
EFEITO DO ÁCIDO ASCÓRBICO SOBRE A ESPERMATOGÊNESE DE ... · Distrofia Muscular de Duchenne....
84
UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE CENTRO DE BIOCIÊNCIAS JANINE KARLA FRANÇA DA SILVA BRAZ EFEITO DO ÁCIDO ASCÓRBICO SOBRE A ESPERMATOGÊNESE DE CAMUNDONGOS COM DISTROFIA MUSCULAR DE DUCHENNE Natal 2015
Transcript of EFEITO DO ÁCIDO ASCÓRBICO SOBRE A ESPERMATOGÊNESE DE ... · Distrofia Muscular de Duchenne....

0
UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE
CENTRO DE BIOCIÊNCIAS
JANINE KARLA FRANÇA DA SILVA BRAZ
EFEITO DO ÁCIDO ASCÓRBICO SOBRE A ESPERMATOGÊNESE DE
CAMUNDONGOS COM DISTROFIA MUSCULAR DE DUCHENNE
Natal
2015

1
JANINE KARLA FRANÇA DA SILVA BRAZ
Efeito do ácido ascórbico sobre a espermatogênese de camundongos com Distrofia
Muscular de Duchenne
Dissertação apresentada ao Programa de Pós-
Graduação em Biologia Estrutural e Funcional da
Universidade Federal do Rio Grande do Norte para
obtenção do título de Mestre em Biologia Estrutural
e Funcional.
Área de concentração:
Ciências morfológicas
Orientador
Prof. Dr.: Carlos Eduardo Bezerra de Moura
Co-orientador
Profa. Dra.: Danielle Barbosa Morais
Natal
2015

2
Autorizo a reprodução e divulgação total ou parcial deste trabalho, por qualquer meio convencional
ou eletrônico, para fins de estudo e pesquisa, desde que citada a fonte.
Catalogação da Publicação na Fonte. UFRN / Biblioteca Setorial do Centro de
Biociências
Braz, Janine Karla França da Silva.
Efeito do ácido ascórbico sobre a espermatogênese de camundongos
com Distrofia Muscular de Duchenne / Janine Karla França da Silva Braz.
– Natal, RN, 2015.
84 f.: il.
Orientador: Prof. Dr. Carlos Eduardo Bezerra de Moura.
Coorientadora: Profa. Dra. Danielle Barbosa Morais
Dissertação (Mestrado) – Universidade Federal do Rio Grande do
Norte. Centro de Biociências. Programa de Pós-Graduação em Biologia
Estrutural e Funcional.
1. Camundongo Mdx. – Dissertação. 2. Estresse oxidativo. – Dissertação. 3. Morfologia. – Dissertação. I. Moura, Carlos Eduardo Bezerra de. II.
Morais, Danielle Barbosa. III. Universidade Federal do Rio Grande do
Norte. IV. Título.
RN/UF/BSE-CB CDU 599.323

3 Nome: BRAZ, Janine Karla França da Silva
Título: Efeito do ácido ascórbico sobre a espermatogênese de camundongos com Distrofia
Muscular de Duchenne
Dissertação apresentada ao Programa de Pós-
Graduação em Biologia Estrutural e Funcional da
Universidade Federal do Rio Grande do Norte para
obtenção do título de Mestre em Biologia Estrutural
e Funcional
Aprovado em:
Banca Examinadora
Prof. Dr. Carlos Eduardo Bezerra de Moura Instituição: UFERSA
Julgamento:____________________________ Assinatura:_________________________
Profa. Dra. Danielle Barbosa Morais Instituição: UFRN
Julgamento:____________________________ Assinatura:_________________________
Prof. Dr. Marcelo Barbosa Bezerra Instituição: UFERSA
Julgamento:____________________________ Assinatura:_________________________

4
Dedico este trabalho aos portadores da Distrofia
Muscular de Duchenne e aos familiares que lutam
diariamente pela vida.

5
AGRADECIMENTOS
À Deus por me permitir crescer através dos percalços e escolhas da vida. Esses dois últimos anos
definitivamente foram de crescimento e ficarão para sempre na memória;
Ao meu marido, João Paulo Araújo Braz, agradeço pela força, bela bondade, pelo carinho, enfim,
por tudo. Sem você eu não chegaria até aqui e não almejaria ir mais longe – PS.: Você já sabe;
Aos meus pais, Francisco Gilberto da Silva e Solange Araújo França, e ao meu irmão Rômulo
França, por acreditarem que esse é apenas o início para fazer grandes transformações;
Ao professor orientador (pai, amigo, irmão), Carlos Eduardo Bezerra de Moura que me lançou um
grande desafio e não me desamparou nos momentos de incertezas, de insegurança e de receios. À
você o meu eterno agradecimento por me tornar o que sou hoje;
À professora co-orientadora, Danielle Barbosa Morais pela paciência e o cuidado com os detalhes
neste trabalho. Com você aprendi que a perfeição é necessária em todos os momentos de nossa
vida;
Às alunas de iniciação científica e amigas, Vilessa Gomes e Nayara Rocha por aceitarem o desafio
matemático que esse trabalho exigiu com competência, ânimo e coragem. À vocês o meu
imensurável obrigada;
À toda equipe do Laboratório Microcelt, em especial, à professora Naianne K. Clebis pela chance
de trabalhar com a DMD e, claro, pela ajuda com as coletas, materiais de laboratório, pela conversa
amiga e científica;
Ao Prof. Fernando L. Ladd pelo auxílio com as lâminas de imunohistoquímica;
Ao Prof. Moacir F. Oliveira pelas imagens de microscopia eletrônica de transmissão;
À toda equipe do Prof. Sérgio Luís P. Matta que gentilmente nos auxiliou com a confecção das
lâminas em resina;

6
À Pró-reitoria de Pós-graduação da UFRN pelo auxílio financeiro com a bolsa de pós-graduação
modalidade demanda social;
Aos amigos que de alguma forma participaram deste trabalho: Maria Cecília Santos e Carlos
Eduardo (Caju) pelo auxílio com a estatística, Anderson Muniz, Schirley Barbosa e Julliane Tamara
Melo (entusiasmo), Maria de Lourdes Freitas e Melyna Solto (técnicas do laboratório de histologia -
DMOR), Nayra Resende e Karoline Vasconcelos (amizade eterna), aos colegas da primeira turma
do Programa de Biologia Estrutural e Funcional (união, alegria e conhecimento).

7
“Vós sois a luz do mundo”.
(Mateus 5:14)

8
RESUMO
BRAZ, J. K. F. S. Efeito do ácido ascórbico sobre a espermatogênese de camundongos com
Distrofia Muscular de Duchenne. 2015. 84f. Dissertação (Mestrado) – Centro de Biociências,
Universidade Federal do Rio Grande do Norte, Natal, 2015.
Objetivou-se avaliar as alterações na espermatogênese provocadas pela Distrofia Muscular de
Duchenne (DMD) e o efeito do tratamento com ácido ascórbico na prevenção dessas injúrias. Neste
trabalho foram utilizados 24 camundongos, sendo 12 da linhagem C57BL/10 (não-distróficos) e 12
da linhagem C57BL/10Mdx (distróficos), divididos em seis grupos com 4 animais cada: C30 =
Controle de 30 dias; D30 = Distrófico com 30 dias; C60 = Controle com 60 dias; D60 = Distrófico
com 60 dias; CS = Controle com 60 dias suplementados com ácido ascórbico e DS60 = Distrófico
com 60 dias suplementados com ácido ascórbico. Os animais dos grupos C30 e D30 foram
eutanasiados aos trinta dias de idade, enquanto que os animais dos demais grupos aos 60 dias de
idade. A suplementação com ácido ascórbico foi ministrada na água na dosagem de 0,005g/dia
durante 30 dias. Após eutanasiados, os testículos (direito e esquerdo) foram coletados, pesados e
seccionados transversalmente, fixados em solução de Karnovysky, incluídos em resina histológica
para análises morfológicas e morfométricas, submetidos à técnica de imunohistoquímica para
caspase-3 e análise ultraestrutural. A distrofia alterou a túnica própria (%), o epitélio (%), lúmen
(%), o volume individual das células de Leydig e o índice Leydigossomático. As análises
ultraestruturais indicaram que a suplementação não foi eficaz na reversão da apoptose mitocondrial
das células de Sertoli. Contudo, o tratamento apresentou resultado satisfatório no estresse oxidativo
em distróficos púberes, reduzindo a densidade de volume das células apoptóticas positivas para
Caspase–3. Além disso, reverteu o quadro de hipertrofia das células de Leydig em distróficos. A
distrofia na pré-puberdade apresentou vesículas lipídicas nas células de Leydig. Dessa forma, a
DMD afetou a organização dos túbulos seminíferos e intertúbulo, no entanto, a suplementação com
ácido ascórbico nas condições experimentais utilizadas para o tratamento da DMD foi suficiente
apenas para reduzir o estresse oxidativo na região basal do compartimento tubular e a hipertrofia
das células de Leydig.
Palavras-chave: Vitamina C; camundongo Mdx; Estresse oxidativo; Morfologia; Testículo.

9
ABSTRACT
BRAZ, J. K. F. S. Effect of ascorbic acid on the spermatogenesis of mice with Duchenne
Muscular Dystrophy. 2015. 84f. Dissertação (Mestrado) – Centro de Biociências, Universidade
Federal do Rio Grande do Norte, Natal, 2015.
The objective was to evaluate alteration in spermatogenenic caused by Duchenne Muscular
Dystrophy (DMD) and the effect of treatment with ascorbic acid to prevent these injuries. In this
study we were used 24 mice, 12 of C57BL / 10 strain (non-dystrophic) and 12 of the strain C57BL /
10Mdx (dystrophic), divided into six groups with four animals each: C30 = 30 days Control; D30 =
Dystrophic 30 days; C60 = Control with 60 days; D60 = Dystrophic 60 days; CS = Control with 60
days supplemented with ascorbic acid and DS60 = Dystrophic 60 days supplemented with ascorbic
acid. The acid ascorbic supplementation was given in water and it was administered at the dosage of
0.005g/day for 30 days. After euthanized, the testicles (right and left) were collected, weighed and
sectioned, fixed in Karnovysky solution, embedded in historesin for histological studies
morphological and morphometric analyzes and submitted to immunohistochemistry for caspase-3
and ultrastructural analysis. The dystrophy changed the tunica propria (%),epithelium (%), lumen
(%), the individual volume of Leydig cells and the index Leydigosomatic. The ultrastructural
analysis showed that supplementation was not effective in reversing the mitochondrial apoptosis of
Sertoli cells. However, the treatment had satisfactory result in oxidative stress in dystrophic
pubertal, reducing the volume density of apoptotic cells positive for Caspase-3. In addition,
reversed hypertrophy above Leydig cells in dystrophic pubertal. The dystrophy prepubertal had
lipid vesicles in the Leydig cells. Thus, the DMD affected the organization of the seminiferous
tubules and intertubule, however, the ascorbic acid supplementation in the experimental conditions
used for the treatment of DMD has been enough to reduce the oxidative stress in the basal region
tubular and hypertrophy of Leydig cells.
Keywords: Vitamin C; Mdx mice; Oxidative stress; Morphology; Testis.

10
SUMÁRIO
1 INTRODUÇÃO .................................................................................................................... 12
2 OBJETIVOS......................................................................................................................... 14
2.1 OBJETIVO GERAL ........................................................................................................... 14
2.2 OBJETIVOS ESPECÍFICOS.............................................................................................. 14
3 REVISÃO DE LITERATURA ............................................................................................ 15
3.1 DISTROFIA MUSCULAR DE DUCHENNE (DMD) ........................................................ 15
3.1.1 A Distrofia Muscular de Duchenne e a reprodução ........................................................ 16
3.2 ÁCIDO ASCÓRBICO ........................................................................................................ 17
3.3 ANATOMIA E FISIOLOGIA DO SISTEMA REPRODUTOR MASCULINO .................. 18
3.3.1 Testículo ....................................................................................................................... 18
3.3.2 Espermatogênese........................................................................................................... 18
3.3.3 Célula de Sertoli ............................................................................................................ 19
3.3.4 Célula de Leydig ........................................................................................................... 19
3.3.5 Ciclo do epitélio seminífero (CES) ................................................................................ 20
4 MATERIAL E MÉTODOS ................................................................................................. 21
4.1 ANIMAIS .......................................................................................................................... 21
4.2 DIETA SUPLEMENTAR COM ÁCIDO ASCÓRBICO .................................................... 21
4.3 EUTANÁSIA E COLETA DO MATERIAL ...................................................................... 21
4.4 PROCESSAMENTO HISTOLÓGICO ............................................................................... 22
4.5 ANÁLISE IMUNOHISTOQUÍMICA PARA CASPASE-3 ................................................ 22
4.6 DENSIDADE DE VOLUME (VV) DAS CÉLULAS CASPASE 3 - POSITIVAS .............. 22
4.7 ESTEREOLOGIA TESTICULAR ..................................................................................... 23
4.8 FREQUÊNCIA DOS ESTÁDIOS DO CICLO DO EPITÉLIO SEMINÍFERO ................... 25
4.9 QUANTIFICAÇÃO DA ESPERMATOGÊNESE .............................................................. 25
4.10 ANÁLISE DA ULTRAESTRUTURA ............................................................................... 26
4.11 ANÁLISE ESTATÍSTICA ................................................................................................. 26
ARTIGO 1: Histomorfometria da espermatogênese de camundongos mdx portadores da
Distrofia Muscular de Duchenne (DMD) suplementados com ácido ascórbico ........................ 32
Resumo ...................................................................................................................................... 32
1. Introdução ........................................................................................................................ 33
2. Material e métodos ........................................................................................................... 34
2.1. Animais ......................................................................................................................... 34
2.2. Dieta suplementar com ácido ascórbico ........................................................................ 34
2.3. Eutanásia e coleta do material ...................................................................................... 34
2.4. Processamento histológico ............................................................................................ 35
2.5. Estereologia testicular .................................................................................................. 35

11
2.6. Frequência relativa dos estádios do ciclo do epitélio seminífero ................................... 36
2.7. Quantificação da espermatogênese ............................................................................... 36
2.8. Análise da ultraestrutura ............................................................................................... 36
2.9. Imunohistoquímica para caspase-3 ............................................................................... 37
2.10. Densidade de Volume (Vv) das células caspase 3 - positivas ......................................... 38
2.11. Análise estatística ......................................................................................................... 38
3. Resultados ......................................................................................................................... 38
3.1. Estereologia testicular .................................................................................................. 38
3.2. Frequência dos estádios do ciclo do epitélio seminífero ................................................ 40
3.3. Quantificação da espermatogênese ............................................................................... 41
3.4. Análise ultraestrutural .................................................................................................. 43
3.5. Contagem de células apoptóticas .................................................................................. 48
4. Discussão ........................................................................................................................... 50
Referências ................................................................................................................................ 55
ARTIGO 2: Histomorfometria do compartimento intertubular em testículos de camundongos
mdx portadores da Distrofia Muscular de Duchenne, tratados com ácido ascórbico .............. 62
Resumo ...................................................................................................................................... 62
1. Introdução ........................................................................................................................ 63
2. Material e Métodos ........................................................................................................... 64
2.1. Animais ......................................................................................................................... 64
2.2. Dieta suplementar com ácido ascórbico ........................................................................ 64
2.3. Eutanásia e coleta do material ...................................................................................... 64
2.4. Processamento histológico ............................................................................................ 65
2.5. Estereologia testicular .................................................................................................. 65
2.6. Análise estatística ......................................................................................................... 66
3. Resultados ......................................................................................................................... 67
4. Discussão ........................................................................................................................... 72
Referências ................................................................................................................................ 75
5. CONCLUSÕES .................................................................................................................... 81
ANEXO A – Certificado Comitê de Ética no Uso dos Animais ICB/USP ................................. 82
ANEXO B – Declaração da Comissão de Ética no Uso de Animais – CEUA/UFRN................ 83

12
1 INTRODUÇÃO
A Distrofia Muscular de Duchenne (DMD) é uma distrofinopatia que afeta 1: 3500 meninos
nascidos (DALTON e SAMPSON, 2015). Os portadores apresentam os primeiros sinais clínicos
dessa distrofia a partir dos 3 anos de idade, demostrando disfunção nos músculos associados ao
joelho e quadril (manobra de Gower), dificultando a mobilidade (EMERY, 2002). Esses sintomas
aparecem em virtude da degeneração muscular progressiva decorrente de mutação no cromossomo
X, no locus gênico Xp21, o que afeta a função da proteína de membrana chamada distrofina,
ausente nos portadores (HOFFMAN et al., 1987; BRADLEY e PARSONS, 1998; TERRIL et al.,
2013). Portadores da DMD podem apresentar variações na gravidade da doença associadas à mesma
mutação, o que dificulta e limita os métodos de intervenção (BRINKMEYER-LANGFORD e
KORNEGAY, 2013).
A degeneração muscular é causada inicialmente por danos nas fibras musculares com
participação de células no processo inflamatório (neutrófilos e macrófagos) que podem perdurar
durante dias em virtude das injúrias causadas à membrana, passando a produzirem EROs (Espécies
Reativos Oxigênio) ao realizarem fagocitose, agravando a resposta inflamatória (TIDBALL, 2005;
WHITEHEAD et al., 2006). O estresse oxidativo é relevante em pacientes com DMD e
camundongos mdx, uma vez que na ausência de distrofina, as espécies reativas de oxigênio estão
elevadas nos músculos e permitem a oxidação de lipídios da membrana, proteínas, ou DNA
(CANTON et al., 2014). Estudos do mecanismo de inflamação e da degeneração muscular de DMD
em humanos são compreendidos com a utilização de cães da raça Golden Retriever, que apresentam
homologia genética com a DMD em humanos, e de camundongos mdx como modelos
experimentais, por possuírem uma mutação que gera o defeito bioquímico semelhante ao
encontrado na DMD (HOFFMAN et al., 1987; WHITEHEAD et al., 2006; MARTINS-JÚNIOR et
al., 2015).
Para reduzir as lesões musculares em DMD, pesquisadores têm realizado estudos com
terapias antioxidantes a base de chá verde (DORCHIES et al., 2006), N-acetilcisteína
(WHITEHEAD et al., 2008) e melatonina (HIBAOUI et al., 2011). O ácido ascórbico é um
antioxidante que protege os testículos de danos decorrentes do estresse oxidativo (SÖNMEZ et al.,
2005). Além disso, em homens com baixo nível de ácido ascórbico foi detectado redução no
número de espermatozoides, na motilidade espermática e aumento no número de espermatozoides
anormais (EBESUNUN et al., 2004).
Não obstante, alguns autores apontam que o hipogonadismo e a atrofia testiscular podem
ocorrer em homens com distrofia muscular miotônica tipo 1 (DM1), uma forma de distrofia

13
muscular que promove a deterioração muscular progressiva e desordem sistêmica, como a atrofia
testicular (CARTER e STEINBECK, 1985; AL-HARBI et al., 2008). Apesar de Askeland et al.
(2013) afirmarem que pacientes portadores da DMD apresentaram morbidades urológicas à medida
em que a distrofia era agravada, estudos relacionados à caracterização da espermatogênese em
indivíduos portadores desse tipo de distrofia ainda não são relatados na literatura. Do mesmo modo,
não há registros da ação antioxidante do ácido ascórbico nos túbulos seminíferos dos portadores da
DMD, tornando relevante a aplicação deste trabalho.
De acordo com Ferlini et al. (2010), a abordagem terapêutica na DMD deve envolver
medidas que possuam resultados adequados e sensatos. Logo, fazendo uso de novas terapias os
pacientes com DMD têm aumentado a sua expectativa de vida para valores acima dos 40 anos nas
últimas décadas (KIENY et al., 2013). Aos pacientes que almejam constituir família é recomendado
o diagnóstico genético pré-implantação para posterior aplicação da técnica de fertilização in vitro
(COCO, 2014). Logo, a necessidade de caracterizar os efeitos dessa distrofia na espermatogênese,
bem como os resultados decorrentes da intervenção pelo uso de ácido ascórbico são fundamentais
para estudos na área clínica médica e farmacológica, visando melhorar o bem estar do paciente
portador da DMD.

14
2 OBJETIVOS
2.1 OBJETIVO GERAL
Avaliar as alterações na espermatogênese provocadas pela Distrofia Muscular de Duchenne
(DMD) e o efeito do tratamento com ácido ascórbico na prevenção dessas injúrias.
2.2 OBJETIVOS ESPECÍFICOS
2.2.1 Avaliar os parâmetros morfométricos testiculares do compartimento tubular e intertubular em
camundongos distróficos suplementados ou não com ácido ascórbico;
2.2.2 Averiguar as alterações ultraestruturais nas células de Sertoli, células de Leydig e túnica
própria de camundongos distróficos tratados ou não com ácido ascórbico;
2.2.3 Quantificar a densidade de volume de células em apoptose positivas para caspase-3 no epitélio
seminífero.

15
3 REVISÃO DE LITERATURA
3.1 DISTROFIA MUSCULAR DE DUCHENNE (DMD)
A Distrofia Muscular de Duchenne (DMD) é caracterizada pela perda muscular progressiva
e fraqueza precoce, podendo levar o paciente a óbito ainda no final da adolescência. Essa descrição
foi feita inicialmente em humanos em 1851 pelo médico inglês Edward Meryon, que postulou que a
distrofia teria caráter hereditário, acometendo somente meninos, com completa degeneração do
sarcolema. Apesar da relevância de suas descrições, os achados de Meryon permaneceram
negligenciados por muito tempo até que o médio neurologista Guillaume Benjamin Amand
Duchenne, em 1868, detalhou a clínica e a histopatologia muscular da distrofia (EMERY, 2002).
Somente em 1986 foi constatado que os pacientes com DMD apresentavam uma deleção do
cromossomo Xp21, o gene da distrofina responsável pela produção da proteína de 427 Da, e sua
ausência desencadeia a distrofia muscular. Esse gene é de caráter recessivo localizado no
cromossomo X materno. Dessa forma, as meninas raramente manifestam a doença por serem
heterozigotas, podendo ainda apresentar sinais clínicos brandos. Já os meninos podem receber o
alelo recessivo do cromossomo X materno e manifestarem a doença (MUKHERJEE et al., 2003).
Os sinais clínicos da DMD se manifestam por volta dos 3 a 5 anos de idade, pela fraqueza muscular
dos joelhos e dos quadris fazendo com que a criança utilize as mãos como se estivesse escalando.
Esse é o sinal característico da manobra de Gower. Aos 12 anos de idade a criança poderá utilizar
cadeira de rodas em virtude da progressão da fraqueza muscular (MORRISON, 2011).
Um complexo distrofina-glicoproteína é formado pela distrofina e por outras proteínas,
permitindo que a actina se conecte a matriz extracelular (integrante da estrutura responsável pela
contratilidade muscular). A distrofina, juntamente com as sarcoglicanas e outra glicoproteínas,
podem estar associadas a tecidos musculares e não-musculares, enfatizando a necessidade de
conhecer a doença em diferentes aspectos e propor formas alternativas de intervenção nas diferentes
áreas teciduais comprometidas pela ausência de distrofina (DAVIS, 1997; HOFFMAN E
DRESSMAN, 2001).
Há alguns anos a expectativa de vida dos portadores da DMD era de 20 anos de idade
podendo alcançar pouco mais de 30 anos, sendo a principal causa da morte relacionada a problemas
cardiorrespiratórios (DAVIS, 1997; EMERY, 2002). Todavia, nos últimos anos tem-se observado a
evolução da expectativa de vida dos portadores de DMD em virtude da eficiência da técnica de
ventilação por traqueostomia, um método seguro e eficaz (KIENY et al., 2013). Esse aumento, na
expectativa de vida, gira em torno de 40 a 50 anos de idade, um avanço significativo e pode trazer

16
consigo o interesse reprodutivo, que deve ser acompanhado do aconselhamento genético (DALTON
e SAMPSON, 2015; PEDLOW et al., 2015).
Para o estudo da distrofia são utilizados frequentemente modelos animais, sendo o
camundongo mdx o mais adequado para avaliar os mecanismos de mionecrose e regeneração na
distrofia muscular de Duchenne, a fim de se desenvolver estratégias terapêuticas. Esse camundongo
desenvolve a distrofia muscular recessiva associada ao cromossomo X (locus Xp21), e não possui a
proteína distrofina. No desenvolvimento de sua miopatia é possível observar alterações musculares
associadas à mionecrose e presença de infiltrados de linfócitos que sugerem a participação do
sistema linfocitário, bem como a presença de moduladores da expressão de componentes da matriz
extracelular que auxiliam no direcionamento dos linfócitos ao local da lesão (SEIXAS et al., 1997).
A ausência da distrofina na DMD eleva os nível de espécies reativas de oxigênio (EROs)
nos músculos, levando a oxidação de lipídios, proteínas ou do DNA (RENJINI et al., 2012). Em
murinos a degeneração muscular inicia no período pré-púbere, com um processo inflamatório, e
posteriormente ocorre a progressão necrótica (DISATNIK et al., 1998). Esse processo desencadeia
a degeneração, a fibrose e o déficit funcional da musculatura do diafragma, musculatura esquelética
e cardíaca em camundongos mdx e em pacientes com DMD (STEDMAN et al., 1991). Todavia,
antioxidantes naturais podem oferecer proteção aos danos decorrentes do estresse oxidativo em
camundongos mdx, através da restauração dos níveis de glutationa (DORCHIES et al., 2009).
3.1.1 A Distrofia Muscular de Duchenne e a reprodução
Os estudos relacionados à espermatogênese na distrofia muscular estão descritos para a
distrofia miotônica, obtidos por meio da medição dos níveis de testosterona sérica livre e total, bem
como descrição de hipogonadismo nos indivíduos portadores (AL –HARBI, 2008).
Alguns trabalhos com cães Golden Retriever, outro modelo comumente utilizado para o
estudo da Distrofia Muscular de Duchenne (GRMD), relatam alterações como criptorquidísmo,
hérnia inguinoescrotal, exposição constante da glande na ausência de ereção, aspermia, maior taxa
de anormalidade espermática, degeneração testicular com fibrose, hialinização e retração das fibras
do músculo cremáster, o que pode comprometer a aptidão reprodutiva dos cães afetados (LUPPI,
2006).
Através de estudos com o camundongo mutante mdx3cv
, um novo modelo para estudos
funcionais da região C-terminal da distrofina em tecidos musculares e não musculares, foi
observado que na ausência da proteína Dp71 os flagelos dos espermatozoides estavam alterados,
levando a redução de sua motilidade e da fecundidade, em virtude da redução ou ausência da β-

17
distroglican e superexpressão e relocalização de α-sintrofina, proteínas essas associadas ao
complexo de distrofina (HERNÁNDEZ-GONZÁLES et al., 2005).
3.2 ÁCIDO ASCÓRBICO
O ácido ascórbico (AA) ou vitamina C foi isolado pela primeira vez 1928 por Szent-
Gyorgyi, sendo atualmente considerada como um antioxidante solúvel em água e insolúvel na maior
parte dos solventes orgânicos. Os seres humanos não o sintetizam, mas adquirem através da
ingestão de frutas cítricas, morangos, tomates, brócolis, nabo e outros vegetais folhosos. Quando
essa vitamina não é consumida, cerca de 3% das reservas permanecem reduzidas diariamente e o
indivíduo começa a apresentar sintomas clínicos de escorbuto (GONÇALVES, 2008). Dessa forma,
o AA é necessário para a realização de funções fisiológicas importantes, como o controle do
colesterol sanguíneo, além de contribuir para a síntese de catecolaminas que regulam o sistema
nervoso central, auxilia na absorção de ferro intestinal, e possui função imunológica e antioxidante,
protegendo o DNA das células da ação dos radicais livres (IQBAL et al., 2004). Tendo em vista as
propriedades antioxidantes da vitamina C, pesquisadores conseguiram reforçar a defesa
antioxidante e consequentemente reduzir o estresse oxidativo na distrofia muscular do tipo
facioescapulohumeral (PASSERIEUX et al., 2014).
Essa vitamina é conhecida pela fórmula molecular C6H8O6, ou pelo nome químico 2-oxi-L-
treohexônio-1,4-lactona-2,3-enediol. Pode ser encontrada na forma reduzida e ativa (ácido L-
ascórbico) e na forma oxidada (ácido L-deidroascórbico) (MORÁN et al., 2006). O ácido L-
ascórbico apresenta atividade vitamínica total quando comparado ao ácido L-deidroascórbico, o que
corresponde a cerca de 75 a 80% de atividade de vitamina C (PINEDO, 2007). Após a ingestão de
alimentos ricos em vitamina C ocorre sua absorção pela mucosa oral, estômago e intestino, para ser
transportado sob a forma aniônica livre em direção ao interior dos leucócitos e eritrócitos, via
difusão simples (IQBAL et al., 2004). Assim é possível que ocorra a ampla distribuição nos tecidos,
embora alguns órgãos como a glândula suprarrenal, a hipófise e a retina, são grandes armazenadores
de AA (ARANHA et al., 2000).
Além disso, a vitamina C também está presente nos testículos, sendo capaz de ativar
enzimas de hidroxilação que participam dos processos metabólicos (HUSSAIN et al., 2010). A
vitamina C também é encontrada em altas concentrações no plasma seminal. A literatura descreve
que os baixos níveis de ácido ascórbico seminal estão associados com aumento de danos do DNA
espermático, síntese de espermatozoides anormais e infertilidade nos homens (SONG et al., 2006;
COLAGAR e MARZONY, 2009). Logo, estudos apontam que a suplementação com ácido

18
ascórbico pode melhorar os parâmetros espermáticos com aumento significativo na concentração,
motilidade e morfologia dos espermatozoides (AKMAL et al., 2006).
3.3 ANATOMIA E FISIOLOGIA DO SISTEMA REPRODUTOR MASCULINO
3.3.1 Testículo
Os testículos são órgãos pares, de formato oval e estão revestidos pela túnica vaginal e
albugínea. A estrutura glandular dos testículos está dividida em lobos e lóbulos, esses por sua vez
são formados por túbulos seminíferos contorcidos. Os testículos produzem os gametas masculinos
(espermatogênese) e os hormônios sexuais masculinos através de reações enzimáticas, que
conduzem à produção de hormônios esteroides (esteroidegênese) (RUSSELL et al., 1990). A
produção espermática ocorre no compartimento tubular, que consiste nos túbulos seminíferos, e a
síntese de esteroides no compartimento intersticial entre os túbulos seminíferos. Apesar dos
compartimentos apresentarem morfologia e função distinguíveis, eles estão intimamente ligados
entre si para garantir a integridade funcional testicular (WEINBAUER et al., 2010).
Os túbulos seminíferos são avasculares e não possuem inervação sendo constituídos por uma
lâmina própria, lúmen e o epitélio seminífero. A lâmina própria, que constitui a membrana basal, é
uma camada de colágeno com células peritubulares (células mióides). As células mióides são
contráteis que promovem a circulação de fluídos e a propulsão espermática através do lúmen dos
túbulos seminíferos. O epitélio seminífero pode ser classificado como estratificado e nele são
encontradas as células germinativas e as células de Sertoli (RUSSELL et al., 1990).
3.3.2 Espermatogênese
A espermatogênese é um processo que se divide funcionalmente em três fases promovendo
mudanças moleculares e celulares (PARVINEN, 1982). A primeira fase é a proliferativa com a
expansão clonal decorrente de mitoses sucessivas das espermatogônias até a formação dos
espermatócitos. A segunda fase é a meiótica que consiste na divisão por meiose I e II dos
espermatócitos para formação das espermátides. A última fase é a diferenciação das espermátides
arredondas e alongadas para formação do espermatozoide (RUSSELL et al., 1990). Contudo, de
acordo com Cheng et al. (2010) a espermatogênese está dividida em quatro fases distintas: a mitose,
a meiose, a espermiogênese e a espermiação.
Na puberdade as espermatogônias começam a divisão mitótica gerações de células-filhas
que podem manter-se como células-tronco (Espermatogônia tipo A) ou se diferenciarem em
espermatogônia tipo B. A partir destas surgem os espermatócitos primários, que são as maiores
células do epitélio seminífero com 46 cromossomos que entram em prófase e prosseguem a divisão
meiótica formando duas células filhas, os espermatócitos secundários com 23 cromossomos, são de

19
difícil visualização na microscopia em virtude do curto tempo em intérfase, e prosseguem com a
segunda divisão meiótica. As espermátides arredondadas surgem como resultado da segunda
divisão meiótica dos espermatócitos secundários. A partir das espermátides inicia o processo de
espermiogênese, que corresponde a fase final de produção de espermatozoides. Esse é um processo
complexo que envolve a formação do acrossomo, condensação e alongamento do núcleo,
desenvolvimento do flagelo e a perda da maior parte do citoplasma resultando em espermatozoides
no lúmen do túbulo seminífero (WEINBAUER et al., 2010).
3.3.3 Célula de Sertoli
As células de Sertoli foram descritas pelo cientista italiano Enrico Sertoli em 1865, são
cilíndricas e apresentam núcleo e nucléolo tripartido com heterocromatina associada revestindo os
túbulos seminíferos após a puberdade, atuando como células de ligação entre o espaço intertubular e
o lúmen do túbulo seminífero. O citoesqueleto das células de Sertoli é formado por microtúbulos,
filamentos de actina e filamentos intermediários de vimentina. Essa organização permite a
manutenção da forma e polaridade da célula, a movimentação ou posicionamento das organelas, a
formação de pseudópodes e a ancoragem das células germinativas. O domínio apical e lateral dessa
célula é irregular para sustentar as células do epitélio germinativo. Já o domínio basolateral
apresenta junções oclusivas entre as células de Sertoli adjacentes formando o compartimento
adlumial e basal (RUSSELL, 1990; WEINBAUER et al., 2010).
Essas células possuem papel importante na sustentação, proteção e nutrição das células
espermatogênicas em desenvolvimento. Além de fagocitar os corpos residuais, facilitam a liberação
das espermátides maduras para o lúmen dos túbulos seminíferos (RUSSELL, 1990). A proliferação
dessas células ocorre apenas durante o período pré-púbere, dessa forma o bom desenvolvimento e
nessa fase são importantes para a saúde reprodutiva masculina na puberdade. Esse processo é
regulado por fatores hormonais parácrinos e autócrinos (ESCOTT et al., 2014). Além de ser
influenciada pelo hormônio hipofisário folículo estimulante (FSH) para síntese e secreção da
proteína ligante ao andrógeno (ABP) (GRISWOLD, 1998).
3.3.4 Célula de Leydig
As células de Leydig foram descritas pela primeira vez em 1850 pelo zoólogo e anatomista
comparativo alemão Franz Leydig (WEINBAUER et al., 2010). Essas células possuem retículo
endoplasmático liso e mitocôndrias com enzimas associadas à síntese de esteroides, citoplasma
eosinófilo, núcleo arredondado podendo ocasionalmente ser alongado ou poliédrico (MORI e
CHRISTENSEN, 1980). As células de Leydig estão no espaço intertubular e sofrem estimulação do
hormônio luteinizante (LH) mediado pelo monofosfato cíclico de adenosina (AMPc) produzem

20
testosterona, que pode ser convertida em di-hidrotestosterona pela ação enzimática de 5α-redutase.
As altas concentrações de testosterona durante o desenvolvimento das espermatogônias são
mantidas pela ABP produzida pelas células de Sertoli em resposta ao estímulo do FSH
(WEINBAUER et al., 2010).
3.3.5 Ciclo do epitélio seminífero (CES)
A espermatogênese é um processo dinâmico e ordenado formado por associações celulares
em fases específicas do desenvolvimento em secções transversais dos túbulos seminíferos
denominadas de estádios. Essas células evoluem de um estádio anterior para um seguinte
permitindo que cada célula do epitélio seminífero seja maturada em determinado tempo e espaço,
isso faz com que a nova geração celular seja sincronizada. Essa organização celular foi designada
de “ciclo espermatogênico” (REGAUD, 1901). Mas, atualmente a literatura denomina de ciclo do
epitélio seminífero (CES) (PEREY et al., 1961; LEBLONT e CLERMONT, 1962; RUSSELL et al.,
1990). O ciclo é regulado por fatores intrínsecos (células de Sertoli e células germinativas), fatores
extrínsecos (andrógeno e ácido retinóico) e pelo fator espécie-específico (HESS e FRANÇA, 2008).
O CES pode ser classificado de acordo com o método da morfologia tubular
(BERNDTSON, 1977) e o método acrossômico (NEVES et al., 2002). O método da morfologia
tubular identifica 8 estádios para as associações de células germinativas dentro de muitas gerações
em uma área do túbulo, levando em consideração a forma do núcleo e o posicionamento das
espermátides, presença de divisões meióticas e a composição do túbulo seminífero (BERNDTSON,
1977; AMANN, 1986). Já o método acrossômico é baseado no desenvolvimento do sistema
acrossômico ao longo da maturação das espermátides arredondas em alongadas e constitui estádios
desde 6 estádios para o homem a 12 para camundongos (CLERMONT, 1963; RUSSELL, 1990).
O processo espermatogênico completo é determinado pela duração do CES, em mamíferos
são necessários em média 4,5 ciclos, que seria o tempo necessário desde a espermatogônia do tipo
A até a liberação do espermatozoide no lúmen tubular (RUSSELL, 1990). A regulação da duração
do CES é realizada pelas células de Sertoli, uma vez que regulam as fases do epitélio seminífero
(HESS e FRANÇA, 2008).

21
4 MATERIAL E MÉTODOS
4.1 ANIMAIS
Foram utilizados 24 camundongos machos, sendo 12 da linhagem C57BL/10 (animais
controle) e 12 da linhagem C57BL/10Mdx (animais distróficos), provenientes do Biotério da
FIOCRUZ/ Rio de Janeiro e do Biotério do Instituto de Ciências Biomédicas da Universidade de
São Paulo/ICB – USP, respectivamente. O projeto foi aprovado pelo comitê de ética do ICB/USP e
chancelado pelo CEUA/UFRN (Parecer nº 164/2011 CEUA/ICB/USP e Parecer nº 064/2013
CEUA/UFRN) (Anexo A-B). Os animais foram divididos em seis grupos com 4 animais em
cada: Grupo controle com 30 dias de idade (C30); Grupo distrófico com 30 dias de idade (D30);
Grupo controle com 60 dias de idade (C60); Grupo distrófico com 60 dias de idade (D60); Grupo
controle com 60 dias de idade suplementado com ácido ascórbico (CS60) e Grupo distrófico com
60 dias de idade suplementado com ácido ascórbico (DS60). Os animais foram mantidos no
Biotério do Departamento de Anatomia, ICB/USP, em caixas de polietileno, contendo bebedouro
(água) e comedouro (ração), sob temperatura controlada de aproximadamente 22ºC e iluminação
com ciclo de 12 horas claro/escuro.
4.2 DIETA SUPLEMENTAR COM ÁCIDO ASCÓRBICO
Após o desmame (21 dias de idade), os animais receberam ração comercial para roedores
Nuvilab®
(Nuvital, São Paulo, Brasil). A partir dos 30 dias de idade os animais dos grupos
suplementados com ácido ascórbico (CS60 e DS60) receberam diariamente 0,005g de ácido
ascórbico (Sigma-Aldrich, St. Louis, EUA) por quilograma do peso corporal do animal, diluído em
água e administrado por meio de gavagem. A concentração foi calculada de acordo com o protocolo
estabelecido na literatura para modelo de camundongo com distrofia muscular, que se baseia em
Guido et al. (2010) e Tonon et al. (2012).
4.3 EUTANÁSIA E COLETA DO MATERIAL
Os animais foram eutanasiados em câmara de dióxido de carbono hermeticamente fechada,
com entrada de gás na parte superior. Após a eutanásia, os animais foram pesados e os testículos
coletados para serem pesados, seccionados transversalmente e imersos em solução fixadora de
acordo com a técnica histológica a ser empregada: Karnovsky para morfometria testicular sob
microscopia de luz e análise ultraestrutural sob microscopia eletrônica de transmissão, e
Paraformaldeído 4% em solução tampão de PBS para os procedimentos de imunohistoquímica
(KARNOVSKY, 1965; OLIVEIRA, 2004; MORAIS et al., 2014).

22
4.4 PROCESSAMENTO HISTOLÓGICO
Após fixação em Karnovsky por 24h, seguindo-se a desidratação os fragmentos testiculares
destinados à morfometria foram transferidos para álcool 70%. Os fragmentos foram desidratados
em concentrações crescentes de álcool etílico, para posterior inclusão em glicol-metacrilato
(Historesin®, Leica Mycrosistems, Heidelberg, Alemanha). O material foi seccionado a 3µm de
espessura através da técnica semi-seriada utilizando-se micrótomo rotativo Leica RM2255 (Leica
Microsystems, Heidelberg, Alemanha), com intervalo de 40µm entre os cortes, as lâminas
histológicas foram coradas com azul de toluidina/borato de sódio 1% (MORAIS et al., 2014). As
lâminas permanentes confeccionadas foram observadas ao microscópio de luz Motic BA410
(Motic, Causeway Bay, Hong Kong) e microfotogradas para análises e comparações com câmera
digital Moticam 5.0 MP (Motic Instruments Inc, Richmond, Canada). As imagens foram analisadas
ao software Image-Pro Plus®
(Media Cybernetics Inc., Rochville, USA).
4.5 ANÁLISE IMUNOHISTOQUÍMICA PARA CASPASE-3
Para identificar células em apoptose no interior dos túbulos seminíferos foi utilizada a
técnica de imunohistoquímica para caspase 3. Os cortes foram desparafinizados em uma estufa a
60°C durante 30 minutos, colocados em xilol por 5 minutos, reidratados em concentrações
decrescentes de álcool etílico (absoluto, 90, 70 e 50%) e lavados em água destilada. Para
desmascarar os epítopos, a recuperação antigênica foi realizada com solução de citrato de sódio 10
mM (pH 6,0 95°). Esta recuperação foi realizada em panela de pressão de micro-ondas com
potência máxima de 750W, em dois ciclos de 5 minutos. Em seguida as amostras foram arrefecidas
até a temperatura ambiente e lavadas com água destilada. O bloqueio da peroxidase endógena foi
realizado com peróxido de hidrogênio 0,9%, seguindo-se a lavagem com água destilada e solução
salina tamponada com fosfato de sódio (PBS, pH 7,4). O anticorpo primário anti-caspase 3 (diluição
1: 400, Abcam, Cambridge, Inglaterra) foi incubado durante 12h. Em seguida, os antígenos
imunomarcados foram visualizados através do sistema Dako Envision+ (Dako, Carpinteria, EUA),
utilizando-se 3'3-tetra-hidrocloreto de diaminobenzidina (DAB) como cromógeno. As seções foram
finalmente contrastadas com hematoxilina de Harry.
4.6 DENSIDADE DE VOLUME (VV) DAS CÉLULAS CASPASE 3 - POSITIVAS
A densidade de volume (Vv) refere-se a fração de volume ocupada pelas células caspase 3-
positivas nos respectivos espaços referencia, ou seja, o testículo. Para a estimativa de Vv um
sistema teste foi sobreposto sobre o espaço referência (Figura 1), e foram contados o número total
de pontos que tocaram as células de interesse (Pint) e o número total de pontos sobre o espaço
referência (Pref) para obtenção da densidade de volume (Vv) (Vv = ∑Pint / ∑Pref) seguindo o
protocolo proposto por Ladd et al. (2012).

23
Figura 1- Sistema teste aplicado para cálculo da densidade de volume de células apoptóticas
positivas para caspase-3 (ponta da seta).
4.7 ESTEREOLOGIA TESTICULAR
As análises morfométricas foram realizadas de acordo com os protocolos propostos por
Morais et al. (2014a). As proporções volumétricas entre túbulos seminíferos e intertúbulo foram
estimadas pela contagem de 266 pontos projetados sobre 10 imagens obtidas das preparações
histológicas de cada animal, em objetiva de 10x, para quantificar os túbulos seminíferos,
intertúbulo, túnica própria, epitélio seminífero e lúmen (Figura 2). Já o volume dos túbulos
seminíferos foi estimado a partir do percentual ocupado pelo mesmo no testículo e do volume do
parênquima testicular (valor conhecido) (Vol. Túbulo = peso gonadal líquido x Túbulo %/ 100). O
volume do epitélio seminífero foi o produto do percentual do epitélio seminífero vezes o volume do
parênquima testicular, dividido por 100. O peso da túnica albugínea foi estimado a partir da
densidade de volume, pela contagem de 266 pontos projetados sobre 10 imagens obtidas das
preparações histológicas de cada animal, em objetiva de 10x (Vv = No de pontos contados sobre a
túnica albugínea/ Total de pontos contados x 100). O volume absoluto da albugínea foi resultado do
produto da densidade de volume com o volume testicular, levando-se em consideração que sua
densidade é próxima a 1 (JOHNSON et al., 1981; SPRANDO et al., 1998; COSTA et al., 2011).

24
Figura 2 – Modelo de sistema teste para calcular as proporções volumétricas dos componentes
testiculares. Azul de toluidina. Barra = 50µm.
O índice tubulossomático (ITS) foi obtido através da divisão do volume tubular pelo peso
corporal, multiplicado por 100. Já o índice gonadossomático (IGS) foi obtido como quociente da
divisão do peso dos testículos pelo peso corporal, sendo o valor, multiplicado por 100. O diâmetro
dos túbulos seminíferos e altura do epitélio seminífero (mensurada da túnica própria até o lúmen
tubular) foram obtidos pela mensuração de 20 secções transversais de túbulos/animal, escolhendo-
se os túbulos que apresentaram contorno o mais circular possível.
O comprimento dos túbulos seminíferos (CT), em metros, foi estimado a partir da fórmula
CT = VTS/πR2, onde VTS é o volume dos túbulos seminíferos, πR
2 é a área da secção transversal
dos túbulos seminíferos, e R é o diâmetro tubular/2. O CT por grama de testículo foi obtido
dividindo-se este valor pelo peso gonadal.
O percentual de cada elemento do compartimento intertubular foi quantificado a partir da
projeção de 1.000 pontos sobre o intertúbulo de cada animal, quantificando os pontos coincidentes
sobre o núcleo e o citoplasma das células de Leydig, vaso sanguíneo, espaço linfático e tecido
conjuntivo. Os percentuais de cada um destes componentes no intertúbulo e nos testículos foram
estimados, respectivamente, a partir das equações: (Nº de pontos contados sobre o elemento x 100 /
1000) e (% de intertúbulo x % do elemento no intertúbulo / 100), enquanto seus volumes foram

25
estimados pela equação: (% do elemento no testículo x peso do parênquima testicular / 100)
(MORAIS et al., 2014b). Todas essas contagens foram feitas utilizando-se o software Image-Pro
Plus® (Media Cybernetics Inc., Rochville, USA).
Os diâmetros nucleares das células de Leydig foram mensurados quando apresentaram
contorno circular, cromatina perinuclear e nucléolos evidentes, quantificando-se 30 núcleos por
animal. O volume nuclear (VN) e volume citoplasmático (VC) das células de Leydig por animal
foram expressos em µm3, através das respectivas equações: VN = 4/3 πR
3, sendo R o raio nuclear;
VC = % citoplasma x VN / % núcleo. Somando-se VN + VC obteve-se o volume celular.
O cálculo do número total de células de Leydig (TCL) foi obtido pela divisão do volume
total dessas células por parênquima testicular (µm3) pelo volume de cada célula de Leydig (µm
3). O
número de células por grama de testículo foi obtido pela divisão do TCL pelo peso gonadal total. O
índice Leydigossomático (ILS), que quantifica o investimento em células de Leydig em relação à
massa corporal, foi obtido através da equação: ILS = volume total de célula de Leydig por
parênquima testicular / PC x 100, onde PC = peso corporal.
4.8 FREQUÊNCIA DOS ESTÁDIOS DO CICLO DO EPITÉLIO SEMINÍFERO
Os estádios que compõem o ciclo do epitélio seminífero (CES) foram descritos utilizando-se
o método de morfologia tubular que descreve 8 estádios para todas as espécies, baseando-se em
Berndtson (1977). A frequência relativa de cada estádio foi descrita pela caracterização e contagem
de 50 túbulos por animal em secções transversal de forma randomizada.
4.9 QUANTIFICAÇÃO DA ESPERMATOGÊNESE
De acordo com Russell et al. (1990) deve ser escolhido o estádio mais frequente do ciclo
do epitélio seminífero para determinar as taxas celulares. Dessa forma, nessa pesquisa foram
quantificadas a população de cada tipo celular encontrada no estádio 8 do CES, através da contagem
dos núcleos das células germinativas e do nucléolo das células de Sertoli. Foram utilizadas 10
secções transversais circulares de túbulos seminíferos por animal para, então, mensurar os
diâmetros nucleares de espermatogônias do tipo A (A), espermatócitos primários na transição pré-
leptótenos para leptóteno (PL-L), espermatócitos primários em paquíteno (PQ), espermátides
arredondadas (Ar) e nucléolos de células de Sertoli (Se) (RUSSELL et al., 1990). As contagens
foram corrigidas de acordo com a espessura do corte histológico e o diâmetro nuclear ou nucleolar
das células germinativas e células de Sertoli, respectivamente, de acordo com o método de
Abercrombie (1946) modificado por Amann e Almquist (1962).

26
A partir desses dados o índice das células de Sertoli foi obtido com a finalidade de indicar a
capacidade de suporte dessas células por total de células germinativas. Esse índice foi determinado
através do resultado da razão entre os números corrigidos de células germinativas e o número
corrigido de células de Sertoli, utilizando-se a fórmula: (A+PL-L+PQ+Ar) / Se. Os índices mitótico
(PL-L / A), meiótico (Ar / PQ) e o rendimento geral da espermatogênese (Ar / A) também foram
quantificados.
4.10 ANÁLISE DA ULTRAESTRUTURA
Após fixação em solução de Karnovsky por 24h, os fragmentos testiculares foram
submetidos à microscopia eletrônica de transmissão (MET), seguindo o protocolo proposto por
Oliveira (2004). Os fragmentos foram imersos em glutaraldeído a 2,5%, tamponado com fosfato de
sódio a 0,1M, pH 7,4. Após a fixação o material foi lavado três vezes durante 10 minutos cada no
mesmo tampão. Em seguida ocorreu a pós-fixação com tetróxido de ósmio 2% por duas horas. Os
fragmentos foram lavados com solução tampão por três vezes durante dez minutos cada. Após esse
procedimento o material foi imerso em acetato de uranila a 3% overnight, lavado com tampão
fosfato e desidratado com série crescente de álcool etílico (50-100%) durante 10 minutos em cada.
Após desidratação, os fragmentos foram imersos em óxido de propileno por 10 minutos para
garantir total desidratação do tecido. Em seguida o material foi embebido em resina araldite 502
(Polysciences Inc, California, EUA), seguido da confecção dos blocos. Foram então obtidos cortes
semi-finos com ultramicrótomo, os quais foram corados com solução aquosa azul de toluidina a 1%
para a identificação dos locais para a realização de cortes ultrafinos. Cortes de 70nm de espessura
foram confeccionados com navalha de diamante e colocados em telas de cobre com 200 mesh para
posterior contraste com acetato de uranila saturado a 2%, por sete a dez minutos. Em seguida
utilizou-se citrato de chumbo a 0,5%, durante o mesmo período. O material foi analisado em
Microscópio Eletrônico de Transmissão no Laboratório de MEV (Jeol® 100 CX II, Tóquio, Japão)
da Faculdade de Medicina Veterinária e Zootecnia da USP.
4.11 ANÁLISE ESTATÍSTICA
Os parâmetros testiculares quantitativos foram expressos em média ± desvio padrão. Estes
foram submetidos à análise de variância (ANOVA) e teste de múltiplas comparações de Kruskall-
Wallis, seguido do teste de Mann-Whitney com correção de Bonferroni, utilizando-se o software
PAST® versão 2.17 (HAMMER et al., 2001). Admitiu-se o nível de significância de P<0,05.

27
REFERÊNCIAS
ABERCROMBIE, M. Estimation of nuclear populations from microtome sections. The
Anatomical Records, v. 94, n. 2, p. 239-247, 1946.
AKMAL, M. et al. Improvement in human semen quality after oral supplementation of vitamin C.
Journal of Medicinal Food, v. 9, n. 3, p. 440-2, 2006.
AL-HARBI, T. M. et al. Hypogonadism is common in men with myopathies. Journal of Clinical
Neuromuscular Disease, v. 9, n. 4, p. 397–401, 2008.
AMANN, R. P.; ALMQUIST, J. O. Reproductive capacity of dairy bulls. VIII. Dand indirect
measurement of testicular sperm production. Journal of Dairy Science, v. 45, n. 6, p. 774-781,
1962.
AMANN, R. P. Detection of alterations in testicular and epididymal function in laboratory animals.
Environmental Health Perspectives, v. 70, p. 149-158, 1986
ARANHA, F. Q. et al. O papel da vitamina C sobre as alterações orgânicas no idoso. Revista de
Nutrição da PUC-Campinas, v. 13, n. 2, p. 89-97, 2000.
ASKELAND, E. J. et al. Urological manifestations of duchenne muscular dystrophy. Journal of
Urology, v. 190, n. 4, p. 1523-1528, 2013.
BERNDTSON, W. E. Methods for quantifying mammalian spermatogenesis: a review. Journal of
Animal Science, v. 44, n. 5, p. 818-883, 1977.
BRADLEY, D.; PARSONS, E. Newborn screening for Duchenne muscular dystrophy. Seminars in
Neonatology, v. 3, n. 1, p. 27-34, 1998.
BRINKMEYER-LANGFORD, C.; KORNEGAY, J. N. Comparative genomics of X-linked
muscular dystrophies: The golden retriever model. Current Genomics, v. 14 n. 5, p. 330-342,
2013.
CANTON, M.; MENAZZA, S.; DI LISA, F. Oxidative stress in muscular dystrophy: from generic
evidence to specific sources and targets. Journal of Muscle Research and Cell Motility, v. 35,
n.1, p. 23-36, 2014.
CARTER, J. N.; STEINBECK, K. S. Reduced adrenal androgens in patients with myotonic
dystrophy. The Journal of Clinical Endocrinology and Metabolism, v. 60, n. 3, p. 611-614, 1985.
CHENG, C. Y. et al. Regulation of spermatogenesis in the microenvironment of the seminiferous
epithelium: new insights and advances. Molecular and Cellular Endocrinology, v. 315, n. 1, p.
49-56, 2010.
CLERMONT, Y. The cycle of the seminiferous epithelium in man. American Journal of
Anatomy, v. 112, n. 1, p. 35-51, 1963.
COCO, R. Reprogenetics: preimplantational genetics diagnosis. Genetics and Molecular Biology,
v. 37, n. 1, p. 271-284, 2014.

28
COLAGAR, A. H.; MARZONY, E. T. Ascorbic Acid in human seminal plasma: determination and
its relationship to sperm quality. Journal of Clinical Biochemistry and Nutrition, v. 45, n.2, p.
144-9, 2009.
COSTA, D. S. et al. Intrinsic rate of spermatogenesis in free-ranging feral pigs (Sus scrofa sp).
Animal Reproduction Science, v. 125, n. 1-4, p. 165-169, 2011.
DALTON, J.; SAMPSON, J. B. Overview of Adult Muscular Dystrophies. In: GOLDMAN, J.
(Org.). Genetic Counseling for Adult Neurogenetic Disease. Nova York: Springer, 2015. p. 233-
237.
DAVIS, K. E. Challenges in duchenne muscular dystrophy. Neuromuscular Disorders, v. 7, n.8,
p. 482-486, 1997.
DISATNIK, M. H. et al. Evidence of oxidative stress in mdx mouse muscle: studies of the pre-
necrotic state. Journal of the Neurological Sciences, v. 161, n. 1, p. 77-84, 1998.
DORCHIES, O. M. et al. Green tea extract and its major polyphenol (-)- epigallocatechin gallate.
The American Journal of Physiology - Cell Physiology, v. 290, n. 2, p. 616 –625, 2006.
EBESUNUN, M. O. et al. Plasma and semen ascorbic levels in spermatogenesis. The West
African Journal of Medicine, v. 23, n. 4, 290–293, 2004.
EMERY, A. E. H. The muscular dystrophies. Lancet, v. 359, n. 9397, p. 687-695, 2002.
ESCOTT, M. G. et al. Mechanisms of hormonal regulation of sertoli cell development and
proliferation: a key process for spermatogenesis. Current Molecular Pharmacology, v. 7, n. 2, p.
96-108, 2014.
FERLINI, A. et al. Dystrophin restoration in skeletal, heart and skin arrectorpili smooth muscle of
mdx mice by ZM2 NP-AON complexes. Gene Therapy, v. 17, n. 3, p. 432-438, 2010.
GONÇALVES, A. A. S. S. Avaliação da capacidade antioxidante de frutas e polpas de frutas
nativas e determinação dos teores de flavonóides e vitamina C. 2008. 88p . Tese (Mestrado em
Ciência dos Alimentos). Universidade de São Paulo, São Paulo. 2008.
GRISWOLD, M. D. The central role of Sertoli cells in spermatogenesis. Seminars in Cell &
Developmental Biology, v. 9, p. 411-416, 1998.
GUIDO, N. A. et al. Fiber type composition of the sternomastoid and diaphragm muscles of
dystrophin‐deficient mdx mice. The Anatomical Record, v. 293, n.10, p. 1722-1728, 2010.
HAMMER, O; HARPER, D. A. T.; RYAN, P. D. PAST: Paleontological statistics software
package for education and data analysis. Palaeontologia Electronica. v. 4, n. 1, p. 1-9, 2001.
HERNÁNDEZ-GONZÁLEZ, E. O. et al. Absence of dp71 in mdx3cv mouse spermatozoa alters
flagellar morphology and the distribution of ion channel and nNos. Journal of Cell Science, v. 118,
n. 1, p. 137-145, 2005.
HIBAOUI, Y. et al. Melatonin improves muscle function of the dystrophic mdx5Cv mouse, a
model for Duchenne muscular dystrophy. Journal of Pineal Research, v. 51, n. 2, p. 163–171,
2011.

29
HOFFMAN, E. P.; DRESSMAN, D. Molecular pathophysiology and targeted therapeutics for
muscular dystrophy. Trends in Pharmacological Sciences, v. 22, n. 9, p. 465-470, 2001.
HOFFMAN, E. P.; BROWN, R. H.; JR. KUNKEL, L. M. Dystrophin: the protein product of the
duchenne muscular dystrophy locus. Cell, v. 51, n. 6, p. 919-928, 1987.
HUSSAIN, J.; HUSSAIN, J.; SHARMA, K. C. Fluoride and health hazards: community perception
in a fluorotic area of central Rajasthan (India) an arid environment. Environmental Monitoring
and Assessment, v. 162, n. 1, p. 1-14, 2010.
IQBAL, K.; KHAN, A.; KHATTAK, M. M. A. K. Biological significance of ascorbic acid
(Vitamin C) in human health–a review. Pakistan Journal of Nutrition, v. 3, n. 1, p. 5-13, 2004.
JOHNSON, L.; PETTY, C. S.; NEAVES, W. B. A new approach to qualification of
spermatogenesis and its application to germinal cell attrition during human spermatogenesis.
Biology of Reproduction, v. 25, n. 1, p. 217–226, 1981.
KARNOVSKY, M. J. A formaldehyde-glutaraldehyde fixative of high osmolarity for use in
electron microscopy. The Journal of Cell Biology, v. 27, p. 137-138, 1965.
KIENY, P. et al. Evolution of life expectancy of patients with Duchenne muscular dystrophy at
AFM Yolaine de Kepper centre between 1981 and 2011. Annals of Physical and Rehabilitation
Medicine, v. 56, n. 6, p. 443–454, 2013.
LADD, A. A. L. et al. SCG postnatal remodelling–hypertrophy and neuron number stability–in
Spix's Yellow-toothed Cavies (Galea spixii). International Journal of Developmental
Neuroscience, v. 30, n. 2, p. 129-137, 2012.
LEBLOND, C. P.; CLERMONT, Y. Definition of the stages of the cycle of the seminiferous
epithelium in the rat. Annals of the New York Academy of Sciences, v. 55, n. 4, p. 548-573, 1952.
LUPPI, M. M. C. P. Avaliação andrológica de cães da raça Golden Retriever sadios e afetados
pela distrofia muscular. 2006. 109f. Dissertação (Mestrado em Ciências) - Faculdade de Medicina
Veterinária, Universidade de São Paulo, São Paulo, 2006.
MARTINS-JÚNIOR, H. A. et al. Profiles of Steroid Hormones in Canine X-Linked Muscular
Dystrophy via Stable Isotope Dilution LC-MS/MS. PloS one, v. 10, n. 5, e0126585-e0126585,
2015.
MORAIS, D. B. et al. Evaluation of the cell population of the seminiferous epithelium and
spermatic indexes of the bat Sturnira lilium (Chiroptera: Phyllostomidae). PloS one. v. 9, p.
e101759, 2014a.
MORAIS, D. B. et al. Histomorphometric characterization of the intertubular compartment in the
testes of the bat Sturnira lilium. Animal Reproduction Science, v. 147, n. 3, p. 180-186, 2014b.
MORÁN, G. A. G. et al. Aspectos bioclínicos y patobiológicos de la vitamina C en la especie
humana. Revista CES Medicina, v. 20, n. 2, p. 53-72, 2006.
MORI, H., CHRISTENSEN, A.K. Morphometric analysis of Leydig cells in the normal rat testis.
The Journal of Cell Biology, v. 84, n. 2, p. 340-354, 1980.

30
MORRISON, L. A. Dystrophinopathies. In: AMATO, A. A.; GRIGGS, R. C. (eds.). Handbook of
Clinical Neurology. Amsterdan: Elsevier, 2011. p. 11–39.
MUKHERJEE, M. et al. Denovo mutations in sporadic deletional Duchenne muscular dystrophy
(DMD) cases. Experimental and Molecular Medicine, v. 35, n. 2, p. 113–117, 2003.
NEVES, E. S., CHIARINI-GARCIA, H., FRANÇA, L. R. Comparative testis morphometry and
seminiferous epithelium cycle length in donkeys and mules. Biology of Reproduction, v. 67, n. 1,
p. 247-255, 2002.
OLIVEIRA, M. F. Placentação em mocós, Kerodon rupestre Wied 1920. 2004. 209f. Tese
(Doutorado em Anatomia dos Animais Domésticos e Silvestres) - Faculdade de Medicina
Veterinária e Zootecnia da Universidade Federal de São Paulo, São Paulo, 2004.
PARVINEN, M. Regulation of the seminiferous epithelium. Endocrine Reviews, v. 3, n. 4, p. 404-
417, 1982.
PASSERIEUX, E. et al. Effects of vitamin C, vitamin E, zinc gluconate, and selenomethionine
supplementation on muscle function and oxidative stress biomarkers in patients with
facioscapulohumeral dystrophy: a double-blind randomized controlled clinical trial. Journal of
Free Radicals in Biology and Medicine, v. 81, p. 158-169, 2014.
PINEDO, R. A. Estudo da estabilização da polpa de camu-camu (Myciaria dubia (N.B.K)
Vc.Vaugh) congelada visando a manutenção de ácido ascórbico e antocianinas. 2007. 108p.
Tese (Doutorado em Engenharia Química). Universidade Estadual de Campinas, Campinas, 2007.
PEDLOW, K. et al. Assisted standing for Duchenne muscular dystrophy. Cochrane Librar,. v.3, n.
2, p. 1-9, 2015.
PEREY, B. et al. The wave of the seminiferous epithelium in the rat. American Journal of
Anatomy, v. 108, n. 1, p. 47-77, 1961.
REGAUD, C. Etudes sur la structure des tubes seminiferes et sur la spermatogenese chez les
mammiferes. Archives d'Anatomie Microscopique et de Morphologie Expérimentale, v. 4, n.
101, p. 231, 1901.
RENJINI, R. et al. Oxidative damage in muscular dystrophy correlates with the severity of the
pathology: role of glutathione metabolism. Neurochemical Research, v. 37, n. 4, p. 885-898, 2012
RUSSELL, L. D. et al. Histological and histopathological evaluation of the testis. Bolesta: Cache
River Press, 1990. 280p.
SEIXAS, S. L. et al. Importância do camundongo mdx na fisiopatologia da distrofia muscular de
duchenne. Arquivos de Neuro-Psiquiatria, v. 55, n. 3B, p. 610-617, 1997.
SONG, G. J.; NORKUS, E. P.; LEWIS, V. Relationship between seminal ascorbic acid and sperm
DNA integrity in infertile men. International Journal of Andrology, v. 29, n. 6, p. 569-75, 2006.
SÖNMEZ, M.; TURK, G.; YUCE, A. The effect of ascorbic acid supplementation on sperm
quality, lipid peroxidation and testosterone levels of male Wistar rats. Theriogenology, v. 63, n. 7,
p. 2063–2072, 2005.

31
SPRANDO, R. L. et al. Testing the potential of sodium fluoride to affect spermatogenesis: a
morphometric study. Food and Chemical Toxicology, v. 36, n. 12, p. 1117-1124, 1998.
STEDMAN, H. H. et al. The mdx mouse diaphragm reproduces the degenerative changes of
Duchenne muscular dystrophy. Nature, v. 352, n. 6335, p. 536-539, 1991.
TERRIL, J. R. et al. Oxidative stress and pathology in muscular dystrophiefocus on protein thiol
oxidation and dysferlinopathies. FEBS Journal, v. 280, n.17, p. 4149–4164, 2013.
TIDBALL, J. G. Inflammatory processes in muscle injury and repair. American Journal of
Physiology. Regulatory, Integrative and Comparative Physiology, v. 288, n.2, p. 345–353,
2005.
TONON, E. Ascorbic acid protects the diaphragm muscle against myonecrosis in mdx mice.
Nutrition. v. 28, n. 6, p. 686-690, 2012
WEINBAUER, G. F. et al. Physiology of testicular function. In: NIESCHLAG E. et al. (eds.).
Andrology. Heidelberg: Springer Berlin, 2010.p. 11-59.
WHITEHEAD, N. P. et al. N-Acetylcysteine ameliorates skeletal muscle pathophysiology in mdx
mice. The Journal of Physiology, v. 586, n.7, p. 2003–2014, 2008.
WHITEHEAD, N. P.; YEUNG, E. W.; ALLEN, D. G. Muscle damage in mdx (dystrophic) mice:
role of calcium and reactive oxygen species. Clinical and Experimental Pharmacology and
Physiology, v. 33, n. 7, p. 657–662, 2006.

32
ARTIGO 1: Histomorfometria da espermatogênese de camundongos mdx portadores da
Distrofia Muscular de Duchenne (DMD) suplementados com ácido ascórbico
Resumo
Objetivou-se avaliar as alterações na espermatogênese provocadas pela Distrofia Muscular de
Duchenne (DMD) e o efeito do ácido ascórbico (AA) na prevenção dessas injúrias. Foram
utilizados 12 camundongos da linhagem C57BL/10 (não-distróficos) e 12 da linhagem
C57BL/10Mdx (distróficos) distribuídos nos grupos: Controle com 30 dias (C30); Distrófico com
30 dias (D30); Controle com 60 dias (C60); Distrófico com 60 dias (D60); Controle com 60 dias
suplementados com ácido ascórbico (CS60) e Distrófico com 60 dias suplementados (DS60). Os
animais foram suplementados com ácido ascórbico 0,005g/dia. Após eutanásia, os testículos foram
coletados, pesados e seccionados transversalmente. O material foi fixado e processado
histologicamente para análises morfológicas, morfométricas, ultraestruturais e imunohistoquímica
para caspase-3. A DMD reduziu o percentual de epitélio seminífero em D60 quando comparando a
D30 (P<0,05). Verificou-se maior percentual de túnica própria em D30 em relação a C30 e D60
(P<0,05). A distrofia não alterou a morfometria das células do epitélio germinativo, logo elevou
rendimento geral da espermatogênese em D60 (P<0,05). As células de Sertoli dos camundongos
distróficos púberes apresentaram vacuolizações citoplasmáticas e alteração da conformação
mitocondrial em relação a C60. A suplementação reduziu o número de células positivas para
caspase-3 em distróficos quando comparado a CS60. Dessa forma, a DMD promoveu os danos
testiculares ainda na pré-puberdade de camundongos, mas não inviabilizou a espermatogênese.
Além disso, a concentração de AA utilizada reverteu o quadro apoptótico no compartimento tubular
em camundongos distróficos.
Palavras-chave: Espermatogênese; epitélio seminífero; apoptose; camundongo mdx.

33
1. Introdução
A Distrofia Muscular de Duchenne (DMD) é a forma mais comum das distrofias
musculares, sendo uma doença autossômica recessiva ligada à mutação no cromossomo X, que
afeta aproximadamente 1:3600-6000 meninos nascidos (Hoffman et al., 1987; Bradley & Parsons,
1998). O gene da DMD afeta a função da proteína de membrana chamada distrofina, que por estar
ausente nos portadores da doença, acarreta degeneração muscular progressiva (Hoffman et al.,
1987; Terriel et al., 2013). A distrofina funcional está associada à actina e ao complexo de
glicoproteínas de membrana, protegendo a membrana de danos mecânicos durante a contração.
Assim a ausência dessa proteína deixa o sarcolema mais susceptível a danos após a contração
muscular, podendo induzir a necrose das fibras musculares cardíacas e esqueléticas, ocasionando
inflamação com produção de Espécies Reativas do Oxigênio (ERO's) (Vandebrouck et al., 2002;
Lapidos et al., 2004; Tidball, 2005; Whitehead et al., 2006). Há evidências de que os níveis
elevados dos ERO's podem agravar as distrofias musculares, além de desencadear a apoptose nas
células germinativas testiculares (Rao & Shaha, 2000; Tidball & Wehling-Henricks, 2007).
Para reduzir a necrose muscular promovida pelo estresse oxidativo em camundongos mdx,
estudos tem utilizado antioxidantes, como o extrato de chá verde. A dieta suplementar com chá
verde em camundongos distróficos protegeu o tecido muscular contra a necrose e estimulou a
adaptação do músculo tornando-o mais forte e mais resistente (Dorchies et al., 2006). De acordo
com Ângulo et al. (2011), a vitamina C (ácido ascórbico) é um micronutriente associado à
fertilidade e sua deficiência pode ocasionar distúrbios na espermatogênese. Em um estudo realizado
com camundongos que apresentavam déficit de vitamina C foi observado maior frequência de
apoptose nos espermatozoides, demonstrado através da marcação de fragmentação do DNA por
meio do ensaio de TUNEL (Yazama et al., 2006).
De acordo com Drucker et al. (1963) a produção e maturação dos espermatozoides nos
túbulos seminíferos podem ser alteradas em portadores da distrofia muscular miotônica (DM1).
Esses autores relataram desorganização na estrutura testicular, marcada pela redução progressiva da
atividade espermatogênica, degeneração das células germinativas e células de Sertoli, espessamento
da lâmina própria e esclerose tubular. Contudo, não há relatos na literatura que abordem
quantitativamente a morfologia do compartimento tubular, bem como os efeitos decorrentes da
intervenção pelo uso de ácido ascórbico na espermatogênese de pacientes portadores da DMD,
tornando esse estudo relevante para a área da clínica médica e farmacológica.
Além disso, com os avanços no tratamento da DMD, os portadores tem aumentado sua
expectativa de vida para 40 a 50 anos de idade, um avanço significativo levando-se em

34
consideração que esta expectativa estava em torno de 20 anos de idade (Pedlow et al., 2015). De
acordo com Abbott et al. (2015) um paciente britânico do sexo masculino com 30 anos de idade
possui uma vida familiar estável, casado e conseguiu ser pai, ou seja, o aumento da expectativa de
vida elevou o número de portadores sexualmente ativos e a possibilidade de serem pais.
Diante do exposto, este trabalho teve como objetivo avaliar as alterações do compartimento
tubular provocadas pela DMD em camundongos mdx, bem como a eficácia da suplementação com
ácido ascórbico sobre estas possíveis injúrias.
2. Material e métodos
2.1. Animais
Foram utilizados 24 camundongos machos, sendo 12 da linhagem C57BL/10 (animais
controle) e 12 da linhagem C57BL/10Mdx (animais distróficos), provenientes do Biotério da
FIOCRUZ/ Rio de Janeiro e do Biotério do Instituto de Ciências Biomédicas da Universidade de
São Paulo/ICB – USP, respectivamente. O projeto foi aprovado pelo comitê de ética do ICB/USP e
chancelado pelo CEUA/UFRN (Parecer nº 164/2011 CEUA/ICB/USP e Parecer nº 064/2013
CEUA/UFRN) (Anexo A-B). Os animais foram divididos em seis grupos com 4 animais em
cada: Grupo controle com 30 dias de idade (C30); Grupo distrófico com 30 dias de idade (D30);
Grupo controle com 60 dias de idade (C60); Grupo distrófico com 60 dias de idade (D60); Grupo
controle com 60 dias de idade suplementado com ácido ascórbico (CS60) e Grupo distrófico com
60 dias de idade suplementado com ácido ascórbico (DS60). Os animais foram mantidos no
Biotério do Departamento de Anatomia, ICB/USP, em caixas de polietileno, contendo bebedouro
(água) e comedouro (ração), sob temperatura controlada de aproximadamente 22ºC e iluminação
com ciclo de 12 horas claro/escuro.
2.2. Dieta suplementar com ácido ascórbico
Após o desmame (21 dias de idade), os animais receberam ração comercial para roedores
Nuvilab®
(Nuvital, São Paulo, Brasil). A partir dos 30 dias de idade os animais dos grupos
suplementados com ácido ascórbico (CS60 e DS60) receberam diariamente 0,005g de ácido
ascórbico (Sigma-Aldrich, St. Louis, EUA) por quilograma do animal, diluído em água e
administrado por meio de gavagem. A concentração foi calculada com base no protocolo
estabelecido na literatura (Guido et al., 2010; Tonon et al., 2012).
2.3. Eutanásia e coleta do material
Os animais foram eutanasiados em câmara de dióxido de carbono hermeticamente fechada,
com entrada de gás na parte superior. Após a eutanásia, os animais foram pesados e os testículos
coletados para serem pesados, seccionados transversalmente e imersos em solução fixadora de

35
acordo com a técnica histológica a ser empregada: Karnovsky para morfometria testicular sob
microscopia de luz e análise ultraestrutural sob microscopia eletrônica de transmissão, e
Paraformaldeído 4% em solução tampão de PBS para os procedimentos de imunohistoquímica
(Karnovsky, 1965; Oliveira, 2004; Morais et al., 2014a).
2.4. Processamento histológico
Após fixação em Karnovsky por 24h, os fragmentos testiculares destinados à morfometria
foram transferidos para álcool 70%. Os fragmentos foram desidratados em concentrações crescentes
de álcool etílico, para posterior inclusão em glicol-metacrilato (Historesin®, Leica Mycrosistems,
Heidelberg, Alemanha). O material foi seccionado a 3µm de espessura através da técnica semi-
seriada com micrótomo rotativo Leica RM2255 (Leica Microsystems, Heidelberg, Alemanha), com
intervalo de 40µm entre os cortes, e as lâminas histológicas destinadas à morfometria foram coradas
com azul de toluidina/borato de sódio 1% (Morais et al., 2014a). As lâminas permanentes
confeccionadas foram observadas ao microscópio de luz Motic BA410 (Motic, Causeway Bay,
Hong Kong) e microfotogradas para análises e comparações com câmera digital Moticam 5.0 MP
(Motic Instruments Inc, Richmond, Canada). As imagens foram analisadas ao software Image-Pro
Plus®(Media Cybernetics Inc., Rochville, USA).
2.5. Estereologia testicular
As análises morfométricas foram realizadas de acordo com os protocolos propostos por
Morais et al. (2014a). As proporções volumétricas entre túbulos seminíferos e intertúbulo foram
estimadas pela contagem de 266 pontos projetados sobre 10 imagens obtidas das preparações
histológicas de cada animal, em objetiva de 10x, para quantificar os túbulos seminíferos,
intertúbulo, túnica própria, epitélio seminífero e lúmen. Já o volume dos túbulos seminíferos foi
estimado a partir do percentual ocupado pelos mesmos no testículo e do volume do parênquima
testicular (Vol. Túbulo = peso gonadal líquido x Túbulo %/ 100). O volume do epitélio seminífero
foi o produto do percentual do epitélio seminífero com o volume do parênquima testicular, dividido
por 100. O peso da túnica albugínea foi estimado a partir da densidade de volume pela contagem de
266 pontos projetados sobre 10 imagens obtidas das preparações histológicas de cada animal, em
objetiva de 10x (Vv = No de pontos contados sobre a túnica albugínea / Total de pontos contados x
100). O volume absoluto da albugínea foi resultado do produto da densidade de volume com o
volume testicular, levando-se em consideração que a densidade testicular em mamíferos é
aproximadamente 1 (Johnson et al., 1981; Sprando et al., 1998; Costa et al., 2011).
O índice tubulossomático (ITS) foi obtido através da divisão do volume tubular pelo peso
corporal multiplicado, por 100. Já o índice gonadossomático (IGS) foi obtido como quociente da

36
divisão do peso dos testículos pelo peso corporal, sendo o valor, multiplicado por 100. O diâmetro
dos túbulos seminíferos e altura do epitélio seminífero (mensurada da túnica própria até o lúmen
tubular) foram obtidos pela mensuração de 20 secções transversais de túbulos/animal, escolhendo-
se os túbulos que apresentaram contorno o mais circular possível.
O comprimento dos túbulos seminíferos (CT), em metros, foi estimado a partir da fórmula
CT = VTS/πR2, onde VTS é o volume dos túbulos seminíferos, πR
2 é a área da secção transversal
dos túbulos seminíferos, e R é o diâmetro tubular/2. O CT por grama de testículo foi obtido
dividindo-se este valor pelo peso gonadal.
2.6. Frequência relativa dos estádios do ciclo do epitélio seminífero
Os estádios que compõem o ciclo do epitélio seminífero (CES) foram descritos utilizando-se
o método de morfologia tubular (Berndtson, 1977). A frequência relativa de cada estádio foi
descrita pela caracterização e contagem de 50 túbulos por animal em secções transversal de forma
randomizada.
2.7. Quantificação da espermatogênese
Quantificou-se a população de cada tipo celular encontrada no estádio 8 do CES, através da
contagem dos núcleos das células germinativas e do nucléolo das células de Sertoli. Foram
utilizadas 10 secções transversais circulares de túbulos seminíferos por animal para, então,
mensurar os diâmetros nucleares de espermatogônias do tipo A (A), espermatócitos primários na
transição pré-leptótenos para leptóteno (PL-L), espermatócitos primários em paquíteno (PQ),
espermátides arredondadas (Ar) e nucléolos de células de Sertoli (Se) (Russell et al., 1990). As
contagens foram corrigidas de acordo com a espessura do corte histológico e o diâmetro nuclear ou
nucleolar das células germinativas e células de Sertoli, respectivamente, de acordo com o método de
Abercrombie (1946) modificado por Amann & Almquist (1962).
A partir desses dados o índice das células de Sertoli foi obtido com a finalidade de indicar a
capacidade de suporte dessas células por total de células germinativas. Esse índice foi determinado
através do resultado da razão entre os números corrigidos de células germinativas e o número
corrigido de células de Sertoli, utilizando-se a fórmula: (A+PL-L+PQ+Ar) / Se. Os índices mitótico
(PL-L / A), meiótico (Ar / PQ) e o rendimento geral da espermatogênese (Ar / A) também foram
quantificados.
2.8. Análise da ultraestrutura
Após fixação em solução de Karnovsky por 24h, os fragmentos testiculares foram
submetidos à microscopia eletrônica de transmissão (MET). Os fragmentos foram imersos em

37
glutaraldeído a 2,5%, tamponado com fosfato de sódio a 0,1M, pH 7,4. Após a fixação o material
foi lavado três vezes durante 10 minutos cada no mesmo tampão. Em seguida ocorreu a pós-fixação
com tetróxido de ósmio tamponado a 2% por duas horas (tamponado em fosfato de sódio 0,1M e a
pH 7,4). Os fragmentos foram lavados com solução tampão por três vezes durante dez minutos
cada. Após esse procedimento o material foi imerso em acetato de uranila a 3% overnight, lavado
com tampão fosfato e desidratado com série crescente de álcool etílico (50-100%) durante 10
minutos em cada.
Após desidratação, os fragmentos foram imersos em óxido de propileno por 10 minutos para
garantir total desidratação do tecido. Em seguida o material foi embebido em resina araldite 502
(Polysciences Inc, California, EUA), para a confecção dos blocos. Foram então obtidos cortes semi-
finos com ultramicrótomo, os quais foram corados com solução aquosa de azul de toluidina a 1%
para a identificação dos locais para a realização de cortes ultrafinos. Cortes de 70nm de espessura
foram confeccionados com navalha de diamante e colocados em telas de cobre com 200 mesh para
posterior contraste com acetato de uranila saturado a 2%, por sete a dez minutos e o citrato de
chumbo a 0,5%, durante o mesmo período. O material foi analisado em Microscópio Eletrônico de
Transmissão (Jeol® 100 CX II, Tóquio, Japão).
2.9. Imunohistoquímica para caspase-3
Para identificar células em apoptose no interior dos túbulos seminíferos foi utilizada a
técnica de imunohistoquímica para marcação da caspase-3. Os cortes foram desparafinizados em
uma estufa a 60°C durante 30 minutos e colocados em xilol por 5 minutos, reidratados em
concentrações decrescentes de álcool etílico (100%, 90%, 70% e 50%) e lavados em água destilada.
Para desmascarar os epítopos, a recuperação antigênica foi realizada com solução de citrato de
sódio 10 mM (pH 6,0 95°). Esta recuperação foi realizada em panela de pressão de micro-ondas
com potência máxima de 750W, em dois ciclos de 5 minutos. Em seguida as amostras foram
arrefecidas até a temperatura ambiente e lavadas com água destilada. O bloqueio da peroxidase
endógena foi realizado com peróxido de hidrogênio 0,9%, seguindo-se a lavagem com água
destilada e solução salina tamponada com fosfato de sódio (PBS, pH 7,4). O anticorpo primário
anti-caspase 3 (diluição 1: 400, Abcam, Cambridge, Inglaterra) foi incubado durante a noite (12h).
Em seguida, os antígenos imunomarcados foram visualizados através do sistema Dako Envision+
(Dako, Carpinteria, EUA), usando 3'3-tetra-hidrocloreto de diaminobenzidina (DAB) como
cromógeno. As seções foram finalmente contrastadas com hematoxilina de Harry e montadas para
análise em microscopia de luz.

38
2.10. Densidade de Volume (Vv) das células caspase 3 - positivas
A densidade de volume (Vv) refere-se a fração de volume ocupada pelas células caspase 3-
positivas no respectivo espaço referência, ou seja, o testículo. Para a estimativa de Vv foram obtidos
3 campos por animal e aplicado um sistema teste sobre o espaço referência. Foram contados o
número total de pontos que tocam as células de interesse (Pint) os quais foram divididos pelo
número total de pontos sobre o espaço referência (Pref) (Ladd et al., 2012).
2.11. Análise estatística
Os parâmetros testiculares quantitativos foram expressos em média ± desvio padrão. Estes
foram submetidos à análise de variância (ANOVA) e teste de múltiplas comparações de Kruskall-
Wallis, seguido do teste de Mann-Whitney com correção de Bonferroni, utilizando-se o software
PAST® versão 2.17 (Hammer et al., 2001). Admitiu-se o nível de significância de P<0,05.
3. Resultados
3.1. Estereologia testicular
Na tabela 1 observam-se os resultados biométricos dos grupos estudados. O peso corporal
dos grupos distróficos pré-púbere (D30) e púbere (D60) se comportaram semelhante ao controle, ou
seja, aumentando na puberdade. A suplementação aumentou significativamente o peso corporal e
da gônada dos animais distróficos quando comparado CS60, P<0,05. O IGS foi significativamente
menor nos animais distróficos pré-púberes quando comparado C30. Já o ITS foi maior nos animais
de 30 dias, tanto controle quanto distrófico, quando comparados, respectivamente a C60 e D60.
Tabela 1 – Médias da biométrica e índices morfométricos dos testículos de camundongos
portadores de Distrofia Muscular de Duchenne, nos diferentes grupos experimentais.
Médias com pares de letras maiúsculas diferentes na mesma linha (A-B; C-D; E-F) comparação entre grupos Controle e
Distrófico (P<0,05); Médias com pares de letras minúsculas diferentes na mesma linha (a-b; c-d) comparação entre
Controle (C30-C60; C60-CS60) e Distrófico (D30-D60; D60-DS60) (P<0,05). Não foram consideradas as comparações
(C30-CS60; C30-D60; C30-DS60; C60-D30; C60-DS60; CS60-D30; CS60-D60; D30-DS60). Teste de Kruskal-Wallis
seguido de Mann Whitney com correção de Bonferroni. Dados expressos em média ± D.P. C30: Controle com 30 dias,
C60: Controle com 60 dias, CS60: Controle com 60 dias suplementado com ácido ascórbico, D30: Distrófico com 30
dias, D60: Distrófico com 60 dias, DS60: Distrófico com 60 dias suplementado com ácido ascórbico. IGS = índice
gonadossomático e ITS = índice tubulossomático.
Com relação à proporção volumétrica do compartimento tubular, a distrofia ainda no
período pré-púbere (D30) elevou o percentual de túnica própria comparado a C30 e D60, bem como

39
a suplementação elevou significativamente esse parâmetro em CS60 em relação a C60 (Tabela 2).
Contudo, o percentual de lúmen foi reduzido pela distrofia tanto no período pré-púbere quanto no
púbere quando comparando os animais dos grupos C30 e C60, respectivamente. O diâmetro tubular
foi elevado em distróficos D60 em relação a D30, esse comportamento também foi evidente entre
os controles C60 e C30. O percentual de epitélio foi significativamente elevado pela distrofia em
D30 em relação aos distróficos 60 dias não suplementado. Os distróficos pré-púberes apresentaram
redução do percentual da albugínea em relação a C30 e D60.
O comprimento tubular (CT) por testículo foi significativamente maior em pré-púberes
quando comparado a púberes nos grupos distróficos, bem como o comprimento tubular por grama
de testículo de D30 quando comparado a C30 e D60 (Tabela 2).

40
Tabela 2 – Médias da morfometria do compartimento tubular dos testículos de camundongos
portadores de Distrofia Muscular de Duchenne, nos diferentes grupos experimentais.
Médias com pares de letras maiúsculas diferentes na mesma linha (A-B; C-D; E-F) comparação entre grupos Controle e
Distrófico (P<0,05); Médias com pares de letras minúsculas diferentes na mesma linha (a-b; c-d) comparação entre
Controle (C30-C60; C60-CS60) e Distrófico (D30-D60; D60-DS60) (P<0,05). Não foram consideradas as comparações
(C30-CS60; C30-D60; C30-DS60; C60-D30; C60-DS60; CS60-D30; CS60-D60; D30-DS60). Teste de Kruskal-Wallis
seguido de Mann Whitney com correção de Bonferroni. Dados expressos em média ± D.P. C30: Controle com 30 dias,
C60: Controle com 60 dias, CS60: Controle com 60 dias suplementado com ácido ascórbico, D30: Distrófico com 30
dias, D60: Distrófico com 60 dias, DS60: Distrófico com60 dias suplementado com ácido ascórbico. Proporção
volumétrica (%) e volume (mL) dos compartimentos testiculares, diâmetro tubular (µm), altura do epitélio (µm),
comprimento tubular por testículo (CT) (m), comprimento tubular por grama de testículo (CT/g de T) (m).
3.2. Frequência dos estádios do ciclo do epitélio seminífero
De acordo com o método da morfologia tubular, o epitélio seminífero dos camundongos
distrófico foi descrito com 8 estádios, semelhante ao grupos controle. As médias das frequências
relativas de cada estádio encontram-se na Figura 1. O estádio 4 foi o menos frequente (0,58%),
enquanto que o estádio 8 foi o mais frequente (30,05%). Apesar de estar presente no grupo C30 e de
ser o estádio mais frequente nos demais grupos, a distrofia em pré-púberes apenas não apresentou o
estádio 8 em seu ciclo,. As fases pré-meiótica (estádios 1-3), meiótica (estádio 4) e pós-meiótica
(estádios 5-8) representaram respectivamente 36,17%, 0,58% e 61,09% do ciclo do epitélio
seminífero.
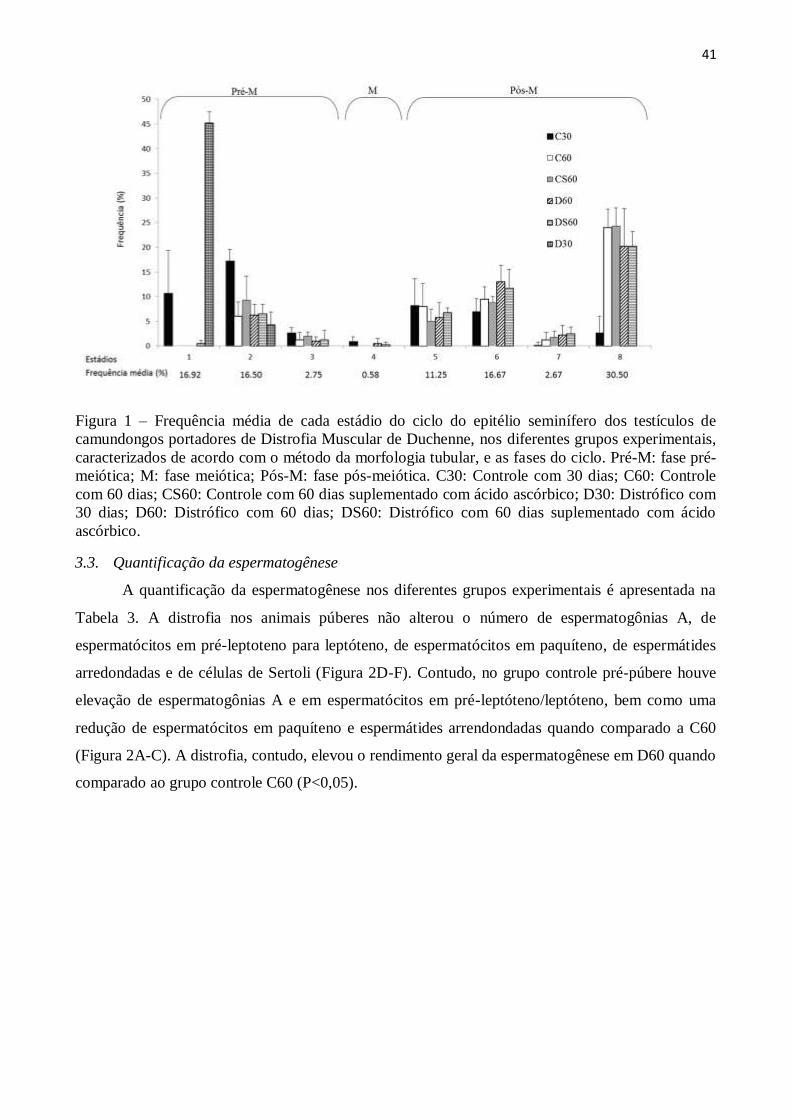
41
Figura 1 – Frequência média de cada estádio do ciclo do epitélio seminífero dos testículos de
camundongos portadores de Distrofia Muscular de Duchenne, nos diferentes grupos experimentais,
caracterizados de acordo com o método da morfologia tubular, e as fases do ciclo. Pré-M: fase pré-
meiótica; M: fase meiótica; Pós-M: fase pós-meiótica. C30: Controle com 30 dias; C60: Controle
com 60 dias; CS60: Controle com 60 dias suplementado com ácido ascórbico; D30: Distrófico com
30 dias; D60: Distrófico com 60 dias; DS60: Distrófico com 60 dias suplementado com ácido
ascórbico.
3.3. Quantificação da espermatogênese
A quantificação da espermatogênese nos diferentes grupos experimentais é apresentada na
Tabela 3. A distrofia nos animais púberes não alterou o número de espermatogônias A, de
espermatócitos em pré-leptoteno para leptóteno, de espermatócitos em paquíteno, de espermátides
arredondadas e de células de Sertoli (Figura 2D-F). Contudo, no grupo controle pré-púbere houve
elevação de espermatogônias A e em espermatócitos em pré-leptóteno/leptóteno, bem como uma
redução de espermatócitos em paquíteno e espermátides arrendondadas quando comparado a C60
(Figura 2A-C). A distrofia, contudo, elevou o rendimento geral da espermatogênese em D60 quando
comparado ao grupo controle C60 (P<0,05).
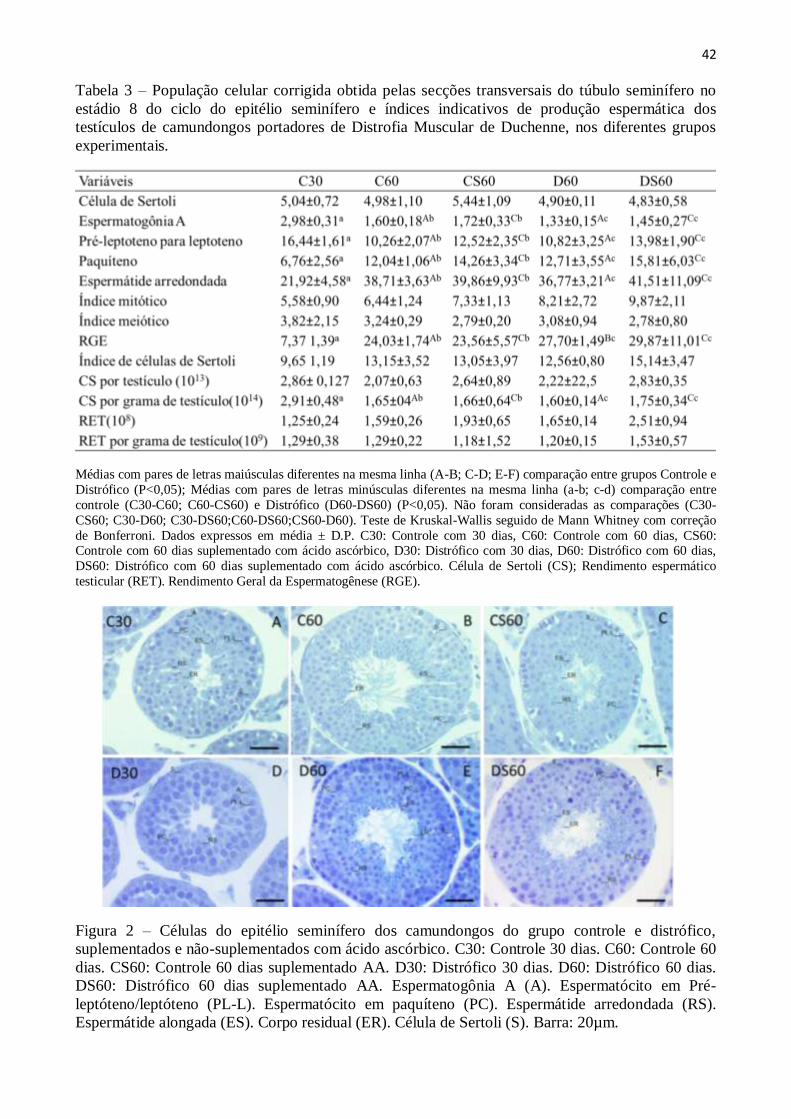
42
Tabela 3 – População celular corrigida obtida pelas secções transversais do túbulo seminífero no
estádio 8 do ciclo do epitélio seminífero e índices indicativos de produção espermática dos
testículos de camundongos portadores de Distrofia Muscular de Duchenne, nos diferentes grupos
experimentais.
Médias com pares de letras maiúsculas diferentes na mesma linha (A-B; C-D; E-F) comparação entre grupos Controle e
Distrófico (P<0,05); Médias com pares de letras minúsculas diferentes na mesma linha (a-b; c-d) comparação entre
controle (C30-C60; C60-CS60) e Distrófico (D60-DS60) (P<0,05). Não foram consideradas as comparações (C30-
CS60; C30-D60; C30-DS60;C60-DS60;CS60-D60). Teste de Kruskal-Wallis seguido de Mann Whitney com correção
de Bonferroni. Dados expressos em média ± D.P. C30: Controle com 30 dias, C60: Controle com 60 dias, CS60: Controle com 60 dias suplementado com ácido ascórbico, D30: Distrófico com 30 dias, D60: Distrófico com 60 dias,
DS60: Distrófico com 60 dias suplementado com ácido ascórbico. Célula de Sertoli (CS); Rendimento espermático
testicular (RET). Rendimento Geral da Espermatogênese (RGE).
Figura 2 – Células do epitélio seminífero dos camundongos do grupo controle e distrófico,
suplementados e não-suplementados com ácido ascórbico. C30: Controle 30 dias. C60: Controle 60
dias. CS60: Controle 60 dias suplementado AA. D30: Distrófico 30 dias. D60: Distrófico 60 dias.
DS60: Distrófico 60 dias suplementado AA. Espermatogônia A (A). Espermatócito em Pré-
leptóteno/leptóteno (PL-L). Espermatócito em paquíteno (PC). Espermátide arredondada (RS).
Espermátide alongada (ES). Corpo residual (ER). Célula de Sertoli (S). Barra: 20µm.

43
3.4. Análise ultraestrutural
A microscopia eletrônica de transmissão revelou que a túnica própria possui células mióides
com usual formato fusiforme aderido à membrana basal nos grupos controle ou em variados graus
de desorganização nos grupos distróficos. Nos animais C30 a túnica própria foi formada por uma
camada de células mióides e uma camada de fibras colágenas (Figura 3A). Já em D30 as fibras
colágenas estavam dispostas separando a camada de células mióides da membrana basal (Figura
3B). O grupo C60 apresentou célula mióide alongada e núcleo mais arredondado (Figura 3C).
Todavia, em D60 as células mióides apresentaram formato oval e núcleo irregular com uma camada
de fibras colágenas separando-as da membrana basal (Figura 3D). Em CS60 a célula mióide era
fusiforme, a camada de fibras colágenas além de possuir vacúolos e estava desarticulada da lâmina
própria (Figura 3E). Em DS60 houve perda da integridade da célula mióide descaracterizando a
túnica própria (Figura 3F).
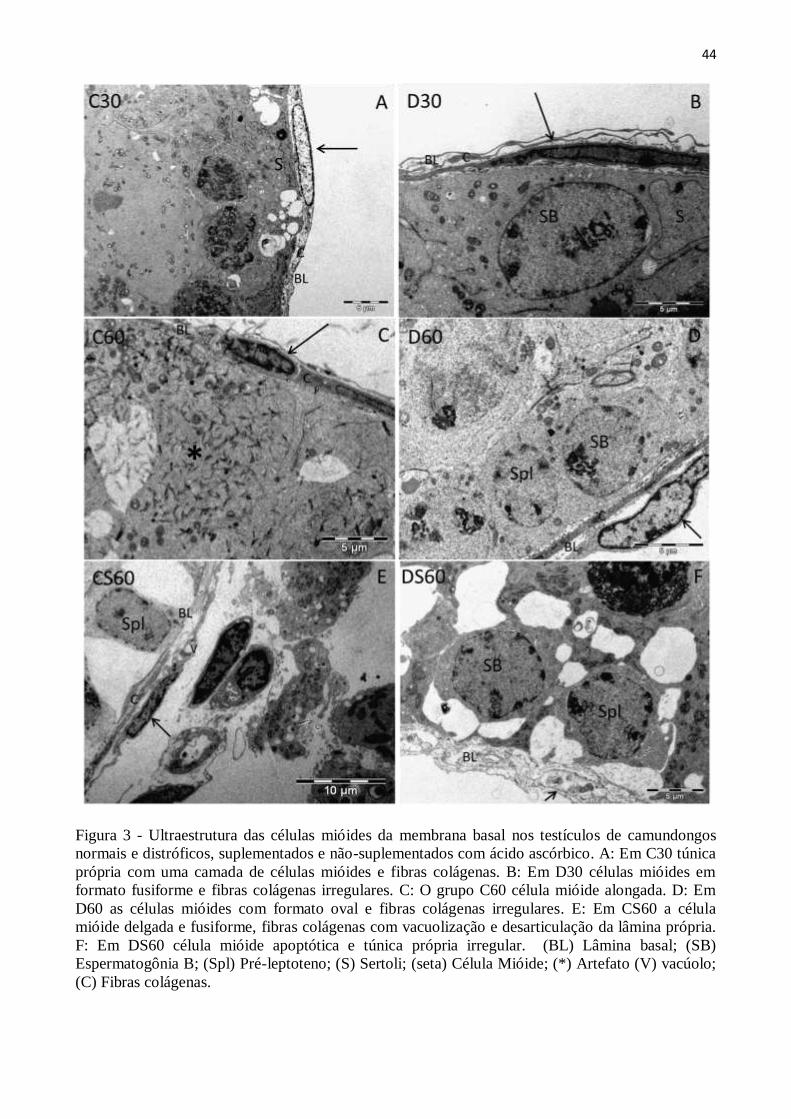
44
Figura 3 - Ultraestrutura das células mióides da membrana basal nos testículos de camundongos
normais e distróficos, suplementados e não-suplementados com ácido ascórbico. A: Em C30 túnica
própria com uma camada de células mióides e fibras colágenas. B: Em D30 células mióides em
formato fusiforme e fibras colágenas irregulares. C: O grupo C60 célula mióide alongada. D: Em
D60 as células mióides com formato oval e fibras colágenas irregulares. E: Em CS60 a célula
mióide delgada e fusiforme, fibras colágenas com vacuolização e desarticulação da lâmina própria.
F: Em DS60 célula mióide apoptótica e túnica própria irregular. (BL) Lâmina basal; (SB)
Espermatogônia B; (Spl) Pré-leptoteno; (S) Sertoli; (seta) Célula Mióide; (*) Artefato (V) vacúolo;
(C) Fibras colágenas.

45
As células de Sertoli apresentaram típicas variações no formato nuclear e do nucléolo
proeminente nos grupos controle (Figura 4A, 4D, 4G). Nos grupos distróficos as células de Sertoli
possuem núcleo que varia de arredonda a alongado com irregularidades, nucléolos proeminentes e
vacuolização citoplasmática (Figura 4C, 4E, 4H). As junções dessas células foram do tipo gap ou
oclusão (Figura 4A-I).
A morfologia mitocondrial variou nos grupos estudados. Na Figura 5A-B observa-se, que as
mitocôndrias encontradas nas espermátides em distróficos 30 foram semelhante às encontradas em
C30, tipo ortodoxa (formato oval com cristas lamelares interpostas por matriz eletrodensa) e
condensada (formato alongado ou oval com dilatações e espaços entre as cristas). As células de
Sertoli do grupo C60 apresentaram mitocôndrias do tipo ortodoxa (Figura 5C). Todavia, o grupo
D60 a predominância foi a do tipo condensada (Figura 5D). Nos grupos suplementados
apresentaram características de apoptose mitocondrial nas espermátides, como a presença de
núcleos eletrodensos e clusters mitocondriais (Figura 5E-F).
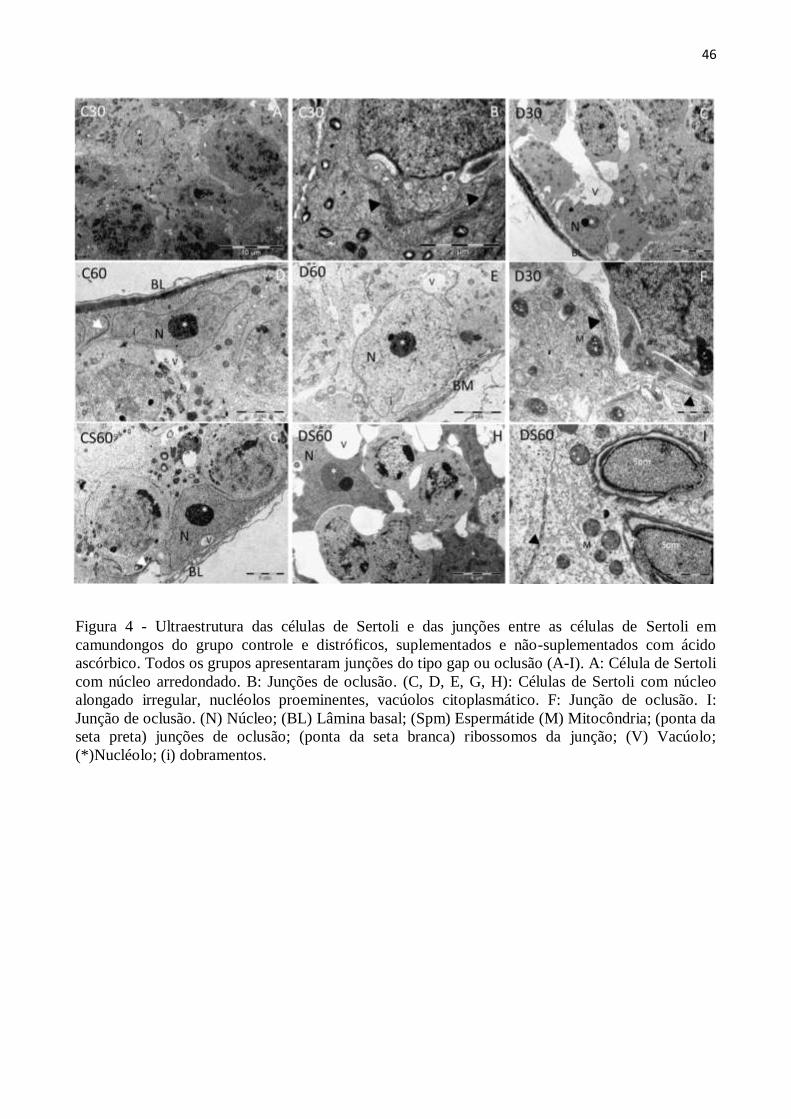
46
Figura 4 - Ultraestrutura das células de Sertoli e das junções entre as células de Sertoli em
camundongos do grupo controle e distróficos, suplementados e não-suplementados com ácido
ascórbico. Todos os grupos apresentaram junções do tipo gap ou oclusão (A-I). A: Célula de Sertoli
com núcleo arredondado. B: Junções de oclusão. (C, D, E, G, H): Células de Sertoli com núcleo
alongado irregular, nucléolos proeminentes, vacúolos citoplasmático. F: Junção de oclusão. I:
Junção de oclusão. (N) Núcleo; (BL) Lâmina basal; (Spm) Espermátide (M) Mitocôndria; (ponta da
seta preta) junções de oclusão; (ponta da seta branca) ribossomos da junção; (V) Vacúolo;
(*)Nucléolo; (i) dobramentos.
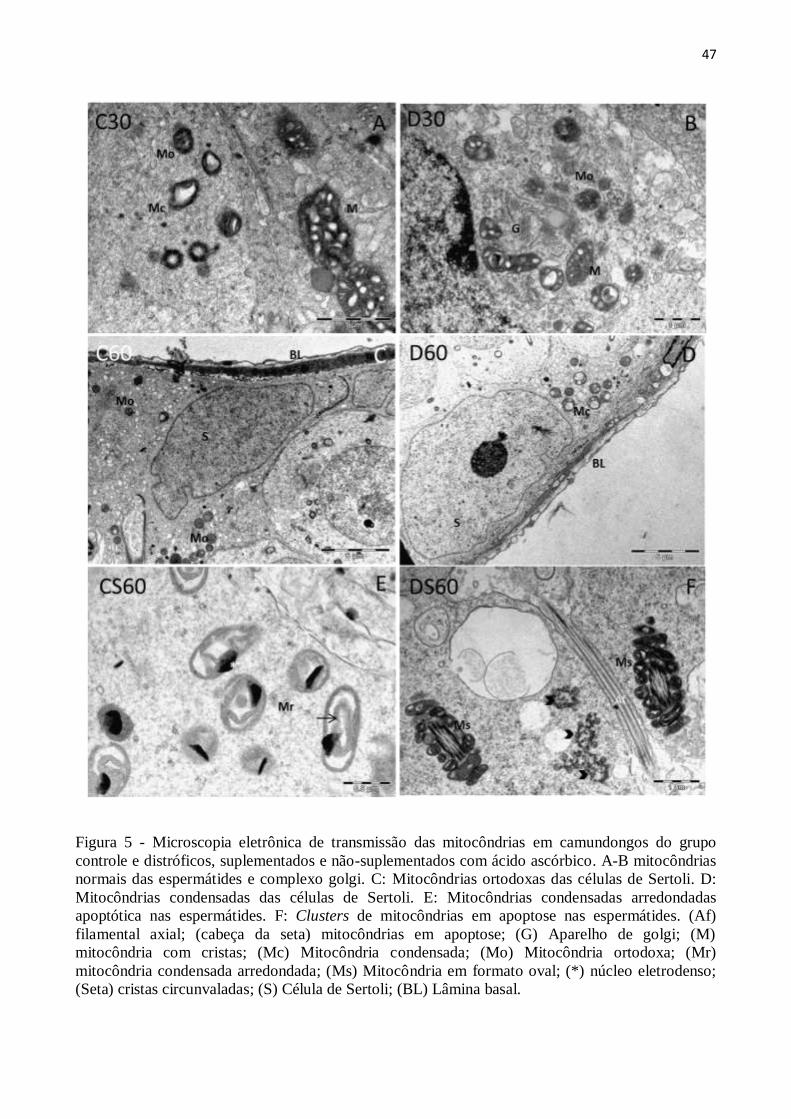
47
Figura 5 - Microscopia eletrônica de transmissão das mitocôndrias em camundongos do grupo
controle e distróficos, suplementados e não-suplementados com ácido ascórbico. A-B mitocôndrias
normais das espermátides e complexo golgi. C: Mitocôndrias ortodoxas das células de Sertoli. D:
Mitocôndrias condensadas das células de Sertoli. E: Mitocôndrias condensadas arredondadas
apoptótica nas espermátides. F: Clusters de mitocôndrias em apoptose nas espermátides. (Af)
filamental axial; (cabeça da seta) mitocôndrias em apoptose; (G) Aparelho de golgi; (M)
mitocôndria com cristas; (Mc) Mitocôndria condensada; (Mo) Mitocôndria ortodoxa; (Mr)
mitocôndria condensada arredondada; (Ms) Mitocôndria em formato oval; (*) núcleo eletrodenso;
(Seta) cristas circunvaladas; (S) Célula de Sertoli; (BL) Lâmina basal.

48
3.5. Contagem de células apoptóticas
Através da densidade de volume (Vv) das células apoptóticas em secções transversais do
túbulo seminífero é possível observar que a suplementação com ácido ascórbico nos distróficos
(DS60: 0,78±0,0009%) reduziu a expressão de células positivas para caspase-3 quando comparado
ao grupo CS60 (2,39±0,009%). A distrofia apresentou redução de células apoptóticas em D60
(0,19±0,0003%) quando comparado ao grupo distrófico pré-púbere D30 (2,01±0,0048%). Isso
também foi observado entre os grupos controle C60 (0.0129±0.0006%) e C30 (0,145±0,0004%).
(Figura 6A-F).
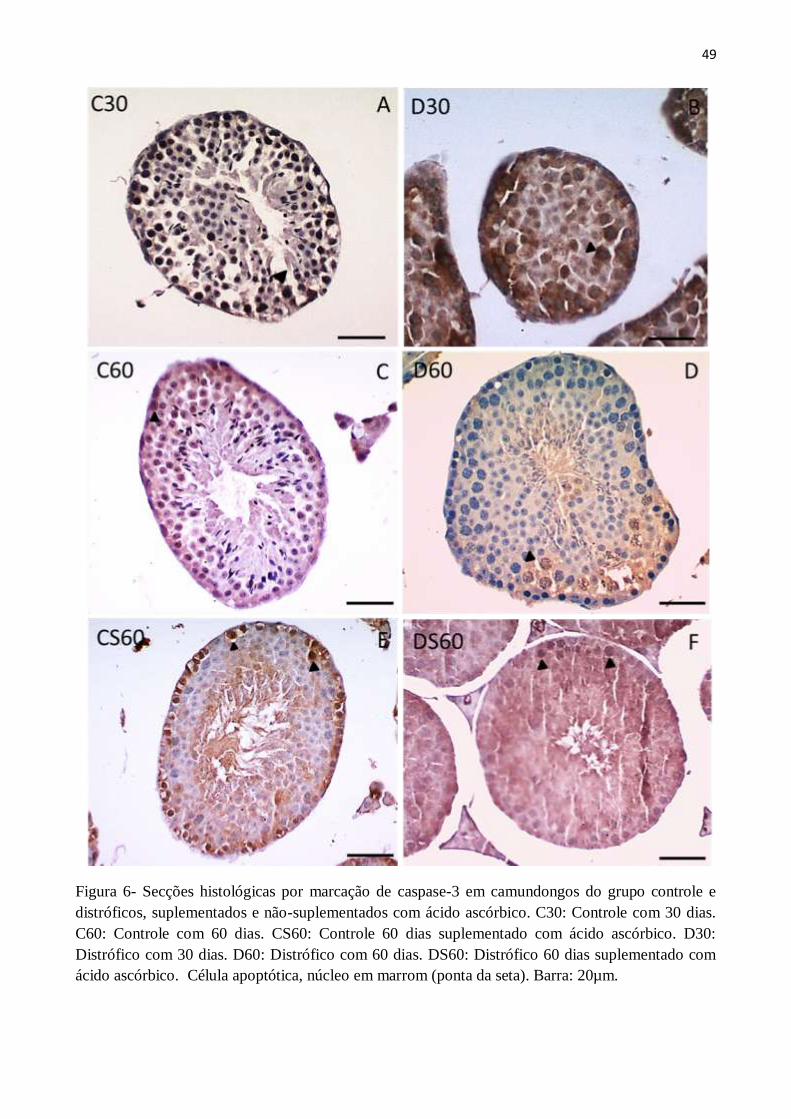
49
Figura 6- Secções histológicas por marcação de caspase-3 em camundongos do grupo controle e
distróficos, suplementados e não-suplementados com ácido ascórbico. C30: Controle com 30 dias.
C60: Controle com 60 dias. CS60: Controle 60 dias suplementado com ácido ascórbico. D30:
Distrófico com 30 dias. D60: Distrófico com 60 dias. DS60: Distrófico 60 dias suplementado com
ácido ascórbico. Célula apoptótica, núcleo em marrom (ponta da seta). Barra: 20µm.

50
4. Discussão
A morfologia e a morfometria do compartimento tubular na Distrofia Muscular de Duchenne
ainda não está esclarecida na literatura científica, assim como o efeito da suplementação a base de
ácido ascórbico nessa patologia. Dessa forma, esse trabalho avaliou se a morfometria tubular e os
índices de produção espermáticos em camundongos mdx podem ser alterados pela DMD, bem
como, se há influência da suplementação com ácido ascórbico sobre esses parâmetros. De acordo
com França & Russell (1998), tanto o peso como o tamanho testicular são indicadores quantitativos
da produção espermática. A elevação do peso da gônada pode implicar em alterações do epitélio
seminífero ou edema intersticial (Sellers et al., 2007). Todavia, nesse trabalho a suplementação com
ácido ascórbico em distróficos púberes elevou o peso testicular sem alterar significativamente o
epitélio seminífero e o intertúbulo. Isso possivelmente ocorreu em virtude da intervenção do ácido
ascórbico na manutenção da integridade fisiológica nos testículos, visto que diversas funções
enzimáticas da vitamina C são essenciais para a síntese, desenvolvimento e manutenção testicular
(Yousef, 2005).
A suplementação com ácido ascórbico ao elevar o peso corporal em distróficos púberes pode
ter influenciado, positivamente, o comportamento reprodutivo. Essa variável é necessária para
determinação do índice gonadossomático (IGS), que representa a percentagem de massa corpórea
que corresponde ao testículo, e, do índice tubulussomático (ITS), que quantifica a proporção de
túbulos seminíferos em relação à massa corporal (Paula et al., 2002). O IGS mostrou-se reduzido
em camundongos distróficos na pré-puberdade quando comparado ao controle C30, isso implica
que é requerido menor investimento de massa corporal para formação da gônada em virtude do
maior peso corporal (Kenagy & Trombulak, 1986). Nessa pesquisa os valores médios do ITS nos
camundongos controle foram próximos ao descrito Melo et al. (2010) para ratos, 0,50%, no entanto,
no presente trabalho observa-se uma redução no ITS na transição da pré-puberdade para puberdade
nos grupos distróficos e controle. Estes resultados indicam que na distrofia, semelhante ao controle,
o maior investimento em túbulos seminíferos pode estar diretamente relacionado com a maior
quantidade de energia requerida para a produção espermática no período pré-púbere.
A túnica albugínea e o mediastino testicular são constituintes morfológicos dos testículos,
apesar de não estarem diretamente relacionados à espermatogênese pode alterar o peso da gônada e
interferir em alguns parâmetros morfométricos (Azevedo et al., 2006). O percentual testicular
ocupado pela túnica albugínea foi abaixo da média encontrada para ratos (6,5%, Russell & França,
1995). Contudo, a distrofia reduziu esse percentual ainda no período pré-púbere, assim como, em
testículos de camundongos expostos a fluoreto de sódio, cujos percentuais de túnica albugínea
foram diferentes para o grupo controle e menores em relação ao padrão para ratos, com percentual

51
de 1,81%, e para animais expostos a diferentes concentrações de fluoreto de sódio, com 1,63-2,01%
(Sprando et al., 1998).
Ao remover a túnica albugínea é obtido o parênquima testicular, que é dividido em
compartimento tubular e compartimento intertubular, sendo o compartimento tubular o principal
componente do testículo. A organização do compartimento tubular foi semelhante às descrições
prévias para a maioria dos mamíferos, com túbulos seminíferos compostos pela túnica própria com
células mióides, epitélio seminífero e lúmen (Russell et al., 1990; França & Russell, 1998; França &
Godinho, 2003; Costa et al., 2006; Beguelini et al., 2009; Costa et al., 2011). Na túnica própria dos
animais de todos os grupos observou-se apenas uma camada de células mióides, assim como
descrito na literatura para outros roedores (Bustos-Obregón, 1976). Essas células são contráteis e a
distrofia alterou sua morfologia mostrando núcleo irregular, conforme observado na análise
ultraestrutural, isso sugere perda de suas características funcionais, segundo afirmam Martin et al.
(1992) em estudo realizado em pacientes com a síndrome de Klynefelter. Nesse caso os
camundongos distróficos podem apresentar comprometimento funcional, como ao elevar a força
para o movimento dos fluidos e da propulsão dos espermatozoides no túbulo seminífero (Clermont,
1958; Ross, 1967; Suvanto & Cormano, 1970; Russell et al., 1989a).
De acordo com Russell & França (1995) a túnica própria, ou tecido peritubular, compreende
1,2% do testículo em ratos. Na pré-puberdade, o percentual de túnica própria foi maior nos
distróficos em relação aos animais controle, indicando uma maior espessura dessa túnica. Isso
certamente foi decorrente do aumento de deposição de fibras colágenas (Bustos-Obregón, 1976).
Essa elevação pode inibir a gametogênese em virtude do desequilíbrio das vias parácrinas de
comunicação entre o túbulo e intertúbulo (Adam et al., 2011, 2012). A suplementação com ácido
ascórbico no grupo controle aumentou significativamente o percentual de túnica própria, que na
análise ultraestrutural revelou aumento de fibras colágenas que descaracterizaram a integridade da
lâmina basal, possivelmente decorrente da participação crucial do ácido ascórbico na síntese de
colágeno (Khalid et al., 2004). No entanto, nos distróficos suplementados não foi observado esse
aumento no percentual da túnica própria, entretanto, verificou-se comportamento semelhante das
fibras colágenas na MET. Esses achados ultraestruturais foram semelhantes ao descrito na literatura
para ratos tratados com nicotina e segundo Aydos et al. (2001) podem prejudicar a progressão
normal da espermatogênese.
O percentual de lúmen foi reduzido pela distrofia e pode ocasionar uma restrição parcial do
refluxo de fluido testicular (Russell et al., 1989b). O lúmen está diretamente relacionado ao
diâmetro tubular, um indicador da atividade espermatogênica testicular (Attal et al., 1963; Sinha-

52
Hikim et al., 1988; França & Cardoso, 1998). Os valores encontrados para o diâmetro tubular em
todos os grupos desse trabalho estavam abaixo do descrito para a maioria dos amniotas, 180-300µm
(Roosen-Runge, 1977). Porém, de acordo com Roosen-Runge (1961) os ratos adultos possuem
redução do diâmetro tubular ao logo da rede testicular, podendo alcançar valor igual ou menor a
100µm. Dessa forma, nesse trabalho o diâmetro tubular em distróficos comportou-se de forma
semelhante aos animais dos grupos controle, e a suplementação não afetou significativamente essa
variável em distróficos, assim como em ratos após a suplementação com extratos vegetais de
Tynnanthus fasciculatus (Melo et al., 2010).
O diâmetro tubular ao longo de todo o testículo de ratos pode sofrer diferenças significativas
em virtude das variações ao longo do ciclo do epitélio seminífero, estando relacionado com o
aumento do lúmen tubular que antecede a espermiação (Wing & Christensen, 1982). Logo, medir a
altura do epitélio seminífero é uma ferramenta que permite investigar efetivamente as variações da
produção espermática ao longo do ciclo do epitélio seminífero, uma vez que o epitélio seminífero
reflete diretamente o processo espermatogênico (Azevedo et al., 2006). Apesar de não apresentar
variação significativa a altura do epitélio dos grupos estudados foi abaixo da média encontrada para
animais domésticos, 60-100µm (França & Russell, 1998). Contudo, é possível que ocorra redução
da altura do epitélio sem alterar severamente o processo espermatogênico (França et al., 2000).
A distrofia em animais pré-púberes elevou o comprimento tubular por testículo a valores
superiores ao descrito para ratos Sprague-Dawley adultos com 21,8±0,6 metros de túbulo
seminífero (Ghosh et al., 1992). Isso também foi observado no comprimento tubular por grama de
testículo, um parâmetro produtivo, logo é provável que a morfologia testicular da distrofia tenha se
preparado para a produção espermática na pré-puberdade, mas fisiologicamente a puberdade
precoce não acontece. Já que os pacientes portadores da DMD possuem puberdade tardia decorrente
do número de doenças crônicas associadas, problemas nutricionais secundários ou aumento da
demanda energética (Ezri et al., 2012).
O epitélio seminífero é constituído por células germinativas e somáticas organizadas em 8
estádios celulares de acordo com o método da morfometria tubular (Berndtson, 1977; Russell et al.,
1990). Através dessa organização é possível obter o CES, uma ferramenta utilizada para
compreender a espermatogênese e identificar possíveis erros (Russell et al., 1990). Além disso, a
análise da frequência relativa do CES é fundamental para determinar a duração de cada estádio,
sendo os estádios menos frequentes aqueles de curta duração, enquanto os estádios mais frequentes
são aqueles de maior duração (França et al., 1998; Costa et al., 2011). Nesse trabalho a distrofia
apresentou 8 estádios, sendo o estádio 8 o mais frequente, e o 4 o menos frequente. Resultados

53
semelhantes foram encontrados para outras espécies de roedores (Melo et al., 2013; Morais et al.,
2014b). O processo espermatogênico pode ser divido nas fases pré-meiótica, meiótica e pós-
meiótica e, para fins de análise comparativa, as frequências são agrupadas e distribuídas entre as
fases (Courot et al., 1970). Assim como descrito para outros roedores, a fase pós-meiótica foi a mais
representativa do ciclo nesse trabalho, implicando em alta diferenciação celular no estádio 8
(Morais et al. 2014b). A distribuição das fases nos grupos estudados foi semelhante, contudo foi
possível observar que o estádio 8 no controle pré-púbere e sua ausência no grupo distrófico de 30
dias de idade. Esse estádio é caracterizado pela presença de espermatozoides no lúmen,
confirmando a baixa atividade espermatogênica no período pré-púbere de animais distróficos.
Ao longo do ciclo do epitélio seminífero os estádios possuem diferentes linhagens celulares
que ao serem quantificadas permitem a análise funcional do parênquima testicular e estimam a
produção espermática (França, 1991). Ainda que a distrofia não tenha alterado a morfometria das
células do epitélio seminífero durante a puberdade, ocorreu aumento do rendimento geral da
espermatogênese, mas possivelmente prejudicou a espermiogênese. Isso pode ser decorrente da
redução da frequência do estádio 8 em distróficos púberes quando comparado ao controle de mesma
idade, visto que é nesse estádio que encontramos espermátides alongadas prontas para a
espermiação no lúmen, e que não são avaliadas quantitativamente pelo rendimento geral da
espermatogênese. Dessa forma, pode haver implicações na qualidade seminal em distróficos. De
acordo com Hernández-González et al. (2005) camundongos distróficos mutantes, que não
expressam produtos do gene da distrofina, possuíram alteração morfológica flagelar, causando
redução progressiva da motilidade espermática, na ausência da proteína Dp71, uma isoforma da
proteína distrofina de baixo peso molecular. Todavia, segundo Peres et al. (2014) cães distróficos
Golden Retriever apresentam volume de ejaculação, concentração espermática, motilidade e
morfologia espermática dentro dos padrões normais, mas é possível que ocorra teratozoospermia
(mais de 20% dos espermatozoides totais com anormalidades) e redução do volume seminal
decorrente da ineficiente contração do músculo prostático. Dessa forma, apesar da morfometria
favorecer o aumento do rendimento da espermatogênese, a qualidade seminal dependerá da
gravidade da DMD.
Além das células germinativas, o número de células de Sertoli é um fator determinante da
capacidade espermática e do tamanho testicular em mamíferos, visto que são reguladoras da
espermatogênese (Orth et al., 1988; Hess et al., 1993; Weinbauer et al., 2010). Além disso, cada
célula de Sertoli é capaz de suportar um número de células germinativas, conhecido como índice da
célula de Sertoli (Russell & Peterson, 1984). Apesar de não haver alteração nos índices de célula de
Sertoli, os animais distróficos pré-púberes apresentaram vacuolizações nessas células. Esse achado

54
antecede as alterações nas células do epitélio germinativo, sendo considerado por alguns autores
como o primeiro sinal morfológico dos danos tubulares e que acontece em virtude de dilatações no
retículo endoplasmático (Creasy, 2001; Boekelheide, 2005). O aumento da densidade de volume de
células positivas para caspase-3, nos túbulos dos animais distróficos, pode está relacionada com as
alterações das células de Sertoli e consequentemente maior apoptose do epitélio germinativo nesses
animais.
As mitocôndrias testiculares modificam sua morfologia, localização e metabolismo ao longo
da espermatogênese (Ramalho-Santos et al., 2009). Segundo De Martino et al. (1979) as
mitocôndrias se classificam em três tipos: tipo ortodoxa predominam em células de Sertoli,
espermatogônias e em espermatócitos pré-leptóteno/leptóteno; tipo intermediária em espermatócito
em zigóteno; e o tipo condensada em espermatócito em paquíteno e espermátides. Contudo, as
células de Sertoli podem alterar a morfologia de suas mitocôndrias através da liberação de fatores.
A liberação de Activina A pelas células de Sertoli induz a mudança para a forma condensada, a fim
de aumentar a eficiência metabólica e contribuir para a regulação da diferenciação de células
germinativas (Meinhardt et al., 2000). Dessa forma, a distrofia alterou a morfologia mitocondrial
no período púbere para o tipo condensada para melhorar o metabolismo energético de forma
compensatória.
A ativação da apoptose pela via de caspase-3 em distróficos foi semelhante ao controle.
Logo, é possível afirmar que ocorreu apoptose fisiológica, visto que esse evento é necessário para
manter a homeostasia das proporções de células de Sertoli e de células germinativas durante a
primeira onda da espermatogênese (Rodriguez et al., 1997; Pentikäinen et al., 2003). A apoptose
durante esse período pode ser elevada pela redução dos níveis de FSH, e inibida pelo aumento de
testosterona (Rodriguez et al., 1997; Jahnukainen et al., 2004). A concentração utilizada na
suplementação com ácido ascórbico para o grupo controle não foi suficiente para reduzir a apoptose
fisiológica na puberdade, visto que a apoptose via caspase-3 em espermatogônia A e espermatócito
em paquíteno é um controle preventivo da produção exacerbada de gametas, durante a primeira
onda da espermatogênese na puberdade (Jahnukainen et al., 2004). Contudo, em distróficos
suplementados houve uma redução na Vv de células positivas para caspase-3. Portanto a
suplementação atuou reduzindo o estresse oxidativo na região basal do compartimento tubular, uma
vez que o tratamento com antioxidantes como a vitamina E, o ácido ascórbico e o selênio atuam na
apoptose desencadeada pela via das caspases (Koyuturk et al., 2006).
Com isso é possível inferir que a Distrofia Muscular de Duchenne afeta alguns parâmetros
da morfologia testicular e, consequentemente, o processo da espermatogênese ainda no período pré-

55
púbere do indivíduo, mas não impede a reprodução. Além disso, a concentração de ácido ascórbico
utilizada foi eficaz para reverter os processos apoptóticos na região basal do compartimento tubular
em camundongos distróficos.
Referências
Abbott D, Jepson M, Hastie J. (2015) Men living with long‐term conditions: exploring gender and
improving social care. Health Soc Care Community 1:8.
Abercrombie M. (1946) Estimation of nuclear populations from microtome sections. Anat Rec
94, 239-247.
Adam M, Urbanski HF, Garyfallou VT, et al. (2012) High levels of the extracellular matrix
proteoglycan decorin are associated with inhibition of testicular function. Int J Androl 25, 1-12.
Adam M, Schwarzer JU, Kohn FM, et al. (2011) Mast cell tryptase stimulates production of
decorin by human testicular peritubular cells: possible role of decorin in male infertility by
interfering with growth factor signaling. Human Rep 26, 2613-2625.
Amann RP, Almquist JO (1962) Reproductive capacity of dairy bulls. VIII. Dand indirect
measurement of testicular sperm production. J Dairy Sci 45, 774-781.
Angulo C, Maldonado R, Pulgar E, et al. (2011) Vitamin C and oxidative stress in the
seminiferous epithelium. Biol Res 44,169-180.
Attal J, Courot M, Richetin C, et al. (1963) Développement testiculaire et établissement de la
spermatogénèse chez le taureau. Ann Biol Anim Biochim Biophys 3, 219-241.
Aydos K, Güven MC, Can B, et al. (2001) Nicotine toxicity to the ultrastructure of the testis in
rats. BJU Int 88, 622-626.
Azevedo MHF, Paula TAR, Matta SLP, et al. (2006) Morfometria testicular e o túbulo
seminífero da onça-pintada (Panthera onca) adulta. Revista CERES 53, 374-381.
Beguelini MR, Moreira PR, Faria KC, et al. (2009) Morphological characterization of the
testicular cells and seminiferous epithelium cycle in six species of Neotropical bats. J Morpho 270,
943-953.

56
Berndtson WE (1977) Methods for quantifying mammalian spermatogenesis: a review. J Anim Sci
44, 818-883.
Boekelheide K (2005) Sertoli cell toxicantes. In: Sertoli Cell Biology, ed. (eds Skinner MK,
Griswold MD), pp. 345-382, San Diego: Elsevier.
Bradley D, Parsons E (1998) Newborn screening for Duchenne muscular dystrophy. Semin
Neonatol 3, 27-34.
Bustos‐Obregon E (1976) Ultrastructure and function of the lamina propria of mammalian
seminiferous tubules. Andrologia 8, 179-185.
Clermont Y (1958) Contractile elements in the limiting membrane of the seminiferous tubules of
the rat. Exp Cell Res 15, 438-440.
Costa DS, Macedo DB, Santos MD, et al. (2011) Intrinsic rate of spermatogenesis in free-ranging
feral pigs (Sus scrofa sp). Anim Reprod Sci 125, 165-169.
Costa DS, Paula TAR, Matta SLP (2006) The intertubular compartment morphometry in
capybaras (Hydrochoerus hydrochaeris) testis. Anim Reprod Sci 91, 173-179.
Courot M, Hochereau-de Riviers MT, Ortavant R (1970) Spermatogenesis. In: The Testis, ed
(eds Johnson AD, Gomes WR, Vandemark NL.), pp. 339-432, New: York: Academic Press.
Creasy DM (2001) Pathogenesis of male reproductive toxicity. Toxic Pathol 29, 64-76.
De Martino C, Floridi A, Marcante ML, et al. (1979) Morphological, histochemical and
biochemical studies on germ cell mitochondria of normal rats. Cell Tissue Res 196, 1-22.
Dorchies OM, Wagner S, Vuadens O, et al. (2006) Green tea extract and its major polyphenol (-)-
epigallocatechin gallate. Am J Physiol Cell Physiol 290, 616 –625.
Drucher WD, Blanc WA, Rowland LP, et al. (1963) The testis in myotonic muscular dystrophy: a
clinical and pathologic study with a comparison with the Klinefelterm syndrome. J Clin Endocrinol
Metab 23, 59-75.
Ezri J, Marques-Vidal P, Nydegger A (2012). Impact of disease and treatments on growth and
puberty of pediatric patients with inflammatory bowel disease. Digestion 85, 308-319.

57
França LR, Godinho CL (2003) Testis morphometry, seminiferous epithelium cycle length,
and daily sperm production in domestic cats (Felis catus). Biol Reprod 68, 1554–1561
França LR, Leal MC, Sasso-Cerri E, et al. (2000) Cimetidine (Tagamet) is a reproductive
toxicant in male rats affecting peritubular cells. Biol Repro 63, 1403-1412.
França LR, Cardoso FM (1998) Duration of spermatogenesis and sperm transit time through the
epididymis in the Piau boar. Tissue Cell 30, 573-582.
França LR, Ogawa T, Avarbock MR, et al. (1998) Germ cell genotype controls cell cycle during
spermatogenesis in the rat. Biol Reprod 59, 1371-1377.
França LR, Russell LD (1998) The testis of domestic mammals. In: Male Reproduction: A
Multidisciplinary Overview, 1ª ed. (eds Martinez-Garcia F, Regadera J), pp. 197–219, Madrid:
Churchill Livingstone.
França LR (1991) Análise morfofuncional da espermatogênese de suínos adultos da raça Piau
(Tese). Belo Horizonte (MG): Universidade Federal de Minas Gerais. 180p.
Ghosh S, Bartke A, Grasso P, et al. (1992) Structural manifestations of the rat Sertoli cell to
hypophysectomy: a correlative morphometric and endocrine study. Endocrinology 131, 485-49.
Guido AN, Campos GER, Neto HS, et al. (2010) Fiber type composition of the sternomastoid and
diaphragm muscles of dystrophin‐deficient mdx mice. Anat Rec 293, 1722-1728.
Hammer O, Harper DAT, Ryan PD (2001) PAST: Paleontological statistics software package for
education and data analysis. Palaeontol Electron 4, 1-9.
Hernández-González, EO et al. (2005) Absence of dp71 in mdx3cv mouse spermatozoa alters
flagellar morphology and the distribution of ion channel and nNos. J Cell Sci 118, 137-145, 2005.
Hess RA, Cooke PS, Bunick D, et al. (1993) Adult testicular enlargement induced by neonatal
hypothyroidism is accompanied by increased Sertoli and germ cell numbers. Endocrinology 132,
2607-2613.
Hoffman EP, Brown RH, Jr Kunkel LM (1987) Dystrophin: the protein product of the Duchenne
muscular dystrophy locus. Cell 51, 919-928.

58
Jahnukainen K, Chrysis D, Hou M, et al. (2004) Increased apoptosis occurring during the first
wave of spermatogenesis is stage-specific and primarily affects midpachytene spermatocytes in the
rat testis. Biol Reprod 70, 290-296.
Johnson L, Petty CS, Neaves WB (1981) A new approach to qualification of spermatogenesis
and its application to germinal cell attrition during human spermatogenesis. Biol Reprod 25,
217–226.
Karnovsky MJ (1965) A formaldehyde-glutaraldehyde fixative of high osmolarity for use in
electron microscopy. J Cell Biol 27, 137.
Kenagy GJ, Trombulak SC (1986) Size and function of mammalian testis in relation to
body size. J Mammal 67, 1–22.
Khalid I, Alam K, Khattak MMAK (2004) Biological significance of ascorbic acid (vitamin C)
in human health-a review. Pak J Nutr 3, 5-13.
Koyuturk M, Yanardag R, Bolkent S, et al. (2006). Influence of combined antioxidants against
cadmium induced testicular damage. Environ Toxicol Pharmacol 21, 235-240.
Ladd AAL, Ladd FVL, Silva AA, et al. (2012) SCG postnatal remodelling–hypertrophy and
neuron number stability–in Spix's Yellow-toothed Cavies (Galea spixii). Int J Dev Neurosci 30,
129-137.
Lapidos KA, Kakkar R, McNally ME (2004) The dystrophin glycoprotein complex: signaling
strength and integrity for the sarcolemma. Circ Res 94, 1023-1031.
Martin R, Santamaria L, Nistal M, et al. (1992). The peritubular myofibroblasts in the testes
from normal men and men with Klinefelter's syndrome. A quantitative, ultrastructural, and
immunohistochemical study. J Pathol 168, 59-66.
Meinhardt A, McFarlane JR, Seitz J, et al. (2000) Activin maintains the condensed type of
mitochondria in germ cells. Mol Cell Endocrinol 168, 111-117.
Melo FCSA, Sousa TP, Costa KLC, et al. (2013) Descriptive morphometry and stereology of the
tubular compartment in the wild rodent Hylaeamys megacephalus (Rodentia: Cricetidae) from
Central Brazil. Anim Reprod Sci 138, 110-117.

59
Melo F, Matta SL, Paula TA, et al. (2010) The effects of Tynnanthus fasciculatus (Bignoniaceae)
infusion on testicular parenchyma of adult Wistar rats. Biol Res 43, 445-450.
Morais DB, Barros MS, Paula TA, et al. (2014a) Evaluation of the cell population of the
seminiferous epithelium and spermatic indexes of the bat Sturnira lilium (Chiroptera:
Phyllostomidae). PloS one 9, e101759.
Morais ACT, Balarini MK, Lopes EO, et al. (2014b) The tubular compartment and the
spermatogenic dynamics of the wild rodent Oxymycterus nasutus (Rodentia: Cricetidae). Anim
Reprod Sci 149, 249-258.
Orth JM, Gunsalus GL, Lamperti AA (1988) Evidence from sertoli cell-depleted rats indicates
that spermatid number in adults depends on numbers of Sertoli cells produced during perinatal
development. Endocrinology 122, 787-794.
Paula TAR, Costa DS, Matta SLP (2002) Avaliação histológica quantitativa do testículo de
capivaras (Hydrochoerus hydrochaeris) adultas. Biosci J 18, 121–136.
Pedlow K, McDonough S, Lennon S (2015) Assisted standing for Duchenne muscular dystrophy.
Cochrane Libr 3, 1-9.
Pentikäinen V, Dunkel L, Erkkilä K (2003) Male germ cell apoptosis. Endocr Dev 5, 56-80
Peres MA, Rocha AM, Vannucchi CI, et al. (2014) Semen analysis of Golden Retriever healthy
dogs and those affected by muscular dystrophy. Andrologia 46, 277-282.
Rao AV, Shaha C (2000) Role of glutathione S-transferasesinoxidative stress-induced male germ
cell apoptosis. Free Radic Biol Med 29, 1015–27.
Ramalho-Santos J, Varum S, Amaral S, et al. (2009) Mitochondrial functionality in
reproduction: from gonads and gametes to embryos and embryonic stem cells. Hum Reprod Update
15, 553-572.
Rodriguez I, Ody C, Araki K, et al. (1997). An early and massive wave of germinal cell apoptosis
is required for the development of functional spermatogenesis. EMBO J 16, 2262-2270.
Ross MH (1967) The fine structure and development of the peritubular contractile cell component
in the seminiferous tubules of the mouse. Am J Anat 121, 523-557.

60
Roosen-Runge EC (1977) The process of spermatogenesis in animals. Massachusetts: The Murray
Printing Company. 213p.
Roosen-Runge EC (1961) The rete testis in the albino rat: its structure, development and
morphological significance. Cells Tissues Organs 45, 1-30.
Russell LD, França LR (1995) Building a testis. Tissue Cell 27, 129 -147.
Russell LD, Ettlin RA, Sinha-Hikim AP, et al. (1990) Histological and histopathological
evaluation of the testis. Bolesta: Cache River Press. 280p.
Russell LD, Saxena NK, Turner TT (1989a). Cytoskeletal involvement in spermiation and sperm
transport. Tissue Cell 21, 361-379.
Russell LD, Bartke A, Goh JC (1989b) Postnatal development of the Sertoli cell barrier, tubular
lumen, and cytoskeleton of Sertoli and myoid cells in the rat, and their relationship to tubular fluid
secretion and flow. Am J Ana 184, 179-189.
Russell LD, Peterson RN (1984) Determination of the elongate spermatid—Sertoli cell ratio in
various mammals. J Reprod Fertil 70, 635-641.
Sellers RS, Mortan D, Michael B, et al. (2007) Society of Toxicologic Pathology position paper:
organ weight recommendations for toxicology studies. Toxicol Pathol 35, 751-755.
Sinha-Hikim A, Bartke A, Russell LD (1988) Morphometric studies on hamster testes in
gonadally active and inactive states: light microscope findings. Biol Reprod 39, 1225-1237.
Sprando RL, Collins TFX, Black T, et al. (1998) Testing the potential of sodium fluoride to
affect spermatogenesis: a morphometric study. Food Chem Toxicol 36, 1117-1124.
Suvanto O, Kormano M (1970) The relation between in vitro contractions of the rat seminiferous
tubules and the cyclic stage of the seminiferous epithelium. J Reprod Fertil 21, 227-232.
Terrill JR, Radley-Crabb HA, Iwasaki T, et al. (2013) Oxidative stress and pathology in
muscular dystrophiefocus on protein thiol oxidation and dysferlinopathies. FEBS J 280, 4149–4164.
Tidball JG, Wehling-Henricks M (2007) The role of free radicals in the pathophysiology of
muscular dystrophy. J Appl Physiol 102, 1677–1686.

61
Tidball JG (2005) Inflammatory processes in muscle injury and repair. Am J Physiol Regul Integr
Comp Physiol 288, 345–353.
Tonon E, Ferretti R, Shiratori JH, et al. (2012) Ascorbic acid protects the diaphragm muscle
against myonecrosis in mdx mice. Nutrition 28, 686-690.
Vandebrouck C, Martin D, Schoor MC, et al. (2002) Involvement of TRPC in the abnormal
calcium influx observed in dystrophic (mdx) mouse skeletal muscle fibers. J Cell Biol 158, 1089–
1096.
Yazama F, Furuta K, Fujimoto M, et al. (2006) Abnormal spermatogenesis in mice unable to
synthesize ascorbic acid. Anat Sci Int 81, 115–125.
Yousef MI (2005). Protective role of ascorbic acid to enhance reproductive performance of male
rabbits treated with stannous chloride. Toxicology 207, 81-89.
Weinbauer GF, Luetjens CM, Simoni M, et al. (2010) Physiology of testicular function. In:
Andrology: Male Reproductive Health and Dysfunction (eds Neischlag E, Behre HM, Nieschlag S),
pp. 11-59, New York: Springer
Whitehead NP, Yeung EW, Allen DG (2006) Muscle damage in mdx (dystrophic) mice: role of
calcium and reactive oxygen species. Clin Exp Pharmacol Physiol 33, 657–662.
Wing TY, Christensen AK (1982) Morphometric studies on rat seminiferous tubules. Am J Anat
165, 13-25.

62
ARTIGO 2: Histomorfometria do compartimento intertubular em testículos de camundongos
mdx portadores da Distrofia Muscular de Duchenne, tratados com ácido ascórbico
Resumo
Objetivou-se caracterizar as alterações no compartimento intertubular testicular (CIT) provocadas
pela Distrofia Muscular de Duchenne (DMD) e o efeito protetor do ácido ascórbico (AA). Foram
utilizados 24 camundongos, 12 da linhagem C57BL/10 (não-distróficos) e 12 da linhagem
C57BL/10Mdx (distróficos), divididos em grupos: C30 e C60 = Controle (não-distrófico) 30 e 60
dias de idade; D30 e D60 = Distrófico 30 e 60 dias de idade; CS60 e DS60 = Controle e distrófico
com 60 dias de idade suplementado com AA. A suplementação com 0,005g/dia de AA foi
ministrada por 30 dias. Após eutanásia, os testículos foram coletados, fixados e incluídos em
historesina para análises morfométricas e análise ultraestrutural A suplementação no grupo DS60
aumentou o volume do intertúbulo em relação ao CS60 (média ± desvio padrão, 40,3 ± 16,7µL
versus 24,2 ± 1,4µL, p<0,05). A distrofia em D30 reduziu a porcentagem de células de Leydig no
intertúbulo e no volume do parênquima testicular (4,85±3,83%; 0,70±0,40µL, respectivamente) em
relação a C30 (17,25±5,75%; 4,20±1,40µL respectivamente) (p<0,05). Na ultraestrutura das células
de Leydig verificou-se, precocemente, a presença de vesículas lipídicas nos distróficos pré-púberes
30 dias (D30). Houve hipertrofia das células de Leydig quando comparado D60 com C60, sendo
revertida pela suplementação com AA. Pode-se concluir que a DMD afeta a organização do CIT e a
suplementação de AA reverteu a hipertrofia células de Leydig no intertúbulo.
Palavras-chave: Distrofina; Antioxidantes; Vitamina C; Célula de Leydig; Espermatogênese.

63
1. Introdução
A Distrofia Muscular de Duchenne (DMD) é a forma mais comum das distrofias musculares,
sendo uma doença genética recessiva letal ligada ao cromossomo X, que acarreta a degeneração
muscular progressiva, em virtude da ausência da proteína distrofina. A DMD afeta 1 a cada 3.500
meninos nascidos e seus sinais clínicos geralmente se desenvolvem ainda na primeira infância, onde
os portadores apresentam dificuldade em correr, subir escadas e caem com frequência ao caminhar
(Emery, 2002; Kieny et. al., 2013). Além disso, podem se desenvolver complicações cardíacas,
respiratórias, gastrointestinais e ortopédicas com a progressão da distrofia (Bushby et. al., 2010). Há
relatos que portadores de outra modalidade de distrofia, a Distrofia Muscular Miotônica (DM1),
apresentavam ainda hipogonadismo, afetando a função tubular e intersticial dos testículos, variação
no nível normal de produção de testosterona e disfunção erétil (Peric et. al., 2013). Os níveis de
testosterona livre e total no soro foram mensurados em diferentes distrofinopatias, dentre elas a
DMD, revelando que 54% dos portadores tinham baixo nível de testosterona total, enquanto 39%
apresentaram baixo nível de testosterona total e livre e 8% apresentaram baixo nível de testosterona
livre com níveis normais de testosterona total (Al-Harbi et. al., 2008).
Há evidências de que níveis elevados de espécies reativas de oxigênio (EROs) podem agravar
as distrofias musculares, além de desencadear a apoptose nas células germinativas testiculares (Rao
e Shaha, 2000; Tidball e Wehling-Henricks, 2007). Logo, o uso de antioxidantes, como o ácido
ascórbico, além de reduzir a necrose muscular promovida pelo estresse oxidativo (Dorchies et. al.,
2006), poderia exercer um efeito similar no parênquima testicular. De fato, segundo Sönmez et. al.
(2005), o uso de ácido ascórbico elevou significativamente os índices de testosterona e aumentou a
fertilidade de ratos Wistar.
A síntese de testosterona é realizada pelas células de Leydig, localizadas no compartimento
intertubular (Fawcett et. al., 1973; Setchell, 2004). Assim, a quantificação dessas células, através da
aplicação da morfometria intertubular, pode fornecer indicativos importantes quanto à atividade
gonadal e a fertilidade (Russell et. al., 1990; Paula et. al., 2002; Morais et. al., 2014). Todavia, não
foram encontrados trabalhos relatando a atuação do ácido ascórbico nas células de Leydig, bem
como a quantificação morfométrica dessas células em portadores da DMD.
Dessa forma, este estudo serve de suporte para novos tratamentos com sucesso em
distrofinopatias, garantindo o aumento da expectativa de vida dos portadores ao longo dos anos.
Isso é evidenciado quando se compara a expectativa de vida de portadores da DMD na década de
60, de 14,4 anos, com os dados de estudos atuais afirmando que os portadores podem viver por mais
de 40 anos (Kieny et. al., 2013; Saito et. al., 2014; Dalton et. al., 2015). Esses fatores contribuem
para aumentar a busca pelos portadores da DMD por alternativas de intervenção reprodutiva por

64
meio do aconselhamento genético e aplicação de biotecnologias (Wallace et. al., 2005; Dalton et.
al., 2015).
Neste sentido, esse trabalho teve por objetivo caracterizar as alterações no compartimento
intertubular provocadas pela DMD e o efeito do tratamento a base de ácido ascórbico.
2. Material e Métodos
2.1. Animais
Foram utilizados 24 camundongos machos, sendo 12 da linhagem C57BL/10 (controle, não
distróficos) e 12 da linhagem C57BL/10Mdx (animais distróficos), provenientes do Biotério da
FIOCRUZ/Rio de Janeiro e do Biotério do Instituto de Ciências Biomédicas da Universidade de
São Paulo/ICB/USP, respectivamente. O projeto foi aprovado pelo comitê de ética do ICB/USP e
chancelado pelo CEUA/UFRN (Parecer nº 164/2011-CEUA/ICB/USP e Parecer nº 064/2013-
CEUA/UFRN). Os animais foram divididos em seis grupos com 4 animais cada: Grupo Controle
com 30 dias de idade (C30); Grupo Distrófico com 30 dias de idade (D30); Grupo Controle com 60
dias de idade (C60); Grupo Distrófico com 60 dias de idade (D60); (CS60) Grupo Controle
suplementado com ácido ascórbico com 60 dias de idade e (DS60) Grupo Distrófico suplementado
com ácido ascórbico com 60 dias de idade. Os animais foram mantidos no biotério do
Departamento de Anatomia ICB/USP em caixas de polietileno, contendo bebedouro e comedouro,
sob temperatura controlada de 22ºC e iluminação com ciclo de 12 horas claro/escuro.
2.2. Dieta suplementar com ácido ascórbico
Após o desmame (21 dias de idade), os animais receberam ração comercial para roedores
Nuvilab®
(Nuvital, São Paulo, Brasil). A partir dos 30 dias de idade os animais dos grupos
suplementados com ácido ascórbico – AA (CS60 e DS60) receberam diariamente 0,005g de ácido
ascórbico (Sigma-Aldrich, St. Louis, EUA) diluído em água e administrado por meio de gavagem
por 30 dias. A concentração foi calculada levando em consideração o protocolo estabelecido na
literatura por Guido et. al. (2010) e Tonon et. al. (2012) para camundongos distróficos.
2.3. Eutanásia e coleta do material
Após o período experimental, os animais foram eutanasiados em câmara de dióxido de
carbono hermeticamente fechada, com entrada de gás na parte superior. Após a eutanásia os animais
foram pesados e os testículos coletados, pesados, seccionados transversalmente e fixados por
imersão em solução de Karnovsky por 24h para morfometria testicular sob microscopia de luz e
para análise ultraestrutural sob Microscopia Eletrônica de Transmissão (MET) (Karnovsky, 1965;
Oliveira, 2004; Morais et. al., 2014).

65
2.4. Processamento histológico
Os fragmentos testiculares destinados às análises morfométricas foram desidratados em série
etanólica crescente (70 a 100%) e incluídos em glicol-metacrilato (Historesin®
LeicaMycrosistems,
Heidelberg, Alemanha). Foram obtidas secções histológicas semiseriadas a 3µm de espessura
utilizando-se micrótomo rotativo Leica RM2255 (Leica Microsystems, Heidelberg, Alemanha),
com intervalo de 40µm entre os cortes, sendo as lâminas coradas com azul de toluidina/borato de
sódio 1%. As preparações foram analisadas ao microscópio Motic BA410 (Motic, CausewayBay,
Hong Kong) e microfotogradas para análises com câmera digital Moticam 5.0 MP (Motic
Instruments Inc, Richmond, Canada), com auxílio do Software Motic Images Plus 2.0 OML (Motic,
Xiamen, China).
Já os fragmentos fixados em Karnovsky após 24h destinados a análise da ultraestrutura
celular foram imersos em solução de Glutaraldeído 2,5% tamponado com fosfato de sódio a 0,1M,
pH 7,4. Após fixação o material foi lavado três vezes durante 10 minutos cada, no mesmo tampão.
Em seguida ocorreu a pós-fixação com tetróxido de ósmio 2% por duas horas (tamponado em
fosfato de sódio 0,1M, pH 7,4). Os fragmentos foram novamente lavados com solução tampão por
três vezes durante dez minutos cada. A seguir o material foi imerso em acetato de uranila a 3%
overnight, lavado em tampão fosfato, desidratado em série etanólica crescente (50 a 100%), durante
10 minutos e imerso em óxido de propileno por 10 minutos para garantir total desidratação do
tecido. Em seguida o material foi embebido em resina araldite 502 (PolysciencesInc, California,
EUA) para confecção dos blocos. Cortes semi-finos foram obtidos com ultramicrótomo, os quais
foram corados com solução aquosa de azul de toluidina a 1% para a identificação dos locais para a
realização de cortes ultrafinos. Cortes a 70nm de espessura foram confeccionados com navalha de
diamante e colocados em telas de cobre com 200 mesh para posterior contraste com acetato de
uranila saturado a 2%, por sete a dez minutos, seguido de citrato de chumbo a 0,5%, pelo mesmo
período. O material foi analisado em Microscópio Eletrônico de Transmissão (Jeol®
100 CX II,
Tóquio, Japão).
2.5. Estereologia testicular
O peso da túnica albugínea foi estimado a partir da densidade de volume (Vv), pela
contagem de 266 pontos projetados sobre 10 imagens obtidas das preparações histológicas de cada
animal, em objetiva de 10x (Vv = No de pontos contados sobre a túnica albugínea/ Total de pontos
contados x 100). O volume absoluto da albugínea foi o resultado do produto da densidade de
volume com o volume testicular, levando-se em consideração que a densidade do testículo é
aproximadamente a 1 (Johnson et. al., 1981; Sprando et. al., 1998; Costa et. al., 2011).

66
As proporções volumétricas entre os túbulos seminíferos e o intertúbulo foram estimadas
contando-se 266 pontos projetados sobre 10 imagens para cada animal, utilizando-se a equação: (No
de pontos contados sobre túbulo ou intertúbulo / Total de pontos contados x 100). O percentual de
cada elemento do compartimento intertubular foi quantificado a partir da projeção de 1.000 pontos
sobre o intertúbulo de cada animal, quantificando os pontos coincidentes sobre o núcleo e o
citoplasma das células de Leydig, vasos sanguíneos, espaço linfático e tecido conjuntivo. Os
percentuais de cada um destes componentes no intertúbulo e nos testículos foram estimados,
respectivamente, a partir das equações: (Nº de pontos contados sobre o elemento x 100 / 1000) e (%
de intertúbulo x % do elemento no intertúbulo / 100), enquanto seus volumes foram estimados pela
equação: (% do elemento no testículo x peso do parênquima testicular / 100), e expressos em µL
(Morais et. al., 2014). Todas essas contagens foram feitas utilizando-se o software Image-Pro Plus®
(Media Cybernetics Inc., Rochville, USA).
Os diâmetros nucleares das células de Leydig foram mensurados quando apresentaram
contorno circular, cromatina perinuclear e nucléolos evidentes, quantificando-se 30 núcleos por
animal. O volume nuclear (VN) e volume citoplasmático (VC) das células de Leydig por animal
foram expressos em µm3, através das respectivas equações: VN = 4/3 πR
3, sendo R o raio nuclear;
VC = % citoplasma x VN / % núcleo. Somando-se VN + VC obteve-se o volume celular.
O cálculo do número total de células de Leydig (TCL) foi obtido pela divisão do volume
total dessas células por parênquima testicular (µm3) pelo volume de cada célula de Leydig (µm
3). O
número de células por grama de testículo foi obtido pela divisão do TCL pelo peso gonadal total. O
índice Leydigossomático (ILS), que quantifica o investimento em células de Leydig em relação à
massa corporal, foi obtido através da equação: ILS = volume total de célula de Leydig por
parênquima testicular / PC x 100, onde PC = peso corporal.
2.6. Análise estatística
Os parâmetros testiculares quantitativos foram expressos em média ± desvio padrão e
submetidos à análise de variância (ANOVA), com teste de múltiplas comparações Kruskall-Wallis
seguido do teste de Mann-Whitney com correção de Bonferroni, utilizando-se o software PAST®
versão 2.17 (Hammer et. al., 2001). Admitiu-se o nível de significância de p < 0,05.

67
3. Resultados
A Tabela 1 contém as médias para cada grupo controle e distróficos, referentes aos pesos
corporal e testicular, percentuais e volumes de túbulos seminíferos e intertúbulo, bem como quanto
às proporções volumétricas dos componentes do intertúbulo. O intertúbulo apresentou-se composto
por células de Leydig, vasos sanguíneos, tecido conjuntivo e espaço linfático (Figura 1A-F).
No grupo distrófico pré-púbere as células de Leydig reduziram seu percentual no intertúbulo
(4,85 ± 3,83% versus 17,25 ± 5,75%) (p < 0,05), o percentual no parênquima testicular (0,74 ±
0,51% versus 4,16 ± 1,42%) (p < 0,05) e o volume no parênquima testicular (0,7±0,4µL versus 4,2
±1,4µL) em relação ao controle pré-púbere. Contudo, o grupo púbere distrófico (D60) elevou o
percentual de células de Leydig no parênquima testicular em relação a D30 (2,65 ± 1,02% versus
0,74 ± 0,51%) (Tabela 1).
O volume do intertúbulo, apesar de não sofrer alteração significativa entre os grupos
distróficos e controle, ocorreu uma elevação no grupo distrófico que recebeu a suplementação com
ácido ascórbico (DS60) em relação a CS60 (40,3 ± 16,7µL versus 24,2 ± 1,4µL, p<0,05). Porém, a
distrofia no período púbere (D60) aumentou o volume do espaço linfático no parênquima em
relação ao controle de 60 dias de idade (C60) (9,8 ± 1,3µL versus 4,3 ± 1,2µL). Além disso, os
animais do grupo D60 quando comparado aos distróficos pré-púberes (D30), apresentaram volume
(9,8 ± 01,3µL versus 2,8 ± 0,9µL, p<0,05) e percentual do espaço linfático elevado no parênquima
(7,20±1,83% versus 2,95 ± 1,29%, p<0,05) (Tabela 1).
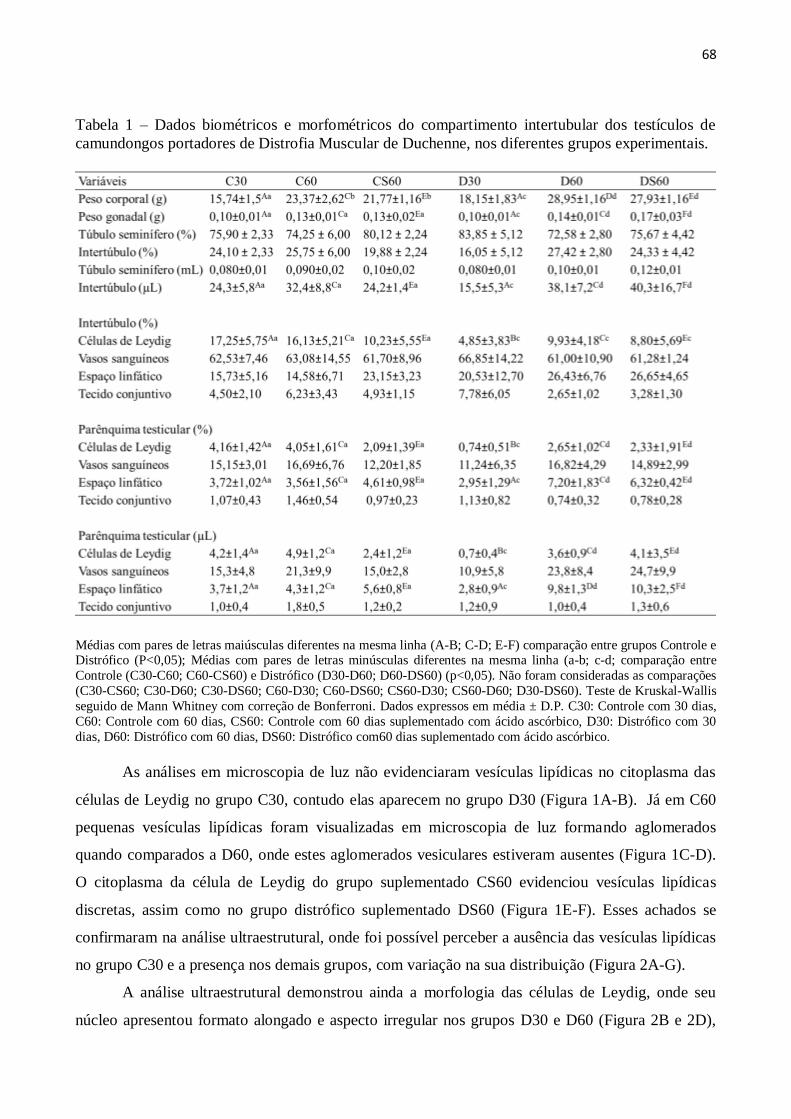
68
Tabela 1 – Dados biométricos e morfométricos do compartimento intertubular dos testículos de
camundongos portadores de Distrofia Muscular de Duchenne, nos diferentes grupos experimentais.
Médias com pares de letras maiúsculas diferentes na mesma linha (A-B; C-D; E-F) comparação entre grupos Controle e Distrófico (P<0,05); Médias com pares de letras minúsculas diferentes na mesma linha (a-b; c-d; comparação entre
Controle (C30-C60; C60-CS60) e Distrófico (D30-D60; D60-DS60) (p<0,05). Não foram consideradas as comparações
(C30-CS60; C30-D60; C30-DS60; C60-D30; C60-DS60; CS60-D30; CS60-D60; D30-DS60). Teste de Kruskal-Wallis
seguido de Mann Whitney com correção de Bonferroni. Dados expressos em média ± D.P. C30: Controle com 30 dias,
C60: Controle com 60 dias, CS60: Controle com 60 dias suplementado com ácido ascórbico, D30: Distrófico com 30
dias, D60: Distrófico com 60 dias, DS60: Distrófico com60 dias suplementado com ácido ascórbico.
As análises em microscopia de luz não evidenciaram vesículas lipídicas no citoplasma das
células de Leydig no grupo C30, contudo elas aparecem no grupo D30 (Figura 1A-B). Já em C60
pequenas vesículas lipídicas foram visualizadas em microscopia de luz formando aglomerados
quando comparados a D60, onde estes aglomerados vesiculares estiveram ausentes (Figura 1C-D).
O citoplasma da célula de Leydig do grupo suplementado CS60 evidenciou vesículas lipídicas
discretas, assim como no grupo distrófico suplementado DS60 (Figura 1E-F). Esses achados se
confirmaram na análise ultraestrutural, onde foi possível perceber a ausência das vesículas lipídicas
no grupo C30 e a presença nos demais grupos, com variação na sua distribuição (Figura 2A-G).
A análise ultraestrutural demonstrou ainda a morfologia das células de Leydig, onde seu
núcleo apresentou formato alongado e aspecto irregular nos grupos D30 e D60 (Figura 2B e 2D),
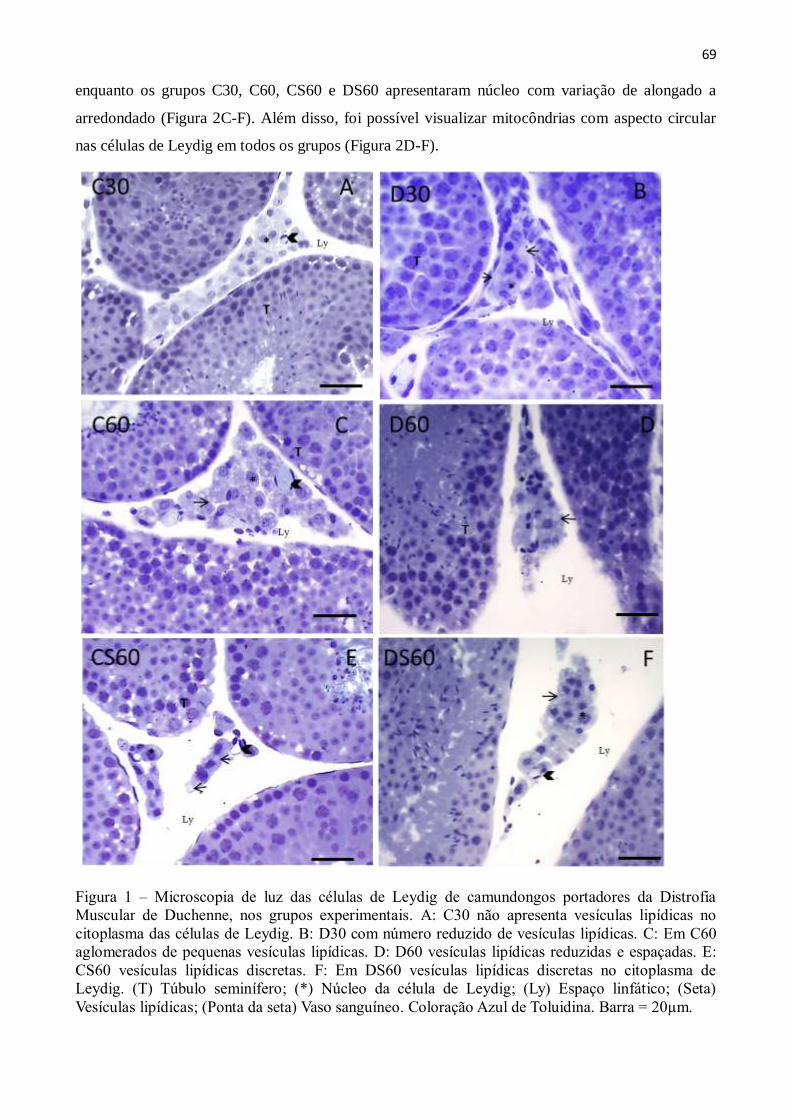
69
enquanto os grupos C30, C60, CS60 e DS60 apresentaram núcleo com variação de alongado a
arredondado (Figura 2C-F). Além disso, foi possível visualizar mitocôndrias com aspecto circular
nas células de Leydig em todos os grupos (Figura 2D-F).
Figura 1 – Microscopia de luz das células de Leydig de camundongos portadores da Distrofia
Muscular de Duchenne, nos grupos experimentais. A: C30 não apresenta vesículas lipídicas no
citoplasma das células de Leydig. B: D30 com número reduzido de vesículas lipídicas. C: Em C60
aglomerados de pequenas vesículas lipídicas. D: D60 vesículas lipídicas reduzidas e espaçadas. E:
CS60 vesículas lipídicas discretas. F: Em DS60 vesículas lipídicas discretas no citoplasma de
Leydig. (T) Túbulo seminífero; (*) Núcleo da célula de Leydig; (Ly) Espaço linfático; (Seta)
Vesículas lipídicas; (Ponta da seta) Vaso sanguíneo. Coloração Azul de Toluidina. Barra = 20µm.
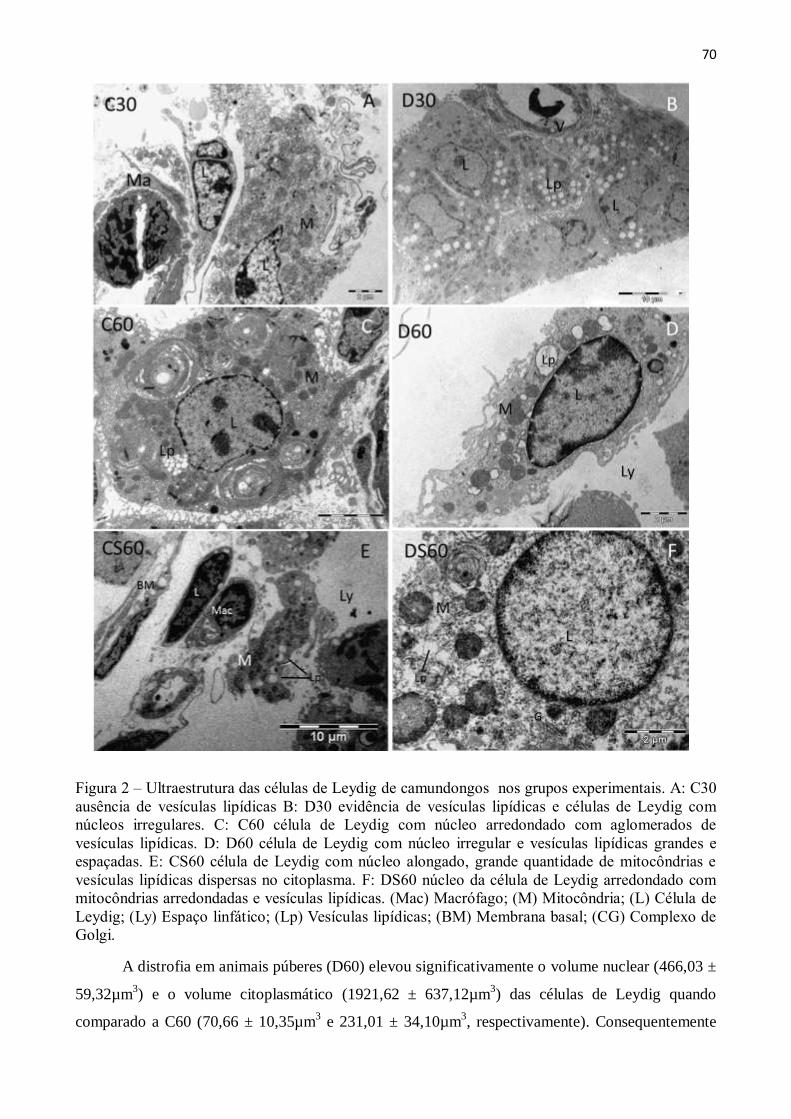
70
Figura 2 – Ultraestrutura das células de Leydig de camundongos nos grupos experimentais. A: C30
ausência de vesículas lipídicas B: D30 evidência de vesículas lipídicas e células de Leydig com
núcleos irregulares. C: C60 célula de Leydig com núcleo arredondado com aglomerados de
vesículas lipídicas. D: D60 célula de Leydig com núcleo irregular e vesículas lipídicas grandes e
espaçadas. E: CS60 célula de Leydig com núcleo alongado, grande quantidade de mitocôndrias e
vesículas lipídicas dispersas no citoplasma. F: DS60 núcleo da célula de Leydig arredondado com
mitocôndrias arredondadas e vesículas lipídicas. (Mac) Macrófago; (M) Mitocôndria; (L) Célula de
Leydig; (Ly) Espaço linfático; (Lp) Vesículas lipídicas; (BM) Membrana basal; (CG) Complexo de
Golgi.
A distrofia em animais púberes (D60) elevou significativamente o volume nuclear (466,03 ±
59,32µm3) e o volume citoplasmático (1921,62 ± 637,12µm
3) das células de Leydig quando
comparado a C60 (70,66 ± 10,35µm3 e 231,01 ± 34,10µm
3, respectivamente). Consequentemente

71
elevando o volume individual das células de Leydig em distróficos na puberdade (D60) em relação
ao grupo C60 (2387,66 ± 665,54µm3
versus 301,68 ±40,33µm3). A suplementação com ácido
ascórbico no grupo distrófico (DS60) reduziu significativamente o volume nuclear (62,08 ±
17,43µm3), volume citoplasmático (224,90 ± 117,06µm
3) e volume individual das células de Leydig
(286,98 ± 131,80µm3) em relação a D60 (Tabela 2).
A distrofia elevou o diâmetro nuclear das células de Leydig na puberdade (D60) quando
comparado ao período pré-púbere (D30) (9,61 ±0,39µm versus 4,50±0,23µm), resultado semelhante
foi observado entre os controles púbere e pré-púbere. Contudo, a suplementação, em distróficos
(DS60), atuou reduzindo significativamente essa variável (4,88 ± 0,50µm) quando comparado a
D60. Já o percentual do núcleo foi reduzido na puberdade em distróficos quando comparado a D30
(20,41 ± 4,54% versus 36,93 ± 11,67%). Essa tendência também foi observada entre os grupos C30
e C60 (Tabela 2).
A distrofia reduziu significativamente o índice Leydigossomático tanto em D30 em relação
a C30 (0,0040 ± 0,0027% versus 0,026 ± 0,009%) quanto em D60 em relação a C60 (0,0123 ±
0,0030% versus 0,022 ± 0,009%). Essa redução também aconteceu quanto ao número de células de
Leydig por grama de testículo, em D30 em relação a C30 (5,81 ± 4,24 x107 versus 37,72 ± 13,56
x107) e em D60 em relação a C60 (1,11 ± 0,26x10
7versus 13,41 ± 4,65x10
7) (Tabela 2).
Tabela 2– Morfometria das células de Leydig e índice Leydigossomático do compartimento
intertubular dos testículos de camundongos portadores de Distrofia Muscular de Duchenne, nos
diferentes grupos experimentais.
Médias com pares de letras maiúsculas diferentes na mesma linha (A-B; C-D; E-F) comparação entre grupos Controle e
Distrófico (P<0,05); Médias com pares de letras minúsculas diferentes na mesma linha (a-b; c-d) comparação entre
Controle (C30-C60; C60-CS60) e Distrófico (D30-D60; D60-DS60) (p<0,05). Não foram consideradas as comparações
(C30-CS60; C30-D60; C30-DS60; C60-D30; C60-DS60; CS60-D30; CS60-D60; D30-DS60).Teste de Kruskal-Wallis
seguida de Mann Whitney com correção de Bonferroni. Dados expressos em média ± D.P. C30: Controle com 30 dias,
C60: Controle com 60 dias, CS60: Controle com 60 dias suplementado com ácido ascórbico, D30: Distrófico com 30
dias, D60: Distrófico com 60 dias, DS60: Distrófico com60 dias suplementado com ácido ascórbico.

72
4. Discussão
São escassos os estudos sobre a organização e quantificação dos componentes do intertúbulo,
bem como, a influência do ácido ascórbico na Distrofia Muscular de Duchenne. Todavia, é
fundamental compreender as alterações reprodutivas dessa patologia, bem como a influência do
tratamento de antioxidantes, através da quantificação dos percentuais e volume dos componentes
testiculares, uma vez que podem oferecer subsídios para o desenvolvimento de novas terapias no
sistema reprodutor masculino de pacientes portadores dessa alteração genética.
Os testículos dos mamíferos podem ser divididos funcionalmente em dois compartimentos: o
compartimento tubular ou espermatogênico e o compartimento intertubular ou androgênico. O
percentual destes compartimentos pode variar de acordo com a espécie, sendo considerado um dos
fatores responsáveis pela diferença na eficiência da produção espermática (Russell et. al., 1990;
França e Russell, 1998). Nesse trabalho, o intertúbulo foi morfologicamente semelhante às
descrições para os demais mamíferos, tanto nos grupos controle quanto nos distróficos, sendo esse
compartimento composto por células de Leydig, vasos sanguíneos, tecido conjuntivo e espaço
linfático (França e Russell, 1998). De acordo com Fawcett et. al. (1973) a organização do
intertúbulo de camundongos e de ratos é descrita como tipo 1, onde as células de Leydig ocupam
uma área reduzida contrastando com a maior extensão ocupada pelos espaços linfáticos. Todavia,
essas proporções podem variar, como foi descrito que para um mesmo gênero de roedores é
possível encontrar predominância de espaço linfático em virtude da síntese de outros produtos
esteroides, como feromônio, ou de células de Leydig, que pode estar relacionada à síntese de outros
esteroides além da testosterona (Taylor e Horner, 1970; Fawcett et. al., 1973). Dessa forma, a
variação da categoria de intertúbulo encontrada nesse trabalho entre animais controle e distróficos
pode ser justificada.
O volume do compartimento intertubular foi elevado pela suplementação em distróficos e pode
estar relacionado ao aumento do espaço linfático, permitindo aumento de fluidos sanguíneos no
interstício, como observado em ratos tratados com baixas doses de cádmio (Predes et. al., 2010). Os
valores encontrados no intertúbulo para a proporção e volume de vasos sanguíneos e tecido
conjuntivo não diferiram entre as idades dos distróficos, distróficos tratados e não-tratados.
Todavia, o aumento do volume do espaço linfático no parênquima foi mais acentuado à medida que
o camundongo alcançou a puberdade. Isso implica em aumento da capacidade dos vasos linfáticos
em eliminar dos testículos os materiais vascularmente segregados, bem como, manter as
concentrações adequadas de andrógeno em ambos os testículos e nos vasos sanguíneos (Fawcett et.
al., 1973).

73
Embora as proporções volumétricas dos componentes intertubulares possam variar entre as
espécies, a maioria dos mamíferos apresenta como importante constituinte funcional do
compartimento intertubular, as células de Leydig, que secretam esteroides e feromônios essenciais
para o comportamento sexual de machos e o desenvolvimento da espermatogênese (Evans et. al.,
1996; França e Russell, 1998; Paula et. al., 2002). A literatura descreve que nos mamíferos, de
forma geral, ocorre aumento das proporções das células de Leydig no parênquima testicular na fase
pré-púbere e segue em regressão até a puberdade (Setchell, 1978). Contudo, no presente estudo a
distrofia provocou a redução dessas proporções ainda no período pré-púbere acentuando na
puberdade. Isso pode estar relacionado ao estresse oxidativo decorrente da distrofia, assim como
observado em animais expostos ao cádmio (Blanco et. al., 2007).
A análise ultraestrutural revelou que morfologicamente o núcleo das células de Leydig era
arredondado com heterocromatina nos grupos controle e distrófico suplementado com 60 dias,
semelhante ao padrão observado para outros mamíferos (Christensen, 1965; França e Russell, 1998;
Costa et. al., 2006; Paula et. al., 2007). Todavia, os animais dos grupos distróficos apresentaram
núcleos irregulares. Apesar de Hooker (1970) descrever como normal os núcleos das células de
Leydig com formas elipsoides ou poligonais, estudos com coelhos apresentaram núcleos irregulares
na célula de Leydig após exposição a inseticidas (Almasiova et. al., 2014). De acordo com as
observações em ratos tratados com herbicida esse pleomorfismo nuclear e outras mudanças
estruturais nas células de Leydig podem reduzir o nível de testosterona plasmática (Victor-Costa et.
al., 2010).
A morfologia da ultraestrutura citoplasmática das células de Leydig comumente apresenta
vesículas lipídicas, formadas por ésteres de colesterol, substrato para biossíntese da testosterona que
é catalisada por enzimas localizadas na membrana do retículo endoplasmático liso e na membrana
das mitocôndrias (Mori e Christensen, 1980). Apesar desses autores afirmarem que em ratos pré-
púberes as vesículas lipídicas nas células de Leydig estão ausentes, no presente estudo a distrofia
apresentou essas estruturas ainda na pré-puberdade. Essa estocagem precoce de substrato para
esteroidogênese deve ser compensatória, em virtude da desregulação do eixo hipotalâmico-
hipofisário-gonadal que acontece em portadores da DMD. Nesse caso é provável que exista
comprometimento funcional das células de Leydig devido à redução da atividade da enzima
superóxido dismutase que altera as enzimas esteroidegênicas e aumenta os danos oxidativos nas
células de Leydig (Sarkar et. al., 2004; Ricci et. al., 2009). Visto que, apesar de apresentarem níveis
elevados de LH, os pacientes portadores da distrofia miotônica tipo 1 e de outros tipos de
distrofinopatias possuem níveis de testosterona total abaixo dos níveis descritos para homens
púberes não-distróficos, confirmando a existência de insuficiência androgênica (Lou et. al., 1994;
Al-Harbi et. al., 2008; Orngreen et. al., 2012).

74
A literatura descreve que para portadores da distrofia muscular miotônica do tipo 1 (DM1)
ocorre hiperplasia (aumento do número de células de Leydig) como um mecanismo compensatório
decorrente do aumento dos hormônios gonadotrópicos e de falhas funcionais nas células de Leydig
(Febres et. al., 1975; Roses e Adams, 1996; Ashizawa e Sarkar, 2011). Contudo no presente
trabalho, observou-se que a DMD atuou elevando consideravelmente o volume nuclear e
citoplasmático das células de Leydig e, consequentemente, o seu volume individual na puberdade,
caracterizando uma hipertrofia. Essa alteração quando relatada em ratos expostos a vapor de
formaldeído demostrou redução dos índices de testosterona resultantes de falha funcional das
células de Leydig (Razi et. al., 2013). No entanto percebe-se que a suplementação de ácido
ascórbico utilizada em camundongos distróficos reverteu esse quadro provavelmente em virtude da
elevação de glutationa, que reduz os radicais livres que promovem estresse oxidativo testicular,
semelhante ao descrito para o tratamento combinado de ácido ascórbico com uma droga
quimioterápica em camundongos com tumor (Longchar e Prasad, 2015).
Na maioria dos mamíferos domésticos a média é de 20-60 milhões de células de Leydig por
grama de testículo (França e Russell, 1998). Em ratos e camundongos as médias estão entre 10,5
milhões e 25 milhões, respectivamente (Zirkin e Ewing, 1987). Segundo Castro et. al. (2002) esse
parâmetro pode ser utilizado para monitorar os níveis de testosterona, uma vez que há uma
correlação positiva entre as células de Leydig por grama de testículo e os níveis tecidual e
plasmático da testosterona em coelhos normais. Logo, a distrofia ao reduzir consideravelmente o
número de células de Leydig por grama de testículo pode ter reduzido os níveis de testosterona
plasmático tanto no período pré-púbere quanto na puberdade. A concentração plasmática de
testosterona é um parâmetro importante que necessita ser avaliada em trabalhos futuros para
confirmar essa possível correlação.
A redução do peso gonadal e o aumento do volume das células de Leydig em animais
distroficos implicou no aumento da proporção de massa corporal alocada nas células de Leydig, o
índice Leydigossomático, em relação aos animais controle. Tendo em vista que esse índice permite
a comparação com espécies de diferentes tamanhos (Melo et. al., 2015) e que cães da raça Golden
Retriever são comumente utilizados como modelo para a DMD (Martins-Júnior et. al., 2015), em
cães normais sem raça definida observa-se que em esse índice varia com a idade do animal,
aumentando até a puberdade onde há maior produção espermática, e reduzindo no período pós-
púbere (Mascarenhas et. al., 2006), esse comportamento difere dos achados descritos para animais
distróficos no presente trabalho, certamente esse maior investimento de massa corporal em células
de Leydig é mais uma ação compensatória para garantir maior suporte hormonal para
espermatogênese.

75
Pode-se concluir que a Distrofia Muscular de Duchenne afetou as proporções morfométricas
dos componentes do compartimento intertubular, especialmente o número e morfologia da célula de
Leydig a depender da idade. No entanto, a suplementação de ácido ascórbico foi capaz de reverter,
a hipertrofia acentuada das células de Leydig observada em camundongos distróficos púberes.
Referências
Al-Harbi, T.M., Bainbridge, L.J., McQueen, M.J., Tarnopolsky, M.A., 2008. Hypogonadism is
common in men with myopathies. J. Clin. Neuromuscul. Dis. 9, 397–401.
Almasiova, V., Holovska, K.,Tarabova, L., Cigankova, V., Lukacinova, A., Nistiar, F., 2014.
Structural and ultrastructural study of the rabbit testes exposed to carbamate insecticide. J. Environ.
Sci. Health. A. 47, 1319-1328.
Ashizawa T.E.T.S.U.O., Sarkar, P.S., 2011. Myotonic dystrophy types 1 and 2. Handb. Clin.
Neurol. 101, 193-237.
Blanco, A., Moyano, R., Vivo, J., Flores-Acuna, R., Molina, A., Blanco, C., Agüera, E., Monterde,
J.G., 2007. Quantitative changes in the testicular structure in mice exposed to low doses of
cadmium. Environ. Toxicol. Pharmacol. 23, 96-101.
Bushby, K., Finkel, R., Birnkrant, D.J., Case, L.E., Clemens, P.R., Cripe, L., Kaul, A., Kinnett, K.,
McDonald, C., Pandya, S., Poysky, J., Shapiro, F., Tomezsko, J., Constantin, C., 2010. Diagnosis
and management of Duchenne muscular dystrophy, part 1: diagnosis, and pharmacological and
psychosocial management. Lancet Neurol. 9, 77-93.
Castro, A.C.S., Berndtson, W.E., Cardoso, F.M., 2002, Plasma and testicular testosterone levels,
volume density and number of Leydig cells and spermatogenic efficiency of rabbits. Braz. J. Med.
Biol. Res. 35, 493-498.
Christensen, A.K., 1965. The fine structure of testicular interstitial cells in guinea pigs. J. Cell. Biol.
26, 911-935.
Costa, K.L.C., Matta, S.L.P., Gomes, M.L.M., Paula, T.A.R., Freitas, K.M., Carvalho, F.A.R.,
Silveira, J.A., Dolder, H., Mendis-Handagama, S.L.M.C., 2011. Histomorphometric evaluation of

76
The neotropical brown brocket deer Mazama gouazoubira testis, with an emphasis on cell
population indexes of spermatogenic yield. Anim. Reprod. Sci. 127, 202–212.
Costa, D.S., Paula, T.A.R., Matta, S.L.P., 2006. The intertubular compartment morphometry in
capybaras (Hydrochoerus hydrochaeris) testis. Anim. Reprod. Sci. 91, 171-179.
Dalton, J., Goldman, J.S., Sampson, J.B., 2015. The Muscular Dystrophies, In: Goldman, J.S.
(Eds.), Genetic Counseling for Adult Neurogenetic Disease. Springer, New York, pp. 251-262.
Dorchies, O.M., Wagner, S., Vuadens, O., Waldhauser, K., Buetler, T.M.,Kucera, P., Ruegg, U.T.,
2006. Green tea extract and its major polyphenol (-)-epigallocatechingallate. Am. J. Physiol. Cell.
Physiol. 290, 616 –625.
Emery, A.E.H., 2002.The muscular dystrophies.Lancet. 359, 687-695.
Evans, A.C.O., Pierso, R.A., Garcia, A., Mcdougall, L.M., Hrudka, F., Rawlings, N.C.,
1996.Changes in the circulating hormone concentrations, testes histology and testes
ultrasonography during sexual maturation in beef bulls. Theriogenology. 46, 345-347.
Fawcett, D.W., Neaves, W.B., Flores, M.N., 1973. Comparative observations on intertubular
lymphatics and the organization of the interstitial tissue of the mammalian testis. Biol.Reprod. 9,
500–532.
Febres, F., Scaglia, H., Lisker, R., Espinosa, J., Morato, T., Shkurovich, M., Pérez-palacios, G.,
1975. Hypothalamic-pituitary-gonad function in patients with myotonic dystrophy. J. Clin.
Endocrinol. Metab. 41, 833-840.
França, L.R., Russell, L.D., 1998. The testis of domestic animals, in: Martinez-Garcia, F.,
Regadera, J., (Eds.), Male Reproduction: A Multidisciplinary Overview. Churchill Livingstone
Madrid, pp. 197–219.
Guido, A.N., Campos, G.E.R., Neto, H.S., Marques, M.J., Minatel, E., 2010. Fiber type
composition of the sternomastoid and diaphragm muscles of dystrophin‐deficient mdx mice. Anat.
Rec. 293, 1722-17.

77
Hammer, O., Harper, D.A.T., Ryan, P.D., 2001. PAST: Paleontological statistics software package
for education and data analysis. Palaeontol.Electron. 4, 1-9.
Hooker, C.W., 1970. The intertubular tissue of the testis, in: Johnson, A.D., Gomes, W.R.,
Vandemark, N.L. (Eds.), The Testis. Academic Press, New York, pp. 483–550.
Johnson, L., Petty, C.S., Neves, W.B., 1981. A new approach to qualification of spermatogenesis
and its application to germinal cell attrition during human spermatogenesis. Biol. Reprod. 25, 217–
226.
Karnovsky, M.J., 1965. A formaldehyde-glutaraldehyde fixative of high osmolarity for use in
electron microscopy. J. Cell. Biol. 27, 137.
Kieny, P., Chollet, S., Delalande, P., Le Fort, M., Magot, A., Pereon, Y., Verbe, B.P., 2013.
Evolution of life expectancy of patients with Duchenne muscular dystrophy at AFM Yolaine de
Kepper centre between 1981 and 2011. Ann. Phys. Rehabil. Med. 56, 443-454.
Longchar, A., Prasad, S.B. 2015. Biochemical changes associated with ascorbic acid–cisplatin
combination therapeutic efficacy and protective effect on cisplatin-induced toxicity in tumor-
bearing mice. Toxicol. Rep. 2, 489-503.
Lou, X.Y., Nishi, Y., Haji, M., Antoku, Y., Tanaka, S., Ikuyama, S., Yanase, T., Takayanagi, R.,
Nawata, H., 1994. Reserved Sertoli cell function in the hypogonadic male patients with myotonic
dystrophy.Fukuoka IgakuZasshi. 85, 168-174.
Martins-Júnior, H.A., Simas, R.C., Brolio, M.P., Ferreira, C.R., Perecin, F., Nogueira, G.P.,
Miglino, M.A., Martins, D.S., Eberlin, M.N., Ambrósio, C.E., 2014. Profiles of Steroid Hormones
in Canine X-Linked Muscular Dystrophy via Stable Isotope Dilution LC-MS/MS. PloS one. 10,
e0126585-e0126585.
Mascarenhas, R.M., Paula, T.A.R., Matta, S.L.P., Lanna, L.L.E., Fonseca, C.C., Neves, M.T.D.D.,
2006. Morfometria macro e microscópica e índices somáticos dos componentes testículares de cães
sem raça definida, da puberdade à senilidade/macro. Ceres. 53, 113-119.

78
Melo, F.C.S.A., Sousa, T.P., Lugate, K., Costa, C., Matta, S.L.P., Melo, F.R., 2015. Morphometry
and stereology of the interstitial compartment in the wild rodent Hylaeamys megacephalus
(rodentia: cricetidae) from central Brazil. Encicl. 11, 2331-2339.
Morais, D.B., Barros, M. S., Freitas, M.B., Paula, T. A., Matta, S.L. 2014. Histomorphometric
characterization of the intertubular compartment in the testes of the bat Sturnira lilium. Anim.
Reprod. Sci. 147, 180-186.
Mori, H., Christensen, A.K., 1980. Morphometric analysis of Leydig cells in the normal rat testis. J.
Cell. Biol. 84, 340-354.
Oliveira M.F. 2004. Placentação em mocós, Kerodon rupestris (Wied 1920) (Tese). São Paulo (SP):
Faculdade de Medicina Veterinária e Zootecnia da Universidade Federal de São Paulo. 208p.
Orngreen, M.C., Arlien-Søborg, P., Duno, M., Hertz, J. M., Vissing, J. 2012. Endocrine function in
97 patients with myotonic dystrophy type 1. J. Neurol. 259, 912-920.
Paula, T.A R., Matta, S.L.P., Silva Jr, V.A., Costa, D.S., Fonseca, C.C., Neves, M.T.D., 2007.
Interbular space characterization in adult capybara (Hydrochoerus hydrochaeris) testis. Braz. Arch.
Biol. Technol. 50, 289-297.
Paula, T.A.R., Costa, D.S., Matta, S.L.P., 2002. Avaliacão histológica quantitativa do testículo de
capivaras (Hydrochoerus hydrochaeris) adultas. Biosci. J. 18, 121–136.
Peric, S., Nisic, T., Milicev, M., Basta, I.,Marjanovic, I., Peric, M., Lavrnic, D., Stojanovic, V.R.,
2013. Hypogonadism and erectile dysfunction in myotonic dystrophy type 1. Acta. Myol. 32, 106-
109.
Predes, F.S., Diamante, M.A.S., Dolder, H., 2010. Testis response to low doses of cadmium in
Wistar rats. Int. J. Clin. Exp. Pathol. 91, 125-131.
Rao, A.V., Shaha, C., 2000. Role of glutathione S-transferasesinoxidative stress-induced male germ
cell apoptosis. Free. Radic. Biol. Med. 29, 1015–27.

79
Razi, M., Malekinejad, H., Sayrafi, R., Hosseinchi, M. R., Feyzi, S., Moshtagion, S. M., Janbaz, H.,
2013. Adverse effects of long-time exposure to formaldehyde vapour on testicular tissue and sperm
parameters in rats. In Veterinary research forum: an international quarterly journal. Faculty of
Veterinary Medicine, Urmia, pp. 213.
Ricci, G., Catizone, A., Esposito, R., Pisanti, F. A., Vietri, M.T., Galdieri, M., 2009.Diabetic rat
testes: morphological and functional alterations. Andrologia. 41, 361-368.
Roses, A.D., Adams, C., 1996. Myotonic dystrophy. Neurogenetics. 495, 913-930.
Russell, L.D., Ettlin, R.A., Sinha-Hikim, A.P., Clegg, E.D., 1990. Mammalian spermatogenesis,
in: Russell, L.D., Ettlin, R.A., Sinha-Hikim, A.P., Clegg, E.D. (Eds.), Histological and
Histopathological Evaluation of the Testis. Cache River Press, Clearwater, pp. 1–40.
Saito, T., Kawai, M., Matsumura, T., Fujimura, H., Sakoda, S., 2014. GP 163: Clinical features of
Duchenne muscular dystrophy aged over 40 years. Neuromuscular Disord.24, 854.
Sarkar, P.S., Paul, S., Han, J., Reddy, S., 2004. Six5 is required for spermatogenic cell survival and
spermiogenesis. Hum. Mol. Gen. 13, 1421-1431.
Setchell, B.P., 2004. Hormones: what thetestis really sees. Reprod. Fertil.Dev. 16, 535–545.
Setchell, B.P., 1978. The mammalian testies, first ed. Paul Elek, London.
Sönmez, M., Türk, G., Yüce, A., 2005. The effect of ascorbic acid supplementation on sperm
quality, lipid peroxidation and testosterone levels of male Wistar rats. Theriogenology. 63, 2063-
2072.
Sprando, R.L., Collins, T. F. X., Black, T., Olejnik, N., Rorie, J., 1998. Testing the potential of
sodium fluoride to affect spermatogenesis: a morphometric study.Food Chem. Toxicol. 36, 1117-
1124.
Taylor, J.M., Horner, B.E., 1970. Observations on reproduction in Leggadina (Rodentia, Muridae).
J. Mammal. 51, 10-17.

80
Tidball, J.G., Wehling-Henricks, M., 2007. The role of free radicals in the pathophysiology of
muscular dystrophy. J. Appl. Physiol. 102, 1677–1686.
Tonon, E., Ferretti, R., Shiratori, J.H., Santo Neto, H., Marques, M.J., Minatel, E., 2012. Ascorbic
acid protects the diaphragm muscle against myonecrosis in mdx mice. Nutrition. 28, 686-690.
Victor-Costa, A.B., Bandeira, S.M.C., Oliveira, A G., Mahecha, G.A.B., Oliveira, C.A., 2010.
Changes in testicular morphology and steroidogenesis in adult rats exposed to Atrazine.
Reprod.Toxicol. 29, 323-331.
Wallace, W.H.B., Anderson, R.A., Irvine, D.S., 2005. Fertility preservation for young patients with
cancer: who is at risk and what can be offered?.Lancet Oncol. 6, 209-218.
Zirkin, B.R., Ewing, L.L., 1987. Leydig cell differentiation during maturation of the rat testis: A
stereological study of cell number and ultrastructure. Anat. Rec. 219, 157–163.

81
5. CONCLUSÕES
Portanto, pode-se afirmar que a DMD provocou alterações significativas na morfologia do
compartimento tubular e intertubular dos testículos de camundongos distróficos, mas não impediu a
espermatogênese em camundongos púberes. Já a suplementação com ácido ascórbico, nas
condições experimentais aplicadas, foi capaz de reduzir a apoptose celular provocada por estresse
oxidativo nos compartimentos tubular e reverteu a hipertrofia das células de Leydig observada em
camundongos distróficos púberes.

82
ANEXO A – Certificado Comitê de Ética no Uso dos Animais ICB/USP

83
ANEXO B – Declaração da Comissão de Ética no Uso de Animais – CEUA/UFRN


















