
Carlos A. Eslava Campos Ulises ... - La Medicina es Así
24
Unidad VII - Ecología médica Bacteriología Carlos A. Eslava Campos Secretaría de Salud, Hospital Infantil de México Federico Gómez, Unidad de Hemato-Oncología e Investigación, Laboratorio de Patogenicidad Bacteriana. Ciudad de México, México. Ulises Hernández Chiñas Secretaría de Salud, Hospital Infantil de México Federico Gómez, Unidad de Hemato-Oncología e Investigación, Laboratorio de Patogenicidad Bacteriana. Ciudad de México, México. José Molina López Secretaría de Salud, Hospital Infantil de México Federico Gómez, Unidad de Hemato-Oncología e Investigación, Laboratorio de Patogenicidad Bacteriana. Ciudad de México, México. María E. Chávez Berrocal Secretaría de Salud, Hospital Infantil de México Federico Gómez, Unidad de Hemato-Oncología e Investigación, Laboratorio de Patogenicidad Bacteriana. Ciudad de México, México. Silvia Giono Cerezo Instituto Politécnico Nacional, Escuela Nacional de Ciencias Biológicas, Departamento de Microbiología, Laboratorio de Bacteriología Médica. Ciudad de México, México. Antecedentes Theodor Escherich ,en 1885, identificó y describió una bacteria a la que inicialmente llamó Bacterium coli commune var neopolitanum, debido a que la aisló de las heces de niños con diarrea, y propuso la participación de este microorganismo como agente causal del padecimiento. Sin embargo, ya que la bacteria también era aislada de niños sanos, se descartó su participación en la patogénesis. Tuvo que transcurrir más de medio siglo para que la bacteria fuera reconocida como patógeno intestinal, en 1945 cuando Bray 1 generó la hipótesis de la existencia de subtipos de E. coli que podrían explicar la diarrea infantil de etiología desconocida. Fue Neter 2 , quién utilizando un esquema de tipificación con sueros para identificar antígenos de la bacteria (O:H), propuesto por Kauffmann, probo la hipótesis de Bray y confirmó que ciertas serovariedades de E. coli presentaban asociación con diarrea en niños. Finalmente, la participación de E. coli en la patogénesis de las diarreas fue confirmado por M. Levin 3 al desafiar voluntarios con la bacteria. H. coli es una bacteria gramnegativa no esporulada, anaerobia facultativa que habita el intestino y las heces tanto de animales de sangre caliente como de reptiles. Es un
Transcript of Carlos A. Eslava Campos Ulises ... - La Medicina es Así

Unidad VII - Ecología médicaBacteriología
Carlos A. Eslava Campos
Secretaría de Salud, Hospital Infantil de México Federico Gómez, Unidad de Hemato-Oncología e Investigación, Laboratorio dePatogenicidad Bacteriana. Ciudad de México, México.
Ulises Hernández Chiñas
Secretaría de Salud, Hospital Infantil de México Federico Gómez, Unidad de Hemato-Oncología e Investigación, Laboratorio dePatogenicidad Bacteriana. Ciudad de México, México.
José Molina López
Secretaría de Salud, Hospital Infantil de México Federico Gómez, Unidad de Hemato-Oncología e Investigación, Laboratorio dePatogenicidad Bacteriana. Ciudad de México, México.
María E. Chávez Berrocal
Secretaría de Salud, Hospital Infantil de México Federico Gómez, Unidad de Hemato-Oncología e Investigación, Laboratorio dePatogenicidad Bacteriana. Ciudad de México, México.
Silvia Giono Cerezo
Instituto Politécnico Nacional, Escuela Nacional de Ciencias Biológicas, Departamento de Microbiología, Laboratorio deBacteriología Médica. Ciudad de México, México.
Antecedentes
Theodor Escherich ,en 1885, identificó y describió una bacteria a la que inicialmente llamóBacterium coli commune var neopolitanum, debido a que la aisló de las heces de niños condiarrea, y propuso la participación de este microorganismo como agente causal delpadecimiento. Sin embargo, ya que la bacteria también era aislada de niños sanos, sedescartó su participación en la patogénesis.Tuvo que transcurrir más de medio siglo para que la bacteria fuera reconocida comopatógeno intestinal, en 1945 cuando Bray1 generó la hipótesis de la existencia de subtipos deE. coli que podrían explicar la diarrea infantil de etiología desconocida. Fue Neter 2, quiénutilizando un esquema de tipificación con sueros para identificar antígenos de la bacteria(O:H), propuesto por Kauffmann, probo la hipótesis de Bray y confirmó que ciertasserovariedades de E. coli presentaban asociación con diarrea en niños. Finalmente, laparticipación de E. coli en la patogénesis de las diarreas fue confirmado por M. Levin3 aldesafiar voluntarios con la bacteria.H. coli es una bacteria gramnegativa no esporulada, anaerobia facultativa que habita elintestino y las heces tanto de animales de sangre caliente como de reptiles. Es un

Unidad VII - Ecología médicaBacteriología
microorganismo importante ya que junto a más de 500 especies bacterianas convive ensimbiosis con el hospedero y constituye la microbiota intestinal del tracto digestivo.4 E. coli seencuentra en el intestino grueso, especialmente en el ciego y el colon y reside en la capa democo que cubre las células epiteliales del tracto intestinal, lo que representa un nichoecológico nutricional al que la bacteria se adaptó por su alto contenido de carbohidratos,metabolito requerido para la adquisición de energía.5
Existe una gran diversidad de clonas de E. coli en el hospedero, debido a factores como eltamaño del cuerpo, la morfología del intestino, los hábitos alimenticios, tiempo de retencióndigestiva y el resto de microorganismos integrantes de la microbiota.6 E. coli es una de lasprimeras especies bacterianas que colonizan el intestino, las cepas iniciales pueden originarsede la biota fecal materna y puede alcanzar una densidad mayor a 10 UFC por gramo deheces, antes de la expansión de las bacterias anaerobias.9
Después de los dos años de edad, la densidad de las bacterias se estabiliza y permanece enconcentraciones de aproximadamente 10 UFC, para posteriormente disminuir gradualmente.8
La relación entre E. coli y el hospedero podría ser definida como comensalismo ya que labacteria provee beneficios como interferir en la colonización por microorganismos patógenos,elaborar compuestos como vitaminas y enzimas necesarios para el buen funcionamiento delorganismo5 y, por su parte, la bacteria recibe nutrientes importantes para su sobrevivencia.La plasticidad del genoma de E. coli le ha permitido evolucionar de tal manera que además deexistir las variedades que viven como parte de la biota del intestino, han emergido clonasvirulentas, algunas que colonizan el tracto digestivo y otras que en su evolución adquirieronla capacidad de establecerse fuera de este aparato (Tabla 1).
Tabla 1.Grupos clonales y características generales de E. coli.
Grupo
Comportamiento
Colonización intestinal Diarrea Infección extraintestinal
Comensal +++ – +
DEC +++ +++ –
ExPEC + – +++

Unidad VII - Ecología médicaBacteriología
(-) ausente(+) poco(+++) máximo.
Las propiedades de virulencia que ha desarrollado E. coli se deben a la adquisición horizontalde elementos genéticos móviles, como son los plásmidos, las islas de patogenicidad y losvirus de bacterias conocidos como fagos.7 (Cuadro 1)

Unidad VII - Ecología médicaBacteriología
Aunque en la actualidad existen diferentes procedimientos para definir la identidad de losmicroorganismos en los inicios de la bacteriología, fue mediante ensayos de aglutinación conel suero y cepas de la bacteria aisladas de niños con cuadros de diarrea, que pudoestablecerse que existían variedades de E. coli responsables del padecimiento. Con dichosantecedentes, Kauffman desarrollo un sistema para la obtención en conejos, de sueros con

Unidad VII - Ecología médicaBacteriología
anticuerpos específicos para la identificación de las variedades de la bacteria asociadas con laproducción de diarrea en niños.7
Estructura antigénica de E. coli.
Para la caracterización de la bacteria se realiza la identificación de los antígenos somáticos(O), capsulares (K) y flagelares (H). El antígeno O forma parte de las cadenas laterales decarbohidratos del lipopolisacárido, hasta el momento existen 186 variedades reportadasreconocidas internacionalmente (O1-O186). Este es un antígeno termoestable, resiste elcalentamiento a 121°C.Los antígenos K son polisacáridos termolábiles que rodean la célula a manera de envoltura ocomo cápsula rudimentaria. Se conocen tres variedades de este antígeno y, considerandoalgunas características físicas, para reconocerlas se les han asignado las letras L, B y A. Seconocen 91 variedades del este antígeno y se reportan de K1 a K91. La composición delantígeno flagelar H es de naturaleza proteica y por lo mismo son termolábiles, las cepas de E.coli no móviles no los presentan, aunque algunas cepas poseen los genes asociados a suexpresión, hasta el momento se reconocen 56 variedades del antígeno. Al realizar lacaracterización solo del antígeno somático se establece el serogrupo de la bacteria, cuandose identifica la variedad flagelar entonces se define el serotipo, y la fórmula antigénicacompleta se establece cuando se caracterizan los tres antígenos.La participación de E. coli en la etiología de cuadros de diarrea en niños que reportó Bray en1945,1 fue confirmada en México por un grupo de investigadores del Hospital Infantil (1946)que reportaron una cepa a la que llamaron Escherichia coli Gómez, iniciando una etapa decrecimiento explosivo en el estudio de la bacteria a nivel mundial, en la que ha participado ungran número de investigadores mexicanos.8 Con el empleo de la serología pudo definirse queexistían variedades intestinales patógenas y no patógenas de la bacteria, sin embargo, elmismo T. Escherich señaló que este microorganismo era responsable de las infecciones devías urinarias de niños y niñas, lo que vislumbró la existencia de variedades responsables deinfecciones extraintestinales.7
La serología por un largo tiempo fue de gran utilidad para identificar al microorganismo,posteriormente, sabiendo que la bacteria podía aceptar material genético externo y que lascaracterísticas de virulencia de cada grupo se asociaban a genes diferentes, surgió lapregunta en relación con el origen filogenético de los diferentes grupos patógenos de labacteria.Evolución de Escherichia coli
La adaptación evolutiva de E. coli sucede a través de una selección positiva o de cambiosgenéticos al azar, afectado con frecuencia por la colonización de un nuevo sitio o durante latransmisión a nuevos hospederos, llevando a E. coli a la evolución de su virulencia y la
Unidad VII - Ecología médicaBacteriología
aparición de linajes patogénicos de la bacteria. Dependiendo de la transmisión y del sitio deinfección, estos pueden ser subdivididos en tres categorías denominados patógenosoportunistas, accidentales y profesionales.Las cepas de E. coli comensales pueden ser oportunistas, cuando salen del intestino yocasionan una infección en un hospedero que presenta barreras de defensa o sistemasinmunocomprometidos. Una característica importante de los patógenos oportunistas es quepueden retornar a su hábitat original durante la infección, un ejemplo es E. coli uropatógena(UPEC). Los accidentales, son patógenos que no circulan entre humanos sino que sonadquiridos por contacto con otras especies o a través del ambiente. Un ejemplo son las cepasde E. coli enterohemorrágica por ser un patógeno zoonótico. Los patógenos profesionalescausan una enfermedad para poder continuar su circulación en la naturaleza y puedentransmitirse durante una infección activa, un ejemplo es E. coli enteroinvasiva y E. colienterotoxigénica.H. coli por sus propiedades como bacteria patógena se integró a ocho tipos o variedadespatógenas, dos que pueden colonizar estructuras fuera del intestino denominadas E. coliextraintestinales (ExPEC por sus siglas en inglés) grupo que incluye las cepas de E. coliuropatógena (UPEC) y las que producen meningitis neonatal (NMEC). (Tabla 2)
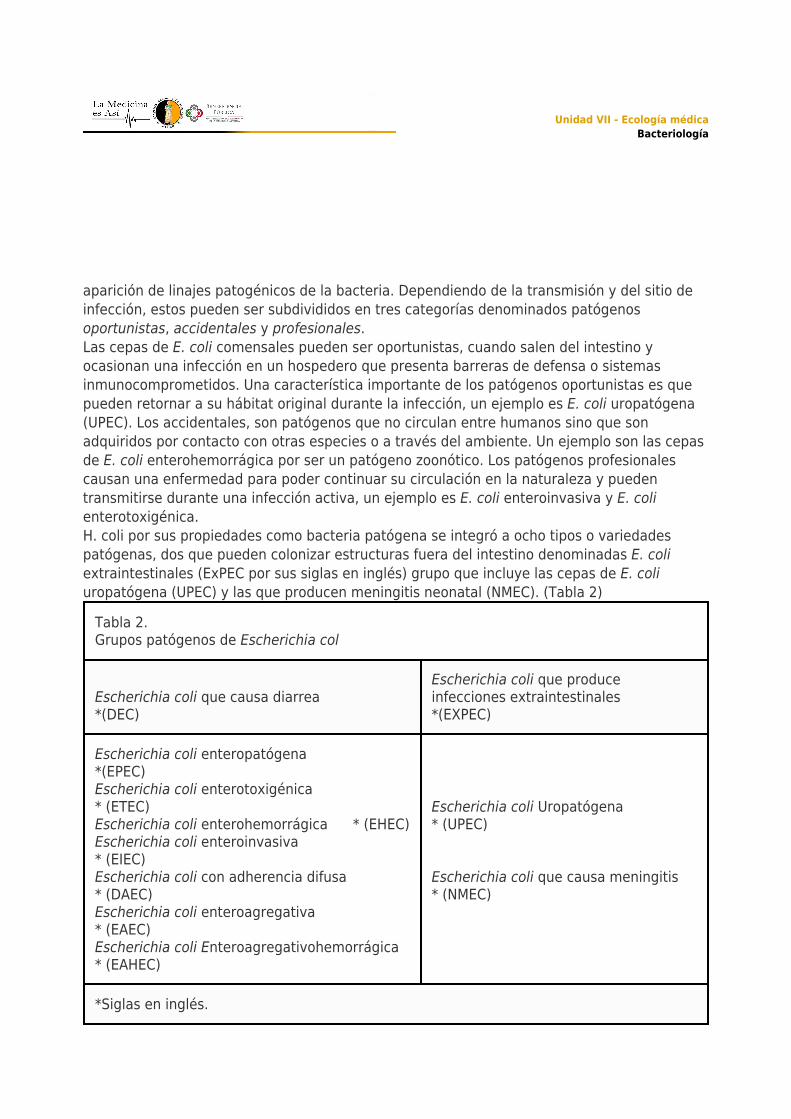
Tabla 2.Grupos patógenos de Escherichia col
Escherichia coli que causa diarrea*(DEC)
Escherichia coli que produceinfecciones extraintestinales*(EXPEC)
Escherichia coli enteropatógena*(EPEC)Escherichia coli enterotoxigénica* (ETEC)Escherichia coli enterohemorrágica * (EHEC)Escherichia coli enteroinvasiva* (EIEC)Escherichia coli con adherencia difusa* (DAEC)Escherichia coli enteroagregativa* (EAEC)Escherichia coli Enteroagregativohemorrágica* (EAHEC)
Escherichia coli Uropatógena* (UPEC) Escherichia coli que causa meningitis* (NMEC)
*Siglas en inglés.

Unidad VII - Ecología médicaBacteriología
En relación con las variedades que afectan el tracto digestivo, se han reportado seisvariedades que se conocen como E. coli productoras de diarrea (DEC). Las cepas DEC seclasifican de acuerdo con su mecanismo de patogenicidad y el cuadro clínico que producen,integrando los patotipos: E. coli enteropatógena (EPEC), E. coli enterotoxigenica (ETEC), E. colienteroinvasiva (EIEC), E. coli enterohemorragica (EHEC), E. coli enteroagregativa (EAEC) y E.
coli de adherencia difusa (DAEC).7 En el año 2011 en el continente europeo se presentó unbrote de diarrea con cuadros de colitis hemorrágica y casos de síndrome urémico hemolítico,asociados todos éstos a una cepa con características de los grupos EAEC y EHEC. Por sucomportamiento híbrido se le asignó el nombre de E. coli enteroagregativohemorrágica(EAHEC), sin embargo, al momento no se han reportado nuevos brotes producidos por esta ocepas similares.Grupos filogenéticos
El avance en la tecnología ha sido un factor decisivo en el desarrollo del conocimiento en labacteriología, particularmente en el caso de E. coli., al respecto se han utilizado diferentesprocedimientos para definir la identidad de la bacteria. En 1974 Millikan propuso el uso delsistema de enzimas multilocus (MLEE; multilocus enzyme electrophoresis), que consiste enanalizar los cambios presentados en diferentes enzimas elementales para la sobrevivencia dela bacteria. Posteriormente se implementa el procedimiento de análisis de secuencias deaminoácidos de las enzimas multilocus. Este sistema es utilizado para analizar la distribuciónde los grupos patógenos de la bacteria.9 Con la información generada por los diferentesinvestigadores, se definieron los grupos filogenéticos o filogrupos A, B1, B2, D de E. coli, que,por la similitud génica, a su vez se integraron en los subgrupos A0, A1, B1, B21, B22, D1, D2.10
Los grupos filogenéticos A y B1 están conformados por cepas de E. coli comensalesprovenientes de la microbiota “normal” del intestino grueso de humanos y animales; hayestudios que demostraron que estas cepas no presentan factores de virulencia importantes.En contraste, los grupos filogenéticos B2 y D están conformados principalmente por cepascon alto potencial patogénico caracterizado por la expresión de diversos factores devirulencia asociados a enfermedades extraintestinales.11
Para la clasificación en grupos filogenéticos se utiliza una prueba de PCR múltiple (reacción encadena de la polimerasa), en esta se amplifican los genes chuA que codifican para unaproteína fijadora de grupo Hemo, yjaA presente en el genoma de E. coli str. K-12 y unfragmento de ADN designado TspE4.C2, obtenido a partir de una biblioteca genómica encepas secuenciadas de los diferentes grupos filogenéticos.12

Unidad VII - Ecología médicaBacteriología
Grupos patógenos de Escherichia coli que producen trastornosintestinales
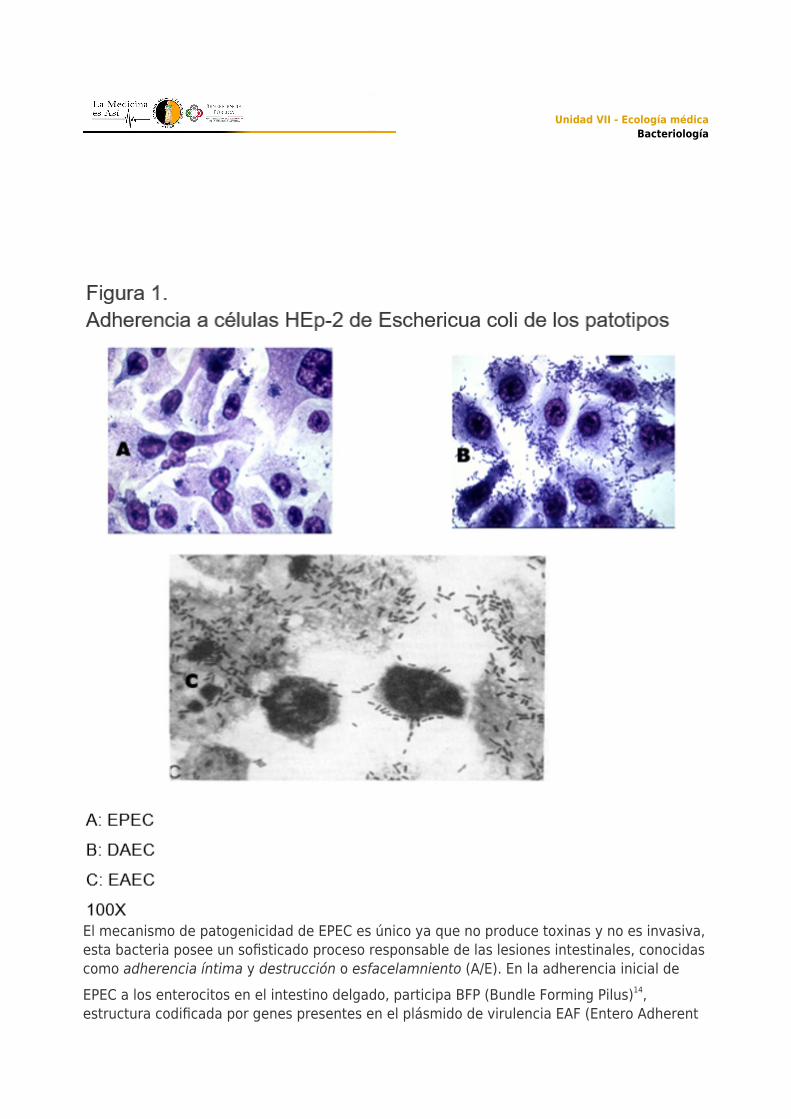
Escherichia coli enteropatógena (EPEC)Este fue el primer grupo patógeno de la bacteria que se relacionó con casos de diarrea agudaen niños menores de cinco años que asistían a guarderías, EPEC es uno de los principalesagentes asociados a diarrea aguda que puede ir acompañado por fiebre baja y vomito.7 Noobstante que con el empleo de la serología se podía identificar a las cepas EPEC,transcurrieron más de tres décadas para que en 1979,13 se describiera una de las propiedadesrelacionadas con la patogenicidad de la bacteria, que consistía en su capacidad deadherencia. (Figura 1)
Unidad VII - Ecología médicaBacteriología
El mecanismo de patogenicidad de EPEC es único ya que no produce toxinas y no es invasiva,esta bacteria posee un sofisticado proceso responsable de las lesiones intestinales, conocidascomo adherencia íntima y destrucción o esfacelamniento (A/E). En la adherencia inicial deEPEC a los enterocitos en el intestino delgado, participa BFP (Bundle Forming Pilus)14,estructura codificada por genes presentes en el plásmido de virulencia EAF (Entero Adherent

Unidad VII - Ecología médicaBacteriología
Factor). BFP forma largos haces de fimbrias que permiten la interacción entre las bacteriaspara formar microcolonias (adherencia localizada), así como con la N-acetil-lactosamina,receptor ubicado en la superficie de la célula huésped.En el evento de adherencia y destrucción de microvellosidades, también participa el sistemade secreción de tipo III (T3SS). Dicho sistema permite a la bacteria introducir al enterocito delhospedero al receptor (Tir) de la intimina proteína de superficie de la bacteria. Además de Tir,se introducen otros compuestos que activan señales intracelulares e inducen arreglos delcitoesqueleto del enterocito, como es el caso de la polimerización de la actina.15 Una vez quela bacteria se adhiere íntimamente, se inicia la formación de un pedestal con un proceso finalque induce la destrucción de las vellosidades intestinales (A/F). La expresión de los elementosantes referidos se asocia a un conjunto de genes codificados en la isla de patogenicidad LEE(locus of enetrocyte effacement).7
Los cambios en las células del epitelio intestinal contribuyen al tipo de diarrea que sepresenta debido a la perdida de la superficie de adsorción. Diferentes reportes sugieren queEPEC tiene un efecto directo sobre el proceso de transporte de iones, en particular losrelacionados con solutos, electrolitos, serotoninergicos y en el transporte de ácidos grasos.16
Clasificación de EPECOriginalmente, algunos serogrupos de EPEC fueron asociados a la diarrea infantil, sinembargo, en la actualidad su clasificación está basada en la presencia específica de genes devirulencia, utilizando técnicas moleculares, se han encontrado algunos en cepas EPEC que nose encuentran dentro de los serogrupos o serotipos clásicos. Los genes eae (intimina) y bfpA(pili en forma de manojo) han sido utilizados para la clasificación de EPEC y para susubdivisión en cepas EPEC típicas (eae +, bfpA+) y cepas EPEC atípicas (eae+, bfpA-).7,14 Pordefinición, todas las cepas EPEC carecen de genes para la producción de la toxina de Shiga(stx-).EpidemiologíaEPEC es uno de los principales agentes patógenos de diarrea en niños menores de 2 años enpaíses en desarrollo. Estudios epidemiológicos que utilizan métodos moleculares asocian aEPEC en entre 5 y 10 % de los episodios de diarrea en el mundo, pero cuando el diagnósticose basa en el patrón de adherencia localizado sobre células HEp-2 o por serotipificación, suprevalencia aumenta en promedio de 10 a 20 % con una gran variabilidad en los diferentesestudios.17,18,19
En México, las autoridades de salud han comunicado que EPEC se presenta endémicamentehasta en 6 % de la población.20 Sin embargo, estudios previos realizados en niños de la ciudadde Guadalajara, Jalisco y en una comunidad rural del estado de Morelos, refieren entre 17.5 y19 % la incidencia de diarrea por cepas EPEC.21,22

Unidad VII - Ecología médicaBacteriología
Datos similares publicados en México refieren la importancia de cepas de EPEC atípicas enniños menores con cuadros de diarrea aguda, internados en hospitales de diferentes ciudadesdel país.23
Escherichia coli enterotoxigenica (ETEC)Este grupo patógeno de la bacteria ocasiona cuadros de diarrea secretora caracterizados porabundante secreción del fluido intestinal,24 afecta a niños menores con consecuencias fatalesy a individuos que viajan de países industrializados a otros en vías de desarrollo, por lo que aeste síndrome se le conoce como diarrea del viajero. La virulencia atribuida a ETEC incluye laadherencia al epitelio intestinal por factores de colonización (CFA) y la elaboración deenterotoxinas de tipo termo lábil (TL) y termo estable (TE).ETEC posee diferentes fimbrias, cuyo receptor es un manosido y se conocen como manosasensibles, similares a las que tienen otros tipos de E. coli no enterotoxigénicos, a estas se lesdenomina fimbrias tipo 1 (F1). Existe otro grupo de fimbrias resistentes a la manosa, estas seencuentran presentes en 70 % de las cepas ETEC. Se definen como antígenos K cuando setrata de cepas de origen animal y de estas se reconocen los antígenos K88 (F2), k99 (F3)K987 (F4).En humanos los factores antigénicos de colonización (CFA) han sido los más estudiados, deellos se han descrito CFA/I, CFA/II, E8775 y PCFO59. La toxina termolábil (TL) de elevado pesomolecular (de 85,000 a 90,000 Da) tiene una estructura muy similar a la toxina del cólera y aligual que en ésta el sitio primario de unión es el monosialogangliósido GM-1. Al unirse a lascélulas de la mucosa intestinal e internalizarse induce ribosilación del ADP, estimula laadenilatociclasa e incrementa los niveles de AMPc con una cinética similar a la que induce la
toxina del cólera; el evento se relaciona con la salida de Cl–, HCO3– y agua que persiste
durante horas después que la toxina se ha fijado a la célula.Se conocen dos variedades de la toxina TLI y TLII que no cruzan antigénicamente, la primeraes expresada por cepas de E. coli aisladas de animales y humanos, sin embargo, TLIIraramente se identifica en bacterias aisladas de humanos. TLI es una toxina del grupo ABcompuesta por las subunidades A (dimérica) con actividad de enzima, y B (pentamérica) quese une a ganglosidos GM1. La toxina termoestable (TE) tiene un peso molecular bajo (2,000Da), presenta un principio de acción rápido y no se une a los gangliósidos de la membranacelular de la pared. En su estado natural no es antigénica, actúa por estimulación de laguanilatociclasa con la acumulación de GMP cíclico en células de la mucosa.Los niveles intracelulares elevados de guanilciclasa dan lugar a la secreción de cloruros porlas células de las criptas de forma similar a la toxina colérica, pero TE no altera la reabsorciónde cloruro de sodio neutro por el borde en cepillo de la vellosidad intestinal. Su efectofisiológico también incluye aumento del movimiento de líquidos de la sangre hacia elintestino, lo que da como resultado una diarrea semilíquida que puede ser indistinguible de la

Unidad VII - Ecología médicaBacteriología
diarrea producida por Vibrio cholerae.Algunas cepas de ETEC pueden presentar otros factores de virulencia como la enterotoxinatermo estable EAST1 (Entero Agregative Stable Toxin), EastA una serín proteasaautosecretada por enterobacterias (SPATE) que rompe catepsina G, acelerando la salida defluido. Otro factor de virulencia es CylA, una citotoxina formadora de poro.25
EpidemiologíaDesde que ETEC se reconoció como bacteria asociada a la diarrea aguda en viajeros y en lapoblación de países en desarrollo, diferentes investigadores se han dado a la tarea dedocumentar estudios epidemiológicos asociados a ETEC en diarreas, principalmente enpoblación pediátrica de todo el mundo.Un reporte de 136 estudios con 17,205 aislamientos de ETEC refiere que la mitad de los casos(49 %) se presentó en la población de áreas endémicas y 17 % en personas que viajaron apaíses en desarrollo. Englobando los resultados referidos los autores encontraron que de60 % de las cepas ETEC aisladas, solo 17 % expresa LT o en combinación con ST (33 %). Laexpresión de CFA fue la más común en todas las regiones del mundo [CFA/I (17 %), CFA/II(9 %), CFAIV (18 %).]Estudios epidemiológicos realizados en países en vías de desarrollo, incluidos Latinoamérica yMéxico, han demostrado que ETEC y EPEC son dos de los principales patógenos aislados enlos casos de diarrea infantil, y muchas de estas clonas son las causantes de diarrea de losextranjeros que visitan nuestro país.27,28
Escherichia coli enterohemorrágica (EHEC)Los bovinos son un reservorio clave para la transmisión de EHEC a los humanos, por logeneral ocurre a través de alimentos contaminados y de agua.29 Ha sido referido que EHECO157 es descendiente de una cepa EPEC O55, pues presenta la isla de patogenicidad LEE yproduce el evento A/E a las células del epitelio intestinal que colonizan (la región distal delíleon y el intestino grueso en los seres humanos).A diferencia de EPEC, la infección por EHEC puede causar síntomas más severos como colitishemorrágica (CH), afecciones potencialmente mortales, como el síndrome urémico hemolítico(SUH) y púrpura trombocitopénica (PT). Estos síntomas son en gran parte debidos a la acciónde las toxinas Shiga (Stx1 y/Stx2) elaboradas por la bacteria, la producción de estas toxinasdio lugar a una serie de discusiones respecto a cómo debería llamarse a este grupo debacterias; la conclusión fue que el término EHEC se utilice solo para cepas del serogrupoO157 y el de STEC (Shiga Toxin Escherichia coli) en cepas de otros serogrupos productores dela toxina.30 La familia de toxinas tipo Shiga contiene principalmente las variedades Stx1 yStx2, estas difieren en su secuencia de aminoácidos por lo que son antigénicamentediferentes.

Unidad VII - Ecología médicaBacteriología
La expresión de ambas se relaciona con bacteriófagos lisogénicos insertados en elcromosoma de la bacteria. Stx1 es similar a la toxina Shiga de Shigella dysenteriae tipo 1, dela misma se conocen las variantes stx/vtx1a, vtx1c, vtx1d. De Stx2 se han descrito lasvariantes Stx2a, Stx2b, Stx2c, Stx2d, Stx2e, Stx2f y Stx2g, de estas toxinas solo el tipo Stx2es el que se ha asociado a la producción de SUH.31
Las toxinas son del tipo A-B compuestas por una subunidad A de 32 kDa unida de forma nocovalente al anillo pentamérico de la subunidad B. La subunidad A tiene una actividad de N-glicosidasa que cataliza la despurinización de un solo residuo de adenina del ribosoma 60S,haciéndolos inactivos e inhibiendo la síntesis de proteínas.La subunidad B guía la unión de la toxina al globotriasilceramido (Gb-3), glicolípido receptorpresente en la membrana plasmática de ciertas células eucariotas. Gb-3 se encuentra en lascélulas glomerulares de riñón en humanos (CD77/Gb3) y un nuevo receptor de Stx se hadescrito en las criptas del intestino delgado de las células de Panteth. En este grupo debacterias se han identificado otros factores asociados a su virulencia, como es el caso del piliHCP (Hemorrhagic Coli Pilus), al respecto de este pili se ha reportado que activa la expresiónde citocinas proinflamatorias.En relación con las cepas STEC, estas pertenecen a una gran variedad de serotipos y puedenpresentar los genes para la expresión de toxinas tipo shiga (SLTX), se aíslan de animales y sehan relacionado con cuadros de diarrea en humanos.32
EpidemiologiaEHEC es el agente responsable de brotes de gastroenteritis grave en los países en desarrollode América del Norte, Japón y partes de Europa; la mayoría de los brotes de gastroenteritispor EHEC se deben al serotipo O157:H7, mientras que otros serotipos son de importanciamédica en países en desarrollo. En México, aunque existen reportes del aislamiento de cepasO157:H7 de animales, su frecuencia en humanos es baja.33
En un estudio34,35 se observó la presencia de anticuerpos contra el LPS O157 en 20 % de unamuestra de sueros de humanos y animales, frecuencia similar a la reportada en estudiosrealizados en EUA y Canadá en áreas en las que la bacteria es endémica, pero en las que lafrecuencia de casos de CH, SUH y PT es muy baja.Los datos referidos sugieren que los anticuerpos contra el LPS O157 son de tipo protector.Existen reportes del cruce antigénico entre el LPS O157 y el de otras bacterias incluidasalgunas Salmonelas, esta podría ser una explicación de la baja incidencia de aislamiento de labacteria en nuestro país.Escherichia coli enteroinvasiva (EIEC)Las cepas EIEC afectan la mucosa del colon y producen un cuadro disentérico similar, aunquemenos severo del que produce Shigella dysenteriae tipo 1. EIEC difiere de otros patovares de

Unidad VII - Ecología médicaBacteriología
E. coli, entre otros aspectos, por ser una bacteria intracelular obligada que no presentanflagelos ni factores de adherencia y por no fermentar la lactosa. La virulencia de EIEC seasocia a un plásmido de 220 kb que codifica para un sistema de secreción tipo III (T3SS) y unlocus Mxi-Spa, requerido para la invasión, sobrevivencia celular y apoptosis de macrófagos.33
El primer paso del proceso de patogénesis de dichas cepas es la adherencia de las bacterias alas microvellosidades de la mucosa intestinal y posteriormente al borde en cepillo delenterocito. La bacteria es capturada e internalizada por las células M, estas células laspresentan a los macrófagos en los que la bacteria estimula la apoptosis, por lo que al morir elmacrófago son liberadas.Las bacterias interaccionan con la célula intestinal que forma una vesícula en su membrana,lo anterior da lugar a que se facilite la penetración de la bacteria establecida y multiplicadaen el interior de la célula intestinal. Por efecto de eventos de señalización intracelular, seactiva la expresión en la bacteria de un sistema de secreción tipo III, este induce lapolimerización de actina celular, lo que favorece el movimiento de la bacteria, para que através de su migración por el citoesqueleto se internalice a las células adyacentes.EpidemiologíaEIEC presenta una distribución mundial y se ha reportado como causa frecuente de diarrea enBrasil, Estados Unidos y Europa. Estudios epidemiológicos realizados en México23,27 muestranque las cepas EIEC son poco frecuentes como causales de diarrea, identificándosepreferentemente después del sexto mes de vida.Escherichia coli enteroagregativa EAECEn países tanto en desarrollo como desarrollados, EAEC es considerado un patógenoemergente que se reconoce como la causa más común de diarrea del viajero después deETEC.32,36 EAEC puede colonizar los intestinos delgado y grueso y el paciente puede presentaruna leve inflamación del colon. La diarrea causada por EAEC es aguda y en algunos casospersistente, acompañada de moco y sangre.La característica fenotípica de EAEC es la adherencia llamada agregativa, en la cual estáninvolucrados genes que se encuentran en el plásmido de virulencia (pAA).7,33 Estos genescodifican para fimbrias de adherencia agregativa (AAF), que presentan adhesinas agrupadasen la familia Dr y median la adherencia a la mucosa intestinal. En ensayos in vitro con célulasHEp-2 (Figura 1) forman un patrón de adherencia en forma de ladrillos apilados, llamadoagregativo.Las variantes AAF/I, AAF/II, AAF/III, Hda han sido identificadas en las cepas EAEC37 aunque sedesconoce su receptor, se sabe que presentan una carga positiva que une las fimbrias allipopolisacárido, cuya carga es negativa. Por otro lado, la presencia de una proteína llamadadispersina que se asocia al lipopolisicarido, enmascara la carga negativa de este y promuevela dispersión de EAEC a través de la mucosa intestinal, contrarrestando la agregación entre

Unidad VII - Ecología médicaBacteriología
las bacterias.Después de adherirse a la capa de moco intestinal, EAEC es capaz de penetrarla gracias a laactividad mucolitica de Pic, serina proteasa de la familia SPATE.38 El fenotipo de adherenciaagregativo contribuye a la formación de una biopelícula sobre los enterocitos, lo que dificultala eliminación de la bacteria. El daño a la mucosa ocasionado por las cepas EAEC se debe a lasecreción de toxinas como Pet (Plasmid-encoded toxin), SPATE, cuyo blanco es la fodrina;39,40
la toxina es internalizada a la célula eucariota por endocitosis a través de un mecanismomediado por clatrina, posteriormente Pet viaja hasta el retículo endoplásmico para despuésencontrarlo en el citoplasma. Otras tres toxinas han sido encontradas en el sobrenadante deEAEC, enterotoxina de Shigella 1 (ShET1), EAEST1 y Sat (SPATE identificado originalmente enUPEC), sin embargo, su papel en la patogénesis de EAEC no está claro.EpidemiologíaNumerosos estudios epidemiológicos de EAEC han sido realizados en varios países, un análisisglobal de los resultados muestra que EAEC es la causa de diarrea en 15 % de los niños queviven en países en desarrollo y en 4 % de los residentes en países industrializados. Este yotros estudios han determinado el papel de EAEC en enfermedades diarreicas en niños,adultos mayores y personas infectadas con VIH que viven en países tanto en desarrollo ycomo desarrollados.32
El tipo patógeno de EAEC, ha sido relacionado con diferentes tipos de diarrea. Estudiosepidemiológicos en Perú, México, Brasil, India, Kenia e Israel asocian a esta bacteria concuadros de diarrea persistente. Sin embargo, otros estudios en Bangladesh, Perú y Tailandiaasocian a la misma bacteria con casos de diarrea aguda. A la fecha existen diferentesestudios realizados en ciudadanos americanos, españoles y canadienses que viajaron a paísesen desarrollo y presentaron diarrea. Estos y otros resultados ubican a EAEC como unpatógeno asociado a diarrea del viajero después de ETEC.32,36
EAEC también es la causa de brotes de diarrea, en México se reportaron dos brotes de diarreaen los cuneros de un hospital pediátrico y en otras partes del mundo como la india, Serbia,Japón e Inglaterra.36
Escheichia coli de adherencia difusa (DAEC)Este grupo en ensayos de adherencia en células HeLa y HEp-2 genera un patrón definidocomo adherencia difusa (Figura 1). Este patrón esta mediado por fimbrias (de la familia Dr yF1845) y adhesinas no fimbriales (Afa), colectivamente llamadas adhesinas Afa/Dr, estas dosmoléculas ayudan a DAEC a colonizar el intestino delgado y han sido implicadas en diarrea enniños menores de 5 años, así como en infecciones del tracto urinario en adultos.41
Las adhesinas Afa-Dr interaccionan con el factor de aceleración de caída (DAF) ubicado sobrela superficie de las células epiteliales del intestino y del tracto urinario. La interacción Afa/DRcon DAF dispara una cascada de señalizaciones intracelulares dependientes del calcio, cuyo

Unidad VII - Ecología médicaBacteriología
resultado es la elongación y daño a las microvellocidades del borde de cepillo a través de ladesorganización de componentes del citoesqueleto. Además, la interacción Afa/DR-DAFinduce la secreción de IL-8 promoviendo la transmigración de polimorfonucleares (PMN) pormedio del epitelio y la interacción de Afa/DR con PMN acelera la apoptosis celular y undecrecimiento de la fagocitosis por PMN (42). Algunas subclases de las adhesinas Afa-Dr pueden interaccionar con CECAM (moléculas deadhesión relacionados con antígenos carcinoembrionario), familia de receptores encontradosen la superficie de membranas y en particular en acúmulos de lípidos. Esta interacción (Afa-Dr-CECAM) degrada enzimas involucradas en la absorción y secreción intestinal lo cualcontribuye a la diarrea.41
A diferencia de otras E. coli patógenas, la patogénesis de DAEC se debe predominantementea la interacción de las adhesinas Afa‑Dr con las células del hospedero. Existen reportes encepas DAEC que refieren la presencia de Sat, otra toxina de la familia SPATE, con lesionesintercelulares que incrementan la permeabilidad celular. Hasta la fecha no existen otrossistemas de secreción u otros factores de virulencia identificados en cepas DAEC Afa-Dr.EpidemiologíaExisten pocos estudios que relacionen el patotipo DAEC con enfermedades gastrointestinales,observando que la susceptibilidad es dependiente de la edad, ya que se ha asociado a niñosmayores de 5 años. En México se reportó la participación de este patotipo en cuadros dediarrea en niños mayas; estudios más recientes en Brasil con niños menores de 2 añosencuentran una cierta prevalencia de DAEC en cuadros de diarrea. Las razones de estefenómeno relacionado con la edad se desconocen, así como los factores involucrados en lainfección por DAEC.32
Escherichia coli asociada a infecciones extra intestinales (ExPEC)
El grupo ExPEC ha conservado la capacidad de mantenerse en la mucosa del intestino sinocasionar trastornos al hospedero, tiene la habilidad de diseminarse y colonizar otros hábitatsy de manera consecuente ocasionar infecciones extraintestinales.43
Escherichia coli Uropatógena (UPEC)UPEC es responsable de entre 70 y 80 % de las infecciones del tracto urinario (ITUs)adquiridas en la comunidad y de 40 a 50 % de las ITUs de origen nosocomial. UPEC se agrupadentro de las E. coli asociadas a infecciones extraintestinales (ExPEC) y comparte estaclasificación con los patotipos de E. coli que causan meningitis neonatal (NMEC). UPEC seconsidera un patógeno oportunista con capacidad de afectar la susceptibilidad del hospedero,a través de la producción de factores de virulencia que le permiten adherirse, colonizar ydañar al uroepitelio.44
En la actualidad, está bien documentado que, aunque las cepas UPEC se localizan en tracto

Unidad VII - Ecología médicaBacteriología
gastrointestinal, son diferentes a las cepas comensales que forman parte de la biotaintestinal. El análisis por serología en diferentes estudios refiere que los serogrupos O1, O2,O4, O6, O7, O15, O18, O25, O75, O83 y O175 son los que con mayor frecuencia se relacionancon la etiología de las ITU.UPEC puede diseminarse por vía ascendente desde un foco infeccioso en la uretra anterior,mediante la producción de factores de virulencia diversos que contribuyen a su patogénesis.Además, produce una gran variedad de fimbrias o pili (tipo 1, fimbria P, fimbria S, fimbria F1Cy Curli), estas fimbrias se unen eficientemente al uroepitelio y están codificadas por losoperones fim, pap, sfa, foc y csg, respectivamente.Las fimbrias producidas por cepas UPEC presentan variación de fase, es decir, se expresan ose reprimen dependiendo de los cambios en las condiciones ambientales de temperatura,osmolaridad, pH y disponibilidad de nutrientes. Estas características confieren a la bacteriauna ventaja adaptativa, que permite mantener poblaciones mixtas de bacterias productoras yno productoras de fimbrias funcionales, y es así que incrementa la adherencia, coloniza einvade a los tejidos del hospedero de forma eficiente.44.Las fimbrias requieren de la interacción de un receptor específico en la membrana delhospedero, favorecen el proceso de adherencia y el inicio de un mecanismo de invasión quepermite la internalización de la bacteria dentro de compartimentos unidos a la membrana,similares a endosomas tardíos en células superficiales en sombrilla. El crecimiento de UPECen el interior de las células, se restringe y la bacteria entra en un estado de quiescencia; sinembargo, puede salir de la célula, entrar a otras e iniciar una multiplicación descontrolada enel citosol. En UPEC se favorece la interacción entre bacterias y células, lo que representa unevento esencial para el desarrollo de comunidades bacterianas intracelulares (IBCs)formadoras de biopelículas en el uroepitelio.46
UPEC secreta toxinas como la α-hemolisina (HlyA), factor necrotizante citotóxico-1 (Cnf1) y elfactor de invasión del epitelio (IbeA), que se exportan a través de sistemas de secreción tipo I,II y III; lo que facilita la translocación de moléculas efectoras que actúan en sistemas deseñalización celular durante la activación en la producción de citocinas pro yantiinflamatorias; asimismo, participan en el ensamble de fimbrias o pilis, en la activación depéptidos antimicrobianos y en la secreción de proteínas enzimáticas.Otras toxinas que se secretan como proteínas autotransportadoras son las proteínas Sat, quese translocan a través del sistema secreción tipo V y poseen habilidad para inducir un efectocitopático en la célula hospedera. La expresión de Sat se asocia con daño renal en modelosmurinos, se caracteriza por la disolución de la membrana del glomérulo, pérdida de lascélulas epiteliales tubulares y vacuolización en el tejido renal.44,45,46
La proteína Tos (Type one secretion A) codificada por tosA es un miembro de la familia deproteínas RTX (Repeat-in-toxin), que están distribuidas en bacterias gramnegativas y secaracteriza por presentar regiones repetidas ricas en glicina y aspartato, cercanas al carboxilo

Unidad VII - Ecología médicaBacteriología
terminal de la proteína y por contener un sistema de secreción tipo 1 (Type 1 secretionsystem, T1SS), esto permite la exportación de la proteína dentro del medio ambienteextracelular.tosA (originalmente upxA) se encuentra en la isla de patogenicidad (PAI-CFT073-aspV) junto a losgenes homólogos tolC, hlyB y hlyD. La presencia del captador de fierro (chuA, E. coli haem-utilization-gene) es característica en aislados tosA+ y sugiere su asociación con el linajefilogenético B2. Estos estudios apoyan a tosA como un marcador de virulencia en las cepasclínicas UPEC y representan un aporte a la investigación en la clínica médica.En un estudio en el que se analizó la relación entre cepas UPEC y DEC, se identificó lapresencia de genes de cepas de Escherichia coli enteroagregativa (EAEC), lo que les permitióa los autores proponer que este grupo de cepas DEC pudieran considerarse potenciales UPEC.En el Hospital Infantil de México, Federico Gómez, se realizó un estudio prospectivo en el quese analizó la presencia de genes de virulencia de E. coli que causa diarrea en cepas aisladasde niños con IVU. Los resultados mostraron un gran número de cepas que presentaron genesde EPEC, ETEC, STEC y EAEC; en el mismo estudio se analizaron los serotipos de las bacteriasque correlacionaron con los serogrupos UPEC de importancia epidemiológica (datos nopublicados).El objetivo fundamental del tratamiento de las ITU consiste en utilizar un antimicrobiano quegarantice la erradicación del microorganismo responsable. En la actualidad se considera quelo que más beneficia a los pacientes es indicar el antibiótico más potente durante el periodomás breve, para lograr el objetivo de controlar la infección. La selección del antimicrobianodependerá del agente causal, de los patrones de sensibilidad en la comunidad y en el mediohospitalario, y de las características del paciente (edad, género, embarazo, localizaciónanatómica de la infección y condiciones de comorbilidad). Los factores relacionados con elantimicrobiano a utilizar incluyen su farmacodinamia, perfil de efectos adversos y facilidad enla administración.En cepas clínicas UPEC de pacientes pediátricos, aproximadamente 80 % presentamultirresistencia a antibióticos que se utilizan para el tratamiento (Hernández et al.manuscrito en preparación). Los mecanismos involucrados en la multirresistencia sondiversos y se encuentran en función de las características del antibiótico utilizado; así comode las características de las cepas bacterianas.Se conocen varios mecanismos que confieren resistencia a cepas de UPEC, uno de ellos serelaciona con la pérdida de porinas pues reduce el paso del antibiótico a través de lamembrana celular; también puede deberse a eventos de inactivación enzimática por laproducción de β-lactamasas, alteración de la permeabilidad por expulsión activa delantibiótico del interior de la bacteria antes de que esta pueda tener un efecto; pormodificación enzimática del antibiótico que cambia su topología y evita que interactúe con susitio blanco, por mutaciones del sitio blanco que impiden que el antibiótico se una a su sitiode acción, por modificaciones ribosómicas que impiden al antibiótico unirse a este e inhibir la

Unidad VII - Ecología médicaBacteriología
síntesis de proteínas o por efecto de la derivación metabólica al utilizar una enzimaalternativa resistente para evitar el efecto inhibidor del antibiótico (inhibidores de folatos).Los antibióticos β-lactámicos (penicilinas, cefalosporinas, carbapenémicos y monobactámicos)se utilizan como primera opción para el tratamiento de las ITU y su mecanismo de resistenciaestá asociado con frecuencia a la producción de enzimas [β-lactamasas-AmpC, metalo-β-lactamasas (MBL) y carbapenemasa (KPC)] que hidrolizan el anillo β-lactámico y lo inactivan.El surgimiento de cefalosporinas de tercera y cuarta generación resistentes a β-lactamasasdio origen al desarrollo de β-lactamasas de espectro extendido (BLEEs) como un mecanismode resistencia bacteriana.Las BLEEs más representativas en E. coli están codificadas por blaTEM, blaSHV y blaCTX y ademásampCCMT, de las β-lactamasas-AmpC del tipo CMT. La resistencia mediada por BLEEs yenzimas modificadoras se facilita por la adquisición de plásmidos y transposones quepromueven la expansión horizontal epidémica de genes de resistencia entre comunidadesbacteriana.47,48
Los aminoglucósidos (gentamicina, kanamicina, trobamicina) se utilizan como segunda opciónpara el tratamiento de las ITU, las bacterias inactivan estos antibióticos porque sintetizanenzimas modificadoras que N-adenilan, N-acetilan los grupos amino y O-fosforilan los gruposhidroxilo que utilizan como cofactores donadores acetil-CoA y ATP respectivamente. Entre lasprincipales enzimas modificadoras están la acetil-transferasa (codificada por los genes aacA),fosfatidil-transferasa (aph), adenil-transferasa (ant, aad), con afinidad específica por susustrato (48).Epidemiología de infecciones del tracto urinario (ITU)Las infecciones del tracto o vías urinarias representan un serio problema de salud pública,anualmente se reportan 150 millones de casos en el mundo, aproximadamente 40 % de lasmujeres y 12 % de los hombres presentan durante el transcurso de su vida, al menos unevento de ITU con síntomas. Desde hace varios años las infecciones de vías o tracto urinario,han ocupado los primeros lugares dentro de las 20 principales causas de morbilidad que sereportan en la República Mexicana.En el 2016 se notificaron 4 106 966 casos con incidencia de 3,358.84, ambas por 100,000habitantes. Las infecciones recurrentes (persistentes) del tracto urinario (IRTU) se presentandesde la infancia, lo anterior por el hecho de que hasta 8 % de los niños en edades desde unoy hasta doce meses presentan por lo menos una infección de vías urinarias.49 Por otro lado,diferentes estudios epidemiológicos señalan que aproximadamente la cuarta parte de lasmujeres que se ven afectadas por ITU puede presentar una infección recurrente en un periodode seis a doce meses. Una consecuencia importante de las ITU recurrentes, es que puedenconducir a alteraciones de la función renal.Escherichia coli productora de meningitis neonatal(NMEC)La meningitis neonatal presenta altas tasas de morbilidad y mortalidad y se ha comprobado

Unidad VII - Ecología médicaBacteriología
que después de Streptococcus agalactiae (Grupo B), Escherichia coli es el segundomicroorganismo relacionado con el padecimiento. Durante el embarazo en la mujer seproduce mayor colonización por cepas con antígeno capsular K1. Estas cepas son lasprincipalmente implicadas en la meningitis neonatal.H. coli K1 es la variedad más común relacionada con meningitis neonatal (NMEC), perteneceprincipalmente al grupo filogenético B2 y en menor proporción al filogrupo D. La clonaO18:K1:H7 se ha reportado a nivel mundial y otras como O83:K1 y O45:K1 solo en algunospaíses. En relación con los factores de virulencia de ECNM, se ha identificado que estáncodificados en el cromosoma y por material genético externo (plásmidos, Islas dePatogenicidad). Un hecho relevante respecto a este grupo de bacterias es el escasoconocimiento que se tiene en relación con la patogénesis de la enfermedad, lo que en granmedida influye en las altas tasas de morbilidad y mortalidad.50
Para entender el paso de las bacterias por la barrera hematoencefálica (BBB), se handesarrollado modelos in vitro con células endoteliales microvasculares del cerebro humano(HBMEC) y en animales de laboratorio. Al respecto se observó que para que esto ocurra serequiere de una bacteriemia severa, la adherencia e invasión de las bacterias a las célulasendoteliales y que la bacteria viable cruce la barrera hematoencefálica.Estudios con diferentes cepas de E. coli K1 mostraron que componentes como la proteína demembrana externa (OmpA), las proteínas Ibe, AslA, TraJ y CNF1 interactúan con receptoresespecíficos de HBMEC, lo que contribuye a la translocación de la bacteria por la BBB. Lainvasión de E. coli K1 al sistema nervioso central (CNC) ocasiona inflamación y alteracionesintracraneales como pleocitosis, ruptura de la BBB y daño neuronal. El tratamiento de lospacientes con meningitis neonatal debe realizarse con antimicrobianos, al respecto algunoscomités recomiendan no utilizar como terapia empírica cefalosporinas de tercera generación,aztreonam o quinolonas con el objetivo de disminuir la resistencia bacteriana, así como laemergencia de enterobacterias productoras de betalactamasas de espectro extendido(BLEEs).Las infecciones por E. coli con BLEE han experimentado importantes cambios epidemiológicosen los últimos tiempos.47,48 Estas enzimas confieren resistencia a un gran número deantibióticos de uso común, como penicilina, ampicilina, cefalosporinas de cualquiergeneración (excepto cefamicinas), aztreonam y en un porcentaje no desdeñable de casos,también a los betalactámicos asociados a inhibidores de betalactamasas, aminoglucósidos,tetraciclinas y cotrimoxazol. Este patrón de multirresistencia supone una dificultadterapéutica, que explica su asociación en numerosos estudios con mayor mortalidad, duraciónde la estancia hospitalaria y costo económico.
Referencias:1.- Bray J. Isolation of antigenically homogeneous strains of Bact. coli neapolitanum fromsummer diarrhea of infants. J Pathol Bacteriol 1945;57:239‑47.

Unidad VII - Ecología médicaBacteriología
2.- Neter E, Westphal O, Luderitz O, Needell M. Demonstration of antibodies againstenteropathogenic Escherichia coli in sera of children of various ages. Pediatrics1955;16:801-8.3.- Levine MM, Bergquist EJ, Nalin DR, et al. Escherichia coli strains that cause diarrhoea butdo not produce heat- labile or heat-stable enterotoxins and are non-invasive. Lancet1978;1:1119-22.4.- Holmes E, Li JV, Marchesi JR, Nicholson JK. Gut microbiota composition and activity inrelation to host metabolic phenotype and disease risk. Cell Metab. 2012;16:559-645.- Chang DE, Smalley DJ, Conway T, et al. Carbon nutrition of Escherichia coli in the mouseintestine. Proc Natl Acad Sci U S A 2004;101:7427-32.6.- Tenaillon O, Skurnik D, Picard B, Denamur E. The population genetics of commensalEscherichia coli. Nat Rev Microbiol. 2010;8:207-17.7.- Kaper JB, Nataro JP, Mobley HL. Pathogenic Escherichia coli. Nat Rev Microbiol.2004;2:123-40.8.- Olarte J, Varela G. A complete somatic antigen common to Salmonella adelaide,Escherichia coli-gomez, and Escherichia coli 0111:B4. J Lab Clin Med. 1952;40:252‑49.- Escobar-Páramo P, Clermont O, Bui H, et al. A Specific Genetic Background Is Required forAcquisition and Expression of Virulence Factors in Escherichia coli. Mol. Biol. Evol.2004;21:1085–94.10.- Chaudhuri RR, Henderson IR. The evolution of the Escherichia coli phylogeny. Infect, GenEvol. 2012;12:214–26.11.- Chakraborty A, Saralaya V, Adhikari P, Shenoy S, Baliga S, Hegde A. Characterizationof Escherichia coli Phylogenetic Groups Associated with Extraintestinal Infections in SouthIndian Population. Ann Med Heal Scien Res. 2015;5:241-6.12.- Clermont O, Bonacorsi S, Bingen E. Rapid and simple determination of the Escherichiacoli phylogenetic group. Appl Environ Microbiol. 2000;66:4555-8.13.- Cravioto A, Gross RJ, Scotland SM, Rowe B. An adhesive factor found in Escherichia colibelonging to the traditional infantile enteropathogenic serogroups. Microbiology1979;6:3427-37.14.- Giron JA, Donnenberg MS, Martin WC, Jarvis KG, Kaper JB. Distribution of the bundle-forming pilus structuralgene (bfpA) among enteropathogenic Escherichia coli. J Infect Dis1993;168:1037–4115.- Croxen MA, Finlay B. Molecular mechanisms of Escherichia coli pathogenicity. Nat RevMicrobiol 2010;8:26-38.16.-Gill RK, Borthakur A, Hodges K, et al. Mechanism underlying inhibition of intestinal apicalCl/OH exchange following infection with enteropathogenic E. coli. J Clin Invest2007;17:428‑37.17.- Croxen MA, Finlay B. Molecular mechanisms of Escherichia coli pathogenicity. Nat RevMicrobiol. 2010;8:26-38.18.- Fischer-Walker CL, Black RE. Diarrhoea morbidity and mortality in older children,

Unidad VII - Ecología médicaBacteriología
adolescents, and adults. Epidemiol Infect 2010;138:1215–26.19.- Ochoa TJ, Mercado EH, Ruiz J, et al. Frecuencia y patotipos de Escherichia colidiarrogénica en niños peruanos con y sin diarrea. Rev Peru Med Exp Salud Publica2011;28:13-20.20.- Flisser A, Velasco-Villa A, Martínez-Campos C, et al. Infectious diseases in Mexico. Asurvey from 1995-2000. Arch Med Res 2002;33:343-50.21.- Mathewson JJ, Oberhelman RA, Dupont HL, Javier de la Cabada F, Garibay EV.Enteroadherent Escherichia coli as a cause of diarrhea among children in Mexico. J ClinMicrobiol 1987;25:1917-19.22.-Cravioto A, Tello A, Navarro A, et al. Association of Escherichia coli HEp-2 adherencepatterns with type and duration of diarrhoea. Lancet 1991;337:262‑4.23.- Estrada‐Garcia T, Lopez‐Saucedo C, Thompson‐Bonilla R, et al. Association ofdiarrheagenic Escherichia coli Pathotypes with infection and diarrhea among Mexican childrenand association of atypical enteropathogenic E. coli with acute diarrhea. J ClinMicrobiol 2009;47:93–8.24.- Dubreuil J. D. Enterotoxigenic Escherichia coli targeting intestinal epithelial tightjunctions: An effective way to alter the barrier integrity. Microbial Pathogenesis2017;113:129–34.25.-Croxen MA, Finlay BB. Molecular mechanisms of Escherichia coli pathogenicitywww.nature.com/reviews/micro 2010.26.- Gupta S, Keck J, Ram P, Crump J, Miller M, Mintz E. Part III. Analysis of data gapspertaining to enterotoxigenic Escherichia coli infections in low and medium humandevelopment index countries, 1984-2005. Epidemiol Infect 2008;136:721–38.27.- Cravioto A, Reyes RE, Ortega R, Fernández G, Hernández R, López D. Prospective study ofdiarrhoeal disease in a cohort of rural Mexican children: incidence and isolated pathogensduring the first two years of life. Epidemiol Infect 1988;101:123-34.28.- Estrada-García T, Cerna JF, Paheco-Gil L, et al. Drug-resistant diarrheogenic Escherichiacoli, Mexico. Emerg Infect Dis 2005;11:1306-8.29.- Gyles CL. Shiga toxin-producing Escherichia coli: an overview. J Anim Sci2007;85(13):E45-E62.30.- Quilliam RS, Chalmers RM, Jones DL, et al. Seroprevalence and risk factors associatedwith Escherichia coli O157 in a farming population. Zoon Pub Health 2012;59:83-8.31.- Tarr PI, Gordon CA, Chandler WL. Shiga-toxin-producing Escherichia coli and haemolyticuraemic syndrome. Lancet 2005;365:1073–86.32.- Croxen MA, Law RJ, Finlay BB, et al. Recent Advances in Understanding EntericPathogenic Escherichia coli. Clin Microbiol Rev 2013;26:822–80.33.- Callaway TR, Anderson RC, Tellez G, et al. Prevalence of Escherichia coli O157 in cattleand swine in central Mexico. J. Food Prot 2004;67:2274-6.34.- Navarro A, Eslava C, Cravioto A, et al. Antibody response to Escherichia coli O157 andother lipopolysaccharides in healthy children and adults. Clin Diag Lab Immunol

Unidad VII - Ecología médicaBacteriología
2003 ;10:797-01.35.- Navarro A, Eslava C, Cravioto A, et al. Common epitopes in lipopolysaccharides ofdifferent Enterobacteriaceae are associated with an immune response against Escherichia coliO157 in bovine serum samples. J Med Microbiol 2007;56:1447-54.36.- Villaseca JM, Hernández U, Sainz-Espuñes TR, Rosario C, Eslava C. EnteroaggregativeEscherichia coli an Emergent Pathogen with Different Virulence Properties; Rev. Latin.Microbiol 2005;47:140-59.37.- Boisen N, Struve C, Scheutz F, Krogfelt KA, Nataro JP. New adhesin of enteroaggregativeEscherichia coli related to the Afa/Dr/AAF family. Infect. Immun 2008;76:3281–92.38.- Henderson IR, Czeczulin J, Eslava C, Noriega F, Nataro JP. Characterization of Pic, aSecreted Protease of Shigella flexneri and Enteroaggregative Escherichia coli. Infect Immun1999;67:5587-96.39.- Eslava C, Navarro-García F, Czeczulin JR, Henderson IR, Cravioto A, and Nataro JP. Pet, anAutotransporter Enterotoxin from Enteroaggregative Escherichia coli. Infect Immun1998;66:3155-63.40.- Villaseca JM, Navarro-García F, Mendoza-Hernández G, Nataro JP, Cravioto A, Eslava C. PetToxin from Enteroaggregative Escherichia coli Produces Cellular Damage Associated WithFodrin Disruption. Infect Immun 2000;68:5920-7.41.- Servin AL. Pathogenesis of Afa/Dr diffusely adhering Escherichia coli. Clin. Microbiol. Rev2005;18:264–92.42.- Le Bouguénec C, Servin AL. Diffusely adherent Escherichia coli strains expressing Afa/Dradhesins (Afa/Dr DAEC): hitherto unrecognized pathogens. FEMS Microbiol Lett2006;256:185-94.43.- Katouli M. Population structure of gut Escherichia coli and its role in development ofextra-intestinal infections. Iran J Microbiol 2010;2:59-72.44.- Flores-Mireles AL, Walker JN, Caparon M, Hultgren SJ. Urinary tract infections:epidemiology, mechanisms of infection and treatment options. Nat Rev Microbiol2015;13:269–84.45.- Lewis AJ, Richards AC, Mulvey MA. Invasion of host cells and tissues by uropathogenicbacteria. Microbiol spectr 2016;4:1-2946.- Agarwal J, Srivastava S, Singh M. Pathogenomics of uropathogenic Escherichia coli. IndianJ Med Microbiol 2012;30:141-9.47.- Peleg AY, Hooper DC. Hospital-acquired infections due to gram-negative bacteria. N Engl JMed 2010;362:1804‑13.48.- Franiczek R, Sobieszczanska B, Turniak M, et al. ESBL-producing Escherichia coli isolatedfrom children with acute diarrhea – antimicrobial susceptibility, adherence patterns andphylogenetic background. Adv Clin Exp Med 2012;21:187–92.49.- Anuario de Morbilidad 1984 -2016 Dirección General de Epidemiología DIE –INFORMACIÓN EPIDEMIOLÓGICA Secretaria de Salud.50.- Xie Y, Kolisnychenko V, Paul-Satyaseela M, et al. Identification and Characterization of

Unidad VII - Ecología médicaBacteriología
Escherichia coli RS218–Derived Islands in the Pathogenesis of E. coli Meningitis J Infect Dis2006;194:358–64.Temas relacionados
Infecciones Asociadas a Cuidados de la Salud y la ResistenciaAntimicrobiana en Bacterias del Grupo Eskape en MéxicoBacteriología, Unidad VII - Ecología médicabacterias, ESKAPE, IAAS, infección


















