
ESTUDO DA BIOGÊNESE DOS MELANOSSOMAS E … · colegiado de ciÊncias biolÓgicas e da saÚde –...
31
COLEGIADO DE CIÊNCIAS BIOLÓGICAS E DA SAÚDE – CCBS CURSO DE BIOTECNOLOGIA ESTUDO DA BIOGÊNESE DOS MELANOSSOMAS E O CONTROLE DA SÍNTESE DA MELANINA PELO INIBIDOR TRICICLAZOL NOS FUNGOS DEMÁCEOS FONSECAEA PEDROSOI E WANGIELLA DERMATITIDIS. Akio Kosaka Kajishima Rio de Janeiro 2009
Transcript of ESTUDO DA BIOGÊNESE DOS MELANOSSOMAS E … · colegiado de ciÊncias biolÓgicas e da saÚde –...

COLEGIADO DE CIÊNCIAS BIOLÓGICAS E DA SAÚDE – CCBS
CURSO DE BIOTECNOLOGIA
ESTUDO DA BIOGÊNESE DOS MELANOSSOMAS E O
CONTROLE DA SÍNTESE DA MELANINA PELO INIBIDOR
TRICICLAZOL NOS FUNGOS DEMÁCEOS FONSECAEA
PEDROSOI E WANGIELLA DERMATITIDIS.
Akio Kosaka Kajishima
Rio de Janeiro
2009

Akio Kosaka Kajishima
Aluno do curso de Biotecnologia
Matricula: 0613800280
ESTUDO DA BIOGÊNESE DOS MELANOSSOMAS E O
CONTROLE DA SÍNTESE DA MELANINA PELO INIBIDOR
TRICICLAZOL NOS FUNGOS DEMÁCEOS FONSECAEA
PEDROSOI E WANGIELLA DERMATITIDIS.
Trabalho de Conclusão de Curso, TCC,
apresentado ao Curso de Graduação em
Biotecnologia, da UEZO como parte dos
requisitos para a obtenção do grau de
Tecnólogo em Biotecnologia, sob a
orientação do Prof. Anderson Jack Franzen.
Rio de Janeiro
Agosto de 2009

ii
ESTUDO DA BIOGÊNESE DOS MELANOSSOMAS E O CONTROLE DA
SÍNTESE DA MELANINA PELO INIBIDOR TRICICLAZOL NOS FUNGOS
DEMÁCEOS FONSECAEA PEDROSOI E WANGIELLA DERMATITIDIS.
Elaborado por Akio Kosaka Kajishima
Aluno do curso de Biotecnologia da UEZO
Este trabalho de Graduação foi analisado e aprovado com
Grau:____________________
Rio de Janeiro, 25 de Agosto de 2009.
_________________________________________________________
Jessica Manya Bittencourt Dias Vieira, Doutora em Ciências Biológicas (Microbiologia)
Membro da Banca
_________________________________________________________
Sergio Henrique Seabra, Doutor em Ciências Biológicas (Biofísica)
Membro da Banca
_________________________________________________________
Rafael Berrelho Bernini, Mestre em Ciências (Físico-Química)
Professor de TCC
_________________________________________________________
Anderson Jack Franzen, Doutor em Ciências Biológicas (Biofísica)
Professor Orientador e presidente da Banca
Rio de Janeiro, RJ – Brasil
Agosto de 2009.

iii
Dedico este trabalho à minha noiva, Vanessa,
por me apoiar em todos os momentos.
Meus pais, meus irmãos e a todos meus
familiares, por sempre estarem ao meu lado.
E principalmente a Deus por todas as bênçãos.

iv
Agradeço ao meu orientador, Anderson Franzen,
pelos ensinamentos e broncas que me puseram no
caminho certo, e por todo seu apoio;
Ao Sergio Seabra, por me orientar nos momentos
que sempre precisava;
Aos colegas de curso, que sofremos e alegramos
juntos durante toda graduação;
Aos professores, por me passarem todo meu
conhecimento;
Aos companheiros de laboratório, por todos os
momentos que passamos quebrando a cabeça em
nossa pesquisa;
Aos técnicos de nossos laboratórios, sem o apoio
de vocês meu trabalho nunca sairia;
A todos que em algum momento me apoiaram e
tornaram isso possível.

v
RESUMO
Com o aumento de grupos de pacientes gravemente doentes e/ou
imunocomprometidos como na AIDS e diabetes, aumento do número de pacientes
submetidos à quimioterapia de câncer ou terapia imunossupressiva para transplante de
órgãos e tratamento com antibióticos de largo espectro ou glicocorticóides, a freqüência de
infecções fúngicas tem aumentado, assim como o número de espécies potencialmente
invasivas. Duas infecções se destacam, sendo elas a Cromoblastomicose, tendo como seu
principal agente etiológico o fungo Fonsecaea pedrosoi, e a Feohifomicose, que tem como
um dos principais agentes etiológico a Wangiella dermatitidis. São infecções cutâneas e
subcutâneas de alta relevância ao homem e de grande ocorrência no Brasil e no exterior.
Uma das principais características dos fungos causadores destas infecções é a
produção de melanina. Esta melanina tem como função a proteção dos fungos, agindo
como um seqüestrador de radicais oxidativos do oxigênio tornando estes fungos mais
resistentes aos mecanismos de defesa do hospedeiro e, também, a drogas antifúngicas.
O uso do triciclazol como fungicida em plantações de arroz é muito comum no
controle do patógeno Pyricularia oryzae, por ter a característica de inibir a biossíntese da
melanina, este tem sido alvo de estudos em fungos demáceos causadores de infecções
como a Cromoblastomicose e Feohifomicose. A diminuição da produção de melanina
demonstrou a diminuição da resistência dos fungos aos antifúngicos comercialmente
usados.
A inibição na produção da melanina tem se mostrado um possível alvo na síntese
de novos fármacos antifúngicos.
Neste estudo foi visto a inibição na produção de melanina do fungo demáceo
Wangiella dermatitidis em uma concentração de 16µg/ml, tornando possível futuros
estudos em associação a antifúngicos.
Palavras Chave: Wangiella dermatitidis, Fonsecaea pedrosoi, Melanina,
Cromoblastomicose, Feohifomicose, fungos, triciclazol.

vi
ABSTRACT
With the increase in groups of severely ill patients and / or immunocompromised
patients such as in AIDS and diabetes, increased number of patients undergoing cancer
chemotherapy or immunosuppressive therapy for organ transplantation and treatment with
broad spectrum antibiotics or glucocorticoids, the frequency of fungal infections has
increased, as well as the number of potentially invasive species. Two infections stand out,
these being the chromoblastomycosis, having as its main agent fungus Fonsecaea
pedrosoi, and phaeohyphomycosis, which has as one of the main etiological agents of
Wangiella dermatitidis. Are cutaneous and subcutaneous infections and disorders of high
relevance to man, and a high incidence in Brazil and abroad.
One of the main characteristics of the fungi causing these infections is the
production of melanin. This has the function of melanin protection from fungi, acting as a
kidnapper of oxidative radicals of oxygen making them more resistant to fungal defense
mechanisms of the host and also the antifungal drugs.
The use of tricyclazole with a fungicide on rice plantations is very common in the
control of pathogen Pyricularia oryzae, having the characteristic of inhibiting melanin
biosynthesis, this has been investigated in dematiaceous fungi that cause infections such as
chromoblastomycosis and phaeohyphomycosis. The decrease in melanin production
showed a decrease in resistance of fungi to antifungal agents used commercially.
The inhibition of melanin production has shown a possible target in the synthesis of
new antifungal drugs.
This study was seen to inhibit the production of melanin dematiaceous fungus
Wangiella dermatitidis in a concentration of 16μg/ml, making possible future studies in
combination with antifungals.
Keywords: Wangiella dermatitidis, Fonsecaea pedrosoi, melanin, chromoblastomycosis,
phaeohyphomycosis, fungus, tricyclazole.

vii
LISTA DE FIGURAS
FIGURA 1: A PLANTA, ESPINHOS, EXAMES MICROSCÓPIOS DIRETOS DOS ESPINHOS E CULTURA. ........................ 4
FIGURA 2: MACRO E MICROMORFOLOGIA DO FONSECAEA PEDROSOI................................................................ 5
FIGURA 3: CORPOS ESCLERÓTICOS OU CÉLULAS MURIFORMES DO FONSECAEA PEDROSOI ................................. 5
FIGURA 4: MACRO E MICROMORFOLOGIA DO FUNGO WANGIELLA DERMATITIDIS ............................................... 7
FIGURA 5: ESQUEMA DAS VIAS DE PRODUÇÃO DE MELANINA EM FUNGOS ....................................................... 9
FIGURA 6: ESQUEMA DA VIA BIOSSINTÉTICA DA MELANINA DESCRITA PARA WANGIELLA DERMATITIDIS. ......... 10
FIGURA 7: WANGIELLA DERMATITIDIS CRESCIDO EM MEIO DEXTROSE DE SABOURAUD LÍQUIDO NA AUSÊNCIA
DO INIBIDOR TRICICLAZOL ................................................................................................................. 14
FIGURA 8: MICROGRAFIA DO MICÉLIO DO FUNGO WANGIELLA DERMATITIDIS NA AUSÊNCIA DO INIBIDOR
TRICICLAZOL ..................................................................................................................................... 15
FIGURA 9: MICROGRAFIA DO MICÉLIO DO FUNGO WANGIELLA DERMATITIDIS NA PRESENÇA DO INIBIDOR
TRICICLAZOL ..................................................................................................................................... 15
FIGURA 10: MICROGRAFIA DO CONÍDIO DO FUNGO WANGIELLA DERMATITIDIS NA AUSÊNCIA DO INIBIDOR
TRICICLAZOL ..................................................................................................................................... 16
FIGURA 11: MICROGRAFIA DO CONÍDIO DO FUNGO WANGIELLA DERMATITIDIS NA PRESENÇA DO INIBIDOR
TRICICLAZOL. .................................................................................................................................... 16

viii
SUMÁRIO
RESUMO .................................................................................................................................................. V
ABSTRACT ............................................................................................................................................ VI
LISTA DE FIGURAS ............................................................................................................................ VII
SUMÁRIO ........................................................................................................................................... VIII
1. INTRODUÇÃO ................................................................................................................................ 1
2. OBJETIVO ..................................................................................................................................... 12
3. METODOLOGIA ........................................................................................................................... 13
4. RESULTADOS ............................................................................................................................... 14
4.1. INIBIÇÃO DA BIOSSÍNTESE DA MELANINA NO FUNGO DEMÁCEO WANGIELLA
DERMATITIDIS PELO INIBIDOR TRICICLAZOL. ............................................................................ 14
4.2. MICROSCOPIA ELETRÔNICA DE VARREDURA DO MICÉLIO DO FUNGO WANGIELLA
DERMATITIDIS NA AUSÊNCIA DO TRICICLAZOL. ....................................................................... 14
4.3. MICROSCOPIA ELETRÔNICA DE VARREDURA DO MICÉLIO DO FUNGO WANGIELLA
DERMATITIDIS NA PRESENÇA DO TRICICLAZOL. ....................................................................... 15
4.4. MICROSCOPIA ELETRÔNICA DE VARREDURA DO CONÍDIO DO FUNGO WANGIELLA
DERMATITIDIS NA AUSÊNCIA DO TRICICLAZOL. ....................................................................... 16
4.5. MICROSCOPIA ELETRÔNICA DE VARREDURA DO CONÍDIO DO FUNGO WANGIELLA
DERMATITIDIS NA PRESENÇA DE TRICICLAZOL......................................................................... 16
5. DISCUSSÃO ................................................................................................................................... 17
6. CONCLUSÃO ................................................................................................................................ 18
7. REFERENCIAS BIBLIOGRÁFICAS ........................................................................................... 19

1. INTRODUÇÃO
O estudo dos fungos tem grande importância, pois são espécies envolvidas na
agricultura, na indústria e na medicina. Na agricultura, algumas espécies fúngicas são
potencialmente perigosas às plantações por colonizarem as mesmas trazendo prejuízo
financeiro, como é o caso do fungo demáceo Magnaphorte grisea que para colonizar a
planta necessita da produção da melanina para formar a apressoria, estrutura usada para
penetrar na folha (Kawamura et al, 1997). Na indústria alimentícia, existem espécies de
grande importância, como as que entram na fabricação de alimentos (incluindo bebidas),
de ácidos orgânicos, de queijos e inúmeros outros. Na indústria farmacêutica, a produção
de fármacos a partir de substâncias fúngicas como antibióticos ou o próprio fungo usado
como regulador intestinal além de outras funções. Além de serem usados nos processos de
fabricação, eles podem ser usados para controlar a poluição do meio ambiente provocado
pelo descarte das indústrias, processo chamado de biorremediação, como é o caso do
Phanerochaete chrysosporium que vem sendo pesquisado por sua grande atuação
degradativa sobre vários materiais, como exemplos os corantes usados na indústria têxtil
(Kunz et al, 2002). Na medicina, durante as duas últimas décadas, a freqüência de
infecções fúngicas tem aumentado dramaticamente, assim como o número de espécies
potencialmente invasivas. Os fatores que contribuem para este aumento incluem: (a) o
aumento de grupos de pacientes gravemente doentes e/ou imunocomprometidos como na
AIDS, (b) aumento do número de pacientes submetidos à quimioterapia de câncer ou
terapia imunossupressiva para transplante de órgãos, (c) procedimentos médicos mais
agressivos como cirurgia extensiva e uso de catéteres, (d) nutrição parenteral, (e)
tratamento com antibióticos de largo espectro ou glicocorticóides. O aumento do número
de pacientes propensos a infecções fúngicas oportunistas tem sido descrita em tal escala
que basicamente qualquer fungo conhecido pode causar infecção nestes indivíduos (Perfect
& Schell, 1996). Este fato, associado à escassez de estudos epidemiológicos em países em
desenvolvimento, acaba comprometendo o diagnóstico micológico destas infecções e
piorando os quadros clínicos das doenças.
Duas infecções fúngicas de interesse ocorrem no mundo, são estas a
Cromoblastomicose e Feohifomicose.
A Cromoblastomicose é infecção fúngica que envolve tipicamente a pele e tecidos
subcutâneos, sendo que tecidos profundos e ossos não estão envolvidos, tendo os membros

2
inferiores os mais afetados devido à proximidade do solo, e são causadas por severas
espécies de fungos pigmentados da família Dematiaceae. Normalmente, a inoculação
ocorre com um trauma causado por ferramentas, farpas ou espinhos (Fader & Mc
Ginnis,1988; Silva, Souza & Rozental, 1999). A doença tem um desenvolvimento lento,
podendo levar anos até o aparecimento da lesão. A lesão inicia-se por uma área
avermelhada no sítio da inoculação, que leva a descamação da pele, conhecida como lesões
tipo placa, e que evoluem para grandes verrugas e úlceras, de aspecto semelhante à de
couve flor (McGinnis, 1983). Estas lesões são secas, não provocam dor, não têm mau
cheiro e, portanto torna-se muito comum observarmos casos de lavradores que apresentam
a lesão por muitos anos (Caligiorne, 1999). Podendo levar quatorze anos em média, da
inoculação ao diagnóstico, tempo suficiente para a doença estar em um estágio avançado,
prejudicando a locomoção do paciente, e dificultando assim o tratamento (Minotto, 2001).
As camadas epidérmicas podem ser infiltradas por leucócitos polimorfonucleares e
células gigantes, ocasionalmente formando microabcessos. Na derme, ocorre resposta
tecidual granulomatosa, com infiltrado celular, que pode ser focal ou difuso, composto de
linfócitos, leucócitos polimorfonucleares, abundantes leucócitos mononucleares, células
plasmáticas, eosinófilos e ocasionalmente células de Langerhans. São observados, também,
abscessos de variável grau de necrose e supuração. Em lesões antigas, o tecido fibroso
forma o maior componente das lesões e granulomas podem estar presentes. Células
escleróticas (ou corpos muriformes) podem ser vistas dentro de macrófagos, de células
gigantes ou extracelularmente (Silva et al.,2002). A análise histopatológica da
Cromoblastomicose inclui hiperplasia pseudoepiteliomatosa, dermo granuloma, formação
de microabcessos epidermal e fibrose (McGinnis, 1983). Com relação à imunologia é
demonstrada uma supressão parcial na resposta celular de pacientes com
Cromoblastomicose (Fuchs & Pecher, 1992), quanto à resposta humoral ocorre uma grande
variação individual a antígenos de fungos agentes de Cromoblastomicose, sendo que, a
persistência in situ do fungo é o principal fator patológico na reação granulomatosa e na
remodelação da lesão cutânea, enquanto que a presença de enzimas como a neutrófilo
elastase, colagenase intersticial e importantes citocinas como a fatores-β de transformação
e crescimento (TGF-β), fator-α de necrose tumoral (TNF-α) e interferon-γ (IFN-γ),
sugerem uma explicação para esta reação tecidual (Esterre et al, 2000). O papel de certas
citocinas no controle de infecções invasivas por fungos demáceos representa importante

3
desafio para a pesquisa nesta área e oferece novas abordagens para o tratamento de
infecções fúngicas, principalmente em organismos imunocomprometidos.
Diferentes fungos demáceos podem causar a patologia, dentre as principais espécies
estão Rhinocladiella aquaspersa, Cladophialophora carrionii, Fonsecaea compactum,
Phialophora verrucosum, Wangiella dermatitidis e Fonsecaea pedrosoi (Iwatsu et al.,
1979; Gimenes et al., 2005), sendo este último o principal agente etiológico da
Cromoblastomicose (Rippon, 1988; Bonifaz, Paredes-Solis & Saul, 2004).
Foram estudados 1.323 casos em Madagascar, acompanhados de 1985 a 1997,
descrevendo dois ecossistemas bem definidos: um no norte do país, causado por
Fonsecaea pedrosoi e, outro no sul, por Cladophialophora carrionii (Esterre ET AL.,
2000).
Na Colômbia, Fonsecaea pedrosoi demostrou ser o principal agente etiológico da
Cromoblastomicose (Cardona-Castro & Agudelo-Flórez, 1999).
No Japão, também foram isolados fungos dematíaceos como Fonsecaea pedrosoi,
Cladophialophora batiana, Wangiella dermatitidis e Exophiala jeanselmei, sendo os
principais agentes etiológicos da Cromoblastomicose no país (Iwatsu et al., 1979; Iwatsu et
al., 1980).
No Brasil, a infecção ocorre em várias regiões, sendo a região amazônica
considerada a principal área endêmica da doença, sendo os trabalhadores rurais do sexo
masculino os principais atingidos (Granja, et al., 2000; Silva, et al., 1999; Oliveira, 1972).
Outra região estudada é o estado do Rio Grande do Sul, foi realizado um estudo de
prevalência de Cromoblastomicose por um período de 29 anos, encontrando 100 casos da
doença, tendo uma predominância da infecção em pessoas do sexo masculino (4:1) e de
agricultores brancos com idade variando entre 50-59 anos, com lesões de membros
inferiores, a maioria proveniente da região do norte do estado (Minotto et al., 2001).
Sendo o principal agente etiológico da Cromoblastomicose, o Fonsecaea pedrosoi é
de grande interesse em estudo. Ele é um fungo polimórfico que vive na natureza como
saprófita, sendo encontrado no solo, nos vegetais e em troncos de madeiras apodrecidos
(Nimrichter et al., 2004; Lacaz et al., 2002). O fungo é isolado de algumas fontes como os
espinhos da planta dormideira (Mimosa pudica) (Salgado et al., 2004) e do coco Babaçu
(Orbignya phalerata Martius) (Marces et al., 2006).

4
Figura 1: A planta, espinhos, exames microscópios diretos dos espinhos e cultura. Mimosa pudica, também
conhecida como dormideira (A) e uma imagem com maior aumento dos espinhos (B). O isolamento do
espinho (C) e examinado, onde é possível ver a presença de dois tipos de hifas demáceos (D, setas pretas),
uma hifa septada, e outra com uma porção terminal globular (D, seta brancas) ou pequenas (3 a 4 células)
hifas com uma extremidade com fim brusco e outra extremidade com forma de copo (E). Em cultura, após 14
dias em estufa, colônias de fungos filamentosos pretos crescem na superfície do espinho (F) (Salgado et al.,
2004). Escala das barras: A: 6 cm, B: 2 cm, C e F: 1 cm, D: 50 μm, E: 30 μm.
O Fonsecaea pedrosoi é considerado fungo polimórfico por apresentar mais de
duas formas de desenvolvimento, como os conídios, hifas ou micélios considerados as
formas saprofíticas, e os corpos escleróticos, ou células muriformes, considerados como a
forma patogênica (Silva et al., 2002).

5
Figura 2: Macro e micromorfologia do Fonsecaea pedrosoi. Cultura e microcultura do fungo obtido da
Mimosa pudica (A e C) e de lesões depacientes (B e D). Hifas apresnetando conidióforos terminais (setas
pretas) e conídios (setas brancas) (Salgado et al. 2004). Escala das barras: A e B: 3cm, C e D: 12μm
Figura 3: Corpos escleróticos ou células muriformes do Fonsecaea pedrosoi (setas pretas), forma patogênica
(Salgado et al., 2004). Escala da barra: B:15 μm
Fonsecaea pedrosoi cresce em meios de cultura ideais como sabouraud, butterfield
e czapeck dox, na forma de hifas escuras e conídios - propágulos que facilitam a dispersão

6
do fungo no ambiente, apresentando escura-pigmentação, variando de verde oliva a preto,
devido à deposição de melanina na parede celular do fungo (Alviano et al., 1990).
Outra infecção de grande importância é a Feohifomicose, esta expressão foi usada,
primeiramente, por AJELLO em 1974 para designar micoses subcutâneas e sistêmicas,
tanto em homens, quanto em animais, no qual o agente etiológico é um fungo demáceo
que, nos tecidos se apresenta sob a forma de células leveduriformes, ao lado de hifas
curtas, regulares ou alongadas, às vezes distorcidas ou dilatadas ou, então, em formas
combinadas. A Feohifomicose é quase sempre infecção oportunística, provocando lesões
superficiais ou profundas, sendo seus agentes mais freqüentes a Exophiala jeanselmei e
Wangiella dermatitidis. No passado essas infecções eram referidas como
Cromoblastomicose sistêmica, cromomicose cística, ceratocromomicose, demaciomicose
cerebral e cromohifomicose, por vários investigadores (Lacaz et al, 1984). A infecção se
inicia após a implantação traumática do fungo na pele (Hohl et al, 1983), que com o
neurotropismo apresentado por esta espécie, o sistema nervoso central é o local mais
acometido (Chang et al, 2000). A Feohifomicose pode ser classificada de acordo com a via
de penetração do fungo no organismo, o grau de envolvimento tecidual e o estado
imunitário do hospedeiro, em cinco modalidades, sendo estas, superficial, cutânea,
subcutânea, sistêmica e oportunista.
Feohifomicose superficial é causada por fungos demáceos capazes de invadir as
camadas superficiais do extrato córneo da epiderme, sem provocar reação tecidual. A
Feohifomicose cutânea é resultante da invasão da camada córnea da epiderme por um
fungo que provoca destruição tecidual e reação inflamatória. A Feohifomicose subcutânea
é decorrente da inoculação do fungo, por ocasião de um traumatismo, resultando lesão
localizada e com caráter supurativo. A Feohifomicose sistêmica é resultado da inalação de
propágulos por hospedeiro imunocompetente, consequentemente inicia por lesão pulmonar
e, posteriormente, se dissemina por via hematogênica a uma víscera ou sistema. A
Feohifomicose oportunista é infecção fúngica que ocorre em hospedeiro imunossuprimido,
pode resultar da inoculação ou de inalação do agente, com posterior disseminação
hematogênica a um ou vários órgãos ou sistemas (Londero, 1987).
Um dos principais agentes etiológico é o fungo dimórfico e demáceo conhecido
como Wangiella (Exophiala) dermatitidis. Além desta, outras patologias como
conjuntivite, otites, pneumonia e endocardites podem ser causadas por esta espécie
(Vartian et al, 1985), em alguns casos pode causar a Cromoblastomicose (Rippon et al.,

7
1988). A Wangiella dermatitidis também está relacionada com aborto de bovinos
(Knudtson & Kirkbride, 1992).
Figura 4: Macro (A) e micromorfologia (B) do fungo Wangiella dermatitidis
O fungo é basicamente saprófito, normalmente cresce no solo ou em madeiras em
decomposição (Kazanas, 1985). Examinando morcegos da Amazônia, REIS & MOK
(1979), isolaram, do macerado do fígado, baço, pulmões, e conteúdo intestinal desses
animais, amostras de Wangiella dermatitidis, sendo os mesmos eventuais reservatórios
desse fungo na natureza (Lacaz et al, 1984). Em cultura, as colônias macroscópicas deste
fungo crescem lentamente em ágar batata a temperatura de 25°C e após 3 a 4 semanas a
cor se torna preta, devido à produção de melanina, porém estes fungos podem crescer em
temperaturas elevadas até 42°C e não assimilam nitrato de potássio (Larone, 1995). No
local da infecção a 37º C, estas espécies dependem da quitina para crescer, pois mutantes
(WdChs5p) que não sintetizam a quitinase, enzima que degrada a quitina, tornaram-se
menos virulentos, o que será revertido com a reposição do gene (Liu et al, 2004). A
Wangiella dermatitidis produz melanina a partir de um substrato endógeno pela ação de
outra classe de enzima, a policetídio sintase (Langfelder et al, 2003) e mutantes deficientes
na capacidade de síntese da melanina apresentaram expressiva diminuição na mortalidade
dos ratos infectados, quando comparados com o tipo selvagem, fato devido à diminuição
da formação de hifas invasivas pelos mutantes (Dixon et al, 1992). A quebra no gene da
policetídeo sintase, enzima chave na síntese da DHN-melanina, produziu cepas albinas e
bem menos virulentas (Feng et al, 2001).
Uma das principais características dos fungos demáceos como Fonsecaea pedrosoi
e Wangiella dermatitidis é a sua capacidade de produzir pigmento in vivo e in vitro, já
caracterizado como melanina (Alviano et al 1991; Langfelder et al 2003). A produção de

8
pigmento tem sido relacionada com a dificuldade de se combater estas infecções nos
hospedeiros. A melanina pode agir como um seqüestrador de radicais oxidativos do
oxigênio tornando estes fungos mais resistentes aos mecanismos de defesa do hospedeiro
e, também, a drogas antifúngicas (Rozental et al, 1994 e 1996). A melanina pode ser
definida como um pigmento que varia de verde a preto, com alto peso molecular e
densidade de ampla distribuição, sendo encontrado em bactérias, fungos, plantas e animais
(Wheeler & Bell, 1988; Bell & Wheeler, 1995). A biossíntese da melanina ocorre a partir
da polimerização oxidativa de compostos fenólicos e indólicos (Vidal-Cros et al, 1994,
Wheeler & Bell, 1988) Nas plantas e nos animais, a enzima responsável pela formação da
melanina é a tirosinase, uma monofenolase multifuncional (3,4-dihidroxifenilalanina
oxigênio oxidoredutase) contendo cobre (Seiji et al., 1963). A tirosinase catalisa a
hidroxilação e a oxidação da tirosina (atividade monofenolásica), e a oxidação de DOPA
(atividade orto-difenolásica), que na seqüência através de reações enzimáticas e de
autopolimerização levam até a formação da melanina. A atividade monofenolásica é o
passo limitante na biossíntese da melanina (Seiji, 1963 & Raposo et al, 2001). A tirosinase
é sintetizada no retículo endoplasmático, glicosilada no lúmen do Golgi e transportada
ainda sob a forma inativa através de vesículas até a organela responsável pela síntese da
melanina (Mishina, 1992).
A melanogênese é o processo de biossíntese da melanina e a biogênese dos
melanossomas nos animais se inicia com os estágios pré-melanossomais I e II. No estágio
I, as organelas não contêm melanina, mas contém internamente vesículas membranosas
com material irregular e fibroso e se apresentam elétronluscente em micrografias
eletrônica, lembrando os endossomas tardios. No estágio II, as fibras formam internamente
um feixe alongado e paralelo por toda a extensão da organela, que ganha formato elíptico,
dando a ela um aspecto estriado bem característico por microscopia eletrônica de
transmissão. No estágio III, a melanina começa a ser depositada sobre o feixe o que é
demonstrado pelo aumento da elétrondensidade da organela e o desaparecimento do feixe.
Finalmente, no estágio IV, as lâminas não são mais visualizadas, sendo a organela
altamente elétrondensa nas micrografias eletrônicas devido ao seu completo preenchimento
com a melanina (Seiji, et al, 1963; Raposo et al, 2001). A organela parece ter uma
linhagem lisossomal, pois apresenta acúmulo da Pmel 17, uma proteína da matriz fibrosa
do estágio II dos premelanossoma, em endossomas tardios (Raposo et al, 2001) e ainda
pelo fato de que a melanização ocorre em pH ácido e que a modificação do pH da organela

9
modula a quantidade e qualidade da melanina (Orlow, 1995). Os melanócitos apresentam
um importante papel na proteção do corpo contra a radiação ultravioleta, onde o número de
células, o grau de melanização e o modo de distribuição dos melanócitos entre as células
epidermais estão diretamente relacionados ao bom desempenho desta função (Weis, 1977).
Nos fungos, a biossíntese da melanina tem vários precursores naturais incluindo
1,8-dihidroxi-naftaleno (DHN), gama-glutaminil-4-hidroxibenzeno, catecol, catecolaminas
e tirosina (Blois, 1978), sendo a via da DOPA-melanina e da DHN-melanina as principais.
A primeira tem como substrato inicial o aminoácido dihidroxifenilalanina (DOPA) e a
segunda o acetato que se transforma em dihidroxinaftaleno (DHN) (Polak, 1990) (Fig. 4).
Figura 5: Esquema das vias de produção de melanina em fungos (Adaptado de Polak Polak,1990)
Na DOPA-melanina, as enzimas principais são fenoloxidases, que são divididas em
lacase, que catalisa somente um passo oxidativo atuando a partir da DOPA e a que catalisa
dois passos oxidativos, a tirosinase, atuando tanto a partir da tirosina quanto a partir da

10
DOPA. Na DHN-melanina, o acetil-coenzima A é necessário para iniciar a via, pois será
convertido a acetato, porém malonil-coenzima A pode ser usado para ser convertido a
1,3,6,8-tetrahidroxinaftaleno (THN), ambos os substratos iniciais dependem da ação da
policetídeo sintase (PKS). A enzima responsável do próximo passo, que é a transformação
do THN a scitalona, é a THN redutase que é alvo de inibição do triciclazol, levando a
formação de flaviolina. A desidratação de scitalona promove a formação de 1,3,8-
trihidroxinaftaleno que sofre uma segunda redução catalisada por uma redutase, que
também é inibida na presença de triciclazol, acumulando assim a hidroxijugalona. A partir
da vermelona ocorre uma desidratação que forma o 1,8-Dihidroximelanina (DHN), o qual
dá o nome à via biossintética (Bell & Wheeler, 1986) (Fig. 5). Para formação da melanina
a partir do composto DHN passos de dimerização e polimerização acontecem catalisados
por uma lacase (Edens et al, 1999).
Figura 6: Esquema da via biossintética da melanina descrita para Wangiella dermatitidis. [H]= reações de
redução, [O]= reações de oxidação, Tr= sítio de inibição do triciclazol (Taylor et al., 1987; Wang et al.,
1995)
O 5-metil-1,2,4-triazol(3,4-b)benzotiazol ou triciclazol é usado na agricultura como
fungicida do patógeno de arroz Pyricularia oryzae. O triciclazol é pulverizado sobre a
planta e passa ter sua ação no fungo que infecta a folha, não é volátil e tem relativa
solubilidade em água e pode ser recuperado do solo (Krieger et al, 2000). Os fungos
demáceos como Fonsecaea pedrosoi, Cladosporium carrioni e Exophiala (Wangiella)
dermatitidis sintetizam a melanina pela DHN-melanina, pois na presença do triciclazol
sofrem mudanças na coloração da cultura passando de preta para avermelhada (Taylor et

11
al, 1987). Em Sporothrix schenckii patógeno humano causador da sporotricose, o
triciclazol bloqueou a biossíntese da melanina tornando o meio marrom-avermelhado
representando o acúmulo de 2-hidroxijulgone (2-HJ), onde o passo enzimático inibido será
do 1,3,8-THN a vermelona, acumulando 1,3,8-THN que instável e auto-oxida 2-HJ, porém
não será capaz de inibir o passo anterior de 1,3,6,8-THN a scitalona, pois não será
detectado o produto proveniente da transformação do 1,3,6,8-THN, a flaviolina (Romero-
Martinez et al, 2000).
As células controle de F. pedrosoi apresentam coloração que varia do verde oliva a
preto, enquanto que os fungos cultivados na presença do triciclazol demonstram uma
coloração avermelhada, seja em meio sólido ou líquido, significando distinção na formação
da melanina pela mudança do padrão de refringência da célula analisada por microscopia
óptica. A viabilidade do fungo não sofre alterações consideráveis na presença do
triciclazol, pois os resultados da citometria de fluxo com o marcador FUN-1, que indica a
atividade metabólica celular, demonstram que na presença do triciclazol os conídios de F.
pedrosoi são 7 % menos viáveis do que as células controle (Franzen et al, 2006), o que
pode ser explicado pela menor captação da glicose, do acúmulo de fenóis e ainda na
inibição da deposição de melanina na célula (Costa et al., 2003). A viabilidade de células
de células epiteliais LLC-MK2 também se mostrou com pequena variação na presença do
triciclazol, em uma concentração oito vezes maior, apenas foram inviabilizadas 5% das
células (Franzen et al, 2006). A ultraestrutura das células crescidas com triciclazol não
demonstra diferenças topográficas, porém no citoplasma destas células podem ser
visualizados melanossomas vazios ou pouco preenchidos indicando não só um possível
comprometimento na formação do pigmento, quanto no transporte do mesmo (Franzen et
al, 2006) para a parede celular (Franzen et al, 1999). Ainda, com técnicas refinadas de
preparo de amostra para microscopia eletrônica, demonstram-se detalhes da estrutura
interna do Fonsecaea pedrosoi jamais visualizadas e a diferença no tamanho da parede do
triciclazol para a célula controle, o que pode estar relacionado com a diminuição da
proteção dos conídios crescidos com triciclazol em relação ao controle (Franzen et al,
2006). A via de biossintética da produção de melanina no fungo Fonsecaea pedrosoi
parece ser mesmo a via da DHN-melanina, pois o fungo demonstra alteração na coloração
das culturas na presença de triciclazol (Franzen et al, 2006).

12
2. OBJETIVO
Analisar o efeito da droga triciclazol sobre a melanização do fungo demáceo Wangiella
dermatitidis e comparar, similaridade e diferença, com o já estudado processo de biogênese
do fungo Fonsecaea pedrosoi na síntese de melanina.

13
3. METODOLOGIA
Cultivo dos fungos
Os fungos demáceos utilizados foram Fonsecaea pedrosoi e Wangiella
dermatitidis, isolados de pacientes humanos com Cromoblastomicose, no Hospital
Universitário Clementino Fraga Filho, da Universidade Federal do Rio de Janeiro, e no
Laboratório de dermato-imunologia UEPA/MC – Marituba, Pará. Os fungos foram
mantidos em meio definido Agar Dextrose de Sabouraud (Peptona bacteriológica 5,0 g/L;
hidrolisado enzimático de caseína 5,0 g/L; dextrose 40,0g/L; agar 15,0 g/L), a 28ºC. O
cultivo foi feito em meio Dextrose de Sabouraud líquido (Peptona bacteriológica 5,0 g/L;
hidrolisado enzimático de caseína 5,0 g/L; dextrose 40,0g/L) em Erlenmeyers de 500 ml,
sob agitação para obtenção de conídios, a 28ºC por 5 dias. A obtenção dos conídios foi
feita pela filtração à vácuo da suspensão de fungos em um funil de Büchner com filtro de
vidro sinterizado com poros de 40-60G e quitassato.
Inibição da formação da melanina
A inibição da formação da melanina foi analisada no fungo Exophiala (Wangiella)
dermatitidis, na presença do triciclazol, o inibidor da síntese da via DHN-melanina. O
triciclazol foi adicionado ao meio de cultivo em concentrações de 16µg/ml da droga,
dissolvida em DMSO. A concentração final de DMSO no meio de cultivo não ultrapassou
0,2%, concentração esta sem efeito citotóxico sobre as espécies fúngicas. Como controle, o
fungo foi cultivado na ausência do triciclazol.
Processamento para Microscopia Eletrônica de Varredura
As amostras foram fixadas por uma hora, em temperatura ambiente, e por 24hs a
10ºC com uma solução contendo 2,5% de glutaraldeido, 4% de paraformaldeido, em
tampão cacodilato de sódio 0,1M, pH 7,2. As células, então, foram lavadas em tampão
cacodilato de sódio, pós-fixadas em tetróxido de ósmio 1% e ferrocianeto de potássio 0,8%,
com conseguinte lavagem em tampão cacodilato de sódio 0,1M, desidratadas em
concentrações crescente de etanol, secas em ponto crítico de CO2, e montadas em stub de
latão. Sendo, em seguida, recobertas com uma fina camada de ouro para observação em
microscópio eletrônico de varredura JEOL JSM-6490LV.

14
4. RESULTADOS
4.1. INIBIÇÃO DA BIOSSÍNTESE DA MELANINA NO FUNGO DEMÁCEO
WANGIELLA DERMATITIDIS PELO INIBIDOR TRICICLAZOL.
Observam-se na figura 5, tubos contendo o fungo Wangiella dermatitidis crescido
somente em meio Dextrose de Sabouraud líquido (tubos A e C), amostra controle. E
crescido em meio Dextrose de Sabouraud líquido contendo o inibidor triciclazol na
concentração de 16µg/ml (tubos B e D), amostra TC. Em cultura contendo triciclazol, o
fungo Wangiella dermatitidis apresenta coloração alterada (figura 5, B e D) em relação à
cultura controle (Figura 5, A e C)
Figura 7: Wangiella dermatitidis crescido em meio Dextrose de Sabouraud líquido na ausência do inibidor
triciclazol, amostra controle (tubos A e C), e na presença de 16µg/ml de triciclazol, amostra TC (tubos B e
D). Tubos divididos em micélios (A e B) e conídios (C e D)
4.2. MICROSCOPIA ELETRÔNICA DE VARREDURA DO MICÉLIO DO
FUNGO WANGIELLA DERMATITIDIS NA AUSÊNCIA DO TRICICLAZOL.
A micrografia demonstra o micélio dividido em hifa e seus conidióforos (A) e o
conidióforo (B).
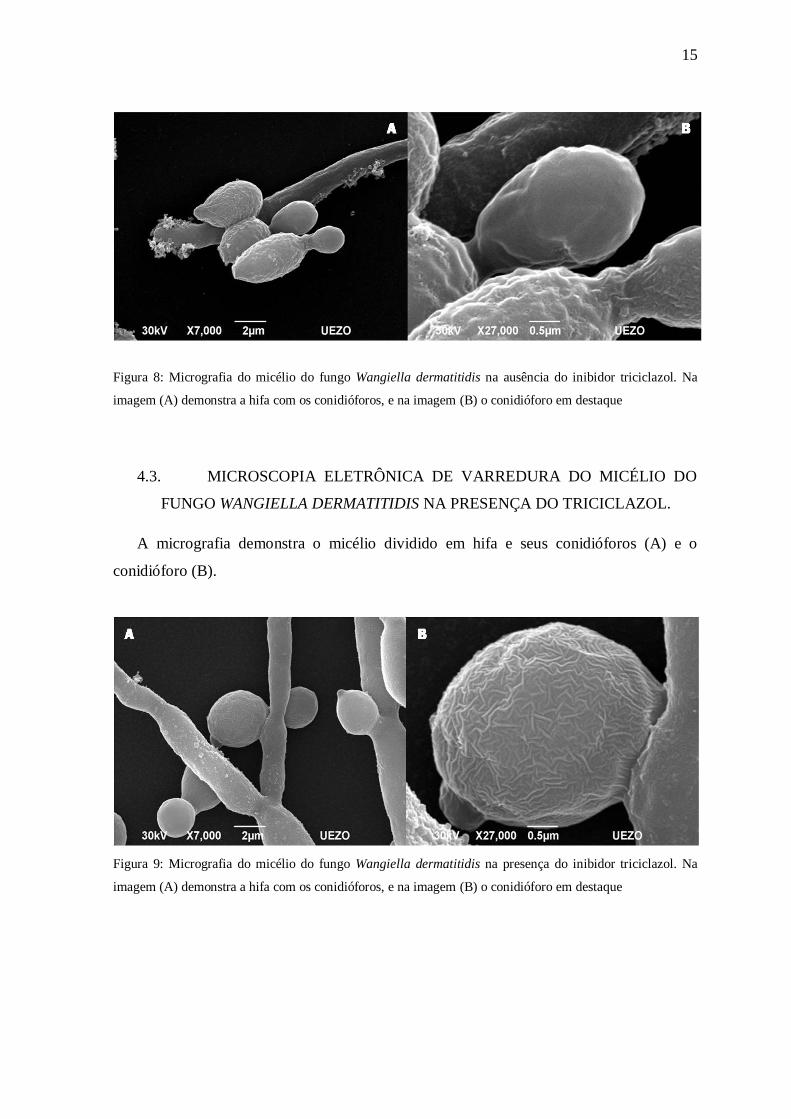
15
Figura 8: Micrografia do micélio do fungo Wangiella dermatitidis na ausência do inibidor triciclazol. Na
imagem (A) demonstra a hifa com os conidióforos, e na imagem (B) o conidióforo em destaque
4.3. MICROSCOPIA ELETRÔNICA DE VARREDURA DO MICÉLIO DO
FUNGO WANGIELLA DERMATITIDIS NA PRESENÇA DO TRICICLAZOL.
A micrografia demonstra o micélio dividido em hifa e seus conidióforos (A) e o
conidióforo (B).
Figura 9: Micrografia do micélio do fungo Wangiella dermatitidis na presença do inibidor triciclazol. Na
imagem (A) demonstra a hifa com os conidióforos, e na imagem (B) o conidióforo em destaque
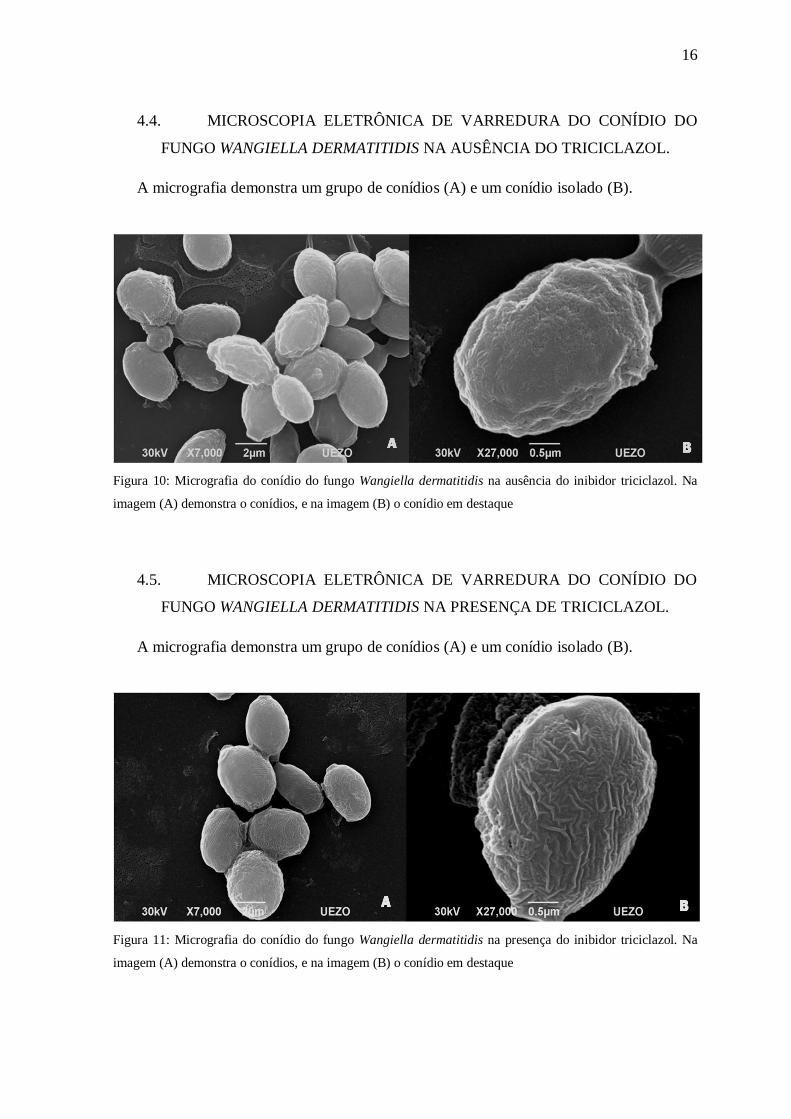
16
4.4. MICROSCOPIA ELETRÔNICA DE VARREDURA DO CONÍDIO DO
FUNGO WANGIELLA DERMATITIDIS NA AUSÊNCIA DO TRICICLAZOL.
A micrografia demonstra um grupo de conídios (A) e um conídio isolado (B).
Figura 10: Micrografia do conídio do fungo Wangiella dermatitidis na ausência do inibidor triciclazol. Na
imagem (A) demonstra o conídios, e na imagem (B) o conídio em destaque
4.5. MICROSCOPIA ELETRÔNICA DE VARREDURA DO CONÍDIO DO
FUNGO WANGIELLA DERMATITIDIS NA PRESENÇA DE TRICICLAZOL.
A micrografia demonstra um grupo de conídios (A) e um conídio isolado (B).
Figura 11: Micrografia do conídio do fungo Wangiella dermatitidis na presença do inibidor triciclazol. Na
imagem (A) demonstra o conídios, e na imagem (B) o conídio em destaque

17
5. DISCUSSÃO
As micoses subcutâneas são infecções causadas por grupo diversificado de fungos que
atacam o homem e os animais. As lesões aparecem inicialmente a partir de um ponto de
inoculação do fungo, através de traumas. Podem permanecer localizados ou se espalhar
pelos tecidos adjacentes, por via linfática ou hematogênica. A maioria dos fungos
envolvidos são sapróbios habituais do solo e vegetais em decomposição. Outra
característica comum aos fungos causadores destas infecções é a produção de melanina.
Esta melanina tem sido relacionada à resistência dos fungos a resposta imunológica do
hospedeiro, e ao uso de antifúngicos (Rozental et al, 1994 e 1996).
Duas infecções tem se destacado, a Cromoblastomicose e a Feohifomicose, ambas
causadas por fungos demáceos produtores de melanina, tendo como agentes etiológicos os
fungos Fonsecaea pedrosoi e Wangiella dermatitidis (Alviano et al 1991; Langfelder et al
2003).
O uso de triciclazol, um inibidor da via DHN-melanina, tem sido muito estudada em
fungos produtor de melanina. Em cultura contendo triciclazol, o fungo Wangiella
dermatitidis apresenta coloração alterada em relação à cultura controle, o mesmo já foi
descrito em Fonsecaea pedrosoi (Franzen et al, 2006) e Sporothrix schenkii (Romero-
Martinez, 2000). Essa coloração é devido a acumulação de intermediários da via
melanogênica, como 2-hidroxijuglona, 3- hidroxijuglona e flaviolina (Taylor, Wheeler &
Szaniszlo, 1987). Demonstrando a via DHN-melanina como a via de biossíntese da
melanina no fungo Wangiella dermatitidis (Taylor et al, 1987).
A ultraestrutura das células crescidas com triciclazol não demonstra diferenças
topográficas, assim como no fungo Fonsecaea pedrosoi (Franzen et al, 2006).
Em Wangiella dermatitidis e Fonsecaea pedrosoi a melanina está relacionada à
capacidade invasiva (Brush & Money, 1999; Farbiarz et al., 1992), tornando a melanina
importante na disseminação do fungo pelos tecidos, estendendo assim a infecção.
Diversos trabalhos indicam a inibição da formação de melanina como uma estratégia
de controle biológico de um patógeno (Nosanchuk & Casadevall, 2003), tornando a
inibição da síntese de melanina um alvo na produção de fármacos.

18
6. CONCLUSÃO
Com os resultados obtidos, conclui-se que, em uma concentração de 16µg/ml do
inibidor triciclazol, ocorre aparente inibição da síntese de melanina no fungo Wangiella
dermatitidis, sendo necessários novos estudos pra comprovar essa inibição.
Devido à baixa toxicidade apresentada pelo triciclazol, uma possível associação com
antifúngicos já utilizados, tornaria mais eficaz o tratamento destas infecções, diminuindo
assim o custo e tempo de tratamento.
Mais experimentos são necessários no desenvolvimento de novos fármacos, mas o uso
de inibidores de biossíntese da melanina, como triciclazol, para que torna-se possível o
tratamento destas infecções.

19
7. REFERENCIAS BIBLIOGRÁFICAS
ALVIANO, C. S., ROZENTAL S., DE SOUZA, W., ANGLUSTER, J & TRAVASSOS,
L. R. Characterization of Fonsecaea pedrosoi melanin. J Gen Microbiol. v.137, p. 837-844. 1991.
BELL, A. A. & WHEELER, M. H. Biosynthesis and Functions of Fungal Melanins. Ann.
Rev Phytopathol, v. 24, p.411- 451, 1986.
BLOIS, M. S. The melanins: Their structure and synthesis. Photochem Photobiol. v. 3 p. 115-134, 1978.
BONIFAZ, A., PAREDES-SOLIS, V. & SAUL, A. Treating chromoblastomycosis with
systemic antifungals. Expert Opin Pharmacother. v. 5, n. 2, p. 247-254. Review. 2004.
BRUSH, L. & MONEY, N. P. Invasive hyphal growth in Wangiella dermatitidis is
induced by stab inoculation and shows dependence upon melanin biosynthesis. Fungal
Genet. Biol. v. 28, n. 3, p. 190-200. 1999.
CALIGIORNE, R. B., RESENDE, M. A., OLIVEIRA, R. C. B. W., VALÉRIO, H. M.,
CORDEIRO, R. A., AZEVEDO, V. FUNGOS DEMATIÁCEOS: Fungos negros que
afetam animais, plantas e o homem. Biotecnologia Ciência & Desenvolvimento. v. 11, p. 22-25. 1999.
CARDONA-CASTRO, N, AGUDELO-FLÓREZ, P. Development of a chronic
chromoblastomycosis model in immunocompetent mice. Med. Mycology. v. 37, n. 2, p. 81-83. 1999.
CHANG, C. L., D. S. KIM, D. J. PARK, H. J. KIM, C. H. LEE, AND J. H. SHIN. Acute
Cerebral Phaeohyphomycosis due to Wangiella dermatitidis Accompanied by Cerebrospinal Fluid Eosinophilia. J Clin Microbiol. v. 38, p. 1965-1966. 2000.
COSTA, J.M., SCROFERNEKER, M.L., CORBELLINI, V.A. Study of different nitrogen
sources on glucose uptake and production of melanin precursors and fungal mass of Fonsecaea pedrosoi culture in tricyclazole. Process Biochemistry. v. 39, p. 633-636. 2004.
DIXON, D. M., J. MIGLIOZZI, C. R. COOPER, JR., O. SOLIS, B. BRESLIN, AND P. J.
SZANISZLO. Melanized and non-melanized multicellular form mutants of Wangiella
dermatitidis in mice: mortality and histopathological studies. Mycoses. v.35, p. 17–21.
1992.
EDENS WA, GOINS TQ, DOOLEY D, HENSON M. Purification and characterization of
a secreted laccase of Gaeumannomyces graminis var. tritici. Appl Environ Microbiol. v.
65, p. 3071-3074. 1999.
ESTERRE, P., M. JAHEVITRA, AND A. ANDRIANTSIMAHAVANDY. Humoral
immune response in chromoblastomycosis during and after therapy. Clin. Diagn. Lab.
Immunol. v. 7, p. 497–500. 2000.
FADER, RC, AND M. R. MCGINNIS. Infections caused by dematiaceous fungi:
chromoblastomycosis and phaeohyphomycosis. Infect. Dis. Clin. North Am. v. 2, p. 925–
938. 1988.

20
FARBIARZ, S. R., DE CARVALHO, T. U., ALVIANO, C. & DE SOUZA W. Inhibitory
effect of melanin on the interaction of Fonsecaea pedrosoi with mammalian cells in vitro. J. Med. Vet. Mycol. v. 30, n. 4, p. 265-73. 1992.
FENG, B., WANG, X., HAUSER, M., KAUFMANN, S., JENTSCH, S., HAASE, G.,
BECKER, J. M. & SZANISZLO, P. J. Molecular cloning and characterization of
WdPKS1, a gene involved in dihydroxynaphthalene melanin biosynthesis and virulence in
Wangiella (Exophiala) dermatitidis. Infection and Immunity. v. 69, n. 3, p.1781-1794. 2001.
FRANZEN A.J., DE SOUZA W., FARINA M., ALVIANO C.S. & ROZENTAL S.
Morphometric and densitometric study of the biogenesis of electron-dense granules in Fonsecaea pedrosoi. FEMS Microbiol. Lett. v. 173(2) p. 395-402. 1999.
FRANZEN, A.J., CUNHA, M.L., BATISTA, E.J.O., SEABRA, S.H., DE SOUZA, W. &
ROZENTAL, S. Effects of tricyclazole (5-methyl-1,2,4-triazol[3,4] benzothiazole), a
specific DHN-melanin inhibitor, on the morphology of Fonsecaea pedrosoi conidia and
sclerotic cells. Microscopy Research Technics. v. 69 n. 9, p. 729 – 737. 2006
FUCHS, J. & PECHER, S. Partial supperssion of cell mediated immunity in chromoblastomycosis. Mycophatologia. v. 119, p. 73-76. 1992.
GRANJA, A. N., SILV. C. N. P., COSTA, J. M. L., SILVA, R. R. & MARQUES, S. G.
Correlação da apresentação clínica e histopatológica de 60 casos de Cromoblastomicose, São Luís-MA. Rev. Soc. Bras. Med. Trop. v. 33, p. 232. 2000.
HOHL, P. E., H. P. HOLLEY, JR., E. PREVOST, L. AJELLO, & A. A. PADHYE.
Infections due to Wangiella dermatitidis in humans: report of the first documented case from the United States and a review of the literature. Rev Infect Dis. v. 5, p. 854-64. 1983.
IWATSU, T., MIYAJI, M., TAGUCHI, H., OKAMOTO, S. & KURITA, N. Skin test-
active substance prepared from culture filtrate of Fonsecaea pedrosoi. Mycopathologia. V. 67, p. 101-105. 1979.
IWATSU, T., MIYAJI, M. & OKAMOTO, S. Isolation of Phialophora verrucosa and
Fonsecaea pedrosoi from nature in Japan. Mycopathologia. v. 75, p. 149-158. 1981.
KAZANAS, N., Pathogenicity of a fungus resembling Wangiella dermatitidis isolated from edible mushrooms. Appl Environ Microbiol. v. 51, n. 2, p. 261-267. 1985.
KRIEGER, M.S., COOK, W.L., KENNARD, L.M. Extraction of Tricyclazole from Soil
and Sediment with Subcritical Water. J Agric Food Chem. v. 48, p. 2178-83. 2000.
KNUDTSON, W. U., AND C. A. KIRKBRIDE. Fungi associated with bovine abortion in the northern plains states (USA). J Vet Diagn Invest. 4:181-5. 1992.
KUNZ, A., PERALTA-ZAMORA, P., MORAES, S. G. Novas tendências no tratamento
de efluentes têxteis. Quím. Nova. v.25, n.1, p.78-82. 2002.
LACAZ, C. S., PORTO, E. ANDRADE, J. G. & QUEIROZ, F. T. F. Feohifomicose disseminada por Exophiala spinifera. An bras Dermatol, v. 59, n. 5, p. 238-243. 1984.

21
LANGFELDER K., STREIBEL M., JAHN B., HAASE G. & BRAKHAGE A.A.
Biosynthesis of fungal melanins and their importance for human pathogenic fungi. Fungal
Genet. Biol. v. 38, n. 2, p. 143-58. 2003.
LARONE, D. H. Medically Important Fungi - A Guide to Identification, 3rd ed. ASM
Press, Washington, D.C. 1995.
LIU H, KAUFFMAN S, BECKER JM, SZANISZLO PJ. Wangiella (Exophiala)
dermatitidis WdChs5p, a Class V Chitin Synthase, Is Essential for Sustained Cell Growth
at Temperature of Infection. Eukaryot. Cell. v. 3, p. 40-51. 2004.
LONDERO, A. T. Feo-hifomicose Interesse em dermatologia. An bras Dermatol. v. 62, p. 327-331. 1987
MCGINNIS M.R. Chromoblastomycosis and phaeohyphomycosis: new concepts,
diagnosis, and mycology. J. Am. Acad. Dermatol. v. 8, n. 1, p. 1-16. 1983.
MINOTTO R., BERNARDI C.D., MALLMANN L.F., EDELWEISS M.I. &
SCROFERNEKER M.L. Chromoblastomycosis: a review of 100 cases in the state of Rio
Grande do Sul, Brazil. J. Am. Acad. Dermatol. v. 44, n. 4, p. 585-592. 2001.
MOLL, H. The role of chemokines and accessory cells in the immunoregulation of cutaneous leishmaniasis. Behring Inst. Mitt. v. 99 p. 73–78. 1997.
NOSANCHUK J.D., CASADEVALL A. The contribution of melanin to microbial
pathogenesis. Cell. Microbiol. v. 5, n. 4, p. 203-23. 2003.
OLIVEIRA, L.G. - Isolamento e identificação de agentes da cromomicose em Belo Horizonte, Rev. Soc. Bras. Med. Trop. v. 7, n. 1, p. 7-10; 1973.
ORLOW SJ. Melanosomes are specialized members of the lysosomal lineage of
organelles. J Invest Dermatol. v. 105, p. 3-7. 1995.
PERFECT J.R. AND SCHELL W.A. The new fungal opportunists are coming. Clin.
Infect. Dis. v. 22, p. 112-121. 1996.
POLAK A. Melanin as a virulence factor in pathogenic fungi. Mycoses. v. 33, n. 5, p. 215-
24. 1990.
RAPOSO G, TENZA D, MURPHY DM, BERSON JF, MARKS MS. Distinct protein
sorting and localization to premelanosomes, Melanosomes, and lysosomes in pigmented
melanocytic cells. J Cell Biol. v. 19, p. 809-24. 2001.
RIPPON, J.W. Medical Micology: The Pathogenic Fungi and the Pathogenic Actinomycetes., 3
rd ed. W.B. Sounders, Philadelphia. p. 276-296. 1988.
ROMERO-MARTINEZ R., WHEELER M., GUERRERO-PLATA A., RICO G. &
TORRES-GUERRERO H. Biosynthesis and functions of melanin in Sporothrix schenckii. Infect. Immun. v. 68, n. 6, p. 3696-703. 2000.
ROZENTAL S., ALVIANO C.S. & DE SOUZA W. The in vitro susceptibility of
Fonsecaea pedrosoi to activated macrophages. Mycopathologia. V. 126, n. 2, p. 85-91. 1994.

22
ROZENTAL S, ALVIANO C.S. AND DE SOUZA W. Fine structure and cytochemical
study of the interaction between Fonsecaea pedrosoi and rat polymorphonuclear leukocyte. J. Med. Vet. Mycol. v. 34, p. 323-330. 1996.
SALGADO, C. G., SILVA, J. P. DINIZ, J. A., SILVA, M. B., COSTA, P. F., TEXEIRA,
C. & SALGADO, U. I. Isolation of fonsecaea pedrosoi from thorns of mimosa pudica, a
probable natural source of chromoblastomycosis. Rev. Inst. Med. trop. S. Paulo. v. 46, p.
33-36, 2004.
SEIJI, M. SHIMAO, K. BIRBECK, M.S.C. & FITSPATRICK, T.B. Subcellular
localization of melanin biosynthesis. Ann. New York Acad. Sci. v. 100, p. 497-533. 1963.
SHARON, N. & LIS, H. Lectins as cell recognition molecules. Science. v. 240, p. 227-33.
1989.
SILVA J.P., DE SOUZA W. & ROZENTAL S. Chromoblastomycosis: a retrospective
study of 325 cases on Amazonic Region (Brazil). Mycopathologia. v. 143, n. 3, p. 171-
175. 1999.
SILVA J.P., ALVIANO, D.S, ALVIANO, C.S, DESOUZA W. TRAVASSOS, L.R.
DINIZ, J.A.P. & ROZENTAL S. Comparison of fonsecaea pedrosoi sclerotic cells
obtained in vivo and in vitro: ultrastructure and antigenicity. FEMS Immunol Med
Microbiol. v. 33, p. 63-69. 2002.
TAYLOR, B.E., WHEELER, M.H. & SZANISZLO, P.J. Evidence for pentaketide melanin
biosynthesis in dematiaceous human pathogenic fungi. Mycologia. v. 79, n. 2, p. 320-322. 1987.
WEISS LW, ZELICKSON AS. Giant melanosomes in multiple lentigines syndrome. Arch
Dermatol. v. 113, p. 491-4. 1977.
WHEELER, M. H. AND BELL, A. A. Melanins and their importance in pathogenic fungi. Curr Top Med Mycol. v. 2, p. 338-387. 1988.
VARTIAN, C. V., D. M. SHLAES, A. A. PADHYE, AND L. AJELLO. Wangiella
dermatitidis endocarditis in an intravenous drug user. Am J Med. v. 78, p. 703-7. 1985.
VIDAL-CROS, A., VIVIANI, F., LABESSE, G., BOCCARA, M. AND GAUDRY, M.
Polyhydroxynaphthalene reductase involved in melanin biosynthesis in Magnaporthe
grisea. Purification, cDNA cloning and sequencing. Eur J Biochem. v. 219, p. 985-992. 1994.
YU, R.J., GRAPPEL, S.F. & BLANK, F. Inhibition of Keratinases by α2-Macroglobulin.
Experientia, v. 28, p. 886. 1972.


















