
Deriva genética - docs.ufpr.brlehtonen/populacoes/03aDerivagenetica.pdf · DERIVA GENÉTICA...
55
Deriva genética
Transcript of Deriva genética - docs.ufpr.brlehtonen/populacoes/03aDerivagenetica.pdf · DERIVA GENÉTICA...

Deriva genética

Introdução • Em qualquer organismo vivo, formam-se mais
gametas do que aqueles que poderiam sobreviver.
• Esse é um dos princípios básicos da teoria darwiniana de seleção natural.
• Quais desses gametas sobreviverão e quais não são determinados, em parte, pelo acaso: como se participassem de um sorteio.
• Como esse processo de amostragem não altera as frequências alélicas em nenhuma direção predeterminada, esse processo é conhecido como deriva genética aleatória.

DERIVA GENÉTICA ALEATÓRIA E AMOSTRAGEM BINOMIAL
• Considere uma população grande, em equilíbrio de H-W com os alelos A e a em frequências iguais p = q = 1/2.
• Nessa população, as frequências genotípicas são: ¼ AA, ½ Aa e ¼ aa.
• Suponha que essa população estivesse a ponto de colapsar e que apenas quatro indivíduos escolhidos ao acaso sobreviveriam para perpetuar o grupo.
• Seria possível, apenas pelo acaso que os sobreviventes consistissem apenas em 4 indivíduos AA: essa possibilidade tem uma probabilidade (1/4)4 = 1/256

DERIVA GENÉTICA ALEATÓRIA E AMOSTRAGEM BINOMIAL
• Da mesma forma, seria possível que os quatro indivíduos fossem aa.
• Qualquer outra combinação de genótipos poderia ocorrer, e não seria difícil obter a probabilidade para cada combinação.
• Se o tamanho da nova colônia permanecer em apenas quatro indivíduos em cada geração, esse tipo de amostragem aleatória ocorre a cada geração.

DERIVA GENÉTICA ALEATÓRIA E AMOSTRAGEM BINOMIAL
• Em cada ciclo reprodutivo, haverá a possibilidade de uma grande mudança nas frequências gênicas causada unicamente por um processo de sorteio.
• Uma consequência da deriva aleatória torna-se logo clara: em algum momento, a população terá todos os seus alelos A ou todos os alelos a.
• A razão para isso é que, uma vez que a população atinge um estado de "fixação", ela congela.
• Apenas novos mutantes ou migrantes podem reintroduzir variação.

DERIVA GENÉTICA ALEATÓRIA E AMOSTRAGEM BINOMIAL
• Se não há cruzamento preferencial, amostrar quatro indivíduos diploides é perfeitamente igual a amostrar oito gametas haploides.
• A probabilidade de cada um dessas combinações é dada por uma distribuição binomial, a qual corresponde aos sucessivos termos da expansão (1/2 A + 1/2 a)8.

DERIVA GENÉTICA ALEATÓRIA E AMOSTRAGEM BINOMIAL
• O processo de sortear gametas a partir de uma população finita tem os mesmos pressupostos daqueles que resultaram nas frequências de Hardy-Weinberg.
• Mas, nesse caso, as frequências alélicas podem mudar de geração em geração por causa da variação aleatória devida ao tamanho finito da população.

DERIVA GENÉTICA ALEATÓRIA E AMOSTRAGEM BINOMIAL
• Nesse modelo os adultos reprodutores em cada geração somam N indivíduos diploides. Esses indivíduos produzem um conjunto infinito de gametas no qual as frequências alélicas são as mesmas dos adultos.
• Desse conjunto infinito gametas, 2N são sorteados e unidos ao acaso para formar os zigotos da próxima geração.
• Esse modelo do processo de amostragem resulta na distribuição binomial de todas as combinações possíveis de A e a.

DERIVA GENÉTICA ALEATÓRIA E AMOSTRAGEM BINOMIAL
• Embora as mudanças nas frequências alélicas em virtude da deriva genética aleatória desafiem a nossa capacidade de prevê-Ias, o comportamento médio das frequências alélicas em um grande número de populações pode ser predito.

DERIVA GENÉTICA ALEATÓRIA E AMOSTRAGEM BINOMIAL
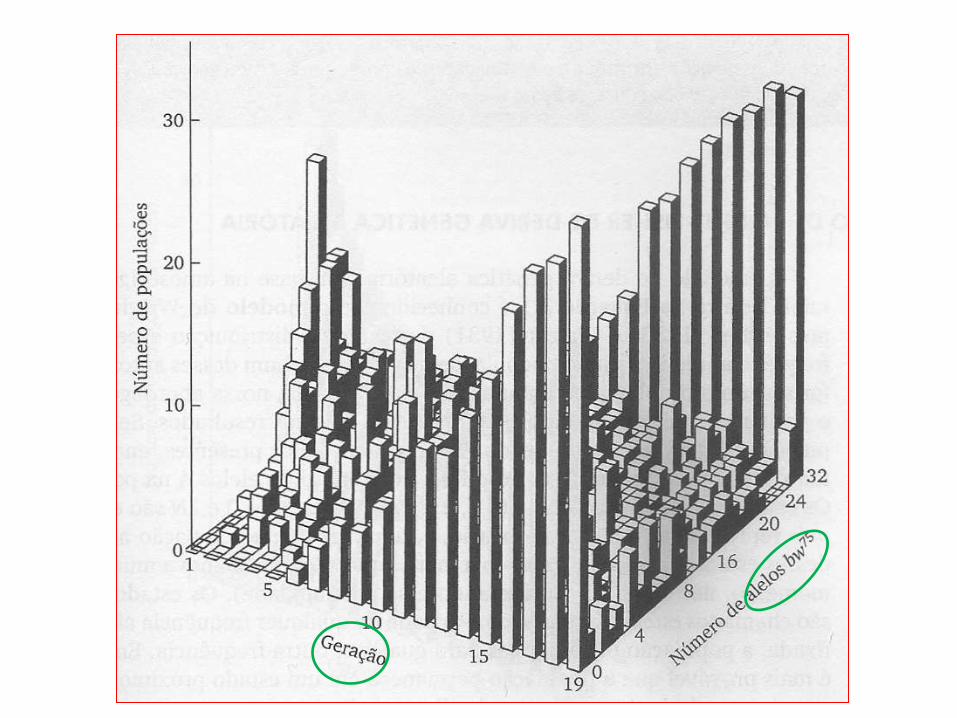
• Um experimento real forneceu resultados apresentados na figura a seguir.
• O gráfico mostra a história de 19 gerações de deriva genética aleatória em 107 subpopulações de Drosophila melanogaster.
• Cada subpopulação foi formada com 16 moscas heterozigotas bw (brown eyes) e mantida a um tamanho constante de 16 indivíduos, escolhendo-se ao acaso oito machos e oito fêmeas para produzir a próxima geração.

MODELO DE WRIGHT-FISHER DE DERIVA GENÉTICA ALEATÓRIA
• Esse modelo de deriva genética aleatória com base na amostragem binomial é conhecido como modelo de Wright-Fisher
• Fisher (1930) e Wright (1931) derivaram a distribuição esperada das frequências alélicas nas subpopulações.

MODELO DE WRIGHT-FISHER DE DERIVA GENÉTICA ALEATÓRIA
• Se uma população contém 2N alelos, sendo A e a os dois alelos presentes, então o “estado” da população pode ser descrito pelo número de alelos A na população.
• Os estados possíveis são então 0, 1, 2, ... 2N.
• Os estados 0 e 2N são especiais pois representam estados de fixação, e, uma vez que a população atinge um desses estados, não pode deixá-lo a menos que ocorra uma nova mutação.

MODELO DE WRIGHT-FISHER DE DERIVA GENÉTICA ALEATÓRIA
• Os estados 0 e 2N são chamados estados de absorção.
• Partindo de qualquer frequência alélica não fixada, a população pode derivar para qualquer outra frequência.
• Entretanto, é mais provável que a população permaneça em um estado próximo do anterior do que derive para um estado distante.

MODELO DE WRIGHT-FISHER DE DERIVA GENÉTICA ALEATÓRIA
• Por exemplo, se 2N = 32, então a chance de ir por deriva de 30 cópias do alelo A para 29 cópias em uma geração é 0,186, enquanto a chance de ir por deriva até 27 cópias é de 0,033.
• A probabilidade de que uma população derive de estado que contém i cópias para um com j cópias do alelo A é conhecida como probabilidade de transição.

MODELO DE WRIGHT-FISHER DE DERIVA GENÉTICA ALEATÓRIA
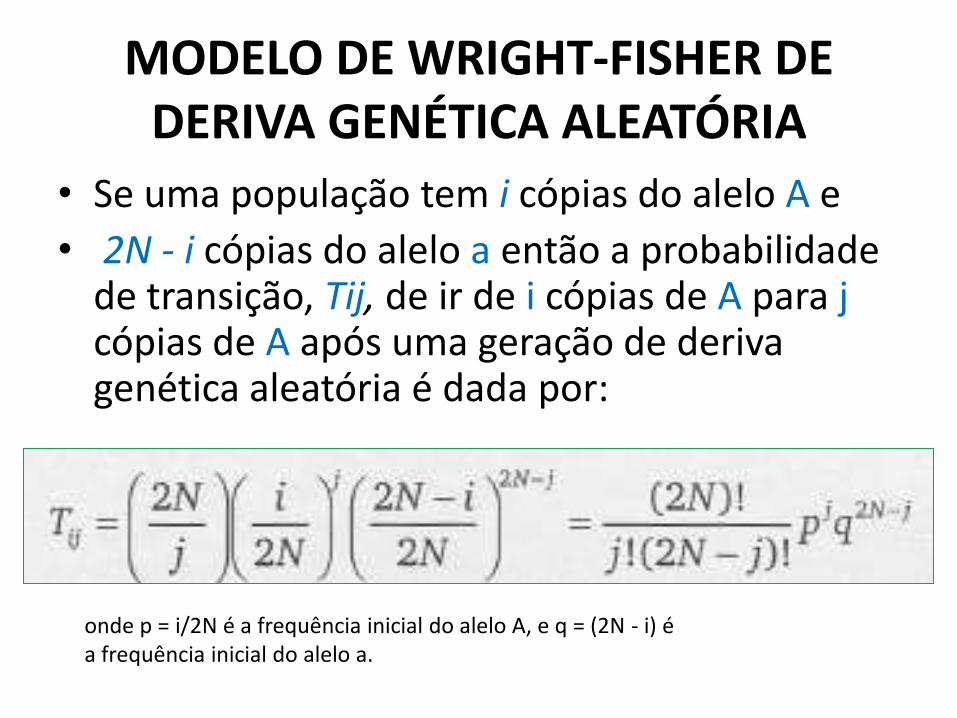
• A probabilidade de transição para o modelo Wright-Fisher é obtida diretamente pela distribuição binomial:
MODELO DE WRIGHT-FISHER DE DERIVA GENÉTICA ALEATÓRIA
• Se uma população tem i cópias do alelo A e
• 2N - i cópias do alelo a então a probabilidade de transição, Tij, de ir de i cópias de A para j cópias de A após uma geração de deriva genética aleatória é dada por:
onde p = i/2N é a frequência inicial do alelo A, e q = (2N - i) é a frequência inicial do alelo a.

MODELO DE WRIGHT-FISHER DE DERIVA GENÉTICA ALEATÓRIA
• As probabilidades de transição podem ser colocadas em uma matriz quadrada T, com os Tij fornecendo a probabilidade de transição do estado i para o estado j para i, j = 0, 1, 2 ... 2N.
• A matriz T contém todo o necessário para prever a distribuição esperada para as populações ao longo de uma série de gerações.
• Esse tipo de modelo, expresso em termos • de estados discretos com probabilidades fixas de
ir de um estado a outro, é conhecido como cadeia de Markov

MODELO DE WRIGHT-FISHER DE DERIVA GENÉTICA ALEATÓRIA

MODELO DE WRIGHT-FISHER DE DERIVA GENÉTICA ALEATÓRIA
• O modelo de Wright-Fisher incorpora uma característica importante da deriva genética aleatória:
• É que a magnitude da mudança ao acaso é maior quando as frequências alélicas são 1/2 do que quando as frequências alélicas são mais desequilibradas.
• A mudança é maior porque a variância na distribuição das amostragens é maior quando p = 1/2.
• No modelo de Wright-Fisher, a variância nas frequências alélicas em uma geração de deriva genética aleatória é dada por pq/(2N),
• que corresponde à variância da proporção em uma distribuição binomial.
• A variância cai a zero em p = 0 e p = 1.

DERIVA ALEATÓRIA EM UMA POPULAÇÃO SUBDIVIDIDA
• A maioria das populações reais se encontra subdividida em unidades menores: – os humanos se concentram em cidades, povoados e
vilarejos; – animais formam manadas, rebanhos e cardumes; – plantas se agregam em bosques.
• Esse tipo de subdivisão é remanescente da estrutura populacional na natureza
• As subpopulações não estão isoladas geneticamente umas das outras em virtude de alguma migração de indivíduos entre as subpopulações, o que resulta em fluxo gênico, ou troca de genes, entre elas.

DERIVA ALEATÓRIA EM UMA POPULAÇÃO SUBDIVIDIDA
• Entretanto, a deriva genética aleatória tenderá a causar diferenciação das frequências alélicas entre as subpopulações, mesmo diante de algum fluxo gênico.

Estrutura populacional
• Pode-se pensar que a deriva genética seja importante apenas em poucas espécies.
• Entretanto, a maioria das espécies é formada por demes pequenos e com localização estável
• Essas populações podem divergir substancialmente através da deriva,dando origem à variação geográfica nas espécies.
• Até mesmo em espécies como o pinheiro Pinus ponderosa, que tem distribuição contínua sobre vastas extensões, o acasalamento ocorrerá entre os indivíduos mais próximos.

• O número de indivíduos nas proximidades com os quais um indivíduo têm maior probabilidade de se reproduzir aleatoriamente é 4πs2D,
• onde D é a densidade da população (o número de indivíduos por unidade de área) e s é o desvio padrão das distâncias entre os locais de nascimento dos indivíduos e os dos descendentes.
• Essas distâncias são frequentemente pequenas.

População reprodutora
• O contingente global de uma população (N) não tem valor algum em genética.
• As populações se subdividem em isolados
• Dentro desse isolado o importante é a população reprodutora (Nr)
• Nr é o conjunto de indivíduos que está contribuindo para formar a geração seguinte.

População reprodutora • Pode acontecer que a variância do número de filhos
por casal é muito grande
• Considere 2 populações com Nr = 1000
• Em uma das populações a fertilidade tem uma distribuição normal
• Na outra, a fertilidade tem uma distribuição de Poisson
• Esses casais que compõem a Nr não tem a mesma importância genética, logo sua simples contagem não oferece uma visão realista da estrutura genética da população.

População reprodutora
• Outros problemas com o uso de Nr:
• Ciclos regulares de variação numérica
• Número diferente de homens e mulheres
• Elevada taxa de endocruzamento

População efetiva
• Sewall Wright parâmetro fundamental para a genética de populações é a população efetiva (Ne)
• Existem diversas fórmulas para calcular Ne
• (ver Freire-Maia, 1974, pg 135-137)
• As estimativas de Ne para populações humanas variam em torno de ¼ de N

• Se apenas metade dos indivíduos de uma população de N = 50 se reproduzir, a perda de heterozigosidade por geração (1/2N) será de 1/50 ao invés de 1/100.
• A população possuirá um TAMANHO EFETIVO Ne de 25;
• é Ne que determina a taxa de deriva genética.
• Um fator que pode reduzir o tamanho efetivo da população é uma proporção sexual desbalanceada.

• Se, por exemplo, os machos protegem haréns de fêmeas contra outros machos, os poucos machos que se reproduzem contribuem desproporcionalmente às gerações subsequentes, podendo ocorrer aumento da deriva genética.
• Se a população reprodutiva consiste de Nm machos e Nf fêmeas, o tamanho efetivo da população é
• Ne= (4NmNf)/(Nm+Nf)
• Assim, 100 tribos de camundongos, cada uma com um macho e quatro fêmeas, constituem uma população efetiva de 320 ao invés de 500.

• À medida que alguns indivíduos deixam mais descendentes do que outros, a futura população irá descender de relativamente poucos indivíduos.
• Se k e σk2 são, respectivamente, a média e a variância no número de descendentes por pai que sobrevive até a época da reprodução e se σk2 > k, o tamanho efetivo da população é menor do que o número de indivíduos na população.

• Qualquer fator que cause tal variação reduz o tamanho efetivo, incluindo a seleção, a qual, é por definição, a desigualdade do sucesso reprodutivo.
• Eisen (1975), por exemplo, descobriu que a variação genética da taxa de crescimento nos camundongos era virtualmente reduzida a zero após 14 gerações de forte seleção artificial, quando se reduziu o tamanho populacional a 4 ou 8 pares; entretanto, ainda existia variação em linhagens selecionadas mantidas com 16 pares.

• O tamanho efetivo da população será ainda mais reduzido se as gerações se sobrepuserem de modo que os descendentes possam se acasalar com seus pais (Felsenstein 1971, Giesel 1971) ou se o tamanho da população flutuar ao longo das gerações.
• As flutuações no tamanho populacional colocam-nas em GARGALOS (afunilamento ou "bottenecks") durante os quais a variação genética é reduzida.

• Uma aproximação do tamanho efetivo Ne, neste caso, é encontrada através da média harmônica do tamanho populacional,
• 1/Ne= (1/t)Σ(1/N)
• Por exemplo, se em cinco gerações sucessivas uma população consistir de 100, 150, 25, 150 e 125 indivíduos, Ne seria aproximadamente igual a 70 ao invés da média aritmética, 110;
• Ne=1/((1/5)*(1/100+1/150+1/25+1/150+1/125)) • as populações de pequeno tamanho exercem maior
influência sobre o valor de Ne que as de grande tamanho.

• Normalmente é bastante difícil a obtenção de informações suficientes sobre uma população natural - distâncias de dispersão, proporção sexual efetiva, frequência de acasalamentos através das gerações, variância do sucesso reprodutivo - para que se possa medir o tamanho efetivo da população.
• Mesmo quando apenas um ou poucos desses fatores são avaliados, tal como a variância do sucesso reprodutivo, é comum encontrar em populações humanas e de Drosophila em laboratório um Ne menor que 75 por cento do número real de indivíduos (Crow e Kimura 1970).

• Merrell (1968) contou o número de oviposições de rãs-leopardo (Rana pipiens) em lagoas cujo número total de rãs havia sido estimado;
• assumindo-se que cada oviposição resultou de um casal diferente, ele verificou que apenas de 1 a 67 por cento das rãs havia se acasalado com sucesso.

MUTAÇÕES EM POPULAÇÕES FINITAS
• Em uma população muito grande, a taxa de mudança da frequência gênica provocada apenas por mutação é provavelmente baixa, porque a taxa de mutação em um loco provavelmente estará por volta de u = 10-5 ou 10-6 por gameta por geração.
• Se aceitarmos que o alelo A (com frequência p) muta para A' (com freqüência q = 1 - p) numa taxa u e que a taxa de mutação de A' para A é v, a mudança na frequência de A' será q = u(1 - q) – vq por geração.

• Assim, se q = 0,5, u = 10-5 e v = 10-7, q passará para 0,50000495 na próxima geração. Nesta taxa, serão necessárias aproximadamente 70.000 gerações para se chegar a meio caminho da frequência gênica de equilíbrio.
• A frequência gênica de equilíbrio, é encontrada colocando-se q = 0 e procurando-se a solução para q, é
• q = /( + v)
• Assim, se u = 10-5 e v = 10-7, a frequência de equilíbrio estável seria q = 0,99, mas levaria um tempo muito longo para ser alcançada; a mutação, por si só, é uma força muito fraca.

• A derivação anterior pressupõe que existam apenas dois alelos distinguíveis, como ocorre algumas vezes com características fenotípicas grosseiras.
• Se, entretanto, reconhecermos como alelos quaisquer diferenças na sequência de nucleotídeos de um gene grande, uma dada sequência pode mutar para qualquer uma de milhares de outras sequências e a taxa de reversão à sequência de nucleotídeos anterior será desprezível.
• Deste modo, por mutação somente, qualquer alelo se tornará extremamente raro e a população irá conter um vasto número de sequências de nucleotídeos diferentes.

• Em uma população finita de tamanho efetivo Ne, os alelos são perdidos por deriva genética e a mutação é, portanto, importante por restabelecer a variação genética.
• Se as taxas de mutação e o tamanho populacional permanecerem constantes por um tempo suficientemente longo, ocorrerá eventualmente um balanceamento entre as taxas de perda e ganho de alelos por mutação
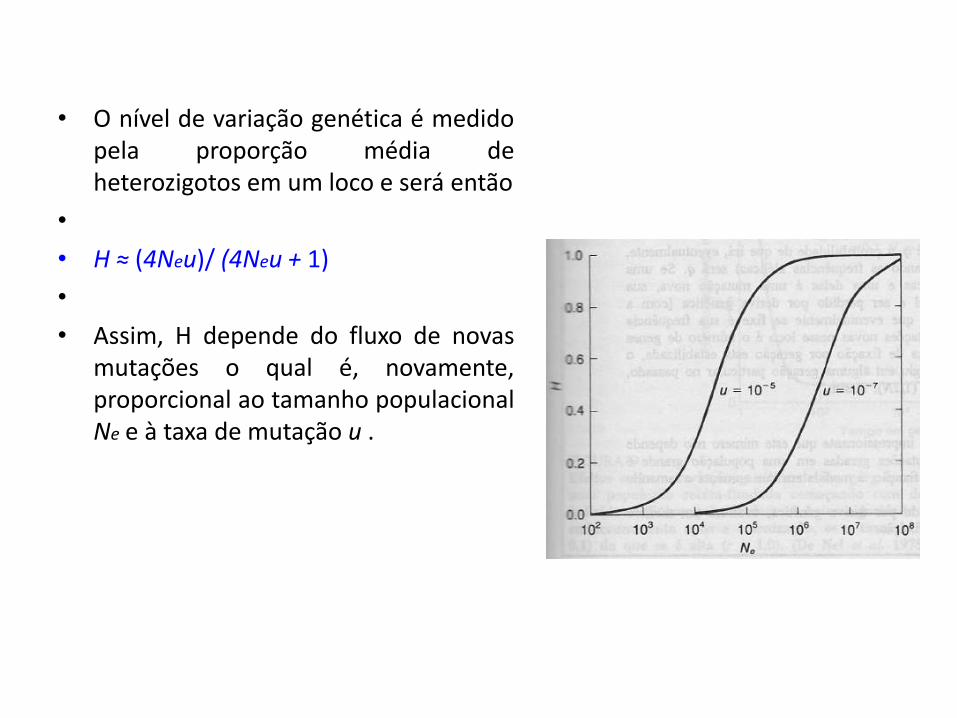
• O nível de variação genética é medido pela proporção média de heterozigotos em um loco e será então
•
• H ≈ (4Neu)/ (4Neu + 1)
•
• Assim, H depende do fluxo de novas mutações o qual é, novamente, proporcional ao tamanho populacional Ne e à taxa de mutação u .

• Por exemplo, se u = 10-6 e Ne = 30.000 , H será, aproximadamente, 0,11.
• Tenha em mente que Ne é o tamanho populacional efetivo e que estamos considerando apenas alelos hipotéticos com efeitos precisamente equivalentes sobre a sobrevivência e a reprodução.

EFEITO DO FUNDADOR
• Os acidentes genéticos inerentes às populações de tamanho pequeno podem ser importantes durante a colonização.
• Se ocorre a colonização, por um ou poucos indivíduos, de uma ilha ou fragmento de um novo hábitat não ocupado anteriormente por uma espécie, todos os genes da população originada derivarão dos poucos portados pelos fundadores e de mutações e imigrantes subsequentes.

• Uma colônia fundada por um par de indivíduos diplóides pode ter, no máximo, quatro alelos por loco, embora possa existir um número muito maior de alelos na população de onde vieram.
• Entretanto, ainda que o número de alelos seja reduzido, o grau de heterozigosidade, e portanto a variância genética, é quase tão alto quanto o da população de onde vieram os colonizadores.

• Na média é (1 - 1/2N)Ho, onde Ho é a proporção de heterozigotos na população originadora e N é o número de colonizadores.
• Isto ocorre simplesmente porque os alelos raros pouco contribuem para o nível de heterozigosidade da população originadora e são esses mesmos alelos que provavelmente estarão ausentes da colônia.
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,9
0 1 2 3 4 5 6 7 8
Série1

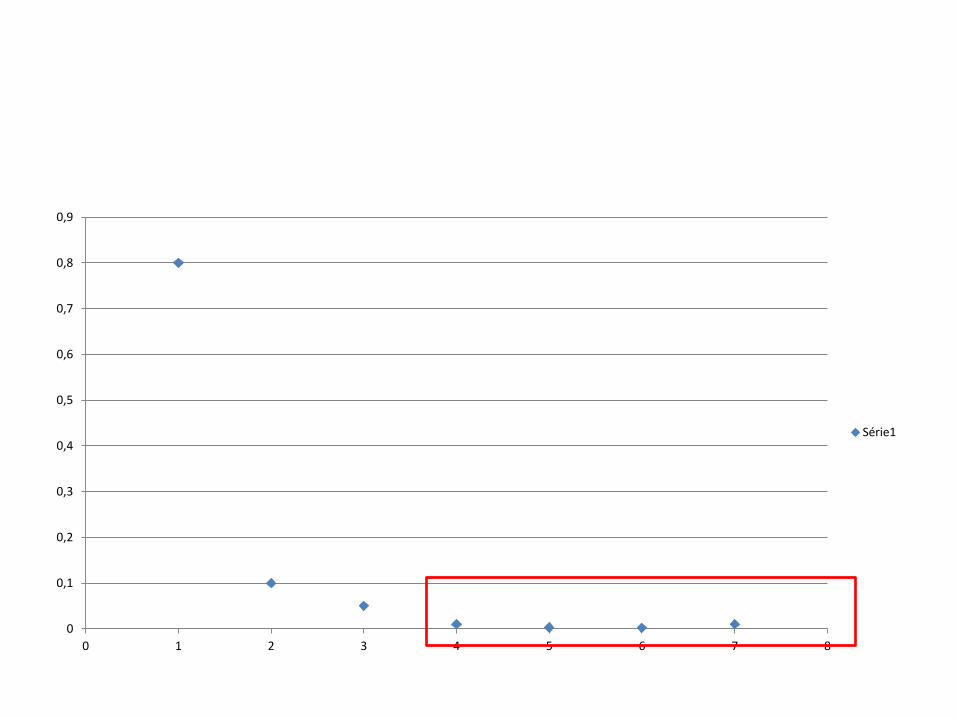
• Entretanto, se a colônia permanecer pequena, a heterozigosidade da colônia diminuirá rapidamente por deriva genética e apenas lentamente haverá sua recomposição por mutação e deriva; a menos que a seleção natural aumente a freqüência dos alelos raros.
• Se, entretanto, a colônia crescer rapidamente, não haverá grande redução na quantidade de variação genética.
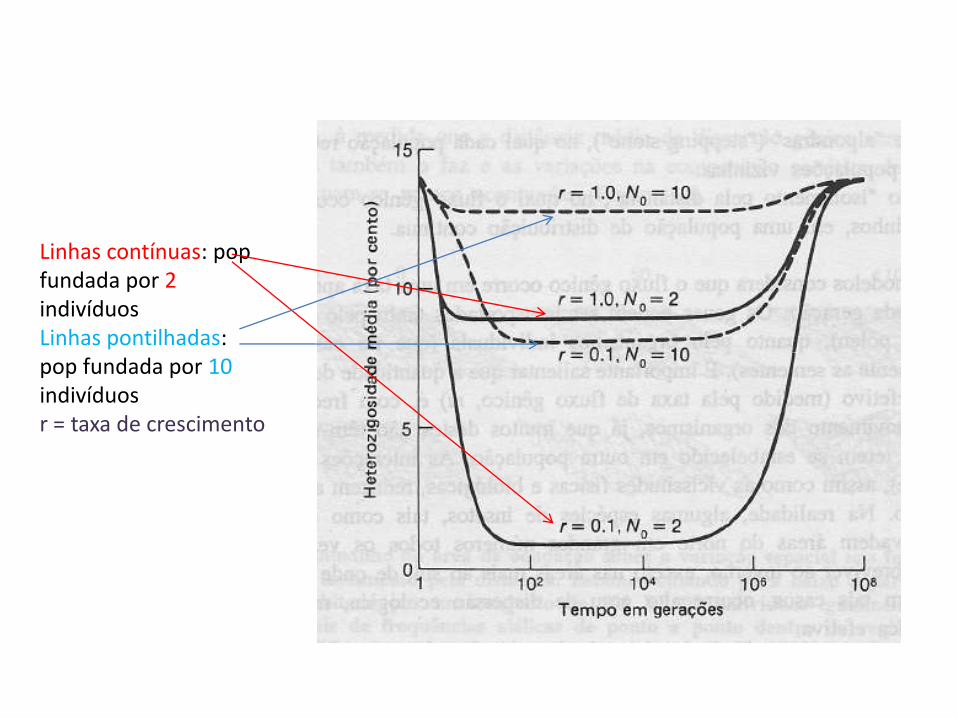
Linhas contínuas: pop fundada por 2 indivíduos Linhas pontilhadas: pop fundada por 10 indivíduos r = taxa de crescimento

• O mesmo princípio aplica-se a qualquer população que passe por um acentuado gargalo.
• Mayr (1954, 1963) argumentou que o EFEITO DO FUNDADOR afetaria de tal forma a constituição genética de uma população que haveria uma antecipação de seu desenvolvimento em uma nova espécie.

• Um exemplo extremo do efeito de gargalo é provido pelo elefante-marinho-do-norte (Mirounga angustirostris).
• A caça reduziu esta espécie a aproximadamente 20 animais por volta de 1890; desde então a população cresceu para mais de 30.000.

• O tamanho efetivo da população deve ter sido menor que 20 no seu ponto mais crítico, uma vez que a espécie é poligínica - menos de 20% dos machos copulam com todas fêmeas.

•Bonnell e Selander (1974), através de eletroforese, não descobriram nenhuma variação genética em uma amostra de 24 locos eletroforéticos nessa espécie. •Já no caso do elefante-marinho-do-sul (M. leonina), que nunca sofreu redução tão drástica no número de indivíduos, têm sido observada variação genética.

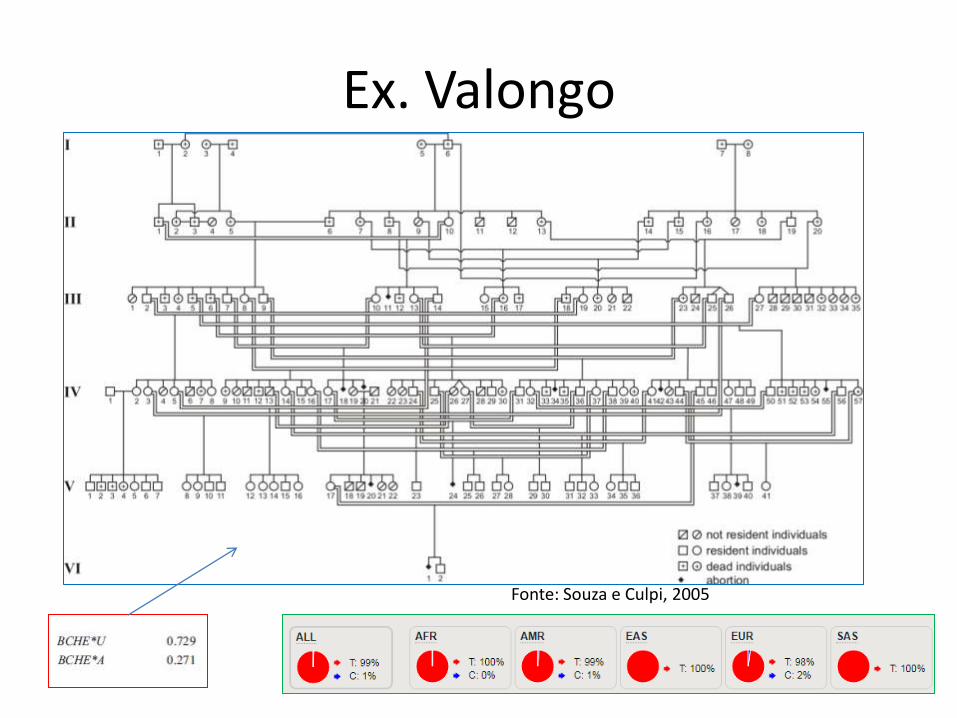
Ex. Valongo
Fonte: Souza e Culpi, 2005

Referências
• Freire-Maia, N. Genética de Populações Humanas. Hucitec. 1974. Capítulo 5
• Futuyma, D. Biologia Evolutiva. 2a ed. SBG. 1992. Capítulo 5, pg 139 a 144
• Hartl, D.L. e Clark, A. G. Princípios de genética de populações. 4ª ed. Artmed. 2010. Capítulo 3
• Souza, I.R. e Culpi, L.. Valongo, genetic studies on an isolated Afro-Brazilian community. Genetics and Molecular Biology, 28, 3, 402-406 (2005)
• Bonnell, M.L. e Selander, R.K. Elephant Seals: Genetic Variation and Near Extinction. Science. 24;184(4139):908-9. 1974


















