Biofísica - Universidade de São Paulo | Escola Superior ... · A principal função da membrana...
60
azevedolab.net © 2012 Dr. Walter F. de Azevedo Jr. Biofísica Membrana celular Prof. Dr. Walter F. de Azevedo Jr. 1
Transcript of Biofísica - Universidade de São Paulo | Escola Superior ... · A principal função da membrana...
azevedolab.net
© 2
01
2 D
r. W
alter
F.
de
Aze
ve
do
Jr.
Biofísica
Membrana celular
Prof. Dr. Walter F. de Azevedo Jr.
1
Como é a estrutura da membrana celular?
Como funcionam os venenos de vespas?
Como funcionam os venenos de serpentes?
O que é o potencial de membrana?
Fonte: http://www.kurzweilai.net/ azevedolab.net 2

Membrana
celular
Interação com
a membrana
Bomba de
Na+ / K+
Testes
clínicos Material
adicional
azevedolab.net 3

A principal função da membrana celular é
manter moléculas, tão diversas como
proteínas e pequenos solutos, no interior
da célula. A membrana funciona para
regular seletivamente sua permeabilidade,
ou seja, a facilidade com a qual moléculas
e íons atravessam a membrana. O
estudo da composição da membrana faz
uso de diversas técnicas e metodologias
físicas, discutiremos a seguir os principais
modelos da membrana e iniciaremos o
estudo de fenômenos elétricos na célula.
No livro de Oparin, “A Origem da Vida”,
ele propôs que para qualquer forma de
vida é necessária a presença de uma
barreira, que separe a parte “viva” do
meio que a cerca. Esse trabalho destaca
a necessidade de uma membrana para
isolar, até mesmo as formas de vida mais
simples, do meio exterior.
Modelo computacional da bicamada fosfolipídica
da membrana celular.
4
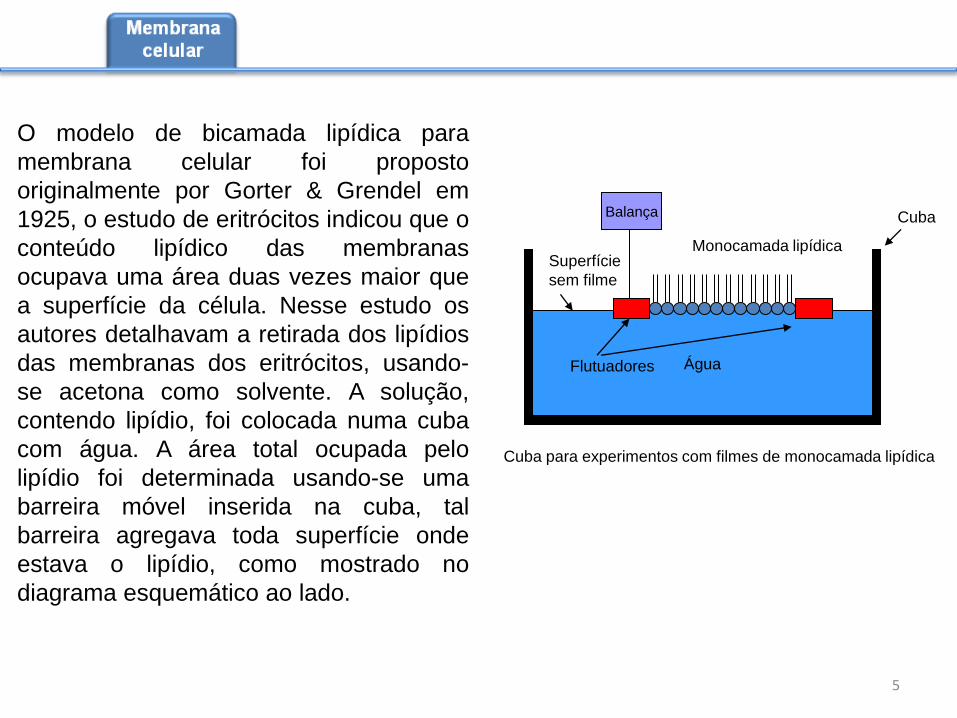
O modelo de bicamada lipídica para
membrana celular foi proposto
originalmente por Gorter & Grendel em
1925, o estudo de eritrócitos indicou que o
conteúdo lipídico das membranas
ocupava uma área duas vezes maior que
a superfície da célula. Nesse estudo os
autores detalhavam a retirada dos lipídios
das membranas dos eritrócitos, usando-
se acetona como solvente. A solução,
contendo lipídio, foi colocada numa cuba
com água. A área total ocupada pelo
lipídio foi determinada usando-se uma
barreira móvel inserida na cuba, tal
barreira agregava toda superfície onde
estava o lipídio, como mostrado no
diagrama esquemático ao lado.
Água
Monocamada lipídica
Balança
Superfície
sem filme
Flutuadores
Cuba
Cuba para experimentos com filmes de monocamada lipídica
5

A monocamada lipídica é formada por
fosfolipídios. Cada um apresenta uma
cabeça polar (esfera azul), que fica em
contato com a água, e duas caudas
hidrofóbicas, que não entram em contato
com a água. A área total do filme de
lipídio, sobre a superfície da água, foi
determinado com sendo
aproximadamente igual ao dobro da área
do eritrócito. Tal observação levou à
hipótese da bicamada lipídica (indicada
abaixo), com uma parte polar voltada para
os meios intracelular e extracelular e a
parte hidrofóbica voltada para o interior da
membrana, escondida do solvente.
Podemos pensar na bicamada lipídica
como um cobertor dobrado, com as
caudas hidrofóbicas no meio do cobertor,
e as cabeças polares formando a parte
externa.
Água
Monocamada lipídica
Balança
Superfície
sem filme
Flutuadores
Cuba
Cuba para experimentos com filmes de monocamada lipídica
Bicamada lipídica
Cabeças
polares Caudas
hidrofóbicas
6
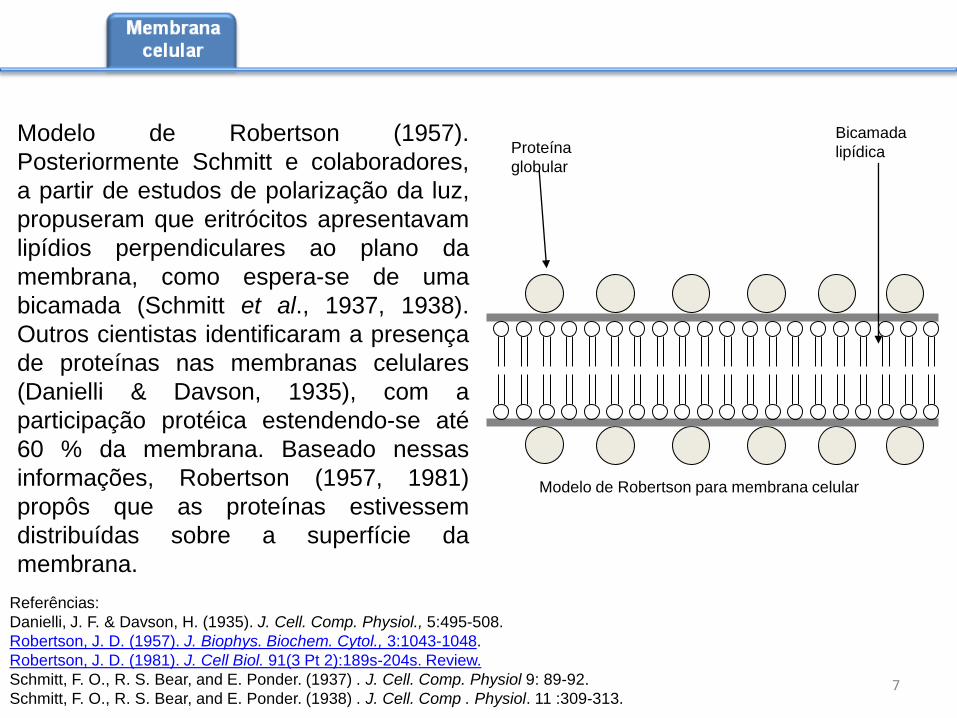
Modelo de Robertson (1957).
Posteriormente Schmitt e colaboradores,
a partir de estudos de polarização da luz,
propuseram que eritrócitos apresentavam
lipídios perpendiculares ao plano da
membrana, como espera-se de uma
bicamada (Schmitt et al., 1937, 1938).
Outros cientistas identificaram a presença
de proteínas nas membranas celulares
(Danielli & Davson, 1935), com a
participação protéica estendendo-se até
60 % da membrana. Baseado nessas
informações, Robertson (1957, 1981)
propôs que as proteínas estivessem
distribuídas sobre a superfície da
membrana.
Referências:
Danielli, J. F. & Davson, H. (1935). J. Cell. Comp. Physiol., 5:495-508.
Robertson, J. D. (1957). J. Biophys. Biochem. Cytol., 3:1043-1048.
Robertson, J. D. (1981). J. Cell Biol. 91(3 Pt 2):189s-204s. Review.
Schmitt, F. O., R. S. Bear, and E. Ponder. (1937) . J. Cell. Comp. Physiol 9: 89-92.
Schmitt, F. O., R. S. Bear, and E. Ponder. (1938) . J. Cell. Comp . Physiol. 11 :309-313.
Proteína
globular
Bicamada
lipídica
Modelo de Robertson para membrana celular
7
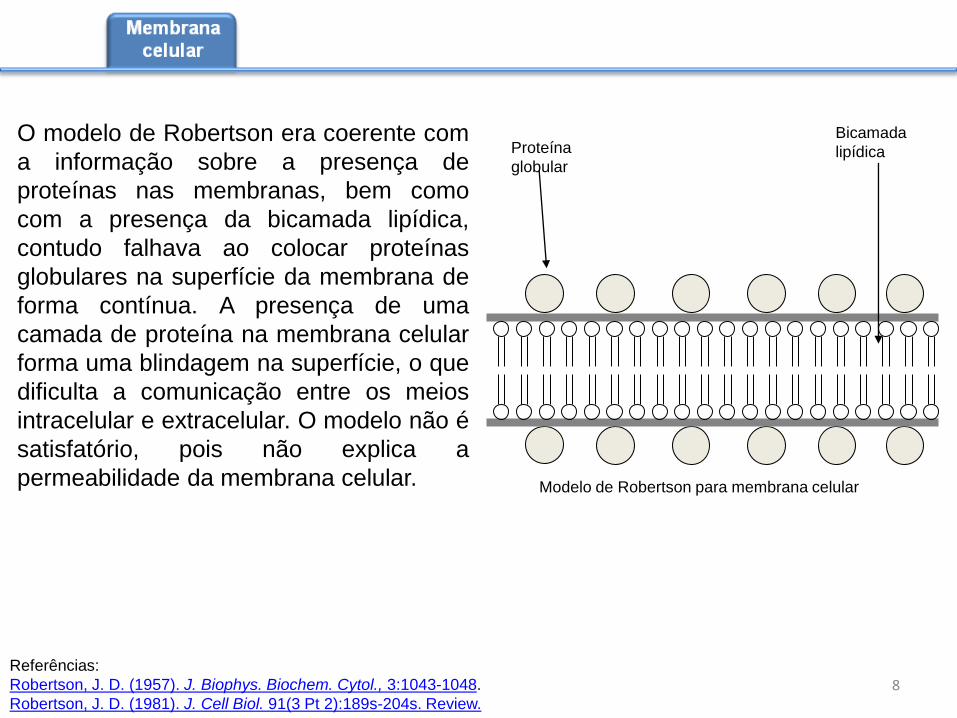
O modelo de Robertson era coerente com
a informação sobre a presença de
proteínas nas membranas, bem como
com a presença da bicamada lipídica,
contudo falhava ao colocar proteínas
globulares na superfície da membrana de
forma contínua. A presença de uma
camada de proteína na membrana celular
forma uma blindagem na superfície, o que
dificulta a comunicação entre os meios
intracelular e extracelular. O modelo não é
satisfatório, pois não explica a
permeabilidade da membrana celular.
Proteína
globular
Bicamada
lipídica
Modelo de Robertson para membrana celular
Referências:
Robertson, J. D. (1957). J. Biophys. Biochem. Cytol., 3:1043-1048.
Robertson, J. D. (1981). J. Cell Biol. 91(3 Pt 2):189s-204s. Review.
8

Proteína intrínseca, ou
transmembrana
Proteína extrínseca
O modelo de mosaico fluido indica duas proteínas inseridas
na bicamada lipídica (elipsóides cinzas). A proteína da
esquerda é uma proteína extrínseca e a da direita uma
proteína intrínseca. Os fosfolipídios são indicados com a
cabeça polar em preto e a cauda hidrofóbica pelas linhas que
saem da esfera preta.
Referência : Singer SJ, Nicolson GL. Science. 1972 ;175(23):720-
31.
Modelo de mosaico fluido. Experimentos
mais detalhados mostraram deficiências
nos diversos modelos de membrana. Em
1972 Singer e Nicolson propuseram um
novo modelo para a membrana, chamado
de modelo de mosaico fluido. Nesse
modelo temos a bicamada lipídica, onde
encontram-se inseridas proteínas. O
modelo prevê duas formas de proteínas
inseridas na membrana, uma que
atravessa toda a membrana, chamada
proteína intrínseca, ou transmembranar. A
segunda forma de proteína localiza-se
sobre a membrana, sendo encontrada
tanto no exterior como voltada para o
citoplasma. Tal forma de proteína é
chamada extrínseca.
9
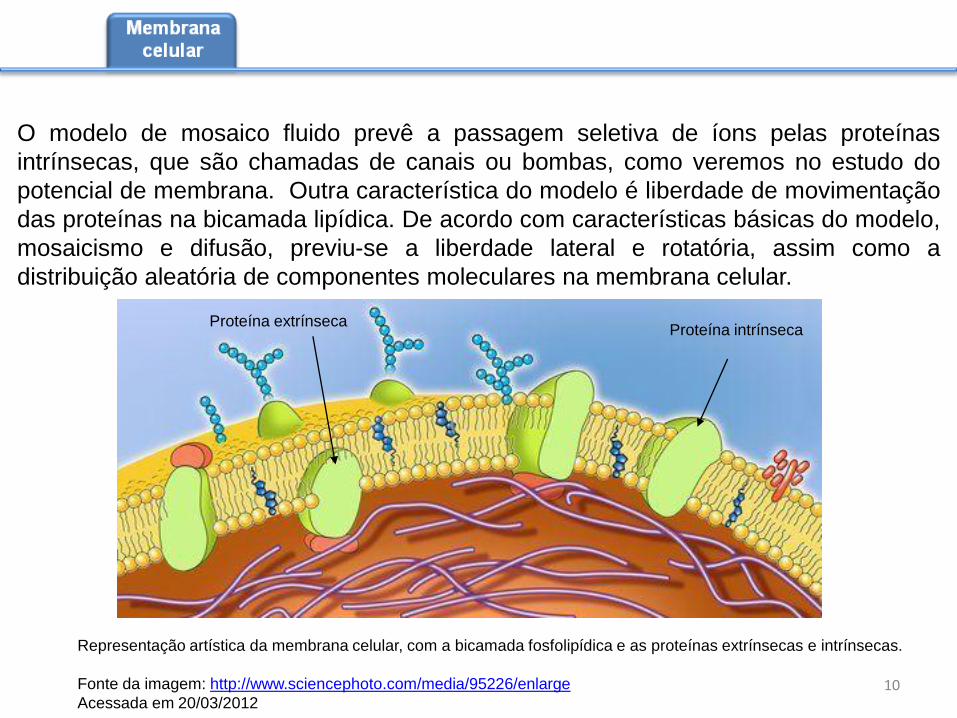
Representação artística da membrana celular, com a bicamada fosfolipídica e as proteínas extrínsecas e intrínsecas.
Fonte da imagem: http://www.sciencephoto.com/media/95226/enlarge
Acessada em 20/03/2012
Proteína extrínseca
O modelo de mosaico fluido prevê a passagem seletiva de íons pelas proteínas
intrínsecas, que são chamadas de canais ou bombas, como veremos no estudo do
potencial de membrana. Outra característica do modelo é liberdade de movimentação
das proteínas na bicamada lipídica. De acordo com características básicas do modelo,
mosaicismo e difusão, previu-se a liberdade lateral e rotatória, assim como a
distribuição aleatória de componentes moleculares na membrana celular.
Proteína intrínseca
10

CH2 CH CH2 O P O X
O
O-O
C
R1
O
O
C
R2
O
Biomembranas são baseadas
principalmente em lipídios, com
predominância de fosfolipídios. A estrutura
química geral de uma molécula de
fosfolipídio é mostrada ao lado (figura a).
Tal molécula é basicamente um glicerol
(figura b), sobre a qual foram ligadas as
cadeias de ácidos graxos (R1 e R2). O
grupo fosfato permite a ligação de
qualquer molécula, designada na figura
por pelo grupo X. Um dos ácidos graxos
típicos, encontrados nos fosfolipídios, é
chamado ácido palmítico (figura c) .
a)
b)
HO CH2 C CH2 OH
OH
H
Fosfolipídio
c) Glicerol
Ácido palmítico
11

A molécula de ácido palmítico (ou ácido
hexadecanóico) apresenta 16 carbonos e
31 hidrogênios. Tal ácido graxo é dito
saturado, pois apresenta o maior número
de possível de hidrogênios ligados. A
presença de ligações duplas na cadeia de
ácido graxo indica que o mesmo é não
saturado. As duas cadeias R1 e R2 não
precisam ser homogêneas, ou seja,
podem apresentar cadeias de tamanhos
distintos. Nos fosfolipídios, uma parte da
molécula é polar, a cabeça hidrofílica, e a
parte apolar é composta pelas duas
cadeias de ácidos graxos. O diagrama
esquemático ao lado ilustra uma molécula
de fosfolipídio. Moléculas que apresentam
parte polar e parte hidrofóbica são
chamadas anfipáticas. Na figura da
abaixo temos a representação CPK do
fosfolipídio.
Cabeça polar
Caudas hidrofóbicas
Caudas hidrofóbicas
Cabeça polar
12

Diversos modelos computacionais de
biomembranas foram construídos e
submetidos à simulação de dinâmica
molecular. Na dinâmica molecular,
aspectos sobre a mobilidade dos sistemas
moleculares são simulados, de forma que
a interação da bicamada fosfolipídica com
moléculas de água pode ser verificada.
Tais modelos usam diferentes
componentes para a formação da
bicamada, no exemplo ao lado foram
usadas de moléculas de 1-palmitoil-2-
oleoil-sn-glicerol-3-fosfatidilcolina,
formando uma caixa retangular, onde
temos moléculas de água interagindo com
a parte polar da bicamada. No modelo
computacional, vemos claramente que as
caudas hidrofóbicas não interagem com
as moléculas d’água.
Hid
rofílic
a H
idro
fóbic
a H
idro
fílic
a
Modelo computacional da membrana
Diagrama esquemático da membrana
Hid
rofílic
a H
idro
fóbic
a H
idro
fílic
a
13

Podemos pensar que o modelo
computacional representa um fatia da
membrana celular, sem a presença de
proteínas. É como se tivéssemos cortado
uma fatia em formato de cubo da
biomembrana. Na simulação
computacional da membrana, as
moléculas de água posicionam-se nas
regiões polares, como era de se esperar. A
vantagem da simulação computacional é
que uma vez confirmada a capacidade de
simulação de aspectos conhecidos, como
a interação com água, podemos adicionar
moléculas aos sistemas e prever seu
comportamento. Por exemplo, podemos
prever qual o comportamento de fármacos
com a bicamada. Tal simulação é de
interesse no desenvolvimento de
fármacos, visto que para que possam agir
a maioria dos fármacos têm que
atravessar a membrana.
Hid
rofílic
a H
idro
fóbic
a H
idro
fílic
a
Modelo computacional da membrana
Diagrama esquemático da membrana
Hid
rofílic
a H
idro
fóbic
a H
idro
fílic
a
14
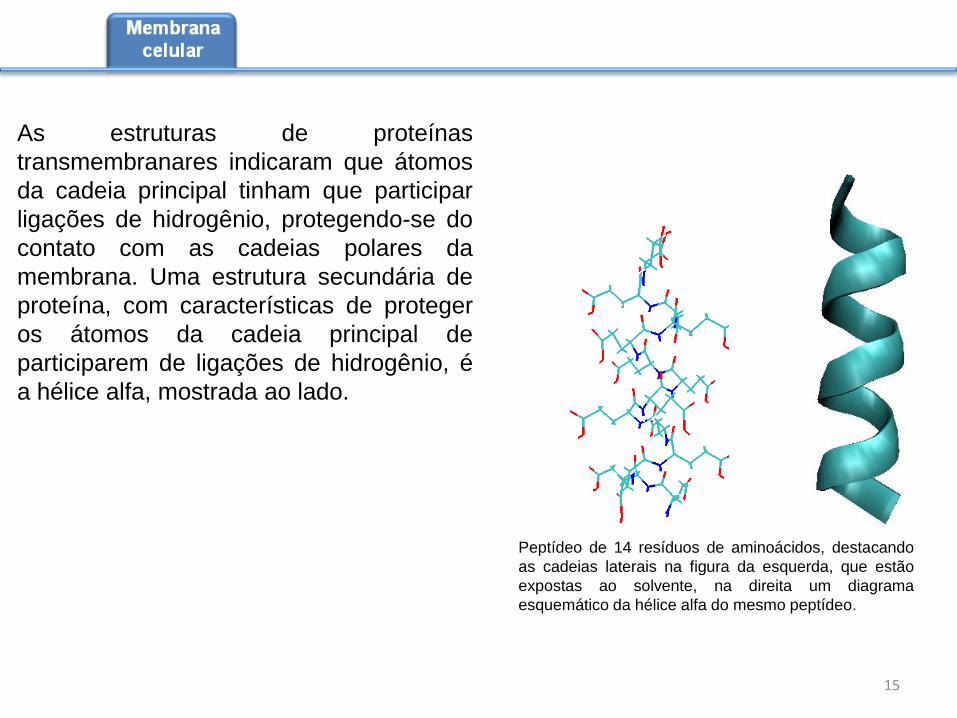
As estruturas de proteínas
transmembranares indicaram que átomos
da cadeia principal tinham que participar
ligações de hidrogênio, protegendo-se do
contato com as cadeias polares da
membrana. Uma estrutura secundária de
proteína, com características de proteger
os átomos da cadeia principal de
participarem de ligações de hidrogênio, é
a hélice alfa, mostrada ao lado.
Peptídeo de 14 resíduos de aminoácidos, destacando
as cadeias laterais na figura da esquerda, que estão
expostas ao solvente, na direita um diagrama
esquemático da hélice alfa do mesmo peptídeo.
15

Como exemplo de proteína
transmembranar, temos o complexo
proteico, centro de reação fotossintético
da bactéria púrpura R. viridis, que é o
local da etapa inicial da captura de
energia luminosa na fotossíntese. Tal
complexo proteico é composto de quatro
cadeias polipeptídicas (tetrâmero),
indicadas na figura ao lado, nas cores
azul, vermelha, cinza e dourado. Há
também quatorze cofatores de baixo peso
molecular, indicados em amarelo. Entres
os cofatores temos cromóforos, que
absorvem a energia de excitação, que é
convertida em potencial eletroquímico,
através da membrana.
Referência: Deisenhofer, J. & Michel, H. (1989) EMBO J.
8:2149-2170.
16

Nesta representação, os resíduos de
aminoácidos hidrofóbicos estão em cinza,
os polares em verde, os ácidos em
vermelho e os básicos em azul. Átomos
pertencentes aos cofatores estão em
ciano. Vemos claramente uma
heterogeneidade na distribuição de
cargas na superfície das proteínas. No
centro temos uma região
preponderantemente hidrofóbica, e nas
partes superior e inferior uma
concentração de resíduos de aminoácidos
carregados e polares, o que caracteriza
uma região hidrofílica.
Referência: Deisenhofer, J. & Michel, H. (1989) EMBO J.
8:2149-2170.
132 Å
72 Å
Hid
rofí
lica
Hid
rofó
bic
a
Hid
rofí
lica
17
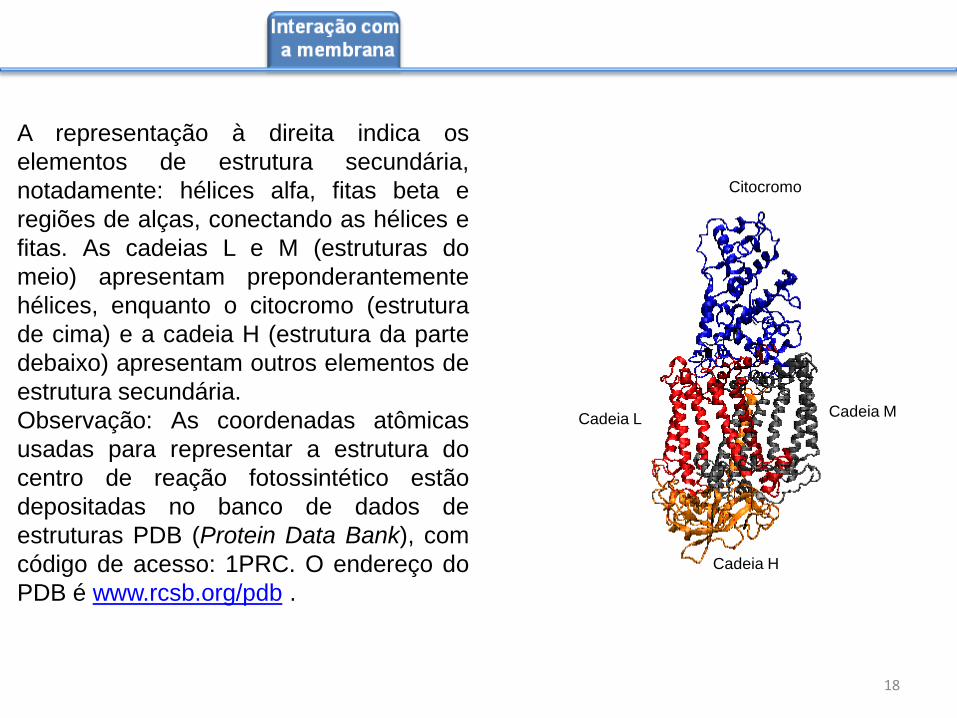
A representação à direita indica os
elementos de estrutura secundária,
notadamente: hélices alfa, fitas beta e
regiões de alças, conectando as hélices e
fitas. As cadeias L e M (estruturas do
meio) apresentam preponderantemente
hélices, enquanto o citocromo (estrutura
de cima) e a cadeia H (estrutura da parte
debaixo) apresentam outros elementos de
estrutura secundária.
Observação: As coordenadas atômicas
usadas para representar a estrutura do
centro de reação fotossintético estão
depositadas no banco de dados de
estruturas PDB (Protein Data Bank), com
código de acesso: 1PRC. O endereço do
PDB é www.rcsb.org/pdb .
Cadeia H
Cadeia M Cadeia L
Citocromo
18

Citoplasma
Interação das cadeias L e M (vermelho e cinza) do
centro de reação fotossintético com a bicamada lipídica.
Hélice alfa é a estrutura secundária
comumente encontrada em segmentos
transmembranares de proteínas de
membranas. Normalmente temos a
combinação de vários segmentos de
hélice, formando feixes de hélices
transmembranares, como nas estruturas
das cadeias L e M da estrutura do
centro de reação fotossintético mostrada
ao lado. A partir da análise da estrutura
do centro de reação fotossintético,
podemos propor um modelo para
interação da proteína com a membrana.
No modelo temos que a parte
hidrofóbica, formada por hélices alfa,
interage com a membrana celular,
conforme indicado no diagrama
esquemático ao lado.
Meio extracelular
19

Muitas das proteínas, que interagem com
a membrana celular, apresentam regiões
em hélice alfa. Para entendermos esta
interação proteína-membrana, vamos
olhar alguns detalhes da estrutura da
hélice alfa. Uma hélice alfa, como
mostrada na figura a, tem as cadeias
laterais apontando para fora da hélice
(indicadas com setas). Tal hélice alfa pode
ser representada como um cilindro (figura
b). Nas figuras c e d temos a visão de
cima de cada uma das representações da
hélice alfa. As visões facilitam a
identificação das regiões hidrofóbicas e
hidrofílicas das hélices alfa, como nas
figuras c e d.
a) b)
c) d)
Cadeias laterais
Cadeias laterais
20

Nas estruturas de feixes de hélices
transmembranares, verifica-se que a parte
da hélice, que toca os lipídios, é
relativamente mais hidrofóbica que a
parte que participa do contato hélice-
hélice. No diagrama esquemático à
direita, temos um feixe de 4 hélices (figura
b), onde vemos que a região entre as
hélices é mais hidrofílica que a região em
contato com a bicamada lipídica. A região
das hélices alfa, em contato com as
caudas hidrofóbicas da bicamada
fosfolipídica, estão indicadas por setas.
a) Hélice transmembranar
b) 4 hélices transmembranares
Superfície mais hidrofóbica
Superfície mais hidrofílica 21

Outra estrutura possível, para proteínas
transmembranares, é a folha beta. Um
arranjo onde a folha beta fecha-se sobre
si forma um arranjo similar a um barril,
sendo denominado barril beta. A estrutura
em barril proporciona as características
físico-químicas desejáveis para uma
proteína transmembranar, tal como
blindagem dos átomos da cadeia principal
que participam de ligações de hidrogênio.
A figura ao lado mostra a estrutura de
uma porina. As porinas criam canais na
membrana permitindo a passagem de
moléculas de baixa massa molecular
(inferior a 700 D), tais como molécuolas
de água, íons, glucose e outros
nutrientes.
Barril beta da porina de Rhodopseudomonas blastica.
Código de acesso PDB: 8PRN
22

Toxinas da vespa solitária Anterhynchium
flavormarginatum micado. Essa vespa
injeta seu veneno em lagartas e deposita
seus ovos próximos à vítima. Devido à
ação tóxica de seu veneno, a lagarta fica
paralisada, mas viva. Ao eclodirem, as
jovens vespas terão um banquete fresco e
vivo. Tal comportamento indicou que o
veneno da A. flavormarginatum micado,
poderia ser uma fonte rica de moléculas
com atividades antimicrobianas. Podemos
imaginar o veneno das vespas como um
coquetel de moléculas. A questão é: qual
ou quais moléculas apresentam as
atividades biológicas de interesse? Foto: Cortesia do Dr. K. Konno.
23

Referência: Sforça, M. L., Oyama, S. Jr., Canduri, F., Lorenzi,
C. C., Pertinhez, T. A., Konno, K., Souza, B. M., Palma, M. S.,
Ruggiero Neto, J., Azevedo, W. F. Jr., Spisni, A. (2004).
Biochemistry 43:5608-5617.
Foto: Cortesia do Dr. K. Konno.
Mastoparanos são peptídeos isolados em
vespas e abelhas, muitos deles
apresentam atividade biológicas, tais
como, degranulação de mastócitos,
atividades antimicrobiana e hemolítica.
Essas moléculas atuam principalmente na
membrana celular, provavelmente
rompendo a integridade da bicamada
lipídica. O mastoparano, identificado no
veneno da A. flavormarginatum micado
(EMP-AF), apresenta atividade hemolítica
e de degranulação de mastócitos (Sforça
et al., 2004).
24

Abaixo temos as estruturas primárias (sequência de resíduos de aminoácidos) de um
mastoparano típico e do EMP-AF, os resíduos Asn, Lys e Ser são polares. Ambos
peptídeos apresentam 14 resíduos de aminoácido. Uma das características de tais
peptídeos é que eles apresentam o terminal C amidado, como pode ser visto no final
da estrutura primária. A numeração acima da sequência indica o número do resíduo de
aminoácido. As sequências são indicadas com o código de três letras, com o código de
uma letra entre parênteses abaixo do código de três letras.
Referência: Sforça, M. L., Oyama, S. Jr., Canduri, F., Lorenzi, C. C., Pertinhez, T. A., Konno, K., Souza, B. M., Palma, M. S.,
Ruggiero Neto, J., Azevedo, W. F. Jr., Spisni, A. (2004). Biochemistry 43:5608-5617.
Mastoparano típico
EMP-AF
25

Abaixo temos as estruturas secundárias de um mastoparano típico e do EMP-AF, os
resíduos de asparagina Asn (N) e serina Ser (S) são indicados em rosa. Os resíduos
de lisina Lys (K) indicados em vermelho. Os peptídeos apresentam estrutura em hélice
alfa, com o resíduos polares de um lado da hélice e resíduos apolares do lado oposto,
o que dá um caráter anfipático ao peptídeo.
Referência: Sforça, M. L., Oyama, S. Jr., Canduri, F., Lorenzi, C. C., Pertinhez, T. A., Konno, K., Souza, B. M., Palma, M. S.,
Ruggiero Neto, J., Azevedo, W. F. Jr., Spisni, A. (2004). Biochemistry 43:5608-5617.
Mastoparano típico EMP-AF
26

Toxinas da vespa solitária Anoplius
samariensis. Essa vesta injeta seu
veneno em aranhas e deposita seus ovos
próximos à vítima, da mesma forma que a
A. flavormarginatum micado. A ação
tóxica de seu veneno paralisa a aranha,
que será comida viva pelas vespas ao
eclodirem.
Referência: Konno, K., Hisada, M., Fontana, R., Lorenzi, C. C., Naoki, H., Itagaki, Y., Miwa, A., Kawai, N., Nakata, Y.,
Yasuhara, T., Ruggiero Neto, J., de Azevedo, W. F. Jr., Palma, M. S., Nakajima, T. (2001). Bioch. Biophys. Acta. 1550:70-80.
Foto: Cortesia do Dr. K. Konno.
27

Anoplin. O veneno da A. samariensis é
uma possível fonte de moléculas com
atividades biológicas. O peptídeo Anoplin,
um decapeptídeo (10 resíduos de
aminoácido) com o terminal C amidado e
sequência, GLLKRIKTLL, foi identificado
no veneno da A. samariensis. Testes de
atividade biológica mostraram ação
antimicrobiana desse peptídeo, sendo o
menor peptídeo que se tem notícia a
apresentar tal ação. A versão com o
terminal C não-amidado apresenta
atividade antimicrobiana reduzida, quando
comparada com o Anoplin-NH2. Tal
observação pode ser usada como guia
para o desenho de peptídeos
antimicrobianos, visto que a atividade está
relacionada com a presença do grupo
amida no terminal C.
Foto: Cortesia do Dr. K. Konno.
28
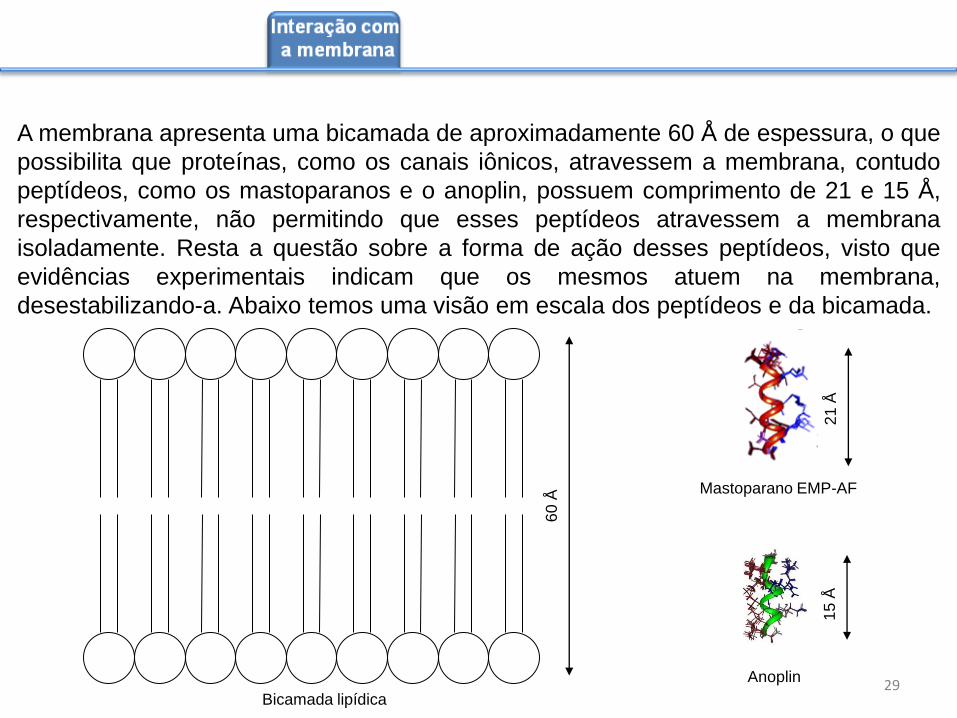
A membrana apresenta uma bicamada de aproximadamente 60 Å de espessura, o que
possibilita que proteínas, como os canais iônicos, atravessem a membrana, contudo
peptídeos, como os mastoparanos e o anoplin, possuem comprimento de 21 e 15 Å,
respectivamente, não permitindo que esses peptídeos atravessem a membrana
isoladamente. Resta a questão sobre a forma de ação desses peptídeos, visto que
evidências experimentais indicam que os mesmos atuem na membrana,
desestabilizando-a. Abaixo temos uma visão em escala dos peptídeos e da bicamada.
60
Å Mastoparano EMP-AF
Anoplin
21
Å
15
Å
Bicamada lipídica 29

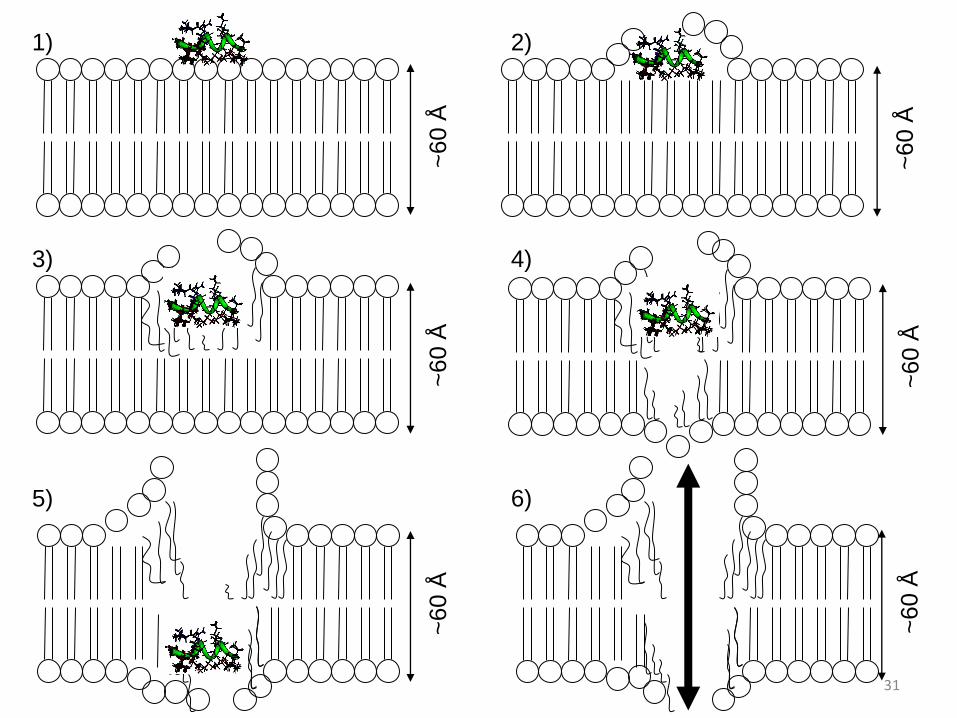
Uma possível forma de ação dos peptídeos, é por meio do desmonte da camada
externa da membrana, o que levaria à sua desestruturação e consequente quebra da
membrana celular. A interação do peptídeo com a membrana é um fenômeno
complexo, que não é possível acessarmos a partir exclusivamente da estrutura 3D.
Contudo, baseado nas estruturas 3D dos peptídeos e da membrana, foi possível
propor diversos modelos para interação do peptídeo mastoparano com a membrana.
Um dos modelos propõe que esta interação é um processo de diversas etapas
(indicadas abaixo e no esquema do slide seguinte).
1) Ancoragem do peptídeo na membrana celular;
2) Início da desestabilização da membrana celular;
3) Início da desmontagem da primeira camada lipídica;
4) Início da desmontagem da segunda camada lipídica;
5) Desmontagem da bicamada lipídica;
6) Fluxo de substâncias para o interior e exterior da célula leva à morte da célula.
30
~60 Å
~60 Å
~60 Å
~60 Å
~60 Å
~6
0 Å
1) 2)
3) 4)
5) 6)
31

Fosfolipase A2 de venenos de
serpentes. Venenos de serpentes podem
ser pensados como coquetéis de
proteínas e peptídeos, que apresentam
um vasto espectro de atividades
biológicas, entre elas muitas de interesse
farmacológico. Há dois objetivos
principais nos estudos dos componentes
dos venenos de serpentes. Um focado
nos soros e possíveis inibidores para os
acidentes ofídicos, que ainda ocorrem no
Brasil e no resto do mundo. Outro foco
está centrado nas propriedades
farmacêuticas dos componentes dos
venenos.
Fonte da imagem: http://www.cobrasbrasileiras.com.br/bothrops_jararacussu.html
Acessado em 19/03/2012 32

Entre as proteínas, identificadas nos
venenos, destacam-se as
metaloproteases e as fosfolipases A2
(discutidas aqui). As fosfolipases A2 são
enzimas que catalisam a clivagem de
fosfolipídios, componentes da membrana.
Tal enzima foi intensivamente estudada e
está presente no veneno de diversas
serpentes. Entres suas atividades,
destacam-se a atividade miotóxica,
apresentada pelo veneno das serpentes
do gênero Bothrops. A ação miotóxica é
responsável por danos extensos no tecido
muscular, se não tratada. A fosfolipase A2
apresenta 122 resíduos de aminoácidos
em sua estrutura primária, e sua
estrutura 3D foi elucidada por técnicas de
difração de raios X. A estrutura apresenta
6 hélices principais, indicadas por
cilindros azuis e duas fitas em amarelo.
C
N
Estrutura da fosfolipase A2 de Bothrops jararacussu.
33

Estudos estruturais das fosfolipases A2
lançaram as bases moleculares para o
entendimento da atividade miotóxica. O
modelo mais aceito propõe que, a
clivagem dos fosfolipídios da membrana
celular levam à ação miotóxica. Modelos
computacionais da interação da
fosfolipases A2 com a membrana
identificaram detalhes da interação, como
mostrado ao lado. A fosfolipase A2 , uma
proteína rica em hélices (indicadas como
cilindros azuis), apresenta interações
eletrostática e hidrofóbica com a
membrana, o que possibilita a
desestruturação desta e a consequente
atividade tóxica. A simulação
computacional indicou que a ligação da
fosfolipase A2 à membrana, desloca
moléculas de água, o que pode facilitar a
interação da enzima com os fosfolipídios.
Fonte da imagem:
http://www.ks.uiuc.edu/Research/smd_imd/pla2/
Acessada em 19/03/2012
Modelo computacional da interação da fosfolipase A2 com a
monocamada da membrana. Os fosfolipídios são indicados
com caudas hidrofóbicas em verde.
34
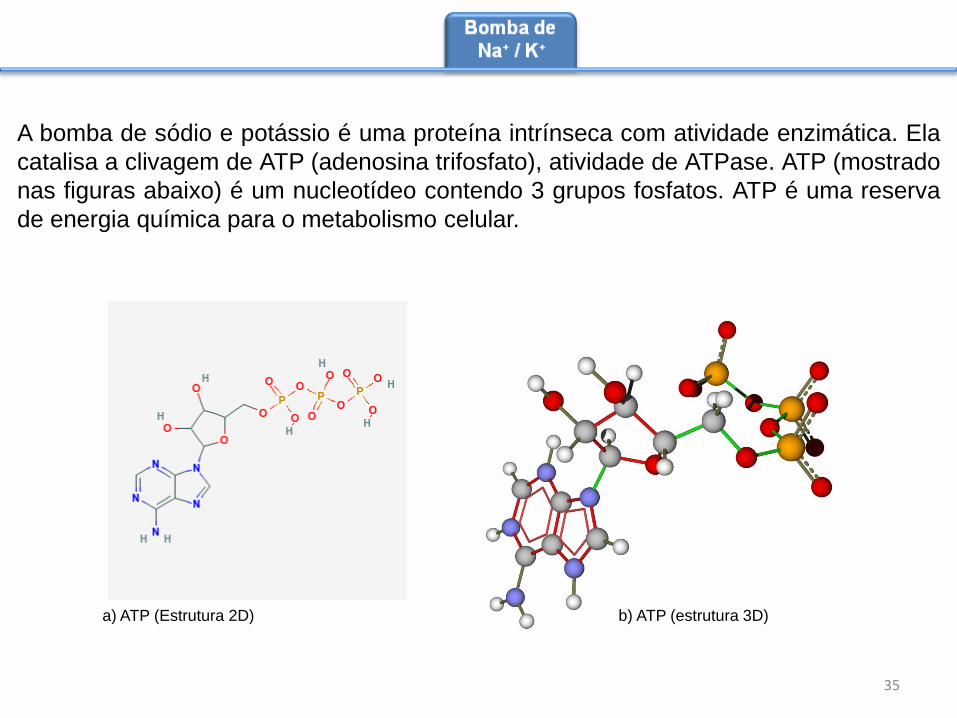
A bomba de sódio e potássio é uma proteína intrínseca com atividade enzimática. Ela
catalisa a clivagem de ATP (adenosina trifosfato), atividade de ATPase. ATP (mostrado
nas figuras abaixo) é um nucleotídeo contendo 3 grupos fosfatos. ATP é uma reserva
de energia química para o metabolismo celular.
a) ATP (Estrutura 2D) b) ATP (estrutura 3D)
35

A ação da bomba de Na+/K+ segue as seguintes etapas:
1) A bomba de Na+/K+ , com ATP ligado, liga-se a 3 íons de Na+ intracelulares.
2) ATP é hidrolisado, causando a fosforilação de um resíduo de Asp, da bomba de
Na+/K+ com a liberação de ADP.
3) A mudança conformacional, da bomba de Na+/K+ , expõe os íons de Na+, que por
apresentarem baixa afinidade pela bomba de Na+/K+, são liberados para o meio
extracelular.
4) A bomba de Na+/K+ liga-se a 2 íons de K+ extracelulares, isto causa a
desfosforilação da bomba, trazendo-a de volta à sua conformação anterior,
transportando K+ para dentro da célula.
5) A forma desfosforilada da bomba de Na+/K+ apresenta afinidade mais alta por íons
de Na+, os íons de K+ são liberados, a molécula de ATP liga-se à bomba.
6) O sistema está pronto para um novo ciclo.
36

ATP PI ADP
PI
PI
PI
ATP
ATP
Na+
Na+
K+
1 2
3
4 5
6
Meio intracelular
Meio extracelular Meio extracelular Meio extracelular
Meio extracelular Meio extracelular Meio extracelular
Meio intracelular Meio intracelular
Meio intracelular Meio intracelular Meio intracelular
37

PI
Na+
Meio extracelular
Meio intracelular
K+
Canais de K+ são os canais usualmente abertos na membrana plasmática de
neurônios em repouso. Assim, ocorre saída de íons K+, o que deixa um excesso de
carga negativa no interior da célula e, como resultado, um potencial negativo. A ação
conjunta da bomba de Na+/K+ e do canal de K+ leva a um acúmulo de carga positiva
no meio extracelular. Tal situação, tem como conseqüência, uma diferença de
potencial negativa do meio intracelular com relação ao meio extracelular. Se
colocarmos um eletrodo no interior de um neurônio em repouso, teremos um potencial
elétrico de algumas dezenas de milivolts negativos, tal potencial é chamado potencial
de repouso.
Bomba de Na+/K+ canal de K+
38

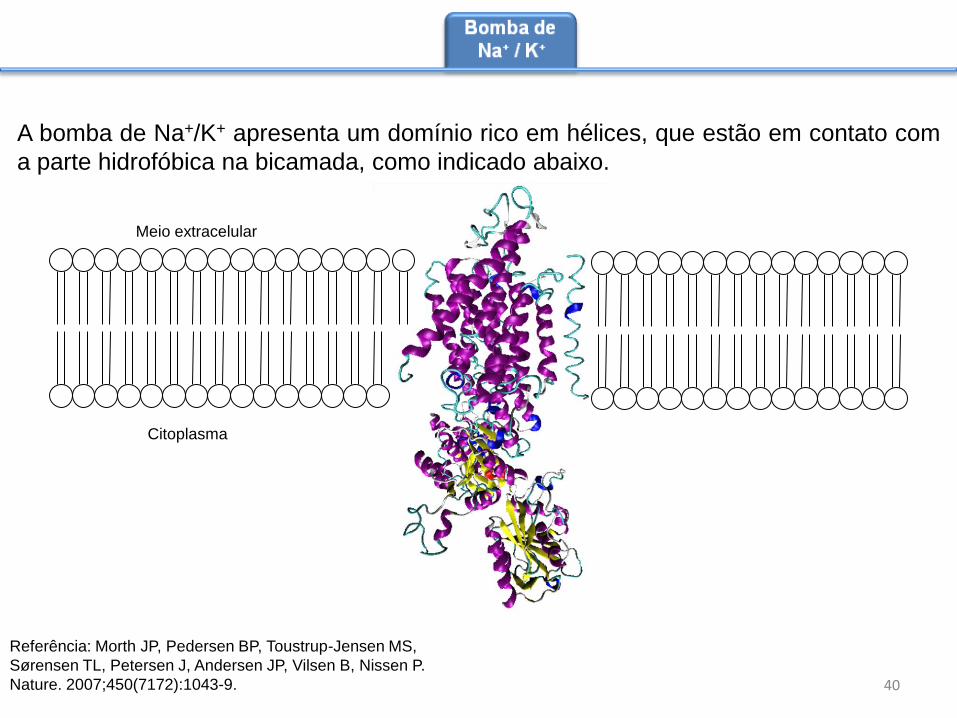
Aproximadamente um terço de todo ATP
da célula é usado para o funcionamento
da bomba de Na+/K+, o que indica a
importância para o metabolismo celular da
bomba de Na+/K+. Em 2007 foi elucidada
a estrutura tridimensional da Na+/K+,
mostrada na figura ao lado (Morth et al.,
2007). A análise da estrutura indicou uma
divisão clara de regiões hidrofóbicas e
hidrofílicas, que sugerem a inserção na
bomba de Na+/K+ na membrana,
conforme o modelo mostrado no próximo
slide.
Referência: Morth JP, Pedersen BP, Toustrup-Jensen MS,
Sørensen TL, Petersen J, Andersen JP, Vilsen B, Nissen P.
Nature. 2007;450(7172):1043-9.
39
Meio extracelular
Citoplasma
Referência: Morth JP, Pedersen BP, Toustrup-Jensen MS,
Sørensen TL, Petersen J, Andersen JP, Vilsen B, Nissen P.
Nature. 2007;450(7172):1043-9.
A bomba de Na+/K+ apresenta um domínio rico em hélices, que estão em contato com
a parte hidrofóbica na bicamada, como indicado abaixo.
40

Diferentes formas de estudo de sistemas
biológicos.
In silico. Usa simulação computacional
para o estudo de sistemas biológicos. Tem
como principal vantagem o baixo custo, e
a possibilidade de testarmos diversos
sistemas diferentes em computador. O
principal problema é a confiabilidade dos
sistemas simulados. Nem sempre é
possível obtermos modelos
computacionais realísticos para um
sistema biológico. A abordagem in silico é
comum no estudo de novos fármacos. É
uma forma de acelerar o processo de
descoberta e desenvolvimento de
fármacos, adicionando inteligência e ética
ao desenvolvimento de novos fármacos. Fármaco contra tuberculose em estudo in silico.
Fonte: Bio-Inspired Algorithms Applied to Molecular Docking
Simulations. Heberlé G, De Azevedo WF Jr. Curr Med Chem
2011; 18 (9): 1339-1352. 41

Adicionamos inteligência, pois podemos
testar milhares de sistemas em
computador. No caso específico do
desenho de fármacos in silico, podemos
testar em computador milhares, até
mesmos milhões de moléculas, que
apresentam potencial de se tornarem
fármacos.
Adicionamos uma abordagem ética ao
desenvolvimento de fármacos, pois ao
invés de sacrificarmos milhões de cobaias
em testes pré-clínicos, focamos os testes
pré-clínicos nas moléculas mais
promissoras.
Muitos testes pré-clínicos são feitos em camundongos e em
cachorros. 42

In silico (uma história de sucesso).
Algumas décadas atrás ocorreu a primeira
grande epidemia de AIDS. O HIV foi
identificado como o causador da AIDS. No
estudo da biologia do vírus foi identificado
que o vírus apresenta uma poliproteína,
que precisa ser clivada para que o HIV
seja ativo. As partes clivadas da
poliproteína formam o vírus maduro que
infecta as células. Se a clivagem for
inibida, o vírus é impedido de atingir sua
forma madura. A elucidação de estrutura
tridimensional da protease do HIV
permitiu que, o modelo chave fechadura
fosse usado para investigar in silico o
potencial de novos fármacos contra o HIV.
Estrutura tridimensional da protease do HIV resolvida por
cristalografia por difração de raios X. Código PDB: 7HVP
43

In vitro. Usa experimentos de laboratório
sem envolvimento direto de cobaias
(testes em tubos de ensaios). Tem um
custo relativamente mais alto que a
simulação computacional, mas é mais
realista, visto que os experimentos são
realizados em situações próximas às
encontradas no ser vivo.
In vivo. É o experimento mais realista,
visto que é feito em seres vivos. Tem
como principais problemas o custo
elevado, quando comparado com as
outras abordagens, e os aspectos éticos
envolvidos.
Fonte da imagem: http://www.vivopharm.com.au/us/in_vitro.php
Acessada em 08/04/2012
Cientistas demonstram a eficácia de nanopartículas para
entrega de droga anticancerígena (doxorubicin) nas células
alvos.
Fonte da imagem:
http://newsroom.ucla.edu/portal/ucla/srpview.aspx?id=138683
44

O teste mais avançado e realista dos
estudos in vivo é o testes clínico. Em tais
testes, moléculas com potencial
farmacológico são aplicados em
humanos. Os testes visam verificar se o
fármaco em potencial apresenta eficácia e
toxicidade tolerável. Normalmente, são
realizados testes pré-clínicos, onde os
fármacos em potencial são testados in
vitro e in vivo, com animais não humanos.
Classicamente os testes clínicos são
divididos nas seguintes fases: Fase 0. Tal
denominação é recente ("Guidance for
Industry, Investigators, and Reviewers".
Food and Drug Administration).
Consultado em 04/09/ 2011.) e visa testar
o fármaco em potencial em humanos em
doses sub-terapêuticas, onde é testado se
o fármaco em potencial comporta-se
como esperado.
Pílulas de óleo de Salvia officinalis (ou placebo) usada em
testes clínicos como remédio para memória. Fonte da
imagem:http://www.sciencephoto.com/media/281222/enlarg
e . Acessada em 08/04/2012 45
Fase I. Nesta fase um grupo
relativamente pequeno de indivíduos
(entre 100 e 200) são testados. A fase I
testa a tolerância da droga e usa
indivíduos saudáveis. As regras para a
participação nos testes variam de país
para país, nos EUA pagam para
indivíduos participarem de tais testes.
Fase II. Nesta fase é testada a eficácia da
droga, bem como sua toxidade. O número
de indivíduos pode chegar a 300.
Fase III. Nesta fase é realizado um
estudo multicentro e com um número
maior de pacientes, até 3000.
Fase IV. É a fase pós-mercado, ocorre
depois do fármaco ter sido liberado para
comercialização.
Seriado Two and half men. Alan foi convencido a participar
de um teste clínico.
Fonte da imagem:
http://www.youtube.com/watch?v=8ypYeK8L-DQ
Acessada em 08/04/2012 46

As regras de Lipinski (Lipinski et al., 1997,
2001) são parâmetros usados para
avaliação do potencial, que uma molécula
apresenta de ser absorvida pelo
organismo humano. Apesar de terem sido
publicadas em 1997, essas regras já eram
usadas há muito tempo pelo laboratório
farmacêutico Pfizer. O seu sucesso na
avaliação do potencial farmacológico
deve-se às seguintes razões:
Fácil aplicação;
Fácil memorização;
São evidentes do ponto de vista físico-
químico;
São baseadas em pesquisa básica
sólida e funcionam efetivamente. Lipinski, C.A., Lombardo, F., Dominy, B.W., Feeney, P.J.. Adv
Drug Deliv Rev. 1997, 23(1-3):3-25.
Lipinski, C.A., Lombardo, F., Dominy, B.W., Feeney, P.J.. Adv
Drug Deliv Rev. 2001, 46(1-3):3-26
Fonte da imagem:
http://www.sciencephoto.com/media/268423/view
Acessada em 04/05/2012)
47

As regras de Lipinski são baseadas nas
propriedades físico-químicas de
compostos que formam a base de dados
USAN (United States Adopted Names
Directory), que apresentam moléculas que
passaram a fase 1 de testes clínicos.
Estudos de fase 1 envolvem
determinação de aspectos relacionados à
toxidade e farmacocinética. Compostos
que chegam à fase 2 são aqueles que
apresentam absorção e toxidade
tolerável, o que sinaliza que investimentos
podem ser realizados em mais testes. As
regras de Lipinski analisam :
Doadores e aceitadores de ligação de
hidrogênio;
LogP;
Massa molecular.
Fonte da imagem:
http://www.sciencephoto.com/media/268488/view
Acessada em 04/05/2012)
48

As regras de Lipinski baseiam-se em
critérios físico-químicos que as drogas
devem apresentar para serem absorvidas
pelo organismo humano. A absorção está
relacionada à solubilidade e à presença
de ligações de hidrogênio. Para um dado
composto, é avaliado o número total de
potenciais átomos que podem estar
envolvidos nessas ligações, aqui
separados em duas classes na análise.
1) Número de doadores de ligação de
hidrogênio (hydrogen-bond donors)(HBD).
Indica o número de hidrogênios presentes
na molécula, que podem fazer ligação de
hidrogênio, principalmente ligados aos
oxigênio e nitrogênio.
2) Aceitadores de ligação de hidrogênio
(hydrogen-bond acceptors)(HBA). Indica o
número de aceitadores de ligação de
hidrogênio, na maioria das vezes átomos
de oxigênio e nitrogênio.
Fonte da imagem:
http://www.sciencephoto.com/media/350658/view
Acessada em 04/05/2012)
49

A aplicação das regras de Lipinski baseia-se na avaliação da solubilidade do composto
em água, o aumento do número de ligações de hidrogênio contribui para esta
solubilidade, mas dificulta a passagem pela membrana, visto que aumenta o número
de ligações de hidrogênio que devem ser quebradas para a passagem da bicamada
fosfolipídica. Vamos ver um exemplo de como contar os doadores e aceitadores de
ligação de hidrogênio. Usaremos a molécula código CID2244, disponível no site:
http://pubchem.ncbi.nlm.nih.gov/summary/summary.cgi?cid=2244&loc=ec_rcs.
Indicaremos o número de doadores como HBD e o de aceitadores como HBA. Na
molécula usaremos a seguinte notação:
Aceitador de ligação de hidrogênio
Doador de ligação de hidrogênio
50
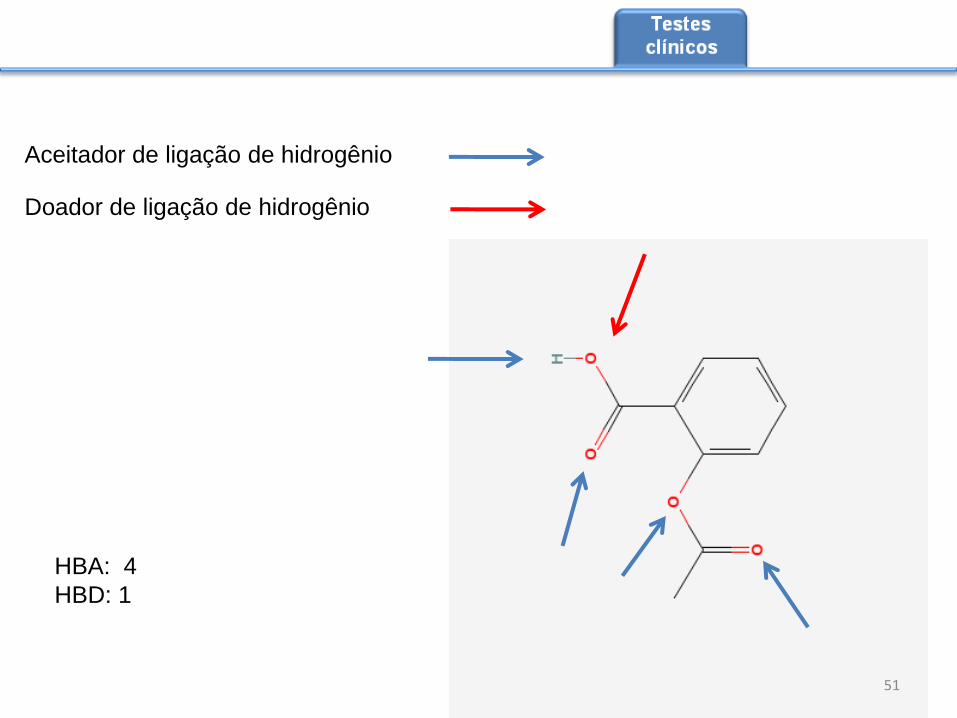
Aceitador de ligação de hidrogênio
Doador de ligação de hidrogênio
HBA: 4
HBD: 1
51

Definimos a lipofilicidade de um composto, como a tendência deste para uma
partição numa matriz lipídica apolar contra uma matriz aquosa. Uma forma de
determinarmos experimentalmente a lipofilicidade de um composto é a partir da
partição deste entre fases imiscíveis, tais como octanol (fase apolar) e um tampão
aquoso (fase polar). Os valores de partição podem ser expressos em termos de Log P,
que é o coeficiente de partição de um composto entre octanol e uma fase aquosa
(tampão onde a molécula está neutra),
onde [Composto]octanol é a concentração do composto na fase apolar, e
[Composto]aquosa é a concentração na fase aquosa. Há algoritmos estabelecidos para
determinarmos o Log P de um composto (Hansch et al., 1995; Cheng et al., 2007).
aquosa
octanol
Composto
CompostologP Log
Hansch, C., Leo, A. and Hoekman, D. (1995) Exploring QSAR. Fundamental and applications in chemistry and biology, volume
I. Hydrophobic, electronic and steric constants, volume 2. New York: Oxford University Press.
Cheng, T.; Zhao, Y.; Li, X.; Lin, F.; Xu, Y.; Zhang, X.; Li, Y.; Wang, R.*; Lai, L. "Computation of Octanol-Water Partition
Coefficients by Guiding an Additive Model with Knowledge", J. Chem. Inf. Model. 2007, 47, 2140-2148.
52

Um guia geral para absorção gastrointestinal, após administração oral da droga,
propõe que o Log P deve estar na faixa de 0 a 3 para uma situação ótima, como
indicado na figura abaixo.
Bio
dis
ponib
ilida
de o
ral
Log P
Baixa
Alta
-4 -3 -2 -1 0 1 2 3 4 5 6 7
Faixa ótima
Baixa permeabilidade
da bicamada lipídica
Baixa solubilidade aquosa
Figura baseada em Kerns, E. H. e Di, L. Drug-like properties; Concepts, structure, design and methods. Academic Press,
Elsevier, London, 2008. (pg. 45)
53

Regras de Lipinski. A partir da estrutura
de um composto podemos avaliar
características da estrutura, tais como
ligações de hidrogênio, Log P e MW. Uma
absorção e permeabilidade adequadas
são esperadas para moléculas que
apresentem as seguintes características:
HBD 5
HBA 10 (5x2)
Log P 5
MW 500 (5x100)
Todos os aspectos considerados são
múltiplos de 5, por isso tal regra é
também chamada de regra dos 5 (rule of
five).
Fonte da imagem:
http://www.sciencephoto.com/media/350657/view
Acessada em 04/05/2012)
54

Relação com as outras disciplinas
Na aula de hoje vimos detalhes sobre as
membranas celulares, um tópico
relacionado com as disciplinas Biologia
Celular e Tecidual e Biologia molecular.
Vimos a importância farmacológica de
peptídeos tóxicos e dos venenos de
serpentes, as fases dos testes clínicos e
as regras de Lipinski, aspectos
relacionados com a disciplina
Farmacologia, do sétimo semestre do
curso de Biologia. Ao analisarmos os
comportamentos das vespas, temos uma
relação clara com as disciplinas de
Zoologia I e II, do quinto e sexto
semestres, respectivamente. Obviamente,
estamos estudando as bases moleculares
da membrana celular, um tópico de
estudo da Química. O potencial de
repouso da célula é objeto da Física.
Aula de
hoje
Química
Física
Biologia
Molecular
Biologia
Celular e
Tecidual
Zoologia I e II
Farmacologia
55

Artigos indicados
Selecionei 3 artigos que considero trazem resultados interessantes sobre moléculas
que interagem com a membrana.
1) How C-terminal Carboxyamidation Alters the Biological Activity of Peptides from
the Venom of the Eumenine Solitary Wasp. Sforça ML Oyama Jr, S, Canduri F,
Lorenzi CCB, Pertinhez TA, Konno K, Souza BM, Palma MS, Ruggiero Neto J,
Azevedo Jr. WF, Spisni A. Biochemistry 2004; 43(19): 5608-5617. Este artigo
descreve as bases estruturais para a atividade biológicas dos venenos das
vespas.
2) Conformation and lytic activity of eumenine mastoparan: a new antimicrobial
peptide from wasp venom. Cabrera MPS, Souza BM, Fontana R, Konno K, Palma
MS, Azevedo Jr WF, Ruggiero Neto J. J Pept Res 2004; 64(3): 95-103. Este artigo
descreve a atividade antimicrobiana de peptídeos isolados das vespas.
3) Crystallographic and Spectroscopy characterization of a molecular hinge:
conformation change in Bothropstoxin I, a dimeric Lys49-Phospholipase
homologue. Giotto, MTS, Garratt, RC, Oliva, G, Mascarenhas, YP, Giglio, JR,
Cintra, AC, De Azevedo Jr., WF, Arni, RK, Ward, RJ. Proteins. 30(4):442-454,
1998. Este artigo descreve a estrutura da fosfolipase A2 extraída do veneno da
Bothrops jararacussu.
56

Filme indicado
Não conheço um bom filme que possa ter
relação com a aula de hoje, mas há um
filme de TV chamado de “Abelhas
Assassinas”. No filme ocorre a chegada
de abelhas agressivas numa pequena
cidade no estado de Washington-EUA.
Uma nota de interesse. Na década de
1950, na cidade de Rio Claro-SP, estudos
de cruzamentos de abelhas africanas com
abelhas europeias foram realizados, para
o aprimoramento da produção de mel. O
descuido no manejo das abelhas
africanas (muito agressivas) permitiu a
fuga de um enxame. Décadas depois,
abelhas africanizadas chegaram aos
EUA, aumentado o número de acidentes
com picadas de abelhas, o que foi
inspiração para o filme citado acima. Fonte da imagem: http://www.imdb.com/title/tt0353625/
Acessado em 19/03/2012 57

Notícia relacionada
Sempre que possível gosto de destacar a
importância do pensamento inovador no
estudo de biologia. A notícia ao lado
descreve uma empresa de Piracicaba-SP,
que vende enxames de vespa para
controle biológico de pragas em lavouras.
Essa empresa foi considerada entre as
mais inovadoras do mundo.
Fonte da notícia:
http://economia.uol.com.br/ultimas-
noticias/redacao/2012/03/02/empresa-
brasileira-de-agricultura-esta-entre-as-
mais-inovadoras-do-mundo.jhtm
58
Sites indicados
Segue uma breve descrição de dois sites relacionados à aula de hoje. Se você tiver
alguma sugestão envie-me ([email protected]).
1) http://www.susanahalpine.com/anim/Life/memb.htm . Este site mostra animações
flash do modelo de mosaico fluido. O site é em inglês.
2) http://www.brookscole.com/chemistry_d/templates/student_resources/shared_reso
urces/animations/ion_pump/ionpump.html . Este site mostra uma animação com o
funcionamento da bomba de sódio e potássio. O site é em inglês.
59

Referências
Danielli, J. F. & Davson, H. (1935). J. Cell. Comp. Physiol., 5:495-508.
Garcia, E. A. C. Biofísica. Editora Savier, 2000.
Gorter, E. & Grendel, F. (1925). J. Exp. Med. 41:439-443.
Kerns, E. H. e Di, L. Drug-like properties; Concepts, structure, design and methods.
Academic Press, Elsevier, London, 2008.
Lipinski, C.A., Lombardo, F., Dominy, B.W., Feeney, P.J.. Adv Drug Deliv Rev. 1997,
23(1-3):3-25.
Lipinski, C.A., Lombardo, F., Dominy, B.W., Feeney, P.J.. Adv Drug Deliv Rev. 2001,
46(1-3):3-26
Purves, W. K., Sadava, D., Orians, G. H., Heller, H. G. Vida. A Ciência da Biologia. 6a
ed. Artmed editora. 2002.
Schmitt, F. O., R. S. Bear, and E. Ponder. (1937) . J. Cell. Comp. Physiol 9: 89-92.
Schmitt, F. O., R. S. Bear, and E. Ponder. (1938) . J. Cell. Comp . Physiol. 11 :309-313.
Schneider, G. & Baringhaus, K. –H. Molecular Design. Concepts and Applications.
WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim, 2008
http://www1.umn.edu/ships/9-2/membrane.htm
60