
Ecologia de fêmeas de ouriço-preto Chaetomys subspinosus...
89
Pontifícia Universidade Católica de Minas Gerais Programa de Pós-graduação em Zoologia de Vertebrados Ecologia de fêmeas de ouriço-preto Chaetomys subspinosus (Olfers 1818) (Rodentia: Erethizontidae) nas florestas de restinga do Parque Estadual Paulo César Vinha, Guaraparí, Espírito Santo PEDRO AMARAL DE OLIVEIRA Belo Horizonte, MG 2006
-
Upload
nguyendien -
Category
Documents
-
view
213 -
download
0
Transcript of Ecologia de fêmeas de ouriço-preto Chaetomys subspinosus...

Pontifícia Universidade Católica de Minas Gerais
Programa de Pós-graduação em Zoologia de Vertebrados
Ecologia de fêmeas de ouriço-preto Chaetomys subspinosus
(Olfers 1818) (Rodentia: Erethizontidae) nas florestas de
restinga do Parque Estadual Paulo César Vinha,
Guaraparí, Espírito Santo
PEDRO AMARAL DE OLIVEIRA
Belo Horizonte, MG
2006

Pedro Amaral de Oliveira
Ecologia de fêmeas de ouriço-preto Chaetomys subspinosus (Olfers
1818) (Rodentia: Erethizontidae) nas florestas de restinga do
Parque Estadual Paulo César Vinha, Guaraparí, Espírito Santo
Dissertação apresentada ao Programa de Pós-graduação em Zoologia de Vertebrados da Pontifícia Universidade Católica de Minas Gerais, como requisito para obtenção do título de Mestre em Zoologia de Vertebrados.
Orientador: Prof. Dr. Adriano G. Chiarello
Belo Horizonte, MG
2006

FICHA CATALOGRÁFICA Elaborada pela Biblioteca da Pontifícia Universidade Católica de Minas Gerais
Oliveira, Pedro Amaral de O48e Ecologia de fêmeas de ouriço-preto Chaetomys subspinosus (Olfers 1818)
(Rodentia: Erethizontidae) nas florestas de restinga do Parque Estadual Paulo César Vinha, Guaraparí, Espírito Santo / Pedro Amaral de Oliveira. Belo Horizonte, 2010.
87f.: il. Orientador: Adriano Garcia Chiarello Dissertação (Mestrado) – Pontifícia Universidade Católica de Minas Gerais.
Programa de Pós-Graduação em Zoologia de Vertebrados. 1. Mamíferos. 2. Erethizontidae. 3. Porco espinho. 4. Habitat. 5. Ecologia. 6.
Dieta. 7. Movimentação. 8. Mata Atlântica. 9. Setiba, Restinga (ES). I. Dias, Ronaldo Brêtas de Carvalho. II. Pontifícia Universidade Católica de Minas Gerais. Programa de Pós-Graduação em Zoologia de Vertebrados. III. Título.
CDU: 599.324

4
AGRADECIMENTOS
Sem nenhuma dúvida, este trabalho seria impossível sem a orientação e apoio de
várias pessoas envolvidas e agradece-las é apenas uma maneira de reconhecer que sem elas
tudo ficaria muito mais difícil.
• Agradeço, primeiramente, ao orientador deste trabalho Adriano Chiarello, pelo
exemplo profissional, pela oportunidade de aprendizado e todos os valiosos
ensinamentos. Pelas críticas, sugestões e apoio. E acima de tudo, pelos inesquecíveis
momentos compartilhados ao longo de quatro anos no interior deste “mundo paralelo”
conhecido como Mata Atlântica.
• Ao grande amigo e colega Rodrigo Barata, pela força de vontade, pela paciência nos
piores momentos e pela excelente companhia, imprescindível durante todos os mais de
dois anos de trabalhos de campo. Faltam palavras para descrever o quanto foi
fundamental sua participação no projeto. Os ouriços agradecem.
• Especialmente ao Antonio Oliveira de Jesus, o Toninho. Sem nenhuma dúvida, a
participação desta pessoa extraordinária literalmente “salvou” este projeto, assim
como todo o “Plano de Manejo” da espécie no nível nacional. Nós e os ouriços-pretos
te agradecemos de coração.
• Ao amigo e escalador arvorista Oscar E. Yepes, pela participação eficiente e bastante
divertida quando os estudos ainda eram em Santa Teresa. Obrigado pela ajuda na
captura dos “Fodomys subsacaneosus”.
• A todo o pessoal do Parque Estadual Paulo César Vinha, que nos receberam tão bem,
com verdadeira hospitalidade e bom humor. Em especial ao Michel e a Carolina, pelo
apoio incondicional ao projeto. Aos estagiários do parque, a toda a galera da
“vigilância”, a Juliana, Janine, Jailton, Rogério, Américo e o “Setiba”. Cuidem bem
deste local, pois ele é verdadeiramente muito especial.
• Ao João Henrique, amigo sempre prestativo e atual responsável pelo PEPCV,
obrigado pela imagem de satélite cedida gentilmente, foi uma ajuda que enriqueceu
muito o trabalho.

• A todas as pessoas de Santa Teresa, obrigado ao Hélio Fernandes, diretor do Museu de
Biologia Mello Leitão. E a todos os responsáveis pela Estação Biológica de Santa
Lúcia, à Rose e em especial ao casal Lourdes e José Molino. Muita felicidade e saúde
para todos os seus.
• Ao Marcello Passamani, que gentilmente emprestou o equipamento utilizado bem no
início do projeto, quando ainda estávamos na região serrana e que permitiu um
aprendizado inicial fundamental para o transcorrer do trabalho.
• Um agradecimento especial para todas as pessoas do “Instituto Dríades de Pesquisa e
Conservação da Biodiversidade” e todas as demais envolvidas de alguma maneira
neste projeto, obrigado pelo apoio e confiança, muito bom trabalhar junto com vocês e
fazer parte de uma iniciativa tão importante.
• Ao colega de “perrengue” Gaston Giné, obrigado pelas várias discussões ao longo do
projeto, pela ajuda em dias de muita chuva e pelo dia de surf. Boa sorte no seu
doutorado, conte comigo para o que for preciso.
• À Déborah Faria, valeu toda a paciência durante o projeto, obrigado pela confiança no
meu trabalho. Tenho certeza que valeu a pena todo o trabalho enorme que fizemos
juntos.
• Ao Rivelino Galvão, o primeiro a chamar a atenção para a importância dos ouriços no
Parque de Setiba e por ter sido quem nos apresentou para o Toninho. Obrigado pela
hospitalidade, pelo apoio quando tudo ainda era “só o começo”. Um abraço em toda a
família. Valeu pelos vários artigos, eles já rodaram o Brasil.
• Ao amigo Thiago Soares, valeu pela ajuda em Setiba, tenho certeza que foi uma
experiência gratificante para todos nós.
• Aos colegas que acompanharam os trabalhos em algumas campanhas no PEPCV,
Laura Albert, Óscar Yepes e Maria Cecília “Ciça”, valeu por quebrarem um pouco da
nossa rotina, vocês são uma excelente companhia.
• Ao amigo e médico veterinário Nelson Curi, pelo auxílio fundamental nos momentos
de manejo direto com os ouriços e pela companhia sempre bem humorada.
• A todos os colegas do mestrado, aos demais professores e funcionários, obrigado pela
excelente convivência. Com certeza um período muito especial para mim.

• Obrigado também ao Prof. Dartagnan Lino Viana, do Centro de Geoprocessamento da
PUC /Arquidiocese (CEGIPAR) na PUC-Minas, pela ajuda fundamental com os
programas utilizados para as análises de área de vida e movimentação. Valeu a
paciência.
• A Tudy e ao Léo, meus primeiros orientadores, por terem feito nascer uma verdadeira
paixão pelos “bichos de pêlo”, obrigado pelo apoio incondicional e por serem pessoas
tão especiais. Um abraço enorme em toda a galera “cipoeira” da maaastuuu!!
• Em especial aos meus pais, fonte de força e amor. Simplesmente vocês são o Máximo,
exemplos de vida para uma vida inteira, ou mais...
• Aos meus dois irmãos, amo vocês demais, se cuidem.
• Especialmente a Cláudia, a minha razão de sempre voltar pra casa, você mais do que
ninguém sabe o custo de encarar um trabalho como este, obrigado pela paciência.
• À CAPES pelo financiamento da bolsa de mestrado.
• Gostaria de agradecer ao Ministério do Meio Ambiente, que através do Fundo
Nacional do Meio Ambiente (Edital FNMA/PROBIO 01/2003) financiou o Projeto
“Elaboração do Plano de Manejo do Ouriço-preto Chaetomys subspinosus (Olfers,
1818)” do qual esta dissertação é um dos muitos frutos que ainda virão.
• Ao IBAMA, através da Diretoria de Fauna e Recursos Pesqueiros e da Coordenação
Geral de Fauna, pela concessão da licença que permitiu o desenvolvimento desta
pesquisa.
• Ao Fundo de Incentivo a Pesquisa da PUC-Minas que financiou parcialmente o
presente trabalho com a aquisição de alguns equipamentos e logística para as
campanhas no Espírito Santo a partir do segundo semestre de 2005 (FIP/PUC
2005/36-TLE).
“Que a paz seja convosco”.

Nossa tarefa deve ser libertar-nos da nossa
prisão individual pela ampliação do nosso
circulo de compaixão, para abranger todas as
criaturas vivas e toda a natureza em sua beleza.
Albert Einstein

RESUMO
O ouriço-preto (Chaetomys subspinosus) é um roedor arborícola endêmico da Mata Atlântica
e ainda pouco estudado, sendo considerado atualmente como uma espécie ameaçada de
extinção. Para o estudo apresentado nesta dissertação, três fêmeas adultas foram marcadas
com rádio-colar e monitoradas de março de 2005 a fevereiro de 2006, em porções de floresta
de restinga presentes no Parque Estadual Paulo César Vinha, Espírito Santo. Foram coletadas
informações básicas a respeito da ecologia da espécie como o seu padrão de uso do espaço
(área de vida), sua dieta, atividade e movimentação. Os pontos de localização foram coletados
durante o dia e a noite (18:00 - 23:00 h), quando foram também coletados dados sobre a
atividade noturna e a dieta dos animais. As áreas de vida encontradas foram relativamente
pequenas (média de 2,14 ha pelo Mínimo Polígono Convexo ou 1,09 ha pelo método Kernel)
e os ouriços-pretos percorreram em média 147 metros nas noites observadas. A espécie exibiu
hábito estritamente arbóreo e noturno, descendo ao chão apenas para defecar em latrinas. Em
conjunto, as atividades de movimentação e alimentação foram as mais freqüentes (médias de
32,46 % e 20,34 % respectivamente). No entanto, mesmo sendo monitorados no seu período
principal de atividade, os animais repousaram boa parte do tempo observado (média de 34,84
%). Quanto à sua dieta, os ouriços-pretos foram bastante seletivos em relação às espécies
vegetais utilizadas e as partes consumidas, podendo ser considerado um folívoro quase
restrito. Um total de 57 árvores foram utilizadas na sua alimentação, correspondendo a 14
espécies vegetais, com cada indivíduo usando um número ainda menor (4, 6 e 10). Pera
glabrata, Tapirira guianensis e Pera leandri foram as três espécies notadamente mais
importantes na dieta dos ouriços-pretos. Assim como já foi observado em outros mamíferos
folívoros arborícolas, o ouriço-preto também adota uma estratégia comportamental
nitidamente baseada na economia de energia, possivelmente relacionada com a baixa
capacidade nutricional da sua dieta constituída predominantemente de folhas.
Palavras-chave: Mamíferos. Erethizontidae. Ouriço-preto. Chaetomys. Área de vida.
Ecologia. Mata Atlântica. Restinga. Endêmica. Parque Estadual Paulo César Vinha.

ABSTRACT
The ecology of female thin-spined porcupine Chaetomys subspinosus (Olfers 1818)
(Rodentia: Erethizontidae) in restinga forests at the Parque Estadual Paulo César
Vinha, southeast Brazil.
The thin-spined porcupine (Chaetomys subspinosus) is a poorly known arboreal rodent,
endemic to a distinct portion of the Brazilian Atlantic Forest and regarded as an endangered
species. Three adult female porcupines were monitored from March 2005 to February 2006 at
the Parque Estadual Paulo César Vinha, in areas of restinga forest of Espírito Santo state. Data
on home range, activity budgets, diet and movements were collected. Home ranges were very
small (average of 2,14 ha with Minimum Convex Polygon and 1,09 with Kernel method) and
the animals traveled an average of 147 m during the nights of observation (18:00 - 22:00 h).
Chaetomys subspinosus exhibited an arboreal and strictly nocturnal habit, descending to the
forest floor only to defecate. Together, movement and feeding were the activities registered
more often (averages of 32,46 % e 20,34 % respectively). However, the animals were also
lethargic, resting an average of 34,84 % of time during monitoring time. The porcupines were
highly selective regarding diet and plant parts consumed, being almost strictly folivore. A
total of 57 trees were used as food resource, corresponding a total of 14 plant species, but
each individual made up its diet with an even smaller number of species (4 – 12). The three
most important plant species present in the porcupine’s diet were Pera glabrata, Tapirira
guianensis and Pera leandri. Thus, like other mammals with arboreal and folivorous habits,
C. subspinosus adopts a behavioral strategy based on energy economy, possibly related to its
nutritional-poor diet, composed mainly of leaves.
Key-words: Mammals. Erethizontidae. Thin-spined Porcupine. Chaetomys. Home
range. Ecology. Atlantic Forest. Restinga. Endemic. Paulo Cesar Vinha Park.

L ISTA DE FIGURAS
Figura 1. Fêmea de Chaetomys subspinosus (Ch-160) capturada e monitorada no Parque
Estadual Paulo César Vinha, Guaraparí, ES. 18
Figura 2. Indivíduo de C. subspinosus (Ch-EBSL) capturado e acompanhado no estudo piloto
em Santa Teresa. 25
Figura 3. Localização geral do Parque Estadual Paulo César Vinha (PEPCV), no município
de Guaraparí, Espírito Santo. 26
Figura 4. Interior da formação de floresta de restinga típica da área de estudo no PEPCV,
Guaraparí, Espírito Santo. 27
Figura 5. C. subspinosus (Ch-160) marcado com colar equipado com rádio-transmissor,
mostrando o reflexo dos adesivos refletores e dos olhos do animal. 29
Figura 6. Imagem de satélite mostrando o contorno da área do PEPCV e a localização geral
dos três indivíduos estudados. 30
Figura 7. Momento de captura de um ouriço-preto para retirada do radio-transmissor, feita
manualmente retirando o animal de seu local de repouso. 31
Figura 8. Áreas de vida calculadas para Ch-970 pelo MPC, usando todos os pontos (linha
contínua) ou apenas os pontos diurnos (linha tracejada). 41
Figura 9. Áreas de vida calculadas para Ch-274 pelo MPC, usando todos os pontos (linha
contínua) ou apenas os pontos diurnos (linha tracejada). 42

Figura 10. Áreas de vida calculadas para Ch-160 pelo MPC, usando todos os pontos (linha
contínua) ou apenas os pontos diurnos (linha tracejada). 43
Figura 11. Área de vida da fêmea Ch-970, calculada através dos métodos MPC (linhas) e
Kernel (contornos). Os contornos indicam as diferentes probabilidades de ocorrência dos
mesmos em suas áreas de uso. 45
Figura 12. Área de vida da fêmea Ch-274, calculada através dos métodos MPC (linhas) e
Kernel (contornos). Os contornos indicam as diferentes probabilidades de ocorrência dos
mesmos em suas áreas de uso. 46
Figura 13. Área de vida da fêmea Ch-160, calculada através dos métodos MPC (linhas) e
Kernel (contornos). Os contornos indicam as diferentes probabilidades de ocorrência dos
mesmos em suas áreas de uso. 47
Figura 14. Área de vida acumulada por mês (MPC), ao longo monitoramento de cada ouriço-
preto no PEPCV, no período de março de 2005 a fevereiro de 2006. Os valores acima das
linhas correspondem ao número de registros de localização acumulados até o respectivo mês,
para cada indivíduo. 48
Figura 15. Áreas de vida sazonais da fêmea Ch-970, calculadas pelo MPC. 50
Figura 16. Áreas de vida sazonais da fêmea Ch-274, calculadas pelo MPC . 50
Figura 17. Áreas de vida sazonais da fêmea Ch-160, calculadas pelo MPC. 51
Figura 18. Médias sazonais das porcentagens de tempo alocados na categoria “repouso” para
os ouriços-pretos estudados no PEPCV. Barras verticais indicam erro padrão. Resultados
significativos são mostrados sobre cada par de barras (p<0,05; teste Mann-Whitney). 53

Figura 19. Médias sazonais das porcentagens de tempo alocados na categoria
“movimentando” para os ouriços-pretos estudados no PEPCV. Barras verticais indicam erro
padrão. Resultados significativos (p<0,05) ou marginalmente significativos (0,05<p<0,10)
são mostrados sobre cada par de barras (teste Mann-Whitney). 54
Figura 20. Médias sazonais das porcentagens de tempo alocados na categoria “alimentando”
para os ouriços-pretos estudados no PEPCV. Barras verticais indicam erro padrão. Resultados
significativos (p<0,05) ou marginalmente significativos (0,05<p<0,10) são mostrados sobre
cada par de barras (teste Mann-Whitney). 54
Figura 21. Taxa de utilização de árvores de alimentação (árvores/hora) observadas entre as
estações seca e chuvosa (teste de Mann-Whitney). 57
Figura 22. Média individual e total dos “distâncias mínimas” percorridas pelos ouriços-pretos
nas estações seca e chuvosa. 58
Figura 23. Média individual e total das “distâncias completas” percorridas pelos ouriços-
pretos nas estações seca e chuvosa. O valor marginalmente significativo do teste-t
(0,05<p<0,10) é mostrado para o animal Ch-166; para os demais não houve diferença
significativa. 59
Figura 24. Sítio de repouso diurno (Ch-160), mostrando os referidos emaranhados de cipós,
folhas secas e trepadeiras. 60
Figura 25. Detalhe da latrina utilizada pela fêmea Ch-160, mostrando o acúmulo de fezes
recentes e mais antigas. 61
Figura 26. Individuo de ouriço-preto durante a captura (já no chão), em postura corporal
típica de intimidação. 63

L ISTA DE TABELAS
Tabela 1. Identificação, peso, tipo de habitat dominante e detalhes do monitoramento noturno
dos animais marcados com rádio-colar no PEPCV, no período de março de 2005 a fevereiro
de 2006. 28
Tabela 2. Número de árvores utilizadas (pontos), de registros nestes pontos e valores de áreas
de vida estimadas pelos métodos MPC e Kernel para cada animal monitorado no PEPCV no
período de março de 2005 a fevereiro de 2006. Os resultados são mostrados para todos os
pontos ("total") e somente com os registros obtidos nos sítios de repouso diurnos usados
pelos ouriços-pretos ("diurno"). 39
Tabela 3. Áreas de vida e respectivo tamanho amostral (n) dos três ouriços-pretos
monitorados no PEPCV, estimadas pelo MPC para os períodos seco e chuvoso, entre abril de
2005 a fevereiro de 2006. 49
Tabela 4. Porcentagem média de tempo alocado nas atividades principais anotadas para cada
ouriço-preto em 33 noites de monitoramento, no período de março de 2005 a fevereiro de
2006 no PEPCV. 52
Tabela 5. Número de registros de alimentação e de espécies vegetais presentes na dieta dos
ouriços-pretos monitorados no período de março de 2005 a fevereiro de 2006. Também são
mostradas as três espécies principais da dieta (em ordem decrescente de importância) e suas
respectivas porcentagens relativas de registros. 55
Tabela 6. Valores médios (metros), erro padrão (EP), tamanho amostral e valores mínimos e
máximos observados para os animais estudados no PEPCV, no período de março de 2005 a
fevereiro de 2006. Duas medidas são apresentadas, uma usando apenas os dois pontos nos
sítios de repouso diurnos (distância mínima) e outra considerando também os pontos
marcados durante a noite (distância completa). 58

SUMÁRIO
1 INTRODUÇÃO 14
1.1 Mata Atlântica 14
1.2 As restingas 15
1.3 Os mamíferos da Mata Atlântica 15
1.4 Os ouriços-cacheiros (Erethizontidae) 16
1.5 O ouriço-preto 18
1.6 Uso do espaço e definição de área de vida 20
1.7 Rádio-telemetria com mamíferos 21
1.8 Contexto conservacionista do estudo 21
2 OBJETIVOS 23
2.2 Objetivo principal 23
3 MATERIAL E MÉTODOS 24
3.1 Estudo piloto 24
3.2 Área de estudo 25
3.3 Captura e marcação dos ouriços-pretos 28
3.4 Coleta dos dados 32
3.5 Área de vida e uso do habitat 33
3.5.1 Mínimo polígono convexo 34
3.5.2 Área de vida acumulada 34
3.5.3 Método Kernel 35
3.6 Padrão de atividade 36
3.7 Espécies vegetais utilizadas na dieta 37
3.8 Movimentação 37
3.9 Análises estatísticas 38

4 RESULTADOS 39
4.1 Uso do espaço 39
4.1.1 Mínimo polígono convexo 40
4.1.2 Método Kernel 43
4.1.3 Área de vida acumulada 47
4.1.4 Variações sazonais das áreas de vida 48
4.2 Padrão de atividade 51
4.2.1 Variações sazonais no padrão de atividade 52
4.3 Espécies vegetais utilizadas na dieta 55
4.4 Movimentação 57
4.4.1 Variações sazonais na movimentação 58
4.5 Sítios de repouso diurno 59
4.6 Utilização de latrinas 61
5 DISCUSSÃO 63
5.1 Captura e monitoramento por rádio-telemetria 65
5.2 Sobre as estimativas de área de vida 65
5.3 Áreas de vida e uso do habitat 66
5.4 Padrão de atividade e movimentação 69
5.5 Dieta dos ouriços-pretos 71
5.6 Sítios de repouso diurno 73
5.7 Utilização de latrinas 74
5.8 Observações sobre cuidados parentais 75
5.9 Implicações conservacionistas 75
6 REFERÊNCIAS 79

14
1. INTRODUÇÃO
1.1 Mata Atlântica
Desde o período Colonial, se estabeleceram dentro do Bioma da Mata Atlântica os
principais centros urbanos, industriais e silviculturais do Brasil, conseqüentemente a região
abriga atualmente grande parte da população do país. Este processo de ocupação ocorreu
historicamente de forma desordenada, tendo como conseqüência um enorme preço ambiental:
a área original da Mata Atlântica era de 1.227.600 km2 e atualmente existem apenas cerca de
91.930 km2, ou seja, entre 7,5% e 8% da cobertura original (Myers et al., 2000; Galino-Leal
& Câmara, 2003). O pouco que restou se encontra distribuído na forma de fragmentos
isolados de vegetação (predominantemente secundária), imersos em uma matriz dominada por
pastagens e áreas agrícolas de diversos tipos de monocultura, sendo que destes remanescentes,
uma porcentagem ainda menor (menos de 2%) se encontra legalmente protegida como
Unidade de Conservação (Tabarelli et al., 2005). O processo de fragmentação, além de
diminuir as populações de espécies vegetais e animais, promove o isolamento daquelas que
permaneceram nas “ilhas” remanescentes, colocando-as em alto risco de extinção local
(MacArthur & Wilson, 1967; Shafer, 1981; Chiarello, 1999). Mesmo em grandes
remanescentes, a superexploração principalmente pela caça ilegal, pode levar a extinção das
populações locais (Redford, 1992).
A Mata Atlântica brasileira é um dos ecossistemas mais ricos em diversidade e
endemismos de espécies vegetais e animais no mundo, sendo que em conjunto, somente os
mamíferos, aves, répteis e anfíbios que ocorrem neste Bioma somam cerca 1.807 espécies, das
quais 389 são endêmicas (MMA, 2000). Em virtude dessa imensa riqueza biológica, seu
estágio avançado de degradação e permanente ameaça, é atualmente reconhecida como uma
das 34 “áreas prioritárias para a conservação da biodiversidade” em todo o mundo, sendo
considerada uma das oito mais importantes (Mittermeier et al., 1998, Myers et al., 2000;
Mittermeier et al., 2004).
No estado do Espírito Santo, a situação da Mata Atlântica é pouco melhor que a
encontrada no restante do país, restando cerca de 30 % da cobertura original de florestas (SOS
mata atlântica & INPE, 2002). No entanto, as áreas de preservação existentes são, de uma

15
forma geral, ecologicamente pequenas e a maioria dos remanescentes são fragmentos isolados
dentro de propriedades particulares. Mesmo assim, o estado ainda abriga uma enorme
biodiversidade, principalmente na sua região central, uma área reconhecida como um centro
de endemismo (“Rio Doce”) na Mata Atlântica (Aguiar et al., 2003).
1.2 As Restingas
Ao longo da costa brasileira encontramos planícies litorâneas arenosas de idade
quaternária, resultantes da ação conjunta de fatores como fontes de areia, correntes de deriva
litorânea, variações do nível relativo do mar e armadilhas para retenção de sedimentos
(Suguio & Tessler, 1984). Essas planícies de cordões arenosos são, geralmente, ocupadas por
comunidades vegetais características e diversificadas, resultado de fatores ambientais locais,
como topografia, proximidade do mar, condições do solo, profundidade do lençol freático,
bem como pelas diferentes inter-relações, aumentando em complexidade na medida em que
estão mais distantes do oceano (Lacerda et al., 1993).
Além dessas feições associadas aos diferentes tipos de vegetação, deve ser
mencionada a presença de inúmeras lagoas, normalmente associadas às depressões entre
cordões, à chegada de pequenos córregos ou à localização superficial do lençol freático. A
este complexo fitogeográfico tem-se dado o nome de Restinga, embora o termo “restinga”
também apresente outros significados (Suguio & Tessler, 1984). Assim, as restingas são
grandes mosaicos de ecossistemas definidos e interligados funcionalmente (Araújo & Lacerda
1987) considerados em seu conjunto um ecossistema associado à Mata Atlântica. Devido ao
caráter único da formação de cada porção de restinga ao longo do litoral, relacionados a seus
aspectos geomorfológicos, cada uma tem suas peculiaridades florísticas próprias (Cerqueira,
2000; Assis et al., 2004a). Dentre as comunidades vegetais presentes nas restingas brasileiras
estão as florestas de restinga, que podem ou não sofrer inundações durante o ano (Araújo &
Henriques, 1984). Estas possuem um porte arbóreo relativamente baixo e uma distribuição
muitas vezes relacionada à proximidade de corpos d’água ou do lençol freático.
1.3 Os mamíferos da Mata Atlântica
A fauna de mamíferos brasileira é uma das maiores do mundo, na primeira lista
dedicada exclusivamente ao grupo foram listadas 524 espécies (Fonseca et al., 1996), um
número que vem aumentando à medida que novas espécies são descritas e revisões são feitas

16
dentro de grupos ou espécies com grande distribuição geográfica. Segundo Reis et al. (2006),
atualmente são reconhecidas 652 espécies (24,61% a mais). Considerando somente o bioma
da Mata Atlântica, ocorrem 261 espécies de mamíferos, das quais 73 são endêmicas (Myers et
al., 2000). Este alto endemismo deve-se principalmente aos roedores, marsupiais e primatas
(Fonseca & Kierulff, 1989; Costa et al., 2000). No entanto, contrário a este cenário de riqueza
e endemismos, a situação crítica de redução de habitat faz com que a Mata Atlântica abrigue
inúmeras espécies de mamíferos “globalmente ameaçadas”, devido ao grau de ameaça que
sofrem suas populações, como por exemplo: Leontopithecus rosalia (mico-leão-dourado), L.
chrysomelas (mico-leão-da-cara-dourada), Cebus xanthosternos (macaco-prego-do-peito
amarelo), Brachyteles hypoxanthus e B. arachnoides (muriqui ou mono-carvoeiro), Bradypus
torquatus (preguiça-de-coleira) e o ouriço-preto (Chaetomys subspinosus). Assim como em
outras partes do mundo, essas espécies possuem um importante papel na conservação dos
ecossistemas onde sobrevivem, servindo como símbolos para a conservação regional, como
espécies indicadoras e como ponto focal para programas de conscientização pública e
educacional, permitindo melhores perspectivas para os esforços de conservação no país.
1.4 Os ouriços-cacheiros (Erethizontidae)
Atualmente, estão listados 71 gêneros e 235 espécies de roedores no Brasil (Oliveira &
Bonvicino, 2006). Os roedores neotropicais pertencentes à família Erethizontidae, são
conhecidos popularmente como ouriços-cacheiros, ouriços-comuns, luiz-cacheiros ou porcos-
espinhos. Apresentam pêlos modificados em espinhos distribuídos pelo corpo e não possuem
os primeiros dígitos (hálux e pólex), que são substituídos funcionalmente por uma calosidade
óssea. São animais noturnos, predominantemente solitários e arborícolas especializados
(Charles-Dominique et al., 1981), representando uma das diversas radiações de mamíferos em
hábitos arbóreos nas florestas tropicais (Voss & Angermann, 1997). Os trabalhos enfocando a
biologia do grupo são ainda escassos, porém acredita-se que tenham todos uma dieta
herbívora e que, assim como outros mamíferos arborícolas também herbívoros, apresentem
baixos níveis de atividade (Moojen, 1952; Chiarello et al., 1997; Emmons & Feer, 1997;
Santos Junior, 1998; Eisemberg & Redford, 1999).
O número de espécies e gêneros neotropicais incluídos nesta família ainda é
controverso, principalmente como conseqüência dos poucos exemplares depositados em
coleções científicas e a falta de mais estudos de revisão taxonômica dentro do grupo
(Concepcion & Molinari, 1991), com espécies sendo ainda descritas recentemente (Voss &

17
Silva, 2001). Segundo Oliveira & Bonvicino (2006), são reconhecidos para o Brasil três
gêneros divididos em duas subfamílias: Erethizontinae, com os gêneros Coendou Lacépède,
1799 e Sphiggurus Cuvier, 1823; e a subfamília Chaetomyinae, que compreende apenas o
monotípico C. subspinosus, representando uma linhagem evolutiva única e por isso destacado
por Oliver & Santos (1991) como um dos mais distintos e importantes mamíferos endêmicos
da Mata Atlântica.
Embora Voss & Angermann (1997) não reconheçam Sphiggurus, como um gênero
distinto, a separação de Sphiggurus e Coendou sugerida por Husson (1978) foi aceita por
outros autores (Honacki et al., 1982; Woods, 1993) e recentemente confirmada através de
dados genéticos (Concepcion & Molinari, 1991; Bonvicino et al., 2000; 2002). O quinto
gênero, Chaetomys, não é considerado por alguns pesquisadores como um membro dos
Erethizontidae (Patterson & Wood, 1982), que propuseram seu posicionamento junto aos
“ratos-de-espinho” da família Echimyidae, opinião seguida na “Lista Anotada dos Mamíferos
do Brasil” (Fonseca et al., 1996). Por outro lado, outros pesquisadores já posicionavam
Chaetomys como um gênero dentro dos Erethizontidae (Moojen, 1952; Cabrera, 1957; Nowak
& Paradiso, 1983), e mais recentemente, através de características morfológicas cranianas
(Martin, 1994; Carvalho, 2000), reforçaram a sua permanência na família dos ouriços
verdadeiros. De fato, através de dados moleculares Vilela et al. (2009) não só confirmam
como posicionam Chaetomys como uma forma basal dentro da família. Sendo assim, o
presente trabalho acompanha Oliveira & Bonvicino (2006) e considera um gênero incluído
nos Erethizontidae.
Apesar de possuírem uma das mais amplas distribuições geográficas entre as famílias
de roedores das Américas, o conhecimento atual a respeito da biologia do grupo se deve
principalmente aos estudos com o também monotípico Erethizon dorsatum (porco-espinho-
norte-americano) ((Dodge & Barnes, 1975; Smith, 1979; Rose, 1987; Snyder & Linhart,
1997; Griesemer et al., 1998; Sweitzer & Berger, 1998; Griesemer et al., 1999; Olson &
Lewis, 1999; Ilse & Hellgren, 2001), sendo escassos os trabalhos de campo sobre os gêneros
neotropicais de ouriços. Estes estão também concentrados em uma única espécie de ouriço-
cacheiro (Coendou prehensilis), destacando-se o clássico de Charles-Dominique et al. (1981)
e o trabalho de Montgomery & Lubin (1978) sobre sua movimentação nos Lhanos da
Venezuela. Recentemente, no Brasil, Santos Junior (1998) estudou indivíduos translocados de
C. prehensilis nos Cerrados do Triângulo Mineiro. Outras informações relevantes a respeito
da biologia desta mesma espécie foram fornecidas por Roberts et al. (1985) através do estudo
de longo prazo de um pequeno grupo de ouriços em cativeiro.

18
1.5 O ouriço-preto
O ouriço-preto Chaetomys subspinosus (Olfers, 1818) é um roedor arborícola com
cerca de 2 kg de peso, cuja localidade tipo é Ilhéus (Ávila-Pires, 1967), no estado da Bahia. É
uma espécie endêmica da Mata Atlântica (Figura 1), sendo considerada atualmente como
ameaçada de extinção na categoria “vulnerável” (MMA, 2006; IUCN, 2004; Chiarello et al.,
2008). Na região de estudo é conhecido como jaú-torino, possuindo vários nomes locais ao
longo de sua distribuição, como guandú, borê, ouriço-de-espinho-mole, luiz-cacheiro-preto,
luiz-preto, espeta-mangaba-preto, entre outros. Ao contrário do que o nome indica, apresenta
pelagem de cor marrom-acinzentada e difere das outras espécies de ouriços-cacheiros quanto
à forma e distribuição dos seus espinhos, apresentando uma gradação que vai de espinhos
mais curtos e duros, encontrados apenas ao redor da cabeça, para espinhos cada vez mais
longos, ondulados e pouco rijos seguindo na direção caudal.
Figura 1. Fêmea de Chaetomys subspinosus (Ch-160) capturada e monitorada no Parque Estadual Paulo César Vinha, Guaraparí, ES.
Principalmente devido a seus hábitos crípticos e sua pelagem escura, indivíduos de C.
subspinosus são extremamente difíceis de serem encontrados na natureza. Este fato, aliado a
sua aparente baixa densidade populacional, contribuíram para o baixo número de espécimes

19
depositados em coleções científicas e para que esta espécie permanecesse por mais de 30 anos
sem nenhum registro científico, até ser “redescoberta” nas matas do sul da Bahia (Santos et
al., 1987). Conseqüentemente o conhecimento a respeito da distribuição original de C.
subspinosus e sua situação atual ainda é escasso. Segundo Faria & Giné (2005), atualmente a
distribuição da espécie se estende do sul estado de Sergipe, (com o limite norte no rio
Sergipe), até a região sul do Espírito Santo, tendo como limite oeste as florestas deciduais, a
Caatinga no estado da Bahia e as florestas estacionais semideciduais do oeste de Minas
Gerais. No entanto, devido à redução da Mata Atlântica nestas regiões os fragmentos mais
importantes e que podem ainda conter populações significativas estão localizados entre o sul
da Bahia e o Espírito Santo (Santos et al., 1987; Oliver & Santos, 1991; Faria & Giné, 2005).
Historicamente a espécie também ocorria no norte do Rio de Janeiro, porém devido ao estado
avançado de degradação da região, não existem mais relatos para este estado (Gastón Giné,
com. pess.).
Em 2004, durante um inventário realizado em áreas prioritárias para a conservação da
Mata Atlântica no vale do rio Jequitinhonha (Probio/MMA), foi confirmado o primeiro
registro da espécie para Minas Gerais, no município de Bandeira, extremo nordeste do estado
(Fabiano R. Melo, com. pess. e Pedro A. Oliveira, obs. pess.).
Anterior a conclusão deste trabalho, eram praticamente inexistentes quaisquer estudos
de longa duração com esta espécie, conseqüentemente, muito pouco se conhecia a respeito da
sua história natural básica. A informação disponível na literatura se restringia a alguns
comentários encontrados em Moojen (1952) e Coimbra-Filho (1972), anteriores a lacuna
histórica de registros científicos. No entanto, após o trabalho desenvolvido por Santos et al.
(1987), outros pesquisadores retomaram as pesquisas com esta espécie. O primeiro estudo
ecológico foi conduzido por Chiarello et al. (1997) na Estação Biológica de Santa Lúcia, ES.
Posteriormente, outras informações foram fornecidas por Galvão (2003) e Galvão & Leite
(2005) nas florestas de restinga do Parque Estadual Paulo César Vinha, ES. Recentemente,
concomitantes ao presente trabalho, estudos também foram feitos no sul da Bahia com
indivíduos de C. subspinosus observados na natureza (Giné et al., 2006a / 2006b) e alguns
mantidos em cativeiro (Giné et al., 2005a / 2005b).
1.6 Uso do espaço e definição de área de vida
Uma das questões centrais abordadas em estudos ecológicos é a compreensão do uso
do espaço pelos animais (Kernohan et al., 2001), relacionado com outros aspectos como seu

20
padrão de forrageamento, a distribuição dos recursos e seleção de habitat (Harris et al., 1990).
Normalmente, estes estudos são feitos através da estimativa das “áreas de vida” ou “áreas de
uso” dos animais estudados - quando estas realmente existem - a partir das localizações
pontuais observadas por um determinado período e com o auxílio de diversos estimadores de
contorno e distribuição de utilização (Jacob & Rudran, 2003). Estas informações podem ser
complementadas com dados a respeito de seu padrão de atividade e da movimentação dos
animais dentro deste espaço definido, sendo estas importantes não somente no nível
individual, como também tendo implicações na dinâmica das populações e estruturação da
comunidade, podendo também (em alguns casos) indicar o sistema social da espécie (Komers
& Brotherton, 1997).
Dificilmente observa-se na natureza animais que se movem ao acaso, estando o
deslocamento normalmente relacionado com a procura de alimentos, sofrendo influencia de
sua localização e disponibilidade, bem como a presença de rotas conhecidas e/ou seguras, a
procura de parceiros e cuidados com a prole. Embora existam exceções, a maioria dos animais
passam a maior parte do tempo em uma área definida, dentro da qual estão familiarizados e
onde podem se movimentar com segurança e eficiência. Esta área é reconhecida como sua
“área de vida” (Bergallo, 1990; Powell, 2000).
Burt (1943) forneceu a primeira definição conceitual de área de vida (“home range”)
para mamíferos, como sendo “a área atravessada por um individuo em suas atividades
normais em busca de alimento, acasalamento e cuidado com a prole. Saídas ocasionais, talvez
de natureza exploratória, não devem ser consideradas parte da área de vida”. Embora esta seja
uma definição clara conceitualmente, existem alguns problemas inerentes, como foi destacado
por Powell (2000) e Kernohan et al. (2001). Incluindo o uso de termos ambíguos como
“normais” e a falta de um componente temporal especificado, Burt também não se deteve em
como quantificar as saídas ocasionais (“outliers”) e como definir a área onde são feitas as
saídas. Desta forma, ainda não existe um consenso geral a respeito da chamada “área de
vida”. Uma das definições, que leva em consideração os estimadores mais atuais, foi sugerida
por Kernoham et al. (2001) como sendo a “área com uma probabilidade definida de
ocorrência de um animal, em um período de tempo especifico”.
1.7 Rádio-telemetria com mamíferos
A rádio-telemetria é uma técnica que permite ao pesquisador, com o auxilio de um
receptor e uma antena direcional, determinar a direção ou aproximar-se da origem do sinal do

21
transmissor, a fim de registrar dados sobre a posição, movimentos e atividade, através da
localização direta ou indireta (triangulação) do animal marcado. A partir destes dados, se
forem coletados ao longo de um gradiente temporal, pode-se determinar padrões de uso do
espaço, de atividade, movimentação e utilização de recursos (Harris et al., 1990; Jacob &
Rudran, 2003). Avanços tecnológicos como a miniaturização dos componentes eletrônicos
tem permitido que pesquisas de campo sejam realizadas sem grande interferência no
comportamento, atividade e movimentação natural destes animais. O uso integrado de
ferramentas como sistemas de posicionamento global (GPS), sistemas de informação
geográfica (SIG) e programas de computador com diversos estimadores tem permitido
avanços nas análises de dados provenientes de rádio-telemetria (Kernohan et al., 2001).
Na região neotropical, esta técnica vem sendo utilizada com sucesso em estudos com
outras espécies de ouriços-cacheiros (Montgomery & Lubin, 1978; Santos Junior, 1998) e
ainda outros mamíferos arborícolas, como por exemplo, as preguiças (Montgomery &
Sunquist, 1975; Chiarello, 1998a, 1998b; Cassano, 2006), o jupará (Julien-Laferrière, 1993,
1999; Kays & Gittleman, 2001) e algumas espécies de marsupiais (Moraes, 2003; Lira, 2005).
Também foram estudados a ecologia e comportamento de outros roedores também noturnos,
herbívoros e arborícolas como o rato-da-taquara (Kannabateomys amblyonyx) (Silva,1993;
Silva, 2005) e o rato-do-cacau (Callistomys pictus) (Moura & Fonseca, 2005) todos utilizando
a localização dos animais por rádio-telemetria como apoio fundamental.
1.8 Contexto conservacionista do estudo
A falta de dados biológicos é um dos maiores problemas para o esforço de
conservação atualmente empreendido em todo mundo, em especial nas regiões tropicais,
prejudicando o delineamento de estratégias que sejam efetivas para conservação da
biodiversidade. Estudos a respeito da ecologia de uma espécie, que permitam uma melhor
compreensão das necessidades e do papel desempenhado por ela, são aspectos fundamentais
para o bom planejamento de sua estratégia conservacionista (Cullen & Valadares-Pádua,
1997). Recentemente, diversas iniciativas por parte do governo brasileiro e de organizações
não-governamentais tem procurado incrementar a conservação dos mamíferos no Brasil,
incluindo melhorias na legislação, a criação de novas unidades de conservação, a definição de
áreas prioritárias para sua conservação e a elaboração de planos de manejo para espécies
consideradas ameaçadas de extinção (Costa et al., 2005).

22
Dentro deste contexto, este trabalho faz parte de um projeto mais amplo: “Elaboração
do Plano de Manejo do Ouriço-preto, Chaetomys subspinosus (Olfers, 1818)”, financiado pelo
MMA (Edital FNMA/ PROBIO 01/ 2003). Este plano de manejo teve a coordenação do
“Instituto Dríades de Pesquisa e Conservação da Biodiversidade”, sediado em Ilhéus, com a
parceria de outras instituições como a PUC-Minas, Centro de Pesquisas do Cacau - CEPLAC
e Universidade Federal do Espírito Santo. Este projeto foi desenvolvido principalmente em
áreas do sul da Bahia e no Espírito Santo e procurou definir uma estratégia de manejo
conservacionista para o ouriço-preto. As ações propostas serão embasadas em um conjunto
inédito de informações biológicas obtidas por uma equipe multidisciplinar de pesquisadores,
durante 24 meses de estudo, que incluíram pesquisas nas áreas de ecologia, genética,
comportamento (in situ e ex situ) e distribuição da espécie.
A escassez de estudos anteriores significa que os dados apresentados neste trabalho
serão comparados com estudos feitos com outros Erethizontidae e com outros mamíferos
arborícolas. Os resultados apresentados não somente fornecem dados sobre a biologia de C.
subspinosus especificamente, como também adicionam informações a respeito dos mamíferos
folívoros arborícolas, com baixos níveis de metabolismo e atividade. Desta forma,
colaborando no entendimento do papel que desempenham estes “podadores (browsers) das
copas” em ecossistemas tropicais.

23
2. OBJETIVOS
2.2 Objetivo principal
Através do acompanhamento na natureza de três indivíduos marcados com colares
equipados com rádio-transmissores, este estudo teve como objetivo principal estudar a
ecologia do ouriço-preto quanto ao seu padrão de uso do espaço, de atividade, movimentação
e dieta nas florestas de restinga do Parque Estadual Paulo César Vinha (PEPCV).
Apresentando também outros aspectos relacionados da sua biologia como a utilização de
latrinas, uma observação estrutural dos sítios de repouso diurnos utilizados pelos animais e
algumas observações sobre interações mãe/filhote. Os objetivos específicos foram:
• Calcular a área de vida dos ouriços-pretos com o auxilio de dois estimadores de
contorno e distribuição de utilização, observando também o contorno obtido
somente com as árvores utilizadas como sítios de repouso diurnos, em
comparação com sua área de vida;
• Estudar o padrão de atividade da espécie, discutindo seu orçamento temporal nas
primeiras horas da noite, bem como as implicações para a atividade geral da
espécie ao longo do dia;
• Identificar quais são as principais espécies vegetais das florestas de restinga
presentes na dieta do ouriço-preto, abordando alguns aspectos do seu padrão de
forrageamento;
• Caracterizar a movimentação dos ouriços-pretos quanto aos seus deslocamentos
entre os locais de repouso diurno (distância mínima) e as distâncias percorridas nas
noites de acompanhamento (distância completa);
• Observar as variações sazonais no tamanho e contorno das áreas de vida estimadas,
no padrão de atividade e na movimentação dos ouriços-pretos, comparando
também o número de árvores de alimentação visitadas nas noites das estações seca
e chuvosa.

24
3. MATERIAL E MÉTODOS
3.3 Estudo piloto
É importante mencionar que anteriormente ao trabalho conduzido no PEPCV,
desenvolveu-se o que foi considerado um “estudo piloto” do mesmo, no período de julho a
dezembro de 2004. Neste período, foram realizadas campanhas mensais à região de Santa
Teresa, ES, mais especificamente na Estação Biológica de Santa Lúcia (EBSL), visando
encontrar e capturar indivíduos de C. subspinosus para realizar pesquisas sobre sua ecologia.
No dia 22/09/2004, uma fêmea adulta de ouriço-preto (Ch-EBSL; Figura 2) foi
capturada em vegetação ombrófila densa, com o auxílio de um escalador profissional (Oscar
Echevery Yepes). Esta fêmea foi marcada com rádio-transmissor (Telonics Inc. modelo 205),
sendo posteriormente monitorada por três meses (seis noites) até ser encontrada morta no
chão da mata em dezembro de 2004, aparentemente sem sinais de predação e sem apresentar
ferimentos em decorrência do colar (espécime depositado na coleção do Museu de Biologia
Mello Leitão, Santa Teresa). Durante as noites em que foi monitorada foram identificadas
diversas possibilidades e as dificuldades de se acompanhar no campo as atividades noturnas
desta espécie, que normalmente se movimenta no estrato mais alto da vegetação, de forma
silenciosa e diversas vezes encoberta pela vegetação, principalmente nos eventos de
alimentação.
Após a perda do animal em Santa Teresa e devido à dificuldade de obter novos
encontros na EBSL, em março de 2005 o estudo foi redirecionado para a floresta de restinga
do PEPCV, onde outros estudos com ouriços (C. subspinosus e Sphiggurus villosus) já
haviam sido feitos por Galvão (2003) e onde atuava no auxilio dos pesquisadores um
conhecido morador da região (Antônio Oliveira de Jesus) experiente no encontro e captura de
ouriços (Rivelino Galvão, com. pess.). Outro motivo que determinou a mudança da área de
estudo para o PEPCV foi a própria estrutura da mata ali presente, uma vez que as formações
florestais presentes nas restingas são reconhecidamente mais baixas que outras formações
como a ombrófila densa, o que facilitaria sobremaneira o acompanhamento das atividades dos
ouriços-pretos marcados. Permitindo a obtenção de informações precisas e de forma mais
sistemática, principalmente se tratando de uma coleta feita à noite.

25
Figura 2. Indivíduo de C. subspinosus (Ch-EBSL) capturado e acompanhado no estudo piloto em Santa Teresa.
Devido ao relativo pouco tempo de observação, aliado a dificuldade em acompanhar o
deslocamento e a atividade do individuo Ch-EBSL em Santa Teresa, apenas alguns
comentários serão feitos a seu respeito, quando pertinentes, concentrando as análises
efetuadas nos ouriços-pretos estudados nas florestas de restinga do PEPCV.
3.2 Área de estudo
O Parque Estadual Paulo César Vinha (PEPCV), antigo Parque Estadual de Setiba
(Decreto Estadual no 4903/94), está localizado no município de Guaraparí, ES (Figura 3),
possuindo cerca de 1500 ha de área e 25 km de perímetro, estendendo-se da Rodovia do Sol
(“Rodosol”, rodovia estadual ES-160) para leste até o Oceano Atlântico, confrontando-se ao
norte com a Ponta da Fruta e ao sul com o Recreio de Setiba. O clima da região, segundo a
classificação Koëpen, é Aw do tipo tropical com verão chuvoso e inverno seco. A temperatura
média anual é de 23o C, com uma precipitação anual média de 1.307 mm e a umidade relativa
média anual é de 80% (Fabris & Cesar 1996). De acordo com Pereira (1990), a presença de
lagos, o desnível do solo (distância entre o lençol freático e a superfície), a direção dos ventos,
a distância ao mar e a formação geológica são os principais fatores que contribuem para a
presença de um mosaico de formações florestais no PEPCV, que é típico dos ambientes de
restinga.

26
Figura 3. Localização geral do Parque Estadual Paulo César Vinha (PEPCV), no município de Guaraparí, Espírito Santo.
O PEPCV representa a mais extensa área de vegetação de restinga existente no litoral
sul do Espírito Santo, sendo uma das poucas unidades de conservação localizadas na zona
costeira do estado (Venturini et al., 1996; MMA, 1996). A formação de restinga presente no
PEPCV é considerada atualmente como uma das “áreas prioritária para a conservação da
biodiversidade da Mata Atlântica e Campos Sulinos”, posicionada na categoria de “alta
importância biológica” (área 234; MMA, 2000).
Um estudo florístico na área do parque, conduzido por Assis et al. (2004a) num trecho
de floresta de restinga imediatamente após a formação pós-praia, encontrou 172 espécies
pertencentes a 54 famílias, com predomínio de Myrtaceae (25 espécies), incluindo espécies
endêmicas ao PEPCV e outras ameaçadas de extinção, destacando sua importância para a
conservação da flora da restinga. Esta formação de floresta de restinga (Figura 4) presente no
PEPCV sofreu grande influência da Mata Atlântica na sua composição, no entanto apresenta
baixa similaridade em relação às outras áreas de restinga arbórea do litoral brasileiro (Assis et
al., 2004b).

27
Figura 4. Interior da formação de floresta de restinga típica da área de estudo no PEPCV, Guaraparí, Espírito Santo.
Segundo Venturini et al. (1996), a fauna de mamíferos presente no PEPCV é
composta por cerca de 43 espécies, sendo três delas marinhas. De maneira sintópica com
Chaetomys, uma segunda espécie de ouriço-cacheiro (Sphiggurus villosus), conhecido
localmente como ouriço-amarelo ou ouriço-verdadeiro, ocorre na mata de restinga do PEPCV,
porém com uma densidade populacional aparentemente bem maior (Galvão, 2003). No
Parque estão presentes também outras espécies de mamíferos como o sagui-da-cara-branca
(Callithrix geoffroyi), único primata da reserva, e marsupiais como a cuíca-lanosa (Caluromys
philander), a cuíca-de-quatro-olhos (Philander frenata), o jupati (Metachirus nudicaudatus)
entre outras. Devido ao tamanho relativamente pequeno da reserva e a proximidade com áreas
urbanas de Guaraparí, a presença de grandes predadores como a onça-parda (Puma concolor)
e a onça-pintada (Panthera onca) é pouco provável, o que torna outros mamíferos como a
irara (Eira barbara), o cachorro-do-mato (Cerdocyon thous), o mão-pelada (Procyon
cancrivorus), o jaguarundi (Puma yagouaroundi) e até mesmo cães domésticos, os potenciais
predadores de ouriços na área do PEPCV.
No litoral sul do Espírito Santo, a urbanização não planejada e a pecuária extensiva
ainda são importantes fatores de desmatamento. Na área de entorno do PEPCV - Área de
Proteção Ambiental (APA) de Setiba - o turismo, a especulação imobiliária, os loteamentos e
a extração ilegal de areia são as principais causas de degradação ambiental (MMA, 1996).
Segundo informações da própria administração do trecho sob concessão (entre Guaraparí e
28
Vila Velha), o tráfego intenso de veículos na Rodosol (limite imediato do parque e que corta a
APA de Setiba ao meio), além do ruído constante, causa diversos atropelamentos de animais
silvestres.
3.3 Captura e marcação dos ouriços-pretos
O trabalho se baseou em dados coletados através da localização e acompanhamento
visual de indivíduos adultos de Chaetomys, no período de março de 2005 a fevereiro de 2006,
em vegetação de floresta de restinga no PEPCV. Para tanto, foram feitas 12 campanhas
mensais com quatro dias de duração cada, totalizando 33 noites de monitoramento e com 150
horas e 50 minutos de observação noturna. Outras informações a respeito dos animais
capturados e monitorados durante o estudo se encontram na Tabela 1.
Tabela 1. Identificação, peso, tipo de habitat dominante e detalhes do monitoramento noturno dos animais
marcados com rádio-colar no PEPCV, no período de março de 2005 a fevereiro de 2006.
Monitoramento noturno
Animal * Peso (g) Captura habitat período No noites horas
Ch-970 2700 04/03/05 Floresta de Restinga 25/03/05 a 13/11/05 10 47:20
Ch-274 2200 04/03/05 Floresta de Restinga 25/03/05 a 21/02/06 12 52:20
Ch-160 2100 05/03/05 Floresta de Restinga 27/03/05 a 22/02/06 11 51:10
Total 33 150:50
* Os números referem-se às freqüências individuais dos radio-transmissores colocados nos animais monitorados.
Foi realizada uma campanha de três dias para procura e captura da espécie alvo no
início do mês de março, onde foram encontradas e capturadas três fêmeas adultas da espécie,
todas em áreas de floresta de restinga. Estas foram marcadas com rádio-transmissores de
diferentes freqüências presos a colares do tipo “ball-chain” (Telonics Inc.-modelo 205), de
maneira semelhante ao estudo realizado por Chiarello et al. (1997) e Giné et al. (2006a) com
esta mesma espécie e a outros estudos feitos com Coendou prehensilis (Santos Junior, 1998;
Charles-Dominique et al., 1981). Os transmissores utilizados pesavam cerca de 50 gramas,
representando menos de 3,0 % do peso corporal dos animais, abaixo do limite de 5,0 %
recomendado por Cochran (1980) para não influenciar o comportamento de mamíferos.

29
Seguindo a recomendação feita por Gaston Giné (com. pess.), que desenvolve
pesquisas sobre a ecologia de C. subspinosus no sul da Bahia, adesivos refletores (3M) foram
colados nos radio-transmissores, reduzindo consideravelmente a dificuldade de acompanhar a
movimentação dos animais à noite, uma vez que os olhos do ouriço-preto possuem apenas um
reflexo pálido quando iluminados com fontes de luz artificiais (Figura 5).
Figura 5. C. subspinosus (Ch-160) marcado com colar equipado com rádio-transmissor, mostrando o reflexo dos adesivos refletores e dos olhos do animal.
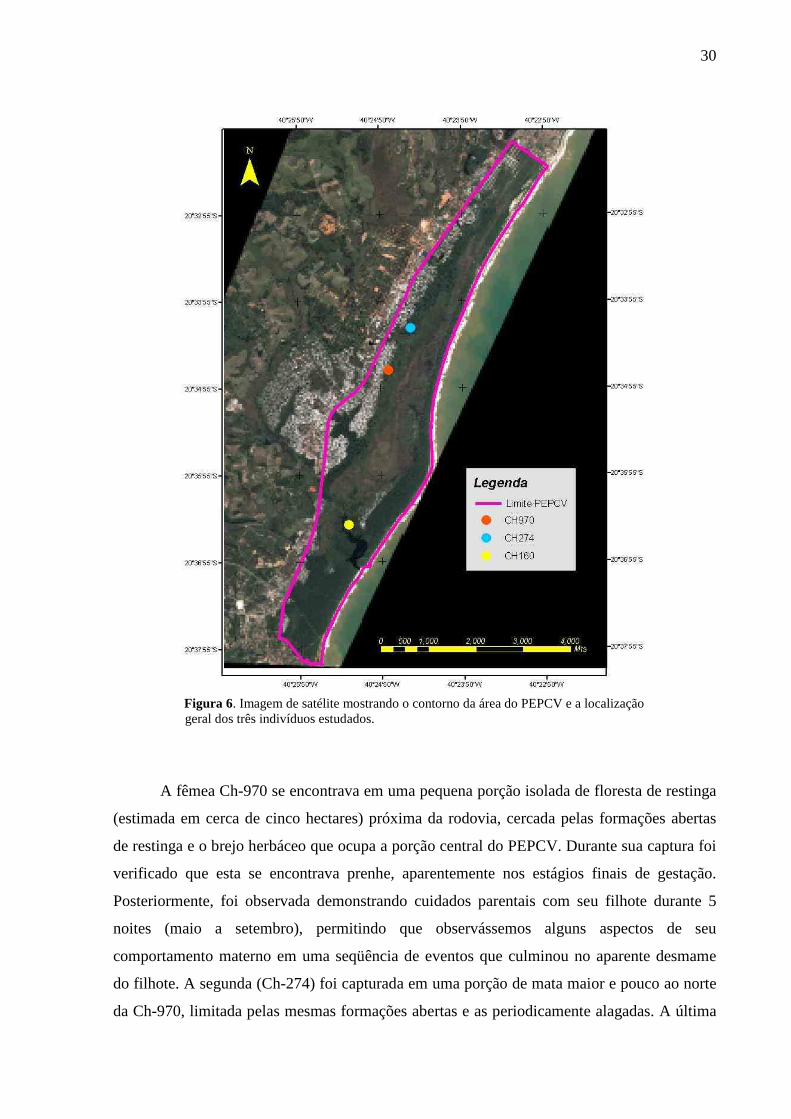
O contorno do PEPCV e a localização geral de cada uma das três fêmeas estudadas
são mostrados na Figura 6. O limite oeste do parque coincide com o traçado da Rodosol, os
extremos norte e sul fazem limite com áreas urbanas e condomínios particulares e a leste com
a faixa litorânea (praia).
30
Figura 6. Imagem de satélite mostrando o contorno da área do PEPCV e a localização geral dos três indivíduos estudados.
A fêmea Ch-970 se encontrava em uma pequena porção isolada de floresta de restinga
(estimada em cerca de cinco hectares) próxima da rodovia, cercada pelas formações abertas
de restinga e o brejo herbáceo que ocupa a porção central do PEPCV. Durante sua captura foi
verificado que esta se encontrava prenhe, aparentemente nos estágios finais de gestação.
Posteriormente, foi observada demonstrando cuidados parentais com seu filhote durante 5
noites (maio a setembro), permitindo que observássemos alguns aspectos de seu
comportamento materno em uma seqüência de eventos que culminou no aparente desmame
do filhote. A segunda (Ch-274) foi capturada em uma porção de mata maior e pouco ao norte
da Ch-970, limitada pelas mesmas formações abertas e as periodicamente alagadas. A última

31
fêmea capturada (Ch-160) estava distante das outras duas cerca de 4 km, em uma área
periodicamente alagada, dentro da faixa de floresta de restinga (Mata Esclerófila Litorânea)
presente após a formação pós-praia e próxima a Lagoa de Caraís, uma das principais atrações
turísticas do PEPCV.
Todas foram encontradas durante o dia, através de procura ativa (cerca de 28 horas e
30 minutos no total) nos seus respectivos sítios de repouso diurnos. A procura e captura foi
realizada com o auxilio de um morador do entorno da reserva, mencionado anteriormente,
extremamente competente na localização e captura de ouriços. Esta foi realizada
manualmente após subir cautelosamente na árvore onde se encontrava o animal (Figura 7),
com a apreensão sendo feita segurando pelo terço final da cauda e retirando o animal da
árvore.
Figura 7. Momento de captura de um ouriço-preto para retirada do radio-transmissor, feita manualmente retirando o animal de seu local de repouso.
Os animais foram então trazidos ao solo, contidos fisicamente pelo tórax com o auxílio
de uma forquilha de madeira, pesados (dinamômetro Pesola, precisão de 100g) e
posteriormente sedados com uma associação de Cloridrato de Xilazina (2 mg/kg de peso vivo)
e Cloridrato de Quetamina (5 mg/kg de peso vivo), injetada intramuscularmente. O
monitoramento anestésico foi realizado durante todo o tempo de sedação. A dose utilizada
resultou em sedação moderada, com média de 46 minutos até a recuperação, tempo suficiente
para que os animais fossem manipulados e para a colocação dos rádio-transmissores. Um

32
exame clínico completo foi realizado no local por um Médico Veterinário especializado
(Nelson H. A. Curi, CRMV/MG 6454) e coletadas amostras de sangue total, sangue de ponta
de orelha e algumas fezes. Os três indivíduos foram soltos no mesmo local da captura, e
monitorados por cerca de uma hora para o diagnóstico de sua adaptação ao equipamento de
rádio transmissão e até que estivessem recuperados dos efeitos dos sedativos.
A fêmea Ch-970 foi monitorada da sua captura em março até dezembro (8 meses), em
janeiro de 2006 foi encontrada morta em uma área aberta próxima a sua latrina, localizada na
borda da mata. Embora não confirmado, esta foi possivelmente predada, uma vez que os
eventos de defecação observados estão entre os raros momentos onde os animais descem ao
chão, ficando expostos a predadores terrestres. As outras duas foram monitoradas durante
todo o período de estudo, sendo recapturadas em maio de 2006 quando foram retirados os
colares.
3.4 Coleta dos dados
Durante cada dia de campo, no período claro do dia, com o auxilio de um receptor
(Titley Eletronics - modelo Regal 2000) e uma antena direcional “Yagi” de três elementos
(Titley Eletronics), duas das três fêmeas marcadas foram encontradas em seus sítios de
repouso diurno através do método do “rastreamento terrestre” ou “homing in on the animal”
(White & Garrott, 1990), que consiste em seguir a direção de maior intensidade do sinal até a
confirmação visual da posição do animal. Estas localizações diurnas foram utilizadas para a
análise do uso do espaço e para a descrição e caracterização dos abrigos diurnos, procurando
investigar quais variáveis poderiam estar influenciando na escolha de tais locais.
No período noturno, uma das três fêmeas marcadas foi monitorada durante seu período
de maior atividade, que segundo Chiarello et al. (1997) e Giné et al. (2005a) acontece nas
primeiras horas da noite. Nestas observações (entre 18:00 e 23:00) foram coletados dados a
respeito de suas principais atividades em amostragem “animal focal”, utilizando registro
“instantâneo” (Altmann, 1974), com um período amostral de 1 minuto e um intervalo de 9
minutos entre os registros. As localizações obtidas para cada um dos animais durante os
acompanhamentos noturnos foram também utilizadas para as estimativas das áreas de vida.
As observações noturnas foram conduzidas por pelo menos dois observadores, tendo o
auxílio de dois binóculos (Nikon e Minolta 8x40) e quatro lanternas de diferentes intensidades
de luz (duas lanternas Maglight recarregáveis de 12 watts e duas lanternas de cabeça Petzl

33
modelo Duo de 3 watts), utilizadas alternadamente dependendo da distância observador /
animal.
3.5 Área de vida e uso do habitat
Para o estudo da área de vida e uso do habitat foram utilizadas as localizações nos
sítios de repouso diurnos, assim como as obtidas durante os acompanhamentos noturnos,
marcadas nas árvores (pontos) onde os animais efetuaram suas principais atividades como:
alimentação, defecação (utilização de latrinas) e interações sociais (ex: cuidado parental).
Cada vez que o indivíduo foi localizado durante o período do dia ou durante a noite nos
pontos referidos acima, o tronco da árvore ou o cipó onde o mesmo foi observado foi
identificado com fita e numerado para posterior mapeamento.
Devido à proximidade recorrente das árvores utilizadas e os valores de acuracidade
obtidos por aparelho de GPS (em torno de 15 metros), o posicionamento espacial foi feito
utilizando uma bússola de visada (Suunto KB-14) e trena métrica (20 m), tendo como
referência inicial em cada área um ou dois pontos próximos da borda, marcados em GPS e
com melhor acuracidade, conferindo maior precisão a todas as localizações obtidas.
Posteriormente, utilizando o programa “AutoCAD Map-2000”, as coordenadas em UTM
(projeção Universal Transversa de Mercator) para cada árvore marcada foram definidas e
estes pontos utilizados nas análises de uso do espaço e movimentação dos ouriços. A partir
destes pontos de localização, para cada indivíduo marcado, foi calculada sua área de vida com
base em dois métodos usuais e complementares, o Mínimo Polígono Convexo (MPC) (Hayne,
1949) e o método Kernel (Worton, 1989). O primeiro deles reconhecidamente superestima a
área de vida incluindo áreas não utilizadas, enquanto o segundo tende a produzir uma
estimativa mais conservadora, incluindo informações sobre a estrutura interna da área.
Para o cálculo do MPC foi utilizado o mesmo programa “AutoCAD Map-2000” e para
o estimados Kernel a extensão HRE (“Home Range Extension”) para “ArcView” (Rodgers &
Carr, 1998), utilizando o programa “ArcGIS 8.3”. Todas as análises espaciais foram feitas no
Centro de Geoprocessamento da PUC/Arquidiocese (CIGEPAR) na PUC-Minas, com o
auxilio do Geógrafo Dartagnan Lino Viana. Uma imagem de satélite (IKONUS, ano 2005) foi
utilizada para o posicionamento real das áreas de vida dentro do mosaico de formações
vegetais de Restinga.

34
3.5.1 Mínimo polígono convexo
Dentre todos os estimadores disponíveis, o MPC pode ser considerado o mais antigo e
comum, ainda muito utilizado em função de sua simplicidade e comparabilidade entre estudos
(Powell, 2000). Sendo recomendada sua inclusão como um de dois ou mais métodos de
estimativa de área de vida (Harris et al., 1990). Consiste na união dos pontos mais externos da
distribuição das localizações formando um polígono convexo e então calcula-se a área do
polígono com 100% dos pontos ou outra porcentagem (normalmente 95%), se for de interesse
do estudo eliminar os pontos mais extremos da amostra (“outliers”) ou considerados de
natureza exploratória.
Todos os registros de localização utilizados neste trabalho foram obtidos mediante
observações diretas dos indivíduos estudados, sendo anotados nos pontos (árvores) onde os
indivíduos realizaram efetivamente suas principais atividades. Desta forma, foi assumido no
presente estudo que a porcentagem mais adequada seria com 100% dos pontos (MPC Total),
sem que nenhum destes fosse excluído como extremo ou exploratório, sob pena de
descartarmos certamente locais importantes da área de vida como repouso diurno,
alimentação e latrinas, localizados na periferia da área do polígono.
Procurando investigar se somente as localizações feitas durante o dia nos sítios de
repouso produziriam uma estimativa correspondente da área de vida, em comparação com a
obtida com todos os registros, bem como se os pontos utilizados estavam distribuídos em toda
a área de vida ou concentrados, o mesmo MPC Total foi feito utilizando somente os pontos
obtidos durante o período do dia (MPC Diurno).
Para observar possíveis variações entre as estações (seca e chuvosa) no tamanho e no
posicionamento das áreas utilizadas, foram estimadas pelo MPC as áreas para cada animal
estudado, considerando como período seco os meses entre abril a setembro e o chuvoso entre
outubro e fevereiro. O primeiro mês amostrado (março) não entrou nesta análise sazonal de
áreas, pois foi anterior ao período seco amostrado, tornando-o inadequado para o
agrupamento posterior de localizações feitos na estação chuvosa.
3.5.2 Área de vida acumulada
Áreas de vida calculadas através do MPC são dependentes diretamente do número de
localizações registradas e à medida que se acumulam ao longo do tempo para um mesmo
indivíduo, estas tendem a crescer cada vez mais lentamente, até chegarem próximas a uma

35
assíntota, ou seja, um valor onde localizações adicionais resultam em acréscimos mínimos na
área estimada (Harris et al., 1990; Kernohan et al., 2001). Assim, visando confirmar se a
estimativa feita ao longo dos meses de estudo resultou em um valor final estável, bem como
observar quais seriam os valores mínimos de localizações para uma aproximação real da área
de vida utilizada pelos ouriços-pretos de maneira geral, foram calculadas as áreas (MPC) com
os registros acumulados a cada mês de estudo para cada um dos animais. Valores mínimos de
localizações foram obtidos nos meses em que pontos adicionais não representaram mais
aumentos relevantes nas áreas estimadas.
3.5.3 Método Kernel
Proposto por Worton, (1989), o estimador não-paramétrico de Kernel foi um grande
aperfeiçoamento das funções probabilísticas de densidade, sendo considerado atualmente o
melhor método disponível para análises de área de vida através da “distribuição de utilização”
(Kernohan et al., 2001; Powell, 2000). A importância do conceito de “distribuição de
utilização” está na transformação de uma informação bidimensional de área em uma de
volume, onde o terceiro eixo representa a intensidade de utilização, ou em alguns casos,
representando a importância de determinadas regiões de sua área de vida. Este conceito pode
também ser entendido como a probabilidade de um animal ser encontrado em cada ponto do
espaço.
Este método utiliza um grupo particular de funções de probabilidade de densidade,
chamadas de “kernels” e representadas graficamente por pequenos “morros” arredondados,
associados a cada uma das localizações amostradas. A partir da combinação dos “kernels” de
cada localização, obtém-se uma estimativa da real função de densidade da distribuição de
utilização, sendo a estimativa da densidade em qualquer ponto da distribuição equivalente à
média de todos os kernels que nele se sobrepõem. Pode-se então, descrever contornos de
probabilidade de densidade e estimar a área de vida com base na porcentagem desejada desta
distribuição de utilização (Worton, 1989; Seaman & Powell, 1996; Powell, 2000; Kernohan et
al., 2001).
O método Kernel está dividido em “fixo” e “adaptativo”, sendo o primeiro
considerado de melhor performance especificamente para estimativas de área de vida
(Seaman & Powell, 1996; Seaman et al., 1999; Powell, 2000; Kernohan et al., 2001). Uma
das questões mais críticas neste método é a definição da “largura da banda” (H) conhecida

36
também como “parâmetro de suavização” (“smoothing parameter”), existindo diversos meios
estatísticos possíveis para uma escolha não-arbitrária (Silverman, 1986). No entanto, segundo
Kernohan et al. (2001) existem alguns problemas relacionados aos métodos mais utilizados
para escolha do H como o da validação cruzada dos quadrados mínimos (“least square cross
validation”) (Hlscv) e o valor de referência (Href). Quando os registros de localização estão no
mesmo ponto ou muito próximos entre si, o valor escolhido no Hlscv aparece muito pequeno,
quebrando os contornos em diversos picos em torno dos pontos amostrados (“undersmooth”).
Já nos casos em que a distribuição é multimodal, o valor obtido pelo Href resulta em contornos
super suavizados (“oversmooth”), perdendo detalhes da estrutura interna da distribuição de
utilização (Silverman, 1986).
Estes dois métodos estatísticos de escolha do H, presentes no programa HRE (Hlscv e
Href), como previsto anteriormente, produziram nas análises preliminares estimativas ora
suavizando demais o contorno, ora para menos. Desta forma, foi utilizado para este trabalho
com os ouriços-pretos o Kernel fixo, com largura da banda H escolhida pelo usuário (Huser)
utilizando valores intermediários aos obtidos estatisticamente pelos métodos referidos acima
(Huser=0.2 para Ch-274 e Ch-160; Huser=0.3 para Ch-970) e que forneceram contornos
considerados adequados e comparáveis entre os animais. Também não foi utilizada a opção de
variância x/y do HRE, mantendo a forma dos “contornos” circulares. Seguindo o padrão
consensual entre pesquisadores (embora arbitrário), a área de vida estimada pelo método
Kernel foi definida como sendo a menor área contendo 95% da distribuição de utilização
(Worton, 1989; Seaman & Powell, 1996; Powell, 2000). Os centros de atividade para cada
animal foram definidos como sendo os contornos de até 50 % da distribuição de utilização
(Powell, 2000).
3.6 Padrão de atividade
Cada fêmea marcada foi monitorada uma noite por mês, as categorias de atividade
amostradas começaram a ser anotadas a partir do início de suas atividades, ou a partir do
horário do fim do pôr-do-sol “twilight end” (“escuridão astronômica”, calculada através do
programa “Earthsun - versão 4.6”). Terminando em torno de 23:00 h, ou mais cedo nos casos
de chuvas e/ou de pouca visibilidade.
Quatro categorias principais de atividade foram amostradas: “repouso” (Rep.) quando
estava dormindo (repouso noturno), ou acordado, mas sem nenhuma movimentação
horizontal ou vertical; “movimentando” (Mov.) foi anotada quando houve deslocamentos

37
verticais ou horizontais do corpo nos galhos e troncos; “alimentando” (Alim.) foi anotada
quando um item alimentar estava sendo engolido, mastigado ou sendo trazido para a boca.
Todas as demais atividades menos freqüentes como coçar o corpo (auto-catação), a utilização
das latrinas (defecação) e as interações sociais entre adultos e entre a mãe (Ch-970) e seu
filhote foram agrupados na categoria “outros” (Out.). Nos registros onde o animal monitorado
estava encoberto pela vegetação ou onde não foi possível determinar a categoria de atividade
no intervalo do período amostral, esta foi anotada como “indeterminada” (Indet.).
Nas comparações feitas no padrão de atividade entre estações, para efeito de análise, a
estação seca compreendeu o período de abril a setembro e a chuvosa de outubro a março,
conforme divisão adotada em trabalhos anteriores (ex: Chiarello, 1998a) e de acordo com a
média histórica de precipitação e temperatura da cidade de Vitória, ES, (INCAPER, 2006).
3.7 Espécies vegetais utilizadas na dieta
Foram coletadas amostras de material vegetal de 50 árvores marcadas durante o
estudo, para posterior identificação das espécies de plantas que foram utilizadas na sua
alimentação. Estas foram analisadas e identificadas pelo Prof. Dr. Oberdan José Pereira,
botânico especialista em vegetação de restinga e profundo conhecedor da região do PEPCV,
com diversos trabalhos a respeito da flora do Parque (ex: Pereira, 1990). Estas espécies foram
então classificadas em ordem de importância individual e total, de acordo com o número de
registros instantâneos da atividade de “alimentação” anotados para cada uma delas durante as
noites de monitoramento.
3.8 Movimentação
Para o estudo da movimentação da espécie foram calculados as “distâncias mínimas” e
as respectivas “distâncias completas” em cada noite de monitoramento. A primeira medida foi
considerada como a distância horizontal entre o local do início da atividade e a posição de
repouso diurno subseqüente (repouso diurno - repouso diurno), anotada no dia seguinte.
As “distâncias completas” foram consideradas como a distância total percorrida por
cada animal durante uma noite de monitoramento, sendo utilizado a distância em linha reta do
início de suas atividades noturnas, passando pelos pontos (árvores) onde efetuou suas
principais atividades (referidas acima) até sua localização na manhã seguinte. Ambas as
distâncias foram calculadas através do programa “AutoCAD Map 2000”.

38
3.9 Análises estatísticas
Para comparar a proporção de tempo gasto nas principais atividades nas noites de
monitoramento entre os indivíduos, o número de registros em cada uma foi transformado em
porcentagem e então utilizado nas comparações feitas através do teste não paramétrico de
Kruskal-Wallis (Siegel, 1956), uma vez que os dados de porcentagem não possuem
distribuição normal. Como apenas parte da noite foi usada para a amostragem das atividades e
o período observado variou entre as noites de observação (média de 4,58 horas/noite, EP =
0,13 horas/noite, amplitude de 2,17 a 5,67 horas/noite) foi testado a possibilidade desta
variação ter interferido no tempo gasto nas atividades. Para isto foi usado a Correlação de
Spearman (Siegel, 1956). Para a comparação das variações sazonais (estação seca x estação
chuvosa) no tempo gasto nas atividades, foi usado o teste não paramétrico de Mann-Whitney
(U) (Siegel, 1956).
As distâncias médias medidas para os animais foram comparadas entre os indivíduos
através de uma Análise de Variância seguida de um teste de Tukey (post hoc). Para tanto, os
dados foram checados quanto à normalidade através do teste de Kolmogorov-Smirnov
(p>0,05 em todos os casos) e quanto à homogeneidade de variâncias através do teste de
Levene (p>0,05 em todos os casos). Para a comparação sazonal destas distâncias foi usado o
teste t de student (t-test) para duas amostras independentes.
Análises adicionais foram feitas para testar se houve variação significativa no número
de árvores visitadas para alimentação/noite entre as estações. Para isso dividiu-se o número de
árvores visitadas pelo total de horas observadas em cada noite e comparou-se o índice
resultante (árvores visitadas/hora) através do teste não paramétrico de Mann-Whitney. Em
seguida, calculou-se o tempo gasto alimentando em cada árvore dividindo-se o tempo total
gasto em alimentação pelo número total de arvores visitadas por noite. O índice resultante
(horas de alimentação/arvore) foi comparado utilizando o também o teste de Mann-Whitney.
Todos os testes estatísticos foram feitos no programa “Statistical Package for the Social
Science” (SPSS) versão 8.
39
4. RESULTADOS
4.1 Uso do espaço
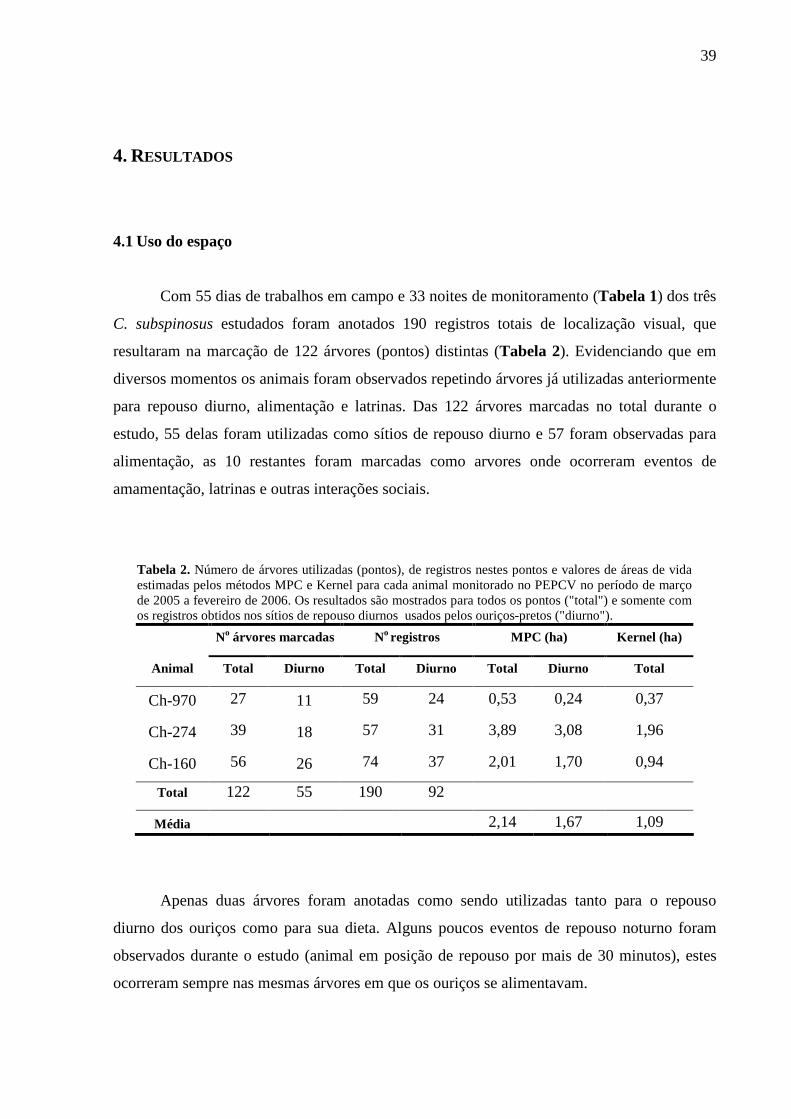
Com 55 dias de trabalhos em campo e 33 noites de monitoramento (Tabela 1) dos três
C. subspinosus estudados foram anotados 190 registros totais de localização visual, que
resultaram na marcação de 122 árvores (pontos) distintas (Tabela 2). Evidenciando que em
diversos momentos os animais foram observados repetindo árvores já utilizadas anteriormente
para repouso diurno, alimentação e latrinas. Das 122 árvores marcadas no total durante o
estudo, 55 delas foram utilizadas como sítios de repouso diurno e 57 foram observadas para
alimentação, as 10 restantes foram marcadas como arvores onde ocorreram eventos de
amamentação, latrinas e outras interações sociais.
Tabela 2. Número de árvores utilizadas (pontos), de registros nestes pontos e valores de áreas de vida estimadas pelos métodos MPC e Kernel para cada animal monitorado no PEPCV no período de março de 2005 a fevereiro de 2006. Os resultados são mostrados para todos os pontos ("total") e somente com os registros obtidos nos sítios de repouso diurnos usados pelos ouriços-pretos ("diurno").
No árvores marcadas No registros MPC (ha) Kernel (ha)
Animal Total Diurno Total Diurno Total Diurno Tota l
Ch-970 27 11 59 24 0,53 0,24 0,37
Ch-274 39 18 57 31 3,89 3,08 1,96
Ch-160 56 26 74 37 2,01 1,70 0,94
Total 122 55 190 92
Média 2,14 1,67 1,09
Apenas duas árvores foram anotadas como sendo utilizadas tanto para o repouso
diurno dos ouriços como para sua dieta. Alguns poucos eventos de repouso noturno foram
observados durante o estudo (animal em posição de repouso por mais de 30 minutos), estes
ocorreram sempre nas mesmas árvores em que os ouriços se alimentavam.

40
4.1.1 Mínimo polígono convexo
As estimativas de tamanho de área de vida calculadas através do MPC utilizando todos
os pontos amostrados para as três fêmeas produziram resultados marcadamente diferentes,
com um valor médio de 2,14 ha, mas com uma variação de cerca de 7 vezes entre a menor
(0,53 ha; Ch-970) e a maior área de vida (3,89 ha; Ch-274). De maneira geral, as três áreas
“diurnas” acompanham o contorno da borda da floresta de restinga, com uma concentração de
pontos de repouso nas porções próximas ao brejo herbáceo, em áreas periodicamente
inundadas (principalmente Ch-160).
Todas as três áreas estimadas utilizando somente as localizações nos sítios de repouso
diurno foram menores para todas as fêmeas quando comparadas às áreas totais (média = 1,67
ha; variando de 0,24 a 3,08 ha/individuo) (Tabela 2). Como pode ser observado nas Figuras
8, 9 e 10, os polígonos desenhados utilizando somente estes pontos diurnos se encontram
incluídos dentro do polígono total, chegando a acompanhar a forma deste último (Ch-160 e
Ch-274).
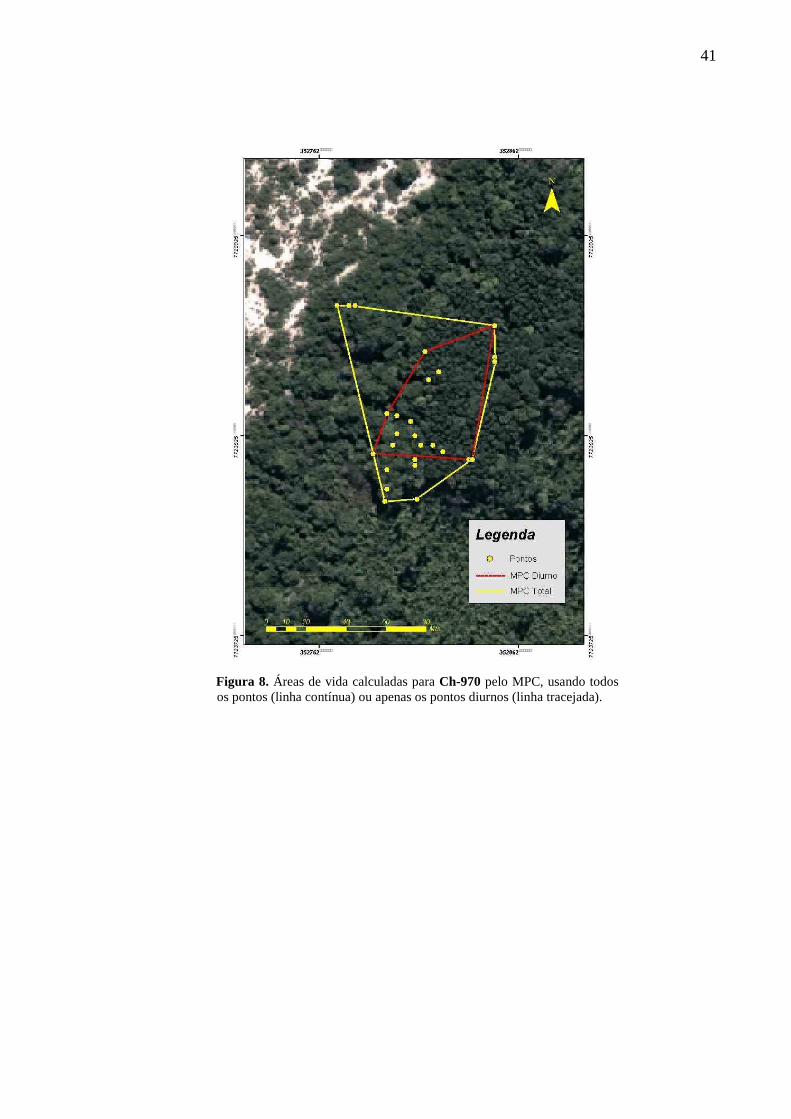
41
Figura 8. Áreas de vida calculadas para Ch-970 pelo MPC, usando todos os pontos (linha contínua) ou apenas os pontos diurnos (linha tracejada).
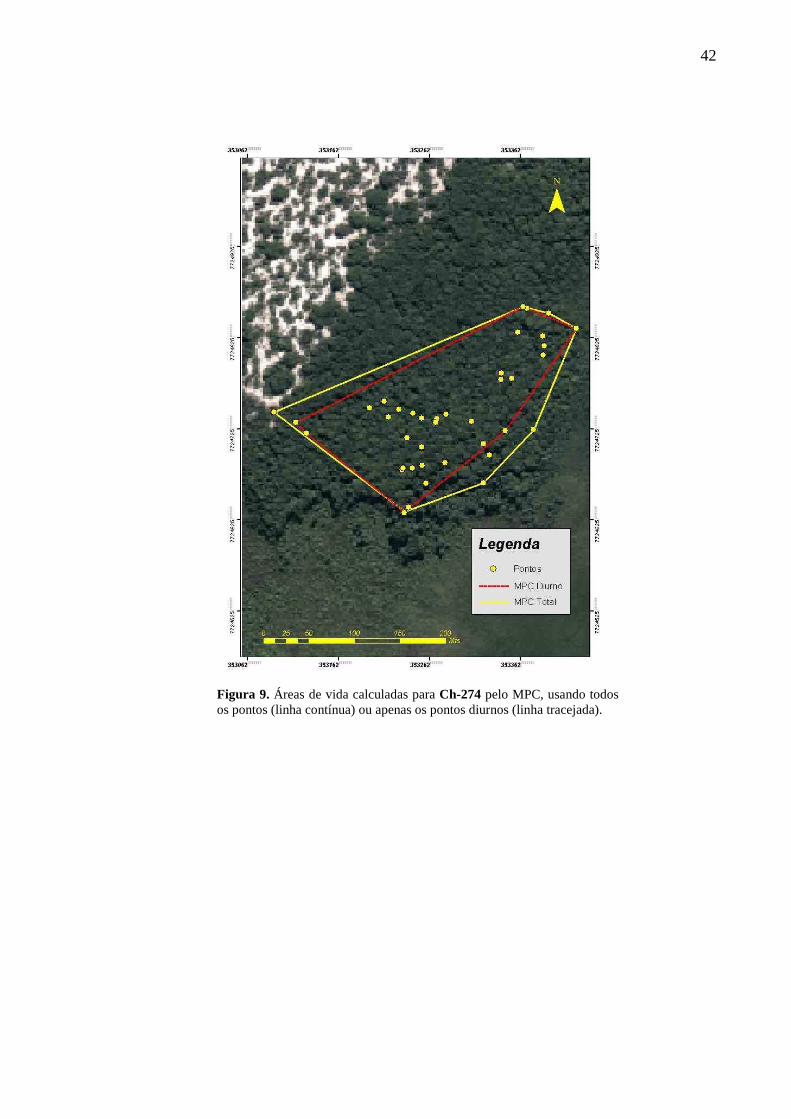
42
Figura 9. Áreas de vida calculadas para Ch-274 pelo MPC, usando todos os pontos (linha contínua) ou apenas os pontos diurnos (linha tracejada).
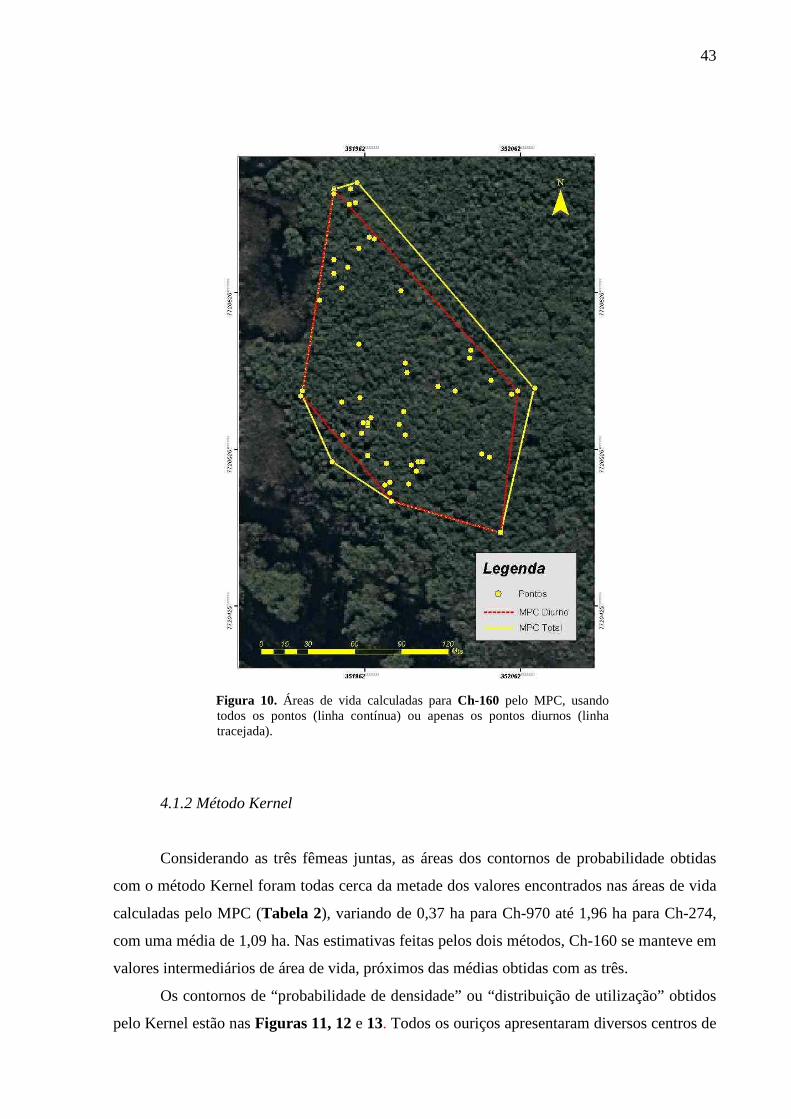
43
Figura 10. Áreas de vida calculadas para Ch-160 pelo MPC, usando todos os pontos (linha contínua) ou apenas os pontos diurnos (linha tracejada).
4.1.2 Método Kernel
Considerando as três fêmeas juntas, as áreas dos contornos de probabilidade obtidas
com o método Kernel foram todas cerca da metade dos valores encontrados nas áreas de vida
calculadas pelo MPC (Tabela 2), variando de 0,37 ha para Ch-970 até 1,96 ha para Ch-274,
com uma média de 1,09 ha. Nas estimativas feitas pelos dois métodos, Ch-160 se manteve em
valores intermediários de área de vida, próximos das médias obtidas com as três.
Os contornos de “probabilidade de densidade” ou “distribuição de utilização” obtidos
pelo Kernel estão nas Figuras 11, 12 e 13. Todos os ouriços apresentaram diversos centros de

44
atividade (Kernel 50 %), ou seja, as atividades estavam concentradas em várias “porções”
específicas da área de vida, localmente relacionadas com a concentração de árvores utilizadas
na sua dieta e sítios de repouso diurnos próximos a estas. De maneira geral, os ouriços-pretos
concentraram suas atividades próximas aos limites das formações florestais, em porções que
sofrem influência das inundações no brejo herbáceo, porém não deixaram de utilizar também
pontos mais internos da floresta de restinga, normalmente em locais de repouso diurno.
A fêmea Ch-970 utilizou uma área de vida muito reduzida (Figura 11; Tabela 2),
apresentando 5 centros de atividade, sendo o principal na porção sul, onde se encontrava seu
filhote e onde havia uma concentração de árvores utilizadas para alimentação (principalmente
da espécie Tapirira guianensis), mais alguns sítios diurnos utilizados recorrentemente e a
latrina localizada no limite sul da sua área, onde foi observada em seis oportunidades, em
noites diferentes. Os três pontos encontrados na periferia noroeste de sua área de vida
correspondem a duas árvores de alimentação (Pera glabrata e Pera leandri) e uma onde
amamentou seu filhote. O ponto com contorno de probabilidade 10% isolado na ponta
nordeste corresponde a um sítio de repouso utilizado 6 vezes, próximo a uma T. guianensis
usada na dieta.
Ch-274 obteve a maior área de vida em ambas estimativas (Tabela 2), apresentando 9
centros de atividade, principalmente no centro da restinga arbórea e na borda com o brejo
herbáceo (Figura 12). O contorno central corresponde a um sítio diurno próximo de várias
árvores de alimentação das espécies T. guianensis e P. glabrata, sendo as duas espécies
principais da dieta dos ouriços-pretos no PEPCV (ver adiante). A mancha na porção nordeste
da área de vida também corresponde a pontos de repouso e a presença das duas mesmas
espécies de árvores. O ponto mais extremo a oeste da área é a localização da latrina usada por
este indivíduo, no limite com a formação aberta de restinga.
Para Ch-160, a porção mais utilizada de sua área (sul) corresponde a uma
concentração de seis árvores da sua alimentação e de três sítios de repouso próximos a estes
pontos, bem no limite com o brejo herbáceo (Figura 13). Esta fêmea apresentou 12 centros de
atividade, sendo os 7 centros localizados a oeste posicionados sobre porções periodicamente
inundadas. A “mancha” mais ao norte reflete uma concentração de árvores de P. glabrata,
além de uma árvore de repouso diurno próxima.
Em 16 oportunidades foram observados durante o dia outros indivíduos tanto de C.
subspinosus (n=11) como de S. villosus (n=5) em sítios de repouso diurnos já marcados
anteriormente para os animais estudados, bem como sozinhos em arvores próximas ou no

45
interior das áreas de vida calculadas. Durante a noite também foram registradas interações
com outros ouriços-pretos (filhotes, jovens e adultos).
Figura 11. Área de vida da fêmea Ch-970, calculada através dos métodos MPC (linhas) e Kernel (contornos). Os contornos indicam as diferentes probabilidades de ocorrência dos mesmos em suas áreas de uso.
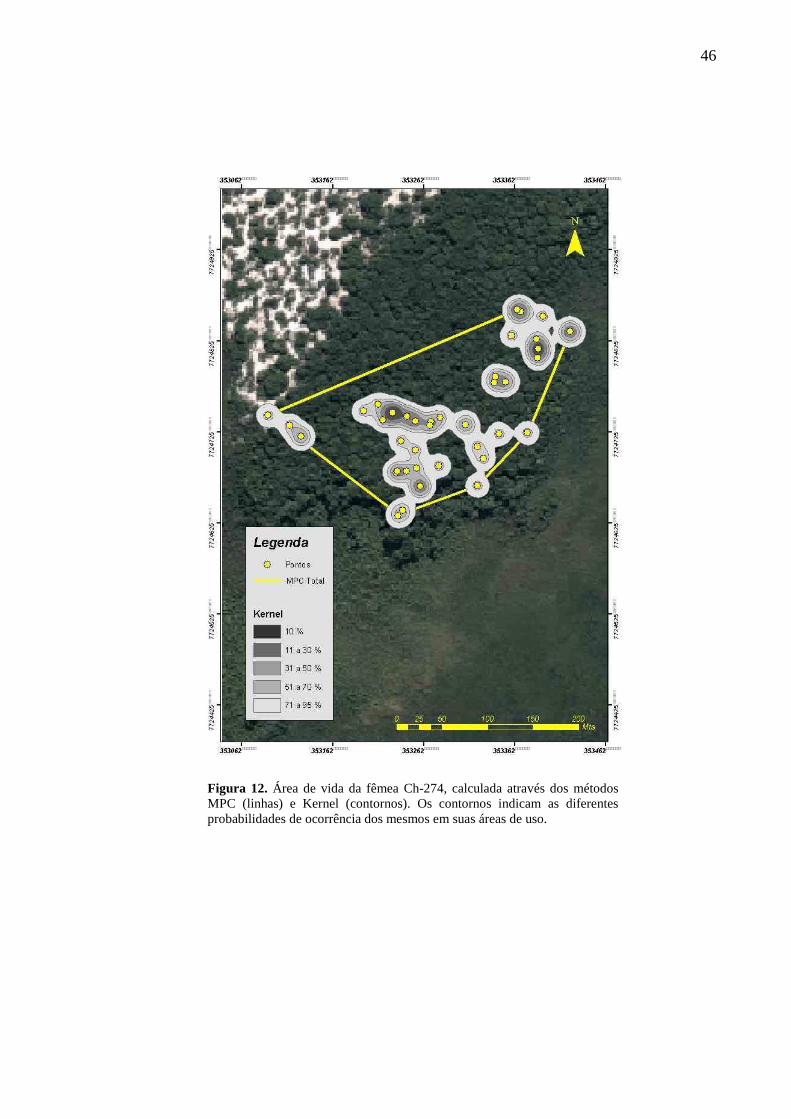
46
Figura 12. Área de vida da fêmea Ch-274, calculada através dos métodos MPC (linhas) e Kernel (contornos). Os contornos indicam as diferentes probabilidades de ocorrência dos mesmos em suas áreas de uso.

47
Figura 13. Área de vida da fêmea Ch-160, calculada através dos métodos MPC (linhas) e Kernel (contornos). Os contornos indicam as diferentes probabilidades de ocorrência dos mesmos em suas áreas de uso.
4.1.3 Área de vida acumulada
Como mostrado pela Figura 14, a área acumulada para os três animais aumentou
substancialmente nos primeiros três a quatro meses de estudo, ou seja, em torno de 20
registros de localização para cada um. Com pouco mais do que três meses de monitoramento
a área de vida da fêmea Ch-970 se estabilizou em apenas 0,5 ha. Com as fêmeas Ch-274 e Ch-
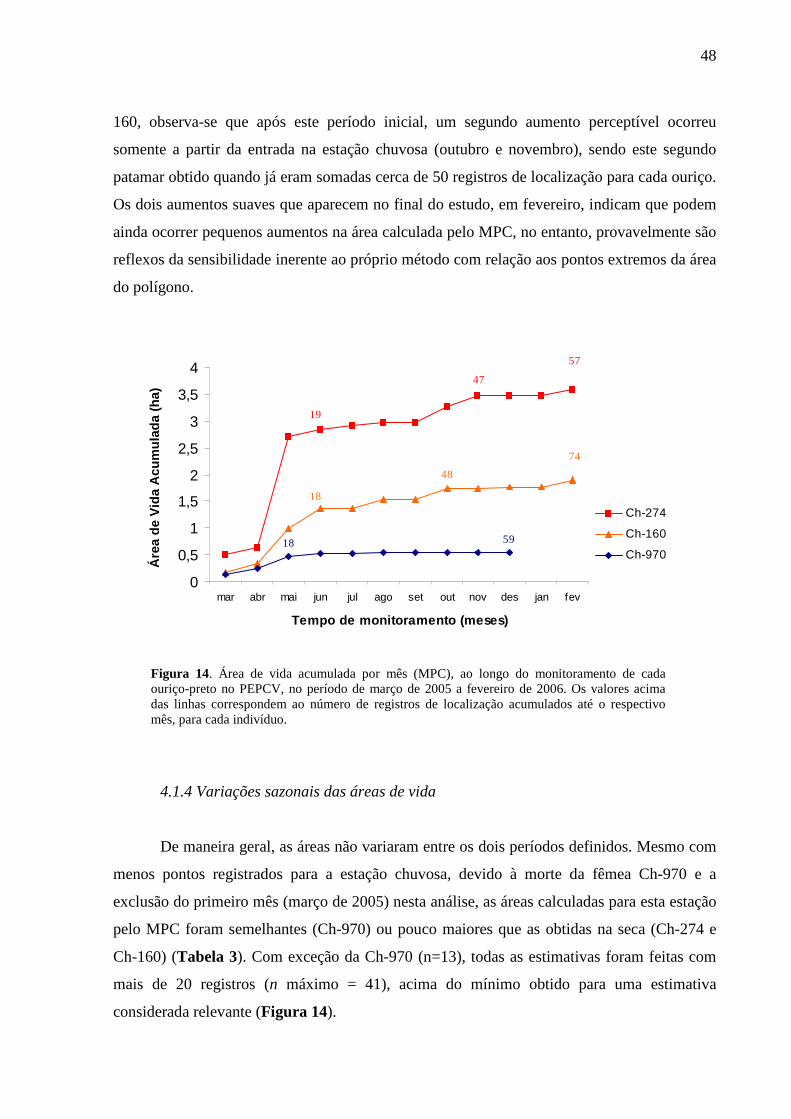
48
160, observa-se que após este período inicial, um segundo aumento perceptível ocorreu
somente a partir da entrada na estação chuvosa (outubro e novembro), sendo este segundo
patamar obtido quando já eram somadas cerca de 50 registros de localização para cada ouriço.
Os dois aumentos suaves que aparecem no final do estudo, em fevereiro, indicam que podem
ainda ocorrer pequenos aumentos na área calculada pelo MPC, no entanto, provavelmente são
reflexos da sensibilidade inerente ao próprio método com relação aos pontos extremos da área
do polígono.
0
0,5
1
1,5
2
2,5
3
3,5
4
mar abr mai jun jul ago set out nov des jan fev
Tempo de monitoramento (meses)
Áre
a de
Vid
a A
cum
ulad
a (h
a)
Ch-274
Ch-160
Ch-970
48
47
18
18
19
57
74
59
Figura 14. Área de vida acumulada por mês (MPC), ao longo do monitoramento de cada ouriço-preto no PEPCV, no período de março de 2005 a fevereiro de 2006. Os valores acima das linhas correspondem ao número de registros de localização acumulados até o respectivo mês, para cada indivíduo.
4.1.4 Variações sazonais das áreas de vida
De maneira geral, as áreas não variaram entre os dois períodos definidos. Mesmo com
menos pontos registrados para a estação chuvosa, devido à morte da fêmea Ch-970 e a
exclusão do primeiro mês (março de 2005) nesta análise, as áreas calculadas para esta estação
pelo MPC foram semelhantes (Ch-970) ou pouco maiores que as obtidas na seca (Ch-274 e
Ch-160) (Tabela 3). Com exceção da Ch-970 (n=13), todas as estimativas foram feitas com
mais de 20 registros (n máximo = 41), acima do mínimo obtido para uma estimativa
considerada relevante (Figura 14).

49
Tabela 3. Áreas de vida e respectivo tamanho amostral (n) dos três ouriços-pretos monitorados no PEPCV, estimadas pelo MPC para os períodos seco e chuvoso, entre abril de 2005 a fevereiro de 2006.
Área de Vida MPC (ha)
Estação Ch-970 (n) Ch-274 (n) Ch-160 (n)
Seca 0,43 (41) 2,48 (27) 1,05 (38)
Chuva 0,42 (13) 3,14 (23) 1,28 (31)
(n) = número de localizações registradas em cada estação.
Os contornos obtidos com o MPC para os registros agrupados por estação (Figuras
15, 16 e 17) ilustram, além dos tamanhos semelhantes das áreas, que os ouriços-pretos
demonstraram o que pode ser interpretado como “fidelidade territorial” às porções de mata
ocupadas, mantendo-se praticamente na mesma região, mesmo quando os pontos foram
separados na primeira e segunda metade do estudo.
Para Ch-160, a partir da estação chuvosa, de maneira geral ocorreu uma concentração
dos pontos a oeste da área, assim como uma expansão para o norte da área do polígono
observado na seca. Provavelmente um reflexo da utilização de uma porção periodicamente
inundada da borda da formação florestal e que possui diversas árvores de alimentação
visitadas por este indivíduo, principalmente das espécies P. glabrata e Inga laurina e Inga
fagifolia.
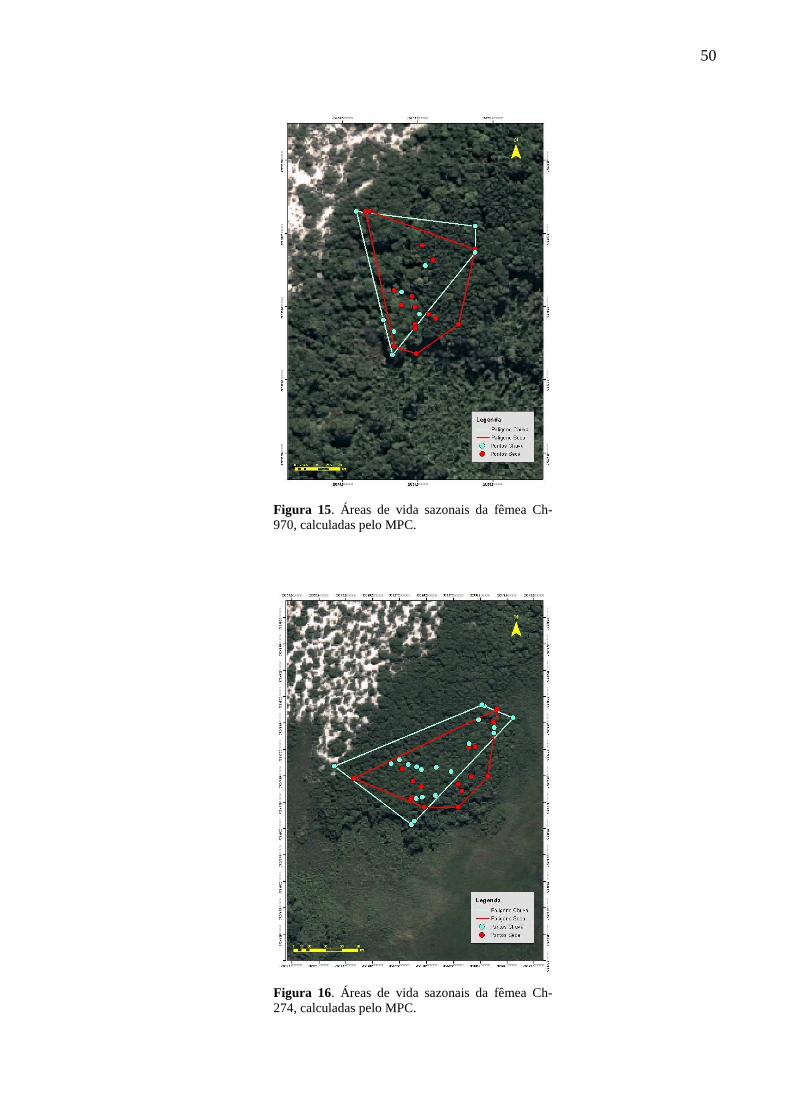
50
Figura 15. Áreas de vida sazonais da fêmea Ch-970, calculadas pelo MPC.
Figura 16. Áreas de vida sazonais da fêmea Ch-274, calculadas pelo MPC.

51
Figura 17. Áreas de vida sazonais da fêmea Ch-160, calculadas pelo MPC.
4.2 Padrão de atividade
As 150 horas e 50 minutos de observação noturna das três fêmeas de ouriço-preto
monitoradas (Tabela 1), resultaram em 932 registros instantâneos, assim divididos: 291 para
Ch-970; 323 registros para Ch-274 e 318 para Ch-160. Quanto a sua atividade diária de
maneira geral, observa-se que durante todo o período claro do dia os ouriços permanecem em
repouso em um de seus abrigos diurnos, praticamente não se movimentam e raramente
mudam de posição, excetuando-se alguns poucos eventos de auto-catação e pequenas
mudanças na posição do corpo. A espécie pode ser considerada estritamente noturna,
iniciando suas atividades no crepúsculo, após o pôr-do-sol (em média 26,5 minutos) e antes
dos horários de escuridão astronômica.
A partir de então, pelo menos nas cinco primeiras horas da noite, os ouriços-pretos
concentraram suas atividades em alimentação e movimentação (Tabela 4), esta última feita
normalmente nos próprios galhos das árvores onde se alimentam ou no deslocamento entre
estas. A alimentação foi menor que as outras duas atividades principais para todos os animais
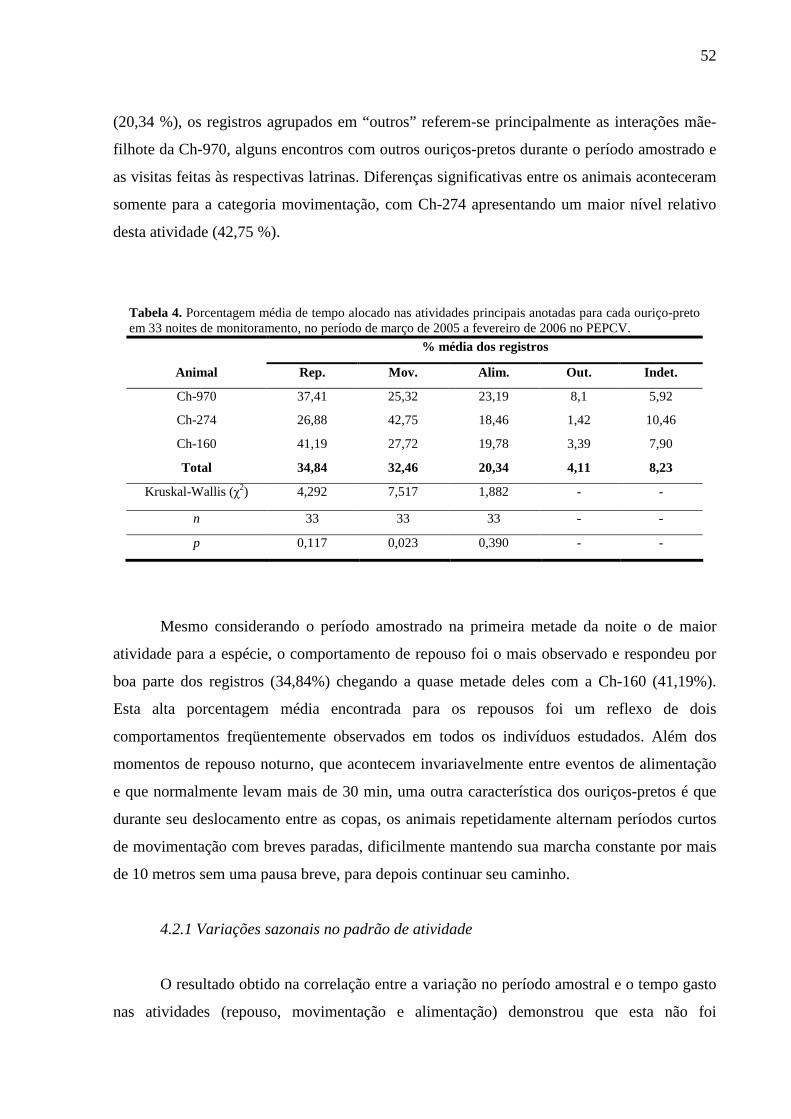
52
(20,34 %), os registros agrupados em “outros” referem-se principalmente as interações mãe-
filhote da Ch-970, alguns encontros com outros ouriços-pretos durante o período amostrado e
as visitas feitas às respectivas latrinas. Diferenças significativas entre os animais aconteceram
somente para a categoria movimentação, com Ch-274 apresentando um maior nível relativo
desta atividade (42,75 %).
Tabela 4. Porcentagem média de tempo alocado nas atividades principais anotadas para cada ouriço-preto em 33 noites de monitoramento, no período de março de 2005 a fevereiro de 2006 no PEPCV.
% média dos registros
Animal Rep. Mov. Alim. Out. Indet.
Ch-970 37,41 25,32 23,19 8,1 5,92
Ch-274 26,88 42,75 18,46 1,42 10,46
Ch-160 41,19 27,72 19,78 3,39 7,90
Total 34,84 32,46 20,34 4,11 8,23
Kruskal-Wallis (χ2) 4,292 7,517 1,882 - -
n 33 33 33 - -
p 0,117 0,023 0,390 - -
Mesmo considerando o período amostrado na primeira metade da noite o de maior
atividade para a espécie, o comportamento de repouso foi o mais observado e respondeu por
boa parte dos registros (34,84%) chegando a quase metade deles com a Ch-160 (41,19%).
Esta alta porcentagem média encontrada para os repousos foi um reflexo de dois
comportamentos freqüentemente observados em todos os indivíduos estudados. Além dos
momentos de repouso noturno, que acontecem invariavelmente entre eventos de alimentação
e que normalmente levam mais de 30 min, uma outra característica dos ouriços-pretos é que
durante seu deslocamento entre as copas, os animais repetidamente alternam períodos curtos
de movimentação com breves paradas, dificilmente mantendo sua marcha constante por mais
de 10 metros sem uma pausa breve, para depois continuar seu caminho.
4.2.1 Variações sazonais no padrão de atividade
O resultado obtido na correlação entre a variação no período amostral e o tempo gasto
nas atividades (repouso, movimentação e alimentação) demonstrou que esta não foi
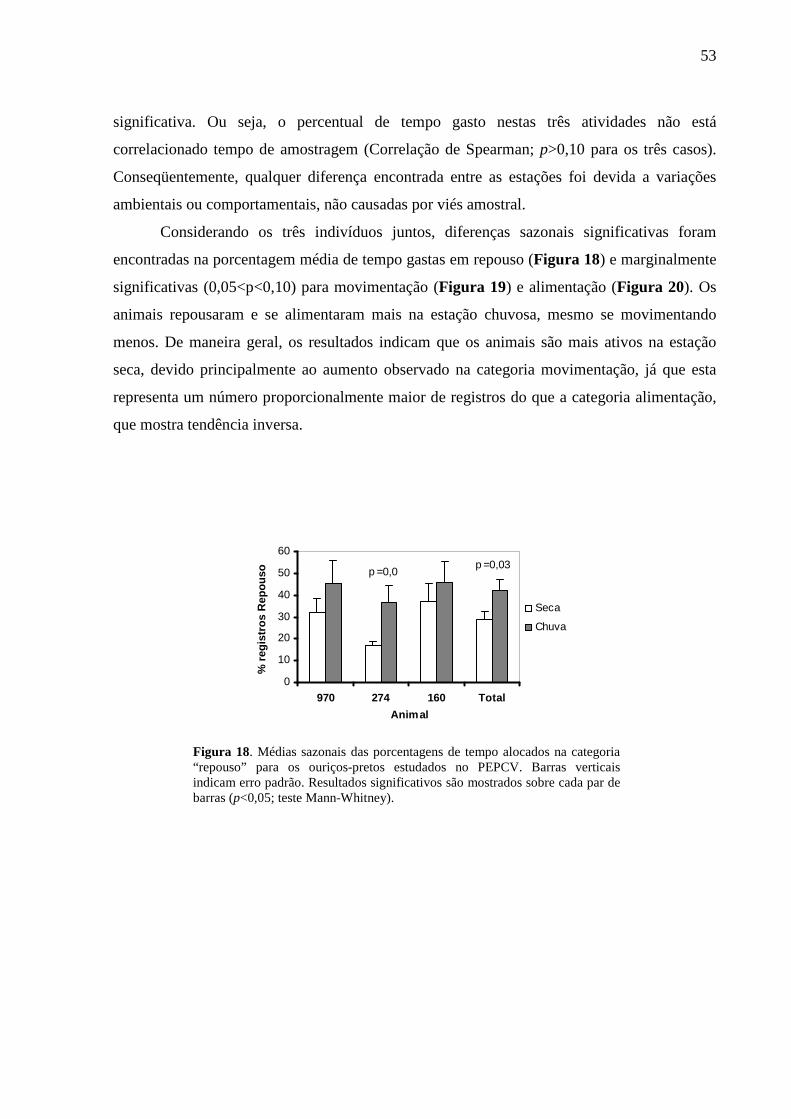
53
significativa. Ou seja, o percentual de tempo gasto nestas três atividades não está
correlacionado tempo de amostragem (Correlação de Spearman; p>0,10 para os três casos).
Conseqüentemente, qualquer diferença encontrada entre as estações foi devida a variações
ambientais ou comportamentais, não causadas por viés amostral.
Considerando os três indivíduos juntos, diferenças sazonais significativas foram
encontradas na porcentagem média de tempo gastas em repouso (Figura 18) e marginalmente
significativas (0,05<p<0,10) para movimentação (Figura 19) e alimentação (Figura 20). Os
animais repousaram e se alimentaram mais na estação chuvosa, mesmo se movimentando
menos. De maneira geral, os resultados indicam que os animais são mais ativos na estação
seca, devido principalmente ao aumento observado na categoria movimentação, já que esta
representa um número proporcionalmente maior de registros do que a categoria alimentação,
que mostra tendência inversa.
0
10
20
30
40
50
60
970 274 160 Total
Animal
% r
egis
tros
Rep
ouso
Seca
Chuva
p =0,03p =0,0
Figura 18. Médias sazonais das porcentagens de tempo alocados na categoria “repouso” para os ouriços-pretos estudados no PEPCV. Barras verticais indicam erro padrão. Resultados significativos são mostrados sobre cada par de barras (p<0,05; teste Mann-Whitney).
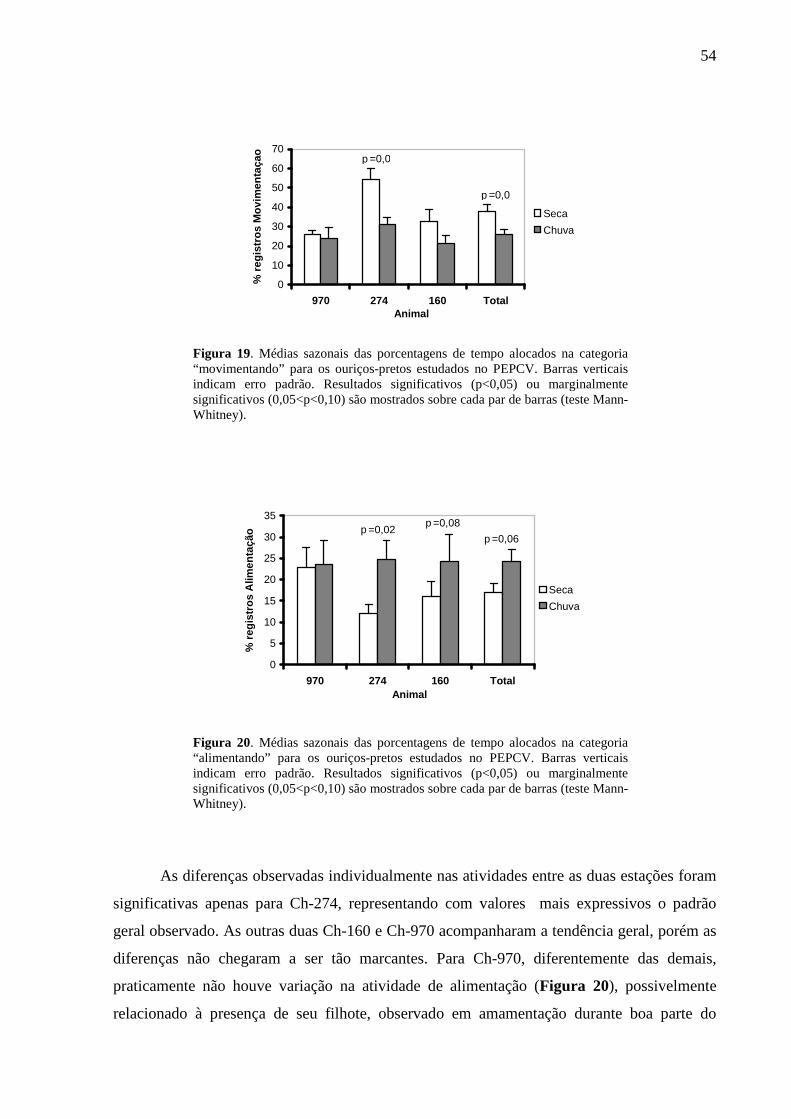
54
0
10
20
30
40
50
60
70
970 274 160 TotalAnimal
% r
egis
tros
Mov
imen
taça
o
Seca
Chuva
p =0,0
p =0,0
Figura 19. Médias sazonais das porcentagens de tempo alocados na categoria “movimentando” para os ouriços-pretos estudados no PEPCV. Barras verticais indicam erro padrão. Resultados significativos (p<0,05) ou marginalmente significativos (0,05<p<0,10) são mostrados sobre cada par de barras (teste Mann-Whitney).
0
5
10
15
20
25
30
35
970 274 160 TotalAnimal
% r
egis
tros
Alim
enta
ção
Seca
Chuva
p =0,08
p =0,06p =0,02
Figura 20. Médias sazonais das porcentagens de tempo alocados na categoria “alimentando” para os ouriços-pretos estudados no PEPCV. Barras verticais indicam erro padrão. Resultados significativos (p<0,05) ou marginalmente significativos (0,05<p<0,10) são mostrados sobre cada par de barras (teste Mann-Whitney).
As diferenças observadas individualmente nas atividades entre as duas estações foram
significativas apenas para Ch-274, representando com valores mais expressivos o padrão
geral observado. As outras duas Ch-160 e Ch-970 acompanharam a tendência geral, porém as
diferenças não chegaram a ser tão marcantes. Para Ch-970, diferentemente das demais,
praticamente não houve variação na atividade de alimentação (Figura 20), possivelmente
relacionado à presença de seu filhote, observado em amamentação durante boa parte do
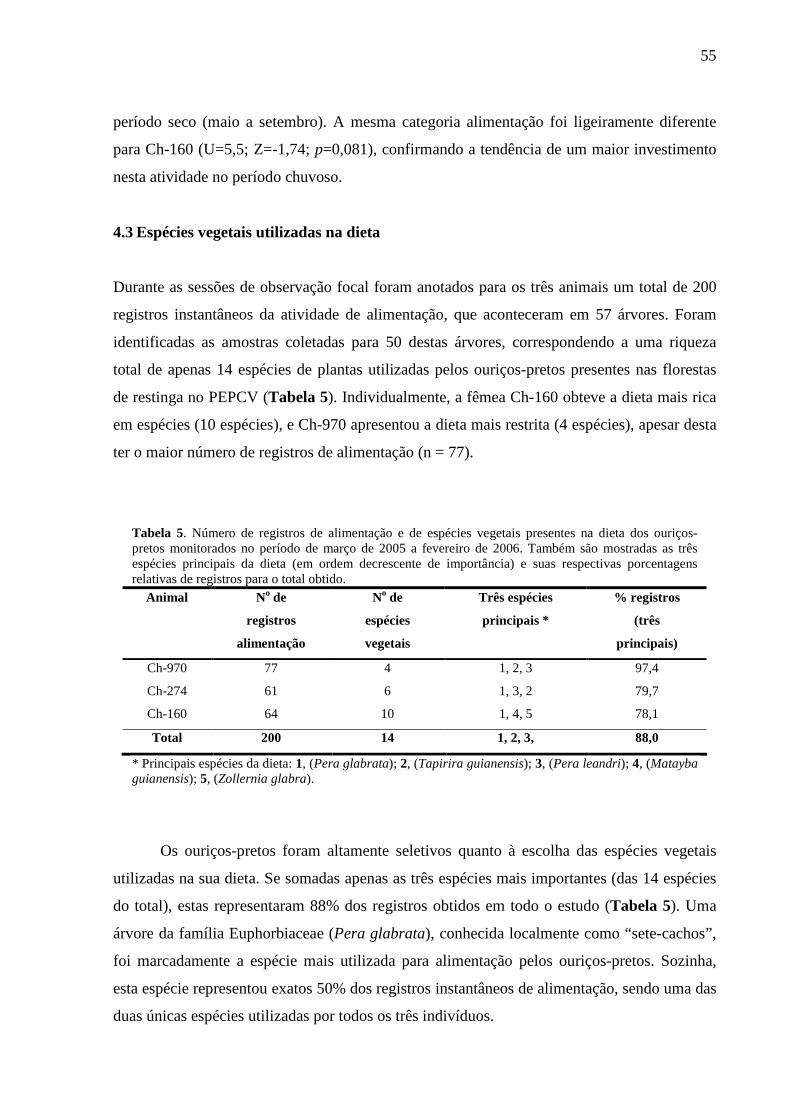
55
período seco (maio a setembro). A mesma categoria alimentação foi ligeiramente diferente
para Ch-160 (U=5,5; Z=-1,74; p=0,081), confirmando a tendência de um maior investimento
nesta atividade no período chuvoso.
4.3 Espécies vegetais utilizadas na dieta
Durante as sessões de observação focal foram anotados para os três animais um total de 200
registros instantâneos da atividade de alimentação, que aconteceram em 57 árvores. Foram
identificadas as amostras coletadas para 50 destas árvores, correspondendo a uma riqueza
total de apenas 14 espécies de plantas utilizadas pelos ouriços-pretos presentes nas florestas
de restinga no PEPCV (Tabela 5). Individualmente, a fêmea Ch-160 obteve a dieta mais rica
em espécies (10 espécies), e Ch-970 apresentou a dieta mais restrita (4 espécies), apesar desta
ter o maior número de registros de alimentação (n = 77).
Tabela 5. Número de registros de alimentação e de espécies vegetais presentes na dieta dos ouriços-pretos monitorados no período de março de 2005 a fevereiro de 2006. Também são mostradas as três espécies principais da dieta (em ordem decrescente de importância) e suas respectivas porcentagens relativas de registros para o total obtido.
Animal No de
registros
alimentação
No de
espécies
vegetais
Três espécies
principais *
% registros
(três
principais)
Ch-970 77 4 1, 2, 3 97,4
Ch-274 61 6 1, 3, 2 79,7
Ch-160 64 10 1, 4, 5 78,1
Total 200 14 1, 2, 3, 88,0
* Principais espécies da dieta: 1, (Pera glabrata); 2, (Tapirira guianensis); 3, (Pera leandri); 4, (Matayba guianensis); 5, (Zollernia glabra).
Os ouriços-pretos foram altamente seletivos quanto à escolha das espécies vegetais
utilizadas na sua dieta. Se somadas apenas as três espécies mais importantes (das 14 espécies
do total), estas representaram 88% dos registros obtidos em todo o estudo (Tabela 5). Uma
árvore da família Euphorbiaceae (Pera glabrata), conhecida localmente como “sete-cachos”,
foi marcadamente a espécie mais utilizada para alimentação pelos ouriços-pretos. Sozinha,
esta espécie representou exatos 50% dos registros instantâneos de alimentação, sendo uma das
duas únicas espécies utilizadas por todos os três indivíduos.

56
A segunda mais importante foi Tapirira guianensis da família Anacardiaceae,
representando 21,5% dos registros obtidos. Todas as outras que aparecem como principais
(Pera leandri, Matayba guianensis e Zollernia glabra) respondem por menos de 10% dos
registros. Além da seletividade encontrada para as espécies vegetais presentes na dieta dos
animais, as árvores também foram individualmente selecionadas (14 indivíduos foram
visitados repetidas vezes ao longo do trabalho). A fêmea Ch-970 foi o caso mais extremo de
seletividade observado, com as três espécies principais da sua dieta correspondendo a 97,4%
dos registros. Mesmo para Ch-160, com a maior riqueza relativa de espécies, as três mais
importantes responderam por 78,1 % dos registros.
Os ouriços-pretos possuíram uma dieta composta predominantemente de folhas (97 %
considerando brotos, folhas novas e maduras), selecionando preferencialmente as folhas mais
novas (59 % dos registros totais de alimentação para folhas) presentes nas extremidades dos
galhos. Outras partes vegetais como invólucros florais e alguns poucos frutos de P. glabrata
(em setembro e outubro), bem como inflorescências de M. guianensis, foram registrados
relativamente poucas vezes durante os eventos de alimentação, aparentemente se tratando de
um recurso explorado oportunísticamente pelos animais.
Ao longo de uma noite de observação, os animais normalmente se alimentam em mais
de uma árvore e em mais de uma espécie. Quando pretendem se alimentar, os ouriços-pretos
assumem um comportamento estereotipado, eles vão até as copas nas porções finais dos
galhos, prendem os membros posteriores em galhos “seguros” e posicionam a cauda para
conferir ainda mais mobilidade aos membros anteriores e a cabeça. A partir de então, se
mostram igualmente seletivos quanto às partes vegetais consumidas, ativamente selecionando
as folhas das pontas (mais novas).
A diferença encontrada na taxa de visitação de árvores de alimentação (árvores
alim./hora) entre as estações foi marginalmente significativa (Mann-Whitney; U=77,5;
W=248,5; p=0,065), sendo que o número de árvores/hora foi maior na estação chuvosa
(média de 0,59 árvores/hora) do que na estação seca (Figura 21) (média de 0,44
árvores/hora).

57
00,10,20,30,40,50,60,7
Seca Chuva Estação
Árv
ores
/hor
a
Figura 21. Taxa de utilização de árvores de alimentação (árvores/hora) observadas entre as estações seca e chuvosa (Teste de Mann-Whitney).
Já o tempo despendido em alimentação em cada árvore não variou significativamente
entre as estações seca e chuvosa (Teste de Mann-Whitney; U=97; W=250; p=0,382). Ou seja,
os ouriços visitaram um número ligeiramente superior de árvores na estação chuvosa, mas
sem mudanças significativas no tempo gasto em cada uma delas entre as estações.
4.4 Movimentação
As médias totais e individuais das duas distâncias calculadas para os ouriços-pretos se
encontram na Tabela 6. O número de distâncias mínimas usando apenas dois pontos/noite
(sitio diurno a sitio diurno) utilizados nas análises (n=51) foi maior que o utilizado para as
distâncias completas (n=31) obtidas usando todos os pontos disponíveis/noite, pois foram
considerados também os registros onde mesmo sem monitoramento noturno de determinado
animal, foram coletados dois pontos diurnos consecutivos nos seus locais de repouso. De
maneira geral, entre pontos de repouso diurnos consecutivos (distancia mínima), os ouriços-
pretos se deslocaram ao longo do ano em média 72,58 metros (máximo de 218,9 metros).
Diferenças significativas entre os indivíduos foram encontradas apenas nas médias de
distância mínima para a fêmea Ch-274 (Tabela 6), que possuiu também a maior área de vida
estimada e menor número de repousos (ver acima).
Utilizando todos os pontos obtidos em cada noite de monitoramento, as distâncias
completas foram em média de 147 metros (máximo de 324 metros e mínimo de 21,2 metros).
Durante o dia os ouriços-pretos raramente se movimentam de um local de repouso para outro,
sendo assim as distâncias completas percorridas no período da noite podem ser consideradas
um valor representativo para todas as 24 horas do dia. No entanto, deve ser mencionado que
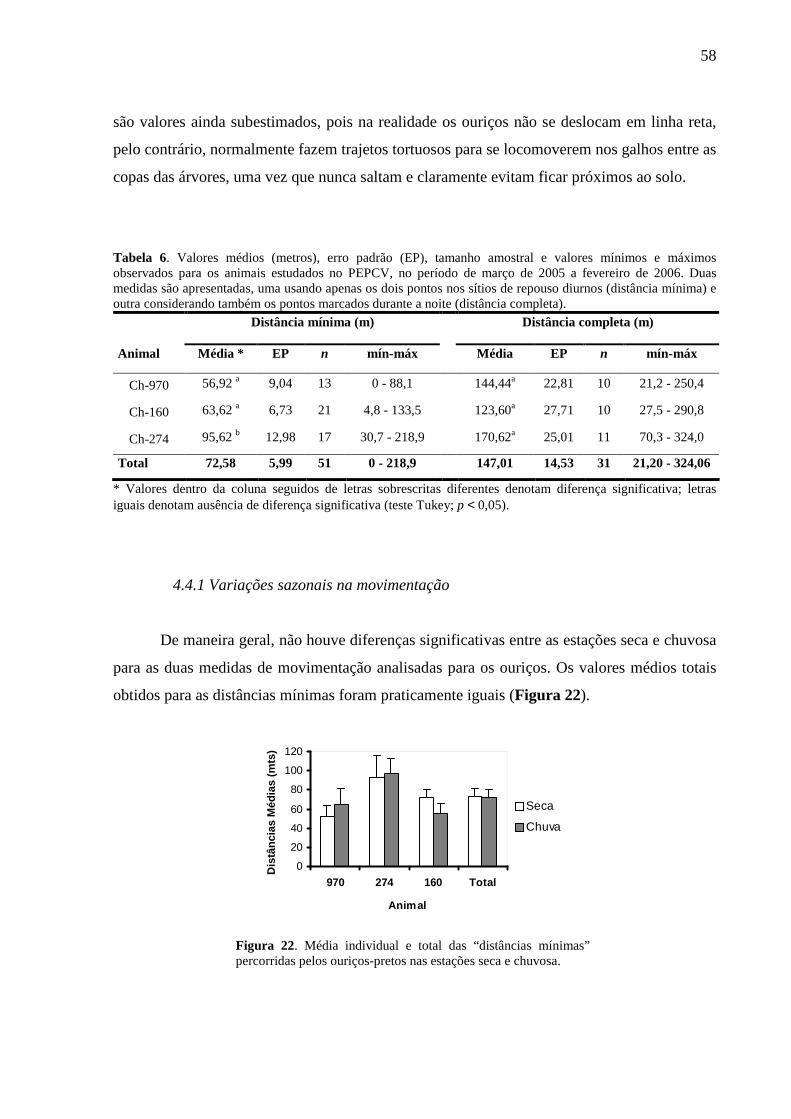
58
são valores ainda subestimados, pois na realidade os ouriços não se deslocam em linha reta,
pelo contrário, normalmente fazem trajetos tortuosos para se locomoverem nos galhos entre as
copas das árvores, uma vez que nunca saltam e claramente evitam ficar próximos ao solo.
Tabela 6. Valores médios (metros), erro padrão (EP), tamanho amostral e valores mínimos e máximos observados para os animais estudados no PEPCV, no período de março de 2005 a fevereiro de 2006. Duas medidas são apresentadas, uma usando apenas os dois pontos nos sítios de repouso diurnos (distância mínima) e outra considerando também os pontos marcados durante a noite (distância completa).
Distância mínima (m) Distância completa (m)
Animal Média * EP n mín-máx Média EP n mín-máx
Ch-970 56,92 a 9,04 13 0 - 88,1 144,44a 22,81 10 21,2 - 250,4
Ch-160 63,62 a 6,73 21 4,8 - 133,5 123,60a 27,71 10 27,5 - 290,8
Ch-274 95,62 b 12,98 17 30,7 - 218,9 170,62a 25,01 11 70,3 - 324,0
Total 72,58 5,99 51 0 - 218,9 147,01 14,53 31 21,20 - 324,06
* Valores dentro da coluna seguidos de letras sobrescritas diferentes denotam diferença significativa; letras iguais denotam ausência de diferença significativa (teste Tukey; p < 0,05).
4.4.1 Variações sazonais na movimentação
De maneira geral, não houve diferenças significativas entre as estações seca e chuvosa
para as duas medidas de movimentação analisadas para os ouriços. Os valores médios totais
obtidos para as distâncias mínimas foram praticamente iguais (Figura 22).
0
20
40
60
80
100
120
970 274 160 Total
Animal
Dis
tânc
ias
Méd
ias
(mts
)
Seca
Chuva
Figura 22. Média individual e total das “distâncias mínimas” percorridas pelos ouriços-pretos nas estações seca e chuvosa.

59
A diferença entre as distâncias completas totais também não foi expressiva, embora
tenha apresentado uma tendência para um maior percurso no período seco, uma constatação
reforçada pelo único indivíduo que variou de maneira marginalmente significativa suas
médias sazonais (Ch-160), percorrendo na seca mais do que o dobro da distância média
observada no período chuvoso (Figura 23).
0
50
100
150
200
250
970 274 160 Total
Animal
Dis
tânc
ias
Méd
ias
(mts
)
Seca
Chuva
p =0,066
Figura 23. Média individual e total das “distâncias completas” percorridas pelos ouriços-pretos nas estações seca e chuvosa. O valor marginalmente significativo do teste-t (0,05<p<0,10) é mostrado para o animal Ch-166; para os demais não houve diferença significativa.
A distância mínima média entre os locais de repouso diurno para Ch-970 na estação
seca foi a menor das três fêmeas, em conformidade com o menor MPC obtido somente com
os sítios diurnos de repouso (Tabela 2), o que possivelmente também está relacionado com a
presença de seu filhote.
4.5 Sítios de repouso diurno
Em 92 registros de localização diurna foram utilizados 55 sítios diferentes para
repouso diurno pelos animais estudados no PEPCV (Tabela 2). A altura média dos locais
exatos onde os animais se encontravam foi de 6,4 m (amplitude: 0,5-17 m), em árvores com
DAP médio de 12,5 cm e altura média de 8 m (amplitude: 1-19 m). Na maioria dos locais
utilizados (85,7%) os animais estavam em galhos de pequeno diâmetro (< 5 cm) e
ligeiramente inclinados (ângulo entre 0° e 30°) em 57,7% dos registros. Os ouriços-pretos
foram encontrados em 65,2% das localizações em locais sombreados, associados a

60
emaranhados de cipós, trepadeiras e folhas secas (chamadas popularmente de “baceiros” ou
“cofeiros”) (Figura 24). Todos os ouriços-pretos repetiram locais utilizados em meses
anteriores, em alguns casos chegando a 7 repetições de um mesmo sítio (Ch-970). Na
realidade, boa parte dos registros anotados em árvores já marcadas anteriormente possuíam
estes emaranhados de vegetação, típicos de ambientes de borda.
Figura 24. Sítio de repouso diurno (Ch-160), mostrando os referidos emaranhados de cipós, folhas secas e trepadeiras.
Sem que uma busca específica fosse feita sistematicamente, foram observados 12
encontros de outros C. subspinosus (não marcados) em sítios de repouso diurnos já utilizados
pelas fêmeas estudadas. Destes, em quatro deles havia outro ouriço-preto utilizando a mesma
árvore no mesmo momento, bem próximos dos animais marcados (em alguns casos quase se
tocando), sem demonstrar nenhum comportamento agressivo aparente. Em três oportunidades
diferentes ocorreram encontros com S. villosus em locais também já utilizados, sendo que em
uma delas foi observado um adulto desta espécie na mesma árvore que a fêmea Ch-274,
separados por poucos centímetros e em um local de repouso utilizado freqüentemente. Em
outras 6 oportunidades encontramos indivíduos de S. villosus em repouso diurno, sozinhos e
em árvores não utilizadas pelos ouriços-pretos marcados, mas próximas ou no interior das
áreas de vida delimitadas pelo MPC.
Como foi exposto anteriormente, em apenas duas árvores de repouso foram também
observados eventos de alimentação, sendo uma da espécie Manilkara subsericea e um

61
individuo de Inga fagifolia. As espécies Protiun icicariba, Fícus sp., Emmotun nitens e T.
guianensis foram as mais utilizadas como sítios de repouso pelos animais. A espécie mais
importante da dieta dos ouriços-pretos no PEPCV (P. glabrata) não foi usada em nenhum
momento como sitio de repouso diurno pelos indivíduos monitorados.
4.6 Utilização de latrinas
Todos os três animais monitorados foram observados utilizando “latrinas” (locais de
defecação recorrente), todas elas situadas em sítios estruturalmente complexos como
aglomerados de bromélias no chão da mata ou em ocos de arvore, todas posicionadas na
periferia de suas áreas de vida. Os animais Ch-970 e Ch-160 foram observados
individualmente fazendo uso de apenas uma latrina e Ch-274 utilizando duas. A fêmea Ch-
970 foi observada utilizando a mesma latrina 6 vezes durante o estudo, em noites diferentes,
em todos os eventos descendo quase ao chão, entrando em uma copa “fechada” (trepadeiras)
de uma árvore caída localizada bem na borda, sempre nas primeiras horas da noite e aí
permanecendo por 13 a 23 minutos. Ch-160 foi observada em apenas uma oportunidade
utilizando sua latrina, esta também desceu quase no chão (30 cm de altura) em um local bem
protegido no meio de um aglomerado de bromélias “gravatás”, permaneceu lá por cerca de 9
minutos. Posteriormente foi confirmada a grande quantidade de fezes no local (Figura 25).
Figura 25. Detalhe da latrina utilizada pela fêmea Ch-160, mostrando o acúmulo de fezes recentes e mais antigas.

62
A fêmea Ch-274 foi registrada utilizando suas latrinas cada uma em uma
oportunidade, uma também estava em um aglomerado de bromélias no chão, no limite entre a
restinga arbórea e as formações abertas de restinga (extremo oeste do MPC; Figura 12) e a
outra se tratava de uma árvore de maior porte já morta, que apresentava um grande oco na sua
extremidade superior, abaixo de uma bromélia.

63
5. DISCUSSÃO
5.1 Captura e monitoramento por rádio-telemetria
Nos momentos de captura dos animais no PEPCV (e na EBSL em Santa Teresa)
alguns comportamentos interessantes foram observados. Os animais permaneceram nos
respectivos locais de repouso, encarando sempre frontalmente quando o seu captor estava
próximo. Uma vez no solo, ao contrário do que acontece com S. villosus que foge
prontamente pelo chão (obs. pess.), os ouriços-pretos permanecem de frente para o “predador”
(pesquisador) quando acuados, assumindo uma postura típica que procura aumentar seu
tamanho corporal, eriçando todos os pêlos e assentado nos membros inferiores (Figura 26),
algumas vezes chegando a balançar o galho em que se encontrava.
Figura 26. Individuo de ouriço-preto durante a captura (já no chão), em postura corporal típica de intimidação.
Também diferentemente dos outros ouriços-cacheiros (S. villosus), que liberam um
forte odor repulsivo e “chocalham” seus espinhos produzindo um som chiado alto e

64
claramente ameaçador, os ouriços-pretos por sua vez batem os dentes rápida e repetidamente,
produzindo também um barulho característico em um “display” também intimidador para
possíveis predadores. As diferenças entre os dois comportamentos característicos dos ouriços-
pretos e demais Erethizontidae neotropicais provavelmente estão relacionadas com a estrutura
e distribuição dos espinhos nos primeiros, que são moles na maior parte do corpo e rijos
somente na cabeça. Ou seja, para esta espécie oferecer o dorso para o predador no caso de
fuga seria oferecer a parte menos protegida do corpo. Por isso quando acuados, os ouriços-
pretos permanecem “de frente”, mostrando os espinhos da cabeça e acentuando a mensagem
com os dentes, os quais, quando chegam a ser utilizados são capazes de causar ferimentos
profundos (Rivelino Galvão, com. pess.).
O campo para estudos utilizando a radio-telemetria no Brasil é enorme e ainda pouco
explorado, apenas na última década esta técnica passou efetivamente a figurar como um dos
métodos à disposição dos pesquisadores brasileiros para a obtenção de importantes
informações sobre a ecologia e comportamento de espécies silvestres (Jacob & Rudran,
2003). De fato, é uma ferramenta particularmente atraente para estudos com animais noturnos
e/ou que não são facilmente localizados em seu ambiente natural, o que normalmente é o caso
para boa parte dos mamíferos neotropicais, incluindo o ouriço-preto.
Embora outros trabalhos com C. prehensilis (Santos Junior, 1998) e com E. dorsatum
(Griesemer et al., 1999) relatem ferimentos no pescoço dos ouriços em detrimento do uso do
colar, nenhum dos indivíduos marcados na EBSL e no PEPCV apresentaram ferimentos,
somente uma moderada queda local de pêlos (espinhos). Contribuíram para isto o tamanho e
peso proporcionalmente pequenos dos transmissores utilizados (veja Métodos) e o tipo de
coleira adotado (corrente de bolas de aço). Entretanto, considerando a pequena extensão das
áreas de vida ocupadas pelos ouriços-pretos, recomenda-se que rádio-transmissores ainda
mais leves (menores) devam ser usados preferencialmente em trabalhos futuros com esta
espécie, uma vez que não são necessários dispositivos com grande alcance do sinal emitido.
Surpreendentemente, os ouriços-pretos se habituaram com relativa facilidade tanto à
presença do rádio-colar quanto à dos pesquisadores (barulho e lanternas) já nas primeiras
noites (meses) de acompanhamento, passando diversas vezes bem próximos dos observadores
e aparentemente permanecendo com todas as suas atividades normais durante todo o período
de estudo. Desta forma, as suas atividades puderam ser anotadas a uma distância que variou
de cerca de 1,5 a 15 metros do observador, permitindo visualização acurada dos indivíduos e
de suas respectivas atividades. A rádio-telemetria foi certamente fundamental para encontrar
os indivíduos, mas uma vez localizados, o próprio comportamento característico da espécie

65
(lentidão de movimentos, a rápida habituação à presença dos observadores e à iluminação
artificial), bem como sua pequena área de vida e a estatura relativamente baixa da mata de
restinga, permitiram um acompanhamento visual detalhado e sistemático da movimentação e
atividade dos animais estudados, possibilitando a marcação exata da sua localização em todos
os registros. Esta pode ser considerada uma situação não muito comum para estudos
ecológicos na natureza feitos com radio-telemetria, onde de maneira geral são obtidas as
localizações apenas através da técnica da triangulação (Jacob & Rudran, 2003), o que limita o
acesso a outras informações biológicas.
5.2 Sobre as estimativas das áreas de vida
O efeito da auto-correlação (dependência estatística entre pontos consecutivos) das
localizações sucessivas em estimativas de área de vida já foi abordado em alguns estudos,
porém os resultados são ainda contraditórios. Segundo Harris et al. (1990) e White & Garrott
(1990), a autocorrelação possui um efeito negativo em algumas análises. Por outro lado, Solla
et al. (1999) afirmam que a autocorrelação é positiva do ponto de vista da obtenção de
informações mais relevantes da própria área de vida e que sua eliminação teria sim um efeito
negativo. Nos casos em que a pesquisa estiver interessada nas distâncias diárias percorridas e
outras medidas de curto prazo, a independência estatística pode torna-se irrelevante (Reynolds
& Laundré, 1990). Segundo Kernohan et al. (2001), mesmo em estimativas de área de vida,
uma amostragem adequada das localizações ao longo do período de estudo, onde estejam
incluídos fatos biologicamente importantes para cada espécie, é muitas vezes mais importante
que a determinação de um intervalo de tempo independente. Em concordância com esta visão,
no presente trabalho a abordagem utilizada privilegiou o registro de locais onde ocorreram
atividades biologicamente importantes para os ouriços, independentemente da possível
existência de autocorrelação entre os pontos, visando coletar a maior quantidade de dados
ecológicos possíveis.
Existem diversas limitações inerentes ao método MPC, pois este fornece apenas linhas
de contorno da área, sendo muito sensível a pontos externos, superestimando a medida da área
ao incorporar partes nunca usadas (White & Garrott, 1990). O MPC ainda desconsidera a
estrutura interna de utilização, assumindo que a área é usada uniformemente, o que raramente
é observado, principalmente em mamíferos. Uma aproximação mais acurada da realidade de
uso do espaço pelos animais poderia ser representada através dos contornos de utilização
fornecidos pelo Kernel 95% (Figuras 11, 12 e 13). Estes, quando sobrepostos aos contornos

66
do MPC confirmam seu aspecto mais conservador, deixando de incluir porções muitas vezes
não utilizadas pelos animais, reduzindo esta que é a principal deficiência do método MPC.
Entretanto, mesmo sendo considerado um dos melhores estimadores de área de vida
disponível atualmente, o Kernel também compartilha falhas com outros estimadores. A
premissa de independência estatística entre localizações consecutivas na amostra, por
exemplo, não considera a própria variável sendo medida, uma vez que animais não se movem
no ambiente ao acaso (Powell, 2000). Procurando satisfazer esta pressuposição dos
estimadores, o caráter temporal da amostra é então desconsiderado, perdendo informações
biológicas importantes (Solla et al., 1999). Isto pode ser exemplificado no presente estudo
comparando as áreas estimadas com todos os pontos com aquelas onde apenas os pontos de
repouso diurnos são usados. Os registros dos sítios de repousos diurnos são considerados
independentes entre si, já que representam os extremos de deslocamento de uma noite (início
e fim), quando comparado aos que incluem os demais pontos anotados ao longo da noite.
Entretanto, quando a área de vida é estimada utilizado apenas estes registros nos repousos
diurnos, ela tem uma extensão inferior àquela na qual pontos não independentes são utilizados
(Figuras 8, 9 e 10). Ou seja, se o critério de independência entre os pontos for usado
indiscriminadamente, pode-se obter uma estimativa robusta do ponto de vista estatístico, mas
pouco acurada do ponto de vista ecológico. Adicionalmente, a maneira como foram coletados
os pontos de localização, nos locais onde os animais realizaram suas principais atividades,
permitiu que os contornos de distribuição de utilização obtidos no método Kernel refletissem
biologicamente pontos considerados particularmente importantes dentro das áreas de vida
estimadas.
5.3 Áreas de vida e uso do habitat
As estimativas feitas através do MPC produziram valores semelhantes aos encontrados
por Giné et al. (2006a) estudando esta mesma espécie no sul da Bahia, onde os ouriços
também ocuparam áreas em torno de 2 hectares. Outros folívoros arborícolas também
apresentam áreas de vida parecidas, como é o caso de preguiças do gênero Bradypus. Cassano
(2006) observou áreas entre 0,44 e 5,73 ha e Chiarello (1998a) entre 2,8 e 5,9 ha para B.
torquatus em estudos realizados na Bahia e no Espírito Santo, respectivamente. Montgomery
& Sunquist (1975) encontraram para B. variegatus valores entre 0,5 e 3,7 ha, muito
semelhantes aos observados com os ouriços-pretos no PEPCV. A pequena extensão destas
áreas está de acordo com o hábito essencialmente folívoro destas espécies, já que folívoros

67
arborícolas tendem a apresentar áreas de vida menores do que outras espécies de mamíferos
de porte semelhante, mas com dieta energeticamente mais rica (ex: frutos) (McNab, 1963). Os
valores estimados para os ouriços-pretos são muito menores do que as áreas de vida do
ouriço-cacheiro (C. prehensilis; média de 24,61 ha, variando de 17,17 a 35,55 ha; Santos
Junior, 1998). O ouriço-cacheiro é um animal relativamente mais ativo e que inclui maiores
proporções de frutos na sua dieta (Charles-Dominique et al., 1981; Santos Junior, 1998). Isto
reflete diferentes estratégias de forrageamento por parte destes dois erethizontídeos,
principalmente no que se refere a comportamentos relacionados com uma economia
energética e que são mais presentes em animais com dieta acentuadamente folívora (MacNab,
1978).
Assim como foi observado na fêmea Ch-970, que possuía um filhote em fase de
amamentação e usou a menor área de vida dentre as três fêmeas estudadas no PEPCV, no
estudo conduzido no sul da Bahia a menor área de vida também foi registrada para uma fêmea
com filhote em amamentação (Gastón Giné, com. pess.). Isto sugere que os animais
restringem sua área de uso de modo a estarem sempre próximos de seus filhotes, uma vez que
estes raramente passam o dia juntos no mesmo sítio diurno. Adicionalmente, é possível que
fêmeas lactantes não se afastem muito de seus filhotes devido a limitações metabólicas, já que
a fase de lactação tem um alto custo energético (Robbins, 1993). Áreas de vida menores para
fêmeas acompanhadas de filhotes já foram observadas em preguiças-de-coleira (B. torquatus;
Chiarello, 1998), sugerindo que este comportamento pode ser comum entre mamíferos
folívoros arborícolas.
Ainda são poucos os trabalhos que utilizaram o método Kernel para estimativas das
áreas de vida de erethizontídeos neotropicais (Giné, 2006a). No entanto, os resultados
encontrados demonstram que o método realmente permite uma melhor compreensão da
estrutura ou do uso interno da área de vida. Os principais centros de atividade encontrados
estão relacionados a um ou mais locais de repouso diurno, próximos de várias árvores de
alimentação. Cassano (2006) utilizando o Kernel com B. torquatus encontrou que esta espécie
também concentra suas atividades em árvores especificas, normalmente relacionadas à
alimentação e que foram freqüentadas em seguidas oportunidades. Tais locais foram referidos
em Montgomery & Sunquist (1975) como “árvores modais”. Moraes (2003), estudando
marsupiais da espécie Micureus demerarae, observou que os abrigos freqüentados por estes
pequenos mamíferos estavam localizados dentro ou próximo às áreas mais utilizadas,
sugerindo que tais abrigos poderiam fornecer proteção contra predadores, ou estar mais
próximos das fontes de recursos alimentares.

68
Mas porque os ouriços-pretos possuem uma área de vida? Stamps (1995) argumenta
que animais possuem áreas de vida porque os benefícios de permanecer em locais definidos,
onde é possível se movimentar através de rotas conhecidas e assim também evitar ou escapar
de predadores são simplesmente maiores do que os custos. Para animais como os ouriços-
pretos, que possuem dieta energeticamente pobre e que são presas potenciais de vários
predadores de médio e grande porte, a hipótese de Stamps possivelmente se aplica. No
entanto, como lembrado por Powell (2000), as razões para que animais mantenham área de
vida certamente são ainda mais amplas, como as relacionadas com a distribuição dos recursos
mais importantes e com sua reprodução. Todos os ouriços-pretos monitorados incluíram parte
de sua área de vida acima de porções periodicamente inundadas das florestas de restinga,
concentrando suas atividades próximas ou acima destas áreas, o que está provavelmente
relacionado com a densidade das duas espécies mais importantes da sua dieta (Pera glabrata
e Tapirira guianensis), uma vez que estas duas espécies estão normalmente associadas a solos
encharcados, próximas ao lençol freático ou a corpos d’água (Oberdan Pereira, com. pess.).
Embora fosse esperado, quanto ao uso do habitat é importante mencionar que em
nenhum momento os ouriços-pretos foram vistos em formações que não a formação florestal
de restinga, chegando diversas vezes até árvores literalmente na borda com o brejo herbáceo e
com a restinga aberta. A espécie pode, desta forma, ser considerada como arborícola e de
hábitos predominantemente florestais. Mesmo nos eventos de defecação os ouriços evitam
tocar o chão, ficando extremamente alertas e escolhendo locais protegidos para as suas
latrinas. Somente em uma ocasião uma das fêmeas monitoradas (Ch-274) foi observada
chegando próxima ao solo, saindo de uma árvore de repouso diurno exatamente na borda para
o interior da mata, onde não havia conexão pelas copas. Outros trabalhos também
confirmaram o hábito arborícola quase restrito (Chiarello et al., 1997; Giné et al., 2005c; Giné
et al., 2006a), no entanto, deslocamentos pelo chão foram observados nestes trabalhos e
existem alguns relatos de animais observados cruzando estradas (Rivelino Galvão, com.
pess.). Provavelmente os ouriços-pretos utilizam o chão somente quando o seu deslocamento
pelas copas e galhos não for possível.
Foram obtidos diversos registros de outros indivíduos de C. subspinosus e de S.
villosus, utilizando sítios diurnos já marcados anteriormente, e outras vezes sozinhos em
árvores localizadas no interior das áreas de vida estimadas. O compartilhamento de sítios de
repouso e provavelmente das próprias “trilhas de odor” (ver adiante) existentes entre as
árvores de interesse para sua alimentação, sugerem que possivelmente ocorrem extensas
sobreposições de áreas de vida entre os indivíduos de ouriço-preto, talvez se tratando de

69
indivíduos proximamente relacionados como irmãos, ex-filhotes e parceiros sexuais. Esta
hipótese é corroborada pelas observações feitas com indivíduos de C. subspinosus em
cativeiro (Gastón Giné, com. pess.), onde animais adultos se mostraram agressivos somente
na presença de outros indivíduos não relacionados, o que não ocorria com relação a seus
filhotes desmamados ou jovens. Roberts et al. (1985) também observou em C. prehensilis
mantidos em cativeiro que nenhum dos pais demonstrou agressividade direcionada a seus
filhotes desmamados, tolerando sua presença inclusive na presença do próximo infante.
Quanto à área de vida acumulada ao longo do tempo e o número de localizações, Giné
et al., (2006a) observou que já nas primeiras noites de observação dos ouriços-pretos foi
possível determinar sua área de vida, sem aumentos posteriores relevantes, devido
principalmente à capacidade dos animais de percorrerem boa parte de suas pequenas áreas em
uma noite de atividade (ver adiante). Os resultados encontrados com os ouriços-pretos no
PEPCV foram semelhantes ao encontrado por Santos Junior (1998) com C. prehensilis, onde
foram observados também dois patamares, obtidos próximos a 20 e 50 localizações, embora
estes tenham sido valores obtidos com animais translocados. Possivelmente, exceto para as
áreas menores registradas, um número superior a 5 noites ou mais de 50 localizações seriam
suficientes para uma estimativa relativamente estável das áreas ocupadas por indivíduos desta
espécie.
Segundo Powell (2000), a fidelidade territorial é necessária para a manutenção de uma
área de vida e pode ser usada para indicar se um animal ali se estabeleceu ou não. Embora não
tenha sido testada estatísticamente, os três animais estudados apresentaram o que pode ser
considerado como fidelidade territorial, pois permaneceram praticamente na mesma porção da
mata durante todo o ano de monitoramento, o que pode ser exemplificado pelos dois
contornos obtidos para as estações seca e chuvosa (Figuras 15, 16 e 17). Giné et al. (2006a)
também aponta para esta utilização constante de uma área de vida bem definida para os
ouriços-pretos.
5.4 Padrão de atividade e movimentação
O ouriço-preto é uma espécie noturna, assim como outros roedores Erethizontidae
como C. prehensilis (Montgomery & Lubin, 1978; Charles-Dominique et al., 1981; Santos
Junior, 1998) e Echimyidae arborícolas como o rato-do-bambú (Kannabateomys amblyonyx)
(Silva, 1993; Silva, 2005) e o rato-do-cacau (Callistomys pictus) (Moura & Fonseca, 2005).
Estudos realizados com C. subspinosus encontraram dois picos de atividade, concentrados no

70
início e final da noite, intercalados com um período de repouso noturno mais prolongado
(Chiarello et al., 1997; Giné et al., 2005a). Outros roedores também folívoros como K.
amblyonyx, apresenta este mesmo padrão geral de atividade (Silva, 1993; Santos Junior,
1998), normalmente com o primeiro pico de atividade sendo mais expressivo que o segundo.
De maneira geral, os ouriços-pretos estudados no PEPCV passaram boa parte do
tempo parados ou em repouso. O mesmo foi registrado por Chiarello (1997) e Giné et al.
(2005a) observando os animais ao longo de toda a noite, onde a categoria repouso foi a mais
freqüente (superior a 60 %), seguida também pela movimentação e alimentação. Portanto,
além de permanecerem em repouso durante todo o período claro do dia, os ouriços ainda
ficam boa parte da noite parados, mesmo no seu período de maior atividade (primeiras horas
da noite). Esta baixa atividade exibida pela espécie está de acordo com o esperado para
mamíferos com dietas altamente folívoras e com baixas taxas de metabolismo (McNab, 1963;
Mendes, 1989; Queiroz, 1995; Chiarello, 1998a; Cassano, 2006). Através dos aspectos
hematológicos encontrados para as amostras de sangue coletadas no PEPCV (Nelson Curi,
dados não publicados), foi confirmado o relativo baixo metabolismo dos ouriços-pretos em
comparação com outros mamíferos de porte semelhante.
A distância completa percorrida por noite observada no presente trabalho (média de
147,0 m) está próxima do valor registrado para a mesma espécie no sul da Bahia (média de
201, 4 m; Giné et al., 2006a). Os valores ligeiramente superiores observados no sul da Bahia
talvez se devam ao fato de que lá os animais foram monitorados ao longo de todo o período
da noite, onde foram anotadas suas posições a cada 10 minutos. Quanto às distâncias
mínimas, Montgomery & Lubin (1978) relatam para C. prehensilis valores de deslocamento
noturno entre pontos de repouso diurno consecutivos que chegam a 700 metros, com uma
média de 254 metros, bem superiores aos valores encontrados no PEPCV para os C.
subspinosus (média de 72 metros; Tabela 6).
De maneira geral, as distâncias calculadas podem ser consideradas relativamente
pequenas para um animal deste porte e reforçam o observado com relação às áreas de vida,
indicando que C. subspinosus é uma espécie pouco ativa, adotando possivelmente uma
estratégia de forrageamento que minimiza o seu gasto energético. No entanto, em comparação
com outros folívoros arborícolas especialistas como as preguiças-de-coleira Bradypus
torquatus (Chiarello, 1998a; Cassano, 2006), os ouriços-pretos percorrem distâncias muito
maiores em um período de 24 horas, podendo percorrer toda a extensão de sua área de vida
em apenas uma noite de atividade.

71
Ao contrário do que foi relatado em Nowak & Paradiso (1983), que afirmam que
Chaetomys é um animal que se move rapidamente e realiza saltos, foi observado que ao se
locomoverem os ouriços-pretos são extremamente cuidadosos, não saltam, andam
vagarosamente, pausadamente e raramente fazem barulho, assim como observado nos estudos
de Chiarello et al., (1997). As fêmeas estudadas no PEPCV passaram repetidas vezes pelos
mesmos galhos de uma árvore de interesse para outra, sempre posicionando sua cauda
preênsil de forma a atuar como segurança ou mesmo um quinto membro em algumas
situações. Enquanto se movimentam, o que fazem sempre utilizando bastante suas longas
vibrissas, os animais freqüentemente cheiram os galhos a sua frente antes de continuar, o que
indica um provável sistema de marcação de “trilhas de odor” (durante a captura e manuseio é
evidente o cheiro forte e característico dos ouriços). Estas trilhas são possivelmente
compartilhadas com outros indivíduos da mesma espécie e até mesmo outras espécies de
ouriços como o sintópico S. villosus.
Outros indivíduos destas espécies foram encontrados durante o estudo dentro das três
áreas de vida dos animais marcados, bem como utilizando sítios diurnos já marcados
anteriormente. Em diversas oportunidades observamos outros ouriços-pretos adultos se
aproximando e até mesmo seguindo as fêmeas marcadas pelos mesmos galhos que elas
estavam utilizando. (Gastón Giné, com. pess.) também sugere a provável marcação ativa dos
galhos utilizados formando uma malha de trajetos reconhecidos também por outros ouriços.
Reforçando esta hipótese, Martinez et al. (2006) relatou a presença de duas estruturas
glandulares na base do ânus, tanto em fêmeas quanto em machos de Chaetomys,
possivelmente relacionadas com a produção de odores utilizados na marcação destas trilhas.
Adicionalmente, Felicioli et al. (1993) encontrou em Histrix cristata evidências que apontam
para uma impressionante função discriminativa no olfato de porcos-espinhos. E Sackett
(1913) relata que E. dorsatum apresentou habilidade extraordinária para memorizar labirintos
e ainda se lembrar deles por centenas de dias.
5.5 Dieta dos ouriços-pretos
As árvores utilizadas para repouso diurno estavam distribuídas em toda a área de vida
dos ouriços-pretos, invariavelmente próximas de árvores de alimentação. De forma geral, os
resultados indicam claramente que C. subspinosus apresenta uma dieta especializada e
estritamente folívora, com um reduzido número de espécies vegetais, muito inferior ao
registrado para outros mamíferos arborícolas herbívoros e primariamente folívoros como as

72
preguiças do gênero Bradypus (Chiarello, 1998b; Montgomery & Sunquist, 1975) e barbados
como Alouatta guariba (Chiarello, 1994; Mendes, 1989). Em nenhum momento foram
observados itens animais na sua dieta, sendo que as outras partes vegetais consumidas (brotos,
inflorescências e frutos) responderam por menos de 3 % da dieta ao longo do ano. A fêmea
Ch-970 se alimentou praticamente apenas de P. glabrata e T. guianensis ao longo de todo o
ano, representando cerca de 90 % dos registros obtidos. Aparentemente, a distribuição local
destas duas espécies (especialmente P. glabrata) são determinantes da localização das áreas
de vida ocupadas pelos animais, próximas ou acima das porções periodicamente alagadas das
florestas de restinga, como mencionado anteriormente.
Dados sobre dieta de ouriços-cacheiros neotropicais são ainda escassos. A análise do
conteúdo estomacal de dois C. subspinosus capturados no município de Salvador, Bahia,
revelou presença de flores e frutos, incluindo fibras de frutos da palmeira dendê (Elaeais
guineensis) (Kuniy et al., 2005). Estes resultados porém se referem a uma amostra muito
pequena (dois indivíduos) e de espécimes capturados em uma região muito alterada
antrópicamente. Um estudo sobre Coendou prehensilis reporta uma dieta concentrada em
cerca de apenas 3 espécies (Santos Junior, 1998), lembrando, entretanto que os indivíduos
estudados eram translocados e duas das três espécies mais consumidas eram plantas exóticas
como a manga (Mangifera indica) e o milho (Zea mays).
Os ouriços-pretos mostraram uma clara preferência por folhas novas, embora tenham
também consumido folhas maduras em menor quantidade. Esta preferência possivelmente
está relacionada com a maior digestibilidade e valor nutricional encontrado nestas quando
comparadas com as folhas maduras (Milton, 1980; Montgomery & Sunquist, 1978). Da
mesma forma, uma preferência acentuada por folhas novas foi demonstrada para preguiças
(Chiarello, 1998b) e primatas como os barbados (Chiarello, 1994) e muriquís (Nogueira,
1996).
As espécies consumidas por Chaetomys representam uma parcela mínima das espécies
vegetais potencialmente disponíveis para estes animais nas florestas de restinga, uma vez que
Assis et al. (2004a) encontrou 172 espécies pertencentes a 54 famílias amostrando cerca de
apenas 4 hectares. As duas famílias mais importantes na dieta dos animais de estudo,
Euphorbiaceae e Anacardiaceae, ocuparam a 33a e a 36a posição, respectivamente, em termos
de valor de importância em um estudo florístico realizado no PEPCV (Assis et al., 2004b).
Estes dois trabalhos, porém, inventariaram as espécies presentes em uma área amostral
relativamente restrita do parque. Portanto, generalizações a partir do mesmo devem ser feitas
tendo em vista esta limitação.

73
Assim como para outros folívoros arborícolas, provavelmente esta especialização
alimentar é um resultado do desenvolvimento de adaptações fisiológicas que permitem tolerar
uma gama pequena de compostos secundários (Freeland, 1991). No caso especifico dos
ouriços-pretos, estas adaptações incluem seu baixo metabolismo e atividade, que permitem
sua sobrevivência mesmo com itens pobres energeticamente, assim como uma anatomia
interna que aponta para um fermentador cecal diferenciado, funcional e desenvolvido
(Martinez et al., 2006). Adicionalmente, através destas adaptações, os folívoros podem
utilizar recursos não disponíveis para animais generalistas, diminuindo ou até mesmo
evitando competição inter-específica (Moss, 1991).
Com relação à variação sazonal dos recursos alimentares, de maneira geral, os dados
indicam pouca variação nos parâmetros estudados. Apenas a taxa de visitação foi ligeiramente
superior na estação chuvosa. Este aumento parece estar relacionado a um ligeiro aumento do
número de árvores usadas na alimentação nesta estação. Entretanto esta variação não implicou
em aumentos significativos nas tanto na forma como na extensão das áreas de vida entre as
estações. Estes resultados sugerem que a floresta de Restinga estudada apresentou pequena
variação sazonal na oferta dos itens mais consumidos pelo ouriço-preto (folhas de árvores).
Infelizmente a área de estudo (PEPCV) ainda não foi objeto de estudo desta natureza e
portanto não há como analisar mais detalhadamente as causas da pequena variação encontrada
na dinâmica de uso da área entre as estações.
5.6 Sítios de repouso diurno
Assim como outros mamíferos, os Erethizontidae normalmente não dormem em
qualquer lugar da sua área de vida, pelo contrario, são bastante seletivos quanto às
características estruturais do seu local de repouso, como foi demonstrado em outros estudos
com o ouriço-cacheiro (Roberts et al., 1985; Santos Junior, 1998) e o próprio ouriço-preto
(Barata et al., 2005; Giné et al., 2006b). Sem nenhuma exceção, todos os sítios de repouso se
localizavam acima do solo em copas de árvores, um comportamento também apresentado por
outros erethizontídeos neotropicais como C. prehensilis (Santos Junior, 1998).
De maneira geral, os sítios de repouso ocupados pelos ouriços contam quase sempre
com a presença dos emaranhados de cipós e trepadeiras, principalmente os que foram
repetidamente utilizados, sugerindo certa preferência e fidelidade por estes locais para seu
repouso diurno. Giné et al. (2006b) também observou o uso dos sítios diurnos feito de
maneira não uniforme pelos ouriços-pretos, com os locais mais sombreados reconhecidos

74
como “baceiros” sendo igualmente os mais freqüentados. Este comportamento pode estar
relacionado a uma estratégia para evitar ou dificultar o acesso de predadores, além de proteger
os ouriços de fatores climáticos como o sol forte e chuvas. Certamente, o uso destes
emaranhados dificulta sobremaneira o avistamento da espécie, principalmente em áreas de
florestas com porte mais alto, sendo possivelmente um dos motivos pelos quais são animais
dificilmente registrados em inventários faunísticos.
Como os ouriços-pretos foram observados em raras oportunidades se alimentando em
árvores utilizadas também para repouso diurno, pode-se concluir que a escolha destes sítios
parece estar mais relacionada à estrutura física destes locais do que ao seu papel como fonte
alimentar. Porém, podem estar espacialmente relacionadas com a distribuição destas últimas.
Nos trabalhos realizados inicialmente na Estação Biológica de Santa Lúcia, a fêmea Ch-EBSL
foi localizada apenas 12 vezes (somente em sítios de repouso diurno), sendo 7 destes registros
anotados para o mesmo local, um grande “oco de árvore” situado à cerca de 20 metros de
altura. Um comportamento também observado nos ouriços-pretos estudados em mata
ombrófila (Gastón Giné, com. pess.). Possivelmente, os animais estudados no PEPCV não
foram observados dentro de ocos devido à quase ausência destes locais nas áreas de floresta
de restinga ocupadas pelos indivíduos.
5.7 Utilização de latrinas
O comportamento de descer ao chão para defecar já havia sido registrado por Chiarello et al.
(1997) com o próprio ouriço-preto e foi também observado em C. prehensilis (Roberts et al.,
1985; Santos Junior, 1998). Outros folívoros arborícolas como as preguiças dos gêneros
Bradypus e Choloepus chegam também até o solo para depositar suas fezes (Montgomery &
Sunquist, 1975). Pela quantidade de fezes encontradas, mesmo com poucos registros para Ch-
160 e Ch-274, fica claro que são locais utilizados freqüentemente pelos ouriços-pretos
marcados. Observou-se uma preferência estrutural por locais protegidos por bromélias ou
outra vegetação fechada, uma estratégia possivelmente relacionada à defesa contra
predadores, uma vez que a atividade de defecação é um dos poucos momentos onde o animal
se expõe para predadores terrestres. Latrinas únicas foram também observadas no sul da
Bahia, sendo utilizadas uma vez a cada noite, porem situadas em locais mais altos como em
palmeiras e ocos de troncos (Gastón Giné, com. pess.). Importante mencionar que são quase
ausentes palmeiras de maior porte no PEPCV, bem como árvores com ocos grandes o
suficiente para os animais entrarem. Deste modo, a escolha diferenciada dos tipos de latrinas

75
em diferentes localidades pode estar relacionada à estrutura da vegetação e a oferta e
disponibilidade local de tais micro-ambientes.
5.8 Observações sobre cuidados parentais
Devido ao tamanho reduzido da área de vida para Ch-970, seu o filhote estava sempre
por perto do local do repouso diurno, sendo que algumas vezes foi encontrado em abrigos já
marcados anteriormente. Pelas informações obtidas, um único filhote foi amamentado por no
mínimo 5 meses (maio a setembro). Em maio e junho a fêmea foi observada indo até a árvore
onde se encontrava o filhote e o amamentando por cerca de 20 minutos. Em julho, após
mamar por alguns minutos, o filhote foi observado mastigando poucas folhas de P. glabrata
ao lado da mãe, possivelmente aprendendo de quais espécies deve se alimentar. A partir de
setembro este estava muito ativo, se movendo com mais velocidade até onde a fêmea se
encontrava. Em outubro observamos que os mamilos da mãe não estavam mais aparentes e
que o filhote se aproximou do seu ventre, mas esta ativamente impediu que este tivesse acesso
às mamas. A partir de novembro não foi observado mais a aproximação do filhote.
De maneira geral, mesmo sendo observados poucos eventos, o filhote saiu de um
período inicial onde ficava passivo e a espera da mãe, até um período em que este ia
ativamente de encontro à mesma, terminando com sua independência. Uma seqüência
semelhante à relatada por Roberts et al. (1985) para C. prehensilis observados em cativeiro,
que indicam três fases principais de desenvolvimento dos filhotes: um período de total
dependência nutricional da mãe; um período de nutrição misturada, onde o filhote ainda
depende do leite materno, mas já consome sólidos; e um período de independência
nutricional. Durante os eventos de amamentação, enquanto o filhote se encontrava próximo à
mãe, esta conduziu uma limpeza cuidadosa do mesmo, utilizando tanto as garras anteriores
quanto os dentes.
5.9 Implicações conservacionistas
Como ficou demonstrado, o uso da rádio-telemetria para o estudo dos ouriços-pretos
foi perfeitamente aplicável, sendo que estudos futuros podem ser realizados sem problemas,
principalmente se forem seguidas às recomendações feitas a respeito do peso e forma dos
dispositivos utilizados. O tipo característico dos locais escolhidos pelos animais para repouso
diurno, em locais sombreados, com a presença de folhas secas e trepadeiras, além da
coloração dos próprios ouriços-pretos, pode ser vista como um dos principais fatores para que

76
esta espécie tenha até o momento tão poucos registros e espécimes coletados. Por outro lado,
esta utilização de locais conspícuos, bem como o comportamento de defecar em latrinas,
podem auxiliar no encontro dos indivíduos na natureza. Adicionalmente, a participação de
pessoas que conheçam o local de estudo e principalmente que saibam encontrar os animais é
fundamental para se trabalhar com os ouriços, uma vez que devido ao próprio habito dos
animais, estes são dificilmente capturados utilizando armadilhas.
As formações florestais de restinga presentes no PEPCV são reconhecidas atualmente
como “áreas prioritárias para a conservação da Mata Atlântica” (MMA, 2000). Com base em
dados de entrevistas feitas ao longo de toda a distribuição tida como original para esta
espécie, as evidencias indicam que o ouriço-preto é mais abundante nas florestas de restinga
dos estados do Sergipe, Bahia e Espírito Santo do que nas florestas ombrófilas densas e semi-
decíduas do interior (Gastón Giné, com. pess.). Demonstrando a importância desta formação
florestal presente nas restingas para a conservação da espécie, podendo ser visto também
como um alerta, uma vez que nestes estados as restingas são ambientes que sofrem
historicamente uma grande pressão de exploração humana, principalmente por parte do setor
imobiliário e plantações de eucalipto, devido a sua localização privilegiada em baixadas
litorâneas e a relativa facilidade de ocupação em comparação, por exemplo, com os ambientes
de mangue. Especificamente em áreas de vegetação de restinga, quase não existem Unidades
de Conservação e quando estão presentes são relativamente pequenas para pretensões
conservacionistas. Desta forma, como será recomendado no plano de manejo da espécie, a
criação de novas UC’s em áreas de restinga, bem como assegurar a necessidade de avaliações
mais eficientes sobre a presença desta espécie em locais sujeitos ao licenciamento ambiental,
seriam ações que incrementariam sobremaneira os esforços de conservação do ouriço-preto a
nível nacional.
Ainda não existem trabalhos a respeito do tamanho das populações remanescentes da
espécie, mas aparentemente estas se encontram em processo de declínio (Santos et al., 1987;
Oliver & Santos, 1991; Faria & Giné, 2005). O único trabalho que aborda aspectos a respeito
da situação genética da espécie concluiu que embora a espécie ainda esteja espacialmente
distribuída na maior parte da sua área original, o histórico de exploração e fragmentação das
populações em remanescentes cada vez menores e isolados acarretou em perdas genéticas
detectáveis e significativas, com um índice considerado elevado de endogamia (Gonçalves,
2005; Gonsalves et al. 2005). Um quadro preocupante, pois além dos possíveis problemas
adicionais devido a continua depressão endogâmica que provavelmente aparecerá nas
próximas gerações, sabe-se que populações que se encontram isoladas e com poucos

77
indivíduos estão mais vulneráveis a eventos que produzem variação demográfica (Shafer,
1981). Ou seja, à medida que as populações de uma espécie diminuem, aumenta a
probabilidade de extinção da mesma. Devido a esta situação geral de crescente ameaça esta
espécie está atualmente listada como ameaçada de extinção (categoria “Vulnerável”) na Lista
Nacional das Espécies da Fauna Brasileira Ameaçadas de Extinção (MMA, 2006) e na Lista
Vermelha da IUCN (IUCN, 2004).
Pelos dados coletados no PEPCV, a dieta seletiva dos animais, porém concentrada em
espécies consideradas comuns e amplamente distribuídas, bem como a pequena área de vida
ocupada, sugerem que a espécie não é extremamente exigente em termos de habitat. Desta
forma, as principais causas para este aparente declínio populacional seriam principalmente a
perda e alteração do seu hábitat natural - uma vez que se trata de uma espécie restrita a uma
pequena e já completamente alterada porção da Mata Atlântica - juntamente com os fatores
genéticos.
Em conjunto com estes fatores, a caça é atualmente outra pressão que afeta as
populações de ouriço-preto, sendo uma atividade reconhecidamente ilegal e praticada em
ampla escala pelas populações rurais ao longo da sua área de distribuição (Faria & Giné,
2005). Estabelecer com que intensidade esta prática está afetando as populações, bem como a
simples aplicação da Lei de Crimes Ambientais seriam maneiras de diminuir esta pressão
negativa.
Como o ouriço-preto não havia sido alvo de estudos de longo prazo anteriormente a
realização deste trabalho, diversas informações a respeito da sua biologia ainda não foram
investigadas, como por exemplo, a sua densidade na natureza em diferentes formações
florestais, a estrutura social das populações e os demais aspectos básicos de sua reprodução.
Obter estas informações seria importante para estimar o tamanho das populações
remanescentes e a possível necessidade ou não de interliga-las através de fluxo gênico (com o
uso de translocações, por exemplo). Adicionalmente, seria muito interessante realizar uma
comparação entre os ouriços-pretos e os ouriços-cacheiros-amarelos (S. villosus). Esta espécie
filogeneticamente próxima e de ocorrência sintópica aparentemente ocorre em densidades
locais sempre superiores aos primeiros. Investigar o grau de sobreposição dos nichos
ocupados pelas duas espécies e como se desenvolve a provável competição interespecífica
seria um dos meios para avaliar as causas desta disparidade demográfica, procurando assim
compreender melhor o aparente declínio das populações de ouriço-preto.

78
Chaetomys subspinosus é uma espécie ainda pouco familiar para a população de uma
forma geral e até no meio científico, mesmo entre mastozoólogos. No entanto, devido a suas
características físicas e seu comportamento inofensivo, pode ser considerada uma espécie
carismática e que tem potencial para ser utilizada como uma espécie símbolo ou “bandeira”
para a Mata Atlântica, aparecendo como um dos importantes representantes deste ecossistema
ameaçado e de imensa biodiversidade.

79
6 REFERÊNCIAS
Aguiar, A. P.; Chiarello, A. G.; Mendes, S. L. & Matos, E. N. 2003. The Central and Serra do Mar Corridors in the Brazilian Atlantic Forest. In: The Atlantic Forest of South America, biodiversity status, threats, and outlook. (eds) Garlindo-Leal, C. & Câmara, I. G. Island Press. 118-132. Altmann, J. 1974. Observational study of behavior: sampling methods. Behavior. n. 49: p. 227-267.
Araújo, D. S. D. & Henriques, R. P. B. 1984. Análise florística das restingas do estado do Rio de Janeiro. In: Restingas: Origem, estrutura e processos (L.D. Lacerda, D.S.D. Araújo, R. Cerqueira & B. Turcq, orgs.). Universidade Federal Fluminense / CEUFF, Niterói, p.159-193.
Araújo, D.S.D. & Lacerda, L.D. 1987. A natureza das restingas. Ciência Hoje n 6 (33): 42-48.
Assis, A. M.; Thomaz, L. D. & Pereira, O. J. 2004a. Florística de um Floresta de Restinga no Município de Guaraparí, Espírito Santo, Brasil. Acta Bot. Bras. 18 (1): 191-201.
Assis, A. M.; Pereira, O. J. & Tomaz, L. D. 2004b. Fitossociologia de uma floresta de Restinga no Parque Estadual Paulo César Vinha, Setiba, município de Guarapari (ES). Revista Brasil. Bot., V.27, n.2, p.349-361.
Ávila-Pires, F. D. 1967. The type-locality of “Chaetomys subspinosus” (Olfers, 1818) (Rodentia, Caviomorpha). Museu Nacional, Rio de Janeiro. Rev. Brasil. Biol. 27 (2) : p. 177-179.
Barata, R. S. L.; Oliveira, P. A. & Chiarello, A. G. 2005. Abrigos diurnos utilizados pelo ouriço-preto, Chaetomys subspinosus (Rodentia, Erethizontidae), no Parque Estadual Paulo César Vinha, Guaraparí, ES. In: Livro de Resumos do III Congresso Brasileiro de Mastozoologia. Vitória: UFES/Sociedade Brasileira de Mastozoologia.
Bergallo, H. G. 1990. Fatores Determinantes do Tamanho da Área de Vida em Mamíferos. Ciência e Cultura, n. 42 (12): 1067-1072.
Bonvicino, C. R.; Almeida, F. C. & Cerqueira, R. 2000. The karyotype of Sphiggurus villosus (Rodentia: Eretizontidae) from Brazil. Stud. Neotrop. Fauna & Environm 35: 81-83.
Bonvicino, C. R.; Penna-Firme, V.; Braggio, E. 2002. : Molecular and karyologic evidence of the taxonomic status of Coendou and Sphiggurus (Rodentia: Hystricognathi). J. Mamm. 83 (4): 1071-1076.
Burt, W. H. 1943. Territoriality and home range concepts as applied to mammals. J. Mamm. 24: 346-352.
Cabrera, A. 1961. Catálogo de los mamíferos de América del Sur. II. Rev. Museo Argentino Cienc. Nat. Bernardino Rivadavia, Zool, 4 (2): 309-732.

80
Carvalho, G. 2000. Substitution of the deciduous premolar in Chaetomys subspinosus (Olfers, 1818) (Hystricognathi, Rodentia) and its taxonomic implications. International Journal of Mammalian Biology. 65: 187-190.
Cassano, C. R. 2006. Ecologia e conservação da preguiça-de-coleira (Bradypus torquatus Illiger, 1811) no sul da Bahia. Dissertação de Mestrado. Universidade estadual de Santa Cruz, Ilhéus, BA. 93pp.
Cerqueira, R. 2000. Biogeografia das restingas. In: Esteves, FA, Lacerda, LD (eds). Ecologia de restingas e lagoas costeiras. NUPEM/UFRJ, Rio de Janeiro. p. 65-75.
Charles-Dominique, P.; Atramentowicz, M.; Charles-Dominique, G. H.; Hladick, A.; Hladick, C. M. & Prévost, M. F. 1981. Les mammiferes frugivores arboricoles nocturnes d’une forêt guyanaise: inter-relations plantes-animaux. Revue d'Écologie (Terre et Vie). 35: 341-435.
Chiarello, A.G. 1994. Diet of the brown Houler Monkey Alouatta fusca in a Semi-deciduos Forest Fragment of Southeastern Brazil. Primates, 35 (1): 25-34.
Chiarello, A.G. 1998a. Activity budgets and ranging patterns of the Atlantic forest maned sloth Bradypus torquatus (Xenarthra: Bradypodidae). J. Zool. Lond. 246, 1-10.
Chiarello, A. G. 1998b. Diet of the Atlantic forest maned sloth Bradypus torquatus (Xenarthra: Bradypodidae). J. Zool. Lond. 246, 11-19.
Chiarello, A. G.; Passamani, M. & Zortéa, M. 1997. Field observations on the thin-spined porcupine, Chaetomys subspinosus (Rodentia; Echimyidae). Mammalia, t. 61, n.1: p. 29-36.
Chiarello, A. G.; Aguiar, L. M. S.; Cerqueira, R.; Melo, F. R.; Rodrigues, F. H. G. & Silva, V. M. F. 2008. Mamíferos Ameaçados de Extinção no Brasil. In: Machado, A. M. B.; Drummond, G. M. & Paglia, A. (Ed.). 2008. Livro Vermelho da Fauna Brasileira Ameaçada de Extinção. MMA – Ministério do Meio Ambiente; Belo Horizonte MG: Fundação Biodiversitas. 2v. 1420 p.
Cochran, W.W. 1980. Wildlife telemetry. In: Wildlife Management Techniques Manual, 4th edition (S.D. Schemnitz, ed.), Wildlife Society, Washington. Pp. 507-520.
Coimbra-Filho, A. F. 1972. Mamíferos ameaçados de extinção no Brasil. In: Espécies da Fauna Brasileira Ameaçadas de Extinção. Ed. Academia Brasileira de Ciências, Rio de Janeiro.
Concepcion, J. L. & Molinari, J. 1991. Sphiggurus vestitus pruinosus (Mammalia, Rodentia, Erethizontidae): The Karyotype and its Phylogenetic Implications, Descriptive Notes. Studies on Neotropical Fauna and Environment. Vol. 26, n 4. p. 237-241.
Costa, C. M. R., Leite, Y. R., Fonseca, G. A. B. & Fonseca, M. T. 2000. Biogeography of South Americam Forest Mammals: Endemism and Diversity in the Atlantic Forest. Biotropica 32 (4b): 872-881.
Costa, L. P. ; Leite, Y. R. ; Mendes, S. L. & Ditchfield, A. D. 2005. Mammal Conservation in Brazil. Conservation Biology, Vol. 19. n. 3: p. 672-679.

81
Cullen Jr. L. & Valladares-Pádua. 1997. Métodos para estudos de ecologia, manejo e conservação de primatas na natureza. In: Manejo e conservação de vida silvestre no Brasil. (orgs) Valladares-Padua, C. & Bodmer, R. E. Cap. 15: p. 239-269.
Dodge, W. E. & Barnes, V. G. 1975. Movements, home range, and control of porcupines in western Washington (Wildlife Leaflet No. 507). U. S. Department of Interior, Fish and Wildlife Service.
Eisenberg, J.F. & Redford, K.H. 1999. Mammals of the Neotropics. The Central Neotropics, Vol. 3, Ecuador, Peru, Bolivia, Brazil. The University of Chicago Press, Chicago.
Emmons, L.H. & Feer, F. 1997. Neotropical Rainfores Mammals. A Field Guide, Second Edition. The University of Chicago Press, Chicago.
Fabris, L. C. & O. Cesar, 1996. - Estudos florísticos em uma mata litorânea no sul do Estado do Espírito Santo, Brasil. Bol. Mus. iol. Mello Leitão (N. Sér.), 5: 15-46.
Faria, D. & Giné, G. 2005. Distribuição atual do ouriço-preto (Chaetomys subspinosus, Olfers 18180 nos estados de Sergipe e Bahia, Brasil. In: Livro de Resumos do III Congresso Brasileiro de Mastozoologia. Vitória: UFES/Sociedade Brasileira de Mastozoologia.
Felicioli, A.; Ganni, M.; Garibotti, M. Pelosi, P. 1993. Multiple types and forms of odorant-binding proteins in the Old-World porcupine Hystrix cristata. Comp. Biochem. Physiol. 105 (3-4):775-84.
Freeland, W. J. 1991. Plant secondary metabolities: biochemical coevolution with herbivores. In: (eds.) Palo, R. T. & Robbins, C. T. Plant Defenses against mammalian herbivory. Boca Raton, CRC Press: 61-81.
Fonseca, G. A. B.; Herrmann, G.; Leite, Y.; Mittermeier, R.; Rylands, A. & Patton, J. 1996. Lista anotada dos mamíferos do Brasil. Conservation International & Fundação Biodiversitas. Occasional papers in Conservation Biology, v. 4: p. 1-38.
Garlindo-Leal, C. & Câmara, I. G. 2003. Atlantic Forest Hotspot Status: An Overview. In: Garlindo-Leal, C. & Câmara, I. G. (orgs) The Atlantic Forest of South America, biodiversity status, threats, and outlook. Island Press, Washington, DC. 488p.
Galvão, R. 2003. Informações biológicas sobre Sphiggurus villosus e Chaetomys subspinosus (Rodentia, Erethizontidae) no Parque Estadual Paulo César Vinha, Guaraparí, ES. Monografia de Graduação. Universidade Federal do Espírito Santo. Vitória.
Galvão, R. & Leite, Y. R. 2005. Uso do carretel de rastreamento “spool in line” em ouriços (Rodentia, Erethizontidae). In: Livro de Resumos do III Congresso Brasileiro de Mastozoologia. Vitória: UFES/Sociedade Brasileira de Mastozoologia.
Giné, G. A. F.; Le Pendu, Y.; Filadelfo, V.; Cunha, S. N. S. & Faria, D. 2005a. Estudo do padrão de atividade do ouriço-preto (Chaetomys subspinosus, Olfers 1818) em cativeiro. In: Livro de Resumos do III Congresso Brasileiro de Mastozoologia. Vitória: UFES/Sociedade Brasileira de Mastozoologia.

82
Giné, G. F.; Le Pendu, Y.; Nogueira, S. S. C.; Barros, K. S. & Faria, D. 2005b. Preferência alimentar e locais de repouso utilizados pelo ouriço-preto (Chaetomys subspinosus, Olfers 1818) em cativeiro. In: Livro de Resumos do III Congresso Brasileiro de Mastozoologia. Vitória: UFES/Sociedade Brasileira de Mastozoologia.
Giné, G. A. F.; Le Pendu, Y.; Cunha, S. N. S.; Filadelfo, V. & Faria, D. 2005c. Padrão de atividade, preferência alimentar e uso do espaço pelo ouriço-preto (Chaetomys subspinosus, Olfers 1818) e ouriço-amarelo (Sphiggurus insidiosus, Lichtenstein 1818) em cativeiro. In: VII Congresso de Ecologia do Brasil, 2005, Caxambu. Anais do VII Congresso de Ecologia do Brasil.
Giné, G. A. F.; Le Pendu, Y.; Duarte, J. M. B.; Faria, S. & Faria, D. 2006a. Uso do espaço pelo ouriço-preto (Chaetomys Subspinosus) na região cacaueira do sul da Bahia. In: VII Congresso Internacional sobre Manejo de Fauna Silvestre na Amazônia e América Latina, 2006, Ilhéus. Anais do VII Congresso Internacional sobre Manejo de Fauna Silvestre na Amazônia e América Latina.
Giné, G. A. F.; Faria, D.; Le Pendu, Y. & Duarte, J. M. B. 2006b. Locais de repouso diurno utilizados pelo ouriço-preto (Chaetomys Subspinosus, Olfers 1818) na natureza. In: VII Congresso Internacional sobre Manejo de Fauna Silvestre na Amazônia e América Latina, 2006, Ilhéus. Anais do VII Congresso Internacional sobre Manejo de Fauna Silvestre na Amazônia e América Latina.
Gonsalves, C. 2005. Diversidade Genética da espécie de roedor Chaetomys suspinosus (ouriço-preto) para auxiliar na elaboração de seu plano de manejo. Dissertação de Mestrado em Genética e Biologia Molecular. Universidade Estadual de Santa Cruz, Fundação Para Amparo à Pesquisa do Estado da Bahia.
Gonsalves, C.; Martínez, R. A. & Gaiotto, F. A. 2005. DNA da espécie de roedor Chaetomys subspinosus (ouriço-preto) auxilia na elaboração do seu plano de manejo. In: 51o Congresso Brasileiro de genética, 2005, Águas de Lindóia, SP. Resumos do 51º Congresso Brasileiro de genética. São Paulo : Sociedade Brasileira de Genética
Griesemer, S. J.; Fuller, T. K. & Degraaf, R. M. 1998. Habitat use by porcupines (Erethizon dorsatun) in central Massachusetts: effects of topography and forest composition. 1998. Am. Midl. Nat . 140: p. 271-279.
Griesemer, S. J.; Hale, M. O.; Rose, U. & Fuller, T. K. 1999. Capturing and marking adult North Americam porcupines. Wildlife Society Bulletin. 27 (2): p. 310-313.
Harris, S.; Cresswell, W. J.; Forde, P. G.; Trewhella, W. J. Woollard, T. & Wrays, S. 1990. Home-range analysis using radio-tracking data – a review of problems and techniques particularly as applied to the study of mammals. Mammal. Rev. 20, 2/3: 97-123.
Hayne, D. W. 1949. Calculation of size of home range. J. Mammalogy. n. 30: p.1-18.
Honacki, J. H.; Kinman, K. E. & Koeppl, J. W. 1982. (eds.) Mammal species of the world: taxonomic and geographic reference. Allem Press, Inc., The Assoc. Syst. Coll., Lawrence, Kansas. 694 pp.
Husson, A. M. 1978. The mammals of Suriname. E. J. Brill, Leiden. 569 pp.

83
Ilse, L. M. & Hellgren, E. C. 2001. Demographic and behavioral characteristics of North American Porcupines (Erethizon dorsatum) in Pinyon-Juniper Woodlands of Texas. Am. Midl. Nat . n. 146: p.329-338.
INCAPER. 2006. Sistema de Informações Agrometeorológicas. Disponível em página da Web: http:// www.incaper.es.gov.br/clima/index/.
IUCN. 2004. The IUCN 2004 Red List of Threatened Species. IUCN, Switzerland. Disponível em página da Web: http://www.redlist.org/.
Jacob, A. A. & Rudran, R. 2003. Radio-telemetria em estudos populacionais. In: Métodos de Estudos em Biologia da Conservação e Manejo de Vida Silvestre. (orgs) Cullen, L.; Rudran, R. & Valladares-Pádua, C. UFPR. p. 285-342.
Julien-Laferriere, D. 1993. Radio-tracking observations on ranging and foraging patterns by Kinkajous (Potos flavus) in French Guiana. J. Trop. Ecology. 9: p. 19-32.
Julien-Laferriere, D. 1999. Foraging strategies and food partitioning in the neotropical frugivorous mammals Caluromys philander and Potos flavus. J. Zool. Lond. n. 247: 71-80.
Kays, R. W. & Gittleman, J. L. 2001. The social organization of the kinkajou Potos flavus (Procyonidae). J. Zool. Lond. 253: 491-504.
Kernoham, B. J.; Gitlen, R. A. & Millspaugh, J. J. 2001. Analysis of animal space use and movements. In: Radio Tracking and Animal Populations (Millspaugh, J.J. & Marzluff, J.M., eds.) Academic Press, San Diego. Pp. 125-166.
Komers, P. E. & Brotherton, P. N. M. 1997. Female space use is the best predictor of monogamy in mammals. Proc. R. Soc. Lond. 264: 1261-1270.
Lacerda, L.D.; Araújo, D.S.D. & Maciel, N.C. 1993. Dry coastal ecosystems of the tropical Brazilian coast. In: van der Maarel E. (ed). Dry coastal ecosystems: Africa, America, Asia, Oceania. Elsevier, Amsterdam. p. 477-493.
Lira, P. K. 2005. Padrões espaciais de marsupiais em uma paisagem fragmentada de Mata Atlântica no Estado do Rio de Janeiro. Dissertação de Mestrado. Universidade Federal do Rio de Janeiro. Rio de Janeiro. 90 pp.
Martin, T. 1994. On the systematic position of Chaetomys subspinosus (Rodentia: Caviomorpha) based on evidence from the incisor enamel microstructure. J. Mamm. Evol. Vol. 2, n. 2. p. 117 – 131.
Martinez, R. A.; Giné, G.; Le Pendu, Y.; Alvarez, M. & Faria, D. 2005. Algumas considerações sobre a anatomia interna e externa do ouriço-preto Chaetomys subspinosus Olfers, 1818 (Rodentia, Erethizontidae). In: Livro de Resumos do III Congresso Brasileiro de Mastozoologia. Vitória: UFES/Sociedade Brasileira de Mastozoologia.
McNab, B. K. 1963. Bioenergetics and the determination of home range size. Am. Nat. 97: 133-140.

84
McNab, B. K. 1978. Energetics of Arboreal folivores: physiological problems and ecological consequences of feeding on an ubiquitous food suply. Pages 153-162. In: G. G. Montgomery, editor. The Ecology of Arboreal Folivores. Smithsonian Institution Press, Washington,DC.
Myers, N.; Mittermeier, R. A.; Mittermeier, C. G.; Fonseca, G. A. & Kent, J. 2000. Biodiversity hotspots for conservation priorities. Nature 403: 853-858.
Mendes, S. L. 1989. Estudo Ecológico de Alouatta fusca (Primates: Cebidae) na Estaçao Ecológica de Caratinga, MG. Rev. Nord. Biol. 6: 71-104.
Mendes, S. & Padovan, M. P. 2000. A Estação Biológica de Santa Lúcia, Santa Teresa, Espírito Santo. Bol. Mus. Biol. Mello Leitão. 11/12: p. 7-34.
Milton, K. 1980. The Foraging Strategy of Howler Monkeys. Columbia Univ. Press, New York.
Mittermeier, R. A.; Myers, N.; Thonsen, J. B.; Fonseca, G. A. B. & Olivieri, S. 1998. Biodiversity hotspots and major tropical wilderness areas: approaches to setting conservation priorities. Conservation Biology 12. 516-520.
Mittermeier, R. A; Gil, P. R.; Hoffman, M.; Pilgrin, J.; Brooks, T.; Mittermeier, C. G.; Lamoreux, J. & Fonseca, G. A. B. 2004. Hotspots revisited: Eath’s Biologically Richest and Most Threatened Terrestrial Ecoregions. Conservation International.
MMA - Ministério do Meio Ambiente. 1996. Perfil dos Estados Litorâneos do Brasil: subsídios À implantação do Programa de Gerenciamento Costeiro. Brasília: Programa Nacional do Meio Anbiente.
MMA - Ministério do Meio Ambiente. 2000. Avaliação e Ações Prioritárias para a Conservação da Biodiversidade da Mata Atlântica e Campos Sulinos. Conservation International do Brasil; Fundação SOS Mata Atlântica; Fundação Biodivérsitas; Instituto de Pesquisas Ecológicas, Secretaria do Meio Ambiente do Estado de São Paulo, SEMAD & Instituto Estadual de Florestas–MG. 40 p.
MMA - Ministério do Meio Ambiente. 2006. Lista nacional das espécies da fauna brasileira ameaçadas de extinção. Disponível em página da Web: http // www.mma.gov.br.
Montgomery, G. G. & Lubin, Y. D. 1978. Movements of Coendou prehensilis in the Venezuelan Ilanos. J. Mammalogy. 59 (4): p. 877 – 888.
Montgomery, G. G. & Sunquist, M. E. 1975. Impact of sloths on neotropical forest energy flow and nutrient cycling. In: Tropical ecology systems, trends in terrestrial and aquatic research. Golley, F.B. & Medin, E. (Eds). Berlin, Springer-Verlag.
Moss, R. 1991. Diet selection – an ecological perspective. Proc. Nutr. Soc. 50: 71-75.
Moojen, J. 1952. Os roedores do Brasil. Vol. 1. Intit. Nacion. do livro, Rio de Janeiro.
Moraes, E. M. 2003. Área de Uso, deslocamento e padrão de atividade de Micureus demerarae (Thomas, 1905) (Mammalia: Didelphidae) na reserva Biológica União, estado do Rio de Janeiro, Brasil. Dissertação de Mestrado. Pontifícia Universidade Católica de Minas Gerais. 61 pp.

85
Moura, R. T. & Fonseca, G. A. B. 2005. Observações preliminares sobre ocorrência, distribuição, biologia e ecologia do rato-do-cacau (Callistomys pictus), na Mata Atlântica, sul da Bahia. In: Livro de Resumos do III Congresso Brasileiro de Mastozoologia. Vitória: UFES/Sociedade Brasileira de Mastozoologia.
Nogueira,C. P. 1996. The diet of the muriqui females, Brachyteles arachnoids in different reproductive conditions. Neotropical Primates, 4: 156.
Novak, R. M. & Paradiso. 1983. Walker’s mammals of the world. The Johns Hopkins University Press, Baltimore and London.
Oliveira, J. A. & Bonvicino, C. B. 2006. Ordem Rodentia. In: Mamíferos do Brasil. (eds) Reis, N. R.; Peracchi, A. L.; Pedro, W. A. & Lima, I. P. Londrina. p. 347-406.
Oliver, W. L. R & Santos, I. B 1991. Threatened endemic mammals of the Atlantic forest region of south-eastern Brazil. Wildl. Preserv. Trust Special Sci. Rep. 4, 1-126.
Olson, R. & Lewis, A. M. 1999. Porcupine ecology and damage management techniques for rural homeowners. University of Wyoming.
Passamani, M.; Mendes, S. & Chiarello, A. G. 2000. Non-volant mammals of the Estação Biológica de Santa Lúcia and adjacent areas of Santa Teresa, Espírito Santo, Brazil. Bol. Mus. Biol. Mello Leitão. 11/12: p. 201-214.
Pereira, O. J., 1990. - Caracterização fitofisionômica da restinga de Setiba - Guaraparí - Espírito Santo. in: II Simpósio de Ecossistemas da Costa Sul e Sudeste Brasileira - Estrutura, Função e Manejo. ACIESP, São Paulo. p. 207-209
Patterson, B. & Wood, A. E. 1982. Rodents from the Deseaden Oligocene of Bolivia and the relationships of the Caviomorpha. Bull. Mus. Comp. Zool. 149: p. 371-573.
Powell, R. A. 2000. Animal home ranges and territories and home range estimators. In: Research techniques in animal ecology: controversies and consequences. (eds) Boitani, L. & Fuller, T. K. Columbia University Press, New York: p.65-110.
Queiroz, H. L. 1995. Preguiças e Guaribas, os mamíferos folívoros do Mamirauá. Brasília: CNPq e Sociedade Civil do Mamirauá.
Redford, K. H. 1992. The Empty Forest. BioScience, 42 (6).
Reis, N. R.; Shibatta, O. A.; Peracchi, A. L.; Pedro, W. A. & Lima, I. P. 2006. Sobre os mamíferos do Brasil. In: Mamíferos do Brasil (eds) Reis, N. R.; Peracchi, A. L.; Pedro, W. A. & Lima, I. P. Londrina. p. 17-25.
Reynolds, R. D. & Laundrè, J. W. 1990. Time intervals for estimating pronghorn and coyote home ranges and daily movements. Journal of Wildlife Management 54: 316-322.
Roberts, M.; Brand, S. & Maliniak, E. 1985. The biology of captive prehensile-tailed porcupines, Coendou prehensilis. J. Mammalogy. 66 (3): p. 476-482.
Robbins, C.T., 1993. Wildlife Feeding and Nutrition . Academic Press, San Diego, 2nd edition.

86
Rodgers, A. R. & Carr, A. P. 1998. The Home Range Extension for ArcView. Ontario Ministry of Natural Resources, Center for Northern Forest Ecosystem Research, Thunder Bay, Ontario, Canada.
Rose, U. 1987. Denning and winter range of the porcupine. Can. J. Zool. 65: 981-986.
Sackett, L. W. 1913. The Canada Porcupine: A study of the learning process. Behav. Monogr. 2: 1-84.
Santos Junior, T. S. 1998. Monitoramento de Coendou Prehensilis (Rodentia: Erethizonthidae) resgatados no reservatório da Usina Hidrelétrica de Miranda (MG) e translocados para a Reserva do Jacob, Nova Ponte – MG . Dissertação de Mestrado. Universidade Federal de Brasília, Brasília. 56 pp.
Santos, I. B.; Oliver, W. L. & Rylands, A. B. 1987. Distribution and status of two of tree porcupines, Chaetomys subspinosus and Sphiggurus insidiosus in south-east Brazil. Dodo, J. Jersey. Wildl. Preserv. Trust. 24 : 43-60.
Seaman, D. E. & Powell, R. A. 1996. An avaluation of the accuracy of Kernel density estimators for home range analysis. Ecology. n. 77: p. 2075-2085.
Seaman, D. E.; Millspaugh, J. J.; Kernohan, B. J.; Brundige, G. C.; Readeke, K. L. & Gitzen, R. A. 1999. Effects of sample size on kernel home range estimates. Journal of Wildlife Management 63: p. 739-747.
Shafer, M. L. 1981. Minimum population sizes for species conservation. BioCience, 31:131-134.
Siegel, S. 1956. Nonparametric statistics for the behavioral sciences. MacGraw-Hill Book Company, New York.
Silva, L. F. B. M. 1993. Ecologia do rato de bambu, Kannabaetomys amblyonyx (Wagner, 1845) na Reserva Biológica de Poço das Antas, Rio de Janeiro. Dissertação de Mestrado. Universidade Federal de Minas Gerais, Belo Horizonte.
Silva, R. B. 2005. Ecologia do Rato-da-taquara (Kannabateomys amblyonyx) no Parque Estadual de Itapuã. Mastozoologia Neotropical. Vol. 12, n. 1. p. 98-99.
Silverman, B. W. 1986. Density estimation for statistics and data analysis. London: Chapman & Hall.
Smith, G. W. 1979. Movements and home range of the porcupine in northeastern Oregon. Northwest Sci. 53: 277-282.
Snyder, M. A.; Linhart, Y. B. 1997. Porcupine feeding patterns: Selectivity by a generalist herbivore? Can. J. Zool. vol. 75, no. 12, pp. 2107-2111.
Solla, S. R.; Borduriansky, R. & Brooks, R. J. 1999. Eliminating autocorrelation reduces biological relevance of home range estimates. Journal of Animal Ecology 68: p. 221-234.
SOS Mata Atlântica & INPE. 2002. Atlas dos Remanescentes Florestais da Mata Atlântica no Período 1995-2000. Relatório final. Fundação SOS Mata Atlântica, São Paulo.

87
Suguio, K. & Tessler, M.G. 1984. Planícies de cordões litorâneos quartenários do Brasil: origem e nomenclatura. In: Lacerda LD, Araújo DSD, Cerqueira R, Turq B (eds). Restingas: Origem, estrutura e processos. CEUFF, Niterói. p. 15-26.
Stamps, J. 1995. Motor learning and the value of familiar space. American Naturalist 146: 41-58.
Sweitzer, R. A. & Berger, J. 1998. Evidence for female biased dispersal in North American Porcupines (Erethizon dorsatum). J. Zool. Lond. n. 244: p. 159-166.
Tabarelli, M.; Pinto, L. P.; Silva, J. C.; Hirota, M. & Bedê, L. 2005. Challenges and opportunities for biodiversity conservation in the Brasilian Atlantic Forest. Conservation Biology 19 (3): 695-700.
Venturini, A.C., A. M. S. Ofranti, J. B. M. Varejão & P.R. Paz, 1996. - Aves e Mamíferos na restinga. Parque Estadual Paulo César Vinha. Setiba – Guaraparí – Espírito Santo. Secretaria de Estado do Desenvolvimento Sustentável. Governo do Estado do Espírito Santo. Vitória.
Vilela, R. V.; Machado, T.; Ventura, K.; Fagundes, V.; Silva, M. J. J. & Yonenaga-Yassuda, Y. 2009. The taxonomic status of the endangered thin-spined porcupine, Chaetomys subspinosus (Olfers, 1818), based on molecular and karyologic data. Evolutionary Biology, 9:29.
Voss, R. S. & Angermann, R. 1997. Revisionary notes on neotropical porcupines (Rodentia: Erethizontidae). 1. Type material described by Olfers (1818) and Kuhl (1820) in the Berlin Zoological Museum. Am. Mus. Nov. (3214): p. 1-44.
Voss, R. & da Silva, M. N. F. 2001. Revisionary notes on neotropical porcupines (Rodentia: Erethizontidae). 2. A review of the Coendou vestitus group with descriptions of two new species from Amazonia. Am. Mus. Nov. (3351): p. 1-36.
White, G. C. & Garrott, R. A. 1990. Analysis of Wildlife Radio-tracking Data. Academic Press, London. 383 pp.
Worton, B. J. 1989. Kernel methods for estimating the utilization distribution in home range studies. Ecology. n. 70: p.164-168.
Woods, C. A. 1993. Suborder Hystricognathi. In: Mammal Species of the World. (Ed.) Wilson, D. E. & Reeder, D. A. M. 2a ediçao. London: Smithsonian Inst. Press. pp. 789.


















