
Filogeografia de Pipa carvalhoi (Miranda-Ribeiro,...
68
1 UNIVERSIDADE FEDERAL DO ESPÍRITO SANTO CENTRO DE CIÊNCIAS HUMANAS E NATURAIS PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS BIOLÓGICAS Filogeografia de Pipa carvalhoi (Miranda-Ribeiro, 1937) Paulo Victor Scherrer de Araújo Vitória, ES 2016
Transcript of Filogeografia de Pipa carvalhoi (Miranda-Ribeiro,...

! 1!
UNIVERSIDADE FEDERAL DO ESPÍRITO SANTO
CENTRO DE CIÊNCIAS HUMANAS E NATURAIS
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS BIOLÓGICAS
Filogeografia de Pipa carvalhoi (Miranda-Ribeiro, 1937)
Paulo Victor Scherrer de Araújo
Vitória, ES
2016

! 2!
UNIVERSIDADE FEDERAL DO ESPÍRITO SANTO
CENTRO DE CIÊNCIAS HUMANAS E NATURAIS
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIAS BIOLÓGICAS
Filogeografia de Pipa carvalhoi (Miranda-Ribeiro, 1937)
Paulo Victor Scherrer de Araújo
Orientadora: Leonora Pires Costa
Dissertação submetida ao Programa de Pós-Graduação em Ciências Biológicas (Biologia Animal) da Universidade Federal do Espírito Santo como requisito parcial para a obtenção do grau de Mestre em Biologia Animal
Vitória, ES
Agosto, 2016

! 3!
AGRADECIMENTOS
Agradeço primeiramente à minha esposa, Rebecca Pereira Cruz, por todo amor, carinho e
apoio emocional durante esse processo. Você foi muito mais do que esposa, foi também
assistente de laboratório, parceira de campo e revisora. Dedico essa conquista a você.
A meus pais, meu irmão e minha cunhada por toda motivação e apoio. Vocês são meus
principais professores e dedico não só essa, mas todas as conquistas a tudo que
investiram em mim.
A minha orientadora Leonora Pires Costa por mais uma vez ter aceitado orientar um
projeto com animais de pele lisa. Sua ajuda foi fundamental para minha formação como
biólogo e agora, como mestre em biologia animal. Obrigado por todo apoio e pela
confiança depositada em mim.
Um agradecimento especial a Eduardo Loyola Muhl, meu grande parceiro desde os
tempos de graduação, sempre disposto a ajudar, ser ajudado e virar madrugadas
trabalhando. Precisamos dormir mais.
J. F. R. Tonini, C. S. Lima, J. Justino e E. L. Muhl pelos ensinamentos das técnicas
laboratoriais. Agradeço também a todos os colegas do Núcleo de Genética aplicada à
Conservação da Biodiversidade, UFES, por me socorrerem em diversos momentos
durante esses anos de convivência. Eu nunca conseguiria gerar uma sequência de DNA
sem a ajuda de vocês.
Aos companheiros LaMabianos (Laboratório de Mastozoologia e Biogeografia – UFES)
por todos os momentos de debate, discussão e descontração que tornam o dia-a-dia de
trabalho muito mais agradável. Trabalhar ao lado de vocês é uma honra e esse trabalho
contou com a contribuição de cada um de vocês.
Konrad Radzikowski pelo companherismo nos campos e toda parceria durante o período
que esteve no Brasil. Danke Schön!

! 4!
A. Lima, A. Garda, R. Ávila, M. Napoli, M. Trefaut, D. Santana e C. Haddad, demais
curadores e técnicos em curadoria pela doação de alíquotas de tecidos para a realização
das análises moleculares. A. Fouquet, D. Massemin e R. Ernst por toda ajuda em relação
aos Pipidae.
Aos professores do PPGBAN por todos os ensinamentos e trocas de ideias. Vocês são
todos grande inspiração na minha vida profissional.
Aos professores componentes da banca pela disponibilidade de avaliar e contribuir para o
enriquecimento desse trabalho.
Ao Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) e à
Fundação de Amparo a Pesquisa e Inovação do Espírito Santo (FAPES) pelos recursos
aplicados ao Núcleo de Genética Aplicado à Conservação da Biodiversidade. À FAPES e
à Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) pela bolsa de
mestrado.
Por fim, a todos os demais envolvidos de alguma forma na minha vida profissional e
acadêmica. Muito obrigado aos meus queridos alunos, aos funcionários da UFES e aos
meus colegas do PPGBAN.

! 5!
Resumo
Pipa carvalhoi (Anura: Pipidae) é um anfíbio aquático primitivo que se distribui ao
longo das drenagens costeiras do leste do Brasil. Sua distribuição é marcada por uma
disjunção superior a 500 km onde não existem registros da ocorrência do animal. Suas várias
adaptações ao hábito de vida subaquático, que incluem a retenção de linha lateral, ausência
de língua e alimentação por sucção sugerem baixa capacidade de dispersão por terra e
consequentemente, alta estruturação genética entre suas populações. O objetivo do presente
estudo foi, então, compreender os padrões de diversificação genética de Pipa carvalhoi ao
longo de sua distribuição geográfica e a influência de seu hábito de vida nos processos
históricos que moldaram sua diversidade. Foram sequenciados os marcadores mitocondriais
12S, 16S e NADH desydrogenase (ND2) assim como os marcadores nucleares Seventh in
Absentia (SIA) e Tyrosinase (TYR) para 79 amostras de tecidos de P. carvalhoi, provenientes
de grande parte de sua distribuição geográfica, além de outras duas espécies do gênero. Foi
revelada uma forte estruturação genética associada a divisão das principais regiões
hidrográficas. Sua diversificação data do Paleogeno e parece estar associada ao surgimento
das principais drenagens costeiras do Brasil. A alta estruturação genética, a ausência de
indícios recentes de fluxo gênico e a divergência profunda revelam diversidade críptica ao longo
da distribuição de Pipa carvalhoi.
Palavras-chaves: Filogenética; Biogeografia; Anura; Mata Atlântica; Pipidae.

! 6!
SUMÁRIO
Introdução ................................................................................................................ 5
Objetivos .................................................................................................................. 8
Objetivo Geral ......................................................................................................... 8
Objetivos Específicos .............................................................................................. 9
Materiais e Métodos ................................................................................................ 9
Amostras Populacionais .......................................................................................... 9
Métodos Moleculares e Filogenéticos ..................................................................... 10
Resultados ............................................................................................................... 12
Discussão ................................................................................................................. 17
Conclusões .............................................................................................................. 29
Referências Bibliográficas ...................................................................................... 31
TABELAS .................................................................................................................. 40
FIGURAS ...................................................................................................................
APÊNDICES ..............................................................................................................
45
56

! 7!
1. Introdução
Estudos de genética molecular apontam que a diversidade de espécies
reconhecidas está subestimada em grande parte dos animais e plantas (Oliver et al.
2013). Em anfíbios, essa subestimação é evidente, uma vez que nos últimos 20 anos,
aproximadamente 3000 espécies foram descritas (Köhler et al. 2005; Glaw & Köhler,
1998; Köhler et al. 2005; AmphibiaWeb, 2016). Dessa forma, observa-se um aumento de
mais de 35% no número absoluto de espécies, sendo este decorrente de dois fatores: i:
acesso a regiões tropicais anteriormente não exploradas, aumentando o número de
espécies endêmicas) e ii: abordagens teóricas associadas de novas metodologias, como
a filogeografia e a bioacústica, que aliadas à taxonomia clássica, revelam a diversidade
de espécies crípticas (Köhler et al. 2005; Fouquet et al. 2007).
Das 7528 espécies de anfíbios reconhecidas atualmente, 88% pertencem à Ordem
Anura (AmphibiaWeb, 2016), distribuídas, principalmente, na região Neotropical (Köhler
et al. 2005), na qual o Brasil possui o maior índice de riqueza de anuros em âmbito
global, sendo 982 espécies registradas até o momento no total. Dessas, mais de 500
(Haddad et al. 2013) ocorrem na Mata Atlântica (MA), um dos biomas mais ameaçados
do mundo em decorrência da ocupação humana. Dos seus mais de 1.300.000 km² de
mata originais, a MA está hoje reduzida a menos de 8% de sua extensão, representada
por pequenos fragmentos dispersos ao longo da costa brasileira e em Missiones,
Argentina (Galindo-Leal & Câmara, 2005). Sua relevância ecológica, atrelada à ameaça
de destruição que vem sofrendo, evidenciam a importância de se estudar os processos
causadores dessa diversidade.
Estudos filogeográficos têm sido desenvolvidos, com a utilização de diferentes
organismos como modelo, como anfíbios e mamíferos, na busca por padrões que
buscam investigar a diversidade de espécies e os processos históricos ligados à

! 8!
especiação neste bioma (e.g. Carnaval, 2002; Pellegrino et al., 2005; Grazziotin et al.,
2006; Carnaval et al., 2009; Fitzpatrick et al., 2009; Costa & Leite, 2012). Observa-se
diferentes propostas para explicar a origem da alta diversidade de espécies encontrada
na MA, como as hipótese dos refúgios pleistocênicos (Haffer, 1969; Carnaval et al.
2009), a hipótese de isolamento por rios (Wallace, 1952; Sick, 1967), hipótese dos
gradientes ecológicos (Endler, 1982), hipótese paleogeográfica (Chapman, 1917; Bates,
2001) e, mais recentemente, hipótese da Mata Atlântida (Leite et al., 2016).
Apesar dos muitos esforços empregados na busca por um padrão geral que
compreendesse toda a diversidade registrada na MA, os estudos indicam que espécies
diferentes, mesmo aquelas com distribuição geográfica similar, podem ter sido
influenciadas por processos históricos distintos (Batalha-Filho & Miyaki, 2011; Tonini et
al. 2013). Sendo assim, para compreender corretamente o padrão filogeográfico de uma
espécie, além dos aspectos ambientais associados, deve ser levado em conta também
sua história natural, já que vários fatores como filopatria, capacidade de dispersão,
competição com espécies próximas e presença de parasitas e patógenos podem
influenciar diretamente na estruturação geográfica das populações (Martins &
Domingues, 2011; Tonini et al. 2013). Em seus estudos sobre anfíbios amazônicos,
Fouquet et al. 2007 menciona que espécies de anuros que são associadas a áreas
abertas, como Leptodactylus fuscus, Adenomera hylaedactyla e Scinax ruber, bem como
as que estão associadas a rios e grandes pântanos, como Lithobates palmipes, Pipa
pipa, Sphaenorhynchus lacteus, Hypsiboas ranipes e Dendropsophus nanus, tendem a
apresentar menor estruturação filogeográfica.
Os Pipidae são anuros primitivos altamente adaptados à vida aquática, que se
distribuem ao longo da África subsaariana (gêneros Xenopus, Hymenochirus e
Pseudhymenochirus) e América do Sul e Central (gênero Pipa). Adaptações para o
hábito subaquático incluem retenção de linha lateral no adulto, ausência de língua,

! 9!
alimentação por sucção, ausência de tímpano, ausência de cordas vocais e
membranas interdigitais (Trueb & Cannatella, 1986; Tobias et al. 2011). Suas
espécies habitam corpos d’água permanentes e temporários, bem como riachos
lênticos e remanso de córregos, onde camuflam-se na lama e em folhas depositadas
no substrato (Carvalho, 1937; Trueb &.Cannatella, 1986). Xenopus corresponde ao
gênero mais estudado da família, sendo também o mais diverso, com 29 espécies
atualmente reconhecidas. Sabe-se que esses organismos apresentam uma alta
capacidade de migração entre habitats, tanto entre rios diferentes, no período seco,
quanto entre poços, no período úmido (Rödel, 2000).
Diferentes estudos apontam que, em espécies aquáticas, a estruturação
genética pode ocorrer devido à separação geográfica entre bacias hidrográficas
(Lovejoy & de Araujo 2000; Turner et al. 2004; Hubert et al. 2007; Sistrom et al. 2009;
Cooke et al. 2009;Pearse et al. 2006). Em Xenopus, porém, observa-se que
determinadas espécies possuem ampla distribuição, com alta conexão entre
populações de diferentes bacias hidrográficas, sugerindo que a divisão entre bacias
não deve ter sido o fator primordial na diversificação desse gênero (Evans et al. 2004;
2015; Furman et al. 2015). A separação das espécies Xenopus ocorreu em períodos
anteriores ao Pleistoceno, com os processos de especiação relacionados a eventos
alopoliploidia independentes, em que duas espécies se hibridizam e o zigoto é
estabilizado pela duplicação cromossômica (Kobel, 1996; Evans et al. 2004; 2015).
Pipa é o gênero endêmico da região Neotropical, composto por sete espécies,
seis das quais ocorrem ao longo e acima das bacias dos rios Amazonas e Orinoco (P.
arrabali, P. aspera, P. myersi, P. parva, P. pipa e P. snethlageae) e uma espécie (P.
carvalhoi) com ocorrência na costa leste do brasil, abrangendo os biomas da MA e
Caatinga, com limite sul em Vitória, Espírito Santo (Silva et al. 2010; Santana et al.
2014). Dentre as características que distinguem o gênero Pipa dos demais Pipidae

! 10!
africanos está o formato das pontas dos dedos e sua reprodução, em que os ovos
são carregados nas costas da fêmea durante o desenvolvimento (Trueb & Cannatella,
1986; Azrabe, 1999; Canedo, 2006). P. carvalhoi pode ocupar ambientes
relativamente impactados, sendo frequentemente encontrado em meios antropizados,
incluindo lagos artificiais (Silva et al. 2010; Santana et al. 2014). Possui distribuição
disjunta no leste do Brasil, com um primeiro agrupamento de populações no Nordeste
(Ceará, Paraíba, Pernambuco, Rio Grande do Norte e Sergipe), e um segundo ao
longo do Sul da Bahia, Espírito Santo e Minas Gerais (Silva et al. 2010; Santana et al.
2014).
Trabalhos sobre filogeografia de anfíbios na MA já levaram em conta espécies
com diferentes modos de vida, como arborícolas (Carnaval et al., 2009; Tonini et al.
2013), terrestres (Thomé et al. 2010; Fouquet et al. 2012) e semi-fossoriais (Tonini et
al., 2013). Porém, segundo Tonini et al. 2013, a diferença no modo de vida das
espécies pode acarretar respostas diferenciadas aos eventos biogeográficos e
paleoclimáticos comuns à área de distribuição.
Para Pipa carvalhoi há um desconhecimento das relações filogeográficas, dos
processos históricos relacionados à sua distribuição e da influência de seu hábito de
vida nos padrões de segregação das populações. Por tratar-se da única espécie
subaquática dentro da diversidade de anuros que abarca sua distribuição, questiona-
se se P. carvalhoi pode revelar padrões filogeográficos completamente distintos
àqueles conhecidos para outras espécies de hábitos distintos. Dessa forma,
objetivamos i: investigar a distribuição da diversidade genética de P. carvalhoi ao
longo de sua distribuição geográfica; ii: compreender se há segregação entre as
populações associadas à diferentes bacias hidrográficas.
2. Objetivos:

! 11!
2.1. Objetivo Geral:
Compreender os padrões de diversificação genética de Pipa carvalhoi ao longo
de sua distribuição geográfica.
2.2. Objetivos Específicos:
i. Mapear a diversidade genética e relação filogenética entre as diferentes
populações de P. carvalhoi;
ii. Avaliar e datar os processos históricos ligados a diversificação genética de P.
carvalhoi;
iii. Estudar a ligação entre P. carvalhoi e as espécies amazônicas de Pipa, além
de reconstruir potenciais rotas de colonização da Mata Atlântica por esta espécie.
3. Material e Métodos
3.1 Amostras Populacionais
Para as análises genéticas foram utilizados tecidos muscular e hepático,
conservados em etanol 95%.. Parte do material foi obtido por doação das coleções “Célio
Fernando Batista Haddad” (CFBH) da Universidade Federal de São Paulo (UNESP),
Coleção Herpetológica da Universidade Federal do Rio Grande do Norte (CHBEZ),
Coleção de Anfíbios e Répteis da Universidade Rural do Cariri (URCA-G), Coleção de
Anfíbios da Universidade Federal da Bahia (UFBA), Coleção de Anfíbios e Répteis do
Instituto Nacional de Pesquisas da Amazônica (CHINPA), por amostras cedidas pelos
pesquisadores Dr. Miguel Trefaut Rodrigues da Universidade de São Paulo (USP) e Dr.
Diego José Santana da Universidade Federal de Mato Grosso do Sul (UFMS). O restante
das amostras foram coletadas durante este estudo nos municípios de Santa Teresa (ES),
Santa Maria de Jetibá (ES), Colatina (ES), e em Poté (MG) (Figura 1). A captura dos

! 12!
espécimes foi realizada por meio de busca ativa em corpos d’água, tanto temporários
quanto permanentes, com auxílio de um puçá de pesca de malha fina. Os exemplares
coletados foram eutanasiados em etanol 20% para retirada de tecido hepático, fixados em
formalina 10% por 48 horas e transferidos para conservação em etanol 70%.
Foram obtidos um total de 79 amostras de tecido hepático ou muscular, sendo
destas, 55 amostras de Pipa carvalhoi distribuidas ao longo de toda a distribuição
geográfica conhecida para a espécie (Figura 1), além de 21 amostras de P. pipa e 2
amostras de P. arrabali. Adicionalmente, foram utilizadas sete amostras de outras
espécies da família Pipidae, disponíveis no Genbank nas análises do marcador molecular
12S (Apêndice 1). A lista de amostras e suas respectivas localidades estão representadas
na Tabela 1.
3.2 Métodos Moleculares e Filogeográficos
As amostras de tecido foram submetidas ao protocolo de extração com sal descrito
por Bruford et al. (1992) e posteriormente submetidas à eletroforese em gel de agarose
1% para checagem do sucesso da extração de DNA.
Os marcadores mitocondriais NADH dehydrogenase 2 (ND2), 16S, 12S, e os
nucleares Seventh in Absentia (SIA) e Tyrosinase (TYR) foram amplificados através da
clonagem in vitro pela reação em cadeia da polimerase (PCR) em um termociclador. Os
primers utilizados e os perfis de PCR encontram-se na Tabela 2.
Os produtos de PCR foram purificados com ExoSAP-IT (GE Healthcare Life
Sciences) e sequenciados com o kit BigDye Terminator 3.1 (Applied Biosystems). As
sequências foram obtidas por meio do sequenciador automático ABI 3500 (Applied
Biosystems) no Núcleo de Genética Aplicada à Biodiversidade da Universidade Federal
do Espírito Santo – UFES.

! 13!
As sequências foram alinhadas pelo método MUSCLE (Edgar, 2004) implementado
no software MEGA 6 (Tamura et al. 2013) e, posteriormente, conferidas manualmente. O
melhor modelo mutacional foi calculado para cada marcador molecular utilizado pelo
software JModelTest (Posada, 2008). A partir das sequências alinhadas foram geradas
árvores de afinidade filogenética pelos algoritmos de Máxima Verossimilhança (MV) e
Inferência Bayesiana (IB) utilizando-se, respectivamente, os softwares MEGA 6 (Tamura
et al., 2013) e BEAST v1.6.2 (Drummond & Rambaut, 2007). Para cada algoritmo, 6
cladogramas foram gerados: um para cada marcador molecular individual e os demais
para os cinco marcadores concatenados. Os valores para suporte dos clados da MV foi
gerado por meio de 1000 replicações de bootstrap. Para a IB foi utilizado o algoritmo de
integração de Monte Carlo via cadeias de Markov, percorrendo 10 milhões de gerações,
com árvores amostradas a cada 1000 gerações, descartando-se as 1000 primeiras como
burn-in. Como grupo externo utilizado para as análises filogenéticas foram alocados os
táxons Pseudopaludicola sp., Pipa pipa e P. arrabali.
As distâncias genéticas foram geradas pelo software MEGA 6 (Tamura et al. 2013)
utilizando o modelo de Kimura-2-parâmetros (K2P). Dados de dinâmica populacional
foram estimados por meio de testes de neutralidade D de Tajima (Tajima, 1989), Fs de Fu
(Fu, 1997), diversidade nucleotídica (π) e haplotípica (h). Foi testada a correlação entre
distância genética e distância geográfica através do teste de Mantel pelo software Past
3.11 (Hammer et al. 2001). Para avaliar a conexão entre os haplótipos foram geradas
redes de haplótipos pelo software PopArt (Bandelt et al., 1999).
O relógio molecular foi calculado de forma a estimar, em escala temporal, a
separação das linhagens de P. carvalhoi e também da separação de P. carvalhoi das
demais espécies de Pipa. Os tempos de divergência foram calculados a partir de duas
diferentes matrizes de dados. A primeira matriz contava com seis espécies de Pipidae,
incluindo três espécies de Pipa (P. carvalhoi, P. parva e P. pipa) e três espécies africanas

! 14!
(Hymenochirus sp., Silurana tropicalis e Xenopus borealis). Tal matriz foi utilizada para
calcular, a partir do marcador 12S, a origem do gênero Pipa e da separação entre as
linhagens das três principais linhagens do gênero. A segunda matriz contou com os cinco
marcadores concatenados, utilizando amostras de P. carvalhoi dos três clados
recuperados pelas análises filogenéticas, além de P. pipa e P. arrabali. Em ambas as
análises foi utilizado relógio molecular relaxado log-normal. O ponto de calibração foi, na
primeira análise, baseado na hipótese de que a separação entre Pipa e os outros gêneros
de Pipidae foi desencadeada pela quebra da supercontinente Gondwana, onde América
do Sul e África eram terras contínuas, separando esses continentes. Diante disso,
calibramos a base do gênero Pipa com uma distribuição normal a priori tendo média de
102 Ma e desvio padrão de 7 Ma.
O número de haplótipos (H) e de sítios polimórficos (S), testes de neutralidade (D
de Tajima (1989); Fs de Fu (1997)) e índice de divergência genética (Fst) foram
estimados por meio do software DnaSP v.5 (Librado e Rozas, 2009). A correlação entre
distâncias genéticas e geográficas foi verificado por meio do Teste de Mantel
implementado pelo software Arlequin 3.5 (Excoffier e Lischer, 2010).
4. Resultados
Foram obtidas sequências de DNA mitocondrial de Pipa carvalhoi de 41 amostras
para o marcador 12S, 50 amostras para o marcador 16S, 40 amostras para o marcador
ND2, além de sequências de DNA nuclear de 54 amostras para o marcador SIA e 21
amostras para o marcardor TYR. Foram obtidas, ainda, 18 sequências de P. pipa e duas
de P. arrabali para cada um dos 5 marcadores genéticos, com exceção de ND2, em que
não foi possível o sequenciamento de P. arrabali e somente 6 sequências de P. pipa
foram obtidas com sucesso. No alinhamento, incluindo as três espécies de Pipa, as
matrizes de dados apresentaram 329 pares de bases (pb) para 12S, 370 pb para 16S,

! 15!
867 pb,para ND2, 376 pb para SIA e 300 pb para TYR. Os marcadores nucleares foram
os mais conservados. Dentre eles, SIA apresentou apenas 4% de sítios variáveis (15
sítios) e TYR apresentou 11,3% de sítios variáveis (34 sítios). Quanto aos marcadores
mitocondriais, uma maior porcentagem de sítios polimórficos foi recuperado, sendo eles
de 29,5% para 16S (109 sítios), 45,5% para 12S (143 sítios) e 66,6% para ND2 (577
sítios).
Os conjuntos de dados não apresentaram saturação de substituições de bases
significativa pelo índice de saturação do método de Xia e Xie (2001). O melhor modelo
evolutivo selecionado para as conjuntos de dados foi HKY para SIA, HKY +G para 16S,
HKY +I para 12S e TYR e GTR +G para ND2.
Pipa carvalhoi foi recuperado monofilético em todas as topologias com alto suporte,
independente do algorítmo utilizado. O monofiletismo de P. carvalhoi foi recuperado
também nas análises concatenadas com todos os marcadores. P. pipa e P. arrabali foram
sempre mais próximas entre si quando comparadas a P. carvalhoi nas análises
filogenéticas. Nas árvores de IB e MV para as matrizes de 12S, 16S e concatenadas, foi
recuperado o monofiletismo das três espécies (Apêndice 2 a 5, 12 e 13). Para as matrizes
de TYR e SIA, porém, P. arrabali e P. pipa formaram uma grande politomia, tanto na IB
quanto na MV (Apêndice 8 a 11). A matriz de dados de ND2 não apresentou amostras de
P. arrabali, portanto, nas árvores de IB e MV para este gene só foi recuperado o
monofiletismo de P. carvalhoi e P. pipa (Apêndice 6 e 7). A matriz de 12S recuperou a
espécie P. parva como grupo mais basal dentro do gênero Pipa (Figura 2).
Na árvore de espécies, assim como dentre as árvores de genes, para os
marcadores mitocondriais (12S, 16S e ND2), P. carvalhoi está estruturado em três clados,
nomeados aqui como Atlântico Sudeste, Atlântico Leste e Atlântico Nordeste (Figura 3),
em referência às bacias hidrográficas nas quais estão concentrados a maior parte dos
indivíduos agrupados. As amostras de Pernambuco, na região da Bacia do São Francisco

! 16!
e de Minas Gerais, nas regiões das Bacias Atlântico Leste e Atlântico Sudeste, ficaram
espalhadas pelos três clados. Os clados foram também separados com alto suporte nas
árvores de genes concatenadas. Os marcadores nucleares (SIA e TYR), porém,
recuperaram P. carvalhoi como uma grande politomia, sem evidenciar a divisão interna de
grupos com alto suporte. Somente a árvore de IB da matriz de dados de SIA separou o
clado Atlântico Nordeste das demais localidades de P. carvalhoi (Apêndice 8).
O clado Atlântico Sudeste reuniu amostras de todas as localidades do Espírito
Santo, hidrograficamente localizadas na bacia Atlântico Sudeste, e as amostras de Poté,
Minas Gerais, localizadas na bacia Atlântico Leste, porém, próximas à divisa com a bacia
Atlântico Sudeste. Esse clado foi monofilético em todas as filogenias de genes
mitocondriais e, também, de genes concatenados. As amostras do Espírito Santo sempre
formaram subclado separado das amostras de Minas Gerais com alto suporte estatístico
na IB e MV (Apêndice 2 a 7, 12 e 13).
O clado Atlântico Leste agrupou amostras de todas as localidades no estado da
Bahia, localizados na bacia Atlântico Leste, as amostras de Ferros, Minas Gerais,
localizada na bacia Atlântico Sudeste e Buíque, além das amostras de Pernambuco,
localizada na bacia do São Francisco. Nas filogenias de ND2, as amostras de
Pernambuco foram agrupadas em um clado à distinção dos demais, na base de P.
carvalhoi, sem qualquer relação de monofiletismo com os representantes da bacia
Atlântico Leste (Apêndice 6 e 7).
O clado Atlântico Nordeste agrupou as amostras do Ceará e Rio Grande do Norte,
localizadas na bacia Atlântico Nordeste, além de amostras de Araçuaí, Minas Gerais,
cidade localizada na bacia Atlântico Leste, na região do Vale do Jequitinhonha. Esse
clado foi recuperado tanto nas filogenias de genes mitocondriais e concatenados, quando
na IB para o marcador nuclear SIA. As amostras de Minas Gerais e Rio Grande do Norte

! 17!
formaram um subclado separado das amostras do Ceará nas árvores de matriz de dados
mitocondriais e concatenados.
As redes de haplótipos dos marcadores mitocondriais também recuperaram os três
clados de P. carvalhoi (Figura 4, 5 e 6). As localidades de Minas Gerais estiveram
representadas pelos três agrupamentos, sendo Araçuaí sempre mais próxima das
amostras da Bacia Atlântico Nordeste, enquanto as amostras de Ferros sempre mais
próximas das amostras da Bacia Atlântico Leste e a de Poté mais próxima das amostras
da Bacia Atlântico Sudeste. Para a rede de haplótipos de SIA foram encontrados apenas
dois haplótipos, um somente com as amostras do clado Atlântico Nordeste, e outro com
as amostras dos clados Atlântico Leste e Atlântico Sudeste. Todos os indivíduos
amostrados compartilham o mesmo haplótipo de TYR.
O clado com maior número de haplótipos foi sempre o Atlântico Leste (12 de 12S, 5
de 16S), onde poucas localidades diferentes compartilharam haplótipos únicos. No clado
Atlântico Sudeste, a localidade de Poté, Minas Gerais, sempre segregou-se como
haplótipo distinto das amostras das localidades do Espírito Santo, que, por sua vez,
formou um haplótipo único de 12S e dois haplótipos diferentes de 16S. O clado Atlântico
Nordeste formou três haplótipos tanto de 12S quanto de 16S, representados pelas duas
localidades do Ceará, e um terceiro haplótipo reunindo amostras do Rio Grande do Norte
e de Araçuaí, Minas Gerais. A rede de ND2 recuperou haplótipos únicos apenas dentro de
mesmas localidades, porém, a maioria das amostras formaram haplótipos únicos.
Pipa carvalhoi apresentou alta divergência entre clados para os marcadores
mitocondriais e seus haplótipos mais próximos estão separados por números de passos
mutacionais semelhantes aos que separaram P. arrabali de P. pipa (Tabela 3). Os
haplótipos de P. carvalhoi foram sempre separados dos haplótipos das espécies
amazônicas por um alto número de passos mutacionais. P. carvalhoi apresentou sempre
alta divergência genética em relação às espécies amazônicas P. pipa e P. arrabali (17% ±

! 18!
3% no 12S; 16% ± 3% no 16S; 54% ± 5% no ND2; 3% ± 1% no SIA; 10% ± 4% no TYR).
A divergência entre P. pipa e P. arrabali, porém, foi baixa para todos os marcadores (6% ±
2% no 12S; 3% ± 1% no 16S; >1% em SIA e TYR). Considerando as divergências
intraespecíficas, P. carvalhoi (5% no 12S; 3% no 16S; 19% no ND2) apresentou sempre
valores maiores que P. pipa (1,5% no 12S; 2% no 16S; 5% no ND2) (Tabela 3). Pipa.
arrabali, por apresentar apenas duas amostras, apresentou divergência sempre menor
que 1%.
A divergência genética identificada entre os clados de P. carvalhoi foi alta para os
marcadores mitocondriais, sempre maior ou igual a divergência entre P. pipa e P. arrabali.
Para os marcadores nucleares, a divergência foi sempre menor que 1%, assim como a
divergência entre as espécies amazônicas. O padrão de proximidade genética entre os
diferentes clados, entretanto, variou entre os marcadores. Para 12S, o clado Atl.SE foi
mais distante de seu clado vizinho Atl.E (8% ± 2%) do que quando comparado ao clado
Atl.NE (7% ± 2%), e a menor divergência genética foi apresentada entre Atl.NE e Atl.E
(6% ± 1%). Já para os marcadores 16S e ND2, Atl.NE e Atl.E apresentaram a maior
distância genética entre clados (5% ± 1% no 16S e 26% ± 3% no ND2), enquanto as
distâncias entre os clados Atl.NE e Atl.SE foi a mesma que entre Atl.SE e Atl.E (4% ± 1%
no 16S e 23% ± 2% no ND2). Em nenhuma das análises os clados Atl.NE e Atl.SE, que
são mais distantes geograficamente, apresentaram a maior distância genética. As
amostras de Pernambuco, que nas análises de ND2 formaram um clado bastante distinto
dos demais, apresentaram uma alta divergência em relação aos clados predefinidos por
este trabalho (41% ± 4% em relação a Atl.E e Atl.SE, e 46% ± 4% em relação ao clado
Atl.NE). Quanto às divergências internas de cada clado, Atl.E apresentou a maior
divergência em 12S (2%), Atl.NE teve a maior divergência em 16S (2%) e Atl.SE foi o
mais divergente em ND2 (6%) (Tabela 3). O teste de Mantel não recuperou relação entre
distância genética e distância geográfica (p=0,74).

! 19!
O relógio molecular calculado a partir das matrizes concatenadas datou o
surgimento de Pipa carvalhoi durante o período Paleogeno entre 75,5 – 33,4 milhões de
anos atrás e a divergência das linhagens de P. pipa e P. arrabali bem mais recente, no
Mioceno, entre 16,5 – 4 milhões de anos atrás. Os três clados identificados para P.
carvalhoi divergiram também durante o Mioceno, sendo que a primeira divisão que levou
ao clado Atlântico Leste foi estimada entre 23,5 – 6,8 milhões de anos atrás, enquanto a
segunda divisão, entre os clados Atlântico Sudeste e Atlântico Nordeste foi estimada entre
20,2 – 6,2 milhões de anos atrás. O relógio molecular calculado a partir da matriz de 12S
datou o surgimento de P. parva entre 87,6 – 33,9 milhões de anos atrás e a divisão entre
P. carvalhoi e P. pipa entre 77,2 – 24,8 milhões de anos atrás (Figura 7 e 8).
Grande parte dos valores encontrados nos testes de neutralidade não foram
significativos (Tabela 4). Valores significativos indicando expansão populacional foram
encontrados apenas para o clado Atlântico Sudeste, segundo o marcador 12S, tanto pelo
teste D de Tajima quanto pelo Fs de Fu. Os valores de Fst indicaram diferenciação
genética muito significativa (Fst > 0,25; Wright 1978, Hartl e Clark 1997) entre os clados
de Pipa carvalhoi (Tabela 3). Os valores de diferenciação genética entre os clados de P.
carvalhoi foram sempre próximos ou maiores do que os valores de diferenciação entre P.
pipa e P. arrabali. De acordo com o marcador 12S, os maiores valores de Fst ocorreram
entre os clados Atlântico Sudeste e Atlântico Nordeste (0,85), entre o Atlântico Leste e
Atlântico Nordeste (0,93) segundo o marcador 16S e os identifacados para o Atlântico
Leste e Atlântico Sudeste pelo marcador ND2 (0,78).
5. Discussão
A relação evolutiva entre as três espécies de Pipa utilizadas no trabalho sustentam
a proximidade não apenas geográfica mas também genética entre P. pipa e P. arrabali.

! 20!
Os dados corroboram com a hipótese filogenética de Trueb e Cannatella (1986) que diz
que P. carvalhoi teria surgido de um evento de vicariância que resultou em sua separação
com o ancestral comum à linhagem que deu origem, posteriormente, a P. arrabali e P.
pipa. Dados osteológicos descritos em Trueb e Cannatella (1986) e Cannatella e Trueb
(1988), além da falta de estágio larval no desenvolvimento, também suportam a monofilia
do grupo P. arrabali e P. pipa, juntamente com as espécies P. aspera e P. snethlageae,
não incluídas nas análises do presente trabalho. A falta de modificações dérmicas bucais
e o corpo pouco achatado, característica presente em P. pipa e P. snethlageae, traz
grande semelhança externa entre P. carvalhoi e P. Arrabali, Porém, tal característica foi
parafilética em todas as análises evolutivas realizadas pelo presente trabalho.
Nesse estudo, a distância genética entre P. pipa e P. arrabali nunca ultrapassou
significativamente as divergências entre os clados internos a P. carvalhoi, e as amostras
das duas espécies exibiram haplótipo único nas análises de DNA nuclear, que possui
taxas evolutivas lentas em relação aos marcadores mitocondriais (Avise, 2000).
As análises de 12S, que incluíram P. parva, recuperaram a espécie como basal
dentro do gênero, corroborando a hipótese filogenética de Trueb e Cannatella (1986) e
Cannatella e Trueb (1988), que posiciona P. parva, juntamente com P. myersi como grupo
basal dentro da evolução do gênero pela presença de 5 sinapomorfias compartilhadas
com Hymenochirus, grupo-irmão de Pipa. O posicionamento do grupo Pipa parva como
primitivo na evolução do gênero foi considerado possível de estar errada pelo próprio
trabalho de Trueb e Cannatella (1986), devido à presença de características convergentes
entre o grupo e P. arrabali, P. snethlageae e P. pipa. Os dados gerados pelo presente
estudo suportam o posicionamento de P. parva como grupo basal dentro do gênero,
assim como representado nas análises de Trueb e Cannatella (1986) e Cannatella e
Trueb (1988).

! 21!
Estudos sobre a sistemática e a relação de parentesco das espécies de Pipa são
escassos e antigos. O primeiro estudo comparativo sobre o grupo foi feito por Noble
(1925), onde o autor separa Pipa em dois gêneros com base no nível de achatamento
dorso-ventral do corpo e presença de modificações dérmicas bucais: Pipa, que
corresponderia às atuais P. pipa e P. snethlageae, e Protopipa, um gênero que agrupou
as espécies P. aspera e P. parva, espécie pouco estudada até então. Pipa carvalhoi foi
posteriormente descrita como Protopipa carvalhoi por Miranda-Ribeiro (1937a), nome
utilizado também por Carvalho (1937). No mesmo ano, porém, Miranda-Ribeiro (1937b)
descreveu o gênero Hemipipa para a inclusão de P. carvalhoi, baseado na presença de
estágio larval. Hemipipa carvalhoi foi utilizado por uma série de autores nos anos que
sucederam sua descrição (Carvalho, 1939; Myers & Carvalhoi, 1945; Paterson, 1955;
Weigoldt, 1976). Dunn (1948) propôs a inclusão de todas as espécies em um único
gênero Pipa, forma que foi suportada pelas análises filogenéticas posteriores (Trueb &
Cannatella, 1986; Cannatella & Trueb, 1988; Trueb & Massemin, 2001).
Dunn (1948) caracterizou como difícil de organizar filogeneticamente as cinco
espécies de Pipa descritas até o dado momento, mas descreveu P. carvalhoi como mais
primitiva e P. pipa como mais especializada na maioria dos caracteres analisados, exceto
pela especialização das pontas dos dedos, onde P. parva apresentava a forma mais
primitiva. Trabalhos posteriores (Trueb & Cannatella, 1986; Cannatella & Trueb, 1988;
Trueb & Massemin, 2001) suportam o grupo P. parva e P. myersi como basais ao gênero,
e P. arrabali, P. aspera, P. pipa e P. snethlageae como grupo mais especializado dentro
da filogenia (Figura 9).
O presente trabalho representa o primeiro estudo molecular utilizando mais de
duas espécies de Pipa e, ainda, o primeiro estudo a visitar a relação de parentesco entre
as espécies do gênero desde o trabalho de Trueb e Massemin (2001). Apesar de não
incluirem amostras das espécies P. aspera, P. myersi e P. snethlageae na matriz de

! 22!
dados analisada, o parentesco evolutivo das mesmas com, respectivamente, P. arrabali,
P. parva e P. pipa tem alto suporte (Trueb & Cannatella, 1986; Cannatella & Trueb, 1988;
Trueb & Massemin, 2001). Esse resultado corrobora a homologia do desenvolvimento
direto dentro do gênero e posiciona essa característica como derivada dentro de sua
história evolutiva, sendo presente apenas em P. arrabali, P. aspera, P. pipa e P.
snethlageae. Características como presença de dentes maxilares são, porém,
convergências entre P. parva, P. pipa e P. snethlageae e representam homoplasias da
característica derivada (Trueb & Cannatella, 1986).
Não existem evidências plausíveis para o reconhecimento de Hemipipa e Protopipa
como gêneros separados de Pipa. Os níveis de divergência encontrados entre as
espécies são altos em relação aos encontrados em outras espécies de Anuros (Vences et
al. 2005a; Vences et al. 2005b), porém, os altos níveis de divergência e o alto número de
passos mutacionais na separação entre os haplogrupos de cada espécie podem ser fruto
da separação muito antiga sofrida pelas principais linhagens dentro do gênero e o
isolamento gerado pela separação das principais bacias hidrográficas que abrigam as
espécies.
As relações filogenéticas entre as espécies de Pipa, bem como a divisão de
haplótipos corroboram a hipótese biogeográfica de Trueb e Cannatella (1986) que sugere
que o ancestral do gênero era, após a separação dos continentes Africano e Sul
Americano, espalhado por toda América do Sul. Esse ancestral sofreu,
subsequentemente, uma vicariância que o dividiu em duas espécies. A primeira delas
restrita à região ao Noroeste da América do Sul, dando origem a P. parva e P. myersi,
hoje restritas às regiões da bacia do rio Maracaibo, na Venezuela e no noroeste da
Colombia, respectivamente. A outra que ocupava a região hidrográfica dos rios Orinoco e
Amazonas e ainda áreas do Sudeste e Nordeste do Brasil. Em seguida, um novo evento
vicariante resultou em uma espécie isolada na costa leste do Brasil, que deu origem à P.

! 23!
carvalhoi e a linhagem que deu origem a P. arrabali , P. aspera, P. snethlageae e P. pipa
teria se diversificado nas regiões Amazônicas e do rio Orinoco.
A divisão do supercontinente Gondwana parece ter tipo papel crucial na origem da
diversidade atual de Pipidae. Estes (1975) propôs que a quebra da Gondwana foi o
evento responsável pela divisão entre Pipa e os gêneros africanos Hymenochirus e
Pseudhymenochirus. Tal hipótese foi utilizada como ponto de calibração das análises de
relógio molecular no presente estudo, já que não existem indícios da presença de fósseis
de Pipa na África e nem de fósseis de Hymenochirus ou Pseudhymenochirus na América
do Sul. A ideia de que anfíbios podem se dispersar sobre uma barreira marinha já foi
utilizada como base para explicação sobre a distribuição de diversas espécies,
principalmente relacionados a eventos de colonização de ilhas oceânicas (Evans et al.
2003; Vences et al. 2003), porém, é muito menos frequente do que a dispersão por meio
de ambientes terrestres (Bewick et al. 2012).
Estes (1975a,b) descreveu “Xenopus” romeri do Paleoceno tardio na Bacia de
Itaboraí. A presença de um fóssil de Xenopus na América do Sul tornaria pouco provável
a hipótese de diversificação de Xenopus durante o Paleoceno/Eoceno. Este fóssil foi
reanalisado posteriormente por Cannatella e Trueb (1988) e Báez e Harrison (2005) que
posicionaram ”Xenopus” romeri anterior ao surgimento do gênero Xenopus. A família
Pipidae, favorecida pelo hábito de vida aquático, é uma das mais numerosas em termos
de registros fósseis dentre todos os anfíbios. Em sua grande maioria os registros datam
do Cretáceo e Terciário e estão localizados por várias regiões da África e América do Sul.
Tais registros dão suporte às hipóteses biogeográficas de diversificação da família. A
presença de Singidella e Pachycentrata, grupo-irmão dos atuais Hymenochirus e
Pseudhymenochirus, somente na África dá suporte à hipótese de divisão da linhagem
Pipinae (Hymenochirus, Pseudhymenochirus e Pipa) pela quebra do supercontinente
Gondwana. Não existe, porém, nenhum registro fóssil de espécies de Pipa, nem mesmo

! 24!
de espécies próximas a este táxon, para ser utilizado como ponto de calibração para as
análises de relógio molecular.
A datação mostrou que a separação entre as principais linhagens de Pipa
aconteceu pouco depois da quebra da Gondwana, ainda durante as primeiras fases do
Período Terciário, cerca de 61 Ma para o primeiro evento de vicariância, que separou o
grupo P. parva das demais espécies e 53 Ma para o segundo evento vicariante, isolando
a linhagem de P. carvalhoi na costa leste. O Paleoceno e Eoceno parecem ter sido
importantes também na formação da diversidade atual das linhagens Africanas de
Pipidae. Evans et al. (2004) estimaram que todos os Xenopus atuais originaram-se entre
50.5 e 81.3 Ma.
Diferente das espécies de Pipa, determinados Xenopus possuem distribuição
muito ampla e com alta conexão entre populações de diferentes bacias hidrográficas,
sugerindo que esse não deve ter sido o fator primordial na diversificação desse
gênero (Evans et al. 2004; 2015; Furman et al. 2015). O histórico de especiação
desse gênero africano, porém, parece estar relacionado a diversos processos de
alopoliploidia independentes, com a presença de espécies tetraploides, octoploides e
até dodecaploides (Kobel, 1996; Evans et al. 2004; 2015). Em Pipa, as bacias
hidrográficas parecem ter atuado no isolamento das principais linhagens e, ainda,
pontualmente na divisão de espécies-irmãs, como entre P. parva e P. myersi, em que
P. parva está restrita a áreas da bacia do Rio Maracaibo, onde P. myersi não ocorre. As
espécies das regiões do Orinoco e Amazônica possuem ampla distribuição geográfica,
incluindo diversos pontos de simpatria, onde os níveis de divisão interna de redes
hidrográficas não parecem ter tido papel importante nos eventos de especiação.
As análises intraespecíficas recuperaram, em P. carvalhoi, três clados distintos,
nomeados aqui de acordo com as bacias hidrográficas a qual fazem parte a maioria de

! 25!
suas amostras. A composição de cada clado foi similar em todas as análises filogenéticas,
independente do marcador utilizado.
O clado Atlântico Nordeste é composto por amostras dos estados do Ceará e Rio
Grande do Norte, juntamente com as amostras de Araçuaí, Minas Gerais. O clado
Atlântico Leste é composto pelas amostras situadas na Bahia, Pernambuco e,
adicionalmente, as amostras de Ferros, Minas Gerais. O clado Atlântico Sudeste foi
composto pelas amostras de todo o Espírito Santo, além de Poté, Minas Gerais.
As medidas de distância genética recuperaram, para genes mitocondriais, alta
divergência entre clados e baixa divergência interna aos clados. As distâncias, em geral,
são superiores ao padrão de distâncias intraespecíficas presentes na literatura. Vences
(2005a,b), revela que a variação intrapopulacional em 16S varia de 0 a 3%, enquanto
entre diferentes populações da mesma espécie a variação é de 0 a 5,1% e entre espécies
diferentes, entre 1 e 16,5%, com valores mais baixos entre espécies de distribuição
alopátrica. Sugere-se (Vences 2005a,b) que valores em torno de 5% para 16S sejam
utilizados como limiar para a definição de espécies em Anfíbios, enquanto Fouquet et al.
(2007) sugere o valor mínimo de 3% entre espécies diferentes para o mesmo marcador.
Os valores de divergência recuperados no presente estudo para o marcador 16S entre os
clados de P. carvalhoi foram de 4% ± 1% entre os clados Atl.NE e Atl.SE e entre Atl.SE e
Atl.E, e de 5% ± 1% entre os clados Atl.NE e Atl.E. Em adição, foram recuperados valores
baixos de divergência dentro dos clados, sendo 0% para Atl.E e 1% para Atl.SE. A
divergência entre as espécies P. pipa e P. arrabali para o gene 16S foi também de 5% ±
1%.
Para ND2, tem sido considerado o limiar de 10% para a divergência entre espécies
de anuros (Crawford, 2003). A distância recuperada entre os clados de P. carvalhoi foi de
22% a 29%, muito acima do limiar utilizado para outras espécies de anuros. Não foi
possível amplificar nenhuma sequência de ND2 para P. arrabali, mas a distância entre P.

! 26!
pipa e P. carvalhoi foi, em média, 54%. As divergências dentro dos clados foi, porém, de
2% para os clados Atl.E e Atl.NE, e de 6% para o clado Atl.SE.
Não existe limite de divergência interespecífica determinado para o gene 12S,
porém, Nogueira et al. (2016) recuperou valores de 4% a 10% entre espécies do grupo
Scinax catharinae e entre 7% e 17% entre espécies do grupo Scinax ruber. Os valores de
divergência entre os clados de P. carvalhoi variaram entre 6% e 8% no presente estudo,
enquanto entre as espécies P. pipa e P. arrabali a distância foi de 6%. As divergências
dentro dos clados foi sempre menor de 2% para o mesmo marcador.
Uma alta estruturação genética foi identificada para todos os clados de P.
carvalhoi. Os valores de divergência genética entre os clados foram sempre iguais, ou
maiores, do que os valores de divergência entre P. pipa e P. arrabali. A separação de P.
pipa e P. arrabali é suportada também por diversos caracteres morfológicos e
osteológicos (Noble, 1925; Dunn, 1948; Trueb & Cannatella, 1986; Cannatella & Trueb,
1988) e cada uma dessas espécies possuem ainda, uma espécie-irmã mais próxima que
não foi incluída no presente estudo (P. aspera para P. arrabali e P. snethlageae para P.
pipa). Os valores altos de Fst sugerem ausência de fluxo gênico entre os diferentes
clados.
Além de altos valores de distância genética, o relógio molecular recuperou tempo
de divergência entre os clados de P. carvalhoi mais antigo que a separação de P. arrabali
com P. pipa. A separação entre os clados de P. carvalhoi ocorreu durante o Mioceno, há
cerca de 14,4 Ma, com intervalo de 95% de confiança entre 6,8 e 23,5 Ma. A separação
entre P. pipa e P. arrabali foi também durante o Mioceno, por volta de 9,5 Ma, com
intervalo de 95% de confiança entre 4,0 e 16,5 Ma.
Grande parte dos pesquisadores sistematas concordam hoje que o melhor futuro
possível para a tomada de decisões taxonômicas é a abordagem integrativa. Devido à
sua complexidade, a diversidade de espécies precisa ser analisada por meio de múltiplas

! 27!
perspectivas, já que a evolução das diferentes características acontece em taxas
diferentes e podem variar entre os diferentes grupos de seres vivos (Padial et al. 2010).
Aspectos morfológicos, ecológicos e comportamentais devem ser analisados, juntamente
com os dados moleculares, para que seja definido com maior precisão se Pipa carvalhoi
corresponde a um complexo de diferentes espécies.
A relação de parentesco entre os três clados internos a P. carvalhoi variou entre os
diferentes marcadores. As árvores de 16S e ND2 separaram os clados como Atl.SE e
Atl.NE como um grupo monofilético separado do clado Atl.E, embora para o último
marcador a IB tenha tido suporte moderado (91%). Em SIA, porém, foi recuperada a
monofilia dos clados Atl.SE e Atl.E com moderado suporte filogenético (94%). Para as
árvores de TYR e 12S foi recuperada uma tricotomia entre os três clados. As análises
concatenadas corroboram a monofilia dos clados Atl.SE e Atl.NE, que possuem a maior
distância genética entre si. Tal relação se torna possível se assumirmos que, após a
segregação dos indivíduos da bacia Atlântico Leste, os demais indivíduos das bacias
Atlântico Nordeste e Sudeste mantiveram contato por vias interioranas, mantendo, assim,
o fluxo gênico. O fato de Minas Gerais apresentar amostras pertencentes a ambos os
clados, Nordeste e Sudeste, sugere que pode ter sido esse o ponto de contato entre eles.
As amostras de Pernambuco estão localizadas na bacia do São Francisco. Apesar
de serem as únicas amostras localizadas nessa bacia, tais amostras só formaram um
clado em separado dos demais representantes das demais bacias hidrográficas nas
análises de ND2, onde se configuram como o grupo mais basal da espécie. Para outros
marcadores, incluindo a análise concatenada, as amostras da bacia do São Francisco
foram sempre internas ao clado Atlântico Leste, sendo monofiléticas apenas em 12S. Os
níveis de divergência par a par e a posição filogenética das amostras de tal localidade
sugerem fluxo gênico recente. Porém, existe uma disjunção entre a localidade de
Pernambuco e as demais localidades da Bahia de pelo menos 500 km, onde nunca foi

! 28!
registrada a presença da espécie (Santana et al. 2014). A coleta de novos espécimes e
inclusão de novas sequências provenientes de Pernambuco, Paraíba, Ceará e Sergipe,
estado onde a ocorrência de P. carvalhoi foi recém registrada, ajudarão a definir com
melhor precisão a relação filogenética da região em relação às outras localidades e a
investigar uma quebra filogenética entre os dois clados. Vale pontuar que a disjunção na
distribuição geográfica de P. carvalhoi era de 1500 km quando registrada por Myers e
Carvalho (1945). Com o registro de novas localidades (Santana et al. 2014) a disjunção
conhecida hoje não ultrapassa os 500 km. Portanto, essa faixa onde a ocorrência da
espécie não é registrada por ser resultado do menor esforço de coleta local, ou ainda de
eventos de extinção local e recente.
P. carvalhoi já foi estudada sobre diferentes aspectos estruturais e
comportamentais. Estudos osteológicos foram extremamente importantes para a
descrição de toda a diversidade do gênero (Paterson, 1955; Trueb & Cannatella, 1986;
Cannatella & Trueb, 1988; Trueb & Massemin, 2001), assim como aspectos morfológicos
de todas as espécies (Noble, 1925; Dunn, 1948), aspectos histológicos do dorso de
fêmeas foram investigados por Fernandes et al. (2011), características bioacústicas e
reprodutivas foram realizadas por Heuwinkel e Bunten (1996) e a composição da dieta
estudada por Canedo et al. (2006). Em nenhum dos trabalhos citados, porém, o foco foi a
investigação da variação geográfica ou as divergências intraespecíficas. Dunn (1948) cita
as localidades conhecidas para P. carvalhoi estavam divididas em dois grupos: um grupo
entre Pernambuco e Ceará, ocorrendo em altutides de 590 a 1000 metros e um outro
grupo em terras baixas, representado por outro grupo formado por, até então, uma única
localidade a 1500 km de distância do primeiro grupo, no Espírito Santo. O presente
estudo representa, então, a primeira tentativa de organização intraespecífica das
linhagens de P. carvalhoi.

! 29!
Apesar de muitas novas localidades terem sido descobertas para P. carvalhoi após
os estudos morfológicos de Dunn (1948), a distribuição geográfica da espécie continua a
apresentar uma disjunção entre as amostras do Nordeste e do Sudeste. A disjunção
corresponde hoje a uma faixa de aproximadamente 500 km entre a localidade de Poço
Redondo, em Sergipe e Jequié, na Bahia que são, respectivamente, a localidade mais ao
sul conhecida para o primeiro grupo e aquela mais ao norte conhecida para o segundo
grupo (Santana et al. 2014). Apesar do padrão de disjunção existente geograficamente
para a espécie, os haplótipos de P. carvalhoi não apresentaram padrão de parentesco
evolutivo semelhante ao observado em sua distribuição geográfica em nenhuma das
análises realizadas.
Grande parte das espécies de anfíbios estão altamente ligadas a ambientes
úmidos e corpos d’água devido principalmente à presença de larva aquática,
chamada de girino, e também da alta suscetibilidade à desidratação (Duellman e
Trueb, 1994). Pipa, porém, é o único gênero Neotropical a apresentar adultos
adaptados à vida subaquática, e se utilizar de tais ambientes aquáticos para todas as
suas funções biológicas, tais como alimentação, reprodução, desenvolvimento e
forrageamento, de forma que era esperado uma associação muito maior com o
padrão de dispersão apresentado por peixes de água doce, onde a diversidade está
associada principalmente aos processos físicos relacionados ao surgimento e
diferenciação das bacias hidrográficas (Costa & Bockmann, 1994; Armbruster, 1998;
Quevedo & Reis, 2002), do que de outras espécies de anfíbios, explicados por
padrões ligados à fauna terrestre como a Hipótese dos Refúgios (Haffer, 1969; Carnaval
et al. 2009; Menezes et al. 2016).
A relação evolutiva recuperada para as amostras costeiras, entretanto, pode ser
explicada pela divisão das bacias hidrográficas. Amostras das diferentes bacias
hidrográficas formaram clados separados nas análises filogenéticas, haplótipos separados

! 30!
e apresentaram alta estruturação genética entre elas. Associado a isso, amostras de uma
mesma bacia apresentaram sempre alta proximidade filogenética e haplotípica, além de
baixa divergência genética entre elas. Nossos dados contavam com amostras de apenas
uma localidade da bacia do São Francisco, porém, tais amostras só formaram um clado
único em ND2. Nas demais análises, as amostras da bacia do São Francisco estiveram
inclusas ao clado Atlântico Leste.
A separação entre as bacias hidrográficas não explica, porém, todas as relações
filogenéticas encontradas para P. carvalhoi, falhando em explicar principalmente o padrão
encontrado para as amostras de Minas Gerais, localizadas mais ao interior do continente.
Amostras de Araçuaí, Minas Gerais, área localizada na Bacia Atlântico Leste,
representam o mesmo haplótipo ou um haplótipo muito próximo a amostras do Rio
Grande do Norte e Ceará, localizados na Bacia Atlântico Nordeste, apesar de estarem
separados por mais de 1000 km. Amostras de Ferros, Minas Gerais, apesar de estarem
localizadas na Bacia Atlântico Sudeste, foram sempre mais próximas filogeneticamente às
amostras da Bacia Atlântico Leste, localizadas principalmente na Bahia. Amostras de
Poté, Minas Gerais, localizadas na Bacia Atlântico Leste, foram sempre mais próximas
filogeneticamente às amostras do Espírito Santo, localizadas na Bacia Atlântico Sudeste.
Porém, é conhecida a habilidade de P. carvalhoi e de outras espécies de Pipidae de
movimentação, inclusive para migração, e forrageamento fora da água (Myers &
Carvalho, 1945; Dunn, 1948; Tinsley et al. 1996; Lobos & Jaksic, 2005), principalmente
após chuvas torrenciais, explicando a presença de indivíduos dessas espécies em lagos
isolados. É possível que a proximidade maior entre rios e afluentes de diferentes bacias
hidrográficas nas regiões do interior do sudeste do Brasil tenha favorecido eventos
migratórios por meio terrestre durante as estações chuvosas, resultando em um padrão
de distribuição local que independe da divisão de bacias hidrográficas. Costa (2001)

! 31!
defende a origem mista de bacias hidrográficas e propõe que cabeceiras de rios são
áreas onde ocorrem complexas trocas de fauna entre áreas adjacentes.
É conhecido também que as transgressões oceânicas podem ter moldado a
diversificação de espécies durante o Quaternário. Durante períodos de glaciação, o nível
do mar chegou a ficar em média 100 metros abaixo do nível atual, expondo a plataforma
continental (Ribeiro, 2006; Leite et al. 2016). O aumento do curso dos rios, combinados a
uma série de movimentos verticais de blocos continentais gerados pela diminuição dos
níveis dos mares favoreceu novas conexões entre rios de diferentes regiões hidrográficas
durante o Pleistoceno (Ribeiro, 2006).
O baixo número de amostras interioranas em nossos dados não permite uma
conclusão precisa sobre uma maior influência de processos migratórios ou vicariantes nos
padrões de parentesco encontrados para as amostras de Minas Gerais. Porém,
independente do processo que tenha influenciado o padrão encontrado, o mesmo foi
gerado por eventos recentes, provavelmente do final do Terciário e começo do
Quaternário, devido ao nível de semelhança genética encontrado entre amostras dessas
populações e aquelas provenientes da costa leste do país.
6. Conclusões
Pipa surgiu a partir da segregação entre América do Sul e África pela quebra do
supercontinente Gondwana no Cretáceo. O Paleoceno e Eoceno foram de grande
importância para a diversificação do gênero, assim como foi para os gêneros africanos
Xenopus e Silurana. A diversificação inicial de Pipa foi influenciada por dois eventos de
vicariância dividindo as espécies por regiões hidrográficas. As espécies P. parva e P.
myersi formam o primeiro grupo a se separar, ficando restritas à região da Bacia do rio
Maracaibo. Posteriormente um segundo evento vicariante separou P. carvalhoi nas bacias

! 32!
da costa Leste brasileira e a linhagem que deu origem às espécies P. pipa, P. arrabali, P.
aspera e P. snethlageae nas regiões Amazônica e do Orinoco.
Pipa carvalhoi apresenta alta estruturação filogenética, com três clados compostos
por indivíduos associados a diferentes regiões hidrográficas (Atlântico Leste, Atlântico
Nordeste e Atlântico Sudeste). A segregação entre as diferentes regiões hidrográficas
mostra-se como uma barreira ao fluxo gênico e parece ter sido responsável pela
estruturação genética da espécie, principalmente das populações costeiras. Para as
populações de Minas Gerais, mais ao interior, a hidrografia não explica o padrão
filogenético, indicando que outros fatores podem ter atuado na estruturação genética das
populações. A relação filogenética entre os três clados variou entre os diferentes
marcadores moleculares. A análise concatenada sugere que os clados Atlântico Nordeste
e Atlântico Sudeste sejam monofiléticos, apesar de serem os clados mais distantes
geograficamente. Para que isso seja possível, a faixa do interior, onde existem
representantes de todos os três clados pode ter servido como zona de contato entre as
populações distantes.
As amostras da bacia do São Francisco foram internas ao clado Atlântico Leste,
com exceção das análises de ND2, onde esse grupo formou um clado separado e basal
em relação aos outros grupos da espécie. Apesar da proximidade genética, P. carvalhoi
apresenta uma disjução na distribuição geográfica de cerca de 500 km entre as
populações da bacia Atlântico Leste e da bacia do São Francisco, onde não existem
registros de ocorrência para a espécie que pode ser devido à subamostragem regional ou
mesmo eventos de extinção recentes no local.
As distâncias genéticas, valores de Fst e o tempo de divergência estimado pelo
relógio molecular mostram que a separação encontrada entre os clados internos a P.
carvalhoi supera os valores assumidos como limiares para a divisão de espécies em
anfíbios. Os valores são, ainda, sempre maiores ou iguais àqueles que separam P.

! 33!
arrabali e P. pipa, espécies largamente sustentadas por caracteres moleculares,
morfológicos e comportamentais.
Pipa carvalhoi é, em suma, uma linhagem de origem muito antiga que teve sua
diversificação influenciada pelo surgimento das principais drenagens costeiras do Brasil.
Tais eventos não servem, porém, para explicar toda a diversidade da espécie. A alta
estruturação genética, a ausência de indícios recentes de fluxo gênico e a divergência
profunda revelam a possibilidade de diversidade críptica ao longo da distribuição de Pipa
carvalhoi.
!
!
Referências Bibliográficas:
Armbruster, J.W. Phylogenetic relationships of the suckermouth armored catfishes os the Rhinelepis group
(Loricariidae: Hypostominae). Copeia 3, 620-636, 1998.
Arzabe, C. Reproductive activity patterns of anurans in two different altitudinal sites within the Brazilian
Caatinga. Revista Brasileira de Zoologia 16(3), 851-864, 1999.
Avise, J.C. Phylogeography: The History and Formation of Species. Harvard University Press, Cambridge,
MA. 447 p, 2000.
Báez, A.M. & Harrison, T. A new pipine frog from an Eocene crater lake in north-central Tanzania.
Palaeontology 48, 723–737, 2005.
Bandelt, H., Forster, P., Röhl, A. Median-joining networks for inferring intraspecific hylogenies. Mol. Biol.
Evol. 16, 37-48, 1999.

! 34!
Batalha-Filho, H. & Miyaki, C.Y. Filogeografia da Mata Atlântica. Revista da Biologia Especial B, 31–34,
2011.
Bates, J.M. Avian diversification in Amazonia: evidence for historical complexity and a vicariance model for a
basic pattern of diversification. In: Vieira I, D'Incão M.A, Silva J.M.C, Oren, D. (eds) Diversidade Biológica
e Cultural da Amazônia. Belém. Museu Paraense Emílio Goeldi 119-138, 2001.
Bewick, A.J, Chain, F.J.J, Heled, J., Evans, B.J. The pipid root. Systematic Biology 61(6): 913-916, 2012.
Bruford, M.W., Hanotte, O., Brookfield, J.F.Y., Burke, T. Single-locus and DNA fingerprinting In: Hoelzel AR
(ed) Molecular genetic analyses of populations. A Pratical Approach. Oxford. IRL Press 225–269, 1992.
Cannatella, D. C. & Trueb, L. Evolution of pipoid frogs: Intergeneric relationships of the aquatic frog family
Pipidae (Anura). Zoological Journal of the Linnean Society 94, 1–38, 1988.
Canedo, C., Garcia, J.P., Fernandes, R., Pombal, J.P. Diet of Pipa carvalhoi (Amphibia, Pipidae) is not
influenced by female parental care. Herpetological Review 37(1), 44-45, 2006
Carnaval, A.C., Moritz, C., Hickerson, M., Haddad, C. & Rodrigues, M. Stability predicts diversity in the
Brazilian Atlantic Forest hotspot. Science 323: 785-789, 2009.
Carnaval, A.C.O.Q. Phylogeography of four frog species in forest fragments of northeastern Brazil – a
preliminary study. Integrative and Comparative Biology 42: 913-921, 2002.
Carvalho, A.L. Notas ecológicas e zoogeográficas sobre vertebrados no Nordeste Brasileiro. Revista O
Campo, 12-13, 1937.
Carvalho, A.L. Notas sobre Hemipipa carvalhoi Mir.-Rib. (Batrachia Anura: Pipidae). 1ª parte. Boletim
Biológico (Nova Série) 4(3), 394- 414, 1939.
Cooke, G.M., Chao, N.L., Beheregaray, L.B. Phylogeography of a flooded forest specialist fish from central
Amazonia based on intron DNA: the cardinal tetra Paracheirodon axelrodi. Freshw. Biol. 54, 1216- 1232,
2009.

! 35!
Costa, L.P. The historical bridge between the Amazon and the Atlantic Forest of Brazil: a study of molecular
phylogeography with small mammals. Journal of Biogeography 30: 71-86, 2003.
Costa, L.P., Leite, Y.R. Historical fragmentation shaping vertebrate diversification in the Atlantic forest
biodiversity hotspot. Bones, clones, and biomes: the history and geography of recent neotropical mammals.
Edited by: Patterson BD, Costa LP. 2012, Chicago: University of Chicago Press
Costa, W.J.E.M. & Bockmann, F.A. A new genus and species of Sarcoglanidinae (Siluriformes:
Trichomycteridae) from southeastern Brazil, with a re-examination of subfamilial phylogeny. Journal of
Natural History 28, 715-730, 1994.
Crawford, A.J., Bermingham, E., Polania, C. The role of tropical dry forest as a long-term barrier to dispersal:
a comparative phylogeographic analysis of dry forest tolerant and intolerant frogs. Molecular Ecology 16:
4789-4807, 2007.
Drummond, A. J. & Rambaut, A. "BEAST: Bayesian evolutionary analysis by sampling trees." BMC
Evolutionary Biology 7: 214, 2007.
Duellman, W.E. & TRUEB, L. Biology of Amphibians. Baltimore, The Johns Hopkins University Press
670p, 1994.
Dunn, E.R. American frogs of the family Pipidae. American Museum Novitates 1348, 1-13, 1948.
Edgar, R.C. MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids
Res 32: 1792-1797, 2004.
Endler, J.A. Problems in distinguishing historical from ecological factors in biogeography. American
Zoologist 22: 441–452, 1982.
Estes, R. Xenopus from the Paleocene of Brazil and its zoogeographic importance. Nature 254, 48-40,
1975a.
Estes, R. Fossil Xenopus from the Paleocene of South America and zoogeography of pipid frogs.
Herpetologica 31, 263-278, 1975b.

! 36!
Evans, B.J., Brown, R.M., McGuire, J.A., Supriatna, J., Andayani, N., Diesmos, A., Melnick, D.J., Cannatella,
D.C. Phylogenetics of fanged frogs: testing biogeographical hypotheses at the interface of the Asian and
Australian faunal zones. Systematic Biology 52, 1-29, 2003.
Evans, B.J., Kelley, D.B., Tinsley, R.C., Melnick, D.J., Cannatella, D.C. A mitochondrial DNA phylogeny of
African clawed frogs: phylogeography and implications for polyploid evolution. Molecular Phylogenetics and Evolution 33, 197–213, 2004.
Evans, B.J., Carter, T.F., Greenbaum, E., Gvoždík, V., Kelley, D.B., McLaughlin, P.J., Pauwels, O.S., Portik,
D.M., Stanley, E.L., Tinsley, R.C., Tobias, M.L., Blackburn, D.C. Genetics, morphology, advertisement calls,
and historical records distinguish six new polyploid species of African clawed frog (Xenopus, Pipidae) from
west and Central Africa. PLoS One 10, 2015.
Excoffier, L., Lischer, H.E.L. Arlequin suite ver 3.5: a new series of programs to perform population genetics
analyses under Linux and Windows. Mol Ecol Resour 10, 564–567, 2010.
Fernandes, T.L., Antoniazzi, M.M., Sasso-Cerri, E., Egami, M.I., Lima, C., Rodrigues, M.T., Jared,
C. 2011. Carrying progeny on the back: Reproduction in the Brazilian aquatic frog Pipa carvalhoi. South Am
J Herpetol 6, 161–176, 2011.
Fitzpatrick, S.W., Brasileiro, C.A., Haddad C.F.B. & Zamudio K.R. Geographical variation in genetic structure
of an Atlantic Coastal Forest frog reveals regional differences in habitat stability. Molecular Ecology 18(13):
2877-2896, 2009.
Fouquet, A., Gilles, A., Vences, M., Marty, C., Blanc, M., Gemmell, N.J. Underestimation of Species
Richness in Neotropical Frogs Revealed by mtDNA Analyses. PLoS ONE 2(10), 2007.
Fouquet, A., Recoder, R., Teixeira, M., Cassimiro, J., Amaro, R.C., Camacho, A., Damasceno, R., Carnaval,
A.C., Moritz, C. & Rodrigues, M.T. Molecular phylogeny and morphometric analyses reveal deep divergence
between Amazonia and Atlantic Forest species of Dendrophryniscus. Molecular Phylogenetics and
Evolution 62, 823–838, 2012.
Fu, Y.X. Statistical tests of neutrality of mutations against population growth, hitchhiking and background
selection. Genetics 147, 915–925, 1997.

! 37!
Furman, B.L.S., Bewick, A.J., Harrison, T.L., Greenbaum, E., Gvoždík, V. et al. Pan-African
phylogeography of a model organism, the African clawed frog Xenopus laevis. Mol Ecol 24, 909-925,
2015.
Galindo-Leal, C. e Câmara, I. G. Status do hotspot Mata Atlântica: uma síntese. Capítulo 1. In: Galindo-
Leal, C. e Câmara, I.G. (Eds). Mata Atlântica: Biodiversidade, Ameaças e Perspectivas. Belo Horizonte:
Conservação Internacional, 3-11 pp, 2005.
Glaw, F., Köhler, J. Amphibian species diversity exceeds that of mammals. Herpetological Review 29, 11–
12, 1998.
Grazziotin, F.G., Monzel, M., Echeverrigaray, S. & S.L. Bonatto. Phylogeography of the Bothrops jararaca
Complex (Serpentes:Viperidae): past fragmentation and island colonization in the Brazilian Atlantic Forest.
Molecular Ecology 15: 3969-3982, 2006.
Haddad, C.F.B., Toledo, L.F., Prado, C.P.A., Loebmann, D., Gasparini, J.L., Sazima, I. Guia dos Anfíbios
da Mata Atlântica. Anolis Books Editora, 1ª Edição, 544p, 2013.
Haffer, J. Speciation in Amazonian birds. Science 165, 131–131, 1969.
Hammer, Ø., Harper, D.A.T., Ryan, P.D. Past: paleontological statistics software package for education and
data analysis. Palaeontologia Electronica 4, 1, 2001.
Hartl, D.L., Clark, A.G. Principles of populations genetics, 3th ed. Sunderland, Massachusetts:
Sinauer. 542p., 1997.
Heuwinkel, H. & Bunten, G. Hydroacoustical investigations on the frog Pipa carvalhoi. Bioacoustics 6(4),
317, 1996.
Hubert, N., Torrico, J.P., Bonhomme, F., Renno, J.F. Species polyphyly and mtDNA introgression among
three Serrasalmus sister-species. Molecular Phylogenetics and Evolution 46, 375–381, 2007.
Kobel, H.R. Reproductive capacity of experimental Xenopus gilli x X. l. laevis hybrids. In: Tinsley, R.C.,
Kobel, H.R. (Eds.), The Biology of Xenopus, Oxford: Clarendon Press. 73-80, 1996.

! 38!
Köhler, J., Vieites, D.R., Bonett, R.M., Garcia, F.H., Glaw, F., Steinke, D., Vences, M. New amphibians and
global conservation: A boost in species discoveries in a highly endangered vertebrate group. Bioscience 55,
693–696, 2005.
Leite, Y.L.R., Costa, L.P., Loss, A.C., Rocha, R.G., Batalha-Filho, H., Bastos, A.C., Quaresma, V.S.,
Fagundes, V., Paresque, R., Passamani, M., Pardini, R. Neotropical forest expansion during the last glacial
period challenges refuge hypothesis. Proc. Natl. Acad. Sci. 113, 1008–1013, 2016.
Librado, P., Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data.
Bioinformatics 25, 1451–1452, 2009.
Lobos, G. & Jaksic, F.M. The ongoing invasion of African clawed frogs (Xenopus laevis) in Chile: causes of
concern. Biodivers. Conserv. 14, 429-439, 2005
Lovejoy, N.R. & Araujo, M.L.G. Molecular systematics, biogeography and population structure of Neotropical
freshwater needlefishes of the genus Potamorrhaphis. Molecular Ecology 9, 259-268, 2000.
Martins, F.M. Historical biogeography of the Brazilian Atlantic forest and the Carnaval–Moritz model of
Pleistocene refugia: what do phylogeographical studies tell us? Biological Journal of the Linnean Society
104 (3): 499-509, 2011.
Menezes, L., Canedo, C., Batalha-Filho, H., Garda, A.A., Gehara, M., Napoli, M.F. Multilocus
Phylogeography of the Treefrog Scinax eurydice (Anura, Hylidae) Reveals a Plio-Pleistocene Diversification
in the Atlantic Forest. PloS ONE 11 (6), 2016.
Miranda-Ribeiro, A. Sobre uma collecção de vertebrados do nordeste brasileiro. Primeira parte: peixes e
batrachios. Revista O Campo, 54-56, 1937a.
Miranda-Ribeiro, A. Ainda os batrachios do nordeste. Revista O Campo 26, 1937b.
Myers, G.S. & Carvalho, A.L. Notes on some new or little known Brazilian amphibians, with an examination
of the history of the Plata salamander, Ensatina platensis. Boletim do Museu Nacional do Rio de Janeiro,
Nova Série Zoologia 35, 1-24, 1945.
Noble, G. K. A new genus of Surinam Toads (Pipidae). American Museum Novitates 164, 1–3, 1925.

! 39!
Nogueira, L., Solé, M., Siqueira, S., Affonso, P.R.A.M., Strüssmann, C., Sampaio, I. Genetic analysis reveals
candidate species in the Scinax catharinae clade (Amphibia: Anura) from Central Brazil. Genetics and
Molecular Biology 39(1), 49-53, 2016.
Oliver, L.A., Rittmeyer, E.N., Kraus, F., Richards, S.J., Austin, C.A. Phylogeny and phylogeography of
Mantophryne (Anura: Microhylidae) reveals cryptic diversity in New Guinea. Molecular Phylogenetics and
Evolution 67, 600–607, 2013.
Padial, J.M., Miralles, A., De la Riva, I., Vences, M. The integrative future of taxonomy. Front. Zool. 7, 16,
2010.
Paterson, N. The skull of the toad, Hemipipa carvalhoi Mir.-Rib. with remarks on other Pipidae. Proc. Zool.
Soc. 125, 223-252, 1955.
Pearse, D.E., Arndt, A.D., Valenzuela, N., Miller, B.A., Vitor Cantarelli, J.W., Sites, J.R. Estimating population
structure under non-equilibrium conditions in a conservation context: continent- wide population genetics of
the giant Amazon river turtle Podocnemis expansa (Chelonia; Podocnemidae). Molecular Ecology 15, 985-
1006, 2006.
Pellegrino, K.C.M., Rodrigues, M.T., Waite, A.N., Morando, M., Yassuda, Y.Y. & Sites, J.W. Phylogeography
and specieslimits in the Gymnodactylus darwinii complex (Gekkonidae, Squamata): genetic structure
coincides with river system in the Brazilian Atlantic Forest. Biological Journal of the Linnean Society 85,
13–26, 2005.
Posada, D. JModelTest: Phylogenetic Model Averaging. Molecular Biology and Evolution 25: 1253-1256,
2008.
Quevedo, R. & Reis, R.E. Pogonopoma obscurum: a new species of loricariid catfish (Siluriformes:
Loricariidae) from southern Brazil, with comments on the genus Pogonopoma. Copeia 2002 (2), 402-
410, 2002.
Ribeiro, A.C. Tectonic history and the biogeography of the freshwater fishes from the coastal drainages of
eastern Brazil: an example of faunal evolution associated with a divergent continental margin. Neotropical
Ichthyology 4(2), 225-246, 2006.

! 40!
Rödel, M.O. Herpetofauna of West Africa, Vol. I. Amphibians of the West African Savanna. – Edition
Chimaira, Frankfurt, Germany, 2000.
Santana, D.O., Franco, S.C., Rocha, S.M., Freitas, E.B., De-Carvalho, C.B., Faria, R.G. First record of Pipa
carvalhoi (Miranda-Ribeiro, 1937) (Anura: Pipidae) in the state of Sergipe, northeastern Brazil. Check List 10(2): 407-408, 2014
Silva, E.T., Fernandes, V.D., Santana, D.J., Feio, R.N. Amphibia, Anura, Pipidae, Pipa carvalhoi (Miranda-
Ribeiro, 1937): Distribution extension and geographic distribution map in the southeast of Brazil. Check List
(UNESP) 6(3): 451-453, 2010.
Sistrom, M.J., Chao, N.L., Beheregaray, L.B. Population history of the Amazonian one-lined pencilfish based
on intron DNA data. J Zool 4, 287–298, 2009.
Tajima, F. Statistical method for testing the neutral mutation hypothesis by DNA popymorphism. Genetics
123, 585-595, 1989.
Tamura, K., Stecher, G., Peterson, D., Filipski, A., Kumar, S. Molecular Biology and Evolution 30, 2725-
2729, 2013.
Tinsley, R. C., Loumont, C., Kobel, H. R. Geographical distribution and ecology; pp 35-59. In: TINSLEY, R. C. &
KOBEL, H. R. (eds.): The biology of Xenopus; Oxford (Oxford Univ. Press), 440 pp. 1996
Tobias, M.L., Evans, B.J., Kelley, D.B. Evolution of advertisement call in African clawed frogs. Behaviour
148, 519–549, 2011.
Thomé, M.T.C., Zamudio, K.R., Giovanelli, J.G.R., Haddad, C.F.B., Baldissera, J.F.A. & Alexandrino, J.
Phylogeography of endemic toads and post-Pliocene persistence of the Brazilian Atlantic Forest. Molecular Phylogenetics and Evolution 55, 1018–1031, 2010.
Tonini, J., Costa, L.P. & Carnaval, A. Phylogeographic structure is strong in the Atlantic Forest; predictive
power of correlative paleodistribution models, not always. Journal Of Zoological Systematics &
Evolutionary Research 51(2): 114-121, 2013.

! 41!
Trueb, L. & Cannatella, D.C. Systematics, morphology, and phylogeny of the genus Pipa (Anura:Pipidae).
Herpetologica 42(4):412–449, 1986.
Trueb, L. & Massemin, D. The osteology and relationships of Pipa aspera (Amphibia: Anura: Pipidae), with
notes on its natural history in French Guiana. Amphib-Reptilia 22, 33-54, 2001.
Turner, T.F., McPhee, M.V., Campbel, P., Winemiller, K.O. Phylogeography and interespecific genetic
variation of prochilodontid fishes endemic to rivers in North South America. J. of Fish Bio. 64, 186-201,
2004.
Vences, M., Vieites, D.R., Glaw, F., Brinkmann, H., Kosuch, J., Vieth, M., Meyer, A. Multiple overseas
dispersal in amphibians. Proceedings of the Royal Society of London Series B, Biological Sciences
270, 2435-2442, 2003.
Vences, M., Thomas, M., Bonett, R.M., Vieites, D.R., Deciphering amphibian diversity through DNA
barcoding: chances and challenges. Philosophical Transactions of the Royal Society London Series B,
Biological Sciences 360, 1859– 1868, 2005a.
Vences, M., Thomas, M., der Meijden, A., Chiari, Y., Vieites, D.R. Comparative performance of the 16S
rRNA gene in DNA barcoding of amphibians. Frontiers Zool. 2, 5. 2005b.
Wallace, A.R. On the monkeys of the Amazon. Proceedings Zoologist Society London 20: 107-110, 1852.
Weigoldt, P. Beobachtungen zur Biologie und Ethologie von Pipa (Hemipipa) carvalhoi Mir. Rib. 1937
(Anura, Pipidae). Z. Tierpsychol. 40, 80-99, 1976.
Wright, S. Evolution and Genetics of Populations: Variability within and among Natural Populations.
University of Chicago Press, Chicago, 465 pp, 1978.
Xia, X., Xie, Z. DAMBE: Software package for data analysis in molecular biology and evolution. J Hered 92,
371–373, 2001.
!
!!!
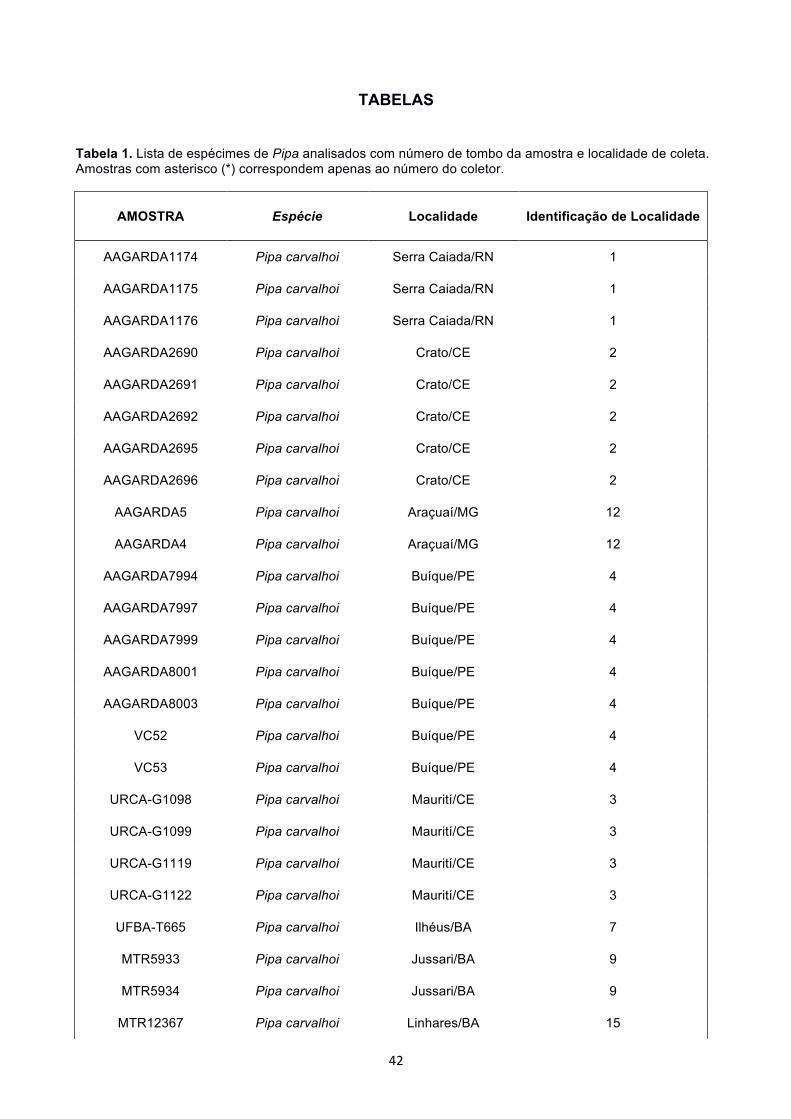
! 42!
!TABELAS
Tabela 1. Lista de espécimes de Pipa analisados com número de tombo da amostra e localidade de coleta. Amostras com asterisco (*) correspondem apenas ao número do coletor.
AMOSTRA Espécie Localidade Identificação de Localidade
AAGARDA1174 Pipa carvalhoi Serra Caiada/RN 1
AAGARDA1175 Pipa carvalhoi Serra Caiada/RN 1
AAGARDA1176 Pipa carvalhoi Serra Caiada/RN 1
AAGARDA2690 Pipa carvalhoi Crato/CE 2
AAGARDA2691 Pipa carvalhoi Crato/CE 2
AAGARDA2692 Pipa carvalhoi Crato/CE 2
AAGARDA2695 Pipa carvalhoi Crato/CE 2
AAGARDA2696 Pipa carvalhoi Crato/CE 2
AAGARDA5 Pipa carvalhoi Araçuaí/MG 12
AAGARDA4 Pipa carvalhoi Araçuaí/MG 12
AAGARDA7994 Pipa carvalhoi Buíque/PE 4
AAGARDA7997 Pipa carvalhoi Buíque/PE 4
AAGARDA7999 Pipa carvalhoi Buíque/PE 4
AAGARDA8001 Pipa carvalhoi Buíque/PE 4
AAGARDA8003 Pipa carvalhoi Buíque/PE 4
VC52 Pipa carvalhoi Buíque/PE 4
VC53 Pipa carvalhoi Buíque/PE 4
URCA-G1098 Pipa carvalhoi Maurití/CE 3
URCA-G1099 Pipa carvalhoi Maurití/CE 3
URCA-G1119 Pipa carvalhoi Maurití/CE 3
URCA-G1122 Pipa carvalhoi Maurití/CE 3
UFBA-T665 Pipa carvalhoi Ilhéus/BA 7
MTR5933 Pipa carvalhoi Jussari/BA 9
MTR5934 Pipa carvalhoi Jussari/BA 9
MTR12367 Pipa carvalhoi Linhares/BA 15

! 43!
MTR15988 Pipa carvalhoi Camacan/BA 10
MTR16082 Pipa carvalhoi Camacan/BA 10
DS35 Pipa carvalhoi Ferros/MG 14
DS36 Pipa carvalhoi Ferros/MG 14
DS42 Pipa carvalhoi Ferros/MG 14
DS43 Pipa carvalhoi Ferros/MG 14
PVS108 Pipa carvalhoi Santa Maria de Jetibá/ES 18
PVS109 Pipa carvalhoi Santa Maria de Jetibá/ES 18
PVS110 Pipa carvalhoi Santa Teresa/ES 17
PVS111 Pipa carvalhoi Santa Teresa/ES 17
PVS112 Pipa carvalhoi Santa Teresa/ES 17
PVS113 Pipa carvalhoi Colatina/ES 16
PVS114 Pipa carvalhoi Colatina/ES 16
PVS115 Pipa carvalhoi Colatina/ES 16
PVS116 Pipa carvalhoi Colatina/ES 16
PVS56 Pipa carvalhoi Poté/MG 13
PVS57 Pipa carvalhoi Poté/MG 13
CH2225 Pipa carvalhoi Porto Seguro/BA 11
CH2297 Pipa carvalhoi Porto Seguro/BA 11
CH3973 Pipa carvalhoi Buararema/BA 8
CH3974 Pipa carvalhoi Buararema/BA 8
CH3975 Pipa carvalhoi Buararema/BA 8
CH3976 Pipa carvalhoi Buararema/BA 8
CH3977 Pipa carvalhoi Buararema/BA 8
CH4248 Pipa carvalhoi Una/Ilhéus/BA 7
CH12862 Pipa carvalhoi Jequié/BA 5
CH16157 Pipa carvalhoi Uruçuca/BA 6
CH16158 Pipa carvalhoi Uruçuca/BA 6
CH16159 Pipa carvalhoi Uruçuca/BA 6
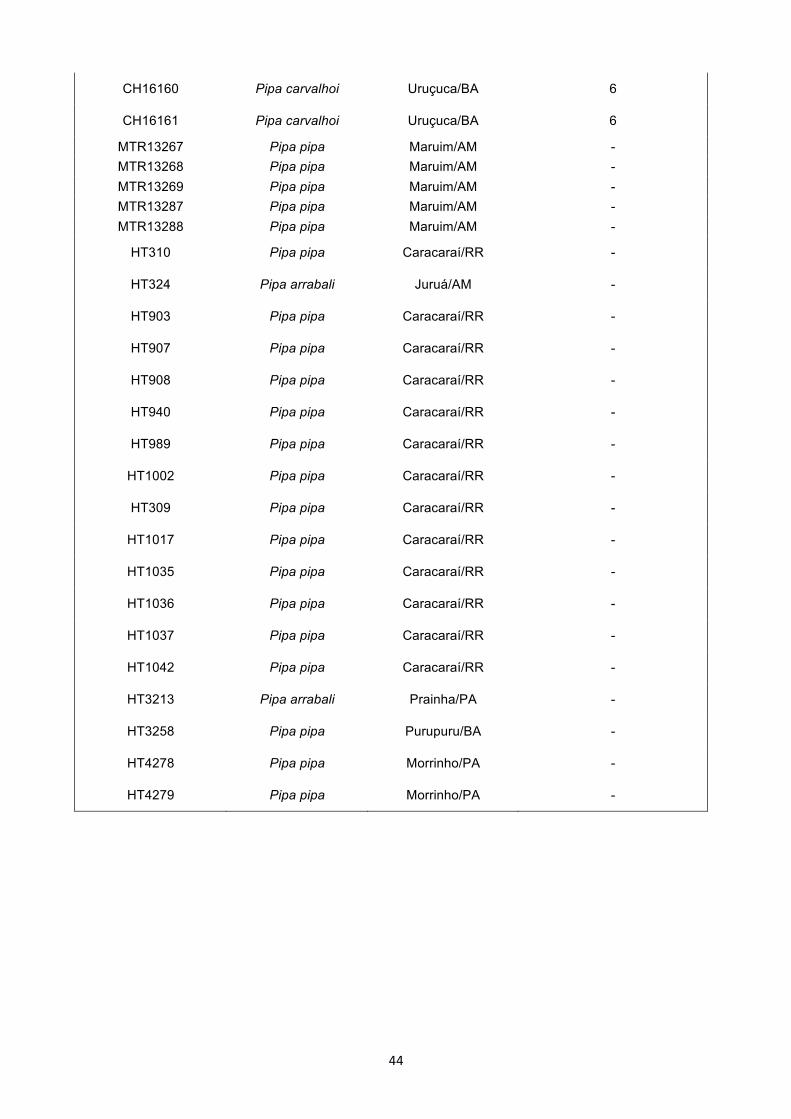
! 44!
CH16160 Pipa carvalhoi Uruçuca/BA 6
CH16161 Pipa carvalhoi Uruçuca/BA 6
MTR13267 Pipa pipa Maruim/AM - MTR13268 Pipa pipa Maruim/AM - MTR13269 Pipa pipa Maruim/AM - MTR13287 Pipa pipa Maruim/AM - MTR13288 Pipa pipa Maruim/AM -
HT310 Pipa pipa Caracaraí/RR -
HT324 Pipa arrabali Juruá/AM -
HT903 Pipa pipa Caracaraí/RR -
HT907 Pipa pipa Caracaraí/RR -
HT908 Pipa pipa Caracaraí/RR -
HT940 Pipa pipa Caracaraí/RR -
HT989 Pipa pipa Caracaraí/RR -
HT1002 Pipa pipa Caracaraí/RR -
HT309 Pipa pipa Caracaraí/RR -
HT1017 Pipa pipa Caracaraí/RR -
HT1035 Pipa pipa Caracaraí/RR -
HT1036 Pipa pipa Caracaraí/RR -
HT1037 Pipa pipa Caracaraí/RR -
HT1042 Pipa pipa Caracaraí/RR -
HT3213 Pipa arrabali Prainha/PA -
HT3258 Pipa pipa Purupuru/BA -
HT4278 Pipa pipa Morrinho/PA -
HT4279 Pipa pipa Morrinho/PA -
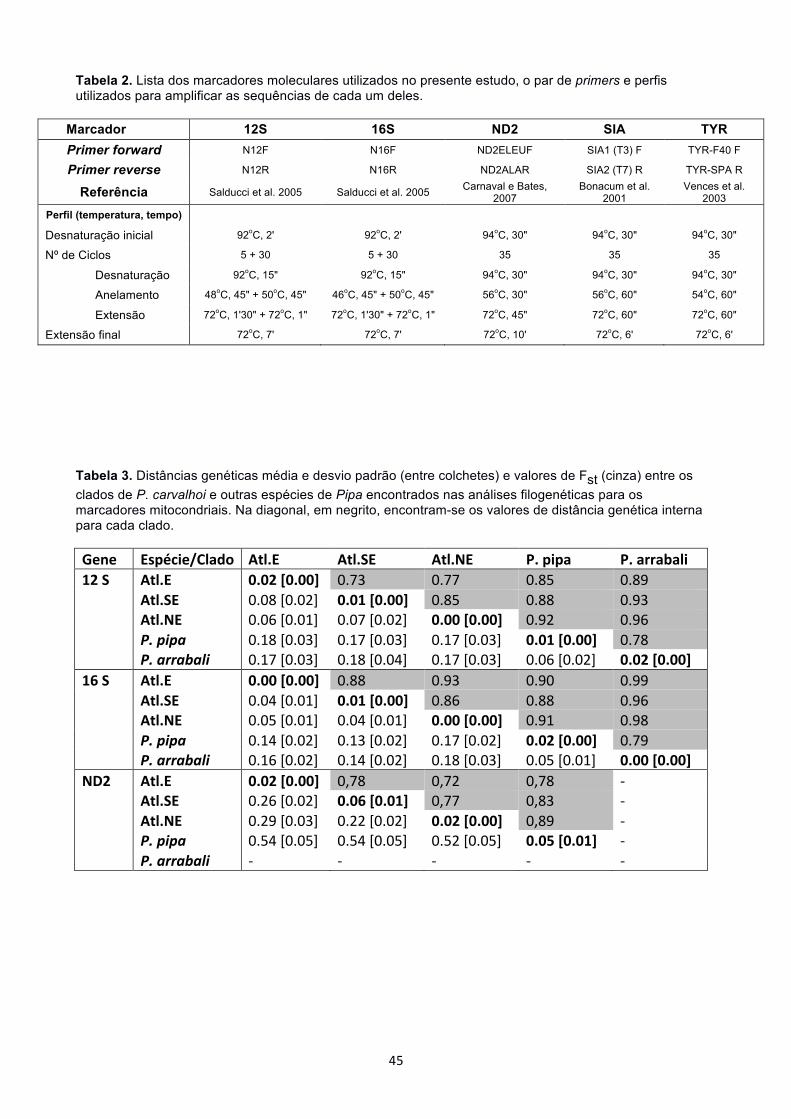
! 45!
Tabela 2. Lista dos marcadores moleculares utilizados no presente estudo, o par de primers e perfis utilizados para amplificar as sequências de cada um deles.
Marcador 12S 16S ND2 SIA TYR Primer forward N12F N16F ND2ELEUF SIA1 (T3) F TYR-F40 F
Primer reverse N12R N16R ND2ALAR SIA2 (T7) R TYR-SPA R
Referência Salducci et al. 2005 Salducci et al. 2005 Carnaval e Bates, 2007
Bonacum et al. 2001
Vences et al. 2003
Perfil (temperatura, tempo)
Desnaturação inicial 92oC, 2' 92oC, 2' 94oC, 30" 94oC, 30" 94oC, 30"
Nº de Ciclos 5 + 30 5 + 30 35 35 35
Desnaturação 92oC, 15" 92oC, 15" 94oC, 30" 94oC, 30" 94oC, 30"
Anelamento 48oC, 45" + 50oC, 45" 46oC, 45" + 50oC, 45" 56oC, 30" 56oC, 60" 54oC, 60"
Extensão 72oC, 1'30" + 72oC, 1" 72oC, 1'30" + 72oC, 1" 72oC, 45" 72oC, 60" 72oC, 60"
Extensão final 72oC, 7' 72oC, 7' 72oC, 10' 72oC, 6' 72oC, 6'
Tabela 3. Distâncias genéticas média e desvio padrão (entre colchetes) e valores de Fst (cinza) entre os clados de P. carvalhoi e outras espécies de Pipa encontrados nas análises filogenéticas para os marcadores mitocondriais. Na diagonal, em negrito, encontram-se os valores de distância genética interna para cada clado. Gene$ Espécie/Clado$ Atl.E$ Atl.SE$ Atl.NE$ P.$pipa$ P.$arrabali$12$S$ Atl.E$ 0.02$[0.00]$ 0.73! 0.77! 0.85! 0.89!$$ Atl.SE$ 0.08![0.02]! 0.01$[0.00]$ 0.85! 0.88! 0.93!$$ Atl.NE$ 0.06![0.01]! 0.07![0.02]! 0.00$[0.00]$ 0.92! 0.96!$$ P.#pipa# 0.18![0.03]! 0.17![0.03]! 0.17![0.03]! 0.01$[0.00]$ 0.78!$$ P.#arrabali# 0.17![0.03]! 0.18![0.04]! 0.17![0.03]! 0.06![0.02]! 0.02$[0.00]$16$S$ Atl.E$ 0.00$[0.00]$ 0.88! 0.93! 0.90! 0.99!$$ Atl.SE$ 0.04![0.01]! 0.01$[0.00]$ 0.86! 0.88! 0.96!$$ Atl.NE$ 0.05![0.01]! 0.04![0.01]! 0.00$[0.00]$ 0.91! 0.98!$$ P.#pipa# 0.14![0.02]! 0.13![0.02]! 0.17![0.02]! 0.02$[0.00]$ 0.79!$$ P.#arrabali# 0.16![0.02]! 0.14![0.02]! 0.18![0.03]! 0.05![0.01]! 0.00$[0.00]$ND2$ Atl.E$ 0.02$[0.00]$ 0,78! 0,72! 0,78! 0!$$ Atl.SE$ 0.26![0.02]! 0.06$[0.01]$ 0,77! 0,83! 0!$$ Atl.NE$ 0.29![0.03]! 0.22![0.02]! 0.02$[0.00]$ 0,89! 0!$$ P.#pipa# 0.54![0.05]! 0.54![0.05]! 0.52![0.05]! 0.05$[0.01]$ 0!$$ P.#arrabali# 0! 0! 0! 0! 0!
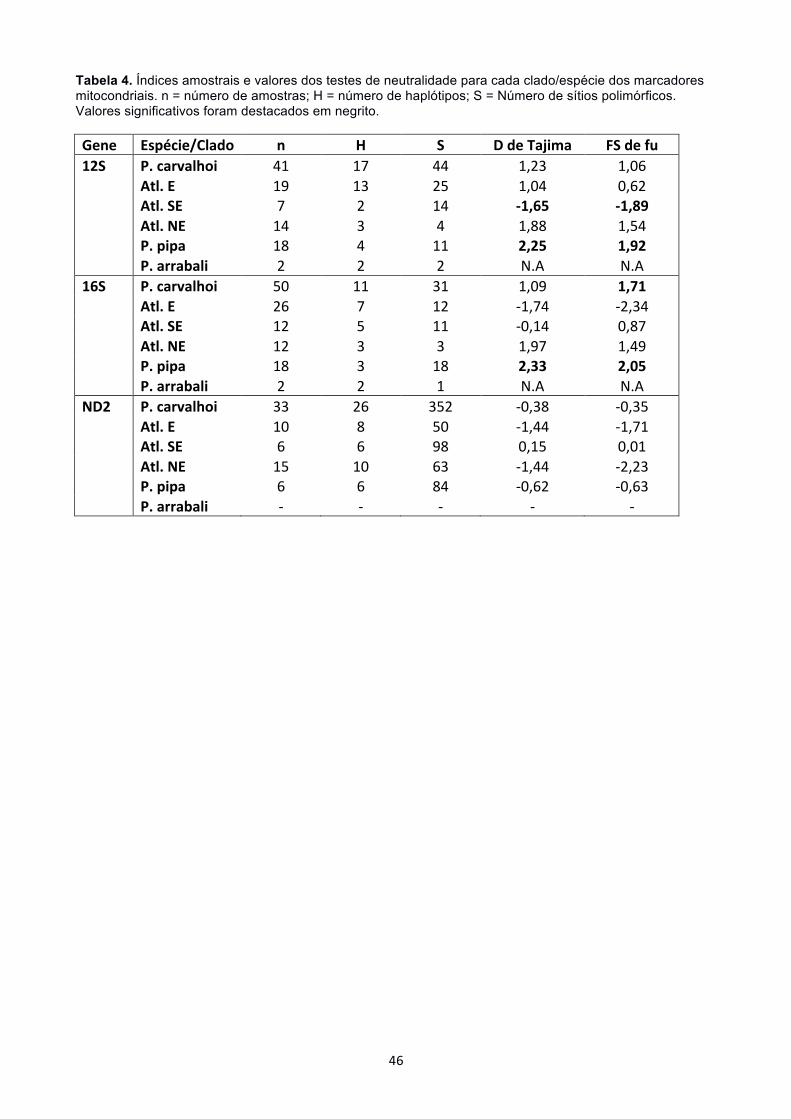
! 46!
Tabela 4. Índices amostrais e valores dos testes de neutralidade para cada clado/espécie dos marcadores mitocondriais. n = número de amostras; H = número de haplótipos; S = Número de sítios polimórficos. Valores significativos foram destacados em negrito. Gene$ Espécie/Clado$ n$ H$ S$ D$de$Tajima$ FS$de$fu$12S$ P.$carvalhoi$ 41! 17! 44! 1,23! 1,06!$$ Atl.$E$ 19! 13! 25! 1,04! 0,62!$$ Atl.$SE$ 7! 2! 14! J1,65$ J1,89$$$ Atl.$NE$ 14! 3! 4! 1,88! 1,54!$$ P.$pipa$ 18! 4! 11! 2,25$ 1,92$$$ P.$arrabali$ 2! 2! 2! N.A! N.A!16S$ P.$carvalhoi$ 50! 11! 31! 1,09! 1,71$$$ Atl.$E$ 26! 7! 12! 01,74! 02,34!$$ Atl.$SE$ 12! 5! 11! 00,14! 0,87!$$ Atl.$NE$ 12! 3! 3! 1,97! 1,49!$$ P.$pipa$ 18! 3! 18! 2,33$ 2,05$$$ P.$arrabali$ 2! 2! 1! N.A! N.A!ND2$ P.$carvalhoi$ 33! 26! 352! 00,38! 00,35!$$ Atl.$E$ 10! 8! 50! 01,44! 01,71!$$ Atl.$SE$ 6! 6! 98! 0,15! 0,01!$$ Atl.$NE$ 15! 10! 63! 01,44! 02,23!$$ P.$pipa$ 6! 6! 84! 00,62! 00,63!$$ P.$arrabali$ 0! 0! 0! 0! 0!
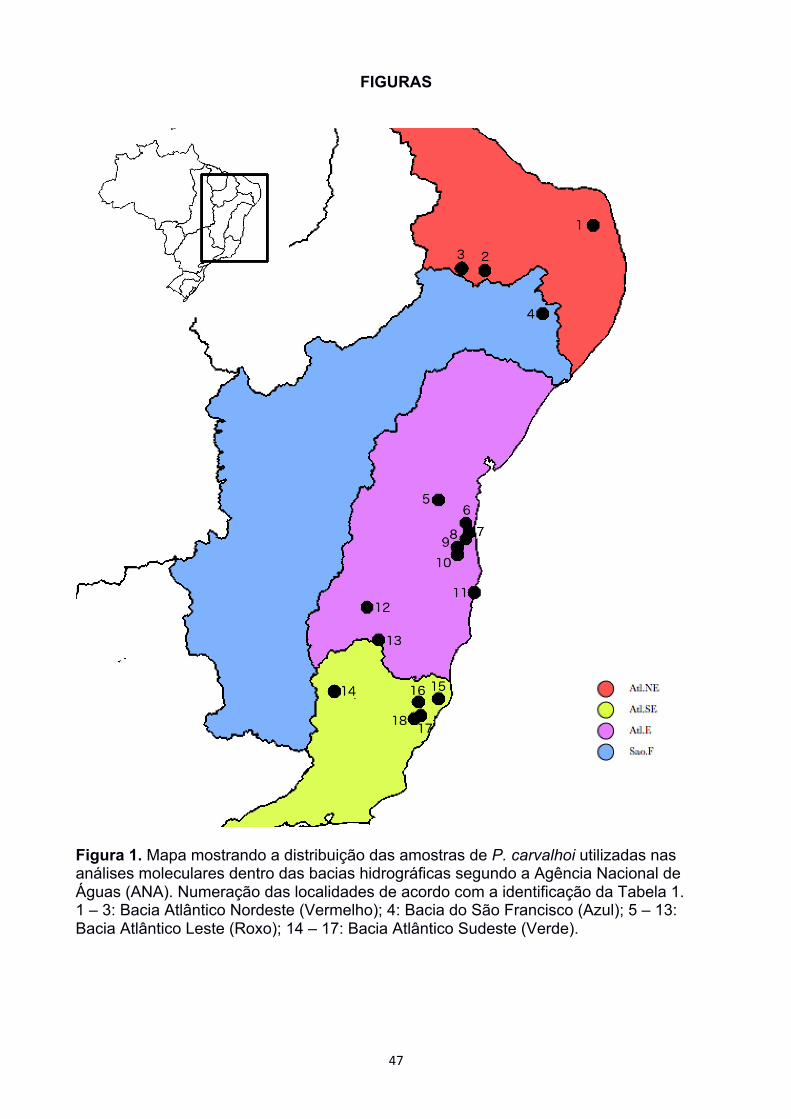
! 47!
FIGURAS
Figura 1. Mapa mostrando a distribuição das amostras de P. carvalhoi utilizadas nas análises moleculares dentro das bacias hidrográficas segundo a Agência Nacional de Águas (ANA). Numeração das localidades de acordo com a identificação da Tabela 1. 1 – 3: Bacia Atlântico Nordeste (Vermelho); 4: Bacia do São Francisco (Azul); 5 – 13: Bacia Atlântico Leste (Roxo); 14 – 17: Bacia Atlântico Sudeste (Verde).

! 48!
Figura 2. Árvore Filogenética de genes concatenados para as espécies de Pipa. Valores ao lado dos ramos representam, respectivamente, o valor de bootstrap da Máxima Verossimilhança (ML) e probabilidade posterior da Inferência Bayesiana (IB).

! 49!
Figura 3. Árvore Filogenética de genes concatenados para os clados de Pipa carvalhoi. Valores ao lado dos ramos representam, respectivamente, o valor de bootstrap da Máxima Verossimilhança (ML) e probabilidade posterior da Inferência Bayesiana (IB).
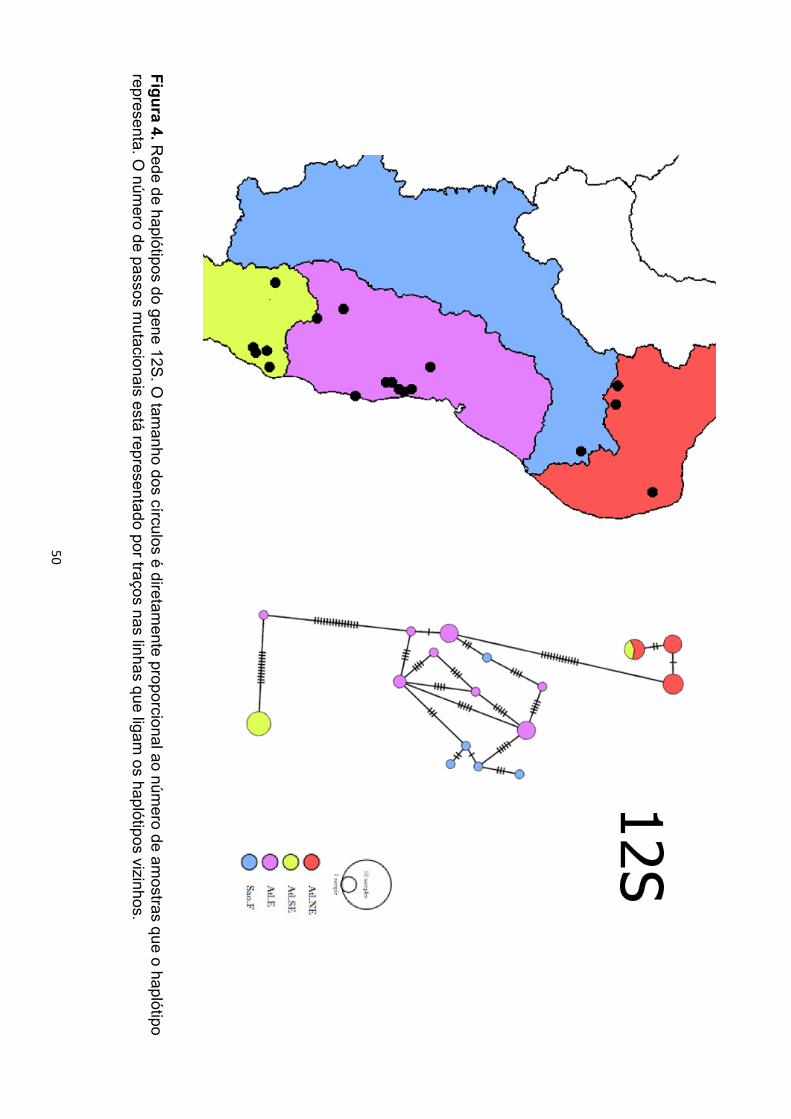
!50!
Figura 4. R
ede de haplótipos do gene 12S. O
tamanho dos círculos é diretam
ente proporcional ao número de am
ostras que o haplótipo representa. O
número de passos m
utacionais está representado por traços nas linhas que ligam os haplótipos vizinhos.
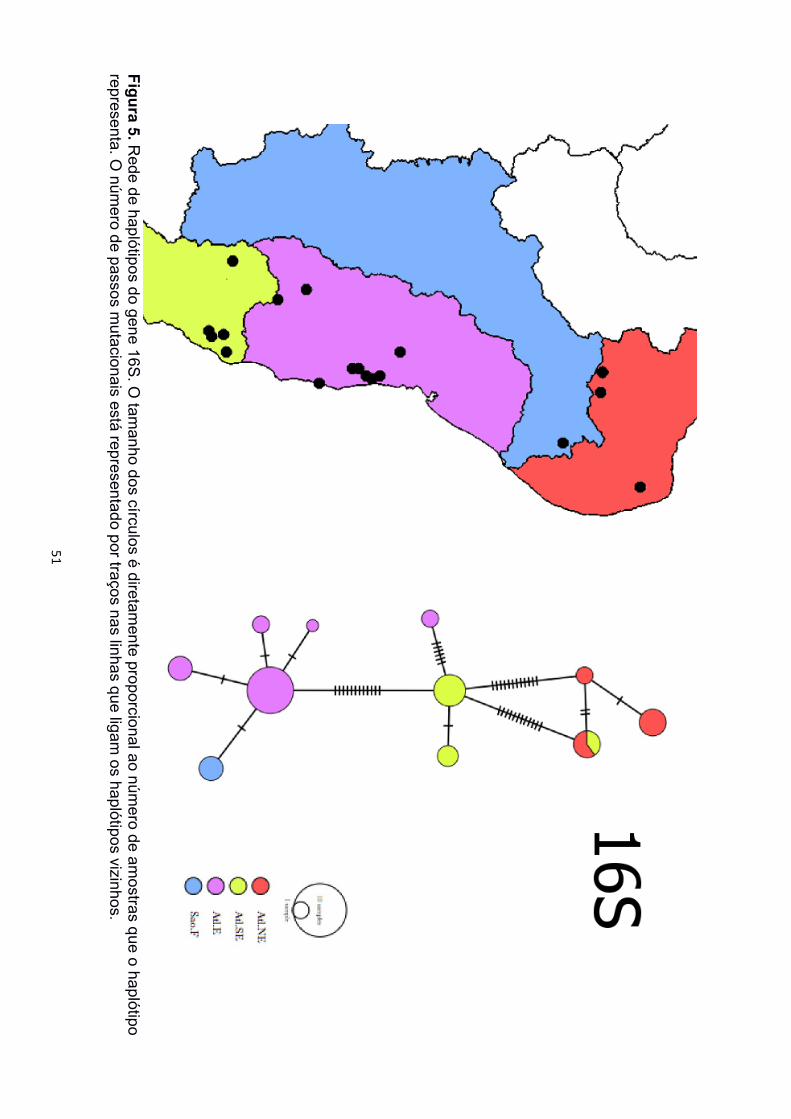
!51!
Figura 5. R
ede de haplótipos do gene 16S. O
tamanho dos círculos é diretam
ente proporcional ao número de am
ostras que o haplótipo representa. O
número de passos m
utacionais está representado por traços nas linhas que ligam os haplótipos vizinhos.
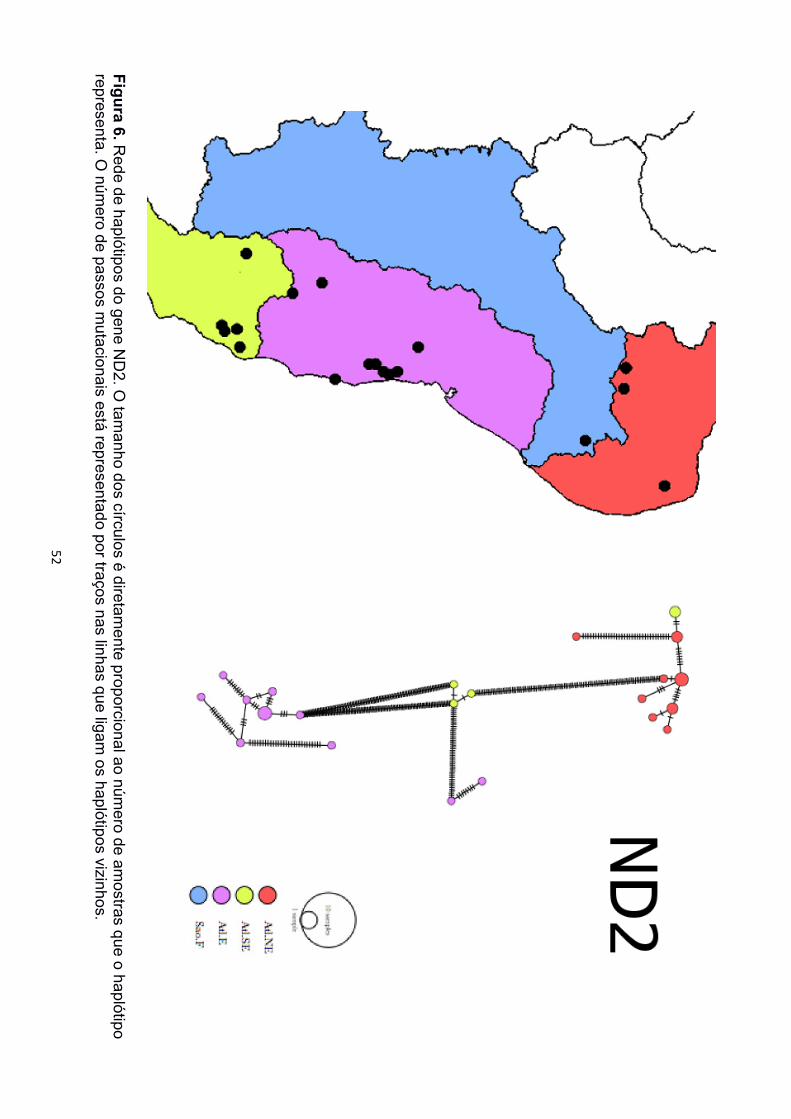
!52!
Figura 6. Rede de haplótipos do gene N
D2. O
tamanho dos círculos é diretam
ente proporcional ao número de am
ostras que o haplótipo representa. O
número de passos m
utacionais está representado por traços nas linhas que ligam os haplótipos vizinhos.

!53!
Figura 7. Árvore com
datação por Relógio M
olecular baseados nas sequências de 12S. O
s representam o tem
po para cada ramo em
m
ilhões de anos. As barras azuis representam
intervalo de confiança de 95%.
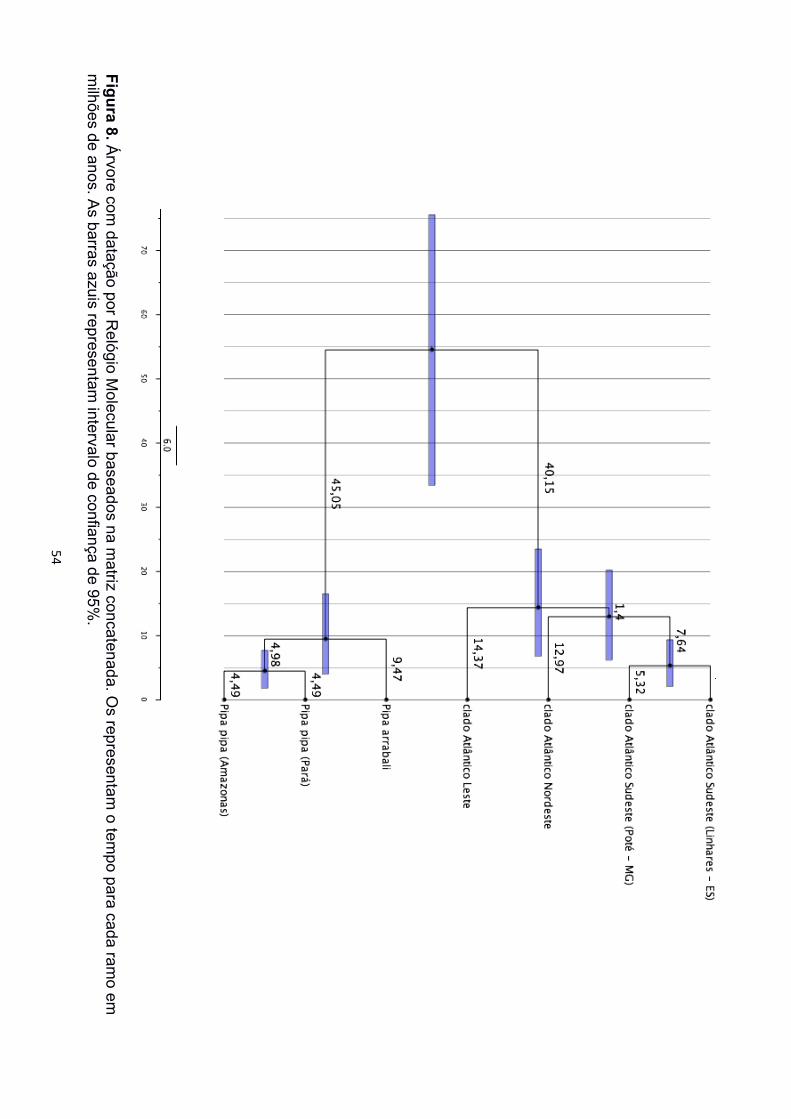
!54!
Figura 8. Árvore com
datação por Relógio M
olecular baseados na matriz concatenada. O
s representam o tem
po para cada ramo em
m
ilhões de anos. As barras azuis representam
intervalo de confiança de 95%.

!55!
Figura 9. Relação de parentesco entre espécies de P
ipa proposta por Trueb & C
annatella, 1986.

!56!
AP
ÊN
DIC
ES
AP
ÊN
DIC
E 1: Lista sequências de 12S
obtidas no GenB
ank para as análises de relógio molecular. A
s siglas correspondem aos
números de acesso do G
enBank.
E
spécie A
cesso Genbank
Pseudhym
enochirus merlini
NC
_015618.1 H
ymenochirus boettgeri
NC
_015615.1 S
ilurana tropicalis N
C_006839.1
Xenopus borealis
NC
_018776.1 P
ipa pipa G
Q244477.1
Pipa carvalhoi
NC
_015617.1 P
ipa parva A
Y581622.1
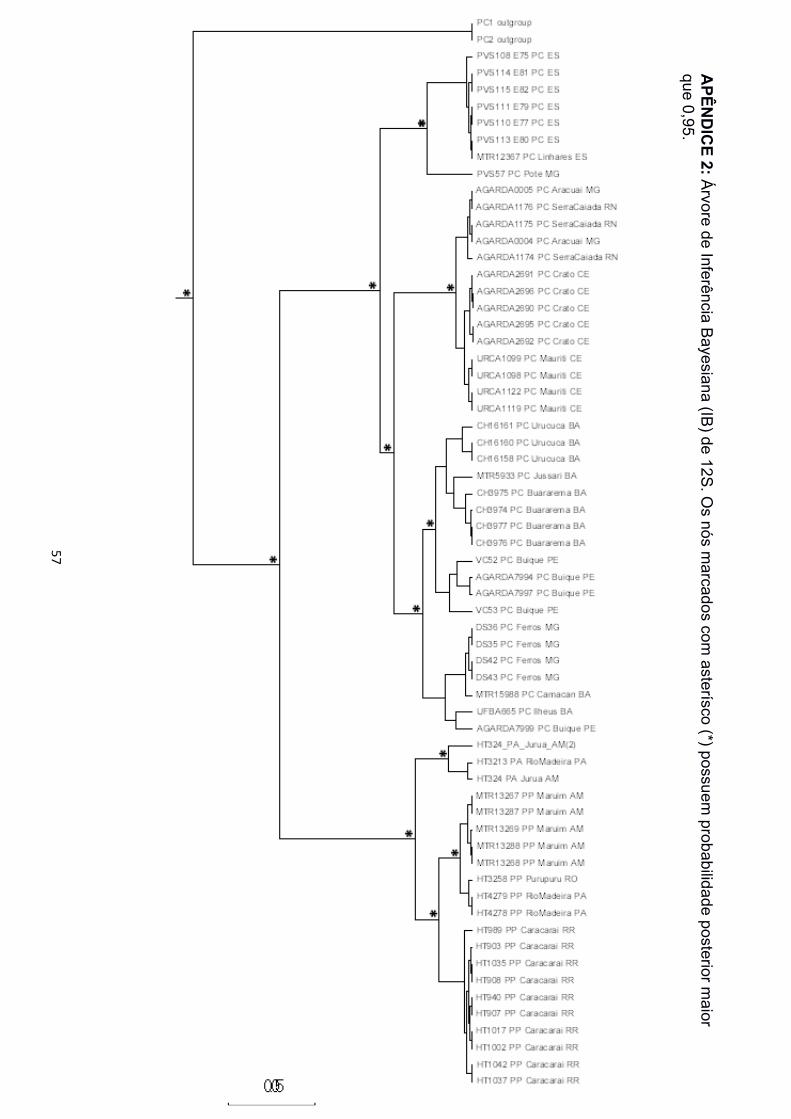
!57!
AP
ÊN
DIC
E 2: Á
rvore de Inferência Bayesiana (IB
) de 12S. O
s nós marcados com
asterísco (*) possuem probabilidade posterior m
aior que 0,95.

!58!
AP
ÊN
DIC
E 3: Á
rvore de Máxim
a Verossim
ilhança (MV
) de 12S. O
s valores dos nós correspondem ao bootstrap.
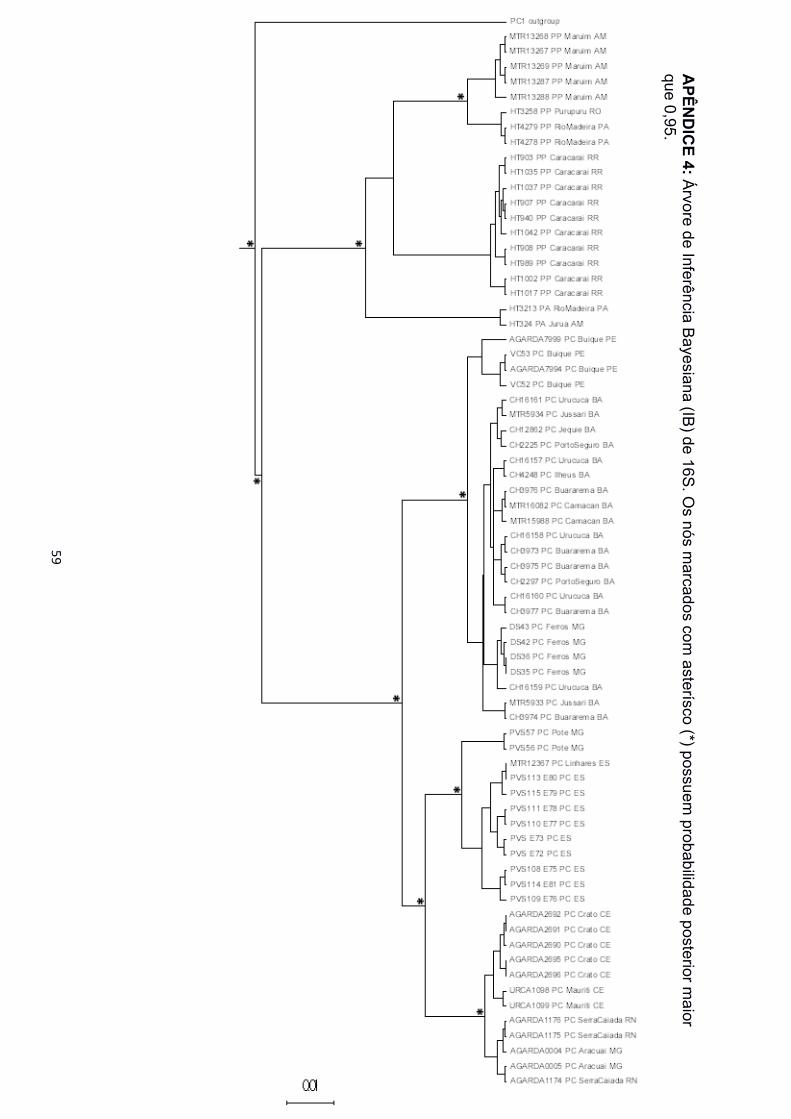
!59!
AP
ÊN
DIC
E 4: Á
rvore de Inferência Bayesiana (IB
) de 16S. O
s nós marcados com
asterísco (*) possuem probabilidade posterior m
aior que 0,95.

!60!
AP
ÊN
DIC
E 5: Á
rvore de Máxim
a Verossim
ilhança (MV
) de 16S. O
s valores dos nós correspondem ao bootstrap.
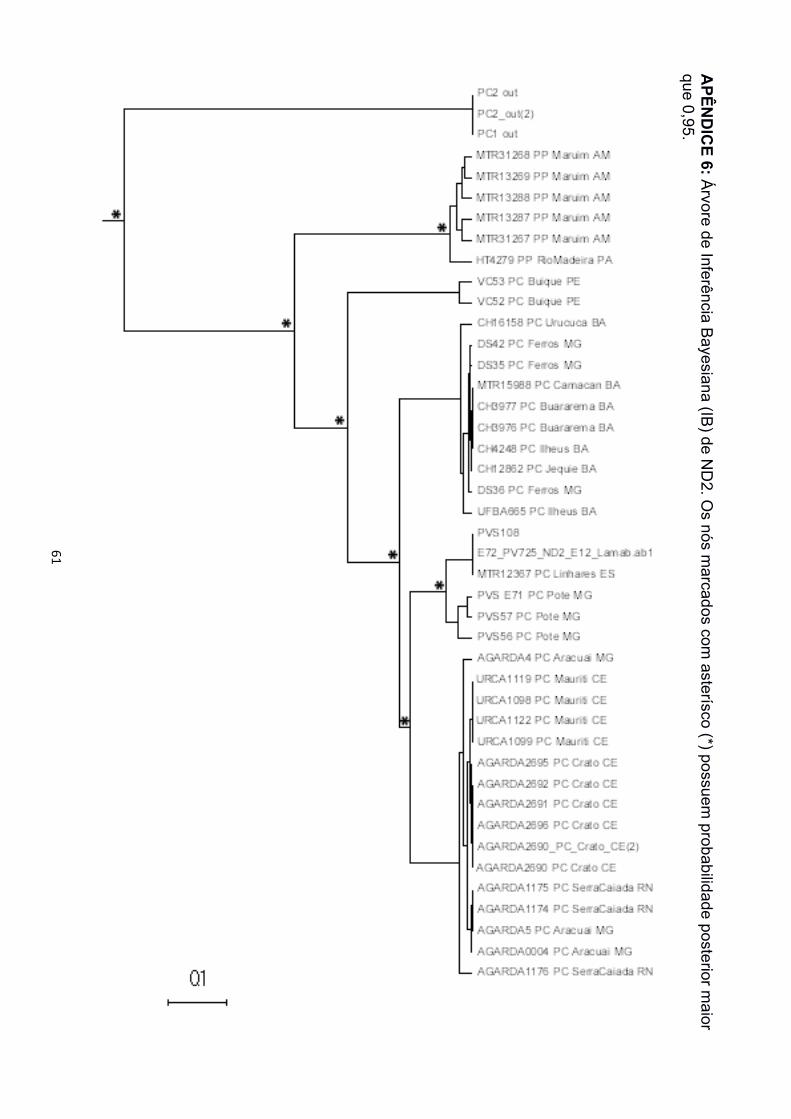
!61!
AP
ÊN
DIC
E 6: Á
rvore de Inferência Bayesiana (IB
) de ND
2. Os nós m
arcados com asterísco (*) possuem
probabilidade posterior maior
que 0,95.
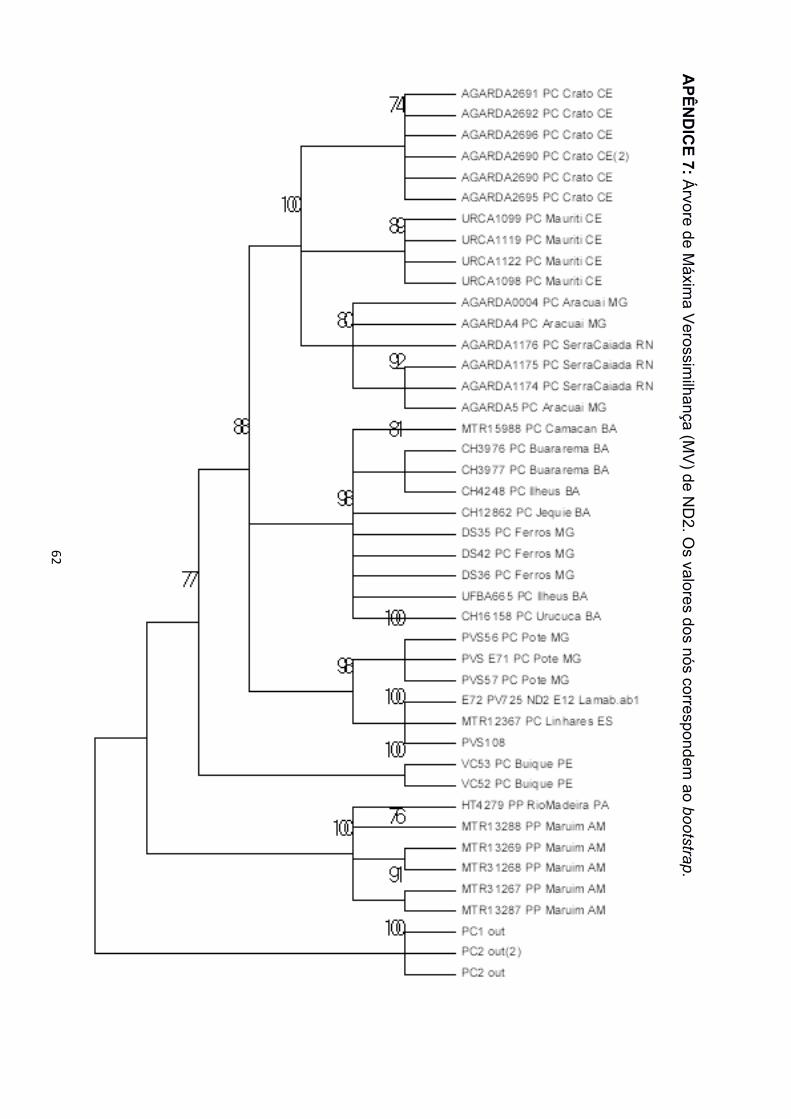
!62!
AP
ÊN
DIC
E 7: Á
rvore de Máxim
a Verossim
ilhança (MV
) de ND
2. Os valores dos nós correspondem
ao bootstrap.
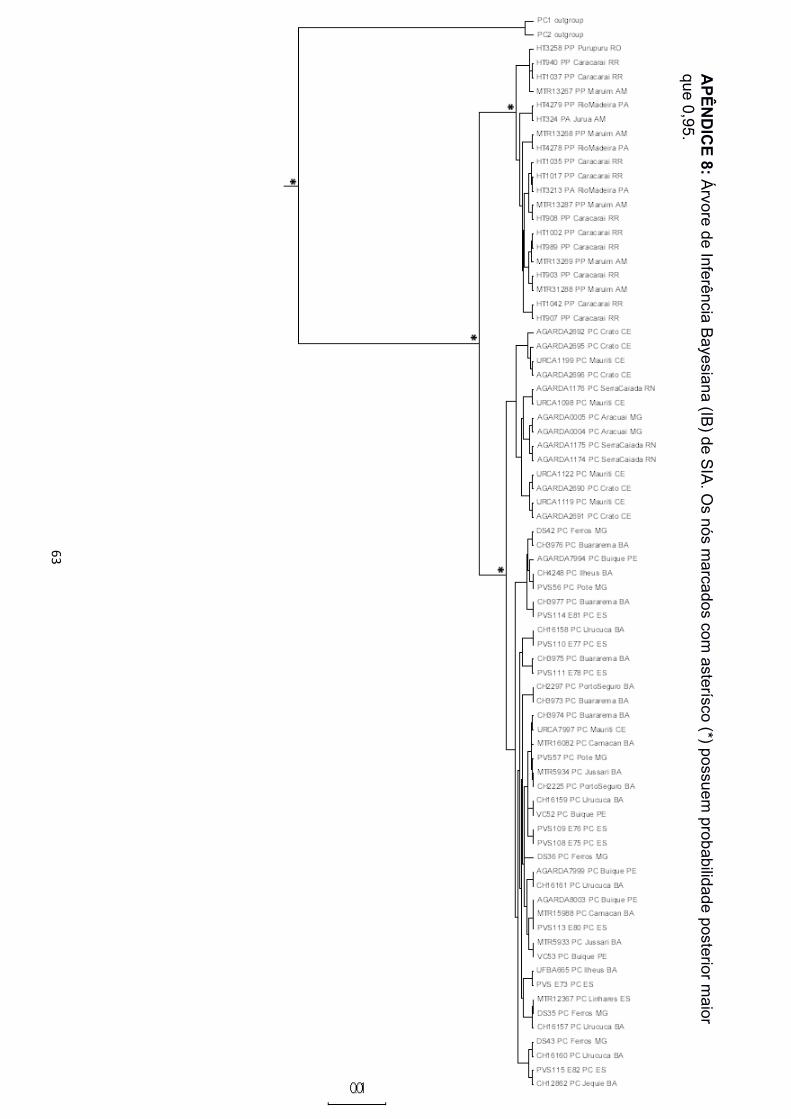
!63!
AP
ÊN
DIC
E 8: Á
rvore de Inferência Bayesiana (IB
) de SIA
. Os nós m
arcados com asterísco (*) possuem
probabilidade posterior maior
que 0,95.

!64!
AP
ÊN
DIC
E 9: Á
rvore de Máxim
a Verossim
ilhança (MV
) de SIA
. Os valores dos nós correspondem
ao bootstrap.
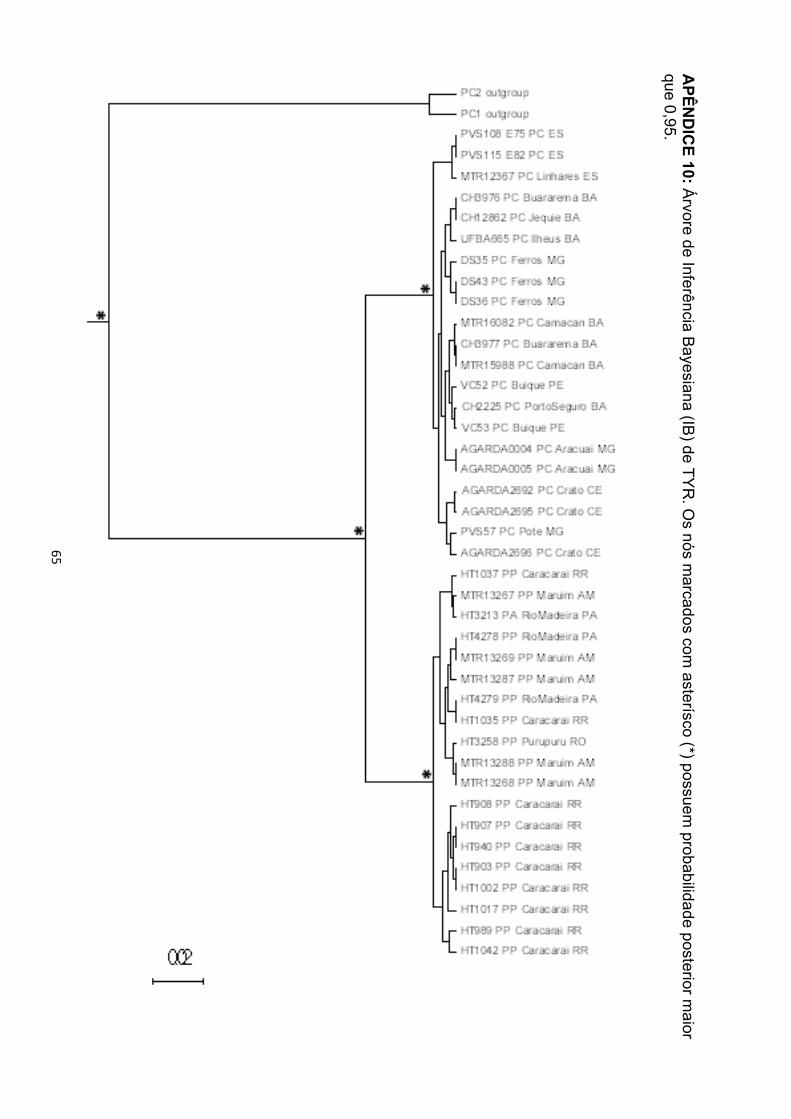
!65!
AP
ÊN
DIC
E 10: Á
rvore de Inferência Bayesiana (IB
) de TYR
. Os nós m
arcados com asterísco (*) possuem
probabilidade posterior maior
que 0,95.

!66!
AP
ÊN
DIC
E 11: Á
rvore de Máxim
a Verossim
ilhança (MV
) de TYR
. Os valores dos nós correspondem
ao bootstrap.

!67!
AP
ÊN
DIC
E 12: Á
rvore de Inferência Bayesiana (IB
) dos dados concatenados. Os nós m
arcados com asterísco (*) possuem
probabilidade posterior m
aior que 0,95.

!68!
AP
ÊN
DIC
E 13: Á
rvore de Máxim
a Verossim
ilhança (MV
) dos dados concatenados. Os valores dos nós correspondem
ao bootstrap.


















