
FORAMINÍFEROS COMO FERRAMENTA DE ESTUDO NA GEOLOGIA AMBIENTAL … · 2017-11-03 · universidade...
161
Universidade Federal do Rio Grande do Norte Centro de Ciências Exatas e da Terra Programa de Pós-Graduação em Geodinâmica DISSERTAÇÃO DE MESTRADO FORAMINÍFEROS COMO FERRAMENTA DE ESTUDO NA GEOLOGIA AMBIENTAL DO ESTUÁRIO DO RIO POTENGI E DA PLATAFORMA INTERNA, RN, BRASIL Autora: CRISTIANE LEÃO CORDEIRO DE FARIAS Orientadora: Dra. PATRÍCIA PINHEIRO BECK EICHLER Co – Orientadora: DRA. HELENICE VITAL Dissertação n o 159 Natal-RN, 16 de novembro de 2015
Transcript of FORAMINÍFEROS COMO FERRAMENTA DE ESTUDO NA GEOLOGIA AMBIENTAL … · 2017-11-03 · universidade...

Universidade Federal do Rio Grande do Norte
Centro de Ciências Exatas e da Terra
Programa de Pós-Graduação em Geodinâmica
DISSERTAÇÃO DE MESTRADO
FORAMINÍFEROS COMO FERRAMENTA DE ESTUDO
NA GEOLOGIA AMBIENTAL DO ESTUÁRIO DO RIO
POTENGI E DA PLATAFORMA INTERNA, RN, BRASIL
Autora:
CRISTIANE LEÃO CORDEIRO DE FARIAS
Orientadora:
Dra. PATRÍCIA PINHEIRO BECK EICHLER
Co – Orientadora:
DRA. HELENICE VITAL
Dissertação no 159
Natal-RN, 16 de novembro de 2015

UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE
CENTRO DE CIÊNCIAS EXATAS E DA TERRA
PROGRAMA DE PÓS-GRADUAÇÃO EM GEODINÂMICA
DISSERTAÇÃO DE MESTRADO
FORAMINÍFEROS COMO FERRAMENTA DE ESTUDO
NA GEOLOGIA AMBIENTAL DO ESTUÁRIO DO RIO
POTENGI E DA PLATAFORMA INTERNA, RN, BRASIL
Autora:
Cristiane Leão Cordeiro de Farias
Dissertação apresentada em 16 de novembro
de 2015, ao Programa de Pós-Graduação em
Geodinâmica e Geofísica – PPGG, da
Universidade Federal do Rio Grande do Norte
- UFRN como requisito à obtenção do Título
de Mestre em Geodinâmica e Geofísica, com
área de concentração em Geodinâmica.
Comissão Examinadora:
Dra. Patrícia Pinheiro Beck Eichler (orientadora)
Dr. Moab Praxedes Gomes (membro interno)
Dra. Silvia Helena de Melo e Sousa (membro externo)
Natal-RN, 16 de novembro de 2015.

i

ii
RESUMO
O estuário do rio Potengi tem sido afetado por diversos fatores antrópicos ao
longo dos anos, como dragagens periódicas, dejetos industriais e domésticos,
intenso tráfego e entre outros fatores, ocasionando vários desastres ambientais,
entre eles o notório acidente ecológico ocorrido em Julho de 2007, que abrangeu
os municípios de São Gonçalo do Amarante, Macaíba e Natal. Os foraminíferos
atuam como ferramentas de estudo viáveis nesses ambientes, pois são capazes de
indicar ambientes ecologicamente estressados, apontando as alterações
hidrográficas e os ambientes deposicionais em estuários. Verificar gradientes
ambientais em locais antropicamente impactados no rio Potengi e em sua
plataforma interna através de espécies de foraminíferos, e sua resposta aos
fatores físicos, químicos e geológicos são importantes no diagnostico de
ambientes. Os resultados mostram a dominância de foraminíferos oportunistas
Ammonia tepida, Bolivina striatula, Quinqueloculina patagonica e Q. miletti
especialmente nas regiões próximas às fazendas de carcinicultura e ao esgoto do
Canal do Baldo em ambientes de granulometria fina; e Q. lamarckiana indica
penetração da cunha salina no rio Potengi. Ocorreram algumas espécies
características de manguezais, tolerantes à baixa salinidade como Trochammina.
inflata e T. squamata no Canal do Rio Potengi, sugerindo provável transporte do
material sedimentológico para as regiões de plataforma interna e que o
contribuinte fluvial é capaz de exportar organismos mixohalinos, tolerantes à
baixa salinidade, para a plataforma interna. Pode-se concluir nesse estudo que o
estuário é regido por dois ambientes distintos, onde a área mais externa sofre
forte influência marinha e a área mais interna sofre fraca influência salina. O rio
Potengi é dominado por espécies calcárias e aglutinantes, enquanto que a foz e a
plataforma interna são dominadas pelas espécies calcárias, aglutinantes e
porcelanáceas, as quais são típicas de ambientes com alta salinidade e
hidrodinâmica. A distribuição dos foraminíferos está condicionada às correntes
fluviais e marinhas, a granulometria e a disponibilidade de CaCO3 e de matéria
orgânica. Palavras chave: rio Potengi, estuário, plataforma interna, foraminíferos
bentônicos, granulometria.

iii
ABSTRACT
The Potengi River estuary has been affected by various anthropogenic factors
over the years, as periodic dredging, industrial and domestic waste, traffic and
other factors, causing various environmental disasters, including the notorious
ecological accident in July 2007, which covered the municipalities of São
Gonçalo do Amarante, Macaíba and Natal. Foraminifera serve as viable study
tools in these environments; they are able to identify ecologically stressed
environments, pointing out hydrographic changes and depositional environments
in estuaries. The necessity to check the differences in environmental gradients in
places anthropically impacted in Potengi River and adjacent inner shelf through
species of foraminifera, and, the responses of these organisms to physical,
chemical and geological factors is to provide baseline in the diagnosis of
environments. The results show the dominance of opportunistic Ammonia tepida,
Bolivina striatula, Quinqueloculina patagonica and Q. miletti especially in
regions close to shrimp farms and Baldo Channel sewage in fine grain
environments; and Q. lamarckiana indicates penetration of the saline waters in
Potengi River. The occurrence of low-salinity tolerant foraminiferal species
typical of mangrove environments as Trochammina inflata and T. squamata in
Potengi River Channel suggest they probably could have been transported from
mangrove area near the Potengi river mouth to the inner shelf regions. These
findings suggest Potengi River is able to export mixohaline and mangrove
organisms to inner shelf. Two distinct environments were observed, the
outermost area is more influenced by marine influence and the innermost area is
less influenced. Calcareous and agglutinated species dominate Potengi River,
while mouth and inner shelf areas are dominated by calcareous, agglutinated and
porcelaneous species, which are typical of highly saline and hydrodynamic
environments and the contributive factors that controls foraminiferal distribution
were balance of marine and freshwater currents, grain size, availability of CaCO3
and organic matter.
Key words: Potengi River, estuary, inner shelf, benthic foraminifera, grain size.

iv
AGRADECIMENTOS
Agradeço à Universidade Federal do Rio Grande do Norte pelo apoio logístico e
por toda a estrutura necessária para o desenvolvimento do trabalho.
Agradeço aos seguintes projetos que viabilizaram meu mestrado: CNPq INCT
AmbTropic/WG 2.1, CAPES–Ciências do Mar 207-10; PRH22-
ANP/MCT;PLAT N-NE Rede 05 / FINEP / CTPETRO; CNPq; PROBRAL 337-
10 (CAPES/DAAD), CAPES PVE 151/2012 (Ciências sem Fronteiras),
MCTI/CNPq Nº 23/2011 - Apoio Técnico para Fortalecimento da Paleontologia
Nacional.(552976/2011-3), CAPES através do Edital Ciências do Mar
(207/2010) para a bolsa de Post Doc no Laboratório de Geologia e Geofísica
Marinha e Monitoramento Ambiental da Universidade Federal do Rio Grande do
Norte (GGEMMA-UFRN-Brazil). Agradeço a todos os professores e
funcionários do Departamento de Geologia da UFRN pelos apoios prestados e
em especial à Nildinha, que me ajudou em todos os contratempos e estresses, que
não foram poucos. Muito obrigada!
Sou grata ao Laboratório de Sedimentologia do Departamento de Geologia da
UFRN e à técnica/chefe do laboratório Fátima e dos seus monitores pelo
fornecimento dos produtos químicos e dos equipamentos necessários para a
realização dos procedimentos analíticos.
Com muita gratidão ao Museu Câmara Cascudo pelo fornecimento do laboratório
para a realização dos procedimentos granulométricos; à chefe do departamento
Iracema Miranda da Silveira e a Janny e Fiética pela ajuda prestada, pelas
risadas e pela amizade.
É com muita gratidão ao Departamento de Geoquímica da UFRN pelo
fornecimento do laboratório para a realização dos procedimentos biológicos; e
agradeço também a professora Raquel Franco De Souza e aos monitores
Emerson, Robson Rafael e Ingrid de Castro dos Santos deste departamento pela
ajuda nesta etapa.
Agradeço ao Departamento de Química da UFRN e à professora Dra. Rosangela
de Carvalho Balaban a utilização do laboratório para a realização dos

v
procedimentos granulométricos. Agradeço também às monitoras Luciana e
Sthepanie pela ajuda prestada e pelo seu tempo em ajudar nas análises.
Meus agradecimentos também ao Departamento de Geografia da UFRN pelo
fornecimento do laboratório para a realização dos procedimentos, em especial à
professora Zuleide Maria Carvalho Lima e a Joyce e Ivaniza não só pela ajuda
prestada, mas também pelas risadas e pelos momentos musicais ecléticos (MPB,
samba, rock, etc). Foi muito divertido trabalhar com vocês!
Serei eternamente grata à minha orientadora, mentora e amiga, profa. Dra.
Patrícia Pinheiro Beck Eichler por todo o apoio, paciência e suporte não só no
desenvolvimento desse trabalho, mas também por ter me aberto as portas e me
dado a oportunidade de trabalhar como estagiária por dois anos. A você, querida
mestra, minha eterna gratidão!
É com muita satisfação que agradeço à minha co-orientadora profa. Dra.
Helenice Vital que me recebeu no GEMMA e auxiliou no meu aprimoramento
acadêmico e desse trabalho, através do fornecimento de materiais bibliográficos
sobre geologia, oceanografia e enviando declarações para vários departamentos
da UFRN e agradeço, ainda, pela oportunidade no GEMMA. A você, professora
e amiga, muitíssimo obrigada!
Sou totalmente grata ao professor Dr. Moab Praxedes Gomes pelo
esclarecimento de dúvidas e sugestões de grande valia no meu trabalho.
Também agradeço ao professor Dr. Narendra Kumar Srivastava por informações,
sugestões e bibliografias.
É com muita honra e gratidão que dedico essa vitória ao professor Dr. Barun Sen
Gupta (professor visitante da Lousiana State University) pelo suporte, apoio e
paciência na identificação de espécies de foraminíferos e suas características
singulares. Muito obrigada por tudo!
É com muita satisfação e consideração que agradeço ao IBAMA pela ajuda
prestada, em especial ao superintendente Alvamar Costa de Queiroz e o
coordenador de gabinete Luiz Eduardo Carvalho Bonilha pelo fornecimento do
mapa da área afetada e dos pareceres técnicos; ao analista ambiental Airton De
Grande por ceder um horário de seu tempo para assistir ao filme – documentário

vi
“Rio Contado”; e à recepcionista do gabinete da superintendência Francisca
Batista Mendes, pela cortesia e atenção.
Agradeço carinhosamente aos meus pais Paulo Roberto Cordeiro de Farias e
Maria Goretti Leão Cordeiro de Farias; às minhas irmãs Danielle e Caroline Leão
Cordeiro de Farias e a minha querida sobrinha e anjinha Isabelinha pelo amor e
carinho incondicionais e me ajudaram a vencer todas as dificuldades da vida;
principalmente ao meu pai, que me ajudou nos estudos com geologia geral para a
prova de mestrado e nos módulos necessários para o mestrado, me tirando as
dúvidas sempre que era necessário.
Sou grata a todos os meus colegas e amigos do laboratório de Geologia e
Geofísica Marinha e Monitoramento Ambiental (GEMMA), em especial ao
André, que com MUITA paciência me ajudou a confeccionar os mapas da área
de estudo e de contorno das espécies de foraminíferos; à Isabelle Barros que me
explicou as diferentes granulometrias existentes; à Isabelle e ao Cordeiro que me
explicaram a metodologia granulométrica aplicada no museu; e ao Canindé, que
coletou as amostras do rio Potengi.
Às minhas amigas de longa data Andrea “Adrenalina” e Rízia que, mesmo tendo
vidas muito corridas e ocupadas, sempre acharam um jeitinho de sairmos e
jogarmos conversa fora. Adoro vocês!
Agradeço principalmente a Deus pela oportunidade que me foi dada, pela
família, pelos amigos que adquiri ao longo da vida e pelas pessoas que me
ajudaram a vencer em todas as dificuldades que surgiram no caminho e por ter
vencido esta nova etapa que se chama mestrado, que era um sonho que há tanto
tempo estava sendo pelejado arduamente e agora foi superado com muito
trabalho duro e muito suor.

vii
ÍNDICE
RESUMO ii
ABSTRACT iii
AGRADECIMENTOS iv
ÍNDICE vii
ÍNDICE DE FIGURAS ix
ÍNDICE DE TABELAS xiv
LISTA DE SÍMBOLOS E ABREVIATURAS xvi
1 – Introdução 18
2 – Área de estudo 22
2.1 – Definição de estuários, classificação, características sedimentares e
importância
22
2.1.1 Definições de estuários 22
2.1.2 Hidrodinâmica do fluxo e dispersão dos sedimentos 25
2.1.3 Importância dos estuários 26
2.1.4 Ameaças aos estuários 27
2.2 – Definição e importância da geologia ambiental 38
2.3 – O estuário do rio Potengi 29
2.4 – Aspectos fisiográficos 30
2.5 – O acidente de Julho de 2007 31
2.6 – Aspectos sociais 33
2.7 – Aspectos geológicos regionais 35
3 – Materiais e métodos 41
3.1 – Amostragem 41
3.2 – Parâmetros biológicos 44
3.3 – Parâmetros sedimentológicos 46
3.4 – Constituintes orgânicos 52
3.5 – Análise estatística 53
3.5.1 Análise univariada 53
3.5.2 Análise multivariada 54
4 – Resultados 56
4.1 – Descrição dos dados abióticos 58
4.1.1 Amostragem de Outubro de 2011 58
4.1.2 Amostragem de Janeiro de 2012 62
4.1.3 Análise comparativa dos mapas de georreferenciamento da
granulometria de Outubro de 2011 e Janeiro de 2012
66
4.2 – agrupamentos e similaridades das comunidades de foraminíferos 72
4.2.1 Amostragem de Outubro de 2011 73
4.2.2 Amostragem de Janeiro de 2012 84
4.2.3 Análise comparativa dos mapas de georreferenciamento dos

viii
foraminíferos de Outubro de 2011 e Janeiro de 2012 94
5 – Discussões 101
6 – Conclusões 109
REFERÊNCIAS 110
APÊNDICE 117
8 – Artigo 119
8.1 – Introdução 123
8.2 – Caracterização da área de estudo 123
8.2.1 O rio Potengi 123
8.2.2 Aspectos fisiográficos 123
8.2.3 Aspectos geologicos 124
8.3 – Materiais e métodos 124
8.3.1 Parâmetros biológicos 126
8.3.2 Parâmetros sedimentológicos 127
8.3.3 Análises estatísticas 127
8.4 – Resultados 128
8.5 – Discussões 149
8.6 – Conclusões 155
8.7 – Agradecimentos 156
Referências 157

ix
ÍNDICE DE FIGURAS
Figura 2.1: Representação esquemática da morfologia de um estuário e dos
processos nele atuantes, comparando os limites destes de acordo com as
definições de Pritchard (1967) e Dalrymple et al. (1992). (B) Distribuição
esquemática e intensidade relativa dos processos físicos operando no
estuário, indicando o zoneamento faciológico tripartite resultante.
Observar os sentidos de fornecimento de sedimento e a bidirecionalidade
dos processos de onda e de marés, que afetarão também as estruturas
sedimentares primárias. Compilado de Boyd et al. (2006).
23
Figura 2.2: Estuário dominado por ondas. Distribuição de (A) Tipos de
energia; (B) componentes morfológicos do estuário em planta e (C)
Distribuição em seção longitudinal das fácies sedimentares no interior de
um estuário dominado por ondas idealizado. NMM = Nível médio do mar.
Washover = depósitos de arrombamento da barreira. Adaptado de
Dalrymple et al. (1992 por Boyd et al. (2006). Compilado de Boyd et al.
(2006).
24
Figura 2.3: Estuário ideal dominado por marés. Distribuição de: (A) Tipos
de energia e: (B) Elementos morfológicos em planta no interior do
estuário. Compilado de Boyd et al. (2006).
25
Figura 2.4: Modelo de circulação estuarina e rotas dos sedimentos de
acordo com a distribuição de fluxos fluvial e marinho (ondas e correntes
de marés). Compilado de Rossetti (2008).
26
Figura 2.5: Peixes e crustáceos mortos no rio Potengi, Natal, RN. Fonte:
http://www.meioambiente.ufrn.br/wp-content/uploads/2015/08/0-CAPA-
Mortandade-de-peixes-POTENGINATAL.jpg.
32
Figura 2.6: Vista geral do estuário Potengí-Jundiaí, incluindo o retângulo
amarelo com a área diretamente afetada pelo acidente, medida em
extensão do rio de 3 km na data de 29 de julho de 2007, por meio de
estimativa visual (elevada concentração de peixes mortos). O trecho do rio
com destaque em azul mostra a extensão de distribuição de peixes mortos
no dia 30 de julho de 2007, por cerca de 10 km, até a ponte do Guarapes.
(Compilado do Parecer Técnico IBAMA n.º 02021.000032/2015-87,
2015).
32
Figura 2.7: (A) Calha do rio Potengi; vista para jusante, da ponte da BR-
304/RN, km 269, vendo-se antigos terraços fluviais entalhados pelo curso
atual. (B) Fotos: Farias (janeiro de 2016).
34
Figura 2.8: Calha do rio Potengi, vista para montante, da ponte da BR-
304/RN, km 269, vendo-se a draga e os montes de areia extraída do rio,
acumulados na margem direita aguardando transporte. Foto: Farias
(janeiro de 2016).
34
Figura 2.9: Vista aérea oblíqua do estuário do rio Potengi, mostrando a
ocupação humana de suas margens. Em primeiro plano, delimitado pelo
quadrado amarelo, o local do acidente ecológico de 2007. Ao fundo, a
cidade de Natal. Foto: Farias (2011).
35

x
Figura 2.10: Mapa geológico simplificado da região em torno de Natal.
Fonte: Frazão (20103, modificado de Bezerra et al., 2001).
36
Figura 2.11: Mapa geológico simplificado da região da foz do estuário
Potengi. Compilado de Cunha (1982).
36
Figura 2.12: (A) Praia do Y, margem direita, próximo à foz do rio Potengi,
vendo-se ao fundo (montante do rio) bancos lamosos colonizados pela
vegetação de mangue. (B) Ponte Antenor Navarro e a praia da Redinha
Velha ao fundo (margem esquerda do rio Potengi). Fotos: Farias (2013).
37
Figura 2.13: Vista aérea oblíqua do estuário do rio Potengi em direção à
sua foz, mostrando os bancos lamosos colonizados por vegetação de
mangue e em parte ocupados pela carcinicultura e pela área urbana.
38
Figura 2.14: Mapa de distribuição de sedimentos no estuário do rio
Potengi. Compilado de Frazão e Vital (2007).
39
Figura 3.1: Mapa da área de estudo das duas amostragens realizadas. (A)
Outubro de 2011 e (B) Janeiro de 2012.
42
Figura 3.2: Draga van veen usada para a coleta das amostras. 43
Figura 3.3: Coleta do material sedimentológico e biológico. 43
Figura 3.4: A sonda CTD. 43
Figura 3.5: (A) Procedimento de peneiramento em via úmida utilizando-se
duas peneiras de 0,500 mm e 0,062 mm; (B) As amostras sendo colocadas
dentro da estufa para serem secadas dentro da estufa a 60º C e; (C) A
utilização do tricloroetileno para separar as carapaças dos foraminíferos
das frações mais pesadas, as quais se depositam no fundo.
45
Figura 3.6: À esquerda, os foraminíferos sendo transferidos com o pincel
da placa de análise para as lâminas de fundo preto (triagem); e à direita, a
identificação dos foraminíferos baseado no livro de Boltovskoy et al.
(1980).
46
Figura 3.7: (A) Eliminação de água contendo sais que possam afetar as
análises; (B) secagem dos sedimentos nas chapas sob a temperatura de 60º
C e; (C) processo de quarteamento dos sedimentos para as análises de
matéria orgânica, CaCO3 e granulometria.
47
Figura 3.8: Em (A) e (B) as amostras sendo tratadas com o peróxido
(H2O2); e em (C) as amostras sendo lavadas após o tratamento.
49
Figura 3.9: A primeira etapa da análise granulométrica. Em (A) a
sequência de peneiras no Agitador de Peneiras RO-TAP Modelo “T”; (B)
a foto ilustrativa da peneira 1.00 mm mostrando seu conteúdo; (C) as
placas de metal enumeradas de 1 a 8 de acordo com a ordem da sequência
de peneiras e seu conteúdo já retirado; (D) as amostras sendo pesadas e
catalogadas no bloco de notas; (E) as amostras sendo guardadas
separadamente em saquinhos para que em (F) sejam guardados para
arquivo.
50
Figura 3.10: (A) Cadinhos de porcelana contendo os sedimentos dentro da
mufla; (B) mufla sendo ligada para queimar a matéria orgânica (M.O); (C)
após a queima da MO, os cadinhos são colocados dentro da estufa para
resfriar e; (D) pesagem dos sedimentos para mensurar a perda de M.O.
51

xi
Figura 4.1: Dados pluviométricos em (A) Outubro de 2011 e (B) Janeiro
de 2012.
56
Figura 4.2 Análise de PCA de Outubro de 2011, mostrando gráfico com os
agrupamentos das estações conforme os valores positivos e negativos do
PC1 e PC2, além da disposição dos parâmetros abióticos em relação a
todos os quadrantes.
60
Figura 4.3: Análise de PCA de Janeiro de 2012, mostrando gráfico com os
agrupamentos das estações conforme os valores positivos e negativos do
PC1 e PC2, além da disposição dos parâmetros abióticos em relação a
todos os quadrantes.
64
Figura 4.4: Porcentagem de cascalho no rio Potengi e na plataforma
interna em Outubro de 2011 e Janeiro de 2012.
67
Figura 4.5: Porcentagem de areia grossa no rio Potengi e na plataforma
interna em Outubro de 2011 e Janeiro de 2012.
68
Figura 4.6: Porcentagem de areia muito grossa no rio Potengi e na
plataforma interna em Outubro de 2011 e Janeiro de 2012.
68
Figura 4.7: Porcentagem de areia média no rio Potengi e na plataforma
interna em Outubro de 2011 e Janeiro de 2012.
69
Figura 4.8: Porcentagem de areia fina no rio Potengi e na plataforma
interna em Outubro de 2011 e Janeiro de 2012.
70
Figura 4.9: Porcentagem de areia muito fina no rio Potengi e na plataforma
interna em Outubro de 2011 e Janeiro de 2012.
71
Figura 4.10: Porcentagem de argila e silte no rio Potengi e na plataforma
interna em Outubro de 2011 e Janeiro de 2012.
72
Figura 4.11: Dendograma formado pelo CLUSTER mostrando o
agrupamento das estações conforme a similaridade das espécies de
foraminíferos encontradas entre elas em Outubro de 2011.
82
Figura 4.12: Grupos formados a partir da aplicação de MDS nos dados
bióticos de Outubro de 2011.
84
Figura 4.13: Dendograma formado pelo CLUSTER mostrando o
agrupamento das estações conforme a similaridade das espécies de
foraminíferos encontradas entre elas em Janeiro de 2012.
93
Figura 4.13: Grupos formados a partir da aplicação de MDS nos dados
bióticos de Janeiro de 2012.
94
Figura 4.14: Abundância relativa do gênero Ammonia spp. em Outubro de
2011 e Janeiro de 2012.
95
Figura 4.15: Abundância relativa do gênero Bolivina spp. em Outubro de
2011 e Janeiro de 2012.
96
Figura 4.16: Abundância relativa da espécie Caronia exilis em Outubro de
2011 e Janeiro de 2012.
97
Figura 4.17: Abundância relativa do gênero Elphidium spp. em Outubro de
2011 e Janeiro de 2012.
98
Figura 4.18: Abundância relativa do gênero Quinqueloculina spp. em
Outubro de 2011 e Janeiro de 2012.
98
Figura 4.19: Abundância relativa do gênero Textularia spp. em Outubro de
2011 e Janeiro de 2012.
99

xii
Prancha 1: Espécies principais ocorrentes no rio Potengi e na plataforma
interna. 1: Ammonia tepida; 2: Bolivina striatula; 3: Caronia exilis; 4:
Cibicides spp; 5: Elphidium spp; 6: Hanzawaia boueana; 7: Pseudononium
atlanticum; 8 – 10: Quinqueloculina spp; 11: Textularia spp.
117
Figura 8.1: Mapa da área de estudo das duas amostragens realizadas, a
primeira em outubro de 2011 (triângulos) e a segunda em janeiro de 2012
(círculos).
125
Figura 8.2: Prancha ilustrativa das espécies dominantes selecionadas. 1:
Ammonia spp; 2: Bolivina striatula; 3: Caronia exilis; 4: Cibicides spp; 5:
Elphidium spp; 6: Hanzawaia boueana; 7: Pseudononium atlanticum; 8 –
10: Quinqueloculina spp; 11: Textularia spp.
127
Figura 8.3: Dados pluviométricos em Outubro de 2011 (A) e em Janeiro de
2012 (B).
128
Figura 8.4: Gráfico com os agrupamentos das estações conforme os
valores do PC1 e PC2, além da disposição dos parâmetros abióticos em
relação aos quadrantes da primeira coleta.
134
Figura 8.5: Dados abióticos de PCA de Janeiro de 2012 135
Figura 8.6: Dendograma mostrando o agrupamento das estações conforme
a similaridade das espécies de foraminíferos encontradas entre elas da
primeira coleta.
136
Figura 8.7: As análises de CLUSTER das amostras demonstrando o nível
de similaridade entre as estações, dividindo-as em três grupos: Plataforma
Interna (5 – 16), Canal do Rio Potengi (17 – 24) e Foz do Rio Potengi (1 –
4).
138
Figura 8.8: Abundância relativa do gênero Ammonia spp. em Outubro de
2011 e em Janeiro de 2012.
141
Figura 8.9: Abundância relativa do gênero Bolivina spp. em Outubro de
2011 e em Janeiro de 2012.
142
Figura 8.10: Abundância relativa da espécie Caronia exilis em Outubro de
2011 e em Janeiro de 2012.
142
Figura 8.11: Abundância relativa do gênero Quinqueloculina spp. em
Outubro de 2011 e em Janeiro de 2012.
143
Figura 8.12: Abundância relativa do gênero Textularia em Outubro de
2011 e em Janeiro de 2012.
143
Figura 8.13: Porcentagem de cascalho no Rio Potengi e na plataforma
continental em Outubro de 2011 e Janeiro de 2012.
144
Figura 8.14: Porcentagem de areia grossa no Rio Potengi e na plataforma
continental em Outubro de 2011 e Janeiro de 2012.
145
Figura 8.15: Porcentagem de areia muito grossa no Rio Potengi e na
plataforma continental em Outubro de 2011 e Janeiro de 2012.
145
Figura 8.16: Porcentagem de areia média no Rio Potengi e na plataforma
continental em Outubro de 2011 e Janeiro de 2012.
146
Figura 8.17: Porcentagem de areia fina no Rio Potengi e na plataforma
continental em Outubro de 2011 e Janeiro de 2012.
147

xiii
Figura 8.18: Porcentagem de areia muito fina no Rio Potengi e na
plataforma continental em Outubro de 2011 e Janeiro de 2012.
147
Figura 8.19: Porcentagem de argila e silte no Rio Potengi e na plataforma
continental em Outubro de 2011 e Janeiro de 2012.
158

xiv
ÍNDICE DE TABELAS
Tabela 4.1: Quadro gerado pelo PCA, com destaque para a boa variação
cumulativa encontrada no PC2.
58
Tabela 4.2: Valores do PC1 e PC2 para os parâmetros abióticos usados no
PCA. Os valores positivos e negativos do PC1 e PC2 desta tabela estão
correspondentes aos eixos PC1 e PC2 da figura 24, os quais possuem
valores positivos e negativos para determinado componente abiótico.
59
Tabela 4.3: Parâmetros abióticos da coleta de Outubro de 2011. *Nota: a
estação 3 veio pouco material sedimentológico e foi insuficiente para as
porcentagens.
61
Tabela 4.4: Dados fornecidos pela análise BIOENV para Outubro de 2011,
mostrando os principais fatores que se relacionam com a biota da região.
62
Tabela 4.5: Parâmetros abióticos de Janeiro de 2012. 65
Tabela 4.6: Dados fornecidos pela análise BIOENV para Janeiro de 2012,
mostrando os principais fatores que se relacionam com a biota da região.
65
Tabela 4.7: Relação das espécies quanto à composição da carapaça na
tabela à esquerda; e relação das espécies quanto ao hábitat: marinho,
mixohalino, mangue ou cosmopolita na tabela à direita em Outubro de
2011.
74
Tabela 4.8: Relação das espécies identificadas vivas e mortas de cada
estação na amostragem de Outubro de 2011.
76
Tabela 4.9: Frequência absoluta das espécies identificadas na amostragem
de Outubro de 2011.
77
Tabela 4.10: Frequência relativa das espécies identificadas na amostragem
de Outubro de 2011.
78
Tabela 4.11: Relação das espécies quanto à composição da carapaça na
tabela à esquerda; e relação das espécies quanto ao hábitat: marinho,
mixohalino, mangue ou cosmopolita na tabela à direita em Janeiro de 2012.
85
Tabela 4.12: Relação das espécies identificadas vivas e mortas de cada
estação na amostragem de Janeiro de 2012.
87
Tabela 4.13: Frequência absoluta das espécies identificadas na amostragem
de Janeiro de 2012.
89
Tabela 4.14: Frequência relativa das espécies identificadas na amostragem
de Janeiro de 2012.
91
Tabela 8.1: Parâmetros abióticos da coleta de Outubro de 2011. 129
Tabela 8.2: Análise granulométrica por estação de Outubro de 2011. 130
Tabela 8.3: Parâmetros abióticos da coleta de Janeiro de 2012. 131
Tabela 8.4: Análise granulométrica por estação de Janeiro de 2012. 132
Tabela 8.5: Quadro gerado pelo PCA, com destaque para a variação
cumulativa encontrada na somatória do PC1 e PC2.
133
Tabela 8.6: Valores do PC1 e PC2 para os parâmetros abióticos usados no
PCA.
133
Tabela 8.7: Frequência relativa das espécies encontradas na coleta de
Outubro de 2011.
139

xv
Tabela 8.8: Frequência relativa das espécies encontradas na coleta de
janeiro de 2012.
140

xvi
LISTA DE SÍMBOLOS E ABREVIATURAS
a.C Antes de Cristo
BIOENV Matriz de dados biológicos e abióticos
CaCO3 Carbonato de cálcio
cm3 Centímetros cúbicos
CLUSTER Análise de agrupamentos para as estações e/ou espécies
cm Centímetros
C org. Medido EA Carbono orgânico medido em analisador elementar
C org. Total Carbono orgânico total
CTD Aparelho que mede a profundidade, condutividade e temperatura oC Graus Celsius
C2HCl3
Tricloroetileno
g/ml Grama por mililitro
g/cm3 Grama por centímetro cúbico
HCl Ácido clorídrico
H2O2 Peróxido de hidrogênio
km Quilômetros
km2 Quilômetros quadrados
Ltda. Limitada
m Metros
M.a Milhões de anos
MDS Escala multidimensional não – métrica
mg Miligramas
ml Mililitros
mm Milímetros
M.O Matéria orgânica
N total Nitrogênio total
No Número
PC Componente principal
PCA Análise do componente principal
Ppm Partes por milhão
spp Espécies
vpm Voltas por minuto
vs Versus
δ Isótopo
δ13
C Isótopo de carbono 13
δ15
N Isótopo de nitrogênio 15

17
Capítulo 1

18
1 – Introdução
O rio Potengi é considerado um dos maiores estuários da costa brasileira e
faz fronteira com os municípios de Macaíba, São Gonçalo do Amarante e Natal.
Cunha (2004) considera que o estuário do Potengi exerce um papel fundamental
de sustentação das atividades urbanas e turísticas, com significativo reflexo no
equilíbrio socioeconômico da região de Natal. Além da população que tem
atividades de subsistência diretamente ligadas ao estuário, como é o caso
específico da comunidade de pescadores, o estuário tem um importante papel no
abastecimento de mercadorias, combustíveis e no escoamento da produção
primária e secundária do Estado do Rio Grande do Norte, através do Porto de
Natal, situado na margem direita.
Contudo, este importante estuário tem sido afetado pela ação antrópica,
como foi mostrado nos trabalhos de Ramos e Silva et al. (2006), Silva et al.
(2001), Pereira (1999), Frazão (2003) e Cunha (2004), envolvendo dragagens
periódicas da calha do rio, descargas de efluentes sem tratamento dos esgotos e
industriais de vários setores, crescente expansão demográfica e industrial da
cidade, atividades de turismo e lazer, construção de margens artificiais,
construção de um guia de corrente na região da Boca da Barra e a
superurbanização da região costeira.
A geologia ambiental tem a função de analisar as interações humanas com
o ambiente físico, onde são estudados os processos naturais da Terra, como os
processos erosivos e os potenciais riscos naturais que afetam a população, como
as inundações, terremotos, entre outros (Keller, 2012). Essa ciência tem também
importância na análise dos processos geológicos, como a deposição dos
sedimentos de fundo marinho (Suguio, 1999).
Tais efeitos antrópicos afetam o ecossistema, causando processos erosivos
provocadas pelas correntes fluviais das marés, formando bancos de areia nas
margens côncavas e remobilização dos sedimentos erodidos nas margens
convexas (Pereira, 1999), contaminação por metais pesados lançados pelas
fábricas e esgotos (Ramos e Silva et al., 2006; Silva et al., 2001; De Souza et al.,
2010). Como resultado mais danoso desses fatores, ocorreu o acidente ecológico

19
de grandes proporções em Julho de 2007, com a morte de 40 toneladas de peixes,
além de aves e animais mortos na área que abrangeu os municípios de São
Gonçalo do Amarante, Macaíba e Natal (Parecer Técnico no
02021.000145/2014-00 do IBAMA, 2014).
Os foraminíferos são protistas bentônicos ou planctônicos que surgiram no
Cambriano (Era Paleozóica, há aproximadamente 570 M.a) e desde então eles
proliferaram, evoluíram e existem até o Recente, principalmente em ambientes
marinhos (Vilela, 2004). Os foraminíferos do início do Paleozóico possuíam
apenas tecas aglutinantes (que continuam até o Recente), porém no decorrer do
Paleozóico, surgiram as tecas microgranulares e finalmente os foraminíferos com
tecas calcárias lamelares ou porcelanosas (Boersma, 1978). Até o presente,
existem 60.000 espécies de fósseis e recentes validamente catalogadas e 10.000
espécies estimadas (incluindo apenas 40 – 50 espécies planctônicas) viventes,
constituindo o grupo mais diverso de microrganismos com carapaça nos mares
modernos (Debenay, 2012). Os foraminíferos podem ser utilizados em vários
setores das ciências da terra, como a paleoecologia, paleogeografia,
bioestratigrafia, exploração de petróleo, cronologia dos depósitos do quaternário
e estudos evolutivos (Yassini & Jones, 1995).
A dinâmica populacional dos foraminíferos responde bem à contaminação
e poluição, como foi mostrado por Eichler et al. (2014), em que o teor de acidez
dos sedimentos provocado por acidente de óleo na Baía de Guanabara resultou na
dissolução das carapaças de carbonato de cálcio de várias espécies de
foraminíferos. Assim, dependendo da tolerância dos foraminíferos uma
determinada situação, Eichler et al. (2003) determinaram que uma mesma região
pode apresentar aumento exagerado de algumas espécies, mas por outro lado
pode apresentar também ausência de espécies que não toleram esta mesma
condição.
O estudo de indicadores ambientais, como os foraminíferos, pode ser
ainda relacionado com a análise dos aspectos físicos, geológicos e químicos da
região, sendo esta uma boa oportunidade para estudos ambientais, segundo a
Comissão do Registro Geológico da Dinâmica da Biosfera (Committee on the

20
Geologic Record of Biosphere Dynamics, 2005). Isto pode ser justificado, pois
são capazes de sintetizar as características gerais do ambiente, ressaltar as
variações ambientais de curtos períodos de tempo e reagir sensivelmente às
variações sazonais e aos efeitos antrópicos, reconhecendo a problemática local
através de avaliações e diagnósticos ambientais. As análises granulométricas são
importantes no estudo dos sedimentos, pois sua distribuição pode ser
característica de ambientes deposicionais fornecendo também informações sobre
os processos físicos atuantes durante a sedimentação (processos hidrodinâmicos),
como foi citado por Suguio (1980).
Os foraminíferos se correlacionam a granulometria como foi mostrado nos
trabalhos de Eichler et al. (2007), Mahiques et al. (2009), Ruiz et al. (2005),
Sarita et al. (2015) e Wang & Chappell (2001), onde as espécies oportunistas
geralmente são encontradas em sedimentos finos, nos quais a matéria orgânica é
facilmente depositada (areia fina, muito fina, silte e argila).
O presente trabalho desenvolvido no estuário do rio Potengi e em sua
plataforma interna tem como objetivo analisar as espécies de foraminíferos
bentônicos e as respostas destes aos fatores físicos, químicos e geológicos, além
de identificar as possíveis áreas afetadas por poluição e contaminação,
fornecendo dados para avaliações de impactos ambientais. Dados abióticos
(salinidade, densidade, temperatura, profundidade, granulometria, teor dos
constituintes orgânicos e teor de CaCO3) foram correlacionados às espécies de
foraminíferos, evidenciando os locais de deposição em sedimentos mixohalinos e
marinhos.

21
Capítulo 2

22
2 – Área de Estudo
Neste capítulo serão apresentadas a definição, classificação, características
sedimentares e importância dos estuários; definição e importância da geologia
ambiental quanto à relação Homem – Ambiente; o estuário do rio Potengi,
contendo suas descrições gerais, aspectos fisiográficos, o acidente ocorrido em
Julho de 2007, os aspectos sociais dos municípios por onde esse rio percorre e os
aspectos geológicos da área d estudo e do rio Potengi.
2.1 Definição de estuários, classificação, características sedimentares e
importância
Neste tópico será apresentada a definição dos estuários, sob a ótica
geológica e oceanógrafa, a hidrodinâmica do fluxo e dispersão dos sedimentos,
sua importância e as ameaças existentes nesses ambientes.
2.1.1 Definições de estuário
Existem inúmeras definições para os estuários, entre elas Pritchard (1967)
e Fairbridge (1968), que definem esse ambiente como “um corpo aquoso
litorâneo semi-fechado em que a salinidade é diluída pela ação da descarga
fluvial”; e também de Rahmani (1988), definindo-o como “um corpo de água
costeiro que possui conexão com o mar aberto contendo uma mistura de água
marinha de origem fluvial”. Garrison (2002) também definiu o estuário como um
corpo de água rodeado por terra, onde a água doce se mistura com a água
salgada, propondo origens e um padrão de circulação de águas, propondo origens
e um padrão de circulação de águas. Todavia, conforme observado por
Dalrymple (1992) e Boyd et al. (2006), estas definições possuíam enfoque
oceanográfico, sendo ambíguas num contexto geológico e difíceis de aplicar a
sedimentos e rochas sedimentares antigas.
A morfologia idealizada de um estuário representa um corpo d’água de
forma em forma de funil, cujo lado maior abre-se para o oceano. O estuário é o
palco das interações entre os processos marinhos (ondas e correntes) e os
fluviais. O predomínio de um processo sobre o outro depende ao longo do tempo
de controles autocíclicos (inerentes ao próprio sistema deposicional) e alocíclicos
(externos ao sistema deposicional e de abrangência regional a mundial, como as

23
variações eustáticas do nível do mar, clima e tectonismo). A Figura 2.1 ilustra a
morfologia, os limites de fácies e os processos estuarinos atuantes.
Figura 2.1: (A) Representação esquemática da morfologia de um estuário e dos processos nele atuantes,
comparando os limites destes de acordo com as definições de Pritchard (1967) e Dalrymple et al. (1992).
(B) Distribuição esquemática e intensidade relativa dos processos físicos operando no estuário, indicando
o zoneamento faciológico tripartite resultante. Observar os sentidos de fornecimento de sedimento e a
bidirecionalidade dos processos de onda e de marés, que afetarão também as estruturas sedimentares
primárias. Compilado de Boyd et al. (2006).
Os estuários foram definidos geologicamente por Dalrymple et al. (1992)
e Boyd et al. (2006) como feições de natureza transgressiva, representando o
preenchimento de depressões por sedimentos durante a subida relativa do nível
do mar. Os estuários atuais são originários do aumento do nível do mar nos
últimos 18.000 anos (120 m), devido ao intenso derretimento das geleiras
continentais durante a Época do Pleistoceno (1.6 – 10 M.a), mais comumente
conhecida como a Era do Gelo (Thurman & Trujillo, 1999).
Nos níveis de mar baixo, ocorre a incisão de vales fluviais. Quando o nível
do mar começa a subir novamente, as porções mais distais destes vales são
alagadas por águas salgadas e submetidas à ação das marés. Os estuários assim
formados aprisionam sedimentos fornecidos pelos rios e, consequentemente,
pouco sedimento consegue chegar na plataforma. Fortes correntes de maré
podem causar erosão no fundo marinho do material sedimentar pré-existente. O
material lamoso normalmente migra em mar aberto para águas mais profundas,

24
enquanto que as areias ou são transportadas em direção a terra ou são deixadas
no mesmo local da plataforma à medida que aumenta a profundidade das águas
do mar (Dalrymple, 1992).
Rahmani (1988) propôs um modelo “tripartite” de preenchimento dos
estuários para rochas sedimentares do Cretáceo Superior na Província de Alberta,
Canadá e sugeriu processos deposicionais atuantes neste tipo de estuário.
Ressaltou também a importância do modelo tripartite para a paleogeografia e
Exploração de petróleo. A classificação de estuários mais atualizada é a de Boyd
et al. (2006), na qual os autores dividem este importante sistema deposicional
transicional em dois tipos fundamentais, com base no poder relativo dos
processos de ondas e marés: (a) estuários dominados por ondas (Figura 2.2) e
(b) estuários dominados por marés (Figura 2.3). De acordo com estes autores,
a maior parte dos estuários (incluindo o rio Potengi) se encaixa na sefiniçao de
estuários dominados por marés em ambiente de macromarés (definidas por > 4 m
de amplitude, de acordo com Davies (1964, 1972), porém também pode haver
dominância da maré em amplitudes muito menores de maré se a ação das ondas
for limitada e/ou se o prisma de maré é grande.
Figura 2.2 Estuário dominado por ondas. Distribuição de (A) Tipos de energia; (B) componentes
morfológicos do estuário em planta e (C) Distribuição em seção longitudinal das fácies sedimentares no
interior de um estuário dominado por ondas idealizado. NMM = Nível médio do mar. Washover =
depósitos de arrombamento da barreira. Adaptado de Dalrymple et al. (1992 por Boyd et al. (2006).
Compilado de Boyd et al. (2006).

25
Figura 2.3: Estuário ideal dominado por marés. Distribuição de: (A) Tipos de energia e: (B) Elementos
morfológicos em planta no interior do estuário. Compilado de Boyd et al. (2006).
2.1.2 Hidrodinâmica do fluxo e dispersão dos sedimentos
Os sedimentos trazidos para dentro dos estuários derivam principalmente
do influxo fluvial, da plataforma continental, da erosão da margem e fundo do
estuário e de atividades biológicas. A combinação da ação das correntes de maré,
descarga fluvial e ondas, resulta numa grande variedade de fácies (Clifton, 1982).
Os sedimentos podem ser transportados por um longo tempo ao longo do
eixo do estuário pela interação complexa entre o fluxo fluvial, as correntes de
maré cheia e vazante e a circulação estuarina. Podem ser novamente
ressuspensos, passando por novos ciclos de deposição e erosão, conforme
mostrado na Figura 2.4 (Rossetti, 2008). O influxo fluvial tem grande efeito na
hidrodinâmica dos estuários (dependendo da descarga do rio), por trazer grandes
quantidades de sedimentos e por influenciar diretamente a salinidade. Ao entrar
no estuário, o sedimento de fundo é transportado em direção ao mar, sendo
depositado na zona de convergência, a qual é formada pelo encontro da água
doce com a cunha salina que flui em direção à cabeceira do estuário (Figura 2.4).

26
Figura 2.4: Modelo de circulação estuarina e rotas dos sedimentos de acordo com a distribuição de fluxos
fluvial e marinho (ondas e correntes de marés). Compilado de Rossetti (2008).
O sedimento estuarino é formado por areia bem selecionada e lama. A
areia geralmente possui origem marinha e a lama é de origem fluvial.
Normalmente estes dois tipos de sedimentos ocorrem interlaminados, podendo
estar intensamente bioturbados, a ponto de apagar estruturas sedimentares pré-
existentes. A tendência é que os sedimentos se tornem progressivamente mais
finos em direção a montante do estuário, de forma que a parte central do estuário
geralmente é lamosa (Figuras 2.2 e 2.3). Na base dos canais estuarinos ocorrem
clastos grossos, incluindo fragmentos de plantas, conchas, intraclastos argilosos e
seixos. Nas planícies de maré ocorrem sedimentos de suspensão (silte e argila),
que também geralmente estão intensamente bioturbados pelas raízes da
vegetação, crustáceos e vermes (Clifton, 1982).
2.1.3 Importância dos estuários
Os estuários são importantes sob diversos pontos de vista. Os estuários
servem de abrigo a inúmeros organismos, podendo servir também não só como
abrigo e de berçários naturais para animais marinhos e para a comunidade
aquática em geral, como também fornecem valiosos frutos do mar para a
população, como recurso de alimentos de valor. Servem como fazendas naturais,
onde a vegetação aquática sustenta uma complexa cadeia alimentar; controlam

27
ações erosivas, alagamentos e as inundações, absorvendo o choque de ondas
provenientes de tempestades antes que cheguem às áreas internas e causem
destruição das propriedades ribeirinhas. Constituem um sistema de filtragem
natural da poluição, em que ajudam a limpar a água dos efluentes industriais e
domésticos dos estuários para os rios e também um sistema de purificação do ar,
pela presença de plantas que absorvem o dióxido de carbono do ar e libera
oxigênio.
É um centro de educação ambiental, funcionando como “laboratórios
locais” para escolas, centros recreativos e tratamento de saúde mental; centros de
trabalhos para os pescadores, ecologistas e guias turísticos, sendo ainda atração
turística, onde atraem observadores de pássaros, pescadores à linha e anzol,
caçadores e barqueiros (Klee, 1999; Garrison, 2002). Boyd et al. (2006)
consideram que os estuários e os vales incisos por eles ocupados durante os
níveis de mar alto são importantes ambientes deposicionais, porque são locais de
habitação humana, portos, além de representarem significativas acumulações de
petróleo formado em rochas geradoras estuarinas ricas em matéria orgânica e
reservatórios de hidrocarbonetos. São ainda repositórios de informações
importantes sobre a sedimentação em ambientes de mar baixo e transgressiva
inicial nas configurações da rampa e da plataforma. Constituem ambientes
complexos, possivelmente um grupo único de ambientes sedimentares.
2.1.4 Ameaças aos estuários
Apesar de sua importância, Klee (1999) mencionou que os estuários são
anualmente degradados pela ação antrópica, onde são dragados para tornarem
fundos os canais para a navegação e depositar seus sedimentos nas terras de
agricultura ou construções de residências. De acordo com Garrison (2002), os
estuários continuam sendo atrativos para o homem, onde são construídos portos,
marinas, recursos recreativos (natação, transportes marinhos como lanchas e
barcos), além de servir de habitação da população ribeirinha e construção de
campos para a agricultura e carcinicultura, ocasionando poluição de suas águas.

28
Para Thurman & Trujillo (1999), os principais contribuintes para a
poluição são os esgotos (hospitalares e domésticos), derramamento de petróleo e
produtos químicos provenientes dos pesticidas e mercúrio, que são provenientes
das indústrias. No que diz respeito ao derramamento de petróleo, Pinet (2003)
cita que dependendo do ecossistema em que este produto foi lançado, os níveis
dos impactos iniciais e a recuperação variam bastante.
2.2 – Definição e importância da geologia ambiental
O Homem está diretamente ligado ao ambiente em que vive, tirando deste
os recursos necessários para sua sobrevivência. Mas ao mesmo tempo está
propenso aos efeitos naturais, como inundações, maremotos, tempestades e
outros (Klee, 1999; Garrison, 2002). Para que se possa estudar essa relação
Homem – Ambiente, é necessário conhecer os fundamentos da geologia
ambiental e aplicar o planejamento das atividades humanas, antecipando
eventuais riscos naturais.
Para Keller (2012) essa ciência é considerada interdisciplinar, pois ela
depende dos aspectos da química (composição dos materiais da Terra), física
(leis naturais) e biologia (entendimento das formas de vida). Este autor considera,
com base em várias aplicações, que a geologia ambiental pode ser definida como
um ramo da ciência da Terra que estuda todo o espectro das interações humanas
com o ambiente físico. Sendo assim, a geologia ambiental é um ramo da Ciência
Ambiental, isto é, a ciência das migrações, entre as ligações entre os processos
físicos, biológicos e sociais, no estudo do ambiente.
A geologia ambiental, no sentido estrito, é a geologia aplicada ao uso da
informação geológica para nos ajudar a resolver os conflitos da utilização da
terra, para minimizar a degradação ambiental e para maximizar os resultados
benéficos de nossos ambientes naturais modificados. Já Suguio (1999) descreveu
como mais complexas essas interações, onde os ambientes naturais afetam
também a atmosfera, a hidrosfera e a criosfera da Terra. Os processos naturais da
Terra, como as ondas marinhas, as condições meteorológicas e os processos

29
erosivos afetam as atividades humanas, especialmente onde houver potenciais
riscos geológicos relacionados, como inundações, terremotos e vulcanismos.
2.3 O Estuário do rio Potengi
O estuário do rio Potengi é considerado um dos maiores da costa brasileira e
faz fronteira com os municípios de Macaíba, São Gonçalo do Amarante e Natal
(De Souza et al., 2010). O canal principal apresenta em média 170 m de largura e
profundidade entre 8 e 10 m, recebendo as águas dos rios Jundiaí, Doce e
Potengi, sendo este último com a maior área de drenagem (De Souza et al., 2010;
Frazão, 2003; Cunha, 2004). A dinâmica do complexo estuarino rio
Potengi/Jundiaí tem sua dinâmica regida pelas correntes de maré, devido à baixa
descarga fluvial nos altos cursos e a atuação da maré tem intensa penetração no
rio, fato constatado pela presença de manguezais em locais distantes da Boca da
Barra (Pereira, 1999).
O estuário do rio Potengi foi classificado por Frazão (2003) como do tipo
planície costeira, segundo critério de Fairbridge (1980). Quanto aos processos
físicos dominantes, este autor considerou este estuário como dominado por marés
ou de maré, de acordo com critérios de Dalrymple (1992) e Reinson (1992).
Justificou esta classificação com base no fato das correntes de maré exercerem
um papel fundamental na costa leste potiguar, apesar da amplitude de maré em
torno de 2,8 m ser considerada como mesomaré elevada por Davies (1964).
Sua nascente está localizada no município de Cerro Corá, localizada na
microrregião da Serra de Santana, no Seridó Potiguar, a 180 km da capital Natal
e as suas águas banham os municípios de Cerro Corá, São Tomé, Barcelona, São
Paulo do Potengi, São Pedro, Ielmo Marinho, São Gonçalo do Amarante e Natal,
sendo que o rio Potengi é perene a partir de São Gonçalo do Amarante.
Contudo, este importante estuário tem sido afetado pela ação antrópica,
como foi mostrado nos trabalhos de Frazão (2003) e Cunha (2004). De acordo
com o primeiro autor, 60% do esgoto da cidade de Natal é lançado diretamente
em seu curso sem tratamento algum; há várias fontes de matéria orgânica, além
da presença das atividades bem desenvolvidas das indústrias e do Porto que

30
também contribuem para o impacto, como dragagens periódicas e construção de
estruturas defletoras de corrente, as quais provocaram efeitos contrários aos
desejados, como os citados por Frazão (2003) e Cunha (2004).
Silva et al. (2001) determinaram em ostras nas proximidades do canal do
Baldo que o estuário está contaminado com metais pesados como Mn, Fe, Cu, Ni
e Pb e que os níveis de Zn, Cd e Cr apresentaram concentrações maiores do que
os índices permitidos. Pereira (1999) citou como impactos antrópicos a crescente
expansão demográfica e industrial da cidade, com o aterramento de grandes
extensões da margem direita do estuário, o despejo de dejetos urbanos e
industriais, atividades de turismo e lazer, dragagem da calha do rio (ampliação da
capacidade portuária), construção de margens artificiais, construção de um guia
de corrente na região da Boca da Barra, a superurbanização da região costeira,
impedindo a ação natural dos ventos e da maré no transporte de sedimentos e
esculpindo uma paisagem artificial, entre outras.
2.4 Aspectos fisiográficos
O clima predominante é do tipo "As", ou seja, clima tropical com estação
seca no período em que o sol está mais alto e os dias são mais longos. O clima é
quente e úmido, sendo a estação seca do verão e chuvosa do inverno, segundo a
classificação de Köppen (RADAMBRASIL, 1981). A pequena precipitação
pluviométrica é determinada pelo Anticiclone do Atlântico Sul (localizado sobre
o Oceano Atlântico), que possui altas pressões e impõe a região, sob seu
domínio, um tempo estável e sem chuvas. Durante o período do fim do verão e
todo o outono, este Anticiclone perde força (nas regiões Norte e Nordeste do
Brasil), proporcionando a ação da Zona de Convergência Intertropical (ZCIT),
responsável pelos ventos alísios provenientes dos hemisférios Norte e Sul
(Nimer, 1972).
As marés que ocorrem no estuário Potengi e áreas adjacentes são de
natureza semi-diurna, com a variação média das marés de sizígias de cerca de
2.30m e das marés de quadratura, de cerca de 0,85 m e amplitude máxima em
torno de 2,83 m, caracterizando um regime de mesomaré. De acordo com Frazão

31
(2003) comparando-se os valores das velocidades medidas com a amplitude da
maré, durante a maré de sizígia, verifica-se que no estuário Potengi as
velocidades em enchente são maiores do que em vazante. Ainda segundo este
autor os valores máximos atingidos na situação de enchente são de cerca de 1,08
m/s. A média da altura significativa das ondas é de aproximadamente 0,8 m e
apresentam um período de 13 s. De acordo com a Estação Climatológica da
UFRN - Natal, os ventos mais frequentes são predominantes do quadrante Leste,
com frequência Leste-Sudeste e velocidades médias mensais sempre em torno de
4 e 4,5 m/s.
2.5 O Acidente de Julho de 2007
Em julho de 2007 ocorreu um desastre ambiental de grandes proporções,
onde foram encontradas 40 toneladas de peixes, além de aves e animais mortos
na área que abrangeu São Gonçalo do Amarante, Macaíba e Natal (Figura 2.5). O
Parecer Técnico n.º 02021.000145/2014-00 (2014) do IBAMA (Instituto
Brasileiro do Meio Ambiente) cita que ocorreu grande mortalidade de peixes e
invertebrados estuarinos e marinhos no estuário Potengi – Jundiaí (Figura 2.6)
em quatro períodos: Setembro de 2004; Novembro de 2005; Julho de 2007 e
Julho de 2010.
Trabalhos realizados no rio Potengi com ostras Crassostrea rhizophorae
que atuaram como bioindicadores mostrou que, antes dos incidentes
mencionados acima, o rio Potengi já sofre forte influência de metais pesados
provenientes das dragagens periódicas, agricultura (fertilizantes e pesticidas),
emissão dos esgotos não tratados, lixo hospitalar e industriais (Silva et al., 2001).
O mesmo foi observado no trabalho de Ramos e Silva et al. (2006), que fizeram
um estudo com as árvores dos manguezais vermelhos e sua resposta aos metais
pesados. Como foi observado, todos esses trabalhos evidenciaram formas de
contaminação no rio Potengi.

32
Figura 2.5: Peixes e crustáceos mortos no rio Potengi, Natal, RN. Fonte:
http://www.meioambiente.ufrn.br/wp-content/uploads/2015/08/0-CAPA-Mortandade-de-peixes-
POTENGINATAL.jpg
Figura 2.6: Vista geral do estuário Potengí-Jundiaí, incluindo o retângulo amarelo com a área
diretamente afetada pelo acidente, medida em extensão do rio de 3 km na data de 29 de julho de 2007, por
meio de estimativa visual (elevada concentração de peixes mortos). O trecho do rio com destaque em azul
mostra a extensão de distribuição de peixes mortos no dia 30 de julho de 2007, por cerca de 10 km, até a
ponte do Guarapes. (Compilado do Parecer Técnico IBAMA n.º 02021.000032/2015-87, 2015).
Corpos d´água confinados (lagoas costeiras e lagunas estranguladas) e
também ambientes aquáticos de pequeno tamanho (açudes, pequenos lagos e
tanques de cultivo) são especialmente sensíveis ou vulneráveis às mortandades
maciças, pois apresentam baixíssima capacidade de diluir poluentes ou oferecer
pontos de refúgio aos organismos, potencializando os fenômenos fisiológicos e

33
toxicológicos, até mesmo de efluentes domésticos. Por outro lado, ambientes
aquáticos amplos ou abertos, com maior circulação de água, permitem escape ou
fuga dos organismos para outros refúgios, o que faz com que mortandades
maciças nestes ambientes sejam menos frequentes ou associadas ao processo de
elevada rapidez de ação, como substâncias químicas de elevado efeito
toxicológico ou de Demanda Química de Oxigênio (DQO), normalmente
presentes em efluentes industriais (Parecer Técnico IBAMA no.
02021.000145/2014-00, 2014).
2.6 Aspectos Sociais
O filme “Rio Contado” (2015) mostra a pouca informação por parte da
população de Cerro Corá a respeito da nascente do rio Potengi e sua importância.
Em São Tomé e Barcelona, na microrregião da Borborema Potiguar, o rio
Potengi é seco em torno de nove meses e durante a seca suas águas localizam-se
no lençol freático. Em Barcelona ocorre a deposição de esgotos e abatedouros
que despejam carcaças, sangue e vísceras dos animais em uma vala e na época da
cheia são transportadas para o rio Potengi.
Em São Paulo do Potengi, na microrregião do Agreste Potiguar, a seca
ocorre durante nove meses e foi construída a Barragem Campo Grande com a
intenção de acumular água e favorecer a agricultura e a criação de gado; porém o
rio recebe efluentes domésticos e hospitalares, poluindo dessa forma as suas
águas. Em São Pedro e Ielmo Marinho, também no Agreste Potiguar, ocorrem
períodos de seca durante quase todo o ano e chove por três meses. A Figura 2.7
mostra a calha parcialmente seca do rio Potengi no município de São Pedro.

34
Figura 2.7: (A) Calha do rio Potengi; vista para jusante, da ponte da BR-304/RN, km 269, vendo-se
antigos terraços fluviais entalhados pelo curso atual. (B) Fotos: Farias (janeiro de 2016).
Em Ielmo Marinho e São Pedro ocorrem dragagens periódicas do leito do
rio Potengi (Figura 2.8), onde a extração da areia é muito intensa, provocando o
rebaixamento da calha do rio e, consequentemente, o rebaixamento do lençol
freático. A água baixou tanto que tornou impraticável a agricultura, pois além
dos solos secarem, os terrenos à beira do rio estão diminuindo, porque a extração
de areia provoca o desbarrancamento das propriedades, as quais vão perdendo
terreno, enquanto o rio fica mais largo e prejudica as atividades agropecuárias e
agrícolas.
Figura 2.8: Calha do rio Potengi, vista para montante, da ponte da BR-304/RN, km 269, vendo-se a
draga e os montes de areia extraída do rio, acumulados na margem direita aguardando transporte. Foto:
Farias (janeiro de 2016).

35
Nos municípios de São Gonçalo do Amarante, Macaíba e Natal, o rio
Potengi, já com feições estuarinas, é perene e nestes três municípios recebe a
contaminação de esgotos domésticos, hospitalares e industriais, além do lixo
jogado em suas águas. O manguezal em suas margens foi parcialmente destruído
para a instalação de fazendas de carcinicultura e pela ocupação humana (Figura
2.9).
Figura 2.9: Vista aérea oblíqua do estuário do rio Potengi, mostrando a ocupação humana de suas
margens. Em primeiro plano, delimitado pelo quadrado amarelo, o local do acidente ecológico de 2007.
Ao fundo, a cidade de Natal. Foto: Farias (2011).
2.7 Aspectos geológicos regionais
A área de estudo está localizada no litoral oriental do Estado do Rio
Grande do Norte, inserida na bacia litorânea denominada Bacia Sedimentar
Costeira Pernambuco-Paraíba-Rio Grande do Norte (Frazão, 2003). Estudos
recentes realizados por Lima Filho (2013), a região estudada situa-se
estruturalmente sobre a plataforma de Natal, localizada entre o Alto de Touros e
a Sub-Bacia de Canguaretama. Possui sedimentos quaternários recentes como
dunas fixas ou móveis, aluviões, terraços fluviais e mangues (Figura 2.10) e são
encontradas, ainda, uma sequência “Tércio – Quaternária” representada pela
Formação Barreiras, a qual é observada em toda a faixa litorânea brasileira, desde
o Rio de Janeiro até o Pará (Frazão, 2003).
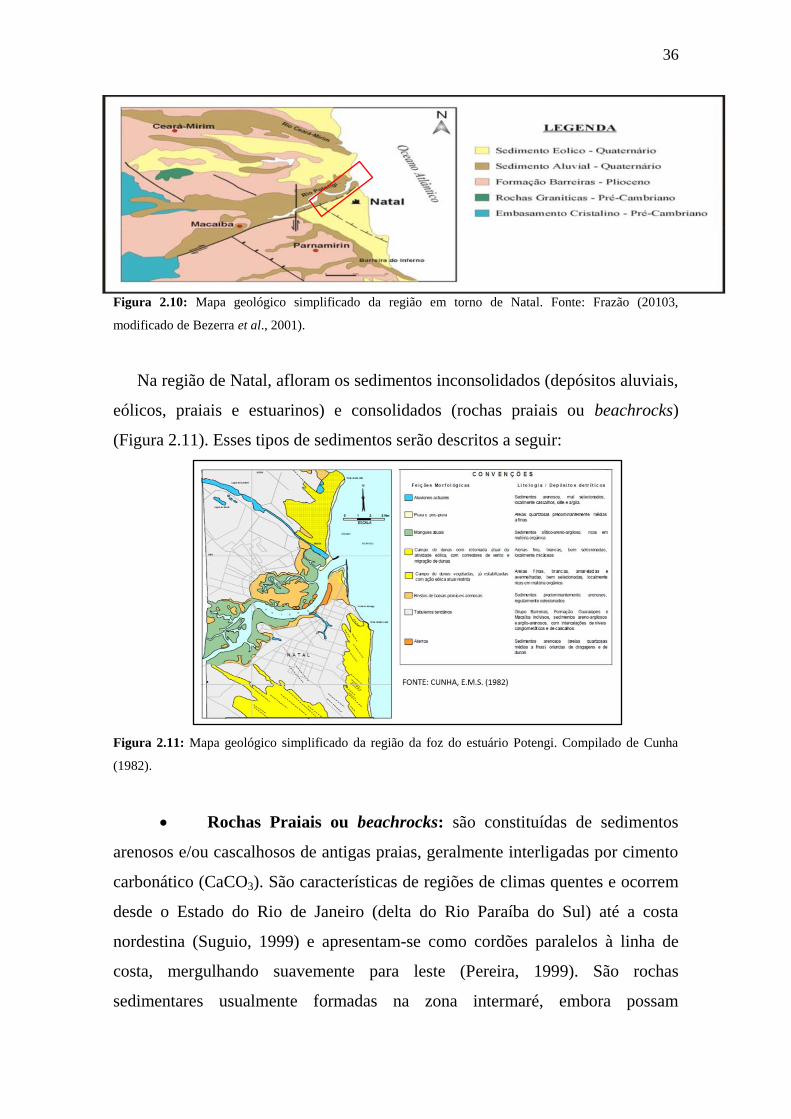
36
Figura 2.10: Mapa geológico simplificado da região em torno de Natal. Fonte: Frazão (20103,
modificado de Bezerra et al., 2001).
Na região de Natal, afloram os sedimentos inconsolidados (depósitos aluviais,
eólicos, praiais e estuarinos) e consolidados (rochas praiais ou beachrocks)
(Figura 2.11). Esses tipos de sedimentos serão descritos a seguir:
Figura 2.11: Mapa geológico simplificado da região da foz do estuário Potengi. Compilado de Cunha
(1982).
Rochas Praiais ou beachrocks: são constituídas de sedimentos
arenosos e/ou cascalhosos de antigas praias, geralmente interligadas por cimento
carbonático (CaCO3). São características de regiões de climas quentes e ocorrem
desde o Estado do Rio de Janeiro (delta do Rio Paraíba do Sul) até a costa
nordestina (Suguio, 1999) e apresentam-se como cordões paralelos à linha de
costa, mergulhando suavemente para leste (Pereira, 1999). São rochas
sedimentares usualmente formadas na zona intermaré, embora possam

37
desenvolver-se em zona sublitorânea, podendo ser úteis como indicadoras na
variação do nível do mar.
Depósitos eólicos: representados por dunas fixas e móveis,
recobrem toda a região, embora estejam ocupadas em grande parte pela área
urbana de Natal. Elas capeiam a Formação Barreiras e os sedimentos recentes,
apresentando direção SE – NW fornecida pelos ventos. São observadas duas
gerações de dunas: paleodunas, de coloração avermelhada e as dunas mais
recentes, de coloração branca;
Depósitos aluviais: ocorrem na calha do rio Potengi e são
constituídos por sedimentos de canais, terraços e planícies fluviais, que podem
ser relacionadas às planícies de inundação. São mais representativos nos cursos
médio e alto do rio Potengi (Figuras 2.7 e 2.8);
Depósitos praiais: são sedimentos arenosos submetidos à ação da
hidrodinâmica costeira e retrabalhamento constante (Figura 2.12), sendo
transportados para outras áreas pela ação das ondas, correntes marinhas e marés,
produzindo diferenças na morfologia praial por processos fluviais e,
principalmente, por ação marinha e eólica, com aporte continuo de sedimentos.
Na parte distal do estuário do rio Potengi são observadas praias arenosas próximo
à Ponte Antenor Navarro (Iate Clube de Natal - praia do Y e Redinha Velha
(Figura 2.12);
Figura 2.12: (A) Praia do Y, margem direita, próximo à foz do rio Potengi, vendo-se ao fundo (montante
do rio) bancos lamosos colonizados pela vegetação de mangue. (B) Ponte Antenor Navarro e a praia da
Redinha Velha ao fundo (margem esquerda do rio Potengi). Fotos: Farias (2013).

38
Depósitos estuarinos: presentes no baixo curso do rio Potengi, são
influenciados principalmente pelas marés. Na baixamar, ficam expostos bancos
lamosos das margens estuarinas, colonizados pela vegetação de mangue (Figura
2.13).
Os sedimentos estuarinos holocênicos do fundo do rio Potengi e litorâneos
da sua foz foram estudados por Frazão (2003). São predominantemente arenosos,
variando de selecionados a bem selecionados, por vezes siltosos, sendo
reconhecidas duas fácies texturais importantes: Fácie Lamosa e Fácie Arenosa
(Figura 2.14). A distribuição destas fácies aparentemente oscila em função da
periodicidade das marés e intensidade das correntes. Frazão e Vital (2007)
identificaram, por meio de dados hidroacústicos, duas zonas de exsudação de gás
metano na área de estudo deste trabalho, provavelmente por efeito da
neotectônica atuante na região.
Figura 2.13: Vista aérea oblíqua do estuário do rio Potengi em direção à sua foz, mostrando os bancos
lamosos colonizados por vegetação de mangue e em parte ocupados pela carcinicultura e pela área
urbana.
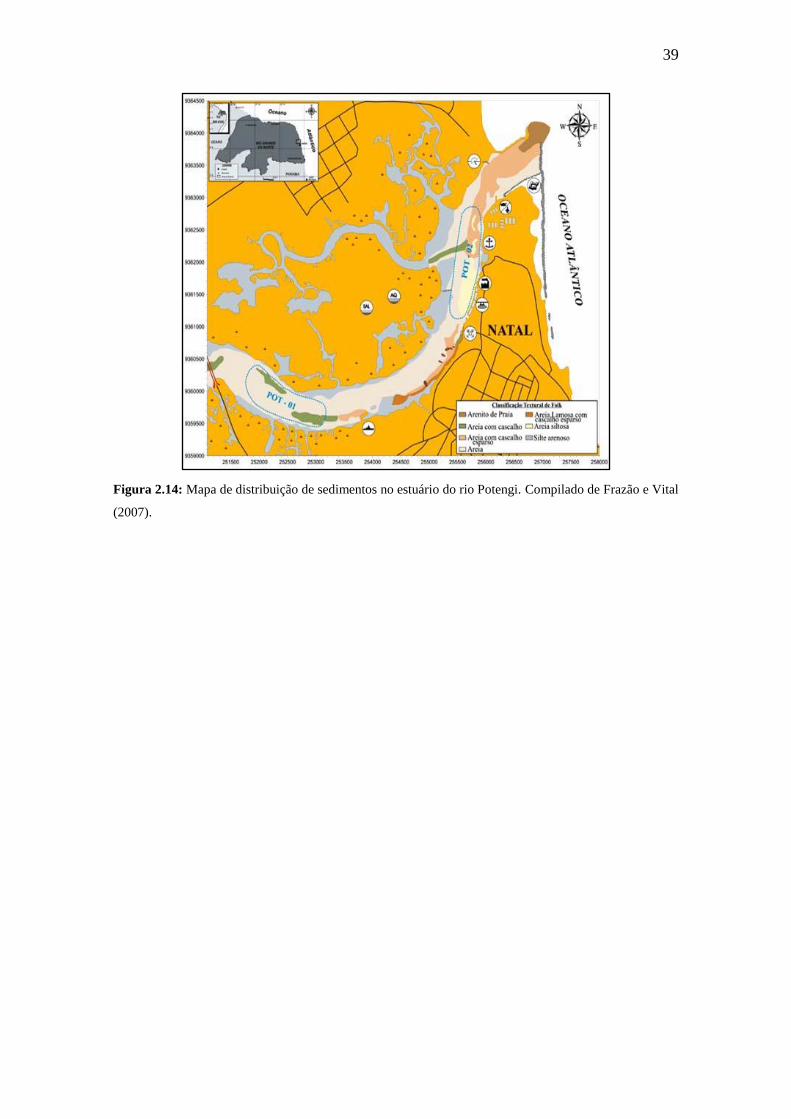
39
Figura 2.14: Mapa de distribuição de sedimentos no estuário do rio Potengi. Compilado de Frazão e Vital
(2007).

40
Capítulo 3

41
3 – Materiais e métodos
Neste capítulo serão detalhados os procedimentos de campo, laboratoriais
e a utilização de programas estatísticos.
3.1 Amostragem
Sedimentos superficiais de 18 estações foram coletadas em outubro de
2011 e 24 estações em Janeiro de 2012 (Figura 3.1), totalizando 42 amostras. As
coletas foram realizadas no curso superior do canal do rio Potengi, que possui
estreita comunicação com o local dos acidentes mencionados e descritos
anteriormente (Figura 2.14). O propósito de se fazer a coleta em ambientes
diferentes (rio e plataforma interna) é verificar as diferenças dos gradientes de
salinidade, temperatura e profundidade, característicos de estuários.
Foi utilizado o amostrador pontual Van Veen (Figura 3.2) e de cada
amostra coletada foi retirada uma fina camada superior delas (Figura 3.3).
O primeiro centímetro retirado foi colocado em frascos com o corante
Rosa de Bengala (1g/1000 ml de álcool), cuja função é corar o protoplasma dos
organismos que estiverem vivos, distinguindo-os dos mortos, no momento da
coleta. O objetivo de separar os indivíduos vivos dos mortos é verificar quais
indivíduos estavam vivos na hora da coleta. A função do álcool é impedir o
ataque bacteriano. Para a medida dos dados físicos (salinidade, condutividade,
densidade, temperatura e profundidade) foi utilizado o aparelho CTD
(Conductivity, Temperature and Depth) da marca Seabird (Figura 3.4).
Na segunda amostragem foram obtidos dados de carbono orgânico e
nitrogênio onde as amostras foram submetidas a análise elementar e isotópica no
analisador elementar Costec Instruments Elemental Combustion System acoplado
ao detector de espectrometria de massas com razão isotópica Thermo Scientific
Delta V Advantage Isotope Ratio MS (EAI – RMS).

42
Figura 3.1: Mapa da área de estudo das duas amostragens realizadas. (A) Outubro de 2011 e (B) Janeiro
de 2012.
A B

43
Figura 3.2: Draga van veen usada para a coleta das amostras.
Figura 3.3: Coleta do material sedimentológico e biológico.
Figura 3.4: A sonda CTD.

44
3.2 Parâmetros biológicos
Em laboratório, o sedimento (50 cm3) foi peneirado em via úmida usando-
se duas peneiras sucessivas de 0,500 e 0,062 mm e posteriormente foi colocado
em papel filtro (Figura 3.5A), para em seguida ser submetido à secagem na estufa
a 60º C (Figura 3.5B). Os foraminíferos foram separados do sedimento por
flotação em tricloroetileno (C2HCl3) dentro de uma capela (Figura 3.5C), pois
suas carapaças são mais leves do que o C2HCl3 e flotam para a superfície,
enquanto que as frações mais pesadas depositam-se no fundo, de acordo com
Boltovskoy (1965).
Após a separação, os foraminíferos foram transferidos com pincel placa de
análise para lâminas especiais de fundo preto, processo denominado de triagem,
para então serem analisados, possibilitando a identificação das espécies, a qual
foi feita utilizando-se microscópio binocular de luz refletida (Figura 3.6). Tal
identificação foi baseada principalmente em Boltovskoy et al. (1980). Uma
prancha ilustrativa de fotos feita em Microscópio Eletrônico de Varredura
(MEV) das principais espécies encontra-se em Anexos.
Os mapas de contorno foram gerados baseados nas tabelas de frequencia
relativa de espécies de foraminíferos através do programa Surfer 9. Foram
selecionadas 6 espécies dominantes para discussão dos resultados.

45
Figura 3.5: (A) Procedimento de peneiramento em via úmida utilizando-se duas peneiras de 0,500 mm e
0,062 mm; (B) As amostras sendo colocadas dentro da estufa para serem secadas dentro da estufa a 60º C
e; (C) A utilização do tricloroetileno para separar as carapaças dos foraminíferos das frações mais
pesadas, as quais se depositam no fundo.
A B
C

46
Figura 3.6: À esquerda, os foraminíferos sendo transferidos com o pincel da placa de análise para as
lâminas de fundo preto (triagem); e à direita, a identificação dos foraminíferos baseado no livro de
Boltovskoy et al. (1980).
3.3 Parâmetros Sedimentológicos
O material coletado foi lavado três vezes sucessivamente para a
eliminação de sais que pudessem afetar as análises realizadas (Figura 3.7A). Em
seguida, foi levado para placas de aquecimento, onde ficou sob temperatura de
60 C até sua secagem completa (figura 3.7B). Posteriormente, todo o material
foi quarteado para as análises de matéria orgânica (10g), granulometria (50g) e
CaCO3 (10g) (Figura 3.7C), constituindo o peso inicial, e o restante foi
arquivado.
47
Figura 3.7: (A) Eliminação de água contendo sais que possam afetar as análises; (B) secagem dos
sedimentos nas chapas sob a temperatura de 60º C e; (C) processo de quarteamento dos sedimentos para
as análises de matéria orgânica, CaCO3 e granulometria.
Para a análise de carbonato de cálcio, as amostras foram tratadas com
ácido clorídrico (HCl) diluído a 10% para eliminar o CaCO3 e a seguir o material
foi lavado para eliminar o HCl. Então, após a secagem completa à 60º C nas
placas de aquecimento, foi feita uma nova pesagem para mensurar quanto de
massa a amostra perdeu, ou seja, quanto em massa havia de CaCO3. Para a
análise granulométrica, as amostras foram tratadas com peróxido de hidrogênio
(H2O2), diluído em água destilada (100 ml de H2O2 para 900 ml de água
destilada), para eliminar a matéria orgânica. Após o ataque, as amostras foram
A B
C

48
lavadas para eliminar o H2O2 e então foram secas nas placas de aquecimento em
temperatura de 60º C (Figura 3.8).
Com a matéria orgânica eliminada, foi feita a análise granulométrica que
consistiu em duas etapas: na primeira etapa foi realizado o peneiramento em uma
sequência de sete peneiras (4,00 mm; 2,00 mm; 1,00 mm; 0,500 mm; 0,250 mm;
0,125 mm e 0,063 mm) e uma base colocadas em um agitador de peneiras RO-
TAP (Figura 3.9A) durante 10 minutos em uma frequência de cinco vpm (voltas
por minuto). Antes desse procedimento, as amostras foram colocadas em placa
de metal e aquecidas por 10 minutos para retirar a umidade dos sedimentos,
pesadas inicialmente e depositadas na sequência de peneiras.
Após o término do tempo estipulado, as peneiras foram retiradas e os
sedimentos retidos nas peneiras (Figura 3.9B) foram retirados através de escovas
com cerdas grossas e agulhas de seringas (0.500 mm e 0.250 mm) e depositados
separadamente em placas de metal enumerados de 1 a 8, de acordo com a ordem
das peneiras, como mostrado na figura abaixo (Figura 3.9C). Após verificar que
nenhum sedimento ficou nas peneiras, estes foram pesados e catalogados em um
bloco de notas (Figura 3.9D), depois guardados separadamente em um saquinho
com identificação de cada amostra (Figura 3.9E) e finalmente são guardados
todos juntos para arquivo (Figura 3.9F).

49
Figura 3.8: Em (A) e (B) as amostras sendo tratadas com o peróxido (H2O2); e em (C) as amostras sendo
lavadas após o tratamento.
A
C
B

50
Figura 3.9: A primeira etapa da análise granulométrica. Em (A) a sequência de peneiras no Agitador de
Peneiras RO-TAP Modelo “T”; (B) a foto ilustrativa da peneira 1.00 mm mostrando seu conteúdo; (C) as
placas de metal enumeradas de 1 a 8 de acordo com a ordem da sequência de peneiras e seu conteúdo já
retirado; (D) as amostras sendo pesadas e catalogadas no bloco de notas; (E) as amostras sendo guardadas
separadamente em saquinhos para que em (F) sejam guardados para arquivo.
C D
E F
A B

51
Para a análise de matéria orgânica (MO), um total de 10g de cada amostra
pesada inicialmente foi depositada em cadinhos de porcelana e colocados na
mufla Modelo F2 - DM Monofásico (Figura 3.10A) inicialmente a 300º C por 20
– 30 minutos e posteriormente a 600º C por 5 horas (Figura 3.10B). No dia
seguinte, estes foram retirados com pinça e colocados em estufa (Figura 3.10C),
deixando em temperatura ambiente. Após este procedimento, houve nova
pesagem das amostras para mensurar quanto de massa de matéria orgânica foi
perdida (Figura 3.10D).
Figura 3.10: (A) Cadinhos de porcelana contendo os sedimentos dentro da mufla; (B) mufla sendo ligada
para queimar a matéria orgânica (M.O); (C) após a queima da MO, os cadinhos são colocados dentro da
estufa para resfriar e; (D) pesagem dos sedimentos para mensurar a perda de M.O.
A B
C D

52
3.4 Constituintes orgânicos
Para a determinação do teor de carbono orgânico total e da razão isotópica
de carbono orgânico, o CaCO3 foi previamente retirado das amostras. Em
seguida, aproximadamente 10 mg de sedimento foram acondicionados em
cartuchos de estanho (5 mm x 9 mm) e submetidos a análise elementar e
isotópica no analisador elementar Costec Instruments Elemental Combustion
System acoplado ao detector de espectrometria de massas com razão isotópica
Thermo Scientific Delta V Advantage Isotope Ratio MS (EAI – RMS).
O teor de nitrogênio total e sua razão isotópica foram determinados em
alíquota de 10 mg de sedimento sem a remoção de CaCO3 acondicionados em
cápsulas de estanho, sendo submetidas a análise elementar e isotópica no EAI-
RMS. Os teores de carbono orgânico e nitrogênio total foram expressos em
porcentagem (%). As razões isotópicas do carbono orgânico e nitrogênio foram
expressas na notação δ (‰):
Onde, “R amostra” é a razão isotópica medida na amostra e “R padrão” é a
razão isotópica medida no padrão de referência. A proporção em massa (%) de
carbono e nitrogênio foi obtida através da relação entre as proporções do padrão
e das áreas correspondentes aos picos do padrão e da amostra:
O padrão adotado na quantificação em massa foi o Solo LECO 502-309 –
LECO Corporation. Os teores de carbono e nitrogênio são, respectivamente,
13,77% e 0,092%. A calibração para as análises isotópicas do carbono orgânico e
nitrogênio total foram feitas através dos padrões certificados USGS-40 (ácido
glutamínico: δ13
C = -26,389‰ vs PDB; δ15
N = -4,5‰ vs ar) e IAEA-600
(cafeína: δ13
C = -27,771‰ vs PDB; δ15
N = +1,0‰ vs ar).

53
A avaliação da repetitividade das análises foi realizada através da análise
em replicata (n = 10) de uma amostra de sedimento adotado como padrão
secundário. A repetitividade para a análise de carbono orgânico total foi: δ13
C =
0,11‰; Corg = 0,06%. Para nitrogênio total, a repetitividade foi de: δ15
N =
0,13‰; Ntotal = 0,002%.
3.5 Análise estatística
A análise estatística foi realizada com o Programa Primer 5 e 6, que
consiste nas análises univariadas (permite a análise de cada variável, podendo
esta ser medida para uma ou mais amostras independentes) e multivariadas
(inclui os métodos de análise das relações de múltiplas variáveis dependentes
e/ou múltiplas variáveis independentes, quer se estabeleçam ou não relações de
causa/efeito entre estes dois grupos) dos dados bióticos e abióticos.
As análises univariadas (índices de diversidade, dominância e
equitatividade) e multivariadas (PCA, CLUSTER, MDS e BIOENV) foram
baseadas na matriz de dados de frequência absoluta de espécies de foraminíferos
e de dados ambientais (profundidade, temperatura, salinidades, porcentagens de
areia, silte, argila, carbono orgânico e enxofre).
A matriz de dados ambientais foi padronizada e submetida à análise de
PCA para identificar tendências biológicas e ambientais. Nos dados biológicos
foram aplicadas as análises de MDS e CLUSTER. O BIOENV correlaciona as
matrizes biológicas e abióticas para verificar qual o fator abiótico mais
importante na distribuição de espécies.
Para a visualização espacial dos dados foi utilizado o programa Surfer 9,
onde a matriz de dados bióticos e granulométricos foram utilizados para
ilustração dos resultados obtidos.
3.5.1 Análise univariada
A análise univariada engloba os índices de dominância (espécies que
dominam determinada área), diversidade (variedade de espécies) e equitatividade
(estabilidade do ambiente) e mostra boa efetividade quando utilizadas em
conjunto para avaliar mudanças na estrutura de comunidade.

54
Essa análise visa apontar quais as espécies dominantes em uma área, quais
as espécies existentes e se o ambiente é favorável, não favorável ou em
equilíbrio.
3.5.2 Análise multivariada
Os dados de profundidade, salinidade, condutividade, densidade,
temperatura, teor de CaCO3 e distribuição de tamanhos de partículas do
sedimento (granulometria), além da distribuição das espécies de foraminíferos
foram estudados especialmente através de PCA (Principal Components Analysis)
que tem a função de analisar quais os componentes principais da amostra de
estudo (temperatura, granulometria fina, salinidade, etc); CLUSTER (análise de
agrupamentos para as estações e/ou espécies, correlacionando a matriz dos dados
biológicos e abióticos, apontando a variável ambiental mais influente na
dinâmica de espécies) tem a função de analisar quais os agrupamentos das
espécies em comum encontradas nas estações; BIOENV (Environmental
Biology), para determinar qual o fator abiótico mais importante na distribuição
das espécies; e MDS (non metric Multi Dimensional Scaling) que tem a função
de formar agrupamentos através das estações que são influenciadas pelos dados
bióticos.
Os dendrogramas do CLUSTER foram realizados para as estações (Modo
Q) e espécies (Modo R). A matriz de dados ambientais (profundidade,
temperatura, salinidades e porcentagens de areia, silte, argila, carbono orgânico e
enxofre) foi padronizada e submetida à análise de PCA para identificar
tendências ambientais, como dominância e diversidade. Nos dados biológicos
foram aplicadas as análises de MDS e CLUSTER. O programa ArcMap 10
confeccionou os mapas da área de estudo de Outubro de 2011 e Janeiro de 2012;
e o Surfer 9 com o método de interpolação “Krigging” utilizou os dados de
abundância relativa dos gêneros de foraminíferos e da granulometria que
tivessem porcentagens relativas significativas para gerar mapas de
georeferenciamento, comparando as duas amostragens.

55
Capítulo 4
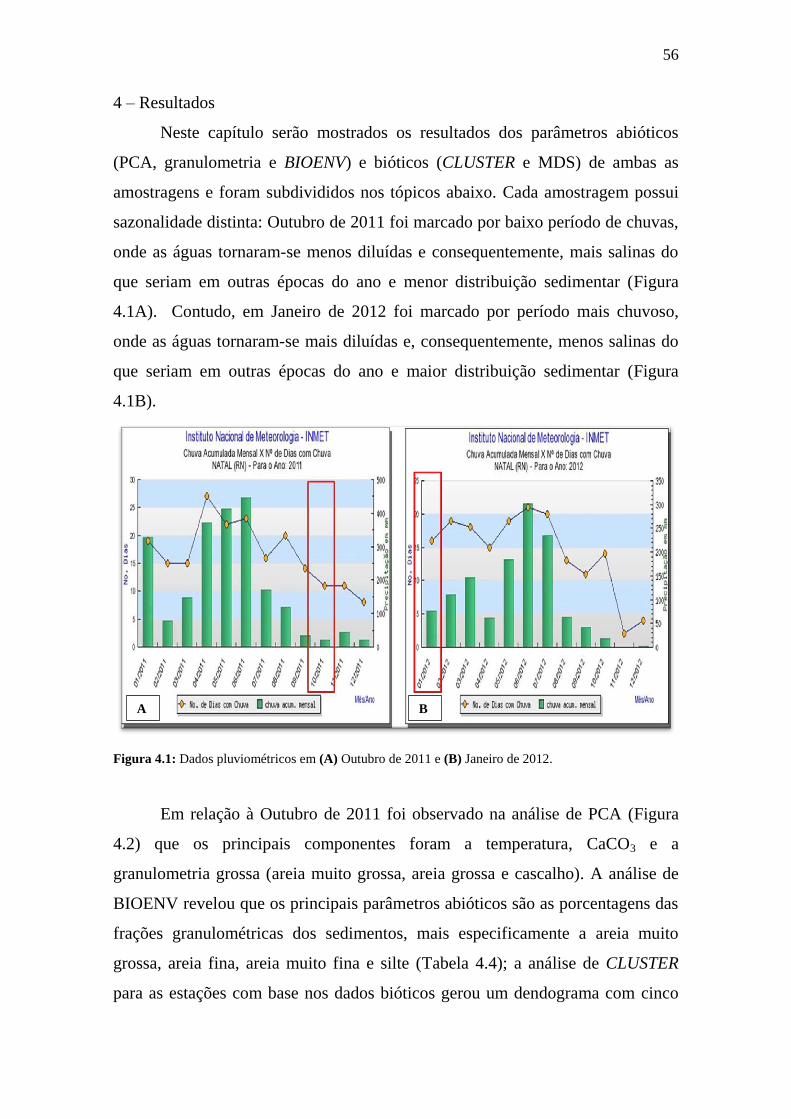
56
4 – Resultados
Neste capítulo serão mostrados os resultados dos parâmetros abióticos
(PCA, granulometria e BIOENV) e bióticos (CLUSTER e MDS) de ambas as
amostragens e foram subdivididos nos tópicos abaixo. Cada amostragem possui
sazonalidade distinta: Outubro de 2011 foi marcado por baixo período de chuvas,
onde as águas tornaram-se menos diluídas e consequentemente, mais salinas do
que seriam em outras épocas do ano e menor distribuição sedimentar (Figura
4.1A). Contudo, em Janeiro de 2012 foi marcado por período mais chuvoso,
onde as águas tornaram-se mais diluídas e, consequentemente, menos salinas do
que seriam em outras épocas do ano e maior distribuição sedimentar (Figura
4.1B).
Figura 4.1: Dados pluviométricos em (A) Outubro de 2011 e (B) Janeiro de 2012.
Em relação à Outubro de 2011 foi observado na análise de PCA (Figura
4.2) que os principais componentes foram a temperatura, CaCO3 e a
granulometria grossa (areia muito grossa, areia grossa e cascalho). A análise de
BIOENV revelou que os principais parâmetros abióticos são as porcentagens das
frações granulométricas dos sedimentos, mais especificamente a areia muito
grossa, areia fina, areia muito fina e silte (Tabela 4.4); a análise de CLUSTER
para as estações com base nos dados bióticos gerou um dendograma com cinco
A B

57
grupos distintos (C1, C2, C3, C4 e C5), onde os três primeiros grupos estão
relacionados à foz do rio e os dois últimos estão relacionados à região interna do
estuário (Figura 4.19).
Foram identificadas 31 espécies, das quais as espécies calcárias foram as
dominantes, seguidas das espécies aglutinantes e porcelanosas (Tabela 4.7); as
espécies dominantes foram A. tepida, B. striatula, Q. miletti, Q. patagonica e Q.
lamarckiana, sendo as primeiras oportunistas (tolerantes a ambientes poluídos e
estressados) e a última proveniente de ambiente marinho, mostrando dessa forma
que o rio Potengi sofre estresse ambiental e há penetração da cunha marinha no
canal do rio de estudo até a estação 10, observada na tabela de frequência relativa
(Tabela 4.10).
Em relação à Janeiro de 2012 a granulometria fina (areia fina, areia muito
fina, silte e argila), carbono orgânico, nitrogênio e CaCO3 foram os componentes
principais a dominarem o PCA (Figura 4.11); a análise de BIOENV revelou o
CaCO3 como o fator mais evidente, seguido pela porcentagem de carbono
orgânico (Tabela 4.6); a análise de CLUSTER para as estações com base nas
espécies revelaram a formação de três grupos: Foz do Rio Potengi (1 – 4),
Plataforma Interna (estações 5 – 16), Canal do Rio Potengi (17 – 24) (figura
4.29); foi observado aumento na diversidade das espécies (41 espécies), mas
baixa dominância de 6 espécies, como A. tepida, B. striatula, Q. lamarckiana, Q.
miletti, Q. patagonica e T. gramen, sendo essa última encontrada principalmente
na plataforma interna (Tabela 4.27).
Ambas as amostragens foram dominadas principalmente por espécies
calcárias, seguida por espécies porcelanosas e aglutinantes. De modo geral,
houve domínio de espécies marinhas, como Q. lamarckiana e o gênero
Textularia, seguida por espécies mixohalinas, cosmopolitas e a presença de
algumas espécies oportunistas, principalmente das espécies A. tepida, Q. miletti e
Q. patagonica (Tabela 4.11). Essas análises serão apresentadas nos tópicos a
seguir contendo as descrições dos dados abióticos de ambas as campanhas
mostrando suas variações, a descrição da variação dos sedimentos em ambas as

58
amostragens, descrição das comunidades dos foraminíferos e será feita uma
análise comparativa de ambas as amostragens.
4.1. Descrição dos dados abióticos
Serão fornecidos a seguir os resultados dos dados abióticos de 2011 e
2012, com base nas análises de PCA para Outubro de 2011(Figuras 4.2) e Janeiro
de 2012 (Figura 4.11) e os mapas de georreferenciamento da granulometria da
primeira amostragem (Figuras 4.3 – 4.10) e da segunda amostragem (Figuras
4.12 – 4.18).
4.1.1 Amostragem de Outubro de 2011
Através do aparelho CTD, foi possível adquirir os dados físicos da
profundidade, CaCO3, temperatura, salinidade, densidade e condutividade. Por
meio desses dados, foi possível fazer as análises estatísticas através do Programa
Primer 5 e 6, onde foi realizada a somatória dos componentes principais dos
dados abióticos (PC) e foram selecionados aqueles que apresentaram maior
porcentagem de variação (PC1: 50.1 e PC2: 24.6) e a somatória de ambas
resultou em 74.6% das amostras (Tabela 4.1).
Tabela 4.1: Quadro gerado pelo PCA, com destaque para a boa variação cumulativa encontrada no PC2.
Após determinar essa somatória e componentes principais (PC1 e PC2),
foram analisados os valores abióticos obtidos em campo juntamente com os
valores de PC1 e PC2 no programa Primer 5 e 6, que determinou a formação de
quatro grupos distintos, cujos valores positivos e negativos dos eixos x (PC1) e y
(PC2) correspondem à tabela 4.2.
PC Autovalor % Variação % Variação Cumulativa
1 7.01 50.1 50.1
2 3.44 24.6 74.6
3 1.03 7.4 82.0
4 0.99 7.0 89.0
5 0.56 4.0 93.1

59
Tabela 4.2: Valores do PC1 e PC2 para os parâmetros abióticos usados no PCA. Os valores positivos e
negativos do PC1 e PC2 desta tabela estão correspondentes aos eixos PC1 e PC2 da figura 24, os quais
possuem valores positivos e negativos para determinado componente abiótico.
A análise de Componente Principal (figura 4.2) para os dados abióticos
em função das estações (Figura 3.1A) determinou a formação de quatro grupos:
O grupo P1 localizado nos valores negativos para o eixo x do PC1 e
positivos no eixo y do PC2 é influenciado pelos parâmetros de temperatura,
CaCO3 e areia muito grossa, representados pelas estações 14 e 17, localizadas na
região central do rio Potengi, próximos às fazendas de carcinicultura e das salinas
(figura 11A), onde ambas são influenciadas pela temperatura, a estação 17 é
fortemente influenciada pelo CaCO3 e pela areia muito grossa e a estação 14 tem
fraca influencia de CaCO3 e areia muito grossa;
O grupo P2 localizado nos valores positivos dos eixos x de PC1 e y de
PC2 é influenciado pelos parâmetros granulométricos de cascalho e areia grossa,
representados pelas estações 2, 5, 11, 12 e 18, indicando que são típicos de
ambientes hidrodinâmicos mais intensos, localizados próximo à desembocadura
(estação 2), na foz (5) e no canal principal (11, 12 e 18), próximos ao Canal do
Baldo e do Porto de Natal;
O grupo P3 localizado com valores negativos no eixo x para PC1 e eixo y
para PC2 é influenciado pela granulometria fina (areia fina, muito fina, silte e
argila), representado pelas estações 1, 13, 15 e 16, localizados no canal principal
do rio Potengi, típicos de ambientes de baixa hidrodinâmica;
Variáveis PC1 PC2
Profundidade 0.177 -0.104
Temperatura -0.320 0.171
Salinidade 0.337 -0.198
Densidade 0.340 -0.198
Condutividade 0.322 -0.196
CaCO3 (%) -0.301 0.165
Cascalho (%) 0.003 0.490
Areia muito grossa (%) -0.004 0.491
Areia grossa (%) 0.237 0.197
Areia média (%) 0.026 -0.278
Areia fina (%) -0.280 -0.296
Areia muito fina (%) -0.319 -0.246
Silte (%) -0.329 -0.158
Argila (%) -0.314 -0.216

60
O grupo P4 localizados nos valores positivos para o eixo x para PC1 e
valores negativos para o eixo y para PC2 é influenciado pelos parâmetros de
profundidade, salinidade, condutividade, densidade e areia média, representados
pelas estações 3, 4, 6, 7, 8, 9 e 10, localizados na foz do rio Potengi (3, 4, 6, 7, 8
e 9), que é tipicamente salina, densa e condutora, possuindo granulometria média
que é tipicamente hidrodinâmica, e na foz do Rio Doce (10).
Figura 4.2: Análise de PCA de Outubro de 2011, mostrando gráfico com os agrupamentos das estações
conforme os valores positivos e negativos do PC1 e PC2, além da disposição dos parâmetros abióticos em
relação a todos os quadrantes.
Observando a Tabela 4.3 em conjunto com o mapa da figura 3.1A pode-se
observar que o local mais profundo da amostragem é a estação 10, com 14,21 m
de profundidade, localizada na foz do rio Doce. Além dela, as estações 2, 3, 4, 5,
6 e 7 também apresentaram maiores profundidades, sendo que a estação 2
localiza-se próximo à desembocadura e as estações 3, 4, 5, 6 e 7 situam-se na foz,
sendo estas últimas com valores de profundidade superiores a 10 m. É importante
observar que estas são profundidades absolutas, sem correção do nível do mar.
A variação da temperatura não foi tão significativa (27.8 – 28.3º C), já que
a área de trabalho se encontra em um clima quente e úmido, mas pôde-se
perceber que as maiores temperaturas pertenceram às estações nas regiões mais
internas do rio e rasas (estações 14 – 18) e os menores valores pertenceram às
estações nas regiões localizadas próximos à foz (2), na foz (3 – 8) e nas
PC1
PC
2

61
proximidades do Rio Doce (9 – 10). É importante ressaltar que ocorrem
dragagens periódicas no rio Potengi para facilitar a passagem de barcos em seu
canal (Cunha, 2004). A salinidade é maior nas estações próximas à foz (2 e 9), na
foz (3 – 8) e mais baixa nas estações localizadas no canal do rio (1, 14 – 18), com
exceção das estações 10 – 13 que apresentaram maiores valores, porém estão
próximas às salinas.
Densidade e condutividade apresentaram maiores valores nas estações
próximas à foz (2 e 9), na foz (3 – 8) e em algumas estações na foz do Rio Doce
(10) e no canal principal (11 – 13 e 18), com menores valores nas estações
localizadas no curso superior (14 – 17), provavelmente isso ocorreu pela baixa
quantidade de salinidade existente nesse trecho.
Tabela 4.3: Parâmetros abióticos da coleta de Outubro de 2011. *Nota: a estação 3 veio pouco material
sedimentológico e foi insuficiente para as porcentagens.
A análise de BIOENV aponta a areia muito grossa como o parâmetro
principal mais influente na distribuição dos foraminíferos para Outubro de 2011
(Tabela 4.4).
Estações Profundidade (m) Temperatura (oC) Salinidade (ppm) Densidade (g/cm3) Condutividade (ppm) CaCO3 (%) Cascalho (%) Areia muito grossa (%) Areia grossa (%) Areia média (%) Areia fina (%) Areia muito fina (%) Silte (%) Argila (%)
1 8,28 27,98 35,56 22,86 57,01 5,00 0,00 0,00 14,05 39,80 31,44 7,67 6,02 1,02
2 12,61 27,80 36,02 23,28 57,46 0,82 7,56 3,75 67,78 15,79 3,31 1,02 0,38 0,41
3 13,25 27,80 36,28 23,48 57,82 0,00 0,00 0,00 0,00 0,00 0,00 0,00 0,00 0,00
4 10,82 27,77 36,34 23,52 57,89 0,00 0,43 0,00 46,57 43,99 5,75 1,98 0,67 0,62
5 13,71 27,80 36,42 23,59 58,03 0,00 7,38 16,48 58,07 13,40 3,17 0,99 0,19 0,32
6 12,19 27,83 36,11 23,34 57,63 3,31 2,60 0,00 43,93 42,79 7,15 2,07 0,77 0,70
7 11,36 27,85 36,18 23,38 57,75 1,60 4,90 0,00 46,96 38,60 6,29 1,88 0,63 0,74
8 5,55 27,89 36,24 23,39 57,86 2,50 0,00 0,00 23,41 68,20 4,60 2,37 0,72 0,70
9 9,06 27,90 36,27 23,42 57,92 0,00 0,00 0,00 44,56 45,02 6,65 2,19 0,93 0,65
10 14,21 27,90 36,23 23,41 57,87 0,83 0,00 0,00 24,10 50,69 18,57 3,68 2,16 0,80
11 3,52 28,10 36,14 23,23 57,95 0,83 0,15 19,08 59,71 15,41 3,66 1,15 0,45 0,39
12 6,86 27,93 36,31 23,44 58,02 1,67 6,59 14,74 61,63 12,59 2,44 1,05 0,62 0,35
13 6,45 27,93 36,15 23,31 57,79 0,00 0,00 0,00 11,90 41,75 33,12 7,00 5,40 0,83
14 4,40 28,20 35,50 22,72 57,16 1,63 11,31 14,57 41,30 25,40 3,96 1,80 1,28 0,38
15 4,87 28,27 35,22 22,49 56,84 5,79 0,00 0,00 0,05 12,25 39,35 19,26 25,89 3,20
16 9,58 28,32 35,02 22,34 56,60 4,07 0,00 0,00 16,32 40,05 26,75 7,89 7,60 1,39
17 7,53 28,19 35,03 22,39 56,48 7,50 17,50 36,99 24,78 13,62 2,37 1,03 3,18 0,54
18 8,87 27,98 35,94 23,14 57,54 0,83 7,05 20,70 38,78 26,13 4,14 1,25 1,50 0,44
Parâmetros abióticos da coleta em Outubro 2011

62
Tabela 4.4: Dados fornecidos pela análise BIOENV para Outubro de 2011, mostrando os principais
fatores que se relacionam com a biota da região.
4.1.2 Amostragem de Janeiro de 2012
Os valores expressos para PC1 e PC2 nas tabelas 4.1 e 4.2 para os dados
abióticos da primeira amostragem também se aplicaram para a análise de PCA
aplicada nos dados abióticos (Figura 4.3), onde observou-se a formação de quatro
grupos distintos: P1, P2, P3 e P4, com base no mapa da figura 3.1B.
O grupo P1, localizado nos valores negativos para o eixo x do PC1 e
positivos no eixo y para PC2, é influenciado pelos parâmetros de granulometria
fina (areia fina, areia muito fina, silte e argila), representadas pelas estações 1, 4,
10, 14, 17, 21, 22 e 23, onde a estação 14 está situada na plataforma interna sul
em Ponta Negra, sendo influenciada pela areia muito fina. As estações 1 e 23 são
influenciadas pelo silte e argila, as quais estão localizadas no canal principal do
rio próximos à Marina Iate Clube; e as estações 4, 10, 17, 21 e 22 são
influenciadas pela areia fina, sendo que as estações 4 e 10 encontram-se na
plataforma interna norte e as três últimas encontram-se no canal principal do rio,
onde a estação 17 situa-se no curso superior. A estação 21 encontra-se próximo
ao canal do Baldo e a estação 22 está próximo ao Porto de Natal,
respectivamente.
O grupo P2, localizado nos valores positivos para o eixo x do PC1 e y do
PC2, é influenciado pelos parâmetros de porcentagem de carbono orgânico,
nitrogênio e CaCO3, representado pelas estações 6, 7, 8, 15 e 20, onde as estações
7, 8 e 20 são influenciadas pelas porcentagens de carbono orgânico e nitrogênio,
que indicam ambientes com altas concentrações de matéria orgânica. As estações
7 e 8 situam-se na plataforma interna norte e provavelmente ocorre a deposição
8 Areia muito grossa (%) 8 Areia muito grossa (%)
11 Areia fina (%) 11 Areia fina (%)
12 Areia muito fina (%) 13 Silte (%)
13 Silte (%)
Correlação: 0.156 Correlação: 0.155
Seleção de variáveis (Outubro/2011)

63
natural de matéria orgânica, indicando ambiente anóxico, já a estação 20 situa-se
no canal principal do rio próximo às salinas e as fazendas de carcinicultura. As
estações 6, 9 e 15, as quais situam-se na plataforma interna, são influenciadas
pelo CaCO3, sugerindo que existe grande contribuição biológica na plataforma
interna.
O grupo P3, localizado nos valores negativos para o eixo x do PC1 e y do
PC2, é influenciado pelo parâmetro da profundidade representado pelas estações
2, 5 e 24, onde as estações 2 e 24 encontram-se próximas à desembocadura e a
estação 5 situa-se na plataforma interna e de acordo com a Tabela 4.4 dos dados
abióticos a estação 24 foi a mais profunda desse grupo.
O grupo P4, localizado no valor positivo para o eixo x do PC1 e negativo
para o eixo y do PC2, é influenciado pela granulometria grossa (areia grossa,
areia muito grossa e areia média) representado pelas estações 3, 9, 11, 12, 13, 16,
18 e 19, onde as estações 3, 11, 12, 13 e 16 situam-se na plataforma interna e as
estações 18 e 19 estão localizadas próximas às fazendas de carcinicultura e às
salinas, no canal principal do rio Potengi. A porcentagem de cascalho não foi
acrescentada na análise de PCA. A porcentagem de areia grossa, areia muito
grossa e areia média são típicas de ambientes hidrodinâmicos mais intensos.

64
Figura 4.3: Análise de PCA de Janeiro de 2012, mostrando gráfico com os agrupamentos das estações
conforme os valores positivos e negativos do PC1 e PC2, além da disposição dos parâmetros abióticos em
relação a todos os quadrantes.
Analisando a tabela 4.5 dos dados abióticos em função do mapa na figura
3.1B os locais mais profundos se encontram na plataforma interna (estações 5, 11
e 15), mas também existem alguns trechos profundos no rio Potengi,
provavelmente pelas dragagens periódicas (estações 1, 2, 22, 23 e 24). O CaCO3
possui maiores frequências na plataforma interna (estações 3, 5, 6, 7, 8, 9, 10, 11,
13, 14, 15 e 16) e mais baixas no canal do rio Potengi (estações 1, 2, 4, 12, 17,
18, 19, 20, 21, 22, 23 e 24). Isso pode ser explicado pela grande disposição de
materiais sedimentológicos de origem bioclástica na plataforma interna, ao
contrário do que acontece no rio Potengi. O desenvolvimento de organismos
marinhos (principalmente os moluscos, corais, algas calcárias, espongiários e
briozoários, por exemplo) é mais favorecido em condições de águas do mar
claras, limpas, mornas e ricas em nutrientes, o que não ocorre no interior do
estuário do rio Potengi.
Os teores muito baixos de carbono orgânico (porcentagens de carbono
orgânico medido em analisador elementar e carbono orgânico total) sugerem
P1 P2
P3 P4

65
ambiente oxidante, ou seja, é inadequado à preservação da matéria orgânica,
como é o caso das estações 6, 7, 15, 20 e 22, ao contrário da estação 8.
Os teores de nitrogênio mostraram-se elevados na estação 7, a qual está
situada na plataforma interna, indicando grande contribuição de matéria orgânica
proveniente das atividades biológicas na plataforma; a estação 20 também
apresentou valores altos, a qual situa-se próxima às salinas e às fazendas de
carcinicultura, indicando que esse ambiente pode estar sob efeito de nitrificação,
que é um processo químico-biológico na formação de nitrito no solo pela ação
conjunta de bactérias quimiossintetizantes nitrificantes, pela ação de conversão
da amônia em nitrato.
Os baixos valores de δ13C/
δ 12C e
δ15N/
δ 14N indicam altos conteúdos
de proteínas (razão C: estações 6, 7, 8 e 15 situadas na plataforma interna;
estações 20 e 22 no canal principal do rio, onde a estação 20 está próxima às
salinas e fazendas de carcinicultura e a estação 22 está próximo ao Porto de
Natal; razão N: 10, 14 e 21, onde as estações 10 e 14 situam-se na plataforma
interna – praia dos artistas e Ponta Negra, respectivamente; estação 21 situa-se no
canal principal próximo ao Canal do Baldo) e, portanto, indicam fonte derivada
de fitoplâncton e zooplâncton. Há probabilidade dos valores mais elevados de
δ15N poderem indicar fonte de fitoplâncton e os valores menores de
zooplâncton.
Tabela 4.5: Parâmetros abióticos de Janeiro de 2012.
Estações Profundidade (m) % CaCO3 % C org medido EA % C org total % N total δ13C/12C δ15N/14N Cascalho (%) Areia muito grossa (%) Areia Grossa (%) Areia média (%) Areia fina (%) Areia muito fina (%) Silte + Argila (%)
1 10.7 2,61 0,01 2,72 0,00 0,04 1,13 6,10 34,59 36,91 21,22
2 13 2,17 0,00 3,65 6,63 34,02 48,98 5,98 0,73
3 11 15,64 0,02 1,97 0,07 4,92 25,84 50,34 14,46 4,29 0,07
4 5.1 1,83 0,00 0,39 14,39 13,90 67,75 3,53 0,04
5 13 22,13 0,02 2,46 0,27 25,35 35,31 7,79 22,32 8,74 0,23
6 5 84,37 0,12 0,02 0,02 -10,42 2,02 0,00 9,91 41,61 16,15 30,32 1,94 0,08
7 8 78,15 0,29 0,06 0,08 -11,39 3,19 0,04 6,94 51,71 15,11 25,54 0,63 0,02
8 9 93,03 4,07 0,28 0,03 -18,17 2,26 0,00 0,12 16,20 16,13 59,83 7,63 0,09
9 8 61,36 0,04 2,25 0,00 1,40 10,29 71,30 14,90 1,95 0,16
10 9 16,30 0,01 -2,16 0,00 0,36 19,17 10,01 67,73 1,98 0,75
11 12.5 15,61 0,00 1,30 42,23 48,46 6,67 1,24 0,10
12 10 1,72 0,00 1,79 75,63 22,56 0,02 0,00 0,00
13 11 45,98 0,18 52,07 30,55 13,55 3,29 0,33 0,03
14 12 22,45 0,02 -0,37 0,00 0,18 0,26 1,48 9,26 74,64 14,20
15 13.8 70,08 0,30 0,09 0,02 -14,49 0,61 0,03 2,04 11,18 53,83 30,81 1,98 0,13
16 11 21,69 0,02 3,00 0,00 0,95 13,75 53,08 26,80 3,61 1,81
17 9 5,77 0,01 3,66 0,00 3,49 12,91 10,33 71,82 1,20 0,26
18 5.7 2,49 0,00 1,07 15,36 52,18 30,69 0,64 0,06
19 4.7 9,77 0,02 0,99 4,69 61,00 32,67 0,47 0,15
20 5.6 6,60 0,27 0,26 0,05 -17,67 3,68 0,00 0,44 1,94 54,26 42,05 1,19 0,13
21 5.9 7,40 0,02 -0,65 0,06 4,69 5,72 13,00 53,06 20,20 3,26
22 13.9 5,52 0,12 0,11 0,03 -11,78 1,53 0,00 3,26 10,12 43,30 34,20 4,52 4,60
23 13 3,08 0,00 0,15 0,57 1,67 45,14 47,93 4,55
24 13.7 2,30 0,01 3,28 0,00 3,11 13,21 53,00 27,80 2,70 0,17
Parâmetros abióticos da coleta em Janeiro/2012

66
A análise de BIOENV revelou que o CaCO3 é o fator mais importante na
distribuição da comunidade de foraminíferos seguido pela porcentagem de
carbono orgânico (Tabela 4.6), indicando que o carbonato de cálcio presente nos
sedimentos é o principal colaborador na formação das carapaças dos
foraminiferos e o carbono orgânico é o fator secundário para a formação das
comunidades dos foraminiferos.
Tabela 4.6: Dados fornecidos pela análise BIOENV para Janeiro de 2012, mostrando os principais fatores
que se relacionam com a biota da região.
4.1.3 Análise comparativa dos mapas de granulometria de Outubro de
2011 e Janeiro de 2012
A seguir serão comentados os dados granulométricos nos mapas de
georreferenciamento em função dos dados granulométricos da Tabela 4.5.
Em Outubro de 2011 a fração de cascalho apresentou maior valor na
estação 17 (17%), localizada próxima às fazendas de carcinicultura e das salinas.
Sua menor frequência situou-se nas estações localizadas na foz (estações 5 e 6) e
em alguns trechos do canal principal (estações 12 e 18), desaparecendo na foz do
Rio Doce (estação 10), na foz do rio Potengi (estações 4, 7 e 8) e grande parte do
canal principal (estações 1, 9, 11, 13 – 16). Em Janeiro de 2012, apesar do
decréscimo significativo na estação 5 (0.28%), a maior concentração ocorre na
plataforma interna, enquanto que na foz e no canal principal do rio é ausente
(Figura 4.4).
2 CaCO3 (%) 2 CaCO3 (%)
3 C org medido EA (%)
Correlação: 0.247 Correlação: 0.223
Seleção de variáveis (Janeiro/2012)

67
Figura 4.4: Porcentagem de cascalho no rio Potengi e na plataforma interna em Outubro de 2011 e
Janeiro de 2012.
Em Outubro de 2011, areia grossa ocorre em maiores frequências (45 –
70%) próximo à foz (estação 2), na foz (estações 5, 6 e 7) e no canal central do
rio Potengi (estações 11 e 12). As menores porcentagens (15%) ocorrem no curso
superior do canal (estação 16) e próximo ao rio Doce (estação 1), desaparecendo
próximo à estação 15, localizada no curso superior do rio. Em Janeiro de 2012
foram observadas maiores porcentagens na estação 12, localizada na plataforma
interna (75%) e também na plataforma interna norte, na estação 7 (50%).
Menores frequências (5 - 10%) de areia grossa nas estações 2, 9, 21 e 22, onde a
estação 9 encontra-se na plataforma interna, a estação 2 está próxima à foz e as
estações 21 e 22 estão no curso principal do rio. Essa fração desaparece nas
estações 14 e 23, as quais situam-se na plataforma interna sul (Ponta Negra) e no
rio Potengi (Marina Iate Clube) (Figura 4.5).
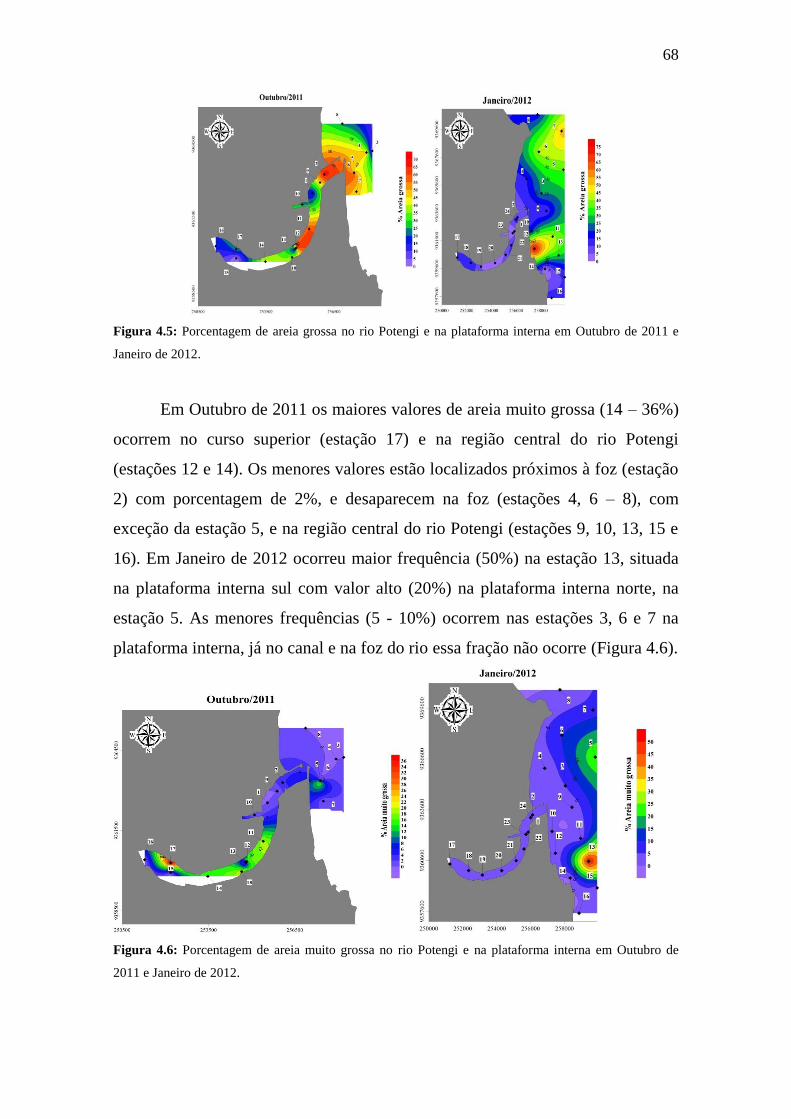
68
Figura 4.5: Porcentagem de areia grossa no rio Potengi e na plataforma interna em Outubro de 2011 e
Janeiro de 2012.
Em Outubro de 2011 os maiores valores de areia muito grossa (14 – 36%)
ocorrem no curso superior (estação 17) e na região central do rio Potengi
(estações 12 e 14). Os menores valores estão localizados próximos à foz (estação
2) com porcentagem de 2%, e desaparecem na foz (estações 4, 6 – 8), com
exceção da estação 5, e na região central do rio Potengi (estações 9, 10, 13, 15 e
16). Em Janeiro de 2012 ocorreu maior frequência (50%) na estação 13, situada
na plataforma interna sul com valor alto (20%) na plataforma interna norte, na
estação 5. As menores frequências (5 - 10%) ocorrem nas estações 3, 6 e 7 na
plataforma interna, já no canal e na foz do rio essa fração não ocorre (Figura 4.6).
Figura 4.6: Porcentagem de areia muito grossa no rio Potengi e na plataforma interna em Outubro de
2011 e Janeiro de 2012.
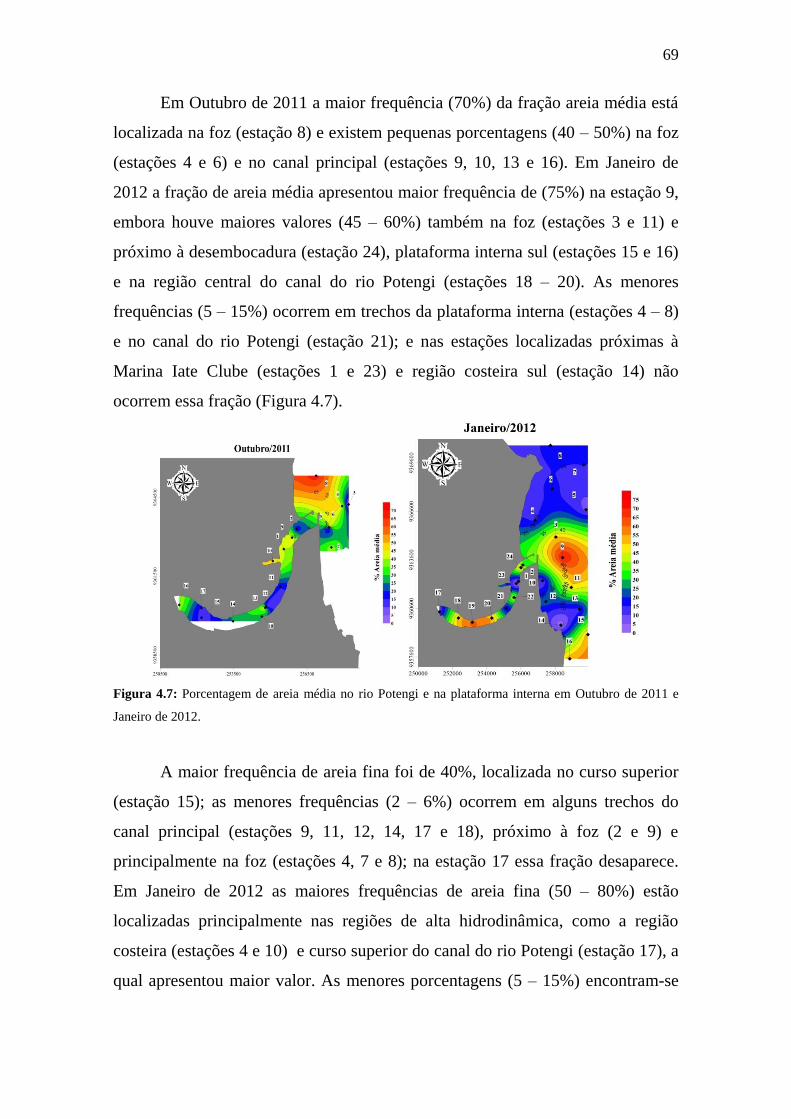
69
Em Outubro de 2011 a maior frequência (70%) da fração areia média está
localizada na foz (estação 8) e existem pequenas porcentagens (40 – 50%) na foz
(estações 4 e 6) e no canal principal (estações 9, 10, 13 e 16). Em Janeiro de
2012 a fração de areia média apresentou maior frequência de (75%) na estação 9,
embora houve maiores valores (45 – 60%) também na foz (estações 3 e 11) e
próximo à desembocadura (estação 24), plataforma interna sul (estações 15 e 16)
e na região central do canal do rio Potengi (estações 18 – 20). As menores
frequências (5 – 15%) ocorrem em trechos da plataforma interna (estações 4 – 8)
e no canal do rio Potengi (estação 21); e nas estações localizadas próximas à
Marina Iate Clube (estações 1 e 23) e região costeira sul (estação 14) não
ocorrem essa fração (Figura 4.7).
Figura 4.7: Porcentagem de areia média no rio Potengi e na plataforma interna em Outubro de 2011 e
Janeiro de 2012.
A maior frequência de areia fina foi de 40%, localizada no curso superior
(estação 15); as menores frequências (2 – 6%) ocorrem em alguns trechos do
canal principal (estações 9, 11, 12, 14, 17 e 18), próximo à foz (2 e 9) e
principalmente na foz (estações 4, 7 e 8); na estação 17 essa fração desaparece.
Em Janeiro de 2012 as maiores frequências de areia fina (50 – 80%) estão
localizadas principalmente nas regiões de alta hidrodinâmica, como a região
costeira (estações 4 e 10) e curso superior do canal do rio Potengi (estação 17), a
qual apresentou maior valor. As menores porcentagens (5 – 15%) encontram-se

70
na plataforma interna (estações 3, 5, 9 e 14), desaparecendo na região sul
(estações 12 e 13) (figura 4.8).
Figura 4.8: Porcentagem de areia fina no rio Potengi e na plataforma interna em Outubro de 2011 e
Janeiro de 2012.
Em Outubro de 2011 a maior porcentagem de areia muito fina é de 20%
no curso superior (estação 15) seguida pelas estações 16 e 9, no curso superior e
próxima ao rio Doce respectivamente, ambas com 8%. Os valores mais baixos (1
– 4%) ocorrem ao longo do canal principal (9 – 12 e 14, 17 e 18), e na foz do rio
Potengi (2 – 8). Em Janeiro de 2012 a maior frequência de areia muito fina foi na
estação 14 (75%), situada na plataforma interna sul, em Ponta Negra; existe
porcentagem significativa no canal principal do rio Potengi (35 – 40%) nas
estações 1 e 23, situadas próximas à Marina Iate Clube. A menor frequência (5%)
ocorre em alguns trechos do Rio (estações 2, 21, 22e 24,) e também na
plataforma interna (estações 5 e 8) e não ocorre no curso superior (estações 17 –
20) e na plataforma interna (estações 3, 4, 6, 7, 9 – 13, 15 e 16) (Figura 4.9).
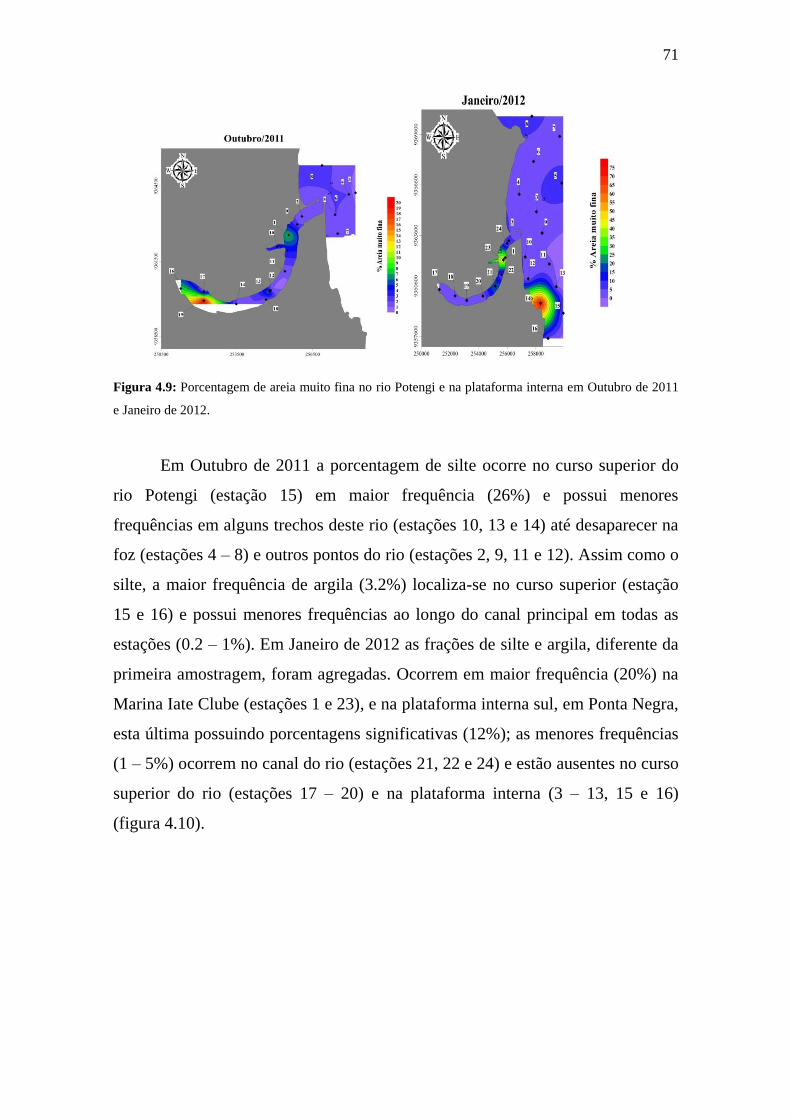
71
Figura 4.9: Porcentagem de areia muito fina no rio Potengi e na plataforma interna em Outubro de 2011
e Janeiro de 2012.
Em Outubro de 2011 a porcentagem de silte ocorre no curso superior do
rio Potengi (estação 15) em maior frequência (26%) e possui menores
frequências em alguns trechos deste rio (estações 10, 13 e 14) até desaparecer na
foz (estações 4 – 8) e outros pontos do rio (estações 2, 9, 11 e 12). Assim como o
silte, a maior frequência de argila (3.2%) localiza-se no curso superior (estação
15 e 16) e possui menores frequências ao longo do canal principal em todas as
estações (0.2 – 1%). Em Janeiro de 2012 as frações de silte e argila, diferente da
primeira amostragem, foram agregadas. Ocorrem em maior frequência (20%) na
Marina Iate Clube (estações 1 e 23), e na plataforma interna sul, em Ponta Negra,
esta última possuindo porcentagens significativas (12%); as menores frequências
(1 – 5%) ocorrem no canal do rio (estações 21, 22 e 24) e estão ausentes no curso
superior do rio (estações 17 – 20) e na plataforma interna (3 – 13, 15 e 16)
(figura 4.10).

72
Figura 4.10: Porcentagem de silte e argila e silte no rio Potengi e na plataforma interna em Outubro de
2011 e Janeiro de 2012.
4.2 Agrupamentos e similaridades das comunidades de foraminíferos
Neste tópico serão explanadas as análises de CLUSTER, MDS e os mapas
da disposição espacial das espécies que apareceram em maior frequência e
daquelas que sofreram algum tipo de alteração espacial, em conjunto com as
tabelas absoluta e relativa de ambas as amostragens. Serão mostradas também
quais as espécies atuantes nos ambientes marinhos, fluviais e de mangue e quais
são as espécies oportunistas, ou seja, são capazes de sobreviver em ambientes
poluídos e estressados, podendo inibir a presença de outras espécies através da
competição por nutrientes e matéria orgânica, e quais as espécies cosmopolitas,
ou seja, podem ser encontrados em vários ambientes diferentes.

73
4.2.1 Outubro de 2011
Na primeira amostragem foram identificadas 31 espécies, onde foi
observada a quantidade de indivíduos vivos e mortos (Tabela 4.8) para verificar
se as condições ambientais são favoráveis (ambiente fornece nutrientes
suficientes para a comunidade, sem existir a necessidade de competição),
desfavoráveis (ambiente não fornece nutrientes suficientes, ocasionando
competição intraespecífica (indivíduos da mesma espécie) ou interespecífica
(indivíduos de espécies diferentes, onde uma inibe a presença de outras) ou em
equilíbrio (o número de indivíduos vivos e mortos é igual ou muito próximos um
do outro) às comunidades de foraminíferos. Depois disso, foi confeccionada a
tabela de frequência absoluta (Tabela 4.9), que consiste na soma dos indivíduos
vivos e mortos, para dessa forma elaborar a tabela de frequência relativa (Tabelas
4.10), a qual serviu para as análises de CLUSTER (Figura 4.11) e MDS (Figura
4.12).
Das espécies identificadas foi possível observar que o rio Potengi é
dominado principalmente por foraminíferos que possuem carapaça calcária,
seguida por foraminíferos de carapaças aglutinantes e porcelanáceas (Tabela 4.7
à esquerda), onde estas últimas, na maioria das vezes, possuem características
marinhas e de ambientes altamente hidrodinâmicos, como a espécie porcelanácea
Quinqueloculina lamarckiana e o gênero aglutinante Textularia spp, salvo
exceções das espécies Quinqueloculina miletti e Quinqueloculina patagonica,
que preferem ambientes mixohalinas, além de se adaptarem em ambientes
ecologicamente estressados (oportunistas) e a espécie aglutinante Caronia exilis
que é tipicamente mixohalina (Tabela 4.7 à direita).

74
Tabela 4.7: Relação das espécies quanto à composição da carapaça na tabela à esquerda; e relação das
espécies quanto ao hábitat: marinho, mixohalino, mangue ou cosmopolita na tabela à direita em Outubro
de 2011.
Será discutida a seguir a tabela dos indivíduos vivos e mortos que
aparecem em maior frequência nas estações, como Ammonia tepida, Bolivina
striatula e Quinqueloculina lamarckiana (Tabela 4.8) em função da disposição
espacial das amostras na figura 3.1A.
A. tepida apresentou maiores valores de indivíduos vivos (>500
indivíduos) em relação aos mortos na estação 2 (820 indivíduos), a qual está
situada na desembocadura do rio, na estação 10 (760 indivíduos) situada na foz
do Rio Doce, e na estação 14 (600 indivíduos) que se encontra próximo às
fazendas de carcinicultura e das salinas, indicando que esses ambientes são
favoráveis à sobrevivência da espécie. As menores frequências (<500 indivíduos)
foram nas estações 9 (390 indivíduos), 12 (92 indivíduos) 17 (95 indivíduos)e 18
(60 indivíduos), indicando que esses ambientes não possuem condições para a
espécie.
Relação das espécies quanto ao tipo de carapaça Outubro/2011
Calcárias
Ammonia tepida
Bolivina pulchella
Bolivina striatula
Bulimina elongata
Cibicides sp
Cibicides variabilis
Discorbis bertheloti
Discorbis peruvianus
Discorbis sp
Discorbis williamsoni
Elphidium discoidale
Elphidium galvestonense
Elphidium poeyanum
Fissurina laevigata
Hanzawaia boueana
Lagena striata
Oolina globosa
Pseudononium atlanticum
Aglutinantes
Ammotium salsum
Caronia exilis
Reophax sp
Textularia agglutinans
Textularia earlandi
Textularia gramen
Porcelanosas
Cornuspira involvens
Pyrgo peruviana
Quinqueloculina intricata
Quinqueloculina gregaria
Quinqueloculina lamarckiana
Quinqueloculina miletti
Quinqueloculina patagonica
Relação das espécies marinhas/dulcícolas/mangue/cosmopolitas Outubro/2011 *O = oportunista
Cosmopolitas
Ammonia tepida (O)
Mixohalinas
Bolivina pulchella
Bolivina striatula (O)
Elphidium discoidale (O)
Elphidium galvestonense
Elphidium poeyanum
Quinqueloculina miletti (O)
Quinqueloculina patagonica (O)
Mangue
Ammotium salsum
Caronia exilis
Marinhas
Bulimina elongata
Cibicides sp
Cibicides variabilis
Cornuspira involvens
Discorbis bertheloti
Discorbis peruvianus
Discordis sp
Discorbis williamsoni
Fissurina laevigata
Hanzawaia boueana
Lagena striata
Oolina globosa
Pyrgo peruviana
Pseudononion atlanticum
Quinqueloculina intricata
Quinqueloculina gregaria
Quinqueloculina lamarckiana
Reophax sp
Textularia agglutinans
Textularia earlandi
Textularia gramen

75
B. striatula apresentou maior número de indivíduos vivos em relação aos
mortos (>100 indivíduos) na estação 16 (450 indivíduos), localizada no curso
superior do Rio localizada próxima as salinas e das fazendas de carcinicultura.
Os indivíduos que apresentaram menores valores (<100 indivíduos) foram as
estações 10 e 13 (90 indivíduos), localizadas na foz do Rio Doce. Quanto aos
indivíduos mortos em relação aos vivos a espécie apresentou maior número
(>100 indivíduos) na estação 5 (120 indivíduos), a qual está situada na foz. Os
menores valores (<100 indivíduos) situam-se nas estações 4 (40 indivíduos) e 6
(40 indivíduos), dispostas também na foz, indicando que esse ambiente não
possui condições de sustento desse organismo. As estações que apresentaram
ambiente em equilíbrio foram as amostras 9 e 17 pois disponibilizam nutrientes
suficientes para os organismos, situadas respectivamente próximo à foz e
próximo às fazendas de carcinicultura e às salinas. As amostras 3, 7 e 8 não
apresentaram indivíduos, provavelmente por estarem mais afastados da foz e essa
espécie é mixohalina.
Q. lamarckiana apresentou maior número de indivíduos vivos em relação
aos mortos (>100 indivíduos) na estação 5 (840 indivíduos). O menor valor
(<100 indivíduos) foi da estação 7, situada ao sul da foz, que apresentou 60
indivíduos. Quanto ao número de indivíduos mortos em relação aos vivos os
maiores valores (>100 indivíduos) a estação 1 apresentou maior valor (300
indivíduos), localizada próxima à foz do Rio Doce. O menor valor (<100
indivíduos) ocorre na estação estação 6 (90 indivíduos) situada na foz. O
ambiente parece estar em equilíbrio na estação 2 e as amostras 13 e 15 – 18
apresentam ausência da espécie por não apresentarem características marinhas e
no caso da estação 13 provavelmente o ambiente indica ausência dos nutrientes
necessários.

76
Tabela 4.8: Relação das espécies identificadas vivas e mortas de cada estação na amostragem de Outubro
de 2011.
Observando a tabela de frequência absoluta das espécies de foraminíferos
(Tabela 4.9) em conjunto com o mapa da figura 3.1A foi constatado que a
espécie A. tepida possui maior número de indivíduos em relação às demais
espécies principalmente na estação 15 (24.660 indivíduos), localizada próxima à
Estação de Tratamento de Água de Quintas, com exceção da estação 8 (6
indivíduos), a qual está localizada ao norte da foz. Q. lamarckiana também
possui maior número de indivíduos, principalmente na estação 5 (1.290
indivíduos), localizada na foz e possui menor número na estação 12 (4
indivíduos), localizada na região central do rio Potengi e desaparece no curso
superior. Q. miletti possui maior número de indivíduos na estação 15 (1800) e
menor na estação 8 (1 indivíduo) e desaparece na foz e em alguns pontos no rio.
Q. patagonica ocorre em maior número nas estações 1 (380 indivíduos) e 5 (300
indivíduos), localizadas no canal do rio e na foz respectivamente, e parece existir
competição interespecífica entre Q. patagonica e Q. miletti. Textularia earlandi
apresenta maior número de indivíduos na estação 5 (210 indivíduos), a qual está
Espécies\estações
Vivos Mortos Vivos Mortos Vivos Mortos Vivos Mortos Vivos Mortos Vivos Mortos Vivos Mortos Vivos Mortos Vivos Mortos Vivos Mortos Vivos Mortos Vivos Mortos Vivos Mortos Vivos Mortos Vivos Mortos Vivos Mortos Vivos Mortos Vivos Mortos
Ammonia tepida 380 2420 820 780 200 440 30 280 120 1350 30 220 0 150 1 5 390 130 760 140 540 1140 92 54 360 1290 600 420 1980 22680 1080 1620 95 15 60 22
Ammotium salsum 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 60 0 90 0 0 0 180 0 0 0 0
Bulimina elongata 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 10 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Bolivina pulchella 0 0 0 0 0 0 0 0 0 30 0 0 0 10 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Bolivina striatula 40 0 140 100 0 0 10 40 30 120 0 40 0 0 0 0 20 20 90 10 30 60 12 0 90 0 120 0 180 0 450 0 10 10 0 2
Cornuspira involvens 0 0 0 40 0 0 0 20 0 0 0 30 0 0 0 0 0 0 0 10 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 1
Cibicides sp 0 0 0 0 0 0 0 0 0 0 0 40 0 10 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Cibicides variabilis 0 0 0 0 0 0 0 10 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Discorbis bertheloti 0 0 0 0 0 0 0 10 0 30 0 0 0 0 0 0 0 10 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Discorbis peruvianus 0 0 0 20 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 10 0 30 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Discorbis sp 0 20 0 0 0 0 0 0 0 0 0 20 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Discorbis williamsoni 0 0 0 0 100 0 0 10 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Elphidium discoidale 0 60 20 60 0 0 0 10 0 120 0 10 0 0 0 0 0 0 0 30 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Elphidium galvestonense 0 80 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Elphidium poeyanum 0 0 0 0 0 0 0 0 0 0 0 0 0 30 0 2 0 0 0 0 0 0 0 0 0 0 0 0 0 0 90 0 0 0 0 0
Fissurina laevigata 0 20 0 0 0 0 0 10 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Caronia exilis 0 20 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 10 0 30 0 0 0 30 60 60 0 0 0 0 0 5 0 0
Hanzawaia boueana 0 0 0 0 0 0 0 10 0 30 0 10 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Lagena striata 0 0 0 0 0 0 0 0 0 0 0 10 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Oolina globosa 0 20 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Pyrgo peruviana 0 40 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 - 0 0 0 0 0
Pseudononion atlanticum 0 60 0 0 0 0 0 0 0 0 0 10 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Quinqueloculina intricata 0 20 0 0 0 20 0 0 0 0 10 40 0 0 0 0 0 0 0 20 0 0 0 0 0 0 - 0 0 0 0 0 0 0 0 0
Quinqueloculina gregaria 0 80 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Quinqueloculina lamarckiana 100 300 20 20 0 140 130 100 840 450 10 90 60 40 6 12 0 100 30 70 0 30 0 4 0 0 0 30 0 0 0 0 0 0 0 0
Quinqueloculina miletti 0 0 140 160 20 80 0 0 0 120 10 0 0 0 0 1 0 0 40 20 60 150 0 0 30 60 0 60 0 1800 180 360 0 0 0 0
Quinqueloculina patagonica 40 340 20 20 0 20 30 60 0 300 0 170 0 0 0 0 70 20 30 60 0 30 4 2 0 0 0 90 0 0 0 0 0 10 4 2
Reophax sp 0 20 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Textularia agglutinans 0 20 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Textularia earlandi 0 60 0 0 0 0 0 50 0 210 0 0 0 50 0 3 0 0 0 30 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Textularia gramen 40 20 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Relação de espécies vivas/mortas por estação para Outubro/2011
7 8 9 10 11 121 2 3 4 5 6 16 17 1813 14 15

77
localizada na foz e é inexistente ao longo do canal por tratar-se de uma espécie
marinha.
Tabela 4.9: Frequência absoluta das espécies identificadas na amostragem de Outubro de 2011.
De acordo com a tabela de frequência relativa das espécies de
foraminíferos (Tabela 4.10) em função do mapa da figura 3.1A, a espécie A.
tepida apresenta maior dominância (estações 12, 13, 15 e 18) no canal do rio
Potengi, próximo às fazendas de carcinicultura, às salinas, à Estação de
Tratamento de Água de Quintas e no canal do baldo; e as menores frequências
ocorrem na foz. Q. lamarckiana dominou na foz e próximo à esta, mas possuiu
menor frequência ao longo do canal do rio Potengi; as oportunistas Q. miletti e
Q. patagonica apresentaram baixas frequências, mas parece haver competição
entre essas espécies por nutrientes e matéria orgânica. Textularia earlandi obteve
maiores frequências na foz e próximo a esta e é inexistente ao longo do canal por
tratar-se de uma espécie marinha.
ESPÉCIES/ESTAÇÕES 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18
Ammonia tepida 2800 1600 640 310 1470 250 150 6 520 900 1680 146 1650 1020 24660 2700 110 82
Ammotium salsum 0 0 0 0 0 0 0 0 0 0 0 0 60 90 0 180 0 0
Bulimina elongata 0 0 0 0 0 0 0 0 0 10 0 0 0 0 0 0 0 0
Bolivina pulchella 0 0 0 0 30 0 10 0 0 0 0 0 0 0 0 0 0 0
Bolivina striatula 40 240 0 50 150 40 0 0 40 100 90 12 90 120 180 450 20 2
Caronia exilis 0 40 0 20 0 30 0 0 0 10 0 0 0 0 0 0 0 1
Cornuspira involvens 0 0 0 0 0 40 10 0 0 0 0 0 0 0 0 0 0 0
Cibicides sp 0 0 0 10 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Cibicides variabilis 0 0 0 10 30 0 0 0 10 0 0 0 0 0 0 0 0 0
Discorbis bertheloti 0 20 0 0 0 0 0 0 0 10 30 0 0 0 0 0 0 0
Discorbis peruvianus 20 0 0 0 0 20 0 0 0 0 0 0 0 0 0 0 0 0
Discordis sp 0 0 100 10 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Discorbis williamsoni 60 80 0 10 120 10 0 0 0 30 0 0 0 0 0 0 0 0
Elphidium discoidale 80 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Elphidium galvestonense 0 0 0 0 0 0 30 2 0 0 0 0 0 0 0 90 0 0
Elphidium poeyanum 20 0 0 10 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Fissurina laevigata 20 0 0 0 0 0 0 0 0 10 30 0 30 120 0 0 5 0
Hanzawaia boueana 0 0 0 10 30 10 0 0 0 0 0 0 0 0 0 0 0 0
Lagena striata 0 0 0 0 0 10 0 0 0 0 0 0 0 0 0 0 0 0
Oolina globosa 20 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Pyrgo peruviana 40 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Pseudononion atlanticum 60 0 0 0 0 10 0 1 0 0 0 0 0 0 0 0 0 0
Quinqueloculina intricata 20 0 20 0 0 50 0 0 0 20 0 0 0 0 0 0 0 0
Quinqueloculina gregaria 80 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Quinqueloculina lamarckiana 400 40 140 230 1290 100 100 18 100 100 30 4 0 30 0 0 0 0
Quinqueloculina miletti 0 300 100 0 120 10 0 1 0 60 210 0 90 60 1800 540 0 0
Quinqueloculina patagonica 380 40 20 90 300 170 0 0 90 90 30 6 0 90 0 0 10 6
Reophax sp 20 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Textularia agglutinans 20 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Textularia earlandi 60 0 0 50 210 0 50 3 0 30 0 0 0 0 0 0 0 0
Textularia gramen 60 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Frequência absoluta das espécies de foraminíferos encontradas em Outubro/2011

78
Tabela 4.10: Frequência relativa das espécies identificadas na amostragem de Outubro de 2011.
Serão discutidas a seguir as análises de CLUSTER (Figura 4.11) e MDS
(Figura 4.12). Para a análise de CLUSTER e MDS foi utilizado o programa
Primer 5 e 6, onde foi realizada a análise dos agrupamentos das espécies em
função da sua disposição espacial (mapa da Figura 3.1A) e dos parâmetros
abióticos, como a temperatura, profundidade, salinidade, densidade,
condutividade e granulometria (Tabela 4.4) que atuam nesses ambientes.
A análise de CLUSTER para as estações (Modo Q) em função dos dados
bióticos e abióticos gerou um dendograma com cinco grupos distintos (Figura
4.11) e nível de similaridade de 45%, em que analisando a disposição das
amostras (Figura 3.1A), dos dados abióticos (Tabela 4.4) e da frequência relativa
das espécies (Tabela 4.10) desse agrupamento, podem-se constatar dois
ambientes distintos, em que um está relacionado à região da foz do rio Potengi e
à plataforma interna, representado pelas estações de 1 a 11 dentro dos grupos C1,
C2 e C3; e outro ambiente corresponde à região mais interna do estuário,
representado pelas estações 12 a 18, as quais são constituintes dos grupos C4 e
C5.
ESPÉCIES/ESTAÇÕES 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18
Ammonia tepida 66,67 67,80 62,75 38,27 39,20 33,33 42,86 19,35 68,42 65,69 80,00 86,90 85,94 66,67 92,57 68,18 75,86 90,11
Ammotium salsum 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 3,13 5,88 0,0 4,55 0,0 0,0
Bulimina elongata 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,73 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Bolivina pulchella 0,0 0,0 0,0 0,0 0,80 0,0 2,86 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Bolivina striatula 0,95 10,17 0,0 6,17 4,00 5,33 0,0 0,0 5,62 7,30 4,29 7,14 4,69 7,84 0,68 11,36 13,79 2,20
Caronia exilis 0,48 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,73 1,43 0,0 1,56 7,84 0,0 0,0 3,45 0,0
Cornuspira involvens 0,0 1,69 0,0 2,47 0,0 4,00 0,0 0,0 0,0 0,73 0,0 0,0 0,0 0,0 0,0 0,0 0,0 1,10
Cibicides sp 0,0 0,0 0,0 0,0 0,0 5,33 2,86 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Cibicides variabilis 0,0 0,0 0,0 1,23 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Discorbis bertheloti 0,0 0,0 0,0 1,23 0,80 0,0 0,0 0,0 1,32 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Discorbis peruvianus 0,0 0,85 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,73 1,43 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Discordis sp 0,48 0,0 0,0 0,0 0,0 2,67 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Discorbis williamsoni 0,0 0,0 9,80 1,23 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Elphidium discoidale 1,43 3,39 0,0 1,23 3,20 1,33 0,0 0,0 0,0 2,19 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Elphidium galvestonense 1,90 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Elphidium poeyanum 0,0 0,0 0,0 0,0 0,0 0,0 8,57 6,45 0,0 0,0 0,0 0,0 0,0 0,0 0,0 2,27 0,0 0,0
Fissurina laevigata 0,48 0,0 0,0 1,23 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Hanzawaia boueana 0,0 0,0 0,0 1,23 0,80 1,33 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Lagena striata 0,0 0,0 0,0 0,0 0,0 1,33 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Oolina globosa 0,48 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Pyrgo peruviana 0,95 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Pseudononion atlanticum 1,43 0,0 0,0 0,0 0,0 1,33 0,0 3,23 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Quinqueloculina intricata 0,48 0,0 1,96 0,0 0,0 6,67 0,0 0,0 0,0 1,46 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Quinqueloculina gregaria 1,90 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Quinqueloculina lamarckiana 9,52 1,69 13,73 28,40 34,40 13,33 28,57 58,06 13,16 7,30 1,43 2,38 0,0 1,96 0,0 0,0 0,0 0,0
Quinqueloculina miletti 0,0 12,71 9,80 0,0 3,20 1,33 0,0 3,23 0,0 4,38 10,00 0,0 4,69 3,92 6,76 13,64 0,0 0,0
Quinqueloculina patagonica 9,05 1,69 1,96 11,11 8,00 22,67 0,0 0,0 11,84 6,57 1,43 3,57 0,0 5,88 0,0 0,0 6,90 6,59
Reophax sp 0,48 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Textularia agglutinans 0,48 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Textularia earlandi 1,43 0,0 0,0 6,17 5,60 0,0 14,29 9,68 0,0 2,19 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Textularia gramen 1,43 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Frequência relativa das espécies de foraminíferos encontradas em Outubro 2011

79
O grupo C1 é constituído pelas estações 7 e 8, que apresentam como
principal característica a dominância de Ammonia tepida, Quinqueloculina
lamarckiana e, em menor frequência, Textularia earlandi. Ambas as estações são
encontradas na foz e apresentaram altos teores de salinidade, densidade e
densidade, principalmente a estação 8 (salinidade: 36,24 ppm; densidade: 23,39
g/cm3 e condutividade: 57,86 ppm), enquanto que a estação 7 apresentou valores
um pouco menores (salinidade: 36,18 ppm; densidade: 23,38 g/cm3 e
condutividade: 57,75 ppm). Quanto à granulometria, as frações de areia grossa
(estação 7: 46,96%) e areia média (estação 8: 68,20%) foram as dominantes,
indicando dessa forma que o grupo representa ambientes hidrodinâmicos e com
características marinhas e as frações finas apresentaram-se menos expressivas.
O grupo C2 apresentou-se menos homogêneo quanto às espécies em
relação aos grupos, podendo até mesmo dividi-lo em dois subgrupos: um
formado pelas estações 1, 3 e 10, cujas espécies dominantes são basicamente
Ammonia tepida e a Quinqueloculina lamarckiana; enquanto o outro subgrupo é
composto pelas estações 2 e 11, com A. tepida e Q. miletti. As estações 2, 3 e 10
apresentaram maior profundidade em relação às outras, com 12,6 m (estação 2),
13.3 m (estação 3) e 14.2 m (estação 10), sendo esta a mais profunda e a estação
11 foi coletada em profundidade rasa (3,5 m). A estação 3 obteve maiores valores
quanto à salinidade (36,28 ppm) e densidade (23,48 g/cm3), a qual situa-se na
foz. A estação 10, a qual está situada na foz do Rio Doce, apresentou valores
altos de salinidade (36,23 ppm), provavelmente devido à sua proximidade com as
salinas.
A estação 1 apresentou menores valores de salinidade (35,56 ppm),
densidade (22,86 g/cm3) e condutividade (57,01 ppm), a qual está localizada no
canal principal do rio e mais afastada das salinas. A estação 11, localizada
próximo ao Porto de Natal, apresentou valor alto na condutividade (57,95 ppm)
seguido da estação 10 (57,87 ppm), onde na maioria dos demais parâmetros a
estação 11 apresentou valores menores. Quanto à granulometria a estação 1
obteve valores altos de areia média (39,80%) e areia fina (31,44%) e de CaCO3
(5%), a estação 2 possui valores altos de cascalho (7,56%) e areia grossa

80
(67,78%), a estação 10 possui valor alto de areia média (50,69%) e a estação 11
possui valor alto de areia grossa (59,71%). Foi observado que a fração grossa foi
dominante nesse grupo, indicando ambiente de alta hidrodinâmica e com
características mixohalinas. As frações finas apresentaram-se em menor
frequência.
Já o Grupo C3, formado pelas estações 4, 5, 6 e 9, é composto por
Ammonia tepida, Quinqueloculina lamarckiana e Q. patagonica. Todas as
estações apresentaram valores similares de temperatura (27,8 – 27,9º C), a
estação 5 apresentou valor alto de salinidade (36,42 ppm) e densidade (23,59
g/cm3), enquanto que a estação 9 obteve valor alto de condutividade (57,92 ppm)
e a estação 6 apresentou teor de CaCO3 mediano (3,31%), apesar de ter obtido
valores menores para os demais parâmetros. Quanto à granulometria a estação 9,
situada próximo à foz, obteve valor alto de areia média (45,02%) seguida da
estação 4 (43,99%), situada na foz; a estação 5 obteve valores altos de cascalho
(7,38%), areia grossa (16,48%) e areia muito grossa (58,07%), indicando dessa
forma que esse grupo pertence a um ambiente altamente hidrodinâmico e de
domínio marinho, com baixa presença de frações finas.
O grupo C4 formado pelas estações 12, 17 e 18, onde as estações 12 e 18
encontram-se próximas ao Canal do Baldo e a estação 17 está próxima às
fazendas de carcinicultura e das salinas, apresentou baixa variedade de espécies
com predominância de Ammonia tepida, Bolivina striatula e Quinqueloculina
patagonica. As estações 17 e 18 apresentaram valor médio similar de
temperatura, já a estação 17 apresenta valor mais elevado (28,2º C) e índice alto
de CaCO3 (7,50%), em contrapartida apresentou menores valores de salinidade,
densidade e condutividade em relação à estação 12, onde apresentou esses
valores mais altos (salinidade: 36,31 ppm; densidade: 23,44 g/cm3 e
condutividade: 58,02 ppm).
Quanto à granulometria a estação 12 apresentou alta frequência de areia
grossa (61,63%), a estação 18 apresentou alta frequência de areia média
(26,13%) e a estação 17 dominou principalmente nas frações grossas, como
cascalho (17,50%) e areia muito grossa (36,99%) e apresentou silte (3,18%) em

81
sua composição sedimentar, indicando dessa forma que esse grupo é de ambiente
tipicamente estuarino e hidrodinâmico moderado, com presença de granulometria
fina significativa.
O grupo C5 composto pelas estações 13 a 16 apresenta as seguintes
espécies como sendo dominantes: Ammonia tepida, Bolivina striatula e
Quinqueloculina miletti, com exceção da estação 14 que apresentou Caronia
exilis como dominante. Quanto aos parâmetros abióticos as estações de 14 a 16
apresentaram profundidades rasas (estação 14: 4,4 m – estação 16: 9,6 m) e
salinidades baixas, com exceção da estação 13 (36,15 ppm), o qual apresentou
densidade (23,31 g/cm3) e condutividade (57,79 ppm) altas, enquanto que a
estação 15 apresentou altos índices de CaCO3 (5,79%).
Quanto à granulometria foi observada a presença das granulometrias
grossas (cascalho, areia grossa, areia muito grossa e areia média) e finas (areia
fina, areia muito fina, silte e argila). As estações 13 e 16 obtiveram frequências
similares de areia média (40,05 - 41,75%), enquanto que a estação 14 obteve
valor alto de cascalho (11,31%), areia grossa (41,30%) e areia muito grossa
(14,57%) e a estação 15 obteve valor alto de areia fina (39,35%), areia muito fina
(19,26%), silte (25,89%) e argila (3,20%). Foi observado que esse grupo é típico
de ambiente estuarino com hidrodinâmica moderada, com a presença de ambas
as granulometrias.

82
Figura 4.11: Dendograma formado pelo CLUSTER mostrando o agrupamento das estações conforme a
similaridade das espécies de foraminíferos encontradas entre elas em Outubro de 2011.
Através da tabela relativa dos foraminíferos (Tabela 4.10) a análise de
MDS gerou quatro grupos distintos: M1, M2, M3 e M4, onde M1 é representada
pelas estações 13, 15 e 16; M2 é representada pelas estações 12, 17 e 18; M3 é
representada pelas estações 1 – 6, 9 – 11 e 14; e M4 é representada pelas estações
7 e 8 (Figura 4.12).
O grupo M1 obteve como espécies dominantes A. tepida e Q. miletti,
onde a primeira apresentou alta frequência na estação 15 (92,57%), localizada
próxima à Estação de Tratamento de Água de Quintas, e a segunda apresentou
menor frequência na estação 16 (13,64%), localizada no curso superior do rio.
Esse grupo é dominado por espécies oportunistas e indicaram as prováveis áreas
afetadas, que são as proximidades da Estação de Tratamento de Água de Quintas
e no curso superior do rio.
O grupo M2 apresentou dominância de A. tepida, B. striatula e Q.
patagonica, onde a primeira apresentou maior frequência na estação 18
(90,11%), situada próximo ao Canal do Baldo, enquanto que a segunda e a
terceira apresentaram menores frequências, respectivamente, na estação 17
Ambiente marinho (Foz
e proximidades no Rio
Potengi)
Ambiente estuarino
(Região interna do Rio
Potengi)

83
(13,79%) e (6,90%), próxima às salinas e fazendas de carcinicultura. Esse grupo
também apresentou espécies oportunistas, como A. tepida, B. striatula e Q.
patagonica, indicando como prováveis locais poluídos as proximidades do Canal
do Baldo e das salinas e fazendas de carcinicultura.
O grupo M3 apresentou dominância de A. tepida e Q. lamarckiana, onde
a primeira apresentou alta frequência na estação 11 (80%), situada próxima ao
Porto de Natal, e a segunda apresentou menor frequência na estação 5 (34,40%),
localizada na foz. A dominância de A. tepida indica probabilidade de estresse
ambiental e poluição, enquanto Q. lamarckiana, que apresentou baixas
frequências na foz, indica o fraco aporte da cunha salina na entrada do rio
Potengi nesse período.
O grupo M4 apresentou maior diversidade em relação aos demais grupos
com a dominância de A. tepida, E. poeyanum, Q. lamarckiana e T. earlandi,
onde Q. lamarckiana obteve frequência mais alta na estação 8 (58,06%), situada
na foz ao norte, seguida da A. tepida na estação 7 (estação 7: 42,86%), também
situada na foz mas ao sul, enquanto que E. poeyanum e T. earlandi apresentaram
frequências baixas, respectivamente, na estação 7 (8,57% e 14,29%). O grupo
apresentou espécies mixohalinas (A. tepida e E. poyeanum) quanto marinhas (Q.
lamarckiana e T. earlandi), evidenciando altas salinidades nas regiões distais da
foz (Q. lamarckiana).
Fazendo uma comparação entre esta análise e a do CLUSTER, constatou-
se que os grupos M2 e M4 se mostraram idênticos, respectivamente, aos grupos
C4 e C1 da análise de CLUSTER, enquanto que o grupo M1 se mostrou
semelhante ao grupo C5, diferenciando apenas pela ausência da estação 14, pois
esta não apresentou similaridade das espécies em relação às estações presentes no
grupo M1. Quanto ao grupo M3 dessa análise, ele engloba os grupos C2 e C3 da
análise de CLUSTER, além da estação 14, pois como foi observada na descrição
acima, essa estação apresenta similaridade de espécies em relação a esse grupo.

84
Figura 4.12: Grupos formados a partir da aplicação de MDS nos dados bióticos de Outubro de 2011.
4.2.2 Amostragem de Janeiro de 2012
Na segunda amostragem foi observado aumento na diversidade de
foraminíferos identificados (41 espécies), onde foi observada a quantidade de
indivíduos vivos e mortos (Tabela 4.13). Depois disso, foi confeccionada as
tabelas de frequência absoluta (Tabela 4.14) e relativa (Tabela 4.15), a qual
serviu para as análises de CLUSTER (Figura 4.21) e MDS (Figura 4.22). Das
espécies identificadas foi possível observar que o rio Potengi é dominado
principalmente por foraminíferos que possuem carapaça calcária e a plataforma
interna é dominada por espécies aglutinantes e porcelanosas (Tabela 4.11 à
esquerda), salvo exceções das espécies porcelanosas Quinqueloculina miletti e
Quinqueloculina patagonica, que preferem ambientes mixohalinos, além de
serem adaptadas a ambientes ecologicamente estressados (oportunistas) e as
espécies aglutinantes Caronia exilis e o gênero Trochammina spp. que são
tipicamente mixohalinos (Tabela 4.11 à direita).

85
Tabela 4.11: Relação das espécies quanto à composição da carapaça na tabela à esquerda; e relação das
espécies quanto ao hábitat: marinho, mixohalino, mangue ou cosmopolita na tabela à direita em Janeiro
de 2012.
Será discutida a seguir a tabela dos indivíduos vivos e mortos que
aparecem em maior frequência nas estações, como Ammonia tepida, Bolivina
striatula e Quinqueloculina lamarckiana (tabela 4.12) em função da disposição
espacial das amostras na figura 3.1B.
A. tepida apresentou maiores valores (>1.000 indivíduos) de indivíduos
vivos em relação aos mortos nas estações 17 – 20, localizadas no curso superior
do rio, onde as estações 18 e 19 estão situadas próximas às fazendas de
carcinicultura e a estação 20 está próxima ao Canal do Baldo, sendo que a
estação 20 obteve valores elevados (1200 indivíduos), enquanto que o menor
valor (<1.000) foi na estação 19 (110 indivíduos). Quanto aos mortos em relação
aos vivos a espécie apresentou altas frequências (> 1.000 indivíduos) em quase
todas as amostras, principalmente na estação 14 (8.730 indivíduos), situado em
Ponta Negra na plataforma interna sul; enquanto que a menor frequência (<1.000
indivíduos) foi na estação 8 (24 indivíduos).
Relação das espécies quanto ao tipo de carapaça Janeiro/2012
Calcárias
Ammonia tepida
Amphicoryna scalaris
Bolivina striatula
Buccella peruviana
Bulimina marginata
Buliminella elegantissima
Cassidulina subglobosa
Cibicides fletcheri
Cibicides valvulanatus
Cibicides variabilis
Discorbis flordana
Discorbis peruvianus
Discorbis valvulatus
Discorbis williamsoni
Elphidium articulatum
Elphidium discoidale
Fissurina laevigata
Fissurina lucida
Foraminífero bentônico jovem
Hanzawaia boueana
Hopkinsina pacifica
Lagena striata
Lenticulina sp
Orthomorphina sp
Poroeponides lateralis
Pseudononium atlanticum
Uvigerina peregrina
Aglutinantes
Caronia exilis
Textularia earlandi
Textularia gramen
Trochammina discorbis
Trochammina inflata
Trochammina squamata
Porcelanosas
Cornuspira involvens
Miliolinella subrotunda
Pyrgo nasuta
Quinqueloculina intricata
Quinqueloculina lamarckiana
Quinqueloculina miletti
Quinqueloculina patagonica
Relação das espécies marinhas/dulcícolas/mangue/cosmopolitas Janeiro/2012 *O = oportunista
Cosmopolitas
Ammonia tepida (O)
Mixohalinas
Bolivina striatula (O)
Buccella peruviana
Buliminella elegantissima (O)
Elphidium articulatum
Elphidium discoidale (O)
Fissurina lucida
Milliolinella subrotunda
Quinqueloculina milletti (O)
Quinqueloculina patagonica (O)
Mangue
Caronia exilis
Trochammina discorbis
Trochammina inflata
Trochammina squamata
Marinhas
Amphicoryna scalaris
Bulimina marginata
Cassidulina subglobosa
Cibicides fletcheri
Cibicides valvulanatus
Cibicides variabilis
Cornuspira involvens
Discorbis flordana
Discorbis peruvianus
Discorbis valvulatus
Discorbis williamsoni
Fissurina laevigata
Foraminífero bentônico jovem
Hanzawaia boueana
Hopkinsina pacifica
Lagena striata
Lenticulina sp
Orthomorphina sp
Poroeponides lateralis
Pseudononium atlanticum
Pyrgo nasuta
Quinqueloculina intricata
Quinqueloculina lamarckiana
Textularia earlandi
Textularia gramen
Uvigerina peregrina

86
B. striatula apresentou maiores valores (> 100 indivíduos) de indivíduos
vivos em relação aos mortos nas estações 7, 15 e 16 (plataforma interna), 17
(curso superior), 18 (próximo às fazendas de carcinicultura e salinas), 20
(próximo ao Canal do Baldo), 22 e 24 (próximos ao Porto de Natal e Marina Iate
Clube respectivamente), sendo a estação 24 a mais numerosa (540 indivíduos), e
o menor valor (<100 indivíduos) foi na estação 17, que apresentou apenas 7
indivíduos vivos seguido da estação 7 que apresentou 10 indivíduos. Quanto ao
nímero de mortos em relação aos vivos o maior valor (>100 indivíduos)
apresentou-se nas estações 1, 2, 3 e 14, sendo a estação 1 (marina Iate Clube) a
mais numerosa (630 indivíduos) e o menor valor (<100) apresentou-se nas
estações 4 e 6, situada na plataforma interna norte (4 indivíduos cada uma), da
mesma forma que a estação 8 apresenta-se ausente, provavelmente por estar bem
afastada da costa e essa espécie é mixohalina com pouca tolerância à baixas
salinidades.
Q. lamarckiana apresentou maiores valores (>100 indivíduos) em relação
ao número de vivos sobre os mortos em apenas três estações (2, 8 e 11) todas
localizadas na plataforma interna, onde a estação 11 apresentou maior valor (370
indivíduos), indicando ambiente ideal ao seu desenvolvimento, enquanto que o
menor valor (< 100 indivíduos) foi na estação 8 (17 indivíduos), provavelmente
indicando baixo índice de nutrientes e salinidade. Quanto ao numero de mortos
em relação aos vivos o maior valor (>800 indivíduos) foi na estação 14 (2.460
indivíduos) e o menor valor foi na estação 17 (2 indivíduos).

87
Tabela 4.12: Relação das espécies identificadas vivas e mortas de cada estação na amostragem de Janeiro
de 2012.
Espé
cies\e
staç
ões
Indi
vídu
os v
ivos
/mor
tos
Vivo
sM
orto
sVi
vos
Mor
tos
Vivo
sM
orto
sVi
vos
Mor
tos
Vivo
sM
orto
sVi
vos
Mor
tos
Vivo
sM
orto
sVi
vos
Mor
tos
Vivo
sM
orto
sVi
vos
Mor
tos
Vivo
sM
orto
sVi
vos
Mor
tos
Vivo
sM
orto
sVi
vos
Mor
tos
Vivo
sM
orto
sVi
vos
Mor
tos
Vivo
sM
orto
sVi
vos
Mor
tos
Vivo
sM
orto
sVi
vos
Mor
tos
Vivo
sM
orto
sVi
vos
Mor
tos
Vivo
sM
orto
sVi
vos
Mor
tos
Amm
onia
rolsh
ause
ni0
00
00
00
00
00
00
00
00
00
00
00
00
00
300
00
00
00
00
00
00
00
00
00
0
Amm
onia
tepi
da36
045
0034
588
567
523
4010
3931
016
0018
129
027
05
2412
010
1090
430
6039
011
010
4012
036
075
087
3010
013
7597
536
1511
149
131
5511
079
1200
900
500
1575
2280
2920
810
6720
4050
3600
Amm
otiu
m sa
lsum
00
150
00
00
00
00
00
00
00
100
00
00
00
00
00
00
00
00
00
00
00
00
00
00
Amph
icory
na sc
alar
is0
00
00
00
00
100
00
00
00
00
00
200
00
00
00
015
150
00
00
00
00
00
00
00
0
Boliv
ina
sp0
00
00
00
10
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
0
Boliv
ina
stria
tula
9063
015
105
4549
51
410
800
410
00
00
9010
500
9010
4040
6030
048
025
012
075
70
222
21
340
025
5044
028
030
9054
012
0
Boliv
ina
varia
bilis
00
00
00
01
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
Bucc
ella
peru
vian
a0
450
045
450
00
800
40
200
10
100
100
00
200
800
210
025
00
00
00
10
00
00
00
00
00
Bucc
ella
sp0
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
0
Bulim
ina
mar
gina
ta0
00
00
00
00
100
20
00
00
100
010
100
00
00
300
015
00
00
00
00
00
00
00
00
0
Bulim
ina
pata
goni
ca0
00
00
01
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
0
Bulim
inell
a ele
gant
issim
a45
180
00
090
00
010
02
00
00
00
010
00
00
00
00
00
00
00
00
00
00
00
00
090
00
Cass
idul
ina
subg
lobo
sa0
900
00
450
10
100
10
100
00
200
00
00
00
00
00
00
150
03
30
00
050
00
8030
300
0
Cibi
cides
flet
cher
i0
450
00
00
30
100
00
00
00
00
00
00
00
00
150
2550
00
00
00
00
00
00
00
00
00
Cibi
cides
val
vula
natu
s0
00
00
450
00
800
20
200
00
00
00
400
00
200
00
00
00
00
00
00
00
00
00
00
0
Cibi
cides
var
iabi
lis0
015
150
00
00
300
00
00
10
100
00
100
00
00
00
00
00
00
00
00
00
00
00
00
0
Corn
uspi
ra in
volv
ens
00
015
013
50
00
100
20
00
00
010
00
00
00
00
900
00
00
00
00
00
00
040
800
00
60
Disc
orbi
s ber
thelo
ti0
00
00
00
00
00
00
00
010
00
00
00
00
200
00
00
00
00
00
00
00
00
00
00
0
Disc
orbi
s flo
rdan
a0
00
00
00
00
00
00
00
00
00
00
00
00
060
025
045
150
00
00
020
00
00
030
300
0
Disc
orbi
s per
uvia
nus
00
00
00
00
00
01
100
00
2020
00
00
100
00
00
00
00
00
00
00
00
00
00
00
00
Disc
orbi
s val
vula
tus
00
00
00
00
00
00
00
01
00
00
1070
010
010
00
00
030
600
00
00
00
00
00
120
00
060
Disc
orbi
s var
iabi
lis0
00
00
00
00
300
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
0
Disc
orbi
s will
iam
soni
00
030
013
51
110
300
30
00
00
00
00
020
00
2090
00
500
00
00
00
00
200
00
00
00
60
Elph
idiu
m sp
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
Elph
idiu
m a
rticu
latu
m0
00
00
00
00
00
00
00
00
00
00
100
00
400
600
250
00
00
00
00
00
00
00
00
0
Elph
idiu
m d
iscoi
dale
013
50
00
00
310
301
10
100
00
400
1010
400
300
00
180
050
075
01
00
01
200
00
017
600
300
3060
Elph
idiu
m p
oyea
nun
00
00
00
01
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
300
00
Fiss
urin
a la
evig
ata
090
00
00
00
00
00
00
00
010
00
00
00
00
00
00
00
00
00
00
00
00
00
030
00
Fiss
urin
a lu
cida
00
00
045
00
010
00
010
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
Fiss
urin
a sp
00
015
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
Fora
minif
ero
bento
nico
jove
m0
00
00
00
010
00
00
00
00
00
00
00
00
400
00
250
600
00
00
00
00
00
00
00
0
Furs
enko
ina
pont
oni
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
015
00
00
00
00
00
00
00
00
Gau
dryin
a ex
ilis
00
015
045
00
030
00
00
00
00
00
020
00
00
00
00
00
00
00
00
00
00
00
00
300
Hanz
awai
a bo
uean
a0
00
00
450
10
00
030
00
010
600
200
300
400
060
00
2515
600
00
00
10
00
00
00
00
30
Hopk
insin
a pa
cifica
00
00
013
50
00
00
00
00
00
00
00
00
100
00
00
00
00
00
00
00
00
00
00
00
0
Lage
na la
evis
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
030
Lage
na st
riata
00
00
00
00
00
00
00
00
00
00
00
010
00
060
00
00
00
00
00
00
00
040
00
00
Lent
iculin
a sp
00
00
00
00
00
00
00
00
00
00
100
00
040
00
00
015
00
00
00
00
00
00
00
00
Mill
iolin
ella
subr
otun
da0
00
150
450
00
00
00
00
00
00
00
00
500
200
900
250
150
00
00
00
00
00
030
300
30
Noni
onell
a pu
lchell
a0
00
00
00
00
00
00
00
00
100
00
00
00
00
00
00
00
00
00
00
00
00
00
00
0
Orth
omor
phin
a fil
iform
is0
00
00
450
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
0
Plan
ctonic
a0
00
00
00
00
100
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
0
Poro
epon
ides
late
ralis
00
150
00
00
00
00
00
05
00
00
1020
00
020
00
00
4515
00
00
00
00
00
00
00
00
Pseu
dono
nium
atla
nticu
m0
450
150
450
10
100
13
010
02
00
010
020
040
00
3018
025
00
150
00
00
10
00
00
00
600
60
Pyrg
o na
suta
045
00
00
00
020
00
00
01
020
00
00
00
8020
030
5025
300
00
01
00
00
00
00
00
00
Quin
quelo
culin
a gr
egar
ia0
00
00
450
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
0
Quin
quelo
culin
a in
trica
ta0
900
00
00
010
100
10
401
010
100
1010
600
00
2030
900
5015
00
00
00
00
200
00
060
180
00
Quin
quelo
culin
a la
mar
ckia
na90
1035
7560
495
900
817
570
570
739
6010
017
1112
024
060
140
370
360
190
260
700
800
1740
2460
350
450
1200
1605
02
15
56
100
100
2510
024
072
039
011
4048
010
80
Quin
quelo
culin
a m
illet
ti45
495
150
4590
52
2020
01
00
00
00
00
200
010
00
090
00
3030
00
00
00
00
00
400
030
3021
0
Quin
quelo
culin
a pa
tago
nica
135
2115
120
120
180
1035
18
100
260
648
3050
32
7048
020
230
130
160
120
320
014
011
1024
0022
522
531
554
02
31
40
20
6010
012
528
092
042
093
045
075
0
Quin
quelo
culin
a sp
00
150
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
Rosa
lina
sp0
00
00
00
00
00
00
00
00
00
00
00
00
200
00
250
00
00
00
00
00
00
00
00
0
Trilo
culin
a sp
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
Text
ular
ia ea
rland
i0
00
00
180
00
00
00
00
00
00
00
1010
050
160
120
021
00
5015
600
00
00
00
00
00
00
00
0
Text
ular
ia g
ram
en0
00
045
01
344
054
06
460
900
1020
700
023
038
00
6014
052
015
087
075
400
3030
00
00
00
20
00
00
00
300
0
Text
ular
ia sp
00
015
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
Troc
ham
min
a di
scor
bis
00
00
00
01
00
00
00
00
00
00
00
00
00
00
00
030
00
00
00
00
00
00
00
00
Troc
ham
min
a in
flata
045
00
450
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
030
00
Troc
ham
min
a sq
uam
ata
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
00
800
00
00
Uvi
gerin
a pe
regr
ina
00
00
00
01
040
00
00
00
030
00
100
020
00
00
00
150
00
00
00
00
00
00
00
090
2223
2415
1617
1819
2010
1112
1314
21
Rel
ação
de
espé
cies v
ivas
/mor
tas p
or e
staç
ão p
ara
Jane
iro/2
012
12
34
56
78
9

88
Observando a tabela de frequência absoluta das espécies de foraminíferos
(Tabela 4.13) em conjunto com o mapa da figura 3.1B foi constatado que a
espécie A. tepida possui maior número de indivíduos na estação 14 (9.480
indivíduos) na região costeira sul, seguida pelas estações 23 (7.530 indivíduos) e
24 (7.650 indivíduos), localizados no canal do rio Potengi, com exceção da
estação 8 (29 indivíduos), a qual está localizada próxima à região costeira norte.
Q. lamarckiana também possui maior número de indivíduos,
principalmente na estação 14 (4200 indivíduos), localizada na região costeira sul
e menor quantidade na estação 17, localizada no curso superior do rio Potengi. Q.
miletti possui maior número na estação 1 (585 indivíduos) e menor na estação 6
(1 indivíduo), localizada na plataforma interna norte, desaparecendo no curso
superior do rio e na plataforma interna. Q. patagonica possui maior número na
estação 14 (3.510 indivíduos), localizada na região costeira sul, e menor na
estação 19 (2 indivíduos). Textularia gramen apresenta maior número de
indivíduos na estação 14 (1.020 indivíduos), a qual está localizada na região
costeira sul, e menor número na estação 19 (2 indivíduos), localizada no canal do
rio Potengi e é inexistente no curso superior, na foz e em alguns trechos da
plataforma interna.
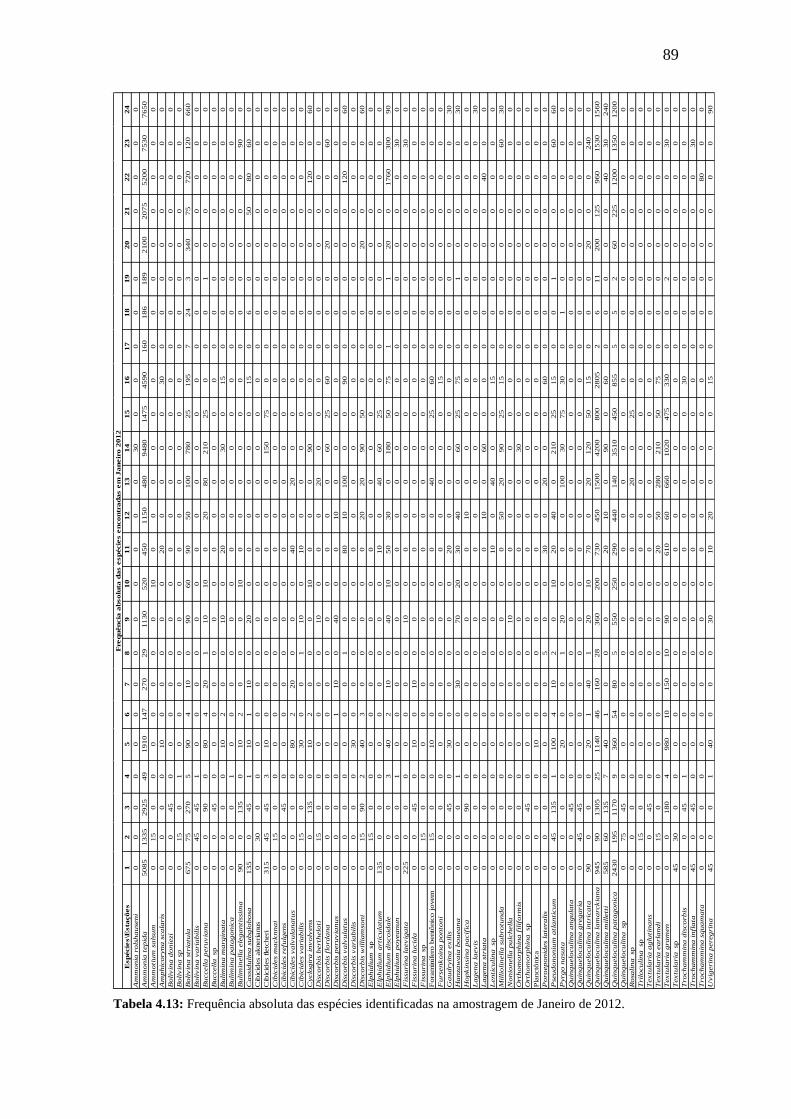
89
Tabela 4.13: Frequência absoluta das espécies identificadas na amostragem de Janeiro de 2012.
Espécie
s\E
sta
ções
12
34
56
78
910
11
12
13
14
15
16
17
18
19
20
21
22
23
24
Am
monia
rols
hauseni
00
00
00
00
00
00
030
00
00
00
00
00
Am
monia
tepid
a5085
1335
2925
49
1910
147
270
29
1130
520
450
1150
480
9480
14
75
459
016
0186
189
2100
20
75
5200
75
30
76
50
Am
moti
um
sals
um
015
00
00
00
010
00
00
00
00
00
00
00
Am
phic
oryna s
cala
ris
00
00
10
00
00
020
00
00
30
00
00
00
00
Bolivin
a d
onie
zi
00
45
00
00
00
00
00
00
00
00
00
00
0
Bolivin
a s
p0
15
01
00
00
00
00
00
00
00
00
00
00
Bolivin
a s
tria
tula
675
75
270
590
410
090
60
90
50
100
780
25
19
57
24
33
40
75
720
120
66
0
Bolivin
a v
aria
bilis
045
45
10
00
00
00
00
00
00
00
00
00
0
Buccella p
eruvia
na
00
90
080
420
110
10
020
80
210
25
00
01
00
00
0
Buccella sp
00
45
00
00
00
00
00
00
00
00
00
00
0
Bulim
ina m
argin
ata
00
00
10
20
010
020
00
30
015
00
00
00
00
Bulim
ina p
ata
gonic
a0
00
10
00
00
00
00
00
00
00
00
00
0
Bulim
inella e
leganti
ssim
a90
0135
010
20
00
10
00
00
00
00
00
00
90
0
Cassid
ulina s
ubglo
bosa
135
045
110
110
020
00
00
00
15
06
00
50
80
60
0
Cib
icid
es a
knerianus
030
00
00
00
00
00
00
00
00
00
00
00
Cib
icid
es fle
tcheri
315
45
45
310
00
00
00
00
150
75
00
00
00
00
0
Cib
icid
es m
acknnai
015
00
00
00
00
00
00
00
00
00
00
00
Cib
icid
es r
efu
lgens
00
45
00
00
00
00
00
00
00
00
00
00
0
Cib
icid
es v
alv
ula
natu
s0
00
080
220
00
040
020
00
00
00
00
00
0
Cib
icid
es v
aria
bilis
015
00
30
00
110
010
00
00
00
00
00
00
0
Cyclo
gir
a involv
ens
00
135
010
20
00
10
00
090
00
00
00
0120
06
0
Dis
corbis
berth
elo
ti0
15
00
00
00
10
00
020
00
00
00
00
00
0
Dis
corbis
flo
rdana
00
00
00
00
00
00
060
25
60
00
020
00
60
0
Dis
corbis
peruvia
nus
00
00
01
10
040
00
10
00
00
00
00
00
00
Dis
corbis
valv
ula
tus
00
00
00
01
00
80
10
100
00
90
00
00
0120
06
0
Dis
corbis
varia
bilis
00
00
30
00
00
00
00
00
00
00
00
00
0
Dis
corbis
william
soni
015
90
240
30
00
00
20
20
90
50
00
00
20
00
06
0
Elp
hid
ium
sp
015
00
00
00
00
00
00
00
00
00
00
00
Elp
hid
ium
arti
cula
tum
135
00
00
00
00
010
040
60
25
00
00
00
00
0
Elp
hid
ium
dis
coid
ale
00
03
40
210
040
10
50
30
0180
50
75
10
120
01760
300
90
Elp
hid
ium
poyeanun
00
01
00
00
00
00
00
00
00
00
00
30
0
Fis
surin
a laevig
ata
225
00
00
00
010
00
00
00
00
00
00
03
00
Fis
surin
a lucid
a0
045
010
010
00
00
00
00
00
00
00
00
0
Fis
surin
a sp
015
00
00
00
00
00
00
00
00
00
00
00
Fora
min
ífero
bentô
nic
o jovem
015
00
10
00
00
00
040
02
560
00
00
00
00
Fursenkoin
a p
onto
ni
00
00
00
00
00
00
00
015
00
00
00
00
Gaudryin
a e
xilis
00
45
030
00
00
020
00
00
00
00
00
00
30
Hanzaw
aia
boueana
00
01
00
30
070
20
30
40
060
25
75
00
10
00
03
0
Hopkin
sin
a p
acif
ica
00
90
00
00
00
00
10
00
00
00
00
00
00
Lagena laevis
00
00
00
00
00
00
00
00
00
00
00
03
0
Lagena s
tria
ta0
00
00
00
00
00
10
060
00
00
00
040
00
Lenti
culina
sp
00
00
00
00
00
10
040
00
15
00
00
00
00
Milliolinella s
ubrotu
nda
00
00
00
00
00
050
20
90
25
15
00
00
00
60
30
Nonio
nella p
ulc
hella
00
00
00
00
10
00
00
00
00
00
00
00
0
Orth
om
orphin
a f
ilif
orm
is0
00
00
00
00
00
00
30
00
00
00
00
00
Orth
om
orphin
a sp
00
45
00
00
00
00
00
00
00
00
00
00
0
Pla
nctô
nic
a0
00
010
00
00
00
00
00
00
00
00
00
0
Poroeponid
es late
ralis
00
00
00
05
00
30
020
00
60
00
00
00
00
Pseudononiu
m a
tlanti
cum
045
135
1100
410
20
10
20
40
0210
25
15
00
10
00
60
60
Pyrgo n
asuta
00
00
20
00
120
00
0100
30
75
30
01
00
00
00
Quin
quelo
culina a
ngula
ta0
045
00
00
00
00
00
00
00
00
00
00
0
Quin
quelo
culina g
regaria
045
45
00
00
00
00
00
00
00
00
00
00
0
Quin
quelo
culina intr
icata
90
00
020
140
120
10
70
020
120
50
15
00
020
00
240
0
Quin
quelo
culina lam
arckia
na
945
90
1305
25
1140
46
160
28
360
200
730
450
1500
4200
800
280
52
61
12
00
12
5960
15
30
15
60
Quin
quelo
culina m
ille
tti
585
60
135
740
10
00
020
10
090
060
00
00
040
30
24
0
Quin
quelo
culina p
ata
gonic
a2430
195
1170
9360
54
80
5550
250
290
440
140
3510
450
85
55
52
60
22
51200
13
50
12
00
Quin
quelo
culina
sp
075
45
00
00
00
00
00
00
00
00
00
00
0
Rosalina sp
00
00
00
00
00
00
20
02
50
00
00
00
00
Triloculina sp
015
00
00
00
00
00
00
00
00
00
00
00
Textu
laria
aglu
tinans
00
45
00
00
00
00
00
00
00
00
00
00
0
Textu
laria
earla
ndi
015
00
00
00
00
20
50
280
210
50
75
00
00
00
00
Textu
laria
gram
en
00
180
4980
10
150
10
90
0610
60
660
1020
475
33
00
02
00
03
00
Textu
laria
sp
45
30
00
00
00
00
00
00
00
00
00
00
00
Trocham
min
a d
iscorbis
00
45
10
00
00
00
00
00
30
00
00
00
00
Trocham
min
a infl
ata
45
045
00
00
00
00
00
00
00
00
00
03
00
Trocham
min
a s
quam
ata
00
00
00
00
00
00
00
00
00
00
080
00
Uvig
erin
a p
eregrin
a45
00
140
00
030
010
20
00
015
00
00
00
09
0
Frequência
absolu
ta d
as e
spécie
s e
ncontr
adas e
m J
aneir
o 2
012

90
De acordo com a tabela de frequência relativa das espécies de
foraminíferos (tabela 4.14) em função do mapa da figura 3.1B observou-se que a
espécie dominante foi A. tepida principalmente nas estações 17, 18 e 19
localizadas no curso superior e próximas às fazendas de carcinicultura. A espécie
Q. lamarckiana dominou a plataforma interna em alta porcentagem,
principalmente na estação 13 (40, 8%) e possui baixas frequências no canal do
rio Potengi e diminui em direção ao curso superior. A espécie T. gramen ocorre
nas estações 5, 7, 11 e 13, localizadas na plataforma interna e sua frequência é
baixa ao longo do rio, desaparecendo no curso superior.

91
Tabela 4.14: Frequência relativa das espécies identificadas na amostragem de Janeiro de 2012.
Esp
écie
s\E
staç
ões
12
34
56
78
910
1112
1314
1516
1718
1920
2122
2324
Am
mon
ia te
pida
47,1
65,9
41,1
42,2
37,4
51,4
32,5
34,5
45,0
46,8
17,1
46,6
13,0
45,7
39,3
48,2
91,4
81,6
89,6
75,5
81,4
50,4
65,2
64,6
Am
phic
oryn
a sc
alar
is0,
00,
00,
00,
00,
20,
00,
00,
00,
00,
00,
80,
00,
00,
00,
00,
30,
00,
00,
00,
00,
00,
00,
00,
0
Boliv
ina
stri
atul
a6,
36,
74,
46,
01,
81,
41,
20,
03,
65,
43,
42,
02,
73,
80,
72,
04,
010
,51,
412
,22,
97,
01,
05,
6
Bucc
ella
per
uvia
na0,
00,
01,
90,
01,
61,
42,
41,
20,
40,
90,
00,
82,
21,
00,
70,
00,
00,
00,
50,
00,
00,
00,
00,
0
Bulim
ina
mar
gina
ta0,
00,
00,
00,
90,
20,
70,
00,
00,
40,
00,
80,
00,
00,
10,
00,
20,
00,
00,
00,
00,
00,
00,
00,
0
Bulim
inel
la e
lega
ntis
sim
a0,
80,
01,
90,
00,
20,
70,
00,
00,
00,
90,
00,
00,
00,
00,
00,
00,
00,
00,
00,
00,
00,
00,
80,
0
Cas
sidu
lina
subg
lobo
sa1,
30,
00,
60,
90,
20,
31,
20,
00,
80,
00,
00,
00,
00,
00,
00,
20,
02,
60,
00,
02,
00,
80,
50,
0
Cib
icid
es fl
etch
eri
2,9
4,4
1,3
2,6
0,2
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,7
2,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
Cib
icid
es v
alvu
lana
tus
0,0
0,0
0,0
0,0
1,6
0,7
2,4
0,0
0,0
0,0
1,5
0,0
0,5
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
Cib
icid
es v
aria
bilis
0,0
0,7
0,0
0,0
0,6
0,0
0,0
1,2
0,4
0,0
0,4
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
Cyc
logi
ra in
volv
ens
0,0
0,0
1,9
0,0
0,2
0,7
0,0
0,0
0,0
0,9
0,0
0,0
0,0
0,4
0,0
0,0
0,0
0,0
0,0
0,0
0,0
1,2
0,0
0,5
Dis
corb
is fl
orda
na0,
00,
70,
00,
00,
00,
00,
00,
00,
40,
00,
00,
00,
50,
30,
70,
60,
00,
00,
00,
70,
00,
00,
50,
0
Dis
corb
is p
eruv
ianu
s0,
00,
00,
00,
00,
00,
31,
20,
01,
60,
00,
00,
40,
00,
00,
00,
00,
00,
00,
00,
00,
00,
00,
00,
0
Dis
corb
is v
alvu
latu
s0,
00,
00,
00,
00,
00,
00,
01,
20,
00,
03,
00,
42,
70,
00,
00,
90,
00,
00,
00,
00,
01,
20,
00,
5
Dis
corb
is w
illia
mso
ni0,
00,
71,
31,
71,
41,
00,
00,
00,
00,
00,
00,
80,
50,
41,
30,
00,
00,
00,
00,
70,
00,
00,
00,
5
Elp
hidi
um a
rtic
ulat
um1,
30,
00,
00,
00,
00,
00,
00,
00,
00,
00,
40,
01,
10,
30,
70,
00,
00,
00,
00,
00,
00,
00,
00,
0
Elp
hidi
um d
isco
idal
e0,
00,
00,
03,
40,
80,
71,
20,
01,
60,
91,
91,
20,
00,
91,
30,
80,
60,
00,
50,
70,
017
,12,
90,
8
Fis
suri
na la
evig
ata
2,1
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,4
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,3
0,0
Fis
suri
na lu
cida
0,0
0,0
0,6
0,0
0,2
0,0
1,2
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
Fora
min
ífero
ben
tôni
co jo
vem
0,0
0,7
0,0
0,0
0,2
0,0
0,0
0,0
0,0
0,0
0,0
0,0
1,1
0,0
0,7
0,6
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
Gau
dryi
na e
xilis
0,0
0,0
0,6
0,0
0,6
0,0
0,0
0,0
0,0
0,0
0,8
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,3
Han
zaw
aia
boue
ana
0,0
0,0
0,0
0,9
0,0
0,0
3,6
0,0
2,8
1,8
1,1
1,6
0,0
0,3
0,7
0,8
0,0
0,0
0,5
0,0
0,0
0,0
0,0
0,3
Hop
kins
ina
paci
fica
0,0
0,0
1,3
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,4
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
Lage
na s
tria
ta0,
00,
00,
00,
00,
00,
00,
00,
00,
00,
00,
00,
40,
00,
30,
00,
00,
00,
00,
00,
00,
00,
40,
00,
3
Lent
icul
ina
sp0,
00,
00,
00,
00,
00,
00,
00,
00,
00,
00,
40,
01,
10,
00,
00,
20,
00,
00,
00,
00,
00,
00,
00,
0
Mill
iolin
ella
sub
rotu
nda
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
2,0
0,5
0,4
0,7
0,2
0,0
0,0
0,0
0,0
0,0
0,0
0,5
0,3
Ort
hom
orph
ina
sp0,
00,
00,
60,
00,
00,
00,
00,
00,
00,
00,
00,
00,
00,
10,
00,
00,
00,
00,
00,
00,
00,
00,
00,
0
Por
oepo
nide
s la
tera
lis0,
00,
00,
00,
00,
00,
00,
06,
00,
00,
01,
10,
00,
50,
00,
00,
60,
00,
00,
00,
00,
00,
00,
00,
0
Pse
udon
oniu
m a
tlant
icum
0,0
2,2
1,9
0,9
2,0
1,4
1,2
2,4
0,0
0,9
0,8
1,6
0,0
1,0
0,7
0,2
0,0
0,0
0,5
0,0
0,0
0,0
0,5
0,5
Pyr
go n
asut
a0,
00,
00,
00,
00,
40,
00,
01,
20,
80,
00,
00,
02,
70,
12,
00,
30,
00,
40,
00,
00,
00,
00,
00,
0
Qui
nque
locu
lina
intr
icat
a0,
80,
00,
00,
00,
40,
34,
81,
20,
80,
92,
70,
00,
50,
61,
30,
20,
00,
00,
00,
70,
00,
02,
10,
0
Qui
nque
locu
lina
lam
arck
iana
8,8
4,4
18,4
21,6
22,3
16,1
19,3
33,3
14,3
18,0
27,8
18,2
40,8
20,2
21,3
29,4
1,1
2,6
5,2
7,2
4,9
9,3
13,2
13,2
Qui
nque
locu
lina
mill
etti
5,4
3,0
1,9
6,0
0,8
0,3
0,0
0,0
0,0
0,0
0,8
0,4
0,0
0,4
0,0
0,6
0,0
0,0
0,0
0,0
0,0
0,4
0,3
2,0
Qui
nque
locu
lina
pata
goni
ca22
,59,
616
,57,
87,
018
,99,
66,
021
,922
,511
,017
,83,
816
,912
,09,
02,
92,
20,
92,
28,
811
,611
,710
,1
Text
ular
ia e
arla
ndi
0,0
0,7
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,8
2,0
7,6
1,0
1,3
0,8
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
Text
ular
ia g
ram
en0,
00,
02,
53,
419
,23,
518
,111
,93,
60,
023
,22,
417
,94,
912
,73,
50,
00,
00,
90,
00,
00,
00,
30,
0
Troc
ham
min
a di
scor
bis
0,0
0,0
0,6
0,9
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,3
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
Troc
ham
min
a in
flata
0,4
0,0
0,6
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,3
0,0
Troc
ham
min
a sq
uam
ata
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,8
0,0
0,0
Uvi
geri
na p
ereg
rina
0,4
0,0
0,0
0,9
0,8
0,0
0,0
0,0
1,2
0,0
0,4
0,8
0,0
0,0
0,0
0,2
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,8
Freq
uênc
ia r
elat
iva
das
espé
cies
de
fora
min
ífero
s en
cont
rada
s em
Jan
eiro
201
2

92
Serão discutidas a seguir as análises de CLUSTER (Figura 4.13) e MDS
(Figura 4.14) com base na disposição espacial das estações (mapa da Figura
3.1B) e dos parâmetros abióticos como profundidade, CaCO3, carbono orgânico,
nitrogênio e granulometria (Tabela 4.4) que atuam nesses ambientes.
A análise de CLUSTER (Figura 4.21) juntamente com a tabela de
frequência relativa das espécies (tabela 4.4) revelaram a formação de três grupos
de acordo com as espécies de foraminíferos encontradas: Plataforma Interna (5 –
16), Canal do Rio Potengi (17 – 24) e Foz do Rio Potengi (1 – 4). As espécies
oportunistas Ammonia tepida e Bolivina striatula dominaram nos três grupos
formados tanto no CLUSTER como no MDS, onde A. tepida apresentou altas
porcentagens, principalmente no canal do rio Potengi; enquanto que a espécie
Bolivina striatula aparece em baixas porcentagens, especialmente no rio Potengi,
indicando provavelmente ambiente ecologicamente estressado e poluído.
Cibicides fletcheri, o qual indica ambientes costeiros de alta energia, geralmente
anexada a rochas ou plantas, também apareceu em maiores quantidades na Foz
do rio Potengi, o que explica a presença dessa espécie.
As espécies tipicamente marinhas Hanzawaia boueana, Elphidium
discoidale, Cibicides valvulatanus, C. variabilis e Discorbis peruvianus
aparecem com maior frequência na Plataforma Interna. A espécie de habitat
marinho e de ambientes com alta hidrodinâmica Quinqueloculina lamarckiana
dominou os três grupos, mas em maior porcentagem na Plataforma Interna e
menor na Foz do Rio Potengi. Q. patagonica e Cassidulina subglobosa aparecem
em menor densidade no Canal do Rio Potengi. A espécie oportunista e
mixohalina Q. miletti apresenta frequências mais altas na Foz do Rio Potengi,
indicando deposiçao de matéria organica e outros nutrientes nesse ambiente.
Pseudononion atlanticum, Textularia earlandi, T. gramen e Trochammina inflata
aparecem na Foz do Rio Potengi e na Plataforma Interna. A presença dessas
espécies de foraminíferos tolerantes à baixa salinidade e tipicamente
mixohalinas, como T. inflata, T. squamata e Caronia exilis podem sugerir que o
contribuinte fluvial é capaz de exportar organismos de ambientes mixohalinos
em direção à foz e à plataforma interna.

93
Figura 4.13: Dendograma formado pelo CLUSTER mostrando o agrupamento das estações conforme a
similaridade das espécies de foraminíferos encontradas entre elas em Janeiro de 2012.
A análise de MDS (Figura 4.14) diferenciou três grupos, onde as espécies
de foraminíferos indicadores foram Ammonia tepida, Quinqueloculina
patagonica, Cassidulina subglobosa, Trochamina inflata e T. squamata, as quais
ocorrem principalmente no canal do rio Potengi. Bolivina striatula, Cibicides
fletcheri, Quinqueloculina lamarckiana, Q. miletti, Pseudononion atlanticum,
Textularia earlandi, T. gramen e Trochamina discorbis ocorrem principalmente
na foz do rio Potengi. Hanzawaia boueana, Elphidium discoidale, Cibicides
valvulatanus, C. variabilis, Discorbis peruvianus, Quinqueloculina lamarckiana,
Pseudononion atlanticum, Textularia earlandi e T. gramen aparecem com maior
frequência na plataforma interna. Isso indica que o grupo Canal do Rio Potengi é
dominado por espécies mixohalinas e de manguezais além da presença de
espécies oportunistas como A. tepida e Q. patagonica, indicando ambiente
ecologicamente estressado e provavelmente poluído. O grupo da Foz do Rio
Potengi apresentou espécies mixohalinas e de manguezais (B. striatula, C.
fletcheri, Q. miletti e T. discorbis) quanto marinhas (Q. lamarckiana, P.
atlanticum, T. gramen e T. earlandi), indicando mistura das águas marinhas e

94
fluviais. O grupo Plataforma Interna apresentou a maioria das espécies típicas
marinhas, com exceção de T. discorbis, E. discoidale, C. valvulatanus e C.
variabilis, que são mixohalinas e de manguezais, indicando que o aporte fluvial
pode tê-las transportado para a desembocadura e, através das correntes marinhas,
foram levadas para a plataforma interna.
Figura 4.14: Grupos formados a partir da aplicação de MDS nos dados bióticos de Janeiro de 2012.
4.2.3 Análise comparativa dos mapas de georreferenciamento dos
foraminíferos de Outubro de 2011 e Janeiro de 2012
Serão descritos a seguir os mapas de contorno dos seis foraminíferos
selecionados para estudo: Ammonia spp, Bolivina spp, Caronia exilis, Elphidium
spp, Quinqueloculina spp e Textularia spp. que foram dominantes nas duas
amostragens (Figuras 4.15 – 4.20).
Em Outubro de 2011 Ammonia spp. apresentou maior frequência na
estação 15 (95%), a qual está próxima à Estação de Tratamento de Água de
Quintas e também apresentou maior frequência nas estações 12, 13 e 18, situadas
respectivamente próximas às salinas e fazendas de carcinicultura e ao Canal do
Baldo. A menor frequência ocorre na foz com 15% na estação 8, situada a norte
da foz. Em Janeiro de 2012 esse gênero ocupou a metade do canal principal,
apresentando maior frequência nas estações 17 e 19 (85%) e foi observado
Plataforma
Interna
Canal do
Rio Potengi
Foz do Rio
Potengi

95
também domínio desse gênero na plataforma interna e na foz, mas em baixas
frequências (40%), representados pelas estações 4, 6, 9, 10, 12, 14 e 16. As
menores frequências estão localizadas na plataforma interna, com 10%
representadas pela estação 13 (Figura 4.15).
Figura 4.15: Abundância relativa do gênero Ammonia spp. em Outubro de 2011 e Janeiro de 2012.
Em Outubro de 2011 Bolivina spp. obteve maior frequência na estação 17,
situada próxima às fazendas de carcinicultura (14%) e também na foz (8%),
possuindo menor frequência na região mais afastada da desembocadura na
estação 3 (1%), desaparecendo ao norte da foz (estação 8) e em alguns trechos do
rio (estação 1 e 15). Em Janeiro de 2012 esse gênero apresentou maior frequência
na estação 20 (12%), situada próximo à área do Canal do Baldo e às fazendas de
carcinicultura e das salinas. Assim como no mapa de Ammonia spp, verificou-se
a formação de uma “pluma” na foz do rio Potengi, representado pelas estações 4
e 10 (5%). A menor frequência encontrada está na estação 8, localizada na
plataforma interna norte (1%), desaparecendo na plataforma interna sul,
representada pela estação 15 (Figura 4.16).

96
Figura 4.16: Abundância relativa do gênero Bolivina spp. em Outubro de 2011 e Janeiro de 2012.
Em Outubro de 2011 Caronia exilis localiza-se principalmente na região
central do canal do rio Potengi na estação 14 (8%) e sua frequência diminui
próxima às fazendas de carcinicultura nas estações 12 e 13 (1%), desaparecendo
no curso superior (estação 16), próxima à Estação de Tratamento de Água de
Quintas (estação 15), esgoto do Canal do Baldo (estação 18) e ao se aproximar da
foz (estações 1, 2 e 9) e na foz (3 – 8). Em Janeiro de 2012 além de apresentar
porcentagem muito baixa, essa espécie ocorre na estação 11 (0.8%), na
plataforma interna sul, embora existam também pequenas colônias na plataforma
interna norte, representadas pelas estações 3 e 5 (0.5%) e próxima à foz do rio
Potengi, representada pela estação 24 (0.3%). Desaparece nos extremos norte e
sul da plataforma interna e ao canal do rio Potengi (Figura 4.17).

97
Figura 4.17: Abundância relativa da espécie Caronia exilis em Outubro de 2011 e Janeiro de 2012.
Em Outubro de 2011 Elphidium spp. é mais frequente na estação 7, ao sul
da foz (9%) e também na estação 8, a norte da foz (6%). A frequência é baixa em
alguns trechos do rio nas estações 10 e 16 (2%) e também na região mais
afastada da foz, nas estações 4 a 6 com o mesmo valor, enquanto que na área
mais afastada da foz na estação 3 e no canal principal nas estações 9, 11 a 15, 17
e 18 esse gênero desaparece. Em Janeiro de 2012 esse gênero apresentou maior
frequência na estação 22 (16%), situada próximo ao Porto de Natal, apresenta
baixas frequências na plataforma interna representada pelas estações 9 e 12 – 15
(1%), desaparecendo ao longo do canal do rio Potengi (estações 17 – 21), na foz
(estações 2 e 24) e na plataforma interna (estações 3, 5, 6 e 8) (Figura 4.18).

98
Figura 4.18: Abundância relativa do gênero Elphidium spp. em Outubro de 2011 e Janeiro de 2012.
Em Outubro de 2011 Quinqueloculina spp. obteve maior frequência na
estação 8 (65%), localizada à norte da foz enquanto as menores frequências (5%)
estão localizadas próximas às fazendas de carcinicultura e das salinas (estações
12, 13 e 17), a estação 15 está próxima à Estação de Tratamento de Água de
Quintas e a estação 18 está próximo ao Canal do Baldo. Em Janeiro de 2012 esse
gênero apresentou-se dominante na plataforma interna, principalmente na estação
13, situada na plataforma interna sul (46%), porém existe porcentagem maior na
estação 1, situada na Marina Iate Clube (34%) e as menores frequências
encontram-se no curso superior do canal principal (2%) representado pelas
estações 17 – 19 (Figura 4.19).
Figura 4.19: Abundância relativa do gênero Quinqueloculina spp. em Outubro de 2011 e Janeiro de
2012.
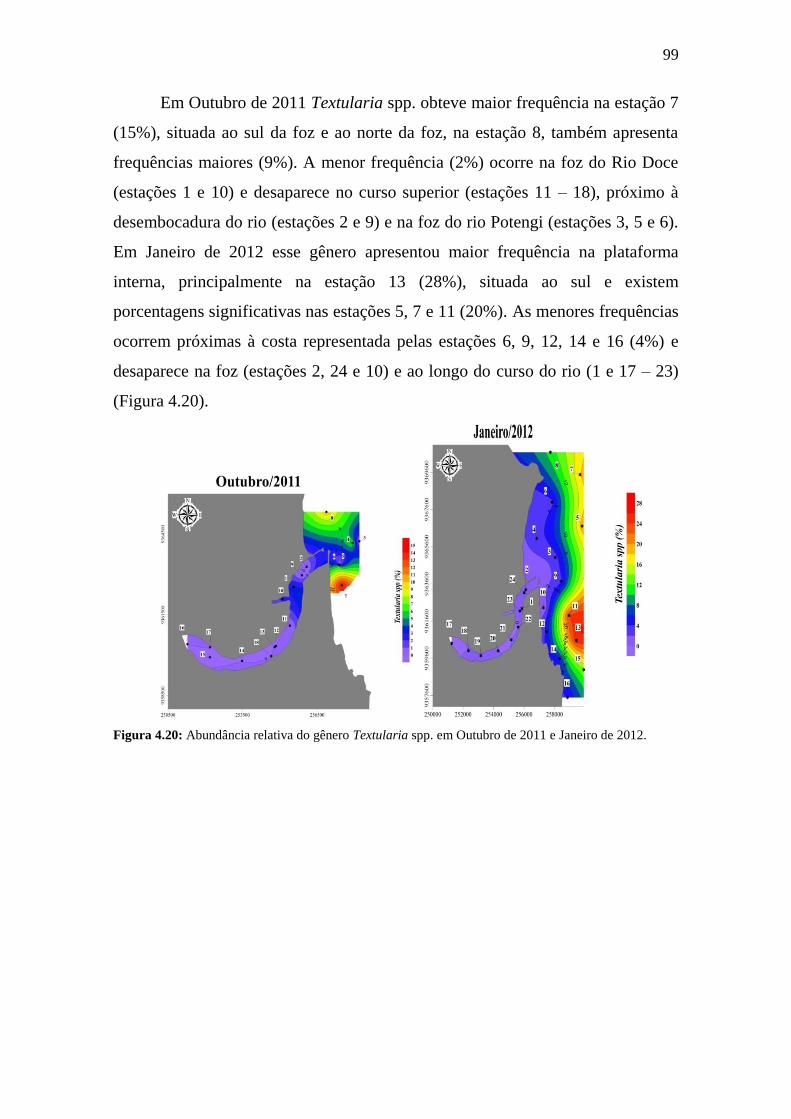
99
Em Outubro de 2011 Textularia spp. obteve maior frequência na estação 7
(15%), situada ao sul da foz e ao norte da foz, na estação 8, também apresenta
frequências maiores (9%). A menor frequência (2%) ocorre na foz do Rio Doce
(estações 1 e 10) e desaparece no curso superior (estações 11 – 18), próximo à
desembocadura do rio (estações 2 e 9) e na foz do rio Potengi (estações 3, 5 e 6).
Em Janeiro de 2012 esse gênero apresentou maior frequência na plataforma
interna, principalmente na estação 13 (28%), situada ao sul e existem
porcentagens significativas nas estações 5, 7 e 11 (20%). As menores frequências
ocorrem próximas à costa representada pelas estações 6, 9, 12, 14 e 16 (4%) e
desaparece na foz (estações 2, 24 e 10) e ao longo do curso do rio (1 e 17 – 23)
(Figura 4.20).
Figura 4.20: Abundância relativa do gênero Textularia spp. em Outubro de 2011 e Janeiro de 2012.

100
Capítulo 5

101
5 – Discussões
Ambas amostragens revelaram que a composição da carapaça dos
foraminíferos do rio Potengi e da plataforma interna é dominado por espécies
calcárias, seguida de espécies porcelanáceas. No entanto, na segunda amostragem
verificou-se que a plataforma interna é dominada também por espécies
aglutinantes e porcelanáceas, o que pode ser explicado pela granulometria
característica nesses ambientes. Trabalhos de Wang e Chappell (2001), Ruiz et
al. (2005), Eichler et al. (2007), Mahiques et al. (2009) e Sarita et al. (2015)
discutem aspectos e relações de foraminíferos com a granulometria, onde
espécies de aglutinantes selecionam grãos finos e grossos para a construção de
suas carapaças, e espécies porcelanáceas são geralmente relacionadas a
granulometria mais grossa.
O valor elevado de indivíduos vivos para A. tepida nas proximidades das
fazendas de carcinicultura, das salinas e do Canal do Baldo, reforçam sua
característica cosmopolita e indicadora de ambiente caracterizado por baix
salinidade, teor de argila e alto teor de carbono orgânico (Mleis e Covelli, 2013).
B. striatula também dominou na proximidade das fazendas de carcinicultura e
das salinas e na foz na primeira amostragem, enquanto que na segunda
amostragem houve domínio nas proximidades do Canal do Baldo.
De acordo com Eichler et al. (2003, 2007), Ruiz et al. (2005), Uehara et
al. (2007), Berkeley et al. (2008), Mahiques et al. (2009) e Debenay (2012), o
gênero Bolivina costuma viver em lagoas costeiras, estuários e baías e são
também tolerantes a ambientes com baixos teores de oxigênio dissolvido e alto
teor de carbono orgânico indicando talvez um local impactado em ambientes de
granulometria fina. A presença de provável competição interespecífica de Q.
miletti e Q. patagonica observada no presente estudo necessitam ainda de estudo
para comprovar essa hipótese. De acordo com Debenay (2012) o gênero
Quinqueloculina pode ser encontrado em lagoas costeiras, pântanos salgados,
estuários e baías, sendo também considerados oportunistas e tolerantes à
ambientes poluídos, como as espécies Quinqueloculina miletti e Q. patagonica.
Essa maior concentração das espécies oportunistas no estuário pode ser explicada

102
pela grande quantidade de efluentes provenientes do Canal do Baldo, indústrias
de vários setores e da agricultura, principalmente o Esgoto do Canal do Baldo,
que lança seus efluentes não tratados diretamente no leito do rio Potengi (mais de
60%) diariamente, como foi mostrado nos trabalhos de Ramos e Silva et al.
(2006) e Silva et al. (2001), onde o primeiro autor estudou as variações de metais
pesados através das raízes das árvores dos manguezais vermelhos e o segundo
autor utilizou ostras Crassostrea rhizophorae como bioindicadores, onde ambos
apontaram discutiram os efeitos antrópicos, como dragagens periódicas, uso de
pesticidas e fertilizantes na agricultura e a emissão dos esgotos não tratados
combinados com o despejo de efluentes industriais e farmacêuticos no estuário.
Algumas espécies encontradas no Rio Potengi e plataforma interna podem
indicar ambientes ecologicamente estressados (A. tepida, B. striatula, Q.
patagonica e Q. miletti), apontar as presenças da influência marinha no estuário
(Q. lamarckiana) e aporte fluvial para a foz (Trochammina spp), além de serem
utilizados como indicadores em estudos paleoambientais e subambientes
passados (Comissão do Registro Geológico da Dinâmica da Biosfera, 2005;
Mahiques et al., 2009; Uehara et al., 2007; Ruiz et al., 2005; Wang e Chappell,
2001).
Algumas espécies de foraminíferos oportunistas foram encontrados nos
trabalhos de Eichler et al. (2007), realizado no Canal Bertioga (SP); De Souza et
al. (2010), realizado no rio Potengi; Eichler et al (2014), realizado na Baía de
Guanabara (RJ), Eichler – Coelho et al. (1997), Uehara et al. (2007) e Mahiques
et al. (2009), realizados no sistema estuarino Cananéia – Iguape (SP), Ruiz et al.
(2005), realizado em três estuários: Guadiana, Piedras e Tinto – Odiel,
localizados ao longo da Costa de Huelva, Sudoeste da Espanha, e Berkeley et al.
(2008), realizado na linha de costa adjacente ao Rio Cocoa, Queensland,
Austrália.
Além das espécies oportunistas, foram encontradas também espécies
marinhas, como T. gramen, T. earlandi, Q. lamarckiana, H. boueana, Cibicides
valvulatanus, Cibicides variabilis, U. peregrina e P. atlanticum, sendo que U.
peregrina aparece somente em Janeiro de 2012. A maior frequência de Q.

103
lamarckiana fica restrita na foz e são encontradas baixas frequências no curso
superior; já em Janeiro de 2012 foi observado que as maiores frequências situam-
se na plataforma interna e percebem-se porcentagens significativa até a estação
22, situada nas proximidades do Porto de Natal. Os autores Eichler et al. (2007) e
Wang & Chappell (2001) descrevem o gênero Quinqueloculina como
foraminífero típico de ambientes de alta hidrodinâmica da plataforma interna.
Apesar das espécies H. boueana e P. atlanticum aparecerem em baixas
frequências restritas à foz P. atlanticum, H. boueana, T. gramen e T. earlandi
aparecem em maior frequência na plataforma interna e nas proximidades da foz.
Segundo Eichler et al. (2012), a espécie Hanzawaia boueana está relacionada à
Água Tropical sob a influência da Corrente Brasileira, e sua presença no rio
Potengi e na plataforma indica ambientes de alta salinidade e nesse estudo pode
revelar a penetração da cunha salina no rio Potengi, enquanto que P. atlanticum e
U. peregrina estão associados à plataforma interna, juntamente com Q.
lamarckiana. O gênero Cibicides é típico de ambientes costeiros de alta energia,
geralmente anexado a rochas ou plantas (Scott et al., 2004; Debenay, 2012), o
que explica a sua localização na plataforma e na foz do rio Potengi, nunca no
interior do mesmo. Apesar de não obter o dado de salinidade na segunda
amostragem, o Rio Potengi parece possuir gradiente de influência marinha até as
proximidades do Porto de Natal.
Em ambas as amostragens foi identificada a espécie mixohalina e típica de
manguezais C. exilis, onde em Outubro de 2011 ocorreu no canal central do Rio
Potengi próxima às salinas e as fazendas de carcinicultura. Já na segunda
amostragem foi observado que, além do desaparecimento dessa espécie no rio e
seu aparecimento na plataforma interna, sua frequência está muito baixa,
situando-se na plataforma interna. As espécies do gênero Trochammina, típicas
de áreas de manguezal (Eichler et al., 2007), também foram encontradas na
plataforma, apenas na segunda amostragem, indicando a presença da pluma
estuarina de água doce na plataforma interna revelando a alta competência do Rio
Potengi de exportar material para a plataforma.

104
As análises de PCA revelaram que os principais parâmetros que atuaram
na distribuição dos foraminíferos foram a granulometria, o CaCO3, carbono
orgânico e o nitrogênio, seguidas de profundidade, salinidade, condutividade e
densidade. O BIOENV revelou tambem que a granulometria foi o fator que
definiu a distribuição dos foraminíferos em Outubro de 2011, enquanto que em
Janeiro de 2012 foi o carbonato de cálcio. Pode-se concluir que altas
porcentagens de areia grossa e areia muito grossa revelam ambientes de alta
hidrodinâmica, e baixa frequência de cascalho, areia média, areia fina, areia
muito fina, silte e argila, típicas de ambientes moderadamente hidrodinâmicos
(areia média e areia fina) e baixa hidrodinâmica (frações finas). Comparando as
análises de MDS foi observado que tanto em Outubro de 2011 e Janeiro de 2012
o Canal do Rio Potengi é dominado por espécies oportunistas (Outubro de 2011:
A. tepida, B. striatula e Q. patagonica; Janeiro de 2012: A. tepida e Q.
patagonica) evidenciando os prováveis locais de contaminação, como as
proximidades da Estação de Tratamento de Água de Quintas, Canal do Baldo,
salinas e fazendas de carcinicultura.
O rio Potengi apresenta frações de areia média e areia fina, indicando
ambiente moderadamente hidrodinâmico, embora tenha apresentado
porcentagem significativa de silte e argila na estação 1, situada próxima à Marina
Iate Clube, indicando ser um ambiente deposicional e de baixa hidrodinâmica.
Na plataforma interna as frações de areia grossa e areia média foram dominantes,
enquanto que a fração de areia fina ficou concentrada na plataforma interna
norte, provavelmente por causa das correntes marinhas, e as frações de areia
muito fina e silte e argila apresentaram porcentagens significativas na estação 14,
situada em Ponta Negra, indicando ambientes deposicionais e de baixa
hidrodinâmica. Segundo Frazão et al. (2003) a dispersão do material
sedimentológico se dá pelas marés, quase sempre acompanhados por correntezas
horizontais. As correntes de maré predominam na circulação do estuário Potengi
e na plataforma interna, junto ao litoral. As correntes induzidas pelo fluxo do rio
Potengi são restritas à região estuarina e são induzidas pelos ventos alísios na
direção SE e E, transportando grandes quantidades de sedimentos na plataforma

105
interna, em direção à costa e são responsáveis pela orientação das ondas, as quais
atingem a linha de costa, gerando correntes de deriva litorânea que transportam
os sedimentos no sentido S – N (Pereira, 1999; Suguio, 1999).
No estudo de Frazão & Vital (2007), o canal do rio Potengi é formado
predominantemente pela fração arenosa, com a presença de barras alongadas de
areia com cascalho esparso no curso superior e na foz do rio Doce, apresenta
areia siltosa na área das indústrias, no Porto de Natal, no Iate Clube, as margens
do canal são compostas por silte arenoso, nas proximidades da foz apresenta as
feições areia lamosa com cascalho esparso e arenito de praia. Essa informação
concorda com a análise de Outubro de 2011, mas em Janeiro de 2012 foi
observado que o cascalho não aparece nos locais citados pelo autor, podendo
indicar que houve forte influência fluvial que provavelmente deslocou essas
frações para outros ambientes.
Segundo a tabela dos dados abióticos, as porcentagens de CaCO3 foram
muito altas na plataforma interna da segunda amostragem, que indicam maior
contribuição biológica, ao contrário do constatado no estuário. Os teores muito
baixos de carbono orgânico sugerem um ambiente oxidante, inadequado à
preservação da matéria orgânica. Para Cunha (2004), a distribuição do CaCO3 no
estuário do rio Potengi se mostrou menor (5%), ocupando as zonas mais
profundas no estuário médio. Os valores maiores estão associados aos
sedimentos mais finos, situados nas zonas menos profundas junto às margens, em
resposta à ação seletiva das correntes de marés e, além disso, alguns locais do
estuário são ricos em conchas calcárias, devido às ações de drenagem como é o
caso dos canais que interligam o rio Potengi.
Quanto à matéria orgânica, esse mesmo autor afirmou nos seus estudos
que a matéria orgânica é derivada dos manguezais característicos dos ambientes
com alta produtividade primária (plantas e árvores), onde há enriquecimento por
decomposição da matéria orgânica de origem próxima aos dejetos domésticos.
Neste estudo os locais que apresentaram maiores valores de matéria orgânica
apresentaram maior predominância de foraminíferos oportunistas, como B.
striatula, A. tepida, Q. patagonica e Q. miletti.

106
A temperatura variou de acordo com a disposição espacial das estações,
onde as amostras coletadas no canal principal apresentaram temperaturas maiores
e as amostras coletadas próximas à desembocadura e na foz apresentaram
temperaturas mais baixas, salientando que o parâmetro de temperatura não está
no grupo P1 provavelmente pela existência das dragagens periódicas no rio e
estas alteram a configuração do fundo. Na segunda amostragem a profundidade
variou principalmente próximo da foz do rio e na plataforma interna, tendo esta
última a profundidade esperada. A temperatura não foi o fator determinante na
distribuição dos foraminíferos, pois a área de estudo localiza-se em área tropical
e as águas são quentes, cuja variação é mínima, como foi mostrado no estudo de
Frazão et al. (2003).
Nos estudos de Pereira (1999) e Cunha (2004) realizados no rio Potengi,
onde nesse trecho as zonas mais profundas (entre 15 a 12,5 m) encontram-se no
canal principal, entre o Iate Clube e a Ponte de Igapó (ponte antiga) e à medida
que se afasta do trecho a profundidade diminui. Segundo a autora, esse fato é
explicado pela ação antrópica que faz as dragagens do rio para facilitar a
navegação e o aporte de navios, além das ações erosivas provocadas pelas
correntes fluviais das marés, formando bancos de areia nas margens côncavas e
remobilização dos sedimentos erodidos nas margens convexas. Esses estudos
confirmaram as variações de profundidade no canal do Rio no atual trabalho, em
particular na área do Porto e do Iate Clube.
Os parâmetros de salinidade, condutividade e densidade obtiveram
maiores valores nas estações coletadas próximas da foz e na foz, já que é um
ambiente que sofre influência constante do mar e obtiveram menores valores nas
estações coletadas nas regiões mais internas. Segundo Cunha (2004), a influência
estuarina na circulação costeira se restringe às zonas próximas da saída do
estuário e em pequena escala, sendo que a influência é maior nas condições de
maré alta. A linha de recifes situada na entrada do estuário é responsável pela
penetração das correntes costeiras no interior do rio.

107
Como foi observado acima, os fatores que contribuem para a distribuição
dos foraminíferos são os aportes fluviais e marinhos, granulometria e a
disposição de CaCO3 e matéria orgânica.

108
Capítulo 6

109
6 – Conclusões
A análise sistemática dos foraminíferos presentes em sedimentos do
rio Potengi e plataforma interna adjacente indica a presença de dois
ambientes: a área mais externa é regida pela forte influência marinha e a
área mais interna é mixohalina;
A forte influência do mar ao longo do rio Potengi é revelada através
da presença da espécie Quinqueloculina lamarckiana. O gênero
Trochammina indica a presença da pluma estuarina de água doce na
plataforma interna revelando a alta competência do rio Potengi de exportar
material da região de manguezal para a plataforma;
O rio Potengi é dominado por espécies calcárias e aglutinantes,
indicadoras de aporte continental, enquanto que a foz e a plataforma
interna são dominadas pelas espécies aglutinantes e porcelanáceas, as
quais são típicas de ambientes com alta salinidade e hidrodinâmica;
A presença das espécies Ammonia tepida, Quinqueloculina
patagonica e Q. miletti, juntamente com as porcentagens elevadas de areia
muito fina, silte e argila, revelaram que os principais locais de deposição
provavelmente indicam sedimentos poluídos ou contaminados;
Os fatores que contribuíram para a distribuição dos foraminíferos
foram as correntes fluviais e marinhas, a granulometria, porcentagem de
CaCO3 e matéria orgânica;
A ocorrência de B. striatula nas estações próximas à foz, às salinas
e às fazendas de carcinicultura reforçam sua característica;

110
Referências
BERKELEY A., PERRY C.T., SMITHERS S.G., HORTON B.P. 2008.
The spatial and vertical distribution of living (stained) benthic
foraminifera from a tropical, intertidal environment, north Queensland,
Australia. Revista Marine Micropaleontology. 69: 204 – 261.
BOERSMA A. 1978. Foraminifera. In: HAQ, B.U.; BOERSMA, A. (eds.)
Introduction to marine micropaleontology. New York: Elsevier, p. 19-77.
BOLTOVSKOY E. 1965. Los foraminiferos recientes. Eudeba, Buenos
Aires, 510 pp.
BOLTOVSKOY E., GIUSSANI G., WATANABE S., WRIGHT R. 1980.
Atlas of benthic shelf foraminifera of southwest Atlantic. Dr. W. Junk
(ed.), Hague, 154 pp.
BOYD R., DALRYMPLE R.W., ZAITLIN B.A. 2006. Estuarine and
incised valley facies models In: POSAMENTIER, W.; WALKER, R.G.
(ed.) Facies Models Revisited. SEPM (Society for Sedimentary Geology),
Special Publication n. 84, ISBN 1-56576-121-9, p. 171–235.
CLIFTON H.E. 1982. Estuarine deposits. In: Sandstone depositional
environments – AAPG Memoir 31. AAPG: Tulsa, p. 179-189.
COMMITTEE ON THE GEOLOGIC RECORD OF BIOSPHERE
DYNAMICS. 2005. The Geological Record of Ecological dynamics:
Understanding the Biotic Effects of Future environmental Change. The
National Academic Press, Washington, 216 pp.
CUNHA E.M.S. 2004. Evolución actual del litoral de Natal – RN (Brasil)
y sus aplicaciones a la gestión integrada. Evolução atual do litoral de
Natal – RN (Brasil) e suas aplicações a gestão integrada. Tese de
doutorado. Universitat de Barcelona. Department d´Ecologia, Facultat de
Biologia. Programa Doctorado de Ciencias del Mar.
DALRYMPLE R.W., ZAITLIN B.A., BOYD R. 1992. Estuarine facies
models: Conceptual basis and stratigraphic implications: Journal of
Sedimentary Petrology, Tulsa, v. 62, p. 1130–1146.,

111
DAVIES J.L. (ed.). 1964, A morphogenic approach to world shorelines.
Zeits. Geomorph. Stuttgart, v. 8 (Special Number), p. 172-142.
DAVIES J.L. 1972. Geographical Variation in Coastal Development.
Longman, London, 204 pp.
DEBENAY J.P. 2012. A guide to 1.000 foraminifera from Southwestern
Pacific New Caledonia. IRD Éditions, Publications Scientifiques du
Muséum, Paris, 385 pp.
DE GRANDE A.M. 2015. Rio Contado. [Filme – Documentário].
Produção de Airton Miguel De Grande. Direção de Airton Miguel de
Grande. Natal. IBAMA. Vídeo digital, inédito, 95 min. Color. Son.
DE SOUZA V.M., LAUT L.L.M., DA SILVA F.S., FIGUEIREDO JR
A.G., VITAL H., FRAZÃO E. 2010. Benthic foraminífera and bacterial
activity as a proxy for environmental Characterization in Potengi Estuary,
Rio Grande do Norte, Brazil. Rio de Janeiro. Anuário de geociências. 33 –
1: 20 – 34.
EICHLER – COELHO P.B., DULEBA W., EICHLER B.B., COELHO –
JUNIOR C. 1997. Determinação do impacto ecológico do Valo Grande
(Iguape, SP) a partir das associações de foraminíferos e tecamebas.
Revista Brasileira de Biologia. 57: 463 – 477.
EICHLER P.P.B., EICHLER B.B., MIRANDA L.B., PEREIRA E.R.M.,
KFOURI P.B.P., PIMENTA F.M., BÉRGAMO A.L., VILELA C.G.
2003. Benthic foraminiferal response to variations in temperature, salinity,
dissolved oxygen and organic carbon in the Guanabara Bay, Rio de
Janeiro, Brazil. Anuário do Instituto de Geociências. Rio de Janeiro. 26:
36-51.
EICHLER P.P.B., EICHLER B.B., MIRANDA L.B., RODRIGUES A.R.
2007. Modern foraminiferal facies in a subtropical estuarine channel,
Bertioga, São Paulo, Brazil. Journal of Foraminiferal Research. 37: 234-
247.
EICHLER P.P.B., EICHLER B.B., SENGUPTA G.B., RODRIGUES
A.R. 2012. Foraminifera as indicators of marine pollutant contamination

112
on the inner continental shelf of Southern Brazil. Marine Pollution
Bulletin. 64:22 – 30.
EICHLER P.P.B., EICHLER B.B., PEREIRA E.R.M., VITAL, H. 2014.
Evaluation of Environmental and ecological effects due to the accident in
an oil pipe from Petrobrás in Guanabara Bay, RJ, Brazil. Open Journal of
Marine Science. 4: 298 - 315.
FAIRBRIDGE R.W. 1968. The encyclopedia of geomorphology. Van
Nostrand Reinhold, New York. 1295 pp.
FRAZÃO E.P. 2003. Caracterização hidrodinâmica e morfo – sedimentar
do estuário Potengi e áreas adjacentes: subsídios para controle e
recuperação ambiental no caso de derrames de hidrocarbonetos.
Dissertação (Mestrado). Centro de Ciências Exatas e da Terra,
Universidade Federal do Rio Grande do Norte. 145 p.
FRAZÃO E. & VITAL H. 2007. Estruturas rasas de gás em sedimentos no
estuario Potengi (Nordeste do Brasil). Brazilian Journal of Geophysics.
Rio de Janeiro. 25. 17 – 26.
GARRISON T. 2002. Oceanography: an invitation to marine science.
Thomson Learning Inc, United Estates of America, 554 pp.
IBAMA. 2014. Parecer Técnico n.o 02021.000145/2014-00. Natal,
NUPAEM-DICAF/SUPES/IBAMA/RN, 107 p.
IBAMA. 2015. Parecer Técnico n.o 02021.000032/2015-87. Natal,
NUPAEM-DITEC/SUPES/IBAMA/RN, 51 p.
KELLER E.A. 2012. Introduction to environmental geology. Prentice
Hall, USA. 768 pp.
KLEE G.A. 1999. The coastal environment: toward integrated coastal and
marine sanctuary management. Prentice – Hall Inc, United Estates of
America, 281 pp.
LIMA FILHO M. L. 2013. Bacia Pernambuco e Paraíba. Rio de Janeiro.
Agência Nacional do Petróleo, Gás Natural e Biocombustíveis, Rodada
11.

113
MAHIQUES M.M., BURONE L., FIGUEIRA R.C.L., LAVENÉRE -
WANDERLEY A.A.O., CAPELLARI B., ROGACHESKI C.E.,
BARROSO C.P., DOS SANTOS L.A.S., CORDERO L.M., CUSSIOLI
M.C. 2009. Anthropogenic influences in a lagoonal environment: a
multiproxy approach at the Valo Grande mouth, Cananéia-Iguape system
(SE Brazil). Brazilian Journal of Oceanography. 57: 325-337.
MELIS R. & COVELLI S. 2013. Distribution and morphological
abnormalities of recent foraminifera in the Marano and Grado Lagoon
(North Adriatic Sea, Italy). Mediterranean Marine Science. 14: 432 – 450.
NIMER E. 1972. Climatologia da Região Nordeste do Brasil. Revista
Brasileira de Geografia, 34: 3 – 51.
PEREIRA L.B.F. 1999. Mapeamento geológico, estratigrafía cenozoica e
estruturaçao neotectonica da regiao costeira entre Ponta Negra e Redinha e
Estuario do Rio Potengi. Relatório (Graduação). Centro de Ciências
Exatas e da Terra, Universidade Federal do Rio Grande do Norte. 80 p.
RADAM BRASIL. 1981. Série Levantamento de Recursos Naturais do
Projeto RADAM BRASIL.
RAMOS E SILVA, C.A., DA SILVA A.P., DE OLIVEIRA S.R. 2006.
Concentration, stock and transport rate of heavy metals in a tropical red
mangrove, Natal, Brazil. Marine Chemistry. 99. 2 – 11.
ROSSETTI D.F. 2008.Ambientes estuarinos. In: A.J.C.L.P. SILVA,
M.A.N.F. ARAGÃO, A.J.C. MAGALHÃES (Org). Ambientes de
sedimentação siliciclástica do Brasil. Beca – BALL Edições. São Paulo,
Brasil. P.: 194 – 211. REINSON G.E. 1992. Transgressive barrier islands
and estuarine systems. In: WALKER R.G., JAMES N.P. Facies models
response to sea level changes. Ontario: Geological Association of Canada,
Geotext 1, p. 179-194.
PINET P. R. 2003. Invitation to oceanography. Jones and Bartlett
Publishers Inc, London, UK. 556 pp.

114
PRITCHARD D.W. 1967. What is an estuary: Physical viewpoint? In:
LAUFF G.H. (Ed.) Estuaries. Washington: Am. Assoc. for Adv. Sci.,
Publ. n. 83, p. 3-5.
RAHMANI R.A. 1988. Estuarine tidal cannel and nearshore
sedimentation of a late cretaceous epicontinental sea, Drumheller, Alberta,
Canada. In De Bouer P.L, Van Gelder A. and Nio S.D. (Eds). Tide
influenced sedimentary environments and facies. Reidel. Boston. P 433 –
474.
RUIZ F., GONZÁLEZ – REGALADO M.L., PENDÓN J.G., ABAD M.,
OLÍAS M., MUÑOZ J.M. 2005. Correlation between foraminifera and
sedimentary environments in recent estuaries of Southwestern Spain:
applications to Holocene reconstructions. Quaternary International. 140 –
141: 21 – 36.
SARITA C., DELMINDA M., SIMON, C., DAVID S., TOMASZ B.
2015. Ecological zonation of benthic foraminifera in the lower Guadiana
Estuary (southeastern Portugal). Marine micropaleontology. 114: 1 – 18.
SCOTT D.B., SCHAFER C.T., MEDIOLI F.S. 2004. Monitoring in
Coastal Environments Using Foraminifera and Thecamoebian Indicators.
Cambridge University Press. Cambridge, United Kingdom.
SILVA C.A.R., RAINBOW P.S., SMITH B.D. and SANTO Z.L. 2001.
Biomonitoring of trace metal contamination in the Potengi Estuary, Natal
(Brazil), using the oyster Crassostrea rhizophorae, a local food source.
Water Research. 35: 4072 – 4078.
SUGUIO K. 1980. Rochas sedimentares: Propriedades, gênese
importância ecológica. Ed. Edgard Blücher Ltda, São Paulo, 500 pp.
SUGUIO K. 1999. Geologia do Quaternário e mudanças ambientais
(passado + presente = futuro?). Paulo´s Comunicação e Artes Gráficas.
São Paulo, 366 pp.
THURMAN H.V & TRUJILLO A.P. 1999. Essentials of oceanography.
Prentice – Hall, Inc. USA.

115
UEHARA R.S., DULEBA W., PETRI S., MAHIQUES M.M.,
RODRIGUES M. 2007. Micropaleontologia e sedimentologia aplicadas à
análise paleoambiental: um estudo de caso em Cananéia, São Paulo,
Brasil. Revista Brasileira de Paleontologia. 10: 137-150.
VILELA C.G. 2004. Foraminíferos. In: I.S Carvalho (Ed). Paleontologia.
Ed. Interciência. Rio de Janeiro. P.: 269 – 282.
WANG P. & CHAPPELL J. 2001. Foraminifera as Holocene
environmental indicators in the South Alligator River, Northern Australia.
Quaternary International. 83–85: 47–62.
YASSINI I. & JONES B.G. 1995. Foraminiferida and ostracoda from
estuarine and shelf environments on the southeastern coast of Australia.
University of Wollongong Press. Wollongong. Austrália.

116
Apêndice

117
Apêndice 1
Prancha 1: Espécies principais ocorrentes no rio Potengi e na plataforma interna. 1: Ammonia
tepida; 2: Bolivina striatula; 3: Caronia exilis; 4: Cibicides spp; 5: Elphidium spp; 6: Hanzawaia
boueana; 7: Pseudononium atlanticum; 8 – 10: Quinqueloculina spp; 11: Textularia spp.

118
8 – Artigo

119
Artigo “Geologia ambiental do estuário do Rio Potengi e plataforma
continental adjacente, RN, Brasil”
Submetido à revista de Geociências
Cristiane Leão Cordeiro de Farias*, Patrícia Pinheiro Beck Eichler e Helenice
Vital
Programa de Pós-graduação em Geodinâmica e Geofísica, Universidade Federal
do Rio Grande do Norte, 59072-970, Natal, RN, Brazil.
*Corresponding author: [email protected], [email protected],
[email protected]. Telephone (55) 84 98849 – 1949

120
Resumo
Rio Potengi, receptor de vários poluentes e contaminantes, foi avaliado através
de 42 amostras ao longo do estuário, na foz e plataforma continental interna.
Análises univariadas e multivariadas foram aplicadas à matriz dos dados bióticos
e abióticos dessas áreas. Os resultados mostram a dominância de foraminíferos
oportunistas A. tepida, B. striatula, Q. patagonica e Q. miletti especialmente nas
regiões próximas às fazendas de carcinicultura e ao esgoto do Canal do Baldo em
ambientes de granulometria fina, e Q. lamarckiana indicadora da penetração da
cunha salina e ambientes de alta hidrodinâmica associada a sedimentos de areia
grossa a muito fina. A presença de espécies características marinhas H. boueana,
E. discoidale, P. atlanticum, T. earlandi e T. gramen na Foz do Rio Potengi e na
plataforma interna indicam ambientes de altas salinidades. A ocorrência de
algumas espécies tolerantes à baixa salinidade como T. inflata e T. squamata no
Canal do Rio Potengi sugerem que provavelmente devem ter sido transportados
do manguezal próximo à foz do Rio Potengi para as regiões de plataforma
interna, sugerindo que o contribuinte fluvial é capaz de exportar organismos de
água doce preferivelmente em direção sul do que em direção norte.
Palavras chave: Rio Potengi, estuário, plataforma interna, foraminíferos
bentônicos, granulometria.

121
Abstract
Potengi River, which has been receiving pollutants and contaminants, was
evaluated through 42 samples along the estuary, mouth and adjacent continental
inner shelf. Univariate and multivariate analysis were applied to the matrix of
biotic and abiotic data of those areas. The results show the dominance of
opportunistic forams like A. tepida, B. striatula, Q. patagonica and Q. miletti in
carciniculture and sewage from Canal do Baldo in fine sediments, and Q.
lamarckiana indicative of saline penetration in high hydrodynamic environments
associated to coarse sand to very fine sand substrate. The presence of marine
characteristic species H. boueana, E. discoidale, P. atlanticum, T. earlandi and T.
gramen in Potengi River´s mouth and inner shelf indicates high salinity
environments. However, occurrence of low-salinity tolerant foraminiferal species
typical of mangrove environments as T. inflata and T. squamata in Potengi
River´s Channel suggest they probably could have been transported from
mangrove area near the Potengi River mouth to the inner shelf regions. These
findings also suggest Potengi River is able to export mixohaline organisms
preferably towards south than north.
Key words: Potengi River, estuary, inner shelf, benthonic foraminifera, grain
size.

122
8.1 – Introdução
Os foraminíferos são microrganismos pertencentes ao reino protista,
podendo ser bentônicos infaunais (enterrados no sedimento), epifaunais (sobre o
sedimento) e fixos ao substrato; ou planctônicos. Surgiram no Cambriano (570
milhões de anos) e vivem até o Recente, sobretudo em ambientes marinhos, mas
existem espécies dulcícolas. Possuem carapaça (teca) de formas variadas,
composta por carbonato de cálcio (CaCO3), aglutinada por partículas do
sedimento do meio em que vivem ou, mais raramente, orgânica ou silicosa. A
teca é formada por uma (unilocular) ou várias (multilocular) câmaras que se
intercomunicam através de uma ou mais aberturas, chamada de forâmen (Vilela,
2004).
São extremamente valorizados na Indústria do Petróleo, devido a sua
utilização na geologia para fins bioestratigráficos e na datação de rochas. No que
diz respeito a estudos ecológicos, a dinâmica populacional dos foraminíferos
responde bem à poluição, devido às alterações por ela impostas no teor de acidez
dos sedimentos, o qual afetará facilmente a distribuição de determinadas
espécies, por exemplo. Assim, dependendo da tolerância dos foraminíferos à
determinada situação, uma mesma região pode apresentar aumento exagerado de
algumas espécies oportunistas, mas por outro lado pode apresentar também
ausência de espécies que não toleram esta mesma condição (Eichler et al., 2003).
De acordo com a Comissão do Registro Geológico da Dinâmica da
Biosfera (Committee on the Geologic Record of Biosphere Dynamics, 2005), o
estudo de indicadores ambientais, como os foraminíferos, pode ser relacionado à
aspectos físicos, geológicos e químicos da região, sendo esta uma boa
oportunidade para estudos ambientais. Isto pode ser justificado, pois os
foraminíferos são capazes de sintetizar as características gerais do ambiente,
ressaltar as variações ambientais de curtos períodos de tempo e reagir
sensivelmente às variações sazonais e aos efeitos antrópicos, reconhecendo a
problemática local através de avaliações e diagnósticos ambientais. De Souza et
al. (2010) realizam a primeira caracterização de foraminíferos no Potengi e
apontam a sua utilização como proxy ambiental.

123
Neste contexto o presente trabalho foi desenvolvido no estuário do Rio
Potengi e na sua plataforma continental adjacente. Dados abióticos (salinidade,
densidade, temperatura, profundidade, granulometria, teor dos constituintes
orgânicos e teor de CaCO3) foram correlacionados aos dados de foraminíferos. O
objetivo principal é analisar as espécies de foraminíferos bentônicos e as
respostas destes aos fatores físicos, químicos e geológicos da região de estudo,
além de identificar as possíveis áreas afetadas. Especificamente o presente
trabalho apresenta a dinâmica da população de foraminíferos correlacionada à
granulometria e ao conteúdo de matéria orgânica dos sedimentos, tentando dessa
forma evidenciar os locais de deposição e sedimentação.
8.2 – Caracterização da área de estudo
8.2.1 O rio Potengi
O estuário do Rio Potengi localiza-se na cidade de Natal, no Estado do
Rio Grande do Norte, situado na região nordeste do Brasil. Sua nascente está
localizada no município de Cerro Corá, localizada na microrregião da Serra de
Santana, no Seridó Potiguar, a 180 km da capital Natal e as suas águas banham
os municípios de Cerro Corá, São Tomé, Barcelona, São Paulo do Potengi, São
Pedro, Ielmo Marinho, São Gonçalo do Amarante e Natal, sendo que o Rio
Potengi é perene a partir de São Gonçalo do Amarante.
Neste estuário existe o Porto de Natal, onde ocorre intenso tráfego
marítimo de embarcações de diversos portes e, para que os maiores possam
adentrar no canal, são necessárias dragagens periódicas e o alargamento. Além
do Porto, o estuário do Rio Potengi é afetado pela carcinicultura, agricultura,
fábricas de vários setores e esgotos domésticos, tendo como principal receptor
destes últimos o Canal do Baldo (De Souza et al., 2010).
8.2.2 Aspectos fisiográficos
O "clima" predominante é do tipo "AS", quente e úmido, sendo a estação
seca do verão e chuvosa do inverno, segundo a classificação de Köppen
(RADAMBRASIL, 1981). A pequena precipitação pluviométrica é determinada

124
pelo Anticiclone do Atlântico Sul (localizado sobre o Oceano Atlântico), que
possui altas pressões e impõe a região, sob seu domínio, um tempo estável e sem
chuvas. Durante o período do fim do verão e todo o outono, este Anticiclone
perde força (nas regiões Norte e Nordeste do Brasil), proporcionando a ação da
Zona de Convergência Intertropical (ZCIT), responsável pelos ventos alísios
provenientes dos hemisférios Norte e Sul (Nimer, 1972).
As marés que ocorrem no estuário do Potengi e áreas adjacentes são de
natureza semi-diurna, com a variação média das marés de sizígias de cerca de
2.30m e das marés de quadratura, de cerca de 0.85 m e amplitude máxima em
torno de 2.83 m, caracterizando um regime de mesomaré. De acordo com a
Estação Climatológica da UFRN - Natal, os ventos mais frequentes são
predominantes do quadrante leste, com frequência Leste-Sudeste, com
velocidades médias mensais sempre em torno de 4 e 4.5 m/s.
8.2.3 Aspectos geológicos
A região de estudo localiza-se no domínio da Bacia Pernambuco –
Paraíba, mais especificamente na Sub – bacia da Paraíba, que é a mais
setentrional das bacias sedimentares da costa leste brasileira, situando-se entre os
paralelos 6º S e 9º S. Foi subdividida por Córdoba et al. (2007) em duas Sub –
bacias: Paraíba, a norte; e Pernambuco (também conhecida como Bacia do Cabo,
ao sul). De acordo com Lima Filho (2013), a região estudada situa-se
estruturalmente sobre a plataforma de Natal, que fica entre as Bacias Alto de
Touros e a Sub – bacia Canguaretama, com um total de 33.000 km2, sendo 9.000
km2 em sua porção emersa e 24.000 km
2 na sua porção imersa.
8.3 Materiais e métodos
Foram realizadas duas coletas em estações diferentes: a primeira ocorreu
em outubro de 2011, com a coleta de 18 estações e a segunda ocorreu em janeiro
de 2012, com a coleta de 24 estações (Figura 8.1).
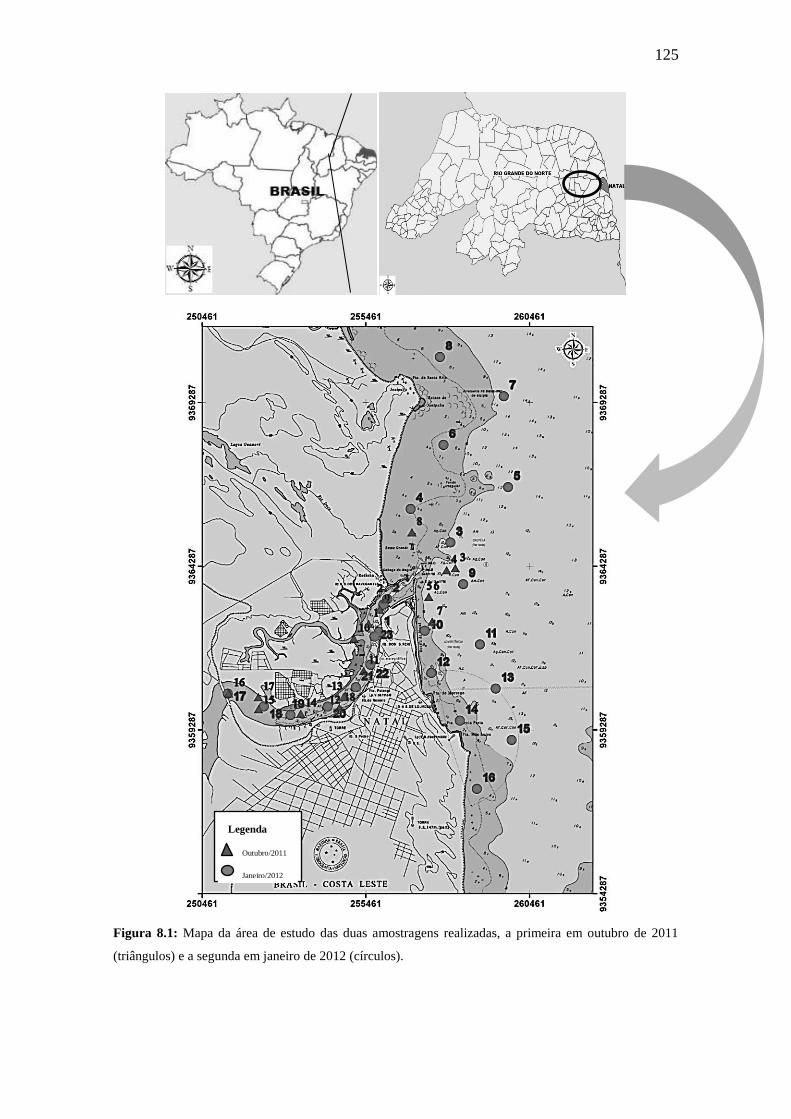
125
Figura 8.1: Mapa da área de estudo das duas amostragens realizadas, a primeira em outubro de 2011
(triângulos) e a segunda em janeiro de 2012 (círculos).
Legenda
Outubro/2011
Janeiro/2012

126
O objetivo da coleta em ambientes diferentes (rio e plataforma) é verificar
as diferenças de gradientes de salinidade, temperatura e profundidade. Foi
utilizado o amostrador pontual Van Veen e de cada amostra coletada foi retirada
uma fina camada superior, o primeiro centímetro, a qual foi colocada em frascos
com o corante Rosa de Bengala (1g/1000 ml de álcool), cuja função é corar o
protoplasma dos organismos que estiverem vivos, distinguindo-os dos mortos, no
momento da coleta. A função do álcool é impedir o ataque bacteriano. Para a
medida dos dados físicos (salinidade, condutividade, densidade, temperatura e
profundidade) foi utilizado o aparelho CTD (Conductivity, Temperature and
Depth) da marca Seabird.
8.3.1 Parâmetros biológicos
Para obtenção dos dados biológicos, as amostras foram peneiradas em via
úmida utilizando duas peneiras sucessivas de 0,500 e 0,062 mm e posteriormente
colocadas em papel filtro e secas na estufa à 60o C. Em seguida as amostras
foram tratadas com tricloroetileno (C2HCL3) dentro de uma capela. Nesse
processo, as carapaças dos foraminíferos sendo mais leves que o produto flotam
para a superfície e as frações mais pesadas depositam-se no fundo (Boltovskoy et
al. 1980). Após a separação, os foraminíferos foram transferidos com pincel para
lâminas especiais de fundo preto (triagem), para então serem analisados,
possibilitando a identificação das espécies, a qual foi feita utilizando-se
microscópio binocular de luz refletida.
Após identificadas as espécies, os organismos vivos e mortos foram
somados e feita a tabela de abundância total de foraminíferos. Essa somatória é a
frequência absoluta das espécies, a partir da qual foi calculada a porcentagem
relativa. A partir da frequência absoluta foram aplicadas as análises univariadas e
multivariadas com o Programa Primer 5 e 6, e a partir da porcentagem relativa
das espécies foram confeccionados mapas de contorno através do programa
Surfer 9. As espécies selecionadas para discussão dos resultados são as
dominantes na área de estudo (Figura 8.2).
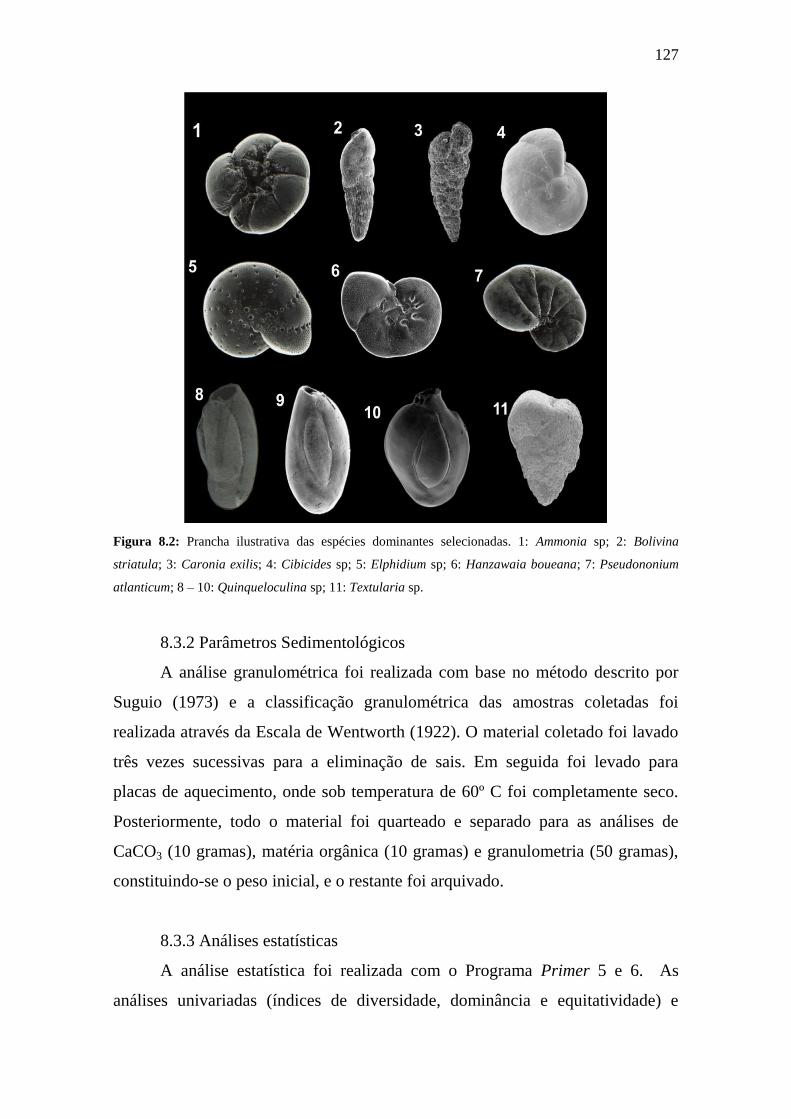
127
Figura 8.2: Prancha ilustrativa das espécies dominantes selecionadas. 1: Ammonia sp; 2: Bolivina
striatula; 3: Caronia exilis; 4: Cibicides sp; 5: Elphidium sp; 6: Hanzawaia boueana; 7: Pseudononium
atlanticum; 8 – 10: Quinqueloculina sp; 11: Textularia sp.
8.3.2 Parâmetros Sedimentológicos
A análise granulométrica foi realizada com base no método descrito por
Suguio (1973) e a classificação granulométrica das amostras coletadas foi
realizada através da Escala de Wentworth (1922). O material coletado foi lavado
três vezes sucessivas para a eliminação de sais. Em seguida foi levado para
placas de aquecimento, onde sob temperatura de 60º C foi completamente seco.
Posteriormente, todo o material foi quarteado e separado para as análises de
CaCO3 (10 gramas), matéria orgânica (10 gramas) e granulometria (50 gramas),
constituindo-se o peso inicial, e o restante foi arquivado.
8.3.3 Análises estatísticas
A análise estatística foi realizada com o Programa Primer 5 e 6. As
análises univariadas (índices de diversidade, dominância e equitatividade) e

128
multivariadas (PCA e CLUSTER) foram baseadas na matriz de dados de
frequência absoluta de foraminíferos e de dados ambientais (profundidade,
temperatura, salinidade, porcentagens de areia, silte, argila, carbono orgânico e
enxofre). A matriz de dados ambientais foi padronizada e submetida à análise de
PCA para identificar tendências ambientais. Nos dados biológicos foram
aplicadas as análises de CLUSTER. Para a confecção dos mapas de
georeferenciamento foi utilizado o programa Surfer 9, onde a matriz de dados
bióticos e granulométricos foram submetidos para contornar os valores de
abundância.
8. 4 – Resultados
A coleta de Outubro de 2011 foi marcada por um período baixo de chuvas,
onde as águas tornaram-se menos diluídas e, consequentemente, mais salinas do
que seriam em outras épocas do ano e menor distribuição sedimentar (figura
5.3A). Janeiro de 2012 foi marcada por um período com mais chuva, onde as
águas tornaram-se mais diluídas e menos salinas do que seriam em outras épocas
do ano e uma maior distribuição sedimentar (Figura 8.3B).
Figura 8.3: Dados pluviométricos em Outubro de 2011 (A) e em Janeiro de 2012 (B).
Os parâmetros abióticos (profundidade, temperatura, salinidade,
densidade, condutividade e CaCO3) de Outubro de 2011 podem ser visualizados
na figura 4 e a porcentagem das frações granulométricas na figura 5; e para
A B

129
Janeiro de 2012 os parâmetros abióticos (profundidade e CaCO3) podem ser
visualizados na figura 6 e a porcentagem das frações granulométricas na figura 7,
formando a base de dados abióticos. De acordo com a tabela de parâmetros
abióticos (Tabela 8.1), as estações coletadas em trechos mais profundos
apresentam temperaturas mais baixas, enquanto que as estações coletadas em
trechos mais rasos apresentaram temperaturas altas. As estações 2, 9 e 10
provavelmente são os locais que sofrem processos erosivos, como a dragagem.
A salinidade é mais alta na foz ou nas estações próximas a esta e mais
baixa nas estações localizadas no curso superior e na região central, com exceção
da estação 12 que apresentou salinidade alta, que se localiza próximo às salinas.
O CaCO3 apresentou maiores leituras nas estações próximas à Marina Iate clube
e no curso superior, mais baixas e inexistentes nas estações próximas à foz, na
foz e em alguns trechos do rio. Os parâmetros de densidade e condutividade são
mais altos na foz e próximas a ela e mais baixas no curso superior do Rio
Potengi.
Tabela 8.1: Parâmetros abióticos da coleta de Outubro de 2011.
Estações Profundidade (m) Temperatura (oC) Salinidade Densidade Condutividade CaCO3 (%)
1 8,3 28,0 35,56 22,86 57,01 5,00
2 12,6 27,8 36,02 23,28 57,46 0,82
3 13,3 27,8 36,28 23,48 57,82 0,00
4 10,8 27,8 36,34 23,52 57,89 0,00
5 13,7 27,8 36,42 23,59 58,03 0,00
6 12,2 27,8 36,11 23,34 57,63 3,31
7 11,4 27,9 36,18 23,38 57,75 1,60
8 5,6 27,9 36,24 23,39 57,86 2,50
9 9,1 27,9 36,27 23,42 57,92 0,00
10 14,2 27,9 36,23 23,41 57,87 0,83
11 3,5 28,1 36,14 23,23 57,95 0,83
12 6,9 27,9 36,31 23,44 58,02 1,67
13 6,5 27,9 36,15 23,31 57,79 0,00
14 4,4 28,2 35,50 22,72 57,16 1,63
15 4,9 28,3 35,22 22,49 56,84 5,79
16 9,6 28,3 35,02 22,34 56,60 4,07
17 7,5 28,2 35,03 22,39 56,48 7,50
18 8,9 28,0 35,94 23,14 57,54 0,83
Parâmetros abióticos da coleta em Outubro 2011

130
De acordo com a tabela de granulometria (Tabela 8.2) o cascalho
apresentou maiores frequências nas estações localizadas na foz e também em
alguns trechos do canal do Rio Potengi, mas a fração arenosa foi dominante,
principalmente areia grossa e areia média, que correspondem a ambientes de alta
hidrodinâmica, como o canal do rio Potengi e na foz respectivamente. As frações
de silte e argila apresentam maiores valores no curso superior do canal e baixa
frequência no canal e na foz do Rio Potengi.
Tabela 8.2: Análise granulométrica por estação de Outubro de 2011.
A análise de PCA aplicada nos dados abióticos de Janeiro de 2012
(Figuras 8.3 e 8.4) apresentam valores similares de PC1 e PC2 de Outubro de
2011 (Figura 8.4). Os locais mais profundos se encontram na plataforma interna,
mas também existem alguns trechos profundos no Rio Potengi, provavelmente
pelas dragagens periódicas.
Atraves da tabela dos dados abiotciso de Janeiro de 2012 (Tabela 5.3),
observou-se que as estações próximas à região costeira e a região norte possuem
baixa profundidade, já na região sul ocorre variação de profundidade entre as
amostras, provavelmente por ações erosivas que podem ser naturais
(hidrodinâmica marinha) ou antrópicas (dragagem), localizadas na Praia dos
Artistas, Ponta Negra e Barreira do Inferno.
Estações Cascalho (%) Areia muito grossa (%) Areia grossa (%) Areia média (%) Areia fina (%) Areia muito fina (%) Silte (%) Argila (%)
1 0,0 0,0 14,05 39,80 31,44 7,67 6,02 1,02
2 7,56 3,75 67,78 15,79 3,31 1,02 0,38 0,41
3 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
4 0,43 0,0 46,57 43,99 5,75 1,98 0,67 0,62
5 7,38 16,48 58,07 13,40 3,17 0,99 0,19 0,32
6 2,60 0,00 43,93 42,79 7,15 2,07 0,77 0,70
7 4,90 0,00 46,96 38,60 6,29 1,88 0,63 0,74
8 0,0 0,0 23,41 68,20 4,60 2,37 0,72 0,70
9 0,0 0,0 44,56 45,02 6,65 2,19 0,93 0,65
10 0,0 0,0 24,10 50,69 18,57 3,68 2,16 0,80
11 0,15 19,08 59,71 15,41 3,66 1,15 0,45 0,39
12 6,59 14,74 61,63 12,59 2,44 1,05 0,62 0,35
13 0,0 0,0 11,90 41,75 33,12 7,00 5,40 0,83
14 11,31 14,57 41,30 25,40 3,96 1,80 1,28 0,38
15 0,0 0,0 0,05 12,25 39,35 19,26 25,89 3,20
16 0,0 0,0 16,32 40,05 26,75 7,89 7,60 1,39
17 17,50 36,99 24,78 13,62 2,37 1,03 3,18 0,54
18 7,05 20,70 38,78 26,13 4,14 1,25 1,50 0,44
Porcentagem granulométrica da coleta em Outubro 2011

131
Se compararmos a profundidade entre as estações coletadas nos pontos em
comum (Outubro de 2011: estações 2, 8, 7, 16, 15, 18 e 9; Janeiro de 2012: 2, 4,
10, 17, 18, 20 e 24, respectivamente), pôde-se verificar que houve aumento na
profundidade na estação 24, próximo à desembocadura, enquanto que houve
diminuição na plataforma interna e na região central do Rio Potengi,
respectivamente. O CaCO3 possui maiores frequências na plataforma interna e
mais baixas no canal do Rio Potengi. As estações com baixas frequências são
aquelas que se encontram ao longo do canal do Rio Potengi. Se compararmos os
dados de CaCO3 entre as duas amostragens nas estações em comum, pode-se
verificar que houve aumentos sigificativos desse parâmetro na regiao costeira sul.
A menor frequência de ganho foi na estação próxima à foz. Houve diminuição de
CaCO3 nas estações localizadas no canal central e na região costeira norte,
respectivamente.
Tabela 8.3: Parâmetros abióticos da coleta de Janeiro de 2012.
Estações Profundidade (m) CaCO3 (%)
1 10.7 2,61
2 13 2,17
3 11 15,64
4 5.1 1,83
5 13 22,13
6 5 84,37
7 8 78,15
8 9 93,03
9 8 61,36
10 9 16,30
11 12.5 15,61
12 10 1,72
13 11 45,98
14 12 22,45
15 13.8 70,08
16 11 21,69
17 9 5,77
18 5.7 2,49
19 4.7 9,77
20 5.6 6,60
21 5.9 7,40
22 13.9 5,52
23 13 3,08
24 13.7 2,30
Parâmetros abióticos da coleta em Janeiro 2012

132
De acordo com a tabela de granulometria (Tabela 8.4) verificou-se
aumento das porcentagens das frações arenosas, e a frequência de cascalho foi
muito baixa comparada à campanha de Outubro de 2011 e a maior frequência
localiza-se na plataforma interna. A fração dominante foi a arenosa,
principalmente areia grossa, areia fina e areia muito fina, localizadas,
respectivamente na região costeira, curso superior e região costeira sul. As
frações de areia muito grossa e areia média também foram altas.
Silte e argila obtiveram frequências altas nas estações 1 e 14, onde a
estação 1 está localizada no canal do Rio Potengi próximo à Marina Iate Clube e
a estação 14 localiza-se na região costeira sul e ao longo do canal; porém na foz e
na plataforma interna possuem baixas frequências em alguns desses trechos e em
outros essa fração é inexistente.
Tabela 8.4: Análise granulométrica por estação de janeiro de 2012.
Estações Cascalho (%) Areia muito grossa (%) Areia grossa (%) Areia média (%) Areia fina (%) Areia muito fina (%) Silte + Argila (%)
1 0,00 0,04 1,13 6,10 34,59 36,91 21,22
2 0,00 3,65 6,63 34,02 48,98 5,98 0,73
3 0,07 4,92 25,84 50,34 14,46 4,29 0,07
4 0,00 0,39 14,39 13,90 67,75 3,53 0,04
5 0,27 25,35 35,31 7,79 22,32 8,74 0,23
6 0,00 9,91 41,61 16,15 30,32 1,94 0,08
7 0,04 6,94 51,71 15,11 25,54 0,63 0,02
8 0,00 0,12 16,20 16,13 59,83 7,63 0,09
9 0,00 1,40 10,29 71,30 14,90 1,95 0,16
10 0,00 0,36 19,17 10,01 67,73 1,98 0,75
11 0,00 1,30 42,23 48,46 6,67 1,24 0,10
12 0,00 1,79 75,63 22,56 0,02 0,00 0,00
13 0,18 52,07 30,55 13,55 3,29 0,33 0,03
14 0,00 0,18 0,26 1,48 9,26 74,64 14,20
15 0,03 2,04 11,18 53,83 30,81 1,98 0,13
16 0,00 0,95 13,75 53,08 26,80 3,61 1,81
17 0,00 3,49 12,91 10,33 71,82 1,20 0,26
18 0,00 1,07 15,36 52,18 30,69 0,64 0,06
19 0,02 0,99 4,69 61,00 32,67 0,47 0,15
20 0,00 0,44 1,94 54,26 42,05 1,19 0,13
21 0,06 4,69 5,72 13,00 53,06 20,20 3,26
22 0,00 3,26 10,12 43,30 34,20 4,52 4,60
23 0,00 0,15 0,57 1,67 45,14 47,93 4,55
24 0,00 3,11 13,21 53,00 27,80 2,70 0,17
Porcentagem granulométrica da coleta em Janeiro 2012

133
A análise de componente principal (PCA) que foi aplicada nos dados
abióticos de Outubro de 2011 revelou que a somatória do PC1 e PC2 explicam
74,6% da variação cumulativa das amostras (Tabela 8.5).
Tabela 8.5: Quadro gerado pelo PCA, com destaque para a variação cumulativa encontrada na somatória
do PC1 e PC2.
Os parâmetros abióticos (profundidade, temperatura, salinidade,) e os
valores de PC1 e PC2 (figura 8.9) que determinaram a formação de quatro
grupos distintos são apresentados na tabela 8.6.
Tabela 8.6: Valores do PC1 e PC2 para os parâmetros abióticos usados no PCA.
Observou-se que no gráfico da figura 8.4, o grupo P1 é semelhante com
relação à temperatura, teor de CaCO3 e de areia muito grossa sendo formado
pelas estações 14 e 17, localizadas no estuário. Já o grupo P2, formado pelas
estações 2, 5, 11, 12 e 18, apresentam porcentagens similares e elevadas de
cascalho e areia grossa, correspondentes à ambientes com alta hidrodinâmica. O
grupo P3 se caracteriza por porcentagens elevadas de areia fina, areia muito fina,
PC Autovalor % Variação % Variação Cumulativa
1 7.01 50.1 50.1
2 3.44 24.6 74.6
3 1.03 7.4 82.0
4 0.99 7.0 89.0
5 0.56 4.0 93.1
Variáveis PC1 PC2
Profundidade 0.177 -0.104
Temperatura -0.320 0.171
Salinidade 0.337 -0.198
Densidade 0.340 -0.198
Condutividade 0.322 -0.196
CaCO3 (%) -0.301 0.165
Cascalho (%) 0.003 0.490
Areia muito grossa (%) -0.004 0.491
Areia grossa (%) 0.237 0.197
Areia média (%) 0.026 -0.278
Areia fina (%) -0.280 -0.296
Areia muito fina (%) -0.319 -0.246
Silte (%) -0.329 -0.158
Argila (%) -0.314 -0.216

134
silte e argila, sendo formado pelas estações 1, 13, 15 e 16, sendo as estações 1 e
13 caracterizadas por ambientes de baixa hidrodinâmica e as estações 15 e 16
com alta hidrodinâmica. O grupo P4 é composto pelas estações 3, 4, 6, 7, 8, 9 e
10 e apresenta como principal característica os valores elevados de salinidade,
condutividade, densidade e porcentagem de areia média.
Figura 8.4: Gráfico com os agrupamentos das estações conforme os valores do PC1 e PC2, além da
disposição dos parâmetros abióticos em relação aos quadrantes da primeira coleta.
A análise de PCA aplicada nos dados abióticos de Janeiro de 2012
(Tabelas 8.3 e 8.4) apresentam valores similares de PC1 e PC2 de Outubro de
2011 (Tabelas 8.1 e 8.2). De acordo com o PCA (Figura 8.5), as estações 1, 14 e
23 são influenciadas pela granulometria, sendo que a 14 é influenciada pela areia
muito fina e a 1 e 23 são influenciadas pelo silte e argila. As estações 4, 10, 17,
21 e 22 são influenciadas pela areia fina, enquanto que 7, 8 e 20 são
influenciadas pelos parâmetros de carbono orgânico e nitrogênio, que indicam
ambientes com altas concentrações de matéria orgânica.
As estações 6, 9 e 15 são influenciadas pelo CaCO3, que provavelmente
foram coletadas próximas às áreas de recifes de corais. As estações 3, 11, 12, 13,
16, 18 e 19 são influenciadas por areia média e grossa, correspondentes à
ambientes de alta hidrodinâmica, sendo que os pontos 18 e 19 estão localizados

135
no Rio Potengi. E as estações 2 e 5 são influenciadas pela profundidade, sendo a
estação 2 localizada próxima à foz do Rio Potengi e a estação 5 localizada na
plataforma.
Figura 8.5: Dados abióticos de PCA de Janeiro de 2012.
A análise de CLUSTER para as estações (Modo Q) baseada nos dados
biológicos para a primeira coleta, gerou um dendograma com cinco grupos
distintos (Figura 5.6) e nível de similaridade de 45%. Analisando a disposição
das amostras nesse agrupamento e com base no posicionamento espacial delas
(Figura 5.1), podem-se constatar dois ambientes distintos, em que um está
relacionado à região da foz do Rio Potengi e à plataforma interna, representado
pelas estações de 1 a 11 dentro dos grupos C1, C2 E C3; e outro ambiente
corresponde à região mais interna do estuário, representado pelas estações 12 a
18, as quais são constituintes dos grupos C4 e C5.
O grupo C1 é constituído pelas estações 7 e 8, que apresentam como
principal característica dominância de Ammonia tepida, Quinqueloculina
lamarckiana e, em menor frequência, Textularia earlandi. Essas estações
também apresentaram valores similares quanto à temperatura, salinidade,
densidade e condutividade. O grupo C2 apresenta menor homogeneidade dos

136
grupos, podendo até mesmo dividi-lo em dois subgrupos: um formado pelas
estações 1, 3 e 10, cujas espécies dominantes são basicamente Ammonia tepida e
a Quinqueloculina lamarckiana; enquanto o outro subgrupo é composto pelas
estações 2 e 11, com A. tepida e Q. miletti.
Já o Grupo C3, formado pelas estações 4, 5, 6 e 9, foi composto por A.
tepida, Q. lamarckiana e Q. patagonica. Todas as estações apresentaram valores
similares de temperatura, salinidade, densidade e condutividade e teores nulos de
carbonato, com exceção da estação 6 que apresentou um teor de CaCO3 mediano
e valores menores para os demais parâmetros. Já na parte mais interna do Rio
Potengi há o grupo C4 formado pelas estações 12, 17 e 18 com baixa variedade
de espécies, apenas 4, com predominância de Ammonia tepida, Bolivina striatula
e Quinqueloculina patagonica. As estações 12 e 18 apresentaram valor mediano
e similar de temperatura, já a estação 17 apresentou valor mais elevado. Também
nessa região do rio, há o grupo C5 composto pelas estações 13 a 16, as quais não
apresentaram boa similaridade quanto aos parâmetros abióticos. Esse grupo
apresenta as seguintes espécies como dominantes: Ammonia tepida, Bolivina
striatula e Quinqueloculina miletti, com exceção da estação 14 que apresentou a
Caronia exilis como dominante.
Figura 8.6: Dendograma mostrando o agrupamento das estações conforme a similaridade das espécies de
foraminíferos encontradas entre elas da primeira coleta.
A análise de CLUSTER (figura 8.7) para Janeiro de 2012 revelou a
formação de três grupos de acordo com as espécies de foraminíferos encontradas:

137
Plataforma Interna (5 – 16), Canal do Rio Potengi (17 – 24) e Foz do Rio Potengi
(1 – 4) (figura 8.13). A espécie Ammonia tepida dominou os três grupos
formados no CLUSTER, principalmente no Canal do Rio Potengi.
Em menores porcentagens a espécie Bolivina striatula dominou nos três
grupos, especialmente no Canal do Rio Potengi. Cibicides fletcheri também
apareceu em maiores quantidades na Foz do Rio Potengi. Hanzawaia boueana,
Elphidium discoidale, Cibicides valvulatanus, C. variabilis e Discorbis
peruvianus aparecem com maior frequência na plataforma interna.
Quinqueloculina lamarckiana dominou os três grupos, mas em grande
quantidade na plataforma interna e em menor porcentagem na Foz do Rio
Potengi.
Quinqueloculina patagonica e Cassidulina subglobosa aparecem com
menor densidade no Canal do Rio Potengi. Quinqueloculina miletti apresenta
frequências mais altas na Foz do Rio Potengi. Pseudononion atlanticum,
Textularia earlandi, T. gramen e Trochamina discorbis aparecem na Foz do Rio
Potengi e na plataforma interna. A ocorrência de algumas espécies de
foraminíferos tolerantes à baixa salinidade – típicos de ambientes de manguezais
como Trochamina inflata e T. squamata no canal do Rio Potengi – sugere que o
contribuinte fluvial é capaz de exportar organismos de água doce preferivelmente
em direção sul do que em direção norte. Esses organismos provavelmente devem
ter sido transportados do manguezal próximo à foz do Rio Potengi para as
regiões de plataforma interna.
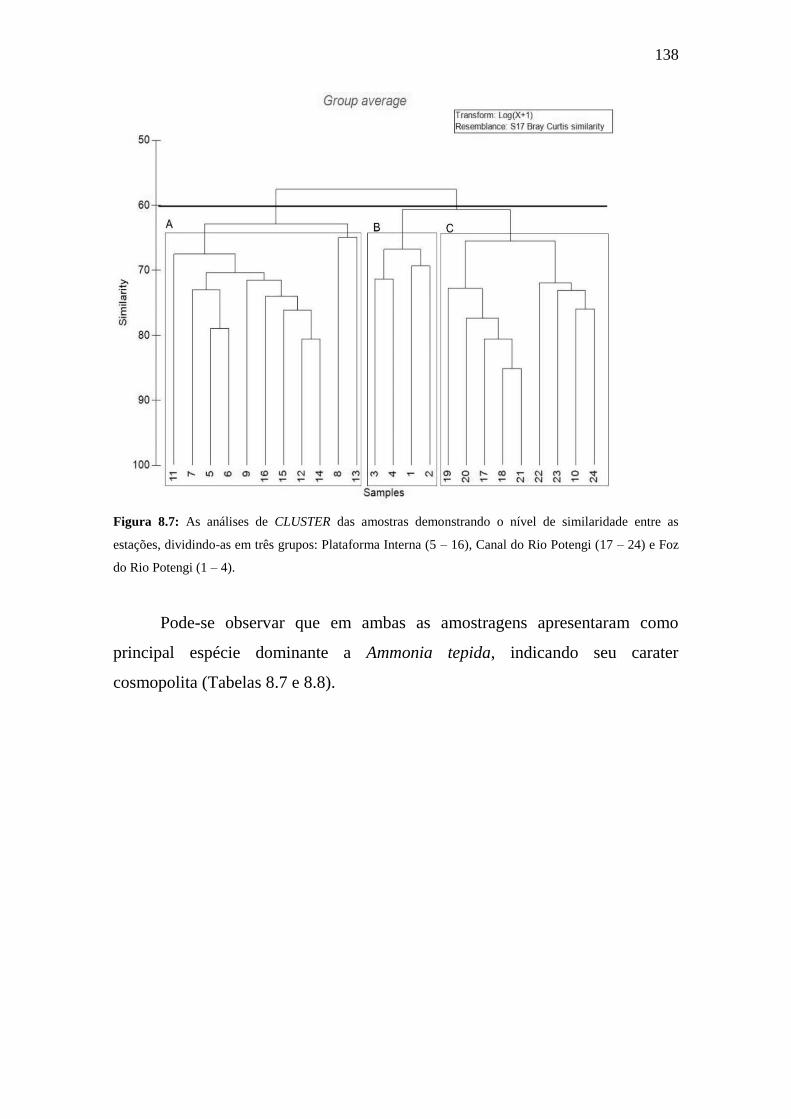
138
Figura 8.7: As análises de CLUSTER das amostras demonstrando o nível de similaridade entre as
estações, dividindo-as em três grupos: Plataforma Interna (5 – 16), Canal do Rio Potengi (17 – 24) e Foz
do Rio Potengi (1 – 4).
Pode-se observar que em ambas as amostragens apresentaram como
principal espécie dominante a Ammonia tepida, indicando seu carater
cosmopolita (Tabelas 8.7 e 8.8).
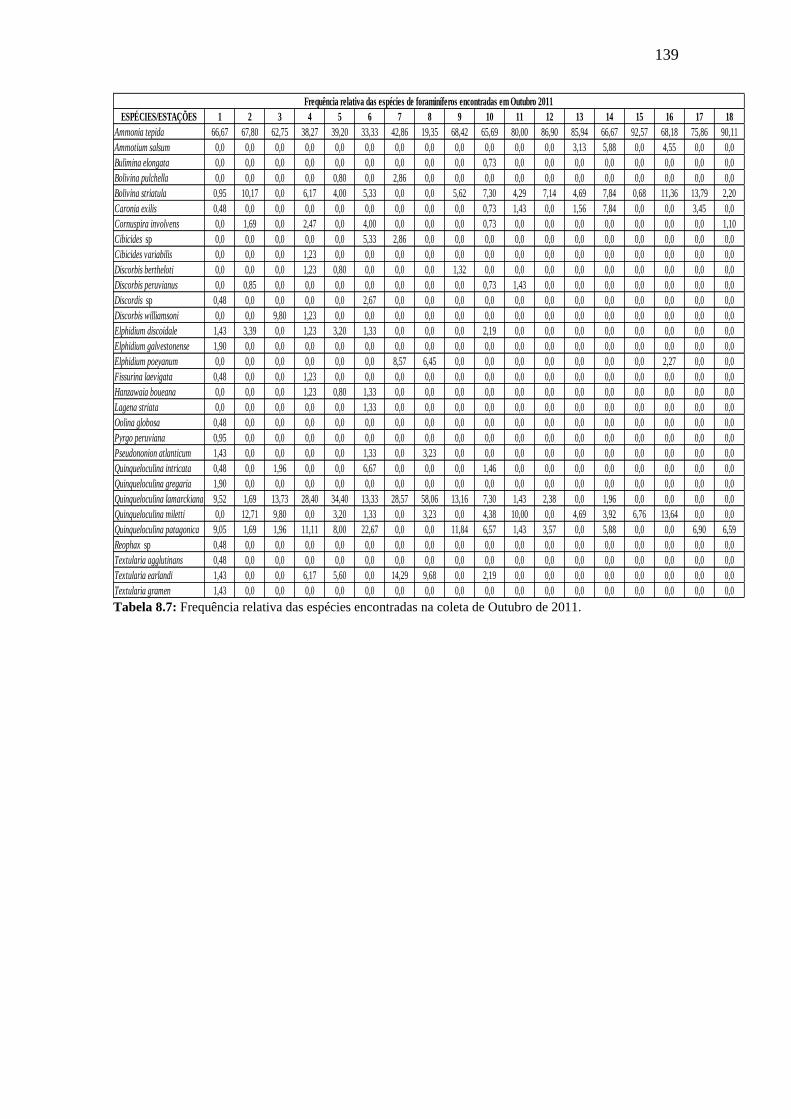
139
Tabela 8.7: Frequência relativa das espécies encontradas na coleta de Outubro de 2011.
ESPÉCIES/ESTAÇÕES 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18
Ammonia tepida 66,67 67,80 62,75 38,27 39,20 33,33 42,86 19,35 68,42 65,69 80,00 86,90 85,94 66,67 92,57 68,18 75,86 90,11
Ammotium salsum 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 3,13 5,88 0,0 4,55 0,0 0,0
Bulimina elongata 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,73 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Bolivina pulchella 0,0 0,0 0,0 0,0 0,80 0,0 2,86 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Bolivina striatula 0,95 10,17 0,0 6,17 4,00 5,33 0,0 0,0 5,62 7,30 4,29 7,14 4,69 7,84 0,68 11,36 13,79 2,20
Caronia exilis 0,48 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,73 1,43 0,0 1,56 7,84 0,0 0,0 3,45 0,0
Cornuspira involvens 0,0 1,69 0,0 2,47 0,0 4,00 0,0 0,0 0,0 0,73 0,0 0,0 0,0 0,0 0,0 0,0 0,0 1,10
Cibicides sp 0,0 0,0 0,0 0,0 0,0 5,33 2,86 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Cibicides variabilis 0,0 0,0 0,0 1,23 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Discorbis bertheloti 0,0 0,0 0,0 1,23 0,80 0,0 0,0 0,0 1,32 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Discorbis peruvianus 0,0 0,85 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,73 1,43 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Discordis sp 0,48 0,0 0,0 0,0 0,0 2,67 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Discorbis williamsoni 0,0 0,0 9,80 1,23 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Elphidium discoidale 1,43 3,39 0,0 1,23 3,20 1,33 0,0 0,0 0,0 2,19 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Elphidium galvestonense 1,90 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Elphidium poeyanum 0,0 0,0 0,0 0,0 0,0 0,0 8,57 6,45 0,0 0,0 0,0 0,0 0,0 0,0 0,0 2,27 0,0 0,0
Fissurina laevigata 0,48 0,0 0,0 1,23 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Hanzawaia boueana 0,0 0,0 0,0 1,23 0,80 1,33 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Lagena striata 0,0 0,0 0,0 0,0 0,0 1,33 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Oolina globosa 0,48 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Pyrgo peruviana 0,95 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Pseudononion atlanticum 1,43 0,0 0,0 0,0 0,0 1,33 0,0 3,23 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Quinqueloculina intricata 0,48 0,0 1,96 0,0 0,0 6,67 0,0 0,0 0,0 1,46 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Quinqueloculina gregaria 1,90 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Quinqueloculina lamarckiana 9,52 1,69 13,73 28,40 34,40 13,33 28,57 58,06 13,16 7,30 1,43 2,38 0,0 1,96 0,0 0,0 0,0 0,0
Quinqueloculina miletti 0,0 12,71 9,80 0,0 3,20 1,33 0,0 3,23 0,0 4,38 10,00 0,0 4,69 3,92 6,76 13,64 0,0 0,0
Quinqueloculina patagonica 9,05 1,69 1,96 11,11 8,00 22,67 0,0 0,0 11,84 6,57 1,43 3,57 0,0 5,88 0,0 0,0 6,90 6,59
Reophax sp 0,48 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Textularia agglutinans 0,48 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Textularia earlandi 1,43 0,0 0,0 6,17 5,60 0,0 14,29 9,68 0,0 2,19 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Textularia gramen 1,43 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0 0,0
Frequência relativa das espécies de foraminíferos encontradas em Outubro 2011

140
Tabela 8.8: Frequência relativa das espécies encontradas na coleta de janeiro de 2012.
Esp
écie
s\E
staç
ões
12
34
56
78
910
1112
1314
1516
1718
1920
2122
2324
Am
mon
ia te
pida
47,1
65,9
41,1
42,2
37,4
51,4
32,5
34,5
45,0
46,8
17,1
46,6
13,0
45,7
39,3
48,2
91,4
81,6
89,6
75,5
81,4
50,4
65,2
64,6
Am
phic
oryn
a sc
alar
is0,
00,
00,
00,
00,
20,
00,
00,
00,
00,
00,
80,
00,
00,
00,
00,
30,
00,
00,
00,
00,
00,
00,
00,
0
Boliv
ina
stri
atul
a6,
36,
74,
46,
01,
81,
41,
20,
03,
65,
43,
42,
02,
73,
80,
72,
04,
010
,51,
412
,22,
97,
01,
05,
6
Bucc
ella
per
uvia
na0,
00,
01,
90,
01,
61,
42,
41,
20,
40,
90,
00,
82,
21,
00,
70,
00,
00,
00,
50,
00,
00,
00,
00,
0
Bulim
ina
mar
gina
ta0,
00,
00,
00,
90,
20,
70,
00,
00,
40,
00,
80,
00,
00,
10,
00,
20,
00,
00,
00,
00,
00,
00,
00,
0
Bulim
inel
la e
lega
ntis
sim
a0,
80,
01,
90,
00,
20,
70,
00,
00,
00,
90,
00,
00,
00,
00,
00,
00,
00,
00,
00,
00,
00,
00,
80,
0
Cas
sidu
lina
subg
lobo
sa1,
30,
00,
60,
90,
20,
31,
20,
00,
80,
00,
00,
00,
00,
00,
00,
20,
02,
60,
00,
02,
00,
80,
50,
0
Cib
icid
es fl
etch
eri
2,9
4,4
1,3
2,6
0,2
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,7
2,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
Cib
icid
es v
alvu
lana
tus
0,0
0,0
0,0
0,0
1,6
0,7
2,4
0,0
0,0
0,0
1,5
0,0
0,5
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
Cib
icid
es v
aria
bilis
0,0
0,7
0,0
0,0
0,6
0,0
0,0
1,2
0,4
0,0
0,4
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
Cyc
logi
ra in
volv
ens
0,0
0,0
1,9
0,0
0,2
0,7
0,0
0,0
0,0
0,9
0,0
0,0
0,0
0,4
0,0
0,0
0,0
0,0
0,0
0,0
0,0
1,2
0,0
0,5
Dis
corb
is fl
orda
na0,
00,
70,
00,
00,
00,
00,
00,
00,
40,
00,
00,
00,
50,
30,
70,
60,
00,
00,
00,
70,
00,
00,
50,
0
Dis
corb
is p
eruv
ianu
s0,
00,
00,
00,
00,
00,
31,
20,
01,
60,
00,
00,
40,
00,
00,
00,
00,
00,
00,
00,
00,
00,
00,
00,
0
Dis
corb
is v
alvu
latu
s0,
00,
00,
00,
00,
00,
00,
01,
20,
00,
03,
00,
42,
70,
00,
00,
90,
00,
00,
00,
00,
01,
20,
00,
5
Dis
corb
is w
illia
mso
ni0,
00,
71,
31,
71,
41,
00,
00,
00,
00,
00,
00,
80,
50,
41,
30,
00,
00,
00,
00,
70,
00,
00,
00,
5
Elp
hidi
um a
rtic
ulat
um1,
30,
00,
00,
00,
00,
00,
00,
00,
00,
00,
40,
01,
10,
30,
70,
00,
00,
00,
00,
00,
00,
00,
00,
0
Elp
hidi
um d
isco
idal
e0,
00,
00,
03,
40,
80,
71,
20,
01,
60,
91,
91,
20,
00,
91,
30,
80,
60,
00,
50,
70,
017
,12,
90,
8
Fis
suri
na la
evig
ata
2,1
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,4
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,3
0,0
Fis
suri
na lu
cida
0,0
0,0
0,6
0,0
0,2
0,0
1,2
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
Fora
min
ífero
ben
tôni
co jo
vem
0,0
0,7
0,0
0,0
0,2
0,0
0,0
0,0
0,0
0,0
0,0
0,0
1,1
0,0
0,7
0,6
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
Gau
dryi
na e
xilis
0,0
0,0
0,6
0,0
0,6
0,0
0,0
0,0
0,0
0,0
0,8
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,3
Han
zaw
aia
boue
ana
0,0
0,0
0,0
0,9
0,0
0,0
3,6
0,0
2,8
1,8
1,1
1,6
0,0
0,3
0,7
0,8
0,0
0,0
0,5
0,0
0,0
0,0
0,0
0,3
Hop
kins
ina
paci
fica
0,0
0,0
1,3
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,4
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
Lage
na s
tria
ta0,
00,
00,
00,
00,
00,
00,
00,
00,
00,
00,
00,
40,
00,
30,
00,
00,
00,
00,
00,
00,
00,
40,
00,
3
Lent
icul
ina
sp0,
00,
00,
00,
00,
00,
00,
00,
00,
00,
00,
40,
01,
10,
00,
00,
20,
00,
00,
00,
00,
00,
00,
00,
0
Mill
iolin
ella
sub
rotu
nda
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
2,0
0,5
0,4
0,7
0,2
0,0
0,0
0,0
0,0
0,0
0,0
0,5
0,3
Ort
hom
orph
ina
sp0,
00,
00,
60,
00,
00,
00,
00,
00,
00,
00,
00,
00,
00,
10,
00,
00,
00,
00,
00,
00,
00,
00,
00,
0
Por
oepo
nide
s la
tera
lis0,
00,
00,
00,
00,
00,
00,
06,
00,
00,
01,
10,
00,
50,
00,
00,
60,
00,
00,
00,
00,
00,
00,
00,
0
Pse
udon
oniu
m a
tlant
icum
0,0
2,2
1,9
0,9
2,0
1,4
1,2
2,4
0,0
0,9
0,8
1,6
0,0
1,0
0,7
0,2
0,0
0,0
0,5
0,0
0,0
0,0
0,5
0,5
Pyr
go n
asut
a0,
00,
00,
00,
00,
40,
00,
01,
20,
80,
00,
00,
02,
70,
12,
00,
30,
00,
40,
00,
00,
00,
00,
00,
0
Qui
nque
locu
lina
intr
icat
a0,
80,
00,
00,
00,
40,
34,
81,
20,
80,
92,
70,
00,
50,
61,
30,
20,
00,
00,
00,
70,
00,
02,
10,
0
Qui
nque
locu
lina
lam
arck
iana
8,8
4,4
18,4
21,6
22,3
16,1
19,3
33,3
14,3
18,0
27,8
18,2
40,8
20,2
21,3
29,4
1,1
2,6
5,2
7,2
4,9
9,3
13,2
13,2
Qui
nque
locu
lina
mill
etti
5,4
3,0
1,9
6,0
0,8
0,3
0,0
0,0
0,0
0,0
0,8
0,4
0,0
0,4
0,0
0,6
0,0
0,0
0,0
0,0
0,0
0,4
0,3
2,0
Qui
nque
locu
lina
pata
goni
ca22
,59,
616
,57,
87,
018
,99,
66,
021
,922
,511
,017
,83,
816
,912
,09,
02,
92,
20,
92,
28,
811
,611
,710
,1
Text
ular
ia e
arla
ndi
0,0
0,7
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,8
2,0
7,6
1,0
1,3
0,8
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
Text
ular
ia g
ram
en0,
00,
02,
53,
419
,23,
518
,111
,93,
60,
023
,22,
417
,94,
912
,73,
50,
00,
00,
90,
00,
00,
00,
30,
0
Troc
ham
min
a di
scor
bis
0,0
0,0
0,6
0,9
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,3
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
Troc
ham
min
a in
flata
0,4
0,0
0,6
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,3
0,0
Troc
ham
min
a sq
uam
ata
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,8
0,0
0,0
Uvi
geri
na p
ereg
rina
0,4
0,0
0,0
0,9
0,8
0,0
0,0
0,0
1,2
0,0
0,4
0,8
0,0
0,0
0,0
0,2
0,0
0,0
0,0
0,0
0,0
0,0
0,0
0,8
Freq
uênc
ia r
elat
iva
das
espé
cies
de
fora
min
ífero
s en
cont
rada
s em
Jan
eiro
201
2

141
Os dados obtidos da tabela relativa (Tabela 8.8) para a comparação de
gêneros de foraminíferos (Figuras 8.8 a 8.12) e porcentagem relativa da
granulometria (Figuras 8.13 e 8.19) encontrada entre as duas coletas serão
descritos a seguir.
Em Outubro de 2011, verificou-se maior abundância do gênero Ammonia
sp. (75 – 95%) próxima às fazendas de carcinicultura, às salinas, à Estação de
Tratamento de Água de Quintas e ao Canal do Baldo. A menor frequência ocorre
na foz com 15%. Em Janeiro de 2012, observa-se que, além do pequeno
decréscimo de sua frequência (85% de frequência), houve aumento da área de
domínio. As menores frequências estão localizadas na plataforma interna, com
10% (Figura 8.8).
Figura 8.8: Abundância relativa do gênero Ammonia sp. em Outubro de 2011 e em Janeiro de 2012.
Em Outubro de 2011, Bolivina sp. foi encontrada no canal superior (14%
de frequência) próxima às fazendas de carcinicultura, entretanto na foz apresenta
menor frequência (1%). Na amostragem de Janeiro de 2012, houve decréscimo
de sua frequência próximo à área do Esgoto do Baldo (12%) e às fazendas de
carcinicultura (8%). A menor frequência encontrada está na plataforma interna
norte (1%) e desaparece na plataforma interna sul (Figura 8.9).
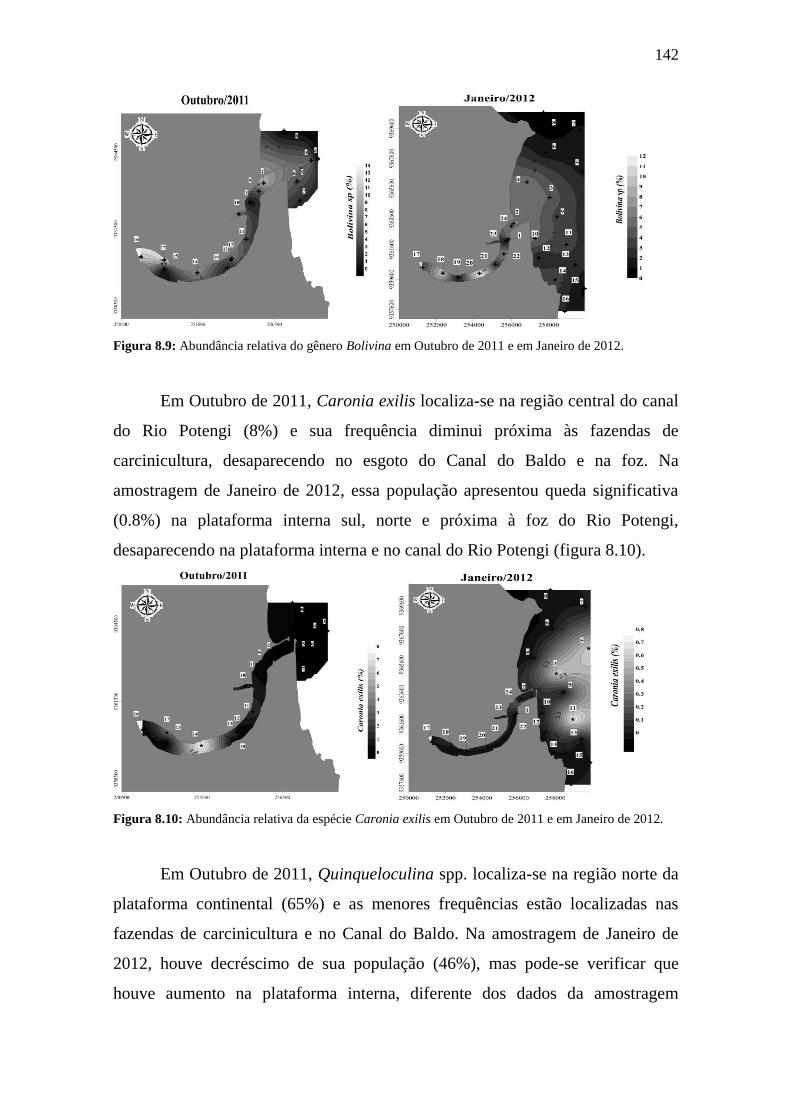
142
Figura 8.9: Abundância relativa do gênero Bolivina em Outubro de 2011 e em Janeiro de 2012.
Em Outubro de 2011, Caronia exilis localiza-se na região central do canal
do Rio Potengi (8%) e sua frequência diminui próxima às fazendas de
carcinicultura, desaparecendo no esgoto do Canal do Baldo e na foz. Na
amostragem de Janeiro de 2012, essa população apresentou queda significativa
(0.8%) na plataforma interna sul, norte e próxima à foz do Rio Potengi,
desaparecendo na plataforma interna e no canal do Rio Potengi (figura 8.10).
Figura 8.10: Abundância relativa da espécie Caronia exilis em Outubro de 2011 e em Janeiro de 2012.
Em Outubro de 2011, Quinqueloculina spp. localiza-se na região norte da
plataforma continental (65%) e as menores frequências estão localizadas nas
fazendas de carcinicultura e no Canal do Baldo. Na amostragem de Janeiro de
2012, houve decréscimo de sua população (46%), mas pode-se verificar que
houve aumento na plataforma interna, diferente dos dados da amostragem
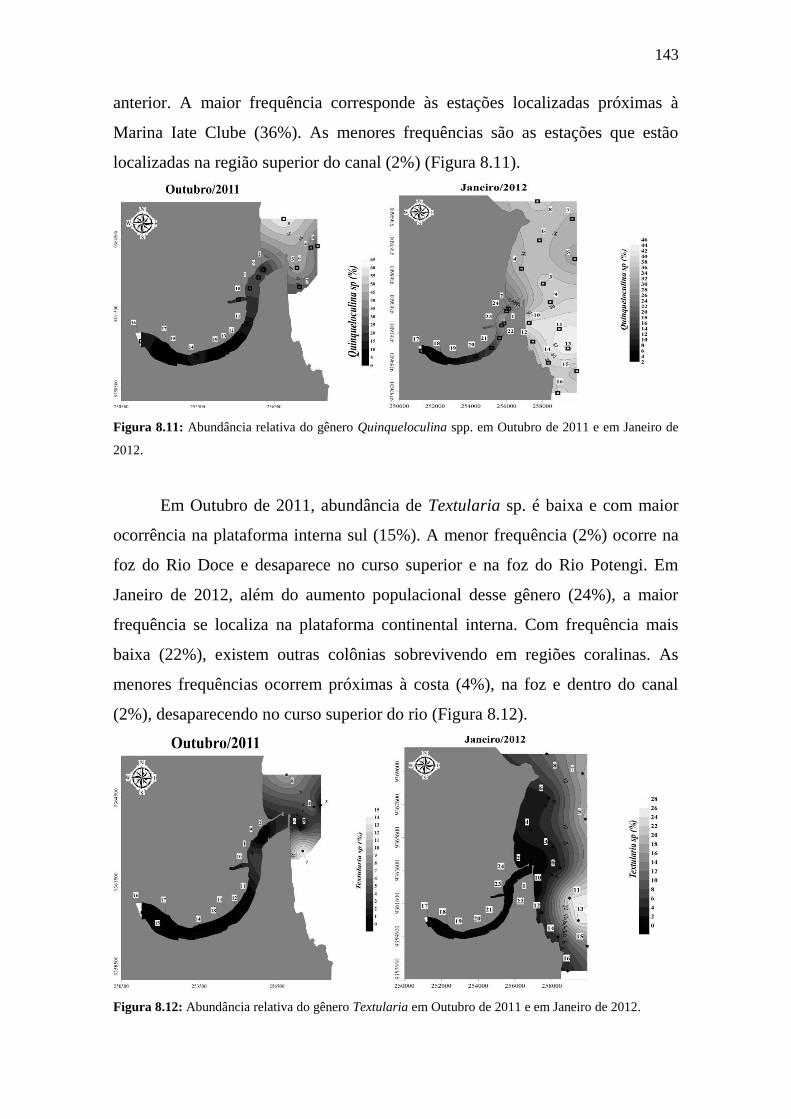
143
anterior. A maior frequência corresponde às estações localizadas próximas à
Marina Iate Clube (36%). As menores frequências são as estações que estão
localizadas na região superior do canal (2%) (Figura 8.11).
Figura 8.11: Abundância relativa do gênero Quinqueloculina spp. em Outubro de 2011 e em Janeiro de
2012.
Em Outubro de 2011, abundância de Textularia sp. é baixa e com maior
ocorrência na plataforma interna sul (15%). A menor frequência (2%) ocorre na
foz do Rio Doce e desaparece no curso superior e na foz do Rio Potengi. Em
Janeiro de 2012, além do aumento populacional desse gênero (24%), a maior
frequência se localiza na plataforma continental interna. Com frequência mais
baixa (22%), existem outras colônias sobrevivendo em regiões coralinas. As
menores frequências ocorrem próximas à costa (4%), na foz e dentro do canal
(2%), desaparecendo no curso superior do rio (Figura 8.12).
Figura 8.12: Abundância relativa do gênero Textularia em Outubro de 2011 e em Janeiro de 2012.

144
Em Outubro de 2011, a fração mais alta é a de cascalho (17%) que ocorre
na estação 17 e na foz; e a mais baixa são nas estações distantes da foz e em
alguns trechos do canal, desaparecendo na foz do Rio Doce. Em Janeiro de 2012,
sua porcentagem decaiu (0.28%) na plataforma interna, enquanto que na foz do
Rio Potengi essa fração é inexistente; no canal do Rio Potengi e na plataforma
interna essa fração ocorre em menores porcentagens (Figura 8.13).
Figura 8.13: Porcentagem de cascalho no Rio Potengi e na plataforma continental em Outubro de 2011 e
Janeiro de 2012.
Em Outubro de 2011, areia grossa ocorre em maiores frequências (50 –
70%) próxima à foz e ao trecho central do Rio Potengi. As menores porcentagens
(15%) se encontram na região superior do canal, região central e próxima ao rio
Doce; desaparecendo próximo às estações 15 e 3. Em Janeiro de 2012, a
porcentagem de areia grossa é maior (75%) na região costeira sul, enquanto que
na plataforma interna norte a menor frequência é de 5%, desaparecendo em
alguns pontos do Rio Potengi (Figura 8.14).

145
Figura 8.14: Porcentagem de areia grossa no Rio Potengi e na plataforma continental em Outubro de
2011 e Janeiro de 2012.
Em Outubro de 2011, areia muito grossa (14 – 36%) ocorre no curso
superior e região central do Rio Potengi; menores frações estão localizadas
próximo à foz com porcentagem de 2%, e desaparecem na região mais afastada
dela e em pequena área da região central do Rio Potengi. Em Janeiro de 2012,
essa frequência é maior do que na primeira amostragem; as maiores frequências
(20 – 50%) ocorrem na plataforma interna, e as menores frequências (5%) na
plataforma interna, no canal e na foz do rio, enquanto que na região costeira não
ocorre areia muito grossa (Figura 8.15).
Figura 8.15: Porcentagem de areia muito grossa no Rio Potengi e na plataforma continental em Outubro
de 2011 e Janeiro de 2012.

146
Em Outubro de 2011 a maior frequência (70%) da fração de areia média
está localizada na plataforma interna (estação 8) e existem pequenas
porcentagens (25 – 45%) na foz e no canal do Rio Potengi; a menor frequência é
de 10%. Em Janeiro de 2012 a frequência ficou 5% maior do que na amostragem
anterior. As maiores frequências são entre 55 – 75%, localizadas na plataforma
interna e na região central do canal do Rio Potengi. As menores frequências estão
localizadas em alguns trechos da plataforma interna e no canal do Rio Potengi
(10 – 15%) e nas estações localizadas próximas à Marina Iate Clube; e na região
costeira sul não apresentaram areia média (figura 8.16).
Figura 8.16: Porcentagem de areia média no Rio Potengi e na plataforma continental em Outubro de
2011 e Janeiro de 2012.
Em Outubro de 2011, a maior frequência de areia fina foi de 40%,
localizada no curso superior; as menores frequências (4 – 6%) ocorrem em
alguns trechos do canal e principalmente na foz e na estação 17 essa fração
desaparece. Em Janeiro de 2012 pode-se perceber que a frequência dobrou na
porcentagem da fração da amostragem anterior. As maiores frequências (55 –
80%) estão localizadas principalmente nas regiões de maior hidrodinâmica, como
a região costeira e no curso superior do canal do Rio Potengi; as menores
porcentagens (5 – 25%) encontram-se na plataforma continental interna,
principalmente na região sul (Figura 8.17).

147
Figura 8.17: Porcentagem de areia fina no Rio Potengi e na plataforma continental em Outubro de 2011 e
Janeiro de 2012.
Em Outubro de 2011, a maior frequência de areia muito fina é de 20% e
está localizada no curso superior; outro local com frequência significativa é
próxima ao rio Doce, com 8%; essa fração tem menores frequências em alguns
trechos do rio e na foz do Rio Potengi (1 – 6%). Em Janeiro de 2012 houve
aumento da frequência (75%), localizada na região costeira sul e existe um trecho
do canal do Rio Potengi com porcentagens significativas (35 – 40%); a menor
frequência (5%) está presente em alguns trechos do Rio e na plataforma interna e
ausente no curso superior e na plataforma interna (Figura 8.18).
Figura 8.18: Porcentagem de areia muito fina no Rio Potengi e na plataforma continental em Outubro de
2011 e Janeiro de 2012.

148
Em Outubro de 2011, frequência mais alta de argila (3.2%) localiza-se no
curso superior; e na região superior do canal do Rio Potengi e à medida que se
aproxima da foz a sua frequência torna-se mínima (0.6%), desaparecendo nas
regiões afastadas mais longes da foz. Porcentagem de silte ocorre no curso
superior do Rio Potengi, em maior frequência (26%) e menores frequências em
trechos no canal do Rio Potengi, e desaparecendo na foz e outros pontos do rio.
Em Janeiro de 2012, a fração de silte e argila ocorre em maior frequência (20%)
na Marina Iate Clube, e na região costeira sul, com porcentagens significativas (7
– 12%); as menores frequências ocorrem próximas à foz e estão ausentes no
canal superior do Rio Potengi e na plataforma interna (Figura 8.19).
Figura 8.19: Porcentagem de argila e silte no Rio Potengi e na plataforma continental em Outubro de
2011 e Janeiro de 2012.

149
8.5 – Discussões
No sudeste do país foram realizados estudos no Canal Bertioga (Eichler et
al., 2007), no Sistema Estuarino – Lagunar Cananéia – Iguape (SP) (Eichler –
Coelho et al., 1997; Uehara et al., 2007 e Mahiques et al., 2009) e na Baía de
Guanabara (RJ) (Eichler et al., 2014), e servem como modelo de comparação ao
do Rio Potengi, pois são muito similares na questão dos impactos ambientais
antrópicos e na oceanografia onde o ambiente aquático apresenta diferença em
gradientes físico-químicos e biológicos.
Em Eichler – Coelho (1997) foram encontrados foraminíferos das espécies
Ammobaculites sp, Difflugia prottaeiformis (indicadores de água doce) e
Elphidium excavatum antes do rompimento da Barragem do Valo Grande, um rio
no Estado de São Paulo. Após o rompimento, foram encontradas as espécies
calcárias da subordem Rotaliina e foraminíferos aglutinantes da subordem
Textulariina; além disso, foram encontradas também carapaças deformadas da
espécie E. excavatum, que mostra a relação brusca da mudança das condições
ambientais da zona estuarina, decorrente do rompimento. Nenhuma dessas
espécies aparece no Rio Potengi, revelando o alto teor salino desse rio.
Os grupos formados pela análise de CLUSTER em outubro de 2011
revelaram Ammonia tepida como espécie dominante indicando sua característica
cosmopolita, seguida pela Quinqueloculina lamarckiana e Textularia earlandi.
Ruiz et al. (2005) desenvolveu um trabalho em três estuários: Guadiana, Piedras
e Tinto – Odiel, os quais estão localizados ao longo da Costa de Huelva,
Sudoeste da Espanha, onde a espécie Ammonia tepida foi também mais
representativa nesse estudo.
Nossos dados mostram ainda uma diferenciação de dois subgrupos, onde
um está dominado por Ammonia tepida e Quinqueloculina lamarckiana; e o
outro está dominado por Ammonia tepida e Quinqueloculina miletti. Os
foraminíferos dominantes encontrados no Canal Bertioga (Eichler et al., 2007)
em comum com o Rio Potengi são os gêneros Ammonia, Bolivina e
Quinqueloculina.

150
No trabalho de Mahiques et al. (2009), foram encontradas espécies
calcárias mixohalinas de foraminíferos, dominadas geralmente por Ammonia
tepida, Elphidium excavatum e E. gunteri na zona basal (200 – 130 cm) no
testemunho; a zona intermediária (130 – 50 cm) foi caracterizada pela presença
de poucas espécies aglutinantes e tecamebas; já a zona do topo (50 – 0 cm) foi
marcada por um leve aumento na densidade e riqueza totais das espécies
aglutinantes (Miliammina fusca) e das tecamebas. A única espécie em comum
que aparece no rio Potengi é A. tepida.
Já no trabalho de Uehara et al. (2007), a qual também utilizou
testemunhos, a espécie dominante foi Pararotalia cananeiaensis, a qual é
endêmica dessa região. Nesses testemunhos foram encontradas espécies típicas
de ambiente de plataforma interna (Hanzawaia boueana e Pseudononium
atlanticum), inclusive Globigerina bulloides e Uvigerina bifurcata, próprias de
meios mais distais (288 – 178 cm). A partir de 153 cm de profundidade, os
foraminíferos calcários mixohalinos (Cribroelphidium excavatum e Ammonia
tepida) e os aglutinantes (Arenoparrela mexicana e Haplophragmoides wilberti)
tornam-se mais abundantes que os marinhos. De 143 a 12 cm de profundidade, os
aglutinantes aumentam progressivamente rumo ao topo, sugerindo a presença de
manguezais através da espécie Miliammina fusca, a qual indica maior aporte de
água doce entrando no sistema. No topo do testemunho, a presença de calcários
mixohalinos em associação com aglutinantes, volta a ser abundante, sugerindo
ambiente de borda de manguezal com influência marinha moderada. As espécies
apresentadas pela autora que tem em comum com o Rio Potengi são Ammonia
tepida, Hanzawaia boueana e Pseudononium atlanticum.
No trabalho de Wang & Chappell (2001) realizado no rio Alligator do Sul,
Darwin, Austrália, foram identificadas espécies coradas onde a maioria das
espécies mencionadas pelos autores é desconhecida, exceto o gênero
Arenoparrella e um integrante da família dos lagenídeos (Lagena striata) que
foram encontradas na segunda amostragem no Rio Potengi. No trabalho de
Berkeley et al. (2008) no rio Cocoa, Queensland, Austrália, foram encontradas
espécies aglutinantes e calcárias, das quais algumas espécies não ocorrem no rio

151
Potengi. No trabalho desenvolvido por Sarita et al. (2015), os autores analisaram
a distribuição espacial e sazonal das comunidades de foraminíferos bentônicos
vivos no estuário de Guadiana. Foram identificadas várias espécies, das quais a
grande maioria também não é encontrada em nossa costa ou no rio Potengi, com
exceção da espécie Trochammina inflata.
No nosso estudo Quinqueloculina lamarckiana indica a presença de cunha
salina que penetra pela foz do rio Potengi até alguns locais rio adentro. O gênero
Quinqueloculina é também típico de ambientes de alta hidrodinâmica (Eichler et
al., 2007; Wang & Chappell, 2001) da plataforma continental e no nosso estudo
sua presença indica o alto teor salino do Rio Potengi. De acordo com Debenay
(2012) pode ser encontrado em lagoas costeiras, pântanos salgados, estuários e
baías, sendo também considerados oportunistas e tolerantes à ambientes
poluídos, como as espécies Quinqueloculina miletti e Q. patagonica.
Observamos uma diferenciação nos nossos dados, onde uma espécie está
indicando provavelmente a contaminação por esgotos e a outra por detritos
provenientes da carcinicultura; além disso, parece existir competição entre ambas
as espécies por nutrientes e matéria orgânica, onde uma espécie inibe a outra e
vice-versa, principalmente nos ambientes prováveis de serem poluídos.
A formação de três grupos distintos pelo CLUSTER em Janeiro de 2012:
Foz do Rio Potengi (1 – 4), Plataforma Interna (5 – 16 e Canal do Rio Potengi
(17 – 24) apresentaram as espécies Ammonia tepida e Quinqueloculina
lamarckiana dominando em altas porcentagens, sendo que Ammonia tepida
dominou principalmente no Canal do Rio Potengi e em menor frequência na
plataforma interna; enquanto que Quinqueloculina lamarckiana dominou
principalmente na plataforma interna e em menores frequências no Canal do Rio
Potengi. As mesmas espécies de foraminíferos foram observadas no trabalho de
De Souza et al. (2010), no Rio Potengi, onde seis amostras em pontos de
poluição aparentes no rio estabeleceram modelo de zonação para avaliar o
impacto ambiental da área através da distribuição dos agrupamentos dos
foraminíferos associados à atividade bacteriana e aos parâmetros físicos –
químicos.

152
No trabalho de Eichler et al. (2007) Quinqueloculina milletti aparece
exclusivamente no estuário em baixa frequência associada às salinidades
marinhas normais, à alta claridade da água e à boa circulação. As maiores
frequências de Quinqueloculina lamarckiana ocorrem na região de plataforma
interna e no Canal do Rio Potengi.
Em baixas porcentagens a espécie Bolivina striatula dominou nos três
grupos, especialmente na Foz do Rio Potengi, mas nas estações próximas às
fazendas de carcinicultura e ao esgoto do Canal do Baldo ocorrem frequências
mais altas. De acordo com Debenay (2012) e Eichler et al. (2003), o gênero
Bolivina é característico de lagoas costeiras, estuários e baías, tolerantes a
ambientes com baixos teores de oxigênio dissolvido e alto teor de carbono
orgânico indicando talvez um local impactado em ambientes de granulometria
fina (Eichler et al., 2007). O gênero Bolivina aparece também na região de
manguezal onde existe abundância de nutrientes e sedimentos finos em Bertioga
(Eichler et al., 2007).
As espécies Hanzawaia boueana, Elphidium discoidale, Cibicides
valvulatanus, Cibicides variabilis e Discorbis peruvianus aparecem em maiores
frequências na plataforma interna ao Rio Potengi. Se compararmos nossos dados
aos dados de Eichler et al. (2014), na Baía de Guanabara (RJ), onde os autores
encontraram maiores frequências de Cassidulina subglobosa, Discorbis
williamsoni, Quinqueloculina seminulum, Buliminella elegantissima e Bolivina
elongata, observamos que apenas a espécie Quinqueloculina seminulum não
ocorre no rio Potengi. Quinqueloculina patagonica e Cassidulina subglobosa
aparecem em baixa frequência no Canal do Rio Potengi.
A espécie Cassidulina subglobosa foi encontrada no trabalho de Eichler et
al. (2003), na entrada da Baía de Guanabara, em maiores frequências indicando
salinidades mais altas. A espécie Quinqueloculina miletti apresenta frequências
mais altas na Foz do Rio Potengi, enquanto que Textularia earlandi, T. gramen e
Trochammina discorbis aparecem na plataforma interna. Essa distribuição revela
a zonação relacionada à influência da penetração de cunha salina no Rio Potengi
e a ocorrência de Trochammina discorbis na plataforma indica a presença da

153
pluma estuarina de água doce na plataforma continental adjacente revelando a
alta competência do Rio Potengi de exportar material para a plataforma.
Espécies de Trochammina inflata e T. squamata ocorreram em
pouquíssimas frequências no Canal do Rio Potengi e a espécie Trochammina
inflata também foi encontrada no trabalho de Sarita et al. (2015), que foi
realizada no Estuário Guadiana em Portugal, onde foi identificada em quase
todas as amostras, mas encontra-se afiliada a Jadammina macrescens.
Assim como a espécie Caronia exilis, o gênero Textularia sp é uma
espécie aglutinante e costuma viver em ambientes de granulometria grossa, mas a
diferença entre ambas as espécies é que esta é adaptada para ambientes altamente
hidrodinâmicos e salinos, enquanto que o gênero Caronia é típico de ambientes
lagunares (Debenay, 2012).
A análise de PCA revelou que a granulometria grossa (areia muito grossa,
areia grossa e cascalho), temperatura e CaCO3 exerceram maior influência nas
estações em Outubro de 2011, enquanto que os parâmetros que exerceram menor
influência nas estações foram a granulometria fina (areia média, areia fina, areia
muito fina, silte e argila), profundidade, salinidade, condutividade e densidade. A
distribuição granulométrica pode ser utilizada para detectar os ambientes de
deposição e cada ambiente tem seu nível energético característico com parâmetro
granulométrico condicionado à hidro dinamicidade.
No canal do rio a granulometria é arenosa, principalmente areia grossa e
areia média, que se revela como ambiente altamente hidrodinâmico. A pequena
porcentagem de argila, a qual é depositada nas margens do Rio Potengi,
representam, no entanto, a presença de alguns ambientes de menor energia.
Quanto à salinidade, o Rio Potengi apresenta basicamente dois ambientes
distintos: um ambiente mais interno, o qual não é influenciado pela alta
salinidade; e outro ambiente que é a área próxima à foz que é influenciada pela
alta salinidade. A presença de CaCO3 encontra-se associada à presença de
bioclastos, principalmente nas proximidades com a foz dos rios Doce e Potengi.

154
A análise de PCA em Janeiro de 2012 determinou que os principais
fatores reguladores da fauna de foraminíferos são a porcentagem de carbono
orgânico, nitrogênio, CaCO3 e granulometria fina (areia fina, areia muito fina,
silte e argila); e os parâmetros que são menos determinantes são a profundidade e
a granulometria grossa (areia média e areia grossa). As porcentagens de CaCO3
foram muito altas nas estações que se encontram na plataforma interna,
provavelmente essas áreas onde foram coletadas possuem recifes de corais em
suas proximidades. Os teores muito baixos de Carbono orgânico sugerem um
ambiente oxidante, inadequado à preservação da matéria orgânica.
De acordo com as análises granulométricas e o PCA, a granulometria
dominante é a arenosa, composta pelas frações areia grossa, areia muito fina e
areia fina, revelando ambiente com alta hidrodinâmica. Pouquíssimas estações
são influenciadas pelo silte e argila, que indicam ambientes de baixa
hidrodinâmica. A porcentagem de cascalho variou entre as duas coletas: na
primeira amostragem essa fração apresentou maior frequência, enquanto que na
segunda amostragem houve um decréscimo significativo. Uma possível
explicação para esse fato é que as frações de cascalho tenham sido deslocadas
para outros locais de deposição pelas correntezas fluviais e também pelo
constante retrabalhamento do leito marinho.
Em Janeiro de 2012 a frequência de areia grossa apresentou-se mais alta
que Outubro de 2011 próximo à foz e no trecho central do Rio Potengi. O
aumento dessa fração se deve pelo constante retrabalhamento hidrodinâmico do
leito marinho e o input de sedimentos através dos rios e pelo transporte das águas
pluviais. De acordo com Klee (1999), a zona costeira é afetada por fatores
marinhos, como marés, ondas, correntes, ressurgência e aumento do nível do
mar, além da influência dos rios costeiros, que tem a função crucial de depositar
os sedimentos no mar e também de renovar o suprimento sedimentar das praias.
Em Outubro de 2011 a maior frequência de areia fina encontra-se em
alguns trechos com pequenas porcentagens no canal do Rio Potengi; já em
Janeiro de 2012 a frequência dessa fração dobrou e encontra-se no canal superior
do Rio Potengi e existem também frequências relativamente maiores localizadas

155
em regiões de maior hidrodinâmica, como a região costeira. Essa fração mostrou-
se típica de local deposicional principalmente na região norte da região costeira
norte e parte da região costeira sul, próximo ao Rio Potengi, provavelmente pela
grande descarga das águas fluviais juntamente com as correntezas marinhas que
vem pelo sul, mostrando que ocorre o retrabalhamento do material
sedimentológico. Areia muito fina apresentou maior frequência na primeira
amostragem e aumento, principalmente no canal do Rio Potengi, indicando
ambientes deposicionais de baixa hidrodinâmica e provavelmente poluídos ou
contaminados.
De acordo com Lisitzin (1996), a granulometria mais grossa é proveniente
de regiões topográficas mais altas, apesar do material mais fino ser depositado no
fundo oceânico. As maiores concentrações das frações arenosas e siltosas são
encontradas próximas à costa. Para Garrison (2002), apesar das frações maiores
(matacões, pedregulhos e seixos) ocorrerem no mar, a maioria dos sedimentos
marinhos é de partículas finas (areia, silte e argila).
Altas frequências de silte e argila estão localizadas na região superior do
Rio Potengi próxima às fazendas de carcinicultura e às salinas, enquanto que as
menores frequências se encontram nas áreas mais próximas da foz, com alta
hidrodinâmica. Apesar de não ter sido possível fazer a comparação de
porcentagem entre as frações de silte e argila da segunda amostragem, foi
possível verificar que ambas as frações ocorrem nos mesmos locais de deposição
em ambas as amostragens e estas foram deslocadas por ação hidrodinâmica
fluvial da região superior para outro setor do rio Potengi. Essas frações são
típicas de ambientes deposicionais e de baixa hidrodinâmica e em geral
relacionadas a locais com sedimentos poluídos ou contaminados.
8.6 – Conclusões
A análise sistemática dos foraminíferos presentes em sedimentos do Rio
Potengi e plataforma interna adjacente indica a presença de dois ambientes: um
influenciado pela ação das marés e outro, mais interno, menos influenciado pela
ação marinha;

156
A forte influência do mar ao longo do Rio Potengi através da presença da
espécie Quinqueloculina lamarckiana. Por sua vez a Trochammina discorbis
indica a presença da pluma estuarina de água doce na plataforma continental
adjacente revelando a alta competência do Rio Potengi de exportar material para
a plataforma;
A presença das espécies Ammonia tepida, Quinqueloculina patagonica e
Quinqueloculina miletti, juntamente com as porcentagens elevadas das frações
granulométricas de areia muito fina, silte e argila, revelaram os principais locais
de deposição que provavelmente contém sedimentos poluídos ou contaminados;
Existe possibilidade de haver competição entre as espécies
Quinqueloculina patagonica e Quinqueloculina miletti por nutrientes e matéria
orgânica.
8.7 – Agradecimentos
As autoras gostariam de agradecer a Universidade Federal pelo espaço
físico para a realização dos procedimentos laboratoriais; aos departamentos de
geologia, geografia, química e ao Museu Câmara Cascudo pelo fornecimento do
laboratório e materiais para a realização dos procedimentos laboratoriais;
agradecemos ao CNPq INCT AmbTropic/WG 2.1, CAPES-Ciências do Mar 207-
10; PRH22-ANP/MCT; PLAT N-NE Rede 05 / FINEP / CTPETRO; CNPq;
PROBRAL 337-10 (CAPES/DAAD), CAPES PVE 151/2012 (Ciências sem
fronteiras), PE 0242/2013 (Ciências sem Fronteiras), MCTI/CNPq Nº 23/2011 -
Apoio Técnico para Fortalecimento da Paleontologia Nacional.(552976/2011-3),
Capes através do Edital Ciências do Mar (207/2010) para a bolsa de Post Doc no
Laboratório de Geologia e Geofísica Marinha e Monitoramento Ambiental da
Universidade Federal do Rio Grande do Norte (GGEMMA-UFRN-Brazil) e
CNPq (303481/09-9) pelo suporte financeiro da realização desse trabalho.

157
Referências
Berkeley A., Perry C.T., Smithers S.G and Horton B.P. 2008. The spatial and
vertical distribution of living (stained) benthic foraminifera from a tropical,
intertidal environment, north Queensland, Australia. Revista Marine
Micropaleontology. 69: 204 – 261.
Boltovskoy E. 1965. Los foraminiferos recientes. Buenos Aires: Eudeba, 510 p.
Committee on the Geologic Record of Biosphere Dynamics. 2005. The
Geological Record of Ecological dynamics: Understanding the Biotic Effects of
Future environmental Change. Washington: The National Academic Press. p 30
– 74.
Córdoba V.C., Jardim de Sá E.F., Souza D.C. and Antunes A.F. 2007. Bacia
Pernambuco-Paraíba. Boletim de Geociências da Petrobras. Rio de Janeiro. 15:
391-403.
Debenay J.P. 2012. A guide to 1.000 foraminifera from Southwestern Pacific
New Caledonia. Paris: IRD Éditions. Publications Scientifiques du Muséum. 385
p.
De Souza V.M., Laut L.L.M., Da Silva F.S., Figueiredo Jr A.G., Vital H. and
Frazão E. 2010. Benthic foraminífera and bacterial activity as a proxy for
environmental Characterization in Potengi Estuary, Rio Grande do Norte, Brazil.
Anuário de geociências. UFRJ. 33 – 1: 20 – 34.
Eichler – Coelho P.B., Duleba W., Eichler B.B. and Coelho – Junior C. 1997.
Determinação do impacto ecológico do Valo Grande (Iguape, SP) a partir das
associações de foraminíferos e tecamebas. Revista Brasileira de Biologia. 57:
463 – 477.
Eichler P.P.B., Eichler B.B, Miranda L.B., Pereira E.R.M., Kfouri P.B.P.,
Pimenta F.M., Bérgamo A.L. and Vilela C.G. 2003. Benthic foraminiferal
response to variations in temperature, salinity, dissolved oxygen and organic
carbon in the Guanabara Bay, Rio de Janeiro, Brazil. Anuário do Instituto de
Geociências – UFRJ. Rio de Janeiro. 26: 36-51;

158
Eichler P.P.B., Eichler B.B., Miranda L.B. and Rodrigues A.R. 2007. Modern
foraminiferal facies in a subtropical estuarine channel, Bertioga, São Paulo,
Brazil. Journal of Foraminiferal Research. 37: 234-247.
Eichler P.P.B., Eichler B.B, Pereira E.R.M. and Vital H. 2014. Evaluation of
Environmental and ecological effects due to the accident in na oil pipe from
Petrobrás in Guanabara Bay, RJ, Brazil. Open Journal of Marine Science. 4: 298
- 315.
Garrison T. 2002. Oceanography: an invitation to marine science, 4th
ed., United
Estates of America, Thomson Learning Inc. 554 pp.
IBAMA. 2014. Parecer Técnico n.o 02021.000145/2014-00. NUPAEM-
DICAF/SUPES/IBAMA/RN. Natal. 107 pp.
IBAMA. 2015. Parecer Técnico n.o 02021.000032/2015-87. NUPAEM-
DITEC/SUPES/IBAMA/RN. Natal. 51 pp.
Klee G.A. 1999. The coastal environment: toward integrated coastal and marine
sanctuary management. United Estates of America. Prentice – Hall Inc. 281 pp.
Lima Filho M.L. 2013. Bacia Pernambuco e Paraíba. Agência Nacional do
Petróleo, Gás Natural e Biocombustíveis, Rodada 11. Rio de Janeiro.
Lisitzin A.P. 1996. Oceanic sedimentation: lithology and geochemistry.
American Geophysical Union, Washington, DC. 1996. P. 54 – 55. Translated
from the Russian by C. Woolhiser.
Mahiques M.M., Burone L., Figueira R.C.L., Lavenére - Wanderley A.A.O.,
Capellari B., Rogacheski C.E, Barroso C.P., dos Santos L.A.S., Cordero L.M.
and Cussioli M.C. 2009. Anthropogenic influences in a lagoonal environment: a
multiproxy approach at the Valo Grande mouth, Cananéia-Iguape system (SE
Brazil). Brazilian Journal of Oceanography. 57: 325-337.
Nimer E. 1972. Climatologia da Região Nordeste do Brasil. Revista Brasileira de
Geografia, 34: 3 – 51.
RADAM BRASIL. 1981. Série Levantamento de Recursos Naturais do Projeto
RADAM BRASIL.

159
Ruiz F., González – Regalado M.L., Pendón J.G, Abad M., Olías M. and Muñoz
J.M. 2005. Correlation between foraminífera and sedimentary environments in
recent estuaries of Southwestern Spain: applications to Holocene reconstructions.
Quaternary International. 140 – 141: 21 – 36.
Sarita C., Delminda M., Simon C., David S. and Tomasz B. 2015. Ecological
zonation of benthic foraminifera in the lower Guadiana Estuary (southeastern
Portugal). Marine micropaleontology. 114:1 – 18.
Silva C.A.R., Rainbow P.S, Smith B.D and Santo Z.L. 2001. Biomonitoring of
trace metal contamination in the Potengi Estuary, Natal (Brazil), using the oyster
Crassostrea rhizophorae, a local food source. Water Research. 35: 4072 – 4078.
Vilela C.G. 2004. Foraminíferos. In: Carvalho I.S (Ed). Paleontologia. Rio de
Janeiro. Ed. Interciência. p 269 – 282.
Wang P. and Chappell J. 2001. Foraminifera as Holocene environmental
indicators in the South Alligator River, Northern Australia. Revista Quaternary
International. 83–85: 47–62.
Uehara R.S., Duleba W., Petri S., Mahiques M.M and Rodrigues M. 2007.
Micropaleontologia e sedimentologia aplicadas à análise paleoambiental: um
estudo de caso em Cananéia, São Paulo, Brasil. Revista Brasileira de
Paleontologia. 10: 137-150.


















