Comportamento Exploratório de Ratos no Labirinto em Cruz...
111
UNIVERSIDADE DE SÃO PAULO Faculdade de Filosofia, Ciências e Letras de Ribeirão Preto Departamento de Psicologia e Educação Programa de Pós-graduação em Psicobiologia Comportamento Exploratório de Ratos no Labirinto em Cruz Elevado, Ingestão de Açúcar e Manuseio Diogo Antonio Blóes Chagas Tese de Doutorado apresentada à Faculdade de Filosofia, Ciências e Letras de Ribeirão Preto – USP para a obtenção do título de Doutor em Ciências, Área de Psicobiologia. Ribeirão Preto – SP 2007
Transcript of Comportamento Exploratório de Ratos no Labirinto em Cruz...

UNIVERSIDADE DE SÃO PAULO Faculdade de Filosofia, Ciências e Letras de Ribeirão Preto
Departamento de Psicologia e Educação Programa de Pós-graduação em Psicobiologia
Comportamento Exploratório de Ratos no Labirinto em Cruz Elevado, Ingestão de Açúcar e Manuseio
Diogo Antonio Blóes Chagas
Tese de Doutorado apresentada à Faculdade de Filosofia, Ciências e Letras de Ribeirão Preto – USP para a obtenção do título de Doutor em Ciências, Área de Psicobiologia.
Ribeirão Preto – SP 2007

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

FICHA CATALOGRÁFICA
Blóes Chagas, Diogo Antonio Comportamento exploratório de ratos no labirinto em cruz elevado,
ingestão de açúcar e manuseio. – Ribeirão Preto, 2007. 108 pp ; il.; 30 cm.
Tese de Doutorado, apresentada à Faculdade de Filosofia, Ciências e Letras de Ribeirão Preto/USP – Área de concentração: Psicobiologia.
Orientador: Morato, Silvio.
1. Ingestão de açúcar. 2. Manuseio experimental. 3. Comportamento exploratório. 4. Ansiedade. 5. Labirinto em cruz elevado. 6. Campo aberto. 7. Ratos.

UNIVERSIDADE DE SÃO PAULO
Faculdade de Filosofia, Ciências e Letras de Ribeirão Preto Departamento de Psicologia e Educação
Programa de Pós-graduação em Psicobiologia
Comportamento Exploratório de Ratos no Labirinto em Cruz Elevado, Ingestão de Açúcar e Manuseio
Diogo Antonio Blóes Chagas
Orientador: Prof. Dr. Silvio Morato
Ribeirão Preto – SP 2007


Agradecimentos À minha mãe, Márcia, que foi e continuará sendo a pessoa mais importante da minha vida, me apoiando desde o início. Sem você com certeza teria sido muito difícil! Não há maneiras de transformar em palavras o que sinto por você e o quanto sou grato por tudo que me fez! Te amo! Aos meus irmãos Pedro e Patrícia por sempre estarem ao meu lado incondicionalmente. Vocês são muito importante! Penso em vocês todos os dias! À minha avó, Henriqueta, que lutou muito na vida para poder educar seus filhos e netos. Sua força e persistência é um modelo para todos! Ao professor Dr. Silvio Morato, pela amizade, orientação, formação e apoio dispensados durante a realização deste trabalho. Também o agradeço por me incentivar a trabalhar paralelamente ao doutoramento. Poucos professores deste programa teriam feito dessa forma.Obrigado por ser tolerante para com meus atrasos frente aos prazos do programa! Ao amigo e professor Dr. Ari Bassi do Nascimento, pelas sugestões sobre o trabalho e pelas conversas nos bares. Você teve e continua tendo grande participação na minha formação profissional! Aos colegas Paulinho, Milena, Célio, Ana, Javier pelo companheirismo. Aos amigos de Poços de Caldas: Regiane, Gustavo (purga) e Francine (M..!). Gosto muito de vocês! À Reni e família, por terem me acolhido maravilhosamente em Poços. Obrigado por ter me tratado como um de seus filhos! Não é a toa que a chamo de mãe 2. Aos professores do programa de Psicobiologia que colaboraram de maneira direta ou indireta na produção deste trabalho. Aos colegas da Psicobiologia. Ao CNPq pelo apoio financeiro durante o primeiro ano, sem o qual a realização deste trabalho não seria possível.
i

ii

Índice Agradecimentos ................................................................................................... i Resumo ................................................................................................................ 1 Abstract ................................................................................................................ 2 Introdução ............................................................................................................ 3 Ingestão de substâncias doces e ansiedade ..................................................................... 3 Ingestão de substâncias doces e outros comportamentos apetitivos ............................... 5 Comportamento exploratório e ansiedade ou medo .......................................................... 7 Manuseio e o comportamento de ratos ........................................................................... 12 Problemas e objetivos ..................................................................................................... 13 Experimento I ..................................................................................................... 16 Método ............................................................................................................................. 18 Sujeitos .................................................................................................................... 18 Equipamento ............................................................................................................ 18 Procedimento ........................................................................................................... 19 Medida da ingestão de açúcar ......................................................................... 19 Teste no labirinto em cruz elevado .................................................................. 20 Medida da ingestão de açúcar como uma função do peso dos animais ......... 20 Análise estatística .................................................................................................... 20 Resultados ....................................................................................................................... 21 Discussão ........................................................................................................................ 29 Experimento II .................................................................................................... 38 Método ............................................................................................................................. 40 Sujeitos .................................................................................................................... 40 Equipamento ............................................................................................................ 40 Procedimento ........................................................................................................... 41 Medida de ingestão .......................................................................................... 41 Labirinto em cruz elevado ................................................................................ 41 Análise estatística .................................................................................................... 42 Resultados ....................................................................................................................... 42 Discussão ........................................................................................................................ 45 Experimento III ................................................................................................... 51 Método ............................................................................................................................. 53 Sujeitos .................................................................................................................... 53 Equipamento ............................................................................................................ 53 Procedimento ........................................................................................................... 54 Campo aberto .................................................................................................. 54 Labirinto em cruz elevado ................................................................................ 55 Ingestão de açúcar ........................................................................................... 55 Análise estatística .................................................................................................... 55 Resultados ....................................................................................................................... 56 Discussão ........................................................................................................................ 64 Experimento IV .................................................................................................. 69 Método ............................................................................................................................. 70 Sujeitos .................................................................................................................... 70 Equipamento ............................................................................................................ 71
iii

Procedimento .......................................................................................................... 71 Sessões de consumo ...................................................................................... 71 Labirinto em cruz elevado ............................................................................... 71 Drogas.............................................................................................................. 71 Análise estatística ................................................................................................... 72 Resultados ...................................................................................................................... 72 Discussão ........................................................................................................................ 76 Discussão Geral ............................................................................................... 81 Preferência por substâncias de gosto doce ............................................................ 81 Ingestão de substâncias palatáveis e modelos de ansiedade ................................ 83 Considerações finais ............................................................................................... 88 Referências Bibliográficas ............................................................................... 91
iv

RESUMO Vários estudos mostraram que existem diferenças individuais no consumo de substâncias doces, um fenômeno observado em roedores e humanos. Além disso, alguns trabalhos mostraram que essa variabilidade individual no consumo das substâncias doces prediz diferenças individuais em outros comportamentos. Dessa forma, o presente trabalho teve o objetivo de verificar se existe uma relação entre ingestão de substâncias doces e comportamentos relacionados à ansiedade ou à atividade locomotora em ratos. Para isso, quatro experimentos foram realizados. No Experimento I, ratos tiveram acesso a açúcar granulado ou ração moída durante uma hora por oito dias consecutivos. Ratos bons e maus consumidores de açúcar foram classificados com base na média de ingestão de açúcar e, posteriormente, foram testados no labirinto em cruz elevado. Os ratos que ingeriram maiores quantidades de açúcar apresentaram maiores freqüências de comportamentos relacionados à ansiedade ou à atividade locomotora. Esse efeito pode ser função da ingestão de açúcar ou do manuseio durante o período de ingestão. Baseado nisso, o objetivo do Experimento II foi o de verificar se o procedimento de manuseio e a ingestão de açúcar tinham efeitos independentes sobre as medidas comportamentais registradas no labirinto em cruz elevado. Para isso, ratos foram submetidos a diferentes períodos de manuseio (0, 1, 5 e 10 min) durante um período de 14 dias. Esse manuseio consistiu em alisar o pelo do dorso do rato com o dedo. Metade desses ratos teve acesso a açúcar e ração logo após o manuseio, enquanto a outra metade não. Um outro grupo de animais apenas permaneceu no biotério durante esse período. Os resultados mostraram que à medida que aumentou o tempo de manuseio diminuiu a quantidade de açúcar ingerida, sem alteração da quantidade de ração ingerida. Os registros comportamentais no labirinto foram inconsistentes, pois o procedimento de manuseio e a ingestão de açúcar afetaram tanto as medidas relacionadas à ansiedade como a atividade locomotora no labirinto. Pode ser que o procedimento de manusear os animais por 14 dias os tenha tornado menos ansiosos no labirinto. Em função disso, o objetivo do Experimento III foi o de minimizar o efeito do manuseio da fase de ingestão e verificar se existe correlação entre ingestão de açúcar e comportamentos relacionados à ansiedade ou à atividade locomotora. Para isso, utilizaram-se dois modelos animais, o campo aberto e o labirinto em cruz elevado. Somente após realizados os testes no campo aberto e no labirinto, realizou-se o procedimento de sete dias de ingestão. Os animais que ingeriram maiores quantidades de açúcar comportaram-se como menos ansiosos no campo aberto e locomoveram-se mais no labirinto. Isso mostrou que essas correlações são independes do fator manuseio. Baseado nisso, o objetivo do Experimento IV foi o de verificar se esses efeitos deveram-se à percepção do gosto doce ou ao aumento de açúcar no sangue. Para isso os animais foram distribuídos em quatro grupos: ratos que tiveram acesso a açúcar e ração por uma hora diária durante sete dias, ratos que tiveram acesso a açúcar e ração por somente uma hora, ratos que receberam uma única injeção de glicose (1 M, 1 ml/kg, i.p.) 30 minutos antes do teste no labirinto, e ratos que apenas permaneceram no biotério por sete dias. Após as sessões de ingestão dos respectivos grupos, os ratos foram imediatamente testados no labirinto. Tanto os ratos do grupo que tiveram acesso ao açúcar por sete dias como os do grupo que recebeu injeção de glicose comportaram-se como menos ansiosos e mais ativos no labirinto. Concluiu-se que ingerir açúcar torna os ratos menos ansiosos, mas os procedimentos não permitiram deslindar quais os mecanismo subjacentes a esse efeito. Palavras-chave: Ingestão de açúcar; manuseio experimental; comportamento exploratório; ansiedade; labirinto em cruz elevado; campo aberto; ratos.
1

ABSTRACT Several studies have demonstrated the existence of individual differences in the consumption of sweet substances, a phenomenon observed in rodents and humans. Besides, some reports have shown this individual variability in the consumption of sweet substances to predict individual differences in other behaviors. Thus, the present work aimed at investigating whether there is a relationship between the ingestion of sweet substances and behaviors related to anxiety or locomotor activity in rats. For this, four experiments were carried out. In Experiment I, rats had access to commercial sugar or ground rat chow during one hour for eight consecutive days. Good and bad sugar consumer rats, thus classified on the basis of the average sugar ingestion, were later tested in the elevated plus-maze. Rats that ingested larger amounts of sugar exhibited larger frequencies of behaviors related to anxiety and locomotor activity. Such an effect could have been due to sugar ingestion or to the handling during the consumption period. Based on this, the aim of Experiment II was to investigate whether handling procedure and sugar ingestion had independent effects on behavioral measures recorded in the elevated plus-maze. For this, rats were submitted to different handling periods (0, 1, 5 and 10 min) for a period of 14 days. Handling consisted of combing a rat’s back fur with the finger. Half of the subjects had access to sugar and chow immediately after handling while the other half did not. Another group of rats just remained in the animal room during this period. Results showed that the longer the handling, less sugar was ingested without alterations in chow consumption. Behavioral records in the plus-maze were inconsistent, since both handling and sugar ingestion affected both anxiety- and locomotion-related measures. It could be the case that handling the animals for 14 days has rendered them less anxious inn the plus-maze. Because of this, the objective of Experiment III was to attempt at minimizing the effect of handling during the consumption period and investigate whether there is a correlation between sugar ingestion and anxiety- and locomotor-related behaviors. Two animal models were used: the open-field and the elevated plus-maze. The rats were tested in the open-field and, 24 h later, in the elevated plus-maze. Only after both tests were the animals submitted to the 7-day consumption period. Rats that ingested larger amounts of sugar behaved as less anxious and perambulated more in the plus-maze. This showed that these correlations are independent of handling. Based on this, Experiment IV aimed at investigating whether these effects were due to the perception of sweet taste or to the increase in blood sugar. For this, the animals were divided into four groups: rats that had daily 1-h periods of access to sugar and chow for seven days, rats that had access only to a 1-h session with sugar and chow available, rats that received a single i.p. glucose injection (1 M, 1 ml/kg) 30 min before being tested in the plus-maze, and rats that just remained in the animal room for seven days. All animals were tested in the elevated plus-maze after the consumption session(s) or injection. Both the rats that had access to sugar for seven days and the ones that were injected with glucose behaved in a less anxious manner and were more active in the plus-maze. It was concluded that the more sugar the rats ingested the less anxious they behaved and the move they moved about the plus-maze but the procedures did not allow uncovering the mechanisms underlying this effect. Keywords: Sugar ingestion; experimental handling; exploratory behavior; anxiety; elevated plus-maze; open-field; rats.
2

INTRODUÇÃO
Ingestão de substâncias doces e ansiedade
Um dos primeiros efeitos documentados das drogas ansiolíticas, tais como os
benzodiazepínicos, foi sua habilidade para aumentar a ingestão de comida e de água em
ratos e cães. Randall et al., (1960) observaram que a administração aguda de
clordiazepóxido aumentou a quantidade de comida ingerida por ratos. Os mesmos autores
também examinaram o efeito da administração crônica dessa droga sobre o comportamento
de ingestão em cachorros. Eles mostraram que a quantidade de comida ingerida dobrou
quando a droga foi administrada por um período de cinco dias. Com base nessa observação,
os autores concluíram que a droga estimula o apetite nesses animais.
Além de se observar que as drogas benzodiazepínicas aumentavam a quantidade de
comida ingerida por ratos saciados em uma condição de livre acesso á comida (Poschel,
1971), alguns trabalhos também mostraram que essas drogas também aumentavam a
ingestão de comida na presença de condições com caráter aversivo. O clordiazepóxido
aumentou a freqüência do comportamento de pressão à barra, contingente ao recebimento
de comida e choque, e a ingestão suprimida por aversão condicionada ao gosto (Margules e
Stein, 1967). Esses mesmos autores também observaram que o oxazepam aumentou a
freqüência do comportamento de pressão à barra suprimido por um procedimento de
extinção além de também aumentar a ingestão suprimida por gosto amargo.
Esse efeito facilitador da ingestão de comida e de líquidos foi observado em
animais saciados e privados (Cole, 1983), com comida familiar (Xenakis e Sclafani, 1981)
e não familiar (Shepard e Estall, 1984), em ambiente familiar (Soubrié et al., 1975) ou
novo (Shepard e Estall, 1984), e em grande variedade de espécies, tais como o rato
3

(Cooper e Francis, 1979a e b), gato (Della-Fera et al., 1980), cavalos (Brown et al., 1976),
macaco Rhesus (Foltin e Schuster, 1983) e humanos (Ayd, 1962).
Digno de nota é o fato de alguns trabalhos mostrarem que as drogas
benzodiazepínicas parecem ter o efeito de aumentar a quantidade ingerida de comida
palatável em uma condição em que há escolha. Animais que receberam drogas
benzodiazepínicas aumentaram significativamente o consumo de comida ou de líquidos
palatáveis (comida doce ou água com sacarina) comparado ao consumo de ração de
laboratório ou de água (Cooper, 1987; Cooper e McClelland, 1980; Cooper e Yerbury,
1988; Parker, 1991). Esse mesmo efeito de aumentar a ingestão de substâncias palatáveis
em ratos também foi observado em estudos nos quais uma fístula gástrica permitia a
retirada do que era ingerido, anulando assim a modulação da ingestão por eventos pós-
ingestivos (Cooper e Yerbury, 1988). Bainbridge (1968) mostrou que fêmeas de rato que
receberam clordiazepóxido aumentaram a freqüência da resposta de pressão à barra
mantida pela apresentação de leite condensado adocicado. Esses resultados sugerem que as
drogas benzodiazepínicas afetam a percepção do gosto palatável. Poschel (1971) relatou
que diazepam, nitrazepam, oxizapem e medazepam aumentaram o consumo de leite
condensado adocicado somente na primeira exposição dos ratos a essa comida. Dessa
forma, ele sugeriu que esse aumento na ingestão seja função de um efeito da droga sobre a
resposta neofóbica inicial ao leite adocicado.
Esse aumento na ingestão induzida por drogas benzodiazepínicas ainda não é bem
compreendido. Uma sugestão encontrada na literatura é que o aumento na ingestão resulta
de uma influência emocional inibitória atenuada pela droga. Tal influência emocional
poderia estar suprimindo a ingestão ou comportamento relacionado à ingestão (Margules e
Stein, 1967), uma hipótese baseada na redução no estado de ansiedade. Uma segunda
interpretação encontrada na literatura é que as drogas benzodiazepínicas têm uma ação
direta sobre algum mecanismo específico de ingestão, isto é, pode inibir a ação de algum
4

mecanismo de saciedade ou estimular algum mecanismo de fome (Margules e Stein, 1967).
Diferente da hipótese anterior, esta não considera relevante o estado de ansiedade.
Margules e Stein (1967) sugeriram que ambas as hipóteses devem ser consideradas.
Quando a ingestão é emocionalmente suprimida, os benzodiazepínicos agiriam sobre o
mecanismo emocional; quando suprimida apenas por fatores de saciedade, agiriam sobre o
mecanismo de saciedade.
Ingestão de substâncias doces e outros comportamentos apetitivos
Ha na literatura, além de trabalhos mostrando que as drogas benzodiazepínicas
aumentam o consumo de comida e induzem preferência por comida palatável, outros
mostrando ainda que os ratos exibem considerável variabilidade em sua propensão a
ingerir uma substância doce (De Cabo et al., 1995; Sills e Crawley, 1996; DeSouza et al.,
1998). Adicionalmente, essa variabilidade individual na ingestão de substância doce pode
predizer diferenças individuais em resposta a aspectos apetitivos, tais como exploração,
aproximação e reforço (DeSouza et al., 1998). Por exemplo, ingestão elevada de sacarina
correlaciona-se com ingestão elevada de etanol (Kampov-Polevoy et al., 1990; Gosnell e
Krahn, 1992; Overstreet et al., 1993) e ratos seletivamente criados como bons ou maus
consumidores de etanol consumiram, respectivamente, quantidades elevadas ou baixas de
sacarina (Sinclair et al., 1992). Gosnell et al., (1995) mostraram que ratos com alta
preferência por sacarina auto-administraram mais morfina que ratos com baixa preferência.
Além disso, ratos que ingeriram quantidades elevadas de sacarose auto-administraram
psicoestimulantes intravenosamente mais prontamente que ratos que ingeriram baixas
quantidades de sacarose (DeSouza et al., 2000).
Paralelamente às observações de que os ratos apresentam diferenças individuais no
consumo de substâncias doces, observou-se ainda que os ratos exibem padrões diferentes
5

de atividade locomotora em ambientes novos. Alguns ratos apresentam altas taxas de
atividade locomotora enquanto outros apresentaram baixas taxas da mesma (Piazza et al.,
1989). Interessante é o fato de que alguns pesquisadores mostraram que a taxa de atividade
locomotora induzida por ambientes novos pode predizer a resposta a drogas de abuso
(Piazza et al., 1989), como se observou nas diferenças individuais de consumo de
substâncias doces e nas respostas a essas drogas.
Assim, animais que apresentaram altas taxas de atividade locomotora em ambientes
novos auto-administraram anfetamina mais rapidamente e foram mais sensíveis aos efeitos
estimulantes da anfetamina sobre a atividade locomotora do que os animais que
apresentaram baixas taxas de atividade locomotora em ambientes novos. A atividade
locomotora induzida por um ambiente novo também permitiu prever: (1) a atividade
locomotora induzida por cocaína e cafeína (Hooks et al., 1992), (2) atividade locomotora e
auto-administração induzida por etanol (Gingras e Cools, 1995), e (3) hiperatividade
locomotora ao estímulo contextual condicionado em resposta aos efeitos da anfetamina
(Jodogne et al., 1994).
Estudos de microdiálise in vivo contribuíram para ressaltar ainda mais as
semelhanças entre ratos com altas taxas de atividade locomotora em ambiente novo e ratos
que consomem quantidades elevadas de açúcar. Ratos que apresentaram os níveis mais
altos de atividade locomotora induzida por anfetamina, assim como ratos que consumiram
quantidades elevadas de açúcar, mostraram atividade dopaminérgica elevada no núcleo
acumbente, quando comparados, respectivamente, a ratos com níveis mais baixos de
atividade locomotora induzido por anfetamina (Piazza et al., 1990; Hooks et at., 1992) e a
ratos que consumiram quantidades baixas de açúcar (Sills e Vaccarino, 1991; Sills et
al.,1993).
Esses dados sugerem que, quando selecionamos ratos que consomem quantidades
elevadas de açúcar livremente disponível, também estamos selecionando ratos que
6

apresentam atividade locomotora elevada em ambientes novos. Além disso, Sills e
Vaccarino (1994) relataram que níveis elevados de ingestão de açúcar predizem atividade
locomotora elevada em resposta à administração de anfetamina. Os mesmo autores
também sugeriram que os mecanismos que regulam a ingestão de açúcar e atividade
locomotora induzida por anfetamina se sobrepõem.
É interessante notar que alguns trabalhos mostram que essa variabilidade
locomotora em resposta a ambientes novos pode servir para prever comportamentos de
ratos em modelos animais de exploração livre. Quando comparados a ratos com baixa taxa
de atividade locomotora em ambientes novos, ratos com alta taxa de atividade locomotora
em ambientes novos apresentam menor freqüência de comportamentos relacionados à
ansiedade em testes tais como a caixa claro-escuro e o labirinto em cruz elevado (Kabbaj et
al., 2000). Um fato interessante é que os ratos com alta taxa de atividade locomotora em
ambientes novos mostraram os mais altos níveis de corticosterona na corrente sangüínea,
quando comparado a ratos com baixa taxa de atividade locomotora em ambientes novos
(Piazza et al., 1991; Dellu et al., 1996; Kabbj et al., 2000). Adicionalmente, DeSouza et al.,
(1998) relataram que ratos bons consumidores de sacarose mostraram resposta de
sobressalto a um estímulo acústico de magnitude menor, em comparação com ratos maus
consumidores de sacarose. Os mesmos autores também relataram que os ratos que
preferiram sacarose apresentaram menor freqüência de comportamentos relacionados à
ansiedade no labirinto em cruz elevado quando comparados a ratos que não demonstraram
preferência por sacarose.
Comportamento exploratório e ansiedade ou medo
O comportamento exploratório é estudado em ratos selvagens e de laboratório.
Entretanto, Halliday (1966) relatou que situações que induzem movimentos exploratórios
7

também induzem comportamentos tais como defecação e congelamento (freezing). Dessa
maneira, sugeriu que, ao menos em ratos, a resposta à novidade pode ser interpretado em
termos de duas tendências opostas: exploração e medo. Como estados de medo não são
acessíveis ao experimentador, utilizar registros de comportamentos relacionados à
fisiologia ajudam a inferir sobre esse estado. Ratos selvagens nem sempre respondem a um
estímulo novo aproximando-se, apesar de apresentarem comportamento exploratório.
Nessa situação, é mais provável observá-los imóveis (freezing) ou simplesmente parados
por um período de tempo quando colocados frente a algo estranho (Bindra e Spinner,
1958). Esses comportamentos também podem ser observados em ratos de laboratório
quando arranjadas as devidas condições. Essas respostas à novidade são comumente
chamadas de medo (Barnett, 1975). Vários autores já discutiram os aspectos aversivos de
objetos ou ambientes não familiares sobre o comportamento de ratos (Halliday, 1966;
Berlyne, 1950). A esquiva de objetos ou ambientes estranhos e também de animais
estranhos de espécie igual ou diferente é comum no reino animal. Esse comportamento de
esquiva é aprendido na vida precoce de muitas espécies de aves e mamíferos durante o
período em que há investimento parental (Barnett, 1975). A combinação dos
comportamentos de explorar e de esquivar-se é provavelmente o resultado de pressões
seletivas que maximizaram a maneira desses animais interagirem com os recursos e
perigos de seu ambiente (Barnett, 1975).
Um dos modelos mais utilizados no estudo da ansiedade, o labirinto em cruz
elevado, foi desenvolvido na década de 50 a partir de trabalhos sobre comportamento
exploratório. Em um estudo clássico, Montgomery (1951), que já havia demonstrado
que, devido à habituação, o comportamento exploratório do rato diminui com o tempo
de exposição ao aparelho, estudou de que maneira ratos exploram um labirinto em
forma de Y com dois braços abertos e um fechado, com paredes baixas e elevado do
chão (Montgomery, 1955). Nessa ocasião, ele propôs que a estimulação oriunda da
8

novidade promove tanto o comportamento de medo como o de exploração, gerando um
conflito do tipo aproximação-esquiva. Considerou que tanto os braços abertos como os
fechados evocam o comportamento exploratório, mas que os primeiros evocam também
o medo com maior intensidade do que os últimos, resultando numa maior exploração
dos braços fechados pelo animal quando colocado nesse tipo de labirinto. Relatou ainda,
que a força do medo evocado pela situação nova diminuia com o tempo de exposição
direta à estimulação e que, depois de um período sem exposição, a re-exposição ao
mesmo estímulo resulta em recuperação espontânea do medo.
A partir desses resultados, Morrison e Stephenson (1970) sugeriram que o número
de entradas e o tempo de permanência do rato no braço aberto do labirinto em Y
elevado poderiam ser aumentados com drogas que diminuíssem a ansiedade. Handley e
Mithani (1984) utilizaram pela primeira vez um labirinto em cruz elevado, constituído
de dois braços fechados por paredes baixas (10 cm), cruzados por dois braços abertos.
Nele elas observaram que os ratos apresentavam uma preferência pela exploração dos
braços fechados e que drogas ansiolíticas aumentavam o número de entradas nos braços
abertos enquanto as ansiogênicas o diminuíam. Em 1985, Pellow e colaboradores
validaram o labirinto em cruz elevado como modelo de ansiedade dos pontos de vista
farmacológico, fisiológico e comportamental., Seu aparato era constituído por dois
braços abertos e dois fechados com paredes altas (desta vez com os clássicos 40 cm de
altura) e sua funcionalidade como modelo animal de ansiedade baseava-se na atividade
exploratória e na já descrita aversão natural dos ratos pelos braços abertos. Esta aversão
aos braços abertos parece ser desencadeada pelo espaço aberto dos braços sem paredes e
parece estar ligada a uma tendência ao tigmotatismo e não desencadeada pela altura
(Treit et al., 1993).
9

Neste modelo, os indicadores de ansiedade são calculados a partir do número de
entradas e do tempo gasto nos braços abertos do labirinto, correlacionando-se
negativamente com a ansiedade. Esses indicadores são freqüentemente aumentados
pelas drogas ansiolíticas clássicas e diminuídos pelas drogas ansiogênicas (Handley e
Mithani, 1984; Pellow et al., 1985; Pellow e File, 1986; Guimarães et al., 1990; Cruz et
al., 1994). Pellow et al., (1985) mostraram que a freqüência de entradas, mas
principalmente a porcentagem de entradas, nos braços abertos do labirinto são boa
medida de ansiedade enquanto o número total de entradas relaciona-se com as
tendências exploratórias e locomotoras do animal., Recentemente, alguns estudos
mostraram que a atividade motora geral estaria mais relacionada com o número de
entradas nos braços fechados do labirinto em cruz elevado (File, 1992; Cruz et al.,
1994) mas o tempo gasto nesses braços não reflete a atividade geral sendo, ao invés, um
indicador de ansiedade da mesma forma que as medidas nos braços abertos (File, 1992;
Cruz et al., 1994).
Alguns trabalhos têm mostrado que a suscetibilidade dos ratos para explorar os
braços abertos do labirinto é influenciada por muitos fatores. Morato e Castrechini
(1989) mostraram que quanto maior o nível de luminosidade ambiental, menor a
exploração dos braços abertos. Maisonnette et al., (1993) mostraram que o alojamento
individual torna os ratos mais sensíveis à aversividade dos braços abertos, reduzindo sua
exploração. Da mesma forma, o número de animais alojados por caixa (Botelho et al.,
2007), as condições de alojamento e transporte (Morato e Brandão, 1996, 1997), a idade
dos animais testados (Frussa-Filho et al., 1991; Wright et al., 1991), a cepa (Rex et al.,
1996) e o sexo dos mesmos (Steenbergen et al., 1990) também modificaram a
exploração dos braços abertos.
10

Em 1987, Lister, usando camundongos suíços NIH, demonstrou a generalidade de
espécies que o modelo é capaz de abranger. Empregando uma versão do labirinto em
escala proporcional ao tamanho dos camundongos e a mesma duração de sessão, ele
encontrou resultados comportamentais e farmacológicos comparáveis aos de ratos e
concluiu que essa versão modificada do labirinto em cruz elevado para camundongos
também era um teste útil para o estudo de compostos ansiolíticos e ansiogênicos.
Embora o efeito ansiolítico obtido com drogas que agem no complexo receptor
GABA-benzodiazepina seja consistente no labirinto em cruz elevado (Rodgers e Cole,
1994), outros tipos de droga geralmente produzem resultados mais variáveis. Resultados
inconsistentes foram obtidos com drogas serotonérgicas usando os indicadores clássicos
de ansiedade no labirinto em cruz elevado. Por exemplo, foi relataram-se efeitos
ansiolítico (Almeida et al., 1991; Critchley et al., 1992), ansiogênico (Critchley e
Handley, 1987; Pellow et al., 1987) e ausência de efeito (Critchley et al., 1992; Pellow e
File, 1986) de tratamentos com alguns agonistas 5-HT1A. Resultados inconsistentes
também foram relatados para antagonistas de receptores 5-HT2 (Pellow et al., 1987) e 5-
HT3 (File e Johnston, 1989).
Quando os animais exploram um labirinto em cruz elevado, além dos indicadores
convencionais de ansiedade, é possível observar ainda uma variedade de
comportamentos passíveis de análise mais detalhada. Lorenz (1981) salienta que a
principal característica do estudo dessa variedade de comportamentos é investigar sob
quais circunstâncias um dado animal de uma determinada espécie, idade e estado
emocional exibe um comportamento funcional sem necessidade de treinamentos
específicos. Comportamentos desse tipo foram investigados há algum tempo no
labirinto em cruz elevado. Cruz e colaboradores (1994), utilizando uma análise do
comportamento do rato neste modelo, chamada por eles de etofarmacológica,
11

demonstraram que algumas categorias comportamentais podiam se somar aos índices
tradicionais de ansiedade, já que foram alteradas da mesma maneira pela administração
de dois ansiolíticos (nitrazepam e diazepam) e de maneira oposta pela administração de
duas drogas ansiogênicas (pentilenotetrazol e FG 7142). Por exemplo, a exploração das
extremidades dos braços abertos do labirinto em cruz elevado também pode ser
considerado um bom indicador de ansiedade (Cruz et al., 1994). Neste estudo, o tempo
gasto nas extremidades foi menor em ratos tratados com pentilenotetrazol e FG 7142 e
maior nos animais tratados com midazolam e nitrazepan, em relação aos respectivos
grupos controle. Outro comportamento, o de levantar-se nos quartos traseiros (rearing),
foi relacionado à atividade motora (Cole e Rodgers, 1993), já que sua expressão foi
diminuída pelo FG 7142 e pelo pentilenotetrazol (Cruz et al., 1994) mas não foi alterada
pelos ansiolíticos benzodiazepínicos (Cole e Rodgers, 1993; Cruz et al., 1994).
Manuseio e o comportamento de ratos
Sabe-se que o manuseio de ratos, precoce ou tardio, produz alterações
comportamentais e fisiológicas (Andrews e File, 1993; Clausing et al., 1997; Fernández-
Teruel et al., 2002). Na fase neonatal, a manipulação de ratos altera a produção de
corticosteróides, aumenta a arborização dentrítica na região do hipocampo e melhora a
memória (Meaney et al., 1990), neutraliza os efeitos da desnutrição (Levitsky e Barnes,
1972) e aumenta a quantidade de receptores de glicocorticóides no hipocampo, de GABAA
em núcleos do sistema noradrenérgico ascendente e de benzodiazepinas no córtex frontal e
em alguns núcleos do complexo amigdalóide e do sistema noradrenérgico ascendente
(Anisman et al., 1998).
No rato adulto, faz com que algumas drogas ansiogênicas percam seus efeitos
(Brett e Pratt, 1990; Andrews e File, 1993; Hogg, 1996). Também se observou um
12

aumento de atividade e de comportamentos exploratórios específicos, aumento esse
normalmente associado com uma diminuição na defecação em uma variedade de testes
envolvendo diferentes graus de novidade, como o teste do campo aberto (Denenberg et al.,
1978), teste de exposição a caixas conhecidas e não conhecidas (Ferré et al., 1995), teste da
tábua de buracos (File 1978; Fernández-Teruel et al., 1992a) e labirinto em túnel
(Fernández-Teruel et al., 1991). Entretanto, quando foram usados testes mais específicos
medindo ansiedade ou medo (incondicionado ou condicionado), observou-se um aumento
na exploração dos braços abertos do labirinto em cruz elevado (Fernández-Teruel et al.,
1990; Andrews e File, 1993; McIntosh et al., 1999) e no número de visitas e tempo gasto
nos compartimentos abertos e iluminados no labirinto em túnel e na caixa claro-escuro
(Fernández-Teruel et al., 1992b). Além disso, observou-se que, imediatamente após a
exposição a um único choque, ratos manuseados mostraram menor supressão de um
comportamento apetitivo do que ratos não manuseados (Bodnoff et al., 1987). Esses efeitos
descritos acima são caracterizados como de tipo ansiolítico.
Alguns autores relataram que esse efeito do manuseio sobre comportamentos
relacionados à ansiedade ou atividade locomotora pode ser mediado pela alteração no
sistema GABA-benzodiazepina (Biggio, 1983; File et al., 1990; File et al., 1992). Essas
diferenças são similares àquelas associadas ao tratamento com benzodiazepínicos, que
reduz a supressão de um comportamento apetitivo mantido pela exposição a um ambiente
novo ou a um evento aversivo (Bodnoff et al., 1987).
Vários trabalhos relataram que o tratamento com benzodiazepínicos alterou o
comportamento de ingerir comida. Os resultados descritos acima sugerem que o manuseio
altera o comportamento exploratório de ratos e também o sistema GABA-benzodiazepina.
Dessa forma, o manuseio pode ser uma variável que afeta o comportamento de ingerir
comida ou líquidos palatáveis. Quando realizada uma busca na literatura não se encontrou
13

quaisquer trabalhos que tenham investigado o efeito do manuseio sobre o comportamento
de ingerir comida ou líquidos palatáveis.
Problemas e objetivos
O aumento do apetite e da ação sedativa do clordiazepóxido foram descritos juntos
nos primeiros relatos lidando com o 1,4-benzodiazepina (Randall et al., 1960).
Adicionalmente, esses efeitos foram demonstrados utilizando-se outros compostos
benzodiazepínicos. Entretanto, o interesse clínico e experimental dessas drogas ocorreu em
função de seus efeitos ansiolítico, sedativos e anti-punição. Apesar do pouco interesse no
efeito dessas drogas sobre a ingestão de alimentos e líquidos, alguns trabalhos começaram
a descrever a relação entre essas drogas e ingestão. Além de mostrarem que essas drogas
afetam a ingestão de alimentos, um efeito mais específico foi observado: sempre que havia
disponibilidade de dois ou mais alimentos com diferenças de palatabilidade, drogas que
agem no sistema GABA-benzodiazepina aumentaram a quantidade ingerida de substâncias
palatáveis, sobretudo quando comparada à quantidade ingerida de ração de laboratório
(Margules e Stein, 1967). A literatura apresenta duas hipóteses para a relação entre
benzodiazepínicos e ingestão de comida palatável: essas drogas podem agir sobre estados
de ansiedade ou sobre mecanismos relacionados à fome e saciedade. Ainda não há dados
suficientes que permitam sair desse impasse.
Paralela a essas observações, muitos trabalhos mostraram que os ratos variam
quanto à ingestão livre de substâncias palatáveis (De Cabo et al., 1995; DeSouza et al.,
1998). Quando se classificam ratos pelas quantidades consumidas de substâncias palatáveis,
de forma a podermos ter bons e maus consumidores, podemos predizer alguns
comportamentos como: auto administração de drogas, atividade locomotora em ambientes
novos em resposta ao tratamento com anfetamina, e atividade dopaminérgica no núcleo
acumbente. Adicionalmente, a atividade locomotora de ratos em ambientes novos também
14

foi correlacionada aos comportamentos descritos acima (Piazza et al., 1990; Hooks et at.,
1992). Esses trabalhos mostraram que se pode usar uma classe de comportamentos para
predizer outras.
É interessante o fato de que ratos que ingeriram muita comida palatável, em
detrimento de ratos que ingeriram pouca, também auto-administraram com maior
freqüência algumas drogas (Gosnell et al., 1995; DeSouza et al., 2000). Levando-se em
consideração que drogas benzodiazepínicas alteram a ingestão de substâncias palatáveis,
podemos utilizar a ingestão das mesmas para prever comportamentos relacionados à
ansiedade em modelos animais de ansiedade?
Baseado nessas colocações, torna-se pertinente verificar se ingestão de comida
palatável pode funcionar como um evento preditivo de comportamentos relacionados à
ansiedade em ratos. Pois, se os benzodiazepínicos agem de acordo com a primeira hipótese,
pode-se concluir que os ratos respondem de maneira diferente (mais ou menos ansiosos)
quando exposto em ambientes aversivos, sendo que a quantidade ingerida de comida
palatável pode ser um indicador dessa maneira de responder. O Experimento I verificou
como ratos classificados como bons ou maus consumidores de açúcar se comportam em
modelos animais de ansiedade. Sabendo que o manuseio afeta o comportamento de ratos
no labirinto em cruz elevado de maneira semelhante ao tratamento com benzodiazepínicos
(Bodnoff et al., 1987), o Experimento II verificou se o manuseio durante o procedimento
de ingestão pode afetar a ingestão ao mesmo tempo que o comportamento no labirinto em
cruz elevado. O Experimento III teve o objetivo de tentar anular os efeitos do manuseio
durante o procedimento de ingestão para verificar com mais especificidade se há
correlação entre ingestão e comportamento exploratório, em condições de exploração livre.
Finalmente, o Experimento IV teve o objetivo de verificar se ingestão prévia de açúcar
altera o comportamento de ratos no labirinto em cruz elevado.
15

Ações independentes dos benzodiazepínicos sobre a ingestão ou ansiedade têm
importantes implicações clínicas. Complicações podem surgir quando pessoas ingerem
essas drogas com finalidade ansiolítica, pois pode ocorrer aumento de ingestão com
conseqüente aumento de peso. Adicionalmente, membros da família dos benzodiazepínicos
podem ser descritos ou desenvolvidos com função relativamente específica para o
comportamento de ingestão, de forma a poder se tornar uma ferramenta valiosa no
tratamento de alguns casos de distúrbio alimentar.
Experimento I Vários estudos mostraram que existem diferenças individuais no consumo de
substâncias doces, um fenômeno notado em roedores e humanos (Brennan et al., 2001).
Assim, quando açúcar granulado está disponível, a quantidade consumida varia
amplamente entre os ratos (DeSouza et al., 1998; De Cabo et al., 1995; Sills e Crawley,
1996). Alguns trabalhos mostraram que essa variabilidade individual no consumo de
substâncias doces prediz diferenças individuais em outros comportamentos com perfil
apetitivo. Por exemplo, alta ingestão de sacarina correlaciona-se com alta ingestão de
etanol (Kampov-Polevoy et al., 1990; Gosnell e Krahn 1992; Overstreet et al., 1993) e
ratos seletivamente criados como bons ou maus consumidores de etanol consumiram,
respectivamente, altas ou baixas quantidades de sacarina (Sinclair et al., Li, 1992). Gosnell
et al. (1995) mostraram que ratos com alta preferência por sacarina auto-administram mais
morfina que ratos com baixa preferência. Além disso, ratos que ingerem quantidades
elevadas de sacarose auto-administram psicoestimulantes intravenosamente mais
prontamente que ratos que ingerem baixas quantidades de sacarose (DeSouza et al., 2000).
Paralelamente às observações de que ratos apresentam diferenças individuais no
consumo de substâncias doces, observou-se que ratos exibem padrões diferentes de
atividade locomotora em ambientes novos. Alguns ratos apresentam altas taxas de
16

atividade locomotora enquanto outros apresentam baixas taxas de atividade locomotora
(Piazza et al., 1989). Interessante é o fato de que alguns pesquisadores mostraram que a
taxa de atividade locomotora induzida por ambientes novos correlaciona-se com respostas
a drogas de abuso (Piazza et al, 1989), como observado em relação às diferenças
individuais no consumo de substâncias doces e resposta a essas mesmas drogas.
Segundo Dess e Minor (1996), por outro lado, o comportamento de ingestão está
relacionado com comportamentos afetivos tais como medo e ansiedade. Ratos
seletivamente criados para exibir um fenótipo de emocionalidade diferem quanto à
ingestão de sacarina (Gray, 1987; Overstreet et al., 1993). As linhagens que apresentaram
maior freqüência de comportamentos relacionados à ansiedade tomaram menor quantidade
de sacarina que as linhagens que apresentaram menor freqüência desses comportamentos.
DeSouza et al (1998) relataram que ratos tidos como bons consumidores de sacarose
mostraram resposta de sobressalto de magnitude menor diante de um estímulo acústico, em
comparação com ratos tidos como maus consumidores de sacarose. Os mesmos autores
também relataram que ratos que preferiram sacarose apresentaram freqüência menor de
comportamentos relacionados à ansiedade no labirinto em cruz elevado quando
comparados a ratos que não demonstraram preferência por sacarose.
Se ratos que preferem substâncias doces comportam-se como ratos que apresentam
atividade locomotora elevada em ambientes novos, pode ser que os dados relatados por
DeSouza et al (1998) não estejam relacionados somente à ansiedade. Além disso, em seu
experimento, esses autores não registraram medidas relacionadas à atividade locomotora
no labirinto em cruz elevado. Em função disso, o presente experimento teve dois objetivos.
Primeiro, fazer um procedimento semelhante ao protocolo experimental de DeSouza et al.
(1998) para verificar se ratos classificados como bons consumidores de açúcar granulado
apresentam menor freqüência de comportamentos relacionados à ansiedade no labirinto em
cruz elevado, quando comparados a ratos classificados como maus consumidores de açúcar
17

granulado. Segundo, verificar se os ratos que preferem açúcar granulado à ração de
laboratório moída apresentam atividade locomotora aumentada no labirinto em cruz
elevado em relação aos ratos que não demonstram preferencia por açúcar granulado á
ração moída de laboratório.
Método
Sujeitos.
Foram utilizados 108 ratos machos de uma derivação Wistar provenientes do
biotério central da Universidade de São Paulo em Ribeirão Preto, pesando 200 ± 10 g. Os
sujeitos foram alojados em caixas de polipropileno (40 x 34 x 17 cm) em grupos de seis,
em um ciclo claro/escuro de 12 horas (luzes acesa as 7:00 h) e temperatura mantida entre
24 e 27° C, com livre acesso a ração comercial durante todo o experimento. Todos foram
submetidos a um período de habituação de 72 horas antes de qualquer manipulação
experimental. Quarenta sujeitos tiveram seu peso corporal medido diariamente enquanto os
demais foram pesados no início e no final do experimento.
Equipamento.
Utilizou-se um labirinto de madeira com dois braços abertos e dois fechados por
paredes laterais de 40 cm de altura. Os braços têm 50 cm de comprimento e 10 cm de
largura e estão arranjados de modo que os do mesmo tipo localizam-se em lados opostos,
unidos por um quadrado central de 10 x 10 cm. Os braços abertos são providos de bordas
de acrílico de 0,5 cm de altura. O labirinto é elevado 50 cm do chão e é iluminado por uma
lâmpada de 100 W, localizada 1,75 m acima do quadrado central. No intervalo entre as
sessões, o labirinto era limpo com uma solução de 5 % de etanol e, em seguida, seco com
uma flanela. As sessões experimentais eram gravadas com uma câmara de vídeo
18

posicionada ao lado da lâmpada e sobre o centro do labirinto. As sessões foram gravadas
em fitas VHS com um videocassete e observadas em um aparelho televisor. O
comportamento dos ratos no labirinto foi registrado em um computador com um programa
(X-Plo-Rat) desenvolvido em nosso laboratório para essa finalidade e em seguida foi
analisado observador treinado. Para avaliação do deslocamento dos animais no labirinto e
da distância percorrida no mesmo, o piso do labirinto foi dividido em quadrados de 10 cm
de lado em uma transparência de plástico colocada sobre a tela do monitor da televisão
usada para as observações.
Procedimento.
Nas sessões, o rato era gentilmente colocado no centro do labirinto com o focinho
voltado para um dos braços fechados. Cinco minutos depois encerrava-se a sessão e o
animal era retirado. Avaliaram-se as categorias comportamentais que se seguem.: (1)
entradas e tempo gasto em cada braço. Uma entrada era definida como a passagem das
quatro patas do animal de um braço para outro. (2) Entradas e tempo gasto nas
extremidades (dois últimos quadrados) dos braços abertos. (3) Distância percorrida nos
braços — estimada pelo total de cruzamento nos quadrados. (4) Tempo gasto levantando-
se, registrado sempre que o animal se colocava sobre as patas traseiras, independentemente
de apoiar-se nas paredes. (5) Tempo gasto esticando-se, registrado sempre que o animal
mantinha as patas traseiras em um lugar e esticava o tronco deslocando-se para frente com
movimentos das patas traseiras e retornando à posição original.; (6) Tempo gasto
mergulhando a cabeça, registrados sempre que a cabeça se projetava além do limite das
bordas do piso do labirinto e abaixo de seu nível, de modo que o eixo rostro-caudal da
cabeça colocava-se em uma posição quase vertical.
MEDIDA DA INGESTÃO DE AÇÚCAR. Durante esta fase utilizou-se açúcar comercial
granulado. Depois da habituação de 72 h, iniciou-se a fase da medida de ingestão de
19

açúcar. Por oito dias, em uma sessão diária de 60 min, os animais eram colocados
individualmente em uma caixa análoga àquela onde estavam alojados, com dois
comedouros de aço inoxidável contendo, respectivamente, açúcar comercial e ração moída.
Os comedouros eram pesados no início e final de cada sessão. A seguir, os animais eram
colocados de volta em suas gaiolas. Esta fase permitiu classificar os ratos conforme o
consumo de açúcar e distribuí-los em dois grupos com base na ingestão dos dois últimos
dias. No grupo de alto consumo, estavam os animais com consumo superior a um desvio
padrão acima da média de todos os ratos; no grupo de baixo consumo, estavam os animais
com consumo inferior a um desvio padrão abaixo da média de todos os ratos. Os animais
que não preencheram nenhum desses critérios foram utilizados apenas em parte da análise.
TESTE NO LABIRINTO EM CRUZ ELEVADO. Vinte e quatro horas após a última sessão de
ingestão, os animais foram testados no Labirinto em Cruz Elevado. Por ocasião do teste, os
ratos eram colocados gentilmente no quadrado central do labirinto com o focinho voltado
para um dos braços fechados e permitiu-se que explorassem livremente por cinco minutos.
Durante esses cinco minutos de sessão, foram registrados a freqüência de entradas em cada
tipo de braço, o tempo de permanência nos braços abertos, a distância percorrida nos dois
tipos de braços (estimada a partir do número de quadrados atravessados), as entradas e
tempo de permanência nas extremidades dos braços abertos, o tempo gasto nos
comportamentos de esticar-se, mergulhar a cabeça e levantar-se.
MEDIDA DA INGESTÃO DE AÇÚCAR COMO UMA FUNÇÃO DO PESO DOS ANIMAIS. Para verificar
se a ingestão de açúcar ou ração variava com o peso dos animais calculou-se quantos
gramas de açúcar ou ração foram ingeridos por quilo de rato, tanto para os animais
classificados como alto ou baixo consumidores. Os animais foram submetidos às mesmas
condições descritas na fase de ingestão de açúcar, sendo pesados a cada dois dias, em um
total de 4 pesagens. Para cada dia de pesagem foi calculada a ingestão em gramas por quilo
de rato. Em seguida foi calculada a média dessas quatro medidas.
20

Análise estatística.
Os dados coletados foram analisados com o teste t de Student e com o teste de
correlação de Pearson, ambos com nível de significância P < 0,05.
Resultados
A Figura 1 mostra o consumo de açúcar e ração dos animais extraídos de um grupo
de 68 ratos e classificados como bons (N = 7) ou maus (N = 7) consumidores de açúcar.
Como esperado, os bons consumidores de açúcar ingeriram quantidades significativamente
maiores de açúcar em comparação ao consumo de ração de laboratório (t[14] = 5,85, P <
0,001). Os maus consumidores ingeriram quantidades significativamente menores de
açúcar em comparação ao consumo da ração de laboratório (t[12] = -2,30, P = 0,04). Além
disso, os bons consumidores ingeriram quantidades significativamente maiores de açúcar
que os maus consumidores (t[14] = 5,85,; P < 0,001). A preferência por açúcar não afetou o
consumo da ração de laboratório, já que não houve diferença no consumo de ração entre
bons e maus consumidores (t[13] = -0,14,; P = 0,88).
A
*0
1
2
3
4
5
Bons Maus Bons MausTipo de seleção
Cons
umo
de a
çúca
r (g)
Açúcar Ração
B
*0
1
2
3
4
Bons Maus Bons MausTipo de seleção
Cons
umo
de ra
ção
(g)
Açúcar Ração
Figura 1. Média da ingestão de açúcar granulado e de ração de laboratório moída por ratosclassificados como bons e maus consumidores de açúcar (A) ou bons e maus consumidores deração (B). *, Diferente dos bons consumidores (t de Student, P < 0,05).
21
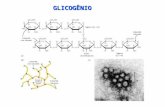
A Figura 2A mostra a ingestão de açúcar e ração dos animais extraídos de um
grupo de 40 ratos pesados diariamente e classificados como bons (N = 8) ou maus (N = 6)
consumidores de açúcar. Novamente, como esperado, os bons consumidores de açúcar
ingeriram quantidades significativamente maiores de açúcar em comparação ao consumo
de ração de laboratório (t[14] = 6,54, P < 0,001). Para os animais do grupo de baixo
consumo de açúcar não houve diferença entre o consumo de açúcar e ração de laboratório
(t[10] = -0,85, P = 0,42). Os bons consumidores de açúcar ingeriram quantidades
significativamente maiores de açúcar que os maus consumidores de açúcar (t[12] = 6,33, P <
0,001). O consumo de ração não diferiu entre os bons e os maus consumidores (t[12] = -1,15,
P = 0,27). A Figura 2B mostra o consumo de açúcar e ração normalizado para o peso
corporal do bons e maus consumidores de açúcar. Igual ao consumo absoluto de açúcar
mostrado nas Figuras 1 e 2A, a ingestão dos bons consumidores normalizada pelo peso
também foi significativamente maior que o consumo de ração (t[14] = 6,77,; P < 0,001).
Igualmente, a ingestão normalizada de açúcar dos bons consumidores foi maior que a dos
maus consumidores (t[12] = 3,20,; P = 0,008). Os dados da Figura 2B mostram que o
elevado consumo de açúcar não está relacionado ao peso dos animais. Em outras palavras,
A B
*
0
1
2
3
A R A R
Inges
tão de
açúc
ar (g
)
Bons consumidores
°
01
23
45
6
A R A R
Inges
tão de
açúc
ar (g
/Kg)
Maus consumidores
Grupos
Figura 2. Ingestão de açúcar granulado (A) e de ração de laboratório moída (R) de animais classificados como bons e maus consumidores de açúcar. A, Média (+ E. P. M.) da ingestão absoluta, em gramas. B, Média (+ E. P. M.) da ingestão normalizada pelo peso corporal dos animais, em gramas por quilo de peso corporal. *, Diferente do consumo de açúcar; °, diferente dos bons consumidores de açúcar (t de Student, P < 0,05).
22
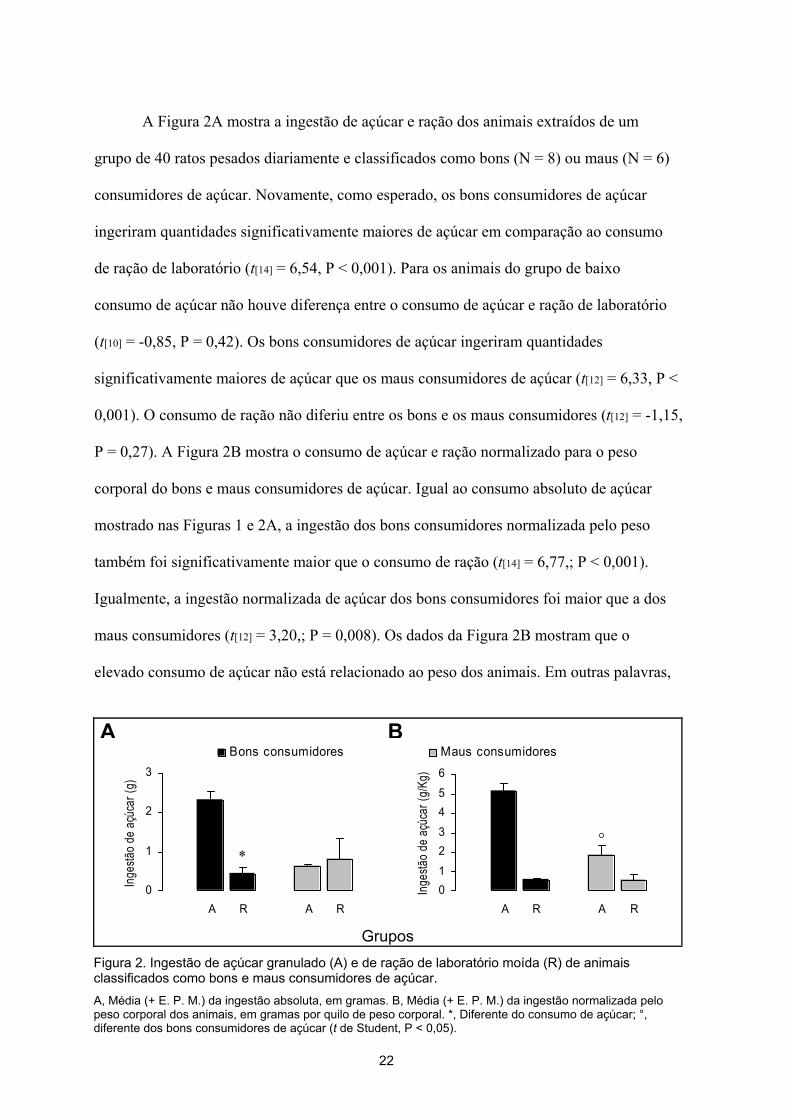
os ratos que consumiram mais não o fizeram simplesmente porque eram mais pesados mas
sim porque comeram proporcionalmente mais.
A Figura 3 mostra as medidas obtidas no labirinto em cruz elevado relacionadas à
atividade exploratória. Pode-se observar na figura que, quando os ratos são classificados
pelo consumo de açúcar, os bons consumidores entram mais nos braços fechados (t[13] =
3,126, P = 0,008), percorrem distâncias maiores tanto nos braços fechados (t[13] = 3,086, P
= 0,009) quanto nos abertos (t[13] = 3,018, P = 0,010), além de apresentarem um maior total
de entradas que os maus consumidores (t[13] = 3,263, P = 0,006). No entanto, quando os
ratos são classificados pelo consumo de ração, não há diferenças estatisticamente
significativas entre os bons e maus consumidores quanto à freqüência de entradas nos
A
*
0
2
4
6
8
10
12
Bons Maus Bons MausTipo de seleção
Entra
das n
os b
raço
s fec
hado
Açúcar Ração
B
*
0
2
4
6
8
10
Bons Maus Bons MausTipo de seleção
Distâ
ncia
perco
rrida
nos b
raço
sfe
chad
os
Açúcar Ração
C
*
0
1
2
3
4
5
Bons Maus Bons Maus
Tipo de seleção
Distâ
ncia
perco
rrida
nos b
raço
sab
erto
s (m)
Açúcar Ração
D
*
02468
1012141618
Bons Maus Bons MausTipo de seleção
Tota
l de
entra
da
Açúcar Ração
Figura 3. Medidas relacionadas à atividade. Freqüência de entradas nos braços fechados (A), distância percorrida nos braços fechados (B) e abertos (C), e total de entradas (D). *, Diferente dos bons consumidores de açúcar (t de Student, P < 0,05).
23
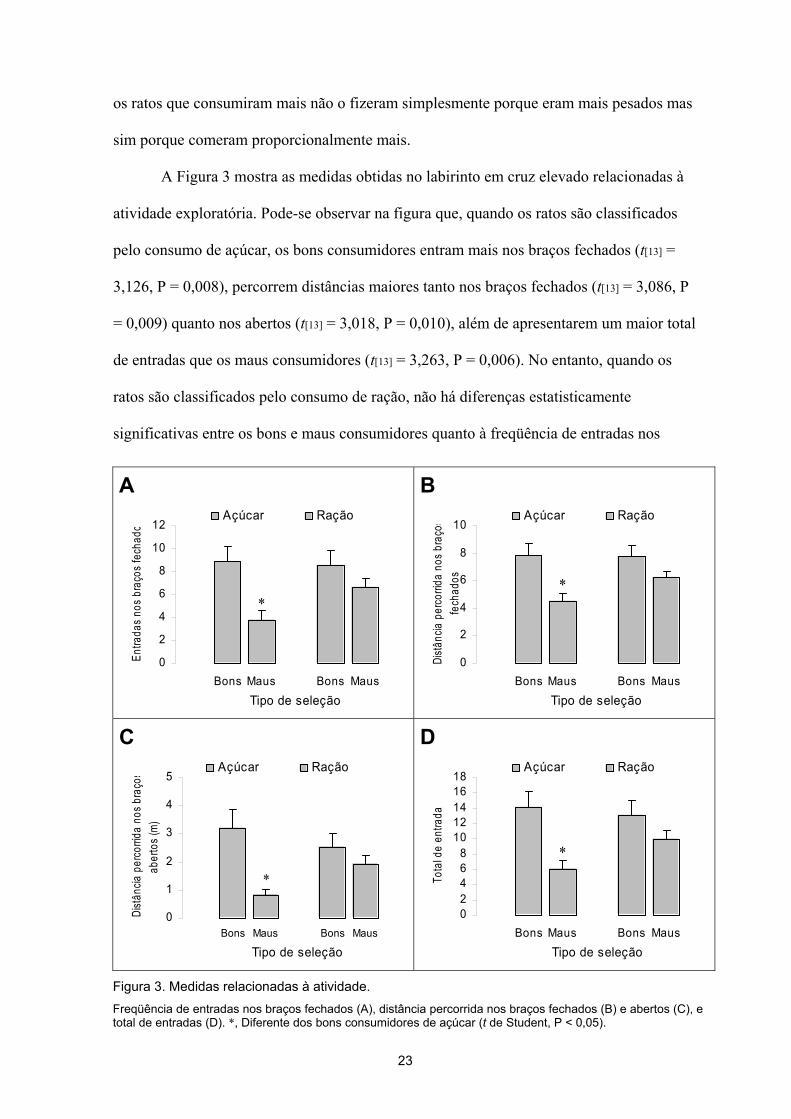
braços fechados (t[23] = 1,353, P = 0,189), distâncias percorridas nos braços fechados (t[23]
= 1,753, P = 0,093) ou nos abertos (t[23] = 1,039, P = 0,310), e ao total de entradas nos dois
tipos de braços (t[23] = 1,446, P = 0,162).
A Figura 4 mostra medidas obtidas no labirinto em cruz elevado mais relacionadas
à ansiedade. Pode-se observar na figura que os bons consumidores não gastam mais tempo
nos braços abertos (t[13] = 1,857, P = 0,086) que os maus consumidores, mas entram neles
com maior freqüência (t[13] = 2,487, P = 0,027). A porcentagem de entradas nos braços
abertos dos bons (36,17 ± 4,12%) e maus consumidores (40,14 ± 5,67%) não foi
estatisticamente significativa (t[13] = -0,575, P = 0,575). Além disso, os bons consumidores
A
01020304050607080
Bons Maus Bons MausTipo de seleção
Temp
o no
s bra
ços a
berto
s (s
Açúcar Ração
B
*
0
12
34
56
7
Bons Maus Bons MausTipo de seleção
Entra
das n
os b
raço
s abe
rto
Açúcar Ração
C
*0
510
1520
2530
35
Bons Maus Bons MausTipo de seleção
Temp
o na
extr
em a
berta
(s
Açúcar Ração
D
*
0
1
2
3
4
5
Bons Maus Bons MausTipo de seleção
Entra
das:
extre
mida
des a
berta
Açúcar Ração
Figura 4. Medidas relacionadas à ansiedade. Tempo gasto nos braços abertos (A), porcentagem de entradas nos braços abertos (B), tempo gasto nas extremidades abertas (C), e freqüência de entradas nas extremidades abertas (D). *, Diferente dos bons consumidores de açúcar (t de Student, P < 0,05).
24

de açúcar entram mais (t[13] = 3,018, P = 0,010) e passam mais tempo (t[13] = 2,574, P =
0,023) nas extremidades dos braços abertos. No entanto, quando os ratos são classificados
pelo consumo de ração, não há diferenças significativas entre bons e maus consumidores
quanto a tempo gasto (t[23] = 0,142, P = 0,888) nem quanto à freqüência de entradas nos
braços abertos (t[23] = 1,242, P = 0,227). Como no caso da classificação pelo consumo de
açúcar, quando os ratos são classificados pelo consumo de ração a porcentagem de
entradas (não mostrada na figura) nos braços abertos dos bons (33,06 ± 3,14%) e maus
consumidores (32,21 ± 4,40%) não foi estatisticamente significativa (t[23] = -0,575, P =
0,575) . Igualmente, os bons consumidores de ração não diferiram quanto à freqüência de
entradas (t[23] = 0,712, P = 0,484) nem quanto ao tempo gasto (t[23] = -0,415, P = 0,682) nas
extremidades dos braços abertos.
A Figura 5 mostra o tempo gasto nos comportamentos de esticar-se, mergulhar a
cabeça e levantar-se. Pode-se observar que o tempo gasto esticando-se foi o mesmo tanto
entre os bons como entre os maus consumidores, seja de açúcar (t[13] = 0,091, P = 0,929)
seja de ração (t[23] = 1,449, P = 0,161). Por outro lado, os bons consumidores de açúcar
mergulharam a cabeça por um tempo significativamente maior que os maus consumidores
(t[13] = 2,626, P = 0,021), enquanto os bons e maus consumidores de ração não se
A
02468
10121416
Bons Maus Bons MausTipo de seleção
Tem
po e
stic
ando
-se
(s)
Açúcar Ração
B
*
0
2
4
6
8
10
Bons Maus Bons MausTipo de seleção
Tem
po m
ergu
lhan
do a
ca
beça
(s)
Açúcar Ração
C
*
05
1015
202530
35
Bons Maus Bons MausTipo de seleção
Tem
po le
vant
ando
-se
(s)
Açúcar Ração
Figura 5. Tempo gasto nos comportamentos de esticar-se (A), mergulhar a cabeça (B), e levantar-se (C). *, Diferente dos bons consumidores de açúcar (t de Student, P < 0,05).
25

diferenciaram (t[23] = -0,015, P = 0,998). Igualmente, os bons consumidores de açúcar
levantaram-se por mais tempo que os maus consumidores (t[13] = 2,579, P = 0,021),
enquanto os bons e maus consumidores de ração novamente não se diferenciaram (t[23] =
1,111, P = 0,278).
A Figura 6 e a Tabela 1 mostram as correlações entre a ingestão de açúcar e as
medidas indicadoras de atividade exploratória obtidas no labirinto em cruz elevado dos
bons e maus consumidores de açúcar. Há uma correlação positiva estatisticamente
significativa entre ingestão de açúcar e freqüência de entradas nos braços fechados,
distância percorrida nos braços fechados, distância percorrida nos braços abertos e
freqüência total de entradas nos braços abertos e fechados. Não ocorreram correlações
significativas entre a ingestão de ração e as medidas de atividade exploratória (Tabela 1).
A
01234567
0 5 10 15 20Entradas nos braços fechados
Inge
stão
de
açúc
ar (g
)
B
01234567
0 5 10Distância percorrida nos braços fechados (m)
Inge
stão
de
açúc
ar (g
)
15
C
01234567
0 2 4 6 8Distância percorrida nos braços abertos (m)
Inge
stão
de
açúc
ar (g
)
D
01234567
0 10 20 30Total de entradas
Inge
stão
de
açúc
ar (g
)
Figura 6. Correlações entre a ingestão de açúcar e freqüência de entradas nos braços fechados (A), distância percorrida nos braços fechados (B) e abertos (C), e total de entradas (D) dos melhores e piores consumidores de açúcar..
26

Tabela 1. Correlação entre as quantidades ingeridas de açúcar e de ração e o comportamento dos ratos bons e maus consumidores de açúcar no labirinto em cruz elevado. r, Coeficiente de correlação de Pearson; P, probabilidade.
Melhores e piores consumidores de açúcar
Açúcar Ração
Comportamentos r P r P Entradas nos braços abertos 0.712 0.003 0.062 0.826 Entradas nos braços fechados 0.760 0.001 0.174 0.536 Total de entradas nos braços 0.815 0.000 0.140 0.618 Tempo gasto nos braços abertos (s) 0.624 0.013 0.034 0.903 Porcentagem de entradas nos braços abertos 0.070 0.804 -0.183 0.514 Entrada média nos braços abertos (s) 0.110 0.696 0.225 0.420 Distância percorrida nos braços abertos (m) 0.780 0.001 0.102 0.716 Distância percorrida nos braços fechados (m) 0.722 0.002 0.256 0.358 Tempo gasto no quadrado central (s) 0.452 0.090 0.389 0.152 Entradas nas extremidades dos braços abertos 0.784 0.001 0.091 0.748 Tempo nas extremidades braços abertos (s) 0.783 0.001 0.061 0.830 Freqüência de mergulhos de cabeça 0.745 0.001 0.116 0.680 Tempo gasto mergulhando a cabeça (s) 0.656 0.008 0.016 0.956 Freqüência de levantadas 0.471 0.077 0.365 0.181 Tempo gasto levantando-se (s) 0.307 0.267 0.020 0.944 Freqüência de esticadas -0.113 0.689 0.524 0.045 Tempo gasto esticando-se (s) -0.011 0.970 0.380 0.162 A Figura 7 e a Tabela 1 mostram as correlações entre a ingestão de açúcar e as
medidas indicadoras de atividade exploratória obtidas no labirinto em cruz elevado mais
relacionadas à ansiedade dos bons e maus consumidores de açúcar. Houve correlação
positiva estatisticamente significativa entre ingestão de açúcar e tempo gasto nos braços
abertos, tempo gasto nas extremidades dos braços abertos e freqüência de entradas nas
extremidades dos braços abertos mas não entre ingestão de açúcar e porcentagem de
entradas nos braços abertos. Não ocorreram correlações significativas entre a ingestão de
ração e as medidas de ansiedade (Tabela 2).
A Figura 8 e a Tabela 1 mostram as correlações entre a ingestão de açúcar e outras
medidas registradas no labirinto em cruz elevado dos bons e maus consumidores de açúcar.
Observou-se uma correlação positiva entre a ingestão de açúcar e o tempo gasto
mergulhando a cabeça mas não entre ingestão de açúcar e tempo gasto esticando-se e
levantando-se.
27

A
01234567
0 50 100 150 200Tempo gasto nos braços aberto (s)
Inge
stão
de
açúc
ar (g
)
B
012345678
0 1 2 3 4 5 6Entradas nos braços abertos (% )
Inge
stão
de
açúc
ar (g
)
C
01234567
0 20 40Tempo gasto nas extremidades abertas (s)
Inge
stão
de
açúc
ar (g
)
60
D
01234567
0 2 4 6 8 1Entradas nas extremidades abertas
Inge
stão
de
açúc
ar (g
)
0
Figura 7. Correlações entre a ingestão de açúcar e tempo gasto nos braços abertos (A), porcentagem de entradas nos braços abertos (B), Tempo gasto nas extremidades abertas (C), e freqüência de entradas nas extremidades abertas (D) dos melhores e piores consumidores de açúcar..
A
01234567
0 10 20 30 40 50Tem po es ticando (s )
Inge
stão
de
açúc
ar (g
)
B
012345678
0 5 10 15 20 25Tempo mergulhando a cabeça (s)
Inge
stão
de
açúc
ar (g
)
C
01234567
0 10 20 30 40 50Tempo levantando-se (s)
Inge
stão
de
açúc
ar (g
)
Figura 8. Correlações entre a ingestão de açúcar e tempo gasto esticando-se (A), mergulhando a cabeça (B), e levantando-se (C) dos melhores e piores consumidores de açúcar.
A Tabela 2 mostra as correlações entre os consumos de açúcar e de ração e
comportamentos exibidos no labirinto em cruz elevado pelos bons e maus consumidores de
ração. Pode-se notar que ocorreu uma única correlação significativa entre o consumo de
açúcar e a duração média das entradas nos braços abertos. Ocorreram, ainda, duas
correlações significativas entre o consumo de ração e as freqüências de levantar-se e
esticar-se.
28
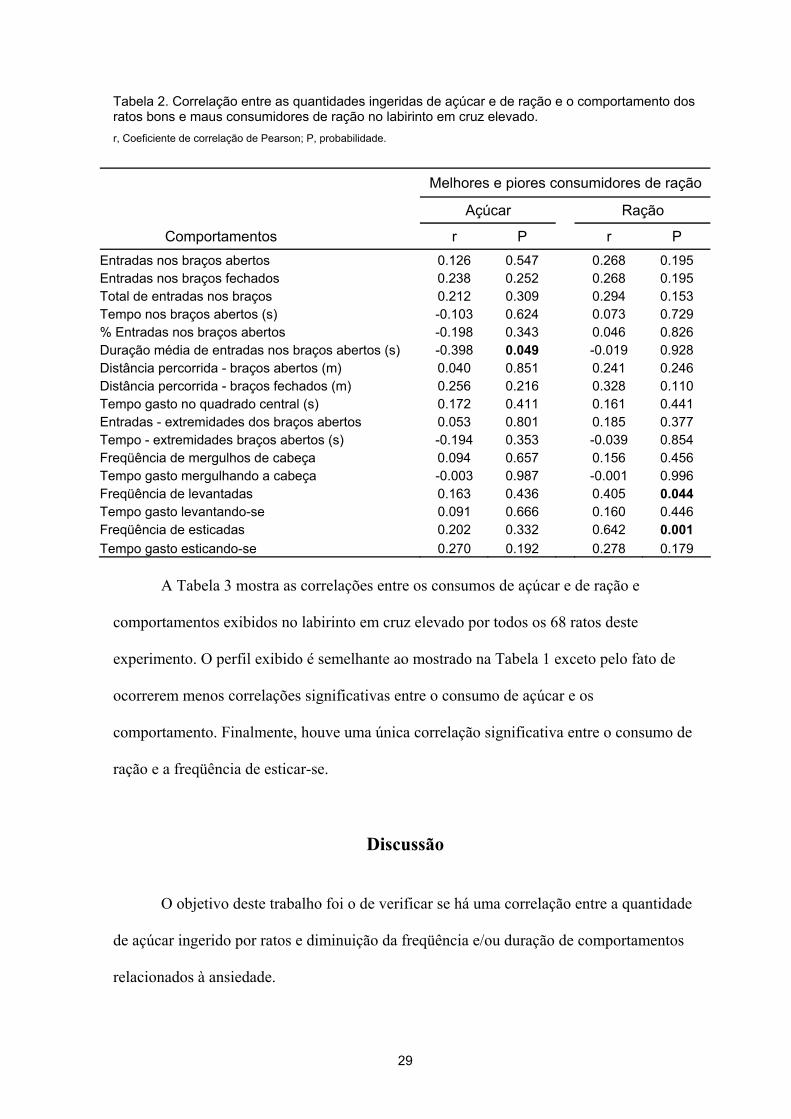
Tabela 2. Correlação entre as quantidades ingeridas de açúcar e de ração e o comportamento dos ratos bons e maus consumidores de ração no labirinto em cruz elevado. r, Coeficiente de correlação de Pearson; P, probabilidade.
Melhores e piores consumidores de ração
Açúcar Ração
Comportamentos r P r P Entradas nos braços abertos 0.126 0.547 0.268 0.195 Entradas nos braços fechados 0.238 0.252 0.268 0.195 Total de entradas nos braços 0.212 0.309 0.294 0.153 Tempo nos braços abertos (s) -0.103 0.624 0.073 0.729 % Entradas nos braços abertos -0.198 0.343 0.046 0.826 Duração média de entradas nos braços abertos (s) -0.398 0.049 -0.019 0.928 Distância percorrida - braços abertos (m) 0.040 0.851 0.241 0.246 Distância percorrida - braços fechados (m) 0.256 0.216 0.328 0.110 Tempo gasto no quadrado central (s) 0.172 0.411 0.161 0.441 Entradas - extremidades dos braços abertos 0.053 0.801 0.185 0.377 Tempo - extremidades braços abertos (s) -0.194 0.353 -0.039 0.854 Freqüência de mergulhos de cabeça 0.094 0.657 0.156 0.456 Tempo gasto mergulhando a cabeça -0.003 0.987 -0.001 0.996 Freqüência de levantadas 0.163 0.436 0.405 0.044 Tempo gasto levantando-se 0.091 0.666 0.160 0.446 Freqüência de esticadas 0.202 0.332 0.642 0.001 Tempo gasto esticando-se 0.270 0.192 0.278 0.179
A Tabela 3 mostra as correlações entre os consumos de açúcar e de ração e
comportamentos exibidos no labirinto em cruz elevado por todos os 68 ratos deste
experimento. O perfil exibido é semelhante ao mostrado na Tabela 1 exceto pelo fato de
ocorrerem menos correlações significativas entre o consumo de açúcar e os
comportamento. Finalmente, houve uma única correlação significativa entre o consumo de
ração e a freqüência de esticar-se.
Discussão
O objetivo deste trabalho foi o de verificar se há uma correlação entre a quantidade
de açúcar ingerido por ratos e diminuição da freqüência e/ou duração de comportamentos
relacionados à ansiedade.
29

Tabela 3. Correlação entre as quantidades ingeridas de açúcar e de ração e o comportamento de todos os ratos no labirinto em cruz elevado. r, Coeficiente de correlação de Pearson; P, probabilidade.
Todos os sujeitos (N = 68)
Açúcar Ração
Comportamentos r P r P Entradas nos braços abertos 0.266 0.028 0.128 0.297 Entradas nos braços fechados 0.330 0.006 0.111 0.367 Total de entradas nos braços 0.341 0.004 0.134 0.277 Tempo nos braços abertos (s) 0.194 0.113 0.029 0.813 % Entradas nos braços abertos -0.046 0.712 0.045 0.717 Entrada média nos braços abertos (s) -0.070 0.570 -0.021 0.863 Distância percorrida - braços abertos (m) 0.236 0.053 0.108 0.382 Distância percorrida - braços fechados (m) 0.316 0.009 0.120 0.329 Tempo gasto no quadrado central (s) 0.207 0.090 0.103 0.405 Entradas - extremidades dos braços abertos 0.260 0.033 0.087 0.483 Tempo - extremidades braços abertos (s) 0.088 0.477 -0.001 0.992 Freqüência de mergulhos de cabeça 0.179 0.143 0.119 0.332 Tempo gasto mergulhando a cabeça 0.187 0.127 0.009 0.941 Freqüência de levantadas 0.125 0.309 0.190 0.120 Tempo gasto levantando-se 0.088 0.476 0.034 0.785 Freqüência de esticadas -0.036 0.772 0.349 0.004 Tempo gasto esticando-se 0.009 0.941 0.113 0.361
As análises dos ratos bons e maus consumidores de açúcar mostraram que, de um
modo geral, a tendência a consumir mais açúcar é indicadora de comportamento
exploratório no labirinto em cruz elevado característico de animais com baixos níveis de
medo ou ansiedade. Além disso, os efeitos observados foram específicos do consumo de
açúcar, uma vez que a análise dos ratos quando classificados em bons e maus
consumidores de ração não mostrou quaisquer efeitos do consumo da mesma sobre os
comportamentos exibidos no labirinto. Mais que isso, a análise do comportamento desses
animais mostrou correlações positivas entre o consumo de açúcar e diversos dos
comportamentos registrados. De um modo geral, essas correlações não aconteceram com o
consumo de ração. Parte das correlações se manteve quando todos os animais foram
analisados.
Os dados obtidos sobre as diferenças de consumo de açúcar, desvinculados do
consumo de ração, estão de acordo com vários trabalhos da literatura (De Cabo et al. 1995;
30

Brennan et al. 2001; Sills e Crawley, 1996; Sills e Vaccarino, 1996, Desousa et al., 1998).
Quando submetemos ratos à ingestão livre de dois tipos de comida, açúcar granulado e
ração moída de laboratório, pudemos classificá-los e agrupá-los em bons e maus
consumidores de açúcar sem que os grupos diferissem no consumo da ração de laboratório
moída. Em outras palavras, os animais de ambos os grupos não consumiram nem mais nem
menos ração por conta da quantidade de açúcar ingerida no mesmo período.
Adicionalmente, o consumo de açúcar não estava relacionado ao ganho de peso do animal.
Podemos encontrar vários trabalhos na literatura que mostraram que maior ingestão
de substâncias doces se correlaciona com: maior atividade locomotora quando os animais
são submetidos a tratamento agudo e crônico com anfetamina (Sills e Crawley, 1996), alta
ingestão de etanol (Kampov-Polevoy et al., 1990; Gosnell e Krahn 1992; Overstreet et al.,
1993); maior freqüência de auto-administração de morfina (Gosnell et at, 1995) e de
psicoestimulantes (DeSouza et al., 2000). Além dessas correlações, alguns autores
sugeriram que maior ingestão de açúcar se correlaciona negativamente com
comportamentos relacionados à ansiedade.
Segundo Dess e Minor (1996), a ingestão de qualquer tipo de comida interage com
processos afetivos, tais como medo e ansiedade. Ratos submetidos a choques consomem
menor quantidade de substâncias adulteradas por sacarina ou quinino, mas aumentam a
ingestão de açúcar (Dess e Chapman, 1990; Dess e Choe, 1994). Esses dados apóiam a
hipótese de que animais ansiosos ingerem mais açúcar. Em função disso, Dess e Minor
(1996) sugeriram que os estados de medo e ansiedade poderiam estar relacionados com a
ingestão de substâncias doces.
Quando utilizamos o labirinto em cruz elevado observamos que bons consumidores
de açúcar apresentaram alterações nas medidas comportamentais, tanto relacionadas à
ansiedade quanto à atividade locomotora, quando comparados aos maus consumidores de
açúcar. Os bons consumidores de açúcar apresentaram maior freqüência de entradas nos
31

braços fechados, maior freqüência de entradas totais nos braços fechados e abertos e maior
distância percorrida nos braços fechados. Como essas medidas estão mais relacionadas à
atividade locomotora podemos dizer que bons consumidores foram mais ativos que os
maus consumidores no labirinto em cruz elevado.
Em relação às medidas comportamentais relacionadas à ansiedade, observamos que
os bons consumidores de açúcar apresentaram maior freqüência de entradas nos braços
abertos e nas extremidades dos mesmos e maior tempo de permanência nos braços abertos.
Baseado nestas medidas relacionadas à ansiedade, diríamos que os bons consumidores de
açúcar apresentaram menos comportamentos relacionados à ansiedade que os maus
consumidores. Entretanto, não observamos diferenças entre bons e maus consumidores de
açúcar em uma medida comumente relacionada à ansiedade, que é a porcentagem de
entradas nos braços abertos. Geralmente, a porcentagem de entradas nos braços abertos é
mais sujeita a diminuição que as outras medidas relacionadas à ansiedade quando os
animais estão ansiosos. Como houve diferenças entre os bons e maus consumidores de
açúcar tanto em medidas relacionadas à ansiedade quanto à atividade locomotora, e não
houve diferença entre esses grupos para a porcentagem de entradas nos braços abertos,
torna-se difícil dizer se os animais estão se locomovendo mais ou estão menos ansiosos.
Outras medidas registradas no labirinto em cruz elevado podem ajudar nesse tipo
de situação. Segundo Cruz et al. (1994), o comportamento de esticar-se está relacionado às
condições aversivas que geram ansiedade. Entretanto, também não observamos diferenças
entre bons e maus consumidores de açúcar quanto ao tempo que os animais gastaram
comportando-se desta maneira. Segundo os mesmos autores (Cruz et al., 1994), os
comportamentos de mergulhar a cabeça para fora da plataforma dos braços abertos e
levantar-se de modo a apoiar-se nas patas dianteiras estão, respectivamente, mais
relacionados à ansiedade e à atividade locomotora e/ou exploratória. Assim, os bons
consumidores gastaram mais tempo que maus consumidores mergulhando a cabeça
32

(indicador de ansiedade diminuída) e levantando-se (indicador de maior atividade geral).
Considerando-se essas três medidas, pode-se dizer que elas indicam que os bons
consumidores de açúcar estavam menos ansiosos e mais ativos que os maus consumidores.
Qualquer uma dessas diferenças comportamentais citadas acima não foram
observadas quando comparamos bons e maus consumidores de ração. Isso sugere que não
é a ingestão de qualquer alimento que pode estar relacionada a essas medidas registradas
no labirinto em cruz elevado, mas parece que a percepção do gosto doce de substâncias
palatáveis é uma variável relevante nesta condição.
Além disso, quando realizamos testes de correlação, observamos que não existe
apenas uma diferença nas medidas registradas no labirinto em cruz elevado entre bons e
maus consumidores, mas também uma correlação significativa e positiva entre ingestão de
açúcar e a freqüência de alguns comportamentos registrados nesse modelo. Quando
selecionamos grupos de ratos e os classificamos como bons e maus consumidores de
açúcar, observamos que quanto maior a quantidade de açúcar ingerido maior a: freqüência
de entradas nos braços fechados, a freqüência total de entradas nos braços fechados e
abertos, e a distância percorrida em cada tipo de braço. Para o mesmo grupo de ratos, a
mesma correlação significativa e positiva foi observada entre maior ingestão de açúcar e
algumas medidas comportamentais relacionadas à ansiedade como: o tempo de
permanência nos braços abertos e nas extremidades dos mesmos e a freqüência de entradas
nas extremidades dos braços abertos. Isso mostra que quanto maior a quantidade de açúcar
ingerido maior será a freqüência das categorias comportamentais citadas acima.
Quando fizemos o teste de correlação para todos os animais independentemente de
os classificarmos como bons ou maus consumidores de açúcar, também observamos uma
correlação significativa e positiva para algumas medidas comportamentais. Quanto maior o
consumo de açúcar maior a: freqüência de entradas nos braços abertos e fechados, a
freqüência de entradas totais nos dois tipos de braços, a distância percorrida nos braços
33

fechados e a freqüência de entradas nas extremidades dos braços abertos. Ainda nesta
condição, temos alterações tanto em medidas relacionadas à ansiedade como à atividade
locomotora. Isso nos permite supor que a quantidade de açúcar ingerida pode funcionar
como um previsor para alguns comportamentos no labirinto em cruz elevado. Alem disso,
quando se levou em conta o consumo de ração, não observamos qualquer correlação entre
ingestão ração e medidas registradas no labirinto em cruz elevado. Essa ausência de
correlação foi observada tanto quando se analisaram apenas os bons e maus consumidores
de ração como quando se analisaram todos os sujeitos.
Apesar de os resultados do presente experimento não oferecerem condições
suficientes para separarmos um efeito ansiolítico de um efeito locomotor no labirinto em
cruz elevado, os resultados de outros trabalhos permitem levantar a hipótese de que bons
consumidores de açúcar locomovem-se mais que maus consumidores de açúcar. Desouza
et al. (1998) mostraram que ratos classificados como bons consumidores de açúcar
apresentaram porcentagem de entradas e tempo de permanência maiores nos braços abertos
em relação aos ratos classificados como maus consumidores de açúcar. Entretanto, esses
autores não fizeram registro de medidas comportamentais relacionadas à atividade
locomotora. Baseado nesses resultados os autores acima sugeriram que diferenças
comportamentais no consumo de sacarose podem predizer respostas relacionadas à
ansiedade no labirinto em cruz elevado.
Por outro lado, Piazza et al. (1989) e Hooks et al. (1991) mostraram que ratos que
exibiram altos níveis de atividade locomotora em resposta a ambientes novos mostraram
também locomoção mais exacerbada em resposta à anfetamina e à cocaína do que ratos
que exibiram baixo nível de atividade locomotora em resposta a ambientes novos.
Interessante é que Sills e Vaccarino (1994) mostraram que ratos que ingeriram altas
quantidades de açúcar também apresentaram locomoção mais exacerbada em resposta ao
34

tratamento agudo e crônico com anfetamina do que ratos que consumiram baixa
quantidade de açúcar.
As semelhanças entre ratos classificados como bons consumidores de açúcar e ratos
que exibem altos níveis de atividade locomotora em ambientes novos não se restringem
apenas à resposta locomotora ao tratamento com anfetamina. Estudos de microdiálise in
vivo mostraram que ratos que apresentaram os mais altos níveis de atividade locomotora
induzida por anfetamina também mostraram maior atividade dopaminérgica no núcleo
acumbente, quando comparados a ratos com níveis mais baixos de atividade locomotora
induzida por anfetamina, tanto sob condições de linha de base quanto em resposta a
tratamentos com psicoestimulantes (Hooks et at. 1992; Piazza et al. 1990). No mesmo
sentido, Sills e Vaccarino (1991) e Sills et al. (1993) relataram que ratos bons
consumidores de açúcar mostraram maior atividade dopaminérgica no núcleo acumbente
que ratos maus consumidores de açúcar.
Uma hipótese na literatura é a de que o sistema dopaminérgico esteja implicado na
atividade locomotora e na ingestão de substâncias doces. Vários trabalhos mostraram que o
tratamento com anfetamina afeta a locomoção (Kelly et al., 1975; Vaccarino et al., 1986;
Jones e Robbins, 1992; Weissenborn e Winn 1992) e a ingestão (Colle e Wise, 1988; Sills
e Vaccarino 1991; Sills et al. 1993). Esses trabalhos parecem indicar que, quando
selecionamos ratos que exibem atividade locomotora elevada frente a ambientes novos,
também selecionamos ratos bons consumidores de açúcar.
Abercrombie et al. (1989) e Imperato et al. (1989) mostraram que ratos submetidos
a choques na cauda apresentaram níveis elevados de dopamina no núcleo acumbente. Na
medida em que eventos estressantes afetam o sistema dopaminérgico, pode ser que ratos
bons e maus consumidores de açúcar apresentem variações diferentes neste sistema e uma
outra maneira de responder diferente pode ser a atividade locomotora. Dessa forma, ao
serem colocados no labirinto em cruz elevado, um modelo que envolve conflito, ratos bons
35

consumidores de açúcar locomovem-se mais que ratos maus consumidores tanto nos
braços abertos como nos fechados, e segundo Kabbaj et al. (2000), a atividade locomotora
elevada está associada à novidade e não ao ambiente familiar. Há vários trabalhos na
literatura relatando que ratos preferem ambientes familiares à ambientes novos, o que
mostra que um ambiente novo pode ser aversivo. No presente experimento (assim como na
maioria esmagadora dos relatos na literatura), o labirinto em cruz elevado é um ambiente
novo e pode ter favorecido o aumento na atividade locomotora de ratos classificados como
bons consumidores de açúcar mais que no caso dos maus consumidores.
Os dados do presente trabalho podem ser interpretados de maneira diferente da
interpretação que Desouza et al. (1998) fizeram sobre seus resultados, de que ratos bons
consumidores de açúcar sejam menos ansiosos que ratos maus consumidores no labirinto
em cruz elevado. Esses autores mostraram alterações em apenas duas medidas relacionadas
à ansiedade e não mostraram registro de medidas relacionadas à atividade locomotora.
Inclusive, se ingestão alta ou baixa de açúcar podem estar relacionadas a comportamentos
relacionados à ansiedade, outros trabalhos sugerem uma hipótese diferente da relatada por
Desouza et al. (1998).
Quando ratos foram expostos a um ambiente novo, mostraram um aumento nos
níveis de corticosterona em relação a controles não expostos (Oitzl et al., 1997).
Interessantemente, quando comparado a ratos com baixa resposta locomotora a um
ambiente novo, ratos com alta resposta locomotora mostraram níveis mais elevados de
corticosterona por um período mais prolongado quando foram exposto a um ambiente novo
inescapável (Piazza et al., 1989) ou submetidos ao modelo de restrição comportamental
(Dellu et al., 1996). Isso sugere que tomar como medida apenas o comportamento de
locomover-se não é suficiente para se inferir estados de ansiedade.
De maneira semelhante, Kabbaj et al. (2000) classificaram animais com alta e baixa
resposta locomotora frente a um ambiente novo e posteriormente testaram esses animais na
36

caixa claro/escuro e no labirinto em cruz elevado. Verificaram que os animais com alta
resposta locomotora frente a um ambiente novo exibiram maior freqüência de entradas no
ambiente claro e permaneceram mais tempo nesse ambiente que os animais com baixa
resposta locomotora frente a um ambiente novo. Adicionalmente, os ratos com alta
resposta locomotora frente a um ambiente novo permaneceram mais tempo nos braços
aberto do labirinto em cruz elevado e mostraram maior atividade locomotora nos mesmos
quando comparados aos animais com baixa resposta locomotora frente a um ambiente
novo. Esses resultados parecem indicar que, quando testamos na gaiola de transição
claro/escuro ou no labirinto em cruz elevado, ratos classificados como tendo alta resposta
locomotora frente a um ambiente novo exibem comportamentos menos relacionados à
ansiedade que ratos classificados como tendo baixa resposta locomotora em um ambiente
novo. Entretanto, os animais com alta resposta locomotora exibiram maiores níveis de
corticosterona no plasma quando comparados aos animais com baixa resposta locomotora
(Dellu et al., 1996; Piazza et al., Simon 1991). Isso sugere que ratos com alta resposta
locomotora frente a um ambiente novo são mais ansiosos que ratos com baixa resposta
locomotora frente a um ambiente novo. Como mostrado acima, há semelhanças
comportamentais entre ratos bons consumidores de açúcar e ratos com alta atividade
locomotora em ambientes novos. Assim, pode ser que o fato de os bons consumidores de
açúcar se locomoverem mais no labirinto em cruz elevado seja um indício de que esse
modelo é mais aversivo para estes do que para maus consumidores.
Há um outro problema que potencialmente poderia dificultar a interpretação dos
resultados do presente experimento. Durante a execução do protocolo experimental deste
experimento, o manuseio crônico e diário dos animais, pelo experimentador pode ter
afetado o comportamento tanto no labirinto em cruz elevado como o comportamento de
consumir açúcar durante a fase de ingestão. Alguns trabalhos mostraram que o manuseio
afeta o comportamento de ratos no labirinto em cruz elevado de modo a aumentar a
37

freqüência de entradas e tempo de permanência nos braços abertos (Andrews e File, 1993).
Adicionalmente, há trabalhos mostrando que os benzodiazepínicos facilitaram a ingestão
de comida e líquido palatáveis em uma variedade de espécies (por exemplo, Birk e Noble,
1982). As diferenças nas medidas comportamentais observadas no labirinto em cruz
elevado e no comportamento de ingerir de açúcar poderiam ser influenciadas pelo
manuseio. Da mesma maneira que ratos respondem de modo diferente quando em
ambientes novos, é possível que as diferenças no comportamento de consumir açúcar e nas
medidas comportamentais registradas no labirinto em cruz elevado sejam uma resposta ao
manuseio. Dessa forma, o experimento a seguir teve como objetivo verificar se o manuseio
crônico e/ou a ingestão de açúcar afetam o comportamento no labirinto em cruz elevado.
Experimento II As alterações nas medidas comportamentais relacionados à ansiedade e à atividade
locomotora no labirinto em cruz elevado observadas no Experimento I podem ser efeito: (1)
da ingestão de açúcar por um período prolongado, (2) da manipulação crônica dos animais
ou (3) de ambos os procedimentos.
Nos procedimentos experimentais realizados com animais, são várias as condições
às quais podem ser submetidos. Uma delas é o manuseio que o experimentador realiza,
conceito nem sempre bem definido. Um manuseio pouco ou nada definido é o contato
ocasional e rápido do ser humano com o animal em procedimentos rotineiros em
laboratórios que utilizam modelos animais. Dentre os exemplos desse manuseio rápido e
rotineiro, incluem-se a pesagem dos animais, injeções de drogas, transporte para a situação
de teste, troca de gaiolas durante procedimentos de limpeza etc
Geralmente, a manipulação experimental consiste em (a) retirar o animal da gaiola,
colocá-lo na palma da mão e retorná-lo, ou (b) manipulá-lo na própria gaiola, ou ainda (c)
retirá-lo da gaiola, mantê-lo seguro na mão, onde é realizada a manipulação. A
38

manipulação pode consistir em alisar suavemente os animais com o polegar no sentido
céfalo-caudal ou massagear os pêlos das costas e do pescoço, sendo que este tipo de
manipulação apresenta as mais variadas formas em relação a como é realizada e à duração.
Por exemplo, para Meinrath e Flaherty (1987), manipulação foi separar a mãe da ninhada e
colocar os filhotes um por vez em um compartimento individual por 3 minutos, do segundo
ao décimo quinto dia após o desmame. Abel (1971) chamou de manipulação a remoção
individual dos filhotes da ninhada e carregá-los por 3 minutos no corredor adjacente à sala
do laboratório, sendo que neste período o animal foi repetidamente colocado na palma da
mão do experimentador. Clausing et al. (1997) chamaram de manipulação à retirada do
filhote da ninhada e segurá-lo na mão por cinco minutos. Andrews e File (1993) chamaram
de manipulação pegar o animal (170 g), pesá-lo e aplicar uma injeção intraperitoneal de
água, por 7 dias consecutivos. Brett e Pratt (1990) relataram que os ratos foram
manuseados por dois dias, mas não descreveram as condições de manipulação. Quando
buscamos na literatura resultados referentes aos efeitos da manipulação constatamos que
não há um consenso do que chamar de manipulação.
Sabe-se que o manuseio precoce ou tardio produz alterações comportamentais e
fisiológicas (Andrews e File, 1993; Clausing et al. 1997; Fernández-Teruel et al. 2002) e
um sistema envolvido é o GABA-benzodiazepínico (Biggio, 1983; File et al., 1990a, 1992).
Em função do sistema envolvido e dos resultados comportamentais observados em
modelos animais de ansiedade, essas alterações produzidas em ratos pelo manuseio foram
caracterizada como sendo de tipo ansiolítica. Paralelamente, alguns trabalhos mostraram
que os benzodiazepínicos facilitaram a ingestão de comida e de líquidos em uma variedade
de espécies (Britton e Britton, 1981; Birk e Noble, 1982), embora não se conheça(m) o(s)
mecanismo(s) subjacente(s) a esse efeito. O efeito facilitador dessas drogas sobre a
ingestão de comida ou de líquido é interpretado de duas maneiras: (a) como uma redução
nos componentes aversivos da situação de teste, que normalmente suprimem o consumo
39

(Britton e Britton, 1981) ou (b) como uma modificação direta do mecanismo biológico que
controla a ingestão de comida e líquido (Cooper, 1980). Dessa maneira, o manuseio pode
ter um efeito semelhante ao efeito de drogas benzodiazepínicas sobre o comportamento de
ingestão de comida ou líquidos palatáveis.
Baseado na primeira hipótese mencionada acima, os ratos podem responder
diferentemente frente à manipulação repetida do experimentador. Isso pode ter efeitos
sobre a ingestão de açúcar ou ração durante a fase de ingestão e sobre as medidas
comportamentais registradas no labirinto em cruz elevado. No experimento anterior não
tivemos como saber se as medidas comportamentais registradas no labirinto em cruz
elevado foram afetadas pela ingestão prévia de açúcar ou pela manipulação prolongada do
experimentador ou se esses dois procedimentos têm efeitos no labirinto. Em função do
exposto acima, o objetivo deste experimento foi verificar se a manipulação experimental
pode afetar (a) o consumo de substâncias palatáveis em ratos e (b) os comportamentos no
labirinto em cruz elevado. Adicionalmente, verificar se ingestão prévia de açúcar e
manipulação experimental prolongada têm efeitos aditivos quando ratos são testados no
labirinto em cruz elevado.
Método
Sujeitos.
Foram utilizados 81 ratos Wistar machos (170 ± 10 g) provenientes do biotério
Geral do Campus da Universidade de São Paulo em Ribeirão Preto e mantidos nas mesmas
condições descritas no Experimento I.
Equipamento.
O mesmo utilizado no Experimento I
40

Procedimento.
Após o período de habituação os animais foram distribuídos em grupos (N = 9)
como se descreve a seguir. (1) Grupo Biotério— permaneceu em caixas no biotério por 14
dias sendo manipulado apenas para troca de maravalha; (2) Grupo Estimulação —
subdividido nos grupos 0, 1, 5 e 10, referência aos respectivos tempos de estimulação
diário (0, 1, 5 ou 10 minutos). Após o período de estimulação, os animais eram colocados
por 1 hora em uma caixa igual à que viviam no alojamento, na qual estavam disponíveis
dois comedouros de aço inoxidável vazios. O grupo 0 apenas era colocado na caixa por 1
hora. Passado esse tempo, os animais eram devolvidos às suas caixas de alojamento. Este
procedimento durou 14 dias. (3) Grupo Estimulação e Consumo — durante 14 dias os
animais foram submetidos aos mesmos procedimentos que os animais do grupo
Estimulação, mas, que após a estimulação eram colocados em uma caixa igual à de
alojamento por 1 hora, na qual estavam disponíveis em comedouros de aço inoxidável
açúcar comercial granulado e ração moída. Após 1 hora os animais eram devolvidos às
suas caixas de alojamento. Os comedouros foram pesados antes e depois do período de
ingestão. A estimulação foi realizada da seguinte forma: o rato foi colocado em uma gaiola
(30 x 18 x 13 cm) com maravalha e recebeu dois tipos de estimulação sem ser seguro:
alisamento da parte superior do corpo (dorso) e massagem breve (pressão ligeira e curta do
corpo) até completar o período correspondente a seu grupo.
MEDIDA DE INGESTÃO. Para os animais do grupo Estimulação e Consumo, mediu-se a
ingestão das duas substâncias. A média de ingestão de cada grupo foi calculada usando-se
os registros de ingestão das duas últimas sessões de cada rato.
LABIRINTO EM CRUZ ELEVADO. No décimo quinto dia, os ratos foram testados no labirinto
em cruz elevado por um período de cinco minutos de modo análogo ao realizado no
Experimento I, analisando-se os mesmos comportamentos.
41

Análise estatística.
A ingestão de açúcar e de ração foi analisada por uma análise de variância (Anova)
de duas vias, com tempo de manuseio como um dos fatores (quatro níveis: 0, 12, 5 e 10
minutos de manipulação) e substância consumida como um fator de medidas repetidas
(dois níveis: açúcar e ração). Sempre que apropriado, as médias de grupo foram
comparadas com o teste post-hoc de Duncan. Os dados comportamentais foram submetidos
a uma Anova de uma via, seguida pelo teste post-hoc de Dunnett, de comparações com um
grupo controle. Em todos os casos o nível de significância foi fixado em P < 0,05.
Resultados
A Figura 9 mostra o consumo de açúcar granulado e ração de laboratório moída dos
grupos de animais submetidos aos diferentes períodos de manuseio. A Anova mostrou uma
diferença geral significativa no fator consumo (F[1,28] = 3,428; P = 0,031) e o teste de
Duncan indicou que o consumo de açúcar foi maior que o de ração moída. Como outro
efeito geral, a Anova mostrou ainda que a manipulação influenciou o consumo (F[3,28] =
55,920; P < 0,001) e o teste de Duncan apontou que os sujeitos manipulados por 10 min
consumiram significativamente menos que os manipulados por 0 e 5 min. Finalmente, a
Anova mostrou uma interação significativa entre os dois fatores (F[3,28] = 6,091; P = 0,003).
O teste de Duncan evidenciou que os ratos manipulados por 0, 1 e 5 min consumiram
significativamente mais açúcar que ração. Evidenciou também que o consumo de açúcar
dos ratos manipulados por 10 min foi significativamente menor que os manipulados por 0,
1 e 5 min. Finalmente, mostrou que o consumo de açúcar dos ratos manipulados por 1 e 5
min foi significativamente menor que o consumo dos ratos não manipulados.
A Figura 10 e a Tabela 4 mostram as medidas correlacionadas com ansiedade. A
Anova mostrou um efeito do manuseio na porcentagem de entradas nos braços abertos
42

● ●
°* * *
0
1
2
3
4
0 1 5 10Tem po de m anuseio (m in)
Inge
stão
(g)
Açúcar Ração
Figura 9. Consumo de açúcar e de ração nas duas últimas sessões de medida de consumo pelos ratos do grupo Estimulação e Consumo. *, Deferente do consumo de açúcar; °, diferente do açúcar 0, 1 e 5; ●, diferente do açúcar 0. (Duncan, P < 0,05)
(F[8,72] = 2,081; P = 0,049) e o teste de Dunnett mostrou que os animais sem acesso ao
açúcar e manuseados por 10 min apresentaram porcentagens maiores que os do grupo
controle (Figura 1A). Também houve um efeito do manuseio na freqüência de entradas nos
braços abertos (F[8,72] = 2,684; P = 0,012), no tempo gasto nos braços abertos (F[8,72] =
2,824; P = 0,009) e na freqüência de entradas nas extremidades abertas (F[8,72] = 2,956; P =
0,006) e o teste de Dunnett mostrou que tempos e freqüências significativamente maiores
que os dos animais do grupo controle ocorreram nos animais que consumiram açúcar e
foram manuseados por 1 min e nos que não tiveram acesso ao açúcar e foram manuseados
por 10 min (Figura 1B e C; Tabela 4). Finalmente, observou-se um efeito do manuseio
sobre o tempo gasto nas extremidades abertas (F[8,72] = 3,003; P = 0,006) e o teste de
Dunnett mostrou que os animais que consumiram açúcar e foram manuseados por 1 min
permaneceram mais tempo nessas extremidades do que os controles (Figura 1D). Não se
observaram efeitos do manuseio sobre o tempo gasto mergulhando a cabeça (F[8,72] = 1,124;
P = 0,358; Tabela 4) e esticando-se (F[8,72] = 1,202; P = 0,310; Tabela 4).
A Figura 11 e a Tabela 4 mostram as medidas relacionadas à atividade locomotora
e/ou exploratória. A Anova não mostrou efeitos significativos do manuseio sobre a
freqüência de entradas nos braços fechados (F[8,72] = 0,579; P = 0,792; Figura 11A),
43

A
*
05
101520253035404550
C 0 1 5 10 0 1 5 10Tempo de manuseio
Entra
das n
os br
aços
aber
tos (%
)
Biotério Com ingestão Sem ingestão
B
* *
0102030405060708090
C 0 1 5 10 0 1 5 10Tempo de manuseio
Temp
o nos
braç
os ab
ertos
(s)
Biotério Com ingestão Sem ingestão
C *
*
0
1
2
3
4
5
6
C 0 1 5 10 0 1 5 10
Tempo de manuseio
Entra
das n
as ex
tremi
dade
s abe
rtas
Biotério Com ingestão Sem ingestão
D *
05
101520253035404550
C 0 1 5 10 0 1 5 10Tempo de manuseio
Temp
o nas
extre
mida
des a
berta
s (s)
Biotério Com ingestão Sem ingestão
Figura 10. Medidas relacionadas à ansiedade. Porcentagem de entradas nos braços abertos (A), tempo gasto nos braços abertos (B), freqüência de entradas nas extremidades dos braços abertos (C), e tempo gasto nas extremidades abertas (D). C, grupo controle; *, diferente dos controles (Duncan, P < 0,05).
Tabela 4. Medidas (média ± E. P. M.) de diversos comportamentos exibidos pelos ratos do grupo controle e os manipulados, com acesso ou não ao açúcar em sessões de 60 min. *, Diferente do grupo controle, C (Duncan, P < 0,05).
Estimulado e Consumo Estimulado
Parâmetro C 0 1 5 10 0 1 5 10
Freqüência de entradas nos braços abertos
3,00 ±0,41
3,44 ±0,63
6,56 ±0,69*
4,11 ±0,72
4,89 ±0,72
4,44 ±0,65
4,11 ±0,54
4,22 ±0,91
5,78 ±0,70*
Total de entradas 11,00 ±0,94
12,44 ±0,85
16,67 ±0,96
13,33 ±1,41
14,44±1,34
13,56 ±1,18
13,33 ±1,20
12,67 ±1,76
14,00 ±1,31
Distância percorrida nos braços abertos (m)
7,32 ±3,05
7,02 ±2,08
17,12 ±6,78
13,18 ±6,28
15,73±5,32
13,53 ±5,75
9,52 ±3,18
8,50 ±2,59
18,54 ±8,44
Tempo gasto mergulhando a cabeça (s)
4,45 ±0,82
6,67 ±1,31
10,34 ±2,08
5,61 ±0,55
8,48 ±1,84
7,18 ±2,0
6,96 ±1,56
5,71 ±1,79
8,53 ±2,40
Tempo gasto esticando-se (s) 10,52 ±2,10
10,70 ±3,12
4,59 ±1,32
6,38 ±1,47
7,02 ±1,70
7,12 ±2,23
11,43 ±4,27
10,81 ±3,20
4,79 ±1,13
Tempo gasto levantando-se (s) 36,17 ±3,06
34,87 ±3,28
37,54 ±2,34
40,11 ±3,31
34,12±4,01
32,08 ±3,51
36,05 ±2,60
36,35 ±3,69
36,53 ±3,54
44

freqüência total de entradas nos dois tipos de braços (F[8,72] = 1,549; P = 0,156; Tabela 4),
distância percorrida nos braços fechados (F[8,72] = 0,480; P = 0,866; Figura 11B) e abertos
(F[8,72] = 0,684; P = 0,704; Tabela 4) e Tempo gasto levantando-se (F[8,72] = 0,458; P =
0,881; Tabela 4).
A
0
2
4
6
8
10
12
C 0 1 5 10 0 1 5 10Tempo de manuseio
Entra
das n
os br
aços
fech
ados
Biotério Com ingestão Sem ingestão
B
0123456789
10
C 0 1 5 10 0 1 5 10
Tempo de manuseio
Ditân
cia no
s bra
ços f
echa
dos (
m)
Biotério Com ingestão Sem ingestão
Figura 11. Medidas relacionadas à atividade locomotora e/ou exploratória. Freqüência de entradas nos braços fechados (A) e distância percorrida nos braços fechados (B).
Discussão
No experimento I, observamos que animais que ingeriram maiores quantidades de
açúcar, contrapostos aos que ingeriram menores quantidades, apresentaram maiores
freqüências de comportamentos tanto relacionados à ansiedade como à atividade
locomotora no labirinto em cruz elevado. Entretanto, não tínhamos como saber se essas
diferenças são efeito da ingestão de açúcar ou do procedimento de manuseio executado no
protocolo da fase de ingestão. O presente experimento foi uma tentativa de controlar essas
duas condições para verificar seus efeitos sobre as medidas comportamentais registradas
no labirinto em cruz elevado.
As diferentes quantidades de manuseio a que os animais foram expostos não
alteraram a ingestão de ração moída de laboratório. Entretanto, a ingestão de açúcar
diminuiu conforme aumentava o tempo de manuseio. Willner et al. (1987) mostraram que
45

o consumo de sacarose diluída em água diminuiu durante um período de 3 semanas em que
ratos foram expostos a diferentes condições de estresse. Em contrapartida, o consumo de
sacarose aumentou nos animais não expostos as condições de estresse durante esse mesmo
período. Além disso, Katz (1982) relatou que ratos submetidos a um período de 3 semanas
de exposição a diferentes condições estressantes diminuíram o consumo de sacarose e de
sacarina sem que houvesse alteração no consumo de água.
No presente experimento, a maneira de manusear os animais foi semelhante para
todos os grupos, mas a duração da sessão de manuseio foi a variável que realmente afetou
a ingestão de açúcar. Considerando os resultados de Willner et al. (1987) e de Katz (1981),
podemos supor que sessões mais longas de manuseio deixaram os animais mais estressados,
a julgar pelo fato de terem ingerido menores quantidades de açúcar. Se esse foi o caso, a
duração do manuseio foi a condição relevante e não a freqüência de contatos com o
experimentador, pois, com exceção dos controles, todos os animais deste experimento
tiveram a mesma freqüência de contato com o experimentador. Ainda, os resultados deste
experimento, dos experimentos de Willner et al. (1987) e de Katz (1981) sugerem que as
manipulações realizadas nos animais alteraram somente a ingestão de substâncias
palatáveis.
Apesar do exposto acima, quando os animais deste experimento foram testados em
um modelo animal de ansiedade — labirinto em cruz elevado — observamos que a
condição de manuseio e a ingestão de açúcar produziram efeitos sobre o comportamento
exploratório dos ratos que são difíceis de serem explicados.
Em comparação com os ratos do grupo controle, os animais que foram expostos às
sessões de 10 min de manuseio sem posterior ingestão de açúcar aumentaram a
porcentagem de entradas, tempo e freqüência de entradas nos braços abertos bem como a
freqüência de entradas nas extremidades dos braços abertos. Entretanto, os animais
expostos às sessões de manuseio de 0, 1 e 5 min mas sem ingestão posterior de açúcar não
46

diferiram dos animais do grupo controle. Parece que manusear os animais por 10 minutos
durante 14 dias afeta seu comportamento no labirinto em cruz elevado. Esses mesmos 10
min de manuseio, no entanto, não produziram qualquer efeito sobre o comportamento dos
animais que receberam a ingestão posterior de açúcar. Talvez a ingestão de açúcar tenha
anulado os efeitos do manuseio por 10 min. Mesmo assim, o grupo de animais manuseado
por 10 min com ingestão posterior de açúcar consumiu significativamente menos açúcar
que os grupos de animais com ingestão posterior de açúcar e manuseados por 0, 1 e 5 min.
Isso sugere que a ingestão de açúcar pode ter tido pouca relevância para explicar o fato de
o grupo de animais manuseados por 10 min não diferirem dos ratos do grupo controle.
Nesse caso, por que os animais manuseados por 10 min com ingestão posterior de açúcar
não se comportaram de modo semelhante aos animais manuseados por 10 min sem
ingestão de açúcar?
Também observamos que, comparados aos controles, os animais manuseados por 1
min e com ingestão posterior de açúcar aumentaram o tempo gasto nos braços abertos, a
freqüência de entradas e o tempo gasto nas extremidades desses braços no labirinto em
cruz elevado. Entretanto, não observamos qualquer efeito sobre o comportamento
exploratório dos animais manuseados por 0, 5 e 10 min e com ingestão posterior de açúcar.
Os animais expostos às sessões de manuseio por 1 ou 5 min ingeriram quantidades
semelhantes de açúcar. Isso sugere que o tempo de manuseio talvez seja relevante para
justificar o efeito observado no grupo manuseado por 1 min. Entretanto, os animais
expostos às sessões de 1 min de manuseio sem ingestão posterior de açúcar não diferiram
dos animais do grupo controle. Adicionalmente, se considerarmos que a ingestão de açúcar
é um fator de impacto sobre o desempenho no labirinto em cruz elevado, o grupo que não
foi manuseado teria que se destacar dos outros, pois ingeriu uma quantidade
significativamente maior de açúcar quando comparado aos outros grupos.
47

Alguns trabalhos mostraram que a ingestão prévia de substância palatável pode
afetar o comportamento em situações posteriores. Por exemplo, a ingestão prévia de
substância palatável aumenta a latência de retirada da pata no teste da chapa quente, efeito
interpretado como analgesia (Segato et al., 1997). Entretanto, não encontramos trabalhos
na literatura mostrando que a ingestão prévia de substâncias palatáveis afeta
comportamento de ratos em modelos de exploração livre.
Em relação ao manuseio, vários trabalhos mostraram seus efeitos em situações de
exploração livre ou que exijam um operante específico. Levine et al. (1956) foram os
primeiros a relatar que o manuseio logo após o nascimento melhora o desempenho de ratos
na tarefa de esquiva ativa de duas vias. Posteriormente, vários estudos confirmaram esses
resultados, estendendo esse efeito de melhora para várias linhagens de ratos.
Adicionalmente, verificou-se que o manuseio precoce aumenta a atividade e
comportamento exploratório específico de ratos em vários modelos, tais como campo
aberto (Fernández-Teruel et al., 1992b), exposição a caixas conhecidas e desconhecidas
(Ferré et al., 1995) e tábua de buracos (Fernández Teruel et al., 1992a). Quando foram
utilizados testes mais relacionados à ansiedade verificou-se que o procedimento de
manuseio precoce tornou os animais menos ansiosos, visto aumentarem a exploração nos
braços abertos de um labirinto em cruz elevado (Fernandez-Teruel et al., 1990) e o número
de visitas e tempo de permanência em compartimentos iluminados e abertos em um
labirinto em túnel e na caixa claro-escuro (Fernández-Terual et al., 1991b).
No presente experimento os animais foram manuseados quando adultos (cerca de
200 g). Andrews e File (1993) relataram que, em comparação com ratos não manuseados,
ratos adultos (170 g) expostos a um período de 7 dias de manuseio (pegar o rato
diariamente, pesá-lo e aplicar uma injeção intraperitoneal de salina) apresentaram maiores
porcentagens de tempo gasto e de entradas nos braços abertos de um labirinto em cruz
elevado. Além disso, os autores salientaram que a história de manuseio prévia à chegada
48

dos animais no laboratório também é um fator importante a ser investigado, pois pode ter
efeitos relevantes nesse tipo de teste.
Meinrath e Flaherty (1987) também relataram efeitos do manuseio em ratos adultos.
Um grupo de animais com 60 dias de idade foi exposto ao procedimento de contraste de
ingestão de sacarose e 4 dias depois ao teste no campo aberto. Outro grupo foi exposto ao
procedimento na ordem inversa: campo aberto e quatro dias depois contraste de ingestão
de sacarose. Esse protocolo experimental de contraste de ingestão de sacarose durou 17
dias e nesse período os animais eram deslocados de suas caixas de alojamento para caixas
onde poderiam ingerir 4% ou 32% de sacarose, individualmente. Esses autores
observaram que houve efeito da ordem de execução dos protocolos: (1) contraste seguido
de campo aberto ou (2) campo aberto seguido de contraste. Os animais da primeira ordem
locomoveram-se mais no campo aberto em relação aos animais da segunda ordem.
Adicionalmente, esses animais mostraram maior freqüência de levantadas (ficar em pé nas
patas traseiras) e menor número de bolos fecais. Esses autores sugeriram que os
procedimentos repetidos (contato com o experimentador, pesagem, remoção e
deslocamento da caixa de alojamento) afetaram os animais de modo a torná-los menos
ansiosos no campo aberto.
Um ponto interessante a ser mencionado foi que os animais que se locomoveram
mais no campo aberto no experimento de Meinrath e Flaherty (1987), além de serem
manipulados mais vezes pelo experimentador também ingeriram sacarose antes do teste no
campo aberto. No entanto, os autores não fizeram qualquer sugestão de que a ingestão de
sacarose, de alguma forma, tenha sido relevante para justificar o aumento nas medidas
comportamentais registradas nesse modelo.
Brett e Pratt (1990) também relataram efeitos interessantes do manuseio. Esses
autores não encontraram diferenças significativas no comportamento exploratório no
labirinto em cruz elevado entre animais manuseados e não manuseados. Essa diferença
49

surgia apenas quando a manipulação era crônica. Em um procedimento subagudo, grupos
independentes de ratos receberam injeções de salina ou 2,5 mg/Kg de diazepam por três
dias consecutivos e posteriormente eram testados no labirinto em cruz elevado. Os animais
que receberam diazepam aumentaram a porcentagem de entradas e o tempo gasto nos
braços abertos quando comparados aos controles. Entretanto, quando um procedimento
crônico (28 dias) foi realizado não houve diferença entre os grupos controle e experimental
quanto às mesmas medidas do comportamento exploratório no labirinto em cruz elevado.
Como a única diferença entre os protocolos subagudo e crônico foi a freqüência com que
os animais foram manuseados, os autores apontaram o manuseio como a variável ser
considerada.
Esses resultados sugerem que o manuseio tem efeitos sobre medidas
comportamentais registradas em modelos de exploração livre. Entretanto, no presente
experimento não temos como dizer se os efeitos que observamos estão mais relacionados
ao manuseio, à ingestão de açúcar, a uma interação entre manuseio e ingestão de açúcar ou
se até mesmo se é possível dizer que são idiossincrasias.
No experimento I observamos diferenças entre bons e maus consumidores de
açúcar nas medidas registradas no labirinto em cruz elevado, entretanto, não conseguimos
fazer distinção se essas diferenças estavam mais relacionadas à ansiedade ou à atividade
locomotora. Os resultados do presente experimento também são difíceis de serem
compreendidos e também não ajudaram a explicar os resultados do Experimento I.
Os resultados do Experimento I parecem sugerir que existe uma relação entre
ingestão de açúcar e atividade locomotora no labirinto em cruz elevado. Entretanto, o
labirinto em cruz elevado é um modelo freqüentemente utilizado para registrar
comportamentos relacionados à ansiedade, sendo que outros modelos como o campo
aberto são mais utilizados quando se pretende investigar atividade locomotora. Para tentar
identificar efeitos sobre atividade locomotora ou sobre comportamentos relacionados à
50

ansiedade, torna-se necessário adicionar o teste no campo aberto. Com essa finalidade,
realizou-se o experimento a seguir.
Experimento III Os resultados obtidos nos experimentos anteriores sugeriram, além da utilização de
outro modelo animal, que parece relevante um maior controle das condições de manuseio
anteriores aos testes, principalmente no do campo aberto, pois muitos trabalhos mostraram
que o manuseio prévio pode alterar a atividade locomotora neste modelo de teste. Nesse
sentido, primeiro programaram-se os testes no campo aberto e no labirinto em cruz elevado
e em seguida as sessões de ingestão de açúcar. Nos experimentos anteriores era ao
contrário: primeiro a fase de ingestão de açúcar e depois o teste no labirinto em cruz
elevado. Dessa forma, de um modo geral, aqui vamos visar verificar se a ingestão de
açúcar está relacionada à atividade locomotora ou a comportamentos indicadores de
ansiedade. Com esse fim, utilizaram-se dois modelos, o campo aberto e o labirinto em cruz
elevado, na seguinte seqüência experimental: testes no campo aberto, no labirinto em cruz
elevado e a fase de ingestão de açúcar. A intenção foi a de deixar fase de ingestão de
açúcar por último em uma tentativa de amenizar os efeitos do manuseio.
O teste no campo aberto consiste em medir os comportamentos que ocorrem em um
ambiente novo e sem lugares protegidos. Os comportamentos observados dependem da
interação do rato com muitos fatores tais como: (1) experiência da remoção de um
ambiente familiar; transferência para o campo aberto, (2) características do campo aberto e
de seu ambiente circundante e (3) o protocolo experimental programado (Walsh e
Cummins, 1976).
Apesar de ser um modelo bastante utilizado não existe um padrão entre os
pesquisadores, tanto na forma como no uso deste modelo. Seu tamanho (Broadhurst, 1957;
Blizard, 1971), forma e cor (Delbarre et al., 1970; Oldham e Morlock, 1970), subdivisões
51

(Morrison e Thatcher, 1969), textura do chão (Satinder, 1968), odores (McCall, 1969) e
localização da área de início (Clark et. al. 1970; King, 1970) têm sido variados. Do mesmo
modo encontramos variações no ambiente circundante, tais como (1) a natureza,
intensidade e posição da luz, (2) odores e a visibilidade e (3) posição do observador (Walsh
e Cummins, 1976). De fato, é difícil pensar em uma condição desse modelo que já não
tenha sido modificada. Esta dificuldade de padronização pode ser explicada pelo fato de os
próprios pesquisadores não descreverem os detalhes de suas variáveis de procedimento.
Além das variações citadas acima, também observamos que há variações nas
variáveis dependentes registradas nesse modelo, como por exemplo, medidas fisiológicas,
endocrinológicas e comportamentais (Walsh e Cummins, 1976). Apesar disso, o modelo é
bem conhecido por ser utilizado para investigar atividade locomotora. A atividade
locomotora tem sido registrada pelo número de subdivisões atravessadas. Adicionalmente,
atividade locomotora também tem sido registrada pela sua localização, sendo que as
subdivisões do aparato podem ser compreendidas nas áreas central ou periférica (Ivinskis,
1970). Dessa forma, a atividade locomotora sobre ocorrida em uma ou outra área também
tem sido usada como uma medida de ansiedade (Morrison e thatcher, 1969; Valle, 1970).
Diferentes espécies foram testadas no campo aberto, com diferentes tamanhos. Os
resultados mostram que alterações no tamanho do campo aberto tem efeitos sobre a
atividade locomotora, quanto maior o tamanho do campo aberto maiores os níveis de
atividade locomotora em ratos (Montgomery, 1951; Broadhurst, 1957).
Outra condição que também afeta a atividade locomotora nesse modelo é o nível de
iluminação. Em condições com altos níveis de iluminação os ratos mostram atividade
locomotora reduzida. Blizard (1971) relatou que quanto maior o campo aberto, mais os
animais se locomovem sob condição de baixa iluminação. Valle (1970) relatou que
atividade locomotora, o comportamento de ficar em pé sobre as patas traseiras e o
comportamento conhecido como tigmotaxia — ficar próximo a superfícies verticais — são
52

menos freqüentes em condições de alta iluminação. Livesey e Egger (1970) relataram que
a atividade locomotora era menor nas condições de iluminação 9,3 lux do que na de 0,005
lux em ratos de 47 ou 115 dias.
Levando em consideração os resultados obtidos no Experimento I sobre ingestão de
açúcar e comportamentos relacionados à ansiedade ou atividade locomotora no labirinto
em cruz elevado, o objetivo deste experimento foi o de verificar se a ingestão de açúcar
está relacionado à atividade locomotora ou aos comportamentos indicadores de ansiedade.
Para isso, comparamos o comportamento exploratório de ratos em dois modelos: o campo
aberto e o labirinto em cruz elevado. Os animais foram submetidos às seguintes condições
experimentais: teste no campo aberto, teste no labirinto em cruz elevado e sessões de
ingestão de açúcar. A fase de ingestão de açúcar foi realizada por último com a finalidade
de amenizar possíveis efeitos do manuseio.
Método
Sujeitos.
Foram utilizados 26 ratos Wistar machos (300 ± 10 g) provenientes do biotério
Geral do Campus da Universidade de São Paulo em Ribeirão Preto e mantidos nas mesmas
condições descritas no Experimento I.
Equipamento.
Utilizou-se o mesmo labirinto em cruz elevado do Experimento I. Além desse
aparato, utilizou-se também um campo aberto. Este era constituído por caixa quadrada de
madeira (120 x 120 x 40 cm) com piso de fórmica marrom escuro, com paredes laterais de
madeira. Suspensa acima da região central do campo aberto havia uma câmera de vídeo
acoplada a um vídeo cassete e a um aparelho de televisão localizados em uma sala
53

adjacente à sala adjacente à sala de teste, como no caso do labirinto em cruz elevado. Por
ocasião do registro, também como foi feito para o labirinto em cruz elevado, colocava-se
um gabarito transparente sobre a tela do aparelho de televisão com a imagem do campo
aberto com o piso dividido em 36 quadrados (cada um com o equivalente a 20 cm de lado
no piso do aparato). Isso permitiu a localização precisa dos deslocamentos do animal e dos
comportamentos emitidos em cada um dos quadrados do campo aberto.
Procedimento.
CAMPO ABERTO. Após o período de habituação de três dias, os animais foram testados no
campo aberto por cinco minutos. O animal era gentilmente colocado na área central do
piso do campo aberto podendo explorá-la livremente durante 5 minutos. Durante o teste, a
luz da sala permaneceu desligada. Foram registradas a distância total percorrida (estimada
a partir do número de quadrados nos quais o animal entrava) e a distância percorrida em
cada região do campo aberto (Figura 12): área periférica (20 quadrados com pelo menos
um dos lados com parede), área intermediária (12 quadrados adjacentes internamente à
área periférica) e área central (4 quadrados no centro do campo aberto). Para o registro,
utilizou-se uma câmera equipada com lâmpada infravermelha. O registro comportamental
foi feito com o mesmo programa utilizado nos Experimentos I e II.
Figura 12. Diagrama do campo aberto dividido em 36 quadrados. Área periférica, quadrados hachurados; área intermediária, quadrados em branco; área central, quadrados pontilhados. Cada quadrado tem 20 cm de lado. A divisão em quadrados refere-se à divisão sobre o gabarito transparente; o piso do aparato não continha quaisquer divisões.
54

LABIRINTO EM CRUZ ELEVADO. Vinte e quatro horas após o teste no campo aberto, os
animais foram testados no labirinto em cruz elevado por cinco minutos do mesmo modo
que nos Experimentos I e II, analisando-se os mesmos comportamentos. Durante os cinco
minutos de teste, ao contrário do que ocorreu nos Experimentos I e II, a luz da sala
permaneceu desligada.
INGESTÃO DE AÇÚCAR. Vinte e quatro horas após o teste no labirinto em cruz elevado teve
início a fase de ingestão. Durante oito dias, em sessões diárias de 30 min, os animais foram
colocados individualmente em uma caixa semelhante à de alojamento, na qual estavam
disponíveis ração moída em comedouros de aço inoxidável, e dois frascos com bicos de
aço inoxidável contendo açúcar dissolvido em água de torneira (240 g/l) e apenas a água de
torneira. Os comedouros foram pesados no início e final de cada sessão, permitindo obter a
quantidade consumida. Os frascos continham divisões que permitiam medir o nível de
líquido, também permitindo obter a quantidade consumida durante a sessão. Em seguida,
os animais eram devolvidos às suas respectivas gaiolas-viveiro, com ração e água a
vontade.
Análise estatística.
A ingestão de açúcar e de ração foi analisada pelo teste de Mann-Whitney enquanto
a exploração do campo aberto foi analisada por Anova de uma via com medidas repetidas
(três níveis: áreas periférica, intermediária e central) seguida, sempre que apropriado, pelo
teste post-hoc de comparações de médias de grupos de Duncan. As correlações entre o
consumo de açúcar e de ração e os comportamentos no campo aberto e no labirinto em
cruz elevado foram feitas com o teste de correlações de Pearson. Em todos os casos o nível
de significância foi fixado em P < 0,05.
55

Resultados
A Figura 13 mostra a média da ingestão de açúcar e de ração nas duas últimas
sessões de consumo. Pode-se observar que o açúcar foi preferido pelos ratos, tanto em
relação à água quanto em relação à ração.
**
0
2
4
6
8
Açúcar Água Açúcar RaçãoSubstância
Con
sum
o (m
l)
0.0
0.5
1.0
1.5
2.0
Con
sum
o (g
)
Líquidos Sólidos
Figura 13. Ingestão média de açúcar e ração nas duas últimas sessões do período de consumo. *, Significativamente diferente do consumo de açúcar (Mann-Whitney, P < 0,05).
A Figura 14 mostra o padrão de exploração do campo aberto. A Anova mostrou
diferenças na quantidade de quadrados explorados em cada área (F[2,50] = 96,004; P <
0,001), no tempo gasto em cada área (F[2,50] = 6219,017; P < 0,001) e na distância nelas
percorrida (F[2,50] = 684,550; P < 0,001). O teste de Duncan mostrou que os animais
atravessaram mais quadrados na área intermediária do que nas outras duas e mais na
periférica do que na central. No entanto gastaram muito mais tempo e percorreram
distâncias maiores na área periférica do que nas áreas intermediária e central.
°
*
0
2
4
6
8
10
12
P I C
Freq
üênc
ia de
entra
das
° *0
50
100
150
200
250
300
P I C
Temp
o gas
to (s)
* *0
5
10
15
20
25
30
P I C
Distâ
ncia
perco
rrida (
m)
Áreas do campo aberto
Figura 14. Parâmetros de ocupação do campo aberto. P, Área periférica; I, área intermediária; C, área central. *, Diferente de P e I; °, diferente de P (Mann-Whitney, P < 0,05).
56

A Tabela 5 mostra as correlações entre o consumo de açúcar e a exploração do
campo aberto. Mostra também a correlação entre esta exploração e o consumo de ração.
Pode-se observar que houve uma correlação significativa entre o consumo de açúcar e a
distância percorrida na área central, isto é, ratos que consumiram mais açúcar
perambularam pela área central mais que os que consumiram menores quantidades de
açúcar. Além disso, houve também uma correlação positiva entre o consumo de ração e o
tempo gasto na área periférica, isto é, ratos que consumiram mais ração permaneceram
mais tempo na área periférica do que os que consumiram menos ração.
Tabela 5. Correlação entre o consumo (g) de açúcar e de ração e os comportamentos no campo aberto. P, Área periférica; I, área intermediária; C, área central. *, Correlação de Pearson estatisticamente significativa, P < 0,05.
Parâmetro
Freqüência de entradas Tempo gasto (s) Distância percorrida (m)
Substância consumida P I C P I C P I C
Açúcar 0.341 0.342 0.315 -0.368 0.291 0.348 0.134 0.332 0.391*Ração -0.176 -0.174 -0.135 0.390* -0.380 -0.295 0.266 -0.194 -0.203
A Figura 15 mostra as correlações individuais entre consumo tanto de açúcar como
de ração e freqüência de entradas, tempo gasto e distância percorrida campo aberto sempre
que a correlação apresentou uma probabilidade menor que 0,10. Pode-se notar que o
consumo de açúcar tendeu a se correlacionar com a freqüência de entradas nas áreas
periféricas e intermediária, com o tempo gasto nas áreas periférica e central e com a
distância percorrida nas áreas intermediária e central. Também se pode notar que houve
uma tendência à correlação entre o consumo de ração e o tempo gasto nas áreas periférica e
intermediária. As correlações com probabilidade entre 0,05 e 0,10, no entanto, foram
consideradas apenas como indicativas.
A Tabela 6 mostra as médias (± E. P. M.) dos comportamentos exibidos pelos ratos
nos braços do labirinto em cruz elevado. Não se analisaram as diferenças devido ao fato de
os animais se constituírem em um grupo único. Destaca-se. no entanto, o tempo maior
57

gasto nos braços fechados do labirinto, o que nem sempre se observou com os outros
comportamentos.
Área periférica Área intermediária Área central
A (Açúcar, P < 0,10)
0
2
4
6
8
10
12
14
0 1 2 3 4
Ent
rada
s
B (Açúcar, P < 0,10)
02
468
101214
1618
0 1 2 3 4
Ent
rada
s
C (Açúcar, P < 0,10; ração, P < 0,05)
250
260
270
280
290
300
0 1 2 3 4
Tem
po
D (Ração P < 0,10)
0
5
10
15
20
25
30
0 1 2 3 4
Tem
po (s
)
E (Açúcar, P < 0,10)
0
5
10
15
20
25
30
35
0 1 2 3 4
Tem
po (s
)
F (Açúcar, P < 0,10)
0
1
2
3
4
5
6
0 1 2 3 4
Dis
tânc
ia p
erco
rrida
(m)
G (Açúcar, P < 0,05)
0
1
2
3
4
0 1 2 3 4
Dis
tânc
ia p
erco
rrid
a (m
)
Consumo (g)
Figura 15. Correlação entre consumo de açúcar e ração e parâmetros de ocupação do campo aberto.
São mostradas apenas as correlações com P < 0,10. ●, Açúcar (linha de tendência pontilhada); °, ração (linha de tendência cheia).
58

Tabela 6. Comportamentos exibidos nos braços do labirinto em cruz elevado (média ± E. P. M.).
Área do labirinto
Medidas Braços abertos Braços fechados Centro Total
Freqüência Entradas 10.27±0.65 10.62±0.53 - 20.88±0.96 Porcentagem do total 48.77±1.66 51.23±1.66 - - Distância percorrida (m) 6.91±0.53 8.76±0.40 - 15.67±0.73 Levantadas 0.15±0.07 14.58±0.87 1.54±0.47 16.27±1.15 Esticadas 2.92±0.46 2.19±0.35 1.54±0.28 6.65±0.68 Mergulhos de cabeça 6.04±0.78 0.04±0.04 1.38±0.24 7.46±0.83 Limpadas 0.23±0.08 5.12±0.46 0.12±0.06 5.46±0.47
Tempo (s) Entradas 94.93±6.24 157.44±6.65 47.63±2.44 - Porcentagem do total 31.64±2.08 52.48±2.22 15.88±0.81 - Levantadas 0.14±0.09 22.21±1.76 1.97±0.66 24.32±2.17 Esticadas 3.56±0.66 3.04±0.65 1.76±0.31 8.35±1.15 Mergulhos de cabeça 6.62±0.98 0.03±0.03 2.15±0.41 8.80±1.18 Limpadas 0.12±0.06 24.79±4.18 0.05±0.04 24.96±4.21
A Tabela 7 contém as médias (± E. P. M.) dos comportamentos ocorridos nas
extremidades dos braços do labirinto em cruz elevado.
Tabela 7. Comportamentos exibidos nas extremidades dos braços do labirinto em cruz elevado (média ± E. P. M.). Extremidades
Freqüência Tempo
Medidas Braços abertos
Braços fechados
Braços abertos
Braços fechados
Entradas 8.19±0.70 10.19±0.48 43.70±4.45 84.65±5.49 Levantadas 0.00±0.00 12.44±0.76 0.00±0.00 20.32±1.68 Esticadas 1.65±0.38 0.08±0.05 2.01±0.53 0.06±0.04 Mergulhos de cabeça 2.31±0.43 0.00±0.00 2.56±0.53 0.00±0.00 Limpadas 0.04±0.04 3.42±0.40 0.03±0.03 14.93±3.15
A Tabela 8 mostra as correlações entre a ingestão de açúcar e de ração nas duas
últimas sessões do período de consumo e os comportamentos exibidos nos braços abertos e
fechados e no centro do labirinto em cruz elevado. Mostra também, sempre que pertinente,
os totais de cada um dos parâmetros considerados. Pode-se notar que houve uma
correlação positiva entre a ingestão de açúcar e a freqüência de entradas e a distância
59
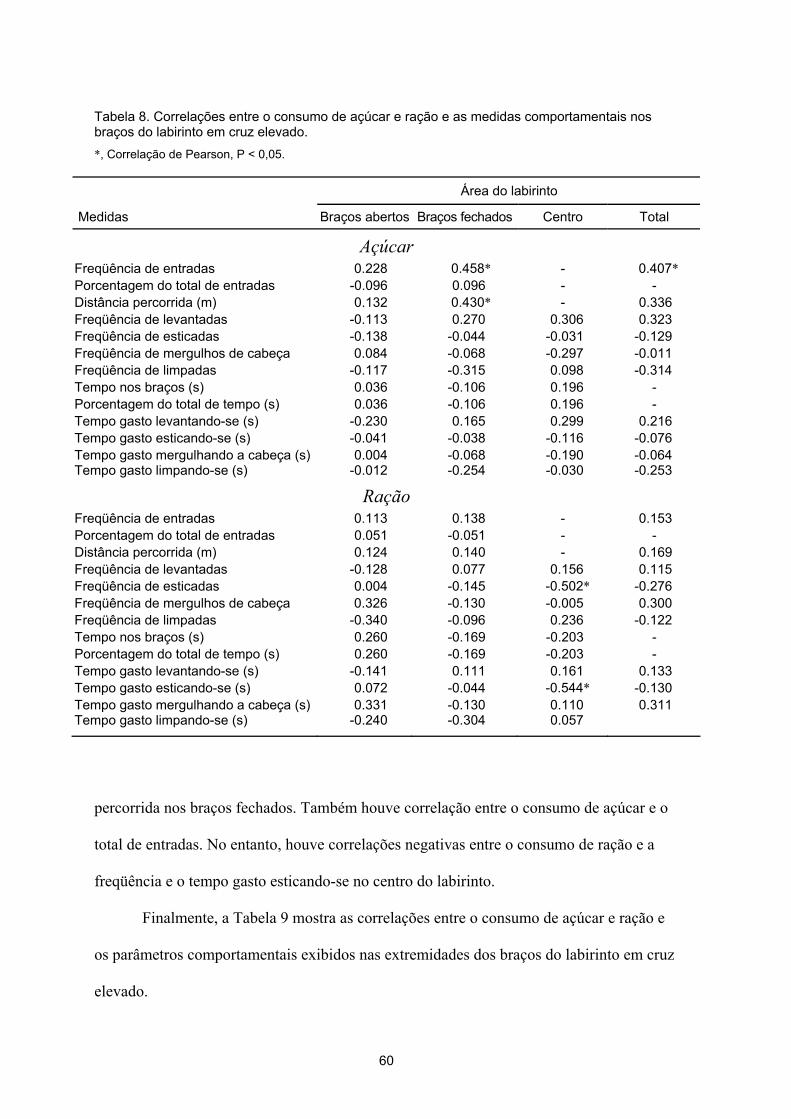
Tabela 8. Correlações entre o consumo de açúcar e ração e as medidas comportamentais nos braços do labirinto em cruz elevado. *, Correlação de Pearson, P < 0,05.
Área do labirinto
Medidas Braços abertos Braços fechados Centro Total
Açúcar Freqüência de entradas 0.228 0.458* - 0.407* Porcentagem do total de entradas -0.096 0.096 - - Distância percorrida (m) 0.132 0.430* - 0.336 Freqüência de levantadas -0.113 0.270 0.306 0.323 Freqüência de esticadas -0.138 -0.044 -0.031 -0.129 Freqüência de mergulhos de cabeça 0.084 -0.068 -0.297 -0.011 Freqüência de limpadas -0.117 -0.315 0.098 -0.314 Tempo nos braços (s) 0.036 -0.106 0.196 - Porcentagem do total de tempo (s) 0.036 -0.106 0.196 - Tempo gasto levantando-se (s) -0.230 0.165 0.299 0.216 Tempo gasto esticando-se (s) -0.041 -0.038 -0.116 -0.076 Tempo gasto mergulhando a cabeça (s) 0.004 -0.068 -0.190 -0.064 Tempo gasto limpando-se (s) -0.012 -0.254 -0.030 -0.253
Ração Freqüência de entradas 0.113 0.138 - 0.153 Porcentagem do total de entradas 0.051 -0.051 - - Distância percorrida (m) 0.124 0.140 - 0.169 Freqüência de levantadas -0.128 0.077 0.156 0.115 Freqüência de esticadas 0.004 -0.145 -0.502* -0.276 Freqüência de mergulhos de cabeça 0.326 -0.130 -0.005 0.300 Freqüência de limpadas -0.340 -0.096 0.236 -0.122 Tempo nos braços (s) 0.260 -0.169 -0.203 - Porcentagem do total de tempo (s) 0.260 -0.169 -0.203 - Tempo gasto levantando-se (s) -0.141 0.111 0.161 0.133 Tempo gasto esticando-se (s) 0.072 -0.044 -0.544* -0.130 Tempo gasto mergulhando a cabeça (s) 0.331 -0.130 0.110 0.311 Tempo gasto limpando-se (s) -0.240 -0.304 0.057
percorrida nos braços fechados. Também houve correlação entre o consumo de açúcar e o
total de entradas. No entanto, houve correlações negativas entre o consumo de ração e a
freqüência e o tempo gasto esticando-se no centro do labirinto.
Finalmente, a Tabela 9 mostra as correlações entre o consumo de açúcar e ração e
os parâmetros comportamentais exibidos nas extremidades dos braços do labirinto em cruz
elevado.
60

Tabela 9. Correlações entre consumo de açúcar e ração e medidas comportamentais nas extremidades dos braços do labirinto em cruz elevado.
Extremidades dos braços
Freqüência Tempo
Medidas Abertos Fechados Abertos Fechados
Açúcar Entradas 0.105 0.317 -0.125 -0.328 Levantadas -0.038 0.194 -0.038 0.134 Esticadas -0.222 0.056 -0.138 0.047 Mergulhos de cabeça -0.027 -0.038 -0.082 -0.038 Limpadas -0.221 -0.336 -0.221 -0.416*
Ração Entradas 0.236 0.092 0.364 -0.057 Levantadas -0.147 0.084 -0.147 0.125 Esticadas 0.101 0.129 0.100 0.139 Mergulhos de cabeça 0.456* -0.147 0.384 -0.147 Limpadas -0.124 -0.140 -0.124 -0.179
Pode-se notar uma correlação negativa entre a ingestão de açúcar e o tempo gasto se
limpando nas extremidades dos braços fechados e uma correlação positiva entre a ingestão
de ração e a freqüência de mergulhos de cabeça nas extremidades dos braços abertos.
A Figura 16 mostra as correlações individuais com probabilidade menor que 0,10
entre o consumo tanto de açúcar como de ração e os seguintes comportamentos ligados à
exploração: distância percorrida nos braços fechados, distância percorrida total, entradas
nos braços fechados, total de entradas nos dois tipos de braços, e tempo gasto nas
extremidades dos braços abertos. As correlações com probabilidade entre 0,05 e 0,10, no
entanto, foram consideradas apenas como indicativas.
A Figura 17 mostra correlações individuais significativas entre o consumo de ração
e o comportamento de levantar-se (freqüência e tempo no centro do labirinto). A Figura 18
mostra as correlações individuais com probabilidade menor que 0,10 entre consumo de
ração e parâmetros do comportamento de mergulhar a cabeça (tempo nos braços abertos,
freqüência e tempo nas extremidades abertas). E, por último, a Figura 19 mostra as
correlações individuais com probabilidade menor que 0,10 entre consumo de açúcar e
61

parâmetros do comportamento de se limpar (freqüência e tempo nas extremidades dos
braços fechados).
A (açúcar, P < 0,05)
02468
10121416
0 1 2 3
Consumo (g)
Distâ
ncia
no fe
chad
o (m)
4
B (açúcar, P < 0,10)
0
5
10
15
20
25
0 1 2 3
Consumor (g)
Distâ
ncia
total
(m)
4
C (Açúcar, P < 0,05)
02468
1012141618
0 1 2 3
Consumo (g)
Entra
das n
o fec
hado
4
D (Açúcar, P < 0,05)
0
5
10
15
20
25
30
35
0 1 2 3
Consumo (g)
Total
de en
trada
s
4
E (Ração, P < 0,10)
0
20
40
60
80
100
120
0 1 2 3 4Consumo (g)
Temp
o extr
emida
de ab
erta
Figura 16. Correlação entre consumo de açúcar e ração e parâmetros de entradas nos braços do labirinto em cruz elevado. São mostradas apenas as correlações com P < 0,10.●, Açúcar (linha de tendência pontilhada); °, ração (linha de tendência cheia).
62
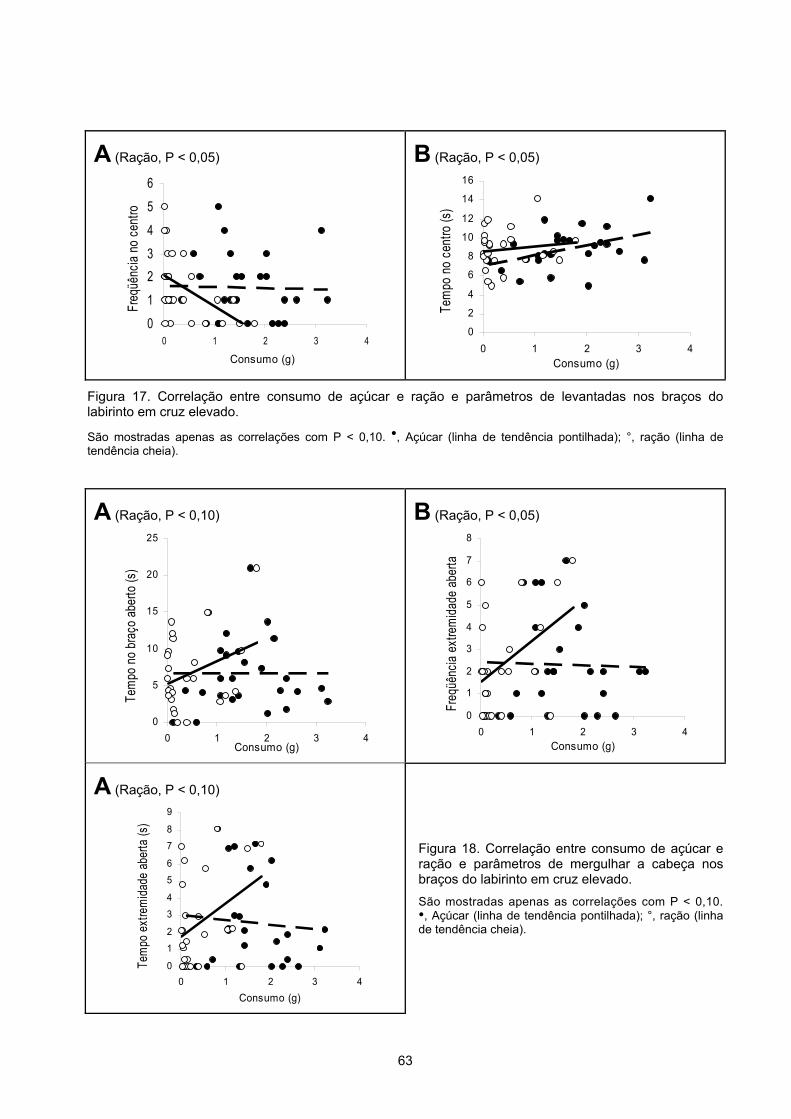
A (Ração, P < 0,05)
0123456
0 1 2 3 4
Consumo (g)
Freq
üênc
ia no
centr
o
B (Ração, P < 0,05)
0
2
4
6
8
10
12
14
16
0 1 2 3 4Consumo (g)
Temp
o no c
entro
(s)
Figura 17. Correlação entre consumo de açúcar e ração e parâmetros de levantadas nos braços do labirinto em cruz elevado.
São mostradas apenas as correlações com P < 0,10. ●, Açúcar (linha de tendência pontilhada); °, ração (linha de tendência cheia).
A (Ração, P < 0,10)
0
5
10
15
20
25
0 1 2 3 4Consumo (g)
Temp
o no b
raço a
berto
(s)
B (Ração, P < 0,05)
0
1
2
3
4
5
6
7
8
0 1 2 3Consumo (g)
Freq
üênc
ia ex
tremi
dade
abert
a
4
A (Ração, P < 0,10)
0
1
2
3
4
5
6
78
9
0 1 2 3 4Consumo (g)
Temp
o extr
emida
de ab
erta (
s)
Figura 18. Correlação entre consumo de açúcar e ração e parâmetros de mergulhar a cabeça nos braços do labirinto em cruz elevado. São mostradas apenas as correlações com P < 0,10.●, Açúcar (linha de tendência pontilhada); °, ração (linha de tendência cheia).
63

A (Açúcar, P < 0,10)
0
1
2
3
4
5
6
7
8
9
0 1 2 3 4
Consumo (g)
Freq
üênc
ia ex
tremi
dade
fec
hada
B (Açúcar, P < 0,05)
0
10
20
30
40
50
60
70
0 1 2 3Consumo (g)
Tem
po e
xtre
mid
ade
fech
ada
(s)
4
Figura 19. Correlação entre consumo de açúcar e ração e parâmetros de limpar-se nos braços do labirinto em cruz elevado.
São mostradas apenas as correlações com P < 0,10. ●, Açúcar (linha de tendência pontilhada); °, ração (linha de tendência cheia).
Como mencionado no caso das figuras anteriores, as correlações com probabilidade
entre 0,05 e 0,10, foram consideradas apenas como indicativas.
Discussão
No Experimento I observamos que, em comparação com os ratos que ingeriram
quantidades menores, os animais que ingeriram maiores quantidades de açúcar
apresentaram freqüências maiores de comportamentos relacionados tanto à ansiedade
como à atividade locomotora no labirinto em cruz elevado. Entretanto, não tínhamos como
saber se essas diferenças eram efeito da ingestão de açúcar ou do procedimento de
manuseio executado no protocolo da fase de ingestão. No Experimento II, tentamos
verificar se a condição de manuseio acarretava efeitos sobre a ingestão de açúcar ou
afetava as medidas registradas no labirinto em cruz elevado. Os resultados foram
inconsistentes e também não conseguimos verificar se as alterações observadas nas
64

medidas registradas no labirinto em cruz elevado eram função do manuseio ou da ingestão
de açúcar.
No presente experimento utilizaram-se dois modelos animais: a arena e o labirinto
em cruz elevado. Adicionalmente, tentamos amenizar os efeitos do manuseio, ao qual os
ratos foram expostos durante a fase de ingestão, sobre as medidas comportamentais
registradas no campo aberto e no labirinto em cruz elevado. Dessa forma, os ratos foram
expostos aos procedimentos da fase de ingestão depois dos testes no campo aberto e no
labirinto em cruz elevado.
Mais atividade locomotora no labirinto em cruz elevado parece funcionar como um
previsor de maior ingestão de açúcar, pois os animais que mostraram maior freqüência de
entradas nos braços fechados, maior freqüência total de entradas nos dois tipos de braços e
maior distância percorrida nos braços fechados ingeriram, posteriormente, maiores
quantidades de açúcar. Não houve correlação entre medidas comportamentais relacionadas
á ansiedade e quantidade de ingestão de açúcar. Também não houve correlação entre
qualquer das medidas comportamentais registradas no labirinto em cruz elevado e a
ingestão de ração de laboratório. Os resultados deste experimento sugerem que atividade
locomotora registrada no labirinto em cruz elevado varia da mesma forma que a ingestão
de açúcar registrada em sessões posteriores. No experimento I, observamos as mesmas
correlações observadas neste experimento, com a diferença de que também houve
correlação entre medidas comportamentais relacionadas à ansiedade e à quantidade de
ingestão de açúcar. Essa diferença pode ter ocorrido porque os animais do Experimento I
foram manuseados antes do teste no labirinto em cruz elevado, o que não ocorreu neste
experimento. Dessa forma, o efeito de tipo ansiolítico do Experimento I parece ter sido
acarretado pelo manuseio.
No presente experimento, os resultados observados no campo aberto sugerem uma
análise diferente da análise realizada sobre os resultados observados no labirinto em cruz
65

elevado. No campo aberto os animais que percorreram maior distância na área central
ingeriram maiores quantidades de açúcar em sessões posteriores. É relevante mencionar
que houve correlação positiva (com índice de significância 0,08, indicativo, portanto) entre
o tempo gasto na área central e a quantidade de ingestão de açúcar em sessões posteriores.
Entretanto, não houve correlação entre freqüência de entradas na área central e a
quantidade de açúcar ingerido. Segundo Prut e Belzung (2003), o tempo gasto, a
freqüência de entradas e a porcentagem da distância percorrida na área central são medidas
relacionadas à ansiedade. Apesar de não observarmos correlações significativas entre todas
as medidas comportamentais relacionadas à ansiedade e à quantidade de açúcar ingerido,
os resultados sugerem que comportamentos relacionados à ansiedade no campo aberto
sejam um previsor da ingestão de açúcar em sessões posteriores.
Entretanto temos que tomar cuidado quando tentamos analisar esses
comportamentos registrados no campo aberto. Roth e Katz (1979) submeteram um grupo
de ratos a um período de uma hora de estresse por luz fluorescente (70 W) e som (95 dB),
enquanto outro grupo permaneceu no biotério. Após esse período os animais foram
testados no campo aberto. Os animais que foram estressados apresentaram maior
freqüência de cruzamentos nos quadrados centrais do campo aberto, quando comparados
aos animais controle. Utilizando-se somente esse registro de comportamento pode-se
pensar que os animais do grupo estressado comportaram-se como menos ansiosos no
campo aberto em relação aos animais do grupo controle. Entretanto, quando se utiliza o
registro dos níveis de corticosterona, observa-se que os níveis dos animais estressados são
maiores que os dos animais controle. Os registros dos níveis de corticosterona sugeriram
que os animais submetidos à luz e ao som comportaram-se como mais ansiosos, embora
tenham se locomovido mais na área central do campo aberto. Utilizar medidas
comportamentais e endocrinológicas diminui as chances de se fazer uma análise incorreta.
66

No presente experimento não houve registro de medidas endocrinológicas e, em função
disso, as análises foram realizadas com precaução.
Em relação às medidas relacionadas à atividade locomotora os resultados foram
inconsistentes. Segundo Prut e Belzung (2003), a freqüência de entradas e a porcentagem
de distância percorrida na área periférica são medidas comportamentais relacionadas à
atividade locomotora. Houve uma correlação positiva (embora com índice de significância
0,08) entre a freqüência de entradas na área periférica e a quantidade ingerida de açúcar.
Isso sugere uma tendência de medidas como freqüência de entradas na área periférica e
quantidade ingerida de açúcar a variarem da mesma forma. Apesar de medidas como
freqüência de entradas na área periférica e quantidade de açúcar ingerido terem variado da
mesma forma, ambas aumentaram. Dessa forma, não temos condições de dizer se uma
medida afetou a outra.
Em relação à ingestão de ração de laboratório houve uma correlação positiva com o
tempo gasto na área periférica. Nenhuma outra medida registrada no campo aberto se
correlacionou com a ingestão de ração de laboratório. De um modo geral, tanto as medidas
comportamentais registradas no labirinto em cruz elevado assim como no campo aberto
não se correlacionaram com a quantidade ingerida de ração de laboratório. Isso sugere que,
quando se observa que certas medidas comportamentais registradas no labirinto em cruz
elevado ou no campo aberto correlacionam-se com a ingestão de açúcar, um fator a
considerar é a percepção do gosto doce. Os resultados do Experimento I e do presente
experimento apresentaram resultados parecidos aos de outros pesquisadores. Alguns
trabalhos mostraram que a variabilidade no consumo de substâncias doces permite predizer
diferenças comportamentais com perfil apetitivo. Por exemplo, alta ingestão de sacarina
correlaciona-se com alta ingestão de etanol (Kampov-Polevoy et al., 1990; Gosnell e
Krahn, 1992; Overstreet et al., 1993) e ratos seletivamente criados como bons e maus
consumidores de etanol consumiram, respectivamente, altas e baixas quantidades de
67

sacarina (Sinclair et al., 1992). Gosnell et al. (1995) mostraram que ratos com alta
preferência por sacarina auto-administram mais morfina que ratos com baixa preferência.
Além disso, ratos que ingerem quantidades elevadas de sacarose auto-administram
psicoestimulantes intravenosamente mais prontamente que ratos que ingerem baixas
quantidades de sacarose (DeSouza et al., 2000).
Outro fator importante é que tanto as medidas registradas no labirinto em cruz
elevado assim e no campo aberto como o registro da ingestão de açúcar são variáveis
dependentes do protocolo experimental e pode não haver relação entre elas. Muitos autores
defendem uma relação direta entre algumas variáveis dependentes, por exemplo, ingestão
de etanol e atividade locomotora em ambiente novo (Gingras e Cools, 1995, 1996),
ingestão de açúcar e comportamentos relacionados à ansiedade no labirinto em cruz
elevado (DeSouza et. al., 1998), alta ingestão de sacarina e alta ingestão de etanol
(Kampov-Polevoy et al., 1990; Gosnell e Krahn, 1992; Overstreet et al., 1993), e alta
ingestão de sacarina e auto-administração de morfina (Gosnell, et. al 1995). Sugerir que
existe uma relação entre os registros comportamentais no labirinto em cruz elevado ou no
campo aberto e a ingestão de açúcar em sessões posteriores pode ser precipitada. O
presente experimento não apresenta condições para identificar as variáveis que
favoreceram as correlações entre ingestão de açúcar e comportamentos registrados no
campo aberto ou no labirinto em cruz elevado. O teste de Pearson, utilizado neste
experimento, apenas nos mostra se a ingestão de açúcar e as medidas comportamentais
registradas no campo aberto ou no labirinto em cruz elevado variam da mesma forma ou
não. Entretanto, dizer que existe uma relação entre esses eventos requer estudos mais
detalhados. Com base nesses resultados, o experimento a seguir teve o objetivo de verificar
se ingestão de açúcar afeta o comportamento de ratos no labirinto em cruz elevado.
68

Experimento IV Os resultados do Experimento III mostraram que os ratos que ingeriram maiores
quantidades de açúcar apresentaram maior freqüência de comportamentos relacionados à
atividade locomotora no labirinto em cruz elevado e maior freqüência de comportamentos
relacionados à ansiedade no campo aberto. Em relação à ingestão de ração moída, não
ocorreram quaisquer correlações com as medidas comportamentais registradas no labirinto
em cruz elevado ou no campo aberto. Os resultados do Experimento I também mostraram
correlações entre maior quantidade de açúcar ingerida e maior freqüência de
comportamentos relacionados à ansiedade ou à atividade locomotora registrados no
labirinto em cruz elevado. Os resultados dos Experimentos I e III apenas mostraram que
quantidade ingerida de açúcar correlacionou-se positivamente com a freqüência de
comportamentos relacionados à ansiedade e à atividade locomotora no labirinto em cruz
elevado e no campo aberto.
Alguns trabalhos também mostraram que maior consumo de substâncias doces
correlacionou-se com outros comportamentos em diferentes modelos animais (Kampov-
Polevoy et al., 1990; Gosnell e Krahn 1992; Sinclair et al., 1992; Overstreet et al., 1993;
Gosnell et al., 1995). DeSouza et. al (1998) relataram que ratos bons consumidores de
sacarose mostraram resposta de sobressalto de menor magnitude diante de um estímulo
acústico, quando comparados a ratos maus consumidores de sacarose. Os mesmos autores
também relataram que os ratos que preferiram sacarose apresentaram menor freqüência de
comportamentos relacionados à ansiedade no labirinto em cruz elevado quando
comparados a ratos que não demonstraram preferência por sacarose.
Para qualquer dos resultados apresentados nos experimentos deste trabalho não se
pode afirmar que a quantidade ingerida de açúcar afetou o comportamento registrado no
labirinto em cruz elevado, embora tenham ocorrido correlações significativas. Em função
69

disso, o presente experimento teve o objetivo de verificar se a ingestão prévia de açúcar
afeta o comportamento de ratos no labirinto em cruz elevado.
Método
Sujeitos
Foram utilizados 32 ratos Wistar machos (170 ± 10 g) provenientes do biotério
Geral do Campus da Universidade de São Paulo em Ribeirão Preto e mantidos nas mesmas
condições descritas no Experimento I.
Após o período de habituação às condições do biotério, os animais foram
distribuídos em quatro Grupos (N = 8), três deles baseados na quantidade de sessões de
consumo (ver abaixo) a que seriam submetidos antes de serem testados no labirinto em
cruz elevado e um outro que receberia injeção de glicose. Para isso, os animais dos três
primeiros Grupos foram privados de comida por 24 horas uma única vez e em seguida
submetidos a uma sessão de habituação ao consumo, para que as substâncias oferecidas
não fossem novidade. Depois dessa sessão, um primeiro Grupo (0) permaneceu no biotério
por sete dias e os sujeitos foram manipulados apenas durante a limpeza da gaiola e troca de
maravalha. No oitavo dia, os animais foram testados no labirinto em cruz elevado e
imediatamente após, foram submetidos a uma sessão de consumo sem qualquer privação.
Não foram, exceto pela sessão de treino com privação, submetidos a nenhuma sessão de
consumo antes do teste no labirinto. Um segundo Grupo (1) permaneceu no biotério por
sete dias e, igual aos do primeiro Grupo, foram manipulados apenas durante a limpeza da
gaiola e troca de maravalha. No oitavo dia foram submetidos à sessão de consumo sem
privação. Imediatamente após, esses animais foram testados no labirinto em cruz elevado.
Foram, excetuando-se a sessão de treino com privação, submetidos a uma única sessão de
consumo antes do teste no labirinto. Um terceiro Grupo (7) foi submetido a sete sessões de
70

consumo sem qualquer privação, iniciadas 24 horas depois da sessão de habituação ao
consumo com privação. No oitavo dia, depois das sete sessões de consumo, foram
submetidos ao teste no labirinto. Exceto pela sessão de treino de consumo, esses animais
não foram mais submetidos a qualquer privação. Finalmente, para testar o efeito de açúcar
no metabolismo, um quarto Grupo (G) permaneceu no biotério por sete dias e, igual aos
sujeitos do primeiro e segundo Grupos, foi manipulados apenas durante a limpeza da
gaiola e troca de maravalha. No oitavo dia, os animais receberam injeção intraperitoneal de
glicose 30 minutos antes de serem testado no labirinto em cruz elevado. Esses animais
nunca foram submetidos a privação.
Equipamento
Labirinto em cruz elevado. idem ao do Experimento I
Procedimento
SESSÕES DE CONSUMO. Os animais foram transportados para uma sala adjacente em uma
caixa pequena e colocados individualmente em gaiolas idênticas às usadas para o
alojamento no biotério, para uma sessão de consumo de uma hora sem qualquer privação.
Durante esse período, estavam disponíveis nessa gaiola um recipiente com ração moída, e
dois frascos contendo água de torneira e solução de açúcar (240 g/l) respectivamente. Os
comedouros e os frascos com líquidos foram pesados antes e depois do período de
consumo. A diferença entre esses registros indicava o consumo de cada animal.
LABIRINTO EM CRUZ ELEVADO. Os animais foram testados no labirinto em cruz elevado
por cinco minutos do mesmo modo que nos Experimentos I e II, analisando-se os mesmos
comportamentos.
DROGAS. Os animais receberam injeções intraperitoniais de glicose. A droga (Drogasil,
Brasil) foi usada em concentração de 1 M e injetada em um volume de l ml/kg. As
soluções de açúcar foram preparadas usando-se açúcar comercial e água de torneira.
71

Análise estatística
Os dados coletados foram submetidos a Anova de uma via e, sempre que
apropriado, seguida do teste de Duncan de comparações entre médias de Grupo. Em todos
os casos, usou-se nível de significância P < 0,05.
Resultados
A Figura 20 mostra o ingestão durante as sessões de consumo. A Anova não
mostrou quaisquer diferenças no consumo de água (F[2,21] = 0,505, P = 0,610) ou de
solução de açúcar (F[2,21] = 0,526, P = 0,598). O consumo de ração, no entanto, foi alterado
pela exposição às
**
012345678
0 1 7 0 1 7 0 1 7
Sessões de consumo antes do teste no labirinto em cruz elevado
Inge
stão
(ml)
0.0
0.5
1.0
1.5
2.0
2.5
3.0
Inge
stão
(g)
Água Solução Ração
Figura 20. Consumo de água, solução de açúcar (240 g/l) e ração nos períodos de ingestão. No submetido a sete dias de consumo, foram utilizadas as médias das últimas duas sessões. *, Significativamente diferente do Grupo 0 (Duncan, P < 0,05).
diferentes quantidades de sessões de consumo (F[2,21] = 4,053, P = 0,031). O teste de
Duncan mostrou que os animais submetidos a 1 e 7 sessões prévias ao teste no labirinto em
cruz elevado ingeriram quantidade de ração significativamente menor que os sujeitos do
Grupo submetido ao teste labirinto após o período de sete dias e ã sessão de consumo
depois do teste comportamental (Grupo 0).
A Figura 21A mostra a freqüência de entradas nos braços abertos do labirinto. A
Anova mostrou um efeito do número de sessões de consumo e da injeção de glicose (F[3,28]
= 4,810, P = 0,008) e o teste de Duncan indicou que os animais submetidos a sessões de
72

consumo prévias ao teste comportamental e os que receberam injeção de glicose entraram
mais freqüentemente nos braços abertos do que os animais não submetidos à sessão prévia.
No entanto, a porcentagem de entradas nesses braços (Figura 21B) não foi afetada seja
pelas sessões prévias seja pela injeção de glicose (F[3,28] = 2,148, P = 0,117). A Figura 21C
mostra que o tempo gasto nos braços abertos, da mesma forma que a freqüência de
entradas nos mesmos, também foi alterado pela quantidade de sessões prévias de consumo
e pela injeção de glicose (F[3,28] = 4,861, P = 0,008). O teste de Duncan mostrou todos os
grupos gastaram mais tempo nos braços abertos do que o Grupo 0. Da mesma forma, a
Anova mostrou que a quantidade de sessões prévias de consumo e a injeção de glicose
alteraram tanto a freqüência de entradas (F[3,28] = 3,540, P = 0,027) quanto o tempo gasto
(F[3,28] = 3,113, P = 0,042) nas extremidades dos braços abertos (Figuras 21D e 21E). O
teste de Duncan mostrou que a exposição a uma sessão prévia e à injeção de glicose
aumentaram a freqüência e tempo gasto explorando as extremidades, em comparação com
o Grupo 0.
A Figura 22 (A e B) mostra as medidas ligadas à motricidade e atividade
exploratória propriamente dita. A Anova mostrou que a quantidade de sessões prévias de
consumo e a injeção de glicose alteraram tanto a freqüência de entradas nos braços
fechados (F[3,28] = 5,138, P = 0,006) quanto a distância percorrida neles (F[3,28] = 3,067, P =
0,044). O teste de Duncan mostrou que a exposição a uma sessão prévia e à injeção de
glicose aumentaram tanto a freqüência de entradas nos braços fechados quanto a distância
neles percorrida, quando comparados com o Grupo 0. A Figura 22C mostra a distância
percorrida nos braços abertos do labirinto. A Anova mostrou um efeito tanto do número de
sessões de consumo quanto da injeção de glicose (F[3,28] = 3,315, P = 0,034) e o teste de
Duncan de comparações entre médias de grupos indicou que os animais dos grupos
submetidos a sessões de consumo prévias ao teste comportamental e os que receberam
injeção de glicose percorreram maiores distâncias nos braços abertos do que os animais
não submetidos à sessão prévia (Grupo 0).
73

A *
**
0
1
2
3
4
5
6
7
0 1 7 G
Entra
das n
os br
aços
aber
tos
B
0
10
20
30
40
50
0 1 7 G
Entra
das n
os br
aços
aber
tos (%
)
C
* **
0
10
20
30
40
50
60
70
0 1 7 G
Temp
o nos
braç
os ab
ertos
(s)
D
**
0
1
2
3
4
5
6
0 1 7 G
Entra
das n
as ex
tremi
dade
s abe
rtas
E
**
0
5
10
15
20
25
30
35
0 1 7 G
Temp
o nas
extre
mida
des a
berta
s (s)
Figura 21. Exploração dos braços abertos do labirinto em cruz elevado.
0, 1 e 7, Quantidade de sessões de consumo antes do teste no labirinto em cruz elevado; G, injeção de glicose (1 M, 1 ml/kg). *, Diferente do Grupo 0 (Duncan, P < 0,05).
74

A
* *
0
3
6
9
12
15
0 1 7 G
Entra
das n
os br
aços
fech
ados
B
**
0
2
4
6
8
10
12
0 1 7 GDistâ
ncia
perco
rrida
no fe
chad
o (m)
C
**
*
0
1
2
3
4
5
0 1 7 G
Distâ
ncia
perco
rrida
no ab
erto
(m)
Figura 22. Medidas ligadas à atividade motora e exploratória.
0, 1 e 7, Quantidade de sessões de consumo antes do teste no labirinto em cruz elevado; G, injeção de glicose (1 M, 1 ml/kg). *, Diferente do Grupo 0 (Duncan, P < 0,05).
Finalmente, a Figura 23 mostra outros comportamentos exibidos no labirinto em
cruz elevado. A Anova não mostrou diferenças devidas a sessões de consumo nem às
injeções de glicose sobre os comportamentos de levantar (F[3,28] = 2,471, P = 0,082),esticar
(F[3,28] = 1,482, P = 0,241) e mergulhar a cabeça (F[3,28] = 1,934, P = 0,147). Mostrou, no
entanto, efeitos sobre o comportamento de limpar (F[3,28] = 1,934, P = 0,147). O teste de
Duncan indicou que os sujeitos submetidos a 1 e 7 sessões prévias de consumo, mas não os
que receberam a injeção de glicose, limparam-se significativamente menos que os sujeitos
do Grupo 0, que não foram submetidos a sessões prévias de consumo.
75

A
0
5
10
15
20
25
30
35
40
0 1 7 G
Temp
o lev
antan
do-se
(s)
B
0
5
10
15
20
25
0 1 7 G
Temp
o esti
cand
o-se
(s)
C
0
2
4
6
8
10
12
14
16
18
0 1 7 G
Temp
o mer
gulha
ndo a
cabe
ça (s
)
D
**
0
10
20
30
40
50
60
70
0 1 7 G
Temp
o lim
pand
o-se
(s)
Figura 23. Outros comportamentos exibidos no labirinto em cruz elevado.
0, 1 e 7, Quantidade de sessões de consumo antes do teste no labirinto em cruz elevado; G, injeção de glicose (1 M, 1 ml/kg). *, Diferente do Grupo 0 (Duncan, P < 0,05).
Discussão
O presente experimento teve o objetivo de verificar se a ingestão prévia de
substâncias doces afeta o comportamento de ratos no labirinto em cruz elevado. Os dados
obtidos sobre a ingestão das soluções de açúcar mostraram que não houve diferença na
média diária de ingestão dessas substâncias em função do número de dias de ingestão para
os diferentes grupos de ratos. Adicionalmente, a exposição ao labirinto em cruz elevado
não teve efeitos sobre a ingestão de açúcar. Entretanto, a média diária de ingestão dos ratos
que tiveram apenas uma sessão de ingestão antes do teste no labirinto em cruz elevado foi
76

significativamente maior que a dos ratos que tiveram uma sessão de ingestão após o teste
ou 7 sessões de ingestão antes do teste. Essa diferença provavelmente se deu pelo fato de
um dos ratos desse grupo (que tiveram uma única sessão de ingestão antes do teste no
labirinto em cruz elevado) ter ingerido uma enorme quantidade de solução, o que elevou a
média de ingestão desse grupo. Levando em consideração esse fator podemos dizer que
não houve diferença significativa na média diária de ingestão de água, solução de açúcar e
ração de laboratório entre os diferentes grupos de ratos. Adicionalmente, podemos verificar
que, quando têm livre acesso a uma substância de gosto doce, os ratos a preferem em
detrimento de substâncias que não produzam essa sensação. Como já observado nos
experimentos anteriores, os ratos ingeriram quantidades de solução de açúcar maiores do
que água ou ração de laboratório.
Em relação ao teste no labirinto em cruz elevado, houve um efeito da ingestão
prévia de açúcar sobre as medidas comportamentais relacionadas à atividade locomotora.
Os ratos que tiveram uma única sessão de ingestão de solução de açúcar antes do teste no
labirinto aumentaram a freqüência de entradas nos braços fechados, a freqüência total de
entradas nos dois tipos de braços e a distância total percorrida nos braços fechados, em
comparação com os ratos que apenas permaneceram no biotério. Esses resultados sugerem
que o acesso livre à solução de açúcar por uma hora aumenta a atividade locomotora
quando os ratos são testados no labirinto em cruz elevado imediatamente após a ingestão.
Também houve um aumento na freqüência total de entradas nos dois tipos de braços para o
grupo de ratos que tiveram sete sessões de ingestão de açúcar. Quanto a esse resultado,
entretanto, é difícil separar o que é efeito da ingestão de açúcar e o que é efeito do
manuseio obrigatório durante as sessões de ingestão. Como discutido no Experimento II,
essa condição de manuseio (pegar o rato freqüentemente e colocá-lo em uma caixa
semelhante à que fica alojado) pode afetar o comportamento de ratos em modelos que
medem atividade locomotora. Alguns trabalhos mostraram esse efeito do manuseio sobre a
77

atividade locomotora em ratos (Andrews e File, 1993; Meinrath e Flaherty, 1987; Brett e
Pratt, 1990). Os ratos que tiveram apenas uma sessão de ingestão prévia ao teste no
labirinto em cruz elevado tiveram menos contato com o experimentador e
conseqüentemente não foram tão manuseados quanto os animais que tiveram sete sessões
de ingestão. Dessa forma, pode-se sugerir que o aumento na atividade locomotora exibido
pelos animais que tiveram apenas uma sessão de ingestão está menos relacionado ao
manuseio.
Um resultado interessante foi que os ratos que receberam a injeção intraperitoneal
de glicose também aumentaram a atividade locomotora no labirinto em cruz elevado:
mostraram aumentos na freqüência de entradas nos braços fechados, freqüência total de
entradas nos dois tipos de braços e distância percorrida nos braços fechados. Esses
resultados mostram que conseqüências pós-ingestivas da administração de glicose podem
ter afetado a atividade locomotora dos ratos no labirinto em cruz elevado. Dessa forma, a
sensação oral produzida pela ingestão da solução de açúcar quando os animais tiveram
uma ou sete sessões de ingestão parece não ser o principal fator afetando as medidas
comportamentais registradas no labirinto.
Uma variável que pode ter afetado o comportamento dos ratos que receberam
glicose intraperitoneal foi a picada da agulha. Alguns trabalhos mostraram que situações
como administração de choques elétricos ( Anderson et al, 1976), medo condicionado
(Baron, 1964; Cappell e Rodin, 1969), isolamento (Ader e Friedman, 1964; Goldsmith et al,
1978), barulho excessivo (Cox e Lee, 1976) e iluminação intensa (Dixon e DeFries, 1968)
tiveram efeitos de suprimir a atividade locomotora e/ou exploratória. Entretanto, Roth e
Katz (1979) mostraram que uma condição aversiva aumentou a atividade locomotora de
ratos em um campo aberto. Ratos expostos a uma luz fluorescente de 70 W e a um ruído
branco de 95 dB por um período de uma hora aumentaram a freqüência de cruzamentos
nos quadrados do centro e nos quadrados da periferia do campo aberto comparado a ratos
78

que não foram expostos a essa condição. Adicionalmente, a latência para iniciar o
movimento, quando colocado no campo aberto, foi significativamente menor nos animais
estressados do que nos controles. Dessa forma, o aumento na atividade locomotora dos
ratos que no presente experimento receberam glicose pode ser mais um efeito da picada da
agulha que da administração de glicose intraperitoneal. Para verificar esta possibilidade
deveria existir um grupo de ratos que tivesse recebido injeção intraperitoneal de salina.
Em relação às medidas comportamentais relacionadas à ansiedade também
observamos um efeito da ingestão de açúcar. Os ratos que tiveram uma única sessão de
ingestão de açúcar percorreram distâncias maiores nos braços abertos, permaneceram lá
mais tempo, e entraram mais e permaneceram mais tempo nas extremidades dos braços
abertos do que os ratos que ficaram no biotério. Os ratos que tiveram sete sessões de
ingestão apenas gastaram mais tempo gasto nos braços abertos do que os ratos que
permaneceram no biotério. Para este grupo (sete sessões de ingestão) vale a mesma análise
feita acima sobre as medidas comportamentais relacionadas à atividade locomotora, isto é,
o aumento pode ser função do manuseio. Apesar de várias medidas comportamentais
relacionadas à ansiedade terem aumentado por causa da ingestão de solução de açúcar, não
houve qualquer efeito sobre a porcentagem de entradas nos braços abertos. Contudo, houve
alterações significativas em várias medidas comportamentais relacionadas à ansiedade, o
que permite sugerir que a ingestão prévia de solução de açúcar teve um efeito ansiolítico
em ratos quando testados no labirinto em cruz elevado.
Novamente, os ratos que receberam injeções intraperitoniais de glicose também se
comportaram como se estivessem menos ansiosos: mostraram aumentos na distância
percorrida e no tempo gasto nos braços abertos, na freqüência de entradas e tempo gasto
nas extremidades desses braços. Como já discutido acima, é difícil dizer se esse efeito foi
causado pela picada ou pela administração intraperitoneal de glicose. Adicionalmente, os
resultados desse grupo fazem pensar que os ratos estavam menos ansiosos. Mas, como
79

houve alterações concomitantes tanto nas medidas relacionadas à atividade locomotora
quanto nas relacionadas à ansiedade, além de um provável estresse da picada, pode ser que
na realidade os ratos estivessem mais ansiosos. No Experimento I, salientou-se que tomar
como medida apenas o comportamento de locomover-se no labirinto não é parâmetro
suficiente para inferir estados de ansiedade. Seria interessante também utilizar medidas
fisiológicas, o que não ocorreu neste experimento.
Alguns trabalhos mostraram que a ingestão prévia de substâncias palatáveis afetou
o comportamento de ratos em situações posteriores. Por exemplo, a ingestão prévia de
substâncias palatáveis aumentou a latência da retirada da pata no teste da chapa quente, ao
aumentar a analgesia (Segato et. al, 1997). Entretanto, não encontramos trabalhos na
literatura mostrando que a ingestão prévia de substâncias palatáveis tenha afetado o
comportamento de ratos em modelos de exploração livre. Em função disso, os resultados
do presente trabalho levantam algumas questões. Por que os animais que tiveram sete
sessões de ingestão de açúcar comportaram-se diferente dos animais que tiveram apenas
uma única sessão de ingestão de açúcar? Lembrando que a média diária de açúcar foi
semelhante para todos os grupos, a única diferença foi a freqüência de contatos com o
protocolo da sessão de ingestão, isto é, os contatos com o açúcar, com o experimentador, a
retirada da caixa de alojamento, o próprio período de ingestão, no qual o animal ficava
separado de outros animais etc. A condição de repetição pode ter tornado esses animais
mais habituados, quando comparados aos animais que tiveram apenas uma sessão de
ingestão antes do teste no labirinto em cruz elevado. Além disso, o protocolo da sessão de
ingestão pode ter funcionado como uma condição de estresse para esses animais por ter
ocorrido apenas uma vez, sem dar condições para que se habituassem. Se de fato ocorreu,
essa condição pode ter favorecido o aumento na freqüência de comportamentos
relacionados à locomoção e o aumento na freqüência de comportamentos não relacionados
a um estado de ansiedade.
80

Discussão geral
Preferência por substâncias de gosto doce
Vários estudos mostraram que existem diferenças individuais no consumo de substâncias
doce, um fenômeno recorrente em roedores e humanos (Brennan et al., 2001). Os
resultados do presente trabalho sobre o consumo de açúcar e ração vão na mesma direção
de vários trabalhos na literatura (De Cabo et al., 1995; Sills e Crawley, 1996; Sills e
Vaccarino, 1996, DeSousa et al., 1998). Quando submetemos nossos ratos à ingestão livre
de dois tipos de comida, açúcar granulado e ração moída de laboratório, pudemos
classificá-los como bons ou maus consumidores de açúcar sem que qualquer dos dois
grupos apresentasse alteração no consumo da ração moída de laboratório. Em outras
palavras, os animais de ambos os grupos não consumiram mais (ou menos) ração por conta
da quantidade de açúcar ingerida no mesmo período. Além disso, o consumo mais elevado
de açúcar não se correlacionou com o peso dos ratos.
A literatura já mostra há muito que moléculas que servem como carregadores
energéticos, como açúcares, são preferidas por alguns animais (Pfaffman, 1975; Scott e
Giza, 1987). Em parte, isso pode explicar o fato de alguns animais mostrarem maior
preferência pelo açúcar á ração moída de laboratório. Entretanto, a escolha por uma
comida não se restringe apenas ao valor energético, pois há um intervalo desde a ingestão
até a metabolização de moléculas que funcionam como carregadores energéticos, e a
indicação de o que ingerir parece ocorrer no momento de escolha do alimento e não no
período de metabolização das moléculas em açúcares. Desse modo, alguns eventos que
afetam a escolha de alimentos, para maximizar a probabilidade do armazenamento de
energia, tornaram-se evolutivamente importantes (Galef, 1996). A percepção do gosto doce
81

sinaliza uma fonte energética para certos animais (Bassi, 1996). Dessa forma, alguns
animais apresentam diferenças na preferência por fontes diferentes de alimento em função
da percepção de gosto e não de energia adquirida. Essa percepção do gosto varia entre as
espécies em função da disponibilidade do que há para comer (Galef, 1996). Por exemplo,
herbívoros parecem ser pouco sensíveis á percepção do gosto amargo (Glendining, 1994),
pois a mesma não restringe sua dieta.
Há trabalhos na literatura que fundamentam a hipótese acima. Mook et. al. (1993)
mostraram que ratos continuam ingerindo sacarose mesmo que o líquido fosse drenado
através de uma fístula esofágica, tendo portanto pouca ou nenhuma conseqüência nutritiva.
Ratos também beberam soluções de sacarina ainda que a mesma não tivesse calorias e
fosse eliminada sem quebras metabólicas (Smith e Foster, 1980). Esses dados mostram que
para alguns animais a percepção do gosto doce é um dos fatores que afetam a ingestão.
Além da percepção do gosto doce, o custo para acessar um ou outro alimento
também interfere na escolha do quê comer. Quando beija-flores podiam escolher entre
comedouros com uma alta taxa de ingestão de energia instantânea, porém com solução de
sacarose de baixa concentração e comedouros com baixa taxa de energia instantânea porém
com solução de sacarose hiperconcentrada, o custo de acesso, ou seja a distância do ninho
aos comedouros revelou-se importante para a escolha. Quando o ninho estava próximo aos
comedouros os beija-flores preferiram a solução de sacarose pouco concentrada; entretanto,
quando estava longe eles preferiram a solução hiperconcentrada (Tamm, 1989).
Bassi (1996) também mostrou que o custo de um alimento afeta a escolha do quê
comer. Em seu estudo, ratos tiveram disponível sacarina, água e ração. À medida que o
custo para acessar a sacarina aumentou os animais deixaram de pressionar a barra e
preferiram ingerir água, uma vez que a água era um recurso livremente disponível.
Adicionalmente, Collier e Johnson (2000) mostraram que, quando ratos podiam ingerir
sacarose ou ração de laboratório livremente, preferiram a sacarose. Quando estipulado um
82

custo para ingerir a sacarose, os ratos passaram a ingerir ração de laboratório. Quanto
maior o custo para ingerir a sacarose maior foi a preferência pela ração. No presente
experimento não houve custo tanto para o açúcar granulado como para a ração moída de
laboratório e, por isso, pudemos observar que grande parte dos animais preferiu o açúcar
granulado.
Ingestão de substâncias palatáveis e modelos de ansiedade
Segundo Dess e Minor (1996) comportamento de ingerir interage com processos
tais como medo e ansiedade. Em alguns estudos, a exposição a choques elétricos
intermitentes alterou a taxa do comportamento de comer, sugerindo um persistente estado
de medo ou ansiedade (Helmstetter e Fanselow, 1993; Dess e VanderWeele, 1994). Ratos
seletivamente criados para exibir um fenótipo de emocionalidade diferiram quanto à
ingestão de sacarina (Gray, 1987 apud Dess e Minor, 1996). Ratos das linhagens que
apresentaram maior freqüência de comportamentos relacionados à ansiedade tomaram
menor quantidade de sacarina e de quinino quando comparados aos das linhagens que
apresentaram menor freqüência de comportamentos relacionados à ansiedade. DeSouza et
al. (1998) relataram que ratos tidos como bons consumidores de sacarose mostraram
resposta de sobressalto de menor magnitude frente de um estímulo acústico, em
comparação com ratos maus consumidores de sacarose. Os mesmos autores também
relataram que ratos que preferiram sacarose apresentaram menor freqüência de
comportamentos relacionados à ansiedade no labirinto em cruz elevado do que ratos que
não demonstraram preferência por sacarose. Os resultados relatados acima sugerem que
ratos que apresentaram maior freqüência de comportamentos relacionados á ansiedade
também ingeriram menores quantidades de substância doce.
83

Esses dados sugerem que, quando selecionamos ratos em função da quantidade de
açúcar ingerida, o fato de alguns deles ingerirem quantidades de açúcar maiores do que
outros pode nos indicar como eles se comportarão quando submetidos a alguns modelos
animais de ansiedade. Os resultados do Experimento I também mostraram que a ingestão
de substâncias doces está correlacionada com comportamentos relacionados à ansiedade no
labirinto em cruz elevado. Os resultados do Experimento III mostraram que, quando se fez
o inverso, isto é, registraram-se as medidas comportamentais no labirinto em cruz elevado
ou na arena, e posteriormente, registrou-se a quantidade de açúcar ingerida obtivemos uma
conclusão semelhante.
Os resultados dos Experimentos I e III, bem como de outros trabalhos na literatura,
apenas apontam uma correlação entre duas medidas comportamentais, sendo que uma
delas é a quantidade ingerida de açúcar e a outra pode variar de acordo com os objetivos do
respectivo trabalho: medidas comportamentais registradas no labirinto em cruz elevado ou
arena (Experimentos I, II e III; DeSouza et al., 1998), quantidade de ingestão de etanol
(Kampov-Polevoy et al., 1990; Gosnell e Krahn 1992; Overstreet et al 1993), auto-
administração de morfina (Sinclair et al., 1992) ou auto-administração de um
psicoestimulante (Gosnell et al., 1995).
Apesar de haver uma correlação entre quantidade ingerida de açúcar e algumas
medidas comportamentais exibidas no labirinto em cruz elevado ou na arena, não podemos
dizer que uma dessas medidas interfere na outra. Entretanto, alguns trabalhos mostraram
que a ingestão prévia de uma substância palatável aumentou a latência de retirada da pata
no teste da chapa quente, um efeito interpretado como analgesia (Segato et. at, 1997).
Além disso, Blass et al. (1987) mostraram que a infusão intra-oral de solução de sacarose
em ratos com 10 dias de idade aumentou rapidamente a latência de retirada da pata no teste
da placa quente e diminuiu o número de vocalizações provocadas pelo isolamento social.
Esse efeito sobre as vocalizações sugere que a infusão de sacarose teve um efeito
84

ansiolítico. Dessa forma, teria a ingestão de açúcar efeitos sobre comportamento
exploratório? Mais especificamente, afetaria o comportamento de ratos na arena ou
labirinto em cruz elevado?
Os resultados do experimento IV mostraram que a ingestão prévia de açúcar afetou
o comportamento de ratos no labirinto em cruz elevado, mas somente quando esteve
disponível por uma hora antes do teste nesse aparato. Um período de uma hora de ingestão
de açúcar aumentou a freqüência de comportamentos tanto relacionados à ansiedade como
à atividade locomotora. Não há trabalhos na literatura relatando o efeito de ingestão de
substâncias doces sobre comportamento de animais em modelos de exploração livre.
Adicionalmente, também se observou que a injeção intraperitoneal de glicose meia hora
antes do teste no labirinto teve efeitos semelhantes à condição de disponibilizar açúcar por
meia hora antes do teste. Entretanto, é preciso cautela para discutir os resultados do grupo
de ratos que recebeu injeção intraperitoneal de glicose, pois o efeito sobre as medidas
registradas no labirinto em cruz elevado podem ser em função da picada da agulha (ver
discussão Experimento IV).
Embora os resultados do presente trabalho corroborem outros trabalhos na literatura,
isto é, mostrem que há uma correlação entre ingestão de substâncias doces e certos
comportamentos registrados no labirinto em cruz elevado ou na arena (Experimentos I e
III), ainda seria interessante especular que condições favoreceram essas correlações.
Alguns trabalhos mostraram que eventos ocorridos na vida precoce de ratos afetaram o
desenvolvimento dos mesmos e esse efeito teve duração até a vida adulta (Levine, 1956,
1957, 1962; Haltmeyer et al., 1967; Núñez et al., 1995). Em ratos, uma condição que tem
efeitos sobre o modo como responderão frente a situações estressantes é o grau de cuidado
materno recebido no início da vida (Ogawa et al., 1994; Penke et al., 2001; Champagne et
al., 2003), definido como freqüência de lambidas (licking) e limpadas (grooming) da mãe
para os filhotes.
85

Assim, quando comparados a ratos criados por mães que apresentavam maior
freqüência de cuidados maternos, ratos criados por mães que apresentaram menor
freqüência de cuidados maternos durante a primeira semana de vida exibiram maiores
níveis de resposta fisiológica ao estresse, medido pelos hormônios de estresse circulantes
no sangue(Liu et al., 1997). Esses animais também apresentaram maior freqüência de
comportamentos relacionados à ansiedade quando testados em uma variedade de modelos
animais de ansiedade (Francis et al., 1999). Interessante é que esses mesmos ratos, quando
expostos a um evento estressante (quatro choques de 0,6 mA com três segundos de duração
em intervalos de 15 segundos), aumentaram a ingestão de uma substância palatável
(salgados de manteiga de amendoim) em detrimento da ração de laboratório, quando
comparados a ratos criados por mães que apresentaram maior freqüência de cuidados
maternos (Hancock et al., 2005). Esses dados apóiam os resultados dos Experimentos I e
III pois, quando os ratos foram testados no labirinto em cruz elevado e na arena, houve
correlações positivas significativas entre quantidade ingerida de açúcar e comportamentos
relacionados à ansiedade e à atividade locomotora.
Segundo Hancock et al. (2005), ratos criados por mães que apresentaram menor
freqüência de cuidados maternos mostraram-se mais ansiosos na vida adulta e ingeriram
maior quantidade de substâncias palatáveis. Na discussão do Experimento I foi levantada a
hipótese de que os ratos que consumiram maior quantidade de açúcar comportaram-se
como mais ansiosos no labirinto em cruz elevado. Esses resultados contrastam com os
relatados por Gray (1987), pois estes relataram que maior freqüência de comportamentos
relacionados à ansiedade está relacionada com menor quantidade ingerida de sacarina.
Entretanto, este autor utilizou ingestão de sacarina e, em função disso, é importante
ressaltar que Bassi (1996) relatou que ratos Sprague-Dawley preferiram água à sacarina,
sugerindo que a sacarina pode ser percebida como mais aversiva (gosto amargo) para essa
linhagem. Adicionalmente, ratos submetidos a choques inescapáveis consumiram menor
86

quantidade de substância adulterada por sacarina ou quinino, mas aumentaram a ingestão
de açúcares (Dess e Chapman, 1990; Dess e Choe, 1994). Esses dados suportam a idéia de
que a percepção do gosto amargo da sacarina pode estar relacionada às condições genéticas
e/ou exposição a eventos aversivos, de modo que essas condições podem salientar as
propriedades de gosto amargo da sacarina. Assim, a percepção do gosto amargo da
sacarina parece ser um fator importante quando se discute os resultados de Gray (1987),
comparando-os aos do presente trabalho.
Segundo DeSouza et al. (1998), ratos menos ansiosos consumiram maior
quantidade de sacarose, o que contrasta com os resultados de Hancock et al. (2005).
Quanto aos resultados de DeSouza et al. (1998), discutiu-se no Experimento I que utilizar
apenas as medidas comportamentais registradas no labirinto em cruz elevado não é um
bom parâmetro para discutir estados de medo e/ou ansiedade; medidas fisiológicas
ajudariam a discutir melhor esses estados. Adicionalmente, DeSouza et al. (1998)
apresentaram os registros de apenas duas categorias comportamentais bastante
relacionadas à ansiedade e não apresentaram quaisquer medidas relacionadas à atividade
locomotora. Em função disso, sugeriu-se no Experimento I que os ratos desses autores
estivessem mais ansiosos e não menos ansiosos, como sugerido pelos autores, concordando
com a hipótese de Hancock et al. (2005).
Um outro fator que reforça a relação entre ingestão de substâncias de gosto doce e
comportamentos relacionados à ansiedade é que não houve diferenças na ingestão de ração
de laboratório entre ratos criados por mães que apresentaram maior ou menor freqüência
de cuidados maternos (Hancock et al., 2005). Isto é, ratos mais e menos ansiosos não
ingeriram quantidades diferentes de ração de laboratório. Esse resultado é pertinente para
os resultados dos quatro experimentos deste trabalho. Quando realizado o teste de
correlação entre medidas registradas no labirinto em cruz elevado e arena e quantidade
87

ingerida de ração de laboratório, não houve correlação entre ingestão e as medidas
comportamentais.
Considerações finais
Um aspecto a ser considerado, tanto nos resultados deste experimento como no de
outros, é qual condição funciona como condição controle quando o objetivo é estudar os
efeitos da manipulação precoce ou tardia. Embora haja muita inconsistência no que se
chama de manuseio, ou “handling” (ver introdução do Experimento I), a condição de
controle é aquela em que o animal permanece no biotério durante todo o experimento,
sendo manuseado o mínimo possível. Entretanto, alguns trabalhos mostraram resultados
interessantes sobre os efeitos da manipulação e da ausência da manipulação sobre o
desempenho em atividades na vida adulta (Levine et al., 1955, 1967; Levine, 1962). Levine
(1960) submeteu um grupo de ratos jovens a choques elétricos leves programados sempre
para o mesmo período do dia. Em um segundo grupo os ratos foram colocados em uma
caixa igual por um período de tempo igual, mas não receberam choques. Um terceiro grupo
de ratos permaneceu no ninho e não foi manuseado durante todo o tempo de duração do
experimento. Quando os ratos chegaram à idade adulta, os experimentadores registraram
comportamentos relacionados à ansiedade. Para surpresa dos experimentadores, foi o
terceiro grupo quem apresentou comportamentos diferentes. O comportamento dos ratos
que receberam choques não foi diferente do comportamento dos ratos que foram apenas
manipulados, porém não receberam choques.
Esse protocolo experimental foi repetido expondo ratos a uma variedade de
condições estressantes e à diferentes formas de manipulações (Levine et al., 1955, 1967;
Levine, 1960, 1962) . Observou-se que os ratos não manipulados sempre se comportaram
de uma maneira diferente dos ratos que receberam choques ou dos que foram apenas
88

manipulados. Quando os ratos foram colocados em um ambiente não familiar (caixa de
plástico transparente), os não manipulados se encolhiam em um canto da caixa por um
período maior de tempo, enquanto os dos outros dois grupos exploraram todo o espaço da
caixa. Resultados semelhantes foram observados quando utilizaram o campo aberto: os
ratos que receberam choques ou que foram apenas manipulados locomoveram-se mais
(número de quadrados atravessados) do que os não manipulados. Nas duas condições de
teste, os ratos não manipulados se escondiam em um dos cantos ou se arrastavam
timidamente, tendendo a defecar e urinar com maior freqüência do que os que receberam
choques ou simplesmente foram manipulados. Tomando estes comportamentos em
conjunto, pode-se sugerir que os ratos não manipulados apresentaram freqüência de
comportamentos relacionados à ansiedade maior do que a dos ratos que receberam choques
ou que simplesmente foram manipulados.
Adicionalmente, alguns trabalhos também mostraram que ratos manipulados na
infância apresentaram resposta fisiológica a eventos estressantes diferente da de ratos não
manipulados (Levine,1962; Levine et al., 1967). Esses autores registraram os níveis de
esteróides na corrente sangüínea de ratos estimulados e não estimulados imediatamente
após receberem choques elétricos. Antes de receberem os choques ambos os grupos de
ratos mostraram níveis semelhantes de esteróides. Após os choques, entretanto, os ratos
que foram expostos ao estresse na infância mostraram níveis de esteróides nos primeiros
15 minutos mais altos que os dos ratos que não foram expostos ao estresse na infância. Os
ratos que não foram expostos ao estresse na infância demoraram mais tempo para
apresentar níveis de esteróides semelhantes aos dos estressados na infância. Os ratos não
estressados na infância também apresentaram níveis altos de esteróides por um período
mais longo quando comparados aos estressados na infância. Esses resultados sugerem que
há um padrão distinto de resposta ao estresse nos dois tipos de ratos. Considerando o
comportamento como um ajuste econômico às pressões a que o organismo está exposto, a
89

velocidade e o tempo de duração da resposta às pressões ambientais parecem otimizar a
relação custo/benefício nos ratos estressados na infância e os somente manipulados. O
atraso e o prolongamento da resposta de estresse nos ratos não manipulados em
comparação com os estressados na infância ou apenas manipulados sugerem um
comportamento que minimiza a relação custo/benefício, isto é, tem pouco valor de
sobrevivência.
Esses dados comentados acima sugerem que os animais do presente experimento
sujeitados à situação que foi denominada de controle, talvez sejam os animais que
devessem ser tratados como pertencentes à condição experimental. Em muitos laboratórios,
o animal considerado “normal ” é aquele que permanece no biotério com comida e água à
disposição, sem qualquer tipo de pressão seletiva sobre sua maneira de se comportar.
Parece um tanto evidente que, dessa maneira, os animais estariam despreparados para um
ambiente que não se mantém constante (ambiente natural). Muitos trabalhos utilizando
animais mostraram que tornar o ambiente variável, isto é, sujeitar o comportamento à
seleção tem efeitos sobre o desempenho na vida adulta. Em outras palavras, esses animais
melhoraram sua habilidade para resolver certos problemas impostos pelo experimentador
(Levine, 1956; Denenberg e Karas, 1961; Núñez et al., 1995). Além disso, podemos citar
as condições em que os animais são criados e mantidos em cativeiro. Quando chegam à
vida adulta, suas chances de sobreviver em um ambiente natural são mínimas. Se o
objetivo de se fazer pesquisa é entender o comportamento levando-se em consideração a
filogênese, e não vejo como não fazer isso, deveríamos pensar sobre a condição que
chamamos de controle. Manter o animal em um biotério sem que ele precise se empenhar
para conseguir comida ou água e até mesmo livrando-o de entrar em contato com eventos
aversivos é a condição mais bizarra que se possa imaginar: não há evidências da existência
de nenhuma espécie animal que foi selecionada nessas condições.
90

Referências Bibliográficas
Abel, E. L. (1971). Habituation as a factor in early handling. Journal of Comparative and Physiological
Psychology, 74 (2): 219-221.
Abercrombie, E. D., Keefe, K. A., DiFrischia, D. S. e Zigmond, M. J. (1989). Differential effects of stress on
in vivo dopamine release in striatum, nucleus accumbens and medial frontal cortex. Journal of
neurochemistry, 52: 1655-1658.
Ader, R. e Friedman, S. B. (1964). Social factors affecting emotionality and resistance to disease in animals.
Psychological Reports, 15: 535-541.
Almeida, S. S.: De Oliveira, L M.; Graeff, F. G. (1991). Early life protein malnutrition changes exploration
of the elevated plus-maze and reactivity to anxiolytics. Psychopharmacology, 103: 513-518.
Anderson, D. C., Koehn, C., Crowell, C., Lupo, J. (1976). Different intensities of unsignalled inescapable
shock treatments as determinants of non-shock motivated open field behavior: A resolution of disparate
results. Physiology and Behavior, 17: 391-394.
Andrews, N. e File, S. E. (1993). Handling history of rats modifies behavioural effects of drugs in the
elevated plus-maze test of anxiety. European Journal of Pharmacology, 235: 109-112.
Anisman, H., Zaharia, M. D., Meaney, M. J. e Merali, Z. (1998). Do early life events permanently alter
behavioral and hormonal responses to stressors ? International Journal of Developmental Neuroscience,
16: 149-64.
Ayd, F. J. (1962). A critical appraisal of chlordiazepoxide. Journal of Neuropsychiatry, 3: 117-180.
Bainbridge, J. G. (1968). The effect of psychotropic drugs on food reinforced behaviour and on food
consumption. Psychopharmacologia, 12 (3): 204-213.
Barnett, S. A. (1975). Movement in the living space. In. The rat: a study behavior. The University of Chicago
Press, Chicago e Londres.
Baron, A. (1964). Suppression of exploratory behavior by aversive stimulation. Journal of Comparative and
Physiological Psychology, 57: 299-301.
Bassi, A. N. (1996). Efeitos dos custos do acesso e do consume sobre a ingestão de soluções doces, calórica e
não calóricas, num sistema de economia fechada. Tese de doutorado apresentada ao Departamento de
Psicologia e Educação da Faculdade de Filosofia, Ciências e Letras de Ribeirão Preto – Universidade
de São Paulo.
Berlyne, D. E. (1950). Novelty and curiosity as determinants of exploratory behavior. British Journal of
Psychology, 41: 68-80.
Biggio, G. (1983). The action of stress B-carbolines, diazepam and Ro 15-1788 on GABA receptors in the rat
brain, benzodiazepine recognition site ligands. In. Biochemistry and Pharmacology (Eds.) Biggio, G. e
Costa, A. Raven Press New York, 105.
Bindra, D. e Spinner, N. (1958). Response to different degrees of novelty. Journal of Experimental Analysis
of Behavior, 1: 241-350.
Birk, J. e Noble, R. G. (1981). Naloxone antagonism of diazepam-induced feeding in the Syrian-hamster. Life
Sciences, 29 (11): 1125-1131.
91

Birk, J. e Noble, R. G. (1982). Bicuculine blocks diazepam-induced feeding in Syrian hamsters. Life
Sciences, 30 (4): 321-325.
Blass, E., Fitzgerald, E. e Kehoe, P. (1987). Interaction between sucrose, pain and isolation distress.
Pharmacology, Biochemistry and Behavior, 26: 483-489.
Blizard, D. A. (1971). Situational determinants of open field behavior in mus musculus. British Journal of
Psychology, 62: 245-252.
Bodnoff, S. R., SuranyVadotte, B., Quirion, R. e Meaney, M. J. (1987). Postnatal handling reduces novelty-
induced fear and increase (3H) flunitrazepam binding in rat brain. European Journal of Pharmacology,
144: 105.
Botelho, S., Estanislau, C. e Morato, S. (2007). Effects of under- and overcrowding on exploratory behavior
in the elevated plus-maze. Behavioural Processes, 74: 357–362.
Brennan, K., Roberts, D. C. S. e Anisman, H. (2001). Individual differences in sucrose consumption in the rat:
motivational and neurochemical correlates of hedonia. Psychopharmacology, 157: 269-276.
Britton, D. R. e Britton K. (1981). A sensitive open field measure of anxiolytic drug activity. Pharmacology,
Biochemistry and Behavior, 15: 577-582.
Brett, R. R. e Pratt, J. A. (1990). Chronic handling modifies the anxiolytic effect of diazepam in the elevated
plus-maze. European Journal of Pharmacology, 178: 135–138.
Broadhurst, P. L. (1957). Determinants of emotionality in the rat: situational factors. British Journal of
Psychology, 48: 1-12.
Brown, D. F., Houpt, K. A. e Schryver H. F. (1976). Stimulation of food intake in horses by diazepam and
promazine. Pharmacology, Biochemistry and Behavior, 5: 495-497.
Cappell, H. e Rodin, J. (1969). Fear increasing effects of experience with shock. Psychonomic Science, 17:
267-269.
Champagne, F. A., Francis, D. D., Mar, A. e Meaney, M. J. (2003). Variations in maternal care in the rat as a
mediating influence for the effects of environment on development. Physiology and Behavior, 79: 359-
371.
Clark, C., Gorman, D. e Vernadakis, A. (1970). Effects of prenatal administration of psychotropic drugs on
behavior of developing rats. Developmental Psychology, 3: 225-235.
Clausing, P., Mothes, H. H., Opitz, B. e Kormann, S. (1997). Differential effects of communal rearing and
preweaning handling on open-field behavior and hot plate latencies in mice. Behavioural Brain
Research, 82: 179-184.
Cole, S. O. (1983). Combined effects of chlordiazepoxide treatment and food deprivation on concurrent
measure of feeding and activity. Pharmacology, Biochemistry and Behavior, 18: 369-372.
Cole, J. C. e Rodgers, R. (1993). An ethological analysis of the effects chlordiazepoxide and bretazenil (Ro
16-6028) in the murine elevated plus-maze test. Behavioral Pharmacology, 4: 573-580.
Colle, L. e Wise, R. A. (1988). Concurrent facilitatory and inhibitory effects of amphetamine on stimulation-
induced eating. Brain Research, 459: 356-360.
Collier, G. e Johnson, D. F. (2000). Sucrose intake as a function of its cost and the cost of chow. Physiology
and Behavior, 70: 477-487.
Cooper, S. J. (1980). Benzodiazepines as appetite-enhancing compounds. Appetite, 1: 7-19.
92

Cooper, S. J. (1987). Chlordiazepoxide-induced selection of saccharin-flavoured food in the foodeprivet rat.
Physiology and Behavior, 41: 539-542.
Cooper, S. J. e Francis, R. L. (1979a). Effects of acute or chronic administration of chlordiazepoxide on
feeding parameters using two food textures in the rat. Journal of Pharmacy and Pharmacology, 31 (11):
743-746.
Cooper, S. J. e Francis, R. L. (1979b). Water intake and time course of drinking after single or repeated
chlordiazepoxide injections. Psychopharmacology (Berlim), 65 (2): 191-195.
Cooper, S. J. e McClelland, A. (1980). Effects of chlordiazepoxide, food familiarizarion, and prior shock
experience on food choice in rats. Pharmacology, Biochemistry and Behavior, 12: 23-28.
Cooper, S. J. e Yerbury, R. E. (1988). Clonazepam selectively increases saccharin ingestion in a two-choice
test. Brain Research, 456: 173-176.
Cox, T. e Lee, J. (1976). Noise stress and the effects of viloxazine (vivalan), a new antidepressant, on open
field activity in rats. Pharmacology, Biochemistry and Behavior, 4: 729-730.
Critchley, M. A. E. e Handley, S. L. (1987). 5-HT1A ligands effects in the X-maze anxiety test. British
Journal of Pharmacology, 92: 660.
Critchley, M. A. E., Njung’e, K. e Handley, S. L. (1992). Action and some interactions of 5-HT1A ligands in
the elevated X-maze and effects of dorsal raphe lesions. Psychopharmacology, 106: 484-490.
Cruz, A. P. M., Frei, F. e Graeff, F. G. (1994). Ethopharmacological analysis of rat behavior on the elevated
plus-maze. Pharmacology, Biochemistry and Behavior, 49: 171-179.
De Cabo, C., De Souza, N. J., Traubici, M. E. e Vaccarino, F. J. (1995). Anxiety-like response in the plus-
maze is predicted by combined individual differences in sugar feeding and novel exploration. Society
for Neuroscience Abstracts, 21: 448.
Delbarre, B., Dumas, G. e Guionniere, M. (1970). An automated open field method. Psychopharmacology,
18: 227-230.
Della-Fera, M. A., Baile, C. A. e McLaughlin, C. L. (1980). Feeding elicited by benzodiazepine-like
chemicals in puppies and cats: Structure-activity relationships. Pharmacology, Biochemistry and
Behavior, 12 (2): 195-200.
Dellu, P. F., Piazza, P. V., Mayo, W., Le Moal, M. e Simon, H. (1996). Novelty-seeking in rats —
biobehavioral characteristics and possible relationship with the sensation-seeking trait in man.
Neuropsychobiology, 34 (3): 136-145.
Denenberg, V. H. e Karas, G. G. (1961). Interactive effects of age and duration of infantile experiences upon
weight gain and mortality in the rat. Journal of Comparative and Physiological Psychology, 54: 685-
689.
Denenberg, V. H., Garbanati, J., Sherman, G., Yutzey, D. A. e Kaplan, R. (1978). Infantile stimulation
induces brain lateralization in rats. Science, 201 (4361): 1150-1152.
DeSouza, N. J., Bush, D. E. e Vaccarino, F. J. (2000). Self-administration of intravenous amphetamine is
predicted by individual differences in sucrose feeding in rats. Psychopharmacology, 148: 52-48.
DeSouza, N. J., Wunderlich, G. R., De Cabo, C. e Vaccarino, F. J. (1998). Individual differences in sucrose
intake predict behavioral reactivity in rodent models of anxiety. Pharmacology, Biochemistry and
Behavior, 60 (4): 841-846.
93

Dess, N. K. e Chapman, C. D. (1990). Individual differences in taste, body weight, and depression in the
helplessness rat model and in humans. Brain Research Bulletin, 24: 669-676.
Dess, N. K. e Choe, S. (1994). Stress selectively reduces sugar + saccharin mixture intake but increases
proportion of calories consumed as sugar by rats. Psychobiology, 22: 77-84.
Dess, N. K. e Minor, T. R. (1996). Taste and emotionality in rats selectively bred for high versus low
saccharin intake. Animal Learning and Behavior, 24 (1): 105-115.
Dess, N. K. e VanderWeele, D. A. (1994). Lithium chloride and inescapable, unsignaled tail shock
differentially affect meal patterns of rats. Physiology and Behavior, 56: 203-207.
Dixon, L. K. e DeFries, J. C. (1968). Effects of illumination on open field behavior in mice. Journal of
Physiological and Comparative Psychology, 66: 803-805.
Fernández-Teruel, A., Escorihueta, R. M., Driscoll, P., Tobeña, A. e Bãttig, K. (1992a). Differential effects
of early simulation and/or perinatal flumazenil treatment in young low-and high-avoidance rats.
Psychopharmacology, 108 (1-2): 170-176.
Fernández-Teruel, A., Escorihueta, R. M., Jiménez, P. e Tobeña, A. (1990). Infantile stimulation and
perinatal administration of Ro 15-1788: additive anxiety-reducing effects in rats. European Journal of
Pharmacology, 191: 111-114.
Fernández-Teruel, A., Escorihueta, R. M., Núñez, J. F., Gomà, M., Driscoll, P. e Tobeña, A. (1992b). Early
stimulation effects on novelty-induced behavior in two psychogenetically-selected rat lines with
divergent emotionality profiles. Neuroscience Letters, 137 (2):185-188.
Fernández-Teruel, A., Escorihueta, R. M., Núñez, J. F., Zapata, A., Boix, F., Salazar, W. e Tobeña, A. (1991).
The early acquisition of two-way (shuttle-box) avoidance as an anxiety-mediated behavior:
psychopharmacological validation. Brain Research Bulletin, 26: 173-176.
Fernández-Teruel, A., Gimenez-Lirt, L., Escoriheuta, R. M., Gil, L., Aquilar, R., Steimer, T. e Tobeña, A.
(2002). Early life handling stimulation and environmental enrichment: are some of their effects
mediated by similar neuron mechanisms? Pharmacology, Biochemistry and Behavior, 73 (1): 233-245.
Ferré, P., Nüñez, J. F., Garcia, E., Tobeña, A., Escorihueta, R. M. e Fernández-Teruel, A. (1995). Postnatal
handling reduces anxiety as measured by emotionality rating and hyponeophagia tests in female rats.
Pharmacology, Biochemistry and Behavior, 51: 199-203.
File, S. (1978). The ontogeny of exploration in the rat: habituation and effects of handling. Developmental
Psychobiology, 11: 321-8, 1978.
File, S. E. (1992). Behavioral detection of anxiolytic action. In. J. M. Elliot, D. Heal e C. A. Marsden (Orgs.),
Experimental Approaches to Anxiety and Depression. John Wiley & Sons Ltd, Chichester.
File, S. E. e Johnston, A. L. (1989). Lack of effects of 5-HT3 receptor antagonists in the social interaction and
elevated plus-maze tests of anxiety in the rat. Psychopharmacology, 99: 248-251.
File, S. E., Andrews, N., Wu: Y., Zharkovsky, A. e Zangrossi Jr., H. (1992). Modification of
chlordiazepoxide’s behavioural and neurochemical effects by handling and plus-maze experience.
European Journal of Pharmacology, 218: 9-14.
File, S., Andrews, N. e Zharkovsky, A. (1990). Handling habituation and chlordiazepoxide have different
effects on GABA and 5-HT function in the frontal cortex and hippocampus. European Journal of
Pharmacology, 190: 229.
94

Foltin, R. W. e Schuster, C. R. (1983). Interaction between the effects of intragastric meals and drugs on
feeding in rhesus monkeys. Journal of Pharmacology and Experimental Therapeutics, 226: 405-411.
Francis, D., Caldji, C., Champagne, F. A., Plotsky, P. M. e Meaney, M. J. (1999). The role of corticotrophin-
releasing factor-norepinephrine systems in mediating the effects of early experience on the
development of behavioral and endocrine response to stress. Biological Psychiatry, 46: 1153-1166.
Frussa-Filho, R., Otoboni, J.R.., Uema, F.T. e Sá-Rocha, L.C. (1991). Evalution of memory and anxiety in
rats observed in the elevated plus-maze: effects of age and isolation. Brazilian Journal of Medical and
Biological Research, 24: 725-728.
Galef, B. G. (1996). Food selection: Problems in understanding how we choose foods to eat. Neuroscience
and Biobehavioral Reviews, 20 (1): 67-73.
Gingras, M. A. e Cools, A. R. (1995). Differential ethanol intake in high and low responder to novelty.
Behavioural Pharmacology, 6 (7): 718-723.
Gingras, M. A. e Cools, A. R. (1995). Differential ethanol intake in high and low responders to novelty.
Behavioural Pharmacology, 6: 718-723.
Gingras, M. A. e Cools, A. R. (1996). Analysis of the biphasic locomotor response to ethanol in high and low
responders to novelty: a study in Nijmegen Wistar rats. Psychopharmacology, 125: 258-264.
Glendining, J. I. (1994). Is the bitter rejection response always adaptive? Physiology and Behavior, 56: 1217-
1227. (Apud Bassi, 1996)
Goldsmith, J. F., Brain, P. F. e Bento, D. (1978). Effects of the duration of individual or group housing on
behavioral and adrenocortical reactivity in male mice. Physiology and Behavior, 21: 757-760.
Gosnell, B. A. e Krahn, D. D. (1992). The relationship between saccharin and alcohol intake in rats. Alcohol,
9: 203-206.
Gosnell, B. A., Lane, K. E., Bell, S. M. e Krahn, D. D. (1995). Intravenous morphine self-administration by
rats with low versus high saccharin preferences. Psychopharmacology, 117: 248-252.
Gray, J. (1987). The Psychology of Fear and Stress. Cambridge University Press, Cambridge: 92-114. (Apud
Dess e Minor, 1996)
Guimarães, F. S., Chiaretti, T. M., Graeff, F. G. e Zuardi, A. W. (1990). Antianxiety effect of cannabidol in
the elevated plus-maze. Psychopharmacology, 100: 558-559.
Halliday, M. S. (1966). Exploration and fear in the rat. Symposium of the Zoological Society of London, 18:
45-59.
Haltmeyer, G. C., Denenberg, V. H. e Zarrow, M. X. (1967). Modification of the plasma corticosterone
response as a function of infantile stimulation and electric shock parameters. Physiology and Behavior,
2: 61-63.
Hancock, S. D., Menard, J. L. e Olmstead, M. C. (2005). Variations in maternal care influence vulnerability
to stress-induced binge eating in female rats. Physiology and Behavior, 85 (4): 430-439.
Handley, S L. e Mithani, S. (1984). Effects of alpha-adrenoceptor agonists in a maze-exploration model of
“fear”-motivated behavior. Naunyn-Schmiedeberg’s Archives of Pharmacology, 327: 1-5.
Helmstetter, F. J. e Fanselow, M. S. (1993). Aversively motivated changes in meal patterns of rats in a closed
economy: the effects of shock density. Animal Learning and Behavior, 21: 168-175.
95

Hogg, S. (1996). A review of the validity and variability of the elevated plus-maze as an animal model of
anxiety. Pharmacology, Biochemistry and Behavior, 54: 21-30.
Hooks, M. S., Jones, G. H., Liem, B. J. e Justice, J. B. (1992). Sensitization and individual differences to IP
amphetamine, cocaine, or caffeine following repeated intracranial amphetamine infusion.
Pharmacology, Biochemistry, and Behavior, 43: 815-823.
Hooks, M. S., Jones, G. H., Smith, A. D., Neill, D. B. e Justice, J. B. (1991). Individual differences in
locomotor activity and sensitization. Pharmacology, Biochemistry, and Behavior, 38: 467-470.
Imperato, A., PuglisAllegra, S., Casolinin, P., Zocchi, A. e Angelucci, L. (1989). Stress-induced
enhancement of dopamine and acetylcholine release in limbic structures: role of corticosterone.
European Journal of Pharmacology, 165 (2-3): 337-338.
Ivinskis, A. (1970). A study of validity of open field measures. Australian Journal of Psychology, 22: 175-
183.
Jodogne, C., Marinelli, M., Le Moal M. e Piazza, P. V. (1994). Animals predisposed to develop amphetamine
self-administration show higher susceptibility to develop contextual conditioning of both amphetamine-
induced hyperlocomotion and sensitization. Brain Research. 657(1-2): 236-244.
Jones, G. H. e Robbins, T. W. (1992). Differential effects of mesocortical, mesolimbic and mesostriatal
dopamine depletion on spontaneous, conditioned and drug-induced locomotor activity. Pharmacology,
Biochemistry, and Behavior, 43: 887-895.
Kabbaj, M., Devine, D. P., Savage, V. R. e Akil, H. (2000). Neurobiological correlates of individual
differences in novelty-seeking behavior in the rat: differential expression of stress-related molecules.
Journal Neuroscience, 20: 6983-6988.
Kampov-Polevoy, A. B, Kasheffskaya, R. P. e Sinclair, J. D. (1990). Initial acceptance of ethanol: gustatory
factors and patterns of alcohol drinking. Alcohol, 7: 115-120.
Katz, R. J. (1982). Animal model of depression: Pharmacological sensitivity of a hedonic deficit.
Pharmacology, Biochemistry, and Behavior, 16: 965-968.
Kelly, P. H., Seviour, P. W. e Iversen, S. D. (1975). Amphetamine and apomorphine responses in the rat
following 6-OHDA lesions of the nucleus accumbens, septi and corpus striatum. Brain Research, 94:
507-522.
King, D. L. (1970). Comparison between two methods of demonstrating relatedness of emotionality variables
in rats. Journal of Comparative and Physiological Psychology, 72: 238-243.
Levine, S. (1956). A further study of infantile handling and adult avoidance learning. Journal of Personality,
25 (1): 70-80.
Levine, S. (1957). Infantile experience and consummatory behavior in adulthood. Journal of Comparative
and Physiological Psychology, 50 (6): 609-612.
Levine, S. (1960). Stimulation in infancy. Scientific American, 202 (5): 80-86.
Levine, S. (1962). Plasma-free corticosteroid response to electric shock in rats stimulated in infancy. Science,
135 (3506): 795-796.
Levine, S., Chevalier, J. A. e Korchin, S. L. (1955). The effects of early shock and handling on later
avoidance learning. Journal of Personality, 24: 475-493.
Levine, S., Haltmeyer, G. C., KAras, G. G. e Denenberg, V. H. (1967). Physiological and Behavioral effects
of Infantile Stimulation. Physiology and Behavior, 2: 55-59.
96

Levitsky, D. A. e Barnes, R. H. (1972). Nutritional and environmental interactions in the behavioral
development of the rat: Long-term effects. Science, 176: 68-71.
Lister, R. G. (1987). The use of a plus-maze to measure anxiety in the mouse. Psychopharmacology, 92: 180-
185.
Liu, D., Diorio, J., Tannenbaum, B., Caldji, C., Francis, D. e Freedman, A. (1997). Maternal care,
hippocampal glucocorticoid receptors, and hypothalamic-pituitary-adrenal response to stress. Science,
277: 1659-1662.
Livesey, P. J. e Egger, G. J. (1970). Age as a factor in open field responsiveness in the white rat. Journal of
Comparative and Physiological Psychology , 73: 93-99.
Lorenz, K. Z. (1981). The Foundations of Ethology. Simon and Schuster, Nova York.
McCall, R. (1969). Caretaker effect in rats. Developmental Psychology, 1: 771.
Maisonnette, S., Morato, S. e Brandão, M. L. (1993). Modulation by 5-HT1A receptors of the anxiogenic
effects induced by isolation in the elevated plus maze test. Physiology and Behavior, 54: 753-758.
Margules, D. L. e Stein, L. (1967). Neuroleptics vs tranquilizers: evidence from animals of mode and site of
action. In. H. Brill, Neuropsychopharmacology. Exerpta Medica Foundation, Amsterdã.
Mclntosh, J., Anisman, H. e Merali, Z. (1999). Short-and long-periods of neonatal maternal separation
differentially affect anxiety and feeding in adults rats: gender-dependent effects. Developmental Brain
Research, 113: 97-106.
Meaney, M. J., Aitken, D. H., Bhatnagar, M., Bodnoff, S.R., Mitchell, J. B. e Sarrieau, A. (1990). Neonatal
handling and the development of the adrenocortical response to stress. Em N. Gunzenhauser, T. B.
Brazelton e T. Field (Org.), Advances in Touch: New Implications in Human Development. Johnson &
Johnson Consumer Products, Inc., Skill-man, NJ: 11-22.
Meinrath, A. B. e Flaherty, C. F. (1987). Preweanling handling influences open-field behavior, but not
negative contrast or sucrose neophobia. Animal Learning and Behavior, 15 (1): 83-92.
Montgomery, K. C. (1951). The relation between exploratory behavior and spontaneous alternation in the
white rat. Journal of Comparative and Physiological Psychology, 44: 582-589.
Montgomery, K. C. (1955). The relationship between fear induced by novel stimulation and exploratory
behavior. Journal of Comparative and Physiological Psychology, 48: 254-260.
Mook, D. G., Oberlin, A., Tyndall, E. e Wagner, S. (1993). Persistence of sham feeding after intragastric
meals in the rat: concentration-intake relation. Psychobiology, 21: 55-59.
Morato, S. e Castrechini: (1989). Effects of floor surface and environmental illumination on exploratory
activity in the elevated plus maze. Brazilian Journal of Medical and Biological Research, 22: 707-710.
Morato, S., Brandão, M.L. (1996). Transporting rats to the test situation and exploratory behavior in the
elevated plus-maze. Psychobiology, 24, (3): 247-252, 1996.
Morato, S., Brandão, M.L. (1997). Paradoxical increase of exploratory behavior in the elevated plus maze by
rats exposed to two kinds of aversive stimuli. Brazilian Journal of Medical And Biological Research,
30: 1113-1120.
Morrison, C. F. e Stephenson, J. A. (1970). Drugs effects on a measure of unconditioned avoidance in the rat.
Psychopharmacologia, 18 (2): 133-143.
Morrison, B. J. e Thatcher, K. (1969). Overpopulation effects on social reduction of emotionality in the
albino rat. Journal of Comparative and Physiological Psychology, 69: 658-662.
97

Núñez, J. F., Ferré, P., Garcia, E., Escorihuela, R. M., Fernández-Teruel, A. e Tobeña, A. (1995). Postnatal
handling reduces emotionality ratings and accelerates two-way active avoidance in female rats.
Physiology and Behavior, 57: 831-835.
Ogawa, T., MIkume, M., Kuroda, , Y., Muncoka, K., Mori, K. J. e Takahashi, K. (1994). Periodic maternal
deprivation alters stress response in adult offspring: potentiates the negative feedback regulation of
restraint stress-induced adrenocortical response and reduces the frequencies of open field induced
behaviors. Pharmacology, Biochemistry and Behavior, 49: 961-967.
Oitzl, M. S., vanHaarst, A. D., e deKloet, E. R. (1997). Behavioral and neuroendocrine responses controlled
by the concerted action of central mineralocorticoid (MRS) and glucocorticoid receptors (GRS).
Psychoneuroendocrinology, 22: 87-93.
Oldham, J. e Morlock, H. (1970). The effects of open field size on activity in the Mongolian gerbil.
Psychonomic Science, 20: 290.
Overstreet, D. H., Kampov-Polevoy, A. B., Rezvani, A. H., Murrelle, L., Halikas, J. A. e Janowsky, D. S.
(1993). Saccharin intake predicts ethanol intake in genetically heterogeneous rats as well as different rat
strains. Alcoholism: Clinical and Experimental Research, 17: 366-369.
Parker, L A. (1991). Chlordiazepoxide nonspecifically enhance consumption of saccharin solution.
Pharmacology, Biochemistry and Behavior, 38: 375-377.
Pellow, S. e File, S. E. (1986). Anxiolytic and anxiogenic drug effects on exploratory activity in elevated
plus-maze: a novel test of anxiety in the rat. Pharmacology, Biochemistry and Behavior, 24: 525-529.
Pellow, S., Chopin:, File, S. e riley, M. (1985). Validation of open: closed arm entries in an elevated plus-
maze as a measure of anxiety in the rat. Journal of Neuroscience Methods, 14: 149-167.
Pellow, S., Johnston, A. L. e File, S. E. (1987). Selective agonists and antagonists for 5-hydroxytriptamine
receptor subtypes, and interactions with yohimbine and FG 7142 using the elevated plus-maze test in
rats. Journal of Pharmacy and Pharmacology, 39: 917-928.
Penke, Z., Felszeghy, K., Fernette, B., Sage, D., Nyakas, C. e Burlet, A. (2001). Postnatal maternal
deprivation produces long-lasting modifications of the stress response, feeding and stress-related
behavior in the rat. European Journal of Neuroscience, 14: 747-755.
Pfaffman, C. (1975). Phylogenetic origins of sweet sensitivity. In: Denton, D. A. e Coghlan, J. P. (Orgs.),
Olfaction and Taste. Academic Press, Nova York. (Apud Galef, 1996)
Piazza, P. V., Deminiere, J. M., Le Moal, M. e Simon, H. (1989). Factors that predict individual vulnerability
to amphetamine self-administration. Science, 245: 1511-1513.
Piazza, P. V., Deminiere, J. M., LeMoal, M e Simon, H. (1990). Stress-and pharmacologically-induced
behavioral sensitization increases vulnerability to acquisition of amphetamine self-administration.
Brain Research, 514: 22-26.
Piazza, P. V., Maccari, S., Deminiere, J. M., Le Moal, M., Mormede, P. e Simon, H. (1991). Corticosterone
levels determine individual vulnerability to amphetamine self-administration. Proceedings of the
National Academy of Sciences of the United States of America, 88: 2088-2092.
Poschel, B. P. H. (1971). A simple and specific screen for benzodiazepine-like drugs. Psychopharmacologia,
19: 193-198.
Prut, L. e Belzung, C. (2003). The open field as a paradigm to measure the effects of drugs on anxiety-like
behaviors: a review. European Journal of Pharmacology, 463: 3-33.
98

Randall, L. O., Schallek, W., Heise, G. a., Keith, E. F. e Bagdon, R. E. (1960). The psychosedative properties
of methaminodiazepoxide. Journal of Pharmacology and Experimental Therapeutics, 129: 163-197.
Rex, A., Sondern, U., Voigt, J. P., Franck, S. e Fink, H. (1996). Strain differences in fear-motivated behavior
of rats. Pharmacology, Biochemistry and Behavior, 54 (1): 107-111.
Rodgers, R. J. e Cole, J. C. (1994). The elevated plus-maze: pharmacology, methodology and ethology. In. S.
J. Cooper e C. A. Hendrie (Orgs.), Ethology and Psychopharmacology. John Wiley and Sons,
Chichester.
Roth, K. A. e Katz, R. J. (1979). Stress, behavioral arousal, and open field activity – A reexamination of
emotionality in the rat. Neuroscience and Biobehavioral Reviews, 3: 247-263.
Roth, K. A. e Richard, J. K. (1979). Stress, behavioral arousal, and open field activity - A reexamination of
emotionality in the rat. Neuroscience and Biobehavioral Reviews, 3: 247-263.
Satinder, K. P. (1968). A note on the correlation between open field and escape avoidance behavior in the rat.
Journal of Psychology, 69: 3-6.
Scott, T.R. e Giza, B. K. (1987). Neurophysiological aspects of sweetness. In: Dobbing, J. (Org.). Sweetness.
Spring-Verlag, Londres: 15-31.
Segato, F. N., Castro-Souza, C., Segato, E. N., Morato, S. e Coimbra, N. C. (1997). Sucrose ingestion causes
opioid analgesia. Brazilian Journal of Medical and Biological Research, 30: 981-984.
Shepard, R. A. e Estall, L. B. (1984). Anxiolytic actions of chlordiazepoxide determine its effects on
hyponeophagia in rats. Psychopharmacology ( Berlim) 82: 343-347.
Sills, T. L. e Crawley, J. N. (1996). Individual differences in sugar consumption predict amphetamine-
induced dopamine overflow in nucleus accumbens. European Journal Pharmacology, 303: 177-181.
Sills, T. L. e Vaccarino, F. J. (1991). Facilitation and inhibition of feeding by a single dose of amphetamine:
relationship to baseline intake and accumbens cholecystokinin. Psychopharmacology, 105: 329-334.
Sills, T. e Vaccarino, F. J. (1994). Individual differences in sugar intake predict the locomotor response to
acute and repeated amphetamine administration. Psychopharmacology, 116: 1-8.
Sills, T. e Vaccarino, F. J. (1996). Individual differences in sugar consumption following systemic or
intraaccumbens administration of low doses of amphetamine in nondeprived rats. Pharmacology
Biochemistry and Behavior, 54: 665-670.
Sills, T., Baird, J. P. e Vaccarino, F. J. (1993). Individual differences in the feeding effects of amphetamine:
role of nucleus accumbens dopamine and circadian factors. Psychopharmacology, 112: 211-218.
Sinclair, J. D., Kampov-Polevoy, A., Stewart, R. e Li, T. K. (1992). Taste preference in rat lines selected for
low and high alcohol consumption. Alcohol, 9: 155-160.
Smith, J. C. e Forster, D. F. (1980). Some determinants of intake of glucose+saccharin solutions. Physiology
and Behavior, 25: 127-133. (Apud Bassi, 1996)
Soubrié, P., Kulkarni, S., Simon, P. e Bossier, J. R. (1975). Effets des anxiolytiques sur la prise de nourriture
de rats et de souris placés en situation nouvelle ou familière. Psychopharmacology, 45 (2): 203-210.
Steenbergen, H. L., Heinsbroek, R. P. W., Van Hest, A. e Van de Pol, N. E. (1990). Sex-dependent effects of
inescapable sock administration on shuttlebox-escape performance and elevated plus-maze behavior.
Physiology and Behavior, 48: 571-576.
Tamm, S. (1989). Importance of energy costs in central place foraging by hummingbirds. Ecology, 70: 195-
205. (Apud Bassi, 1996)
99

Treit, D., Menard, J. e Royan, C. (1993). Anxiogenic stimuli in the elevated plus-maze. Pharmacology,
Biochemistry and Behavior, 44: 463-469Vaccarino, F. J., Amalric, M. Swerdlow, N. R. e Koob, G. F.
(1986). Blockade of amphetamine but not opiate-induced locomotion following antagonism of
dopamine function in the rat. Pharmacology Biochemistry Behavior, 24, 61-65.
Valle, F. P. (1970). Effects of strain, sex, and illumination on open field behavior of rats. American Journal
of Psychology, 83: 103-111.
Walsh, R. e Cummins, R. A. (1976). The open field test: A critical review. Psychological Bulletin, 83 (3):
482-504.
Weissenborn, R. e Winn, P. (1992). Regulatory behaviour, exploration and locomotion following NMDA or
6-OHDA lesions in the rat nucleus accumbens. Behavioural Brain Research, 51, 127-137.
Wright, I. K., Upton, N. e Marsden, A. (1991). Resocialization of isolation-rearing rats does not alter their
anxiogenic profile on the elevated x-maze model of anxiety. Physiology and Behavior, 50: 1129-1132.
Xenakis, S. and A. Sclafani. (1981). The effects of pimozide on the consumption of a palatable saccharin-
glucose solution in the rat. Pharmacology, Biochemistry and Behavior, 15: 435-442.
100

Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas

Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo