
Comparação de protótipos de armadilhas e atraentes de...
107
UNIVERSIDADE FEDERAL DE MINAS GERAIS INSTITUTO DE CIÊNCIAS BIOLÓGICAS PROGRAMA DE PÓS GRADUAÇÃO EM PARASITOLOGIA CARLA MARIA DIAS LOPES Comparação de protótipos de armadilhas e atraentes de oviposição para captura de fêmeas grávidas de Culex quinquefasciatus Say, (1823) (Diptera: Culicidae) em campo BELO HORIZONTE MINAS GERAIS – BRASIL 2007
Transcript of Comparação de protótipos de armadilhas e atraentes de...

UNIVERSIDADE FEDERAL DE MINAS GERAIS
INSTITUTO DE CIÊNCIAS BIOLÓGICAS
PROGRAMA DE PÓS GRADUAÇÃO EM PARASITOLOGIA
CARLA MARIA DIAS LOPES
Comparação de protótipos de armadilhas e atraentes de
oviposição para captura de fêmeas grávidas de
Culex quinquefasciatus Say, (1823) (Diptera: Culicidae) em campo
BELO HORIZONTE
MINAS GERAIS – BRASIL
2007

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

II
CARLA MARIA DIAS LOPES
Comparação de protótipos de armadilhas e atraentes de
oviposição para captura de fêmeas grávidas de
Culex quinquefasciatus Say, (1823) (Diptera: Culicidae) em campo
ORIENTADOR: Dr. ÁLVARO EDUARDO EIRAS
BELO HORIZONTE MINAS GERAIS – BRASIL
2007
Dissertação apresentada ao Departamento de Parasitologia do Instituto de Ciências Biológicas da Universidade Federal de Minas Gerais, como requisito final para a obtenção do título de Mestre em Parasitologia. Área de concentração: Entomologia

III
Aos meus pais e irmã pela presença sempre constante.

IV
AGRADECIMENTOS
Ao Dr. Álvaro Eduardo Eiras, pela orientação, estímulo, paciência, confiança e amizade,
meu agradecimento especial.
À Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), pela
concessão de bolsa de mestrado.
À coordenação da Pós-Graduação em Parasitologia da Universidade Federal de Minas
Gerais (UFMG), aos professores e funcionários do Departamento, pela estrutura oferecida
ao bom desenvolvimento deste trabalho.
Ao Laboratório de Ecologia Química de Insetos Vetores, que me acolheu, pela estrutura
física e pelo corpo de pesquisadores, sempre disponíveis e estimuladores incansáveis.
À minha querida Turma de Mestrado “Família Mexicana” (Ana Flávia, Ceres Luciana,
Fernanda Regina, Kelly Key, Priscila Helena, Renata Cristina e Sydnei Leonardo), pelos
bons e maus momentos compartilhados durante o curso.
Às regionais Venda Nova, Norte e Pampulha e seus funcionários Francisco (Venda Nova),
Nerice e Adriana (Norte) e Cristiano (Pampulha) pelo auxílio fundamental na busca e
contato com residentes de áreas dos trabalhos em campo.
Aos proprietários e funcionários do clube Lareira, da Ilumef e da oficina do Nem, que
sempre estiveram de portas abertas para realização de qualquer experimento ao qual me
propusesse.
À Maria Cristina, pelo auxílio contínuo na realização da análise da paridade.
Ao Tiago, que com muito carinho, sempre se dispunha a me levar ao campo.

V
Ao João, pelo auxílio fundamental na tradução.
Á Rosemary Roque pelo importante auxílio nas análises estatísticas.
À Ione, por tornar possível o contato com os funcionários das regionais.
Ao Dr. Ivan, do Departamento de Estatística da Escola de Veterinária da UFMG, meu
agradecimento pelo auxílio no tratamento estatístico dos dados biológicos.
À todas as instituições e pessoas que contribuíram diretamente para a realização deste
trabalho, o meu muitíssimo obrigada!
AGRADECIMENTOS PESSOAIS
À minha família querida, tenho muito a agradecer: ao meu pai pelos bons conselhos e
suporte financeiro nos momentos de aperto, a minha mãezinha querida pelo colo naqueles
dias em que parecia que iria fraquejar, e a minha irmã pela admiração e confiança.
Ao Tiago, pelo relacionamento que tanto cresceu e se consolidou nos últimos meses,
agradeço pelos momentos de aconchego e pelo seu amor.
À todos os amigos que estiveram comigo durante esses dois anos e meio e mesmo que me
incentivaram a ingressar no Mestrado, agradeço pelo apoio, carinho e pelos bons momentos
vividos. Vocês foram fundamentais para que eu passasse por cada etapa desta minha
empreitada com um pouco mais de tranqüilidade e felicidade.

VI
RESUMO Populações adultas de Culex quinquefasciatus são monitoradas principalmente através de armadilhas luminosas ou iscadas com CO2 e animais, sendo estas dispendiosas, o que dificulta o monitoramento em áreas com baixo poder econômico. Recentemente, foi desenvolvida uma armadilha adesiva, denominada P1T1, de baixo custo, capaz de capturar fêmeas grávidas deste mosquito. A armadilha P1T1 consiste em um recipiente de polietileno de cor preta, fosco (20 x 30 x 9 cm), com tampa com abertura retangular (8 x 16 cm), ao qual é acrescido 1L de água desclorada. Acima do nível d’água é colocado um cartão adesivo responsável pela captura dos insetos. Neste trabalho, objetivou-se aumentar o potencial de captura desta armadilha, testando algumas modificações no seu desenho original e no atraente sintético de oviposição utilizado em conjunto com esta. Inicialmente, foram desenvolvidos quatro novos protótipos, construídos tendo como base um balde de cor preta, que também capturariam os mosquitos através de cartão adesivo. Estes novos modelos diferenciavam-se entre si quanto ao número e posição de aberturas, em forma de funis, que serviam como entradas de acesso do mosquito no interior da armadilha e saída da pluma de odor gerada pelo atraente químico. Todos os modelos continham no interior um cartão adesivo responsável pela captura dos insetos. Através de teste de fumaça, pôde-se observar que a armadilha P1T1 foi responsável pela formação de uma pluma de odor mais concentrada, que dispersou-se homogeneamente pela área adjacente, sofrendo menores influências das correntes de ar, quando comparada às armadilhas em forma de balde. A armadilha P1T1 mostrou-se mais eficaz na captura de adultos de C. quinquefasciatus, em comparação aos outros protótipos desenvolvidos no presente trabalho. Dentre os 225 C. quinquefasciatus (145 machos e 80 fêmeas) capturados, a armadilha P1T1 capturou cerca de 46% de machos, sendo a média de captura significativamente maior que a obtida por três dos novos protótipos (Mann Whitney, p<0.05). A armadilha P1T1 capturou 50% do total de fêmeas, sendo estatisticamente mais eficiente que os demais modelos (Mann Whitney, p<0.05). A armadilha P1T1 foi usada para avaliar combinações binárias dos atraentes sintéticos de oviposição, onde o escatol foi combinado com o indol, nonanal ou p-cresol. O escatol demonstrou ser mais atrativo do que as combinações binárias (Mann Whitney, p<0.05). Os resultados da avaliação de combinações terciárias e quaternárias dos referidos atraentes sintéticos de oviposição foram inconclusivos, devido à baixa captura de adultos de C. quinquefasciatus durante o experimento. A armadilha P1T1 mostrou-se específica para captura de fêmeas grávidas de C. quinquefasciatus, quando comparada à armadilha MosquiTRAP® contendo o AtrAedes® (atraente sintético de oviposição específico para fêmeas de Aedes aegypti) (Mann Whitney, p=0.02). No entanto, sua captura não foi diferente da obtida pela MosquiTRAP® iscada com escatol (Mann Whitney, p= 0.19). Os resultados confirmam a viabilidade do modelo P1T1 como armadilha para captura de fêmeas grávidas de C. quinquefasciatus e demonstram que as combinações de atraentes avaliadas não aumentaram o seu potencial de captura, pelo menos nas concentrações avaliadas.

VII
ABSTRACT Culex quinquefasciatus adult populations have been mainly monitored by light traps or baited with CO2 or animals using such equipment is costly, thus, it is difficult to monitor in areas with low dispensible income. Recently, a new sticky trap capable of capturing gravid females of C. quinquefasciatus, called P1T1, was developed with low cost. The P1T1 consists of a black polyethylene container (20 x 30 x 9 cm) with a cover with a central rectangular opening (8 x 16 cm), to which 1L of tap water is added. A sticky card is placed above the water level to capture mosquitoes. The aim of this work was to increase the potential of P1T1 to catch mosquitoes, through modifications of the trap design and different combinations of synthetic oviposition attractants. Four new models of traps (buckets A, B, C and D) were developed using a black bucket as the main body and a sticky card. These new models differed by the number and position of openings, and the addition of funnels, that served as entrances for mosquitoes and exits for the odor plume. By smoke tests, it could be observed that the P1T1 trap allowed formation of a more concentrated odor plume that was dispersed more homogeneously into adjacent areas and was less influenced by wind currents than the other trap designs. The P1T1 was also more efficient in the capture of adult C. quinquefasciatus, in comparison to the other prototypes developed and evaluated in the present work. Among the 225 C. quinquefasciatus (145 males and 80 females) captured, P1T1 caught about 46% of the total males, where the mean number of caught mosquitoes was signigicantly higher than buckets A, B and C (Mann Whitney, p <0.05). The P1T1 trap captured 50% of the total females and it was significantly more efficient than the other models (Mann Whitney, p <0.05). When the P1T1 trap was used to evaluate binary combinations of skatole with indol, nonanal or p-cresol, skatole alone demonstrated to be significantly more attractive than the binary combinations (Mann Whitney, p <0.05). The results of tertiary and quaternary combinations of the synthetic oviposition attractants were inconclusive, due to low capture of adult C. quinquefasciatus during experiments. The P1T1 trap was shown to be specific to the capture of gravid female C. quinquefasciatus, when compared to the MosquiTRAP containing AtrAedes® (synthetic oviposition attractant specific for Aedes aegypti females) (Mann Whitney, p=0.02). However, P1T1 was not significantly more efficient than the MosquiTRAP® baited with skatole (Mann Whitney, p = 0.19). The results confirm the ability of P1T1 trap as a trap to capture gravid females of C. quinquefasciatus and demonstrated that skatole alone is more efficient than the other combinations of attractants tested, at least in the concentrations evaluated.

VIII
LISTA DE FIGURAS
Figura 1 - Protótipos de armadilhas desenvolvidos tendo como base um balde: (a)
balde A, (b) balde B, (c) balde C e (d) balde D. Ver texto para detalhes.
47
Figura 2 - Protótipos de armadilhas testados. Da esquerda para direita: balde A, balde
D, balde C e balde B. Ao centro, P1T1.
48
Figura 3 - (a) Confecção dos furos nos cartões adesivos, com o auxílio de chave de
fenda e martelo; (b) suporte sobre o qual se apóia o cartão adesivo e (c)
cartão adesivo da armadilha P1T1, confeccionado a partir de aparas de
cartão emendadas.
49
Figura 4 – Esquema da armadilha P1T1 mostrando a evaporação da água localizada
abaixo do cartão, representada pelas setas, através de perfurações no
cartão.
49
Figura 5 – (a) Mangueira de PVC (4,5 cm de largura por 5cm de comprimento),
utilizada como liberador do escatol; (b) suporte para o liberador contendo
o atraente, confeccionado com um clipe médio fixado ao centro do cartão
adesivo de cada protótipo de armadilha: vista lateral e (c) vista superior.
52
Figura 6 – Esquema de ovários de culicídeos mostrando as características observadas
no estudo da paridade: (a) ovário de fêmea nulípara e (b) ovário de fêmea
onípara. Fonte: FORATTINI, 2002, p. 121.
53
Figura 7 - Área experimental situada no bairro Jaqueline: (a, b, c e d) possíveis
criadouros de Culex quinquefasciatus, constituídos por represamentos de
escape da rede de esgoto e áreas de instalação das armadilhas: (e) Oficina
do Nem e (f) Ilumef.
55

IX
Figura 8 - Liberadores: (a) tubos de polietileno, utilizados para os atraentes líquidos
e (b) mangueira de PVC, para atraentes sólidos. Suporte dos atraentes,
confeccionado com fragmento de caixa de ovo de codorna, posicionado
ao centro do cartão adesivo da armadilha P1T1: (c) vista lateral e (d) vista
superior.
58
Figura 9 – Armadilha para captura de adultos de Aedes aegypti (MosquiTRAP)
utilizada no experimento de comparação entre o melhor protótipo de
armadilha para captura de fêmeas grávidas de Culex quinquefasciatus
com MosquiTRAP em campo. (a) Parte inferior contendo tela de
proteção; (b) parte superior (tampa); (c) AtrAedes (atraente de
oviposição sintético) e (d) cartão adesivo.
63
Figura 10 – Dispersão da pluma de odor produzida por cloreto de amônia na
armadilha P1T1.
67
Figura 11 – Dispersão da pluma de odor produzida por cloreto de amônia no (a)
balde C e no (b) balde D.
68
Figura 12 – Pluma de odor produzida por cloreto de amônia liberada pelos (a) baldes
A e (b) B.
69

X
LISTA DE QUADROS
Quadro 1: Principais armadilhas utilizadas para captura de mosquitos adultos e suas
aplicações.
39
Quadro 2: Disposição das armadilhas em campo, nas posições definidas dentro de cada
uma das três regiões (1, 2 e 3), durante a realização do experimento de avaliação de
protótipos de armadilhas para captura de fêmeas grávidas de Culex quinquefasciatus.
56
Quadro 3: Tratamentos do experimento de avaliação de combinações binárias de atraentes
de oviposição para captura de fêmeas grávidas de Culex quinquefasciatus, acrescidos
às armadilhas P1T1 contendo 1L de água desclorada.
59
Quadro 4: Disposição dos diferentes tratamentos em campo, nas posições definidas dentro
de cada região (1, 2 e 3), durante a realização do experimento de avaliação de
combinações binárias de atraentes sintéticos de oviposição para captura de fêmeas
grávidas de Culex quinquefasciatus. E: escatol, I: indol, N: nonanal e P: p-cresol.
60
Quadro 5: Tratamentos do experimento de avaliação de combinações terciárias e
quaternária de atraentes de oviposição para captura de fêmeas grávidas de Culex
quinquefasciatus, acrescidos às armadilhas P1T1 contendo 1L de água desclorada.
61
Quadro 6: Disposição dos diferentes tratamentos em campo, nas posições definidas dentro
de cada região (1 e 2), durante a realização do experimento de avaliação de
combinações terciárias e quaternária de atraentes sintéticos de oviposição para
captura de fêmeas grávidas de Culex quinquefasciatus. E: escatol; I: indol; N:
nonanal e P: p-cresol.
62
Quadro 7: Disposição dos diferentes tratamentos em campo, nas posições definidas dentro
de cada região (1, 2 e 3), durante a realização do experimento de comparação do
melhor protótipo de armadilha para captura de fêmeas grávidas de Culex
quinquefasciatus com MosquiTRAP.
64

XI
LISTA DE TABELAS
Tabela 1. Período de utilização de atraentes sintéticos de oviposição em liberadores,
de acordo com o período de estabilização das taxas de evaporação (LIMA,
2005).
38
Tabela 2. Captura total e média de fêmeas e machos de Culex quinquefasciatus pelos
protótipos de armadilhas em experimento de campo no bairro Jaqueline,
Belo Horizonte-MG, no período de 25 a 30/09/06. N = 15.
71
Tabela 3. Número e porcentagem de fêmeas de Culex quinquefasciatus capturadas,
por estágio fisiológico, por cada um dos cinco protótipos de armadilhas
avaliados em experimento de campo no bairro Jaqueline, Belo Horizonte-
MG, no período de 25 a 30/09/06. N = 15.
72
Tabela 4. Captura total e média de machos e de fêmeas de Culex quinquefasciatus
por cada um dos cinco tratamentos, no experimento de avaliação de
combinações binárias de atraentes sintéticos de oviposição realizado no
bairro Jaqueline, Belo Horizonte-MG, realizado no período de 03 a
13/11/06. N = 15.
74
Tabela 5. Número e porcentagem de fêmeas de Culex quinquefasciatus capturadas,
por estágio fisiológico, por cada um dos tratamentos do experimento de
avaliação de combinações binárias de atraentes sintéticos de oviposição
realizado no bairro Jaqueline, Belo Horizonte-MG, no período de 03 a
13/11/06. N = 15.
75

XII
Tabela 6. Captura total e média de machos e de fêmeas de Culex quinquefasciatus
por cada um dos tratamentos do experimento de avaliação de combinações
terciárias e quaternária de atraentes sintéticos de oviposição no bairro
Jaqueline, Belo Horizonte-MG, no período de 19 a 31/01/07. N =12. E:
escatol, I: indol, N: nonanal e P: p-cresol.
77
Tabela 7. Número e porcentagem de fêmeas de Culex quinquefasciatus capturadas,
por estágio fisiológico, por cada um dos tratamentos do experimento de
avaliação de combinações terciárias e quaternária de atraentes sintéticos de
oviposição no bairro Jaqueline, Belo Horizonte-MG, no período de 19 a
31/01/07. N = 12.
78
Tabela 8. Captura total e média de machos e de fêmeas de Culex quinquefasciatus
por tratamento do experimento de comparação entre o melhor protótipo de
armadilha para captura de fêmeas grávidas de Culex quinquefasciatus com
MosquiTRAP no bairro Jaqueline e no ICB/UFMG, Belo Horizonte-MG,
nos períodos de 22 a 30/03/07 e de 16 a 24/04/07. N = 24.
80
Tabela 9. Captura total de machos e fêmeas de Aedes aegypti por tratamento do
experimento de comparação entre o melhor protótipo de armadilha para
captura de fêmeas grávidas de Culex quinquefasciatus com MosquiTRAP
no bairro Jaqueline e no ICB/UFMG, Belo Horizonte-MG, nos períodos de
22 a 30/03/07 e de 16 a 24/04/07. N = 24.
81
Tabela 10. Captura de outros Insecta e Aracnida por cada tratamento do experimento
de comparação entre o melhor protótipo de armadilha para captura de
fêmeas grávidas de Culex quinquefasciatus com MosquiTRAP no bairro
Jaqueline e no ICB/UFMG, Belo Horizonte-MG, nos períodos de 22 a
30/03/07 e de 16 a 24/04/07. N = 24.
81

XIII
SUMÁRIO
RESUMO VI ABSTRACT VII LISTA DE FIGURAS VIII LISTA DE QUADROS X LISTA DE TABELAS XI 1. INTRODUÇÃO 15 2. REVISÃO DE LITERATURA 20
2.1 Biologia de Culex quinquefasciatus 20 2.2 Postura de ovos 22 2.3 Principais métodos de controle e monitoramento de mosquitos 24 2.4 A utilização de semioquímicos para o monitoramento e/ou controle de
mosquitos 28
2.5 Atraentes de oviposição para Culex quinquefasciatus 34 2.6 A utilização de armadilhas e atraentes para a captura de mosquitos adultos 36 2.7 A armadilha P1T1 41
3. OBJETIVOS 44 3.1 Geral 44 3.2 Específicos 44
4. MATERIAL E MÉTODOS 46 4.1 Desenvolvimento e avaliação de novos protótipos de armadilhas para
captura de fêmeas grávidas de Culex quinquefasciatus 46
4.1.1 Protótipos de armadilhas avaliados 46 4.1.2 Avaliação da dispersão de odores nos diferentes protótipos de
armadilha desenvolvidos 49
4.1.3 Atraente Sintético de Oviposição (ASO) 51 4.1.4 Paridade das fêmeas coletadas em campo 52 4.1.5 Área Experimental 54
4.1.5.1 Área 1: Regional Venda Nova 54 4.1.5.1.1 Avaliação da presença de Culex
quinquefasciatus no Clube Lareira e em suas proximidades
54
4.1.5.2 Área 2: Regional Norte 55 4.1.6 Experimento 1: Avaliação dos protótipos de armadilha de
oviposição para captura de fêmeas grávidas de Culex quinquefasciatus
56
4.2 Experimento 2: Avaliação de combinações binárias de atraentes sintéticos de oviposição
58
4.3 Experimento 3: Avaliação de combinações terciárias e quaternária de atraentes sintéticos de oviposição
61
4.4 Experimento 4: Comparação do melhor protótipo de armadilha para captura de fêmeas grávidas de Culex quinquefasciatus com MosquiTRAP
63
4.5 Análises estatísticas 65 5. RESULTADOS 66

XIV
5.1 Desenvolvimento e avaliação de novos protótipos de armadilhas para captura de fêmeas grávidas de Culex quinquefasciatus
66
5.1.1 Avaliação da dispersão de odores nos diferentes protótipos de armadilha desenvolvidos
66
5.1.2 Experimento 1: Comparação entre os diferentes protótipos desenvolvidos de armadilhas para captura de fêmeas grávidas de Culex quinquefasciatus
70
5.1.2.1 Área 1: Regional Venda Nova 70 5.1.2.2 Área 2: Regional Norte 70
5.2 Experimento 2: Avaliação de combinações binárias de atraentes sintéticos de oviposição
73
5.3 Experimento 3: Avaliação de combinações terciárias e quaternária de atraentes sintéticos de oviposição
76
5.4 Experimento 4: Comparação do melhor protótipo de armadilha para captura de fêmeas grávidas de Culex quinquefasciatus com MosquiTRAP
79
6. DISCUSSÃO 82 7. CONCLUSÕES 95 8. REFERÊNCIAS BIBLIOGRÁFICAS 97

15
1. INTRODUÇÃO
O mosquito Culex quinquefasciatus Say, 1823 (Diptera: Culicidae) é o principal
vetor de filariose linfática no mundo, doença causada pelos helmintos Wuchereria
bancrofti, Brugia malayi e B. timori (OMS, 1998). O vetor C. quinquefasciatus também foi
incriminado como vetor de encefalites que ocasionalmente podem acometer o homem
(CONSOLI e OLIVEIRA, 1994). No Brasil, a filariose linfática é causada apenas pela W.
bancrofti, sendo conhecida como filariose bancroftiana, cujo vetor é a fêmea de C.
quinquefasciatus, a qual ingere as microfilárias do parasito durante repasto sanguíneo
realizado em pessoas infectadas (FORATTINI, 2002). A endemia tem distribuição
geográfica focal, atingindo a periferia de núcleos urbanos situados em regiões úmidas e
quentes, como Recife (PE), Maceió (AL) e Belém (PA) (FONTES, 2000; FORATTINI,
2002). No mundo, estima-se em 120 milhões o número de pessoas parasitadas e em 1,2
bilhão os indivíduos que vivem em áreas endêmicas com o risco de contrair a doença,
sendo que a maioria dos infectados vive na África, seguido de Índia, Sul da Ásia, Regiões
do pacífico e nas Américas (WHO, 2000).
O programa de controle da doença preconiza o tratamento em massa da população
de áreas de risco com o medicamento dietilcarbamazina, bem como a redução do contato
do homem com o mosquito, através da redução da densidade populacional do vetor nas
áreas endêmicas (FONTES, 2000; WHO, 1988).
O grande incômodo causado pelos ruídos do mosquito C. quinquefasciatus e suas
picadas à noite, que podem causar reações alérgicas, associados à abundância do inseto, são
fatores que dificultam a vida das populações atingidas, constituindo-se em causa de

16
diminuição da qualidade de vida, desvalorização econômica de imóveis situados em regiões
com alta densidade do mosquito, prejuízos à criação de animais, bem como redução na
eficiência do trabalho (CONSOLI e OLIVEIRA, 1994; FORATTINI, 2002).
O controle de C. quinquefasciatus deve ser focado fundamentalmente na redução
dos criadouros potenciais desse mosquito, mediante obras de saneamento. O uso de
substâncias tóxicas (controle químico) reveste-se sempre de aspecto emergencial, pois tem
a impropriedade de poluir o ambiente, e de existirem populações de insetos resistentes
(FORATTINI, 2002).
Armadilhas utilizadas para captura de fêmeas grávidas de C. quinquefasciatus tais
como a armadilha de Reiter “gravid trap” (REITER, 1983; 1987), a armadilha CDC
(Centers for Disease Control and Prevention) com luz ultravioleta, acrescida de atraentes de
oviposição (RITCHIE, 1984) e a armadilha CFG (conterflow geometry), também possuindo
atraentes de oviposição, como infusões de gramíneas (MBOERA et al., 2000a), podem
obter uma captura eficiente, porém possuem um custo elevado, o que limita a sua utilização
em áreas de poder econômico baixo. Todas as armadilhas citadas utilizam baterias, o que
além de aumentar os custos, dificulta sua operacionalidade em campo, visto que as baterias
só funcionam continuamente entre 24-48 horas, devendo ser constantemente trocadas e
recarregadas. Em uma cidade com alta densidade populacional de mosquitos e baixo poder
econômico, associação que ocorre com freqüência, o monitoramento através de armadilhas
deste tipo se tornaria inviável, visto que seria necessário um grande número das mesmas.
No Brasil, seria de grande importância o emprego de armadilha e atraente de
oviposição de baixo custo no monitoramento e/ou controle de populações de C.
quinquefasciatus por meio de coleta massal de fêmeas grávidas. Tal metodologia consiste
em coletar seletivamente o maior número possível de insetos-alvo para reduzir sua

17
população a um nível que não cause danos (BENTO, 2001). Uma armadilha de baixo custo,
com atraente de oviposição para captura de fêmeas grávidas, poderia monitorar densidades
de populações e funcionar como ferramenta adicional no controle integrado, removendo
mosquitos do ambiente.
Um modelo de armadilha desenvolvido recentemente para fêmeas grávidas de C.
quinquefasciatus, denominada P1T1, utiliza um recurso que elimina a necessidade de fonte
de energia: um cartão de polietileno impregnado com cola entomológica e posicionado na
armadilha acima da lâmina d’água ou infusão, perfurado para permitir a emanação dos
voláteis responsáveis pela atração dos mosquitos, é responsável pela captura de fêmeas de
C. quinquefasciatus. A armadilha foi desenvolvida com base nos comportamentos de pré-
oviposição e oviposição do mosquito, registrados em filmagens através de microcâmeras e
vídeo cassete (LIMA, 2005).
Lima (2005) testou nesta armadilha diversos atraentes de oviposição sintéticos em
liberadores de polietileno (para atraentes líquidos) e de silicone (para atraentes sólidos, em
pó). O escatol se mostrou como o atraente de oviposição sintético, dentre os testados (fenol,
p-cresol, nonanal, escatol e indol), com a maior porcentagem de captura de fêmeas e, assim
como o p-cresol, não apresentou diferença significativa de captura, em relação ao seu
controle (P1T1 contendo infusão de gramínea Panicum maximum a 75%). Com relação à
captura de fêmeas grávidas, os atraentes escatol, indol e p-cresol obtiveram
significativamente maior percentual que os seus controles (infusão a 75%). Não foram
realizados testes de combinações dos referidos atraentes de oviposição sintéticos, o que
poderia aumentar o potencial de captura da armadilha.
Em testes de comparação entre as armadilhas P1T1 iscadas com escatol e CDC, a
primeira se mostrou específica para fêmeas grávidas e ingurgitadas de C. quinquefasciatus,

18
visto que apresentou um percentual de captura maior de tais categorias do inseto, em
comparação à CDC (LIMA, 2005). Sendo assim, a armadilha P1T1, acrescida de atraentes
de oviposição sintéticos, se constitui em uma importante e potencial ferramenta no
monitoramento das populações de C. quinquefasciatus, pois se mostra específica e eficaz na
captura de fêmeas grávidas do mosquito, é de fácil operacionalidade e de baixo custo,
fatores relevantes especialmente no contexto sócio-econômico do país. No entanto, o seu
potencial de captura relativamente baixo impede a sua utilização como ferramenta de
controle do inseto. Portanto, a armadilha P1T1 necessita de maiores estudos visando
aumentar a sua eficiência de captura de fêmeas grávidas de C. quinquefasciatus.
O presente trabalho visa desenvolver e avaliar novos protótipos de armadilhas para
captura de fêmeas grávidas de C. quinquefasciatus, que diferenciem da armadilha P1T1 em
aspectos visuais, como tamanho e forma. Também foram relevantes fatores olfativos dos
protótipos desenvolvidos, como a diferença na formação da pluma de odor obtida por cada
protótipo desenvolvido, o que dificulta ou facilita o encontro da fonte do odor pelo inseto.
Os protótipos construídos possuíam diferentes formas de acesso do inseto ao interior da
armadilha, onde se situa o cartão adesivo responsável pela captura do mesmo, representadas
por diferentes números e posições de aberturas dos protótipos desenvolvidos. A associação
destes fatores resulta no potencial de captura de cada protótipo de armadilha, o que foi
avaliado em campo.
Ainda com o intuito de aumentar o potencial de captura da armadilha para captura
de fêmeas grávidas de C. quinquefasciatus, o presente trabalho avaliou diferentes
combinações binárias, terciárias e quaternárias de atraentes sintéticos de oviposição em
polímeros liberadores, acrescidos ao melhor protótipo de armadilha obtido, visando
aumentar a atratividade obtida pela utilização dos atraentes isolados.

19
Também foram realizados experimentos em campo para avaliar a especificidade do
melhor protótipo de armadilha desenvolvido e da melhor combinação de atraentes de
oviposição sintéticos na captura de fêmeas grávidas de C. quinquefasciatus, em
comparação à MosquiTRAP, utilizando AtrAedes (atraente utilizado para captura de
fêmeas de Aedes aegypti) ou a melhor combinação obtida para fêmeas grávidas de C.
quinquefasciatus, pois a MosquiTRAP, apesar de ser específica para fêmeas grávidas de
A. aegypti, freqüentemente captura fêmeas do gênero Culex.

20
2. REVISÃO DE LITERATURA
2.1 Biologia de Culex quinquefasciatus
A espécie C. quinquefasciatus é considerada cosmopolita e distribui-se pelas regiões
tropicais e subtropicais, sendo comumente encontrado no ambiente humano, mantendo-se
relacionada com o homem em todas as fases de sua vida (FORATTINI et al., 1993). A
espécie está presente em todos os estados brasileiros, sendo conhecida popularmente como
muriçoca, pernilongo, mosquito ou carapanã. A ineficiência ou ausência de saneamento
básico pode contribuir para a abundância e prevalência deste mosquito, visto que suas
formas imaturas se desenvolvem em coleções de água altamente poluídas, dotadas de
abundante matéria orgânica e detritos, relativamente protegidas de prolongada exposição
solar (CONSOLI e OLIVEIRA, 1994; FORATTINI, 1963). Através de monitoramento com
armadilhas luminosas “New Jersey 50” na cidade de São Paulo, pôde-se associar a maior
prevalência de espécies de Culex em áreas com abastecimento de água, porém sem rede de
esgoto (FORATTINI et al., 1973). Este mosquito é mais freqüente nos meses quentes e
chuvosos, devido à ampliação de seus criadouros, no entanto é coletado durante todo o ano,
ininterruptamente (CONSOLI e OLIVEIRA, 1994).
O mosquito C. quinquefasciatus possui desenvolvimento pós-embrionário do tipo
holometábolo, ou seja, possui metamorfose completa, passando pelos estágios de ovo
(depositados juntos, sob a forma de jangada), larva (L1-L4), pupa e adulto. O ciclo tem
duração de cerca de 13 dias, podendo sofrer alterações de acordo com variações na
temperatura e na disponibilidade de alimento nos criadouros (FORATTINI, 1963). A

21
temperatura ótima para o desenvolvimento da maioria dos mosquitos tropicais encontra-se
entre 24 e 28° C (CONSOLI e OLIVEIRA, 1994).
Os adultos de C. quinquefasciatus têm hábito essencialmente noturno e são
altamente endofílicos; sendo que as fêmeas procuram no domicílio abrigo e fontes para o
exercício da hematofagia (CONSOLI e OLIVEIRA, 1994; EIRAS, 2000; FORATTINI,
2002). As fêmeas possuem apreciável antropofilia, embora possam procurar outros
mamíferos, aves e até mesmo répteis como fontes para o repasto sanguíneo (CONSOLI e
OLIVEIRA, 1994; FORATTINI, 1963). As fêmeas de C. quinquefasciatus são ditas
anautógenas, pois é necessário que ocorra uma primeira alimentação sanguínea para o
desenvolvimento de seus ovos e sua deposição na água (FORATTINI, 1963). Existe
concordância gonotrófica nesta espécie, pois cada oviposição é precedida por um repasto
sanguíneo (CONSOLI e OLIVEIRA, 1994; FORATTINI, 1963).
Na região neotropical e no Brasil, C. quinquefasciatus é considerado como o mais
eficiente vetor da filariose bancroftiana ao homem (CONSOLI e OLIVEIRA, 1994;
FORATTINI, 2002). São fatores da espécie de mosquito que facilitam a veiculação deste
patógeno o hábito preferencialmente antropofílico, visto que o parasito Wuchereria
bancrofti não possui outro reservatório além do homem, bem como o hábito hematofágico
noturno, pois é neste momento em que são encontradas microfilárias em abundância
circulando pelo sangue periférico (CONSOLI e OLIVEIRA, 1994).
A febre do Nilo Ocidental é uma encefalite viral causada por um flavivírus que
acomete principalmente aves e ocasionalmente o homem, podendo ser transmitida por C.
quinquefasciatus e que, desde o seu primeiro registro em Uganda, em 1937, vem sendo
observada em diversas regiões do planeta (ARAÚJO, 2004). Nas Américas, a primeira
epidemia ocorreu na região metropolitana de Nova York, se espalhando por 12 estados

22
(SALGADO, 2000). Johnson et al. (2003) constataram que o vírus atenuado, utilizado na
fabricação de vacina contra o vírus do Nilo Ocidental, não pode ser transmitido por seus
vetores, como C. quinquefasciatus, o que o torna um bom candidato para o controle da
doença.
2.2 Postura de ovos
O gênero Culex, assim como os gêneros, Culiseta, Uranotaenia, Armigeres e
Coquillettidia depositam seus ovos na lâmina d’água em conjunto, unidos um ao outro com
o auxílio de uma mucilagem; estes ficam flutuando no meio aquático, formando, então,
uma jangada (CLEMENTS, 1999; FORATTINI, 2002).
Geralmente, as fêmeas de C. quinquefasciatus depositam seus ovos durante a
escotofase (FORATTINI, 1963), sendo esta atividade cíclica e obedecendo ao ritmo
circadiano (CLEMENTS, 1999). Segundo Mboera et al. (2000b), em estudos em campo
utilizando o feromônio sintético de oviposição (FOS) (5R, 6S)-6-acetoxi-5-hexadecanolide,
foram verificados dois picos de oviposição: entre a 1ª e a 3ª h da escotofase ocorre a maior
deposição de ovos, seguido do intervalo entre a 1ª e a 3ª h da fotofase. Em condições de
laboratório, Lima (2005) observou dois picos de postura, o primeiro entre a 1ª e 2ª hora de
escotofase e o segundo entre a 8ª e 10ª hora da escotofase, não tendo sido observada
nenhuma deposição de jangada durante a fotofase. Em laboratório, as fêmeas de C.
quinquefasciatus depositam seus ovos no período entre o 4º e o 8º dias após a alimentação
sanguínea, não havendo diferença significativa entre as médias de jangadas depositadas
neste período (LIMA, 2005).

23
As fêmeas de C. quinquefasciatus podem apresentar vários ciclos gonotróficos,
distinguindo-se, assim, nulíparas, como aquelas que ainda não ovipositaram e oníparas as
que o fizeram ao menos uma vez. Dentre as oníparas, diz-se paucíparas as que desovaram
uma ou duas vezes e multíparas as que produziram mais do que três cargas de ovos
(FORATTINI, 1963).
A deposição de ovos em mosquitos pode ser dividida em dois estágios: o
comportamento de pré-oviposição, caracterizado pelo comportamento de procura das
fêmeas grávidas por um possível criadouro, seguido do comportamento de forrageamento,
ou seja, avaliação do ambiente em que será realizada a postura de ovos; e a oviposição, que
consiste no ato de deposição de ovos (CLEMENTS, 1999). Através de filmagens e em
condições de laboratório, Lima (2005) observou que as fêmeas grávidas de C.
quinquefasciatus apresentam seqüências de comportamentos de explorar o criadouro antes
da oviposição, gastando mais tempo neste comportamento do que quando comparada ao
comportamento de oviposição. As fêmeas grávidas desta espécie realizam a oviposição
geralmente seguindo as etapas de pouso nas paredes internas do criadouro, deslocamento na
interface reservatório/água, posicionamento do primeiro par de pernas nas paredes e os
outros dois na superfície líquida, oviposição das jangadas e vôo de saída do criadouro
(LIMA, 2005).
Geralmente, o comportamento de oviposição de fêmeas grávidas de mosquitos
obedece à seguinte seqüência: vôo, orientação, encontro e aceitação, podendo ocorrer pouso
e avaliação da superfície do criadouro. Em cada estágio, diferentes sinais serão percebidos
pelas fêmeas: durante o vôo, são importantes os estímulos visuais e olfativos, enquanto que
para os estágios em pouso, os estímulos incluem fatores físicos e químicos envolvidos no
contato com as superfícies (CLEMENTS, 1999).

24
As fêmeas de C. quinquefasciatus se tornam mais sensíveis às diversas substâncias
isoladas de criadouros quando grávidas, o que pode ser visto através de estudos
eletrofisiológicos com antenas de fêmeas grávidas (BLACKWELL et al., 1993; DU e
MILLAR, 1999; MILLAR et al., 1992). Por exemplo, fêmeas grávidas apresentam maiores
amplitudes de respostas das antenas aos compostos atraentes de oviposição, como 3-metil-
indol, enquanto os machos são praticamente insensíveis ao mesmo composto
(BLACKWELL et al., 1993).
Os comportamentos de pré-oviposição e oviposição são difíceis de serem
observados naturalmente, porém filmagens podem fornecer dados consistentes, como foi
feito para fêmeas grávidas de Aedes aegypti (GOMES, 2003) e de C. quinquefasciatus
(LIMA, 2005), nos quais foram observados aspectos da exploração do criadouro e
comportamentos de oviposição.
2.3 Principais métodos de controle e monitoramento de mosquitos
O controle de insetos vetores de doenças pode ser obtido através de medidas de
controle químico, físico e biológico, direcionado de acordo com o inseto alvo. Ainda,
medidas de proteção individual podem ser utilizadas, visando evitar o hábito hematofágico
dos mosquitos, protegendo o homem através do uso de repelentes ou cortinados
(FORATTINI, 2002).
O controle de culicídeos tem-se voltado preferencialmente para as formas imaturas,
mediante adequada manipulação do ambiente, através do controle físico, mecânico ou
ambiental, que trata de atividades de planejamento, organização, execução e vigilância das

25
modificações do ambiente que objetivam torná-lo pouco propício à sobrevivência da
população de mosquitos (FORATTINI, 2002). Para tanto, podem ser adotadas medidas
como canalização do esgoto, destinação correta ao lixo, com a construção de aterros
sanitários, eliminação de recipientes que possam acumular água em áreas urbanas, como
garrafas, pneus, recipientes plásticos e bromélias (CONSOLI e OLIVEIRA, 1994, EIRAS,
2000; FORATTINI, 2002).
O controle químico, realizado através da utilização de inseticidas para o controle de
mosquitos, é amplamente utilizado em vários países, sendo o principal suporte dos
programas de combate e controle de insetos vetores de doenças (CONSOLI e OLIVEIRA,
1994). A dose única e subletal de determinados inseticidas tem efeito significativo na
reprodução de mosquitos, por diminuir a longevidade das fêmeas, o que diminui o número
de cópulas e de oviposições por fêmea durante sua vida, e por diminuir o número de ovos
depositados pelas fêmeas tratadas, em relação ao controle (ROBERT e OLSON, 1989).
Até 1995, o DDT (Dicloro-difenil-tri-cloro-etano, organoclorado) foi o inseticida
químico mais empregado, por ser relativamente barato, porém seu alto poder residual, e o
fato de não ser biodegradável, sendo acumulativo nas gorduras de animais de sangue
quente, fez com que o mesmo deixasse de ser utilizado no Brasil, medida preconizada pela
OMS, mas que em alguns países ainda não foi estabelecida (CONSOLI e OLIVEIRA,
1994; FORATTINI, 2002).
Inseticidas à base de piretróides são utilizados no Brasil através da técnica de
aplicação denominada “ultra baixo volume”, popularmente conhecido como fumacê.
Apesar da eficiência deste método estar condicionada às condições climáticas, devido ao
aceleramento operacional e diminuição dos custos ainda é amplamente utilizado

26
(FORATTINI, 2002). Populações de C. quinquefasciatus do Oeste da África apresentam
resistência a piretróides (CHANDRE et al., 1998).
Populações de C. quinquefasciatus de São Paulo apresentam resistência ao
Malathion, inseticida fosforado utilizado nos programas de controle locais, e ao
Fenitrothion, nunca usado no local, o que sugere existência de resistência cruzada
(BRACCO et al., 1999). Em uma avaliação de seis espécies de mosquitos vetores, a
linhagem de campo de C. quinquefasciatus se mostrou a menos susceptível ao Malathion, o
que demonstra seu potencial de desenvolver resistência a este inseticida (DORTA et al.,
1993).
O temefós (organofosforado), amplamente utilizado no Brasil na formulação
granulada impregnada em areia, para a eliminação de larvas de mosquitos, é de baixa
toxicidade aos mamíferos, no entanto atualmente tem efeito residual no ambiente e já se
depara com populações resistentes (FORATTINI, 2002). A resistência de espécies de
Culex, como C. quinquefasciatus, do Sri Lanka, se deve ao uso em larga escala de
inseticidas organofosforados (KARUNARATNE e HEMINGWAY, 2001).
As principais restrições à utilização de inseticidas químicos no controle de insetos
vetores são sua inespecificidade, a existência de resistência aos mesmos e seu alto poder
residual (CONSOLI e OLIVEIRA, 1994).
Diferentes inibidores do desenvolvimento de insetos podem ser utilizados em
programas de controle das quatro espécies de mosquitos: Culex pipiens fatigans, Anopheles
gambiae, A. aegypti e Anopheles quadrimaculatus (BUSVINE et al., 1976). Tidos como
inseticidas químicos alternativos, podem ser empregados no controle de larvas de
mosquitos. Estes reguladores do crescimento atuam como análogos do hormônio juvenil,
que se utilizados na água inibem a metamorfose, sendo o mais conhecido o Methoprene, ou

27
inibem a formação de quitina, impedindo a muda do inseto (CONSOLI e OLIVEIRA,
1994).
Produtos de origem vegetal podem ser utilizados como inibidores de crescimento,
reprodução e oviposição ou como repelentes (CONSOLI e OLIVEIRA, 1994). A
combinação de cipermetrina e extratos de Solanum xanthocarpum mostrou sinergismo na
atividade larvicida contra C. quinquefasciatus, porém devem ser feitos estudos para avaliar
a toxicidade em humanos e animais, especialmente para formas aquáticas (MOHAN et al.,
2006). O resíduo do desfibramento de folhas de Agave sisalana (resíduo do sisal) pode ser
usado no controle de C. quinquefasciatus, por sua atividade larvicida (PIZARRO et al.,
1999), porém não há registros de sua utilização no controle do mosquito.
Como controle biológico entende-se a medida que visa à redução da densidade
populacional de determinado vetor, utilizando agentes biológicos, como predadores
naturais e agentes entomopatológicos (FORATTINI, 2002). Dentre os predadores
vertebrados destacam-se os peixes larvíparos, como Gambusia affinis e Poecilis reticulata
(CONSOLI e OLIVEIRA, 1994). Em relação aos invertebrados, é grande a lista de
predadores, dentre eles hidrozoários, planárias, crustáceos e insetos (FORATTINI, 2002).
Entre os insetos, o grupo que mais se destaca são os culicídeos pertencentes ao gênero
Toxorhynchites, cujas formas larvárias são predadoras de espécies de Aedes, mas as formas
adultas não são hematófagas, sendo, portanto, inócuos sob esse ponto de vista (CONSOLI e
OLIVEIRA, 1994; FORATTINI, 2002). Um problema desta modalidade de controle é a
habilidade de algumas espécies de mosquitos para detectar a presença de predadores nos
sítios de oviposição, evitando-os (AVIEITAM e BLAUSTEIN, 2004).
Como bactérias entomopatogênicas destaca-se Bacillus thurigiensis, bactéria
comumente encontrada no solo, formadora de esporos que produzem toxinas de elevado

28
poder inseticida, provocando a lise de células gástricas do inseto, como Aedes, Culex,
Psorophora e Anopheles. Outro agente é o Bacillus sphaericus, com especificidade para
Culex, principalmente C. quinquefasciatus (FORATTINI, 2002). A atividade larvicida de
B. sphaericus contra C. quinquefasciatus não varia de acordo com a densidade larvária de
mosquitos, mas é diretamente proporcional à temperatura (entre 15º C e 25º C), a
mortalidade de larvas aumenta quando há evaporação em pequenos criadouros, e é
inversamente proporcional ao período de incidência da luz solar direta sobre o criadouro
artificial (PINTO, 1995).
O fungo Metarhizium anisopliae causa mortalidade em larvas de C.
quinquefasciatus e em adultos de A. gambiae e C. quinquefasciatus submetidos à
formulação oleosa contendo conídia deste fungo, podendo ser utilizado no controle de
insetos endofílicos (ALVES et al., 2002; SCHOLTE et al., 2003).
O controle integrado representa a orientação mais adequada nas atividades de
combate aos mosquitos de interesse epidemiológico (FORATTINI, 2002). Neste contexto,
armadilhas para captura de mosquitos podem auxiliar no controle destes, desde que
possuam um alto potencial de captura, auxiliando na redução da densidade populacional do
mosquito-alvo a um nível que evite a transmissão de doenças (BENTO, 2001).
2.4 A utilização de semioquímicos para o monitoramento e/ou controle de mosquitos
As substâncias químicas estão envolvidas na mediação de um grande número de
comportamentos de insetos, desde a comunicação entre indivíduos de uma mesma espécie
ao reconhecimento de fatores ambientais e procura de alimento (comunicação

29
intraespecífica e interespecífica) (CARDÉ e BELL, 1996). Tais substâncias químicas são
ditas semioquímicos, que significa sinal químico. Os compostos responsáveis pela
comunicação interespecífica são ditos aleloquímicos; substâncias responsáveis pela
comunicação entre indivíduos de uma mesma espécie são ditas feromônios (VILELA e
DELLA LUCIA, 2001). Dentre os aleloquímicos, podemos distinguir:
Alomônios: substâncias favoráveis ao emissor e desvantajosas ao receptor (DICKE e
SABELIS, 1992), como as substâncias de defesa dos artrópodes (VILELA e DELLA
LUCIA, 2001);
Cairomônios: substâncias favoráveis ao receptor, mas desvantajosas ao emissor (DICKE e
SABELIS, 1992), como o odor do hospedeiro, utilizado por insetos hematófagos para a sua
localização (EIRAS, 2001);
Sinomônios: substâncias favoráveis a ambos, tanto receptor quanto emissor (DICKE e
SABELIS, 1992), como os odores florais que atraem os insetos polinizadores (VILELA e
DELLA LUCIA, 2001);
Apneumônios: substâncias que beneficiam o receptor e que são emitidas por material inerte
(NORDLUND e LEWIS, 1976), como os voláteis de infusão de gramínea, atrativos para
fêmeas grávidas de C. quinquefasciatus (HAZARD et al., 1967; ISOE et al., 1995b;
MILLAR et al., 1992).
Os feromônios são considerados substâncias mensageiras entre indivíduos de uma
mesma espécie, não sendo confundidos com os hormônios por serem secretados para o
exterior do corpo do emissor (VILELA e DELLA LUCIA, 2001). Um exemplo é o
feromônio de oviposição de C. quinquefasciatus, produzido pelos ovos após o contato com

30
a água e que atrai outras fêmeas grávidas, exercendo um fenômeno de agregação das
mesmas, as quais depositam suas jangadas no mesmo criadouro (CLEMENTS, 1999).
O comportamento de oviposição é mediado por sinais físicos, químicos e biológicos
e, na ausência de um sinal (como privação de água ou luz), uma série inteira de
comportamentos que termina na deposição de ovos pode ser ativada pelos sinais restantes
(ISOE e MILLAR, 1996), sendo a escolha dos locais de oviposição de fêmeas de C.
quinquefasciatus baseada em informações recebidas durante fases pré-adultas (L4 e pupa)
e/ou logo após a emergência, sugerindo alguma forma de condicionamento (McCALL e
EATON, 2001).
Segundo Clements (1999), os compostos químicos podem gerar em fêmeas grávidas
de mosquitos respostas, segundo as quais serão classificados em:
Atraentes, quando fazem com que as fêmeas grávidas orientem seu vôo em direção à fonte;
Arrestantes, quando fazem com que as fêmeas grávidas permaneçam onde estão, por
pararem sua locomoção, não responderem ao estímulo ou por mudarem sua rota;
Repelentes, quando faz com que as fêmeas grávidas voem em direção contrária à fonte do
odor;
Deterrentes, uma substância não volátil que inibe a oviposição, a qual ocorreria na ausência
da mesma;
Estimulantes, quando incita a oviposição.
Isoe e Millar (1995) procuraram caracterizar os fatores que mediam a escolha do
local de oviposição por fêmeas grávidas de Culex tarsalis, obtidos de infusão fermentada e
matéria orgânica, e sugeriram que a oviposição pode ocorrer em resposta a fatores como
estimulantes de oviposição de contato (não-voláteis) no lugar de atraentes voláteis.

31
Na procura de novas técnicas para o controle de insetos vetores de doenças por
métodos seletivos, os semioquímicos têm potencial promissor no controle comportamental,
podendo ser utilizados como atraentes em armadilhas para o monitoramento e/ou controle
destes insetos, porém, ainda assim, são escassos os estudos relacionados aos insetos vetores
de doenças humanas e veterinárias (EIRAS, 2001). Os feromônios são semioquímicos
geralmente atóxicos, não causam poluição ambiental e são específicos (JUTSUM e
GORDON, 1989). Além disso, as chances de resistência dos insetos aos feromônios
sintéticos são remotas (EIRAS, 2001).
A combinação de testes comportamentais e eletrofisiológicos pode resultar em
importantes informações sobre quais sinais químicos um mosquito utiliza e em quais
concentrações estes são efetivos. Compostos químicos de hospedeiros utilizados como
fonte de repasto sanguíneo, plantas e sítios de oviposição podem ser utilizados como
atrativos e repelentes, podendo ser utilizados como ferramenta complementar no controle
de mosquitos (DAVIS e BOWEN, 1994).
O primeiro trabalho de identificação de feromônio de oviposição foi realizado com
C. quinquefasciatus. A molécula identificada como eritro-6-acetoxi-4-hexadecanolide
(ADH) é secretada na região apical dos ovos e estimula a oviposição de fêmeas grávidas
(LAURENCE e PICKETT, 1982). A variabilidade nas respostas ao feromônio foi
evidenciada por vários autores (LAURENCE e PICKETT, 1985; MORDUE et al., 1992;
MILLAR et al., 1994; OTIENO et al., 1988; PILE et al., 1991). Para se obter uma resposta
significante de coleta de jangadas depositadas por fêmeas grávidas de C. quinquefasciatus
em gaiolas, foi necessária uma dose de 0,02 µg do feromônio acrescido a 250 mL de água
desclorada, equivalente a 1/15 da quantidade produzida por uma jangada (0,3 µg)

32
(LAURENCE e PICKETT, 1985). Enquanto Millar et al. (1994) observou, em testes em
gaiola, que uma quantia variando entre 2-15 jangadas coespecíficas (em 80 mL de água
destilada) aparentemente não teve efeito significativo na oviposição de fêmeas grávidas de
C. quinquefasciatus, comparado à oviposição obtida no controle contendo apenas água
destilada. Otieno et al. (1988) avaliaram o isômero do feromônio natural, feromônio de
oviposição sintético (FOS) (5R, 6S)-6-acetoxi-5-hexadecanolide, em campo e só obtiveram
uma deposição de jangadas significativa, em comparação a águas limpas, após 18 dias de
experimento. Estudos em túnel de vento e com equipamentos de vídeo demonstraram a
atratividade do FOS para fêmeas grávidas de C. quinquefasciatus (PILE et al., 1991).
Estudos comportamentais e eletrofisiológicos demonstraram a atração de fêmeas grávidas
de C. quinquefasciatus em resposta ao FOS e à água poluída. Foi demonstrado ainda que a
combinação dos dois tem um efeito aditivo, tanto na sensibilidade das antenas (estudo
eletrofisiológico), quanto na oviposição (estudo comportamental) (MORDUE et al., 1992).
Outros semioquímicos, ditos apneumônios, têm mostrado atratividade para fêmeas
grávidas de A. aegypti e Aedes albopictus, como infusões de gramíneas (ALLAN e KLINE,
1995; CHADEE et al., 1993; HAZARD et al., 1967; ROQUE, 2002), bem como
substâncias isoladas de infusões (ALLAN e KLINE, 1998) e águas contendo formas
imaturas de mosquitos da mesma espécie (ALLAN e KLINE, 1995; ALLAN e KLINE,
1998). A atratividade também foi demonstrada para fêmeas grávidas de C. quinquefasciatus
e C. tarsalis por apneumônios como infusão de gramíneas (HAZARD et al., 1967; ISOE et
al., 1995a, b; LIMA, 2005; MILLAR et al., 1992), águas poluídas (BLACKWELL et al.,
1993) e locais com ou que previamente tinham larvas co-específicas (MOKANY e SHINE,
2003).

33
As infusões de gramíneas são utilizadas em armadilhas de oviposição nos
programas de controle e monitoramento de mosquitos como A. aegypti e C.
quinquefasciatus em todo o mundo, inclusive no Brasil (ALLAN e KLINE, 1995; ISOE et
al., 1995a; JAYANETTI et al., 1988; KLINE et al., 2006; LIMA, 2005; MBOERA et al.,
2000a; ROQUE, 2002). A espécie de gramínea utilizada, a concentração utilizada e a idade
da mesma podem influenciar na sua atratividade, pois a microfauna presente nesses meios
sofre constantes alterações, o que resulta na liberação de diferentes voláteis (SANT’ANA,
2006).
O número de jangadas depositadas por Culex pipiens e Culex restuans em
armadilhas de oviposição iscadas com infusão varia de acordo com o tipo de substrato
utilizado para o preparo da infusão, com a idade da infusão, com o modo de preparo da
infusão e de acordo com a época do ano (LAMPMAN e NOVAK, 1996). Ao avaliar
diferentes concentrações de infusão da gramínea Panicum maximum (capim colonião) para
o uso em armadilhas de oviposição, foi verificado que as armadilhas contendo infusão a
75% capturaram significativamente mais fêmeas grávidas de C. quinquefasciatus, em testes
em campo e laboratório (LIMA, 2005). Diferentes concentrações de infusão da gramínea
Sclerica bractaeda, acrescidas a ovitrampas, foram avaliadas em campo, quanto à
atratividade de A. aegypti, porém não houve diferença significativa na deposição de ovos
entre as diferentes concentrações da infusão avaliadas (CHADEE et al., 1993). A atração de
fêmeas de C. tarsalis não variou entre infusão da gramínea Cynadon dactylon (capim
Bermuda) a 10% e a 100%, porém a deposição de ovos foi significativamente maior em
resposta à infusão a 100% (ISOE et al., 1995b). Hazard et al. (1967) descreveram que
infusão de alfafa diluída a 10% resultou em uma deposição de jangadas significativamente
maior, quando comparada à água destilada. Já Du e Millar (1999) observaram que infusões

34
de capim Bermuda a 100% foram repelentes quando comparadas à água destilada (DU e
MILLAR, 1999). Por outro lado, a idade da infusão pode alterar a espécie de mosquito
atraída pela mesma. Para a atração de fêmeas de C. quinquefasciatus, os melhores
resultados foram obtidos com infusões entre 10 e 21 dias (ISOE et al., 1995b).
Reiter (1983) padronizou uma metodologia para o preparo e acondicionamento de
infusões de gramíneas, utilizando capim Bermuda, com o intuito de reduzir as alterações na
microfauna; o preparo conta com, além da gramínea, levedura de cerveja e lactoalbumina
hidrolisada e a infusão deve ser mantida em recipiente fechado, em geladeira até o
momento de sua utilização.
Ainda assim, a utilização de infusão apresenta desvantagens tais como o tempo
demandado em seu preparo ou alterações das concentrações dos voláteis emitidos, o que
torna essencial a identificação dos compostos presentes na mesma.
2.5 Atraentes de oviposição para Culex quinquefasciatus
Através de cromatografia gasosa foram identificados vários compostos da infusão
de gramínea capazes de atrair e/ou estimular a oviposição de C. quinquefasciatus, como
fenol, 4-metil-fenol (p-cresol), 4-etil-fenol, indol e 3-metil-indol (escatol) (MILLAR et al.,
1992).
Experimento em campo evidenciou que 3-metil-indol (escatol) é o componente
atrativo primário para fêmeas grávidas de C. quinquefasciatus, pois não houve diferença
significativa na captura entre armadilhas de Reiter (1983) contendo apenas escatol e

35
armadilhas com uma solução atraente, contendo escatol, 4-metil-fenol, 4-etil-fenol, fenol e
indol (BEEHLER et al., 1994).
A técnica de eletroantenografia (EAG) pode ser utilizada na triagem de compostos
presentes em infusões de gramíneas, verificando a quais as fêmeas grávidas respondem,
porém não exclui os testes de comportamento, visto que não se sabe de que modo a
resposta gerada vai modificar seu comportamento. As antenas de fêmeas grávidas de C.
quinquefasciatus apresentaram respostas significativas aos compostos 4-metil-fenol,
naftaleno, indol associado a 2-undecanona e 3-metil-indol e respostas fracas ou
intermediárias ao fenol, nonanal, 4-etil-fenol e 2-tridecanona; tais estudos foram avaliados
em diferentes dosagens, sendo que o 3-metil-indol foi atrativo em concentrações baixas
como 10 ng/L (DU e MILLAR, 1999).
Testes de campo com combinações de compostos atrativos foram realizados visando
o monitoramento de espécies de Culex (MBOERA et al., 1999; 2000 a, b). Em comparação
entre o FOS, infusão de gramínea e voláteis de possível criadouro, ambos os tratamentos
receberam mais jangadas que apenas água destilada, e a combinação de FOS e voláteis do
criadouro sugeriram sinergismo, visto que a captura foi maior do que a de cada tratamento
utilizado isoladamente (MBOERA et al., 1999). A atratividade para fêmeas de C.
quinquefasciatus é dose-dependente; o 3-metil-indol capturou significativamente mais
jangadas nos recipientes de oviposição nas concentrações de 10-6 e 10-5 µg/L (MBOERA et
al., 2000b).
Testes comportamentais e de eletrofisiologia com C. quinquefasciatus mostraram
que água poluída, obtida através da fermentação de fezes de coelho, diluída a 1% e
acrescida de 0,05 µg de FOS mostraram um efeito sinérgico sobre a oviposição das fêmeas,

36
enquanto que os mesmos tratamentos, isolados ou combinados, quando não-diluídos,
apresentavam captura semelhante ao controle (água destilada) (BLACKWELL et al., 1993).
A adição de culturas de bactérias em águas para oviposição pode interferir na
atratividade de fêmeas grávidas de mosquitos, pois a atratividade para fêmeas grávidas de
C. quinquefasciatus de agentes biológicos utilizados no controle de larvas desta espécie,
como B. sphaericus e B. thurigiensis var. israelensis foi avaliada, demonstrando que ambos
foram responsáveis pelo aumento significativo na oviposição, quando comparado ao
estimulante de oviposição 4-metil-fenol (p-cresol) (POONAM et al., 2002).
2.6 A utilização de armadilhas e atraentes para a captura de mosquitos adultos
A combinação de fatores olfativos e visuais apresentados pelas armadilhas é
fundamental, por torná-las mais eficazes (EIRAS, 2001). A forma como os atraentes se
difundem na armadilha através da formação de plumas de odores e a constituição das
substâncias são fatores responsáveis por alterações na eficácia das mesmas quanto à captura
dos mosquitos (DU e MILLAR, 1999). Quanto à dispersão dos odores, estes podem se
disseminar em várias direções e possuírem estrutura diversa, modulada pela turbulência do
vento (EIRAS e MAFRA-NETO, 2001).
Alguns locais da pluma podem conter uma concentração maior ou menor do
composto, o que tanto pode facilitar quanto confundir os insetos para o encontro do
composto. Em insetos voadores, o vôo mediado pelo odor da fonte é o resultado da junção
da anemotaxia (resposta em direção ao vento) e um movimento de ziguezague. Como a
pluma de odor é formada por filamentos dos compostos, estes insetos interceptarão os
filamentos em diferentes momentos do vôo, através da antena e, quando os insetos não

37
localizarem os filamentos, poderão começar a voar em ziguezague e contra o vento, na
tentativa de encontrarem novamente a pluma de odor; caso não encontrem o composto, ou
se este estiver em altas concentrações, os insetos podem desistir da procura e voarem para
outras direções. Para que o comportamento do inseto em resposta à pluma seja elucidado, é
necessário a intercepção e não intercepção dos filamentos de odor em vários momentos
(EIRAS e MAFRA-NETO, 2001).
As armadilhas para captura de insetos, principalmente pragas agrícolas, podem se
beneficiar da utilização de liberadores, que são polímeros capazes de liberar as substâncias
atrativas na armadilha a uma taxa relativamente constante, durante todo o período de
captura dos mosquitos. Os liberadores são artefatos que têm em sua estrutura (septos de
borracha, polietilenos, fibras ocas, grânulos de PVC) moléculas dispostas com espaço entre
elas, pelos quais possa ocorrer a difusão dos gases atrativos. Além da constância na taxa de
liberação, são vantagens da utilização de liberadores para os atraentes a proteção dos
componentes químicos ativos contra a degradação pelo oxigênio do ar ou pela luz e a
aplicabilidade em diferentes situações de manejo.
Lima (2005) avaliou atraentes identificados de infusões de gramínea que
apresentaram respostas significativas para fêmeas grávidas de C. quinquefasciatus,
baseando-se em dados de literatura (BEEHLER et al., 1994; DU e MILLAR, 1999;
MBOERA et al., 2000b; MILLAR et al., 1992). Foram avaliados as taxas de evaporação, o
equilíbrio do sistema e duração de atraentes de oviposição sintéticos (TAB. 1), em
diferentes liberadores. Assim, o período de utilização de cada atraente, após o preparo do
liberador, deve variar de acordo com o período de liberação constante do mesmo.O
liberador contendo escatol, apesar de permanecer liberando o atraente até 110 dias após o

38
seu preparo, deve ser utilizado até 80 dias após o seu prepao, pois a partir desta data ocorre
mudança de cor na mangueira de PVC, utilizada como liberador.
TABELA 1 - Período de utilização de atraentes sintéticos de oviposição em liberadores, de acordo
com o período de estabilização das taxas de evaporação (LIMA, 2005).
Composto Tipo de liberador
Tempo de
equilíbrio
(dias)
Peso médio de
liberação
(mg/dia)
Tempo de
liberação
(dias)
Fenol Tubo de polietileno 7 6 a 7 60
P-cresol Tubo de polietileno 15 4 a 7 90
Nonanal Tubo de polietileno 5 2,3 a 3,4 110
Escatol Tubos de PVC 11 2,3 a 4,5 110
Indol Tubos de PVC 15 3 a 4 90
O QUADRO 1 apresenta um sumário de estudos utilizando armadilhas para captura
de mosquitos adultos com diferentes objetivos, como pesquisa das populações locais de
mosquitos, monitoramento ou ferramenta auxiliar no controle das espécies.

39
QUADRO 1: Principais armadilhas utilizadas para captura de mosquitos adultos e suas aplicações.
Armadilha(s) utilizada(s) Autor (es) Objetivo do trabalho
Protótipos elaborados pelos autores, iscados com infusão de alfafa.
Lewis et al (1974)
Avaliar, quanto às variações nas taxas de captura de mosquitos da espécie C. quinquefasciatus, pequenas modificações nas entradas e formas de retenção dos mosquitos nos protótipos elaborados.
CDC gravid trap.
Reiter (1985); Service (1977)
Desenvolvimento de armadilha por Paul Reiter que utiliza infusão de gramínea como atrativo para fêmeas grávidas de Culex.
CDC gravid trap.
Reiter (1987)
Corrigir problemas operacionais, como captura de formigas que ingeriam as fêmeas de C. quinquefasciatus capturadas, do modelo anterior.
CDC gravid trap.
Jayanetti et al. (1988)
Avaliar a presença de vetores de filarioses no peridomicílio no Sri Lanka, obtendo maior captura de fêmeas grávidas de C. quinquefasciatus.
Falcão.
Teodoro et al. (1994)
Monitoramento de mosquitos no sul do Brasil.
CDC (Center for Disease Control and Prevention) e EVS (Encephalitis Vírus Surveillance).
Ritchie e Kline (1995)
Estudo em campo na Austrália comparando a eficácia das duas armadilhas constatou que a armadilha luminosa CDC capturou mais mosquitos que a armadilha EVS, e que octenol e dióxido de carbono aumentaram a coleta de três espécies.
Armadilha de Shannon, CDC e New Jersey.
Gomes (2002)
Revisão que propõe a utilização das armadilhas citadas para a vigilância entomológica de culicídeos, especialmente am ambiente extradomiciliar.
Armadilha P1T1
Lima (2005)
Desenvolvimento de armadilha específica para captura de fêmeas grávidas de C. quinquefasciatus que não utiliza baterias ou energia elétrica.

40
New Jersey.
Kim et al. (2006)
Monitoramento de mosquitos na Coréia, visando determinar o período ideal para a aplicação de inseticidas.
EVS, MMF (Mosquito Magnet Freedom), MML (Mosquito Magnet Liberty) e MMX (Mosquito Magnet-X).
Cooperband e Carde (2006)
Filmagens de fêmeas de C. quinquefasciatus e C. tarsalis à procura de repasto sanguíneo avalia a eficiência de captura das quatro armadilhas iscadas com dióxido de carbono, em túnel de vento. A MMX obteve maior captura das duas espécies avaliadas.
Armadilha luminosa CDC, CDC gravid trap, MMX e MM-Pro (Mosquito Magnet Professional).
Kline et al. (2006)
Avaliação das quatro armadilhas quanto à captura de espécies de Culex na Flórida, sendo C. quinquefasciatus a espécie mais coletada por todos os modelos de armadilhas. A armadilha MM-Pro é a responsável pela maior captura, em comparação às outras armadilhas avaliadas.
CFG (conterflow geometry) e CDC, iscadas com FOS e infusão de gramínea.
Mboera et al. (2000a)
Avaliar a eficácia das armadilhas, quanto à captura de fêmeas grávidas de C. quinquefasciatus, em testes de campo na Tanzânia. A CFG capturou significativamente mais fêmeas do que a CDC, ambas iscadas com FOS; e a CFG iscada com FOS + infusão de gramínea foi superior aos tratamentos utilizados isoladamente, sendo que os últimos foram diferentes à CFG contendo apenas água.

41
A maioria das armadilhas citadas no quadro 1 necessita de energia elétrica ou
baterias para seu funcionamento e são, em sua grande maioria, inespecíficas, porém,
quando acrescidas de atraentes de oviposição podem capturar fêmeas grávidas de C.
quinquefasciatus. Um modelo recente de armadilha, conhecida como MosquiTRAP® utiliza
um recurso que elimina a necessidade de energia elétrica: um cartão adesivo de polietileno
fixado em suas paredes internas é responsável pela captura de fêmeas de A. aegypti
(EIRAS, 2002). Este modelo foi elaborado baseado em observações do comportamento de
oviposição de fêmeas grávidas de A. aegypti, através de filmagens (GOMES, 2003) e
consiste em um recipiente plástico na cor preta e com formato cilíndrico, utilizando água e
um atraente de oviposição sintético contendo nonanal (AtrAedes®), desenvolvido a partir de
voláteis de infusões de gramínea (Panicum maximum). A armadilha foi especialmente
desenvolvida para coletar fêmeas grávidas de A. aegypti, mas também é capaz de capturar
fêmeas do gênero Culex (LIMA, 2005). A armadilha captura principalmente fêmeas
grávidas e é baseada na estimulação visual (cor preta) e na olfativa (atraente de oviposição).
Os insetos capturados pela MosquiTRAP® são retirados com pinça entomológica e
identificados ainda em campo, no momento da vistoria da armadilha, com auxílio de uma
lupa manual (10X). A possibilidade de identificação dos exemplares capturados ainda em
campo dispensa a rotina de laboratório, agilizando a obtenção de dados (GAMA et al.,
2006).
2.7 A armadilha P1T1

42
A armadilha P1T1 (LIMA, 2005) foi desenvolvida no Laboratório de Ecologia
Química de Insetos Vetores (Instituto de Ciências Biológicas (ICB), Universidade Federal
de Minas Gerais (UFMG)) e consiste em um recipiente de polietileno de cor preta fosca (20
x 30 x 9 cm) com tampa com abertura retangular (8 x 16 cm), ao qual é acrescido 1 L de
água desclorada e acima do nível d’água é construído um suporte para o cartão adesivo. A
posição do cartão adesivo na armadilha P1T1, localizado acima da superfície líquida,
impede a deposição de jangadas na superfície líquida, evitando que a armadilha se
transforme em um criadouro. O material adesivo empregado nas armadilhas possibilita a
fácil retirada e identificação dos mosquitos capturados, além de não danificá-los,
possibilitando que estes sejam dissecados para a determinação de seus estágios fisiológicos.
Os atraentes de oviposição sintéticos (AOS) em polímeros (mangueira de PVC e tubos de
polietileno) apresentam atratividade para fêmeas de C. quinquefasciatus e equilíbrio nas
taxas de evaporação, podendo ser utilizados continuamente em campo ou laboratório por
um período médio de 60 dias. A seletividade da armadilha P1T1 contendo liberador com
escatol a fêmeas grávidas e ingurgitadas de C. quinquefasciatus (quando comparada à
armadilha iscada com outros atraentes) é útil em trabalhos de campo para amostragens
visando à observação de índices de infectividade das fêmeas de parasitos que utilizam esta
espécie de mosquito como vetor (LIMA, 2005).
Visando aumentar o potencial de captura da armadilha P1T1, o escatol pode ser
utilizado em combinações binárias (quando utilizado em combinação com outro AOS),
terciárias (quando o escatol for utilizado combinado a outros dois AOS) e quaternárias
(resultante da combinação entre escatol e os três atraentes restantes), podendo ser utilizados
os atraentes p-cresol, nonanal e indol, cada qual em seu liberador (de polietileno, para
atraentes líquidos ou de silicone, para atraentes sólidos), acrescidos à armadilha P1T1, de

43
acordo com a combinação a ser utilizada. A utilização de diferentes tipos de liberadores
para atraentes sólidos e líquidos deve-se à diferença da porosidade entre os dois materiais
utilizados, o que torna o tubo de polietileno ideal para ser utilizado como liberador de
atraentes líquidos e os tubos de PVC para atraentes sólidos.
Atualmente, a única armadilha existente específica para a captura de fêmeas
grávidas de C. quinquefasciatus é a Reiter’s Gravid Trap, que utiliza baterias, o que
dificulta sua operacionalidade. Além disso, é utilizado nesta armadilha, como atraente de
oviposição, a infusão de gramínea, cujos inconvenientes no preparo e utilização foram
descritos anteriormente.
Em testes de comparação entre as armadilhas P1T1 iscadas com escatol e CDC,
realizados por Lima (2005), a primeira se mostrou específica para fêmeas grávidas e
ingurgitadas de sangue de C. quinquefasciatus, visto que apresentou um percentual de
captura maior de tal categoria, em comparação à CDC. Dentre os adultos de C.
quinquefasciatus capturados durante o experimento, as fêmeas grávidas e ingurgitadas
representaram 56,2% do total capturado pela P1T1 e somente 16,5% dos adultos capturados
pela CDC.
A P1T1, desenvolvida na UFMG, tem como vantagens não utilizar baterias,
substituída na armadilha pela utilização de cartão adesivo responsável pela captura de
insetos, ser altamente específica e poder ser utilizada com liberadores contendo atraentes
sintéticos de oviposição. Porém, seu potencial de captura deve ser aumentado, para que
torne possível a sua utilização em programas de controle de C. quinquefasciatus (LIMA,
2005).

44
3. OBJETIVOS
3.1 Geral:
Desenvolver e avaliar novos protótipos e formulações de atraentes sintéticos de oviposição
para fêmeas grávidas de Culex quinquefasciatus.
3.2 Específicos:
1- Elaborar diferentes protótipos de armadilhas de oviposição;
2- Avaliar a dispersão da pluma de odor nos protótipos de armadilhas desenvolvidos em
comparação à armadilha P1T1;
3- Avaliar em campo a captura de fêmeas grávidas de C. quinquefasciatus pelos protótipos
de armadilhas desenvolvidos em comparação à armadilha P1T1;
4- Testar diferentes combinações binárias, terciárias e quaternária de atraentes sintéticos
de oviposição para fêmeas grávidas de C. quinquefasciatus em campo;
5- Avaliar a especificidade do melhor protótipo de armadilha desenvolvido e da melhor
combinação de atraentes de oviposição sintéticos na captura de fêmeas grávidas de C.

45
quinquefasciatus, em comparação à MosquiTRAP®, utilizando AtrAedes® (atraente
utilizado para captura de fêmeas grávidas de Aedes aegypti) ou a melhor combinação
obtida para fêmeas grávidas de C. quinquefasciatus em campo.

46
4. MATERIAL E MÉTODOS
4.1 Desenvolvimento e avaliação de novos protótipos de armadilhas para captura de
fêmeas grávidas de Culex quinquefasciatus
4.1.1 Protótipos de armadilhas avaliados
As armadilhas para captura de fêmeas grávidas de C. quinquefasciatus foram
desenvolvidas baseadas em observações de comportamento de pré-oviposição e oviposição
(LIMA, 2005).
Um total de cinco protótipos de armadilhas para captura de fêmeas grávidas de C.
quinquefasciatus foi avaliado em campo (FIG. 2). Todos os protótipos de armadilhas
desenvolvidos foram construídos usando como base um balde plástico com tampa (com
cerca de 10 L de capacidade, diâmetros de 26 cm na base e 30 cm na abertura, e 39 cm de
altura) de cor preto fosco (FIG. 1 e FIG. 2). Quatro dos protótipos desenvolvidos se
diferenciam quanto ao número e posicionamento de aberturas circulares (oito cm de
diâmetro cada abertura), cada qual com um funil interno (8 cm de comprimento e 3 cm de
diâmetro na abertura final) de tela de malha fina de plástico preto (0,5 mm). O funil foi
acrescido às aberturas com o objetivo de evitar o escape de mosquitos após sua entrada na
armadilha, evitando que houvesse diferença entre o número de insetos que entrassem nos
protótipos e o número de insetos capturados pelos cartões adesivos posicionados no interior
das armadilhas. Os códigos e a caracterização dos quatro protótipos são listados a seguir:

47
Balde A= balde preto com uma abertura na tampa superior;
Balde B = balde preto com quatro aberturas na tampa superior;
Balde C = balde preto com uma abertura na tampa e quatro aberturas laterais;
Balde D = balde preto com quatro aberturas na tampa e quatro aberturas laterais.
FIGURA 1 - Protótipos de armadilhas desenvolvidos tendo como base um balde: (a) balde A, (b)
balde B, (c) balde C e (d) balde D. Ver texto para detalhes.
a) b)
c) d)
10 cm

48
A armadilha P1T1 (LIMA, 2005) consiste em um recipiente retangular de
polietileno de cor preto fosco (20 x 30 x 9 cm) com tampa com abertura retangular (8 x 16
cm) (FIG. 2).
FIGURA 2 - Protótipos de armadilhas testados. Da esquerda para direita: balde A, balde D, balde C
e balde B. Ao centro, P1T1.
Todos protótipos receberam 1 L de água desclorada, acima da qual se situava um
suporte interno construído com palito abaixador de língua (10 X 1,5 cm) (FIG. 3a), fixado
às paredes da armadilha com o auxílio de cola quente. Um cartão adesivo inodoro de cor
preta, circular, com diâmetro de cerca de 25 cm foi colocado sobre o suporte interno para as
armadilhas construídas tendo como base o balde. Para a armadilha P1T1 foi usado o mesmo
cartão adesivo, porém retangular (20 X 30 cm). Os cartões foram produzidos a partir de
aparas de cartão emendadas e recortadas no formato desejado (FIG. 3b). O cartão continha
furos com tamanho aproximado de 0,5 cm, feitos com o auxílio de chave de fenda com
ponta fina e martelo (FIG. 3c), distribuídos ao longo do cartão aleatoriamente, para permitir
a evaporação da água localizada abaixo do cartão (FIG. 4).

49
FIGURA 3 - (a) Confecção dos furos nos cartões adesivos, com o auxílio de chave de fenda e
martelo; (b) suporte sobre o qual se apóia o cartão adesivo e (c) cartão adesivo da armadilha P1T1,
confeccionado a partir de aparas de cartão emendadas.
FIGURA 4 – Esquema da armadilha P1T1 mostrando a evaporação da água localizada abaixo do
cartão, representada pelas setas, através de perfurações no cartão.
4.1.2 Avaliação da dispersão de odores nos diferentes protótipos de armadilha
desenvolvidos
As armadilhas foram testadas em campo, na área externa do ICB da UFMG. Cada
protótipo foi testado isoladamente, em momentos diferenciados, a fim de permitir o registro
a) b) c)
Cartão furado
Água desclorada

50
completo dos dados da dispersão da fumaça liberada em cada protótipo. A dispersão da
fumaça por cada um dos protótipos de armadilhas avaliados foi observada nas mesmas
condições de vento, sendo observada a pluma de odor em situações de calmaria e de
correntes de vento para todos os protótipos.
No centro da base interna de cada protótipo de armadilha foi posicionada uma placa
de Petri contendo dois chumaços de algodão, um embebido com HCl (ácido clorídrico) e o
outro com NH4OH (hidróxido de amônio). A reação do ácido clorídrico (HCl) com o
hidróxido de amônio (NH4OH) produz cloreto de amônio (NH4Cl), o qual se apresenta na
forma de uma fumaça branca, facilmente visualizada em um contraste escuro, composto por
um pano preto posicionado abaixo da armadilha (chão) e atrás da mesma, em uma parede
ao seu fundo.
A fumaça liberada por cada armadilha foi filmada e fotografada com o auxílio de
uma máquina fotográfica digital em ambiente fechado (sem vento), dentro de um aquário
de acrílico sem tampa, cúbico, com arestas de 1 m, e em ambiente aberto (com vento), no
exterior do prédio do ICB. Para a comparação da dispersão da fumaça em cada protótipo de
armadilha foram utilizados os recursos fotográficos, através da análise de cada fotografia
obtida e da visualização quadro a quadro dos vídeos produzidos. Também foram
desenhados os contornos da fumaça em uma folha de acetato fixada ao monitor do
microcomputador. Baseado nestes dados seguiu-se a análise da dispersão de odores em
cada protótipo testado.
A velocidade do vento foi medida com o auxílio de uma trena e um cronômetro. A
trena foi posicionada aberta até a marca de 1 m, acima de um fundo escuro (obtido através
de um pano preto). Ao lado do ponto zero da trena estava uma placa de Petri fechada
contendo dois chumaços de algodão, um embebido com HCl (ácido clorídrico) e o outro

51
com NH4OH (hidróxido de amônio). No momento da abertura da placa de Petri, o
cronômetro tinha o seu contador de tempo disparado, o qual era zerado quando a fumaça
chegasse à marca de 1 m da trena. Tal procedimento nos permitiu calcular a velocidade do
vento (obtida através da distância percorrida por unidade de tempo) Foram realizadas dez
repetições consecutivas da medição e a velocidade do vento foi obtida através da média dos
resultados obtidos.
Os dados de temperatura e umidade relativa do ar foram obtidos com o auxílio de
um aparelho termo-higrômetro.
4.1.3 Atraente Sintético de Oviposição (ASO)
Em capela de exaustão, cerca de 200mg de 3-metil-indol (escatol) (Acros, 99% de
pureza) foram acondicionados em mangueiras de PVC (5 x 4,5 cm) (FIG. 5a). Com o
auxílio de uma seladora (Everest®, modelo X), os liberadores foram lacrados e o atraente
pôde ser utilizado para os testes em campo no período entre 11 e 80 dias após ter sido
preparado o liberador contendo o atraente (LIMA, 2005). Os liberadores contendo os
atraentes foram posicionados em um clipe médio fixo em pé, com o auxílio de cola quente,
sempre ao centro do cartão adesivo de cada protótipo de armadilha (FIG. 5b e 5c), de modo
que todas as superfícies do liberador estivessem livres.

52
FIGURA 5 – (a) Mangueira de PVC (4,5 cm de largura por 5cm de comprimento), utilizada como
liberador do escatol; (b) suporte para o liberador contendo o atraente, confeccionado com um clipe
médio fixado ao centro do cartão adesivo de cada protótipo de armadilha: vista lateral e (c) vista
superior.
4.1.4 Paridade das fêmeas coletadas em campo
Todas as fêmeas de C. quinquefasciatus coletadas nas armadilhas foram dissecadas
para a observação de sua paridade, baseando-se na técnica de DETINOVA (1962), na qual
se disseca os ovários e ovaríolos. Tal técnica permite a classificação das fêmeas de
mosquitos, sendo nulíparas aquelas que apresentam as extremidades traqueolares
enoveladas (FIG. 6a) e oníparas as fêmeas com as mesmas traquéolas distendidas (FIG. 6b)
(CONSOLI e OLIVEIRA, 1994).
a) b) c)

53
FIGURA 6 – Esquema de ovários de culicídeos mostrando as características observadas no estudo
da paridade: (a) ovário de fêmea nulípara e (b) ovário de fêmea onípara. Fonte: FORATTINI, 2002,
p. 121.
As fêmeas que se apresentaram ingurgitadas de sangue foram classificadas como
recém alimentadas (RA) e as que possuíam ovos como grávidas (GR). Dentre as grávidas,
estágios de desenvolvimento também foram classificados como: fêmeas com ovos e sangue
na porção superior do intestino; fêmeas com ovos e sangue na parte inferior do intestino e
fêmeas com ovos completamente desenvolvidos. Quando o estágio fisiológico não pôde ser
observado, a fêmea foi classificada como de estágio indeterminado.
a) b)

54
4.1.5 Área Experimental
4.1.5.1 Área 1: Regional Venda Nova
As armadilhas foram testadas em campo, inicialmente no Clube Lareira, situado à
rua Visconde de Taunay, bairro São João Batista, regional Venda Nova de Belo Horizonte
(MG). A área apresenta reclamações constantes dos moradores relatadas aos agentes de
saúde da região, devido ao incômodo causado pela hematofagia noturna por pernilongos.
No interior do Clube Lareira, existe um represamento de água rica em matéria orgânica, o
qual poderia constituir um criadouro de C. quinquefasciatus.
4.1.5.1.1 Avaliação da presença de Culex quinquefasciatus no Clube Lareira e em suas
proximidades
Três armadilhas CDC foram instaladas em três áreas distintas, uma no vestiário dos
funcionários do clube, outra na varanda de uma residência situada a um quarteirão do clube
e outra numa área coberta existente nos fundos de outra residência, esta situada a dois
quarteirões do clube. As armadilhas foram instaladas no final da tarde de 15/08/2006 e
retiradas na manhã seguinte, na qual se procedeu a leitura das mesmas. Os exemplares de
C. quinquefasciatus capturados foram colocados em potes de acrílico, com etiqueta
identificadora do local de instalação da armadilha e foram levados ao laboratório para
identificação.

55
4.1.5.2 Área 2: Regional Norte
No bairro Jaqueline, situado na regional Norte de BH, foram encontrados criadouros
de C. quinquefasciatus, onde pôde-se observar facilmente abundantes formas imaturas,
sendo estes criadouros a céu aberto constituídos por três represamentos resultantes do
escape de conteúdos da rede de esgoto da Copasa (FIG. 7a, 7b, 7c e 7d), formados a partir
de Maio de 2006, e que continuam sem solução até o presente momento.
Através de conversas com os moradores e trabalhadores, bem como com os agentes
de saúde da região, foi constatado o incômodo noturno causado pela hematofagia. Em
visitas a dois endereços na região (FIG 7e e 7f) também pôde-se visualizar a presença de
insetos adultos em locais escuros e abrigados (como em cabines de caminhões, caixas de
madeira utilizadas para guardar ferramentas, dentre outros).
FIGURA 7 - Área experimental situada no bairro Jaqueline: (a, b, c e d) possíveis criadouros de
Culex quinquefasciatus, constituídos por represamentos de escape da rede de esgoto e áreas de
instalação das armadilhas: (e) Oficina do Nem e (f) Ilumef.
a) b) c)
f) e) d)

56
4.1.6 Experimento 1: Avaliação dos protótipos de armadilhas para captura de fêmeas
grávidas de Culex quinquefasciatus
No Clube, foram selecionados três locais (1, 2 e 3), 100 m eqüidistantes. Em cada
área no clube Lareira, foi definido cinco posições (A, B, C, D e E), nas quais foram
colocadas as armadilhas a serem testadas para comparação. O experimento contou com
cinco tratamentos a serem testados, cada qual representado por um dos protótipos de
armadilhas anteriormente descritos (P1T1, balde A, balde B, balde C e balde D). Todas as
armadilhas possuíram como atraente sintético 3-metil-indol (escatol) acondicionado em
mangueira de PVC. Houve, diariamente, troca de posição das armadilhas entre as cinco
posições escolhidas, dentro de cada uma das três regiões (QUADRO 2). Para disposição
das armadilhas em campo foi utilizado o quadrado latino (5x5), e a normalidade dos dados
de captura em campo foi testada através do teste de Kolmogorov Smirnov (Z). O
experimento foi realizado de 7 a 12/08/2006.
QUADRO 2: Disposição das armadilhas em campo, nas posições definidas dentro de cada uma das
três regiões (1, 2 e 3), durante a realização do experimento de avaliação de protótipos de armadilhas
para captura de fêmeas grávidas de Culex quinquefasciatus.
Dia de leitura Posição
1 2 3 4 5
A P1T1 Balde A Balde B Balde C Balde D
B Balde A Balde B Balde C Balde D P1T1
C Balde B Balde C Balde D P1T1 Balde A
D Balde C Balde D P1T1 Balde A Balde B
E Balde D P1T1 Balde A Balde B Balde C

57
No bairro Jaqueline, foram escolhidas três regiões (1, 2 e 3), distantes umas das
outras no mínimo 100 m, uma situada em uma oficina mecânica de automóveis pesados
(Oficina do Nem) (região 1) (FIG. 6e) e as outras duas (regiões 2 e 3) situadas em uma
fábrica de postes de alumínio (Ilumef) (FIG. 6f). Em cada região, foram definidas cinco
posições (A, B, C, D e E), nas quais foram colocadas as armadilhas a serem testadas para
comparação. O delineamento experimental (QUADRO 2), os tratamentos e os
procedimentos de leitura foram semelhantes aos utilizados no clube Lareira.
A vistoria das armadilhas foi realizada a cada 24 horas e o número de C.
quinquefasciatus foi contabilizado, incluindo machos e fêmeas. As fêmeas capturadas
tiveram determinados seus estágios fisiológicos, através de dissecção de seus ovários
(DETINOVA, 1962). Além disso, outros Artrópodes, quando capturados, também foram
contados e identificados, ao nível de Ordem. Durante a vistoria das armadilhas, todos os
mosquitos capturados no interior de cada protótipo, foram retirados com auxílio de uma
pinça e colocados em tubo de plástico com etiqueta identificadora do tipo de armadilha,
posição da mesma, dia de leitura e número de espécimes coletados.
O protótipo de armadilha que capturou o maior número de C. quinquefasciatus,
principalmente fêmeas grávidas, foi utilizado nos experimentos posteriores.

58
4.2 Experimento 2: Avaliação de combinações binárias de atraentes sintéticos de
oviposição
Foram testadas, inicialmente, combinações binárias dos seguintes atraentes: 4-metil-
fenol (p-cresol) (Sigma Aldrich, 98% de pureza), nonanal (Acros, 95% de pureza), indol
(Acros, 99% de pureza) e 3-metil-indol (escatol) (Acros, 99% de pureza), sendo os dois
primeiros líquidos e os dois últimos em forma de pó. Em capela de exaustão, 200
microlitros dos atraentes líquidos foram distribuídos em tubos de polietileno com 5 cm de
comprimento e 0,5 cm de espessura (FIG. 8a), enquanto que os atraentes em forma de pó,
cerca de 200 mg, foram acondicionados em mangueiras de PVC (5 x 4,5 cm) (FIG. 8b).
Todos os liberadores foram lacrados com o auxílio de uma seladora (Everest®, modelo X).
Os liberadores contendo os atraentes foram posicionados em suportes confeccionados com
fragmentos de caixa de ovo de codorna situada no centro do cartão adesivo da armadilha
utilizada (P1T1), de modo que todas as superfícies do liberador estivessem livres (FIG. 8c e
8d).
FIGURA 8 - Liberadores: (a) tubos de polietileno, utilizados para os atraentes líquidos e (b)
mangueira de PVC, para atraentes sólidos. Suporte dos atraentes, confeccionado com fragmentos de
caixa de ovo de codorna, posicionado ao centro do cartão adesivo da armadilha P1T1: (c) vista
lateral e (d) vista superior.
a) b) c) d)

59
Todos os atraentes sintéticos foram colocados no interior da armadilha P1T1 que
continha 1 L de água desclorada abaixo do cartão adesivo. O escatol foi usado como
composto principal da P1T1 devido ao fato de ser o mais atrativo para C. quinquefasciatus
dentre os atraentes utilizados (escatol, indol, p-cresol e nonanal). Armadilhas apenas com
escatol como atraente foram os controles do experimento (QUADRO 3). Um quinto
tratamento, também utilizado como controle do experimento, consistia em armadilha do
mesmo tipo (P1T1) contendo apenas água desclorada sem nenhum liberador contendo
atraente de oviposição sintético (QUADRO 3).
QUADRO 3: Tratamentos do experimento de avaliação de combinações binárias de
atraentes de oviposição para captura de fêmeas grávidas de Culex quinquefasciatus,
acrescidos às armadilhas P1T1 contendo 1 L de água desclorada.
Tratamento Escatol Indol P-Cresol Nonanal
1 X - - -
2 X X - -
3 X - X -
4 X - - X
5 - - - -
As combinações binárias foram testadas em campo, na fábrica de postes de alumínio
(Ilumef) utilizada no experimento anterior, situada no bairro Jaqueline, regional Norte de
Belo Horizonte. Na área, foram escolhidas três áreas (1, 2 e 3), distantes uma da outra ao
menos 100 m. Em cada região, foram definidas cinco posições (A, B, C, D e E), nas quais
foram colocadas as armadilhas com os tratamentos a serem testados para comparação.
Houve, a cada dois dias, troca de lugar das armadilhas entre as cinco posições escolhidas,

60
dentro de cada uma das três regiões (QUADRO 4). Para disposição das armadilhas em
campo foi utilizado o quadrado latino (5x5).
QUADRO 4: Disposição dos diferentes tratamentos em campo, nas posições definidas dentro de
cada região (1, 2 e 3), durante a realização do experimento de avaliação de combinações binárias de
atraentes sintéticos de oviposição para captura de fêmeas grávidas de Culex quinquefasciatus. E:
escatol; I: indol; N: nonanal e P: p-cresol.
Dia de leitura Posição
1 2 3 4 5
A Água Escatol E + I E + N E + P
B Escatol E + I E + N E + P Água
C E + I E + N E + P Água Escatol
D E + N E + P Água Escatol E + I
E E + P Água Escatol E + I E + N
A vistoria a cada dois dias das armadilhas e a dissecação de fêmeas foi realizada
segundo a mesma metodologia do experimento anterior.
O melhor tratamento dentre os testados foi utilizado nos experimentos de
combinações terciárias e quaternárias.

61
4.3 Experimento 3: Avaliação de combinações terciárias e quaternária de atraentes
sintéticos de oviposição
Como não foi obtida uma combinação binária com melhor captura (experimento 2),
foram testadas todas as combinações terciárias possíveis entre os atraentes, sendo o
controle escatol utilizado isoladamente (QUADRO 5). A combinação quaternária também
foi testada (QUADRO 5), além da armadilha contendo apenas água.
QUADRO 5: Tratamentos do experimento de avaliação de combinações terciárias e quaternária de
atraentes sintéticos de oviposição para captura de fêmeas grávidas de Culex quinquefasciatus,
acrescidos às armadilhas P1T1 contendo 1 L de água desclorada.
Tratamento Escatol Indol P-cresol Nonanal
1 X - - -
2 X X X -
3 X X - X
4 X - X X
5 X X X X
6 - - - -
Houve, a cada dois dias, troca de lugar das armadilhas entre as seis posições
escolhidas (A, B, C, D, E e F), dentro de cada uma das duas regiões (1 e 2) (QUADRO 6).
Para disposição das armadilhas em campo foi utilizado o quadrado latino (6x6), e a
normalidade dos dados de captura em campo também foi testada através do teste de
Kolmogorov Smirnov (Z).

62
QUADRO 6: Disposição dos diferentes tratamentos em campo, nas posições definidas dentro de
cada região (1 e 2), durante a realização do experimento de avaliação de combinações terciárias e
quaternárias de atraentes sintéticos de oviposição para fêmeas grávidas de Culex quinquefasciatus.
E: escatol; I: indol; N: nonanal e P: p-cresol.
Dia de leitura Posição
1 2 3 4 5
A Água Escatol E + I + N E + I + P E + N + P
B Escatol E + I + N E + I + P E + N + P E + I + N + P
C E + I + N E + I + P E + N + P E + I + N + P Água
D E + I + P E + N + P E + I + N + P Água Escatol
E E + N + P E + I + N + P Água Escatol E + I +N
F E + I + N + P Água Escatol E + I +N E + I + P
A vistoria foi realizada a cada dois dias das armadilhas e a dissecação de fêmeas foi
realizada segundo a mesma metodologia dos experimentos anteriores.
A melhor formulação foi utilizada nos experimentos de comparação do melhor
protótipo com outra armadilha para fêmeas grávidas.

63
4.4 Experimento 4: Comparação do melhor protótipo de armadilha para captura de
fêmeas grávidas de Culex quinquefasciatus com MosquiTRAP
O melhor protótipo de armadilha (P1T1) e o melhor tratamento dos testes de
combinação de atraentes de oviposição sintéticos (escatol utilizado isoladamente) foram
avaliados quanto à seletividade de captura de fêmeas grávidas de C. quinquefasciatus em
comparação com a armadilha MosquiTRAP® (FIG. 9).
FIGURA 9 – Armadilha para captura de adultos de Aedes aegypti (MosquiTRAP) utilizada no
experimento de comparação entre o melhor protótipo de armadilha para captura de fêmeas grávidas
de Culex quinquefasciatus com MosquiTRAP em campo. (a) Parte inferior contendo tela de
proteção; (b) parte superior (tampa); (c) AtrAedes (atraente de oviposição sintético) e (d) cartão
adesivo.
O experimento contou com os seguintes tratamentos:
Tratamento 1: P1T1 + Escatol.
Tratamento 2: MosquiTRAP + Escatol.
Tratamento 3: MosquiTRAP + AtrAedes.
Fonte: Álvaro Eiras
Fonte: Rosemary Roque

64
À cada armadilha MosquiTRAP® foram acrescidos cerca de 275 mL de água
desclorada e o atraente AtrAedes® foi posicionado no cartão adesivo da armadilha, fixado
com o auxílio de um percevejo.
As armadilhas foram testadas em campo, inicialmente na mesma área da região
metropolitana de Belo Horizonte utilizada nos experimentos anteriores (bairro Jaqueline).
Foram escolhidas três áreas (1, 2 e 3), distantes no mínimo 100 m uma da outra, cada uma
representando uma repetição do experimento. Em cada região, foram definidas três
posições (A, B e C), nas quais foram colocadas as armadilhas a serem testadas para
comparação. Houve, a cada quatro dias, troca de posição das armadilhas entre as três
posições escolhidas, dentro de cada uma das três áreas (QUADRO 7). Para disposição das
armadilhas em campo foi utilizado o quadrado latino (3X3).
QUADRO 7: Disposição dos diferentes tratamentos em campo, nas posições definidas dentro de
cada região (1, 2 e 3), durante a realização do experimento de comparação do melhor protótipo de
armadilha para captura de fêmeas grávidas de Culex quinquefasciatus com MosquiTRAP.
Dia de leitura Posição
1 2 3
A P1T1 + Escatol MosquiTRAP + Escatol MosquiTRAP + AtrAedes
B MosquiTRAP + Escatol MosquiTRAP + AtrAedes P1T1 + Escatol
C MosquiTRAP + AtrAedes P1T1 + Escatol MosquiTRAP + Escatol
A vistoria a cada quatro dias das armadilhas e a dissecação de fêmeas foi realizada
segundo a mesma metodologia dos experimentos anteriores.
Em um segundo momento, foram realizadas outras cinco repetições do experimento
no ICB da UFMG. Na área, foram escolhidas cinco regiões (1, 2, 3, 4 e 5), distantes no
mínimo 100 m uma da outra. Em cada região, foram definidas três posições (A, B e C), nas

65
quais foram colocadas as armadilhas a serem testadas para comparação. Houve, a cada
quatro dias, troca de posição das armadilhas entre as três posições escolhidas, dentro de
cada uma das cinco áreas (QUADRO 7).
A vistoria a cada quatro dias das armadilhas e a dissecação de fêmeas foi realizada
segundo a mesma metodologia dos experimentos anteriores.
4.5 Análises estatísticas
Para a análise estatística da quantidade de machos, fêmeas e fêmeas grávidas de C.
quinquefasciatus coletadas em campo foi feito o teste de normalidade Kolmogorov
Smirnov (Z). Como os dados não apresentaram distribuição normal (p< 0,05, Kolmogorov
Smirnov), a comparação entre os tratamentos avaliados em cada experimento foi feita com
o teste de Kruskal-Wallis. Diferenças significativas entre as médias foram comparadas pelo
teste de Mann-Whitney em nível de 5%.

66
5. RESULTADOS
5.1 Desenvolvimento e avaliação de novos protótipos de armadilhas para captura de
fêmeas grávidas de Culex quinquefasciatus
5.1.1 Avaliação da dispersão de odores nos diferentes protótipos de armadilha
desenvolvidos
Durante a realização do experimento, a temperatura oscilou entre 26° e 29° C
(mínima e máxima), a umidade relativa do ar permaneceu em torno de 67% e a velocidade
média do vento obtida foi de 0,23 ± 0,06 m/s.
A fumaça liberada pela armadilha P1T1 se mostrou mais concentrada próximo à
mesma do que a liberada pelos protótipos que utilizam como base o balde, e sua dispersão
na área adjacente também se mostrou mais constante, o que pôde ser observado através de
uma pluma mais concentrada e mais facilmente visualizada (FIG. 10). A fumaça liberada
pela P1T1 demorava mais tempo para se dispersar na corrente de vento, sofrendo menor
influência das condições climáticas e atingindo, assim, maior área em uma concentração
maior.

67
FIGURA 10 – Dispersão da pluma de odor produzida por cloreto de amônio na armadilha P1T1.
Dentre os protótipos em forma de balde, pôde-se notar que os baldes C e D (FIG.
11), por possuírem aberturas laterais, permitiram uma circulação do ar no interior das
armadilhas, o que fez com que a fumaça liberada se dispersasse em várias direções.
Entretanto, nos mesmos protótipos, a fumaça foi liberada de uma maneira dispersa, pouco
concentrada. Ainda, dependendo da direção do vento, freqüentemente a pluma de odor foi
interrompida, o que pode dificultar a chegada do mosquito à fonte de odor.

68
FIGURA 11 – Dispersão da pluma de odor produzida por cloreto de amônio no (a) balde C e no (b)
balde D.
Nos baldes A e B (FIG. 12), a pluma de odor formada era liberada mais
continuamente do que nos baldes C e D, sendo mais concentrada no balde A (com abertura
única) e mais dispersa no balde B (com quatro aberturas), porém em situações de calmaria,
freqüentemente, a liberação da pluma de odor se interrompia, sendo necessário o retorno de
uma corrente de ar mais forte para que a pluma de odor voltasse a ser liberada, o que
acontecia especialmente no balde A.
a) b)

69
FIGURA 12 – Pluma de odor produzida por cloreto de amônio liberada pelos (a) baldes A e (b) B.
Entretanto, devem ser considerados testes de captura de insetos para avaliar, além da
dispersão de odores, a influência das diferenças do número de aberturas e de suas
localizações no acesso ao interior da armadilha. A entrada dos mosquitos ocorre nas
aberturas dos protótipos, bem como a saída dos mesmos, a qual resulta na captura do inseto
ou não pelos cartões adesivos situados no interior da armadilha.
a) b)

70
5.1.2 Experimento 1: Comparação entre os diferentes protótipos desenvolvidos de
armadilhas para captura de fêmeas grávidas de Culex quinquefasciatus
5.1.2.1 Área 1: Regional Venda Nova
Durante a realização do experimento de avaliação de protótipos de armadilhas para
captura de fêmeas grávidas de Culex quinquefasciatus, nenhum mosquito foi capturado
pelos tratamentos avaliados. Na investigação da área e suas adjacências, através da
utilização da armadilha CDC, apenas uma fêmea de C. quinquefasciatus foi capturada a um
quarteirão do local dos testes (Clube Lareira), portanto, a área foi descartada.
5.1.2.2 Área 2: Regional Norte
O total de 225 C. quinquefasciatus foi capturado nas armadilhas, destes 145 machos
e 80 fêmeas, durante 5 dias de experimento. Das 15 observações (5 armadilhas x 3
monitoramentos) realizadas durante o período, o balde A obteve captura média de machos
significativamente diferente dos baldes C e D (Mann Whitney, p= 0,003 e p= 0,001,
respectivamente), tendo apresentado a menor captura média dentre todas as armadilhas. A
média de captura de machos da P1T1 não foi significativamente maior que a do balde D
(Mann Whitney, p= 0,054), porém a P1T1 capturou o triplo de machos que o balde D.
Foram capturados pela armadilha P1T1 79 machos, perfazendo uma média de 5,3 ± 2,35
machos/armadilha/noite, que foi significativamente maior do que a observada com os
baldes A, B e C (Mann Whitney, p= 0,000, p=0,005 e p= 0,04, respectivamente). Entre as
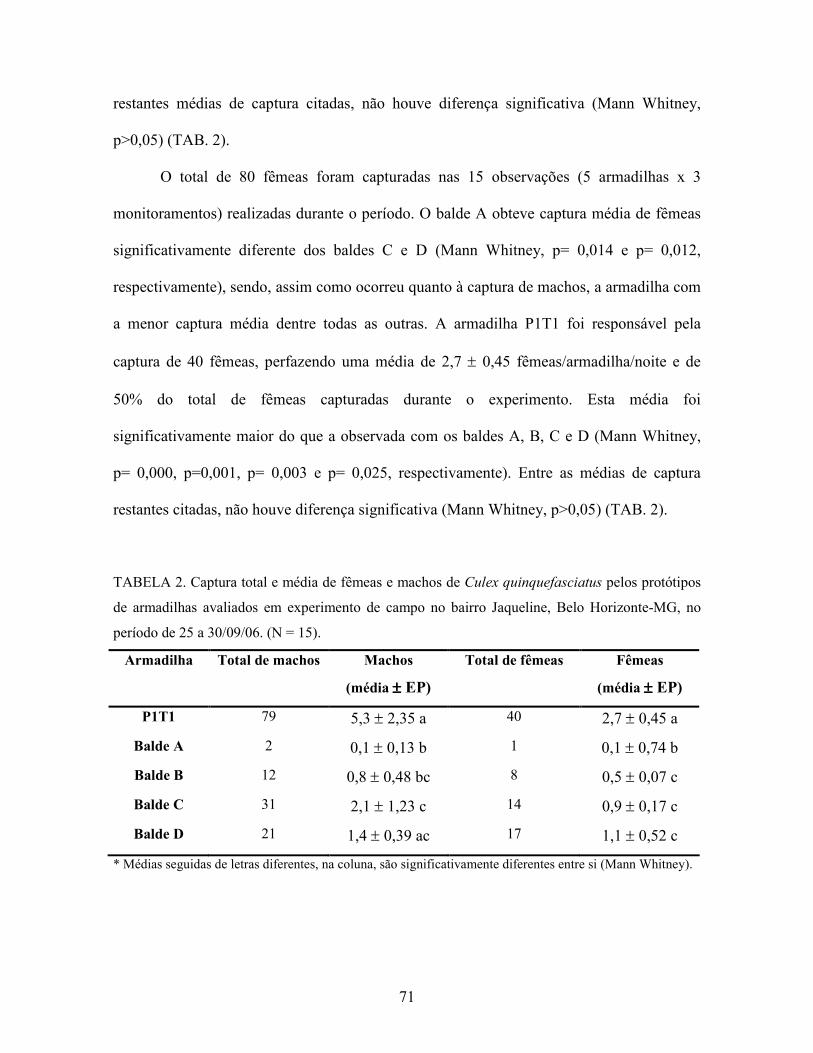
71
restantes médias de captura citadas, não houve diferença significativa (Mann Whitney,
p>0,05) (TAB. 2).
O total de 80 fêmeas foram capturadas nas 15 observações (5 armadilhas x 3
monitoramentos) realizadas durante o período. O balde A obteve captura média de fêmeas
significativamente diferente dos baldes C e D (Mann Whitney, p= 0,014 e p= 0,012,
respectivamente), sendo, assim como ocorreu quanto à captura de machos, a armadilha com
a menor captura média dentre todas as outras. A armadilha P1T1 foi responsável pela
captura de 40 fêmeas, perfazendo uma média de 2,7 ± 0,45 fêmeas/armadilha/noite e de
50% do total de fêmeas capturadas durante o experimento. Esta média foi
significativamente maior do que a observada com os baldes A, B, C e D (Mann Whitney,
p= 0,000, p=0,001, p= 0,003 e p= 0,025, respectivamente). Entre as médias de captura
restantes citadas, não houve diferença significativa (Mann Whitney, p>0,05) (TAB. 2).
TABELA 2. Captura total e média de fêmeas e machos de Culex quinquefasciatus pelos protótipos
de armadilhas avaliados em experimento de campo no bairro Jaqueline, Belo Horizonte-MG, no
período de 25 a 30/09/06. (N = 15).
Armadilha Total de machos Machos
(média ±±±± EP)
Total de fêmeas Fêmeas
(média ±±±± EP)
P1T1 79 5,3 ± 2,35 a 40 2,7 ± 0,45 a
Balde A 2 0,1 ± 0,13 b 1 0,1 ± 0,74 b
Balde B 12 0,8 ± 0,48 bc 8 0,5 ± 0,07 c
Balde C 31 2,1 ± 1,23 c 14 0,9 ± 0,17 c
Balde D 21 1,4 ± 0,39 ac 17 1,1 ± 0,52 c
* Médias seguidas de letras diferentes, na coluna, são significativamente diferentes entre si (Mann Whitney).
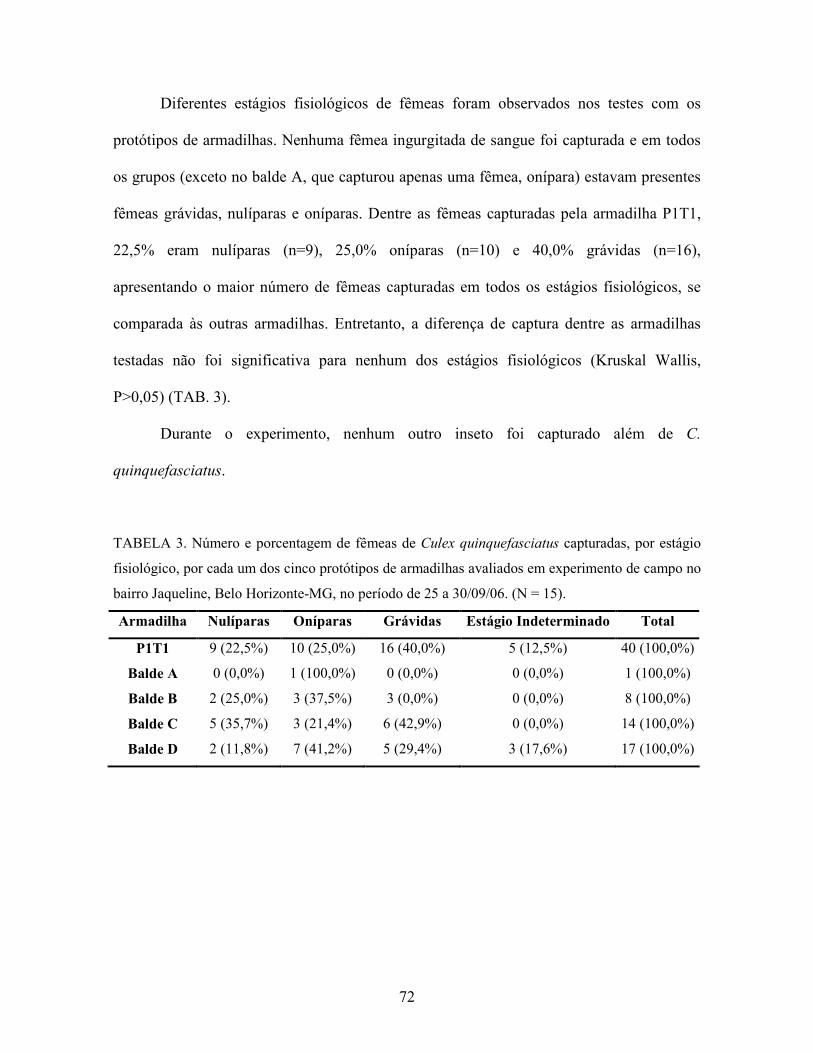
72
Diferentes estágios fisiológicos de fêmeas foram observados nos testes com os
protótipos de armadilhas. Nenhuma fêmea ingurgitada de sangue foi capturada e em todos
os grupos (exceto no balde A, que capturou apenas uma fêmea, onípara) estavam presentes
fêmeas grávidas, nulíparas e oníparas. Dentre as fêmeas capturadas pela armadilha P1T1,
22,5% eram nulíparas (n=9), 25,0% oníparas (n=10) e 40,0% grávidas (n=16),
apresentando o maior número de fêmeas capturadas em todos os estágios fisiológicos, se
comparada às outras armadilhas. Entretanto, a diferença de captura dentre as armadilhas
testadas não foi significativa para nenhum dos estágios fisiológicos (Kruskal Wallis,
P>0,05) (TAB. 3).
Durante o experimento, nenhum outro inseto foi capturado além de C.
quinquefasciatus.
TABELA 3. Número e porcentagem de fêmeas de Culex quinquefasciatus capturadas, por estágio
fisiológico, por cada um dos cinco protótipos de armadilhas avaliados em experimento de campo no
bairro Jaqueline, Belo Horizonte-MG, no período de 25 a 30/09/06. (N = 15).
Armadilha Nulíparas Oníparas Grávidas Estágio Indeterminado Total
P1T1 9 (22,5%) 10 (25,0%) 16 (40,0%) 5 (12,5%) 40 (100,0%)
Balde A 0 (0,0%) 1 (100,0%) 0 (0,0%) 0 (0,0%) 1 (100,0%)
Balde B 2 (25,0%) 3 (37,5%) 3 (0,0%) 0 (0,0%) 8 (100,0%)
Balde C 5 (35,7%) 3 (21,4%) 6 (42,9%) 0 (0,0%) 14 (100,0%)
Balde D 2 (11,8%) 7 (41,2%) 5 (29,4%) 3 (17,6%) 17 (100,0%)

73
5.2 Experimento 2: Avaliação de combinações binárias de atraentes sintéticos de
oviposição
O total de 525 C. quinquefasciatus foi capturado na armadilha P1T1, contendo
diferentes combinações binárias de atraentes e os controles, durante 10 dias de
experimento. Destes, 285 machos e 240 fêmeas. Das 15 observações (5 armadilhas x 3
monitoramentos) realizadas durante o período, a armadilha contendo escatol e nonanal
obteve captura média significativamente diferente da armadilha contendo apenas água
(Mann Whitney, p= 0,008), tendo apresentado a segunda maior captura média dentre todos
os tratamentos. A armadilha contendo apenas escatol capturou 160 machos, perfazendo
uma média de 10,7 ± 3,97 machos/armadilha/leitura. Esta média foi significativamente
maior do que a observada com as armadilhas contendo apenas água, contendo escatol e
indol e contendo escatol e p-cresol (Mann Whitney, p= 0,000, p=0,012 e p= 0,006,
respectivamente). A média de captura da armadilha contendo apenas escatol só não foi
significativamente maior do que a da armadilha contendo escatol e nonanal (3,3 ± 0,84,
n=50) (Mann Whitney, p= 0,122), porém a armadilha contendo apenas escatol capturou
mais do que o triplo de machos que a armadilha contendo escatol e nonanal. Entre as
restantes médias de captura citadas, não houve diferença significativa (Mann Whitney,
p>0,05) (TAB. 4).
Dentre as 240 fêmeas capturadas nas 15 observações realizadas durante o
experimento, a armadilha contendo escatol e nonanal obteve captura média
significativamente diferente da armadilha contendo apenas água (Mann Whitney, p=
0,003), tendo apresentado a segunda maior captura média dentre todos os tratamentos. As

74
armadilhas contendo apenas escatol capturaram 127 fêmeas (53% do total de fêmeas),
perfazendo uma média de 8,5 ± 2,75 fêmeas/armadilha/leitura. Esta média foi
significativamente maior do que a observada nas armadilhas contendo apenas água,
contendo escatol e indol e contendo escatol e p-cresol (Mann Whitney, p= 0,000, p= 0,012
e p= 0,006, respectivamente). A média de captura da armadilha contendo apenas escatol só
não foi significativamente maior que a da armadilha contendo escatol e nonanal (3,1 ± 0,67,
n=46) (Mann Whitney, p= 0,122), porém a armadilha contendo apenas escatol capturou
mais do que o dobro de fêmeas que a armadilha contendo escatol e nonanal. Entre as
restantes médias de captura citadas, não houve diferença significativa (Mann Whitney,
p>0,05) (TAB. 4). A presença de diferença significativa de captura média entre os cinco
tratamentos foi semelhante para machos e fêmeas.
TABELA 4 - Captura total e média de machos e de fêmeas de Culex quinquefasciatus por cada um
dos cinco tratamentos, no experimento de avaliação de combinações binárias de atraentes sintéticos
de oviposição realizado no bairro Jaqueline, Belo Horizonte-MG, realizado no período de 03 a
13/11/06. (N = 15).
Tratamento Total de
machos
Machos
(média ±±±± EP)
Total de
fêmeas
Fêmeas
(média ±±±± EP)
Água 12 0,8 ± 0,26 b 17 1,1 ± 0,40 b
Escatol 160 10,7 ± 3,97 a 127 8,5 ± 2,75 a
Escatol + Indol 36 2,4 ± 0,74 bc 28 1,9 ± 0,47 bc
Escatol + Nonanal 50 3,3 ± 0,84 ac 46 3,1 ± 0,67 ac
Escatol + p-cresol 27 1,8 ± 0,52 bc 22 1,5 ± 0,16 bc
* Médias seguidas de letras diferentes, na coluna, são significativamente diferentes entre si (Mann Whitney).
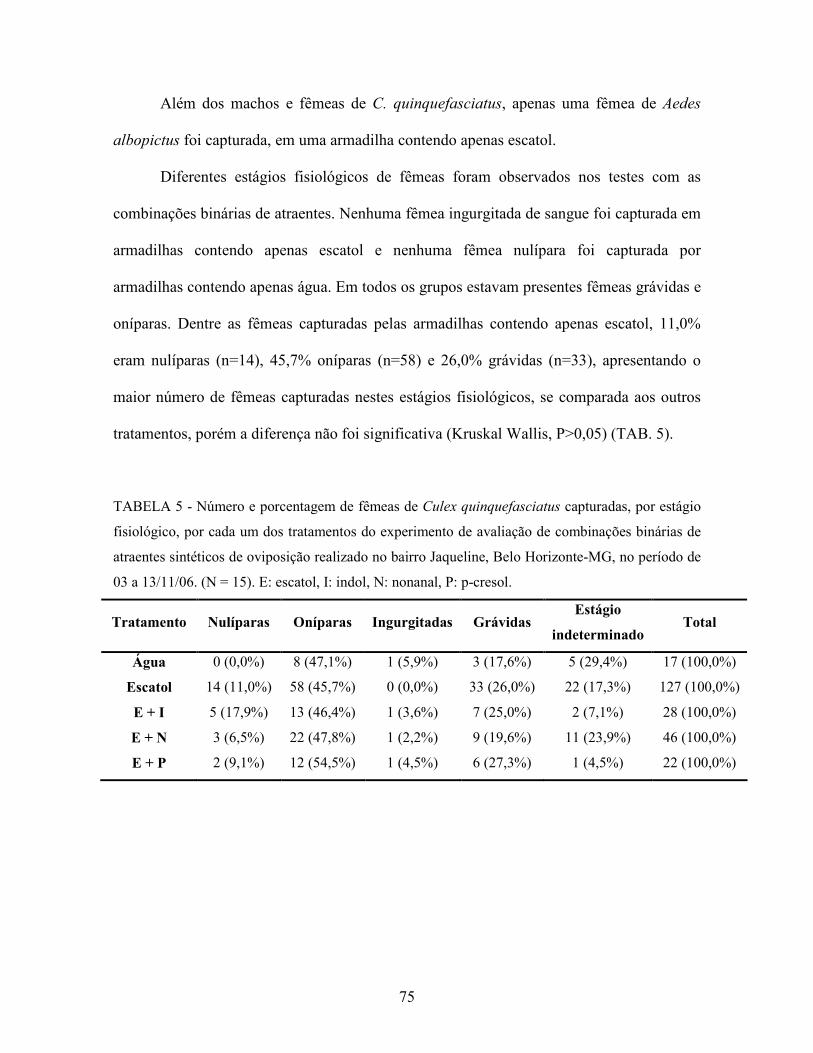
75
Além dos machos e fêmeas de C. quinquefasciatus, apenas uma fêmea de Aedes
albopictus foi capturada, em uma armadilha contendo apenas escatol.
Diferentes estágios fisiológicos de fêmeas foram observados nos testes com as
combinações binárias de atraentes. Nenhuma fêmea ingurgitada de sangue foi capturada em
armadilhas contendo apenas escatol e nenhuma fêmea nulípara foi capturada por
armadilhas contendo apenas água. Em todos os grupos estavam presentes fêmeas grávidas e
oníparas. Dentre as fêmeas capturadas pelas armadilhas contendo apenas escatol, 11,0%
eram nulíparas (n=14), 45,7% oníparas (n=58) e 26,0% grávidas (n=33), apresentando o
maior número de fêmeas capturadas nestes estágios fisiológicos, se comparada aos outros
tratamentos, porém a diferença não foi significativa (Kruskal Wallis, P>0,05) (TAB. 5).
TABELA 5 - Número e porcentagem de fêmeas de Culex quinquefasciatus capturadas, por estágio
fisiológico, por cada um dos tratamentos do experimento de avaliação de combinações binárias de
atraentes sintéticos de oviposição realizado no bairro Jaqueline, Belo Horizonte-MG, no período de
03 a 13/11/06. (N = 15). E: escatol, I: indol, N: nonanal, P: p-cresol.
Tratamento Nulíparas Oníparas Ingurgitadas Grávidas Estágio
indeterminado Total
Água 0 (0,0%) 8 (47,1%) 1 (5,9%) 3 (17,6%) 5 (29,4%) 17 (100,0%)
Escatol 14 (11,0%) 58 (45,7%) 0 (0,0%) 33 (26,0%) 22 (17,3%) 127 (100,0%)
E + I 5 (17,9%) 13 (46,4%) 1 (3,6%) 7 (25,0%) 2 (7,1%) 28 (100,0%)
E + N 3 (6,5%) 22 (47,8%) 1 (2,2%) 9 (19,6%) 11 (23,9%) 46 (100,0%)
E + P 2 (9,1%) 12 (54,5%) 1 (4,5%) 6 (27,3%) 1 (4,5%) 22 (100,0%)

76
5.3 Experimento 3: Avaliação de combinações terciárias e quaternária de atraentes
sintéticos de oviposição
Durante a realização deste experimento observou-se uma população reduzida na
área. Somente o total de apenas 21 C. quinquefasciatus foi capturado em todas armadilhas
contendo diferentes combinações terciárias e quaternária de atraentes e os controles. Destes
nove machos e 12 fêmeas, durante 12 dias de experimento (leituras feitas a cada dois dias).
Das 12 observações (6 armadilhas x 2 monitoramentos) realizadas durante o período, cinco
machos (55,6% do total) foram capturados pelas armadilhas contendo apenas escatol,
perfazendo uma média de 0,4 ± 0,26 machos/armadilha/leitura. Entre os machos capturados
pelos diferentes tratamentos, não houve diferença significativa (Kruskal Wallis, p= 0,256).
Os outros tratamentos obtiveram uma captura menor que o escatol e nas armadilhas
contendo as combinações terciárias escatol + indol + nonanal e escatol + indol + p-cresol
não foi capturado nenhum macho durante a realização do experimento (TAB. 6).
Dentre as 12 fêmeas, as armadilhas contendo as combinações terciárias escatol +
indol + nonanal e escatol + indol + p-cresol capturaram uma fêmea cada tratamento, ao
longo de experimento, e a armadilha contendo apenas água capturou duas fêmeas. As
armadilhas contendo apenas escatol capturaram oito fêmeas (66,7% do total), perfazendo
uma média de 0,7 ± 0,26 fêmeas/armadilha/leitura. Esta média foi significativamente maior
do que a observada nas armadilhas contendo a combinação terciária escatol + p-cresol +
nonanal e contendo a combinação quaternária (escatol + indol + p-cresol + nonanal) (Mann
Whitney, p= 0,015). Entre as restantes médias de captura citadas, não houve diferença
significativa (Mann Whitney, p>0,05) (TAB. 6).

77
TABELA 6 - Captura total e média de machos e de fêmeas de Culex quinquefasciatus por cada um
dos tratamentos do experimento de avaliação de combinações terciárias e quaternárias de atraentes
sintéticos de oviposição no bairro Jaqueline, Belo Horizonte-MG, no período de 19 a 31/01/07. (N =
12). E: escatol, I: indol, N: nonanal e P: p-cresol.
Tratamento Total
de machos
Machos
(média ±±±± EP)
Total de fêmeas Fêmeas
(média ±±±± EP)
Água 1 0,1 ± 0,08a 2 0,2 ± 0,17 abc
Escatol 5 0,4 ± 0,26a 8 0,7 ± 0,26 a
E + I + N 0 0,0 ± 0,00a 1 0,1 ± 0,08 abc
E + I + P 0 0,0 ±0,00a 1 0,1 ±0,08 abc
E + P + N 2 0,2 ± 0,11a 0 0,0 ± 0,00 bc
E + I + N + P 1 0,1 ± 0,08a 0 0,0 ± 0,00 c
* Médias seguidas de letras diferentes, na coluna, são significativamente diferentes entre si (Mann Whitney).
Além dos machos e fêmeas de C. quinquefasciatus, foram capturadas três fêmeas de
Aedes aegypti, destas duas foram capturadas em armadilhas contendo apenas escatol e a
outra em uma armadilha contendo a combinação quaternária dos atraentes.
Apenas dois estágios fisiológicos de fêmeas foram observados nos testes com as
combinações terciárias e quaternária de atraentes. Nenhuma fêmea ingurgitada de sangue
tampouco nulípara foi capturada pelos tratamentos avaliados. O estágio fisiológico mais
observado foi o de fêmeas grávidas, presentes em todos os tratamentos positivos para
fêmeas de C. quinquefasciatus. Porém, o estágio fisiológico que inclui as fêmeas oníparas
contou com maior número de captura (sete das 12 fêmeas capturadas eram oníparas).
Dentre as fêmeas capturadas pelas armadilhas contendo apenas escatol, 75,0% eram
oníparas (n=6) e 12,5% grávidas (n=1). As armadilhas contendo apenas escatol capturaram,
quanto às fêmeas oníparas, maior número que as demais, porém a diferença não foi
significativa (Kruskal Wallis, P>0,05) (TAB. 7).
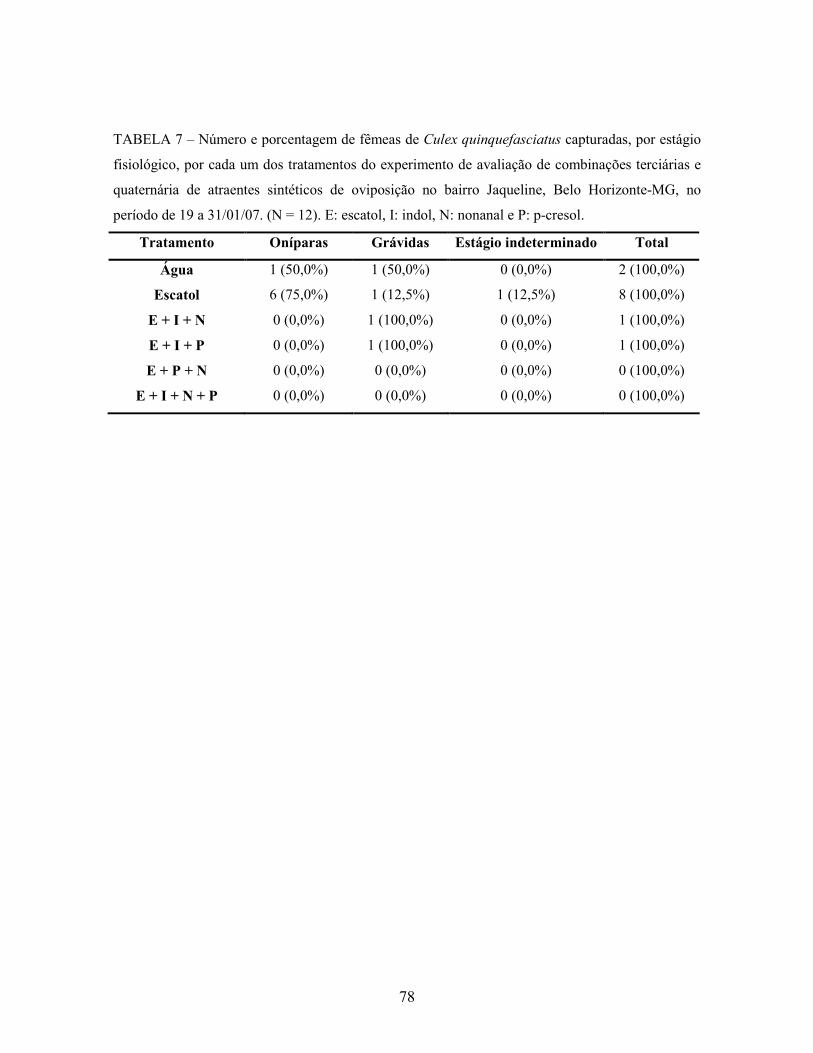
78
TABELA 7 – Número e porcentagem de fêmeas de Culex quinquefasciatus capturadas, por estágio
fisiológico, por cada um dos tratamentos do experimento de avaliação de combinações terciárias e
quaternária de atraentes sintéticos de oviposição no bairro Jaqueline, Belo Horizonte-MG, no
período de 19 a 31/01/07. (N = 12). E: escatol, I: indol, N: nonanal e P: p-cresol.
Tratamento Oníparas Grávidas Estágio indeterminado Total
Água 1 (50,0%) 1 (50,0%) 0 (0,0%) 2 (100,0%)
Escatol 6 (75,0%) 1 (12,5%) 1 (12,5%) 8 (100,0%)
E + I + N 0 (0,0%) 1 (100,0%) 0 (0,0%) 1 (100,0%)
E + I + P 0 (0,0%) 1 (100,0%) 0 (0,0%) 1 (100,0%)
E + P + N 0 (0,0%) 0 (0,0%) 0 (0,0%) 0 (100,0%)
E + I + N + P 0 (0,0%) 0 (0,0%) 0 (0,0%) 0 (100,0%)

79
5.4 Experimento 4: Comparação do melhor protótipo de armadilha para captura de
fêmeas grávidas de Culex quinquefasciatus com MosquiTRAP
Apenas 16 C. quinquefasciatus foram capturados nos tratamentos avaliados, destes
seis machos e 10 fêmeas, durante 24 dias de experimento. Das 24 observações (3
armadilhas x 8 monitoramentos) realizadas durante o período, cinco machos (83,3% do
total) foram capturados pelas armadilhas P1T1 contendo escatol, perfazendo uma média de
0,2 ± 0,10 machos/armadilha/leitura, seguida da armadilha MosquiTRAP contendo
escatol, que capturou um macho (16,7%); a armadilha MosquiTRAP contendo AtrAedes
não capturou nenhum macho de C. quinquefasciatus. Entre os machos capturados pelos
diferentes tratamentos, não houve diferença significativa (Kruskal Wallis, p= 0,062) (TAB.
8).
Dentre as 10 fêmeas capturadas nas 24 observações realizadas, as armadilhas
MosquiTRAP contendo escatol capturaram duas fêmeas, enquanto oito foram capturadas
pelas armadilhas P1T1 contendo escatol, perfazendo uma média de 0,3 ± 0,14
fêmeas/armadilha/leitura. Esta média foi significativamente maior do que a observada nas
armadilhas MosquiTRAP contendo AtrAedes (Mann Whitney, p= 0,020), sendo
responsável pela captura de 80% do total de fêmeas capturadas durante o experimento.
Entre as restantes médias de captura citadas, não houve diferença significativa (Mann
Whitney, p>0,05) (TAB. 8).

80
TABELA 8. Captura total e média de machos e de fêmeas de Culex quinquefasciatus por tratamento
do experimento de comparação entre o melhor protótipo de armadilha para captura de fêmeas
grávidas de Culex quinquefasciatus com MosquiTRAP no bairro Jaqueline e no ICB/UFMG, Belo
Horizonte-MG, nos períodos de 22 a 30/03/07 e de 16 a 24/04/07. (N = 24).
Tratamento Total de
machos
Machos
(média ±±±± EP)
Total de
fêmeas
Fêmeas
(média ±±±± EP)
P1T1 + Escatol 5 0,2 ± 0,10a 8 0,3 ± 0,14 a
MosquiTRAP + Escatol 1 0,0 ± 0,04a 2 0,1 ± 0,06 ac
MosquiTRAP + AtrAedes 0 0,0 ± 0,00a 0 0,0 ± 0,0 bc
* Médias seguidas de letras diferentes, na coluna, são significativamente diferentes entre si (Mann Whitney).
Todas as fêmeas de C. quinquefasciatus capturadas durante a realização dos
experimentos, por todos os tratamentos avaliados, eram grávidas.
Durante a realização do experimento, foram capturados 12 A. aegypti, destes apenas
um macho (capturado pela armadilha P1T1 contendo Escatol) e 11 fêmeas. Não houve
diferença significativa entre os machos capturados pelos diferentes tratamentos (Kruskal
Wallis, p= 0,368). Dentre as 11 fêmeas de A. aegypti capturadas, uma foi capturada por
uma armadilha MosquiTRAP contendo escatol, quatro em armadilhas P1T1 contendo
escatol e seis em MosquiTRAP contendo AtrAedes, porém a diferença não foi
significativa (Kruskal Wallis, p= 0,123) (TAB. 9).

81
TABELA 9 - Captura total de machos e de fêmeas de Aedes aegypti por tratamento do experimento
de comparação entre o melhor protótipo de armadilha para captura de fêmeas grávidas de Culex
quinquefasciatus com MosquiTRAP no bairro Jaqueline e no ICB/UFMG, Belo Horizonte-MG,
nos períodos de 22 a 30/03/07 e de 16 a 24/04/07. (N = 24).
Tratamento Total de machos Total de fêmeas
P1T1 + Escatol 1 4
MosquiTRAP + Escatol 0 1
MosquiTRAP + AtrAedes 0 6
Além dos Culicídeos citados, foram capturados outros dípteros, 305 Psychodidae,
do gênero Clogmyia, antigo Telmatoscopus (“mosca de banheiro"), dois Muscidae, além de
outros insetos (cinco Formicidae e 10 Himenópteros). Também foram encontrados três
Aracnida durante a realização do experimento (TAB 10).
TABELA 10. Captura de outros Insecta e Aracnida por tratamento do experimento de comparação
entre o melhor protótipo de armadilha para captura de fêmeas grávidas de Culex. quinquefasciatus
com MosquiTRAP no bairro Jaqueline e no ICB/UFMG, Belo Horizonte-MG, nos períodos de 22
a 30/03/07 e de 16 a 24/04/07. (N = 24).
Número de Artrópodes capturados Categorias
P1T1 + Escatol MosquiTRAP + Escatol MosquiTRAP + AtrAedes
Psychodidae 226 35 44
Muscidae 1 1 0
Formicidae 3 2 0
Himenoptera 9 1 0
Aracnida 3 0 0

82
6. DISCUSSÃO
Os resultados deste trabalho mostraram o potencial da armadilha P1T1 na captura
de Culex quinquefasciatus, especialmente de fêmeas, utilizando atraentes sintéticos de
oviposição. Para isto, conhecimentos básicos sobre comportamento de pré-oviposição e
oviposição da espécie-alvo de mosquito forneceram subsídios importantes para avaliação
de protótipos de armadilhas e para a avaliação do comportamento dos mosquitos frente a
diferentes combinações dos atraentes.
Com o objetivo de aumentar o potencial de captura obtido pela P1T1, foram
desenvolvidos diferentes protótipos de armadilhas. Lima (2005) observou a especificidade
da armadilha P1T1 quanto à captura de fêmeas grávidas de C. quinquefasciatus, porém o
seu potencial de captura deveria ser aumentado para que esta constituísse um instrumento
de monitoramento eficaz utilizado como medida adicional em programas de controle do
vetor. Para tal uso, as armadilhas devem possuir um alto potencial de captura, auxiliando na
redução da densidade populacional do mosquito-alvo a um nível que evite a transmissão de
doenças (BENTO, 2001).
Os protótipos construídos usando balde como parte principal, tinham formato e
tamanho diferenciados da P1T1, bem como apresentavam número e posições diferentes de
aberturas. As variações de número e posições de aberturas alteram a forma da dispersão do
odor pela armadilha e as possibilidades de entrada e escape do inseto. Um funil de tela foi
acrescido a cada abertura dos protótipos em balde com o objetivo de evitar o escape de
mosquitos após sua entrada na armadilha, evitando que houvesse diferença entre o número
de insetos que entrassem nos protótipos e o número de insetos capturados pelos cartões

83
adesivos posicionados no interior das armadilhas. O escape de mosquitos antes de sua
captura foi um problema observado na armadilha P1T1 durante a realização dos
experimentos. Durante a leitura dos experimentos, no momento em que a tampa da
armadilha P1T1 era retirada, escapavam alguns insetos.
Visando avaliar a dispersão da pluma de odor nos protótipos de armadilha
desenvolvidos (tendo como base um balde), em comparação à P1T1, foi realizado o teste de
fumaça, que forneceu subsídios importantes para, inicialmente especular o potencial de
captura de cada protótipo. Posteriormente este pôde ser confirmado através da avaliação da
eficácia na captura de insetos-alvo em campo. A armadilha P1T1 foi responsável pela
formação de uma pluma de odor mais concentrada, que se dispersou mais homogeneamente
pela área adjacente, sofrendo menores influências das correntes de vento, quando
comparada aos protótipos de armadilhas em forma de balde. Dentre os protótipos em forma
de balde, pôde-se notar que a presença de aberturas laterais permitiu uma circulação do ar
no interior das armadilhas, o que fez com que a fumaça liberada se dispersasse em várias
direções, apesar de se apresentar mais dispersa, pouco concentrada e sendo interrompida de
acordo com as rajadas de vento. Apesar da maior continuidade de liberação da pluma de
odor pelos protótipos sem aberturas laterais, em situações de ar parado a pluma facilmente
se interrompia, permanecendo desta maneira por um longo tempo, o que poderia atrapalhar
a orientação do mosquito para o sítio de oviposição.
A forma como os atraentes se difundem na armadilha através da formação de
plumas de odores e a constituição das substâncias são fatores responsáveis por alterações na
eficácia das mesmas quanto à captura dos mosquitos (DU e MILLAR, 1999). A dispersão
dos odores é modulada pela turbulência do vento e a concentração da pluma de odor pode
tanto facilitar quanto confundir os insetos para o encontro do composto. Para que o inseto

84
se oriente pela pluma de odor, é necessário o contato intermitente com o composto,
resultante da intercepção e não intercepção dos filamentos de odor em vários momentos
pelo inseto (EIRAS e MAFRA-NETO, 2001). Sendo assim, a avaliação do potencial de
captura das armadilhas em campo se torna essencial para a interpretação da influência da
dispersão dos odores em sua eficácia.
Devido à ausência total de captura do inseto de interesse (fêmeas grávidas de C.
quinquefasciatus) no Clube Lareira, durante a realização do experimento de avaliação de
protótipos de armadilhas para captura de fêmeas grávidas, foi realizado um experimento
curto com a armadilha CDC para analisar se realmente a área não possuía insetos e
investigar novas possíveis áreas experimentais. A ausência de insetos-alvo durante a
realização dos experimentos e a captura de apenas uma fêmea a um quarteirão do local de
testes (Clube Lareira) fez com que a área fosse descartada, sendo repetido o experimento no
bairro Jaqueline.
Um total de 225 C. quinquefasciatus foi capturado nas armadilhas avaliadas (P1T1,
Baldes A, B, C e D), sendo 145 machos e 80 fêmeas. Dentre estes, a armadilha P1T1
capturou cerca de 46% do total de machos, sendo significativamente maior sua captura que
a obtida pelos baldes A, B e C. Para as fêmeas, a P1T1 capturou 50% do total, sendo a
captura significativamente maior que os demais protótipos. Quanto aos estágios fisiológicos
das fêmeas capturadas, apesar de não ter sido observada diferença significativa entre os
protótipos avaliados dentre de nenhuma das categorias, a armadilha P1T1 capturou mais
exemplares de grávidas, nulíparas e oníparas dentre os protótipos avaliados. Através destes
resultados, aliados à avaliação da dispersão da pluma de odor nos protótipos de armadilhas,
pudemos constatar que a P1T1 facilitou o encontro do sítio de oviposição para fêmeas. Os
experimentos de avaliação da dispersão dos odores e do potencial de captura foram

85
realizados em campo, em ambiente sujeito à turbulência do vento. Nestas condições, a
estrutura das plumas nas armadilhas P1T1 se apresentou de maneira mais homogênea,
situação na qual os odores podem ser distribuídos igualmente, o que fez com que,
conseqüentemente, este sítio atraísse maior percentual de fêmeas, especialmente de fêmeas
grávidas.
Os protótipos tipo balde com aberturas laterais capturaram mais insetos que os sem
estas aberturas, provavelmente devido à interrupção da dispersão da pluma de odor nas
armadilhas sem aberturas laterais em condições de ar parado ou pela presença de mais
acessos de entrada dos insetos nas armadilhas devido ao maior número de aberturas nos
protótipos com aberturas laterais. A maior captura de fêmeas e machos de C.
quinquefasciatus do balde D em relação aos outros baldes permite excluir a possibilidade
de a saída dos mosquitos pelas aberturas diminuir a eficácia de captura das mesmas, visto
que, caso isto ocorresse, quanto mais aberturas possuísse um protótipo, mais insetos
escapariam, deixando, assim, de ser capturados. Maiores estudos quanto ao comportamento
do inseto em resposta às modificações nas armadilhas, tanto em resposta à forma de
dispersão de odores, quanto ao desenho do protótipo, poderiam ser realizados através de
filmagem da dispersão do odor e dos padrões comportamentais dos insetos aos protótipos,
porém a filmagem do experimento foi descartada em virtude do pior desempenho dos
protótipos desenvolvidos frente à P1T1, que foi avaliada por Lima (2005).
Dentre as três áreas experimentais em que foram realizados os experimentos de
avaliação de protótipos de armadilhas (quadrados latinos diferentes), apenas uma obteve
menor captura que as outras duas (região 1, situada na Oficina do Nem). Portanto, os
próximos experimentos não foram realizados nesta região.

86
Sabe-se que não apenas a estrutura da pluma de odor será responsável pela maior
atratividade ao sítio, mas também, a composição de odores, suas concentrações e o tamanho
da superfície do criadouro (CLEMENTS, 1999). Assim, a utilização de atraentes sintéticos
de oviposição pode aumentar a atratividade da armadilha P1T1. A utilização de infusão de
gramínea como atraente aumenta significativamente a captura de fêmeas pela P1T1 (LIMA,
2005). Contudo, seu uso apresenta desvantagens tais como o tempo de preparo ou
alterações das concentrações dos voláteis emitidos, o que torna essencial o uso de atraentes
sintéticos de oviposição. Através de cromatografia gasosa, Millar et al. (1992) identificaram
vários compostos da infusão de gramínea capazes de atrair e/ou estimular a oviposição de
C. quinquefasciatus, tais como fenol, 4-metil-fenol (p-cresol), 4-etil-fenol, indol e 3-metil-
indol (escatol). Em experimentos de campo, quando foi avaliada uma mistura de compostos
(escatol, p-cresol, 4-etil-fenol, fenol e indol) em comparação com o escatol, ambos
dissolvidos em água, não houve diferença significativa entre o número de jangadas
ovipositadas entre os grupos. Portanto, o escatol isoladamente é o responsável pela
atração/estímulo ao sítio de oviposição (BEEHLER et al., 1994).
As armadilhas para captura de insetos, como ocorre para pragas agrícolas, podem se
beneficiar da utilização de liberadores, que são polímeros capazes de liberar as substâncias
atrativas na armadilha a uma taxa relativamente constante, durante todo o período de
captura dos mosquitos. Além da constância na taxa de liberação, são vantagens da
utilização de liberadores para os atraentes a proteção dos componentes químicos ativos
contra a degradação pelo oxigênio do ar ou pela luz e a aplicabilidade em diferentes
situações de manejo (BENTO, 2001). Lima (2005) avaliou as perdas médias de massa dos
compostos isolados de infusão de gramínea fenol, p-cresol, nonanal, escatol e indol em
liberadores, fornecendo dados importante quanto ao período de utilização destes liberadores

87
contendo atraentes sintéticos de oviposição em campo. Nos experimentos com os
liberadores contendo atraentes sintéticos de oviposição, usando a armadilha P1T1, a infusão
a 100% capturou maior média de fêmeas, mas, quando cada grupo foi observado
individualmente, não houve diferença significativa na captura média de fêmeas usando
escatol e p-cresol. Estes resultados sugerem que as taxas médias de evaporação destes
atraentes nos liberadores apresentam atratividade similar ao da infusão a 100%, o que não
ocorreu com os outros atraentes como indol, nonanal e fenol (LIMA, 2005). Lima (2005)
observou que o escatol isoladamente liberado em polímeros pode ser o responsável pela
atratividade de fêmeas grávidas às P1T1, ou seja, é responsável pelo encontro do sítio de
oviposição. Porém, diferentes combinações de atraentes sintéticos podem aumentar a
atratividade, visto que os testes demonstraram que a infusão capturou em média maior
número de fêmeas que todos os atraentes.
No experimento de avaliação de combinações binárias de atraentes de oviposição
sintéticos do presente trabalho, o total de 525 C. quinquefasciatus foi capturado nas
armadilhas, destes 285 machos e 240 fêmeas. O escatol isoladamente foi responsável pela
captura de cerca de 56% dos machos, sendo a média de captura significativamente maior do
que a observada com as armadilhas contendo apenas água, contendo escatol e indol e
contendo escatol e p-cresol. A média de captura da armadilha contendo apenas escatol só
não foi significativamente maior que a da armadilha contendo escatol e nonanal, porém a
armadilha contendo apenas escatol capturou mais do que o triplo de machos que a
armadilha contendo escatol e nonanal. As armadilhas contendo apenas escatol capturaram
cerca de 53% do total de fêmeas capturadas durante o experimento e todos os estágios
fisiológicos.

88
Os resultados do experimento de combinação binária de atraentes de oviposição
sintéticos do presente trabalho corroboram com os de Beehler et al. (1994), que
observaram, em experimentos de campo utilizando armadilhas de Reiter (gravid trap), que
armadilhas contendo escatol obtém uma captura de fêmeas de C. quinquefasciatus
significativamente diferente de armadilhas contendo apenas água. Estes autores também
observaram que a adição de outros atraentes, em combinação ao escatol, não resultam em
um aumento da atratividade, pois não houve diferença significativa de captura entre a
combinação de atraentes e o escatol utilizado isolado.
Provavelmente, os machos e as fêmeas de outros estágios fisiológicos (não
grávidas) de C. quinquefasciatus capturados, foram atraídos pela cor escura das armadilhas,
em busca de um local para abrigar-se. Os machos, ainda, podem ter sido atraídos pelas
fêmeas capturadas, visando à cópula. Durante os experimentos de comparação entre os
diferentes protótipos desenvolvidos de armadilhas para captura de fêmeas grávidas de C.
quinquefasciatus e de avaliação de combinações binárias de atraentes sintéticos de
oviposição, realizados no bairro Jaqueline, houve uma maior captura de machos da espécie-
alvo, em comparação à captura de fêmeas, o que provavelmente ocorreu devido à maior
densidade populacional deste sexo na área experimental.
Os resultados sugerem que, dentre as combinações testadas e semelhantes às
concentrações utilizadas em avaliações dos atraentes segundo Lima (2005), a utilização do
escatol isolado possui maior potencial de atratividade dos insetos-alvo do que quando
associado aos outros atraentes sintéticos. A combinação do escatol com os demais atraentes
exerce um efeito antagonista, pois o potencial de captura é diminuído. O experimento
corrobora dados da literatura, pois o escatol capturou significativamente mais fêmeas que a
água (DU e MILLAR, 1999) e, dependendo da concentração do escatol, há maior

89
atratividade de fêmeas por este tratamento, se comparado à água, demonstrada pela
deposição de jangadas (BEEHLER et al., 1994; BLACKWELL et al., 1993; MBOERA et
al., 2000b).
Provavelmente, alterações na concentração dos compostos secundários resultem, se
não em maior captura do que o escatol utilizado isoladamente, no mesmo potencial de
captura, pois estes compostos estão presentes em infusões de gramínea, que atraem
igualmente fêmeas grávidas, se comparadas ao escatol isoladamente (LIMA, 2005). Ainda,
uma combinação de atraentes (dentre os quais estavam escatol, p-cresol e indol, testados no
presente trabalho) foi tão atrativa quando o escatol utilizado isoladamente (BEEHLER et
al., 1994).
Como não houve uma combinação binária mais atrativa que o escatol isolado, os
mesmos atraentes utilizados no experimento de avaliação de combinações binárias foram
avaliados em todas combinações terciárias possíveis, também sendo testada a combinação
quaternária dos atraentes, acrescidos à armadilha P1T1 (cada atraente em seu liberador, na
mesma concentração anterior), em comparação aos controles representados pela armadilha
P1T1 contendo apenas água e contendo água e escatol isolado.
No entanto, houve uma reduzida captura de C. quinquefasciatus durante todo o
experimento, onde apenas o total de 21 insetos foi capturado, sendo nove machos e 12
fêmeas. Os experimentos foram realizados em um período chuvoso (Janeiro de 2007), que
provavelmente contribuiu para uma baixa densidade de insetos-alvo no local de realização
dos experimentos. O aumento do nível e da velocidade de vazão da água nos criadouros
diminui a densidade das formas imaturas nestes locais durante o período chuvoso.
Conseqüentemente, a captura obtida pela P1T1 iscada com escatol não foi
significativamente diferente da captura obtida pelo controle (P1T1 contendo apenas água).

90
A diferença de atratividade de fêmeas entre água e escatol já foi observada no presente
trabalho, no experimento de avaliação de combinações binárias e em outros relatos
(BEEHLER et al., 1994; BLACKWELL et al., 1993; MBOERA et al., 2000b).
Dentre as 12 fêmeas capturadas, oito foram capturadas pelas armadilhas contendo
apenas escatol (66,7% do total de fêmeas capturadas durante o experimento), média
significativamente maior do que a observada nas armadilhas contendo a combinação
terciária escatol + p-cresol + nonanal e contendo a combinação quaternária (escatol + indol
+ p-cresol + nonanal). Contudo, os experimentos não foram repetidos porque, considerando
os dados do experimento de avaliação das combinações binárias (cujos resultados foram
válidos) com o presente experimento, concluiu-se que a combinação dos atraentes, cada um
em seu liberador e nas concentrações utilizadas não resultaria em maior atratividade de
fêmeas, aumentando o potencial de captura da armadilha P1T1, como era o objetivo inicial.
A melhoria da atratividade deste instrumento de monitoramento poderia ser obtida
através da avaliação de diferentes combinações entre os atraentes utilizados, em diferentes
concentrações de cada um, sendo utilizados em liberadores próprios para cada atraente ou
todos os atraentes num mesmo liberador. Neste caso, uma base com óleos minerais ou
outros emulsionáveis (excipientes) que não alterem as propriedades químicas dos atraentes
e que não possua odor poderá ser testada.
È importante salientar que os trabalhadores dos locais de realização dos testes
constantemente reclamavam do odor liberado pelo escatol, inclusive queixando-se de fortes
dores de cabeça provavelmente associadas ao mau cheiro. Assim, o forte odor do escatol
poderia ser motivo de reclamações e até rejeição da armadilha por parte de moradores,
dificultando sua utilização em ambiente intradomiciliar.

91
A baixa densidade de insetos na área de realização dos experimentos pôde ser
constatada através dos resultados obtidos nos experimentos de avaliação de combinações
terciárias e quaternária de atraentes sintéticos de oviposição e de comparação entre a
MosquiTRAP e a P1T1. Moradores da região confirmaram a diminuição da população de
C. quinquefasciatus, pois pararam de ser importunados durante as noites pelas picadas
destes insetos. Ainda, ao final das três repetições do experimento de comparação entre
MosquiTRAP e AtrAedes, no bairro Jaqueline, devido à ocorrência de casos confirmados
de dengue, ocorreu a aplicação de inseticidas no local, então se fez necessária a busca de
novo local para a realização de mais repetições do experimento.
Foram realizadas visitas a locais na região da Pampulha, acompanhadas de
profissionais da Regional Pampulha, com o objetivo de encontrar uma área com alta
densidade dos insetos-alvo para a realização de repetições do experimento de comparação
entre a P1T1 e a MosquiTRAP. Os locais visitados sabidamente funcionam como
criadouros da espécie-alvo em épocas de alta infestação, nos quais foram procedidas a
busca ativa por formas imaturas do inseto, através de conchas, e de adultos, através de
inspeção, dentro de residências, de locais normalmente utilizados como abrigo dos
mosquitos durante o dia (locais escuros), além de perguntas aos moradores das
proximidades quanto à existência de incômodos causados pelas picadas noturnas. Não foi
observada uma área com alta densidade de formas imaturas tampouco de adultos de C.
quinquefasciatus, portanto o experimento não foi realizado nas áreas visitadas.
A realização de um experimento por outro grupo de pesquisadores do Laboratório
de Ecologia Química de Insetos Vetores da UFMG, utilizando a armadilha BG-Sentinel, na
região externa do ICB/UFMG, demonstrou a presença de insetos adultos na região (R.,

92
MOTA, Laboratório de Ecologia Química de Insetos Vetores, comunicação pessoal), então,
foram realizadas outras cinco repetições do último experimento neste local.
A armadilha MosquiTRAP apresenta-se com baixo custo e fácil operacionalidade e
já é utilizada com atraentes sintéticos de oviposição para captura de Aedes sp. (EIRAS,
2003), mas freqüentemente captura também C. quinquefasciatus. Apenas 16 C.
quinquefasciatus foram capturados no experimento de comparação entre MosquiTRAP e
P1T1, sendo seis machos e 10 fêmeas. Entre os machos capturados pelos diferentes
tratamentos, não houve diferença significativa. Dentre as 10 fêmeas capturadas, oito (80%
do total) foram capturadas pelas armadilhas P1T1 contendo escatol, cuja média de captura
foi significativamente maior do que a observada nas armadilhas MosquiTRAP contendo
AtrAedes, que não capturou nenhuma fêmea de C. quinquefasciatus. Todas as fêmeas de
C. quinquefasciatus capturadas durante o experimento eram grávidas.
Em experimentos em campo realizados por Lima (2005) utilizando infusão de
gramínea seca com lactoalbumina hidrolisada e levedo de cerveja (75%), não houve
diferença significativa entre os números médios de fêmeas grávidas na P1T1 e na
MosquiTRAP. No entanto, ao trocar a infusão anteriormente utilizada por infusão fresca a
100% nas duas armadilhas, houve um aumento tanto no número total quanto na média de
fêmeas grávidas capturadas na MosquiTRAP. O aumento na captura pode ter ocorrido
devido ao maior poder de atratividade da infusão fresca a 100%, pela possibilidade de as
plumas de odores emanadas na armadilha MosquiTRAP facilitarem o encontro deste sítio
de postura ou por as fêmeas em campo apresentarem o comportamento preferencial de
toque nas paredes internas do sítio, local onde se situa o cartão adesivo na MosquiTRAP.
No entanto, deve-se considerar que durante o experimento citado de comparação entre a

93
P1T1 e a MosquiTRAP, a armadilha P1T1 utilizava, para a captura dos insetos, um cartão
impregnado com cola entomológica, e a autora observou que esta cola exercia uma
repelência dos insetos, fator determinante para a diferença de captura entre a P1T1 e a
MosquiTRAP.
Os resultados da avaliação em campo corroboram os dados de Lima (2005),
mostrando que a P1T1 atinge o objetivo para o qual foi concebida, capturar com
seletividade fêmeas alimentadas e grávidas de C. quinquefasciatus. Ainda, pôde-se
confirmar que as condições da armadilha permitem que os indivíduos capturados sejam
facilmente contados, identificados e transferidos para o laboratório para eventuais exames
de infectividade. Contudo, os resultados continuam indicando que a atratividade da
armadilha P1T1 deve ser melhorada, visando aumentar o seu potencial de captura para uso
em larga escala, como uma maneira de monitorar as variações na densidade populacional
do inseto-alvo.
Uma possibilidade para a melhoria da armadilha poderá vir da utilização de várias
combinações de atraentes de oviposição sintéticos, em diferentes concentrações e
formulações, ou seja, o desenvolvimento de uma mistura atrativa para fêmeas grávidas de
C. quinquefasciatus, visto que as combinações avaliadas no presente trabalho não
cumpriram o objetivo de aumentar a atratividade da armadilha.
Ainda, a mistura de atraentes sintéticos pode ser combinada ao feromônio sintético
de oviposição (FOS), em diferentes concentrações no intuito de obter-se um efeito aditivo à
mistura, o que não foi realizado no presente trabalho devido à indisponibilidade de FOS, o
qual não é produzido em larga escala, e sua síntese ou extração para a utilização como
atrativo utilizado na P1T1 demandaria um tempo demasiadamente longo para a realização

94
de uma dissertação de mestrado. O presente trabalho continuou mostrando que a armadilha
possui a especificidade desejada, porém que o seu potencial de captura deve ser melhorado,
permitindo, assim sua utilização em programas de monitoramento de C. quinquefasciatus.

95
7. CONCLUSÕES
Os resultados deste estudo sobre o desenvolvimento de armadilha adesiva em
avaliações com diferentes combinações de atraentes sintéticos de oviposição permitem
concluir que:
1- A armadilha P1T1 foi responsável pela formação de uma pluma de odor mais
concentrada, que se dispersou mais homogeneamente pela área adjacente, sofrendo
menores influências das correntes de vento, quando comparada aos protótipos em forma de
balde.
2- A armadilha P1T1 se mostrou mais eficaz na captura de adultos de Culex
quinquefasciatus, em comparação aos outros protótipos desenvolvidos no presente trabalho.
3- A armadilha P1T1 contendo apenas escatol apresentou maior potencial de captura, em
comparação às combinações binárias de atraentes sintéticos de oviposição e à armadilha
contendo apenas água.
4- Os resultados da avaliação de combinações terciárias e quaternária de atraentes sintéticos
de oviposição foram inconclusivos.

96
5- A armadilha MosquiTRAP contendo o AtrAedes não se mostrou específica à captura
de fêmeas grávidas de C. quinquefasciatus, quando comparada à armadilha P1T1.

97
8. REFERÊNCIAS BIBLIOGRÁFICAS ALLAN, S. A.; KLINE, D. L. Evaluation of organic infusions and synthetic compounds mediant oviposition in Aedes albopictus and Aedes aegypti (Diptera: Culicidae). Journal of Chemical Ecology, v. 21, p. 1847-1860, 1995. ALLAN, S. A.; KLINE, D. L. Larval rearing water and preexisting eggs influence oviposition by Aedes aegypti and Ae. albopictus (Diptera: Culicidae). Journal of Medical Entomology, v. 35, p. 943-947, 1998. ALVES, S. B. et al. Potential of some Metarhizium anisopliae isolates for control of Culex quinquefasciatus (Dipt., Culicidae). Journal of Applied Entomology, v. 126, p. 504-509, 2002. ARAÚJO, F. A. A. et al. Inquérito sorológico em aves migratórias e residentes de Galinhos/RN para detecção do vírus da Febre do Nilo Ocidental e outros vírus. SVS-Boletim Eletrônico epidemiológico, v. 2, p. 1-12, 19/02/2004. Disponível em <www.saude.gov.br/svs>. AVIETAM; BLAUSTEIN, L. Oviposition habitat selection by mosquitoes in response to predator (Notonecta maculata) density. Physiological Entomology, v. 29, p. 188-191, 2004. BEEHLER, J. W.; MILLAR, J. G.; MULLA, M. S. Field evaluation of synthetic compounds mediating oviposition in Culex mosquitoes (Diptera: Culicidae). Journal of Chemical Ecology, v. 20, p. 281-291, 1994. BENTO, J. M. S. 2001. Fundamentos do monitoramento, da coleta massal e do confundimento de insetos-praga. In: VILELA, E. F.; DELLA LUCIA, M. T. Feromônios de Insetos: Biología, química e emprego no manejo de pragas. Ribeirão Preto: Holos, 2001. p. 135-144. BLACKWELL, A. et al. A behavioural and eletrophysiological study of oviposition cues for Culex quinquefasciatus. Physiological Entomology, v. 18, p. 343-348, 1993. BRACCO, J. E.; BARATA, J. M. S.; MARINOTTI, O. Evaluation of inseticide resistance and biochemical mechanisms in a population of Culex quinquefasciatus (Diptera: Culicidae) from São Paulo, Brazil. Memórias do Instituto Oswaldo Cruz, v. 94, p. 115-120, 1999. BUSVINE, J. R.; RONGSRIYAM, Y.; BRUNO, D. Effects of some insect development inhibitors on mosquito larvae. Pestic. Sci., v. 7, p. 153-160, 1976. CARDÉ, R. T.; BELL, W. Chemical ecology of insects 2. New York: Publisher, 1996.

98
CHADEE, D. D. et al. Oviposition response of Aedes aegypti mosquitoes to different concentrations of hay infusion in Trinidad, West Indies. Journal of the American Mosquito Control Association, v. 9, p. 346-348, 1993. CHANDRE, F. et al. Pyrethroid resistance in Culex quinquefasciatus from West Africa. Medical and Veterinary Entomology, v. 12, p. 359-366, 1998. CLEMENTS, A. N. Adult circadian rhythms. In:_______. The Biology of Mosquitoes, Sensory Reception and Behavior. New York: CABI Publishing, 1999. p. 207-238. CLEMENTS, A. N. The regulation of adult behavior. In:_______. The Biology of Mosquitoes, Sensory Reception and Behavior. New York: CABI Publishing, 1999. p. 239-262. CLEMENTS, A. N. The modification of adult behavior by geophysical and climatic factors. In:_______. The Biology of Mosquitoes, Sensory Reception and Behavior. New York: CABI Publishing, 1999. p. 263-286. CLEMENTS, A. N. The source and characteristics of host cues. In:_______. The Biology of Mosquitoes, Sensory Reception and Behavior. New York: CABI Publishing, 1999. p. 433-457. CLEMENTS, A. Host finding. In:_______. The Biology of Mosquitoes, Sensory Reception and Behavior. New York: CABI Publishing, 1999. p. 458-479. CLEMENTS, A. N. Egg laying. In:_______. The Biology of Mosquitoes, Sensory Reception and Behavior. New York: CABI Publishing, 1999. p. 552-643. CONSOLI, R. A. G. B.; OLIVEIRA, R. L. Principais mosquitos de importância sanitária no Brasil. Rio de Janeiro: Ed. Fiocruz, 1994. COOPERBAND, M. F.; CARDÉ, R. T. Orientation of Culex mosquitoes to carbon dioxide-baited traps: flight manouvres and trapping efficiency. Medical and Veterinary Entomology, v. 20, p. 11-26, 2006. DAVIS, E. E.; BOWEN, M. F. Sensory physiological basis for attraction in mosquitoes. Journal of the American Mosquito Control Association, v. 10, p. 316-325, 1994. DETINOVA, T. S. Age-grouping methods in diptera of medical importance with special reference to some vectors of malaria. Monograph Series. World Health Organization (WHO), Geneva, 47, 216 p. 1962. DICKE, M.; SABELIS, M. W. Cost and benefits of chemical information conveyance: proximate and ultimate factors. In: ROLTBERG, B. D.; ISMAN, M. B. Insect chemical ecology: an evolutionary approach. New York: Chapman and Hall, 1992. p. 122-155.

99
DORTA, D. M.; VASUKI, V.; RAJAVEL, A. Evaluation of organophosphorus and synthetic pyrethroid inseticides against six vector mosquitoe species. Revista de saúde Pública, v. 27, p. 391-397, 1993. DU, Y.; MILLAR, J. G. Eletroantennogram and oviposition bioassay responses of Culex quinquefasciatus and Culex tarsalis (Diptera: Culicidae) to chemicals in odors from bermuda grass infusions. Journal of Medical Entomology, v. 36, p. 158-166, 1999. EIRAS, A. E. Culicidae. In: NEVES, D. P. Parasitologia Humana. 10ª edição. São Paulo: Editora Atheneu, 2000. p.320-333. EIRAS, A. E.; MAFRA-NETO, A. Olfatometria aplicada ao estudo do comportamento de insetos. In: VILELA, E. F.; DELLA LUCIA, T. M. C. Feromônios de Insetos: Biología, química e emprego no manejo de pragas. Ribeirão Preto: Holos, 2001. p. 99-112. EIRAS, A. E. Mediadores químicos entre hospedeiros e insetos vetores de doenças médico-veterinárias. In: VILELA, E. F.; DELLA LUCIA, T. M. C. Feromônios de Insetos: Biologia, química e emprego no manejo de pragas. Ribeirão Preto: Holos, 2001. p. 99-112. EIRAS, A. E. Armadilha para captura do mosquito Aedes sp. INPI 0203907-9 (Depósito de Patente), 2002. EIRAS, A. E. Proposta de novo método de monitoramento e de novos índices de vigilância entomológica usando MosquiTRAP, uma nova armadilha para captura de adultos do mosquito Aedes aegypti. Relatório Técnico de Pesquisa submetido à FUNASA. 36 p. 2003. FONTES, G. Filariidae: Wuchereria bancrofti – Filariose Linfática. In: NEVES, D. P. Parasitologia Humana. 10ª edição. São Paulo: Editora Atheneu, 2000. p.269-277. FORATTINI, O. P. Culicidae. Morfologia e classificação. In:_______. Entomologia Médica. São Paulo: Editora da Universidade de São Paulo, 1963. p. 123-184. FORATTINI, O. P. Culicidae. Biologia. In:_______. Entomologia Médica. São Paulo: Editora da Universidade de São Paulo, 1963. p. 185-303. FORATTINI, O. P. et al. Observações sobre os mosquitos Culex da cidade de São Paulo, Brasil. Revista de Saúde Pública, v. 7, p. 315-330, 1973. FORATTINI, O. P. et al. Studies on mosquitoes (Diptera: Culicidae) and anthropic environment. 4- Survey of resting adults and synanthropic behaviour and South-Eastern, Brazil. Revista de Saúde Pública, v. 27, p. 398-411, 1993. FORATTINI, O. P. Culicidologia Médica: Identificação, Biologia, Epidemiologia. São Paulo: Editora da Universidade de São Paulo, 2002.

100
GAMA, R. A.; SILVA, I. M.; RESENDE, M. C.; EIRAS, A. E. Evaluation of the sticky MosquiTRAP for monitoring Aedes aegypti (Diptera: Culicidae) in the district of Itapoã, Belo Horizonte, Minas Gerais, Brazil. Memórias do Instituto Oswaldo Cruz. 2006 GOMES, A. C. Vigilância Entomológica. Informe Epidemiológico do SUS, v. 11, p. 79-90, 2002. GOMES, A. Seqüência do comportamento de oviposição do mosquito Aedes aegypti (Diptera: Culicidae) em condições de laboratório. 2003. 85f. Dissertação (Mestrado em Parasitologia) - Instituto de Ciências Biológicas, Universidade Federal de Minas Gerais, Belo Horizonte, 2003. HARZARD, E. I.; MAYER, M. S.; SAVAGE, K. E. Attraction and oviposition stimulation of gravid female mosquitoes by bacteria isolated from hay infusions. Mosquito News, v. 27, p. 133-136, 1967. ISOE, J. et al. Oviposition responses of Culex tarsalis and Culex quinquefasciatus to aged Bermuda grass infusions. Journal of the American Mosquito Control Association, v. 11, p. 39-44, 1995 b. ISOE, J.; MILLAR, J. G.; BEEHLER, J. W. Bioassays for Culex (Diptera: Culicidae) mosquito oviposition attractants and stimulants. Journal of Medical Entomology, v. 32, p. 475-482, 1995 a. ISOE, J.; MILLAR, J. G. Characterization of factors mediating oviposition site choice by Culex tarsalis. Journal of the American Mosquito Control Association, v. 11, p. 21-28, 1995. ISOE, J.; MILLAR, J. G. Water deprivation and light affect responses of Culex tarsalis (Diptera: Culicidae) to oviposition site cues. Environmental Entomology, v. 25, p. 1383-1390, 1996. JAYANETTI, S. R.; PERERA, H. A. S.; WIJESUNDARA, M. S. Evaluation of the CDC gravid mosquito trap for sampling periodomestic mosquito filarial vectors. Mosquito-Borne Diseases Bulletin, v.5, p. 18-21, 1988. JOHNSON, B. W. et al. Growth characteristics of the veterinary vaccine candidate ChimeriVaxTM-West Nile (WN) virus in Aedes and Culex mosquitoes. Medical and Veterinary Entomology, v. 17, p. 235-243, 2003. JUTSUM, A. R.; GORDON, R. F. S. Introduction. Pheromones: importance to insects and role in pest management. In:______. Insect Pheromones in Plant Protection. London: John Wiley e Sons, 1989. p. 1-13. KARUNARATNE, S. H. P. P.; HEMINGWAY, J. Malathion resistance and prevalence of the malathion carboxylesterase mechanism in populations on mosquito vectors of disease in Sri Lanka. Bulletin of the World Health Organization, v. 79, p. 1060-1064, 2001.

101
KIM, H. C. et al. Seasonal prevalence of mosquitoes collected from light traps in the Republic of Korea in 2003. Entomological Research, v. 36, p. 139-148, 2006. KLINE, D. L.; PATNAUDE, M.; BARNARD, D. R. efficacy of four trap types for detecting and monitoring Culex spp. In North Central Florida. Journal of Medical Entomology, v. 43, p. 1121-1128, 2006. LAMPMAN, R. L.; NOVAK, R. J. Oviposition preferences of Culex pipiens and Culex restuans for infusion-baited traps. Journal of the American Mosquito Control Association, v. 12, p. 23-32, 1996. LAURENCE, B. R.; PICKET, J. A. Erythro-6-Acetoxy-hexadecanolide, the major component of a mosquito oviposition attractant pheromone. Journal of the Chemical Society, Chemistry Communication, p. 59-60, 1982. LAURENCE, B. R.; PICKET, J. A. An oviposition attractant pheromone in Culex quinquefasciatus Say (Diptera: Culicidae). Bulletin of Entomology Research, v. 75, p. 283-290, 1985. LEWIS, L. F. et al. Collecting ovigerous Culex pipiens quinquefasciatus Say near favorable resting sites with louvered traps baited with infusions of alfalfa pellets. Mosquito News, v. 34, p. 436-439, 1974. LIMA, C. A. Comportamento de oviposição e desenvolvimento de armadilha iscada com atraentes de oviposição natural e sintético para captura de fêmeas grávidas de Culex quinquefasciatus Say, 1823 (Díptera: Culicidae). 2005. 187 f. Tese (Doutorado em Parasitologia) – Instituto de Ciências Biológicas, Universidade Federal de Minas Gerais, Belo Horizonte, 2005. MBOERA, L. E. G. et al. Influence of synthetic oviposition pheromone and volatiles from soakage pits and grass infusions upon oviposition site-selection of Culex mosquitoes in Tanzania. Journal of Chemical Ecology, v. 25, p. 1855-1865, 1999. MBOERA, L. E. G et al. Sampling gravid Culex quinquefasciatus (Diptera: Culicidae) in Tanzania with traps baited with synthetic oviposition pheromone and grass infusions. Journal of Medical Entomology, v. 37, p. 172-176, 2000a. MBOERA, L. E. G. et al. Oviposition and behavioral responses of Culex quinquefasciatus to skatole and synthetic oviposition pheromone in Tanzania. Journal of Chemical Ecology, v. 26, 1193-1203, 2000b. McCALL, P. J.; EATON, G. Olfactory memory in the mosquito Culex quinquefasciatus. Medical end Veterinary Entomology, v. 15, p. 197-203, 2001. MILLAR, J. G.; CHANEY, J. D.; MULLA, M. R. Identification of oviposition attractants for Culex quinquefasciatus from fermented Bermuda grass infusions. Journal of the American Mosquito Control Association, v. 8, p. 11-17, 1992.

102
MILLAR, J. G. et al. Interaction of the Culex quinquefasciatus egg raft pheromone with a natural chemical associated with oviposition sites. Journal of the American Mosquito Control Association, v. 10, p. 374-379, 1994. MOHAN, L.; SHARMA, P.; SRIVASTAVA, C. N. Evaluation of Solanum xanthocarpum extract for cypermethrin against larvae of the filarial vector Culex quinquefasciatus (Say). Entomological Research, v. 36, p. 220-225, 2006. MOKANY. P. J.; SHINE, R. Oviposition site selection by mosquitoes is affected by cues from conspecific larvae and anuran tadpoles. Austral Ecology, v. 28, p. 33-37, 2003. MORDUE, A. J. et al. Behavioural and electrophysiological evaluation of oviposition attractants for Culex quinquefasciatus Say (Diptera: Culicidae). Experientia, v. 48, p. 1109-1111, 1992. NORDLUND, D. A.; LEWIS, W. J. Terminology of chemical-releasing stimuli in intraespecific and interspecific interaction. Journal of Chemical Ecology, v. 2, p. 211-220, 1976. ORGANIZATIÓN MUNDIAL DE LA SALUD. Lucha contra la filariasis linfática. Manual para personal sanitario. Geneva, 1998. OTIENO, W. A. et al. A field trial of the synthetic oviposition pheromone with Culex quinquefasciatus Say (Diptera: Culicidae) in Kenya. Bulletin of Entomological Research, v. 78, p. 463-470, 1988. PILE, M. M.; SIMMONDS, S. J.; BLANEY, W. M. Odour-mediated upwind flight of Culex quinquefasciatus mosquitoes elicited by a synthetic attractant. Physiological Entomology, v. 16, p. 77-85, 1991. PINTO, C. J. C. Eficácia larvicida de Bacillus sphaericus Neide, 1902 contra Culex quinquefasciatus Say, 1823 (díptera: Culicidae) frente a alguns fatores ambientais. 1995. 102 f. Dissertação (Mestrado em Parasitologia) – Instituto de Ciências Biológicas, Universidade Federal de Minas Gerais, Belo Horizonte, 1995. PIZARRO, A. P. B. et al. O aproveitamento do resíduo da indústria do sisal no controle de larvas de mosquitos. Revista da Sociedade Brasileira de Medicina Tropical, v. 36, p.23-29, 1999. POONAM, S.; PAILY, K. P.; BALARAMAN, K. Oviposition attractancy of bacterial culture filtrates – responses of Culex quinquefasciatus. Memórias do Instituto Oswaldo Cruz, v. 97, p. 359-362, 2002. REITER, P. A. A portable battery-powered trap for collecting gravid Culex mosquitoes. Mosquito News, v. 43, p. 496-508, 1983.

103
REITER, P. A. A field trial of expanded polystyrene balls for the control of Culex mosquitoes in pit latrines. Journal of the American Mosquito Control Association, v. 1, p. 519-524, 1985. REITER, P. A revised version of the CDC gravid mosquito trap. Journal of the American Mosquito Control Association, v. 3, p. 325-327, 1987. RITCHIE, S. A. Hay infusion and isopropyl alcohol-baited CDC light trap; a simple, effective trap for gravid Culex mosquitoes. Mosquito News, v. 44, p. 404-407, 1984. RITCHIE, S. A.; KLINE, D. L. Comparison of CDC and EVS light traps baited with carbon dioxide and octenol for trapping mosquitoes in Brisbane, Queensland (Diptera: Culicidae). Journal of Australian Entomology Society, v. 34, p. 215-218, 1995. ROBERT, L. L.; OLSON, J. K. Effects of sublethal dosages of inseticides on Culex quinquefasciatus. Journal of the American Mosquito Control Association, v. 5, p. 239-246, 1989. ROQUE, R. A Avaliação de armadilhas iscadas com infusões de gramíneas como atraentes e/ou estimulantes de oviposição do mosquito Aedes (Stegomyia) sp (Díptera: Culicidae). 97p. Dissertação (Mestrado em Parasitologia): Instituto de Ciências Biológicas, Universidade Federal de Minas Gerais, Belo Horizonte, 2002. SALGADO, C. C. Análisis de situación de salud em las Américas, 1999-2000. Boletín Epidemiológico/OPS, v. 21, p. 1-16, 2000. SANT’ANA, A. D.; ROQUE, R. A.; EIRAS, A. E. Characteristics of grass infusions as oviposition attractants to Aedes (Stegomyia) (Diptera: Culicidae). Journal of Medical Entomology, v. 43, p. 214-220, 2006. SCHOLTE, E. J. et al. Infection of malaria (Anopheles gambiae s. s.) and filariasis (Culex quinquefasciatus) vectors with the entomopathogenic fungus Metarhizium anisopliae. Malaria Journal, v. 2, p. 1-8, 2003. SERVICE, M. W. Sampling adults with carbon dioxide traps, light-traps, visual attraction traps and sound traps. In:______. Mosquito Ecology – Field Sampling Methods. New York: John Wiley e Sons, 1977. TEODORO, U. et al. Mosquitos de ambientes peri e extradomiciliares na região sul do Brasil. Revista de Saúde Pública, v. 28, p. 107-115, 1994. VILELA, E. F.; DELLA LUCIA, T. M. C. Introdução aos semioquímicos e terminologia. In: VILELA, E. F.; DELLA LUCIA, T. M. C.. Feromônios de Insetos: biologia, química e emprego no manejo de pragas. Ribeirão Preto: Holos, 2001. p. 9-12. WORLD HEALTH ORGANIZATION. Control of Lymphatic Filariasis (A Manual for Health Personnel). Geneva, 1988.

104
WORLD HEALTH ORGANIZATION. Control of Lymphatic Filariasis (A Manual for Health Personnel). Geneva, 2000.

Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas
Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo


















