
EQUILÍBRIO IÓNICO E POTENCIAIS DE REPOUSO DAS MEMBRANAS
33
M Filomena Botelho EQUILÍBRIO IÓNICO E POTENCIAIS DE REPOUSO DAS MEMBRANAS
Transcript of EQUILÍBRIO IÓNICO E POTENCIAIS DE REPOUSO DAS MEMBRANAS

M Filomena Botelho
EQUILÍBRIO IÓNICO E
POTENCIAIS DE REPOUSO DAS MEMBRANAS

Potenciais de repouso das membranas
Potenciais de repouso das membranas
O equilíbrio de Donnan não explica o potencial de repouso das células nervosas e musculares, mas pode contudo explicar a:
- Diferença de potencial existente através das paredes dos capilares, que são :
-impermeáveis aos iões de proteinato negativos-permeáveis aos pequenos iões
A razão pela qual os modelos de membrana propostos não explicam o potencial de repouso, é que a condição que foi imposta era a de equilíbrio, a qual não é verdadeira

A experiência demonstrou que o potencial de repouso é um- potencial de difusão,
isto é:Há uma diferença de potencial entre as duas fases, que é produzido por uma interdifusão dos iões.
Célula nervosaA célula nervosa pode encontrar-se, sob o ponto de vista eléctrico, em dois estados:
- De repouso – praticamente não passa corrente eléctrica através da membrana (do axónio)
- Activo – ocorre um potencial de acção o qual é acompanhado por correntes de solutos iónicos através da membrana (de dentro para fora e de fora para dentro), que são detectáveis

Para que haja mudança de um estado eléctrico para outro, é necessário que a célula seja estimulada
- eléctrico- químico- mecânico
Repouso → nervo (axónio) de lula
Consideremos três iões:- Na+
- K+
- Cl-
[K+]i = 400 mM/l [K+]e = 10 mM/lK+
Estudos feitos com traçadores, mostraram que a membrana celular, no estado de repouso é:
- permeável ao ião K+

Repouso → nervo (axónio) de lula
[Cl-]i = 40 mM/l [Cl-]e = 540 mM/lCl-
A permeabilidade da membrana celular das células nervosas ao ião Cl-, no repouso é elevada
[Na+]i = 41 mM/l [Na+]e = 460 mM/lNa+
A permeabilidade da membrana celular das células nervosas ao ião Na+, no repouso é praticamente nula
Por outro lado:No estado de repouso, o interior do axónio é negativo em relação ao exteriorMedidas efectuadas por microeléctrodos, o potencial do interior da célula em relação ao exterior, é de –61 mV, no estado de repouso

Mas, apesar daquela diferença de concentração que se verifica para qualquer dos iões, ela mantém-se desde que não actue nenhum estimulo.
Isto é:Desde que não actue nenhum estímulo, mantêm-se os gradientes de
- concentração- potencial eléctrico
De facto:
As correntes eléctricas resultantes dos movimentos dos iões, através da membrana, dependem de:
- gradientes – forçam os iões através das paredes celulares• potencial químico• potencial eléctrico
- permeabilidades

No caso exemplo do axónio da lula, como deverão actuar as forças de difusão e as forças eléctricas, para os três iões referidos, no estado de repouso, durante o qual a corrente eléctrica iónica através da membrana é nula
esquema:
Intracelular-61 mV
Extracelular0 mV
K+K+ D
E
Cl-Cl-E
D
Na+Na+D
E
Intr
acel
ula
r-6
1 m
V
Extracelular0 mV
K+K+ D
E
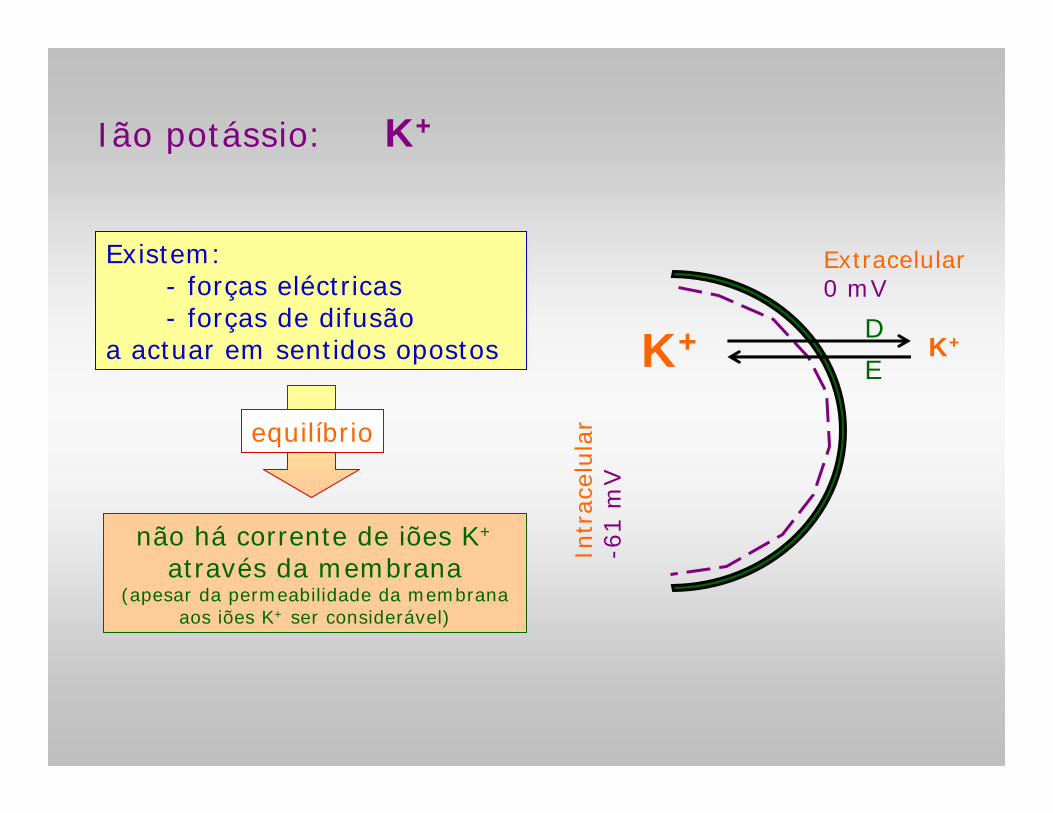
Ião potássio: K+
Existem:- forças eléctricas- forças de difusão
a actuar em sentidos opostos
não há corrente de iões K+
através da membrana(apesar da permeabilidade da membrana
aos iões K+ ser considerável)
equilíbrio

Intr
acel
ula
r-6
1 m
V
Extracelular0 mV
Ião cloro: Cl-
Existem:- forças eléctricas- forças de difusão
a actuar em sentidos opostos
não há corrente de iões Cl-através da membrana
(apesar da permeabilidade da membrana aos iões Cl- ser considerável)
equilíbrio Cl-Cl-E
D

Intr
acel
ula
r-6
1 m
V
Extracelular0 mV
Ião sódio: Na+
Existem:- forças eléctricas- forças de difusão
a actuar no sentido do interior da célula
No repouso, mantêm-se as diferenças de concentrações de Na+ através da membrana(a permeabilidade da membrana aos iões Na+ no repouso é extremamente baixa)
Na+Na+D
E
equilíbrio
equação de Nernst a cada ião

No repouso, podemos pois aplicar a equação de Nernst a cada ião, pois não havendo corrente eléctrica através da membrana, podemos pensar que pode existir equilíbrio
Generalizando para qualquer ião i, podemos então dizer que:
µii = µi
e~ ~
∆ψ = ψii – ψi
e = - lnRTFZi
Cii
Cie
∆ψ – traduz a diferença de potencial eléctrico que terá que existir através da membrana para que a relação
para cada espécie se mantenha constante
Cii
Cie

Se aplicarmos a equação de Nernst para o caso do axónio da lula, dado como exemplo, aos três iões considerados (K+, Cl-, Na+) deveremos obter
valores para a diferença de potencial eléctrico que deverão ser iguais à diferença de potencial eléctrico obtida quando a medimos
A aplicação da equação de Nernst aos três iões em causa (K+, Cl-, Na+) dá os seguintes resultados
K+ ∆ψ K+ = -93 mV
Cl- ∆ψ Cl- = -66 mV
Na+ ∆ψ Na+ = +61 mV
Como se poderão explicar estes resultados tão díspares?
?

A diferença entre os resultados obtidos por medições e os obtidos com a aplicação da equação de Nernst, é porque:
o equilíbrio não é um pressuposto válido pois corresponde à realidade
A experiência provou que o:
potencial de repouso é um potencial de difusão, isto é, a diferença de potencial entre duas fases da membrana, resulta de uma
- difusão bidireccional dos iõesmas não é um estadio de equilíbrio

M Filomena Botelho
Modelo de membrana com campo eléctrico
constante no interior ou modelo de campo
eléctrico constante –Equação de Goldman

Modelo de campo eléctrico constante
Equação de GoldmanModelo de campo eléctrico constante
Equação de Goldman
Partindo da equação de Nernst-Planck chegou-se a uma relação capaz de reproduzir fielmente os potenciais das células excitáveis em função:
- das concentrações iónicas- dos coeficientes de permeabilidade
Goldman considerou:- campo eléctrico constante no interior da membrana
(gradiente de potencial eléctrico constante na fase da amostra)
- coeficientes de partição constantes e iguais nas interfaces da membrana

Assim, as condições são:
dVdxE = -⃗ V – potencial eléctrico
C1’ = β1 C1
C2’ = β2 C2
β1 = β2
β1β2
coeficientes de partição
C1C2
Concentrações nas interfaces membrana-solução
Interior da célula
Exterior da célula
∆x
V2
V1
C1
C2
C1’
C2’

Partindo então da equação de Nernst-Planck:
e impondo as condições de Goldman, a resolução da equação vem:
Ji = -u Zi∣Zi∣
RT + u Ci F ∣Zi∣dVdx( )
dCi
dx
Ji = ∆Vβ ui F ∣Zi∣
∆xC1 – C2 . e
FRT- ∆V Zi
1 – eF
RT- ∆V Zi
Para o:• ião i• valência Zi• mobilidade ui
Campo eléctrico constanteβ1 = β2 = β

Ji = ∆Vβ ui F ∣Zi∣
∆xC1 – C2 . e
FRT- ∆V Zi
1 – eF
RT- ∆V Zi
Na ausência de campo eléctrico e quando se trata de moléculas ionizadas esta equação deve transformar-se na equação que foi deduzida para a densidade de corrente de um soluto através da membrana originam exclusivamente por difusão
Porém quando fazemos tender ∆V → 0, a equação anterior transforma-se numa indeterminação
Como levantar a indeterminação ?

Peguemos outra vez na equação, e multipliquemos por Para podermos simplificar alguma coisa
RTRT
Ji = ∆Vβ ui∣Zi∣F
∆xC1 – C2 . e
FRT- ∆V Zi
1 – eF
RT- ∆V Zi
RTRT
y y
y
Se:
Zi = +1
β ui RT∆x = K
A equação tomará a seguinte forma:
Ji = K yC1 – C2 . e
-y
1 – e-y

Ji = K yC1 – C2 . e
-y
1 – e-y
lim Ji = K (C1 – C2. e )-y
1 – e-y
y∆V → 0
Uma vez que:
y = F ∆VRT
tende para 0 quando ∆V → 0, a indeterminação surge
através do cociente 1 – e
-yy
Esta indeterminação pode ser levantada, desenvolvendo em
série e-y

Levantar a indeterminação, desenvolvendo e-y em série.1 – e
-yy
Assim:
e-y = 1 – y + - + - LLy2
2!y3
3!y4
4!
Desta maneira:
lim =1 – e- y
yy → 0
= limy → 0 y y2
1 - + LL2! 3!
1
y → 0 y2 y3
1 - 1 + y - + LL2! 3!
ylim =
= 1

assim: lim Ji = K (C1 – C2. e )-y
1 – e-y
y∆V → 0
β ui RT∆xJi = x 1 x (C1 – C2)
β ui RT∆xJi = x (C1 – C2)
Permeabilidade P da membrana para a espécie iónica i
como:∆Cs = C1 – C2
Podemos então dizer que, quando ∆V = 0:
Ji = P . ∆CsEquação formalmente idêntica à equação da difusão para moléculas neutras – Equação de Fick

Uma outra situação que a equação de Nernst-Planck, integrada nas condições atrás indicadas, deve contemplar é a situação de equilíbriosituação de equilíbriono caso de uma membrana permeável à espécie iónica i, isto é, a situação em que Ji = 0
Continuando a considerar o ião com valência Z = +1, vem:
1 – e RT
Ji = 0 = ∆Vβ ui F
∆xC1 – C2 . e
FRT- ∆V
F- ∆V
0 = C1 – C2 . eF
RT- ∆V
FRT- ∆V= eC1
C2

Aplicando logaritmos, vem para ∆V:
FRT- ∆V= eC1
C2
∆V = lnRTF
C2C1
Equação de Nernst(iões positivos e monovalentes)
Traduz a diferença de potencial existente através da membrana quando há equilíbrio
Se considerarmos iões de valência Zi, a equação toma a forma:
∆V = lnRTF Zi
C2C1

Pi =β ui RT∆x F
Retomemos a equação de Nernst-Planck
Como o coeficiente de permeabilidade, vamos considerar:
Ji = ∆Vβ ui F ∣Zi∣
∆xC1 – C2 . e
FRT- ∆V Zi
1 – eF
RT- ∆V Zi
vem:
Ji = ∆VPi ∣Zi∣ F2
RTC1 – C2 . e
FRT- ∆V Zi
1 – eF
RT- ∆V Zi

Aplicando esta equação aos três principais iões (K+, Na+, Cl-) que criam correntes através da parede do axónio, vem
JCl- = PCl- ∆VRT
[Cl-]1 – [Cl-]2 . eF
RT+ ∆V
1 – eF
RT ∆V
F2Cl-
JK+ = PK+ ∆VRT
[K+]1 – [K+]2 . eF
RT- ∆V
1 – eF
RT- ∆V
F2
K+
JNa+ = PNa+ ∆VRT
[Na+]1 – [Na+]2 . eF
RT- ∆V
1 – eF
RT- ∆V
F2Na+

Como a corrente total através da membrana é nula:
JK+ = PK+ ∆VRT
[K+]1 – [K+]2 . eF
RT- ∆V
1 – eF
RT- ∆V
F2
K+
JNa+ = PNa+ ∆VRT
[Na+]1 – [Na+]2 . eF
RT- ∆V
1 – eF
RT- ∆V
F2Na+
JK+ + JNa+ + JCl- = 0
Para simplificar ao máximo, vejamos:
JCl- = PCl- ∆VRT
[Cl-]1 – [Cl-]2 . eF
RT+ ∆V
1 – eF
RT ∆V
F2Cl-F
RT ∆V
FRT ∆V
e
e
=

JCl- = PCl- ∆VRT
[Cl-]1 – [Cl-]2 . eF
RT+ ∆V
1 – eF
RT ∆V
F2Cl-F
RT ∆V
FRT ∆V
e
e
=
= PCl- ∆VRT
[Cl-]1. e – [Cl-]2
FRT- ∆V
e – 1F
RT- ∆V
F2 -1-1
=
= PCl- ∆VRT
[Cl-]2 – [Cl-]1 . eF
RT- ∆V
1 – eF
RT- ∆V
F2
Somando agora, vem:

JK+ + JNa+ + JCl- = 0
+ PCl- {[Cl-]2 – [Cl-]2 . e } = 0F
RT- ∆V
PNa+ {[Na+]1 – [Na+]2 . e } + PK+ {[K+]1 – [K+]2 . e } +F
RT- ∆VF
RT- ∆V
PNa+[Na+]1 + PK+[K+]1 + PCl-[Cl-]2 = PNa+[Na+]2 + PK+[K+]2 + PCl-[Cl-]1 e F
RT- ∆V
e =F
RT- ∆V PNa+[Na+]1 + PK+[K+]1 + PCl-[Cl-]2
PNa+[Na+]2 + PK+[K+]2 + PCl-[Cl-]1
Somando agora, vem:

Logaritmizando vem:
FRT- ∆V = ln
PNa+[Na+]1 + PK+[K+]1 + PCl-[Cl-]2
PNa+[Na+]2 + PK+[K+]2 + PCl-[Cl-]1
e resolvendo em ordem a ∆V, vem:
RTF∆V = ln
PNa+[Na+]2 + PK+[K+]2 + PCl-[Cl-]1PNa+[Na+]1 + PK+[K+]1 + PCl-[Cl-]2
Equaçãode
Goldman
foi aplicada a inúmeras situações, a células de diferentes naturezas, dando valores de ∆V praticamente coincidentes com os valores experimentais

Esta equação de Goldman pode ser expressa em termos de permeabilidade:
- relativos- absolutos
SePK+αK+ = PNa+
PCl-αCl- = PNa+
as permeabilidades podem ser expressas em relação à permeabilidade do Na+.
A equação de Goldman toma a seguinte forma:
RTF∆V = ln
[Na+]2 + αK+[K+]2 + αCl-[Cl-]1[Na+]1 + αK+[K+]1 + αCl-[Cl-]2

Os coeficientes de permeabilidade- obtém-se a partir de medições de potencial- são supostamente independentes da concentração
Para o caso do axónio, pode obter-se o valor correcto para a diferença de potencial entre os dois lados da membrana, se entrarmos com a seguinte relação para as concentrações:
PK : PNa : PCl = 1 : 0,04 : 0,45
∴
αK+ = 25 αCl- = 11,25

Caso do axónio da lula:
No repouso:
PK : PNa : PCl 1 : 0,04 : 0,45
No potencial de acção:
PK : PNa : PCl 1 : 20 : 0,45
Na célula em repouso, se considerarmos as permeabilidades do sódio e do cloro em relação à do potássio, à qual era atribuído o valor de 1, verificamos:
- menos permeável ao cloro- muitíssimo pouco permeável ao sódio
Na célula durante o potencial de acção, contrariamente, se considerarmos as mesmas condições, verificamos:
- permeabilidade ao cloro se mantém- enorme subida da permeabilidade ao sódio


















