
Efeitos do beta caroteno e do urucum sobre a expressão de ...
115
UNIVERSIDADE FEDERAL DE OURO PRETO INSTITUTO DE CIÊNCIAS E BIOLÓGICAS NÚCLEO DE PESQUISA EM CIÊNCIAS BIOLÓGICAS PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS BIOLÓGICAS LORENA SOUZA E SILVA Efeitos do beta caroteno e do urucum sobre a expressão de genes hepáticos do metabolismo do colesterol em ratas hipercolesterolêmicas Ouro Preto 2013
Transcript of Efeitos do beta caroteno e do urucum sobre a expressão de ...

UNIVERSIDADE FEDERAL DE OURO PRETO
INSTITUTO DE CIÊNCIAS E BIOLÓGICAS
NÚCLEO DE PESQUISA EM CIÊNCIAS BIOLÓGICAS
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS BIOLÓGICAS
LORENA SOUZA E SILVA
Efeitos do beta caroteno e do urucum sobre a
expressão de genes hepáticos do metabolismo
do colesterol em ratas hipercolesterolêmicas
Ouro Preto
2013

LORENA SOUZA E SILVA
Efeitos do beta caroteno e do urucum sobre a
expressão de genes hepáticos do metabolismo
do colesterol em ratas hipercolesterolêmicas
Tese apresentada ao Programa de Pós-
Graduação em Ciências Biológicas da
Universidade Federal de Ouro Preto como
parte integrante dos requisitos para a obtenção
do título de Doutor em Ciências Biológicas,
área de Concentração: Bioquímica Estrutural e
Fisiológica
Orientador: Prof. Dr. Marcelo Eustáquio Silva
Ouro Preto
2013

Catalogação: [email protected]
S586e Silva, Lorena Souza e.
Efeitos do beta caroteno e do urucum sobre a expressão de genes hepáticos
do metabolismo do colesterol em ratas hipercolesterolêmicas [manuscrito] /
Lorena Souza e Silva - 2013.
xv, 97f.: il. color.; graf., tab.
Orientador: Prof. Dr. Marcelo Eustáquio Silva.
Tese (Doutorado) - Universidade Federal de Ouro Preto.
Instituto de Ciências Exatas e Biológicas. Núcleo de Pesquisas em Ciências
Biológicas. Programa de Pós-Graduação em Ciências Biológicas.
Área de concentração: Bioquímica Estrutural e Fisiológica.
1. Colesterol - Teses. 2. Expressão gênica - Teses. 3. Hipercolesterolemia -
Teses. 4. Rato como animal de laboratório - Teses. I. Silva, Marcelo Eustáquio.
II. Universidade Federal de Ouro Preto. III. Título.
CDU: 612.397:633.863


SILVA, L.S. Dedicatória
ii
DEDICATÓRIA
Aos meus pais, Zilda e
Geraldo, e à minha irmã
Renata, com muito amor.

SILVA, L.S. Agradecimento Especial
iii
AGRADECIMENTO ESPECIAL
Agradeço ao Prof. Dr. Marcelo Eustáquio Silva, a orientação e os
ensinamentos imprescindíveis para a realização deste trabalho. Agradeço a confiança
creditada a mim e principalmente a oportunidade de aprender com você durante todos
esses anos.
MUITO OBRIGADA!

SILVA, L.S. Agradecimentos
iv
AGRADECIMENTOS
A Deus pelo dom da vida e pela dádiva do saber, por estar sempre comigo guiando
meus passos e me iluminando.
Aos meus pais e minha irmã, POR TUDO. Por acreditarem junto comigo nos meus
sonhos e não medirem esforços para que eu os realizasse. Pelo amor incondicional e
pelo apoio em todas as minhas decisões.
A minha família, que mesmo a distância torceu por mim.
Ao André, pelo tão dedicado amor e por fazer parte de todos os momentos dessa
trajetória. Obrigada pela paciência e serenidade com que me ouvia e principalmente por
nunca duvidar de que tudo iria dar certo!
As professoras Maria Lúcia Pedrosa e Daniela Caldeira Costa, pela parceria,
ensinamentos e oportunidades oferecidas para a realização deste trabalho. Muito
obrigada!
Ao Professor Rinaldo Cardoso dos Santos, pelo exemplo como mestre e profissional.
Agradeço por sempre me auxiliar tão prontamente, e por todas as sugestões dadas no
aprimoramento da escrita da tese e dos artigos.
A Professora Cíntia Lopes de Brito Magalhães, que de uma maneira tão solícita e
paciente me ensinou e ajudou com a técnica de qPCR. A sua ajuda foi extremamente
importante para a realização deste trabalho.
Aos colegas dos Laboratórios de Nutrição Experimental (LABNEX) e Bioquímica
Metabólica (LBM), Aline Mayrink de Miranda, Ana Flávia Santos Sampaio,
Bárbara Moreira Quirino, Bianca Figueiredo, Bruno da Cruz Pádua, Carolina
Morais Araújo, Danielle de Lima Ávila, Emerson Cruz de Oliveira, Fabiano Kenji

SILVA, L.S. Agradecimentos
v
Haraguchi, Fernanda Camini, Glaucy Rodrigues de Araújo, Isabel Emerich,
Joamyr Victor Rossoni Júnior, Joyce Ferreira Guerra, Kely Raspante, Maísa
Silva, Natália Nogueira do Nascimento, Pedro Henrique Amorim Miranda, Poliane
Maciel, Rogério Tadeu Ferreira, Sandra Camilo, Simone de Fátima Viana Cunha,
Waleska Claudia Dornas Amaral, a minha gratidão e reconhecimento de que a ajuda
de todos vocês foi fundamental para a realização deste trabalho.
Aos amigos que se tornaram irmãos. Obrigada Aline, Bruno, Carolina, Glaucy,
Joamyr, Melina, Natália, Pedro e Simone pela amizade, cumplicidade, sorrisos e
abraços. Como foi bom e divertido cada dia com vocês! A vocês eu dedico todo o meu
carinho!
Aos colegas da Pós Graduação e aos secretários e secretárias do NUPEB. Em especial à
Maria Aparecida Trópia pelo atendimento e auxilio sempre prestados com muita
simpatia e boa vontade.
Aos técnicos do Laboratório de Nutrição Experimental e Bioquímica Metabólica, Jair
Pastor Mota, Clodoaldo e Renata Rebeca, pela disponibilidade com que sempre me
atenderam.
Aos Laboratórios pertencentes ao Núcleo de Pesquisas em Ciências Biológicas, em
especial ao Laboratório de Biologia Molecular e Celular, Laboratório de Doença de
Chagas, Laboratório de Microbiologia e Laboratório de Imunoparasitologia pelo suporte
necessário a realização deste trabalho.
A querida e eterna República Tchu Tchu Tchu, que tanto amo! Obrigada á todas por
fazerem parte da minha história e por dividirem comigo os momentos mais incríveis que
passei em Ouro Preto. Vocês são parte de mim!

SILVA, L.S. Resumo
vi
RESUMO
Diversas pesquisas indicam que os carotenóides podem modificar o metabolismo
de lipídios e neste contexto, o beta caroteno e o urucum, um corante natural rico em
carotenóides, foram descritos em apresentar efeitos hipolidêmicos em diversos modelos
experimentais. Uma vez que evidências científicas sobre os mecanismos associados aos
efeitos hipolipidêmicos do beta caroteno e urucum ainda não estão bem estabelecidas, o
objetivo do presente trabalho foi investigar os efeitos destes compostos sobre a
expressão de genes hepáticos do metabolismo do colesterol em um modelo de rato com
hipercolesterolemia induzida por dieta. Quarenta ratas albinas da linhagem Fisher,
foram divididas em cinco grupos de 8 animais de acordo com o tratamento recebido. O
grupo C recebeu dieta padrão AIN-93M (4% óleo de soja), o grupo H recebeu dieta
hipercolesterolemiante (25% de óleo de soja e 1% de colesterol), o grupo HBC recebeu
a dieta hipercolesterolemiante suplementada com 0,2% beta caroteno, o grupo HU 0,1%
recebeu a dieta hipercolesterolemiante suplementada com 0,1% de semente de urucum
em pó e o grupo HU 0,2% recebeu a dieta hipercolesterolemiante suplementada com
0,2% de semente de urucum em pó. Após oito semanas de experimentação, os animais
foram anestesiados e eutanasiados. O sangue foi coletado para a determinação do perfil
lipídico e o fígado foi removido para determinação da expressão de genes envolvidos no
metabolismo do colesterol. Para melhor entendimento dos resultados, os grupos foram
divididos como experimento 1 (grupos C, H e HBC) e experimento 2 (grupos C, H, HU
0,1% e HU 0,2%), onde foram avaliados os efeitos do beta caroteno e do urucum
separadamente e respectivamente. A suplementação do beta caroteno à dieta
hipercolesterolemiante melhorou significativamente o perfil lipídico sérico dos animais,
promovendo reduções no colesterol total, nas lipoproteínas aterogênicas, nos
triglicérides e no índice aterogênico. Além disso, o beta caroteno reduziu o conteúdo de
gordura e colesterol no fígado, bem como aumentou a excreção de gordura e colesterol
nas fezes. Nenhuma alteração na expressão dos genes SREBP-2, HMG-CoA R, LDLR,
PPARα, CYP7A1, ABCG5 ou ABCG8 foi obtida pela suplementação com beta
caroteno, sugerindo que seu efeito hipocolesterolemiante possa estar relacionado à
diminuição da absorção intestinal do colesterol. A suplementação do urucum à dieta

SILVA, L.S. Resumo
vii
hipercolesterolemiante promoveu melhoras no perfil lipídico sérico dos animais, como a
redução do colesterol total, das lipoproteínas aterogênicas e do índice aterogênico, bem
como promoveu alterações na expressão de genes do metabolismo de colesterol. A
suplementação do urucum em ambas as concentrações elevou as expressões de
CYP7A1, SHP e ABCB11. Estes resultados sugerem que o efeito hipocolesterolemiante
do urucum estaria relacionado à via de catabolismo do colesterol através da up
regulation da CYP7A1, a enzima responsável pela conversão de colesterol á ácidos
biliares. A partir desses resultados observamos que embora por mecanismos diferentes,
tanto o beta caroteno quanto urucum exerceram efeitos hipolipidêmicos importantes.
Palavras-chaves: Beta caroteno, urucum, metabolismo do colesterol, expressão gênica,
hipercolesterolemia, ratos.

SILVA, L.S. Abstract
viii
ABSTRACT
Several studies indicate that carotenoids may modify lipid metabolism and in
this context, beta-carotene and annatto, a natural colorant rich in carotenoids, have been
reported to present hypolipidemics effects in various experimental models. Since
scientific evidence on the mechanisms associated with hypolipidemic effects of beta-
carotene and annatto are not well established, the objective of this study was to
investigate the effects of these compounds on the expression of liver genes of
cholesterol metabolism in a rat model with diet-induced hypercholesterolemia. Forty
albino rats of Fisher strain were divided into five groups of 8 animals according to the
received treatment. The C group received the standard AIN-93M diet (4% soybean oil),
the H group received a hypercholesterolemic diet (25% of soybean oil and 1%
cholesterol), the HBC group received the hypercholesterolemic diet supplemented with
0.2% beta-carotene, the HU 0.1% group received the hypercholesterolemic diet
supplemented with 0.1% annatto seed powder and the HU 0.2% group received the
hypercholesterolemic diet supplemented with 0.2% annatto seed powder. After eight
weeks of experiment, animals were anesthetized and euthanized. Blood was collected
for determination of the lipid profile and the liver was removed for determination of the
expression of genes involved in cholesterol metabolism. To better understand the
results, groups were divided as experiment 1 (C, H and HBC groups) and experiment 2
(C, H, HU 0.1% and HU 0.2 % groups), to evaluate the effects of beta-carotene and
annato separately and respectively. The supplementation of beta-carotene of the
hypercholesterolemic diet significantly improved the serum lipid profile of the animals,
promoting reductions in total cholesterol, atherogenic lipoproteins, triglycerides and in
the atherogenic index. In addition, beta-carotene reduced the fat and cholesterol content
in the liver, as well as increased fat and cholesterol excretion in the feces. No changes in
the expression of genes SREBP-2, HMG-CoA R, LDLR, PPARα, CYP7A1, ABCG5 or
ABCG8 were obtained by supplementation with beta-carotene, suggesting that its
hypocholesterolemic effect may be related to decreased intestinal absorption of
cholesterol. The supplementation of annatto in the hypercholesterolemic diet promoted
improvements in serum lipid profile of the animals, such as reduction in total
cholesterol, atherogenic lipoproteins and atherogenic index, and promoted changes in

SILVA, L.S. Abstract
ix
the expression of genes of cholesterol metabolism.The addition of annatto in both
concentrations increased the expression of CYP7A1, SHP and ABCB11. These results
suggest that the cholesterol lowering effect of annatto is related to the route of
catabolism of cholesterol by up regulation of CYP7A1, the enzyme responsible for
conversion of cholesterol to bile acids. From these results we observe that although by
different mechanisms, both beta-carotene as annatto exerted important hypolipidemic
effects.
Keywords: Beta-carotene, annatto, cholesterol metabolism, gene expression,
hypercholesterolemia, rats.

SILVA, L.S. Sumário
x
SUMÁRIO
LISTA DE ABREVIATURAS ....................................................................................... xii LISTA DE TABELAS ................................................................................................... xiv
LISTA DE FIGURAS ..................................................................................................... xv CAPÍTULO 1 – INTRODUÇÃO ..................................................................................... 2 CAPÍTULO 2 – REVISÃO BIBLIOGRÁFICA .............................................................. 5
2.1 - Dislipidemias e Mecanismos de Regulação do Metabolismo do Colesterol ........ 5 2.2 - Carotenóides e sua Importância na Dieta ........................................................... 14 2.3 - Beta Caroteno ..................................................................................................... 19 2.4 - Urucum (Bixa orellana L.) e Urucum ................................................................ 22
CAPÍTULO 3 – OBJETIVOS ........................................................................................ 29 3.1 – Objetivo Geral .................................................................................................... 29 3.2 – Objetivos Específicos ........................................................................................ 29
CAPÍTULO 4 – MATERIAL E MÉTODOS ................................................................. 31
4.1 - Animais ............................................................................................................... 31 4.2 - Delineamento Experimental ............................................................................... 31
4.3 - Dietas .................................................................................................................. 32
4.4 - Eutanásia ............................................................................................................. 33
4.5 - Dosagens dos Parâmetros Bioquímicos .............................................................. 33 4.6 - Extração e Dosagem da Gordura Hepática ......................................................... 34
4.6.1 - DOSAGEM DE COLESTEROL TOTAL NA GORDURA HEPÁTICA ....................... 34 4.7 - Extração e Dosagem da Gordura Fecal .............................................................. 35
4.7.1 - DOSAGEM DE COLESTEROL TOTAL NA GORDURA FECAL ............................. 35
4.8 - Ensaio de RT-PCR Quantitativa em Tempo Real .............................................. 36 4.8.1 - EXTRAÇÃO DO RNA TOTAL ......................................................................... 36
4.8.2 - SÍNTESE DO CDNA ....................................................................................... 37
4.7.3 - DESENHO DOS OLIGONUCLEOTÍDEOS INICIADORES ...................................... 37 4.8.4 - CURVA DE EFICIÊNCIA DO OLIGONUCLEOTÍDEOS INICIADORES.................... 39
4.8.5 - RT-PCR QUANTITAIVA EM TEMPO REAL..................................................... 39
4.9 - Análise Estatística .............................................................................................. 40
CAPÍTULO 5 – RESULTADOS .................................................................................... 42 5.1 - Experimento 1 .................................................................................................... 42
5.1.1 - INGESTÃO ALIMENTAR, GANHO DE PESO E EFICIÊNCIA ALIMENTAR ........... 42
5.1.2 - PERFIL LIPÍDICO E ÍNDICE ATEROGÊNICO ..................................................... 43 5.1.3 - PESO RELATIVO DO FÍGADO E QUANTIDADES DE GORDURA E COLESTEROL
TOTAL HEPÁTICO..................................................................................................... 44 5.1.4 - EXCREÇÃO FECAL E QUANTIDADES DE GORDURA E COLESTERL TOTAL
FECAL ...................................................................................................................... 45
5.1.5 - EXPRESSÃO DE GENES DO METABOLISMO HEPÁTICO DO COLESTEROL ...... 46 5.2 - Experimento 2 .................................................................................................... 48
5.2.1 - INGESTÃO ALIMENTAR, GANHO DE PESO E EFICIÊNCIA ALIMENTAR ........... 48 5.2.2 - PERFIL LIPÍDICO E ÍNDICE ATEROGÊNICO ..................................................... 49

SILVA, L.S. Sumário
xi
5.2.3 - PESO RELATIVO DO FÍGADO E QUANTIDADES DE GORDURA E COLESTEROL
TOTAL HEPÁTICO..................................................................................................... 50
5.2.4 - EXCREÇÃO FECAL E QUANTIDADES DE GORDURA E COLESTERL TOTAL
FECAL ...................................................................................................................... 51 5.2.5 - EXPRESSÃO DE GENES DO METABOLISMO HEPÁTICO DO COLESTEROL ........ 52
CAPÍTULO 6 – DISCUSSÃO ....................................................................................... 57 6.1- Experimento 1 ..................................................................................................... 57
6.2 - Experimento 2 .................................................................................................... 65 CAPÍTULO 7 – CONCLUSÕES ................................................................................... 72 REFERÊNCIAS BIBLIOGRÁFICAS ........................................................................... 74
ANEXOS.........................................................................................................................92

SILVA, L.S. Lista de Abreviaturas
xii
LISTA DE ABREVIATURAS
ABCGs - do inglês, ATP-binding cassette subfamily G transporters
ACAT - Acil-CoA- colesterol aciltransferase
AIN - do inglês- American Institute of Nutrition
ANOVA - Análise de variância
apoA1 - Apolipoproteína AI
apoAII - Apolipoproteína AII
ApoAV - Apolipoproteína AV
apoCIII - Apolipoproteína CIII
BARE - Elemento de resposta á ácidos biliares
C - Grupo controle
CT - do inglês, threshold cycle
CYP7A1 - Colesterol-7α -hidroxilase
DCV - Doenças Cardiovasculares
FXR - Receptor X farnesóide
FXRE - Elemento de resposta a FXR
GAPDH - Gliceraldeído 3-fosfato desidrogenase
H - Grupo que recebeu dieta hipercolesterolemiante
HBC - Grupo que recebeu dieta hipercolesterolemiante suplementada com beta
caroteno
HDL - Lipoproteína de alta densidade
HMG-CoA R - 3-hidróxi-3-metil-glutaril CoA redutase
HU 0,1% - Grupo que recebeu dieta hipercolesterolemiante suplementada com urucum
à 0,1%
HU 0,2% - Grupo que recebeu dieta hipercolesterolemiante suplementada com urucum
à 0,2%
INSIG -1 - Gene 1 induzido por insulina
JNK - c-Jun N-terminal quinase
LDL - Lipoproteína de baixa densidade
LDLR - Receptor de LDL

SILVA, L.S. Lista de Abreviaturas
xiii
LRH-1 - do inglês, liver receptor homolog-1
LXR - Receptor X hepático
LXRE - Elemento de resposta á LXR
PPAR - Receptores ativados por proliferadores de peroxisomos
qPCR- Reacão em cadeia da polimerase quantitativa
RXR - Receptor X retinóide
SCAP - Proteína ativadora da clivagem de SREBP
SHP - do inglês, small heterodimer partner
SRE - Elemento de resposta á esterol
SREBP-2 - Proteína ligadora aos elementos de resposta a esterol - 2
VLDL - Lipoproteína de densidade muito baixa

SILVA, L.S. Lista de Tabelas
xiv
LISTA DE TABELAS
Tabela I - Carotenóides de Bixa orellana.......................................................................26
Tabela II - Composição das dietas experimentais (g/Kg de dieta).................................32
TABELA III - Oligonucleotídeos iniciadores utilizados para a análise qRT-PCR........38
Tabela IV - Ingestão alimentar diária, ganho de peso corporal, e eficiência alimentar
em ratas tratadas com dieta padrão, dieta hipercolesterolemiante e dieta
hipercolesterolemiante suplementada com e beta caroteno.............................................42
Tabela V - Perfil lipídico sérico e índice aterogênico de ratas tratadas com dieta padrão,
dieta hipercolesterolemiante e dieta hipercolesterolemiante suplementada com e beta
caroteno...........................................................................................................................44
Tabela VI – Peso relativo do fígado e conteúdos totais de gordura e colesterol hepático
em ratas tratadas com dieta padrão, dieta hipercolesterolemiante e dieta
hipercolesterolemiante suplementada com e beta caroteno.............................................45
Tabela VII – Excreção fecal diária e conteúdos totais de gordura e colesterol fecal em
ratas tratadas com dieta padrão, dieta hipercolesterolemiante e dieta
hipercolesterolemiante suplementada com beta caroteno...............................................46
Tabela VIII - Ingestão alimentar diária, ganho de peso corporal, e eficiência alimentar
em ratas tratadas com dieta padrão, dieta hipercolesterolemiante e dieta
hipercolesterolemiante suplementada com urucum.........................................................48
Tabela IX - Perfil lipídico sérico e índice aterogênico de ratas tratadas com dieta
padrão, dieta hipercolesterolemiante e dieta hipercolesterolemiante suplementada com
urucum............................................................................................................................50
Tabela X – Peso relativo do fígado e conteúdos totais de gordura e colesterol hepático
em ratas tratadas com dieta padrão, dieta hipercolesterolemiante e dieta
hipercolesterolemiante suplementada com urucum.........................................................51
Tabela XI – Excreção fecal diária e conteúdos totais de gordura e colesterol fecal em
ratas tratadas com dieta padrão, dieta hipercolesterolemiante e dieta
hipercolesterolemiante suplementada com urucum.........................................................52

SILVA, L.S. Lista de Figuras
xv
LISTA DE FIGURAS
Figura 1 - Ativação de SREBP por clivagem proteolítica................................................8
Figura 2- Modelo simplificado da regulação do metabolismo de colesterol e ácidos
biliares por LXR e FXR...................................................................................................10
Figura 3 - Modelo de regulação da expressão de CYP7A1 por sinalização em cascata através de
FXR e JNK...................................................................................................................................12
Figura 4 - Efeitos biológicos do PPARα........................................................................14
Figura 5 - Estruturas da -ionona e dos principais carotenóides dietéticos...................16
Figura 6- Papel dos carotenóides na prevenção de doenças crônicas............................18
Figura 7 - Fotos de urucum............................................................................................22
Figura 8- Estruturas moleculares da cis e trans bixina e cis e trans norbixina..............24
Figura 9 – Curva de crescimento semanal dos grupos de animais do experimento 1....43
Figura 10- Expressão do mRNA de genes hepáticos relacionados ao metabolismo de
lipídios (SREBP-2, HMG CoA R, LDLR, PPAR α, CYP7A1, ABCG5 e ABCG8)......47
Figura 11 - Curva de crescimento semanal dos grupos de animais do experimento 2...49
Figura 12 - Expressão do mRNA de genes hepáticos relacionados ao metabolismo de
lipídios (SREBP-2, HMG CoA R, LDLR, PPAR α).......................................................53
Figura 13 - Expressão do mRNA de genes hepáticos relacionados ao metabolismo de
lipídios (CYP7A1, LXR, FXR, ABCG5, ABCG8, SHP, ABCG11)..............................55
Figura 14 - Resumo dos principais efeitos do beta caroteno no soro,
nas fezes e no fígado.....................................................................................................................64
Figura 15 – Proposta do efeito hipocolesterolêmico do urucum.................................................70


SILVA, L.S. Capítulo 1 - Introdução
2
CAPÍTULO 1 – INTRODUÇÃO
As doenças cardiovasculares (DCV) são as principais causas de morte em países
economicamente desenvolvidos, bem como em países de economia emergente (Leifert e
Abeywardena, 2008). De acordo com a Organização Mundial da Saúde (OMS), todo
ano mais de 17 milhões de pessoas morrem vítimas de DCVs, as quais estão entre as
principais responsáveis pelo aumento da morbi-mortalidade dentre as afecções não
transmissíveis.
Idade, sexo, predisposição genética, estilo de vida e dieta são reconhecidos como
os principais fatores de risco modificáveis para DCV. Os efeitos negativos da ingestão
excessiva de gorduras saturadas e colesterol, por exemplo, já estão bem estabelecidos
no desenvolvimento de doenças cardiovasculares (Leifert e Abeywardena, 2008).
Para reduzir as DCV por meio da redução das concentrações de colesterol
circulante, muita atenção tem sido focada na pesquisa por intervenções dietéticas para a
hipercolesterolemia (Park et al, 2009). Tem sido demonstrado que alguns fitoquímicos
de plantas medicinais tradicionais têm efeitos hipolipidêmicos, por modularem a
expressão de genes envolvidos no metabolismo de lipídios e lipoproteínas (Chung et al.,
2007, 2008). Também tem sido demonstrado que os suplementos antioxidantes podem
efetivamente reduzir o perfil lipoproteico aterogênico em doentes com hiperlipidemia e
aterosclerose (Diaz et al., 1997). Estas observações sugerem que a atividade
antioxidante e os efeitos de redução do colesterol de vários extratos vegetais podem
reduzir o desenvolvimento e/ou progressão da aterosclerose e doenças cardiovasculares
(Chung et al., 2007; Chung et al., 2008).
Os carotenóides são um grande grupo de pigmentos presentes na natureza,
identificados em plantas superiores, algas, fungos, bactérias e em alguns animais. Eles
são responsáveis pelas cores do amarelo ao vermelho de frutas, vegetais, fungos e
flores, e são utilizados comercialmente como corantes alimentícios e em suplementos
nutricionais. A produção mundial de carotenóides é calculada em 100 milhões de
toneladas por ano e seu mercado global em 2005 foi estimado em US$ 935 milhões com
projeções de elevação para US$ 1 bilhão até antes do ano de 2009 (Guzman, 2005;
Uenojo et al., 2007). Estes compostos, além do poder corante, possuem importante
função nutricional na dieta humana. São precursores de vitamina A, além de outras

SILVA, L.S. Capítulo 1 - Introdução
3
ações benéficas como proteção contra certos tipos de câncer, doenças cardiovasculares,
cataratas, degeneração macular e fortalecimento do sistema imunológico (Krinsky,
1994; Olson, 1999).
Por muitos anos, o representante mais proeminente dos carotenóides, o beta
caroteno, foi usado como corante de alimentos. O beta caroteno é provavelmente o mais
estudado, e um dos principais presentes na dieta, sangue e tecidos (Krinsky e Johnson,
2005). Evidências experimentais indicam que a ingestão deste carotenóide está
associada à diminuição do risco para doença arterial coronariana (Gaziano e Hennekens,
1993) e sua administração foi responsável por efeitos hipolipidêmicos em ratos e
coelhos hipercolesterolêmicos (Amen e Lachance 1974; Erdman e Lachance, 1974; Tsai
e Mazeedi, 1988; Tsai et al., 1992; Shaish et al., 1995).
O urucuzeiro (Bixa orellana L.) é uma planta nativa da Floresta Tropical da
América Central e do Sul, suas sementes são uma rica fonte de pigmentos vermelhos
alaranjados que têm sido amplamente usados pela indústria de corantes de alimentos, os
quais se constituem numa mistura de muitos carotenóides, tais como bixina, norbixina,
fitoenos e δ-carotenos (Mercadante et al., 1996; Fernandes et al., 2002; Felicissimo et
al, 2004). Esta planta é conhecida por estar associada ao tratamento de várias doenças
na medicina popular e o consumo dos extratos do urucum tem mostrado várias
propriedades bioativas, incluindo propriedades antioxidantes, através da modulação da
produção de espécies reativas de oxigênio, propriedades antitumorais (Junior et al.,
2005; Agner et al., 2005; Rossoni Jr. et al., 2011), bem como efeitos hipolipidêmicos
em ratos, camundongos e coelhos (Lima et al., 2001; Lima et al, 2003; de Paula et al.,
2009; Ferreira et al., 2012).
Muitas linhas de pesquisa tem sugerido que os carotenóides podem modificar o
metabolismo de lipídios (Shih et al., 2008). Entretanto, ainda faltam informações de
como o beta caroteno e o urucum exercem seus efeitos hipolipidêmicos, e se os fatores
que regulam o metabolismo do colesterol podem ser influenciados pelo seu consumo.
Neste contexto, o objetivo deste trabalho foi investigar os efeitos do urucum e do beta
caroteno sobre a expressão de genes hepáticos do metabolismo do colesterol em um
modelo de rato com hipercolesterolemia induzida por dieta.


SILVA, L.S. Capítulo 2 - Revisão Bibliográfica
5
CAPÍTULO 2 – REVISÃO BIBLIOGRÁFICA
2.1 - DISLIPIDEMIAS E MECANISMOS DE REGULAÇÃO DO METABOLISMO
DO COLESTEROL
O colesterol é um componente estrutural essencial da membrana celular e um
importante precursor dos hormônios esteróides, oxisteróis e ácidos biliares (Rezen et al,
2011). Apesar de sua importância, quantidades excessivas de colesterol nas células
podem ocasionar patologias, incluindo a perda da função da membrana, a precipitação
na forma de cristais, ou resultar em danos ateroscleróticos (Weber et al, 2004; Oram,
2002).
Existe um delicado equilíbrio no organismo para cobrir as necessidades de
lipídios das células e, ao mesmo tempo, evitar seu acúmulo excessivo. Por razões não
muito claras, esse equilíbrio pode romper-se, elevando o nível de um ou mais
componentes lipídicos na corrente sanguínea, ao que se denomina dislipidemia
(Gonçalves et al., 2006).
A apresentação laboratorial das dislipidemias compreende quatro situações bem
definidas: hipercolesterolemia isolada (valores de aumentados de colesterol total);
hipertrigliceridemia isolada (valores aumentados de triglicérides); hiperlipidemia mista
(valores aumentados do colesterol total e dos triglicérides) e diminuição isolada do
colesterol HDL ou em associação com aumento do colesterol LDL e/ou dos triglicérides
(Sociedade Brasileira de Cardiologia, 2001).
A hipercolesterolemia é um fator de risco dominante que contribui para o
desenvolvimento e progressão da aterosclerose e subsequente doença cardiovascular,
uma das doenças mais sérias em humanos (Deppa e Varalakshmi, 2005).
As doenças cardiovasculares são a principal causa de mortalidade global, sendo
responsável por quase 17 milhões de mortes anualmente; a aterosclerose em particular,
é a principal contribuinte para a patogênese de infarto cerebral e do miocárdio. Por
muitos anos, tem sido mostrado que o excesso do colesterol LDL é o principal fator de

SILVA, L.S. Capítulo 2 - Revisão Bibliográfica
6
risco para a aterosclerose. As lipoproteínas de baixa densidade (LDLs) são as principais
carreadoras do colesterol no plasma e aparecem envolvidas no desenvolvimento de
várias doenças degenerativas tais como aterosclerose, carcinogênese, envelhecimento e
diabetes mellitus. Muitos estudos têm sugerido que modificações peroxidativas da LDL
é um importante fator nas mudanças ateroscleróticas (Nader et al., 2010).
Os ácidos graxos poliinsaturados nas lipoproteínas são quimicamente
vulneráveis ás espécies reativas de oxigênio. No caso da LDL, elas são atacadas pelos
radicais livres e então modificadas oxidativamente. A LDL oxidada (ox-LDL) é
responsável pela liberação de fatores que iniciam o recrutamento de monócitos e
promovem sua diferenciação em macrófagos. Ox-LDLs são reconhecidas por receptores
scavengers e internalizadas em macrófagos, células endoteliais e células musculares
lisas, levando á formação de células espumosas, o marcador do início das lesões
ateroscleróticas (Kaliora et al., 2009 ; Nader et al., 2010).
A manutenção da homeostase do colesterol é muito complexa e hermeticamente
controlada por mudanças nas concentrações do mRNA de múltiplas enzimas envolvidas
nas vias de biossíntese, captação e efluxo do colesterol. A 3-Hidróxi-3-metilglutaril
CoA redutase (HMG-CoA redutase) é a enzima limitante na biossíntese do colesterol. A
oxidação do colesterol pela colesterol-7α-hidroxilase (CYP7A1), a enzima limitante da
síntese de ácidos biliares, é a principal via de eliminação do colesterol do corpo e os
receptores de LDL (LDLR) desempenham um papel vital na captação e remoção do
colesterol plasmático (Reena et al., 2011).
Para que a homeostase do colesterol seja alcançada, o organismo deve integrar a
síntese de novo de colesterol, a captação do colesterol da circulação mediada por
receptores, a deposição de colesterol nas membranas e a secreção de lipoproteínas, bem
como o armazenamento intracelular de ésteres de colesterol e o metabolismo de
oxisteróis, ácidos biliares e hormônios esteróides. Estes processos são regulados por
vários fatores endógenos e exógenos e são dependentes do tipo de tecido. Proteínas
envolvidas na homeostase do colesterol são reguladas á níveis transcricionais e
traducionais e pós-transcricionalmente por modificações e degradação de proteínas
(Rezen et al, 2011).
A rápida regulação dos teores de colesterol livre na célula é conseguida por meio
modificação covalente de enzimas. Estudos mostram que a fosforilação inativa a HMG-

SILVA, L.S. Capítulo 2 - Revisão Bibliográfica
7
CoA redutase e ativa a CYP7A1 e a ACAT (Acil-CoA- colesterol aciltransferase),
enzima presente no fígado que catalisa a formação de ésteres de colesterol a partir de
colesterol (Scallen e Sanghvi, 1983). Como uma ação rápida, um aumento da
concentração de colesterol, leva a fosforilação que diminui a biossíntese e aumenta a
esterificação do colesterol e o metabolismo de ácidos biliares. Se esta regulação não é
suficiente, segue a regulação ao nível transcricional. Se o conteúdo de colesterol no
fígado é baixo, a expressão gênica é modulada, resultando em maior captação de
colesterol do plasma e menor efluxo, maior biossíntese de colesterol, menor síntese de
ácidos biliares e maior liberação do armazenamento de colesterol esterificado. Quando o
colesterol é abundante, há uma diminuição na biossíntese de colesterol, um aumento do
efluxo de colesterol para o plasma e para a bile e um aumento da síntese de ácidos
biliares e esterificação. Estes processos são regulados principalmente por dois fatores de
transcrição, que funcionam como sensores de esteróis celulares, o SREBP-2 (proteína
ligadora de elementos de resposta a esteróis) e o LXR (receptor X hepático) (Rezen et
al., 2011).
SREBP-2 é um fator de transcrição que reside no retículo endoplasmático. Na
presença de colesterol, SREBP-2 está ligado a duas outras proteínas: SCAP (proteína
ativadora da clivagem de SREBP) e INSIG-1 (gene 1 induzido por insulina). Quando o
nível de colesterol da membrana do retículo endoplasmático diminui, SCAP altera a sua
conformação e facilita a dissociação de INSIG-1 do complexo SREBP-2-SCAP,
permitindo o complexo de migrar para o Complexo de Golgi, onde SREBP-2 é clivado
pelas proteases S1P e S2P (proteases site-1 e site 2 respectivamente). O SREBP clivado
migra então para o núcleo, onde se liga ao elemento de resposta á esterol (SRE) e altera
a transcrição de mais de trinta genes envolvidos na síntese de colesterol e lipídios,
incluindo o receptor de LDL e a HMG-CoA redutase (Horton et al, 2002; Redinger,
2003; Gent e Braakman, 2004; Weber et al., 2004) (Figura 1). Quando os níveis de
colesterol intracelular estão elevados, SCAP interage fortemente com INSIG, o
transporte de SREBP do retículo endoplasmático é inibido e não há síntese de colesterol
e nem de receptores de LDL (Gent e Braakman, 2004; Weber et al., 2004; Eberlé et al,
2004).

SILVA, L.S. Capítulo 2 - Revisão Bibliográfica
8
Figura 1 - Ativação de SREBP por clivagem proteolítica. SREBP: Proteína ligadora aos
elementos de resposta a esterol, SCAP: Proteína ativadora de clivagem, INSIG: Gene 1 induzido
por insulina, S1P: Protease site-1, S2P: Protease site-2, RE: Retículo endoplasmático. Fonte:
Adaptado de Eberlé et al., 2004.
O catabolismo do colesterol em ácidos biliares e sua subsequente excreção nas
fezes é principal meio de eliminação do colesterol do corpo. A síntese dos ácidos
biliares é limitada aos hepatócitos e ocorre através de duas vias distintas: a via clássica
ou neutra, e a via alternativa ou ácida. Uma vez sintetizados, os ácidos biliares são
conjugados a aminoácidos (glicina ou taurina) antes de serem secretados nos canalículos
biliares. Os ácidos biliares, colesterol, fosfolipídios, pequenas quantidades de pigmentos
biliares e proteínas constituem a bile, que é armazenada na vesícula biliar até que seja
segregada para o intestino delgado em resposta a gorduras da dieta. A bile está
envolvida na emulsificação e absorção de lipídios dietéticos e vitaminas lipossolúveis
do intestino delgado. Como os ácidos biliares atravessam o intestino delgado até
atingirem o íleo terminal, aproximadamente 95% são reabsorvidos e transportados de
volta para o fígado através da circulação enterohepática. Devido á aproximadamente 5%
dos ácidos biliares serem eliminados em cada ciclo enterohepático, o fígado deve

SILVA, L.S. Capítulo 2 - Revisão Bibliográfica
9
sintetizar uma quantidade equivalente para manter um pool constante de ácidos biliares.
Assim, o metabolismo de ácidos biliares é um processo hermeticamente regulado, e a
sua perturbação afeta toda a homeostase do colesterol no corpo (Lee et al., 2006).
A CYP7A1 é uma enzima específica do fígado, que cataliza a primeira e
limitante etapa da via clássica da síntese de ácidos biliares. A enzima converte
colesterol em 7α-hidroxicolesterol, e etapas enzimáticas subsequentes levam
principalmente a conversão de 7 α- hidroxicolesterol a ácido cólico. Devido ao fato de a
síntese de ácidos biliares ser a principal via responsável pela manutenção da homeostase
do colesterol no corpo, a regulação da atividade CYP7A1 se demonstra importante
(Ando et al., 2005).
Tem sido demonstrado que a expressão do gene da CYP7A1 é controlada por
fatores transcricionais da superfamília dos receptores nucleares. O LXR, cujos ligantes
endógenos são os oxisteróis, regula positivamente a transcrição da CYP7A1, enquanto o
FXR, (receptor X farnesóide), cujos ligantes são o ácidos biliares, regula negativamente
a CYP7A1 (Ando et al., 2005).
O LXR, originalmente encontrado no fígado, é dimerizado a outro receptor
nuclear, o receptor X retinóide (RXR), formando o heterodímero LXR/RXR. Uma vez
sinalizado pelos oxisteróis, o complexo LXR/RXR se liga ao elemento de resposta a
LXR (LXRE) na região promotora de genes alvo como a CYP7A1, promovendo a
regulação positiva da enzima e diminuindo os níveis de colesterol intracelular
(Redinger, 2003; Wójcicka et al, 2007; Rezen et al 2011).
Estudos usando genéticos e agonistas sintéticos tem definido outros importantes
papéis para o LXR no controle do metabolismo de colesterol. O tratamento de animais
com agonistas de LXR resulta em mudanças na expressão de genes, promovendo o
efluxo de colesterol de células periféricas tais como os macrófagos, a excreção de
colesterol do fígado e a inibição da absorção do colesterol no intestino (Calkin e
Tontonoz, 2010).
No nível molecular, o LXR controla o efluxo de colesterol por regular a
expressão de genes que codificam os transportadores ABCA1 e ABCG1
(transportadores cassete de ligação a ATP) (Figura 2). Upregulation de ABCA1 e
ABCG1 resulta em aumentada transferência de colesterol intracelular para as partículas
de HDL. O acúmulo de colesterol oxidado e outras formas modificadas de colesterol por

SILVA, L.S. Capítulo 2 - Revisão Bibliográfica
10
macrófagos presentes nas paredes dos vasos sanguíneos é um evento crítico na
patogênese da aterosclerose, e a capacidade dos agonistas de LXR em aumentar o
efluxo de colesterol nos macrófagos, estimulou um grande interesse no potencial
terapêutico destes compostos (Schulman e Heyman , 2006).
A ativação de LXR também regula a expressão dos transportadores ABCG5 e
ABCG8 (Figura 2). A expressão desses transportadores é bastante restrita ao fígado e
ao intestino, onde estas proteínas funcionam para promover a excreção do colesterol
(fígado) e limitar a absorção do colesterol (intestino) (Berge et al., 2000; Repa et al.,
2002; Yu et al., 2003).
Assim, pela mobilização do colesterol em células periféricas, promoção da
excreção hepática do colesterol e limitação da sua absorção no intestino, a ativação do
LXR resulta em uma perda líquida de colesterol. Este processo de tráfico de colesterol
para HDL e, finalmente, para fora do corpo tem sido denominado transporte reverso de
colesterol. É importante notar que os agonistas de LXR diminuem a aterosclerose em
modelos animais, e tem sido sugerido que o aumento do transporte reverso de colesterol
desempenha um papel importante nesta atividade (Levin et al., 2005).
Figura 2 - Modelo simplificado da regulação do metabolismo de colesterol e ácidos biliares
por LXR e FXR. Oxisteróis aumentam a transcrição de ABCA1, ABCG5/G8, e CYP7A1
através da ativação de LXR. Ácidos biliares aumentam a transcrição de SHP e ABCB11 através
da ativação de FXR. SHP inibe a transcrição de CYP7A1 através de um mecanismo indireto.
Fonte: Adaptado de Shibata et al., 2007.
Os ácidos biliares são detergentes biológicos com uma série de funções
importantes incluindo a geração do fluxo biliar hepático, a excreção biliar de colesterol,

SILVA, L.S. Capítulo 2 - Revisão Bibliográfica
11
a emulsificação de lipídios no intestino e a captação de vitaminas solúveis em lipídios.
Entretanto, eles são altamente citotóxicos e o desequilíbrio de sua síntese e excreção
resulta em vários processos patológicos. Ácidos biliares têm sido designados como
reguladores cruciais, como ligantes de receptores nucleares importantes na homeostase
de ácidos biliares e colesterol e como iniciadores de cascatas de sinalização importantes
na regulação metabólica e função hepática (Eloranta e Kullak-Ublick, 2005).
Os ácidos biliares hidrofóbicos, ácido quenodesoxicólico, litólico e deoxicólico,
são os ligantes ativadores de FXR mais potentes. Estudos forneceram evidências de que
a repressão da CYP7A1 mediada por FXR ocorre através de um mecanismo indireto
envolvendo dois receptores nucleares adicionais, o LRH-1 (liver receptor homolog-1),
também chamado de CPF, FTF ou NR5A2, e o SHP (small heterodimer partner). O
LRH-1 é um fator de transcrição positivo requerido para a transcrição máxima da
CYP7A1 e também para a expressão do SHP (Gupta et al., 2001; Gupta et al., 2002).
Os ácidos biliares ativam a transcrição de SHP através da ligação do
heterodímero FXR/RXR ao elemento de resposta a FXR (IR1-motif) na região
promotora de SHP. Elevados níveis da proteína SHP inibem a transcrição da CYP7A
através da inibição da atividade de LRH-1, o qual se liga a BARE (elemento de resposta
á ácidos biliares) na região promotora de CYP7A1 (Figura 3). Dessa maneira, o FXR
não se liga diretamente a região promotora da CYP7A1, mas regula a transcrição dessa
enzima indiretamente pela ativação do SHP, o qual interage com LRH-1, que é fator de
transcrição de CYP7A1 (Gupta et al., 2001; Gupta et al., 2002, Xu et al., 2004).
Além desse mecanismo, outro mecanismo adicional para a repressão da
CYP7A1 mediada pelos ácidos biliares envolve a ativação da via da c-Jun N-terminal
quinase (JNK). Foi demonstrado, em culturas primárias de hepatócitos de ratos, que os
ácidos biliares ativam a via da JNK, que por sua vez, ativa o fator de transcrição c-Jun.
c-Jun ativado liga-se então ao elemento AP1 no promotor de SHP e up regulates sua
atividade transcricional (Gupta et al., 2002). Gupta et al., 2001 também sugeriu que c-
Jun possa também interagir diretamente com LRH-1 para reprimir a transcrição da
CYP7A1 (Figura 3).

SILVA, L.S. Capítulo 2 - Revisão Bibliográfica
12
Figura 3 - Modelo de regulação da expressão de CYP7A1 por sinalização em cascata
através de FXR e JNK. Este modelo prediz que os ácidos biliares presentes nos hepatócitos,
ativam tanto o FXR quanto a cascata de sinalização da JNK. FXR e c-Jun ativados, aumentam a
transcrição de SHP-1 através da ligação aos elementos IR-1 e AP-1 no promotor de SHP-1,
respectivamente. Elevados níveis de SHP-1, reprimem a transcrição de CYP7A1 por interação
com LRH-1, um fator de transcrição positivo que se liga à região BARE-II no promotor da
CYP7A1. Sugere-se que c-Jun também possa interagir diretamente com LRH-1 para reprimir
transcrição de CYP7A1. Fonte: Adaptado de Gupta et al., 2001.
Os ácidos biliares, através da ativação do FXR, não só aumentam a transcrição
de SHP, mas também aumentam a transcrição de ABCB11, o principal transportador de
efluxo biliar dos ácidos biliares, dessa forma o pool de ácidos biliares é garantido
(Shibata et al., 2007; Li et al, 2011). (Figura 2).
Os receptores ativados por proliferadores de peroxisomos (PPARs) são proteínas
transdutoras pertencentes à superfamília de receptores nucleares. Estes fatores de
transcrição ativados por ligantes catalisam e coordenam diferentes eventos bioquímicos
a fim de se obter a homeostase celular de lipídios e açúcares, além de orquestrarem
respostas inflamatórias a estímulos (Chinetti-Gbaguidi et al., 2005; Kota et al., 2005;
Perrone et al., 2005).
Depois da ativação pelos ligantes, os PPARs formam heterodímeros com o
RXR; esses heterodímeros tornam-se ativados, e se ligam aos elementos de resposta a
proliferadores de peroxisomos (PPRE) localizados nas regiões promotoras dos genes

SILVA, L.S. Capítulo 2 - Revisão Bibliográfica
13
alvo, causando aumento ou diminuição da transcrição de muitos genes envolvidos
nestes processos biológicos (Figura 4) (Chinetti-Gbaguidi et al., 2005; Kota et al.,
2005; Perrone et al., 2005).
Entre os PPARs três isotipos tem sido identificados: PPAR, PPAR, e PPAR,
sendo o primeiro, o principal receptor envolvido no metabolismo de lipídios Chinetti-
Gbaguidi et al., 2005; Kota et al., 2005; Perrone et al., 2005).
O PPARα é um receptor nuclear controlador do metabolismo lipídico (Cornwell
et al., 2004; Chinetti-Gbaguidi et al., 2005; Kota et al., 2005; Perrone et al., 2005). É
expresso em numerosos tecidos de roedores e humanos incluindo fígado, rins, coração,
músculo esquelético e tecido adiposo marrom e também em uma série de células
vasculares tais como as células endoteliais, monócitos e macrófagos (Cornwell et al.,
2004; Kota et al., 2005). Além disso, é ativado por ligantes naturais endógenos
(agonistas), incluindo ácidos graxos, eicosanóides, fosfolípides oxidados e produtos
lipolíticos derivados de lipoproteínas, bem como por drogas como os fibratos (Cornwell
et al., 2004;Chinetti-Gbaguidi et al., 2005; Kota et al., 2005; Perrone et al., 2005).
O papel crítico dos agonistas de PPARα na -oxidação de ácidos graxos tem
sido bem documentado; em adição eles também estimulam a absorção de ácidos graxos,
por aumentar a expressão de proteínas transportadoras de ácidos graxos e translocases
de ácidos graxos (Kota et al., 2005).
A ativação de PPAR α, por seus ligantes, resulta em aumento da lipólise e do
clearance de lipoproteínas aterogênicas ricas em triglicérides a partir da ativação da
lipase lipoproteica e da apolipoproteína AV (apoAV) e redução da produção do inibidor
da lipase lipoproteica: a apolipoproteína CIII (apoCIII) (Chapman, 2006).
A terapia com ativadores de PPAR α também promove a β-oxidação de ácidos
graxos, reduzindo a biodisponibilidade destes para a síntese de triglicérides, inibe a
síntese de novo de ácidos graxos a partir da redução da atividade da acetil-CoA
carboxilase e da ácido graxo sintase e aumenta a síntese das apolipoproteínas AI e AII
(apoAI e apoAII) (Schoonjans et al., 1996).
Em particular, as vias de metabolismo de lipídios têm revelado o mecanismo de
ação de muitas drogas naturais e sintéticas que têm sido vitais no tratamento de várias
desordens metabólicas (Kota et al., 2005). PPARs apresentam-se como bons alvos de

SILVA, L.S. Capítulo 2 - Revisão Bibliográfica
14
drogas para a correção do perfil global de riscos que predispõem um indivíduo a
doenças cardiovasculares (Chinetti-Gbaguidi et al., 2005).
Figura 4 - Efeitos biológicos do PPARα. FA: ácído graxo, FFA: ácido graxo livre, LPL:
lipoproteína lipase; TG: triglicérides; ABCA1: transportador cassete de ligação a ATP.Fonte:
Adaptado de Kota et al., 2005.
2.2 - CAROTENÓIDES E SUA IMPORTÂNCIA NA DIETA
Os carotenóides são uma família de compostos pigmentados que são sintetizados
por plantas e microorganismos, mas não em animais. Nas plantas, eles contribuem para
a maquinaria fotossintética e as protegem contra os danos da luz. Frutas e vegetais
constituem as principais fontes de carotenóides na dieta humana (Mangels et al.,1993;
Agarwal e Rao, 2000; Johnson, 2002).
Os carotenóides estão presentes como micronutrientes em frutas e vegetais e são
os responsáveis por suas cores amarelo, laranja e vermelha. As propriedades
antioxidantes dos carotenóides tem sido foco de inúmeras pesquisas, eles são os
responsáveis pelas propriedades benéficas de frutas e vegetais na prevenção de doenças
humanas, incluindo doenças cardiovasculares, câncer e outras doenças crônicas, além de

SILVA, L.S. Capítulo 2 - Revisão Bibliográfica
15
serem importantes fontes dietéticas de vitamina A. (Astrog et al., 1997; Paiva e Russell,
1999).
Mais de 600 carotenóides têm sido até agora identificados na natureza.
Entretanto, somente cerca de 40 estão presentes em uma dieta humana típica. Destes 40,
cerca de 20 foram identificados no sangue e tecidos humanos. Cerca de 90% dos
carotenóides no corpo e dieta humana são representados por beta caroteno, α-caroteno,
licopeno, luteína e criptoxantina (Gerster, 1997).
Todos os carotenóides possuem estrutura poliisoprenóide e uma longa cadeia de
duplas ligações conjugadas (Britton, 1995). Eles são divididos em duas classes:
carotenos não polares (ex. beta caroteno e licopeno) contendo somente carbono e
hidrogênio e xantofilas polares (ex. cantaxantina e zeaxantina) que possuem pelo menos
um átomo de oxigênio (Shih et al., 2008).
A sequencia de biossíntese dos carotenóides nas plantas é a seguinte: fitoeno
fitoflueno ζ -caroteno neurosporeno licopeno - caroteno beta caroteno.
Cada etapa enzimática de fitoeno para licopeno adiciona uma dupla ligação á molécula,
resultando em licopeno, o qual é uma molécula simétrica contendo 13 duplas ligações.
A etapa biossintética após o licopeno envolve ciclização enzimática dos grupos finais,
que resulta em - caroteno (uma anel -ionona) e beta caroteno (dois anéis -ionona). A
adição de oxigênio á molécula leva á formação de xantofilas (Goodwin, 1980).
A concentração de cada carotenóide na fruta ou no vegetal sugere que uma
enzima ou um conjunto de enzimas possam ser a etapa limitante na cascata
biossintética. Por exemplo: uma alta concentração de licopeno em tomates vermelhos
sugere uma falta de atividade enzimática suficiente para converter licopeno a -
caroteno (ou seja, atividade insuficiente da ciclase) (Beecher, 1998).
Os diferentes carotenóides são derivados essencialmente por modificações na
base estrutural por ciclização dos grupos finais e por introdução de oxigênio, resultando
assim nas suas cores e propriedades antioxidantes características (Rao e Rao 2007). A
representação das estruturas da -ionona e dos principais carotenóides dietéticos são
mostradas na Figura 5.

SILVA, L.S. Capítulo 2 - Revisão Bibliográfica
16
Figura 5 - Estruturas da -ionona e dos principais carotenóides dietéticos. Fonte: Adaptado
de Uenojo et al., 2007 e Rao e Rao, 2007.
A biodisponibilidade dos carotenóides parece ser dependente de vários fatores
(Parker et al., 1999). Em geral, a absorção dos carotenóides depende da sua
biodisponibilidade na matriz alimentar e da sua solubilidade em micelas (Krinsky,
1993; Boileau et al, 2002). Muitos carotenóides são absorvidos melhor na presença de
gorduras na dieta e a partir de alimentos processados do que a partir de fontes não
transformadas (Stahl et al., 1992; Bohm e Bitsch, 1999). A natureza das isoformas dos
carotenóides também afeta a sua biodisponibilidade e absorção, (isto é, trans-isômeros
de licopeno são menos absorvidos que os isômeros cis) (Bohm e Bitsch, 1999). Outros
fatores que influenciam a absorção dos carotenóides incluem a presença de fibra
dietética, o estado de saúde do indivíduo, e a forma física do carotenóide (van het Hof et
al., 2000).

SILVA, L.S. Capítulo 2 - Revisão Bibliográfica
17
Os carotenóides são absorvidos nas células da mucosa gastrointestinal e
aparecem inalterados na circulação e tecidos (Erdman et al., 1993; Parker, 1996). No
intestino os carotenóides são absorvidos por difusão passiva após serem incorporados
nas micelas, que são formadas por gordura dietética e ácidos biliares. Os carotenóides
micelares são então incorporados nos quilomícrons e lançados no sistema linfático. Eles
são, então, incorporados em lipoproteínas no fígado e liberados para a corrente
sanguínea. Os carotenóides são absorvidos diferencialmente por diferentes tecidos.
Pouco se sabe sobre os mecanismos de absorção dos carotenóides nos tecidos até o
momento. O principal local de armazenamento de carotenóides é o tecido adiposo
(Parker,1988; Erdman et al., 1993).
Com base em estudos epidemiológicos uma relação positiva é sugerida entre a
maior ingestão alimentar e concentrações teciduais de carotenóides e menor risco de
doenças crônicas (Agarwal e Rao, 2000, Johnson 2002; Elliott, 2005). Beta caroteno e
licopeno têm mostrado estar inversamente relacionados ao risco de doenças
cardiovasculares e certos tipos de câncer (Johnson 2002; Ribaya-Mercado et al., 2004).
As propriedades antioxidantes dos carotenóides têm sido sugeridas como o principal
mecanismo pelo qual eles proporcionam seus efeitos benéficos. Estudos recentes têm
mostrando também que os carotenóides podem mediar seus efeitos através de outros
mecanismos, tais como comunicação por junções do tipo gap, regulação do crescimento
celular, modulação da expressão gênica e resposta imune e como moduladores de
enzimas metabolizadoras de fármacos (Astrog, 1997; Paiva e Russell, 1999; Jewell e
O’Brien, 1999; Bertram, 1999). No entanto, os carotenóides tais como e α e beta
caroteno e a -criptoxantina têm a vantagem adicional de serem capazes de ser
convertidos em vitamina A e estarem relacionados ao papel de prevenção e
desenvolvimento de doenças crônicas (Rao e Rao, 2007). O papel dos carotenóides na
prevenção de doenças crônicas e suas ações biológicas encontram-se resumidas na
Figura 6.

SILVA, L.S. Capítulo 2 - Revisão Bibliográfica
18
Figura 6 - Papel dos carotenóides na prevenção de doenças crônicas. Fonte: Adaptado de
Rao e Rao, 2007.
As ações antioxidantes dos carotenóides são baseadas em suas propriedades de
quelar o oxigênio singleto e de interceptar radicais peroxila (Stahl e Sies, 1996), sendo a
habilidade em quelar o oxigênio singleto, a melhor ação antioxidante documentada
pelos carotenóides (Paiva e Russell, 1999). O mecanismo de desativação do oxigênio
singleto pode ocorrer pela transferência física da energia de excitação deste para o
carotenóide ou pela reação química do carotenóide com ele. Em condições normais no
organismo, 95% da desativação do oxigênio singleto é física, restando somente 5% para
reagir quimicamente (Barreiros e David, 2006).
Em adição à capacidade de quelar o oxigênio singleto, tem sido muito
demonstrada a habilidade de vários carotenóides em interferir nas reações em cadeia
iniciadas por radicais, particularmente com aquelas que resultam em peroxidação
lipídica. Os carotenóides sequestram os radicais peroxila formados na peroxidação

SILVA, L.S. Capítulo 2 - Revisão Bibliográfica
19
lipídica, interrompendo, assim, a sucessão de reações que podem danificar
compartimentos lipofílicos (Krinsy, 1998).
Tudo indica que as propriedades e funções dos carotenóides são diretamente
dependentes de sua conformação estrutural básica (Britton, 1995) e modulada pelos
diferentes grupos funcionais, peculiares a cada grupo de carotenóides (Di Mascio et al.,
1990). A atividade quelante dos carotenóides depende principalmente do número de
duplas ligações conjugadas da molécula e é influenciada em uma menor extensão pelos
grupos finais dos carotenóides (cíclicos ou acíclicos) ou a natureza dos substituintes nos
carotenóides contendo grupos finais cíclicos. O licopeno (onze duplas ligações
conjugadas e duas duplas ligações não conjugadas) está entre os mais eficientes
queladores de oxigênio singleto dos carotenóides naturais (Krinsky, 1998).
2.3 - BETA CAROTENO
Dentre os carotenóides, o beta caroteno é provavelmente o mais estudado, e um
dos principais presentes em nossa dieta, sangue e tecidos. As principais fontes dietéticas
de beta caroteno incluem folhas e vegetais verdes bem como frutas e vegetais laranja e
amarelo. Dos 50 diferentes carotenóides que podem ser metabolizados em vitamina A, o
beta caroteno é o que possui a maior atividade pró-vitamina A (Krinsky e Johnson, 2005).
As funções biológicas do beta caroteno incluem transferência de energia na
fotossíntese; transferência de energia para fotoproteção, e conversão metabólica a
retinóides, em animais com ingestão inadequada de vitamina A pré-formada (Krinsky,
1994). Esta última constitui até o momento a única função biológica comprovada do
carotenóide em humanos (Olson, 1996).
A conversão metabólica do beta caroteno à vitamina A é quimicamente possível
devido á sua estrutura molecular que contém anéis -ionona não substituídos, ligados à
cadeia lateral poliênica. Sendo assim, o carotenóide pode, teoricamente, gerar duas
moléculas de vitamina A (Rodriguez-Amaya, 1997).
A maior parte do beta caroteno absorvido pela mucosa duodenal é convertida a
retinol (60 a 70%) que em seguida é metabolizado a ésteres de retinila no enterócito.
Estes, juntamente com a molécula intacta do carotenóide, atingem o fígado através da
linfa, veiculados por quilomicrons remanescentes. No fígado, os ésteres de retinila são

SILVA, L.S. Capítulo 2 - Revisão Bibliográfica
20
convertidos a retinol e se ligam à proteína ligante do retinol, os quais circulam no
plasma associados à outra proteína transportadora, a transtiretina, que direciona o retinol
para os tecidos, onde o retinol pode ser oxidado à retinal e, posteriormente em ácido
retinóico. O beta caroteno é transportado no plasma por lipoproteínas e estocado
principalmente no tecido adiposo. Os carotenóides também podem ser transportados por
outras lipoproteínas como a HDL, LDL e VLDL e a conversão metabólica do
carotenóide a retinóides pode ocorrer também em tecidos de diferentes órgãos, tais
como o pulmão e os rins (Olson, 1994; Wang, 1994; Van Vliet, 1996, Parker et al.,
1996, Ross, 2003).
O beta caroteno é principal fonte de vitamina A na dieta dos Estados Unidos.
Esta pró-vitamina e a vitamina A e seus derivados, os retinóides, tem sido considerados
como protetores e terapêuticos nos processos de carcinogênese (Blakely et al, 1998).
Inicialmente, existiam dúvidas sobre o poder quimiopreventivo do beta caroteno,
se este estaria, realmente, associado exclusivamente ao beta caroteno, ou se deveria ser
atribuído à sua conversão em vitamina A que, por sua vez, promoveria a ação.
Posteriormente, os estudos revelaram que o beta caroteno não só possuía uma ação
exclusiva, como esta era mais potente que a promovida pela vitamina A (Willis e
Wians, 2003).
A pressão de oxigênio (pO2) nos tecidos é um dos interferentes da atuação
antioxidante do beta caroteno. Inicialmente foi atribuída ao beta caroteno uma atividade
pró-oxidante, ou seja, de promoção da oxidação, ao invés de proteção contra essa
oxidação, quando esse atuava em tecidos sob tensões de oxigênio muito elevadas
(Palozza, 1998), entretanto, investigações posteriores, mais detalhadas, observaram que
ocorre apenas um decréscimo da ação antioxidante, pelo processo de auto-oxidação do
beta caroteno (Krinsky, 2001). A partir dessas observações, inclusive, pôde ser
concluído que seria importante, nesses casos, utilizar a vitamina E em associação, uma
vez que essa atua eficazmente em tecidos sob altas tensões de oxigênio (Zhang e
Omaye, 2000).
A concentração de beta caroteno também influencia sua ação antioxidante, de
modo que concentrações que superam os 4-5 μM prejudicam sua habilidade protetora
e/ou a revertem em pró-oxidativa (Lowe et al., 1999; Krinsky, 2001), neste último caso
podendo, inclusive, promover uma lesão de material genético (Woods et al., 1999). A

SILVA, L.S. Capítulo 2 - Revisão Bibliográfica
21
demonstração desse efeito reverso por diversos estudos (Paolini et al., 1999; Lowe et
al., 1999), indica que suplementações com altas doses de micronutrientes devem ser
atentamente acompanhadas pelos estudos, como em qualquer intervenção farmacológica
(Goodman et al., 2003).
Por outro lado, sob condições fisiológicas de pO2 e adequadas concentrações
plasmáticas (1 a 4-5 μM) (Lowe et al., 1999; Zhang e Omaye, 2000, Krinsky, 2001), o
beta caroteno é capaz de prevenir danos celulares (Lowe et al., 1999); diminuir os níveis
de espécies reativas de oxigênio no meio intracelular (Bestwick e Milne, 200),
reduzindo os riscos de lesão de material genético; e promover ação antioxidante em
células pulmonares expostas a nitrosaminas específicas do tabaco (Weitberg e Corvese,
1997).
Estudos em ratos e camundongos indicaram uma supressão ou redução da
peroxidação de lipídios com a administração do beta carotene e uma melhora no status
antioxidante (Iyama et al, 1996; Whittaker et al,1996; Kraus et al, 1997 e Riondel et al,
2002).
A estrutura em duplas ligações conjugadas é primariamente a responsável pela
excelente habilidade do beta caroteno em quelar fisicamente o oxigênio singleto sem
degradação e pela reatividade química do beta caroteno com radicais livres tais como o
radical peroxil (ROO•), radical hidroxil (•OH), e radical superóxido (O2-•). Em geral,
quanto maior a cadeia poliênica, maior a habilidade de estabilização do radical peroxil.
Foi demonstrado que o radical peroxil foi cerca de 100-1000 vezes mais reativo com
carotenóides do que com o hidrogênio alílico em ácidos graxos poliinsaturados. Portanto,
nas concentrações suficientes, os carotenóides podem proteger lipídios de danos
peroxidativos (Valko et al., 2004).
O beta caroteno tem sido relatado em diminuir significativamente lesões
ateroscleróticas em coelhos hipercolesterolêmicos, assim como o α-tocoferol. Beta
caroteno é mais efetivo quelante do oxigênio singleto em baixas pressões de oxigênio
enquanto a vitamina E é mais efetiva em altas pressões de oxigênio. Em resumo,
antioxidantes dietéticos, como o beta caroteno, podem proteger o colesterol LDL contra a
oxidação e ajudar a prevenir a aterosclerose experimental. Evidências epidemiológicas
sugerem que os antioxidantes também podem ajudar a prevenir manifestações clínicas de
doenças artérias coronárias (Sulli et al., 1998).

SILVA, L.S. Capítulo 2 - Revisão Bibliográfica
22
2.4 - URUCUM (BIXA ORELLANA L.) E URUCUM
O urucuzeiro (Bixa orellana L.) é uma planta nativa da Floresta Tropical da
América Central e do Sul (Felicissimo et al, 2004) é um arbusto que pode alcançar de 2
a 9 m de altura, sendo uma planta ornamental, pela beleza e colorido de suas flores e
frutos (Barbosa-Filho, 2006). Suas flores possuem cor rosa claro e surgem nas
extremidades dos ramos, formando fascículos onde nascem cápsulas ovoides com dois
carpelos cobertos de espinhos flexíveis de coloração variável entre o verde, vermelho-
pálido e roxo. No interior destes fascículos são encontradas, em média, 40 sementes. As
sementes são grosseiramente arredondadas, revestidas por uma polpa mole de coloração
avermelhada, as quais se tornam secas, duras e de coloração escura com o
amadurecimento (Franco et al., 2006). (Figura 7).
Figura 7 - Fotos de urucum. A: Urucuzeiro, B: Floração e frutificação, C: Folhas (posições
dorsal e ventral), D: Raiz, E: Fruto, F: Fruto aberto com sementes expostas. Fonte: Barbosa-
Filho, J.M. (2006).
A B C
D E F

SILVA, L.S. Capítulo 2 - Revisão Bibliográfica
23
Do urucum são fabricados os corantes naturais mais difundidos na indústria de
alimentos, ou seja, os produtos de urucum representam aproximadamente 70% (em
quantidade) de todos os corantes naturais e 50% de todos os ingredientes naturais que
têm função corante nos alimentos (Ghiraldini, 1994).
De suas sementes podem se obter corantes com grande variação de tons, que vão
desde o amarelo-laranja ao vermelho (Nielsen, 1990; Ghiraldini, 1994). Na indústria de
alimentos são utilizadas para dar cor em manteiga, margarina, maionese, molhos,
mostarda, salsichas, sopa, sucos, sorvetes, produtos de panificação, macarrão e queijo,
sendo também bastante empregado na indústria da impressão e na tintura (Franco et al.,
2006).
Hoje, o urucum é cultivado em muitos países tropicais como: Bolívia, Brasil, Sri
Lanka, República Dominicana, Equador, Guiana, índia, Jamaica, México, Peru e
Suriname e em menor escala na África, como por exemplo, na Angola, Quênia, Nigéria,
Tanzânia e no Pacífico como as Filipinas e Hawaii (Preston e Richard, 1980).
Estimativas de 2006 mostravam que o Peru era o maior produtor mundial de urucum,
seguido do Brasil e do Quênia (Balaswamy et al., 2006).
No Brasil, cerca de 70% dos grãos produzidos destinam-se ao processamento de
colorau (corante doméstico), 20% são utilizados na produção do corante e 10% são
exportados (Batista, 1994). Em 2000, foi estimado em 1.200 t o consumo de colorífico
no Brasil. Para o segmento de condimentos, especiarias e temperos, o colorífico
representa 44,6%, seguido da pimenta-do-reino com 35,4%, a canela 4,1%, o cominho
4,0% e 11,9% para a pimenta com cominho, bicarbonato, orégano, louro, erva doce,
cravo, camomila e outros. (Franco et al., 2006).
O urucum é uma mistura de muitos carotenóides, tais como bixina, norbixina,
fitoenos e δ-carotenos (Mercadante et al., 1996), sendo a bixina e a norbixina os dois
principais produtos processados do urucum, representando 90% do total de pigmentos
encontrados em suas sementes (Preston e Richard, 1980).
Dos carotenóides presentes nas sementes do urucum, 80% consistem do
apocarotenóide bixina, o qual tem sido encontrado até o momento somente em Bixa
orellana (Mercadante et al., 1996).
A bixina (C25H30O4) é um carotenóide que não possui atividade pró-vitamina A,
contendo dois grupos carboxílicos, sendo que um é um éster monometílico (De Oliveira

SILVA, L.S. Capítulo 2 - Revisão Bibliográfica
24
et al., 2003). Embora menor que os carotenóides usuais, a bixina é também formada por
uma cadeia de duplas ligações conjugadas alternadas, os quais permitem algumas
características especiais. Este pigmento ocorre na forma não estável cis (metil-
hidrogênio 9’-cis-6,6’diapocaroteno 6,6’dioato) na fruta da planta, tornando-se logo na
forma estável trans sobre a extração por organosolventes e/ou calor (Lyng et al, 2005).
A norbixina, forma desmetilada da bixina, (C24H27O4), tem forte poder de
coloração, é um apocarotenóide hidrossolúvel, extraído via saponificação da polpa da
semente e posterior acidificação para precipitar um pó contendo 30-40% de norbixina
(Giuliano et al., 2003).
A conversão de bixina em norbixina é feita pela diluição da bixina em meio
alcalino, a bixina perde um grupo metil produzindo a norbixina, pigmento de cor
vermelho intenso que após a separação e secagem é comercializada na forma de pó ou
pasta (Carvalho e Hein, 1989). (Figura 8).
Figura 8 - Estruturas moleculares da cis e trans bixina e cis e trans norbixina. Fonte:
Adaptado: Scotter, M. J. (1995).
O teor de bixina presente nas sementes de urucum varia de acordo com a
variedade da cultura, do solo, do clima e dos tratos culturais, podendo encontrar
sementes com menos de 1% e até com mais de 4% de bixina. Na semente bruta sua
concentração pode chegar até 5,0%. Contudo, diferentes variedades de sementes
apresentam teores às vezes inferiores a 2,0%. O valor comercial da semente é baseado

SILVA, L.S. Capítulo 2 - Revisão Bibliográfica
25
no percentual de bixina. Teores mínimos de 2,5% são normalmente exigidos para
exportação (Franco et al, 2002).
Como a demanda de urucum e outros corantes naturais aumentaram,
especificações mais rigorosas são impostas e melhores entendimentos de sua química e
bioquímica são garantidos (Ghiraldini, 1994).
Prévias investigações fitoquímicas têm revelado a presença de muitos
carotenóides derivados, alguns terpenóides, tocotrienóis, arenos e flavonóides
(incluindo luteonina e apigenina) nas sementes do urucum. Investigações sobre suas
folhas têm revelado a presença de flavonóides bisulfatos e nas raízes foi encontrado o
triterpeno ácido tomentósico (Shilpi et al., 2006).
Outros carotenóides, em quantidades muito pequenas, foram isolados das
sementes de urucum e suas estruturas foram elucidadas através de técnicas
espectroscópicas como espectrometria de massas e ressonância magnética nuclear. Estes
novos carotenóides puderam ser divididos em três grupos: ésteres metílicos de
apocarotenóides (C30 e C32) (Mercadante et al.,1996, Mercadante et al.,1997a),
diapocarotenóides com grupos terminais éster metílico, cetona e aldeído (C19, C22,
C24 e C25) (Jondiko e Pattenden, 1989; Mercadante et al.,1997b) e ácidos de
diapocarotenóides (C22 e C24) esterificados com grupos metil e geranilgeraniol
(Mercadante et al.,1999). Além desses carotenóides, um carotenóide derivativo com 14
carbonos foi isolado (Mercadante et al.,1997b) e a presença de fitoeno, fitoflueno, ζ-
caroteno e neurosporeno, todos com 40 carbonos, foi confirmada (Mercadante et
al.,1996).
Muitas outras informações referentes à composição química da semente, raiz e
folhas do urucum, são encontradas na literatura. Contudo, talvez pelas diferentes
procedências das plantas, métodos analíticos empregados, ou ainda a custa da própria
instabilidade apresentada por alguns de seus componentes, os percentuais variam
bastante (Oliveira, 2005).
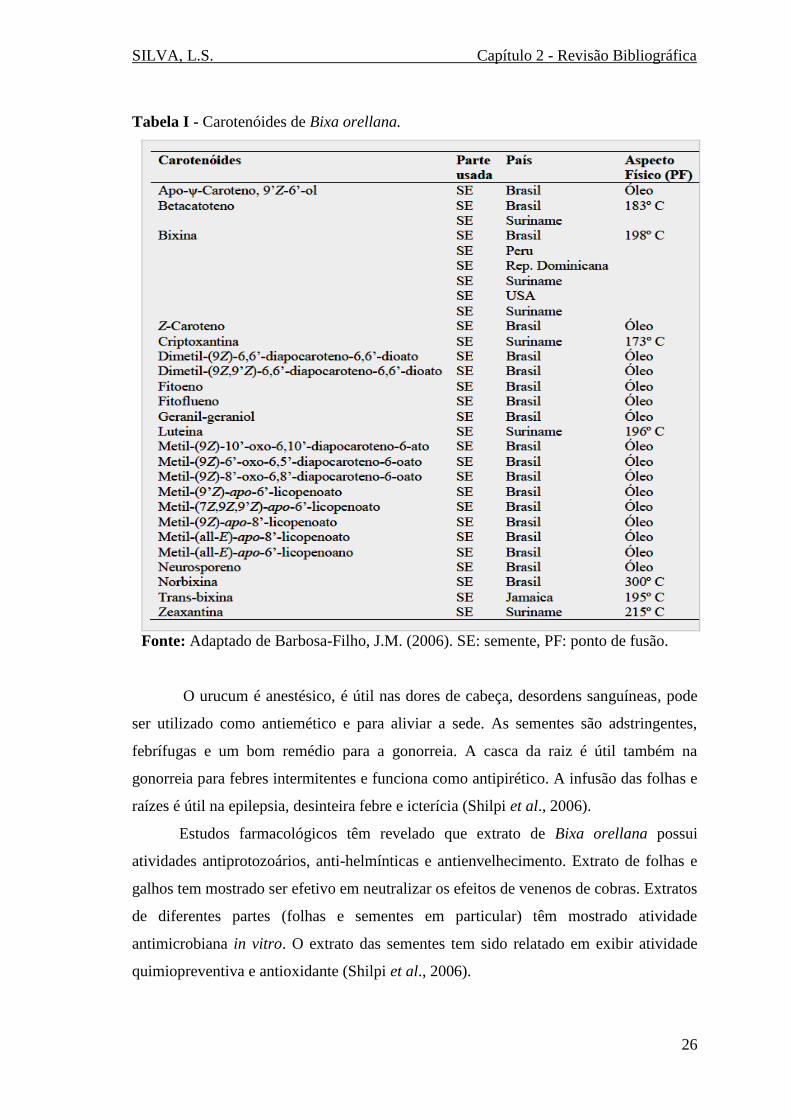
Atualmente mais de duas dezenas de substâncias foram isoladas das sementes de
Bixa orellana. A Tabela I mostra os principais carotenóides encontrados nas sementes
desta planta.
SILVA, L.S. Capítulo 2 - Revisão Bibliográfica
26
Tabela I - Carotenóides de Bixa orellana.
Fonte: Adaptado de Barbosa-Filho, J.M. (2006). SE: semente, PF: ponto de fusão.
O urucum é anestésico, é útil nas dores de cabeça, desordens sanguíneas, pode
ser utilizado como antiemético e para aliviar a sede. As sementes são adstringentes,
febrífugas e um bom remédio para a gonorreia. A casca da raiz é útil também na
gonorreia para febres intermitentes e funciona como antipirético. A infusão das folhas e
raízes é útil na epilepsia, desinteira febre e icterícia (Shilpi et al., 2006).
Estudos farmacológicos têm revelado que extrato de Bixa orellana possui
atividades antiprotozoários, anti-helmínticas e antienvelhecimento. Extrato de folhas e
galhos tem mostrado ser efetivo em neutralizar os efeitos de venenos de cobras. Extratos
de diferentes partes (folhas e sementes em particular) têm mostrado atividade
antimicrobiana in vitro. O extrato das sementes tem sido relatado em exibir atividade
quimiopreventiva e antioxidante (Shilpi et al., 2006).

SILVA, L.S. Capítulo 2 - Revisão Bibliográfica
27
Entre os carotenóides naturais, bixina é um dos mais efetivos supressores
biológicos de moléculas de oxigênio singleto e pode contribuir para a proteção das
células e tecidos contra os efeitos deletérios dos radicais livres, tendo também um
efetivo inibidor da peroxidação de lipídios (Di Mascio et al., 1990; Zhang et al., 1991;
Silva et al., 2001).
Existem trabalhos relacionando a atuação da bixina sobre a hipercolesterolemia
e estresse oxidativo (Zhang et al., 1991; Silva et al., 2001, Lima et al., 2001; Lima et al,
2003) e estudos toxicológicos têm demonstrado que ela não apresenta efeitos tóxicos
significativos em ratos e camundongos (Fernandes et al.,2002; Bautista et al., 2004).
Neste contexto, o urucum, por suas propriedades e peculiaridades, se torna um atrativo
alvo de pesquisa, e a investigação de suas atuações sobre o metabolismo lipídico
justifica a necessidade novos experimentos.


SILVA, L.S. Capítulo 3 - Objetivos
29
CAPÍTULO 3 – OBJETIVOS
3.1 – OBJETIVO GERAL
Investigar o efeito do beta caroteno e da semente do urucum sobre a expressão
de genes do metabolismo hepático do colesterol em ratos com hipercolesterolemia
induzida por dieta.
3.2 – OBJETIVOS ESPECÍFICOS
Avaliar os efeitos da dieta hipercolesterolemiante, do beta caroteno e do urucum
sobre:
I - Parâmetros biométricos dos animais;
II - Perfil lipídico sérico;
III – Armazenamento de gorduras e colesterol total no fígado;
IV – Excreção de gorduras e colesterol total nas fezes;
V- Expressão do mRNA dos principais receptores, enzimas e transportadores
envolvidos no metabolismo hepático do colesterol.


SILVA, L.S. Capítulo 4 - Material e Métodos
31
CAPÍTULO 4 – MATERIAL E MÉTODOS
4.1 - ANIMAIS
Foram utilizadas ratas albinas da linhagem Fisher, com idade de 80 a 95 dias e
peso médio de 195 g, provenientes do Laboratório de Nutrição Experimental da Escola
de Nutrição (ENUT) da Universidade Federal de Ouro Preto. Todos os animais foram
mantidos em gaiolas individuais dispostas em ambiente com temperatura, luminosidade
e umidade controladas e receberam água “ad libitum”. O protocolo experimental
relativo ao uso dos animais foi aprovado pela Comissão de Ética no Uso de Animais da
Universidade Federal de Ouro Preto (CEUA-UFOP, certificado n° 2009/13).
4.2 - DELINEAMENTO EXPERIMENTAL
Quarenta ratas Fisher albinas, foram divididas em 5 grupos de oito animais:
Grupo 1- Grupo controle (C), que recebia dieta padrão seguindo recomendações do
American Institute of Nutrition - AIN-93 (Reeves et al., 1993) ;
Grupo 2- Grupo hipercolesterolêmico (H) que recebia dieta hipercolesterolemiante
composta por 25% de óleo de soja e 1% de colesterol (Matos et al, 2005) ;
Grupo 3- Grupo hipercolesterolêmico e beta caroteno 0,2% (HBC) (dieta
hipercolesterolemiante acrescida de beta caroteno em pó na concentração de 0,2%);
Grupo 4- Grupo hipercolesterolêmico e urucum 0,1% (HU 0,1%) (dieta
hipercolesterolemiante acrescida de semente de urucum em pó na concentração de
0,1%);
Grupo 5- Grupo hipercolesterolêmico e urucum 0,2% (HU 0,2%) (dieta
hipercolesterolemiante acrescida de semente de urucum em pó na concentração de
0,2%).
Antes do início do experimento, os animais passaram por um período de 2
semanas para adaptação às dietas, onde os animais do grupo C receberam dieta controle
e os animais dos demais grupos receberam a dieta hiperlipídica. Após este período, os
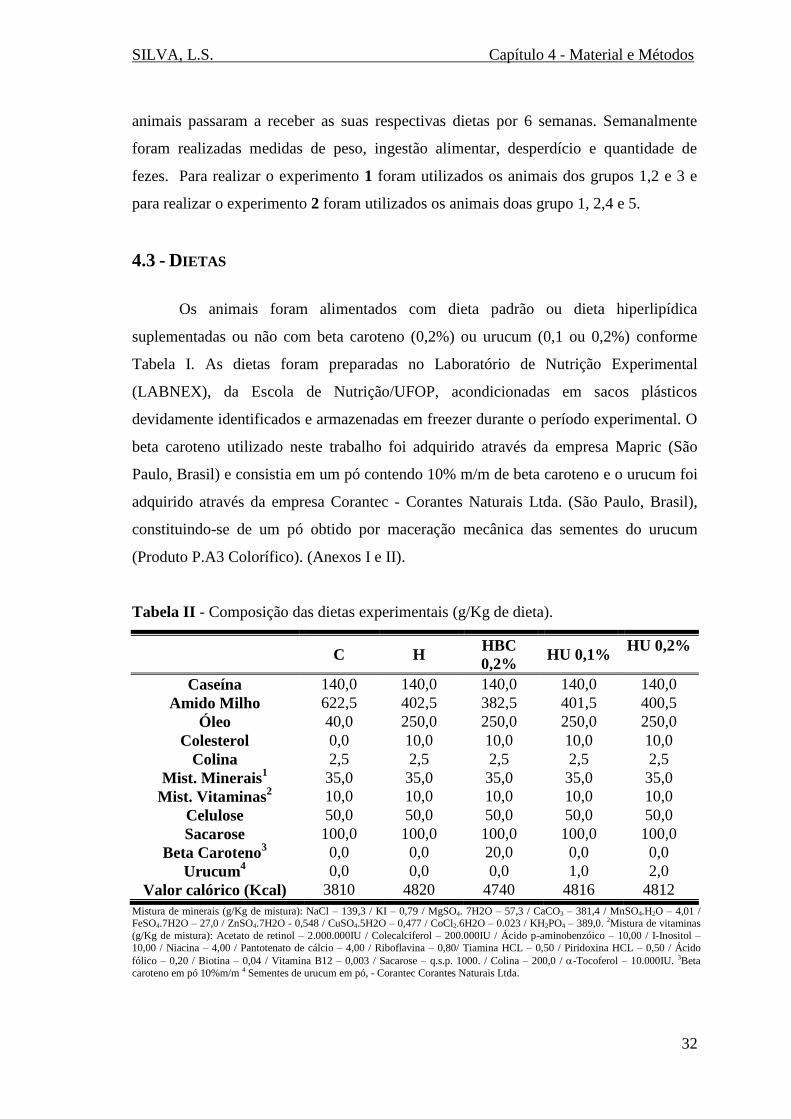
SILVA, L.S. Capítulo 4 - Material e Métodos
32
animais passaram a receber as suas respectivas dietas por 6 semanas. Semanalmente
foram realizadas medidas de peso, ingestão alimentar, desperdício e quantidade de
fezes. Para realizar o experimento 1 foram utilizados os animais dos grupos 1,2 e 3 e
para realizar o experimento 2 foram utilizados os animais doas grupo 1, 2,4 e 5.
4.3 - DIETAS
Os animais foram alimentados com dieta padrão ou dieta hiperlipídica
suplementadas ou não com beta caroteno (0,2%) ou urucum (0,1 ou 0,2%) conforme
Tabela I. As dietas foram preparadas no Laboratório de Nutrição Experimental
(LABNEX), da Escola de Nutrição/UFOP, acondicionadas em sacos plásticos
devidamente identificados e armazenadas em freezer durante o período experimental. O
beta caroteno utilizado neste trabalho foi adquirido através da empresa Mapric (São
Paulo, Brasil) e consistia em um pó contendo 10% m/m de beta caroteno e o urucum foi
adquirido através da empresa Corantec - Corantes Naturais Ltda. (São Paulo, Brasil),
constituindo-se de um pó obtido por maceração mecânica das sementes do urucum
(Produto P.A3 Colorífico). (Anexos I e II).
Tabela II - Composição das dietas experimentais (g/Kg de dieta).
C H
HBC
0,2% HU 0,1%
HU 0,2%
Caseína 140,0 140,0 140,0 140,0 140,0
Amido Milho 622,5 402,5 382,5 401,5 400,5
Óleo 40,0 250,0 250,0 250,0 250,0
Colesterol 0,0 10,0 10,0 10,0 10,0
Colina 2,5 2,5 2,5 2,5 2,5
Mist. Minerais1 35,0 35,0 35,0 35,0 35,0
Mist. Vitaminas2 10,0 10,0 10,0 10,0 10,0
Celulose 50,0 50,0 50,0 50,0 50,0
Sacarose 100,0 100,0 100,0 100,0 100,0
Beta Caroteno3 0,0 0,0 20,0 0,0 0,0
Urucum4 0,0 0,0 0,0 1,0 2,0
Valor calórico (Kcal) 3810 4820 4740 4816 4812
Mistura de minerais (g/Kg de mistura): NaCl – 139,3 / KI – 0,79 / MgSO4. 7H2O – 57,3 / CaCO3 – 381,4 / MnSO4.H2O – 4,01 / FeSO4.7H2O – 27,0 / ZnSO4.7H2O - 0,548 / CuSO4.5H2O – 0,477 / CoCl2.6H2O – 0.023 / KH2PO4 – 389,0. 2Mistura de vitaminas
(g/Kg de mistura): Acetato de retinol – 2.000.000IU / Colecalciferol – 200.000IU / Ácido p-aminobenzóico – 10,00 / I-Inositol –
10,00 / Niacina – 4,00 / Pantotenato de cálcio – 4,00 / Riboflavina – 0,80/ Tiamina HCL – 0,50 / Piridoxina HCL – 0,50 / Ácido
fólico – 0,20 / Biotina – 0,04 / Vitamina B12 – 0,003 / Sacarose – q.s.p. 1000. / Colina – 200,0 / -Tocoferol – 10.000IU. 3Beta
caroteno em pó 10%m/m 4 Sementes de urucum em pó, - Corantec Corantes Naturais Ltda.

SILVA, L.S. Capítulo 4 - Material e Métodos
33
4.4 - EUTANÁSIA
Ao fim de cada experimento os animais foram deixados oito horas em jejum e
após anestesia com isoflurano, foram eutanaziados. O sangue foi coletado através de
secção do plexo braquial, recolhido em tubos de polipropileno (com 2,0 mL de
capacidade), centrifugado a 1500 g por 15 minutos e o soro coletado. O soro foi
devidamente aliquotado e estocado a -80°C para a posterior realização das dosagens
bioquímicas.
Os fígados dos animais foram extraídos, lavados em solução salina, pesados,
imersos em nitrogênio líquido e imediatamente estocado a -80°C para posteriores
análises. O conteúdo fecal foi removido do ceco dos animais, seco em estufa ventilada a
60°C, macerado, pesado e utilizado para extração da gordura total.
4.5 - DOSAGENS DOS PARÂMETROS BIOQUÍMICOS
As dosagens das concentrações séricas de colesterol total, colesterol HDL e
triglicérides foram realizadas de acordo com as orientações do fabricante dos kits
comerciais Labtest Diagnóstica S.A. Os princípios dos métodos e seus respectivos
protocolos estão descritos no Anexo III.
Para a determinação da concentração do colesterol não-HDL foi feita a subtração
da concentração do colesterol total pela concentração do colesterol HDL.
O colesterol não-HDL é usado para estimar a concentração de partículas
potencialmente aterogênicas no soro, tais como LDL, IDL e VLDL. Para obtenção do
índice aterogênico foi calculada a razão colesterol não-HDL /colesterol HDL.
Para a garantia do controle de qualidade das determinações bioquímicas foi
utilizado o soro Qualitrol 1H da Labtest Diagnóstica S.A.
Colesterol não-HDL (mmol/L) = Colesterol Total (mmol/L) – Colesterol HDL (mmol/L)

SILVA, L.S. Capítulo 4 - Material e Métodos
34
4.6 - EXTRAÇÃO E DOSAGEM DA GORDURA HEPÁTICA
A gordura hepática foi extraída usando uma mistura de clorofórmio e metanol
(2:1, v/v), conforme descrito por Folch et al. (1957). Posteriormente, foi quantificada
gravimetricamente e o colesterol total foi mensurado no extrato lipídico.
Procedimento de extração da gordura hepática:
Inicialmente, 400 mg de fígado foram homogeneizados com 8 mL da mistura
clorofórmio-metanol (2:1v/v) e o homogenato vertido em tubos de vidro pré-lavados
com éter de petróleo. Os tubos foram centrifugados a 1000 g por 10 minutos. O
sobrenadante foi retirado e colocado em outro tubo de vidro previamente pesado e
identificado. Foram acrescentados 2 mL da solução de NaCl 0,73%, nova centrifugação
foi realizada e a fase superior desprezada. Posteriormente, a parede interna do tubo de
vidro foi lavada com 3 mL da solução de Folch - clorofórmio/metanol/água (3: 48: 47).
A fase superior foi desprezada e os tubos com os extratos lipídicos foram secos em
estufa semiaberta a 40ºC para evaporação total do solvente. Após a evaporação, os
tubos foram resfriados no dessecador e pesados novamente.
Cálculo:
4.6.1 - DOSAGEM DE COLESTEROL TOTAL NA GORDURA HEPÁTICA
Anteriormente ao procedimento de dosagem do colesterol total, a gordura
hepática extraída foi diluída em isopropanol. Dessa forma, em um tubo de polipropileno
diluições de 1:50 foram preparadas a partir da homogeneização de 10 μL de gordura em
490 μL em isopropanol.
Em seguida, a concentração de colesterol total nessa amostra diluída foi
determinada, utilizando o kit para colesterol total da Labtest Diagnóstica S.A. (Lagoa
Quantidade de Gordura no Fígado (mg) = [peso do tubo final (g) – peso do tubo inicial (g)] x 1000

SILVA, L.S. Capítulo 4 - Material e Métodos
35
Santa, MG, Brasil) e os resultados foram expressos em mg de colesterol total/ grama de
fígado.
4.7 - EXTRAÇÃO E DOSAGEM DA GORDURA FECAL
A gordura fecal foi extraída usando uma mistura de clorofórmio e metanol (2:1,
v/v), conforme descrito por Folch et al. (1957). Posteriormente, foi quantificada
gravimetricamente e o colesterol total foi mensurado no extrato lipídico.
Procedimento de extração da gordura fecal:
Em tubos de vidro pré-lavados com éter de petróleo, 100 mg de fezes maceradas
foram homogeneizadas com 1,9 mL da mistura clorofórmio-metanol (2:1v/v) e 0,4 mL
de metanol. Os tubos foram centrifugados a 1000 g por 10 minutos. O sobrenadante foi
retirado e colocado em outro tubo de vidro previamente pesado e identificado. Foram
acrescentados 0,8 mL de clorofórmio puro e 0,64 mL da solução de NaCl 0,73%, nova
centrifugação foi realizada e a fase superior desprezada. Posteriormente, a parede
interna do tubo de vidro foi lavada com 0,9 mL da solução de Folch -
clorofórmio/metanol/água (3: 48: 47). A fase superior foi desprezada e os tubos com os
extratos lipídicos foram colocados em estufa semiaberta a 40ºC para evaporação total do
solvente. Após a evaporação, os tubos foram resfriados no dessecador e pesados
novamente.
Cálculo:
4.7.1 - DOSAGEM DE COLESTEROL TOTAL NA GORDURA FECAL
Anteriormente ao procedimento de dosagem do colesterol total, a gordura fecal
extraída foi diluída em isopropanol. Dessa forma, em um tubo de polipropileno
Quantidade de Gordura nas Fezes (mg) = [peso do tubo final (g) – peso do tubo inicial (g)] x 1000

SILVA, L.S. Capítulo 4 - Material e Métodos
36
diluições de 1:50 foram preparadas a partir da homogeneização de 10 μL de gordura em
490 μL em isopropanol.
Em seguida, a concentração de colesterol total nessa amostra diluída foi
determinada, utilizando o kit para colesterol total da Labtest Diagnóstica S.A. (Lagoa
Santa, MG, Brasil) e os resultados foram expressos em mg de colesterol total/ grama de
fezes.
4.8 - ENSAIO DE RT-PCR QUANTITATIVA EM TEMPO REAL
4.8.1 - EXTRAÇÃO DO RNA TOTAL
O RNA total do tecido hepático de ratos foi isolado utilizando o sistema SV
Total RNA Isolation System (Promega Corporation, Madison, USA) de acordo com as
instruções do fabricante. Resumidamente, 60 mg do tecido hepático foram
homogeneizados com 350μL do tampão de lise (RNA Lysis Buffer). Uma alíquota de
175 μL do homogenato obtido foi transferida para um novo tubo, onde foram
adicionados 350 μL do tampão de diluição (RNA Diltion Buffer). A mistura foi
colocada em banho-maria por exatamente 3 minutos e centrifugada a 1500 g por 10
minutos. Ao sobrenadante, foram adicionados 200 μL de etanol 95% e o volume total
foi transferido para a membrana da coluna do Spin Basket Assembly e centrifugados á
1500 g por 1 minuto. À membrana dessa coluna foram adicionados 600μL de RNA
Wash Solution, seguida por centrifugação a 1500 g por 1 minuto. Posteriormente foram
adicionados 50 μL do mix de incubação de DNAse o qual foi incubado a temperatura
ambiente por 15 minutos. Após este período, 200μL de DNAse Stop Solution foram
adicionados a coluna e centrifugados a 1500 g por 1 minuto. Foram realizadas duas
lavagens com RNA Wash Solution, (600 μL a 1500 g por 1 minuto e 250 μL a 1500 g
por 2 minutos respectivamente). A coluna foi transferida para o tubo de eluição e sobre
sua membrana foram adicionados 100 μL de Nuclease Free Water, que foram
posteriormente centrifugados a 1500 g por 1 minuto. Por último, a concentração e
pureza do RNA total foram determinadas em espectrofotômetro NanoVue (GE
Healthcare, UK).

SILVA, L.S. Capítulo 4 - Material e Métodos
37
4.8.2 - SÍNTESE DO CDNA
O ácido desoxirribonucleico complementar (cDNA) foi sintetizado a partir de
1µg de RNA total utilizando o kit Hight-Capacity cDNA Reverse Transcription da
Applied Biosystems, (Foster City, CA) de acordo com as instruções do fabricante. O
meio de reação continha 2 μL de tampão 10x (500 mM de KCl, 100 mM deTris-HCl, 25
mM de MgCl2, pH 8,3), 0,8 μL da mistura de desoxiribonucleotídeos trifosfato
(dNTPs) 100 mM, 2 μL de primers randômicos e 1 μL da enzima transcriptase reversa
MultiScribe (50 U/μL). A reação foi realizada nas seguintes condições, 10 minutos a
25°C, seguido de 120 minutos a 37°C e 5 minutos a 85°C no termociclador Biocycler
modelo MJ96+.
4.7.3 - DESENHO DOS OLIGONUCLEOTÍDEOS INICIADORES
Os oligonucleotídeos iniciadores utilizados para a amplificação dos transcritos
de interesse foram construídos de acordo com sequências de nucleotídeos publicadas na
literatura (tabela III).

SILVA, L.S. Capítulo 4 - Material e Métodos
38
TABELA III – Oligonucleotídeos iniciadores utilizados para a análise qRT-PCR.
Gene Oligonucleotídeos Iniciadores
(sequência 5’ para 3’) Referência
SREBP-2 F - TGGGCTTCTTGGCTAGCTACTT
R - TTCGCTCCATGAAAAACTTCTG Reena et al. (2011)
HMG CoA F - AGATACTGGAGAGTGCCGAGAAA
R - TTTGTAGGCTGGGATGTGCTT Li et al. (2009)
LDLR F - CCAACCTGAAGAATGTGGTG
R - CAGGTCCTCACTGATGATGG Li et al. (2009)
PPAR α
F - TGTCGAATATGTGGGGACAA
R - AAACGGATTGCATTGTGTGA
Palou et al. (2008)
CYP7A1 F - TACTTCTGCGAAGGCATTTGG
R- GAACACAGAGCATCTCCCTGG Reena et al. (2011)
LXR F - GGCCCTGCATGCCTATGT
R- CATTAGCATCCGTGGGAACA Shibata et al. (2007)
FXR F - TGACAAAGAAGCCGCGAAT
R- CACACAGCTCATCCCCTTTTATT Shibata et al. (2007)
SHP F - GGAGCAGCCCTCGTCTCA
R- ACACTGTATGCAAACCGAGGAA Shibata et al. (2007)
ABCG5 F - AGGCTCAGTTACAGGCTCAGAG
R - GTCCCACTTCTGCTGGCATGAT
Scoogan et al. (2009)
ABCG8 F - CGTCAGATTTCCAATGACTTCCG
R - TCCGTCCTCCAGTTCATAGTACA Scoogan et al. (2009)
ABCB11 F - GCACCCTTGAGAGGACCTTTT
R - AGCGTTGCCGGATCGA
Shibata et al. (2007)
GAPDH F - ATGGAGAAGGCTGGGGCTCACCT
R - AGCCCTTCCACGATGCCAAAGTTGT Xiong et al. (2010)

SILVA, L.S. Capítulo 4 - Material e Métodos
39
4.8.4 - CURVA DE EFICIÊNCIA DO OLIGONUCLEOTÍDEOS INICIADORES
Para determinar a eficiência da amplificação dos genes alvo e do gene de
referência endógena foram construídas curvas padrões para cada amplificado, a partir de
diluições seriadas do cDNA de uma mesma amostra. A análise de regressão linear
dos valores de “Threshold Cycle” (CT) em função do logaritmo da respectiva diluição
forneceu o coeficiente angular da reta (a, em y = ax+b) que foi utilizado para o cálculo
da eficiência de amplificação do produto pelos iniciadores, conforme equação abaixo.
4.8.5 - RT-PCR QUANTITAIVA EM TEMPO REAL
A análise da expressão gênica foi realizada através da técnica de PCR em tempo
real (qPCR). A quantificação dos produtos formados durante os ciclos de amplificação
foi realizada com o reagente Power SYBR® Green PCR Master Mix (Applied
Biosystems, Foster City, CA, USA). As reações foram realizadas em placas de 96
poços, sendo adicionados em cada poço 2 μL de cDNA (100 ng), 0,48 μL de cada
primer (forward e reverse, 10 μM), 6 μL Power de SYBR®
Green Master Mix e volume
final de água livre de DNAse para 12 μL. As reações foram realizadas nas seguintes
condições, 50°C por 2min, 95° C por 10 min e então 40 ciclos de 95°C por 15s
(desnaturação) e 60°C por 1min (anelamento dos iniciadores e extensão dos produtos)
no termociclador ABI 7300 (Applied Biosystems, CA, USA). O gerenciamento do
termociclador e a coleta dos dados gerados durante a amplificação foram realizados pelo
programa 7000 System SDS Software (Applied Biosystems). Todas as análises foram
realizadas em triplicata técnica. A especificidade dos produtos obtidos foi confirmada
pela análise das curvas de dissociação do produto amplificado ao final de cada reação.
Os dados obtidos foram analisados utilizando o método de quantificação relativa
da expressão gênica (CT comparativo ou ∆∆CT), que permite quantificar diferenças no
nível de expressão de um gene específico entre as diferentes amostras. A expressão dos
genes alvo foi determinada em função da expressão do gene de referência endógena

SILVA, L.S. Capítulo 4 - Material e Métodos
40
gliceraldeído 3- fosfato desidrogenase (GAPDH) e uma amostra normalizadora (grupo C)
foi utilizada como base para os resultados de expressão comparativa. De posse dos
valores de CT, que corresponde ao número do ciclo na fase exponencial da PCR em que
a fluorescência ultrapassa o valor basal, foi calculado o ∆CT de cada amostra, na qual o
valor do CT do gene de referência endógena (GAPDH) foi subtraído do CT do gene alvo.
Em seguida foram calculados os valores de ∆∆CT, na qual o valor do ∆CT da
amostra normalizadora (grupo C) foi subtraído do ∆CT das amostras teste (demais
grupos experimentais).
Os valores do ∆∆CT obtidos foram utilizados em uma fórmula aritmética para o
cálculo final da diferença de expressão dos genes entre as amostras analisadas, dada por
2-∆∆CT
.
4.9 - ANÁLISE ESTATÍSTICA
Todos os dados foram submetidos ao teste de normalidade Kolmogorov-
Smirnov. Os parâmetros avaliados apresentaram distribuição normal e foram analisados
pelo teste de análise de variância univariada (ANOVA-one way) seguido pelo teste de
Bonferroni para determinação das diferenças entre os grupos. Para as análises da
expressão gênica foi utilizado teste-t de Student.
Os dados foram expressos como média ± desvio padrão e as diferenças foram
consideradas significativas para p <0,05. Todas as análises foram realizadas utilizando o
software GraphPad Prism versão 5.00 para Windows (San Diego, California, USA).
∆CT = CT do gene alvo - CT do gene de referência endógena
∆∆CT = ∆CT da amostra teste - ∆CT da amostra normalizadora


SILVA, L.S. Capítulo 5 - Resultados
42
CAPÍTULO 5 – RESULTADOS
5.1 - EXPERIMENTO 1
5.1.1 - INGESTÃO ALIMENTAR, GANHO DE PESO E EFICIÊNCIA ALIMENTAR
Os parâmetros ingestão alimentar diária, ganho de peso corporal e eficiência
alimentar foram avaliados nos grupos experimentais e estão apresentadas na tabela IV.
Os animais que receberam as dietas hipercolesterolemiantes (H e HBC)
apresentaram menor ingestão alimentar em relação ao controle, entretanto foram os que
apresentaram maior ganho de peso corporal e consequentemente aqueles com maior
eficiência alimentar.
Tabela IV - Ingestão alimentar diária, ganho de peso corporal, e eficiência alimentar
em ratas tratadas com dieta padrão, dieta hipercolesterolemiante e dieta
hipercolesterolemiante suplementada com e beta caroteno.
Grupos Experimentais
Variáveis C H HBC
Ingestão Alimentar (g/dia) 15,38 2,00a
12,14 1,79b 11,20 0,60
b
Ganho de peso corporal (g) 32,04 9,45b
69,31 13,22a
56,16 15,45a
Eficiência Alimentar (%)* 5,03 1,62
b 13,69 2,49
a 11,85 2,96
a
Os resultados estão apresentados como média desvio padrão (n=8). Letras sobrescritas diferentes na
mesma linha indicam diferenças estatisticamente significativas entre os grupos. Os dados foram
analisados por análise de variância univariada (one way ANOVA) seguido pelo Teste de Bonferroni
(p ≤ 0,05). C: grupo controle; H: dieta hipercolesterolemiante; HBC: dieta hipercolesterolemiante + beta
caroteno 0,2%. * (Ganho de peso corporal)/ (ingestão alimentar) x 100.
A Figura 9 demonstra a evolução do ganho de peso dos animais ao longo do
período experimental. Durante todo o período foi observado que os animais do grupo H
obtiveram os maiores pesos e que os animais do grupo HBC se mantiveram com pesos
intermediários aos pesos obtidos pelos animais dos grupos C e H.
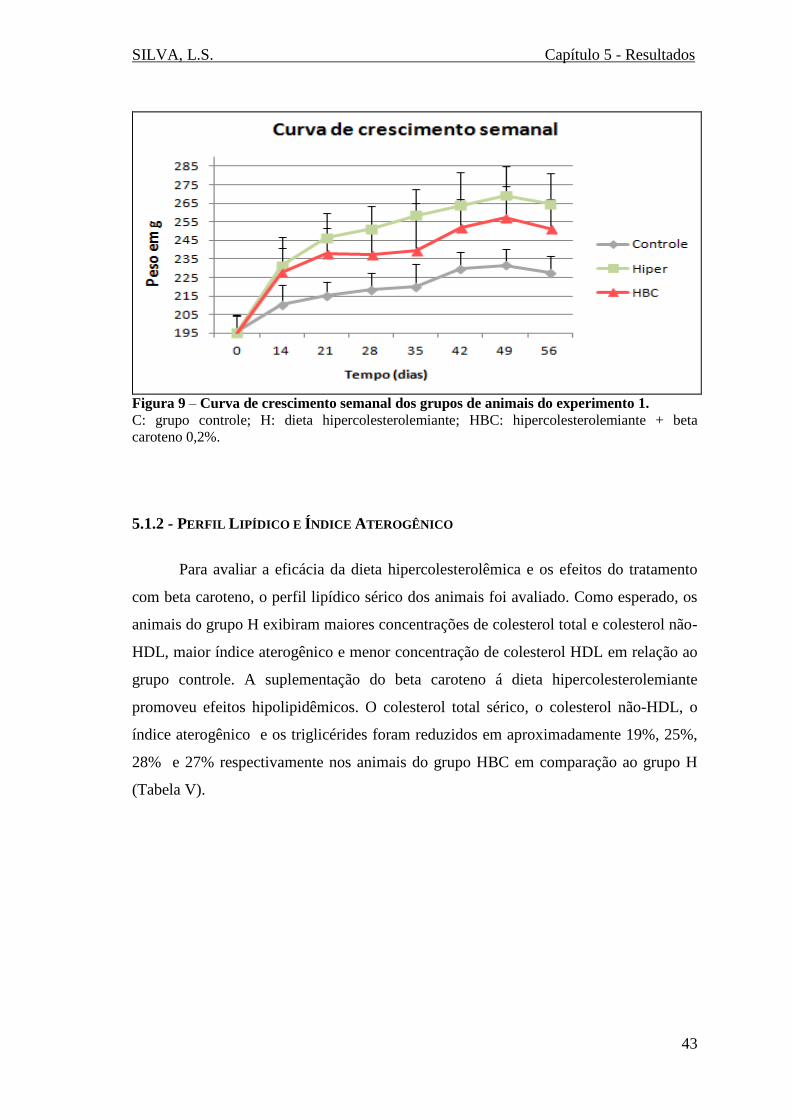
SILVA, L.S. Capítulo 5 - Resultados
43
Figura 9 – Curva de crescimento semanal dos grupos de animais do experimento 1.
C: grupo controle; H: dieta hipercolesterolemiante; HBC: hipercolesterolemiante + beta
caroteno 0,2%.
5.1.2 - PERFIL LIPÍDICO E ÍNDICE ATEROGÊNICO
Para avaliar a eficácia da dieta hipercolesterolêmica e os efeitos do tratamento
com beta caroteno, o perfil lipídico sérico dos animais foi avaliado. Como esperado, os
animais do grupo H exibiram maiores concentrações de colesterol total e colesterol não-
HDL, maior índice aterogênico e menor concentração de colesterol HDL em relação ao
grupo controle. A suplementação do beta caroteno á dieta hipercolesterolemiante
promoveu efeitos hipolipidêmicos. O colesterol total sérico, o colesterol não-HDL, o
índice aterogênico e os triglicérides foram reduzidos em aproximadamente 19%, 25%,
28% e 27% respectivamente nos animais do grupo HBC em comparação ao grupo H
(Tabela V).
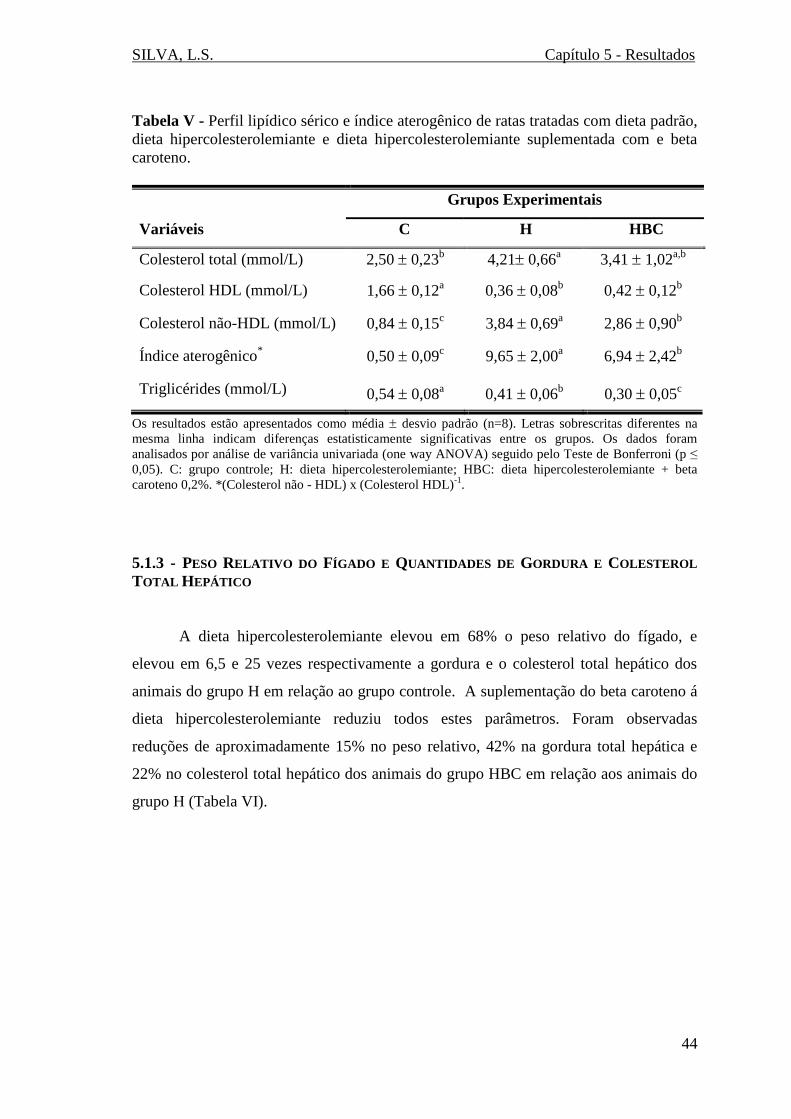
SILVA, L.S. Capítulo 5 - Resultados
44
Tabela V - Perfil lipídico sérico e índice aterogênico de ratas tratadas com dieta padrão,
dieta hipercolesterolemiante e dieta hipercolesterolemiante suplementada com e beta
caroteno.
Grupos Experimentais
Variáveis C H HBC
Colesterol total (mmol/L) 2,50 0,23b
4,21 0,66a
3,41 1,02a,b
Colesterol HDL (mmol/L) 1,66 0,12a
0,36 0,08b
0,42 0,12b
Colesterol não-HDL (mmol/L) 0,84 0,15c
3,84 0,69a
2,86 0,90b
Índice aterogênico* 0,50 0,09
c 9,65 2,00
a 6,94 2,42
b
Triglicérides (mmol/L) 0,54 0,08a 0,41 0,06
b 0,30 0,05
c
Os resultados estão apresentados como média desvio padrão (n=8). Letras sobrescritas diferentes na
mesma linha indicam diferenças estatisticamente significativas entre os grupos. Os dados foram
analisados por análise de variância univariada (one way ANOVA) seguido pelo Teste de Bonferroni (p ≤
0,05). C: grupo controle; H: dieta hipercolesterolemiante; HBC: dieta hipercolesterolemiante + beta
caroteno 0,2%. *(Colesterol não - HDL) x (Colesterol HDL)-1
.
5.1.3 - PESO RELATIVO DO FÍGADO E QUANTIDADES DE GORDURA E COLESTEROL
TOTAL HEPÁTICO
A dieta hipercolesterolemiante elevou em 68% o peso relativo do fígado, e
elevou em 6,5 e 25 vezes respectivamente a gordura e o colesterol total hepático dos
animais do grupo H em relação ao grupo controle. A suplementação do beta caroteno á
dieta hipercolesterolemiante reduziu todos estes parâmetros. Foram observadas
reduções de aproximadamente 15% no peso relativo, 42% na gordura total hepática e
22% no colesterol total hepático dos animais do grupo HBC em relação aos animais do
grupo H (Tabela VI).

SILVA, L.S. Capítulo 5 - Resultados
45
Tabela VI – Peso relativo do fígado e conteúdos totais de gordura e colesterol hepático
em ratas tratadas com dieta padrão, dieta hipercolesterolemiante e dieta
hipercolesterolemiante suplementada com e beta caroteno.
Grupos Experimentais
Variáveis C H HBC
Peso relativo do fígado (%)* 2,59 0,10
c 4,37 0,15
a 3,69 0,30
b
Gordura total hepática
(mg de gordura/g de fígado) 57,56
6,78
c 377,30
20,77
a 216,00 42,03
b
Colesterol total hepático
(mg de colesterol/g de fígado) 3,87
0,49
c 99,13
8,58
a 77,42 12,17
b
Os resultados estão apresentados como média desvio padrão (n=8). Letras sobrescritas diferentes na
mesma linha indicam diferenças estatisticamente significativas entre os grupos. Os dados foram
analisados por análise de variância univariada (one way ANOVA) seguido pelo Teste de Bonferroni (p ≤
0,05). C: grupo controle; H: dieta hipercolesterolemiante; HBC: dieta hipercolesterolemiante + beta
caroteno 0,2%. *(Peso do fígado) / (Peso corporal) x 100.
5.1.4 - EXCREÇÃO FECAL E QUANTIDADES DE GORDURA E COLESTERL TOTAL FECAL
A tabela VII indica que os animais do grupo H apresentaram elevações nos
conteúdos totais de gordura e colesterol fecal em relação aos animais controle. A
suplementação do beta caroteno á dieta hipercolesterolemiante promoveu uma elevação
de 52% e 46% respectivamente na gordura e colesterol total fecal no grupo HBC
comparado ao grupo H. A excreção fecal diária não foi afetada nem pela dieta
hipercolesterolemiante e nem pelo beta caroteno

SILVA, L.S. Capítulo 5 - Resultados
46
Tabela VII – Excreção fecal diária e conteúdos totais de gordura e colesterol fecal em
ratas tratadas com dieta padrão, dieta hipercolesterolemiante e dieta
hipercolesterolemiante suplementada com beta caroteno.
Grupos Experimentais
Variáveis C H HBC
Excreção fecal (g/dia) 0,93 0,09
0,90 0,03
0,93 0,07
Gordura total fecal (mg de
gordura/g de fezes) 44,71 12,03
c 129,90 27,79
b 197,50 20,78
a
Colesterol total fecal (mg
de colesterol/ g de fezes) 2,34 0,53
c 80,41 7,17
b 117,30 22,36
a
Os resultados estão apresentados como média desvio padrão (n=8). Letras sobrescritas diferentes na
mesma linha indicam diferenças estatisticamente significativas entre os grupos. Os dados foram
analisados por análise de variância univariada (one way ANOVA) seguido pelo Teste de Bonferroni (p ≤
0,05). C: grupo controle; H: dieta hipercolesterolemiante; HBC: dieta hipercolesterolemiante + beta
caroteno 0,2%.
5.1.5 - EXPRESSÃO DE GENES DO METABOLISMO HEPÁTICO DO COLESTEROL
Para investigar os mecanismos moleculares envolvidos no efeito
hipocolesterolêmico do beta caroteno, a expressão de importantes genes relacionados ao
metabolismo do colesterol foi avaliada por qRT-PCR, incluindo o SREBP-2, HMG
CoA-R, LDLR, PPAR α, ABCG5, ABCG8 e CYP7A1. Assim, o efeito da dieta
hipercolesterolemiante foi avaliado comparando-se o grupo C com o grupo H e o efeito
do beta caroteno foi avaliado comparando-se o grupo H com o grupo HBC.
A Figura 10 mostra que os animais do grupo H apresentaram reduções na
expressão dos genes SREBP-2, HMG CoA-R e LDLR e aumentos nas expressões de
PPAR α e CYP7A1 ( 0,5 e 7,3 vezes respectivamente) quando comparados aos animais
do grupo C. A suplementação de beta caroteno á dieta hipercolesterolemiante não afetou
a expressão de nenhum destes genes, sendo os animais dos grupos H e HBC
estatisticamente iguais. A expressão dos transportadores ABCG5 e ABCG8 entre os
grupos C e H e entre os grupos H e HBC foram também estatisticamente iguais.
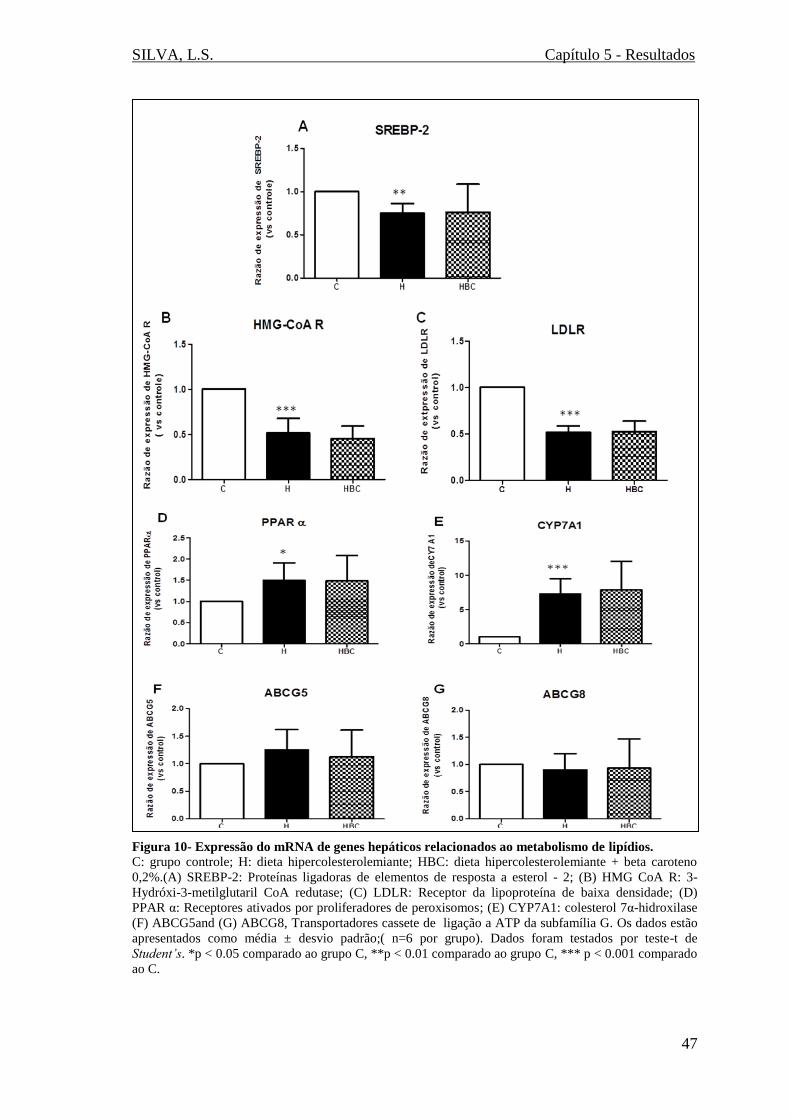
SILVA, L.S. Capítulo 5 - Resultados
47
Figura 10- Expressão do mRNA de genes hepáticos relacionados ao metabolismo de lipídios.
C: grupo controle; H: dieta hipercolesterolemiante; HBC: dieta hipercolesterolemiante + beta caroteno
0,2%.(A) SREBP-2: Proteínas ligadoras de elementos de resposta a esterol - 2; (B) HMG CoA R: 3-
Hydróxi-3-metilglutaril CoA redutase; (C) LDLR: Receptor da lipoproteína de baixa densidade; (D)
PPAR α: Receptores ativados por proliferadores de peroxisomos; (E) CYP7A1: colesterol 7α-hidroxilase
(F) ABCG5and (G) ABCG8, Transportadores cassete de ligação a ATP da subfamília G. Os dados estão
apresentados como média ± desvio padrão;( n=6 por grupo). Dados foram testados por teste-t de
Student’s. *p < 0.05 comparado ao grupo C, **p < 0.01 comparado ao grupo C, *** p < 0.001 comparado
ao C.
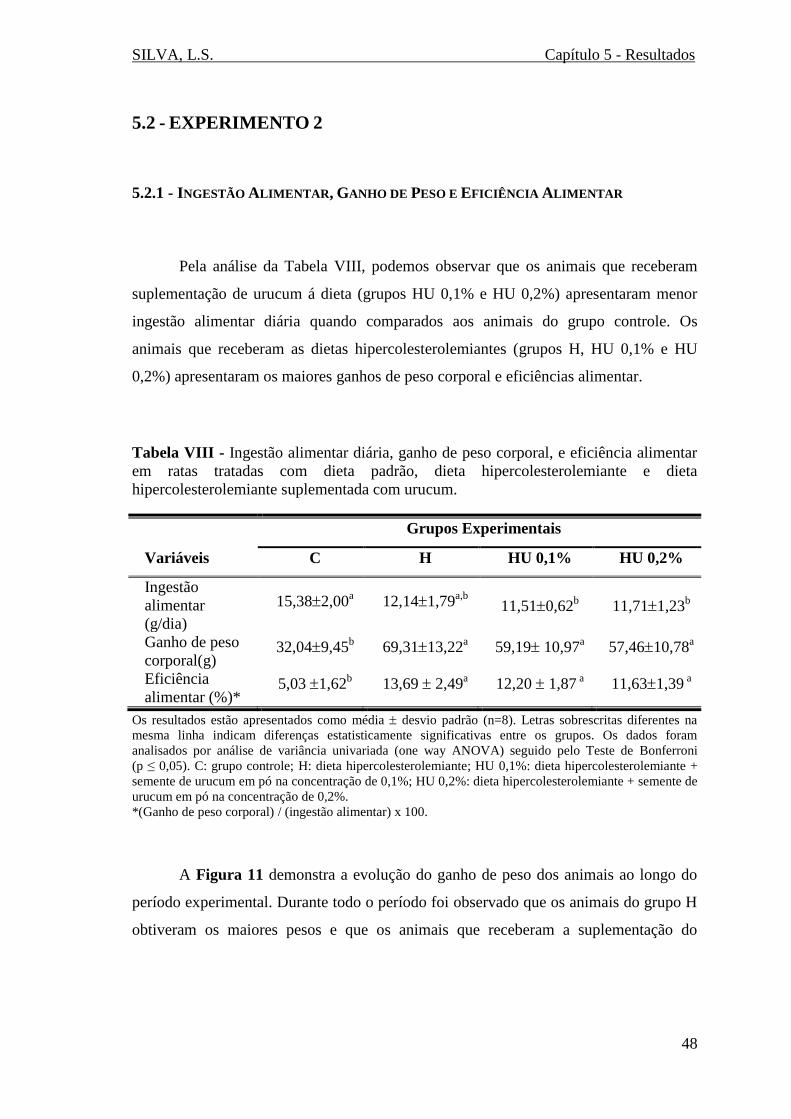
SILVA, L.S. Capítulo 5 - Resultados
48
5.2 - EXPERIMENTO 2
5.2.1 - INGESTÃO ALIMENTAR, GANHO DE PESO E EFICIÊNCIA ALIMENTAR
Pela análise da Tabela VIII, podemos observar que os animais que receberam
suplementação de urucum á dieta (grupos HU 0,1% e HU 0,2%) apresentaram menor
ingestão alimentar diária quando comparados aos animais do grupo controle. Os
animais que receberam as dietas hipercolesterolemiantes (grupos H, HU 0,1% e HU
0,2%) apresentaram os maiores ganhos de peso corporal e eficiências alimentar.
Tabela VIII - Ingestão alimentar diária, ganho de peso corporal, e eficiência alimentar
em ratas tratadas com dieta padrão, dieta hipercolesterolemiante e dieta
hipercolesterolemiante suplementada com urucum.
Grupos Experimentais
Variáveis C H HU 0,1% HU 0,2%
Ingestão
alimentar
(g/dia)
15,382,00a
12,141,79a,b
11,510,62b 11,711,23
b
Ganho de peso
corporal(g) 32,049,45
b 69,3113,22
a 59,19 10,97
a 57,4610,78
a
Eficiência
alimentar (%)* 5,03 1,62
b 13,69 2,49
a 12,20 1,87
a 11,631,39
a
Os resultados estão apresentados como média desvio padrão (n=8). Letras sobrescritas diferentes na
mesma linha indicam diferenças estatisticamente significativas entre os grupos. Os dados foram
analisados por análise de variância univariada (one way ANOVA) seguido pelo Teste de Bonferroni
(p ≤ 0,05). C: grupo controle; H: dieta hipercolesterolemiante; HU 0,1%: dieta hipercolesterolemiante +
semente de urucum em pó na concentração de 0,1%; HU 0,2%: dieta hipercolesterolemiante + semente de
urucum em pó na concentração de 0,2%.
*(Ganho de peso corporal) / (ingestão alimentar) x 100.
A Figura 11 demonstra a evolução do ganho de peso dos animais ao longo do
período experimental. Durante todo o período foi observado que os animais do grupo H
obtiveram os maiores pesos e que os animais que receberam a suplementação do
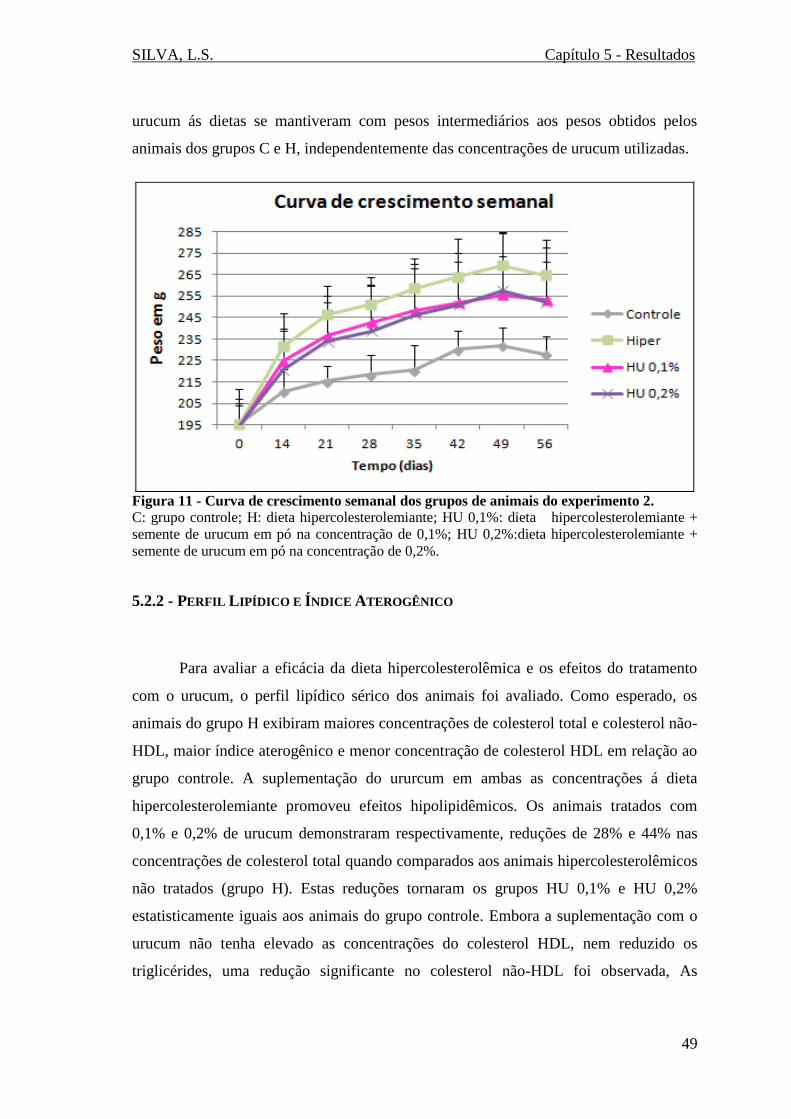
SILVA, L.S. Capítulo 5 - Resultados
49
urucum ás dietas se mantiveram com pesos intermediários aos pesos obtidos pelos
animais dos grupos C e H, independentemente das concentrações de urucum utilizadas.
Figura 11 - Curva de crescimento semanal dos grupos de animais do experimento 2. C: grupo controle; H: dieta hipercolesterolemiante; HU 0,1%: dieta hipercolesterolemiante +
semente de urucum em pó na concentração de 0,1%; HU 0,2%:dieta hipercolesterolemiante +
semente de urucum em pó na concentração de 0,2%.
5.2.2 - PERFIL LIPÍDICO E ÍNDICE ATEROGÊNICO
Para avaliar a eficácia da dieta hipercolesterolêmica e os efeitos do tratamento
com o urucum, o perfil lipídico sérico dos animais foi avaliado. Como esperado, os
animais do grupo H exibiram maiores concentrações de colesterol total e colesterol não-
HDL, maior índice aterogênico e menor concentração de colesterol HDL em relação ao
grupo controle. A suplementação do ururcum em ambas as concentrações á dieta
hipercolesterolemiante promoveu efeitos hipolipidêmicos. Os animais tratados com
0,1% e 0,2% de urucum demonstraram respectivamente, reduções de 28% e 44% nas
concentrações de colesterol total quando comparados aos animais hipercolesterolêmicos
não tratados (grupo H). Estas reduções tornaram os grupos HU 0,1% e HU 0,2%
estatisticamente iguais aos animais do grupo controle. Embora a suplementação com o
urucum não tenha elevado as concentrações do colesterol HDL, nem reduzido os
triglicérides, uma redução significante no colesterol não-HDL foi observada, As
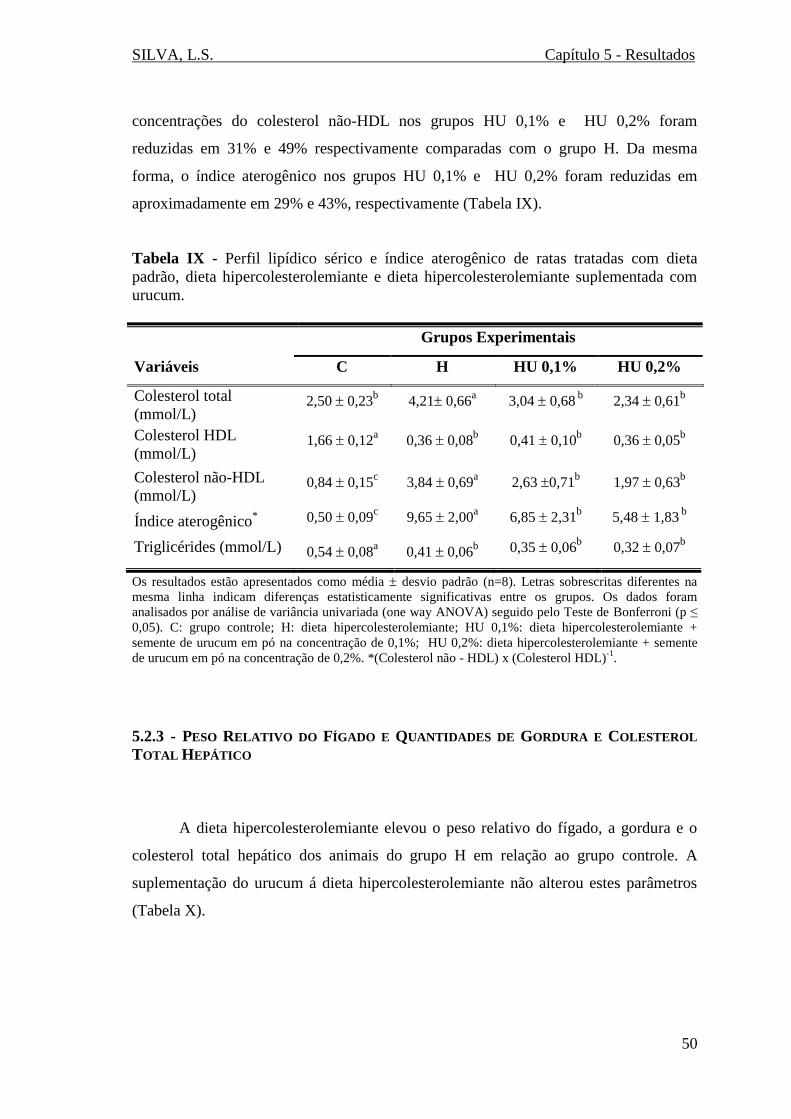
SILVA, L.S. Capítulo 5 - Resultados
50
concentrações do colesterol não-HDL nos grupos HU 0,1% e HU 0,2% foram
reduzidas em 31% e 49% respectivamente comparadas com o grupo H. Da mesma
forma, o índice aterogênico nos grupos HU 0,1% e HU 0,2% foram reduzidas em
aproximadamente em 29% e 43%, respectivamente (Tabela IX).
Tabela IX - Perfil lipídico sérico e índice aterogênico de ratas tratadas com dieta
padrão, dieta hipercolesterolemiante e dieta hipercolesterolemiante suplementada com
urucum.
Grupos Experimentais
Variáveis C H HU 0,1% HU 0,2%
Colesterol total
(mmol/L) 2,50 0,23
b 4,21 0,66
a 3,04 0,68
b 2,34 0,61b
Colesterol HDL
(mmol/L) 1,66 0,12
a 0,36 0,08
b 0,41 0,10
b 0,36 0,05
b
Colesterol não-HDL
(mmol/L) 0,84 0,15
c 3,84 0,69
a 2,63 0,71
b 1,97 0,63
b
Índice aterogênico* 0,50 0,09
c 9,65 2,00
a 6,85 2,31
b 5,48 1,83
b
Triglicérides (mmol/L) 0,54 0,08a 0,41 0,06
b 0,35 0,06
b 0,32 0,07
b
Os resultados estão apresentados como média desvio padrão (n=8). Letras sobrescritas diferentes na
mesma linha indicam diferenças estatisticamente significativas entre os grupos. Os dados foram
analisados por análise de variância univariada (one way ANOVA) seguido pelo Teste de Bonferroni (p ≤
0,05). C: grupo controle; H: dieta hipercolesterolemiante; HU 0,1%: dieta hipercolesterolemiante +
semente de urucum em pó na concentração de 0,1%; HU 0,2%: dieta hipercolesterolemiante + semente
de urucum em pó na concentração de 0,2%. *(Colesterol não - HDL) x (Colesterol HDL)-1
.
5.2.3 - PESO RELATIVO DO FÍGADO E QUANTIDADES DE GORDURA E COLESTEROL
TOTAL HEPÁTICO
A dieta hipercolesterolemiante elevou o peso relativo do fígado, a gordura e o
colesterol total hepático dos animais do grupo H em relação ao grupo controle. A
suplementação do urucum á dieta hipercolesterolemiante não alterou estes parâmetros
(Tabela X).
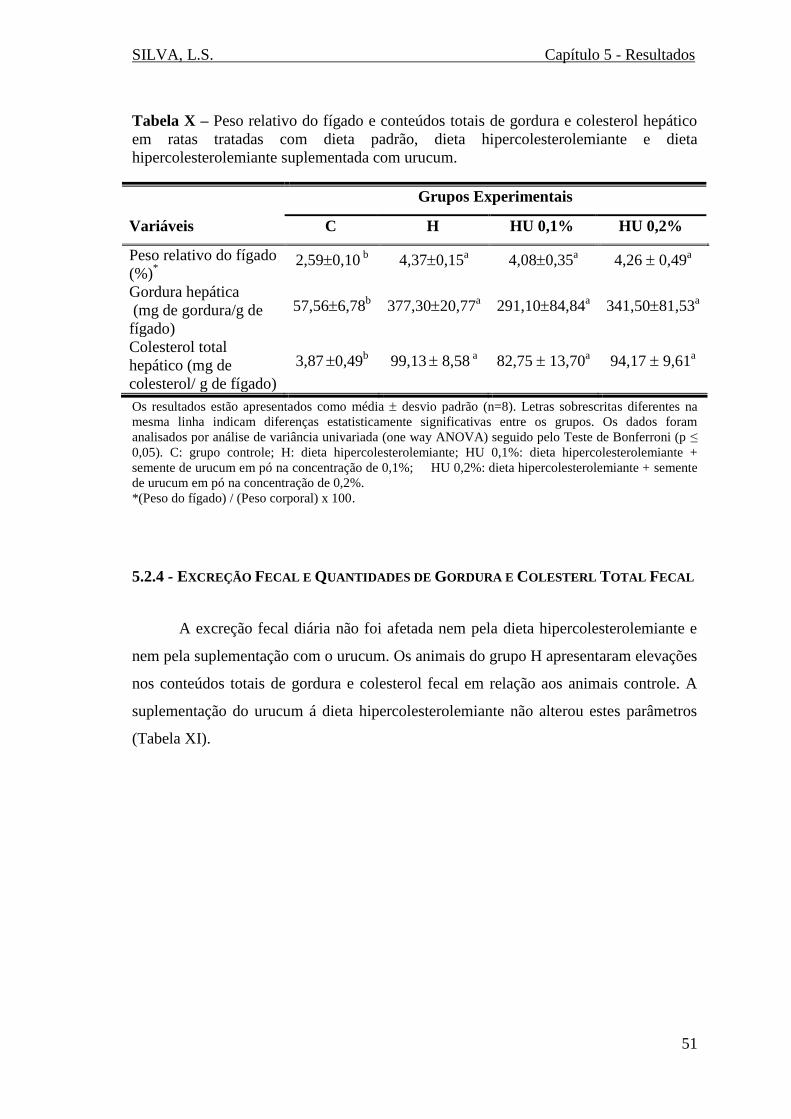
SILVA, L.S. Capítulo 5 - Resultados
51
Tabela X – Peso relativo do fígado e conteúdos totais de gordura e colesterol hepático
em ratas tratadas com dieta padrão, dieta hipercolesterolemiante e dieta
hipercolesterolemiante suplementada com urucum.
Grupos Experimentais
Variáveis C H HU 0,1% HU 0,2%
Peso relativo do fígado
(%)*
2,590,10 b
4,370,15a
4,080,35a
4,26 0,49a
Gordura hepática
(mg de gordura/g de
fígado)
57,566,78b
377,3020,77a
291,1084,84a
341,5081,53a
Colesterol total
hepático (mg de
colesterol/ g de fígado)
3,87 0,49
b 99,13
8,58
a 82,75 13,70
a 94,17 9,61
a
Os resultados estão apresentados como média desvio padrão (n=8). Letras sobrescritas diferentes na
mesma linha indicam diferenças estatisticamente significativas entre os grupos. Os dados foram
analisados por análise de variância univariada (one way ANOVA) seguido pelo Teste de Bonferroni (p ≤
0,05). C: grupo controle; H: dieta hipercolesterolemiante; HU 0,1%: dieta hipercolesterolemiante +
semente de urucum em pó na concentração de 0,1%; HU 0,2%: dieta hipercolesterolemiante + semente
de urucum em pó na concentração de 0,2%.
*(Peso do fígado) / (Peso corporal) x 100.
5.2.4 - EXCREÇÃO FECAL E QUANTIDADES DE GORDURA E COLESTERL TOTAL FECAL
A excreção fecal diária não foi afetada nem pela dieta hipercolesterolemiante e
nem pela suplementação com o urucum. Os animais do grupo H apresentaram elevações
nos conteúdos totais de gordura e colesterol fecal em relação aos animais controle. A
suplementação do urucum á dieta hipercolesterolemiante não alterou estes parâmetros
(Tabela XI).
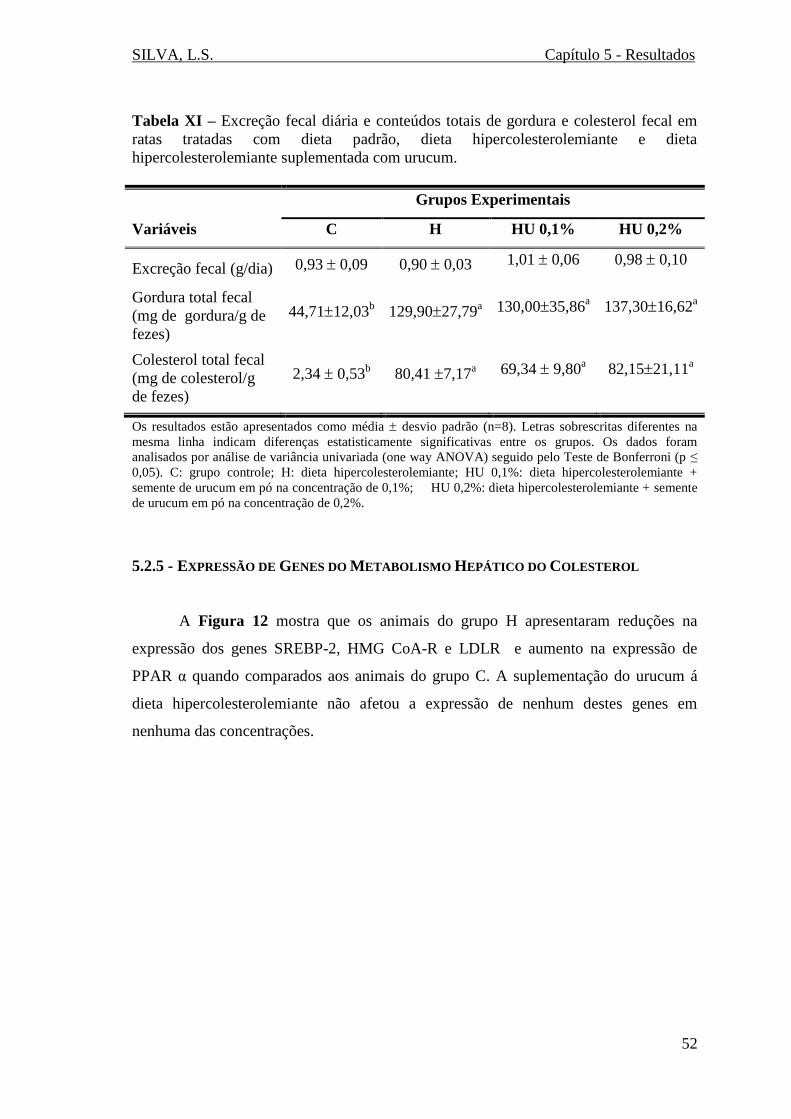
SILVA, L.S. Capítulo 5 - Resultados
52
Tabela XI – Excreção fecal diária e conteúdos totais de gordura e colesterol fecal em
ratas tratadas com dieta padrão, dieta hipercolesterolemiante e dieta
hipercolesterolemiante suplementada com urucum.
Grupos Experimentais
Variáveis C H HU 0,1% HU 0,2%
Excreção fecal (g/dia) 0,93 0,09
0,90 0,03 1,01 0,06 0,98 0,10
Gordura total fecal
(mg de gordura/g de
fezes)
44,7112,03b 129,9027,79
a 130,0035,86
a 137,3016,62
a
Colesterol total fecal
(mg de colesterol/g
de fezes)
2,34 0,53b 80,41 7,17
a 69,34 9,80
a 82,1521,11
a
Os resultados estão apresentados como média desvio padrão (n=8). Letras sobrescritas diferentes na
mesma linha indicam diferenças estatisticamente significativas entre os grupos. Os dados foram
analisados por análise de variância univariada (one way ANOVA) seguido pelo Teste de Bonferroni (p ≤
0,05). C: grupo controle; H: dieta hipercolesterolemiante; HU 0,1%: dieta hipercolesterolemiante +
semente de urucum em pó na concentração de 0,1%; HU 0,2%: dieta hipercolesterolemiante + semente
de urucum em pó na concentração de 0,2%.
5.2.5 - EXPRESSÃO DE GENES DO METABOLISMO HEPÁTICO DO COLESTEROL
A Figura 12 mostra que os animais do grupo H apresentaram reduções na
expressão dos genes SREBP-2, HMG CoA-R e LDLR e aumento na expressão de
PPAR α quando comparados aos animais do grupo C. A suplementação do urucum á
dieta hipercolesterolemiante não afetou a expressão de nenhum destes genes em
nenhuma das concentrações.
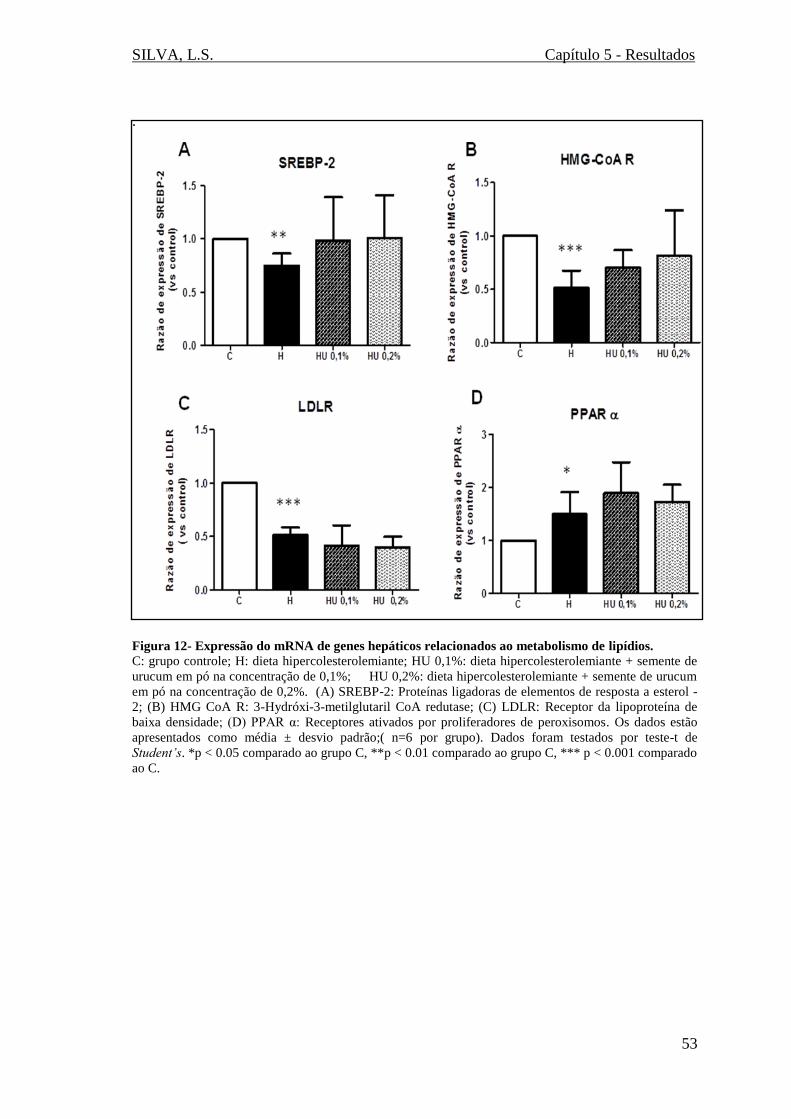
SILVA, L.S. Capítulo 5 - Resultados
53
.
Figura 12- Expressão do mRNA de genes hepáticos relacionados ao metabolismo de lipídios.
C: grupo controle; H: dieta hipercolesterolemiante; HU 0,1%: dieta hipercolesterolemiante + semente de
urucum em pó na concentração de 0,1%; HU 0,2%: dieta hipercolesterolemiante + semente de urucum
em pó na concentração de 0,2%. (A) SREBP-2: Proteínas ligadoras de elementos de resposta a esterol -
2; (B) HMG CoA R: 3-Hydróxi-3-metilglutaril CoA redutase; (C) LDLR: Receptor da lipoproteína de
baixa densidade; (D) PPAR α: Receptores ativados por proliferadores de peroxisomos. Os dados estão
apresentados como média ± desvio padrão;( n=6 por grupo). Dados foram testados por teste-t de
Student’s. *p < 0.05 comparado ao grupo C, **p < 0.01 comparado ao grupo C, *** p < 0.001 comparado
ao C.

SILVA, L.S. Capítulo 5 - Resultados
54
Os níveis de mRNA da CYP7A1 foram elevados em 7,3 vezes no grupo H em
comparação ao grupo C. A suplementação com urucum elevou significativamente os
níveis de mRNA dessa enzima. O aumento na expressão da CYP7A1 no grupo HU
0,1% foi de 1,6 vezes e no grupo HU 0,2 foi de 1,5 vezes, ambos em relação ao grupo
H.
A expressão dos receptores LXR e FXR, bem como a expressão dos
transportadores ABCG5 e ABCG8 não foram alteradas nem pela dieta
hipercolesterolemiante e nem pela suplementação com urucum. Assim, não houveram
diferenças nas comparações (C x H ), ( H x HU 0,1%) e (H x HU 0,2%).
A dieta hipercolesterolemiante não afetou a expressão de SHP e ABCB11, mas a
suplementação do urucum a esta dieta promoveu elevações nos níveis de mRNA para
SHP e ABCB11. O aumento na expressão de SHP nos grupos HU 0,1% e HU 0,2% foi
de aproximadamente 1,8 e 1,9 vezes respectivamente relação ao grupo H e o aumento
na expressão de ABCB11 nos grupos HU 0,1% e HU 0,2% foi de aproximadamente 2,0
e 1,8 vezes respectivamente (Figura 13).
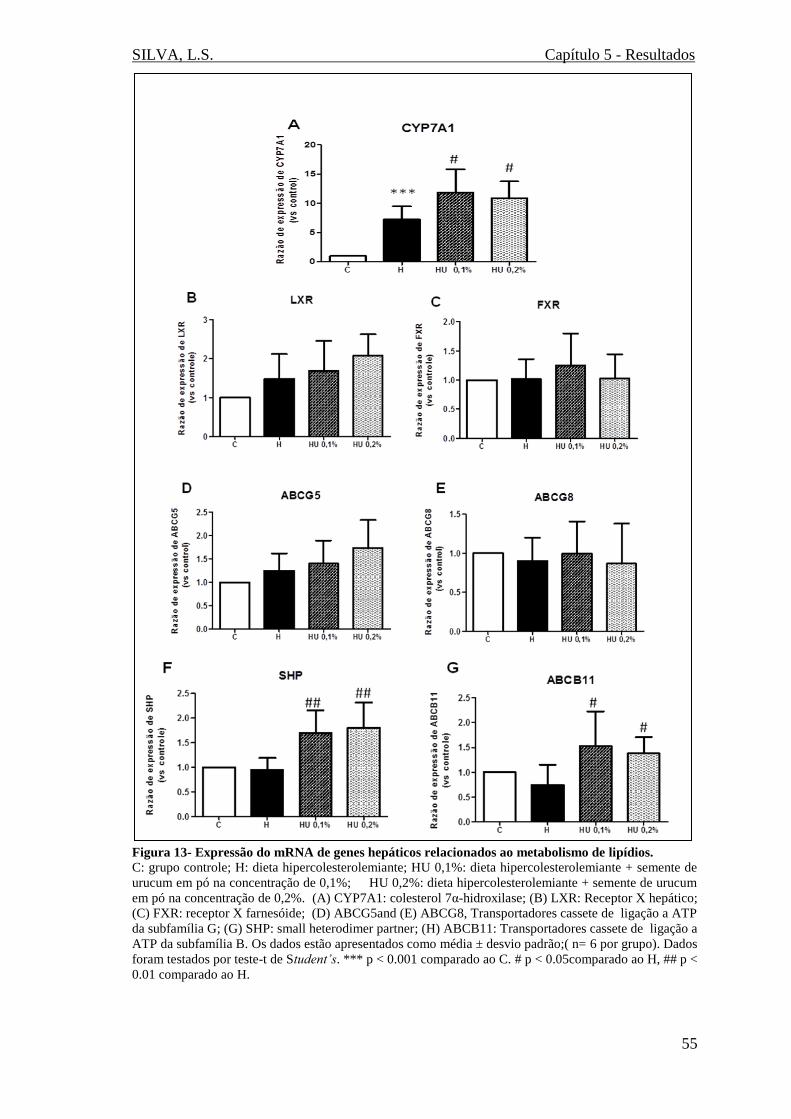
SILVA, L.S. Capítulo 5 - Resultados
55
Figura 13- Expressão do mRNA de genes hepáticos relacionados ao metabolismo de lipídios. C: grupo controle; H: dieta hipercolesterolemiante; HU 0,1%: dieta hipercolesterolemiante + semente de
urucum em pó na concentração de 0,1%; HU 0,2%: dieta hipercolesterolemiante + semente de urucum
em pó na concentração de 0,2%. (A) CYP7A1: colesterol 7α-hidroxilase; (B) LXR: Receptor X hepático;
(C) FXR: receptor X farnesóide; (D) ABCG5and (E) ABCG8, Transportadores cassete de ligação a ATP
da subfamília G; (G) SHP: small heterodimer partner; (H) ABCB11: Transportadores cassete de ligação a
ATP da subfamília B. Os dados estão apresentados como média ± desvio padrão;( n= 6 por grupo). Dados
foram testados por teste-t de Student’s. *** p < 0.001 comparado ao C. # p < 0.05comparado ao H, ## p <
0.01 comparado ao H.


SILVA, L.S Capítulo 6 - Discussão
57
CAPÍTULO 6 – DISCUSSÃO
6.1- EXPERIMENTO 1
A hiperlipidemia, comum na população mundial, é considerada como um fator
de risco altamente modificável para as doenças cardiovasculares, tais como a doença
coronária cardíaca e doenças arteriais periféricas. Níveis elevados de lipídios no sangue,
especialmente aumentados níveis séricos de LDL, podem acelerar a aterosclerose e a
redução dos níveis lipídicos, portanto, tem sido considerada uma abordagem importante
para prevenir ou retardar a progressão da aterosclerose (Durrington, 2003; Arsenault et
al, 2009).
Dado o interesse público na prevenção e/ou tratamento das DCV e outras
doenças metabólicas, estudos têm sido realizados para identificar produtos naturais que
possam regular a dislipidemia (Yeon et al., 2012) e nessa perspectiva, estudos
epidemiológicos têm encontrado associações inversas entre os níveis de beta caroteno
no soro ou tecido adiposo e as doenças cardiovasculares (Kritchevsky, 1999).
O beta caroteno demonstrou diminuir o colesterol sérico e as lesões
ateroscleróticas aórticas em ratos e coelhos hipercolesterolêmicos respectivamente
(Amen e Lachance, 1974; Erdman e Lachance, 1974; Shaish et al., 1995). A alta
ingestão de beta caroteno e a suplementação dietética de vitamina E foram associados a
uma diminuição do risco de doença coronariana arterial (Gey et al., 1991; Gaziano e
Hennekens, 1993; Stampfer et al., 1993). O beta caroteno mostrou efeitos anti-
hiperlipidêmicos em ratos espontaneamente hipertensos (Tsai e Mazeedi, 1988; Tsai et
al., 1992), e estudos em ratos e camundongos indicaram a supressão ou a redução da
peroxidação lipídica e uma melhora no status antioxidante após a administração de beta
caroteno (Iyama et al., 1996; Whittaker et al., 1996; Kraus et al., 1997; Riondel et al.,
2002).
O experimento 1 do nosso trabalho, avaliou os efeitos do beta caroteno na
concentração de 0,2%, sobre parâmetros relacionados ao metabolismo do colesterol
utilizando ratos com hiepercolesterolemia como modelo animal.

SILVA, L.S Capítulo 6 - Discussão
58
Sabe-se que a concentração de beta caroteno tem influência na sua ação
antioxidante, podendo prejudicar sua habilidade protetora e/ou a reverter em pró-
oxidativa. Além disso, sabe-se que o colesterol dietético exerce efeito pró-oxidativo
sobre a peroxidação de lipídios e oxidação de lipoproteínas. A escolha da dose utilizada
neste estudo, foi baseada no trabalho de Shih et al. (2008), no qual a suplementação de
beta caroteno na dose de 0,2%, á dieta hipercolesterolemiante proporcionou em ratos,
efeitos hipocolesterolemiantes sem comprometimento da suas capacidade antioxidante.
Os resultados da Tabela IV indicam que apesar de os animais tratados com as
dietas hipercolesterolemiantes (grupos H e HBC) apresentarem menor ingestão
alimentar em relação ao controle eles foram os que apresentaram maior ganho de peso
corporal e maior eficiência alimentar.
Os ratos ajustam sua ingestão alimentar em função de seus requerimentos
energéticos (Baker et al., 1979), sendo assim, quanto mais calorias por grama uma dieta
apresentar, menor será a quantidade ingerida pelos animais para cobrir suas
necessidades calóricas. As dietas oferecidas aos animais dos grupos H e HBC
acondicionavam maior valor calórico, justificando assim, a menor ingestão apresentada
pelos animais destes grupos (Valor energético: C = 3810 Kcal; H= 4820 Kcal e HBC =
4740 Kcal).
Os nossos resultados de perfil lipídico sérico, demonstraram que a
suplementação de beta caroteno à dieta hipercolesterolêmica foi efetiva em promover
efeitos hipolipidêmicos, sendo capaz de reduzir as concentrações séricas de colesterol
total e colesterol não-HDL, bem como o índice aterogênico dos animais do grupo HBC
em 19%, 25% e 28%, respectivamente. (Tabela V).
As funções fisiológicas e os efeitos colaterais do colesterol têm sido amplamente
documentados. Ratos alimentados com um nível elevado de colesterol geralmente
apresentam fígado gorduroso caracterizado por um maior peso de tecido hepático e uma
aparência macroscópica anormal (Yuan et al., 1998; Yuan e Kitts, 2002). O presente
estudo mostrou que de fato, a administração da dieta hipercolesterolemiante promoveu
uma elevação no peso do fígado dos animais do grupo H, bem como um maior estoque
de gorduras totais, o qual pode ser atribuído ao maior acúmulo de colesterol encontrado
no fígado (Tabela VI). A suplementação de beta caroteno reduziu o peso do fígado, a
quantidade de gordura e o colesterol total hepático, demonstrando que a suplementação

SILVA, L.S Capítulo 6 - Discussão
59
de beta caroteno pode melhorar os níveis de lipídios tanto no soro quanto no fígado dos
animais. Semelhantes aos resultados encontrados neste estudo, Shih et al., (2008),
relataram que o colesterol dietético elevou significativamente as concentrações do
colesterol plasmático e hepático em ratos, e que a suplementação de beta caroteno
também a 0,2% melhoraram estas elevações de concentração induzidas pelo colesterol.
Quanto aos triglicérides, observamos que os animais que receberam dieta
hipercolesterolemiante apresentaram menores concentrações se comparados aos animais
que receberam a dieta controle e que o tratamento com beta caroteno reduziu as
concentrações séricas deste lipídio (Tabela V). Uma redução similar dos triglicérides
séricos em animais alimentados com dietas ricas em colesterol e gordura insaturada
também foram observadas por Turbino-Ribeiro et al., (2003), Thomas-Moyá et al.,
(2006) e Souza et al., (2010). Estes resultados podem ser explicados pela alta
quantidade de gorduras insaturadas usadas nessas dietas. É bem conhecido que ácidos
graxos insaturados inibem importantes enzimas relacionadas ao metabolismo da VLDL
resultando assim, em menores níveis de triglicérides no plasma (Connor et al., 1993).
O consumo de beta caroteno está associado com diversos efeitos benéficos á
saúde, entretanto os mecanismos adjacentes á sua atividade hipocolesterolemiante ainda
não estão claros. O presente estudo examinou também os efeitos do beta caroteno sobre
a excreção fecal de colesterol e caracterizou seus efeitos sobre a expressão hepática dos
genes SREBP-2, HMG-CoA redutase, LDLR , CYP7A1, PPARα, ABCG5 e ABCG 8.
A homeostase hepática do colesterol é mantida pelo balanço da síntese de novo,
captação e eliminação do colesterol (Li et al., 2011). A biossíntese e a captação do
colesterol são altamente reguladas ao nível transcricional através de mecanismos de
feedback negativo, os quais são controlados pelo SREBP-2 (Sato,2010).
SREBP-2 é um fator de transcrição que regula a homeostase do colesterol em
virtude da sua capacidade para se ligar e ativar os promotores de genes que codificam a
HMG-CoA redutase e o LDLR (Miserez et al., 2002). Tanto a HMG-CoA redutase
quanto o LDLR têm um elemento de regulação de esterol nas suas regiões promotoras e
são, portanto, geralmente regulados por SREBP-2 (Sato et al., 1999).
A análise de qRT-PCR para este experimento, revelou que os animais do grupo
de H apresentaram expressão de SREBP-2 significativamente menor em relação ao
grupo controle, e que a expressão dos genes HMG CoA-R e LDLR seguiu o mesmo

SILVA, L.S Capítulo 6 - Discussão
60
padrão de expressão do SREBP-2 (Figura 10). Tem sido sugerido que uma dieta
hipercolesterolêmica pode levar a um estado de saturação de colesterol nos hepatócitos,
o que resulta na diminuição da regulação da síntese de novo de colesterol e na menor
absorção de LDL através de receptor, com a diminuição da expressão de SREBP- 2.
Os nossos resultados sugerem que a saturação dos hepatócitos pelo colesterol foi
a responsável pela redução da expressão do SREBP-2, isto por sua vez refletiu em
menores expressões de HMG-CoA redutase e LDLR. Resultados de outros estudos
indicam que a expressão hepática dos genes HMG-CoA-R e LDLR em ratos
alimentados com dietas ricas em lipídios são reguladas negativamente em comparação
com a expressão obtida em ratos controle, corroborando então com os resultados
obtidos neste estudo (Rudling et al., 1992; Dhingra e Bansal 2006; Shibata et al., 2007;
Yiu et al., 2011; Gao et al., 2013). A suplementação de beta caroteno á dieta
hipercolesterolemiante não afetou a expressão hepática dos genes SREBP-2, HMG-
CoA-R e LDL-R.
A suplementação com beta caroteno também não afetou a expressão dos genes
PPARα e CYP7A1. Sendo as expressões destes genes aumentadas no grupo H em
comparação com o controle (Figura 10).
Os PPARs são fatores de transcrição pertencentes à superfamília dos receptores
nucleares. Os PPARs, com três isoformas (α, e ), estão envolvidos na regulação a
longo prazo do metabolismo lipídico, e suas atividades são moduladas por ligantes
derivados de lipídios endógenos. O PPARα é expresso no fígado e regula o catabolismo
dos ácidos graxos. Ele desempenha um papel importante na regulação da homeostase de
ácidos graxos através da regulação dos genes alvo que codificam enzimas da oxidação e
transporte de ácidos graxos (Ferré, 2004; Evans et al., 2009). Quando o PPARα é
ativado, ele promove a oxidação dos ácidos graxos, a síntese de corpos cetônicos, e
poupam glicose (Park et al., 2009).
O experimento 1, demonstra que a expressão do PPARα foi aumentada pela
dieta hipercolesterolêmica.Com base em estudos moleculares, é aceitável que a
sinalização do PPARα in vivo depende da disponibilidade de ligantes e da expressão do
receptor. Uma dieta rica em gorduras, presumivelmente, fornece mais ácidos graxos
como ligantes de PPARα e, portanto, pode ser capaz de up-regulate genes alvo do
PPARα e melhorar o metabolismo dos ácidos graxos através da via de sinalização do

SILVA, L.S Capítulo 6 - Discussão
61
PPARα. Isto, por sua vez, pode reduzir a acúmulo de gordura corporal (Hsu e Huanga,
2007).
Bonila et al., (2000) observaram que ratos Wistar alimentados com dietas ricas
em gordura (250 g de óleo vegetal / kg de dieta), exibiram aumentadas expressões
hepáticas de PPARα e acil-CoA oxidase, além de uma maior ativação de PPARα.
Vários autores (Gottlicher et al., 2002; Forman et al., 1997; Krey et al., 1997; Kliewer
et al., 1997; Murakami et al., 1999; Lin et al., 1999) demonstraram em estudos in vitro,
que ácidos graxos de cadeia longa não apenas se ligam como também ativam o PPARα.
Banner et al., (1993) identificaram o ácido palmítico, ácido linoleico, ácido oleico e o
ácido araquidônico como ativadores endógenos PPARα. A potência relativa de cada
ácido graxo varia com o tipo de medição e tipo de PPAR avaliado. Em experimentos
avaliando o PPARα de camundongos, ratos e humanos, a potência de ligação e de
ativação do ácido oleico foram comparáveis à do ácido linoleico (Gottlicher et al., 1992;
Murakami et al., 1999; Lin et al., 1999).
O fígado é o principal órgão envolvido na regulação do metabolismo do
colesterol, sendo a conversão do colesterol a ácidos biliares o caminho principal do
catabolismo deste esterol. A CYP7A1 é a enzima limitante na via biossintética hepática
dos ácidos biliares, e deste modo controla a homeostase do colesterol e ácidos biliares
(Li et al., 2011)
A deficiência de CYP7A1 em seres humanos está associada à
hipercolesterolemia e aterosclerose prematura (Pullinger et al., 2002). Miyake et al.,
(2002) relataram que a expressão transgênica de CYP7A1 no fígado de camundongos
preveniu a aterosclerose induzida por dieta litogênica e posteriormente, Li et al., (2010)
relataram que camundongos overexpressing CYP7A1 se mostraram protegidos contra a
hipercolesterolemia, obesidade e resistência à insulina induzidas por dietas ricas em
lipídios. O efeito de redução do colesterol pelo estímulo da síntese de ácidos biliares
tem sido atribuído ao aumento da conversão de colesterol em ácidos biliares e ao
estímulo de captação de colesterol hepático mediado por LDLR.
Em nosso experimento, os animais do grupo H exibiram um aumento
significativo de 7,3 vezes na expressão de CYP7A em comparação com o grupo
controle, corroborando com outros estudos que têm demonstrado que a atividade e a
expressão do mRNA da enzima CYP7A1 são aumentadas pela presença de colesterol

SILVA, L.S Capítulo 6 - Discussão
62
dietético (Guo et al., 2011; Yiu et al., 2011; Boone et al., 2011). A expressão hepática
da CYP7A1 não foi afetada pela suplementação do beta caroteno.
Ao avaliar as vias de biossíntese, captação e catabolismo do colesterol pôde-se
observar que a expressão de importantes genes dessas vias não foi afetada pela
administração do beta caroteno. Assim, foi investigado se os efeitos hipolipidêmicos
obtidos pelo beta caroteno estariam relacionados a alterações nas vias de efluxo do
colesterol como por exemplo: o transporte do colesterol do fígado para a bile, mediado
por transportadores, ou a eliminação direta do colesterol nas fezes.
A administração de beta caroteno aumentou significativamente a excreção de
gorduras e colesterol total nas fezes (Tabela VII). A diminuição dos níveis de
colesterol sérico e a elevação do colesterol fecal obtido pela administração do beta
caroteno sugerem que ele pode diminuir a absorção do colesterol no intestino e
aumentar sua excreção nas fezes, ou ainda apresentar alterações metabólicas nas células
intestinais.
A absorção do colesterol desempenha um papel importante na regulação de sua
homeostase no corpo inteiro (Grundy, 1983; Wilson e Rudel, 1994) e vários
transportadores têm sido sugeridos em afetar direta ou indiretamente a absorção do
colesterol intestinal. Neste contexto, os transportadores ABCG5 ABCG8, desempenham
papéis significativos na excreção direta do colesterol via bie (Rudel, 1994) e a sua
expressão foi avaliada.
O colesterol hepático é secretado para a bile através dos transportadores de
efluxo de colesterol ABCCG5 e ABCG8, presentes na membrana canalicular dos
hepatócitos (Graf et al., 2003; Yu et al., 2005). No intestino, ABCG5/G8 promove o
efluxo de esteróis vegetais e de colesterol para o lúmen, prevenindo a absorção de
esteróis vegetais e limitando a absorção do colesterol dietético. Mutações nos genes
ABCG5 e / ou ABCG8 causam sitosterolemia em seres humanos (Berge et al., 2000) e
camundongos knockout para os genes ABCG5/G8 exibem uma secreção biliar de
colesterol acentuadamente diminuída e um aumento na absorção intestinal do colesterol
fracionado (Yu et al., 2002).
Yu et al., (2002) mostraram que camundongos com aumento na expressão
desses transportadores no fígado e no intestino delgado apresentaram alterações
importantes no metabolismo do colesterol como, aumento na concentração de colesterol

SILVA, L.S Capítulo 6 - Discussão
63
na bile, redução na absorção do colesterol e aumento na síntese hepática deste esterol.
Nishimoto et al., (2009) demonstraram que a suplementação dietética com óleo de
peixe, rico em ácidos graxos poliinsaturados, aumentou significativamente a expressão
hepática dos transportadores ABCG5 e ABCG8 e a quantidade de colesterol excretada
nas fezes em camundongos.
As análises de qRT-PCR revelaram que nem a dieta hipercolesterolemiante e
nem o beta caroteno afetaram a expressão de genes ABCG5 e ABCG8 (Figura 10).
Embora não possamos excluir a possibilidade de uma regulação pós-transcricional na
expressão desses genes, estes resultados sugerem que a elevação da excreção fecal do
colesterol promovida pelo beta caroteno está mais relacionada com a diminuição da
absorção intestinal do colesterol do que com o aumento da sua eliminação na bile pelos
transportadores ABCG5 e ABCG8.
Rasmussen et al., (2009), avaliaram em camundongos C57BL/6J o efeito
hipocolesterolêmico da alga comestível Nostoc comune e observaram que sua
administração, reduziu o colesterol total sérico, reduziu a absorção intestinal e elevou a
excreção fecal de esteróis, sem afetar a expressão intestinal de ABCG5.
Uma vez que o beta caroteno não exerceu um efeito direto sobre a expressão dos
transportadores de colesterol, a redução da absorção do colesterol parece estar atribuída
ao seu efeito no interior do lúmen intestinal. Os passos principais na absorção de
colesterol são a emulsificação no estômago, a hidrólise da ligação éster com uma
esterase pancreática específica, a solubilização micelar, a absorção no intestino delgado
e o transporte para o sistema linfático (Ros, 2000). Devido à insolubilidade do colesterol
em água, a solubilização do colesterol em micelas mistas é um requisito para a
eficiência da sua absorção. O colesterol solubilizado é então transferido das micelas
para a membrana das células da borda em escova intestinal.
Park et al., (2009), observaram em ratos hipercolesterolêmicos tratados com
extratos de milho uma redução do colesterol sérico e uma elevação na excreção fecal de
colesterol, e propuseram, que os componentes ativos do milho poderiam estar
interferindo tanto na afinidade das micelas com a membrana intestinal quanto na
afinidade do colesterol com as micelas. Além disso, os mesmos autores relataram que
essas alterações no lúmen intestinal afetariam o metabolismo hepático do colesterol e
poderiam afetar a síntese e o catabolismo das lipoproteínas.

SILVA, L.S Capítulo 6 - Discussão
64
De maneira semelhante ao observado por Park et al.,(2009), podemos sugerir
que o beta caroteno possa estar agindo de maneira semelhante, uma vez que o aumento
da excreção fecal do colesterol não pode ser atribuído ao maior efluxo de colesterol do
fígado para a bile mediado pelos transportadores ABCG5 e ABCG8.
Em conclusão o beta caroteno não afetou a expressão hepática dos principais
genes envolvidos na homeostase do colesterol e os nossos resultados sugerem que a
diminuição da absorção intestinal do colesterol e o subsequente aumento da sua
excreção nas fezes foram os fatores primários para o efeito hipocolesterolemiante obtido
pelo beta caroteno (Figura 14).
Figura14: Resumo dos principais efeitos do beta caroteno no soro, nas fezes e no fígado.

SILVA, L.S Capítulo 6 - Discussão
65
6.2 - EXPERIMENTO 2
As sementes do urucum são uma importante fonte de corante natural usada na
indústria de alimentos, encontrando aplicações em produtos têxteis, cosméticos e
farmacêuticos. O urucum é conhecido por estar associado ao tratamento de várias
doenças na medicina popular e alguns estudos têm demonstrado efeitos antioxidantes e
hipocolesterolêmicos dos extratos de urucum.
O consumo do extrato de urucum demonstrou exibir várias propriedades
bioativas, incluindo propriedades antioxidantes e antitumorais (Júnior et al., 2005;
Agner et al., 2005). A suplementação de 0,09% de extrato de urucum modulou a
produção de espécies reativas de oxigênio e de óxido nítrico em neutrófilos de ratos
diabéticos (Rossoni Jr. et al., 2011). O tratamento com bixina e norbixina, os
componentes principais das sementes de urucum, promoveram efeitos positivos na
síndrome metabólica, melhorando a sensibilidade à insulina em adipócitos, através da
activação de PPAR (Takahashi et al.,2009). Em camundongos, o extrato aquoso das
sementes de urucun foi capaz de reverter a hipertrigliceridemia induzida por Triton,
frutose e etanol, demonstrando um efeito hipolipemiante (Ferreira et al., 2012), e em
ratos que foram alimentados com uma dieta rica em gorduras, o tratamento com extrato
aquoso de urucum reduziu o e colesterol total e LDL e elevou o colesterol HDL ( de
Paula et al., 2009).
O segundo experimento foi realizado para investigar a atividade hipolipemiante
do urucum utilizando ratos hipercolesterolêmicos induzidos por dieta como modelo
experimental.
Os nossos resultados mostraram que a suplementação de urucum a dieta
hipercolesterolemiante reduziu os níveis de colesterol total e colesterol não-HDL, bem
como o índice aterogênico (Tabela IX), demonstrando que o consumo de urucum
melhora o perfil de lipídios no soro e pode exercer um efeito ateroprotetor.
Tem sido demonstrado que fitoquímicos de algumas plantas medicinais
tradicionais têm efeitos hipolipidêmicos através da modulação de genes envolvidos no
metabolismo de lipídios e lipoproteínas (Chung et al., 2007; Chung et al., 2008). Assim,

SILVA, L.S Capítulo 6 - Discussão
66
nós focamos na caracterização dos efeitos do urucum sobre a transcrição de genes
hepáticos envolvidos no metabolismo do colesterol.
A Figura 12 demonstra que a suplementação de urucum a dieta
hipercolesterolêmica não alterou a expressão de SREBP-2 HMG-CoA-R, LDLR e
PPARα. Dessa forma, avaliamos se o efeito hipocolesterolemiante promovido pelo
urucum poderia estar relacionado á via de catabolismo do colesterol ou relacionado ás
vias de efluxo do colesterol como a sua eliminação através da bile mediada pelos
receptores ABCG5 e ABCG8, ou sua eliminação direta nas fezes.
Nossos resultados mostraram que a suplementação com urucum não alterou a
expressão de ABCG5 e ABCG8 (Figura 13), nem as quantidades de gordura e
colesterol total no fígado e nas fezes (Tabela X e XI), e dessa forma podemos assumir
que a secreção hepática do colesterol na bile mediada por estes transportadores não foi
estimulada e nem o efluxo de colesterol diretamente nas fezes.
A secreção biliar de colesterol quer sob a forma de colesterol livre ou ácido
biliar, é a única rota significativa na eliminação do colesterol em mamíferos. CYP7A1 é
a enzima limitante na via de biossíntese hepática dos ácidos biliares e deste modo
controla a homeostase do colesterol e ácido biliar (Li et al., 2011). Em algumas
espécies, o consumo de colesterol, em quantidades que excedem as exigências
nutricionais resulta em uma resposta homeostática que inclui a indução da expressão de
CYP7A1 e o aumento da síntese de ácidos biliares (Kern, 1991; Dueland et al., 1993)
Por outro lado, hamsters (Horton et al, 1995) e camundongos knockout para o receptor
LXR (Peet et al., 1998) exibem uma incapacidade em induzir a CYP7A1 em resposta
ao colesterol dietético e uma marcante susceptibilidade em acumular colesterol em
lipoproteínas no plasma e no fígado. Estes resultados sugerem que a modulação das vias
sintéticas de colesterol/ácidos biliares contribui para a manutenção da homeostase do
colesterol.
A estimulação gênica da expressão de CYP7A1, por dietas ricas em colesterol,
varia entre as espécies. Ratos e camundongos exibem uma marcante up-regulation
enquanto hamsters, humanos, macacos, cobaias e coelhos não estimulam ou até mesmo
reprimem a expressão de CYP7A1. Os ratos são, portanto, capazes de se adaptarem a
grandes flutuações de entrada de esteróis e são resistentes a hipercolesterolemia
induzida por dieta, enquanto que os coelhos e hamsters desenvolvem

SILVA, L.S Capítulo 6 - Discussão
67
hipercolesterolemia em uma dieta rica colesterol (Dueland et al., 1993; Rudel et al.,
1994; Horton et al., 1995; Nguyen et al., 1999).
Estudos de Horton et al.,(1995) mostraram que as diferenças de sensibilidade ao
colesterol dietético no rato e no hamster são principalmente devidas a diferenças na
regulação da CYP7A1. Um baixo nível basal de expressão da CYP7A1, acoplado a uma
incapacidade na indução da expressão e atividade da enzima, torna o hamster mais
sensível ao colesterol dietético do que o rato.
Outra particularidade observada em ratos e camundongos é que estes animais
possuem o ácido muricólico, um ácido biliar natural trihidroxilado, como seu principal
ácido biliar. Em humanos, este ácido biliar não ocorre em quantidades substanciais e
também não pode ser metabolizado á partir do ácido quenodesoxicólico como ocorre
em fígados de ratos (Denk et al., 2002).
Elevados níveis de ácidos biliares hidrofóbicos no fígado e outros
compartimentos do corpo são encontrados em doenças colestáticas primárias mais
comuns, como a cirrose biliar primária e a colangite esclerosante primaria. O ácido
muricólico, devido à presença de um grupo hidroxila adicional na posição 6 do anel
esteróide, é mais hidrofílico que o ácido ursodeoxicólico, o qual é utilizado como
tratamento médico para cirrose biliar primária (Maillette et al., 2010).
Tem sido demosntrado, a efetividade do ácido muricólico na dissolução de
cálculos biliares de colesterol in vitro (Montet et al., 1987), e em um recente estudo, o
ácido muricólico mostrou ser mais efetivo que o ácido ursodeoxicóclico em prevenir e
dissolver cálculos biliares em camundongos (Wang e Tazuma, 2002) . Adicionalmente,
o ácido muricólico promoveu efeitos benéficos tanto na colestase induzida por ácido
taurolitocólico em hepatócitos primários de ratos (Milkiewicz et al., 2002) quanto em
colestase induzida por ácido taurocólico em fígados perfundidos de ratos tratados com
colchicina (Katagiri et al., 1992).
A Figura 13 demonstra que a dieta hipercolesterolemiante promoveu um
aumento na expressão de CYP7A1 em 7,3 vezes no grupo H em comparação com o
grupo controle, e que a suplementação de urucum a esta dieta, promoveu elevações
significativas na expressão de CYP7A1 nos grupos HU 0,1% e HU 0,2%, em
comparação com o grupo H. Assim, estes resultados sugerem, que o urucum possa estar
interferindo na modulação hepática da síntese de ácidos biliares e que seu efeito

SILVA, L.S Capítulo 6 - Discussão
68
hipocolesterelemiante pode ser principalmente atribuído ao aumento do catabolismo do
colesterol pela up-regulation da CYP7A1.
Nós reconhecemos a limitação de se considerar apenas os níveis de expressão de
RNAm como uma medida da via de fluxo, bem como consideramos a importância de se
confirmar se alterações na expressão dos genes encontram-se refletidas nos níveis de
proteínas. Entretanto como suporte da nossa hipótese, os estudos de Buhman et al.,
2000; Yang et al., 2003 e Shibata et al., 2007a, demonstraram uma relação linear entre o
aumento da expressão gênica de CYP7A1, a maior atividade da enzima e a redução do
colesterol sérico.
A transcrição da CYP7A1 é regulada positivamente pelo LXR, que tem como
ligantes os oxisteróis, e negativamente pelo FXR, que tem como ligantes os ácidos
biliares.
Os oxisteróis aumentam a transcrição de ABCA1, ABCG5/G8, e CYP7A1
através da ativação de LXR e os ácidos biliares aumentam a transcrição de SHP e
ABCB11 através da ativação de FXR (Shibata et al., 2007). Com base em tais
informações, as alterações na expressão de CYP7A1 e ABCG5/G8, genes alvos de
LXR, foram verificadas para indicar se houve ativação do LXR, assim como as
alterações na expressão de SHP e ABCB11, genes alvo de FXR, foram verificadas para
indicar se houve ativação do FXR.
A Figura 13 demonstra que as expressões de LXR, FXR, ABCG5/G8, SHP e
ABCB11 não foram alteradas pela dieta hipercolesterolemiante e que as expressões de
SHP e ABCB11 foram elevadas pela suplementação do urucum á essa dieta.
Corroborando com os nossos resultados, Xu et al., (2004) observaram que a
expressão e a atividade da CYP7A1 de ratos Sprague-Dawley alimentados com 2% de
colesterol foram elevadas, e que as expressões de LXR, FXR, SHP e ABCB11 não
foram alteradas pelo colesterol dietético.
Além dos efeitos do colesterol dietético, os autores avaliaram também os efeitos
do ácido cólico sobre a expressão destes genes e os resultados indicaram que os animais
tratados com ácido cólico apresentaram maiores expressões de SHP e ABCB11 em
comparação aos animais não tratados. Segundo os autores, esta elevação foi devida á
ativação do FXR, uma vez que o pool dos ácidos biliares, os quais são ligantes de FXR,
foi expandido.

SILVA, L.S Capítulo 6 - Discussão
69
Os nossos resultados demonstram que os animais tratados com urucum
apresentaram maiores expressões de CYP7A1, SHP e ABCB11. A partir daí podemos
sugerir que a maior expressão da CYP7A1, promovida pelo urucum, estaria levando a
uma maior conversão do colesterol á ácidos biliares, e que assim como os resultados
obtidos por Xu et al., (2004) a expansão do pool de ácidos biliares estaria
proporcionando uma maior ativação do FXR e consequentemente uma maior
transcrição de SHP e ABCB11, explicando assim, as elevações de suas expressões nos
grupos HU 0,1% e HU 0,2%.
Apesar de SHP, ter sua expressão aumentada e ser um fator inibidor da
expressão da CYP7A1, os animais dos grupos HU 0,1% e HU 0,2%, apresentaram
mesmo assim, elevações na expressão da CYP7A1. A explicação para este fato é que, a
nível molecular, embora a CYP7A1 seja up-regated por LRX e down-regulated por
FXR, o mecanismo estimulatório de LXR predomina sobre o mecanismo inibitório de
FXR, quando ratos são expostos a uma dieta rica em colesterol (Wójcicka et al., 2007).
Kerr et al., (2002) ralataram que a expressão de CYP7A1 em camundongos
knockout para SHP foi apenas 1,3 vezes mais abundante que em camundongos
selvagens.
Gupta et al., (2002) observaram que ratos alimentados com colesterol 2%
exibiram aumentos nas expressões de SHP e CYP7A1 e que a expressão de FXR não
foi alterada. Ao suplementar a dieta de colesterol com ácido cólico a 1%, os autores
observaram uma marcante elevação do SHP e uma concomitante redução da CYP7A1.
Entretanto, foi observado também, que mesmo com a presença do ácido cólico na dieta,
o nível do RNAm da CYP7A1 ainda foi 300% maior do que o obtido nos animais
controle.
Os autores concluíram então, que sob condições onde, tanto o SHP (via
estimulação de FXR), quanto o LXR são ativados, o efeito estimulatório de LXR
substitui o efeito inibitório de FXR, o que resulta na indução da expressão de CYP7A1
em ratos. Além disso, os autores sugeriram que os ácidos biliares não são reguladores
transcricionais de FXR e que eles apenas ativam este receptor através de uma maneira
ligante dependente.
Os nossos resultados indicam que mesmo não havendo aumento na transcrição
de LXR e FXR, estes receptores nucleares foram ativados, o que pôde ser verificado

SILVA, L.S Capítulo 6 - Discussão
70
pela maior transcrição da CYP7A1 e de SHP e ABCB11 respectivamente. Além disso,
foi observado que mesmo com a ativação simultânea de LXR e FXR, a expressão da
CYP7A1 foi estimulada, o que corrobora com os resultados descritos anteriormente, que
demonstram a predominância do efeito estimulatório de LXR sobre o efeito inibitório
de FXR na indução da expressão da CYP7A1.
Em resumo, nossos resultados demonstraram que a suplementação com urucum
não afetou a expressão dos genes hepáticos envolvidos na síntese e captação do
colesterol, mas aumentou significativamente a expressão da CYP7A1, a principal
enzima envolvida no catabolismo do colesterol. Diante destes resultados, podemos
concluir que o efeito hipocolesterolemiante do urucum envolve a up-regulation da
CYP7A1 que aumenta a conversão do colesterol á ácidos biliares, os quais foram os
responsáveis pela elevação da expressão de SHP e ABCB11 via ativação do FXR.
A Figura 15 demonstra uma possível hipótese dos mecanismos associados ao
efeito hipocolesterolemiante do urucum em ratos com hipercolesterolemia induzida por
dieta.
Figura 15 – Proposta do efeito hipocolesterolêmico do urucum. Animais tratados com urucum
apresentaram menor concentração sérica de colesterol total, e maior expressão hepática dos genes
CYP7A1, SHP e ABCB11. O mecanismo sugerido é que talvez o urucum possa ter agido como um
ativador de LXR e este receptor, uma vez ativado estimulou a transcrição da CYP7A1. A elevação da
CYP7A1 levou a uma maior conversão de colesterol á ácidos biliares, o que resultou em: 1) redução da
concentração do colesterol total sérico e 2) aumento dos ácidos biliares. Com o maior pool de ácidos
biliares, o FXR tornou-se ativado e promoveu uma maior transcrição de SHP e ABCB11.


SILVA, L.S. Capítulo 7 - Conclusões
72
CAPÍTULO 7 – CONCLUSÕES
De acordo com os resultados obtidos e sob as condições experimentais
empregadas no presente estudo, podemos concluir que a suplementação tanto do beta
caroteno quanto do urucum á dieta hipercolesterolemiante contribuiu para a melhora no
perfil lipídico.
Os animais que receberam dieta hipercolesterolemiante e beta caroteno
apresentaram:
- redução do colesterol total sérico, colesterol não HDL, triglicérides e índice
aterogênico;
- redução da gordura e colesterol total hepático;
- aumento na excreção de gordura e de colesterol nas fezes.
Os animais que receberam dieta hipercolesterolemiante e urucum apresentaram:
- redução do colesterol total sérico, colesterol não HDL e índice aterogênico;
- aumento na expressão da CYP7A1;
- aumento na expressão de SHP e ABCB11
A partir desses resultados podemos observar que o beta caroteno e o urucum
apresentaram efeitos hipocolesterolemiantes. Entretanto, seus mecanismos de atuação
foram diferentes:
O mecanismo associado ao efeito hipocolesterolêmico do beta caroteno não
envolve alterações na expressão de genes hepáticos do metabolismo do
colesterol e pode estar relacionado á menor absorção intestinal de colesterol, que
foi evidenciada pelas maiores concentrações de colesterol nas fezes ou á
alterações do metabolismo do colesterol nas células intestinais.
O mecanismo associado ao efeito hipocolesterolêmico do urucum está
relacionado à conversão do colesterol á ácidos biliares, através da up regulation
da CYP7A1, que por sua vez foi responsável pelo desencadeamento uma série
de eventos que culminaram em maiores expressões de SHP e ABCB11.


SILVA, L.S. Referências Bibliográficas
74
REFERÊNCIAS BIBLIOGRÁFICAS
Agarwal, S.; Rao, A. V. (2000). Carotenoids and chronic diseases. Drug Metab Drug
Interact. 17(1–4): 189–210.
Agner, A. R.; Bazo, A. P.; ,Ribeiro L. R.; Salvadori, D. M. (2005). DNA damage and
aberrant crypt foci as putative biomarkers to evaluate the chemopreventive effect of
annatto (Bixa orellana L.) in rat colon carcinogenesis. Mutat Res. 582: 146-154.
Amen, R.J.; Lachance, P. A. (1974). The effects of beta-carotene and canthaxanthin on
serum cholesterol levels in the rats. Nutr Rep Int. 10: 269-276.
Ando, H.; Tsuruoka, S.; Yamamoto, H.; Takamura, T.; Kaneko, S.; Fujimura A. (2005).
Regulation of cholesterol 7α-hydroxylase mRNA expression in C57BL/6 mice fed an
atherogenic diet. Atherosclerosis 178: 265-269.
Arsenault, B.J.; Rana, J.S.; Stroes, E.S.; Despres, J.P.; Shah, P.K.; Kastelein, J.J.;
Wareham, N.J.; Boekholdt, S.M.; Khaw, K.T. (2009). Beyond low-density lipoprotein
cholesterol: respective contributions of non-high-density lipoprotein cholesterol levels,
triglycerides, and the total cholesterol/high-density lipoprotein cholesterol ratio to
coronary heart disease risk in apparently healthy men and women. J Am Coll Cardiol.
55: 35-41.
Astrog P. (1997). Food carotenoids and cancer prevention: an overview of current
research. Trends Food Sci Technol. 8: 406-413.
Astrog, P.; Gradele,t S.; Berges, R.; Suschetet, M. (1997). Dietary lycopene decreases
initiation of liver preneoplastic foci by diethylnitrosamine in rat. Nutr Cancer. 29: 60-
68.
Baker, H. J. Lindsey, J.R. Weisbroth, S. H. (1979). The laboratory rat. American
College of Laboratory. Animal Medice Series. v. 1 Biology and Diseases, p.128,
Academic Press, Inc, Florida.
Balaswamy, K., Prabhakara Rao, P.G., Satyanarayana, A., Rao, D.G. (2006). Stability
of bixin in annatto oleoresin and dye powder during storage. Food Science and
Technology, 39: 952-956.
Banner, C. D.; Gottlicher, M.; Widmark, E.; Sjovall, J.; Rafter, J. J.; Gustafsson, J. A.
(1993). A systematic analytical chemistry/cell assay approach to isolate activators of
orphan nuclear receptors from biological extracts: characterization of peroxisome
proliferator-activated receptor activators in plasma. J Lipid Res. 34: 1583-15 91.

SILVA, L.S. Referências Bibliográficas
75
Barbosa-Filho, J.M. (2006). Bixa orellana: Retrospectiva de usos populares, atividades
biológicas, fitoquímicas e emprego na fitocosmética, no Continente Americano. In:
Simpósio Brasileiro do Urucum, p.1-14. João Pessoa
Barreiros, A. L. B. S.; David J. M. (2006). Estresse oxidativo: relação entre geração de
espécies reativas e defesas do organismo. Química Nova. 29(1): 113-123.
Batista, C. L.L.C. (1994). Produção e avaliação da estabilidade de corante hidrossolúvel
de urucum. Universidade Federal de Viçosa, p 71. Viçosa.
Bautista, A. R. P. L.; Miranda, M. S.; Batista, M. S.; Moreira; E. L.T. Silva, I. M.M.;
Gomes, I. C. S. (2004). Avaliação da toxicidade oral subcrônica da bixina para ratos.
Brazilian Journal of Pharmaceutical Sciences. 40(2): 229-233.
Beecher, G. R. (1998). Nutrient content of tomatoes and tomatoes products. Proc Soc
Exp Biol Med. 218: 98–100.
Berge,K. E.; Tian, H.; Graf, G.A. Yu, L.; Grishin, N. V.; Schultz,J.; Kwiterovich, P.;
Shan,B.; Barnes,R.; Hobbs, H.H. (2000). Accumulation of dietary cholesterol in
sitosterolemia caused by mutations in adjacent ABC transporters. Science.
290(5497):1771-1775.
Bertram, J. S. (1999). Carotenoids and gene regulation. Nutr Rev. 57: 182-191.
Bestwick, C. S.; Milne, L. (2000). Effects of beta-carotene on antioxidant enzyme
activity, intracellular reactive oxygen and membrane integrity within post confluent
Caco-2 intestinal cells. Biochim Biophys Acta. 1474(1): 47-55.
Blakely, S.R.; Lynnard,S.; Adkins,J.; Knight, E.V. (1988). Effects of -Carotene and
Retinyl Palmitate on Corn Oil-Induced Superoxide Dismutase and Catatase in Rats. J.
Nutr. 118: 152-158.
Bohm, V.; Bitsch, R. (1999). Intestinal absorption of lycopene from different matrices
and interactions to other carotenoids, the lipid status, and the antioxidant capacity of
human plasma. Eur J Nutr. 38: 118 -125.
Boileau, T. W.; Boileau, A.C.; Erdman, J. W. Jr. (2002). Bioavailability of all-trans and
cis-isomers of lycopene. Exp Biol Med. 227: 914 -919.
Bonilla, S.; Redonnet, A.; Noel-Suberville, C.; Pallet, V.; Garcin, H.; Higueret, P.
(2000). High-fat diets affect the expression of nuclear retinoic acid receptor in rat liver.
Br J Nutr. 83: 665-671.
Boone, L. R.; Brooks, P. A.; Niesen, M. I.; Ness, G. C. (2011). Mechanism of resistance
to dietary cholesterol. J Lipids. 2011: 9 pages.
Britton G. (1995). Structure and properties of carotenoids in relation to function. FASEB
J. 9:1551-1558.

SILVA, L.S. Referências Bibliográficas
76
Buhman, K. K.; Furumoto, E. J.; Donkin, S. S.; Story, J. A. (2000). Dietary psyllium
increases expression of ileal apical sodium-dependent bile acid transporter mRNA
coordinately with dose-responsive changes in bile acid metabolism in rats. J Nutr. 130:
2137-2142.
Calkin, A. C.; Tontonoz, P. (2010). Liver x receptor signaling pathways and
atherosclerosis. Arterioscler Thromb Vasc Biol. 30(8): 1513-1518.
Carvalho, P. R. N., Hein, M. (1989). Urucum – Uma fonte de corante natural. Coletânea
ITAL, v.19 (1), p. 25-33. Campinas
Chapman, M.J. (2006). Fibrates: therapeutic review. Br. J. Diab. Vasc. Dis. 6: 11-19.
Chinetti-Gbaguidi, G.; Fruchart, Jean C.; Staels, B. (2005). Role of the PPAR family of
nuclear receptors in the regulation of metabolic and cardiovascular homeostasis: new
approaches to therapy. Current Opinion in Pharmacology. 5(2): 177-183.
Chung, M. J.; Kang, A. Y.; Park, S. O.; Park, K. W.; Jun, H. J.; Lee, S. J. (2007). The
effect of essential oils of dietary wormwood (Artemisia princeps), with and without
added vitamin E, on oxidative stress and some genes involved in cholesterol
metabolism. Food ChemToxicol. 45: 1400-1409.
Chung, M. J.; Sung, N. J.; Park, C. S.; Kweon, D. K.; Mantovan,i A.; Moon, T. W.; Lee,
S. J.; Park, K. H. (2008). Antioxidative and hypocholesterolaemic activities of water-
soluble puerarin glycosides in HepG2 cells and in C57 BL/6J mice. Eur J Pharmacol.
578: 159-170.
Connor, W. E,; De Francesco, C. A.; Connor, S. L.(1993). N-3 fatty acids from fish oil.
Effects on plasma lipoproteins an hypertriglyceridemic patients. Ann N Y Acad Sci. 683:
16-34.
Cornwell, P. D.; de Souza, A. T.; Ulrich, R. G. (2004). Profiling of hepatic gene
expression in rats treated with fibric acid analogs. Mutat.Res. 549 (1-2): 131-145.
Denk, G. U.; Kleiss, C. P.; Wimmera, R.; Vennegeerts, T.; Reiter, F. P.; Schulz
S.;.Zischka, H.; Rust, C.(2012). Tauro--muricholic acid restricts bile acid-induced
hepatocellular apoptosis by preserving the mitochondrial membrane potential.
Biochemical and Biophysical Research Communications. 424:758-764.
de Oliveira, A. C.; Silva, I. B.; Manhaes-Rocha, D.A.; Paumgartten, F. J. (2003).
Induction of liver monooxygenases by annatto and bixin in female rats. Braz. J. Med.
Biol. Res. 36(1): 113-118.
de Paula, H.; Pedrosa, M. L.; Rossoni Jr, J.V.; Haraguchi, F. K.; Santos, R. C.; Silva,
M. E. (2009). Effect of an aqueous extract of annatto (Bixa orellana) seeds on lipid
profile and biochemical markers of renal and hepatic function in hipercholesterolemic
rats. Braz Arch Biol Technol. 52: 1373-1378.

SILVA, L.S. Referências Bibliográficas
77
Deppa, P.R.; Varalakshmi, P. (2005). Atheroprotective effect of exogenous heparin-
derivative treatment on the aortic disturbances and lipoprotein oxidation in
hypercholesterolaemic diet fed rats. Clinica Chimica Acta. 355: 119-130.
Dhingra, S.; Bansal, M.P. (2006). Hypercholesterolemia and LDL receptor mRNA
expression, modulation by selenium supplementation. Biometals. 19: 493-501.
Di Mascio, P.; Devasagayam, T. P.; Kaiser, S.; SIES, H. (1990). Carotenoids,
tocopherols and thiols as biological singlet molecular oxygen quenchers. Biochem. Soc.
Trans. 18(6):1054-1056.
Diaz, M. N.; Frei, B.; Vita, J.A.; Keaney, J. F. JR. (1997). Antioxidants and
atherosclerotic heart disease. New Eng J of Med. 33: 408-416.
Dueland, S.; Drisko, J.; Graf, L.; Machleder, D.; Lusis, A. J.; Davis, R. A. (1993).
Effect of dietary cholesterol and taurocholate on cholesterol-7α-hydroxylase and hepatic
LDL receptors in inbred mice. J Lipid Res. 34: 923–931.
Durrington, P. (2003). Dyslipidaemia. Lancet. 362: 717-731.
Eberlé, D.; Hegarty, B.; Bossard, P., Ferré, P.; Foufelle, F. (2004). SREBP transcription
factors: master regulators of lipid homeostasis. Biochimie. 86: 839-848.
Elliott, R. (2005). Mechanisms of genomic and non-genomic actions of carotenoids.
Biochim Biophys Acta. 1740: 147-54.
Eloranta, J.J.; Kullak-Ublick, G.A. (2005). Coordinate transcriptional regulation of bile
acid homeostasis and drug metabolism, Arch. Biochem. Biophys. 433: 397-412.
Erdman Jr, J. W.; Bierer, T. L.; Gugger, E. T. (1993). Absorption and transport of
carotenoids. Ann NY Acad Sci.691: 76-85.
Erdman, J.W.; Lachance, P.A. (1974). Failure of the non-vitamin A active carotenoid
lycopene to act as an antihypercholesterolemic agent in rats. Nutr. Rep. Int. 10: 277-
284.
Evans, J. L.; Goldfine, I.D.; Maddux, B. A.; Grodsky, G. M. (2009). Oxidative stress
and stress-activated signaling pathways: a unifying hypothesis of type 2 diabetes.
Endocr Rev. 23: 599-622.
Felicissimo, M. P.; Bittencourt, C.; Houssiau, L.E.; Pireaux, J. J. (2004). Time-of-flight
secondary ion mass spectrometry and X-ray photoelectron spectroscopy analyses of
Bixa orellana seeds. J.Agric.Food Chem. 52(7): 1810-1814.
Fernandes, A. C. S.; Almeida, C. A.; Albano, F.; Laranja, G. A. T.; Felzenszwalb, I.;
Lage, C. L. S.; Sá, C. C. N. F.; Moura, A. S.; Kovary, K. (2002). Norbixin ingestion did
not induce any detectable DNA breakage in liver and kidney but caused a considerable

SILVA, L.S. Referências Bibliográficas
78
impairment in plasma glucose levels of rats and mice. Journal of Nutritional
Biochemistry. 13: 411- 420.
Ferré, P. (2004). The biology of peroxisome proliferator-activated receptors:
relationship with lipid metabolism and insulin sensitivity. Diabetes. 53(Suppl1): 43-50.
Ferreira, J. M.; Sousa, D. F.; Dantas, M. B.; Fonseca, S. G. C.; Menezes, D. B.; Martins,
A.M.C.; Queiroz, M. G. R. (2012). Effects of Bixa orellana L. Seeds on
Hyperlipidemia. Phytother Res. DOI: 10.1002/ptr.4675.
Folch, J.; Lees, M.; Sloane Stanley, G. H. (1957). A simple method for the isolation and
purification of total lipides from animal tissues. J Biol Chem. 226: 497-509.
Forman, B. M.; Chen, J.; Evans, R. M. (1997). Hypolipidemic drugs, polyunsaturated
fatty acids, and eicosanoids are ligands for peroxisome proliferator-activated receptors
alpha and delta. Proc Natl Acad Sci USA. 94: 4312-4317.
Franco, C. F. O.; Silva, F. C. P.; Filho, J. C.; Neto, M. B.; José, A. R. S.; Rebouças, T.
N. H.; Fontinélli, I. S. C. (2002). Urucuzeiro: agronegócio de corantes naturais.
EMEPA/SAIA, João Pessoa, p.120.
Franco, C.F.O.; Silva, F.C.P.; Cazé Filho, J.; Barreiro Neto, M.; José, A.R.S.; Rebouças,
T.N.H.; Fontinélli, I.S.C. (2006). Etnobotânica e Taxonomia do Urucuzeiro.Simpósio
Brasileiro do Urucum. João Pessoa.
Gao, H.; Liu, Z, Qu, X.; Zhao, Y. (2013). Effects of Yerba Mate tea (Ilex
paraguariensis) on vascular endothelial function and liver lipoprotein receptor gene
expression in hyperlipidemic rats. Fitoterapia. 84: 264-272.
Gaziano, J. M.; Hennekens, C. H. (1993). The role of beta-carotene in the prevention of
cardiovascular disease. In: L. M.Canfield, N. I. Krinsky, J. A. Olso, (ed) Carotenoids in
Human Health, New York Academy of Sciences, p. 148-155, New York.
Gent, J.; Braakman, I. (2004). Low-density lipoprotein receptor structure and folding.
Cell Mol.Life Sci. 61(19-20): 2461-2470.
Gerster, H. (1997). The potential role of lycopene for human health. J Am Coll
Nutr.16:109-26.
Gey K. F.; Puska, P.; Jordan, P.; Moser, U. K. (1991). Inverse correlation between
plasma vitamin E and mortality from ischemic heart disease in crosscultural
epidemiology. Am J Clin Nutr. 53(1): 326S-334S.
Ghiraldini, E. (1994). Corantes naturais mais comumente usados na indústria de
alimentos. Revista Brasileira de Corantes Naturais, 2(1): 83-87.
Giuliano, G.; Rosati, C. E.; Bramley, P. M. (2003). To dye or not to dye: biochemistry
of annatto unveiled. Trends Biotechnol. 21(12): 513-516.

SILVA, L.S. Referências Bibliográficas
79
Gonçalves, M. C. R.; Diniz, M. F. F. M.; Borba, J. D. C.; Nunes, X. P.; BARBOSA-
Filho, J. M. (2006). Beringela (Solanum melongena L.) – mito ou realidade no combate
as dislipidemias? Revista Brasileira de Farmacognosia. 16(2): 252-257.
Goodman, G. E.; Schaffer, S.; Omenn, G.S.; Chen, C.; King, I. (2003). The association
between lung and prostate cancer risk, and serum micronutrients: results and lessons
learned from beta-carotene and retinol efficacy trial. Cancer Epidemiol Biomarkers
Prev. 12(6): 518-526.
Goodwin, T. W. (1980). The Biochemistry of the Carotenoids. Vol. 1:“Plants.” New
York: Chapman and Hall, p 203.
Gottlicher, M.; Widmark, E.; Li, Q.; Gustafsson, J. A. (1992). Fatty acids activate a
chimera of the clofibric acid-activated receptor and the glucocorticoid receptor. Proc
Natl Acad Sci USA. 89: 4653-4657.
Graf, G. A.; Yu, L.; Li, W. P.; Gerard, R.; Tuma, P. L.; Cohen, J. C.; Hobbs H.H .
(2003) ABCG5 and ABCG8 are obligate heterodimers for protein trafficking and biliary
cholesterol excretion. J Biol Chem. 278: 48275-48282.
Grundy, S. M. (1983). Absorption and metabolism of dietary cholesterol. Annu Rev Nutr.
3: 71-96.
Guo, J.; Be,i W.; Hu, Y.; Tang, C.; He, W.; Liu, X.; Huang, L.; Cao, Y.; Hu, X.; Zhong,
X.; Cao, L. (2011). A new TCM formula FTZ lowers serum cholesterol by regulating
HMG-CoA reductase and CYP7A1 in hyperlipidemic rats. J Ethnopharmacol. 135:
299-307.
Gupta, S.; Stravitz, R. T.; Dent, P.; Hylemon P. B. (2001). Down-regulation of
cholesterol 7a-hydroxylase (CYP7A1) gene expression by bile acids in primary rat
hepatocytes is mediated by the c-Jun N-terminal Kinase pathway. J Biol Chem. 276(19):
15816-582.
Gupta, S.; Pandak, W. M.; Hylemona, P. B. (2002). LXRα is the dominant regulator of
CYP7A1 transcription. Bioch Biophys Res Commun. 293: 338-343.
Guzman, D. (2005). Value of carotenóides sector to cross billion dollar mark. Chemical
Market Reporter. 268(1): 33.
Horton, J. D.; Cuthbert, J. A.; Spady, D. K. (1995). Regulation of hepatic 7α-
hydroxylase expression and response to dietary cholesterol in the rat and hamster. J.
Biol. Chem. 270(10): 5381-5387.
Horton, J. D.; Goldstein, J. L.; Brown, M. S. (2002). SREBPs: activators of the
complete program of cholesterol and fatty acid synthesis in the liver. J Clin
Invest. 109(9): 1125–1131.

SILVA, L.S. Referências Bibliográficas
80
Hsu, S. C.; Huanga, C. (2007). Changes in liver PPARa mRNA expression in response
to two levels of high-safflower-oil diets correlate with changes in adiposity and serum
leptin in rats and mice. J Nutr Biochem. 18: 86-96.
Iyama, T.; Takasuga, A.; Azuma, M. (1996). Beta-carotene accumulation in mouse
tissues and a protective role against lipid peroxidation. Internat J Vit Nutr Res. 66: 301-
305.
Jewell, C.; O’Brien, N. M. (1999). Effect of dietary supplementation with carotenoids
on xenobiotic metabolizing enzymes in the liver, lung, kidney and small intestine of the
rat. Br J Nutr. 81: 235-242.
Johnson, E. J. (2002). The role of carotenoids in human health. Nutr Clin Care. 5(2):
47-49.
Jondiko, I. J. O.; Pattenden, G. (1989). Terpenoids and an apocarotenoid from seeds of
Bixa orellana. Phytochem., 28: 3159-3162.
Júnior, A. C.; Asad, L. M.; Oliveira, E. B.; Kovary, K.; Asad, N. R.; Felzenszwalb, I.
(2005). Antigenotoxic and antimutagenic potential of an annatto pigment (norbixin)
against oxidative stress. Genet Mol Res. 4: 94-99.
Kaliora, A. C.; Kountouri, A. M.; Karathanos, V. T. (2009). Antioxidant Properties of
Raisins (Vitis vinifera L.). J Med Food. 12(6): 1302–1309.
Katagiri, K.; Nakai, T.; Hoshino, M.; Hayakawa, T.; Ohnishi, H.; Okayama, Y.;
Yamada, T.; Ohiwa, T.; Miyaji, M.; Takeuchi, T. (1992). Tauro-beta-muricholate
preserves choleresis and prevents taurocholate-induced cholestasis in colchicine-treated
rat liver. Gastroenterology. 102: 1660-1667.
Kern, F. Jr. (1991). Normal plasma cholesterol in an 88 year old man who eats 25 eggs
a day: mechanisms of adaptation. N Engl J Med. 324: 896-99.
Kerr, T. A.; Saeki, S.; Schneider, M.; Schaefer, K.; Berdy, S.; Redder, T.; Shan, B.
David W. Russell,1,4 and Margrit Schwarz. (2002). Loss of nuclear receptor shp
impairs but does not eliminate negative feedback regulation of bile acid synthesis.
Developmental Cell. 2: 713-720.
Kliewer, S. A. Sundseth, S. S.; Jones, S.A.; Brown, P.J.; Wisely, G.B.; Koble, C. S.;
Devchand, P.; .Wahli, W.; Willson, T. M. Lenhard, J. M.; Lehmann, J. M. (1997).
Fatty acids and eicosanoids regulate gene expression through direct interactions with
peroxisome proliferator-activated receptors alpha and gamma. Proc Natl Acad Sci USA.
94: 4318-4323.
Kota, B. P.; Huang, T. H.; Roufogalis, B. D. (2005). An overview on biological
mechanisms of PPARs. Pharmacol.Res. 51(2): 85-94.

SILVA, L.S. Referências Bibliográficas
81
Kraus, A.; Roth, H. P.; Kirchgessner, M. (1997). Supplementation with vitamin C,
vitamin E or beta-carotene influences osmotic fragility and oxidative damage of
erythrocytes of zinc-deficient rats. J Nutr. 127:1290–1296.
Krey, G.; Braissant,.; O, L’Horset, F.; Kalkhoven, E.; Perroud, M.; Parker, M. G.;
Wahli, W. (1997). Fatty acids, eicosanoids, and hypolipidemic agents identified as
ligands of peroxisome proliferator-activated receptors by coactivator-dependent
receptor ligand assay. Mol Endocrinol 11: 779-791.
Krinsky, N. I. (1993). Actions of carotenoids in biological systems. Annu Ver Nutr.13:
561- 187.
Krinsky, N. I. (1994). The biological properties of carotenoids. Pure Appl Chem. 66:
1003-1010.
Krinsky, N. I. (1998). Overview of lycopene, carotenoids, and disease prevention. Proc
Soc Exp Biol Med. 218: 95-97.
Krinsky, N. I. (2001). Carotenoids as antioxidants. Nutrition. 17(10): 815-817.
Krinsky,N. I; Johnson.E.J. (2005).Carotenoid actions and their relation to health and
disease. Mol Aspects Medicin. 26:459-516.
Kritchevsky, S.B. (1999). -Carotene, Carotenoids and the Prevention of Coronary
Heart Disease. J. Nutr. 129: 5-8.
Lee, F. Y.; Lee, H.; Hubbert, M. L.; Edwards, P.A.; Zhang, Y. (2006). FXR, a
multipurpose nuclear receptor. Trends Biochem Sciences. 31(10): 572-580.
Leifert, W. R.; Abeywardena, E. M. Y. (2008). Cardioprotective actions of grape
polyphenols. Nutrition Research., 28:729-737.
Levin, N.; Bischoff, E. D.; Daige, C. L.; Thomas, D.; Vu, C. T.; Heyman, R.
A.; Tangirala, R. K.; Schulman, I. G. (2005). Macrophage liver X receptor is required
for antiatherogenic activity of LXR agonists. Arterioscler Thromb Vasc Biol.
25(1):135–142.
Li, G.; Liu, X.; Zhu, H.; Huang, L.; Liu, Y.; Ma, C.; Qin, C. (2009). Insulin resistance
in insulin-resistant and diabetic hamsters (Mesocricetus auratus) is associated with
abnormal hepatic expression of genes involved in lipid and glucose metabolism.
Comparative Medicine. . 59(5): 449-458.
Li, T.; Matozel, M.; Boehme, S.; Kong, B.; Nilsson, L. M.; Guo, G.; Ellis, E.; Chiang, J.
Y. L. (2011). Overexpression of cholesterol 7α-hydroxylase promotes hepatic bile acid
synthesis and secretion and maintains cholesterol homeostasis. Hepatology 53: 996-
1006.

SILVA, L.S. Referências Bibliográficas
82
Li, T.; Owsley, E.; Matozel, M.;, Hsu, P.; Novak, C. M.; Chiang, J. Y. (2010).
Transgenic expression of cholesterol 7α-hydroxylase in the liver prevents high-fat diet-
induced obesity and insulin resistance in mice. Hepatology. 52: 678-690.
Lima, L. R. P.; Oliveira, T. T.; Nagem, T. J.; Pinto, A. S.; Stringheta, P. C.; Tinoco, A.
L. A.; Silva, J. F. (2001). Bixina, Norbixina e Quercetina e seus efeitos no metabolismo
lipídico de coelhos.Braz. J. Vet. Res. anim. Sci.38:(4):196-200.
Lima, L. R. P.; Oliveira, T. T.; Nagem, T. J. (2003). Efeitos do flavonóide quercetina e
dos corantes bixina e norbixina sobre parâmetros sanguíneos de coelhos. Rev.
Nutr.16(3): 305-314.
Lin, Q.; Ruuska, S. E.; Shaw, N. S.; Dong, D.; Noy, N. (1999). Ligand selectivity of the
peroxisome proliferator-activated receptor alpha. Biochemistry. 38: 185-190.
Lowe, G. M.; Booth, L. A.; Young. A. J.; Bilton, R. F. (1999). Lycopene and beta-
carotene protect against oxidative damage in HT29 cells at low concentrations but
rapidly lose this capacity at higher doses. Free Radic Res. 30(2): 141-151.
Lyng, S.M.O.; Passos, M.; Fontana, J.D. (2005). Bixin and -cyclodextrin inclusion
complex and stability tests. Process Biochemistry. 40:.865–872.
Mangels, A. R.; Holden, J. M.; Beecher, G. R.; Forman, M. R. Lanza E. (1993).
Carotenoid contents of fruits and vegetables: an evaluation of analytical data. J Am Diet
Assoc. 93: 284-296.
Matos, S.L.; de Paula, H., Pedrosa; M.L.; Santos, R.C.; Oliveira, E.L.;Chianca Júnior,
D.A.; Silva, M.E. (2005). Dietary models for inducing hypercholesterolemia in rats.
Brazilian Archives of Biology and Technology. 48: 203-209.
Mercadante, A. Z..; Steck, A.; Rodriguez-Amaya, D. B.; Pfander, H.; Britton, G.
(1996). Isolation of methyl 9’Z-apo- 6’-lycopenoate from Bixa orellana. Phytochem.
41: 1201-1203.
a) Mercadante, A .Z.; Steck, A.; Pfander, H. (1997). Isolation and identification of new
apocarotenoids from annatto (Bixa orellana L.) seeds. J. Agric. Food Chem. 45: 1050-
1054.
b) Mercadante, A. Z.; Steck, A.; Pfander, H. (1997). Isolation and structure elucidation
of minor carotenoids from annatto (Bixa orellana L.) seeds. Phytochem. 46: 1379-1383.
Mercadante, A. Z.; Steck, A.; Pfander, H. (1999). Three minor carotenoids from annatto
(Bixa orellana L.) seeds. Phytochem. 52: 135-139.
Miserez, A. R.; Muller, P. Y.; Barella, L.; Barella, S.; Staehelin, H. B.; Leitersdorf, E.;
Kark, J. D.; Friedlander, Y. (2002). Sterol regulatory element binding protein (SREBP-
2) contributes to polygenic hypercholesterolemia. Atherosclerosis 164: 15-26.

SILVA, L.S. Referências Bibliográficas
83
Miyake, J. H.; Duong-Polk, X. T.; Taylor, J.M.; Du, E. Z.; Castellani, L. W.; Lusis, A.
J.; Davis, R. A.(2002) Transgenic expression of cholesterol 7α-hydroxylase prevents
atherosclerosis in C57BL/6J mice. Arterioscler Thromb Vasc Biol. 22: 121-126.
Montet, J. C.; Parquet, M.; Sacquet, E.; Montet, A. M.; Infante, R.; Amic, J. (1987).
Betamuricholic acid; potentiometric and cholesterol-dissolving properties. Biochim.
Biophys. Acta. 918: 1-10.
Milkiewicz, P.; Roma, M. G.; Elias, E.; Coleman, R. (2002). Hepatoprotection with
tauroursodeoxycholate and beta muricholate against taurolithocholate induced
cholestasis: involvement of signal transduction pathways. Gut. 51: 113-119.
Murakami, K.; Ide, T.; Suzuki, M.; Mochizuki, T.; Kadowaki, T. (1999). Evidence for
direct binding of fatty acids and eicosanoids to human peroxisome proliferators-
activated receptor alpha. Biochem Biophys Res Commun. 260: 609-613.
Nader, M. A.; El-Agamy, D. S.; Suddek G. M. (2010). Protective Effects of Propolis
and Thymoquinone on Development of Atherosclerosis in Cholesterol-Fed Rabbits.
Arch. Pharm. Res. 33(4): 637-643.
Nguyen, L. B.; Xu, G.; Shefer, S.; Tint, G. S.; Batta, A.; Salen, G. (1999). Comparative
regulation of hepatic sterol 27-hydroxylase and cholesterol 7alpha-hydroxylase
activities in the rat, guinea pig, and rabbit: effects of cholesterol and bile acids,
Metabolism. 48(12): 1542-1548.
Nielsen, M.R. (1990). Natural colors for ice-cream. Scandinavian Dairy Information.
4(4): 56-58.
Nishimoto, T.; Pellizzon, M. A. Aihara, M. Stylianou, I.M.; Billheimer, J. T.; Rothblat,
G.; Rader, D. J. (2009). Fish oil promotes magrofage reverse cholesterol transport in
mice. Arteriocler Thromb Vasc Biol. 29: 1502-1508.
Oliveira, J.S. (2005). Caracterização, extração e purificação por cromatografia de
compostos de urucum (Bixa orellana l.). Universidade Federal de Santa Catarina.
Centro Tecnológico. Departamento de Engenharia Química e Engenharia de Alimentos.
Olson, J. A. (1994). Absorption, transport, and metabolism of carotenoids in huma
Olson, J. A. (1996). Benefits and liabilities of vitamin A and carotenoids. Journal of
Nutrition.126: 1208S-1212S.
Olson, J. A. (1999). Carotenoids and human health. Arch Lat Am Nutr. 49: 7-11.
Oram, J. F. (2002). Molecular basis of cholesterol homeostasis: lessons from Tangier
disease and ABCA1. Trends Mol.Med. 8(4): 168-173.

SILVA, L.S. Referências Bibliográficas
84
Paiva, S.; Russell, R. (1999). Beta carotene and other carotenoids as antioxidants. J Am
Coll Nutr.18:426-433.
Palou, M.; Priego, T.; Sánchez, J.; Villegas E.; Rodríguez, A. M.; Palou, A.; Picó, C.
(2008). Sequential changes in the expression of genes involved in lipid metabolism in
adipose tissue and liver in response to fasting. Pflugers Arch Eur J Physiol. 456: 825–
836.
Palozza, P. (1998). Prooxidant actions of carotenoids in biologic systems. Nutr Rev.
56(9): 257-265.
Paolini, M.; Cantelli-Forti, G.; Perocco, P.; Pedulli, G. F.; Abdel-Rahman S. Z.;
Legator, M. S. (1999). Co-carcinogenic effect of beta-carotene. Nature. 398(6730): 760-
761.
Park, C. H.; Cho, E. J.; Yokozawa, T. (2009). Protection Against Hypercholesterolemia
by Corni Fructus Extract and Its Related Protective Mechanism. J Med Food, 12: 973-
981.
Parker, R. S. (1988). Carotenoid and tocopherol composition in human adipose tissue.
Am J Clin Nutr. 47:-33-36.
Parker, R. S. (1996). Absorption, metabolism and transport of carotenoids. FASEB J.
10: 542-551.
Parker, R. S.; Swanson, J.E.; You, C.S.; Edwards, A. J.; Huang, T. (1999).
Bioavailability of carotenoids in human subjects. Proc Nutr Soc. 58: 155-162.
Peet, D. J.; Turley, S. D.; Ma, W.; Janowski, B. A.; Lobaccaro, J. M.; Hammer, R. E.;
Mangelsdorf, D. J. (1998). Cholesterol and bile acid metabolism are impaired in mice
lacking the nuclear oxysterol receptor LXR. Cell. 93: 693-704.
Perrone, M. G.; Santandrea, E.; Dell'uomo, N.; Giannessi, F.; Milazzo, F. M., Sciarroni,
A. F.; Scilimati, A.; Tortorella, V. (2005). Synthesis and biological evaluation of new
clofibrate analogues as potential PPARalpha agonists. Eur.J.Med.Chem. 40(2):143-154.
Preston, H.D.; Rickard, M.D. (1980). Extraction and chemistry of annatto. Food
Chemistry. 5(1): 47-56.
Pullinger, C. R.; Eng, C.; Salen, G.; Shefer, S.; Batta, A. K.; Erickson, S. K.;. Verhagen,
A.; Rivera, C. R.; Mulvihill, S. J.; Malloy, M. J.; Kane, J. P. (2002). Human cholesterol
7α-hydroxylase (CYP7A1) deficiency has a hypercholesterolemic phenotype. J Clin
Invest. 110: 109-117.
Rao, A.V.; Rao, L.G. (2007). Carotenoids and human health. Pharmacological
Research. 55:207-216.

SILVA, L.S. Referências Bibliográficas
85
Rasmussen, H.E.; Blobaum, K. R.; Jesch, E. D.; Ku, C. S.; Park, Y.K.; Lu, F.; Carr, T.
P.; Lee, J. Y. (2009). Hypocholesterolemic effect of Nostoc commune var. sphaeroides
Ku¨tzing, an edible blue-green alga Eur J Nutr. 48: 387–394.
Redinger, R. N. (2003). The coming of age of our understanding of the enterohepatic
circulation of bile salts. Am J Surg. 185(2): 168-172.
Reena, M. B.; Gowda, L. R.; Lokesh, B. R. (2011). Enhanced hypocholesterolemic
effects of interesterified oils are mediated by upregulating ldl receptor and cholesterol
7-α- hydroxylase gene expression in ratsj. Nutr. 141: 24-30.
Reeves, P.G., Nielsen, F. H., Fahey, G. C. (1993). AIN-93 purified diets for laboratory
rodents: Final report of the American Institute of Nutrition ad hoc Writing Committee
on the reformulation of the AIN-76A rodent diet. J. Nutrition.123(11): 467-472.
Repa, J. J.; Berge, K. E.; Pomajzl, C.; Richardson, J. A.; Hobbs, H.; Mangelsdorf, D. J.
(2002). Regulation of ATP-binding cassette sterol transporters ABCG5 and ABCG8 by
the liver X receptors alpha and beta. J Biol Chem. 277(21): 18793-18800.
Rezen, T.; Rozman, D.; Pascussi, J.M.; Monostory, K. (2011). Biochimica et Biophysica
Acta. 1814: 146-160.
Ribaya-Mercado, J. D.; Blumberg, J. B. (2004). Lutein and zeaxanthin and their
potential roles in disease prevention. J Am Coll Nutr. 23(6): 567S–5687S.
Riondel, J.; Wong, H. K.; Glise, D.; Ducros, V.; Favier, A. (2002). The effect of a
water-dispersible beta-carotene formulation on the prevention of age-related lymphoid
neoplasms in mice. Anticancer Res. 22: 883–888.
Rodriguez-Amaya, D. B. (1997). Carotenoids and food preparation: the retention of
provitamin a carotenóides in prepared, processed, and stored foods. Arlington, U.S.
Agency for International Development, p. 88.
Ros, E. (2000). Intestinal absorption of triglyceride and cholesterol. Dietary and
pharmacological inhibition to reduce cardiovascular risk. Atherosclerosis. 151: 357-379.
Ross, A. C. (2003). Retinoid prodution and catabolism: role of diet in regulating retinal
esterefication and retinoic acid oxidation. Journal of Nutrition. 133: 291-296.
Rossoni Jr, J. V.; Araújo, G. R.; Pádua, B. C.; Chaves, M. M.; Pedrosa, M. L.; Silva,
M. E.; Costa, D. C. (2011). Annatto extract and -carotene modulate the production of
reactive oxygen species/nitric oxide in neutrophils from diabetic rats. J Clin Biochem
Nutr. 50(3): 177-183.
Rudel, L.; Deckelman, C.; Wilson, M.; Scobey, M.; Anderson, R. (1994). Dietary
cholesterol and downregulation of cholesterol 7a-hydrox- ylase and cholesterol
absorption in African green monkeys, J. Clin. Invest. 93: 2463-2472.

SILVA, L.S. Referências Bibliográficas
86
Rudling, M. (1992). Hepatic mRNA levels for the LDL receptor and HMG CoA
reductase show coordinate regulation in vivo. J Lipid Res. 33:493-501.
Santos, R. D.; Pereira, A.; Neto, A. A.; Souza, A. D.; Avezum A.; Faludi, A. A.;
Spósito, A. C.; Loures-Vale A. A.; Lottenberg, A. M.; Chagas, A. C. P.; Mansur, A. P.;
Guimarães, A.; Duncan, B.; Caramelli, B.; Carvalho, C. C.; Polanczyk, C.; Scherr, C. ;
Armaganijan, D. ; Dante, M.G.; Quintão, E.; Mello, E. P.; Nakandakare, E.; Oppi, E.;
Cotta, E. F.; Janovich, E.; Moriguchi, E.; Fonseca, F. H.; Picheth, G. ; Dioguardi, G.;
Rosito, G.; Xavier, H. T.; Lopes, I. L.; Cruz, I.; Issa, J. S.; Diament, J.; Ramires, J. F.
A.; Basques, J. C.; Lima, J. C.; Nicolau, C.; Krieger, J. E.; Santos, J. E.; Galvão, J. J.;
Aldrighi, J. A; Novazzi, J. P.; Almeida, L. P.; Bricarello, L. P.; Rabelo, L.; Uint, L.;
César, L. M. C.; Bodanese, L. C.; Introcaso, L.; Izar, M. C.; Magalhães, M. C.; Catelli,
M. H.; Scartezini, M.; Bertolami, M. C.; Malachias, M. C.; Batlouni, M.; Ghorayeb, N.;
Clausell, N.; Forti, N.; Garcia, F. S.; Gebara, O.; Coelho, O. C.; Luz, P. L.; Maranhão,
R. C.; Stein, R.; Perim, R. C.; Giannini, S. D.; Kaiser, S.; Ihara, S.; Martinez, T.;
Carvalho, T.; Filho, W. S. (2001). III Diretrizes brasileiras sobre dislipidemias e diretriz
de prevenção da aterosclerose do departamento de aterosclerose da Sociedade Brasileira
de Cardiologia. Arquivos Brasileiros de Cardiologia. 77: 1-48.
Sato, R. (2010). Sterol metabolism and SREBP activation. Arch Biochem Biophys. 501:
177-181.
Sato, R.; Miyamoto, W.; Inoue, J.; Terada, T.; Imanaka, T.; Maeda, M. (1999). Sterol
regulatory element-binding protein negatively regulates microsomal triglyceride transfer
protein gene transcription. J Biol Chem. 274: 24714-24720.
Scallen, T. J.; Sanghvi A. (1983). Regulation of three key enzymes in cholesterol
metabolism by phosphorylation/dephosphorylation (3-hydroxy-3-methylglutaryl-
coenzyme A reductase/acyl-coenzyme A: cholesterol acyltransferase/cholesterol 7α-
hydroxylase/homeostasis). Proc. Nati. Acad. Sci. USA. 80: 2477-2480.
Schoonjans, K.; Staels, B.; Auwerx, J. J. (1996). Role of the peroxisome proliferator-
activated receptor (PPAR) in mediating the effects of fibrates and fatty acids on gene
expression. Lipid Res. 37(5): 907-925.
Schulman, I. G.; Heyman, R. A. (2006). LXR as a therapeutic target for atherosclerosis.
In: Packard CJ, Rader DJ, eds. Lipids and Atherosclerosis. 93-100.
Scoggan, K. A.; Gruber, H.; Chen, Q.; Plouffe, L. J.; Lefebvre, J. M.; Wang, B.;
Bertinato, J.; L'Abbé, M. R.; Hayward, S.; Ratnayake, W.M. N. (2009). Increased
incorporation of dietary plant sterols and cholesterol correlates with decreased
expression of hepatic and intestinal Abcg5 and Abcg8 in diabetic BB rats Journal of
Nutritional Biochemistry. 20: 177-186.
Scotter, M. J. (1995). Characterization of the Coloured Thermal Degradation Products
of Bixin from Annatto and a Revised Mechanism for their Formation. Food Chemistry.
53(.2): 177-185.

SILVA, L.S. Referências Bibliográficas
87
Shaish, A.; Daugherty, A.; O’Sullivan, F.; Schonfeld, G.; Heinecke, J.W. (1995). Beta-
carotene inhibits atherosclerosis in hypercholesterolemic rabbits. J. Clin. Invest.
96:2075-2082.
Shibata S.; Hayakawa K.; Egashira, Y.; Sanada, H. (2007). Roles of nuclear receptors in
the up-regulation of hepatic cholesterol 7α-hydroxylase by cholestyramine in rats. Life
Sciences. 80: 546-553.
a) Shibata, S.; Hayakawa, K.; Egashira, Y.; Sanada, H. (2007). Hypocholesterolemic
mechanism of chlorella: chlorella and its indigestible fraction enhance hepatic
cholesterol catabolism through up-regulation of cholesterol 7alpha- hydroxylase in rats.
Biosci Biotechnol Biochem.71: 916-25.
Shih, C. K.; Chang, J. H.; Yang, S. H.; Chou, T. W.; Cheng, H. H. (2008). -Carotene
and canthaxanthin alter the pro-oxidation and antioxidation balance in rats fed a high-
cholesterol and high-fat diet. Br J Nutr. 99:59-66.
Shilpi, J.A.; Rahman, T.U.; Uddin, S.J.; Shahanur, A.; Sadhu, S.K.; Véronique, S.
(2006). Preliminary pharmacological screening of Bixa orellana L. leaves. Journal of
Ethnopharmacology. 108: 264-271.
Silva, C. R.; Antunes, L. M. G.; Bianchi, M. L. (2001). Antioxidant action of bixin
against cisplatin-induced chromosome aberrations and lipid peroxidation in rats.
Pharmacol. Res. 43(6): 561-566.
Souza, M. O.; Silva, M.; Silva, M. E. Oliveira, R.P.; Pedrosa, M. L. (2010). Diet
supplementation with acai (Euterpe oleracea Mart.) pulp improves biomarkers of
oxidative stress and the serum lipid profile in rats. Nutrition. 26: 804-810.
Stahl, W.; Schwarz, W.; Sundquist, A. R.; Sies, H. (1992). Cis-trans isomers of
lycopene and beta-carotene in human serum and tissues. Arch Biochem Biophys. 294:
173-177.
Stahl, W.; Sies, H. (1996). Lycopene: a biologically important carotenoid for humans?
Arch Biochem Biophys. 336: 1–9.
Stampfer, M. J.; Hennekens, C. H.; Manson, J. E.; Colditz, G. A.; Rosner, B.; Willett,
W. C. (1993) Vitamin E consumption and the risk of coronary disease in women. N
Engl J Med. 328: 1444-1449.
Sulli, K. C.; Sun, J.; Giraud, D. W.; Moxley, R. A.;.Driskell, J. A. (1998). Effects of b-
carotene and a-tocopherol on the levels of tissue cholesterol and triglyceride in
hypercholesterolemic rabbits. J. Nutr. Biochem. 9: 344–350.
Takahashi, N,; Goto, T.; Taimatsu, A.; Egawa, K.; Katoh, S.; Kusudo, T.; Sakamoto, T.;
Ohyane, C.; Lee, J. Y.; Kim, Y.; Uemura, T.; Hirai, S.; Kawada, T. (2009). Bixin
regulates mRNA expression involved in adipogenesis and enhances insulin sensitivity

SILVA, L.S. Referências Bibliográficas
88
in 3T3-L1 adipocytes through PPAR activation. Biochem Biophys Res Commun. 390:
1372-1376.
Thomás-Moyá, E.; Gianotti, M.; Lladó, I.; Proenza, A.M. (2006). Effects of caloric
restriction and gender on rat serum paraoxonase 1 activity. Journal of Nutritional
Biochemistry, 17: 197-203.
Tsai, A. C.; Mazeedi, H. A. (1988) Effects of beta-carotene supplementation on serum
lipid levels in spontaneously hypertensive (SH) rats. FASEB J .2: A854.
Tsai, A. C.; Mazeedi, H. A.; Mameesh, M. S. (1992). Dietary betacarotene reduces
serum lipid concentration in spontaneously hypertensive rats fed a vitamin A-fortified
and cholesterolenriched diet. J Nutr. 122: 1768–1771.
Turbino-Ribeiro, S. M. L.; Silva, M. E.; Chianca, D. A. Jr.; de Paula, H.; Cardoso, L.
M.; Colombari, E.; Pedrosa, M. L. (2003). Iron overload in hypercholesterolemic rats
affects iron homeostasis and serum lipids but no blood pressure. J Nutr.133: 15-20.
Uenojo, M.; Maróstica Jr., M. R.; Pastore G. M. (2007). Carotenóides:propriedades,
aplicações e biotransformação para formação de compostos de aroma. Quim Nova. 3:
616-622.
Valko, M.; Izakovic, M.; Mazur, M.; Rhodes, C. J.; Telser, J. (2004). Role of oxygen
radicals in DNA damage and cancer incidence. Molecular and Cellular Biochemistry.
266:37–56.
Van Het Hof, K. H.; West, C. E.; Weststrate, J. A.; Hautvast, J. G. (2000). Dietary
factors that affect the bioavailability of carotenoids. J Nutr. 130: 503-506.
Van Vliet, T. (1996). Absorption of -carotene and other carotenoids in humans and
animal models. Eur J Clin Nutr. 50 (3): 32S-37S.
Wang, D. Q.; Tazuma, S. (2002). Effect of beta-muricholic acid on the prevention and
dissolution of cholesterol gallstones in C57L/J mice. J. Lipid Res. 43: 1960–1968.
Wang, X. D. (1994). Review: absorption and metabolism of - carotene. J Am Coll Nut.
13(4): 314-325.
Weber, L. W., Boll, M.E Stampfl, A. (2004). Maintaining cholesterol homeostasis:
sterol regulatory element-binding proteins. World J.Gastroenterol. 10(21): 3081-3087.
Weitberg, A. B.; Corvese, D. (1997). Effect of vitamin E and beta-carotene on DNA
strand breakage induced by tobacco-specific nitrosamines and stimulated human
phagocytes. J Exp Clin Cancer Res. 16(1): 11-14.
Wenniger, L. M. B.; Beuers, U. (2010). Bile salts and cholestasis. Dig. Liver Dis. 42:
409-418.

SILVA, L.S. Referências Bibliográficas
89
Whittaker, P.; Wamer, W. G.; Chanderbhan, R. F.; Dunkel, V. C. (1996). Effects of
alpha-tocopherol and beta- carotene on hepatic lipid peroxidation and blood lipids in
rats with dietary iron overload. Nutr Cancer. 25: 119-128.
Willis, M.S.; Wians, F. H. Jr. (2003). The role of nutrition in preventing prostate
cancer: a review of the proposed mechanism of action of various dietary substances.
Clin Chim Acta. 330(1-2): 57-83.
Wilson, M. D.; Rudel, L. L. (1994). Review of cholesterol absorption with emphasis on
dietary and biliary cholesterol. J Lipid Res. 35: 943-955.
Wójcicka, G.; Wiśniewska, A. J.; Horoszewicz, K.; Bełtowski, J. (2007). Liver X
receptors (LXRs). Part I: Structure, function, regulation of activity, and role in lipid
metabolism. Postepy Hig Med Dosw. 61: 736-759.
Woods, J. A.; Bilton, R. F.; Young, A. J. (1999). Beta-carotene enhances hydrogen
peroxide-induced DNA damage in human hepatocellular HepG2 cells. FEBS Lett.
449(2-3): 255-258.
Xiong, Q.; Xie, P.; Li, H.; Hao, L.; Li, G.; Qiu, T.; Liu, Y. (2010). Acute effects of
microcystins exposure on the transcription of antioxidant enzyme genes in three organs
(liver, kidney, and testis) of male wistar rats. J Biochem Mol Toxicol. 24: 361-367.
Xu, G.; Pan, L.; Li, H.; Shang, Q.; Honda, A.; Shefer, S.; Bollineni, J.; Matsuzaki, Y.;
Tint, G. S.; Salen, G. (2004). Dietary cholesterol stimulates CYP7A1 in rats because
farnesoid X receptor is not activated. Am J Physiol Gastrointest Liver Physiol. 286:
730-735.
Yang, J. L.; Kim, Y. H.; Lee, H. S.;Lee, M. S.; Moon, Y. K.( 2003). Barley β-glucan
lowers serum cholesterol based on the up-regulation of cholesterol 7α-hydroxylase
activity and mRNA abundance in cholesterol- fed rats. J. Nutr. Sci. Vitaminol. 49: 381-
387.
Yeon, J.Y.; Kim, H.; Sung, M. (2012). Diets rich in fruits and vegetables suppress blood
biomarkers of metabolic stress in overweight women. Prev. Med.
doi:10.1016/j.ypmed.2011.12.026
Yiu, W. F.; Kwan, P. L.; Wong, C. H.; Kam, T. S.; Chiu, S. M.; Chan, S. W.; Chan, R.
(2011). Attenuation of fatty liver and prevention of hypercholesterolemia by extract of
Curcuma longa through regulating the expression of CYP7A1, LDL-receptor, HO-1,
and HMG-CoA reductase. J Food Sci. 76(3): 80-89.
Yu L.; Hammer, R. E.; Li-Hawkins, J.; Von Bergmann, K.; Lutjohann, D.; Cohen, J. C.
(2002). Disruption of Abcg5 and Abcg8 in mice reveals their crucial role in biliary
cholesterol secretion. Proc Natl Acad Sci USA. 99: .16237-16242.
Yu, L.; Gupta, S.; Xu, F.; Liverman, A. D.; Moschetta, A.; Mangelsdorf, D. J. (2005).
Expression of ABCG5 and ABCG8 is required for regulation of biliary cholesterol
secretion. J Biol Chem. 280: 8742-8747.

SILVA, L.S. Referências Bibliográficas
90
Yu, L.; York, J.; von Bergmann, K.; Lutjohann, D.; Cohen, J. C.; Hobbs, H. H. (2003).
Stimulation of cholesterol excretion by the liver X receptor agonist requires ATP-
binding cassette transporters G5 and G8. J Biol Chem. 278(18): 15565-15570.
Yuan, Y V.; Kitts, D. D. (2002). Dietary source and cholesterol interactions alter plasma
lipids and tissue susceptibility to oxidation in spontaneously hypertensive (SHR) and
normotensive Wistar Kyoto (WKY) rats. Mol Cell Biochem. 232: 33-47.
Yuan, Y. V.; Kitts, D. D.; Godin, D. V. (1998.) Variations in dietary fat and cholesterol
intakes modify antioxidant status of SHR and WKY rats. J Nutr 128: 1620-1630.
Zhang, L. X.; Cooney, R. V.; Bertram, J. S. (1991). Carotenoids enhance gap junctional
communication and inhibit lipid peroxidation in C3H/10T1/2 cells: relationship to their
cancer chemopreventive action. Carcinogenesis. 12: 2109-2114.
Zhang, P.; Omaye, S. T. (2000). Beta-carotene and protein oxidation: effects of ascorbic
acid and alpha- tocopherol. Toxicology. 146(1): 37-47.
Zhang, Y.; Breevoort, S. R.; Angdisen, J.; Fu, M.;. Schmidt, D. R; Holmstrom, S. R.;
Kliewer, S. A. Mangelsdorf, D. J.; Schulman, I. G. (2012). Liver LXRα expression is
crucial for whole body cholesterol homeostasis and reverse cholesterol transport in
mice. J Clin Invest. 122(5): 1688-1699.


SILVA, L.S. Anexos
92
ANEXO I – Certificado de Análise – Beta Caroteno 10%
SILVA, L.S. Anexos
93
ANEXO II – Fluxograma de Produção – Produto PA3 Colorífico

SILVA, L.S. Anexos
94
ANEXO III – Protocolo das Dosagens Bioquímicas
Colesterol HDL
Princípio
As lipoproteínas de muita baixa densidade (VLDL) e as lipoproteínas de baixa
densidade (LDL) são quantitativamente precipitadas e, após centrifugação, o colesterol
ligado às lipoproteínas de alta densidade (Colesterol HDL) é determinado no
sobrenadante.
Amostra
O jejum não é imprescindível para a dosagem de colesterol HDL. Usar soro. O analito é
estável por 3 dias entre 2 – 8ºC.
Produto Utilizado
Colesterol HDL, Catálogo 13 - ANVISA – 10009010026
Labtest Diagnóstica: Av. Paulo Ferreira da Costa, 600. Lagoa Santa, MG, 33400-000
Precipitante: Armazenar entre 2 – 8°C. Contém ácido fosfotúngstico 1,5 mmol/L e
cloreto de magnésio 54 mmol/L.
Padrão - 20 mg/dL: Armazenar entre 2 – 8°C. Armazenar bem vedado para evitar
evaporação.
Equipamentos
1. Centrífuga para tubos.
2. Fotômetro capaz de medir com exatidão a absorbância em 500 nm ou filtro verde
(490 a 510 nm).
3. Banho-maria mantido à temperatura constante (37 ºC).
4. Pipetas para medir amostras e reagentes.
5. Cronômetro.
Procedimento
Precipitação das VLDL e LDL
Em um tubo 12 x 75 colocar 0,25mL de soro e 0,25mL de precipitante. Agitar
vigorosamente durante 30 segundos. A agitação sugerida é fundamental para obtenção
de resultados consistentes. Centrifugar a 3.500 rpm por pelo menos 15 minutos para
obter um sobrenadante límpido. Pipetar o sobrenadante límpido imediatamente após a
centrifugação, tomando o cuidado para não ressuspender o precipitado, a fim de evitar
resultados falsamente elevados.
Utilizar com o Reagente 1 - Colesterol Liquiform Labtest Cat. 76. Tomar 3 tubos de
ensaio e proceder como a seguir:

SILVA, L.S. Anexos
95
Branco Teste Padrão
Sobrenadante ----- 0,1 mL -----
Padrão (nº 2) ----- ----- 0,1 mL
Reagente 1 1,0 mL 1,0 mL 1,0 mL
Misturar e colocar no banho-maria a 37 ºC durante 10 minutos. O nível da água no
banho deve ser superior ao nível do reagente nos tubos de ensaio. Determinar as
absorbâncias do teste e padrão em 500 nm ou filtro verde (490 a 540 nm) acertando o
zero com o branco. A cor é estável por 60 minutos.
Cálculos
HDL mg/dL =
Conversão de mg/dL para Unidade SI: mmol/L = mg/dL x 0,026
Colesterol Total
Princípio
O colesterol total é determinado de acordo com as seguintes reações:
Ésteres de colesterol Colesterol + Ácidos graxos
Colesterol + O2 Colest-4-en-ona + H2O2
2H2O2 + Fenol + 4-Aminoantipirina Antipirilquinonimina + 4H2O
A intensidade da cor vermelha formada é diretamente proporcional à concentração de
colesterol na amostra.
Amostra
O jejum não é imprescindível para a dosagem de colesterol total. Usar soro.
Anticoagulantes como citrato, oxalato ou EDTA produzem resultados falsamente
diminuídos. O analito é estável por 7 dias entre 2 – 8 ºC e vários meses a 10 ºC
negativos.
Produto Utilizado
Colesterol Liquiform, Catálogo 76-2/100 - ANVISA - 10009010068
Labtest Diagnóstica: Av. Paulo Ferreira da Costa, 600. Lagoa Santa, MG, 33400-000
Reagente 1: Armazenar entre 2 – 8°C. Contém tampão 50 mmol, pH 7,0, fenol 24,0
mmol/L, colato de sódio 500 µmol/L, azida sódica 15 mmol/L, 4 aminoantipirina 500
µmol/L, colesterol esterase ≥250 U/L, colesterol oxidase ≥250 U/L e peroxidase ≥1000
U/L.

SILVA, L.S. Anexos
96
Padrão 200 mg/dL: Armazenar entre 2 – 8°C. Após o manuseio armazenar bem vedado
para evitar evaporação. Contém azida sódica 15 mmol/L.
Equipamentos
1. Fotômetro capaz de medir com exatidão a absorbância em 500 nm ou filtro verde
(490 a 510 nm).
2. Banho-maria mantido à temperatura constante (37 ºC).
3. Pipetas para medir amostras e reagentes.
4. Cronômetro.
Procedimento
Tomar 3 tubos de ensaio e proceder como a seguir:
Branco Teste Padrão
Amostra ----- 0,01 mL -----
Padrão (nº 2) ----- ----- 0,01 mL
Reagente 1 1,0 mL 1,0 mL 1,0 mL
Misturar e colocar em banho-maria 37 ºC 10 minutos. O nível de água no banho deve
ser superior ao nível de reagentes nos tubos de ensaio. Determinar as absorbâncias do
teste e padrão em 500 nm ou filtro verde (490 a 510 nm), acertando o zero com o
branco. A cor é estável por 60 minutos.
Cálculos
Colesterol mg/dL=
Conversão de mg/dL para Unidade SI: mmol/L = mg/dL x 0,026
Triglicérides
Princípio
Os triglicérides são determinados de acordo com as seguintes reações:
Triglicérides Glicerol + Ácidos Graxos
Glicerol + ATP Glicerol-3-Fosfato + ADP
Glicerol-3-Fosfato + O2 Dihidroxiacetona + H2O2
2 H2O2 + 4 Aminoantipirina + 4-Clorofenol Quinoneimina + 4 H2O

SILVA, L.S. Anexos
97
A intensidade da cor vermelha formada é diretamente proporcional à concentração de
triglicérides na amostra.
Amostra
Jejum de 12 a 14 horas. Usar soro ou plasma com EDTA. O analito é estável por dois
dias entre 2 – 8°C. Armazenamento prolongado da amostra não é recomendado, porque
várias substâncias podem ser hidrolizadas liberando glicerol, levando à obtenção de
resultados falsamente elevados. A heparina promove a ativação in vivo ou in vitro da
lipase da lipoproteína, fazendo com que a concentração dos triglicérides se reduza
gradativamente em amostras contendo heparina.
Produto Utilizado
Triglicérides Liquiform, Catálogo 87 - ANVISA - 10009010070
Labtest Diagnóstica: Av. Paulo Ferreira da Costa, 600. Lagoa Santa, MG, 33400-000
Reagente 1 - Armazenar entre 2 - 8 ºC. Contem tampão 50 mmol/L, pH 6,9; acetato de
magnésio 4 mmol/L; 4-clorofenol 5 mmol/L; 4-aminoantipirina 300 mol/L ATP 1,0
mmol/L; lipase da lipoproteína 1400 U/L; glicerolquinase 1000 U/L e azida sódica 7
mmol/L.
Padrão - 200 mg/dL - Armazenar entre 2 - 30 ºC. Contém triglicérides 200 mg/dL e
azida sódica 7 mmol/L.Após o manuseio sugere-se armazenar bem vedado, para evitar
evaporação.
Equipamentos
1. Fotômetro capaz de medir com exatidão a absorbância em 505 nm ou filtro verde
(490 a 520nm).
2. Banho-maria mantido à temperatura constante (37°C).
3. Pipetas para medir amostras e reagentes.
4. Cronômetro.
Procedimento
Tomar 3 tubos de ensaio e proceder como a seguir:
Branco Teste Padrão
Amostra ----- 0,01 mL -----
Padrão (nº 2) ----- ----- 0,01 mL
Reagente 1 (nº 1) 1,0 mL 1,0 mL 1,0 mL
Misturar e colocar no banho-maria 37 ºC por 10 minutos. O nível da água no banho
deve ser superior ao nível do reagente nos tubos de ensaio. Determinar as absorbâncias
do teste e padrão em 505 nm ou filtro verde (490 a 520 nm) acertando o zero com o
branco. A cor é estável por 60 minutos.
Cálculos
Triglicérides (mg/dL) =
Conversão de mg/dL para Unidade SI: mmol/L = mg/dL x 0,0113
![Pathfinder - Classes [Beta]](https://static.fdocumentos.com/doc/165x107/5571f33449795947648da99a/pathfinder-classes-beta.jpg)



![Pathfinder - Combate [Beta]](https://static.fdocumentos.com/doc/165x107/5571f33449795947648da99d/pathfinder-combate-beta.jpg)













