
Detecção de genes de virulência em diferentes fagotipos e ...€¦ · CASTILLA, K. S. Detecção...
78
KARINA SALVAGNI CASTILLA Detecção de genes de virulência em diferentes fagotipos e ribotipos de Salmonella Enteritidis utilizando a Reação em Cadeia da Polimerase (PCR) São Paulo 2003
Transcript of Detecção de genes de virulência em diferentes fagotipos e ...€¦ · CASTILLA, K. S. Detecção...

KARINA SALVAGNI CASTILLA
Detecção de genes de virulência em diferentes fagotipos e ribotipos de Salmonella Enteritidis utilizando a Reação em
Cadeia da Polimerase (PCR)
São Paulo 2003

KARINA SALVAGNI CASTILLA
Detecção de genes de virulência em diferentes fagotipos e ribotipos de Salmonella Enteritidis utilizando a Reação em
Cadeia da Polimerase (PCR)
São Paulo 2003
Dissertação apresentada paraobtenção do título de Mestre,junto à Faculdade de MedicinaVeterinária e Zootecnia daUniversidade de São Paulo. Departamento: Medicina Veterinária Preventiva eSaúde Animal Área de concentração: Epidemiologia Experimental eAplicada às Zoonoses. Orientador: Antonio José Piantino Ferreira



FOLHA DE APROVAÇÃO
Nome do autor: CASTILLA, Karina Salvagni
Título: Detecção de genes de virulência em diferentes fagotipos e ribotipos de
Salmonella Enteritidis utilizando a Reação em Cadeia da Polimerase (PCR)
Aprovado em:
Banca Examinadora
Prof. Dr. ___________________ Instituição: __________________________
Julgamento: ___________________ Assinatura: __________________________
Prof. Dr. ___________________ Instituição: __________________________
Julgamento: ___________________ Assinatura: __________________________
Prof. Dr. ___________________ Instituição: __________________________
Julgamento: ___________________ Assinatura: __________________________
Dissertação apresentada àFaculdade de MedicinaVeterinária e Zootecnia daUniversidade de São Paulo,para obtenção do título deMestre.

AGRADECIMENTOS
À Deus, pelo dom da vida, pela minha saúde física e mental, e pelas oportunidades
que tem me oferecido.
Aos meus pais, pelo amor, dedicação e ensinamentos durante a minha educação e
pela oportunidade de eu poder ter escolhido esta profissão.
Ao meu marido pelo apoio nas horas difíceis, pela sua ajuda e paciência incansável
durante a elaboração desta dissertação.
À CAPES, pela bolsa que me foi concedida.
À Faculdade de Veterinária e Zootecnia na Universidade de São Paulo, pela
oportunidade de aperfeiçoar os meus conhecimentos.
Ao meu orientador Prof. Dr. Antonio José Piantino Ferreira pela oportunidade,
orientação, amizade e confiança sobre mim depositada.
À Prof. Dra. Claudete Serrano Astolfi Ferreira, que me ajudou na condução deste
trabalho, sempre me ensinando e mostrando o caminho mais correto.
À Prof. Dra. Iolanda A Nunes por ter gentilmente cedidos as amostras de
Samonellas.
Ao laboratório Biovet pelo fornecimento das aves para a realização do experimento.
À Prof. Dra. Maria Regina Baccaro pela utilização do laboratório de Sanidade Suína.
À laboratorista Maria José Aparecida Louro, grande amiga, que com seu carinho e
ajuda tornaram as atividades agradáveis no laboratório.
À Dra. Andréa Micke Moreno, as colegas Renata Paixão e Daniela Sabatini Doto,
pela dedicação, paciência e, sobretudo, pela amizade e ajuda durante a elaboração
do PCR, possibilitando ótimos e agradáveis momentos de convivência.

Aos meus colegas de laboratório, Amarílis Novaes Délboux, Laura Villareal Buitrago,
Marcelo Ribeiro Silva Pequini, Marco Aurélio Elmer Lopes, Rofson Falcão, Ronaldo
Santos de Paço e Terezinha Knöbl que sempre me ajudaram, principalmente nas
colheitas de material.
Ao funcionário Paulo Sérgio de Souza pela ajuda no manejo das aves.
OBRIGADA

CASTILLA, K. S. Detecção de genes de virulência em diferentes fagotipos e ribotipos de Salmonella Enteritidis utilizando a Reação em Cadeia da Polimerase (PCR). [Virulence genes detection in different phage types and ribotypes of Salmonella Enteritidis by Polimerase Chain Reaction (PCR)]. 2003. 77 f. Dissertação (Mestrado em Epidemiologia Experimental e Aplicada às Zoonoses) – Faculdade de Medicina Veterinária e Zootecnia, Universidade de São Paulo, São Paulo, 2003.
RESUMO
O objetivo deste estudo foi o de verificar a ocorrência de quatro genes de virulência
em Salmonella Enteritidis de acordo com o fagotipo e ribotipo, assim como a
virulência “in vivo”. Os genes estudados foram invA, spvC, sefC e pefA em 120
amostras isoladas de várias fontes, pertencentes a diferentes fagotipos e ribotipos
provenientes de sete estados brasileiros. Para a verificação da presença dos genes,
as amostras foram examinadas pela técnica de Reação em Cadeia da Polimerase
(PCR) individualmente e em multiplex. A ocorrência para o gene invA foi de 100%
nas amostras (120/120), spvC em 94% (113/120), sefC em 97,5% (117/120) e pefA
em 97% (116/120) das amostras. Foi possível caracterizar as amostras em cinco
diferentes perfis. O primeiro, P1, foi positivo para os genes invA, spvC, sefC e pefA,
P2 para invA, spvC e pefA, P3 para invA, sefC e pefA, P4 para invA e sefC e o
último P5 para os genes invA e spvC. Para as amostras PT4RT1 obteve-se o perfil
P1 em 94% (64/68), P2 em 1,5% (1/68), P3 em 3% (2/68) e P4 em 1,5% das
amostras (1/68). Para as amostras PT4RT2, 86% (18/21) pertenceram ao perfil P1,
5% (1/21) ao P3 e 9% (2/21) das amostras ao perfil P4. Nas amostras PT4RT3 80%
(4/5) pertenceram ao perfil P1 e 20% (1/5) ao P2. Nas amostras PT4TR9, 91%
(10/11) pertenceram ao perfil P1 e 9% (1/11) ao P3. Todas as amostras PT4RT5,

PT7RT1 e PT4RT10 pertenceram ao perfil P1 e apenas a amostra PT1 apresemtou
o perfil P5. A virulência foi avaliada desafiando as aves via oral e subcutânea,
através da colonização do ceco e invasão do fígado e baço. O gene spvC está
relacionado com a sobrevivência e aumento da média de crescimento da bactéria no
fígado e baço, mas neste estudo não demonstrou diferença na porcentagem de aves
com reisolamento positivo para a bactéria nestes orgãos. Os genes sefC e pefA
foram importantes para promover a colonização cecal, pois somente quando estes
dois genes simultaneamente estiveram presentes no perfil, obteve-se o
reisolamento cecal quando as aves foram desafiadas oralmente sendo esta via a
principal rota de infecção deste patógeno.
Palavras chaves: Salmonella. Genes. Virulência. PCR.

CASTILLA, K. S. Virulence genes detection in different phage types and ribotypes of Salmonella Enteritidis by Polimerase Chain Reaction (PCR). [Detecção de genes de virulência em diferentes fagotipos e ribotipos de Salmonella Enteritidis utilizando a Reação em Cadeia da Polimerase (PCR)]. 2003. 77 f. Dissertação (Mestrado em Epidemiologia Experimental e Aplicada às Zoonoses) – Faculdade de Medicina Veterinária e Zootecnia, Universidade de São Paulo, São Paulo, 2003.
ABSTRACT
This study has the intention of verify the four virulence genes event in Salmonella
Enteritidis according with phage type and ribotype and the virulence “in vivo”. The
genes studied were invA, spvC, sefC and pefA in 120 Salmonella Enteritidis isolates
from different origins, belongs at different ribotypes and phage types of seven
Brazilian states. To verify the genes presence the strains were examined by
Polimerase Chain Reaction (PCR) technique, singly and multiplex. The event of invA
gene was in 100% (120/120) of strains, the spvC gene was in 94% (113/120) of
strains, the sefC gene was in 97.5% (117/120) of strains and the pefA gene was in
97% (116/120) of strains. There were discovered five different profiles. The pattern
one, P1, was positive for invA, spvC, sefC e pefA. The P2 was positive for invA, spvC
and pefA. The P3 was positive for invA, sefC e pefA. The P4 was positive for invA
and sefC and the last one, P5, was positive for invA and spvC. For the PT4RT1
strains, the P1 profile was present in 94% (64/68) of strains; P2 profile was in 1.5%
(1/68); P3 in 3% (2/68); and P4 in 1.5% (1/68) of strains. For the PT4RT2 strains, the
P1 profile was present in 86% (18/21) of strains; P3 profile was in 5% (1/21); and P4
in 9% (2/21) of strains. For the PT4RT3 strains, the P1 profile was present in 80%
(4/5) of strains, and P2 in 20% (1/5) of strains. For the PT4RT9 strains, the P1 profile

was present in 91% (10/11) of strains, and P3 in 9% (1/11) of strains. The strains:
PT4RT5, PT7RT1 e PT4RT10 belongs at the P1 profile, and only the PT1 strain
belongs at the P5’s profile. The virulence was evaluated challenging the birds orally
and subcutaneous, through the colonization of the caecum, liver and spleen invasion.
The gene spvC was related with the survivance and the increase of the average grow
of the bacteria in the liver and spleen, but this study doesn’t demonstrate the
difference in the percentage of positive birds for the bacteria in their organs. The sefC
and pefA genes were important to promote the caecum colonization, because only
when both genes were present simultaneously in the profile, obtain the caecum
isolation when the birds were been by orally challenged this way the main route of
the infection of this pathogen.
Key words: Salmonella. Genes. Virulence. PCR.

LISTA DE FIGURAS
Figura 1 - Eletroforese em gel de agarose 1,5% corado com
brometo de etídeo, mostrando os produtos de
amplificação para os genes invA, spvC, sefC e pefA
....................................................................................... 52

LISTA DE QUADROS
Quadro 1 - Descrição das amostras de Salmonella Enteritidis,
segundo o fagotipo, ribotipo, estado, ano e fonte de
isolamento..................................................................... 43
Quadro 2 - Descrição dos primers utilizados para os genes invA,
spvC, sefC e pefA ........................................................ 47

LISTA DE TABELAS
Tabela 1 - Aves SPF desafiadas pela via oral e via subcutânea, com
os cinco perfis de Salmonella Enteritidis relacionados aos
genes invA, spvC, sefC e pefA .................................... 49
Tabela 2 - Cinco perfis de Salmonella Enteritidis relacionando o
fagotipo, ribotipo e fonte de isolamento quanto a
presença ou ausência dos genes invA, spvC, sefC e pefA
....................................................................................... 51
Tabela 3 - Resultado do reisolamento de Salmonella Enteritidis em
fígado, baço e ceco de pintos SPF desafiados por via
oral................................................................................. 53
Tabela 4 - Resultado do reisolamento de Salmonella Enteritidis em
fígado, baço e ceco de pintos SPF desafiados por via
subcutânea.................................................................... 55

LISTA DE ABREVIATURAS
DNA Ácido desoxirribonucléico
DnTP Deoxirribonucleotídeo
EDTA Ácido etilenodiaminotetracético
kb Kilobase
kDa KiloDalton
M Molar
MDa MegaDalton
mg Miligrama (10-3 grama)
ml Mililitro (10-3 grama)
mM Milimolar (10-3 grama)
pb Pares de base
PCR Reação em Cadeia da Polimerase
pH Potencial hidrogeniônico
Primers Oligonucleotídeos
PT Fagotipo
RT Ribotipo
UFC Unidades Formadoras de Colônias
V Volt
Xg Força centrífuga
µm Micrômetro (10-6 metro)
µl Microlitro (10-6 metro)

SUMÁRIO
1 REVISÃO DA LITERATURA............................................................................17
1.1 Caracterização do gênero Salmonella.............................................................17
1.2 Salmonelose....................................................................................................18
1.3 Salmonella Enteritidis em Saúde Pública........................................................21
1.4 Fatores de Virulência de Salmonella Enteritidis..............................................25
1.4.1 Gene invA........................................................................................................26
1.4.2 Gene spvC.......................................................................................................30
1.4.3 Gene sefC........................................................................................................35
1.4.4 Gene pefA........................................................................................................39
1.5 Detecção dos genes de virulência pela Reação em Cadeia da Polimerase
(PCR) ........................................................................................................................ 40
2 OBJETIVOS.....................................................................................................43
3 MATERIAIS E MÉTODOS ..............................................................................44
3.1 Cultivo Bacteriano............................................................................................44
3.2 Extração do DNA Bacteriano...........................................................................47
3.3 Amplificação do DNA bacteriano.....................................................................48
3.4 Detecção do produto amplificado....................................................................49
3.5 Infecção experimental em aves SPF...............................................................50
4 RESULTADOS.................................................................................................52
4.1 Amplificação do DNA bacteriano utilizando a PCR.........................................52
4.2 Infecção Experimental em aves SPF desafiadas pela via oral........................54
4.3 Infecção Experimental em aves SPF desafiadas pela via subcutânea...........55
5 DISCUSSÃO ....................................................................................................57

6 CONCLUSÃO..................................................................................................64
REFERÊNCIAS...............................................................................................65

17
1 REVISÃO DA LITERATURA
1.1 Caracterização do gênero Salmonella
As bactérias do gênero Salmonella pertencem à família Enterobacteriaceae,
são bastonetes gram negativos, anaeróbios facultativos medindo de 0,7 a 1,5 µm por
2,0 a 5,0 µm. A maioria dos sorotipos é móvel devido à presença de flagelos
peritríquios, crescem em temperatura ótima de 37°C, mas já foram observados
crescimentos em temperaturas entre 5 e 45°C, sendo o pH 7 ideal, mas podendo
variar entre pH 4 e pH 9. Produzem gás a partir da fermentação da glicose e gás
sulfídrico, são capazes de descarboxilar os aminoácidos lisina e ornitina, reduzir
nitratos a nitritos e utilizar citrato como fonte de carbono (GAST, 1997).
Segundo Popoff e colaboradores (1998) este gênero divide-se em duas
espécies, Salmonella enterica e Salmonella bongori. Salmonella enterica por sua vez
subdivide-se em seis subespécies, ou seja, enterica, salamae, arizonae, diarizonae,
houtenae e indica, enquanto Salmonella bongori tem como única subespécie
bongori.
Taxonomicamente a classificação proposta por Popoff e Le Minor (1997) é:
gênero, espécie, subespécie e sorotipo. Desta forma a grafia para o sorotipo
Enteritidis fica descrita como: Salmonella enterica subespécie enterica sorotipo
Enteritidis, mas deste ponto em diante esta denominação será abreviada para
gênero e sorotipo, ou seja, o agente será descrito como Salmonella Enteritidis.
A identificação da espécie ou sorotipo de Salmonella é realizada através de
métodos como: perfil bioquímico e sorológico, resistência a antimicrobianos,

18
produção ou sensibilidade a bactericinas, fagotipagem, perfil plasmidial e perfil de
restrição a endonucleases (DOYLE, 1990).
1.2 Salmonelose
Salmonellas são patógenos intracelulares que habitam o trato gastrointestinal
de várias espécies animais e do homem, resultando em diferentes manifestações da
doença, incluindo febre entérica, bacteremia e gastroenterites (HENDERSON et al.,
1999).
A doença causada por Salmonella é genericamente chamada de salmonelose
(DOYLE, 1990). Alguns sorotipos estão associados com maior freqüência ou mais
adaptados a determinados hospedeiros. É o caso da Salmonella Cholerasuis que
causa septicemia, pneumonia e enterocolite em suínos; Salmonella Dublin causa
enterite mucohemorrágica, septicemia, pneumonia e aborto em bovinos e
septicemia, enterite necrótica, pneumonia e mortalidade em bezerros; Salmonella
Abortusequi causa aborto em eqüinos; Salmonella Abortusovis causa aborto em
ovinos; Salmonella Gallinarum causa febre tifóide em galinhas e Salmonella
Pullorum pulorose em galinhas e perus. Sorotipos adaptados ao homem incluem
Salmonella Typhi causando febre tifóide e Salmonella Paratyphi causando febre
paratifóide. Salmonella Enteritidis faz parte dos mais de 2300 sorotipos de
Salmonellas que não são adaptadas a hospedeiros específicos chamadas
paratíficas, causando salmonelose em humanos e em uma ampla variedade de
animais (POPPE, 1999).
Em humanos Salmmonella Enteritidis possui um período de incubação
variando de poucas há 72 horas e a duração da doença é entre 4 a 10 dias.

19
Normalmente em surtos desse sorotipo são observados sinais e sintomas como
diarréia com cólicas, cefaléia, calafrios, dor abdominal, náusea, febre e vômito
(INFORME TÉCNICO, 1996; PASSARO et al., 1996).
Segundo Bryan e Doyle (1995) a ingestão de 10 UFC dessa bactéria podem
causar a doença em humanos adultos.
As aves comerciais constituem um dos mais importantes reservatórios de
Salmonellas, e são responsáveis pela introdução da mesma na alimentação
humana, através da contaminação da carne e dos ovos, principalmente por
Salmonella Enteritidis (GAST, 1997). Nas granjas a bactéria é introduzida através de
rações contaminadas por matérias primas principalmente as de origem animal, como
farinhas de carne, ossos, peixe, penas e vísceras (HOFER et al., 1998), pela água,
contato com aves ou animais portadores, vetores como insetos, roedores, pessoas e
equipamentos (BORLAND, 1975; GAST, 1998). Entre os anos de 1979 a 1991 o
Laboratório de Enterobactérias do Instituto Oswaldo Cruz (IOC) diagnosticou 2.293
amostras de Salmonella isoladas de rações e seus insumos (HOFER et al., 1998).
A bactéria pode sobreviver no feno e meio ambiente por poucos dias a várias
semanas e sua sobrevivência na água é maior na presença de matéria orgânica
(BRYAN e DOYLE, 1995).
Segundo Nakamura et al. (1997), existem três mecanismos de transmissão
horizontal de Salmonella: através da água, comida e aerossóis. Estes autores
sugerem que a rapidez da transmissão horizontal de Salmonella Enteritidis pode ser
afetada por padrões de fluxo de ar. Gast e colaboradores (1998) verificaram a
transmissão horizontal de Salmonella Enteritidis através do ar e observaram que o
isolamento da bactéria foi maior nas penas das aves (77%) do que nos órgãos
(33%).

20
A contaminação do ovo ocorre através da contaminação da ave no ovário
pela presença da bactéria nos folículos ovarianos contaminando a gema, através da
presença no oviduto resultando na presença da bactéria na albumina e
conseqüentemente no ovo durante a sua formação, ou pode ocorrer à possibilidade
do ovo ser contaminado após a formação da casca, onde a bactéria presente nas
fezes penetra através da casca quando ocorrer o contato ao passar pela cloaca ou
no meio ambiente (BARROW, LOVELL,1991; MC ILROY, 1989; SHIVAPRASAD et
al., 1990; STOKES, 1956).
Durante a matança e o processamento dos frangos, a presença de
Salmonella (incluindo o sorotipo Enteritidis) no conteúdo intestinal, pele e penas das
aves resulta em conseqüente contaminação da carne e seus subprodutos (BRYAN,
DOYLE, 1995). A contaminação cruzada pode ocorrer principalmente em mercados
e cozinhas industriais e a refrigeração imprópria do produto leva a bactéria a
multiplicar-se rapidamente (DUGUID, NORTH, 1991).
Costa et al. (1997) analisaram amostras de carcaças durante o ano de 1996
colhidas de abatedouros com controle sanitário permanente, abatedouro sem
controle permanente e no mercado varejista no estado de São Paulo, sendo que
eles encontraram a seguinte positividade para Salmonella: 8,3%, 13,3% e 35%
respectivamente.
A detecção de Salmonella Enteritidis em aves adultas através da
sintomatologia torna-se difícil, pois geralmente mesmo contaminadas as aves são
clinicamente normais e tornam-se portadoras disseminando a doença. Este sorotipo
atinge mais aves até duas semanas de idade, podendo causar uma septicemia
aguda. Quando atinge clinicamente aves adultas pode causar anorexia, diarréia,
diminuição na produção e eclosão devido à mortalidade embrionária e

21
ocasionalmente pericardite, hepatite e esplenomegalia. (GAST, 1997; POPPE et al.,
1993)
1.3 Salmonella Enteritidis em Saúde Pública
Até meados da década de oitenta, Salmonella Enteritidis era isolada de
humanos e animais com uma freqüência considerada de baixa a média na maioria
dos países. No final dos anos oitenta ocorreu um dramático aumento na prevalência
deste sorotipo isolado de infecções em humanos ultrapassando Salmonella
Typhimurium que, até então, ocupava o primeiro lugar. Esta mudança ocorreu
inicialmente em países da Europa e depois em países das Américas (POPPE, 1999;
RODRIGUE et al., 1990).
A susceptibilidade e a severidade da infecção em humanos depende de
fatores como a dose infectante do patógeno, o estado imune e idade do hospedeiro.
A doença não é considerada grave em indivíduos imunocompetentes, porém
pacientes imunodeficientes principalmente aqueles com Síndrome da
Imunodeficiência Adquirida (AIDS), desnutridos, crianças e idosos são mais
susceptíveis devido ao sistema imune estar menos efetivo, podendo em alguns
casos evoluir para óbito (GRUENEWALD et al., 1994; MILDVAN et al., 1982). A
infecção pode evoluir para uma septicemia e causar várias seqüelas como
pericardite, doenças neurológicas e neuromusculares, artrites, espondilites e
osteomielites (POPPE, 1999).
As infecções foram freqüentemente associadas com o consumo de carne de
frango e ovos, nesse último caso principalmente quando consumidos crus ou
insuficientemente cozidos na composição de alimentos como, por exemplo, em

22
doces (BICHILER et al., 1994; DUGUID, NORTH, 1991; SCHIMID,
BAUMGARTNER, 1999; SCUDERI, 1999; THIEL, 1999). Salmonella Enteritidis é
encontrada também em outras fontes de alimentação humana, porém em menor
freqüência, como nas carnes suína e bovina (ÂNGULO, SWERLOW, 1999), no leite
cru (WOOD et al., 1991), na carne de peru e produtos lácteos (PERALES,
AUDICANA 1989).
Estudos controlados de casos esporádicos de Salmonella Enteritidis
constataram que ovos crus ou semicrus são os mais importantes fatores de risco de
infecção em humanos (HELDBERG et al., 1993; MORSE et al., 1994; PASSARO et
al., 1996). Segundo Perales e Audicana (1989) a maioria dos surtos de Salmonella
ocorridos na Espanha são transmitidos por ovos.
Na Holanda as aves comerciais constituem o principal reservatório de
Salmonella Enteritidis, apesar dela já ter sido isolada de outras espécies animais
como suínos, ovinos, eqüinos, bovinos, cães, gatos, pássaros, roedores e répteis, no
entanto estas espécies representam somente 1% do total dos isolamentos. O
aumento deste sorotipo em aves comerciais desde 1988 coincide com o aumento de
isolamentos do mesmo em humanos, evidenciando serem as aves a principal fonte
de infecção para humanos, sendo a prevalência de setecentos casos para cada cem
mil habitantes, principalmente entre crianças de zero a cinco anos de idade (VAN DE
GIESSEN et al., 1999).
Na Inglaterra e País de Gales ocorreu um aumento na incidência do sorotipo
Enteritidis isolado em aves a partir do ano de 1985, onde a ocorrência, que era de
3,3%, foi para 48,3% em 1989. Neste período ocorreu também um aumento no
número de isolamentos de toxiinfecções humanas por este sorotipo, sendo o
fagotipo 4 o principal responsável pelas infecções (WALL, WARD, 1999)

23
Na Dinamarca os sorotipos encontrados em pacientes predominam em aves
comerciais e estão praticamente ausentes nas carnes suína e bovina. Na última
década houve um aumento da incidência de salmonelose humana, em 1994 foram
isolados em torno de oitenta e dois casos para cada cem mil habitantes, sendo que
em 1995 somente Salmonella Enteritidis teve um número de casos de quarenta e um
para cada cem mil habitantes e em 1996 a salmonelose foi a zoonose mais
reportada (GERNER-SMIDT, WEGENER 1999).
Na Alemanha a maioria dos surtos e casos esporádicos de Salmonellla
Enteritidis têm sido associados com o consumo de ovos de galinha. Neste país, em
1984, houve um aumento significativo no número de surtos causados por Salmonella
Enteritidis, porém desde 1992 este número vem diminuindo. Ainda assim estima-se
que os casos de infecção em humanos variem entre um a dois milhões por ano
(TSCHÄPE et al., 1999).
Na Escócia o Scottish Centre for Infecction and Environmrntal Health (SCIEH)
demonstrou a importância de Salmonella Enteritidis isolada numa investigação feita
como parte do programa de toxiinfecções de origem alimentar na Europa durante o
período de 1990 a 1994 (MUNRO et al., 1999).
A salmonelose foi a doença mais comum de origem alimentar nos Estados
Unidos no final da década de oitenta, sendo que em 1990 Salmonella Enteritidis foi o
sorotipo mais isolado (RODRIGUE et al., 1990). Há uma estimativa de que ocorram
oitocentas mil a quatro milhões de infecções humanas por Salmonella (trezentos a
mil e quinhentos casos para cada cem mil pessoas) por ano, sendo que em 1996
somente pelo sorotipo Enteritidis ocorreram duzentas e cinquenta mortes. A bactéria
foi isolada com maior freqüência em pessoas idosas e especialmente em crianças
menores de cinco anos de idade (ÂNGULO, SHERDLOW 1999).

24
A ocorrência de Salmonella Enteritidis cresceu nas últimas duas décadas na
França, o número de casos praticamente dobraram de 1986 para 1996, sendo que a
incidência da doença foi maior em crianças com menos de cinco anos de idade do
que em pacientes mais velhos (GRIMONT et al., 1999).
Na década de oitenta a prevalência deste sorotipo também aumentou na
Suíça, ocorrendo um pico epidêmico em 1992 com setenta e nove casos da doença
para cada cem mil habitantes, porém o número de casos diminuiu até o final dos
anos noventa (SCHIMID, BAUMGARTNER, 1999).
Na Áustria, Salmonella Enteritidis teve proporções epidêmicas a partir do ano
de 1988, com um pico da doença em 1992 e um decréscimo da mesma entre 1993 a
1995, sendo que até o final da década de noventa permaneceu em um nível estável
(THIEL,1999).
Salmonella Enteritidis foi isolada em 3 a 4% de casos humanos nos anos
oitenta e aumentou para mais de 30% na década de noventa na Itália (BINKIN et al.,
1993).
Segundo Wong e pesquisadores (1994) a prevalência de Salmonella em
Hong Kong foi estimada em dois mil casos por ano e um aumento significante do
sorotipo Enteritidis foi observado no final dos anos oitenta e início dos anos noventa.
Taunay e colaboradores (1996) identificaram em São Paulo, no período de
1950 a 1990, um total de 45.862 cepas de Salmonella, 31.517 isoladas de infecções
humanas e 14.345 isoladas de material não humano, sendo que, Salmonella
Enteritidis correspondeu a um percentual de 0,4% dos isolamentos.
No período de 1991 a 1995 Tavechio et al. (1996) avaliaram 5.490 amostras
de Salmonellas isoladas no estado de São Paulo. Foram isoladas 2.254 cepas de
origem humana e 3.236 cepas de origem não humana de diferentes sorotipos, sendo

25
que Salmonella Enteritidis correspondeu a 1,2% dos isolados em 1991, 2% em
1992, 43,3% em 1994 e 64,5% em 1995.
Lírio e colaboradores (1998) verificaram que a presença de Salmonella
Enteritidis isolada de alimentos pelo Laboratório de Controle de Alimentos de São
Paulo foi de 70,6%, sendo que o maior número de isolamentos foi em frango “in
natura” e lingüiça crua.
Segundo Hofer e pesquisadores (1997), os dados no Brasil que abordam a
ocorrência de sorotipos de Salmonella são exíguos, tanto em relação às situações
clínicas, quanto às epidemiológicas. Eles analisaram no período de 1962 a 1991,
2.123 amostras de Salmonellas, onde 87,21% das mesmas foram isoladas de Gallus
gallus de diversos estados brasileiros, sendo que o sorotipo Enteritidis passou de
freqüente no período de 1972 a 1981 para muito freqüente no período de 1982 a
1991.
Entre os anos oitenta e noventa o Centro de Referência Nacional de Cólera e
outras Enteroinfecções Bacterianas (FIOCRUZ) mostrou um aumento na incidência
de Salmonella Enteritidis isolada de origem humana (QUINTAES et al., 1997)
1.4 Fatores de Virulência de Salmonella Enteritidis
Durante o período em que Salmonella leva para entrar no trato gastroinstestinal
até ser fagocitada por macrófagos ela enfrenta uma variedade de defesas do
organismo hospedeiro. Os fatores de virulência expressados por bactérias
patogênicas freqüentemente ajudam as mesmas a sobreviver em condições hostis
como temperatura elevada, pH ácido, diminuição da tensão de oxigênio e privação
de nutrientes (VALONE et al., 1993)

26
A virulência da Salmonella é altamente complexa, necessitando a expressão
de numerosos genes, os quais codificam toxinas, adesinas, invasinas ou outros
fatores de virulência. Estes fatores podem estar localizados em elementos genéticos
transmissíveis, como, os “transposons”, plasmídeos ou bacteriófagos, assim como
podem fazer parte de regiões especificas particulares no cromossoma da bactéria,
chamadas de Ilhas de patogenicidade. Estas Ilhas estão presentes no genoma de
sorotipos patogênicos de determinadas espécies, mas ausentes ou raramente
presentes em variantes não patogênicas da mesma espécie (HACKER et al., 1997).
1.4.1 Gene invA
Um dos primeiros passos no ciclo de patogenicidade da bactéria intracelular
facultativa Salmonella sp. é a invasão de células do epitélio intestinal, como um fator
essencial de virulência no processo de infecção. As bases da genética molecular
para esta invasão parecem ser complexas, e muitos genes estão envolvidos neste
processo. A maioria destes genes está localizada entre 58 e 60 minutos no
cromossoma da bactéria, em um local denominado de SPI-1 (Salmonella
Pathogenicity Islands - 1) (GINOCHIO et al, 1994; HACKER et al., 1997; PORTER,
CURTISS III, 1997).
Ilhas de patogenicidade são formadas por seqüências específicas de DNA
bacteriano, que codificam determinantes de virulência responsáveis por estabelecer
interações específicas com o hospedeiro (MARCUS et al., 2000). Um total de cinco
Ilhas tem sido descrito para Salmonella enterica: SPI-1 onde os genes inv, spa e hil
são necessários para auxiliar na invasão da bactéria em células não fagociticas,
como as células epiteliais da mucosa intestinal (HACKER et al, 1997), SPI-2, SPI-3 e

27
SPI-4 onde seu genes são primariamente requeridos para o crescimento e
sobrevivência da bactéria com o hospedeiro manifestando a fase sistêmica da
doença e por último a SPI-5 que codifica fatores de virulência envolvidos na
inflamação e secreção de cloretos caracterizando a fase entérica da doença
(MARCUS et al., 2000).
As Ilhas de patogenicidade SPI-1 e SPI-2 codificam um sistema de secreção
tipo III que são estruturais e funcionalmente distintos (MARCUS et al., 2000). SPI-1
transporta determinantes de virulência chamados de proteínas efetoras, as quais são
secretadas dentro ou fora da ilha, junto às células eucarióticas suceptíveis do
hospedeiro modulando suas funções e permitindo a entrada nas mesmas (ZHOU,
GÁLAN, 2001). As proteínas efetoras subvertem as funções normais da célula
hospedeira, beneficiando assim a invasão pela bactéria (MECSAS, STRAUSS
1996). As proteínas efetoras de SPI-2 são necessárias para a multiplicação e
sobrevivência em macrófagos e causar infecção sistêmica (MARCUS et al., 2000).
Gálan e Curtiss (1989) identificaram uma região com quatro genes para
Salmonella Typhimurium, invA, invB, invC e invD, que possuem a capacidade de
complementar a invasão no trato intestinal, demonstrando que mutações nesses
genes afetaram a virulência desse sorotipo, pois quando administrados oralmente
(rota natural de entrada da bactéria) impediram a invasão da bactéria diminuindo a
capacidade de colonizar as placas de Peyer, a parede do intestino delgado, o baço e
aumentaram a dose letal (LD50) para camundongos. Quando este mesmo mutante
foi administrado intraperitonealmente não demonstrou nenhuma alteração na DL50
ou na capacidade de colonizar o baço.
Gálan e Curtis III (1991) concluíram em trabalho realizado com diferentes
sorotipos de Salmonella, incluindo Salmonella Enteritidis, que os sorotipos que não

28
possuíam o gene invA foram incapazes de expressar invABC e foram deficientes
para invadir células de mamíferos (Henle-407), indicando que o “operon” invABC
teve um papel importante na invasão em células epiteliais. O gene invA é o primeiro
no “operon”, onde os genes invA, -B e -C estão arranjados na mesma unidade
transcricional e o gene invD está localizado em uma unidade transcricional diferente
(GALÁN et al., 1992).
Galán e colaboradores (1992) conceituaram que a função do gene invA foi a
de aumentar a capacidade de invasão de Salmonella Typhimurium em células
epitelias. Os autores concluiram que ele foi essencial para a invasão fenotípica, pois
mutantes deste gene foram deficientes na invasão de células epiteliais,
demonstrando que a proteína InvA faz parte de uma família de proteínas, as quais
possuem a função de transportar proteínas específicas através da membrana
celular bacteriana.
Rahn e pesquisadores (1992) utilizaram o gene invA na reação da PCR, e
constataram que ele pode ser usado para diferenciar Salmonella sp. de outros
microorganismos, pois a seqüência de nucleotídeos contida neste gene estava
presente e era única para as 630 amostras Salmonella pertencentes a 100
diferentes sorotipos. Porém duas amostras de Salmonella Litchfield e duas de
Salmonella Sentfenberg foram negativas para a presença deste gene.
Amostras de Salmonella Typhimurium mutantes para os genes invA e invG
diminuiram a capacidade destas amostras em invadir células MDCK (Madin-Darby
Canine Kidney) (CLARK et al., 1996). Porter e Curtis III (1997) realizaram mutações
nos genes invA, invB e invC de Salmonella Enteritidis e Salmonella Typhimurium e
constataram uma redução na virulência oral e na colonização da parede do íleo e
baço em pintinhos de um dia de vida. Eles também testaram estes mutantes

29
intraperitonealmente e não obtiveram efeito na virulência, suportando a idéia de que
Salmonella tenha múltiplos fatores de virulência, alguns dos quais funcionando em
nível de pós-invasão.
Um nível de regulação na invasão de células eucarióticas que não requer
síntese de proteínas e sim contato do organismo com a célula do hospedeiro foi
demontrado por Ginochio et al. (1994). O contato celular resultou em estruturas
supramoleculares na superfície da membrana celular de Salmonella Typhimurium,
as quais foram diferentes dos flagelos ou “pilli”. Os autores demonstraram que a
agregação, a perda ou retração dessas estruturas foi necessária para a
internalização da bactéria em cultura de células epiteliais, pois mutantes de
Salmonella deficientes na formação transitória dessas estruturas foram incapazes de
invadir as mesmas, demonstrando que estas estruturas tiveram um papel na
interação entre a bactéria e as células do organismo hospedeiro. Eles encontraram
dois fenótipos: o primeiro onde mutações nos genes invG e invC resultaram em
organismos que falharam na agregação dos apêndices, e o segundo no qual cepas
mutantes no invA e invE, resultaram em organismos que agregaram as estruturas,
porém falharam na hora de recolher ou desprender as mesmas em incubação com
células epiteliais. Mais de vinte proteínas do sistema de secreção tipo III codificado
pela SPI-1 fazem parte da formação dessas estruturas de superfície da membrana
celular, as quais são essenciais para invadir tanto células M quanto células epiteliais
(PLANO et al., 2001; ZHOU, GALÁN 2001). A proteína InvA faz parte do sistema
funcional de secreção tipo III (MECSAS, STRAUSS, 1996; PLANO et al., 2001)
Células M possuem um papel fundamental na patogenicidade de Salmonella
Typhimurium uma vez que durante os primeiros estágios da infecção, elas são o
primeiro local de invasão do intestino de camundongos e em testes realizados “in

30
vitro” (JONES et al., 1994; PENHEITER et al., 1997). Clark e colaboradores (1998)
demonstraram que é necessário a presença do gene invA para uma eficiente
invasão nas células M .
1.4.2 Gene spvC
Plasmídeos são segmentos de DNA extracromossomal da bactéria que
carreiam informação genética adicional e se replicam de forma independente
(BICHER et al., 1994).
Plasmídeos virulentos estão envolvidos na sobrevivência e crescimento da
bactéria em células de hospedeiros, o conceito é de que eles não estão envolvidos
na interação inicial entre Salmonella e a mucosa intestinal nem são requeridos para
a invasão, pois não são necessários para uma eficiente colonização das placas de
Peyer. Contudo eles habilitam o organismo a persistir nas células do sistema
reticuloendotelial, como o baço e fígado (GULIG, CURTISS III, 1987; SWAMY et al.,
1996).
Nakamura et al. (1985) demonstraram que cepas de Salmonella Enteritidis
que não possuíam um plasmídeo de 36–Mda, foram menos virulentas quando
comparadas a cepas que possuíam o plasmídeo, sugerindo a associação deste com
virulência em camundongos.
Um plasmídeo de 100-kb presente em sorotipos de Salmonella Typhimurium
foi importante na virulência, pois as amostras foram mais eficientes quando
administradas oralmente em infectar nódulos linfáticos mesentéricos e baço de
camundongos quando possuíam o plasmideo, sugerindo que o papel

31
desempenhado por ele, foi o de realizar uma complexa interação com células do
sistema reticuloendotelial (GULIG, CURTISS III, 1987).
Barrow e pesquisadores (1987) relacionaram o plasmídeo de 85-kb presente
em Salmonella Gallinarum à virulência em galinhas, pois quando ele foi eliminado
houve uma redução na virulência da amostra quando administrada via oral e
intraperitoneal. Estes autores demonstraram que ele aumentou a sobrevivência e o
crescimento da bactéria em células do sistema reticuloendotelial. Em Salmonella
Pullorum, o plasmídeo de 85-kb foi associado com uma completa perda de virulência
quando eliminado do sorotipo e inoculado oralmente para galinhas, sendo esta a
rota natural de infecção. Quando a via de administração utilizada foi a parenteral,
houve um decréscimo na virulência, sendo que a reintrodução do plasmídeo
restaurou a virulência nas duas vias de inoculação (BARROW, LOVEL, 1988).
Um plasmídeo de virulência de 50-MDa presente em Salmonella Dublin
esteve também presente em 678 das 1021 amostras de Salmonella de diferentes
sorotipos testados por hibridização, sendo que a maioria dos sorotipos positivos foi
Typhimurium, Enteritidis e Dublin (WOODWARD et al., 1989).
Chart e colaboradores (1989) constataram que a virulência de Salmonella
Enteritidis fagotipo 4 para camundongo BALB\c dependeu da presença de um
plasmídeo de 38-MDa.
Mutações em um plasmídeo de 60-MDa de amostras virulentas de Salmonella
Typhimurium resultaram em uma redução na capacidade adesiva e na eliminação da
capacidade em invadir células HeLa, sendo que a reintrodução deste plasmídeo
restaurou tanto a capacidade adesiva quanto a invasiva. Estas cepas mutantes
também tiveram a virulència reduzida quando inoculadas oralmente em
camundongos (JONES et al., 1992).

32
Fierer e colaboradores (1992) demonstraram que 76% das 79 amostras de
Salmonellas isoladas de sangue humano e 42% das 33 amostras de fezes humanas
possuíam plasmídeos. Salmonellas são isoladas de infecções naturais sistêmicas
mais freqüentemente que em infecções entericas, tanto em animais quanto em
humanos. A presença de plasmídeos indica que eles possuem um papel importante
na patogenicidade de Salmonellas não tifóides (FIERER et al, 1993).
Sorotipos de Salmonellas não tifóides associados com doença extra-intestinal
possuem plasmídeos que codificam importantes genes para a indução de uma
infecção sistêmica e letal (GUINEY et al., 1995; ROUDIER et al., 1992).
Foi demonstrado em Salmonella Typhimurium que plasmídeos de virulência
aumentaram a média de crescimento da bactéria em camundongos (GULIG,
DOYLE, 1993).
Halavatkar e Barrow (1993) pesquisaram a virulência para o plasmídeo de 54-
kb presente em Salmonella Enteritidis PT4, e verificaram que ele foi essencial para
total expressão de virulência em camundongos. No entanto não observaram nenhum
papel do plasmídeo na produção da doença e mortalidade em galinhas poedeiras
jovens ou na sua contribuição na localização da bactéria em órgãos viscerais,
incluindo os ovários. Segundo estes autores, um resultado semelhante ocorreu
quando um plasmídeo de 90-kb de Salmonella Typhimurium foi eliminado da
bactéria.
Não foram necessários plasmídeos de virulência de Salmonellla em
camundongos para o deslocamento da bactéria através da mucosa intestinal,
demonstrando que cepas de Salmonella sem os plasmídeos foram aptas a colonizar
e persistir no baço e fígado, mas na fase sistêmica da infecção não houve um
crescimento adequado (GUILLOREAU et al, 1996).

33
Salmonella Typhimurium, Salmonella Enteritidis, Salmonella Dublin,
Salmonella Cholerasuis, Salmonella Abortusovis, Salmonella Pullorum e Salmonella
Gallinarum possuem plasmideos de virulência de diferentes tamanhos e composição
genética, mas, no entanto, todos contêm uma região altamente conservada de 7,8-
kb (GUINEY et al., 1995; LIBBY et al., 1997; ROTGER, CSADÉSUS, 1999). A
análise da seqüência de nucleotídeos desta região em comum para estes sorotipos
revelou cinco genes designados de genes spv (Salmonella plasmide virulence):
spvR, spvA, spvB, spvC e spvD que codificam as proteínas SpvR, SpvA, SpvB e
SpvC respectivamente (GULIG et al., 1993).
Uma parte na virulência de Salmonella tem sido relacionada a estes genes.
Hefferman et al. (1987) concluíram que o “operon” spv promoveu a sobrevivência e
aumentou a média de crescimento da bactéria no fígado, baço e nódulos linfáticos
de camundongos infectados oralmente. Gulig et al. (1993) demonstraram que estes
genes restauraram a virulência em cepas curadas de plasmideos em camundongos.
Testes de virulência indicaram que a proteína SpvB foi essencial para
Salmonella Dublin causar letalidade em camundongos. A abolição de spvA não teve
efeito na virulência. Os genes spvC e spvD foram instáveis na inoculação “in vivo”,
mas estes mutantes necessitaram de inóculos em concentrções mais altas que o
normal para matar camundongos. A proteína SpvB foi necessária para a expressão
do fenótipo de virulência (ROUDIER et al, 1992).
Os quatro genes estruturais, spvA, spvB, spvC e spvD, são transcritos como
um “operon” e ativados pelo produto do gene spvR, o qual é produzido na fase
estacionária de crescimento provavelmente devido ao acúmulo nessa fase de
produtos metabólicos (VALONE et al., 1993).

34
Fierer et al. (1993) demonstraram que o “operon” spv contribuiu para o
crescimento intracelular de Salmonella, pois os genes spv foram expressos dentro
das células do hospedeiro. Eles estudaram a expressão do gene spvB e
demonstraram que ele foi necessário para a virulência em camundongos e que o
gene spvA não foi, sugerindo que spvB foi ativado na resposta a diferentes
estímulos dentro da célula do hospedeiro.
Segundo Gulig e colaboradores (1993), o gene spvC aumenta a média de
crescimento nas células do hospedeiro afetando a interação com o sistema. Valone
et al. (1993) constataram que nenhuma proteína Spv foi expressa quando
Salmonella Dublin teve mutações no gene spvC, sendo que a perda na expressão
das proteínas SpvA e SpvB pode ter ocorrido, segundo os autores, devido ao
rompimento da estabilidade de transcrição ou pode indicar que spvC é requerido
para expressão de outras proteínas.
Nos resultados encontrados por Libby e colaboradores (1997), os genes spv
de Salmonella Dublin promoveram um aumento na proliferação da bactéria em
tecidos intestinais locais e extraintestinais de hospedeiros naturais, demonstrando
que este sorotipo em bezerros causou mortalidade maior quando possuía estes
genes plasmideais.
A localização no organismo bacteriano da proteína spvA é na membrana
externa; SpvB no citoplasma e membrana interna; SpvC no citoplasma e a proteína
SpvD foi exportada para fora da célula no sobrenadante mas também foi encontrada
no citoplasma e nas membranas segundo pesquisas realizadas por El-Gedayle et al.
(1997). Estes mesmos autores sugerem que o papel das proteínas intracelulares
SpvB e SpvC é o de aumentar a replicação da bactéria no meio intracelular

35
intrafagossomal, onde o crescimento bacteriano é normalmente restrito por um
limitado suplemento de nutrientes essenciais.
1.4.3 Gene sefC
Muitas bactérias patogênicas são capazes de invadir as células do hospedeiro
e conseqüentemente residir intracelularmente. Este processo complexo acontece
devido a uma série de eventos controlados bioquimicamente. O primeiro evento é a
fixação na superfície da célula do hospedeiro, sendo este essencial na
patogenicidade (FINLAY, FALKOW, 1989). Existem vários mecanismos para esta
fixação, sendo que um dos principais é mediado por fímbrias (CLOUTHIER et al.,
1993).
Fímbrias também chamadas de “pilli” são filamentos proteicos produzidos na
superfície estrutural da bactéria. São compostas de arranjamentos helicoidais
idênticos e possuem subunidades proteicas chamadas fibrinas (COLLINSON et al.,
1991). Estes filamentos auxiliam na fixação da bactéria na superfície da célula
eucariótica do hospedeiro contribuindo para a patogenicidade (FINLAY, FALKOW,
1989). Muitas proteínas parecem estar envolvidas na estrutura, biossíntese e
construção de um dado tipo de fímbria. As fímbrias são classificadas de acordo com
sua morfologia e sua capacidade de aglutinar eritrócitos em: tipo 1 que aglutinam
eritrócitos na ausência de manose; tipo 2 que não aglutinam eritrócitos e fímbrias
tipo 3 as quais aglutinam eritrócitos tratados com ácido tânico na presença de
manose (DUGUID et al., 1966).
Ernest et al. (1990) descreveram que a fímbria tipo 1 foi um dos fatores que
contribuiu para a invasão bacteriana em células Hep–2 em Salmonella Typhimurium,

36
pois a invasão foi muito mais efetiva com bactérias fimbriadas do que com as não
fimbriadas.
Finlay e Falkow (1990) observaram em testes de aderência de Salmonella em
células intestinais epiteliais de humano (Caco-2) a existência de muitas estruturas
como fimbrias entre a bactéria e a superfície da célula hospedeira.
Através de testes realizados de hemaglutinação, foi constatado que
Salmonella Enteritidis utilizou fímbrias tipo 1 e tipo 3 de para aderir as células
epiteliais bucais humanas e células epiteliais de intestino de camundongo
(ASLANZADEH, PAULISSEN, 1992).
A presença da fímbria tipo 1 aumentou a invasão de Salmonella Typhimurium
e Salmonella Braenderup em células HeLa, demonstrando que esta fímbria teve um
papel acessório na virulência (HORIUCHI et al.,1992).
Salmonella Enteritidis tem sido descrita produzindo cinco tipos de fímbrias,
referidas como SEF14 (FEULTRIER et al., 1988), SEF21 (MÜLLER et al., 1991),
SEF17 (COLLINSON et al., 1991), as quais são compostas por proteínas de peso
molecular de 14, 21 e 17 KDa respectivamente, fímbria tipo 3 (ASLANZADEH,
PAULISSEN, 1992) e BFP (“bundle-forming pili”) (SOHEL et al., 1993). Sendo estas
morfologicamente, bioquimicamente e imunologicamente distintas (COLLINSON et
al., 1991).
Naugthon e colaboradores (2001) verificaram a importância da fimbria SEF21
como fator de virulência nos estágios iniciais da infecção, onde o número de
Salmonellas encontrado no estômago e intestino de ratos foi maior nas primeiras
horas após a inoculação quando a bactéria expressou esta fímbria.
Feultrier et al. (1986) reportaram a fímbria SEF14 como do tipo 1, porém
Thorns et al. (1990) demontraram que esta fímbria tem uma estrutura muito menor

37
que a tipo 1 comumente reportada, sugerindo um tipo de fimbria até então não
descrita.
Salmonella Enteritidis produz delgados filamentos de fímbrias designados de
SEF14. O gene de 5,3 Kb que codifica a fímbria SEF14 foi cortado com a enzima de
restrição Hind-III e um fragmento de 3,9 Kb obtido foi seqüenciado e subdividido em
três genes: sefA, sefB e sefC. Os genes sefABC fazem parte de um complexo
“operon”, sendo responsáveis pela estrutura e expressão da fímbria SEF14 . A
seqüência dos nucleotídeos destes três genes em Salmonella Enteritidis representou
a primeira seqüência de um “operon” fimbrial de importância invasiva na
enteropatogenicidade desta bactéria (CLOUTHIER et al., 1993). Em 2001 Collinghan
e Woodward identificaram mais dois genes no “operon” de SEF14, sefD e sefE.
Clouthier e pesquisadores (1993) após sequenciarem os três genes sefABC,
verificaram que sefA codifica uma única fímbria, sefB e sefC codificam proteínas que
possuem similaridade com proteínas acessórias de Escherichia coli e Klebsiela
pneumoniae e sefC e com outras proteínas da membrana externa. Estes autores
também demontraram que sefB e sefC não se expressam na ausência de sefA, pois
suas transcrições iniciam na região promovida por sefA.
Em um estudo realizado em 1993, Turcotte e Woodward estudaram a
distribuição da fímbria SEF14 e constataram que ela foi limitada ao grupo D das
Salmonellas, sendo que a expressão destes genes foi limitada aos sorotipos
Enteritidis, Dublin, Moscow e Blegdam.
Peralta et al. (1994) demonstraram que a fímbria SEF14 contribuiu na
aderência de Salmonella Enteritidis em células epiteliais e que anticorpos
provenientes de gemas de ovos de galinhas imunizadas diretamente contra esta

38
fímbria, contribuiram na proteção, pois diminuiram a colonização e a invasão durante
os estágios iniciais da infecção.
Thiagarajan e colaboradores (1996-b) inocularam experimentalmente três
sorotipos de Salmonella Enteritidis em galinhas, sendo que o primeiro expressou
dois tipos de fímbrias, SEF14 e SEF21, o segundo sorotipo expressou somente
SEF21 e o terceiro não expressou fímbrias. A menor intensidade de colonização
cecal e excreção fecal nas aves foi obtida com o sorotipo que não expressou
fímbrias, porém o isolamento da bactéria no fígado e baço, não diferiu entre os três
sorotipos testados. Este resultado confirma o papel da fímbria na ligação de
Salmonella Enteritidis em células epiteliais.
A fímbria SEF14 de Salmonella Enteritidis esteve envolvida no ligamento da
bactéria em células granulosas do ovário de galinhas, pois este sorotipo interagiu
melhor com fibronectina, colágeno e laminina na presença desta proteína fimbrial
(THIAGARAJAN et al., 1996-a)
Thorns e pesquisadores (1996) inocularam aves pela via oral com amostras
de Salmonella Enteritidis negativas para a fímbria SEF14 e amostras positivas para
a mesma. Como resultado, 18% das aves inoculadas com o fenótipo negativo para a
fímbria eliminaram a bactéria enquanto que 35% das aves inoculadas com o fenótipo
positivo eliminaram o agente.
Segundo Ogunniyi et al. (1997), a fímbria SEF14 pode ser importante na
colonização das placas de Peyer de camundongos BALB/c e contribuir no processo
de infecção.
Segundo Bäumler et al. (1997) mutações no gene sefA de Salmonella
Enteritidis reduziu a capacidade de colonização em células de galinha, durante a
primeira semana da infecção.

39
Edwards e colaboradores (2000) reportaram que uma suposta subunidade
adesina da fímbria SEF14 de Salmonella Enteritidis, SefD, foi essencial para a
virulência em camundongos. A fímbria SEF14 foi considerada essencial para
eficiente internalização e sobrevivência do agente em macrófagos na infecção
intraperitoneal, relacionando este sistema fimbrial na infecção sistêmica, além de
sua participação no estágio inicial da colonização em células epiteliais.
1.4.4 Gene pefA
Segundo Friedrich et al. (1993) um plasmídeo de 90-kb descrito em
Salmonella Typhimurium conferiu aumento da virulência em camundongos
promovendo a disseminação da infecção após a invasão intestinal. A seqüência de
nucleotídeos de 13,9 Kb de um segmento desse plasmídeo codifica proteínas de
membrana externa envolvidas na biossíntese fimbrial em bactérias entéricas. Nesta
região encontra-se o “operon” pef (Plasmid encoded fimbriae) que contém quatro
genes, pefA, pefB, pefC e pefD e adicionais ORFs (janelas abertas de leitura)
responsáveis pela formação da fímbria PEF (ROTGER et al., 1999). Os autores
Friedrich e colaboradores (1993) realizaram mutações nos genes pefA, pefC, e nos
orf5 e orf6 concluindo que estas mutações reduziram ou bloquearam a apresentação
da fímbria PEF, indicando que as proteínas codificadas por estes genes foram
transportadas através da membrana citoplasmática e contribuiram para a formação
estrutural da fímbria.
Bäumler et al. (1996) investigaram o papel do “operon” fimbrial pef e
evidenciaram que fímbrias codificadas por plasmídeo de Salmonella Typhimurium
mediaramm a adesão no intestino delgado de camundongos e foram necessárias

40
para acúmulo de fluidos em camundongos jovens, porém este não é um resultado
conclusivo para outras espécies, pois nas linhagens de células epiteliais T84 do
intestino de humanos, a fímbria PEF não mediou a adesão e em coelhos não causou
acúmulo de fluídos.
O “operon” fimbrial pef foi relacionado com sorotipos isolados freqüentemente
de animais domésticos. A distribuição filogenética entre linhas clonais de Salmonella,
evidenciou que “operon” foi introduzido recentemente dentro de duas linhagens de
Salmonella enterica subespécie I (a primeira incluindo os sorotipos Enteritidis e
Cholerasuis e a segunda linhagem o sorotipo Typhimurium) predominantemente
isoladas de animais de sangue quente (BÄUMLER et al., 1997).
Van der Velden et al. (1998) realizaram mutações em quatro “operons”
fimbriais de Salmonella Typhimurium, os quais foram lpf, pef, fim e agf onde
verificaram que esta mutação simultânea reduziu a capacidade de colonizar o
intestino de camundongos, atenuando a virulência.
1.5 Detecção dos genes de virulência pela Reação em Cadeia da Polimerase
(PCR)
O DNA presente no cromossoma é composto por uma fita dupla, enrolada em
forma de hélice. Cada fita é constituída de bases nucleotídicas conhecidas pelas
letras A, C, G e T que são adenina, citosina, guanina e timina respectivamente.
Estas bases estão ligadas entre si em uma mesma fita de DNA através de um
esqueleto formado por moléculas de deoxirribose. A base A de uma fita sempre
pareia com a base T na outra fita, enquanto que a base C sempre pareia com a base

41
G da outra fita de DNA, através de ligações de pontes de hidrogênio sendo deste
modo as duas fitas de DNA chamadas de complementares (JOHNSON, 1985).
Como os genes possuem seqüências de DNA específicas, seqüências de DNA
de determinadas regiões são específicas e particulares de determinado gene,
portanto não são encontradas em nenhum outro gene de um mesmo ou diferentes
organismos (JOHNSON, 1985). Estas regiões são o alvo exato para amplificação
numa reação de PCR.
A amplificação do DNA pela PCR ocorre através da repetição cíclica de três
etapas: a primeira onde o DNA é aquecido a 94 ou 95°C, causando a separação da
dupla fita de DNA quebrando as pontes de hidrogênio. Na segunda etapa a
temperatura é reduzida para 50 a 60°C permitindo que a dupla fita de DNA se forme
novamente. Nesta fase, o excesso de iniciadores irá parear com as duas fitas de
DNA nas regiões complementares, este processo é chamado de anelamento. Na
terceira etapa chamada de extensão a temperatura é aumentada para 72°C, que é a
temperatura ótima para a ação da enzima Taq DNA polimerase, ela sintetiza novas
fitas de DNA a partir da extremidade 3’ do iniciador. Os nucleotídeos são
adicionados a nova fita de DNA pela Taq DNA polimerase de forma complementar,
seqüencial, com base na seqüência do DNA molde, sintetizando, a enzima realiza a
extensão do fragmento hibridizado. A PCR aproveita os produtos sintetizados (novas
cadeias) como matéria prima nas próximas reações de extensão, daí o nome reação
em cadeia da polimerase, que amplifica exponencialmente os novos produtos de
DNA (ATLAS, BEJ, 1994; TAYLOR et al., 1991).
O segmento de DNA amplificado então pode ser visualizado sob luz ultra violeta
em gel de agarose corado com brometo de etídeo e comparado a um marcador

42
padrão devendo corresponder ao tamanho da banda esperado, conforme o par de
iniciadores utilizado (BOOM et al., 1990; TAYLOR, 1991).

43
2 OBJETIVOS
Caracterizar as amostras de Salmonella Enteritidis, já fagotipadas e ribotipadas,
quanto a presença dos genes invA, spvC, sefC e pefA.
Verificar a invasão e a colonização das amostras de Salmonella Enteritidis, cor-
relacionando a presença ou ausência dos genes invA, spvC, sefC e pefA, através da
infecção experimental em aves SPF, pela via oral e subcutânea.

44
3 MATERIAIS E MÉTODOS
3.1 Cultivo Bacteriano
Foram utilizadas 120 amostras de Salmonella Enteritidis gentilmente cedidas
pela Profa. doutora Iolanda A Nunes já fagotipadas (NUNES et al., 2002) e
ribotipadas (NUNES et al., 1999) conforme mostra o quadro 1.
Continua
DESIGNAÇÃO DA AMOSTRA FAGOTIPO RIBOTIPO ESTADO ANO FONTE
317 PT4 RT1 PR 1997 Ovos Bicados 301 PT1 - SC 1996 Galinha - Coração 265 PT4 RT1 SC 1996 Galinha 192 PT4 RT1 SC 1996 Galinha 469 PT4 RT1 SC 1996 Galinha 472 PT4 RT1 SC 1996 Galinha 194 PT4 RT1 SC 1996 Galinha 254 PT4 RT1 SC 1996 Galinha 197 PT4 RT1 SC 1996 Galinha - Coração 276 PT4 RT1 SC 1996 Galinha - Cavidade Abdominal 256 PT4 RT1 SC 1996 Galinha - Cavidade Abdominal 199 PT4 RT1 SC 1996 Galinha - Intestino 466 PT4 RT1 SC 1996 Galinha - Ceco 204 PT4 RT1 SC 1996 Galinha 482 PT4 RT1 SP 1995 Postura Comercial 488 PT4 RT1 SP 1995 Postura Comercial 104 PT4 RT1 PR 1996 Fezes de Homem 501 PT4 RT1 MT 1997 Swab de Arrasto - Matriz 67 sem. 492 PT4 RT1 SC 1996 Cama de Frango 470 PT4 RT1 SC 1996 Cama de Frango 458 PT4 RT1 SC 1996 Cama de Frango 496 PT4 RT1 PR 1997 Forro de Caixa - Matriz de Corte 489 PT4 RT1 SP 1995 Ovário - Postura Comercial 460 PT4 RT1 SP 1995 Postura Comercial - Fígado, Baço e Bile 484 PT4 RT1 SP 1995 Postura Comercial 457 PT4 RT1 SP 1997 Pintos de Corte 467 PT4 RT1 SC 1996 Saco da Gema - Pintos de 1 dia 299 PT4 RT1 SC 1996 Saco da Gema - Pintos de 1 dia 281 PT4 RT1 PR 1996 Fezes de Homem 282 PT4 RT1 PR 1996 Sangue de Homem 248 PT4 RT1 PR 1996 Fezes de Homem
45
Continuação
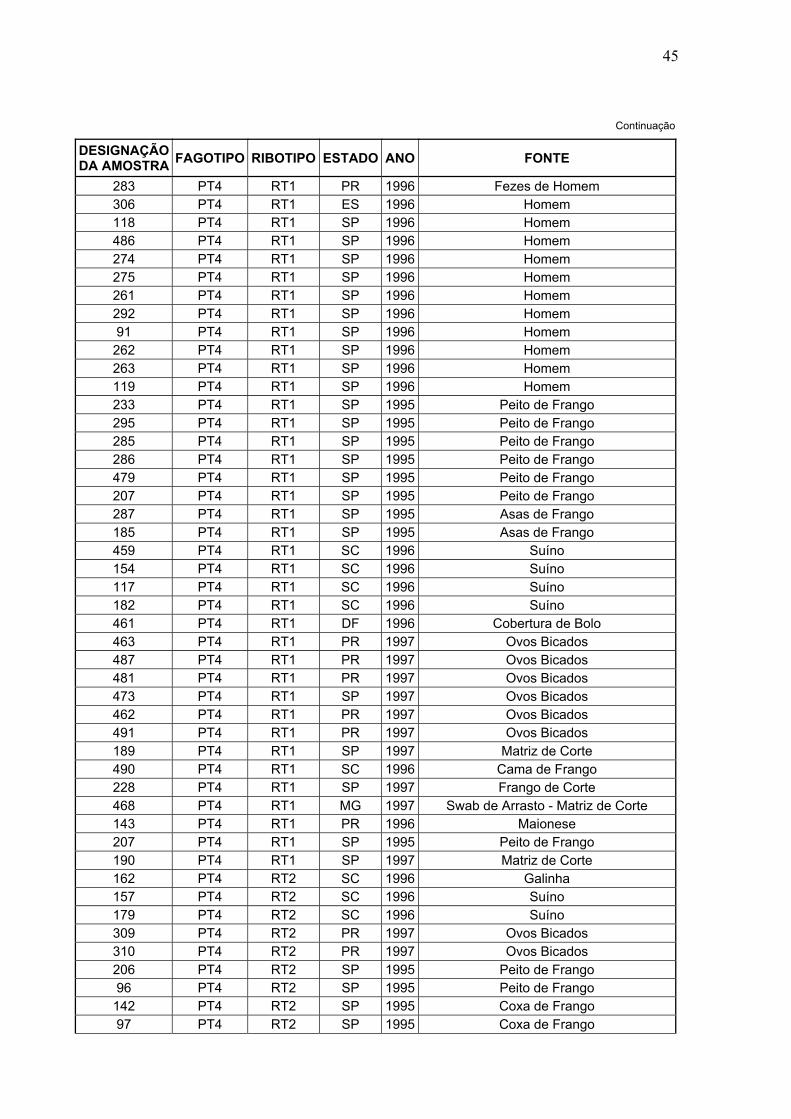
DESIGNAÇÃO DA AMOSTRA FAGOTIPO RIBOTIPO ESTADO ANO FONTE
283 PT4 RT1 PR 1996 Fezes de Homem 306 PT4 RT1 ES 1996 Homem 118 PT4 RT1 SP 1996 Homem 486 PT4 RT1 SP 1996 Homem 274 PT4 RT1 SP 1996 Homem 275 PT4 RT1 SP 1996 Homem 261 PT4 RT1 SP 1996 Homem 292 PT4 RT1 SP 1996 Homem 91 PT4 RT1 SP 1996 Homem 262 PT4 RT1 SP 1996 Homem 263 PT4 RT1 SP 1996 Homem 119 PT4 RT1 SP 1996 Homem 233 PT4 RT1 SP 1995 Peito de Frango 295 PT4 RT1 SP 1995 Peito de Frango 285 PT4 RT1 SP 1995 Peito de Frango 286 PT4 RT1 SP 1995 Peito de Frango 479 PT4 RT1 SP 1995 Peito de Frango 207 PT4 RT1 SP 1995 Peito de Frango 287 PT4 RT1 SP 1995 Asas de Frango 185 PT4 RT1 SP 1995 Asas de Frango 459 PT4 RT1 SC 1996 Suíno 154 PT4 RT1 SC 1996 Suíno 117 PT4 RT1 SC 1996 Suíno 182 PT4 RT1 SC 1996 Suíno 461 PT4 RT1 DF 1996 Cobertura de Bolo 463 PT4 RT1 PR 1997 Ovos Bicados 487 PT4 RT1 PR 1997 Ovos Bicados 481 PT4 RT1 PR 1997 Ovos Bicados 473 PT4 RT1 SP 1997 Ovos Bicados 462 PT4 RT1 PR 1997 Ovos Bicados 491 PT4 RT1 PR 1997 Ovos Bicados 189 PT4 RT1 SP 1997 Matriz de Corte 490 PT4 RT1 SC 1996 Cama de Frango 228 PT4 RT1 SP 1997 Frango de Corte 468 PT4 RT1 MG 1997 Swab de Arrasto - Matriz de Corte 143 PT4 RT1 PR 1996 Maionese 207 PT4 RT1 SP 1995 Peito de Frango 190 PT4 RT1 SP 1997 Matriz de Corte 162 PT4 RT2 SC 1996 Galinha 157 PT4 RT2 SC 1996 Suíno 179 PT4 RT2 SC 1996 Suíno 309 PT4 RT2 PR 1997 Ovos Bicados 310 PT4 RT2 PR 1997 Ovos Bicados 206 PT4 RT2 SP 1995 Peito de Frango 96 PT4 RT2 SP 1995 Peito de Frango 142 PT4 RT2 SP 1995 Coxa de Frango 97 PT4 RT2 SP 1995 Coxa de Frango

46
Continuação
DESIGNAÇÃO DA AMOSTRA FAGOTIPO RIBOTIPO ESTADO ANO FONTE
212 PT4 RT2 SP 1995 Asas de Frango 208 PT4 RT2 SP 1995 Asas de Frango 213 PT4 RT2 SP 1995 Asas de Frango 214 PT4 RT2 SP 1995 Asas de Frango 100 PT4 RT2 MG 1996 Carcaça 101 PT4 RT2 SC 1996 Frango de Corte 102 PT4 RT2 SC 1996 Frango de Corte 314 PT4 RT2 MT 1997 Forro de Caixa - Pintos de Corte 198 PT4 RT2 SC 1996 Galinha - Cavidade Abdominal 219 PT4 RT2 RJ 1996 Fezes de Homem 220 PT4 RT2 DF 1996 Fezes de Homem 225 PT4 RT2 SC 1996 Ceco de Galinha 293 PT4 RT3 SP 1996 Homem 205 PT4 RT3 SP 1995 Peito de Frango 145 PT4 RT3 PR 1996 Maionese 215 PT4 RT3 DF 1996 Massa de Bolo 304 PT4 RT3 PR 1996 Fezes de Homem 311 PT4 RT5 PR 1997 Ovos Bicados 268 PT4 RT5 SC 1996 Galinha 307 PT4 RT5 SC 1996 Pele 152 PT4 RT5 SC 1996 Suíno 210 PT4 RT9 SP 1995 Peito de Frango 95 PT4 RT9 SP 1995 Peito de Frango 153 PT4 RT9 SC 1996 Suíno 160 PT4 RT9 SC 1996 Suíno 103 PT4 RT9 SC 1996 Fezes de Homem 183 PT4 RT9 SP 1996 Fezes de Homem 92 PT4 RT9 SP 1996 Fezes de Homem 227 PT4 RT9 SP 1996 Homem 294 PT4 RT9 SP 1996 Homem 94 PT4 RT9 SP 1996 Homem 209 PT4 RT9 SC 1997 Suíno 221 PT4 RT10 SC 1997 Fezes de Homem 270 PT7 RT1 SC 1996 Galinha - Fígado 303 PT7 RT1 SC 1996 Galinha - Fígado 201 PT7 RT1 SC 1996 Galinha - Fígado 202 PT7 RT1 SC 1996 Galinha - Ceco 204 PT7 RT1 SC 1996 Galinha - Fígado 305 PT7 RT1 RJ 1996 Homem 186 PT7 RT1 SP 1995 Asas de Frango 113 PT7 RT1 SC 1996 Fígado de Galinha 174 PT7 RT5 SC 1996 Fígado de Galinha
Quadro 1 - Descrição das amostras de Salmonella Enteritidis, segundo o fagotipo,
ribotipo, estado, ano e fonte de isolamento.

47
As amostras foram semeadas em meio McConkey e uma colônia de cada
amostra foi cultivada em 3 ml de caldo Luria Bertani (LB) por 24 horas a 37°C.
Como controle positivo para os genes invA e spvC foi utilizada uma amostra de
Salmonella typhimurium ATCC 14028 (Laboratório de Ornitopatologia da
Universidade de São Paulo), para os genes sefC e pefA a amostra de Salmonella
Enteritidis 248, e para o controle negativo foi utilizada uma amostra de Escherichia
coli K12 (Laboratório de Ornitopatologia da Universidade de São Paulo).
3.2 Extração do DNA Bacteriano
O protocolo para extração do DNA bacteriano foi realizado segundo o método
descrito por Boom et al. (1990).
Uma alíquota de 200 µl do crescimento bacteriano de cada amostra cultivada
em LB foi adicionada a um microtubo contendo 1 ml de tampão de lise (120 gramas
de Tiocianato de Guanidina; 1 ml de Triton 100x; 111,2 ml de Tris-HCl 0,1 M pH 6,4
e 8,8 ml de EDTA 0,5 M) e 40 µl da suspensão carreadora (1 grama de Terra
Diatomácea; 50 µl de HCl e 5 ml de água ultrapura). O microtubo foi agitado durante
1 minuto e deixado sobre a bancada durante 10 minutos. Posteriormente foi
centrifugado em alta velocidade a 10.000 Xg durante 90 segundos. O sobrenadante
foi descartado e foi adicionado ao pélete formado 0,5 ml de tampão de lavagem (120
gramas de Tiocianato de Guanidina, e 100 ml de Tris-HCl 1M pH 6,4) agitando
durante 15 segundos. Após este último procedimento o tubo foi centrifugado em alta
velocidade durante 90 segundos, o sobrenadante foi descartado, sendo esta etapa
repetida mais uma vez com tampão de lavagem, duas vezes com etanol 70% e uma
última vez com acetona, sendo que nesta última a centrifugação durou 2 minutos. O

48
pélete obtido no microtubo foi seco em estufa a 37°C durante 20 minutos. Após a
completa secagem do pélete foi adicionado ao mesmo 150 µl de tampão de eluição
(10 mM Tris-HCl e 1mM EDTA pH 8.0) , agitado por 60 segundos e o microtubo foi
mantido à 52-56°C durante 10 minutos. Por último o microtubo foi centrifugado
durante 5 minutos a 10.000 Xg, onde após o sobrenadante que continha o DNA
bacteriano foi removido para novos tubos e armazenado a -20°C até ser utilizado na
reação da PCR. Este mesmo procedimento foi utilizado para os controles positivos e
negativos.
3.3 Amplificação do DNA bacteriano
Os oligonucleotídeos iniciadores (primers) para os genes invA e spvC (SWAY
et al., 1996), sefC (BÄUMLER et al., 1997) e pefA (BÄUMLER et al., 1996), foram
sintetizados pela Invitrogen Brasil Ltda e estão descritos no Quadro 2.
GENE SEQUÊNCIA DE NUCLEOTÍDEOS PARES DE BASE (pb)
F: TTGTTACGGCTATTTTGACCA invA R: CTGACTGCTACCTTGCTGATG
521
F: CGGAAATACCATCTACAAATA spvC R: CCCAAACCCATACTTACTCTG
669
F: GCGAAAACCAATGCGACTGTAG sefC R: CCCACCAGAAACATTCATCCC
1103
F: AGGGAATTCTTCTTGCTTCCATTCCATTATTGCACTGGG pefA R: TCTGTCGACGGGGGATTATTTGTAAGCCACT
520
Quadro 2 - Descrição dos primers utilizados para os genes invA, spvC, sefC e pefA
Para a realização da PCR 0,5µl do DNA de cada amostra foi adicionado a um
microtubo contendo 15,15 µl de água ultrapura, 2,5 µl de tampão de PCR 10x (100
mM Tris-HCl pH 8.3; 500 mM KCl; 15mM MgCl2 e 0,01% [w/v] gelatina), 0,75 µl de
MgCl2, 0,5 µl de dNTP (deoxynucleotideos triphosphates), 0,5 µl do primer-F e 0,5 µl

49
do primer-R (40 pmoles), 0,1 µl de Taq-polimerase, totalizando um volume de 25 µl.
Antes de colocar os microtubos no termociclador foi adicionada uma gota de óleo
mineral em cada um.
A reação da PCR foi testada também simultaneamente para dois genes sob a
forma de multiplex, os quais foram: invA e spvC juntos e pefA e sefC. O
procedimento para a amplificação foi o mesmo.
A amplificação foi realizada no termociclador (PT200 MJ. Research) com os
seguintes programas para a desnaturação, anelamento e extensão das fitas de
DNA:
Para os genes invA e spvC: 30 ciclos de 93°C durante 1 minuto, 42°C
durante 1 minuto e 72°C durante 2 minutos.
Para os genes pefA e sefC foram realizados 30 ciclos de 95°C durante 1
minuto , 50°C durante 1 minuto e 72°C durante 2 minutos.
Após o término do programa os tubos foram retirados do aparelho e guardados sob
refrigeração.
3.4 Detecção do produto amplificado
Para cada 10 µl do produto da PCR de cada amostra foi adicionados 3µl de
tampão de corrida (loading buffer 6x), após a homogeneização foram colocados 10
µl em cada poço do gel de agarose a 1,5% (para cada 100ml de TBE 0,5x foram
colocadas 1,5 gramas de agarose). Os produtos da PCR juntamente com o
marcador de pares de base 100bp DNA ladder (Invitrogen Brasil Ltda) foram então
separados por eletroforese em corrida de 80 V durante 1 hora e 30 minutos. O gel foi

50
então corado com uma solução de Brometo de Etídeo (10mg/ml) durante 10
minutos, descorado por mais 10 minutos em água destilada e fotografado sob luz
ultravioleta.
3.5 Infecção experimental em aves SPF
De acordo com os resultados, cinco amostras representando os cinco perfis
genéticos relacionados aos genes foram inoculados em pintinhos SPF (Specific
Pathogen Free) de um dia de idade, pertencentes à linhagem Legorhn, cedidos pelo
laboratório Biovet.
As concentrações que foram utilizadas para desafiar as aves via oral e via
subcutânea encontram-se na tabela 1.
Tabela 1 - Aves SPF desafiadas pela via oral e via subcutânea, com os cinco
perfis de Salmonella Enteritidis relacionados aos genes invA, spvC,
sefC e pefA
GENES CONCENTRAÇÃO DE DESAFIO PERFIL GENÉTICO invA spvC sefC pefA Log 10 UFC/0,1ml
VIA ORAL2 Log 10 UFC/0,1ml VIA
SUBCUTÂNEA2
1º Perfil (P1) + + + + 7,5 x 101 3,2 x 101
2º Perfil (P2) + + - + 1 x 101 6,6 x 101
3º Perfil (P3) + - + + 5 3,5 x 101
4º Perfil (P4) + - + - 8,3 x 101 4,1 x 101
5º Perfil (P5) + + - - 1 x 101 2,5 x 101

51
Cada perfil genético e sua respectiva concentração foi administrado antes do
alojamento das aves, na dose de 0,1 ml, através de uma sonda oral e também via
subcutânea em um grupo de 5 pintinhos para cada concentração. O grupo controle
foi de 5 aves para cada via e recebeu solução fisiológica estéril 85% como placebo.
Cada grupo de 5 aves foi alojado em uma bateria de 1,7m2. O aviário foi
previamente lavado e desinfetado respeitando o vazio sanitário, sendo que a cama
utilizada foi maravalha nova. Cada bateria manteve seu próprio comedouro e
bebedouro até o final do experimento. Ração e água foram dadas “ad libidum”.
As aves que não morreram até o décimo dia pós-inóculo, foram sacrificadas
por deslocamento cervical. Após a morte ou sacrifício de cada ave, foram removidos
assepticamente o fígado, baço e o ceco. Os órgãos foram colocados em sacos
plásticos esterelizados (Inlab), pesados, macerados e foi acrescentada água
peptonada a 0,1% (Merk) para pré-enriquecimento a cada saco. Após ser feita uma
homogeneização, 0,1 ml do tecido homogeneizado foi plaqueado em meio XLT4
(Xylosine-Lysine-Tergitol-4/Difco). As placas foram mantidas em estufa a 37°C e a
leitura foi realizada em 24, 48, 72 e 96 horas.
Uma colônia negra de cada placa, sugestiva de Salmonella, foi testada com
soro polivalente anti-Salmonella (Promicro-lot: 02839) durante 2 minutos sob
agitação para a certificação da aglutinação e identificação como Salmonella.

52
4 RESULTADOS
4.1 Amplificação do DNA bacteriano utilizando a PCR
As amostras analisadas para os genes spvC, sefC, pefA e invA apresentaram
o seguinte resultado: o gene invA esteve presente em 100% (120/120) das
amostras; spvC em 94% (113/120), sefC em 97,5% (117/120) e o gene pefA em
97% (116/120) das amostras.
As amostras foram caracterizadas em cinco diferentes perfis conforme mostra
a tabela 2.
Tabela 2 - Cinco perfis de Salmonella Enteritidis relacionando o fagotipo,
ribotipo e fonte de isolamento quanto a presença ou ausência dos
genes invA, spvC, sefC e pefA
GENES DESIGNAÇÃO DA AMOSTRA
FAGOTIPO / RIBOTIPO FONTE PERFIL
GENÉTICO invA spvC sefC pefA 306 PT4RT1 Homem P1 + + + + 119 PT4RT1 Homem 102 PT4RT3 Frango
P2 + + - +
459 PT4RT1 Suíno 182 PT4RT1 Suíno 212 PT4RT2 Frango 94 PT4RT9 Homem
P3 + - + +
287 PT4RT1 Frango 219 PT4RT2 Homem 213 PT4RT2 Frango
P4 + - + -
301 PT1 Galinha - Coração P5 + + - -
Dentre as amostras PT4RT1 94% (64/68) pertenceram ao perfil P1, 1,5%
(1/68) ao perfil P2, 3% (2/68) ao P3 e 1,5% (1/68) das amostras pertenceram ao
perfil P4. Para as amostras PT4RT2, 86% (18/21) pertenceram ao perfil P1, 5%
(1/21) ao P3 e 9% (2/21) ao perfil P4. Nas amostras PT4RT3, 80% (4/5)

53
apresentaram o perfil P1 e 20% (1/5) o perfil P2. Sendo que para as amostras
PT4RT9 o resultado foi de 91% (10/11) pertencentes ao perfil P1 e 9% (1/11) ao P3.
As amostras PT4RT5, PT7RT1 e PT4RT10 apresentaram o perfil P1 e apenas a
amostra PT1 apresentou o perfil 5. Todas as amostras pertencentes ao quadro 1 que
não se encontram relacionadas na tabela 2, pertenceram ao perfil P1.
A figura 1 mostra os diferentes resultados encontrados para os quatro genes
estudados por eletroforese em gel de agarose.
Figura 1 - Eletroforese em gel de agarose 1,5% corado com brometo de etídeo
mostrando os produtos de amplificação para os genes invA, spvC, sefC e pefA.
M-marcador de peso molecular 100 pb; 1- Salmonella Enteritidis 248 controle
positivo para os genes sefC e pefA; 2-Salmonella Enteritidis 119 negativa para
sefC e positiva para pefA; 3- Salmonella Enteritidis 182 positiva para sefC e pefA;
4- Salmonella Enteritidis 287 positiva para sefC e negativa pefA; 5- Escherichia
coli K12 controle negativo; M1- marcador de peso molecular 100 pb; 6-
Salmonella Typhimurium controle positivo para os genes invA e spvC; 7-
Salmonella Enteritidis 94 positiva para invA e negativa para spvC; 8- Salmonella

54
Enteritidis 102 positiva para invA e spvC; e 9- Escherichia coli K12 controle
negativo.
4.2 Infecção Experimental em aves SPF desafiadas pela via oral
A tabela 3 mostra o resultado obtido do reisolamento no fígado, baço e ceco,
do total de aves desafiadas com os cinco perfis da bactéria, pela via oral.
Tabela 3 - Resultado do reisolamento de Salmonella Enteritidis, em fígado, baço
e ceco de pintos SPF desafiados por via oral.
GENES CONCENTRAÇÃO DO DESAFIO
Nº DE AVES REISOLADAS/ Nº DE AVES
DESAFIADAS
DESIGNAÇÃO DAS AMOSTRAS DE SALMONELLA ENTERITIDIS
FAGOTIPO/ RIBOTIPO
PERFIL GENÉTICO
invA spvC sefC pefA Log 10 UFC/0,1 ml Fígado/ Baço Ceco
Amostra 306 PT4 RT1 P1 + + + + 7,5 x 101 2 / 5 1 / 5
Amostra 102 PT4 RT3 P2 + + - + 1 x 101 2 / 5 0 / 5
Amostra 459 PT4 RT1 P3 + - + + 5 5 / 5 1 / 5
Amostra 287 PT4 RT1 P4 + - + - 8,3 x 101 1 / 5 0 / 5
Amostra 301 PT1 P5 + + - - 1 x 101 2 / 5 0 / 5
Conforme mostra a tabela 3, o reisolamento cecal somente foi possível na
presença simultânea dos genes sefC e pefA nos perfis P1 e P3. Quando houve a
presença de apenas um destes genes não foi possível obter o reisolamento pela via
oral.
No reisolamento do fígado e baço, a presença destes genes não se mostrou
importante. Quando os genes sefC e pefA estiveram presentes nos perfis P1 e P3, o

55
reisolamento nestes órgãos foi de 40% e 60% respectivamente, sendo que no perfil
P5, onde eles estavam ausentes o reisolamento foi de 40%. Nos perfis negativos
para apenas um dos genes, P2 e P4, o reisolamento foram de 40 e 20%
respectivamente.
A presença ou ausência do gene spvC não interferiu na taxa de reisolamento
cecal quando as aves foram desafiadas via oral. No reisolamento do fígado e baço
quando este gene esteve ausente a porcentagem para o perfil P3 foi de 100% e para
P4 de 20%. Nos perfis P1, P2 e P5 onde ele esteve presente, foi obtido um
reisolamento de 40%.
Não houve reisolamento bacteriano no fígado, baço e ceco, nas cinco aves do
grupo controle.
4.3 Infecção Experimental em aves SPF desafiadas pela via subcutânea
A presença simultânea dos genes sefC e pefA nos perfis P1 e P3, resultou
uma taxa de reisolamento das aves no fígado e baço de 80 e 60% respectivamente.
Na amostra negativa para estes dois genes, P5, o reisolamento foi de 100% e nos
perfis P2 e P4 negativos para apenas um deles foi de 60 e 80% respectivamente.
Também não houve correlação entre a presença ou ausência de um ou do dois
genes sefC e pefA com a taxa de reisolamento a partir do ceco pela via de desafio
subcutânea, conforme a tabela 4.
Quando o gene spvC esteve presente no perfis P1, P2 e P5, a porcentagem
de reisolamento nas aves do fígado e baço foi de 80, 60 e 100% respectivamente. A
porcentagem de reisolamento encontrada nas amostras negativas para os genes, P3

56
e P4, foi de 60 e 80%, respectivamente. O resultado do reisolamento dos perfis P1,
P2, P3 e P5 foi de 60%, e para o perfil P4 foi 20% (Tabela 4).
Não houve reisolamento bacteriano no fígado, baço e ceco, nas cinco aves do
grupo controle.
No desafio pela via subcutânea, a mortalidade das aves iniciou 24 horas
depois da inoculação, porém não houve uma correlação entre os genes dos cinco
perfis estudados e a mortalidade.
Tabela 4 - Resultado do reisolamento de Salmonella Enteritidis em fígado, baço e
ceco de pintos SPF desafiados por via subcutânea.
GENES CONCENTRAÇÃO DO DESAFIO
Nº DE AVES REISOLADAS/ Nº DE AVES
DESAFIADAS
DESIGNAÇÃO DAS AMOSTRAS DE SALMONELLA ENTERITIDIS
FAGOTIPO/ RIBOTIPO
PERFIL GENÉTICO
invA spvC sefC pefA Log 10 UFC/0,1 ml Fígado/ Baço Ceco
Amostra 306 PT4 RT1 P1 + + + + 3,2 x 101 4 / 5 3 / 5
Amostra 102 PT4 RT3 P2 + + - + 6,6 x 101 3 / 5 3 / 5
Amostra 459 PT4 RT1 P3 + - + + 3,5 x 101 3 / 5 3 / 5
Amostra 287 PT4 RT1 P4 + - + - 4,1 x 101 4 / 5 1 / 5
Amostra 301 PT1 P5 + + - - 2,5 x 101 5 / 5 3 / 5

57
5 DISCUSSÃO
O gene pefA faz parte do “operon” pef que codifica a fímbria PEF e localiza-se
num plasmídeo de virulência de 90 kb de algumas Salmonellas como os sorotipos
Enteritidis e Typhimurium segundo Woodward et al. (1996). Estes mesmos autores
hibridizaram algumas amostras de Salmonellas para os gene pefA e encontraram
68% de positividade para o sorotipo Typhimurium, sendo que isolados individuais
dos sorotipos Cholerasuis e Bovismorbificans hibridizaram para o gene e apenas
uma das amostras hibridizadas do sorotipo Enteritidis foi negativa. Em nosso estudo,
a positividade foi em 3,3% (4/120) das amostras testadas, sendo que 96,7%
(116/120) foram negativas para o gene pefA através da PCR.
No total das amostras negativas para o gene pefA, 50% (2/4) pertenciam ao
PT4RT2, 25% (1/4) ao PT4RT1 e 25% (1/4) ao PT1 conforme a tabela 2.
Os genes sefA, sefB e sefC fazem parte da fímbria SEF14, sendo que sefB e
sefC não são expressos na ausência de sefA, pois a transcrição dos genes sefB e
sefC inicia na região promotora sefA. Esses três genes fazem parte de um complexo
“operon” responsável pela expressão e construção da fímbria SEF14 (CLOUTHIER
et al., 1993). Dentre as amostras de Salmonella Enteritidis analisadas para a
presença do gene sefC, 2,5% (3/20) das amostras foram negativas. Destas, 33,3%
(1/3) eram PT4RT1, 33,3% (1/3) PT4RT3 e o restante PT1.
Segundo Clouthier et al. (1993), o gene sefC codifica uma proteína que tem
homologia com proteínas de membrana externa e possui elevado peso molecular.
Segundo estes autores, os sistemas fimbriais caracterizados que codificam proteínas
de membrana externa de alto peso molecular são presumidamente essenciais para a

58
construção e localização da fímbria. Partindo deste princípio, a ausência deste gene
implicaria na ausência da fímbria. Como proposto por Rajashekara et al. (1998), o
diagnóstico usando o antígeno fimbrial rSEF14 para Salmonella Enteritidis como
uma ferramenta específica e sensível não poderia ser utilizado na ausência de
spvC.
As fímbrias possuem um papel importante na adesão e invasão da bactéria
em células epiteliais intestinais e também na persistência em ceco (CLOUTHIER et
al., 1993). Thorns et al. (1996) demonstraram que a colonização cecal em aves foi
maior na primeira semana após desafio quando Salmonellla Enteritidis possuía a
fímbria SEF14. Os resultados encontrados neste estudo são semelhantes ao
trabalho de Thiagarajan et al. (1996-b) que demonstraram que sorotipos de
Salmonella Enteritidis inoculados oralmente expressando duas fímbrias SEF21 e
SEF14 tiveram maior colonização cecal em galinhas do que os sorotipos que
expressaram somente uma ou nenhuma das fímbrias. Semelhantemente em nosso
estudo, o reisolamento cecal de Salmonela Enteritidis quando o desafio foi realizado
pela via oral, somente foi possível na presença simultânea dos genes sefC e pefA no
mesmo perfil (tabela 3). Como este dois genes são formadores de fímbrias,
provavelmente pode haver uma relação sinérgica entre ambos. Isto justifica o nosso
resultado de reisolamento cecal negativo quando o gene sefC foi avaliado
isoladamente. Rajashera et al. (2000) encontraram resultado semelhante quando
estudaram o papel da patogenicidade da fímbria SEF14 em infecções em galinhas
através de cepas mutantes para o gene sefC, e constataram não haver diferença
significativa entre as cepas mutantes e não mutantes em relação a capacidade de
colonizar os cecos e na excreção fecal,

59
Segundo Allen-Vercoe et al. (1999), a colonização cecal é um importante
passo na patogenia de Salmonella Enteritidis em animais e no homem e um aspecto
relevante na contaminação do ovo via cecal. Dados encontrados por outros autores
sugerem que a fímbria SEF14 tenha um papel importante na patogenicidade como
um determinante de virulência. Thiagarajam et al. (1996-a) demonstraram que a
fímbria SEF14 influenciou na aderência da bactéria às células da granulosa do
ovário de galinha. Peralta et al. (1994) demonstraram que anticorpos anti-SEF14
derivados da gema do ovo diminuíram a colonização da bactéria durante os estágios
iniciais da infecção.
Para a avaliação do reisolamento do fígado, baço e ceco nas aves desafiadas
via subcutânea e também para o reisolamento do fígado e baço quando o desafio foi
via oral, tanto a presença quanto a ausência dos genes sefC e pefA nos perfis, não
demonstrou interferir na capacidade de colonização e invasão neste estudo
conforme tabelas 3 e 4. Estes resultados estão de acordo com Thiagarajam et al.
(1996-b) que estudaram o papel da fímbria SEF14 quando inoculada oralmente em
galinhas e verificaram que a presença dela não interferiu nos reisolamentos obtidos
do fígado, baço e órgãos reprodutivos. Ogumiyi et al. (1997) também constataram
que a fímbria SEF14 não teve papel na infecção sistêmica, sendo que ela foi
avaliada pela colonização da bactéria no baço de camundongos infectados
intraperitonealmente, indicando que a expressão desta fímbria não é crítica após ter
atravessado a barreira epitelial.
As amostras de Salmonella Enteritidis foram 100% (120/120) positivas para o
gene invA, o qual faz parte da ilha de patogenicidade–1 (SPI-1). Segundo Gálan e
Curtis III (1989), os genes de invasão de SPI-1 são requeridos na infecção oral para
causar doença em camundongos, pois mutações na bactéria para o gene invA de

60
Salmonella Typhimurium aumentaram a dose letal para camundongos e reduziram a
capacidade de colonizar as placas de Peyer após inoculação. Estas mesmas
amostras não atenuaram a virulência sistêmica quando inoculados
intraperitonealmente em camundongos. Em 1997 Porter e Curtiss III realizaram
mutações nos genes invA, invB e invC de Salmonella Typhimurium e constataram
uma redução na virulência oral e na colonização em pintinhos de um dia de idade,
porém quando estas mesmas amostras foram testadas intraperitonealmente estes
genes não interferiram na virulência. Estes resultados sugerem que a
patogenicidade do gene invA possui função apenas nos passos iniciais da infecção,
a qual ocorre naturalmente através da infecção oral.
A associação entre sorotipos virulentos de Salmonella que rotineiramente
produzem doença sistêmica e a presença de plasmídeos de alto peso molecular tem
sido reportada por vários autores (BAIRD et al.,1985; BARROW, LOVEL, 1988;
GULIG, CURTISS III 1987; HEFFERMAN et al.,1987).
Swamy et al. (1996) estudaram 245 amostras de Salmonella isoladas de
origem avícola, humana e água para a hibridização com o gene spvC e constataram
que somente 15,1% dos isolados foram positivos, sendo que 100% dos ovos, 3,8%
dos cecos de galinhas e 5,9% de isolados humanos foram positivos e todas os
isolados de água foram negativos para o gene. Em nosso estudo, 94% (113/120)
das amostras foram positivas para o gene sefC, sendo que destas, 65% (73/113)
dos isolados de origem avícola, 75% (6/8) de origem suína, 92% (24/26) de fonte
humana e 100% (4/4) de alimentos diversos foram positivos.
Para Bichler et al. (1994), o perfil plasmideal de uma bactéria para uma
determinada cepa pode ser específico durante um intervalo de tempo e uma área
limitada. Em seus estudos concluíram que dentro de um mesmo perfil plasmideal

61
podem existir diferentes fagotipos ou que para o mesmo fagotipo podem existir
diferentes perfis plasmideais. Em nosso estudo foi constatado que para o PT4 de
diferentes ribotipos, RT1, RT2 e RT9, ocorreram diferenças em relação à
positividade para o gene spvC e para PT4RT1 e PT4RT2 em relação ao gene pefA,
ou seja, houve uma diferença quanto a presença destes genes plasmideais,
demonstrando haver diferenças no perfil para o mesmo fagotipo e ribotipo e
diferentes ribotipos com o mesmo perfil.
Gulig e Curtiss III (1987), concluíram que cepas de Salmonella Typhimurium
deficientes em plasmídeos não possuíram defeito na capacidade de aderência ou
invasão de células epiteliais e segundo Hefferman et al. (1987), os genes spv são
requisitados somente após a invasão do intestino para a sobrevivência no sistema
reticuloendotelial. Os resultados encontrados neste estudo estão de acordo com
estes autores, onde a presença ou ausência do gene spvC não interferiu no
reisolamento cecal nas duas vias desafio utilizadas conforme tabelas 3 e 4. Este
gene está inserido em uma região de 8-kb que é altamente conservada entre os
plasmídeos de virulência de várias Salmonellas (GUINEY et al., 1995).
Valone et al. (1993) realizaram uma inserção no gene spvC de Salmonella
Dublin onde constataram que houve um rompimento na estabilidade de transcrição,
pois não houve expressão das proteínas SpvA e SpvB. As proteínas que foram
expressas pelo “operon” spv, incluindo SpvC, promoveram um aumento da
proliferação intracelular da bactéria em hospedeiros naturais, disseminado a bactéria
além do intestino para os nódulos linfáticos mesentéricos, baço e fígado,
aumentando desta maneira a severidade da infecção e produzindo doença letal ( El-
GEDAILY et al., 1997, HEFFERMAN et al., 1987; LIBBY et al., 1997). Em nossos
resultados, porém, quando o desafio foi via oral, os perfis P1, P2 e P5 (tabela 3),

62
positivos para o gene spvC apresentaram uma porcentagem de reisolamento nas
aves do fígado e baço de 40%. Já nos perfis P3 e P4, negativos para o gene, a
porcentagem foi de 100 e 20% respectivamente. Neste estudo obtivemos resultados
semelhantes aos de Hinton et al. (1990) que estudaram um plasmídeo de 38-MDa e
relataram que ele não teve nenhum efeito na capacidade das cepas de Salmonella
Enteritidis PT4 invadirem o fígado ou colonizar o ceco de galinhas de um dia de
idade após a infecção oral, sugerindo que para galinhas ele não foi necessário para
além das barreiras epiteliais intestinais.
Amostras de Salmonella Enteritidis livres de plasmídeos permaneceram
totalmente virulentas para galinhas (HALAVATKAR, BARROW, 1993). Salmonellas
tifóides causam doença invasiva e não possuem os genes spv (ROTGER et al.
1999). No entanto, segundo Fierer et al. (1993), Salmonellas que possuem
plasmídeos são isoladas de infecções sistêmicas naturais em animais e humanos
indicando que eles possuem um papel importante na patogenicidade desta doença.
Para Barrow et al. (1989), genes cromossomais também são importantes como
determinantes de virulência, particularmente na capacidade para sobreviver e
multiplicar em células do sistema reiculoendotelial. Roudier et al. (1990) em um
estudo, introduziram um plasmídeo de virulência pertecente aos sorotipos Dublin,
Typhimurium e Clolerasuis em sorotipos isentos como Heidelburg e Newport,
conferindo para estas últimas a capacidade de causar virulência em camundongos,
porém quando introduzido para outros sorotipos como o Derby e Sainpaul não.
Wallis et al. (1995) identificaram um plasmídeo de virulência em Salmonella
Dublin e concluíram que ele apenas afetou a persistência da bactéria em locais
sistêmicos, não influenciando na fase entérica da infecção ou na disseminação
sistêmica da doença. Em nossso estudo quando o desafio foi realizado pela via

63
subcutânea, os perfis P1, P2 e P5, positivos para o gene spvC, não demonstraram
diferença no reisolamento das aves no fígado e baço em relação aos perfis P3 e P4
negativos para o gene (tabela 4).
O conhecimento do genótipo e fenótipo da bactéria e o uso de um modelo de
infecção “in vivo” podem ajudar no melhor entendimento para a prevenção da
infecção ou doença em humanos. A importância do uso de aves para a infecção “in
vivo” é que elas são o principal veículo desta zoonose. Devido ao elevado número
de genes envolvidos na virulência deste patógeno, são necessários mais estudos
sobre outros genes de virulência para determinar a importância de cada um na
patogenicidade da doença.

64
6 CONCLUSÃO
1- Foram observados cinco perfis genéticos em relação aos quatro genes
estudados através da técnica de PCR. Perfil P1 positivo para os genes invA,
spvC, sefC e pefA; P2 positivo para invA, spvC e pefA; P3 positivo para invA,
sefC e pefA; P4 positivo para invA e sefC e o perfil P5 positivo para os genes
invA e spvC.
2- As amostras de Salmonella Enteritidis que possuem o mesmo fagotipo e
ribotipo possuem diferentes perfis genéticos e amostras de diferentes
ribotipos apresentam o mesmo perfil genético.
3- A presença simultânea dos genes sefC e pefA permitiu o reisolamento cecal
nos animais estudados desafiados pela via oral.
4- Todas as amostras estudadas de Salmonella Enteritidis foram positivas para
o gene invA através da PCR.
5- O gene spvC não influenciou a capacidade de invasão no fígado e baço das
aves desafiadas tanto pela via oral quanto pela via subcutânea.

65
REFERÊNCIAS
ALLEN-VERCOE, E. E.; WOODWARD, M. J. Colonisation of the chicken caecum by afimbriate and aflagellate derivatives of Salmonella enterica serotype Enteritidis. Veterinary Microbiology, v. 69, p. 265-275, 1999.
ANGULO, F. J.; SWERDLOW, D. L. Epydemiology of Human Salmonella enterica Serovar Enteritidis. Infections in the united States. In: SAEED, A. M.; GAST, R. K.; POTTER, M. E.; WALL, P. G. Salmonella enterica serovar Enteritidis in humans and animals. Ames: Iowa State University Press, 1999. p. 3-18.
ASLANZADEH, J.; PAULISSEN, L. J. Adherence and Pathogenesis of Salmonella enteritidis in Mice. Microbiology and Immunology, v. 36, p. 885-893, 1990.
ASLANZADEH, J.; PAULISSEN, L. J. Role of type 1 and type 3 fimbrial on the adherence and pathogenesis of Salmonella enterica in mice. Microbiology and Immunology, v. 36, p. 351-359, 1992.
ATLAS, R. M.; BEJ, A. K. Polymerase Chain Reaction. In: GERHARDT, P; MURRAU, R. G. E; WOOD, W. A.; KRIEG, N. R. Methods for General and Molecular Bacteriology. Washington, DC; American Society for Microbiology, 1994. p. 418-435. BARROW, P. A.; LOVELL, M. A. Experimental infection of egg-layng hens with Salmonella enterica phage type 4. Avian Pathology, v. 20, p. 335-348, 1991.
BARROW, P. A.; LOVELL, M. A. Funcional Homology of Virulence Plasmids in Salmonella gallinarum, Salmonella pullorum and Salmonella typhimurium. Infection and Immunity, v. 57, n, 10, p. 3136-3141, 1989.
BARROW, P. A.; LOVELL, M. A. The association between a large molecular mass plasmid and virulence in a strain of Salmonella pullorum. Journal of General Microbiology, v. 134, p. 2307-2316, 1988.
BARROW, P. A.; SIMPSON, J. M.; LOVELL. M. A. Intestinal colonization in the chicken by food-poisoning Salmonella serotypes, microbial characteristics associated with faecal excretion. Avian Pathology, v. 17, p. 571-588, 1988.

66
BARROW, P. A.; SIMPSON, J. M.; LOVELL, M. A.; BINNS, M. Contribuition of Salmonella gallinarum Large Plasmid toward Virulence in Fowl Typhoid. Infection and Immunity, v. 55, n. 2, p. 388-392, 1987.
BÄUMLER, A. J.; GILDE, A. J.; TSOLIS, R. M. van der VELDEN, A. W. M.; AHMER, B. M. M.; HEFFRON, F. Contribuition of horizontal gene transfer and deletion events to development of distinctive patterns of fimbrial operons during evolution of Salmonella Serotypes. Journal of Bacteriology, v. 179, n. 2, p. 317-322, 1997.
BÄUMLER, A. J.; TSOLIS, R. M.; BOWE, F. A.; KUSTERS, J. G.; HOFFMANN, S.; HEFFRON, F. The pef fimbrial operon of Salmonella typhimurium mediates adhesion to murine samll intestine and is necessary for fluid accumulation in the infant mouse. Infecction and Immunity, v. 64, n. 1, p. 61-68, 1996.
BICHLER, L. A.; NAGARAJA, K. V.; POMEROY, B. Plasmid Diversity in Salmonella enteritidis of Animal, Poultry, and Human Origin. Journal of Food Protection, v. 57, n. 1, p. 4-11, 1994.
BINKIN, N.; SCUDERI, G.; NOVACO,F.; GIOVANARDI, G. L.; PAGANELLI, G.; FERRARI, G.; CAPELLI, O.; RAVAGLIA, l.; ZILIOLI, F.; AMADEI, V.; MAGLIANI, W.; VIANI, E.; RICCO, D.; BONINI, B., MAGRI, M.; ALESSANDRINI, A; BURSI, G.; BARIGAZZI, G.; FANTASIA, M.; FILETICI, E.; SALMASO, S. Egg-related Salmonella enteritidis, Italy, 1991. Epidemiology and Infection. v. 110, p. 227-237, 1993.
BOOM, R.; SOL, C. J. A.; SALIMANS, M. M. M.; JANSEN, C. L.; WERTHEIM-van DILLEN, P. M. E.; VAN DER NOORDAA, J. Rapid and Simple Method for Purification of Nucleic Acids. Journal of Clinical Microbiology, v. 28, n. 3, p. 495-503, 1990.
BORLAND, E. D. Salmonella infection in poultry. The Veterinary Records, v. 97, p. 406-408, 1975.
BRASIL. Secretaria Estado da Saúde. Salmonella Enteritidis. Informe Técnico, mar. 1996.
BRYAN, F. L.; DOYLE, M. P. Health Risks and Consequences of Salmonella and Campylobacter jejuni in Raw Poultry. Journal of Food Protection, v. 58, n. 3, p. 326-344, 1995.

67
CHART, H.; THRELFALL, E. J.; ROWE, B. Virulence of Salmonella enteritidis phage type 4 is related to the possesion of a 38 MDa plasmid. Federation of European Microbiological Societie Microbiology Letters, v. 58, p. 299-303, 1989.
CLARK, M. A.; HIRST B. H.; JEPSON, M. A. Inoculum Composition and Salmonella Pathogenicity Island 1 Regulate M-Cell Invasion and Epithelial Destruction by Salmonella typhimurium. Infection and Immunity, v. 66, n. 2, p. 724-731, 1998.
CLARK, M. A.; REED, K. A.; LODGE, J.; STEPHEN, J.; HIRST, B. H.; JEPSON, M. A. Invasion of Murine Intestinal M Cells by Salmonella typhimurium inv Mutants Severely Deficient for Invasion of Cultured Cells. Infection and Immunity, v. 64, n. 10, p. 4363-4368, 1996.
CLOUTHIER, S. C.; MÜLLER, K-H.; DORAN, J. L.; COLLINSON, S. K.; KAY, W. W. Characterization of three fimbrial genes, sefABC, of Salmonella enteritidis. Journal of Bacteriology, v. 175, n. 9, p. 2523-2533, 1993.
COLLIGHAN, R. J.; WOODWARD, M. J. The SEF14 fimbrial antigen of Salmonella enterica serovar Enteritidis is encoded within a pathogenicity islet. Veterinary Microbiology, v. 80, p. 235-245, 2001.
COLLINSON, S. K.; EMÖDY, L MÜLLER, K-H.; TRUST, T. J.; KAY, W. W. Purification and Characterization of Thin, Aggregative Fimbriae from Salmonella enteritidis. Journal of Bacteriology, v. 173, n. 15, p. 4773-4781, 1991.
COSTA, F. N.; ROSSI JUNIOR, O. D.; NADER FILHO, A.; TAVECHIO, A. T. Sorovares de Salmonella isolados de carcaças e cortes de frango obtidos na indústria e no comércio em Jaboticabal, Estado de São Paulo, em 1996. Revista Brasileira de Ciência Veterinária, v. 4, n. 3, p. 97-100, 1997.
DOYLE, M. P. Salmonella. In: CLIVER, D. O. Foodborne diseases. San Diego : Academic Press, 1990. p. 185-204.
DUGUID, J. P.; ANDERSON, E. S.; CAMPBRLL, I. Fimbriae and adhesive properties in Salmonellae. Journal Path. Bact. , v. 92, p. 107-138, 1966.
DUGUID, J. P.; NORTH, R. A. E. Egg and salmonella food-poisoning: an evaluation. Journal of Medical Microbiology, v. 34, p. 65-72, 1991.

68
EDWARDS, R. A.; SCHIFFERLS, D. M; STANLEY, R. M. A role for Salmonella fimbriae in intraperitoneal infections. Proceedings of National Academy of Science from USA. v. 97, n. 3, p. 1258-1262, 2000.
EL-GEDAILY, A.; PAESOLD, G.; KRAUSE, M. EXPRESSION Profile and Subcelullar Location of the Plasmid-Encoded Virulence (Spv) Proteins in Wild-Type Salmonella dublin. Infection and Immunity, v. 65, n. 8, p. 3406-3411, 1997.
ERNEST, R. K.; DOMBROSKI, D. M.; MERRIK, J. M. Anaerobiosis, Type 1 Fimbriae, and Growth Phase Are Factores That Affect Invasion of Hep-2 Cells by Salmonella typhimurium. Infection and Immunity, v. 58, n. 6, p. 2014-2016, 1990.
FEUTRIER, J.; KAY, W. W.; TRUST, T. J. Cloning and Expression of a Salmonella enteritidis Fimbrin Gene in Escherichia coli. Journal of Bacteriology, v. 170, n. 9, p. 4216-4222, 1988.
FEUTRIER, J.; KAY, W. W.; TRUST, T. Purification and Characterization of Fimbriae from Salmonella enteritidis. Journal of Bacteriology, v. 168, n. 1, p. 221-227, 1986.
FIERER, J.; ECKANN, L.; FANG, F.; PFEIFER, C.; FINLAY, B. B.; GUINEY, D. Expression of the Salmonella virulence plasmid gene spvB in cultured macrophages and nonphagocytic cell. Infection and Immunity, v. 61, n. 12, p. 5231-5236, 1993.
FIERER, J.; KRAUSE, M.; TAUXE, R.; GUINEY, D. Salmonella typhimutium Bacteremia: Association with the Virulence Plasmid. The Journal of Infections Diseases, v. 166, p. 639-642, 1992.
FINLAY, B.; FALKOW, S. Common Themes in Microbial Pathogenicity. Microbiological Reviews, v. 53, n. 2, p. 210-230, 1989.
FINLAY, B.; FALKOW, S. Salmonella Interactions whit Polarized Human Intestinal Cells. The Journal of Infecctions Diseases, v. 162, p. 1096-1106. 1990.
FRIEDRICH, M. J.; KINSEY, N. E.; VILA, J.; KADNER, R. J. Nucleotide sequence of a 13,9 kb segment of the 90kb virulence plasmid of Salmonella typhimurium: the presence of fimbrial biosynthetic genes. Molecular Microbiology, v. 8, n. 3, p. 543-558, 1993.
GALÁN, J. E.; CURTISS III, R. Cloning and molecular characterization of genes whose products allow Salmonella typhimurium to penetrate tissue culture cells.

69
Procceedings of Natlonal Academy of Science from USA, v. 86, p. 6383-6387, 1989.
GALÁN, J. E.; CURTISS III, R. Distribuition of the invA, -B, C, and –D Genes of Salmonella typhimurium among Other Salmonella Serovars invA Mutants of Salmonella typhi Are Deficient for Entry into Mammalian Cells. Infection and Immunity, v. 59, n. 9, p. 2901-2908, 1991.
GALÁN, J. E.; GINOCCHIO, C.; COSTEAS. Molecular and Functional Characterization of the Salmonella Invasion Gene invA: Homology of INVA to Members of a New Protein Family. Journal of Bacteriology, v. 174, n.13, p. 4338-4349), 1992.
GAST, R. K. Paratyphoid Infections. In: BARNES, H. J.; BEARD, C. W.; McDOUGALD, L. R.; SAIF, Y. M. Ed. CALNEK, B. W. Diseases of Poultry. 10th. Ames: Iowa State University Press. 1997. p. 97-122. GAST. R. K.; MITCHELL, B. W.; HOLT, P. S. Airbone transmission of Salmonellla enteritidis infection between groups of chicks in controlled-environment isolation cabinets. Avian Diseases, v. 42, n.2, p. 315-320, 1998.
GERNER- SMIDT, P.; WEGENER, H. C. Salmonella enterica serovar Enteritidis in Demmark. In: SAEED, A. M.; GAST, R. K.; POTTER, M. E.; WALL, P. G. Salmonella enterica serovar Enteritidis in humans and animals. Ames: Iowa State University Press 1999. p. 63-69.
GINOCCHIO, C.; OLMSTED, S. B.; GÁLAN, J. E. Contact with Epithelial Induces the Formation of Surface Appendages on Salmonella typhimurium. Cellular, v. 76, p. 717-724, 1994.
GRIMONT, P. A. D.; BOUVET, P.; GRIMONT, F.; DESENCLOS, J. C. Salmonella enterica Serovar Enteritidis in France: Epydemiology, Prevention and Control. In: SAEED, A. M.; GAST, R. K.; POTTER, M. E.; WALL, P. G. Salmonella enterica serovar Enteritidis in humans and animals. Ames: Iowa State University Press, 1999. p. 43-49.
GRUENEWALD, R.; BLUM, S.; CHAN, J. Relationship between human immunodeficiency virus infection and salmonelosis in 20-to 59 –Year-Old Residents of New York City. Clinical Infections Diseases, v. 18, p. 358-363, 1994.

70
GUILOTEAU, L. A.; WALLIS, T. S.; GAUTIER, A. V.; MacINTYRE, S.; PLATT, D. J.; LAX, A. J. The Salmonella virulence plasmid enhaces Salmonella-induced lysis og macrophages and influences inflammatory responses. Infecction and Immunity, v. 68, n. 8, p. 3385-3393, 1996.
GULIG, P. A.; CURTISS III, R. Plasmid-Associated Virulence of Salmonella typhimurium. Infection and Immunity, v. 55, n. 12, p. 2891-2901, 1987.
GULIG, P. A.; DOYLE, T. J. The Salmonella typhimurium Virulence Plasmid Increases the Growt Rate of Salmonellae in Mice. Infection and Immunity, v. 61, n. 2, p. 501-511, 1993.
GULIG, P. A.; DANBARA, H.; GUINEY, D. G.; LAX, A. J.; NOREL, F.; RHEN, M. Molecular anslysis of spv virulence genes of the salmonella virulence plasmids. Molecular Microbiology, v. 7, n. 6, p. 825-830, 1993.
GUINEY, D. G.; LIBBY, S.; FANG, F. C.; KRAUSE, M.; FIERER, J. Growth-phase regulation of plasmid virulence genes in Salmonella. Trends in Microbiology, v. 3, n. 7, p. 275-279, 1995.
HACKER, J.; BLUM-OEHLER, G.; MÜHLDORFER, I.; TSCHÄPE, H. Pathogenicity islands of virulent bacteria: structure, function and impact on microbial evolution. Molecular Microbiology, v. 23, n. .6, p. 1089-1097, 1997.
HALAVATKAR, H.; BARROW, P. A. The role a 54-kb plasmid in the virulence of strains of Salmonella Enteritidis of phage type 4 for chickens and mice. Journal of Medical Microbiology, v. 38, p. 171-176, 1993.
HEFFERNAN, E, J.; FIERER, J.; CHIKAMI, G.; GUINEY, D. Natural History of Oral Salmonella dublin Infection in BALB/c Mice: Effect of na 80-kilobase-pair Plasmid on Virulence. The Journal of Infections Diseases, v. 155, n. 6, 1987.
HELDBERG, C. W.; DAVID, M. J.; WHITE, K. E.; MACDONALD, K. L.; OSTERHOLM, M. T. Role of Egg Consumption in Sporedic Salmonella enteritidis and Salmonella typhimurium Infections in Minnesota. The Journal of Infection Diseases, v. 167, p. 107-111, 1993. HENDERSON, S. C.; BOUNOUS, D. I.; LEE, M. D. Early events in the pathogenesis of avian salmonellosis. Infection and Immunity, v. 67, n. 7, p. 3580-3586, 1999.

71
HINTON, M.; THREFALL, E. J.; ROWE, B. The invasiveness of different strains of Salmonella enteritidis phage type 4 for young chickens. Federation of European Microbiological Societies Microbiology Letters, v. 70, p. 193-196, 1990.
HOFER, E.; SILVA FILHO, S. J.; REIS, E. M. F. Prevalência de Sorovares de Salmonella Isolados de Aves no Brasil. Pesquisa Veterinária Brasileira, v. 17, n. 2, p. 55-62, 1997.
HOFER, E.; SILVA FILHO, S. J.; REIS, E. M. F. Sorovares de Salmonella isolados de matérias-primas e de ração para aves no Brasil .Pesquisa Veterinária Brasileira, v. 18, n. 1, p. 21-27, 1998.
HORIUCHI, S.; INAGAKI, Y.; OKAMURA, N.; NAKAYA, R.; YAMAMMOTO, N. Type 1 Pili Enhance the Invasion of Salmonella braenderup and Salmonella typhimurium to HeLa Cells. Microbiology and Immunology, v. 36, n. 6, p. 593-602, 1992.
JONES, B. D.; GHORI, N.; FALKOW, S. Sallmonella typhimurium Initiates Murine Infection by Penetrating and Destroying the Specialized Epithelial M Cells of the Peyer’s Patches. The Journal of Experimental Medicine, v. 180, p. 15-23, 1994. JONES, G. W.; RABERT, D. K.; SVINARICH, D. M.; WHITFIELD, H. J. Association of Adhesive, Invasive, and Virulent Phenotypes pf Salmonella typhimurium with Autonomous 60-Megadalton Plasmids. Infection and Immunity, v. 38, n.2, p.476-486, 1992.
JOHNSON, J. L. Determination of DNA Base Composition. Methodos in Microbiology. In: Academic Press. Inc. London, v. 18, p. 1-29, 1985.
LIBBY, S.; ADAMS, L. G.; FICHT, T. A.; ALLEN, C.; WHITFORD, H. A.; BUCHMEIER, N. A.; BOSSIE, S.; GUINEY, D. The spv Genes the Salmonella dublin Virulence Plasmid Are Requerid for Severe Enteritis and Systemic Infection in the Natural Host. Infection and Immunity, v. 65, n. 5, p. 1786-1792, 1997.
LÍRIO, V. S.; SILVA, E. A.; STEFONI, S.; CAMARGO, D; RECCO, E. A P.; MALUF, Y. T.; MIYAZAWATT, T. T.; NEVES, D. V. D. A; OLIVEIRA, V. M. R. Freqüência de 17 sorotipos de Salmonella isolados em alimentos. Higiene Alimentar, v. 12, n. 55, p. 36-42, 1998.
MARCUS, S. L; BRUMELL, J. H.; PFEIFER, G, FINLAY, B. B. Salmonella pathogenicity islands: big virulence in small packages. Microbes and Infection, v. 2, p. 145-156, 2000.

72
MECSAS, J.; STRAUSS, E. J. Molecular Mechanisms of Bacterial Virulence: Type III Sacretion and Pathogenicity Islands. Emerging Infections Diseases, v. 2, n. 2, p. 271-285, 1996.
MC ILROY, S. G.; MC CRACKEN, R. M. NEILL, S. D.; O’BRIEN, J. J. Control, prevention and eradication of Salmonella enteritidis infection in broiler and broiler breeder flocks. The Veterinary Record, v. 125, p. 545-548, 1989.
MILDVAN, D.; MATHUR, U.; ENLOW, R. W.; ROMAIN, P. L.; WINCHESTER, R. J.; COLP, C.; SINGMAN, H.; ADELSENBERG, B.; SPIGLAND, I. Opportunistic Infections and Immune in Homosexual Men. Annais of Internacional Medicine, v. 96, n. 1, p. 700-704, 1982.
MORSE, D. L. BIRHEAD, G. S.; GUARDINO, J.; KONDRACKI, S. F.; GUZEWICH, J. J. Outbreak and Sporadic Egg-Associated Cases of Salmonella enteritidis: New York’s Experience. American Journal of Public Health, v. 84, p. 859-860, 1994.
MÜLLER, K. H.; COLLINSON, S. K.; TRUST, T. J.; KAY, W. W. Type 1 Fimbriae of Salmonella enteritidis. Journal of Bacteriology, v. 173, n. 15, p. 4765-4772, 1991. MUNRO, D. S.; GIRDWOOD, R. W. A; REILLY, W. J. Salmonella enterica Serovar Enteritidis in Scotland. In: SAEED, A. M.; GAST, R. K.; POTTER, M. E.; WALL, P. G. Salmonella enterica serovar Enteritidis en humans and animals. Ames: Iowa State University Press, 1999. p. 27-31. NAKAMURA, M.; SATO, S.; OHYA, T.; SUZUKI, S.; IKEDA, S. Possible Relationship of a 36-Megadalton Salmonella enteritidis Plasmid to Virulence in Mice. Infection and Immunity, v. 47, n. 3, p. 831-833, 1985.
NAKAMURA, M.; TAKAGI, M.;TAKAHASHI, T.;SUKUZIS, S.; SOTOS, S.;TAKEHARA, K. The effect of the flow on horizontal transmission of Salmonella enteritidis in chickens. Avian Diseases, v. 41, n. 2, p. 354-360, 1997.
NAUGHTON, P. J.; GRANT, G.; BARDOCZ, S.; ALLEN-VERCOE, E.; WOODWARD, J.; PUSTZAI, A. Expression of type 1 fimbriae (SEF21) of Salmonella enterica serotype Enteritidis in the early colonization of the rat intestine. Journal of Medical Microbiology, v. 50, p. 191-197, 2001.

73
NUNES, A I. Salmonella Enteritidis – fagotipos, susceptibilidade a drogas antimicrobianas e epidemiologia molecular baseada na sonda complementar ao rRNA. 1999. 119 f. Tese (Doutorado). Instituto de Ciências Biomédicas, Universidade de São Paulo, São Paulo, 1999.
NUNES, A. I.; HELMUTH, R.; SCHROETER, A.; MEAD, G. C.; SANTOS, M. A. A.; SOLARI, S. A.; SILVA, O. R.; FERREIRA, A. J. P. Phage Typing of Salmonella Enteritidis from Different Sources in Brazil. Journal of Food Protection, v. 66, n. 2, p. 324-327, 2003.
OGUNNIYI, A. D.; KOTLARSKI, I.; MORONA, R.; MANNING, P. A. Role of SefA Subunit Protein of SEF14 Fimbriae in the Pathogenesis of Salmonella enterica serovar Enteritidis. Infection and Immunity, v. 65, n. 2, p. 708-717, 1997.
PASSARO, D. J.; REPORTER, R.; MASCOLA, L.; KILMAN, L.; MALCOM, G. B.; ROLKA, H.; WERNER, B.; VUGIA, D. J. Epidemic Salmonella enteritidis Infection in Los Angeles Country, California. The Predominance of Phage Type 4. West of Journal Medical, v. 165, p. 126-130, 1996.
PENHEITER, K. L.; MATHUR, N.; GILES, D.; FAHLEN, T.; JONES, B. D. Non-invasive Salmonella typhimurium mutants are avirulent brcause of na inability to enter and destroy M cells of ileal Peye´s patches. Molecular Microbiology, v. 24, n. 4, p. 697-709, 1997.
PERALES, I.; AUDICANA, A. The role of hen’s eggs in outbreaks of salmonellosis in north Spain. Internacional Journal of Food Microbiology, v. 8, n. 175-180, 1989.
PERALTA, R. C.; YOKOYAMA, H.; IKEMORI, Y.; KUROKY, M.: KODOMA, Y. Passive immunisation against experimental salmonellosis in mice by orally administred hen egg-yolk antibodies specific for 14-kDa fimbriae of Salmonella Enteritidis. Journal of Medical Microbiology, v. 41, p. 29-35, 1994.
PLANO, G. V.; DAY, J. B.; FERRACCI, F. Type III export: new uses for na old pathway. Molecular Microbiology, v. 40, n. 2, p. 284-293, 2001.
POPOFF, M. Y.; BOCKEMÜEL, J.; BRENER, F. W. Supplement 1997 (no. 41) to the Kauffmann-White scheme. Research in Microbiology, v. 149, p. 601-604, 1998.
POPOFF, M. Y.; LE MINOR, L. L. Antigenic formulas of Salmonella servars. In: National Salmonella Center by the who collaborating Centre for Reference and Research on Salmonella. Paris: Institute Pasteur, 1997.

74
POPPE, C. Epidemiology of Salmonella enterica serovar Enteritidis In: SAEED, A. M.; GAST, R. K.; POTTER, M. E.; WALL, P. G. Salmonella enterica serovar Enteritidis in humans and animals. Ames: Iowa State University Press, 1999. p. 3-18.
POPPE, C.; DEMCZUK, W., MC FADDEN, K., JOHNSON, R. R. Virulence of Salmonella enteritidis phage types 4, 8 and 13 and other Salmonella spp for day-old-chicks, hens and mice. Canadian Journal Veterinary Research, v.57, p. 281-287, 1993.
PORTER, S. B.; CURTIS III, R. Effect of inv Mutations on Salmonella Virulence and Colonization in 1-Day-Old White Leghorn Chicks. Avian Diseases, v. 41, n.1, p. 45-47, 1997.
QUINTAES, B. R.; REIS, E. M. F.; RODRIGUES, D. P.; HOFER, E. Marcadores epidemiológicos em Salmonella Enteritidis. In: CONGRESSO BRASILEIRO DE MICROBIOLOGIA, Rio de Janeiro, 1997. Anais…, 1997. p.54.
RAHN, K.; De GRANDIS, S. A.; CLARKE, R. C.; McEWEM, S. A.; GÁLAN, J. E; GINOCHIO, C.; CURTISS III, R.; GYLES, C. L. Amplification of an invA sequence of Salmonella typhimurium by polimerase chain reaction as a specific method of detection of Salmonella. Molecular and Cellular Probes, v. 6, p. 271-279, 1992.
RAJASHERA, G.; MUNIR, S.; ALEXEYER, M. F.; HAHVORSON, A. D.; WELLS, C. L.; NAGAJARA, K. V. Pathogenic Role of SEF14, SEF17 and SEF21 Fimbriae in Salmonella enterica Serovar Enteritidis Infection of Chickens. Applied and Environmental Microbiology, v. 66, n. 4, p. 1759-1763, 2000.
RAJASHEKARA, G.; MUNIR, S.; LAMICHHANE, C. M.; BACK, A.; KAPUR, V.; HALVORSON, D. A.; NAGAJARA, K. V. Application of Recombinant Fimbrial Prorein for the Specific Detection of Salmonella enteritidis Infection in Poultry. Diagnostic in Microbioloigy Infection Disease, v .32, p. 147-157, 1998.
RODRIGUE, D. C.; TAUXE, R. V.; ROWE, B. Internacional increase in Salmonella enteritidis: A new pandemic? Epidemiology and Infection, v. 105, p. 12-27, 1990.
ROTGER, R.; CASADÉSUS, J. The viruelnce plasmids of Salmonella. Internacional Microbiology, v. 2, p. 177-184, 1999.

75
ROUDIER, C.; FIERER, J, GUINEY, D. G. Characterization of translation termination mutations in the spv operon of the Salmonella virulence plasmid pSDL2. Journal of Bacteriology, v. 174, n. 20, p. 6418-6423, 1992.
SCHMID, H.; BAUMGARTNER, A. Salmonella enterica Serovar Enteritidis in Switzerland: Recognition, Development, and control of the Epidemic. In: SAEED, A. M.; GAST, R. K.; POTTER, M. E.; WALL, P. G. Salmonella enterica serovar Enteritidis in humans and animals. Ames: Iowa State University Press, 1999. p. 81-91.
SHIVAPRASAD, H. L.; TYMONEY, J. F.; MORALES, B. L., BAKER, R. C. Pathogenesis of Salmonella enteritidis Infection in Laying Chickens. I. Studies on Egg Transmission, Clinical Signes, fecal Shedding, and Serologic Responses. Avian Diseases, v. 34, p. 548-557, 1990.
SCUDERI, G. Epydemiology of Salmonella enterica Serovar Enteritidis Infections in Italy. In: SAEED, A. M.; GAST, R. K.; POTTER, M. E.; WALL, P. G. Salmonella enterica serovar Enteritidis in humans and animals. Ames: Iowa State University Press, 1999. p. 111-115.
SOHEL, I; PUENTE, J. L.; MURRAY, W. J.; VUOPIO-VARKILA, J.; SCHOOLNIK, G. K. Cloning and characterization of the bundle-forming pilin gene of enteropathogenic Escherichia coli and its distribuition in Salmonella serotypes. Molecular Microbiology, v. 7, n. 4, p. 563-575, 1993.
STOKES, J. L.; OSBORNE, W. W. BAYNE, H. G. Penetration and growth of Salmonella in shell eggs. Food Research, v. 21, p. 510-518, 1956.
SWAMY, S. C.; BARNHART, H.; LEE, M. D.; DREESEN. Virulence determinants invA and spvC in Salmonellae isolated from poultry products, Wastewater, and human sources. Applied and Environmental Microbiology, v. 62, n. 10, p. 3768-3771, 1996.
TAVECHIO, A. T.; FERNANDES, S. A.; NEVES, B. C.; DIAS, A. M. G.; IRINO, K. Changing Patterns of Salmonella Serovars: Increase of Salmonella Enteritidis im São Paulo, Brazil. Revista do Instituto de Medicina Tropical de São Paulo, v. 38, n. 5, p. 315-322, 1996.
TAUNAY, A. E.; FERNANDES, S. A.; TAVECHIO, A T.; NEVES, B. C.; DIAS, A. M. G.; IRINO, K. The role of public helth laboratory in the problem of salmonellosis in São Paulo, Brazil. Revista do Instituto de Medicina Tropical de São Paulo, v. 38, n. 2, p. 119-127, 1996.

76
TAYLOR, G. R. Polymerase chain reaction: basic principles and automation. In: Pherson, M. J.; Quirke, P.; Taylor, G. PCR: A Practical approach. Oxford: Oxford University Press,1991. p. 1-14.
THIAGARAJAN, D.; SAEED, M.; TUREK, J.; ASEM, E. K. In vitro attachment and invasion of chicken ovarian granulosa cell by Salmonella enteritidis phage type 8. Infecction and Immunity, v. 64, n. 12, p. 5015-5021, 1996-a.
THIAGARAJAN, D; THACKER, H. L.; SAEED, A. M.. Experimental Infection of Laying Hens with Salmonella enteritidis Strains that Express Different Types of Fimbriae. Poultry Science, v. 75, p. 1365-1372, 1996-b.
THIEL, W. Salmonella enterica Serovar Enteritidis. Phage Type in Austria: From Understanding to Intervention. In: SAEED, A. M.; GAST, R. K, POTTER, M. E.; WALL, P. G. Salmonella enterica serovar Enteritidis in humans and animals. Ames: Iowa State University Press, 1999. p. 91-109.
THORNS, C.; SOJKA, M. G. CHASEY, D. Detection of a Novel Fimbrial Structure on the Surface of Salmonella enteritidis by using a Monoclonal Antibody. Journal of Clinical Microbiology, v. 28, n. 11, p. 2409- 2414, 1990.
THORNS, C. J.; TURCOTTE, C.; GEMMELL, C. G.; WOODWARD, M. J. Studies into the role the SEF14 fimbrial antigen in the pathogenesis of Salmonella enteritidis. Microbial Pathogenesis, v. 20, p. 235-246, 1996.
TSCHÄPE, H.; LIESENGANG, A.; GERICKE, B, PRAGER, R.; RABSCH, W.; HELMUTH, R. Ups and Downs of Salmonella enterica Serovar Enteritidis in Germany. In: SAEED, A. M.; GAST, R. K.; POTTER, M. E.; WALL, P. G. Salmonella enterica serovar Enteritidis in humans and animals. Ames: Iowa State University Press, 1999. p. 51-61.
TURCOTTE, C; WOODWARD, M. J. Cloning DNA nucleotide sequence and distribuition of the gene encoding the SEF14 fimbrial antigen of Salmonella enteritidis. Journal of General Microbiology, v. 139, p. 1477-1485, 1993.
VALONE, S. E.; CHIKAMI, G. K.; MÜLLER, V. L. Stress Induction of the Virulence Proteins (SpvA, -B, -C) from Native Plasmid pSDL2 of Salmonella dublin. Infection and Immunity, v. 61, n. 2, p. 707-713, 1993.

77
VAN DE GIESSEN, A. W.; VAN LEEWWEN, W. J.; VAN PELT, W. Salmonella enterica Serovar Enteritidis in the Netherlands: Epidemiology, Prevention and Control. In: SAEED, A. M.; GAST, R. K.; POTTER, M. E.; WALL, P. G. Salmonella enterica serovar Enteritidis in humans and animals. Ames: Iowa State University Press, 1999. p. 71-79.
VAN DER VELDEN, A. W. M.; BÄUMLER, A. J.; TSOLIS, R. M.; HEFRON, F. Multiple Fimbrial Adhesins Are Requerid for Full Virulence of Salmonella typhimurium in Mice. Infection and Immunity, v. 66, n. 6, p. 2803-2808, 1998.
ZHOU, D.; GALÁN, J. Salmonella entry into host cell: the work in concert of type III secreted effector proteins. Microbes and Infection, v. 3, p. 1293-1298, 2001.
WALL, P. G.; WARD, L. R. Epidemiology of Salmonella enterica Serovar Enteritidis Phage Type 4 in England and Wales. In: SAEED, A. M.; GAST, R. K.; POTTER, M. E.; WALL, P. G. Salmonella enterica serovar Enteritidis in humans and animals. Ames: Iowa State University Pres, 1999. p. 19-25. WALLIS, T. S.; PAULIN, S. M.; PLESTED, J. S.; WATSON, P. R.; JONES, P. W. The Salmonella Dublin Virulence Plasmid Mediates Systemic but Not Enteric Phases of Salmonellosis in Cattle. Infection and Immunity, v. 63, n. 7, p. 2755-2761, 1995. WONG, S. S. Y.; YUEN, K. Y.; YAM, W. C.; LEE, T. Y.; CHAU, P. Y. Changing epidemiology of human salmonellosis in Hong Kong, 1982-93. Epidemiology and Infection, v. 113, 425-434, 1994.
WOOD, J. D.; CHALMERS, G. A.; FENTON, R. A.; PRITCHARD, J.; SCHOONDERWOERD, M.; LICHTEMBERG, W. L. Persistent shedding of Salmonella enteritidis from the udder of a cow. Canadian Veterinary Journal, v. 32, p. 738-741, 1991.
WOODWARD, M. J.; ALLEN-VERCOE, E.; REDSTONE, J. S. Distribuition, gene sequence and expression in vivo of the plasmid encoded fimbrial antigen of Salmonella serotype Enteritidis. Epidemiology and Infection, v. 117, p. 17-28, 1996.
WOODWARD, M. J.; McLAREN, I.; WRAY, C. Distribuition of virulence plasmids within Salmonellae. Journal of General Microbiology, v. 135, p. 503-511, 1989.


















