Estimación del índice de área foliar (LAI) de un pinar ... · Estimación del índice de área...
21
Estimación del índice de área foliar (LAI) de un pinar costero de Pinus halepensis Miller y Pinus pinea L. Emilio Garcia a,b* , Maria José Lledó b , Juan Rafael Sánchez b a* Postgrado Ciencias del Desarrollo (CIDES), Universidad Mayor de San Andrés, La Paz, Bolivia phone/fax 00591 2786169 email: [email protected] b Departmento de Ecología, Universidad de Alicante, Ap. Correos 99 – E-03080, Alicante, España. Phone 0034 96 590 3625 Fax 0034 96 590 9832 email: [email protected] Resumen Se ha estimado el Índice de Área Foliar (LAI) de una ecosistema dunar fijado con Pinus halepensis y Pinus pinea, implantado en los años 1897-1926 a través de una metodología sencilla de cosecha de acículas localizados en un prisma imaginario en copas de árboles casi homogéneos. Los principales resultados nos indican que esta metodología puede aminorar los trabajos arduos de las metodologías destructivas por lo que esta metodología utilizada se lo ha denominado como semidestructiva, ya que afecta solo parte de las copas. El LAI obtenido ha variado desde 1,55 a 5,51 para ambas especies, mostrándonos que los que tienen mas alto LAI son los pinos piñoneros en comparación a los pinos alepos. Esta diferencia básicamente se ha asumido que es debido a la estructura de la copa del árbol, ya que los primeros tienen una copa más compacta y las segundas presentan mas huecos y esto hace que su estructura sea menos homogénea y menos densa. De manera general se puede concluir que el LAI obtenido de forma directa en la parcela de experimentación de Guardamar del Segura es de 3,61±2,00 (una masa forestal de copas mas o menos homogéneas); el índice de corrección para los datos del LAI-2000 (instrumento óptico que se utiliza para medir el LAI) estimado para la parcela de Guardamar del Segura es de 0,77±0,12; si se estima el LAI de la parcela por medio del LAI-2000, se deberá corregir por un segundo factor 2,26 para obtener el valor cercano al obtenido por medición directa. Y finalmente los datos de LAI están altamente correlacionados con el diámetro normal de los árboles, por lo que su utilización potencial en el cálculo del LAI de todo el pinar se puede realizar con la medición de los diámetros normales de una muestra estratégica representativa de este ecosistema Palabras clave: Pinus halepensis, Pinus pinea, LAI, LAI-2000, método destructivo, biomasa. 1. Introducción El índice de área foliar (LAI) para coníferas está definido como la mitad del área de la superficie de las acículas por unidad de área de superficie de suelo (Walter y otros, 2003). El LAI es uno de los índices mas estudiados a la hora de estimaciones de procesos de producción en ecosistemas ya que permite caracterizar la estructura de la cubierta vegetal (Mussche y otros, 2001), relaciona los procesos clave del ecosistema, el balance hidrológico y de energía (Feldkirchner y Gower, 2001, Bolstad y Gower, 1990), y de fundamental importancia, para estimaciones de intercambio de carbono, agua, nutrientes y luz (Thomas y Winner, 2000, Gower y Norman, 1991). Aunque el método directo para estimar el LAI es el mas eficaz, toma mucho tiempo, es muy laborioso (Eklundh y otros, 2001; Nilson y otros, 1999), y evita trabajos de investigación de largo aliento. Comprenden desde la recolecta de acículas de un área determinada (Gower y otros, 1999), aplicación de ecuaciones alométricas a datos de diámetro de un rodal (Law y
Transcript of Estimación del índice de área foliar (LAI) de un pinar ... · Estimación del índice de área...

Estimación del índice de área foliar (LAI) de un pinar costero de Pinus halepensis Miller y Pinus pinea L. Emilio Garciaa,b*, Maria José Lledób, Juan Rafael Sánchezb a*Postgrado Ciencias del Desarrollo (CIDES), Universidad Mayor de San Andrés, La Paz, Bolivia phone/fax 00591 2786169 email: [email protected] bDepartmento de Ecología, Universidad de Alicante, Ap. Correos 99 – E-03080, Alicante, España. Phone 0034 96 590 3625 Fax 0034 96 590 9832 email: [email protected] Resumen Se ha estimado el Índice de Área Foliar (LAI) de una ecosistema dunar fijado con Pinus halepensis y Pinus pinea, implantado en los años 1897-1926 a través de una metodología sencilla de cosecha de acículas localizados en un prisma imaginario en copas de árboles casi homogéneos. Los principales resultados nos indican que esta metodología puede aminorar los trabajos arduos de las metodologías destructivas por lo que esta metodología utilizada se lo ha denominado como semidestructiva, ya que afecta solo parte de las copas. El LAI obtenido ha variado desde 1,55 a 5,51 para ambas especies, mostrándonos que los que tienen mas alto LAI son los pinos piñoneros en comparación a los pinos alepos. Esta diferencia básicamente se ha asumido que es debido a la estructura de la copa del árbol, ya que los primeros tienen una copa más compacta y las segundas presentan mas huecos y esto hace que su estructura sea menos homogénea y menos densa. De manera general se puede concluir que el LAI obtenido de forma directa en la parcela de experimentación de Guardamar del Segura es de 3,61±2,00 (una masa forestal de copas mas o menos homogéneas); el índice de corrección para los datos del LAI-2000 (instrumento óptico que se utiliza para medir el LAI) estimado para la parcela de Guardamar del Segura es de 0,77±0,12; si se estima el LAI de la parcela por medio del LAI-2000, se deberá corregir por un segundo factor 2,26 para obtener el valor cercano al obtenido por medición directa. Y finalmente los datos de LAI están altamente correlacionados con el diámetro normal de los árboles, por lo que su utilización potencial en el cálculo del LAI de todo el pinar se puede realizar con la medición de los diámetros normales de una muestra estratégica representativa de este ecosistema
Palabras clave: Pinus halepensis, Pinus pinea, LAI, LAI-2000, método destructivo, biomasa.
1. Introducción
El índice de área foliar (LAI) para coníferas está definido como la mitad del área de la superficie de las acículas por unidad de área de superficie de suelo (Walter y otros, 2003). El LAI es uno de los índices mas estudiados a la hora de estimaciones de procesos de producción en ecosistemas ya que permite caracterizar la estructura de la cubierta vegetal (Mussche y otros, 2001), relaciona los procesos clave del ecosistema, el balance hidrológico y de energía (Feldkirchner y Gower, 2001, Bolstad y Gower, 1990), y de fundamental importancia, para estimaciones de intercambio de carbono, agua, nutrientes y luz (Thomas y Winner, 2000, Gower y Norman, 1991).
Aunque el método directo para estimar el LAI es el mas eficaz, toma mucho tiempo, es muy laborioso (Eklundh y otros, 2001; Nilson y otros, 1999), y evita trabajos de investigación de largo aliento. Comprenden desde la recolecta de acículas de un área determinada (Gower y otros, 1999), aplicación de ecuaciones alométricas a datos de diámetro de un rodal (Law y

otros, 2001; Cherry y otros, 1998), hasta la recolección de caída de hojas (Law y otros, 2001; Hebert y Jack, 1998). Buscando evitar este esfuerzo, se han desarrollado estimaciones indirectas que comprenden desde la captación de imágenes de la copa de los árboles (Walter y otros, 2003; Leblanc y otros, 2002; Martens y otros, 1993; Bolstad y Gower, 1990) método comúnmente llamado de mediciones ópticas; desarrollo de expresiones empíricas a partir de mediciones de arquitectura de la planta (Barrantes y Gracia, 1989), hasta el análisis de imágenes satelitales (Eklundh y otros, 2001; Nilson y otros, 1999).
Uno de los métodos indirectos mas utilizados en este tipo de estimaciones es el que fue desarrollado a partir de la observación de huecos y la distribución angular de la fracción de éstos en la cubierta vegetal como el que utiliza el LAI-2000. Este aparato mide la atenuación de la luz que penetra los diferentes estratos verticales de una cubierta vegetal utilizando 5 diferentes ángulos cenit, calculando para estos ángulos un índice de existencia de cobertura de la parte de arriba y por debajo de ésta (Law y otros, 2001). Se asume una serie de condiciones: (a) que el follaje es totalmente negro. El aparato tiene filtros que rechazan las radiaciones mayores a 490 nm, pero el follaje refleja y transmite una pequeña radiación en el resto del espectro, (b) los elementos del follaje son pequeños en relación al resto del campo a la visión del sensor, (c) el follaje esta orientado al azar respecto del ángulo acimutal, (d) que el follaje esta distribuido al azar. Por otro lado, se ha evidenciado que este instrumento mide un índice de la totalidad de la planta incluyendo la parte leñosa (Smolander y Stenberg, 1996), las hojas y ramas muertas que queda sostenido entre las ramas vivas (Vose y Swank, 1990). Además, en pinos, las acículas se agrupan en ramillos y no están distribuidos al azar (Eklundh y otros, 2001), y tiene agrupaciones del follaje a escala superiores a la del ramillo (agrupación a nivel de ramas, verticilos, copas, y cubierta total) (Chen y otros, 1997) incurriendo por tanto en una subestimación del valor del LAI (Nilson, 1999; Nilson y otros, 1999; Smith y otros, 1993, citado por Thomas y Winner, 2000; Stenberg y otros, 1994; Deblonde y otros, 1994; Gower y Norman, 1991).
El ceptómetro mide la radiación fotosinteticamente activa (400-700 nm) incidentes en 80 sensores localizados a 1 cm de intervalo a lo largo de una varilla de 80 cm de largo. La transmitancia del PAR que atravieza la copa del árbol y el PAR directo mas el coeficiente de extinción son los factores que intervienen para computar el LAI a través de la ecuación de Beer-Lambert (Vose y Swamk, 1990).
Sin embargo, todos los instrumentos de mediciones ópticas, como el LAI-2000 y el ceptómetro, no distinguen el follaje del material leñoso, por lo tanto miden el índice de área de la totalidad de la copa de la planta (LAI efectivo o LAIe) (Gower y otros, 1999).
Las mediciones se hacen más irreales en casos donde la copa de los árboles no son continuas y existen muchos huecos, no sólo dentro de las copas sino en el conjunto de los árboles, que a la hora de realizar estimaciones dentro del ecosistema, se hacen mas dificultosos ya que encierran errores de medición y errores de criterios de aplicación de método. Otra de las dificultades de la medición del LAI, es la mal formación de las copas de los árboles. La presencia de factores medioambientales adversos al crecimiento de las acículas del pino, como el spray marino, hace que la copa del pino se incline en dirección opuesta a la dirección de circulación del causante de este efecto. Esta reacción de la planta hace que sus acículas se desarrollen sólo en el 50% de la copa del árbol.
Este trabajo se condujo bajo tres objetivos: (1) estimar el LAI del bosque de pino a partir de mediciones ópticas utilizando el LAI-2000 y el ceptómetro, (2) estimar los índices que afectan al LAI efectivo y (3) Comparar los valores de mediciones ópticas con los obtenidos por mediciones directas.

2. Teoría Como consecuencia de los fallos mencionados arriba, inherentes en los instrumentos de mediciones ópticas, los investigadores han aplicado una serie de correcciones a las mediciones obtenidas por éstos. Para el LAI-2000 están descritas extensamente en Chen (1996) y Chen y otros (1997). Otros en cambio han preferido sustentar su corrección basándose en la arquitectura de la planta, como Smolander y Stenberg (1996) que expresaron los resultados del LAI-2000 multiplicando esta información por 2, ya que asumieron que la parte leñosa era la mitad de la totalidad de la copa. También se ha realizado la corrección multiplicando al resultado un índice basado en la relación del área total de proyección de las acículas con el área de la silueta de la copa (Hebert y Jack, 1998); otros autores han incorporado elementos como el índice de agrupación de acículas (Chen, 1996), un índice razón de área de acículas en relación al ramillo (para agrupaciones dentro de la copa) y un índice razón de la parte leñosa en relación al área total de la copa (Stenberg y otros, 2003).
Barclay y Trofymow (2000) y Chen (1996) indican que el LAI-2000 mide el LAI efectivo, el cual es el producto del LAI y el índice de agrupación del follaje.
2.1. Índices de agrupación del follaje.
El índice de agrupación del follaje (Ω) es el más estudiado ya que esta relacionado directamente a los regímenes de luz y la estructura de la vegetación (Walter y otros, 2003), aunque la contribución del material no fotosintetizador es también importante (Lopez-Serrano y otros, 2000), como son las ramas y las hojas secas. Una ordenación de los elementos que concurren en la obtención de un índice de corrección para las mediciones obtenidas con el LICOR 2000, en principio, parte de a) la agrupación de follaje, tanto a nivel ramillo, dentro de la copa de árbol y dentro de la cubierta, b) el grado de contribución del material no fotosinteticamente activo o madera, el cual según Chen y otros (1997), tiene hasta un 30% en proporción en copas de LAIs bajos, implicándose de manera específica la cuantificación de las ramas ya que estas sostienen a las acículas (Kucharik y otros, 1998). Por lo tanto este es un índice de corrección requerido para convertir el LAIe a LAI. Según Chen (1996),
Ω= eLAI
LAI (1)
El Ω es igual a 1 cuando la distribución espacial de las hojas es al azar. Cuando las hojas están distribuidas muy regularmente, toma un valor mayor a la unidad, y cuando las hojas están muy agrupadas, en el extremo caso, las hojas están detenidas unas sobre otras, este índice toma un valor menor a la unidad.
Las acículas de las coníferas tienen diferentes niveles de agrupación, por lo que se ha tomado al ramillo (el agrupamiento básico de acículas distribuido alrededor de la más pequeña rama) como la unidad básica de follaje que afecta la transmitancia de la radiación solar (Fassnacht y otros, 1994). También se ha determinado a través de la distribución de los tamaños de huecos en la copa, que el tamaño de las unidades básicas de follaje es el promedio del ancho de la proyección de la ramilla (Chen, 1997). Esta relación explica porque a una pequeña distancia, los pequeños huecos en la copa desaparecen debido al efecto de la penumbra. Por eso es difícil medir el total del área de las acículas dentro de las ramillas con los instrumentos ópticos. Para esto se separa Ω en dos componentes:

E
E
γΩ
=Ω (2)
Donde γE es la razón área de acículas a ramillo que cuantifica el efecto del agrupamiento del follaje dentro de la ramilla y ΩE es el índice de agrupamiento que cuantifica el efecto de agrupación del follaje a escalas superiores a la del ramillo.
2.1.1. Razón área de acículas a ramillo (γE)
Las confieras tienes sus acículas agrupadas y por lo tanto es difícil medir este índice con los instrumentos ópticos. Chen (1996) desarrolló la siguiente relación:
s
nE A
A=γ (3)
Donde An es el área total en el ramillo y As es la mitad del área total del ramillo (que es el área de la superficie imaginaria de la esfera del ramillo). Y donde As se puede calcular a través de (Lopez-Serrano, 1999):
)75cos()45cos()15cos()75cos()0,90()45cos()0,45()15cos()0(
2°+°+°
°°°+°°°+°°=
xAxAxAA ppp
s (4)
Donde Ap es el área proyectada en tres ángulos (0°, 45° y 90°) de incidencia de la cámara, manteniendo el ramillo horizontal, se puede calcular la mitad de la superficie total del ramillo.
2.1.2. Índice de agrupamiento para escalas superiores a las del ramillo (ΩE)
Cuando los elementos del follaje están agrupados a niveles mayores, es más probable observar mayores huecos en la copa. Por eso, se utiliza la distribución del tamaño de los huecos para cuantificar ΩE. Este índice se expresa de la siguiente forma (Leblanc y otros, 2002; Chen y Chilar, 1995):
[ ] [ ][ ])0(ln
)0(ln)0()0(1
mr
mmrmE F
FFF −+=Ω (5)
Donde Fm(0) es la fracción de huecos medidos en la copa del árbol y Fmr(0) es la fracción de huecos para una copa con elementos posicionados al azar. El primer elemento se puede obtener a través de la medición de la transmitancia de la radiación difusa o directa a determinado ángulo cenit. Mientras que el segundo se obtiene midiendo con el TRAC (Tracing Radiation nad Arquitectura of Canopies) o alternativamente utilizando MVI (Multiband Vegetation Imagen) el cual utiliza una relación diferente (Kucharik, y otros, 1999).

2.1.3. Razón entre el área de la superficie leñosa y el área de la superficie total (α)
El índice de área de la planta toma en cuenta también el índice de área de la parte leñosa.
)1( αλ
−−=−⎟⎟⎠
⎞⎜⎜⎝
⎛Ω
= WLAIWxLAILAI tE
Ee (6)
Donde W es el índice de área de la parte leñosa (fuste, ramas y piñas), y α =W/LAIt
Por lo que la relación para estimar el LAI quedaría como:
⎟⎟⎠
⎞⎜⎜⎝
⎛Ω
−=E
EeLAILAIγ
α )1( (7)
Esta expresión ha sido ampliamente aplicada por diferentes autores como Barclay y Trofymow, 2000; Nilson, 1999; Chen, 1996, entre otros.
2. Materiales y métodos
2.1. Descripción del sitio
El estudio fue conducido en una parcela del pinar de la población de Guardamar del Segura (38°06.128' N 0°38.965' W), a 45 Km al SE de la ciudad de Alicante. El pinar se localiza junto a la desembocadura del río Segura y se implantó en las dunas con el fin de fijarlas y establecer una barrera que detuviera su avance. Dentro del municipio, este conjunto se encuentra a unos 800 m al NE del municipio. Su ancho máximo alcanza unos 1.800 m y su largo máximo unos 6.000 m aproximadamente; el área total de la pinada alcanza unos 134.425 Ha aproximadamente. El área tiene una precipitación variable, como promedio anual alcanza los 300 mm, una temperatura media anual de 17°C. Esta formada principalmente de Pinus halepensis Mill y P. Pinea, y su densidad de plantación varia (de 2x2 hasta de 3x2 m), así como la edad de los árboles (que va desde los 90 hasta los 30 años). El área basal del rodal es de (0,19 ± 0,075 m, n=790).
Entre la mayor presencia de plantas de porte menor están Elymus farctus (Viv) Runemark ex Melderis, Carpobrotus edulis, Helichrysum stoechas, Ononis natrix, Medicago marina, Echinophora spinosa, Silene ramossisima Desf.
2.2. Método directo de obtención del LAI
Partiendo de la definición del LAI para confieras, para obtener información de estimación de área de acículas y por tanto del LAI, se seleccionaron 7 individuos de P. halepensis y otros 7 de P. pinea para un muestreo destructivo.
Una vez ubicados los individuos en el sitio, se determinaron las áreas de las copas de los árboles, los polígonos proyectados en el suelo y los diámetros altura pecho y basal. El área de la copa de los árboles fue medido a través del método de triángulos según:

Ac = T1 +T2 + ….. T8 (8)
Donde T es el área del triángulo escaleno o isósceles, según se haya identificado luego en el proceso de cálculo. La altura de la copa de los árboles fue medido al estar instalando los prismas, antes de la corta, a través de una plomada métrica.
Posteriormente, se ha instalado en las copas de estos árboles un prisma imaginario de 0,5x0,5x2,5 m a través de un dispositivo casero de madera, jalones e hilos. Por razones de estimación de los errores debido a la ubicación de cada prisma, se ha instalado también en dos árboles tres prismas (P1, P2, P3), (1 en cada especie) una a continuación de otra, en línea empezando de la parte mas exterior a la copa. Una vez realizado la cosecha, se ha identificado que el LAI promedio fue la que correspondía al prisma del medio (P2), ubicándose, por tanto, en los demás individuos el prisma en este sitio. Ubicadas en la parte central de la copa del árbol y en todos los árboles a la misma orientación, se ha procedido a medir el LAI de éstos con el LAI-2000 utilizando el tapón de 45° y obtenido luego los datos del anillo a 38°.
Luego se ha procedido a una corta estratificada cada 50 cm de las ramas, ramillos y acículas encontradas dentro del prisma. Los estratos a partir del más alto se nominaron como E1, E2, .... En como el más bajo. Según al tamaño de la copa se alcanzo a corta de 4 a 6 estratos. La corta se realizo cuidadosamente, separando el producto de cada estrato en bolsas de plástico, los cuales fueron etiquetados y sellados, para luego ser trasladarlos al laboratorio donde se procedió al trabajo posterior de pesa y separación.
En el laboratorio, se ha realizado los siguientes trabajos:
a) Obtención de la razón área de acícula a ramillo: se procedió a seleccionar los ramillos completos, medir los diámetros del ramillo entero y de la base de la rama de éste, pesarlos y posteriormente separarlos en acículas y ramas. El peso fresco como el seco, se obtuvo con una balanza eléctrica de 0.05 Kg de precisión.
b) Obtención del área de las acículas: la obtención del área de las acículas se la hizo de las muestras frescas antes de secarlos. Para esto utilizó un escaner EPSON bajo una resolución continua, en tipo de 8 bits para evitar el efecto de las sombras en las figuras. Para procesarlas se utilizó el programa Imagen 1.3. (Nacional Institutes of Health, 2003), el cual permite obtener el área a través de un proceso de calibración. Se escanearon 120 acículas en promedio para cada estrato.
c) Medición de los diámetros de los ramillos: la medición de los diámetros de las bases las ramas o ramillos, y su respectivo peso fresco nos permitió realizar regresiones alométricas con estos dos parámetros. Esto se practicó ya que la separación de las ramas de las acículas no solo nos llevó mucho tiempo, sino que necesitábamos estimar el peso de las acículas en ramas sin acículas. Bajo este método se pudo estimar el total de biomasa foliar del árbol multiplicando la biomasa conocida en la rama de la muestra, infiriendo luego al número total de ramas de la copa. También se correlacionó los datos de área de las acículas con los diámetros. (Tabla 1)
d) Peso seco de las muestras: concluida la obtención del área, se procedió a secarlos en el horno a 65° por 24 horas, para luego volverlos a pesar y obtener así el peso seco de las muestras.

e) Estimación de la razón área parte leñosa al total del área, (α): se extrapolaron los valores de pesos secos con los valores de pesos frescos obtenidos en todo el prisma, por lo cual se obtuvo en un área conocida, un valor estimado.
Finalmente para el cálculo del LAI por medición directa o destructiva (LAID) se obtuvo primeramente, el cociente entre la mitad del área proyectada de las acículas en el prisma y la superficie proyectada por el prisma. Luego de las estimaciones se obtuvo el cociente entre el área proyectada de las acículas en la copa del árbol y la superficie proyectada por el mismo.
2.3. Métodos indirectos
2.3.1. Mediciones con el LAI-2000
Mediciones arriba y por debajo de la copa de los árboles elegidos fueron tomadas con el LAI-2000 previas a los cortes de los mismos. Las mediciones por debajo de la copa, se realizó justo por debajo del prisma utilizando el tapón de lente de 45°. Durante la medición el aparato fue sostenido horizontalmente y en una sola orientación. Para evitar la influencia solar las mediciones se realizaron en la tarde, en la mañana, o en días nublados para minimizar el efecto dispersión del sol . Una vez hecho esto, se procedió al trabajo de gabinete, consistente en recuperar los datos del LAI-2000 al ordenador, verificar los datos, recomputarlos en su caso, utilizando el programa C2000.
Para fines de escalamiento también se tomaron medidas en el pinar a lo largo de un transecto de 700 m, cuasi perpendicular a la costa. Las medidas se realizaron en subtransectos de 100 m y un intervalo de 10 m, utilizando el criterio descrito en el manual del LAI-2000 y el método transecto longitud-finita (Gower, 1996). Las mediciones se realizaron una vez por estación .
2.3.2. Mediciones con el ceptómetro
La radiación fotosinteticamente activa (PAR) fue medido con un radiómetro portátil (Ceptómetro Sunfleck, ®Decagon, Delta-T Devices LTD, Cambridge, England) que cuenta con 80 sensores de radiación, ubicados a 1 cm de intervalo a lo largo de una sonda.
El LAI se calcula el método descrito por Pierce y Running (1988), y el Manual del Usuario del Ceptómetro (1989) usando el cociente adimensional PAR abajo y PAR arriba, el coeficiente de extinsión y la ley de Beer-Lambert, como sigue:
kQQ
LAI o
i
CEPTOMETRO
ln−= (9)
Donde, el Qi es el PAR debajo de la copa del árbol; Qo: PAR arriba del dosel del árbol; k el coeficiente de extinción. Qi y Qo se midieron por debajo y por arriba de la copa del árbol respectivamente, entre las 10:00 y 14:00 del día. Asumiendo que la distribución de los ángulos de las acículas es elipsoidal (Thomas y Winner, 2000, Norman y Campbell, 1989), k se estimó de acuerdo al Manual de ceptómetro, Decagon Devices (1989), como sigue:

733.0
21
22
)182.1(774.1)tan(
−++Θ+
=xx
xk (10)
Donde, x es la relación entre la longitud horizontal y vertical de la copa del árbol, θ el ángulo al zenit, ángulo medido desde la vertical.
x se estimó midiendo el área de la sombra de la copa del árbol proyectada al suelo y el área plana vertical de esta misma copa. θ se estimó a través de la medida directa del ángulo que el sol origina respecto a una línea vertical. Colocando dos piezas de madera unidas en ángulo recto, la vertical de 10 cm y la horizontal de 30 cm, y un nivel de mano en la horizontal. Se midió la sombra hecha por la pieza vertical proyectada en la pieza horizontal, donde θ = (arctan (x/10)) donde x es la longitud de la sombra y 10 el tamaño de la pieza vertical.
3. Resultados y discusión.
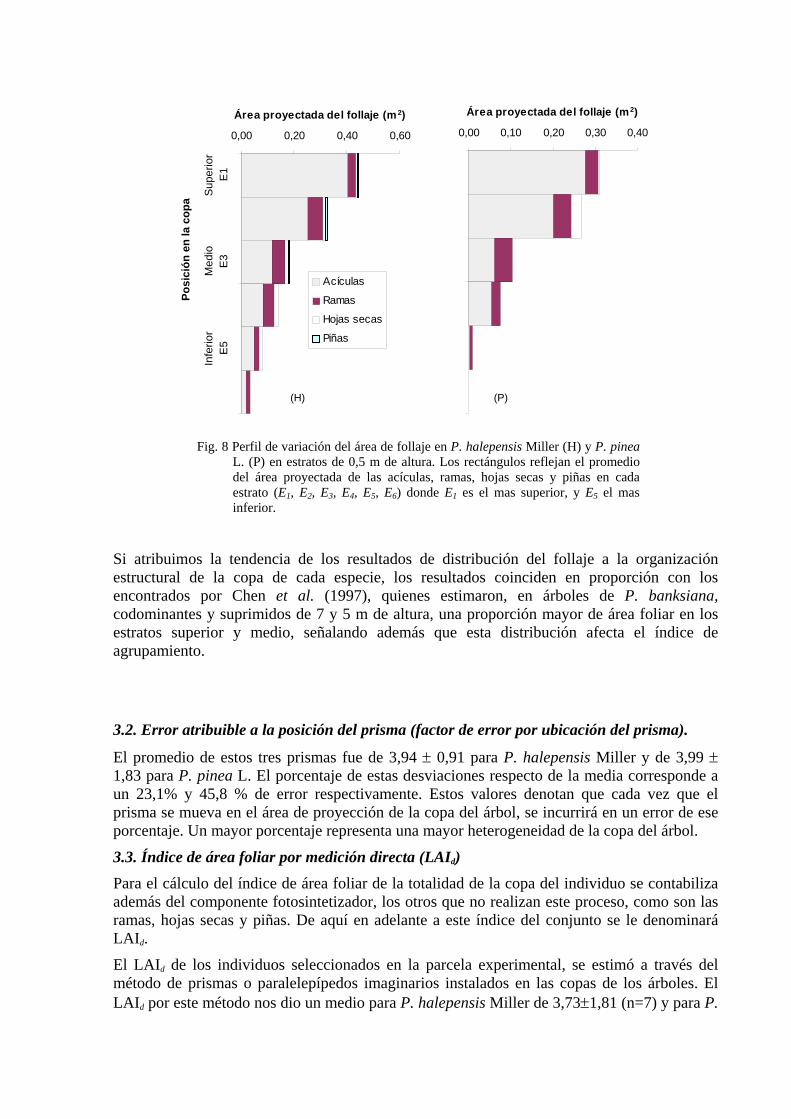
3.1. Área del follaje. En cada prisma, se presentan los datos por estratos. El análisis de los resultados de la cosecha de acículas, ramas, hojas muertas y secas, y piñas en los prismas dieron un promedio del área proyectada del follaje bastante variable entre estratos como puede observarse en la Fig. 8. Por ejemplo, para acículas de las dos especies, se observa una clara diferencia de la superficie media de acículas por prisma (50x50 cm2). Para P. halepensis Miller se ha obtenido de 0,18 ± 0,19 m2 de superficie de acículas en 0,25 m2 de superficie de suelo, y para P. pinea L. 0,14 ± 0,17 m2 en la misma superficie de suelo. Los elevados valores de desviación estándar encontrados, indican la alta variabilidad entre estratos dentro de la copa del árbol.
Sin embargo la mayor proporción de la biomasa de acículas se ha encontrado en los estratos superiores, coincidiendo con el trabajo de Lledó (1983) que trabajo con Pinus halepensis, quien encontró en cuatro árboles de diferentes alturas que el mayor porcentaje de acículas se encuentra en los estratos medios.
El aporte a la proyección total en pino carrasco es el siguiente: 63% del total del follaje es acícula, 27% ramas, 8% hojas muertas y secas y un 2% de piñas; en el pino piñonero el 47% es acícula, 37% son ramas, un 1% son hojas secas y ningún porcentaje de piñas, aunque esto es excepcional.
Esta variación se refleja más en las observaciones dentro de cada estrato. Así vemos que la mayor cantidad de acículas se encuentran en los primeros 3 estratos en un 87% para P. halepensis Miller y en 92% para P. pinea L.. Smolander y Stenberg (1996) trabajando con P. sylvestris L. de 8,7 m de alto y 8,7 cm de diámetro, encontraron que el 88,13% del total de la composición de la copa, es acícula.
0,00 0,20 0,40 0,60
Sup
erio
rE
1M
edio
E3
Infe
rior
E5
Posi
ción
en
la c
opa
Área proyectada del follaje (m 2)
Acículas
Ramas
Hojas secas
Piñas
0,00 0,10 0,20 0,30 0,40
Área proyectada del follaje (m2)
Fig. 8 Perfil de variación del área de follaje en P. halepensis Miller (H) y P. pinea
L. (P) en estratos de 0,5 m de altura. Los rectángulos reflejan el promedio del área proyectada de las acículas, ramas, hojas secas y piñas en cada estrato (E1, E2, E3, E4, E5, E6) donde E1 es el mas superior, y E5 el mas inferior.
Si atribuimos la tendencia de los resultados de distribución del follaje a la organización estructural de la copa de cada especie, los resultados coinciden en proporción con los encontrados por Chen et al. (1997), quienes estimaron, en árboles de P. banksiana, codominantes y suprimidos de 7 y 5 m de altura, una proporción mayor de área foliar en los estratos superior y medio, señalando además que esta distribución afecta el índice de agrupamiento.
3.2. Error atribuible a la posición del prisma (factor de error por ubicación del prisma).
El promedio de estos tres prismas fue de 3,94 ± 0,91 para P. halepensis Miller y de 3,99 ± 1,83 para P. pinea L. El porcentaje de estas desviaciones respecto de la media corresponde a un 23,1% y 45,8 % de error respectivamente. Estos valores denotan que cada vez que el prisma se mueva en el área de proyección de la copa del árbol, se incurrirá en un error de ese porcentaje. Un mayor porcentaje representa una mayor heterogeneidad de la copa del árbol.
3.3. Índice de área foliar por medición directa (LAId) Para el cálculo del índice de área foliar de la totalidad de la copa del individuo se contabiliza además del componente fotosintetizador, los otros que no realizan este proceso, como son las ramas, hojas secas y piñas. De aquí en adelante a este índice del conjunto se le denominará LAId.
El LAId de los individuos seleccionados en la parcela experimental, se estimó a través del método de prismas o paralelepípedos imaginarios instalados en las copas de los árboles. El LAId por este método nos dio un medio para P. halepensis Miller de 3,73±1,81 (n=7) y para P.
(H) (P)
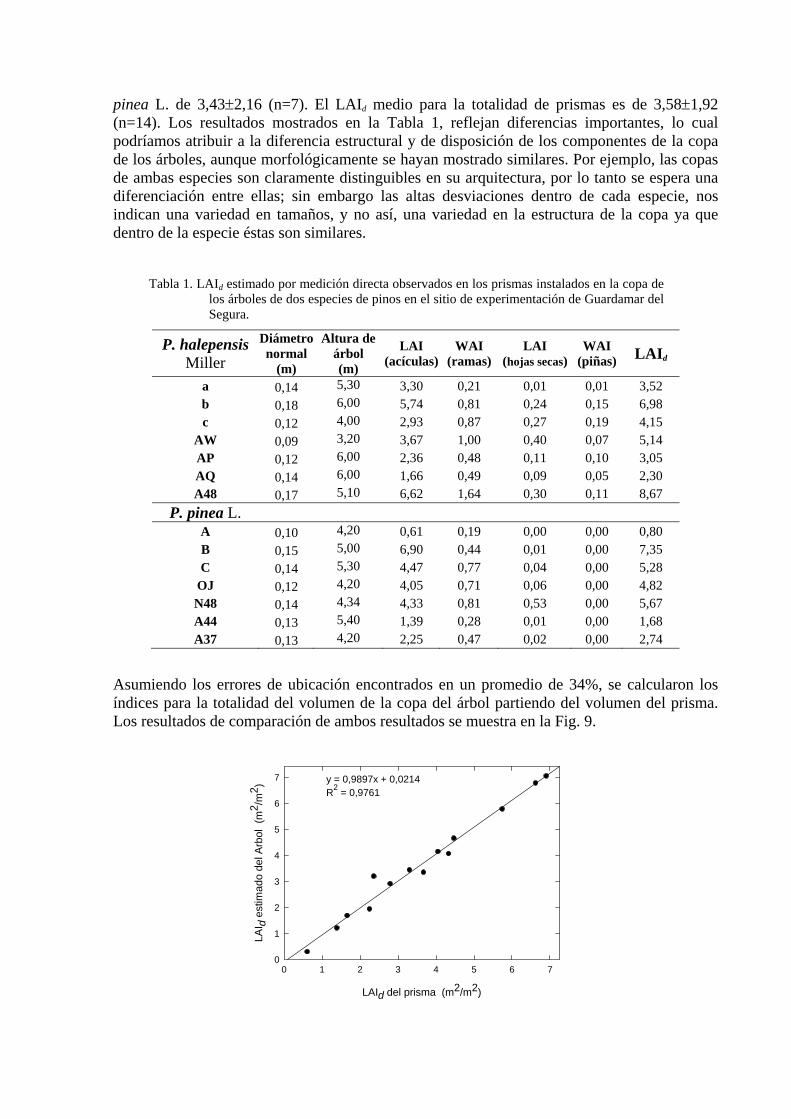
pinea L. de 3,43±2,16 (n=7). El LAId medio para la totalidad de prismas es de 3,58±1,92 (n=14). Los resultados mostrados en la Tabla 1, reflejan diferencias importantes, lo cual podríamos atribuir a la diferencia estructural y de disposición de los componentes de la copa de los árboles, aunque morfológicamente se hayan mostrado similares. Por ejemplo, las copas de ambas especies son claramente distinguibles en su arquitectura, por lo tanto se espera una diferenciación entre ellas; sin embargo las altas desviaciones dentro de cada especie, nos indican una variedad en tamaños, y no así, una variedad en la estructura de la copa ya que dentro de la especie éstas son similares.
Tabla 1. LAId estimado por medición directa observados en los prismas instalados en la copa de
los árboles de dos especies de pinos en el sitio de experimentación de Guardamar del Segura.
P. halepensis Miller
Diámetro normal
(m)
Altura de árbol (m)
LAI (acículas)
WAI (ramas)
LAI (hojas secas)
WAI (piñas) LAId
a 0,14 5,30 3,30 0,21 0,01 0,01 3,52 b 0,18 6,00 5,74 0,81 0,24 0,15 6,98 c 0,12 4,00 2,93 0,87 0,27 0,19 4,15
AW 0,09 3,20 3,67 1,00 0,40 0,07 5,14 AP 0,12 6,00 2,36 0,48 0,11 0,10 3,05 AQ 0,14 6,00 1,66 0,49 0,09 0,05 2,30 A48 0,17 5,10 6,62 1,64 0,30 0,11 8,67
P. pinea L. A 0,10 4,20 0,61 0,19 0,00 0,00 0,80 B 0,15 5,00 6,90 0,44 0,01 0,00 7,35 C 0,14 5,30 4,47 0,77 0,04 0,00 5,28
OJ 0,12 4,20 4,05 0,71 0,06 0,00 4,82 N48 0,14 4,34 4,33 0,81 0,53 0,00 5,67 A44 0,13 5,40 1,39 0,28 0,01 0,00 1,68 A37 0,13 4,20 2,25 0,47 0,02 0,00 2,74
Asumiendo los errores de ubicación encontrados en un promedio de 34%, se calcularon los índices para la totalidad del volumen de la copa del árbol partiendo del volumen del prisma. Los resultados de comparación de ambos resultados se muestra en la Fig. 9.
LAId del prisma (m2/m2)
0 1 2 3 4 5 6 7
LAI d
est
imad
o de
l Arb
ol (
m2 /m
2 )
0
1
2
3
4
5
6
7
y = 0,9897x + 0,0214 R2 = 0,9761

Fig. 9 Relación LAId obtenido en el prisma y LAI estimado para el total de la copa del árbol (n=14).
La evaluación de los resultados muestra un R2 altamente significativo (p< 0,0001). Sin embargo esta relación encierra un sesgo debido a la heterogeneidad de la copa del árbol. La correlación esta basada en la cantidad de elementos encontrados en los prismas y aunque se ha tomado en cuenta el error debido a la posición de éste, solo se la tomo en una orientación.
La relación entre la biomasa estimada de acículas en el total del volumen de la copa del árbol y el LAId ha mostrado tener una alta significancia (R2=0,8169; a=-0,0696; b=0,00708; Dev Stand=0,825; n=14).
3.4. Índice de área foliar efectiva del ceptómetro y LAI-2000 Previo a la obtención de los productos que permitieron el cálculo del LAId, en Julio del 2003 se realizó la obtención de datos con el LAI-2000 y el ceptómetro. El procedimiento siguió las instrucciones del manual de los dos instrumentos (Decagon Devices, 1989; y Li-Cor, 1992).
3.4.1. Mediciones con el ceptómetro. Para el cálculo del LAI estimado con el ceptómetro, que en realidad vendría a ser el índice de área de la planta se utiliza a partir de ahora la denominación de LAIec (LAI efectivo del ceptómetro). Los datos de radiación fotosinteticamente activa (PAR) se tomaron al medio día (de 11:45 a 15:30 horas).
Los valores medios encontrados con este instrumento son para P. halepensis Miller de 3,25±1,21 (n=7) y para P. pinea L. de 2,72±1,53 (n=7). La media para el total de individuos es de 2,99±1,35 (n=14).
Para llegar a los anteriores resultados, se asumió una capacidad de absorbancia de la radiación solar de la acícula del 84%, valor que utiliza Law et al. (2001) para coníferas. Calculamos el coeficiente de extinción (k) bajo diferentes ángulos cenit de incidencia solar (34°, 35°, 37°, 39°, 41°, 44°, 46°, 55° y 57°). Para este cálculo se incluyeron también datos de invierno 2002-2003. La tendencia del k con un x de 0,51 se puede observar en la Fig. 10 (a). x es el parámetro de la relación del área proyectada de un elemento de la copa sobre un plano horizontal y su proyección en un plano vertical (Campbell y Norman, 1990). La figura muestra que los máximos valores de k obtenidos son de 0,89 y 0,94, indicando una posición muy horizontal de las acículas, sesgo de medición que atribuimos a la cercanía del instrumento a las ramas. El valor promedio de k para todos los árboles fue de 0,61 ± 0,12, altos en relación a los valores encontrados por Jarvis y Leverenz, (1983) citado por Bolstad y Gower (1990) de 0,45; 0,42 y 0,28, pero, en promedio, comparables con los valores encontrados por estos últimos (0,85; 0,73 y 0,37).
La regresión no linear de la relación del coeficiente de extinción y el ángulo cenit ajusta a la curva de la forma )exp(bxayy o += (R2= 0,99189, con un Error estándar = 0,0025).
Por otro lado en el análisis de las mediciones por debajo (Qi) y arriba (Qo) de la copa del árbol, se vio una fuerte correlación entre el LAI estimado por el ceptómetro y el logaritmo de la transmitancia de la radiación (Qi/Qo) (R2 = 0,9323; p=0,0001; n=76). La correlación se puede observar en la Fig. 10 (b).
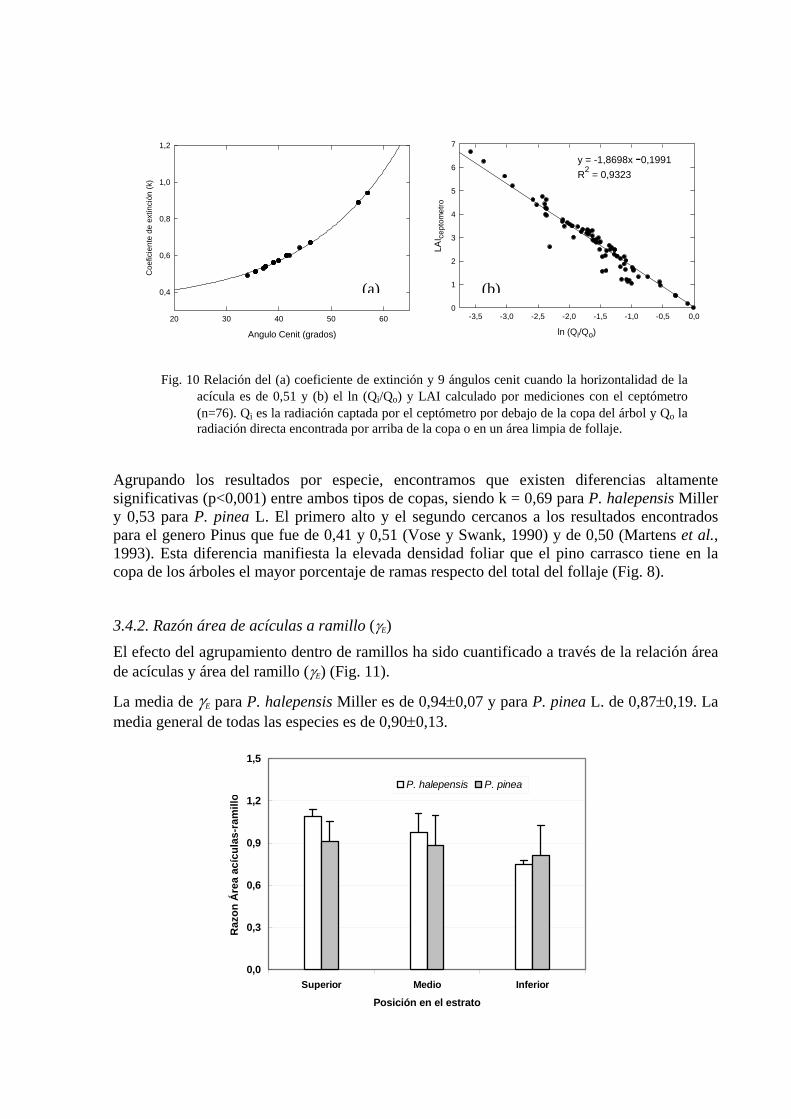
Angulo Cenit (grados)
20 30 40 50 60
Coe
ficie
nte
de e
xtin
ción
(k)
0,4
0,6
0,8
1,0
1,2
ln (Qi/Qo)
-3,5 -3,0 -2,5 -2,0 -1,5 -1,0 -0,5 0,0
LAI c
epto
met
ro
0
1
2
3
4
5
6
7
Fig. 10 Relación del (a) coeficiente de extinción y 9 ángulos cenit cuando la horizontalidad de la acícula es de 0,51 y (b) el ln (Qi/Qo) y LAI calculado por mediciones con el ceptómetro (n=76). Qi es la radiación captada por el ceptómetro por debajo de la copa del árbol y Qo la radiación directa encontrada por arriba de la copa o en un área limpia de follaje.
Agrupando los resultados por especie, encontramos que existen diferencias altamente significativas (p<0,001) entre ambos tipos de copas, siendo k = 0,69 para P. halepensis Miller y 0,53 para P. pinea L. El primero alto y el segundo cercanos a los resultados encontrados para el genero Pinus que fue de 0,41 y 0,51 (Vose y Swank, 1990) y de 0,50 (Martens et al., 1993). Esta diferencia manifiesta la elevada densidad foliar que el pino carrasco tiene en la copa de los árboles el mayor porcentaje de ramas respecto del total del follaje (Fig. 8).
3.4.2. Razón área de acículas a ramillo (γE)
El efecto del agrupamiento dentro de ramillos ha sido cuantificado a través de la relación área de acículas y área del ramillo (γE) (Fig. 11).
La media de γE para P. halepensis Miller es de 0,94±0,07 y para P. pinea L. de 0,87±0,19. La media general de todas las especies es de 0,90±0,13.
y = -1,8698x -0,1991 R2 = 0,9323
(b)(a)
0,0
0,3
0,6
0,9
1,2
1,5
Superior Medio Inferior
Posición en el estrato
Raz
on Á
rea
acíc
ulas
-ram
illo
P. halepensis P. pinea

Fig. 11 Promedio de variación de la relación de área de acículas Vs. área proyectada del ramillo en diferentes ángulos (efecto de agrupación dentro de ramillos). Las medidas fueron estimadas tomando los datos del paralelepípedo imaginario, en estratos de 0,50 m de altura. Las líneas continuas sobre las barras expresan la variación de la desviación estándar.
Los valores de γE varían entre especies y dentro de cada estrato, aunque esta variación no es estadísticamente significativa. Esta variación puede deberse al muestreo temporal de los ramillos, ya que las muestras se tomaron en una época donde los primordios de la última cohorte estaban en su fase de crecimiento, cuantificado en un 22,23% para el pino carrasco y de 24,89% para el pino piñonero. Aunque los valores encontrados encierran un error del 30% aproximadamente, por el número pequeño de muestras (n=23), éstos son muy bajos en comparación a los hallados por Chen (1996) para P. banksiana, en un promedio de la parte superior de 1,61 y 1,71; en la parte media de la copa de 1,40 y 1,44; y en la parte inferior de la copa de 1,22 y 1,37, de entre pinos jóvenes y longevos, respectivamente. Los valores mas bajos que encontramos se hallaron entre los 0,79 y 0,83 en P. halepensis Miller y P. pinea L. respectivamente. Asumimos que en estas dos especies, esto se debió a que la proyección del ramillo (As) fue mayor a la proyección de acículas (An), indicando una mayor separación de acículas que en los otros individuos, causando la aparición de huecos que sesga el tamaño del área de proyección del ramillo. El método de muestreo también puede haber causado un error en el cálculo de este índice. Aunque Gower y Norman (1990) encontraron que los ramillos visualmente son estructuras diferentes, y considerados como las unidades básicas de absorción de radiación fotosinteticamente activa (Fassnacht et al., 1994), se ha observado que este acercamiento difiere mucho de la realidad. Aunque utilizamos un método muy simple para obtener el As, nos dimos cuenta que es muy difícil saber exactamente donde comienza y termina el límite del borde del ramillo. Coincidimos con Chen et al. (1996), cuando él indica que el trabajo de determinación del índice de agrupamiento dentro de ramillos es la que mas ardua.
3.4.3. Razón entre el área de la superficie leñosa y el área de la superficie total (α)
En los árboles del pinar de Guardamar del Segura tienen muchas ramas muertas en la parte mas baja de la copa del árbol. Es lógico pensar que estas ramas también interceptan la radiación solar por debajo de esta estructura. Por lo tanto, las ramas muertas y vivas, también afectan la transmitancia del PAR, y las mediciones de LAI con instrumentos ópticos. Por esta razón se midió el porcentaje de influencia que las ramas aportan cuando se esta midiendo con el LAI-2000.
El material leñoso permanece invariable durante toda la estación, y por tanto solo necesita ser medido una vez. El volumen de influencia de este material fue calculado utilizando la relación (9), donde el área proyectada se calculó como la mitad del diámetro de la base multiplicado por la longitud de la rama. Esta ecuación muestra la proporción de ramas respecto del total de componentes. Los resultados agrupados por estratos se observan en la Fig. 12.
La media de α para P. halepensis Miller es de 0,47±0,14 y para P. pinea L. de 0,56±0,21. La media general de los valores α en conjunto nos da 0,51 ± 0,17.
0,0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
Superior Medio Inferior
Posición en el estrato
Frac
ción
leño
sa ( α
)
P. halepensis P. pinea
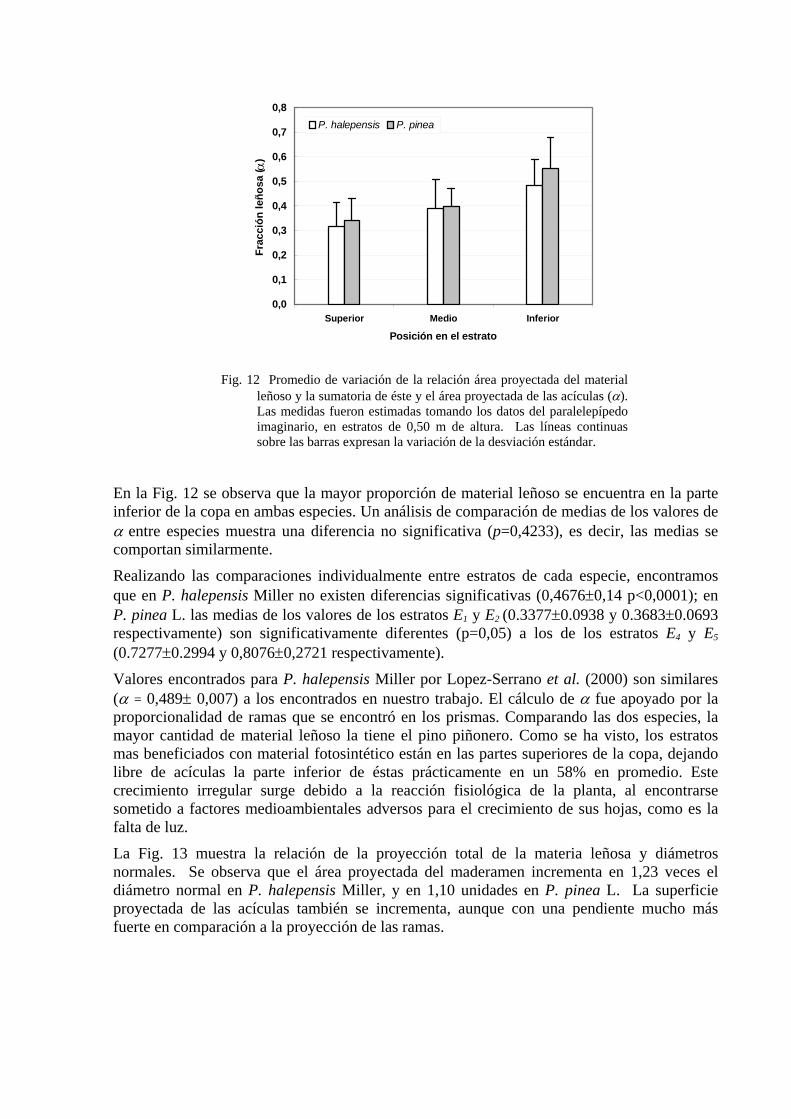
Fig. 12 Promedio de variación de la relación área proyectada del material
leñoso y la sumatoria de éste y el área proyectada de las acículas (α). Las medidas fueron estimadas tomando los datos del paralelepípedo imaginario, en estratos de 0,50 m de altura. Las líneas continuas sobre las barras expresan la variación de la desviación estándar.
En la Fig. 12 se observa que la mayor proporción de material leñoso se encuentra en la parte inferior de la copa en ambas especies. Un análisis de comparación de medias de los valores de α entre especies muestra una diferencia no significativa (p=0,4233), es decir, las medias se comportan similarmente.
Realizando las comparaciones individualmente entre estratos de cada especie, encontramos que en P. halepensis Miller no existen diferencias significativas (0,4676±0,14 p<0,0001); en P. pinea L. las medias de los valores de los estratos E1 y E2 (0.3377±0.0938 y 0.3683±0.0693 respectivamente) son significativamente diferentes (p=0,05) a los de los estratos E4 y E5 (0.7277±0.2994 y 0,8076±0,2721 respectivamente).
Valores encontrados para P. halepensis Miller por Lopez-Serrano et al. (2000) son similares (α = 0,489± 0,007) a los encontrados en nuestro trabajo. El cálculo de α fue apoyado por la proporcionalidad de ramas que se encontró en los prismas. Comparando las dos especies, la mayor cantidad de material leñoso la tiene el pino piñonero. Como se ha visto, los estratos mas beneficiados con material fotosintético están en las partes superiores de la copa, dejando libre de acículas la parte inferior de éstas prácticamente en un 58% en promedio. Este crecimiento irregular surge debido a la reacción fisiológica de la planta, al encontrarse sometido a factores medioambientales adversos para el crecimiento de sus hojas, como es la falta de luz.
La Fig. 13 muestra la relación de la proyección total de la materia leñosa y diámetros normales. Se observa que el área proyectada del maderamen incrementa en 1,23 veces el diámetro normal en P. halepensis Miller, y en 1,10 unidades en P. pinea L. La superficie proyectada de las acículas también se incrementa, aunque con una pendiente mucho más fuerte en comparación a la proyección de las ramas.

0
0,4
0,8
1,2
1,6
2
0 0,05 0,1 0,15 0,2 0,25 0,3
Diametro normal (m)
Áre
a pr
oyec
tada
(m2 )
0 0,05 0,1 0,15 0,2 0,25 0,3
Diametro normal (m)
Acículas
Parte leñosa
Fig. 13 Relación del área proyectada de la hoja y la parte leñosa de 7 árboles de P. halepensis
Miller (a) y 7 árboles de P. pinea L (b) con el diámetro normal (diámetro 1,3 m sobre el nivel del suelo).
Se advierte que la proyección del área de las ramas es mas o menos constante hasta un diámetro normal de 0,26. esta invariabilidad al menos hasta el diámetro señalado evidencia la invariabilidad de la parte leñosa durante cambios estacionales en un período agrícola en especial en nuestra zona de estudio, con sequías de larga duración y temperaturas elevadas.
3.4.4. Índice de área foliar corregido
La Tabla 2 muestra los resultados de la corrección de la medición con el LAI-2000. La aplicación de estos factores se realiza de acuerdo a la ecuación (7). Para todos los valores obtenidos con el LAI-2000 se verificó una sobreestimación de los valores obtenidos por el instrumento utilizado respecto de los valores obtenidos con la corrección. Esta sobrevaloración de la información obtenida por el instrumento es explicada porque el LAI-2000 (y el ceptómetro a su vez), no discrimina entre acículas, ramas, ni otros componentes de la copa del árbol. Los instrumentos ópticos en realidad miden un índice representante de la totalidad de los componentes de la copa del árbol.
Las mediciones en el pino alepo tuvo un 42% en promedio mayor a la del obtenido por medición con el instrumento, en contra de los 30% en promedio del pino piñonero. Sin embargo, la menor diferencia observada fue de un 14% en la primera especie en comparación a los 18% de la segunda especie.
Tabla 2. Resumen de valores del LAI-2000, y los índices de corrección obtenidos en la
copa de los árboles de dos especies de pinos en el sitio de experimentación de Guardamar del Segura. Ω se tomo de Chen et al., (1997) promediando los valores de los pinos longevos.
Especie Código LAI-2000 1-α γE Ω LAI corregido
P. halepensis Miller
a 1,91 0,44 1,13 0,77 1,23 b 2,99 0,57 0,92 0,77 2,04 c 1,67 0,50 0,79 0,77 0,86
AW 2,10 0,64 0,91 0,77 1,59 AP 1,31 0,60 1,13 0,77 1,15
AQL 1,05 0,79 0,84 0,77 0,90 A48 3,20 0,67 1,01 0,77 2,81
(a) (b)

P. pinea L.
A 0,47 0,65 0,84 0,77 0,33 B 3,58 0,51 1,16 0,77 2,75 C 2,48 0,57 0,95 0,77 1,74
AOJ 2,52 0,71 0,97 0,77 2,25 AN48 2,63 0,59 1,24 0,77 2,50 A44 1,00 0,58 0,83 0,77 0,62 A37 1,27 0,65 0,94 0,77 1,01
Se evidencia que la mayor proporción de ramas, expresados en los valores de α, influye de manera importante en los valores tomados por el LAI-2000. Calculando las sobreestimaciones sin tomar el Ω, vemos que la proporción se evidencia más, ya que los valores del LAI-2000 muestran mayor variabilidad. Esto hace innegable la necesidad de la aplicación de los índices de corrección a los datos obtenidos por este instrumento.
De la misma forma y coincidiendo con los trabajos de Lopez-Serrano et al. (2000), cuando se calcula el LAI, sin tomar γE, se observa una gran variación de éstos y una baja correlación entre valores de LAI y γE (R2 =0,44; a=1,201; b=-0,1064; SD=0,11; n=14). Atribuimos esta variación a las características morfológicas de la copa del pino, en especial, la de P. halepensis Miller, ya que en la agrupación de sus acículas también se observa la presencia de hasta una tercera cohorte, proveniente de hace 3 años, causando una sobreestimación del área del ramillo.
En adición, la ausencia o presencia de cohortes, causa pequeños huecos entre las ramas que podrían causar una sobreestimación del As. De la misma forma, el alto número de acículas por ramillo, puede haber introducido un error en los cálculos finales.
Realizando la regresión lineal de los dos grupos de valores, separados en grupos de la misma especie, encontramos una buena correlación para ambas unidades. Para P. halepensis Miller se obtuvo: R2 = 0,76; a=-0,34; b=0,95; SD=0,41; n=7; y para P. pinea L.: R2 = 0,92; a=-0,06; b=0,84; SD=0,25; n=7. Según Gower et al. (1999) los valores del LAI-2000 son considerados como PAI (un indice foliar para la totalidad de la planta), y la diferencia entre estos valores y los obtenidos mediante corrección, nos da un índice de área para la parte leñosa (WAI). Aunque los resultados obtenidos por ellos son bastante diferentes entre pinos, coincidimos con los valores encontrados por Chen y Chilar (1995) mencionados por Chen (1996), en cuya investigación se obtuvo un WAI de 0,32, cercano al valor medio encontrado por nosotros (WAI=0,38) para P. pinea L. El valor obtenido para P. halepensis Miller (WAI=0,51) se acerca más a los obtenidos por Gower et al. (1999) y Chen et al. (1997) para pinos longevos (0,73 y 0,72 respectivamente).
Finalmente, luego de esta corrección, los valores promedios de LAI encontrados en la parcela de experimentación para P. halepensis Miller es de 1,52±0,71 y para P. pinea L. 1,69±0,69. Según estos resultados, el factor que se multiplicaría a los datos obtenidos por el LAI-2000 será para P. halepensis Miller es de 0,75±0,14 y para P. pinea L. 0,78±0,11.
El promedio del LAI de la totalidad de los árboles es de 1,56±0,81. El factor de corrección obtenido para el conjunto es de 0,77±0,12.
3.5. Comparación de los índices de área foliar por mediciones ópticas
Con el ceptómetro y el LAI-2000 se obtienen un LAIe, ya que ambos son instrumentos de medición óptica.
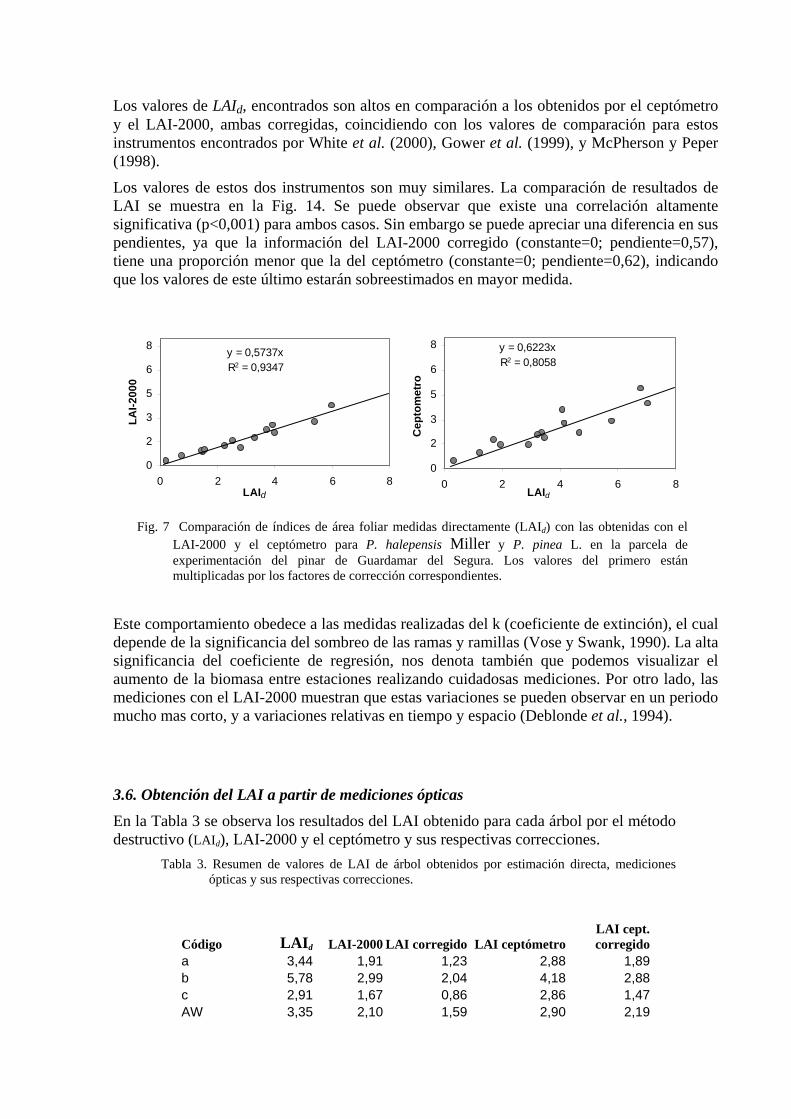
Los valores de LAId, encontrados son altos en comparación a los obtenidos por el ceptómetro y el LAI-2000, ambas corregidas, coincidiendo con los valores de comparación para estos instrumentos encontrados por White et al. (2000), Gower et al. (1999), y McPherson y Peper (1998).
Los valores de estos dos instrumentos son muy similares. La comparación de resultados de LAI se muestra en la Fig. 14. Se puede observar que existe una correlación altamente significativa (p<0,001) para ambos casos. Sin embargo se puede apreciar una diferencia en sus pendientes, ya que la información del LAI-2000 corregido (constante=0; pendiente=0,57), tiene una proporción menor que la del ceptómetro (constante=0; pendiente=0,62), indicando que los valores de este último estarán sobreestimados en mayor medida.
y = 0,5737xR2 = 0,9347
0
2
3
5
6
8
0 2 4 6 8LAId
LAI-2
000
y = 0,6223xR2 = 0,8058
0
2
3
5
6
8
0 2 4 6 8LAId
Cep
tom
etro
Fig. 7 Comparación de índices de área foliar medidas directamente (LAId) con las obtenidas con el
LAI-2000 y el ceptómetro para P. halepensis Miller y P. pinea L. en la parcela de experimentación del pinar de Guardamar del Segura. Los valores del primero están multiplicadas por los factores de corrección correspondientes.
Este comportamiento obedece a las medidas realizadas del k (coeficiente de extinción), el cual depende de la significancia del sombreo de las ramas y ramillas (Vose y Swank, 1990). La alta significancia del coeficiente de regresión, nos denota también que podemos visualizar el aumento de la biomasa entre estaciones realizando cuidadosas mediciones. Por otro lado, las mediciones con el LAI-2000 muestran que estas variaciones se pueden observar en un periodo mucho mas corto, y a variaciones relativas en tiempo y espacio (Deblonde et al., 1994).
3.6. Obtención del LAI a partir de mediciones ópticas
En la Tabla 3 se observa los resultados del LAI obtenido para cada árbol por el método destructivo (LAId), LAI-2000 y el ceptómetro y sus respectivas correcciones.
Tabla 3. Resumen de valores de LAI de árbol obtenidos por estimación directa, mediciones ópticas y sus respectivas correcciones.
Código LAId LAI-2000 LAI corregido LAI ceptómetroLAI cept. corregido
a 3,44 1,91 1,23 2,88 1,89b 5,78 2,99 2,04 4,18 2,88c 2,91 1,67 0,86 2,86 1,47AW 3,35 2,10 1,59 2,90 2,19

AP 3,20 1,31 1,15 2,35 2,07AQL 1,69 1,05 0,90 2,03 1,76A48 6,78 3,20 2,81 5,54 4,88A 0,30 0,47 0,33 0,65 0,47B 7,05 3,58 2,75 5,20 3,98C 4,66 2,48 1,74 3,10 2,19AOJ 4,15 2,52 2,25 3,10 2,78AN48 4,07 2,63 2,50 3,72 3,58A44 1,21 1,00 0,62 1,50 0,95A37 1,94 1,27 1,01 1,80 1,45 3,61 2,01 1,56 2,99 2,32
Los valores medios muestran una similitud entre los instrumentos ópticos, sin embargo, éstos son menores a los obtenidos por el método directo.
6. Conclusiones Con los resultados obtenidos se puede llegar a las siguientes conclusiones:
En la obtención del LAI de forma directa:
En este trabajo se ha comprobado que en la obtención del LAI a partir de la utilización de prismas en copas de árboles de arquitectura heterogénea, se incurre en un error de muestreo.
En la obtención del LAI con el LAI-2000:
Los datos de este instrumento son bastante precisos si se toma cuidado y detalle en su adquisición en el campo.
El Indice de Área Foliar obtenido por el LAI-2000 tienen una sobreestimación de sus valores, ya que no discrimina la parte fotosintetizadora de la no fotosintetizadora. Se ha evidenciado que estos datos necesitan ser corregidos por los índices de agrupamiento que representan la presencia de las ramas, y la forma de agrupación de los componentes de la copa del árbol. El valor de este índice obtenido para la parcela de Guardamar del Segura es de 0,77±0,12.
En la obtención del LAI con el ceptometro:
Los resultados de LAI obtenidos por este instrumento se apoyan en la utilización de constantes y ecuaciones. La información que complementa el cálculo puede variar según la estación en la que se tome.
No es aconsejable la utilización de la una media de k, ya que ésta es representativa de un sitio y una determinada hora.
En la aplicación de los resultados encontrados:
El LAI obtenido de forma directa en la parcela de experimentación de Guardamar del Segura es de 3,61±2,00 (una masa forestal de copas mas o menos homogéneas).
El índice de corrección para los datos del LAI-2000 estimado para la parcela de Guardamar del Segura es de 0,77±0,12.
Si se estima el LAI de la parcela por medio del LAI-2000, se deberá corregir por un segundo factor 2,26 para obtener el valor cercano al obtenido por medición directa.

Los datos de LAI están altamente correlacionados con el diámetro normal de los árboles, por lo que su utilización potencial en el cálculo del LAI de todo el pinar se puede realizar con la medición de los diámetros normales de una muestra estratégica representativa de este ecosistema.
4. Agradecimientos
Al proyecto “Los pinares costeros y la vegetación psamofítica: análisis de sus procesos hidrológicos y propuesta de integración en una restauración en mosaico” por haber prmitido esta investigación.
5. Referencias
Barclay, H.J. and Trofymow, J.A. 2000. Relationship of readings from the LI-COR canopy analyser to total one-side leaf area index and stand structure in immature Douglas-fir. Forest Ecology Management 132 (2000) 121-126.
Barrantes, O. and Gracia, C.A. 1989. Estimación del área foliar a partir de la superficie de albura en Pinus sylvestris. Options Mediterraneennes – Série Séminaires No.3, 1989: 53-56.
Bolstad, P.V., and Gower, S.T. 1990. Estimation of leaf area index in fourteen southern Wisconsin forest stands using a portable radiometer. Tree Physiology 7, 115-124.
Campbell, G.S. and Norman, J.M. 1990. The description and measurement of plant canopy structure. In Plant Canopies: their growth, form and function. Cambridge University Press. 1990. 200 pag.
Chen, J.M. and Cihlar, J. 1995. Quantifying the effect of canopy architecture on optical measurements of leaf area index using two gap size analysis methods. IEEE Transactions on Geoscience and Remote Sensing, Vol. 33 No.3 May, 1995.
Chen, J.M. 1996. Optically-based methods for measuring seasonal variation of leaf area index in boreal conifer stands. Agricultural and Forest Metereology, Vol 80, Issue 2-4, July 1996, pp 135-163.
Chen, J.M., Rich, P.M., Gower, S.T., Norman, J.M., and Plummer, S. 1997. Leaf area index of boreal forests: theory, techniques, and measurements. Journal of Geophysical Research 102 (D24): 29, 429-29, 443.
Cherry, M., Hingston, A., Battaglia, M., and Beadle, C. 1998. Calibrating the LI-COR LAI-2000 for estimating leaf area index in eucalypt plantations. Tasforest Vol. 10: 75-82.
Deblonde, G., Penner, M., and Royer, A. 1994. Measuring leaf area index with the LI-COR LAI-2000 in pine stands. Ecology, Vol. 75, pp 1507-1511.
Decagon Devices, Inc. 1989. Sunfleck Ceptometer, User manual. Delta-T Devices Ltd., Cambridge. 34 pag.
Eklundh, L., Harrie, L., Kuusk, A. 2001. Investigating relationships between Landsat ETM+ sensor data and leaf area index in a boreal conifer forest. Remote Sensing of Environment 78 (2001) 239-251.
Fassnacht, K., Gower, S. T., Norman, J.M., and McMurtric, R.E. 1994. A comparison of optical and direct methods for estimating foliage surface area index in forest. Agric. For. Metereol. Vol 71, 1-2 (183-207).

Feldkirchner, D.C. and Gower, S.T. 2001. Using the LI-COR LAI-2000 to estimate leaf area index and light transmittance in forest canopies. Methodology papers series of the 4th International Conference on ILTER in East and Pacific Region, Ulaanbaata-Hatgal, Mongolia, 2001, pp. 12-14
Gower, S. T. and Norman, J. M., 1990. Rapid estimation of leaf area index in conifer and broad-leaf plantations. Ecology, 72(5): 1896-1900
Gower, S.T., Kucharik, C.J., Norman, J.M. 1999. Direct and indirect estimation of leaf area index, fAPAR, and net primary production of terrestrial ecosystems. Remote Sensing of Environment, 70, 29-51.
Hebert, M. T., & Jack, S. 1998. Leaf area index and site water balance of loblolly pine (Pinus taeda L.) across a precipitation gradient in East Texas. Forest Ecology and Management 105 (1998) 273-282.
Kucharik, C.J., Norman, J.M., and Gower, S.T. 1998. Measurements of branch area and adjusting leaf area index indirect measurements. Agricultural and Forest Metereology, Vol. 91, Issues 1-2, pp. 69-88.
Kucharik, C.J., Norman, J.M., and Gower, S.T. 1998. Characterization of radiation regimes in non-random forest canopies: theory, measurements, and a simplified modelling approach. Tree Physiology 19, 695-706.
Law, B.E., Van Tuyl, S., Cescatti, A., and Baldocchi, D.D. 2001. Estimation of leaf area index in open-canopy ponderosa pine forest at different successional stages and management regimes in Oregon. Agricultural and Forest Meteorology 108 (2001) 1-14.
Leblanc, S.G., Chen, J.M., and Kwong, M. 2002. Tracing Radiation and Architecture of Canopies – TRAC Manual V.2.1.3.
Leblanc, S.G., Fernandes, R., and Chen, J.M. 2002. Recent advancements in optical field leaf area index, foliage heterogeneity, and foliage angular distribution measurements. Proceedings of International Geoscience and Remote Sensing Society, 24-28 June 2002, Toronto.
Leverenz, J.W. and Hinckley, T.M. 1990. Shoot structure, leaf area index and productivity of evergreen conifer stands. Tree Physiology 6, 135-149.
Li-Cor, Inc. 1992. LAI-2000 Plant canopy analyser. Nebraska. 224 pag.
Lledó, M.J. (1982) Datos energéticos, estructurales y químicos de Pinus halepensis Mill. en el pinar de la Sierra Espuña (provincia de Murcia). Tesina de Licenciatura. Universidad de Valencia. 202 pag.
Lopez-Serrano, F.R., Landete-Castillejos, T., Martinez-Millan, J., Cerro-Barja, A. 2000. LAI estimation of natural pine forest using a non-standard sampling technique. Agricultural and Forest Meteorology 101 (2000) 95-111.
Martens, S.N., Ustin, S.L., Rousseau, R.A. 1993. Estimation of tree canopy leaf area index by gap fraction analysis. Forest Ecology and Management 61 (1993) 91-108.
Mussche, S., Samson, R., Nachtergale, L., De Schrijver, A., Lemeur, R., and Lust, N. 2001. A comparison of optical and direct methods for monitoring the seasonal dynamics of leaf area index in deciduous forest. Silva Fennica 35 (4): 373-384.
Nacional Institutes of Health. 2003. Writing ImageJ Plugins–A Tutorial. Upper Austria, University of Applied Sciences, Dept. of Media Technology and Design, Hagenberg, Austria. 50 pag.

Nilson, T. 1999. Inversion of gap frequency data in forest stand. Agricultural and Forest Metereology 98-99 (1999) 437-448.
Nilson, T., Anniste, J., Lang, M., Praks, J. 1999. Determination of needle area indices of coniferous forest canopies in the NOPEX region by ground-based optical measurements and satellite images. Agricultural and Forest Metereology 98-99 (1999) 449-462.
McPherson, E.G. and Peper, P.J. 1998. Comparison of five methods for estimating leaf area index of open deciduous trees. Journal of arboriculture 24(2):98-111. 1998.
Pérez, C., A.J. (1994) Atlas Climàtic de la Comunitat Valenciana. Generalitat Valenciana. Valencia. 1994. 205 pag.
Raventos, J., De Luis, M., Gras, M.J., Cufar, K., Gonzáles-Hidalgo, J.C., Bonet, A., Sanchez, J.R. (2001) Growth of Pinus pinea and Pinus halepensis as affected by dryness, marine spray and land use changes in a Mediterranean semiarid ecosystem. Dendrochronologia 19(2)-2001:211-220.
SIG oleícola español (1997). Cartas de vuelo. Provincia de ALICANTE, Municipio de Guardamar del Seggura, Polígono 4 / Parcela 185.
Superficie 33,273 ha / Olivos 0 / Fecha Vuelo 10/1997
Smith, N.J., Chen, J.M., and Black, T.A. 1993. Effects of clumping on estimates of stand leaf area index using the LI-COR LAI-2000. Canadian Journal of Forest Research, Vol. 23, pp. 1940-1943.
Smolander, H., Stenberg, P. 1996. Response of LAI-2000 estimates to changes in plant area index in a Scots pine stand. Tree Physiology 16, 345-349.
Stenberg, P., Linder, S., Smolander, H., and Flower-Ellis, J. 1994. Performance of the LAI-2000 plant canopy analyzer in estimating leaf area index of some Scot pine stands. Tree Physiology 14, 981-995.
Stenberg, P., Nilson, T., Smolander, H., and Voipio, P. 2003. Gap fraction based estimation of LAI in Scots pine stands subjected to experimental removal of branches and stems. Can. J. Remote Sensing, Vol. 29 No. 3, pp. 363-370, 2003.
Thomas, S.C., and Winner, W.E. 2000. Leaf area index of an old-growth Douglas-fir forest estimated from direct structural measurement in the canopy. Can. J. For. Res. 30: 1922-1930 (2000).
Vose, J.M., and Swank, W.T. 1990. Assessing seasonal leaf area dynamics and vertical leaf area distribution in eastern white pine (Pinus strobes L.) with a portable light meter. Tree Physiology 7, 125-134.
Walter, J.M.N., Fournier, R.A., Soudani, K., Meyer, E. 2003. Integrating clumping effects in forest canopy structure: an assessment through hemispherical photographs. Can. J. Remote Sensing. Vol. 29 No.3, pp 388-410, 2003.
White, M.A., Asner, G.P, Nemani, R.R., Privette, J.L., and Running, S.W. Measuring fractional cover and leaf area index in arid ecosystem: Digital camera, radiation transmittance, and laser altimetry methods. Remote Sen. Environ. 74:45-57(2000)