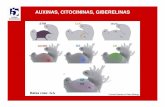
Citocininas
27
Reguladores da Divisão Celular Cristina Martinho da Silva Felipe Augusto Miranda Ramos Guilherme Rodrigues Fernandes Campos
-
Upload
cristina-martinho -
Category
Documents
-
view
543 -
download
1
Transcript of Citocininas

Reguladores da Divisão Celular
Cristina Martinho da Silva
Felipe Augusto Miranda Ramos
Guilherme Rodrigues Fernandes Campos

Sumário:
• Histórico
• Biossíntese, Metabolismo e Transporte
• Percepção e Transdução do Sinal
• Efeitos Fisiológicos
• Conclusão

Histórico
• Década de 1950, Dr. Folke Skoog da
Universidade de Winsconsin (EUA) ao cultivar
a medula da planta do tabaco in vitro,
procurava uma substância que fosse
responsável pela divisão celular.
• O cultivo do tabaco em in vitro, utilizando meios
nutritivos contendo AIA, extrato de levedura e
água de coco, promoveu uma intensa
proliferação de células.

Histórico
• 1955 Isolada por Carlos Miller, bases nitrogenadas do
esperma Arenque.
• Denominada Cinetina, por ser capaz de aumentar a
citocinese

Histórico
• Skoog et al., em 1965 propuseram o termo citocinina
para compostos com atividade biológica igual à
cinetina,ou seja, capazes de promover citocinese em
células vegetais.
• David Letham, em 1973 isolou a
primeira citocinina natural de plantas em
extrato de milho verde (Zea mays),
denominando-a de Zeatina

Biossíntese, Metabolismoe Transporte
• Formuladas de derivados de unidades de
isopropeno.
• Quimicamente:
Isopropeno
Borracha Natural
Carotenóides
GA
BA
Compostos de Defesa Vegetal

Biossíntese
• Principal região de síntese de
Cks da planta, mas é
sintetizada também em
embriões em desenvolvimento

Conjugação eHidrólise
Quebra da cadeia lateral
Importante mecanismo do controle do nivel endógeno.
• Maior parte é encontrada na forma conjugada com
moléculas de açucar.

Transporte
• As Cks são transportadas para a
parte aérea através do xilema na
forma de ribosídeos.
• As Cks são encontradas no
floema durante a translocação
de assimilados de folhas
senescentes (fontes) para as
partes jovens da planta
(drenos) na forma de
glicosídeos.

Percepção e transdução do Sinal
Regulação da Divisão Celular
Diferenciação Celular
Senescência
Maturação de Cloroplastos
Expansão Celular
Indução da Mobilização de Nutrientes
Agente Secreto da Simbiose

(1) A Citocinina Liga-se à porção extra celular da
proteína CRE1, num domínio chamado CHASE.
AHK2 e AHK3 provavelmente também agem como
receptores de Ck.
(2) A Ligação da Ck a esses
receptores promove a sua
histidina quinase. O fosfato é
transferido para um resíduo
aspartato (D) dos domínios
receptores.(3) O Fosfato é então transferido
para uma histidina conservada,
presente em uma proteína AHP.
(4) A fosforilação promove o
movimento da proteína AHP para
o interior do núcleo, onde o
fosfato é tranferido para um
resíduo aspartato localizado no
domínio receptor de uma ARR
tipo-B (Arabidopsis response
regulators)
(5) A fosforilação desta ARR tipo-B ativa o
domínio de saída que induz a transcrição
dos genes que codificam as ARRs tipo-A.
(6) As ARRs tipo-A provavelmente são
também fosforiladas pelas proteínas AHP..
(7) As ARRs tipo-A fosforiladas interagem
com vários efetores, mediando mudanças
na função celular apropriada à Ck.

(1) A Citocinina Liga-se à porção extra celular da
proteína CRE1, num domínio chamado CHASE.
AHK2 e AHK3 provavelmente também agem como
receptores de Ck.
(2) A Ligação da Ck a esses
receptores promove a sua
histidina quinase. O fosfato é
transferido para um resíduo
aspartato (D) dos domínios
receptores.(3) O Fosfato é então transferido
para uma histidina conservada,
presente em uma proteína AHP.
(4) A fosforilação promove o
movimento da proteína AHP para
o interior do núcleo, onde o
fosfato é tranferido para um
resíduo aspartato localizado no
domínio receptor de uma ARR
tipo-B (Arabidopsis response
regulators)
(5) A fosforilação desta ARR tipo-B ativa o
domínio de saída que induz a transcrição
dos genes que codificam as ARRs tipo-A.
(6) As ARRs tipo-A provavelmente são
também fosforiladas pelas proteínas AHP..
(7) As ARRs tipo-A fosforiladas interagem
com vários efetores, mediando mudanças
na função celular apropriada à Ck.

(1) A Citocinina Liga-se à porção extra celular da
proteína CRE1, num domínio chamado CHASE.
AHK2 e AHK3 provavelmente também agem como
receptores de Ck.
(2) A Ligação da Ck a esses
receptores promove a sua
histidina quinase. O fosfato é
transferido para um resíduo
aspartato (D) dos domínios
receptores.(3) O Fosfato é então transferido
para uma histidina conservada,
presente em uma proteína AHP.
(4) A fosforilação promove o
movimento da proteína AHP para
o interior do núcleo, onde o
fosfato é tranferido para um
resíduo aspartato localizado no
domínio receptor de uma ARR
tipo-B (Arabidopsis response
regulators)
(5) A fosforilação desta ARR tipo-B ativa o
domínio de saída que induz a transcrição
dos genes que codificam as ARRs tipo-A.
(6) As ARRs tipo-A provavelmente são
também fosforiladas pelas proteínas AHP..
(7) As ARRs tipo-A fosforiladas interagem
com vários efetores, mediando mudanças
na função celular apropriada à Ck.

(1) A Citocinina Liga-se à porção extra celular da
proteína CRE1, num domínio chamado CHASE.
AHK2 e AHK3 provavelmente também agem como
receptores de Ck.
(2) A Ligação da Ck a esses
receptores promove a sua
histidina quinase. O fosfato é
transferido para um resíduo
aspartato (D) dos domínios
receptores.(3) O Fosfato é então transferido
para uma histidina conservada,
presente em uma proteína AHP.
(4) A fosforilação promove o
movimento da proteína AHP para
o interior do núcleo, onde o
fosfato é tranferido para um
resíduo aspartato localizado no
domínio receptor de uma ARR
tipo-B (Arabidopsis response
regulators)
(5) A fosforilação desta ARR tipo-B ativa o
domínio de saída que induz a transcrição
dos genes que codificam as ARRs tipo-A.
(6) As ARRs tipo-A provavelmente são
também fosforiladas pelas proteínas AHP..
(7) As ARRs tipo-A fosforiladas interagem
com vários efetores, mediando mudanças
na função celular apropriada à Ck.

(1) A Citocinina Liga-se à porção extra celular da
proteína CRE1, num domínio chamado CHASE.
AHK2 e AHK3 provavelmente também agem como
receptores de Ck.
(2) A Ligação da Ck a esses
receptores promove a sua
histidina quinase. O fosfato é
transferido para um resíduo
aspartato (D) dos domínios
receptores.(3) O Fosfato é então transferido
para uma histidina conservada,
presente em uma proteína AHP.
(4) A fosforilação promove o
movimento da proteína AHP para
o interior do núcleo, onde o
fosfato é tranferido para um
resíduo aspartato localizado no
domínio receptor de uma ARR
tipo-B (Arabidopsis response
regulators)
(5) A fosforilação desta ARR tipo-B ativa o
domínio de saída que induz a transcrição
dos genes que codificam as ARRs tipo-A.
(6) As ARRs tipo-A provavelmente são
também fosforiladas pelas proteínas AHP..
(7) As ARRs tipo-A fosforiladas interagem
com vários efetores, mediando mudanças
na função celular apropriada à Ck.

(1) A Citocinina Liga-se à porção extra celular da
proteína CRE1, num domínio chamado CHASE.
AHK2 e AHK3 provavelmente também agem como
receptores de Ck.
(2) A Ligação da Ck a esses
receptores promove a sua
histidina quinase. O fosfato é
transferido para um resíduo
aspartato (D) dos domínios
receptores.(3) O Fosfato é então transferido
para uma histidina conservada,
presente em uma proteína AHP.
(4) A fosforilação promove o
movimento da proteína AHP para
o interior do núcleo, onde o
fosfato é tranferido para um
resíduo aspartato localizado no
domínio receptor de uma ARR
tipo-B (Arabidopsis response
regulators)
(5) A fosforilação desta ARR tipo-B ativa o
domínio de saída que induz a transcrição
dos genes que codificam as ARRs tipo-A.
(6) As ARRs tipo-A provavelmente são
também fosforiladas pelas proteínas AHP.
(7) As ARRs tipo-A fosforiladas interagem
com vários efetores, mediando mudanças
na função celular apropriada à Ck.

(1) A Citocinina Liga-se à porção extra celular da
proteína CRE1, num domínio chamado CHASE.
AHK2 e AHK3 provavelmente também agem como
receptores de Ck.
(2) A Ligação da Ck a esses
receptores promove a sua
histidina quinase. O fosfato é
transferido para um resíduo
aspartato (D) dos domínios
receptores.(3) O Fosfato é então transferido
para uma histidina conservada,
presente em uma proteína AHP.
(4) A fosforilação promove o
movimento da proteína AHP para
o interior do núcleo, onde o
fosfato é tranferido para um
resíduo aspartato localizado no
domínio receptor de uma ARR
tipo-B (Arabidopsis response
regulators)
(5) A fosforilação desta ARR tipo-B ativa o
domínio de saída que induz a transcrição
dos genes que codificam as ARRs tipo-A.
(6) As ARRs tipo-A provavelmente são
também fosforiladas pelas proteínas AHP..
(7) As ARRs tipo-A fosforiladas interagem
com vários efetores, mediando mudanças
na função celular apropriada à Ck.

Efeitos Fisiológicos
Regulação da Divisão Celular
Diferenciação Celular
Senescência
Maturação de Cloroplastos
Expansão Celular
Indução da Mobilização de Nutrientes
Agente Secreto da Simbiose

Efeitos Fisiológicos
Regulação da Divisão Celular
• As citocininas (Cks) atuam em etapas específicas do ciclo
celular, regulando a atividade de proteínas denominadas
ciclinas, que controlam a divisão celular.
CKs Gene cycd3 Ciclinas
Ciclinas
Gene cdc2
Cks + Axs
Cinase
Gene cdc2
Cks

Efeitos Fisiológicos
Diferenciação Celular
• As Cks atuam na diferenciação das células, sobretudo no
processo de formação das gemas caulinares.
• Importante interação com auxinas na regulação do
crescimento em plantas intactas, e na indução de raízes,
calos e caules em cultura de tecidos.

Efeitos Fisiológicos
Senescência
• A aplicação de Ck em uma folha ou parte de uma folha
atrasa o envelhecimento da mesma.
• A ação da Ck seria uma mobilização e acúmulo de solutos
de outras partes da planta, ou da folha, para a região
tratada, aumentando o metabolismo na região e atrasando
sua senescência.

Efeitos Fisiológicos
Maturação dos Cloroplastos
• Plantas crescendo no escuro, após a germinação, não
desenvolvem cloroplastos.
• Se forem transferidas para a luz em tempo, e antes disso
receberem doses adequadas de Ck, o esverdeamento é
mais rápido e mais intenso.
As Cks atuam sobre o fitocromo B (Phy B), através do ARR4 -
regulador de resposta da via de sinalização das citocininas.
Esse impede a reversão da forma ativa do Phy B que absorve luz
na faixa do vermelho extremo, para a forma inativa do Phy B, que
absorve luz na faixa do vermelho.
Desse modo, as Cks mantêm o Phy B na forma ativa e ocasiona a
diferenciação dos cloroplastos.

Efeitos Fisiológicos
Expansão Celular
• A aplicação de Ck em certos tecidos vegetais pode
promover sua expansão sem aumento da matéria seca.
• Isso indica um aumento na plasticidade das paredes
celulares, permitindo maios absorção de água e tamanho
final

Efeitos Fisiológicos
Indução da Mobilização de Nutrientes
• As Ck estão envolvidas no estabelecimento de drenos
• As Cks atuam de modo direto em, pelo menos, duas proteínas
(invertase e transportador de hexose) as quais são necessárias
para o descarregamento apoplástico no floema
• A invertase diminui o potencial químico da sacarose na região do
descarregamento, favorecendo a chegada continua desse
nutriente.
• O transportador de hexose é necessário para que os açúcares
entrem nas células do dreno.

Efeitos Fisiológicos
Agente Secreto da Simbiose
• A interação simbiótica entre a bactéria Rhizobium e as
leguminosas levam a indução de um novo órgão: o nódulo
fixador de nitrogênio.
• Em estudos recentes, Florian
et al. (2008) sugeriram e
discutiram um modelo no
qual a Ck poderia também
mediar a infecção bacteriana
pelo controle da expressão
do gene NIN (Nodule
inception) e regulação de
outros fatores de transcrição
que regulam mecanismos
que operam local e
sistemicamente.


Bibliografia e Sites
• http://www.fisiologiavegetal.ufc.br/Aulas%20em%20PDF%20PG/Unidade%20XVI%20-%20Citocininas.pdf (Acessado em 15/11/2010)
• TAIZ, L. & ZEIGER, E. – Fisiologia Vegetal – 3ª edição; 2004 – Editora Artmed, Porto Alegre/ RS.
• KERBAUY, G.B.; Fisiologia Vegetal – Editora Guanabara Koogan / Rio de Janeiro,2004