
ESTUDO DA ESTIMULAÇÃO ELÉTRICA NERVOSA … · superiores, as duas principais são...
78
Cristiane Magela de Freitas Santos ESTUDO DA ESTIMULAÇÃO ELÉTRICA NERVOSA TRANSCUTÂNEA NA NOCICEPÇÃO E NO EDEMA INDUZIDOS PELA ADMINISTRAÇÃO DE SEROTONINA INTRADÉRMICA. Belo Horizonte Escola de Educação Física, Fisioterapia e Terapia Ocupacional 2010
-
Upload
trinhtuyen -
Category
Documents
-
view
212 -
download
0
Transcript of ESTUDO DA ESTIMULAÇÃO ELÉTRICA NERVOSA … · superiores, as duas principais são...

Cristiane Magela de Freitas Santos
ESTUDO DA ESTIMULAÇÃO ELÉTRICA NERVOSA
TRANSCUTÂNEA NA NOCICEPÇÃO E NO EDEMA INDUZIDOS
PELA ADMINISTRAÇÃO DE SEROTONINA INTRADÉRMICA.
Belo Horizonte
Escola de Educação Física, Fisioterapia e Terapia Ocupacional
2010

Cristiane Magela de Freitas Santos
ESTUDO DA ESTIMULAÇÃO ELÉTRICA NERVOSA TRANSCUTÂNEA NA
NOCICEPÇÃO E NO EDEMA INDUZIDOS PELA ADMINISTRAÇÃO DE
SEROTONINA INTRADÉRMICA
Belo Horizonte
Escola de Educação Física, Fisioterapia e Terapia Ocupacional
2010
Dissertação apresentada ao Programa de Pós-Graduação em
Ciências da Reabilitação, nível mestrado, da Escola de Educação
Física, Fisioterapia e Terapia Ocupacional da Universidade
Federal de Minas Gerais (UFMG), como requisito parcial para
obtenção do Título de Mestre em Ciências da Reabilitação.
Orientador: Prof. Dr. Marcos Antônio de Resende
Co-orientadora: Profª Drª Janetti Nogueira de Francischi

S237e
2010
Santos, Cristiane Magela de Freitas Estudo da estimulação elétrica nervosa transcutânea na nocicepção e no edema induzidos pela administração de serotonina intradérmica.. [manuscrito] / Cristiane Magela de Freitas Santos – 2010. 75f. Orientador: Marcos Antônio de Resende Co-orientadora: Janetti Nogueira de Francischi
Mestrado (dissertação) – Universidade Federal de Minas Gerais, Escola de Educação Física, Fisioterapia e Terapia Ocupacional.
Bibliografia: f. 60-63
1. Serotonina - Teses. 2. Analgesia - Teses. 3. Opióides – Teses. 4. Estimulação elétrica nervosa transcutânea – Teses. I. Resende, Marcos Antônio de. II. Francischi, Janetti Nogueira de. III.Universidade Federal de Minas Gerais. Escola de Educação Física, Fisioterapia e Terapia Ocupacional. IV. Título.
CDU: 612.76 Ficha catalográfica elaborada pela equipe de bibliotecários da Biblioteca da Escola de Educação Física, Fisioterapia e Terapia Ocupacional da Universidade Federal de Minas Gerais.



Dedico esta dissertação aos meus pais e ao grande amor da minha vida,
motivos pelos quais procuro me superar a cada dia...

AGRADECIMENTOS
Agradecimento é a expressão de um ser humano quando sente gratidão por um ato.
É muito satisfatório ter a quem agradecer neste momento. Sou imensamente grata a
todos aqueles, que mesmo na correria do dia-a-dia, reservaram momentos do seu
precioso tempo para me ajudar na elaboração deste trabalho.
Agradeço a Deus por iluminar sempre os meus caminhos e pela força para
ultrapassar barreiras ou optar por novos caminhos. Aos meus pais, aos quais devo a
vida e o incentivo para tornar-me uma pessoa cada vez melhor, em meus atos como
ser humano e em meu desempenho profissional.
Ao Professor Doutor Marcos Antônio de Resende, por acreditar na minha
capacidade de realizar este trabalho e também por me guiar no caminho científico. À
Professora Doutora Janetti Nogueira de Francischi, o meu muito obrigada pelos
esclarecimentos farmacológicos, apoio crítico e construtivo que em muito
engrandeceu minha caminhada. À Professora Doutora Ligia de Loiola Cisneros, por
me direcionar no caminho à docência, ajudando-me a superar limitações e a
acreditar no meu desejo de ser Mestre.
Aos colegas do laboratório, Patrícia, Webster, Rafael, Rinaldo e Débora, pelo apoio
nos experimentos e companheirismo.
E, claro, a você, Joel, pelo apoio, carinho e amor. Por todos os finais de semana e
feriados dos quais você abdicou para estar ao meu lado, seja no laboratório, me
auxiliando nos experimentos, seja em casa analisando dados e elaborando textos.
Sem você, esta jornada teria sido árdua. Amo você. Muito obrigada!

RESUMO
A estimulação elétrica nervosa transcutânea (TENS) é definida como a
aplicação de corrente elétrica por meio de eletrodos na pele para alívio da
dor. Diferentes teorias são propostas para explicar o mecanismo de ação
analgésica da TENS. Estudos recentes mostraram que parte dessa analgesia
pode, também, ser mediada por ação de neurotransmissores em receptores
periféricos. O objetivo desse estudo foi investigar o efeito da TENS de baixa
(BF: 10HZ) e alta (AF: 130HZ) freqüência na nocicepção e no edema
induzidos por serotonina (5-HT) administrada intradermicamente na pata do
rato (10µg/0,1ml). TENS de Baixa e alta frequência foram aplicadas na pata
direita durante 20 minutos e, imediatamente após, a 5-HT foi administrada. A
nocicepção foi medida pelo método Hargreaves e o edema pelo
hidropletismômetro (Ugo Basile). A TENS de BF inibiu em 100% a
hiperalgesia da 5-HT, mas não a de AF. TENS de AF e BF não inibiram o
edema induzido por 5-HT. O pré-tratamento com naltrexona (Nx: 50µg; I.pl.),
reverteu o efeito antinociceptivo da TENS de BF. A TENS de BF ao longo do
tempo foi capaz de inibir a nocicepção induzida por 5-HT por até 30 minutos
após o término de sua aplicação. Nossos resultados confirmaram a ausência
de efeito anti-inflamatório pela TENS e a participação de opioide endógeno
periférico na analgesia da TENS de baixa freqüência.

ABSTRACT
Transcutaneuous electrical nerve stimulation (TENS) is defined as the application of
an electrical current to the skin through electrodes for pain relief. Various theories
have been proposed in order to explain the analgesic mechanism of TENS. Recent
studies have demonstrated that part of this analgesia can also be mediated through
neurotransmitters found in peripheral receptors. The present study aims to
investigate the effects of low frequency (LF: 10 HZ) TENS as well as high
frequency (HF: 130 HZ) TENS in nociception and edema induced by
intradermally administered serotonin (5-HT), injected into rat paw (10µg/0.1
ml). LF and HF TENS were applied to the right paw for 20 min, and 5-TH was
administered immediately after. The Hargreaves method was used to measure
nociception, and the hydroplethysmometer (Ugo Basile) was used to measure
edema. The LF TENS reduced by about 100% 5-TH hyperalgesia, but the same
did not happen with the HF TENS. HF and LF TENS did not inhibit 5-HT-induced
edema. Pre-treatment with naltrexone (Nx: 50µg; I.pl.) reversed the
antinociceptive effects of LF TENS. LF TENS application could inhibit
nociception 5-HT-induced edema for 30 min maximum after the end of its
application. Our results have confirmed the lack of an antiinflammatory effect
through the use of TENS as well as the participation of a peripheral endogenous
opioid in LF TENS analgesia.

LISTA DE ABREVIATURAS E SIGLAS
5-HT – 5-hidroxitriptamina
AF – Alta frequência
ANOVA – analysis of variance
APTA – American Physical Therapy Association
ATP - Adenosina Trifosfato
BF – Baixa frequência
CEBIO - Centro de Bioterismo do Instituto de Ciências Biológicas
CETEA – Comitê de Ética em Experimentação Animal
CGRP – Calcitonin gene related peptide
EPM – erro padrão da média
g – grama
Hz – Hertz
i.d. – intradérmica
i.pl. – intraplantar
Kg - kilogramas
LIF- Leukemia inhibitory factor
m/s – metros por segundo
mg - miligrama
ml – mililitros
NaCl – Cloreto de Sódio

NGF – Nerve growth factor
Nx – Naltrexona
s.c.- subcutânea
SNC – Sistema Nervoso Central
TENS – Estimulação Elétrica Nervosa Transcutânea
TNF-α – Factor de necrose tumoral alfa
µg – microgramas
µl – microlitros

SUMÁRIO
Capítulo 1- INTRODUÇÃO............................................................................... ...... 11
1.1 Considerações iniciais ...................................................................................... 11
1.2 Dor inflamatória ................................................................................................ 13
1.3 Serotonina ........................................................................................................ 15
1.4 Mecanismos endógenos de controle da dor ..................................................... 17
1.5 Estimulação Elétrica Nervosa Transcutânea (TENS) ....................................... 20
1.6 Objetivos do estudo .......................................................................................... 24
1.6.1 Objetivo geral ................................................................................................ 24
1.6.2 Objetivos específicos..................................................................................... 24
Capítulo 2 - MATERIAL E MÉTODOS ................................................................... 25
2.1 Animais............................................................................................................. 25
2.2 Ambientalização dos animais ........................................................................... 26
2.3 Substâncias utilizadas ...................................................................................... 26
2.4 Administração das substâncias ........................................................................ 27
2.5 Teste nocipeptivo – Algesímetro ...................................................................... 27
2.5.1 Descrição do teste de hiperalgesia térmica .................................................. 28
2.5.2 Avaliação do edema inflamatório – Hidropletismômetro ............................... 29
2.6 Aparelho de Estimulação Elétrica Nervosa Transcutânea (TENS) ................... 30
2.7 Procedimento experimental .............................................................................. 32
2.7.1 Curva dose resposta da serotonina injetada perifericamente ........................ 32
2.7.2 Pré-tratamento dos animais com antagonistas de serotonina ....................... 32
2.7.2.1 Mensuração do volume da pata dos ratos após pré-tratamento com
antagonistas de serotonina.......................................................................................33
2.8 Aplicação da TENS .......................................................................................... 33
2.8.1 Efeito da TENS sobre a hiperalgesia e o edema induzidos por serotonina ... 33
2.9 Pré-tratamento intraplantar com naltrexona ..................................................... 34
2.10 Estudo do tempo de efeito da Tens de baixa frequência................................ 35
2.11 Análise estatística .......................................................................................... 35
Capítulo 3 - REFERÊNCIAS .................................................................................. 36 Capítulo 4 – ARTIGO: Estudo da estimulação elétrica nervosa transcutânea na 42 nocicepção e no edema induzidos pela administração de serotonina intradérmica. Capítulo 5 – CONSIDERAÇÃOES FINAIS................................................................74
ANEXOS ................................................................................................................ .75

11
Capítulo 1 - INTRODUÇÃO
1.1 Considerações iniciais
A dor é um fenômeno complexo que envolve não somente a transmissão do
estímulo nocivo da periferia ao córtex, mas, também, todo o processo cognitivo e
emocional que ocorre no cérebro (JULIUS et al., 2001).
A relação entre a percepção da dor e o estímulo nocivo não é proporcional,
segundo Melzack e Wall (1965), isto é, a intensidade da dor percebida não está
diretamente relacionada ao tamanho da lesão sofrida, pois pode ser modulada em
seu trajeto ascendente. Tipicamente ela ocorre em resposta a um estímulo lesivo,
mas, por vezes, também na ausência deste. Assim, a Associação Internacional para
o Estudo da Dor (IASP) definiu a dor como uma experiência sensorial e emocional
desagradável, associada à lesão tecidual real ou potencial, ou descrita em termos de
tal lesão (IASP, 2008). Esta variação na percepção da dor está relacionada aos
aspectos cognitivos, emocionais, motores, bem como aos mecanismos
descendentes, inibitórios da dor, que participam de sua modulação (CASEY, 1999).
Isto explica fenômenos como um soldado com grandes feridas no campo de batalha
não relatar dor (MELZACK et al., 1965; MELZACK, 1999).
A nocicepção é uma forma especializada de sinalização sensorial, que
converte informação sobre o dano tecidual iminente (ou atual) (BARANAUSKAS et
al., 1998). Tal sensação é determinada pela estimulação das fibras aferentes
primárias do tipo Aδ e C, que transmitem o impulso nociceptivo até a medula
espinhal.

12
Existe uma diferença conceitual entre os termos nocicepção e dor. O primeiro
refere-se às manifestações neurofisiológicas geradas pelo estímulo lesivo enquanto
o segundo envolve a percepção de um estímulo aversivo e exige a capacidade de
abstração e elaboração dos impulsos sensoriais (MILLAN, 1999; ALMEIDA et al.,
2004).
A transmissão do estímulo doloroso, assim como sua inibição, envolve
múltiplos mecanismos. Os estímulos nociceptivos chegam de maneira
altamente organizada no corno dorsal da medula espinhal, com as fibras Aδ
terminando principalmente nas lâminas I e V e as fibras C na lâmina II. A
partir destas regiões, são acionados neurônios de projeção e interneurônios
de segunda ordem na medula espinhal, sendo que alguns são ativados
somente por estímulos nocivos (específicos para nocicepção) e outros
respondem a diferentes estímulos de alta ou baixa intensidade (largo
espectro). Esta conexão sináptica entre as fibras aferentes primárias e os
neurônios do corno dorsal envolve neurotransmissores como o glutamato e a
substância P, que são responsáveis pela produção de potenciais pós-
sinápticos excitatórios rápidos e lentos respectivamente (MILLAN, 2002).
A neuroanatomia e a organização das vias ascendentes da dor são
altamente complexas. Neuroanatomicamente dois tipos de vias ascendentes
podem ser identificados, monossinápticas e polissinápticas (MILLAN, 2002).
As primeiras projetam diretamente para centros cerebrais superiores e
incluem os tratos espino-mesencefálico, espino-braquial, espino-talâmico e
espino-reticular. Já as vias polissinápticas possuem uma estação relé de
neurônios de segunda ordem antes de se projetarem aos centros cerebrais

13
superiores, as duas principais são espino-cervical e sistema lemniscal
(MILLAN, 2002). Nos centros superiores do sistema nervoso central a
informação é finalmente processada para produzir a percepção da dor
(GRUBB, 1998).
Durante o processo inflamatório, os nociceptores que estão localizados no
tecido danificado, são excitados por estímulos não dolorosos (alodínia) ou
apresentam uma resposta intensificada ao estímulo nociceptivo (hiperalgesia). Essa
alteração nos nociceptores pode ser produzida por mudanças físicas no tecido
lesado (edema) e por mediadores inflamatórios que alteram o limiar das terminações
nervosas aos estímulos térmicos e mecânico (GRUBB, 1998).
1.2 Dor inflamatória
Ao sofrer uma lesão tecidual, o organismo aciona mecanismos cujo propósito
é limitar o dano e facilitar a regeneração. Esses mecanismos fazem parte da
resposta inflamatória, que é caracterizada pelos quatro sinais cardinais: dor, rubor,
calor, tumor e, também, por um quinto sinal que é a perda da função (GALLIN et al.
1992).
O papel da dor inflamatória é alertar o organismo sobre a presença de um
dano e prevenir traumas futuros ao tecido anteriormente lesado (RIEDEL et al.,
2001).
As células danificadas após o comprometimento tecidual liberam, no local da
lesão, ATP e prótons que, juntamente com a bradicinina derivada do plasma, vão
produzir efeitos excitatórios. Simultaneamente, os mastócitos, que estão

14
normalmente presentes nos tecidos, liberam aminas (5-hidroxitriptamina e histamina)
e metabólitos do ácido araquidônico, incluindo as prostaglandinas (COUTAUX et al.,
2005). Essas substâncias alteram o limiar das terminações nervosas ao estímulo
térmico e mecânico (GRUBB, 1998). Assim como os mastócitos, os macrófagos
também já se encontram nos tecidos e migram para o local da lesão onde liberam
citocinas (Interleucinas 1 e 6 e Fator de Necrose Tumoral alfa – TNF-α), e fatores de
crescimento (NGF – Fator de Crescimento do Nervo e LIF – Fator Inibitório da
Leucemia) (COSTIGAN et al., 2000). Já as fibras aferentes primárias contribuem
com o processo inflamatório liberando neuropeptídeos, tais como a substância P e
peptídeo relacionado ao gene da calcitonina (CGRP) (COSTIGAN et al., 2000;
COUTAUX et al. 2005). Tais mediadores químicos, dentre outros, agem
principalmente através da sensibilização dos nociceptores (JULIUS et al., 2001).
Muitos dos receptores de alto limiar respondem a uma variedade de estímulos
térmico, químico e mecânico e são definidos como nociceptores polimodais (KIDD et
al., 2001).
Os nociceptores distinguem-se de outros tipos de receptores com base em
sua morfologia, velocidade de condução e limiar de ativação (BESSON, 1999). Eles
são terminais diferenciados com velocidade de condução lenta (fibras C < 2,5 m/s e
fibras Aδ 2,5 a 20,0 m/s), e são normalmente ativados por estímulos de intensidade
nociva (GRUBB, 1998). Os estímulos nocivos que resultam em uma sensação de
dor rápida, aguda e bem localizada, em geral refletem a ativação das fibras Aδ, que
são minimamente mielinizadas e conduzem a dor primária; e a sensação de dor
lenta, crônica e em queimação, em geral, é conduzida por fibras do tipo C, que não

15
são mielinizadas, e participam da condução da dor secundária (GALLIN et al. 1992;
BESSON, 1999; JULIUS et al., 2001).
Um dos mediadores químicos liberados no tecido lesado e inflamado é a
serotonina. Ela é capaz de facilitar a saída de proteínas do plasma e de provocar
hiperalgesia em ratos e humanos (OKAMOTO et al., 2002).
1.3 Serotonina
A serotonina (5-HT) é encontrada em neurônios, plaquetas, mastócitos
cutâneos de roedores e basófilos (TOKUNAGA et al., 1998). No sistema nervoso
central (SNC), está associada com o processamento e modulação da dor
(SOMMER, 2004). Vários estudos mostraram que, na medula espinhal, a 5-HT
liberada de estruturas do tronco cerebral exerce uma ação analgésica (MILLAN,
2002; SOMMER, 2004).
As fibras descendentes serotoninérgicas surgem de núcleos localizados no
bulbo ventromedial rostral, sendo que o núcleo magno da rafe é a principal fonte de
fibras serotoninérgicas que se projetam para a medula espinhal (YOSHIMURA et al.,
2006). Já em tecidos periféricos, a aplicação de 5-HT produz dor em humanos e
nocicepção em roedores (SUFKA et al., 1992; TAMBELLI et al., 2006).
Os receptores 5-HT podem ser classificados em, no mínimo, três, e no
máximo sete classes. Elas incluem 5-HT1, 5-HT2 e 5-HT3, bem como o receptor “não
clonado” 5-HT4. Os genes dos receptores 5-HT5, 5-HT6 e 5-HT7 foram clonados
recentemente, mas ainda têm que ser caracterizados operacional e
transducionalmente em tecidos intactos (HOYER et al., 1994).

16
Os efeitos fisiológicos da serotonina nos tecidos, como um agente pro -
inflamatório e pro-nociceptivo, podem ser descritos em termos da heterogeneidade
de seus receptores identificados na periferia (TAIWO et al., 1992; TAMBELLI et al.,
2006). Possíveis mecanismos de ação, diretos e indiretos, foram propostos
(SOMMER, 2004). Uma possibilidade é a de que a 5-HT periférica age nos
receptores 5-HT1 e 5-HT2 para potencializar mudanças vasculares e, de forma
indireta, pode provocar o fluxo de outros elementos pro-nociceptivos e pro-
inflamatórios (bradicinina e eicosanóides) do sangue. Alternativamente, a serotonina
do plasma e/ou derivada dos tecidos pode ter ação direta nos receptores 5-HT3,
localizados nas fibras aferentes Aδ/C, produzindo nocicepção (GIORDANO et al.,
1989; SUFKA et al., 1992; TOKUNAGA et al., 1998; SOMMER, 2004; TAMBELLI et
al., 2006). Dessa forma, a serotonina liberada no tecido lesado ou inflamado
contribui para a sensibilização das fibras nervosas. Estudos realizados para
determinar os mecanismos envolvidos em outros componentes da inflamação, tais
como o edema induzido por 5-HT (extravasamento de plasma), foram focados
primariamente na inflamação cutânea na pata de ratos (PIERSE, 1995). Os subtipos
de receptores que mediam o extravasamento de plasma no tecido cutâneo induzido
por 5-HT não foram determinados (PIERSE,1995).
Tanto a administração local, quanto sistêmica de antagonista 5-HT,
metisergida, inibiu o edema induzido por 5-HT na pata do rato (SUFKA, 1991). A
serotonina injetada por via intra-articular aumentou substancialmente o
extravasamento de plasma no tecido sinovial. O receptor 5-HT2A foi identificado nos
terminais eferentes simpáticos do tecido sinovial e sugerido como principal mediador
do edema nesse tecido (PIERSE, 1995).

17
1.4 Mecanismos endógenos de controle da dor
Em 1965, Melzack e Wall sugeriram a existência de uma espécie de comporta
no corno dorsal da medula. De acordo com essa teoria, alguns neurônios teriam a
capacidade de suprimir a transmissão do sinal doloroso do corno dorsal da medula,
fechando, então, um portão hipotético, e inibindo a passagem do impulso doloroso
(Teoria da Comporta Espinhal) (MELZACK et al., 1965). As células da lâmina II do
corno posterior (substância gelatinosa) são principalmente interneurônios inibitórios
curtos que se projetam às lâminas I e V, e regulam a transmissão na primeira
sinapse da via nociceptiva entre as fibras aferentes primárias e os neurônios de
transmissão do trato espino-talâmico. Assim, as células da substância gelatinosa
respondem tanto à atividade das fibras aferentes que entram na medula quanto à
atividade das vias descendentes (RANG et al., 2008).
A sensação de dor pode ser modificada pelos sistemas endógenos inibitórios
da dor, predominantemente através das vias descendentes de noradrenalina,
serotonina e opióides endógenos, tais como β-endorfina e dinorfina (YOSHIMURA et
al., 2006). Essas vias descendentes constituem um dos mecanismos de controle da
comporta que controlam a transmissão de impulsos dolorosos (relacionados com a
sensação de dor) no corno posterior da medula espinhal. Muitas classes de
neurônios do corno dorsal, as próprias fibras aferentes primárias e as fibras
descendentes do cérebro exercem uma potente influência modulatória sobre a
transferência de informação nociceptiva que chega da periferia para a medula
espinhal e da medula para o cérebro (MILLAN et al., 2002).
As células noradrenérgicas são classificadas de A1 a A7 e estão amplamente
distribuídas no mesencéfalo, das quais um número substancial de neurônios está

18
em A5-A7 (locus coeruleus) e tem uma significante projeção descendente para a
medula espinhal (JONES, 1991; PERTOVAARA, 2006; YOSHIMURA et al., 2006).
Estudos farmacológicos evidenciaram a ação antinociceptiva da noradrenalina no
sistema inibitório descendente (JASMIN et al., 2002; TANABE et al., 2005).
Os receptores de catecolaminas são classicamente divididos em duas
categorias principais: alfa e beta-adrenoceptores, sendo o efeito antinociceptivo da
noradrenalina mediado principalmente pelos α-2-adrenoceptores (FAIRBANKS et al.,
1999).
Muitos estudos têm relacionado a serotonina (5-HT) na modulação
descendente da transmissão nociceptiva (ZHAO et al., 2007). De maneira
semelhante ao sistema noradrenérgico, as fibras descendentes serotoninérgicas
surgem de núcleos localizados no bulbo ventromedial rostral, sendo que o núcleo
magno da rafe é a principal fonte de fibras serotoninérgicas que se projetam para a
medula espinhal (YOSHIMURA et al., 2006; RANG et al., 2007).
Os opióides estão envolvidos tanto em componentes ascendentes quanto
descendentes da modulação da dor (STAMFORD, 1995). Existem três tipos
principais de receptores: opióides µ (mu), κ (kappa) e δ (delta) que estão presentes
no hipotálamo, substância cinzenta periaquedutal, corno dorsal da medula e nos
nervos aferentes periféricos (STAMFORD, 1995; HOLDEN et al., 2005). O receptor µ
opióide é foco nos estudos de dor, pois sua ativação é necessária para a ação da
maioria dos analgésicos potentes (FIELDS, 2004). Os opióides promovem a
analgesia através de diversos mecanismos, dentre eles a ativação das vias
descendentes e a inibição da transmissão no corno dorsal (DELEO, 2006). Tanto os
terminais aferentes primários quanto os neurônios de segunda ordem, no corno
dorsal da medula, apresentam receptores µ e δ opióides. A aplicação de agonistas µ

19
opióides reduz a liberação de neurotransmissores excitatórios pelos terminais
aferentes primários através da inibição dos canais de cálcio voltagem-dependentes
(FIELDS, 2007). Segundo a revisão de Fields (2007) outro controle ocorre através
da ação direta na despolarização de neurônios de segunda ordem, no corno dorsal,
pela abertura de canais de potássio. Essas ações dos opióides no corno dorsal
contribuem para eficácia clínica de agonistas µ opióide tais como a morfina.
Há, também, evidências de que os opióides inibem a descarga de
terminações aferentes nociceptivas na periferia, particularmente sob condições de
inflamação, nas quais aumenta a expressão dos receptores opióides pelos
neurônios sensitivos (OBARA et al., 2007; RANG et al., 2008). Os opióides podem
inibir a liberação de neurotransmissores excitatórios (ex: substância P) das
terminações nervosas sensoriais (MACHELSKA et al., 2002).
Existem diferentes ligantes endógenos, da classe dos peptídeos opióides,
como as endorfinas, encefalinas e dinorfinas, que exibem diferentes afinidades e
seletividades pelos três receptores opióides µ (endorfinas e encefalinas), δ
(encefalinas e endorfinas) e κ (dinorfinas) (MACHELSKA et al., 2002; STEIN et al.,
2009). Esses peptídeos opióides endógenos são identificados na pele, em tecidos
subcutâneos e particularmente em células inflamatórias (STEIN et al., 2009).
Stein et al. (1993) verificaram em seu estudo que a naloxona, um antagonista
opióide, injetada por via intra-articular, intensificou a sensação de dor após cirurgia
de artroscopia no joelho, sugerindo a participação de receptores opióides na
analgesia pós-operatória.
Semelhante aos estudos realizados para verificar a participação dos
receptores opióides na analgesia periférica, Gentili et al. (1996) demonstraram que a

20
administração de clonidina, um agonista α2- adrenérgico, por via intra-articular
reduziu a dor no período pós-operatório de artroscopia do joelho.
É bem descrito na literatura que a interação sinérgica entre receptores
opióides e α2-adrenérgicos, quando co-administradas morfina e clonidina no SNC,
resultam em uma intensificação da analgesia (STEIN et al., 2009). Buerkle et al.
(2000) concluíram que, a co-administração de um opióide e um agonista α2-
adrenérgico, por via intra-articular, resultou em alívio intensificado da dor no pós-
operatório de cirurgia de joelho, quando comparado com cada um administrado
isoladamente, o que sugere que essa sinergia pode ocorrer também na periferia
(BUERKLE et al., 2000).
Além da utilização da farmacologia clássica, outros recursos podem ser
utilizados como coadjuvantes no tratamento da dor e evitar os efeitos colaterais
advindos do uso frequente dessas drogas. Um recurso não farmacológico, não
invasivo, sem efeitos colaterais, com poucas contra-indicações e comum na prática
clínica do fisioterapeuta é a estimulação elétrica nervosa transcutânea (TENS)
(JOHNSON, 2001; SLUKA, 2003).
1.5 Estimulação Elétrica Nervosa Transcutânea (TEN S)
A estimulação elétrica nervosa transcutânea é definida, pela American
Physical Therapy Association (APTA), como a aplicação de corrente elétrica, por
meio de eletrodos de superfície, na pele para alívio da dor (APTA, 1997; SLUKA et
al., 2006). Dois tipos de TENS são usados clinicamente, TENS de baixa frequência

21
(frequência de estimulação < 10 Hz) e TENS de alta frequência (frequência > 50 Hz)
(AINSWORTH et al., 2006; SLUKA et al., 2006).
Diferentes teorias são propostas para explicar o mecanismo de ação da TENS
(RADHAKRISHNAN et al., 2003). A mais conhecida foi descrita no estudo de Wall e
Sweet (1967) pautada na teoria da comporta espinhal da dor introduzida por
Melzack e Wall, em 1965. Essa teoria forneceu uma explicação convincente sobre a
natureza da dor, e forneceu uma base teórica para a efetividade da TENS no
tratamento da dor (KING et al., 2001). Assim, a informação nociceptiva das fibras
aferentes de pequeno diâmetro (Aδ e C) é inibida pela estimulação das fibras de
grande diâmetro Aβ e o estímulo doloroso é impedido de alcançar os centros supra-
espinhais (MELZACK et al., 1965). Então, a TENS utilizada clinicamente seria capaz
de estimular fibras de grande diâmetro e promover o controle da dor (KING et al.,
2001). Além disso, estudos neurofarmacológicos sugeriram a ação de mecanismos
espinhal e supra-espinhal relacionados a neurotransmissores, na analgesia induzida
pela TENS (RADHAKRISHNAN et al., 2003).
Opióides endógenos liberados no sistema nervoso central estão envolvidos
no mecanismo analgésico da TENS (SLUKA et al., 1999) que abrange,
predominantemente, mecanismos centrais (espinhal e supra-espinhal) (SLUKA et
al., 1999, 2003; AINSWORTH et al., 2006) e, mais recentemente, também periféricos
(SABINO et al., 2008).
A naloxona administrada no corno dorsal na medula espinhal, em ratos
artríticos, impediu o efeito antinociceptivo da aplicação da TENS, confirmando a
teoria que a TENS exerce seu efeito através da liberação de opióides a nível
espinhal (SLUKA et al., 1999).

22
Ainda em relação ao envolvimento de neurotransmissores centrais nos
mecanismos de ação da TENS, Radhakrishnan et al. (2003) investigaram o papel
dos receptores serotoninérgicos e noradrenérgicos no mecanismo anti-hiperalgésico
da TENS. Neste estudo eles demonstraram que subtipos de receptor 5-HT2A e 5-HT3
espinhais estão envolvidos na anti-hiperalgesia induzida pela TENS de baixa
frequência, mas não na de alta frequência, sugerindo um importante papel do
sistema serotoninérgico central em mediar a antinocicepção da TENS
(RADHAKRISHNAN et al., 2003). Neste mesmo estudo, a administração intratecal
de antagonista α2-adrenoceptor (yohimbina) não afetou o efeito analgésico da TENS
de baixa e alta frequência, mostrando que o mecanismo analgésico da TENS não
envolve ativação de α2-adrenoceptor espinhal (RADHAKRISHNAN et al., 2003;
KING et al., 2005).
Evidências em estudos recentes sugerem que parte do efeito analgésico da
TENS pode ser mediado por ação de neurotransmissores em receptores periféricos
(SLUKA et al., 2003). A administração periférica de um antagonista seletivo de α2-
adrenoceptor bloqueou o efeito anti-hiperalgésico mediado pela TENS, sugerindo
que esses receptores são ativados pela liberação endógena de noradrenalina em
resposta à TENS, e exógena pela administração de clonidina, um agonista α2-
adrenérgico (KING et al., 2005).
Estudos desenvolvidos por Sabino et al. (2008) mostraram que a TENS de
baixa e alta frequência induzem efeito anti-hiperalgésico através de diferentes
mecanismos de ação: a TENS de baixa frequência através da liberação de opióides
e a TENS de alta frequência por mecanismos não opióides. Esse mesmo estudo
sugere que a antinocicepção induzida pela TENS de baixa frequência é devido à
liberação local de opióides endógenos (SABINO et al., 2008).

23
A literatura mostra que o mecanismo de analgesia da TENS envolve outros
sistemas além dos mencionados na teoria do portal espinhal. Além da teoria do
portão, outras vias anatômicas, neurotransmissores e seus receptores e tipos de
neurônios, estão envolvidos no mecanismo de analgesia induzida pela TENS
(SLUKA et al., 2006) .
Porém, apesar de muitos esforços em elucidar os mecanismos pelos quais a
TENS exerce seu efeito anti-hiperalgésico, pouco se conhece a respeito de suas
ações nos tecidos periféricos.
Poucos estudos na literatura associam o uso da TENS com a atividade
antiinflamatória. Investigações sugerem a não efetividade da TENS de alta e de
baixa frequência em prevenir o edema inflamatório (RESENDE et al., 2004).
Até o presente momento, nas bases de dados consultadas, não foram
encontradas investigações que mostrem a capacidade da TENS em bloquear
estímulo nociceptivo e edema induzido por uma amina biogênica, como a
serotonina, encontrada nos tecidos periféricos em condições inflamatórias agudas.
Assim, há também a necessidade de pesquisa básica para compreender o
mecanismo de analgesia, induzida pela estimulação elétrica nervosa transcutânea,
tem motivado estudos na área.
Os trabalhos em modelos animais minimizam o efeito placebo da TENS,
permitem melhor controle sobre a extensão e tempo de injúria, os parâmetros são
melhor controlados, além de facilitar a análise da posição dos eletrodos e possibilitar
a realização de exames neurobiológicos.
Clinicamente, a TENS é raramente administrada de forma isolada.
Combinações de tratamentos farmacológicos e não-farmacológicos para controle da
dor são frequentes. O entendimento do mecanismo de ação desse recurso permite

24
aos profissionais que o utilizam serem capazes de intensificar os efeitos de diversos
tipos de analgésicos e reduzir os efeitos colaterais dos agentes farmacológicos
produzindo um melhor alívio da dor.
Diante do exposto, a investigação dos mecanismos de ação da TENS se faz
importante, para que sua indicação seja realizada com maior precisão, observando-
se as diferentes situações clínicas e parâmetros para sua aplicação.
1.6 Objetivos do estudo
1.6.1 Objetivo geral
Verificar o efeito da TENS de baixa e alta frequência na nocicepção e edema
induzidos por serotonina administrada perifericamente, na pata de ratos.
1.6.2 Objetivos específicos
• Verificar se a administração periférica por injeção intraplantar de
naltrexona, um antagonista opióide específico, antes da aplicação da
TENS, restaura o efeito nociceptivo da serotonina.
• Verificar o tempo de efeito da TENS aplicada antes da indução da
nocicepção, no pico de hiperalgesia da serotonina injetada
perifericamente.

25
Capítulo 2 - MATERIAIS E MÉTODOS
O presente estudo faz parte de um projeto de pesquisa intitulado “Estudo da
participação da via serotoninérgica e noradrenérgica na modulação da dor
inflamatória após estimulação elétrica nervosa transcutânea de baixa e alta
frequência”, sob orientação do Prof. Dr. Marcos Antonio de Resende foi aprovado
pelo Comitê de Ética em Experimentação Animal da UFMG - CETEA (Parecer n°
237/08) (Anexo 1) e, somente após a sua aprovação, foi iniciado o estudo.
2.1 Animais
Foram utilizados ratos Holtzman machos com peso entre 280 e 310 gramas
(para os experimentos no teste algesimétrico de Hargreaves , e de 160 a 200g (para
experimentos no hidropletismômetro). Os animais foram fornecidos pelo CEBIO –
Centro de Bioterismo do Instituto de Ciências Biológicas da UFMG. A variação de
peso dos animais para os dois testes deveu-se a limitações dos métodos. Em nosso
estudo-piloto, pudemos observar que animais menores apresentavam um
comportamento exploratório duradouro, provavelmente devido às dimensões do
compartimento na caixa de acrílico utilizada para colocar os animais durante o teste
(18 cm x 29 cm x 12.5 cm), o que inviabilizou medidas em intervalos menores de
tempo. Optamos, então, por utilizar animais maiores, segundo Hargreaves et al.
(1988), que utilizaram animais com peso entre 250 e 300g. Os experimentos foram
realizados com grupos de 4 a 10 animais, perfazendo um total de 161 animais
utilizados em todo o estudo.

26
2.2 Ambientalização dos animais
Os animais permaneceram no biotério em caixas de plástico (seis por gaiola), com
forração de maravalha, com livre acesso à água e comida. No dia anterior aos
experimentos, os animais foram privados de alimento e levados para a sala de
experimento, com ciclo claro-escuro de 12 horas e temperatura controlada (de 22 a
24°C). Foi realizada a adaptação ao aparelho algesí metro por 30 minutos, para que
os animais se habituassem às caixas de teste, à manipulação imposta pelo
pesquisador e aos procedimentos de teste. Essa metodologia assemelha-se aos
padrões pré-estabelecidos em estudos similares (RESENDE et al., 2006; SABINO et
al., 2008).
2.3 Substâncias utilizadas
As substâncias utilizadas nesse estudo foram: serotonina (Serotonin, Sigma®,
E.U.A), maleato de metisergida (Methysergide maleate, Sigma®, E.U.A), pizotifeno
(Pizotifen, Sandoz®, Brasil), cloridrato de ondansetrona (Ondansetron, Glaxo®,
Brasil), naltrexona (Naltrexone hydrochloride, Sigma®, E.U.A) e solução salina
fisiológica estéril (NaCl 0,9%).
2.4 Administração das substâncias
Todas as substâncias foram preparadas em solução salina estéril, momentos
antes do experimento. Elas foram administradas após as medidas do limiar de
retirada ou do volume da pata no baseline.

27
Para indução da nocicepção, os animais receberam injeção intradérmica (i.d.)
de serotonina (10µg) ou salina na pata direita. Um volume total de 0,1ml foi
administrado. A pata contralateral desses animais não recebeu nenhuma
administração. A injeção intradérmica de serotonina (10µg) também foi utilizada para
induzir edema. O mesmo volume final (0,1ml) foi administrado na pata direita dos
animais; já a pata contralateral recebeu igual volume de salina.
Injeção subcutânea (s.c.) foi administrada no dorso dos animais próximo à
região cervical para administração dos antagonistas de serotonina (pizotifeno,
metisergida e ondansetrona) ou salina.
Naltrexona (50µg/0,1ml) ou salina (0,1ml) foram injetadas na pata direita por
via intraplantar (i.pl.), sendo que nenhuma substância foi administrada na pata
contralateral durante esses experimentos.
2.5 Teste nociceptivo - Algesímetro
Para a medida do tempo de latência de retirada da pata foi utilizado o
aparelho Plantar test da Ugo Basile® (modelo 7370, Itália), que é um algesímetro
térmico descrito originalmente por Hargreaves et al. (1988). Essa técnica avalia a
resposta nociceptiva a estímulos térmicos em animais não restritos.
2.5.1 Descrição do teste de hiperalgesia térmica
Para o teste, o animal foi cuidadosamente colocado em compartimento de
acrílico individual (18 cm x 29 cm x 12.5 cm), posicionado sobre uma superfície de
vidro especial, que permitiu a passagem de forma homogênea da luz e do calor. Foi

28
realizado um período de ambientalização (± 5 minutos), suficiente para que o animal
terminasse a exploração do ambiente. Após esse tempo, a fonte de luz
infravermelha era colocada sob cada uma das patas traseiras do animal. O teste foi
iniciado por um interruptor que ativou a fonte de calor e disparou o cronômetro
eletrônico. Quando o animal retirava a pata, a fonte de luz e o relógio eram
interrompidos automaticamente, e o tempo de retirada, em segundos, era registrado.
Um tempo máximo de 30 segundos foi utilizado como tempo de corte para impedir
dano tecidual.
A latência de retirada da pata foi considerada como resposta ao calor, quando
o animal realizava movimentos de sacudir, lamber ou erguer a pata traseira. Duas
medidas de cada pata foram realizadas com um curto intervalo de tempo entre as
mesmas. A média das duas medidas foi utilizada para calcular o delta (∆) da
latência de retirada, obtido através da diferença entre a pata direita (inflamada) e a
pata esquerda (controle), em segundos. Todas as medidas foram realizadas pelo
mesmo avaliador e no mesmo período do dia. As aferições foram realizadas no
tempo zero e após 5, 15, 30, 60 e 120 minutos da administração de serotonina
intradérmica na região plantar.
FIGURA 1: Método Hargreaves – Aparelho Ugo Basile® (modelo 7370)

29
2.5.2 Avaliação do edema inflamatório – Hidropleti smômetro
Para aferir o aumento de volume das patas traseiras, causado pelo estímulo
pró-inflamatório, foi utilizado um hidropletismômetro da Ugo Basile® (modelo 1750,
Itália), composto por duas câmaras transparentes, uma contendo o fluido que seria
deslocado e a outra contendo um transdutor de sinal elétrico, ligadas por um sistema
de vasos comunicantes.
O método consistiu na imersão da pata do animal até a articulação tíbio-
társica, em uma solução salina e detergente dentro da cubeta do aparelho. O
deslocamento de fluido, assim provocado, acionou um transdutor cujo sinal foi
transformado em mililitros e pôde ser lido em um mostrador digital, colocado ao lado
na bancada. O sinal elétrico produzido foi proporcional ao volume deslocado pelas
patas avaliadas.
O cálculo do edema da pata foi obtido através dos valores de variação entre
o volume da pata inflamada (direita) e o volume da pata contralateral (esquerda) (∆,
em ml), nos mesmos tempos utilizados para medida do limiar nociceptivo, isto é,
tempo zero e após 5, 15, 30, 60 e 120 minutos do estímulo pró-inflamatório. As
medidas foram realizadas por um mesmo examinador previamente treinado
(Doutoranda Patrícia Paiva Lima) e os valores expressos como média ± EPM, por
grupo, da diferença do volume (ml) entre as patas traseiras, em cada tempo.

30
(A) (B)
FIGURA 2: Medida pletismométrica do volume das patas posteriores dos ratos (Hidropletismômetro Ugo Basile, modelo 1750)
2.6 Aparelho de Estimulação Elétrica Nervosa Trans cutânea (TENS)
O aparelho de TENS utilizado para induzir antinocicepção foi o Neurodyn
III/Ibramed®. O aparelho foi devidamente calibrado e os parâmetros de estimulação
administrados foram: baixa (BF:10 Hz) e alta frequência (AF: 130 Hz), com duração
de pulso de 130 µs. A intensidade para todos os grupos foi considerada no limiar
sensorial, isto é, a intensidade foi aumentada até o limiar motor com contração
muscular visível e, em seguida, reduzida para o nível sensorial, conforme descrito
por Sluka et al. (1998); (Sluka, 2001), e realizado em experimentos prévios do nosso
laboratório (RESENDE et al., 2004; SABINO et al., 2006, 2008).
A intensidade do estímulo foi determinada no início da aplicação da TENS
para cada animal. A duração da estimulação elétrica nervosa transcutânea foi de 20
minutos. A escolha dos parâmetros relativos à frequência, a duração do pulso, o

31
tempo de aplicação e a determinação da intensidade escolhidos para este estudo
experimental foram semelhantes aos utilizados na clínica fisioterapêutica
(JOHNSON, 2001; SLUKA et al., 2003). Eletrodos com 1 cm² foram especialmente
construídos para os experimentos e fixados na face plantar e dorsal da pata
posterior direita do animal com fita adesiva, não sendo utilizado gel acoplador. Antes
de iniciar a estimulação aguardou-se um curto período de tempo para que os
animais cessassem a exploração do ambiente e se adaptassem à condição
experimental. Ao início da passagem de corrente elétrica pela pata dos animais eles
apresentaram comportamento de fuga e agitação que cessou após
aproximadamente 3 minutos da aplicação da TENS.
Experimentos com o aparelho desligado foram conduzidos a fim de controle.
Os eletrodos foram fixados na pata direita tratada com 5-HT da mesma forma que no
grupo que recebeu a estimulação elétrica, porém o aparelho de TENS foi mantido
desligado da tomada, durante os 20 minutos correspondentes à aplicação da TENS.
FIGURA 3: Aparelho de TENS Neurodyn III/Ibramed®

32
FIGURA 4: Técnica de aplicação da TENS
2.7 Procedimento experimental
2.7.1 Curva dose-resposta da serotonina injetada p erifericamente
Para o estudo da serotonina (5-HT), diferentes doses dessa substância (0,1;
1,0 e 10 µg) (OLIVEIRA et al., 2007) foram administradas em grupos distintos de
animais (n=5 em todos os experimentos), na pata traseira direita. Nenhuma
administração foi feita na pata esquerda. As medidas foram obtidas nos tempos zero
e após 5, 15, 30, 60 e 120 minutos da injeção de serotonina. A dose de escolha para
indução da nocicepção foi de 10 µg de serotonina.
2.7.2 Pré-tratamento dos animais com antagonistas de serotonina
Todas as drogas utilizadas foram administradas por injeção subcutânea no
dorso da região cervical dos animais meia hora antes da administração de
serotonina (10µg/0,1ml i.d) (ABBOTT et al., 1997). As medidas foram obtidas nos
mesmos tempos dos experimentos anteriores. Os seguintes antagonistas foram

33
utilizados: metisergida (2mg/kg) (ZARRINDAST et al., 1995), ondansetrona (2mg/Kg)
e pizotifeno (2mg/kg) (FRANCISCHI et al.,1996). Os animais do grupo controle
receberam salina meia hora antes da injeção de serotonina. O efeito nociceptivo da
serotonina após o pré-tratamento com seus antagonistas foi, então, avaliado.
2.7.2.1 Mensuração do volume da pata dos ratos após pré-tratamento com
antagonistas de serotonina
Grupos de animais distintos dos anteriores foram também tratados com as
mesmas concentrações de antagonistas ou salina, meia hora antes da
administração intradérmica de 5-HT, para mensurar seu efeito sobre o edema na
pata traseira direita. Todos os animais receberam salina por injeção intradérmica
(0,1ml) na pata esquerda. As medidas foram obtidas no tempo zero e nos 5, 15, 30,
60 e 120 minutos seguintes à aplicação de serotonina.
2.8 Aplicação da TENS
2.8.1 Efeito da TENS sobre a hiperalgesia e o edem a induzidos por serotonina
TENS de baixa e de alta frequências foram aplicadas na pata direita, em
grupos distintos de 5 a 10 animais, 30 minutos antes de os animais receberem a
injeção de serotonina intradérmica na região plantar (10µg/100µl). Durante a
aplicação da TENS, os ratos moviam-se “livremente”, em caixas especialmente
construídas para experimentos com animais (30x20 cm).

34
A fim de evitar que eles provocassem algum dano aos eletrodos
confeccionou-se, em cartolina, um colar “elizabetano” preso por fita adesiva ao redor
da cabeça dos animais. Os ratos utilizaram o colar durante os 20 minutos de
aplicação da TENS.
Dois estudos em grupos distintos foram realizados: um para avaliar o efeito da
TENS de baixa e alta frequências sobre a hiperalgesia provocada pela serotonina e
o outro experimento para avaliar a ação da TENS sobre o edema induzido pela 5-
HT.
2.9 Pré-tratamento intraplantar com naltrexona
A fim de estudar o mecanismo de ação da TENS de baixa frequência, sobre a
hiperalgesia provocada por serotonina perifericamente, foi utilizada naltrexona, um
antagonista opióide, por via intraplantar (50 µg/100µl), preparado em solução salina.
Essa substância foi administrada 30 minutos antes da aplicação da TENS de baixa
frequência. A dose escolhida foi baseada no estudo de Rodrigues et al. (2005).
Esses autores demonstraram em seu estudo que esta dose de 50 µg injetada na
pata contralateral, não tinha efeito sobre a hiperalgesia da pata inflamada. Sabino et
al. (2008) também utilizaram essa mesma dose em seu estudo que investigou os
mecanismos envolvidos no efeito antinociceptivo da TENS de alta e baixa
frequência.

35
2.10 Estudo do tempo de efeito da TENS de baixa fr equência
Para estudo da duração do tempo de ação da TENS, os animais receberam
TENS de baixa frequência (10 Hz), com tempo de pulso de 130 µs, por 20 minutos.
Ela foi aplicada no tempo zero. Nos ratos controle a TENS foi utilizada desligada por
igual período. Grupos, com quatro animais cada, receberam injeção intradérmica
plantar de serotonina (10µg) nos diferentes tempos após aplicação da TENS de BF,
ou seja, 5, 25, 55 e 115 minutos após a TENS ter sido retirada. As medidas do
tempo de retirada da pata foram realizadas no Hargreaves, sempre cinco minutos
após a injeção de 5-HT, aos 10 ,30, 60 ou 120 minutos após a aplicação da TENS
na pata direita.
2.11 Análise estatística
A preparação do banco de dados, assim como a análise estatística, foram
realizadas por meio do programa GraphPad Prism 4.00, instalado em ambiente
Windows®.
Os resultados estão representados como a média ± erro padrão da média. As
comparações estatísticas das diferenças entre os grupos nos diversos tempos de
medidas foram feitas utilizando-se análise de variância (One-way ANOVA), com
correção post hoc pelo teste de Bonferroni. Para os experimentos com apenas dois
grupos, foi utilizado para verificar diferenças entre dois grupos nos diversos tempos
a Two-way ANOVA. Um valor de p inferior a 0,05 foi considerado estatisticamente
significativo.

36
Capítulo 3 - REFERÊNCIAS ABBOTT, F. V.; HONG, Y.; BLIER, P. Persisting sensitization of the behavioural response to formalin-induced injury in the rat through activation of serotonin receptors. Neuroscience , v. 77, n. 2, p. 575-584, 1997. AINSWORTH, L.; BUDELIER, K.; CLINESMITH, M.; FIEDLER, A.; LANDSTRON, R.; LEEPER, B. J.; MOELLER, L.; MUTCH, S.; O’DELL, K.; ROSS, J.; RADHAKRISHNAN, R.; SLUKA, K. A. Transcutaneous electrical nerve stimulation (tens) reduces chronic hiperalgesia induced by muscle inflammation. Pain , v. 120, p. 182-187, 2006. ALMEIDA, R. F.; ROIZENBLATT, S.; TUFIK, S. Afferent pain pathways: a neuroanatomical review. Brain research, v. 1000, p. 504-510, 2000. APTA. American physical therapy association, guide to physical therapy practice. Physical Therapy , v. 77, p. 1171-1624, 1997. BARANAUSKAS, G.; NISTRI, A. Sensitizations of pain pathways in the spinal cord: cellular mechanisms. Progress in Neurobiology, v. 54, p. 349-365, 1998. BESSON, J. M. The Neurology of pain. The Lancet , v. 353, p. 1610-1615, 1999. BUERKLE, H.; HUGE, V.; WOLFGART, M.; STEINBECK, J.; MERTES, N.; VAN AKEN, H.; PRIEN, T. Intra-articular clonidine analgesia after knee arthroscopy. European Journal of Anaesthesiology , v. 17, p. 295-299, 2000. CASEY, K. L. Forebrain mechanisms of nociception and pain: Analysis through imaging. Proceedings of the National Academy of Sciences of the United States of America , v. 96, p. 7668-7674, 1999. COSTIGAN, M.; WOOLF, C. J. Pain: molecular mechanisms. The Journal of Pain , v. 1, n. 3, supplement 1, p. 35-44, 2000. COUTAUX, A.; ADAM, F.; WILLER, J. C.; BARS, D.L. Hyperalgesia and allodynia: peripheral mechanisms. Joint Bone Spine , v. 72, p. 359-371, 2005. DELEO, J. A. Basic science of pain. The Journal of Bone and Joint Surgery , v. 88-A, supplement 2, p. 58-62, 2006.

37
FAIRBANKS, C. A.; WILCOX, G. L. Moxonidine, a selective α2-adrenergic and imidazoline receptor agonist, produces spinal antinociception in mice. The Journal of Pharmacology and Experimental Therapeutics , v. 290, n. 1, p. 403-412, 1999. FIELDS, H. State-dependent opióide control of pain. Neuroscience , v. 5, p. 565-575, 2004. FIELDS, W. L. Understanding how opioids contribute to reward and analgesia. Regional Anesthesia and Pain Medicine , v. 32, n. 3, p. 242-246, 2007. FRANCISCHI, J. N.; DIAS, M. F.; ROCHA, O. A.; CASTRO, M. S. A.; TATSUO, M. A. K. F.; FARINELLI, P.; PACHECO, C. M. F.; FERREIRA-ALVES, D. L.; SIROIS, P. Pharmacological characterization of sephadex-induced oedema in rat paws: predominant role of serotonin and platelet-activating factor. International archives of allergy and immunology , v. 109, p. 398-406, 1996. GALLIN, J.I.; GOLDSTEIN, I.M.; SNYDREMAN, R. Inflammation: basic principles and clinical correlates. 2. ed. New York: Raven Press , 1992. GENTILI, M.; JUHEL, A.; BONNET, F. Peripheral analgesic effect of intra-articular clonidine. Pain , v. 64, p. 593-596, 1996. GIORDANO, J.; ROGERS L. V. Peripherally administered serotonin 5HT3 receptor antagonists reduce inflammatory pain in rats. European journal of pharmacology, v. 170, p. 83-86, 1989.
GRUBB, B. D. Peripheral and central mechanisms of pain. British Journal Anaesthesia , v. 81, p. 8-11,1998. HARGREAVES, K.; DUBRER, R.; BROWN, F.; FLORES, C.; JORIS, J. A new and sensitive method for measuring thermal nociception in cutaneous hiperalgesia. Pain , v. 32, p. 77-88, 1988. HOLDEN, J. E.; JEONG Y.; FORREST, J. M. The endogenous opioid system and clinical pain management. The American Academy of Clinical Neuropsychology Clinical Issues , v. 16, p. 291-301, 2005. HOYER, D.; CLARKE, D. E.; FOZARD, J. R.; HARTIG, P. R.; MARTIN, G. R.; MYLECHARANE, E. J.; SAXENA, P. R.; HUMPHREY, P. P. A. International union of pharmacology classification of receptors for 5-hydroxytryptamine (serotonin). The American Society for Pharmacology and Experimental Therapeutics , v. 46, n. 2, p. 157-203, 1994.

38
IASP. Internacional association for the study of pain Web site. Pain terminology . Disponível em: <http://www.iasp-pain.org/terms-p.html> Acesso em: 8 ago. 2008. JASMIN, L.; TIEN, D.; WEINSHENKER, D.; PALMITER, R. D.; GREEN, P.G.; JANNI, G.; OHARA, P.T. The NK1 receptor mediates both the hyperalgesia and the resistance to morphine in mice lacking noradrenaline. Proceedings of the National Academy of Sciences of the United States of America , v. 99, n. 2, p. 1029–1034, 2002. JONES, S. L. Descending noradrenergic influences on pain. Progress in brain research , v. 88, p. 381-394, 1991. JULIUS, D.; BASBAUM, A.I. Molecular mechanisms of nociception. Nature , v. 413, p. 203-210, 2001. KIDD, B. L.; URBAN, L. A. Mechanism of inflammatory pain. British Journal of Anaesthesia , v. 87, p. 3-11, 2001. KING, E. W.; AUDETTE, K.; ATHMAN, G. A.; NGUYEN, H. O. X.; SLUKA, K. A.; FAIRBANKS, C. A. Transcutaneous electrical nerve stimulation activates peripherally located alpha-2A adrenergic receptors. Pain , v. 115, p. 364-373, 2005. KING, E. W.; SLUKA, K. A. The effect of varying frequency and intensity of transcutaneous electrical nerve stimulation on secondary mechanical hyperalgesia in an animal model of inflammation. Them Journal of Pain, v. 2, n. 2, p. 128-133, 2001. MACHELSKA, H.; STEIN, C. Immune mechanisms in pain control. Anesthesia & Analgesia , v. 95, p. 1002-1008, 2002. MELZACK, R. From the gate to the neuromatrix. Pain , Supplement 6, p. S 121-126, 1999. MELZACK, R.; WALL, P. D. Pain mechanisms: new theory. Science , v. 150, p. 971-979, 1965. MILLAN, M. J. Descending control of pain. Progress in Neurobiology , v. 66, p. 355-474, 2002. MILLAN, M. J. The induction of pain: an integrative review. Progress in Neurobiology. n. 57, p. 1-164, 1999.

39
OBARA, I.; MAKUCH, W.; SPETEA, M.; SCHÜTZ, J.; SCHMIDHAMMER, H.; PRZEWLOCKI, R.; PRZEWLOCKA, B. Local peripheral antinociceptivo effects of 14-O-methyloxymorphone derivatives in inflammatory and neuropathic pain in the rat. European Journal of Pharmacology , v. 558, p. 60-67, 2007. OKAMOTO, K.; IMBE, H.; MORIKAWA, Y.; ITOH, M.; SEKIMOTO, M.; NEMOTO, K.; SENBA, E. 5-HT2A receptor subtype in the peripheral branch of sensory fibers is involved in the potentiation of inflammatory pain in rats. Pain , v. 99, p. 133-143, 2002. OLIVEIRA, M. C. G.; PELEGRINE-DA-SILVA, A.; PARADA, C. A.; TAMBELI, C. H. 5HT acts on nociceptive primary afferents through an indirect mechanism to induce hiperalgesia in the subcutaneous tissue. Neuroscience , p. 708-714, 2007. PERTOVAARA, A. Noradrenergic pain modulation. Progress in Neurobiology , v. 80, p. 53-83, 2006. RADHAKRISHNAN, R.; KING, E. W.; DICKMAN, J. K.; HEROLD, C. A.; JOHNSTON, N. F.; SPURGIN, M. L.; SLUKA, K. A. Spinal 5-HT2 and 5-HT3 receptors mediate low, but not high, frequency TENS – induced antihyperalgesia in rats. Pain , v. 5, p. 205-213, 2003. RANG, H. P.; DALE, M. M.; RITTER, J. M. Fármacos analgésicos. In: RANG, H. P.; DALE, M. M.; RITTER, J. M. Farmacologia . 6. ed. Rio de Janeiro: Elsevier. 2008. p. 492-507. RESENDE, M. A.; GONÇALVES, H. H.; SABINO, G. S.; PEREIRA, L. S. M.; FRANSCISCHI, J. N. Redução do efeito analgésico da estimulação elétrica nervosa transcutânea em ratos tolerantes à morfina. Revista Brasileira de Fisioterapia , v. 10, n. 3, p. 291-296, 2006. RIEDEL, W.; NEECK, G. Nociception, pain and antinociception: current concepts. Zeitschrift für Rheumatologie , v. 60, n. 6, p. 404-415, 2001. RODRIGUES, A.R.A.; DUARTE, I.D.G. The peripheral antinociceptive effect induced by morphine is associated with ATP-sensitive K+ channels. British Journal of Pharmacology , v. 129, p. 110-114, 2000. SABINO, G. S.; DE SOUZA, M. V. S.; REZENDE, M. A. Estimulação elétrica nervosa transcutânea no pós-operatório de cirurgia torácica ou abdominal. Fisioterapia e Movimento ; v. 19, n. 1, p. 59-72, 2006.

40
SABINO, G. S.; SANTOS, C. M. F.; FRANCISCHI, J. N.; RESENDE, M. A.. Release of endogenous opioids following transcutaneous electric nerves stimulation in an experimental model of acute inflammatory pain. The Journal of Pain , v. 9, n. 2, p. 157-163, 2007. SLUKA, K.A. The basic science mechanisms of tens and clinical implications. American Pain Society , v. 11, n. 2, 2001. SLUKA, K.A.; BAILEY, K.; BOGUSH, J.; OLSON, R.; RICKETTS, A. Treatment with either high or low frequency TENS reduces the secondary hiperalgesia observed after injection of kaolin and carrageenan into the knee joint. Pain , v. 77, p. 97-102, 1998. SLUKA, K.A.; DEACON, M.; STIBAL, A.; STRISSEL, S.; TERPSTRA, A. Spinal blockade of opioid receptors prevents the analgesia produced by tens in arthritic rats. Journal Pharmacology Experimental Therapeutics , v. 289, n. 2, p. 840-846, 1999. SLUKA, K.A.; LISI, T.L.; WESTLUND, K.N. Increased release of serotonin in the spinal cord during low, but not high, frequency transcutaneous electric nerve stimulation in rats with joint inflammation. Archives of physical medicine and Rehabilitation , v. 87, p. 1137-1140, 2006. SLUKA, K.A.; WALSH, D. Transcutaneous electrical nerve stimulation: basic science mechanisms and clinical effectiveness. The Journal of Pain , v. 4, n. 3, p. 109-121, 2003. SOMMER, C. Serotonin in pain and analgesia: actions in the periphery. Molecular neurobiology , v. 30, p. 117-125, 2004. STAMFORD, J.A. Descending control of pain. British Journal of Anaesthesia , v. 75, p. 217-227, 1995. STEIN, C.; CLARK, J.D.; OH, U.; VASKO, M.R.; WILCOX, G.L.; OVERLAND, A.C.; VANDERAH, T.W.; SPENCER, R.H. Peripheral mechanisms of pain and analgesia. Brain Research Reviews , v. 60, p. 90-113, 2009. STEIN, C.; HASSAN, A.H.S. Local analgesic effect of endogenous opioid peptides. Lancet , v. 342, p. 321-327, 1993.

41
SUFKA, K.J.; SCHOMBURG, F.M.; GIORDANO, J. Receptor mediation of 5-ht-induced inflammation and nociception in rats. Pharmacology Biochemistry & Behavior , v. 41, p. 53-56, 1991. TAIWO, Y.O.; LEVINE, J.D. Serotonin is a directly-acting hyperalgesic agent in the rat. Neuroscience , v. 48, p. 485-490, 1992. TAMBELI, C.H.; OLIVEIRA, M.C.G.; CLEMENTE, J.T.; PELEGRINI-DA-SILVA, A.; PARADA, C.A. A Novel mechanism involved in 5-hydroxytryptamin-induced nociception: the indirect activation of primary afferents. Neuroscience , v. 141, p. 1517-1524, 2006. TANABE, M.; TAKASU, K.; KASUYA, N.; SHIMIZU, S.; HONDA, M.; ONO, H. Role of descending noradrenergic system and spinal a2-adrenergic receptors in the effects of gabapentin on thermal and mechanical nociception after partial nerve injury in the mouse. British Journal of Pharmacology , v. 144, p. 703-714, 2005. TOKUNAGA, A.; SAIKA, M.; SENBA, E. 5-HT2A receptor subtype is involved in the thermal hyperalgesic mechanism of serotonin in the periphery. Pain , v. 76, p. 349-355, 1998. YOSHIMURA, M.; FURUE, H. Mechanisms for the anti-nociceptive actions of the descending noradrenergic and serotoninergic systems in the spinal cord. Journal of Pharmacological Sciences , v. 101, p. 107-117, 2006. ZARRINDAST, M.R.; SAJEDIAN, M.; REZAYAT, M.; GHAZI-KHANSARI, M. Effects of 5HT receptor antagonists on morphine-induced tolerance in mice. European Journal of Pharmacology , v. 273, p. 203-207, 1995. ZHAO, Z.Q.; CHIECHIO, S.; SUN, Y.G.; ZHANG, K.H.; ZHAO, C.S.; SCOTT, M.; JOHNSON, R.L.; DENERIS, E.S.; RENNER, K.J. ; GEREAU IV, R.W.; CHEN, Z.F. Mice lacking central serotonergic neurons show enhanced inflammatory pain and an impaired analgesic response to antidepressant drugs. The Journal of Neuroscience , v. 27, n. 22, p. 6045– 6053, 2007.

42
Capítulo 4 – ARTIGO: Estudo da estimulação elétrica nervosa transcutânea na
nocicepção e no edema induzidos pela administração de serotonina
intradérmica
Cristiane Magela de Freitas Santos1, Marcos Antônio de Resende2, Patrícia Paiva
Lima3, Janetti Nogueira de Francischi4
1Mestranda em Ciências da Reabilitação, Universidade Federal de Minas Gerais –
UFMG, Belo Horizonte, MG, Brasil.
2Professor Doutor, Departamento de Fisioterapia, Universidade Federal de Minas
Gerais – UFMG, Belo Horizonte, MG, Brasil.
3Doutoranda, Departamento de farmacologia do instituto de ciências biológicas,
Universidade Federal de Minas Gerais – UFMG, Belo Horizonte, MG, Brasil.
4Professora Doutora, Departamento de farmacologia do instituto de ciências
biológicas, Universidade Federal de Minas Gerais – UFMG, Belo Horizonte, MG,
Brasil.
Correspondência para Marcos Antônio de Resende: Avenida Antônio Carlos, 6637.
Departamento de Fisioterapia. CEP: 31.270-901. Belo Horizonte - MG. Tel:
(31)34097412. [email protected]
Título para as páginas do artigo: Estudo da TENS na nocicepção e edema da 5-HT.
Palavras-chave: TENS, Sistema nervoso periférico, Serotonina, Opióides
endógenos, Nocicepção,

43
RESUMO
A estimulação elétrica nervosa transcutânea (TENS) é definida como a
aplicação de corrente elétrica por meio de eletrodos na pele, para alívio da
dor. Diferentes teorias são propostas para explicar o mecanismo de ação
analgésica da TENS. Estudos recentes mostraram que parte dessa analgesia
pode, também, ser mediada por ação de neurotransmissores localizados em
receptores periféricos. O objetivo deste estudo foi investigar o efeito da TENS
de baixa (BF: 10 Hz) e alta (AF: 130 Hz) frequência na nocicepção e no
edema induzidos por serotonina (5-HT) administrada intradermicamente na
pata do rato (10µg/0,1ml). TENS de BF e de AF foram aplicadas na pata
direita durante 20 minutos e, imediatamente após, a 5-HT foi administrada. A
nocicepção foi medida pelo método Hargreaves e o edema, pelo
hidropletismômetro (Ugo Basile). A TENS de BF inibiu em 100% a
hiperalgesia da 5-HT, mas não a de AF. TENS de AF e BF não inibiram o
edema induzido por 5-HT. O pré-tratamento com naltrexona (Nx: 50µg; I.pl.)
reverteu o efeito antinociceptivo da TENS de BF. A TENS de BF foi capaz de
inibir a nocicepção induzida por 5-HT por até 30 minutos após o término de
sua aplicação. Nossos resultados confirmaram a ausência de efeito anti-
inflamatório pela TENS e a participação de opióides endógenos periféricos na
analgesia da TENS de baixa frequência.
Perspectivas : Este estudo fornece informações relevantes sobre os
mecanismos de ação analgésicos da TENS nos tecidos periféricos. Uma
melhor compreensão a respeito desse recurso não invasivo, sem efeitos

44
colaterais e de baixo custo para alívio da dor, é de grande importância para
fisioterapeutas, cientistas e população em geral.
ABSTRACT
Transcutaneuous electrical nerve stimulation (TENS) is defined as the application of
an electrical current to the skin through electrodes for pain relief. Various theories
have been proposed in order to explain the analgesic mechanism of TENS. Recent
studies have demonstrated that part of this analgesia can also be mediated through
neurotransmitters found in peripheral receptors. The present study aims to
investigate the effects of low frequency (LF: 10 HZ) TENS as well as high
frequency (HF: 130 HZ) TENS in nociception and edema induced by
intradermally administered serotonin (5-HT), injected into rat paw (10µg/0.1
ml). LF and HF TENS were applied to the right paw for 20 min, and 5-TH was
administered immediately after. The Hargreaves method was used to measure
nociception, and the hydroplethysmometer (Ugo Basile) was used to measure
edema. The LF TENS reduced by about 100% 5-TH hyperalgesia, but the same
did not happen with the HF TENS. HF and LF TENS did not inhibit 5-HT-induced
edema. Pre-treatment with naltrexone (Nx: 50µg; I.pl.) reversed the
antinociceptive effects of LF TENS. LF TENS application could inhibit
nociception 5-HT-induced edema for 30 min maximum after the end of its
application. Our results have confirmed the lack of an antiinflammatory effect
through the use of TENS as well as the participation of a peripheral endogenous
opioid in LF TENS analgesia. Perspectives : This study provides relevant
information about the analgesic mechanisms of TENS in peripheral tissues. A
better understanding of this noninvasive treatment, which has low costs and

45
no side-effects for the relief of pain, represents an important issue to physical
therapists, to scientists and to the general population.
Short title: Study of TENS on nociception and edema induced by 5-HT
Key words: TENS, Pheripheral nervous system, Serotonin, Endogenous
opioids, Nociception, Analgesia.

46
INTRODUÇÃO
A estimulação elétrica nervosa transcutânea (TENS) é definida, pela American
Physical Therapy Association (APTA), como a aplicação de corrente elétrica por
meio de eletrodos de superfície na pele para alívio da dor.2,28 O tratamento da dor
aguda normalmente é realizado com drogas e alguns agentes não farmacológicos,
sendo um deles, a TENS, utilizada frequentemente na clínica de Fisioterapia.40 Dois
tipos de TENS são usadas clinicamente, TENS de baixa (< 10 Hz) frequência (TENS
de BF) e TENS de alta (> 50 Hz) frequência (TENS de AF).1,28
Diferentes mecanismos são propostos para explicar a ação analgésica da TENS.21
Estudos neurofarmacológicos sugerem a participação de mecanismos espinhais e
supraespinhais relacionados a neurotransmissores para explicar a analgesia
provocada pela TENS.21,28,29 Alguns estudos têm verificado o envolvimento de
opióides endógenos liberados no sistema nervoso central após aplicação da
TENS1,27,29 e, mais recentemente, nosso grupo mostrou evidências da participação
de vias opioidérgicas periféricas.25 Além disso, subtipos de receptor 5-HT2A e 5-HT3
espinhais parecem estar envolvidos na anti-hiperalgesia induzida pela TENS de BF,
mas não na de AF, sugerindo um importante papel do sistema serotoninérgico
central em mediar a antinocicepção da TENS21. No entanto, outros estudos
mostraram que o mecanismo analgésico da TENS não envolve a participação de α2-
adrenoceptor espinhal13,21 e que parte da analgesia da TENS pode ser mediada por
ação de neurotransmissores em receptores periféricos.29
A 5-HT liberada de estruturas do tronco cerebral exerce uma ação analgésica15,30 e,
em tecidos periféricos, a aplicação dessa substância produz edema e dor em

47
humanos e nocicepção em roedores.36,38 Provavelmente, a 5-HT periférica age nos
receptores 5-HT1 e 5-HT2 provocando mudanças vasculares e, de forma indireta,
alterar o fluxo de outros elementos pro-nociceptivos e pró-inflamatórios como a
bradicinina e eicosanóides do sangue. Alternativamente, a serotonina do plasma
e/ou derivada dos tecidos pode ter ação direta sobre os receptores 5-HT3,
localizados nas fibras aferentes Aδ/C, produzindo nocicepção.30,36,38,39
Na literatura consultada, os estudos não mostraram a efetividade da TENS em
bloquear a dor e edema induzidos por uma amina biogênica, como a serotonina, que
pode ser encontrada nos tecidos periféricos em condições inflamatórias agudas.
Além disso, poucos estudos associaram o uso da TENS como agente não
farmacológico para reduzir o edema inflamatório.23 No presente estudo, foi
investigado o mecanismo analgésico da TENS de BF e de AF na hiperalgesia e
edema induzidos por serotonina administrada perifericamente. Investigação do
tempo de efeito da TENS, no pico de hiperalgesia da serotonina injetada
perifericamente, também foi conduzida.

48
MATERIAIS E MÉTODOS
Animais
Foram utilizados ratos Holtzman machos com peso entre 280 e 310 gramas para os
experimentos no teste algesimétrico de Hargreaves (1988)6 e 160 a 200g para
experimentos no instrumento hidropletismômetro. Os animais foram fornecidos pelo
CEBIO – Centro de Bioterismo do Instituto de Ciências Biológicas da UFMG. Os
experimentos foram realizados em grupos de 4 a 10 animais, perfazendo um total de
161 animais utilizados em todo o estudo. O desenho experimental foi previamente
aprovado pelo Comitê de Ética em Experimentação Animal da UFMG - CETEA
(Parecer n° 237/08).
Ambientalização dos animais
Os animais permaneceram no biotério em caixas de plástico (seis por gaiola) com
forração de maravalha com livre acesso à água e comida. No dia anterior aos
experimentos, os animais foram privados de alimento e levados para a sala de
experimento, com ciclo claro-escuro de 12 horas e temperatura controlada (22 a
24°C). Foi realizada a adaptação ao aparelho algesí metro por 30 minutos, para que
os animais se habituassem às caixas de teste, à manipulação imposta pelo
pesquisador e aos procedimentos de teste.22,25
Substâncias utilizadas
As substâncias utilizadas neste estudo foram: serotonina (Serotonin, Sigma®, E.U.A),
maleato de metisergida (Methysergide maleate, Sigma®, E.U.A), pizotifeno (Pizotifen,

49
Sandoz®, Brasil), cloridrato de ondansetrona (Ondansetron, Glaxo®, Brasil),
naltrexona (Naltrexone hydrochloride, Sigma®, E.U.A) e solução salina fisiológica
estéril (NaCl 0,9%).
Administração das substâncias
Todas as substâncias foram preparadas em solução salina estéril momentos antes
do experimento. Elas foram administradas após as medidas do limiar de retirada ou
do volume da pata no baseline. Para indução da nocicepção um volume de 0,1 ml de
serotonina (10µg) por via intradérmica (i.d.), foi administrada na pata direita. A pata
contralateral desses animais não recebeu nenhuma substância. A injeção
intradérmica de serotonina (10µg) também foi utilizada para induzir edema e o
mesmo volume final (0,1ml) foi administrado na pata direita dos animais; já a pata
contralateral recebeu igual volume de salina. Injeção subcutânea (s.c.) dos
antagonistas de serotonina - pizotifeno (2 mg/Kg), metisergida (2 mg/Kg) e
ondansetrona (2 mg/Kg) ou salina (0,1 ml) foram administradas no dorso dos
animais próximo à região cervical. Naltrexona (50µg/0,1ml) ou salina (0,1ml) foram
injetadas na pata direita por via intraplantar (i.pl.), sendo que nenhuma substância foi
administrada na pata contralateral durante esses experimentos.
Teste nociceptivo – Algesímetro
Para a medida do tempo de latência de retirada da pata, foi utilizado o aparelho
Plantar test da Ugo Basile® (modelo 7370, Itália), que permite o registro de medidas
de hiperalgesia ao calor na pata de ratos, descrito originalmente por Hargreaves, K.
(1988).6 Para o teste, o animal foi colocado em compartimento de acrílico individual

50
(18 cm x 29 cm x 12.5 cm), posicionado sobre uma superfície de vidro especial, que
permitiu a passagem de forma homogênea da luz e do calor. Foi realizado um
período de ambientalização (± 5 minutos). Após esse tempo, a fonte de luz
infravermelha foi colocada sob cada uma das patas traseiras do animal. Um tempo
máximo de 30 segundos de aplicação foi utilizado como tempo de corte para impedir
dano tecidual. Duas medidas de cada pata foram obtidas. A média das duas
medidas foi utilizada para calcular o delta (∆) da latência de retirada da pata, obtido
através da diferença entre a pata direita (inflamada) e a pata esquerda (controle), em
segundos. Todas as medidas foram realizadas pelo mesmo avaliador e no mesmo
período do dia. As aferições foram realizadas no tempo zero e após 5, 15, 30, 60 e
120 minutos da administração de serotonina intradérmica na região plantar.
Avaliação do edema inflamatório – Hidropletismômetr o
Para aferir o aumento de volume das patas traseiras causado pela serotonina, foi
utilizado um hidropletismômetro da Ugo Basile® (modelo 1750, Itália). O cálculo do
edema da pata foi obtido através dos valores de variação entre o volume da pata
inflamada (direita) e o volume da pata contralateral (esquerda) (∆, em ml) nos
mesmos tempos utilizados para medida do limiar nociceptivo, isto é, tempo zero e
após 5, 15, 30, 60 e 120 minutos do estímulo pró-inflamatório. As medidas foram
realizadas por um mesmo examinador, previamente treinado, e os valores expressos
como média ± EPM, por grupo, da diferença do volume (ml) entre as patas traseiras,
em cada tempo.

51
Aparelho de Estimulação Elétrica Nervosa Transcutân ea (TENS)
O aparelho de TENS utilizado para induzir antinocicepção foi o Neurodyn
III/Ibramed®. O aparelho foi devidamente calibrado e os parâmetros de estimulação
administrados foram: baixa (BF: 10 Hz) e alta frequência (AF: 130 Hz), com duração
de pulso de 130 µs e tempo de aplicação de 20 minutos. O limite de intensidade
sensorial foi considerado imediatamente abaixo do limiar motor. Eletrodos com 1 cm²
foram especialmente construídos para os experimentos e fixados na face plantar e
dorsal da pata posterior direita do animal com fita adesiva. A fim de evitar que eles
provocassem algum dano aos eletrodos, confeccionou-se, em cartolina, um colar
Elizabetano, que foi preso por fita adesiva ao redor da cabeça dos animais.
Protocolo experimental
Para a escolha da dose de serotonina a ser utilizada no estudo, diferentes doses
dessa substância (0,1; 1,0 e 10 µg)19 foram administradas em grupos distintos de
animais na pata traseira direita. Em seguida, outro estudo foi realizado com a
administração por via subcutânea no dorso dos animais dos antagonistas
metisergida,5,41 ondansetrona e pizotifeno5 meia hora antes da administração de
serotonina (10µg/0,1ml i.d). Para verificar o efeito da TENS de BF e de AF na
hiperalgesia e edema provocados pela serotonina, a estimulação elétrica de baixa e
alta frequência foi aplicada na pata direita 30 minutos antes dos animais receberem
a injeção de serotonina intradérmica na região plantar (10µg/100µl). Foi realizado um
experimento como controle com a TENS desligada para avaliar o nível de analgesia
provocado pela manipulação dos animais. Em um outro experimento, naltrexona (50
µg/100µl)24, um antagonista opióide, foi administrado por via intraplantar 30 minutos

52
antes da aplicação da TENS de BF. Para verificar duração do tempo de efeito da
TENS, outro experimento foi conduzido. Grupos com 4 animais cada receberam
injeção intradérmica plantar de serotonina (10µg) nos diferentes tempos após
aplicação da TENS de BF. As medidas do tempo de retirada da pata foram
realizadas no Hargreaves, sempre 5 minutos após a injeção de 10 µg de 5-HT, aos
10 ou 30 ou 60 ou 120 minutos após a aplicação da TENS na pata direita.
Análise estatística
A preparação do banco de dados, assim como a análise estatística, foram
realizadas por meio do programa GraphPad Prism 4.00, instalado em ambiente
Windows®.
Os resultados estão representados como a média ± erro padrão da média. As
comparações estatísticas das diferenças entre os grupos nos diversos tempos de
medidas foram feitas utilizando-se análise de variância (One-way ANOVA), com
correção post hoc pelo teste de Bonferroni. Para os experimentos com apenas dois
grupos, foi utilizado para verificar diferenças entre dois grupos nos diversos tempos
a Two-way ANOVA. Um valor de p inferior a 0,05 foi considerado estatisticamente
significativo.

53
RESULTADOS
Nocicepção induzida por serotonina
Conforme mostrado na FIG. 1A, a injeção intradérmica (i.d.) de 5-HT (0,1; 1,0 e 10
µg), na pata traseira de ratos, provocou uma diminuição na latência de retirada da
pata nas três doses empregadas quando comparadas com o controle (salina). O
efeito e duração máximos da 5-HT ocorreram aos 5 e 30 minutos, respectivamente,
com a maior dose utilizada de 5-HT.
Edema induzido por serotonina
De acordo com a FIG. 1B, a injeção intraplantar de 5-HT (0,1; 1,0 e 10 µg), na
pata traseira de ratos, provocou um aumento no volume da pata nas três doses
empregadas quando comparadas com o controle (salina). A maior dose utilizada (10
µg) foi a dose de escolha
Administração de antagonistas da 5-HT na nocicepção e edema
Para identificar os principais receptores envolvidos, nos efeitos nociceptivo e
edematogênico da 5-HT, antagonistas específicos de receptores da 5-HT foram
administrados. As FIG. 2A e 2B mostraram que, nos animais pré-tratados com
pizotifeno e metisergida, respectivamente, ocorreu um completo bloqueio do efeito
nociceptivo e do edema induzidos pela maior dose de serotonina utilizada (10µg). Já
o pré-tratamento com ondansetrona não alterou os efeitos nociceptivo e
edematogênico induzidos pela 5-HT.

54
Efeito da aplicação da TENS de baixa (BF) e alta fr equência (AF) na nocicepção
e edema provocados por serotonina
O pré-tratamento do animal com a TENS de BF reverteu completamente o efeito
nociceptivo provocado pela injeção local de serotonina quando comparado ao
controle (FIG. 3A). Já na FIG. 3B, podemos observar que a TENS de AF não
reverteu o efeito da 5-HT. Entretanto, nem a TENS de BF nem a de AF modificaram
a resposta de edema provocado pela serotonina, quando comparadas ao controle,
conforme mostrado na FIG. 3C.
Efeito da naltrexona na antinocicepção induzida pel a TENS de baixa frequência
no modelo de administração periférica da serotonina
Como o tratamento com a TENS foi efetivo apenas quando a baixa frequência foi
utilizada, procuraram-se verificar possíveis mecanismos endógenos envolvidos.
Conforme mostrado na FIG. 4, o pré-tratamento com naltrexona, um antagonista
opióide, reverteu o efeito analgésico da TENS de BF na hiperalgesia da serotonina.
Tempo de duração do efeito analgésico da TENS de BF
Para verificar a duração do efeito antinociceptivo da TENS de BF, variou-se o
período após o término de aplicação da mesma antes da administração da
serotonina. A TENS de BF mostrou-se eficaz em inibir o efeito da serotonina por até
30 minutos após o término de sua aplicação (FIG. 5).

55
DISCUSSÃO
Nossos dados confirmam estudos prévios sobre o efeito hiperalgésico da serotonina
em tecidos periféricos.17,36 Inclusive, a injeção de 5-HT por via intradérmica plantar
provocou nocicepção térmica dose-dependente, resultado que está de acordo com
dados de Oliveira et al. (2007)19, que verificaram nocicepção mecânica induzida pela
5-HT na pata de ratos. De forma semelhante a 5-HT administrada intraplantarmente
induziu edema dose-dependente, resultado que concorda com os obtidos por Sufka
et al., (1991)36.
Os efeitos dos antagonistas de receptor 5-HT na resposta de retirada ao estímulo
térmico e no edema foram analisados utilizando-se injeção intradérmica plantar de
serotonina. Como mostrado nas FIG. 2A e 2B, a metisergida, um antagonista de
receptores 5-HT2/5-HT1, e o pizotifeno, antagonista 5-HT2,3,8,35,36 foram capazes de
antagonizar o efeito nociceptivo e edematogênico da serotonina. Já a ondansetrona,
antagonista 5-HT3,8,26,36 não inibiu os efeitos da 5-HT. Embora estudos tenham
demonstrado a participação de receptores 5-HT1, 5HT2 e 5-HT3 no efeito
hiperalgésico da serotonina na periferia.17,36,37,42 , nossos resultados confirmam os
obtidos por Tokunaga et al. (1998),39 nos quais a administração de antagonista de 5-
HT2, mas não de 5-HT3, antagonizou a hiperalgesia induzida pela injeção de 5-HT. A
variação encontrada na ativação de receptores 5 HT3 pode ser devido a diferenças
de métodos de avaliação, espécies de animais e doses das drogas utilizadas nos
diferentes estudos. Entretanto, outros estudos sugerem o não envolvimento do
receptor 5-HT3 no edema obtido pela 5-HT.5,11,20,42 . Tal achado foi confirmado neste
estudo, uma vez que a ondansetrona falhou em suprimir o edema induzido pela 5-
HT. Por outro lado, o receptor 5-HT2 parece ser o principal responsável por tal

56
efeito,11,20 o que está de acordo com os nossos dados, pois a metisergida e
pizotifeno inibiram em quase 100% a formação de edema pela serotonina na pata
dos ratos.
A utilização da TENS como coadjuvante eficaz no tratamento de condições
inflamatórias agudas e crônicas ainda não está totalmente confirmada. Nossos
resultados demonstraram uma completa inibição do efeito nociceptivo da serotonina
administrada perifericamente pela TENS de BF (10 Hz), mas não pela de AF (130
Hz). No entanto, nenhuma das duas frequências foi capaz de inibir o edema induzido
pela 5-HT, confirmando achados prévios de nosso laboratório.23 nos quais não foi
encontrada atividade antiedematogênica da TENS em modelo de inflamação
induzido por carragenina. A sequência de mecanismos pelos quais a 5-HT influencia
a inflamação e a nocicepção não é clara. Os diversos tipos de receptores podem
diferentemente mediar as ações inflamatórias e nociceptivas da serotonina nos
tecidos periféricos.36
É descrito na literatura que a TENS utiliza diferentes mecanismos para exercer seu
efeito analgésico.22,25,28 e que a analgesia da TENS de BF e de AF envolvem a
liberação endógena de opióides,9,12 sendo a analgesia da TENS de BF, mas não da
AF, revertida por uma baixa dose de naloxona23, um antagonista opioide
específico.4 Além disso, em estudo prévio realizado por Rodrigues et al. (2000),24 foi
demonstrado que a dose de 50 µg de naltrexona, administrada na pata do rato,
antagonizou o efeito antinociceptivo induzido pela morfina, no local da lesão, quando
administrado ipsilateralmente, mas não contralateralmente, confirmando sua ação
em receptores opióides locais. A partir dessas informações, um experimento foi
conduzido para verificar a ação analgésica periférica da TENS. Conforme foi

57
verificado, o pré-tratamento com a TENS de BF provavelmente resultou na liberação
local de opióides e impediu o efeito nociceptivo induzido pela posterior administração
de 5-HT. É possível que os agonistas de receptores µ opióides ajam por inibir a
ativação da adenilato ciclase por mediadores inflamatórios, tais como a serotonina e
prostaglandina E2 em neurônios aferentes periféricos, enquanto os agonistas de
receptor δ e κ opióides inibem a secreção de substâncias pró-inflamatorias por
neurônios eferentes simpáticos.7
Alguns estudos mostraram que os opióides podem produzir potente analgesia por
ativação de receptores em neurônios sensoriais periféricos.18,32,33 Esses receptores
opióides estão localizados em neurônios sensoriais na periferia e centralmente33, em
animais33,34 e humanos.16 A ativação desses receptores por opióides pode inibir os
canais iônicos excitatórios como TRPV1, canais de Ca2+ e também a liberação de
neuropeptídeos pró-inflamatórios como substância P e peptídeo relacionado ao gene
da calcitonina (CGRP), das terminações nervosas sensoriais periféricas, resultando
em efeito antinociceptivo.33 A administração de 50 µg de naltrexona por via
intraplantar, 30 minutos antes da TENS de BF, reverteu o efeito analgésico da TENS
no modelo de nocicepção da serotonina.
Os dados desta pesquisa estão de acordo com os obtidos por Sabino et al. (2008),25
que sugeriram a participação opióide local na analgesia da TENS de BF no modelo
da carragenina. De acordo com Stein (1993)32, os agonistas µ opióides são mais
potentes que os agonistas δ e κ em induzir efeito antinociceptivo periférico. Um
estudo recente descreveu a presença de β-endorfina imunoreativa em queratinócitos
na pele de ratos.10 A liberação de β-endorfina pelos queratinócitos produziu
antinocicepção por agir nos receptores µ opióides nos neurônios aferentes

58
primários.10 Alterações nas membranas dos queratinócitos pela aplicação da TENS
de BF podem induzir à liberação de β-endorfina e contribuir para o efeito anti-
hiperalgesico.25 Estudos imunohistoquímicos posteriores são necessários para
investigar essa contribuição.
O tempo de analgesia obtido com a TENS de BF, aplicada anteriormente à
administração da serotonina, foi de 30 minutos. No estudo de Resende et al.
(2004)23, a TENS foi aplicada após a carragenina, obtendo uma analgesia de uma
hora e 30 minutos. Essa diferença na duração da analgesia provavelmente se deu
pela diferença no momento de aplicação da TENS, indicando que a analgesia obtida
com a aplicação da TENS após a instalação do processo inflamatório torna-se mais
duradoura. É provável que, após a indução da inflamação periférica, o transporte
axonal de receptores opióides para as terminações nervosas sensitivas seja
intensificado.14 Células imunes presentes no tecido inflamado ou que migram até o
local da lesão como granulócitos, monócitos/macrófagos e linfócitos de roedores e
humanos são também uma fonte de peptídeos opióides, que podem induzir
analgesia no tecido lesado.31 Esses fatores podem ter contribuído para uma
analgesia mais duradoura quando a TENS foi aplicada após a indução da
inflamação pela carragenina do que pela serotonina na pata do rato.23
Uma limitação deste estudo foi provavelmente a indução de algum estresse
provocado pela manipulação dos animais, uma vez que os experimentos foram
conduzidos com os animais acordados. No entanto, para controlar a interferência do
estresse nos resultados, um grupo controle de animais foi exposto a todas as
manipulações, bem como ao TENS desligado. Além disso, algumas pequenas
diferenças no índice da latência de retirada da pata foram encontradas em alguns

59
experimentos, o que se justifica pelas diferentes épocas do ano em que os
experimentos foram conduzidos, mesmo com temperatura ambiente e outros
parâmetros do estudo mantidos constantes.
Uma vez que a TENS de BF inibiu a nocicepção induzida pela serotonina através de
seu efeito analgésico local, clinicamente ela pode ser útil para estimular mecanismos
analgésicos de ação local, atuando de forma periférica e central no controle da dor
aguda e/ou crônica. Mais estudos são necessários para esclarecer os mecanismos
de ação analgésica da TENS e fundamentar seu uso como uma opção de
tratamento não invasivo e não farmacológico da dor, resguardando o paciente dos
efeitos colaterais advindos do uso frequente de drogas analgésicas.

60
REFERÊNCIAS 1. AINSWORTH, L.; BUDELIER, K.; CLINESMITH, M.; FIEDLER, A.; LANDSTRON, R.; LEEPER, B. J.; MOELLER, L.; MUTCH, S.; O’DELL, K.; ROSS, J.; RADHAKRISHNAN, R.; SLUKA, K. A. Trancutaneous electrical nerve stimulation (TENS) reduces chronic hiperalgesia induced by muscle inflammation. Pain , v. 120, p. 182-187, 2006. 2. APTA, American Physical Therapy Association, Guide to Physical Therapy Practice. Physical Therapy , v. 77, p. 1171-1624, 1997. 3. CENTURION, D.; MEHOTRA, S.; SÁNCHES-LÓPEZ, A.; GUPTA, S.; VANDENBRINK, A. M.; VILLALÓN, C. M. Potential vascular α1-adrenoceptor blocking properties of an array of 5-HT receptor ligands in the rat. European Journal of Pharmacology, v. 535, p. 234-242, 2006. 4. DHAWAN, B. N.; CESSELIN, F.; RAGHUBIR, R.; REISINE, T.; BRADLEY, P. B.; PORTOGHESE, P. S.; HAMON, M. International union of pharmacology. XII. classification of opioid receptors. Pharmacological reviews, v. 48, n. 4, p. 567-592, 1996.
5. FRANCISCHI, J. N.; DIAS, M. F.; ROCHA, O. A.; CASTRO, M. S. A.; TATSUO, M. A. K. F.; FARINELLI, P.; PACHECO, C. M. F.; FERREIRA-ALVES, D. L.; SIROIS, P. Pharmacological characterization of sephadex-induced oedema in rat paws: predominant role of serotonin and platelet-activating factor. International archives of allergy and immunology , v. 109, p. 398-406, 1996. 6. HARGREAVES, K.; DUBRER, R.; BROWN, F.; FLORES, C.; JORIS, J. A new and sensitive method for measuring thermal nociception in cutaneous hiperalgesia. Pain , v. 32, p. 77-88, 1988. 7. HONG, Y.; ABBOTT, F. V. Peripheral opióide modulation of pain and inflammation in the formalin test. European Journal of Pharmacology, v. 227, p. 21-28, 1995. 8. HOYER, D.; CLARKE, D. E.; FOZARD, J. R.; HARTIG, P. R.; MARTIN, G. R.; MYLECHARANE, E. J.; SAXENA, P. R.; HUMPHREY, P. P. A. International union of pharmacology classification of receptors for 5-hydroxytryptamine (serotonin). The American Society for Pharmacology and Experimental Therapeutics , v. 46, n. 2, p. 157-203, 1994.
9. HUGHES JR., G. S.; LICHSTEIN, P. R.; WHITLOCK, D.; HARKER, C. Response of plasma beta-endorphins to transcutaneous electrical nerve stimulation in healthy subjects. Physical Therapy , v. 64, n. 7, p. 1062-1066, 1984.

61
10. IBRAHIM, M. M.; PORRECA, F.; LAI, J.; ALBRECHT, P. J.; RICE, F. L.; KHODOROVA, A.; DAVAR, G.; MAKRIYANNIS, A.; VANDERAH, R. W.; MATA, H. P.; MALAN JR., P. CB2 cannabinoid receptor activation produces antinociception by stimulating peripheral release of endogenous opioids. Procedings of the National Academy of Sciences, v. 102, n. 8, p. 3093-3098, 2005. 11. INOUE, H.; NAGATA, N.; KOSHIHARA, Y. Participation of serotonin in capsaicin-induced mouse ear edema. The Japanese Journal of Pharmacology, v. 69, p. 61-68, 1995. 12. KALRA, A.; URBAN, M. O.; SLUKA, K. A. Blockade of opioid receptors in rostral ventral medulla prevents antihyperalgesia produced by transcutaneous electrical nerve stimulation (tens). The Journal of Phamacology and Experimental Therapeutics, v. 298, p. 257-263, 2001. 13. KING, E. W.; AUDETTE, K.; ATHMAN, G. A.; NGUYEN, H. O. X.; SLUKA, K. A.; FAIRBANKS, C. A. Transcutaneous electrical nerve stimulation activates peripherally located alpha-2A adrenergic receptors. Pain , v. 115, p. 364-373, 2005. 14. MACHELSKA, H.; STEIN, C. Immune mechanisms in pain control. Anesthesia & Analgesia , v. 95, p. 1002-1008, 2002. 15. MILLAN, M. J. Descending control of pain. Progress in Neurobiology , v. 66, p. 355-474, 2002. 16. MOUSA, S. A.; STRAUB, R. H.; SCHÄFER, M.; STEIN, C. β-Endorphin, met-enkephalin and corresponding opióide receptors within synovium of patients with joint trauma, osteoarthritis and rheumatoid arthristis. Annals of the Rheumatic Diseases, v. 66, p. 871-879, 2007. 17. NITANDA, A.; YASUNAMI, N.; TOKUMO, K.; FUJII, H.; HIRAI, T.; NISHIO, H. Contribution of the peripheral 5-HT2A receptor to mechanical hyperalgesia in a rat model of neuropathic pain. Neurochemistry International, v. 47, p. 394-400, 2005. 18. OBARA, I.; MAKUCH, W.; SPETEA, M.; SCHÜTZ, J.; SCHMIDHAMMER, H.; PRZEWLOCKI, R.; PRZEWLOCKA, B. Local peripheral antinociceptive effects of 14-o-methyloxymorphone derivatives in inflammatory and neuropathic pain in the rat. European Journal of Pharmacology , v. 558, p. 60-67, 2007. 19. OLIVEIRA, M. C. G.; PELEGRINE-DA-SILVA, A.; PARADA, C. A.; TAMBELI, C. H. 5HT acts on nociceptive primary afferents through an indirect mechanism to induce hiperalgesia in the subcutaneous tissue. Neuroscience , p. 708-714, 2007. 20. PIERCE, P. A.; XIE, G.; PEROUTKA, S. J.; GREEN, P. G., LEVINE, J. D. 5-hydroxytryptamine-induced synovial plasma extravasation is mediated via 5-hydroxytryptamine2a receptors on sympathetic efferent terminals. The Journal of Pharmacology and Experimental Therapeutics, v. 275, n. 1, p. 502-508, 1995.

62
21. RADHAKRISHNAN, R.; KING, E. W.; DICKMAN, J. K.; HEROLD, C. A.; JOHNSTON, N. F.; SPURGIN, M. L.; SLUKA, K. A. Spinal 5-HT2 and 5-HT3 receptors mediate low, but not high, frequency TENS – induced antihyperalgesia in rats. Pain , v. 5, p. 205-213, 2003. 22. RESENDE, M. A.; GONÇALVES, H. H.; SABINO, G. S.; PEREIRA, L. S. M.; FRANSCISCHI, J. N. Redução do efeito analgésico da estimulação elétrica nervosa transcutânea em ratos tolerantes à morfina. Revista Brasileira de Fisioterapia , v. 10, n. 3, p. 291-296, 2006. 23. RESENDE, M. A.; SABINO, G. G.; CÂNDIDO, C. R. M.; PEREIRA, L. S. M.; FRANCISCHI, J. N. Local transcutaneous electrical stimulation (TENS) effects in experimental inflammatory edema and pain. European Journal of Pharmacology, v. 504, p. 217-222, 2004. 24. RODRIGUES A. R. A.; DUARTE, I. D. G. The peripheral antinociceptive effect induced by morphine is associated with ATP-sensitive K+ channels. British Journal of Pharmacology , v. 129, p. 110-114, 2000. 25. SABINO, G. S.; SANTOS, C. M. F.; FRANCISCHI, J. N.; RESENDE, M. A.; Release of endogenous opioids following transcutaneous electric nerves stimulation in an experimental model of acute inflammatory pain. The Journal of Pain , v. 9, n. 2, p. 157-163, 2008. 26. SCOTT, J. A.; WOOD, M.; FLOOD, P. The pronociceptive effect of ondansetron in the setting of p-glycoprotein inhibition. International Anesthesia Research Society, v. 103, n. 3, p. 742-746, 2006. 27. SLUKA, K. A.; DEACON, M.; STIBAL, A.; STRISSEL, S.; TERPSTRA, A. Spinal blockade of opioid receptors prevents the analgesia produced by tens in arthritic rats. Journal Pharmacology Experimental Therapeutics , v. 289, n. 2, p. 840-846, 1999. 28. SLUKA, K. A.; LISI, T. L.; WESTLUND, K. N. Increased release of serotonin in the spinal cord during low, but not high, frequency transcutaneous electric nerve stimulation in rats with joint inflammation. Archives of physical medicine and Rehabilitation , v. 87, p. 1137-1140, 2006. 29. SLUKA, K. A.; WALSH, D. Transcutaneous electrical nerve stimulation: basic science mechanisms and clinical effectiveness. The Journal of Pain , v. 4, n. 3, p. 109-121, 2003. 30. SOMMER, C. Serotonin in pain and analgesia: actions in the periphery. Molecular neurobiology , v. 30, p. 117-125, 2004. 31. STEIN, C.; CLARK, J. D.; OH, U.; VASKO, M. R.; WILCOX, G. L.; OVERLAND, A. C.; VANDERAH, T. W.; SPENCER, R. H. Peripheral mechanisms of pain and analgesia. Brain Research Reviews , v. 60, p. 90-113, 2009.

63
32. STEIN, C.; HASSAN, A. H. S. Local analgesic effect of endogenous opioid peptides. Lancet , v. 342, p. 321-327, 1993. 33. STEIN, C.; LANG, L. J. Peripheral mechanisms of opioid analgesia. Current Opinion in Pharmacology, v. 9, p. 3-8, 2009. 34. STEIN, C.; SCHÄFER, MACHELSKA, H. Attacking pain at its source: new perspectives on opioids. Nature Medicine, v. 9, n. 8, 2003. 35. STRACHAN, A. T.; LEIPER, J. B.; MAUGHAN, R. J. Serotonin2C receptor blockade and thermoregulation during exercise in the heat. Medicine & Science in Sports & Exercise, v. 37, n. 3, p. 389-394, 2005. 36. SUFKA, K. J.; SCHOMBURG, F. M.; GIORDANO, J. Receptor mediation of 5-HT-induced inflammation and nociception in rats. Pharmacology Biochemistry & Behavior , v. 41, p. 53-56, 1991. 37. TAIWO, Y. O.; LEVINE, J. D. Serotonin is a directly-acting hyperalgesic agent in the rat. Neuroscience , v. 48, p. 485-490, 1992. 38. TAMBELI, C. H.; OLIVEIRA, M. C. G.; CLEMENTE, J. T.; PELEGRINI-DA-SILVA, A.; PARADA, C. A. A Novel mechanism involved in 5-hydroxytryptamin-induced nociception: the indirect activation of primary afferents. Neuroscience , v. 141, p. 1517-1524, 2006. 39. TOKUNAGA, A.; SAIKA, M.; SENBA, E. 5-HT2A receptor subtype is involved in the thermal hyperalgesic mechanism of serotonin in the periphery. Pain , v. 76, p. 349-355, 1998. 40. WALSH, D. M.; HOWE, T. E.; JOHNSON, M. I.; SLUKA, K. A. Transcutaneous electrical nerve stimulation for acute pain. Cochrane Database of Systematic Reviews, v. 3, 2009. 41. ZARRINDAST, M. R.; SAJEDIAN, M.; REZAYAT, M.; GHAZI-KHANSARI, M. effects of 5HT receptor antagonists on morphine-induced tolerance in mice. European Journal of Pharmacology , v. 273, p. 203-207, 1995. 42. ZEITZ, D. P.; GUY, N.; MALMBERG, A. B.; DIRAJLAL, S.; MARTIN, W. J.; SUN, L.; BONHAUS, D. W.; STUCKY, C. L.; JULIUS, D.; BASBAUM, A. The 5-HT3 subtype of serotonin receptor contributes to nociceptive processing via a novel subset of myelinated and unmyelinated nociceptors. The Journal of Neuroscience, v. 22, n. 3, p. 1010-1019, 2002.
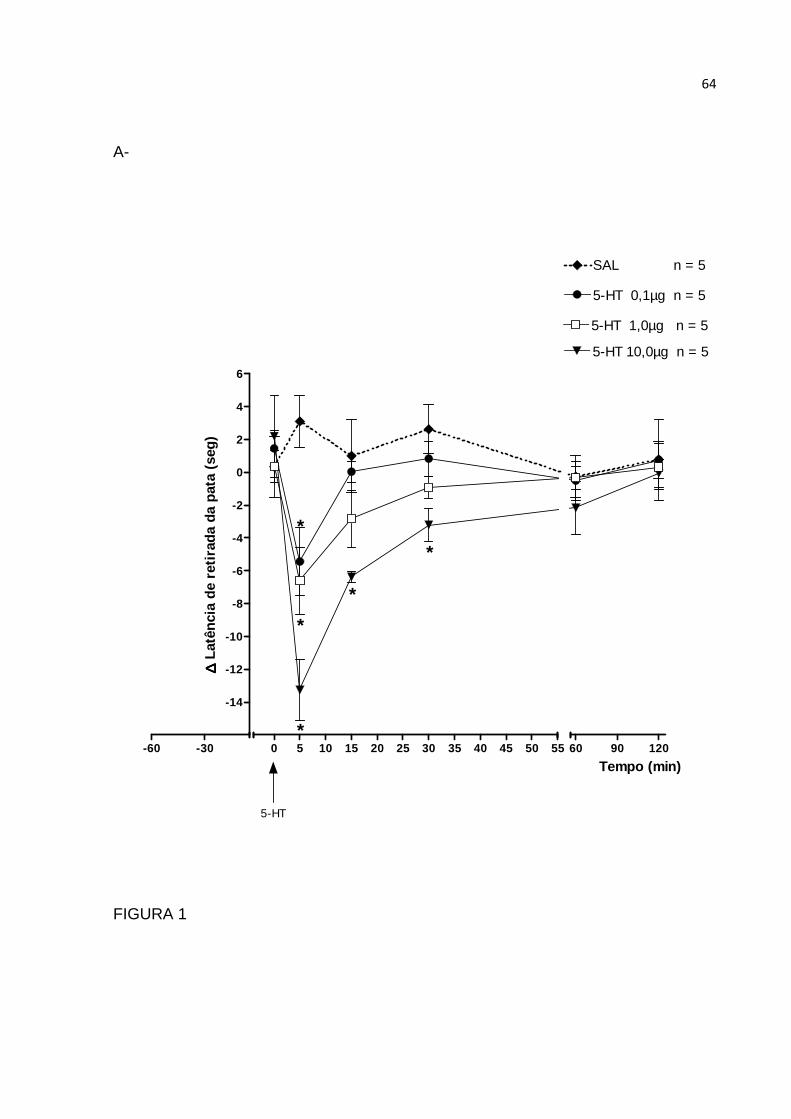
64
A-
-60 -30
-14
-12
-10
-8
-6
-4
-2
0
2
4
6
5-HT 1,0µg n = 5
5-HT 10,0µg n = 5
SAL n = 5
0 5 10 15 20 25 30 35 40 45 50 55
5-HT 0,1µg n = 5
Tempo (min)
*
*
*
*
*
60 90 120
5-HT
∆∆ ∆∆ L
atên
cia
de r
etir
ada
da p
ata
(seg
)
FIGURA 1

65
B-
-5 0 5 10 15 20 25 30 35 40 45 50 550.00
0.25
0.50
0.75
1.00
1.25
1.50
1.75
2.00 SAL n = 50.1mg 5-HT n = 51.0mg 5-HT n = 510.0mg 5-HT n = 5
60 90 120
**
*
**
*
**
5-HT
Time (min)
∆∆ ∆∆ H
ind
paw
vol
ume
(ml)
FIGURA 1

66
A-
-60 -30
-14
-12
-10
-8
-6
-4
-2
0
2
4
6
SAL + 5HT n = 10
PIZO + 5HT n = 5
METIS + 5HT n = 5
0 5 10 15 20 25 30 35 40 45 50 55
ONDA + 5HT n = 5
SAL + SAL n = 4
***
60 90 120
PIZOONDAMETIS
5-HT
Tempo (min)
∆∆ ∆∆ L
atên
cia
de r
etir
ada
da p
ata
(seg
)
FIGURA 2

67
B-
-60 -30
0.25
0.50
0.75
1.00
1.25
1.50
1.75
2.00
SAL + 5-HT n = 7PIZO + 5-HT n = 5
METIS + 5-HT n = 5
0 5 10 15 20 25 30 35 40 45 50 55
ONDA + 5-HT n = 5
** * **
60 90 120
PIZOMETISONDA
5-HT
Tempo (min)
∆∆ ∆∆ v
olum
e da
s pa
tas
tras
eira
s (m
l)
FIGURA 2

68
A-
-60 -30
-14
-12
-10
-8
-6
-4
-2
0
2
4
6
TENS de BF + 5-HT n = 8TENS de BF + SAL n = 6TENS desligado + 5-HT n = 10
0 5 10 15 20 25 30 35
**
*
*
40 50 60 70
TENS 5-HT
Tempo ( min)
∆∆ ∆∆ L
aten
cia
de r
etir
ada
da p
ata
(seg
)
FIGURA 3

69
B-
-60 -30
-14
-12
-10
-8
-6
-4
-2
0
2
4
6
TENS de AF +5-HT n = 8TENS de AF + SAL n = 8TENS desligado + 5-HT n = 10
0 5 10 15 20 25 30 35 40 50 60 70
TENS 5-HT
Tempo (min)
∆∆ ∆∆ L
atên
cia
de r
etir
ada
da p
ata
(seg
)
FIGURA 3

70
C-
-60 -30
0.25
0.50
0.75
1.00
1.25
1.50
1.75
2.00
TENS desligado + 5-HT n = 10
TENS de BF + 5-HT n = 8TENS de AF + 5-HT n = 8
0 5 10 15 20 25 30 35 40 45 50 55
5-HT
60 90 120
TENS
Tempo (min)
∆∆ ∆∆
volu
me
das
pata
s tr
asei
ras
(ml)
FIGURA 3
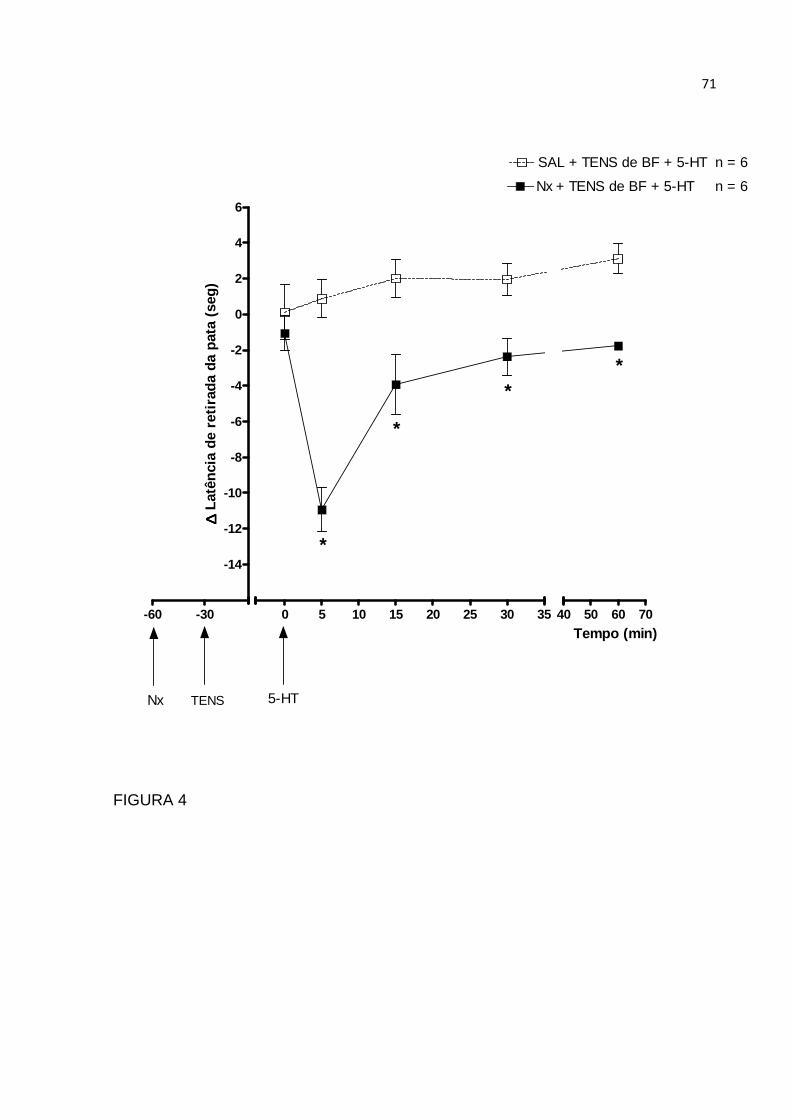
71
-60 -30
-14
-12
-10
-8
-6
-4
-2
0
2
4
6
SAL + TENS de BF + 5-HT n = 6
Nx + TENS de BF + 5-HT n = 6
0 5 10 15 20 25 30 35
*
*
**
40 50 60 70
Nx TENS 5-HT
Tempo (min)
∆∆ ∆∆ L
atên
cia
de r
etir
ada
da p
ata
(seg
)
FIGURA 4
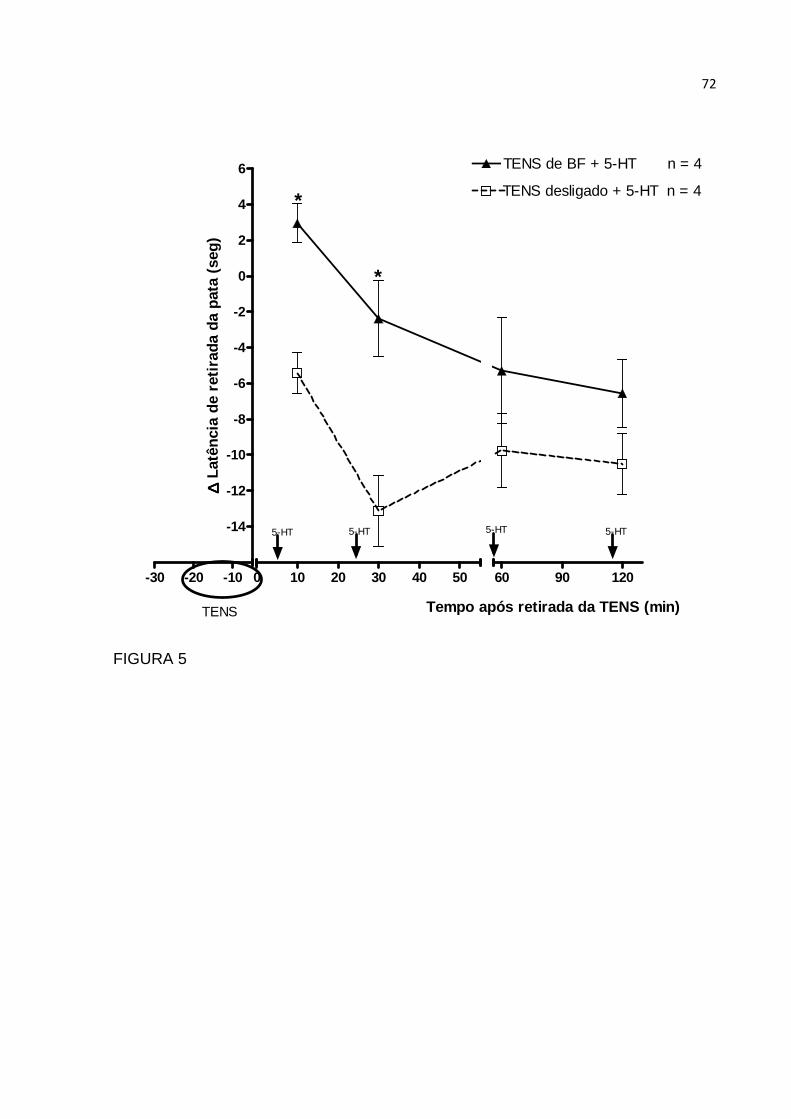
72
-30 -20 -10
-14
-12
-10
-8
-6
-4
-2
0
2
4
6 TENS de BF + 5-HT n = 4
TENS desligado + 5-HT n = 4
0 10 20 30 40 50
*
*
TENS Tempo após retirada da TENS (mi n)
60 90 120
5-HT 5-HT 5-HT 5-HT
∆∆ ∆∆ L
atên
cia
de r
etir
ada
da p
ata
(seg
)
FIGURA 5

73
LEGENDAS DAS FIGURAS
Figura 1A- Efeito dose-dependente de 5-HT na latência de retir ada da pata
seguido a um estímulo térmico. A serotonina (5-HT) foi diluída em salina e injetada
intradermicamente na superfície plantar da pata traseira direita dos animais, sendo
que a pata contralateral não foi tratada. Um volume final de 0,1ml foi administrado.
Cada ponto indica média ± EPM de 5 animais/grupo * indica uma diferença
estatisticamente significativa (p < 0,05).
Figura 1A- Edema dose-dependente de 5-HT intraplantar. Administração
intraplantar de serotonina (5-HT) foi realizada na superfície plantar da pata traseira
direita dos animais, sendo que a pata contralateral foi tratada com o mesmo volume
de salina. Um volume final de 0,1ml foi administrado. Cada ponto indica média ±
EPM de 5 animais/grupo * indica uma diferença estatisticamente significativa (p <
0,05).
Figura 2 – Efeito dos antagonistas de receptores 5-HT, pizotif eno, metisergida,
ondansetrona, na latência de retirada da pata (2A) e no edema (2B) induzidos
por serotonina. Os antagonistas de receptores 5-HT ou salina foram administrados
por injeção subcutânea (s.c.), no dorso dos animais, meia hora antes da
administração intradérmica (i.d.) de serotonina.
A 5-HT (10µg/0,1ml) ou a salina (0,1ml) foram injetadas intradermicamente na
superfície plantar da pata traseira direita dos animais no tempo zero. Todas as
substâncias foram diluídas em salina. As patas contralaterais não foram tratadas na
nocicepção, nem nos experimentos de edema. Cada ponto indica média ± EPM de 4
a 10 animais/grupo. * indica uma diferença estatisticamente significativa (p < 0,05).

74
Figura 3 – Efeitos da TENS de BF e de AF na nocicep ção (A e B) e no edema
(C) induzidos por 5-HT.
A TENS de BF (10 Hz) foi aplicada, na pata traseira direita dos animais, meia hora
antes da injeção intradérmica de 5-HT, por um período de 20 minutos. A pata
contralateral não foi tratada (na nocicepção).
A serotonina foi diluída em salina e administrada em um volume total de 0,1 ml.
Cada ponto indica média ± EPM de 6 a 12 animais/grupo. * indica uma diferença
estatisticamente significativa (p < 0,05).
Figura 4 – Efeito da naltrexona na antinocicepção i nduzida pela TENS de BF.
Naltrexona (50µg/0,1ml) ou a salina (0,1 ml) foram administradas 30 minutos antes
da aplicação da TENS de BF, por injeção intraplantar na pata direita traseira dos
animais. A TENS de BF foi aplicada por 20 minutos e a serotonina, injetada no
tempo zero (30 minutos após início da aplicação da TENS). A pata contralateral não
recebeu tratamento. Cada ponto indica média ± EPM de 6 animais/grupo. * indica
uma diferença estatisticamente significativa (p < 0,05).
Figura 5 - Duração do efeito analgésico da TENS de BF. Grupos com 4 animais
cada receberam injeção intradérmica plantar de serotonina (10µg) nos
diferentes tempos após aplicação da TENS de BF. As medidas da latência de
retirada da pata foram realizadas sempre 5 minutos após a injeção de 5-HT,
nos tempos indicados após a aplicação da TENS na pata direita. A pata
contralateral não recebeu tratamento. * indica uma diferença estatisticamente
significativa (p < 0,05).

75
Capítulo 5- CONSIDERAÇÕES FINAIS
A TENS é um recurso não farmacológico, não invasivo, sem efeitos colaterais,
com poucas contra-indicações e comum na prática clínica do fisioterapeuta.
Clinicamente a TENS é raramente administrada de forma isolada. Combinações de
tratamentos farmacológicos e não-farmacológicos para controle da dor são
freqüentes. O entendimento do mecanismo de ação deste recurso permite aos
profissionais que o utilizam serem capazes de intensificar os efeitos de diversos
tipos de analgésicos e reduzir os efeitos colaterais dos agentes farmacológicos
produzindo um melhor alívio da dor.
Nossos resultados demonstraram uma completa inibição do efeito nociceptivo
da serotonina pela TENS de baixa (10 Hz), mas não pela de alta (130 Hz)
frequência. No entanto, nenhuma das duas freqüências foi capaz de inibir o edema
induzido pela 5-HT, sugerindo que a TENS apresenta eficácia decorrente de ação
analgésica e não anti-inflamatória. Nossos dados sugerem a participação opióide
local na analgesia da TENS de BF também em modelo de nocicepção induzido por
serotonina.
Uma vez que a TENS de BF inibe a nocicepção induzida por serotonina
através de seu efeito analgésico local, clinicamente ela pode ser utilizada para tratar
dor de origem periférica de forma mais eficaz.
Finalmente, esperamos que o nosso estudo tenha contribuído para uma maior
compreensão dos mecanismos analgésicos da TENS e assim tornar fundamentado
seu uso no tratamento da dor inflamatória aguda e crônica.

76
ANEXO 1: Aprovação do estudo pelo Comitê de Ética e m Experimentação Animal da UFMG


















