DE TOXICIDADE GENÉTICA COM TRAÚESèMmA · potência) foram comparados para indução de quebras...
59
ISSN 1516-46911 Ministério ^ rd a Agricultura e do Abastecimento DE TOXICIDADE GENÉTICA COM TRAÚESèMmA % # -• % •V j » /
Transcript of DE TOXICIDADE GENÉTICA COM TRAÚESèMmA · potência) foram comparados para indução de quebras...

ISSN 1516-46911M inistério
^ r d a Agricultura e do Abastecimento
DE TOXICIDADE GENÉTICA COM TRAÚESèMmA
%
# - •
%• V j »
/

REPÚBLICA FEDERATIVA DO BRASIL
Presidente: Fernando Henrique Cardoso
Ministro da Agricultura e do Abastecimento: Marcus Vinícius Pratini de iVIoraes
Empresa Brasileira de Pesquisa Agropecuária - Embrapa
Presidente: Alberto Duque Portugal
Diretores: Dante Daniel Giacomelli Scolari
José Roberto Rodrigues Peres
Elza Angela Battaglia Brito da Cunha
Embrapa Meio Ambiente
Chefe Geral: Bernardo van Raij
Chefe Adjunto de Pesquisa e Desenvolvimento: Deise Maria Fontana Capalbo
Chefe Adjunto Administrativo: Vander Roberto Bisinoto
Empresa Brasileira de Pesquisa Agropecuária Embrapa Meio AmbienteM in istério da A g ricu ltu ra e do A bastecim en to
ISSN 1516-4691
m m m K X m toxiodade geneíica COM mADE^ANIlA
Geraldo Stachetti Rodrigues
Jaguariúna, SP - 1999

EMBRAPA MEIO AMBIENTE - Documentos 14
Exemplares dessa publicação podem ser solicitados à:
Embrapa Meio AmbienteRodovia SP 340 - km 127,5 - Tanquinho VelhoCaixa Postal 69 13820-000, Jaguariúna, SPFone; (19) 3867-8750 Fax: (19) [email protected]
Comitê de Publicações: Aldemir ChaimCélia M. M. de S. Silva Franco Luchini Júlio F. de Queiroz Magda A. de Lima Maria Cristina Tordin
Revisão: Denise Moraes de Oliveira
Normatização: Maria Amélia de Toledo Leme
Produção gráfica: Regina Lúcia Siewert Rodrigues e Franco Ferreira de Moraes
Tiragem: 500 exemplares
RODRIGUES, G.S. Bioensaios de toxicidade genética com
Tradescantia. Jaguariúna: Embrapa Meio A m b ien te , 1999.
56p. (Embrapa Meio Ambiente, Documentos 14).
CDD- 581.15
© Embrapa Meio Ambiente, 1999

SUMARIO
INTRODUÇÃO................................................................................................. 5
TRADESCANTIA E O BIOENSAIO DO MICRONÚCLEO............................. 6
Trad-MCN; fundamentos e desenvolvimento do bloensaio................... 6
O ENSAIO TRAD-MCN COMO UM SISTEMA DE MONITORAMENTOPARA GENOTOXICIDADE NO AMBIENTE.................................................. 10
Poluição do ar..................................................................................................... 10
Poluição aquática................................................................................. 12Contaminantes e acondicionantes do solo........................................ 15Pesticidas, agentes químicos e estresses fisiológicos......................... 17Raios cósmicos e campos eletromagnéticos..................................... 20
TRADESCANTIA E O BIOENSAIO DO PÊLO ESTAMINAL......................... 21
Trad-SHM: Fundamentos e desenvolvimento do bloensaio............... 21
O ensaio Trad-SHIVI como um sistema de monitoramentopara genotoxicidade no ambiente...................................................... 22
CONCLUSÃO............................................................................................... 2 6
REFERÊNCIAS BIBLIOGRÁFICAS................................................................ 27
ANEXOS (Tabelas)..................................................................................... 41


BIOENSAIOS DE TOXICIDADi GENÉTICA COM TRADESCANTIA’
Geraldo S tachettí Rodrigues^
INTRODUÇÃO
Desde os primórdios dos estudos da atividade genética de compostos
químicos e agentes físicos, várias espécies e clones do gênero Tradescantia têm sido utilizados como organismos experimentais, em virtude de uma série de características genéticas favoráveis. Apresentando apenas seis pares de cromossomos grandes e facilmente observáveis, células de quase todas as partes da planta, da ponta da raiz ao tubo polínico em desenvolvim ento, fornecem material excelente para estudos citogenéticos (Ma & Grant, 1982).
Como conseqüência do uso intenso de Tradescantia em estudos
genéticos, encontrou-se uma série de características que permitem a detecção
de agentes que afetam a estabilidade do genoma. Pelo menos quatro dessas características foram selecionadas como indicadores em bloensaios de avaliação
de toxicidade genética. Dois desses, o da mitose em ponta de raiz e o do tubo polínico, são ensaios de aberração cromossômica, nos quais se observam deformações morfológicas visíveis nos cromossomos (Ma, 1982). Um terceiro,
o ensaio da mutação para célula cor-de-rosa em pêlo estaminal (Trad-SHM) (Underbrink et al., 1 973b), é um teste de mutação m itótica pontual que se baseia na expressão de um gene recessivo para cor da flo r em plantas
' A p o i o CNPq, P r ocesso n ú m e r o 5 2 0 6 9 3 - 9 8 - 1 .E c ó l o g o , P h . D . , E m b r a p a M e i o A m b i e n t e . C a i xa p o s t a l 6 9 - C e p 1 3 8 2 0 - 0 0 0 ,
J a g u a r i ú n a , SP,5

heterozigotas. 0 quarto ensaio é um teste c ito ge né tico que se baseia na formação de micronúcleos (Trad-MCN) que resultam da quebra cromossômica
nas células meióticas geradoras do pólen (Ma, 1 979b).
No presente texto, examinam-se as características e fundam entos
dos ensaios Trad-MCN e Trad-SHM e revisam -se os resu ltados até hoje obtidos com estes sistemas na avaliação de g eno tox ican tes am bientais, Ad ic ionalm ente, apresentam-se os proced im entos para a execução de experimentos de avaliação de genotoxicidade ambiental, utilizando-se esses dois bioensaios vegetais. A descrição deta lhada da m etodo log ia para realização desses ensaios, bem como dos ensaios que empregam linhagens
de m ilho e soja, está disponível na C ircular Técn ica da Embrapa Meio
Ambiente, "Bioensaios de toxicidade genética com plantas superiores".
TRADESCANTIA E O BIOENSAIO DO MICRONÚCLEO
Trad-MCN: fundamentos e desenvolvimento do bioensaioEstudos sobre o genoma de Tradescantia iniciaram-se com os
trabalhos pioneiros de Sax & Edmonds (1933) sobre o gam etófito masculino de T. reflexa Raf., quando foram descritas as várias fases do desenvolvimento do micrósporo e determinados os períodos e o ritm o dos eventos meióticos.
Observações im portantes foram feitas sobre os e fe itos de raios-X nos micrósporos dessa espécie (Sax & Edmonds, 1 933). Primeiramente, observou- se que cromossomos meióticos eram mais suscetíveis a quebra ("breakage") que cromossomos mitóticos e, mais importante, cromossomos em divisão eram ao menos dez vezes mais susceptíveis que aqueles em repouso.
Em segundo lugar, quebras ("b re a ks ") não se d is tribu íam aleatoriamente nos cromossomos. Loci posicionados proxim am ente aos
centrômeros apresentavam maior probabilidade de sofrer ruptura sob efeito de radiação. Estas observações levaram à conclusão de que as tensões mecânicas resultantes da espiralação dos cromossomos durante a replicação aum entariam a ocorrência de quebras, in flu en c ian do fo rte m e n te a suscetibilidade a aberrações cromossômicas. Essas inferências seriam mais tarde confirmadas em um estudo dos efeitos da radiação gama gerada por “ Co em T. paludosa And. and Woods. (Sparrow & Singleton, 1953). Os conceitos de temporalidade e sensibilidade qiie emergiram desses estudos tornaram-se extremamente importantes na seleção de bioindicadores para

mutagênese, uma vez que sincronia no desenvolvimento celular e precisão nos períodos de recuperação após os tratamentos mostraram-se dois fatores
decisivos para a performance dos bioensaios.
A m aior suscetibilidade dos cromossomos meióticos, quando
comparados com os m itó ticos, foi mais tarde confirmada em um estudo da
influência da falta de oxigênio na meiose em T. paludosa (Steinitz, 1944). Esta
pesquisa representou a primeira tentativa de observação de micronúcleos nas cé lu las m ães do pólen, com o ind icadores d ire tos de fragm entação crom ossôm ica. Uma taxa espontânea de 0 ,87 % de células contendo
micronúcleos foi definida para T. paludosa, aumentando para 8,0% em células expostas à anaerobiose nos estágios iniciais da prófase.
O ritm o dos estágios da meiose foi ainda melhor caracterizado em um estudo da d iferenciação das anteras de T. paludosa (Taylor, 1950). O
ciclo m eiótico envolve aproximadamente 24h, período no qual deve ocorrer a
exposição e um tem po de recuperação apropriado para que aquelas inflorescências expostas aos agentes tóxicos sejam fixadas para análise do número de micronúcleos, conseqüentemente da atividade genotóxica.
0 c re s c e n te in te re sse nas ca pac idades ra d io m im é tic a s
(especialmente genotóxicas) de substâncias químicas, nos anos 50, sugeriram a utilização de Tradescantia como um bioindicador. Um ensaio de mitose no tubo polínico fo i primeiramente empregado em um estudo comparativo de
agentes quím icos simples em T. paludosa (Smith & Lofty, 1954). Óxido de etileno (um agente m utagênico conhecido), queteno (um composto com resultados inconclusivos), e cloreto de metila (um agente alquilante de baixa potência) foram comparados para indução de quebras em cromátides e aberrações cromossômicas. O ensaio do tubo polínico mostrou-se efetivo na
detecção de genotoxicidade e os resultados revelaram que os compostos mais
ativos (óxido de etileno e queteno) causavam mais numerosas e extensivas aberrações cromossômicas. A propícia seleção de compostos a serem testados nesta pesquisa permitiu a demonstração da sensibilidade da Tradescantia e sua
capacidade para diferenciar precisamente efeitos comparativamente similares.
Numa série de artigos a respeito do papel de determinados nutrientes
na meiose, a produção de m icronúcleos nos micrósporos de T. paludosa foitom ada com o ind icativa de quebra cromossômica (Steffensen, 1953;
Steffensen, 1 954; Steffensen, 1955). Estudando os efeitos da deficiência em7

magnésio (Mg) na meiose, o autor notou uma m aior sensibilidade dos micrósporos quando comparados com pontas radiculares, de acordo com as evidências prévias de maior suscetibilidade de células meióticas que mitóticas. Micronúcleos apareciam em maior número em plantas deficientes em magnésio (Mg), cálcio (Ca) e enxofre (S). Note-se que os dois primeiros nutrientes são responsáveis pela ligação de macromoléculas no núcleo, contribuindo para a estabilidade de proteínas e DNA. Uma taxa espontânea de m icronúcleos de 0 ,84 % foi registrada, aumentando para 3 ,89% em plantas cultivadas em meio deficiente em Ca (Steffensen, 1955). Esses números vieram corroborar observações prévias de produção de micronúcleos em T. paludosa (Steinitz, 1944, anteriormente citado).
Mais de 30 anos após a observação de micronúcleos para detecção de danos à meiose feita por Steinitz, Ma e colaboradores (1978) no Laboratório Nacional Brookhaven desenvolveram o ensaio do micronúcleo-na-tétrade (Trad- MCN). Empregando o clone híbrido 4430 {T. h isutiflora Bush x T. subacauHs Bush), eles compararam a produção de m icronúcleos nas células mães do pólen com mutações para células cor-de-rosa nos pêlos estam inais de Tradescantia expostas ao conhecido agente m utagênico 1,2-dibrom oetano
(DBE).
Já naquele tempo, o ensaio da mutação em pêlos estam inais (Underbrink et al., 1 973b) vinha sendo extensivamente empregado e era um teste reconhecido para mutagênese radiobiológica e química. 0 ensaio do micronúcieo, contudo, exibiu uma eficiência aproximadamente 36 vezes maior. Esta sensibilidade extraordinária foi creditada à especificidade m uito menor do dano necessário para produzir micronúcleos, quando comparada à mutação nos pêlos estaminais. De acordo, poderia assumir-se que numerosos loci em quaisquer dos 1 2 cromossomos de Tradescantia estariam sujeitos a danos que resultariam em quebra dos cromossomos e, conseqüentem ente, em micronúcleos. Em contraste, somente um locus em um único crom ossomo poderia sofrer mutação para produção de células cor-de-rosa em pêlos estaminais (Ma et al., 1 978).
A enorme sensibilidade e simplicidade do ensaio Trad-MCN foi ainda demonstrada em experimentos, nos quais baixas doses de raios-X eram comparadas com dois agentes mutagênicos conhecidos, metanosulfonato de etila (EMS) e azida sódica (NaN.,) em ambas formas, gasosa e líquida (Ma,1 979a), Uma dose de raios-.X de apenas 20-rad induzuj altas freqüências de
micronúcleos (23 MCN,/100 tétrades), enquanto apenas 1,8 M CN/100 células

foram induzidos por 50-rad de raios-X em linfócitos humanos (Countryman e
Heddie, 1976 citado em Ma, 1979a), ou 2,5 M CN /100 eritroblastos de camundongo em cultura de medula óssea expostas a 35-rad de raios-X (Janssen e Ramel, 1976 citado em Ma, 1 979a). Enquanto ocorria 0 ,2% de m utações
por rad no ensaio do pêlo estaminal (Trad-SHM), ocorria 1,6% MCN por rad no ensaio Trad-MCN. A relação entre dose e efeito no ensaio Trad-MCN com
raios-X resultou num coeficiente de correlação de 0,99. Os resultados obtidos com os agentes quím icos confirmaram estes dados, tanto em relação à sensibilidade quanto em relação ao efeito pela dose (Ma, 1979a).
Uma vantagem adicional do ensaio Trad-MCN é o curto período de exposição necessário para se completar um teste - apenas 6h, seguidas de uma recuperação de 24h, para perm itir que as células tratadas na prófase atinjam o estágio de tétrade apropriado para contagem de micronúcleos. Esta periodicidade da meiose foi testada em um estudo da sensibilidade por estágios,
usando exposição a raios-X em T. paludosa (Ma et al., 1980). Grupos de inflorescências receberam uma dose singular de 35 rad de raios-X, após o que inflorescências foram removidas e fixadas em intervalos de 3h, durante 48h pós-irradiação. Um pico de sensibilidade ocorreu depois de 24h, concordando com as observações de Taylor (1 950). Um segundo pico apareceu após cerca de 39h, sugerindo que o início da prófase I e estágios pré-meióticos são também
muito sensíveis.
O emprego do ensaio Trad-MCN para o monitoramento de agentes clastogênicos ambientais foi primeiramente proposto após estudos envolvendo agentes pró-mutagênicos e localidades poluídas (Ma, 1979b; Ma, 1981). Uma
grande vantagem percebida nestes estudos é que nenhuma atividade enzimática externa é necessária para ativar os agentes pró-mutagênicos, dado que o aparato enzimático continuava totalmente funcional nas inflorescências
extraídas das plantas e expostas nos tratamentos.
Por outro lado, várias limitações do ensaio Trad-MCN têm sido apresentadas. O teste, obviamente, oferece apenas um índice relativo de danos genéticos. Translocações, inversões e outros tipos de rearranjos nos cromossomos e cromátides não são revelados como micronúcleos, não sendo,
portanto, detectados pelo ensaio. Não é possível tampouco extrapolar
facilmente os resultados de freqijência de micronúcleos para carcinogenicidade, e os passos metabólicos de agentes mutagênicos e promutagênicos podem ser
9

bastante d iferentes em Tradescantia e outros organismos (especialmente
mamíferos). A lém disso, a alta sensibilidade do sistema resulta em variações
consideráveis na freqüência espontânea de m icronúcleos em diferentes
experim entos, requerendo um sempre cuidadoso controle das condições experimentais e a utilização simultânea de arnosíras controle (Ma, 1 981).
Os proced im entos laboriosos e demorados para contagem de micronúcleos nastétrades consistem em uma desvantagem adicional do teste Trad-MCN (Ma, 1990). Para sanar esta lim itação, facilitando e padronizando
o processo de contagem, um sistema de análise de imagens para micronúcleos
foi desenvolvido (Ma et al., 1 992b). 0 sistema computadorizado é capaz de realizar contagens a uma velocidade 3,5 vezes maior que a contagem manual,
e com uma congruência de 90% nas freqüências observadas.
Uma revisão re lativam ente recente do m onitoram ento in s itu de agentes clastogênicos no ambiente (Ma, 1 990) revelou que até 1990 cerca
de 300 testes haviam sido conduzidos com o ensaio Trad-MCN, numa ampla va riedade de s itu a ç õ e s . C erca de 5 0 % desses te s te s a pon ta ram genotoxicidade. Amostras de águas e solos apresentaram resultados positivos
em cerca de 60% das ocasiões estudadas. Na próxima seção, os estudos de
m onitoramento ambiental com Trad-MCN são apresentados e discutidos.
0 ENSAIO TRAD-MCN COMO UM SISTEMA DE MONITORAMENTO PARA GENOTOXICIDADE NO AMBIENTE
Poluição do ar
Tradescantia foi exposta a vários locais poluídos no estado de Illinois
(EUA) e a gases com um ente encontrados em atmosferas poluídas numa
combinação de testes in situ, no ambiente e in vivo, em laboratório (Ma et al.,1 982). Ensaios in s itu expondo as plantas por 2 a 6h em estacionamentos,
áreas industriais, fazendas e laboratórios apresentaram resultados positivos, especialmente quando emissões de agroquímicos estavam envolvidas. Plantas
expostas em um escritório , uma fazenda de criação animal e uma área
residencial não apresentaram aumentos na freqüência de micronúcleos. ÚLiando fumigadas com os poluentes atmosféricos NO , SO , e 0 , assiin conio ácido
hidrazóico gasoso (HN.j o EMS, as plantas também indicaram clastogênese, 10

sugerindo serem estes gases alguns dos possíveis agentes genotóxicos da atmosfera (Ma et al., 1 982).
Como conseqüência de sua versatilidade, Tradescantia é indicada como um biomonitor para poluição de ambientes fechados, sendo que vários
estudos têm avaliado sua proficiência para os baixos níveis de contam inação que costumeiramente ocorrem em ambientes residenciais. Dentre os resultados
positivos já encontrados nessas situações, citam -se diversos odorizantes comerciais, fumaça de tabaco, p-diclorobenzeno (bolas de naftalina) e outros inseticidas indicados para uso doméstico, além de gases de combustão de óleo diesel (Ma & Harris, 1987a; Ma & Harris, 1 9 8 7 b ).
Um grupo relativam ente incom um de poluentes a tm osféricos estudados in s itu para clastogênese com o ensaio Trad-MCN foi o das fumaças quím icas empregadas pelo exército norte-am ericano. Estes experim entos envolveram outros ensaios incluindo aberrações crom ossôm icas e trocas de cromátides irmãs ("sister chromatid exchange" - SCE) em um roedor (Schaeffer et al., 1987). As fumaças eram geradas a partir de diesel para tanques, "fogoil", e hexacloroetano. Todos estes compostos induziram eventos genotóxicos em ao menos uma das doses. Houve um alto grau de variabilidade (expressa como maiores desvios padrão na produção de micronúcleos) em todos os tratamentos realizados in situ, em relação aos controles de laboratório. Embora este efeito estatístico tenha sido discutido meramente como obscurecendo a relação dose-
efeito dos resultados, isto pode ser indicativo de uma característica inerente deste bioensaio. Os botões menores das inflorescências jovens de Tradescantia
estão sempre encobertos sob botões maiores e folhas, o que pode resultar em sua proteção contra exposição direta, especialmente em experim entos com poluentes atmosféricos, um efeito descrito por Ma (1 979b).
Em um estudo realizado no México, avaliaram-se ao longo do ano os picos de freqüência de m icronúcleos em Tradescantia exposta a uma área pesadamente industrializada, uma área residencial e uma área de ocupação m ista (Ruiz et al., 1992). Plantas expostas à área industria l sempre apresentavam mais micronúcleos que as plantas controle ao longo de todo o ano, enquanto plantas expostas à área residencial tend iam a apresentar incrementos na freqüência de m icronúcleos somente em meses específicos. 0 ensaio Trad-MCN permitiu a detecção de riscos de genotoxicidade causados por emissões de incineradores de lixo municipal (Ma et al., 1993b) e dos drenos de gases de aterros sanitários (Ma et al., 1 993a; Ma et al., 1 996).
11

Dos estudos acima m encionados, pode-se concluir que o ensaio Trad-M CN é adequado para a avaliação de contaminação atmosférica, seja de áreas pesadam ente poluídas, industria is ou urbanas, ou sob condições
normais de ambientes residenciais. Condições atmosféricas, como variações na velocidade e direção dos ventos, normalmente levam a variações estatísticas re la tivam ente altas nos dados. Um resumo dos resultados obtidos com
Tradescantia na avaliação da poluição atm osférica e agentes gasosos está
disponível na Tabela 1.
Poluição aquática
Praticamente todo estudo que envolva a avaliação da presença de agentes m utagênicos em águas naturais deve incorporar um passo de co nce n tra ção dos possíve is agentes tó x ic o s nas am ostras a serem quimicamente analisadas. Isto ocorre devido à intrínseca baixa mutagenicidade dos agentes mais freqüentem ente presentes nas águas, ou devido à sua
concentração m uito baixa, ou ambos.
A necessidade de concentração das amostras foi claramente dem onstrada numa avaliação da probabilidade de se detectar um agente mutagênico em água, utilizando-se o teste de Ames (Johnston & Hopke, 1980). Neste estudo, propôs-se uma variável que ponderava a potência mutagênica e a concentração média de agentes m utagênicos orgânicos com um ente encontrados em águas e a quantidade necessária do agente para induzir uma
duplicação no número de células mutantes no teste de Ames. Considerando- se que: a) geralm ente apenas alíquotas de 1 ml são aplicadas por placa de te s te , b) co m p o s to s o rgân icos tip ica m e n te ocorrem em águas em concentrações na faixa de |ig/L, e c) 95% dos agentes testados até o momento apresentavam uma dose de duplicação de pelo menos 1 500).lg, concluiu-se que uma amostra ambiental média que permitisse a detecção de contaminantes com uma confiança de 95% deveria conter 1 500L. Isto significa que um fator médio de concentração de seis ordens de magnitude seria necessário para reduzir o volume de uma tal amostra para os testes. Deduziu-se que a exposição a agentes mutagênicos presentes em água potável que ocorre no decorrer da
vida de uma pessoa pode ser apreciável, mesmo que não seja possível detectar estes agentes nas amostras (Johnston & Hopke, 1 980).
Talvez a qua lidade m ais im p o rta n te do onsaio T rad-M C N , assim com o
de a lguns o u tro s b ioensa ios vege ta is , é sua capac idade cie de tec ta r ba ix íss im os12

níveis de tox ic idade genética tan to em exposições de cu rto te rm o in s itu ,
quanto em testes in vivo com amostras não concentradas. Isto foi demonstrado em um estudo de dois anos sobre a genotoxicidade das águas de um reservatório para coleta de água de abastecimento municipal e da água tratada proveniente desse reservatório (Ma et al., 1985). Am ostras de água eram coletadas semanalmente e testadas para genotoxicidade e para presença de nutnentes e metais. Os resultados mais im portantes desse estudo dem onstraram uma recorrência sazonal na expressão de picos de freqüência de m icronúcleos, que coincidia com períodos de intensa precipitação e carreamento de solo e água dos campos de soja e milho ao redor do reservatório. A produção de micronúcleos nas amostras de água tratada seguia a tendência observada para as amostras
do reservatório, mas os picos de freqüência eram menores.
Em uma investigação com plem entar, os ensaios Trad-MCN foram conduzidos conjuntam ente com o teste do m icronúcleo em e ritróc ito de cam undongo e amostras adicionais de um poço raso e outro de média profundidade existentes na mesma área do reservatório foram analisadas (Ma et al., 1987). Um padrão similar de freqüência de micronúcleos seguindo-se a precipitação pesada ou derretimento de neve na bacia do lago fo i detectado pelo ensaio Trad-MCN. Análises das amostras de água dos poços mostraram níveis detectáveis de com postos orgânicos, com o cloreto de m etileno, diclorobromoetano, tricloroetileno e tetracloroetileno. Os testes com eritrócitos de camundongo confirmaram os resultados, embora exposições por 6 meses às amostras tivessem sido necessárias, enquanto o ensaio Trad-MCN requisesse
apenas exposições de 30h.
Estes experimentos acima descritos de avaliação da genotoxicidade de águas não podem ser qualificados como in situ, já que em todos os casos as amostras eram trazidas ao laboratório e ensaiadas sob condições controladas.
A avaliação propriamente in s itu de mutagenicidade em ambientes aquáticos
som ente tornou-se possível com a introdução do "aqua toon", um aparato flu tuante desenvolvido especificamente para sustentar material vegetal para exposição a corpos de água. 0 aquatoon foi empregado com sucesso em um estudo in s itu da genotoxicidade dos efluentes de uma fábrica de papel e celulose na margem norte do Lago Superior, no Canadá (Grant et al., 1 992). Os ensaios do micronúcleo e do pêlo estaminal em Tradescantia e o ensaio de aberração cromossômica em ponta radicularem Vicia faba L, foram aplicados
no córrego que recebia os efluentes brutos e na baía do lago na qual este13

córrego desaguava. O ensaio Trad-MCN e o ensaio com \/. faba apresentaram resultados positivos após 24h de exposição em ambos os locais, enquanto o ensaio Trad-SHM produziu resposta inconclusiva. Além de serem mais sensíveis,
os dois ensaios que apresentaram melhores resultados melhor se adaptaram
às cor.d içõcs de experim.entação in s/tu. Isso se deve ao fato de o material desses ensaios poder ser fixado imediatam ente após a exposição, ao passo que o ensaio Trad-SHM requer longos períodos de recuperação em condições controladas antes da análise, o que traz dificuldades em condições de campo
e durante o transporte das amostras.
Um estudo da genotoxic idade de efluentes industriais no M éxico demonstrou incremento na freqijência de micronúcleos no ensaio Trad-MCN,
mesmo após diluição tripla dos efluentes (Ruiz et al., 1992). Da mesma maneira, as lixívias de um aterro sanitário abandonado há 20 anos apresentavam genotoxicidade após diluições de até 20 vezes, enquanto diluições menores que 10 vezes resultavam em efeitos tóxicos (Ma et al., 1 993a).
A genotoxic idade de águas subterrâneas contam inadas com hidrocarbonetos policlorados (PAHs) tratadas em uma usina de purificação, implantada com o objetivo de descontaminar um dos mais importantes aqüíferos da Áustria, foi avaliada numa série de experimentos com o ensaio Trad-MCN (Helma et al., 1993; Helma et al., 1994). Os métodos de purificação consistiam de filtração em carvão ativado e irradiação com luz ultra-violeta (UV). Amostras
coletadas antes de qualquer tra tam ento exibiam atividade clastogênica e
dependente da dose, após exposições por 24h. Quando tratadas no laboratório com quantidades crescentes de UV (até 1 500J/rn^), estas amostras exibiam aumento na freqüência de micronúcleos de forma dose-dependente em relação à irradiação com UV, enquanto os resultados para o controle de água limpa irradiada com UV eram negativos. Os parâm etros quím icos m edidos rotineiram ente na usina de purificação indicavam que as amostras filtradas em carvão ativado apresentavam qualidade de água potável. Em muitos casos,
contudo, maiores freqüências de m icronúcleos foram registradas nessas
amostras, tan to antes quanto após irradiação com UV. 0 mecanismo responsável por esses efeitos era provavelmente a ativação, pela luz UV, de poluentes da água a com postos genotóxicos. Tal conclusão se baseia no fato
de a clastogenicidade das amostras irradiadas ciecrescer com a estocagem, com iima meia vida de aproxim adam ente 1 dia. Os aiitores sugeriram que tratamentos similares com UV para águas dc abastecimento poderiam produzir14

compostos perigosos que pudessem passar sem serem detectados nas estações
de tratam ento (Helma et ai., 1993; Helma et al., 1 994).
Estudos sobre poluição aquática demonstram as aptidões e vantagens
do ensaio Trad-MCN para a avaliação in s itu de agentes genotóxicos no am biente. A capacidade de de tec ta r e fe itos b io lóg icos em am ostras consideradas puras em análises químicas e a possibilidade de se evitarem os
tediosos procedimentos de concentração de am ostras, que podem resultar em perdas ou alteração nos com postos presentes, são características especialmente convenientes do método.
A Tabela 2 apresenta um resumo dos resultados obtidos com o
ensaio Trad-MCN na avaliação de poluentes do ambiente aquático.
Contaminantes e acondicionantes do solo
Vários estudos têm avaliado a m utagenicidade de solos in situ, de extratos de solos de áreas contaminadas, antes e depois da aplicação de medidas de remediação e de materiais acondicionantes de solos propriam ente ditos.
Talvez o acondicionante edáfico produzido em maiores volum es no mundo seja 0 lodo de esgoto municipal. Em geral, o impacto desse material no ambiente se re laciona à sua contam inação por m eta is pesados (L 'H e rm ite & Dehandtschttler, 1980), mas compostos orgânicos complexos são muitas vezes introduzidos nos sistemas de tratamento de esgotos. A possível genotoxicidade desses Iodos foi avaliada em Chicago (EUA), utilizando dois bioensaios vegetais
(pólen ceroso de milho e Trad-MCN) e duas linhagens de Salmonella no teste de Am es (Hopke et al., 1982). Todos esses ensaios indicaram que o lodo continha agentes capazes de induzir genotoxicidade, enquanto o ensaio Trad-
MCN apontava que mesmo diluições quádruplas de lodo eram clastogênicas,
aumentando significativamente a freqüência de micronúcleos em Tradescantia.
A clastogenicidade de vários com postos quím icos com um enteencontrados em depósitos de resíduos perigosos fo i estudada com o ensaioTrad-MCN, numa série de experimentos cujo objetivo era elucidar as possíveis
interações sinergísticas ou antagonísticas dos compostos, quando ocorrendoem misturas (Sandhu et al., 1989). Inicialmente, sete compostos selecionados
da Lista de Compostos Prioritários da Environmental Protection Agency (EPA)
dos EUA (Waters et al., 1 987) foram testados para determ inar suas doses
efetivas mínimas (DEMs). Cinco dos compostos testados produziram resultados15

positivos e foram listados em ordem decrescente de potência no ensaio Trad-
MCN, como segue: tetracetato de chumbo em dim etilsuifóxido (DMSO) (0,4
ppm), heptacloro em DMSO (2,0 ppm), dieldrin em DMSO (3,8 ppm), trióxido
de arsênico em NaOH (4,0 ppm) e 1,2-benz[a,/7|antraceno em etanol (12,5 ppm). Tetracloro etileno (TCE) e aldrin foram imiscíveis em água, impedindo
exposição adequada em solução. Quando expostos na forma gasosa a 30ppm por 2h, TCE apresentou resposta positiva, mas aldrin não.
De posse desses resultados, Ma et al. (1992a) avaliaram a clastogenicidade dos compostos em misturas. Todas as misturas de TCE (um agente não clastogênico) e dieldrin (em concentrações abaixo da DEM) resultaram positivas, sugerindo uma interação sinergística. Surpreendentemente, todas
as misturas de tetracetato de chumbo e trióxido de arsênico (ambos potentes agentes clastogênicos) foram negativas, sugerindo uma interação antagônica.
As outras combinações eram em geral levemente antagônicas, enquanto algumas misturas eram tóxicas, impedindo o desenvolvim ento normal das
tétrades. Essas complexas e, freqüentemente, imprevisíveis respostas induzidas por misturas de compostos químicos levaram os autores a concluir que avaliações
in situ são necessárias quando vários compostos interagem, como normalmente acontece em depósitos de resíduos perigosos.
Gill & Sandhu (1992) expandiram esses estudos testando os mesmos
compostos após incorporação no solo, o que permitiu o uso de Tradescantia
intacta (inclusive raízes) e não apenas das inflorescências. A maioria dos
resultados concordavam com os descritos acima, mas em alguns casos as interações no solo alteraram a expressão da clastogenicidade. Por exemplo, trióxido de arsênico e tetracetato de chumbo, embora não induzissem aumentos
na freqüência de m icronúcleos em solução (como em Ma et al., 1 992a), o
fizeram no solo. Em geral, plantas enraizadas produziram maiores freqüências
de micronúcleos que apenas inflorescências tratadas em solução. Possíveis
razões para esse efeito são aumento de eficiência na ativação metabólica das misturas no sistema radicular das plantas e por microrganismos do solo. Estes
resultados demonstraram novamente que predizer as atividades genotóxicas
de misturas de compostos a partir da análise de seus componentes pode ser uma falácia, enfatizando o valot das avaliações in situ.
A im p ortân c ia dessas co ric lusõ es foi acenU iada pela d e m o n s tra ç ã o
que ácidos tãn icos podem agir com o sinerg is tas na indução de c las togen ic idade
16

em Tradescantia (Knasmuller et al., 1 992). Exposição de Tradescantia por
24h a quantidades crescentes de ácidos tânicos causou um aumento dramático e dependente da dose, nos efeitos clastogênicos de raios-X (35rad), enquanto ácidos tânicos por si só mostraram apenas genotoxicidade moderada. Resultados similares foram obtidos com ácidos tânicos em combinação com outros compostos quím icos. Este resultado pode se revestir de grande importância, pois ácidos tânicos estão presentes na maioria dos alimentos e bebidas, assim com o em águas e solos naturais. Conseqüentemente, praticam ente todo
produto quím ico liberado no ambiente pode interagir e ser potencializado por ácidos tânicos.
O valor do ensaio Trad-M CN como uma ferramenta na avaliação ambiental foi novamente enfatizado num estudo das medidas de biorremediação num depósito de resíduos perigosos (Baud-Grasset et al., 1 993a; Baud-Grasset et al., 1993b). Solos pesadamente contaminados com creosoto (mais de 5 .000ppm de PAHs) foram incubados com o fungo degradador de lignina Phanerochaete chrysosporium Burdsall, por 8 semanas, e os extratos aquosos desses solos foram estudados com o ensaio Trad-MCN. Extratos dos solos pré-incubação eram altamente clastogênicos. P. chrysosporium causou uma dim inuição na contam inação, duplicando a concentração de extrato de solo necessária para induzir uma freqüência de m icronúcleos similar àquela previam ente á incubação (de 1 para 2% ). Novamente o ensaio Trad-MCN mostrou-se extraordinariamente sensível, permitindo a detecção de diferenças
entre amostras proximamente comparáveis.
Os resu lta d o s o b tid o s com Tradescantia na ava liação de
contam inantes do solo estão resumidos na Tabela 3.
Pesticidas, agentes químicos e estresses fisiológicos
Plantas são os receptores biológicos diretos dos pesticidas aplicados no campo. Conseqüentem ente, não surpreende a atenção devotada aos estudos de toxicologia genética de pesticidas em plantas. Uma extensa revisão da g e n o to x ic id a d e de pes tic idas em plantas superiores (Sharma &
Panneerselvan, 1 990) listou um total de 178 ingredientes ativos testados em
pelo menos uma de 31 espécies vegetais diferentes, empregando uma variedade de órgãos e parâm etros genéticos. Aproximadamente 30% dos compostos foram considerados geno tóx icos , enquanto apenas 6% puderam ser considerados inócuos nesses termos.
17

Tradescantia aparece somente uma vez nessa revisão, indicando que esse ensaio não está entre os preferidos para avaliação da toxic idade genética de pestic idas, a despeito de sua sensibilidade e propriedade para avaliações em campo. As primeiras referências ao uso de Tradescantia para testar a genotoxicidade de pesticidas envolveram as atividades citológicas do inseticida m evinfós e do herbicida cianazina (Ahmed & Grant, 1 972b) e do fung ic ida para tra tam en to de sem entes Panogen 15® (m etilm ercúrio diciandiamida) (Ahmed & Grant, 1972a). Nesses casos, testou-se a aberração crom ossôm ica em m itose de pontas radiculares. Ahmed & Grant (1 972b)
mostraram que m evin fós e cianazina induziam aumentos na freqüência de aberrações, mas as doses estudadas foram elevadas (200 a 600ppm ),
consideradas m uito extrem as em term os de contam inação ambiental. Por outro lado, doses de apenas 10ppm de Panogen 1 50<) causavam citotoxicidade, enquanto genotoxicidade óbvia era notada com apenas 1 ppm (Ahmed & Grant, 1972a). Estes resultados apontam para possíveis riscos significativos à saúde nas operações de preparo de calda e aplicação, nos tem pos em que este produto era comercialmente disponível.
Reconhecendo o mérito especial de se estudar agroquímicos in situ, G rant (1982) opinou que nenhum outro organismo seria tão útil quanto
Tradescantia e não haveria qualquer teste tão adequado quanto o ensaio Trad- MCN para a avaliação dos riscos genéticos in s itu . Estas afirm ações foram verificadas em um estudo da genotoxicidade do inseticida malation usado no contro le de pragas em uma casa de vegetação (Ma et al., 1 983). Em um tra tam ento , vasos com plantas intactas foram pulverizados, simulando o contro le convenciona lm ente usado. Tratam entos adicionais envolveram absorção de m alation pelas inflorescências (com ou sem prévia dissolução
com DMSO ou tratamento com a fração microssõmica S-9 de fígado macerado
de cam undongo induzido com A ro c lo r" ') e exposição de plantas intactas a
vapores de malation gerados por aquecimento, em câmaras hermeticamente fechadas. Todas as exposições a soluções de malation, seja pulverizado ou
absorvido pela inflorescência, foram negativas. Já as inflorescências expostas
a vapores de m alation sofreram um sign ificativo aumento na freqüência de m icronúcleos, sugerindo que form as gasosas do pesticida podem ser
particularm ente efetivas (Ma et a!., 1 983).
As clastogenicidacies do Benlate {benonnD e do tio fanato, dois fungicidas usados na conservação de fru lo s para aririazenagern, foram

estudadas com o ensaio Trad-MCN em concentrações de 0 ,0 5 e 0 ,0 7 % , respectivam ente (Huang & Chen, 1993b ; Huang & Chen, 1993c). Am bos
agentes induziram altas freqüências de micronúcleos.
Os resultados obtidos até o presente da genotoxicidade de pesticidas em Tradescantia são resumidos na Tabela 4.
Ma et al, (1984) apresentaram os resultados de 140 ensaios Trad-
MCN realizados com uma variedade de agentes físicos e químicos. Os agentes foram classificados em 9 categorias (números entre colchetes indicam o número
de agen tes te s ta d o s na ca te g o ria ): (a) agen tes re co n h e c id a m e n te carcinogênicos/mutagênicos [15], (b) bebidas comuns 18], (c) produtos químicos comuns [30], (d) drogas comuns [32], (e) pesticidas |1 8], (f) produtos químicos de uso dom éstico [16], (g) radiação ionizante e rad io isótopos [3 ], (h) m onitoram ento in s itu [13[ e (i) m isturas quím icas com plexas [8 ]. A lguns resultados positivos nesses grupos foram ; (a) benzo[a ]p ireno (50|J,M), EMS
(50mM) e azida sódica (0,2m M); (b) etanol (5% ), café descafeinizado (25% ) e cola (50% ); (c) vapores de formaldeído, óxido nitroso e dióxido de enxofre; (d) sacarina e aspirina; (e) B ladex® (cianazina), d icam ba, vapores de diclorvós,hidrazida maléica, p-diclorobenzeno e Tordon® (picloran); (f) alguns odorizantes e cosm éticos; (g) todas as form as de radiação; (h) vários locais poluídos; e (i) vários tipos de gases de com bustão e am ostras de água não
concentradas. Entre os resultados negativos encontravam -se: (a) 1,2-
benzantraceno, metil metanosulfonato e dinitrotolueno; (d) m itom icina C; e (e)
atrazina, simazina e 2,4-D. De 39 agentes testados com o ensaio Trad-MCN e
para os quais havia resultados disponíveis de testes de Ames, 26 forneceram os m esm os resultados, representando uma congruência de 6 7 % . Para pesticidas, 11 de 18 agentes testados resultaram positivos. De 8 pesticidas testados, tan to em Trad-MCN quanto no teste de Am es, som ente simazina forneceu resultados diferentes nos dois testes (Ma et al., 1 984).
Os resu ltados d ispon íve is de ava liações das p ropriedades clastogênicas de agentes químicos selecionados e estresses fis io lógicos em
Tradescantia são apresentados na Tabela 5.
19

Raios cósmicos e campos eletromagnéticos
T. paludosa foi utilizada para estudar os efeitos potenciais de fatores associados a vôo espacial, como aceleração, vibração, falta de gravidade e radiação ionizante (Delone et al., 1 986). .As inflorescências foram fixadas quim icamente a vários intervalos de tempo desde o lançamento da nave, até após a aterrissagem e as figuras mitóticas dos micrósporos foram analisadas
em termos de aberrações. Um tipo especial de aberração foi observado nesse material, consistindo de translocações não-recíprocas complexas e fragmentos
esféricos. 0 surgimento de tais aberrações não estava associado com a duração
do vôo, ou com lançamento ou aterragem. Especulou-se que o agente causativo
seria o pesado bombardeio por radiação cósmica (Delone et al., 1986).
A sensibilidade de Tradescantia à radiação tem sido demonstrada em relação a raios-X, fontes radioisotópicas internas e externas e raios cósmicos. Similarmente, ondas longas de rádio e campos magnéticos de ondas
curtas, presentes nas vizinhanças de antenas de transmissão, têm sido
constatados como danosos a cromossomos em replicação. Em uma série de
experim entos in situ (Haider et al., 1 994), inflorescências de Tradescantia
foram expostas em cinco distâncias de uma antena e em gaiolas plásticas (não isolante) e de Faraday (isolante eletromagnético), distribuídas ao redor de
locais que excediam os padrões para campos elétricos da Associação
Internacional de Proteção a Radiação. Todos os tratamentos resultaram em increm entos na freqijência de micronúcleos quando comparados com os controles de laboratório e, mais importante, comparações entre amostras de gaiolas isolantes e não isolantes resultaram em diferenças altam ente
s ign ifican tes (Haider et al., 1994). Este resultado é particu larm ente
interessante, uma vez que ambos os grupos foram expostos a exatamente as
m esm as cond ições am bienta is, exceto pela in fluência da radiação
eletromagnética. Uma relação dependente da dose com respeito à distância de exposição também suporta a conclusão de que os efeitos observados eram
realmente devidos ao campo eletromagnético.
As atividades clastogênicas de raios-X e outras formas de radiação
ionizante om Tradescantia estão resumidas na Tabela 6,
Em um e s tu d o rece n te , ap o iado pe lo FVograma In te rn a c io n a l de
S eg ura nça Q uím ica, a u tilidade do ensaio Trad M C N (jun tam ente com o u tro s
20

três ensaios vegetais) foi avaliada com quatro compostos genotóxicos conhecidos (Grant & Salamone, 1994) em cinco diferentes laboratórios (Sandhu
et al., 1994a; Sandhu et al., 1994b). Embora os resultados não tenham sido
idênticos,houve boa concordância entre todos laboratórios, sugerindo que o
ensaio Trad-MCN é um bioensaio confiável para detecção de agentes
clastogênicos (Ma et al., 1994b).
Os estudos ora revisados dem onstram que Tradescantia, e em
particu la r o ensaio Trad-M CN, oferece um sistema m uito sensível e
fa c ilm e n te m anuseáve l para ava liações de to x ic id a d e g e n é tic a , especialmente para condições in situ indispensáveis em pesquisas ambientais.
TRADESCANTIA E O BIOENSAIO DO PÊLO ESTAMINAL
Trad-SHM: Fundamentos e desenvolvimento do bioensaio
0 ensaio da mutação em pêlo estaminal (Trad-SHM) baseia-se em m utação pontual (mitótica) na qual é suprimida a expressão do caráter azul dominante em flores de plantas heterozigotas, resultando no aparecimento da
cor rosa recessiva (Emmerling-Thompson & Nawrocky, 1982; Mericle & Mericle,
1967; Mericle & Mericle, 1971; Nayar & Sparrow, 1967). Os estudos iniciais
com esse sistema centraram-se na avaliação dos efeitos genotóxicos e
citotóxicos de radiação ionizante e empregaram as células meristemáticas dos pêlos estaminais do clone 02 de Tradescantia, em substituição a culturas m icrobianas, Nesse ensaio, crescimento normal de pêlos era considerado
equivalente à formação de colônias, enquanto pêlos atrofiados correspondia a eliminação celular nas culturas, devido a efeitos severos, altamente deletérios ou letais. Em adição á utilização de mutação (alteração de cor) como indicador
final das avaliações, alterações genotóxicas como expressão de células gigantes,
duplas ou triplas, bifurcação e outras anomalias de crescimento eram também consideradas juntamente com a perda da integridade reprodutiva das células
dos pêlos como indicadores de genotoxicidade (Nayar & Sparrow, 1 967).
A base genética para expressão de células cor-de-rosa nos pêlos estam inais do clone 4430 de Tradescantia foi estabelecida por meio de cruzamentos recíprocos com a linhagem parental rosa e branca T. subacaulis
Bush (Emmerling-Thompson & Nawrocky, 1980). Determ inou-se que21

pigmentação rosa era dependente de um par de alelos em um único locus, sendo azul (B) dom inante sobre rosa (b) e demonstrou-se que o clone 4 43 0 é homozigoto dominante para o locus branco. A identidade espectrofotométrica de ambos pigmentos, rosa e azul, de quatro clones diferentes de Tradescantia está determinada (Sanda-Kamigawara & Ichikawa, 1 993).
M utação para rosa e perda de integridade reprodutiva nos pêlos
estaminais de várias espécies e híbridos de Tradescantia (Ichikawa & Sparrow, 1967a; Ichikawa & Sparrow, 1 967b; Ichikawa & Sparrow, 1 968; Ichikawa & Sparrow, 1969; Ichikawa et al., 1969; Sparrow & Ichikawa, 1967) tornaram-
se importantes indicadores para o estudo da atividade genotóxica das radiações (Alvarez & Sparrow, 1965; Kappas et al., 1972; Nauman et al., 1976; Nauman et al., 1974; Sparrow et al., 1 973 ; Underbrink et al., 1 973a; Underbrink et al., 1971).
O ensaio Trad-SHM como urn sistema de monitoramento para genotoxicidade no ambiente
Sparrow e colaboradores (1 972) estudaram os efeitos de neutrons e raios-X com o ensaio Trad-SHM (clone 02), estabelecendo uma relação
linear entre dose-resposta para ambos os agentes e uma dose duplicadora da taxa de mutação de apenas 1 rad para raios-X. As freqüências espontâneas de mutação de várias espécies e híbridos de Tradescantia foram definidas com
base em m uitos anos de investigação no Brookhaven National Laboratory (Sparrow & Sparrow, 1 976). Híbridos (i.e., clone 4430) e híbridos putativos (i.e., clone 02) apresentaram freqüências e variação menores na m utação
espontânea, quando comparados com clones de espécies puras, e foram considerados mais adequados para experimentação.
Os efeitos da radioatividade natural sobre a expressão de mutações foram estudados em vôos orbitais (Sparrow et al., 1 968) e pelo cu ltivo de Tradescantia em areia monazítica (Nayar et al., 1970). Notou-se que a taxa de
mLitação aum entava com todas as amostras e os radionuclídeos absorvidos pelas plantas eram muito mais efetivos que a simples radiação externa. Estes resultaaos foram mais tarde confirmados pela exposição de plantas a solos
coletados no canifjo experimental de explosões atômicas das Ilhas Bikini (Ichikawa & Ishii, 1991). Solos que causavam aumentos significativos nas freqüências de m utaçio continham '‘ 'Cs v, '' -'Co, entro ouiros ratíionuclídeos. Outros estudos
22

envolvendo a absorção de radionuclídeos incluem compostos de trítio (Nauman
et al., 1 979; Tano et al., 1984) e '^'1 (Tano & Yamaguchi, 1 979).
0 ensaio Trad-SHM fo i empregado in s itu para m onitorar radiação
ionizante nas vizinhanças de usinas nucleares em um estudo de larga escala realizado no Japão (Ichikawa, 1981). Aum entos significativos nas freqüências de mutação estavam correlacionados tan to com a direção preferencial dos
ventos quanto com os períodos de efetivo funcionamento das usinas. Cebulska- W asilewska (1 992) observou increm ento nas freqüências de m utação em Trad-SHM correlacionadas com contam inação causada em Cracóvia pela
explosão do reator de Chernobyl (a uma distância de 700km ). Incrementos similares na taxa de m utação foram tam bém notificadas de maio a junho de 1986 no Japão (mais de 800km de distância) (Ichikawa et al., 1 996). Nesse caso, variações na taxa espontânea de m utação registradas em um período de 10 anos {1982 a 1992) sempre podiam ser explicadas pelo fator temperatura, ao contrário desse increm ento em 1986. Exposições a radiação tam bém perm itiram a padronização do ensaio Trad-SHM em relação a variações de temperatura (Nauman et al., 1977a; Nauman et al., 1977b), dose (Nauman et al., 1 977c; Nauman et al., 1975) e outras variáveis comuns das condições de experim entação (Underbrink & Sparrow , 1974; Underbrink et al., 1975a;
Underbrink et al., 1975b).
A aplicabilidade do ensaio Trad-SHM a estudos de mutagênese
química foi proposta por Underbrink et al. (1973b) e testada em comparação aos efeitos de radiação e EMS e DBE gaseificados (Nauman et al., 1976). As respostas a agentes químicos apresentaram características similares àquelas dos raios-X (increm ento exponencial seguido de saturação na freqüência de células mutantes) e o clone 4 4 3 0 fo i mais sensível que o clone 02 . Estes
resultados fo ram poste rio rm ente confirm ados com uma variedade de
com postos quím icos e radionuclídeos (Tano, 1987; Tano, 1990; Tano &
Yamaguchi, 1985). Nesses estudos, os agentes mutagênicos foram aplicados
topicam ente, d iretam ente sobre as inflorescências. Doses dim inutas de 5 a
20pg de A/-nitroso-A/-metiluréia e A/-nitroso-A/-etiluréia e 10Opg de EMS foram efetivas e detectáveis no teste Trad-SHM. 0 limite de detecção para radiação
externa estava abaixo de 1 rad.
Essa alta sensibilidade do ensaio Trad-SHM a agentes mutagênicos
químicos foi primeiramente demonstrada após exposição acidental das plantas
23

(clone 02) a vapores que contaminavam o suprimento de ar de um laboratório de Brookhaven. Um incremento súbito na freqüência espontânea de mutações levantou a suspeita que levou à descoberta da contaminação (Sparrow & Schairer, 1 971). Estudos adicionais com Trad-SHM na avaliação de agentes químicos envolvem hidrazida maléica (Gichner et al , 1 982b), M M S, EMS,
DMS (dimetil sulfato) (Ichikawa et al., 1 990 ; Ichikawa & Takahashi, 1 978; Sanda-Kamigawara et al., 1 991 ), com postos A/-nitroso e vários solventes orgânicos, entre outros, assim com o avaliações de ação sinergística entre agentes químicos e entre estes e radiações (Badaev et al., 1 989 ; Gichner et al., 1994; Gichner et al., 1982a; Gichner et al., 1988; Ichikawa, 1992; Ichikawa et al., 1990; Ichikawa et al., 1993; Kuglik et al., 1994 ; Sanda- Kamigawara et al., 1991; Shima & Ichikawa, 1 994; Shima & Ichikawa, 1995a; Shima & Ichikawa, 1 995b; Velem insky et al., 1 987; Vilialobos-Pietrini et al., 1986).
Dem onstrou-se que o ensaio Trad-SHM é ainda capaz de ativar agentes promutagênicos em agentes mutagênicos de ação direta (Gichner et
al., 1980). Benzo-a-pireno, atrazina e vários compostos A/-nitroso foram m utagên icos quando te s ta do s sem tra ta m e n to prévio com frações microssomais (Veleminsky & Gichner, 1988). Xiao e Ichikawa (1995; 1996) relataram a ativação de hidrazida maléica (MH) em um agente m utagênico por ação de peroxidase e dem onstraram que MH agia sinergisticam ente (Cebuiska-Wasilewska et al., 1981) e antagonisticamente com raios-X quando esses eram aplicados antes e depois da MH, respectivamente. Os raios-X suprimiam a ativação da MH quando aplicados a posteriori. Uma revisão da mutagenicidade de radiações ionizantes e agentes químicos estudada com Trad-SHM foi oferecida por Ichikawa (1992).
Talvez a contribu ição mais im portante do ensaio Trad-SHM tenha sido uma série de estudos sobre poluição atmosférica realizados com um laboratório móvel, nos EUA (Schairer, 1979; Schairer & Sautkulis, 1982;
Schairer et al., 1 982 ; Schairer et al., 1 979; Schairer et al., 1 983). A r obtido
em locais p o lu íd os induziam maiores freqüências de mutações que ar filtrado das m esm a s localidades ou ar coletado em áreas controle, com o o Grand Canyon. A mutagenicidade de atmosferas poluídas (Sparrow' & Schairer, 1974) te m sido la m b e m d e m o n s tra d a nas v iz in h a n ç a s de re fin a ria s de ó leo e
coiTi|:i(;xo.s |) f ít io fi i líi n icos ÍL ü w e r i-;l a l. , 1 9 8 3 a ) , fund ições de c h u m b o (Low er
et al., 1 9 78, L o w e r e t al., 1 9 8 3 b l, fabnc.'i de rem éd ios (C ebu Iska -W as ilew ska
24

& Guminska, 1 987) e incinerador m unicipal de resíduos sólidos (Ma, 1 994 ;
Ma et al., 1 993b; Ma et al., 1 996).
A atividade mutagênica de fumaças químicas usadas pelo exército
Norte-Americano também foi avaliada com Trad-SHM . Respostas positivas
foram obtidas com "fogo il" e diesel para tanques, bem como para sua com b inação (S chae ffe r et a l., 1 9 8 7 ). O zôn io , em co n c e n tra ç õ e s ocasionalmente encontradas em áreas poluídas (300 a SOOppb), não apresentou
mutagenicidade no ensaio Trad-SHM (Gichner et al., 1 992 ; Rodrigues et al.,
1 996), embora tenha sido positivo em concentrações mais altas (Schairer,
1 979). Uma revisão do Trad-SHM como um ensaio para agentes mutagênicos gasosos foi publicada sob os auspícios do Programa Gene-Tox, da "USEPA"
(Van'thof & Schairer, 1982).
Em adição a estudos de agentes gasosos, Trad-SHM tem sido usado
para avaliação de ambientes aquáticos (Lower et al., 1 985 ; Tano, 1 989). Episódios de mutagenicidade nas águas de um reservatório no estado Norte-
Americano do Missouri apresentavam correlação com eventos que promoviam
a transferência dos agentes mutagênicos dos sedimentos para a coluna d'água (Lower et al., 1985). Grant e colaboradores (1992) analisaram /n s itu a
genotoxicidade das águas em uma área do Lago Superior nas vizinhanças de
uma fábrica de papel e celulose, usando Trad-SHM, Trad-MCN e o ensaio de aberração crom ossôm ica com Vicia faba . Embora Trad-S H M fosse
sufic ientem ente sensível para detectar atividade m utagênica, o ensaio foi
considerado inferior aos outros em termos de facilidade para manipulação em
campo, sendo que áreas remotas trazem dificuldade para o cultivo das plantas no longo (14 dias) período de recuperação necessário para esse teste.
Há escassêz de in form ações sobre a tiv idade m utagênica de
pesticidas em Trad-SHM (Mohammad & Ma, 1 983). Tomkins & Grant (1972)
estudaram os efeitos de menazon (um inseticida do grupo s-triazina), metobromuron (herbicida do grupo das uréias substituídas) e Daconil 2 7 8 7 ®
(clorotalonil, fungicida do grupo dos hidrocarbonetos aromáticos clorados), embrulhando inflorescências de Tradescantia (clone 02) com algodão embebido
em soluções dos pesticidas (1.500ppm). Nenhuma resposta positiva foi obtida.
A mutagenicidade do herbicida e regulador de crescimento hidrazida maléica
já foi demonstrada em ensaios Trad-SHM empregando o clone 4430 (Gichner
et al., 1 982b; Xiao & Ichikawa, 1 995 ; Xiao & Ichikawa, 1 996). 0 fungicida25

derivado de benzimidazol Benlate® (benomil) foi testado em Trad-SHIVI (clone KU 20) em doses normalmente usadas na agricultura (0,5 a 4,0g/l) (Sakamoto & Takahashi, 1 981 ). Novamente não houve resposta positiva. Em contraste com esses resultados, sete de nove inseticidas foram positivos quando testados com Trad-SHM empregando o clone 4430 (Huang & Chen, 1993a). Diclorvós
(0,1% ), om etoato (0,04% ), metamidofós (9,05% ), M eobal® (3 ,4 -xy ly l metilcarbamato) (0,05% ), mevinfós (0,006% ), Am obem ® (cloramben, em verdade um herbicida) (0,045%) e metil tiofanato (0,07%) resultaram positivos, enquanto triclorfon (0,1 %) e Bassa® (2-sec-butilfenil metilcarbamato) (0,02%
- efeito tóxico registrado) foram negativos. Atrazina resultou mutagênica após exposição crônica do clone 4430 (Schairer & Sautkulls, 1 982) e cianazina também foi notificada como mutagênica em Trad-SHM (sem referência ao clone utilizado) (Veleminsky & Gichner, 1988).
Um resumo dos resultados até hoje obtidos com Trad-SHM na avaliação de mutagênese ambiental está apresentado na tabela 7.
CONCLUSÃO
As abundantes in form ações básicas sobre a genética e o
desenvolvimento de Tradescantia oferecem uma sólida estrutura de suporte
para seu uso como um bioindicador para ensaios de toxicidade genética
ambiental (Ma & Grant, 1 982). Micronúcleos nas células mães dos grãos de pólen são facilmente observáveis, permitindo um baixo grau de incerteza nas
avaliações e diminuindo a subjetividade presente no reconhecimento de aberrações cromossómicas, enquanto a indução de mutações para rosa em células dos pêlos estaminais oferece um indicador somático sensível de
mutagênese. Adicionalmente, os ensaios com Tradescantia têm se provado valiosos nos estudos de sinergismo e antagonismo entre agentes químicos e
entre estes e outros agentes genotóxicos, como radiações, uma propriedade
valiosa para a avaliação de riscos genotóxicos em situações ambientais
complexas (Shima & Ichikawa, 1995b).
Grant (1 994) analisou recentemente a situação atual dos ensaios
com plantas superiores para detecção de agentes mutagênicos ambientais, indn;ando a$ vantagens desses sistemas (í.ti relação a possibilidade de se
reali7art;ni avaliíições m Si tu. Em um estudtj recenterriente patrocinado pelo
26

Programa Internacional de Seguridade Química, a utilidade dos dois ensaios com Tradescantia aqui discutidos (bem como outros três ensaios vegetais)
foi avaliada com quatro reconhecidos agentes químicos genotóxicos (Grant & Salamone, 1994) em cinco laboratórios diferentes (Sandhu e t a l., 1 994a; Sandhu et a l., 1994b). Os resultados obtidos nesse estudo consubstanciaram
o ensaio Trad-SHM como um sistema confiável para a averiguação das propriedades m utagênicas de agentes quím icos (Ma e? a /., 1994a). Em relação ao ensaio Trad-MCN, embora os resultados obtidos para os quatro
agentes não tenham sido idênticos, houve uma boa concordância entre todos os la b o ra tó r io s , suge rindo ser este um b ioensa io c o n fiá v e l para clastogenicidade (Ma e ta l., 1 994b), e como indicado em recente revisão bibliográfica (Rodrigues e ta l., 1 997), Trad-MCN é especialmente apropriado para a monitoração de agentes genotóxicos in situ.
Em conclusão, os estudos aqui revisados dem onstram que os ensaios baseados em Tradescantia o ferecem s is tem as fa c ilm e n te
m anipuláveis e m uito sensíveis para o estudo de tox ic idade genética, especia lm ente para as condições in s itu indispensáveis em estudos
ambientais.
REFERÊNCIAS BIBLIOGRAFICASAHMED, M.; GRANT, W . F. Cytological effects of the mercurial fungic ide Panogen
1 5 on Tradescantia and Vida faba root tips. Mutation Research, v. 1 4, p. 39 V 396 , 1972a.
AHM ED, M .; GRANT, W . F. Cytological effects of the pesticides phosdrin and bladex on Tradescantia and Vicia faba. Canadian Journal of Genetics and Cytology, v. 14, p. 1 57 -165 , 1972b.
ALVAREZ, M . R.; SPARROW, A, H. Comparison of reproductive in te g rity in the stamen hair and root meristem of Tradescantia paludosa fo llow ing acute gamma irradiation. Radiation Botany, v. 5, p. 42 3 -4 30 , 1965.
BADAEV, S. A .; GICHNER, T.; POSPISIL, F.; VELEMINSKY, J. Humic acids inhibit the form ation but not the mutagenicity of N-methyl-N-nitrosourea. Mutation Research, v. 210, p. 9 -13, 1989.
BAUD-GRASSET, F.; BAUD-GRASSET, S.; BIFULCO, J. M.; MEIER, J, M .; M A, T. H. Tradescantia m icronucleus test on the genotoxic ity of PAH-contam inated soil a fte r fungal treatm ent. Ecotoxicology and environmental chemistry - a global perspective. Lisbon: Society of Environmental Toxicology and Chem istry, V 1993a. Abstracts, p. 303.
27

BAUD GRASSET, S.; BAUD-GRASSET, F.; BIFULCO, J. M .: MEIER, J. R.; M A , T, H. Reduction o f g e n o to x ic ity o f a c reosote-contam inated soil a fte r fungal treatm ent determ ined by the Tradescantia m icronucleus test. Mutation Research, V . 303, p. 7 7 -8 2 , 19 93b .
CEBULSKA-W ASILEW SKA, A. Tradescantia s tam en-ha ir m uta tion bioassay on the m utagenicity of radio isotope-contam inated air fo llow ing the Chernobyl nuclear accident and one year later. Mutation Research, v. 27 0 , p. 23 -2 9 , 1992 .
CEBU LSKA-W ASILEW SKA, A .: G UM INSKA, M. The app lica tion o f som atic m utation frequency in Tradescantia to m easurem ents of m utagen ic a c tiv ity of polluted air. Folia Medica Cracovlensia, v. 28 , n. 1-2, p. 1 3 1 -1 3 8 , 1987 .
CEBULSKA-W ASILEW SKA, A .; LEENHOUTS, H. P.; CHADW ICK, K. H. Synergism between EMS and X-rays for the induction o f som atic m utations in Tradescantia. International Journal of Radiation Biology, v. 40 , p. 1 6 3 -1 7 3 , 1981 .
DELONE, N. L.; ANTIPO V, V. V .; PARFENOV, G. P. New type of chrom osom al m utation observed in Tradcscantia patudosa m icrospores dur ing experim ents in space sate llites. Doklady Akademii Nauk Sssr, v. 29 0 , n. 4, p. 9 7 9 -9 8 1 , 1 986 .
EMMERLING-THOMPSON, M .; NAW RO CKY, M. M. Genetic basis fo r using Tradescantia clone 4 4 3 0 as an environm enta l m on ito r of m utagens. The Journal of Heredity, v. 71 , p. 2 6 1 -2 6 5 , 1980 .
EMMERLING-THOMPSON, M .; NAW ROCKY, M. M. Evidence of gam etic m utation fo r flo w e r color in Tradescantia. Environmental and Experimental Botany, v.22, p. 4 0 3 -4 0 8 , 1982 .
GICHNER, T .; LANGEBARTELS, C.; SA N D E R M A N N J r., H. O zone is no t m utagen ic in the Tradescantia and tobacco m utagen ic ity assays. Mutation Research, v. 28 1 , p. 2 0 3 -2 0 6 , 1992 .
GICHNER, T.: LOPEZ, G. C.; W AGNER, E. D.; PLEWA, M. J. Induction o f som atic m utations in Tradescantia clone 4 4 3 0 by three phenylenediam ine isom ers and the antim utagen ic m echanism s of d ie thy ld ith iocarbam ate and am m onium m etavanadate. Mutation Research, v. 306, n. 2, p. 1 6 5 -1 7 2 , 1994 .
GICHNER, T.; VELEM INSKY, J.; PANKO VA, K. D iffe rentia l response to three alkylating n itrosocom pounds and three agricu ltura l chem ica ls in the Salm onella (Ames) and in the Tradescantia, A rab idopsis and barley m utagen ic ity assays. B iologisches Z en tra lb la tt, v. 101, p. 375 38 3 , 1982a.
GICHNER, T.; VELEM INSKY, J.; POKORNY, V. Som atic m uta tion induced by maleic hydra?ide and its po tassium and d ie tliy lam ine salts in the Tradescantia m iita tiiin assay. M u ta tio n Research, v. 103, |5 . 289 293 , 1982b.
G IC H N lR , T .; VEI..EMINSK''i', J .; RIEGER, R. A n tim u ta g e n ic e ffe c ts o f c lie tiiv ld ith ioca rba iiia lc tow a rds iiia ie ic hvclra/ido and N n;trosodie thylam ine- :iHu,i-.iii) m u la iiiM i'c ily 'n tin: ft.'idi'.scaiitia m iilagon ^iiiv asrtay. Biologia Plantarum,V, 3j. n. i , :t , 14 19. ' 98S.
28

GICHNER, T.: VELEM INSKY, J .; UNDERBRINK, A . G, Induction o f som a tic m uta tions by the prom utagen d im e thy l n itrosam ine in hairs of Tradescantia stam en. Mutation Research, v. 78 , p. 3 8 1 -3 8 4 , 19 80 .
GILL, B. S.; SAND HU, S. S. A pp lica tion o f the Tradescantia m icronucleus assay fo r the genetic eva lua tion o f chem ica l m ix tu res in soil and aqueous m edia. Mutation Research, v. 2 7 0 , p. 6 5 -6 9 , 1992 .
GRANT, W . F. C ytogenetic stud ies o f ag ricu ltu ra l chem ica ls in plants. In: FLECK, R.A.; HOLLAENDER, A ., ed.. Genetic toxicology: an agricu ltu ra l perspective. New York: Plenum Press, 19 82 . p. 3 5 3 -3 7 8 . (Basic Life Sciences, 21).
GRANT, W . F. The present s ta tus o f h igher p lan t b ioassay fo r the de tec tion of environm enta l m utagens. Mutation Research, v. 3 1 0 , n. 2, p. 1 75-1 85 , 1 9 9 4 .
GRANT, W . F.; LEE, H. G.; LOGAN, D. M .; SALAM O N E, M . F. The use of Tradescantia and Vicia faba b ioassays fo r the in s itu de tection o f m utagens in an aquatic env ironm en t. Mutation Research, v. 2 7 0 , p. 53 -6 4 , 1992 .
GRANT, W . F.; SALAM ONE, M. F. Com parative m utagen ic ity of chemicals selected for te s t in the International Program on Chemical S a fe ty 's co llabora tive s tudy on plant system s for the de tection o f environm enta l m utagens. Mutation Research, V . 31 0 , n. 2, p, 1 8 7 -2 0 9 , 1994 .
HAIDER, T .; KNASM ULLER, S.; KUNDI, M .; HAIDER, M . C lastogenic e ffe c ts o f rad io frequency radiations on chrom osom es of Tradescantia. Mutation Research, v. 3 2 4 , p. 65 -6 8 , 19 94 .
HELMA, C .; KNASMULLER, S.; SAN YA L, R.; SOMMER, R.; SCHULTEHERMAN, R. The e ffec t o f UV-irradiation on the geno tox ic ity o f contam inated g roundw ater detected by the Tradescantia m icronucleus test. Ecotoxicology and environmental chemistry - A global perspective Lisbon: S ociety o f Environm ental T ox ico logy and C hem istry, 1 9 9 3 . A bs tra c ts , p. 30 3 .
HELMA, C.; SOMMER, R.; SCHULTE-HERMANN, R.; KNASMULLER, S. Enhanced c las togen ic ity of con tam ina ted g ro undw a te r fo llo w in g UV irrad ia tion de tected by the Tradescantia m icronuc leus assay. Mutation Research, v. 3 2 3 , p. 93- 98 , 19 94 .
HOPKE, P. K,; PLEWA, M . J.; JO HNSTO N, J. B.; W EAVER, D.; W OO D, S. G.; LARSON, R. A .; HINESLY, T. M u ltitechn ique screening o f Chicago m unicipal sew age sludge fo r m utagen ic ac tiv ity . Environmental Science and Technology, V . le ', p. 1 4 0 -1 4 7 , 1982 .
HUANG, N.; CHEN, R. The report o f using Tradescantia S tam en Hair m uta tion to tes t 9 insectic ides. Environmental and Molecular Mutagenesis, v. 21 , n. suppl.22, p. 30, 1993a.
HUANG, N.; CHEN, R. Tradescantia m icronucleus (Trad-MCN) te s t on tw o agents used in fru it storage. Environmental and Molecular Mutagenesis, v. 21 , n. suppl. 22, p. 30, 1993b.
29

HUANG, N.; CHEN, R. Use o f Tradescantia m icronucleus assay in de tecting the m utagen ic ity o f tw o agents used in storing fresh fru it. Ecotoxicology and environmental chemistry - A global perspective Lisbon:Society of Environmental Toxico logy and C hem istry , 1993c . Abstracts,, p. 302.
ICHIKAW A, S. In s itu m on ito ring w ith Tradescantia around nuclear pow er plants. Environmental Health Perspectives, v. 3 7, p. I4 b -1 6 4 , I 981.
ICHIKAW A, S. Tradescantia stam en-hair system as an excellent botanical tes te r of m utagen ic ity : its response to ionizing radiations and chemical m utagens, and some synerg is tic e ffe c ts found. Mutation Research, v. 270, p. 3 -2 2 , 1992 .
ICHIKAW A, S.; ISHII, C. Som atic m utation frequencies in the stam en hairs of Tradescantia g ro w n in soil sam ples from the Bikini Island. Japanese Journal Of Genetics, v. 6 6 , n. 1, p. 27 -40 , 1991.
ICH IKAW A, S.; KA N A I, H.; HARADA, H. Somatic m utation frequencies in T ra d e s c a n tia s ta m e n h a irs tre a te d w ith aqueous s o lu t io n s o f e th y l m ethanesulfonate and m ethyl m ethanesulfonate. Japanese Journal of Genetics, V . 65, p . 30 9 , 1990 .
ICHIKAW A, S.; N A K A N O , A ,: KENMOCHI, M.; YAM AM O TO , I.; M URAI, M .; T A K A H A S H I, E.: Y A M A G U C H I, a . ; W A TAN ABE, K.; T O M IY A M A , M .; SUG IYAM A, K.; YOGO, A .; Y AZA K I, T.: OKOMURA, M .: SHIMA, N.; SATOH, M .; YO SHIM O TO , M .; X IAO , L. Z. Yearly variation of spontaneous som atic m utation frequency in the stam en hairs of Tradescantia clone KU 9 grow n outdoors, w h ich show ed a s ign ificant increase after the Chernobyl accident. Mutation Research, v. 3 4 9 , p. 24 9 -2 59 , 1996.
ICHIKAW A, S.: SPARROW , A. H. Radiation-induced loss of reproductive in tegrity in the stam en hair o f Tradescantia blossfeldians M ildbr., a tw e lve-p lo id species. Radiation Botany, v. 7, p. 3 3 3 -3 4 5 , 1967a.
ICHIKAW A, S.: SPARROW , A. H. Radiation-induced loss of reproductive in tegrity in the stam en hairs o f a po lyp lo id series of Tradescantia species. Radiation Botany, v. 7, p. 4 2 9 -4 4 1 , 1967b.
ICHIKAW A, S.; SPARROW , A. H. The use of induced somatic m utations to study cell division rates in irradiated stamen hairs of Tradescantia virginiana L. Japanese Journal of Genetics, v. 43 , p. 57-63 , 1968.
ICHIKAWA, S.; SPARROW, A. H. Analyses of radiation-induced loss of reproductive in tegrity in Tradescantia stam en hairs, an essentially single m eristem atic-ce ll system , Japanese Journa l o f Genetics, v. 44, p. 23 24, 1969.
ICHIKAW A, S.; SPARROW, A. H.; THOMPSON, K. H. M orphologically abnormal cells, somatic m utation and loss of reproductive integrity in iriadiated Tradescantia stamen hairs. Radiation Botany, v. 9. p. 195-211, 1969.
ICHII-AVVA. S.; TA K A H A S H I, C, S. Soniatic nu,tn!ioiKS in Tiadescantia stamen I ici.'S expused to eti lyl rneU lanesuiioi lait;. Enviroiiniental and Experimental Botany,V, 18, |j, 1 9-35, 1978,
30

ICHIKAW A, S.: YAM AG UCHI, A ,; O KUM URA, M, S ynerg istic e ffec ts o f m ethy l m ethanesulfonate and X-rays in inducing som atic m uta tions in the stam en hairs of Tradescantia clones, KU 27 and BNL 4 4 3 0 . Japanese Journal of Genetics, V . 6 8 , n. 4, p. 27 7 -2 92 , 1993.
JOHNSTON, J. B.; HOPKE, P. K. Estim ation o f the w e igh t-dependen t probab ility of de tecting a m utagen w ith the Am es assay. Environmental Mutagenesis, v. 2, p . 4 1 9 -4 2 4 , 1980.
KAPPAS, A .; SPARROW , A. H.; N AW R O C K Y, IVI. M . R e lative b io lo g ica l effectiveness (RBE) of 0 .43 -M ev neutrons and 250 -K vp X-rays fo r som atic aberrations in Tradescantia subacauHs Bush. Radiation Botany, v. 12, p. 2 7 1 - 28 1 , 1972 ,
KNASMULLER, S.; KIM, T. W .; M A, T. H. S ynerg istic e ffe c t be tw een tann ic acid and X-rays detected by the Tradescantia m icronucleus assay. Mutation Research, V . 270, p . 31-37 , 1992.
KUGLIK, P.; VESELSKA, R.; RELICHOVA, J. S en s itiv ity o f p lan t cy tog ene tic and genetic short-te rm assays for evaluating gene tic dam age induced by chem ical m utagens. Cell Biology International, v. 18, n. 5, p. 54 3 , 19 94 .
L'HERMITE, P.; DEHANDTSCHTTLER, J., ed. Copper in animal wastes and sewage sludge. London: D. Reidel 1980. 378 p.
LOWER, W . R.; DROBNEY, V. K.; AHOLT, 8 . J .; POLITTE, R. M u tagen ic ity o f the environm ents in the v ic in ity of an oil re fine ry and a pe trochem ica l com plex. Teratogenesis, Carcinogenesis, and Mutagenesis, v. 3, p. 6 5 -7 3 , 1983a.
LOWER, W . R.; ROSE, P. S.; DROBNEY, V. K. In s itu m utagen ic and o ther e ffec ts associated w ith lead sm elting. Mutation Research, v. 54 , p. 8 3 -9 3 , 1978 .
LOWER, W . R.; THOM PSON, W . A .; DROBNEY, V . K.; YAND ERS, A. F. M utagen ic ity in the v ic in ity o f a lead sm elter. Teratogenesis, Carcinogenesis, and Mutagenesis, v. 3, p. 23 1 -2 53 , 1983b.
LOWER, W . R.; YANDERS, A. F.; MARRERO, T. R.; UNDERBRINK, A . G.; DROBNEY, V. K.; COLLINS, M. D. M u tag en ic ity of bo ttom sed im ent from a w a te r reservoir. Environmental Toxicology and Chemistry, v. 4, p. 13-19 , 1 985 .
M A , T. H. M icronucle i induced by X-rays and chem ical m utagens in m eiotic pollen m other cells of Tradescantia - a prom ising m utagen te s t system . Mutation Research, v. 64, p. 30 7 -3 13 , 1979a.
M A, T. H. Tradescantia m icronuclei (Trad-MCN) tes t fo r environm enta l clastogens. In: KOLBER, A .R.; WONG, T.K.; GRANT, L.D.; DEW OSKIN, R.S.; HUGHES, T .J., ed. In vitro toxicity testing of environmental agents, current and futurep. Part A: Survey of test systems. New York: Plenum Press, 1 979b , p. 19 1 -2 14 .
M A, T. H. Tradescantia m icronucleus bioassay and pollen tube chrom atid aberration tes t fo r in s itu m on ito ring and m utagen screening. Environmental Health Perspectives, v. 37, p. 85-90, 1981.
31

M A, T. H. Tradescantia cy tog ene tic tests (root-tip m itosis, pollen m itos is , pollen m other-cell meiosis). A report o f the U. S. Environmental P rotection A gency Gene-Tox Program. Mutation Research, v, 99, p. 29 3 -3 02 , 1982 .
M A , T. H. Tradescantia m icronucleus test on clastogens and in s itu m on ito ring . In: MENDELSOHN, M .L .; ALBERTINI, R.J., ed. Mutation and the environment. New York: W iley-Liss, 1990 . p. 83-90. (Progress in Clinical and Biological Research, 340).
M A, T. H. Landfill or inc ineration - w h ich is the better w ay to trea t our solid wastes? Environmental and Molecular Mutagenesis, v. 23, suppl. 23 , p. 40, 1 9 9 4 .
M A, T. H.; ANDERSON, V. A .; AHMED, I. Environmental clastogens de tec ted by m e io tic pollen m o the r cells o f Tradescantia. In: TICE, R.R.; CO STA, D.L.; SCHAICH, K .M ., ed. Genotoxic effects of airborne agents. New York: Plenum Press, 1982 . p. 1 4 1 -1 5 7 . (Environmental Science Research, 25).
M A , T. H.: ANDERSON, V. A .; HARRIS, M. M.; BARE, J. L. Tradescantia- M icronucleus (Trad-M CN) tes t on the genotoxic ity of m alathion. Environmental Mutagenesis, v. 5, p. 1 2 7 -1 3 7 , 1983.
M A , T. H,; ANDERSO N, V. A .; HARRIS, M. M .; NEAS, R. E.; LEE, T . S. M utagen ic ity of drinking w a te r detected by the Tradescantia m icronucleus test. Canadian Journal of Genetics and Cytology, v. 27, p. 143 -1 50 , 1985 .
M A, T. H.: CABRERA, G. L,; CEBULSKA-W ASILEW SKA, A.; CHEN, R.: LOARCA, F.; VANDENBERG, A. L.: SALAMONE, M. F. Tradescantia stamen hair m utation bionssay. Mutation Research, v. 310, n. 2, p. 21 1-220, 1994a.
M A , T. H .; C A BR E R A , G. L.; CHEN, R.; GILL, B. S .; S A N D H U , S. S .: VANDENBERG, A. L.; SALAM ONE, M, F. Tradescantia m icronucleus bioassay. Mutation Research, v. 31 0 , n. 2, p. 221-230 , 1994b.
M A, T. H.; GRANT, W . F. The Tradescantias - adventurous plants. The Herbarist, v. 48 , p. 36 -44 , 1982 .
M A, H.: HARRIS, M . 7rac/csca/?r/a micronucleus (Trad-MCN) assay - a potentia l indoor po llu tion m on ito r. Environmental Mutagenesis, v. 9, suppl. 8 , p. 65,1 9 8 7 a .
M A , H.; HARRIS, M. M. Tradescantia m icronucleus (Trad-MCN) bioassay - a pro iiiis ing indoor air po lk ition m onitoring system. In: SEIFERT, B.; ESDORN,H,: FISCHER, M .; RUDEN, H.; WEGNER, J., ed. 4th International Conference on Indoor Air Quality and Climate. Berlin: institu te for W ater, Soil, and Air H '.giene, 1987b. v . l . p , 24 3 -2 47 .
M A H.; HARRIS, M . M .: ANDERSON, V. A ; AHMED, I.; M O H A M M A D , K.;BARL, L.: LIN, G, 7/i;;/cV>c-.';//f/c3-ni:ciOnUL oub i'^rcid M C N ! te s ts ot; 1 40I . i ' i i th iijia ltrcl ruit;;-!!.':;. M uta tion Researcii, v. 138, 1 b7 167, 1984.
32

M A , T. H.; KONTOS, G. J., JR.; ANDERSON, V. A . Stage se n s itiv ity and dose response o f m eio tic chrom osom es o f pollen m othe r cells o f Tradescantia to X- rays. Environmental and Experimental Botany, v. 2 0 , p. 1 6 9 -1 7 4 , 1 9 8 0 .
M A , T. H.; NEAS, R. E.; HARRIS, M. M .; XU, Z.; COOK, C.; SW OFFORD, D. in vivo tests {Tradescantia- and m ouse-m icronucleus) and chem ical analyses on drinking w a te r o f rural com m unities. In: SAND HU, S.S.; DEM ARINI, D .M .; M ASS, M .J .; MOORE, M .M .; MUMFORD, J .L ., ed. Short-term bioassays in the analysis of complex environmental mixtures V . N ew York: Plenum Press, 1987 . p. 18 9 -2 05 .
M A , T. H.; SANDHU, S. S.; PENG, Y.; CHEN, T. D.; KIM , T. W . S ynerg is tic and antagonistic e ffec ts on genotoxic ity o f chem icals com m on ly found in hazardous w aste sites. Mutation Research, v. 2 7 0 , p. 71 -7 7 , 1992a.
M A , T. H.; SPARROW, A. H.; SCHAIRER, L. A .; N A U M A N , A . F. E ffec t o f 1,2- d ib rom oethane (DBE) on m eiotic ch rom osom es o f Tradescantia. M utation Research, v. 58 , p. 25 1 -2 58 , 1978.
M A, T. H.; XU, C.; LIAO, S.; JEONG, B. S. G en o tox ic ity o f landfill gaseous e m iss ion and leacha tes de tec ted by T rade scan tia p la n t b io a ssa ys . In: Ecotoxicology and environmental chemistry - A global perspective. Lisbon: Society of Environm ental Toxico logy and C hem istry , 1993a . p. 304 .
M A, T. H.: XU, C.; LIAO, S.; JEONG, B. S.; LEATHERWOOD, R. //i s/Ya m onitoring of gaseous emission from a municipal incinerator using Tradescantia m icronucleus and Tradescan tia stam en hair m u ta tio n b ioa ssays .In : Ecotoxicology and environmental chemistry - A global perspective. Lisbon: Society o f Environmental T ox ico logy and Chem istry, 1993b. p. 304 .
M A, T. H.; XU, C.; LIAO, S.; MCCONNELL, H.; JEONG, B. S.; W O N, C. D. in s itu m on itoring w ith the Tradescantia bioassays on the ge no to x ic ity o f gaseous em issions from a closed landfill site and an inc inerator. Mutation Research, v. 359 , p. 3 9 -5 2 , 1996.
M A, T. H.; XU, J .; XIA, W .; JONG, X.; SUN, W .; LIN, G. P ro fic iency o f the Tradescantia m icronucleus image analysis system fo r scoring m icronucleus frequencies and data analysis. Mutation Research, v. 2 7 0 , p. 39 -4 4 , 1992b .
MERICLE, L. W .; MERICLE, R. P. M echanistn o f som atic m u ta tio n fo r flo w e rs of hybrid Tradescantia (clone 02). Genetics, v. 56, p. 5 7 6 -5 7 7 , 1967 .
MERICLE, L. W .; MERICLE, R. P. Somatic m uta tions in c lone 0 2 Tradescantia: A search for genetic iden tity . Journal of Heredity, v. 62 , p. 3 2 3 -3 2 8 , 1 97 1 .
M O H A M M A D , K.; M A , T. H. T radescan tia -m ic ronuc leus (Trad-M C N) and Tradescantia-Stamen Hair (Trad-SHM) tests on com m on pesticides. Environmental and Molecular Mutagenesis, v. 5, p. 3 7 0 -3 7 1 , 1983 .
N A U M A N , C. H.; KLOTZ, P. J.; SCHAIRER, L. A. U ptake o f tr it ia te d 1,2- d ibrom oethane by Tradescantia floral tissues: re lation to induced m uta tion frequency in stam en hair cells. Environmental Health Perspectives, v. 19, p. 201-21 5, 1 9 7 9 .
33

NAUM AN, C. H,; SCHAIRER, L, A ,; SAUTKULIS, R, C.; KLUG, E. E. Influence of hypertherm ia on the spontaneous, radiation- and chem ica l-induced m uta tion frequency in Tradescantia stamen hairs. Radiation Botany, v. 70, p. 632, 1 977a.
NAUM AN, C. H.; SCHAIRER, L. A .; SPARROW, A. H. Influence of tem pera ture on spontaneous and radiation-induced som atic m utation in Tradescantia stamen hairs. Mutation Research, v. 50 , p. 2 0 7 -2 1 8 , 1977b.
NAUM AN, C. H.; SPARROW, A. H.; SCHAIRER, L. A. C om parative e ffec ts of ionizing radiation and tw o gaseous chem ical m utagens on som atic m uta tion induction in one m utable and tw o non-m utable clones of Tradescantia. Mutation Research, v. 38, p. 53-70 , 1976 .
NAUM AN, C. H.; SPARROW, A. H.; SCHAIRER, L. A . Low dose m uta tion response relationships in Tradescantia: Principle and comparison to m utagenesis fo llow ing low dose gaseous chem ical m utagen exposures. In: Radiobiological Protection, First European Symposium on Rad-equivalence: proceedings. Luxemburg: Commission of the European C om m unity , 1977c. v. EUR 5725e, p. 13-23.
NAUM AN, C. H.; SPARROW, A. H.; SCHAIRER, L. A .; KLUG, E. E. C om parative effects of ionizing radiation and gaseous chem ical m utagens on m uta tion induction in a m utable clone o f Tradescantia. Radiation Research, v. 59, p. 153 -154 , 1974 .
NAUM AN, C. H.; UNDERBRINK, A . G.; SPARROW, A. H. Influence o f radiation dose rate on somatic m utation induction in Tradescantia stamen hairs. Radiation Research, v. 62, p. 79 -96 , 1975 .
NAYAR, G. G.; GEORGE, K. P.; GOPAL-AYENGAR, A. R. On the biological effects of high background rad ioactiv ity : studies on Tradescantia g ro w n in radioactive m onazite sand. Radiation Botany, v. 10, p. 2 8 7 -2 9 2 , 1970 .
NAYAR, G. G.; s p a r r o w , A. H. Radiation-induced som atic m uta tions and the loss of reproductive in tegrity in Tradescantia stamen hairs. Radiation Botany, v. 7, p. 257 -267 , 1967.
RODRIGUES, G. S.; M A, T. H.; PIMENTEL, D.; WEINSTEIN, L. H. Tradescantia bioassays as m onitoring system s fo r environm enta l m utagenesis - a rev iew . Critical Reviews in Plant Sciences, v. 16, n. 4, p. 3 2 5 -3 5 9 , 1997.
RODRIGUES, G. S.; MADKOUR, S. A .; WEINSTEIN, L. H. G enotoxic a c tiv ity of ozone in Tradescantia. Environmental and Experimental Botany, v. 36, n. 1, p.45-50 , 1996,
RUIZ, E. P.; RABAGO, V. M. E.; LECONA, S. U.; PEREZ, A. B.; M A , T. H. Tradescantia m icronucleus (Trad-MCN) bioassay on clastogenicity of w astew ater anri in s in i m onitoring. Mutation Research, v. 270, p 45 -51 , 1992.
SAKAMOTO, E. T.: TAKAHASHI, C, S. Action of beniate fungicide on Tradescantia iUaniet'i hairs and AHium ccfja \ oo t -tip cells. Revista Brasileira de Genetica, v. 4, IÍ 3, p. 367-381, 1981,
34

SAN D A-KAM IG AW AR A, M .; ICH IKAW A, S. Identity of normal and m utan t flow er- color p igm ents in fou r d iffe re n t Tradescantia clones confirm ed by m eans o f m icroespectropho tom etry . Japanese Journal of Genetics, v. 6 8 , p. 137, 1 993 .
S AN D A -K A M IG A W A R A , M .; IC H IK AW A , S.; W ATANABE, K. Spontaneous, radiation- and EM S-induced som atic pink m uta tion frequencies in tine stam en hair and petals o f a dip lo id c lone o f Tradescantia, KU 27. Environmental and Experimental Botany, v. 31, n. 4, p. 4 1 3 -4 2 1 , 1991.
SANDHU, S. S .; DE SERRES, F. J .; G O PALAN, H. N. B.; GRANT, W . R.; VELEMINSKY, J .; BECKING, G. C. Environm ental m onitoring fo r g e n o to x ic ity w ith plant sys tem s: an in tro du c tion and s tud y design. Mutation Research, v. 310 , n. 2, p. 1 6 9 -1 7 3 , 1994a.
SANDHU, S. S .; DE-SERRES, F. J .; G O PALAN , H. N. B.; GRANT, W . R.; SVENDSG AARD, D .; VELEM IN SKY, J .; BECKING, G. C. Environm enta l m on itoring fo r g e n o to x ic ity w ith p lan t system s: results and recom m endations. Mutation Research, v. 31 0 , n. 2, p. 2 5 7 -2 6 3 , 1994b.
SANDHU, S. S.; M A , T. H.; PENG, Y.; ZHOU, X. C lastogenic ity eva luation of seven chem icals com m on ly found at hazardous industrial w aste sites. Mutation Research, v. 2 2 4 , n. 4, p. 4 3 7 -4 4 6 , 1989 .
SAX, K.; EDMONDS, H. W . Developm ent o f the male gam etophyte in Tradescantia. Botanical Gazette, v. 95 , p. 1 5 6 -1 6 3 , 1933 .
SCHAEFFER, D. J .; N O VAK, E. W .; LOWER, W . R.; YANDERS, A .; KAPILA, S.: W ANG , R. E ffec ts o f chem ical sm okes on flora and fauna under fie ld and laboratory exposures. Ecotoxicology and Environmental Safety, v. 13, p. 301 -3 1 5 , 1987 .
SCHAIRER, L. A . M u tag en ic ity o f am bient air at selected sites in the United States using Tradescantia as a m on ito r. In: KOLBER, T.K.; GRANT, L.D.; DEWOSKIN, R.S.; HUGHES, T .J ., ed. In s itu toxicity testing of environmental agents. Current and future possibilities - Part A: Survey of tests. New York: Plenum Press, 1979 . p. 1 6 7 -1 9 0 .
SCHAIRER, L. A .; SAUTKULIS, R. C. D e tection o f am bient levels o f m utagenic atm ospheric po llu tan ts w ith the higher p lant Tradescantia. In: KLEKOWSKI Jr.,E.J., ed. Environmental mutagenesis, carcinogenesis, and plant biology. New York: Praeger, 19 82 . p. 15 5 -1 94 .
SCHAIRER, L. A .; SAUTKULIS, R. C.; TEMPEL, N. R. M onitoring am bient air for m utagen ic ity using the higher p lant Tradescantia. In: TICE, R.R.; COSTA, D.L.; SCHAICH, K .M ., ed. Genotoxic effects of airborne agents. New York: Plenum Press, 1982 . p. 1 2 3 -1 4 0 (Environm ental Science Research, 25).
SCHAIRER, L. A ,; V A N T HOF, J.; HAYES, C. C.; BURTON, M. R.; DE SERRES,F. J. M easurem ent o f b iological a c tiv ity o f am bient air m ixtures using a mobile laboratory fo r in s itu exposure, pre lim inary results from the Tradescantia plant te s t sys te m . In: W A TE R S , M .; NESN O W ; HUISING H; SAN D H U , S .S .;
35

CLAXTON, ed. Application of short-term bioassays in the fractionation and analysis of complex environmental mixtures. New York: Plenum Press, 1 979 . p. 4 1 9 -4 4 0 .
SCHAIRER, L. A .; V A N T HOF, J.; HAYES, C. C.; BURTON, R, M .; DE SERRES,F. J, Exploratory m on ito ring o f air po llu tants fo r ge no to x ic ity a c tiv ity w ith Tradescantia stam en hair system . Environmental Health Perspectives, \/. 27 , p. 51 -60 , 1983.
SHARIVA, C. B. S. R.; PANNEERSELVAN, N. Genetic tox ico lo gy o f pestic ides in higher plant system s. Critical Reviews in Plant Sciences, v. 9, n. 5, p. 40 9 - 4 4 2 1990 .
SHIM A, N.; ICHIKAW A, S. Synergism detected among m ethyl m ethanesulfonate, ethy- m ethanesulfonate and X-rays in inducing som atic m utations in the stam en hairs o f Tradescantia c lone BNL 4 4 3 0 . Environmental and Experimental Botany,V , 34, p . 3 9 3 -4 0 8 , 1994 .
SHIMA, N.: ICHIKAW A, S. Detection o f synergism among d iffe rent m onofunctional alkylating agents w ith the stam en-hair system of Tradescantia clone BNL 4 4 3 0 . Japanese Journal of Genetics, v. 70, p. 72 4 , 1995a.
SHIM A, N.; ICHIKAW A, S. M utagen ic synergism de tected betw een d im ethy l sulfate and X-rays bu t no t found be tw een A/-methyl-/V-nitrosourea and X-rays in the stam en hairs o f Tradescantia clone BNL 4 4 3 0 . Mutation Research, v. 331 p. 79-87 , 1995b.
SM ITH , S. S.; LOFTY, T. A . C om para tive e ffe c ts o f certa in chem ica ls on Tradescantia chrom osom es as observed at pollen tube m itosis. American Journal of Botany, v. 41 , p. 5 8 9 -5 9 3 , 1954 .
SPARROW, A. H,; IC H IK A W A , S. Com parison o f rad ia tion-induced loss of reproductive in tegrity in the stam en hairs of a po lyp lo id series in Tradescantia. Radiation Botany, v. 31 , p. 63 6 , 1967 .
SPARROW, A. H.: SCHAIRER, L. A . M uta tiona l response in Tradescantia a fte r accidental exposure to a chem ical m utagen. Environmental Mutagen Society Newsletter, v. 5, p. 16 -19 , 1971 .
SPARROW, A. H.; SCHAIRER, L. A, The e ffec ts o f chem ical m utagens (EMS, DBE and specific air po llu tan ts (O3 , SO 2 , N O 2 , N 2 O) on som atic m uta tion rate in Tradescantia . In: BUBININ, N.P., ed. Geneticheskie Poaledstviya Zagryazneniya Okruzhayuschchei Sredy (Genetic effects of pollution in the environment). M oscow : Ins titu te Obshchei Genetiki, 1974 . p. 50 -61 .
SPARROW, A. H.; SCHAIRER, L. A .: M ARIM UTHU, K. M. Genetic and cy to log ic stud es of Tradescantia irradiated during orb ita l flig h t. BioScience, v. 18, p. 582 59 0 , 1 968 .
SPARROW, A. H.; SCHAIRER, L. A .; VILLALOBOS, R. Com parison of som atic m u t jl io i i rates induced in Tiadosc<intia by cliem ica l and [jhysical m utagens,I
36

SPARROW, A. H.; SINGLETON, W . R. The use o f rad iocoba lt as a source of gam m a rays and som e e ffe c ts o f chronic irrad ia tion on g row ing plants. The American Naturalist, v. 87 , n. 8 3 2 , p. 29 -48 , 19 53 .
SPARROW , A. H.: SPARROW , R. C. Spontaneous som a tic m uta tion frequencies fo r flo w e r color in several Tradescantia species and hybrids. Environmental and Experimental Botany, v. 16, p. 2 3 -4 3 , 1976 .
SPARROW , A. H.; UNDERBRINK, A . G.; ROSSI, H. H. M uta tions induced in Tradescantia by small doses o f X -rays and neutrons: analysis o f dose-response curves. Science, v. 176 , p. 9 1 6 -9 1 8 , 1972 .
STEFFENSEN, D. Induction o f chrom osom e breakage at m eiosis by a m agnesium de fic iency in Tradescantia. Proceedings of the National Academy of Sciences,V, 39, p. 6 1 3 -6 2 0 , 19 53 .
STEFFENSEN, D. Irregularities o f chrom osom e d iv is ions in Tradescantia g row n on low sulfa te. Experimental Cell Research, v. 6 , p. 5 5 4 -5 5 6 , 1954 .
STEFFENSEN, D. Breakage o f ch rom osom es in T radescan tia w ith ca lc ium de fic iency. Proceedings of the National Academy of Sciences, v. 41 , p. 155- 1 6 0 , 1955 .
STEINITZ, L. M. The e ffec t o f lack o f oxygen on m eiosis in Tradescantia. American Journal of Botany, v. 31 , p. 4 2 8 -4 4 3 , 1944 .
T A N O , S. Induced som atic m u ta tions by rad ia tion and chem ica ls in Tradescantia. Mutation Research, v. 181 , n. 1, p. 2 0 9 -2 1 4 , 1987 .
TA N O , S. in s itu de tection o f induced m uta tions w ith chem ica ls by Tradescantia. Environmental and Molecular Mutagenesis, v. 1 4, n. suppl. 1 5, p. 1 97, 1 989 .
TAN O , S. In s itu de tection o f induced m uta tions w ith chem ica ls by Tradescantia. In: MENDELSOHN, M .L .; ALBERTINI, R.J., ed. Mutation and the environment. N ew York: W iley-Liss, 1990 . p. 5 7 -6 6 . (Progress in C linical and Biological Research, 340).
TAN O , S.; YAM A G U C H I, H. E ffects o f low dose irrad ia tion from ^^^1 on the induction o f som atic m u ta tions in Tradescantia. Radiation Research, v. 80 , p. 5 4 9 -5 5 5 , 1 9 7 9 .
TANO , S.; YAM AG UCHI, H. E ffects o f several n itroso com pounds on the induction o f som atic m uta tions in Tradescantia w ith special regard to the dose response and threshold dose. Mutation Research, v. 148, p. 5 9 -6 4 , 1985.
T A N O , S.; Y AM AG U C H I, H.; UEDA, S. E ffects o f low dose tritium labeled thym id ine and uridine on the induction o f som atic m u ta tions in Tradescantia. Environmental and Experimental Botany, v. 24, p. 1 7 3 -1 7 7 , 1984.
TAYLOR, J. H. The duration of d iffe ren tia tion in excised anthers. American Journal of Botany, v. 37 , p. 1 3 7 -1 4 3 , 1950 .
TO M KINS, D. J.; GRANT, W . F. Com parative cy to log ica l e ffec ts of the pesticides m enazon, m e tob rom uron and te tra ch lo ro iso p h th a lo n itr ile in Hordeum and
37

Tradescantia. Canadian Journal of Genetics and Cytology, v 14, p. 2 4 5 -2 5 6 , 1972 .
UNDERBRINK, A. G.; CHAIRER, L. A .; SPARROW, A. H. The biological properties of 3 ,9-G eV n itrogen ions V. Determ ination of relative biological e ffec tiveness for som atic m uta tions in Tradescantia. Radiation Research, v. 55, p. 4 3 7 -4 4 4 , 1973a.
UNDERBRINK, A. G.; SCHAIRER, L. A .; SPARROW, A. H. Tradescantia stam en hairs: a rad iobio log ica l tes t system applicable to chemical m utagenesis. In: HOLLAENDER, A ., ed. Chemical mutagens - principles and methods for their detection. New York: Plenum Press, 1973b. p. 171-207.
UNDERBRINK, A. G.; SCHAIRER, L. A .; SPARROW, A. H.; ROSSI, H. H. Relative biological e ffec tiveness of 0 .4 3 -M ev and lower energy neutrons on som atic aberrations and hair length in Tradescantia stamen hairs. International Journal of Radiation Biology, v. 19, p. 215-228 , 1971.
UNDERBRINK, A. G.; SPARROW, A. H. The influence of experimental endpoints, dose, dose rate, neutron energy, nitrogen ions, hypoxia, chrom osom e volum e and oloidy level on RBE in Tradescantia stamen hairs and pollen. In: Biological effects o f neutron Irradiation. Vienna: International A tom ic Energy Agency, 1 974.p, 13b 214 ,
UNDERBRINK, A. G.; SPARROW, A. H.; SAUTKULIS, D.: MILLS, R. E. An elusive fac to r a ffec tin g m uta tion frequency in Tradescantia stam en hairs: its influence on RBE. International Journal of Radiation Biology, v. 28 , p. 527- 538, 1975a.
UNDERBRINK, A . G.; SPARROW, A. H.; SAUTKULIS, D.; MILLS, R. E. Oxygen enhancement ratios (OERs) fo r somatic m utations in Tradescantia stam en hairs. Radiation Botany, v. 15, p. 161 -168 , 1975b.
V A N T HOF, J .; SCHAIRER, L. A . Tradescantia assay system fo r gaseous mutagens. A report o f the U.S. Environmental Protection Agency Gene-Tox Program. Mutation Research, v. 99, p. 303 315, 1982.
VELEMINSKY, J.; BRIZA, J.; GICHNER, T. Benzamide increases the frequency of m utations induced by N-m ethyl-n-nitrosourea in higher plants: Tradescantia, Nicotiana tabacum and Arab idopsis thaliana. Biologisches Zentralblatt, v. 106, n, 1 p. 6 7 -7 1 , 1987 .
VELEMINSKY, J.; GICHNER, T. Mutagenic activ ity of promutagens in plants: indirect evidence o f the ir activa tion . Mutation Research, v. 197, n. 2, p, 221- 242 19 88 .
VILLALOBOS-PIETRINI,, R.; HERNANDES, R,; GUADARRAM A, M, D. L, A .; GOMEZ-ARROYO, S. Cytological detection of somatic mutations in Tradescantia induced by ethanol. Cytologia, v. 51, p, 211-218, 1986,
WATERS, M, D,; STACK, H, P.; BRADY, A, L. Genetic activ ity profiles o f some chemicals found in hazardous wastes. Research Triangle Park: Health E ffects
38

Research Laboratory, Genetic Toxicology D ivision, 19 87 , (Internal Report. P B 8 8 -1 0 7 5 3 7 /X A B ).
XIAO, L, Z,; ICHIKAW A, S. M utagenic in teractions be tw een maleic hydrazide and X-rays in the stamen hairs of Tradescantia c lone BNL 4 4 3 0 . Japanese Journal of Genetics, v, 70 , p. 47 3 , 1995.
XIAO, L. Z.; ICHIKAW A, S. Peroxidase activities in the floral tissues of Tradescantia clone BNI 4 4 3 0 treated w ith maleic hydrazide alone. X -rays alone, or in com binations. Genes and Genetic Systems, v. 71, p. 151, 1 996.
39

40

ANEXOS
41

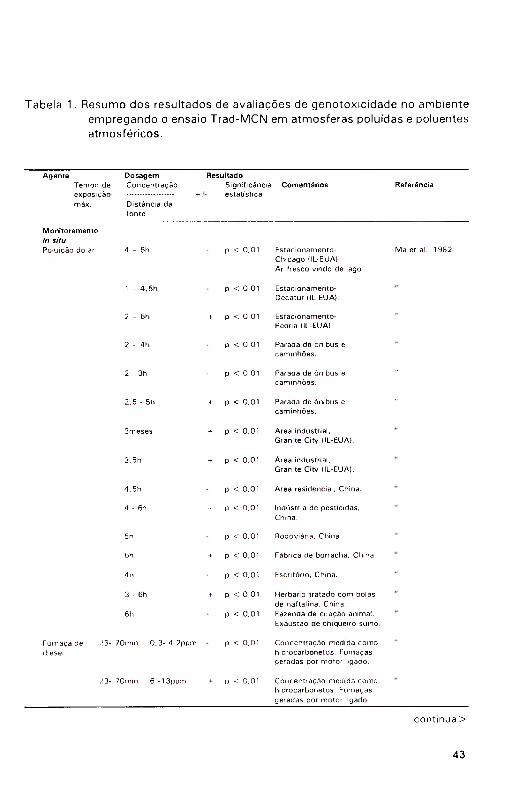
Tabe la 1. Resum o dos resu ltados de ava liações de g e n o to x ic id a d e no am biente em pregando o ensaio T rad-M C N em a tm osfe ras po lu ídas e po luentes a tm osfé ricos.
AgenteTempo de exposição- máx.
DosagemC oncentração.......................... + ,D istância da fon te
ResultadoSignificânciaesta tís tica
Comentários Referência
Monitoramento In s ituPoluição do ar 4 - 6h p < 0,01 Estacíonam ento-
C h icago (IL-EUA)Ar fresco vtndo de lago.
Ma et al.. 1982
1 - 4 ,5 h p < 0,01 Estacionament.0- D ecatur {IL-EUA).
2 - 6h 4 p < 0,01 Estacionam ento- Peona (IL-EUA).
2 - 4h p < 0,01 Parada de ônibus e cam inhões.
2 - 3h p < 0.01 Parada de ônibus e cam inhões.
”
2 ,5 - 5h 4 p < 0 ,01 Parada de ônibus e cam inhões.
"
Bmeses + p < 0,01 Área industria l, G ranite C ity (IL-EUA).
3 .5h -f p < 0,01 Área industria l, G ranite C ity (IL-EUA),
4 ,5 h p < 0,01 Área residencia l. China,
4 - 6h + p < 0,01 Indústria de pestic idas. China.
5h p < 0,01 Rodoviária, China.
6 h + p < 0,01 Fábrica de borracha. Chma.
4h p < 0 ,01 Escritó rio , China,
3 - 6h
6h
+ p < 0.01
p < 0,01
Herbário tra tado com bolas de nafta lina , China,Fazenda de criação animaf. Exaustão de ch ique iro suíno.
Futnaça de /!3- diesel
70m in 0 ,3 - 4 .2ppm p < 0 ,01 C oncentração m edida com o h idrocarbone tos. Fumaças geradas por m o tor ligado.
;^3- 70m in 6 -1 3ppm p < 0,01 C oncentração meclidn como h idrocarbone tos Fumaças geracías por m o to i ligado.
c o n t i n u a >
43
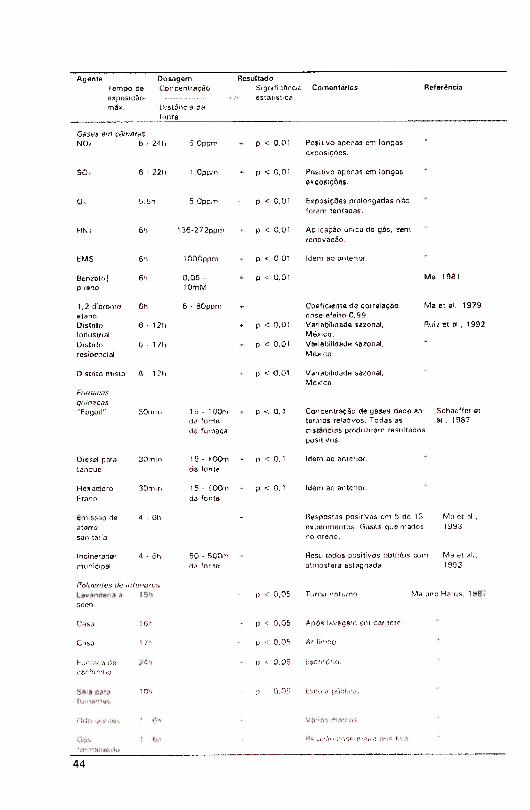
Agente DosagemTem po de ( 'o n cen tra çãoexposfçáo- ....................... ■*máx. L^fsiância da
fiin re
ResultadoS tgn ificáncia Comentários Referênciae s ia tis tica
Gases em cStnarnsNO; B ' 24h G.Oppm p < 0 ,01 P os ilivo apenas em lQr>gas "
exposições.
SO? 6 ■ 2 2 ii 1 -Oppm * p < 0 ,01 P ositjvo apenas em longas exposições.
0:. 5 .5h S.Oppcn p < 0,01 Ejíposições p ro longadas não foram tentadas.
HNn 6h I3 6 '2 7 2 p p m p < 0.01 A plicação única do yas. sem renOvâçáO.
tM S 6h lOOOppm p < 0 ,01 Id«ni ao anterior.
Ben;íOlt4Í pi rsno
6h 0 ,0 5 ■ lO m M
p <■ 0,01 M a, 1981
1,2-cíibfotTio e iono
6h 5 • 80ppm 4- C oefic ien te de corre lação M a et al.. 1979 dose-a fô ito 0 .9 9
D is la loindus ifia l
6 ■ M2U + p < 0 ,0 ) V ariab ilidade sazonai, R u i^ e ta i. , 1992 M éxico .
D is tr itoresidencia l
6 ■ l? h p < 0 .01 V anôbilidade iia io na i. " Méj<ico
D is tr ito m isto
fíf^oaçàsqoinvcàs
6 - 12h fj < ü .ü í '/anabitictade sazonal, M éxico.
"F og o il" 3 0n iin t 5 - 1OOm da fon te d íi fum aça
p < 0 . Î Concen iraçàü de gases dada am S chaeffer et torm os re la tivos. Todas as Sl.. 1987 d is tanc 'as prod 'i? iram rftsnltados positivos.
Diesel pora l3 ftqüe
30m ir\ 1 5 - 1 OOm óa fonte
- p < 0,1 Idem ao anterior.
Hex adoro Eíâno
cm issão deaterrosanitãrírt
30m irv
■4 - 6h
15 - JOOíTv da fon ts
-
p < 0, t Idem ao anterior.
Respostas positivas em 5 cie 13 M íi e t al.. e iípE rim entas. Gases ciueimarios 1993 no dreno.
IncineradormLioiCipai
4 - 6h 50 - B00j>i ds fonte
Resultados pos itivos nb tidos com Ma et al.. iitm os íe ra esl.'^poada, 1 993
PolLiofítçs cív in te rio res
seco
Cas3
Cc»s;i
hun-isçn cie c?.c;ívr':bü
3 6 h
! 7h
1C;>
n •- 0 ,0 5 Turnu nofu /no
p •■'. 0>05 A pôs lí^ví*9en^ &ni c:\fpftlef.
p < O.Oh Ar lurno.
[3 i. O.Ü55 r s c f i iõ d o ,
P O-Or. LL-ooici
M a s n d Hacn s. 15
.»'.‘ .V.' ilor.f--P ‘ '?U-T l,v
44
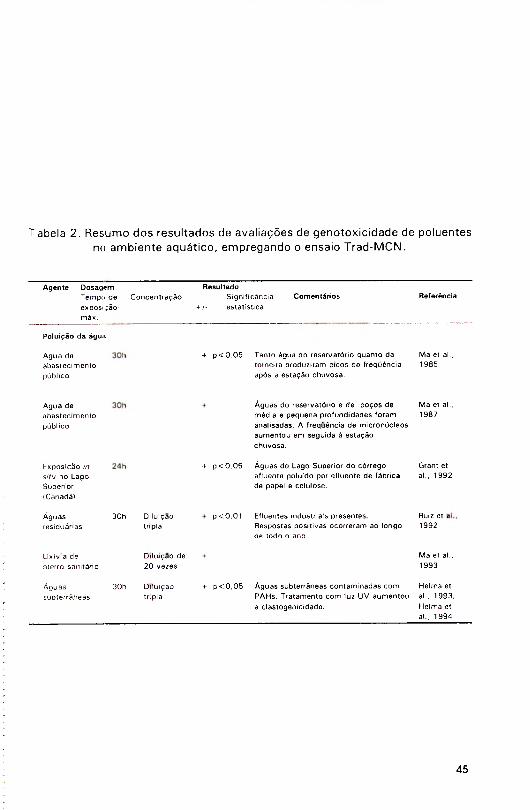
Tabela 2. Resumo dos resultados de avaliações de genotoxicidade de poluentes no ambiente aquático, empregando o ensaio Trad-MCN.
Agente DosagemTempo de C oncen traçãoexposição-máx.
ResultadoS ign ificânc ia
f / - esta tís ticaComentários
Poluição da água
Referência
Água deabastec im entop úb lico
+ p < 0 ,0 5 Tanto água do rese rva tó rio q uan to da Ma et al., to rne ira produziram p icos de freqüênc ia 1985 após a estação chuvosa.
Água deabastec im entopúb lico
Á guas do reservató rio e de poços de m édia e pequena p ro fund idades fo ram analisadas. A freqüência de m icronúcleos aum entou em seguida à estação chuvosa.
M a et al-, 1987
Exposição in s itu no Lago Superio r (Canadá)
4 p < 0 ,0 5 Á guas do Lago Superior do córrego G rant eta fluente po lu ído por e fluen te de fáb rica a l,, 1992 de papel e celulose.
Á guas 30hresiduártas
D ilu içãotrip la
p < 0 ,0 1 E fluentes indus tria is presentes- Ruiz et í
Respostas p os itiva s ocorreram ao longo 1992 de todo o ano.
L ix ív ia de a terro sanità/’io
D ilu iça o de 20 vezes
M a et a l. . 1993
Águas 30hsubterrâneas
D ilu içã o + p < 0 ,0 5 Á guas subterrâneas con tam inadas comtnp la PAHs, T ra tam ento com luz U V aum entou
a c lastogenic idade.
Helm a et al-, 1993; Heim a et al-, 1994
45

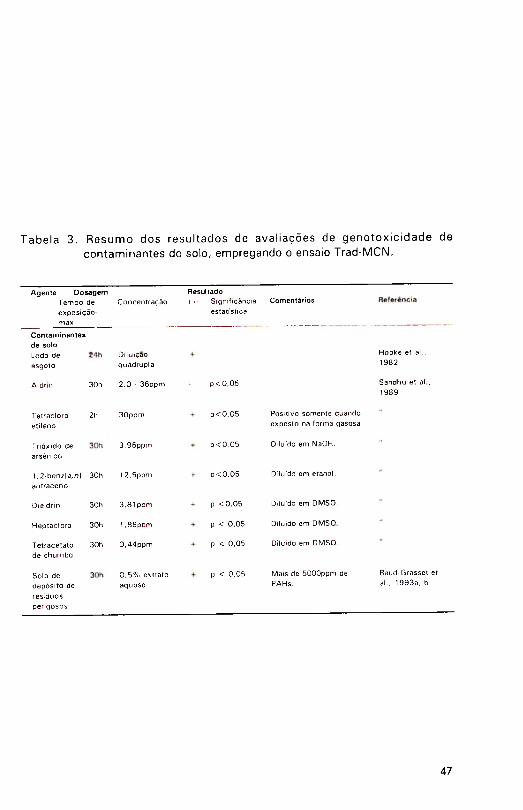
T a be la 3. R e s u m o dos re s u l tad os de ava l iações de g e n o to x ic id a d e decon tam inan tes do solo, empregando o ensaio Trad-MCN.
Agente DosagemTem po de Concentraçãoexposiçãomáx.
Contaminantes de soloLodo de iesgoto
A ldrm 30h
2liTetracloroe tileno
ír ió x id o de arsênico
) ,2 -be n z |a ,/íl 30h antraceno
D ie ld rin
Heptaclo ro
Te trace ta to de chum bo
Solo de depós ito de resíduos perigosos
3üh
30h
30h
Resultado+ /- S ignificância
estatísticaComentários
Dilu içãoquádrup la
2 ,0 ■ 36ppm
30p pm
3.9 6 pp m
1 2.5ppm
3.8 1 pp m
1,88ppm
0 .4 4 p p m
0 ,5 % extra to aquoso
p < 0 .0 5
p < 0 ,0 5
p < 0 ,0 5
p < 0 ,0 5
p < 0 ,0 5
p < 0 ,0 5
p < 0 ,0 5
p < 0 ,0 5
Positivo somente quando exposto na forma gasosa.
Diluído em NaOH.
Diiuído em etanol.
Diluído em DMSO.
Diluído em DMSO-
Diluído em DMSO.
Mais rie 5000ppm de PAHs.
Hopke et al.. 1982
Sandhu et a l. . 1989
Baud-Grasset et al., 1993a , b
47

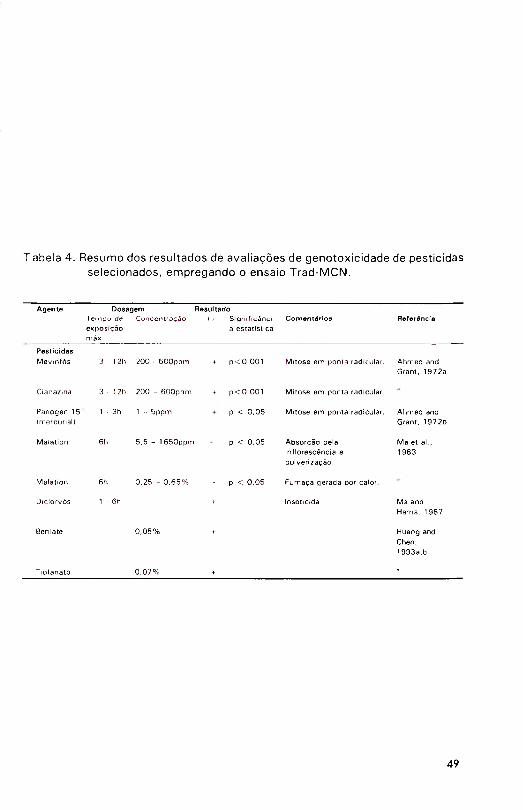
Tabe la 4. Resumo dos resu ltados de avaliações de geno tox ic idade de pestic idasselecionados, em pregando o ensaio T rad-M C N.
Agente DosagemTetnpo de Coticeritraçãoexposição-máx.
Resultado+ /• S ig n ificé n c i
a esta tís ticaComentários Referência
PesticidasM e vin fós 3 - 1 2U 200 • eOOpprn + p < 0 .0 0 1 M ito se em pon ta rad icular. Ahm ed and
Grant. 1972a
Cianazina 3 - 12h 200 * eOOppm + p < 0 ,0 0 1 M ito se em pon ta rad icular.
Panogen 1 5 ' (m ercuria l)
1 - 3h 1 - 5ppm p < 0 ,0 5 M ito se em pon ta rad icular, Ahm ed and Grant, 1972b
M ala tion 6ti 5 ,5 - 1 6 50ppm p < 0 ,0 5 A bsorção pela in flo rescênc ia e pulverização.
Ma et al., 1983
M a la tio ti 6h 0 ,2 5 - 0 ,6 5 % * p < 0 ,0 5 Fum aça gerada por ca lor.
D ic lorvós 1 - 6h + Insetic ida Ma and Harris, 1987
Benlate 0 ,0 5 % + Huang and Chen. 1993a,b
T io fana to 0 ,0 7 % + "
49

50
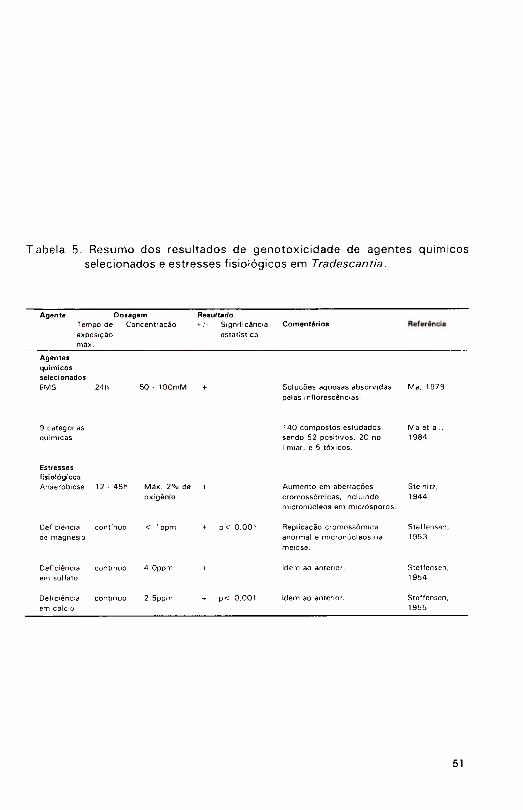
T abe la 5. Resumo dos resu ltados de ge n o to x ic id a d e de agen tes quím icosselecionados e estresses f is io lóg icos em Tradescantia.
Agente DosagemTempo de Concentraçãoexposição-máx.
Resultado+ /- S ig n ificá n c ia C om entários
es ta tís tica
A gentesquím icosselecionadosEMS 24h 50 - lO O m M + S oluções aquosas absorv idas
peias in flo rescências.Ma, 1979
9 categorias quím ícas
1 40 com postos estudados, sendo 52 pos itivo s , 20 no lim ia r, e 5 tóx icos .
M a et al., 1984
Estressesfis io ló g ico sAnaerobiose 12 • 48h M áx. 2% de +
o x igên ioA um en to em aberrações c rom ossôm icas. Inc lu indo m ic ron ú c leo s em m icróspo ros .
S te in it? ,1944
D efic iência ae magnésio
contínuo < 1 ppm + p < 0 ,0 0 1 Replicação crom ossõm ica anorm al e m ic ron ú c leo s na meiose.
S te f fensen, 1953
D efic iência em su lfa to
contínuo 4 ,0 p p m + Idem ao anterior. S te ffensen,1954
Defic iência em cá lc io
contínuo 2 ,5 p p m + p < 0 ,0 0 1 Idem ao anterior. S te ffensen,1955
51

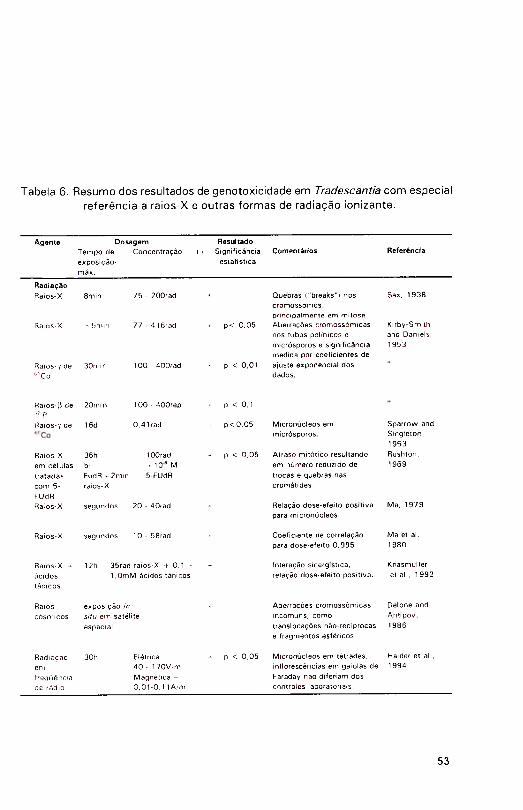
Tabela 6 . Resumo dos resultados de genotoxicidade em Tradescantia com especial re ferência a raios-X e outras fo rm as de radiação ionizante.
Agente DosagemTem po de Concentraçãoexposição-máx.
Resultado+ /- S ignificância
esta tísticaComentários Referência
RadiaçãoRaios-X 8m in 75 - 200rad + Quebras ("breaks") nos
cromossom os, p rincipalm ente em m itose
Sax, 1938
Raios-X - 5m in 77 4 16rad . p < 0 ,0 5 Aberrações crom ossõm icas nos tubos polín icos e m icrósporos e significância medida por coefic ien tes de
K irby-S rn ith and Daniels, 1953
Raios-y de"^'Co
30tTiin 100 - 400rad ^ p < 0,01 ajuste exponencia l dos dados.
Raios-(i dei 7 L3
20m in 100 • 400rep + P < 0.1r
Raios-y de 16d 0 ,41rad -1- p < 0 ,0 5 M icronúcleos em m icrósporos.
S parrow andSingíeton.1953
Raios-X 36h lOOrad + p < 0 ,0 5 A traso m itó tico resultando Rushton,
em cé lulas tra tadas corn 5- FUdR
5- ♦ IO " M F u dR -f2m m 5-FUdR raíos-X
em núm ero reduzido de trocas e quebras nas crom átides.
1969
Raios-X segundos 20 • 40rad + Relação dose-efeito pos itiva para m icronúcleos.
Ma. 1979
Raios-X segundos 10 - 58rad + Coeficiente de corre lação para dose-efe ito 0 ,9 9 5 .
Ma et al. 1980
Raios-X 4- 12h 35rad raios-X + 0,1 - + Interação sinergistica. Knasmuller
ácidostân icos
1 ,0m M ácidos tân icos relação dose-efe ito positiva . et a l.. 1992
Raios exposição ir) + Aberrações crom ossõm icas Delone and
cósm icos Situ ent saté lite espacial
incom uns, como translocações não-recíprocas e fragm entos esféricos.
A n tipov ,1986
Radiação 30h Elétnca •f p < 0 ,0 5 M icronúcleos em tétrades, Haider e t a l, ,
emfreqüência c)e rádio
4 0 - 1 70V,'m M agnética - 0 .0 1 -0 ,1 1 A /m
inflorescências em gaiolas de Faraday não d iferiam dos contro les laboratoria is.
1994
53


Tabela 7. Resumo dos resultados de avaliações de mutagênese ambiental obtidos com o ensaio do pêlo estaminal em Tradescantia (Trad-SHM).
A g e n t e D o s a g e m
T e m p o d e C o n c e n t r a ç ã o
E x p o s i ç ã o
máx
R e s u l t a d o
S i g n i f i c â n c i a C o m e n t á r i o s
e s t a t i s t i c f i
R e f e r ê n c i a
P o l u i ç ã o a t m o s f é r i c a
M o n i t o r a m e n t o in V á n a s 1 0 d i i
l o c a l i d a d e s p o l u í d a s n o s E U A
M e t a l u r g i a E x p o s i ç ã o c r ó
n i c a
E x p o s i
ç ã o c r ó n i c a
0 , 3 -1 1km da fonte1 0 0 - 5 0 0 n
da fonte
E ni i s s õ e s
g a s o s a s d eu m . U e r r o s a n i l á n o I n c i n e r a d o r
in in ic ip a l F u m a ç a s
() II í i n í c a s G a s e s s e l e c n
N O
NO .■
SO .•Ü B E
S E M
C l o r e t o d e V i n i l a
0 !0 ,1
• 6 h
3 0 n 15 - 1 0 0 m da fonte
n a d o s , e m c ü m o r a s
6 h 2 5 0 p p m
6 h 6 h
6 h 6h 6 h
6h 1 1 h ’d6 f i / d
5 0 p p n í
4 0 p p m
1 p \i in5 p p m
7 5 p p m
5 p p m
8 0 0 p p h
P o l u i ç ã o d a á g u a M o n i t o r a ç ã o i n s i t u
Á g u a d e 2 4 f i
u m l a g o
p < 0 , 0 5
S e d I m G o t o de fundo
E x p o s i ç ã o
c i ò n i c a
C a f e i n a
N a N <B e n /T i d I n a
S E M
C r ó II i c 03 h2 - l h
6h
1 0 J M
1 0 ■'5 , 4 x 1 0 ' 1 0 PP m
(j < 0,001
A g e n ! e s q u i n i i c o s
s e i f i c i o n a r í n s . e m s o l u ç ã o
B e n z o l f / ) 2 4 h 3 , 9 x 1 0 " M
p i r e n 0
T a x a s d e m u t a ç ã o m a i s a l t a s , a s s o c i a d a s c o m p r o c e s s a m e n t o
d e p e t r ó l e o
N ã o o c o r r e u c o r r e l a ç ã o e n t r e t a x a d e m u t a ç ã o e d i s t â n c i a d a
m e t a l ú r g i c a
T o d o s o s t e s t e s f o r a n \ e s t a t i s t i c a m e n t e p o s i t i v o s p a r a
u m a l o c a l i d a d e n o T e x a s , e m
r e l a ç ã o a o c o n t r o l e d a c a s a d e
vegetaçãoR e s p o s t a s p o s i t i v a s o l i t i d a s en \
1 d e 1 3 e x p o s i ç õ e s G a s e st) u e I m ,'i d o s ' i a f o ti t r i1 o m i s s õ o
R e s u l t a d o s p o s i t i v o s o b t i d o s c o m a t n i o s f e r a e s t a g n a d a
p < 0 . 0 5
p < 0 , 0 1D< 0,01p < 0 , 0 1
p < 0 , 0 2
p < 0 , 0 2
p = 0 , 6 8
p > 0 , 0 5
p < 0 , 0 5
p < 0 , 0 5
p < 0 , 0 1
p > 0 , 0 5
p < 0 , 0 1 )•) < 0,01
E x p o s i ç ã o c u m u l a t i v a , p o r t r ê s
d i a s
L a g o p o l u í d o c o m e f l u e n t e s d e f a b r i c a ç ã o d e p a p e l e c e l u l o s e . R e s u l t a d o s l o r a m i n c o n c l u s i v o s T a x a s d e m u t a ç ã o t o r a m a l t a s e m t o d o s o s 9 1 t i i a s d o p e r i o d o
experimenta'.
( S c h a i r e r . 1 9 7 9 ;
S c h a i r e r &
S a u t k u l i s . 1 9 8 2 ; S c h a i r e r e t a l . ,
1 9 8 2 )( L o v k í e r e t a l . , 1 9 7 8 ; L o w e r e t a l . , 1 9 8 3 b )
( L o w e r e t a I . ,
1 9 8 3 a I
( M a e t a l , , 1 9 9 3 a )
i M a e t a l . , 1 9 9 3 b )
( S c h a e l f e r e t al . ,
1 9 8 7 )
( S c h a i r e r & S a u t k u l i s , 1 9 8 2 ;
V a n ■ t M o t h S c h a i r e r . 1 9 8 ? i
( G i c h n e r e t a l . ,
1 9 9 2 )( R o d r i g u e s e t a l . ,
1 9 9 6 )
( G r a n i e t a l . , 1 9 9 2 )
e t al , 1 9 8 5 )
R e l a ç ã o d o se - r e s p o s t a p o s i t i v a
d e 1 0 a S O O p p m ,
( S c h a i r e r & S a u t k u l i s ,
1 9 8 2 ; V a n ' t H o f &
S c h a i r e r , 1 9 8 2 }
( N a u m a n e t a 1 , 1 9 7 6 )
DRf: E M S
6 h
A y u d 01 0 p n ni
1 OOnçjI d e m .Soluções químicas . d i r e t a m e n i e s o f i r e ; i n f l o re s c ê l u: la s
t l c l i i k . i w a e t al .1 9 9 0 , I c h I k a vv .) & F a i ^ a h a s h i , 1 9 / 6
S H n d a - K a ni Hj a w a r a
a l . , 1 9 9 1 : I a n o .
1 9 8 7 ; l a n o , 1 9 9 0 , T a n o & Y a r n a c j u c h i ,
1 9 8 5 )
c o n t i n u a > 55

Agente DosagemTempo de ConcentraçãoExposiçãomáx
Resultado + S ignificância Comentários /- estatística
Referência
EMS Agudo lOOng + Soluções químicas aplicadas diretam ente sobre as inflorescéncias.
(Ich ikawa e t al.. 1990, lchikav\/a & Takahashi, 1978; Sanda- Kamigawara et al.. 1991; Tano, 1987; Tano, 1990; Tano & Yamaguchi, 1985)
CompostosA/-nitrosos
Agudo 1 OOng +
Hidrazidamaleica
Agudo + Ocorreu sinergismo com raios-X aplicados antes de hidrazida e supressão dos efeitos com raios-X aplicados a posteriori.
(Xiao & Ichikawa, 1995; Xiao & Ichikawa, 1996}
com postosA/-nitrososPesticidas
24h Vários (Gichner et al., 1982b)
Atrazina Crónico 0,045m g p < .0 1 Dose aplicada por vaso (V an 't Hof & Sdiairer, 1982)
Menazon 24h 1500ppm - p > , 0 5 Inflorescéncias em brdhadas com algodão embebido no pesticida.
(Tomkins & Grant, 1972)
M etobronxjron 1500ppm "Tetracloroiso fta lonitrilo
1500ppm
Benomil 2h 4g/L - p > 0,05 Dose equivalente à recomendada para uso.
{Sakamoto &Takahashi, 1981)
Noveinseticidas
Radiação
Vários -( /- Resposta positiva em sete de nove compostos
(Huang & Chen, 1993a)
raios-7 de ‘■̂’Co
16h 500 R Frequência de mutações diininuíram acima de 300R.
(Nayar & Sparrow. 1967)
Areiaitionazítica
Crônico 1 .SmR/li f p < 0.05 (Nayar et al., 1970)
R aios-X 1 min 160R Relação dose-resposta positiva entre 10 e 160rad.
(Nauman et al., 1976)
Raios-X 1 min 100R + p < 0,01 Relação dose-resposta positiva entre 0,25 e 5rad.
(Sparrow et al., 1972)
Nêutrons 1 min TOR + p < 0,01 Relação dose-resposta positiva entre 0,01 e 8rad.
Radiaçãoendógena
Agudo lOOnCi Compostos marcados com Trítio e 1. Relações dose- resposta positivas.
(Tano, 1987)
Exposição in s/tu a usinas nucleares
Crônico . p < 0 .05 Aum entos s ignificativos nas taxas de mutação estavam relacionados com períodos de operação das usinas e direção dos ventos.
(Ichikawa, 1981)
Solos das ilhas B.kini
76d 1 OOfiR/h 1 p < 0,01 Radioixiclídeos principais eram ’ '^Cs e '’ ’Co.
(Ichikawa & Ishii, 1991)
Ex[)osição in siU/ n atmosfera corctrifi-.inada Exposição i i i s itu ;i '-;Tir!cS'trci ' OI itan-iií iadõ
Crónico
Crônico
I- p < 0,05 Logo após o acidente de Chernobyl. Plantas expostas em Cfacóvin, 700km da fonte de radiação.1 ogo após 0 aciciente cltí Cliernobyl. Plaiitas exno.-^tas iiü ^.OOOkin r j j 'onu: dí; rõdiaçfK.'
(Cebulska-W asilewska, 1992)
(Ic iiikaw a et al., 1996)
56