
COMPOSIÇÃO QUÍMICA E ATIVIDADE ALELOPÁTICA DE … · mesmo tempo, os avanços da Fisiologia...
105
UNESP - UNIVERSIDADE ESTADUAL PAULISTA CÂMPUS DE BOTUCATU INSTITUTO DE BIOCIÊNCIAS COMPOSIÇÃO QUÍMICA E ATIVIDADE ALELOPÁTICA DE EXTRATOS FOLIARES DE Leonurus sibiricus L. (Lamiaceae) LUIZ FERNANDO ROLIM DE ALMEIDA Tese apresentada ao Instituto de Biociências, Câmpus de Botucatu, UNESP, para obtenção do título de Doutor em Ciências Biológicas (Botânica), AC: Fisiologia Vegetal BOTUCATU - SP - 2006 -
Transcript of COMPOSIÇÃO QUÍMICA E ATIVIDADE ALELOPÁTICA DE … · mesmo tempo, os avanços da Fisiologia...

UNESP - UNIVERSIDADE ESTADUAL PAULISTA
CÂMPUS DE BOTUCATU
INSTITUTO DE BIOCIÊNCIAS
COMPOSIÇÃO QUÍMICA E ATIVIDADE ALELOPÁTICA DE
EXTRATOS FOLIARES DE Leonurus sibiricus L. (Lamiaceae)
LUIZ FERNANDO ROLIM DE ALMEIDA
Tese apresentada ao Instituto de
Biociências, Câmpus de Botucatu,
UNESP, para obtenção do título de
Doutor em Ciências Biológicas
(Botânica), AC: Fisiologia Vegetal
BOTUCATU - SP - 2006 -

UNESP - UNIVERSIDADE ESTADUAL PAULISTA
CÂMPUS DE BOTUCATU
INSTITUTO DE BIOCIÊNCIAS
COMPOSIÇÃO QUÍMICA E ATIVIDADE ALELOPÁTICA DE
EXTRATOS FOLIARES DE Leonurus sibiricus L. (Lamiaceae)
LUIZ FERNANDO ROLIM DE ALMEIDA
Profª Drª Maria Elena Aparecida Delachiave
Orientadora
Prof Dr Wagner Vilegas
Co-Orientador
Tese apresentada ao Instituto de
Biociências, Câmpus de Botucatu,
UNESP, para obtenção do título de
Doutor em Ciências Biológicas
(Botânica), AC: Fisiologia Vegetal
BOTUCATU - SP - 2006 -

FICHA CATALOGRÁFICA ELABORADA PELA SEÇÃO TÉCNICA DE AQUISIÇÃO E TRATAMENTO
DA INFORMAÇÃO DIVISÃO TÉCNICA DE BIBLIOTECA E DOCUMENTAÇÃO - CAMPUS DE BOTUCATU - UNESP
BIBLIOTECÁRIA RESPONSÁVEL: SELMA MARIA DE JESUS Almeida, Luiz Fernando Rolim. Composição química e atividade alelopática de extratos foliares de Leonurus sibiricus L. (Lamiaceae) – Luiz Fernando Rolim de Almeida, 2006. 107p. Tese (doutorado) – Universidade Estadual Paulista, Instituto de Biociências de Botucatu, 2004. Orientadora: Maria Elena Aparecida Delachiave Co-orientador: Wagner Vilegas Assunto CAPES: 20303009 1. Fisiologia vegetal CDD 581.1 Palavras-chave: alelopatia, Leonurus; plantas medicinais; óleo essencial; flavonóides, fitoquímica

Dedico
A DEUS.

EFÍGRAFE
“ (....) Essa é a oportunidade histórica da docência, oportunidade que
não existe fora de nós próprios, num certo compartimento do tempo, à espera
que vamos a seu encalço, mas nas relações entre nós e o tempo, na intimidade
dos acontecimentos, no jogo das contradições. Oportunidade que vamos
criando, fazendo na própria história nossa biografia. A história nos castiga
quando não aproveitamos a oportunidade de termos algum compromisso com
as teias sociais que criam e recriam a ciência e a educação. (....)”
Paulo Freire
Pedagogia da Esperança, 1992.

Agradecimentos
Á Profa. Maria Elena Aparecida Delachiave pela confiança e paciência nesses
anos onde pude imaginar e depois vislumbrar os possíveis caminhos a seguir. Obrigado
minha amiga e mestre.
Ao Prof. Wagner Vilegas por ter dado a oportunidade de aprender a fitoquímica
e ter possibilitado a realização de um sonho...saber o que as plantas produzem e como
estudar tudo isso.
Ao Prof. Vincenzo De Feo, pelo apoio e paternalismo ao me receber num país
desconhecido até então, apaixonante por natureza e por conseqüência, obrigado por sua
paixão a botânica e por sua atenção ao brasiliano.
Á Profa. Lourdes Campaner dos Santos, á Dra. Miriam Sannomiya e á Dra.
Emilia Quaranta pela orientação de bancada e pela sinceridade espontânea
imprescindíveis para uma boa Ciência.
Ao Clenílson Rodrigues e Daniel Rinaldi, amigos novos e irmãos de ciência, por
terem me ensinado os passos de um bom fracionamento inerentes de qualquer
fitoquímico.
Aos meus pais, Mario e Mariângela, que ensinaram a olhar pra frente, a
agradecer a Deus e a confiar que meu caminho estava ali do meu lado. Obrigado por
todos esses anos de aprendizado. Amo Vocês...
Aos meus irmãos, Telma, Marinho e Ado, pela lição que deram ao irmão mais
velho de que a construção de uma família vale á pena. Deus abençoe vocês as suas
famílias recém nascidas.
A Bruna, minha noiva e mulher, agradeço pela compreensão, afeto e carinho
nesses anos.

Aos amigos de laboratório. Em Botucatu, João, Yara, Mônica, Camilla, Andréa,
Rodrigo...Em Araraquara, Roberto, Tamara, Juliana Severi, Marcelo, Marcio, Maria
Eloísa, Ana Elisa, Juliana Rodrigues,...Em Salerno, Francesco, Antonio, Angeline,
Carolina, Nillia, Emilia e Laura.
Aos amigos de Botucatu, das repúblicas e dos bares....seria irresponsabilidade
citar todos aqui....obrigado pelas conversas, dúvidas e desafios vividos.
E a todos que de certa maneira me ajudaram a ser mais lúcido, mais sincero,
mais amigo...

PREFÁCIO
Grande número de substâncias químicas oriundas de vegetais foi catalogado
e relacionado com atividades biológicas, em decorrência do aumento de estudos com
plantas medicinais. A Química de Produtos Naturais, como linha de pesquisa, colaborou
para o avanço do conhecimento da flora brasileira, desenvolvendo importantes
metodologias e tecnologias, principalmente nos estudos das plantas medicinais. Ao
mesmo tempo, os avanços da Fisiologia Vegetal, especialmente na ecofisiologia,
levantaram aspectos importantes sobre as relações entre as plantas e as implicações com
fatores bióticos e abióticos. Também, o desenvolvimento de abordagens
interdisciplinares na área de Botânica colaboraram para o reconhecido status científico
da área de ecofisiologia, fato esse que facilitou o acesso a recursos financeiros e
intercâmbio entre pesquisadores de outros países. A alelopatia, interação química entre
plantas e microorganismos, é uma área relativamente nova da ecofisiologia e requer
abordagem interdisciplinar, possibilitando que os trabalhos científicos sobre o assunto
estejam presentes nos mais diferentes periódicos, mostrando a importância e o
crescimento desta área de estudo.
Os estudos interdisciplinares sobre os potenciais alelopáticos vegetais, têm
possibilitado a inclusão de novas teorias químico-ecológicas nas linhas de pesquisa em
Fisiologia Vegetal. As interações entre as plantas e o solo, os mecanismos de defesa
contra patógenos e herbívoros e os fatores ambientais que modulam a produção das
substâncias do metabolismo secundário reforçam a necessidade da abordagem
interdisciplinar.
O presente trabalho procurou colocar em prática os ensinamentos postulados
na literatura pelos demais pesquisadores e buscou aprender a investigar as interações
químicas entre plantas. Ao tentar entender tais relações, a pesquisa nos levou aos

universos desconhecidos da ciência e nos trouxe velhas inquietações sobre novas
dúvidas inerentes do processo de investigação científica. É pela evolução da química de
produtos naturais, aumento de trabalhos publicados em alelopatia, interação entre as
áreas de conhecimentos, entre outros, que este autor agradece aos pesquisadores e
professores que auxiliaram, mesmo às vezes sem tomar conhecimento, na busca
inquieta pelas respostas. Ás tentativas e aos acertos dos que escreveram sobre alelopatia
ofereço esta investigação inicial em Fisiologia Vegetal. Que as velhas moléculas
oriundas de milhões de anos atrás nos permitam conhecer novos caminhos para algumas
áreas da ecofisiologia.
Luiz Fernando Rolim de Almeida

1
S U M Á R I O
Resumo ........................................................................................................... 2
Abstract .......................................................................................................... 4
Introdução ........................................................................................................ 6
Revisão Bibliográfica ..................................................................................... 8
Leonurus sibiricus ..................................................................................... 8
Alelopatia ................................................................................................. 9
Flavonóides ............................................................................................... 12
Capítulo 1- Composição do óleo essencial de Leonurus sibiricus L. ............. Resumo .......................................................................................................Abstract ...................................................................................................... Introdução ...................................................................................................Material e Método ..................................................................................... Resultado e Discussão ................................................................................ Referência Bibliográfica.............................................................................
16171718191922
Capítulo 2- Potencial alelopático de folhas de Leonurus sibiricus L. .............Resumo .......................................................................................................Abstract ...................................................................................................... Introdução ...................................................................................................Material e Método ..................................................................................... Resultado e Discussão ................................................................................ Referência Bibliográfica ............................................................................
24252729303450
Considerações Finais ...................................................................................... 54
Referências Bibliográficas ............................................................................. 56
Anexo .............................................................................................................. 63

2
ALMEIDA, LUIZ FERNANDO ROLIM. COMPOSIÇÃO QUÍMICA E
ATIVIDADE ALELOPÁTICA DE EXTRATOS FOLIARES DE Leonurus
sibiricus L. (LAMIACEAE) 2006. 96p. TESE (DOUTORADO) –
INSTITUTO DE BIOCIÊNCIAS, UNESP - UNIVERSIDADE ESTADUAL
PAULISTA, BOTUCATU.
Resumo: A erva-macaé ou rubim (Leonurus sibiricus L.) pertence à família Lamiaceae da ordem Lamiales. A planta é considerada invasora em lavouras agrícolas, presente em áreas abandonadas, com crescimento espontâneo em quase todo território brasileiro. Exudatos radiculares dessa espécie aumentam a germinação de arroz, trigo e mostarda, mas o extrato aquoso de folhas inibe a germinação de milho e o crescimento de plântulas de tomate. A utilização como erva medicinal é indicada, com base na tradição popular, para resfriado, bronquite e reumatismo; utilizada também na forma de chás, nos casos de sangramento pós-parto, menstruação excessiva, bem como, edema, abscessos e problemas renais. As folhas e flores, em infusão, são capazes de combater vômitos e diarréias. Os extratos metanólicos de folhas demonstram atividade antiinflamatória e combatem células neoplásicas de glândulas mamárias, o que justifica diversos estudos etnofarmacológicos realizados com esta espécie no Brasil. Em relação à composição química, a literatura relata que L. sibiricus apresenta alcalóides, mono, sesqui e diterpenos, e furano-lactonas. Os extratos polares apresentam majoritariamente rutina, derivados e flavonas metoxiladas, além disso, o gênero apresenta as flavonas como marcadores taxonômicos. Outras espécies como L. cardiaca, L. persicus e L. glaucescens também apresentam iridóides e flavonóides que demonstram expressivo potencial medicinal. Os objetivos deste trabalho foram determinar os compostos majoritários das folhas de Leonurus sibiricus; estudar o efeito alelopático dos extratos por infusão e metanólico, das frações flavonoídicas e substâncias isoladas presentes no extrato metanólico. Para determinação do perfil químico do óleo essencial foi utilizado Cromatógrafo Gasoso acoplado a Espectrômetro de Massas (CG-MS) e para a separação das substâncias presentes em extratos e frações foi utilizada Cromatografia Líquida de Alta Eficiência (CLAE). Os terpenos identificados por CG-MS foram sesquiterpenos trans-cariofileno (33,43%), alfa-humuleno (21,49%) e germacreno-D (24,95%) que são responsáveis por, aproximadamente, 70% da composição do óleo. O rendimento deste óleo foi baixo, fato que impossibilitou a avaliação alelopática. Os compostos majoritários encontrados nos extratos (infusão e metanólico) foram flavonóides biglicosilados e monoglicosilados derivados da quercetina. Nos ensaios biológicos foram realizadas avaliações alelopáticas dos extratos (por infusão e metanólico), frações (fracionadas em coluna Sephadex) e substâncias isoladas. Para a avaliação da germinação e crescimento inicial de raiz, os extratos por infusão e metanólico foram preparados nas concentrações de 50, 100, 200, 400 e 800mg L-1; as 12 frações obtidas por separação em Sephadex na concentração de 1g L-1 e os flavonóides isolados (7) nas concentrações de 10-4, 10-5, 10-6 e 10-7M. O controle do extrato por infusão foi água destilada e para o extrato metanólico água destilada com 0,01% de acetona. Para verificar o possível efeito alelopático dos extratos e das frações foram utilizadas sementes de Raphanus sativum, Lactuca sativa e Lepidium sativum e, para as substâncias puras isoladas, sementes de Lactuca sativa. As sementes (10 de cada espécie) foram colocadas para germinar em condição de luz contínua por 4 dias em placas de Petri, com 3 repetições e tendo como substrato papel de

3
filtro umedecido com 5mL de solução; sendo a mesma metodologia para extratos, frações e substâncias isoladas. Os resultados mostraram que os extratos por infusão e metanólico reduziram significativamente apenas a germinação das sementes de Lactuca sativa, enquanto o crescimento inicial das raízes de Raphanus sativum, Lactuca sativa e Lepidium sativum foi afetado pelos extratos, principalmente, nas concentrações mais altas, sendo que o extrato metanólico acarretou redução mais drástica. As frações de Sephadex consideradas flavonoídicas (5, 6, 7, 10, 11 e 12) mostraram atividade, principalmente, no crescimento inicial de raiz das espécies estudadas enquanto a germinação foi pouco afetada. Os compostos majoritários encontrados nos extratos e frações ativas foram os flavonóides quercetina-3-O-β-D-galactopiranosil (1-6)-O-α-L-rhamnopiranosideo; quercetina-3-O-β-D-glucopiranosil (1-6)-O-α-L-rhamnopiranosideo (Rutina); quercetina-3-O-β-D- galactopiranosideo (Hiperina) e quercetina-3-O-β-D-glucopiranosideo (Isoquercetrina), além das agliconas genkanina-3´-hidroxi, genkanina e quercetina que também estão presentes, porém em menor quantidade. Pelos resultados, pode-se concluir que o uso de extratos com perfil químico semelhante podem propiciar respostas alelopáticas diferentes, pois existe forte correlação com a concentração utilizada e estrutura das moléculas; as agliconas dos flavonóides demonstraram maior atividade alelopática (inibição ou promoção) em relação aos glicosídeos, talvez devido a relação com o número de moléculas de açúcares presentes nos esqueletos flavonoídicos.
Palavras-chave: alelopatia, Leonurus, plantas medicinais, fitoquímica, flavonóides, terpenos.

4
Abstract: Chemical compounds and allelopathic activities of Leonurus sibiricus leaves The medicinal plant motherwort or rubim (Leonurus sibiricus L.) belongs to the family Lamiaceae and the order Lamiales; it is considered invasive in crops and is present in abandoned areas, it presents spontaneous growth in great part of Brazil. This vegetal group produces terpenoids and phenolic substances with allelopathic effects, what explains its invasive behaviour in coffee plantations and orchards. Radicular exudates of this species increase the germination of rice, wheat, and mustard, but the aqueous extract inhibits the germination of corn and the growth of tomato plantules. Its use as medicinal herb (medicinal plant) is indicated, based on the folk tradition, for colds, bronchitis, and rheumatism. This plant is used also for teas in cases of after-birth bleeding, excessive menstruation as well as oedema, abscesses, and renal problems. The leaves and flowers are also indicated to combat vomits and diarrhoea when used through infusions. The leaf methanolic extract has also demonstrated anti-inflammatory activity and it combats neoplasic cells of mammary glands, what justifies several ethnopharmacological studies accomplished with this species in Brazil. With regard to the chemical composition, the literature indicates that L. sibiricus presents alkaloids, mono, sesquiterpenes, and diterpenes, and furano-lactones. The polar extract present mainly rutin and its derivates, besides the methoxylated flavones; furthermore, the genus is rich in flavones that are considered taxonomic markers. The phenylpropanoids are present in L. heterophyllus mainly as bi-glycosylated flavonoids. Other genera as L. cardíaca, L. persicus, and L. glaucescens also present iridoids and flavonoids that have demonstrated expressive medicinal potential of this genus. The work aimed at the investigation of the chemical composition of the majority compounds of the Leonurus sibiricus leaves using chromatographic techniques (CG-MS and HPLC) to identify and isolate such substances. Hence, there was performed the essential oil extraction through the method of Clevenger and used the gaseous chromatograph connected with mass spectrometer for the separation of substances. The identification was done through the calculus of indexes of eluid compound retention in the chromatographic system, caparison with the mass spectra. The identified terpenes were the trans-caryophyllene sesquiterpenes (33.43%), alpha-humulene (21.49%), and germacrene-D (24.95%), which are responsible for, approximately 70% of the oil composition. The performance of this oil was low, fact that has made impossible its allelopathic evaluation. On the other hand, the phenylpropiods were extracted through infusion and percolation (in this case, using methanol as solvent). Additionally, there was fractioned, through Sephadex LH-20, the metanolic extract obtaining, therefore, 12 fractions. The isolated flavonoids from the active fractions of Sephadex were mono and bi-glycosylated, besides the aglycone with methyl radicals. The flavonoids are the followings: quercetin-3-O-β-D-galactopyranosyl (1-6)-O-α-L-rhamnopyranoside; quercetin-3-O-β-D-glucopyranosyl (1-6)-O-α-L-rhamnopyranoside (Rutin); quercetin-3-O-β-D-galactopyranoside (Hyperine) and quercetin-3-O-β-D-glucopyranoside (Isoquercetrin), genkanin-3´-hydroxy, genkanin, and quercetin. There were performed biological assays for the allelopathic evaluation of extracts, fractions, and isolated substances. With regard to the germinantion and initial root growth evaluations, there were prepared the extracts through infusion and methanolic in the following concentrations: 50, 100, 200, 400, and 800mg L-1; for the fractions of Sephadex in the concentration of 1g L-1, and the isolated flavonoids in the concentration 10-4, 10-5, 10-6, and 10-7 M. Distilled water was used as check for the extract through infusion, and for the methanolic extract, there was used distilled water with 0.01% of acetone. Seeds of Raphanus sativum, Lactuca sativa, and Lepidium sativum were used for the verification of the allelopahic effect of the extract and flavonoid fractions; for the isolated, pure substances, there was used Lactuca sativa for being considered a sensitive species as well as a indicator of allelopathic activity. The seeds (10 of each species) were placed to germinate under continuous light for 4 days on Petri dishes having as substrate filter paper moisten with 5mL

5
of solution, the same methodology used for extracts and isolated substances.The results demonstrate that the germination was affected only in the first day by the aglycones. With regard to the root initial growth, the activity of tested substances has presented relation with the used concentration, where in the highest concentration (10-4M) the aglycones were responsible for the reduction in the growth and in the other tested concentrations (10-5M, 10-6 and 10-7M), the glycosylated flavonoids have demonstrated their stimulation on the root growth. Similarly, the substances of the secondary methabolism of the leaves of L. sibiricus have presented relation with the biological activity (inibition or promotion), where the release and concentration of theses compounds might be related with the allelopathic effect.
Key words: allelopathy, Leonurus, medicinal plants, phytochemistry, flavonoids, terpenes.

6
INTRODUÇÃO
As propriedades medicinais de L. sibiricus estão presentes em diferentes órgãos, tais
como folhas, que combatem reumatismo crônico e possuem atividade antibacteriana, evitando
dermatites e outros problemas dermatológicos. Na medicina chinesa, as sementes são
consideradas afrodisíacas e a planta seca é prescrita como tonificante e usada em disfunções
menstruais. As folhas e flores são capazes de combater vômitos e diarréias, além de serem
eficientes contra resfriado, bronquite e reumatismo.
A alelopatia é importante fenômeno presente nos agrossistemas, o qual implica em
ampla influência e interação das plantas nas comunidades bióticas. Os efeitos alelopáticos
podem ser diretos ou indiretos nos vegetais, incluindo microrganismos, através da liberação
de produtos naturais para o ambiente (Rice, 1984). Os aleloquímicos sintetizados pelas rotas
do metabolismo secundário de plantas superiores influenciam o estabelecimento e
desenvolvimento de culturas agrícolas e comunidades vegetais (Torres et al., 1996).
Pesquisadores e agricultores reconhecem os aleloquímicos como alternativa viável para
substituir os pesticidas sintéticos, podendo reduzir a poluição ambiental e aumentar a
produtividade agrícola (Qasem & Foy, 2001; Duke et al., 2002).
A riqueza química dos extratos de plantas aromáticas e medicinais faz com que sejam
reconhecidos como fontes de compostos biologicamente ativos, sendo que as substâncias
alelopáticas podem apresentar características químicas semelhantes às das substâncias com
efeitos terapêuticos (Mathela, 1991; Cutler & Cutler, 1999).
A germinação de sementes e o desenvolvimento inicial de plântulas são utilizados para
avaliar o efeito alelopático, sendo freqüentemente usados extratos vegetais ou aleloquímicos
isolados (Friedman, 1995). A sensibilidade a estes compostos varia de acordo com a
concentração aplicada e composição química do extrato (Perez, 1990; Rimando et al., 2001).
Essas substâncias com efeitos alelopáticos podem estar presentes nos diferentes órgãos
vegetais tais como folhas, flores, frutos e gemas (May & Ash, 1990; Larcher, 2000), podendo
ser flavonóides, fenóis, terpenóides, alcalóides, poliacetilenos, ácidos graxos, peptídeos e
outros (Rice, 1984).
Assim, o estudo das plantas medicinais, e respectivos aleloquímicos com a atividade
alelopática, podem representar significativo potencial para a busca de produtos naturais. O
conhecimento dessas substâncias do metabolismo secundário, portanto, é importante para
diferentes áreas do conhecimento científico, dentre elas destacam-se agronômica, taxonômica,
etnobotânica, farmacológica e fitoquímica.

7
Portanto, os objetivos deste trabalho foram investigar a composição química dos
compostos majoritários das folhas de Leonurus sibiricus; estudar o efeito alelopático dos
extratos aquoso (infusão) e metanólico, e também das frações flavonoídicas presentes no
extrato metanólico; e, de acordo com as atividades biológicas em sementes de Lepidium
sativum, Lactuca sativa e Raphanus sativus, identificar e testar as substâncias puras presentes
nas frações ativas em sementes de Lactuca sativa.

8
REVISÃO BIBLIOGRÁFICA
Leonurus sibiricus (L.)
Leonurus sibiricus é planta arbustiva, apresenta flores labiadas e inflorescência
axilares fasciculadas. Pertence a família Lamiaceae e a ordem Lamiales, originária da Índia e
ocorre em regiões tropicais da Ásia, África e América (Lorenzi, 2002). As plantas desta
família produzem terpenóides e substâncias fenólicas com efeitos alelopáticos (Larcher,
2000), o que talvez explique o comportamento de ser invasora (não desejável) em pomares e
cafezais (Lorenzi, 2002). Exudatos radiculares dessa espécie aumentam a germinação de
arroz, trigo e mostarda (Mandal, 2001), mas o extrato aquoso foliar inibe a germinação de
milho e o crescimento de plântulas de tomate (Almeida et al., 2003).
A utilização medicinal das folhas dessa espécie pode prevenir reumatismo crônico,
apresenta atividade antibacteriana e evita dermatites e outros problemas dermatológicos. Na
medicina chinesa, as sementes são consideradas afrodisíacas e a planta seca é prescrita como
tonificante e usada em disfunções menstruais (Bown, 1995). Folhas e flores são capazes de
combater vômitos e diarréias (Hoehne, 1939), além de serem eficientes contra resfriado,
bronquite e reumatismo (Castellucci et al., 2000). O extrato metanólico de folhas, também,
demonstra atividade antiinflamatória (Islam et al., 2005) e combate células neoplásicas de
glândulas mamárias (Nagasawa et al., 1990), o que justifica diversos estudos
etnofarmacológicos realizados com esta espécie no Brasil (Souza et al., 2004).
Em relação à composição química, a literatura relata que L. sibiricus apresenta
alcalóides (Murakami, 1943), mono, sesqui e diterpenos (Savona et al., 1982; Almeida et al.,
2005) e furano-lactonas (Satoh et al., 2003). Os extratos polares apresentam majoritariamente
rutina e seus derivados, além de flavonas metoxiladas (Almeida et al., 2006), as quais são
consideradas marcadores taxonômicos (Giang et al., 2005).
Os fenilpropanóides estão presentes em L. heterophyllus principalmente na forma de
flavonóides glicosilados e recentemente foi isolado a quercetina-3-O-[3-(4-hidroxi-3,5-
dimetoxibenzil) - α - L - rhamnopiranosil] - (1-6) - β - D - galactopiranosideo, um composto
biglicosilado (Cong et al., 2005). Outras espécies como L. cardiaca, L. persicus e L.
glaucescens também apresentam iridóides e flavonóides, o que demonstra expressivo
potencial medicinal deste gênero (Karting et al., 1985; Tasdemir et al., 1999; Milkowska-
Leyck et al., 2002). Estes dados mostram a importância do estudo químico de L. sibiricus,
associando o efeito de extrato e substância isolada em espécies vegetais sensíveis.

9
ALELOPATIA
Os primeiros relatos sobre a capacidade que certas espécies vegetais possuem de
interferir na fisiologia de outras foram feitos por Theophrastus (300 a.C.), seguindo os
trabalhos de Plínio (1 d.C.), Culpeper (1633), Young (1804), De Candolle (1832), Beobachter
(1845), Stickney & Hoy (1881), citados por Rice (1984).
Alelopatia pode ser definida como o efeito de uma planta no crescimento e
estabelecimento de outra (incluindo microrganismos) através da liberação de compostos
químicos para o ambiente (Rice, 1984). Essa interação acarreta resposta, positiva ou negativa,
da planta sensível a tais substâncias chamadas de aleloquímico, as quais são produzidas no
metabolismo secundário (Torres et al., 1996; Inderjit & Nilsen, 2003). Ainda, segundo
Gottlieb (1982), os aleloquímicos são sinais químicos transmitidos ao ambiente, geralmente
em pequena quantidade, e são responsáveis pelas múltiplas interações químicas entre os
diferentes organismos. As interações positivas (promoção, atração, etc) e negativas (inibição,
repelência, etc) indicam funções ecológicas dessas substâncias, importantíssimas para a
evolução e co-evolução das espécies vegetais e animais.
No âmbito da ecologia, o fenômeno da alelopatia pode explicar os mecanismos da
sucessão vegetal, onde espécies invasoras podem excluir espécies nativas a partir de resíduos
e substâncias liberados para o ambiente (Hierro & Callaway, 2003). Neste caso, a atividade
dos aleloquímicos é apenas inibitória e pouco relacionada com a competição por recursos
abióticos (Callaway, 2002). Portanto, a presença dessas espécies numa área de plantio pode
apresentar significativa influência no desenvolvimento de culturas (Batish et al., 2001; Singh
et al., 2001; Qasem & Foy, 2001), como também, no padrão natural de formação e sucessão
das populações e comunidades vegetais (Ferreira & Aquila, 2000).
Desta forma, a alelopatia é importante na resposta ecológica, pois pode interferir nas
populações vegetais tanto em ecossistemas agrícolas, quanto nos naturais. Uma das técnicas
mais utilizadas para estudar a alelopatia envolve o preparo de extratos aquosos foliares ou de
sistema radicular, observando a influência desses extratos na germinação e no crescimento da
raiz (Putnam, 1985; Inderjit & Dakshini, 1990).
Estudos comprovam a ação de aleloquímicos presentes nos extratos de diferentes
espécies vegetais, como de folhas de Triticum sativus, que afeta o crescimento de Gossypium
hirsutum (Hicks et al., 1989) e Oriza sativa (Alsaadawi, 1999), além de inibir a germinação
das próprias cariopses (Kalburtji, 1999). Extratos de Hordeum vulgare afetam a germinação e
crescimento de Triticum durum (Ben-Hammouda et al. 2001), segundo Oudhia & Tripathi

10
(2000), a planta invasora, Parthenium hysterophorus, tem ação alelopática na germinação de
Oriza sativa e, de acordo com Delachiave et al. (1999), Cynodon dactylon afeta a germinação
e desenvolvimento inicial de Lactuca sativa e Lycopersicum esculentum. Outras atividades de
extratos vegetais têm sido demonstradas por outros autores (Nakano et al., 2002; Singh et al.,
2001; Mazzafera, 2003; De Feo et al., 2004), onde ocorre promoção ou inibição do
crescimento, dependendo da concentração utilizada.
Segundo Inderjit (1996), a atividade alelopática na natureza envolve fatores bióticos,
como densidade vegetal, estádios de crescimento e microrganismos, e também fatores
abióticos, como solo, disponibilidade de água, temperatura e incidência de raios UV, dentre
outros. Tais fatores, podem afetar a liberação, disponibilidade, composição química e o modo
de ação das substâncias alelopáticas. A liberação dos compostos ocorre através da
volatilização das substâncias alelopáticas para a atmosfera, como também, o acúmulo destes
aleloquímicos nas fases aquosa e sólida do solo (Rizvi & Rizvi, 1992), os quais podem ser
metabolizados por microorganismos e, portanto, estar presentes na matéria orgânica das
camadas de serrapilheira sujeitos aos processos naturais de lixiviação (Inderjit et al., 2001).
O modo de ação dos aleloquímicos pode ser indireto, quando o composto alelopático
altera, primeiramente, algumas propriedades químicas do solo ou direto quando a substância
interfere no metabolismo vegetal (Ferreira & Aquila, 2000). Segundo Rizvi & Rizvi (1992),
os compostos alelopáticos podem afetar a estrutura e ultra-estrutura celulares; concentração e
balanço hormonal; permeabilidade das membranas afetando a absorção de minerais;
movimento dos estômatos influenciando a fotossíntese; síntese de pigmentos e proteínas;
atividade enzimática; relações hídricas e condução de seiva, como também podem alterar
DNA e RNA.
Harbone (1988) afirma que muitos compostos secundários sintetizados pelas plantas
inibem a germinação de sementes e o crescimento inicial de plântulas. A diversidade química
das substâncias produzidas pelas plantas pode ter origem na evolução vegetal, como resposta
à ação de microrganismos, insetos e vírus. Assim, os compostos com atividades alelopáticas
podem ser usados como alternativa ao uso de herbicidas e fungicidas, além de atuarem como
reguladores exógenos no desenvolvimento de plantas (Waller, 1999).
Técnicas como Cromatografia Gasosa de Alta Resolução (CGAR) e Cromatografia
Líquida de Alta Eficiência (CLAE) são ferramentas analíticas eficientes para identificar a
presença de compostos naturais em extratos brutos, possibilitando a obtenção do perfil
cromatográfico de frações ativas isoladas (Branco & Pizzolatti, 2002). Nos estudos de
alelopatia, para conhecer a composição dos extratos e associar os efeitos, é necessário isolar e

11
identificar a(s) substância(s) que acarreta(m) o efeito tóxico (Inderjit & Del Moral, 1997) ou
benéfico (Macias et al., 1997).
Muitas das substâncias envolvidas na alelopatia são metabólitos secundários que tem
origem na via do ácido chiquímico a qual origina os flavonóides, compostos fenólicos e
taninos (Elliott & Cheng, 1987; Einhellig, 1995), sendo que as concentrações podem ser
influenciadas pelo estresse biótico e abiótico. Os fenilpropanóides têm mostrado influenciar a
absorção de nutrientes (Bergmark et al., 1992; Booker et al., 1992), atividades enzimáticas
relacionadas ao metabolismo oxidativo (Devi & Prasad, 1992), relações hídricas (Barkosky &
Einhellig, 1993), fotossíntese e a respiração (Einhellig & Rasmussen, 1993).
Muitos compostos flavonoídicos, em condições naturais, mostram propriedades
alelopáticas e os mais conhecidos são hesperitina 7-O-rutinosideo, taxifolina 7-O-
arabinosideo e formononetina 7-O-glucosideo (Tomita-Yokotani, 2004) presentes em extratos
e resíduos de Pluchea lanceolata (DC) (Inderjit & Dakshini, 1994, Rice-Evans & Packet,
1998). Recentemente, dois flavonóides, a isorhamnetina 3-O-rutinosideo e a isorhamnetina 3-
O-robinobiosideo, foram isolados do pericarpo de cactus (Hylocereus undaus) e
demonstraram atividade alelopática em Celosia cristata e Neurospora crassa (Tomita-
Yokotani et al., 2004).
Os flavonóides possuem importante papel como compostos de defesa e moléculas
sinalizadoras dos processos de reprodução, patogênese e simbiose, que são produzidas pelas
plantas em grande escala, e no ambiente, têm efeito significativo na composição química do
solo, tendo importância nas interações planta-planta e planta-microrganismo (Rice, 1984;
Shirley, 1996).
Os efeitos desses compostos em plantas incluem diminuição da entrada de oxigênio
nas mitocôndrias e, nos cloroplastos, redução do transporte de elétron e eficiência do
fotossistema II (Moreland & Novitzky, 1988). Essas múltiplas atividades resultam em amplo
espectro de fitotoxicidade (Einhellig, 2004) ou, dependendo da concentração dos flavonóides,
estimulam o crescimento de raízes (Webster et al., 1998).
De modo geral, os flavonóides protegem as células do estresse oxidativo pela
quelação, inativação ou captação de radicais livres, produzidos pelo sistema de transporte de
elétrons (Harbone, 1994). Devido as suas características químicas, capazes de captar elétrons,
atuam como catalisadores na fase fotoquímica da fotossíntese e/ou como reguladores de
canais iônicos envolvidos na fosforilação oxidativa do processo de obtenção de energia
fotossintética (Piettá & Simonetti, 1999). Em função dessas características químicas, os
flavonóides estão envolvidos com os processos relacionados à absorção iônica que prejudicam

12
o padrão de despolarização das membranas celulares das raízes, levando a disfunções no
equilíbrio de íons (Glass & Dunlop, 1974). Os danos metabólicos acarretados pelos
flavonóides, segundo Macias et al. (1997), dependem da esterioquímica das moléculas e
concentração utilizada as quais podem promover ou inibir o crescimento de raízes, sendo no
entanto, pouco esclarecidos os mecanismos de ação destes compostos.
Os compostos fenólicos são considerados potentes inibidores da germinação de
sementes, crescimento da parte aérea e alongamento das raízes (Rice, 1984), como por
exemplo os ácidos p-hidroximandélico; 3,4-dihidroxifenilacético e mandélico, que inibem o
crescimento da parte aérea de arroz. Em relação ao metabolismo enzimático, os fenólicos
presentes em extratos de Pinus laricio inibiram as enzimas glicose-6-fosfato dehidrogenase,
glicose-fosfato isomerase e aldolase que estão relacionadas com a síntese de açúcares
(Muscolo et al., 2001). Ainda, segundo Inderjit & Dakshini (1992), os compostos fenólicos
solúveis em água, oriundos da planta daninha Pluchea lanceolata, influenciaram o teor de
clorofila e a taxa fotossintética líquida de folhas de aspargo, em casa de vegetação.
Em extrato de folhas de Triticum aestivum, Perez (1990) isolou os ácidos
hidroxâmicos DIMBOA e DIBOA, os quais a partir de 2mM reduziram a porcentagem de
germinação e o desenvolvimento inicial de Avena fatua. Da mesma maneira, outros estudos
têm demonstrado que ácidos hidroxâmicos cíclicos, do tipo 4-hidróxi-1,4-benzoxazin-3-onas,
presentes em cereais na forma de glicosídeos, atuam inibindo o crescimento de fungos,
bactérias e vegetais (Kruse et al., 2000).
FLAVONÓIDES
Os flavonóides são pigmentos naturais presentes nos vegetais, capazes de conferir
proteção aos organismos em relação aos danos produzidos pelos agentes antioxidantes, tais
como a radiação ultravioleta, poluição ambiental, substâncias químicas presentes no solo ou
nos alimentos, dentre outros fatores. O organismo humano não produz essas substâncias
protetoras e por isso deve obtê-las através de dieta específica, com alimentos que as
apresentem nas folhas, flores e frutos das plantas (Singleton, 1981; Aheme & O´Brien, 2002).
Os flavonóides foram descobertos por Nobel Szent-Gyorgy, em 1930, que isolou da
casca de limão uma substância, a citrina, que regulava a permeabilidade dos capilares
sanguíneos e denominados, a princípio, por vitamina P (devido à ação na permeabilidade) e
também vitamina C2 (porque se comprovou que alguns flavonóides tinham propriedades
similares à da vitamina C). No entanto, por volta de 1950, o efeito dos flavonóides como

13
vitaminas não foram comprovados e tais denominações foram abandonadas (Martinez-Flórez
et al., 2002).
A estrutura química dos flavonóides apresenta número variável de grupos hidroxilas e
fenóis, com excelentes propriedades de quelação de ferro e outros metais, o que lhes conferem
grande capacidade antioxidante (Havsteen, 1983; Bors et al., 1990; Peres, 1994). Portanto,
desempenham papel essencial de proteção aos fenômenos que provocam danos oxidativos às
células, e têm efeitos terapêuticos em número elevado de patologias incluindo cardiopatia,
isquemia, arterioesclerose e diabete (Jang et al., 1997; Jovanovic et al., 1998).
As propriedades antioxidativas captam radicais livres, como grupos hidroxilas e
superóxido, altamente reativos e relacionados com o início da cadeia de peroxidação lipídica
(Pace-Asciak et al., 1995). Além destes efeitos conhecidos de atividade antioxidante, os
flavonóides apresentam propriedades como estimulação da comunicação entre membranas,
regulação do crescimento celular, indução de enzimas de detoxificação, inibição da
germinação e crescimento de plântulas (Macias et al., 1997;Yang et al., 2000; Igura et al.,
2001; Hoagland & Williams, 2004).
Os flavonóides são compostos de baixo peso molecular que apresentam um esqueleto
carbônico básico (C6-C3-C6), formado por dois anéis fenólicos (A e B) ligados por anel
pirano heterocíclico (C). Os átomos de carbono dos anéis C e A são numerados de 2 a 8 e do
anel B de 2´ a 6´ (Kühnau, 1976). A atividade biológica dos flavonóides como antioxidantes
depende da propriedade redox dos grupos hidroxifenólicos e da relação estrutural entre as
diferentes partes da estrutura química (Bors et al., 1990). Esta estrutura básica permite uma
diversidade de padrões de substituições e variações do anel C e em função das características
estruturais podem ser classificados em 4 grupos (Letan, 1966): 1) Flavanóis, como a catequina
com grupo -OH na posição 3 do anel C; 2) Flavonols, representados pela quercetina, que
possuem grupo carbonila na posição 4 e grupo -OH na posição 3 do anel C; 3) Flavonas,
como a diosmetina, que posuem grupo carbonila na posição 4 do anel C e grupo hidroxila na
posição 3; e 4) Antocianidinas, que possuem grupo -OH na posição 3, mas também possuem
uma dupla ligação entre os carbonos 3 e 4 do anel C. Portanto, as três características
estruturais importantes para a função antioxidante são a presença no anel B do grupo catecol
ou orto-dihidroxi; de dupla ligação na posição 2,3 do anel C e de grupos hidroxilas na posição
3 e 5 do anel A.
Muitos autores relatam que o arranjo do anel B dos flavonóides é provavelmente
responsável pela atividade alelopática e a relação estrutura-atividade está relacionada com a
fitotoxidade destes compostos (Macias et al, 1997; Bais et al., 2003; Parvez et al., 2004;

14
Beninger & Hall, 2005). Evidências de que a atividade dos flavonóides depende da estrutura e
dos radicais substituintes do anel B são demonstradas por Bors et al. (1990). Essas
características estruturais contribuem para aumentar a estabilidade do radical livre e portanto,
conferem a atividade antioxidante. Assim, flavonols são mais potentes do que flavonas devido
à presença do grupo hidroxila na posição 3 do anel A.
Por outro lado, a glicosilação dos flavonóides como ocorre na síntese de rutina, reduz
fortemente a capacidade de remoção dos radicais livres. As moléculas de açúcar podem se
unir às estruturas dos flavonóides, preferencialmente na posição 3 do anel C e com menor
freqüência na posição 7 do anel A, de forma que estes compostos são encontrados como orto-
glucosídeos, sendo a glicose o resíduo de açúcar mais freqüente. Outros açúcares também
presentes nos compostos glicosilados são galactose, ramnose, arabinose e xilose. O esqueleto
cabônico básico dos flavonóides, com os três anéis (A, B C), sem a molécula de açúcar é
conhecido como aglicona. Os glicosídeos são mais solúveis em água e menos reativos frente
aos radicais livres que as agliconas. A adição de um grupo hidroxila no anel B aumenta a
capacidade antioxidante como acontece na miricetina. No entanto, a presença apenas de uma
hidroxila no anel B não é suficiente para conferir atividade biológica ao flavonóide (Piettá,
2000).
Fenilpropanóides, entre eles os flavonóides, são produtos naturais derivados do
aminoácido fenilalanina que sofre desaminação pela enzima fenilalanina amonialiase (PAL).
O exemplo mais simples dessas substâncias são aquelas que contêm somente esqueleto
fenilpropano C6-C3, como os ácidos hidroxicinâmicos, bem como o ácido sináptico, e os
monolignols como os coniferil álcool (Heller & Forkmann, 1993). Fenilpropanóis mais
complexos são formados pela condensação das unidades de fenilpropanos com uma unidade
derivada da via acetato, a malonil coenzima A, gerando outros grupos de flavonóides,
isoflavonóides e stilbenos (Wagner & Farkas, 1975; Midleton et al., 2000).
Os compostos fenilpropanóicos não são encontrados em todas as espécies vegetais,
desta forma as classes de acido hidroxi-cinâmico e flavonóides são características de plantas
superiores, sendo que tais compostos apresentam padrões específicos de substituições
químicas que são peculiares em gêneros e espécies vegetais (Gottlieb, 1982).

15
Capítulo 1
Composição do óleo essencial de rubim (Leonurus sibiricus L.)
Publicado na Revista Brasileira de Plantas Medicinais, v. 8, n.1, p.31-35, 2005.

16
Composição do óleo essencial de rubim (Leonurus sibiricus L. – Lamiaceae)
Resumo: Os óleos voláteis são encontrados na maioria das famílias do reino vegetal dentre
elas, Lamiaceae. As flores e folhas de Leonurus sibiricus têm uso medicinal em resfriados,
bronquite e reumatismo, sendo de fundamental importância a caracterização fitoquímica.
Tendo em vista a importância dessa espécie e a escassez de estudos sobre seus componentes
voláteis, neste trabalho foi identificada a composição do óleo essencial de L. sibiricus L.
(rubim). Para tanto, foi utilizada a técnica de cromatografia gasosa acoplada a espectrometria
de massa. Os resultados indicaram que aproximadamente 70% dos compostos voláteis dessa
espécie são constituídos por trans-cariofileno, alfa-humuleno e germacreno – D. Os outros
compostos também identificados no óleo foram gama-cadineno, beta-bourboreno e alfa-
humuleno.
Palavras-chave: Leonurus sibiricus; Lamiaceae; óleo essencial; plantas medicinais; trans-
cariofileno; germacreno-D; alfa-humuleno
Abstract: Composition of the essencial oil of rubim (Leonurus sibiricus L. – Lamiaceae).
The essential oils are found in a great number of Lamiaceae family species, but few
researches were carried out on this subject. This work had as a goal to identify the Leonurus
sibiricus L. (rubim) essential oil composition. Rubim leaves and flowers in infusion are able
to avoid vomit, diarrhea, and are also indicated in cold, cough, bronchitis and rheumatisms
cases. In order to know what are the phytochemical compositions involved, gas
cromatography techniques with mass spectrometry (GC-MS) was the methodology used. The
results showed 70% volatility compound by trans-cariophylene, alpha-humulene and
germacrene-D. Other substances like gama-cadinene, beta-bourborene and alpha-copaene
were found like compounds of this essential oil species.
Key words: Leonurus sibiricus; Lamiaceae; essential oil; medicinal plants; trans-
cariophylene; germacrene-D; alpha-humulene.

17
INTRODUÇÃO
A erva-macaé ou rubim (Leonurus sibiricus L.) pertence à família Lamiaceae e ordem
Lamiales, a qual produz terpenóides e substâncias fenólicas com efeitos alelopáticos (Larcher,
2000). Segundo Lorenzi (2002), a planta é considerada invasora em lavouras agrícolas,
presente em áreas abandonadas e apresenta crescimento espontâneo em quase todo território
brasileiro.
A utilização como erva medicinal é indicada, com base na tradição popular, para casos
de resfriado, bronquite e reumatismo (Castellucci et al., 2000). A planta é usada na medicina
popular, que recomenda as partes verdes na forma de chás, nos casos de sangramento pós-
parto, menstruação excessiva, bem como contra edema, abscessos e problemas renais (Bown,
1995). De acordo com Hoehne (1939), as folhas e flores, em infusão, também são capazes de
combater vômitos e diarréias. Enquanto, as espécies Leonurus cardiaca e L. japonica são
eficazes no controle de disfunções cardíacas e anti-trombóticas (Bown, 1995).
Nos primeiros estudos fitoquímicos de Leonurus sibiricus foi isolado o alcalóide
estaquidrina (Murakami, 1943), posteriormente, diterpenos (Savona et al., 1982) e furano-
lactonas (Satoh et al., 2003) em suas folhas. Os compostos fenólicos, como o ácido cafeíco,
também foram encontrados nas folhas, raízes e sementes (Almeida et al., 2002), que segundo
Mandal (2001), aumenta a germinação de arroz, trigo e mostarda. Da mesma maneira, frações
flavonoídicas de extrato hidroalcoólico reduzem a germinação e inibem o crescimento de
raízes de pepino (Almeida, et al., 2003). Estudos mostram que a diversidade de estruturas
químicas possibilita diferentes atividades biológicas, através do arranjo entre hidroxilas e
carbonilas com esqueletos carbônicos (Bradow, 1991).
Os óleos voláteis que são liberados para o ambiente apresentam importante função no
ecossistema, agindo como substâncias de sinalização, como defesa e inibição de crescimento.
Para o reconhecimento desses compostos, é indispensável a realização de estudos
fitoquímicos, o que possibilita maior compreensão das relações dos vegetais com o ambiente
(Larcher, 2000). Nesse sentido, muitas espécies aromáticas são estudadas quimicamente por
terem importância nas indústrias de perfume, cosmético e condimento (Endenshaw et al.,
2000).
O estudo das substâncias voláteis, da família Lamiaceae pode colaborar para o
conhecimento fitoquímico das plantas, bem como para evidenciar a diversidade biológica e
química presentes nos vegetais. Portanto, o objetivo deste trabalho foi o de identificar a
composição química dos compostos voláteis de folhas de rubim (L. sibiricus).

18
MATERIAL E MÉTODO
O presente estudo foi realizado no Departamento de Horticultura da UNESP, Campus
de Botucatu e no Centro de Genética, Biologia Molecular e Fitoquímica do Instituto
Agronômico de Campinas (IAC).
A espécie Leonurus sibiricus foi coletada e identificada em Botucatu e está depositada
no herbário Botu com número 23-706. As folhas foram separadas, pesadas e colocadas em
estufa de secagem a 40ºC por 48 horas (40g). Esse material foi submetido a hidro-destilação
em aparelho tipo Clevenger, por 60 min.
A extração dos compostos voláteis foi realizada com a adição de 0,5mL de Acetato de
Etila (“Dinâmica”; lote 7705), seguido do acondicionamento em frasco, contendo sulfato de
sódio (P. A.; “Ecibra”; lote 1951).
Para análise cromatográfica dos constituintes identificados foi utilizada amostra de
100μL em cromatógrafo gasoso acoplado a espectrômetro de massas (CG-EM, Shimadzu,
QP-5000), operando em 70 eV, equipado com coluna capilar de sílica fundida DB-5 (J &
Wiley Scientific, 30m x 0,25mm x 0,25μm), injetor e detector em 240ºC e 230ºC, com gás de
arraste Hélio (1,0 mL min-1), modo split 1/20 e gradiente de temperatura 60ºC-240ºC com
3ºC/min. Através do cálculo dos índices de retenção dos compostos eluídos no sistema
cromatográfico, comparação com os espectros de massas da biblioteca Nist 62:librar do
sistema e dados da literatura (Mc Lafferty & Stauffer, 1989; Adams, 1995) foi possível
identificar os constituintes do óleo de L. sibiricus.
RESULTADO E DISCUSSÃO
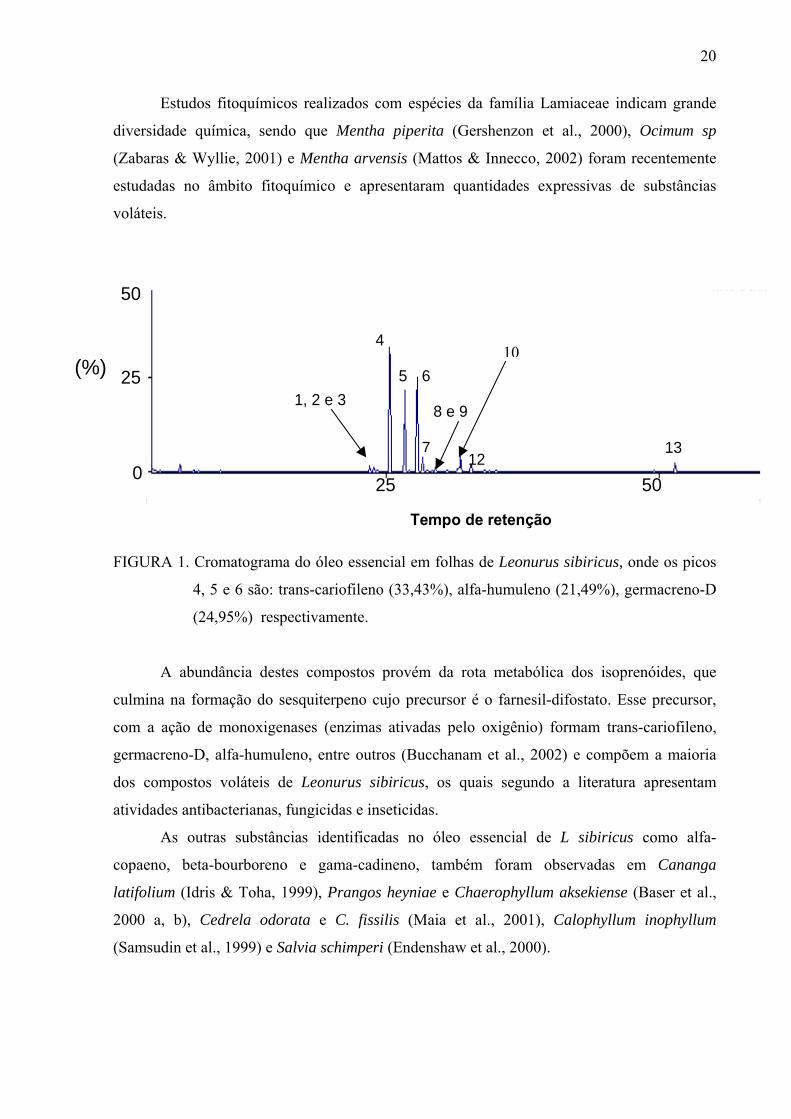
A análise por CG-EM do óleo de Leonurus sibiricus mostrou a presença de 13
compostos, sendo que dentre os identificados pode se observar a predominância dos
sesquiterpenos trans-cariofileno (33,43%), alfa-humuleno (21,49%) e germacreno-D (24,95%)
que são responsáveis por, aproximadamente, 70% da composição do óleo (Tabela 1 e Figura
1). Esses sesquiterpenos também foram encontrados, porém em pequenas quantidades, em
Ocimum selloi em diferentes épocas do ano (Moraes et al., 2002). Essas substâncias
apresentam esqueletos carbônicos com ampla atividade antibacteriana, antifúngica e
inibidores enzimáticos (Abraham, 2001).

19
*N. I. = substância não identificada pelas bibliotecas consultadas.
Ikcal = índice de retenção de Kovats calculado; Iklit = índice de retenção de Kovats da literatura
A presença do trans-cariofileno (pico 4) foi observada anteriormente no óleo essencial
de Phlomis lanata (Couladis et al., 2000), Cannabis sativa (Novak et al., 2001) e Piper duckei
(Abraham, 2001). Estudos comprovam a atividade dessa substância como antimicrobiana,
principalmente em fungos patogênicos e bactérias gram (+/-). A substância alfa-humuleno
(pico 5) encontrada na maioria das espécies aromáticas, apresenta ampla atividade biológica
como inseticida (Yang et al., 2003; Kim et al., 2003), antioxidante (Weel et al., 1999),
antimicrobiano (Bougatsos et al., 2003), como também anticancerígena (Legault et al., 2003).
Germacreno D (pico 6) foi encontrado também em Artemisia parviflora (Rana et al., 2003),
Piper friedrichsthalli e P. pseudollindeni (Vila et al., 2003), Mangifera indica (Lalel et al.,
2003) e em Jupinerus turbinata (Consentino et al., 2003). A literatura relata sua atividade
antibacteriana e fungicida (Ngassapa et al., 2003).
Pico Substância % composição IKcal IKlit
1 tricicleno 1,01 932 926
2 α-copaeno 1,55 1377 1376
3 β-bourboreno 0,80 1386 1384
4 τ-cariofileno 33,43 1422 1418
5 α-humuleno 21,49 1456 1454
6 germacreno D 24,95 1484 1480
7 N. I.* 3,38 X X
8 γ-cadineno 0,99 1516 1513
9 N. I.* 1,28 X X
10 N. I.* 0,87 X X
11 N. I.* 5,67 X X
12 N. I.* 1,87 X X
13 N. I.* 2,22 X X
TABELA 1. Composição do óleo essencial de folhas de Leonurus sibiricus.
20
Estudos fitoquímicos realizados com espécies da família Lamiaceae indicam grande
diversidade química, sendo que Mentha piperita (Gershenzon et al., 2000), Ocimum sp
(Zabaras & Wyllie, 2001) e Mentha arvensis (Mattos & Innecco, 2002) foram recentemente
estudadas no âmbito fitoquímico e apresentaram quantidades expressivas de substâncias
voláteis.
FIGURA 1. Cromatograma do óleo essencial em folhas de Leonurus sibiricus, onde os picos
4, 5 e 6 são: trans-cariofileno (33,43%), alfa-humuleno (21,49%), germacreno-D
(24,95%) respectivamente.
A abundância destes compostos provém da rota metabólica dos isoprenóides, que
culmina na formação do sesquiterpeno cujo precursor é o farnesil-difostato. Esse precursor,
com a ação de monoxigenases (enzimas ativadas pelo oxigênio) formam trans-cariofileno,
germacreno-D, alfa-humuleno, entre outros (Bucchanam et al., 2002) e compõem a maioria
dos compostos voláteis de Leonurus sibiricus, os quais segundo a literatura apresentam
atividades antibacterianas, fungicidas e inseticidas.
As outras substâncias identificadas no óleo essencial de L sibiricus como alfa-
copaeno, beta-bourboreno e gama-cadineno, também foram observadas em Cananga
latifolium (Idris & Toha, 1999), Prangos heyniae e Chaerophyllum aksekiense (Baser et al.,
2000 a, b), Cedrela odorata e C. fissilis (Maia et al., 2001), Calophyllum inophyllum
(Samsudin et al., 1999) e Salvia schimperi (Endenshaw et al., 2000).
25
0
50
(%)
25 50
Tempo de retenção
1, 2 e 3
4
5 6
7
8 e 9
12 13
10

21
REFERÊNCIA BIBLIOGRÁFICA
ABRAHAM, W.R. Bioactive sesquiterpenes produced by fungi: are they useful for humans as
well? Current Medicinal Chemistry, v.8, p.583-606, 2001.
ADAMS, R.P. Identification of essential oil components by gas chromatography / mass
spectroscopy, 4. ed., Illinois: Allured Publishing Corporation, 1995, 659p.
ALMEIDA, L.F.R.; VILEGAS, W.; DELACHIAVE, M.E.A. Investigação química do rubim
(Leonurus sibiricus) In: SIMPÓSIO BRASILEIRO DE PLANTAS MEDICINAIS, 17,
2002, Cuiabá, Anais .... Cuiabá, 2002. p.79
ALMEIDA, L.F.R. et al. Atividade alelopática de extratos e frações de folhas de Leonurus
sibiricus na germinação e desenvolvimento inicial de pepino. Brazilian Journal of Plant
Physiology, v.13 (suplemento), p.351, 2003.
BASER, K.H.C. et al. Composition of the essential oil of Prangos heyniae, a new endemic
from Turkey. Flavour and Fragrance Journal, v.15, p.47-49, 2000a.
BASER, K.H.C. et al. Composition of the essential oil of Chaerophyllum aksekiense, a
recently described endemic from Turkey. Flavour and Fragrance Journal, v.15, p.43-44,
2000b.
BOUGATSOS, C. et al. Composition and antimicrobial activity of the essential oils of
Helichrysum kraussi SCH. BIG. and H. rugulosum LEBS. from South Africa, Flavour and
Fragrance Journal, v.18, p.48-51, 2003.
BOWN, D. The Herb Society of America: Encyclopedia of Herbs and their Uses. 1.ed.,
New York: Darling Kindersley Publ., 1995, 225p.
BRADOW, J.M. Relationships between chemical structure and inhibitory activity of C6
through C9 volatiles emitted by plant residues. Journal of Chemical Ecology, v.17, p.2193-
2212, 1991.
BUCCHANAM, B.B.; GRUISSEN, W.; JONES, R.L. Biochemistry and Molecular Biology
of Plants. 4. ed., Maryland: John Wiley & Sons, 2002, 1367p.
CASTELLUCCI, S. et al. Plantas medicinais relatadas pela comunidade residente na estação
ecológica de Jataí, município de Luís Antônio/SP: uma abordagem etnobotânica. Revista
Brasileira de Plantas Medicinais, v.3, p.51-60, 2000.
CONSENTINO, S. et al. Composition and antimicrobial properties of Sardinian Juniperus
essential oils against foodborne pathogens and spoilage microorganisms. Journal of Food
Protection, v.66, p.1288-1291, 2003.

22
COULADIS, M. et al. Essential oil Plhomis lanata growing in Greece: chemical composition
and antimicrobial activity. Planta Medica, v.66, p.670-672, 2000.
ENDENSHAW, M.M. et al. Volatile oil constituents of the Ethiopian plant Salvia schimperi
Benth. Flavour and Fragrance Journal, v.15, p.27-30, 2000.
GERSHENZON, J.; Mc CONKEY, M.E.; CROTEAU, R.B. Regulation of monoterpene
accumulation in leaves of peppermint. Plant Physiology, v.122, p.205-213, 2000.
HOEHNE, F.C. Plantas e Substâncias Vegetais Tóxicas e Medicinais. 2.ed., São Paulo:
Graphicars, 1939, 355p.
IDRIS, M.S.H.; TOHA, M.Y. Essential oil from flowers of two Cananga species. Review of
Biodiversity and Environmental Conservation, p.1-4, 1999.
KIM, E.H. et al. Acaricidal activity of clave bud oil compounds against Tyrophagus
putrescentiae (Acari-Acaridae). Applied Entomology and Zoology, v.38, p.261-266, 2003.
LALEL, H.J.D.; SINGH, Z.; TAN, B.C. Distribution of aroma volatile compounds in
different parts of mango fruit. Journal Horticultural Science and Biotechnology, v.78,
p.131-138, 2003.
LARCHER, W. Ecofisiologia Vegetal. 2. ed., Stuttgart: Eugen Ulmer, 2000, 531p.
LEGAULT, J. et al. Antitumor activity of balsam oil: production of reactive oxygen species
induced by alpha-humulene as possible mechanism of action. Planta Medica, v.69, p.402-
407, 2003.
LORENZI, H. Plantas Daninhas do Brasil: terrestres, aquáticas, parasitas, tóxicas e
medicinais. 2.ed., São Paulo: Nova Odessa, 2002. 425p.
MAIA, B.H.L. et al. Essential oils of Toona and Cedrela species (Meliaceae): taxonomic and
ecological implications. Journal of Brazilian Chemistry, v.11, p.629-639, 2001.
MATTOS, S.H.; INNECCO, R. Idade ideal de corte de Mentha arvensis como produtora de
óleo essencial de mentol para o Estado de Ceará, Brasil. Revista Brasileira de Plantas
Medicinais, v.5, p.15-18, 2002.
Mc LAFFERTY, F.W.; STAUFFER, D. Registry of spectral data. 3ed., New York: John
Wiley & Sons, 1989, 354p.
MORAES, L.A.S. et al. Phytochemical characterization of essential oil from Ocimum selloi.
Anais da Academia Brasileira de Ciências, v.74, p.183-186, 2002.
MURAKAMI, S. Stachydrim in Leonurus sibiricus L. Acta Phytochemichal, v.13, p.161-
184, 1943.

23
NGASSAPA, O. et al. Composition and antimicrobial activity of essential oils of two
populations of Tanzanian Lippia javanica (Burm. f.) Spreng. (Verbenaceae). Flavour and
Fragance Journal, v.18, p.221-224, 2003.
NOVAK, J. et al. Essential oils of different cultivars of Cannabis sativa L. and their
antimicrobial activity. Flavour and Fragrance Journal, v.16, p.259-262, 2001.
RANA, V.S. et al. Essential oil composition of Artemisia parviflora aerial parts. Flavour and
Fragance Journal, v.14, p.342-344, 2003.
SAMSUDIN, M.W.; IBRAHIM, M.N.; SAID, I. Composition of the steam volatile oil from
Calophyllum inophyllum. Review of Biodiversity and Environmental Conservation, p.1-3,
1999.
SATOH, M. et al. Studies on the constituents of Leonurus sibiricus L. Chemical
Pharmaceutical Bulletin, v.51, p.341-342, 2003.
SAVONA, G. et al. Diterpenoids from Leonurus sibiricus. Phytochemistry, v.21, p.2699-
2701, 1982.
VILA, R. et al. Constituents of the essential oils from Piper friedrichsthalii C.DC. and P.
pseudolindenii C.DC. from Central America. Flavour and Fragrance Journal, v.18, p.198-
201, 2003.
ZABARAS, D.; WYLLIE, S.G. The effect of mechanical wounding of the composition
essential oil from Ocimum minimum L. leaves. Molecules, v.6, p.79-86, 2001.
YANG, Y.C. et al. Ovicidal and adulticidal effects od Eugenia caryophyllata bud and leaf oil
compond on Pedicilus capitatus. Journal of Agricultural and Food Chemistry, v.51,
p.4884-4888, 2003.
WEEL, K.G.C. et al. Antioxidant activity of horehound (Massubium vulgare) grown in
Lithuania. Fett-Lipid, v.101, p.395-400, 1999.

24
Capítulo 2
Potencial alelopático de folhas de Leonurus sibiricus L.
Enviado para Journal of Agricultural and Food Chemistry, 2006.

25
POTENCIAL ALELOPÁTICO DE FOLHAS DE Leonurus sibiricus (Lamiaceae)
Resumo: A interação química entre as plantas é conhecida como alelopatia e está relacionada
com a liberação de substâncias para o ambiente. Os aleloquímicos são produzidos pelo
metabolismo secundário e podem ser, principalmente, terpenos, alcalóides e fenilpropanóides,
substâncias presentes em Leonurus sibiricus, conhecida erva medicinal com ampla atividade
biológica. O presente estudo teve por objetivos avaliar a atividade alelopática de folhas de L.
sibiricus na germinação e desenvolvimento inicial de Raphanus sativus, Lactuca sativa e
Lepidium sativus. Para tanto, foram utilizados extratos por infusão e metanólico, além das
frações do extrato metanólico. As frações com atividade alelopática foram refracionadas em
HPLC com detector de UV, obtendo-se sete flavonóides. Nos extratos por infusão e
metanólico, as concentrações utilizadas nos testes biológicos, foram de 50, 100, 200, 400 e
800 mg L-1; para as 12 frações do Sephadex foram de 1g L-1 e para os flavonóides de 10-4,
10-5, 10-6 e 10-7M. Em todos os testes foi utilizada água destilada como controle e os
flavonóides testados em Lactuca sativa. Os resultados indicaram que as folhas de L. sibiricus
possuem quatro flavonóides majoritários, que são quercetina-3-O-β-D-galactopiranosil(1-6)-
O-α-L-rhamnopiranosideo; quercetina-3-O-β-D-glucopiranosil (1-6)-O-α-L-
rhamnopiranosideo (Rutina); quercetina-3-O-β-D- galactopiranosideo (Hiperina) e quercetina-
3-O-β-D-glucopiranosideo (Isoquercetrina). Além dessas substâncias foram também isoladas
e testadas genkanina-3´-hidroxi, genkanina e quercetina. Os extratos por infusão e metanólico
de folhas de Leonurus sibiricus acarretaram redução significativa apenas na germinação das
sementes de Lactuca sativa, porém não afetaram o processo germinativo de Raphanus sativus
e Lepidium sativum. Por outro lado, o crescimento inicial das raízes de Raphanus sativus,
Lactuca sativa e Lepidium sativum foi mais afetado pelo extrato metanólico, principalmente,
nas concentrações mais altas. As frações de sephadex, consideradas flavonoídicas, mostraram
atividade de redução, principalmente, do crescimento inicial de raiz das espécies estudadas,
sendo que a germinação foi pouco afetada na maioria das concentrações, da mesma forma
quando utilizado extrato. As agliconas foram responsáveis pela inibição da germinação, bem
como do crescimento inicial de raiz na concentração mais elevada (10-4). A genkwanina-
3´hidroxi e quercetina apresentaram maior atividade herbicida na germinação, na
concentração de 10-4M, por outro lado, o comprimento inicial de raiz foi reduzido quando
utilizadas quercetina-3-O-glucose-ramnose, quercetina-3-O-glucose e genkwanina-3´hidroxi.
Nas concentrações de 10-5, 10-6 e 10-7M todas as substâncias isoladas acarretaram estímulo de
crescimento inicial de raiz. Pelos resultados conclui-se que o uso de extratos com perfil

26
químico semelhante pode propiciar respostas alelopáticas diferentes, devido a correlação com
a concentração utilizada e estrutura das moléculas, onde as agliconas demonstraram maior
atividade (inibição ou promoção).
Palavras-chave: alelopatia, Leonurus, plantas medicinais, flavonóides, fitoquímica.

27
Abstract: Allelopathic potential of Leonurus sibiricus leaves. The chemical interaction
between plants is known as allelopathy and it is related to the release of substances to the
environment. The allelochemicals are produced by the secondary metabolism and they can
belong principally to the following classes of substances: terpenes, alkaloid, and
phenylpropanoids; substances present in Leonurus sibiricus, renowned medicinal herb with
wide biological activity. The present study aimed at the evaluation of the allelopathic activity
of the leaves of L. sibiricus in the germination and initial development of Raphanus sativus,
Lactuca sativa, and Lepidium sativus. Thus, there were used extracts through infusion and
methanolic . The fractions with allelopathic activities were refractioned in HPLC with UV
detectors, obtaining seven flavonoids. The concentrations used in the biological tests for the
extracts through infusion and methanolic were the followings: 50, 100, 200, 400, and 800 mg
L-1; for the 12 Dephadex fractions the concentration was 1g L-1; and for the flavonoids, the
concentrations were the followings: 10-4, 10-5, 10-6, and 10-7M. In all of the tests, there were
used distilled water as check and the tested flavonoids in Lactuca sativa. The results indicated
that the leaves of L. sibiricus have four majority flavonoids, which are quercetin-3-O-β-D-
galactopyranosyl(1-6)-O-α-L-rhamnopyranoside; quercetin-3-O-β-D-glucopyranosyl (1-6)-O-
α-L-rhamnopyranoside (Rutin); quercetin-3-O-β-D-galactopyranoside (Hiperine), and
quercetin-3-O-β-D-glucopyranoside (Isoquercetrin). Besides these substances, there were
isolated and tested the genkanin-3´-hydroxy, genkanin, and quercetin. The extracts through
infusion and methanolic of leaves of Leonurus sibiricus caused significative reduction only in
the germination of the seeds of Lactuca sativa, but it has not affected the germinative process
of Raphanus sativus and Lepidium sativum. On the other hand, the initial root growths of
Raphanus sativus, Lactuca sativa, and Lepidium sativum were more affected by the
methanolic extract, principally, in the highest concentrations. The Sephadex fractions, which
are considered flavonoidic have shown activity of reduction, principally, in the initial root
growth of the studied species, being the germination, in the same way when used the extracts,
less affected in the majority of concentrations. The aglycones were responsible for the
inhibition of the germination as well as the root initial growth in the most elevated
concentration (10-4). The genkwanin-3´hydroxy and quercetin have presented the highest
herbicide activity in the germination at the concentration of 10-4M, on the other hand, the root
initial length was reduced when used quercetin-3-O-glucose-rhamnose, quercetin-3-O-glucose
and genkwanin-3´hydroxy. In the concentrations of 10-5, 10-6 and 10-7M, all isolated
substances caused initial root growth stimulus. Based on the results, it concluded that the use
of the extracts with chemical profile similar may propitiate different allelopathic responses

28
due to the correlation to the used concentration and the molecule structure, where the
algycones have demonstrated high activity (inhibition or promotion).
Key words: allelopathy, Leonurus, medicinal plants, flavonoids, phytchemistry.

29
INTRODUÇÃO
Os aleloquímicos sintetizados pelas rotas do metabolismo secundário influenciam
no desenvolvimento e estabelecimento das culturas agrícolas e comunidades vegetais (Torres,
et al., 1996). Pesquisadores e agricultores reconhecem os aleloquímicos como alternativa
viável para substituir os pesticidas sintéticos, visando reduzir a poluição ambiental e aumentar
a produtividade agrícola (Qasem & Foy, 2001; Duke et al., 2002).
A riqueza química dos extratos de plantas aromáticas e medicinais é reconhecida
fonte de compostos biologicamente ativos, sendo que as substâncias alelopáticas podem
apresentar também características semelhantes às das substâncias com efeitos terapêuticos
(Mathela, 1991; Cutler & Cutler, 1999).
A espécie Leonurus sibiricus, originária da Índia, produz terpenóides e substâncias
fenólicas com efeitos alelopáticos (Larcher, 2000) o que explica o comportamento de ser
planta invasora (não desejável) em pomares e cafezais (Lorenzi, 2002). Entretanto, essa
espécie libera exudatos radiculares que aumentam a germinação de arroz, trigo e mostarda
(Mandal, 2001), mas o extrato aquoso de folhas inibe a germinação de milho e o crescimento
de plântulas de tomate (Almeida et al., 2003).
As atividades medicinais de L. sibiricus estão presentes em diferentes órgãos, tais
como folhas, que combatem reumatismo crônico, possuem atividade anti bacteriana, evitam
dermatites e outros problemas dermatológicos. Na medicina chinesa, as sementes são
consideradas afrodisíacas e a planta seca é prescrita como tonificante e usada em disfunções
menstruais (Bown, 1995). Folhas e flores são capazes de combater vômitos e diarréias
(Hoehne, 1939), além de serem eficientes contra resfriado, bronquite e reumatismo
(Castellucci et al., 2000).
O extrato metanólico de folhas, também, demonstrou atividade antiinflamatória
(Islam et al., 2005) e combateu células neoplásicas de glândulas mamárias (Nagasawa et al.,
1990), o que justifica diversos estudos etnofarmacológicos realizados com esta espécie no
Brasil (Souza et al., 2004).
Em relação à composição química, a literatura relata que L. sibiricus apresenta
alcalóides (Murakami, 1943), monoterpenos, sesquiterpenos e diterpenos (Savona et al., 1982;
Boalino et al., 2004; Almeida et al., 2005) e furano-lactonas (Satoh et al., 2003). Os extratos
polares apresentam majoritariamente rutina e seus derivados, além de flavonas metoxiladas
(Almeida et al., 2006).

30
Os fenilpropanóides estão presentes em L. heterophyllus principalmente na forma
de flavonóides glicosilados e recentemente foi isolado a quercetina-3-O-[3-(4-hidroxi-3,5-
dimetoxibenzil)-α-L-rhamnopiranosil]-(1-6)-β-D-galactopiranosideo, um composto
biglicosilado (Cong et al., 2005). Outros gêneros como L. cardíaca, L. persicus e L.
glaucescens também apresentam iridóides e flavonóides que conferem expressivo potencial
medicinal deste gênero (Karting et al., 1985; Tasdemir et al., 1999; Calis et al., 2000;
Milowska-Leyck et al., 2002). Dentro do grupo das substâncias químicas conhecidas como
fenilpropanóide, as flavonas metoxiladas são importantes marcadores quimiotaxonômicos
para o gênero (Giang et al., 2005).
Os flavonóides possuem importante papel como compostos de defesa e são
moléculas sinalizadoras da reprodução, patogênese e simbiose. Os flavonóides são produzidos
pelas plantas em grande escala e quando liberados para o ambiente têm evidente papel na
interação planta-planta e planta-microorganismo (Rice, 1984; Shirley, 1996). Muitos autores
mostram que o arranjo estrutural das moléculas de flavonóides é provavelmente responsável
pela atividade alelopática e esse arranjo está relacionado com a fitotoxidade destes compostos
(Macias et al., 1997; Bais et al., 2003; Parvez et al., 2004; Beninger & Hall, 2005).
Dada a importância medicinal do gênero, os objetivos foram estudar o efeito
alelopático dos extratos por infusão e metanólico; das frações flavonoídicas presentes no
extrato metanólico na germinação de sementes de Raphanus sativus, Lactuca sativa e
Lepidium sativum, além de identificar e testar as substâncias puras presentes nas frações
flavonoídicas ativas, na germinação de sementes de Lactuca sativa.
MATERIAL E MÉTODO
Material vegetal
Folhas de Leonurus sibiricus foram coletadas em Botucatu, Estado de São Paulo,
e identificada por Clemente José Campos, sendo a exsicata depositada no Herbário (BOTU)
do Departamento de Botânica, IB-UNESP com o número 12-706.
Preparo dos extratos
Os extratos foram preparados e analisados no Laboratório de Fitoquímica do
Departamento de Química Orgânica do Instituto de Química da UNESP de Araraquara.

31
Para a extração por infusão, 620g de folhas secas foram colocados em 5 litros de
água em ebulição por 5 minutos, filtrada em papel de filtro e após atingir temperatura
ambiente, a amostra foi submetida a liofilizador, sendo obtido 8,79g de extrato.
Para obtenção do extrato metanólico, 2300g de folhas secas foram submetidos a
extração com metanol (100%) à temperatura ambiente, por percolação durante 5 dias. O
solvente foi evaporado a 40˚C sob pressão reduzida em rotavapor, obtendo-se 389,63g de
extrato.
Quantificação dos compostos majoritários
Extrato por infusão: 1g de folhas secas, com três repetições, foi colocado em
50mL de água fervente por 5 minutos e filtrada em papel de filtro, após atingir temperatura
ambiente. Em seguida, a solução foi submetida a liofilizador, a quantidade de 20mg diluída
em 15mL de metanol: água (1:1), filtrada em filtro de membrana 0,22μm PFE (Millex) e
analisada diretamente em CLAE (Cromatografia Líquida de Alta Eficiência).
Extrato metanólico: Para a preparação de amostras utilizadas na quantificação de
substâncias químicas majoritárias, três repetições com 2g de folhas secas foram
acondicionados com 50mL de clorofórmio e submetidas a banho ultrassônico por 4 horas.
Após esse período foi filtrado e o solvente evaporado a 35˚C, sob pressão reduzida em
rotavapor. Em 1g de resíduo, resultante da primeira filtragem, foi adicionado 25mL de
metanol (100%), também em banho ultrassônico por 4 horas, obtendo-se o extrato seco após
evaporação deste solvente. A seguir, esse extrato foi colocado em cartucho (SPE) SEP PAK
C18 (360g). A sílica desses cartuchos foi ativada com 10mL de metanol e 20mL de água
deionizada, com graus de pureza CLAE. A quantidade de 20mg do extrato seco foi colocado
no cartucho de sílica C18 e a amostra adsorvida foi eluída com 15mL de metanol (100%) e
seca usando N2.
Para quantificar os flavonóides majoritários nos extratos, foi utilizada a curva de
calibração com padrão rutina nas concentrações de 10, 20, 50, 100, 200, 300 e 500 μg mL-1,
relacionando a área dos picos do cromatograma do CLAE com a concentração dos
flavonóides.
Fracionamento
Uma porção (5g) do extrato metanólico foi submetida a coluna de Sephadex LH-
20 de 1,2m x 0,06m com coletor automático, fluxo 250 gotas por minuto com metanol P.A.,
originando 201 frações que foram estudadas em Cromatografia de Camada Delgada (CCD)

32
em placas de sílica gel 20x20 com fase móvel 75:25 (Hexano: Acetato de Etila) e 80:18:2
(Clorofórmio:Metanol:Água) e reveladas com anisaldeído e NP-PEG. Em seguida, foram
reunidas as frações que apresentaram rf (fator de retenção) semelhantes de acordo com as
revelações químicas em CCD, originando 12 novas frações (fluxograma abaixo), sendo as
frações 2, 3 e 4 terpênicas. As frações majoritárias em flavonóides foram submetidas a novo
fracionamento por CLAE para isolamento das substâncias.
Fluxograma para obtenção do extrato metanólico, frações em Sephadex e substâncias puras.
Fr = Frações
* = Frações ativas e re-fracionadas em HPLC
A a G = Substâncias isoladas, onde A e B são flavonóides biglicosídeos; C e D flavonóides
monoglicosídeos e E, F e G agliconas.
Coluna de Sephadex LH-20 (4,6g)
Extrato Metanólico (389,63g)
Fr 12
174 - 201
Fr 11
160 - 173
Fr 10
135 - 159
Fr 9
129 - 134
Fr 8
123 - 128
Fr 7
104 - 122
Fr 6
78 - 103
Fr 5
62-77
Fr 4
39-61
Fr 3
28-38
Fr 2
16-27
Fr 1
1 - 15
* * * * * *
A (7mg) B (8mg)
A, B, C e D
C (10mg) D (6mg)
E (6mg)
F (9mg)
G (9mg)
Folhas secas de Leonurus sibiricus (2300g)

33
Característica do CLAE
Para fracionamento em cromatografia líquida de alta eficiência (CLAE), foi
utilizado filtro de membrana 0,22μm PFE (Millex) na preparação das amostras que foram
submetidas a HPLC Varian Pro-Star equipado com coluna de sílica em fase reversa (250mn x
460 mm i.d., 10μm, Phenomenex Luna). A fase móvel usada foi gradiente linear de aceto
nitrila : água com 28-35% (30min.), aumentando 35-70% (60min), 70-100% (65min) em
fluxo de 1,0mL min-1, e monitorado usando detector ultravioleta com fotodiodo Pro Star 330
em comprimento de onda de 254nm. Os flavonóides isolados foram identificados por análise
de Ressonância Magnética Nuclear (RMN) de 1H e 13C em equipamento Varian INOVA 300
e 500MHz, utilizando como solvente dimetil-sulfoxi deuterado.
Ensaios biológicos
Os extratos por infusão e metanólico, com base na literatura, foram diluídos nas
concentrações de 50, 100, 200, 400 e 800mg L-1; as 12 frações obtidas do Sephadex foram
diluídas na concentração de 1g L-1 e os flavonóides isolados nas concentrações de 10-4, 10-5,
10-6 e 10-7M. Para o controle do extrato por infusão empregou-se água destilada e para o
extrato metanólico água destilada com 0,01% de acetona. Para verificar o possível efeito
alelopático dos extratos e das frações flavonoídicas, foram utilizadas sementes de Raphanus
sativum, Lactuca sativa e Lepidium sativum e, para as substâncias puras isoladas, sementes de
Lactuca sativa por ser considerada espécie sensível e indicadora de atividade alelopática. As
sementes (10 de cada espécie) foram colocadas para germinar em condição de luz contínua
por 4 dias em placas de Petri, tendo como substrato papel de filtro umedecido com 5mL de
cada solução, sendo a mesma metodologia para extratos, frações e substâncias isoladas. Os
parâmetros avaliados foram porcentagem de germinação e crescimento de raiz (medida
direta), com delineamento inteiramente casualizado com 3 repetições. Os dados obtidos foram
submetidos a análise de variância e regressão polinomial.

34
RESULTADO E DISCUSSÃO
Os resultados indicaram que a extração por infusão e metanólica de folhas de
Leonurus sibiricus reduziram de forma marcante apenas a germinação das sementes de
Lactuca sativa, porém não afetaram o processo germinativo de Raphanus sativus e Lepidium
sativum (Tabela 1). Por outro lado, o crescimento inicial das raízes de Raphanus sativus
(Figura 1), Lactuca sativa (Figura 2) e Lepidium sativum (Figura 3) foi mais afetado pelo
extrato metanólico, principalmente, nas concentrações mais altas.
As plântulas de Raphanus sativus apresentaram maior redução no crescimento
inicial de raiz no extrato metanólico (Figura 1B) do que na extração por infusão (Figura 1A),
porém nas concentrações de 50 e 100mg L-1 do extrato metanólico observou-se um aumento
no comprimento de raiz (Figura 1B).
Em Lactuca sativa, as raízes foram afetadas tanto com extração por infusão
(Figura 2A) bem como por extração metanólica (Figura 2B), ocorrendo redução no
crescimento a partir da concentração de 50mg L-1, com maior inibição na concentração de
800mg L-1. Os resultados indicaram que L. sativa foi mais sensível aos compostos presentes
nos extratos utilizados, conforme citado na literatura (Macias et al., 1997; Ferreira & Áquila,
2000; Hoagland & Williams, 2004).
Para as sementes de Lepidium sativum, verificou-se que, a germinação não foi
afetada (Tabela 1), mas a extração por infusão (Figura 3A) e o extrato metanólico (Figura 3B)
acarretaram redução no crescimento inicial das raízes, nas concentrações de 400 e 800mg L-1.
No extrato metanólico, nas concentrações de 50 e 100mg L-1, o crescimento foi,
aproximadamente, 10mm maior em relação ao controle (Figura 3B).
As equações (Figuras 1, 2 e 3) com ajuste linear, obtidas para os experimentos
com extração por infusão, indicam que a atividade dos compostos químicos tem velocidade
constante de inibição, exceto em Lactuca sativa. Por outro lado, as equações de segundo grau
mostram um ponto de estímulo de crescimento, tendo aumentado a atividade inbitória em
concentrações anteriores e posteriores. Os resultados encontrados apresentaram tal
comportamento nas espécies Raphanus sativus e Lepidium sativum, no extrato metanólico,
mostrando pequena atividade estimulatória nas baixas concentrações e em seguida, atividade
inibitória nas concentrações mais elevadas. Para o crescimento inicial das raízes de Lactuca
sativa, espécie comumente conhecida como sensível aos compostos alelopáticos, o ajuste dos
dados observados mostrou função exponencial a qual indica atividade inibitória, tanto do
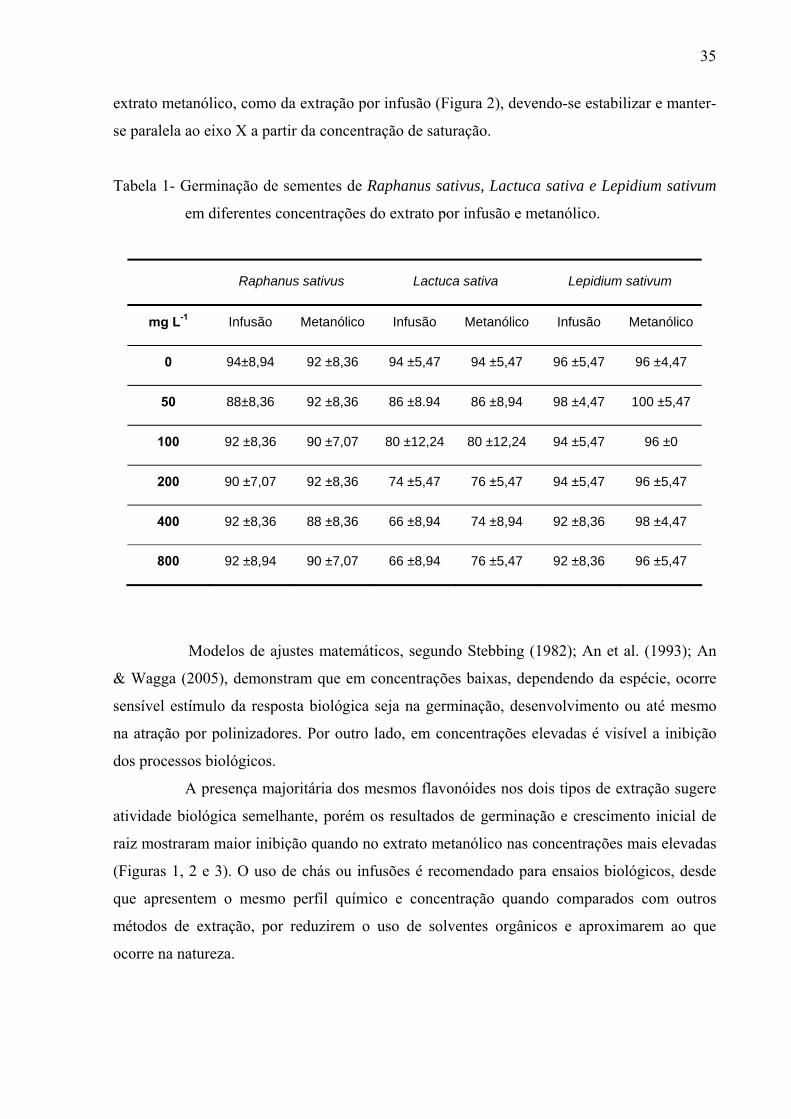
35
extrato metanólico, como da extração por infusão (Figura 2), devendo-se estabilizar e manter-
se paralela ao eixo X a partir da concentração de saturação.
Tabela 1- Germinação de sementes de Raphanus sativus, Lactuca sativa e Lepidium sativum
em diferentes concentrações do extrato por infusão e metanólico.
Raphanus sativus Lactuca sativa Lepidium sativum
mg L-1 Infusão Metanólico Infusão Metanólico Infusão Metanólico
0 94±8,94 92 ±8,36 94 ±5,47 94 ±5,47 96 ±5,47 96 ±4,47
50 88±8,36 92 ±8,36 86 ±8.94 86 ±8,94 98 ±4,47 100 ±5,47
100 92 ±8,36 90 ±7,07 80 ±12,24 80 ±12,24 94 ±5,47 96 ±0
200 90 ±7,07 92 ±8,36 74 ±5,47 76 ±5,47 94 ±5,47 96 ±5,47
400 92 ±8,36 88 ±8,36 66 ±8,94 74 ±8,94 92 ±8,36 98 ±4,47
800 92 ±8,94 90 ±7,07 66 ±8,94 76 ±5,47 92 ±8,36 96 ±5,47
Modelos de ajustes matemáticos, segundo Stebbing (1982); An et al. (1993); An
& Wagga (2005), demonstram que em concentrações baixas, dependendo da espécie, ocorre
sensível estímulo da resposta biológica seja na germinação, desenvolvimento ou até mesmo
na atração por polinizadores. Por outro lado, em concentrações elevadas é visível a inibição
dos processos biológicos.
A presença majoritária dos mesmos flavonóides nos dois tipos de extração sugere
atividade biológica semelhante, porém os resultados de germinação e crescimento inicial de
raiz mostraram maior inibição quando no extrato metanólico nas concentrações mais elevadas
(Figuras 1, 2 e 3). O uso de chás ou infusões é recomendado para ensaios biológicos, desde
que apresentem o mesmo perfil químico e concentração quando comparados com outros
métodos de extração, por reduzirem o uso de solventes orgânicos e aproximarem ao que
ocorre na natureza.
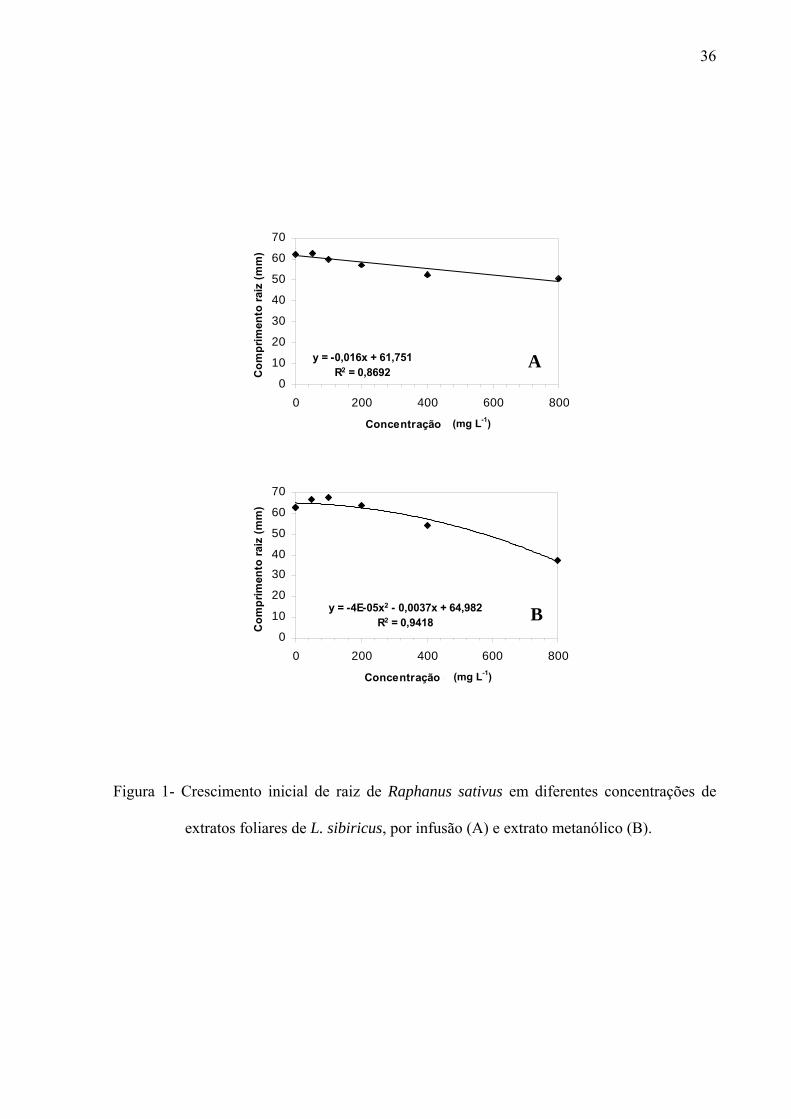
36
Figura 1- Crescimento inicial de raiz de Raphanus sativus em diferentes concentrações de
extratos foliares de L. sibiricus, por infusão (A) e extrato metanólico (B).
(mg L-1)
y = -0,016x + 61,751R2 = 0,8692
0
10
20
30
40
50
60
70
0 200 400 600 800
Concentração (mg L-1)
Com
prim
ento
raiz
(mm
)
y = -4E-05x2 - 0,0037x + 64,982R2 = 0,9418
0
10
20
30
40
50
60
70
0 200 400 600 800
Concentração (mg L-1)
Com
prim
ento
raiz
(mm
)
(mg L-1)
(mg L-1)
A
B
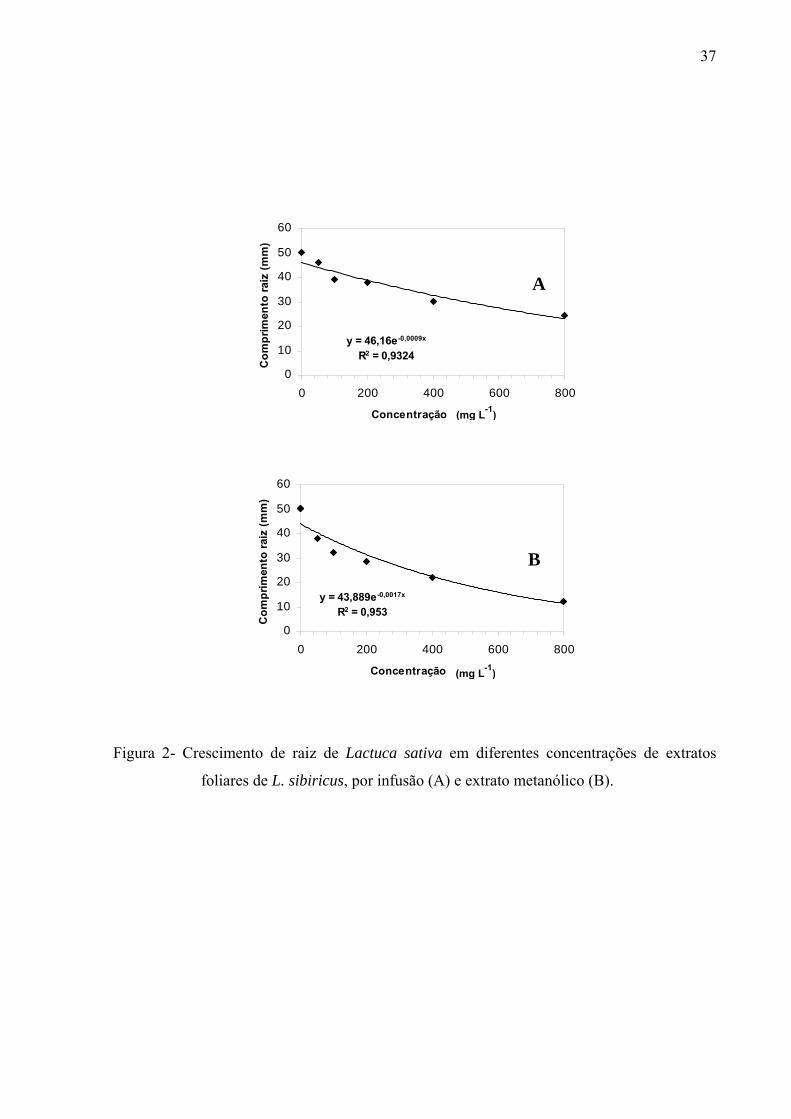
37
Figura 2- Crescimento de raiz de Lactuca sativa em diferentes concentrações de extratos
foliares de L. sibiricus, por infusão (A) e extrato metanólico (B).
y = 46,16e-0,0009x
R2 = 0,93240
10
20
30
40
50
60
0 200 400 600 800
Concentração (mg L-1)
Com
prim
ento
raiz
(mm
)
y = 43,889e-0,0017x
R2 = 0,9530
10
20
30
40
50
60
0 200 400 600 800
Concentração (mg L-1)
Com
prim
ento
raiz
(mm
)
B
A
(mg L-1)
(mg L-1)
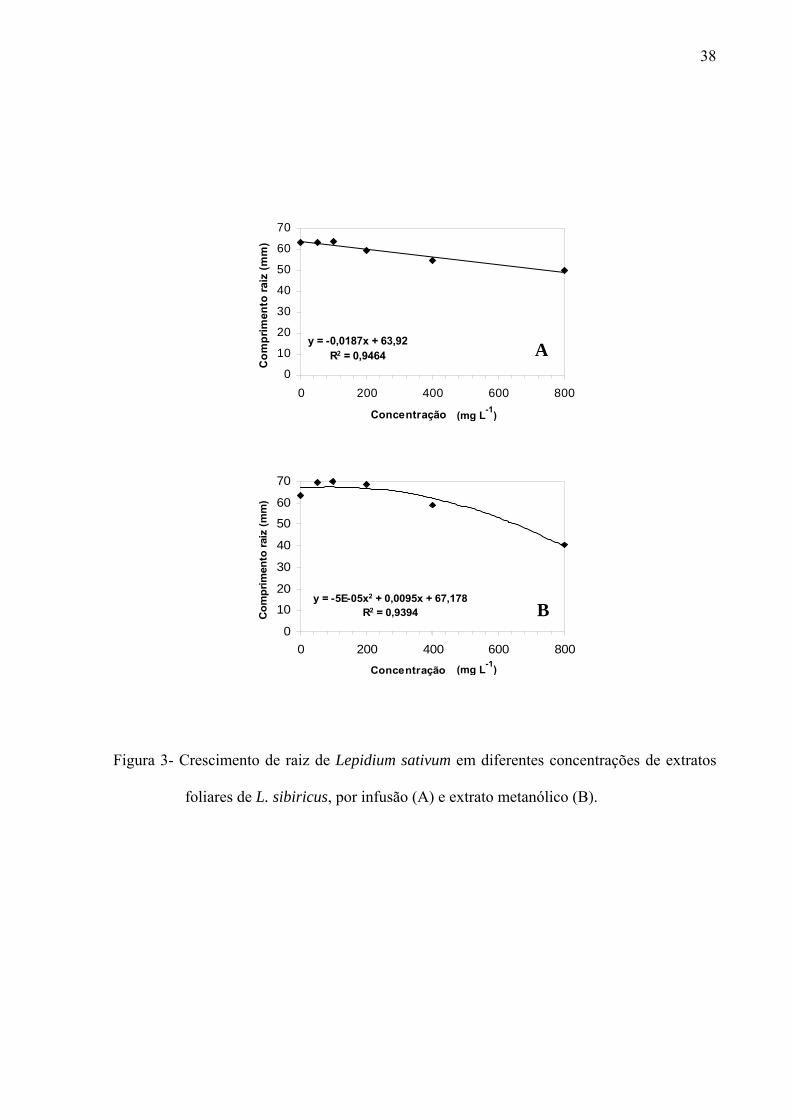
38
Figura 3- Crescimento de raiz de Lepidium sativum em diferentes concentrações de extratos
foliares de L. sibiricus, por infusão (A) e extrato metanólico (B).
y = -0,0187x + 63,92R2 = 0,9464
0
10
20
30
40
50
60
70
0 200 400 600 800
Concentração (mg L-1)
Com
prim
ento
raiz
(mm
)
y = -5E-05x2 + 0,0095x + 67,178R2 = 0,9394
0
10
20
30
40
50
60
70
0 200 400 600 800Concentração (mg L-1)
Com
prim
ento
raiz
(mm
)
(mg L-1)
A
B
(mg L-1)

39
TABELA 2- Quantificação dos flavonóides majoritários da extração por infusão e do extrato
metanólico (± desvio padrão; valores expressos em mg g-1 de folhas secas).
Quercetina 3-O-β-D-galactopiranosil
(1-6)-O-α-L-rhamnopiranosideo
(A)
Quercetina 3-O-β-D- glucopiranosil
(1-6)-O-α-L-rhamnopiranosideo
(B)
Quercetina 3-O-β-D-galactopiranosil
(C)
Quercetina 3-O-β-D- glucopiranosil
(D)
Infusão 4,97 ±0,11 3,47 ±0,30 3,53 ±0,24 4,27 ±0,45
Metanólico 14,72 ±2,86 11,90 ±2,27 13,39 ±2,54 14,71 ±2,78
Os resultados encontrados (Figura 4) mostram que tanto a água quente (infusão)
quanto o metanol (solventes com alta polaridade) são capazes de extrair as mesmas
substâncias majoritárias, mas com diferentes concentrações, que podem acarretar diferentes
níveis de inibição e\ou promoção de crescimento (Macias et al., 1997). Da mesma maneira, as
frações hexânicas e acetato de etila de extratos foliares de Lactuca sativa, acarretaram maior
redução na germinação e crescimento de raiz de Raphanus sativus, quando comparadas com
frações butanólicas e aquosas (Chon et al., 2005). Esses resultados corroboram com os
encontrados, pois sugerem que as substâncias, dependendo do solvente, estão presentes em
diferentes concentrações ativas.
Portanto, as atividades biológicas, além de variarem com a espécie vegetal são
decorrentes da concentração das substâncias encontradas e da variação de esqueletos
carbônicos ativos de flavonóides, que estão relacionados com a atividade alelopática (Figura
5).
As frações consideradas flavonoídicas (5, 6, 7, 10, 11 e 12) mostraram atividade,
principalmente, no crescimento inicial de raiz das espécies estudadas, sendo que a
germinação, da mesma forma quando utilizados extratos (Tabela 1), foi pouco afetada na
maioria das concentrações, nas sementes de Raphanus sativus e Lepidium sativum, embora L.
sativum, nas frações 3 e 4, apresentaram 100% de inibição de germinação (Tabela 4). No
entanto, a germinação de sementes de Lactuca sativa foi afetada mostrando inibição maior
que 50%, nas frações 3, 4, 6, 10, 11 e 12.
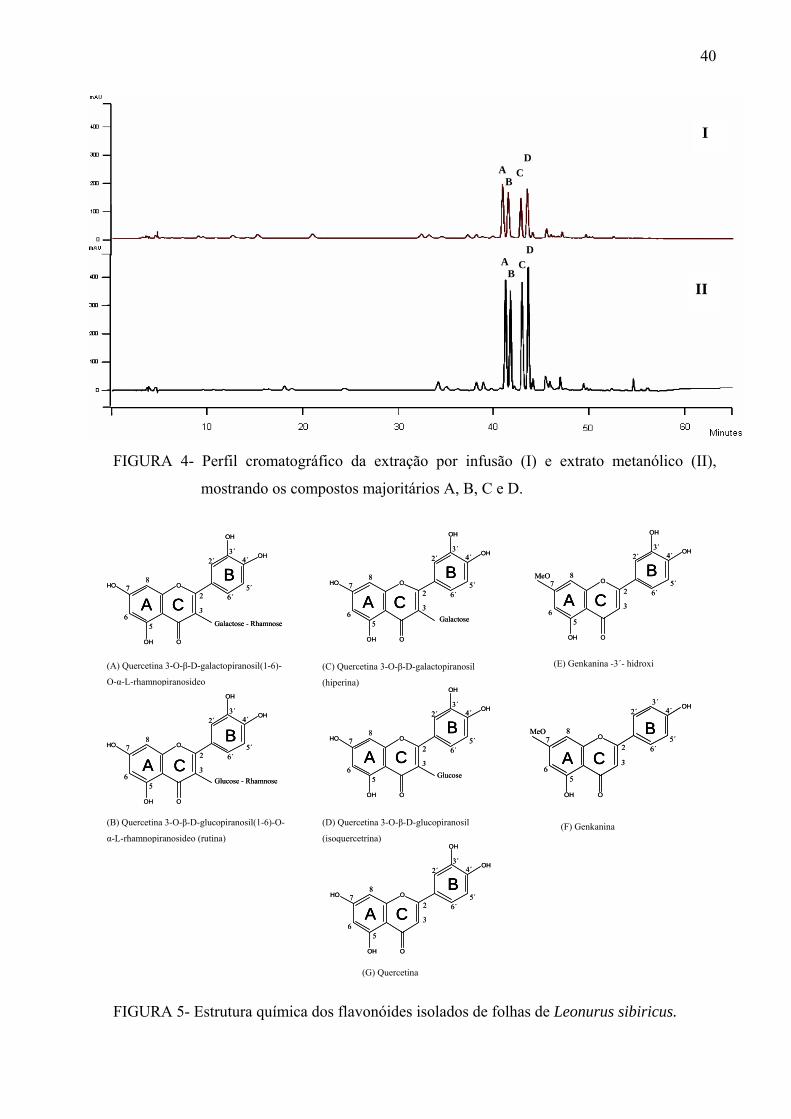
40
FIGURA 4- Perfil cromatográfico da extração por infusão (I) e extrato metanólico (II),
mostrando os compostos majoritários A, B, C e D.
FIGURA 5- Estrutura química dos flavonóides isolados de folhas de Leonurus sibiricus.
I
II
A D
B C
A D
B C
O
O
2´3´
4´
5´6´
3
2
56
78
OH
HO
OH
OH
Galactose - Rhamnose
A C
BO
O
2´3´
4´
5´6´
3
2
56
78
OH
HO
OH
OH
Galactose - Rhamnose
A C
B
A C
B
(A) Quercetina 3-O-β-D-galactopiranosil(1-6)-
O-α-L-rhamnopiranosideo
O
O
2´3´
4´
5´6´
3
2
56
78
OH
HO
OH
OH
Galactose
A C
BO
O
2´3´
4´
5´6´
3
2
56
78
OH
HO
OH
OH
Galactose
A C
B
A C
B
(C) Quercetina 3-O-β-D-galactopiranosil
(hiperina)
O
O
2´3´
4´
5´6´
3
2
56
78
OH
OH
OH
MeO
A C
BO
O
2´3´
4´
5´6´
3
2
56
78
OH
OH
OH
MeO
A C
B
A C
B
(E) Genkanina -3´- hidroxi
O
O
2´3´
4´
5´6´
3
2
56
78
OH
OH
MeO
A C
BO
O
2´3´
4´
5´6´
3
2
56
78
OH
OH
MeO
A C
B
A C
BO
O
2´3´
4´
5´6´
3
2
56
78
OH
HO
OH
OH
Glucose
A C
BO
O
2´3´
4´
5´6´
3
2
56
78
OH
HO
OH
OH
Glucose
A C
B
A C
BO
O
2´3´
4´
5´6´
3
2
56
78
OH
HO
OH
OH
Glucose - Rhamnose
A C
BO
O
2´3´
4´
5´6´
3
2
56
78
OH
HO
OH
OH
Glucose - Rhamnose
A C
B
A C
B
(B) Quercetina 3-O-β-D-glucopiranosil(1-6)-O-
α-L-rhamnopiranosideo (rutina)
(D) Quercetina 3-O-β-D-glucopiranosil
(isoquercetrina)(F) Genkanina
O
O
2´3´
4´
5´6´
3
2
56
78
OH
HO
OH
OH
A C
BO
O
2´3´
4´
5´6´
3
2
56
78
OH
HO
OH
OH
A C
B
A C
B
(G) Quercetina
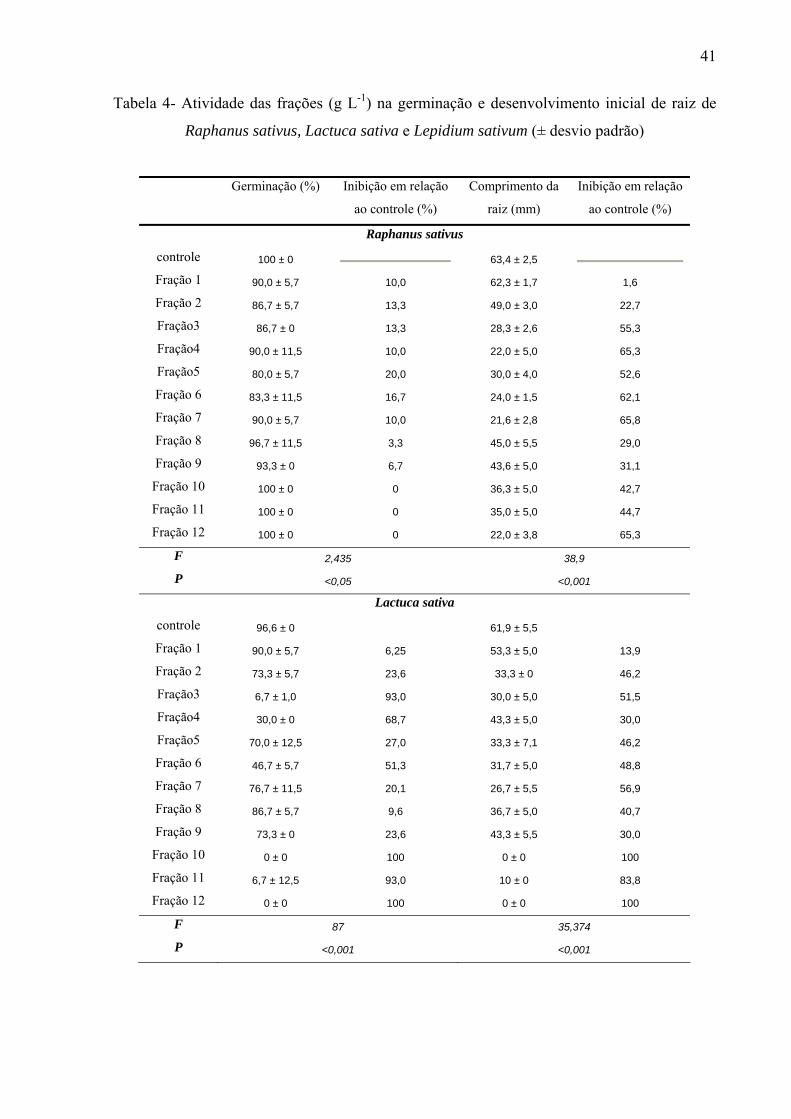
41
Tabela 4- Atividade das frações (g L-1) na germinação e desenvolvimento inicial de raiz de
Raphanus sativus, Lactuca sativa e Lepidium sativum (± desvio padrão)
Germinação (%) Inibição em relação
ao controle (%)
Comprimento da
raiz (mm)
Inibição em relação
ao controle (%)
Raphanus sativus
controle 100 ± 0 63,4 ± 2,5
Fração 1 90,0 ± 5,7 10,0 62,3 ± 1,7 1,6
Fração 2 86,7 ± 5,7 13,3 49,0 ± 3,0 22,7
Fração3 86,7 ± 0 13,3 28,3 ± 2,6 55,3
Fração4 90,0 ± 11,5 10,0 22,0 ± 5,0 65,3
Fração5 80,0 ± 5,7 20,0 30,0 ± 4,0 52,6
Fração 6 83,3 ± 11,5 16,7 24,0 ± 1,5 62,1
Fração 7 90,0 ± 5,7 10,0 21,6 ± 2,8 65,8
Fração 8 96,7 ± 11,5 3,3 45,0 ± 5,5 29,0
Fração 9 93,3 ± 0 6,7 43,6 ± 5,0 31,1
Fração 10 100 ± 0 0 36,3 ± 5,0 42,7
Fração 11 100 ± 0 0 35,0 ± 5,0 44,7
Fração 12 100 ± 0 0 22,0 ± 3,8 65,3
F 2,435 38,9
P <0,05 <0,001
Lactuca sativa
controle 96,6 ± 0 61,9 ± 5,5
Fração 1 90,0 ± 5,7 6,25 53,3 ± 5,0 13,9
Fração 2 73,3 ± 5,7 23,6 33,3 ± 0 46,2
Fração3 6,7 ± 1,0 93,0 30,0 ± 5,0 51,5
Fração4 30,0 ± 0 68,7 43,3 ± 5,0 30,0
Fração5 70,0 ± 12,5 27,0 33,3 ± 7,1 46,2
Fração 6 46,7 ± 5,7 51,3 31,7 ± 5,0 48,8
Fração 7 76,7 ± 11,5 20,1 26,7 ± 5,5 56,9
Fração 8 86,7 ± 5,7 9,6 36,7 ± 5,0 40,7
Fração 9 73,3 ± 0 23,6 43,3 ± 5,5 30,0
Fração 10 0 ± 0 100 0 ± 0 100
Fração 11 6,7 ± 12,5 93,0 10 ± 0 83,8
Fração 12 0 ± 0 100 0 ± 0 100
F 87 35,374 P <0,001 <0,001
42
Tabela 4- continuação
Lepidium sativum
controle 100 ± 0 64,0 ± 2,5 Fração 1 100 ± 5,7 0 62,0 ± 5,7 3,0
Fração 2 70,0 ± 0 26,7 56,0 ± 5,7 12,3
Fração3 0 ± 0 100 0 ± 0 100
Fração4 0 ± 0 100 0 ± 0 100
Fração5 100 ± 0 0 33,1 ± 2,8 48,4
Fração 6 100 ± 5,7 0 31,9 ± 1,1 51,5
Fração 7 96,7 ± 5,7 3,3 30,8 ± 5,7 53,5
Fração 8 100 ± 0 0 36,4 ± 5,0 42,2
Fração 9 100 ± 0 0 53,0 ± 5,7 17,4
Fração 10 100 ± 0 0 26,9 ± 2,8 58,7
Fração 11 100 ± 0 0 22,1 ± 2,0 65,9
Fração 12 100 ± 0 0 38,0 ± 2,8 40,6
F 800,864 85,924 P <0,001 <0,001
Apesar das frações 3 e 4 mostrarem forte atividade alelopática em Raphanus
sativus, Lactuca sativa e Lepidium sativum, principalmente na inibição de crescimento inicial
de raiz, não foram re-fracionadas por apresentarem composição majoritária de terpenos
(dados não mostrados) e não de flavonóides.
As frações flavonoídicas 5 a 7 e 10 a 12 foram re-fracionadas em CLAE e isoladas
sete substâncias (fluxograma e Figura 5), sendo duas biglicosídeos (A e B) nas frações 5 e 6;
duas substâncias com apenas uma molécula de açúcar (C e D) no esqueleto flavonoídico das
frações 6 e 7; duas agliconas metoxiladas (E e F) das frações 10 e 11, além da quercetina (G)
que foi isolada da fração 12. Metodologias de fracionamento auxiliam no estudo guiado de
substâncias ativas e o estudo das frações de Sephadex pode auxiliar no isolamento e
determinação dos aleloquímicos (Hoagland & Williams, 2004), bem como suas atividades
específicas, sendo possível verificar a sensibilidade e seletividade destes compostos. Da
mesma maneira, pesquisas realizadas com Ailanthus altissima determinaram o aleloquímico
alcaloídico ailantona, a partir do estudo prévio das frações do Sephadex do extrato butanólico
e respectivas atividades biológicas (De Feo et al., 2004). Como realizadas neste trabalho,
outras técnicas de cromatografia também foram usadas para verificar o perfil e

43
comportamento cromatográfico dos flavonóides, como por exemplo a CCD, com fases
móveis de diferentes polaridades (Onyikagha & Grotewold, 2004).
Tendo em vista a atividade biológica das frações do Sephadex ter sido mais
evidente nas sementes de Lactuca sativa (Tabela 4), após o re-fracionamento as substâncias
isoladas foram testadas somente em alface, buscando relacionar a estrutura da molécula e
atividade biológica.
Pela Tabela 5, verifica-se que os flavonóides acarretaram maior redução da
porcentagem de germinação de sementes de L. sativa, em relação ao controle, no primeiro dia
nas concentrações mais elevadas. No entanto, a porcentagem final de germinação, com 4 dias,
não foi alterada, em relação ao terceiro dia, independente da concentração e estrutura dos
flavonóides. Portanto, a maior atividade alelopática foi verificada no primeiro e segundo dias,
com a seguinte ordem decrescente genkwanina-3´hidroxi (E) > quercetina (G) > quercetina-3-
O-glucose (D) > quercetina-3-O-glucose-ramnose (B) > genkwanina (F) > quercetina-3-O-
galactose (C) > quercetina-3-O-galactose-ramnose (A).
A genkwanina-3´hidroxi (E) e quercetina (G) apresentaram maior atividade
herbicida na germinação, na concentração de 10-4M, a qual devido a estrutura química, com
grupos catecol nos anéis A, B e C e na posição orto do anel B, respectivamente, podem ter
alterado a permeabilidade das membranas celulares e afetado o alongamento radicular,
necessário a protusão da raiz (Martinez-Flores et al., 2002; Einhellig, 2004; Glass & Dunlop,
1974). Os flavonóides, quercetina-3-O-glucose (D) e quercetina-3-O-glucose-ramnose (B)
reduziram a germinação, devido talvez a presença dos açúcares glucose e glucose-ramnose em
relação as substâncias sem moléculas de açúcar. Por outro lado, a quercetina-3-O-galactose
(C) e quercetina-3-O-galactose-ramnose (A) não afetaram a germinação de forma tão
marcante quanto as substâncias que apresentavam molécula de glucose na estrutura (B e D).
Os resultados encontrados, nos primeiros dias, indicaram que provavelmente os
flavonóides tenham afetado os mecanismos de alongamento celular (Hoagland & Williams,
2004). Enquanto, no final do experimento, possível processo de detoxificação celular, tenha
ocorrido impedindo o efeito de substâncias inibitórias através de ativação de enzimas do
metabolismo oxidativo.
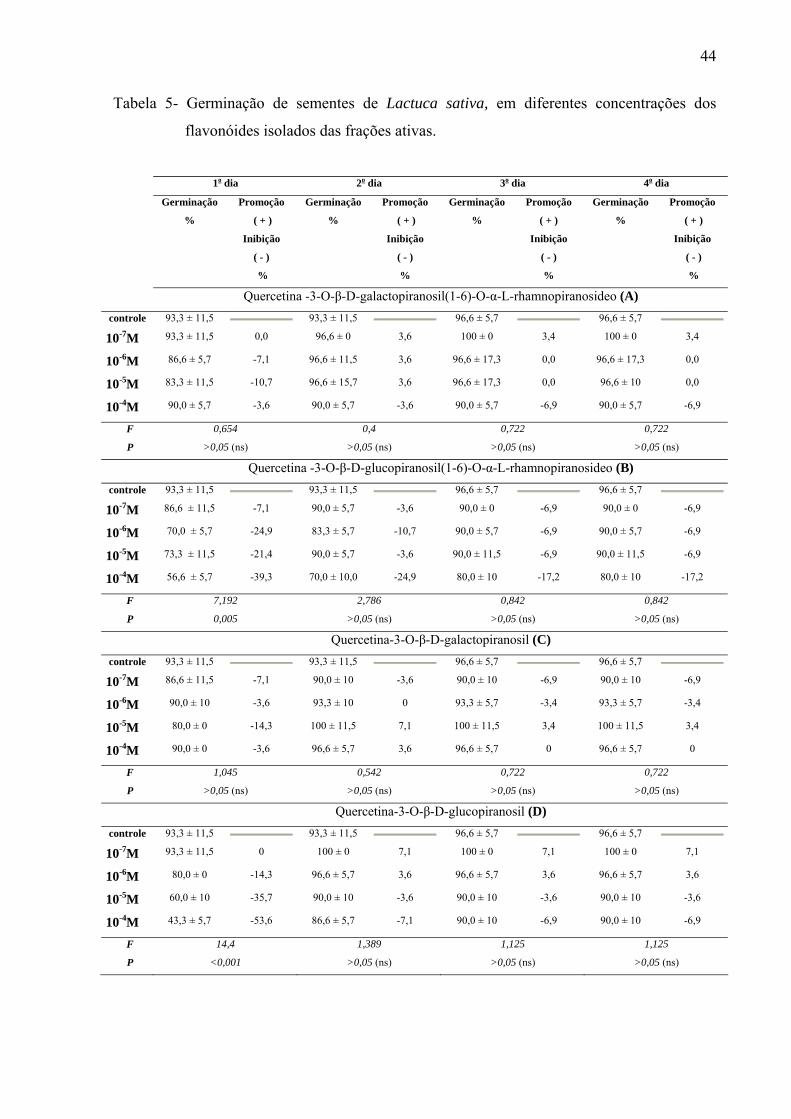
44
Tabela 5- Germinação de sementes de Lactuca sativa, em diferentes concentrações dos
flavonóides isolados das frações ativas.
1o dia 2o dia 3o dia 4o dia
Germinação
%
Promoção
( + )
Inibição
( - )
%
Germinação
%
Promoção
( + )
Inibição
( - )
%
Germinação
%
Promoção
( + )
Inibição
( - )
%
Germinação
%
Promoção
( + )
Inibição
( - )
%
Quercetina -3-O-β-D-galactopiranosil(1-6)-O-α-L-rhamnopiranosideo (A) controle 93,3 ± 11,5 93,3 ± 11,5 96,6 ± 5,7 96,6 ± 5,7
10-7M 93,3 ± 11,5 0,0 96,6 ± 0 3,6 100 ± 0 3,4 100 ± 0 3,4
10-6M 86,6 ± 5,7 -7,1 96,6 ± 11,5 3,6 96,6 ± 17,3 0,0 96,6 ± 17,3 0,0
10-5M 83,3 ± 11,5 -10,7 96,6 ± 15,7 3,6 96,6 ± 17,3 0,0 96,6 ± 10 0,0
10-4M 90,0 ± 5,7 -3,6 90,0 ± 5,7 -3,6 90,0 ± 5,7 -6,9 90,0 ± 5,7 -6,9
F 0,654 0,4 0,722 0,722
P >0,05 (ns) >0,05 (ns) >0,05 (ns) >0,05 (ns)
Quercetina -3-O-β-D-glucopiranosil(1-6)-O-α-L-rhamnopiranosideo (B) controle 93,3 ± 11,5 93,3 ± 11,5 96,6 ± 5,7 96,6 ± 5,7
10-7M 86,6 ± 11,5 -7,1 90,0 ± 5,7 -3,6 90,0 ± 0 -6,9 90,0 ± 0 -6,9
10-6M 70,0 ± 5,7 -24,9 83,3 ± 5,7 -10,7 90,0 ± 5,7 -6,9 90,0 ± 5,7 -6,9
10-5M 73,3 ± 11,5 -21,4 90,0 ± 5,7 -3,6 90,0 ± 11,5 -6,9 90,0 ± 11,5 -6,9
10-4M 56,6 ± 5,7 -39,3 70,0 ± 10,0 -24,9 80,0 ± 10 -17,2 80,0 ± 10 -17,2
F 7,192 2,786 0,842 0,842
P 0,005 >0,05 (ns) >0,05 (ns) >0,05 (ns)
Quercetina-3-O-β-D-galactopiranosil (C) controle 93,3 ± 11,5 93,3 ± 11,5 96,6 ± 5,7 96,6 ± 5,7
10-7M 86,6 ± 11,5 -7,1 90,0 ± 10 -3,6 90,0 ± 10 -6,9 90,0 ± 10 -6,9
10-6M 90,0 ± 10 -3,6 93,3 ± 10 0 93,3 ± 5,7 -3,4 93,3 ± 5,7 -3,4
10-5M 80,0 ± 0 -14,3 100 ± 11,5 7,1 100 ± 11,5 3,4 100 ± 11,5 3,4
10-4M 90,0 ± 0 -3,6 96,6 ± 5,7 3,6 96,6 ± 5,7 0 96,6 ± 5,7 0
F 1,045 0,542 0,722 0,722
P >0,05 (ns) >0,05 (ns) >0,05 (ns) >0,05 (ns)
Quercetina-3-O-β-D-glucopiranosil (D) controle 93,3 ± 11,5 93,3 ± 11,5 96,6 ± 5,7 96,6 ± 5,7
10-7M 93,3 ± 11,5 0 100 ± 0 7,1 100 ± 0 7,1 100 ± 0 7,1
10-6M 80,0 ± 0 -14,3 96,6 ± 5,7 3,6 96,6 ± 5,7 3,6 96,6 ± 5,7 3,6
10-5M 60,0 ± 10 -35,7 90,0 ± 10 -3,6 90,0 ± 10 -3,6 90,0 ± 10 -3,6
10-4M 43,3 ± 5,7 -53,6 86,6 ± 5,7 -7,1 90,0 ± 10 -6,9 90,0 ± 10 -6,9
F 14,4 1,389 1,125 1,125
P <0,001 >0,05 (ns) >0,05 (ns) >0,05 (ns)
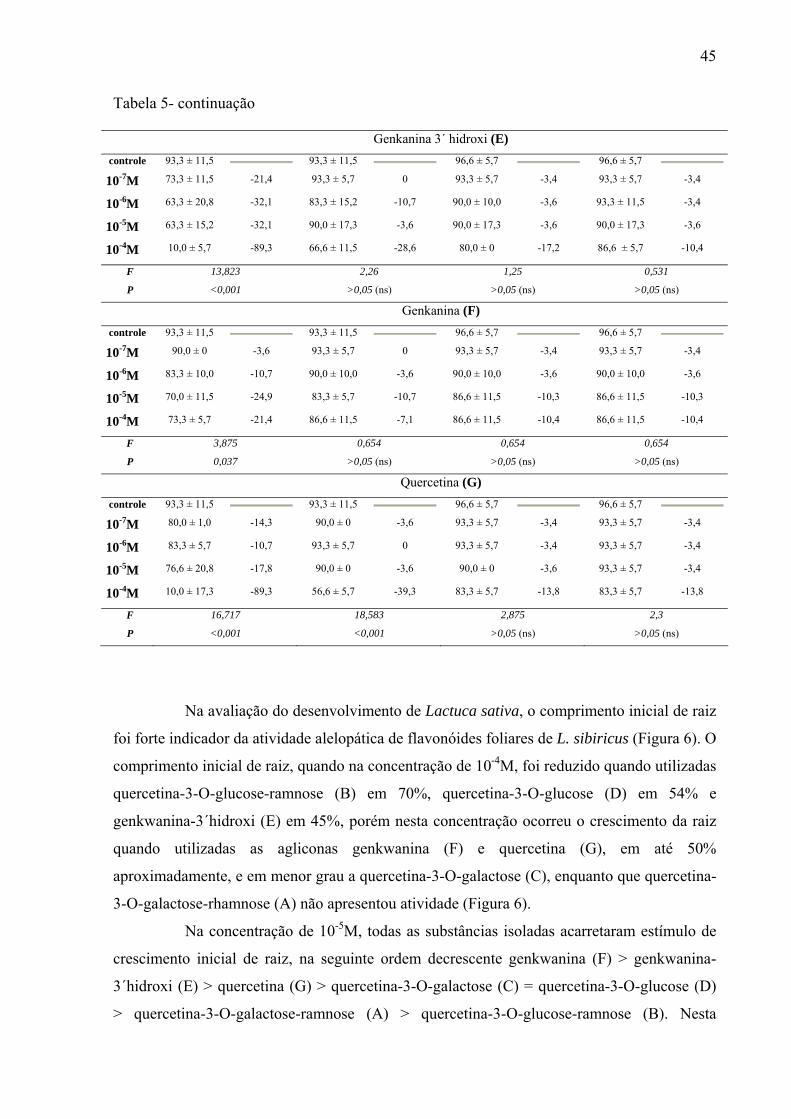
45
Tabela 5- continuação
Genkanina 3´ hidroxi (E) controle 93,3 ± 11,5 93,3 ± 11,5 96,6 ± 5,7 96,6 ± 5,7
10-7M 73,3 ± 11,5 -21,4 93,3 ± 5,7 0 93,3 ± 5,7 -3,4 93,3 ± 5,7 -3,4
10-6M 63,3 ± 20,8 -32,1 83,3 ± 15,2 -10,7 90,0 ± 10,0 -3,6 93,3 ± 11,5 -3,4
10-5M 63,3 ± 15,2 -32,1 90,0 ± 17,3 -3,6 90,0 ± 17,3 -3,6 90,0 ± 17,3 -3,6
10-4M 10,0 ± 5,7 -89,3 66,6 ± 11,5 -28,6 80,0 ± 0 -17,2 86,6 ± 5,7 -10,4
F 13,823 2,26 1,25 0,531
P <0,001 >0,05 (ns) >0,05 (ns) >0,05 (ns)
Genkanina (F) controle 93,3 ± 11,5 93,3 ± 11,5 96,6 ± 5,7 96,6 ± 5,7
10-7M 90,0 ± 0 -3,6 93,3 ± 5,7 0 93,3 ± 5,7 -3,4 93,3 ± 5,7 -3,4
10-6M 83,3 ± 10,0 -10,7 90,0 ± 10,0 -3,6 90,0 ± 10,0 -3,6 90,0 ± 10,0 -3,6
10-5M 70,0 ± 11,5 -24,9 83,3 ± 5,7 -10,7 86,6 ± 11,5 -10,3 86,6 ± 11,5 -10,3
10-4M 73,3 ± 5,7 -21,4 86,6 ± 11,5 -7,1 86,6 ± 11,5 -10,4 86,6 ± 11,5 -10,4
F 3,875 0,654 0,654 0,654
P 0,037 >0,05 (ns) >0,05 (ns) >0,05 (ns)
Quercetina (G) controle 93,3 ± 11,5 93,3 ± 11,5 96,6 ± 5,7 96,6 ± 5,7
10-7M 80,0 ± 1,0 -14,3 90,0 ± 0 -3,6 93,3 ± 5,7 -3,4 93,3 ± 5,7 -3,4
10-6M 83,3 ± 5,7 -10,7 93,3 ± 5,7 0 93,3 ± 5,7 -3,4 93,3 ± 5,7 -3,4
10-5M 76,6 ± 20,8 -17,8 90,0 ± 0 -3,6 90,0 ± 0 -3,6 93,3 ± 5,7 -3,4
10-4M 10,0 ± 17,3 -89,3 56,6 ± 5,7 -39,3 83,3 ± 5,7 -13,8 83,3 ± 5,7 -13,8
F 16,717 18,583 2,875 2,3
P <0,001 <0,001 >0,05 (ns) >0,05 (ns)
Na avaliação do desenvolvimento de Lactuca sativa, o comprimento inicial de raiz
foi forte indicador da atividade alelopática de flavonóides foliares de L. sibiricus (Figura 6). O
comprimento inicial de raiz, quando na concentração de 10-4M, foi reduzido quando utilizadas
quercetina-3-O-glucose-ramnose (B) em 70%, quercetina-3-O-glucose (D) em 54% e
genkwanina-3´hidroxi (E) em 45%, porém nesta concentração ocorreu o crescimento da raiz
quando utilizadas as agliconas genkwanina (F) e quercetina (G), em até 50%
aproximadamente, e em menor grau a quercetina-3-O-galactose (C), enquanto que quercetina-
3-O-galactose-rhamnose (A) não apresentou atividade (Figura 6).
Na concentração de 10-5M, todas as substâncias isoladas acarretaram estímulo de
crescimento inicial de raiz, na seguinte ordem decrescente genkwanina (F) > genkwanina-
3´hidroxi (E) > quercetina (G) > quercetina-3-O-galactose (C) = quercetina-3-O-glucose (D)
> quercetina-3-O-galactose-ramnose (A) > quercetina-3-O-glucose-ramnose (B). Nesta

46
concentração, as agliconas (E, F e G) podem promover o crescimento de raiz mais
efetivamente que os flavonóides glicosilados e dentre estes, talvez, aqueles que apresentam
apenas uma molécula de açúcar (galactose ou glucose) nos esqueletos carbônicos tem maior
atividade.
Atividade semelhante à concentração anterior foi encontrada para 10-6M, com
exceção da ordem decrescente de atividade no crescimento, onde quercetina-3-O-glucose (D)
> genkwanina-3´hidroxi (E) > genkwanina (F) > quercetina-3-O-galactose (C) > quercetina
(G) > quercetina-3-O-galactose-ramnose (A) > quercetina-3-O-glucose-ramnose (B). Nas
concentrações utilizadas, os biglicosídeos (A e B) apresentaram baixa atividade no
crescimento das raízes, quando comparados com as demais substâncias.
As agliconas (F e G) apresentaram menor atividade de promoção de crescimento
na concentração de 10-7M (Figura 6), com seqüência decrescente de quercetina-3-O-galactose
(C) > quercetina-3-O-glucose (D) = genkwanina-3´hidroxi (E) > quercetina-3-O-glucose-
ramnose (B) > quercetina-3-O-galactose-ramnose (A) > genkwanina (F) > quercetina (G).
Comportamento inibitório constante foi verificado nas frações flavonoídicas
(Tabela 4) devido, talvez, as maiores concentrações e efeito sinergístico, quando comparado
com os flavonóides isolados (Tabela 5 e Figura 6), porém quando em menores concentrações
(Figura 6), promoveram o crescimento inicial da raiz (Macias et al., 1997; Hoagland &
Williams, 2004). Esse crescimento, provavelmente, também seja resposta do contato direto
dos extratos, frações e flavonóides com as raízes, por isso mais efetivos neste órgão vegetal
(Nishida et al., 2005).
A quercetina-3-O-glucose-ramnose (B) e o derivado quercetina-3-O-glucose (D)
foram ativos, tanto na germinação (Tabela 5) quanto no crescimento inicial de raiz de
plântulas de L. sativa (Figura 6), inibindo esses processos fisiológicos na maior concentração
testada (10-4M). Resultados semelhantes foram descritos por Basile et al. (2000) e Parvez et
al. (2004) na germinação e crescimento inicial de Raphanus sativus e Arabidospsis thaliana,
respectivamente. Desta forma, os flavonóides glicosilados são conhecidos compostos
alelopáticos e os encontrados em L. sibiricus, também acarretaram estímulo do crescimento
de raízes quando testados isoladamente, nas concentrações de 10-7 a 10-5M (Figura 6).
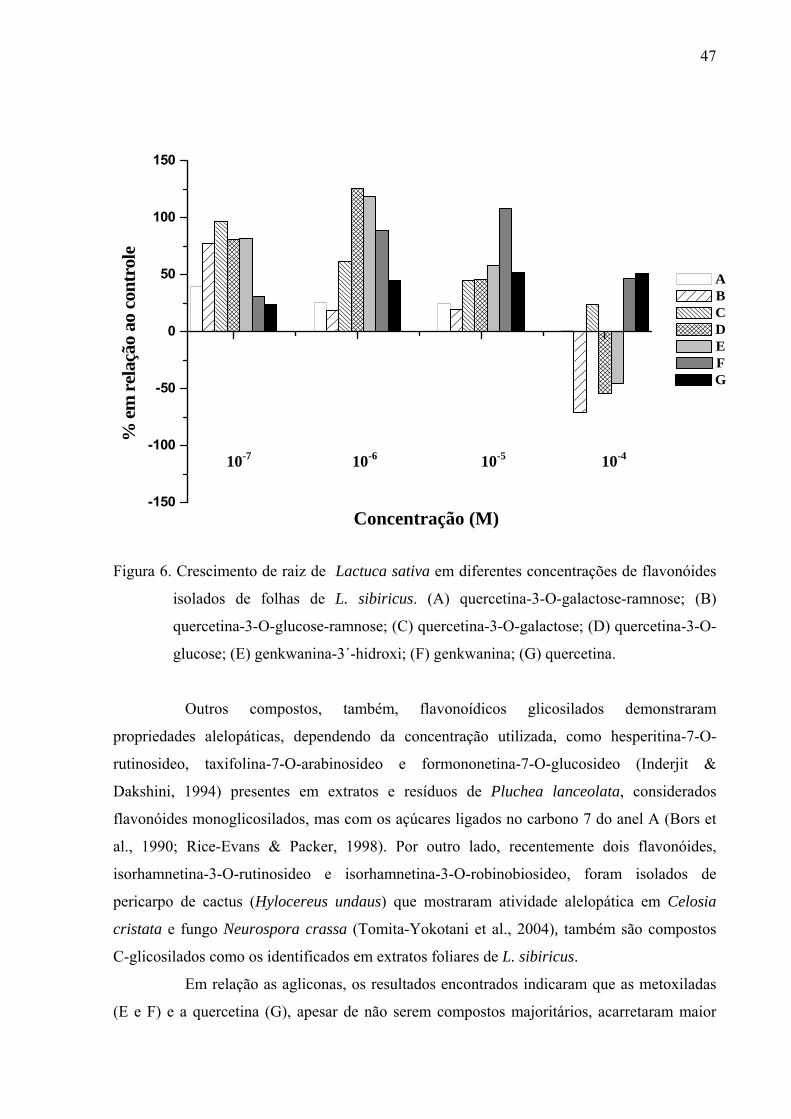
47
Figura 6. Crescimento de raiz de Lactuca sativa em diferentes concentrações de flavonóides
isolados de folhas de L. sibiricus. (A) quercetina-3-O-galactose-ramnose; (B)
quercetina-3-O-glucose-ramnose; (C) quercetina-3-O-galactose; (D) quercetina-3-O-
glucose; (E) genkwanina-3´-hidroxi; (F) genkwanina; (G) quercetina.
Outros compostos, também, flavonoídicos glicosilados demonstraram
propriedades alelopáticas, dependendo da concentração utilizada, como hesperitina-7-O-
rutinosideo, taxifolina-7-O-arabinosideo e formononetina-7-O-glucosideo (Inderjit &
Dakshini, 1994) presentes em extratos e resíduos de Pluchea lanceolata, considerados
flavonóides monoglicosilados, mas com os açúcares ligados no carbono 7 do anel A (Bors et
al., 1990; Rice-Evans & Packer, 1998). Por outro lado, recentemente dois flavonóides,
isorhamnetina-3-O-rutinosideo e isorhamnetina-3-O-robinobiosideo, foram isolados de
pericarpo de cactus (Hylocereus undaus) que mostraram atividade alelopática em Celosia
cristata e fungo Neurospora crassa (Tomita-Yokotani et al., 2004), também são compostos
C-glicosilados como os identificados em extratos foliares de L. sibiricus.
Em relação as agliconas, os resultados encontrados indicaram que as metoxiladas
(E e F) e a quercetina (G), apesar de não serem compostos majoritários, acarretaram maior
10-7 10-6 10-5 10-4
-150
-100
-50
0
50
100
150
% e
m r
elaç
ão a
o co
ntro
le
Concetração (M)
A B C D E F G
10-7 10-6 10-5 10-4
Concentração (M)

48
atividade biológica quanto maior número de grupos catecol (Macias et al., 1997; Einhellig,
2004; Piettá, 2000), além do grupo metoxila no anel A, que também, confere atividade
alelopática (Parvez et al., 2004).
A influência das substâncias isoladas no crescimento inicial das raízes (Figura 6),
talvez seja devido a presença dos radicais no anel B dos flavonóides (Figura 5) o provável
responsável pela atividade alelopática das agliconas (E, F e G), pois a estrutura – atividade
estão relacionadas com a fitotoxicidade destes compostos (Macias et al., 1997; Bais et al.,
2003; Beninger & Hall, 2005; Parvez et al., 2004). Isso porque na substância E tem duas
hidroxilas no anel B, em F apenas uma e em G, embora também apresente duas hidroxilas no
anel B, o anel A deixa de apresetar o radical metoxila presente em E e F.
Especificamente, os fatores determinantes para que esses flavonóides apresentem
atividade antioxidante e alelopática deve-se a um conjunto de fatores relacionados com a a
presença do grupo catecol. O anel B, apresenta características químicas capazes de interagir
com o radical livre (atividade antioxidante), além da presença de dupla ligação na posição 2, 3
conjugadas com o oxigênio na posição 4 do anel C. A presença do grupo hidroxila ou
metoxila nas posições 3, 5 e 7 do anel A, parecem ter menor atividade quando comparada
com os substituintes no anel B e C, no entanto tal atividade pode ser alterada quando outras
substituições estiverem presentes nestes anéis. Essas características estruturais contribuem
para aumentar a estabilidade eletrônica do radical livre e portanto conferirem atividade
antioxidante dos flavonóides (Martinez-Flores et al., 2002; Bors et al., 1990).
Os resultados de crescimento inicial de raiz (Tabela 4 e Figura 6) podem ser
devido às moléculas dos flavonóides apresentarem atividade antioxidante, capaz de capturar
radicais livres, principalmente no meio intracelular onde as moléculas flavonoídicas assumem
cargas negativas em pH neutro (Martinez-Flores et al., 2000) e em baixas concentrações
podem acarretar efeitos promotores de crescimento (Macias et al., 1997; Parvez et al., 2004;
Worsham, 1987), talvez devido ao melhor reaproveitamento de substâncias vitais para a
célula como enzimas, co-enzimas, transportadores de elétrons e proteínas. Altas
concentrações dos flavonóides, por outro lado, podem agir como agente hiperpolarizador de
membrana e alterarem os mecanismos das bombas atepeásicas, tornando os flavonóides
tóxicos para as células, reduzindo o crescimento.
Outra explicação para as mudanças na germinação das sementes e
desenvolvimento inicial das raízes poderia ser devido ao efeito osmótico, em relação às
substâncias presentes nas soluções testadas (Ferreira & Áquila, 2000; Hoagland & Williams,
2004; Bell & Charlwood, 1980). A presença de flavonóides na solução, embora pequena,

49
poderia ter acarretado diminuição do potencial osmótico causando, portanto, diminuição no
crescimento de raiz por dificultar o alongamento celular causado pela entrada de solução.
Além disso, como não ocorreu mudança do Índice Refratométrico da solução inicial e final do
experimento (dados não mostrados), seria pouco provável que os resultados encontrados
sejam devido ao efeito osmótico das soluções.
Pode-se concluir que o uso de extratos com perfil químico semelhante podem
propiciar respostas alelopáticas diferentes, devido a correlação com a concentração utilizada e
estrutura das moléculas, onde as agliconas demonstraram maior atividade (inibição ou
promoção).
Agradecimentos
A Faculdade de Farmácia de Salerno (UNISA) pela disponibilidade do RMN
500MHz e aos pesquisadores Emilia Mancini, Laura Di Martini e Emilia Quaranta, Antonio
Bracci e Francesco Arminante. Ao Instituto de Química da UNESP-Araraquara e os
pesquisadores Daniel Rinaldi e Profa Dra Lourdes Campaner dos Santos.

50
REFERÊNCIA BIBLIOGRÁFICA
ALMEIDA, L.F.R.; DELACHIAVE, M.E.A.; MARQUES, M.O.M. Composição química do
óleo essencial de Leonurus sibiricus. Brasilian Journal of Medicinal Plants, v.8, p.31-35,
2005.
ALMEIDA, L.F.R. et al. Ação alelopática de extratos aquosos de rubim (Leonurus sibiricus)
na germinação e desenvolvimento inicial de alface, tomate e milho. In: IX CONGRESSO
BRASILEIRO DE FISIOLOGIA VEGETAL, 9, 2003, Atibaia-SP. Resumos... Atibaia-SP,
2003. p.56.
ALMEIDA, L.F.R. et al. Constituintes químicos de Leonurus sibiricus (LAMIACEAE). In:
CONGRESSO DA SOCIEDADE BRASILEIRA DE QUÍMICA, 24, 2006, Águas de
Lindóia-SP. Anais ... Águas de Lindóia-SP, 2006. p.94.
AN, M.; WAGGA, W. Mathematical modeling of dose-response relationship (Hormesis) in
allelopathy and its application. Nonlinearity in Biology, Toxicology and Medicine, v.3,
p.153-172, 2005.
AN, M.; JOHNSON, I.R.; LOVETT, J.V. Mathematical modeling of allelopathy: Biological
response to alelochemicals and its interpretation. Journal of Chemical Ecology, v.19,
p.2374-2388, 1993.
BAIS, H.P. et al. Allelopathy and exotic plants: from genes to invasion. Science, v.301,
p.1377–1380, 2003.
BASILE, A. et al. Antibacterial and allelopathic activity of extract from Castanea sativa
leaves, Fitoterapia, v.71, p.8110-8116, 2000.
BELL, E.A.; CHARLWOOD, B.V. Secondary plant products. In: BELL, E.A.;
CHARLWOOD, B.V. Encyclopedia of Plant Physiology, New York: Springer–Verlag,
1980, v.8, 674p.
BENINGER, C.W.; HALL, J.C. Allelopathic activity of luteolin 7-O-β-glucuronide isolated
from Chrysanthemum morifolium L. Biochemical Systematics and Ecology, v.33, p.103–
111, 2005.
BOALINO, D.M. et al. Labdane Diterpenes of Leonurus sibiricus. Journal of Natural
Products, v.67, p.714-717, 2004.
BORS, W. et al. Flavonoids as antioxidants: Determination of radical scavenging efficiencies.
Methods Enzymology, v.186, p.343-355,1990.
BOWN, D. The herb society of America: encyclopedia of herbs and their uses. New York:
Darling Kindersley Publ., 1995, 314p.

51
CALIS, I. et al. Phenylpropanoid glycosides from Leonurus glaucescens. Phytochemistry,
v.61, p.324-329, 2000.
CASTELLUCCI, S. et al. Plantas medicinais relatadas pela comunidade residente na estação
ecológica de Jataí, município de Luís Antônio/SP: uma abordagem etnobotânica. Revista
Brasileira de Plantas Medicinais, v.3, p.51-60, 2000.
CHON, S.U-K. et al. Allelopathic potential in lettuce (Lactuca sativa L.) plants. Scientia
Horticulturae, v.106, p.309–317, 2005.
CONG, Y.; WANG, J.-H.; LI, X. A new flavonoside from Leonurus heterophyllus. Journal
of Asian Natural Products Research, v.7, p.273-277, 2005.
CUTLER, H.G.; CUTLER, S.J. Agrochemicals and Pharmaceuticals: The connection. In:
CUTLER, H.G.; CUTLER, S.J. (Eds) Biologically active natural products: Agrochemicals.
Boca Raton, Florida: CRC Press, 1999, p.1-14.
DE FEO, V. et al. Isolation of phytotoxic compounds from Tree-of-Heaven (Ailanthus
altissima Swingle). Journal of Agricultural and Food Chemistry, v.51, p.1770-1802, 2003.
DUKE, S.O. et al. Strategies for the use of natural products for weed management. Journal of
Pesticide Science, v.27, p.298-306, 2002.
EINHELLIG, F.A. Mode of allelochemical action of phenolic compounds. In: MACIAS, F.A.
et al. (Eds.) Allelopathy: Chemistry and mode of action of allelochemicals. Boca Raton,
Florida: CRC Press, 2004, p.217-238.
FERREIRA, A.G.; AQUILA, M.E.A. Alelopatia: Uma área emergente da ecofisiologia.
Revista Brasileira de Fisiologia Vegetal, v.12 (ed. esp.), p.175-204, 2000.
GIANG, P.M. et al. New Labdane-Type Diterpenoids from Leonurus heterophyllus SW.
Chemical Pharmaceutical Bulletin, v.53, p.938-941, 2005.
GLASS, A.D.M.; DUNLOP, J. Influence of phenolic acids on ion uptake: 4. Depolarization
of membrane potentials. Plant Physiology, v.54, p.855-858, 1974.
HOAGLAND, R.E.; WILLIAMS, R.D. Bioassays-useful tolls of the study of allelopathy. In:
MACIAS, F.A. et al. (Eds.) Allelopathy: Chemistry and mode of action of allelochemicals.
Boca Raton, Florida: CRC Press, 2004, p.315-41.
HOEHNE, F.C. Plantas e Substâncias Vegetais Tóxicas e Medicinais. São Paulo:
Graphicars, 1939, 355p.
INDERJIT; DAKSHINI, K.M.M. Allelopathic potential of the phenolics from the roots of
Pluchea lanceolata. Physiologia Plantarum, v.92, p.571–576, 1994.
ISLAM, M.A. et al. Analgesic and anti-inflammatory activity of Leonurus sibiricus.
Fitoterapia, v.76, p.359-362, 2005.

52
KARTING, T.; GRUBER, A.; MENZINGER, S. Flavonoid-O-Glycosides from the Herbs of
Leonurus cardiaca. Journal of Natural Products, v.48, p.494-495, 1985.
LARCHER, W. Ecofisiologia vegetal. Sttuttgard: Eugen Ulmer, 2000, 315p.
LORENZI, H. Plantas daninhas do Brasil: terrestres, aquáticas, parasitas, tóxicas e
medicinais. São Paulo: Nova Odessa, 2002, 278p.
MACIAS, F.A. et al. Bioactive flavonoids from Helianthus annuus cultivars.
Phytochemistry, v.45, p.683-687, 1997.
MANDAL, S. Allelopathic activity of root exudates from Leonurus sibiricus L. (Raktodrone).
Weed Biology and Management, v.1, p.170-175, 2001.
MARTÍNEZ-FLÓREZ, S.J. et al. Los flavonoides: propiedades y acciones antioxidantes.
Nutricion Hospitalaria, v.6, p.271-278, 2002.
MATHELA, C.S. Allelochemicals in Medicinal and Aromatic Plants. In: NARWAL, S.S.;
TAURO, P. (Eds.) Allelopathy in Agriculture and Forestry. Jodhpur, India: Scientific
Publishers, 1991, p.213-228.
MILKOWSKA-LEYCK, K.; FILIPEK, B.; STRZELECKA, H. Pharmacological effects of
lavandulifolioside from Leonurus cardiaca locomotor activity. Journal of
Ethnopharmacology, v.80, p.85-90, 2002.
MURAKAMI, S. Stachydrim in Leonurus sibiricus L. Acta Phytochemichal, v.13, p.161-
184, 1943.
NAGASAWA, W. et al. Effects of Motherwort (Leonurus sibiricus L) on Preneoplastic and
Neoplastic Mammary-Gland Growth in Multiparous Gr/A Mice Anticancer Research, v.10,
p.203-211, 1990.
NISHIDA, N. et al. Allelopathic effects of volatile monoterpenoids produced by Salvia
leucophylla inhibition of cell proliferation and DNA synthesis in the root apical meristem of
Brassica campestris seedlings. Journal of Chemical Ecology, v.31, p.1187-1203, 2005.
ONYIKAGHA, J.C.; GROTEWOLD, E. The biological and structural distribuition of surface
flavonoids. Recent Research Development Plant Science, v.2, p.1-18, 2004.
PARVEZ M.M. et al. Effects of quercetin and its seven derivatives on the growth of
Arabidopsis thaliana and Neurospora crassa. Biochemical Systematic and Ecology, v.32,
p.631–635, 2004.
PIETTÁ, G.P. Flavonoids as antioxidants. Journal of Natural Products, v.63, p.1035-1042,
2000.
QASEM, J.R.; FOY, C.L. Weed allelopathy, its ecological impacts and future prospects: a
review. Journal of Crop Production, v.4, p.43-119, 2001.

53
RICE, E.L. Allelopathy. New York: Academic Press, 1984, 422p.
RICE-EVANS, C.A.; PACKER, L. Flavonoids in Health and Disease. New York: Marcel
Dekker, 1998, p.447-467.
SATOH, M. et al. Studies of the constituents of Leonurus sibiricus. Chemical
Pharmaceutical Bulletin, v.51, p.341-342, 2003.
SAVONA, G. et al. Diterpenoids from Leonurus sibiricus. Phytochemistry, v.21, p.2699-
2701, 1982.
SHIRLEY, B.W. Flavonoid biosynthesis: new functions for an old pathway. Trends Plant
Science, v.31, p.377-382, 1996.
SOUZA, G.C. et al. Ethnopharmacological studies of antimicrobial remedies in the south of
Brazil. Journal of Ethnopharmacology, v.90, p.135-43, 2004.
STEBBING, A.R.D. Hormesis – the stimulation of growth by low levels of inhibitors.
Science Total Environmental, v.22, p.213-234, 1982.
TASDEMIR, D. et al. Iridoid glycosides of Leonurus persicus. Journal of Natural Products,
v.62, p.811-816, 1999.
TOMITA-YOKOTANI, K. et al. Flavonoids isolated from pericarps of Hylocereus undatus
(Cactaceae) as allelochemicals, Plant and Cell Physiology, v.45, p.181-188, 2004.
TORRES, A. et al. A science of the future. In: First World Congress of Allelopathy, (SAI).
Annals… University of Cadiz, 1996, 278p.
WORSHAM, A.D. Germination of wicthweed seeds. In: MUSSELMAN, L. (ED.) Parasitic
Weeds in Agriculture, Boca Raton, Florida: CRC Press, 1987, p.45-61.

54
CONSIDERAÇÕES FINAIS
As folhas de Leonurus sibiricus apresentaram significativa diversidade química,
tanto de componentes voláteis quanto de flavonóides. Em relação aos terpenos, a presença
majoritária foi de sesquiterpenos, moléculas provavelmente oriundas da rota do ácido
mevalônico que apresentam 15 carbonos na estrutura e que são responsáveis pelo aroma e
atividade biológica do óleo essencial. Por outro lado, os flavonóides majoritários são
derivados da quercetina e apresentam de uma a duas moléculas de açúcar ligadas no carbono
3 do anel heterocíclico; além dessas substâncias majoritárias, as folhas também apresentaram
monoterpenos e agliconas. Essa composição química explica as atividades biológicas
discutidas na literatura e provavelmente, são responsáveis pelas atividades alelopáticas
mencionadas.
Os experimentos de germinação e crescimento inicial em raiz das três diferentes
espécies demonstraram que a atividade alelopática, inicialmente, depende da concentração
utilizada, fato esse verificado nos testes com extratos por infusão e metanólico. Além disso, as
diferentes substâncias isoladas de frações ativas mostraram que a ação alelopática, também,
está relacionada com a estrutura molecular e a presença de grupos químicos específicos,
capazes de alterar o crescimento das raízes. Não foi investigado os locais de ação dessas
substâncias ou delas em conjunto, porém foi observado que a alteração no alongamento e/ou
divisão celular de raízes podem ser em função das características eletrofílicas das moléculas
dos flavonóides. Em outras palavras, a capacidade antioxidante, geralmente benéfica nas
concentrações baixas dos flavonóides, deve ter propiciado melhor funcionamento das células
e portanto, conferido maior crescimento inicial das raízes. Por outro lado, quando em
concentração mais elevada, os flavonóides devem ter alterado os mecanismos celulares de
alongamento e/ou divisão celular, levando a visível inibição do crescimento das raízes.
Essas atividades alelopáticas, positivas (promoção) e negativas (inibição), estão de
acordo com critérios utilizados por muitos autores para definir o termo aleloquímico. Embora
as moléculas encontradas não sejam inéditas, é o primeiro relato da existência e atividade
alelopática de folhas de L. sibiricus, acarretando questionamentos sobre a função e
respectivos sítios de ação dos flavonóides. Não se pode deixar de mencionar que o efeito dos
extratos, os quais apresentam majoritariamente flavonóides glicosilados, pode ser devido ao
efeito de sinergismo destas substâncias, fato que poderá ser comprovado com o estudo em
fatoriais dos sete flavonóides isolados de folhas de L. sibiricus.

55
A espécie L. sibiricus, por estar amplamente distribuída no território brasileiro,
com fácil e rápido crescimento, pode ser um modelo de estudo para observações no âmbito
ecológico, possibilitando a compreensão das interações químicas entre os vegetais. Contudo,
as atividades de extratos e frações ativas, embora distante do que ocorre em condições de
campo, pôde evidenciar e comprovar as observações ocorridas na natureza.

56
REFERÊNCIAS BIBLIOGRÁFICAS
AHERNE, S.A.; O'BRIEN, N.M. Dietary flavonols: chemistry, food content, and metabolism.
Nutrition, v.18, p.75-81 , 2002.
ALMEIDA, L.F.R.; DELACHIAVE, M.E.A.; MARQUES, M.O.M. Composição química do
óleo essencial de Leonurus sibiricus. Brasilian Journal of Medicinal Plants, v.8, p.31-35,
2005.
ALMEIDA, L.F.R.et al. Ação alelopática de extratos aquosos de rubim (Leonurus sibiricus)
na germinação e desenvolvimento inicial de alface, tomate e milho. In: CONGRESSO
BRASILEIRO DE FISIOLOGIA VEGETAL, 9, Resumos... Atibaia-SP, Brasil, p.56, 2003.
ALMEIDA, L.F.R. et al. Constituintes químicos de Leonurus sibiricus (LAMIACEAE). In:
CONGRESSO DA SOCIEDADE BRASILEIRA DE QUÍMICA, SBQ, 24, Anais.... Águas
de Lindóia-SP, p.94, 2006.
ALSAADAWI, I.S. Research on allelopathy in Iraq. In: NARWAL, S.S. (Ed.). Allelopathy
Update. Enfield: Science Publishers, 1999, v.1, p.185-197.
BAIS, H.P. et al. Allelopathy and exotic plants: from genes to invasion. Science, v.301,
p.1377–1380, 2003.
BARKOSKY, R.R.; EINHELLIG, F.A. Effect of a simple phenol on plant water relationships
of soybean Glycine max. Plant Physiology, v.2, p.155-155 (suppl), 1993.
BATISH, D.R. et al., Crop allelopathy and its role in ecological agriculture. Journal of Crop
Production, v.4, p.121-161, 2001.
BEN-HAMMOUDA, M.; GHORBAL, H.; OUESLATI, O. Allelopathic effects of barley
extracts on germination and seedlings growth of bread and durum wheats. Agronomie, v.21,
p.65-71, 2001.
BENINGER, C.W.; HALL, J.C. Allelopathic activity of luteolin 7-O-b-glucuronide isolated
from Chrysanthemum morifolium L. Biochemical Systematics and Ecology, v.33, p.103–
111, 2005.
BERGMARK, C.L.; JACKSON, W.A.; VOLK, R.J. Differential inhibition by ferulic acid of
nitrate and ammonium uptake in Zea mays. Plant Physiology, v.98, p.639-645, 1992.
BOOKER, F.L.; BLUM, U.; FISCUS, E.L. Short-term effects of ferulic acid on ion uptake
and water relations in cucumber seedlings. Journal of Experimental Botany, v.43, p.649-
655, 1992.
BORS, W. et al., Flavonoids as antioxidants: Determination of radical scavenging
efficiencies. Methods Enzymology, v.186, p.343-355, 1990.

57
BOWN, D. The herb society of America: encyclopedia of herbs and their uses. New York:
Darling Kindersley Publ. 1995, 314p.
BRANCO, A.; PIZZOLATTI, M.G. CGAR e CGAR-EM na análise dos constituintes
químicos isolados do extrato hexânico de Sabastiania argutidens (Euphorbiaceae). Química
Nova, v.25, p.15-19, 2002.
CALLAWAY, R.M. The detection of neighbors by plants. Trends in Ecology & Evolution,
v.17, p.104-105, 2002.
CASTELLUCCI, S. et al. Plantas medicinais relatadas pela comunidade residente na estação
ecológica de Jataí, município de Luís Antônio/SP: uma abordagem etnobotânica. Revista
Brasileira de Plantas Medicinais, v.3, p.51-60, 2000.
CONG, Y.; WANG, J.H.; LI, X. A new flavonoside from Leonurus heterophyllus. Journal of
Asian Natural Products Research, v.7, p.273-277, 2005.
CUTLER, H.G.; CUTLER, S.J. Agrochemicals and Pharmaceuticals: The connection. In:
CUTLER, H.G.; CUTLER, S.J. (Eds.). Biologically active natural products:
Agrochemicals, Boca Raton, Florida: CRC Press, 1999, p.1-14.
DE FEO, V. et al. Isolation of Phytotoxic compounds from Tree-of-Heaven (Ailanthus
altissima Swingle). Journal of Agricultural and Food Chemistry, v.51, p.1770-1802, 2004.
DELACHIAVE, M.E.A.; ONO, E.O.; RODRIGUES, J.D. Efeitos alelopáticos de grama-seda
(Cynodon dactylon) na germinação de sementes de pepino, milho, feijão e tomate. Revista
Brasileira de Sementes, v.21, p.194-197, 1999.
DEVI, S.R.; PRASAD, M.N.V. Effect of ferulic acid on growth and hydrolytic enzyme
activities of germinating maize seeds. Journal of Chemical Ecology, v.18, p.1981-1990,
1992.
DUKE, S.O. et al. Strategies for the use of natural products for weed management. Journal of
Pesticide Science, v.27, p.298-306, 2002.
EINHELLIG, F.A. Mode of allelochemical action of phenolic compounds. In: MACIAS, F.A.
et al. (Eds.) Allelopathy: Chemistry and mode of action of allelochemicals. Boca Raton,
Floridaa: CRC Press: 2004, p.217-238.
EINHELLIG, F.A. Allelopathy: Current Status and Future Goals. Acs Symposium Series,
v.582, p.1-24, 1995.
EINHELLIG, F.A.; RASMUSSEN, J.A. Effects of root exudate sorgoleone on
photosynthesis. Journal of Chemical Ecology, v.19, p.369-375, 1993.
ELLIOTT, L.F.; CHENG, H.H. Assessment of allelopathy among microbes and plants. ACS
Symposium Series, v.330, p.504-515, 1987.

58
FERREIRA, A.G.; AQUILA, M.E.A. Alelopatia: Uma área emergente da ecofisiologia.
Revista Brasileira de Fisiologia Vegetal, v.12 (ed. esp.), p.175-204, 2000.
FRIEDMAN, J. Allelopathy, Autotoxicity, and Germination. In: ZIGEL, J.; GALILI, G.
(Eds.) Seed development and germination. New York: Marcel Dekker, 1995, 853p.
GIANG, P.M. et al. New Labdane-Type Diterpenoids from Leonurus heterophyllus SW.
Chemical Pharmaceutical Bulletin, v.53, p.938-941, 2005.
GLASS, A.D.M.; DUNLOP, J. Influence of phenolic acids on ion uptake: 4. Depolarization
of membrane potentials. Plant Physiology, v.54, p.855-858, 1974.
GOTTLIEB, O.R. Micromolecular, evolution, systematics and ecology: an essay into a
novel botanical discipline, Berlin: Springer-Verlag, 1982, 170p.
HARBONE, J.B. Chemotaxonomy of the Leguminosae. In: CODY, V.; MIDDLETON, E.;
HARBONE, J.B. (Eds.), The Flavoniods Advances in Research Since 1980. London:
Chapman & Hall, 1988, p.367-462.
HARBORNE, J.B. The Flavonoids: Advances in Research Since 1986, London: Chapman &
Hall, 1994, p.589-618.
HAVSTEEN, B. Flavonoids: A class of natural products of high pharmacological potency.
Biochemical Pharmacology, v.32, p.1141-1148, 1983.
HELLER, W.; FORKMANN, G. Biosynthesis, In: HARBONE, J.B. (Ed.) The Flavonoids.
Advances in Research since 1986. London: Chapman & Hall, 1993, p.499-535.
HICKS, S.K. et al. Allelopathic effects of wheats straw on cotton germination, emergence and
yield. Crop Science, v.29, p.1057-1061, 1989.
HIERRO, J.L.; CALLAWAY, R.M. Allelopathy and exotic plant invasion. Plant and Soil,
v.256, p.29-39, 2003.
HOAGLAND, R.E.; WILLIAMS, R.D. Bioassays-useful tolls of the study of allelopathy. In:
MACIAS, F.A. et al. (Eds.) Allelopathy: Chemistry and mode of action of allelochemicals.
Boca Raton, Florida: CRC Press, 2004, p.315-41.
HOEHNE, F.C. Plantas e Substâncias Vegetais Tóxicas e Medicinais. São Paulo:
Graphicars, 1939, 355p.
IGURA, K. et al. Resveratrol and quercetin inhibit angiogenesis in vitro. Cancer Letterts,
v.171, p.11-16, 2001.
INDERJIT. Plant phenolics in allelopathy. Botanical Review, v.62, p.186-202, 1996.
INDERJIT; DAKSHINI, K.M.M. The nature of the interference potential of Pluchea
lanceolata (DC) Clarke,C.B. (Asteraceae). Plant and Soil, v.122, p.298-302, 1990.

59
INDERJIT; DAKSHINI, K.M.M. Interference potencial of Pluchea lanceolata (Asteraceae):
growth and physiological responses of asparagus bean, Vigna unguiculata var. sesquipedalis.
American Journal of Botany, v.79, p.977-981, 1992.
INDERJIT; DAKSHINI, K.M.M. Allelopathic potential of the phenolics from the roots of
Pluchea lanceolata. Physiologia Plantarum, v.92, p.571–576, 1994.
INDERJIT; DEL MORAL, R. Is separating resource competition from allelopathy realistic?
Botanical Review, v.63, p.221-230, 1997.
INDERJIT; NILSEN, E.T. Bioassays and field studies for allelopathy in terrestrial plants:
Progress and problems. Critical Reviews in Plant Sciences, v.22, p.221-238, 2003.
INDERJIT; KAUR, M.; FOY, C.L. On the Significance of Field Studies in Allelopathy.
Weed Technology, v.15, p.792–797, 2001.
ISLAM, M.A. et al. Analgesic and anti-inflammatory activity of Leonurus sibiricus.
Fitoterapia, v.76, p.359-362, 2005.
JANG, M.; CAI, L.; UDEANI, G.O. Cancer chemopreventive activity of resveratrol, a natural
product derived from grapes. Science, v.275, p.218-221, 1997.
JOVANOVIC, S.V. et al. Antioxidant properties of flavonoids: reduction potentials and
electron transfer reactions of flavonoid radicals. In: RICE-EVANS C.; PARKER, L. (Eds.)
Flavonoids in health and disease, Nova York: Marcel Dekker, 1998, p.137-161.
KALBURTJI, K.L. Research on allelopathy in Greece. In: NARWAL, S.S. (Ed.) Allelopathy
Update, Enfield: Science Publishers, 1999, v.1, p.37-47.
KARTNIG, T.; GRUBER, A.; MENZINGER, S. Flavonoid-O-Glycosides from the Herbs of
Leonurus cardiaca. Journal of Natural Products, v.48, p.494-495, 1985.
KÜHNAU, J. The Flavonoids: a class of semi-essential food components: their role in human
nutrition. World Revision Nutrition Diet, v.24, p.117-190, 1976.
KRUSE, O. et al. Phosphatidylglycerol is involved in the dimerization of photosystem II.
Journal of Biology Chemistry, v.275, p.6509-6514, 2000.
LARCHER, W. Ecofisiologia vegetal, Sttuttgard: Eugen Ulmer, 2000, 315p.
LETAN, A. The relation of structure to antioxidant activity of quercitin and some of its
derivates. Journal of Food Science, v.31, p.518-523, 1966.
LORENZI, H. Plantas daninhas do Brasil: terrestres, aquáticas, parasitas, tóxicas e
medicinais. São Paulo: Nova Odessa, 2002, 278p.
MACIAS, F.A. et al. Bioactive flavonoids from Helianthus annuus cultivars.
Phytochemistry, v.45, p.683-687, 1997.

60
MANDAL, S. Allelopathic activity of root exudates from Leonurus sibiricus L. (Raktodrone).
Weed Biology and Management, v.1, p.170-175, 2001.
MARTÍNEZ-FLÓREZ, S.J. et al. Los flavonoides: propiedades y acciones antioxidantes.
Nutricion Hospitalaria, v.6, p.271-278, 2002.
MATHELA, C.S. Allelochemicals in medicinal and aromatic plants. In: NARWAL, S. S.;
TAURO, P. (Eds.) Allelopathy in Agriculture and Forestry. Jodhpur, India: Scientific
Publishers, 1991, p.213-228.
MAY, F.E.; ASH, J.E. An assessment of allelopathic potential of Eucalyptus. Australian
Journal of Botany, v.38, p.245-254, 1990.
MAZZAFERA, P. Efeito alelopático do extrato alcoólico do cravo-da-índia e eugenol.
Revista Brasileira de Botânica, v.26, p.231-238, 2003.
MIDDLETON, E. JR.; KANDASWAMI, C.; THEOHARIDES, T.C. The effects of plant
flavonoids on mammalian cells: implications for inflammation, heart disease, and cancer.
Pharmacological Revision, v.52, p.673-751, 2000.
MILKOWSKA-LEYCK, K.; FILIPEK, B.; STRZELECKA, H. Pharmacological effects of
lavandulifolioside from Leonurus cardiaca locomotor activity. Journal of
Ethnopharmacology, v.80, p.85-90, 2002.
MORELAND, D.E.; NOVITZKY, W.P. Interference by flavone and flavonols with
chloroplast-mediated electron transport and phosphorylation. Phytochemistry, v.27, p.3359-
3366, 1988.
MURAKAMI, S. Stachydrim in Leonurus sibiricus L. Acta Phytochemichal, v.13, p.161-
184, 1943.
MUSCOLO, A.; PANUCCIO, M.R.; SIDARI, M. The ascorbate system during the early
stage of germination in Pinus laricio seeds treated with extracts from two different sources of
humus. Seed Science and Technology, v.29, p.275-279, 2001.
NAGASAWA, W. et al. Effects of Motherwort (Leonurus sibiricus L) on Preneoplastic and
Neoplastic Mammary-Gland Growth in Multiparous Gr/A Mice. Anticancer Research, v.10,
p.203-211, 1990.
NAKANO, H. et al. Isolaton and identification of plant growth inhibitors as candidates of
allelopathic effects substances, from aqueous leachate from mesquite (Prosopis juliflora)
leaves. Plant Growth Regulation, v.37, p.113-117, 2002.
ONYILAGHA, J.C.; GROTEWOLD, E. The biological and structural distribuition of surface
flavonoids. Recent Research Development Plant Science, v.2, p.1-18, 2004.

61
OUDHIA, P.; TRIPATHI, R.S. Allelopathic effects of an obnoxious weed Parthenium
hysterophorus on germination and seedling vigour of rice var, Mahamaya. Research on
Crops, v.1, p.111-115, 2000.
PACE-ASCIAK, C.R. et al. The red wine phenolics trans-resveratrol and quercitin block
human platelet aggregation in eicosanoid synthesis implication for protection against coronary
heart disease. Clinical Chemistry Acta, v.235, p.207-219, 1995.
PARVEZ M.M. et al. Effects of quercetin and its seven derivatives on the growth of
Arabidopsis thaliana and Neurospora crassa. Biochemical Systematics and Ecology, v.32,
p.631–635, 2004.
PERES, W. Radicais Livres em níveis biológicos. Pelotas: Ed. Universidade Católica, 1994,
p.49-81.
PEREZ, F.J. Allelopathic effect of hydroxamic acids from cereals on Avena sativa e A. fatua.
Phytochemistry, v.29, p.773-776, 1990.
PIETTÁ, G.P. Flavonoids as antioxidants. Journal of Natural Products, v.63, p.1035-1042,
2000.
PIETTA, P.G.; SIMONETTI, P. Antioxidant Food Supplements in Human Health. San
Diego: Academic Press, 1999, p.283-308.
PUTNAM, A.R. Weed allelopathy. In: DUKE, S.O (Ed.) Weed Physiology: Reproduction
and Ecophysiology. Boca Raton, Florida: CRC Press, 1985, p.131–155.
PUTNAM, A.R.; DUKE, W.B. Allelopathy in agro-ecosystems. Annual Review of
Phytopathology, v.16, p.431-451, 1978.
QASEM, J.R.; FOY, C.L. Weed allelopathy, its ecological impacts and future prospects: a
review. Journal Crop Production, v.4, p.43-119, 2001.
RICE, E.L. Allelopathy. New York: Academic Press, 1984, 422p.
RICE-EVANS, C.A.; PACKER, L. Flavonoids in Health and Disease. New York: Marcel
Dekker, 1998, p.447-467.
RIMANDO, A.M. et al. Searching for rice allelochemicals: an exemple of bioassay-guided
isolation. Agronomy Journal, v.93, p.16-20, 2001.
RIZVI, S.J.H.; RIZVI, V. Exploration of allelochemicals in improving crop productivity. In:
RIZVI, S.J.H.; RIZVI, V. (Eds.). Allelopathy: Basic end applied aspects. London: Chapman
& Hall, 1992, p.443-472.
SATOH, M. et al. Studies of the constituents of Leonurus sibiricus. Chemical
Pharmaceutical Bulletin, v.51, p.341-342, 2003.

62
SAVONA, G. et al. Diterpenoids from Leonurus sibiricus. Phytochemistry, v.21, p.2699-
2701, 1982.
SHIRLEY, B.W. Flavonoid biosynthesis: new functions for an old pathway. Trends Plant
Science, v.31, p.377-382, 1996.
SINGH, H.P.; BATISH, D.R.; KOHLI, R.K. Allelopathy in agroecosystems: a overview.
Journal of Crop Production, v.4, p.1-41, 2001.
SINGLETON, V.L. Flavonoids. In: CHILDESTER, C.O.; MRAK, E.M.; STEWART, G.F.
(Eds.). Advances in Food Research, New York: Academic Press, 1981, p.149-242.
SOUZA, G.C. et al. Ethnopharmacological studies of antimicrobial remedies in the south of
Brazil. Journal of Ethnopharmacology, v.90, p.135-43, 2004.
STOWE, L.G. Allelopathy and its influence on the distribution of plants in an Illinois old
field. Journal of Ecology, v.67, p.1065-1085, 1979.
TASDEMIR, D. et al. Iridoid glycosides of Leonurus persicus. Journal of Natural Products,
v.62, p.811-816, 1999.
TOMITA-YOKOTANI, K. et al. Flavonoids isolated from pericarps of Hylocereus undatus
(Cactaceae) as allelochemicals, Phytochemistry, v.65, p.1247-1253, 2004.
TORRES, A. et al. A science of the future. In: FIRST WORLD CONGRESS OF
ALLELOPATHY. (SAI). Annals… University of Cadiz, 1996, 278p.
WAGNER, H.; FARKAS, L. Síntesis of flavonoids. In: HARBORNE, J.B.; MABRY, T.J.;
MABRY, H. (Eds). The Flavonoids. New York: Academic Press, 1975, p.127-213.
WALLER, G.R. Introduction. In: MACIAS, F.A. et al. (Eds.) Recent advances in
allelopathy. Cadiz: Serv. Publisher Univesity of Cadiz, v.1, 1999, p.231.
WEBSTER, J. et al. Induction of Acclimative Proteolysis of the Light-Harvesting Chlorophyll
a/b Protein of Photosystem II in Response to Elevated Light Intensities. Plant Physiology,
v.118. p.827-834, 1998.
YANG, K.; LAMPRECHT, S.A.; LIU, Y. Chemoprevention studies of the flavonoids
quercetin and rutin in normal and azoxymethane-treated mouse colon. Carcinogenesis, v.21,
p.1655-1660, 2000.

63
A N E X O

64
Capítulo 1
Composição do óleo essencial de Leonurus sibiricus L.

65
A. Espectro de massa em CG-MS de alfa-copaeno

66
B. Espectro de massa em CG-MS de beta-bouboreno.
67
C. Espectro de massa em CG-MS de trans-cariofileno.

68
D. Espectro de massa em CG-MS de alfa-humuleno.
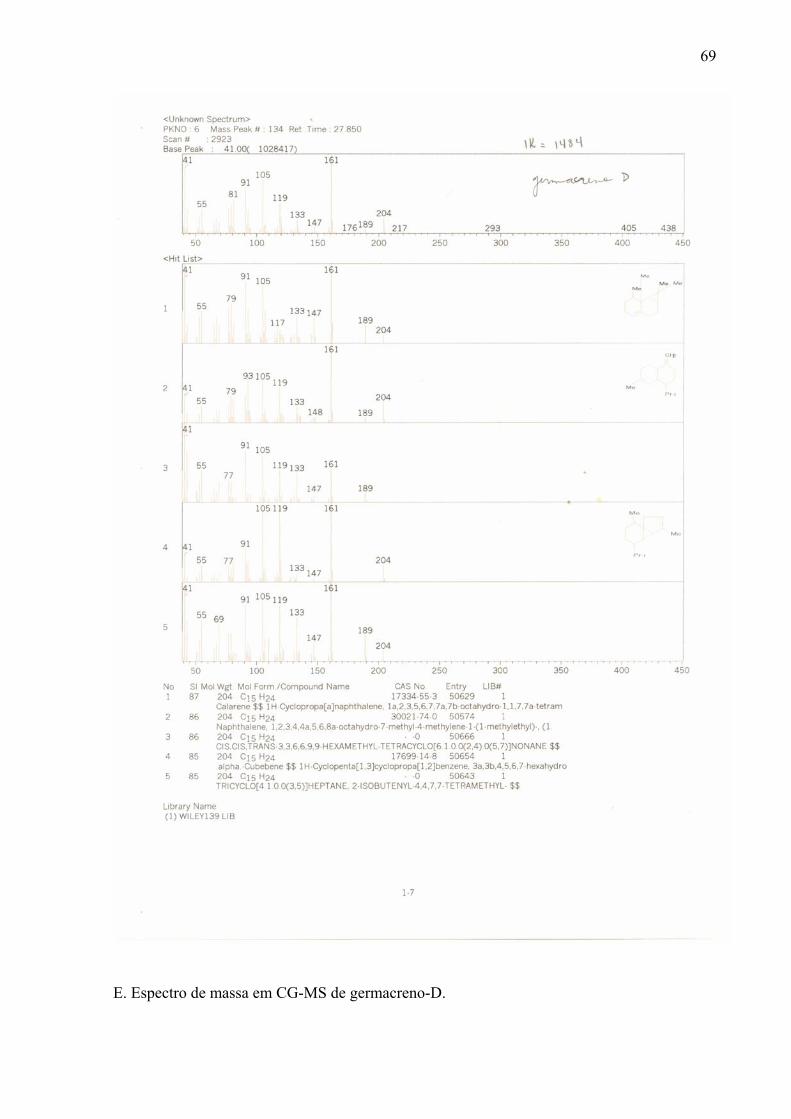
69
E. Espectro de massa em CG-MS de germacreno-D.

70
F. Espectro de massa em CG-MS de gama-cadieno.

71
Capítulo 2
Potencial alelopático de folhas de Leonurus sibiricus L.
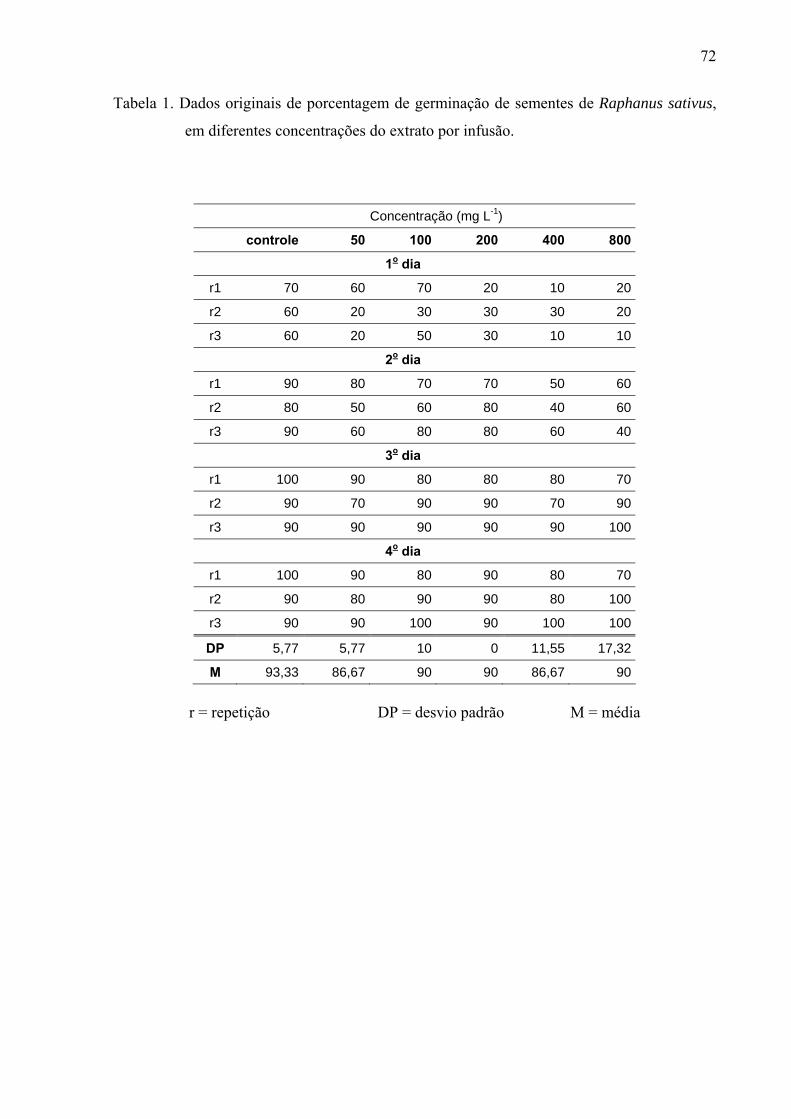
72
Tabela 1. Dados originais de porcentagem de germinação de sementes de Raphanus sativus,
em diferentes concentrações do extrato por infusão.
Concentração (mg L-1)
controle 50 100 200 400 800
1o dia
r1 70 60 70 20 10 20
r2 60 20 30 30 30 20
r3 60 20 50 30 10 10
2o dia
r1 90 80 70 70 50 60
r2 80 50 60 80 40 60
r3 90 60 80 80 60 40
3o dia
r1 100 90 80 80 80 70
r2 90 70 90 90 70 90
r3 90 90 90 90 90 100
4o dia
r1 100 90 80 90 80 70
r2 90 80 90 90 80 100
r3 90 90 100 90 100 100
DP 5,77 5,77 10 0 11,55 17,32
M 93,33 86,67 90 90 86,67 90
r = repetição DP = desvio padrão M = média
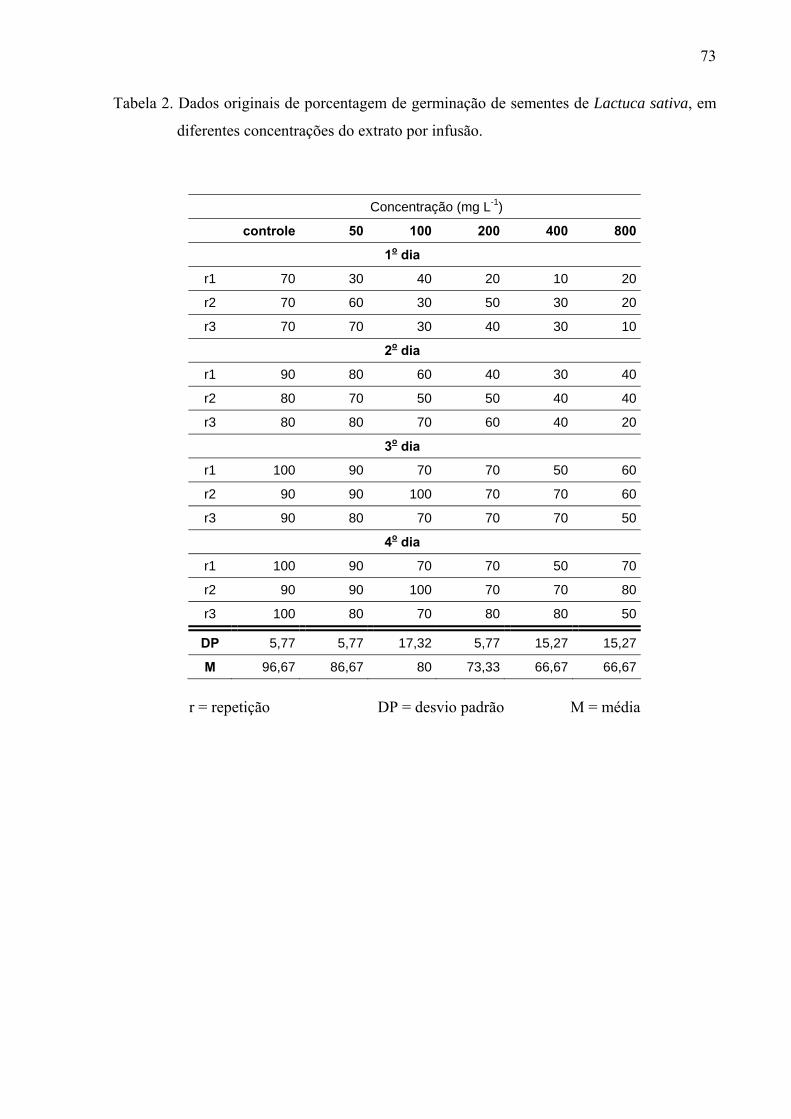
73
Tabela 2. Dados originais de porcentagem de germinação de sementes de Lactuca sativa, em
diferentes concentrações do extrato por infusão.
Concentração (mg L-1)
controle 50 100 200 400 800
1o dia
r1 70 30 40 20 10 20
r2 70 60 30 50 30 20
r3 70 70 30 40 30 10
2o dia
r1 90 80 60 40 30 40
r2 80 70 50 50 40 40
r3 80 80 70 60 40 20
3o dia
r1 100 90 70 70 50 60
r2 90 90 100 70 70 60
r3 90 80 70 70 70 50
4o dia
r1 100 90 70 70 50 70
r2 90 90 100 70 70 80
r3 100 80 70 80 80 50
DP 5,77 5,77 17,32 5,77 15,27 15,27
M 96,67 86,67 80 73,33 66,67 66,67
r = repetição DP = desvio padrão M = média
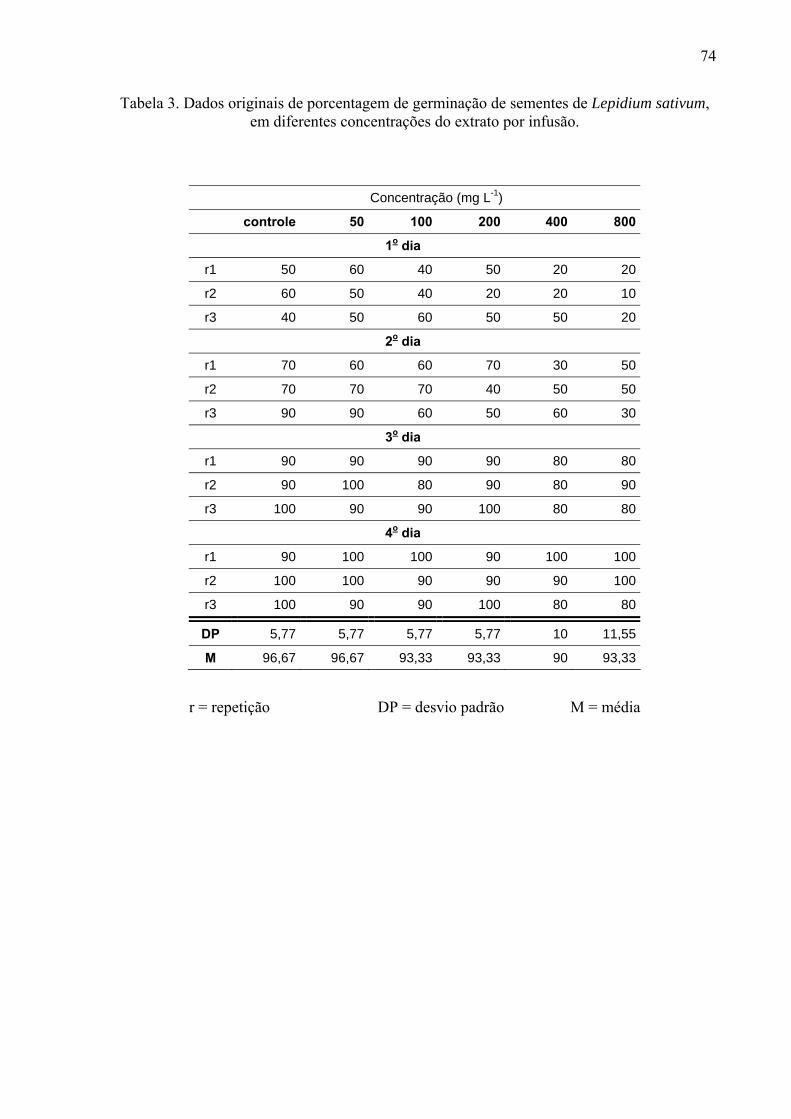
74
Tabela 3. Dados originais de porcentagem de germinação de sementes de Lepidium sativum, em diferentes concentrações do extrato por infusão.
Concentração (mg L-1)
controle 50 100 200 400 800
1o dia
r1 50 60 40 50 20 20
r2 60 50 40 20 20 10
r3 40 50 60 50 50 20
2o dia
r1 70 60 60 70 30 50
r2 70 70 70 40 50 50
r3 90 90 60 50 60 30
3o dia
r1 90 90 90 90 80 80
r2 90 100 80 90 80 90
r3 100 90 90 100 80 80
4o dia
r1 90 100 100 90 100 100
r2 100 100 90 90 90 100
r3 100 90 90 100 80 80
DP 5,77 5,77 5,77 5,77 10 11,55
M 96,67 96,67 93,33 93,33 90 93,33
r = repetição DP = desvio padrão M = média
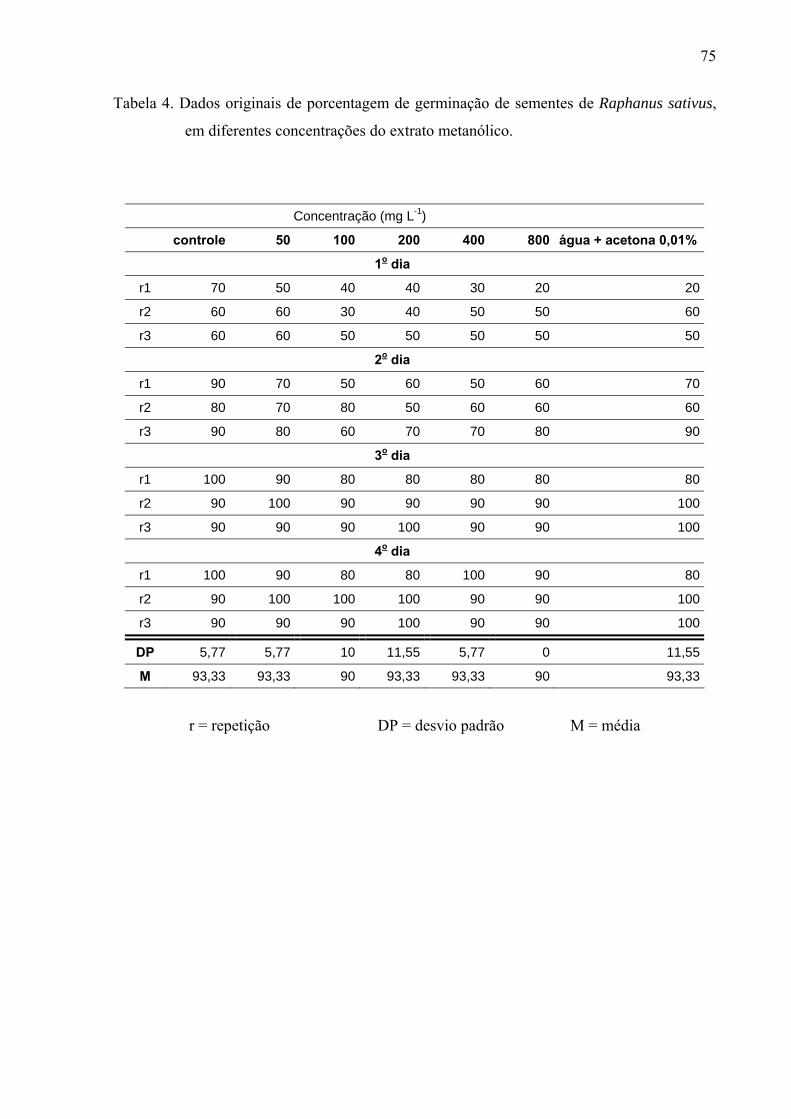
75
Tabela 4. Dados originais de porcentagem de germinação de sementes de Raphanus sativus,
em diferentes concentrações do extrato metanólico.
Concentração (mg L-1)
controle 50 100 200 400 800 água + acetona 0,01%
1o dia
r1 70 50 40 40 30 20 20
r2 60 60 30 40 50 50 60
r3 60 60 50 50 50 50 50
2o dia
r1 90 70 50 60 50 60 70
r2 80 70 80 50 60 60 60
r3 90 80 60 70 70 80 90
3o dia
r1 100 90 80 80 80 80 80
r2 90 100 90 90 90 90 100
r3 90 90 90 100 90 90 100
4o dia
r1 100 90 80 80 100 90 80
r2 90 100 100 100 90 90 100
r3 90 90 90 100 90 90 100
DP 5,77 5,77 10 11,55 5,77 0 11,55
M 93,33 93,33 90 93,33 93,33 90 93,33
r = repetição DP = desvio padrão M = média
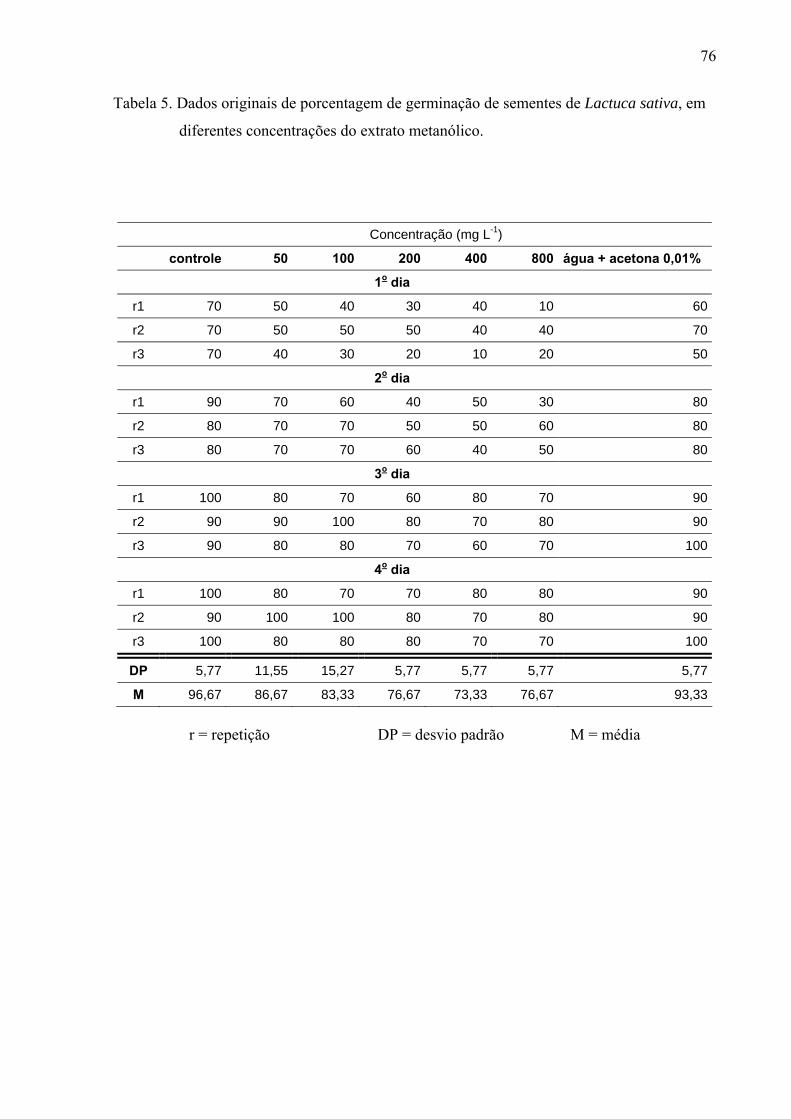
76
Tabela 5. Dados originais de porcentagem de germinação de sementes de Lactuca sativa, em
diferentes concentrações do extrato metanólico.
Concentração (mg L-1)
controle 50 100 200 400 800 água + acetona 0,01%
1o dia
r1 70 50 40 30 40 10 60
r2 70 50 50 50 40 40 70
r3 70 40 30 20 10 20 50
2o dia
r1 90 70 60 40 50 30 80
r2 80 70 70 50 50 60 80
r3 80 70 70 60 40 50 80
3o dia
r1 100 80 70 60 80 70 90
r2 90 90 100 80 70 80 90
r3 90 80 80 70 60 70 100
4o dia
r1 100 80 70 70 80 80 90
r2 90 100 100 80 70 80 90
r3 100 80 80 80 70 70 100
DP 5,77 11,55 15,27 5,77 5,77 5,77 5,77
M 96,67 86,67 83,33 76,67 73,33 76,67 93,33
r = repetição DP = desvio padrão M = média
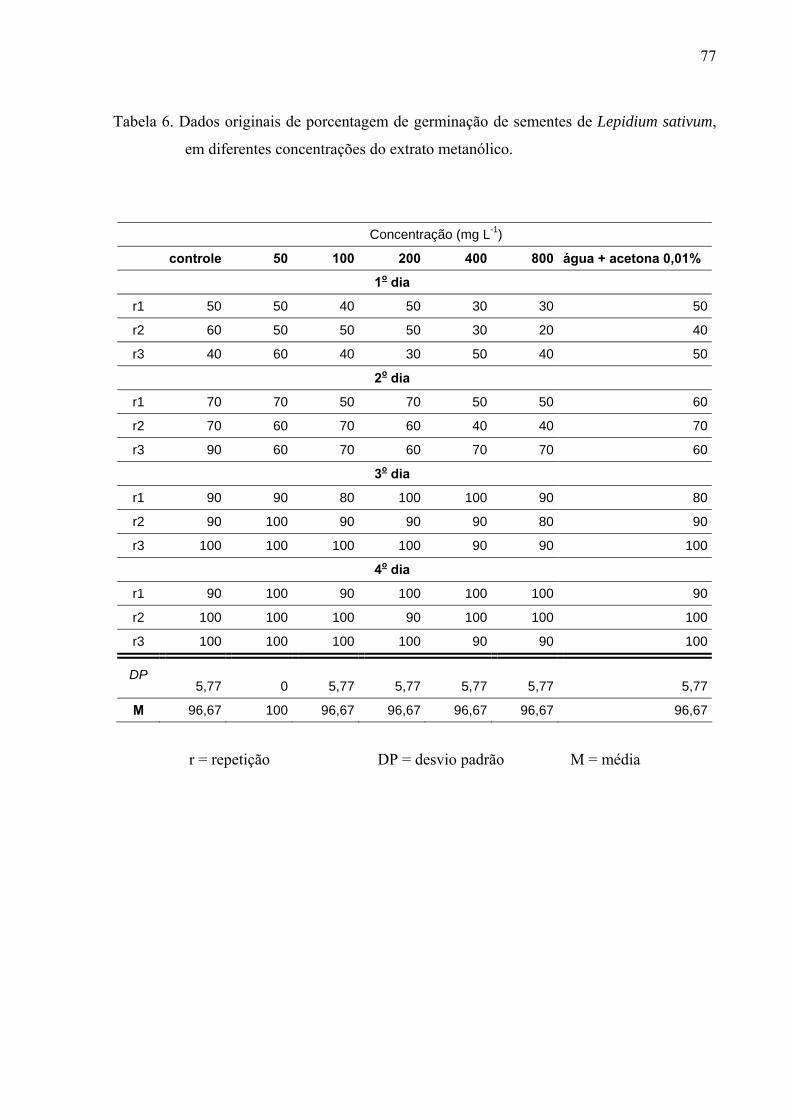
77
Tabela 6. Dados originais de porcentagem de germinação de sementes de Lepidium sativum,
em diferentes concentrações do extrato metanólico.
Concentração (mg L-1)
controle 50 100 200 400 800 água + acetona 0,01%
1o dia
r1 50 50 40 50 30 30 50
r2 60 50 50 50 30 20 40
r3 40 60 40 30 50 40 50
2o dia
r1 70 70 50 70 50 50 60
r2 70 60 70 60 40 40 70
r3 90 60 70 60 70 70 60
3o dia
r1 90 90 80 100 100 90 80
r2 90 100 90 90 90 80 90
r3 100 100 100 100 90 90 100
4o dia
r1 90 100 90 100 100 100 90
r2 100 100 100 90 100 100 100
r3 100 100 100 100 90 90 100
DP 5,77 0 5,77 5,77 5,77 5,77 5,77
M 96,67 100 96,67 96,67 96,67 96,67 96,67
r = repetição DP = desvio padrão M = média
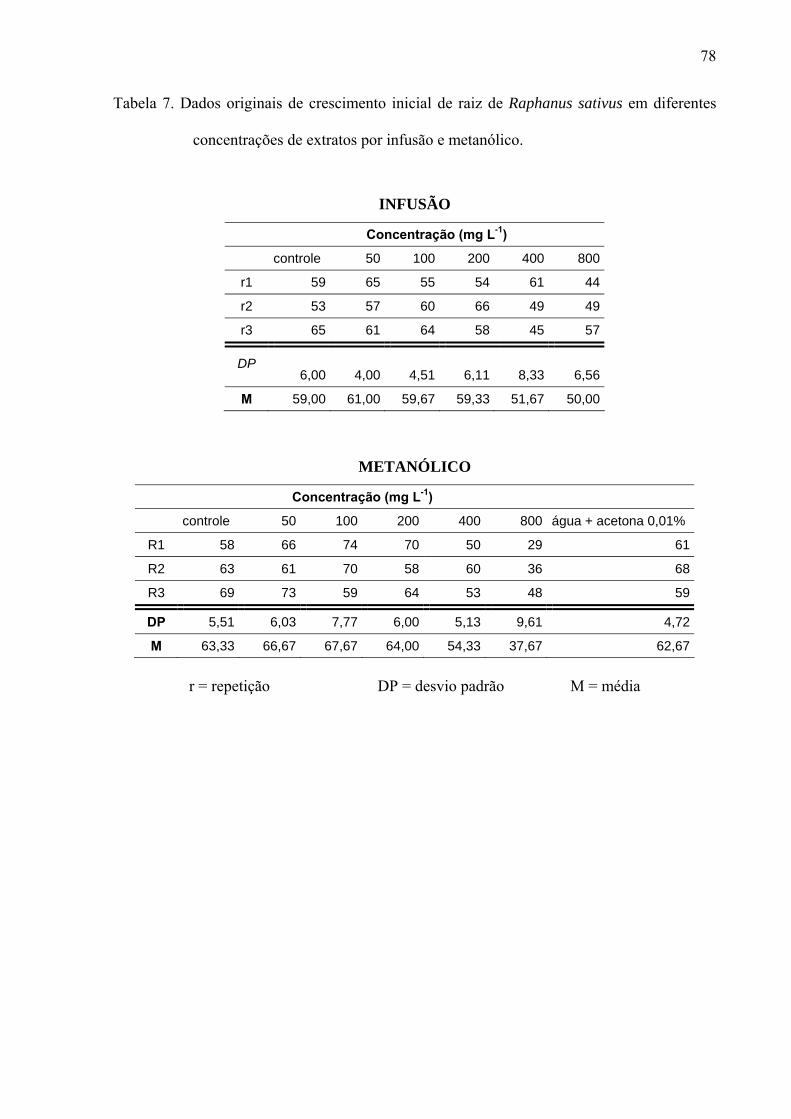
78
Tabela 7. Dados originais de crescimento inicial de raiz de Raphanus sativus em diferentes
concentrações de extratos por infusão e metanólico.
INFUSÃO
Concentração (mg L-1)
controle 50 100 200 400 800
r1 59 65 55 54 61 44
r2 53 57 60 66 49 49
r3 65 61 64 58 45 57
DP 6,00 4,00 4,51 6,11 8,33 6,56
M 59,00 61,00 59,67 59,33 51,67 50,00
METANÓLICO
Concentração (mg L-1)
controle 50 100 200 400 800 água + acetona 0,01%
R1 58 66 74 70 50 29 61
R2 63 61 70 58 60 36 68
R3 69 73 59 64 53 48 59
DP 5,51 6,03 7,77 6,00 5,13 9,61 4,72
M 63,33 66,67 67,67 64,00 54,33 37,67 62,67
r = repetição DP = desvio padrão M = média
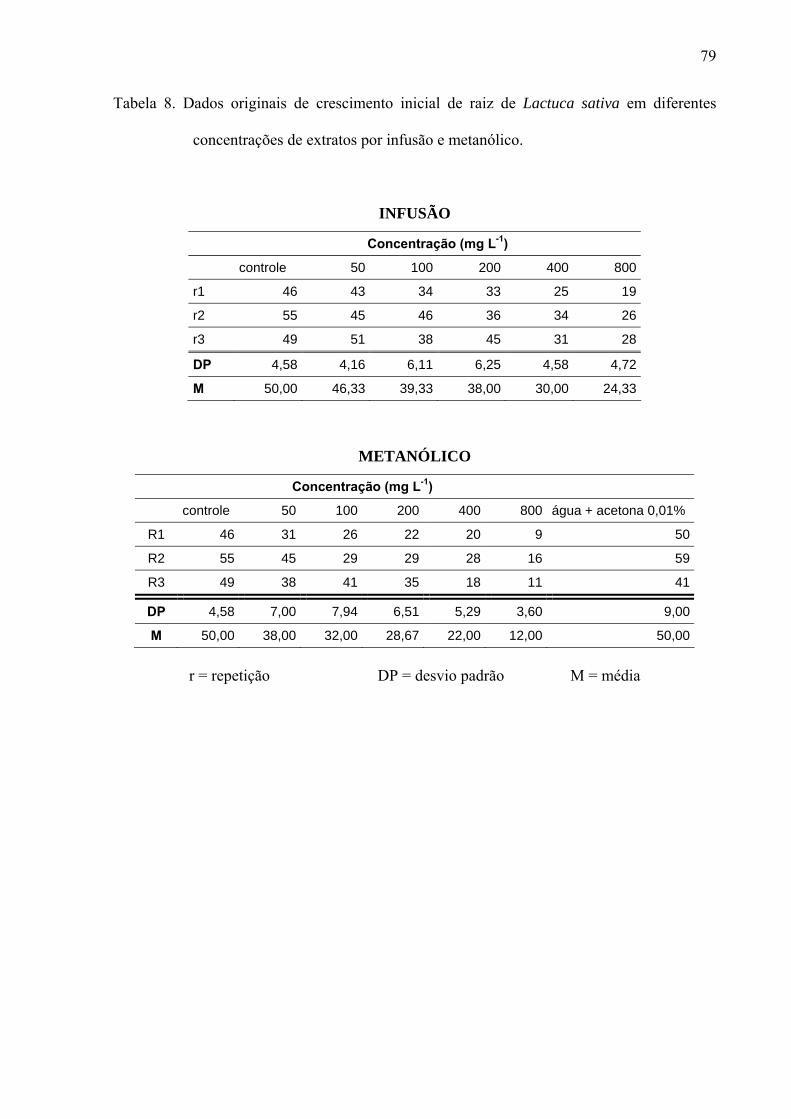
79
Tabela 8. Dados originais de crescimento inicial de raiz de Lactuca sativa em diferentes
concentrações de extratos por infusão e metanólico.
INFUSÃO
Concentração (mg L-1)
controle 50 100 200 400 800
r1 46 43 34 33 25 19
r2 55 45 46 36 34 26
r3 49 51 38 45 31 28
DP 4,58 4,16 6,11 6,25 4,58 4,72
M 50,00 46,33 39,33 38,00 30,00 24,33
METANÓLICO
Concentração (mg L-1)
controle 50 100 200 400 800 água + acetona 0,01%
R1 46 31 26 22 20 9 50
R2 55 45 29 29 28 16 59
R3 49 38 41 35 18 11 41
DP 4,58 7,00 7,94 6,51 5,29 3,60 9,00
M 50,00 38,00 32,00 28,67 22,00 12,00 50,00
r = repetição DP = desvio padrão M = média
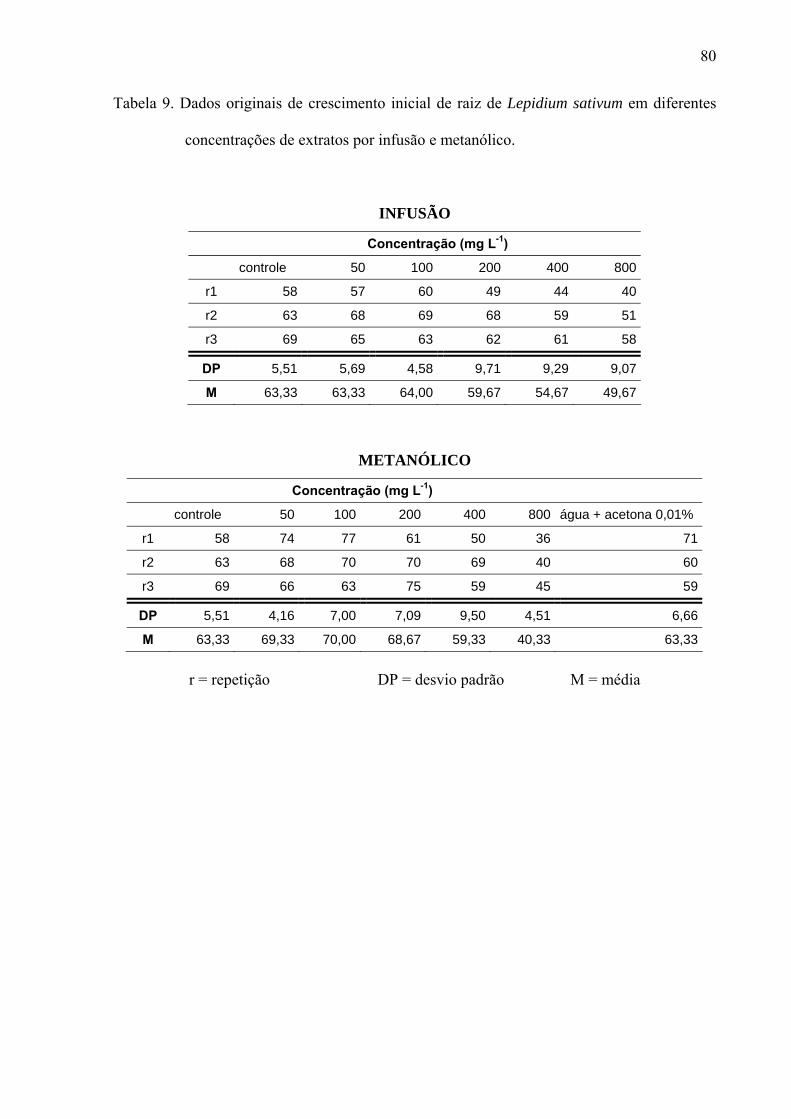
80
Tabela 9. Dados originais de crescimento inicial de raiz de Lepidium sativum em diferentes
concentrações de extratos por infusão e metanólico.
INFUSÃO
Concentração (mg L-1)
controle 50 100 200 400 800
r1 58 57 60 49 44 40
r2 63 68 69 68 59 51
r3 69 65 63 62 61 58
DP 5,51 5,69 4,58 9,71 9,29 9,07
M 63,33 63,33 64,00 59,67 54,67 49,67
METANÓLICO
Concentração (mg L-1)
controle 50 100 200 400 800 água + acetona 0,01%
r1 58 74 77 61 50 36 71
r2 63 68 70 70 69 40 60
r3 69 66 63 75 59 45 59
DP 5,51 4,16 7,00 7,09 9,50 4,51 6,66
M 63,33 69,33 70,00 68,67 59,33 40,33 63,33
r = repetição DP = desvio padrão M = média
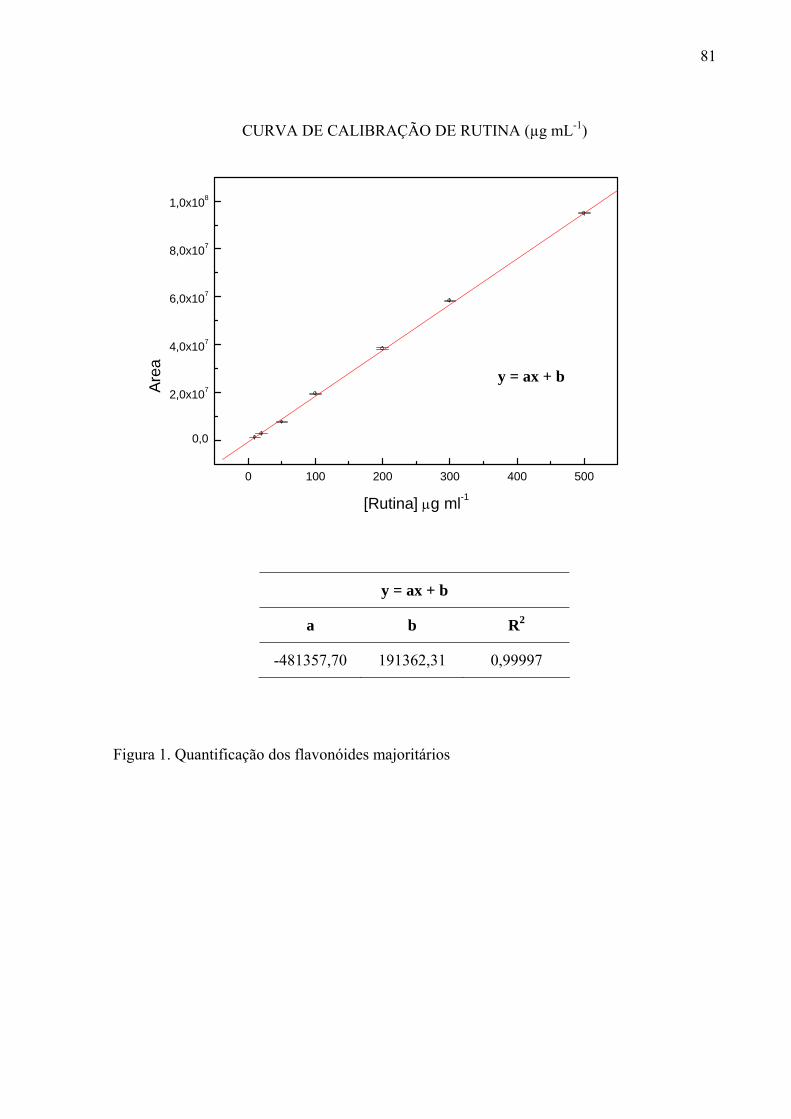
81
CURVA DE CALIBRAÇÃO DE RUTINA (µg mL-1)
Figura 1. Quantificação dos flavonóides majoritários
y = ax + b
a b R2
-481357,70 191362,31 0,99997
0 100 200 300 400 500
0,0
2,0x107
4,0x107
6,0x107
8,0x107
1,0x108
Area
(Con
tage
ns)
[Rutina] μg ml-1
y = ax + b
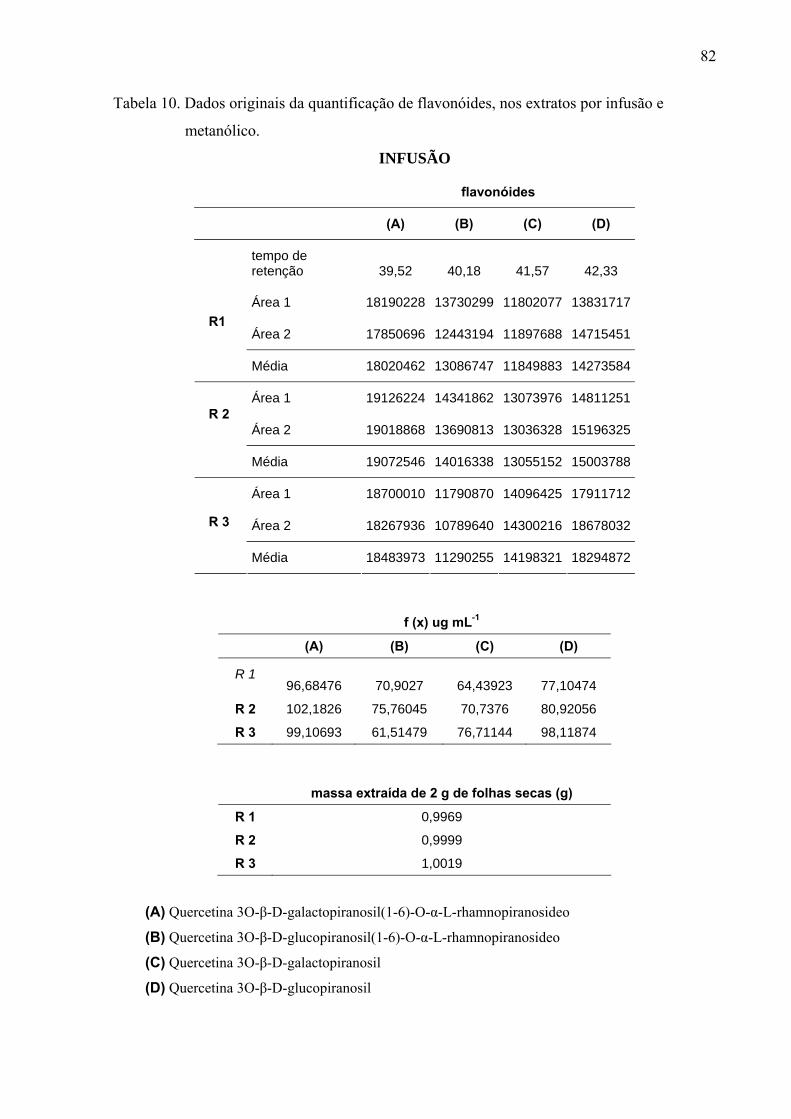
82
Tabela 10. Dados originais da quantificação de flavonóides, nos extratos por infusão e
metanólico.
INFUSÃO
flavonóides
(A) (B) (C) (D)
tempo de retenção 39,52 40,18 41,57 42,33
Área 1 18190228 13730299 11802077 13831717
Área 2 17850696 12443194 11897688 14715451
Média 18020462 13086747 11849883 14273584
Área 1 19126224 14341862 13073976 14811251
Área 2 19018868 13690813 13036328 15196325
Média 19072546 14016338 13055152 15003788
Área 1 18700010 11790870 14096425 17911712
Área 2 18267936 10789640 14300216 18678032
Média 18483973 11290255 14198321 18294872
f (x) ug mL-1
(A) (B) (C) (D)
R 1 96,68476 70,9027 64,43923 77,10474
R 2 102,1826 75,76045 70,7376 80,92056
R 3 99,10693 61,51479 76,71144 98,11874
massa extraída de 2 g de folhas secas (g)
R 1 0,9969
R 2 0,9999
R 3 1,0019
(A) Quercetina 3O-β-D-galactopiranosil(1-6)-O-α-L-rhamnopiranosideo
(B) Quercetina 3O-β-D-glucopiranosil(1-6)-O-α-L-rhamnopiranosideo
(C) Quercetina 3O-β-D-galactopiranosil
(D) Quercetina 3O-β-D-glucopiranosil
R1
R 2
R 3
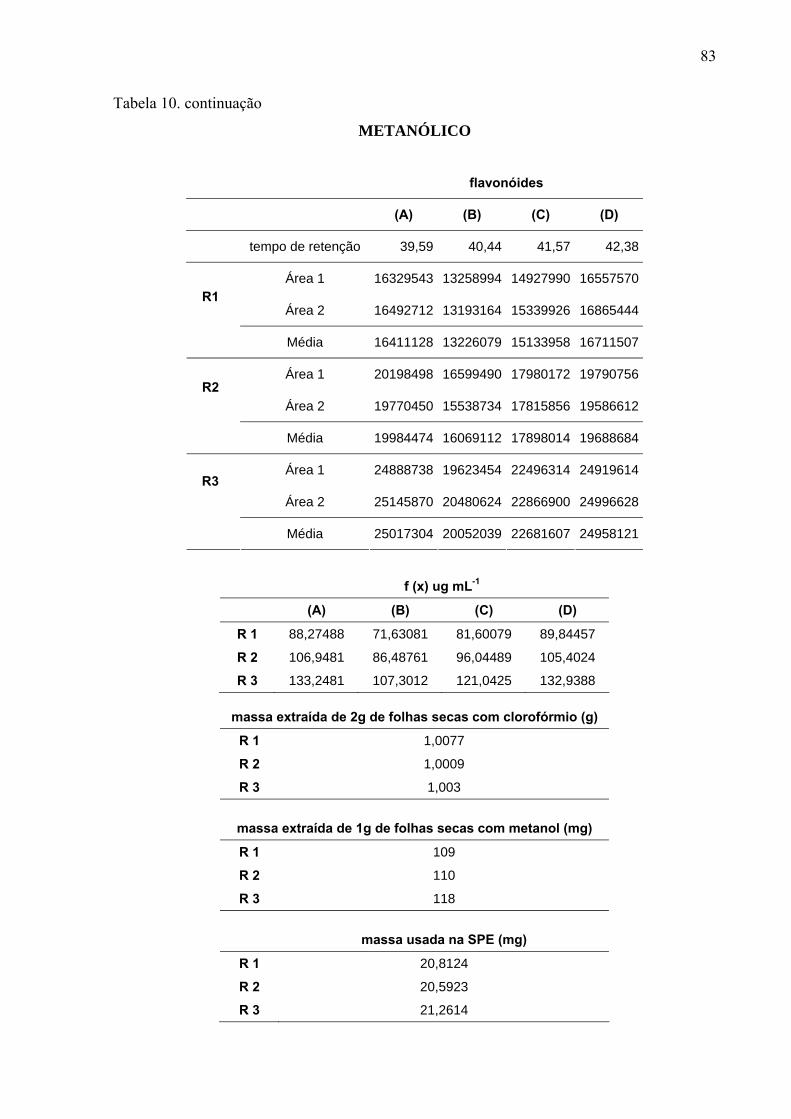
83
Tabela 10. continuação
METANÓLICO
flavonóides
(A) (B) (C) (D)
tempo de retenção 39,59 40,44 41,57 42,38
Área 1 16329543 13258994 14927990 16557570
Área 2 16492712 13193164 15339926 16865444
Média 16411128 13226079 15133958 16711507
Área 1 20198498 16599490 17980172 19790756
Área 2 19770450 15538734 17815856 19586612
Média 19984474 16069112 17898014 19688684
Área 1 24888738 19623454 22496314 24919614
Área 2 25145870 20480624 22866900 24996628
Média 25017304 20052039 22681607 24958121
f (x) ug mL-1
(A) (B) (C) (D)
R 1 88,27488 71,63081 81,60079 89,84457
R 2 106,9481 86,48761 96,04489 105,4024
R 3 133,2481 107,3012 121,0425 132,9388
massa extraída de 2g de folhas secas com clorofórmio (g)
R 1 1,0077
R 2 1,0009
R 3 1,003
massa extraída de 1g de folhas secas com metanol (mg)
R 1 109
R 2 110
R 3 118 massa usada na SPE (mg)
R 1 20,8124
R 2 20,5923
R 3 21,2614
R1
R2
R3

84
Figura 2. Espectro de RMN de 1H do flavonoide isolado quercetina-3-O-galactose-
rhamnose
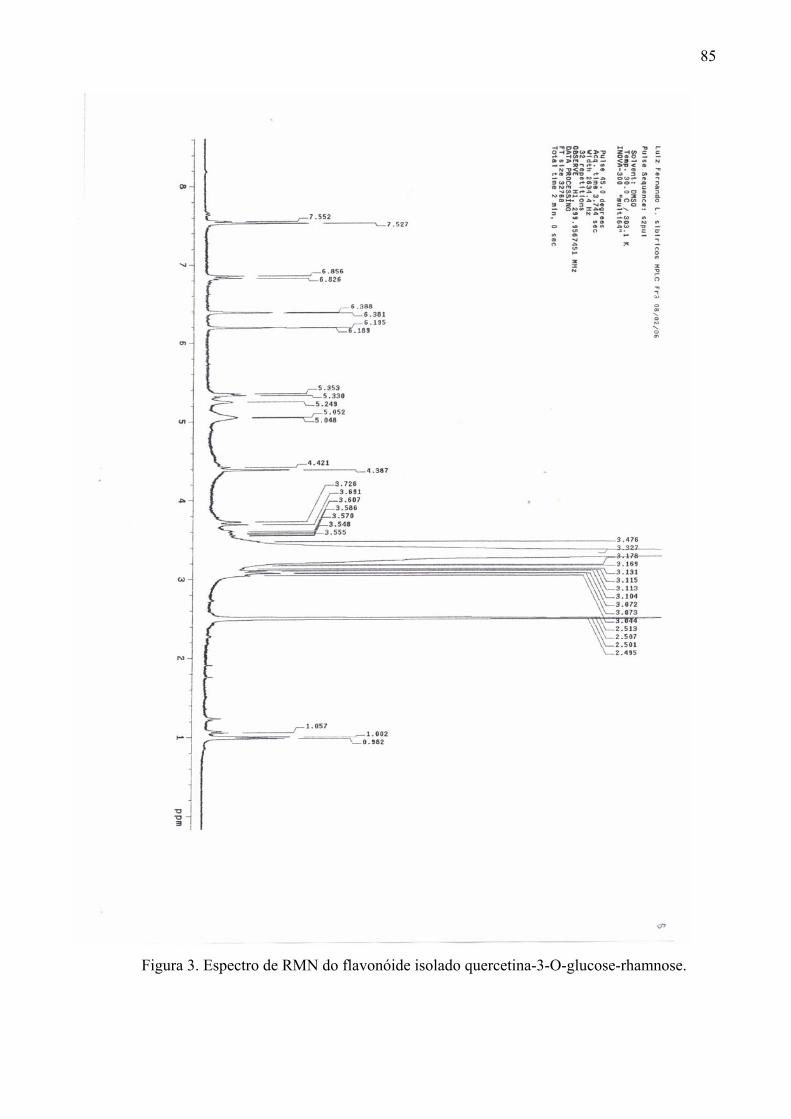
85
Figura 3. Espectro de RMN do flavonóide isolado quercetina-3-O-glucose-rhamnose.

86
Figura 4. Espectro de RMN do flavonóide isolado quercetina-3-O-galactose.

87
Figura 5. Espectro de RMN do flavonóide isolado quercetina-3-O-glucose.

88
Figura 6. Espectro de RMN do flavonóide isolado Genkanina-3-hidroxi.

89
Figura 7. Espectro de RMN do flavonóide isolado Genkanina.
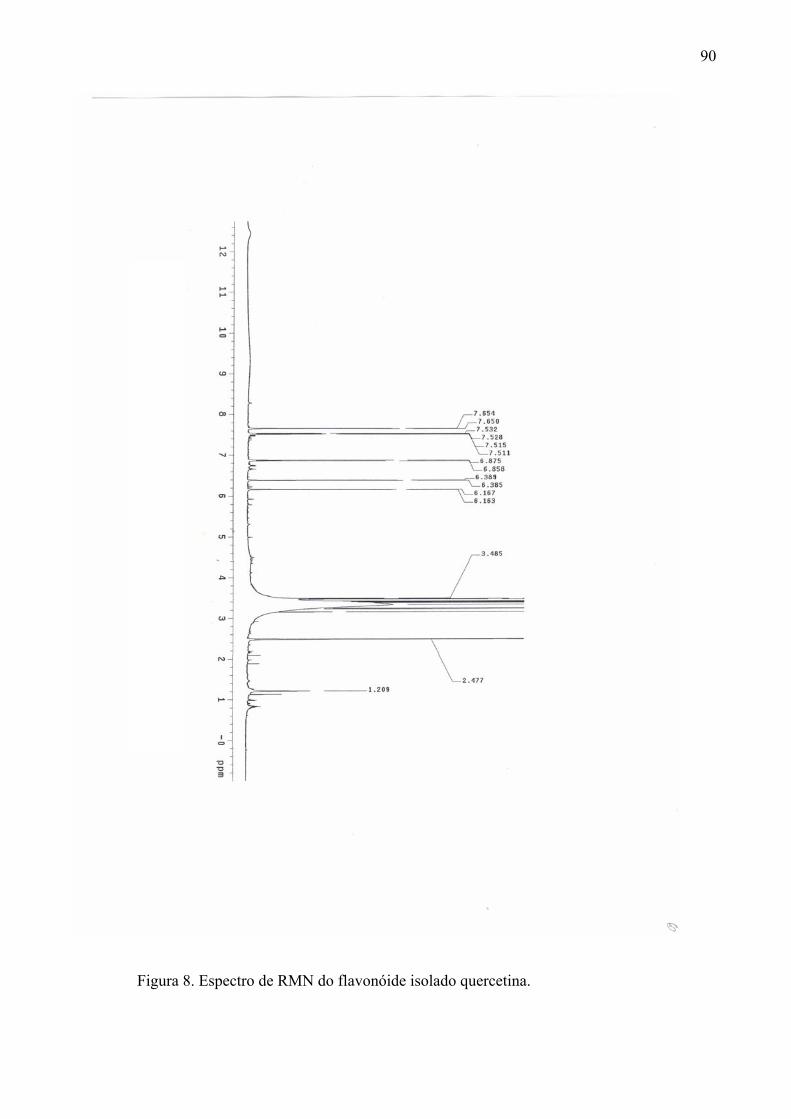
90
Figura 8. Espectro de RMN do flavonóide isolado quercetina.
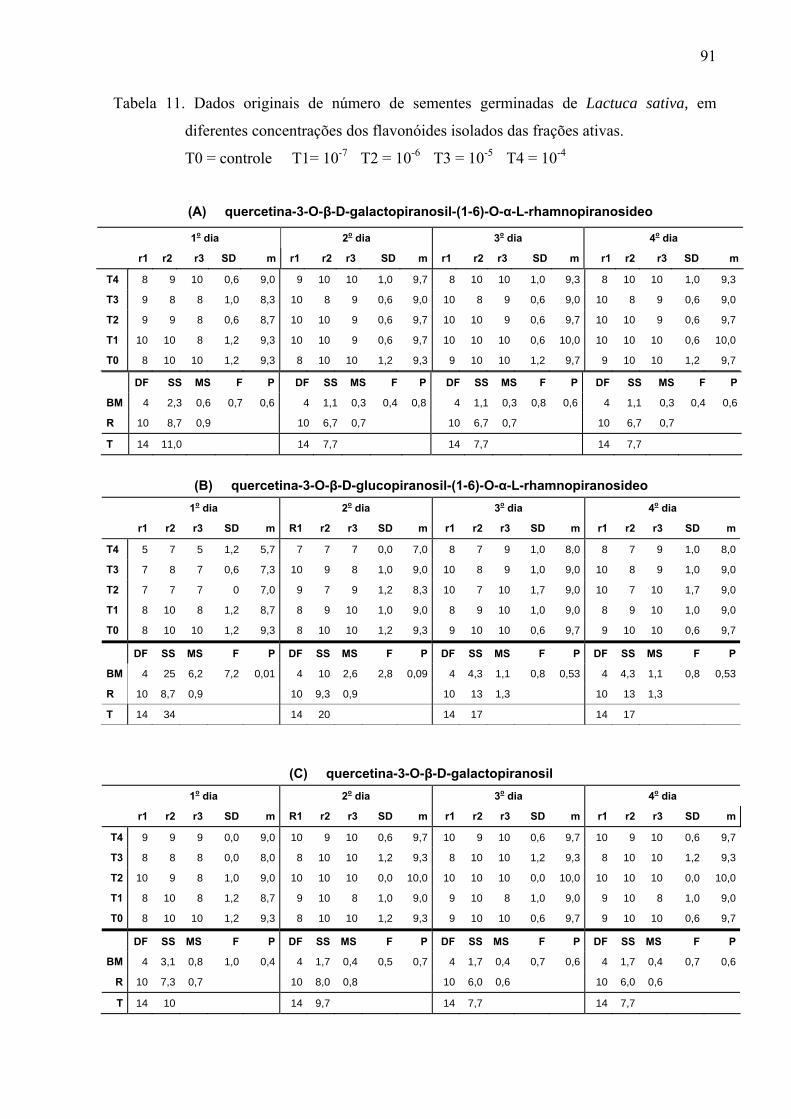
91
Tabela 11. Dados originais de número de sementes germinadas de Lactuca sativa, em
diferentes concentrações dos flavonóides isolados das frações ativas.
T0 = controle T1= 10-7 T2 = 10-6 T3 = 10-5 T4 = 10-4
(A) quercetina-3-O-β-D-galactopiranosil-(1-6)-O-α-L-rhamnopiranosideo
1o dia 2o dia 3o dia 4o dia
r1 r2 r3 SD m r1 r2 r3 SD m r1 r2 r3 SD m r1 r2 r3 SD m
T4 8 9 10 0,6 9,0 9 10 10 1,0 9,7 8 10 10 1,0 9,3 8 10 10 1,0 9,3
T3 9 8 8 1,0 8,3 10 8 9 0,6 9,0 10 8 9 0,6 9,0 10 8 9 0,6 9,0
T2 9 9 8 0,6 8,7 10 10 9 0,6 9,7 10 10 9 0,6 9,7 10 10 9 0,6 9,7
T1 10 10 8 1,2 9,3 10 10 9 0,6 9,7 10 10 10 0,6 10,0 10 10 10 0,6 10,0
T0 8 10 10 1,2 9,3 8 10 10 1,2 9,3 9 10 10 1,2 9,7 9 10 10 1,2 9,7
DF SS MS F P DF SS MS F P DF SS MS F P DF SS MS F P
BM 4 2,3 0,6 0,7 0,6 4 1,1 0,3 0,4 0,8 4 1,1 0,3 0,8 0,6 4 1,1 0,3 0,4 0,6
R 10 8,7 0,9 10 6,7 0,7 10 6,7 0,7 10 6,7 0,7
T 14 11,0 14 7,7 14 7,7 14 7,7
(B) quercetina-3-O-β-D-glucopiranosil-(1-6)-O-α-L-rhamnopiranosideo
1o dia 2o dia 3o dia 4o dia
r1 r2 r3 SD m R1 r2 r3 SD m r1 r2 r3 SD m r1 r2 r3 SD m
T4 5 7 5 1,2 5,7 7 7 7 0,0 7,0 8 7 9 1,0 8,0 8 7 9 1,0 8,0
T3 7 8 7 0,6 7,3 10 9 8 1,0 9,0 10 8 9 1,0 9,0 10 8 9 1,0 9,0
T2 7 7 7 0 7,0 9 7 9 1,2 8,3 10 7 10 1,7 9,0 10 7 10 1,7 9,0
T1 8 10 8 1,2 8,7 8 9 10 1,0 9,0 8 9 10 1,0 9,0 8 9 10 1,0 9,0
T0 8 10 10 1,2 9,3 8 10 10 1,2 9,3 9 10 10 0,6 9,7 9 10 10 0,6 9,7
DF SS MS F P DF SS MS F P DF SS MS F P DF SS MS F P
BM 4 25 6,2 7,2 0,01 4 10 2,6 2,8 0,09 4 4,3 1,1 0,8 0,53 4 4,3 1,1 0,8 0,53
R 10 8,7 0,9 10 9,3 0,9 10 13 1,3 10 13 1,3
T 14 34 14 20 14 17 14 17
(C) quercetina-3-O-β-D-galactopiranosil 1o dia 2o dia 3o dia 4o dia
r1 r2 r3 SD m R1 r2 r3 SD m r1 r2 r3 SD m r1 r2 r3 SD m
T4 9 9 9 0,0 9,0 10 9 10 0,6 9,7 10 9 10 0,6 9,7 10 9 10 0,6 9,7
T3 8 8 8 0,0 8,0 8 10 10 1,2 9,3 8 10 10 1,2 9,3 8 10 10 1,2 9,3
T2 10 9 8 1,0 9,0 10 10 10 0,0 10,0 10 10 10 0,0 10,0 10 10 10 0,0 10,0
T1 8 10 8 1,2 8,7 9 10 8 1,0 9,0 9 10 8 1,0 9,0 9 10 8 1,0 9,0
T0 8 10 10 1,2 9,3 8 10 10 1,2 9,3 9 10 10 0,6 9,7 9 10 10 0,6 9,7
DF SS MS F P DF SS MS F P DF SS MS F P DF SS MS F P
BM 4 3,1 0,8 1,0 0,4 4 1,7 0,4 0,5 0,7 4 1,7 0,4 0,7 0,6 4 1,7 0,4 0,7 0,6
R 10 7,3 0,7 10 8,0 0,8 10 6,0 0,6 10 6,0 0,6
T 14 10 14 9,7 14 7,7 14 7,7
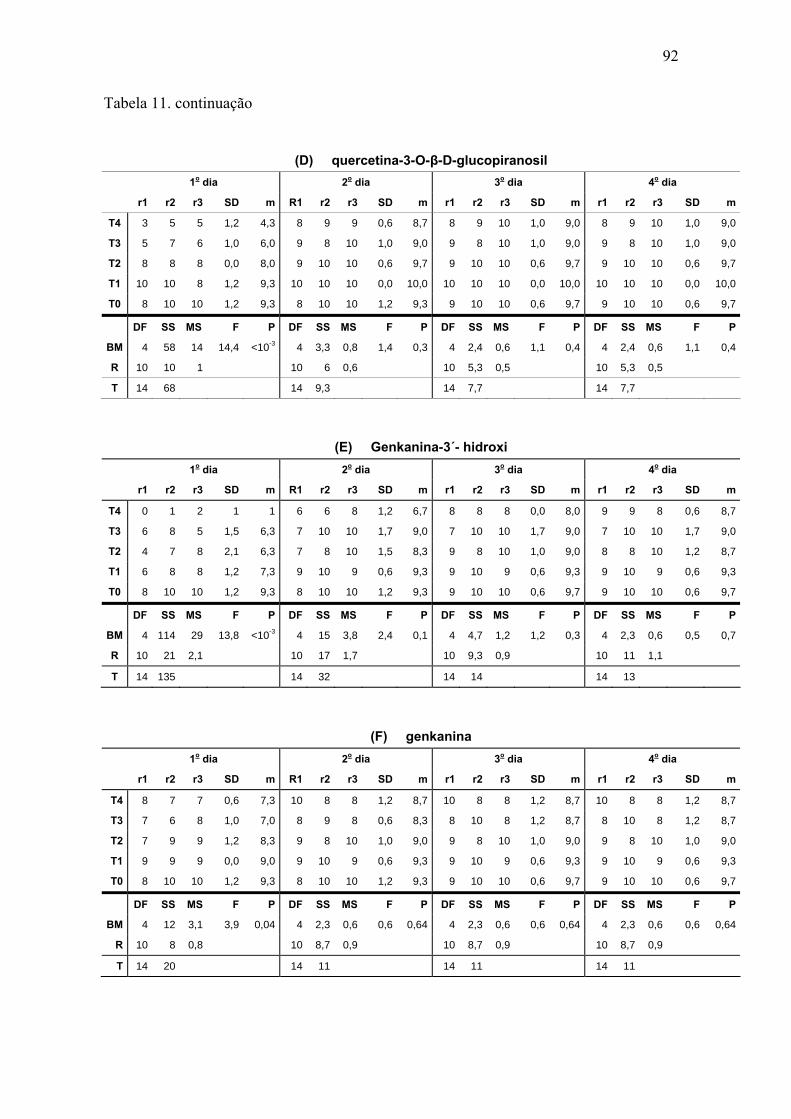
92
Tabela 11. continuação
(D) quercetina-3-O-β-D-glucopiranosil 1o dia 2o dia 3o dia 4o dia
r1 r2 r3 SD m R1 r2 r3 SD m r1 r2 r3 SD m r1 r2 r3 SD m
T4 3 5 5 1,2 4,3 8 9 9 0,6 8,7 8 9 10 1,0 9,0 8 9 10 1,0 9,0
T3 5 7 6 1,0 6,0 9 8 10 1,0 9,0 9 8 10 1,0 9,0 9 8 10 1,0 9,0
T2 8 8 8 0,0 8,0 9 10 10 0,6 9,7 9 10 10 0,6 9,7 9 10 10 0,6 9,7
T1 10 10 8 1,2 9,3 10 10 10 0,0 10,0 10 10 10 0,0 10,0 10 10 10 0,0 10,0
T0 8 10 10 1,2 9,3 8 10 10 1,2 9,3 9 10 10 0,6 9,7 9 10 10 0,6 9,7
DF SS MS F P DF SS MS F P DF SS MS F P DF SS MS F P
BM 4 58 14 14,4 <10-3 4 3,3 0,8 1,4 0,3 4 2,4 0,6 1,1 0,4 4 2,4 0,6 1,1 0,4
R 10 10 1 10 6 0,6 10 5,3 0,5 10 5,3 0,5
T 14 68 14 9,3 14 7,7 14 7,7
(E) Genkanina-3´- hidroxi 1o dia 2o dia 3o dia 4o dia
r1 r2 r3 SD m R1 r2 r3 SD m r1 r2 r3 SD m r1 r2 r3 SD m
T4 0 1 2 1 1 6 6 8 1,2 6,7 8 8 8 0,0 8,0 9 9 8 0,6 8,7
T3 6 8 5 1,5 6,3 7 10 10 1,7 9,0 7 10 10 1,7 9,0 7 10 10 1,7 9,0
T2 4 7 8 2,1 6,3 7 8 10 1,5 8,3 9 8 10 1,0 9,0 8 8 10 1,2 8,7
T1 6 8 8 1,2 7,3 9 10 9 0,6 9,3 9 10 9 0,6 9,3 9 10 9 0,6 9,3
T0 8 10 10 1,2 9,3 8 10 10 1,2 9,3 9 10 10 0,6 9,7 9 10 10 0,6 9,7
DF SS MS F P DF SS MS F P DF SS MS F P DF SS MS F P
BM 4 114 29 13,8 <10-3 4 15 3,8 2,4 0,1 4 4,7 1,2 1,2 0,3 4 2,3 0,6 0,5 0,7
R 10 21 2,1 10 17 1,7 10 9,3 0,9 10 11 1,1
T 14 135 14 32 14 14 14 13
(F) genkanina 1o dia 2o dia 3o dia 4o dia
r1 r2 r3 SD m R1 r2 r3 SD m r1 r2 r3 SD m r1 r2 r3 SD m
T4 8 7 7 0,6 7,3 10 8 8 1,2 8,7 10 8 8 1,2 8,7 10 8 8 1,2 8,7
T3 7 6 8 1,0 7,0 8 9 8 0,6 8,3 8 10 8 1,2 8,7 8 10 8 1,2 8,7
T2 7 9 9 1,2 8,3 9 8 10 1,0 9,0 9 8 10 1,0 9,0 9 8 10 1,0 9,0
T1 9 9 9 0,0 9,0 9 10 9 0,6 9,3 9 10 9 0,6 9,3 9 10 9 0,6 9,3
T0 8 10 10 1,2 9,3 8 10 10 1,2 9,3 9 10 10 0,6 9,7 9 10 10 0,6 9,7
DF SS MS F P DF SS MS F P DF SS MS F P DF SS MS F P
BM 4 12 3,1 3,9 0,04 4 2,3 0,6 0,6 0,64 4 2,3 0,6 0,6 0,64 4 2,3 0,6 0,6 0,64
R 10 8 0,8 10 8,7 0,9 10 8,7 0,9 10 8,7 0,9
T 14 20 14 11 14 11 14 11
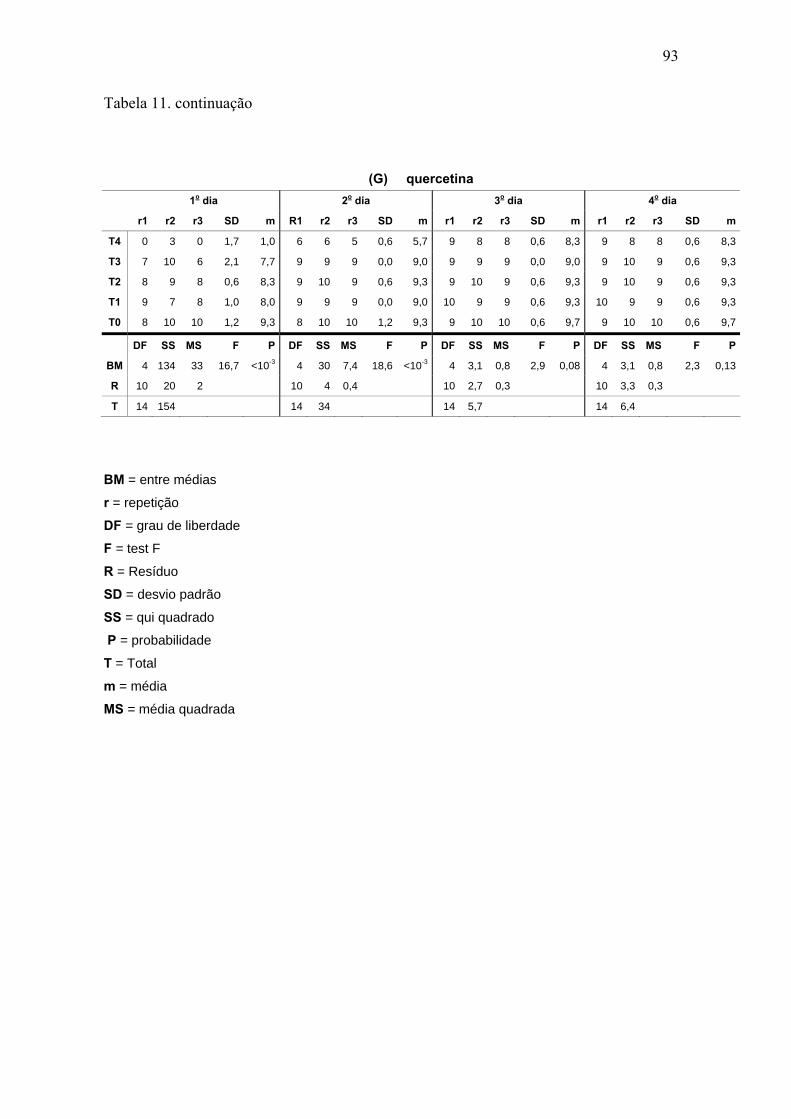
93
Tabela 11. continuação
(G) quercetina 1o dia 2o dia 3o dia 4o dia
r1 r2 r3 SD m R1 r2 r3 SD m r1 r2 r3 SD m r1 r2 r3 SD m
T4 0 3 0 1,7 1,0 6 6 5 0,6 5,7 9 8 8 0,6 8,3 9 8 8 0,6 8,3
T3 7 10 6 2,1 7,7 9 9 9 0,0 9,0 9 9 9 0,0 9,0 9 10 9 0,6 9,3
T2 8 9 8 0,6 8,3 9 10 9 0,6 9,3 9 10 9 0,6 9,3 9 10 9 0,6 9,3
T1 9 7 8 1,0 8,0 9 9 9 0,0 9,0 10 9 9 0,6 9,3 10 9 9 0,6 9,3
T0 8 10 10 1,2 9,3 8 10 10 1,2 9,3 9 10 10 0,6 9,7 9 10 10 0,6 9,7
DF SS MS F P DF SS MS F P DF SS MS F P DF SS MS F P
BM 4 134 33 16,7 <10-3 4 30 7,4 18,6 <10-3 4 3,1 0,8 2,9 0,08 4 3,1 0,8 2,3 0,13
R 10 20 2 10 4 0,4 10 2,7 0,3 10 3,3 0,3
T 14 154 14 34 14 5,7 14 6,4
BM = entre médias
r = repetição
DF = grau de liberdade
F = test F
R = Resíduo
SD = desvio padrão
SS = qui quadrado
P = probabilidade
T = Total
m = média
MS = média quadrada
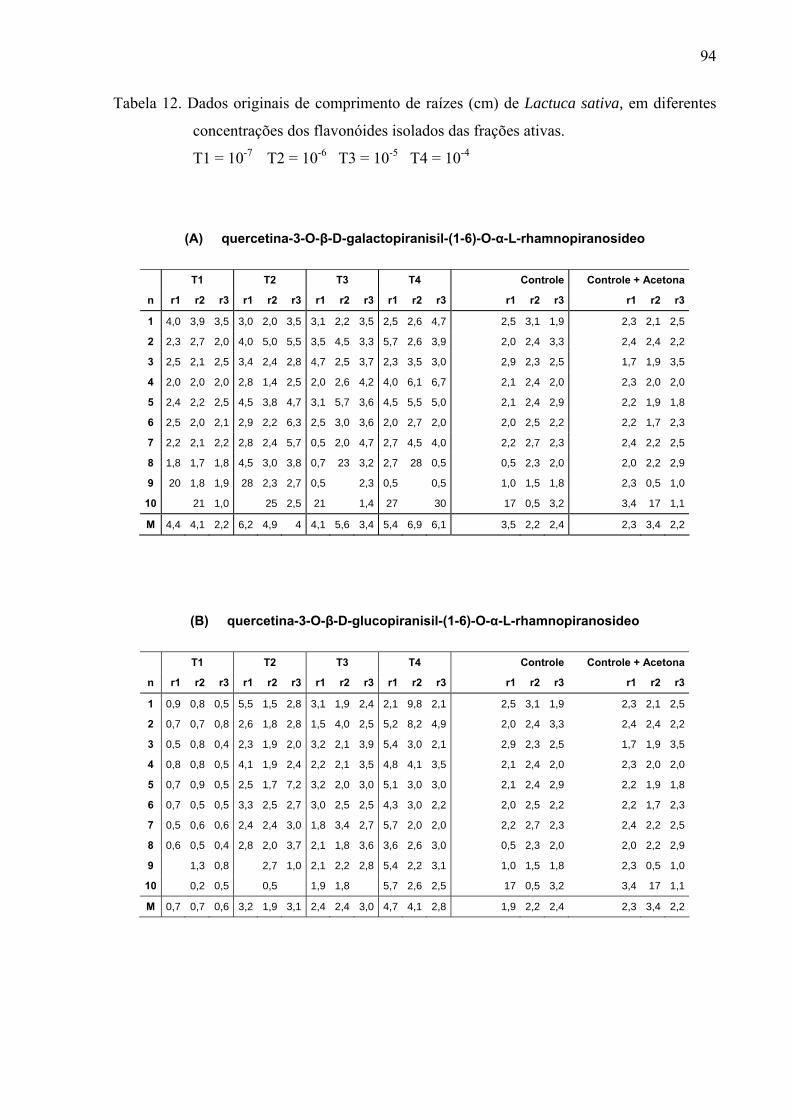
94
Tabela 12. Dados originais de comprimento de raízes (cm) de Lactuca sativa, em diferentes
concentrações dos flavonóides isolados das frações ativas.
T1 = 10-7 T2 = 10-6 T3 = 10-5 T4 = 10-4
(A) quercetina-3-O-β-D-galactopiranisil-(1-6)-O-α-L-rhamnopiranosideo
T1 T2 T3 T4 Controle Controle + Acetona
n r1 r2 r3 r1 r2 r3 r1 r2 r3 r1 r2 r3 r1 r2 r3 r1 r2 r3
1 4,0 3,9 3,5 3,0 2,0 3,5 3,1 2,2 3,5 2,5 2,6 4,7 2,5 3,1 1,9 2,3 2,1 2,5
2 2,3 2,7 2,0 4,0 5,0 5,5 3,5 4,5 3,3 5,7 2,6 3,9 2,0 2,4 3,3 2,4 2,4 2,2
3 2,5 2,1 2,5 3,4 2,4 2,8 4,7 2,5 3,7 2,3 3,5 3,0 2,9 2,3 2,5 1,7 1,9 3,5
4 2,0 2,0 2,0 2,8 1,4 2,5 2,0 2,6 4,2 4,0 6,1 6,7 2,1 2,4 2,0 2,3 2,0 2,0
5 2,4 2,2 2,5 4,5 3,8 4,7 3,1 5,7 3,6 4,5 5,5 5,0 2,1 2,4 2,9 2,2 1,9 1,8
6 2,5 2,0 2,1 2,9 2,2 6,3 2,5 3,0 3,6 2,0 2,7 2,0 2,0 2,5 2,2 2,2 1,7 2,3
7 2,2 2,1 2,2 2,8 2,4 5,7 0,5 2,0 4,7 2,7 4,5 4,0 2,2 2,7 2,3 2,4 2,2 2,5
8 1,8 1,7 1,8 4,5 3,0 3,8 0,7 23 3,2 2,7 28 0,5 0,5 2,3 2,0 2,0 2,2 2,9
9 20 1,8 1,9 28 2,3 2,7 0,5 2,3 0,5 0,5 1,0 1,5 1,8 2,3 0,5 1,0
10 21 1,0 25 2,5 21 1,4 27 30 17 0,5 3,2 3,4 17 1,1
M 4,4 4,1 2,2 6,2 4,9 4 4,1 5,6 3,4 5,4 6,9 6,1 3,5 2,2 2,4 2,3 3,4 2,2
(B) quercetina-3-O-β-D-glucopiranisil-(1-6)-O-α-L-rhamnopiranosideo
T1 T2 T3 T4 Controle Controle + Acetona
n r1 r2 r3 r1 r2 r3 r1 r2 r3 r1 r2 r3 r1 r2 r3 r1 r2 r3
1 0,9 0,8 0,5 5,5 1,5 2,8 3,1 1,9 2,4 2,1 9,8 2,1 2,5 3,1 1,9 2,3 2,1 2,5
2 0,7 0,7 0,8 2,6 1,8 2,8 1,5 4,0 2,5 5,2 8,2 4,9 2,0 2,4 3,3 2,4 2,4 2,2
3 0,5 0,8 0,4 2,3 1,9 2,0 3,2 2,1 3,9 5,4 3,0 2,1 2,9 2,3 2,5 1,7 1,9 3,5
4 0,8 0,8 0,5 4,1 1,9 2,4 2,2 2,1 3,5 4,8 4,1 3,5 2,1 2,4 2,0 2,3 2,0 2,0
5 0,7 0,9 0,5 2,5 1,7 7,2 3,2 2,0 3,0 5,1 3,0 3,0 2,1 2,4 2,9 2,2 1,9 1,8
6 0,7 0,5 0,5 3,3 2,5 2,7 3,0 2,5 2,5 4,3 3,0 2,2 2,0 2,5 2,2 2,2 1,7 2,3
7 0,5 0,6 0,6 2,4 2,4 3,0 1,8 3,4 2,7 5,7 2,0 2,0 2,2 2,7 2,3 2,4 2,2 2,5
8 0,6 0,5 0,4 2,8 2,0 3,7 2,1 1,8 3,6 3,6 2,6 3,0 0,5 2,3 2,0 2,0 2,2 2,9
9 1,3 0,8 2,7 1,0 2,1 2,2 2,8 5,4 2,2 3,1 1,0 1,5 1,8 2,3 0,5 1,0
10 0,2 0,5 0,5 1,9 1,8 5,7 2,6 2,5 17 0,5 3,2 3,4 17 1,1
M 0,7 0,7 0,6 3,2 1,9 3,1 2,4 2,4 3,0 4,7 4,1 2,8 1,9 2,2 2,4 2,3 3,4 2,2
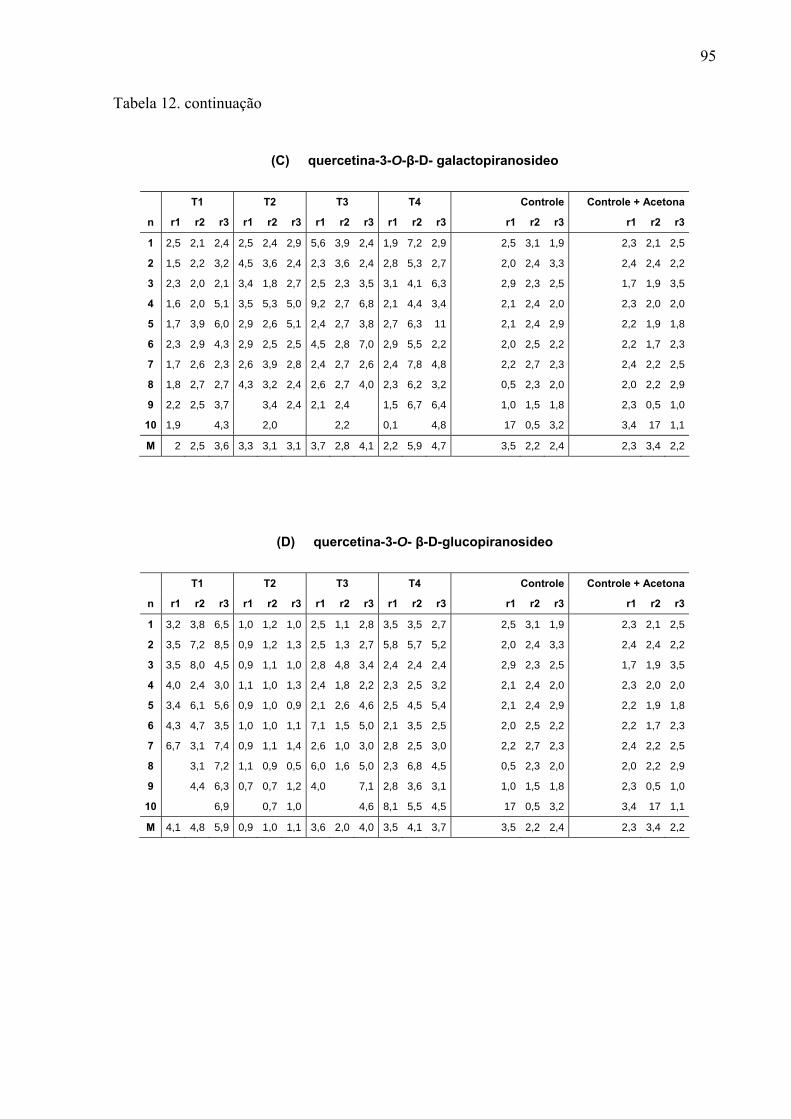
95
Tabela 12. continuação
(C) quercetina-3-O-β-D- galactopiranosideo
T1 T2 T3 T4 Controle Controle + Acetona
n r1 r2 r3 r1 r2 r3 r1 r2 r3 r1 r2 r3 r1 r2 r3 r1 r2 r3
1 2,5 2,1 2,4 2,5 2,4 2,9 5,6 3,9 2,4 1,9 7,2 2,9 2,5 3,1 1,9 2,3 2,1 2,5
2 1,5 2,2 3,2 4,5 3,6 2,4 2,3 3,6 2,4 2,8 5,3 2,7 2,0 2,4 3,3 2,4 2,4 2,2
3 2,3 2,0 2,1 3,4 1,8 2,7 2,5 2,3 3,5 3,1 4,1 6,3 2,9 2,3 2,5 1,7 1,9 3,5
4 1,6 2,0 5,1 3,5 5,3 5,0 9,2 2,7 6,8 2,1 4,4 3,4 2,1 2,4 2,0 2,3 2,0 2,0
5 1,7 3,9 6,0 2,9 2,6 5,1 2,4 2,7 3,8 2,7 6,3 11 2,1 2,4 2,9 2,2 1,9 1,8
6 2,3 2,9 4,3 2,9 2,5 2,5 4,5 2,8 7,0 2,9 5,5 2,2 2,0 2,5 2,2 2,2 1,7 2,3
7 1,7 2,6 2,3 2,6 3,9 2,8 2,4 2,7 2,6 2,4 7,8 4,8 2,2 2,7 2,3 2,4 2,2 2,5
8 1,8 2,7 2,7 4,3 3,2 2,4 2,6 2,7 4,0 2,3 6,2 3,2 0,5 2,3 2,0 2,0 2,2 2,9
9 2,2 2,5 3,7 3,4 2,4 2,1 2,4 1,5 6,7 6,4 1,0 1,5 1,8 2,3 0,5 1,0
10 1,9 4,3 2,0 2,2 0,1 4,8 17 0,5 3,2 3,4 17 1,1
M 2 2,5 3,6 3,3 3,1 3,1 3,7 2,8 4,1 2,2 5,9 4,7 3,5 2,2 2,4 2,3 3,4 2,2
(D) quercetina-3-O- β-D-glucopiranosideo
T1 T2 T3 T4 Controle Controle + Acetona
n r1 r2 r3 r1 r2 r3 r1 r2 r3 r1 r2 r3 r1 r2 r3 r1 r2 r3
1 3,2 3,8 6,5 1,0 1,2 1,0 2,5 1,1 2,8 3,5 3,5 2,7 2,5 3,1 1,9 2,3 2,1 2,5
2 3,5 7,2 8,5 0,9 1,2 1,3 2,5 1,3 2,7 5,8 5,7 5,2 2,0 2,4 3,3 2,4 2,4 2,2
3 3,5 8,0 4,5 0,9 1,1 1,0 2,8 4,8 3,4 2,4 2,4 2,4 2,9 2,3 2,5 1,7 1,9 3,5
4 4,0 2,4 3,0 1,1 1,0 1,3 2,4 1,8 2,2 2,3 2,5 3,2 2,1 2,4 2,0 2,3 2,0 2,0
5 3,4 6,1 5,6 0,9 1,0 0,9 2,1 2,6 4,6 2,5 4,5 5,4 2,1 2,4 2,9 2,2 1,9 1,8
6 4,3 4,7 3,5 1,0 1,0 1,1 7,1 1,5 5,0 2,1 3,5 2,5 2,0 2,5 2,2 2,2 1,7 2,3
7 6,7 3,1 7,4 0,9 1,1 1,4 2,6 1,0 3,0 2,8 2,5 3,0 2,2 2,7 2,3 2,4 2,2 2,5
8 3,1 7,2 1,1 0,9 0,5 6,0 1,6 5,0 2,3 6,8 4,5 0,5 2,3 2,0 2,0 2,2 2,9
9 4,4 6,3 0,7 0,7 1,2 4,0 7,1 2,8 3,6 3,1 1,0 1,5 1,8 2,3 0,5 1,0
10 6,9 0,7 1,0 4,6 8,1 5,5 4,5 17 0,5 3,2 3,4 17 1,1
M 4,1 4,8 5,9 0,9 1,0 1,1 3,6 2,0 4,0 3,5 4,1 3,7 3,5 2,2 2,4 2,3 3,4 2,2
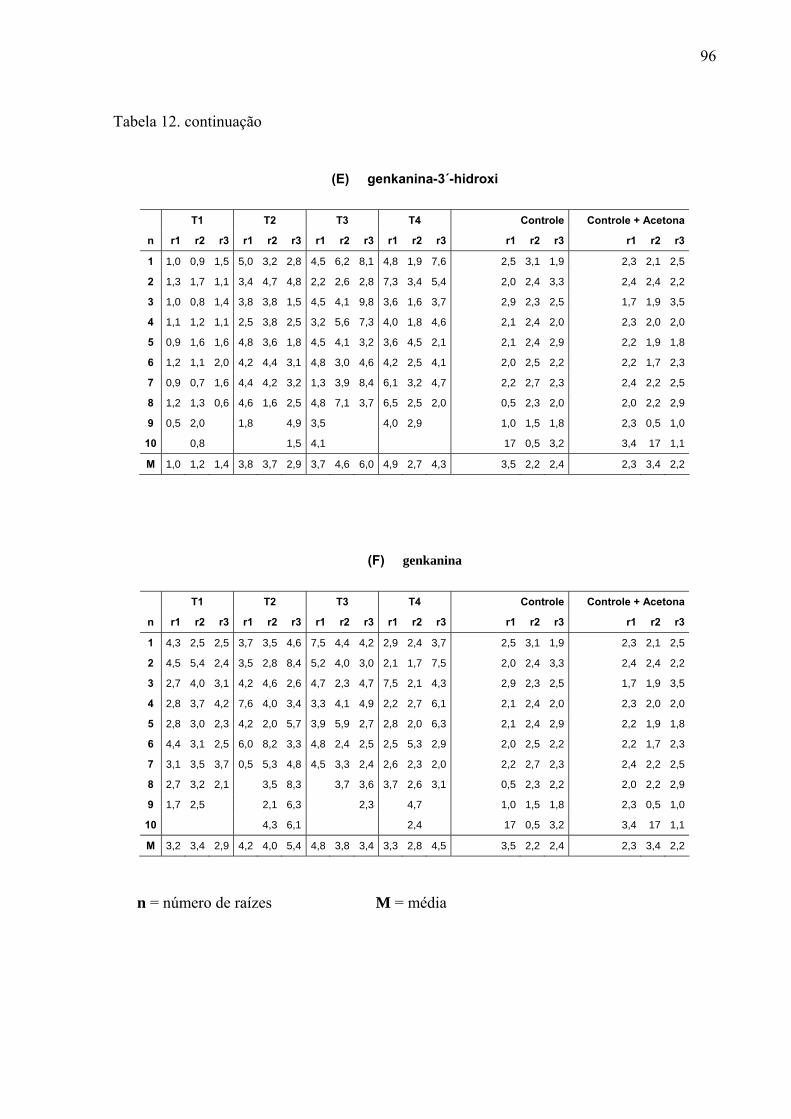
96
Tabela 12. continuação
(E) genkanina-3´-hidroxi
T1 T2 T3 T4 Controle Controle + Acetona
n r1 r2 r3 r1 r2 r3 r1 r2 r3 r1 r2 r3 r1 r2 r3 r1 r2 r3
1 1,0 0,9 1,5 5,0 3,2 2,8 4,5 6,2 8,1 4,8 1,9 7,6 2,5 3,1 1,9 2,3 2,1 2,5
2 1,3 1,7 1,1 3,4 4,7 4,8 2,2 2,6 2,8 7,3 3,4 5,4 2,0 2,4 3,3 2,4 2,4 2,2
3 1,0 0,8 1,4 3,8 3,8 1,5 4,5 4,1 9,8 3,6 1,6 3,7 2,9 2,3 2,5 1,7 1,9 3,5
4 1,1 1,2 1,1 2,5 3,8 2,5 3,2 5,6 7,3 4,0 1,8 4,6 2,1 2,4 2,0 2,3 2,0 2,0
5 0,9 1,6 1,6 4,8 3,6 1,8 4,5 4,1 3,2 3,6 4,5 2,1 2,1 2,4 2,9 2,2 1,9 1,8
6 1,2 1,1 2,0 4,2 4,4 3,1 4,8 3,0 4,6 4,2 2,5 4,1 2,0 2,5 2,2 2,2 1,7 2,3
7 0,9 0,7 1,6 4,4 4,2 3,2 1,3 3,9 8,4 6,1 3,2 4,7 2,2 2,7 2,3 2,4 2,2 2,5
8 1,2 1,3 0,6 4,6 1,6 2,5 4,8 7,1 3,7 6,5 2,5 2,0 0,5 2,3 2,0 2,0 2,2 2,9
9 0,5 2,0 1,8 4,9 3,5 4,0 2,9 1,0 1,5 1,8 2,3 0,5 1,0
10 0,8 1,5 4,1 17 0,5 3,2 3,4 17 1,1
M 1,0 1,2 1,4 3,8 3,7 2,9 3,7 4,6 6,0 4,9 2,7 4,3 3,5 2,2 2,4 2,3 3,4 2,2
(F) genkanina
T1 T2 T3 T4 Controle Controle + Acetona
n r1 r2 r3 r1 r2 r3 r1 r2 r3 r1 r2 r3 r1 r2 r3 r1 r2 r3
1 4,3 2,5 2,5 3,7 3,5 4,6 7,5 4,4 4,2 2,9 2,4 3,7 2,5 3,1 1,9 2,3 2,1 2,5
2 4,5 5,4 2,4 3,5 2,8 8,4 5,2 4,0 3,0 2,1 1,7 7,5 2,0 2,4 3,3 2,4 2,4 2,2
3 2,7 4,0 3,1 4,2 4,6 2,6 4,7 2,3 4,7 7,5 2,1 4,3 2,9 2,3 2,5 1,7 1,9 3,5
4 2,8 3,7 4,2 7,6 4,0 3,4 3,3 4,1 4,9 2,2 2,7 6,1 2,1 2,4 2,0 2,3 2,0 2,0
5 2,8 3,0 2,3 4,2 2,0 5,7 3,9 5,9 2,7 2,8 2,0 6,3 2,1 2,4 2,9 2,2 1,9 1,8
6 4,4 3,1 2,5 6,0 8,2 3,3 4,8 2,4 2,5 2,5 5,3 2,9 2,0 2,5 2,2 2,2 1,7 2,3
7 3,1 3,5 3,7 0,5 5,3 4,8 4,5 3,3 2,4 2,6 2,3 2,0 2,2 2,7 2,3 2,4 2,2 2,5
8 2,7 3,2 2,1 3,5 8,3 3,7 3,6 3,7 2,6 3,1 0,5 2,3 2,2 2,0 2,2 2,9
9 1,7 2,5 2,1 6,3 2,3 4,7 1,0 1,5 1,8 2,3 0,5 1,0
10 4,3 6,1 2,4 17 0,5 3,2 3,4 17 1,1
M 3,2 3,4 2,9 4,2 4,0 5,4 4,8 3,8 3,4 3,3 2,8 4,5 3,5 2,2 2,4 2,3 3,4 2,2
n = número de raízes M = média


















