
Caracterização de populações microbianas em células de ... · o conhecimento da natureza e...
87
Caracterização de populações microbianas em células de combustível para a produção de bioeletricidade Ana Mafalda Marques Cardoso Dissertação para a obtenção do grau de Mestre em Bioengenharia e Nanossistemas Orientador: Prof. Doutor Jorge Humberto Gomes Leitão Júri Presidente: Prof. Doutor Luís Joaquim Pina da Fonseca Orientador: Prof. Doutor Jorge Humberto Gomes Leitão Vogal: Prof. Doutor Gabriel António Amaro Monteiro Novembro 2014
Transcript of Caracterização de populações microbianas em células de ... · o conhecimento da natureza e...

Caracterização de populações microbianas em células de combustível para a produção de bioeletricidade
Ana Mafalda Marques Cardoso
Dissertação para a obtenção do grau de Mestre em
Bioengenharia e Nanossistemas
Orientador: Prof. Doutor Jorge Humberto Gomes Leitão
Júri
Presidente: Prof. Doutor Luís Joaquim Pina da Fonseca
Orientador: Prof. Doutor Jorge Humberto Gomes Leitão
Vogal: Prof. Doutor Gabriel António Amaro Monteiro
Novembro 2014


i
Agradecimentos
Em primeiro lugar, gostaria de agradecer ao Instituto Superior Técnico da Universidade
de Lisboa que facultou o espaço e todas as condições para que a minha formação académica
e científica fosse possível, bem como a todos os docentes que contribuíram para o mesmo.
Ao coordenador do Mestrado em Bioengenharia e Nanossistemas, Professor Doutor
Luís Joaquim Pina da Fonseca, agradeço a oportunidade em frequentar este curso de
Mestrado que contribuiu imenso para o enriquecimento da minha formação académica.
Ao meu orientador, Professor Doutor Jorge Humberto Gomes Leitão, expresso o meu
profundo agradecimento pela orientação, competência científica e disponibilidade, assim como
pelas críticas, correções e pela revisão desta tese de Mestrado. Agradeço também por me ter
recebido no seu grupo de investigação e pela confiança que depositou em mim em todas as
fases do projeto.
Ao doutorando e amigo, André Grilo, agradeço toda a orientação, permanente
disponibilidade e tempo dispensado em ajudar-me passo a passo todas as técnicas
laboratoriais e rotinas do laboratório. O teu acompanhamento e generosidade foram muito
importantes no desenvolvimento deste trabalho.
A todos os meus amigos que me acompanharam neste percurso, não pretendendo
individualizar, um enorme agradecimento por todo o apoio, amizade e partilha dos bons e maus
momentos.
Ao Alexandre, um agradecimento especial pelo apoio incondicional, pelas palavras
certas, por todo o carinho e amizade e especialmente pela paciência e compreensão. Por tudo,
a minha enorme gratidão.
À minha família, um enorme e especial agradecimento, sobretudo aos meus pais e à
minha “mana”. Obrigado por terem acreditado em mim, pelo apoio incondicional, amizade e
motivações diárias. Sem vocês nada disto seria possível e por isso este trabalho é dedicado a
vocês.

ii

iii
Resumo
O crescimento da população mundial e o consequente aumento de resíduos urbanos e
industriais tem suscitado um grande problema ambiental. Os combustíveis fósseis também têm
levantado grandes preocupações uma vez que estes recursos naturais não são renováveis e o
seu uso excessivo levará a um previsível esgotamento das jazidas. Desta forma, procuram-se
desenvolver alternativas que atenuem estes impactos e ainda fazer um aproveitamento das
águas residuais.
As células de combustível microbianas (Microbial Fuel Cells- MFC) constituem uma
nova tecnologia que permite a produção de bioeletricidade de forma sustentável e a partir de
compostos biodegradáveis. Os substratos podem ser hidratos de carbono simples ou, como no
caso em estudo, substratos mais complexos, presentes em águas residuais. De forma a
otimizar e desenvolver a capacidade de produção de bioeletricidade pelas MFCs é fundamental
o conhecimento da natureza e metabolismo dos microrganismos capazes de usar um ânodo
como aceitador de eletrões. O presente trabalho teve como objetivo a caracterização das
populações microbianas presentes em dois reatores anaeróbios distintos, produzindo
eletricidade a partir de acetato como fonte de carbono.
Para tal, recorreu-se a uma metodologia que combina técnicas de biologia molecular, a
análise dos perfis de restrição obtidos após hidrólise com a enzima de restrição Hae III e o uso
de bases de dados contendo genomas microbianos total ou parcialmente sequenciados.
A metodologia utilizada demostrou ser eficaz e identificaram-se 960 clones
correspondentes a um total de 142 perfis de restrição diferentes, resultando na identificação de
58 espécies diferentes. As espécies de bactérias mais abundantes nos reatores pertenciam às
ordens Burkholderiales, Pseudomonadales, Rhodocyclales e Xanthomonadales, pertencentes
ao filo Proteobacterias. No caso dos organismos do domínio Archaea, as espécies mais
abundantes pertenciam à ordem Methanomicrobiales do filo Euryarchaeota.
Palavras-chave: Células de combustível microbianas, bioeletricidade, reatores anaeróbios,
caracterização microbiana, técnicas de biologia molecular.

iv

v
Abstract
World population growth leads to a rapid increase in urban and industrial waste which
renders waste management as a significant environment threat. There’s also a big concern
about the use of fossil fuels because they’re not renewable and its abusive use will end up with
their extinction. Many of the environmental problems our country faces today result from our
fossil fuel dependence. The development of new methods of sustainable energy production
combining wastewater treatment are therefore urgently required.
Microbial fuel cells (MFC) represent a new technology for the production of renewable
energy by direct conversion of organic matter to electricity using bacteria. MFC’s can convert
practically any organic material like simple carbohydrates or, as is the case of the present work,
complex substrates into fuel. For MFC optimization, a complete characterization of the microbial
communities that thrive in the anode compartment is required. The aim of this work is to
characterize the microbial populations of two distinct MFC using acetate as the carbon source
for electricity production.
The methodology used in this study combines molecular biology techniques to obtain of
the 16S rRNA genes and restriction profiles analyses after hydrolysis with the restriction
enzyme Hae III. Databases searches for sequences homologous to the nucleotide sequences
of 16S rRNA encoding genes cloned were subsequently performed.
This methodology has led to the identification of 960 clones belonging to the Bacteria or
Archaea domains, corresponding to a total of 142 distinct restriction profiles, and 58 different
species. The estimative of the species relative abundances showed a large number of bacteria
belonging to Burkolderiales, Pseudomonadales, Rhodocyclales e Xanthomonadales, all of them
orders of Proteobacteria. Archaea exhibiting a high relative abundance belonged to the
Methanomicrobiales, order of the Euryarchaeota.
Keywords: Microbial fuel cells, bioelectricity, anaerobic reactors, microbial characterization,
molecular biology techniques.

vi

vii
Índice
Resumo ................................................................................................................................... iii
Abstract .................................................................................................................................... v
Índice de figuras ...................................................................................................................... ix
Índice de tabelas ..................................................................................................................... xi
Lista de abreviações .............................................................................................................. xiii
1-Introdução............................................................................................................................. 1
1.1. As MFCs (Microbial Fuel Cells) ................................................................................... 1
1.2. Princípio de funcionamento e configuração das MFCs ............................................... 2
1.3. Aplicação das MFCs ................................................................................................... 4
1.4. Transferência eletrónica ............................................................................................ 5
1.5. Processos metabólicos nas MFC................................................................................. 6
1.6. Formação de biofilmes .............................................................................................. 7
1.7. Bactérias electroquimicamente ativas ....................................................................... 8
1.8. Análise da comunidade microbiana ........................................................................... 9
1.9. Aplicação das técnicas de biologia molecular em estudos de biodiversidade. ........... 12
1.10. Metodologias de biologia molecular .................................................................... 13
1.11. Objetivos do trabalho .......................................................................................... 15
2. Material e Métodos............................................................................................................ 17
2.1. Modo de operação e controlo dos bioreatores ............................................................. 17
2.2. Recolha das amostras ................................................................................................. 18
2.3. Avaliação da diversidade das comunidades bacterianas................................................ 19
2.4. Extracção e Purificação de DNA cromossómico ............................................................. 20
2.5. Electroforese em gel de agarose ................................................................................... 20
2.6. Amplificação dos genes rRNA 16S ................................................................................. 20
2.7. Purificação dos produtos de PCR .................................................................................. 22
2.8. Clonagem ..................................................................................................................... 22
2.9. Transformação ............................................................................................................. 23
2.10. Extração de DNA plasmídico ....................................................................................... 24
2.11. Análise dos perfis de restrição .................................................................................... 25
2.12. Análise das sequências dos genes de rRNA 16S ........................................................... 26
3. Resultados e discussão ....................................................................................................... 27
3.1. Recolha das amostras e identificação ........................................................................... 27

viii
3.2. Análise das electroforeses em gel de agarose ............................................................... 27
3.3. Análise dos perfis de restrição dos clones obtidos ........................................................ 30
3.4. Composição das populações microbianas ..................................................................... 32
3.5. Discussão de resultados................................................................................................ 40
3.5.1. Reactor 1 ............................................................................................................... 40
3.5.2. Reactor 2 ............................................................................................................... 43
4. Conclusões ......................................................................................................................... 47
5. Perspectivas futuras ........................................................................................................... 49
6. Bibliografia ......................................................................................................................... 51
Anexo I ................................................................................................................................... 63

ix
Índice de figuras
1. Introdução
Figura 1- Esquematização esquemática dos componentes básicos de uma MFC. 2
Figura 2- Representação esquemática da transferência eletrónica direta. 9
Figura 3: Árvore filogenética universal determinada pela comparação das
sequências rRNA. 14
2. Materiais e Métodos
Figura 4: Fotografia dos bioreatores MFCs donde foram recolhidas as amostras
de biomassa analisados neste trabalho . 17
Figura 5: Representação esquemática dos dois compartimentos dos reatores
MFC, reator 1 e reator 2, com condições de operacionalidade diferentes no
cátodo. 17
Figura 6- Representação esquemática da metodologia usada neste trabalho
para a identificação dos microrganismos presentes nas células microbianas. 18
Figura 7- Representação gráfica, marcas de resistência gene LacZα e
representação esquemática do príncipio para clonagem do fragmento no
vetor PCR2.1 21
3. Resultados e Discussão
Figura 8 – Electroforese em gel de agarose dos amplificados obtidos após reação
de PCR usando DNA total extraído do cátodo e ânodo do reator 1 para o R1 na
fase de menor produção de bioelectricidade. 27
Figura 9 - Electroforese em gel de agarose dos amplificados obtidos após reação
por PCR do DNA total extraído do anôdo do R2 nas duas fases de produção
de bioelectrocidade. 27
Figura 10 – Electroforese em gel de agarose dos amplificados obtidos após
reação por PCR do DNA total extraído do cátodo do reator 2 nas duas fases
de produção de bioelectrocidade. 28
Figura 11: Exemplos de alguns perfis obtidos usando primers para bactérias. 31
Figura 12: Exemplos de alguns perfis obtidos usando primers para archaeas. 31
Figura 13: Comparação da abundância relativa das espécies identificadas no
compartimento anódico do reator 1 no compartimento anódico nas fases de
maior e menor produção de bioeletricidade. 37
Figura 14: Comparação da abundância relativa das espécies identificadas no
compartimento catódico do reator 1 no compartimento anódico nas fases de
maior e menor produção de bioeletricidade. 37
Figura 15: Comparação da abundância relativa das espécies identificadas no
compartimento catódico do reator 2 no compartimento anódico nas fases de

x
maior e menor produção de bioeletricidade 38
Figura 16: Comparação da abundância relativa das espécies identificadas no
compartimento catódico do reator 2 no compartimento anódico nas fases de
maior e menor produção de bioeletricidade 39
Figura 17- Heat map para bactérias e archaea cujas sequências de genes para o
rRNA 16S são mais próximas das sequências obtidas para os dois reatores, nas
duas fases, em ambos os compartimentos. 40
Anexo I
Figura 18: Árvore filogenética correspondente às espécies de bactérias identificadas
no compartimento anódico no reator 1 na fase de maior produção de energia
(R1A_B1). 63
Figura 19: Árvore filogenética correspondente às espécies de bactérias identificadas
no compartimento catódico do reator 1 na fase de menor produção de energia
(R1C_B2). 63
Figura 20: Árvore filogenética correspondente às espécies de archaeas identificadas
no compartimento anódico do reator 1 na fase de maior produção de energia
(R1A_A1). 64
Figura 21: Árvore filogenética correspondente às espécies de bactérias identificadas
no compartimento catódico do reator 1 na fase de maior produção de energia
(R1C_B1). 65
Figura 22: Árvore filogenética correspondente às espécies de bactérias identificadas
no compartimento catódico do reator 1 na fase de menor produção de energia
(R1C_B2). 66
Figura 23: Árvore filogenética correspondente às espécies bacterianas identificadas
no compartimento anódico do reator 2 na fase de maior produção de energia
(R2A_B1). 67
Figura 24: Árvore filogenética correspondente às espécies de Archaea identificadas
no compartimento anódico do reator 2 na fase de maior produção de energia
(R2A_A1). 68
Figura 25: Árvore filogenética correspondente às espécies de bactérias identificadas
no compartimento anódico do reator 2 na fase de menor produção de energia
(R2A_B2). 69
Figura 26: Árvore filogenética correspondente às espécies de bactérias identificadas
no compartimento catódico do reator 2 na fase de maior produção de energia
(R2C_B1). 70
Figura 27: Árvore filogenética correspondente às espécies de bactérias identificadas
no compartimento catódico do reator 2 na fase de menor produção de energia
(R2C_B2). 71

xi
Índice de tabelas
1. Introdução
Tabela 1: Microrganismos capazes de produção de corrente elétrica em cultura
pura numa MFC. 11
2. Materiais e Métodos
Tabela 2 – Sequência e características dos oligonucleótidos iniciadores usados
neste trabalho. 20
Tabela 3- Sumário do programa do PCR (30 ciclos) 20
3. Resultados e Discussão
Tabela 4 - Nomenclatura adotada para designar as amostras de cada reator e
distingui-las quanto ao tipo de organismo, compartimento do reator e fase de
produção de bioeletricidade. 26
Tabela 5: Comparação entre o número total de clones obtido, o número inicial de
perfis diferentes que correspondeu a diferentes clones para sequenciar e o número
efetivo de perfis diferentes. 30
Tabela 6: Espécies microbianas identificadas como as mais próximas das
sequências obtidas por PCR no compartimento anódico do reator 1, na fase de
maior produção de energia. 32
Tabela 7: Espécies microbianas identificadas como as mais próximas das
sequências obtidas por PCR no compartimento anódico do reator 1, na fase de
menor produção de energia. 33
Tabela 8: Espécies microbianas identificadas como as mais próximas das
sequências obtidas por PCR no compartimento catódico do reator 1, na fase de
maior produção de energia. 33
Tabela 9: Espécies microbianas identificadas como as mais próximas das
sequências obtidas por PCR no compartimento catódico do reator 1, na fase de
menor produção de energia. 34
Tabela 10: Espécies microbianas identificadas como as mais próximas das
sequências obtidas por PCR no compartimento anódico do reator 2, na fase de
menor produção de energia. 34
Tabela 11: Espécies microbianas identificadas como as mais próximas das
sequências obtidas por PCR no compartimento anódico do reator 2, na fase de
menor produção de energia. 35
Tabela 12: Espécies microbianas identificadas como as mais próximas das
sequências obtidas por PCR no compartimento catódico do reator 2, na fase de
maior produção de energia. 35

xii
Tabela 13: Espécies microbianas identificadas como as mais próximas das
sequências obtidas por PCR no compartimento catódico do reator 2, na fase de
menor produção de energia. 36

xiii
Lista de abreviações
APS - Persulfato de amónia (Ammonium persulfate)
DNA - Ácido desoxirribonucleico (Deoxyribonucleic acid)
DNTP- Desoxirribonucleotídeos fosfatados (Deoxyribonucleotides)
E. Coli - Escherichia coli
LB - Luria broth
MFC- Células de combustível microbianas (Microbial Fuel Cells)
PCR- Reação em cadeia da polimerase (Polymerase Chain Reaction)
RFLP – Polimorfismo dos fragmentos de restrição (Restriction Fragment Length Polymorphism)
RNA - Ácido ribonucleico (Ribonucleic acid)
TEMED- Tetrametiletilenodiamina (Tetramethylethylenediamine)
TBE- Tris-Borato-EDTA (Tris-borate-EDTA)
TAE- Tris-Acetato-EDTA (Tris-acetate-EDTA)

xiv

1
1-Introdução
Com o aumento da população mundial, a quantidade de resíduos industriais e
domésticos tem aumentando cada vez mais e, embora a população dos países mais
desenvolvidos apresente taxas de crescimento mais baixas, a produção de resíduos nestes
países é extremamente alta. Também os combustíveis fósseis, que têm sustentado o
crescimento industrial e comercial dos países desenvolvidos nos últimos dois séculos, têm
suscitado uma grande preocupação ambiental. Como é evidente, os combustíveis fósseis não
irão conseguir suportar indefinidamente a economia global, além da sua combustão acarretar
vários problemas ambientais, essencialmente devido à emissão de dióxido de carbono.
A exploração de fontes renováveis de energia nos últimos anos tem sido tema de
várias discussões, sendo favorecida face aos combustíveis fósseis, responsáveis em grande
parte pelo aquecimento global e alterações climáticas. A grande preocupação sobre as
recentes alterações climáticas, o aumento da procura pelo petróleo e reservas de gás natural
estão a intensificar a procura de alternativas aos combustíveis fósseis. Atendendo a estes
problemas, o interesse por alternativas energéticas aumentou em todo o mundo.
Recentemente, os investigadores foram também consideravelmente atraídos pelas
características das células combustíveis microbianas ou Microbial Fuel Cells (MFC) que têm a
capacidade de gerar bioeletricidade a partir do metabolismo de compostos orgânicos por
populações microbianas. Esta tecnologia apresenta várias vantagens pois são sistemas limpos,
altamente eficientes, renováveis e com baixa emissão de poluentes. No entanto, a produção de
bioeletricidade a partir de fermentações anaeróbias apresenta ainda várias limitações no que
diz respeito ao seu armazenamento e baixas taxas de produtividade (Logan, 2004).
Atualmente, as MFCs são a melhor opção para a recuperação e conversão de energia
(bioeletricidade), constituindo uma tecnologia futurista capaz de transformar diretamente
energia química em energia elétrica através de reações químicas catalisadas por
microrganismos (Bullen et al., 2006) (Logan, et al., 2006).
1.1. As MFCs (Microbial Fuel Cells)
O primeiro registo de trabalhos relacionados com as MFCs data do ano de 1911,
quando Potter descreveu a produção de energia elétrica a partir de uma cultura mista de
Escherichia coli e Saccharomyces. Só no ano de 1931, é que Cohen foi capaz de produzir uma
voltagem superior a 35V a partir de uma MFC em série. No entanto, existiam ainda muitas
dúvidas em relação ao funcionamento das MFCs e à produção de eletricidade a partir delas.
Em 1980, Allen e Bennetto descobriram que a densidade de corrente e a potência gerada
podiam ser melhoradas usando microrganismos na superfície do ânodo e mediadores de

2
eletrões. A maioria das membranas bacterianas são compostas por lípidos, peptidoglicanos e
lipopolissacáridos, não-condutores, que impedem a transferência de eletrões diretamente para
o ânodo. No entanto, uns anos mais tarde, descobriu-se que certos organismos tinham a
capacidade de fazer a transferência eletrónica diretamente para o ânodo sem a necessidade
de mediadores (Kim, et al., 1999) (Chaudhuri & Lovley, 2003). A partir daí, as MFC foram
consideradas como uma tecnologia viável para gerar energia elétrica e o número de
publicações relacionadas com as MFCs aumentou exponencialmente. Atualmente sabe-se que
a eletricidade gerada é produzida diretamente pela degradação de matéria orgânica (Rabaey &
Verstraete, 2005).
1.2. Princípio de funcionamento e configuração das MFCs
Tal como uma célula electroquímica comum, as MFCs apresentam dois
compartimentos, um contendo o ânodo e outro contendo o cátodo. Nas MFCs, os
microrganismos oxidam compostos orgânicos e transferem os eletrões para o ânodo que
funciona como aceitador de eletrões da cadeia respiratória (Rabaey & Verstraete, 2005). Na
Figura 1 está esquematizada uma célula combustível microbiana constituída pelos
compartimentos anódico e catódico, separados por uma membrana de permuta de protões.
Figura 1- Esquematização esquemática dos componentes básicos de uma MFC (adaptado de Vinay Sharma e Kundu,
2010)

3
Os compostos orgânicos mais utilizados como substratos da reação são glucose,
acetato ou águas residuais. O tipo de compostos utilizados no ânodo é de extrema importância
uma vez que este deve privilegiar o crescimento e fixação de bactérias e ainda manter um
ambiente propício para que estas mantenham a sua atividade eletroquímica adequada à
produção de energia (Liu et al., 2012). Toda a viabilidade dos sistemas MFC prende-se com a
existência de um meio de anaerobiose no ânodo (Kim et al., 2008). Desta forma, os
microrganismos metabolizam a matéria orgânica produzindo eletrões que, por um processo
enzimático, vão resultar em energia para a célula sob a forma de adenosina trifosfato (ATP).
Os eletrões são então libertados para um aceitador de eletrões que pode ser oxigénio, nitrato,
sulfato, entre outros, e os produtos desta reação rapidamente se difundem no meio (Logan,
2008). No entanto sabe-se agora que algumas bactérias são capazes de fazer a transferência
eletrónica de forma exógena à célula, transferindo os eletrões diretamente a um aceitador de
eletrões como um óxido de metal. As bactérias crescem no compartimento anódico e oxidam a
matéria orgânica, libertando eletrões para o ânodo e protões para o meio. A equação (1) é um
exemplo referente a este processo, utilizando acetato como substrato/fonte de carbono.
CH3COO- + 2H2O → 2CO2 + 7H+ +8e- (1)
De seguida os protões resultantes migram para o compartimento catódico através de
uma membrana que permite a troca de protões de forma a igualar a carga transferida pelos
eletrões garantindo a eletroneutralidade de todo o processo. A membrana presente no circuito
é seletiva para protões, impedindo assim a passagem dos eletrões evitando a ocorrência de
um curto-circuito e também impede a passagem de gases intervenientes, um problema
designado por permeabilidade de gás. Esta deve ser ainda resistente às condições geradas
pela redução no cátodo e pela oxidação do ânodo. Por outro lado os eletrões produzidos são
transferidos para o cátodo por um circuito externo. No cátodo, os protões vão reagir com
oxigénio e com os eletrões transferidos pelo circuito externo, formando água – equação (2)
abaixo (Rabaey et al., 2005):
4 H+ + O2 + 4 e- → 2 H2O (2)
O percurso realizado pelos eletrões através do circuito externo vai gerar uma corrente
elétrica que pode ser posteriormente quantificada (Bockris & Srinivasan, 1969).
Resumidamente, o processo global baseia-se na conversão do material orgânico biodegradável
a dióxido de carbono e água, gerando eletricidade. Nestas células as reações ocorrem
controladamente de forma a minimizar ou mesmo eliminar, a presença de subprodutos
indesejáveis.
A transferência direta dos eletrões para o ânodo é muitas vezes dificultada por
sobretensões que causam uma perda da corrente elétrica total obtida pelas MFCs. As
sobretensões são perdas de potencial devido a resistências à transferência de eletrões e a
resistências internas. Segundo Rabaey e Verstraete (2005), este parece ser atualmente o
principal fator limitante da baixa produtividade nas MFCs, sendo provocado pela energia de

4
ativação que as bactérias necessitam para oxidarem os compostos à superfície do ânodo. A
sobretensão pode ser atenuada adicionando, por exemplo, um catalisador (Park & Zeikus,
2003). Também o cátodo pode ser afetado por sobretensões que levam a uma perda
significativa de potencial. Este efeito pode ser atenuado pela adição de soluções de
hexacianoferrato. De forma a tornar o sistema MFC o mais sustentável possível, o cátodo deve
ser exposto ao ar (Rabaey & Verstraete, 2005).
As MFCs destacam-se da maioria dos processos bioenergéticos uma vez que operam
de uma forma eficaz tanto à temperatura ambiente como a baixas temperaturas e também não
é necessário qualquer tratamento dos gases emitidos. Apesar de este ser um processo
altamente eficiente, a grande desvantagem das MFCs continua a ser a sua baixa produção de
energia. Ainda assim, a alta eficiência de produção de eletrões e baixa produção de energia
das MFCs pode ser vista como uma vantagem na alimentação de sistemas de telemetria e
sensores sem fio, que não requerem grandes consumos de energia para transmitir sinais para
recetores situados em locais remotos (Shantaram et al., 2005)
1.3. Aplicação das MFCs
O tratamento de águas residuais é, muito provavelmente, a aplicação mais importante
das MFCs. As águas residuais contêm uma grande quantidade de energia sob a forma de
matéria orgânica biodegradável que pode ser reutilizada para a produção de energia elétrica.
Pelo contrário, as estações de tratamento gastam imensa energia para remover a matéria
orgânica em vez de tentar recuperá-la. Dependendo do conteúdo orgânico das águas
residuais, podem ser aplicados diversos bioprocessos para tratar e reutilizar as águas residuais
de indústrias, agricultura ou resíduos urbanos. A utilização de um sistema MFC teria a
capacidade de tratar as águas residuais através da redução do nível de matéria orgânica.
Moléculas orgânicas como acetato, propionato ou butirato, facilmente seriam quebradas com a
formação de CO2 e H2O. A utilização de certos organismos seria outro benefício das MFCs, por
exemplo a inclusão de microrganismos que apresentam a capacidade de remoção de sulfitos
tal como acontece em estações de tratamento e que poderiam ser incorporados nas MFCs
(Rabaey, 2006). A recuperação de energia das águas residuais iria amortizar o custo do seu
tratamento e reduzir a atual dependência dos combustíveis fósseis, tornando as estações de
tratamento de águas residuais autossustentáveis (Logan, 2008). As MFCs já demonstraram ser
eficazes no tratamento de quase todos os tipos de resíduos gerados, incluindo águas residuais
domésticas, agrícolas, de indústrias como as cervejeiras, refinarias, reciclagem de papel e até
de lixiviados de aterros (Pant D et al., 2010). De forma a atingir este objetivo, as MFCs teriam
que ser otimizadas e dimensionadas à escala industrial e doméstica. Para isso é ainda
necessário compreender a importância de vários fatores que afetam o desempenho das MFCs

5
como a área superficial dos elétrodos, a sua condutividade, concentração de substratos e
ainda, o papel das comunidades microbianas. Idealmente, as MFCs em conjunto com as
energias eólica, solar, geotérmica e das marés, podem contribuir para o cessar do uso de
combustíveis fósseis.
1.4. Transferência eletrónica
De forma a criar condições para que a respiração celular ocorra numa MFC com a
conversão do substrato a CO2, os eletrões têm que ser transferidos extracelularmente para o
elétrodo. As vias metabólicas envolvidas e os mecanismos bioelectroquímicos de conversão de
energia são determinantes na quantidade de energia gerada. A transferência eletrónica que
ocorre no ânodo é apenas devido à necessidade que as bactérias têm de eliminar os eletrões
libertados durante a oxidação do substrato (Schröder, 2007). Certas bactérias conseguem fazer
este processo diretamente mas em certos casos a utilização de mediadores para a
transferência eletrónica é necessária (Schröder, 2007). Diversos trabalhos recentes revelaram
a existência de um terceiro mecanismo de transferência eletrónica, fornecendo evidências de
que as bactérias sintetizam apêndices designados por nanofios (Reguera et al, 2005). Dois
géneros bacterianos capazes de fazer a transferência eletrónica recorrendo a nanofios são
Shewanella e Geobacter (Gorby e Beveridge, 2005). No geral, a maioria das células
bacterianas mantêm-se electroquimicamente inativas devido à estrutura da parede celular
essencialmente composta por material não condutor, tal como lípidos e peptidoglicanos.
Embora a parede celular das bactérias gram-negativas tenha uma superfície eletricamente
carregada sobretudo devido aos lipopolissacáridos presentes na membrana exterior, a
capacidade de permeação para transferência dos eletrões é reduzida. Desta forma, para que a
transferência eletrónica ocorra é necessário a utilização de moléculas mediadoras.
As moléculas mediadoras são intermediários capazes de aceitar eletrões da cadeia
transportadora de eletrões do microrganismo, atravessar a parede e transportar os eletrões até
ao ânodo, onde se oxidam. Os mediadores mais utilizados são a tionina, benzilviologenio, 2.6-
diclorofenolindofenol, 2-hidroxi-1,4-naftoquinona e diversas fenazinas, fenotiazinas,
fenoxoazinas, vermelho neutro, entre outros (Reddy et al., 2010). As principais desvantagens
da utilização de certos tipos de mediadores são a necessidade de reposição constante, o
elevado custo e a formação de resíduos tóxicos para os microrganismos. De entre as bactérias
que requerem mediadores para a transferência eletrónica para o ânodo, encontra-se
Escherichia coli, Actinobacillus succinogenes e Proteus vulgaris (Park & Zeikus, 2000; Kim, et
al., 2000). Por outro lado, as espécies Shewanella putrefaciens, Geobacteraceae
sulfurreducens, Geobacter metallireducens e Rhodoferax ferrireducens são
bioelectroquimicamente ativas e conseguem formar um biofilme na superfície do ânodo e fazer

6
a transferência eletrónica diretamente através da condutância da membrana (Du et al., 2007).
Neste caso o ânodo serve como aceitador final de eletrões da cadeia respiratória dos
microrganismos que compõem o biofilme. Devido a todas as desvantagens económicas e
ambientais do uso de mediadores, a transferência direta de eletrões da cadeia respiratória para
o elétrodo tem sido a opção mais viável (Kim et al., 1999).
O metabolismo energético dos microrganismos do ânodo varia desde anaerobiose a
anaerobiose facultativa (Rabaey e Verstraete, 2005). Os biofilmes também se podem formar na
superfície do cátodo e também podem desempenhar um papel importante na transferência
eletrónica entre os microrganismos. Neste caso, certas espécies como G. metallireducens e G.
sulfurreducens, podem funcionar como aceitadores finais de eletrões, pois recebem os eletrões
do cátodo que funciona como dador de eletrões (Gregory et al, 2004). No cátodo a redução do
oxigénio pode ser catalisada pelos microrganismos, permitindo o seu crescimento usando os
eletrões provenientes do ânodo. As bactérias do cátodo podem obter energia a partir desta
reação porque recebem os eletrões com um potencial superior ao necessário para a redução
do oxigênio (Logan, 2009). Outros mecanismos de redução podem ocorrer no cátodo:
desnitrificação, redução de água (H2O) a peróxido de hidrogénio (H2O2), desidroalogenação,
redução de metais, produção de hidrogénio, produção de metano, redução de perclorato, entre
outros. O facto de os eletrões poderem ser tanto libertados como aceites pelas bactérias,
sugere que a transferência eletrónica entre células é um fenómeno que ocorre naturalmente
em comunidades microbianas (Logan, 2009). Os organismos que populam o cátodo, que
atuam como biocatalisadores, são autotróficos e consomem parte da voltagem para o
crescimento e manutenção celular (Clauwaert et al. 2007).
1.5. Processos metabólicos nas MFC
Rabaey e Verstraete (2005) relacionaram o potencial anódico com as vias metabólicas
utilizadas pelos organismos. De facto, a par dos substratos, o potencial do ânodo vai
determinar o metabolismo bacteriano. Se a corrente da MFC aumenta, o potencial no ânodo
diminui, obrigando as bactérias a entregar os eletrões através de complexos mais reduzidos. O
potencial do ânodo irá influenciar o potencial redox do transportador de eletrões final e assim,
influenciar o metabolismo. Os vários tipos de vias que podem ser distinguidas com base no
potencial do ânodo são: mecanismo oxidativo de elevado potencial redox; mecanismo oxidativo
de potencial redox médio a baixo; e fermentação.
Para potenciais anódicos elevados, as bactérias podem usar a cadeia respiratória do
metabolismo oxidativo. Neste caso os eletrões são transportados através de NADH
desidrogenase, ubiquinona, coenzima Q ou citocromo (Kim et al., 2004). Este processo é
regularmente usado nas MFCs usando culturas mistas com Pseudomonas aeruginosa,

7
Enterococcus faecium e Rhodoferax ferrireducens com uma eficiência estimada de até 65%
(Rabaey et al., 2003). Se a MFC estiver a ser operada com uma resistência externa baixa, a
corrente elétrica durante a formação da biomassa vai ser baixa e por essa razão o potencial da
MFC vai ser também baixo. Por outro lado, o potencial do ânodo vai ser elevado o que resulta
na seleção de organismos aeróbios facultativos e anaeróbios. À medida que a cultura vai
crescendo, a atividade metabólica vai aumentar e portanto, a corrente aumenta, favorecendo o
crescimento de organismos anaeróbios facultativos de baixo potencial redox, devido à
moderação do potencial do ânodo (Rabaey e Verstraete, 2005).
Quando o ânodo apresenta valores mais baixos de potencial mas existem em solução
aceitadores de eletrões como sulfato, os eletrões vão ser entregues preferencialmente a esses
aceitadores. A produção de metano pode ser um indicador de que as bactérias não estão a
usar o ânodo como aceitador de eletrões (Kim, et al., 2004). Por outro lado, se não existirem
aceitadores alternativos de eletrões e no caso de o potencial anódico ser baixo, o processo
usado preferencialmente pelas bactérias é a fermentação. Vários microrganismos dos géneros
Clostridium, Alcaligenes e Enterococcus foram isolados de MFCs e identificados com este tipo
de metabolismo (Rabaey, et al., 2004) (Park, et al., 2001). A fermentação pode resultar numa
produção de acetato ou butirato (Rabaey e Verstraete, 2005). Estes produtos da fermentação
podem ser oxidados por outro tipo de bactérias anaeróbias, como espécies do género
Geobacter, que conseguem produzir eletrões a partir do acetato ou butirato (Vandevivere e
Verstraete, 2001). Se o sistema MFC for operado a uma resistência elevada, o potencial do
ânodo vai também ser baixo mesmo para baixos valores de corrente, levando à seleção de
anaeróbios facultativos de baixo potencial redox e anaeróbios obrigatórios (Rabaey e
Verstraete, 2005).
1.6. Formação de biofilmes
A formação de um biofilme na superfície dos elétrodos é essencial para que a
transferência de eletrões numa MFC seja bem-sucedida. O material dos elétrodos tem ainda
um efeito no tipo de bactérias que aderem e por isso, consoante o tipo de material, a estrutura
microscópica da superfície dos elétrodos vai ser diferente, influenciando as espécies
microbianas aderentes. As bactérias que constituem os biofilmes crescem, reproduzem-se e
produzem substâncias poliméricas extracelulares. Estas substâncias ocupam um volume para
além do volume ocupado pela superfície das células. As células microbianas facilmente se
aderem a superfícies imersas em solução aquosa e a esta estrutura, composta de polímeros e
biomassa aderida, dá-se o nome de biofilme (Liu e Tay, 2000). Biofilmes electroquimicamente
ativos têm uma grande importância, principalmente na oxidação/redução de metais, mas
também na dissolução de minerais, no ciclo do carbono, entre outros (Logan, 2008). Os

8
biofilmes não são estruturas regulares e uniformes, e a maior parte das vezes são constituídos
por mais do que uma espécie de microrganismos. Fatores como a disponibilidade de
nutrientes, caudal, pH, temperatura, entre outros, têm uma grande influência na formação e
estrutura dos biofilmes (Zhang et al., 2011). A resistência externa pode até influenciar a
estrutura dos biofilmes formados nos elétrodos. Como foi dito anteriormente, a resistência
externa, tal como o potencial do ânodo, têm uma grande influência na produção de energia,
controlando as condições de crescimento dos microrganismos bioeletroquimicamente ativos.
De facto, esta é uma questão que tem despertado imenso interesse entre os investigadores e é
possível afirmar que o desempenho de um sistema MFC melhora para valores menores de
resistência externa aplicada (Zhang et al., 2011). Lyon et al. (2010) verificaram grandes
diferenças tanto na energia gerada como na estrutura da comunidade microbiana para
diferentes valores de resistência externa. De facto, se for usada matéria orgânica como
substrato numa MFC com valores de resistência externa baixos, vai haver a formação de mais
eletrões por unidade de tempo a movimentarem-se do biofilme para o elétrodo e por isso, mais
protões a serem produzidos no interior do biofilme. A grande produção de protões leva a uma
diminuição do pH no biofilme, criando um ambiente desfavorável a grande parte dos
microrganismos electroquimicamente ativos. Por outro lado, também se sabe que a estrutura
dos biofilmes facilmente se adapta a alterações do meio em que estão inseridos e por essa
razão, é possível afirmar que a morfologia dos biofilmes é diferente para diferentes valores de
resistência externos (Zhang et al., 2011).
1.7. Bactérias electroquimicamente ativas
A descoberta da existência de bactérias capazes de reduzir ferro para a produção de
energia numa MFC, sem a necessidade de usar mediadores, sugere que esta é uma
característica rara e limitada a um certo número de organismos. No entanto já se verificou que
quatro das cinco classes de Proteobacteria e também alguns organismos dos filos
Acidobacteria e Firmicutes têm a capacidade de gerar energia elétrica sem a utilização de
mediadores (Logan, 2009). Estes microrganismos que são capazes de fazer a transferência
eletrónica extracelularmente são chamados de bactérias exoeletrogénicas ou, mais comumente
designadas, bactérias electroquimicamente ativas. Os eletrões são libertados com a produção
de ATP na cadeia respiratória durante a fosforilação oxidativa. Normalmente, o aceitador final
de eletrões é o oxigénio (respiração aeróbia) mas neste caso as células vão usar outros
compostos como aceitadores finais de eletrões, como por exemplo citocromos, num processo
designado por respiração anaeróbia (Figura 2, A).
Existem pelo menos três mecanismos através dos quais os microrganismos fazem a
transferência eletrónica extracelularmente. O primeiro tipo de mecanismo envolve compostos

9
de ferro, mais especificamente óxidos de Fe (III) que são os aceitadores de eletrões mais
conhecidos. Muitas estirpes de bactérias conseguem libertar os eletrões provenientes da
cadeia respiratória para um óxido de Fe (III) fora da célula, produzindo Fe (II) solúvel (Logan,
2009). Outro mecanismo envolve a transferência eletrónica diretamente de uma célula
microbiana para outra, ou seja, sem a utilização de qualquer intermediário. A espécie
fermentativa Pelotomaculum thermopropionicum liga-se com recurso a filamentos eletricamente
condutores, os nanofios, a células da espécie Methanothermobacter thermautotrophicus
constituindo a primeira evidência de transferência elétrica direta entre diferentes espécies
(Gorby et al., 2006) (Figura 2, B). Várias bactérias utilizam nanofios para a transferência
eletrónica, no entanto apenas foram estudadas com mais detalhe as bactérias Shewanella
oneidensis e Geobacter sulfurreducens. (Malvankar e Lovley, 2014). Malvankar e Lovley (2014)
sugeriram a manipulação genética para aumentar a produção de nanofios e melhorar a sua
condutividade de forma a aumentar a produção de energia elétrica nas MFCs. Uma hipótese
que foi colocada é que a transferência eletrónica entre espécies possa ser uma via de
comunicação entre elas, facto que ainda não foi estudado experimentalmente (Logan, 2009).
Figura 2: Representação esquemática da transferência eletrónica direta via: (A) citocromos, (B) nanofios. (Adaptado de
Arora, 2012).
1.8. Análise da comunidade microbiana
Como foi referido anteriormente, as bactérias redutoras do compartimento anódico
utilizam o ânodo como aceitador final de eletrões. Os mais recentes estudos nas MFC estão
muito centralizados em melhorar a potência gerada de forma a desenvolver sistemas
economicamente fiáveis. O conhecimento das comunidades microbianas que constituem os
biofilmes é muito importante para uma melhor compreensão dos mecanismos de transferência
eletrónica que ocorrem nas MFCs (Rabaey et al., 2005). Como também já foi referido, as MFCs

10
podem ser operadas com culturas puras ou culturas mistas. O estudo de MFCs com culturas
puras é bastante importante para determinar a capacidade de gerar energia e mecanismos de
transferência eletrónica de uma dada estirpe. Existem diversas publicações com a identificação
de muitas espécies com a capacidade de produzir corrente elétrica numa cultura pura,
espécies pertencentes a todas as classes de Proteobacterias. A espécie Geobacter
sulfurreducens tem servido como modelo para clarificar os processos moleculares de
transferência eletrónica. Foi um organismo utilizado em vários estudos e foi até hoje o
organismo que mais eletricidade produziu em cultura pura (Nevin, et al., 2008). A Tabela 1
reúne os organismos capazes de produzir energia elétrica numa MFC a partir de uma cultura
pura.

11
Tabela 1: Microrganismos capazes de produção de corrente elétrica em cultura pura numa MFC.
Microrganismo Grupo taxonómico Referência
Shewanella putrefaciens IR-1 γ-proteobacteria (Kim, et al., 1999)
Proteus vulgaris γ-proteobacteria (Kim, et al., 2000)
Clostridium butyricum Firmicutes (Park, et al., 2001)
Desulfuromonas acetoxidans δ-proteobacteria (Bond, et al., 2002)
Geobacter metallireducens δ-proteobacteria (Bond, et al., 2002)
Rhodoferax ferrireducens β-Proteobacteria (Chaudhuri & Lovley, 2003)
Aeromonas hydrophila A3 δ-proteobacteria (Pham, et al., 2003)
Geobacter sulfurreducens δ-proteobacteria (Bond & Lovley, 2003)
Desulfobulubs propionicus δ-proteobacteria (Holmes, et al., 2004)
Pseudomonas aeruginosa γ-proteobacteria (Rabaey, et al., 2004)
Geopsychrobacter electrodiphilus δ-proteobacteria (Holmes, et al., 2004)
Geothrix fermentans Acidobacteria (Bond & Lovley, 2005)
Saccharomyces cerevisiae Fungi (Walker & Walker, 2006)
Escherichia coli γ-proteobacteria (Zhang, et al., 2006)
Shewanella oneidensis DSP10 γ-proteobacteria (Ringeisen, et al., 2007)
Shewanella oneidensis MR-1 γ-proteobacteria (Bretschger, et al., 2007)
Desulfitobacterium hafniense Firmicutes (Milliken & May, 2007)
Hansenula anómala Fungi (Prasad, et al., 2007)
Acidiphilium sp. 3.2Sup5 α-Proteobacteria (Borole, et al., 2008)
Ochrobactrum antrophi YZ-1 α-Proteobacteria (Zuo, et al., 2008)
Rhodopseudomonas palustris DX-1 α-Proteobacteria (Xing, et al., 2008)
Desulfovibrio desulfuricans δ-proteobacteria (Zhao, et al., 2008)
Therminicola sp. Strain JR Firmicutes (Wrighton, et al., 2008)
Klebsiella pneumoniae L17 γ-proteobacteria (Zhang, et al., 2008)
Arcobacter butzleri ε-proteobacteria (Fedorovich, et al., 2009)
Bacillus subtilis Firmicutes (Nimje, et al., 2009)
Thermincola ferriacetica Z-0001 Firmicutes (Marshall & May, 2009)
Enterobacter cloacae γ-proteobacteria (Rezaei, et al., 2009)
Serratia marcescens γ-proteobacteria (Ghanapriya, et al., 2010)
Corynebacterium sp. strain MFC03 Firmicutes (Liu, et al., 2010)
Nocardiopsis sp. KNU (S strain) Actinobacteria (Hassan, et al., 2012)
Streptomyces enissocaesilis KNU (K
strain) Actinobacteria (Hassan, et al., 2012)
Calditerrivibrio nitroreducens Deferribacteres (Fu, et al., 2013)
Tolumonas osonensis γ-proteobacteria (Luo, et al., 2013)
Cupriavidus basilensis β-Proteobacteria (Friman et al., 2013)

12
No entanto, numa perspetiva mais realista para a aplicação das MFCs ao nível do
tratamento de águas residuais, a utilização de uma cultura mista é mais adequada uma vez
que as culturas puras metabolizam um número limitado de compostos orgânicos. Além disso,
Logan e seus colaboradores (2006) fizeram uma revisão onde se concluiu que as MFCs que
utilizam consórcios de bactérias conseguem produzir substancialmente mais eletricidade do
que culturas puras. Também Ishii et al. (2008) mostrou a partir de uma MFC com apenas uma
câmara (cátodo exposto ao ar), que uma cultura mista produziu 22% mais eletricidade que uma
cultura pura de Geobacter sulfurreducens. Isto pode ser explicado porque geralmente culturas
mistas apresentam maior resistência sob condições de stress, maior taxa de consumo de
substratos e menor especificidade nos substratos (Rabaey et al., 2004). De facto, a utilização
de uma cultura mista de bactérias faz um melhor aproveitamento de toda a matéria orgânica
existente.
Nestes últimos anos foram feitas várias análises ao conteúdo microbiano de biofilmes
de várias MFCs, e verificou-se que não existe nenhuma comunidade bacteriana típica, ou seja,
todas as comunidades analisadas foram diferentes e não existe um organismo mais
proeminente (Aelterman, 2009). Possivelmente também existem ainda bastantes espécies de
bactérias que ainda não foram identificadas como aptas para fazer transferência eletrónica
para um ânodo (Logan et al. 2006). A composição da comunidade microbiana vai depender de
vários fatores como o tipo de inóculo, a configuração da MFC, substratos adicionados e
condições de operacionalidade. Por este motivo torna-se muito importante conhecer as
espécies bacterianas uma vez que, dependendo destes fatores, as bactérias vão usar
diferentes vias metabólicas, sendo por isso um método seletivo e determinante da composição
das populações microbianas. Dado que na MFC os microrganismos vão servir de catalisadores
na transferência de eletrões do substrato para o ânodo, a seleção de bactérias com grande
desempenho é crítica.
1.9. Aplicação das técnicas de biologia molecular em estudos de
biodiversidade
Outrora, a caracterização de populações microbianas baseava-se fundamentalmente
na observação microscópica, nas suas necessidades nutricionais e utilização de diferentes
substratos para a aquisição de carbono e energia e meio de cultivo para o seu crescimento
(métodos ex-situ) (Kennedy, 1999). No entanto, os resultados destas técnicas tornam-se
bastante limitados quando o objetivo é estudar a diversidade de microrganismos que está
associada a um determinado meio ambiente. Os meios de cultivo são seletivos para grupos
específicos o que na maior parte dos casos é extremamente difícil ou mesmo impossível, uma
vez que a maioria dos microrganismos não é sequer cultivável. Por outro lado, muitas vezes

13
não é possível criar as condições para o cultivo de certas espécies cultiváveis, principalmente
devido à falta de informação sobre as necessidades metabólicas e nutricionais de grande parte
dos microrganismos. Com a introdução de técnicas de biologia molecular, houve uma
revolução nas metodologias tradicionais pois não é necessário o cultivo nem o isolamento dos
microrganismos, tornando possível a identificação de microrganismos não cultiváveis,
desconhecidos ou não identificados, e que muitas vezes são determinantes e responsáveis
pelo bom funcionamento de uma cultura. Com base na composição da comunidade microbiana
é ainda possível fazer uma antevisão das funcionalidades de cada espécie, interações
biológicas com o meio e com outros microrganismos e por isso estimar a estabilidade do
processo de tratamento anaeróbio. Normalmente a cooperação entre diferentes
microrganismos oferece um ambiente propício que permite a otimização do processo global
(Wagner & Loy, 2002). Com este conhecimento é possível fazer uma previsão das espécies
que mais contribuem para a produção de bioeletricidade e caracterizar e determinar a
capacidade degradativa da população, indispensável a qualquer biotratamento. O
conhecimento da dinâmica das populações permite ainda o conhecimento das espécies
indesejáveis ou das variações em termos de composição, da estrutura da população ao longo
do processo ou como resultado de variações nas condições ambientais, permitindo ainda o
processo de biotratamento e produção de bioelectricidade.
1.10. Metodologias de biologia molecular
A introdução dos métodos de biologia molecular veio dar suporte às técnicas tradicionais e
embora a atual classificação taxonómica ainda esteja parcialmente baseada nas propriedades
morfológicas e fisiológicas microbianas, cada vez mais se verifica que um grande número de
microrganismos se encontra incorretamente classificado. O método mais utilizado para a
identificação de populações microbianas foi proposto por Pace et al. (1986), que sugeriram o
uso do gene rRNA 16S como marcador molecular ideal para o estudo de populações
microbianas em amostras ambientais. Foi a partir deste método que um ano depois, Carl
Woese e seus colaboradores propuseram uma nova árvore filogenética universal, criada com o
recurso aos métodos moleculares e em particular por recurso à comparação das sequências
nucleotídicas dos genes que codificam o RNA da subunidade menor dos ribossomas, o RNA
16S em procariontes (Figura 3).

14
Figura 3: Árvore filogenética universal determinada pela comparação das sequências rRNA. Uma matriz de
distâncias evolutivas foi calculada a partir dos alinhamentos das sequências de rRNA 16S representativas de cada um
dos três reinos (Carl Woese, 1987).
Com esta proposta, houve um grande incentivo a procurar novas estratégias para
caracterizar o papel dos microrganismos num dado meio e ainda estudar as alterações na
dinâmica das populações. A partir daí, os métodos baseados na análise e sequenciação do
gene rRNA 16S tornaram-se nos mais usados para a análise das comunidades bacterianas
(Dahllöf 2002). Estes métodos foram também bastante bem sucedidos no que diz respeito ao
estudo de comunidades microbianas do solo, mar, rios, lagos, trato gastrointestinal, diversas
amostras clínicas e ainda de reatores anaeróbios para o tratamento de águas residuais (Theron
and Cloete 2000; Narihiro and Sekiguchi 2007).
O conhecimento da composição da comunidade microbiana torna possível antever as
funções de cada espécie, interações biológicas com o meio e com outros microrganismos. A
aquisição de todas estas informações é bastante vantajosa na determinação da estabilidade de
um processo de biotratamento. Normalmente a cooperação entre diferentes microrganismos
oferece um ambiente propício que permite a otimização do processo global (Wagner & Loy,
2002).
Nos últimos anos várias metodologias têm sido desenvolvidas e aperfeiçoadas contribuindo
para o avanço do conhecimento da diversidade genética dos microrganismos, tais como as
análises metagenómicas em que se determinam e analisam as sequências dos genes
codificantes para o rRNA 16S, ou determinando a totalidade das sequências de DNA existentes
na amostra recolhida.

15
1.11. Objetivos do trabalho
A diversidade microbiana em reatores anaeróbios tem sido largamente estudada nos
últimos anos e por isso têm sido obtidas informações muito importantes para a compreensão
da produção de energia nas MFCs. No entanto, existem ainda várias questões que requerem
uma melhor compreensão para compreender a influência da diversidade microbiana na
eficiência das MFCs. Neste contexto, os principais objetivos deste trabalho foram:
i) Caracterizar as populações microbianas existentes em dois reatores anaeróbios
com cátodos distintos (abiótico e biocátodo) com base no conhecimento das
sequências codificantes para o 16S rRNA;
ii) Investigar a diversidade microbiana nos reatores a partir da análise dos perfis dos
fragmentos de restrição dos clones obtidos;
iii) Construção de árvores filogenéticas e comparação da comunidade bacteriana
entre os dois reatores;
iv) Identificação das espécies bioelectroquimicamente ativas por comparação com
literatura disponível;
v) Correlacionar a diversidade microbiana com as respetivas condições de
operacionalidade dos reatores.

16

17
2. Material e Métodos
2.1. Modo de operação e controlo dos bioreatores
As amostras analisadas no presente trabalho foram provenientes de dois bioreatores
MFCs (R1 e R2) com condições de operacionalidade distintas (Figura 4A), operados pelo Dr.
Ramana S. Venkata sob orientação do Prof. Luís Fonseca do Grupo de Bioengenharia do IST.
Os reatores foram inoculados com lamas de uma cultura mista aeróbia, coletadas numa
estação de tratamento de águas residuais (Chelas, Lisboa). O reator 1 foi inoculado com as
lamas apenas no compartimento anódico enquanto o reator 2 foi inoculado nos dois
compartimentos. A alimentação dos reatores foi feita a partir de águas residuais sintéticas.
Como principais fontes de carbono, o ânodo do reator 1 foi enriquecido com acetato de sódio
(10mM) e o ânodo do reator 2 com acetato de sódio (10mM) e carbonato de sódio (3mM).
Foram ainda fornecidas uma solução tampão de fosfato (50mM) e uma solução vitaminada
respetivamente, para o ânodo do reator 1 e para o cátodo do reator 2. No compartimento
catódico do reator 1 o católito usado foi apenas a solução tampão de fosfato (50mM). O pH dos
reatores foi mantido a 7 nos dois compartimentos de ambos os reatores. Os dois
compartimentos apresentavam o mesmo volume e eram separados por uma membrana de
Nafion de condutividade 9.0 ± 5 mS/ cm-1
. Os dois compartimentos catódicos foram
continuamente providos de oxigénio através de uma bomba de ar de forma a manter constante
a quantidade de oxigénio dissolvido (4 ± 0.2 mg/l). Por cada ciclo em modo batch e após a
alimentação e colheita de amostras, os compartimento anódicos foram borbulhados com azoto
gasoso durante 20 minutos, de forma a remover oxigénio dissolvido e a garantir que o meio se
mantinha anaeróbio. Para melhorar a transferência de massa, os compartimentos foram
continuamente agitados com recurso a um agitador magnético. Os meios foram renovados
sempre que a voltagem diminuía até aos 50mV e era nessa altura que a biomassa suspensa
era recolhida, completando assim, um ciclo do reator. Os reatores foram operados durante 28
ciclos à temperatura ambiente (aproximadamente 25°C), correspondendo a 220 dias, enquanto
os valores máximos reprodutíveis de tensão fossem mantidos. Os elétrodos estavam
conectados através de um fio de cobre a uma resistência externa de 1000Ω. A Figura 4B
apresenta uma representação esquemática do modo de operação dos dois reatores.

18
Figura 4: (A) Fotografia dos bioreatores MFCs donde foram recolhidas as amostras de biomassa analisados neste
trabalho, reator 1 à esquerda e reator 2 à direita, com condições de operacionalidade distintas; (B) Representação
esquemática dos dois compartimentos dos reatores MFC, reator 1 e reator 2, com condições de operacionalidade
diferentes no cátodo. (A) cátodo abiótico no R1 e (B) biocátodo no R2. R representa a resistência externa.
2.2. Recolha das amostras
Após o final de um ciclo do reator, as amostras das lamas foram recolhidas por raspagem
dos biofilmes existentes na região dos elétrodos e o material recolhido foi imediatamente
congelado a -80°C. De cada compartimento, de cada reator, foram retiradas amostras de dois
períodos distintos, no período de maior produção de energia e após os 150 e 165 dias para o
reator 2 e reator 1, respetivamente, onde a produção de bioeletricidade era mais baixa (figura
5). A análise da biodiversidade foi investigada para os dois períodos.
(A)
(B)

19
Figura 5: Variação da voltagem (mV) para o reator 1 e reator 2 ao longo do tempo.
2.3. Avaliação da diversidade das comunidades bacterianas
Todo o procedimento experimental para a elaboração deste trabalho foi baseado na
metodologia proposta por Ramos et al. (2008), seguindo uma estratégia que combina a
extração de DNA cromossómico total das amostras dos biofilmes, amplificação dos genes que
codificam para o rRNA 16S por PCR, clonagem e sequenciação dos clones previamente
selecionados a partir de análise do polimorfismo dos fragmentos de restrição (RFLP) obtidos
após hidrólise dos fragmentos de DNA das sequências clonadas. O procedimento está
esquematizado na Figura 6 e é desenvolvido detalhadamente em seguida.
Figura 6- Representação esquemática da metodologia usada neste trabalho para a identificação dos
microrganismos presentes nas células microbianas.
Extração e purificação de DNA cromossómico
Amplificação por PCR
Clonagem
Estabelecimento de perfis de restrição
Sequenciação
Análise bioinformática
Identificação das sequências do rRNA 16S mais próximas

20
2.4. Extração e purificação de DNA cromossómico
A extração e purificação de DNA cromossómico é um processo que se baseia na
utilização de soluções de lise para romper a membrana da célula pela adição de detergentes
para remover lípidos de membrana e ainda promover a lise por adição de proteases e RNAses
para remoção de proteínas e RNA, respetivamente. Após este processo é promovida a
precipitação do DNA pela adição de etanol e acetato de sódio. Devido à insolubilidade do DNA
no etanol, este vai agregar-se e precipitar após centrifugação, facilitando o seu isolamento.
Neste trabalho a extração de DNA cromossómico foi realizada usando o kit comercial “High
Pure PCR Template Preparation Kit” (Roche) segundo as instruções do fabricante, o que
permitiu a purificação de ácidos nucleicos de culturas para posterior processamento.
De modo a analisar o grau de pureza do DNA genómico extraído, este foi analisado
num espectrofotómetro (NanoDrop ® ND-1000 Technologies) por UV/Vis. Através do cálculo
das razões Abs260/Abs280 e Abs260/Abs230 obteve-se uma estimativa do estado de pureza do
DNA (Holler, Skoog, & Crouch).
2.5. Electroforese em gel de agarose
A integridade do DNA genómico purificado foi analisada após electroforese em gel de
agarose (0,8%) em tampão TAE 1x (TAE 50x: 242 g/l Tris Base, 57,1 ml/L acido acético, 100
ml/L de 0.5 EDTA, pH 8) (Sambrook e Russel, 2001). As amostras a analisar foram preparadas
adicionando 10% do volume de corante de aplicação (1% SDS, 50% de Glicerol e 0,05% de
Azul de Bromofenol, 10x concentrado) e submetidas a uma voltagem de 90V durante
aproximadamente 1 hora. Por fim, os géis foram corados por imersão em tampão contendo Gel
Red e visualizados num sistema Gel Doc XR (BioRad, Universal Hood II).
2.6. Amplificação dos genes rRNA 16S
De modo a amplificar os segmentos de DNA dos genes que codificam para o RNA da
subunidade ribossómica 16S, recorreu-se ao PCR usando os oligonucleótidos iniciadores
descritos na Tabela 2. A técnica de PCR permite amplificar pequenos fragmentos do genoma
obtendo-se grandes quantidades de determinada região de DNA. São necessários
oligonucleótidos iniciadores universais ou específicos (sequências iniciadoras),
complementares às sequências localizadas em locais específicos do genoma.

21
Tabela 2 – Sequência e características dos oligonucleótidos iniciadores usados neste trabalho.
Domínio Temperatura
de annealing
(°C)
Forward primer
(5’-3’)
Reverse primer
(3’-5’)
Tamanho
esperado
(bp)
Referências
Bactéria 70 CCAGATCCTACGGGAGGCAG
C
CTTGCTCGGGCCCCGTCAATT
C
605 Rudi et al.,
1997
Archaea 56 ACTGCTCAGTAACACGT CTCCCCCGCCAATTCTTTA 803 Lueders e
Friedrich,
2000
As reações de PCR continham, num volume total de 20 μl, 0,4 μl de cada
oligonucleótido iniciador, 1,5 μl de MgCl2 (1,5 mM), 0,4 μl de DNTPs (0,1 mM), 5 μl de DNA
molde (concentração aproximada de 200 ng), 0,2 μl de Taqmed DNA polimerase (1 U) e 2 μl do
respetivo tampão de reação (10x concentrado) fornecido pelo fabricante da polimerase
(Citomed).
Normalmente a temperatura de annealing é cerca de 5°C abaixo da menor temperatura
de melting do par de oligonucleótidos iniciadores utilizado. Assim, pela fórmula para a
temperatura de melting proposta por Wallace e os seus colaboradores (1979) (Tm (°C) = 2 ( A
+ T ) + 4 ( G + C ) ), tem-se que a temperatura ótima de annealing para qualquer par de primers
pode ser calculada pela seguinte fórmula empírica:
Ta (°C) = 2 ( A + T ) + 4 ( G + C ) - 5°C
em que A, T, C e G da fórmula referem-se ao número de vezes que cada base está presente
na sequência iniciadora. No caso específico deste trabalho, a temperatura ótima de annealing é
de 58,2°C.
A tabela seguinte faz um sumário do programa de PCR que foi usado no trabalho.
Tabela 3- Sumário do programa do PCR (30 ciclos)
Temperatura (°C) Duração (min)
Desnaturação 95 5:00
Annealing
(30 ciclos)
95 0:45
58,2 1:00
72 0:45
Extensão 72 7:00
Fim 4 ∞

22
A obtenção dos produtos de amplificação após a reação de PCR foi confirmada por
electroforese em gel de agarose. Para confirmar o tamanho dos fragmentos amplificados foi
necessário utilizar um marcador de pesos moleculares, neste caso o marcador de origem
comercial 1 Kb Plus (Fermentas). Os géis foram corados e as bandas visualizadas num
transiluminador UV Gel Doc XR (BioRad, Universal Hood II).
2.7. Purificação dos produtos de PCR
Após as reações de amplificação de DNA por PCR, procedeu-se à purificação dos
amplificados de forma a garantir que o DNA se mantinha estável durante os períodos de tempo
de armazenamento. A purificação pode ser feita diretamente a partir dos produtos de PCR mas
também é possível recuperar e purificar DNA contido nos géis de agarose. A recuperação das
amostras em gel foi efetuada no presente trabalho recorrendo ao kit Gel Pure (NZYTech). Este
kit é utilizado para a purificação de fragmentos de DNA entre 100 bp a 10kb, retidos em géis de
agarose. Este método baseia-se na utilização de membranas de sílica-gel que absorvem
seletivamente até 10 μg de fragmentos de DNA na presença de tampões de ligação
específicos. O tampão de ligação vem incluído no kit e contém um pH adequado à ligação do
DNA sem que este interfira com a ligação (ver protocolo do kit) A grande desvantagem é que
neste passo existe o risco de se perderem as extremidades de adenosina dos fragmentos de
DNA e por esta razão, o rendimento do processo poderá ser inferior uma vez que estas
extremidades são indispensáveis para que os produtos de PCR se liguem ao vetor de
clonagem usado.
2.8. Clonagem
Na clonagem, os produtos de PCR purificados são ligados a um vetor de DNA que
passa a ser replicado quando introduzido numa célula hospedeira. A clonagem foi feita
recorrendo ao kit comercial TA Cloning (Invitrogen), que de uma forma rápida e num passo
apenas, permite fazer a inserção dos produtos de PCR no vector. O kit inclui os seguintes
componentes: vector linearizado pCR®2.1, T4 DNA Ligase e respetivo tampão. Este método
baseia-se no facto da Taq polimerase ter uma atividade que adiciona uma única adenosina (A)
ao terminal 3’ dos produtos de PCR (Figura 7). Por outro lado, o vector linearizado fornecido
com o kit apresenta resíduos de timina (T) no terminal 3’, o que vai permitir a ligação eficaz dos
produtos de PCR ao vector linearizado uma vez que as extremidades são coesivas (protocolo
do TA Cloning Kit da Invitrogen). Após as reações de ligação, é promovida a precipitação do
DNA por forma a remover a ligase, bem como o excesso de sais presente no tampão de
ligação, pois estes podem interferir negativamente no processo de eletroporação. A
precipitação foi feita adicionando-se acetato de sódio a 3M (10% do volume total da reação) e

23
etanol (2,5x o volume final da reação), seguido de uma centrifugação de 30 min, 15000 rpm, a
4ºC (Sambrook e Russel, 2001). Após centrifugação, o DNA foi lavado com etanol a 70% e
seco. Por fim, adicionou-se água destilada para ressuspender o DNA das amostras e em
seguida procedeu-se à transformação de células de Escherichia coli.
Figura 7- Representação gráfica, marcas de resistência gene LacZα (esquerda) e representação esquemática do
príncipio para clonagem do fragmento no vetor PCR2.1 (TA Cloning Kit, Invitrogen).
2.9. Transformação
A transformação foi efetuada por dois métodos diferentes, eletroporação ou
transformação clássica, dependendo da disponibilidade do equipamento e das células
disponíveis para o efeito.
No caso da transformação clássica, a suspensão contendo o plasmídeo e as células foi
colocada em gelo durante aproximadamente 30 minutos. De seguida, esta foi colocada
durante 3 minutos num banho a 43°C causando um choque térmico para as células
competentes captarem o DNA e por fim, 5 minutos em gelo. De seguida as amostras
foram colocadas num tubo contendo meio LB líquido e incubado a 37ºC, a 250
rotações num agitador orbital durante 1 hora. As células competentes foram
preparadas a partir de uma colónia de Escherichia coli αDH5 e colocada em 10 ml de
meio LB líquido a 37°C durante a noite de forma a favorecer o seu crescimento.
Seguidamente 4 ml da cultura foram adicionadas a meio LB líquido, durante
aproximadamente 1 hora, até que a densidade ótica inicial (640 nm) de 0,1 aumentasse
até atingir um valor de 0,5. As células foram recolhidas por centrifugação durante 5 min
a 5600 rpm e a 4°C e no final o sobrenadante foi removido. Adicionou-se 100 ml de
MgCl2 0,1M para ressuspender delicadamente o sedimento e centrifugou-se uma vez
mais. De seguida, ressuspenderam-se as células em 100 ml de CaCl2 0,1M e durante
30 minutos foram mantidas em gelo. No final, as células foram recolhidas por
centrifugação e ressuspensas em 22 ml de CaCl2 0,1 M, adicionou-se 3,5 ml de glicerol

24
a 86% e distribuiu-se a suspensão resultante em alíquotas de 300 µl por tubos
eppendorfs. Todo o processo foi realizado em condições de esterilidade e em gelo.
No caso da transformação por electroporação, depois da mistura de ligação de
interesse ter sido incorporada num eppendorf com as células electrocompetentes, a suspensão
resultante foi colocada num sistema de electroporação da BioRad com os seguintes
parâmetros: 2,5 V de tensão e 400 Ω de resistência. Estes valores foram usados para as
células de E. coli. Após a electroporação, as células foram imediatamente colocadas num tubo
com 1 ml de meio LB líquido e incubadas durante aproximadamente 1 hora a 37ºC sob
agitação (250 rpm).
As células electrocompetentes foram preparadas a partir de uma colónia de
Escherichia coli αDH5, colocadas em 10 ml de meio LB líquido e incubados a 37°C durante a
noite. De seguida, um volume adequado da cultura foi transferido para um novo erlenmeyer de
100 ml com LB líquido de modo a obter-se uma densidade óptica (DO) inicial de 0,1 a 640 nm.
A cultura foi incubada até atingir uma DO de 0,8. As células foram depois recolhidas por
centrifugação a 10000 rpm durante 15 minutos a 4°C e o sedimento resultante foi lavado com
água destilada estéril e fria. Foi efetuada novamente uma lavagem mas com 4 ml de glicerol
frio a 10% e por fim ressuspendeu-se em 2 ml de glicerol frio a 10%. No final a suspensão foi
aliquotada em volumes de 110 μl em eppendorfs e conservados a -70°C. Todo o processo foi
efetuado em condições rigorosas de esterilidade e gelo.
Após a transformação espalhou-se a suspensão em meio sólido seletivo para posterior
seleção de clones. A suspensão foi então espalhada em placas de LB sólido com o antibiótico
canamicina, previamente tratadas com X-gal e colocadas na estufa a 37°C durante a noite. O
uso de canamicina em detrimento da ampicilina tem a vantagem de garantir que não se
desenvolvem colónias satélite. As bactérias, ao receberem este DNA, adquirem a capacidade
de crescer em meio de cultura contendo antibiótico assegurando que apenas as células que
expressam o gene de resistência ao antibiótico conseguem crescer na placa. Dado que o
plasmídeo usado com o vetor de clonagem contém o gene lacZ’ que codifica a enzima β-
galactosidase responsável pela clivagem da galactose, a presença de X-gal faz com que as
colónias das bactérias que contenham estes plasmídeos se tornem azuis. A cor azul deve-se à
hidrólise do X-gal pela enzima β-galactosidase formando um produto azul. No entanto, caso
ocorra inserção de DNA no plasmídeo, o gene lacZ’ é interrompido e as colónias que contêm o
DNA exógeno ficam brancas uma vez a enzima β-galactosidase não é produzida. A estirpe
utilizada no trabalho é E.coli αDH5. (Sambrook & Russel, 2001).
2.10. Extração de DNA plasmídico
Os plasmídeos de interesse foram extraídos segundo a técnica de lise alcalina
proposto originalmente por Birnboim e Doly (1979). Este método baseia-se na diferença na
desnaturação, em condições alcalinas (pH ~12), entre o DNA plasmídico e o DNA
cromossómico de alto peso molecular. Plasmídeos de grandes dimensões apresentam

25
propriedades semelhantes ao DNA cromossómico e por essa razão a sua separação é
dificultada. Primeiramente foi efetuada a lise das células com a adição da solução I composta
por Tris-HCl, EDTA (pH=8) e glucose. De seguida adicionou-se uma outra solução de lise,
solução II, que contém SDS para lisar a membrana das células e desnaturar proteínas e NaOH
que eleva o pH para valores alcalinos (pH~12). Nestas condições, ocorre a desnaturação do
DNA cromossómico mas não do DNA plasmídico de menores dimensões. Posteriormente foi
adicionada a solução III, composta por acetato de potássio e ácido acético. A adição de acetato
de potássio tem como objetivo neutralizar a solução devido à elevada concentração de iões H+,
provocando a precipitação do DNA cromossómico. A adição da solução III provoca ainda a
precipitação de proteínas complexadas com SDS, agregadas pela adição da solução II. Após a
adição das três soluções foi efetuada uma centrifugação durante 10 minutos a velocidade
elevada, em que o sobrenadante contendo o DNA plasmídico foi transferido para um novo
tubo. O DNA plasmídico foi então precipitado com a adição de etanol e, no final, a amostra foi
seca sob vácuo. A integridade do DNA foi avaliada por eletroforese em gel de agarose e a
concentração estimada pela absorvância a 260 nm recorrendo ao espectrofotómetro NanoDrop
® (ND-1000 Technologies) (Sambrook & Russel, 2001).
2.11. Análise dos perfis de restrição
A técnica de RFLP (Restriction Fragment Length Polymorphism) baseia-se na
comparação dos perfis de restrição obtidos após hidrólise do DNA com enzimas de restrição e
separação em gel de agarose ou poliacrilamida. O procedimento para obter os perfis de
restrição baseia-se em digestões recorrendo a enzimas de restrição. No presente trabalho
usou-se a enzima Hae III para clivar as ligações fosfodiéster entre G e C nas posições onde a
sequência GGCC é encontrada, originando um grande número de fragmentos de diversos
tamanhos.
Como os fragmentos gerados apresentaram tamanhos muito reduzidos, para uma
melhor resolução foram separados em gel de poliacrilamida (10%) num sistema vertical Mighty
Small (Hoefer). O gel era composto por acrilamida a 30%, tampão TBE (10x) (TBE 10x: 108 g/L
Tris Base, Ácido bórico 55 g/L, 7,44g Na2EDTA•2H2O), água desionizada, persulfato de
amónia (APS) (10%) e tetrametiletilenodiamina (TEMED). Após a polimerização do gel, as
amostras foram adicionadas aos poços do gel e o sistema foi submetido a uma voltagem de
110V. Os géis foram corados por imersão em tampão TAE contendo GelRed e visualizados no
sistema UV Gel Doc XR (BioRad, Universal Hood II). A partir das imagens obtidas, as amostras
foram agrupadas de acordo com o seu perfil de restrição, de maneira a evitar a posterior
sequenciação dos mesmos clones.
Os plamídeos foram sequenciados pela firma Operon MWG Eurofins, usando o primer
universal de sequenciação M13 (5′-CAG GAA ACA GCT ATG ACC-3′).

26
2.12. Análise das sequências dos genes de rRNA 16S
As sequências de DNA obtidas pela sequenciação dos plasmídeos correspondentes
aos diferentes clones de rRNA 16S foram analisadas segundo o programa bioinformático
VecScreen, disponível no portal do NCBI (National Center for Biotecnology Information)
(http://www.ncbi.nlm.nih.gov/). As sequências ao serem analisadas por este programa são
submetidas a um alinhamento local contra o banco de dados de vetores, o Univec. Desta
forma, o programa forneceu diversas informações entre as quais a localização das sequências
contaminantes e os fragmentos suspeitos. Após a remoção das sequências referentes ao vetor,
as sequências restantes foram submetidas à pesquisa de similaridade de nucleótidos através
da ferramenta BLAST (Basic Local Alignment Search Tool). Esta ferramenta permitiu a
comparação entre as sequências de nucleótidos resultantes da sequenciação com sequências
da base de dados. Como resultado, foi possível inferir as relações funcionais e evolucionárias
entre sequências e a identificação dos microrganismos mais similares geneticamente.
2.13. Análise filogenética dos segmentos de rRNA 16S
A construção de árvores filogenéticas é importante de forma a avaliar a comunidade
microbiana presente nos reatores em questão. De forma a obter as árvores filogenéticas, as
sequências dos segmentos de rRNA 16S, clonados e identificados nas bases de dados foram
alinhadas através do software MEGA4. A partir dos alinhamentos foi possível construir árvores
filogenéticas que estimam as divergências temporais, evoluções moleculares e sequências
ancestrais e ainda testar hipóteses evolucionárias. O algoritmo utilizado no MEGA4 foi o
ClustalW, indicado para alinhamentos de várias sequências. Desta forma, as sequências mais
similares, isto é, as que apresentam um melhor score no alinhamento, são as primeiras a ser
alinhadas. Geralmente, as árvores resultantes são próximas do ideal, especialmente quando o
conjunto de dados contém sequências com diferentes graus de divergência (Thompson et al.,
1994).

27
3. Resultados e discussão
3.1. Recolha das amostras e identificação
De forma a facilitar a identificação das amostras obtidas após as reações de PCR
efetuadas sobre o DNA obtido dos reatores, adotou-se uma nomenclatura que reflete o reator,
o compartimento (ânodo ou cátodo), a fase de produção de bioeletricidade e se foram usados
os oligonucleótidos iniciadores para Archaea ou Bacteria. A Tabela 4 sumariza a nomenclatura
adotada.
Tabela 4: Nomenclatura adotada para designar as amostras de cada reator e distingui-las quanto ao tipo de organismo,
compartimento do reator e fase de produção de bioeletricidade. As amostras marcadas com asterisco foram analisadas
por Andreia Fernandes, durante um estágio realizado sob orientação dos professores Luís Fonseca e Jorge Leitão, e
foram incluídos para uma melhor compreensão do trabalho.
Fase de maior produção
de bioeletricidade
Fase de menor produção
de bioeletricidade
Ânodo Cátodo Ânodo Cátodo
Reator 1 Bactérias R1A_B1* R1C_B1 R1A_B2 R1C_B2
Archaeas R1A_A1* R1C_A1 R1A_A2 R1C_A2
Reator 2 Bactérias R2A_B1 R2C_B1 R2A_B2 R2C_B2
Archaeas R2A_A1 R2C_A1 R2A_A2 R2C_A2
3.2. Análise das electroforeses em gel de agarose
Após a extração e purificação do DNA genómico, procedeu-se à eletroforese em gel de
agarose o que permitiu verificar a existência e a integridade do DNA genómico purificado
(resultados não mostrados).
A Figura 8 mostra o resultado da electroforese para o R1 na fase de menor produção
de bioelectricidade, nos dois compartimentos, usando os oligonucleóticos iniciadores para
archaeas e para bactérias. A ausência de bandas para Archaea no R1A (poço 2) foi o
resultado mais relevante e que sugeriu a inexistência de Archaea no compartimento anódico do
R1. Este resultado foi obtido independentemente da confirmação da existência de DNA
extraído do compartimento anódico do reator 1, que não apresentava quaiquer sinais de
degradação quando visualizado após electroforese em gel de agarose.

28
(1) (2) (M) (3) (4)
Figura 8 – Electroforese em gel de agarose dos amplificados obtidos após reação de PCR usando DNA total extraído
do cátodo e ânodo do reator 1 para o R1 na fase de menor produção de bioelectricidade. Pistas: (1)-R1C_A2- Archaea
no compartimento catódico; (2)-R1A_A2- Archaea no compartimento anódico; (M)-marcador de pesos moleculares; (3)-
R1A_B2- bactérias no compartimento anódico; (4) R1C_B2- bactérias no compartimento catódico.
As Figuras 9 e 10 ilustram os resultados da electroforese para o reator 2 nas duas fases de
produção de bioelectricidade para o anôdo, para o anôdo e para o cátodo, respectivamente.
(1) (2) (M) (3) (4)
Figura 9 – Electroforese em gel de agarose dos amplificados obtidos após reação por PCR do DNA total extraído do
anôdo do R2 nas duas fases de produção de bioelectrocidade. Pistas: (1)-R2A_A1- Archaea na fase de maior produção
de bioelectricidade; (2)-R2A-A2- Archaea na fase de menor produção de bioelectricidade; (M)- marcador de pesos
moleculares; (3)-R2A_B1- bactérias na fase de maior produção de bioelectricidade; (4)- R2A_B2- bactérias na fase de
menor produção de bioelectricidade.
No ânodo do reator 2, a ausência de bandas quando se usaram os primers para
Archaea nas reacções de PCR com DNA obtido na fase de menor produção de energia (poço

29
2) sugere a inexistência de Archaea. Apesar da fraca intensidade da banda no poço 1,
correspondente a Archaea no período de maior produção de energia no compartimento
anódico, a quantidade de DNA genómico total foi considerável. Por outro lado, a intensidade
das bandas para bactérias (poço 3 e 4) é maior, o que indica uma boa concentração de DNA
genómico.
(1) (2) (M) (3) (4)
Figura 10 – Electroforese em gel de agarose dos amplificados obtidos após reação por PCR do DNA total extraído do
cátodo do reator 2 nas duas fases de produção de bioelectrocidade. (1)-R2C_A1- Archaea na fase de maior produção
de bioelectricidade; (2)-R2C-A2- Archaea na fase de menor produção de bioelectricidade; (M)- marcador de pesos
moleculares; (3)-R2C_B1- bactérias na fase de maior produção de bioelectricidade; (4)- R2C_B2- bactérias na fase de
menor produção de bioelectricidade.
No cátodo do reator 2 verificou-se a ausência de bandas no poço 2, o que implica a
inexistência de Archaea na fase de menor produção de bioelectricidade. Apesar da banda que
é visivel no poço 1, e que corresponde a Archaea na fase de maior produção de energia, não
foi possível obter o seu DNA genómico, pelo que se considerou a ausência de Archaea no
compartimento catódico do R2. Já com os primers para bactérias, tanto para a fase de maior
produção de bioelectricidade (poço 3) como para a de menor produção (poço 4), obteve-se
uma grande intensidade de bandas. Os resultados descritos foram confirmados pela repetição
das extrações de DNA e das electroforeses e comprovados pela análise estimativa do estado
de pureza do DNA genómico no espectrofotómetro.

30
3.3. Análise dos perfis de restrição dos clones obtidos
Depois de se obter os amplificados por PCR, estes foram sujeitos a purificação para
garantir que eram obtidos apenas os fragmentos amplificados. Posteriormente procedeu-se à
clonagem no vector de clonagem pCR2.1 e de seguida à transformação em células de E. coli
αDH5. Os transformantes foram espalhados na superfície de uma placa contendo meio LB
sólido e com os antibióticos apropriados (descrito na secção Materiais e Métodos). A
transformação resultou numa série de colónias em que os transformantes contendo os
fragmentos de interesse originaram colónias brancas, que foram selecionadas. Foram obtidos
um total de 960 plasmídeos recombinantes a partir do DNA extraído dos dois compartimentos
dos dois reatores, tanto na fase de maior como de menor produção de bioelectricidade (cátodo
e ânodo). De forma a minimizar o número de reações de sequenciação, o DNA foi hidrolisado
com a enzima de restrição Hae III de forma a obter o perfil de restrição para cada um dos
plasmídeos obtidos. Os perfis de restrição obtidos podem ser característicos ao nível das
sequências específicas de cada fragmento. A técnica aproveita-se da possibilidade dessas
diferenças se encontrarem na sequência de reconhecimento da(s) enzima(s) de restrição
utilizada(s) e a sua comparação permite distinguir essas diferenças como diferentes
sequências. A maior vantagem da utilização desta técnica é que minimiza o número de
sequenciações necessárias, uma vez que apenas se sequenciam os clones com perfis de
restrição diferentes, minimizando tempo e recursos. Desta forma, a partir da comparação dos
perfis de restrição, os clones foram agrupados pelo seu perfil. A Tabela 5 resume o número de
clones obtidos em cada ensaio com o número de perfis diferentes obtidos.

31
Tabela 5: Comparação entre o número total de clones obtido, o número inicial de perfis diferentes que correspondeu a
diferentes clones para sequenciar e o número efetivo de perfis diferentes.
Nº total de clones
obtidos
Nº efetivo de perfis
diferentes
R2A_B1 96 8
R2A_A1 119 9
R2C_B1 120 6
R1A_B1 87 8
R1C_B1 126 8
R1A_A1 68 7
R2C_B2 135 20
R2A_B2 41 7
R1C_A2 72 0
R1C_B2 48 8
R1A_B2 48 4
Total 960 85
De facto, no total, o número de reações de sequenciação ficou reduzido de 960 a 142,
o que equivaleu a uma redução de 85% em tempo e recursos. Após a sequenciação verificou-
se que dos 142 perfis iniciais, 85 corresponderam a clones diferentes. As figuras seguintes
ilustram exemplos de perfis de restrição obtidos nos dois reatores, usando primers para o rRNA
16S de bactérias (Figura 11) ou de Archaea (Figura 12). Os perfis aqui ilustrados serviram
como base para agrupar os diferentes clones obtidos.

32
Figura 11: Exemplos de alguns perfis obtidos usando primers para bactérias e sua distribuição pelas ordens. (A)-
Burkholderiales; (B)-Rhodocyclales; (C)-Xanthomonadales; (D)-Rhizobiales; (E)-Nitrosomonadales; (F)-
Enterobacteriales; (G)-Desulfuromonadales; (H)-Sphingomonadales; (I)-Pseudomonadales; (J)-Caulobacterales; (K)-
Flavobacteriales; (L)-Sphingobacteriales.
Figura 12: Exemplos de alguns perfis obtidos usando primers para Archaea. (A)-Methanosarcinales; (B)-
Methanobacteriales; (C)-Methanomicrobiales; (D)-Methanomassiliicoccus; (E)-Nitrososphaerales;
3.4. Composição das populações microbianas
Após a sequenciação, procedeu-se à análise das sequências de DNA obtidas e à
análise filogenética. Para cada uma das sequências obtidas identificou-se o organismo cuja
sequência de rRNA 16S depositada no GenBank é mais próxima e construíram-se as
respetivas árvores filogenéticas de forma a identificar as relações filogenéticas entre os
organismos identificados. As tabelas seguintes (6 a 13) reúnem as espécies de organismos
que foram obtidas nos vários compartimentos e nas diferentes fases de produção de
bioelectricidade dos reatores. As relações filogenéticas foram construídas a partir das
sequências dos clones obtidos e as árvores filogenéticas encontram-se representadas no
Anexo I.
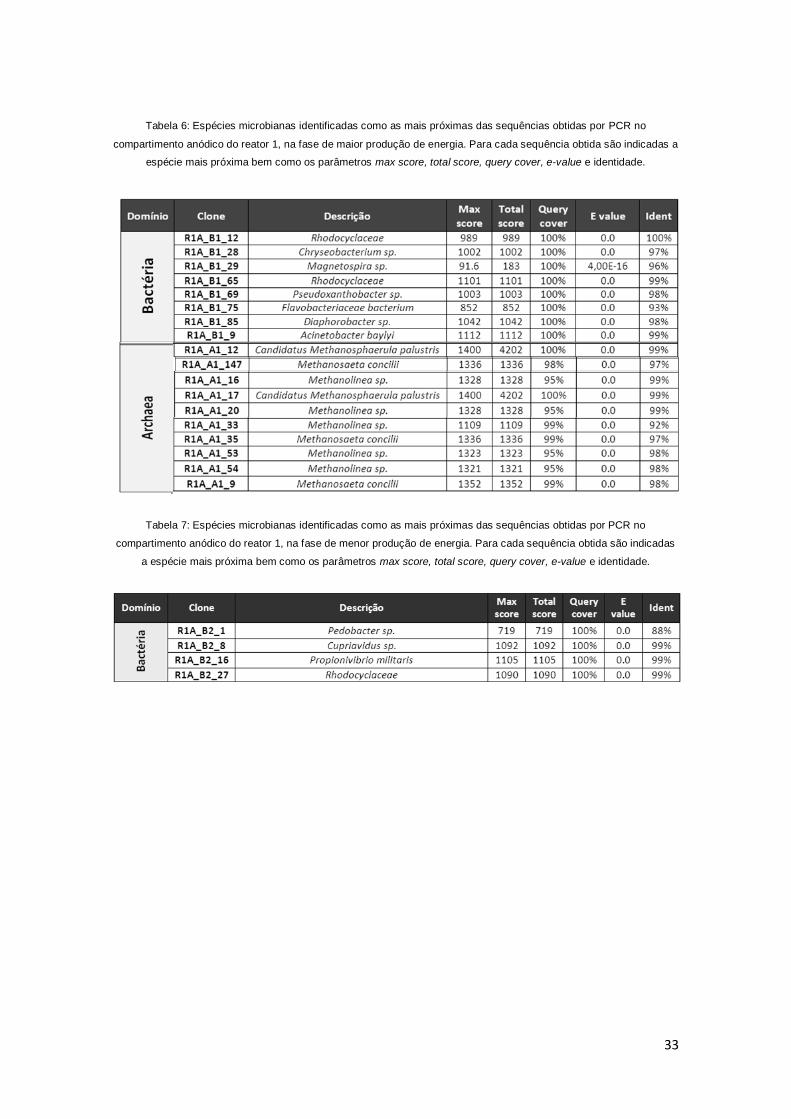
33
Tabela 6: Espécies microbianas identificadas como as mais próximas das sequências obtidas por PCR no
compartimento anódico do reator 1, na fase de maior produção de energia. Para cada sequência obtida são indicadas a
espécie mais próxima bem como os parâmetros max score, total score, query cover, e-value e identidade.
Tabela 7: Espécies microbianas identificadas como as mais próximas das sequências obtidas por PCR no
compartimento anódico do reator 1, na fase de menor produção de energia. Para cada sequência obtida são indicadas
a espécie mais próxima bem como os parâmetros max score, total score, query cover, e-value e identidade.

34
Tabela 8: Espécies microbianas identificadas como as mais próximas das sequências obtidas por PCR no
compartimento catódico do reator 1, na fase de maior produção de energia. Para cada sequência obtida são indicadas
a espécie mais próxima bem como os parâmetros max score, total score, query cover, e-value e identidade.
Tabela 9: Espécies microbianas identificadas como as mais próximas das sequências obtidas por PCR no
compartimento catódico do reator 1, na fase de menor produção de energia. Para cada sequência obtida são indicadas
a espécie mais próxima bem como os parâmetros max score, total score, query cover, e-value e identidade.

35
Tabela 10: Espécies microbianas identificadas como as mais próximas das sequências obtidas por PCR no
compartimento anódico do reator 2, na fase de menor produção de energia. Para cada sequência obtida são indicadas
a espécie mais próxima bem como os parâmetros max score, total score, query cover, e-value e identidade.
Tabela 11: Espécies microbianas identificadas como as mais próximas das sequências obtidas por PCR no
compartimento anódico do reator 2, na fase de menor produção de energia. Para cada sequência obtida são indicadas
a espécie mais próxima bem como os parâmetros max score, total score, query cover, e-value e identidade.

36
Tabela 12: Espécies microbianas identificadas como as mais próximas das sequências obtidas por PCR no
compartimento catódico do reator 2, na fase de maior produção de energia. Para cada sequência obtida são indicadas
a espécie mais próxima bem como os parâmetros max score, total score, query cover, e-value e identidade.
Tabela 13: Espécies microbianas identificadas como as mais próximas das sequências obtidas por PCR no
compartimento catódico do reator 2, na fase de menor produção de energia. Para cada sequência obtida são indicadas
a espécie mais próxima bem como os parâmetros max score, total score, query cover, e-value e identidade.
Com a identificação dos organismos em cada reator e tendo em conta o número de
clones que correspondeu a cada perfil de restrição, foi possível fazer uma análise gráfica que
compara as abundâncias relativas dos organismos nos dois reatores nas duas fases de
produção de bioelectricidade. No entanto, estes resultados poderão ter erros associados
decorrentes de problemas de especificidade das reações de PCR uma vez que, a
especificidade é dada pelos primers e estes podem ter mais afinidade para determinadas
sequências presentes nalguns organismos. As figuras seguintes comparam as abundâncias
relativas dos organismos nas duas fases de produção de bioelectricidade.

37
Figura 11: Comparação da abundância relativa das espécies identificadas no compartimento anódico do reator 1 no
compartimento anódico nas fases de maior e menor produção de bioeletricidade.
Figura 12: Comparação da abundância relativa das espécies identificadas no compartimento catódico do reator 1 no
compartimento anódico nas fases de maior e menor produção de bioeletricidade.
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
R1A
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
R1C
Bacteria Archaea
Bacteria Archaea
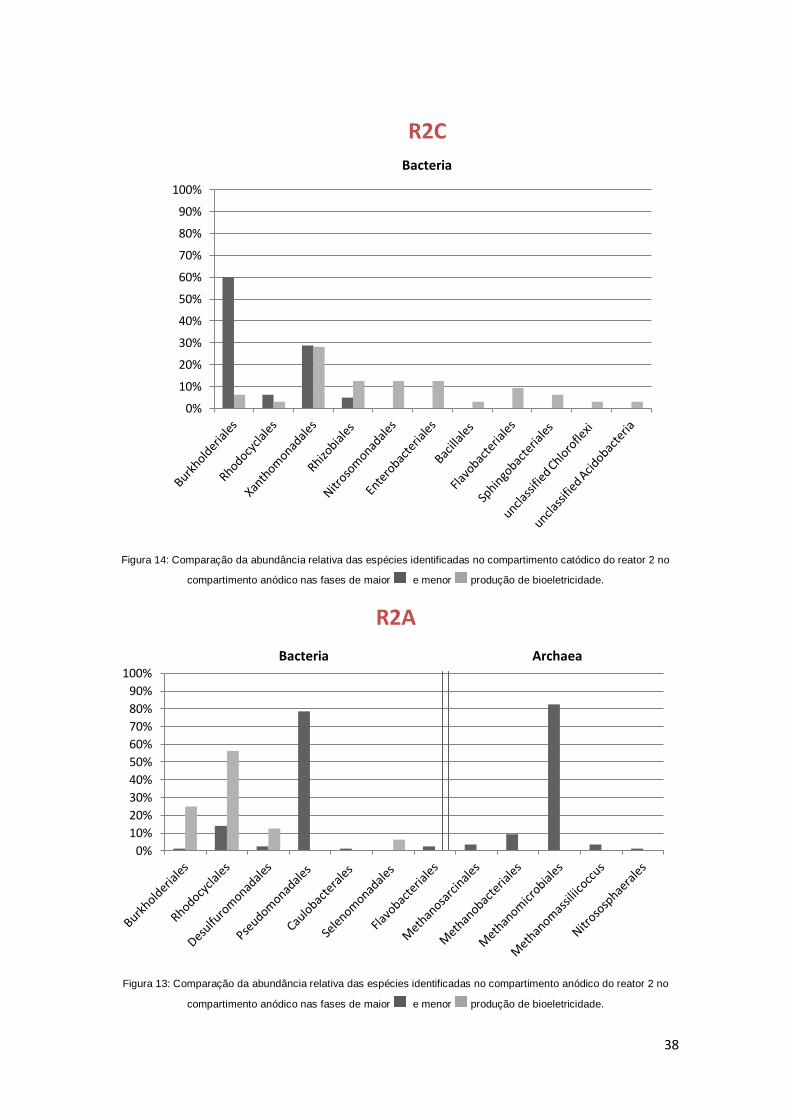
38
Figura 14: Comparação da abundância relativa das espécies identificadas no compartimento catódico do reator 2 no
compartimento anódico nas fases de maior e menor produção de bioeletricidade.
Figura 13: Comparação da abundância relativa das espécies identificadas no compartimento anódico do reator 2 no
compartimento anódico nas fases de maior e menor produção de bioeletricidade.
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
R2C
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
R2A
Bacteria
Bacteria Archaea

39
O esquema da Figura 17 representa um heat map com a distribuição das bactérias e
archaeas obtidos nas duas fases, para os dois reatores nos dois compartimentos. Os
resultados são apresentados ao nível do filo (coluna esquerda) e ao nível da ordem (coluna
direita) com a identificação do número de espécies obtidas em cada ordem (entre parêntesis).
up down Bacteria
R1C R1A R2C R2A R1C R1A R2C R2A
Bac
teri
a
Proteobacteria
60 1 48 1 1 1 2 4 Burkholderiales (8)
1
Rhodospirillales (1)
7 21 5 11
7 1 9 Rhodocyclales (4)
1
23
9
9
Xanthomonadales (3)
7 1 4
6
4
Rhizobiales (10)
4
Nitrosomonadales (1)
26
4
Enterobacteriales (3)
2
2 Desulfuromonadales (1)
1
Sphingomonadales (1)
20 47
62 17
Pseudomonadales (2)
1
Caulobacterales (1)
Firmicutes 1
Bacillales (1)
1 Selenomonadales (1)
Bacteroidetes
1
Cytophagales (1)
4
2
3
Flavobacteriales (3)
1 2
Sphingobacteriales (2)
Chloroflexi
1
unclassified Chloroflexi (1)
Acidobacteria
1
unclassified Acidobacteria (1)
Euryarchaeota
30
3 5
Methanosarcinales (4)
A
rch
aea
8
Methanobacteriales (3)
6
71 53
Methanomicrobiales (4)
3
Methanomassiliicoccus (1)
Thaumarchaeota
1
Nitrososphaerales (1)
Figura 17- Heat map para bactérias e archaea cujas sequências de genes para o rRNA 16S são mais próximas das
sequências obtidas para os dois reatores (R1 e R2), nas duas fases, em ambos os compartimentos (anódico e
catódico). Os organismos são apresentados segundo o filo (coluna da esquerda) e segundo a ordem com a
identificação do número de espécies em cada ordem entre parêntesis (coluna da direita).

40
Com base nas bibliotecas de clones obtidos e na abundância de cada clone avaliado
pelas análises dos perfis de restrição, o filo Proteobacteria aparece como o mais abundante
entre as bactérias, representando mais de 95% dos 250 clones obtidos para as bactérias. De
entre Proteobacterias, Pseudomonadales e Burkholderiales são as ordens mais abundantes,
correspondendo respectivamente a 33% e a 23% do total de Proteobacterias identificadas. No
caso de Archaea, a ordem Methanomicrobiales do filo Euryarchaeota (estritamente redutoras
de dióxido de carbono), é o mais abundante, correspondendo a 90% do total de clones de
Archaea. No total identificaram-se 45 espécies diferentes de bactérias entre as 18 ordens
obtidas e 13 espécies diferentes entre as 5 ordens de Archaea.
3.5. Discussão de resultados
3.5.1. Reator 1
Do compartimento anódico do reator 1 foram obtidos 87 clones contendo sequências
de genes para o rRNA 16S de bactérias e 68 clones para Archaea para a fase de maior
produção de bioelectricidade. Na fase de menor produção de bioelectricidade obtiveram-se 48
clones de bactérias no ânodo (R1A_B2). Por outro lado, no compartimento catódico (R1C_B1)
foram obtidos 126 clones de bactérias para a fase de maior produção de bioelectricidade, 72
clones de Archaea e 48 clones de bactérias para a fase de menor produção de energia.
A Figura 13 mostra que no reator 1, na fase de maior produção de bioelectricidade,
Pseudomanodales e Rhodocyclales são as ordens bacterianas a que pertecem a maior parte
das sequências identificadas no ânodo. Por outro lado, na fase de menor produção de
bioelectricidade, verificou-se a total inexistência de sequências pertencentes a estas classes,
aparecendo espécies pertencentes às ordens das Burkholderiales e Rhodospirillales. Este facto
sugere que embora Burkholderiales e Rhodospirillales sejam vulgarmente identificadas em
MFCs, o seu papel poderá não ter grande relevância na produção de bioelectricidade ou ainda
que, o seu papel na produção de bioelectricidade poderá estar dependende de outras espécies
como por exemplo de Pseudomonadales. As sequências obtidas no cátodo do reator 1 foram
obtidas com uma elevada percentagem para as ordens Burkholderiales, Pseudomonadales e
Enterobacteriales aquando da fase de maior produção de energia (Figura 14). Já na fase de
menor produção de bioeletricidade houve uma clara diminuição de Burkholderiales e total
desaparecimento de Enterobacteriales. Sequências pertencentes à ordem Pseudomonadales
foram também identificadas nesta fase, representando a ordem mais identificada.
Pseudomonadales, Xanthomonadales e Rhizobiales foram as ordens mais representadas no
cátodo na fase de menor produção de energia.
O reator 1 caracterizou-se por uma grande abundância de β-Proteobacterias,
principalmente espécies da ordem Burkholderiales, que foram encontradas em todos
compartimentos nas duas fases de produção de bioelectricidade. Só no compartimento

41
catódico para a fase de maior produção de bioelectricidade (R1C_B1), as sequências
pertencentes à ordem Burkholderiales representaram cerca de 73% dos 126 clones obtidos,
representando apenas 1% dos clones obtidos no compartimento anódico na fase de maior
produção de bioelectricidade (R1A_B1) e 3% e 11%, respetivamente, dos clones obtidos para
o cátodo (R1C_B2) e para o ânodo (R1A_B2) na fase de menor produção de bioelectricidade.
Espécies dos géneros Diaphorobacter e Acidovorax da família Comamonadaceae (ordem
Burkholderiales) foram já identificadas em MFCs enriquecidas com resíduos domésticos
(Lefebvre et al., 2010). Acidovorax poderá estar relacionada com a produção de energia em
MFCs (Sun et al, 2012).
Os membros da família Rhodocyclaceae, pertencentes à ordem das Rhodocyclale, são
também bactérias gram-negativas pertencentes às β-Proteobacterias. Estas só não foram
identificadas no cátodo para a fase de menor produção de energia (R1C_B2). Rhodocyclaceae
representa cerca de 9% dos clones obtidos no cátodo na fase de maior produção de
bioelectricidade (R1C_B1), 30% dos clones obtidos no ânodo na fase de maior produção de
bioelectricidade (R1A_B1) e 78% dos clones obtidos no ânodo na fase de menor produção de
bioelectricidade (R1A_B2). Alguns membros desta família são redutoras de sulfato e clorato,
redutoras de ferro e caracterizam-se pela sua alta tolerância ao oxigénio (Quan et al., 2012).
Pseudomonas é um dos géneros mais representado no reator 1 e tem sido muito
utilizado para abastecer os ânodos das MFCs de forma a otimizar a produção de
bioelectricidade (Yong et al, 2011). Em 2004, Rabaey et al. a partir de uma cultura pura de
Pseudomonas avaliou o potencial eletroquímico de espécies anaeróbias facultativas fazerem a
transferência eletrónica diretamente no ânodo. No presente trabalho 24% dos clones obtidos
no cátodo na fase de maior produção de bioelectricidade (R1C_B1) pertencia ao género
Pseudomonas, representado 60% dos clones obtidos no ânodo na fase de maior produção de
bioelectricidade (R1A_B1) e 50% dos clones obtidos no cátodo na fase de menor produção de
bioelectricidade (R1C_B2). Pseudomonas e Burkholderia são géneros aeróbios podendo
utilizar O2 ou outro aceitador final e, utilizando tanto o oxigénio como o elétrodo do ânodo como
aceitadores de eletrões o que poderá justificar a elevada abundância nos dois compartimentos
do reator.
Outras ordens pertencentes ao filo das Proteobacterias foram identificadas no reator 1.
Várias espécies de Enterobacteriales foram identificadas no cátodo para a fase de maior
produção de energia (R1C_B1). Por exemplo, Serratia é um género que compreende espécies
anaeróbias facultativas, caracterizadas como electroquimicamente ativas pela produção de
bioelectricidade mesmo em culturas puras (Ghanapriya et al., 2010). Shigella boydii foi outro
organismo da ordem das Enterobacteriales identificado no cátodo para a fase de maior
produção (R1C_B1), no entanto, ainda não há informações do seu papel na produção de
bioeletricidade. Dos 126 clones obtidos no cátodo para a fase maior produção de
bioelectricidade (R1C_B1), cerca de 32% corresponderam a sequências homólogas às de
Enterobacteriales.

42
Bacterioidetes é um filo que também está bem representado no reator 1. Foram
identificados vários organismos pertencentes às ordens das Flavobacteriales
(Chryseobacterium) e Cytophagales (Leadbetterella byssophila). Organismos pertencentes ao
filo Bacteroitedes são dos mais importantes no cátodo (Rabaey et al., 2008) e a par das
Proteobacterias são fundamentais para a redução de oxigénio e nitrato no cátodo (Timmers et
al., 2012). Estas representaram cerca de 2% e 6% para o cátodo e ânodo, respectivamente, na
fase de maior produção de bioelectricidade.
No ânodo na fase de maior produção de bioelectricidade (R1A_B1) foi ainda
identificada uma sequência homóloga a Magnetospira, pertencente às Rhodospirillales que são
bactérias fotosintéticas pertencentes às α-Proteobacterias no entanto a sua importância é
desconhecida.
A ordem Rhizobiales foi também bem representada por várias espécies em todos
compartimentos com a exceção do ânodo na fase de menor produção de bioelectricidade.
Rhizobiales representou cerca de 9% de clones obtidos para o cátodo na fase de maior
produção de bioelectricidade (R1C_B1) tendo esta percentagem duplicado (18%) na fase de
menor produção de bioelectricidade (R1C_B2). No ânodo e fase de maior produção de energia
(R1A_B1) foi apenas obtido um clone que correspondia à ordem das Rhizobiales. As espécies
do género Rhizobiales são reconhecidas como aeróbias electroquimicamente ativas (Wang et
al., 2013). As espécies de Rhizobiales obtidas foram Nitrobacter, Mesorhizorbium e
Methylobacterium organo.
Relativamente a Archaea, obtiveram-se organismos deste domínio para o ânodo na
maior fase de produção de bioelectricidade (R1A_A1) e para o cátodo na fase de menor
produção de energia (R1A_A2). No primeiro caso, obtiveram-se duas espécies de
Methanomicrobiales, Methanosphaerula palustris e Methanolinea, e uma pertencente à ordem
das Methanosarcinales, Methanosaeta concilii. Estas são anaeróbias metanogénicas e são
caracterizadas pelo consumo de acetato e produção de metano. No cátodo, para a fase de
menor produção de energia (R1C_A2), identificou-se também Methanosaeta concilii,
Methanolinea mas também Methanosaeta, que faz parte das Archaea da ordem das
Methanosarcinales. Apesar de raramente serem estudadas Archaea em MFCs, é muito comum
encontrá-las, sobretudo espécies metanogénicas (Shehab et al., 2013). Também é comum
encontrá-las em ânodos onde utilizam eletrões para reduzir dióxido de carbono a metano
(Logan, 2012). No cátodo, a existência de Archaea é muito reduzida devido à exposição ao O2
a que este está sujeito e à competição pelo substrato orgânico pelos organismos anaeróbios
facultativos (Park, et al., 2014).
Resumidamente obteve-se uma grande diversidade bacteriana principalmente de
Proteobacterias. Foram identificadas ainda algumas Bacteroidetes no cátodo para a fase de
maior produção de energia (R1C_B1). Em relação às archaeas, foram apenas obtidas
archaeas metanogénicas.

43
3.5.2. Reator 2
No compartimento anódico do reator 2 foram obtidos 96 clones de bactérias e 119
clones de Archaea para a fase de maior produção de bioelectricidade. Na fase de menor
produção de bioelectricidade obtiveram-se 41 clones de bactérias no ânodo (R2A_B2). Por
outro lado, no compartimento catódico foram obtidos 120 clones de bactérias para a fase de
maior produção de bioelectricidade e 135 clones de bactérias para a fase de menor produção
de energia.
Os organismos cuja abundância relativa era mais elevada no ânodo na fase de maior
produção de energia pertenciam, tal como no caso do reator 1, à ordem Pseudomonadales. A
ordem Rhodocyclales representou também uma percentagem elevada. Já na fase de menor
produção de energia, os microrganismos da ordem Pseudomonadales não foram detetados,
tendo-se verificado um aumento bastante significativo na abundância relativa de
Burkholderiales, Rhodocyclales e Desulfuromonadales. Estes resultados sugerem que os
microrganismos da ordem Pseudomonadales serão os organismos que contribuem
significativamente para a produção de bioelectricidade no ânodo. Relativamente ao cátodo
(Figura 15) é possível verificar uma grande abundância de organismos pertencentes às ordens
Burkholderiales e Xanthomonadales na fase de maior produção de bioelectricidade. Na fase de
menor produção de bioeletricidade detetou-se uma grande diversidade de organismos,
praticamente de todas as ordens, sendo as pertencentes à ordem Xanthomonadales as mais
representadas.
Em geral, no reator 2 existia uma grande diversidade de organismos dos filos
Proteobacteria, Firmicutes e Bacteroidetes. Na fase de maior produção de bioelectricidade,
identificou-se para o ânodo uma grande abundância de Pseudomonas que, como já referido, é
das espécies mais utilizadas para aumentar a performance energética de uma MFC. De facto,
estas representaram cerca de 78% dos 96 clones obtidos. Rabaey et al. (2004) referiu que
apesar das condições redox a que os organismos aeróbios, como Pseudomonas, estão
sujeitos, estas tendem a ser os organismos dominantes. Em menor quantidade (14% dos
clones obtidos para esta fase), foram obtidas espécies de Azospira da ordem Rhodocyclacles,
já identificadas em MFCs (Michaelidou et al., 2011). Nesta fase foram ainda identificadas as
espécies Kaisella Koreensis (Flavobacteriales), correspondendo a cerca de 3%, e
Brevundimonas (Caulobacterales), correspondendo a apenas 1% dos clones obtidos.
Foram ainda obtidas no compartimento anódico para as duas fases de produção de
bioeletricidade, sequências para o rRNA 16S semelhantes às das espécies do género
Geobacter, cerca de 3% na fase de maior produção e cerca de 13% na fase de menor
produção. Pertencentes à ordem Desulfuromodales, as espécies do género Geobacter são
consideradas de elevada eficiência na transferência eletrónica para o elétrodo, capazes de
transferir a maioria dos eletrões obtidos a partir do consumo de acetato diretamente para o
elétrodo (Bond e Lovley, 2003).

44
Na fase de menor produção de bioeletricidade, a espécie Propioniovibrio militaris,
pertencente à ordem das Rhodocyclacles, foi a mais abundante no ânodo, representando 56%
do total de clones. No entanto, não foi possível encontrar na literatura nenhuma referência à
espécie e o seu papel em MFCs. Esta espécie é anaeróbia facultativa, não fermentativa e tem
a habilidade de degradar perclorato (Thrash et al., 2009). Desta forma, poderá eventualmente
desempenhar um papel muito importante em MFCs para o tratamento de águas residuais
contaminadas com perclorato, que é um contaminante muito comum em fertilizantes e na água
potável, sendo tóxico para os seres humanos (Butler et al, 2010).
Várias sequências de rRNA 16S semelhantes às dos organismos da ordem
Burkholderiales foram obtidas no ânodo na fase de menor produção (R2A_B2), em particular
com elevada homologia com as das espécies Ottowia, Pelomonas e Cupriavidus. Estas
representaram 25% dos 41 clones obtidos.
Foi ainda obtida uma sequência com homologia com um organismo da família
Veillonellaceae, pertencente à ordem Selenomodales do filo Firmicutes. As bactérias do filo
Firmicutes, gram-positivas, são capazes de fazer a transferência eletrónica diretamente no
ânodo e desempenham um papel importante na geração de bioeletricidade em MFCs
alimentadas a acetato (Wrighton et al, 2008).
Relativamente ao cátodo, na fase de maior produção de energia (R2C_B1) apenas
foram identificados organismos pertencentes ao filo Proteobacteria, sendo 60% pertencentes à
ordem das Burkholderiales. Dos 120 dos clones obtidos, 29% corresponderam a sequências da
ordem Xanthomonadales, representada por Dokdonellas e por Rhodanobacter, géneros que
têm sido encontrados em vários reatores anaeróbios (Quan et al., 2012). Foram ainda
encontradas sequências com homologia para as classes Rhodocyclales (6% dos clones) e
Rhizobiales (5%).
Para a fase de menor produção de bioeletricidade do cátodo (R1C_B2) obteve-se a
mais elevada biodiversidade. Foram encontradas espécies dos filos Proteobacteria, Firmicutes,
Bacteroidetes, Chloroflexi e ainda Acidobacteria. As sequências homólogas ao género
Stenotrophomonas, pertencentes à ordem Xhantomonadales foram as mais abundantes
representando cerca de 28% do total.
Aminobacter sp., Nitrobacter sp., Devosia ginsengisoli e Ochrobactrum sp. foram as
espécies pertencentes à ordem Rhizobiales com maior homologia com as sequências
identificadas (cerca de 13%). Aminobacter tem sido reconhecida em MFCs pelo seu poder
degradativo do ácido nitrilotriacético, um agente quelante industrial muito comum que está a
causar grandes preocupações ambientais (Jang et al, 2006). A presença de Nitrobacter nas
MFCs é também muito importante, uma vez que os membros deste género desempenham um
papel muito importante no processo de nitrificação (Bock e Koops, 1992). A espécie
Ochrobactrum tem sido também identificada em comunidades microbianas do ânodo de MFCs
alimentadas a acetato ou glucose e têm revelado um grande potencial para a produção de
bioelectricidade (Zuo et al., 2008). Tal como outras Rhizobiales, que são α-Proteobacterias,
estas têm a capacidade de fazer a desnitrificação quando sujeitas a condições anaeróbias.

45
Por outro lado, relativamente à ordem Nitrosomonadales, 13% dos clones possuíam
sequências homólogas à espécie Nitrosomonas que é uma proteobactéria nitrificante e que
oxida amónia em nitrito. A presença destas espécies em MFCs é muito útil para o tratamento
de resíduos industriais ou águas residuais e no processo de biorremediação (Kim et al., 2008).
Enterobacteriales foram também identificadas no cátodo com uma abundância relativa
de 13%, como é o caso de Citrobacter freundii. As espécies de Citrobacter, são bactérias
anaeróbias facultativas que conseguem crescer a partir de uma variedade de aceitadores de
eletrões, como o Fe (III), na ausência de oxigénio (George, 2005). Têm sido muito estudadas
nos últimos anos graças à sua capacidade de remoção de metais pesados, redução de
sulfatos, degradação de fenol e degradação de clorofenol (Xu e Liu, 2011). Por estas razões, a
incorporação de Citrobacter em MFCs para o tratamento de águas residuais pode ser bastante
benéfica.
Do filo Proteobacteria foram ainda identificadas Acidovorax e Propiniovibrio militaris,
pertencentes às ordens Burkholderiales (6%) e Rhodocyclacles (3%), respectivamente.
Do filo Firmicutes, foi apenas obtido um clone correspondente a Virgibacillus,
pertencente à ordem Bacillales. Apesar de não haver informações acerca do potencial
electroquímico de Virgibacillus, outras espécies da ordem Bacillales têm sido incluídos em
MFCs com o objectivo de aumentar a sua performance energética (Zhang et al., 2012).
Do filo Bacteroidetes foram identificadas da ordem Flavobacteria as espécies Fluviicola
taffensis e Wandonia haliotis, das quais não há informação de terem sido identificadas em
MFCs nem do seu potencial electroquímico. Do total de clones obtidos, as Flavobacterias
representaram cerca de 9% dos clones. Por outro lado, cerca de 6% correspondeu à ordem
Sphingobacteriales, onde se identificaram sequências homólogas às do género Terrimonas.
Estas são bactérias acidogénicas fermentativas que utilizam vários açúcares e aminas para
produzir ácidos orgânicos que por sua vez são metabolizados pelas bactérias
bioelectroquimicamente ativas para a produção de bioelectricidade (Lu, et al., 2012).
Por fim, obtiveram-se ainda duas sequências com homologia para as espécies não
identificadas dos filos Chloroflexi e Acidobacteria. As Chloroflexi são bactérias verdes não
sulfurosas que obtém energia mediante fotossíntese e as Acidobacteria são bactérias
acidofílicas mas o seu metabolismo ainda não é ainda bem compreendido. São dois filos que
frequentemente têm sido descritos em MFCs mas cuja importância é ainda desconhecida.
Relativamente ao domínio Archaea, apenas foram obtidas sequências homólogas no
ânodo na fase de maior produção de bioelectricidade (R2A_A1). A maioria das espécies
obtidas era do género Methanolinea, pertencentes à ordem Methanomicrobiales com uma
abundancia relativa de 82% (Figura 16). Cerca de 9% dos clones obtidos correspondiam a
algumas espécies da ordem Methanobacteriales como Methanobrevibacter arboriphilus,
Methanobrevibacter smithii e Methanobrevibacter acididurans. Da ordem
Methanomassiliicoccus foi identificada uma espécie denominada por Methanomassiliicoccus
luminyensis. Esta espécie representou cerca de 3% do total de 119 clones obtidos. Todos
estes organismos são metanogénicos, pertencentes ao filo das Euryarchaeotas, caracterizadas

46
pela produção de metano no reator. Por fim, obteve-se uma sequência com homologia com
Candidatus Nitrososphaera gargensis que faz parte do conjunto das Nitrososphaerales do filo
Thaumarchaeota. Organismos deste filo são quimiolitotróficos, oxidam amónia e desempenham
um papel muito importante nos ciclos do azoto e do carbono. O papel das Archaeas em
sistemas MFCs ainda não foi estudado com rigor, no entanto, estão frequentemente presentes
sobretudo archaeas metanogénicas.
Em suma, no ânodo as sequências obtidas mais abundantes apresentaram homologia
com as espécies de Burkholderiales, Rhodocyclacles, Desulfuromodales e Pseudomonadales.
No entanto, para a fase de menor produção de energia as sequências com homologia com
Pseudomodales estão ausentes, o que pode explicar a queda na produção de bioelectricidade
pois, como é conhecido, estas são das espécies mais utilizadas para abastecer as MFCs de
forma a melhorar a produção de bioelectricidade (Pham et al., 2007). No cátodo, apesar da
grande diversidade na fase de menor produção de energia, Burkholderiales e
Xanthomonadales têm uma menor abundância do que tinham na fase de maior produção. As
archaeas apenas foram identificadas no ânodo para a fase de maior produção energética e
eram praticamente todas metanogénicas.

47
4. Conclusões
Neste trabalho, foram analisadas amostras provenientes de dois reatores MFC que
diferiam no facto de um deles possuir um cátodo abiótico e outro um biocátodo. Ambos foram
abastecidos com águas residuais para a produção de bioelectricidade. O objetivo principal foi
caracterizar as comunidades bacterianas combinando metodologias de biologia molecular
(extração de DNA, clonagem e sequenciação) com a análise dos perfis de restrição obtidos
após a hidrólise do DNA pela Hae III seguida da comparação das sequências obtidas com
sequências depositadas em bases de dados. Este tipo de metodologia pode ser adotado para
caracterizar outras comunidades microbianas de qualquer outra natureza.
A diversidade microbiana foi determinada para os dois reatores e houve claramente
uma grande diferença na composição das populações o que sugere que a composição das
comunidades bacterianas em MFCs é muito imprevisível. O método adotado neste trabalho
para a caracterização microbiana (Ramos et al., 2010) demostrou ser eficaz e obteve-se um
número considerável de sequências distintas. Além disso, foi possível obter uma redução de
85% do número de clones a sequenciar para a caracterização microbiana nos dois reatores.
Este tipo de metodologias está sempre sujeito a algum tipo de erros, nomeadamente
durante as reações de PCR ou na distinção dos perfis de restrição. A utilização de mais do que
uma enzima de restrição deve ser considerada, de forma a discriminar clones com perfis de
restrição semelhantes. Por outro lado, é uma técnica que apresenta bastantes vantagens,
como a minimização do número de sequenciações e por ser facilmente reprodutível para outras
caracterizações de populações microbianas.
Dos 960 clones obtidos, identificaram-se 142 perfis diferentes o que resultou na
identificação de 58 espécies diferentes entre bactérias e archaeas. Burkholderiales,
Pseudomonadales, Rhodocyclales e Xanthomonadales, foram as ordens mais abundantes
entre os reatores, todas pertencentes ao filo das Proteobacterias. De entre as Burkholderiales
identificaram-se 8 espécies diferentes, sendo Piscinibacter a mais abundante; das
Pseudomonadales identificaram-se apenas Pseudomonas aeruginosa; das Rhodocyclales
identificaram-se 4 espécies diferentes, sendo Rhodocyclaceae a mais abundante e finalmente
da ordem das Xanthomonadales, onde se identificaram 3 espécies diferentes, a espécie que foi
mais vezes identificada foi a Rhodanobacter. O género Methanolinea, pertencente à ordem das
Methanomicrobiales e ao filo das Euryarchaeota (Archaea) foi o mais representado neste
trabalho.
Existe uma série de fatores que podem influenciar a comunidade bacteriana, como por
exemplo a natureza e composição dos substratos, condições de operacionalidade do reator, o
meio e o tipo de material usado nos elétrodos. Não foi possível estabelecer uma relação entre
a diversidade microbiana com os reatores, nem nas diferentes fases de produção de
bioelectricidade. Como era esperado, a maior parte das espécies identificadas no ânodo eram
facultativas ou anaeróbias. Foram identificadas várias espécies, a maioria já reconhecida como

48
electroquimicamente ativa e outras que desempenham um papel muito relevante no tratamento
de águas residuais. As espécies identificadas e que ainda não foram analisadas no contexto
das MFCs, como era o caso da espécie Propioniovibrio militaris, merecem um estudo mais
aprofundado de forma a compreender o seu papel na comunidade da MFC.
A grande diversidade de espécies que se identificaram neste trabalho, incluindo de
vários filos reflete, por um lado a natureza do inóculo usado (lamas de uma estação de
tratamento de águas residuais), sugerindo que o meio das MFCs passa por um processo de
“auto-optimização” para um melhor desempenho funcional do sistema. Este processo deve
fazer uma seleção das espécies, consoante as interações entre si, quantidade e tipo de
substratos disponíveis, etc. Numa perspetiva a longo prazo, é importante conhecer este tipo de
processos, que demostra a utilidade de sistemas MFCs para o enriquecimento de populações
funcionalmente estáveis, capazes de degradar os compostos orgânicos e ao mesmo tempo
fazerem a transferência de eletrões extracelularmente.
Apesar de não haver comunidades microbianas em MFCs idênticas, estes resultados
contribuem com conhecimentos significativos para a aplicação prática de sistemas MFC em
sistemas de tratamento de águas residuais. O sucesso de uma MFC para o tratamento de
águas residuais depende da concentração e da degradabilidade da matéria orgânica,
temperatura e ausência de substâncias tóxicas entre outras (Logan, 2006).
Embora as MFCs altamente eficazes e em grande escala ainda estejam fora do nosso
alcance, esta é uma tecnologia promissora no qual investigadores e engenheiros estão a
esforçar-se para ultrapassar os vários obstáculos ainda subjacentes. A pressão crescente
sobre o meio ambiente e energias renováveis irão certamente contribuir para o
desenvolvimento desta tecnologia.

49
5. Perspectivas futuras
O estudo da diversidade microbiana e a sua função nas MFCs é um passo muito
importante na compreensão da composição dos biofilmes e na produção de bioelectricidade.
Desta forma, deve ser feito um incentivo a um estudo mais aprofundado de forma a identificar
as espécies que potenciam uma maior produção de bioelectricidade. Um ponto importante é o
estudo da interação (ou competição) entre as diferentes espécies de bactérias e archaeas que
compõe o ânodo. As archaeas também devem ser estudadas com mais rigor de forma a
compreender melhor a sua contribuição nas MFCs.
Para que no futuro as MFCs sejam aplicáveis à larga escala, tem de haver um esforço
no sentido de melhorar o modelo e as condições de operacionalidade das MFCs para que o
custo/eficiência à larga-escala seja compensatório. Eficiência e custo de materiais, arquitetura
física e limitações químicas como a condutividade da solução e o pH, são alguns exemplos de
limitações que as MFCs apresentam (Logan, 2009).
Um maior conhecimento ao nível da transferência eletrónica para a superfície dos
elétrodos seria muito importante de forma a melhorar essa transferência por parte das
bactérias electroquimicamente ativas. Também o potencial das bactérias electroquimicamente
ativas poderia ser potenciado por modificações genéticas sugerido por Arora (2012):
(i) Mutações genéticas nos organismos de forma a formarem pilis mais condutivos
(Yoon et al., 2002);
(ii) Mutações genéticas que melhorem a capacidade dos organismos na formação de
biofilmes;
(iii) Restringir a respiração anaeróbia pelas bactérias desnitrificantes para que os
eletrões fluam apenas para o elétrodo do ânodo e não para outros aceitadores de
eletrões como nitrato ou sulfato em solução;
(iv) Controlar a divisão celular;
(v) Aumentar a habilidade de certos organismos gerarem mediadores.
No futuro, será importante um conhecimento do metagenoma e do transcriptoma dos
biofilmes dos elétrodos, de forma a descrever a expressão genética associada ao seu
metabolismo central e à transferência eletrónica durante a degradação de substratos orgânicos
complexos. A expressão genética vai também elucidar as vias metabólicas e a transdução de
energia em comunidades bacterianas complexas (Ishii et al, 2012).
Para além do tratamento de águas residuais e da produção de bioelectricidade, as
MFCs podem no futuro, ser aplicadas para outros fins. Com a colocação de um elétrodo
anódico em sedimentos marinhos e o elétrodo catódico em água, vai ser possível gerar
bioelectricidade da decomposição bacteriana da matéria orgânica presente nos sedimentos.
Também a energia que atualmente é gerada pelas MFCs, apesar de ser insuficiente para ser
considerada uma fonte de energia renovável praticável, pode ser suficiente para alimentar

50
pequenos dispositivos em locais remotos. As MFCs podem também ser modificadas para
serem usadas como processo de bioremediação ou ainda para a remoção de nitrato
(conversão para nitrito) da água (Logan, 2008)

51
6. Bibliografia
Aelterman, P. (2009). Microbial fuel cells for the treatment of waste streams with energy
recovery. Ph.D. Thesis, Gent University, Belgium.
Allen, R.M.; Bennetto, H.P. (1993). Microbial fuel cells: Electricity production from
carbohydrates. Applied Biochemistry and Biotechnology 39-40: 27–40.
Arora, R. (2013), “The Microbiology of Microbial Electric Systems”. In: Microbial biotechnology:
Energy and Environment, 1st ed. CABI, New Delhi, India, pp. 16-30. ISBN 978-1-84593-
956-4
Bock E, Koops HP (1992). The genus Nitrobacter and related genera. In: The prokaryotes, 2nd
ed., vol 1. Edited by Balows A, Truper HG, Dworkin M, Harder W, Shleifer KH. Springer-
Verlag, New York, pp. 2302-2309.
Bockris, J. O. M. S. S., & Srinivasan, S. (1969). Fuel cells-Their electrochemistry, 1st ed.
McGraw-Hill, New York, pp. 525-531. ISBN 0070063451
Bond, D. R., Holmes, D. E., Tender, L. M., & Lovley, D. R. (2002). Electrode-reducing
microorganisms that harvest energy from marine sediments. Science, 295(5554), 483–5.
doi:10.1126/science.1066771
Bond, D. R., & Lovley, D. R. (2003). Electricity Production by Geobacter sulfurreducens
Attached to Electrodes. Applied and Environmental Microbiology, 69(3), 1548–1555.
doi:10.1128/AEM.69.3.1548-1555.2003
Bond, D. R., & Lovley, D. R. (2003). Electricity production by Geobacter sulfurreducens
attached to electrodes. Applied and Environmental Microbiology, 69(3), 1548–55.
doi:10.1128/AEM.69.3.1548-1555.2003
Bond, D. R., & Lovley, D. R. (2005). Evidence for involvement of an electron shuttle in electricity
generation by Geothrix fermentans. Applied and Environmental Microbiology, 71(4), 2186–
9. doi:10.1128/AEM.71.4.2186-2189.2005
Borole, A. P., O’Neill, H., Tsouris, C., & Cesar, S. (2008). A microbial fuel cell operating at low
pH using the acidophile Acidiphilium cryptum. Biotechnology Letters 30(8), 1367–72.
doi:10.1007/s10529-008-9700-y
Bretschger, O., Obraztsova, A., Sturm, C. A., Chang, I. S., Gorby, Y. A., Reed, S. B., Nealson,
K. H. (2008). Current Production and Metal Oxide Reduction by Shewanella oneidensis

52
MR-1 Wild Type and Mutants. Applied and Environmental Microbiology, 74(2), 553–553.
doi:10.1128/AEM.02560-07
Bullen, R. a, Arnot, T. C., Lakeman, J. B., & Walsh, F. C. (2006). Biofuel cells and their
development. Biosensors & Bioelectronics, 21(11), 2015–45.
doi:10.1016/j.bios.2006.01.030
Butler, C. S., Clauwaert, P., Green, S. J., Verstraete, W., & Nerenberg, R. (2010).
Bioelectrochemical perchlorate reduction in a microbial fuel cell. Environmental Science &
Technology, 44(12), 4685–91. doi:10.1021/es901758z
Chaudhuri, S. K., & Lovley, D. R. (2003). Electricity generation by direct oxidation of glucose in
mediatorless microbial fuel cells. Nature Biotechnology, 21(10), 1229–32.
doi:10.1038/nbt867
Clauwaert, P., Van der Ha, D., Boon, N., Verbeken, K., Verhaege, M., Rabaey, K., & Verstraete,
W. (2007). Open air biocathode enables effective electricity generation with microbial fuel
cells. Environmental Science & Technology, 41(21), 7564–9. doi: 10.1021/es0709831
Cohen, B. (1931). The Bacterial Culture as an Electrical Half-Cell, Journal of Bacteriology, 21,
pp18–19
Dahllöf, I. (2002). Molecular community analysis of microbial diversity. Current Opinion in
Biotechnology, 13(3), 213–7. doi: 10.1016/s0958-1669(02)00314-2
Du, Z., Li, H., & Gu, T. (2007). A state of the art review on microbial fuel cells: A promising
technology for wastewater treatment and bioenergy. Biotechnology Advances, 25(5), 464–
82. doi:10.1016/j.biotechadv.2007.05.004
Friman, H., Schechter, A., Ioffe, Y., Nitzan, Y., & Cahan, R. (2013). Current production in a
microbial fuel cell using a pure culture of Cupriavidus basilensis growing in acetate or
phenol as a carbon source. Microbial Biotechnology, 6(4), 425–34. doi:10.1111/1751-
7915.12026
Fu, Q., Kobayashi, H., Kawaguchi, H., Wakayama, T., Maeda, H., & Sato, K. (2013). A
thermophilic gram-negative nitrate-reducing bacterium, Calditerrivibrio nitroreducens,
exhibiting electricity generation capability. Environmental Science & Technology, 47(21),
12583–90. doi:10.1021/es402749f
George M.G. (2005) Bergey’s Manual of Systematic Bacteriology Volume Two The
Proteobacteria Part B. The Gammaproteobacteria. 2nd Edition. New York,Springer. 651p.
ISBN 978-0-387-28022-6

53
Ghanapriya, K., Rana, S., & Kalaichelvan, P. T. (2010). Electricity generation from Serratia
marcescens isolated from aerobic sludge using Microbial Fuel Cell Technology and its
optimization, 09(10), pp 32–38.
Gorby, Y.A. and Beveridge, T.J. (2005). Composition, reactivity, and regulation of extracellular
metal-reducing structures (bacterial nanowires) produced by dissimilatory metal reducing
bacteria. Warrenton, VA. doi:10.2172/893692
Gorby, Y. A., Yanina, S., McLean, J. S., Rosso, K. M., Moyles, D., Dohnalkova, A., Fredrickson,
J. K. (2006). Electrically conductive bacterial nanowires produced by Shewanella
oneidensis strain MR-1 and other microorganisms. Proceedings of the National Academy
of Sciences of the United States of America, 103(30), 11358–63.
doi:10.1073/pnas.0604517103
Gregory, K. B., Bond, D. R., & Lovley, D. R. (2004). Graphite electrodes as electron donors for
anaerobic respiration. Environmental Microbiology, 6(6), 596–604. doi:10.1111/j.1462-
2920.2004.00593.x
Hassan, S. H. A., Kim, Y. S., & Oh, S.-E. (2012). Power generation from cellulose using mixed
and pure cultures of cellulose-degrading bacteria in a microbial fuel cell. Enzyme and
Microbial Technology, 51(5), 269–73. doi:10.1016/j.enzmictec.2012.07.008
Holmes, D. E., Nicoll, J. S., Bond, D. R., & Lovley, D. R. (2004). Potential role of a novel
psychrotolerant member of the family Geobacteraceae, Geopsychrobacter electrodiphilus
gen. nov., sp. nov., in electricity production by a marine sediment fuel cell. Applied and
Environmental Microbiology, 70(10), 6023–30. doi:10.1128/AEM.70.10.6023-6030.2004
Ishii, S., Watanabe, K., Yabuki, S., Logan, B. E., & Sekiguchi, Y. (2008). Comparison of
electrode reduction activities of Geobacter sulfurreducens and an enriched consortium in
an air-cathode microbial fuel cell. Applied and Environmental Microbiology, 74(23), 7348–
55. doi:10.1128/AEM.01639-08
Jang, J. K., Chang, I. S., Moon, H., Kang, K. H., & Kim, B. H. (2006). Nitrilotriacetic acid
degradation under microbial fuel cell environment. Biotechnology and Bioengineering,
95(4), 772–4. doi:10.1002/bit.20820
Kennedy, A. C. (1999). Microbial diversity in agroecosystem quality. In: Biodiversity in
Agroecosystems (Ed.CRC Press). Florida, pp. 1-19, ISBN: 1-56670-290-9
Kim, B. H., Ikeda, T., Park, H. S., Kim, H. J., Hyun, M. S., Kano, K., Tatsumi, H. (1999).
Electrochemical activity of an Fe(III)-reducing bacterium, Shewanella putrefaciens IR-1, in

54
the presence of alternative electron acceptors. Biotechnology Techniques, 13(7), 475–
478. doi:10.1023/A:1008993029309
Kim, B. H., Park, H. S., Kim, H. J., Kim, G. T., Chang, I. S., Lee, J., & Phung, N. T. (2004).
Enrichment of microbial community generating electricity using a fuel-cell-type
electrochemical cell. Applied Microbiology and Biotechnology, 63(6), 672–81.
doi:10.1007/s00253-003-1412-6
Kim, N., Choi, Y., Jung, S., & Kim, S. (2000). Development of Microbial Fuel Cells Using
Proteus vulgaris, 21(1), 44–48.
Kim, S.-J., Lee, C.-M., Han, B.-R., Kim, M.-Y., Yeo, Y.-S., Yoon, S.-H., Jun, H.-K. (2008).
Characterization of a gene encoding cellulase from uncultured soil bacteria. FEMS
Microbiology Letters, 282(1), 44–51. doi:10.1111/j.1574-6968.2008.01097.x
Lefebvre, O., Nguyen, T. T. H., Al-Mamun, A., Chang, I. S., & Ng, H. Y. (2010). T-RFLP reveals
high β-Proteobacteria diversity in microbial fuel cells enriched with domestic wastewater.
Journal of Applied Microbiology, 109(3), 839–50. doi:10.1111/j.1365-2672.2010.04735.x
Liu, J., Feng, Y., Wang, X., Yang, Q., Shi, X., Qu, Y., & Ren, N. (2012). The effect of water
proofing on the performance of nickel foam cathode in microbial fuel cells. Journal of
Power Sources, 198, 100–104. doi:10.1016/j.jpowsour.2011.09.078
Liu, M., Yuan, Y., Zhang, L., Zhuang, L., Zhou, S., & Ni, J. (2010). Bioelectricity generation by a
Gram-positive Corynebacterium sp. strain MFC03 under alkaline condition in microbial fuel
cells. Bioresource Technology, 101(6), 1807–11. doi:10.1016/j.biortech.2009.10.003
Liu, Y., & Tay, J. H. (2000). A kinetic model for energy spilling-associated product formation in
substrate-sufficient continuous culture. Journal of Applied Microbiology, 88(4), 663–8.
doi:10.1046/j.1365-2672.2000.01009.x
Logan, B. E. (2004). Feature Article: Biologically extracting energy from wastewater:
biohydrogen production and microbial fuel cells. Environment Science and Technology,
38, 160A-167A.
Logan, B. E. (2009). Exoelectrogenic bacteria that power microbial fuel cells. Nature Reviews
Microbiology, 7(5), 375–81. doi:10.1038/nrmicro2113
Logan, B. E. (2010). Scaling up microbial fuel cells and other bioelectrochemical systems.
Applied Microbiology and Biotechnology, 85(6), 1665–71. doi:10.1007/s00253-009-2378-9

55
Logan, B. E. (2012). Essential data and techniques for conducting microbial fuel cell and other
types of bioelectrochemical system experiments. ChemSusChem, 5(6), 988–94.
doi:10.1002/cssc.201100604
Logan, B. E. (2008). Microbial fuel cells. John Wiley & Sons, Inc., Hoboken, New York. ISBN:
978-0-470-23948-3
Logan, B. E., Hamelers, B., Rozendal, R., Schröder, U., Keller, J., Freguia, S., Rabaey, K.
(2006). Microbial fuel cells: methodology and technology. Environmental Science &
Technology, 40(17), 5181–92. doi: 10.1021/es0605016
Lu, L.& Xing, D.F.& Ren, N.Q. (2012), Pyrosequencing reveals highly diverse microbial
communities in microbial electrolysis cells involved in enhanced H2 production from waste
activated sludge. Water Research, 46(7), 2425-2434. Doi:10.1016/j.watres.2012.02.005
Lueders, T., & Friedrich, M. (2000). Archaeal population dynamics during sequential reduction
processes in rice field soil. Applied and Environmental Microbiology, 66(7), 2732–42.
doi:10.1128/AEM.66.7.2732-2742.2000
Luo, J., Yang, J., He, H., Jin, T., Zhou, L., Wang, M., & Zhou, M. (2013). A new
electrochemically active bacterium phylogenetically related to Tolumonas osonensis and
power performance in MFCs. Bioresource Technology, 139, 141–8.
doi:10.1016/j.biortech.2013.04.031
Lyon, D. Y., Buret, F., Vogel, T. M., & Monier, J.-M. (2010). Is resistance futile? Changing
external resistance does not improve microbial fuel cell performance. Bioelectrochemistry
(Amsterdam, Netherlands), 78(1), 2–7. doi:10.1016/j.bioelechem.2009.09.001
Malvankar, N. S., & Lovley, D. R. (2014). Microbial nanowires for bioenergy applications.
Current Opinion in Biotechnology, 27, 88–95. doi:10.1016/j.copbio.2013.12.003
Marshall, C. W., & May, H. D. (2009). Electrochemical evidence of direct electrode reduction by
a thermophilic Gram-positive bacterium, Thermincola ferriacetica. Energy & Environmental
Science, 2(6), 699. doi:10.1039/b823237g
Michaelidou, U., ter Heijne, A., Euverink, G. J. W., Hamelers, H. V. M., Stams, A. J. M., &
Geelhoed, J. S. (2011). Microbial communities and electrochemical performance of
titanium-based anodic electrodes in a microbial fuel cell. Applied and Environmental
Microbiology, 77(3), 1069–75. doi:10.1128/AEM.02912-09

56
Milliken, C. E., & May, H. D. (2007). Sustained generation of electricity by the spore-forming,
Gram-positive, Desulfitobacterium hafniense strain DCB2. Applied Microbiology and
Biotechnology, 73(5), 1180–9. doi:10.1007/s00253-006-0564-6
Moon, H., Chang, I. S., & Kim, B. H. (2006). Continuous electricity production from artificial
wastewater using a mediator-less microbial fuel cell. Bioresource Technology, 97(4), 621–
7. doi:10.1016/j.biortech.2005.03.027
Narihiro, T., & Sekiguchi, Y. (2007). Microbial communities in anaerobic digestion processes for
waste and wastewater treatment: a microbiological update. Current Opinion in
Biotechnology, 18(3), 273–8. doi:10.1016/j.copbio.2007.04.003
Nevin, K. P., Richter, H., Covalla, S. F., Johnson, J. P., Woodard, T. L., Orloff, A. L., Lovley, D.
R. (2008). Power output and columbic efficiencies from biofilms of Geobacter
sulfurreducens comparable to mixed community microbial fuel cells. Environmental
Microbiology, 10(10), 2505–14. doi:10.1111/j.1462-2920.2008.01675.x
Nimje, V. R., Chen, C.-Y., Chen, C.-C., Jean, J.-S., Reddy, A. S., Fan, C.-W., … Chen, J.-L.
(2009). Stable and high energy generation by a strain of Bacillus subtilis in a microbial fuel
cell. Journal of Power Sources, 190(2), 258–263. doi:10.1016/j.jpowsour.2009.01.019
Pace, N. R., Olsen, G. J., & Woese, C. R. (1986). Ribosomal RNA phylogeny and the primary
lines of evolutionary descent. Cell, 45(3), 325–6.
Pant, D., Van Bogaert, G., Diels, L., & Vanbroekhoven, K. (2010). A review of the substrates
used in microbial fuel cells (MFCs) for sustainable energy production. Bioresource
Technology, 101(6), 1533–43. doi:10.1016/j.biortech.2009.10.017
Park, D. H., & Zeikus, J. G. (2000). Electricity generation in microbial fuel cells using neutral red
as an electronophore. Applied and Environmental Microbiology, 66(4), 1292–7. doi:
10.1128/AEM.66.4.1292-1297.2000
Park, D. H., & Zeikus, J. G. (2003). Improved fuel cell and electrode designs for producing
electricity from microbial degradation. Biotechnology and Bioengineering, 81(3), 348–55.
doi:10.1002/bit.10501
Park, H. S., Kim, B. H., Kim, H. S., Kim, H. J., Kim, G. T., Kim, M., … Chang, H. I. (2001). A
Novel Electrochemically Active and Fe(III)-reducing Bacterium Phylogenetically Related to
Clostridium butyricum Isolated from a Microbial Fuel Cell. Anaerobe, 7(6), 297–306.
doi:10.1006/anae.2001.0399

57
Park, T.-J., Ding, W., Cheng, S., Brar, M. S., Ma, A. P. Y., Tun, H. M., & Leung, F. C. (2014).
Microbial community in microbial fuel cell (MFC) medium and effluent enriched with purple
photosynthetic bacterium (Rhodopseudomonas sp.). AMB Express, 4, 22.
doi:10.1186/s13568-014-0022-2
Pham, C. A., Jung, S. J., Phung, N. T., Lee, J., Chang, I. S., Kim, B. H., … Chun, J. (2003). A
novel electrochemically active and Fe(III)-reducing bacterium phylogenetically related to
Aeromonas hydrophila , isolated from a microbial fuel cell. FEMS Microbiology Letters,
223(1), 129–134. doi:10.1016/S0378-1097(03)00354-9
Potter, A. M. C., Character, B., & Sep, N. (2010). Electrical Effects Accompanying the
Decomposition of Organic Compounds. The Royal Society, 84(571), 260–276. doi:
10.1098/rspb.1911.0073
Prasad, D., Arun, S., Murugesan, M., Padmanaban, S., Satyanarayanan, R. S., Berchmans, S.,
& Yegnaraman, V. (2007). Direct electron transfer with yeast cells and construction of a
mediatorless microbial fuel cell. Biosensors & Bioelectronics, 22(11), 2604–10.
doi:10.1016/j.bios.2006.10.028
Quan, X., Quan, Y., Tao, K., & Jiang, X. (2013). Comparative investigation on microbial
community and electricity generation in aerobic and anaerobic enriched MFCs.
Bioresource Technology, 128, 259–65. doi:10.1016/j.biortech.2012.10.001
Quan, X., Quan, Y. & Tao, K. (2012). Effect of anode aeration on the performance and microbial
community of an air–cathode microbial fuel cell. Chemical Engineering Journal, 210, 150–
156. doi:10.1016/j.cej.2012.09.009
Rabaey, K., Boon, N., Siciliano, S. D., Verhaege, M., & Verstraete, W. (2004). Biofuel cells
select for microbial consortia that self-mediate electron transfer. Applied and
Environmental Microbiology, 70(9), 5373–82. doi:10.1128/AEM.70.9.5373-5382.2004
Rabaey, K., Lissenns, G., & Verstraete, W. (2005). Microbial fuel cells: performances and
perspectives (Ed. IWA Publishing), London, pp. 377–399. ISBN: 1843390922
Rabaey, K., Lissens, G., Siciliano, S. D., & Verstraete, W. (2003). A microbial fuel cell capable
of converting glucose to electricity at high rate and efficiency. Biotechnology Letters,
25(18), 1531–5. doi: 10.1023/a:1025484009367
Rabaey, K., Read, S. T., Clauwaert, P., Freguia, S., Bond, P. L., Blackall, L. L., & Keller, J.
(2008). Cathodic oxygen reduction catalyzed by bacteria in microbial fuel cells. The ISME
Journal, 2(5), 519–27. doi:10.1038/ismej.2008.1

58
Rabaey, K., Van de Sompel, K., Maignien, L., Boon, N., Aelterman, P., Clauwaert, P.,
Verstraete, W. (2006). Microbial fuel cells for sulfide removal. Environmental Science &
Technology, 40(17), 5218–24. doi:10.1021/es060382u
Rabaey, K., & Verstraete, W. (2005). Microbial fuel cells: novel biotechnology for energy
generation. Trends in Biotechnology, 23(6), 291–8. doi:10.1016/j.tibtech.2005.04.008
Ramos, C. G., Grilo, A. M., Sousa, S. A., Barbosa, M. L., Nadais, H., & Leitão, J. H. (2010). A
new methodology combining PCR, cloning, and sequencing of clones discriminated by
RFLP for the study of microbial populations: application to an UASB reactor sample.
Applied Microbiology and Biotechnology, 85(3), 801–6. doi:10.1007/s00253-009-2268-1
Reddy, M. V., Srikanth, S., Mohan, S. V., & Sarma, P. N. (2010). Phosphatase and
dehydrogenase activities in anodic chamber of single chamber microbial fuel cell (MFC) at
variable substrate loading conditions. Bioelectrochemistry (Amsterdam, Netherlands),
77(2), 125–32. doi:10.1016/j.bioelechem.2009.07.011
Reguera, G., McCarthy, K. D., Mehta, T., Nicoll, J. S., Tuominen, M. T., & Lovley, D. R. (2005).
Extracellular electron transfer via microbial nanowires. Nature, 435(7045), 1098–101.
doi:10.1038/nature03661
Rezaei, F., Xing, D., Wagner, R., Regan, J. M., Richard, T. L., & Logan, B. E. (2009).
Simultaneous cellulose degradation and electricity production by Enterobacter cloacae in a
microbial fuel cell. Applied and Environmental Microbiology, 75(11), 3673–8.
doi:10.1128/AEM.02600-08
Ringeisen, B. R., Ray, R., & Little, B. (2007). A miniature microbial fuel cell operating with an
aerobic anode chamber. Journal of Power Sources, 165(2), 591–597.
doi:10.1016/j.jpowsour.2006.10.026
Rudi, K., Skulberg, O. M., Larsen, F., & Jakobsen, K. S. (1997). Strain characterization and
classification of oxyphotobacteria in clone cultures on the basis of 16S rRNA sequences
from the variable regions V6, V7, and V8. Applied and Environmental Microbiology, 63(7),
2593–9.
Sambrook, J., Russell, D. W., & Russell, D. W. (2001). Molecular cloning: a laboratory manual
(3-volume set). Cold Spring Harbor, New York: Cold spring harbor laboratory press. ISBN-
10 0-87969-577-3
Skoog, D. A., & West, D. M. (1980). Principles of instrumental analysis (Vol. 158). Philadelphia:
Saunders College.

59
Schröder, U. (2007). Anodic electron transfer mechanisms in microbial fuel cells and their
energy efficiency. Physical Chemistry Chemical Physics : PCCP, 9(21), 2619–29.
doi:10.1039/b703627m
Shantaram, A., Beyenal, H., Raajan, R., Veluchamy, A., & Lewandowski, Z. (2005). Wireless
sensors powered by microbial fuel cells. Environmental Science & Technology, 39(13),
5037–42. doi: 10.1021/es0480668
Sharma, V., & Kundu, P. P. (2010). Biocatalysts in microbial fuel cells. Enzyme and Microbial
Technology, 47(5), 179–188. doi:10.1016/j.enzmictec.2010.07.001
Shehab, N., Li, D., Amy, G. L., Logan, B. E., & Saikaly, P. E. (2013). Characterization of
bacterial and archaeal communities in air-cathode microbial fuel cells, open circuit and
sealed-off reactors. Applied Microbiology and Biotechnology, 97(22), 9885–95.
doi:10.1007/s00253-013-5025-4
Sun, J.-Q., Xu, L., Tang, Y.-Q., Chen, F.-M., Zhao, J.-J., & Wu, X.-L. (2014). Bacterial pyridine
hydroxylation is ubiquitous in environment. Applied Microbiology and Biotechnology, 98(1),
455–64. doi:10.1007/s00253-013-4818-9
Theron, J., & Cloete, T. E. (2000). Molecular techniques for determining microbial diversity and
community structure in natural environments. Critical Reviews in Microbiology, 26(1), 37–
57. doi:10.1080/10408410091154174
Thompson, J. D., Higgins, D. G., & Gibson, T. J. (1994). CLUSTAL W: improving the sensitivity
of progressive multiple sequence alignment through sequence weighting, position-specific
gap penalties and weight matrix choice. Nucleic Acids Research, 22(22), 4673–80. doi:
10.1093/nar/22.22.4673
Thrash, J. C., Pollock, J., Torok, T., & Coates, J. D. (2010). Description of the novel perchlorate-
reducing bacteria Dechlorobacter hydrogenophilus gen. nov., sp. nov.and Propionivibrio
militaris, sp. nov. Applied Microbiology and Biotechnology, 86(1), 335–43.
doi:10.1007/s00253-009-2336-6
Timmers, R. A., Strik, D. P. B. T. B., Hamelers, H. V. M., & Buisman, C. J. N. (2012).
Characterization of the internal resistance of a plant microbial fuel cell. Electrochimica
Acta, 72, 165–171. doi:10.1016/j.electacta.2012.04.023
Vandevivere, P., & Verstraete, W. (2001). Environmental applications. Basic Biotechnology.
Cambridge University Press. ISBN: 9780521549585

60
Wagner, M., & Loy, A. (2002). Bacterial community composition and function in sewage
treatment systems. Current Opinion in Biotechnology, 13(3), 218–27. doi:10.1016/S0958-
1669(02)00315-4
Walker, A. L., & Walker, C. W. (2006). Biological fuel cell and an application as a reserve power
source. Journal of Power Sources, 160(1), 123–129. doi:10.1016/j.jpowsour.2006.01.077
Wallace, R. B., Shaffer, J., Murphy, R. F., Bonner, J., Hirose, T., & Itakura, K. (1979).
Hybridization of synthetic oligodeoxyribonucleotides to phi chi 174 DNA: the effect of
single base pair mismatch. Nucleic Acids Research, 6(11), 3543–57.
Wang, H., Jiang, S. C., Wang, Y., & Xiao, B. (2013). Substrate removal and electricity
generation in a membrane-less microbial fuel cell for biological treatment of wastewater.
Bioresource Technology, 138, 109–16. doi:10.1016/j.biortech.2013.03.172
Woese, C. R. (1987). Bacterial evolution. Microbiological Reviews, 51(2), 221–71.
Wrighton, K. C., Agbo, P., Warnecke, F., Weber, K. A., Brodie, E. L., DeSantis, T. Z., … Coates,
J. D. (2008). A novel ecological role of the Firmicutes identified in thermophilic microbial
fuel cells. The ISME Journal, 2(11), 1146–56. doi:10.1038/ismej.2008.48
Xing, D., Zuo, Y., Cheng, S., Regan, J. M., & Logan, B. E. (2008). Electricity Generation by
Rhodopseudomonas palustris DX-1. Environmental Science & Technology, 42(11), 4146–
4151. doi:10.1021/es800312v
Xu, S., & Liu, H. (2011). New exoelectrogen Citrobacter sp. SX-1 isolated from a microbial fuel
cell. Journal of Applied Microbiology, 111(5), 1108–15. doi:10.1111/j.1365-
2672.2011.05129.x
Yong, Y.-C., Yu, Y.-Y., Li, C.-M., Zhong, J.-J., & Song, H. (2011). Bioelectricity enhancement via
overexpression of quorum sensing system in Pseudomonas aeruginosa-inoculated
microbial fuel cells. Biosensors & Bioelectronics, 30(1), 87–92.
doi:10.1016/j.bios.2011.08.032
Yoon, S. S., Hennigan, R. F., Hilliard, G. M., Ochsner, U. A., Parvatiyar, K., Kamani, M. C.,
Hassett, D. J. (2002). Pseudomonas aeruginosa anaerobic respiration in biofilms:
relationships to cystic fibrosis pathogenesis. Developmental Cell, 3(4), 593–603.
doi:10.1016/S1534-5807(02)00295-2
Zhang, J., Zhang, E., Scott, K., & Burgess, J. G. (2012). Enhanced Electricity Production by Use
of Reconstituted Artificial Consortia of Estuarine Bacteria Grown as Biofilms.
Environmental Science & Technology, 46(5), 2984–2992. doi:10.1021/es2020007

61
Zhang, T., Cui, C., Chen, S., Ai, X., Yang, H., Shen, P., & Peng, Z. (2006). A novel mediatorless
microbial fuel cell based on direct biocatalysis of Escherichia coli. Chemical
Communications (Cambridge, England), (21), 2257–9. doi:10.1039/b600876c
Zhang, W., Sileika, T. S., Chen, C., Liu, Y., Lee, J., & Packman, A. I. (2011). A novel planar flow
cell for studies of biofilm heterogeneity and flow-biofilm interactions. Biotechnology and
Bioengineering, 108(11), 2571–82. doi:10.1002/bit.23234
Zhang, Y., Min, B., Huang, L., & Angelidaki, I. (2009). Generation of electricity and analysis of
microbial communities in wheat straw biomass-powered microbial fuel cells. Applied and
Environmental Microbiology, 75(11), 3389–95. doi:10.1128/AEM.02240-08
Zhao, F., Rahunen, N., Varcoe, J. R., Chandra, A., Avignone-Rossa, C., Thumser, A. E., &
Slade, R. C. T. (2008). Activated Carbon Cloth as Anode for Sulfate Removal in a
Microbial Fuel Cell. Environmental Science & Technology, 42(13), 4971–4976.
doi:10.1021/es8003766
Zuo, Y., Xing, D., Regan, J. M., & Logan, B. E. (2008). Isolation of the exoelectrogenic
bacterium Ochrobactrum anthropi YZ-1 by using a U-tube microbial fuel cell. Applied and
Environmental Microbiology, 74(10), 3130–7. doi:10.1128/AEM.02732-07

62

63
Anexo I
Figura 18: Árvore filogenética correspondente às espécies de bactérias identificadas no compartimento anódico na
fase de maior produção de energia (R1A_B1).
Figura 19: Árvore filogenética correspondente às espécies de bactérias identificadas no compartimento anódico na
fase de menor produção de energia (R1C_B2).
R1A 69
Pseudoxanthobacter
Acinetobacter baylyi
Proteobacterium symbiont of Nilaparva...
R1A 12
Rhodocyclaceae bacterium
Diaphorobacter
Chryseobacterium
Flavobacteriaceae bacterium
R1A 28
R1A 75
R1A 85
R1A 65
R1A 9
0.2

64
Figura 20: Árvore filogenética correspondente às espécies de archaeas identificadas no compartimento anódico na
fase de maior produção de energia (R1A_A1).
A53
A54
A20
A16
Methanolinea sp.
A33
Methanolinea sp.(2)
Methanosphaerula palustris
A12
A17
Methanosaeta concilii
A9
A147
A35
0.02

65
Figura 21: Árvore filogenética correspondente às espécies de bactérias com maior identidade para com as
sequências de RNA 16S obtidas por PCR a partir do DNA total extraído do compartimento catódico do reator 1 na
fase de maior produção de energia (R1C_B1).
R1C 36
R1C 40
R1C 19
R1C 14
R1C 126
R1C 6
R1C 98
Burkholderia
Burkholderia metallica
R1C 57
Rhodocyclaceae bacterium
R1C 22
Acidovorax citrulli
R1C 18
R1C 45
R1C 32
Stenotrophomonas
Pseudomonas 1
Pseudomonas geniculata
Pseudomonas 2
Carbophilus carboxidus
Methylotrophic proteobacterium
Aminobacter
R1C 60
R1C 20
R1C 73
R1C 64
Shigella boydii
Escherichia coli
R1C 104
Serratia
Pseudomonas
R1C 31
R1C 17
Leadbetterella byssophila
0.02

66
Figura 22: Árvore filogenética correspondente às espécies de bactérias com maior identidade para com as
sequências de RNA 16S obtidas por PCR a partir do DNA total extraído no compartimento catódico do reator 1 na
fase de menor produção de energia (R1C_B2).

67
Figura 23: Árvore filogenética correspondente às espécies bacterianas mais próximas das sequências obtidas por
PCR após extração de DNA total do compartimento anódico do reator 2 na fase de maior produção de energia
(R2A_B1).
R2A 31B
R2A 48B
Pseudomonas sp. G1-10
R2A 21B
Pseudomonas sp. X-b2
R2A 1B
Pseudomonas sp. UA-JF2901
R2A 15B
R2A 23B
R2A 22B
Geobacter sulfurreducens
R2A 25B
Brevundimonas sp.
R2A 45B
Rhodocyclaceae sp.
Azospira sp.
Dechlorosoma suillum
R2A 32B
R2A 34B
R2A 12B
Diaphorobacter sp.
R2A 24B
Chryseobacterium koreense
Kaistella koreensis
R2A 62B
0.05

68
Figura 24: Árvore filogenética correspondente às espécies de Archaea cujas sequências de rRNA 16S são mais
próximas das sequências obtidas por PCR após extração de DNA total do compartimento anódico do reator 2 na
fase de maior produção de energia (R2A_A1).
R2A 31A
R2A 71A
Methanolinea sp.
R2A 42A
Methanolinea mesophila
R2A 36A
Methanolinea tarda
R2A 15A
R2A 68A
R2A 35A
Methanomassiliicoccus luminyensis
Methanobrevibacter smithii
R2A 21A
R2A 18A
R2A 46A
R2A 48A
Methanobrevibacter acididurans
Methanobrevibacter arboriphilus
R2A 16A
R2A 17A
R2A 50A
R2A 63A
Candidatus Nitrososphaera gargensis
0.05

69
Figura 25: Árvore filogenética correspondente às espécies de bactérias cujas sequências de rRNA 16S são mais
próximas das sequências obtidas por PCR após extração de DNA total do compartimento anódico do reator 2 na
fase de menor produção de energia (R2A_B2).
R2A B2 10
Ottowia sp.
R2A B2 12
Pelomonas sp.
R2A B2 13
Cupriavidus sp.
R2A B2 29
R2A B2 26
R2A B2 8
Propionivibrio militaris
R2A B2 24
R2A B2 16
R2A B2 11
Veillonellaceae bacterium
Geobacter sp.
R2A B2 30
0.02

70
Figura 26: Árvore filogenética correspondente às espécies de bactérias mais próximas das sequências obtidas por
PCR após extração de DNA total do compartimento catódico do reator 2 na fase de maior produção de energia
(R2C_B1).
R2C 7B
R2C 5B
R2C 32B
Ultramicrobacter hongkongensis
Methyloversatilis universalis
R2C 26B
R2C 29B
Rhizobacter sp.
R2C 21B
Piscinibacter sp.
R2C 44B
R2C 28B
Dokdonella soli
R2C 33B
Rhodanobacter sp.
R2C 8B
R2C 39B
Afipia sp.
R2C 41B
Mesorhizobium sp.
Rhizobium sp.
0.02

71
Figura 27: Árvore filogenética correspondente às espécies de bactérias mais próximas das sequências obtidas por
PCR após extração de DNA total do compartimento catódico do reator 2 na fase de menor produção de energia
(R2C_B2).
R2C B2 75
Nitrobacter sp.
Bradyrhizobium sp.
R2C B2 53
Devosia ginsengisoli
Aminobacter sp.
Ochrobactrum cytisi
Virgibacillus sp.
Chloroflexi
Terrimonas sp.
R2C B2 67
R2C B2 66
Fluviicola taffensis
Wandonia haliotis
Acidobacteria bacterium
R2C B2 44
Stenotrophomonas acidaminiphila
R2C B2 59
Nitrosomonas sp.
R2C B2 74
R2C B2 77
Beta proteobacterium
Propionivibrio militaris
Aquincola sp.
R2C B2 49
R2C B2 45
Acidovorax sp.
Citrobacter freundii
Pseudomonas aeruginosa
R2C B2 55
R2C B2 48
R2C B2 57
R2C B2 8
R2C B2 40
R2C B2 83
R2C B2 47
R2C B2 117
0.2


















