
DESENVOLVIMENTO DE UMA METODOLOGIA PARA …uenf.br/Uenf/Downloads/PGANIMAL_3898_1170089790.pdf · A...
71
DESENVOLVIMENTO DE UMA METODOLOGIA PARA SEXAGEM DE ESPERMATOZÓIDES DE BOVINOS UTILIZANDO ANTICORPOS MONOCLONAIS E COMPLEMENTO Claudia Gomes Fernandes Matta UNIVERSIDADE ESTADUAL DO NORTE FLUMINENSE CAMPOS DOS GOYTACAZES JANEIRO - 2003
Transcript of DESENVOLVIMENTO DE UMA METODOLOGIA PARA …uenf.br/Uenf/Downloads/PGANIMAL_3898_1170089790.pdf · A...

DESENVOLVIMENTO DE UMA METODOLOGIA PARA SEXAGEM DE
ESPERMATOZÓIDES DE BOVINOS UTILIZANDO ANTICORPOS MONOCLONAIS E COMPLEMENTO
Claudia Gomes Fernandes Matta
UNIVERSIDADE ESTADUAL DO NORTE FLUMINENSE CAMPOS DOS GOYTACAZES
JANEIRO - 2003

DESENVOLVIMENTO DE UMA METODOLOGIA PARA SEXAGEM DE ESPERMATOZÓIDES DE BOVINOS UTILIZANDO ANTICORPOS
MONOCLONAIS E COMPLEMENTO
Claudia Gomes Fernandes Matta
Tese Apresentada ao Centro de Ciências e Tecnologias Agropecuárias da Universidade Estadual do Norte Fluminense, como parte das exigências para obtenção do Título de Doutor em Produção Animal
Orientador: Prof. Dr. Marcos Fernando de Resende Matta
CAMPOS DOS GOYTACAZES JANEIRO - 2003

DESENVOLVIMENTO DE UMA METODOLOGIA PARA SEXAGEM DE ESPERMATOZÓIDES DE BOVINOS UTILIZANDO ANTICORPOS
MONOCLONAIS E COMPLEMENTO
Claudia Gomes Fernandes Matta
Tese apresentada ao Centro de Ciências e Tecnologias Agropecuárias da Universidade Estadual do Norte Fluminense, como parte das exigências para obtenção do título de Doutor em Produção Animal
Aprovado em 07 de janeiro de 2003.
Comissão Examinadora:
_________________________________________________________________ Prof. Dr. Gilson Mendes da Cruz (Reprodução Animal) – FACASTELO/FIOCRUZ _________________________________________________________________ Prof. Dr. Clóvis José Pascarelli Souza (Produção Anima) – FIOCRUZ _________________________________________________________________ Prof. Dr. Sergio Aguiar de Barros Vianna (Produção Animal) – UENF
_________________________________________________________________ Prof. Dr. Marcos Fernando de Resende Matta (Imunologia Veterinária) - UENF
(Orientador)

Aos meus pais, pelo incentivo e apoio constante
Ao meu marido Marcos Fernando, e ao meu filho Lucas, pelo amor verdadeiro
Aos meus irmãos, Adriana e Sergio, pelo carinho.
DEDICO

AGRADECIMENTOS
Ao meu orientador, Prof. Dr. Marcos Fernando de Resende Matta, pela
orientação e por ter acreditado em mim.
Ao Prof. Dr. José Frederico Straggiotti Silva pela atenção e companheirismo
Ao Prof. Dr. Clóvis José Pascarelli Souza, presença amiga durante todos
esses anos de UENF.
Aos amigos Guilherme Valente de Souza, Maurício Fraga van Tilburg, Bruno
Fagundes e Carlito Lessa da Silva, companheiros de trabalho.
À Bruna e Juliana, pela ajuda durante os trabalhos no laboratório.
Aos técnicos do LMGA.
À INFIGEN Inc., pela oportunidade de realização dos experimentos.
A todos os professores da UENF que permitiram a realização deste trabalho.
A todos os alunos e professores do Laboratório de Melhoramento Genético
Animal.
À Universidade Estadual do Norte Fluminense (UENF) e ao Centro de
Ciências e Tecnologias Agropecuárias (CCTA), pelo oferecimento deste curso.
À FENORTE, pela concessão de bolsa de estudo.
A Deus, por mais uma conquista.

BIOGRAFIA
CLAUDIA GOMES FERNANDES MATTA, filha de Dolvarino Fernandes de
Aquino e Nadir Gomes Fernandes, nasceu em 22 de junho de 1968, na cidade do
Rio de Janeiro, RJ.
Foi professora do Município do Rio de Janeiro no período de 1988 a 1995.
Formou-se em Ciências Biológicas pela Universidade do Rio de Janeiro no
ano de 1992.
Foi admitida em março de 1995 no curso de Pós-Graduação em Produção
Animal, Mestrado, no Laboratório de Melhoramento Genético Animal da
Universidade Estadual do Norte Fluminense (UENF), em Campos dos Goytacazes -
RJ, tendo defendido a tese em dezembro de 1998.
Em março de 1999, iniciou o curso de Doutorado em Produção Animal, no
Laboratório de Melhoramento genético Animal, submetendo-se à defesa de tese
para conclusão do curso em dezembro de 2002.

CONTEÚDO
RESUMO....................................................................................................................xi
ABSTRACT................................................................................................................xiii
1. INTRODUÇÃO.....................................................................................................1
OBJETIVO GERAL............................................................................................3
OBJETIVOS ESPECÍFICOS.............................................................................3
2. REVISÃO BIBLIOGRÁFICA
2.1 – Sexagem de Espermatozóides de Bovinos................................................4
2.2 - Espermatogênese e Meiose........................................................................5
2.3 - Expressão dos Genes dos Cromossomas X e Y durante a
Espermatogênese...............................................................................................6
2.4 - Metodologias de Sexagem..........................................................................7
2.4.1- Sexagem de Espermatozóides-..............................................................7
2.4.2- Sexagem de Embriões............................................................................9
2.5 - Antígeno HY................................................................................................9
2.6 -O Papel da Proteína HY.............................................................................10
2.7 -A Genética do Antígeno HY........................................................................14
2.8 -Sorologia do Antígeno HY..........................................................................16
2.9 - Purificação do Antígeno HY ..................................................................19

2.10 - Produção de Anticorpos Contra o Antígeno HY....................................20
2.11– Produção de Embriões..........................................................................22
3- MATERIAL E MÉTODOS
3.1–Técnica de imunoafinidade com coluna IMAC.........................................24
3.2–Metodologia de Sexagem de Espermatozóides com anticorpo monoclonal
anti-proteína macho-específica e adição de complemento.............................27
3.2.1-Sêmen..............................................................................................27
3.2.1.1- Tratamentos do Sêmen...........................................................27
3.2.2-Tratamentos para sexagem ............................................................27
3.2.3-Anticorpo monoclonal anti-proteína macho-específica....................28
3.2.4-Fonte de complemento....................................................................28
3.3- Separação pelo gradiente de Percoll................................;......................29
3.4- Imunofluorescência das amostras de células..........................................29
3.5- Eletroforese..............................................................................................29
3.6- Fecundação In Vitro para produção de embriões de bovinos..................30
3.7- Determinação do sexo dos embriões.......................................................30
2.1 - Injeção Intracitoplasmática de espermatozóides.....................................31
3. RESULTADOS E DISCUSSÃO
3.1 –Coluna IMAC............................................................................................31
3.2 – Metodologia de Sexagem pela Ação do Complemento..........................34
4. CONCLUSÕES..........................................................................................46
5. REFERÊNCIAS BIBLIOGRÁFICAS...........................................................48

LISTA DE TABELAS
Tabela 1– Número de embriões analisados por PCR................................................33
Tabela 2 – Análise das diferentes temperaturas de aquecimento do soro objetivando
identificar aquela na qual ocorre inativação da via alternativa do complemento......35
Tabela 3 – Análise da ação do soro sobre os espermatozóides................................35
Tabela 4 – Determinação da temperatura ideal do soro de cobaia. Verificação da
taxa do sexo dos embriões produzidos......................................................................37
Tabela 5 – Comparação da motilidade dos espermatozóides lavados (Lav) e não
lavados (NLav)...........................................................................................................38
Tabela 6 – Motilidade dos espermatozóides nos diferentes tratamentos..................39

Tabela 7 – Análise por PCR, de embriões produzidos com sêmen tratado com 100 µl
anticorpo monoclonal e 100 µl soro de cobaia, e de embriões produzidos sem utilizar
o tratamento...............................................................................................................40
Tabela 8 – Taxa de fertilização usando espermatozóides incubados com anticorpo
monoclonal e soro de cobaia.....................................................................................40
Tabela 9 – Resultados da análise por PCR de 389 embriões resultantes da
fertilização de ovócitos com espermatozóides tratados.............................................41

LISTA DE FIGURAS
Figura 1 – Modelo clássico do cromossoma “Y”........................................................15
Figura 2 - Esquema de fixação dos anticorpos nos espermatozóides.....................25
Figura 3 - Esquema de fixação das células na coluna...............................................25
Figura 4 – Foto da Coluna IMAC, onde observamos a coluna presa ao suporte com
imã, ficando abaixo da mesma um tubo para a coleta da primeira eluição............... 26
Figura 5 - Foto das partes constantes da Coluna Magnética. Para a coleta da
segunda eluição, retira-se a coluna do imã e promove-se nova passagem de tampão
3, podendo se utilizar um êmbulo que se insere na coluna......................................26
Figura 6 – Resultado da análise por PCR de 389 embriões resultantes da fertilização
de ovócitos com espermatozóides tratados, mostrando variações entre os
tratamentos.................................................................................................................41

Figura 7 - Resultados da análise por PCR de 389 embriões resultantes da
fertilização de ovócitos com espermatozóides tratados, mostrando variações entre os
touros..........................................................................................................................42
Figura 8 – Análise computadorizada do gel de eletroforese produzido a partir de
proteínas de membranas de espermatozóides tratados com o pool de anticorpos
monoclonais...............................................................................................................45

RESUMO
MATTA, CLAUDIA GOMES FERNANDES; DS; Universidade Estadual do Norte Fluminense; dezembro de 2002; Desenvolvimento de uma Metodologia para Sexagem de Espermatozóides de Bovinos Utilizando Anticorpos Monoclonais e Complemento; Marcos Fernando de Resende Matta. Neste trabalho, procuramos desenvolver uma metodologia a ser utilizada na
produção de sêmen sexado de bovinos. Utilizamos um anticorpo monoclonal contra
uma proteína macho-específica, associado ao uso de complemento.
Foram testadas diferentes concentrações de líquido ascítico e soro de cobaia
aquecido. Produziram-se embriões e verificou-se a taxa de clivagem e formação de
blastocistos assim como o sexo dos embriões formados.
Observamos que não houve diferença nos resultados quando se utilizaram
variações na quantidade de líquido ascítico e soro. No entanto, observamos
variações entre touros.
Não houve diferenças quando comparadas as taxas de clivagem e fertilização
do tratamento e do controle.

A motilidade dos espermatozóides também não foi alterada quando
submetidos ao tratamento.
Conclui-se que a metodologia onde se utilizam 20µl de líquido ascítico (20µg
/µl de proteína) e 20µl de soro de cobaia aquecido a 52.2°C pode ser adotado, pois
há um menor consumo de reagentes e obtém-se bom resultado. Obteve-se uma
taxa de 80% de embriões fêmeas.
Palavras-chave: imunossexagem; anticorpo monoclonal; bovino

ABSTRACT
MATTA, CLAUDIA GOMES FERNANDES; DS; Universidade Estadual do Norte Fluminense; dezembro de 2002; Development of the Methodology for bovine sexing semen using monoclonal antibody and Complement; Marcos Fernando de Resende Matta.
This work has been tried to develop a methodology for bovine sexing semen.
It was used a monoclonal antibody against a male-specific protein plus guinea
pig serum was tested for sexing semen.
It was used different concentrations of monoclonal antibody and guinea pig
serum in order to verify the complement action on the spermatozoa. The antibody
monoclonal was added to semen and the serum was added too. It was observed a
cytolytic effect on male cells and no effect on female. This way, only the spermatozoa
with X cromossoma will be free for fecundation.
There was not difference between the treatments, but there was between
bulls. There was no trouble with motility spermatozoa.
The results showed almost 80% of female embryos after in vitro fecundation
and PCR analisis.

The treatment with 20µl of the ascitic fluid (20µg /µl of protein) and 20µl guinea
pig serum heated at 52.2°C could be used because the result is good. We observed
a rate of 80% of the female embryos.
Palavras-chave: imuno-sexing; monoclonal antibody; bovine.

1
1 - INTRODUÇÃO
O Brasil possui o maior rebanho bovino comercial do mundo, com cerca de 165
milhões de cabeças. Com este dado, podemos estimar que 40 milhões de fêmeas
entram em idade reprodutiva por ano.
O melhoramento genético do rebanho é a principal base para o
desenvolvimento da atividade. Índices melhores de desfrute são mais rapidamente
obtidos com o melhoramento genético do que com melhorias na nutrição ou no
manejo do rebanho, embora estes sejam fundamentais. Nos últimos anos, devido a
problemas como o da vaca louca, entre outros, o nosso bovino, pelo sistema de
criação a pasto e o surgimento de áreas livres de Febre Aftosa, tornou-se de grande
interesse nos mercados externos. Sem dúvidas, a nossa extensão territorial,
associada ao nosso clima, cria as condições necessárias para que o país seja no
futuro o açougue do mundo. Devido a estes fatos, tornou-se imperativo o
desenvolvimento genético do rebanho de forma a obter um animal ótimo para o
abate, tanto em relação à idade de abate como em relação à qualidade da carne.
Este melhoramento genético é feito utilizando-se touros e vacas geneticamente
superiores. Devido ao fato da criação de bovinos no Brasil ser extensiva, torna
difícil o manejo para inseminação artificial e gera um mercado de tourinhos
geneticamente superiores. A necessidade de tourinhos pode chegar a 400 mil por

2
ano. Para produção destes tourinhos, é fundamental a existência de matrizes de alta
qualidade genética, as quais serão as bases do melhoramento genético do rebanho
nacional. Estas matrizes garantirão 50% do genótipo de nosso plantel de tourinhos,
enquanto que os outros 50% estarão presentes nos touros das centrais de
inseminação artificial. Os tourinhos gerados destas matrizes irão alavancar a
qualidade genética do nosso rebanho e, com isto, o desenvolvimento das
exportações e o superávit do agronegócio nacional, que, no último ano, foi
aproximadamente de 20 bilhões de dólares. (Matta,M.F.R. informações pessoais)
Em relação à produção leiteira, embora não nos traga divisas internacionais,
apresenta outro parâmetro de desenvolvimento de grande importância social, cerca
de 3,5 milhões de empregos diretos e indiretos existem no Brasil diretamente ligados
a esta atividade. A média de produção leiteira, entretanto, é muito baixa, fazendo
com que os produtores tenham poucos resultados financeiros na atividade. A
atividade de produção de leite é financeiramente interessante em escala, o que torna
o melhoramento genético fundamental para que a mesma seja produtiva. Outro fator
importante na atividade é que apenas a fêmea tem interesse, sendo o macho
economicamente inviável.
A possibilidade de se escolher previamente o sexo dos animais vem contribuir
significativamente para dar vantagens aos produtores tanto de gado de leite como
de corte.
Com isto, têm surgido várias técnicas que visam a sexar o sêmen.
A imunossexagem de espermatozóides é uma técnica não-invasiva que busca suprir necessidades da indústria agropecuária. Ela consiste na separação de espermatozóides contendo cromossomo X daqueles contendo o cromossomo Y.
Atualmente, o uso da citometria de fluxo é uma técnica que permite um alto
grau de pureza na amostra final, mas requer um equipamento caro e a quantidade
de células analisadas é limitada. É uma técnica baseada em diferenças físicas entre
a população de espermatozóides, isto é, na quantidade de DNA (o cromossoma X
seria maior do que o cromossoma Y). Métodos imunológicos empregariam
equipamentos mais baratos e uma separação em larga escala. Muitos métodos vêm
sendo pesquisados buscando oferecer uma alternativa para os produtores.
A identificação de proteínas específicas para espermatozóides X ou Y fornece
grandes oportunidades para a separação. Isto aliado ao desenvolvimento de
anticorpos específicos anti-proteína X ou anti-proteína Y para serem utilizados em
técnicas imunológicas.

3
A descoberta de antígenos macho-específicos associada com a tecnologia de
produção de anticorpos monoclonais tornou a imunossexagem de espermatozóides
muito mais promissora.
Trabalhos iniciais foram realizados no Setor de Imunogenética do Laboratório
de Melhoramento Genético Animal da UENF, onde anticorpos monoclonais contra o
antígeno H-Y foram produzidos.
No presente trabalho, desenvolveu-se uma metodologia de sexagem de espermatozóides de bovinos. A metodologia desenvolvida foi de utilização de anticorpos monoclonais antiproteína HY e complemento originário de soro de cobaia, tendo sido, para este objetivo, eliminada a via alternativa. A análise dos espermatozóides, após terem sido tratados com o anticorpo monoclonal e soro de cobaia, foi realizada verificando-se sua motilidade, além da capacidade de fertilidade dos mesmos por meio da observação da taxa de clivagem dos ovócitos fecundados com os espermatozóides imunossexados.
OBJETIVO GERAL
Desenvolvimento de uma metodologia para a produção de embriões de
bovinos utilizando sêmen sexado e complemento.
OBJETIVOS ESPECÍFICOS
- Determinar as concentrações de líquido ascítico e soro de cobaia a serem
utilizados;
- Verificar taxa de clivagem e formação de blastocistos, quando se utiliza
sêmen imuno-sexado;
- Verificar taxa de embriões do sexo feminino, produzidos com a uilização do
sêmen imunossexado.

4
2 - REVISÃO BIBLIOGRÁFICA
2.1 – Sexagem de Espermatozóides em Mamíferos
O principal tema sobre tecnologias reprodutivas é, sem dúvida, o que está
relacionado com o fato de podermos ser capazes de pré-selecionar o sexo dos
descendentes no momento da concepção.
Este fato vai contra a natureza, a qual determina chances iguais para o
nascimento de machos e fêmeas, devido às semelhanças fenotípicas entre os
espermatozóides portadores do cromossoma X e dos portadores do cromossoma Y
(Seidel, 1999).
Nos dias atuais, a situação com relação a pré-seleção é fundamentalmente
diferente do que era há 50 ou 100 anos atrás.
Talvez a mais importante aplicação seja para minimizar doenças ligadas ao
sexo na população humana. Existem mais de 500 doenças conhecidas ligadas ao
cromossoma X, algumas das quais extremamente debilitantes.
Na produção animal, a sexagem se torna particularmente interessante quando
se busca um aumento potencial da eficiência na criação de bovinos, suínos e ovinos
(Hohenboken, 1999; Seidel e Jonhson, 1999).
Outro fator importante é o de se tornar possível o nascimento de animais de
um determinado sexo quando a população se encontrar em problemas de extinção.
Exemplos bem comuns são os de criadores de cavalos, que têm preferência
por determinado sexo e o de produtores de leite que desejam o nascimento de
vacas e o de produtores de corte que preferem o nascimento de animais do sexo
masculino.

5
Todos estes fatores levam ao incremento da seleção do sexo na concepção.
2.2 -Espermatogênese e Meiose
Espermatogênese é o processo de divisão celular e diferenciação que se inicia
nos túbulos seminíferos com espermatogônias indiferenciadas e resulta na produção
de espermatozóides. Parte das espermatogônias não-diferenciadas que se
transformam em espermatogônias diferenciadas, espermatogônias tipo A1. Através
de uma série de divisões mitótica bem controlada, a espermatogônia tipo A1 dá
origem a um grande número de espermatogônias mais avançadas, identificadas em
camundongos como A2, A3, A4, intermediária e espermatogônia Tipo B (OAKBERG,
1956). A espermatogônia Tipo B divide-se para formar o espermatócito primário, que
irá sofrer meiose. Através da primeira divisão meiótica, um espermatócito primário
produz dois espermatócitos secundários, os quais quase que imediatamente entram
na segunda divisão meiótica, resultando em espermátides. A espermátide haplóide
não se divide, mas se diferencia em espermátide alongada, e finalmente, em
espermatozóide. Durante o desenvolvimento das espermatogônias até a formação
de espermatozóides, as células espermáticas migram da periferia em direção ao
centro dos túbulos seminíferos e, finalmente, serão liberadas no lúmen (OAKBERG,
1956; RUSSEL et al., 1990). As células que se originaram de uma mesma
espermatogônia Tipo A1 formam um sincício e estão conectadas umas às outras por
pontes intercelulares. Cada sincício pode conter mais de 50 espermátides, as quais
se mantêm conectadas até o final da espermatogênese (DYM et al., 1971; RUSSEL
et al., 1990).
A meiose se inicia com a replicação do DNA que copia cada cromossoma em
duas cromátides irmãs, após as cópias maternais e paternais de cada par é feita a
recombinação. Durante a primeira divisão meiótica, os cromossomas homólogos se
separam, enquanto que é durante a segunda divisão meiótica que as cromátides
irmãs se separam. Embora o espermatócito primário formado após a primeira divisão
meiótica tenha um DNA contendo 2X, estas células são atualmente haplóides,
contendo o cromossoma X ou Y (ALBERTS et al., 1983; MIYAZAKI et al., 1994).

6
2.3 - Expressão dos Genes do Cromossoma X e Y Durante a Espermatogênese
Após a divisão meiótica, os cromossomas X e Y recém-formados ligados às
espermátides não irão ter diferenças na composição de suas proteínas de
citoplasma e de membrana. A composição protéica dos espermatozóides contendo
os cromossomas X e Y só será diferente quando os genes do cromossomo X e Y
estiverem expressos após a meiose e quando o produto dos genes estiver confinado
nas células haplóides individuais. Assim, eles não são transportados às outras
espermátides por meio de pontes intercelulares. Em camundongos, as pontes
intercelulares se rompem bem ao final da espermiogênese, quando as espermátides
alongadas são liberadas no lúmen dos túbulos seminíferos (DYM et al.,1971;
RUSSEL et al., 1990). A transcrição dos genes cessa num estágio inicial de
espermátide alongada, quando o empacotamento de DNA se torna muito forte e
permite a síntese de novo do RNA.
Durante a espermatogênese, as células germinais sofrem uma divisão meiótica
final e, como resultado deste evento, metade dos espermatozóides contém um
cromossomo “Y” e a outra metade, o cromossomo “X”, determinando, assim, o sexo
genético dos embriões a serem gerados.
Após a divisão meiótica, já no estágio haplóide, os espermatozóides sofrem
modificações morfológicas, desenvolvendo um flagelo, além de outras
características que são capazes de diferenciar as classes de espermatozóides “X” e
“Y”. Estas características fenotípicas são representadas principalmente por proteínas
de superfície de membrana celular que estariam presentes em uma das classes de
espermatozóides e ausentes na outra (EICHWALD, E. J. et al., 1955).
2.4 - Metodologias de Sexagem
Entre os métodos mais estudados está a citometria de fluxo (JOHNSON, 1992),
que possui eficiência em torno de 98% (CUNNINGHAM, 1998), porém necessita de
equipamentos caros, o número de espermatozóides sexados é limitado e é

7
potencialmente invasiva (BLECHER et. al, 1999), devido à utilização de fluorocromos
para marcar o DNA. Outro método é a imunossexagem, com anticorpo monoclonal
para a detecção de antígenos de superfície sexo-específicos nos espermatozóides.
O método é mais simples, mais barato e sem efeitos negativos sobre o
desenvolvimento embrionário, além de não haver limite do número de
espermatozóides para sexagem. Entretanto, este método depende da expressão de
genes do cromossomo X ou Y nas espermátides e de que o produto dessa
expressão não atinja a população de espermatozóides do sexo oposto, através de
pontes intracelulares existentes entre as células durante a espermatogênese
(HENDRICKSEN, 1999). Tem sido reportada a possibilidade real das células
espermáticas X e Y possuírem proteínas para a sexagem espermática (BLECHER et
al., 1999). O antígeno H-Y é um antígeno considerado macho-específico, entretanto,
a sua aplicação na sexagem, principalmente em embriões, não tem atendido às
expectativas, provavelmente devido à baixa afinidade dos anticorpos (VEERHUIS et
al., 1994). Com a identificação de alguns genes que codificam vários epitopos H-Y,
cujos peptídeos são constituintes de moléculas do complexo maior de
histocompatibilidade que está presente na superfície da célula (HENDRIKSEN,
1999), abriu-se uma alternativa de se conseguir diferentes anticorpos monoclonais
para diversos epitopos de proteínas sexo-específicas presentes na membrana.
2.4.1 - Sexagem de Espermatozóides
A sexagem é a separação das duas populações de espermatozóides, em
portadores do cromossomo X e portadores do cromossomo Y, a fim de se utilizar
para a fecundação somente os que darão origem a animais de um sexo pré-
determinado.
A identificação de proteínas sexo-específicas presentes na membrana dos
espermatozóides, associada ao desenvolvimento de um anticorpo específico anti-X
ou anti-Y, permite o estudo de várias técnicas imunológicas objetivando a separação
dos espermatozóides.
Várias abordagens práticas com o intuito de promover a sexagem têm sido
publicadas, tanto na literatura científica, quanto na leiga (BETTERIDGE, 1984).
Segundo alguns autores, a proporção de espermatozóides “X” e “Y” que chegam até

8
a porção superior do trato genital feminino poderia ser alterada e,
conseqüentemente, o sexo do feto. A alteração da taxa de um ou outro sexo do feto
poderia ser conseqüência de vários fatores, como por exemplo, o tempo de
inseminação (WHELAN, 1974 e FRANCE et al., 1984), utilização de métodos de
inseminação, como duchas alcalinas ou ácidas com o intuito de alterar o pH vaginal
(DANIELL, 1983) e, finalmente, a influência da dieta (PAPA et al., 1983;
STOLOWSKI et LORRAIN, 1983). Entretanto, nenhum destes métodos tem
apresentado efeito significativo sobre a taxa do sexo.
A separação das duas populações de espermatozóides baseia-se em suas
diferenças, tais como a massa, motilidade, conteúdo de DNA, carga elétrica de
superfície e determinantes antigênicos da superfície celular.
Até o ano de 1998, de acordo com HOSSAIN e colaboradores, não se
dispunham ainda de método rotineiro e seguro para separação das duas populações
de espermatozóides.
As diferentes metodologias empregadas no fracionamento do sêmen, como a
exploração da diferença da massa de cada população, a motilidade, a carga elétrica
da superfície celular e o conteúdo de DNA, ainda não promoveram, com segurança,
rapidez e de forma econômica, a separação destas duas populações de células.
Atualmente, apenas a citometria de fluxo consegue separar espermatozóides
“X” e “Y” com alta fidelidade. Entretanto, esse método requer equipamentos caros e
tanto o tempo quanto o número de espermatozóides separados por dia é limitado.
A identificação de uma proteína específica na membrana plasmática para
espermatozóides “X” e “Y” pode gerar grandes oportunidades para a escolha do
sexo (HENDRIKSEN,1999). Ela pode permitir o desenvolvimento de um ou mais
anticorpos anti proteína “X” ou anti proteína “Y”, viabilizando a separação através de
técnicas imunológicas.
Método imunológico simples e com equipamentos de baixo custo é proposto, possibilitando uma separação dos espermatozóides em larga escala, com aproveitamento total do ejaculado, evitando o uso de corantes de DNA e excitação por raio laser, com possível efeito negativo no espermatozóide e na viabilidade do embrião (Matta,C.G.F. 2002).
2.4.2. – Sexagem de Embriões
A sexagem de embriões pode ser realizada por métodos invasivos e não-
invasivos.

9
Entre os métodos invasivos, temos a cariotipagem, onde se faz análise na fase de metástase celular para verificação da presença do cromossoma X ou Y. O sucesso desse processo de sexagem raramente excede a 60% dos embriões examinados. Com isto, o método de cariotipagem não é empregado comercialmente. Outro método é o que utiliza uma sonda de DNA específica para o
cromossoma Y. Nesta metodologia, o DNA é ampliado e submetido a uma corrida
eletroforética, corado com brometo de etila e examinado em luz ultravioleta. A
presença de uma banda de 149 pb determina o sexo masculino. Reed et al. (1988)
sexaram embriões com eficácia de 100%. McEvoy (1992) afirmou que a sonda de
DNA é o método mais preciso de sexagem de embriões.
2.5 - Antígeno H-Y
A primeira menção da existência de antígenos H-Y foi feita por EICHWALD e
SILMSER (1955). Em estudos de transplante de pele em camundongos provenientes
de linhagens isogênicas, foi observado que fêmeas normalmente rejeitavam o
transplante de pele de macho. Essa constatação foi observada, mais tarde, em
outras espécies de mamíferos, com alto grau de isogenicidade, como ratos por
BILLINGHAN e SILVERS (1959) e coelhos por CHAI (1968). Essa constatação ficou
conhecida como o fenômeno de Eichwald-Silmser, porque a diferença genética entre
os sexos de animais mamíferos isogênicos foi atribuída ao cromossoma “Y”. Dessa
forma, a rejeição de tecidos de machos deve ser causada por um antígeno macho-
específico, codificado ou regulado pelo cromossoma “Y” (MÜLLER, 1983). Esse
antígeno foi denominado de “antígeno H-Y”, primeiramente por BILLINGHAN e
SILVERS (1960), onde a letra “H” se refere à histocompatibilidade, o “Y” ao
cromossoma “Y”. A presença do antígeno pode ser verificada por meio do teste de
citotoxidade ou pela detecção de anticorpos macho específicos (GOLDBERG et al.,
1971). GOLDBERG e colaboradores (1971) detectaram anticorpos contra
espermatozóides de camundongos, no soro de camundongos fêmeas que
receberam vários transplantes de pele de camundongos machos ou foram
inoculadas com células de baço de machos isogênicos e também no soro de fêmeas
que não apresentaram rejeição aos tecidos de pele de macho (GOLDBERG et al.,
1971). Utilizando o teste de citotoxidade, GOLDBERG e colaboradores (1971)
demonstraram que os espermatozóides, quando expostos ao anticorpo contra a
proteína macho-específica, e, em seguida, a uma suspensão de complemento,

10
apresentavam-se lisados pelo anti-soro antiproteína macho-específica mais
complemento, às vezes em mais de 50%. A lise de tais células foi visualizada em
câmara hemocitométrica através da coloração de Azul de Trypan.
2.6 - O papel da proteína H-Y
O papel crítico do cromossoma "Y" na regulação da morfogênese testicular foi
primeiramente descrito por Ford e colaboradores (1959) e JACOBS e ROSS (1966),
que demonstraram que o cromossoma "Y" estava associado à diferenciação
testicular no homem. O antígeno H-Y tem sido proposto como a chave para o
reconhecimento celular pelo qual o determinante macho ligado ao "Y" atua
(WACHTEL et al., 1975b). O fator determinante da diferenciação testicular foi
proposto por WACHTEL e colaboradores (1975a) como sendo o antígeno
histocompatível macho-específico H-Y e que, por análise cromossomal, este fator
era oriundo de genes situados no braço curto do cromossoma "Y" (MOREIRA FILHO
et al., 1979 e RARY et al., 1979). Assim, um conjunto de gene(s) situado(s) no
cromossoma "Y" promove a evolução da gônada embrionária indiferenciada para
testículo, segundo a hipótese de WELSHONS e RUSSELL ( 1959). Sem a presença
do cromossoma "Y", a gônada embrionária se desenvolverá em ovário (WACHTEL
et al., 1975c). A gônada embrionária indiferenciada, em um indivíduo normal, possui
dois pares de ductos genitais, os ductos Wolffianos e os Mullerianos. Em uma
embriogênese normal, um par regride enquanto o outro persiste, dando origem aos
ductos genitais (HAFEZ, 1982). Assim, nos mamíferos machos são os ductos de
Wolff que persistem, enquanto, nas fêmeas, os ductos Mullerianos permanecem em
função da ausência do cromossoma "Y" (HAFEZ, 1982).
Logo após a fertilização, algumas células mesenquimais do embrião tornam-se túrgidas, diferenciando-se em células de Sertoli, enquanto outras se desenvolvem e se diferenciam em células de Leydig da testis. É nas células de Leydig que ocorrerá a produção e a secreção do hormônio testosterona, que nos machos induzirá ao desenvolvimento dos ductos de Wolff, dando origem ao epidídimo, vasos deferentes, vesícula seminal, ductos ejaculatórios e glândulas acessórias (HAFEZ, 1982).
Na gônada indiferenciada observam-se duas regiões: córtex e medula. Cada
região secreta indutores (hormônios e antígenos de diferenciação) que, de acordo
com HALL e WACHTEL (1980), ditarão o particular padrão de alojamento das
células germinativas e a diferenciação dos demais tecidos gonadais. Se não houver

11
a indução masculina (ausência do cromossoma "Y”), o córtex aumenta, a medula
regride e as células germinativas serão do tipo ovócito, associadas às células
foliculares e a gônada se desenvolverá em ovário. Se o indutor masculino estiver
presente (presença do cromossoma "Y"), a medula se desenvolverá, havendo
correspondente regressão do córtex, as células germinativas serão do tipo
espermatócito, associadas às células tubulares (Sertoli), e a gônada se
desenvolverá em testículo. De acordo com esses dados, o fator chave na
organogênese gonadal seria a regionalização córtico-medular. Assim, o córtex
desenvolverá a arquitetura folicular ovariana, enquanto que a medula seguirá uma
arquitetura tubular de testículo (FRAGOSO, 1994).
O desenvolvimento do embrião em macho está sob o controle de 2 grandes
genes reguladores, um localizado no cromossoma "Y" e outro no cromossoma "X"
(OHNO, 1977b). O gene regulador localizado no cromossoma "Y" é o que
especificará uma proteína plasmática indutora da organização testicular pela gônada
embrionária indiferenciada (OHNO, 1977b). WACHTEL (1983) supôs que o sucesso
do desenvolvimento gonadal era direcionado para a determinação do sexo e
elaboração do fenótipo macho pelo disparo de uma simples molécula. A evidência
de que o antígeno H-Y estava presente em sítios específicos nas células de Sertoli e
células de Leydig quando da exposição in vitro de células ovarianas com o antígeno
H-Y, promovendo a agregação de células, levou WACHTEL (1983) a acreditar que
uma simples molécula pudesse ser o antígeno H-Y.
A atividade biológica do antígeno H-Y e sua capacidade para induzir a organização testicular em gônadas de bovinos indiferenciadas foi testada por OHNO (1979). As gônadas eram possuidoras dos cromossomas "XX" e estavam com idade variando de 30 a 45 dias de idade. O antígeno H-Y foi produzido pelas células de Daudi (linhagem celular derivada do linfoma de Burkitt de homens normais B-46, XY) cultivadas em meio livre de proteína por um período de 16 horas. Assim, as gônadas de bovinos indiferenciadas foram mantidas na solução contendo o antígeno H-Y (sobrenadante de cultura das células de Daudi) por um período de 5 dias, sofrendo uma completa diferenciação testicular, enquanto o controle permaneceu em um estádio indiferenciado.
LUKASA e colaboradores (1992) sugeriram que o papel do antígeno H-Y nas
células germinais primitivas poderia ser para conferir a capacidade de migrar
seletivamente para a medula da gônada indiferenciada e capacidade para iniciar o
desenvolvimento medular. OHNO (1976) propôs que, no estágio de crista gonadal, o
antígeno H-Y poderia mediar o reconhecimento celular, afirmando, então, que a
interação célula-célula e o reconhecimento celular constituem fator chave na
diferenciação e na organogênese. Assim, a presença do antígeno H-Y na superfície
destas células promoveria sua união, crescimento e a característica da diferenciação
que levaria a gônada indiferenciada ao desenvolvimento dos testículos. Por

12
conseguinte, não seria apenas o antígeno H-Y responsável pelo reconhecimento
celular, mas também a presença ou ausência de seu sítio receptor da superfície
celular. OHNO (1976) propôs que a característica distinta das células gonadais, ao
contrário de outras células somáticas, é que aquelas são possuidoras de sítios
receptores para o antígeno H-Y e que somente as células possuidoras do
cromossoma "Y" apresentavam o antígeno H-Y. WACHTEL (1983) mostrou que
diferentes pesquisadores identificaram o antígeno H-Y como sendo idêntico ao fator
de determinação da testi (FDT). MACLAREN e colaboradores (1984) mostraram
evidências definitivas de que o antígeno H-Y e o fator determinador da testi não são
idênticos, quando descrevem Sxr' (a menor seqüência do cromossoma "Y”
conhecida por conter a Tdy do camundongo) que induz a diferenciação sexual
masculina no camundongo, mas não causa expressão do antígeno H-Y. Essa
evidência foi demonstrada por Simpson e colaboradores (1987), que o gene para a
determinação da testi e o gene do H-Y estavam situados separadamente.
Jost e colaboradores (1981) mostraram que os primeiros sinais da
diferenciação testicular em rato ocorrem no 13° dia de gestação. Na ausência de
mitose, algumas células da medula começam a se diferenciar. Essas células, que
são as células de Sertoli iniciais, começam então a se mover juntas e a se
agregarem. Elas circundam as células germinativas primordiais, originando, assim, a
característica da estrutura tubular do testículo. Por intermédio de uma "onda de
diferenciação", aparecem outras células de Sertoli e outros túbulos. As células de
Leydig aparecem entre os elementos indiferenciados do interstício.
Para o desenvolvimento masculino, são necessárias duas secreções do
testículo fetal: substância anti-mulleriana (SAM), produzida pelas células de Sertoli e
testosterona, produzida pelas células de Leydig. A substância anti-mulleriana é um
hormônio protéico que nos indivíduos masculinos causa regressão dos ductos de
Muller, impede o desenvolvimento do útero, dos ductos ovarianos e do terço superior
da vagina. A testosterona é o androgeno principal sintetizado pelo testículo pré e
pós-natal. No embrião ela tem, pelo menos, duas funções: 1°- sua ação local no
testículo promove a maturação da espermatogônia nos túbulos seminíferos; 2°- por
intermédio da circulação sanguínea sistêmica fetal, é induzido o desenvolvimento
das características sexuais secundárias masculinas.
Observações embriológicas e endocrinológicas demonstraram a importância
da testosterona na virilização sexual secundária. A síndrome de feminização

13
testicular, que em ambos, homem e camundongo, é causada por uma mutação de
um gene do cromossoma "X", suprime a recepção de testosterona ao nível de citosol
(OHNO, 1979). Indivíduos afetados por este defeito genético têm fenótipo sexual
feminino, apesar da presença dos testículos perfeitamente diferenciados. Portanto, a
organogênese testicular é um processo complexo, que envolve uma série de sinais
indutores. Este processo pode ser dividido em três etapas, segundo FRAGOSO
(1994):
1°) Estabelecimento do sexo genético, com a união do espermatozóide e do óvulo,
durante a fecundação;
2°) Transformação da gônada inicial indiferenciada em testículo ou ovário, de acordo
com o sexo genético;
3°) Desenvolvimento das características sexuais secundárias, de acordo com o sexo gonadal, num processo mediado por secreções da gônada.
Entretanto, isto não implica que uma diversificação do plano embrionário da
gônada inerentemente feminina ocorra somente no início do desenvolvimento
gonadal. A transformação testicular de gônadas "XX" do bovino "freemartin" (a
fêmea gemelar sincorial de um macho) ocorre muito tarde, após já ter havido uma
considerável diferenciação da gônada feminina em ovário. Parece que o processo
lento da diferenciação de ovário em mamíferos pode sofrer um desvio em direção ao
desenvolvimento testicular, não somente na emergência do desenvolvimento
gonadal como também em períodos mais tardios.
2.7 - A genética do antígeno H-Y
WACHTEL e colaboradores (1975b) demonstraram que o antígeno H-Y nos homens era codificado pelo cromossoma "Y", porque células brancas do sangue de homens com alteração cromossômica possuidoras de 2 cromossomas "Y" absorviam mais anticorpo contra antígeno H-Y do que as normais possuidoras de apenas um cromossoma "Y". KOO e colaboradores (1977) evidenciaram que homens com anomalias
cromossômicas tipo deleção no braço curto do cromossoma "Y” eram negativos
quanto à presença do antígeno H-Y. Entretanto, quando ocorria a deleção no braço
longo desse cromossoma em indivíduos normais, o antígeno H-Y estava presente.
Baseados nessas evidências, KOO e colaboradores (1977) concluíram que o gene
do antígeno H-Y está localizado no braço curto do cromossoma Y.
MOREIRA FILHO e colaboradores (1979) e RARY e colaboradores (1979),
estudando a expressão gênica do antígeno H-Y em pacientes humanos portadores

14
de 2 cromossomas "Y", evidenciaram também que a localização do antígeno H-Y
estava no braço curto do cromossoma "Y".
WOLF (1981) sugeriu a existência de pelo menos 4 genes envolvidos no
sistema H-Y, os quais estariam localizados nos cromossomas "Y", "X" e nos
autossomas. Três genes estruturais e reguladores estariam envolvidos na expressão
das moléculas do antígeno H-Y e o quarto gene, localizado no cromossoma “X”,
codifica um receptor gonadal específico para antigeno H-Y.
WACHTEL (1983) hipotetizou que, em homens normais (46XY), o gene estrutural para antígeno H-Y localizava-se no cromossoma "X" ou em um autossoma, os quais estariam sob o controle de um gene regulador do cromossoma Y, e esse gene regulador seria o responsável pela produção de uma molécula protéica que ativaria o gene estrutural do cromossoma "X" ou do autossoma. WACHTEL (1983) propôs duas explicações para evidenciar que machos com alterações cromossômicas do tipo "XX" e hermafroditas verdadeiros "XX" fossem positivos para antígeno H-Y. A primeira onde o gene estrutural poderia estar no cromossoma "X" ou no autossoma e estaria alterado nesses indivíduos, não necessitando de uma proteína reguladora do cromossoma “Y” para sua expressão. A outra explicação era que a peça do cromossoma “Y”, que incluía o gene regulador, estaria translocada para o cromossoma "X" e, assim, a presença do cromossoma "Y" não se faria necessária para a expressão do antígeno H-Y.
MULLER (1983), em estudos comparativos de aberrações do cromossoma
"Y" em humanos, presumiu que genes nos autossomas e cromossomas sexuais
estivessem envolvidos na expressão do antígeno H-Y. Os genes reguladores
presentes na região paracêntrica dos dois braços, longos e curtos, do cromossoma
"Y", neutralizariam os presumíveis genes repressores do cromossoma "X", e
concluiu que, pelo menos, três genes seriam necessários para a expressão do
antígeno H-Y, sendo os genes reguladores ligados aos cromossomas sexuais e os
gen(es) estruturais ligados aos autossomas.
Por intermédio de pesquisas citogenéticas, diversos pesquisadores têm feito
progresso na análise do cromossoma “Y”. Alguns locus têm sido atribuídos a esse
cromossoma, entretanto, sua localização exata ainda é controvertida em função de
fatores como o pequeno tamanho do cromossoma “Y”, o que dificulta a definição
citogenética e a ausência de recombinações extensivas (LUKASA et al., 1992). LUKASA e colaboradores (1992) apresentam o modelo clássico do cromossoma “Y” tendo 4 sub-regiões como mostrado na figura 1. O cromossoma “Y” é dividido em pelo menos 4 regiões, descritas a seguir:
1 - Uma sub-região de pareamento meiótico homólogo ao cromossoma "X" no braço
curto do "Y" (região pseudoautosomal);
2 - Uma sub-região pericêntrica no braço curto, considerado classicamente como contendo genes de determinação sexual;

15
3- Uma sub-região eucromática do braço longo contendo fatores necessários para a
espermatogênese;
4 - Uma sub-região heterocromática do braço longo geneticamente inerte.
A localização dos fatores determinantes masculinos dentro de qualquer uma
dessas regiões definidas é, usualmente, deduzida com base nas correlações entre
descobertas clínicas e pesquisas citogenéticas em indivíduos com rearranjos
estruturais do cromossoma “Y”.
JACOBS e ROSS (1966) foram os primeiros a propor que o fator determinante
da testis (TDF) no homem estivesse na região pericêntrica do braço curto do
cromossoma “Y”.
Figura 1: Modelo clássico do cromossoma “Y”.
2.8 - Sorologia do antígeno H-Y
A sorologia do antígeno H-Y teve inicio quando GOLDBERG e colaboradores
(1971) utilizaram o teste de citotoxidade, onde as células-alvo eram
espermatozóides de camundongos. Nesse estudo, o anticorpo contra o antígeno H-
Y foi o anti-soro de camundongos fêmeas isogênicas que rejeitaram o transplante de
pele de camundongos machos de mesma linhagem e a fonte de complemento foi

16
soro de coelho. O espermatozóide que apresentava o antígeno H-Y em sua
membrana plasmática era morto após a incubação com anti-soro anti-H-Y e
complemento de coelho. A visualização das células mortas se dava após a adição
de azul de tripan e leitura em microscópio.
KOO e colaboradores (1973), utilizando o vírus mosaico tabaco (VMT) como
marcador visual, puderam observar a localização da molécula do antígeno H-Y em
espermatozóides de camundongo. Nesse experimento, o anti-soro para o antígeno H-Y
foi obtido de fêmeas isogênicas de camundongos da linhagem C57/BL6 após terem
rejeitado o enxerto de pele de macho de mesma linhagem. A seqüência para
evidenciar a localização do antígeno H-Y consistiu em adsorver o anti-soro
previamente com células fêmeas, anticorpos híbridos (anti IgG de camundongo/anti
VMT) e, por último, o VMT. (((Dos espermatozóides corados, três formas
características de coloração foram visualizadas nesse experimento: 1ª) 20 a 40% dos
espermatozóides não apresentaram qualquer coloração ou foram corados a nível
inferior àquele apresentado pelo controle (<20 partículas de VMT/espermatozóide); 2ª)
30 a 40% foram corados com 20/60 partículas de VMT/espermatozóide e 3ª) 20 a 30%
foram corados com 60 a 200 partículas de VMT/espermatozóide. Em 90% dos
espermatozóides corados, o VMT estava localizado principalmente na capa
acrossomal (estrutura de membrana plasmática recobrindo o acrossoma) da cabeça.
Pouca ou nenhuma coloração fora vista em outra parte da cabeça do espermatozóide.
Foi observada, nos 10% dos espermatozóides corados restantes, uma distribuição
mais uniforme do VMT sobre toda a cabeça.
KOO e colaboradores (1978) apresentaram técnica para tipificar o antígeno H-
Y de espermatozóides de camundongo, usando proteína “A” de Staphylococcus
aureus à qual se ligava na porção Fc da IgG. Espermatozóides de camundongos foram
tratados com anti-soro H-Y (produzidos em fêmeas isogênicas de camundongo da
linhagem C57/BL6 inoculadas com células de baço de camundongo macho de mesma
linhagem) e, a seguir, com a proteína A unida com hemácias de carneiro (PA-HC),
analisadas em câmara de Newbauer. Qualquer espermatozóide com três ou mais
ligações PA-HC foi contado como roseta, o que significava ser portador do
cromossoma "Y". O percentual de rosetas foi, então, determinado por ser
significativamente maior para espermatozóides contendo antígeno H-Y do que para os
que não continham.

17
A localização de moléculas do antígeno H-Y, principalmente na capa
acrossomal de espermatozóide de camundongo, também foi mostrada por ZABORSKI
(1979). Nesse experimento, o anti-soro H-Y foi produzido em fêmeas isogênicas de
camundongo da linhagem C57/BL6 inoculadas com células de baço de camundongo
macho de mesma linhagem. Os espermatozóides foram primeiro incubados com igual
volume de anti-soro H-Y por um período de 20 minutos a 37°C e, a seguir, lavados e o
sedimento de espermatozóides ressuspensos em igual volume de conjugado de IgG
de coelho anticamundongo com isotiocianato de fluoresceína (FITC). Finalmente, as
células foram lavadas e incubadas com igual volume de uma suspensão de proteína A
de S. aureus e examinadas em microscópio com luz para fluorescência. Foi verificado
que a ligação da proteína A do S. aureus e a fluorescência do conjugado estavam
confinadas à cabeça do espermatozóide.
BRADLEY e colaboradores (1987) identificaram antígeno macho específico (H-Y) na membrana plasmática do flagelo de espermatozóide epididimário de carneiro, usando anti-soro com alto título para H-Y. O anti-soro foi produzido por inoculações intra-esplênicas de células de rato macho em fêmeas da mesma linhagem isogênica HS. O anti-soro foi adsorvido com células de baço de fêmeas de rato da linhagem HS e com células de macho da mesma linhagem para servir de controle negativo. O antígeno H-Y da membrana plasmática do flagelo do espermatozóide foi analisado por teste de ELISA. Após a purificação de membrana plasmática de flagelo de espermatozóide, o
anti-soro adsorvido com células de machos e fêmeas foi colocado em diferentes
diluições de anticorpo. O anti-soro anti H-Y adsorvido em células de fêmeas
apresentou título de 1:640, enquanto o anti-soro anti H-Y adsorvido em células de
machos apresentou um pico de reação na diluição de 1:40. Quando os soros foram
postos com meio oriundo da cultura de células de Daudi (fonte de H-Y), o anti-soro
adsorvido em células de fêmeas apresentou um título de 1:320, enquanto o anti-soro
adsorvido em células de macho não apresentou nenhuma positividade em qualquer
diluição. A detecção do antígeno H-Y por imunohistoquímica ocorreu por
imunoperoxidase. Quando os espermatozóides foram incubados com anti-soro H-Y
adsorvido com células de fêmeas, ocorreu intensa coloração na membrana
citoplasmática tendo como média percentual 42 positivos para coloração da
imunoperoxidase e 58 negativos. Quando os espermatozóides foram expostos ao anti-
soro H-Y adsorvido com células-macho não ocorreu a coloração marrom,
demonstrando, assim, a especificidade do anti-soro ao antígeno H-Y. Quando os
espermatozóides foram expostos ao anti-soro anti H-Y adsorvidos em células fêmeas
na diluição de 1:200 e, a seguir, a um anticorpo anti-rato conjugado com FITC, esses
apresentaram fluorescência na membrana plasmática da parte posterior da cabeça, na
peça intermediária do flagelo, em, aproximadamente, 50% da população de células.

18
Quando esse procedimento foi realizado com anti-soro H-Y adsorvido em células-
macho, que serviu como controle, ocorreu fraca fluorescência no flagelo.
PETER e colaboradores (1993) conduziram um estudo para a separação de
espermatozóides com base no antígeno de superfície celular H-Y. Esses autores
incubaram espermatozóides de bovinos com anticorpo monoclonal (IgG) contra o
antígeno de superfície celular H-Y associado a esferas de polímeros imantadas
recobertas com um anticorpo contra o isotipo do anticorpo monoclonal. Após um tempo
de incubação, toda a suspensão foi colocada em presença de um magneto. Os
espermatozóides que não estavam ligados às esferas imantadas foram aspirados e
incubados com anticorpo de cabra conjugado com molécula de FITC contra IgG de
camundongo. Analisados para coloração fluorescente em citômetro de fluxo, somente
1,2% destes espermatozóides apresentaram fluorescência. Esses autores, assumindo
que o antígeno de superfície celular H-Y estivesse presente somente em células com o
cromossoma "Y", conseguiram separar um percentual superior a 98% de
espermatozóides possuidores de cromossoma "X".
2.9 - Purificação do antígeno H-Y
GOLDBERG e colaboradores (1971) identificaram o antígeno H-Y pelo teste
de citotoxidade em espermatozóides de camundongos e foi verificado por WACHTEL
(1977) que ue todos os vertebrados heterogaméticos testados expressavam o antígeno
H-Y e que esse antígeno apresentava reação cruzada com outras espécies. OHNO
(1977a) hipotetizou que os antígenos do complexo maior de histocompatibilidade
(MHC) em conjunto com a beta-2-microglobulina serviam como um sítio de ligação
não-específico na superfície celular para a molécula do antígeno H-Y. NILSSON e
colaboradores (1974) verificaram que Células de Daudi (Células do linfoma de Burkitt
humano masculino) são beta-2-microglobulina negativa e OHNO (1979) demonstrou
que elas também são negativas para os antígenos do MHC e que essas células
sintetizavam e secretavam, para o meio de cultura, o antígeno H-Y. OHNO (1979) e
HALL e colaboradores (1981) isolaram o antígeno H-Y de cultura de células de Daudi.
Hall e colaboradores (1981), usando anticorpo monoclonal contra antígeno H-Y fixados
em esferas de Sepharose recobertas com proteína A, foram capazes de isolar uma

19
molécula protéica com peso molecular entre 15.000 e 18.000 Daltons. WACHTEL e
colaboradores (1980) obtiveram o antígeno H-Y, a partir de testículos de machos, após
rápida dissociação e centrifugação. O sobrenadante que continha a molécula do
antígeno H-Y foi filtrado para retirada dos debris celulares e fragmentos de
membranas.
2.10 - Produção de anticorpos contra o antígeno H-Y
Anticorpos monoclonais são proteínas homogêneas específicas que são
formadas em resposta a um antígeno e que reagem especificamente contra esse
antígeno (STRYER, 1992). KOHLER e MILSTEIN (1975) fusionaram células
esplênicas de camundongos imunizados contra hemácias de carneiro com células de
mieloma de camundongo da linhagem BALB/c. As células híbridas obtidas após a
fusão foram chamadas de células híbridas ou de hibridomas. Assim, eles obtiveram
uma célula que possuía a capacidade de secretar grandes quantidades de anticorpo
contra as hemácias de carneiro. As células de hibridomas retinham a característica
das células do mieloma, que era o vigor do crescimento indefinido, e a das células
esplênicas, que era a produção de anticorpos.
GERSHWIN (1981) descreveu o conceito básico envolvido na obtenção de
anticorpo monoclonal pela secreção de células de hibridomas. As células esplênicas
dos camundongos injetados com o antígeno desejado são fusionadas com linhagem
de células de mieloma usando-se polietilenoglicol. KELLEY e LEWIN (1986)
verificaram que, logo após a fusão, surgiam os seguintes híbridos celulares: baço-
baço, baço-mieloma e mieloma-mieloma. OI e HERZENBERG (1980) e KELLEY e
LEWIN (1986) determinaram que, logo após a fusão, as células devem ser transferidas
para um meio seletivo contendo hipoxantina, aminopterina e timidina (HAT). A
aminopterina bloqueia a via normal da síntese de nucleotídeos, forçando a célula para
uma via alternativa dita selvagem que, para isso, utiliza a hipoxantina e a timidina.
GERSHWIN (1981) e KELLEY e LEWIN (1986) verificaram que essa via necessitava
da enzima hipoxantina guanina fosforibosil transferase (HGPRT) e que as células de
mieloma não-fusionadas e as células híbridas mieloma-mieloma não foram capazes de
crescer nesse meio. OI e HERZENBERG (1980) verificaram que as células do baço e
as células fusionadas baço-baço morriam em, no máximo, duas semanas quando em

20
cultura celular. As células híbridas mieloma-baço foram as únicas capazes de
sobreviver e crescer utilizando a HGPRT. Os clones de células híbridas produtoras de
anticorpos específicos para o antígeno, selecionadas por testes imunológicos, são
transferidos para novos poços, de forma que, estatisticamente, seja colocada apenas
uma célula por poço. Dessa forma, as células que advirem serão filhas de uma única
célula ou clones (GERSHWIN, 1981). A especificidade dos vários clones produtores de
anticorpos monoclonais é determinada pela sua reatividade ao antígeno específico em
questão. Após a caracterização dos anticorpos monoclonais produzidos, as culturas
poderão ser produzidas em grandes quantidades in vitro ou os hibridomas poderão ser
injetados na cavidade peritoneal dos animais isogênicos que forneceram as células
esplênicas para a produção de líquido ascítico (OI e HERZENBERG, 1980).
Antes da descoberta do anticorpo monoclonal por KOHLER e MILSTEIN
(1975), anticorpos anti H-Y eram mais comumente obtidos de fêmeas de camundongo
ou rato hiperimunizadas com células de baço de machos da mesma linhagem
isogênica (KOO, 1981). O soro dos animais era testado individualmente para sua
reatividade e os que possuíssem um título alto eram misturados e armazenados a -70o
C até sua utilização (KOO, 1981). Somente 30% das fêmeas de camundongos
imunizadas produziam um soro com alto título de anticorpos anti H-Y (KOO, 1981). Os
anticorpos monoclonais anti H-Y apresentavam a vantagem de poderem ser
produzidos em grande quantidade e com grande especificidade quando comparados
ao soro convencional KOO e colaboradores (1981a).
KOO e colaboradores (1981b) obtiveram dois clones positivos para o antígeno
H-Y após fusionarem células de mieloma NS-1 de camundongo BALB/c e células de
baço de camundongo fêmea da linhagem C57Bl/6 (B6) imunizadas com células de
baço de machos B6. Anticorpos monoclonais contra o antígeno H-Y foram obtidos do
sobrenadante de cultura de células de hibridomas de cada um dos clones ou de líquido
ascítico de fêmea de camundongo inoculado com essas células de hibridomas. A
reatividade dos anticorpos monoclonais foi determinada por ensaio de rosetas de
hemácias, revestidas com proteína A, onde foram acoplados os anticorpos
monoclonais específicos para o antígeno H-Y. Esses preparados imunológicos, quando
colocados em contato com espermatozóides, mostraram que os clones produtores do
anticorpo monoclonal apresentaram grande especificidade para células de machos e
que também possuíam título maior que o anticorpo policlonal anti H-Y. KOO e
GOLDBERG (1978) testaram a especificidade de anticorpos convencionais, pela

21
técnica de formação de rosetas, onde os espermatozóides foram expostos ao
anticorpo contra o antígeno H-Y e, a seguir, adicionada suspensão de hemácias de
carneiro recoberta com anticorpo de coelho específico para o isotipo do primeiro
anticorpo. KOO e colaboradores (1981a) verificaram a especificidade do anticorpo
monoclonal anti H-Y tanto em células de machos quanto em células de fêmeas por
dois diferentes métodos. No primeiro teste, usou o ensaio das rosetas, onde hemácias
de carneiro foram revestidas de proteína A e anticorpo monoclonal anti H-Y e testado
frente a diferentes tecidos, tais como células do baço de camundongo, células brancas
de sangue humano, fibroblasto humano e espermatozóides de camundongo. No
segundo teste, utilizando-se a técnica de citotoxidade celular produzida por células
"natural killer", foi realizado um ensaio onde as células alvo foram os espermatozóides,
colocados em contato com células de linfonodos de camundongos machos e fêmeas e
somente as células de linfonodos de fêmeas reagiram. Os resultados desses
experimentos mostraram que o anticorpo monoclonal anti H-Y era macho específico e
fazia reação cruzada com antígeno H-Y humano. Na determinação do peso molecular
do antígeno H-Y, foi obtido um imunoprecipitado com proteínas de peso molecular de
16.000 Daltons (KOO et al., 1981b), semelhante ao peso molecular do precipitado da
cultura de células de Daudi (HALL et al., 1981). Posteriormente, WACHTEL e
colaboradores (1984) determinaram que a molécula identificada pelo anticorpo contra o
antígeno H-Y não se diferenciava do antígeno H-Y de transplantação.
2.11. Produção de Embriões
A biotecnologia na manipulação de embriões vem sendo empregada desde o
final do século XIX, quando Heape realizou a primeira transferência de embriões
(TE) em coelhos (REICHENBACH, 2002). A primeira TE em bovinos foi realizada por
Willet et al. (1951) através de laparotomia do flanco sob anestesia local
(REICHENBACH, 2002). Entretanto, somente na metade dos anos 70, a TE
dinamizou a comercialização de embriões com o desenvolvimento do método não-
cirúrgico.
O primeiro registro de nascimento de um animal por Produção “in vitro” (PIV)
foi realizado por CHANG em 1955 com coelho. No início da década de 80, nasceu o

22
primeiro bezerro produzido “in vitro”, estando a técnica estabelecida para esta
espécie no início dos anos 90 (GONÇALVES 2002). Hoje, várias equipes realizam
esta técnica no Brasil, estando quase todas estas voltadas para a pesquisa. A
produção “in vitro” de embriões possibilita várias aplicações importantes para a
produção animal, como por exemplo, a diminuição do intervalo entre gerações e a
produção de um grande número de embriões/vaca/ano, permitindo maior eficácia
nos trabalhos de melhoramento genético dos rebanhos. Na pesquisa pode ser
aplicada, na viabilização de projetos de clonagem, injeção intra-citoplasmática de
espermatozóide (ICSI), na sexagem de espermatozóides e de embriões. Esta
técnica é fundamental para a preservação de espécies ou raças de animais em
extinção, entretanto, a utilização da PIV ainda está muito limitada em função da
inconsistência dos resultados referentes às taxas e qualidade de mórulas e
blastocistos e do custo inicial para construção da infraestrutura necessária. A PIV
envolve as técnicas de maturação “in vitro” (MIV), fecundação “in vitro” (FIV) e cultivo
“in vitro” (CIV), as quais são interdependentes.

23
3 - MATERIAL E MÉTODOS
Duas metodologias foram testadas para separação dos espermatozóides
portadores do cromossomo Y, daqueles possuidores do cromossomo X:
a - Técnica de imunoafinidade, com coluna IMAC
b – Técnica de fixação do complemento pela via clássica
3.1.-Técnica de imunoafinidade, com coluna IMAC
A coluna magnética foi desenvolvida para seleção positiva de células
humanas e animais. Ela foi utilizada neste trabalho para separar os espermatozóides
X e Y após a sexagem. Após a lavagem do sêmen, adicionou-se o anticorpo

24
monoclonal produzido em camundongo contra uma proteína macho-específica de
19kDa e incubou-se a amostra por 45 minutos a 37°C. Após este período, a amostra
foi lavada 2 vezes com PBS, centrifugando-se por 5 minutos a 4000 rpm. Adicionou-
se o soro de cobaia, incubando-se por 45 minutos a 37ºC. As células foram
novamente lavadas com PBS, retirando-se o sobrenadante e adicionando-se
anticorpo de cabra anti-camundongo conjugado com ferro (SIGMA®). Este segundo
anticorpo foi utilizado para promover a fixação dos espermatozóides portadores do
cromossoma Y na coluna magnética. Essa suspensão foi incubada por 45
minutos/37°C e, a seguir, lavada com PBS (duas lavagens) e o pellet ressuspendido
em tampão 3 (100 ml de PBS sem Ca+ e Mg+/ EDTA 2 mM/ 0,5% BSA) de acordo
com o protocolo fornecido pelo fabricante. Pela coluna eram passados 3 ml de
tampão 3, no qual estavam diluídas as células. Todo o líquido proveniente da coluna
(1ª eluição) era coletado em um tubo. Depois era separada a coluna do ímã,
adicionados mais 3 ml de tampão 3, fazendo-se, desta forma, uma lavagem da
coluna e retirando-se, assim, as células que estavam fixadas (2ª eluição). Esta
segunda amostra era coletada em um outro tubo. Eram obtidas, então, duas
amostras de células.
Figura 2 : Esquema de fixação dos anticorpos nos espermatozóides
Figura 3: Esquema de fixação das células na coluna
Partícula de Ferro
Molécula de Anticorpo X SPT
Z
Coluna IMAC
Antígeno
Y SPTZ
Anticorpo
anti HY

25
X S
PTZ
Coluna IMAC
IMÃ
Y S
PT
Z
IMÃ
Y SP
TZ
Espermatozóides fixados pelo Ac. Monoclonal e pelo Ac. secundário ligado a
partículas de ferro se fixam ao ímã. Espermatozóides que não reagiram passam
direto pela coluna, sem ficar aderidos a ela (1ª eluição). A segunda eluição é feita,
retirando-se a coluna do equipamento contendo ímã.
Figura 4: Foto da Coluna IMAC, onde observamos a coluna presa ao suporte com
ímã, ficando abaixo da mesma um tubo para a coleta da primeira eluição.
Figura 5 : Foto das partes constantes da Coluna Magnética. Para a coleta da
segunda eluição, retira-se a coluna do ímã e promove-se nova passagem de tampão
3, podendo- se utilizar um êmbulo que se insere na coluna.
Tubo para coleta das eluições
Coluna presa ao imã

26
3.2– Metodologia de Sexagem de Espermatozóides com anticorpo monoclonal
anti-proteína macho-específica e adição de complemento:
A metodologia para sexagem de espermatozóides proposta abrange três etapas:
- Coleta do sêmen
- Adição de anticorpo monoclonal antiproteína macho-específica
- Adição do complemento
Nesta metodologia de sexagem, identificou-se o tratamento que mais se
aproximava de 50% da motilidade das células quando comparado ao grupo controle.
Para utilização desta metodologia, é imperativa a eliminação da via alternativa
do complemento, pela inativação da proteína B.
3.2.1- Sêmen
Utilizou-se sêmen fresco, de touros provenientes do plantel da ABS Global.
3.2.1.1.- Tratamento do Sêmen
O sêmen foi colocado em tubos de 15 ml e centrifugado a 2.000g por 7
minuto,s a fim de que se obtivessem apenas os espermatozóides no fundo do tubo e
Coluna magnética
êmbulo Suprte magnético
imã

27
a partir daí as células fossem utilizadas nos tratamentos de sexagem descritos a
seguir .
3.2.2 -Tratamentos para Sexagem (T)
No tratamento 1 (T1) foram utilizados 107 espermatozóides s/ Plasma Seminal + 20 µµµµl de anticorpo monoclonal + 20 µµµµl de soro de cobaia No tratamento 2 (T2) foram utilizados 107 espermatozóides + 30 µl de anticorpo
monoclonal + 30 µl de soro de cobaia
No tratamento 3 (T3) foram utilizados 107 espermatozóides + 50 µl de anticorpo
monoclonal + 50 µl de soro de cobaia
No tratamento 4 (T4) foram utilizados 107 espermatozóides + 100 µl de anticorpo
monoclonal + 100 µl de soro de cobaia
No tratamento 5 (T5) foram utilizados 107 espermatozóides c/Plasma Seminal + 100
µl de anticorpo monoclonal + 100 µl de soro de cobaia
Controle - 107 espermatozóides sem plasma seminal
Os tratamentos foram realizados em tubos de 2ml.
3.2.3 - Anticorpo Monoclonal AntiProteína Macho-Específica
Utilizaram-se anticorpos monoclonais previamente produzidos pelo Setor de Imunogenética do Laboratório de Melhoramento Genético Animal, contra uma proteína macho-específica (Souza, 1998).
O anticorpo macho-específico produzido foi utilizado para reconhecer os
espermatozóides portadores do cromossoma Y, fixando-se a eles de modo a permitir
a separação das duas populações de espermatozóides (X eY).
3.2.4 - Fonte de complemento

28
Foram testados soros de diferentes espécies animais como fonte de complemento, comparando-se os resultados, considerando-se o espermatozóide como um enxerto xenogeneico, em relação à presença ou ausência de anticorpos que pudessem apresentar reações cruzadas ou inespecíficas.
Foram testadas diferentes temperaturas para verificar qual seria responsável pela inativação da proteína B, sem afetar a via clássica do complemento.
Foram testadas diferentes concentrações de soro, frente à quantidade definida de espermatozóides a fim de se determinar aquela que apresentasse melhor resultado.
3.3 - Separação por gradiente de Percoll
Após a sexagem, os espermatozóides foram separados os vivos dos mortos por gradiente de Percoll para, em seguida, serem utilizados na produção de embriões ou para análise eletroforética comparativa das proteínas de membrana dos espermatozóides vivos com os mortos.
Foram preparados tubos contendo: 0.5 ml de Percoll a 90% e 0.5 ml de
Percoll a 45%. As células foram lentamente colocadas sobre o Percoll a 45% e os
tubos centrifugados a 2000g por 7 minutos. Desta forma, obteve-se a separação
das células vivas, que ficaram no fundo do tubo, e das células mortas, que ficavam
no sobrenadante.
O material resultante foi utilizado para a fecundação “in vitro”, análise das
células após passagem pela coluna magnética e separação das populações das
células para análise eletroforética.
3.4 - Imunofluorescência das amostras de células.
Após a separação por coluna magnética ou pelo Percoll, era feita uma
centrifugação, a 2000 g por 7 minutos, destas células, descartando-se o
sobrenadante e, a seguir, adicionaram-se 35 µl de anticorpo contra IgG de cabra
anti-camundongo conjugado com FITC . Deixava-se em incubação, por 45 minutos,
a 37ºC. As amostras foram lavadas com PBS e o pellet ressupendido em PBS. Por
fim, as amostras eram levadas para o microscópio de fluorescência.
3.5 – Eletroforese

29
Os espermatozóides submetidos ao tratamento de sexagem foram separados
pelo gradiente de percoll. As duas populações de células, vivas e mortas, foram
tratadas com Nonidet P-40 0,5% sob gelo durante uma hora e em seguida
centrifugadas a 14.000g por 20 minutos para a extração das proteínas de
membrana. As proteínas extraídas foram submetidas a uma corrida eletroforética
SDS/PAGE em gel em gradiente de poliacrilamida de 7 a 15%. Realizou-se corrida
eletroforética em 100 Volts. Os géis foram corados com solução de azul de
Coomassie. A descoloração realizou-se por intermédio de várias trocas de solução
de descoloração (40% de ácido acético glacial v/v, 40% de álcool metílico v/v e 20%
de água destilada v/v), até que as bandas protéicas estivessem bem contrastadas
em relação ao fundo do gel.
3.6 - Fecundação “in vitro” para produção de embriões bovinos
A fecundação dos ovócitos seguiu o protocolo do laboratório da INFIGEN.
Foram utilizadas duas metodologias para a fertilização: Injeção
intracitoplasmática de espermatozóides (ICSI) e Fertilização in vitro (FIV).
Após a maturação "in vitro'', levaram-se os oócitos para a fecundação “in vitro”
utilizando-se sêmen imunossexado. A taxa de clivagem foi avaliada 72 h pós-
fecundação e a taxa de blastocistos no oitavo dia.
3.7 - Determinação do sexo dos embriões
A determinação do sexo foi realizada em embriões com oito ou mais células,
através do método de PCR (Polymerase Chain Reaction).
Inicialmente os embriões foram lavados em PBS com 2g/L de BSA. Cada
embrião foi, então, transferido para tubo cônico de PCR contendo 1 µl de solução
tampão (100 Mm Tris, 500 mM KCL e 200 mM MgCl2. 6H2O), 0,75 µl de solução de
proteinase K (20 mg/ml) e 8,25 µl de água ultra-pura (Milli–Q). Os tubos com as
amostras foram levados para o termociclador, onde permaneceram por 1 h a 37ºC,
seguidos de 15 min. a 95ºC (inativar a proteinase K), após o qual foram mantidos a

30
4ºC até serem adicionados de solução contendo primers, dNTP e Taq polimerase
(sol. Mix). Essa solução foi preparada com 2 µl de solução tampão, 200 µM/L de
dNTP, 2 µM/L de primer Y, 2 µM/L de primer bovino (controle), 1 UI de Taq
polimerase e 12,4 µl de água Milli-Q, totalizando 20 µl/tubo. Após a adição da
solução contendo os primers, os tubos foram levados para o termociclador para a
amplificação em 40 ciclos, sendo cada ciclo com 1 min. a 95ºC, seguido de 30 seg. a
56ºC e 1 min. a 72ºC.
Após a amplificação, adicionaram-se a cada tubo 5 µl de uma solução de azul bromofenol 6 x (0,25% de bromofenol, 40% de sacarose e 2% de brometo de etídio) e o conteúdo levado para correr em gel de agarose a 4% e 50 µg/ml de brometo de etídio, por 1 h em 80 volts e 24 mA.
A leitura do gel realizou-se sob luz ultravioleta e considerou-se macho a
amostra que continha as bandas para os primers bovino e macho, fêmea somente
se houvesse a banda para bovino e a ausência de bandas interpretou-se como
problema na amplificação ou ausência de amostra no tubo.
3.8 - Injeção Intracitoplasmática de Espermatozóides
Após os tratamentos de sexagem, escolhiam-se, aleatoriamente,
espermatozóides com bom movimento progressivo e os colocavam diretamente no
interior do ovócito, utilizando-se um micromanipulador para promover a fecundação
dos ovócitos e observar a taxa de clivagem e formação de blastocistos.

31
4 - RESULTADOS E DISCUSSÃO
A imunossexagem de espermatozóides é uma metodologia pesquisada por
vários grupos há dezenas de anos sem que os mesmos obtivessem êxito na sua
aplicação comercial.
Dificuldades como a pouca antigenicidade do antígeno H-Y foram
fundamentais para o insucesso destes pesquisadores. Com o desenvolvimento da
tecnologia dos anticorpos monoclonais, surgiu uma nova perspectiva de
identificação dos antígenos H-Y sendo, portanto, uma poderosa ferramenta para
atingir o objetivo.
4.1 – Coluna IMAC:
Inicialmente foram testadas, com diferentes reagentes, as fixações de
espermatozóides na coluna IMAC. Como anticorpo secundário, portador da
microbilha de ferro, foram utilizados anticorpos monoclonais específicos contra IgG1
32
de camundongo que corresponde à classe do monoclonal usado. Os resultados das
células fixadas variaram bastante, desde 18% a 45% da concentração inicial. Esta
análise se deu após contagem em câmara hematimétrica. Entretanto, foi observado
que todos os espermatozóides fixados na coluna e liberados posteriormente na
segunda eluição estavam mortos. Passamos a utilizar um policlonal, visando a
aumentar a área de ação do anticorpo secundário em relação ao primário na
tentativa de se obter um resultado mais eficiente e constante. Tal fato não ocorreu,
sendo observado que, novamente, os espermatozóides fixados pela coluna e
liberados na segunda eluição estavam mortos.
Teste de imunofluorescência destes espermatozóides foi realizado,
verificando-se que apenas os espermatozóides da segunda eluição fluoresciam,
entretanto além de estarem mortos eram normalmente em número muito menor que
os que da primeira eluição que não fluoresciam e estavam vivos.
Foi realizado um teste de fertilização in vitro com as células provenientes da
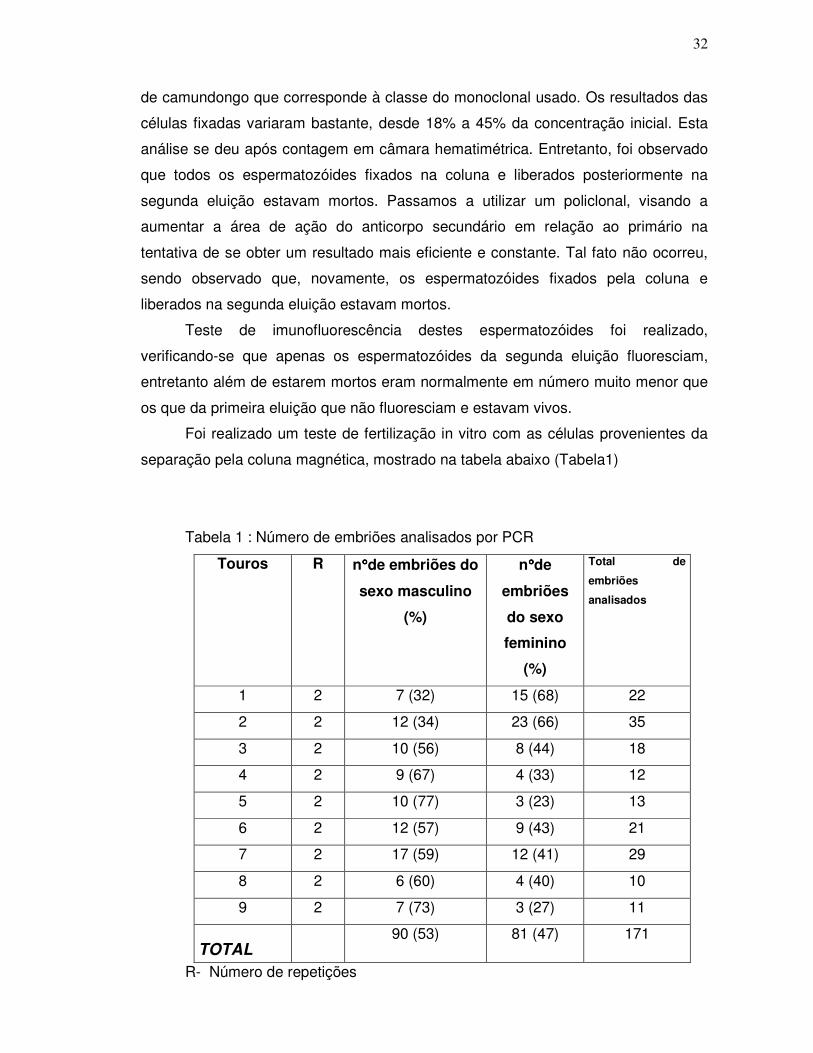
separação pela coluna magnética, mostrado na tabela abaixo (Tabela1)
Tabela 1 : Número de embriões analisados por PCR
Touros R n°°°°de embriões do
sexo masculino
(%)
n°°°°de
embriões
do sexo
feminino
(%)
Total de
embriões
analisados
1 2 7 (32) 15 (68) 22
2 2 12 (34) 23 (66) 35
3 2 10 (56) 8 (44) 18
4 2 9 (67) 4 (33) 12
5 2 10 (77) 3 (23) 13
6 2 12 (57) 9 (43) 21
7 2 17 (59) 12 (41) 29
8 2 6 (60) 4 (40) 10
9 2 7 (73) 3 (27) 11
TOTAL 90 (53) 81 (47) 171
R- Número de repetições

33
Com este resultado, verificamos que a coluna, por si só, não foi eficiente para a sexagem. De acordo com trabalhos desenvolvidos por PETER (1999), foi verificado que
o espermatozóide, ao morrer, apresenta uma reversão de proteínas de membrana,
apresentando uma proteína nova em sua superfície muito imunogênica, ligada ao
complexo maior de histocompatibilidade, que é responsável pela imunofluorescência
observada.
Este fato pode ser claramente evidenciado quando se tenta testar a sexo-
especificidade dos anticorpos monoclonais em espermatozóides vivos e em
espermatozóides mortos. Os espermatozóides vivos ficam sem nenhuma
fluorescência e os mortos apresentam-se sempre muito fluorescentes.
Os resultados obtidos por fluorescência em espermatozóides e em tecidos
são completamente díspares com o observado por SOUZA,1998, quando, em
tecidos de animais do sexo masculino verificava-se grande fluorescência e em
animais do sexo feminino nenhuma ou quase nenhuma fluorescência.
A fluorescência observada nos espermatozóides pode ser devido não só à sensibilidade necessária para reconhecer o antígeno HY, mas também por causa de sua pouca imunogenicidade. A reversão protéica causada pela morte dos espermatozóides faz com que apenas os espermatozóides mortos se aglutinem ou se fixem às colunas. Tal fato tornou inviável a separação dos espermatozóides pelas técnicas de aglutinação, imunoafinidade e coluna IMAC.
A pouca imunogenicidade do antígeno HY faz com que poucos anticorpos se
liguem ao espermatozóide alvo, o que impede a sexagem, tornando-se necessário
para isto que se utilize uma tecnologia muito sensível.
3.2 – Metodologia de Sexagem pela Ação do Complemento
Tal fato nos obrigou a pensar em uma metodologia que fosse suficientemente sensível para reconhecer os anticorpos ligados ao antígeno sexo específico e que não fosse prejudicada pela reversão protéica dos espermatozóides mortos.
O complemento foi selecionado como técnica mais viável para atingir este objetivo, devido a sua alta sensibilidade e grande especificidade para o receptor de C1q no receptor que somente é apresentado no anticorpo ligado ao antígeno.
Entretanto, o complemento possui duas vias de ativação, a clássica e a alternativa. A via clássica é a que interessa para o trabalho, visto só ser ativada na presença do complexo antígeno e anticorpo, enquanto que a via alternativa deve ser eliminada devido ao fato de agir de forma indiscriminada.
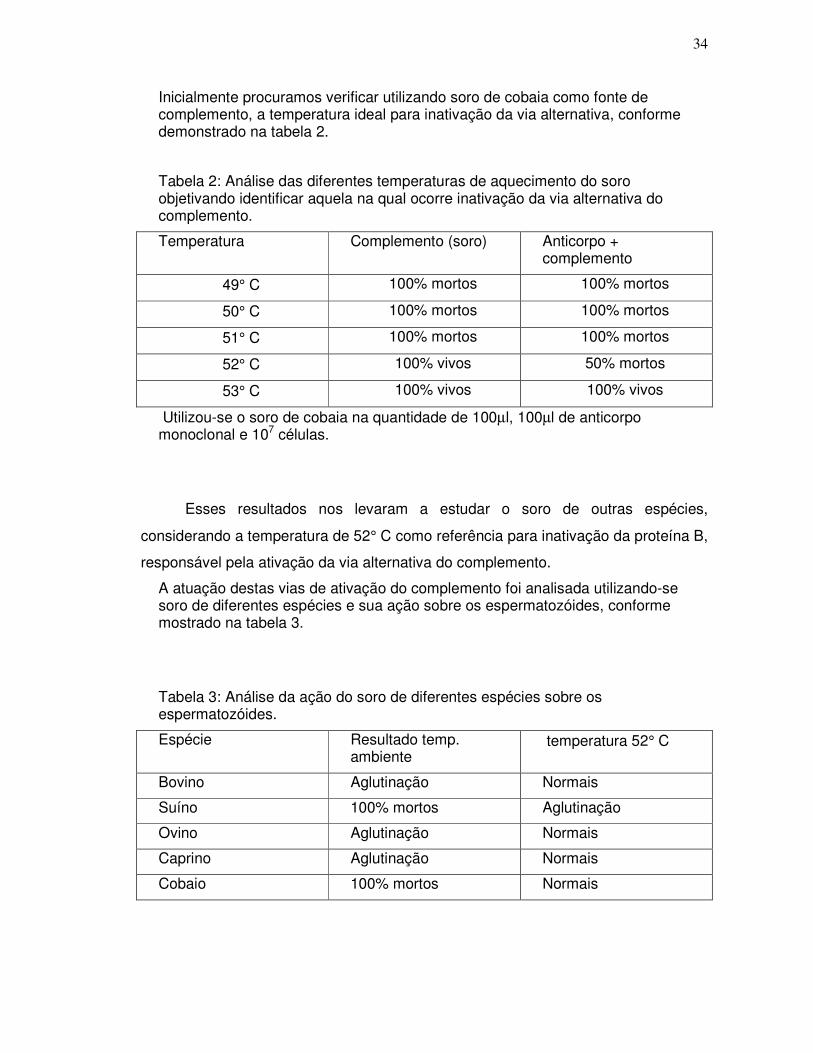
34
Inicialmente procuramos verificar utilizando soro de cobaia como fonte de complemento, a temperatura ideal para inativação da via alternativa, conforme demonstrado na tabela 2.
Tabela 2: Análise das diferentes temperaturas de aquecimento do soro objetivando identificar aquela na qual ocorre inativação da via alternativa do complemento.
Temperatura Complemento (soro) Anticorpo + complemento
49° C 100% mortos 100% mortos
50° C 100% mortos 100% mortos
51° C 100% mortos 100% mortos
52° C 100% vivos 50% mortos
53° C 100% vivos 100% vivos
Utilizou-se o soro de cobaia na quantidade de 100µl, 100µl de anticorpo monoclonal e 107 células.
Esses resultados nos levaram a estudar o soro de outras espécies,
considerando a temperatura de 52° C como referência para inativação da proteína B,
responsável pela ativação da via alternativa do complemento.
A atuação destas vias de ativação do complemento foi analisada utilizando-se soro de diferentes espécies e sua ação sobre os espermatozóides, conforme mostrado na tabela 3.
Tabela 3: Análise da ação do soro de diferentes espécies sobre os espermatozóides.
Espécie Resultado temp. ambiente
temperatura 52° C
Bovino Aglutinação Normais
Suíno 100% mortos Aglutinação
Ovino Aglutinação Normais
Caprino Aglutinação Normais
Cobaio 100% mortos Normais

35
Analisando os resultados obtidos pela ação dos diferentes soros à
temperatura ambiente, verificamos que os soros de bovinos, ovinos e caprinos
aglutinavam os espermatozóides, enquanto que os soros de suíno e cobaia
matavam 100% de todos os espermatozóides.
A aglutinação dos espermatozóides é devida, principalmente, a lesões
superficiais de membrana. Estas lesões de membrana podem ser causadas pela via
clássica ou pela via alternativa do complemento. No caso da via clássic,a isto se
daria principalmente pela reação cruzada de anticorpos que reconheçam alguns
epitopos presentes na superfície de membrana destes espermatozóides que estarão
no caso sofrendo ação semelhante à de transplantes halo ou xenogenêicos. No caso
da via alternativa, esta lesão é causada pelo reconhecimento de algum
oligossacarídio de membrana que tenha ativado a proteína B a iniciar a ativação da
cascata enzimática do complemento. Esta via, entretanto, não foi suficientemente
capaz de matar os espermatozóides nos soros de bovinos, ovinos e caprinos
quando analisadas em temperatura ambiente. Nesta mesma temperatura, os soros
de suíno e de cobaia foram capazes, pela ativação da via alternativa do
complemento, de matar todos os espermatozóides.
Os resultados obtidos a 52° C sugerem que houve a inativação da via
alternativa do complemento no soro de todas as espécies. Em relação ao soro de
suíno, a aglutinação observada de ser devido à presença de anticorpos com reações
cruzadas com epitopos de proteínas de membrana de espermatozóides de bovinos,
os quais ativariam a via clássica do complemento. O fato da via clássica do
complemento, neste caso, não ter apresentado uma reação suficiente para matar os
espermatozóides pode ser devido ao fato de que alguma enzima que faça parte da
via clássica tenha sua ação reduzida pelo aquecimento do soro a 52° C.
A escolha do soro de cobaia como fonte de complemento para trabalhar foi
devido à sua ação tanto à temperatura ambiente quanto a 52° C, além do fato desta
espécie ser possuidoras de forte concentração de complemento no soro.
Devido à grande importância da temperatura ideal de inativação da proteína
B, sem que esta temperatura afete substancialmente outras proteínas que atuam na
cascata enzimática responsável pela ação do complemento, a produção de embriões
foi realizada com o soro de cobaia sendo aquecido a diferentes temperaturas entre
52° C e 53° C, conforme demonstrado na tabela 4.
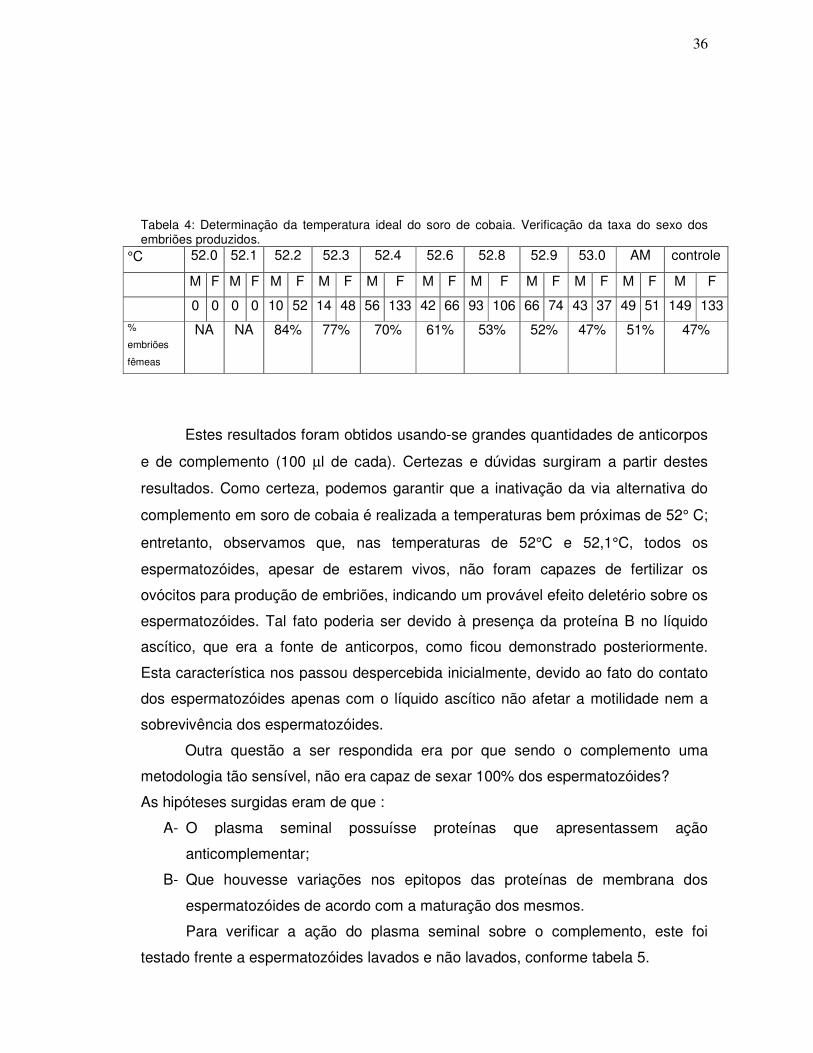
36
Tabela 4: Determinação da temperatura ideal do soro de cobaia. Verificação da taxa do sexo dos embriões produzidos.
°C 52.0 52.1 52.2 52.3 52.4 52.6 52.8 52.9 53.0 AM controle
M F M F M F M F M F M F M F M F M F M F M F
0 0 0 0 10 52 14 48 56 133 42 66 93 106 66 74 43 37 49 51 149 133
%
embriões
fêmeas
NA NA 84% 77% 70% 61% 53% 52% 47% 51% 47%
Estes resultados foram obtidos usando-se grandes quantidades de anticorpos
e de complemento (100 µl de cada). Certezas e dúvidas surgiram a partir destes
resultados. Como certeza, podemos garantir que a inativação da via alternativa do
complemento em soro de cobaia é realizada a temperaturas bem próximas de 52° C;
entretanto, observamos que, nas temperaturas de 52°C e 52,1°C, todos os
espermatozóides, apesar de estarem vivos, não foram capazes de fertilizar os
ovócitos para produção de embriões, indicando um provável efeito deletério sobre os
espermatozóides. Tal fato poderia ser devido à presença da proteína B no líquido
ascítico, que era a fonte de anticorpos, como ficou demonstrado posteriormente.
Esta característica nos passou despercebida inicialmente, devido ao fato do contato
dos espermatozóides apenas com o líquido ascítico não afetar a motilidade nem a
sobrevivência dos espermatozóides.
Outra questão a ser respondida era por que sendo o complemento uma
metodologia tão sensível, não era capaz de sexar 100% dos espermatozóides?
As hipóteses surgidas eram de que :
A- O plasma seminal possuísse proteínas que apresentassem ação
anticomplementar;
B- Que houvesse variações nos epitopos das proteínas de membrana dos
espermatozóides de acordo com a maturação dos mesmos.
Para verificar a ação do plasma seminal sobre o complemento, este foi
testado frente a espermatozóides lavados e não lavados, conforme tabela 5.

37
Tabela 5: Comparação da motilidade dos espermatozóides lavados (Lav) e não
lavados (N. Lav).
temp. 52.0 52.09 52.18 52.27 52.36 52.4 52.6 52.8 52.9 52.9 53.0 controle
Lav. 0% 0% 0% 0% 0% 0% 0% 1% 5% 5% 30% 70%
N.Lav. 30% 30% 30% 40% 40% 40% 50% 50% 60% 60% 60% 70%
Utilizaram-se 100µl de soro de cobaia + 100µl de líquido ascítico + 107
células
Os resultados acima apresentados foram obtidos onde apenas o soro de
cobaia foi aquecido e sugerem, basicamente, que o plasma seminal apresenta
proteínas que atuam de forma a inibir a ação do complemento e que a presença do
plasma seminal é fundamental para sobrevivência dos espermatozóides, visto que
as temperaturas onde o complemento não possuía mais ação, isto é próximo a 53°
C, houve grande mortalidade dos espermatozóides. Devido a estes resultados,
passou-se a retirar o plasma seminal durante o tempo de ação do anticorpo mais
complemento sobre os espermatozóides e readicionou-o logo após. A ação do
complemento foi total em temperaturas próximas a 52°C devido ao líquido ascítico
não ter sido aquecido.
Diferentes concentrações de anticorpo e soro de cobaia foram, então,
utilizados para produção de embriões. Para verificar a ação destas diferentes
concentrações de soro e líquido ascítico sobre os espermatozóides, foram
verificadas a motilidade destes espermatozóides, taxa de fertilização dos ovócitos e
a sexagem promovida. A motilidade dos espermatozóides foi analisada após cada tratamento (antes de ser realizada a separação das
duas populações de células) a fim de se identificar qual tratamento nos daria uma diminuição no número de células vivas que mais se aproximassem da metade do valor do grupo controlem como mostrado na tabela 6.
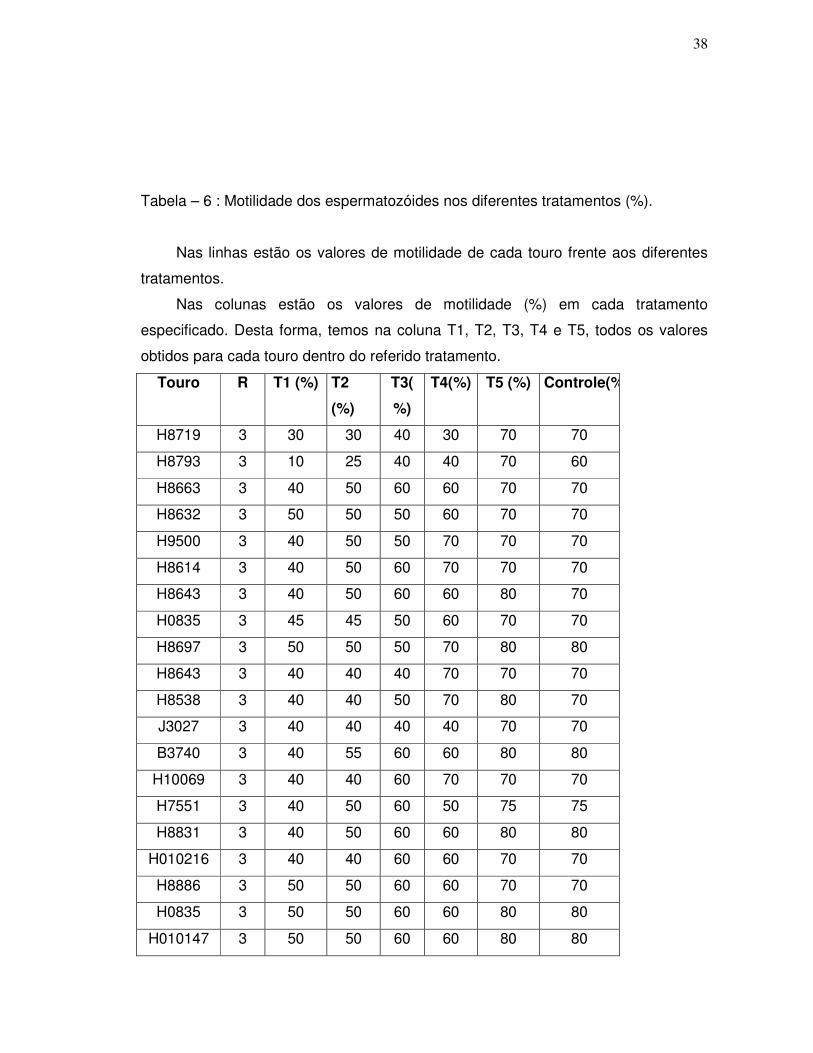
38
Tabela – 6 : Motilidade dos espermatozóides nos diferentes tratamentos (%).
Nas linhas estão os valores de motilidade de cada touro frente aos diferentes
tratamentos.
Nas colunas estão os valores de motilidade (%) em cada tratamento
especificado. Desta forma, temos na coluna T1, T2, T3, T4 e T5, todos os valores
obtidos para cada touro dentro do referido tratamento.
Touro R T1 (%) T2
(%)
T3(
%)
T4(%) T5 (%) Controle(%)
H8719 3 30 30 40 30 70 70
H8793 3 10 25 40 40 70 60
H8663 3 40 50 60 60 70 70
H8632 3 50 50 50 60 70 70
H9500 3 40 50 50 70 70 70
H8614 3 40 50 60 70 70 70
H8643 3 40 50 60 60 80 70
H0835 3 45 45 50 60 70 70
H8697 3 50 50 50 70 80 80
H8643 3 40 40 40 70 70 70
H8538 3 40 40 50 70 80 70
J3027 3 40 40 40 40 70 70
B3740 3 40 55 60 60 80 80
H10069 3 40 40 60 70 70 70
H7551 3 40 50 60 50 75 75
H8831 3 40 50 60 60 80 80
H010216 3 40 40 60 60 70 70
H8886 3 50 50 60 60 70 70
H0835 3 50 50 60 60 80 80
H010147 3 50 50 60 60 80 80
39
Média das
motilidade
s (%)
40.75 45.2 53.5 60 73 72
R – Número de repetições
T – Tratamentos
Realizou-se um teste, onde embriões foram produzidos utilizando-se 100 µl de anticorpo monoclonal antiproteína macho-específica e 100 µl soro de cobaia e embriões produzidos sem que houvesse adição do anticorpo monoclonal e do soro de cobaia. O resultado demonstrou uma taxa maior de embriões do sexo feminino produzidos utilizando-se o tratamento proposto, como mostrado na tabela 7. Tabela 7. Análise por PCR de embriões produzidos com sêmen tratado com 100 µl de anticorpo monoclonal e 100 µl soro de cobaia e de embriões produzidos sem utilizar o tratamento. Anticorpo Monoclonal + soro de cobaia Sem anticorpo monoclonal/ sem soro
Machos Fêmeas % Fêmeas Machos Fêmeas % de Fêmeas
35 139 80 61 105 60
O soro de cobaia foi analisado a fim de se determinar qual a temperatura ideal
a ser utilizada na sexagem dos espermatozóides. Desta forma, observou-se a taxa
de clivagem e formação de blastocistos procurando-se comparar com o grupo
controle.
Observamos que a temperatura de 52.2°C era a mais próxima do controle e,
portanto, aquela a ser definida, conforme mostrado na tabela 8.
Tabela 8 : Taxa de fertilização usando espermatozóides incubados com anticorpo monoclonal e soro de cobaia. Temp. soro 52.2°C 52.3°C 52.4°C 52.6°C 52.8°C
O C B O C B O C B O C B O C B
193 144 72 469 191 69 714 424 187 872 363 126 1149 606 294
Clivagem 75% 41% 59% 42% 53%
Blastocistos 37% 15% 26% 14% 26%
40
Temp.
soro
52.9°C 53.0°C Ac.
Monoclonal Controle
O C B O C B O C B O C B
655 309 131 624 172 54 453 225 75 882 620 270
Clivagem 47% 28% 50% 70%
Blastocistos 20% 9% 17% 31%
Com os resultados de tipo de soro e a temperatura do mesmo definidos, a próxima etapa foi a
de quantificar o anticorpo monoclonal e o soro. 389 embriões foram produzidos utilizando-se os diferentes tratamentos descritos. Observamos uma variação entre os touros, mas não houve variação significativa entre os tratamentos, demonstrando a especificidade do anticorpo monoclonal produzido, como observado na tabela 9. Tabela 9: Resultados da análise por PCR de 389 embriões produzidos a partir da fertilização de ovócitos pelos espermatozóides submetidos aos diferentes tratamentos. Trat. 1 Trat. 2 Trat. 3 Trat. 4 ICSI 20µl/20µl 30µl/30µl 50µl/50µl 100µl/100µl 30µl/30µ Total % F
Touro
M F M F M F M F M F M F
A 2 9 5 9 4 20 4 9 0 13 15 60 80 B 0 1 1 9 2 6 0 1 0 0 3 17 85 C 5 17 3 13 4 9 3 7 0 7 15 53 78 D 2 22 3 27 0 1 0 14 0 0 5 64 93 E 5 8 2 5 2 8 2 4 2 4 13 29 70,7 F 4 6 2 7 5 8 2 6 0 0 13 27 67,5 G 2 21 0 0 3 14 9 26 0 0 14 61 81,3
Total 20 84 16 70 20 66 20 67 2 24 78 311 80 % F 80,77 81,4 76,74 77,0 92,3
M- macho F-Fêmea ICSI-Injeção IntraCitoplasmática de Espermatozóides
Figura 6 : Resultado da análise por PCR de 389 embriões resultantes da fertilização
de ovócitos com espermatozóides tratados, mostrando variações entre os
tratamentos.
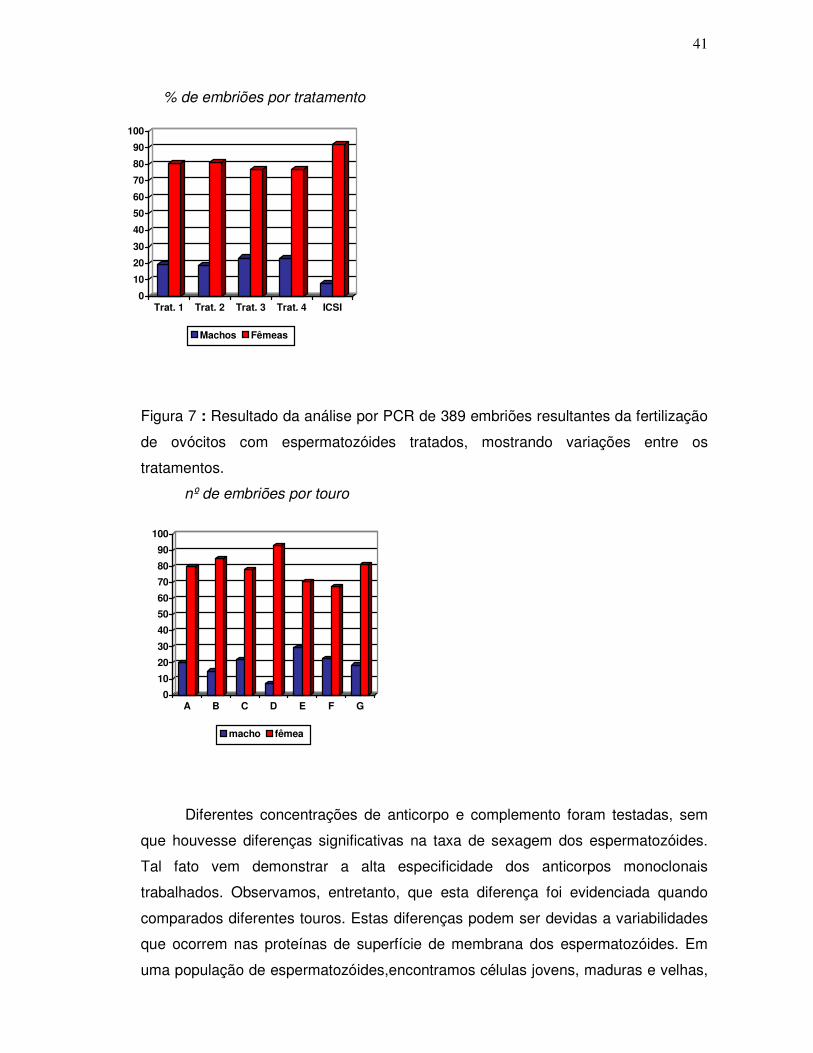
41
% de embriões por tratamento
Figura 7 : Resultado da análise por PCR de 389 embriões resultantes da fertilização
de ovócitos com espermatozóides tratados, mostrando variações entre os
tratamentos.
nº de embriões por touro
Diferentes concentrações de anticorpo e complemento foram testadas, sem
que houvesse diferenças significativas na taxa de sexagem dos espermatozóides.
Tal fato vem demonstrar a alta especificidade dos anticorpos monoclonais
trabalhados. Observamos, entretanto, que esta diferença foi evidenciada quando
comparados diferentes touros. Estas diferenças podem ser devidas a variabilidades
que ocorrem nas proteínas de superfície de membrana dos espermatozóides. Em
uma população de espermatozóides,encontramos células jovens, maduras e velhas,
0
10
20
30
40
50
60
70
80
90
100
Trat. 1 Trat. 2 Trat. 3 Trat. 4 ICSI
Machos Fêmeas
0
10
20
30
40
50
60
70
80
90
100
A B C D E F G
macho fêmea

42
que terão na superfície de suas membranas proteínas apresentando epitopos
diferentes. Estas diferenças, entretanto, podem estar ligadas às diferenças genéticas
entre os touros ou a diferenças entre intervalo de coletas do sêmen destes touros.
Como no laboratório existem outros anticorpos monoclonais, a utilização
concomitante de dois ou mais monoclonais poderia atuar sobre diferentes epitopos
que se encontrem nas diferentes populações de espermatozóides, aumentando o
sucesso da sexagem ou pelo menos tornando mais constante os resultados a serem
obtidos. Um pool destes anticorpos foi empregado como teste para sexagem sem
que, entretanto, se verificassem as concentrações ideais para estes anticorpos.
Após a sexagem, os espermatozóides foram separados por meio de gradiente de
Percoll, as células mortas ficaram entre as concentrações de 45 e 90%, enquanto
que as vivas formaram um pellet. Em princípio, as células vivas deveriam ser as
portadoras do cromossomo X, enquanto que as mortas teriam uma concentração
muito acentuada de espermatozóides possuidores do cromossomo Y. Esta
concentração de espermatozóides contendo cromossomo Y varia com a
percentagem de células mortas no ejaculado antes do trabalho de imunosexagem.
Separação da membrana destas células e análise das proteínas destas
membranas foram realizadas por eletroforese monodimencional, evidenciando
proteínas presentes no perfil protéico das membranas provenientes dos
espermatozóides possuidores do cromossomo Y e ausentes no dos
espermatozóides X. Outros pesquisadores, como Peter et al. (1999) e Hendriks et al.
(1999), pesquisaram esta proteína, utilizando metodologias bastante sensíveis, tais
como separação dos espermatozóides por citometria de fluxo, associada à
eletroforese bidimensional e à coloração pela prata. Esses pesquisadores não
obtiveram resultados satisfatórios devidos, provavelmente, à coloração pela prata,
que não é quantitativa.
Na análise do perfil protéico do gel dos espermatozóides possuidores do cromossomo Y, observou-se uma proteína com peso molecular de 17,18 kDa e outra com peso de 25,81kDa, que estavam ausentes no gel dos espermatozóides possuidores do cromossomo X. Este resultado traz grandes perspectivas para os trabalhos que estamos desenvolvendo, visto que demonstra a possibilidade de se obter uma sexagem melhor utilizando-se mais do que um anticorpo monoclonal, sugerindo que a nossa opinião de haver diferentes epitopos em células com diferentes estágios de maturação é possível. A utilização de diferentes anticorpos monoclonais pode reagir com todas estas formas de apresentação do antígeno H-Y e produzir uma sexagem mais eficiente.

43
A presença de duas proteínas sexo específicas no perfil protéico dos
espermatozóides portadores do cromossomo Y pode ser devido a serem dois
antígenos H-Y diferentes ou a uma forma dimérica da proteína de menor peso
molecular. Neste segundo caso, a proteína poderia ser constituída de peptídeos de
peso molecular próximo a 8,5kDa.
Observou-se, analisando o perfil polipeptidico das proteínas de membrana
dos espermatozóides X, uma proteína com peso molecular que se apresentava com
uma concentração em torno de 4 vezes superior a sua similar observada no
espermatozóide possuidor do cromossomo Y que, pelo presente está sob segredo
industrial, devido à importância econômica do mesmo e ao fato de ainda não ser
patenteado. Tal fato nos levou a acreditar ser esta uma proteína sexo específica
presente nos espermatozóides X. Tais resultados demonstram que a
imunossexagem de espermatozóides para produção de novilhas está totalmente
definida, enquanto que, para produção de tourinhos já possuímos uma perspectiva
bastante interessante. Tais fatos vêm ao encontro de uma necessidade premente do
agronegócio nacional, que é a produção destes reprodutores geneticamente
superiores para a melhoria do rebanho nacional.
Em relação à inseminação artificial, o fato de poder produzir fêmeas em muito
maior escala é fundamental para os produtores de leite. Este fato também é de
grande importância para os pecuaristas produtores de gado de ELITE para corte.
Estes poderão formar suas matrizes das quais surgirão os reprodutores de que o
mercado tanto necessita.

44
Figura 8 - Análise computadorizada do gel produzido a partir de proteínas de
membranas dos espermatozóides tratados pelo pool de anticorpos monoclonais.
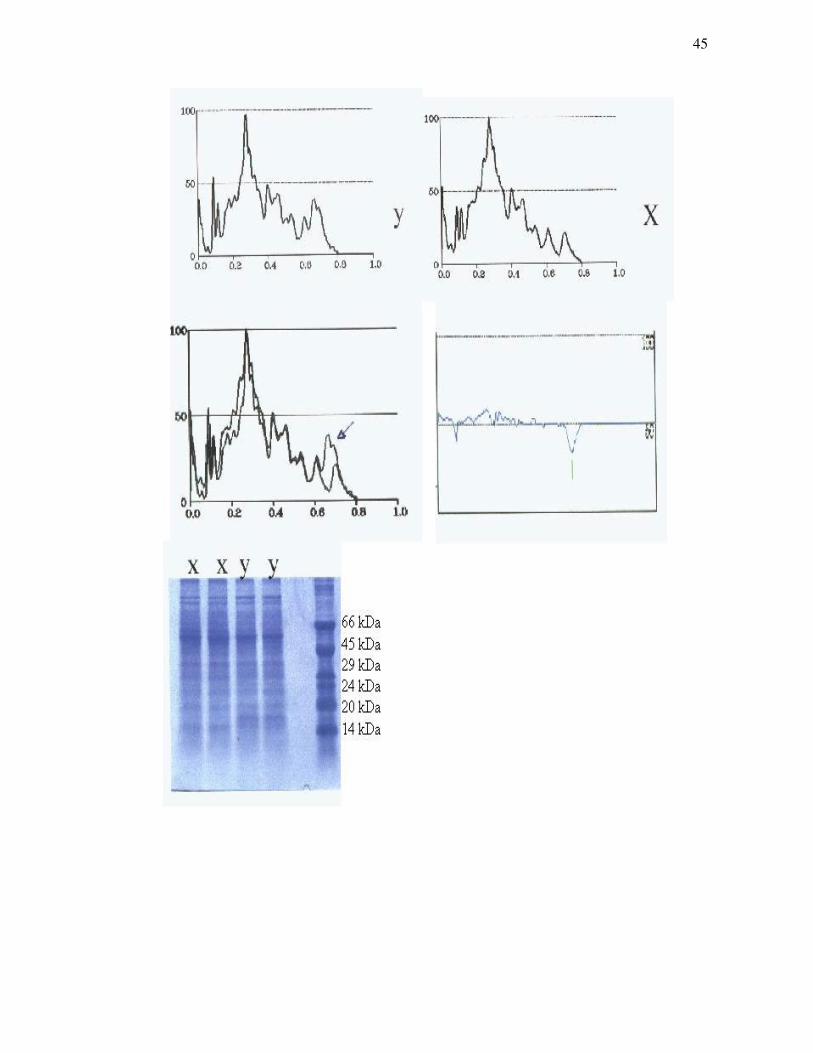
45

46
5 - CONCLUSÕES
Metodologias como a imunoafinidade usando colunas MiniMACS não foram eficientes para separar os
espermatozóides. A análise por PCR dos embriões formados a partir dos espermatozóides separados na coluna serviu para verificarmos que a coluna por si só não é capaz de sexar totalmente as células, provavelmente porque o anticorpo com ferro não foi capaz de se fixar a todas as células que apresentavam o anticorpo monoclonal macho-específico, fixado a elas.
A técnica de imunofluorescência, realizada após passagem pela coluna de afinidade, mostrou que as células da primeira eluição (células vivas) não ficavam fluorescentes enquanto que as células da segunda eluição (células mortas e que ficavam aderidas à coluna) apresentavam fluorescência. Com o objetivo de analisar melhor o efeito da fluorescência, realizou-se um experimento adicional em que todas as células foram mortas por congelamento. Nos questionamos se a fluorescência seria um bom parâmetro de sexagem. Sendo assim, adicionamos o anticorpo monoclonal macho-específico e fez-se a imunofluorescência. Todas as células ficaram fluorescentes, mostrando que a fluorescência, por si só, não é eficiente para a sexagem, pois todas as células mortas apresentavam-se fluorescentes, com maior ou menor intensidade esta fluorescência é devida à reversão de uma proteína do Complexo Maior de Histocompatibilidade, que ocorre quando da morte do espermatozóide, e não está ligada a nenhum fenômeno de apoptose. Desta forma, observamos que o anticorpo monoclonal macho-específico foi capaz de se ligar às duas populações de espermatozóides (X e Y).
Com a separação pelo gradiente de Percoll, obtivemos duas populações
distintas: células vivas e células mortas.
No início dos experimentos, tentamos separar os espermatozóides somente
com a utilização do anticorpo monoclonal macho-específico e coluna magnética. A
percentagem de motilidade era alta, indicando que os espermatozóides Y não
haviam sido totalmente eliminados. Passamos, então, a lavar o sêmen e adicionar
soro de cobaia para matar os espermatozóides Y. Os resultados foram melhores,
mostrando que a utilização do complemento é mais eficaz, sendo esta técnica muito
mais sensível do que a fluorescência, pois, além de observarmos fluorescência nas
células mortas, são necessários vários anticorpos fixados à superfície das células
para que ocorra a fluorescência, enquanto que com o complemento são necessários

47
apenas dois anticorpos fixados ao espermatozóide para que haja toda a reação em
cadeia das proteínas complemento.
Concluímos, então, que a utilização do soro de cobaia associado ao anticorpo
monoclonal com centrifugação pelo gradiente de Percoll foi uma metodologia mais
eficiente para separação dos espermatozóides. Futuramente, utilizaremos o
anticorpo monoclonal antiproteína X associado ao complemento. Desta forma,
esperamos eliminar os espermatozóides portadores do cromossoma X, enquanto
que somente aqueles portadores do cromossoma Y permanecerão vivos para
fecundar os oócitos e dar origem a embriões/bezerros machos.
6 - REFERÊNCIAS BIBLIOGRÁFICAS

48
ALBERTS, B., BRAY, D., LEWIS, J. RAFF, M., ROBERTS, K. WATSON, J.D. (1983)
Molecular Biology of the Cell. New York: Garland Publishing Inc., p.769-811.
AMANN, R. P. (1989) Treatment of sperm to predetermine sex. Theriogenology, v.31,
p.49-60.
AMANN, R. P.; HAMMERSTED, R. H.; VEERAMACHANENI, D. R.N., (1993) The
epididymis and sperm maturation. Reprod. Fertil. Dev., v.5, p.361-381.
BACCHUS, C.; BUSELMAIER, W. (1988) Blastomere karyotyping and transfer of chromosomally selected embryos: implications for the production of specific animal models and human prenatal diagnosis. Human Genetics,v. 80, p.333-336.
BENNETT, D.; BOYSE, E. A. (1973) Sex ratio in progeny of mice inseminated whit
sperm treated with H-Y antiserum. Nature, v.246, p. 308-309.
BETTERIDGE , K. J. (1984) The folklore of sexing. Theriogenology,v. 21 (1), p. 3-6.
BLECHER, S.R., HOWIE, R., LI, S., DETMAR, J., BLAHUT, L.M. (1999) A new
approach to immunological sexing of sperm. Theriogenology, v.52, p. 1309-
1321.
BILLINGHAN, R. E.; SILVERS, W. K. (1959) Inbred animals and tissue transplantation
immunity. Transplantation Bulletin, v.2, p.399-406.
BILLINGHAN, R. E.; SILVERS, W. K. (1960) Studies of tolerance of the Y chromosome
antigen in mice. J. Immunol., v.85, p. 14-16.
BRADLEY, M. P.; FORRESTER, I. T.; HESLOP, B. F. (1987) Identification of a male-
specific (H-Y) antigen on the flagellar plasma membrane of ram epididymal
spermatozoa. Human Genetics, v.75, p.362-367.

49
BREDBACKA, P. (1998) Recent development in embryo sexing and its field
application. Reprod. Nutr. Development, v. 38, p. 605-613
BRYANT B. J. (1980) Method and material for increasing the percentage of mammalian offspring of either sex. U. S. Patent, v.4, p.191.794.
BOOMAN, P. KRUIT, L., VEERHUIS, R., HENGST, A.M., TIEMAN, M., RUCH, F.E.
(1989) Sexing bovine embryos with monoclonal antibodies against the H-Y
antigen. Livestock Prod. Sci. v.23, p.1-16.
CHAI, C. K. (1968) The effect of inbreeding in rabbits. Transplantation, v.6, p.689-693.
CUNNINGHAM, E.P. (1998) The potencial of new reproductive and genetic
technologies. Acta Agric. Scand., Sect. A, Animal Sci., supl. 29, p.67-76.
DANIELL J.F. (1983) Sex-selection procedures. J. Reprod. Med. 1983 Apr; v.28(4),
p. 235-7.
DYM, M., FAWCETT, D.W. (1971) Further observations on the number of
spermatogonia, spermatocytes, and spermatids connected by intercellular
bridges in the mammalian testis. Biol. Reprod. v.28, p.195-215.
EICHWALD, E. J., SILMER,C.R., (1955) Comunicação não titulada, Transplant Bull,
v.2, p.148-149.
FORD, C. E.; JONES, K. W.; POLANI, P. E.; DE ALMEIDA, J. C.; Briggs, J. H.(1959) A
Sex-chromosome anomaly in a case of gonadal dysgenesis (Turner's syndrome).
Lancet, v.1, p.711-713.
FRAGOSO, C. (1994) Anticorpo H-Y e a determinação do sexo no rato. Tese (Mestrado em Ciências) - São Paulo - SP,
Universidade São Paulo - USP, 61p.
FRANCE, J. T.; GRAHAM, F. M.; GOSLING, L.; HAIR, P. I. (1984) A Prospective
study of the preselection of the sex of offspring by timing intercourse relative to
ovulation. Fertility and Sterility, v.41, p. 894-900.

50
GERSHWIN, L. J. (1981) Hybridomas: The production of monoclonal antibodies.
California Veterinarian, v.10, p. 31-33.
GOLDBERG, E.H., BOYSE, E.A., BENNETT, D., SCHEID,M., CARSWELL, E.A.
(1971) Serological demonstration of H-Y (male) antigen on mouse sperm.
Nature, v.232, p.478-480.
GONÇALVES, PB; VISINTIN, JÁ; OLIVEIRA, MA; MONTAGNER, MM; COSTA, LF
(2002) Biotécnicas Aplicadas à Reprodução Animal – Cap.10 pp. 195-226.
HAFEZ, E. S. E. (1982) Reprodução animal. 4a. ed., editora Manole, S. Paulo, p. 720.
HALL, J.L; WACHTEL, S. S. (1980) Primary Sex determination: Genetics and biochemistry. Mol. and Cell Biochem., v.33, p.49-66.
HALL, L.; BUSHKIN, Y.; WACHTEL, S. S. (1981) Immunoprecipitation of human H-Y
antigen. Human Genetics, v.58, p.34-36.
HENDRIKSEN, P.J.M. (1999) Do X and Y spermatozoa differ in proteins?
Theriogenology, v.52, p.1295-1307.
HOHENBOKEN, W. D. Applications of sexed semen in cattle production. Theriogenology, v. 52, p. 1421-1433.
HOLM, P, CALLESEN, H. (1998) In vivo versus in vitro produced bovine ova: similarities and differences relevant for pratical application. Reprod. Nut. Dev., v. 38, p. 579-594
HOPPE, P. C.; KOO, G. C. (1984) Reacting mouse sperm with monoclonal H-Y
antibodies does not influence the sex ratio of eggs fertilized in vitro. J. Reprod.
Immunol., v.6, p. 1-9.
HOSSAIN, A.M., BARIK, S., RIZK, B. (1998) Preconceptional sex selection: past,
present and future. Arch. Andrology, 40(1):3-14.

51
JACOBS P.A.; ROSS A. (1966) Structural abnormalities of the Y chromosome in man.
Nature, 210: 352-354.
JOHNSON, L. A.; FLOOK, J.P.; LOOK, M. V.; Pinkel, D. (1987) Flow sorting of X- and Y- chromosome bearing spermatozoa into two populations. Gamete Research.,v. 16, p. 1-9.
JOHNSON, L.A. (1992) Gender preselection in domestic animals using flow
cytometrically sorted sperm. J. Anim. Sci., v.70, supl.2, p.8-18.
JOHNSON, L. A.; SCHULMAN, J. D. (1994) The safety of sperm selection by flow
cytometry. Human Reproduction, 9: 758-759.
JOST, A: MAGRE, S.; AGELOPOULOU, R. (1981) Early stages of testicular
differentiation in the rat. Human Genetics, v.58, p. 59-63.
KELLEY, K. W.; LEWIN, H. A. (1986) Monoclonal antibodies: Pragmatic application of
immunology and cell biology. Journal Animal Science, v.63, p. 289-309.
KOHLER, G.; MILSTEIN, C. (1975) Continuous culture of fused cells secreting antibody
of predefined specificity. Nature (London), v.256, p. 495-497.
KOO, G. C.; WACHTEL, S. S.; BREG, W. R.; MILLER, O. J. (1977) Mapping the locus
of the H-Y antigen gene on the human Y chromosome. Science, v.198, p 940-942.
KOO, G. C.; GOLDBERG, C. L. (1978) A simplified techniques for H-Y typing. J.
Immuno. Methods, v.23, p.197-201.
KOO, G. C.; TADA, N.; HAMMERLING, U., (1981b) Monoclonal anti H-Y antibodies. In:
Monoclonal antibodies and T cell hybridomas. Perspectives and technical advances.
Amsterdam: Elsevier, p. 143-147.
LUKASA, T.; FRYNS, J. P.; VAN DEN BERGHE (1992) The role of the Y-chromosome
in sex determination. Genetic Counseling, v.3, p.1-11.

52
MCLAREN, A.; SIMPSON, E.; TOMONARI, K.; CHANDLER, P.; HOGG, H. (1984)
Male sexual differentiation in mice lacking H-Y antigen. Nature, 312: 552-555.
MCNUTT TL AND JOHNSON LA. (1996) Flow cytometric of sperm: Influence on
fertilization and embryo/fetal development in the rabbit. Molec Reprod Dev v.43,
p.261-267.
MCEVOY, J. D. (1992) Alteration of the sex ratio. Animal Breeding Abstracts, v. 60,
p. 601-606.
MIYAZACK, W.Y., ORR-WEAVER, T.L. (1994) Sister-chromatid cohesion in mitosis
and meiosis. Ann. Rev. Genet. v.28, p.167-187.
MOREIRA-FILHO, C. A.; OTTO, P. G.; FROTA PESSOA, O. (1979) H-Y antigen
expression in apparent absence of the longain of the Y chromosome. American
Journal Medicine Genetic, v.4, p. 135-139.
MÜLLER, U. (1983) Genetics of H-Y antigen and its gonad-specific receptor. Biblthca
Anat., v.24, p.111-118.
OAKBERG, E.F. (1956) A description of spermatogenesis in the mouse and its use in
analysis of the cycle of the seminiferous epithelium and germ cell renewal. Am.
J. Anal., v.99, p.391-413.
OHNO, S. (1976) Major regulatory genes for mammalian sexual development. Cell, v.7,
p. 315-321.
OHNO, S. (1977a) The Original Function of MHC Antigens as the General Plasma
Membrane Anchorage Site of Organogenesis – directing Proteins. Immunol. Ver.
1977 Jan; v. 33, p.59-69.

53
OHNO, S. (1977b) The Y-Linked H-Y Antigen Locus And The X- Linked Tfm Locus As
Major Regulatory Genes Of The Mammalian Sex Determining Mechanism. J. Steroid
Biochem., v.8, p. 585-592.
OHNO, S. (1979) In: Major Sex Determining Genes. Berlin, Spring-Verlag, p. 81-84.
OI, V. T.; HERZENBERG, L. A. (1980) Immunoglobulin secreting hybrid cell lines. In
Mishell, B. B. and S. M. Shiigi, (eds.) Selected methods in cellular immunology. San
Francisco: Freeman and Co., p. 351-372.
PAPA, F.; HENRION, R.; BREART, G. (1983) Preconceptional selection of fetal sex
using an ionic method. A dietary regime. Results of a prospective clinical study over
2 years. J. de Gynécologie, Obstétrique et Biologie de la Reproduction, v.12, p. 415-
422.
PETER, A. T.; JONES, P. P.; ROBINSON, J. P. (1993) Fraction of Bovine Spermatozoa
for Sex Selection: A Rapid Immunomagnetic Technique to remove spermatozoa that
contain the H-Y Antigen. Theriogenology, 40: 1177-1185.
RARY, J. M.; CUMMINGS, D. K.; JONES, H. W. JR.; ROCK, J. A. (1979) Assignment
of the antigen gene to the short arm of chromosome Y. Hered., 70: 78-80.
REED, K. C., MATTHEWS, M. E., JONES, M. A. (1988) Sex determination of ruminants
using Y-chromossome specific polynucleotides. Published Patent Application. Patent
Cooperative Treaty n°W088/01300.
REICHENBACH, HD; OLIVEIRA, MA; LIMA, PF; SANTOS FILHO, AS; ANDRADE,
JC (2002) Biotécnicas Aplicadas à Reprodução Animal – Cap.8 pp. 127-178.

54
RUSSEL, L.D., ETTLIN, R.A., HIKIM, A.P.S., CLEGG, E.D. (1990) Histological and
histopathological evaluation of the testis. Clearwater F.L.: Cache River Press,
p.49-53.
SEIDEL, G. R. Jr., JONSON, L. A. (1999) Sexing Mammalian Sperm – Overview.
Theriogenology, v.52, p. 1267-1272.
SOUZA, C.J.P (1998) Imunossexagem de Espermatozóides Bovinos. Tese de
doutorado em Produção Animal. Campos dos Goytacazes, R.J., Universidade
Estadual do Norte Fluminense – UENF. 102 p.
STOLOWSKI, J.; LORRAIN, J. (1983) Preconceptional selection of fetal sex.
International Journal of Gynecology and Obstetrics, 18: 440-443.
VEERHUIS, R., HENDRIKSEN, P.J.M., HENGST, A.M., KRUIJT, L., TIEMAN, M.;
BOOMAN, P. (1994)The production of anti-H-Y monoclonal antibodies: their
use in a sex test for bovine embryos. Vet. Immonol. Immunopathol., v.42, p.
317-330.
WACHTEL G.M.; WACHTEL, S.S.; Nakamura, D.;Moreira-Filho, C.A.; Brunner, M.;
Koo, G.C. H-Y antibodies recognize transplantation. 1984; 104:439-41.
WACHTEL, S. S, (1983) In: H-Y antigen and the biology of sex determination. Grune
and Stratton, New York.
WACHTEL, S. S. (1977) H-Y antigen: Genetics and serology. Immunology Review, 33:
3-58.
WACHTEL S. S.; KOO, G. C.; BOYSE E. A. (1975b) Evolutionary conservation of H-Y
(“MALE”) ANTIGEN. NATURE, 254: 270-272.
WACHTELL, S. S; OHNO G. C.; KOO, G. C., BOYSE, E. A. (1975c) Possible role for
H-Y antigen in the primary determination of Sex. Nature, 257: 235-236.

55
WELSONS, W. J. E; RUSSEL, L. B. (1959) The Y-chromosome as the bearer of
male determining factors in the mouse. Proc. Nat. Acad. Sci. (USA), 45: 560-
566.
WHELAN, E. M. (1974) Human sex ratio as a function of the timing of insemination
within the menstrual cycle: A review. Social Biology, 21: 379-384.
WIBERG, U.H. (1987) Revisão. Facts and considerations about Sex-specific
antigens. Human Genet. v.76, p.207-219.
WILD, D. (1994) The immunoassay handbook. New York, Stockton Press: 618 p.
WOLF, U. (1981) Genetic aspects of H-Y antigen. Human Genetics, 58: 25-28.
ZABORSKI, P. (1979) Detection of H-Y antigen on mouse sperm by the use of
Staphylococcus aureus. Transplantation, 27: 348-350.











![transp dinoflagelados - Maio 2010 [Modo de Compatibilidade]uenf.br/Uenf/Downloads/LBT_3660_1273062899.pdf · Filo Dinophyta • causadores das marés vermelhas causadores das marés](https://static.fdocumentos.com/doc/165x107/5a782cd87f8b9aa3618e88fc/transp-dinoflagelados-maio-2010-modo-de-compatibilidadeuenfbruenfdownloadslbt3660.jpg)






