
Estudo e Caracterização da Herpetofauna nas áreas da Duna ... · revisão sistematizada em...
61
Estudo e Caracterização da Herpetofauna nas áreas da Duna da Cresmina, Quinta da Peninha e Quinta do Pisão. Diogo Rama da Silva 2014 - 2015
Transcript of Estudo e Caracterização da Herpetofauna nas áreas da Duna ... · revisão sistematizada em...

Estudo e Caracterização da Herpetofauna nas áreas da Duna da
Cresmina, Quinta da Peninha e Quinta do Pisão.
Diogo Rama da Silva
2014 - 2015


Resumo
Este trabalho teve como objectivo a realização de um estudo de levantamento da
herpetofauna, anfíbios e répteis, com ênfase à riqueza, diversidade e abundância, em
quatro áreas do Parque Natural Sintra-Cascais (PNSC) sob a alçada da Cascais
Ambiente, marca dos serviços prestados pela EMAC – Empresa Municipal de Ambiente
de Cascais, E.M., S.A.: Quinta do Pisão, Quinta da Peninha, Pisão de Baixo e Duna da
Cresmina, num total de aproximadamente 506 hectares.
A amostragem foi realizada entre Outubro de 2014 a Junho de 2015, tendo os
dados sido recolhidos sobre presença e abundâncias, transectos e estações fixas, com
recolha de dados sobre presença e abundâncias, com periodicidade quinzenal. No total
foram prospectadas 19 espécies, das quais 8 são espécies de anfíbios e 11 são espécies
de répteis.
De acordo com os métodos utilizados observou-se que Pelophylax perezi,
Podarcis hispanicus e Psammodromus algirus, uma espécie de anfíbio e duas de
répteis, foram as únicas espécies encontradas em todas as áreas de estudo. Foram
confirmadas as presenças de Discoglossus galganoi e de Acanthodactylus erythrurus,
ambas as espécies, de anfíbio e réptil respectivamente, categorizadas com o estatuto de
Quase Ameaçado – NT pelo Instituto da Conservação da Natureza e das Florestas
(ICNF), espécies estas que apesar de actualmente não serem consideradas como
espécies ameaçadas podem vir a serem-lhes atribuídas categorias de ameaça num futuro
próximo.
Em termos de riqueza específica, as áreas que apresentaram maior número de
espécies foram a Quinta do Pisão e a Peninha, ambas com 13 espécies no total e ambas
com 7 espécies de anfíbios e 6 espécies de répteis. A área com menor riqueza específica
foi a Duna da Cresmina com 5 espécies observadas, 2 de anfíbios e 3 de répteis. No
entanto, foi também nesta área onde ocorreu o maior número de registos para uma única
espécie, Pelodytes punctatus, com 9374 indivíduos observados apenas entre a segunda
quinzena de Outubro e a primeira quinzena de Dezembro. O menor número de registos
ocorreu na área do Pisão de Baixo, para as espécies Natrix maura e Chalcides striatus,
com registo de apenas 1 indivíduo. Os índices de diversidade (H’) e equitabilidade (E)
foram relativamente baixos quando comparados com outros autores, o que sugere
dominância de uma ou mais espécies.
A selecção e utilização de novos programas turísticos de observação da
herpetofauna e a protecção de locais específicos deverá ter em conta os resultados
expostos para um plano de gestão dos recursos na Cascais Ambiente.

Introdução 1 Metodologia 5
1. Áreas de estudo 5
2. Metodologia de campo 7
2.1. Amostragem 7 2.2. Amostragem por varrimento 8 2.3. Amostragem por estações fixas 9
3. Identificação das espécies 11
4. Tratamento dos dados 11
Resultados 13
1. Abordagem geral 13
2. Variação espacial 14 Riqueza específica 14 Diversidade e Equitabilidade 17 Similaridade 17
3. Variação Temporal 18
Riqueza específica 18 Diversidade e Equitabilidade 21
4. Curva de acumulação de espécies 22
5. Curva de elevação 23
Discussão e Conclusão 24 Considerações finais 28 Bibliografia 30 Anexos 34
Anexo 1 - Áreas e locais prospectados 35
Anexo 2 - Biologia das espécies observadas 39
Anfíbios 39
Répteis 47

1
Introdução
O estudo da herpetologia (ramo da zoologia dedicado ao estudo dos répteis e
anfíbios) em Portugal começou em 1864 quando Barbosa du Bocage descreveu uma
nova espécie para a ciência que designou por Chioglossa lusitanica ou salamandra-
lusitânica, como veio a ficar conhecida; foi uma descoberta que teve um grande impacto
junto de naturalistas de diferentes países (Almeida et al., 2001). No entanto, nas dezenas
de anos que se seguiram, existiram poucos registos além de algumas colecções de
museus, realizadas por Paulino de Oliveira (1931) e António Themido (1942), por
exemplo.
Só já nos anos setenta do séc. XX é que todo o material existente sofreria uma
revisão sistematizada em Portugal, nomeadamente com os trabalhos de Crespo (1971,
1972 e 1975) e Malkmus (1979, 1982, 1983, 1984, 1985a, 1985b). É durante os anos
oitenta que muita informação sobre a herpetofauna é produzida em maior escala tendo
como exemplo o lançamento do Atlas de Distribuição dos Anfíbios e Répteis de
Portugal (Crespo e Oliveira, 1989). Pouco depois é publicado o Primeiro Livro
Vermelho dos Vertebrados de Portugal (SNPRCN, 1990) onde é atribuído um estatuto
de conservação a todas as espécies consideradas ameaçadas de extinção.
Desde então, o estudo da herpetologia em Portugal tem evoluído
consideravelmente a nível nacional, repercutindo-se este facto na publicação de
inúmeros trabalhos, dos quais se destacam alguns no âmbito do presente estudo, como
os de Malkmus (1995) e de Godinho et al. (1999) para a formação de atlas de
distribuição e inventariação da herpetofauna regionais, os de Marques (1995) e de Brito
et al. (1996) para estudos de conservação ou ainda outros trabalhos dirigidos a uma
única espécie tais como os de Maymonte (1997), de Brito et al. (1998) e de Brito (2005)
para o lagarto-de-água (Lacerta schreiberi), ou os de Crespo e Sampaio (1994) e de
Brito e Álvares (1996) para a víbora-cornuda (Vipera latastei).
Recentemente foi lançada uma nova edição do Livro Vermelho dos Vertebrados
de Portugal (Cabral et al., 2006). Foi também realizada uma nova revisão ao nível de
classificação das espécies, passando as espécies a serem caracterizadas de acordo com o
seu código genético (ADN) ao invés da análise morfológica, o que veio alterar alguns
dos epítetos genéricos de algumas espécies. Posto isto, muitas espécies foram
posteriormente reclassificadas mantendo, no entanto, os respectivos epítetos específicos
(Speybroeck & Crochet, 2007).
Os anfíbios são componentes integrais de muitos ecossistemas, consistindo
muitas vezes na maior parte da biomassa dos vertebrados (Burton e Likens, 1975). Este
facto, aliado à sua elevada capacidade de digestão e eficiência de produção (Woolbright,
1991), contribui para a manutenção dos ecossistemas em termos energéticos e de fluxo
de carbono (Pearman, 1997), através da manutenção da abundância de artrópodes
(Guyer, 1990) e também da provisão de presas, para cadeias mais complexas de
predadores (Guyer, 1990; Woolbright, 1991 e Duellman e Trueb, 1994).

2
Para além da sua importância para os ecossistemas e de acordo com alguns
autores, os anfíbios têm certas propriedades que os tornam bastante úteis como
bioindicadores ambientais (e.g. Wake e Morowitz, 1991). A sua pele, altamente
permeável a gases e líquidos, resulta numa elevada exposição dos indivíduos a certos
materiais presentes na água, ar e solo, o que os torna sensíveis a alterações no meio
aquático, bem como no meio terrestre (Marques, 1995). Aumentos súbitos na
temperatura ou humidade impele-os a emergir da hibernação e a migrar para planos de
água próximos para se reproduzirem (Araújo et al., 2006). Logo, um possível efeito do
aquecimento global nos anfíbios é uma tendência para uma época reprodutiva cada vez
mais antecipada, à medida que a temperatura média aumenta, apesar de permanecer
incerto qual o impacto que este padrão possa ter na dinâmica populacional (Araújo et
al., 2006).
Por outras palavras, mudanças na temperatura ambiente à escala mundial podem
potencialmente quebrar com os períodos de reprodução e de hibernação e com a
habilidade para encontrar alimento (Blaustein et al., 2001). Por outro lado, a selecção
natural deve favorecer rápidas mudanças nas datas de desova porque uma reprodução
mais antecipada é considerada vantajosa, possibilitando um maior tempo de
desenvolvimento para os juvenis e mais tempo para acumular reservas energéticas antes
da hibernação (Tryjanowski et al., 2003).
Tal como parâmetros relacionados com a temperatura são importantes para
explicar a distribuição das espécies de répteis, também a humidade e a cobertura vegetal
desempenham um papel semelhante para os anfíbios, sendo, no entanto, manifestado
num padrão complexo (Gasc et al., 2004).
Para além destas alterações na sua dinâmica reprodutiva, surge agora uma nova
ameaça que parece empurrar os anfíbios para a extinção, causando um declínio de
populações à escala mundial. Esta ameaça é o Batrachochytrium dendrobatidis
(Longore, Pessier & Nichols DK, 1999), um fungo que causa a quitridiomicose, a
doença que tem vindo a dizimar populações inteiras de anfíbios por todo o mundo. Este
fungo ataca a pele, órgão que é essencial para os anfíbios, sendo então disseminado com
extrema rapidez e facilidade (Negrão, 2009).
Alguns cientistas afirmam que quando o fungo chega a um lugar novo, ele
espalha-se como um incêndio (Lips, 2008). Ainda não se sabe ao certo de onde surgiu
nem como conter o seu crescimento, contudo acredita-se que as mudanças climáticas e
outros factores ambientais estejam a favorecer a proliferação do fungo e os cientistas
correm agora contra o tempo na tentativa de encontrar uma solução para controlar e
exterminar esse problema, que pode extinguir populações inteiras de anfíbios e,
consequentemente, causar um desequilíbrio ecológico irreparável (Negrão, 2009). O
motivo pelo qual as populações de anfíbios sucumbem às infecções por quitrídeos é
ainda desconhecido (Negrão, 2009). No entanto, sabe-se que mudanças inesperadas no
clima podem afectar as relações entre hospedeiros e parasitas, de várias formas,
influenciando um aumento da mortalidade (Juncá, 2001).

3
Devido ao facto de os anfíbios serem componentes chave de muitos
ecossistemas (Burton e Likens, 1975 e Blaustein et al., 2001), mudanças nas suas
populações podem afectar outras espécies dentro das suas comunidades, tais como os
seus predadores (répteis, aves e mamíferos) e presas, mesmo que essas espécies não
tenham sido afectadas por alterações climáticas (Blaustein et al., 2001). Assim, um
declínio nas populações de anfíbios à escala mundial pode ter um impacto prejudicial e
significativo nos ecossistemas e no bem-estar humano (Gardner, 2001).
Em geral, factores como destruição ou modificação de habitat são facilmente
identificáveis como a causa responsável pela perda da biodiversidade local, e como tal,
estas reduções na biodiversidade são frequentemente observadas ao longo da taxonomia
(Gardner, 2001). Além do mais a região Ibérica que engloba Espanha, Portugal e o sul
de França, é considerada uma zona quente da Europa e pensa-se que nesta região a
influência do clima para a distribuição das espécies é secundária, pelo que esta
distribuição ocorre em grande maioria graças a processos históricos, tais como a
diferenciação de espécies nas penínsulas mediterrânicas, que resultam na presença de
espécies endémicas na região, como por exemplo a espécie L. schreiberi (Gasc et al.,
2004).
Ao tomarmos conhecimento dos padrões de distribuição da biodiversidade em
pequenas escalas, subcontinentais a continentais, torna-se mais fácil compreender os
requisitos ecológicos gerais das espécies e das suas comunidades (Omolo, 2006). A
previsão das potenciais distribuições, em locais sem amostragem, pode levar à
descoberta das espécies de répteis e de anfíbios, ou então revelar factores que possam
explicar a sua ausência (Omolo, 2006).
De acordo com Marques (1995) estão presentes no PNSC cerca de 71% das
espécies de anfíbios e 77% das espécies de répteis que existem em território nacional,
confirmando a importância desta zona em termos de diversidade e riqueza. Esta área
apresenta características particulares contendo zonas bastante diversificadas, o que é
reflectido nas preferências demonstradas pelas espécies da herpetofauna que o habitam
(Marques, 1995).
No entanto, a perda extensa de diversidade biológica e as alterações na
distribuição dos organismos (anfíbios e répteis) mostram uma variância considerável
tanto no que respeita à localização geográfica, bem como às características taxonómicas
e ecológicas das espécies envolvidas (Gardner, 2001), o que torna importante a
realização de estudos de inventariação à escala local.
Além disto, as áreas nas quais este estudo foi realizado têm sofrido um aumento
crescente de pressão por parte dos turistas e por algumas intervenções e práticas de
gestão dos habitats que podem ter influenciado as comunidades herpetológicas nos
últimos 30 anos. Face ao exposto, torna-se necessário actualizar os conhecimentos sobre
a ocorrência da herpetofauna nestes locais e a presença/ausência de espécies
importantes para a conservação, como a rã-de-focinho-pontiagudo (Discoglossus
galganoi), o lagarto-de-água (Lacerta schreiberi), a lagartixa-de-dedos-denteados

4
(Acanthodactylus erythrurus), a cobra-de-capuz (Macroprotodon cucullatus) e a víbora-
cornuda (Vipera latastei).
Deste modo, o objectivo principal deste estudo foi o de estudar a distribuição e a
abundância da herpetofauna nas áreas do PNSC sob gestão da Cascais Ambiente. Estas
áreas de estudo constituem apenas uma pequena porção do PNSC e a actualização do
conhecimento sobre estas espécies poderá ser útil para:
A tomada de decisões sobre práticas de gestão através da realização de planos de
gestão de habitats;
A divulgação de uma forma positiva dos aspectos da biologia, importância e
conservação das espécies da herpetofauna aos visitantes destas áreas e,
consequentemente, à sensibilização dos mesmos.

5
Metodologia 1. Áreas de Estudo
As áreas onde este estudo foi realizado localizam-se no PNSC, situado a
noroeste da cidade de Lisboa. Três das áreas, a Quinta do Pisão, o Pisão de Baixo e a Duna da Cresmina, encontram-se a Sul da serra de Sintra, enquanto a última, a Quinta da Peninha, encontra-se em plena serra, na parte mais ocidental da mesma.
A serra de Sintra, complexo montanhoso que domina o PNSC, é uma área com aproximadamente 10km de extensão por 5km de largura, com um perímetro de cerca de 30km e cuja altitude varia entre os 150 e os 529 metros. De acordo com Fabião (1999), esta área pode ser divida em duas regiões, a região ocidental e a central/oriental. A região ocidental apresenta um coberto vegetal maioritariamente arbustivo, dominado sobretudo por carrasco (Quercus coccifera), sargaço (Cistus monspeliensis), roselha (C. crispus), tojo (Ulex densus), murta (Myrtus communis) e rosmaninho (Lavandula pedunculata) (Fabião, 1999). Ocorrem igualmente acácias (Acacia melanoxylon e A. longifolia, principalmente), espécies exóticas infestantes que se estabelecem após a passagem dos incêndios (Fabião, 1999). A região central/oriental apresenta um coberto vegetal predominantemente arbóreo, dominado em grande parte pelo pinheiro bravo (Pinus pinaster), alternando muitas vezes com o eucalipto (Eucalyptus globulus). Existe ainda a presença de carvalho-negral (Quercus pyrenaica), vidoeiro (Betula celtiberica) e teixo (Taxus baccata) (Fabião, 1999).
Figura 1 – Localização das áreas prospectadas (fonte: Google Maps).

6
Nota: As delimitações das áreas na figura 1 não correspondem às suas verdadeiras
fronteiras, servem apenas para dar a entender a sua localização geográfica.
As áreas prospectadas foram as seguintes:
Área 1 (A1) – Quinta do Pisão;
Área 2 (A2) – Quinta da Peninha;
Área 3 (A3) – Pisão de Baixo;
Área 4 (A4) – Duna da Cresmina.
O Pisão de Baixo (área 3) encontra-se inserido na Quinta do Pisão (área 1). No
entanto, como estas duas áreas se encontram separadas fisicamente pela estrada nacional
EN 247-5 torna-se necessário, para efeitos do presente estudo da herpetofauna,
considerá-las como duas áreas distintas.
A área 1, a Quinta do Pisão, está situada a norte do concelho de Cascais e
totalmente inserida no Parque Natural Sintra-Cascais. É uma área com cerca de 355 ha,
dedicada maioritariamente a actividades agro-silvo-pastoris e que, ao encontrar-se numa
zona de transição geológica, apresenta uma ampla heterogeneidade de terreno que,
como tal, compreende uma grande variedade de habitats e nichos ecológicos da zona,
incluindo espécies de fauna e flora de elevado interesse conservacionista. (In:
http://www.cm-cascais.pt/quintadopisao). Está compreendida, aproximadamente, entre
os 100 e os 180m de elevação, apresentando um relevo suave nos planaltos e acentuado
nas dobras de terreno que confinam com ribeiras e linhas de água existentes, com
declives de aproximadamente 35%. É predominantemente dominada pelas áreas agro-
pastoris e as restante por mato, dominado na sua maioria por carrasco (Quercus
coccifera) e floresta, dominada sobretudo por eucalipto (Eucalyptus globulus) e
pinheiro-bravo (Pinus pinaster), com presença de pinheiro-de-Alepo (Pinus halepensis),
sobreiro (Quercus suber), carvalho-cerquinho (Quercus faginea) e medronheiro
(Arbutus unedo), a Norte, e por zambujeiro (Olea europaea) e azinheira (Quercus
rotundifolia) a Sul. Estão registadas várias espécies exóticas infestantes como, por
exemplo, as acácias (Acacia dealbata, A. longifolia, A. mearnsii, A. melanoxylon e A.
metinodes).
A Quinta da Peninha (área 2) compreende uma área com cerca de 60 hectares e
está localizada imediatamente a Sul, adjacente ao Santuário da Peninha. Está
compreendida entre os 340 e os 500m de elevação, aproximadamente. É composta, na
sua maioria, por vegetação rasteira e arbustiva, dominada por giesta (Cytisus striatus),
tojo (Ulex sp.), silvado (Rubus ulmifolius) e conta com algumas manchas de extracto
arbóreo, como eucalipto (Eucalyptos globulus), carvalho-negral (Quercus pyrenaica),
pinheiro (Pinus pinaster), sobreiro (Quercus suber) e cedro-do-buçaco (Cupressus
lusitanica). Destaca-se ainda a presença do feto-de-folha-de-hera (Asplenium
hemionitis), espécie relíquia da floresta subtropical húmida, anterior às glaciações. (In:
http://www.icnf.pt/portal/turnatur/resource/docs-ap/pnsc/pr10-snt-peninh). Na área da
Peninha também se encontram espécies exóticas infestantes, na sua maioria acácias.

7
Localizada a sudoeste da Quinta do Pisão encontra-se o Pisão de Baixo (área 3),
uma área predominantemente de porte arbustivo, dominada pelo carrasco (Quercus
coccifera), com ocorrência de sanguinho-das-sebes (Rhamnus alaternus), tojo (Ulex
sp.), murta (Myrtus communis) e eufórbia (Euphorbia characias). Existe também uma
mancha de pinheiros-de-Alepo (Pinus halepensis) e junto à linha de água, salienta-se a
dominância de silvado (Rubus ulmifolius). Esta área encontra-se sensivelmente entre os
50 e os 100m de elevação.
A Duna da Cresmina (área 4) está localizada junto à faixa costeira, entre as praias da
Cresmina e do Guincho. É uma área com aproximadamente 66 hectares, de substrato
predominantemente arenoso e coberto, na sua maioria, por plantas rasteiras e arbustivas,
como por exemplo o estorno (Ammophila arenaria ssp. arundinacea), o cardo-marítimo
(Eryngium maritimum), a morganheira-das-praias (Euphorbia paralias), o narciso-do-
mar (Pancratium maritimum) e a sabina-das-praias (Juniperus turbinata), e com pouca
ocorrência de extracto arbóreo, maioritariamente de pinheiro-bravo (Pinus pinaster).
Destacam-se ainda algumas espécies endémicas e importantes para a conservação, como
o verbasco-de-flores-grossas (Verbascum litigiosum), a cocleária-menor (Jonopsidium
acaule) e o miosótis-das-praias (Omphalodes kuzinskyanae). Por ser uma zona bastante
exposta, os seus habitats naturais apresentam um delicado equilíbrio ecológico, segundo
um gradiente de solo pobre (areias) e condições climatéricas adversas (ventos fortes
carregados de sal). Para além disso, graças à elevada pressão humana no local, essa
vegetação está muito fragmentada e alterada devido à introdução de espécies sem
interesse conservacionista (In: http://www.cm-cascais.pt/dunadacresmina). Esta área
encontra-se entre os 25 e os 40m de elevação.
2. Metodologia de campo
2.1 Amostragem
Foi feito um reconhecimento inicial às áreas de estudo, mais concretamente para aferir os locais que reunissem as melhores condições para a ocorrência da herpetofauna (tabela 1).
Prospecção
Locais Sigla Nº
Tanque Tq 5
Linha de água Li 3
Transecto T 18
Lagoa L 3
Poço Po 1
Ponto Pt 8
Tabela 1 – Tipo e número de locais prospectados nas áreas de estudo.

8
Área Locais Sigla
Pen
inh
a (á
rea
2)
Transecto 1 T1
Transecto 2 T2
Transecto 3 T3
Transecto 5 T5
Transecto 4 T4
Tanque 3 Tq3
Tanque 2 Tq2
Tanque 1 Tq1
Tabelas 2 e 3 – Locais de amostragem nas áreas 1 e 2.
Tabelas 4 e 5 – Locais de amostragem nas áreas 3 e 4. Para ter uma ideia mais precisa
das suas localizações ver imagens em anexo (anexo 1).
A amostragem de campo foi realizada quinzenalmente, de Outubro de 2014 a
Junho de 2015, com recurso a técnicas directas como prospecção visual, auditiva e
intensiva nos locais escolhidos (Silva, 2007). Para tal, utilizaram-se diferentes tipos de
amostragem, para adultos e larvas, devido em grande parte às especificidades dos
hábitos e habitats das fases larvar e terrestre (Sutherland, 1996).
A amostragem foi idealizada de duas formas: amostragem por varrimento e
amostragem por estações fixas (Marques, 1995):
2.2 Amostragem por varrimento
Esta amostragem teve como propósito o de analisar a presença/ausência de
espécies com recurso a percursos pré-estabelecidos (transectos), tais como linhas e
planos de água e também percursos terrestres (Marques, 1995; Silva, 2007). Este tipo de
amostragem pode ser complementado com recurso a armadilhas tipo pitfall.
Área Locais Sigla
Pis
ão (
área
1)
Tanque 1 Tq1
Linha de água 1 Li1
Transecto 1 T1
Transecto 2 T2
Transecto 3 T3
Transecto 4 T4
Tanque 2 Tq2
Lagoa Grande LG
Transecto 5 T5
Lagoa Pequena LP
Transecto 6 T6
Transecto 7 T7
Poço Po
Lagoa Temporária LT
Linha de água 1/ Transecto 8 Li1/T8
Área Locais Sigla D
un
a C
resm
ina
(áre
a 4
) Ponto 1 Pt1
Ponto 2 Pt2
Ponto 3 Pt3
Ponto 4 Pt4
Ponto 5 Pt5
Ponto 6 Pt6
Ponto 7 Pt7
Ponto 8 Pt8
Área Locais Sigla
Pis
ão d
e B
aixo
(á
rea
3)
Transecto 4 T4
Transecto 3 T3
Transecto 2 T2
Transecto 1 T1
Linha de água/ Transecto 5 Li/T5

9
Amostragem terrestre
Este método envolve a marcação de uma linha imaginária sobre uma dada área,
ao longo da qual os investigadores caminham em intervalos regulares, procurando
sistematicamente por animais (Sutherland, 1996), tendo particular atenção para locais
potenciais de abrigo. Foram utilizados percursos pré-estabelecidos, com um mínimo de
100m de comprimento, recorrendo-se sobretudo à prospecção visual e auditiva, feitos a
uma velocidade média de 2m/s. A extensão dos percursos variou entre áreas e a largura
dos transectos variou consoante o transecto, com um mínimo de 1,5 a 15m. Realizada
em todas as áreas com excepção da área 4.
Figuras 2 e 3 – Transectos T1 e T3 nas áreas 1 e 2, respectivamente.
Amostragem em linha de água
Em tudo semelhante à amostragem terrestre, com excepção para o recurso a
netting (camaroeiro), de modo a recolher alguns dos indivíduos observados e confirmar
a espécie, o estado de desenvolvimento e sexo, se possível. Embora o ideal seja abordar
sempre que possível ambas as margens, tal não foi possível devido à vegetação e ao
terreno. Foram realizadas apenas nas áreas 1 e 3.
Figuras 4 e 5 – Linhas de água Li1 e Li/T5 nas áreas 1 e 3, respectivamente.
2.3 Amostragem por estações fixas
Esta amostragem compreende a prospecção de locais pré-estabelecidos com
elevado potencial para ocorrência de anfíbios, tais como abrigos terrestres e planos de
água, onde foi feita a recolha, registo e posterior reposição de todos os indivíduos
encontrados (Silva, 2007). Este tipo de amostragem visa obter uma estimativa da
abundância das espécies e uma noção da densidade populacional (Marques, 1995),

10
recorrendo também aos dados da amostragem por varrimento (Silva, 2007). No caso da
área 4, cada um dos pontos foi prospectado num raio de 5 a 10m.
Figuras 6 e 7 – Estações fixas Tq2 e Pt3 nas áreas 1 e 4, respectivamente.
Amostragem em planos de água
Neste caso a amostragem dos diferentes planos de água foi realizada por
prospecção visual, auditiva e intensiva com recurso a camaroeiro (netting) (Sutherland,
1996). Neste tipo de abordagem o recurso ao camaroeiro tem como objectivo apanhar o
máximo de animais possíveis (Sutherland, 1996). Todos os esforços foram feitos para
amostrar todas as partes do plano de água de igual modo, tendo no entanto o cuidado de
não danificar a vegetação aquática que serve de abrigo e local de postura para algumas
espécies de anfíbios (Sutherland, 1996).
Figuras 8 e 9 – Planos de água LP e Tq2 nas áreas 1 e 2, respectivamente.
Realizaram-se também amostragens complementares, nomeadamente as
observações ad-hoc, ou seja, observação de espécies fora dos locais e percursos pré-
definidos, mas dentro das áreas de estudo.
Ao longo do esforço de amostragem, no decurso do trabalho de campo, foram
tidas em conta algumas limitações, nomeadamente ao nível das estações fixas, quanto
ao tempo de permanência em cada local, a área a amostrar (só as margens ou todo o
local) e também a frequência da utilização de varrimento ou netting (camaroeiro),
quando confrontado com um local com poucos ou nenhuns indivíduos (Silva, 2007).

11
3. Identificação das espécies
A identificação das espécies foi feita essencialmente in loco com recurso a guias
de campo (Almeida et al., 2001 e Ovenden e Arnold, 2004). Contudo, houve alguns
casos, nomeadamente de larvas de anfíbios, cujas identificações foram confirmadas com
recurso ao guia fotográfico Caldas (2010).
4. Tratamento dos dados
Os dados recolhidos foram colocados numa base de dados em Excel 2007 e
tratados estatisticamente, com o objectivo de determinar diversos parâmetros
populacionais e de comunidade:
Riqueza específica – nº de espécies numa comunidade ou amostra (Olszewski,
2004);
Frequência numérica – Pi:
Frequência de ocorrência:
Diversidade – calculada através do Índice de Shannon-Wienner (Krebs, 1989),
com recurso ao programa informático “Species Diversity & Richness IV”:
H’= i
S
ii pp ln
1
Equitabilidade – medida da distribuição das abundâncias relativas de diferentes
taxa numa comunidade ou amostra, ou uma medida que reflecte a uniformidade
das abundâncias relativas (Olszewski, 2004):
E=max
'
H
H ; Hmax = ln S ; S= riqueza específica

12
Índice de similaridade qualitativo entre áreas através do Coeficiente de Sörensen
(Krebs, 1989):
em que a= nº de espécies na área 1, b= nº espécies na amostra 2 e c= nº de espécies
comuns.
Todos os parâmetros foram analisados de acordo com padrões espaciais e
temporais, em cada uma das áreas de estudo.
Cs= 1002
ba
c

13
Resultados
1. Abordagem Geral
No total deste estudo foram feitas 16.658 observações (tabela 6), das quais
16.213 correspondem aos dados de anfíbios (97,33%), para os três estados de
desenvolvimento (larva, juvenil e adulto) e 445 correspondem aos dados de répteis
(2,67%), para juvenis e adultos.
Anfíbios Nº de
observações
Répteis Nº de
observações
Bufo bufo 1501
Acanthodactylus erythrurus
7
Discoglossus galganoi 3060
Chalcides striatus 1
Lissotriton boscai 129
Hemorrhois hippocrepis 3
Pelodytes punctatus 9374
Lacerta lepida 3
Pelophylax perezi 858
Lacerta schreiberi 4
Pleurodeles waltl 33
Malpolon monspessulanus 2
Salamandra salamandra
1004
Natrix maura 1
Triturus pygmaeus 255
Natrix natrix 4
Total 16214
Podarcis hispanicus 58
Psammodromus algirus 360
Tarentola mauritanica 2
Total de observações 16659
Total 445
Tabela 6 - Observações de anfíbios e répteis no total do período de estudo.
A espécie dos anfíbios que teve o maior número de ocorrências foi Pelodytes
punctatus (sapinho-de-verrugas-verdes), o que equivale a 58% de todas as ocorrências
registadas (tabela 7). Como tal, foi esta a espécie com maior número de registos
observados, com 9374 de um total de 16659 observações, o que corresponde a
aproximadamente 56%. A particularidade é de esta espécie ter sido registada somente
na área 4 (Duna da Cresmina). A espécie Discoglossus galganoi (rã-de-focinho-
pontiagudo) também obteve bons registos, com 3060 observações (19%). Foram
observados indivíduos em todas as fases de desenvolvimento, com excepção das
espécies Pleurodeles waltl (salamandra-de-costelas-salientes) e Triturus pygmaeus
(tritão-pigmeu), para as quais não se observaram juvenis.
Nos répteis o destaque vai para a espécie Psammodromus algirus (lagartixa-do-
mato), com cerca de 80% (tabela 7) dos registos observados para todos os répteis.
Contudo, esta fatia não foi além dos 2,2%, aproximadamente, para o total das
observações. Esta espécie foi também uma, de um total de quatro, para as quais foram
observadas todas as fases de desenvolvimento (juvenil e adulto); as outras três espécies
foram Acanthodactylus erythrurus (lagartixa-de-dedos-dentados), Hemorrhois
hippocrepis (cobra-de-ferradura) e Podarcis hispanicus (lagartixa-ibérica). O menor

14
número de registos ocorreu na área 3, para as espécies Natrix maura (cobra-de-água-
viperina) e Chalcides striatus (cobra-de-pernas-tridáctila), com registo de apenas 1
indivíduo.
O anfíbio Pelophylax perezi (rã-verde) e os répteis Psammodromus algirus e
Podarcis hispanicus foram as únicas espécies observadas em todas as áreas de estudo.
Anfíbios Freq. oc.
Larvas Freq. oc. Juv Freq. oc. Ad
Freq. oc. Total
Bufo bufo 8,71 0,01 0,54 9,26
Discoglossus galganoi 18,42 0,03 0,43 18,88
Lissotriton boscai 0,27 0,03 0,51 0,81
Pelodytes punctatus 57,72 0,07 0,03 57,82
Pelophylax perezi 0,12 0,12 5,06 5,3
Pleurodeles waltl 0,19 0 0,01 0,2
Salamandra salamandra 5,51 0,22 0,46 6,19
Triturus pygmaeus 1,44 0 0,13 1,57
Répteis Freq. oc.
Juv Freq. oc. Ad
Freq. oc. Total
Acanthodactylus erythrurus 0,22 1,35 1,57
Chalcides striatus 0 0,22 0,22
Hemorrhois hippocrepis 0,22 0,45 0,67
Lacerta lepida 0,67 0 0,67
Lacerta schreiberi 0 0,89 0,89
Malpolon monspessulanus 0,45 0 0,45
Natrix maura 0,22 0 0,22
Natrix natrix 0 0,89 0,89
Podarcis hispanicus 0,89 12,14 13,03
Psammodoromus algirus 46,74 34,16 80,9
Tarentola mauritanica 0 0,45 0,45
Tabela 7 – Frequência de ocorrência das espécies (%) por estado de desenvolvimento, no conjunto das áreas, durante todo o período de estudo.
2. Variação Espacial
Riqueza específica (S)
No total foram contabilizadas 19 espécies de herpetofauna, 8 espécies de
anfíbios e 11 espécies de répteis, o que corresponde a 66,7% (12 anfíbios) e 55% (20
répteis) das espécies de herpetofauna do PNSC. Nas figuras abaixo estão representadas
a riqueza específica total para o estudo e a riqueza específica obtida para cada uma das
áreas.
15
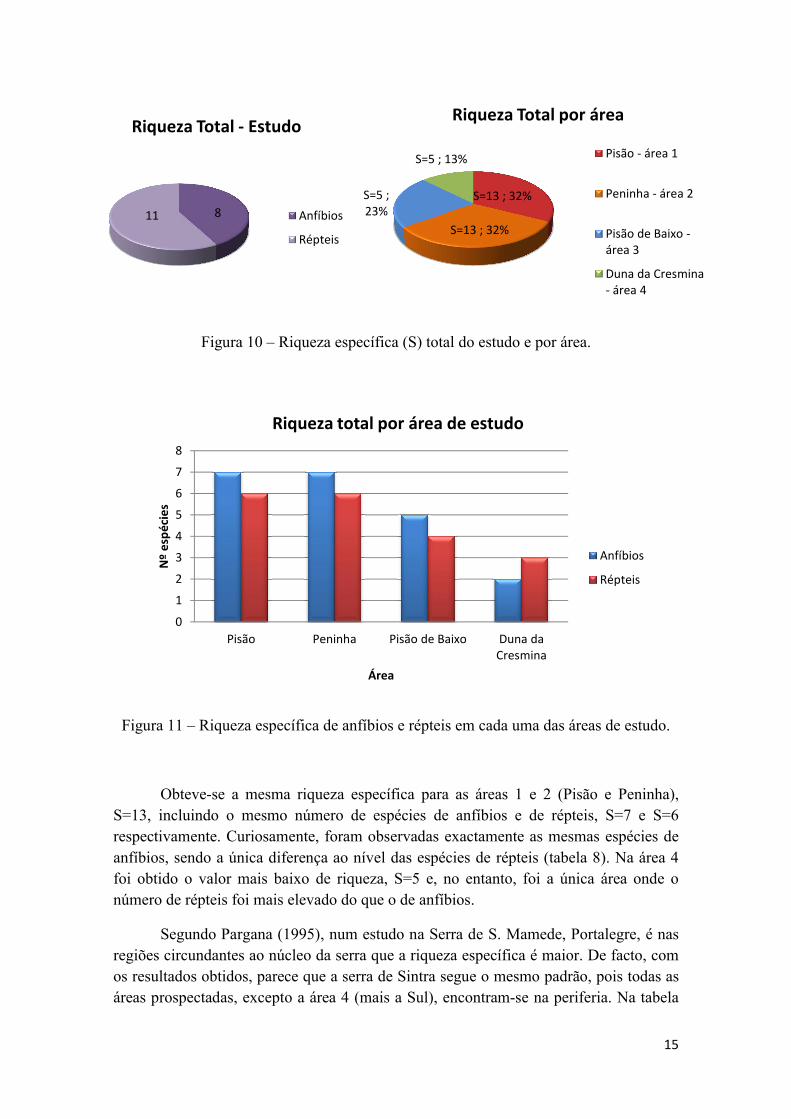
Figura 10 – Riqueza específica (S) total do estudo e por área.
Figura 11 – Riqueza específica de anfíbios e répteis em cada uma das áreas de estudo.
Obteve-se a mesma riqueza específica para as áreas 1 e 2 (Pisão e Peninha),
S=13, incluindo o mesmo número de espécies de anfíbios e de répteis, S=7 e S=6
respectivamente. Curiosamente, foram observadas exactamente as mesmas espécies de
anfíbios, sendo a única diferença ao nível das espécies de répteis (tabela 8). Na área 4
foi obtido o valor mais baixo de riqueza, S=5 e, no entanto, foi a única área onde o
número de répteis foi mais elevado do que o de anfíbios.
Segundo Pargana (1995), num estudo na Serra de S. Mamede, Portalegre, é nas
regiões circundantes ao núcleo da serra que a riqueza específica é maior. De facto, com
os resultados obtidos, parece que a serra de Sintra segue o mesmo padrão, pois todas as
áreas prospectadas, excepto a área 4 (mais a Sul), encontram-se na periferia. Na tabela
811
Riqueza Total - Estudo
Anfíbios
Répteis
S=13 ; 32%
S=13 ; 32%
S=5 ; 23%
S=5 ; 13%
Riqueza Total por área
Pisão - área 1
Peninha - área 2
Pisão de Baixo -área 3
Duna da Cresmina- área 4
0
1
2
3
4
5
6
7
8
Pisão Peninha Pisão de Baixo Duna daCresmina
Nº
esp
éci
es
Área
Riqueza total por área de estudo
Anfíbios
Répteis

16
abaixo (tabela 8) estão assinaladas as riquezas específicas para cada uma das áreas e as
respectivas espécies observadas no decorrer do estudo.
Área 1 Anfíbios Répteis Área 2 Anfíbios Répteis
S=13
B. bufo H. hippocrepis
S=13
B. bufo L. lepida
D. galganoi M. monspessulanus D. galganoi L. schreiberi
L. boscai N. natrix L. boscai N. natrix
P. perezi P. hispanicus P. perezi P.
hispanicus
P. waltl P. algirus P. waltl P. algirus
S. salamandra T. mauritanica S. salamandra T.
mauritanica
T. pygmaeus T. pygmaeus
Área 3 Anfíbios Répteis Área 4 Anfíbios Répteis
S=9
B. bufo C. striatus
S=5
P. punctatus A. erythrurus
D. galganoi N. natrix P. perezi P. hispanicus
L. boscai P. hispanicus P. algirus
P. perezi P. algirus
S. salamandra
Tabela 8 – Riqueza específica para cada um das áreas e respectivas espécies observadas.
Foram igualmente estimados os valores de riqueza dos locais prospectados,
dentro de cada uma das áreas. O esperado seria que as áreas onde se obteve maior
número de espécies fossem aquelas onde se encontrassem os locais com maior riqueza,
o que de facto se veio a verificar. Em relação a estes locais (tabela 9), o que apresentou
maior número de riqueza foi o tanque 1 na área 2 (Peninha), com 11 espécies
observadas, sendo seguido de perto pelo tanque 1 na área 1 (Pisão), com 9 espécies. Os
locais com riqueza mais baixa foram o T1 na área 1 e o Pt2 na área 4 (Duna), nos quais
apenas foi observada uma espécie. O melhor transecto correspondeu ao T5 na área 2,
com 8 espécies observadas.
Área Locais Sigla Número S
Pis
ão
Tanque 1 Tq1 1 9
Linha de água 1 Li1 2 6
Área Locais Sigla Número S
Transecto 1 T1 3 1
Pen
inh
a
Transecto 1 T1 16 3
Transecto 2 T2 4 4
Transecto 2 T2 17 2
Transecto 3 T3 5 2
Transecto 3 T3 18 2
Transecto 4 T4 6 3
Transecto 5 T5 19 8
Tanque 2 Tq2 7 3
Transecto 4 T4 20 2
Lagoa Grande LG 8 4
Tanque 3 Tq3 21 4
Transecto 5 T5 9 3
Tanque 2 Tq2 22 7
Lagoa Pequena LP 10 3
Tanque 1 Tq1 23 11
Transecto 6 T6 11 5

17
Transecto 7 T7 12 3
Área Locais Sigla Número S
Poço Po 13 2
Du
na
Cre
smin
a
Ponto 1 Pt1 29 2
Lagoa Temporária LT 14 7
Ponto 2 Pt2 30 1
Linha de água 2/ transecto 8 Li2/T8 15 2
Ponto 3 Pt3 31 2
Ponto 4 Pt4 32 3
Área Locais Sigla Número S
Ponto 5 Pt5 33 2
Pis
ão d
e B
aixo
Transecto 4 T4 24 2
Ponto 6 Pt6 34 2
Transecto 3 T3 25 2
Ponto 7 Pt7 35 3
Transecto 2 T2 26 3
Ponto 8 Pt8 36 3
Transecto 1 T1 27 3
Linha de água/ Transecto 5 Li/T5 28 6
Tabela 9 – Riqueza específica (S) em cada um dos locais prospectados, em cada uma
das áreas de estudo.
Diversidade e Equitabilidade
Em termos genéricos as áreas com maior diversidade (H’) são aquelas que
apresentam também uma maior equitabilidade (E). No entanto não foi isto o verificado
como se pode comprovar pelos valores obtidos e expostos na tabela 8. A área com
maior diversidade foi a área 1, cujo valor de equitabilidade foi o segundo mais alto. A
maior equitabilidade registou-se na área 3. Isto pode querer dizer que, embora não
sendo a mais diversa das áreas, as espécies nela existentes estão estruturalmente
equilibradas. A área com os valores mais baixos para estes dois parâmetros foi a área 4,
o que sugere pouco equilíbrio das populações e/ou a dominância de uma ou mais
espécies.
Os valores obtidos para a diversidade foram relativamente baixos pois,
idealmente, estes costumam variar entre os 1,5 e os 3,5.
H' E
Área 1 1,383 0,539
Área 2 1,237 0,482
Área 3 1,225 0,558
Área 4 0,052 0,032
Tabela 8 – Valores da diversidade (H’) e equitabilidade (E) para cada uma das áreas de
estudo.
Similaridade
Foi testado o Coeficiente de Sörensen para averiguar a similaridade entre as
áreas estudadas, quer para a herpetofauna quer para o total do estudo. Os resultados
estão representados nas tabelas abaixo (tabelas 9, 10 e 11).

18
De acordo com os resultados, o conjunto de áreas mais semelhante entre si foi o
1 – 2, com aproximadamente 85% de similaridade. Os resultados para os anfíbios e
répteis no mesmo conjunto de áreas foram igualmente os mais altos, com valores de
100% para os anfíbios e cerca de 67% para os répteis. O valor de 100% para os anfíbios
justifica-se pelo que foi referido anteriormente em relação à riqueza específica pois,
para estas áreas, foram observadas exactamente as mesmas espécies de anfíbios.
Áreas Cs
A
nfí
bio
s 1 - 2 100
1 - 3 83,33
1 - 4 22,22
2 - 3 83,33
Áreas Cs
2 - 4 22,22
Tota
l
1 - 2 84,62
3 - 4 28,57
1 - 3 63,64
1 - 4 33,33
Áreas Cs
2 - 3 63,64
Ré
pte
is
1 - 2 66,67
2 - 4 33,33
1 - 3 60
3 - 4 42,86
1 - 4 44,44
2 - 3 40
2 - 4 44,44
3 - 4 57,14
Tabelas 9, 10 e 11 – Similaridade de anfíbios, répteis e total nas áreas de estudo.
Utilizando o mesmo raciocínio, o conjunto de áreas menos semelhante foi o 1 –
4 e o 2 – 4, com um valor de aproximadamente 33% no total, 22% para os anfíbios e
44% para os répteis. Era esperado que a área 4 fosse a que apresentasse maior número
de diferenças já que é a mais inóspita no que diz respeito a habitats, ou seja, todas as
outras áreas apresentam habitats mais variados, maior disponibilidade de abrigos e de
água e, por conseguinte, maior possibilidade para a ocorrência espécies de herpetofauna.
Por seu lado, a área 4 é uma área mais exposta e composta essencialmente por solo
arenoso, mais móvel face a intempéries, o que impede a água de se acumular com
facilidade, dificultando a observação deste tipo de espécies.
3. Variação Temporal
Riqueza específica
A riqueza específica foi estimada com o número de anfíbios e répteis, ao longo
do tempo de estudo (quinzena) e encontra-se sintetizada na figura 12, incluindo curvas
de tendência. Foram também estimadas as riquezas quinzenais para cada uma das áreas
de estudo, esboçadas mais abaixo na figura 13.
19
As curvas de tendência foram adicionadas de modo a verificar se a variação do
número de espécies de herpetofauna ao longo do ano estaria de acordo com a
bibliografia.
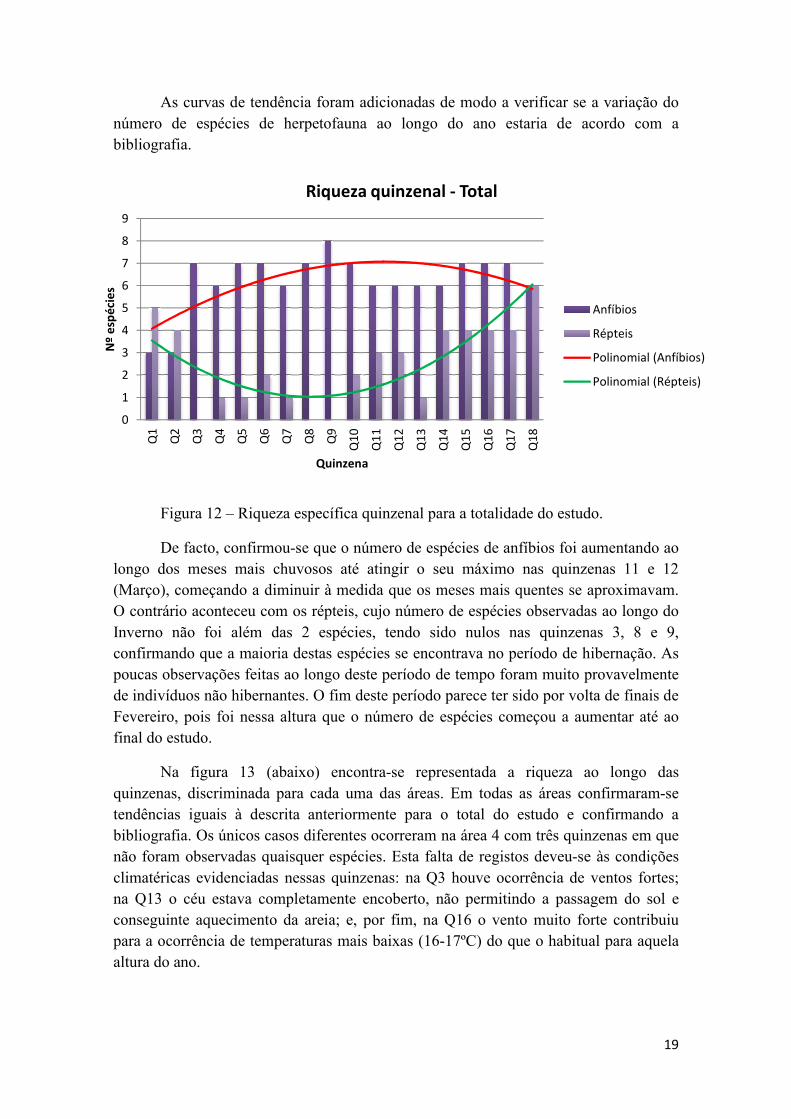
Figura 12 – Riqueza específica quinzenal para a totalidade do estudo.
De facto, confirmou-se que o número de espécies de anfíbios foi aumentando ao
longo dos meses mais chuvosos até atingir o seu máximo nas quinzenas 11 e 12
(Março), começando a diminuir à medida que os meses mais quentes se aproximavam.
O contrário aconteceu com os répteis, cujo número de espécies observadas ao longo do
Inverno não foi além das 2 espécies, tendo sido nulos nas quinzenas 3, 8 e 9,
confirmando que a maioria destas espécies se encontrava no período de hibernação. As
poucas observações feitas ao longo deste período de tempo foram muito provavelmente
de indivíduos não hibernantes. O fim deste período parece ter sido por volta de finais de
Fevereiro, pois foi nessa altura que o número de espécies começou a aumentar até ao
final do estudo.
Na figura 13 (abaixo) encontra-se representada a riqueza ao longo das
quinzenas, discriminada para cada uma das áreas. Em todas as áreas confirmaram-se
tendências iguais à descrita anteriormente para o total do estudo e confirmando a
bibliografia. Os únicos casos diferentes ocorreram na área 4 com três quinzenas em que
não foram observadas quaisquer espécies. Esta falta de registos deveu-se às condições
climatéricas evidenciadas nessas quinzenas: na Q3 houve ocorrência de ventos fortes;
na Q13 o céu estava completamente encoberto, não permitindo a passagem do sol e
conseguinte aquecimento da areia; e, por fim, na Q16 o vento muito forte contribuiu
para a ocorrência de temperaturas mais baixas (16-17ºC) do que o habitual para aquela
altura do ano.
0
1
2
3
4
5
6
7
8
9
Q1
Q2
Q3
Q4
Q5
Q6
Q7
Q8
Q9
Q1
0
Q1
1
Q1
2
Q1
3
Q1
4
Q1
5
Q1
6
Q1
7
Q1
8
Nº
esp
éci
es
Quinzena
Riqueza quinzenal - Total
Anfíbios
Répteis
Polinomial (Anfíbios)
Polinomial (Répteis)

20
Se para os anfíbios a condição essencial para a sua observação é a
disponibilidade de água, para os répteis, é todo um conjunto de condições
(luminosidade, temperatura e ausência de vento, por exemplo) que têm de estar
reunidas, o que pode não facilitar a sua observação.
Figura 13 – Riqueza específica quinzenal em cada uma das áreas analisadas.
0
2
4
6
8
Q1 Q2 Q3 Q4 Q5 Q6 Q7 Q8 Q9 Q10 Q11 Q12 Q13 Q14 Q15 Q16 Q17 Q18
Nº
esp
éci
es
Quinzena
Riqueza quinzenal - Área 1
Anfíbios
Répteis
0
2
4
6
Q1 Q2 Q3 Q4 Q5 Q6 Q7 Q8 Q9 Q10 Q11 Q12 Q13 Q14 Q15 Q16 Q17 Q18
Nº
esp
éci
es
Quinzena
Riqueza quinzenal - Área 2
Anfíbios
Répteis
01
23
4
Q1 Q2 Q3 Q4 Q5 Q6 Q7 Q8 Q9 Q10 Q11 Q12 Q13 Q14 Q15 Q16 Q17 Q18
Nº
esp
éci
es
Quinzena
Riqueza quinzenal - Área 3
Anfíbios
Répteis
0
1
2
3
4
Q1 Q2 Q3 Q4 Q5 Q6 Q7 Q8 Q9 Q10 Q11 Q12 Q13 Q14 Q15 Q16 Q17 Q18
Nº
esp
éci
es
Quinzena
Riqueza quinzenal - Área 4
Anfíbios
Répteis

21
Diversidade e Equitabilidade
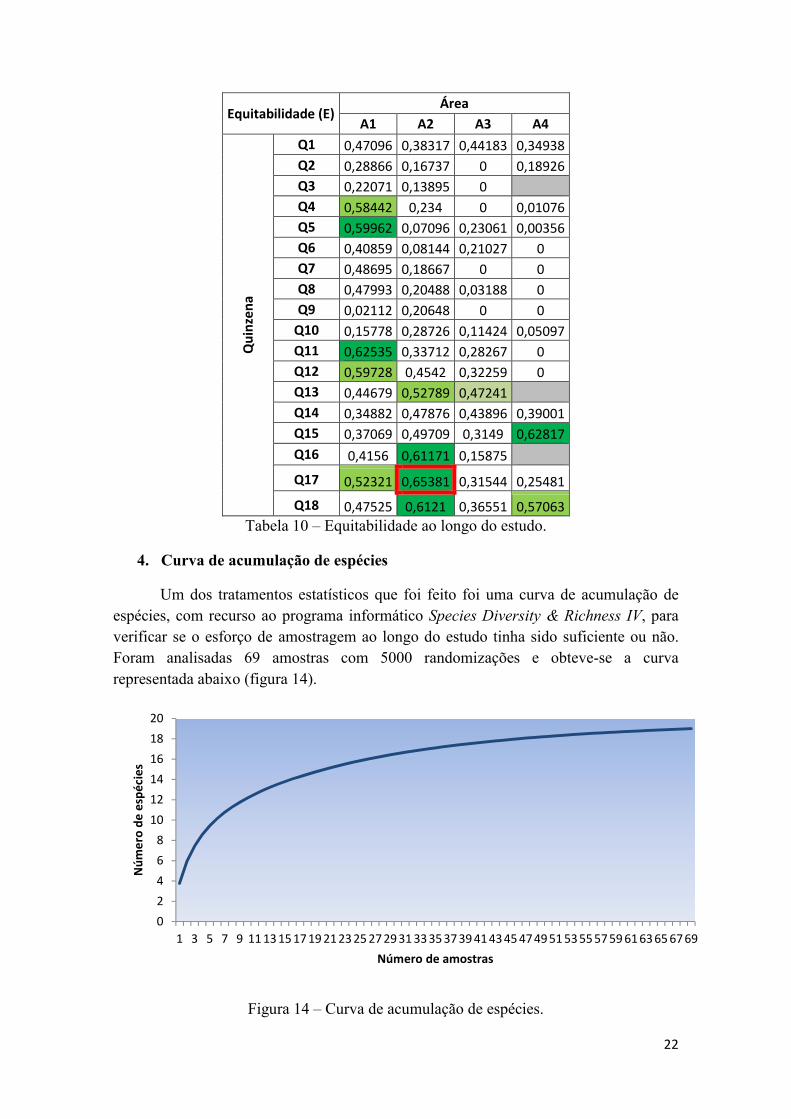
Tal como a riqueza específica, também a diversidade e a equitabilidade foram
estimadas ao longo do período de estudo. Nas tabelas 9 e 10 estão esboçados os valores
de diversidade e de equitabilidade, tendo sido assinalados (vermelho) os valores mais
elevados. Comparativamente à diversidade e equitabilidade da variação espacial houve
algumas diferenças que se destacaram.
Diversidade - H' Área
A1 A2 A3 A4
Qu
inze
na
Q1 1,208 0,983 0,971 0,562
Q2 0,740 0,429 0,000 0,305
Q3 0,566 0,356 0,000
Q4 1,499 0,600 0,000 0,017
Q5 1,538 0,182 0,507 0,006
Q6 1,048 0,209 0,462 0,000
Q7 1,249 0,479 0,000 0,000
Q8 1,231 0,526 0,070 0,000
Q9 0,054 0,530 0,000 0,000
Q10 0,405 0,737 0,251 0,082
Q11 1,604 0,865 0,621 0,000
Q12 1,532 1,165 0,709 0,000
Q13 1,146 1,354 1,038
Q14 0,895 1,228 0,965 0,628
Q15 0,951 1,275 0,692 1,011
Q16 1,066 1,569 0,349
Q17 1,342 1,677 0,693 0,410
Q18 1,219 1,570 0,803 0,918
Tabela 9 – Diversidade ao longo do estudo.
Apesar dos bons valores apresentados, a amostra que obteve o maior valor
ocorreu na área 2, na penúltima quinzena (amostra Q17A2), e não para a área 1, como o
anteriormente obtido. Ocorreram também amostras, somente nas áreas 3 e 4 que
resultaram de frequências de ocorrência muito baixas e, como tal, surgem aqui como
amostras nulas (assinaladas a amarelo). Por essa mesma razão, os valores
correspondentes de equitabilidade foram igualmente nulos.
22
Equitabilidade (E) Área
A1 A2 A3 A4
Qu
inze
na
Q1 0,47096 0,38317 0,44183 0,34938
Q2 0,28866 0,16737 0 0,18926
Q3 0,22071 0,13895 0
Q4 0,58442 0,234 0 0,01076
Q5 0,59962 0,07096 0,23061 0,00356
Q6 0,40859 0,08144 0,21027 0
Q7 0,48695 0,18667 0 0
Q8 0,47993 0,20488 0,03188 0
Q9 0,02112 0,20648 0 0
Q10 0,15778 0,28726 0,11424 0,05097
Q11 0,62535 0,33712 0,28267 0
Q12 0,59728 0,4542 0,32259 0
Q13 0,44679 0,52789 0,47241
Q14 0,34882 0,47876 0,43896 0,39001
Q15 0,37069 0,49709 0,3149 0,62817
Q16 0,4156 0,61171 0,15875
Q17 0,52321 0,65381 0,31544 0,25481
Q18 0,47525 0,6121 0,36551 0,57063
Tabela 10 – Equitabilidade ao longo do estudo.
4. Curva de acumulação de espécies
Um dos tratamentos estatísticos que foi feito foi uma curva de acumulação de
espécies, com recurso ao programa informático Species Diversity & Richness IV, para
verificar se o esforço de amostragem ao longo do estudo tinha sido suficiente ou não.
Foram analisadas 69 amostras com 5000 randomizações e obteve-se a curva
representada abaixo (figura 14).
Figura 14 – Curva de acumulação de espécies.
0
2
4
6
8
10
12
14
16
18
20
1 3 5 7 9 11 13 15 17 19 21 23 25 27 29 31 33 35 37 39 41 43 45 47 49 51 53 55 57 59 61 63 65 67 69
Nú
me
ro d
e e
spé
cie
s
Número de amostras

23
De acordo com o raciocínio, a riqueza encontrada ao longo de um estudo
depende do esforço de amostragem. Observando o gráfico, verifica-se que a curva
começa a atingir um ponto de estabilização, referência para uma suficiência amostral.
5. Curva de elevação
Outro parâmetro analisado foi a variação da riqueza ao longo da elevação. Para
tal, foram determinados os pontos mais baixos e mais altos das áreas em estudo e com
estes calculou-se a elevação média em cada uma das áreas, obtendo-se a curva abaixo
(gráfico y). De acordo com os resultados obtidos, a riqueza específica parece aumentar
com a elevação.
Figura 15 – Curva de elevação – a riqueza específica (S) corresponde às áreas 4,
3, 1 e 2, respectivamente.
32,5m ; S=5
80,5m ; S=9
140m ; S=13
413,5m ; S=13
0
50
100
150
200
250
300
350
400
450
Ele
vaçã
o (
m)

24
Discussão e conclusão
Ao todo estão contabilizadas 32 espécies de herpetofauna para o PNSC, que
corresponde a cerca de 71% das espécies de anfíbios e 77% das espécies de répteis
existentes em território nacional, o que evidencia a importância da zona em termos de
riqueza e diversidade. Todas as áreas prospectadas, com excepção da Peninha,
encontram-se a sul ou na encosta sul da serra de Sintra. São áreas mais expostas ao sol e
por isso mais secas, ao contrário das áreas a norte, com maiores concentrações de
humidade. Seria então espectável que a observação de répteis fosse maior do que a de
anfíbios, facto que se veio a verificar na conclusão deste estudo, no qual foram
observadas 19 espécies de herpetofauna, das quais 8 foram de anfíbios e 11 de répteis.
Estas correspondem a 66,7% das espécies de anfíbios e 55% as espécies de répteis do
PNSC.
Apesar de não obterem as mesmas condições de humidade não quer dizer que
estas áreas não sejam propícias à observação de anfíbios. Com excepção da Duna da
Cresmina, em todas as outras áreas existe a ocorrência e manutenção de água ao longo
de todo o ano, verificando-se apenas alguma escassez e diminuição dos caudais mais
nos meses mais quentes, permitindo alguma estabilidade na dinâmica da manutenção
das populações de herpetofauna. As áreas mais ricas em espécies, e também mais
próximas em termos de similaridade (85% aproximadamente) foram as Quintas do
Pisão e da Peninha. Apesar das diferenças notórias em termos de elevação, quase
trezentos metros, estas áreas albergam uma grande variedade de habitats, facto que
contribui para a ocorrência de um grande número de espécies de herpetofauna. No caso
da Duna da Cresmina também se observaram anfíbios porém apenas nos meses de
Inverno, o que permitiu a manutenção de uma massa de água temporária e a observação
de elevados números de Pelodytes punctatus (sapinho-de-verrugas-verdes).
Infelizmente, cada ano é um caso e o próximo já não terá os valores registados
para esta espécie em particular, devido a ainda não existir essa massa de água (obs.
Pess.), o que vai impedir esta população de se reproduzir e atrasar a sua época de
reprodução. No ano de 2014, esta espécie foi observada no final do mês de Novembro,
altura em que já existia esse pequeno lago temporário. Este é apenas um pequeno
exemplo do impacto que as alterações climáticas têm nestas espécies em particular que,
fazendo parte da base de muitas cadeias alimentar, podem condicionar a sobrevivência
de todas as espécies que dependem delas.
Os locais mais ricos em termos de espécies acabaram por ser estações fixas de
planos de água, como os tanques 1 e 2 na área 2 (Peninha) e o tanque 1 e a lagoa
temporária (LT) na área 1 (Pisão). O melhor transecto foi o T5 na área 2, que incluía
uma mina de água. Era espectável que os melhores registos ocorressem junto a ou em
planos de água, pois os anfíbios dependem directamente de água para a sua
sobrevivência, recorrendo a praticamente todos os locais onde esta exista. Esta procura
de água está, na sua maioria, relacionada com a época de reprodução. Os répteis, por
seu lado, não costumam ser muito observados nestes locais, salvo espécies adaptadas a

25
tal como, por exemplo, as cobras-de-água Natrix maura e Natrix natrix e o lagarto-de-
água Lacerta schreiberi.
No entanto, a presença de répteis junto às massas de água está, em larga medida,
directamente relacionada com a presença de presas que, em muitos casos, pode incluir
anfíbios. Foi observado que é também junto desses locais que cresce a maioria das
plantas silvestres que dão frutos, como é o caso das silvas (Rubus ulmifolius), cujos
frutos são apetecíveis para a maioria das espécies omnívoras dos répteis. Além disso, a
água é também um óptimo chamariz para um sem número de insectos, sobretudo águas
estagnadas, que são presas de ambas as classes de herpetofauna. As aves e mamíferos de
pequenas dimensões, igualmente atraídas pela presença de água e presas, também
podem fazer parte da dieta das espécies da herpetofauna.
Contudo, estas massas de água estão muitas vezes sujeitas a alterações ao nível
de poluição que, caso ocorra algo que altere as condições ideais, pode conduzir à perda
de um elevado número de anfíbios dentro de uma população ou, em casos mais
drásticos, à perda de populações inteiras em larga escala, contribuindo para extinções
locais. Esta sensibilidade deve-se à sua pele extremamente porosa ou, neste caso, à
ausência de uma película de revestimento, permitindo-lhes realizar trocas gasosas
dentro de água mas tornando-os muito mais vulneráveis à poluição. A única excepção é
a espécie Pelophylax perezi (rã-verde) que é mais tolerante à poluição do que as
restantes espécies de anfíbios mas até mesmo essa tolerância é limitada. Deste modo é
seguro concluir que a manutenção da herpetofauna está intimamente ligada às condições
do habitat e, sobretudo, às condições das massas de água.
As espécies que tiveram particular destaque foram os anfíbios Discoglossus
galganoi, com estatuto de quase ameaçada (Cabral et al., 2006) pois desconhece-se o
número dos indivíduos desta população visto não existirem estimativas da densidade
populacional em Portugal (Cabral et al., 2006); Pleurodeles waltl, quase ameaçada (NT)
pelo IUCN, embora ainda não esteja classificada como tal em Portugal, permanecendo
com o estatuto de pouco preocupante (LC); e Triturus pygmaeus, anteriormente
considerada como subespécie de Triturus marmoratus, também categorizada com o
estatuto de quase ameaçada (NT), pois esta espécie encontra-se em declínio,
provavelmente a um ritmo de menos de 30% ao longo de 10 anos, em todo o seu
território, devido à elevada perda de habitats nesse território, fazendo com que esta
espécie se aproxime do estatuto de vulnerável (VU).
No caso dos répteis destaca-se a espécie Acanthodactylus Erythrurus, com
estatuto de quase ameaçada (Cabral et al., 2006) e para a qual também não existem
estimativas da densidade populacional, apesar de poder ser referida como uma espécie
que apresenta localmente efectivos elevados. Além disso, o carácter fragmentado da
distribuição das populações (subpopulações e núcleos populacionais) é explicado
sobretudo por esta ser uma espécie relativamente exigente em termos de habitat
(Camilo-Alves, 1999) e sensível às alterações dos solos para uso agrícola e silvícola
(Cabral et al., 2006).

26
De salientar também a espécie Lacerta schreiberi (Quase Ameaçada – NT –
IUCN, 2009) cuja população terá estado provavelmente em contacto com as populações
do Norte até há relativamente pouco tempo, no entanto, eventualmente devido a
alterações climáticas pós-glaciares e mais recentemente com o elevado crescimento
humano, com a consequente diminuição da área e qualidade de habitat disponível, terá
ocorrido uma fragmentação das populações e mesmo extinção de algumas (Brito et al.,
1998). Estando isolada e num concelho onde a densidade populacional humana é
elevada, surge como área de grande risco e onde a extinção é muito provável a curto
prazo (Brito et al., 1998) ou, como referido por Marques (1995) que caso não se tomem
as precauções necessárias e urgentes de conservação e recuperação da espécie, num
prazo inferior a dez anos poderia ocorrer uma possível extinção local do mesmo.
Felizmente, tal ainda não foi verificado pois foram avistados adultos e juvenis em 2007
(Silva, 2007) e casais de adultos no presente estudo, vinte anos depois.
Por fim, destaque para a espécie Lacerta lepida, uma espécie relativamente
comum em todo o país, porém classificada pelo IUCN como quase ameaçada (NT), pois
esta espécie encontra-se num declínio populacional significativo, a um rácio de menos
de 30% ao longo de uma década, grande parte devido à perda de habitat ao longo de
todo o seu limite de expressão, o que a deixa muito próximo da qualificação de espécie
vulnerável (VU) (IUCN, 2009).
Ao longo do período de amostragem foram tomadas certas precauções para não
causar danos às populações de anfíbios por causa do método de uso do camaroeiro e das
suas desvantagens, apontadas por Sutherland (1996), como o de preservar a vegetação
aquática do fundo, nomeadamente quando eram avistados comportamentos de corte em
Lissotriton boscai. Sutherland (1996) refere ainda a possibilidade de morte das larvas
com guelras externas (Lissotriton boscai, Pleurodeles waltl, Salamandra salamandra e
Triturus pygmaeus) cuja possibilidade destes órgãos ficarem presos na rede é elevada e
pode resultar mesmo na morte do indivíduo. Neste caso, é apenas uma questão do
investigador, pegar com cuidado nas larvas quando capturadas na rede, e pô-las o mais
rapidamente possível num recipiente com água, neste caso um tabuleiro de triagem, para
evitar situações de stress, que felizmente não se verificaram.
Quanto à diversidade é possível aferir, com 95% de certeza (percentagem dada
pelo programa informático Species Diversity & Richness IV que a área da Quinta do
Pisão (área 1) é a área mais diversa, em grande parte graças ao seu tamanho e à grande
variedade de habitats apresentada. De acordo com os valores obtidos, é também possível
confirmar que todas as áreas são mais diversas do que a área 4 (Duna da Cresmina),
sobretudo por causa, uma vez mais, da variedade de habitats, já que nesta área não
existem tantas opções de habitat e, sendo uma área mais exposta aos elementos, com
limitações ao nível de abrigos e disponibilidade de água. Contudo, estes resultados
também servem de indicação quanto à raridade, pois nesta área encontraram-se duas
espécies que não foram encontradas em nenhuma das outras e, como tal, este indício dá
argumentos para que esta área seja considerada de elevado valor ecológico.

27
A última aferição refere-se às restantes áreas, Quinta da Peninha e Pisão de
Baixo (áreas 2 e 3, respectivamente) que, segundo os resultados obtidos, não são
estatisticamente diferentes em termos de diversidade, o que é algo estranho pois são
áreas com diferenças evidentes, tais como, elevação, coberto vegetal, temperatura,
disponibilidade de água ao longo do ano e, até mesmo, em termos de riqueza específica.
No entanto, foi na área 2 que se verificou a amostra com os maiores valores para
diversidade e equitabilidade (amostra Q17A2). Quanto maior a equitabilidade todas as
espécies nessa amostra têm o mesmo número, ou muito semelhante, de indivíduos ou,
por outras palavras, que esta amostra evidencia um equilíbrio no número de indivíduos
por espécie, não sendo forçosamente a amostra com a maior riqueza específica.
Por último, é importante referir as curvas de acumulação de espécies e de
elevação como complemento aos resultados obtidos. A curva de acumulação de espécies
serviu para indicar se o esforço de amostragem ao longo do estudo fora o adequado.
Com efeito, a curva parece atingir um ponto de estabilização o que indica que este
esforço foi suficiente e que a recolha de mais amostras não iria contribuir grandemente
para aumentar o número de espécies observadas.
A curva de elevação serviu para verificar qual o comportamento da riqueza de
espécies face à elevação do terreno. De acordo com a curva obtida, parece que a riqueza
específica aumenta com a elevação, o que de certo modo tem alguma lógica, pois
quanto maior a elevação, maior a ocorrência de pluviosidade e maiores as concentrações
de humidade, condições ideais para os anfíbios. Na sua qualidade de predadores, os
répteis fixam-se, na sua maioria, em locais onde existam números adequados de presas,
logo se existem condições para as presas, os predadores não tardaram a aparecer. De
qualquer modo seria interessante verificar a riqueza no intervalo dos 150 aos 400 metros
de elevação e se esta mantinha a mesma tendência ou apresentaria novos e interessantes
valores.

28
Considerações finais
De acordo com os objectivos preconizados, da caracterização da herpetofauna,
há a salientar a observação de três espécies, encontradas em três das quatro áreas
prospectadas: o anfíbio D. galganoi, e os répteis A. erythrurus e L. schreiberi; as duas
primeiras com o estatuto de quase ameaçada, no Livro Vermelho dos Vertebrados de
Portugal, porém todas com elevado interesse conservacionista. As observações de D.
galganoi, nas suas várias fases de desenvolvimento, vêm realçar a importância destas
áreas como habitats ideais para a sobrevivência desta espécie, face à desconhecida
densidade populacional em Portugal. A. erythrurus e L. schreiberi, por seu lado,
apresentam populações fragmentadas ao longo do país, apesar de a primeira poder
apresentar efectivos mais elevados do que a segunda e ser mais exigente ao nível de
habitats.
As observações de L. schreiberi parecem confirmar a manutenção de um núcleo
populacional já isolado, quando alguns autores já davam como certa a sua extinção local
em menos de uma década. Hoje, ao final de duas décadas, parece que esta população
conseguiu alguma estabilidade, fruto de um esforço de manutenção do seu habitat. É
importante que estes esforços continuem a ser desenvolvidos de modo a reafirmar o
PNSC como um dos mais importantes pólos de biodiversidade, não só para a
herpetofauna, de Portugal.
Infelizmente não foram observadas algumas espécies da herpetofauna em parte
pelo esforço amostral ter sido realizado de uma forma geral e não focado directamente
numa ou outra espécie. No entanto existe sempre a possibilidade de complementar este
estudo com esse tipo de amostragem directa. Dos anfíbios não foram observadas as
espécies Epidalea calamita (sapo-corredor), Alytes obstetricans (sapo-parteiro-comum),
Hyla arborea (rela-arbórea) e Hyla meridionalis (rela-meridional). O sapo-de-unha-
negra (Pelobates cultripes) já está apontado como extinto localmente.
No caso dos répteis, não houve registos das espécies Anguis fragilis (licranço),
Blanus cinereus (cobra-cega), Coronella girondica (cobra-lisa-meridional), Emys
orbicularis (cágado-de-carapaça-estriada), Macroprotodon cucullatus (cobra-de-capuz),
Mauremys leprosa (cágado-mediterrânico), Podarcis carbonelli (lagartixa-de-
Carbonell), Psammodromus hispanicus (lagartixa-do-mato-ibérica), Rhinechis scalaris
(cobra-de-escada) e Vipera latastei (víbora-cornuda). Algumas destas espécies são
muito difíceis de observar por causa dos seus hábitos como, por exemplo, o licranço
(Anguis fragilis) é uma espécie de hábitos preferencialmente subterrâneos.
De futuro, recomenda-se a monitorização e amostragens complementares das
espécies observadas, com especial atenção para as espécies D. galganoi, P. waltl, T.
pygmaeus e A. erythrurus, amostragens dirigidas às espécies não observadas e com
interesse para a conservação como Vipera latastei (víbora-cornuda) e Macroprotodon
cucullatus (cobra-de-capuz) e acções dirigidas à espécie (conservação e sensibilização).
Seria igualmente importante considerar as sugestões feitas por outros autores, neste caso
focando-se no exemplo do lagarto-de-água e passíveis de serem aplicadas a outras

29
espécies de interesse para a conservação, como medidas de conservação ao nível da
protecção e recuperação dos habitats ripícolas propostas por Maymonte (1997), a
recuperação de habitats e gestão adequada dos habitats ainda existentes proposta por
Brito et al. (1998). E ainda a criação em cativeiro para suplementação de populações e
reintrodução de indivíduos no interior do Parque Natural, proposta por Brito et al.
(1998) e por Marques (1995)
Como medidas mais práticas e fáceis de executar sugere-se, para a Quinta do
Pisão, a limpeza da linha de água que abastece a lagoa temporária (LT) e a manutenção
da área do Transecto 6 (T6) o mais intacta possível, pois é um local importante para a
reprodução da espécie D. galganoi, devido às suas várias poças temporárias. Sugere-se
igualmente a limpeza dos tanques, nas áreas da Quinta do Pisão e da Peninha, e a
colocação de rampas, preferencialmente de pedra ou aglomerados de pedras, de modo a
que as espécies de anfíbios e répteis possam circular mais livremente. Sugerem-se
igualmente acções de irradicação de espécies exóticas como o lagostim-da-Louisiana e a
gambúzia, que podem causar graves danos nas populações de anfíbios, pois são
predadores vorazes das suas formas larvares.
Por fim, salienta-se a área da Duna da Cresmina como uma área de elevado valor
ecológico dado a raridade das espécies que lá ocorrem de fauna e de flora e a
potencialidade para a ocorrência de espécies de interesse para a conservação como é o
caso de Vipera latastei (víbora-cornuda) e de Psammodromus hispanicus (lagartixa-do-
mato-ibérica).

30
Bibliografia Almeida N.F., Almeida P.F., Gonçalves H., Sequeira F., Teixeira J. & Almeida F.F.
(2001). Anfíbios e Répteis de Portugal. Guia FAPAS – Fundo para a Protecção dos
Animais Selvagens. Porto.
Araújo M.B., Thuiller W. & Pearson R.G. (2006). Climate warming and the decline of
amphibians and reptiles in Europe. Journal of Biogeography, 33: 1712 – 1728
Blaustein A.R., Belden L.K., Olson D.H., Green D.M., Root T.L., & Kiesecker J.M.
(2001). Amphibian Breeding and Climate Change. Conservation Biology, 15 (6): 1804
– 1809.
Brito J.C. & Álvares F. (1996). Some data on the Distribution and Feeding Habits of
Lataste's Viper (Vipera latastei) and Seoane's Viper (Vipera seoanei) in North-western
Portugal. Abstract IV Congresso Luso-Espanhol / VIII Congresso Espanhol de
Herpetologia. Porto.
Brito J.C., Paulo O.S., Luís C., Godinho R., Rosa H.D., Brito F., Abreu E. & Crespo
E.G. (1996). Bases para uma Estratégia de Conservação de Lacerta schreiberi e seus
Habitats. Relatório Final. Grupo de Conservação Biológica do Centro de Biologia
Ambiental. Faculdade de Ciências da Universidade de Lisboa. 261pp.
Brito J.C., Luís C., Godinho M.R., Paulo O.S. & Crespo E.G. (1998). Bases para a
conservação do Lagarto-de-água (Lacerta schreiberi). Estudos de Biologia e
Conservação da Natureza nº 23. Instituto da Conservação da Natureza. Lisboa.
Brito J.C. (2005). Estratégia de Conservação do Lagarto-de-água em Portugal.
http://www.naturlink.pt
Burton T. M. & Likens G. E. (1975). Salamander populations and biomass in the
Hubbard Brook Experimental Forest. New Hampshire. Copeia, 1975 (3): 541 – 546.
Cabral M.J. (coord.), Almeida J., Almeida P.R., Dellinger T., Ferrand de Almeida N.,
Oliveira M.E., Palmeirim J.M., Queiroz A.I., Rogado L. & Santos-Reis M. (eds.)
(2006). Livro Vermelho dos Vertebrados de Portugal. 2ªed. Instituto da Conservação da
Natureza/Assírio &Alvim. Lisboa
Caldas A. (2010). Anfíbios de Portugal – Guia Fotográfico Quercus. 1ª Edição.
Quercus-ANCN.
Camilo-Alves C. (1999). Análise da distribuição da herpetofauna de Portugal
Continental através dos métodos de regressão logística e análise de sobreposição.
Relatório de Estágio da Licenciatura em Biologia, Faculdade de Ciências da
Universidade de Lisboa, Lisboa. 41 pp.
Crespo E.G. (1971). Anfíbios de Portugal Continental das Colecções do Museu Bocage.
Arquivos do Museu Bocage (2ª série), 3 (8): 203 – 304.

31
Crespo E.G. (1972). Répteis de Portugal Continental das Colecções do Museu Bocage.
Arquivos do Museu Bocage (2ª série), 3 (17): 447 – 612.
Crespo E.G. (1975). Aditamento aos catálogos dos Répteis e Anfíbios de Portugal das
Colecções do Museu Bocage. Arquivos do Museu Bocage (2ª série), 5 (3): 479 – 498.
Crespo E.G. e Oliveira M.E. (1989). Atlas de distribuição dos Anfíbios e Répteis de
Portugal Continental. Serviço Nacional de Parques, Reservas e Conservação da
Natureza. Lisboa.
Crespo E.G. & Sampaio L. (1994). As Serpentes de Portugal. I.C.N. Lisboa.
Duellman W. E. & Trueb L. (1994). Biology of amphibians. John Hopkins University
Press.
Fabião A.M.D. (1999). Caracterização do ambiente físico e da vegetação da Serra de
Sintra e da região envolvente – texto de apoio para visitas de estudo na zona Sintra-
Cascais. Departamento de Engenharia Florestal, Instituto Superior de Agronomia.
Lisboa
Gardner T. (2001). Declining amphibian populations: a global phenomenon in
conservation biology. Animal Biodiversity and Conservation, 24 (2): 1 – 19.
Gasc J.P., Cabela A., Crnobrnja-Isailovic J., Dolmen D., Grossenbacher K., Haffner P.,
Lescure J., Martens H., Martinez Rica J.P., Maurin H., Oliveira M.E., Sofianidou T.S.,
Veith M. & Zuidrewijk A. (Eds.). (2004). Atlas of Amphibians and Reptiles in Europe.
Paris: Museum National d’Histoire Naturelle.
Godinho R., Teixeira J., Rebelo R., Segurado P., Loureiro A., Álvares F., Gomes N.,
Cardoso P., Camilo-Alves C. & Brito J.C. (1999). Atlas of the continental Portuguese
herpetofauna: an assemblage of published and new data. Rev. Esp. Herp, 13: 61 – 82.
Guyer C. (1990). The herpetofauna of La Selva, Costa Rica. In: Four neotropical
rainforests: (A. H. Gentry, Ed.) Yale University Press. 371 – 385.
IUCN. (2009). European Species on the IUCN Red List. Available at:
http://www.iucnredlist.org/europe. (Accessed: 22 June 2009).
Juncá F. A. (2001). Declínio mundial das populações de anfíbios. Sitentibus Série
Ciências Biológicas vol.1. Universidade Estadual de Feira de Santana. Departamento de
Ciências Biológicas. BA. Brasil. (1): 84-87. Maio 2001.
Krebs C.J. (1989). Ecological Methodology. HarperCollins Publishers. University of
British Columbia.
Lips K. R. et al. (2008). Dirigindo a Onda: Reconciliando os papéis de doença e
mudança climática nos declínios de anfíbios. PloS Biologia vol.6. No. 3, e72.

32
Longore J.E., Pessier A.P. & Nichols D.K. (1999). Batrachochytrium dendrobatidis
gen. et sp Nov, a chytrid pathogenic to amphibians. Mycologia 1999; 91: 219 – 27.
Malkmus R. (1979). Zur faunistik und ökologie der amphibien und reptilien in der
Serra de Sintra, Portugal. Nachrichten das Naturwissenschaftlichen Museum.
Aschaffenburg.
Malkmus R. (1982). Beitrag zur Verbreitung der Amphibien und Reptilien in Portugal.
Salamandra, 18 (3/4): 218 – 299.
Malkmus R. (1983). Nachtrag zur Verbreitung der Amphibien und Reptilien Portugals.
Salamandra, 19 (1/2): 71 – 83.
Malkmus R. (1984). Herpetologische Ausflüge in die Serra de Malcata/ Portugal.
Herpetofauna, 31: 6 – 11.
Malkmus R. (1985a). Uber die Herpetofauna im Fátima – Karst Portugal. Natur und
Museum, 115 (6): 174 – 185.
Malkmus R. (1985b). Die Serra da Estrela (Portugal) unter besonderer
Berucksichtigung ihrer Herpetofauna. Bonn. Zool. Beitr., 36 (1/2): 105 – 144.
Malkmus R. (1995). Die Amphibien und Reptilien: Portugals, Madeiras und der
Azoren. Westarp Wissenschaften. Magdeburg.
Marques M.J.A.S. (1995). Determinação de áreas prioritárias para a conservação da
herpetofauna no parque natural Sintra-Cascais. Relatório de estágio para a obtenção de
Licenciatura em Biologia. Faculdade de Ciências da Universidade de Lisboa. Lisboa.
Maymonte M.S.S. (1997). Técnicas de criação em cativeiro, tendo por base a
recuperação do lagarto-de-água (Lacerta schreiberi) na serra de Sintra. Relatório de
estágio para a obtenção de Licenciatura em Biologia. Faculdade de Ciências da
Universidade de Lisboa. Lisboa.
Negrão R. G. (2009). Declínio da população mundial de anfíbios devido a infecção por
Batrachochytrium dendrobatidis (Longore, Pessier & Nichols DK 1999), o fungo
causador da Quitridiomicose. Arte e Ciência. Fevereiro 2009.
Oliveira M.P. (1931). Répteis e anfíbios da Península Ibérica e especialmente de
Portugal. Imprensa da Universidade. Coimbra, 3ª ed. (1ª ed. 1896).
Olszewski T.D. (2004). A unified mathematical framework for the measurement of
richness and evenness within and among multiple communities.
Oikos, 104 (2): 377 – 387.
Omolo DP. 2006. Biodiversity patterns in changing Mediterranean landscapes: a
modelling perspective. Unpublished Msc Thesis, International Institute for Geo-
information Science and Earth Observation, Enschede, The Netherlands.

33
Ovenden D. & Arnold N. (2004). A field guide to the Reptiles and Amphibians of
Britain and Europe. 2ªed. Colins. London.
Pargana J. (1995). Estudo da Herpetofauna no Parque Natural da Serra de S. Mamede.
66 pp.
Pearman P.B. (1997). Correlates of amphibian diversity in an altered landscape of
Amazonian Ecuador. Conservation Biology, 11: 1211 – 1225.
Silva D.A.R.R da (2007). Padrões de distribuição e abundância da Herpetofauna nos
Parques de Sintra – Monte da Lua. Relatório de estágio para a obtenção de Licenciatura
em Biologia. Universidade Lusófona de Humanidades e Tecnologias. Lisboa.
SNPRCN (1990) Livro Vermelho dos Vertebrados de Portugal. Vol. I. Serviço
Nacional de Parques, reservas e conservação da natureza. Lisboa.
Speybroeck J. & Crochet PA. (2007). Species list of the European Herpetofauna – a
tentative update. POD@RCIS 8 (1/2). Netherlands.
Sutherland W.J. (1996) Ecological Census Techniques – a handbook. Cambridge
University Press. New York. USA.
Themido A.A. (1942). Anfíbios e Répteis de Portugal (Catálogo das colecções do
Museu Zoológico de Coimbra). Memórias e Estudos do Museu Zoológico da
Universidade de Coimbra, 133: 1 – 49.
Tryjanowski P., Mariusz R. & Sparks, T.H. (2003) Changes in spawning dates of
common frogs and common toads in western Poland in 1978 – 2002. Annales Zoologica
Fennici, 40: 459 – 464.
Wake D.B. & Morowitz H.J. (1991). Declining amphibian populations – a global
phenomenon?. Findings and recommendations. Alytes, 9 (2): 33 – 42.
Woolbright L.L. (1991). The impact of hurricane Hugo on forest frogs in Puerto Rico.
Biotropica, 23: 462 – 467.

34
ANEXOS

35
Anexo 1 – Áreas e locais prospectados
Figura 16 - Vista aérea da Quinta do Pisão (área 1) e locais prospectados. (Google
Maps)
36
Figura 17 - Vista aérea da Quinta da Peninha (área 2) e locais prospectados. (Google Maps)
37
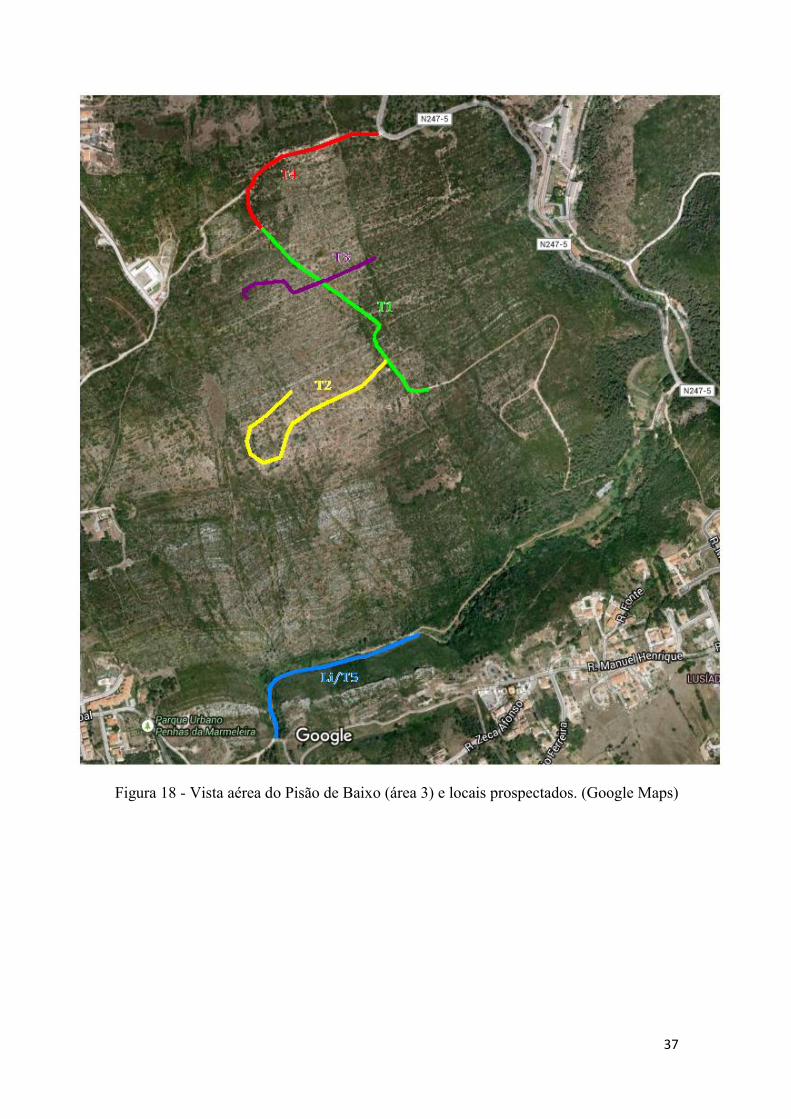
Figura 18 - Vista aérea do Pisão de Baixo (área 3) e locais prospectados. (Google Maps)

38
Figura 19 - Vista aérea da Duna da Cresmina (área 4) e pontos prospectados. (Google Maps)

39
Anexo 2 - Biologia das espécies observadas Anfíbios
1. Salamandra-de-costelas-salientes
Classe: Amphibia; Ordem: Caudata; Família: Salamandridea; Género: Pleurodeles
Espécie: Pleurodeles waltl (Michahelles, 1830)
Descrição: É o maior urodelo ibérico podendo passar dos 30 cm de comprimento. Tem cerca
de 7-9 protuberâncias em cada um dos flancos, amarelas ou alaranjadas. A pele é tipicamente
rugosa e apresenta uma coloração dorsal muito variada. Vivem 10 anos (20 em cativeiro).
Hábitos:
Espécie de hábitos aquáticos encontrando-se activa maioritariamente à noite ou no
crepúsculo. Pode ser encontrada ocasionalmente em terra debaixo de pedras ou
troncos; Pode passar por períodos de estivação entre Novembro e Fevereiro;
Pode migrar para outros locais de água quando os locais onde estão secam ou
começam a secar. Resistente à secura e à poluição;
Habitats: Prefere águas temporárias com fundos de natureza arenosa;
Alimentação: Adultos e larvas – invertebrados aquáticos, insectos, ovos e larvas de outras
espécies de urodelos.
Defesa: Adopta uma postura defensiva tentando espetar as saliências costelares das quais
segrega toxinas.
Reprodução: entre Março e Junho; ovípara
Distribuição: Mais comum no Centro e Sul de Portugal.
Estatuto/Legislação: Estatuto IUCN 2009 – Quase ameaçada (NT). Está incluída no anexo III da
Convenção de Berna.

40
2. Salamandra-de-pintas-amarelas
Classe: Amphibia; Ordem: Caudata; Família: Salamandridea; Género: Salamandra
Espécie: Salamandra salamandra (Linnaeus, 1758)
Descrição: Espécie grande, até 25cm, de cores vivas no dorso com um padrão muito variável
(amarelo, laranja ou vermelho). A pele é lisa e brilhante e está permanentemente húmida.
Hábitos:
Sobretudo nocturna e maioritariamente terrestre; vivem cerca de 20 anos (50 em
cativeiro); os adultos são péssimos nadadores – afogamento de fêmeas;
Utilizam o meio aquático somente para fins reprodutores;
Encontra-se activa todo o ano sendo mais comum entre Setembro e Maio, podendo
ficar inactiva nos meses mais frios nas regiões mais altas.
Habitats: Massas florestais caducifólias (castanheiros e carvalhos, por exemplo) e zonas
agrícolas com oliveiras e gramíneas.
Alimentação:
Adultos – escaravelhos, formigas, caracóis, lesmas, minhocas, centopeias e aranhas;
Larvas – insectos aquáticos, crustáceos, pequenos vermes e larvas de outros anfíbios
incluindo a própria espécie.
Defesa: Secreções tóxicas esbranquiçadas das glândulas parótidas ou adopta uma postura
defensiva baixando a cabeça e arqueando o corpo.
Reprodução:
Entre Setembro e Maio. Possível ocorrência de 2 épocas – Outono e Primavera;
Pouco dimorfismo sexual;
Espécie ovovivípara ou vivípara. As fêmeas dão à luz entre 8-70 larvas, uma vez ou em
várias ocasiões sucessivas ao longo da época.
Distribuição: Um pouco por todo o país.
Estatuto/Legislação: Estatuto IUCN 2009 – Espécie pouco preocupante (LC). Está incluída no
anexo III da Convenção de Berna.

41
3. Tritão-de-barriga-laranja
Classe: Amphibia;
Ordem: Caudata;
Família: Salamandridea;
Género: Lissotriton
Espécie: Lissotriton boscai (Lataste, 1879)
Descrição: Machos mais pequenos que as fêmeas com 6.5-7.5cm e 7-10cm de comprimento,
respectivamente. Coloração dorsal variável do amarelo-acastanhado ao cinzento-acastanhado
escuro e ventre geralmente do laranja-amarelado ao laranja-avermelhado, com manchas
escuras dispersas. Longevidade: 6 anos para os machos e 9 anos para as fêmeas.
Hábitos: Espécie geralmente aquática todo o ano, mas pode ter uma fase terrestre. Actividade
maioritariamente diurna na fase aquática e geralmente nocturna na fase terrestre.
Habitats: Florestas de coníferas e de folhosas caducifólias e meios lênticos de água doce –
pequenas poças, ribeiros, lagoas e poças dentro de cavernas.
Alimentação: Adultos e larvas – invertebrados aquáticos, ovos e larvas da própria espécie e de
outras espécies de urodelos.
Defesa: Fuga e emissão de secreções tóxicas das glândulas cutâneas.
Reprodução:
Dimorfismo sexual – machos mais pequenos com cloaca redonda evidente e com uma
linha branca na extremidade inferior da cauda;
Entre Novembro e Junho, dependendo da região geográfica;
Complexo comportamento de corte dentro de água;
As fêmeas põem 100-250 ovos ao longo da época que são fixados individualmente a
plantas aquáticas ou a outros objectos no fundo da água e eclodem entre 1 a 3
semanas.
Distribuição: Mais comum no norte e centro do país (interior) e junto ao litoral centro e sul.
Estatuto/Legislação: Estatuto IUCN 2009 – Espécie pouco preocupante (LC). Está incluída no
anexo III da Convenção de Berna.

42
4. Tritão-pigmeu
Classe: Amphibia; Ordem: Caudata;
Família: Salamandridea;
Género: Triturus
Espécie: Triturus pygmaeus (Wolterstorff, 1905)
Descrição: Os adultos chegam aos 16cm de comprimento. A coloração dorsal é sobretudo
verde, claro ou escuro, e o ventre é escuro, normalmente preto e com pequenas manchas
brancas. Vivem mais de 10 anos (25 em cativeiro).
Hábitos: Espécie aquática, sobretudo durante a época de reprodução, e terrestre fora da
última. Activa normalmente todo o ano, podendo ficar inactiva nos meses mais frios do
Inverno e nos meses mais quentes do Verão.
Habitats: Meios lênticos de água doce (permanentes ou temporários).
Alimentação:
Adultos – larvas de insectos aquáticos, minhocas, lesmas, caracóis e larvas de outros;
Larvas – pequenos insectos aquáticos e crustáceos – cladóceros e copépodes.
Defesa: Fuga, secreções cutâneas tóxicas e tentativa de morder o agressor.
Reprodução:
Com dimorfismo sexual – as fêmeas e juvenis têm uma risca vertebral alaranjada e os
machos uma crista no dorso;
Entre Outubro-Maio dependendo da região geográfica;
Ocorre à noite e dentro de água;
A fêmea deposita 150-400 ovos que são envolvidos, individualmente, em plantas
aquáticas. A eclosão dá-se 8-14 dias depois da postura;
Metamorfose na Primavera ou até ao final do Verão.
Distribuição: Mais comum a norte e centro do país e também no litoral centro.
Estatuto/Legislação: Estatuto IUCN 2009 – Espécie Quase Ameaçada (NT). Está incluída no
anexo III da Convenção de Berna e no anexo IV da Directiva Habitats.

43
5. Rã-de-focinho-pontiagudo
Classe: Amphibia; Ordem: Anura; Família: Discoglossidae; Género: Discoglossus
Espécie: Discoglossus galganoi (Capula, Nascetti, Lanza, Bullini & Crespo 1985)
Descrição: Pode ter 6-8cm de comprimento. A cabeça é achatada, o focinho pontiagudo, as
pupilas redondas e possuí uma “máscara” na cabeça, entre os olhos e a extremidade do
focinho. A pele é lisa com algumas verrugas e a coloração muito variável (desde o cinza ao
castanho) sendo o ventre esbranquiçado.
Hábitos: Geralmente crepuscular, podendo também ser diurna em dias húmidos e chuvosos.
Permanece activa geralmente todo o ano mas com pouca actividade nos meses mais frios e
também nos mais quentes. Longevidade ronda os 10 anos.
Habitats: Encontra-se nas imediações das massas de água em qualquer tipo de biótopo,
preferindo terrenos encharcados, prados e lameiros.
Alimentação: Adultos – insectos, aranhas, caracóis, lesmas, minhocas ou até mesmo juvenis da
própria espécie; Larvas – matéria vegetal e detritos.
Defesa: Fuga.
Reprodução:
Ocorre no Inverno e na Primavera dentro de água;
O amplexo é inguinal;
A fêmea acasala com vários machos e põe 20-50 ovos de cada vez, podendo pôr cerca
de 1500 num único dia em pequenos grupos no fundo das massas de água ou nas
plantas aquáticas. A eclosão dá-se 2-9 dias depois;
A metamorfose ocorre ao fim de 4-9 semanas.
Distribuição: Ocorre em todo o país mas é mais comum no litoral, o que corresponde a 25% da
área global.
Estatuto/Legislação: Estatuto ICNF 2006 – Quase ameaçada (NT); estatuto IUCN 2009 – Espécie
pouco preocupante (LC). Está incluída no anexo II da Convenção de Berna e nos anexos II e IV
da Directiva Habitats.

44
6. Rã-verde
Classe: Amphibia; Ordem: Anura; Família: Ranidae;
Género: Pelophylax
Espécie: Pelophylax perezi (López-Seoane, 1885)
Descrição: Geralmente com 8,5-10cm de comprimento. A coloração varia de verde, cinzento e
castanho, com manchas irregulares mais escuras sendo muito frequente uma risca vertebral
esverdeada. Tem pregas dorso-laterais castanhas e ventre esbranquiçado com reticulações
cinzentas.
Hábitos: Espécie activa de noite e de dia, altamente aquática e abundante. É boa nadadora e
óptima saltadora, atingindo em salto distâncias até 2 metros. Pode sobreviver em algumas
águas salobras e poluídas. Longevidade ronda os 10 anos.
Habitats: Ocorre numa grande variedade de massas de água, encontrando-se frequentemente
a apanhar sol nas margens.
Alimentação: Espécie muito voraz; Adultos – insectos, aranhas, minhocas, crustáceos,
moluscos, peixes e anfíbios, incluindo os da própria espécie, pequenas rãs e pequenos
roedores; Larvas – matéria vegetal e detritos.
Defesa: Fuga para dentro de água.
Reprodução:
Ocorre na Primavera;
O amplexo é axilar e dá-se sobretudo de noite;
As fêmeas depositam 800-10.000 por época em grandes aglomerados flutuantes;
A eclosão dá-se passados alguns dias e os girinos sofrem a metamorfose 2-4 meses
depois.
Distribuição: Encontra-se uniformemente distribuída por todo o país.
Estatuto/Legislação: Estatuto IUCN 2009 – Espécie pouco preocupante (LC). Está incluída no
anexo III da Convenção de Berna e no anexo V da Directiva Habitats.

45
7. Sapinho-de-verrugas-verdes
Classe: Amphibia;
Ordem: Anura;
Família: Pelodytidae;
Género: Pelodytes;
Espécie: Pelodytes punctatus (Daudin, 1802).
Descrição: Em geral entre 3,5 a 4,5 cm de comprimento (pode alcançar 5 cm), cabeça plana c/
focinho arredondado, s/ glândulas parótidas, pele com numerosas verrugas, frequentemente
esverdeadas ou acastanhadas. Zona ventral esbranquiçada, peito e membros rosados.
Longevidade ronda os 15 anos.
Hábitos: Predominantemente crepuscular ou nocturna, com hábitos diurnos na época de
reprodução. Pode estivar nos meses mais quentes do Verão.
Habitats: Grande variedade, desde dunas costeiras a matos e bosques. Parece demonstrar
preferência por solos calcários. Pode recorrer a grutas, fendas ou concavidades naturais como
refúgio.
Alimentação:
Adultos – pequenos insectos e invertebrados;
Larvas – algas, detritos, fungos e restos de plantas aquáticas.
Defesa: Fuga para a água. Pode segregar muco através da pele.
Reprodução:
Pouco dimorfismo (calosidades nupciais negras nos membros anteriores dos machos);
fêmeas maiores do que os machos;
Amplexo inguinal e dentro de água;
Fêmeas depositam entre 1000 a 1600 ovos, em cordões que fixam a plantas aquáticas.
Distribuição: Mais comum no Centro e Sul de Portugal.
Estatuto/Legislação: Estatuto IUCN 2009 – Espécie pouco preocupante (LC). Está incluída no
anexo III da Convenção de Berna e no anexo V da Directiva Habitats.

46
8. Sapo-comum
Classe: Amphibia; Ordem: Anura; Família: Bufonidae; Género: Bufo
Espécie: Bufo bufo (Linnaeus, 1758)
Descrição: É o maior sapo da Europa podendo atingir os 15cm de comprimento. Apresenta
uma pupila horizontal de cor cobre-dourado e uma pele muito rugosa, normalmente
acastanhada, com glândulas parótidas bastante evidentes e oblíquas.
Hábitos: Espécie de hábitos aquáticos e terrestres, essencialmente crepusculares, mas
também diurnos em dias húmidos e chuvosos. Activa todo o ano, ocorrendo uma diminuição
da actividade no Inverno. Vivem 7 a 10 anos (cerca de 30 em cativeiro).
Habitats: Cursos de água, charcos temporários, florestas, habitats rochosos e arenosos, matos,
terrenos agrícolas e águas estagnadas.
Alimentação: Espécie muito voraz. Adultos – centopeias, escaravelhos, moscas, borboletas,
lesmas, minhocas e ainda outros anfíbios; Larvas – matéria vegetal.
Defesa: Incham o corpo e libertam secreções cutâneas esbranquiçadas das glândulas
parótidas. Também podem expelir urina.
Reprodução:
Apresenta dimorfismo sexual – fêmeas muito maiores que os machos;
Entre Novembro-Abril ou final do Inverno em zonas mais frias;
Migrações até aos locais de reprodução no Outono;
O amplexo é axial ocorrendo competição entre os machos – amplexos de vários
machos e uma só fêmea;
As fêmeas depositam 2000-8000 ovos esféricos em cordões gelatinosos com 5- 10mm
de espessura e até 5m de comprimento. A eclosão dá-se após 2- 3 semanas.
A metamorfose ocorre 2-4 meses depois a eclosão.
Distribuição: Comum em todo o país.
Estatuto/Legislação: Estatuto IUCN 2009 – Espécie pouco preocupante (LC). Está incluída no
anexo III da Convenção de Berna.

47
Répteis
1. Cobra-de-água-de-colar
Classe: Reptilia; Ordem: Squamata;
Família: Colubridae; Género: Natrix
Espécie: Natrix natrix (Linnaeus, 1758)
Descrição: Os machos medem 1,053m
e as fêmeas 1,19m, podendo
ultrapassar os 2m de comprimento. A
coloração dorsal varia do acinzentado
ao verde oliváceo e ao acastanhado,
com manchas escuras dispersas
irregularmente. A cabeça é redonda e
bem definida, as pupilas também são
redondas e apresentam um colar amarelado debruado a preto logo atrás da cabeça o que está
na origem do nome que lhe foi atribuído. A sua longevidade é de 19 anos.
Hábitos: Geralmente de hábitos aquáticos mas também pode ter hábitos terrestres. Está activa
em Março-Outubro. Espécie essencialmente diurna e um pouco nocturna e crepuscular nos
meses mais quentes. É ágil, nada bastante bem e caça frequentemente dentro de água.
Habitats: Biótopos húmidos encontrando-se sobretudo em campos orvalhados e bosques, mas
no sul, ocorre usualmente perto de água.
Alimentação: Sapos, rãs, tritões, girinos, peixes, pequenos mamíferos, aves, outras cobras e
lesmas. Em média ingere uma presa a cada 20 dias.
Defesa: Quando encurralada enrola, incha o corpo e sopra com violência; depois achata a
cabeça e projecta-a como se fosse morder (muito raro). Liberta substâncias nauseabundas pela
cloaca ou vomita o conteúdo estomacal. Pode ainda finge-se de morta permanecendo imóvel
de costas com a boca aberta e a língua de fora.
Reprodução:
Entre a Primavera e o Outono – acasalamentos em Abril- Maio podendo ocorrer
também no final do Verão e do Outono;
Cópulas demoram cerca de três horas e as fêmeas frequentemente só copulam uma
vez;
A fêmea deposita 2-105 ovos, normalmente 30 para uma fêmea madura, 2-5 semanas
após a cópula; Os ovos são depositados em buracos, tocas de mamíferos, debaixo de
troncos e pedras e em montes de estrume e a incubação demora 6-10 semanas.
Distribuição: Um pouco por todo o país mas mais comum no norte e centro.
Estatuto/Legislação: Estatuto IUCN 1996 – Espécie de baixo risco/pouco preocupante (LR/LC).
Está incluída no anexo III da Convenção de Berna.

48
2. Cobra-de-água-viperina
Classe: Reptilia; Ordem: Squamata;
Família: Colubridae; Género: Natrix
Espécie: Natrix maura (Linnaeus,
1758)
Descrição: Cobra aglifa de tamanho
médio com 76,2 cm de comprimento
podendo atingir até 1 metro de
comprimento máximo. Apresenta
uma cabeça triangular com um
focinho aplanado e tem uma
coloração dorsal muito variada sendo
o castanho-escuro a cor
predominante. O desenho dorsal
consiste numa série de manchas escuras de forma e tamanho variáveis ao longo do dorso na
forma de um zig-zag escuro – mimetismo batesiano com as víboras) – o que lhe dá o nome.
Apresenta um padrão em “V” na cabeça e o ventre é normalmente esbranquiçado ou
avermelhado, com manchas escuras quadrangulares. Tem uma longevidade de 20 anos.
Hábitos: Espécie de hábitos aquáticos permanecendo activa de Março a Outubro e inactiva de
Novembro a Fevereiro. Apresenta actividade diurna e nocturna. É ágil em terra e ainda mais na
água, podendo trepar pedras e arbusto. É frequentemente avistada a aquecer-se sobre pedras
nas proximidades de água, semi-submersa ou ainda sobre plantas aquáticas.
Habitats: Encontrada na proximidade de qualquer massa de água – rio, ribeiro, poço, tanque,
charco, etc.
Alimentação: Larvas e adultos de anfíbios, peixes (água doce ou salobra), micromamíferos
(mais raro), anelídeos (minhocas e sanguessugas), répteis – osga ou até mesmo exemplares da
própria espécie.
Defesa: Normalmente foge para a água. Quando encurralada enrosca-se, sopra fortemente e
avança a cabeça para morder, fazendo-o de boca fechada. Quando capturada regurgita a
última vítima e liberta uma substância nauseabunda que esfrega no agressor com movimentos
corporais.
Reprodução: Tem duas épocas de reprodução – Primavera e Outono (no Outono não ocorre
postura – a fêmea só os põe na Primavera seguinte); A fêmea põe 1-14 ovos podendo chegar a
20 ovos (Crespo dependendo do seu tamanho. Estes são depositados na proximidade de água,
sob pedras, entre raízes de arbustos ou entre restos de decomposição. A incubação demora
cerca de 40-45 dias.
Distribuição: É comum em todo o país.
Estatuto/Legislação: Estatuto IUCN 2009 – Espécie pouco preocupante (LC). Está incluída no
anexo III da Convenção de Berna.

49
3. Cobra-de-ferradura
Classe: Reptilia;
Ordem: Squamata;
Família: Colubridae;
Género: Hemorrhois;
Espécie: Hemorrhois hippocrepis
(Linnaeus, 1758)
Descrição: O seu nome deve-se à mancha escura na parte superior da cabeça em forma de
ferradura. Mede entre 80-150cm de comprimento podendo chegar aos 180cm de
comprimento. Cabeça bem diferenciada, pupilas redondas e escamas suaves cujo padrão
dorsal consiste numa série de grandes manchas pouco espaçadas normalmente pretas ou
castanhas escuras. Os flancos têm uma ou mais filas de pequenas manchas pretas e a cor de
fundo varia do castanho ao cinzento com uma pigmentação escura que faz com que pareça
preta. O ventre é amarelo-alaranjado com algumas manchas escuras. Serpente aglifa, ou seja,
não possui colmilhos (dentes venenosos).
Hábitos: Espécie tímida e rápida de hábitos diurnos, podendo apresentar hábitos
crepusculares e nocturnos nas épocas mais quentes do ano. Está activa todo o ano excepto nas
zonas mais frias da sua distribuição, passando por um período de inactividade em Novembro-
Março. Pode escalar bastante bem a arbustos, em declive e em paredes rochosas.
Habitats: Prefere locais secos, muitas vezes rochosos, colinas arbustivas, planícies costais com
pouca vegetação e zonas agrícolas como amendoeiras, alfarrobeiras, oliveiras e vinhas, sendo
frequente em zonas urbanas junto a habitações, edifícios e paredes rochosas.
Alimentação: Pequenos mamíferos, ocasionalmente morcegos, répteis como a osga, lagartixa e
sardão, aves como o pardal, a andorinha e o pintassilgo.
Defesa: Foge e quando encurralada enrosca e dilata ligeiramente a cabeça, incha o corpo,
sopra ruidosamente e morde o agressor caso seja manejada.
Reprodução:
Decorre ao longo da Primavera e início do Verão;
As fêmeas põem até 29 ovos alongados depositados debaixo de pedras ou troncos ou
até mesmo em tocas de roedores. As posturas ocorrem em Junho e a eclosão dá-se
após 1-2 meses de incubação.
Distribuição: Localizada em vários pontos do país mas mais comum no centro e sul.
Estatuto/Legislação: Estatuto IUCN 2009 – Espécie pouco preocupantes (LC). Está incluída no
anexo II da Convenção de Berna e no anexo IV da Directiva Habitats.

50
4. Cobra-de-pernas-tridáctila
Classe: Reptilia;
Ordem: Squamata;
Família: Scincidae;
Género: Chalcides;
Espécie: Chalcides striatus
(Cuvier, 1829).
Descrição: De carácter
esquivo. Espécie de
dimensões médias podendo chegar aos 43,5cm de comprimento. Cabeça pouco diferenciada
do corpo, com uma forma triangular. Dorso acastanhado, podendo também ser acinzentado,
bege ou amarelado, com reflexos metálicos e, geralmente 9 bandas longitudinais escuras (às
vezes 11 ou 13). Corpo coberto por escamas lisas e brilhantes, dando-lhe um aspecto
serpentiforme. Membros de pequenas dimensões, cada um com 3 dedos. Cauda pode ser até
1,5 vezes do comprimento do corpo. Ventre esbranquiçado ou acinzentado.
Hábitos: Espécie de hábitos diurnos. Activa de Fevereiro a Outubro, altura em que entra em
hibernação. Apresentam temperaturas corporais na ordem dos 33°C.
Habitats: habitats com muita humidade, locais expostos com grande insolação e cobertura
herbácea abundante, como prados, campos agrícolas e clareiras de bosques; ocorre também
em zonas de pastagens ou substratos com abundante cobertura de folhas secas.
Alimentação: Consiste na sua maioria em invertebrados: lesmas, grilos, moscas, escaravelhos e
aranhas.
Defesa: Fuga. Pode perder a cauda voluntariamente
Reprodução:
Época inicia-se na Primavera e pode ir até ao Verão.
O acasalamento dá-se entre Abril e Maio;
Espécie ovovivípara.
Fêmea põe 1 a 15 crias totalmente desenvolvidas (eclosão ocorre em Julho e Agosto).
Distribuição: É comum em todo o país.
Estatuto/Legislação: Estatuto IUCN 2009 – Espécie pouco preocupante (LC). Está incluída na
convenção de Berna.

51
5. Cobra-rateira
Classe: Reptilia
Ordem: Squamata
Família: Psammophiidae
Género: Malpolon
Espécie: Malpolon
monspessulanus (Hermann,
1804)
Descrição: Serpente de
grandes dimensões,
ultrapassando facilmente os 2m de comprimento total. Tem uma cabeça relativamente
afunilada e estreita, com um focinho pontiagudo. Dimorfismo sexual ao nível da coloração:
machos – tonalidade de fundo entre os pardos claros e os esverdeados, desenho dorsal
uniforme, têm ainda uma mancha negra junto ao pescoço; fêmeas (e juvenis) – Camuflagem
de negros, brancos, cinzas e castanhos. A sua longevidade pode ultrapassar os 25 anos.
Hábitos: Activa da Primavera ao Outono. Hábitos diurnos e facilmente observada em dias
solarengos. Durante a estivação pode ser crepuscular. Produz um veneno neurotóxico forte
porém inofensivo para humanos pois os dentes inoculadores estão localizados na parte
traseira das mandíbulas.
Habitats: Grande variedade de biótopos, tais como áreas de mato, zonas pedregosas abertas,
ruínas, pedreiras, bosques de carvalhos, soutos, pinhais e montados, jardins e áreas agrícolas.
Alimentação: Varia consoante a idade: juvenis são insectívoros e depois passam a lagartixas,
cobras, pequenos roedores e crias de aves. Mais tarde a sua dieta pode incluir juvenis de
coelho-bravo e sardões adultos.
Defesa: Fuga. Pode optar por uma postura agressiva, projectando a cabeça contra o agressor.
Reprodução:
Época de reprodução na Primavera. Cópulas entre Maio e Junho;
Posturas de 4 a 20 ovos, depositados debaixo de matéria orgânica, pedras ou tocas.
Eclodem 2 meses depois.
Distribuição: É comum em todo o país.
Estatuto/Legislação: Estatuto IUCN 2009 – Espécie pouco preocupante (LC). Está incluída no
anexo III da Convenção de Berna.

52
6. Lagartixa-ibérica
Classe: Reptilia;
Ordem: Squamata;
Família: Lacertidae;
Género: Podarcis
Espécie: Podarcis hispanicus
(Steindachner, 1870)
Descrição: Atinge 6,5cm sem
contar com a cauda e tem
um padrão de desenho e
coloração muito variáveis geralmente castanho ou esverdeado. O ventre pode ser branco,
rosado ou vermelho e ocasionalmente amarelo. O padrão dorsal é geralmente às riscas, em
que a risca vertebral está mais difusa que as dorso-laterais. As fêmeas tem riscas regulares e as
dos machos são mais às manchas. Tem uma longevidade reduzida pois raramente passa os 3
anos.
Hábitos: Espécie ágil, desconfiada e esquiva encontrando-se activa praticamente todo o ano
desde que as temperaturas sejam superiores a 13ºC. É unimodal – observada da parte da
manhã.
Habitats: Rochas, muros, amontoados de pedras, paredes, parapeitos, afloramentos e troncos
de árvores nalguns locais.
Alimentação: Artrópodes tais como moscas, mosquitos, centopeias, aranhas, formigas,
gafanhotos e escaravelhos, indivíduos mais pequenos da própria espécie, detritos e restos de
alimentos em zonas urbanas.
Defesa: Fuga. Emite sons quando capturada tentando morder o agressor.
Reprodução:
Começa em Fevereiro podendo prolongar-se até Abril e as posturas podem ir até
Julho.
Fêmeas põem 1-5 ovos que são chocados em 8 semanas. Podem realizar até 3
posturas por ano sendo o tempo entre cada postura de 22-34 dias.
Distribuição: Por todo o país mas mais abundante no norte e litoral centro.
Estatuto/Legislação: Estatuto IUCN 2009 – Espécie pouco preocupante (LC). Está incluída no
anexo III da Convenção de Berna.

53
7. Lagartixa-do-mato
Classe: Reptilia
Ordem: Squamata
Família: Lacertidae
Género: Psammodromus
Espécie: Psammodromus algirus
(Linnaeus, 1758)
Descrição: Espécie de lagartixa de
tamanho grande e relativamente
larga. Os machos medem, da cabeça ao corpo, 9,1cm e as fêmeas 9,3cm. A cauda mede cerca
de 23,0cm, sendo 2-3 vezes o tamanho do corpo. Tem escamas dorsais grandes e planas –
imbricadas – terminadas em ponta e fortemente aquilhadas. A coloração dorsal é parda
podendo variar do oliváceo ao cobre. Tem duas linhas laterais esbranquiçadas (entre as linhas
apresenta uma tonalidade escura). A zona ventral é sobretudo esbranquiçada com tons
avermelhados ou alaranjados em especial na zona da cauda e apresenta ainda ocelos azulados
atrás dos membros anteriores. As fêmeas têm 1-3 ocelos e machos têm 2-7 ocelos. A sua
longevidade é de 5 a 7 anos.
Hábitos: Espécie extremamente ágil e bastante heliófila – retira-se quando os últimos raios de
sol desaparecerem. Activa desde finais de Fevereiro a Outubro, sendo estritamente diurna
(máximos de actividade durante a manhã e tarde).
Habitats: Zonas de arbustos baixos com grande cobertura de ervas ao nível do solo e evita
substratos arenosos, sendo bastante abundante em pinhais.
Alimentação: Aracnídeos, hemípteros, formicídeos, coleópteros, homópteros, artrópodes e
dípteros.
Defesa: Imobilizam-se por completo ao escutarem ruídos estranhos e se algo se aproxima
fogem velozmente. Quando capturados emitem um grito agudo e curto (audível), tentando
morder o agressor.
Reprodução:
Macho com coloração vermelha (dimorfismo sexual); Machos reproduzem-se com 1-3
fêmeas e as fêmeas com 1-2 machos; O macho escolta à fêmea durante 1-3 dias;
Espécie ovípara – fêmea pode realizar várias posturas por ano. Esta ocorre 30-40 dias
após a cópula, na qual são postos 1-11 ovos, dependendo do tamanho da fêmea. A
incubação demora cerca de 35-45 dias.
Distribuição: É comum em todo o país.
Estatuto/Legislação: Estatuto IUCN 2009 – Espécie pouco preocupantes (LC). Está incluída no
anexo III da Convenção de Berna.

54
8. Lagartixa-de-dedos-dentados
Classe: Reptilia;
Ordem: Squamata;
Família: Lacertidae;
Género:
Acanthodactylus
Espécie:
Acanthodactylus
erythrurus (Schinz,
1833)
Descrição: Tem
aproximadamente 7,8cm de comprimento (cabeça-corpo). O dorso apresenta uma série de
bandas longitudinais castanhas, com manchas escuras intercaladas por riscas mais finas de
tonalidade mais clara.
Hábitos: Atinge o seu pico de actividade quando chega a temperaturas corporais de 25 a 30°C
(actividade directamente relacionada com o seu poder de termorregulação). Hibernam de
acordo com a temperatura ambiental em raízes, galerias, tocas ou pedras. Pode ter actividade
unimodal (contínua) ou bimodal (intercalando um período de interrupção e dois de actividade)
Habitats: Geralmente zonas do litoral em pinhais e áreas de cobertura arbustiva mais ou
menos dispersa.
Alimentação: Composta sobretudo por formigas e escaravelhos. Também podem comer
algumas plantas e flores.
Defesa: Fuga.
Reprodução:
Época de reprodução de Março a Junho;
Fêmeas adquirem tonalidades avermelhadas (dimorfismo sexual);
Posturas de 2 a 5 ovos ocorrendo entre Junho e Agosto; primeiras eclosões em Agosto.
Distribuição: Vários núcleos populacionais isolados de Norte a Sul de Portugal.
Estatuto/Legislação: Estatuto IUCN 2009 – Espécie pouco preocupante (LC); Estatuto ICNF
2006 – Espécie quase ameaçada (NT). Está incluída no anexo III da convenção de Berna.

55
9. Lagarto-de-água
Classe: Reptilia;
Ordem: Squamata;
Família: Lacertidae;
Género: Lacerta
Espécie: Lacerta
schreiberi – Bedriaga,
1878
Descrição: Espécie
com 12-13,5cm de
comprimento do corpo, cauda com 2x o comprimento do corpo. Machos predominantemente
verdes com pequenas manchas pretas, maiores no dorso do que nos flancos e fêmeas
geralmente acastanhadas ou esverdeadas com flancos verdes ou castanhos e com o mesmo
tipo de manchas dos machos. O ventre é às manchas nos machos mas não tão marcado nas
fêmeas, a garganta é branca ou azul nos machos na época de acasalamento e também em
algumas fêmeas, podendo esta cor cobrir toda a cabeça. Os juvenis são acastanhados ou
pretos com manchas redondas ou em barras, brancas ou amareladas debruadas a preto, com a
cauda geralmente laranja ou amarela; estas manchas podem ficar retidas nos sub-adultos. A
longevidade é cerca de 8 anos.
Hábitos: Espécie relativamente ágil mantendo-se activa com hábitos diurnos de Março a
Outubro.
Habitats: Zonas húmidas e frescas, geralmente ao longo de cursos de água com uma certa
corrente e vegetação ripícola densa, como silvas e urtigas podendo surgir no leito ou nas
margens rochosas. Os juvenis podem deslocar-se para meios mais secos.
Alimentação: Moscas, escaravelhos, gafanhotos, abelhas, larvas diversas, aranhas, caracóis e
frutos maduros, como amoras.
Defesa: Geralmente fogem, podendo morder se capturados.
Reprodução:
Acentuado dimorfismo sexual – machos mais coloridos e mais robustos que as fêmeas;
Cópulas em Março-Abril e por vezes violentas (ocorrência de feridas nas fêmeas);
Espécie ovípara. Porém a fêmea pode reter os ovos dentro dela durante 3 semanas
antes da postura. Enterra 5-25 ovos e a incubação demora 65-110 dias.
Distribuição: Todo o país acima do rio Tejo, na Serra de S. Mamede e na Costa Vicentina.
Estatuto/Legislação: Estatuto IUCN 2009 – Espécie quase ameaçada (NT). Está incluída no
anexo II da Convenção de Berna e nos anexos II e IV da Directiva Habitats.

56
10. Osga-comum
Classe: Reptilia;
Ordem: Squamata;
Família: Phyllodactylidae
Género: Tarentola
Espécie: Tarentola mauritanica
(Linnaeus, 1758)
Descrição: Pode atingir os 8,5
cm de comprimento. Tem uma
cabeça grande e bem
diferenciada do corpo. Olhos com pupila vertical. Possui um aspecto aplanado, com quatro
membros pentadáctilos (com cinco dedos). Geralmente a cauda atinge o mesmo comprimento
que o conjunto da cabeça e do corpo. Os dedos têm lamelas (lâminas) digitais não subdivididas
centralmente e as unhas só são visíveis no 3º e 4º dedo. São estas duas características que
permite distingui-la da osga-turca (Hemidactylus turcicus). Coloração muito variada: parte
dorsal esbranquiçada ou acinzentada e com tons de castanho. Chega aos 8-9 anos em
cativeiro.
Hábitos: Com actividade crepuscular e nocturna no verão, podendo ser observados durante o
dia na Primavera e Outono. Hibernam de Novembro-Dezembro a Março. Pode conviver com
várias espécies de lagartos e lagartixas. A sua capacidade de se manter em superfícies verticais
deve-se ao efeito de sucção das lâminas que possui nos dedos e às sedas que cobrem essas
lâminas.
Habitats: Essencialmente habitats rochosos, tais como rochedos, muros, montes de pedras e
paredes de casas, podendo também viver em troncos de árvores.
Alimentação: Sobretudo insectos e aranhas. Podem também ingerir certos vegetais. Os
adultos de maiores dimensões podem ainda predar pequenas lagartixas e outros vertebrados.
Defesa: Pode perder a cauda (esta regenera mas nunca recupera a cor original).
Reprodução:
Pode haver duas épocas de reprodução, de Março a Abril e de Junho a Julho.
Posturas de cerca de 2 ovos brancos de casca dura, depositados em fendas ou debaixo
de pedras. Por vezes, várias fêmeas colocam os ovos no mesmo local. A incubação
pode durar entre 40 e 120 dias.
Distribuição: Amplamente distribuída em Portugal, estando apenas ausente em parte do
quadrante Noroeste.
Estatuto/Legislação: Estatuto IUCN 2009 – Espécie pouco preocupante (LC). Está incluída no
anexo III da Convenção de Berna.

57
11. Sardão
Classe: Reptilia;
Ordem: Squamata;
Família: Lacertidae;
Género: Lacerta
Espécie: Lacerta lepida (Daudin,
1802); Timon lepidus (Daudin,
1802);
Descrição: É o maior lagarto da
Península Ibérica, podendo alcançar 80 cm de comprimento total. De aspecto robusto, possui
uma cabeça proeminente, membros fortes e uma cauda muito comprida. O dorso é verde e
amarelado, com manchas escuras. Nos flancos tem 3 ou 4 filas de manchas azuis (ocelos) por
vezes rodeadas de preto. A cauda tem a mesma coloração do corpo excepto quando se trata
de uma cauda regenerada. O ventre é esbranquiçado ou amarelado. A sua longevidade é
superior a 11 anos (17 a 20 anos em cativeiro).
Hábitos: Os machos são extremamente territoriais, expulsando os competidores com violência.
O período de hibernação inicia-se em Outubro ou Novembro e dura geralmente até Fevereiro
ou Março. Nos meses de Verão permanecem inactivos nas horas de maior calor, apresentando
picos de actividade de manhã e ao fim do dia.
Habitats: Ocorre principalmente em zonas de mato mediterrânico com áreas abertas. Pode
aparecer também na periferia das cidades e em terras de cultivo. Prefere sobretudo locais com
abundância de abrigos, como aglomerados de pedras, muros, arbustos espessos, e com boa
exposição solar. Em geral evita locais húmidos e sombrios e matas densas.
Alimentação: Alimenta-se sobretudo de insectos. Pode também alimentar-se de caracóis,
lesmas, lagartixas, cobras de água e outros répteis, pequenos mamíferos, ovos e crias de aves,
frutas e restos vegetais. Também podem predar os juvenis da própria espécie.
Defesa: Podem soltar a cauda se se sentirem ameaçados (esta regenera-se).
Reprodução:
O macho persegue a fêmea mordiscando-a na parte traseira do corpo. De seguida
imobiliza-a, para evitar ser mordido, e dá-se a cópula.
A gestação dura entre 71 e 102 dias; a fêmea deposita entre 5 e 22 ovos debaixo de
pedras ou troncos, podendo também enterrá-los. A incubação pode durar entre 2 a 3
meses.
Distribuição: Um pouco por todo o país mas mais comum no Centro e Sul.
Estatuto/Legislação: Estatuto IUCN 2009 – Espécie quase ameaçada (NT); estatuto ICNF –
Pouco Preocupante (LC). Está incluída no anexo II da Convenção de Berna.


















