
EFEITOS DO MODAFINIL SOBRE O APRENDIZADO E A … · 2017-12-16 · Efeitos do modafinil sobre o...
77
HELAINE ARRAIS FERNANDES EFEITOS DO MODAFINIL SOBRE O APRENDIZADO E A MEMÓRIA DE CAMUNDONGOS AVALIADOS NA ESQUIVA DISCRIMINATIVA EM LABIRINTO EM CRUZ ELEVADO São Paulo 2011 Tese Apresentada à Universidade Federal de São Paulo – Escola Paulista de Medicina para obtenção do Título de Mestre em Ciências
Transcript of EFEITOS DO MODAFINIL SOBRE O APRENDIZADO E A … · 2017-12-16 · Efeitos do modafinil sobre o...

HELAINE ARRAIS FERNANDES
EFEITOS DO MODAFINIL SOBRE O APRENDIZADO
E A MEMÓRIA DE CAMUNDONGOS AVALIADOS
NA ESQUIVA DISCRIMINATIVA EM LABIRINTO EM
CRUZ ELEVADO
São Paulo
2011
Tese Apresentada à Universidade
Federal de São Paulo – Escola
Paulista de Medicina para obtenção
do Título de Mestre em Ciências

Fernandes, Helaine Arrais Efeitos do modafinil sobre o aprendizado e a memória de camundongos avaliados na esquiva discriminativa em labirinto em cruz elevado/ Helaine Arrais Fernandes. – São Paulo, 2011. x, 67p. Tese (Mestrado) - Universidade Federal de São Paulo. Escola Paulista de Medicina. Programa de Pós-Graduação em Psicobiologia.
Título em inglês: Effects of modafinil on learning and memory of mice evaluated in the plus-maze discriminative avoidance task in mice. 1. Modafinil. 2. Memória. 3. Aprendizado. 4. Ansiedade. 5. Locomoção.

HELAINE ARRAIS FERNANDES
EFEITOS DO MODAFINIL SOBRE O APRENDIZADO
E A MEMÓRIA DE CAMUNDONGOS AVALIADOS
NA ESQUIVA DISCRIMINATIVA EM LABIRINTO EM
CRUZ ELEVADO
Orientador: Prof. Dr. ROBERTO FRUSSA FILHO
Co-orientadora: Dra. LIA RITA AZEREDO BITTENCOURT
São Paulo
2011
Tese Apresentada à Universidade
Federal de São Paulo – Escola
Paulista de Medicina para obtenção
do Título de Mestre em Ciências

UNIVERSIDADE FEDERAL DE SÃO PAULO
ESCOLA PAULISTA DE MEDICINA
PROGRAMA DE PÓS-GRADUAÇÃO EM PSICOBIOLOGIA
Chefe do Departamento de Psicobiologia Profa. Dra. Maria Lucia Oliveira de Souza Formigoni Coordenador do Programa de Pós-Graduação em Psicobiologia Prof. Dr. Marco Túlio de Mello

UNIVERSIDADE FEDERAL DE SÃO PAULO
ESCOLA PAULISTA DE MEDICINA
PROGRAMA DE PÓS-GRADUAÇÃO EM PSICOBIOLOGIA
Prof. Dra. Dalva Lucia Rollemberg Poyares
Prof. Dra. Maria Martha Bernardi
Profa. Dra. Regina Helena da Silva
Profa. Dra. Maria Gabriela Menezes de Oliveira

Esta tese foi realizada nos Departamentos de Farmacologia e Psicobiologia
da Universidade Federal de São Paulo – Escola Paulista de Medicina, com o
apoio financeiro da Associação Fundo de Incentivo à Psicofarmacologia (AFIP) e
do Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq).

v
”Não busques a vaidade de ser melhor que os outros. Contenta-te com a tarefa gloriosa de tentares ser melhor que és. Que tu não
sejas o teu limite de crescimento, mas o teu grande questionamento, o teu grande interrogador”.
(Henrique de Souza Filho - Henfil)

vi
Agradecimentos
Foi há 2 anos que comecei algo... Algo que agora apresento a vocês. E, ao
apresentá-lo sinto um sentimento de alívio, como há muito não sentia!
Senti que brinquei com minha imaginação, meu conhecimento, minha
capacidade... Pode-se dizer que tive muitos momentos difíceis até alcançar meu
objetivo final: a dissertação.
Nesses 2 anos arrisquei-me em uma nova experiência, em um novo
trabalho, em novos amigos. Não sei se o que sinto hoje é diferente de antes...
Continuo assombrada pelo medo do novo, mas gostei e muito!
Gostei até de aprender que existem pessoas que se identificam ou não,
que a pesquisa é difícil, mas surpreendente. Com certeza o que eu mais gostei foi
aprender que o esforço vale à pena! Sim, hoje eu sei!
Hoje eu sei que o mais importante para mim é a aprendizagem. Aprender o
que não sei ainda. Ah! E como existem coisas a serem aprendidas! E por isso
serei sempre uma aprendiz.
Eu creio que esta parte de agradecimentos é uma tarefa difícil, pois muitas
vezes cometemos injustiças e, por esquecimento, não mencionamos nomes de
pessoas que também contribuíram para o trabalho.
Nada na vida conquistamos sozinhos. Sempre precisamos de outras
pessoas para alcançar os nossos objetivos. Muitas vezes um simples gesto pode
mudar a nossa vida e contribuir para o nosso sucesso. E finalizada uma etapa
particularmente importante da minha vida, não poderia deixar de expressar o mais
profundo agradecimento a todos àqueles que me apoiaram nessa longa
caminhada e contribuíram para a realização deste trabalho.
Primeiramente, devo agradecer ao Conselho de Desenvolvimento
Científico e Tecnológico (CNPq) pelo apoio financeiro, o que possibilitou que eu
desenvolvesse um novo trabalho.
Meus agradecimentos ao meu orientador, Prof. Dr. Roberto Frussa Filho,
pela oportunidade oferecida, pela competência científica e acompanhamento do
trabalho, pela disponibilidade e generosidade reveladas ao longo desses anos de
trabalho, assim como pelas críticas, correções e sugestões relevantes feitas
durante a orientação.

vii
À Prof.Dra. Lia Rita Azeredo Bittencourt, minha co-orientadora, pela
competência científica, por ter fornecido o modafinil, e a disponibilidade sempre
manifestada.
Aos membros da banca, por terem aceitado a participar da avaliação deste
trabalho.
Eu fortemente agradeço ao Prof. Dr. Sérgio Tufik e a Profa. Dra. Monica
Andersen, pelo conhecimento adquirido nos cursos e reuniões do sono e pela
oportunidade e apoio nos congressos e eventos científicos.
A Camilla e a Karina por todo apoio em congressos e desenvolvimento da
tese.
Em especial às “Organizações Marinho”: Ao meu pai científico, Prof.
Eduardo Marinho, por ter me apresentado à Pesquisa Científica, por todo apoio e
disposição em decifrar minhas dúvidas, ao Alexandre Justos, um ser humano
como nenhum outro, com um coração enorme e sempre palavras certas em
momentos difíceis, ao Wuo pelas sugestões e apoio bibliográfico. Ao Renan,
Marília, André, Aline, Fran, Thaís e Samira pela alegre convivência e momentos
inovadores, alguns desses foram essenciais no desenvolvimento deste trabalho.
A vocês, muito obrigada!
Um agradecimento especial deve ser feito à Teotila e ao Cleomar, por
possuírem excelente técnica, sempre nos auxiliando nos experimentos e na
obtenção dos mais variados materiais, por sempre se preocuparem com a ordem
do laboratório e pelo ótimo humor.
Às minhas queridas amigas: Liliane e Luciana, pela amizade sólida e
verdadeira por todos os momentos divertidos que passamos juntas. Ao amigo
Luciano, por estar sempre disposto a ajudar e ser um amigo para todas as horas
e à amiga Fernanda pela companhia, apoio e carinho.
Aos meus familiares que sempre me apoiaram. Aos meus pais, Nice e
Raimundo. Aos meus irmãos Andressa, Tatiane e Henrique por sempre torcerem
por mim e ao meu marido Panda por sua extensa paciência, pelo seu amor, por
sempre estar disposto a me ajudar em qualquer situação e principalmente pelo
seu apoio que me conforta e me deixa mais forte para superar meus desafios.
A todos, o meu Eterno Obrigada.

viii
RESUMO
O modafinil, um psicoestimulante utilizado no tratamento da narcolepsia, da
sonolência excessiva durante o dia e da sonolência induzida pela síndrome da
apnéia obstrutiva do sono, parece ser capaz de promover um efeito facilitador de
funções cognitivas. Tal efeito facilitador tem sido buscado pelas sociedades atuais
frente à crescente velocidade do fluxo de informações e à pressão social de
produtividade. Nesse cenário, verificamos os efeitos da administração aguda de
modafinil (32, 64 ou 128 mg/Kg, i.p.) sobre o aprendizado, a consolidação e a
evocação de uma memória discriminativa aversiva, bem como os efeitos da
administração (32, 64 ou 128 mg/Kg por 10 dias, i.p.) sobre a consolidação e a
evocação dessa memória em camundongos. A possível participação do fenômeno
de dependência de estado também foi investigada. Para tanto, utilizamos o
modelo da esquiva discriminativa em labirinto em cruz elevado, que avalia
concomitantemente o aprendizado, a memória, os níveis de ansiedade e a
locomoção de roedores. A administração aguda de modafinil nas doses mais
elevadas (64 ou 128 mg/Kg) prejudicou a consolidação da memória discriminativa
e a habituação ao aparelho, uma memória não-associativa. Por outro lado, tais
déficits parecem ser revertidos com a administração repetida dessa droga. Ainda,
a administração aguda pré-treino ou pré-teste de 32 mg/Kg de modafinil
promoveu déficits de retenção e evocação da tarefa discriminativa,
respectivamente. De importância, tais déficits não foram dependentes de estado.
Ainda, a dose de 64 mg/Kg também promoveu déficits de retenção quando
administrada previamente ao treino. Tais achados não foram relacionados a
alterações nos níveis de emocionalidade dos animais. Tomados em conjunto, os
presentes achados fornecem evidências pré-clínicas do potencial amnéstico do
modafinil.

ix
ABSTRACT
Modafinil, a psychostimulant used in the treatment of narcolepsy, excessive
daytime sleepiness and obstructive-sleep-apnea-induced sleepiness, seems to
induce facilitatory effects on cognition. In this scenario, we investigated the effects
of acute administration of modafinil (32, 64 or 128 mg/Kg, i.p.) on learning,
consolidation and retrieval of a discriminative memory in mice as well as the
effects of chronic administration (32, 64 or 128 mg/Kg for 10 days, i.p.) of this drug
on the consolidation and retrieval of this memory. The possible role of state-
dependency phenomenon was also evaluated. The plus-maze discriminative
avoidance task, an animal model that evaluates concomitantly (but independently)
learning, memory, anxiety-like behavior and motor activity was employed in the
study. In this way, acute administration of the higher doses of modafinil (64 or 128
mg/Kg) induced impairment in the consolidation of the discriminative task and
habituation deficits, a non-associative memory. Conversely, these amnestic effects
were reversed when the drug was administered repeatedly. Moreover, the pre-
training and pre-test acute administration of the 32 mg/kg dose of modafinil
induced retention and retrieval impairments of the discriminative task. Importantly,
these deficits were not state-dependent. Finally, 64 mg/Kg modafinil promoted
retention deficits when administered previous to training. These cognitive effects
were not associated with anxiety alterations. Collectively, our results support pre-
clinical evidence of the potential amnestic effects of modafinil.

x
ÍNDICE
1. INTRODUÇÃO _________________________________________________1
1.1 Memória e ansiedade _______________________________________ 1
1.2 Modafinil _________________________________________________ 7
1.3 Modafinil, Memória e Ansiedade _____________________________ 11
2. OBJETIVOS___________________________________________________14
2.1 Objetivo Geral ___________________________________________ 14
2.2 Objetivos Específicos ______________________________________ 14
3. TRABALHO CIENTÍFICO A SER SUBMETIDO_______________________15
4. CONCLUSÕES_________________________________________________55
4.1 Conclusões Específicas ____________________________________ 55
4.2 Conclusão Geral _________________________________________ 56
5. REFERÊNCIAS BIBLIOGRÁFICAS________________________________57
6. ANEXO_______________________________________________________65

INTRODUÇÃO
1
1. INTRODUÇÃO
1.1 Memória e Ansiedade
Memória se refere ao processo mediante ao qual adquirimos, formamos,
conservamos e evocamos uma informação (Cammarota et al., 2008). A formação da
memória pode ser didaticamente dividida em três fases: o aprendizado, a
consolidação e a evocação. O aprendizado (registro ou aquisição) é o processo pelo
qual a experiência prática ou a observação permitem desenvolver modificações na
conduta que favorecem a adaptação ao meio ambiente. A consolidação (retenção ou
armazenamento) é o processo pelo qual a memória se torna crescentemente
resistente às interferências na ausência de treino, ao longo da simples passagem do
tempo (McGaugh, 2000). A evocação (recordação ou recuperação) refere-se ao
acesso da informação armazenada, podendo ser espontânea ou se dar pela busca
voluntária.
A memória também pode ser definida como a capacidade de reter e manipular
informações adquiridas anteriormente por meio da plasticidade neuronal (Thompson
et al., 2002). Ela pode ser classificada como implícita (ou não-declarativa) ou
explícita (ou declarativa), dependendo de como a informação é armazenada e
evocada; ou memória de curto- ou longo-prazos, dependendo de sua duração
(Thompson et al., 2002).
A memória implícita ou não-declarativa inclui o condicionamento clássico,
habilidades e hábitos, e é em grande parte ou, com freqüência, completamente

INTRODUÇÃO
2
inconsciente. A memória explícita ou declarativa, por outro lado, envolve a
recuperação consciente de eventos ou fatos que tenham ocorrido. Esses tipos de
memória são processados de formas diferentes e em diferentes regiões do cérebro,
embora estejam relacionados por meio de inúmeras conexões neurais (Thompson et
al., 2002).
Assim, o hipocampo é considerado o responsável pela conversão de memória
de curto-prazo em memória de longo-prazo e pela informação espacial, enquanto o
neocórtex, pelo armazenamento da memória de longo-prazo declarativa
(Jerusalinsky et al., 1997). Já o estriado está envolvido com a memória de
procedimento (Mishkin et al., 1984) e a amígdala parece ser especializada no
processo de alerta e informação aversiva e participaria da memória para eventos
com significado emocional (Sarter & Markowitsch, 1985; Davis, 1992; Cahill &
McGaugh, 1990). O septo medial também participa do processo de informação
aversiva, por um mecanismo distinto daquele da amígdala (Izquierdo & Medina,
1991) e o córtex entorrinal, que apresenta conexões mono e polissinápticas com a
amígdala, hipocampo e septo medial (Zola-Morgan et al., 1989; Witter et al., 1989;
Ferreira et al., 1992), processa informação espacial, aversiva, além de outras,
estando particularmente envolvido com a memória declarativa (Zola-morgan et al.,
1989; Witter et al., 1989; Ferreira et al., 1992).
Investigações farmacológicas fornecem base para um importante papel
exercido pela acetilcolina no processo de formação da memória, uma vez que um
efeito amnéstico geralmente é verificado após a administração de antagonistas
colinérgicos como a escopolamina (Weissman, 1967; Glick & Zimmerberg, 1971;

INTRODUÇÃO
3
Wiener & Messer, 1973; Givens & Olton, 1990; Silva et al, 1997). Além disso, o
tratamento com inibidores de colinesterase como a fisostigmina (Straton &
Petrinovich, 1963; Deutsch et al., 1979) é sabiamente estimulador da memória, assim
como o tratamento com agonistas colinérgicos diretos como a nicotina (Garg, 1969;
Battig, 1970; Evangelista et al., 1970; Erickson, 1971;) ou a oxotremonina (Baratti et
al., 1979). Além da relação entre a acetilcolina e a função cognitiva, outros
neurotransmissores como a serotonina (Steckler & Sahgal, 1995) e as catecolaminas
(Dismukes & Rake, 1972; Randt et al., 1971) parecem também participar de forma
relevante do processo de formação da memória.
Dentre os mecanismos celulares e moleculares propostos como base
mecanicística para os processos de aprendizado e memória, destacam-se a
sensibilização e a potenciação de longa duração (LTP – Long Term Potentiation).
A sensibilização consiste em um processo de facilitação pré-sináptica da
transmissão neuronal envolvida nas memórias de curto- e longo-prazos. É um
processo heterossináptico, no qual o aumento da força sináptica é induzido por
interneurônios moduladores ativados pela estimulação. Estes liberam serotonina e
outros neurotransmissores, que se acoplam a receptores transmembrânicos
específicos ligados à proteína Gs, ativando a adenilato ciclase, que catalisa a
conversão de ATP em AMPc, o qual ativa a proteína cinase A (PKA), causando,
aliada à proteína cinase C (PKC), o aumento da liberação de neurotransmissores nos
terminais sinápticos (Kandel, 2003).
A LTP é uma facilitação que consiste em um aumento da amplitude dos
potenciais pós-sinápticos excitatórios devido a uma série de estímulos em alta

INTRODUÇÃO
4
freqüência. Ela possui uma fase precoce transitória (duração de aproximadamente 1
a 3 horas e não requer síntese de novas proteínas e é induzida por apenas uma
série de estimulações) e uma fase tardia de consolidação (ocorre devido a 4 ou mais
séries de estimulações; com duração de, no mínimo 24 horas e síntese de novas
proteínas por meio da via de sinalização AMPc – PKA – MAPK – CREB) (Kandel,
2003).
Assim como a memória é muitas vezes essencial em comportamentos cruciais
para a sobrevivência de uma espécie – tais como busca de alimento, reprodução e
fuga – também o é a ansiedade, que possui suas raízes evolutivas nas reações de
medo dos animais e aparece ligada ao reflexo de fuga, sendo considerada como
uma manifestação do instinto de conservação (Graeff, 1984; 1993; Graeff et al.,
1993).
A ansiedade tem sido descrita como um estado emocional de grande valor
adaptativo, que é experienciado de maneira subjetiva como não prazeroso e
desconfortável e cuja expressão plena envolve alterações comportamentais,
psicofisiológicas e cognitivas (Graeff, 1984; Pratt, 1992).
As bases neurais que envolvem o fenômeno de ansiedade têm sido
amplamente investigadas. A amígdala via mediação serotonérgica, tem sido
apontada como uma das principais estruturas relacionadas ao medo e à ansiedade.
Da amígdala, partem vias que se projetam para o hipotálamo e a matéria cinzenta
periaquedutal. Além disso, o septo medial, o hipocampo e as projeções
noradrenérgicas e serotonérgicas ascendentes, provenientes respectivamente do
locus coeruleus e dos núcleos da rafe, também têm sido implicados nesse processo

INTRODUÇÃO
5
(Gray, 1982; Graeff, 1993). Neuroquimicamente, algumas teorias têm sido propostas
procurando relacionar possíveis disfunções do sistema GABAérgico, tais como níveis
dos ligantes endógenos diminuídos, ou anormalidades no funcionamento do
complexo receptor, com o desenvolvimento do quadro de ansiedade generalizada
(Teicher, 1988; Gorenstein et al., 1994).
A ansiedade pode existir tanto na forma patológica como ser normalmente
experienciada, mas o limite entre o que se denomina ansiedade normal e ansiedade
patológica é difícil de ser determinado (Nutt, 1990; Graeff, 1993). Até um
determinado nível, a ansiedade favorece o desempenho de tarefas motoras e
cognitivas, sendo assim considerada fisiológica, mas passa a ser patológica quando
passa a interferir no comportamento normal do indivíduo (Ashton, 1987; Graeff, 1993;
Silva & Frussa-Filho, 2000).
Muito do conhecimento existente sobre a ansiedade e o medo humano foi
obtido por meio de modelos animais, nos quais se procura reproduzir determinados
aspectos da sintomatologia, da etiologia ou do tratamento da ansiedade. Muitos
modelos de ansiedade estão sendo usados atualmente, sendo a grande maioria
baseada em respostas comportamentais de medo, aprendidas ou espontâneas, que
reproduziriam a ansiedade humana (Rodgers et al., 1997). Dentre os modelos que
envolvem comportamento espontâneo, que são baseados em medos inatos do
animal, podemos ressaltar o labirinto em cruz elevado, um dos modelos animais de
ansiedade mais utilizados (Rodgers et al., 1997). Esse modelo é derivado da
observação de que, quando colocados em labirintos constituídos de braços abertos e
fechados, ratos demonstram claramente preferência pelos braços fechados

INTRODUÇÃO
6
(Montgomery, 1955). A exposição de ratos a situações naturalmente ameaçadoras a
sua espécie, como espaços abertos, explicaria o medo pela exploração dos braços
abertos. O labirinto em cruz elevado foi validado fisiológica, farmacológica e
comportamentalmente como modelo animal de ansiedade. Respostas
comportamentais e fisiológicas indicativas de medo, como imobilidade, defecação e
aumento das concentrações plasmáticas de corticosterona foram observadas após o
confinamento de ratos nos braços abertos do labirinto (Pellow et al., 1985). Os
benzodiazepínicos aumentam seletivamente o número de entradas e o tempo de
permanência dos animais nos braços abertos do aparelho, em relação aos braços
fechados (Handley & Mithany, 1984; Pellow et al., 1985; Pellow & File, 1986).
Paralelamente, drogas ansiogênicas, como por exemplo a cafeína, promovem uma
diminuição desses parâmetros (Lister, 1987).
A memória e a ansiedade estão relacionadas pelo compartilhamento de
estruturas cerebrais (como, por exemplo, a amígdala e o sistema septo-hipocampal),
sendo ambos modulados, pelo menos em parte, pelos receptores GABAA (Izquierdo
& Medina, 1991). Nesse sentido, tem sido sugerido que a ansiedade e a memória
não seriam apenas fenômenos relacionados (Izquierdo & Medina, 1991; Davis et al.,
1997; Silva & Frussa-Filho, 2000), mas que a ansiedade seria, na verdade, um passo
necessário para que ocorresse a formação da memória (Mathews, 1990).
Considerando à estreita relação entre processos mnemônicos e a ansiedade,
bem como a possível interferência de alterações na função motora na quantificação
desses processos, foi desenvolvido, em nosso laboratório, um modelo de estudo
simultâneo de aprendizado/memória, ansiedade e atividade locomotora. Trata-se da

INTRODUÇÃO
7
esquiva discriminativa em labirinto em cruz elevado (Silva et al., 1997; Silva &
Frussa-Filho, 2000). Nesse modelo, realizado em um labirinto em cruz elevado
modificado, no qual o animal deve escolher entre dois braços fechados (um dos
quais aversivo – luz de 100 watts e um jato de ar frio proveniente de um secador de
cabelos) ao mesmo tempo que evita dois braços abertos. Além do estudo da
interação entre memória (quantificada pela preferência pelo braço fechado não-
aversivo em relação ao braço fechado aversivo na sessão de teste) e ansiedade
(avaliada pela evitação aos braços abertos), esse modelo ainda tem a vantagem de
avaliar alterações na locomoção (por meio da quantificação do número de entradas
em todos os braços do aparelho). Tal característica se faz importante tanto para o
estudo de efeitos de drogas (Silva et al., 2002a; Carvalho et al., 2006; Kameda et al.,
2007; Niigaki et al., 2010; Sanday et al., dados submetidos), quanto de
procedimentos experimentais (Castro et al., 2005; Alvarenga et al., 2008) que
conhecidamente alteram a função motora sobre os processos de aprendizado e
memória. Além disso, possibilita a avaliação de drogas com potencial analgésico,
como a morfina sobre os processos cognitivos, como a morfina (Patti et al., 2006),
pois os estímulos aversivos aplicados não parecem promover modificações nos
sistemas de percepção da dor.
1.2 Modafinil
Modafinil [2- (Difenilmetil-sulfinil) acetamida] é um agente que promove um
aumento do estado de alerta. Foi desenvolvido na França por volta de 1990,
especificamente para o tratamento da sonolência diurna excessiva relacionada à

INTRODUÇÃO
8
narcolepsia, para o tratamento do transtorno do sono relacionado à mudança de
turno no trabalho e para a síndrome da apneia/hipopneia obstrutiva do sono (Ferraro
et al, 2001; Silvestri et al., 2002; Ward et al., 2004; Ballon & Feifel, 2006; Minzenberg
& Carter, 2008). Além disso, atualmente o modafinil tem sido utilizado como
tratamento de uma série de outras patologias importantes, como o transtorno de
hiperatividade e déficit de atenção, na sonolência e fadiga relacionadas à doença de
Parkinson e à esclerose múltipla, sedação pós-anestésica, depressão, sintomas
negativos da esquizofrenia e nos déficits cognitivos causados pela doença de
Alzheimer (Ward et al., 2004; Dopheide et al., 2007; Korotkova et al., 2007; Morgan
et al., 2007).
Apesar de suas diversas aplicações, mesmo sendo da mesma classe dos
psicoestimulantes, seus efeitos e suas vias de ação diferem de outras drogas da
mesma classe, como a anfetamina e a cocaína, por exemplo. Dessa forma, o
modafinil parece não promover efeitos colaterais semelhantes a esses
psicoestimulantes, como tolerância e dependência, hipersonolência rebote e efeitos
adversos cardiovasculares (Morgan et al., 2007; Minzenberg & Carter, 2008).
Diversas são as hipóteses que tentam explicar os mecanismos de ação do
modafinil. Estudos têm sugerido que ele pode aumentar a liberação de
catecolaminas (dopamina e noradrenalina), serotonina e glutamato, diminuir a
liberação de GABA em diversas regiões cerebrais, além de ativar neurônios
hipotalâmicos contendo hipocretina/orexina e neurônios do núcleo tuberomamilar
contendo histamina (Minzenberg & Carter, 2008).
A ação do modafinil no sistema dopaminérgico é um tanto controversa. De
fato, diversos estudos demonstram que o modafinil aumenta a atividade motora em

INTRODUÇÃO
9
camundongos, a atividade noturna em macacos e o despertar em gatos, e que esses
efeitos são diminuídos pelo antagonista α1 adrenérgico prazosina, mas não são
afetados pelos antagonistas do receptor D2 sulpiride e haloperidol e nem pelo
antagonista do receptor D1 SCH 23390 (Duteil et al., 1990; Hermant et al., 1991; Lin
et al.; 1992; Tanganelli et al., 1992). Em um estudo realizado por De Séréville e
colaboradores (1994), a análise eletroquímica demonstrou que o modafinil não
interage com o sistema dopaminérgico nigroestriatal em camundongos. Ainda,
parece não alterar o pool citosólico dopaminérgico no estriado de camundongos
(Simon et al.; 1995), além de ter pouca afinidade pelo transportador de dopamina e
pouco afetar a liberação de dopamina no núcleo accumbens (Rush et al., 2002) e no
sistema dopaminérgico nigroestriatal (Ishizuka et al., 2008). Em um estudo realizado
por Lin e colaboradores (1996) foi demonstrado por meio de uma análise imuno-
histoquímica utilizando a proteína c-fos, que o modafinil age principalmente em
neurônios do hipotálamo anterior, mas não em neurônios estriatais ou corticais, nos
quais se encontram os neurônios dopaminérgicos.
Por outro lado, outros estudos têm demonstrado uma maior participação do
sistema dopaminérgico nos efeitos do modafinil. Ferraro e colaboradores (1996)
demonstraram que o modafinil aumenta, de forma dependente de dose, a liberação
de dopamina no núcleo accumbens e que tal aumento é diminuído pelo antagonista
do receptor GABAB baclofen, pelo agonista GABAA muscimol e pelo inibidor da
recaptação do neurotransmissor GABA SKF 89976A, demonstrando haver uma
interação entre o sistema dopaminérgico e GABAérgico nos efeitos do modafinil. De
fato, tais ações do modafinil no sistema dopaminérgico podem ser secundárias à sua

INTRODUÇÃO
10
ação primária na diminuição da liberação do neurotransmissor GABA (Ferraro et al.,
1997).
Além disso, muitas das diversas características da hiperatividade
dopaminérgica, como por exemplo, o aumento da locomoção, são observadas após
o uso do modafinil. Dopheide e colaboradores (2007) demonstraram que o modafinil
aumenta a liberação de dopamina em regiões estriatais e Zolkowska e colaboradores
(2009) demonstraram haver uma correlação entre o aumento de dopamina no
accumbens e um aumento da ambulação e de estereotipia em ratos sob efeito do
modafinil. Também foi demonstrado que o modafinil em altas doses é capaz de inibir
a recaptação de dopamina através da inibição do transportador de dopamina (Mignot
et al., 1994; Ferraro et al., 1996; Madras et al., 2006). De fato, o modafinil tem sido
descrito até mesmo como um agonista de receptores D2 (Korotkova et al., 2007).
Nesse sentido, dados da literatura sugerem que o aumento da liberação de
dopamina parece ser de extrema importância para os efeitos de aumento do estado
de alerta causado pelo modafinil (Qu et al., 2008). Nesse sentido, estudo realizado
por Volkow e colaboradores (2009) demonstrou por meio da análise tomográfica do
cérebro de humanos que o modafinil, assim como o fazem outras drogas de abuso,
também aumenta a liberação de dopamina em áreas cerebrais como o núcleo
accumbens, o que poderia ser um indício de possíveis efeitos abusivos do modafinil.
Ainda, o aumento dos níveis de dopamina no núcleo accumbens e no córtex pré-
frontal parece ser importante para o aumento do estado de alerta em estudos
realizados em ratos (de Saint Hilaire et al., 2001; Murilo-Rodrígues et al., 2007).
De acordo com Wisor & Eriksson (2005) e posteriormente, Morgan e
colaboradores (2007), a interação entre o sistema noradrenérgico e o sistema

INTRODUÇÃO
11
dopaminérgico é necessária para que os efeitos promotores de alerta do modafinil
ocorram. Nesse cenário, Mitchell e colaboradores (2008) propõem que além da
interação noradrenérgica-dopaminérgica ser necessária para o aumento do alerta,
esta também seria necessária para o aumento da atividade locomotora observada
em animais.
A ação do modafinil inibindo a liberação de GABA no núcleo accumbens
aumenta fracamente a liberação de dopamina nessa região, o que também poderia
representar um mecanismo pelo qual ocorreria um aumento na atividade motora em
camundongos e ratos. Além disso, tal mecanismo de ação indireto no sistema
dopaminérgico poderia explicar o baixo potencial de abuso do modafinil, já que
drogas de abuso, como a anfetamina, agem diretamente aumentando a liberação de
dopamina.
1.3 Modafinil, Memória e Ansiedade
A aquisição de memórias de curto-prazo envolve neurônios do córtex pré-
frontal e do córtex parietal, principalmente neurônios dos sistemas dopaminérgicos e
noradrenérgicos (Müller et al., 2004), além de recrutar áreas hipocampais
(Béracochéa et al., 2002). Dessa forma, drogas que agem nessas vias, como os
psicoestimulantes, são efetivos em aumentar a atenção e a aquisição de memória.
Nesse cenário, o modafinil tem se mostrado de grande interesse em estudos de
memória, principalmente por não ter alguns dos efeitos adversos dos
psicoestimulantes clássicos (Turner et al., 2002).

INTRODUÇÃO
12
Estudos que avaliam os efeitos das administrações crônica ou aguda de
modafinil revelam uma melhora no aprendizado, tanto em animais quanto em
humanos (Béracochéa et al., 2007). Além disso, o modafinil parece melhorar a
atenção, o aprendizado e as funções cognitivas de indivíduos privados de sono,
(Pièrard et al., 2007). Esses mesmos autores demonstraram que o modafinil é
também capaz de reverter alterações na arquitetura do ciclo sono-vigília, na memória
e nas emoções, promovidas por 10 horas de privação de sono.
No que se refere aos efeitos do modafinil sobre os processos cognitivos de
roedores, a administração aguda dessa droga aumentou o aprendizado em
camundongos expostos ao labirinto em T quando houve longos intervalos entre as
sessões de aquisição. Além disso, evidências experimentais sugerem que os efeitos
do modafinil sobre a memória são dependentes de tempo e de dose (Morgan et al.,
2007). De fato, alguns estudos sugerem que o modafinil pode aumentar o estado de
atenção em animais, aumentando respostas incipientes e diminuindo a omissão de
erros (Morgan et al., 2007), apesar de outros estudos demonstrarem que o modafinil
não causa tanto efeito nos processos de atenção em humanos (Piérard et al., 2005;
Béracochéa et al., 2007).
Recentemente, tem sido demonstrada uma importante relação entre altos
níveis de corticosterona e os efeitos do modafinil sobre a memória (Piérard et al.,
2005; Béracochéa et al., 2007). Nesse sentido, estudos têm demonstrado que a
aquisição de uma tarefa em roedores estressados e que receberam modafinil é maior
do que em animais nas mesmas condições, porém que receberam solução veículo.
De fato, tem sido demonstrado que o modafinil é mais efetivo em animais que

INTRODUÇÃO
13
apresentam níveis aumentados de estresse. Entretanto, a relação entre modafinil,
liberação de corticosterona e memória permanece desconhecida.
De importância para este trabalho, controvérsias sobre os reais efeitos
benéficos do modafinil na melhora da cognição em humanos ainda são relatadas.
Contudo, parece que o efeito promnéstico induzido por esse fármaco em indivíduos
privados de sono está bem caracterizado (Randall et al., 2004). Nesse sentido, um
estudo realizado por Wesenstein e colaboradores (2002) com uma amostra de 50
pessoas, mostrou que o modafinil aumenta a capacidade cognitiva em doses três
vezes menores quando comparado à cafeína.
Em relação à ansiedade, pouco se sabe sobre os efeitos do modafinil na
emocionalidade. Um estudo realizado por Simon e colaboradores (1994) parece ser
o único que tenta elucidar tal relação. Nesse estudo, esses pesquisadores
compararam os efeitos do modafinil com a dexanfetamina em testes de ansiedade, e
demonstraram que o modafinil, apesar de aumentar a atividade locomotora, não
apresentou efeito ansiogênico quando comparado a dexanfetamina.
Apesar das evidências experimentais apontarem um possível efeito positivo do
modafinil sobre a memória, os efeitos facilitadores dessa droga sobre os processos
cognitivos ainda carecem de experimentação sistemática envolvendo diferentes
modelos de aprendizado e memória. Nesse contexto, a investigação dos efeitos do
modafinil sobre a memória na esquiva discriminativa em labirinto em cruz elevado
desponta como uma interessante hipótese de trabalho. De fato, tal investigação
permitiria verificar a possível participação de alterações na ansiedade e na função
motora sobre os possíveis efeitos promnésticos do modafinil.

OBJETIVOS
14
2. OBJETIVOS
2.1 Objetivo Geral
Investigar os efeitos do modafinil sobre as diferentes fases da memória
(aprendizado, consolidação e evocação) e sobre a ansiedade e a atividade
locomotora de camundongos em um modelo experimental que avalia
simultaneamente memória, atividade locomotora e ansiedade – a esquiva
discriminativa em labirinto em cruz elevado.
2.2 Objetivos Específicos
Avaliar, em camundongos submetidos ao modelo da esquiva discriminativa em
labirinto em cruz elevado, os efeitos da:
administração aguda de diferentes doses de modafinil sobre a consolidação
da memória;
administração aguda de diferentes doses de modafinil sobre o aprendizado e a
evocação da memória;
possível participação do fenômeno de dependência de estado nos possíveis
déficits de memória induzidos pelo modafinil;
administração crônica de diferentes doses de modafinil sobre a consolidação e
evocação da memória.

TRABALHO CIENTÍFICO A SER SUBMETIDO
15
3. INHIBITORY EFFECTS OF MODAFINIL ON MEMORY IN MICE
Helaine A. Fernandesa,b, Camilla L. Pattia,b, Karina A. Zanina,b, Raphael Wuo-Silvaa,
Luciano Fernandes-Santos a,b, Lia R. Bittencourt b, Sergio Tufikb,
Roberto Frussa-Filhoa,b*
aDepartmento de Farmacologia, Universidade Federal de São Paulo, R. Botucatu,
862, Ed. Leal Prado, 1º andar, 04023062, São Paulo, SP, Brasil.
bDepartmento de Psicobiologia, Universidade Federal de São Paulo, R. Napoleão de
Barros, 925, São Paulo, SP, Brasil.
*Correspondence:
Roberto Frussa-Filho
Departamento de Farmacologia – UNIFESP
Rua Botucatu, 862 – Ed. Leal Prado, 1º andar – 04023062 - São Paulo, SP Brasil
Email: [email protected]
Fax (55-11) 5549 4122 x. 222 – Phone – (55-11) 5549 4122 r. 219

TRABALHO CIENTÍFICO A SER SUBMETIDO
16
ABSTRACT
Modafinil, a psychostimulant used to treat narcolepsy, excessive daytime
sleepiness, and sleepiness due to obstructive sleep apnea, appears to promote a
possible facilitatory effect on cognitive function. In this study, we investigated the
effects of the acute administration of 32, 64 or 128 mg/kg of modafinil (MOD) on the
different steps of memory formation and usage (acquisition, consolidation and
retrieval) and the effects of its chronic administration on memory consolidation and
the possible participation of the state-dependency phenomenon in mice using the
plus-maze discriminative avoidance task (PM-DAT), a model that concomitantly
evaluates learning, memory, anxiety levels and locomotion. We observed that the
post-training acute administration of high doses of MOD (64 and 128 mg/kg) impaired
memory consolidation and that these doses also promoted non-associative memory
deficits. After the pre-training treatment, no differences were found when learning was
evaluated, but doses of 32 and 64 mg/kg induced retention deficits in the test session.
In the pre-test administration, only the 32 mg/kg dose promoted retrieval deficits. This
amnestic effect produced by 32 mg/kg of modafinil administered during the pre-test
was not subject to tolerance when this dose was repeated in a previous
administration. Finally, the cognitive deficits produced by modafinil were not due to
the phenomenon of state-dependency. In all, our findings provide pre-clinical
evidence of potential amnesia induced by modafinil.
Key words: modafinil, learning and memory, anxiety, locomotion, mice

TRABALHO CIENTÍFICO A SER SUBMETIDO
17
INTRODUCTION
Modafinil (MOD) is a non-amphetamine type stimulant that acts as a wake-
promoting drug and has been approved for the treatment of excessive daytime
sleepiness in narcolepsy, obstructive sleep apnea, and shift work syndrome. In
addition, MOD shows initial promise for a variety of off-label indications in psychiatry,
including treatment-resistant depression, attention-deficit/hyperactivity disorder and
schizophrenia (Swanson et al., 2006; Minzemberg and Carter, 2008).
There are several hypotheses that attempt to explain the mechanisms of action
of MOD. Studies have suggested that it may increase the release of catecholamines
(dopamine and norepinephrine), serotonin and glutamate, thereby reducing the
release of GABA in various brain regions, and activating hypothalamic neurons
containing hypocretin/orexin neurons of the tuberomammillary nucleus (Minzenberg &
Carter, 2008).
It has been demonstrated that MOD modifies the activity of brain areas
involved with memory, such as the hippocampus and prefrontal cortex (Beracochea,
2003). Within this context, studies indicate that MOD has cognitive-enhancing abilities
in rodents performing a variety of learning/memory exercises in the T-Maze based on
spontaneous alternation behavior (Beracochea et al., 2001), and also based on
enhanced learning (Beracochea, 2003) and memory retrieval (Beracochea et al.,
2007). Still, it was reported that this drug was able to restore the memory impairments
observed after a 10 h sleep deprivation period (Piérard et al., 2007). Concerning its
chronic administration, MOD was reported to improve learning (Beracochea et al.,
2002). Notwithstanding, the facilitative effects of MOD on memory have not always

TRABALHO CIENTÍFICO A SER SUBMETIDO
18
been verified when memory consolidation was specifically investigated. In fact, a
study conducted by Shuman and colleagues (2009) reported that the administration
of MOD immediately after training had no effects on either the Morris water maze task
or the contextual fear memory task.
Although experimental evidence suggests a possible positive effect of MOD on
memory, the facilitatory effects of this drug on cognitive processes still require
systematic experimentation with different models of learning and memory.
Thus, the initial objective of the present study was to investigate the effects of
MOD on the different steps of memory formation in an animal model that
concomitantly evaluates learning, memory, locomotor activity and anxiety, namely the
plus-maze discriminative avoidance task (PM-DAT). This paradigm allows the
concomitant and independent evaluation of the behavioral parameters cited above
(Silva et al., 1997, 1999, 2002a,b, 2004; Silva and Frussa-Filho, 2000; Castro et al.,
2005). In fact, the effects of memory-enhancing (Silva et al., 1997, 1999, 2000; Claro
et al., 1999) or amnesiac procedures (Claro et al., 1999; Silva and Frussa-Filho,
2000, 2002; Silva et al., 1999, 2004; Patti et al., 2006), as well as motor stimulant
(Silva et al., 2002b; Castro et al., 2005) and depressing interventions (Silva et al.,
2002a, 2004; Carvalho et al., 2006), were detected in mice submitted to this
behavioral procedure.

TRABALHO CIENTÍFICO A SER SUBMETIDO
19
MATERIAL AND METHODS
Subjects
Three-month-old Swiss male mice (inbred, raised, and maintained in the
Centre for Development of Experimental Models in Medicine and Biology of
Universidade Federal de São Paulo) were used in the experiments. Animals weighing
30–35 g were housed under controlled temperature (22–23°C) and lighting (12 h light,
12 h dark; lights on at 6:45 a.m.) conditions. Food and water were available ad libitum
throughout the experiments.
The animals used in this study were maintained in accordance with the
National Institute of Health Guide for the Care and Use of Laboratory Animals (NIH
Publications Nº 80-23, revised 1996) and in accordance with the Brazilian Law for
Procedures for Animal Scientific Use (#11794/2008). All experimental procedures
were approved by the Ethics Committee under protocol #1162/08.
Drugs
Modafinil (CEPHALON®) was suspended in a 0.9% NaCl (saline) solution and
administered intraperitoneally at a volume of 10 mL/kg body weight at doses of 32
(MOD32), 64 (MOD64), or 128mg/kg (MOD128). Saline solution (SAL) was used as
the control solution. Behavioral testing started 30 min after the MOD or SAL solutions
injections.

TRABALHO CIENTÍFICO A SER SUBMETIDO
20
Plus-maze Discriminative avoidance task (PM-DAT)
The apparatus employed was a modified elevated plus-maze, made of wood,
containing two enclosed arms with sidewalls and no top (28.5×7×14 cm, 03 lx at the
floor level), opposite two open arms (28.5×7cm, 09 lx at the floor level).
A 100 W lamp was placed exactly over the middle of one of the enclosed arms
(aversive enclosed arm, 660 lx at the floor level). In the training session, each mouse
was placed in the center of the apparatus and, over a period of 10 min, each time the
animal entered the enclosed arm containing the lamp, an aversive situation was
produced until the animal left the arm. The aversive stimuli were the 100 W light and
a cold air blow produced by a hair dryer placed over the aversive enclosed arm. In the
test session (performed in the same room with the observer in the same position), the
mice were again placed in the center of the apparatus and observed for 3 min without
receiving the aversive stimulation. In all experiments, the animals were observed in a
random order, and in a blind manner, and the apparatus was cleaned with a 5%
alcohol solution after each behavioral session.
The total number of entries into any of the arms (an entry was defined as the
entry of all four paws into one arm), the percent time spent in the aversive enclosed
arm (time spent in the aversive enclosed arm divided by the time spent in both
enclosed arms) and the percent time spent in the open arms (time spent in open arms
divided by the time spent in both open and enclosed arms) were calculated. Learning
was evaluated by the percent time spent in the aversive enclosed arm during the
training session. Memory was evaluated by the time spent in the aversive vs. non-
aversive enclosed arms in the test session. The percent time spent in the aversive
enclosed arm among the groups that showed retention of the task in the test session

TRABALHO CIENTÍFICO A SER SUBMETIDO
21
was compared to reveal quantitative differences. Anxiety-like behavior was evaluated
by the percent time spent in the open arms of the apparatus. The total number of
entries into any of the arms was used to evaluate motor activity.
Statistical analysis
The total number of entries into any of the arms, the percent time spent in the
aversive enclosed arm (time spent in aversive enclosed arm divided by the time spent
in both enclosed arms), and percent time spent in open arms (time spent in open
arms divided by the time spent in both open and enclosed arms) were calculated and
compared by the one- or two-way analysis of variance (ANOVA) followed by
Duncan’s test. Memory was evaluated by time spent in the aversive vs. non-aversive
enclosed arms (compared by two-way ANOVA followed by Duncan’s test) and by the
percent time in aversive enclosed arm in the test session. Significance was accepted
at p-values less than 0.05.
Experimental design
Experiment 1: Effects of the acute administration of MOD on memory consolidation of
a discriminative avoidance task in mice
Forty-eight animals were randomly assigned to one of the following groups:
saline (SAL, n=12), modafinil 32 mg/kg (MOD32, n=12), modafinil 64 mg/kg (MOD64,
n=12) or modafinil 128 mg/kg (MOD128, n=12). Mice were trained in the PM-DAT
and, immediately afterwards, received an acute intraperitoneal (i.p.) administration of

TRABALHO CIENTÍFICO A SER SUBMETIDO
22
SAL or MOD at one of the different doses. Ten days later, the animals were submitted
to the test session.
Experiment 2: Effect of the acute administration of MOD on the learning of a
discriminative avoidance task in mice
Forty-eight animals were randomly assigned to one of the following groups:
saline (SAL, n=12), modafinil 32 mg/kg (MOD32, n=12), modafinil 64 mg/kg (MOD64,
n=12) or modafinil 128 mg/kg (MOD128, n=12). Mice received an acute i.p.
administration of SAL or MOD at one of the different doses. Thirty minutes after the
injection, the animals were trained in the PM-DAT. Ten days later, they were
submitted to the test session.
Experiment 3: Effect of the acute administration of MOD on the retrieval of a
discriminative avoidance task in mice
Forty-eight animals were randomly assigned to one of the following groups:
saline (SAL, n=12), modafinil 32 mg/kg (MOD32, n=12), modafinil 64 mg/kg (MOD64,
n=12) or modafinil 128 mg/kg (MOD128, n=12). Mice were trained in the PM-DAT.
Ten days later, the animals received an acute i.p. administration of SAL or MOD at
one of the different doses. Thirty minutes after the injection, the animals were tested.

TRABALHO CIENTÍFICO A SER SUBMETIDO
23
Experiment 4: Role of the state-dependency phenomenon on the cognitive effects of
MOD in mice using the PM-DAT
Forty-eight mice were randomly assigned to one of the following groups: pre-
training/pre-test administration of SAL (SAL-SAL, n=12), pre-test administration of
32mg/kg (SAL-MOD, n=12), pre-training administration of 32 mg/kg of modafinil
(MOD-SAL, n=12) or pre-training/pre-test administration of MOD (MOD-MOD, n=12).
Groups of 24 mice received SAL or 32 mg/kg MOD. Thirty minutes after the injection,
all animals were submitted to a training session in the PM-DAT. Ten days after the
training session, 12 animals from the pre-training SAL group received another SAL
injection, whereas the other 12 mice from the same group received a 32 mg/kg
injection of MOD. Similarly, 12 animals from the pre-training MOD group received a
SAL injection, whereas the other 12 mice received another 32 mg/kg MOD injection.
The test session was performed 30 minutes after the second injection.
Experiment 5: Effects of repeated administration of MOD on memory retention of a
discriminative avoidance task in mice
Forty-eight animals were randomly assigned to one of the following groups:
saline (SAL, n=12), 32mg/kg modafinil (MOD32 n=12), 64mg/kg modafinil (MOD64
n=12) or 128mg/kg modafinil (MOD128 n=12). Mice were trained in the PM-DAT and,
immediately afterwards received an i.p. administration of SAL or MOD at one of the
different doses. On the days following the initial injection, the animals received 9 daily
injections of SAL or MOD. Thirty minutes after the last injection, the animals were
submitted to the test session.

TRABALHO CIENTÍFICO A SER SUBMETIDO
24
RESULTS
Experiment 1: Effects of the acute administration of MOD on memory consolidation of
a discriminative avoidance task in mice
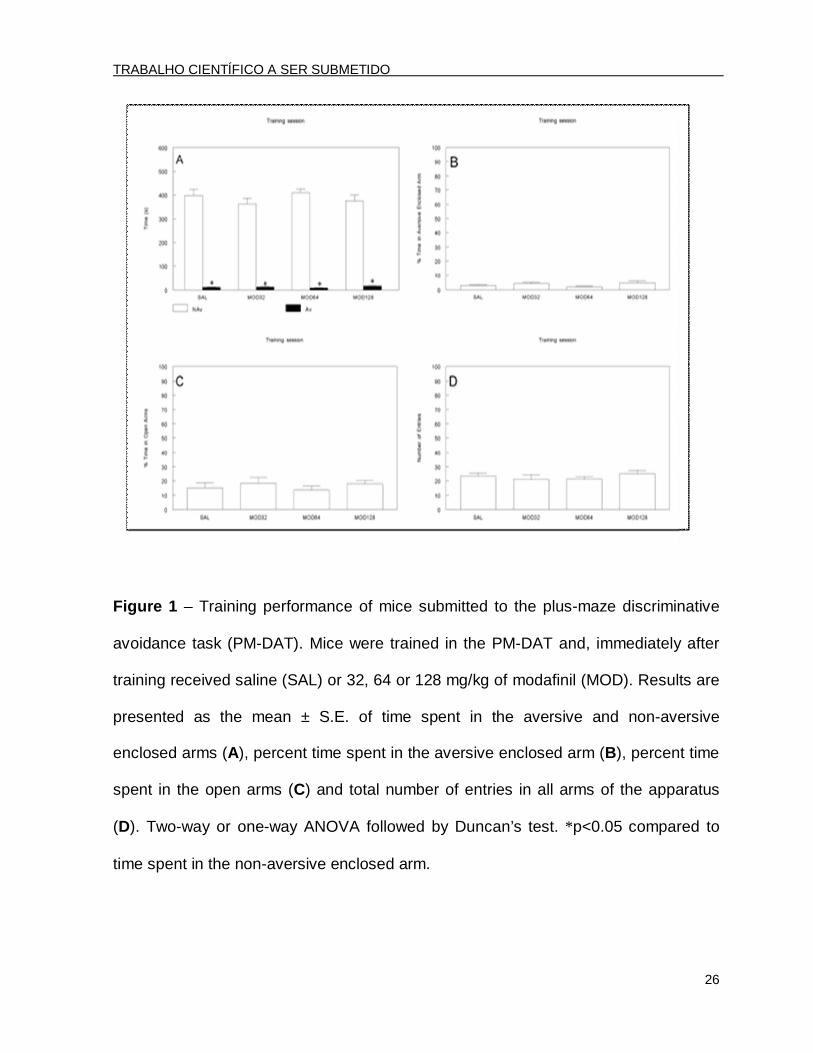
In the training session, a two-way ANOVA revealed significant effects for arm
type [F(1.96)=1152.61; p<0.001]. The post hoc analysis by Duncan’s test revealed
that all of the groups spent significantly less time in the aversive arm than in the non-
aversive enclosed arm (Fig. 1A). During this session, an one-way ANOVA revealed
that there were no significant differences between the groups for the percent time
spent in the aversive enclosed arm F(3.44)=1.88; p>0.05] (Fig. 1B), demonstrating
that all groups showed similar learning levels.
No differences were found in the percent time spent in the open arms in the
training session [F(3.44)=0.49; p>0.05] or in the total number of entries [F(3.44)=0.54;
p>0.05] (ANOVA; Fig. 1C and 1D, respectively). Therefore, all animals presented with
a similar baseline performance.
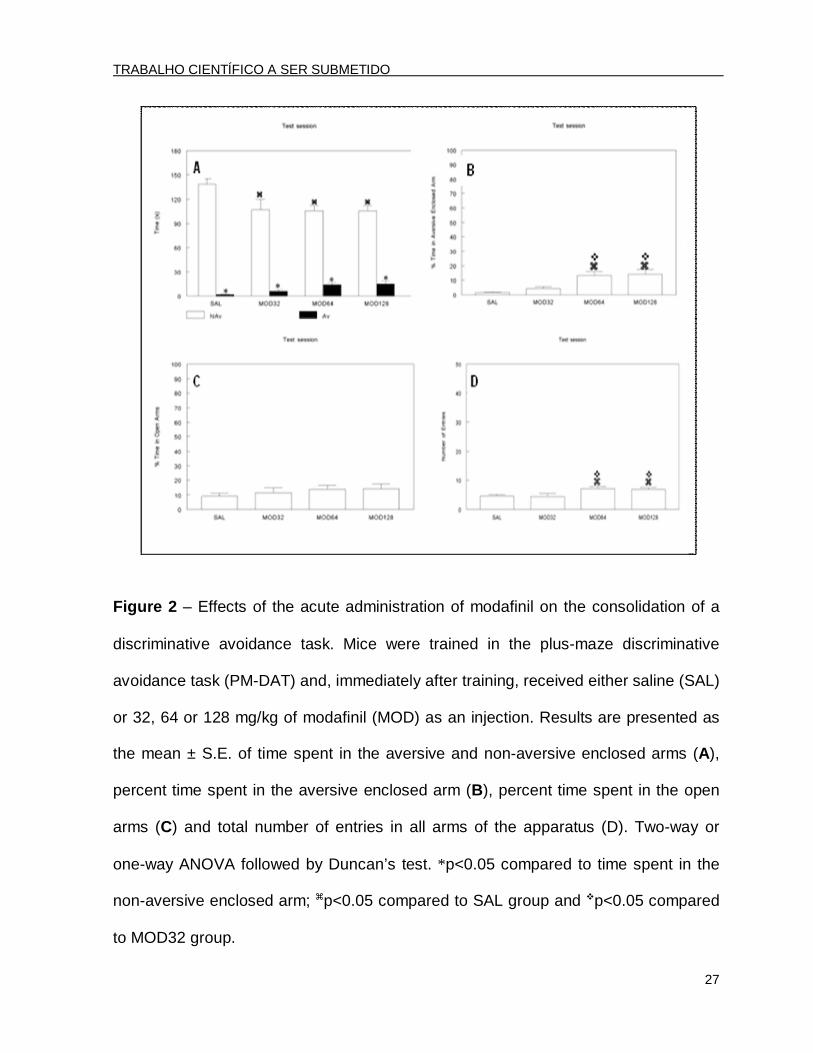
In the test session, performed 10 days after training, a two-way ANOVA
revealed significant arm type [F(1.96)=197.45; p<0.001] and treatment x arm type
interaction effects [F(3.96)=2.87; p<0.05]. The post hoc analysis by Duncan’s test
revealed that all groups spent significantly less time in the aversive enclosed arm
compared to the non-aversive arm (Fig. 2A). The analysis of the percent time spent in
the aversive enclosed arm revealed that the MOD64 and MOD128 groups presented
an improvement in this parameter when compared to the other two groups [(SAL and

TRABALHO CIENTÍFICO A SER SUBMETIDO
25
MOD32) [F(3.44)=5.36; p<0.05] (ANOVA followed by Duncan’s test, Fig. 2B),
showing a retention impairment.
The ANOVA did not reveal significant differences in the percent time spent in
the open arms [F(3.44)=0.66; p>0.05] (Fig. 2C). However, with respect to locomotor
activity, the ANOVA followed by Duncan’s test showed an increased total number of
entries [F(3.44)=3.57; p<0.05] (Fig. 2D) by the animals treated with 64 and 128 mg/kg
of MOD, suggesting habituation deficits.
TRABALHO CIENTÍFICO A SER SUBMETIDO
26
Figure 1 – Training performance of mice submitted to the plus-maze discriminative
avoidance task (PM-DAT). Mice were trained in the PM-DAT and, immediately after
training received saline (SAL) or 32, 64 or 128 mg/kg of modafinil (MOD). Results are
presented as the mean ± S.E. of time spent in the aversive and non-aversive
enclosed arms (A), percent time spent in the aversive enclosed arm (B), percent time
spent in the open arms (C) and total number of entries in all arms of the apparatus
(D). Two-way or one-way ANOVA followed by Duncan’s test. *p<0.05 compared to
time spent in the non-aversive enclosed arm.
TRABALHO CIENTÍFICO A SER SUBMETIDO
27
Figure 2 – Effects of the acute administration of modafinil on the consolidation of a
discriminative avoidance task. Mice were trained in the plus-maze discriminative
avoidance task (PM-DAT) and, immediately after training, received either saline (SAL)
or 32, 64 or 128 mg/kg of modafinil (MOD) as an injection. Results are presented as
the mean ± S.E. of time spent in the aversive and non-aversive enclosed arms (A),
percent time spent in the aversive enclosed arm (B), percent time spent in the open
arms (C) and total number of entries in all arms of the apparatus (D). Two-way or
one-way ANOVA followed by Duncan’s test. *p<0.05 compared to time spent in the
non-aversive enclosed arm; p<0.05 compared to SAL group and p<0.05 compared
to MOD32 group.

TRABALHO CIENTÍFICO A SER SUBMETIDO
28
Experiment 2: Effects of the acute administration of MOD on the learning of a
discriminative avoidance task in mice
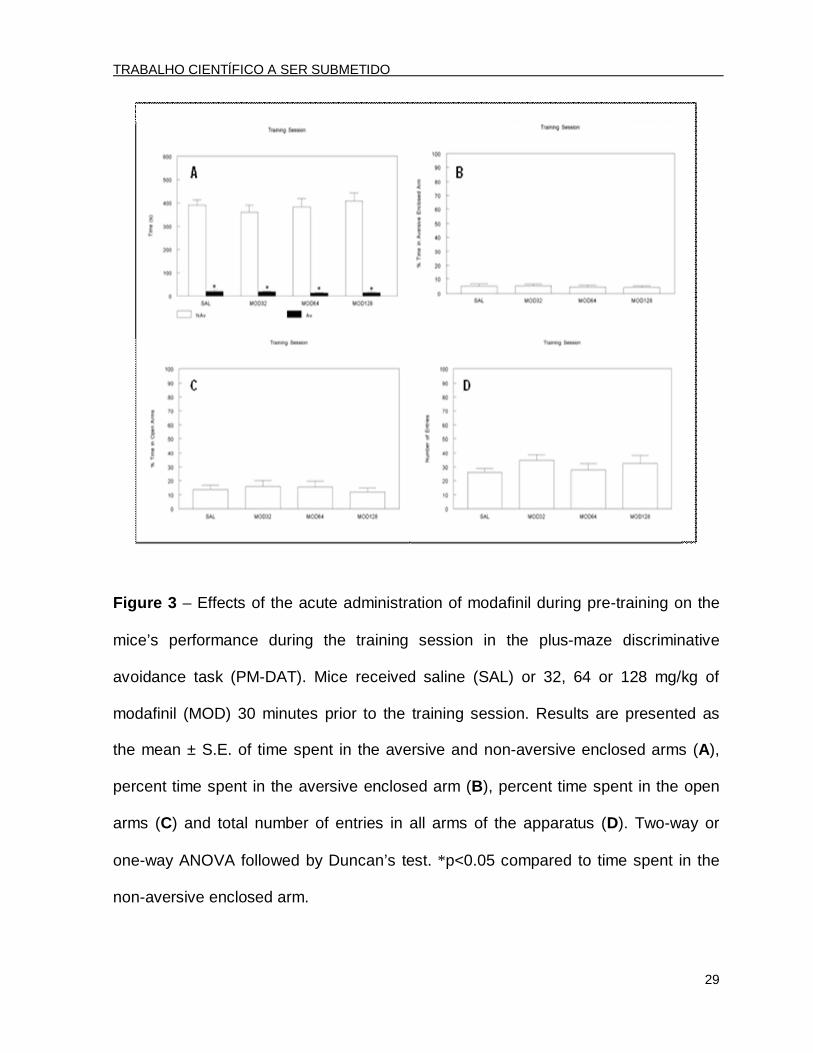
In the training session, the two-way ANOVA revealed significant effects for arm
type [F(1.96)=527.46; p<0.001]. The post hoc analysis by Duncan’s test revealed that
all of the groups spent significantly less time in the aversive arm than in the non-
aversive enclosed arm (Fig. 3A). In this session, the ANOVA revealed that there were
no significant differences among groups in the percent time spent in the aversive
enclosed arm [F(3.47)=0.17; p>0.05] (Fig. 3B), demonstrating that MOD did not
modify learning ability.
During the training session, the ANOVA did not reveal significant differences in
the percent time spent in the open arms [F(3.47)=0.21; p>0,05] (Fig. 3C) or in the
total number of entries [F(3.47)=0.80; p>0.05] (Fig. 3D).
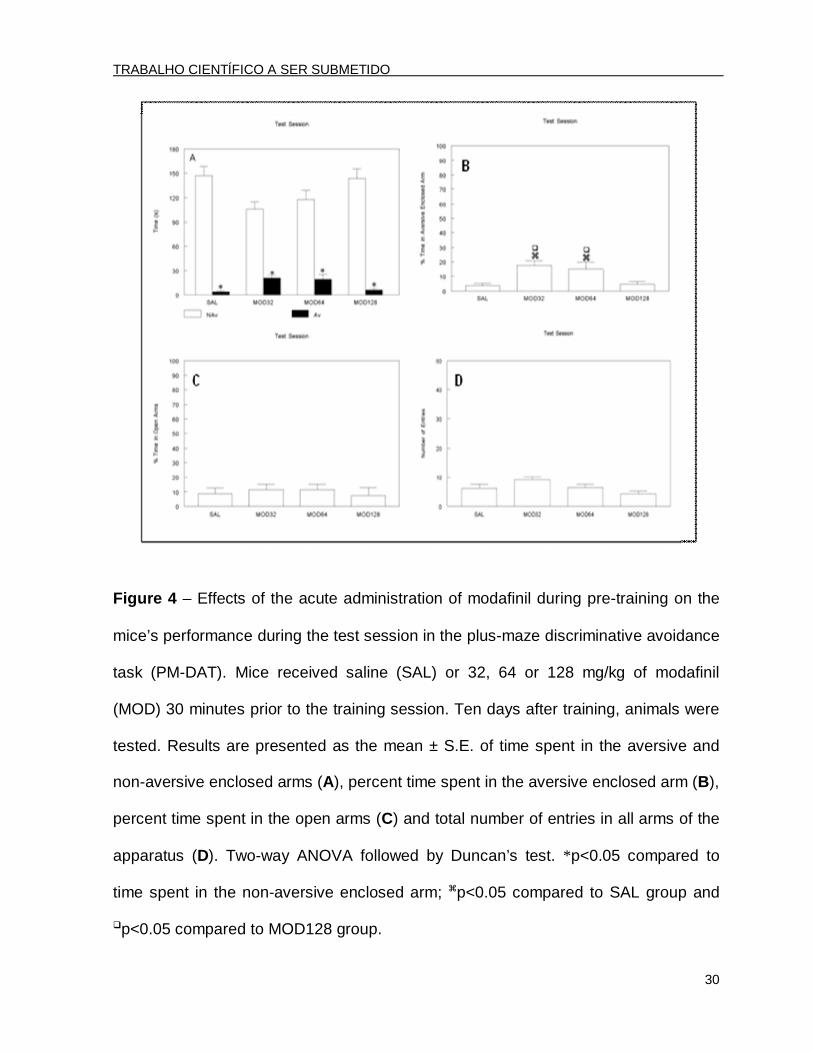
In the test session, performed 10 days after training, the two-way ANOVA
revealed only a significant effect in arm type [F(1.96)=228.91; p<0.001] The post hoc
analysis by Duncan’s test revealed that all of the groups spent significantly less time
in the aversive enclosed arm compared to the non-aversive one (Fig. 4A.
Furthermore, the ANOVA followed by Duncan’s test showed that the MOD32 and
MOD64 groups spent a significantly longer percent time in the aversive enclosed arm
compared to the two other groups (SAL and MOD128) [F(3.47)=6.91; p<0.05]. (Fig.
4B).
The ANOVA did not reveal significant differences among groups in either the
percent time spent in the open arms [F(3.47)=0.24; p>0.05] (Fig. 4C) or in the total
number of entries [F(3.47)=3.25; p<0.05] (Fig. 4D).
TRABALHO CIENTÍFICO A SER SUBMETIDO
29
Figure 3 – Effects of the acute administration of modafinil during pre-training on the
mice’s performance during the training session in the plus-maze discriminative
avoidance task (PM-DAT). Mice received saline (SAL) or 32, 64 or 128 mg/kg of
modafinil (MOD) 30 minutes prior to the training session. Results are presented as
the mean ± S.E. of time spent in the aversive and non-aversive enclosed arms (A),
percent time spent in the aversive enclosed arm (B), percent time spent in the open
arms (C) and total number of entries in all arms of the apparatus (D). Two-way or
one-way ANOVA followed by Duncan’s test. *p<0.05 compared to time spent in the
non-aversive enclosed arm.
TRABALHO CIENTÍFICO A SER SUBMETIDO
30
Figure 4 – Effects of the acute administration of modafinil during pre-training on the
mice’s performance during the test session in the plus-maze discriminative avoidance
task (PM-DAT). Mice received saline (SAL) or 32, 64 or 128 mg/kg of modafinil
(MOD) 30 minutes prior to the training session. Ten days after training, animals were
tested. Results are presented as the mean ± S.E. of time spent in the aversive and
non-aversive enclosed arms (A), percent time spent in the aversive enclosed arm (B),
percent time spent in the open arms (C) and total number of entries in all arms of the
apparatus (D). Two-way ANOVA followed by Duncan’s test. *p<0.05 compared to
time spent in the non-aversive enclosed arm; p<0.05 compared to SAL group and
p<0.05 compared to MOD128 group.

TRABALHO CIENTÍFICO A SER SUBMETIDO
31
Experiment 3: Effects of the acute administration of MOD on the retrieval of a
discriminative avoidance task in mice
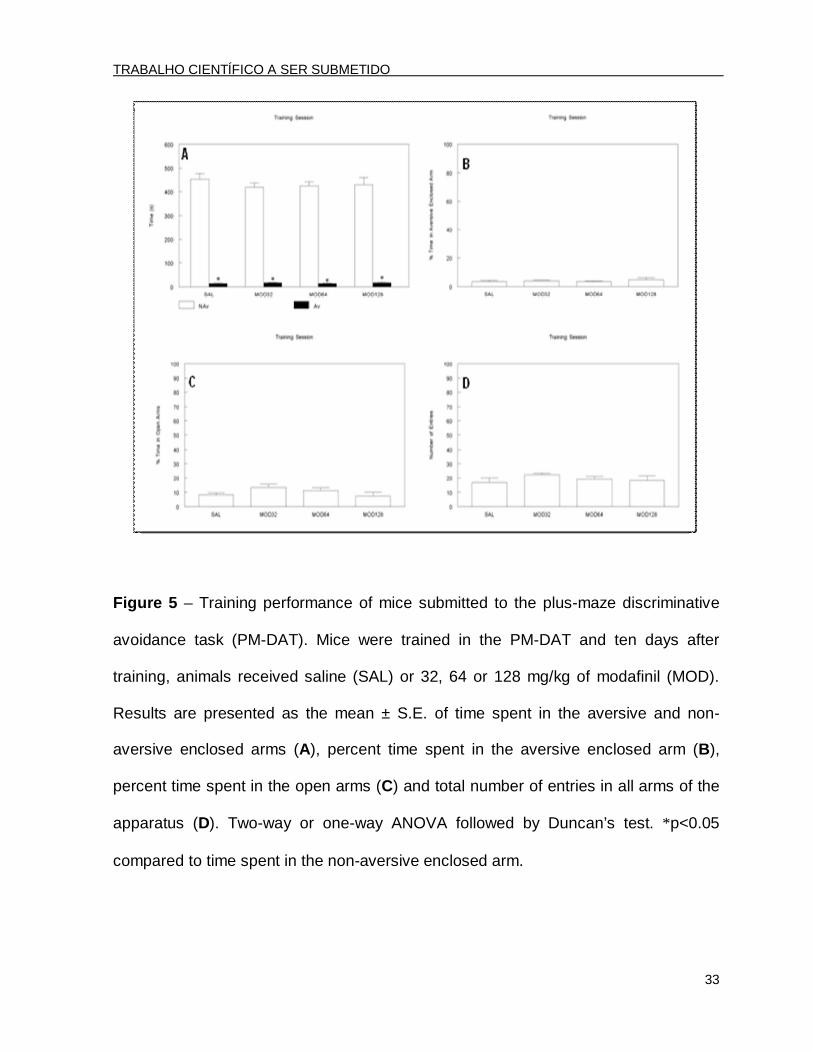
In the training session, the two-way ANOVA revealed significant effects for arm
type [F(1.96)=124.90; p<0.001]. The post hoc analysis by Duncan’s test revealed that
all of the groups spent significantly less time in the aversive arm than in the non-
aversive enclosed arm (Fig. 5A). In this session, the ANOVA revealed that there were
no significant differences among groups for the percent time spent in the aversive
enclosed arm [F(3.44)=0.43; p>0.05] (Fig. 5B), showing a similar learning level.
No differences were found in the percent time spent in the open arms
[F(3.44)=1.63; p>0.05] (Fig. 5C) or in the total number of entries [F(3.44)=0.80;
p>0.05] (Fig. 5D) in the training session, revealing that all of the groups displayed
similar baseline levels of emotionality and locomotion.
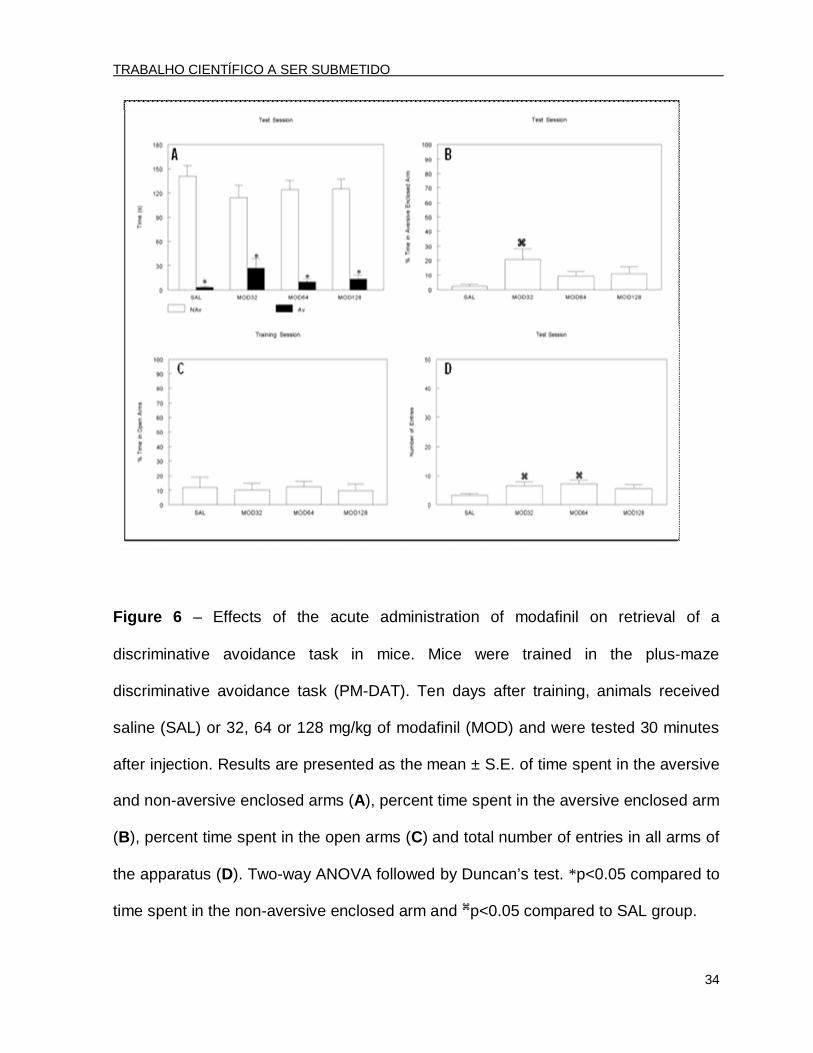
In the test session, performed 10 days after training, the two-way ANOVA
revealed a significant arm type effect [F(1.96)=234.74; p<0.001] (Fig. 6A). The post
hoc analysis by Duncan’s test revealed that all of the groups, irrespective of
treatment, discriminated the enclosed arms, spending significantly more time in the
non-aversive enclosed arm. However, when the percent time spent in the aversive
enclosed arm was analyzed, the ANOVA followed by Duncan’s test revealed that the
animals treated with 32 mg/kg of MOD before testing spent significantly more time in
the aversive enclosed arm (Fig. 6B) compared to the SAL group [F(3.44)=2.37;
p<0.05], indicating a retrieval deficit.
Finally, the ANOVA did not reveal significant differences in the percent time
spent in the open arms during the testing session [F(3.44)=0.05; p>0.05] (Fig. 6C). In

TRABALHO CIENTÍFICO A SER SUBMETIDO
32
contrast, the ANOVA followed by Duncan’s test showed that the animals treated with
32 or 64 mg/kg of MOD had an increase in the total number of entries compared to
the SAL treated group [F(3.44)=3.62; p<0.05] (Fig. 6D).
TRABALHO CIENTÍFICO A SER SUBMETIDO
33
Figure 5 – Training performance of mice submitted to the plus-maze discriminative
avoidance task (PM-DAT). Mice were trained in the PM-DAT and ten days after
training, animals received saline (SAL) or 32, 64 or 128 mg/kg of modafinil (MOD).
Results are presented as the mean ± S.E. of time spent in the aversive and non-
aversive enclosed arms (A), percent time spent in the aversive enclosed arm (B),
percent time spent in the open arms (C) and total number of entries in all arms of the
apparatus (D). Two-way or one-way ANOVA followed by Duncan’s test. *p<0.05
compared to time spent in the non-aversive enclosed arm.
TRABALHO CIENTÍFICO A SER SUBMETIDO
34
Figure 6 – Effects of the acute administration of modafinil on retrieval of a
discriminative avoidance task in mice. Mice were trained in the plus-maze
discriminative avoidance task (PM-DAT). Ten days after training, animals received
saline (SAL) or 32, 64 or 128 mg/kg of modafinil (MOD) and were tested 30 minutes
after injection. Results are presented as the mean ± S.E. of time spent in the aversive
and non-aversive enclosed arms (A), percent time spent in the aversive enclosed arm
(B), percent time spent in the open arms (C) and total number of entries in all arms of
the apparatus (D). Two-way ANOVA followed by Duncan’s test. *p<0.05 compared to
time spent in the non-aversive enclosed arm and p<0.05 compared to SAL group.

TRABALHO CIENTÍFICO A SER SUBMETIDO
35
Experiment 4: Role of the state-dependency phenomenon on the cognitive effects of
MOD using the PM-DAT
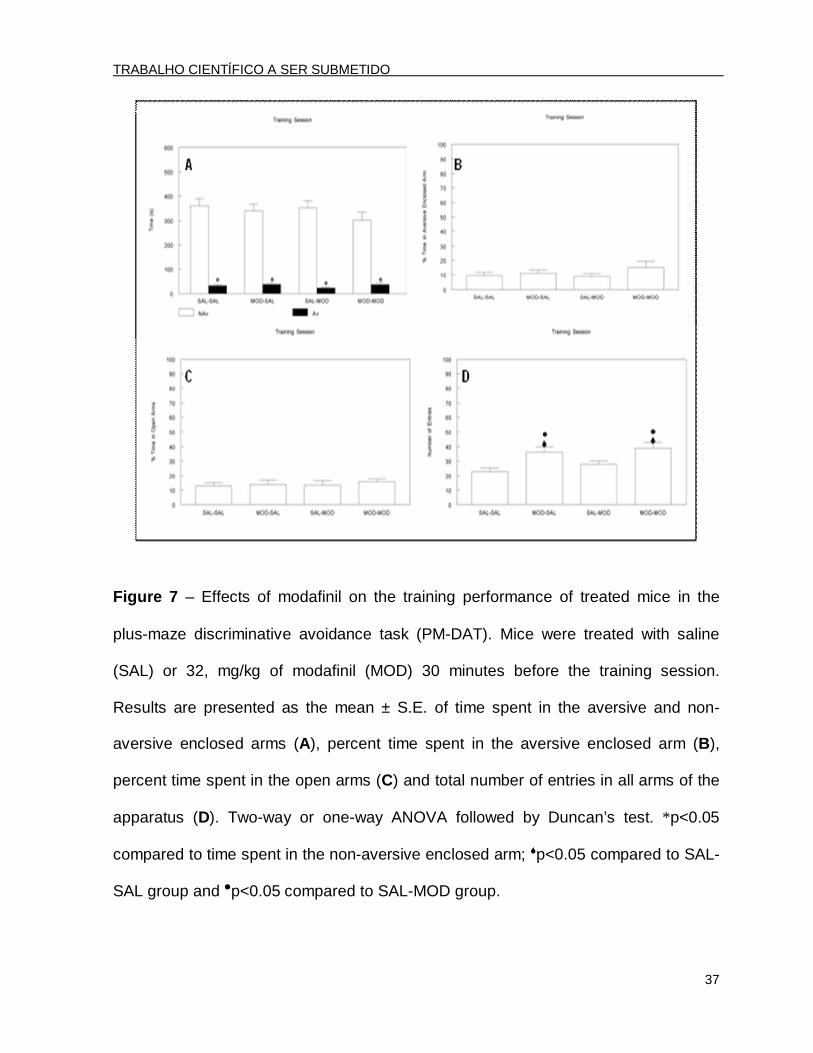
In the training session, the two-way ANOVA revealed only significant effects for
arm type [F(1.96)=5.750; p<0.05]. The post hoc analysis by Duncan’s test revealed
that all of the groups spent significantly less time in the aversive arm than in the non-
aversive enclosed arm (Fig. 7A). In this session, the ANOVA revealed that there were
no significant differences among groups when the percent time spent in the aversive
enclosed arm was analyzed [F(3.44)=1.10; p>0.05] (Fig. 7B). These results
demonstrated that 32 mg/kg of MOD did not modify learning ability, corroborating the
data from Experiment 2 (see Fig. 3A and 3B).
In addition, the ANOVA did not reveal significant differences among groups in
the percent time spent in the open arms [F(3.44)=0.22; p>0.05] (Fig. 7C). On the
other hand, both groups pre-training treated with MOD (MOD-SAL and MOD-MOD
groups) showed increased locomotion compared to the other groups treated with SAL
[F(3.44)=6.21; p<0.05] (Fig. 7D).
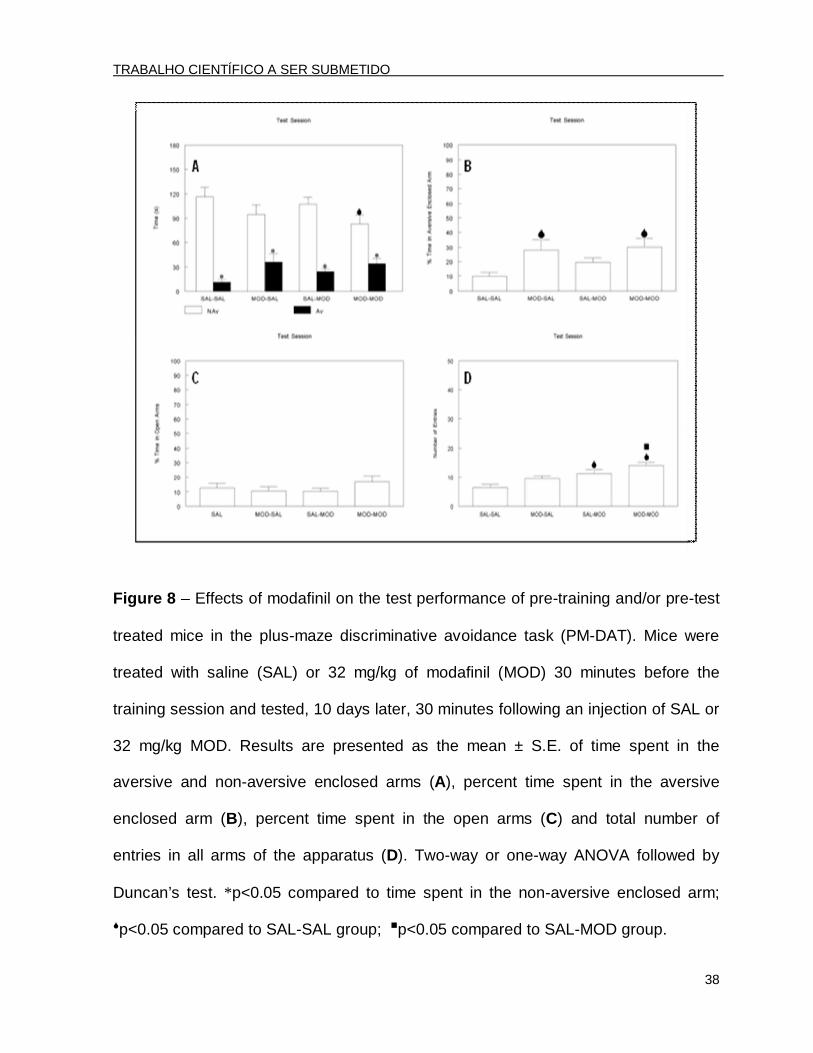
In the test session, performed 10 days after training, the two-way ANOVA
revealed only a significant arm type effect [F(1.96)=133.06; p<0.001]. The post hoc
analysis by Duncan’s test revealed that all of the animals were able to discriminate
the aversive enclosed arm, showing retention of the task (Fig. 8A). Nevertheless, the
ANOVA followed by Duncan’s test showed that the MOD-SAL and MOD-MOD groups
had a significant increase in the percent time in the aversive enclosed arm compared
to the other two groups (SAL-SAL and SAL-MOD) [F(3.44)=3.51; p<0.05] (Fig. 8B),

TRABALHO CIENTÍFICO A SER SUBMETIDO
36
showing that memory impairment induced by pre-training MOD was not related to the
state-dependency phenomenon.
When the percent time spent in the open arms in the testing session was
analyzed, the ANOVA did not reveal significant differences among groups
[F(3.44)=0.91; p>0.05] (Fig. 8C). However, the ANOVA followed by Duncan’s test
showed an increase in the total number of entries in the groups treated with MOD
before testing (the SAL-MOD and MOD-MOD groups) compared to the SAL-treated
groups. Still, the group treated with MOD before training and testing (MOD-MOD) had
increased motor activity compared to the SAL-MOD group [F(3.44)=8.42; p<0.05]
(Fig. 8D).
TRABALHO CIENTÍFICO A SER SUBMETIDO
37
Figure 7 – Effects of modafinil on the training performance of treated mice in the
plus-maze discriminative avoidance task (PM-DAT). Mice were treated with saline
(SAL) or 32, mg/kg of modafinil (MOD) 30 minutes before the training session.
Results are presented as the mean ± S.E. of time spent in the aversive and non-
aversive enclosed arms (A), percent time spent in the aversive enclosed arm (B),
percent time spent in the open arms (C) and total number of entries in all arms of the
apparatus (D). Two-way or one-way ANOVA followed by Duncan’s test. *p<0.05
compared to time spent in the non-aversive enclosed arm; p<0.05 compared to SAL-
SAL group and p<0.05 compared to SAL-MOD group.
TRABALHO CIENTÍFICO A SER SUBMETIDO
38
Figure 8 – Effects of modafinil on the test performance of pre-training and/or pre-test
treated mice in the plus-maze discriminative avoidance task (PM-DAT). Mice were
treated with saline (SAL) or 32 mg/kg of modafinil (MOD) 30 minutes before the
training session and tested, 10 days later, 30 minutes following an injection of SAL or
32 mg/kg MOD. Results are presented as the mean ± S.E. of time spent in the
aversive and non-aversive enclosed arms (A), percent time spent in the aversive
enclosed arm (B), percent time spent in the open arms (C) and total number of
entries in all arms of the apparatus (D). Two-way or one-way ANOVA followed by
Duncan’s test. *p<0.05 compared to time spent in the non-aversive enclosed arm;
p<0.05 compared to SAL-SAL group;p<0.05 compared to SAL-MOD group.

TRABALHO CIENTÍFICO A SER SUBMETIDO
39
Experiment 5: Effects of repeated administration of MOD on memory retention of a
discriminative avoidance task in mice
In the training session, the two-way ANOVA revealed only significant effects for
arm type [F(1.96)=304.48; p<0.001]. The post hoc analysis by Duncan’s test revealed
that all of the groups spent significantly less time in the aversive arm than in the non-
aversive enclosed arm (Fig. 9A). In this session, the ANOVA revealed that there were
no significant differences among groups for the percent time spent in the aversive
enclosed arm F(3.44)=0.35; p>0.05] (Fig. 9B), demonstrating a similar learning of the
task.
The ANOVA did not reveal significant differences in the percent time spent in
the open arms [F(3.44)=0.42; p>0.05] (Fig. 9C) or in the total number of entries
[F(3.44)=0.53; p>0.05] (Fig. 9D). This suggests that all animals had a similar baseline
behavioral performance.
In the test session, performed 10 days after training and 30 min after the end of
the chronic treatment of MOD, the two-way ANOVA revealed a significant effect for
arm type [F(1.96)=99.76; p<0.001] and a significant treatment x arm type interaction
[F(3.96)=2.77; p<0.05] effect. The post hoc analysis by Duncan’s test revealed that all
of the groups were able to discriminate the aversive enclosed arm, irrespective of the
treatment (Fig. 10A). Also, the ANOVA followed by Duncan’s test showed that the
MOD32 group spent a significantly longer percent time in the aversive enclosed arm
compared to the SAL-treated group [F(3.44)=3.61; p<0.05] (Fig. 10B).

TRABALHO CIENTÍFICO A SER SUBMETIDO
40
Finally, the ANOVA did not reveal differences in the percent time spent in the
open arms [F(3.44)=0.29; p>0.05] (Fig. 10C) or in the total number of entries
[F(3.44)=0.80; p<0.05] (Fig. 10D).

TRABALHO CIENTÍFICO A SER SUBMETIDO
41
Figure 9 – Training performance of mice submitted to the plus-maze discriminative
avoidance task (PM-DAT). Mice were trained in the PM-DAT and, immediately after
training, were treated with saline (SAL) or 32, 64 or 128 mg/kg of modafinil (MOD) for
10 consecutive days. Results are presented as the mean ± S.E. of time spent in the
aversive and non-aversive enclosed arms (A), percent time spent in the aversive
enclosed arm (B), percent time spent in the open arms (C) and total number of
entries in all arms of the apparatus (D). Two-way or one-way ANOVA followed by
Duncan’s test. *p<0.05 compared to time spent in the non-aversive enclosed arm.

TRABALHO CIENTÍFICO A SER SUBMETIDO
42
Figure 10 – Effects of chronic administration of modafinil on consolidation of a
discriminative avoidance task in mice. Mice were trained in the plus-maze
discriminative avoidance task (PM-DAT) and, immediately after training, were treated
with saline (SAL) or 32, 64 or 128 mg/kg of modafinil (MOD) for 10 consecutive days
and tested 30 min after the last injection. Results are presented as the mean ± S.E. of
time spent in the aversive and non-aversive enclosed arms (A), percent time spent in
the aversive enclosed arm (B), percent time spent in the open arms (C) and total
number of entries in all arms of the apparatus (D). Two-way or one-way ANOVA
followed by Duncan’s test. *p<0.05 compared to time spent in the non-aversive
enclosed arm and p<0.05 compared to SAL group.

TRABALHO CIENTÍFICO A SER SUBMETIDO
43
DISCUSSION
The main findings of the present study were the following: 1) 64 and 128 mg/kg
of acutely administered MOD, but not 32 mg/kg, impaired the consolidation of a
discriminative avoidance task in mice, subsequently reversed after repeated
administration (10 days); 2) an acute pre-training administration of MOD did not
induce learning modifications nor anxiety-like behavior alteration, but it did impair
retention at doses of 32 and 64 mg/kg; 3) when the drug was given prior to the test,
only the lowest dose (32 mg/kg) was able to impair retrieval—an effect that was
subject to tolerance by repeated administration; 4) finally, the administration of 32
mg/kg was not state-dependent.
Several behavioral changes can influence the processes of acquisition,
processing, storage and retrieval of various memory systems. In this scenario, the
PM-DAT is a behavioral model able to evaluate the interactions among these
mnemonic processes, anxiety and locomotor activity in rodents in an integrative and
concomitant manner. In this paradigm, learning can be assessed by the magnitude of
the avoidance of the enclosed arm during the training session (Patti et al., 2010). The
storage of the task (and, consequently, the processes of consolidation and recall) is
detected during a test session by a subsequent direct comparison between the time
spent in the enclosed arm previously aversive in the training session and in the
enclosed arm that had not previously been aversive. In this way, memory is detected
by the percent time spent in the aversive enclosed arm. In this context, the avoidance
of the aversive enclosed arm upon testing has been validated as a measurement of
retention because amnestic manipulations decrease this effect (Claro et al., 1999;

TRABALHO CIENTÍFICO A SER SUBMETIDO
44
Silva et al., 1999, 2002a, 2004; Silva & Frussa-Filho, 2000, 2002; Kameda et al.,
2007; Alvarenga et al., 2008; Patti et al., 2010). In contrast, memory-improving
treatments increase this effect (Silva et al., 1997, 1999, 2000; Claro et al., 1999).
Furthermore, this behavioral model allows a simultaneous and independent
assessment of anxiety levels (through the avoidance of the open arms of the
apparatus) and locomotor activity (through the number of entries in all arms of the
apparatus). Therefore, the effectiveness of the model at detecting the effects of
anxiolytics agents (Silva & Frussa-Filho, 2000; Calzavara et al., 2004) and anxiogenic
agents (Silva & Frussa-Filho, 2000; Silva et al., 2002a; Silva et al., 2004b) has been
repeatedly demonstrated. At the same time, manipulations known to increase or
decrease the locomotor activity were able to increase (Silva et al., 2002a, Castro et
al., 2005; Kameda et al., 2007) or decrease (Silva et al., 2002b, 2004; Carvalho et al.,
2006) the total number of entries into the arms of the apparatus, respectively.
The results obtained in Experiment 1 show that the post-training acute
administration of 64 and 128 mg/kg of MOD promoted amnesia because the animals
treated at these doses spent an increased percent of their time in the aversive
enclosed arm during the test session. Of note, when MOD was given repeatedly and
subsequently (10 consecutive days; Experiment 5), the amnestic effect is no longer
observed. Although largely speculation, it may be possible that repeated treatment
with these high doses of the drug can enhance memory retrieval, thereby reversing
the post-training consolidation deficit.
To the best of our knowledge, only the study of Shuman and colleagues (2009)
systematically investigated the effects of MOD on memory consolidation. These
authors have reported that MOD was ineffective at modifying the consolidation of

TRABALHO CIENTÍFICO A SER SUBMETIDO
45
context- or cued-conditioned fear tasks in mice. The discrepant findings could be due
to the memory tasks employed in both studies.
These impairing effects on consolidation produced by post-training doses of 64
and 128 mg/kg of MOD were accompanied by hyperlocomotion. Because the drug
was no longer present during the test session (assessed 10 days after the drug’s
administration), impairment in habituation could be an explanation. In this context,
habituation is defined as a decrement in the behavioral response after a repeated
exposure to a certain stimulus (Levitan & Kaczmarek, 1991; Conceição et al., 1994).
In this way, the post-training administration of the drug impaired not only the
consolidation of the discriminative avoidance task but also the consolidation of the
memory of the apparatus as a whole, promoting a non-associative memory deficit.
Regarding learning (Experiment 2), mounting evidence suggests that MOD is
able to facilitate learning depending on the complexity of the task. Accordingly,
Bercacochea and colleagues (2001) found that the administration of 64 mg/kg of
MOD (but not 8 or 32 mg/kg) improved the performance of mice in a T-maze. The
greater the interval between training and testing, which makes the tasks more
complex, the better the performance in animals treated with MOD, suggesting an
interaction between task complexity and the possible cognitive effects of MOD
(Beracochea et al., 2002, 2003). Additionally, the pre-training administration of
75mg/kg of MOD improved performance of rats in the Morris water maze (Shuman et
al., 2009). No learning ability modifications were induced by any of the doses of
modafinil employed in the present study. Accordingly, a possible facilitative effect of
MOD on learning cannot be discarded because the absence of such an effect could
have resulted from a low percent time spent in the aversive enclosed arm at baseline

TRABALHO CIENTÍFICO A SER SUBMETIDO
46
(SAL-treated group), preventing a progressive enhancement of avoidance of that arm
during the training session in MOD-treated mice.
The pre-training administration of 32 and 64 mg/kg (but not 128 mg/kg) of
MOD impaired memory retention. Although these groups were able to discriminate
both enclosed arms (spending more time in the non-aversive enclosed arm, showing
retention of the task), they also spent an increased percent time in the aversive
enclosed arm, suggesting the presence amnestic effects of MOD. Conversely, the
dose of 128 mg/kg of MOD did not affect the pattern of exploration of the aversive
enclosed arm. These findings suggest that MOD promoted memory deficits in an
inverted-U shape fashion.
As for motor activity, a pre-training administration of MOD did not promote
changes in the mice’s locomotion. However, under our laboratory conditions, the
acute administration of MOD was able to promote an increase in the spontaneous
activity of mice exposed to an open-field in a dose-dependent manner. Thus, the
acute administration of doses ranging from about 64 to 128 mg/kg was capable of
promoting hyperlocomotor effects. Still, this stimulant effect was more pronounced
after the administration of 128 mg/kg of the drug (Wuo-Silva et al., 2011). A
speculative explanation for these behavioral differences could be the interaction
between the drug and the experimental environment. In other words, the presence of
the aversive stimuli could represent an unfavorable environment for exploration as
opposed to the open-field, a neutral environment. Thus, this environmental
component could promote an inhibition of exploration, abolishing the hyperlocomotor
effect of the drug. Within context, it should be noted that MOD did not change
animals’ anxiety levels per se at any of the doses tested.

TRABALHO CIENTÍFICO A SER SUBMETIDO
47
In the analysis of a possible effect of MOD on retrieval (Experiment 3), even
though all of the groups were able to discriminate between the enclosed arms of the
apparatus, the group treated with 32 mg/kg of MOD during the pre-test spent a longer
percent time in the aversive enclosed arm, indicating an impaired retrieval of the task.
Conversely, it was shown that under stressful situations (e.g., sleep deprivation),
MOD can restore the performance and activity of sleep-deprived mice, demonstrating
a retrograde effect of MOD on retrieval (Marinelli and Piazza, 2002). This discrepancy
may be due to the low stress involved in the aversive stimuli (no pain) presented in
the PM-DAT compared to that in the passive avoidance task, for example.
Importantly, the inhibitory effect of 32 mg/kg of MOD administration in the pre-test on
the retrieval of the PM-DAT task was not subject to tolerance after a previously
repeated treatment (Experiment 5). These data suggest that chronic treatment with
low doses of modafinil can produce important retrieval impairment.
A pre-test administration of MOD at the doses 32 or 64 mg/kg induced
hyperlocomotion. The enhancement in locomotion could be due to the stimulatory
effects of the drug. Previous exposure to the apparatus could have facilitated the
appearance of such an effect because emotional valence of the environment during
the second exposure seemed to be less aversive, since there was no longer noise
and light stimulation. In line with our hypothesis, MOD did not induce hyperlocomotion
when given prior to training (Experiment 2) when the aversive stimuli were presented.
It could be argued that the impairing effects of MOD on retention and retrieval
are state-dependent. Memory can be state-dependent, in that a response that has
been acquired in a given (i.e., drug-induced) state may not be retrieved when the
organism is in a different state (Colpaert et al., 2001). A pre-test administration of 32

TRABALHO CIENTÍFICO A SER SUBMETIDO
48
mg/kg of MOD was not able to counteract the pre-training-induced amnestic effects of
this drug and vice versa, discounting, at least for this range of doses, the involvement
of the state-dependency phenomenon on MOD-induced memory impairment.
Interestingly, mice in the MOD-MOD group had increased general activity
during the test session. In fact, animals that were treated with MOD before both
sessions had more increased locomotion in the test session compared to animals that
received MOD only before testing. This result agrees with reports that the repeated
treatment with stimulant doses of this drug induces an increase in its locomotor
stimulatory effect in mice, a well-known phenomenon called behavioral sensitization
that was proposed to share neurochemical mechanism with drug dependence (Wuo-
Silva et al., 2011). Specifically, Wuo-Silva and colleagues (2011) have reported that
MOD produced a robust locomotor sensitization, conditioned place preference, and a
cross-sensitization with cocaine. Within this context, although behavioral sensitization
is usually observed after repeated treatment with drugs of abuse, it has been shown
that it is not necessary to repeatedly administer a drug for long periods of time to
produce such a phenomenon. Indeed, a single injection of amphetamine
(Vanderschuren et al., 1999; Costa et al., 2001; Frussa-Filho et al., 2004; Chinen et
al., 2006) or ethanol (Fukushiro et al., 2010) has been reported to enhance locomotor
stimulation produced by a subsequent injection of the drug given hours or weeks
later.
Collectively, our data suggest that MOD has potential amnestic effects
depending on the dose and the timing of administration. Although we did not observe
effects on emotion, modafinil seems to also have an addictive potential. These results
point to the need of caution in prescribing this drug.

TRABALHO CIENTÍFICO A SER SUBMETIDO
49
DISCLOSURE STATEMENT
All authors disclose any actual or potential conflict of interest including any
financial, personal or other relationships with other people or organizations within
three years of beginning the work that could inappropriately influence (bias) the
present work. All authors critically reviewed content and approved final version for
publication.

TRABALHO CIENTÍFICO A SER SUBMETIDO
50
CONTRIBUTORS
HAF, CLP, RWS and RF-F were responsible for the study concept and design.
HAF, LFS and KAZ contributed to the acquisition of animal data. HAF, CLP, KAZ,
RWS, LFS and RF-F assisted with data analysis and interpretation of findings. HAF,
CLP, KAZ and RF-F drafted the manuscript. LRB and ST provided critical revision of
the manuscript for important intellectual content.

TRABALHO CIENTÍFICO A SER SUBMETIDO
51
ACKNOWLEDGMENTS
This research was supported by fellowships from Fundação de Amparo a
Pesquisa do Estado de São Paulo (FAPESP), Conselho Nacional de
Desenvolvimento Científico e Tecnológico (CNPq), Fundação Coordenação de
Aperfeiçoamento de Pessoal de Nível Superior (CAPES) and Associação Fundo de
Pesquisa em Psicobiologia (AFIP). The English language in this manuscript has been
proofread and edited by American Journal Experts (AJE), but we are entirely
responsible for the scientific content of the paper. The authors would like to thank Ms.
Teotila R. R. Amaral, Ms. Marilde Costa, Ms. Claudenice M. dos Santos, Mr. Cleomar
S. Ferreira and Mr. Antonio Rodrigues dos Santos for their capable technical
assistance. R.F.F., S.T. and L.R.B. are recipients of the CNPq fellowship.

TRABALHO CIENTÍFICO A SER SUBMETIDO
52
REFERENCES
Alvarenga TA, Patti CL, Andersen ML, Silva RH, Calzavara MB, Lopez GB, Frussa-Filho R, Tufik S. (2008). Paradoxical sleep deprivation impairs acquisition, consolidation, and retrieval of a discriminative avoidance task in rats. Neurobiol Learn Mem. 90:624-32.
Béracochéa, D; Cagnard, B; Celerier, A; Merrer, J; peres, M; Piérard, C. (2001). First evidence of a delay-dependent working memory – enhancing effect of modafinil in mice. Neuroreport 12: 375-378.
Béracochéa, D; Celerier, A; Borde, N; Valleau, m; Peres, M; Piérard, C., (2002). Improvement of learning processes following chronic systemic administration of modafinil in mice. Pharmacology, Biochemistry and behavior 73: 723-728.
Béracochéa, D; Liscia, P; Tonche, C; Chauveau, F. Jouanin, J. C; Piérard, C. (2007). Stress modulation of the memory retrograde-enhancing effects of the awakening drug modafinil in mice. Psychopharmacology DOI 10.1007/s00213-007-0915-8.
Béracochéa, D; Celerier, A; Peres, M; Piérard, C. (2003). Enhancement of learning processes following na acute modafinil injection in mice. . Pharmacology, Biochemistry and behavior 76: 473-479.
Calzavara MB, Lopez GB, Abílio VC, Silva RH, Frussa-Filho R.(2004) Role of anxiety levels in memory performance of spontaneously hypertensive rats. Behav Pharmacol. 15:545-53.
Carvalho RC, Patti CC, Takatsu-Coleman AL, Kameda SR, Souza CF, Garcez-do-Carmo L, Abílio VC, Frussa-Filho R, Silva RH. (2006) Effects of reserpine on the plus-maze discriminative avoidance task: dissociation between memory and motor impairments. Brain Res. 29:179-83.
Castro, J.P.; Frussa-Filho, R.; Fukushiro, D.F.; Chinen, C.C.; Abílio, V.C.; Silva, R.H.(2005). Effects of long-term continuous exposure to light on memory and anxiety in mice. Physiol. Behav. 86:218-23.
Claro, F.T.; Silva, R.H.; Frussa-Filho, R. (1999). Bovine brain phosphatidylserine attenuates scopolamine-induced amnesia. Physiol. Behav. 67:551-4.
Chinen CC, Faria RR, Frussa-Filho R. (2006) Characterization of the rapid-onset type of behavioral sensitization to amphetamine in mice: role of drug-environment conditioning. Neuropsychopharmacology. 31:151-9.
Colpaert FC, Koek W, Bruins Slot LA. (2001) Evidence that mnesic states govern normal and disordered memory. Behav Pharmacol. 12:575-89.
Conceição IM, Maioline Júnior M, Mattia NF, Chang YH, Smaili S, Frussa-Filho R. (1994) Effect of withdrawal from long-term nifedipine administration on open-field habituation in the rat. Braz J Med Biol Res. 27:1363-7.

TRABALHO CIENTÍFICO A SER SUBMETIDO
53
Costa FG, Frussa-Filho R, Felicio LF. (2001). The neurotensin receptor antagonist, SR48692, attenuates the expression of amphetamine-induced behavioural sensitisation in mice. Eur J Pharmacol. 28:97-103.
Fukushiro DF, Benetti LF, Josino FS, Oliveira GP, Fernandes M, Saito LP, Uehara RA, Wuo-Silva R, Oliveira CS, Frussa-Filho R. (2010). Environmental novelty and illumination modify ethanol-induced open-field behavioral effects in mice. Pharmacol Biochem Behav. 95:13-22.
Kameda SR, Frussa-Filho R, Carvalho RC, Takatsu-Coleman AL, Ricardo VP, Patti CL, Calzavara MB, Lopez GB, Araujo NP, Abílio VC, Ribeiro Rde A, D'Almeida V, Silva RH. (2007). Dissociation of the effects of ethanol on memory, anxiety, and motor behavior in mice tested in the plus-maze discriminative avoidance task. Psychopharmacology (Berl). 192:39-48.
Levitan ES, Hemmick LM, Birnberg NC, Kaczmarek LK. (1991). Dexamethasone increases potassium channel messenger RNA and activity in clonal pituitary cells. Mol Endocrinol. 5:1903-8.
Marinelli M, Piazza PV. (2002). Interaction between glucocorticoid hormones, stress and psychostimulant drugs. Eur J Neurosci. 16:387-94.
McGaugh J. L. (2000). Memory – a century of consolidation. Science 14: 248-251. Minzemberg MJ, Carter CS (2008). Modafinil: a review of neurochemical actions and
effects on cognition. Neuropsychopharmacology 33: 1477-1502.
Morgan, R. E; Crowley, J. M; Smith, R. H; LaRoche, R. B. Dopheide, M. M. (2007). Modafinil improves attention, inhibitory control, and reaction time in healthy, middle-aged rats. Pharmacology, Biochemistry and Behavior 86: 531-541.
Patti CL, Zanin KA, Sanday L, Kameda SR, Fernandes-Santos L, Fernandes HA, Andersen ML, Tufik S, Frussa-Filho R. (2010). Effects of sleep deprivation on memory in mice: role of state-dependent learning. Sleep. 33:1669-79.
Patti, C.L.; Kameda, S.R.; Carvalho, R.C.; Takatsu-Coleman, A.L.; Lopez, G.B.; Niigaki, S.T.; Abílio, V.C.; Frussa-Filho, R.; Silva, R.H. (2006). Effects of morphine on the plus-maze discriminative avoidance task: role of state-dependent learning. Psychopharmacol. 184:1-12.
Piérard, C; Liscia, P; Philippin, J. N; Mons, N; Lafon, T; Chauveau, F; Beers, P. V; Drouet; Serra, A; Jouanin, J. C; Béracochéa, D. (2007). Modafinil restores memory performance and neural activity impaired by sleep deprivation in mice. Pharmacology, Biochemistry and Behavior 88: 55-63.
Piérard, C; Liscia, P; Valleau, M; Drouet, I; Cauveau, F; Huart, B; Bonneau, D; Jouanin, J. C; Beaumont, M. Béracochéa, D. (2006). Modafinil-induced modulation of working memory and plasma corticosterone in chronically-stressed mice. Pharmacology, Biochemistry and Behavior 83: 1-8.
Shuman T, Wood SC, Anagnostaras SG.(2009). Modafinil and memory: effects of modafinil on Morris water maze learning and Pavlovian fear conditioning. Behav Neurosci. 123:257-66.

TRABALHO CIENTÍFICO A SER SUBMETIDO
54
Silva, R.H.; Abílio, V.C.; Takatsu, A.L.; Kameda, S.R.; Grassl, C.; Chehin, A.B.; Medrano, W.A.; Calzavara, M.B.; Registro, S.; Andersen, M.L.; Machado, R.B.; Carvalho, R.C.; Ribeiro, R. De A.; Tufik, S.; Frussa-Filho, R. (2004). Role of hippocampal oxidative stress in memory deficits induced by sleep deprivation in mice. Neuropharmacol. 46:895-903.
Silva, R.H.; Kameda, S.R.; Carvalho, R.C.; Rigo, G.S.; Costa, K.L.; Taricano, I.D.; Frussa-Filho, R. (2002a). Effects of amphetamine on the plus-maze discriminative avoidance task in mice. Psychopharmacol. 160:9-18.
Silva, R.H.; Frussa-Filho, R. (2002). Naltrexone potentiates both amnestic and anxiolytic effects of chlordiazepoxide in mice. Life sci. 72:721-30.
Silva, R.H.; Abílio, V.C.; Torres-Leite, D.; Bergamo, M.; Chinen, C.C.; Claro, F.T.; Carvalho, R. De C.; Frussa-Filho, R. (2002b). Concomitant development of oral dyskinesia and memory deficits in reserpine-treated male and female mice. Behav. Brain res. 132:171-177.
Silva, R.H.; Frussa-Filho, R. (2000). The plus-maze discriminative avoidance task: a new model to study memory-anxiety interactions. Effects of chlordiazepoxide and caffeine. J. Neurosci. Methods. 102:117-25.
Silva, R.H.; Bellot, R.G.; Vital, M.A.; Frussa-Filho, R. (1997). Effects of long-term ganglioside Gm1 administration on a new discriminative avoidance test in normal adult mice. Psychopharmacol. 129:322-8.
Silva, R.H.; Bergamo, M.; Frussa-Filho, R. (2000). Effects of neonatal ganglioside gm1 administration on memory in adult and old rats. Pharmacol. Toxicol. 87:120-5
Silva, R.H.; Felicio,L.F.; Frussa-Filho, R. (1999). Ganglioside Gm1 attenuates scopolamine-induced amnesia in rats and mice. Psychopharmacol. 141:111-7.
Swanson JM, Greenhill LL, Lopez FA, Sedillo A, Earl CQ, Jiang JG, Biederman J (2006). Modafinil film-coated tablets in children and adolescents with attention-deficit/hyperactivity disorder: results of a randomized, double-blind, placebo-controlled, fixed-dose study followed by abrupt discontinuation. J Clin Psychiatry 67: 137-147.
Vanderschuren LJ, Schmidt ED, De Vries TJ, Van Moorsel CA, Tilders FJ, Schoffelmeer AN. (1999). A single exposure to amphetamine is sufficient to induce long-term behavioral, neuroendocrine, and neurochemical sensitization in rats. J Neurosci. 19:9579-86.
Ward, C. P.; Harsh, J. R.; York, K. M.; Stewart, K. L. Mccoy, J. G. (2004). Modafinil facilitates performance on a delayed nonmatching to position swin task in rats. Pharmacology, Biochemistry and Behavior 78: 735-741.
Wuo-Silva, R; Fukushiro, D; Borçoi, A; Fernandes, H.A; Procópio-Souza, R; Hollais, A; Santos, R; Ribeiro, L.T.C; Corrêa, J; Talhati, F; Saito, L; Aramini, T; Kameda, S; Bittencourt, L.R.A.; Tufik, S.; Frussa-Filho, R. (2011). Addictive potential of modafinil and cross-sensitization with cocaine: a preclinical study. Addiction Biology.

CONCLUSÕES
55
4. CONCLUSÕES
4.1. Conclusões Específicas
A administração aguda de 64 ou 128 mg/Kg (mas não de 32 mg/Kg) de
modafinil pós-treino promove déficits na consolidação da memória de uma tarefa de
esquiva discriminativa e de habituação, uma memória não-associativa. Ainda,
quando tal administração é realizada de maneira prolongada (10 dias consecutivos),
há uma reversão dos efeitos amnésticos induzidos por essas maiores doses de
modafinil na tarefa de esquiva discriminativa.
A administração aguda de modafinil não promove modificações na
aquisição da tarefa discriminativa ou nos níveis de ansiedade de camundongos.
Contudo, tal administração pré-treino promove déficits de retenção quando realizada
nas doses de 32 ou 64 mg/Kg.
A administração aguda de 32 mg/Kg modafinil previamente à sessão de
teste promove déficits de evocação da tarefa discriminativa e efeito estimulante nas
doses de 32 ou 64 mg/Kg.
Os déficits de retenção induzidos pela administração pré-treino ou pré-
teste de 32 mg/Kg de modafinil não estão relacionados ao fenômeno de dependência
de estado. Entretanto a administração repetida da droga (antes das sessões de

CONCLUSÕES
56
treino e de teste) parece induzir sensibilização do efeito hiperlocomotor agudo da
droga, um fenômeno comum a todas as drogas de abuso.
4.2. Conclusão Geral
Tomados em conjunto, os presentes achados fornecem evidências pré-
clínicas do potencial amnéstico e de abuso do modafinil.

TRABALHO CIENTÍFICO A SER SUBMETIDO
57
5. REFERÊNCIAS BIBLIOGRÁFICAS
ALVARENGA, T.A.; PATTI, C.L.; ANDERSEN, M.L.; SILVA, R.H.; CALZAVARA, M.B.; LOPEZ, G.B.; FRUSSA-FILHO, R.; TUFIK, S. Paradoxical sleep deprivation impairs acquisition, consolidation, and retrieval of a discriminative avoidance task in rats. Neurobiol. Learn. Mem. 90:624-32, 2008.
ASHTON, H. 1997. Brain systems disorders and psychotropic drugs. Oxford: Oxford university Press.
BALLON, J. S.; FEIFEL, D., 2006. A systematic review of modafinil: potencial clinical uses and mechanisms of action. Journal of Clinical Psychiatry 67 (4): 554-566.
BARATTI, CM; HUYGENS, P; MINO, J; MERLO, A; GARDELLA, 1979.J. Memory facilitation with post-trial injection of oxotremonine and physostigmine in mice. Psycopharmacology. v.64, p. 85-88.
BATTIG, K. 1970. The effect of pre- and post-trial application of nicotine on the 12 problems of Hebb-Willians Test in the rat. Psycopharmacology. v. 18, p. 68-76.
BÉRACOCHÉA, D; CELERIER, A; BORDE, N; VALLEAU, M; PERES, M; PIERARD, C., 2002. Improvement of learning processes following chronic systemic administration of modafinil in mice. Pharmacology, Biochemistry and behavior 73: 723-728.
BÉRACOCHÉA, D; LISCIA, P; TONCHE, C; CHAUVEAU, F. JOUANIN, J. C; PIÉRARD, C. 2007. Stress modulation of the memory retrograde-enhancing effects of the awakening drug modafinil in mice. Psychopharmacology DOI 10.1007/s00213-007-0915-8.
CAHLL, L.; MCGAUGH, J.L., 1990. Amygdaloid complex lesions differentially affect retention of tasks using appetitive and aversive reinforcement. Behavioral Neuroscience 104: 532-543.
CASTRO, J.P.; FRUSSA-FILHO, R.; FUKUSHIRO, D.F.; CHINEN, C.C.; ABÍLIO, V.C.; SILVA, R.H.(2005). Effects of long-term continuous exposure to light on memory and anxiety in mice. Physiol. Behav. 86:218-23.
DAVIS, M., 1992. The role of amygdala in fear-potentiated startle: implications for animal models of anxiety. Trends in Pharmacological Sciences 13: 35-41.
DAVIS, M.; WALKER, D. L.; LEE, Y., 1997. Amygdala and bed nucleus of the strial terminalis: diferential roles in fear and anxiety measured with the acoustic startle reflex. Philosophical Transactions of the Royal Society of London B Biological Sciences 352: 1675-1678.
DEROCHE-GAMONET, V.; DARNAUDÉRY, M.; BRUINS-SLOT, L.; PIAT, F.; LE MOAL, M.; PIAZZA, P.V. Study of the addictive potential of modafinil in naïve

TRABALHO CIENTÍFICO A SER SUBMETIDO
58
and cocaine-experienced rats. Psychopharmacology (Berl). 161: 387-395, 2002.
DEUTSCH, J. A.; ROERS, J. B., 1979. Cholinergic excitability and memory: animal studies and their clinical implications. In: Davis KL; Berger, PA (eds.), Brain Acetilcholine and Neuropsychiatric Disease. Plenum, New York, 175-204.
DISMUKES, R. K.; RAKE, A. V., 1972. Involvement of biogenic amines on memory formation. Psycopharmacology 23: 17-25.
DOPHEINE, M. M.; MORGAN, R. E.; RODVELT, K. R.; SCHACHTMAN, T. R.; MILLER, D. K., 2007. Modafinil evoles striatal [3H] dopamine release and alters the subjective properties of stimulants. European Journal of Pharmacology 568 (1-3): 112-123.
DUTEIL, J.; RAMBERT, F.A.; PESSONNIER, J.; HERMANT, J.F.; GOMBERT, R.; ASSOUS, E. Central alpha 1-adrenergic stimulation in relation to the behaviour stimulating effect of modafinil; studies with experimental animals. Eur J Pharmacol. 180: 49-58, 1990.
ERICKSON, C. K., 1971. Studies on the mechanism of avoidance facilitation by nicotine. Psycopharmacology 22: 357-368.
EVANGELISTA, A. M.; GATTONI, R. C.; IZQUIERDO, I., 1970. Effect of amphetamine, nicotine and hexamethonium on performance of a conditioned response during acquisition and retention trials. Pharmacology 3: 91-96.
FERRARO, L.; TANGANELLI, S.; FUXE, K.; BEBE, B. W.; TOMASINI, M. C.; RAMBERT, F. A.; ANTONELLI, T., 2001. Modafinil does not affect serotonin efflux from rat frontal cortex synaptosomes: comparison with know serotonergic drugs. Brain Research 894: 307-310.
FERRARO, L.; ANTONELLI, T.; O’CONNOR, W. T.; TANGANELLI, S.; RAMBERT, F. A.; FUXE, K. Modafinil: an antinarcoleptic drug with a different neurochemical profile to d-amphetamine and dopamine uptake blockers. Biol Psychiatry. 43: 1181-1183, 1997.
FERRARO, L.; TANGANELLI, S.; O’CONNOR, W. T.; ANTONELLI, T.; RAMBERT, F.;FUXE, K., 1996. The vigilance promoting drug modafinil decreases GABA release in the medail preoptic area and in the posterior hypothalamus of the awake rat: possible involvement of the serotonergic 5-HT3 receptor. Neuroscience Letters 220: 5-8.
FERREIRA, M. B. C.; DA SILVA, R. C.; MEDINA, J. H.; IZQUIERDO, I., 1992. Late posttraining memory processing by entorhinal cortex: involvement of NMDA and GABAergic receptors. Pharmacology, Biochemistry and Behavior 41: 767-771.
GARG, M., 1969. The effect of nicotine on two different types of learning. Psycopharmacology 15: 408-414.

TRABALHO CIENTÍFICO A SER SUBMETIDO
59
GIVENS, B. S.; OLTON, D. S., 1990. Cholinergic and GABAergic modulation of medial septal área: effects on working memory. Behavioral Neuroscience 104: 849-855.
GLICK, S. D.; ZIMMERBERG, B., 1971 Comparative learning impairment and amnesia by scopolamine, phencyclidine and ketamine. Psychonomic Science. 25: 165-166.
GOLD, L. H.; BALSTER, R. L. Evaluating of the cocaine-like discriminative stimulus effects and reinforcing effects of modafinil. Psychopharmacolgy (Berl). 126: 286-292, 1996.
GORENSTEIN, C.; BERNIK, M. A.; GENTIL, V., 1994. Bases biológicas da ansiedade. In: GENTIL, V.; LOTUFEO-NETO. F., eds. Pânico, fobias e obssessões. São Paulo: Edusp, p. 59-71.
GRAEFF, F. G., 1984 Drogas psicotrópicas e seu modo de ação. São Paulo: Edusp.
GRAEFF, F. G., 1993 Ansiedade. In: GRAEFF, F. G.; BRANDÃO, M. L., eds. Neurobiologia das doenças mentais. São Paulo: Editora Lemos. p. 109-144.
GRAEFF, F. G.; VIANA, M. B.; TOMAZ, C., 1993. The elevated T maze, a new experimental model of anxiety and memory, effects of diazepan. Brazilian Journal of Medical and Biological Research 26: 67-70.
GRAY, J. A., 1982. The neuropsycholoogy of anxiety: an inquiry into the functions of the septo-hippocampal system. Oxford: Oxford University Press.
HANDLEY, S. L.; MITHANI, S., 1984. Effects of alpha2-adrenoceptor agonists and antagonists in a maze-exploration model of fear-motivated behavior. Naunyn-Schmiedeberg’s Archives Pharmacology 327: 1-5.
HERMANT, J.F.; RAMBERT, F.A.; DUTEIL, J. Awakening properties of modafinil: effect on nocturnal activity in monkeys (Macaca mulatta) after acute and repeated administration. Psychopharmacology (Berl). 103(1): 28-32, 1991.
HUANG, Q.; ZHANG, L.; TANG, H.; WANG, L.; WANG, Y. Modafinil modulates GABA-activated currents in rat hippocampal pyramidal neurons. Brain Res. 1208: 74-78, 2008.
ISHIZUKA, T.; MURAKAMI, M.; YAMATODANI, A., 2008. Involvement of central histaminergic systems in modafinil-induced but not methylphenidate-induced increases in locomotor activity in rats. European Journal Pharmacology: 1-7
IZQUIERDO, I.; MEDINA, J. H., 1991. GABA-A receptor modulation of memory: the role of endogenous benzodiazepines. Trends in Pharmacological Science 12: 260-265.
JASINSKI, D.R.; KOVACEVIC-RISTANOVIC, R. Evaluation of the abuse liability of modafinil and other drugs for excessive daytime sleepiness associated with narcolepsy. Clin Neuropharmacol. 23(3): 149-156, 2000.

TRABALHO CIENTÍFICO A SER SUBMETIDO
60
JERUSALINSKY, D.; KORNISIUK, E.; IZQUIERDO, I., 1997. Cholinergic neurotransmission and synaptic plasticity concerning memory processing. Neurochemical. Research 22: 507-515.
KANDEL, E. R., 2003. Mecanismos celulares da aprendizagem e as bases biológicas da individualidade In: KANDEL, E. R.; SCHWARTS, J. H.; JESSEL, T. M. eds., Princípios da Neurociência. São Paulo: Manole Editora, p. 1247-1280.
KOROTKOVA, T. M.; KLYUCH, B. P.; PONOMARENKO, A. A.; LIN, J. S.; HAAS, H. L.; SERGEEVA, O. A., 2007. Modafinil inhibits rat midbrain dopaminergic neurons through D2-like receptors. Neuropharmacology 52: 626-633.
LIN, J.S.; HOU, Y.; JOUVET, M. Potential brain neuronal targets for amphetamine-, methylphenidate-, and modafinil-induced wakefulness, evidenced by c-fos immunocytochemistry in the cat. Proc Natl Acad Sci USA., 93(24): 14128-14133, 1996.
LIN, J.S.; ROUSSEL, B.; AKAOKA, H.; FORT, P.; DEBILLY, G.; JOUVET, M. Role of catecholamines in the modafinil and amphetamine induced wakefulness, a comparative pharmacological study in the cat. Brain Res. 591(2): 319-326, 1992.
LISTER, R. G., 1987. The use of a plus-maze to measure anxiety in the mouse. Psychopharmacology 92: 180-185.
MADRAS B.K.; XIE, Z.; LIN, Z.; JASSEN, A.; PANAS, H.; LYNCH, L.; JOHNSON, R.; LIVNI, E.; SPENCER, T.J.; BONAB, A.A.; MILLER, G.M.; FISCHMAN, A.J. Modafinil occupies dopamine and norepinephrine transporters in vivo and modulates the transporters and trace amine activity in vitro. J Pharmacol Exp Ther. 319: 561-569, 2006.
MATHEWS, A., 1990. Why worry? The cognitive function of anxiety. Behaviour Research and Therapy 28: 455-568.
MCGAUGH J. L. (2000). Memory – a century of consolidation. Science 14: 248-251. MIGNOT, E.; NISHINO, S.; GUILLEMINAULT, C.; DEMENT, W.C. Modafinil binds to
the dopamine uptake carrier site with low affinity. Sleep. 17(5): 436-437, 1994.
MITCHELL, H.A.; BOGENPOHL, J.W.; LILES, L.C.; EPSTEIN, M.P.; BOZYCZKO-COYNE, D.; WILLIAMS, M.; WEINSHENKER, D. Behavioral responses of dopamine β-hydroxylase knockout mice to modafinil suggest a dual noradrenergic-dopaminergic mechanism of action. Pharmacol Biochem Behav. 91(2): 217-222, 2008.
MINZENBERG, M. J.; CARTER, C. S., 2008. Modafinil: a review of neurochemical actions and effects on cognition. Neuropsychopharmacology: 1-26.
MISHKIN, M.; MALAMUT, B.; BACHEVALIER, J., 1984. Memories And Habits: Two Neural Systems. In: LYNCH, G.; MCGAUGH, J. L.; WEINBERGER, N. M., eds. Neurobiology of learning and memory. New York, Guilford Press, p. 65-77.

TRABALHO CIENTÍFICO A SER SUBMETIDO
61
MONTGOMERY, K. C., 1955. The relation between fear induced by novel stimulation and exploratory behavior. Journal of Comparative Physiology and Psychology 48: 254-260.
MORGAN, R. E.; CROWLEY, J. M.; SMITH, R. H.; LaROCHE, R. B.; DOPHEIDE, M. M., 2007. Modafinil improves attention, inhibitory control, ande reaction time in healthy, middle-aged rats. Pharmacology, Biochemistry and Behavior 86: 531-541.
MÜLLER, U; STEFFENHAGEN, N; REGENTHAL; BUBLAK P., 2004. Effects of modafinil on working memory processes in humans. Psycopharmacology 177: 161-169.
MURILO-RODRÍGUEZ, E.; HARO, R.; PALOMERO-RIVERO, M.; MILLÁN-ALDACO, D.; DRUCKER-COLÍN, R. Modafinil enhances extracellular levels of dopamine in the nucleus accumbens and increases wakefulness in rats. Behav Brain Res. 176: 353-357, 2007.
NUTT, D. J., 1990. The pharmacology of human anxiety. Pharmacology & Therapeutics 47: 233-266.
PATTI, C.L.; KAMEDA, S.R.; CARVALHO, R.C.; TAKATSU-COLEMAN, A.L.; LOPEZ, G.B.; NIIGAKI, S.T.; ABÍLIO, V.C.; FRUSSA-FILHO, R.; SILVA, R.H. Effects of morphine on the plus-maze discriminative avoidance task: role of state-dependent learning. Psychopharmacol. 184:1-12, 2006.
PIÉRARD, C; LISCIA, P; PHILIPPIN, J. N; MONS, N; LAFON, T; CHAUVEAU, F; BEERS, P. V; DROUET; SERRA, A; JOUANIN, J. C; BÉRACOCHÉA, D. (2007). Modafinil restores memory performance and neural activity impaired by sleep deprivation in mice. Pharmacology, Biochemistry and Behavior 88: 55-63.
PIÉRARD, C; LISCIA, P; VALLEAU, M; DROUET, I; CAUVEAU, F; HUART, B; BONNEAU, D; JOUANIN, J. C; BEAUMONT, M. BÉRACOCHÉA, D. (2006). Modafinil-induced modulation of working memory and plasma corticosterone in chronically-stressed mice. Pharmacology, Biochemistry and Behavior 83: 1-8.
PELLOW, S.; CHOPIN, P.; FILE, S. E.; BRILEY, M., 1985. Validation of open:closed arm entries in an elevated plus-maze as a measure of anxiety in the rat. Journal of Neuroscience Methods 14: 149-167.
PELLOW, S.; FILE, S. E., 1986. Anxyiolitic and anxiogenic drug effects on exploratory activity in an elevated plus-maze: a novel test of anxiety in the rat. Pharmacology, Biochemistry and Behavior 24: 525-529.
PIÉRARD, C; LISCIA, P; PHILIPPIN, J. N; MONS, N; LAFON, T; CHAUVEAU, F; BEERS, P. V; DROUET; SERRA, A; JOUANIN, J. C; BÉRACOCHÉA, D. 2007. Modafinil restores memory performance and neural activity impaired by sleep deprivation in mice. Pharmacology, Biochemistry and Behavior 88: 55-63.
PIÉRARD, C; LISCIA, P; VALLEAU, M; DROUET, I; CAUVEAU, F; HUART, B; BONNEAU, D; JOUANIN, J. C; BEAUMONT, M. BÉRACOCHÉA, D. 2005. Modafinil-induced modulation of working memory and plasma corticosterone in

TRABALHO CIENTÍFICO A SER SUBMETIDO
62
chronically-stressed mice. Pharmacology, Biochemistry and Behavior 83: 1-8.
PRATT, J. A., 1992. The neuroanatomical basis of anxiety. Pharmacology & Therapeutics 55: 149- 181.
QU, W.M.; HUANG, Z.L.; XU, X.H.; MATSUMOTO, N.; URADE, Y. Dopaminergic D1 and D2 receptors are essential for the arousal effect of modafinil. J Neurosci. 28(34): 8462-8469, 2008.
RANDALL, D. C; FLECK, N. L; SHNEERSON, J. M; FILE, S. E. 2004. The cognitive-enhancing properties of modafinil are limited in non-sleep-deprived middle-aged volunteers. Pharmacology, Biochemistry and Behavior 77: 547-555.
RANDT, C. T.; QUARTERMAN, D.; GOLDSTEIN, M.; ANAGNOSTE, B., 1971. Norepinephrine biosyntesis inhibition: effects on memory in mice. Science 172: 498-499.
RODGERS, R. J.; CAO, B. J.; DALVI, A.; HOLMES, A., 1997. Animal models of anxiety: an ethological perspective. Brazilian Journal Medical and Biological Research 30: 289-304.
RUSH, C. R.; KELLY, T. H.; HAYS, L. R.; WOOTEN, A. F., 2002. Discriminative-stimulus effects of modafinil in cocaine-trained humans. Drug and Alcohol Dependence 67: 311-322.
SARTER, M.; MARKOWITSCH, H. J., 1985. The amygdala’s role in human menrmonic processing. Cortex 21: 7-24.
SIEGEL, S., 1956. Non-parametric Statistics for the Behavioral Sciences, New York: McGraw-Hill.
SILVA R. H.; FRUSSA-FILHO, R., 2000. The plus-maze discriminative avoidance task: a new model to study memory- anxiety interactions. Effects of chlordiazepoxide and caffeine. Journal of Neuroscience Methods 102: 117-125.
SILVA, R. H.; ABILIO, V. C.; TORRES-LEITE, D.; BERGAMO, M.; CHINEN, C. C.; CLARO, F. T.; CARVALHO, R. C.; FRUSSA-FILHO, R., 2002 Concomitant development of oral dyskinesia and memory deficits in reserpine-treated male and female mice. Behavioural Brain Research 132 (2): 171-177.
SILVA, R. H.; BELLOT, R.; VITAL, M. A. B. F.; FRUSSA-FILHO, R., 1997. Effects of long-term gklicosideGM1 administration on a new discriminative avoidance test in normal adult mice. Psycopharmacology 129: 322-328.
SILVESTRI, A. J.; SANFORD, L. D.; ROSS, R. J.; MANN, G. L.; PAVLOCK, A.; MORRISON, A. R., 2002. The central nucleos of the amygdale and the wake-promoting effects of modafinil. Brain Research 941: 43-52
SIMON, P; PANISSAUD, C; COSTENTIN, J. 1994. the stimulant effect of modafinil on wakefulnessm is not associated with an increase in anxiety in mice. Psychopharmacology 114: 597-600.
SNEDOCOR, G. W., 1946. Statistical Methods, Iowa State University Press, Ames.

TRABALHO CIENTÍFICO A SER SUBMETIDO
63
STECKLER, T.; SAHGAL, A., 1995. The role of serotoninergic-cholinergic interactions of anticholinesterase drug and maze learning in two strains of rats. Psycopharmacology 5: 47-54.
STOOPS, W.W.; LILE, J.A.; FILLMORE, M.T.; GLASER, P.E.A.; RUSH, C.R. Reinforcing effects of modafinil: influence of dose and behavioral demands following drug administration. Psychopharmacology (Berl). 182: 186-193, 2005.
STRATON, L. D.; PETRINOVICH, L. F., 1963. Post-trial injections of an anticholinesterase drug and maze learning in two strains of rats. Psycopharmacology 5: 47-54.
TANGANELLI, S.; FUXE, K.; FERRARO, L.; JANSON, A. M.; BIANCHI, C., 1992. Inhibitory effects of the psychoative drug modafinil on -aminobutyric acid outflow from the cerebral cortex of the awake freely moving guinea-pig. Archives of Pharmacology 345: 461-465.
TEICHER, M. H., 1988. Biology of anxiety. Medical Clinics of North America 72: 791-814.
THOMPSON, A. M.; GOSNELL, B. A.; WAGNER, J. J., 2002. Enhancement of long-term potentiation in rat hippocampus following cocaine exposure. Neuropharmacology 42: 1039-1042.
TURNER, D. C; ROBBINS, T. W; CLARK, L; ARON, A. R; DOWSON, J; SAHAKIAN, B. J. 2003. Cognitive enhancing effects of modafinil in healthy volunteers. Psychopharmacology 165: 260-269.
VOLKOW, N.D.; FOWLER, J.S.; LOGAN, J.; ALEXOFF, D.; ZHU, W.; TELANG, F.; WANG, G.J.; JAYNE, M.; HOOKER, J.M.; WONG, C.; HUBBARD, B.; CARTER, P.; WARNER, D.; KING, P.; SHEA, C.; XU, Y.; MUENCH, L.; APELSKOG-TORRES, K. Effects of modafinil on dopamine and dopamine transporters in the male human brain. Jama. 301(11): 1148-1159, 2009.
WARD, C. P.; HARSH, J. R.; YORK, K. M.; STEWART, K. L. McCOY, J. G., 2004. Modafinil facilitates performance on a delayed nonmatching to position swin task in rats. Pharmacology, Biochemistry and Behavior 78: 735-741.
WEISSMAN, A., 1967. Drugs and retrograde amnesia. International Review of Neurobiology 10: 167 –168.
WIENER, N. I.; MESSER, J., 1973. Scopolamine-induced impairment of long-term retention in rats. Behavioral Biology 9: 227-234.
WISOR, J.P. ERIKSSON, K.S. Dopaminergic-adrenergic interactions in the wake promoting mechanism of modafinil. Neuroscience. 132: 1027-1034, 2005.
WUO-SILVA, R; FUKUSHIRO, D; BORÇOI, A; FERNANDES, H.A; PROCÓPIO-SOUZA, R; HOLLAIS, A; SANTOS, R; RIBEIRO, L.T.C; CORRÊA, J; TALHATI, F; SAITO, L; ARAMINI, T; KAMEDA, S; BITTENCOURT, L.R.A.; TUFIK, S.; FRUSSA-FILHO, R. (2011). Addictive potential of modafinil and cross-sensitization with cocaine: a preclinical study. Addiction Biology.

REFERÊNCIAS BIBLIOGRÁFICAS
64
ZOLKOWSKA, D.; JAIN, R. ROTHMAN, R.B.; PARTILLA, J.S.; ROTH, B.L.;
SETOLA, V.; PRISINZANO, T.E.; BAUMANN, M.H. Evidence for the involvement of dopamine transporters in behavioral stimulant effects of modafinil. J Pharmacol Exp Ther. 329: 738-746, 2009.
WITTER, M. P.; GROENEWEGEN, H.J.; DA SILVA, F. H.; LOHMAN, A. H. M., 1989. Functional organization of the extrinsic and intrinsic circuitry of the para hippocampal region. Progress in Neurobiology 33: 161-253.
ZOLA-MORGAN, S.; SQUIRE, L. R.; AMARAL, D. G.; SUZUKI, W., 1989. Lesions of perihinal and parahippocampal cortex that spare the amygdada and hippocampal formation produce severe memory impairment. Journal of Neuroscience 9: 4355-4370.
ZOLKOWSKA, D.; JAIN, R. ROTHMAN, R.B.; PARTILLA, J.S.; ROTH, B.L.; SETOLA, V.; PRISINZANO, T.E.; BAUMANN, M.H. Evidence for the involvement of dopamine transporters in behavioral stimulant effects of modafinil. J Pharmacol Exp Ther. 329: 738-746, 2009.

ANEXO
65
6. ANEXO


















