.
A todos os amigos pela paciência que, tenho certeza, tiveram de praticar nestes últimos anos comigo. Neste
agradecimento se incluem os amigos de Belo Horizonte, de Uberaba, de Fortaleza... bem, onde quer que vocês
estejam fica registrado o meu agradecimento.
Aos amigos da Companhia Energética de Minas Gerais e em especial aos funcionários da Estação
Ambiental de Volta Grande que muito me ensinaram neste último ano e à Gerência de Programas e Ações
Ambientais na pessoa de Alberto Zerlotini, que gentilmente me cedeu preciosos dias em Belo Horizonte para a
conclusão deste trabalho.

6
6
SUMÁRIO
1.Introdução . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 8
1.1-Competição por nutrientes em comunidades fitoplanctônicas.. . . . . . . . .
1.2-Nitrogênio e fósforo em ambientes naturais.. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
1.3-Absorção de fósforo e nitrogênio pelo fitoplâncton.. . . . . . . . . . . . . . . . . . . . . . . . . . . .
Objetivos . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
2. Material e Métodos . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
2.1-Área de estudo e escolha das espécies.. . . . . . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
2.2-Descrição das espécies.. .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
2.3-Isolamento e manutenção das culturas estoque.. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
2.4-Delineamento experimental. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
2.4.1-Experimentos de crescimento . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
2.4.2-Experimentos de absorção.. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
2.4.3-Experimentos de competição.. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
2.5-Variação sazonal da comunidade fitoplanctônica e de algumas
variáveis físico químicas no Reservatório da Pampulha.........................
3.Resultados . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
3.1-Crescimento.. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
3.2-Absorção de fósforo e nitrogênio.. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
3.3-Competição entre as espécies.. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
3.4-Variação sazonal das espécies no ambiente natural: Reservatório
da Pampulha.. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
4.Discussão . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
4.1-Crescimento e competição entre as espécies . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
4.2-Absorção de fósforo e nitrogênio . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
4.3-Variação sazonal das espécies no ambiente natural: Reservatório
da Pampulha . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
5.Conclusões ...............................................................................................................................
6.Referências Bibliográficas . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
11
16
18
21
22
22
23
24
25
25
28
29
29
31
31
40
43
47
52
52
58
65
66
67

7
7
RESUMO
Foram avaliadas as capacidades competi t ivas de t rês espécies f i toplanctônicas , Oocyst is
lacustr is (Chlorophyceae) , Microcyst is v ir idis (Cyanobacter ia) e Cryptomonas erosa
(Cryptophyceae) , comuns na composição da comunidade f i toplanctônica do Reservatór io da
Pampulha, um reservatór io urbano eutróf ico, em diferentes razões nit rogênio:fósforo . Foram
real izados exper imentos de crescimento das três espécies em cinco razões N:P,
respect ivamente 4 , 10, 20, 40 e 100. Parâmetros competi t ivos das três espécies , como taxa
máxima de crescimento (µmax) e quant idade mínima de nutr iente requerida para o
crescimento (R*) foram calculados através do modelo de Monod. A capacidade das t rês
espécies de absorver ni t rogênio e fósforo também foi aval iada exper imentalmente . Por f im
foram real izados exper imentos de compet ição com cul turas mistas de pares de espécies em
três razões N:P (4 , 20 e 100) , para que pudesse ser ver if icada a predição obt ida através dos
parâmetros calculados pela curva de crescimento logís t ico. Os dados exper imentais foram
comparados com os dados coletados no Reservatór io da Pampulha a f im de se extrapolar os
resul tados laborator ia is para o ambiente natural . Cryptomonas erosa fo i a espécie que
apresentou, teor icamente, a maior probabi l idade de vencer o processo competi t ivo tanto em
meios l imitados por n i trogênio quanto por fósforo quando cul t ivadas com Oocyst is lacustr is e
Microcyst is v ir idis . Esta previsão foi comprovada pelos exper imentos de competição. Os
exper imentos de absorção demonstraram que esta espécie possui grande probabi l idade de se
tornar dominante em ambientes que não apresentam l imitação de ni t rogênio ao crescimento.
Os dados de var iação sazonal das espécies no Reservatór io da Pampulha corroboraram os
resul tados obt idos em laboratór io mostrando o aumento da densidade desta espécie à medida
que ocorreu o aumento da concentração de fósforo e pr incipalmente, n i t rogênio no
reservatór io.

8
8
1. INTRODUÇÃO
Populações e indivíduos das diferentes espécies não ocorrem isoladamente
em ambientes naturais. Em um contexto ecológico todas as espécies estão
envolvidas em uma rede de complexas interações, pertencentes a um todo
ecológico constituído por partes interdependentes. Desta forma, cada organismo
ou população está ligado a um complexo sistema de interdependência que tem
sua dinâmica influenciada em parte por vários fatores abióticos e em parte pelas
múltiplas atividades dos diversos organismos que o constitui (Putman, 1994).
Este sistema de interdependência é denominado`` Comunidade Ecológica´´
sendo que muitos esforços têm sido despendidos para se desvendar a natureza
das relações que a estruturam e a intensidade em que estas ocorrem na natureza.
Historicamente, tem se observado em estudos ecológicos que uma das
principais forças estruturadoras em comunidades naturais é a competição
interespecífica. Darwin percebeu empiricamente que as populações de diversos
organismos se mantinham em números menores do que seria esperado apenas
contabilizando-se a sua capacidade reprodutiva (Begon et al. , 1995). O ambiente
aonde estas populações se encontravam deveria, assim, estar exercendo alguma
pressão negativa sobre os indivíduos, o que se refletiria em suas taxas de
natalidade, sobrevivência e/ou mortalidade. Gause (1934) postulou que se as
características de utilização de recursos (ex: hábitos alimentares) de duas
espécies fossem similares, estas espécies não poderiam ocupar o mesmo
ambiente ao mesmo tempo, sendo uma delas excluída pela outra. Este axioma
ficou conhecido como ``princípio da exclusão competitiva´´. Os trabalhos de
MacArthur (1958), que observando comunidades de insetos, concluiu que as
diferenças na utilização de diferentes substratos por diferentes espécies
provavelmente se devia a pressões competitivas existentes no passado ( ``the
ghost of competition past´´), e de Hutchinson (1957) que desenvolveu a noção
de nicho ecológico como um hipervolume que engloba todas as características
mensuráveis na história de vida de um organismo, foram de grande utilidade nos

9
9
estudos de interações competitivas entre organismos, pois permitiram aos
pesquisadores uma análise objetiva de padrões encontrados no campo e em
experimentos laboratoriais.
Atualmente, o tema da competição como força estruturadora de
comunidades vem sendo amplamente debatido. A discussão sobre a existência de
nichos vagos em determinados ambientes e a utilização de enfoques neutralistas
na análise de dados obtidos em estudos clássicos têm demonstrado que em
muitos casos, outros processos que não a competição, podem estar envolvidos na
estruturação de comunidades naturais.
Begon et al. (1995) comentam que a essência da competição
interespecífica é a de que indivíduos de uma espécie sofrem redução na
fecundidade, sobrevivência ou crescimento como resultado da exploração de
recursos ou de alguma interferência realizada por indivíduos de uma outra
espécie. Este conceito além de inferir na natureza deletéria da competição, ainda
faz uma importante distinção entre a competição por interferência e a
competição por exploração. Na competição por interferência, a interação entre
indivíduos de espécies diferentes se dá de forma direta, sendo reduzido o acesso
de uma das espécies ao recurso por influência do comportamento agonista de
outra espécie. Na competição por exploração, a utilização do recurso por uma
espécie, é reduzida pelo consumo deste mesmo recurso por uma segunda
espécie, neste caso, entretanto, sem a interação direta entre as duas espécies. A
maioria dos modelos que descrevem interações competitivas tende a se restringir
à competição por exploração. Estes conceitos, no entanto, são arbitrários sendo
util izadas diferentes definições de acordo com o objetivo do autor. Um exemplo
é o trabalho de Schoener (1983) no qual a competição interespecífica é dividida
em seis categorias distintas.
Connell em 1983 revisou trabalhos cujo tema era competição. Sua revisão
englobou 72 estudos publicados em seis importantes revistas (Ecology,
Ecological Monographs, Journal of Ecology, Journal of Animal Ecology, The
American Naturalist e Oecologia).Este autor verificou que em cinco sextos dos
trabalhos que revisou, a competição intra-específica era mais importante para a
estruturação da população da espécie em estudo do que a competição inter-
específica. Esta parece ser uma importante propriedade de populações naturais,
o que permite a coexistência de espécies potencialmente competidoras em

10
10
determinado ambiente. Os estudos revisados por Connell, no entanto, tratavam
principalmente de experimentos realizados com animais terrestres, o que
subestimou a importância da natureza da relação competitiva existente entre
vegetais, tanto terrestres como aquáticos.
Tilman (1982) propôs uma explicação baseada na competição
interespecífica entre espécies e na utilização diferencial de recursos para
explicar a diversidade encontrada entre os vegetais na natureza. Neste modelo,
Tilman considera não só a dinâmica do recurso no ambiente, mas também a
dinâmica de utilização dos recursos pelos organismos observados.
Primeiramente é preciso definir o que será tratado como recurso no
modelo de Tilman. Recursos são, segundo este autor, qualquer substância ou
fator que podem levar a um aumento das taxas de crescimento de determinado
organismo à medida que a sua disponibilidade no ambiente aumenta e que é, ao
mesmo tempo, consumido por este organismo. Desta forma, nutrientes são
considerados recursos neste modelo enquanto temperatura, por não diminuir a
sua disponibilidade em resposta ao seu ‘consumo’ no meio, não pode ser
considerada como tal. Os recursos, segundo Tilman, podem ser classificados
dentro de algumas categorias, sendo os substituíveis e os essenciais os mais
importantes para o entendimento da dinâmica competitiva entre espécies.
Recursos substituíveis são aqueles que podem ser utilizados pelo organismo na
ausência de outros recursos e que os substituem em sua contribuição para o
crescimento de sua população. Organismos podem, por exemplo, utilizar
diferentes fontes de carboidrato para o seu desenvolvimento e consequente
crescimento populacional. Esta categoria é característica de organismos
heterótrofos que consomem nutrientes compostos e que possuem, desta forma,
vários elementos simples incorporados à sua estrutura. Recursos essenciais, por
sua vez, são aqueles sem o qual o organismo não consegue sobreviver e que não
podem ser substituídos por outros. Esta categoria é característica de organismos
autotróficos que utilizam elementos simples em seu metabolismo (ex: Ca, N, P,
Se, Si, etc).
A heterogeneidade espacial e temporal na disponibilidade destes recursos
em determidado ambiente, podem aumentar o número de espécies que podem
coexistir neste ambiente (Tilman et al , 1982). A presença de microhabitats no
ambiente permite a coexistência de um grande número de espécies que de outra

11
11
forma seriam excluídas competitivamente. A variabilidade temporal na
disponibilidade de recursos pode também apresentar consequências na
estruturação de comunidades (Begon et al , 1995). Hutchinson (1961), por
exemplo, observou que as comunidades fitoplanctônicas encontradas em lagos e
oceanos, são em geral muito mais ricas do que se poderia esperar pela escassa
distribuição de recursos nestes ambientes. Este autor percebeu que mudanças
nas condições físicas, como temperatura e intensidade luminosa, e químicas
como as concentrações de nutrientes, que ocorrem em curtos intervalos de tempo
nestes ambientes, poderiam impedir que espécies melhor adaptadas a
determinadas condições ambientais excluíssem as demais. A riqueza encontrada
nestes ambientes, se deveria, desta forma, a repetidas interrupções do processo
de exclusão competitiva das espécies. Este processo permite que espécies de
diferentes estratégias competitivas, pioneiras e clímax, convivam em
determinado ambiente. Esta hipótese ficou conhecida como “ o paradoxo do
plâncton’’.
1.1- Competição por nutrientes em comunidades fitoplanctônicas:
A disponibilidade de nutrientes vem sendo considerada, desde os
primeiros estudos limnológicos, como uma das variáveis que controlam a
estrutura e a biomassa de comunidades fitoplanctônicas bem como a sucessão
sazonal de espécies (Tilman et al , 1982). Diversos estudos têm demonstrado que
não existe limitação nutricional múltipla para o crescimento fitoplanctônico. O
crescimento algal é regulado pelo único nutriente que se apresenta sob a menor
disponibilidade de acordo com a Lei do Mínimo de Liebig (Droop 1973). Razões
nutricionais ótimas para o crescimento algal são espécies-específicas, sendo
determinadas por características fisiológicas específicas de cada espécie
(Bulgarov & Levich, 1999). Estas razões podem, em última instância,
determinar a exclusão ou coexistência das espécies que tomam parte de
interações competitivas (Rhee & Gotham, 1980).
A hipótese da importância relativa das razões nutricionais em relação à
concentração absoluta dos nutrientes nos corpos d'água vem sendo duramente
criticada por alguns autores. Dentre estes Reynolds (1999) comenta que as
espécies fitoplanctônicas não podem ser capazes de ``escolher´´ as razões

12
12
nutricionais que lhes favorecem, e sim, utilizar as concentrações absolutas que
se encontram diluídas no meio. Por outro lado, Droop (1973) e Rhee (1978)
demonstram que as espécies fitoplanctônicas respondem às proporções
nutricionais existentes nos meios em que se encontram e que cada espécie possui
uma razão nutricional ótima para o seu próprio desenvolvimento. Esta razão
seria a quota individual que cada célula necessitaria para efetuar a divisão
celular, ou seja o requisito nutricional mínimo de cada espécie comentado no
modelo de Tilman. Rhee e Gothan (1980) chegaram a calcular a razão ótima
N:P para nove espécies de microalgas e comprovaram que em meios que
possuiam estas razões, as espécies apresentaram o desenvolvimento esperado.
Bulgarov e Levich (1999) comentam que as biomassas relativas das espécies
fi toplanctônicas na comunidade são determinadas pelas razões dos nutrientes
limitantes existentes no meio. A concentração absoluta destes nutrientes
determinaria, desta forma, apenas a biomassa total da comunidade.
A razão nutricional ótima, como comentado acima, é a razão na qual
ocorre a transição da limitação por um nutriente para a l imitação por outro
nutriente. Diferenças na razão N:P atômica de 200 até 1 já foram observadas em
corpos d’água naturais (Stocker & Shortreed, 1985). A transição entre estados
de limitação do crescimento fitoplanctônico por nitrogênio à limitação por
fósforo é comum tanto em ambientes marinhos como em águas continentais
(Sommer, 1989). Estas mudanças são, em geral, sazonais sendo que o afluxo de
dejetos observado em diversos reservatórios urbanos pode contribuir
significativamente neste processo (Giani & Delgado, 1998).
Muitos experimentos têm demonstrado que a habilidade competitiva na
utilização de nitrogênio e fósforo varia entre grupos fitoplanctônicos distintos
(Droop, 1973; Rhee, 1978; Tilman, 1981; Tilman et al 1986, Grover, 1989;
Cerrão et al , 1991; Bulgarov & Levich, 1999). Em períodos de rápido
crescimento as cianobactérias competem mais eficientemente por fósforo do que
as algas verdes. Sob condições de limitação por fósforo, no entanto, sua
capacidade competitiva vem sendo questionada (Bulgarov & Levich 1999).
Smith (1983) observou dominâncias de cianobactérias em 12 lagos diferentes
quando a razão N:P se encontrava abaixo de 29:1. Alguns trabalhos, no entanto,
não corroboram esta generalização. Suttle e Harrison (1988) avaliaram a
capacidade competitiva de uma alga verde, Scenedesmus quadricauda , e de uma

13
13
cianobactéria, Synnechococcus sp , sob condições de limitação por fósforo (altas
razões N:P) utilizando culturas do tipo ``batch´´. Nesta situação foi a
cianobactéria que se mostrou dominante em relação à alga verde.
O enfoque sob as razões nutricionais muda ao se considerar diferentes
formas minerais de nitrogênio e fósforo envolvidas no processo competitivo. De
acordo com McQueen e Lean (1987), variações nas proporções de compostos
nitrogenados também podem influir na estrutura que determinada comunidade
algal apresentará. Segundo estes autores, o aumento da dominância por
cianobactérias pode ser induzido pela diminuição da razão Nitrato: Fósforo
total. Quando esta razão se encontrava acima de 5, a dominância por
cianobactérias nunca foi observada pelos autores.
Estudos que tratam da competição por nutrientes em espécies
fitoplanctônicas são, em geral, realizados com três diferentes enfoques. O
primeiro utiliza comunidades fitoplanctônicas naturais cultivadas in vitro . Neste
caso o experimento de competição utiliza consórcios de espécies algais
coletadas diretamente de reservatórios. Como exemplo, Sommer (1984) cultivou
várias espécies naturais (mais de 30 espécies) variando as razões molares de
sílica e fósforo de 4 a 80. Sob condições de deficiência de sílica (baixas razões
Si:P) apenas Mougeotia thylespora conseguiu se estabelecer, enquanto que sob
condições de deficiência de fósforo as espécies Synedra acus , Asterionella
formosa e Koliella spiculiformis foram as dominantes. Segundo o autor, estes
dados estão de acordo com a ocorrência natural destas espécies no lago.
Outro tipo de experimento é o que utiliza a comunidade fitoplanctônica
natural in situ . Nestes tipos de experimento ocorre a fertilização artificial de
corpos d'água naturais com o acompanhamento das mudanças ocorridas na
estrutura da comunidade algal previamente existente. Tilman (1982) coloca que
experimentos de enriquecimento de todo o lago demonstram claramente que a
adição de fósforo aumentam a biomassa algal e que as proporções dos nutrientes
adicionados (como as razões N:P) têm um efeito dramático na composição das
espécies. Schindler (1977) observou que ao se ferti lizar um pequeno lago com N
e P em razões atômicas de 30 por 1 a espécie dominante seria uma alga verde
(Scenedesmus sp). Quando um lago de características semelhantes era mantido
em uma razão N:P de cerca de 11 o resultado das interações competitivas seria a
dominância por uma cianobactéria. Segundo Schindler ao se fertilizar o lago

14
14
com razões N:P ainda menores (abaixo de 5) outra cianobactéria se tornava
dominante, Anabaena sp, principalmente devido à sua capacidade de fixar N
atmosférico.
O terceiro tipo de experimento utilizado para se avaliar as interações
competitivas entre espécies fitoplanctônicas são os realizados através de
culturas laboratoriais. Experimentos com cultura utilizam gradientes de razões
nutricionais para avaliar a capacidade competitiva das espécies observadas .
Tilman (1976) estudou as interações de duas diatomáceas, Asterionella formosa
e Cyclotella meneghiniana , sob diversas razões N:P. Asterionella. formosa foi a
espécie dominante sempre que a razão se encontrava abaixo de 6:1 enquanto que
C. meneghiniana foi dominante em razões acima de 90:1. Em razões N:P
intermediárias as duas espécies coexitiram em cultura. A utilização deste
enfoque permite que se calcule as razões ótimas de crescimento para cada
espécie em relação a cada nutriente limitante. Rhee (1978), ao realizar um
experimento com culturas isoladas, observou que a razão N:P ótima para o
crescimento de Scenedesmus sp é de 30:1, a mesma observada para a
cianobactéria Anacystis nidulans .
O processo de eutrofização, causado pelo aumento do aporte de matéria
orgânica em reservatórios urbanos, é um problema recorrente em várias partes
do globo atualmente (Krienitz et al , 1996). O crescimento descontrolado dos
organismos planctônicos nestes reservatórios é considerado indesejável do ponto
de vista estético e turístico, especialmente o desenvolvimento de macrófitas
aquáticas e algas filamentosas (Margalef, 1983). No entanto, as consequências
vão além da estética, podendo compreender fenômenos como o esgotamento do
oxigênio da camada inferior da coluna d'água (principalmente em lagos rasos) e
o``bloom´´ de espécies fi toplanctônicas tóxicas, o que pode acarretar uma
mortandade massiva de peixes e mesmo a intoxicação de humanos (Bittencourt-
Oliveira et al , 2001). O fitoplâncton, neste caso, se torna um importante índice
biológico de monitoramento e avaliação da qualidade da água. Desta forma, o
aparecimento de determinadas espécies pode indicar a situação do reservatório
quanto a poluição de suas águas, o que possibilita a proposição de projetos de
manejo e recuperação da área (Wetzel, 1983).
A eutrofização artificial, em geral, leva a uma diminuição das razões N:P
em lagos oligotróficos. Com o aumento do afluxo de dejetos, o fósforo, que é o

15
15
principal elemento limitante ao crescimento de espécies fitoplanctônicas, pode
se acumular causando uma drástica mudança na estrutura e composição destas
comunidades (Henry & Tundisi, 1982). Haarhoff (1992 apud Bulgarov & Levich
1999) observou que as medidas tomadas para reduzir o despejo de dejetos
industriais em um lago hipereutrófico africano causaram a mudança das razões
molares N:P de 4 para 25. Como consequência a biomassa total da comunidade
fitoplanctônica foi reduzida e a antiga cianobactéria dominante, Microcystis
aeruginosa , foi substituída por diversas espécies de algas verdes.
Os estudos realizados no Brasil com a finalidade de avaliar os efeitos da
limitação nutricional em espécies fi toplanctônicas se centraram em sua maioria
em deficiências de fósforo e de nitrogênio. O enfoque principal destes estudos
foi a comunidade como um todo, sendo que espécies foram raramente avaliadas
individualmente. Cerca de metade dos estudos foram baseados em reservatórios,
tanto oligotróficos quanto eutróficos. A outra metade tratou de lagos naturais
principalmente no Vale do Rio Doce e em lagos de várzea amazônicos (Arcifa et
al. , 1995).
Henry & Tundisi (1982, 1983 e 1984) estão entre os primeiros
pesquisadores a utilizar este enfoque experimental em estudos na Represa do
Lobo (SP). Os experimentos consistiram na adição de nutrientes em
concentrações de 10 a 100 vezes superiores do que aquelas encontradas na
coluna d’água do reservatório. As conclusões dos autores foram que tanto o
nitrogênio quanto o fósforo eram limitantes, mas que o último era o fator
principal para a determinação da biomassa algal. Resultados posteriores
demonstraram que o nitrogênio possuía importância igual ou maior do que a de
fósforo na determinação da estrutura fitoplanctônica do reservatório. Não foram
detectadas diferenças na resposta das espécies fitoplanctônicas às diferentes
formas de nitrogênio utilizadas (NH4, NO2, NO3 ou uréia) nos experimentos de
enriquecimento.
A maioria dos estudos em reservatórios tropicais eutróficos, têm
demonstrado que o principal fator limitante para o crescimento e composição da
comunidade fitoplanctônica é o fósforo (Ramos et al. 1986, Henry 1986, Ibañez
1988). Tundisi e Henry (1986) ao realizarem experimentos no Lago Dom
Helvécio no período de circulação da coluna d’água também observaram este
padrão.

16
16
Em experimentos de fertilização foram observadas modificações não só na
estrutura específica da comunidade, mas também em parâmetros fisiológicas das
espécies envolvidas, como concentração de clorofila a, assimilação de nutrientes
e variação da produção primária (Setaro 1984, Henry & Simão 1988, Ibanez
1988). Ibanez (1988) verificou que a adição de nitrogênio e fósforo em amostras
do Lago Paranoá (DF) acarretou uma transição da dominância por uma espécie
de cianobactéria, Cylindrospermopsis brooki , por uma clorofícea, Chlorella sp.
Cerrão et al (1991) observaram que concentrações intermediárias de N e P (300
e 30 µg/l, respectivamente) foram mais eficientes do que as concentrações
máximas (600 e 60 µg/l) no aumento da concentração total de pigmentos de
perifiton em tanques artificiais na Represa do Lobo (SP).
1.2- Nitrogênio e Fósforo em ambientes naturais:
Os organismos precisam de aproximadamente 40 elementos para a
manutenção de sua estrutura e metabolismo. Estes elementos são definidos como
essenciais e podem ser requeridos em grandes (macronutrientes) ou baixas
concentrações (micronutrientes) pelos seres vivos. Nitrogênio e fósforo estão
entre os principais macronutrientes encontrados na composição dos seres vivos,
participando com mais de 1% do peso orgânico seco como definido por Braga et
al (2002). Reynolds (1999) coloca que a dificuldade mais freqüentemente
encontrada por plantas, terrestres ou aquáticas, marinhas ou de água doce, é
satisfazer os seus requerimentos para fósforo e nitrogênio.
Os elementos essenciais fazem parte de ciclos que recebem o nome de
biogeoquímicos. ``Bio´´ porque os organismos vivos interagem no processo de
síntese orgânica e decomposição dos elementos, ``geo´´ porque o meio terrestre
é a fonte dos elementos e químicos porque são ciclos de elementos químicos. A
biogeoquímica é, portanto, a ciência que estuda a troca ou a circulação de
matéria entre os componentes vivos e físicos e químicos da biosfera (Odum,
1971). Pode-se distinguir basicamente dois tipos de ciclos para os elementos
essenciais, os ciclos sedimentares e os gasosos. Nos ciclos sedimentares (tipo do
qual faz parte o fósforo) o reservatório que supre os elementos e os recebe de
volta é a litosfera. Nos ciclos gasosos (tipo do qual faz parte o nitrogênio) o
reservatório é a atmosfera (Esteves, 1998).

17
17
O nitrogênio responde por cerca de 78% da constituição da atmosfera. No
entanto, para ser utilizado pelos seres vivos, o gás nitrogênio (N2) precisa sofrer
uma série de modificações, conhecidas como fixação atmosférica, sendo que
poucos organismos (algumas bactérias e cianofíceas) são capazes de realizá-las.
A principal forma nitrogenada utilizada pelos produtores primários é o nitrato
(NO3-), fruto da decomposição da matéria orgânica ou fixação biológica. Outra
forma utilizada pelos organismos é a amônia (NH3), que pode ser sintetizada
artificialmente a partir do nitrogênio atmosférico a partir de uma técnica
desenvolvida durante a Primeira Guerra Mundial pelo alemão Fritz Haber. Esta
técnica possibilitou o aparecimento dos fertilizantes químicos, utilizados para
aumentar a produtividade de culturas vegetais e atender ao aumento da demanda
de alimentos em conseqüência do crescente aumento da população mundial.
Entretanto, o ciclo equilibrado do nitrogênio depende de um conjunto de fatores
bióticos e abióticos determinados que em conjunto, não estão aptos a assimilar o
excesso sintetizado artificialmente (Braga et al , 2002). Esse excesso de 9x106
t/ano (Kormondy, 1976), carregado para os rios, lagos e lençóis de água
suberrâneos é um dos responsáveis pelo fenômeno da eutrofização, que
compromete a qualidade da água.
O fósforo aparece nos organismos em proporção muito superior aos outros
elementos quando comparado com sua participação nas fontes primárias. Esse
fato justifica a importânica ecológica do fósforo e sugere que este elemento é
provavelmente o fator mais limitante à produtividade primária (Reynolds, 1984).
Por ser um elemento de ciclo fundamentalmente sedimentar, seu pricipal
reservatório é a l itosfera, mais precisamente as rochas fosfatadas e alguns
depósitos formados ao longo de milênios. Por meio de processos erosivos,
ocorre a liberação do fósforo na forma de fosfatos, que serão utilizados pelos
produtores. O fósforo, a exemplo do nitrogênio, é um dos principais fertilizantes
utilizados em agricultura, sendo também despejados na forma de detergentes
fosfatados e o seu excesso vem provocando acentuada deterioração da qualidade
de água dos corpos de água continentais .

18
18
1.3- Absorção de Fósforo e Nitrogênio pelo fitoplâncton.
Antes de se utilizar determinado nutriente para a síntese de estruturas nas
células vivas, é preciso que se obtenha o nutriente do meio externo através de
um processo conhecido como absorção. A absorção dos organismos autotróficos
é limidada a fontes que devem ser solúveis e difusíveis, e quase sempre se
restringe às formas químicas que naturalmente ocorrem no ambiente (Reynolds,
1984).
A absorção nutricional é fator chave no crescimento fitoplanctônico. A
habilidade das espécies em se adaptarem às mudanças na disponibilidade
nutricional no meio em relação aos seus próprios requerimentos de síntese de
matéria pode ser determinante no resultado do processo competitivo. A taxa de
absorção do nutriente cuja concentração é limitante no meio será determinada
externamente pela sua concentração, de acordo com a lei do mínimo, enquanto
que as taxas de absorção dos nutrientes não limitantes serão controladas
internamente para que a célula corresponda à taxa de absorção do nutriente
limitante (Dugdale et al , 1981).
A absorção nutricional é uma função hiperbólica relacionada à
concentração nutricional do meio. Este modelo, uma derivação da equação de
Monod, se baseia na premissa de que o transporte de nutrientes através da
membrana celular deve seguir o modelo de Michaelis-Menten, que descreve a
afinidade enzimática pelo substrato. É necessário, no entanto, que se parta de
alguns requisitos para que se utilize o modelo. O primeiro é que exista apenas
um substrato para a reação, ou, no caso de outros substratos, que as
concentrações se mantenham constantes. O segundo é que apenas as taxas
iniciais de reação são medidas a diferentes concentrações nutricionais, pois após
certo tempo poderá ocorrer a liberação do nutriente pela célula. O terceiro e
último é que o meio é homogêneo o bastante para que não ocorra a depleção do
nutriente disponível às células, ou seja, que na coluna dágua em estudo não
ocorra o processo de estratificação (McCarthy, 1981).
É importante salientar também, que este modelo descreve apenas o
transporte do substrato através da membrana celular, não permitindo a
inferência sobre o metabolismo do mesmo no meio intracelular. Esta definição é
importante, pois as espécies fi toplanctônicas também acumulam reservas

19
19
internas de determinados nutrientes e sua taxa metabólica está intimamente
relacionada à concentração destas reservas, pois, considera-se que são
essenciais para o funcionamento eficiente das enzimas envolvidas no processo
metabólico (McCarthy, 1981).
1.4- Escolha das espécies:
As espécies utilizadas neste experimento são comuns na composição da
comunidade fitoplanctônica do Reservatório da Pampulha, um típico
reservatório eutrófico urbano. Oocystis lacustris , junto a outras espécies de
algas verdes, tem seu pico em biomassa no final do verão e durante grande parte
do outono. Boechat e Giani (2000) observaram picos de densidade deste grupo
de fevereiro a maio no Reservatório da Pampulha. Esta espécie vem
apresentando nos últimos anos um decréscimo em biomassa nas águas do
reservatório, apesar de manter populações estáveis durante a maior parte do ano.
Cryptomonas erosa se mostra abundante durante todo o inverno (junho a início
de setembro), apresentando um aumento acentuado em sua densidade nos
últimos anos. Algas flageladas, em geral, incluindo as criptofíceas, dinofíceas e
algumas crisofíceas costumam apresentar elevada biomassa em reservatórios
urbanos (Rojo & Alvarez-Cobelas, 1993). O fato de poderem movimentar-se é
uma vantagem em ambientes túrbidos com relação à luz e permite, ainda, que
estas algas possam utilizar nutrientes presentes em camadas mais profundas,
podendo em seguida voltar para a região eufótica (Giani et al . 1999). Além
disso, criptofíceas podem apresentar um tipo de alimentação fagotrófica, o que
representa uma fonte adicional de nutrientes, principalmente em reservatórios
com alta concentração de matéria orgânica (Klaveness, 1988). Microcystis
viridis pode apresentar florações em massa no Reservatório da Pampulha da
primavera ao início das chuvas de verão (outubro a janeiro) (Giani, 1994;
Figueredo & Giani, 2000). As cianobactérias, principalmente do grupo
Microcystis , têm um papel muito importante em corpos d'água eutróficos, pela
sua capacidade de se desenvolverem maciçamente (``water blooms´´) e pela sua
potencial toxidade (Paerl, 1996; Giani et al. 1999). No reservatório da Pampulha
têm sido observados eventos desta natureza que causam impactos negativos
sobre o ambiente, desde mau cheiro até mortandade em massa de peixes

20
20
(Goodwin & Giani, 1998). Pinto-Coelho (1992) coloca que em 1980 a estação
de captação de água da Cia. De Saneamento do Estado de Minas Gerais
(COPASA) teve que ser desativada em decorrência dos frequentes blooms de
algas fitoplanctônicas que passaram a ocorrer no reservatório.
Em vista do exposto acima, decidiu-se pela utilização destas três espécies
com características sazonais e fisiológicas distintas e de grande importância na
composição da comunidade fitoplanctônica e na dinâmica ecológica de um típico
reservatório urbano eutrófico.

21
21
OBJETIVOS
O objetivo deste estudo é verificar a capacidade competitiva de Oocystis
lacustris (Chlorophyta-Chlorophyceae), Cryptomonas erosa (Pyrrophyta-
Cryptophyceae) e Microcystis viridis (Cyanobacteria-Cyanophyceae) sob
diferentes razões N:P no meio, visando esclarecer aspectos sobre a dinâmica
competitiva destas espécies em ambientes naturais.
- Objetivos Específicos
• Determinar alguns parâmetros competitivos como taxa de
crescimento em diferentes razões N:P e concentrações mínimas de nitrogênio e
fósforo necessárias para que populações de Oocystis lacustris , Microcystis
viridis e Cryptomonas erosa se mantenham estáveis em cultura.
• Estabelecer a dinâmica de absorção de fósforo e nitrogênio das
três espécies.
• Definir o resultado de interações competitivas entre as três
espécies em diferentes razões N:P.
• Comparar os resultados obtidos em laboratório com os dados de
variação sazonal da comunidade fitoplanctônica e variáveis físico-químicas do
reservatório da Pampulha (MG), um reservatório urbano eutrófico.

22
22
2. MATERIAL E MÉTODOS
2.1- Área de estudo
O reservatório da Pampulha localiza-se na região noroeste do município
de Belo Horizaonte-MG (19o 55´09´´S e 43o 56´ 47´´W). A bacia hidrográfica do
reservatório da Pampulha cobre uma área de 97,57 km2, pertencendo aos
municípios de Belo Horizonte e de Contagem, sendo formada por 40 córregos
dos quais 11 são tributários diretos do reservatório. A área inundada
corresponde a 2,4 km2 e o volume de água a 11,3 milhões de m3. A profundidade
média é de 5 m e a máxima de 16 m (Pinto Coelho, 1998; Tôrres 1999; Heleno,
2002). As principais fontes de poluição do reservatório provêm da sua bacia
hidrográfica, sendo elas o esgoto doméstico e o industrial e os resíduos
transportados pelo escoamento superficial das águas de chuva, além do
crescente assoreamento decorrente da erosão sofrida na bacia de drenagem
(Champs 1992, Sperling 1994, Vignoli 1992). O esgoto doméstico é despejado
diretamente em suas águas e desta forma, o nível de eutrofização do reservatório
vem aumentando consideravelmente nos últimos anos (Freitas, 1994).
2.2- Descrição das espécies
- Oocystis lacustris, Chodat, 1897, (Chlorophyceae): Apresenta células
isoladas ou em colônias de 2, 4, 8 ou 16 células irregularmente distribuídas
numa mucilagem que pode ou não ter nódulos polares. As células são elípticas,
pólos levemente pontiagudos, com ou sem nódulos polares, com comprimento de
9,0 a 29,0 µm e diâmetro de 4,0 a 10,0 µm. Esta espécie pode ter de 1-4
cloroplastos parietais, com ou sem pirenóides. A reprodução é feita por
autósporos, não sendo conhecida reprodução sexuada (Sant’Anna, 1984).
- Cryptomonas erosa, Ehrenberg, 1838, (Cryptophyceae): Células em vista
lateral de contorno elíptico, oblongo a obovado, margem dorsal regular a
irregularmente convexa, margem ventral convexa, reta ou levemente côncava,
raro irregularmente convexa, pólo anterior obliquamente truncado, porção dorsal

23
23
proeminente formando rostro desde pouco a bastante evidente, às vezes sem
rostro, pólo posterior arredondado, às vezes afilado. As medidas celulares são
13,8 a 43,2 µm de comprimento e 6,1 a 21,1 µm de largura. Possui dois
cromatóforos dorsiventrais sem pirenóides nos quais podem ser observados
numerosos grãos de amido. Apresenta dois flagelos subapicais de tamanhos
pouco distintos entre si , aproximadamente tão longos quanto a célula (Castro,
1993).
- Microcystis viridis, Kirchner, 1898, (Cyanophyceae): Colônias
aproximadamente esféricas, elipsoidais ou alongadas de alguma forma. As
células são esféricas variando de 3 a 7 µm de diâmetro. Podem apresentar
vacúolos gasosos em seu interior. A mucilagem em torno das colônias é
indistinta podendo conter um grande número de colônias filhas (Desikachary,
1959).
Figura 1: Fotografias de Oocystis lacustris (A), Cryptomonas erosa (B) e Microcystis viridis (C).
2.3- Isolamento e manutenção das culturas estoques
As espécies foram isoladas a partir de amostras de água coletadas do
Reservatório da Pampulha com redes de 20µm de abertura de poros. Segundo
Kilham (1978) ao se realizar qualquer estudo de fisiologia com espécies
fitoplanctônicas no qual se pretenda comparar os resultados com os encontrados
em determinado corpo d´água, as espécies devem ser isoladas diretamente do
corpo d´água em consideração. A técnica de isolamento consiste na utilização de
pipetas de ponta capilar na separação de uma única célula ou colônia
observando-as através de um microscópio óptico (Rosowski, 1973). As células

24
24
isoladas foram inoculadas em tubos de ensaio contendo meio de cultura WC com
pH de 7.2 (Guillard & Lorenzen, 1972) e incubadas em germinadora à 20oC. O
material utilizado foi esterilizado em autoclave para se evitar a contaminação
por bactérias e outros microrganismos. É sempre melhor evitar a contaminação
por bactérias em experimentos já que os efeitos das bactérias, se em grande
quantidade, podem alterar os resultados (Kilham, 1978). As culturas foram
repicadas a cada 20 dias aproximadamente. Todos os repiques e retiradas de
amostras foram realizados em capela de fluxo laminar estéril . As culturas
estoque foram mantidas em meio WC em câmara germinadora com a temperatura
de 20oC, fotoperíodo 12 h claro: 12h escuro e disponibilidade luminosa média
de 60µmol.m- 1s- 1 .
As culturas de O. lacustris e M. viridis a serem utilizadas neste
experimento foram isoladas em 1994 e vêm sendo mantidas em laboratório nas
condições descritas acima pela equipe do Laboratório de Ficologia,
Departamento de Botânica da Universidade Federal de Minas Gerais.
Cryptomonas erosa foi isolada em 2000 devido a sua crescente importância na
composição fitoplanctônica do Reservatório da Pampulha nos últimos anos.
2.4- Delineamento experimental
O trabalho desenvolvido em laboratório foi dividido em três etapas: os
experimentos de crescimento das espécies, experimentos de competição e
experimentos de absorção dos nutrientes.
-2.4.1 Experimentos de crescimento
As culturas utilizadas no experimento foram do tipo ``batch´´. As espécies
foram cultivadas separadamente em meios WC que tiveram as concentrações
molares de K2HPO4 e NaNO3 modificadas de forma a se atingir as seguintes
razões N:P: 4 e 10 (potencial l imitação por nitrogênio), 20 (sem limitação
nutricional), 40 e 100 (potencial limitação por fósforo) (Giani e Delgado, 1998).
Foram adicionadas soluções de KCl e NaCl de maneira proporcional à retirada
de K2HPO4 e NaNO3 para se manter o equilíbrio iônico dos meios com
deficiência de fósforo e nitrogênio, respectivamente.

25
25
O cultivo se realizou em erlenmeyers de 250ml nos quais foram
adicionados 100 ml de meio de cultura. Os experimentos foram efetuados em
triplicata em câmara germinadora da marca FANEM com fotoperíodo de 12/12
horas, temperatura de 20oC e disponibilidade luminosa média de 60µmol.m- 1s- 1.
Segundo Kilham (1978), em seu estudo com diatomáceas, as condições ideais
para experimentos com culturas do tipo ``batch´´ são temperatura constante,
luminosidade em níveis de saturação e fotoperíodos de 12:12 horas. A autora
enfatiza que muitas espécies não crescerão sob condições de luminosidade
constante, principalmente se estiverem limitadas nutricionalmente.
Foram inoculadas baixas densidades celulares das três espécies no início
dos experimentos, de forma a ser possível acompanhar a sua fase exponencial de
crescimento. A baixa densidade ao início do cultivo é outro fator apontado por
Kilham (1978) como sendo essencial para que se garanta que a dinâmica de
crescimento de espécies cultivadas em meios do tipo ``batch´´ se adeque aos
modelos de crescimento em culturas contínuas e ambientes naturais.
Os experimentos tiveram a duração de 20 dias cada. Foram retiradas
diariamente, sempre no mesmo horário, sub-amostras de 1ml das três réplicas de
cada razão N:P. Estas sub-amostras foram fixadas com lugol acético e duas
réplicas foram contadas em microscópio LEIKA através de hemocitômetro do
tipo Fuchs-Rosenthal, no caso de O. lacustris e C. erosa e hemocitômetro do
tipo Neubawer no caso de M. viridis . Foram contadas no mínimo três lâminas de
cada sub-amostra ou 400 células no caso deste valor ser alcançado antes.
Os valores obtidos em cel/ml foram convertidos para µm3/ml através do
cálculo do biovolume médio das células das três espécies em todos os
tratamentos. Foram utilizadas amostras do início, do meio e do fim de cada
experimento para se medir o biovolume médio de cada espécie em cada
tratamento. A primeira medida foi feita na cultura estoque, utilizada para o
inóculo inicial. A segunda medida foi feita após 10 dias de experimento e a
terceira no último dia de experimento. O cálculo do biovolume de cada espécie
foi realizado através da aproximação do formato das células com formas
geométricas simples, de acordo com Rott (1981). Foram medidas no mínimo 20
células de cada amostra para o cálculo de seu biovolume. As fórmulas utilizadas
foram as seguintes:

26
26
V= π/6.a . b2
Onde o volume (V) é calculado com base na largura média da célula (b) e
no seu comprimento total (a). Esta fórmula é utilizada em espécies que Rott
classificou como tendo forma elipsoidal rotacional com secção circular. Neste
estudo ela foi utilizada para o cálculo do biovolume de O. lacustris e C. erosa .
Para o cálculo do biovolume de M. viridis foi utilizada a seguinte fórmula
que é utilizada para células que Rott classificou como esféricas:
V= π/6.d3
Onde d é o diâmetro da célula.
Foram realizadas comparações entre os valores de biovolume das três
espécies nas diferentes razões N:P, e dentro das mesmas razões N:P em
momentos diferentes do cultivo, através de análises de variância e posteriores
testes de Tukey para amostras com número de casos desiguais (Programa
Statistica 5.0, 1995).
O biovolume médio de cada tratamento para as três espécies ao longo do
experimento foi estimado através do cálculo de uma função polinomial, obtida
pelo programa Excel (Microsoft Office, 1998) que relacionou o volume celular
com o tempo de experimento. A partir destes dados foram construídas as curvas
de crescimento de cada espécie nas diferentes razões N:P em biovolume por
mililitro de cultura (µm3/ml).
As taxas de crescimento para cada amostra foram calculadas através de
regressão linear (program Excel, Microsoft Office 1998), para a fase
exponencial de crescimento das espécies. Foi utilizado o logaritimo natural das
densidades em biovolume de cada espécie. Neste caso µ é o coeficiente de
inclinação da equação da reta em gráfico onde o y é a concentração celular e x é
o tempo de experimento em dias.
Análises de co-variância foram realizadas para comparação da fase
exponencial de crescimento entre tratamentos de uma mesma espécie. Foram
consideradas na análise diferenças significativas (p<0,005) e altamente
significativas (p<0,001)

27
27
Uma regressão não linear foi utilizada para adequar os resultados dos
experimentos de crescimento de cada espécie ao modelo de Monod, utilizando-
se o programa estatístico JMP versão 5.0, seguindo a seguinte equação:
µ= µma x . S/(KS + S)
onde µ e µma x são a taxa de crescimento e a taxa máxima de crescimento
respectivamente, S é a concentração nutricional e KS é a constante de semi-
saturação do crescimento.
Para se obter poder preditivo, sobre os resultados das interações
competitivas entre as três espécies, foi calculada a quantidade mínima de
fósforo e nitrogênio necessária para que a espécie equilibre a sua taxa de
crescimento à sua taxa de perda em cultura:
R*= DKS/(µma x- D)
onde D é a taxa independente de perda de nutrientes (mortalidade)
definida como a taxa hipotética de 0,25d- 1 para culturas do tipo ``batch´´
(Grover, 1989; Giani e Delgado, 1998). De acordo com Tilman (1981) a espécie
com o menor R* para cada nutriente deve, em teoria, apresentar uma melhor
performance competitiva em meios que apresentem concentrações nutricionais
limitantes.
A afinidade pelo substrado, expresso pela curvatura inicial da curva de
saturação, µma x/KS, foi também utilizada como índice de competência
competitiva.
Ao final dos experimentos os meios de cultivo foram filtrados em filtro de
fibra de vidro e foram realizadas análises de clorofila-a seguindo a metodologia
citada por Lorenzen (1967). Com base nos valores de concentração de clorofila-
a medidos, foram calculadas as concentrações de clorofila-a por célula e por
biovolume (µm3) para as três espécies ao final dos experimentos. Foram
realizadas análises de variância (Programa Statistica 5.0, 1995) para se detectar
diferenças estatisticamente significativas entre as concentrações de clorofila nos
meios de cultura, por célula e por biovolume entre as espécies e entre diferentes
razões N:P em uma mesma espécie.

28
28
- 2.4.2 Experimentos de absorção
A complementação dos dados obtidos através da determinação das curvas
de crescimento das três espécies foi obtida através de experimentos de absorção
de fósforo e nitrogênio. Este experimento permitiu que fosse avaliada a
capacidade máxima de absorção de fósforo e nitrogênio das espécies além de
determinar outros parâmetros competitivamente relevantes.
As espécies foram cultivadas por dois dias em meios de cultivo que não
possuiam fósforo e 14 dias em meios de cultivo que não possuíam nitrogênio. As
algas alcançaram, desta forma, o estado conhecido como``starvation´´(Syrett
1981). A partir deste ponto o nutriente foi adicionado em diferentes
concentrações no meio e as espécies foram cultivadas em estufa germinadora
com fotoperíodo de 12:12 horas, temperatura de 20OC e disponibilidade
luminosa média de 60 µmol.m- 1s- 1 .
Nos experimentos de absorção de nitrogênio foi adicionado NaNO3 em 4
diferentes diluições: 3700µg/ml, 370µg/ml, 150µg/ml e 75µg/ml. Amostras
foram coletadas no início do experimento, após 12 horas, 24 horas, 36 horas, 48
horas e 60 horas. As diluições nos experimentos de fósforo foram 1800µg/ml,
180µg/ml, 35µg/ml e 18µg/ml e as coletas foram feitas após 30 minutos, 1 hora,
2 horas, 3 horas e 4 horas.
Foi calculada a taxa de absorção das três espécies para os dois nutrientes
em todas as diluições. O cálculo se baseou na assimilação dos nutrientes pelas
algas, medidos através de sua depleção no meio de cultura. Foi calculado o
biovolume das espécies no meio de cultura e as taxas de absorção foram
calculadas tendo como base a absorção do nutriente por µm3 da cultura.
Uma função hiperbólica relacionando a taxa de absorção de cada espécie
por biovolume com a concentração nutricional, foi calculada, utilizando-se o
programa estatístico JMP versão 5.0, seguindo a seguinte equação, que também
é uma derivação da equação de Monod:
v= Vma x . S/(Km + S)

29
29
onde S é a concentração do substrato, Km é a constante de semi-saturação
e Vma x é a taxa máxima de absorção nutricional. Esta função determina a
capacidade que cada espécie possui de absorver nitrogênio e fósforo em relação
à disponibilidade destes nutrientes no meio.
Foi também calculado o coeficiente de consumo em excesso (R) de
nitrogênio e fósforo para as três espécies. Segundo Tilman (1981) este
coeficiente mede a capacidade da espécie em consumir e armazenar quantidades
de reserva do nutriente quando as condições são favoráveis. O coeficiente foi
calculado dividindo-se a taxa de absorção nutricional nos meios com maior
concentração pela taxa de absorção calculada nos meios com a menor
concentração.
- 2.4.3 Experimentos de competição
Nos experimentos de competição as espécies foram cultivadas juntas, em
culturas mistas de duas espécies, nas razões N:P 4, 20 e 100. Os experimentos
foram realizados em erlenmayers de 250 ml com 100 ml de meio de cultura,
sempre em triplicata. Amostras foram coletadas a cada dois dias e duas réplicas
contadas em microscópio LEIKA com hemocitômetro do tipo Fuchs-Rosenthal.
Foram contadas no mínimo três lâminas de cada sub-amostra ou 400 células da
espécie mais abundante no caso deste valor ser alcançado antes.
Os inóculos iniciais das espécies foram a metade dos utilizados nos
experimentos de crescimento. Desta forma foi possível se acompanhar o
desenvolvimento da interação competitiva entre as espécies desde os estágios
iniciais.
2.5- Variação Sazonal da Comunidade Fitoplanctônica e de algumas
variáveis físico-químicas no Reservatório da Pampulha
Para se comparar os resultados obtidos em laboratório com os encontrados
no Reservatório da Pampulha foram utilizados os dados biológicos e físico-
químicos deste reservatório coletados e analisados no período de janeiro de
1996 a dezembro de 1998 pelos laboratórios de Ficologia, departamento de
Botânica, e Gestão de Reservatórios Tropicais, departamento de Biologia Geral

30
30
da Universidade Federal de Minas Gerais. As coletas ocorreram duas vezes ao
mês em ponto único do reservatório. As amostras foram coletadas através de
garrafa de van Dorn, a um metro de profundidade. As contagens fitoplanctônicas
foram realizadas através de microscópio invertido seguindo método de Uthermöl
(1958). As concentrações de fósforo e nitrogênio foram determinadas em
laboratório. Foram util izados os dados de concentração de fósforo solúvel e
fósforo total. As razões N:P da água do reservatório foram calculadas tendo
como base as concentrações molares de fósforo e nitrogênio (soma de todas as
formas nitrogenadas inorgânicas) durante o período de coleta. Foram calculadas
as razões N:P solúvel e N:P total do reservatório.
Foram realizadas análises de correlação linear (Programa Statistica 5.0,
1995) entre os biovolume das espécies O. lacustris, M. viridis , C. erosa , dos
gêneros Oocystis, Microcystis e Cryptomonas e também da clorofila-a (µg/l),
com as variáveis físico-químicas; temperatura, concentração de nitrogênio
dissolvido (nitrato, nitrito e amônia), fósforo solúvel reativo, fósforo total,
razão N:P solúvel e razão N:P total observados no Reservatório da Pampulha
durante o período de janeiro de 1996 a dezembro de 1998. Foram consideradas
significativas as correlações de P menor ou igual a 0,05.

31
31
3 RESULTADOS
3.1- Crescimento
Serão apresentados inicialmente os resultados de crescimento das espécies
nos meios contendo as diferentes razões N:P. As curvas de crescimento
apresentadas são as médias de duas réplicas, contadas para cada meio com as
cinco razões N:P para cada espécie. A Figura 2 apresenta as curvas de
crescimento das três espécies em logaritmo natural da densidade em células por
milili tro (cel/ml).
Nos experimentos de crescimento de Oocystis lacustris as maiores
densidades foram alcançadas nas razões N:P intermediárias (10, 20 e 40). Nos
meios com as razões N:P 4 e N:P 100 as culturas alcançaram a fase estacionária
de crescimento mais rapidamente e terminaram o experimento em densidades
menores do que nas demais razões. Todos os tratamentos alcançaram a fase
estacionária ao final do experimento. A maior densidade foi alcançada em meios
com razão N:P 10, 4.177.500 cel/ml em média. As menores foram obtidas em
meios com razões N:P 100 que chegaram a 2.255.000 cel/ml em média (Tabela
1).
Microcystis viridis foi a espécie que apresentou maior média final de
densidade com 3.836.500 cel/ml. Oocystis lacustris apresentou 3.347.500 e
Cryptomonas erosa 852.420. Microcystis viridis apresentou densidades finais
muito próximas nos meios com razões N:P 10, 20, 40 e 100 (Figura 2). Os
experimentos realizados em meios de cultivo com razão N:P 4 foram os que
apresentaram menor densidade final. É possível se observar que os cultivos de
M. viridis não alcançaram a fase estacionária de crescimento, à exceção do
cultivo no meio N:P 4. A densidade média nos meios de razão N:P 20 foi a
maior ao final do experimento, com 4.800.000 cel/ml. A menor densidade média
final foi encontrada nos meios de razão N:P 4, 2.097.500 cel/ml (tabela 1).
Cryptomonas erosa , como M. viridis , também apresentou densidades
menores em meios N:P 4 ao final do cultivo. Os meios com razão N:P 20
apresentaram uma densidade média superior aos demais, sendo que em todos as
razões as culturas alcançaram a fase estacionária de crescimento ao final do
experimento. A maior densidade final foi obtida em meios com razão N:P 20,

32
32
com 1.280.000 células por milili tro em média e a menor em meios com razão
N:P 4 com 299.175 em média.
Figura 2: Crescimento de Oocystis lacustris (A), Microcystis viridis (B) e Cryptomonas erosa (C) em ln de cel/ml nas 5 razões N:P.
A)
9
10
11
12
13
14
15
16
0 5 10 15 20 25
Dias
LN c
el/m
l
N:P4N:P10N:P20N:P40N:P100
C)
8
9
10
11
12
13
14
15
0 5 10 15 20
Dias
LN c
el/m
l
N:P4N:P10N:P20N:P40N:P100
B)
9
10
11
12
13
14
15
16
0 5 10 15 20 25
Dias
LN c
el/m
l
N:P4N:P10N:P20N:P40N:P100

33
33
Tabela 1: Médias de densidade inicial e final das três espécies nas cinco razões nutricionais em cel/ml.
Estão representados na tabela 2 os valores médios de biovolume das três
espécies. O biovolume foi calculado em células medidas em três fases distintas
do cultivo das espécies: na cultura estoque a ser inoculada nos meios de cultivo,
na metade do experimento e ao final do experimento. Foram medidas células de
réplicas diferentes (20 em média para cada réplica) sendo estes valores
agrupados posteriormente por razão N:P. Cryptomonas erosa apresentou em
média os maiores valores de biovolume de suas células sendo cerca de 15 vezes
maior do que M. viridis e 6 vezes maior do que O. lacustris . Os valores de
biovolume medidos para as três espécies foram significativamente diferentes
entre si (p<0,005).
A figura 3 mostra a variância entre os valores de biovolume medidos. O
resultado das análises de variância realizadas com os valores de cada espécie
nas diferentes razões e nos diferentes períodos de cultivo demonstrou que as
diferenças foram significativas (p<0,005) apenas para Cryptomonas erosa . Uma
análise de Tukey para números amostrais desiguais separou os valores de
biovolume calculados para C. erosa na cultura estoque e metade do experimento
como sendo iguais entre si e diferentes dos valores obtidos ao final do
experimento, estes últimos significativamente maiores do que os primeiros. Em
M. viridis e O. lacustris não foram encontradas diferenças significativas entre
os tamanhos das células.
O. lacustris C. erosa M. viridisDensidade InicialN:P4 13075 5925 16650N:P10 14070 4210 14500N:P20 12725 4073 25350N:P40 13587 4141 23000N:P100 8675 4773 21500
Densidade FinalN:P4 2327500 299175 2097500N:P10 4177500 802500 3910000N:P20 4022500 1280000 4800000N:P40 3955000 1041250 4400000N:P100 2255000 839175 3975000

34
34
Tabela 2: Valores de biovolume médio das células em µm3, das três espécies nas 5 razões N:P em três períodos distintos dos experimentos.
Figura 3: Valores de biovolume medidos para O. lacustris , M. viridis e C. erosa medidos nas 5 razões N:P utilizadas, na cultura estoque (1), metade do experimento (2 a 6) e final do experimento (7 a 11).
A figura 4 representa as curvas de crescimento das três espécies em
logaritmo natural da densidade em biovolume (µm3/ml). Houve pouca variação
Microcystis
0 1 2 3 4 5 6 7 8 9 10 11 12
Biov
olum
e (u
m3)
0
100
200
300
400
500
600
Oocystis
0 1 2 3 4 5 6 7 8 9 10 11 12
Biov
olum
e (u
m3)
0
200
400
600
800
1000
Cryptomonas
0 1 2 3 4 5 6 7 8 9 10 11 12
Biov
olum
e (u
m3)
0
2000
4000
6000
8000 1- Cultura Estoque
- Metade do experimento2- N:P4 3- N:P104- N:P205- N:P406-N:P100
-Final do experimento7- N:P4 8- N:P109- N:P2010- N:P4011-N:P100
Microcystis
0 1 2 3 4 5 6 7 8 9 10 11 12
Biov
olum
e (u
m3)
0
100
200
300
400
500
600
Oocystis
0 1 2 3 4 5 6 7 8 9 10 11 12
Biov
olum
e (u
m3)
0
200
400
600
800
1000
Cryptomonas
0 1 2 3 4 5 6 7 8 9 10 11 12
Biov
olum
e (u
m3)
0
2000
4000
6000
8000 1- Cultura Estoque
- Metade do experimento2- N:P4 3- N:P104- N:P205- N:P406-N:P100
-Final do experimento7- N:P4 8- N:P109- N:P2010- N:P4011-N:P100
Metade do Experimento Final do ExperimentoCultura Estoque N:P4 N:P10 N:P20 N:P40 N:P100 N:P4 N:P10 N:P20 N:P40 N:P100
O. lacustris 437 325 312 359 399 326 353 355 320 352 392M. viridis 178 195 183 154 144 143 199 192 163 115 159C. erosa 2742 3330 2759 1998 2271 2148 4481 4372 3619 3984 4119

35
35
em relação aos gráficos (Figura 2), já apresentados, em célula por mililitro
(cel/ml) de cultura.
Figura 4: Crescimento de Oocystis lacustris (A), Microcystis viridis (B) e Cryptomonas erosa (C) em Logaritmo na base natural de µm3/ml nas 5 razões N:P.
A)
15
16
17
18
19
20
21
22
0 5 10 15 20 25
Dias
LN u
m3 /m
l
N:P4N:P10N:P20N:P40N:P100
C)
15
16
17
18
19
20
21
22
23
0 5 10 15 20
Dias
LN u
m3 /m
l
N:P4N:P10N:P20N:P40N:P100
B)
15
16
17
18
19
20
21
0 5 10 15 20 25
Dias
LN u
m3 /m
l
N:P4N:P10N:P20N:P40N:P100

36
36
As culturas de C. erosa em todas as razões N:P apresentaram densidade
final superior às de O. lacustris e M. viridis . A densidade média final dos
cultivos com C. erosa foi 2,97.109 µm3/ml contra 1,22.109 µm3/ml de
O.lacustris e 0,64.109 µm3/ml de M.viridis . A tabela 3 apresenta os valores de
densidade inicial e final em biovolume das três espécies em todas as razões N:P
utilizadas nos experimentos. As análises de co-variância realizadas não
mostraram diferenças significativas (p<0,005) entre o crescimento exponencial
de nenhuma das três espécies com relação à razão N:P utilizada.
Tabela 3: Valores de densidade inicial e final das três espécies nas cinco razões nutricionais em biovolume, µm3/ml.106.
A figura 5 apresenta as concentrações de clorofila-a medidas nos meios de
cultura ao final dos experimentos. Cryptomonas erosa apresentou valores de
clorofila-a significativamente superiores (p<0,005) aos valores de O. lacustris e
M. viridis nas 5 razões N:P. A tabela 4 apresenta as concentração de clorofila-a
medidas por célula e por biovolume (µm3) das três espécies em todas as razões
N:P. Cryptomonas erosa apresentou as maiores concentrações por célula e
biovolume em todas as razões N:P, sendo significativamente superiores as
concentrações de O. lacustris e M. viridis que, por sua vez, não diferiram
significativamente entre seus valores.
As concentrações de clorofila-a por célula e por biovolume aumentaram
nas células cultivadas em meios com razões N:P 20, 40 e 100 em O. lacustris e
C. erosa , apesar destas diferenças não serem estatisticamente significativas
O. lacustris C. erosa M. viridisDensidade InicialN:P4 5,71 16,24 2,97N:P10 6,14 11,54 2,58N:P20 5,56 11,17 4,52N:P40 5,93 11,35 4,10N:P100 3,79 13,09 3,83
Densidade FinalN:P4 819,91 1259,78 417,05N:P10 1470,48 3141,03 754,98N:P20 1538,68 3951,43 779,94N:P40 1390,98 3546,23 621,67N:P100 864,09 2942,62 643,31

37
37
(p<0,005). Microcystis viridis não apresentou variação na concentração de
clorofila-a por célula ou por biovolume em relação à razão N:P utilizada. As
maiores concentrações de clorofila-a foram obtidas na razão N:P 40 para M.
viridis e C. erosa , e na razão N:P 20 para O. lacustris .
Figura 5: Concentração de clorofila-a nos meios de cultura em µg/l ao final dos experimentos de crescimento.
Tabela4: Concentração de clorofila-a medida ao final de cada experimento para as três espécies no meio, por célula e por biovolume.
Espécie Razão Conc. Final (ug/L) Conc. Célula (pg/cel) Conc. Biovolume (pg/um3)
O . Lacustris N:P4 42,768 0,0184 5,22.10-6
N:P10 90,882 0,0218 6,18.10-6
N:P20 588,060 0,1462 3,82.10-5
N:P40 272,646 0,0689 1,96.10-5
N:P100 122,958 0,0545 1,42.10-5
M. viridis N:P4 192,456 0,0918 1,53.10-5
N:P10 352,836 0,0902 1,12.10-5
N:P20 486,486 0,1014 1,23.10-5
N:P40 545,292 0,1239 1,57.10-5
N:P100 326,106 0,0820 1,11.10-5
C. erosa N:P4 267,300 0,8935 2,12.10-5
N:P10 775,176 0,9660 2,47.10-5
N:P20 2336,202 1,8252 5,91.10-5
N:P40 2173,322 2,1562 6,26.10-5
N:P100 1512,918 1,8029 5,14.10-5
0
500
1000
1500
2000
2500
4 10 20 40 100
Razão N:P
Clo
rofil
a (u
g/L
)
O. lacustrisM . viridisC. erosa

38
38
A figura 6 apresenta as curvas de crescimento relacionadas à concentração
de fósforo e nitrogênio das três espécies, calculadas a partir dos dados de
densidade em µm3/ml seguindo a equação de Monod. A tabela 5 apresenta os
parâmetros de crescimento calculados a partir destas regressões. Estes
parâmetros servem como índices competitivos das espécies. Segundo Tilman e
Kilham (1976) em um grupo de espécies competindo pelo mesmo nutriente
limitante e que apresentam valores similares de taxas de crescimento máximas, a
espécie com o menor valor de Ks deve sobrepujar as demais. Como as espécies,
em geral, não apresentam valores iguais de taxas máximas de crescimento, a
concentração nutricional requerida por uma população algal em equilíbrio, R*,
ou a curvatura inicial da curva de Monod, µma x/Ks são utilizadas para se prever o
resultado da interação competitiva (Tilman, 1981). A espécie que apresenta o
menor R* ou o maior valor de µma x/Ks deverá vencer o processo competitivo
(Giani e Delgado, 1998).
Tabela 5: Parâmetros de crescimento das três espécies, obtidos a partir de regressões não lineares adaptadas ao modelo de Monod em biovolume. Taxa máxima de crescimento (µma x) , constante média de saturação do crescimento (Ks), concentração nutricional mínima requerida por uma população algal em equilíbrio (R*, calculada para a taxa de mortalidade de 0,25d- 1 e curvatura inicial da curva de Monod (µma x/KS).
Cryptomonas erosa apresentou os parâmetros mais favoráveis nos
experimentos com ambos os nutrientes. O valor de R* calculado para esta
espécie, foi inferior tanto para nitrogênio quanto para fósforo, em relação às
demais espécies. Segundo o modelo de competição diferencial por nutrientes,
esta espécie deveria vencer o processo competitivo quando colocada em culturas
mistas com M. viridis e O. lacustris . Oocystis lacustris e Microcystis viridis
apresentaram valores muito similares calculados para µma x/Ks nos experimentos
Espécie µmax (d-1) Ks (µmol/L) R(µmol/L) µmax/Ks (µmolL-1d-1)
Nitrogênio O. lacustris 0,531 7,880 7,016 0,067M. viridis 0,305 4,650 21,291 0,066C. erosa 0,555 4,950 4,056 0,112
Fósforo O. lacustris 0,543 0,260 0,222 2,088M. viridis 0,307 0,129 0,564 2,381C. erosa 0,648 0,106 0,067 6,113

39
39
com fósforo e nitrogênio mas o valor de R* para O. lacustris foi muito inferior
em ambos.
É importante frisar que apenas O. lacustris apresentou valores crescentes
de taxa de crescimento média com o aumento da concentração de fósforo e
nitrogênio. Cryptomonas erosa apresentou um padrão, no caso das
concentrações de fósforo, inverso, com as maiores taxas de crescimento sendo
obtidas em concentrações menores do nutriente.

40
40
Figura 6: Crescimento das três espécies em relação às concentrações molares de fósforo e nitrogênio. As curvas foram construídas a partir de regressões não lineares dos dados em biovolume.
3.2- Absorção de Fósforo e Nitrogênio
A figura 7 apresenta as curvas de absorção das três espécies nos
experimentos de fósforo e nitrogênio. A tabela 6 apresenta os parâmetros de
absorção calculados para as três espécies: a densidade média em biovolume, a
taxa máxima de absorção em µg do nutriente/µm3/h e o coeficiente de saturação
média de cada espécie, estes dois últimos calculados a partir da equação de
0,000,050,100,150,200,250,300,350,400,450,500,550,60
tx c
resc
0 2 4 6 8 10S
0,000,050,100,150,200,250,300,350,400,450,500,550,60
tx c
resc
0 25 50 75 100 125 150 175S
0,00
0,04
0,08
0,12
0,16
0,20
0,24
0,28
0,32
0,36
tx c
resc
0 2 4 6 8 10S
0,00
0,04
0,08
0,12
0,16
0,20
0,24
0,28
0,32
0,36
tx c
resc
0 25 50 75 100 125 150 175S
0,000
0,075
0,1500,200
0,2750,325
0,4000,450
0,5250,575
Tx c
resc
0 2 4 6 8 10S
0,00
0,10
0,20
0,30
0,40
0,50
0,60
0,70
tx c
resc
0 25 50 75 100 125 150 175s
Oocystis
Microcystis
Cryptomonas
P (µmol/L) N (µmol/L)
0,000,050,100,150,200,250,300,350,400,450,500,550,60
tx c
resc
0 2 4 6 8 10S
0,000,050,100,150,200,250,300,350,400,450,500,550,60
tx c
resc
0 25 50 75 100 125 150 175S
0,00
0,04
0,08
0,12
0,16
0,20
0,24
0,28
0,32
0,36
tx c
resc
0 2 4 6 8 10S
0,00
0,04
0,08
0,12
0,16
0,20
0,24
0,28
0,32
0,36
tx c
resc
0 25 50 75 100 125 150 175S
0,000
0,075
0,1500,200
0,2750,325
0,4000,450
0,5250,575
Tx c
resc
0 2 4 6 8 10S
0,00
0,10
0,20
0,30
0,40
0,50
0,60
0,70
tx c
resc
0 25 50 75 100 125 150 175s
Oocystis
Microcystis
Cryptomonas
P (µmol/L) N (µmol/L)

41
41
Monod. O coeficiente de saturação média é considerado como uma constante
espécie-específica e tem sido reconhecido como uma medida da habilidade
relativa de cada espécie em usar baixas concentrações dos nutrientes. Quando as
concentrações ambientais são baixas, espécies com baixos valores de Km têm
uma vantagem, teórica, sobre espécies com altos Km (Reynolds, 1999).
Tabela 6: Parâmetros de absorção de fósforo (K2HPO4) e nitrogênio (NaNO3) das três espécies. Densidade média de cada espécie nos cultivos em biovolume, taxa máxima de absorção, constante média de saturação da absorção (Km) e coeficiente de consumo em excesso (R).
Nos experimentos de absorção de fósforo, M. viridis apresentou o menor
valor de Km, 44,88 µg/l e a maior taxa de absorção máxima, 1,57.10- 6 µg/µm3/h.
Estes valores foram calculados para uma densidade média de 2,17.108 µm3/ml ao
longo do experimento. Cryptomonas erosa apresentou valores muito próximos a
M. viridis (45,94 µg/l de Km e 1,56. 10- 6µg/µm3/h de absorção máxima) para
uma densidade média de 2,83.108 µm3/ml. Oocystis lacustris foi a espécie cujos
parâmetros de absorção foram menos favoráveis para a absorção de fósforo. Esta
espécie teve 54,95 µg/l de Km e 6,977x10- 7 µg/µm3/h para uma densidade
média de 5,38.108 µm3/ml.
Nos experimentos de absorção de nitrogênio, O. lacustris apresentou o
menor valor de Km, 73,60 µg/l. A sua taxa máxima de absorção foi 5,76.10-6
µg/µm3/dia para uma densidade média de 3,15.108 µm3/ml. Cryptomonas erosa
apresentou a maior taxa de absorção com 2,15.10- 5 µg/µm3/dia, mas em
compensação também possuiu o maior valor de Km, 178,07 µg/l para uma
Fósforo
Densidade Média (um3/ml) Tx máx de absorção (ug/um 3/h) Km (ug/l) R
O . Lacustris 5,38.108 6,98.10-7 54,95 9M. viridis 2,17.108 1,57.10-6 44,88 26C. erosa 2,83.108 1,56.10-6 45,94 33
NitrogênioDensidade Média (um3/ml) Tx máx de absorção (ug/um 3/h) Km (ug/l) R
O . Lacustris 3,15.108 5,76.10-6 73,6 122M. viridis 7,08.107 3,9.10-6 79,31 3C. erosa 1,61.108 2,15.10-5 178,07 18
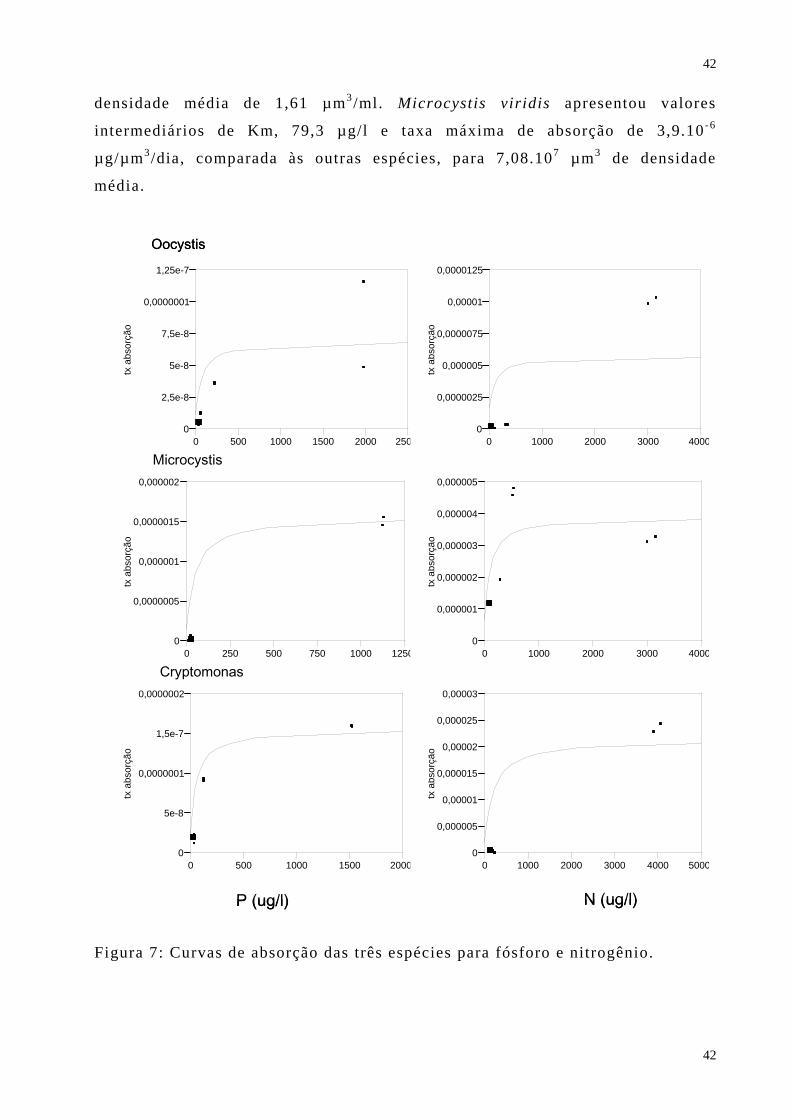
42
42
densidade média de 1,61 µm3/ml. Microcystis viridis apresentou valores
intermediários de Km, 79,3 µg/l e taxa máxima de absorção de 3,9.10- 6
µg/µm3/dia, comparada às outras espécies, para 7,08.107 µm3 de densidade
média.
Figura 7: Curvas de absorção das três espécies para fósforo e nitrogênio.
0
2,5e-8
5e-8
7,5e-8
0,0000001
1,25e-7
tx a
bsor
ção
0 500 1000 1500 2000 2500P
0
0,0000025
0,000005
0,0000075
0,00001
0,0000125
tx a
bsor
ção
0 1000 2000 3000 4000N
0
0,0000005
0,000001
0,0000015
0,000002
tx a
bsor
ção
0 250 500 750 1000 1250P
0
0,000001
0,000002
0,000003
0,000004
0,000005
tx a
bsor
ção
0 1000 2000 3000 4000N
0
5e-8
0,0000001
1,5e-7
0,0000002
tx a
bsor
ção
0 500 1000 1500 2000P
0
0,000005
0,00001
0,000015
0,00002
0,000025
0,00003
tx a
bsor
ção
0 1000 2000 3000 4000 5000N
Oocystis
Microcystis
Cryptomonas
P (ug/l) N (ug/l)
0
2,5e-8
5e-8
7,5e-8
0,0000001
1,25e-7
tx a
bsor
ção
0 500 1000 1500 2000 2500P
0
0,0000025
0,000005
0,0000075
0,00001
0,0000125
tx a
bsor
ção
0 1000 2000 3000 4000N
0
0,0000005
0,000001
0,0000015
0,000002
tx a
bsor
ção
0 250 500 750 1000 1250P
0
0,000001
0,000002
0,000003
0,000004
0,000005
tx a
bsor
ção
0 1000 2000 3000 4000N
0
5e-8
0,0000001
1,5e-7
0,0000002
tx a
bsor
ção
0 500 1000 1500 2000P
0
0,000005
0,00001
0,000015
0,00002
0,000025
0,00003
tx a
bsor
ção
0 1000 2000 3000 4000 5000N
Oocystis
Microcystis
Cryptomonas
P (ug/l) N (ug/l)

43
43
3.3- Competição entre as espécies
Foram realizados três experimentos com culturas mistas para se avaliar o
efeito da razão nutricional na interação competitiva das espécies. Para os
experimentos de competição foram utilizadas apenas as razões nutricionais
extremas dos experimentos de crescimento, N:P 4 e 100, além da razão N:P 20.
O primeiro experimento utilizou O. lacustris e M. viridis e teve a duração de 19
dias (à exceção das réplicas de razão N:P 100 que tiveram 17 dias de duração
devido a inutilização da última coleta). Oocystis lacustris apresentou as maiores
densidades em todas as razões N:P (Figura 8). A maior variação foi observada
na razão N:P 4 e a menor na razão N:P 100. Este resultado está de acordo com o
observado nos parâmetros de crescimento, pois, O. lacustris sempre apresentou
os menores valores de R* em relação a M. viridis , e esta última teve melhor
desempenho nos parâmetros competitivos quando relacionados ao fósforo, o que
explica o seu maior crescimento no meio N:P 100.
O segundo experimento reuniu O. lacustris e C. erosa (Figura 9). Como
comentado anteriormente, C. erosa foi a espécie que apresentou os melhores
índices competitivos. Os experimentos demostraram esta vantagem, na prática
em todas as razões utilizadas com a densidade de C. erosa sendo superior em
todos os experimentos (N:P 4, 20 e 100). O mesmo ocorreu nos experimentos de
competição entre C. erosa e M. viridis (Figura 10).

44
44
Figura 8: Crescimento de Oocystis lacustris e Microcystis viridis em culturas mistas nas razões N:P 4 (A), 20 (B) e 100 (C).
A)
0,00E+00
2,00E+08
4,00E+08
6,00E+08
8,00E+08
1,00E+09
1,20E+09
0 5 10 15 20Dias
Bio
volu
me/
ml
M . viridis
O. lacustris
B)
0,00E+00
2,00E+08
4,00E+08
6,00E+08
8,00E+08
1,00E+09
1,20E+09
1,40E+09
0 5 10 15 20Dias
Bio
volu
me/
ml
C)
0,00E+001,00E+082,00E+083,00E+084,00E+085,00E+086,00E+087,00E+088,00E+08
0 5 10 15 20Dias
Bio
volu
me/
ml

45
45
Figura 9: Crescimento de Oocystis lacustris e Cryptomonas erosa em culturas mistas nas razões N:P 4 (A), 20 (B) e 100 (C).
A)
0,00E+00
1,50E+08
3,00E+08
4,50E+08
6,00E+08
7,50E+08
9,00E+08
0 5 10 15 20Dias
Bio
volu
me/
ml
O. lacustris
C. erosa
B)
0,00E+00
5,00E+08
1,00E+09
1,50E+09
2,00E+09
0 5 10 15 20Dias
Bio
volu
me/
ml
C)
0,00E+00
3,00E+08
6,00E+08
9,00E+08
1,20E+09
1,50E+09
1,80E+09
0 5 10 15 20Dias
Bio
volu
me/
ml

46
46
Figura 10: Crescimento de Microcytis viridis e Cryptomonas erosa em culturas mistas nas razões N:P 4 (A), 20 (B) e 100 (C).
A)
0,00E+00
1,50E+08
3,00E+08
4,50E+08
6,00E+08
7,50E+08
0 5 10 15 20Dias
Bio
volu
me/
ml
M . viridis
C. erosa
B)
0,00E+00
5,00E+08
1,00E+09
1,50E+09
2,00E+09
2,50E+09
0 5 10 15 20
Dias
Bio
volu
me/
ml
C)
0,00E+00
4,00E+08
8,00E+08
1,20E+09
1,60E+09
2,00E+09
0 5 10 15 20
Dias
Bio
volu
me/
ml

47
47
3.4- Variação sazonal das espécies no ambiente natural: Reservatório
da Pampulha
A figura 11 apresenta as variações sazonais de O. lacustris , M. viridis e
C. erosa no período de três anos, janeiro de 1996 a dezembro de 1998, no
Reservatório da Pampulha. No início de 1996 a população de O. lacustris
apresentava a maior densidade nas águas do reservatório. Sua dominânica foi
substituida por M. viridis no início da estação chuvosa do mesmo ano. Esta
espécie apresentou floração em massa em novembro, decaindo em densidade
com o início das chuvas. A densidade de C. erosa começou a ser representativa
no início do outono de 1997 (início da estação seca) e a espécie manteve sua
dominância por praticamente todo o período dos dois anos seguintes, com breves
pico de O. lacustris dividindo esta dominância. Cryptomonas erosa apresentou
dois picos de crescimento máximo, o primeiro em julho de 1996 e o segundo em
junho de 1997, ou seja, sempre no inverno.
Figura 11: Variação da densidade de Cryptomonas erosa , Microcystis viridis e Oocystis lacustris em biovolume (µm3/ml) no reservatório da Pampulha de março de 1997 a abril de 1998.
Para uma análise global das modificações ocorridas na comunidade
fitoplanctônica do reservatório, foram consideradas as variações sazonais dos
gêneros Oocystis , Microcystis e Cryptomonas no mesmo período de três anos, de
janeiro de 1996 a outubro de 1998. Durante estes três anos foram identificadas
0,E+00
2,E+06
4,E+06
6,E+06
8,E+06
1,E+07
1,E+07
1,E+07
2,E+07
out-95 mai-96 dez-96 jun-97 jan-98 jul-98 fev-99
Data
Bio
volu
me
(um
3/l)
M . viridisO . lacustrisC. erosa

48
48
duas espécies pertencentes ao gênero Oocystis , O. lacustris e O. pusilla , três
espécies pertencentes ao gênero Microcystis , M. viridis , M. flos-aquae e M.
aeruginosa , e três de Cryptomonas , C. erosa , C. brasiliensis e C. curvata .
Em linhas gerais, é possível se observar um aumento gradativo da
densidade do gênero Cryptomonas ao longo dos três anos, com picos de
crescimento no inverno. Microcystis apresentou alguns picos de crescimento em
1996 e 1997, sempre no final do ano, início da estação chuvosa. Oocystis , em
comparação às demais, apresentou desidades muito baixas ao longo dos três
anos.
Figura 12: Variação da densidade de espécies dos gêneros Cryptomonas , Microcystis e Oocystis em biovolume (µm3/ml) no Reservatório da Pampulha de janerio de 1996 a novembro de 1998.
A figura 13 apresenta a variação de alguns parâmetros físico-químicos
medidos a um metro de profundidade no Reservatório da Pampulha de janeiro de
1996 a dezembro de 1998. Os valores de temperatura oscilaram sazonalmente ao
longo dos anos. As maiores temperaturas foram medidas no verão (janeiro e
fevereiro), e as menores no inverno (junho e julho). A temperatura máxima
alcançada no período foi 29,5oC e a menor 19,3oC. As concentrações de
nitrogênio também oscilaram sazonalmente no reservatório, sendo os maiores
valores observados no período seco (abril a setembro). A concentração média
deste nutriente aumentou gradualmente ao longo dos três anos no reservatório.
As concentrações de fósforo solúvel não apresentaram padrões de sazonalidade,
sendo observados picos tanto no período chuvoso quanto no seco. As
0,00E+00
1,00E+07
2,00E+07
3,00E+07
4,00E+07
5,00E+07
6,00E+07
out-95 mai-96 dez-96 jun-97 jan-98 jul-98 fev-99
Data
Bio
volu
me
(um
3/m
l)
M icrocystis
Oocystis
Cryptomonas

49
49
concentrações totais de fósforo medidas no reservatório foram maiores no
período seco, a exemplo do nitrogênio. As razões N:P solúvel e N:P total
apresentaram tendências contrárias ao longo dos três anos. Enquanto a razão N:P
solúvel apresentou valores crescentes ao longo dos três anos, a razão N:P total
nas águas do reservatório tendeu a diminuir. A concentração de clorofila-a
medida a um metro de profundidade no reservatório apresentou uma tendência
geral de aumento ao longo dos três anos.
A relação entre a variação das densidades específicas e genéricas além da
concentração de clorofila-a e a variação das características físico-químicas do
reservatório foi testada através de correlações. Os valores obtidos estão
representados na tabela 7.
As densidades de O. lacustris e do gênero Oocystis não se
correlacionaram significativamente com nenhuma das variáveis físico-químicas
do reservatório. A densidade de M. viridis se correlacionou positivamente com a
concentração de fósforo solúvel, apesar desta correlação não ser significativa
para o gênero Microcystis . Cryptomonas erosa teve a sua densidade
correlacionada positivamente com as concentrações de nitrogênio e fósforo
total. As densidades das espécies representantes do gênero Cryptomonas se
correlacionaram negativamente com a temperatura, positivamente com as
concentrações de nitrogênio, positivamente com as concentrações de fósforo
total e positivamente com as razões N:P solúvel. Clorofila se correlacionou
negativamente com fósforo solúvel, positivamente com fósforo total,
positivamente com as razões N:P solúvel e negativamente com as razões N:P
total.

50
50
Figura 13: Variação de temperatura, concentração de nitrogênio (µg/l), concentração de fósforo solúvel (µg/l), fósforo total (µg/l), N:P solúvel, N:P total e clorofila-a, medidos a um metro de profundidade no Reservatório da Pampulha, de janeiro de 1996 a dezembro de 1998.
Temperatura
05
101520253035
dez-95 ago-96 abr-97 jan-98 set-98
oC
Clorofila
0,00
50,00
100,00
150,00
200,00
dez-95 ago-96 abr-97 jan-98 set-98
ug/l
Nitrogênio
0,E+00
2,E+03
4,E+03
6,E+03
8,E+03
dez-95 ago-96 abr-97 jan-98 set-98
ug/l
Fósforo total
0100200300400500600
dez-95 ago-96 abr-97 jan-98 set-98
ug/l
N:P Total
0
20
40
60
80
dez-95 ago-96 abr-97 jan-98 set-98
Raz
ão
Fósforo Solúvel
0
20
40
60
80
out-95 jul-96 mar-97 nov-97 jul-98 mar-99
ug/L
N:P Solúvel
0
100
200300
400
500
600
dez-95 ago-96 abr-97 jan-98 set-98 mai-99
Fós
foro
Sol
úvel
(ug
/l)

51
51
Tabela 7: Valores das correlações entre as densidades das espécies, gêneros e concentração de clorofila do reservatório da Pampulha com algumas variáveis físico-químicas calculadas para o período de janeiro de 1996 a novembro de 1998. Os valores em vermelho são estatisticamente significativos (p<0,005).
Temperatura Nitrogênio Fósforo Solúvel Fósforo Total N:P Solúvel N:P TotalEspécies
O . Lacustris -0,1314 0,1013 0,1715 0,2034 0,0582 -0,0377M. viridis -0,1444 0,1503 0,4816 -0,0267 -0,1821 0,1284C. erosa -0,0993 0,3124 -0,1424 0,2754 0,2376 -0,1338
Gêneros Oocystis -0,12 -0,0921 0,1296 0,2264 0,1222 -0,0644
Microcystis -0,0481 0,1286 0,0928 0,2181 0,0774 -0,1816Cryptomonas -0,256 0,4377 -0,0369 0,2889 0,2606 -0,0785
Clorofila 0,083 0,1328 -0,3776 0,6469 0,4815 -0,4053

52
52
4-DISCUSSÃO
4.1- Crescimento e competição entre as espécies:
Todas as espécies obtiveram maior densidade ao final dos experimentos de
crescimento em meios com razões intermediárias, N:P 10 no caso de O. lacustris
e N:P 20 nos casos de C. erosa e M. viridis . Espera-se, em culturas do tipo
``batch´´, que os nutrientes se tornem gradativamente menos disponíveis para o
crescimento das algas e que a biomassa algal se estabilize e decaia após certo
tempo de cultivo (Kilham, 1978). A biomassa final, desta forma, é regulada pela
concentração dos nutrientes no meio.
Apenas as culturas de M. viridis não apresentaram as suas curvas de
crescimento estabilizadas ao final do experimento, o que pode indicar que para
esta espécie os nutrientes adicionados aos meios não se encontravam
deplecionados ao fim do cultivo. Kilham (1978) argumenta que culturas do tipo
``batch´´ podem ser consideradas como um ``atalho´´ realístico para se obter
informações sobre as habilidades fisiológicas relativas de espécies
fitoplanctônicas como um fator determinante em sua interação na natureza.
Neste caso, baixas densidades iniciais devem ser utilizadas e o experimento
deve ter curta duração para que as concentrações nutricionais não variem
significativamente ao longo do experimento. Estes dois requisitos foram
respeitados nos experimentos de crescimento realizados. Foram adicionadas, em
média, 4.624 células de C. erosa , 12.426 células de O. lacustris e 20.200 de M.
viridis no início dos experimentos. Os cálculos dos parâmetros competitivos se
basearam na fase exponencial de crescimento das espécies. Kilham (1978)
util izou em seus experimentos o período de 4 a 5 dias para o cálculo da taxa de
crescimento das espécies, pois segundo a autora, após este período a
concentração nutricional no meio é reduzida a menos de 25% do total
adicionado. No presente estudo, foram utilizados os dados de crescimento dos
primeiros 5 dias para o cálculo das taxas de crescimento de C. erosa e O.
lacustris e os dados de crescimento dos primeiros 9 dias para o cálculo das taxas
de crescimento de M. viridis .
Tilman et al. (1976) demonstraram que o tamanho da colônia de
Asterionella formosa pode variar, dependendo do nutriente que limita o

53
53
crescimento e do grau desta limitação. Reynolds et al. (1997) comentam que é
largamente reconhecido que o tamanho das células, o formato e a composição
bioquímica do fitoplâncton podem mudar em resposta às mudanças nas
condições ambientais. Lynn et al. (2000) ao estudarem uma espécie de
diatomácea, Stephanodiscus minutus , verificaram que a alga apresentou um
tamanho reduzido quando a disponibilidade de nitrogênio no meio foi l imitada.
Shifrin & Chislom (1981, apud Lynn et al. 2000) afirmam haver um decréscimo
no peso celular para espécies de algas verdes, quando limitadas por nitrogênio.
Para verificar a possibilidade da limitação nutricional interferir no
biovolume celular das espécies, foram feitas medidas de biovolume das três
espécies nas cinco razões N:P em três fases distintas do cultivo, como
especificado na metodologia. Dentre as três espécies C. erosa , apresentou os
maiores valores de biovolume, seguida por O. lacustris e M. viridis . Ao
contrário do esperado, nenhuma espécie apresentou diferenças significativas
(p<0,005) em seu biovolume como conseqüência do cultivo em diferentes razões
N:P.
Outro aspecto verificado neste estudo foi a variação do biovolume celular
em relação ao tempo de cultivo. Esta verificação é fundamental para a análise
dos resultados de crescimento já que todos os índices calculados basearam-se
nos dados em biovolume. Análises de variância realizadas demonstraram que
apenas os valores de biovolume de C. erosa variaram com o tempo de cultivo.
As células medidas após 20 dias de cultivo, ou seja, ao final do experimento,
foram significativamente maiores do que as medidas no início e na metade do
experimento. Esta diferença se explica, para C. erosa , pelo fato desta espécie ter
sido a primeira a estabilizar o seu crescimento em todas as razões N:P, o que
significa que foi a primeira a deplecionar as concentrações nutricionais
adicionadas nos meios a níveis nos quais não poderia mais manter seu
crescimento. Como conseqüência, ao final dos experimentos, esta espécie
apresentava suas células bastante debilitadas, com aparente relaxamento de sua
estrutura celular devido à perda de rigidez de sua parede celular, com
consequente aumento de biovolume. Oocystis lacustris e M. viridis não
apresentaram reflexos tão extremos da escassez de nutrientes A primeira
estabilizou o seu crescimento em período de tempo superior à C. erosa e a

54
54
Segunda não chegou a estabilizar o seu crescimento na maioria dos
experimentos.
Os padrões das curvas de crescimento das espécies em µm3/ml não
diferiram das curvas em cel/ml já que as espécies não apresentaram variações
significativas no biovolume de suas células em diferentes razões N:P. As
análises de co-variância também não demonstraram diferenças significativas
entre o crescimento das espécies em diferentes razões N:P, para os dados em
µm3/ml. Estas análises foram realizadas para o período, considerado neste
estudo, como de crescimento exponencial das espécies. A similaridade entre as
curvas de crescimento das três espécies nas diferentes razões N:P pode ser
explicada, em parte, pela baixa densidade celular inoculada no início do
experimento e pelo pequeno intervalo de tempo utilizado para se calcular as
taxas de crescimento.Como o crescimento exponencial das espécies ocorreu em
períodos curtos, este crescimento nas diferentes razões não parece ter sido
suficientemente diferenciado para ser detectado estatisticamente. Após certo
tempo de cultivo estas diferenças, provalvelmente, se tornariam mais
pronunciadas como conseqüência da diminuição da concentração nutricional
disponível para o crescimento das espécies.
Foi verificado o efeito que as limitações nutricionais exerceram sobre a
composição celular das espécies, através da mensuração de sua concentração de
clorofila-a (Tabela 4). Em todas as espécies, as concentrações de clorofila-a
foram menores nos experimentos de crescimento das espécies em meios com
limitação de nitrogênio ou de fósforo.
Oocystis lacustris e Cryptomonas erosa apresentaram acentuada variação
nas concentrações finais e celulares de clorofila-a, sendo inferiores em meios de
baixas razões N:P. Nestes meios, nos quais a l imitação por nitrogênio é
induzida, era de se esperar uma menor produção das moléculas de clorofila por
parte das células já que o nitrogênio é um dos principais constituintes destas
moléculas (Stryer, 1995). Prezelin (1981) comenta que a concentração de
clorofila pode ser considerada como uma medida indireta da concentração de
nitrogênio intra e extracelular, tanto pela importância que o elemento possui na
composição da clorofila como pela presença de complexos proteínas-clorofila.
Cryptomonas erosa foi a espécie que apresentou os maiores valores totais
e concentração por célula e biovolume de clorofila-a ao fim do experimento de

55
55
crescimento em todas as razões N:P. Por ter atingido maior densidade em todas
as razões e por possuir células significativamente maiores do que as de O.
lacustris e M. viridis , estes resultados eram esperados. Raven & Kubler (2002)
comentam que quanto maior for o volume do organismo fitoplanctônico, maior
terá de ser a sua concentração de clorofila-a por mm3, já que o coeficiente de
absorção específica do pigmento é diminuído. Lynn et al. (2000) afirmam que a
concentração de clorofila tem uma relação positiva com o tamanho da célula,
particularmente com a capacidade em se estocar nitrogênio. A espécie que
apresentou as maiores variações nas concentrações de clorofila-a ao final dos
experimentos foi O. lacustris. Esta variação pode ser explicada por ser esta
espécie uma clorofícea, e, portanto, ter a clorofila-a como principal pigmento
fotossintético. Esta molécula pode, desta forma, funcionar como potencial
indicadora de estresse nutricional para a espécie. As concentrações de clorofila-
a nesta espécie foram menores em meios com baixas concentrações de
nitrogênio e fósforo, sendo a maior concentração obtida em meios com razão
N:P 20. Cryptomonas erosa , que também possui clorofila-a como importante
pigmento fotossintético, apesar de possuir reservas de ficoeritrinas (Sciandra et
al , 2000) também apresentou acentuada variação em sua concentração nas razões
N:P baixas (N:P 4 e 10) em relação às maiores concentrações (N:P 20, 40 e
100). Sciandra et al . (1976), também observaram variação nas concentrações
intracelulares de clorofila-a em Cryptomonas sp. como conseqüência de estresse
causado pela limitação de nitrogênio no meio. Microcystis viridis foi a espécie
que apresentou a menor variação em seu conteúdo intracelular de clorofila-a.
Cianobactérias possuem diversas categorias de pigmentos fotossintetizantes,
dentre os quais se destacam ficocianinas, alo-ficocianinas e ficoeritrinas, que
respondem por grande parte de seu potencial fotossintetizante (Allen et al ,
1990). As variações mínimas no seu conteúdo intracelular de clorofila-a em
diferentes razões N:P pode indicar que nesta espécie a variação na
disponibilidade nutricional pode estar refletida na concentração de outros
pigmentos.
Os resultados obtidos neste estudo estão em consonância com os
observados na literatura (Dillon e Rigler, 1974; Tilman, 1982; Lynn et al. ,
2000), que indicam que as concentrações de clorofila-a estão positivamente
relacionadas às de fósforo e nitrogênio em diversas espécies algais.

56
56
Para o cálculo dos parâmetros que indicam habilidade competitiva das
espécies foram utilizados os dados em biovolume. Grover (1989) coloca que a
existência de uma correlação entre µma x e o volume celular indica a
possibilidade que o tamanho celular seja um preditor útil do resultado da
interação competitiva entre espécies fitoplanctônicas. Neste trabalho foram
utilizadas espécies que diferiam nos valores de biovolume celular, como
comentado acima. A utilização de uma unidade de crescimento como cel/ml
acarretaria em um erro na análise dos resultados, já que não descreveria
corretamente as densidades relativas de cada espécie no meio de cultura.
Os parâmetros de crescimento calculados para as três espécies indicaram
que C. erosa deslocaria competitivamente M. viridis e O.lacustris quando em
meios limitados tanto por fósforo quanto por nitrogênio. Cryptomonas erosa
apresentou os menores valores de R*, que indica a concentração mínima
necessária para manter uma população estável, para os dois nutrientes. Através
destes mesmos experimentos foi previsto que O. lacustris deslocaria
competitivamente M. viridis em culturas mistas, também com limitação de
ambos os nutrientes. Destes resultados presume-se que não é possível, tendo
como base apenas o requerimento por nitrogênio e fósforo, construir um meio no
qual as três espécies possam coexistir. Para que isto ocorresse, as espécies
teriam que diferir em suas habilidades competitivas por fósforo e nitrogênio de
forma a possibilitar que enquanto uma fosse limitada por nitrogênio a outra o
seria por fósforo, ou seja, elas não estariam em competição direta entre si
(Reynolds, 1999). Os experimentos de competição, realizados com culturas
mistas confirmaram esta previsão obtida nos experimentos de crescimento
individual das espécies. Estes resultados demonstram que apesar dos problemas
relacionados à utilização de culturas descontínuas, o modelo de Monod
aparentemente descreve adequadamente os resultados obtidos por experimentos
de curta duração em culturas do tipo ``batch´´ como descrito por Kilham (1978).
A tabela 8 apresenta os valores de R* e µma x/Ks obtidos em alguns
trabalhos realizados com clorofíceas, cianobactérias e criptofíceas. Existe uma
grande variação nos valores obtidos. Esta variação pode ser explicada pela
escolha de diferentes condições de cultivo, como temperatura e luminosidade,
diferente número de diluições nutricionais para o cálculo dos parâmetros e

57
57
utilização de diferentes espécies destes grupos, e mesmo entre espécies iguais
mas de cepas diferentes.
Tabela 8: Parâmetros competitivos de algumas espécies calculados através do modelo de Monod, encontrados na literatura.
* Trabalhos que usaram a concentração nutricional no meio em µmol/l
** Trabalhos que usaram a concentração nutricional no meio em µg/l
Por outro lado a determinação dos parâmetros competitivos calculados
tendo como base o crescimento diferencial das espécies não é suficiente para a
definição dos melhores competidores em diferentes concentrações nutricionais
(Ahlgreen, 1977). É preciso que se conheçam as habilidades que cada espécie
possui em absorver os nutrientes no meio e também a sua estratégia de
armazenamento. Em última instância, são as capacidades de cada espécie em
absorver e responder às concentrações nutricionais no meio que determinarão o
seu desempenho competitivo em ambientes naturais. Muitos trabalhos, desta
forma, focaram as habilidades de absorção nutricional das espécies, pois,
segundo Ahlgreen (1977), a taxa de crescimento pode se dever, muitas vezes, à
Autor Ano de Publicação Nutriente Temperatura (oC) Grupo Espécie R* µmax/Ks
Grover 1989 Fósforo 12 Chlorophyceae Chlamydomonas sp 0,0033* 111*Chlorella sp. 0,01* 36*
Oocystis pusilla 0,011* 44*Scenedesmus quadricauda 0,023* 18*
Cryptophyceae Cryptomonas sp 0,014* 34*
Grover; Sterner & Robinson 1999 Nitrogênio Variável Chlorophyceae Chlorella vulgaris 0,315** 0,13**
Cryptophyceae Cryptomonas sp. 57,29** 0,72**
Cyanobacteria Merismopedia tenuissima 1,49** 0,24**
Fósforo Chlorophyceae Kirchneriella sp. 1,23** 0,27**
Cyanobacteria Merismopedia tenuissima 2,32** 0,2075**
Giani & Delgado 1998 Nitrogênio 20 Chlorophyceae Oocystis lacustris 3,606* 0,146*
Cyanobacteria Synechocystis sp. 2,283* 0,196*
Fósforo Chlorophyceae Oocystis lacustris 0,081* 6,78*
Cyanobacteria Synechocystis sp. 0,188* 2,249*
Este trabalho 2003 Nitrogênio 20 Chlorophyceae Oocystis lacustris 7,016* 0,67*
Cyanobacteria Microcystis viridis 21,291* 0,66*
Cryptophyceae Cryptomonas erosa 4,056* 0,112*
Fósforo Chlorophyceae Oocystis lacustris 0,222* 2,088*
Cyanobacteria Microcystis viridis 0,564* 2,381*
Cryptophyceae Cryptomonas erosa 0,067* 6,113*

58
58
concentração nutricional intracelular e não à concentração de nutrientes no
meios de cultivo.
4.2- Absorção de fósforo e nitrogênio:
Os experimentos de absorção foram realizados para esclarecer o processo
de obtenção de fósforo e nitrogênio pelas três espécies. Reynolds (1999)
comenta que para a construção de biomassa todas as espécies algais necessitam
de aproximadamente os mesmos elementos essenciais na mesma quantidade
relativa, o que realmente varia entre as espécies é a capacidade em absorver
nutrientes e as concentrações externas capazes de saturar a absorção. Segundo
Sommer (1984) existem, teoricamente, três casos de estratégias competitivas
para a absorção nutricional. Espécies adaptadas à velocidade, que possuem uma
alta taxa de absorção e simultânea alta taxa de crescimento. Espécies adaptadas
ao armazenamento que utilizam o pulso de nutrientes adicionados ao meio para
um consumo em excesso (luxury consumption) e espécies com alta afinidade
pelo nutriente, ou seja, com baixos valores de Km. As espécies que se
especializam à velocidade de consumo e divisão celular, são consideradas como
tendo uma estratégia de vida r, ou seja, aproveitam eventos imprevisíveis e
inconstantes para aumentar sua estratégia no ambiente, enquanto as espécies
adaptadas ao armazenamento e à afinidade adotam uma estratégia de vida K,
com a manutenção de populações estáveis por longos períodos de tempo.
Após a adição do nutriente nas culturas são formados reservatórios
provisórios não assimilados cujos tamanhos são determinados pelas condições
anteriores das células (Frenette et al. , 1998; Lomas & Glibert, 2000). Os
resultados dos experimentos de absorção demonstraram que o período de
aclimatação das células em meios sem os nutrientes foram suficientes para que
se alcançasse o estado de ``starvation´´ nas três espécies já que a absorção
nutricional foi sempre positiva em relação ao tempo de experimento para as três
espécies em praticamente todas as diluições.
Os experimentos de absorção de fósforo demonstraram que M. viridis
apresentou as maiores taxas máximas de absorção, ou seja, a maior capacidade
de absorção de fósforo por unidade de tempo comparada às demais. Esta espécie
também foi a que apresentou os menores valores da constante de semi-saturação,

59
59
o que demonstra que para atingir a sua capacidade máxima de absorção necessita
de uma quantidade de fósforo inferior ao necessário para O.lacustris e C. erosa .
Cryptomonas erosa , no entanto, apresentou valores muito próximos aos de M.
viridis , sendo que as duas espécies podem ser consideradas como tendo
habilidades similares de absorção de fósforo. Oocystis lacustris foi a que
apresentou, experimentalmente, o desempenho menos favorável na absorção de
fósforo. Quanto ao nitrogênio, O.lacustris foi a que apresentou o menor valor de
saturação média apesar de C. erosa ter a maior taxa de absorção. A constante de
semi-saturação desta última espécie, no entanto, foi a maior entre as três
espécies, o que indica que necessita de grandes quantidades do nutriente no
meio para alcançar a sua taxa máxima de absorção. Nos experimentos com o
nitrogênio, M. viridis apresentou valores intermediários de taxa de absorção e
constante média de saturação.
Como comentado anteriormente, indivíduos são capazes de adquirir
nutrientes para uso futuro, o que é conhecido como ``consumo em excesso´´
(Tilman, 1982). Um dos parâmetros que pode ser utilizado para se verificar a
capacidade absortiva de determinada espécie é o coeficiente de consumo em
excesso (R) de um determinado nutriente por esta espécie (Tilman, 1981). Este
coeficiente nada mais é do que a razão entre a taxa de absorção de determinado
nutriente pela espécie quando este não é limitante ao seu crescimento e a taxa de
absorção quando o nutriente está em concentração limitante no meio. Ele mede,
desta forma, a capacidade da espécie em consumir e armazenar quantidades de
reserva do nutriente quando as condições são favoráveis, o que aumenta a sua
capacidade de crescimento quando o nutriente se encontra em concentrações
limitantes. Para fósforo, a espécie que possuiu o maior R, maior capacidade de
armazenamento, foi C. erosa . Já para o nitrogênio foi O. lacustris que
apresentou o maior valor de R.
Através do exposto acima, é possível que se trace o perfil de absorção de
fósforo e nitrogênio das três espécies util izadas nos experimentos. O. lacustris ,
como demonstrado pelo experimento de absorção de fósforo, possui baixa
capacidade de absorção deste nutriente. Sua taxa máxima de absorção foi
inferior às das outras duas espécies, a sua constante média de saturação foi a
mais alta, indicando que as concentrações de fósforo necessárias para que a
espécie atinja sua taxa máxima de absorção é alta. A sua afinidade pelo

60
60
substrato foi também a menor encontrada para as três espécies. Cryptomonas
erosa e Microcystis viridis apresentaram comportamento similar em relação à
absorção de fósforo sendo que C. erosa apresentou uma vantagem competitiva
por apresentar maior afinidade pelo substrado. Isvanovics et al (2000)
comentam que cianobactérias são reconhecidamente armazenadoras de fósforo.
Neste estudo verificou-se que C. erosa parece seguir o mesmo comportamento.
Quanto ao nitrogênio, O. lacustris foi a espécie que apresentou o menor
valor de Km e a maior afinidade, demonstrando a sua capacidade de utilizar
baixas concentrações deste nutriente e estocá-lo para uso futuro. Cryptomonas
erosa foi a espécie com maior taxa de absorção demonstrando a sua
superioridade competitiva quando existe uma grande oferta de nitrogênio no
meio. Esta espécie também foi a que apresentou as maiores concentrações de
clorofila-a por biovolume como comentado anteriormente. Esta medida é um
índice da capacidade celular em estocar nitrogênio intracelular (Dortch &
Postel, 1989; Smith et al. 1997; Reynolds, 1999), indicando que a espécie além
de possuir altas taxas de absorção também possui alta capacidade de estocagem
de nitrogênio. Microcystis viridis apresentou uma afinidade muito baixa por
este nutriente.
Para se prever o resultado das interações entre espécies, não só estes
parâmetros de crescimento e absorção devem ser conhecidos. No ambiente
natural as taxas de perda das espécies representadas por mortalidade, predação e
sedimentação exercem grande pressão sobre as populações (Tilman, 1982). A
utilização de taxas baseadas apenas na mortalidade, como a utilizada neste
estudo, é uma simplificação deste processo complexo. Além disso, outros
fatores como temperatura e luminosidade também influenciam dramaticamente o
resultado das interações competitivas e podem ser determinantes no seu
resultado final. Tilman e Kiesling (1984) descobriram que além das baixas
razões N:P, cianobactérias, em geral, necessitam de altas temperaturas para se
tornarem dominantes. Apesar da grande contribuição que estudos experimentais
vêm adicionando ao conhecimento das habilidades fisiológicas das espécies
fitoplanctônicas, é sabido que a sua limitação é enorme se comparados às
diversas variáveis interagentes em ambientes naturais que respondem pela
diversidade de suas comunidades. Como será visto a seguir, uma das formas de

61
61
amenizar esta limitação é a utilização de enfoques experimentais em conjunto
com dados obtidos em corpos d´água naturais.
4.3- Variação sazonal das espécies no ambiente natural: Reservatório
da Pampulha.
Em reservatórios tropicais, oscilações na radiação total e temperatura da
água não são tão altas como as encontradas em reservatórios localizados em
regiões temperadas. No entanto, reservatórios tropicais também sofrem
mudanças sazonais climatológicas, principalmente as relacionadas à
precipitação, que abrangem modificações nas características físicas e químicas
da água (Pinto-Coelho & Giani, 1985; Giani, 1994; Hooker & Hernandez, 1991).
Zevenboom & Mur (1980) comentam que apesar da dificuldade em se extrapolar
os resultados obtidos em laboratório para ambientes naturais, as previsões
qualitativas possibili tam que se prevejam os mecanismos responsáveis pelas
mudanças na comunidade fitoplanctônica. No Reservatório da Pampulha, as
concentrações de nitrogênio e fósforo se encontram em valores não limitantes
para o crescimento das espécies na maior parte do ano. Os dois nutrientes
apresentam uma baixa significativa em sua concentração no início da estação
chuvosa, provavelmente, como conseqüência do aumento do volume de água
existente no reservatório causado pela própria chuva. Tilman (1982) comenta
que a peça central necessária para que seja possível se prever o resultado à
longo prazo da competição entre espécies em ambientes cujas características
fisico-químicas da coluna d´água são instáveis, é o efeito da flutuação dos
recursos nas taxas reprodutivas das espécies a longo prazo. No Reservatório da
Pampulha pode ser observada uma tendência de aumento do fósforo e do
nitrogênio, como demonstrado pelos gráficos da figura 12.
A densidade de C. erosa e do gênero Cryptomonas, constituído no
Reservatório da Pampulha por três espécies, se correlacionam positivamente
(p<0,05) com o aumento da concentração molar de nitrogênio e fósforo total nas
águas do reservatório. Os experimento de crescimento demonstraram que esta
espécie seria capaz de vencer a competição com O. lacustris e M. viridis mesmo
em condições de limitação por fósforo e nitrogênio. No entanto, como
comentado por Kilham (1978) o crescimento das espécies em meios com

62
62
concentrações limitantes de fósforo e nitrogênio pode ser influenciada pelas
reservas destes nutrientes acumuladas nas células, e não, apenas, à concentração
nutricional nos meios de cultivo. Outro aspecto é a quantidade de diluições
utilizadas para se determinar os parâmetros competitivos no modelo de Monod.
Quanto maior o número de diluições maior o poder preditivo do modelo. Para
que seja diminuída a possibilidade de erro ao se inferir na dinâmica de
determinado corpo d´água tendo como base os resultados de experimetos de
crescimento, são realizados experimentos para a identificação dos parâmetros de
absorção nutricional das espécies fitoplanctônicas em diversas concentrações de
nutrientes (Ahlgreen, 1977).
Os experimentos de absorção de nitrogênio e fósforo demonstraram que C.
erosa possui a maior afinidade por fósforo, além de alta taxa máxima de
absorção e baixo valor de constante de semi-saturação para este nutriente, e a
maior capacidade de absorção de nitrogênio. Apesar de possuir a maior taxa
máxima de absorção de nitrogênio, C. erosa possui baixa afinidade e alto valor
de KM para este nutriente. Seria esperado, de acordo com estes resultados, que
C. erosa se tornasse dominante em determinado reservatório, somente se este
reservatório possuísse concentrações não limitantes de nitrogênio. Pode se
observar que as correlações entre a densidade de C. erosa e das demais espécies
do gênero Cryptomonas e as concentrações molares de nitrogênio calculadas
para o Reservatório da Pampulha, foram superiores à correlação com a
concentrações de fósforo total. Huszar & Caraco (1998) não obsevaram nenhuma
correlação entre as concentrações de fósforo em 6 reservatórios temperados e a
densidade de cryptofíceas. Este resultado reforça a suposição de que o aumento
da concentração de nitrogênio é o fator determinante para o aumento da
densidade e dominância de C. erosa e do gênero Cryptomonas no Reservatório
da Pampulha.
Huszar & Caraco (1998) comentam que a concentração de fósforo total
pode ser considerado um bom preditor da biomassa fitoplanctônica em
determinado reservatório e tem sido usado com relativo sucesso na predição de
sua composição taxonômica. A concentração de fósforo total estaria ligada
diretamente à abundância relativa de cianobactérias no reservatório. Neste
estudo não foi observada nenhuma correlação entre a concentração de fósforo
total e as densidades de M. viridis e das espécies constituintes do gênero

63
63
Microcystis . Este fato pode indicar que outras variáveis ambientais estão
influindo na composição taxonômica do reservatório.
A utilização da concentração total de fósforo parece ser mais apropriada
para se descrever a influência deste nutriente na dinâmica da comunidade
fitoplanctônica do Reservatório da Pampulha. A concentração de fósforo solúvel
apresentou picos durante o ano, possuindo correlação negativa com a densidade
fitoplanctônica, medida pela concentração de clorofila-a. Quanto maior a
densidade algal maior seria a absorção deste nutriente essencial para o
crescimento. Deberdt (2002) comenta que a utilização das formas dissolvidas de
nitrogênio e fósforo parece ser a mais usada para experimentos de
enriquecimento, mas que a sua validade para a caracterização de lagos é
questionável. Isto porque as algas possuem mecanismos para seqüestrar e
armazenar fósforo disponível no interior de suas células, o que dificulta julgar a
disponibilidade a partir de concentrações de fósforo solúvel na água. Reynolds
(1977) acha mais apropriado usar a concentração de fósforo total para descrever
a fertilidade de um rio ou lago, contemplando assim o fósforo intracelular
disponível para a célula algal.
As razões N:P, tanto N:P total quanto solúvel, não se correlacionaram
significativamente (p<0,05) com nenhuma das espécies estudadas. A razão N:P
total parece ser a que melhor reflete as mudanças ocorridas no reservatório. A
tendência desta razão é a sua diminuição ao longo dos anos, variando de valores
máximos de 65 a mínimos de 2 sem, no entanto, que isto signifique escassez de
nitrogênio, conceito muitas vezes mal aplicado em estudos limnológicos
(Reynolds, 1999). O aumento da entrada de fósforo, relativamente superior ao
aumento do influxo de nitrogênio, nas águas do reservatório respondem por esta
tendência de diminuição da razão N:P. Giani e Figueiredo (1999) também
observaram esta tendência de aumento da concentração de fósforo total nas
águas do reservatório. A média de concentração durante o ano de 1993 foi de
52,6 µg/l na zona eufótica aumentando para 100,3 µg/l em 1996. Neste estudo,
no entanto, não foi verificado tendência a aumento nas concentrações de nitrato,
nitrito e amônia no decorrer dos anos.
A biomassa da comunidade fitoplanctônica do reservatório se
correlacionou negativamente com a razão N:P total. Esta correlação, no entanto,
parece ser reflexo da alta correlação positiva entre os valores de clorofila-a e

64
64
fósforo total. Pode ser observada uma tendência ao aumento da biomassa algal
no período de estudo e este aumento parce estar ligado diretamente ao aumento
do influxo de fósforo no reservatório (Figura 13), efeito observado em diversos
estudos realizados em ambientes aquáticos ( Schindler, 1977; Reynolds, 1984;
Ramos et al. 1986, Henry 1986, Ibañez 1988; Matveev, 1997).
Diversos fatores, no entanto, poderiam estar influenciando na composição
e biomassa da comunidade fitoplanctônica do reservatório da Pampulha, além
das concentrações de fósforo e nitrogênio. Temperatura, padrões de
estratificação e intensidade de luz são apenas alguns dos fatores que devem
estar contribuindo no metabolismo das espécies e nas suas habilidades de
crescimento e absorção de nutrientes. Ojala et al (1996) comentam que em
ambientes aquáticos o crescimento do fitoplâncton é primariamente determinado
pela disponibilidade luminosa e de nutrientes, cuja utilização e estoques são
modificados pelas variáveis físicas e químicas da água. Figueredo & Giani
(2001) concluíram que no Reservatório da Pampulha, mudanças na diversidade
específica parecem ser controladas basicamente por fatores físico-
climatológicos, já que a concentração nutricional é sempre abundante. Assim, a
instabilidade da lâmina d´água durante o período chuvoso induz a mudanças
rápidas na dominância das espécies. Outro fator que pode determinar o resultado
das interações competitivas entre algas é a predação. Heleno (2002) demonstrou
através de experimentos em laboratório, que a comunidade fitoplanctônica do
Reservatório da Pampulha não é controlada pelo zooplâncton, cuja predação não
afeta sua composição, densidade e diversidade. Um forte indício desta falta de
controle é justamente a alta densidade de cryptofíceas que o reservatório possui,
já que é sabido que este grupo é preferencialmente predado pelo zooplancton por
ser altamente palatável (Giani, 1991).
Pinto-Coelho (1992) comparou estudos realizados no reservatório da
Pampulha nos anos de 84/85 e 90/91. Este autor confirmou o aumento das
concentrações de fósforo e nitrogênio (medida através da concentração de
amônia) nas águas do reservatório entre os dois períodos. A concentração de
fósforo total que nunca ultrapassou 50 µg/l em 1984/85 e 250 µg/l em 1990/91
chegou a ultrapassar 500 µg/l em 1998, como apresentado neste estudo. O autor,
tanto em 1984/85 como em 1990/91 também observou os mesmos padrões
sazonais observados de 1996 a 1998, com as maiores concentrações dos

65
65
nutrientes sendo alcançadas no fim da estação seca (setembro e outubro) nas
águas do reservatório. Quanto às razões nutricionais, foram medidas em 1985/84
razões de 41 a 53 e segundo o autor o fósforo estaria se tornando
progressivamente mais escasso no ecossistema uma vez que sua demanda cresce
dentro do lago. O fósforo, desta forma, seria o elemento chave no controle da
eutrofização do reservatório. No presente estudo foram observados padrões
inversos ao se considerar a razão N:P total, indicando que a tendência da
concentração de fósforo nas águas do reservatório parece seguir um padrão de
aumento, superior ao aumento das concentrações das formas nitrogenadas
(nitrito, nitrato e amônia) em períodos mais recentes.
Os resultados apresentados neste estudo sugerem fortemente que a
composição, densidade e diversidade da comunidade fitoplanctônica do
reservatório da Pampulha, um reservatório eutrófico urbano, parecem ser
determinadas, à longo prazo, pelas concentrações absolutas dos nutrientes em
suas águas e pelas características de crescimento e absorção destes nutrientes
pelas espécies algais que a compõe. Esta afirmação se baseia no estudo das
dinâmicas de crescimento e absorção nutricional de três espécies representativas
da comunidade fitoplanctônica do reservatório e na extrapolação dos dados
obtidos em experimentos laboratoriais aos observatos na variação sazonal desta
comunidade em um período de três anos. Mudanças nas características físico-
climatológicas das águas do reservatório, como comentado por Figueredo e
Giani (2001), parecem responder pelas mudanças instáveis e rapidamente
reversíveis na sucessão de suas espécies fitoplanctônicas.

66
66
5- CONCLUSÕES
- O conhecimento da capacidade de absorção nutricional das espécies
estudadas é de fundamental importância ao se tentar entender a dinâmica
competitiva entre as espécies em ambientes naturais. O conhecimento das
estratégias de absorção das espécies juntamente com os resultados obtidos
através de experimentos de crescimento em diferentes concentrações
nutricionais torna o modelo ainda mais eficiente.
- Os experimentos de crescimento das espécies demonstraram que
Cryptomonas erosa , isolada do reservatório da Pampulha é competitivamente
superior a Oocystis lacustris e Microcystis viridis na utilização de baixas
concentrações de fósforo e nitrogênio.
- Cryptomonas erosa possui alta afinidade por fósforo e em condições de
não limitação de nitrogênio apresenta altas taxas de absorção e armazenamento
deste nutriente. Microcystis viridis possui alta taxa de absorção de fósforo e
baixa constante de semi-saturação para este nutriente. Oocystis lacustris possui
alta afinidade por nitrogênio.
- O aumento da densidade de C. erosa e das outras espécies do gênero
Cryptomonas no Reservatório da Pampulha parece ser conseqüência direta do
aumento da concentração de fósforo e principalmente, de nitrogênio, em suas
águas.
- A composição, densidade e diversidade da comunidade fitoplanctônica
encontrada no Reservatório da Pampulha é provavelmente determinada
primariamente pelas concentrações absolutas de nitrogênio e fósforo em suas
águas e não pelas razões relativas entre estes nutrientes.

67
67
6-REFERÊNCIAS BIBLIOGRÁFICAS
Abrams P. A . 1988. Resouce productivity-consumer species diversity: simple models of
competition in spatially heterogeneous environments. Ecology. 69(5): 1418-1433.
Ahlgren G. 1977. Growth of Oscillatoria agardhii in chemostat culture. 1. Nitrogen and phosphorus
requirements. Oikos 29: 209-224.
Allen M. M.; Law A. & Evans H. 1990. Control of photosynthesis during nitrogen depletion and
recovery in a non-nitrogen-fixing cyanobacterium. Arch. Microbiol. 153: 428-431.
Arcifa M. S.; Starling F. L. R. M.; Sipaúba-Tavares L. H. & Lazzaro X. 1995. Experimental
Limnology in Tundisi J. G.; Bicudo C. E. M. & Tundisi T. M. 1995. Limnology in Brazil.
Brazilian Limnological Society, 1a Edição, Rio de Janeiro, Brazil.
Begon M.; Harper, J. L. & Townsend, C. R. 1995. Ecology: Individuals, Populations and
Communities. 2a edição, Blackwell Science. Victoria, Australia
Berman T., Sherr B. F., Sherr E., Wynne D. & McCarthy J. J. 1984. The characteristics of
ammonium and nitrate uptake by phytolankton in Lake Kinneret. Limnol. Oceanogr. 29(2):
287-297.
Bittencourt-Oliveira M. C., Oliveira M. C. & Yunes J. S. 2001. Cianobactérias tóxicas. Revista de
Biotecnologia 4: 44-47.
Boechat I. & Giani A. 2000. Factors affecting the biochemical composition of seston in a eutrophic
reservoir (Pampulha Reservoir, Belo Horizonte, MG). Rev. Bras. Biol. 60(1): 63-71.
Braga B.; Hespanhol I.; Conejo J. G. L.; Barros M. T. L.; Spencer M.; Porto M.; Nucci N.; Juliano
N. & Eiger S. 2002. Introdução à engenharia ambiental. 1a Edição, Prentice Hall, São Paulo,
Brasil.
Bulgarov N. G. & Levich A . P. 1999. The nitrogen:phosphorus ratio as a factor regulating
phytoplankton community structure. Arch. Hydrobiol. 146(1): 3-22.

68
68
Castro A . A . J. 1993. Cryptophyceae do Estado de São Paulo: Inventário Taxonômico. Tese de
mestrado apresentada ao Instituto de Biociências do Campus de Rio Claro, Universidade
Estadual Paulista, São Paulo.
Champs J. R. B. 1992. Algumas considerações sobre a situação sanitária da Bacia da Pampulha. In:
Anais do Seminário da Bacia Hidrográfica da Pampulha. SEGRAC, Belo Horizonte, 134-
142.
Cerrão G. C.; Carlos V. M.; Santos M. J. & Rigolin O. 1991. Efeito do enriquecimento artificial
sobre a biomassa de perifiton em tanques artificiais na Represa do Lobo (Broa). Rev. Bras.
Biol. 51: 71-78.
Connell, J. H. 1983. On the prevalence and relative importance of interspecific competition:
evidence from field experiments. Amer. Natur. 122 (5): 661-695.
Deberdt G. L. B. 2002. Estudo de cianobacterias em reservatório com elevado grau de trofia
(Reservatório de Salto Grande- Americana- SP). Tese de doutorado defendida no programa
de Engenharia Ambientas da Escola de Engenharia de São Carlos, Universidade de São
Paulo.
Desikachary T. V. 1959. Cyanophyta. 1a Edição, Indian Council of Agricultural Research, New
Delhi.
Dortch Q. & Postel J. R. 1989. Biochemical indicators of N utilization by phytoplankton during
upwelling of the Washington coast. Limnol. Oceanogr. 34 (4): 758-773.
Droop M. R. 1973. Some thoughts on nutrient limitation in algae. J. Phycol. 9: 264-272.
Dugdale R. C.; Jones Jr B. H.; MacIsaac J. J. & Goering J. J. 1981. Adaptation of nutrient
assimilation. In Platt T. Physiological bases of phytoplankton ecology. Otawa, Canada.
Esteves F. A. 1998. Fundamentos de limnologia. 2a edição, Interciência. Rio de Janeiro, Brasil.

69
69
Figueredo C. C. & Giani A. 2001. Seasonal variation in the diversity and species richness of
phytoplankton in a tropical eutrophic reservoir. Hydrobiol. 445: 165-174.
Fong P.l, Zedler J. B. & Donohoe M. 1993. Nitrogen vx. Phosphorus limitation of algal biomass in
shallow coastal lagoons. Limnol. Oceanogr. 38(5): 906-923.
Freitas A.; Santos M. B. L.; Rocha L. A. & Bedê L. C. 1994. Competitive interactionas among
mollusks in urban reservoirs, ponds and lakes. In Ecology and Human Impact on Lakes and
Reservoirs in Minas Gerais. Ed. SEGRAC- Belo Horizonte: 165-188.
Fujimoto N. & Sudo R. 1997. Nutrient-limited growth of Microcystis aeriuginosa and Phormidium
tenue and competition under various N:P supply ratios and temperatures. Limnol. Oceanogr.
42(2): 250-256.
Gause G. F. 1934. The struggle for existence. Reimpresso em 1964, Hafner. Nova York.
Giani A. 1991. Implications of phytoplankton chemical composition for zooplankton production:
experimental evidence. Oecologia 87: 409-16.
Giani A. 1994. Limnology in Pampulha Reservoir: some general observations with enphasis in the
phytoplanktonic community. In: Pinto-Coelho R. M.; Giani A. & Sperling E. Ecology and
human impact on lakes and reservoirs in Minas Gerais. Segrac, Belo Horizonte. p. 141-149.
Giani A. & Delgado P. C. S. 1998. Growth dynamics and competitive ability of a green (Oocystis
lacustris) and a blue green alga (Synechocystis sp.) under different N:P ratios. Verh.
Internat. Verein. Limnol. 26: 1693-1697.
Giani A. & Figueiredo C. C. 1999. Recorrência de padrões sazonais do fitoplâncton num
reservatório eutrófico (Reservatório da Pampulha, MG). In Henry R. Ecologia de
reservatório: estrutura, função e aspectos sociais. Ed FAPESP-Fundbio.
Giani A., Figueredo C. C. & Eterovick P. C. 1999. Algas planctônicas do reservatório da Pampulha
(MG): Euglenophyta, Chrysophyta, Pyrrophyta, Cyanobacteria. Rev. Bras. Biol.22:107-116.

70
70
Goodwin & Giani 1998. Blooms de cianobactérias no reservatório da Pampulha: o efeito das
macrófitas no seu aparecimento. Anais do VIII Seminário Regional de Ecologia. p. 1239-
1245.
Grover J. P. 1989. Phosphorus dependent growth kinetics of 11 species of freshwater algae. Limnol.
Oceanogr. 34(2): 341-348.
Grover J. P.; Sterner R. W. & Robinson J. L. 1999. Algal growth in warm temperate reservoirs:
Nutrient-dependent kinetics of individual taxa and seasonal patterns of dominance. Arch.
Fur Hidrob. 45(1): 1-23.
Guillard R. L. & Lorenzen, C. J. 1972. Yellow-green algae with chlorophyllide c. J. Phycol. 8: 10-14.
Healey F. P. & Hendzel L. L. 1975. Effect of phosphorus deficiency on two algae growing in
chemostats. J. Phycol. 11: 303-309.
Heleno G. von R. 2002. Interação fito-zooplâncton em um reservatório tropical, Lagoa da Pampulha
(MG, Brasil): um estudo experimental. Tese de mestrado defendida no programa de pós-
graduação em Ecologia, Conservação e Manejo da Vida Silvestre da UFMG, Belo Horizonte.
86p.
Henry R. & Tundisi J. G. 1982. Efeitos de enriquecimento artificial por nitrato e fosfato no
crescimento da comunidade fitoplanctônica da Represa do Lobo (Brotas-Itirapina, SP). Ciência
e Cultura, 34: 518-524.
Henry R. & Tundisi J. G. 1983. Responses of the phytoplankton community of a tropical reservoir
(São Paulo, Brazil) to the enrichment with nitrate, phosphate and EDTA. Int. Revue Ges.
Hydrobiol. 68:853-862.
Henry R.; Tundisi J. G. & Curi P. R. 1984. Effects of phosphorus and nitrogen enrichment on the
phytoplankton in a tropical reservoir (Lobo Reservoir, Brazil). Hydrobiologia 118:177-185.

71
71
Henry R. 1986. O crescimento potencial do fitoplâncton da represa de Barra Bonita (Rio Tietê, SP):
uma comparação sazonal dos efeitos de enriquecimento artificial em amostras de seus
tributários. Ciência e Cultura 38:1553-1564.
Henry R. & Simão C. A. 1988. Aspectos sazonais da limitação potencial por N, P e Fe no fitoplâncton
da Represa de Barra Bonita (Rio Tietê, SP). Rev. Bras. Bol. 48: 1-14.
Hooker E. & Hernandez S. 1991. Phytoplankton biomass in Lake Xolotlan (Manágua): its seasonal
and horizontal distribution. Hidobiol. Bull.25 (2): 125-131.
Huszar V. L. M. & Caraco N. F. 1998. The relationship between phytoplankton composition and
physical-chemical variables: a comparison of taxonomic and morphological-functional
descriptors in six temperate lakes. Freshwater Biology 40: 679-696.
Hutchinson, G. E. 1957. Concluding remarks. Cold Spring Harbor Symposium on Quantitative
Biology. 22: 415-427.
Hutchinson G. E. 1961. The paradox of the plankton. Amer. Natur. 95: 137-145.
Ibanez M. S. R. 1988. Response to artificial enrichment with ammonia and phosphate of
phytoplankton from Lake Paranoá (Brasília, DF). Rev Bras. de Biol. 48:453-457.
Isvanovics. V.; Hesham M. S.; Présing M. & Juhos S. V. 2000. Growth and phosphate uptake
kinetics of the cyanobacterium, Cylindrospermopsis raciborskii (Cyanophyceae) in
throughflow cultures. Feshwater Biol. 43:257-275.
Kilham S. S. 1978. Nutrient kinetics of freshwater planktonic algae using batch and semicontinuous
methods. Mitt. Internat. Vereisn. Limnol. 21: 147-157.
Klaveness D. 1988. Ecology of the Cryptomonadida- a first review. In Sandgren C. D. Growth and
reproductive strategies of freshwater phytoplankton. Cambridge University Press. New
York. p. 105-133.
Kormondy E. D. 1976. Concepts of ecology. 2a Edição, Eenglewood Cliffs, Prentice Hall, USA.

72
72
Krienitz L.; Kasprzak P. & Koschel R. 1996. Long term study on the influence of eutrophication,
restoration and biomanipulation on the structure and development of phytoplankton
communities in Feldberg Hausse (Baltic Lake District, Germany). Hydrobiol. 330: 89-110.
Lorenzen C. J. 1967. Determination of clorophyll and pheopigments: spectrophotometric equations.
Limnol. Oceanogr., 12: 343-346.
Lynn S. G.; Kilhan S. S.; Kreeger B. A. & Interlandi S. J. 2000. Effect of nutrient availability on the
biochemical and elemental stoichiometry in the freshwater diatom Stephanodiscus minutus
(Bacillariophyceae). J. Phycol. 36: 510-522.
MacArthur R. H. 1958. Population ecology of some warblers of Northeastern coniferous forests.
Ecology. 39: 599-619.
Margalef R. 1983. Limnología. Ediciones Omega, S. A ., Barcelona.
Matveev V. & Matveeva L. 1997. Grazer control and nutriente limitation of phytoplankton biomass
in two Australian reservoirs. Freshwater Biology 38: 49-65.
McCarthy J. J. 1981. The kinetics of nutrient utilization in Platt T. Physiological bases of
phytoplankton ecology. Otawa, Canada.
McQueen D. J. & Lean D. R. S. 1987. Influence of water temperature and N to P ratios on the
dominance of blue-green algae in lake St. George, Ontario. Can. J. Fish. Aquat. Sci. 44:598-
604.
Nedoma J., Porcalová P., Komãrková J. & Vyhnálek V. 1993. A seasonal study of phosphorus
deficiency in a eutrophic reservoir. Freshwater Biology. 30: 369-376.
Odum E. P. 1971. Fundamentals of ecology. 3a Edição, Saunders.

73
73
Ojala A.; Heaney S. I.; Arbola L. & Barbosa F. 1996. Growth of migrating and non-migrating
cryptophytes in thermally and chemically stratified experimental columns. Freshwater
Biology 35: 599-608.
Paerl H. W. 1996. A comparison fo cyanobacterial bloom dynamics in freswater estuarine and marine
environments. Phycologia 35: 25-35.
Pick F. R. 1989. Species-specific phytoplankton responses to nutriente enrichment in limnetic
enclosures. Arch Hydrobiol. Beih. 3: 177-187.
Pinto-Coelho R. M. 1992. Evolução do grau de eutrofização na lagoa da Pampulha: comparação de
ciclos sazonais de nutrientes (N e P) e organismos planctônicos. In Godinho H. P., Freitas J.
R., Nunes R. L., Souza R. M. V. M., Lombardi W. Anais do seminário da bacia hidrográfica
da Pampulha. Belo Horizonte. SEGRAC, p 1 a 14.
Pinto-Coelho R. M. & Giani A. 1985. Variações sazonais do fitoplâncton e fatores físico-químicos da
água no reservatório Paranoá, Brasília, DF. Cienc. Cult. 37: 2000-2006
Pinto-Coelho R. M. 1998. Effects of eutrophication on seasonal patterns of mesozooplankton in a
tropical reservoir: a 4-year study in Pampulha Lake, Brazil. Feshwater Biol. 40:159-173.
Prezelin B. B. 1981. Light reactions in photosyntesis. In Platt T. Physiological bases of
phytoplankton ecology. Otawa, Canada.
Putman, R. J. 1994. Community Ecology. 1a Edição, Chapman & Hall, USA.
Ramos M. L. L. C.; Goldstain E. G.; Lombardi C. C. & Zagatto P. A. 1986. Estudos sobre a
fertilidade potencial das águas do Reservatório de Barra Bonita. Acta Limnol. Bras. 1:155-
178.
Raven J. A. & Kubler J. E. 2002. New light on the scaling of metabolic rate with the size of algae.
J. Phycol.38: 11-16.

74
74
Reynolds C. S. 1984. The ecology of freshwater phytoplankton. Cambridge University Press,
Cambridge
Reynolds C. S. 1999. Non-determinism to probability, or N:P in the community ecology of
phytoplankton. Arch. Hydrobiol. 146(1): 23-35.
Reynolds R. A.; Stramsky D. & Kiefer D. A. 1997. The effect of nitrogen limitation on the
absorption and scattering properties of the marine diatom Thalasiosira pseudonana. Limnol.
Oceanogr. 42(5): 881-892.
Rhee G. Y. 1973. A continuous culture study of phosphate uptake growth rate and polyphosphate in
Scenedesmus sp. J. Phycol. 9: 495-506.
Rhee G. Y. 1974. Phosphate uptake under nitrate limitation by Scenedesmus sp. and its ecological
implications. J. Phycol. 10: 470-475.
Rhee G. Y. 1978. Effects of N:P atomic ratios and nitrate limitation on algal growth, cell
composition, and nitrate uptake. Limnol. Oceanogr. 23(1): 10-25.
Rhee G. Y. & Gotham I. J. 1980. Optimun N:P ratios and coexistence of planktonic algae. J.
Phycol. 16: 486-489.
Rojo C. M. & Alvarez-Cobelas, 1993. Hypereutrophic phytoplankton and the intermediate
disturbance hypothesis. Hydrobiol. 249: 43-57.
Rosowski J. R.; Vadas R. L. & Kugrens P. 1973. Morphology of lorica of 3 species of euglenoid
Trachelomonas. J. Phycol. 9: 18-28.
Rothhaupt S. O. 1988. Mechanistic resource competition theory applied to laboratory experiments
with zooplankton. Nature. 333: 660-662.
Rott E. 1981. Some results from phytoplankton counting. Schweiz. Z. Hydrol. 43(1):34-62.

75
75
Santanna C. L. 1984. Chlorococcales (Chlorophyceae) do Estado de São Paulo, Brasil. Vaduz. J.
Cramer.
Schindler D. W. 1977. Evolution of P limitation in lakes. Science 195: 260-262.
Schoener T. W. 1983. Field experiments on interspecific competition. Amer. Natur.122(2):241-285.
Setaro F. V. & Melack J. M. 1984. Responses of phytoplankton to experimental nutrient enrichment
in an Amazon floodplain lake. Limnol. Oceanog. 29: 972-984.
Siegel D. A . 1998. Resource competition in a discrete environment: Why are plankton distributions
paradoxical? Limnol. Oceanogr. 43(6): 1133-1146.
Smith V. H. 1983. Low nitrogen to phosphorus ratios favor dominance by Blue-Green algae in lake
Phytoplankton. Science 221: 669-671.
Smith R. E. H.; Grosselin M. & Taguchi S. 1997. The influence of major inorganica nutrients on the
growth and physiology of high arctic ice algae. J. Mar. Sys. 11: 63-70.
Sperling, E. 1994. Corrective measures for the restoration of Pampulha reservoir. In: Pinto-Coelho
R. M.; Giani A. & Sperling E. Ecology and human impact on lakes and reservoirs in Minas
Gerais. Segrac, Belo Horizonte. p. 141-149.
Sommer U. 1984. The paradox of the plankton: Fluctuations of phosphorus availability maintain
diversity of phytoplankton in flow-through cultures. Limnol. Oceanogr. 29(3): 633-636.
Sommer U. 1986. Nitrate and sicicate competition among antarctic phytoplankton. Marine Biology
91, 345-351
Sommer U. 1989. Nutrient status and nutrient competition of phytoplankton in a shallow,
hypertrophic lake. Limnol. Oceanogr. 34: 1162-1173.
Stockner G. & Shortreed S. 1988. Response of Anabaena and Synechococcus to manipulation of
N:P ratios in a lake fertilization experiment. Limnol. Oceanogr. 33: 1348-1361.

76
76
Stryer L. 1995. Biochemistry. 4a edição, W. H. Freeman and Company. New York.
Suttle C.A. & Harrison P.J. 1988. Ammonium and phosphate uptake rates, N:P supply ratios, and
evidence for N and P limitation in some oligotrophic lakes. Limnol. Oceanogr. 33:186-202.
Syrett P. J. 1981. Nitrogen metabolism of microalgae. In Platt T. Physiological bases of
phytoplankton ecology. Otawa, Canada.
Tilman D. 1976. Ecological competition between algae: Experimental confirmation of resource-
based competition theory. Science 192: 463-465.
Tilman D.; Kilham S. S. & Kilham P. 1976. Morphometric changes in Asterionella formosa
colonies under phosphate and silicate limitation. Limnol. Oceanogr. 21: 883-886.
Tilman D. 1981. Tests of resource competition theory using four species of Lake Michigan algae.
Ecology 62(3): 802-815.
Tilman, D. 1982. Resource Competition and Community Structure. 1a Edição, Princeton University
Press, Princeton, USA.
Tilman D.; Kilham S. S. & Kilham P. 1982. Phytoplankton community ecology: the role of limiting
nutrients. Ann Ver. Ecol. Syst. 13: 349-372.
Tilman D.; Kiesling R.; Sterner R.; Kilham S. S. & Johnson F. A. 1986. Green, bluegreen and
diatom algae: Taxonomic differences in competitive ability for phosphorus, silicon and
nitrogen. Arch. Hydrobiol. 106(4): 473-485.
Tôrres, I. C. 1999. Determinação da influência dos tributários na qualidade da água da represa da
Pampulha. Dissertação de mestrado do Programa de Pós-graduação em Ecologia,
Conservação e Manejo da Vida Silvestre da UFMG, Belo Horizonte. 130p.

77
77
Tundisi J. G. & Henry R. 1986. Effects of enrichment on the summer suface phytoplanktonic
community in a stratified tropical lake (Lake Don Elvécio- Parque Florestal do Rio Doce,
Minas Gerais). Rev. Bras. Biol. 46: 231-237.
Utermöhl H. 1958. Zur Vervollkomnung der quantitativen Phytoplankton-Methodik. Mitt. Int. Ver.
Limnol. 9: 1-38.
Vignoli O. 1992. Investigações sedimentológicas na Pampulha- Aevolução do assoreamento e seu
controle. In: Pinto-Coelho R. M.; Giani A. & Sperling E. Ecology and human impact on
lakes and reservoirs in Minas Gerais. Segrac, Belo Horizonte. p. 141-149.
Wetzel R. G. 1983. Limnology. Saunders College Publishing, Michigan.
Zavenboom W. & Mur L. R. 1980. N2 fixing Cyanobacteria: Why they do not become dominant in
Dutch, hypereutrophic lakes. Dev. Hydrobiol. 2: 123-130.


















