DIVERSIDADE GENÉTICA DE Attalea vitrivir (ARECACEAE ... · da bolsa de mestrado. À EMBRAPA...
29
UNIVERSIDADE ESTADUAL DE MONTES CLAROS PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS BIOLÓGICAS – PPGCB 1 BÁRBARA RAYANE RAMOS MUNIZ DIVERSIDADE GENÉTICA DE Attalea vitrivir (ARECACEAE): SUBSÍDIOS PARA CONSERVAÇÃO Montes Claros – MG Fevereiro 2016
Transcript of DIVERSIDADE GENÉTICA DE Attalea vitrivir (ARECACEAE ... · da bolsa de mestrado. À EMBRAPA...

UNIVERSIDADE ESTADUAL DE MONTES CLAROS
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS BIOLÓGICAS – PPGCB
1
BÁRBARA RAYANE RAMOS MUNIZ
DIVERSIDADE GENÉTICA DE Attalea vitrivir (ARECACEAE):
SUBSÍDIOS PARA CONSERVAÇÃO
Montes Claros – MG Fevereiro 2016

0
BÁRBARA RAYANE RAMOS MUNIZ
DIVERSIDADE GENÉTICA DE Attalea vitrivir (ARECACEAE):
SUBSÍDIOS PARA CONSERVAÇÃO
Co-orientador: Dr. Márcio de Carvalho Moretzsohn – EMBRAPA/DF
Co-orientadora: Dra. Vânia Cristina Rennó Azevedo – EMBRAPA/DF
Montes Claros – MG
Fevereiro 2016
Dissertação apresentada ao Programa de Pós-
Graduação em Ciências Biológicas, área de
concentração em Genética da Conservação da
Universidade Estadual de Montes Claros, como parte
dos requisitos para obtenção do título de Mestre em
Ciências Biológicas.
Orientador: Dr. Marcio Antonio Silva Pimenta

1
Dedicatória
Dedico este trabalho à minha mãe Bárbara Soraya e à minha avó Gê que sempre me fizeram
acreditar na concretização dos meus sonhos e colaboraram muito para que eu pudesse realizá-
los; estando sempre ao meu lado, entendendo-me nos momentos de ausência, dando-me
apoio, amor e carinho.

2
Agradecimentos Agradeço, primeiramente, a Deus, por estar comigo em todos os momentos e iluminando-me, sendo meu refúgio e fortaleza durante esta caminhada. A ele, minha eterna gratidão. À diversidade da vida por todas as possibilidades de investigação: a busca por respostas e a formulação de novas perguntas. Agradeço, especialmente, à minha família, afilhados, amigos e incentivadores do meu trabalho; pessoas maravilhosas que sempre estiveram ao meu lado. Ao meu noivo Júnior, cujo, o amor, dedicação e apoio sempre se fizeram presentes nessa trajetória. Ao Professor Marcio Antonio Silva Pimenta pela orientação, dedicação, paciência e amizade, será sempre uma referência de pessoa e profissional. Possibilitou-me aprendizagens únicas, por meio do grande incentivo durante essa jornada. À Universidade Estadual de Montes Claros (UNIMONTES) e ao Programa de Pós-graduação em Ciências Biológicas (PPGCB), através dos professores e funcionários, pela qualidade formação, logística e apoio no decorrer do mestrado. À Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) pela concessão da bolsa de mestrado. À EMBRAPA Recursos Genéticos e Biotecnologia - CENARGEN pelo apoio ao desenvolvimento do projeto nessa Unidade. Em especial ao Márcio de Carvalho Moretzsohn, que não mediu esforços para concretização desse projeto e a Vânia Cristina Rennó Azevedo. Ao Murilo, pela parceria e importante participação na realização desse trabalho.
Ao Elder (Dim), pela amizade e apoio constante em minha vida desde a infância. Aos colegas do Laboratório de Genética Vegetal da Embrapa Recursos Genéticos e Biotecnologia - CENARGEN que colaboraram; em especial a Bruna Jaqueline, Lorena, Pétala, Anadria e Lucineide; vocês foram fundamentais para o desenvolvimento do trabalho. Aos colegas do Laboratório de Genética da Conservação pelo convívio, amizade e apoio incondicional.
A todos vocês, muito obrigada!

3
“A persistência é o caminho do êxito”.
Charles Chaplin

4
SUMÁRIO 1 INTRODUÇÃO .................................................................................................................. 3
2 METODOLOGIA ............................................................................................................... 5
2.1 Área de Estudo e Coleta .................................................................................................. 5
2.2 Extração, Amplificação do DNA e Genotipagem ........................................................... 7
2.3 Análises dos Dados .......................................................................................................... 8
3 RESULTADOS ................................................................................................................. 9
3.1 Diversidade Genética ....................................................................................................... 9
3.2 Estrutura Genética das Populações ................................................................................ 10
3.3 Distância Genética ......................................................................................................... 12
4 DISCUSSÃO .................................................................................................................... 13
5 SUBSÍDEOS PARA CONSERVAÇÃO .......................................................................... 16
6 REFERÊNCIAS ............................................................................................................... 17

1
RESUMO
O babaçu (Attalea sp.) é uma palmeira nativa, pertencente a família Arecaceae, com
ampla distribuição pelo Brasil. A espécie Attalea vitrivir Zona ocorre na região Norte no
estado de Minas Gerais e Sudoeste Baiano, onde o Cerrado é o bioma predominante. O
babaçu é considerado o maior recurso oleífero nativo do mundo e um dos principais produtos
extrativos do Brasil, além de contribuir significativamente para a economia de alguns estados
brasileiros. Diante da importância econômica e social do babaçu, a análise da estrutura e
diversidade genética de populações de A. Vitrivir foi realizada em cinco áreas de duas regiões
distintas, para caracterizar geneticamente a espécie. Foram utilizados 21 pares de primers
microssatélites (SSR) transferidos com sucesso de espécie Orbignya phalerata (sinonímia
Attalea speciosa) para a espécie Attalea vitrivir. Altos valores de diversidade genética foram
encontrados (He: 0,633-0,750), maior dentro das populações do que entre as mesmas,
provavelmente, devido ao isolamento. O software STRUCTURE revelou que as cinco
populações amostradas foram reunidas em dois grupos distintos (K = 2). Coincide com as
regiões norte (maior diferenciação) e sul, e corroborou com os resultados da análise de
variância molecular (AMOVA) que indicou a maior proporção da variabilidade genética entre
indivíduos dentro das populações (93,29%) do que entre populações (6,71%). A alta
diversidade genética nas populações não incluídas na área de proteção ambiental evidencia a
necessidade de novas propostas de ampliação das áreas de conservação, uma vez que, a
ocorrência da espécie é restrita.
Palavras-Chave: Babaçu, Estrutura Genética, SSR, Conservação.

2
ABSTRACT The babassu (Attalea sp.) is a native palm tree belonging to Arecaceae family, with wide
distribution in Brazil. The Attalea vitrivir Zona occurs in the northern region in the state of
Minas Gerais and Bahia Southwest, where the Cerrado is the predominant biome. The
babassu is considered the largest native oil resource in the world and one of the main forest
products in Brazil, and to contribute significantly to the economies of some states. Given the
economic and social importance of babassu, analysis of the structure and genetic diversity of
populations of A. vitrivir was carried out in five areas of two distinct regions, to characterize
genetically the species. Twenty one pairs of microsatellite primers (SSR) were used success
transferred species Orbignya phalerata (synonymy Attalea speciosa) for the species Attalea
vitrivir . High genetic diversity values were found (He: 0.633 to 0.750), most within
populations than between them, probably due to the isolation. The STRUCTURE software
revealed that five populations sampled were grouped into two separate groups (K = 2).
Coincides with the northern regions (greater differentiation) and south, and corroborated the
results of the analysis of molecular variance (AMOVA) indicated that most of the genetic
variability among individuals within populations (93.29%) than among populations (6,71%).
The high genetic diversity in populations not included in the environmental protection area
shows the need for further expansion of proposed conservation areas, since the occurrence of
the species is restricted.
Key-Words: Babassu, Genetic structure, SSR, Conservation.

3
1 INTRODUÇÃO
A família Arecaceae, inicialmente classificada como Palmae, é composta por um
grupo de espécies conhecidas como palmeiras. É um dos principais troncos da evolução das
monocotiledôneas e, atualmente, é constituída por 252 gêneros e aproximadamente 2.600
espécies (Dransfield et al. 2008). A ocorrência natural no Brasil é de 38 gêneros e cerca de
270 espécies (Lorenzi et al. 2010). As palmeiras representam a terceira família botânica mais
importante para o ser humano (Johnson, 1998) e conforme Noblick (1996) depois das
gramíneas é o segundo grupo das monocotiledôneas em importância econômica (cosméticos).
Possuem ampla distribuição, abundância, produtividade e diversidade de usos, grande
importância alimentar, medicinal, sociocultural e econômica para populações locais
(Zambrana et al. 2007). Entretanto, a considerar a relevância, a diversidade e complexidade
sobre a família Arecaceae, estudos ainda são incompletos (Orozco-Segovia et al., 2003; Panza
et al., 2004; Eslabão et al. 2015).
O babaçu (Attalea sp.) é uma palmeira nativa, pertencente a família Arecaceae, com
ampla distribuição pelo Brasil. É considerado o maior recurso oleífero nativo do mundo e um
dos principais produtos extrativos do Brasil, que contribui de maneira significativa para a
economia de alguns Estados da Federação (Alves, 1984; Arrudas et al., 2014). Segundo
Teixeira e Milanez (2003), os babaçuais se distribuem em área de aproximadamente 14,5
milhões de hectares, por sete estados brasileiros (Maranhão, Tocantins, Piauí, Goiás, Minas
Gerais, Mato Grosso e Espírito Santo). O Maranhão é responsável por 90% da produção
nacional do Brasil, local em que se encontra a maior concentração e exploração da espécie
(Lorenzi et al., 1996; Matos et al., 2010). Segundo Frazão (2001), registra-se a ocorrência de
babaçus também no Suriname e na Bolívia (Beni, Pando e Santa Cruz). A palmeira babaçu é
um dos principais recursos naturais não madeireiros no Brasil e é utilizada por milhões de
pequenos agricultores como fonte de renda e alimentação (May et al., 1985; Neves et al.,
2013).
A classificação taxonômica das espécies de babaçu é complexa, uma vez que, foram
realizados poucos estudos do gênero, fato que dificulta a identificação adequada de tais
espécies (Anderson e Balick, 1988). A classificação do babaçu (Attalea sp.) inclui quatro
gêneros: Orbignya, Maximiliana, Scheelea e Attalea (Glassman; 1999). Os gêneros
classificados foram anteriormente considerados correlacionados com Attalea, com diferenças
observadas nas características das flores estaminadas (Henderson, 1995; Pintaud, 2008). No

4
entanto, a partir de 2008, a maioria dos taxonomistas concordou que estes três gêneros
deveriam ser incluídos em Attalea, razão pela qual a nova edição do livro “Genera Palmarum
(2008)” não mais os consideram válidos (Lorenzi et al., 2010). Na região norte do estado de
Minas Gerais, segundo Lorenzi et al. (2010), duas espécies são encontradas, Attalea vitrivir
Zona e Attalea compta Mart. . A Attalea vitrivir Zona é encontrada no norte do estado de
Minas Gerais e sudoeste da Bahia.
O Babaçu (Attalea vitrivir) ocorre na região norte no estado de Minas Gerais e
sudoeste baiano, onde o Cerrado é o bioma predominante (Lorenzi, 1996). Apresenta caule
solitário, pode chegar a 20 m de altura e 50 cm de diâmetro. Os frutos medem de 10 a 14 cm
de comprimento; com mesocarpo branco, farináceo e seco; endocarpo lenhoso, possuem entre
4 e 6 sementes, com endosperma branco e muito oleoso, folhas pinadas, flores pistiladas
(Lorenzi et al, 2010). De acordo com Araújo (2008), o fruto possui amêndoas comestíveis e
alto teor de óleo utilizado na alimentação, empregado em produção de cosmético,
medicamentos e possibilidade de uso para fabricação de biocombustíveis.
Devido à importância econômica e social do Babaçu (A. vitrivir), programas de
melhoramento genético vêm sendo desenvolvidos para aumentar a eficiência do sistema
produtivo (Souza et al., 2008; Silva, 2006). Uma forma eficiente de auxiliar os programas de
melhoramento e conservação da espécie é a análise da variabilidade genética, por meio de
marcadores moleculares, em que é possível detectar dissimilaridades entre diferentes acessos
em nível de DNA.
Os marcadores moleculares podem ser definidos como todo e qualquer fenótipo
molecular oriundo de um gene expresso ou de segmento específico do DNA. É considerado
ferramenta que pode acelerar a obtenção dos índices de diversidade genética (Aguiar, 2012).
Grande variedade de marcadores moleculares está disponível para as diferentes espécies de
vegetais (Borém; Caixeta, 2006). Dentre estes, os microssatélites ou SSR (Simple Sequence
Repeats) são os marcadores moleculares mais ricos em informações e eficientes em análise
genômica minuciosa, uma vez que, são codominantes presentes tanto nas regiões codificadas
como não codificadas distribuídas ao longo do genoma e possuem um alto grau de
polimorfismo, bem como requerem pequena quantidade de DNA pelo uso da técnica PCR
(Polymerase Chain Reaction) (Ritschelet et al., 2004; Agarwal, 2008; Lin et al., 2010; Kalia
et al, 2011).
O emprego da tecnologia de microssatélites envolve algumas limitações, como alto
custo e a grande quantidade de trabalho necessário para o desenvolvimento de primers
específicos para os locos de microssatélites de cada espécie. A presença de sítios

5
microssatélites conservados entre espécies relacionadas, possibilita o aproveitamento de
primers, denominados heterólogos. Dessa forma, primers que foram desenvolvidos para uma
espécie podem ser empregados nas demais espécies do gênero (Ferreira; Grattapaglia, 1998;
Barbará et al., 2007; Rivas et al., 2013).
Muitas transferências de primers microssatélites de plantas tiveram resultados
positivos (Ritschel et al., 2004; Chen et al., 2010; Mnejja et al., 2010; Mistura et al., 2012,
Soares et al., 2013). A transferibilidade de marcadores microssatélites entre espécies
próximas evolutivamente favorece a obtenção rápida de informação genética e permite o
acesso à diversidade da espécie que se deseja estudar. Pode auxiliar no estabelecimento de
estratégias eficientes para conservação tornando-se uma alternativa econômica, além de
rápida para disponibilizar marcadores para a espécie desejada e potencializar estudos da
variabilidade genética (Ritschel et al., 2004; Leite et al., 2007; Peixoto et al., 2009; Rivas et
al., 2013). A transferência de primers da espécie Orbignya phalerata para a espécie A. vitrivir
foi obtida com sucesso, onde 21 pares de primers foram identificados para a espécie.
Assim o presente trabalho busca avaliar a diversidade genética em subpopulações
naturais da espécie A. vitrivir por meio da amplificação de regiões de microssatélites do DNA.
Para isso, serão investigadas as seguintes questões: (i) Qual o nível de variabilidade genética
de A. vitrivir dentro e entre as subpopulações estudadas? (ii) Existe isolamento genético entre
as áreas e/ou regiões? (iii) Os níveis de diversidade genética nas subpopulações localizadas na
Área de Proteção Ambiental do Rio Pandeiros (APA-Pandeiros), localizada no município de
Januária-MG, são suficientes para a conservação da espécie no local?
2 METODOLOGIA
2.1 Área de Estudo e Coleta
Foram selecionados babaçus localizados entre os municípios de Januária (Minas
Gerais) e Cocos (Bahia). A amostragem foi realizada em duas regiões principais, separadas
por aproximadamente 90 km, entre as quais não foi registrada a presença da espécie. As áreas
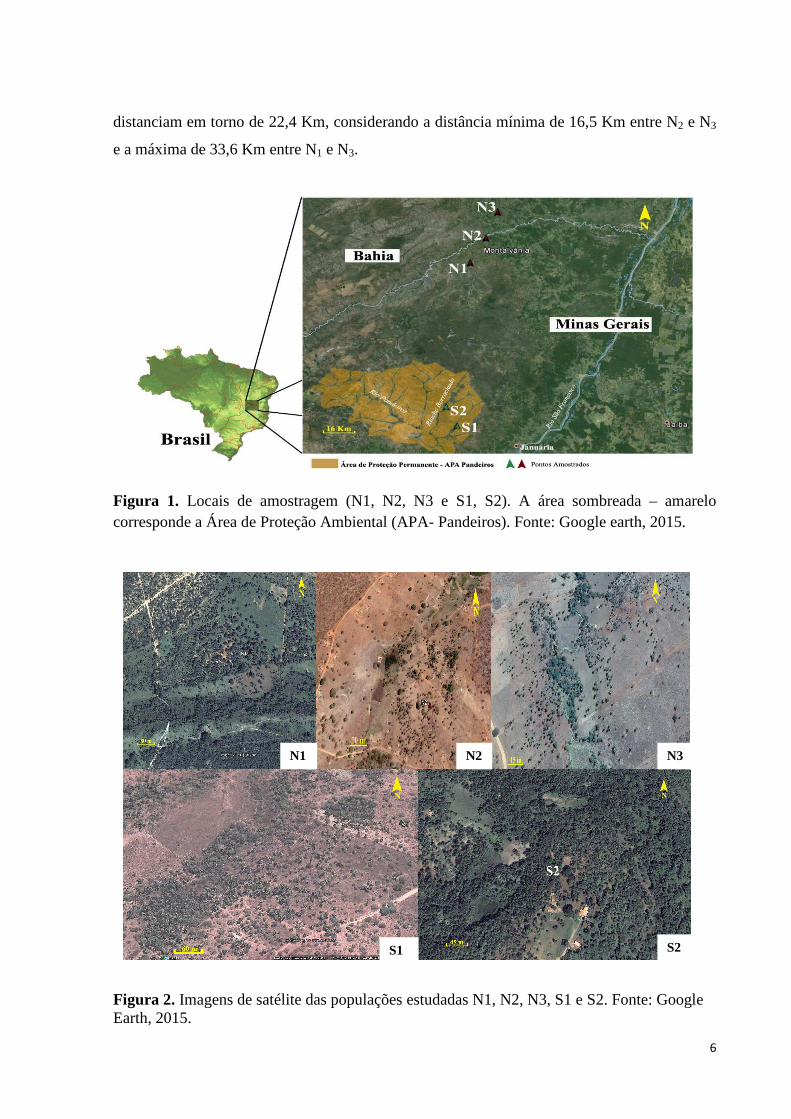
foram denominadas como região Norte (N), e Sul (S) (Figura 1).
Foram amostradas três populações de ocorrência de A. vitrivir na região Norte (N1,
N2, N3) e duas áreas na região Sul (S1 e S2) (Figuras 1 e 2). Na região Norte as áreas
6
distanciam em torno de 22,4 Km, considerando a distância mínima de 16,5 Km entre N2 e N3
e a máxima de 33,6 Km entre N1 e N3.
Figura 1. Locais de amostragem (N1, N2, N3 e S1, S2). A área sombreada – amarelo corresponde a Área de Proteção Ambiental (APA- Pandeiros). Fonte: Google earth, 2015.
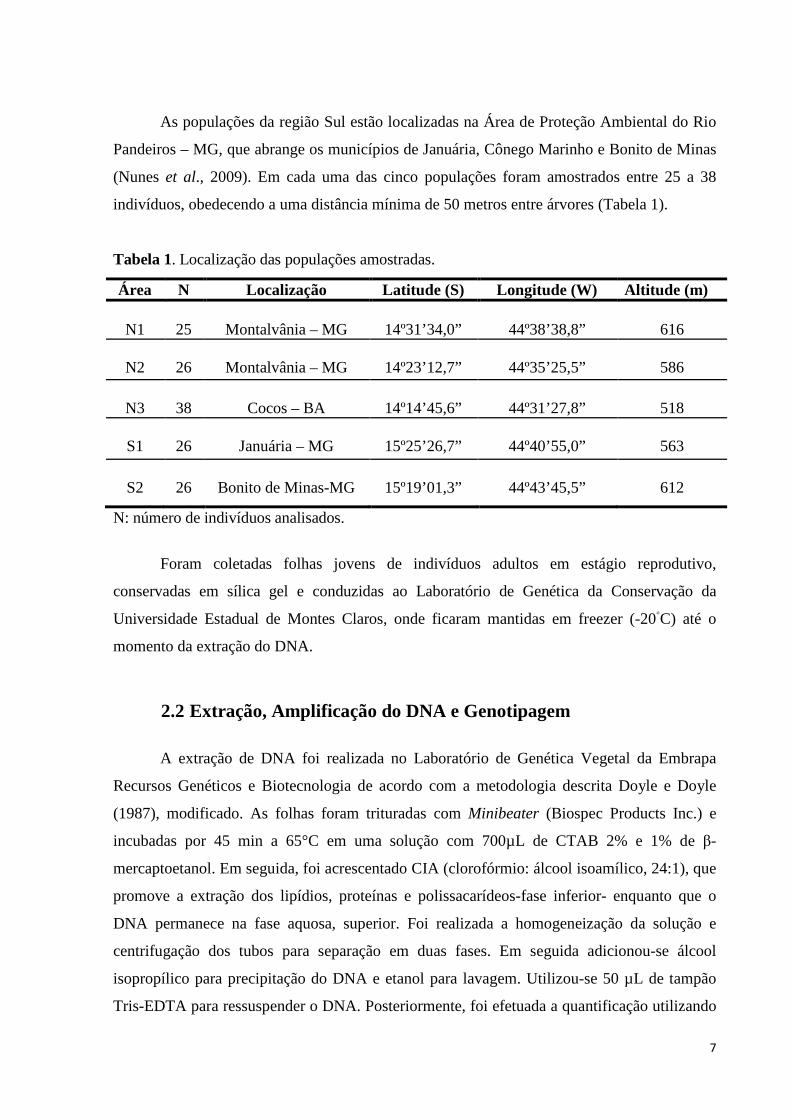
Figura 2. Imagens de satélite das populações estudadas N1, N2, N3, S1 e S2. Fonte: Google Earth, 2015.
N1 N3 N2
S2 S1
7
As populações da região Sul estão localizadas na Área de Proteção Ambiental do Rio
Pandeiros – MG, que abrange os municípios de Januária, Cônego Marinho e Bonito de Minas
(Nunes et al., 2009). Em cada uma das cinco populações foram amostrados entre 25 a 38
indivíduos, obedecendo a uma distância mínima de 50 metros entre árvores (Tabela 1).
Tabela 1. Localização das populações amostradas.
Área N Localização Latitude (S) Longitude (W) Altitude (m)
N1
25
Montalvânia – MG
14º31’34,0”
44º38’38,8”
616
N2 26 Montalvânia – MG 14º23’12,7” 44º35’25,5” 586
N3 38 Cocos – BA 14º14’45,6” 44º31’27,8” 518
S1 26 Januária – MG 15º25’26,7” 44º40’55,0” 563
S2 26 Bonito de Minas-MG 15º19’01,3” 44º43’45,5” 612
N: número de indivíduos analisados.
Foram coletadas folhas jovens de indivíduos adultos em estágio reprodutivo,
conservadas em sílica gel e conduzidas ao Laboratório de Genética da Conservação da
Universidade Estadual de Montes Claros, onde ficaram mantidas em freezer (-20◦C) até o
momento da extração do DNA.
2.2 Extração, Amplificação do DNA e Genotipagem
A extração de DNA foi realizada no Laboratório de Genética Vegetal da Embrapa
Recursos Genéticos e Biotecnologia de acordo com a metodologia descrita Doyle e Doyle
(1987), modificado. As folhas foram trituradas com Minibeater (Biospec Products Inc.) e
incubadas por 45 min a 65°C em uma solução com 700µL de CTAB 2% e 1% de β-
mercaptoetanol. Em seguida, foi acrescentado CIA (clorofórmio: álcool isoamílico, 24:1), que
promove a extração dos lipídios, proteínas e polissacarídeos-fase inferior- enquanto que o
DNA permanece na fase aquosa, superior. Foi realizada a homogeneização da solução e
centrifugação dos tubos para separação em duas fases. Em seguida adicionou-se álcool
isopropílico para precipitação do DNA e etanol para lavagem. Utilizou-se 50 µL de tampão
Tris-EDTA para ressuspender o DNA. Posteriormente, foi efetuada a quantificação utilizando

8
gel de agarose 1% e analise de comparação das amostras em relação à concentração da
amostra conhecida de DNA lambda.
Para caracterização genética A. vitrivir foram utilizados 21 pares de primers
microssatélites, desenvolvidos para a espécie Orbignya phalerata (Attalea speciosa) e
transferidos para a espécie em estudo. No total, foram genotipados 141 indivíduos da espécie,
nas cinco populações estudadas.
As reações de PCR foram realizadas com a utilização de Tampão [(10X Platinum HF
com MgCl2 600 mM, Tris-SO4 (pH 8,9), 180mM (NH4)2SO4, MgSO4)] na concentração de
1X; dNTPs na concentração de 1,3 mM; BSA na concentração de 1,3 mg/mL; primer
forward no concentração de 0,5 µM; primer reverse no concentração de 0,5 µM; Taq
polimerase na concentração de 0,2 u, DNA na concentração de 3 ng e água ultrapura (5,6
µL). O volume total de cada reação foi 13,0 µL; os primers foram marcados com
fluorescências 6-FAM (azul), HEX (verde). As amplificações foram realizadas em
termociclador ABI 9700 (Applied Biosystems), nas seguintes condições: 94ºC por 1 minutos
(1 ciclo), 95ºC por 1 min., 50-60°C por 1 min., 72°C por 1 min. (30 ciclos); e uma extensão
final a 72°C por 20 minutos (1 ciclo). Foram adicionados a 1µL do DNA amplificado, 8,5 µL
de Hi-Di formamida e 0,5 µL do padrão com comprimentos de fragmentos conhecidos,
marcado com fluorescência ROX. Para verificar o êxito da amplificação utilizou gel de
agarose 1% (corado com brometo de etídio) e o DNA lambda visualizados em
transiluminador de luz ultravioleta (UV) modelo Gel logic.
As amostras foram submetidas à eletroforese capilar em sequenciador automático de
DNA ABI-3730 (Applied Biosystems), pertencente ao Laboratório de Genética Vegetal, da
Embrapa Recursos Genéticos e Biotecnologia. Os dados foram analisados pelo programa
GeneMapper (Applied Biosystems).
2.3 Análises dos Dados
Com o auxílio do software GDA (Lewis, 2002) foram estimados o número de alelos
por loco, as frequências alélicas, a heterozigosidade esperada pelo equilíbrio de Hardy-
Weinberg (He) de Nei (1978), a heterozigosidade observada (Ho) e o índice de fixação
intrapopulacional (f). Através do software Arlequin versão 3.1 (Excoffier et al., 2005), foi
realizada a Análise de Variância Molecular (AMOVA) para verificar a distribuição da
diversidade genética entre e dentro das populações. O software STRUCTURE (Prichard et al.,

9
2000) foi utilizado para investigar a estrutura das populações e inferir o número de
agrupamentos geneticamente distintos (K) mais provável de todo o conjunto de dados.
Através do método de Nei (1978) foram obtidas as análises de diversidade genética de A.
vitrivir . O programa HICKORY (Holsinger e Lewis, 2005), foi utilizado para gerar valores de
heterozigosidade genética total (Ht) e heterozigosidade média dentro das populações (Hs),
pelo método Bayesiano; de diferenciação da população (GST-B) (Holsinger et al., 2002) e do
fluxo gênico (Nm) estimado pela fórmula Nm = 0.25(1−GST-B)/GST-B (McDermott, McDonald;
1997). A similaridade genética entre as matrizes de A. vitrivir foi calculada pelo coeficiente
de similaridade de Jaccard com o emprego do software NTSys (Rohlf, 2000) que gerou
dendrograma baseado no agrupamento UPGMA da distância genética θB. O programa Barrier
2.2 (Manni et al., 2004) foi utilizado para analisar a presença de descontinuidades genética
entre as populações no espaço, por meio da triangulação de Delaunay verificadas pelo
algoritmo de Monmonier (Manni et al., 2004) pelas coordenadas geográficas adquiridas pelo
sistema de posicionamento global (GPS) e das distâncias genéticas θB. O Teste de Mantel foi
aplicado com o programa PC-Ord 4.14 (McCune,Mefford; 1997) para verificar a correlação
entre a matriz de distância genética (θB) e a matriz de distância geográfica.
3 RESULTADOS
3.1 Diversidade Genética
Foi observado que a população S1 possui maior índice de diversidade gênica (He:0,750)
e o menor índice (He: 0,633) corresponde à população S2, ambas incluídas na área de
proteção ambiental. As populações N1, N2 e N3 apresentaram maior valor de
heterozigosidade observada (Ho) comparadas com a população S1 que está incluída na área
de proteção. Apesar das populações N1 e N2 não estarem dentro da área de proteção, as
mesmas apresentaram índices de diversidade gênica altos He: 0,735 e 0,714; respectivamente
(Tabela 2).

10
Tabela 2. Estimativa de diversidade genética para cinco áreas de Attalea vitrivir estudadas
com vinte e um locos microssatélites geradas pelo GDA.
População A He Ho f N1 7,333 0,735 0,505 0,318
N2 7,190 0,714 0,540 0,247
N3 7,952 0,684 0,527 0,231
S1 8,476 0,750 0,480 0,367
S2 5,523 0,633 0,537 0,154
Média 7,295 0,703 0,518 0,267
Abreviaturas: A, número médio de alelos por loco; He, diversidade gênica de Nei (1978); Ho, heterozigosidade observada; f, índice de fixação.
O índice de diversidade genética médio dentro das populações (Hs) foi de 0,753; o
índice bayesiano similar ao GST de Nei (GST-B) foi de 0,079; fluxo gênico (Nm) foi de 2,914 e
a heterozigosidade total (Ht) baseado em frequências alélicas médias foi de 0,818.
3.2 Estrutura Genética das Populações
A Análise de Variância Molecular (AMOVA) mostrou que a maior proporção da
variabilidade genética ocorre entre indivíduos dentro das populações (93,29%) do que entre as
populações (Tabela 3). Da diversidade genética total, 6,7% da variação ocorreu entre as
populações.
Tabela 3. Análise da variância molecular (AMOVA) obtida com marcadores microssatélites
em 141 indivíduos de Attalea vitrivir utilizados no estudo.
Fonte de Variação GL SQ VC %VT
Entre populações 4 8,167 0,029 6,7**
Dentro de populações 277 112,500 0,406 93,3**
Total 281 120,667 43,536 100
Graus de liberdade (GL), soma dos quadrados (SQ), componentes de variância (VC), percentagem da variação total (%VT), **significativo ao nível de 1% de probabilidade de erro.
11
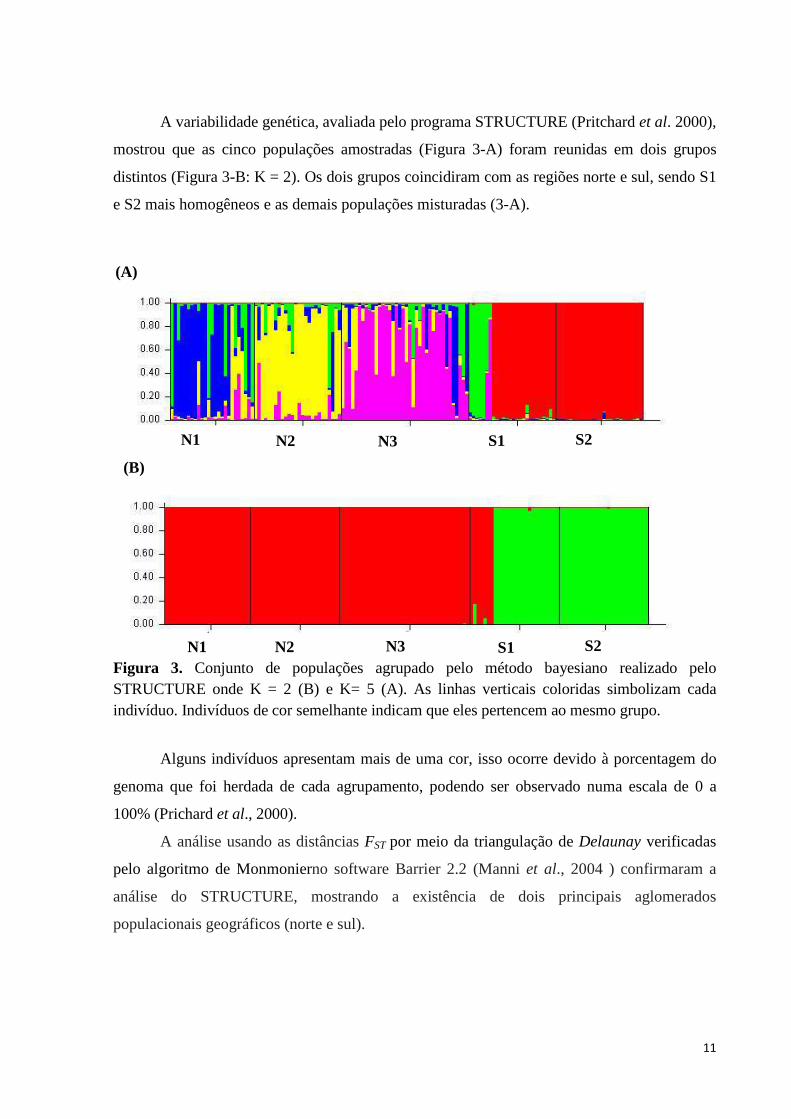
A variabilidade genética, avaliada pelo programa STRUCTURE (Pritchard et al. 2000),
mostrou que as cinco populações amostradas (Figura 3-A) foram reunidas em dois grupos
distintos (Figura 3-B: K = 2). Os dois grupos coincidiram com as regiões norte e sul, sendo S1
e S2 mais homogêneos e as demais populações misturadas (3-A).
Figura 3. Conjunto de populações agrupado pelo método bayesiano realizado pelo STRUCTURE onde K = 2 (B) e K= 5 (A). As linhas verticais coloridas simbolizam cada indivíduo. Indivíduos de cor semelhante indicam que eles pertencem ao mesmo grupo.
Alguns indivíduos apresentam mais de uma cor, isso ocorre devido à porcentagem do
genoma que foi herdada de cada agrupamento, podendo ser observado numa escala de 0 a
100% (Prichard et al., 2000).
A análise usando as distâncias FST por meio da triangulação de Delaunay verificadas
pelo algoritmo de Monmonierno software Barrier 2.2 (Manni et al., 2004 ) confirmaram a
análise do STRUCTURE, mostrando a existência de dois principais aglomerados
populacionais geográficos (norte e sul).
(A)
(B)
N1 S2 S1 N2 N3
N1 N3 N2 S1 S2

12
3.3 Distância Genética
A análise do software Barrier revelou semelhança entre as três populações do norte e as
duas populações situadas ao sul, ou seja, a triangulação de Delaunay indicou a presença de
barreira (Figura 4) que separa as populações do norte (N1, N2 e N3) das populações do sul
(S1 e S2).
Figura 4. Barreira traçada pelo software BARRIER.
Figura 5. A - Mapa físico indicando bacias hidrográficas brasileiras. Fonte: IBGE, 2013.
B - Mapa de unidades de relevo do Brasil. Fonte: IBGE 2006.
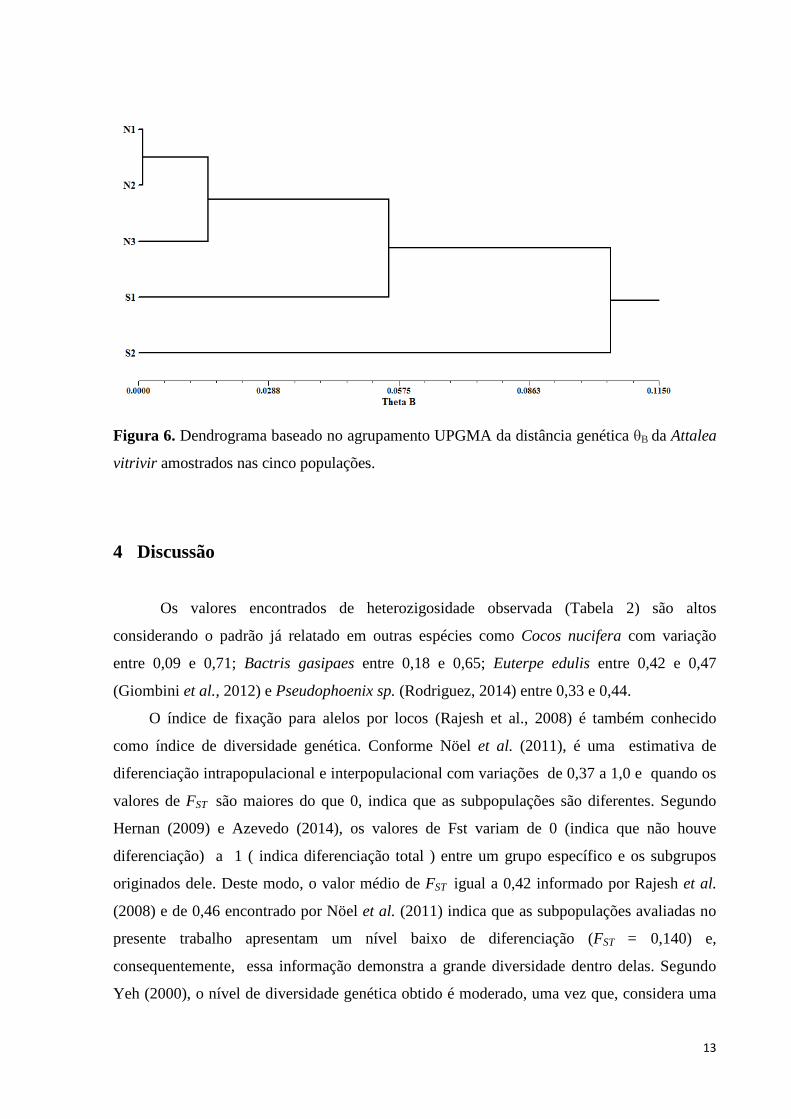
O dendrograma (Figura 6) indica a constituição de dois grandes agrupamentos, um
composto pelas populações situadas na região norte e outro formado pelas demais populações,
corroborando assim com o resultado de K=2 originada pelo STRUCTURE (Prichard et al.
2000), onde o valor de K corresponde ao número real de grupos genéticos.
A B
13
Figura 6. Dendrograma baseado no agrupamento UPGMA da distância genética θB da Attalea
vitrivir amostrados nas cinco populações.
4 Discussão
Os valores encontrados de heterozigosidade observada (Tabela 2) são altos
considerando o padrão já relatado em outras espécies como Cocos nucifera com variação
entre 0,09 e 0,71; Bactris gasipaes entre 0,18 e 0,65; Euterpe edulis entre 0,42 e 0,47
(Giombini et al., 2012) e Pseudophoenix sp. (Rodriguez, 2014) entre 0,33 e 0,44.
O índice de fixação para alelos por locos (Rajesh et al., 2008) é também conhecido
como índice de diversidade genética. Conforme Nöel et al. (2011), é uma estimativa de
diferenciação intrapopulacional e interpopulacional com variações de 0,37 a 1,0 e quando os
valores de FST são maiores do que 0, indica que as subpopulações são diferentes. Segundo
Hernan (2009) e Azevedo (2014), os valores de Fst variam de 0 (indica que não houve
diferenciação) a 1 ( indica diferenciação total ) entre um grupo específico e os subgrupos
originados dele. Deste modo, o valor médio de FST igual a 0,42 informado por Rajesh et al.
(2008) e de 0,46 encontrado por Nöel et al. (2011) indica que as subpopulações avaliadas no
presente trabalho apresentam um nível baixo de diferenciação (FST = 0,140) e,
consequentemente, essa informação demonstra a grande diversidade dentro delas. Segundo
Yeh (2000), o nível de diversidade genética obtido é moderado, uma vez que, considera uma

14
faixa de valores de FST em relação ao nível de diferenciação genética (0,0-0,05 baixo; 0,05-
0,25 moderado; > 0,25 alto).
Altos índices de diversidade genética foram encontrados para A. vitrivir na maior parte
das populações estudadas, exceto nas populações S2 e N3 (He: 0,633; He: 0,684
respectivamente) onde fatores como o isolamento genético, a alta taxa de fragmentação
(Santos et al., 2015) principalmente próximo aos locais estudados; a possível existência de
exploração nesses remanescentes devido a expansão urbana e impactos que alterem as áreas
de ocorrência dessa espécie e/ou comportamento de seus agentes polinizadores podem ser os
responsáveis pela baixa diversidade.
O fluxo gênico (Nm) pode ser considerado como o número absoluto de indivíduos
migrantes que adentram nas subpopulações a cada nova geração (Hartlet. al., 2010).
Conforme Wright (1931), caso o fluxo gênico seja menor que um (Nm<1), a deriva genética
ocasionará em diferenciação populacional substancial, ou seja, a deriva genética tem maior
atuação, resultando em significativa diferenciação genética entre as populações. O fluxo
gênico (Nm) de 2,914 neste trabalho, indica dois migrantes por geração. De acordo com
Wright (1951), valores de Nm maiores que 4,0 migrantes por geração conseguem conter os
efeitos do cruzamento de indivíduos próximos, possíveis de acontecer no interior das
populações.
Baseado na diferenciação genética das regiões estudadas neste trabalho acredita-se que
há duas grandes unidades potenciais para a conservação da espécie: as regiões norte e sul. É
aconselhável que a região Norte (N1, N2 e N3), por apresentar alto índice de diversidade
genética e requerer uma maior atenção, seja incluída em futuros planos de gestão e
conservação da espécie.
Os resultados encontrados para o agrupamento de populações (Tabela 3) assemelham-
se aos encontrados em outras espécies perenes e de fecundação cruzada (Perera et al., 1999;
Lebrun et al., 2005; Mistura et al., 2012; Azevedo, 2014). A alta divergência genética
encontrada entre as populações (Norte e Sul) pode estar associada ao isolamento reprodutivo
das mesmas. Os resultados estão em conformidade com os obtidos por Büttow et al. (2010)
que, em estudo com Butiá capitata, observaram maior variação molecular (83,68%) dentro
das populações. Os mesmos autores também relatam que eficientes mecanismos de dispersão
de pólen e sementes, como se acredita que seja o caso de A. vitrivir, faz com que a
variabilidade dentro das populações seja maior. A distância entre as áreas também deve ser
considerada, uma vez que, favorece o fluxo gênico entre as áreas avaliadas, como relata Boyle
et al. (1990). Silva (2006) estudou a diversidade e estrutura genética da palmeira Geomonas

15
Chottiana Mart. e obteve 86,5% de variabilidade genética dentro de populações,
demonstrando a alta variação genética dentro das populações.
Conforme Lorenzi et al. (2010) A. vitrivir possui ocorrência em uma região geográfica
pequena e fragmentada. A fragmentação e limitação de dispersão de pólen/frutos podem ser
fatores contribuintes para a diferenciação entre as áreas (Figuras 3 e 6). Apesar da ausência de
informações sobre os polinizadores específicos de A. vitrivir, normalmente os besouros da
família Nitidulidae (Mytrops mexicana) atuam como mediadores de polinização em palmeiras
(Henderson, 1995; Gitzendanner et al., 2000; Silberbauer-Gottsberger, 1990; Santos et al.,
2015).
Segundo Silvius (2002), os mamíferos se alimentam dos frutos das palmeiras
(inclusive a do gênero estudado) e os frutos possuem sementes, consideravelmente grandes,
que limitam a dispersão para apenas dispersores maiores (Jordano, 2000). Os roedores são
bastante atraídos pelo mesocarpo de tais frutos (Lorenzi et al., 2004). Duas espécies de
roedores estocadores, Clyomys bishopi e Dasyprocta azarae, são fundamentais na dispersão
de Attalea geraensis (Vieira, 2002). Sendo assim, a atuação de mamíferos e insetos poderia
ter elevado a diferenciação entre as áreas, uma vez que, muitas dessas espécies ficam
confinadas aos fragmentos remanescentes, o que reflete no fluxo gênico entre populações
(Smythe, 1978). A alta diferenciação genética entre os grupos (Figuras 3 e 6) está conforme a
análise realizada pelo software Barrier, o que sugere a existência de barreiras ao fluxo gênico,
como por exemplo, à presença de depressão da bacia hidrográfica do rio São Francisco, de
uma cadeia de montanha ou mesmo limites da fisionomia vegetação de cerrado com a
caatinga, como sugere Brandão (2012) ao trabalhar com Ceiba puiflora.
De acordo com o Teste de Mantel não houve nenhum indício para a correlação entre a
matriz de distância geográfica e estrutura genética (r = 0,212, p = 0,357). As possíveis
barreiras para o fluxo gênico entre as populações devem ser as mesmas citadas conforme a
análise do Barrier.
Segundo o dendrograma gerado pelo software NTSys (Figura 6), há uma tendência de
agrupamento de acordo com a origem das populações. A ausência de correlação entre as
distâncias genéticas e geográficas de A. vitrivir entre fragmentos pode ser explicado pela
baixa divergência genética entre as populações (Tabela 3) que exibem níveis elevados de
fluxo de genes.
As análises com a utilização de marcadores microssatélites (SSR) realizadas revelaram
alta diversidade genética dentro das populações. Conforme os resultados obtidos, infere-se
que as populações remanescentes analisadas conservam variabilidade genética da espécie,

16
sendo relevante para a sobrevivência de A. vitrivir. Os resultados também são pertinentes para
subvencionar medidas de conservação, uma vez que, os níveis de diversidade genética nas
populações localizadas na Área de Proteção Ambiental do Rio Pandeiros (APA-Pandeiros)
não são suficientes para a conservação da espécie.
5 SUBSÍDEOS PARA CONSERVAÇÃO
As análises realizadas indicaram altos níveis de heterozigosidade esperada e
observada, principalmente em indivíduos presentes em áreas não incluídas no ambiente de
proteção ambiental, revelando que parte da variabilidade genética da espécie encontra-se em
áreas fora desse ambiente. As cinco populações avaliadas foram reunidas em dois grupos
(K=2) distintos, coincidindo com as regiões Norte e Sul; sendo que a maior diversidade
genética foi encontrada na população S1 e a menor na população S2. Portanto é necessário
atenção para com a população S2, embora tenha apresentado menor índice de fixação. Os
níveis de diversidade genética encontrada para A. vitrivir foram considerados altos, sendo
maior dentro das populações do que entre as mesmas, provavelmente, devido ao isolamento
reprodutivo.
A alta diversidade genética nas populações não incluídas na área de proteção (N1, N2 e
N3), evidenciam a necessidade de novas propostas de ampliação das áreas de conservação,
uma vez que, sua ocorrência é restrita e as populações S1 e S2 não terem apresentado
diversidade genética suficiente para a conservação da espécie. Além disso, sugere-se para
conservar a variabilidade genética da espécie, a criação e manutenção de bancos de
germoplasma e a dispersão de sementes nas populações como S2 e N3. Sugere-se também o
estabelecimento de programas de educação ambiental nessas áreas (norte e sul) bem como
estudos que descrevam a biologia reprodutiva de A. vitrivir para compreender melhor a
dinâmica dessa espécie.

17
6 REFERÊNCIAS
Agarwal, M.; Shrivastava, N.; Padh, H. 2008. Advances in molecular marker techniques and their applications in plant science. Plant Cell Reports, Heidelberg, v. 27, p. 617-631.
Aguiar, M. S. Os marcadores moleculares como ferramenta no melhoramento genético de plantas. In: I Ciclo de Palestras sobre o Uso de Marcadores Moleculares na Pesquisa Agropecuária.. 2012. Aracajú. Anais... Aracajú: EMBRAPA, 2012, p. 10-14.
Anderson AB, Balick MJ. 1998. Taxonomy of babassu complex. (Orbignya ssp.). Palmae. System Bot. 13:32-50. Arrudas S. R.; Moura, A. C. F.; Salvador, H. F. Salvador; Rocha, S. M. Rocha; Pimenta M. A. S. 2014. Obtenção de Biodiesel de Babaçu (Attalea vitrivir) pelas vias etílica e metílica. FEPEG. Universidade Estadual de Montes Claros.
Azevedo, C. D. O. Diversidade genética de populações de coqueiro (Cocos nucifera L.) via marcadores SSR. 2014. Universidade Estadual do Norte Fluminense Darcy Ribeiro;
Barbará, T.; Palma-Silva, C.; Paggi, Gecele, M.; Bered, F., Fay, M.F.; Lexer, C. 2007. Cross-species transfer of nuclear microsatellite markers: potential and limitations. Molecular Ecology, v.16, n,18, p.3759-3767.
Borém, A. & Caixeta, E. T. 2006. Marcadores moleculares. Viçosa – M.G., 374p..
Boyle, T.; Liengsiri, C.; Piewluang, C. 1990. Genetic structure of black spruce on two contrasting sites. Heredity, Oxford, v.65, n. 3, p.393-399.
Brandão, M. M. 2012. Diversidade genética e filogeografia de Ceiba pubiflora (A. St.-Hil.) K. Schum. (Malvaceae). Tese de Doutorado. Universidade Federal de Lavras - UFLA.
Büttow, M.V.; Castro, C.M.; Schwartz, E.;Tonietto, A.; Barbieri, R.L. 2010. Caracterização molecular de populações de Butia capitata (Arecaceae) do sul do Brasil através de marcadores AFLP. Revista Brasileira de Fruticultura, Jaboticabal, v.32, n.1, p.230-239.
Chen, S.Y.; Lin, Y.Y.; Lin, C.W.; Chen, W.Y.; Yang, C.H.; Ku, H.M. 2010. Transferability of rice SSR markers to bamboo. Euphytica, Wageningen, v.175, p.23-33.
Dransfield, J.; Uhl, N.W.; Asmussen, C.B.; Baker, W.J.; Harley, M.M. & Lewis, C.E. 2008. Genera Palmarum: the evolution and classification of palms. Kew Publishing, Royal Botanical Garden, Londres.732p.

18
Doyle JJ and Doyle JL. 1987. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 19: 11-15. Eslabão, M. P.; Pereira, P. E. E. ; Barbieri, R. L.; Heiden, G. Distribuição geográfica de butia (arecaceae) nos biomas mata atlântica e pampa no rio grande do sul. 2015. XVII Encontro de pós-graduação. UFPEL
Excoffier, L.; Laval, G.; Schneider, S. 2005.Arlequin ver. 3.1: an integrated software package for population genetics data analysis. Berne: University of Berne.
Ferreira, M. E.,Grattapaglia, D. 1998. Introducciónal uso de marcadores moleculares em el Análisis Genético. Brasilia: EMBRAPA-CENARGEN, 220p. Giombini M. I., D. S. Tosto y S. P. Bravo. 2012. Characterization of 20 microsatellites in the Neotropical palm Syagrus romanzoffiana (Arecaceae) identified by cross-amplification from across genera. Molecular Ecology Resources 12: 1196–1197.
Gitzendanner MA and Soltis PS. 2000. Patterns of genetic variation in rare and widespread plant congeners.Am. J. Bot. 87: 783-792
Hartl, D. L.; Clark, A. G. 2010. Princípios de Genética de Populações. 4ª ed. EditoraArtmed: Porto Alegre.
Henderson A, Galeano G, Bernal R. 1995. Field guide to the palms of the Americas. Princeton University Press, Princepton, New Jersey. 352p Hernan, L. 2009. Data analysis for molecular characterization of plant genetic resources. Genet. Resourc. Crop. Ev. 56: 277-292
Holsinger, K.E., Lewis, P.O., Dey, D.K. 2002. Bayesian approach to inferring population structure from dominant markers. Mol. Ecol. 11, 1157–1164.
Holsinger, K.E., Lewis, P.O., 2005. Hickory: A Package for Analysis of Population Genetic Data v.1.0.4. Department of Ecology and Evolutionary Biology. University of Connecticut, Storrs, CT.
IBGE – Instituto Brasileiro de Geografia e Estatística. Disponível em: Acesso em: 15 jan. 2016.
IEPAGRO - Instituto de Estudos e Pesquisa do Agronegócio Rondoniense. 2010. Programa de Ações a Jusante do Médio e Baixo RioMadeira: Unidade de Processamento do Coco de Babaçu. Local: Distrito de Calama. Porto Velho: IEPAGRO, Santo Antônio Energia.

19
Johnson, D.V. 1998. Non-wood forest products 10: tropical palms. [S.l.]. Food and Agriculture Organization of the United Nations (FAO). Disponível em <http://www.fao.org/docrep/x0451e/x0451e00.HTM>. Acesso em 11 Ago 2015.
Jordano P. 2000. Fruits and frugivory. In: Fenner, M. (Ed.). Seeds: The Ecology of Regeneration in Plant Communities (2nd edition). CAB International, Wallingford, UK. pp. 125-165
Jump, A.S., Penuelas, J. 2006. Genetic effects of chronic habitat fragmentation in a wind-pollinated tree. Proc. Natl. Acad. Sci. U. S. A. 103, 8096–8100
Kalia, R. K.; Raí, M. K.; Kalia, S.; Dhawan, A. K. 2011. Microsatellite markers: an overview of the recent progress in plants. Euphytica, Dordrecht, v. 177, p. 309-334,
Lebrun, P., Berguer, A., Hodgkin, T., Baudouin, L. 2005. Biochemical and molecular methods for characterizing coconut diversity. In: Batugal, P., Rao, V.R., Oliver, J. (eds) Coconut Genetic Resources. International Plant Genetic Resources Institute – Regional Office for Asia, the Pacific and Oceania (IPGRIAPO). Serdang, Selangor DE, Malaysia, p. 225-247.
Lin, Z.; Yuan, D.; Zhang, X. 2010. Mapped SSR markers unevenly distributed on the cotton chromosomes. Frontiers of Agriculture in China, China, v. 4, p. 257-264. Leite, T. L.; Diniz, B. T.; Ferreira, M A; Ferreira, M. A. J. F.; Amaral, Z. P. S.; Buso, G S C. 2007. Análise de transferibilidade de primers microssatélites de Cucumis melo para Cucurbita moschata e Luffa cylindrica. Boletim de Pesquisa e Desenvolvimento (Embrapa Recursos Genéticos e Biotecnologia), v. 203, 1-9 p
Lepsh-Cunha, N. Estrutura genética e fenologia de espécies raras de Couratai spp. (Lecythidaceae) na Amazonia. 1996. 147p. Dissertação (Mestrado em Ciências Florestais) - Escola superior de Agricultura Luiz de Queiroz, Piracicaba, SP.
Lewis, P.; Zaykin, D. 2002. Genetic data analysis: computer programe for the analyses of allelic data - version 1.0 (d2). Alburquerque: Universityof New Mexico.
Lewis, P.O. & Zaykin, D. 2000. Genetic Data Analysis: Computer program for the analysis of allelic data. Version 1.0 (d15). Free program distributed by the author over the internet from the GDA. Disponíveem:<http://alleyn.eeb.uconn.edu/gda/,2000>
Lima JRO, Silva RB, Silva CC, Santos LSS, Santos Junior JR, Moura EM, Moura CVR. 2007. Biodisel de babaçu (Orbignyasp.) obtido por via etanólica. Química Nova. 30(3): 600-603.

20
Lorenzi, H. 2010. Geonoma. In: Lorenzi, H.; Noblick, L.R.; Kahn, F. & Ferreira, E. Flora brasileira: Arecaceae (Palmeiras). Instituto Plantarum, Nova Odessa. Pp. 214-255.
Lorenzi, H. et al. 1996. Palmeiras no Brasil: exóticas e nativas. Nova Odessa: Plantarum. 303 p.
Lorenzi, H.; Moreira de Souza, H.; Medeiros-Costa, J.D.; Coelho de Serqueira, L.S. & Ferreira, E. 2004. Palmeiras brasileiras e exóticas cultivadas. Instituto Plantarum, Nova Odessa. 432p.
Lorenzi, H.; Noblick, L.R.; Kahn, F. & Ferreira, E. 2010. Flora brasileira: Arecaceae (Palmeiras). InstitutoPlantarum, Nova Odessa. 382p.
Manni F, Guerard E, Heyer E. 2004. Geographic patterns of (genetic, morphologic, linguistic) variation: how barriers can be detected by using Monmonier’s algorithm. Human Biology 76: 173-190.
Matos DGP, Bernal FSM and Farias Filho MS. 2010. A importancia social e economica do extrativismo do babacu na microrregião de Itapecuru-mirim, estado do maranhão.Anais do XVI Encontro Nacional do Geógrafos. Realizado de 25 a 31 de julho de 2010. Porto Alegre - RS. ISBN 978:85:99907:02:3
May, P.H., Anderson, A.B., Balik, M.J., Frazão, J.M.F., 1985. Subsistence benefits from the babassu palm (Orbignya martiana). Econ. Bot. 39, 113–129
McCune, B., Mefford, M. J. 1997. Multivariate analysis of ecological data. Version 3.0. Gleneden Beach: MjM Software, Software.
McDermott, J. M.; McDonald, B. A. 1993. Gene flow in plant pathosystems. Annual Review of Phytopathology. Palo Alto, v. 31,p. 353-373.
Milach SCK. 1998. Marcadores de DNA. Biotecnologia Ciencia e Desenvolvimento, 5: 14-17.
Mistura, C. C., Barbiere R. L., Castro, C. M., Priore D.,Vilella, J. C. B. 2012. Transferibilidade de marcadores microssatélites de coco (Cocos nucifera) para butiá (Butia odorata).
Mnejja,M. Garcia-mas, J.;Audergon, JM;Arus,P. 2010. Prunus microsatellite marker transferability acrossros aceouscrops. Tree Genetics & Genomes, v.6, n.5, p.689-700.
Ministério do Meio Ambiente –MMA. Promoção Nacional da Cadeia de Valor do Côco Babaçu. Brasilia: MMA, 2009.

21
Nazareno, A.G.; Zucchi, M.I.; Reis, M.S. 2011. Microsatellite markers for Butia eriospatha (Arecaceae), a vulnerable palm species from the Atlantic Rainforest of Brasil.American Journal of Botany, Saint Louis, v.98, n.7, p.198-200.
Nei, M., 1978. Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics 89, 586–590
Neves,S. C., Ribeiro, L. M.,Cunha, I. R. G, Pimenta, M. A. S., Simões, M. O. M, Lopes, P. S. N., 2013. Diaspore structure and germination ecophysiology of the babassu palm (Attalea vitrivir). FLORA-50664; No. Of Pag 11.
Noblick, L. R. 1986. Palmeiras das caatingas da Bahia e as potencialidades econômicas. In: Simpósio Sobre a Caatinga E Sua Exploração Racional, 1986, Brasília, DF, Anais: Simpósio sobre a caatinga e sua exploração racional, Brasília, DF Embrapa, p.99-115
Noblick, L.R. Syagrus. 1996. The Palm Journal, Lawrence, v.126, p. 12-46. Nöel, K.K.J., Edmond, K.K., Louïs, K.K.J. e Eugene, K.K.2011.Microsatellite gene diversity within Philippines dwarf coconut palm (Cocos nucifera L.) resources at Port Bouët, Côte d’Ivoire. Scientific Research and Essays 6 (28): 5986-5992.
Oliveira, A. F. Estrutura genética de populações naturais de Copaifera langsdorffii Desf. a partir de isoenzimas. 2000. 114 p. Dissertação (Mestrado em Engenharia Florestal) - Universidade Federal de Lavras, Lavras, MG.
Peixoto, A. A. P.; Villela, J. L.; Ferreira, M. A. J. F.; Ferreira, M. A.; Amaral, Z. P. de S.; Vieira, R. R. T.; Lira, M. T. R.; Buso, G. S. C.. 2009. Transferibilidade de primers microssatélites de Cucumis melo para Luffa cylindrica. In: Congresso Brasileiro de Melhoramento de Plantas.
Perera, L., Russell, J.R., Provan, J. and Powell, W. 1999. Identification and characterization of microsatellites in coconut (Cocosnucifera L.) and the analysis of coconut populations in Sri Lanka. Mol Ecol 8: 344–346.
Pintaud JC (2008). Laspalmerasen América del Sur. An overview of the taxonomy of Attalea (Arecaceae).Rev. peru. biol. Novembro, 15(supl. 1): 055- 063.
Pritchard, J. K.; Setephens, M. e Donnelly, P. 2000. Inference of population structure using multilocus genotype data.Genetics. 155: 945-959.
Orozco-Segovia, A., Batis, A.I., Rojas-Aréchiga, M., Mendoza, A., 2003. Seedbiologyofpalms: a review. Palms 47, 79–94

22
Transferibilidade de primers microssatélites de Cucumis melo para Luffa cylindrica. In: Congresso brasileiro de melhoramento de plantas, 5., 2009,Guarapari. Anais... Vitória: Incarper, 2009. Disponivel<http://www.alice.cnptia.embrapa.br/handle/doc/662357>. Acesso em: 13 ago. 2015. Rajesh, M.K., Arunachalam, V., Nagarajan, P., Lebrun, P., Samsudeen, K. and Thamban, C. (2008) Genetic survey of 10 Indian coconut landraces by simple sequence repeats (SSRs). Scientia Horticulturae 118: 282-287.
Reis, M. S. Distribuição e dinâmica da variabilidade genética em populações naturais de palmiteiro (Euterpe edulis). 1996. 209p. Tese (Doutorado em Agronomia/Genética e Melhoramento de Plantas) Escola Superior de Agricultura Luiz de Queiroz, Piracicaba, SP.
Ritschel, P. S.; Lins, T. C. L.; Tristan, R. L.; Buso, G. S. C.;Buso, J. A.; Ferreira, M. E. 2004. Development of microsatellite markers from na enriched genomic library for genetic analysis of melon (CucumismeloL.). BMC Plant Biology, v 4, n. 9. p. 2-14. Rivas, l. H.; Giustina, l. D.; Luz, l. N. Karsburg,i. V.; Pereira,t. N. S.; Rossi, a. A. B. Genetic diversity in natural populations of the obroma subincanum mart. In the brazilian amazon. Genetics and molecular research, v. 12, n. 4, p. 4998-5006, 2013
Rodriguez, R. 2014. Genetic diversity and conservation of pseudophoenix (Arecaceae) in Hispaniola. Dissertação de Mestrado, 78p. Florida International University (USA).
Rohlf, F.J., 2000. NTSYS-pc. Numerical taxonomy and multivariate analysis system, Vers. 2.11. Applied Biostatistics, New York.
Rossi, F. S., Rossi, A. A.B., Dardengo, J. F.E., Brauwers,L.R., Silva, M. L. e Sebbenn, A.M. 2014. Genetic diversity in natural populations of Mauritiaflexuosa (Arecaceae) using ISSR markers. ScientiaForestalis. Piracicaba, v. 42, n. 104, p. 631-639, dez.
Santos, M. F. 2011. Variação genética em populações naturais de babaçu (Orbignya phalerata Mart.) por marcadores morfoagronômicos e moleculares. Universidade Federal do Piauí.
Santos, R.R.M; Cavallari M.M.; Pimenta, M.A.S.; Abreu A.G.; Costa M.R.; Guedes M.L. 2015. Population genetic structure of Attalea vitrivir Zona (Arecaceae) in fragmented areas of southeast Brazil. Genetics and Molecular Research 14 (2): 6472-6481.
Silberbauer-Gottsberger I. 1990. Pollination and evolution in palms. Phyton 30: 213-233.

23
Silva, M, S. 2006. Diversidade e estrutura genética em populações naturais de Geonoma schottiana Mart. no Parque Florestal Quedas do Rio Bonito/Mirian de Souza Silva.-Lavras: UFLA, 51p. : il.
Silvius, K. M. 2002. Spatio-temporal patterns of palm endocarp use by three Amazonian forest mammals: granivory or ‘grubivory’, Journal of Tropical Ecology, v. 18, p. 707-723.
Smythe, N. 1978. The Natural History of the Central American Agouti (Dasyproctapunctata). Smithsonian Contributions to Zoology, Washington: Smithsonian Institution Press, v. 257.
Soares, T. N.; Santana, L.; Oliveira, L. K.; Telles, M. P. C.; Collevatti, R. G. 2013. Transferability and characterization of microssatellite loci to in Anacardiumhumile A. St. Hil. (Anacardiaceae). Genetics and Molecular Research, v. 12, n. 3, p. 3146-3149. Souza, I . G. B .; Santos, M. F .; Sittolin , I . M .; Araújo, E. C.E .; Pereira, L.M .; Lima, P . S. 2008. Efficiency in RAPD and ISSR markers for analysis of genetic variability in babassu (Orbignyaphalerata Mart.) . IN.I Genetics Congress of the Midwest .Brasília, 2008.
Teixeira, M.A., 2008. Babassu – A new approach for an ancient Brazilian biomass. Biomass Bioenergy 32, 857–864
Vieira, M. V. 2002. Seasonal Niche Dynamics in Coexisting Rodents of the Brazilian Cerrado. Studies on Neotropical Fauna and Environment, v. 37. 38:7–15
Wright, S. ; 1951. The genetical structure of populations. Annual Eugenics 15:223-354.
Wright, S. 1931. Evolution in Mendelian populations. Genetics, Baltimore, v.16, p.97-159. Yeh, F. C. 2000. Population genetics. In.:Forest conservation genetics: principles and practice. A. Young, D. Boshier e T. Boyle (eds.).pp.21-37. Collingwood: CSIRO Publishing.
Zambrana, N.Y.P.; Byg, A.; Svenning, C.C.; Moraes, M.; Grandez, C. &Balsley, H. 2007. Diversity of palm uses in the western Amazon. Biodiversity and Conservation 16: 2771-2787


















