
DIVERSIDADE GENÉTICA E PADRÕES CROMOSSÔMICOS … · Em memória ao Tio Santo, com o qual aprendi...
50
UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE CENTRO DE BIOCIÊNCIAS PROGRAMA DE PÓS-GRADUAÇÃO EM SISTEMÁTICA E EVOLUÇÃO DIVERSIDADE GENÉTICA E PADRÕES CROMOSSÔMICOS ASSOCIADOS A QUEBRAS DE BARREIRAS PÓS-ZIGÓTICAS EM PEIXES DO GÊNERO Chaetodon (PERCIFORMES) ________________________________________________ Dissertação de Mestrado Natal/RN, fevereiro de 2016 DAVI ZALDER MIGUEL
Transcript of DIVERSIDADE GENÉTICA E PADRÕES CROMOSSÔMICOS … · Em memória ao Tio Santo, com o qual aprendi...

UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE
CENTRO DE BIOCIÊNCIAS
PROGRAMA DE PÓS-GRADUAÇÃO EM SISTEMÁTICA E EVOLUÇÃO
DIVERSIDADE GENÉTICA E PADRÕES CROMOSSÔMICOS
ASSOCIADOS A QUEBRAS DE BARREIRAS PÓS-ZIGÓTICAS
EM PEIXES DO GÊNERO Chaetodon (PERCIFORMES)
________________________________________________
Dissertação de Mestrado
Natal/RN, fevereiro de 2016
DAVI ZALDER MIGUEL

DAVI ZALDER MIGUEL DIVERSIDADE GENÉTICA E PADRÕES CROMOSSÔMICOS ASSOCIADOS
A QUEBRAS DE BARREIRAS PÓS-ZIGÓTICAS EM PEIXES DO GÊNERO
Chaetodon (PERCIFORMES)
Dissertação apresentada ao Programa de
Pós-Graduação em Sistemática e Evolução da
Universidade Federal do Rio Grande do Norte
como parte dos requisitos para obtenção do
título de Mestre em Sistemática e Evolução.
Orientador: Prof. Dr. Wagner Franco Molina
NATAL-RN
2016

Miguel, Davi Zalder. Diversidade genética e padrões cromossômicos associados aquebras de barreira pós-zigóticas em peixes do gênero Chaetodon(Perciformes) / Davi Zalder Miguel. - Natal, 2016. 48 f.: il.
Dissertação (Mestrado) - Universidade Federal do Rio Grandedo Norte. Centro de Biociências. Programa de Pós-Graduação emSistemática e Evolução. Orientador: Prof. Dr. Wagner Franco Molina.
1. Citogenética de Peixes - Dissertação. 2. DistânciaGenética - Dissertação. 3. Peixes Recifais - Dissertação. I.Molina, Wagner Franco. II. Universidade Federal do Rio Grande doNorte. III. Título.
RN/UF/BSE-CB CDU 597.3
Universidade Federal do Rio Grande do Norte - UFRNSistema de Bibliotecas - SISBI
Catalogação de Publicação na Fonte. UFRN - Biblioteca Setorial do Centro de Biociências - CB

DAVI ZALDER MIGUEL DIVERSIDADE GENÉTICA E PADRÕES CROMOSSÔMICOS ASSOCIADOS
A QUEBRAS DE BARREIRAS PÓS-ZIGÓTICAS EM PEIXES DO GÊNERO
Chaetodon (PERCIFORMES)
Dissertação apresentada ao Programa de
Pós-Graduação em Sistemática e Evolução da
Universidade Federal do Rio Grande do Norte
como parte dos requisitos para obtenção do
título de Mestre em Sistemática e Evolução.
Aprovada em 26 de fevereiro de 2016.
Comissão Examinadora:
_____________________________________________ Prof. Dr. Paulo Augusto de Lima Filho – IFRN
_______________________________________________________ Prof. Dr. Gideão Wagner Werneck Félix da Costa – UFRN
_____________________________________________ Prof. Dr. Wagner Franco Molina – UFRN
(Orientador)

Em memória ao Tio Santo, com o qual aprendi a ter fascínio e
amor a natureza.

“The most beautiful thing we can experience is the mysterious. It is the source of all true art and science. He to whom the emotion is a stranger, who can no longer pause to wonder and stand wrapped in awe, is as good as dead —his eyes are closed.”
― Albert Einstein, Living Philosophies

Agradecimentos
Agradeço primeiramente a minha “Namonoiva” Thamires, por estar junto
comigo em todos os momentos, tanto os de felicidade como de tristeza, e
compartilhar comigo esse pequeno instante perdido no universo. Não sei o que
faria sem você!
Agradeço aos colegas do LGRM por todo o apoio, aprendizado e
incentivo durante todos estes dois anos de mestrado. Agradeço ao Prof.
Wagner pela grande oportunidade a mim concedida, pela confiança depositada
em mim e por todo o apoio durante a execução deste trabalho.
Ao Prof. Artoni e todo o pessoal de Ponta Grossa pela estadia e pela
grande oportunidade de aprender coisas novas.
Agradeço a minha mãe e meus familiares por toda a ajuda e suporte
oferecido, mesmo distantes, sempre presentes em pensamento.
Aos meus “filhos” peludos e de quatro pernas, pelo amor incondicional e
por todos os pelos espalhados sobre minhas coisas.
A CAPES pelo apoio financeiro e a coordenação do PPGSE por todo o
suporte oferecido.

SUMÁRIO
Lista de Figuras ............................................................................................... 7
Resumo ............................................................................................................ 8
Abstract ............................................................................................................ 9
Introdução ...................................................................................................... 10
Origem, distribuição e evolução dos peixes-borboleta ..................................... 10
Hibridização e características citogenéticas ..................................................... 12
Objetivos ......................................................................................................... 15
Objetivo Geral .................................................................................................. 15
Objetivos Específicos ....................................................................................... 15
Material e Métodos ......................................................................................... 16
Coleta de material biológico ............................................................................. 16
Técnicas de estimulação mitótica ..................................................................... 16
Obtenção dos cromossomos mitóticos ............................................................. 17
Preparação das lâminas ................................................................................... 18
Hibridação fluorescente in situ (FISH) .............................................................. 18
Microfotografias cromossômicas ..................................................................... 19
Obtenção das sequências gênicas ................................................................... 19
Análise dos dados ............................................................................................ 19
Capítulo 1 - Divergências genéticas entre espécies de peixes-borboleta
capazes de formar híbridos naturais ................................................................ 21
Capítulo 2 - O gênero Chaetodon no Atlântico: Aspectos da diversificação
filética e estase cromossômica ......................................................................... 34
Conclusões ..................................................................................................... 44
Referências Bibliográficas ............................................................................ 45

Lista de Figuras
Tabela 1. Dados citogenéticos de espécies do gênero Chaetodon. 13
Figura 1. Espécies Atlânticas pertencentes ao gênero Chaetodon. Chaetodon
striatus do litoral do Rio Grande do Norte (Brasil) e Key Largo (EUA),
a e b, respectivamente. Chaetodon capistratus, c e C. sedentarius, d,
Barra=1cm. 17
Capítulo 1
Tabela 1. Sequências das 48 espécies de Chaetodon utilizadas nas análises 25
de distância genética, seguidas pelo respectivo número de acesso do
GenBank.
Tabela 2. Média das distâncias genéticas entre espécies do gênero Chaetodon,
formadoras de híbridos naturais e do conjunto total de espécies
analisadas 26
Tabela 3. Distâncias genéticas entre sequências multilocus de pares de
espécies do gênero Chaetodon capazes de formar híbridos naturais 27
Capítulo 2
Figura 1. Mapeamento cromossômico de sítios DNAr 18S (vermelho) e DNAr 39 5S (verde), através do FISH, em C. striatus (a), C. capistratus (b) e
em C. sedentarius (c), e coloração convencional do cariótipo em
Gimsa. Barra = 5 um.

Resumo
A hibridização interespecífica tem sido identificada em diversos grupos animais,
entre estes, de forma recorrente e destacadamente em peixes marinhos da
família Chaetodontidae. O gênero Chaetodon é o mais diverso da família com
cerca de 30 espécies capazes de produzir híbridos viáveis. Todas as
características pertinentes a este fenômeno vêm sendo analisadas, porém
ainda são necessários mais dados sobre a quebra de barreiras reprodutivas
neste gênero. No presente trabalho foram comparadas sequências dos genes
12S, 16S, RAG2 e NADH de espécies do gênero Chaetodon com híbridos
conhecidos, com o objetivo de aferir qual a distância genética entre elas, bem
como comparações citogenéticas por mapeamento cromossômico das regiões
de DNAr 18S e 5S das espécies C. striatus, C. capistratus e C. sedentarius, do
Atlântico Sul e Caribe. Os resultados indicam que as espécies parentais,
mesmo abrigando um alto nível de divergência genética, mantém o potencial
de intercruzamento e produz híbridos viáveis. Um extremo conservadorismo
cariotípico inter e intra-específico foi observado para as espécies. Apesar do
longo tempo decorrido de divergência entre algumas espécies, o potencial
reprodutivo interespecífico, além de fatores ambientais e biológicos, pode ter
sido favorecido pela extensa homogeneidade cariotípica presente na família.
Palavras chave: Citogenética de peixes, distância genética, peixes recifais,
conservadorismo cromossômico, DNAr.

Abstract
Interspecific hybridization has been identified in various animal groups, among
these, on a recurring basis in marine fish, notably in the Chaetodontidae family.
The Chaetodon is the most diverse gender in the family, with about 30 species
able to produce viable hybrids. All relevant features of this phenomenon have
been analyzed, but we still need more data about the break of reproductive
barriers in this genre. In the present study were compared sequences of the
12S gene, 16S, rag2 and NADH of Chaetodon genus with produce known
hybrids, in order to ascertain what the genetic distance between them and
cytogenetic comparisons by chromosomal mapping of regions of rDNA 18S and
5S of C. striatus, C. sedentarius and C. capistratus, from South Atlantic and
Caribbean. The results indicate that parental species, even having a high level
of genetic divergence, maintains the potential intercross and are capable to
produce viable hybrids. An extreme conservatism karyotype inter- and intra-
specific was observed for the species. Despite the long interval of separation
between some species, interspecific reproductive potential as well an
environmental and biological factors may have been favored by extensive
karyotype homogeneity present in the family.
Keywords: Fish cytogenetics, genetic distance, reef fishes, chromosome
conservatism, rDNA.

10
Introdução
Os peixes constituem um vasto grupo parafilético que habita quase
todos os ambientes aquáticos, apresentando grande diversidade morfológica e
comportamental (Nelson, 2006). O grupo dos Perciformes constitui a maior
ordem de vertebrados existentes, com cerca de 40% de todas as espécies de
peixes viventes atualmente e abrange mais de 160 famílias e cerca de 10.000
espécies já descritas (Nelson, 2006). Como parte destas famílias,
Chaetodontidae cujas espécies são conhecidas popularmente como peixes-
borboleta, se destaca como um dos grupos de peixes marinhos com
características peculiares, com cerca de 130 espécies, que habitam
principalmente recifes de corais, apresenta uma ampla diversidade morfológica
e comportamental (Pitss, 1991; Fessler & Westneat, 2007). Devido a suas
características biológicas, Chaetodontidae cuja origem evolutiva é intimamente
ligada a formações de corais é considerado como um dos grupos mais
especializados e adaptados a estes ambientes (Reese, 1975; Gosline 1985;
Hourigan et al., 1988). Dentre os gêneros, Chaetodon é o mais diverso,
possuindo por volta de 90 espécies, com cerca de dois terços destas
relacionadas obrigatoriamente ou de forma facultativa, aos recifes de corais
(Cole & Pratchett, 2008).
Origem, distribuição e evolução dos peixes-borboleta
A origem e evolução da família Chaetodontidae está intimamente ligada
a diversificação e estabelecimento das principais formações recifais modernas.
Os recifes modernos surgiram ao final da grande extinção em massa ocorrida
no final do período Cretáceo, aproximadamente 65 milhões de anos atrás,
cujas características sofreram grandes alterações desde então (Woods, 1998).
Os corais escleractíneos modernos se tornaram a forma dominante no início do
período Cenozoico e estabeleceram seu domínio nos últimos 65 milhões de
anos. Durante esse período Chaetodontidae, vem se adaptando em conjunto,
estabelecendo relações ecológicas importantes com os recifes de corais
(Kauffman & Johnson, 1988; Bellwood & Wainwright, 2002).

11
O registro fóssil mais antigo dos peixes recifais data de cerca de 50
milhões de anos (m.a.), localizado no Monte Bolca, na Itália (Bellwood, 1996).
Diversas espécies da época apresentam características hoje compartilhadas
pelos Chaetodontidae, apesar de revisões e estudos posteriores concluírem
que a origem da família, já com todas as suas características modernas e
atuais ser bem mais recente (Banikov, 2004). De fato, técnicas modernas de
inferência filogenética molecular e evolutiva, sugerem que a origem dos
Chaetodontidae modernos pode ser datada do início do Eoceno, por volta de
40 a 60 m.a., quando o ramo principal deste grupo se diferenciou dos
ancestrais dos Pomacentridae atuais (Cowman & Bellwood, 2011).
O primeiro material a ser considerado como pertencente ao gênero
Chaetodon é Chaetodon penniger, datado do Rupeliano, no Oligoceno, por
volta de 29-35 m.a., encontrado ao sul das montanhas do Cáucaso
(Sytchevskaya et al., 2005). Outra espécie, Chaetodon fisheuri, datado do
Mioceno, encontrada na Algéria, compartilham muitas características com os
peixes-borboleta modernos (Carnevalle, 2006).
Grande parte das filogenias propostas para a família, coloca o gênero
Chaetodon como o mais numeroso em relação a gêneros menores dentro do
grupo. Isso é suportado tanto por filogenias mais antigas (Blum, 1989) como
por estudos mais recentes (Smith et al., 2003; Fessler & Westneat, 2007;
Bellwood et al., 2010). Tais filogenias classificam dois grupos distintos na
família Chaetodontidae, os peixes-borboleta, abrangendo primariamente o
gênero Chaetodon, e os peixes-bandeira, que inclui como principal gênero,
Heniochus, cuja divisão filética se deu por volta de 26 milhões de anos (Fessler
& Westneat, 2007).
O gênero Chaetodon tem sido dividido em quatro principais clados,
agrupados de acordo com ancestralidade comum, local de abrangência e tipo
de alimentação (Fessler & Westneat, 2007). O clado 1 é o menor, contendo
apenas três espécies e todas restritas a regiões de recifes de corais da costa
ocidental da África. O clado 2 agrupa espécies particularmente onívoras, com
abrangência no oceano pacífico e regiões tropicais, contendo diversas
linhagens distintas, com espécies coralívoras e algumas de grandes
profundidades. Há indícios que essa linhagem tenha se originado nos últimos 2
m.a., através de migrações do Oceano índico por meio do Cabo da Boa

12
Esperança (Bellwood et al., 2005). O clado 3, restrito às regiões do Indo-
Pacífico, é composto quase totalmente por indivíduos obrigatoriamente
coralívoros, extremamente dependentes de espécies de corais existentes na
região (Cole et al., 2008). Devido à dependência de determinadas espécies
endêmicas, alguns indivíduos podem ser extremamente sensíveis a mudanças
ambientais e efeitos antrópicos que possam afetar a incidência e frequência
dessas formações de corais (Hixton & Webster, 2002; Jones & McCormick,
2004; Wilson et al., 2006). As espécies pertencentes ao clado 4 constituem-se
grande parte de espécies do Pacífico, com grande incidência de espécies
coralívoras. Porém, diferentemente do clado 3, esse grupo ainda possui
espécies que não necessitam obrigatoriamente de recifes de corais, o que
suporta a hipótese de que peixes de clima temperado estão menos associadas
com ambientes coralinos, onde as espécies do gênero Chaetodon, do Atlântico
temperado tendem a ser mais generalistas do que especializadas (Bellwood &
Wainwright, 2002).
Hibridização e características citogenéticas
Todos os cariótipos descritos das espécies de Chaetodon apresenta a
característica basal comum a grande maioria dos Perciformes, um extremo
conservadorismo cariotípico, com predominância de 48 cromossomos
acrocêntricos em seu complemento cromossômico (Arai, 2011; Motta-Neto et
al., 2011; 2012; Calado et al., 2012), com exceção de C. plebeius e C.
strigangulus que possuem cromossomos metacêntricos e submetacêntricos
(Tabela 1). Tem sido aventado que similaridades cariotípicas poderiam
contribuir para o alto índice de indivíduos híbridos em peixes, sobretudo na
família Chaetodontidae (Molina et al., 2013), cuja incidência de hibridização é
alta, mesmo quando comparado com outras espécies recifais que também
possuem uma grande quantidade de pares interespecíficos (Pyle & Randall,
1994; Domeier et al., 1997; Molina & Galetti, 2002; Yaakub et al., 2006).
As maiores concentrações de híbridos interespecíficos no gênero
Chaetodon estão presentes nas regiões do Índico e Pacífico, locais onde há
grande diversidade de espécies, bem como sobreposição de habitat entre as
espécies (Blum, 1989; Righton et al., 1996; Bellwood & Wainwright, 2002).

13
Os indivíduos híbridos surgem principalmente onde ocorrem
sobreposição da área geográfica de abrangência das espécies, ou quando as
espécies se encontram no limite da abrangência de uma espécie (Brown,
1987). Isso pode ocorrer principalmente devido à baixa densidade que as
espécies de uma dada população apresentam quando estas estão no seu limite
geográfico, o que pode influenciar na formação e estabelecimento de híbridos
(Rhymer & Simberloff, 1996; Rose & Polis, 2000). De fato, diversos fatores
podem ocasionar a ocorrência de pares interespecíficos, pois algumas
espécies de Chaetodon são normalmente monogâmicas (Yabuta, 1997),
facilitando a observação em campo de pares de espécies diferentes e
consequentemente, identificando a ocorrência de espécimes híbridos.
Tabela 1. Dados citogenéticos de espécies do gênero Chaetodon.
Espécie 2n Cariótipo NF Referência
C. auriga 48 48a 48 Arai & Inoue, 1975
C. auripes 48 48a 48 Ojima & Yamamoto, 1990
C. collare 48 48a 48 Nagpure et al., 2006
C. lunula 48 48a 48 Arai & Inoue, 1975
C. plebeius 48 2m+46a 50 Arai & Inoue, 1975
C. sedentarius 48 48a 48 Galetti et al., 2006
C. striatus 48 48a 48 Affonso et al., 2001
C. strigangulus 48 2sm+46a 50 Arai & Inoue, 1975
C. trifasciatus 48 48a 48 Arai & Inoue, 1975
C. vagabundus 48 48a 48 Arai & Inoue, 1975
C. ocellatus 48 48a 48 Molina et al., 2013
Apesar da existência de informações sobre a ocorrência de indivíduos
híbridos, há uma enorme lacuna na compreensão de todos os fenômenos que
podem ocasionar tal fenômeno em ambientes naturais. A raridade com que
esses híbridos são vistos na natureza também contribui para a dificuldade que
se tem a respeito de aspectos morfológicos e comportamentais que esses
animais podem apresentar. Geralmente híbridos apresentam um padrão de
coloração intermediário entre as espécies parentais, o que pode ajudar na

14
localização e identificação de quais indivíduos são realmente híbridos e inferir
indiretamente quais são as duas espécies parentais (Montanari et al., 2012).
Amplos dados estão disponíveis sobre os aspectos reprodutivos e
comportamentais (Fricke, 1986; Driscoll & Driscoll, 1988), biogeográficos
(Blum, 1989; Righton et al., 1996; Bellwood & Wainwright, 2002), filogenéticos
e morfológicos (Blum, 1988; Fessler & Wastneat, 2007) da família
Chaetodontidae. Contudo, os aspectos citogenômicos e estruturais dos
cromossomos e do cariótipo deste grupo de peixes ainda permanecem pouco
estudados. Dados citogenéticos mais resolutivos do cariótipo dos
Chaetodontidae tem surgido recentemente (Molina et al., 2013), contudo,
grande parte do cariótipo e suas características cromossômicas permanece
pouco conhecido para a maioria das espécies.
Diante da possibilidade que um elevado nível de conservadorismo
numérico e estrutural presente nos cromossomos dessas espécies possa
influenciar no fenômeno de hibridização na família Chaetodontidae, faz-se
necessário ampliar as informações citogenéticas de suas espécies.
Adicionalmente o acréscimo de dados citogenéticos propiciará uma maior
compreensão da evolução cromossômica e processos especiativos na família.

15
Objetivos
Objetivo geral
Analisar aspectos da divergência molecular e citogenética, por técnicas
resolutivas, espécies do gênero Chaetodon, visando uma melhor compreensão
dos mecanismos que propiciem a persistência dos processos recorrentes de
hibridização interespecífica em ambientes naturais.
Objetivos específicos
- Utilizar as sequências gênicas disponíveis online de diversas espécies
de peixes pertencentes ao gênero Chaetodon para aferir a distância
genética entre as mesmas.
- Verificar se a distância genética entre as espécies, pode influenciar na
formação e no estabelecimento de híbridos e pares heteroespecíficos.
- Mapear a distribuição de sequências de DNAr 18S e 5S nos
cromossomos de 3 espécies de Chaetodon, C. striatus, C. sedentarius e
C. capistratus, provenientes de diferentes áreas Atlânticas, comparando
ainda aspectos citopopulacionais de C. striatus, entre o Caribe e a
Atlântico Sul.

16
Material e Métodos Coleta de material biológico
Para as análises citogenéticas foram analisados exemplares de 3
espécies do gênero Chaetodon provenientes de duas regiões do Atlântico
(Figura 1). Chaetodon sedentarius (n=1), C. capistratus (n=1) e C. striatus (n=2)
(Figura 2) foram coletados em Key Largo, Flórida, EUA (25°6′23″ N, 80°25′47″
W), C. ocellatus (n=3) e C. striatus (n=5) foram coletados no litoral do Rio
Grande do Norte, nordeste do Brasil. A ilha de Key Largo está localizada ao sul
da Flórida, com cerca de 55 km de extensão, constituindo a ilha mais ao norte
de todas as ilhas da Florida Key. Os espécimes foram coletados utilizando rede
de arrasto e puçás.
Técnicas de Estimulação Mitótica
Após a coleta os espécimes foram submetidos a estimulação por meio
de técnicas de estimulação mitótica, seguindo a metodologia proposta por
Molina et al. 2010, descrita a seguir. Os indivíduos capturados foram
estimulados utilizando medicamentos cujo princípio ativo são lisados de
bactérias e fungos, cujo objetivo visa aumentar a taxa de divisão celular nos
órgãos hematopoiéticos desses animais, para serem posteriormente extraídos.
Foram utilizados 10 mg de lisado de bactérias para 10 ml de água destilada. A
solução foi injetada com o uso de seringas na região dorso-lateral do animal na
proporção de 1 ml para cada 100g de peso. Os indivíduos foram deixados vivos
em seus reservatórios por um período de 24 horas para que a estimulação
ocorra.

17
Figura 1. Espécies Atlânticas pertencentes ao gênero Chaetodon. Chaetodon striatus do litoral do Rio Grande do Norte (Brasil) e Key Largo (EUA), a e b, respectivamente. Chaetodon capistratus, c e C. sedentarius, d, Barra=1cm.
Obtenção de cromossomos mitóticos
Os cromossomos mitóticos foram obtidos mediante a técnica usada e
descrita por Gold et al. (1990), a seguir descritas. Após os indivíduos serem
sacrificados foram removidos fragmentos do rim anterior. Os fragmentos foram
dissociado em 9 ml de meio de cultura RPMI 1640, com o auxílio de pinças e
seringas de vidro desprovidas de agulha, e acrescido de 125µl de colchicina

18
(0,025%). O material foi homogeneizado com o uso de pipetas de Pasteur e
mantido neste meio de cultura por 30 minutos a temperatura ambiente. Ao
término desse tempo, a amostra foi centrifugada por 10 minutos a 900 rpm. O
sobrenadante foi removido e foram acrescentados 9 ml de solução hipotônica
de KCl (0,075M), na qual a suspensão de células, após homogeneização, foi
mantida por 25 minutos à temperatura ambiente. Após esse passo, foram
acrescentados 150µl de fixador (3 metanol; 1 ácido acético) recém preparado e
a solução homogeneizada novamente. O material foi centrifugado por 10
minutos a 900 rpm, descartando o sobrenadante. Um total de 6 ml de fixador foi
adicionado a suspensão, que foi novamente ressuspendida, com o uso de
pipetas de Pasteur. O processo de fixação foi repetido por mais duas vezes, e
após a última centrifugação e eliminação do sobrenadante, foram adicionadas
1,5 ml de fixador. As amostras foram mantidas sobre refrigeração à -20oC em
microtubos de 1,5 ml, para posteriores análises.
Preparação de lâminas
Sobre lâminas cuidadosamente lavadas e recobertas com um filme de
água destilada, aquecido à 60°C, foram gotejados 120µl da suspensão celular.
Após secas, as lâminas foram coradas com corante Giemsa à 5%, por 10
minutos, para análises da morfologia dos cromossomos e do estabelecimento
do valor diploide. As lâminas foram analisadas em microscópio ótico sob
aumento de 1.200X. As melhores metáfases foram utilizadas na montagem dos
cariótipos.
Hibridação fluorescente in situ (FISH)
A hibridação fluorescente in situ (FISH) foi realizada de acordo com a
metodologia descrita por Pinkel et al. 1986, com algumas modificações.
Sequências de DNA in tandem amplificada do DNA genômico de cada espécie
específica foi utilizada para sonda de DNAr 18S e 5S. A sonda 18S foi marcada
por nick translation com digoxigenina-11-dUTP (Roche) e a sonda rDNA 5S por
nick translation com biotina-14-dATP (Roche), de acordo com as instruções do
fabricante. Os cromossomos foram tratados com RNAse livre de DNAse

19
(20mg/mL em 2×SSC) à 37°C por 1 hora, com pepsina (0,005% em HCl 10mM)
à 37°C por 10 minutos e fixados com formaldeído a 1% por 10 minutos, em
seguida desidratados em uma série alcoólica. Os cromossomos foram, então,
desnaturados em 70% formamida/2×SSC à 72°C por 5 minutos. A solução de
hibridização composta por 50% de formamida, 2×SSC, 10% de sulfato de
dextran e a sonda desnaturada (5 ng/µl). Após hibridação overnight à 37°C, as
lâminas foram lavadas em 15% formamida/0.2×SSC à 42°C, por 20 minutos,
0,1×SSC à 60°C, por 15 minutos e Tween20 0,5%/4×SSC, à temperatura
ambiente. O sinal de hibridização da sonda foi detectado usando um conjugado
streptavidina-FITC (Vector) para a sonda DNAr 5S e conjugado anti-
digoxigenina-rodamina (Roche) para a sonda DNAr 18S. Os cromossomos
foram contracorados com Vectashield/DAPI (1,5 µg/ml)(Vector).
Microfotografias cromossômicas
As preparações de FISH e de fluorocromos base específicos foram
analisadas em fotomicroscópio de epifluorescência Olympus BX50, equipado
com sistema digital de captura de imagens Olympus DP73 utilizando o software
cellSens (Olympus Optical Co. Ltd.). Os cromossomos foram classificados de
acordo com a posição do centrômero, em metacêntricos (m), submetacêntricos
(sm), subtelocêntricos (st) e acrocêntricos (a). (Levan et al., 1964).
Obtenção das sequências gênicas
Sequências dos genes NADH, rag2, 12S e 16S de 52 espécies do
gênero Chaetodon, representando 24 pares interespecíficos, foram obtidas do
GenBank. Os genes foram escolhidos pelo seu sólido uso filogenético e por
representar um amplo conjunto de dados disponíveis para os pares
heteroespecíficos neste gênero, formadores de híbridos naturais.
Análise dos dados
Os eletroferogramas obtidos a partir das sequências encontradas no
GenBank foram conferidos com o auxílio do software BioEdit (Hall, 1999). As

20
sequências de cada gene foram separadamente alinhadas através do MUSCLE
(Edgar, 2004), gaps ou áreas não resolvidas, foram removidas por meio do
software MEGA6 (Tamura et al., 2013).
A saturação de transições e transversões nas sequências foram
analisadas com o software DAMBE (Xia & Xie, 2001). O software jModelTest
2.1.4 (Darriba et al., 2012), foi utilizado para selecionar o melhor modelo de
substituição nucleotídica a ser utilizado. As divergências genéticas, baseadas
nas distâncias-P, com 1000 replicações de bootstrap, foram estimadas pelo
software MEGA v6 (Tamura et al., 2013).

21
Capítulo 1
Divergências genéticas entre espécies de peixes-borboleta capazes de
formar híbridos naturais
Miguel, D. Z., Souza, A. S., Molina W. F.
Resumo
O fenômeno da hibridização ocorre nos mais diversos grupos de animais,
porém tem sido identificado de forma acentuada em peixes recifais. O peixes-
borboleta do gênero Chaetodon possuem mais de 30 espécies que se
intercruzam e são capazes de produzir híbridos viáveis. Os aspectos deste
fenômeno vem sendo intensamente estudados, mas ainda não são
inteiramente claros. No presente trabalho foram comparadas sequências dos
genes 12S, 16S, rag2 e NADH de um significativo conjunto de espécies do
gênero Chaetodon que produzem híbridos interespecíficos, com o objetivo de
aferir níveis de distância genética capazes de estabelecer limites à formação
de híbridos. Os resultados indicam que as espécies parentais, mesmo
abrigando um alto nível de divergência genética, relativo a significativos
períodos de tempo desde a divergência evolutiva, ainda podem se intercruzar e
gerar híbridos viáveis.
Palavras-chave: Chaetodon, hibridização, peixes recifais, divergência
genética.

22
Abstract The hybridization phenomenon occurs in all diverse groups of animals, but they
have been identified sharply in reef fish. The butterflyfishes of genus Chaetodon
have more than 30 species that are able to produce viable hybrids. Aspects of
this phenomenon have been thoroughly studied, but are not yet entirely clear. In
the present study were compared sequences of the 12S gene, 16S, rag2 and
NADH of several Chaetodon species that have known hybrids, in order to
ascertain what the genetic distance between these species and ascertain to
what extent this distance can influence the formation of hybrids between the
interspecific pairs. The results indicate that parental strains, even harboring a
high level of genetic diversity also can be crossed and produce viable hybrids
with phylogenetically distant species.
Keywords: Chaetodon, hybridization, reef fish, genetic divergence.
Introdução
A hibridização é um fenômeno amplamente documentado em grande
número de grupos animais e vegetais (Schwenk, 1993; Bierne et al., 2003).
Cerca de 10% de todas as espécies animais possuem a capacidade de
intercruzar e gerar descendentes híbridos, que podem exibir diferentes níveis
de fertilidade e viabilidade (Mallet, 2007). Híbridos naturais se formam em
áreas conhecidas como “zonas de hibridização”, onde espécies distintas,
habitam em simpatria (Barton & Hewitt, 1989). Algumas destas zonas surgem
quando duas espécies já separadas por eventos de vicariância voltam a se
encontrar, e barreiras pré- e pós-zigóticas, ainda não se estabeleceram de
forma completa mesmo decorridos longos períodos de tempo após a
separação (Mayr, 1963).
Os processos de hibridização constituem uma força evolutiva importante
na condução dos processos evolutivos de diversas espécies, ocasionando
tanto a extinção, como o surgimento de novas linhagens e grupos (Hobbs et al,
2013). Do ponto de vista adaptativo, os cruzamentos interespecíficos podem
aumentar o número de genótipos e a diversidade genética de uma população,
permitindo uma maior capacidade de sobrevivência desses indivíduos,
possibilitando que híbridos explorem novos nichos e recursos disponíveis no

23
ambiente que outrora não estavam disponíveis para as espécies parentais
(Seehausen, 2004; Man et al., 2012). De outra forma, a hibridização também
pode levar a processos de especiação reversa (Rhymer & Simberloff, 1996) em
função da diminuição da riqueza do ambiente, causando perda de diversidade
genética e a extinção local ou total das espécies (Seehausen, 2006).
Apesar da ampla documentação da incidência de híbridos, Em geral, tais
indivíduos em ambiente natural são difíceis de serem reconhecidos e
identificados, requerendo analise e observação minuciosa, e pouco se
compreende a respeito das barreiras de fluxo gênico que podem levar a
formação de híbridos em ambiente marinho. Porém, os peixes pertencentes a
família Chaetodontidae, possuem um elevado número de híbridos e
cruzamentos interespecíficos quando em comparação com outras famílias de
peixes marinhos e recifais (Yaakub, 2006). Esses híbridos geralmente ocorrem
em áreas onde as espécies se sobrepõem espacialmente, geralmente na
região entre o oceano Pacífico e Índico (Hobbs et al., 2009). A maioria dos
híbridos é identificável através de seu padrão de coloração, aparentando
mescla entre os padrões das espécies parentais, exibindo características
compartilhadas e comuns a ambos (Montanari et al., 2012).
Dentro da família Chaetodontidae, o gênero Chaetodon possui a maior
parte das espécies de peixe-borboleta descritas, compreendendo mais de 90
espécies conhecidas e identificadas. Cerca de dois terços das espécies do
gênero se encontra nas porções ocidentais do Pacífico e Índico, porém também
são encontradas, em menor número, no Caribe e no Atlântico Sul (Findlay &
Findlay, 2001). Aliado ao nível de especialização de algumas de suas espécies,
que apresentam dependência de recifes de corais (Pitts, 1991), suas
características morfológicas atrativas, as tornam alvos de coleta e
comercialização tornando-as particularmente vulneráveis (Cox, 1994; Pratchett
et al., 2006).
Em geral a viabilidade e a aptidão dos híbridos em peixes diminuem,
conforme as divergências genéticas entre as espécies parentais se acentuam
(Russel, 2003). Neste sentido, as características cromossômicas das espécies
parentais podem influenciar a ocorrência e manutenção de indivíduos híbridos.
De fato, tem sido apontado que cariótipos filogeneticamente mais conservados
em relação ao número de cromossomos ou padrões estruturais, exercem

24
menor restrição pós-zigótica, originando frequentemente híbridos com maior
viabilidade e aptidão (Molina et al., 2013).
Visando determinar as divergências existentes entre pares
heteroespecíficos do gênero Chaetodon, capazes de produzir híbridos viáveis
naturais, foram comparadas as sequências de seus genes mitocondriais (12S e
16S) e nucleares (NADH, rag2), e discutidas as consequências da divergência
genética na manutenção do alto índice de hibridização na família
Chaetodontidae.
Material e Métodos
Obtenção das sequências gênicas
Sequências dos genes NADH, rag2, 12S e 16S de 52 espécies do
gênero Chaetodon, representando 24 pares interespecíficos foram obtidas do
GenBank. Os genes foram escolhidos pelo seu sólido uso filogenético e por
representar um amplo conjunto de dados disponíveis para os pares
heteroespecíficos neste gênero, formadores de híbridos naturais. As
sequencias, bem como os respectivos números de registro do GenBank estão
localizados na Tabela 1.
Análise dos dados
Os eletroferogramas obtidos a partir das sequências encontradas no
GenBank foram conferidos com o auxílio do software BioEdit (Hall, 1999). As
sequências de cada gene foram separadamente alinhadas através do MUSCLE
(Edgar, 2004), gaps ou áreas não resolvidas, foram removidas por meio do
software MEGA6 (Tamura et al., 2013).
A saturação de transições e transversões nas sequências foram
analisadas com o software DAMBE (Xia & Xie, 2001). O software jModelTest
2.1.4 (Darriba et al., 2012), foi utilizado para selecionar o melhor modelo de
substituição nucleotídica a ser utilizado. As divergências genéticas, baseadas
nas distâncias-P, com 1000 replicações de bootstrap, foram estimadas pelo
software MEGA v6 (Tamura et al., 2013).
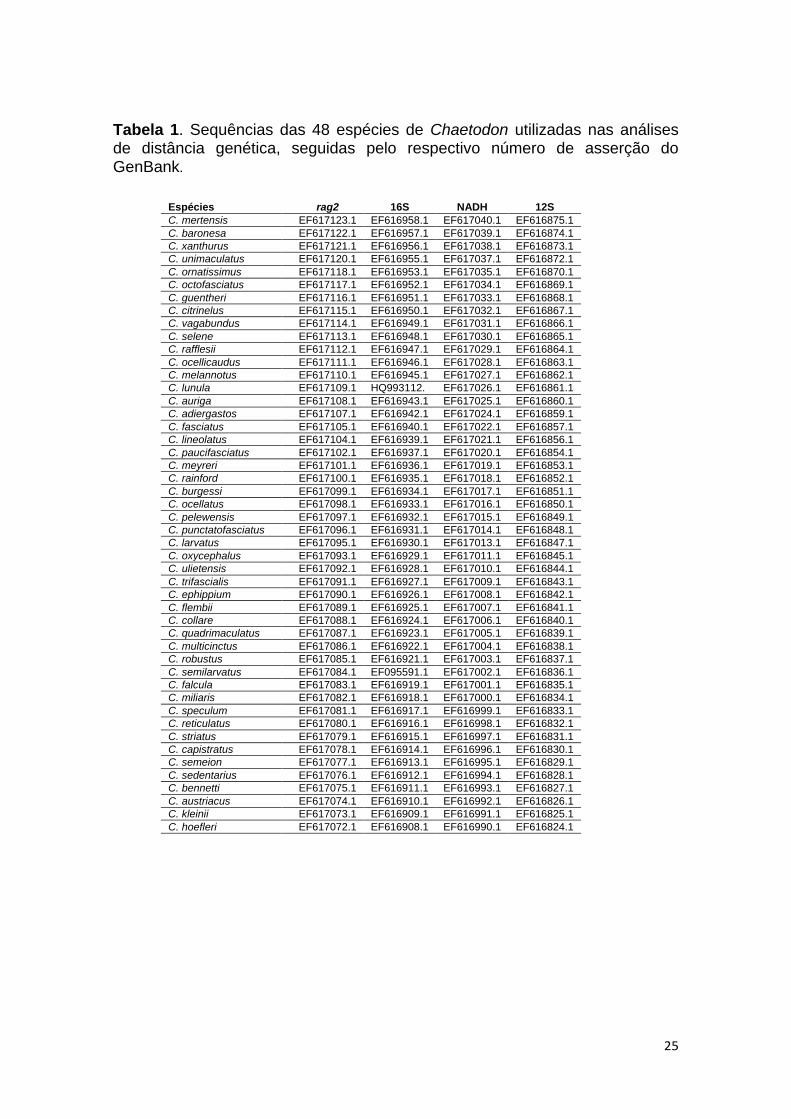
25
Tabela 1. Sequências das 48 espécies de Chaetodon utilizadas nas análises de distância genética, seguidas pelo respectivo número de asserção do GenBank.
Espécies rag2 16S NADH 12S
C. mertensis EF617123.1 EF616958.1 EF617040.1 EF616875.1
C. baronesa EF617122.1 EF616957.1 EF617039.1 EF616874.1
C. xanthurus EF617121.1 EF616956.1 EF617038.1 EF616873.1
C. unimaculatus EF617120.1 EF616955.1 EF617037.1 EF616872.1
C. ornatissimus EF617118.1 EF616953.1 EF617035.1 EF616870.1
C. octofasciatus EF617117.1 EF616952.1 EF617034.1 EF616869.1
C. guentheri EF617116.1 EF616951.1 EF617033.1 EF616868.1
C. citrinelus EF617115.1 EF616950.1 EF617032.1 EF616867.1
C. vagabundus EF617114.1 EF616949.1 EF617031.1 EF616866.1
C. selene EF617113.1 EF616948.1 EF617030.1 EF616865.1
C. rafflesii EF617112.1 EF616947.1 EF617029.1 EF616864.1
C. ocellicaudus EF617111.1 EF616946.1 EF617028.1 EF616863.1
C. melannotus EF617110.1 EF616945.1 EF617027.1 EF616862.1
C. lunula EF617109.1 HQ993112. EF617026.1 EF616861.1
C. auriga EF617108.1 EF616943.1 EF617025.1 EF616860.1
C. adiergastos EF617107.1 EF616942.1 EF617024.1 EF616859.1
C. fasciatus EF617105.1 EF616940.1 EF617022.1 EF616857.1
C. lineolatus EF617104.1 EF616939.1 EF617021.1 EF616856.1
C. paucifasciatus EF617102.1 EF616937.1 EF617020.1 EF616854.1
C. meyreri EF617101.1 EF616936.1 EF617019.1 EF616853.1
C. rainford EF617100.1 EF616935.1 EF617018.1 EF616852.1
C. burgessi EF617099.1 EF616934.1 EF617017.1 EF616851.1
C. ocellatus EF617098.1 EF616933.1 EF617016.1 EF616850.1
C. pelewensis EF617097.1 EF616932.1 EF617015.1 EF616849.1
C. punctatofasciatus EF617096.1 EF616931.1 EF617014.1 EF616848.1
C. larvatus EF617095.1 EF616930.1 EF617013.1 EF616847.1
C. oxycephalus EF617093.1 EF616929.1 EF617011.1 EF616845.1
C. ulietensis EF617092.1 EF616928.1 EF617010.1 EF616844.1
C. trifascialis EF617091.1 EF616927.1 EF617009.1 EF616843.1
C. ephippium EF617090.1 EF616926.1 EF617008.1 EF616842.1
C. flembii EF617089.1 EF616925.1 EF617007.1 EF616841.1
C. collare EF617088.1 EF616924.1 EF617006.1 EF616840.1
C. quadrimaculatus EF617087.1 EF616923.1 EF617005.1 EF616839.1
C. multicinctus EF617086.1 EF616922.1 EF617004.1 EF616838.1
C. robustus EF617085.1 EF616921.1 EF617003.1 EF616837.1
C. semilarvatus EF617084.1 EF095591.1 EF617002.1 EF616836.1
C. falcula EF617083.1 EF616919.1 EF617001.1 EF616835.1
C. miliaris EF617082.1 EF616918.1 EF617000.1 EF616834.1
C. speculum EF617081.1 EF616917.1 EF616999.1 EF616833.1
C. reticulatus EF617080.1 EF616916.1 EF616998.1 EF616832.1
C. striatus EF617079.1 EF616915.1 EF616997.1 EF616831.1
C. capistratus EF617078.1 EF616914.1 EF616996.1 EF616830.1
C. semeion EF617077.1 EF616913.1 EF616995.1 EF616829.1
C. sedentarius EF617076.1 EF616912.1 EF616994.1 EF616828.1
C. bennetti EF617075.1 EF616911.1 EF616993.1 EF616827.1
C. austriacus EF617074.1 EF616910.1 EF616992.1 EF616826.1
C. kleinii EF617073.1 EF616909.1 EF616991.1 EF616825.1
C. hoefleri EF617072.1 EF616908.1 EF616990.1 EF616824.1

26
Resultados
O modelo de substituição GTR foi escolhido para análise das
sequências 12S, 16S, rag2 e NADH, utilizando o Akaike information
criterion (AIC). As divergências genéticas entre os pares interespecíficos e no
gênero foi considerável. Contudo, a maioria das sequências analisadas (12S,
rag2 e NADH), demonstraram menor divergência entre as espécies que
formam híbridos, em relação às demais espécies do gênero (Tabela 1).
Tabela 2. Média das distâncias genéticas entre espécies do gênero
Chaetodon, formadoras de híbridos naturais e do conjunto total de espécies
analisadas.
Comparações interespecíficas Distância Genética
16S 12S rag2 NADH
Médias dos pares interespecíficos 7,82 0,09 0,60 2,11
Médias dentro do gênero 6,00 7,00 2,00 19,00
As distâncias genéticas entre os pares interespecíficos podem ser
observadas na Tabela 2, bem como as respectivas referências sobre a
existência de híbridos. As sequências apresentaram diferentes taxas de
evolução entre si, das quais as mitocondriais 16S e 12S apresentaram
respectivamente as taxas mais elevadas. Os pares mais divergentes quanto as
sequências 16S e NADH foram C. guenteri x C. oxycephalus (18,77% e 6,81%,
respectivamente), enquanto C. pelewensis x C. punctatofaciatus e C.
reticulatus x C. meyeri, não apresentam qualquer divergência entre todas as
sequências das espécies parentais analisadas. As comparações das
sequências rag2 evidenciaram ainda grande divergência entre C. ephipphium x
C. auriga. Em outros pares heteroespecíficos não foram observadas diferenças
entre as espécies, tais como entre C. ulietensis x C. falcula, C. mertensis x C.
xanthurus, C meyeri x C. ornatissimus, e C. multicinctus x C. pelwensis.
A comparação das sequências mitocondriais 16S, que assim como rag2
e NADH, apresentou uma variação na diferença genética relativamente menor
do que a encontrada em 12S. As maiores diferenças observadas foram 7,57%

27
entre C. ocellatus x C. striatus. Os pares C. punctatofasciatus e C. pelewensis,
C. reticulatus x C. meyeri, C. mertensis x C. xanthurus, C. ulietensis x C.
falcula, não apresentaram divergências interespecíficas de sequências.
Tabela 3. Distâncias genéticas entre sequências multilocus de pares de espécies do
gênero Chaetodon capazes de formar híbridos naturais.
Espécies parentais Distâncias Genéticas (%)
16S NADH rag2 12S % Total Referências
C. adiergastos C. collare 8,04 1,76 0,41 2,33 12,54 Hobbs et al., 2013 C. auriga C. ephippium 11,80 3,05 1,65 4,78 21,29 Randall et al., 1977 C. auriga C. vagabundus 6,97 1,41 0,54 3,85 12,56 Hobbs et al., 2013 C. auriga C. ulientensis 10,19 1,29 0,82 3,84 16,14 Hobbs et al., 2013
C. auriga C. fasciatus 10,72 1,29 0,68 5,73 18,42 Randall & Firdman 1981 C. auriga C. lunula 10,72 1,53 0,68 5,41 18,34 Kuiter, 2002 C. auriga C. trifasciatus 17,43 5,05 1,51 6,31 30,03 Schroeder, 2006
C. auriga C. rafflesi 8,58 1,29 0,82 3,24 13,93 Hobbs et al., 2013 C. ephippium C. semeion 5,63 2.93 1,37 2,33 12,26 Randall et al 1977 C. ulietensis C. falcula 0,27 0.35 0,00 0,00 00,62 Hobbs et al., 2013
C. guentheri C. oxycephalus 18,77 6,81 1,65 5,67 32,90 Kuiter, 2002 C. kleinii C. unimaculatus 9,12 0,94 1,10 3,51 14,67 Randall et al., 1977 C. lineolatus C. oxycephalus 2,95 0,94 0,41 1,15 04,45 Hobbs et al., 2013
C. mertensis C. xanthurus 0,27 0,12 0,00 0,00 00,39 Froese & Pauly, 2009 C. meyeri C. ornatissimus 7,24 2,11 0,00 2,94 12,29 Alen et al., 1998 C. miliaris C. multicinctus 7,77 3,64 0,41 4,77 16,59 Hoobs et al., 2013
C. multicinctus C. quadrimaculatus 12,60 4,23 0,54 4,16 21,53 McMillan et al., 1999 C. multicinctus C. pelewensis 0,54 0,47 0,00 0,29 01,30 McMillan et al., 1999 C. ocellatus C. striatus 12,87 5,75 1,10 7,57 27,29 Clavijo, 1987
C. punctatofasciatus C. pelewensis 0,00 0,00 0,00 0,00 00,00 McMillan et al., 1999 C. rafflesi C. vagabundus 10,19 1,53 0,27 2,93 14,92 Kuiter, 2002 C. reticulatus C. ornatissimus 7,24 2,11 0,00 2,04 11,39 Senou et al., 2007
C. reticulatus C. meyeri 0,00 0,00 0,00 0,00 00,00 Senou et al., 2007
Discussão
A origem evolutiva do gênero Chaetodon ocorreu ao final do Oligoceno,
há pouco mais de 20 milhões de anos (Bellwood, 2010). Algumas espécies
como C. guenteri e C. oxychepalus, que divergiram no início do Mioceno,
pouco depois deste período (<20 m.a) surpreendentemente formam híbridos
naturais (Hobbs et al., 2013). Esse par heteroespecífico representa o maior
grau de divergência entre todas as espécies que formam híbridos. Outros pares
como C. auriga x C. trifasciatus também exibem elevada divergência evolutiva,
estabelecida no início do Oligoceno (Fessler & Westneat, 2007).
Em contrapartida, C, meyeri x C. reticulatus e C. pelewensis x C.
punctatofaciatus representam pares heteroespecíficos de espécies
filogeneticamente muito próximas. Portanto, embora a distância genética seja
maior entre espécies filogeneticamente mais distantes, isso não implica

28
necessariamente na ausência de híbridos entre essas espécies. De fato,
apesar da grande maioria dos híbridos ocorrer entre espécies-irmãs, eles
também se mostram viáveis em espécies com uma elevada taxa de
divergência genética
Os resultados mostram que algumas espécies de Chaetodon podem
manter uma alta distância genética entre seus indivíduos e mesmo assim as
barreiras de fluxo gênico permitem que esses animais se intercruzem e formem
pares interespecíficos na natureza. Um exemplo disto ocorre em Chaetodon
striatus e C. ocellatus duas espécies Atlânticas que mantêm um elevado nível
de sobreposição geográfica. Elas ocorrem quase totalmente em simpatria por
todo o litoral sul americano e da América central, abrangendo desde regiões da
Flórida e Caribe, até Santa Catarina e sul do Brasil. Entre elas, existe relato de
hibridização em Porto Rico, no Caribe (Clavijo, 1985), apesar de terem
divergido a cerca de 8,6 m.a (Fessler & Westneat, 2007). Estas espécies
exibem elevado grau de divergência genética em relação a outros pares de
espécies desta família, como demonstrado pela análise de suas sequências.
De fato, essas divergências genéticas são reflexo do elevado período desde a
divisão de suas linhagens (Fessler & Westneat, 2007).
Embora períodos de dezenas a centenas de anos possam ser
suficientes para algumas espécies de peixes alcançarem algum grau
incompatibilidade genética (Behrmann-Godel & Gerlach, 2008), outras podem
não as alcançar mesmo decorridos vários milhões de anos de isolamento
(Russell, 2003).
Do ponto de vista citogenético essas espécies apresentam considerável
grau de similaridade cromossômica (Molina et al., 2013). Essa é uma
característica presente em grande parte dos peixes-borboleta que
compartilham um complemento cromossômico com 2n=48 cromossomos
acrocêntricos (Arai, 2011; Molina et al., 2013). Esse conservadorismo
cromossômico se mostra difundido nas espécies de Chaetodon (Nagpure et al.,
2006; Arai, 2011) sendo também comum para muitos grupos de Perciformes
(Galetti et al., 2000). A ausência de variação numérica e estruturais grosseiras
nos cromossomos, como sinalizam a presença de regiões sintênicas
envolvendo os genes 18S e 5S em C. striatus e C. ocellatus, poderiam reduzir
o papel das barreiras pós-zigóticas entre estas espécies, a partir do qual

29
sugere-se que grande parte do isolamento reprodutivo dessas espécies seriam
mantidas por mecanismos pré-zigóticos (Molina et al., 2013).
Alguns estudos filogenéticos mais recentes caracterizam os peixes-
borboletas do gênero Chaetodon em quatro grandes clados (Fessler &
Westneat, 2007; Bellwood et al., 2010; Cowman & Bellwood, 2011). O clado 4
agrupa a maioria dos indivíduos que formam pares interespecíficos e é
composto principalmente por indivíduos que não são obrigatoriamente
coralívoros. O número de espécies que formam híbridos conhecidos nos
demais clados é relativamente menor, fato que pode ser explicado devido ao
territorialismo que peixes coralívoros em geral possuem, sendo menos
receptivos para a formação de pares interespecíficos (Fricke & Fricke, 1977).
Outro fator igualmente limitante é que esses indivíduos obrigatoriamente
habitam áreas próximas às barreiras de recifes de corais, limitando sua
dispersão e impedindo que o fluxo gênico entre populações isoladas e
distantes se mantenham (Cowman & Bellwood, 2011).
Fenômenos de hibridização entre duas espécies distintas tendem a
ocorrer com relativa frequência em espécies com evolução divergente recente,
ou especiação incipiente. A hibridização nesses casos pode ocorrer devido ao
contato secundário que essas espécies podem manter após algum evento de
vicariância que não perdurou suficientemente para alcançar o isolamento
reprodutivo entre duas espécies (Mayr, 1979; Avise, 2004; Mallet, 2005).
Curiosamente, porém entre os peixes borboletas, espécies mais distantes
filogeneticamente ainda podem manter pares interespecíficos, como no caso
do C. auriga, que é capaz de intercruzar com diversos outras espécies do
gênero, até mesmo com espécies pertencentes a clados diferentes,
filogeneticamente mais distantes (Hobbs et al., 2013). Porém dados mais
específicos sobre como são os hábitos comportamentais, e características
intrínsecas dos híbridos como a capacidade de reprodução e aptidão ao
ambiente são desconhecidas, visto que a maior parte da existência de híbridos
em Chaetodons somente foi relatada através de observação em ambiente
natural.
Grande parte da incidência de pares interespecíficos e indivíduos
híbridos ocorre no Indo-Pacífico onde há o maior hostspot de espécies da
família (Kuriiwa et al., 2007; Hobbs et al., 2009). Os eventos de hibridização

30
ocorrem predominantemente próximo a bordas de barreiras geográficas
conhecidas (Bellwood & Wainwright, 2002).
Provavelmente, a formação de híbridos ocorre quando ambas as
espécies estão em no limite de sua extensão territorial, ou quando uma das
espécies possui baixa frequência em relação a outra (Rhymer & Simberloff,
1996; Rose & Polis, 2000). Dessa forma, uma das espécies parentais é mais
frequente no ambiente do que a outra, levando indivíduos de populações
reduzidas ou com baixa frequência, a se reproduzirem com peixes de outra
espécie (Hobbs et al, 2013). Outro fator que pode contribuir para tal fenômeno
é a ação antrópica, através de reduções populacionais decorrentes da
diminuição gradativa do habitat desses animais, tornando seus indivíduos a
serem mais susceptíveis a cruzamentos com espécies diferentes (Bell et al.,
1984; Dumas et al., 2007).
A incapacidade de observar totalmente os híbridos em ambiente natural
pode contribuir para uma menor estimativa da incidência de híbridos, onde
muitos cruzamentos interespecíficos poderiam passar despercebidos. Como as
espécies de Chaetodon possuem uma coloração bem característica e seus
híbridos apresentam uma variação mesclando as espécies parentais (McMillan
et al., 1999), a observação de híbridos dentro do grupo é mais facilmente
relatada do que em outras famílias. A ausência de estudos aprofundados a
respeito de barreiras de fluxo gênico em ambientes marinhos pode dificultar a
compreensão de como elas podem influenciar na existência e manutenção de
híbridos. Outras famílias como Labridae (Yaakub et al., 2006) e Pomacentridae
(Pyle & Randall, 1994) também possuem uma incidência grande híbridos entre
seus representantes, porém em peixes borboleta essa incidência é
relativamente maior. Outros estudos envolvendo análises citogenéticas mais
detalhadas nessas espécie, poderiam contribuir para o entendimento do papel
dos cromossomos no processo de hibridização em Chaetodontidae.
Referências
Barton, N. H., Hewitt, G. M. (1985). Analysis of hybrid zones. Annual review of Ecology
and Systematics, 113-148.
Allen, G.R., Steene, R. Allen, M. (1998). A guide to angelfishes and butterflyfishes,
OdysseyPublishing, Perth, Australia.

31
Behrmann-Godel, J., & Gerlach, G. (2008). First evidence for postzygotic reproductive
isolation between two populations of Eurasian perch (Perca fluviatilis L.) within
Lake Constance. Frontiers in Zoology, 5:3.
Bell, J. D., & Galzin, R. (1984). Influence of live coral cover on coral-reef fish
communities. Marine Ecology Progress Series, 15(3), 265-274.
Bierne, N., Borsa, P., Daguin, C., Jollivet, D., Viard, F., Bonhomme, F., & David, P.
(2003). Introgression patterns in the mosaic hybrid zone between Mytilus edulis
and M. galloprovincialis. Molecular Ecology, 12, 447-461.
Calado, L.L., Bertollo, L.A.C., Costa, G.W.W.F., & Molina, W.F. (2013). Cytogenetic
studies of Atlantic mojarras (Perciformes-Gerreidae): chromosomal mapping of 5S
and 18S ribosomal genes using double FISH.Aquaculture Research, 44(5), 829-
835.
Clavijo, I.E. 1985. A probable hybrid butterflyfish from the Western Atlantic. Copeia,
1985, 235-238.
Cowman, P. F., & Bellwood, D. R. (2011). Coral reefs as drivers of cladogenesis:
expanding coral reefs, cryptic extinction events, and the development of
biodiversity hotspots. Journal of evolutionary biology, 24, 2543-2562.
Darriba, D., Taboada, G. L., Doallo, R., & Posada, D. (2012). jModelTest 2: more
models, new heuristics and parallel computing. Nature methods, 9, 772-772.
Dumas, P., Kulbicki, M., Chifflet, S., Fichez, R., & Ferraris, J. (2007). Environmental
factors influencing urchin spatial distributions on disturbed coral reefs (New
Caledonia, South Pacific). Journal of Experimental Marine Biology and Ecology,
344(1), 88-100.
Edgar, R. C. (2004), MUSCLE: multiple sequence alignment with high accuracy and
high throughput. Nucleic Acids Research 32, 1792- 1797.
Fessler, J. L., & Westneat, M. W. (2007). Molecular phylogenetics of the butterflyfishes
(Chaetodontidae): taxonomy and biogeography of a global coral reef fish family.
Molecular phylogenetics and evolution, 45, 50-68.
Findley, J. S., & Findley, M. T. (2001). Global, regional, and local patterns in species
richness and abundance of butterflyfishes. Ecological Monographs,71, 69-91.
Fricke, H., Fricke, S. (1977). Monogamy and sex change by aggressive dominance in
coral reef fish. Nature 266, 830 – 832.
Galetti Jr., P. M., Aguilar, C. T., & Molina, W. F. (2000). An overview of marine fish
cytogenetics. Marine Genetics. pp. 55-62.
Hall, T. A. 1999. BioEdit: a user-friendly biological sequence alignment editor and
analysis program for Windows 95/98/NT. Nucleic Acids Symposium Series, 41:95-
98.
Hobbs, J. P. A., Frisch, A. J., Allen, G. R., & Van Herwerden, L. (2009). Marine hybrid
hotspot at Indo-Pacific biogeographic border. Biology Letters,5, 258-261.

32
Hobbs, J. P. A., van Herwerden, L., Pratchett, M. S., & Allen, G. R. (2013).
Hybridisation among butterflyfishes. Biology of Butterflyfish. CRC Press, Florida,
48-69.
Kuiter, R.H. (2002). Butterflyfishes, Bannerfishes and their Relatives. A comprehensive
Guide to Chaetodontidae and Microcanthidae. The Marine Fish Families Series,
TMC Publishing, Chorleywood, UK
Mallet, J. (2007). Hybrid speciation. Nature, 44, 279-283.
McMillan, W. O., Weigt, L. A., & Palumbi, S. R. (1999). Color pattern evolution,
assortative mating, and genetic differentiation in brightly colored butterflyfishes
(Chaetodontidae). Evolution, 247-260.
Man, K. F., Tang, K. S., & Kwong, S. (2012). Genetic algorithms: Concepts and
designs. Springer Science & Business Media.
Mayr, E. (1963). Animal species and evolution. Cambridge, Massachusetts: Harvard
University Press.
Mayr E. (1979). Animal speciation and evolution. In Belknap Press 6th
edn. Cambridge, MA: Belknap Press.
Molina, W. F., Costa, G. W. W. F., Soares, R. X., Affonso, P. R. A.M., Cioffi, M.B.,
Araújo, W. C., & Bertollo, L. A. C. (2013). Extensive chromosome conservatism in
Atlantic butterflyfishes, genus Chaetodon Linnaeus, 1758: Implications for the high
hybridization success. Zoologischer Anzeiger. A Journal of Comparative
Zoology, 253, 137-142.
Montanari, S. R., Van Herwerden, L., Pratchett, M. S., Hobbs, J. P. A., & Fugedi, A.
(2012). Reef fish hybridization: lessons learnt from butterflyfishes (genus
Chaetodon). Ecology and evolution, 2, 310-328.
Motta-Neto, C. C., Cioffi, M. B., Bertollo, L. A. C., & Molina, W. F. (2011). Extensive
chromosomal homologies and evidence of karyotypic stasis in Atlantic grunts of
the genus Haemulon (Perciformes). Journal of Experimental Marine Biology and
Ecology, 401, 75-79.
Motta-Neto, C. C., Lima-Filho, P. A., Araújo, W. C., Bertollo, L. A. C., & Molina, W. F.
(2012). Differentiated evolutionary pathways in Haemulidae (Perciformes):
karyotype stasis versus morphological differentiation. Reviews in Fish Biology and
Fisheries, 22, 457-465.
Pratchett, M.S. 2007. Dietary selection by coral-feeding butterflyfishes
(Chaetodontidae) on the Great Barrier Reef, Australia. Raffles Bulletin of Zoology.
171–176.
Pratchett, M. S., Berumen, M. L., & Kapoor, B. G. (Eds.). (2013). Biology of
butterflyfishes. CRC Press.
Pitts, P. A. (1991). Comparative use of food and space by three Bahamian
butterflyfishes. Bulletin of marine science, 48, 749-756.
Cox, E.F. (1994). Resource use by corallivorous butterflyfishes (Family
Chaetodontidae) in Hawaii. Bulletin of Marine Science, 54, 535-545.

33
Pyle, R.L., & Randall, J.E. (1994). A review of hybridization in marine angelfishes
(Perciformes: Pomacanthidae). Environmental Biology of Fishes,41, 127-145.
Randall, J.E., G.R. Allen, R.C. Steene. (1977). Five probable hybrid butterfl y fi shes of
the genus Chaetodon from the central and western Pacifi c. Records of the
Australian Museum6: 3–26.
Randall, J.E. D. Fridman. (1981). Chaetodon auriga x Chaetodon fasciatus, a hybrid
butterflyfish from the Red Sea. Revue. fr. Aquariol 7: 113–116
Rose, M.D., Polis, G.A. (2000). On the insularity of islands. Ecography,23, 693-701.
Rhymer, J.M., & Simberloff, D. (1996). Extinction by hybridization and introgression.
Annual Review of Ecology and Systematics. 83-109.
Russell, S.T. (2003). Evolution of intrinsic post-zygotic reproductive isolation in fish.
Ann Zool Fennici, 40, 321-329.
Seehausen, O. (2004). Hybridization and adaptive radiation. Trends in ecology &
evolution, 19, 198-207.
Seehausen, O. (2006). Conservation: losing biodiversity by reverse speciation. Current
Biology, 16, 334-337.
Senou, H., Kodato, H., Nomura, T., Yunokawa K. (2006). Coastal fi shes of Ie-jima
Island, the Ryukyu Islands, Okinawa, Japan. Bulletin of the Kanagawa Prefectural
Museum 35: 67–92
Schwenk, K. (1993). Interspecific hybridization in Daphnia: distinction and origin of
hybrid matrilines. Molecular Biology and Evolution, 10, 1289-1302.
Tamura, K., Stecher, G., Peterson, D., Filipski, A., & Kumar, S. (2013). MEGA6:
molecular evolutionary genetics analysis version 6.0. Molecular Biology and
Evolution, 30, 2725-2729.
Yaakub, S.M., Bellwood, D. R., van Herwerden, L., & Walsh, F.M. (2006). Hybridization
in coral reef fishes: introgression and bi-directional gene exchange in Thalassoma
(family Labridae). Molecular Phylogenetics and Evolution, 40, 84-100.
Xia, X., Xie, Z. (2001). DAMBE: software package for data analysis in molecular
biology and evolution. Journal of Heredity, 92, 371-373.

34
Capítulo 2
O gênero Chaetodon no Atlântico: Aspectos da diversificação filética e
estase cromossômica
Miguel, D. Z., Costa G. W. W. F., Soares, R.X., Borges, A. T., Motta-Netto, C.
C., Molina, W.F.
Resumo
Dentre os variados habitantes de recifes de corais, os peixes-borboleta
do gênero Chaetodon se destacam pelo grau de especialização de algumas
espécies e distribuição geográfica, sendo encontrados em praticamente todos
os oceanos do mundo. O gênero possui linhagens inteiras que são
dependentes de recifes de corais, mostrando-se, portanto, susceptíveis a
alterações nestes ambientes. Apesar do vasto conhecimento acumulado sobre
diversos aspectos biológicos, ainda persistem lacunas sobre seus padrões
citogenéticos. O presente estudo analisou aspectos citogenéticos por meio do
mapeamento cromossômico do DNAr 18S e 5S nas espécies Atlânticas C.
striatus, C. capistratus e C. sedentarius. Comparações citopopulacionais foram
realizadas entre indivíduos de C. striatus do Atlântico Sul e litoral do EUA. Os
dados indicam conservadorismo cariotípico entre essas espécies, quanto ao
cariótipo e frequência e distribuição dos sítios ribossomais. Sítios DNAr 18S
estavam localizados exclusivamente em posição pericentromérica no par 10,
enquanto que os sítios DNAr 5S estavam presentes na região pericentromérica
do par 21 para as três espécies analisadas. Um padrão de distribuição similar
dos sítios ribossomais em C. striatus estava presente entre as duas regiões
geográficas analisadas, indicando um cariótipo homogêneo mesmo entre
indivíduos distribuídos uma área geográfica extensa e permeada pela barreira
do rio Amazonas. Os dados citogenéticos significativamente similares nessas
espécies poderiam favorecer a frequente ocorrência de quebras de barreiras
reprodutivas pós-zigoticas no gênero.
Palavras-chave: Peixe-borboleta, citogenética de peixes, conservadorismo
cromossômico, hibridização in situ, Perciformes.

35
Abstract Among the various inhabitants of coral reefs, Chaetodon species stand out for
having a very characteristic and striking coloration, being found in virtually all
the world's oceans. The group has entire groups that are dependent on coral
reefs, and is therefore closely linked to the same ecological, being susceptible
to changes caused by man in the environment. Much is known about various
aspects of belonging to family, such as eating habits, choice of sexual partners,
biogeography and coverage area and staining patterns. But there is a gap in
knowledge of cytogenetic data in most specimens described as belonging to the
family. The present study aimed to map the chromosomally 18S and 5S rDNA
regions of three species of the genus Chaetodon, C. striatus, C. sedentarius
and C. capistratus. The results indicate an extreme conservatism karyotype in
these animals, as both marks were located on the par 21 (18S) and 10 (5S) for
the three species analyzed. Previous cytogenetic studies in C. striatus indicate
that more distant populations remain a homogeneous karyotype even when
they exist in a very large geographical area. The similar karyotype these
species can help in understanding how occur the breaks barriers post-zygotic
gene flow commonly found in these animals.
Keywords: Butterflyfishes, chromosomal conservatism, in situ hybridization,
Perciformes.
Introdução
Peixes da família Chaetodontidae, conhecidos como peixes-borboletas,
são indivíduos importantes na manutenção de recifes e formações recifais de
todo o mundo (Pitss, 1991; Cole & Pratchett, 2008). Possuem uma coloração
bem variada e padrões morfológicos característicos do grupo (McMillan et al.,
1999). Abrangendo cerca de 130 espécies, o gênero Chaetodon é o mais
abundante da família, visto que estão distribuídas em todos os oceanos ao
redor do mundo, com maior concentração de espécies nas regiões próximas ao
oeste do Pacífico, na divisa com o oceano Índico (Blum, 1989; Bellwood &
Wainwright, 2002; Hobs et al., 2009).
Diversas espécies do gênero possuem relações ecológicas íntimas com
corais, de modo que sua capacidade de sobrevivência acaba por ser, em
muitos casos, extremamente dependente da existência desses corais (Reese,
1975; Gosline, 1985; Hourigan et al., 1988). Algumas espécies são ainda mais

36
especialistas, e podem facilmente sofrer influências de alterações ambientais
e/ou atividades antrópicas ser ameaçadas se algum fator antrópico ou
ambiental devido a vulnerabilidade desta relação, já que interferências
negativas podem impactar de forma negativa o crescimento e estabelecimento
dos recifes de corais (Pratchett et al., 2004) e consequentemente a
sobrevivência dos peixes borboleta a eles ligados.
Apesar de dois terços de todas as espécies descritas serem
classificadas como obrigatoriamente coralívoras. Sendo que alguns grupos são
coralívoros facultativos, ou seja, podem se alimentar mesmo os recifes de
corais estando ausentes (Wylie & Paul, 1989). Outros, no entanto, não são
coralívoros, sendo geralmente raspadores de fundo, ou podendo se alimentar
de crustáceos e outros pequenos animais marinhos (Pratchett, 2007; Reavis &
Corpus, 2011). As formas de alimentação desses animais geralmente têm uma
origem evolutiva em comum, sendo o gênero Chaetodon dividido em quatro
grandes táxons cujo habitat e hábitos alimentares são compartilhados pela
maioria dos membros (Fessler & Weastneat, 2007).
Muitos estudos vêm sendo desenvolvidos na área comportamental,
morfológica e biogeográfica desses animais (Fricke, 1986; Cox, 1984; Righton
et al., 1996; Sazima & Sazima, 2001), porém poucos estudos exploram suas
características cromossômicas e seu cariótipo. As poucas espécies
cariotipadas compreendem uma parcela extremamente pequena do grupo,
sendo que esses estudos não se aprofundaram na compreensão de como seu
cariótipo se apresenta e como são suas características cromossômicas (Arai,
2011; Molina et al., 2013). Tais pesquisas realizadas apresentam similaridades
cromossômicas muito evidentes, revelando um extremo conservadorismo
cariotípico que esses indivíduos apresentam. As espécies descritas, em sua
maioria, apresentam 48 cromossomos acrocêntricos em seu cariótipo,
característica essa que é comum a outros grupos de Perciformes. Esse
conservadorismo pode contribuir para que fenômenos como a alta taxa de
hibridização e formação de pares interespecíficos presentes nesse gênero
ocorram.
No presente estudo foram analisadas três espécies diferentes de
Chaetodon: C. striatus, C. capistratus, C. sedentarius. Foram realizados
mapeamentos cromossômicos das regiões DNAr 18S e 5S em indivíduos

37
Chaetodon striatus, cujas características cromossômicas já foram descritas
para populações existentes na região da costa brasileira. Os espécimes
utilizados neste trabalho foram coletados na Flórida, a fim de comparar
biogeograficamente se suas regiões mapeadas apresentam sintenia ou
divergência devido a separação espacial entre essas populações, que são
consideradas distintas.
As duas outras espécies, C. sedentarius e C. capistratus, coletadas
também no mesmo local, foram mapeadas em suas regiões 18S e 5S, que até
o presente momento nunca foram antes analisadas. Este trabalho também
objetivou ajudar na caracterização cariotípica de tais espécies marinhas afim
de ajudar na compreensão de como a evolução e processos de isolamento
reprodutivo podem levar ao surgimento de novas espécies.
Material e Métodos
Indivíduos das espécies de Chaetodon striatus, C. sedentarius e C.
capistratus foram coletados no Oceano Atlântico na região de Key Largo
(25°6′23″N, 80°25′47″O), Flórida, EUA. Amostras de Chaetodon striatus
também foram obtidas no litoral do Rio Grande do Norte. Após a coleta os
espécimes foram submetidos a técnicas de estimulação mitótica (Molina et al.,
2010). Brevemente, nos indivíduos receberam uma injeção intramuscular de
solução de lisados de bactérias e fungos (10 mg do complexo de antígenos /10
ml de água destilada). A solução foi injetada na região dorso-lateral do animal
na proporção de 1 ml para cada 100g de peso. Os indivíduos foram deixados
vivos em seus reservatórios por um período de 24 horas.
Preparação de cromossomos mitóticos
Os indivíduos foram sacrificados com o uso de superdosagem de
Eugenol até que os movimentos branquiais cessassem. Os cromossomos
mitóticos foram obtidos mediante a técnica usada e descrita por Gold et al.
(1990). Um volume de aproximadamente 125µl da suspensão celular foi
gotejado sobre lâmina recoberta com um filme de água destilada aquecido à
60°C. As lâminas após secagem foram coradas com Giemsa 5%, pH 6,8, por
10 minutos e utilizadas nas análises citogenéticas e montagem do cariótipo

38
Hibridação fluorescente in situ (FISH)
A hibridização in situ fluorescente (FISH) foi realizada de acordo com a
metodologia descrita por Pinkel et al. (1986), com algumas modificações. A
sequência de DNA in tandem isolada do DNA genomico de cada espécie
específica foi utilizada para sonda de DNAr 18S e 5S. A sonda 18S foi marcada
por nick translation digoxigenina-11-dUTP (Roche) e a sonda rDNA 5S por nick
translation biotina-14-dATP (Roche) de acordo com as instruções do fabricante.
Os cromossomos foram tratados com RNAse livre de DNAse (20mg / mL em
2×SSC) à 37°C por 1 hora, com pepsina (0,005 % em HCl 10mM) à 37°C por
10 minutos e fixados com formaldeído a 1% por 10 minutos, em seguida
desidratados em uma série alcoólica. Os cromossomos foram, então,
desnaturados em 70% formamida/2×SSC à 72°C por 5 minutos. A solução de
hibridização consistida em 50% de formamida, 2×SSC, 10% de sulfato de
dextran e a sonda desnaturada (5 ng/µl). Após hibridação, overnight à 37°C, as
lâminas foram lavadas em 15% formamida/0.2×SSC à 42°C por 20 minutos,
0,1×SSC à 60°C por 15 minutos e Tween20 0,5%/4×SSC à temperatura
ambiente. O sinal de hibridização da sonda foi detectado usando streptavidina-
FITC conjugadas (Vector) para a sonda DNAr 5S e anti-digoxigenina rodamina
conjugada (Roche) para a sonda DNAr 18S. Os cromossomos foram
contracorados com Vectashield/DAPI (1,5 µg/ml).
As análises em fluorescência foram realizadas em fotomicroscópio de
epifluorescência Olympus BX50, equipado com sistema digital de captura de
imagens Olympus DP73 utilizando o software cellSens (Olympus Optical Co.
Ltd.). Os cromossomos foram classificados de acordo com a posição do
centrômero (Levan et al., 1964).
Resultados
As três espécies analisadas evidenciaram um cariótipo conservado e
idêntico, composto por 2n=48 cromossomos acrocêntricos. Os resultados do
mapeamento dos genes DNAr 18S e 5S nos cromossomos das espécies C.
striatus, C. capistratus e C. sedentarius estão apresentados junto ao cariótipo
na Figura 1.

39
Os sítios se mostraram homeólogos entre as espécies e entre as duas
populações de C. striatus. As sondas DNAr 18S apresentaram marcações em
posição pericentromérica no par 21, enquanto sítios de DNAr 5S, estavam
localizados em posição pericentromérica no par 10.
Figura 1. Cariótipos das espécies C. striatus (a), C. capistratus (b) e C.
sedentarius (c). Mapeamento cromossômico de sítios DNAr 18S (vermelho) e
DNAr 5S (verde), através do FISH, em C. striatus (a), C. capistratus (b) e em C.
sedentarius (c), e coloração convencional do cariótipo em Gimsa. Barra = 5 um.
Discussão
A ausência de variação citogenética entre C. striatus da costa brasileira
e do litoral da Flórida, sugere que o fluxo gênico entre as populações pode
a
b
c

40
ocorrer de forma livre mesmo nas grandes distâncias envolvidas. De fato, esse
resultado pode sugerir um papel menos restritivo do fluxo gênico da barreira
ocasionada pela descarga dos rios Amazonas e Orinoco nesta espécie.
Análises populacionais utilizando marcadores moleculares em C. striatus
apoiam uma baixa estruturação populacional na costa brasileira (Affonso et al.,
2011). Tal condição tem sido encontrada em outras espécies de Chaetodon,
oriundas do Hawaii, que apesar de habitarem uma área geográfica extensa,
não demonstraram diferenciações genéticas evidentes (Craig et al., 2010). A
dispersão larval e hábitos generalistas poderiam atuar para que o fluxo gênico
se mantenha entre essas populações.
Cariótipos com 2n=48 cromossomos acrocêntricos, com sítios
ribossomais localizados em regiões intersticiais dos cromossomos tem sido
considerados caracters basais comuns a um largo número de espécies de
Perciformes (Galetti et al., 2000; Negapture et al., 2006, Arai 2011). Essa
condição ancestral está presente em Pomacanthidae (Affonso et al., 2002),
grupo-irmão dos Chaetodontidae, cuja divergência data do início do Oligoceno,
há proximadamente 30 m.a. (Fessler & Westneat, 2011).
A origem do gênero Chaetodon, na família Chaetodontidae, é um evento
relativamente recente, com uma irradiação a poucos milhões de anos atrás
(Fessler & Westneat, 2011; Hsu et al., 2007). Este curto tempo de divergência
poderia ser um dos fatores a explicar o conservadorismo cariotípico encontrado
no gênero. O pequeno tempo de divergência poderia não ter sido suficiente
para que alterações cromossômicas maiores, tivessem ocorrido de forma mais
intensa no cariótipo.
As altas taxas de hibridização e formação de pares interespecíficos
dentro do gênero Chaetodon (Randall & Fridman, 1981, Randall et al., 1977)
são grandes, mesmo quando comparadas com outros grupos de peixes recifais
que também hibridizam (Montanari & Herwerden, 2012).
As similaridades citogenéticas, demonstrada pelo conservadorismo
estruturais e dos genes ribossomais nas espécies dão suporte sobre o papel
pouco restritivo do cariótipo como efetiva barreira pós-zigótica nas espécies
deste gênero. Dados citogenéticos mais resolutivos envolvendo um maior
número de espécies e decorrente do mapeamento de genes sintênicos são

41
desejáveis para se estabelecer o nível de conservação homeóloga nos
cromossomos das espécies deste gênero.
Referências Affonso, P. R. A. M., Guedes, W., Pauls, E., Galetti Jr., P. M., (2001). Cytogenetic
analysis of coral reef fishes from Brazil (Families Pomacanthidae and
Chaetodontidae). Cytologia (Tokyo) 66, 379–384.
Affonso, P. R A. M., Guedes, W., Pauls, E., Galetti Jr., P. M., (2002). Close
karyotypical and species relationship between two species of marine angelfishes
from South Atlantic: Pomacanthus arcuatus and P. paru (Perciformes,
Pomacanthidae). Caryologia 55, 323–329.
Arai, R., (2011). Fish Karyotypes: A Check List. Springer, Tokyo.
Bellwood, D. R., & Wainwright, P. C., (2002). The history and biogeography of fishes
on coral reefs. Coral reef fishes: dynamics and diversity in a complex ecosystem,
5-32.
Blum, S. D., (1989). Biogeography of the Chaetodontidae: an analysis of allopatry
among closely related species. Environmental Biology of Fishes, 25(1-3), 9-31.
Craig, M. T., Eble, J. A., Bowen, B. W., (2010). Origins, ages and population histories:
comparative phylogeography of endemic Hawaiian butterflyfishes (genus
Chaetodon). J. Biogeogr. 37, 2125–2136.
Fessler, J. L., Westneat, M. W., (2007). Molecular phylogenetics of the butterflyfishes
(Chaetodontidae): taxonomy and biogeography of a global coral reef fish family.
Molecular phylogenetics and evolution, 45(1), 50-68.
Fricke, H. W., (1986). Pair swimming and mutual partner guarding in monogamous
butterflyfish (Pisces, Chaetodontidae): a joint advertisement for territory. Ethology,
73(4), 307-333.
Galetti Jr., P. M., Aguilar, C. T., Molina, W. F., (2000). An overview of marine fish
cytogenetics. Hydrobiologia, 420, 55–62.
Gosline, W. A., (1985). Relationships among some relatively deep-bodied percoid fish
groups. Ichthyology Journal, 31(4), 351-357.
Hobbs, J. P. A., Frisch, A. J., Allen, G. R., Van Herwerden, L., (2009). Marine hybrid
hotspot at Indo-Pacific biogeographic border. Biology Letters, 5(2), 258-261.
Hsu, K. C., Chen, J. P., Shao, K. T., (2007). Molecular phylogeny of Chaetodon
(Teleostei: Chaetodontidae) in the Indo-West Pacific: evolution in geminate
species pairs and species groups. The Raffles Bulletin of Zoology, 14, 77-86.

42
Molina, W. F., da Costa, G. W. W. F., Soares, R. X., Affonso, P. R. A. M., Cioffi, M.B.,
Araújo, W. C., Bertollo, L. A. C., (2013). Extensive chromosome conservatism in
Atlantic butterflyfishes, genus Chaetodon Linnaeus, 1758: Implications for the high
hybridization success. Zoologischer Anzeiger-A Journal of Comparative
Zoology, 253(2), 137-142.
Molina, W. F., Galetti Jr., P. M., (2004). Karyotypic changes associated to the
dispersive potential on Pomacentridae (Pisces, Perciformes). J. Exp. Mar. Biol.
Ecol. 309, 109–119.
Montanari, S. R., Van Herwerden, L., Pratchett, M. S., Hobbs, J. P. A., Fugedi, A.,
(2012). Reef fish hybridization: lessons learnt from butterflyfishes (genus
Chaetodon). Ecology and evolution, 2(2), 310-328.
Nagpure, N. S., Kumar, R., Srivastava, S. K., Kushwaha, B., Gopalakrishnan, A.,
Basheer, V. S., (2006). Cytogenetic characterization of two marine ornamental
fishes, Chaetodon collare and Stegastes insularis. J. Mar. Biol. Ass. India 48, 267–
269.
Nielsen, E. E., Hemmer-Hansen, J., Poulsen, N. A., Loeschcke, V., Moen, T.,
Johansen, T., Mittelholzer, C., Taranger, G. L., Ogden, R., Carvalho, G. R., (2009).
Genomic signatures of local directional selection in a high gene flow marine
organism; the Atlantic cod (Gadus morhua). BMC Evolutionary Biology, 9(1), 276.
Pratchett, M. S., (2007). Dietary selection by coral-feeding butterflyfishes
(Chaetodontidae) on the Great Barrier Reef, Australia. Raffles Bull Zool, 14, 171-
176.
Pratchett, M. S., Wilson, S. K., Berumen, M. L., McCormick, M. I. (2004). Sublethal
effects of coral bleaching on an obligate coral feeding butterflyfish. Coral Reefs,
23(3), 352-356.
Randall, J. E., Allen, G. R., Steen, R. C., (1977). Five probable hybrid butterflyfishes of
the genus Chaetodon from the central and western pacific. Rec. West. Aust. Mus.
6, 3–26.
Randall, J. E., Fridman, D., (1981). Chaetodon auriga × Chaetodon fasciatus, a hybrid
butterflyfish from the Red Sea. Rev. Fr. Aquariol. 7, 113–116
Reavis, R. H., Copus, J. M., (2011). Monogamy in a feeding generalist, Chaetodon
trichrous, the endemic Tahitian butterflyfish. Environmental biology of fishes, 92(2),
167-179.
Reese, E. S., (1975). A comparative field study of the social behavior and related
ecology of reef fishes of the family Chaetodontidae1) 2). Zeitschrift für
Tierpsychologie, 37(1), 37-61.
Righton, D., Kemp, J., Ormond, R., (1996). Biogeography, community structure and
diversity of Red Sea and western Indian Ocean butterflyfishes. Journal of the
Marine Biological Association of the United Kingdom, 76(01), 223-228.

43
Wylie, C. R., Paul, V. J., (1989). Chemical defenses in three species of Sinularia
(Coelenterata, Alcyonacea): effects against generalist predators and the
butterflyfish Chaetodon unimaculatus Bloch. Journal of Experimental Marine
Biology and Ecology, 129(2), 141-160.

44
Conclusões
Diante das análises realizadas foi possível realizar algumas inferências:
- As distâncias genéticas dos genes nucleares e mitocondriais entre as
espécies, indicam que mesmo longos períodos de divergência evolutiva não
são suficientes para efetivo bloqueio pós-zigótico no gênero Chaetodon.
- As espécies C. striatus, C. capistratus e C. sedentarius apresentam
padrão conservativo quanto aos aspectos macroestruturais dos cromossomos
e organização dos genes DNAr 18S e 5S.
- O conservadorismo cromossômico em Chaetodon também é manifesto
no nível populacional. A despeito da vasta extensão oceânica, populações de
C. striatus, do sul do Atlântico Ocidental não apresentam diferenciações
citogenéticas em relação àqueles do litoral sul dos EUA.
- As similaridades cariotípicas interespecíficas encontradas no gênero
Chaetodon não podem ser descartadas como fator importante nos processos
de hibridação em espécies simpátricas de peixes borboleta.
- A despeito de alguns dados disponíveis, a ausência de dados
citogenômicos e taxonômicos mais extensos no gênero Chaetodon, ainda
dificulta a compreensão plena do papel dos cromossomos no processo de
hibridização dos peixes-borboleta. Dessa forma, a compreensão deste
frequente fenômeno, que desafia os padrões de coesão genética de outros
grupos de peixes, deve ser ampliada diante da grande relevância para a
compreensão dos aspectos evolutivos deste carismático grupo de peixes
recifais.

45
5. Referências Bibliográficas
Arai, R. (2011). Fish karyotypes: a check list. Springer Science & Business
Media
Affonso, P.R.A.M., Guedes, W., Pauls, E. and Galetti Jr., P.M. (2001)
Cytogenetic analysis of coral reef fishes from Brazil (families
Pomacanthidae and Chaetodontidae). Cytologia, 66: 379–384.
Arai, R., Inoue, M. (1975) Chromosomes of nine species of Chaetodontidae and
one species of Scorpidae from Japan. Bull. Natn. Sci. Mus. Tokyo, (A), 1:
217–224
Bannikov, A. F. (2004). Fishes from the Eocene of Bolca, northern Italy,
previously classified with the Chaetodontidae (Perciformes). Studi e
Ricerche sui Giacimenti Terziari di Bolca, 10, 57-76.
Bellwood, D. R. (1996). The Eocene fishes of Monte Bolca: the earliest coral
reef fish assemblage. Coral Reefs, 15(1), 11-19.
Bellwood, D. R. (1997). Reef fish biogeography: habitat associations, fossils
and phylogenies. In Proceedings of the 8th International Coral Reef
Symposium (Vol. 1, pp. 379-384).
Bellwood, D. R., & Wainwright, P. C. (2002). The history and biogeography of
fishes on coral reefs. Coral reef fishes: dynamics and diversity in a complex
ecosystem, 5-32.
Bellwood, D. R., Hughes, T. P., Connolly, S. R., & Tanner, J. (2005).
Environmental and geometric constraints on Indo‐Pacific coral reef
biodiversity. Ecology Letters, 8(6), 643-651.
Blum, S. D. (1989). Biogeography of the Chaetodontidae: an analysis of
allopatry among closely related species. Environmental Biology of
Fishes,25(1-3), 9-31.
Brown, J. H. (1984). On the relationship between abundance and distribution of
species. American naturalist, 255-279.
Carnevale, G. (2006). Morphology and biology of the Miocene butterflyfish
Chaetodon ficheuri (Teleostei: Chaetodontidae). Zoological Journal of the
Linnean Society, 146(2), 251-267.
Cole, A. J., Pratchett, M. S., & Jones, G. P. (2008). Diversity and functional
importance of coral‐feeding fishes on tropical coral reefs. Fish and
Fisheries, 9(3), 286-307.
Cowman, P. F., & Bellwood, D. R. (2011). Coral reefs as drivers of
cladogenesis: expanding coral reefs, cryptic extinction events, and the
development of biodiversity hotspots. Journal of evolutionary biology,
24(12), 2543-2562.

46
Driscoll, J. W., & Driscoll, J. L. (1988). Pair behavior and spacing in
butterflyfishes (Chaetodontidae). Environmental biology of fishes, 22(1), 29-
37.
Fessler, J. L., & Westneat, M. W. (2007). Molecular phylogenetics of the
butterflyfishes (Chaetodontidae): taxonomy and biogeography of a global
coral reef fish family. Molecular phylogenetics and evolution, 45(1), 50-68.
Fricke, H. W. (1986). Pair swimming and mutual partner guarding in
monogamous butterflyfish (Pisces, Chaetodontidae): a joint advertisement
for territory. Ethology, 73(4), 307-333.
Galetti Jr, P. M., Aguilar, C. T., & Molina, W. F. (2000). An overview of marine
fish cytogenetics. Marine Genetics. 55-62.
Galetti Jr., P.M., Molina, W.F., Affonso, P.R.A.M. and Aguilar, C.T. (2006)
Assessing genetic diversity of Brazilian reef fishes by chromosomal and
DNA markers. Genetica,126: 161-177
Gosline, W. A. (1985). Relationships among some relatively deep-bodied
percoid fish groups. Japanese Journal of Ichthyology, 31(4), 351-357.
Hixon, M. A., & Webster, M. S. (2002). Density dependence in reef fish
populations. Coral reef fishes: dynamics and diversity in a complex
ecosystem. Academic Press, San Diego, California, USA, 303-325.
Hourigan, T. F., Timothy, C. T., & Reese, E. S. (1988). Coral reef fishes as
indicators of environmental stress in coral reefs. In Marine organisms as
indicators. 107-135. Springer New York.
Jones, G. P., McCormick, M. I., Srinivasan, M., & Eagle, J. V. (2004). Coral
decline threatens fish biodiversity in marine reserves. Proceedings of the
National Academy of Sciences of the United States of America, 101(21),
8251-8253.
Kauffman, E. G., & Johnson, C. C. (1988). The morphological and ecological
evolution of Middle and Upper Cretaceous reef-building rudistids. Palaios,
194-216.
Molina, W. F., & Galetti-Jr, P. M. (2002). Robertsonian rearrangements in the
reef fish Chromis (Perciformes, Pomacentridae) involving chromosomes
bearing 5S rRNA genes. Genetics and Molecular Biology, 25(4), 373-377.
Motta-Neto, C. C., Cioffi, M. B., Bertollo, L. A. C., & Molina, W. F. (2011).
Extensive chromosomal homologies and evidence of karyotypic stasis in
Atlantic grunts of the genus Haemulon (Perciformes). Journal of
Experimental Marine Biology and Ecology, 401(1), 75-79.
Motta-Neto, C. C., Lima-Filho, P. A., Araújo, W. C., Bertollo, L. A. C., & Molina,
W. F. (2012). Differentiated evolutionary pathways in Haemulidae
(Perciformes): karyotype stasis versus morphological differentiation.
Reviews in Fish Biology and Fisheries, 22(2), 457-465.

47
Nagpure, N.S., Kumar, R., Srivastava, S.K., Kushwaha, B., Gopalakrishnan, A.,
Basheer, V.S. (2006) Cytogenetic characterization of two marine
ornamental fifishes, Chaetodon collare and Stegastes insularis. J. Mar. Biol.
Ass. India, 48: 267–269.
Nelson, J. S. (2006). Fishes of the World. John Wiley & Sons.
Ojima, Y., Yamamoto, K. (1990) Cellular DNA contents of fishes determined by
flow cytometry. La Kromosomo, II, 57: 1871–1888.
Pitts, P. A. (1991). Comparative use of food and space by three Bahamian
butterflyfishes. Bulletin of marine science, 48(3), 749-756.
Pyle, R. L., & Randall, J. E. (1994). A review of hybridization in marine
angelfishes (Perciformes: Pomacanthidae). Environmental Biology of
Fishes,41(1-4), 127-145.
Reese, E. S. (1975). A comparative field study of the social behavior and
related ecology of reef fishes of the family Chaetodontidae. Zeitschrift für
Tierpsychologie, 37(1), 37-61.
Rhymer, J. M., & Simberloff, D. (1996). Extinction by hybridization and
introgression. Annual Review of Ecology and Systematics, 83-109.
Righton, D., Kemp, J., & Ormond, R. (1996). Biogeography, community
structure and diversity of Red Sea and western Indian Ocean
butterflyfishes.Journal of the Marine Biological Association of the United
Kingdom, 76(01), 223-228.
Rose, M. D., Polis, G. A. (2000). On the insularity of islands. Ecography,23(6),
693-701.
Smith, W., Webb, J. F., & Blum, S. D. (2003). The evolution of the laterophysic
connection with a revised phylogeny and taxonomy of butterflyfishes
(Teleostei: Chaetodontidae). Cladistics, 19(4), 287-306.
Sytchevskaya, E. K. Late Oligocene-Early Miocene history of the ichthyofauna
of Eastern Paratethys. In Current Research in Vertebrate Palaeontolgy 3rd
Annual Meeting of the European Associa-tion of Vertebrate
Palaeontologists (EAVP) in the Hessisches Landesmuseum Darmstadt 18th
to 23th July 2005. 65.
Wilson, S. K., Graham, N. A., Pratchett, M. S., Jones, G. P., & Polunin, N. V.
(2006). Multiple disturbances and the global degradation of coral reefs: are
reef fishes at risk or resilient? Global Change Biology, 12(11), 2220-2234.
Wood, R. (1998). The ecological evolution of reefs. Annual Review of Ecology
and Systematics, 179-206.
Yaakub, S. M., Bellwood, D. R., van Herwerden, L., & Walsh, F. M. (2006).
Hybridization in coral reef fishes: introgression and bi-directional gene
exchange in Thalassoma (family Labridae). Molecular phylogenetics and
evolution, 40(1), 84-100.

48
Yabuta, S. (1997). Spawning migrations in the monogamous butterflyfish,
Chaetodon trifasciatus. Ichthyological Research, 44(2-3), 177-182.


















