
Function of glutathione peroxidases in legume root nodules
41
Function of glutathione peroxidases in legume root nodules 1 2 Manuel A. Matamoros 1,* , Ana Saiz 1,* , Maria Peñuelas 1 , Pilar Bustos- 3 Sanmamed 2 , Jose M. Mulet 3 , Maria V. Barja 3 , Nicolas Rouhier 4,5 , Marten 4 Moore 6 , Euan K. James 7 , Karl-Josef Dietz 6 and Manuel Becana 1,† 5 6 1 Departamento de Nutrición Vegetal, Estación Experimental de Aula Dei, Consejo Superior 7 de Investigaciones Científicas (CSIC), Apartado 13034, 50080 Zaragoza, Spain 8 2 Institut des Sciences du Végétal, Avenue de la Terrasse, 91198 Gif-sur-Yvette, France 9 3 Instituto de Biología Molecular y Celular de Plantas, Universidad Politécnica de Valencia- 10 CSIC, Camino de Vera, 46022 Valencia, Spain 11 4 Université de Lorraine, Interactions Arbres-Microorganismes, UMR1136, F-54500 12 Vandoeuvre-lès-Nancy, France 13 5 INRA, Interactions Arbres-Microorganismes, UMR1136, F-54280 Champenoux, France 14 6 Biochemistry and Physiology of Plants, W5-134, Bielefeld University, D-33501, Germany 15 7 The James Hutton Institute, Invergowrie, Dundee, DD2 5DA, UK 16 17 * These authors contributed equally to this manuscript. 18 † To whom correspondence should be addressed. E-mail: [email protected] 19 20 Authors' e-mails: M.A.M. ([email protected]), A.S. ([email protected]), M.P. (maria. 21 [email protected]), P.B.-S. ([email protected]), J.M.M. (jmmulet@ 22 ibmcp.upv.es), M.V.B. ([email protected]), N.R. (Nicolas.Rouhier@univ-lorraine. 23 fr), M.M. ([email protected]), E.K.J. ([email protected]), K.-J.D. (karl- 24 josef.dietz@ uni-bielefeld.de), M.B. ([email protected]) 25 26 Author for correspondence: 27 Manuel Becana 28 Estación Experimental de Aula Dei, Consejo Superior de Investigaciones Científicas, 29 Apartado 13034, 50080 Zaragoza, Spain 30 Tel: +34-976-716055, Email: [email protected] 31 32 Received: 30 December 2014 33 Running title: Glutathione peroxidases of nodules 34 35 Total word count: 8526 36 Number of Tables: 2 37 Number of Figures: 7 38 Figures in colour: 2 (print: Figures 4 and 6; online-only: Figure S1) 39 40 Supplementary data Tables: 1, Figures: 3 41
Transcript of Function of glutathione peroxidases in legume root nodules

Function of glutathione peroxidases in legume root nodules 1
2 Manuel A. Matamoros1,*, Ana Saiz1,*, Maria Peñuelas1, Pilar Bustos-3
Sanmamed2, Jose M. Mulet3, Maria V. Barja3, Nicolas Rouhier4,5, Marten 4
Moore6, Euan K. James7, Karl-Josef Dietz6 and Manuel Becana1,† 5 6
1 Departamento de Nutrición Vegetal, Estación Experimental de Aula Dei, Consejo Superior 7
de Investigaciones Científicas (CSIC), Apartado 13034, 50080 Zaragoza, Spain 8 2 Institut des Sciences du Végétal, Avenue de la Terrasse, 91198 Gif-sur-Yvette, France 9 3 Instituto de Biología Molecular y Celular de Plantas, Universidad Politécnica de Valencia-10
CSIC, Camino de Vera, 46022 Valencia, Spain 11 4 Université de Lorraine, Interactions Arbres-Microorganismes, UMR1136, F-54500 12
Vandoeuvre-lès-Nancy, France 13 5 INRA, Interactions Arbres-Microorganismes, UMR1136, F-54280 Champenoux, France 14 6 Biochemistry and Physiology of Plants, W5-134, Bielefeld University, D-33501, Germany 15 7 The James Hutton Institute, Invergowrie, Dundee, DD2 5DA, UK 16
17 * These authors contributed equally to this manuscript. 18 † To whom correspondence should be addressed. E-mail: [email protected] 19
20 Authors' e-mails: M.A.M. ([email protected]), A.S. ([email protected]), M.P. (maria. 21 [email protected]), P.B.-S. ([email protected]), J.M.M. (jmmulet@ 22 ibmcp.upv.es), M.V.B. ([email protected]), N.R. (Nicolas.Rouhier@univ-lorraine. 23 fr), M.M. ([email protected]), E.K.J. ([email protected]), K.-J.D. (karl-24 josef.dietz@ uni-bielefeld.de), M.B. ([email protected]) 25 26 Author for correspondence: 27 Manuel Becana 28 Estación Experimental de Aula Dei, Consejo Superior de Investigaciones Científicas, 29 Apartado 13034, 50080 Zaragoza, Spain 30 Tel: +34-976-716055, Email: [email protected] 31 32 Received: 30 December 2014 33
Running title: Glutathione peroxidases of nodules 34 35 Total word count: 8526 36 Number of Tables: 2 37 Number of Figures: 7 38 Figures in colour: 2 (print: Figures 4 and 6; online-only: Figure S1) 39 40 Supplementary data Tables: 1, Figures: 3 41

2
Short statement. Glutathione peroxidases are antioxidant enzymes localized to different 1
cell compartments, including the nucleus. Transcriptional and post-translational 2
regulation via S-nitrosylation strongly suggest functions in hormonal cascades and nitric 3
oxide redox signaling. 4
5
Abstract 6
7 Glutathione peroxidases (Gpxs) are antioxidant enzymes not studied so far in 8
legume nodules, despite the fact that reactive oxygen species are produced at 9
different steps of the symbiosis. The function of two Gpxs that are highly expressed 10
in nodules of the model legume Lotus japonicus was examined. Gene expression 11
analysis, enzymatic and nitrosylation assays, yeast cell complementation, in situ 12
mRNA hybridization, immunoelectron microscopy, and LjGpx-green fluorescent 13
protein (GFP) fusions were used to characterize the enzymes and to localize each 14
transcript and isoform in nodules. The LjGpx1 and LjGpx3 genes encode 15
thioredoxin-dependent phospholipid hydroperoxidases and are differentially 16
regulated in response to nitric oxide (NO) and hormones. LjGpx1 and LjGpx3 are 17
nitrosylated in vitro or in plants treated with S-nitrosoglutathione (GSNO). 18
Consistent with the modification of the peroxidatic cysteine of LjGpx3, in vitro 19
assays demonstrated that this modification results in enzyme inhibition. The 20
enzymes are highly expressed in the infected zone, but the LjGpx3 mRNA is also 21
detected in the cortex and vascular bundles. LjGpx1 is localized to the plastids and 22
nuclei, and LjGpx3 to the cytosol and endoplasmic reticulum. Based on yeast 23
complementation experiments, both enzymes protect against oxidative stress, salt 24
stress, and membrane damage. It is concluded that both LjGpxs perform major 25
antioxidative functions in nodules, preventing lipid peroxidation and other oxidative 26
processes at different subcellular sites of vascular and infected cells. The enzymes 27
are probably involved in hormone and NO signaling, and may be regulated through 28
nitrosylation of the peroxidatic cysteine essential for catalytic function. 29
30 Key words: Antioxidants, glutathione peroxidases, legume nodules, Lotus japonicus, 31
nitric oxide, reactive oxygen species, S-nitrosylation. 32

3
1 2
Abbreviations: ABA, abscisic acid; ACC, 1-aminocyclopropane-1-carboxylic acid; CK, 3
cytokinin; Gpx, glutathione peroxidase; GSH, glutathione; GSNO, S-nitrosoglutathione; 4
GSSG, glutathione disulfide; JA, jasmonic acid; NO, nitric oxide; ROS, reactive oxygen 5
species; SA, salicylic acid; Trx, thioredoxin. 6
7
Introduction 8 9
Glutathione peroxidases (Gpxs) are ubiquitous enzymes that catalyze the reduction of 10
H2O2 or organic peroxides to water or the corresponding alcohols using glutathione 11
(GSH) or thioredoxins (Trxs) as electron donors (Herbette et al., 2007; Brigelius-Flohé 12
and Maiorino, 2013). These enzymes were initially described in mammals, where eight 13
clades can be distinguished based on amino acid sequences, substrate specificity, and 14
subcellular localization (Herbette et al., 2007; Brigelius-Flohé and Maiorino, 2013). Four 15
groups of Gpxs, termed ‘classical’ or cytosolic (Gpx1), gastrointestinal (Gpx2), plasmatic 16
(Gpx3), and phospholipid hydroperoxidases (Gpx4), contain seleno-Cys instead of Cys at 17
the catalytic site. Gpx6, located in the olfactory system, is a selenoprotein in humans and 18
pigs but not in rodents, whereas an epididymis-specific (Gpx5) and two recently 19
discovered Gpxs associated to the endoplasmic reticulum (Gpx7 and Gpx8) do not 20
contain seleno-Cys (Brigelius-Flohé and Maiorino, 2013). 21
Plant Gpxs are most similar in terms of amino acid sequences to the mammalian 22
Gpx4 enzymes but lack seleno-Cys (Herbette et al., 2007), with the single exception of 23
the Gpx from the unicellular green alga Chlamydomonas reinhardtii (Fu et al., 2002). 24
The fact that Cys is less reactive than seleno-Cys may explain why plant Gpxs are less 25
efficient in scavenging reactive oxygen species (ROS) than their mammalian counterparts 26
(Herbette et al., 2007). Plant Gpxs usually have three Cys residues (Supplementary Fig. 27
S1), but only the first ('peroxidatic') Cys and the third ('resolving') Cys are required for 28
catalysis and Trx regeneration (Navrot et al., 2006; Koh et al., 2007). The Gpxs are 29

4
encoded by small multigene families, comprising five to eight members in the model 1
plants so far examined (Rodriguez Milla et al., 2003; Margis et al., 2008; Navrot et al., 2
2006; Ramos et al., 2009). Many plant Gpxs may protect membranes from peroxidative 3
damage (Gueta-Dahan et al., 1997; Herbette et al., 2002) and some Arabidopsis thaliana 4
Gpx isoforms may play additional roles in redox transduction and stress signaling (Miao 5
et al., 2006; Chang et al., 2009). 6
Legumes establish symbiotic associations with rhizobia forming root nodules, which 7
are unique organs that fix atmospheric N2 into ammonium. Nodules contain O2-sensitive 8
metalloproteins and leghemoglobin that favor ROS production (Dalton, 1995; Becana et 9
al., 2010). However, low steady-state ROS levels are required for critical functions such 10
as plant organ development and stress perception (Foyer and Noctor, 2005; Puppo et al., 11
2005). To offset the potential toxicity of ROS while allowing them to play signaling 12
roles, nodules contain an impressive array of antioxidants, although only the enzymes and 13
metabolites of the ascorbate-GSH pathway have been studied in detail to elucidate their 14
role in peroxide metabolism (Dalton, 1995; Becana et al., 2010). In sharp contrast, the 15
function of Gpxs in nodules has been overlooked, despite early studies showing that Gpx 16
activity is responsive to oxidative stress (Gueta-Dahan et al., 1997) and that ROS and 17
nitric oxide (NO) are involved at different stages of the symbiosis (Puppo et al., 2013). 18
Six Gpx genes have been identified in the model legume L. japonicus and two of them, 19
LjGpx1 and LjGpx3, are highly expressed in nodules (Ramos et al., 2009). Here, a 20
detailed characterization of these two isoforms is provided by combining enzyme activity 21
assays, expression profiles, mRNA and protein localizations in nodules, and functional 22
complementation of a yeast Gpx-deficient mutant. Because Gpx activities rely on critical 23
Cys residues (Jung et al., 2002; Navrot et al., 2006; Herbette et al., 2007) and S-24
nitrosylation is an important post-translational modification underlying NO signaling 25
(Astier et al., 2012), the possible regulation of LjGpx1 and LjGpx3 activities by 26
nitrosylation has been studied by using dedicated mass spectrometry (MS) methods. 27

5
Materials and Methods 1 2
Plant growth and treatments 3 4 Seeds of Lotus japonicus (Regel) Larsen ecotype MG20 were sown, seedlings were 5
inoculated with Mesorhizobium loti strain R7A, and plants were grown in controlled 6
environment cabinets as previously described (Ramos et al., 2009). Plants to be used for 7
biochemical and microscopy studies were grown for 46 d in pots (1 liter) containing 8
vermiculite and were irrigated twice a week with B&D nutrient solution (Broughton & 9
Dilworth, 1971) supplemented with 0.25 mM NH4NO3. 10
Expression profiles of LjGpx1 and LjGpx3 were determined in nodules of plants 11
exposed to stress and hormones. (a) Nitro-oxidative stress. This was induced by treating 12
the plants with cadmium (Cd) or S-nitrosoglutathione (GSNO). Plants grown for 46 d in 13
pots were separated into two groups. One set of plants was treated with 100 M CdCl2 in 14
water and nodules were harvested after 6 h. The other set of plants was transferred to 15
Erlenmeyer flasks containing 250 ml of 1:10 HEN buffer [100 mM HEPES (pH 8.0), 1 16
mM EDTA, 0.1 mM neocuproine] supplemented with either 5 mM GSNO or glutathione 17
disulfide (GSSG; control). The flasks were protected from light and plants were treated 18
for 6 h. (b) Phytohormones. Nodulated plants were grown hydroponically for 44 d 19
(Tovar-Méndez et al., 2011) and treated for 48 h with 50 M abscisic acid (ABA), 20
salicylic acid (SA), jasmonic acid (JA), 1-aminocyclopropane-1-carboxylic acid (ACC), 21
or cytokinin (CK, an equimolar mixture of kinetin and 6-benzyl-aminopurine). Stock 22
solutions (100 mM) were prepared in 2 ml of ethanol (ABA, ACC, SA), 23
dimethylsulfoxide (JA), or 1 M NaOH (CKs), and added to 4 l of the aerated hydroponic 24
solution (1:4 B&D nutrient solution lacking combined nitrogen, pH 6.6). Control plants 25
were treated with the same concentrations of ethanol, dimethylsulfoxide, or NaOH. 26
27
Expression analysis of LjGpx genes 28 29 Total RNA was extracted from nodules and processed as described (Ramos et al., 2009). 30
Quantitative reverse-transcription PCR was performed with the primers listed in 31
Supplementary Table S1 using a 7500 Real-Time PCR System (Applied Biosystems). 32
Transcript levels were normalized with ubiquitin and the relative values of gene 33

6
expression were calculated using the 2exp(-CT) method, where CT is the threshold 1
cycle (Livak and Schmittgen, 2001). The stability of ubiquitin expression during the 2
treatments was confirmed with eIF-4A (eukaryotic initiation factor 4A) and PP2A 3
(subunit of the Ser/Thr protein phosphatase 2A) as additional reference genes. 4
5
Biochemical characterization of LjGpxs 6 7
Expression and purification of recombinant proteins. LjGpx1 and LjGpx3 fragments 8
encoding the predicted mature proteins (Supplementary Fig. S1) were amplified by PCR 9
from nodule cDNA using PfuUltra II DNA polymerase (Agilent) and primers 10
(Supplementary Table S1) compatible with pET200 directional TOPO expression kits 11
(Invitrogen). Protein expression was induced in Escherichia coli BL21 (DE3) by the 12
addition of 1 mM isopropyl--D-thiogalactopyranoside for 4 h at 37ºC. Bacteria were 13
harvested by centrifugation, resuspended in 50 mM potassium phosphate (pH 8.0) 14
containing 300 mM NaCl and 40 mM imidazole, and sonicated 6 x 30 s. Extracts were 15
cleared by centrifugation and supernatants were loaded onto HiTrap chelating HP Ni-16
affinity columns (GE Healthcare Life Sciences). The His-tagged proteins were eluted 17
with buffer supplemented with 250 mM imidazole, desalted, and concentrated by 18
ultrafiltration. 19
20 Biochemical assays. LjGpx1 and LjGpx3 activities were determined by monitoring 21
NADPH oxidation at 340 nm (extinction coefficient = 6.22 mM-1 cm-1) under steady-state 22
conditions. The reaction mixture comprised TE buffer [30 mM Tris-HCl (pH 8.0), 1 mM 23
EDTA], 1 M A. thaliana NADPH-Trx reductase, 20 M poplar (Populus trichocarpa) 24
Trxh1, 150 nM recombinant enzymes, and 0.4 mM NADPH (Navrot et al., 2006). The 25
activities were recorded using 0.5-30 μM phosphatidylcholine hydroperoxide and 5-1000 26
μM H2O2, t-butyl hydroperoxide, and cumene hydroperoxide. Phosphatidylcholine 27
hydroperoxide was synthesized as described by Maiorino et al. (1990) and its 28
concentration standardized by the FOX colorimetric method (Wolff, 1994). The Gpx 29
activity was determined after subtracting the spontaneous reduction rate observed in the 30
absence of Gpx. The apparent Km and Vmax values were calculated by nonlinear 31
regression using a Michaelis-Menten equation. To study the effect of S-nitrosylation on 32

7
enzyme activities, recombinant LjGpx1 and LjGpx3 were treated with 1 mM GSNO or 1
GSSG (control) for 1 h at 37ºC in the dark. Excess reagents were removed by 2
ultrafiltration and enzyme activity was assayed with H2O2 as described above. 3
4 Interaction of LjGpxs with endogenous Trxs. The procedure of Balmer et al. (2003) was 5
followed as shown schematically in Figure 1. The L. japonicus Trxh4 (LjTrxh4) and a 6
mutated derivative (Cys-60-Ser), produced by site-directed mutagenesis, were cloned 7
using specific primers (Supplementary Table S1) as indicated for LjGpxs. Both proteins 8
had an N-terminal poly-His tag and were purified by Ni-affinity chromatography. 9
Purified Trxs (7 mg) were bound to CNBr-activated Sepharose 4B (1.25 g Sepharose) 10
following the manufacturer's instructions (GE Healthcare Life Sciences). Extracts of L. 11
japonicus nodules were prepared in TE + protease inhibitor cocktail (Roche). The 12
extracts were cleared by centrifugation and the supernatants were separated into two 13
fractions, which were passed through the columns containing either the wild-type or the 14
mutated proteins. The columns were previously washed with TE + 2 mM DTT to ensure 15
complete reduction of bound Trxs, and then with TE alone to remove excess DTT. The 16
nodule extracts (25-40 mg of protein) were passed continuously overnight through the 17
columns, which were afterwards washed with five volumes of TE buffer and another five 18
volumes of TE + 500 mM NaCl. The bound proteins were then eluted with TE + 10 mM 19
DTT and identified by liquid chromatography coupled to tandem mass spectrometry (LC-20
MS/MS) in both data-dependent and target acquisition modes. In the latter case, between 21
four and nine tryptic peptides were searched for each LjGpx protein. The MS instrument 22
was a Velos LTQ (Thermo Scientific) equipped with a microelectrospray ionization 23
source. Samples containing 2 g protein were diluted up to 20 l with 5% methanol and 24
1% formic acid, and loaded on the chromatographic system. Details of the 25
chromatography and detection conditions are given in Sainz et al. (2015). 26
27
Complementation of LjGpxs in yeast 28 29 Methods for yeast (Saccharomyces cerevisiae) manipulation and for preparation of rich 30
Yeast extract-Peptone-Dextrose medium (YPD) and minimal Synthetic-Dextrose growth 31
medium (SD) were as described by Guthrie and Fink (1991). YPD was used for 32
experiments and SD for selecting transformant colonies and precultures. 33

8
Complementation with LjGpx1 and LjGpx3 was carried out with the triple deletion 1
mutant gpx1/2/3 (MATa his31 leu2 met15 ura30 gpx1::URA3 2
gpx2::His3MX6 gpx3::KanMX6) derived from the BY4741 strain (Avery and Avery, 3
2001). The constructs encoding the mature LjGpx1 and LjGpx3 proteins were cloned into 4
pENTR/D-TOPO and recombined into the yeast expression vector pAG425GPD-ccdB 5
using Gateway and LR Clonase II. The mutant strain was transformed with the constructs 6
by the lithium acetate-polyethylene glycol method (Gietz and Woods, 2002). Growth 7
assays were performed in solid YPD medium by spotting serial dilutions of saturated 8
cultures onto plates with the concentrations of stress inducers and the exposure times as 9
indicated. The peroxides were added on top of the solidified medium, whereas NaCl and 10
caffeine were added prior to autoclaving. Linolenic acid was prepared from a 11
concentrated stock in YPD medium containing 1% (w/v) tergitol (Avery and Avery, 12
2001) and supplied to the medium after autoclaving but prior to gelification. Control 13
experiments with media supplemented with 1% tergitol alone showed no effect on yeast 14
growth. 15
16
Localization of LjGpx transcripts and proteins in nodules 17 18 In situ RNA hybridization. Antisense and sense digoxigenin-labeled RNA probes based 19
on gene-specific primers (Supplementary Table S1) were synthesized using the DIG 20
RNA Labeling Kit (Roche). The protocols of Bustos-Sanmamed et al. (2013) were 21
followed and the process was fully automated with an InsituPro VSi instrument (Intavis, 22
Germany). Nodule sections were examined with a DMI6000 B inverted microscope 23
(Leica). 24
25 Immunoblots. Antisera were raised in rabbits with c. 1 mg of purified recombinant LjGpx1 26
and LjGpx3 proteins and were used to purify polyclonal monospecific antibodies by 27
chromatography in CNBr-activated Sepharose 4 Fast Flow following conventional 28
protocols (BioGenes, Germany). The antibodies were further purified by 29
immunoadsorption with protein extracts of E. coli. Immunoblots were performed as 30
described (Rubio et al., 2009). The secondary antibody was a goat anti-rabbit IgG 31
horseradish peroxidase conjugate (Sigma). The primary and secondary antibodies were 32
used at dilutions of 1:500 and 1:20000, respectively, and immunoreactive proteins were 33

9
detected by chemiluminescence. 1
2 Immunogold localization. Nodules were fixed in 4% paraformaldehyde and 0.1% 3
glutaraldehyde in 50 mM sodium cacodylate buffer (pH 7.0). Procedures for sample 4
dehydration in ethanol and infiltration in LR White resin at low temperatures were 5
performed in a Leica AFS2 as described (Rubio et al., 2009; Sainz et al., 2013). Ultrathin 6
sections were collected on pyroxylin-coated Ni-grids and incubated for 1 h with each 7
antibody diluted 1:10 in blocking/diluting buffer. The sections were then washed and 8
incubated for 1 h with 15-nm gold particles conjugated to protein A (BB International, 9
UK) diluted 1:100 in the same buffer (Rubio et al., 2009). Serial sections treated with 10
non-immune serum substituting for LjGpx antibodies served as negative controls. 11
Sections were viewed and digitally photographed using a JEM 1400 transmission electron 12
microscope (JEOL, Japan). 13
14 Localization using GFP fusions and protoplast transformation. The open reading frames 15
of LjGpx1 and LjGpx3, bearing the sequences encoding the putative transit peptides, were 16
amplified by PCR, cloned into pENTR/D-TOPO (Invitrogen), and recombined into the 17
Gateway binary vector pGWB5 (Nakagawa et al., 2007) with LR Clonase II (Invitrogen). 18
In these constructs, the green fluorescent protein (GFP) was translationally fused at the 19
C-terminus of the LjGpx proteins and the expression of the fusion protein was driven by 20
the cauliflower mosaic virus 35S promoter. Mesophyll protoplasts were isolated from A. 21
thaliana leaves and 5 g plasmid DNA was delivered into protoplasts by the downsized 22
polyethylene glycol-mediated transfection method (Seidel et al., 2004). Subcellular 23
localization was visualized using a confocal laser scanning microscope (LSM 780, Zeiss, 24
Germany) using excitation at 488 nm (GFP and chlorophyll) and emission at 499-535 nm 25
(GFP) and 650-700 nm (chlorophyll). 26
27
Detection of S-nitrosylation of LjGpx1 and LjGpx3 28
29 This was performed using the biotin (Jaffrey et al., 2001) and His-tag (Camerini et al., 30
2007) switch assays. Biotin switch assay. Recombinant LjGpx1 and LjGpx3 were diluted 31
to 1 mg ml-1 in HEN buffer and incubated with 1 mM GSNO or GSSG (control) for 1 h at 32
37ºC in the dark with shaking. Reagents were removed by acetone precipitation and two 33

10
washes with ice-cold acetone. Free thiols were blocked in HEN buffer with 100 mM N-1
ethylmaleimide (NEM) and 2.5% SDS for 1 h at 37ºC in the dark with shaking. Excess 2
NEM was removed by acetone precipitation/washing and proteins were solubilized in 3
HENS buffer (HEN + 1% SDS). The biotin switch was performed for 1 h at 37ºC in the 4
dark in HENS buffer containing 20 mM ascorbate and 0.25 mg ml-1 HPDP-Biotin 5
(Pierce). Excess reagents were removed by acetone precipitation and washing. Proteins 6
were resuspended in HENS buffer, separated on 15% SDS gels, and transferred onto 7
polyvinylidene fluoride membranes. Anti-biotin antibody (Sigma) was used at 1:10000. 8
His-tag switch assay. Incubation with GSNO and derivatization of free thiols with 100 9
mM NEM were as described for the biotin switch but replacing biotin by the alkylating 10
peptide I-CH2-CO-Gly-Arg-Ala-(His)6. After incubation for 1 h at 37ºC in the dark, 11
proteins were dialyzed overnight in 10 mM NH4HCO3, concentrated, and analyzed by 12
matrix-assisted laser desorption/ionization time-of-flight MS. Affinity purification of 13
biotinylated proteins. Nodulated plants were treated with 5 mM GSNO or GSSG for 6 h. 14
Proteins were extracted in HEN buffer with 0.2% SDS and protease inhibitors, and 15
subjected to the biotin switch. Dry pellets were resuspended in binding buffer consisting 16
of 25 mM HEPES (pH 7.7), 1 mM EDTA, 100 mM NaCl, 0.8% Triton X-100, and 50 l 17
of streptavidin-agarose resin (Sigma). Samples were incubated overnight at 4ºC and then 18
the agarose beads were washed ten times with a buffer comprising 25 mM HEPES (pH 19
7.7), 1 mM EDTA, 600 mM NaCl, and 0.8% Triton X-100. Biotinylated proteins were 20
eluted by boiling the beads for 10 min in SDS loading buffer [50 mM Tris-HCl (pH 6.8), 21
10% glycerol, 1% SDS, 0.01% bromophenol blue, 50 mM DTT]. After centrifugation, 22
proteins were separated on 15% SDS gels and transferred to membranes for immunoblot 23
analysis with LjGpx antibodies. 24
25
26
Results 27
28
LjGpx1 and LjGpx3 are Trx-dependent phospholipid hydroperoxidases 29
30 Previous work had shown that LjGpx1 and LjGpx3 are highly expressed in nodules 31
(Ramos et al., 2009) and that the LjGpx3 mRNA level is 6.8-fold greater in nodules than 32

11
in uninfected roots (Colebatch et al., 2002). These observations prompted us to focus on 1
the function of LjGpx1 and LjGpx3. To this end, recombinant enzymes were produced 2
and their activities assayed toward various hydroperoxides using Trx and GSH as 3
potential electron donors. However, no LjGpx activity was detected with GSH as 4
reductant and with H2O2 or organic peroxides as substrates, and hence further work was 5
done exclusively with Trx. Kinetic analyses indicated that the two LjGpx isoforms 6
catalyze the Trx-dependent reduction of H2O2, t-butyl hydroperoxide, and cumene 7
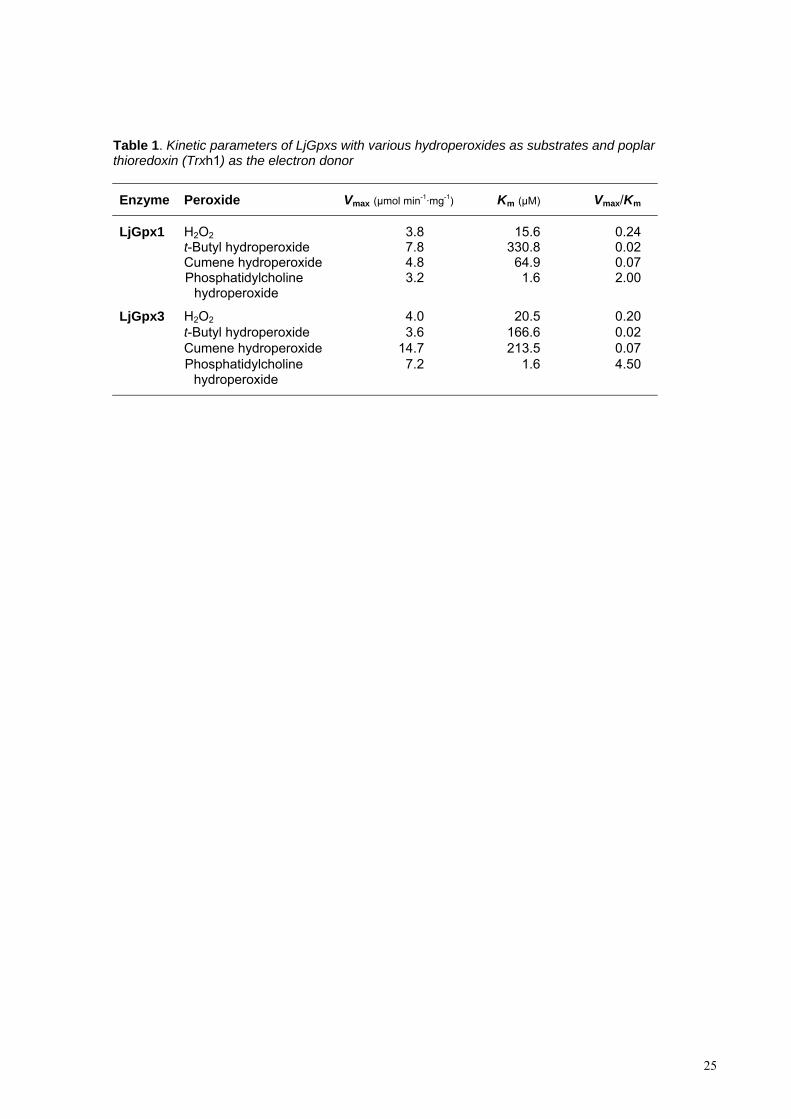
hydroperoxide (Table 1). The apparent affinities of both isoforms for organic peroxides 8
(Km ~60-300 M) were lower than for H2O2 (Km ~20 M). The opposite trend was seen 9
for the maximum velocities (Vmax), with apparent values of ~4 mol min-1 mg-1 protein 10
for H2O2 and 4-15 mol min-1 mg-1 protein for organic peroxides. The apparent affinity 11
of LjGpx1 and LjGpx3 for phospholipid hydroperoxides was much higher (Km ~1.6 M) 12
and the apparent Vmax of LjGpx3 doubled that of LjGpx1. As a result, the Vmax/Km ratios 13
of the two enzymes, which are an indication of their catalytic efficiencies, were very high 14
for lipid peroxides (2-4.5), low for H2O2 (0.2), and very low for organic peroxides (0.02-15
0.07) (Table 1). All these data led us to conclude that LjGpx1 and LjGpx3 are Trx-16
dependent phospholipid hydroperoxidases. 17
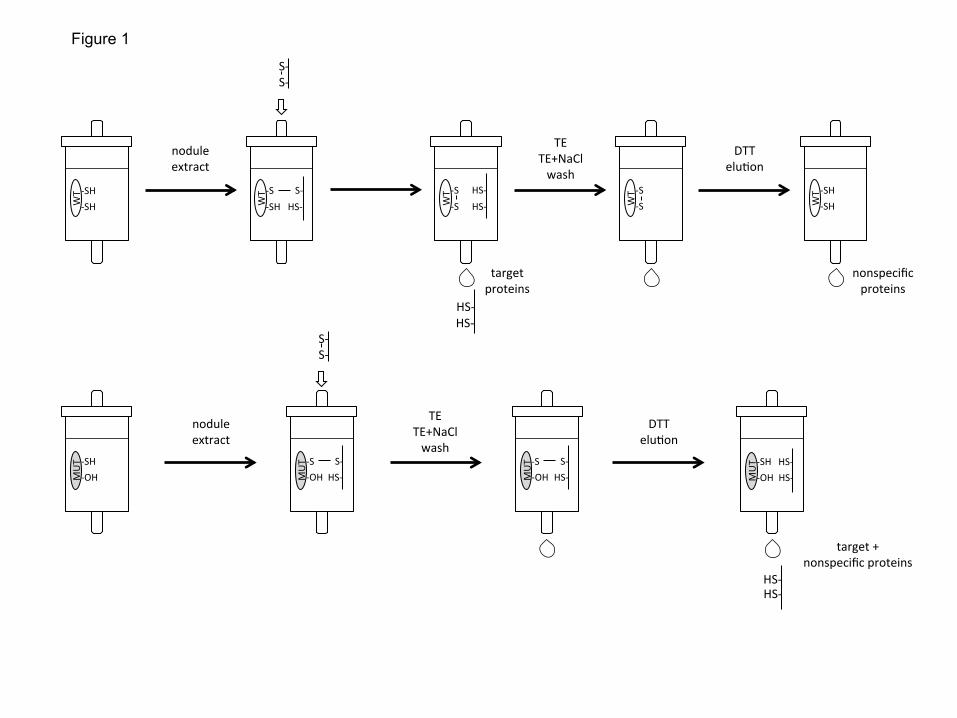
The interaction between LjGpxs and endogenous Trxs was demonstrated using the 18
cytosolic isoform LjTrxh4 that is highly expressed in nodules (Tovar-Méndez et al., 19
2011). The Cys-60-Ser derivative of LjTrxh4 was used for an affinity binding assay 20
based on the formation of a stable heterodisulfide bond between the remaining Cys of the 21
active site and the Cys residues of the target proteins (Fig. 1; Balmer et al., 2003). The 22
wild-type protein served as a control for nonspecific binding. In three independent 23
preparations of nodules, LjGpx1 and LjGpx3 were identified as protein targets because 24
they become covalently bound to mutated LjTrxh4 but not to the wild-type protein (Table 25
2), which indeed supports the thioredoxin-dependency of the two LjGpx isoforms. 26
LjGpx2 was also found to be a target of LjTrxh4 (Table 2), but LjGpx4, LjGpx5, or 27
LjGpx6 (Ramos et al., 2009) were not detected even though the highly sensitive target 28
mode was used in the MS analysis. 29
30

12
LjGpx1 and LjGpx3 are differentially expressed in response to nitro-1
oxidative stress and hormones, and the proteins protect against oxidative 2
damage 3 4 Because both LjGpx isoforms are very active in reducing phospholipid hydroperoxides to 5
innocuous lipid alcohols, they may protect cells from oxidative damage. This hypothesis 6
was tested by two experimental approaches. 7
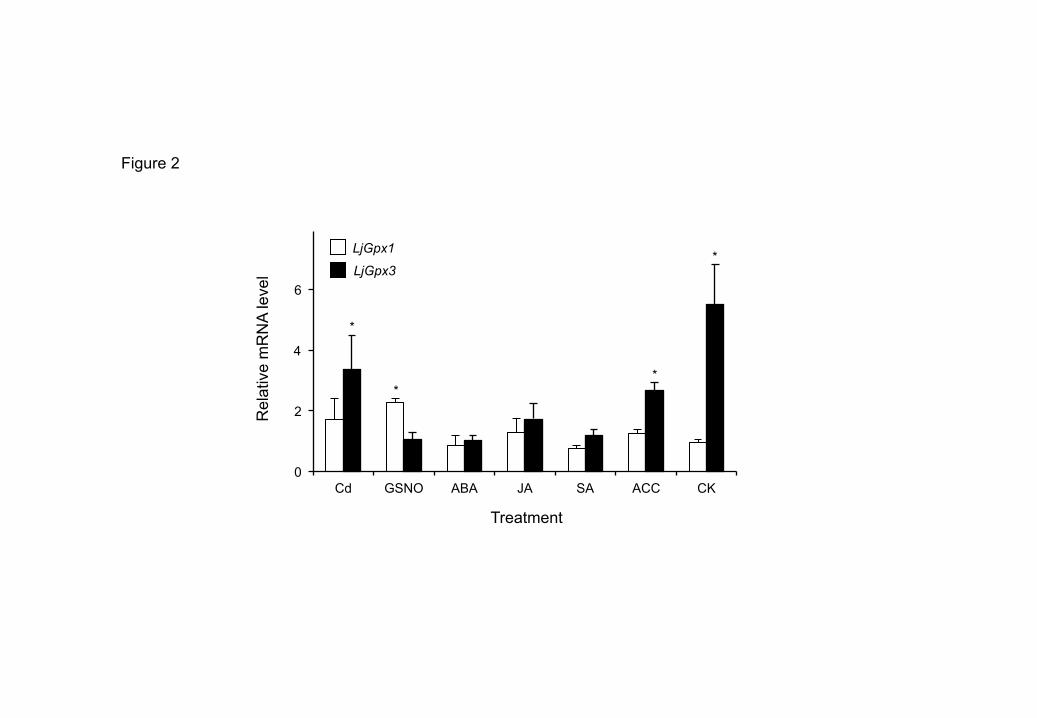
Firstly, the expression of LjGpx1 and LjGpx3 in nodules was analyzed under nitro-8
oxidative stress elicited by Cd, a heavy metal that promotes ROS production (Romero-9
Puertas et al., 2004) or by GSNO, a NO-releasing metabolite implicated in trans-10
nitrosylation reactions (Perazzolli et al., 2004). LjGpx3 was upregulated by Cd whereas 11
LjGpx1 was responsive to GSNO (Fig. 2), which reflects differential gene regulation and 12
strongly suggests an antioxidative role for the proteins. The effects of phytohormones at a 13
physiologically relevant concentration (50 M) on gene expression were also compared 14
because at least some of them are mediated by NO (Bright et al., 2006). LjGpx3 but not 15
LjGpx1 was upregulated in nodules of plants treated with the ethylene precursor ACC or 16
with CK, whereas the expression of the two genes was not affected by the stress signaling 17
compounds ABA, JA, and SA (Fig. 2). 18
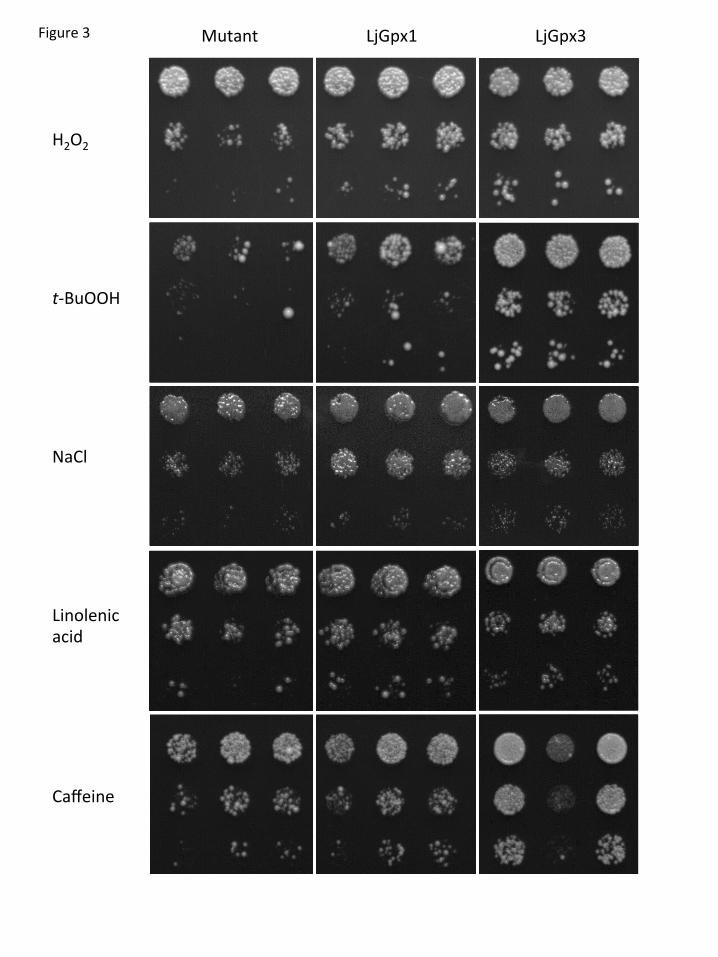
The second approach to assess the antioxidative role of LjGpxs was to perform 19
functional complementation in yeast (Fig. 3). This strategy is extensively used with plant 20
proteins which in most cases are functional in yeast (Serrano et al., 1999). Moreover, the 21
use of yeast enabled us to examine directly the effects of LjGpx expression in a 22
completely Gpx-null background. Thus, the effects of two peroxides, which are LjGpx 23
substrates, on the growth of the yeast Gpx-deficient mutant and the corresponding 24
transformed cells were investigated (Fig. 3). The concentrations of H2O2 and t-butyl 25
hydroperoxide were optimized to maximize differences in phenotype. Both LjGpx1 and 26
LjGpx3 complemented the defective growth of the mutant in the presence of H2O2 or t-27
butyl hydroperoxide (compare growth at the highest dilution, 1:1000). While the 28
protection afforded by LjGpx1 and LjGpx3 against H2O2 was similar, LjGpx3 had a 29
greater protective effect against t-butyl hydroperoxide. Likewise, yeast cells expressing 30
LjGpx1 and LjGpx3 exhibited better growth than the mutant under salt stress induced by 31
NaCl (Fig. 3). To determine whether LjGpx1 and LjGpx3 improve tolerance to stress 32

13
imposed on the plasma membrane and/or the cell wall, yeast cells were treated with 1
linolenic acid and caffeine. Yeast cells are unable to synthesize polyunsaturated fatty 2
acids but incorporate exogenously added linolenic acid into the membranes, making them 3
prone to peroxidation (Avery and Avery, 2001). Also, caffeine induces alteration of the 4
yeast cell wall architecture and may affect membrane integrity (Kuranda et al., 2006). 5
Cells expressing either of the two LjGpx proteins showed greater tolerance to linolenic 6
acid and caffeine than the mutant strain (Fig. 3). 7
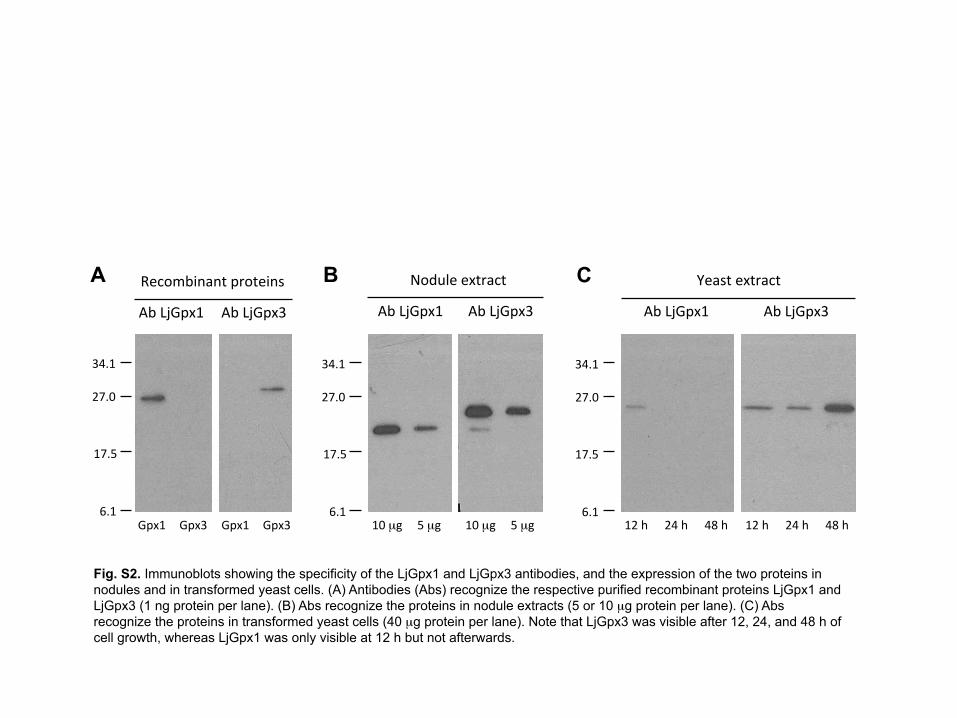
To compare expression levels of LjGpx1 and LjGpx3 in yeast, antibodies were 8
produced and used on immunoblots (Supplementary Fig. S2). Because the antibodies 9
were intended to be employed also for immunolocalization studies of the LjGpxs, which 10
require very high specificity, monospecific antibodies were purified from antisera by 11
affinity chromatography and then repurified by immunoadsorption with E. coli protein 12
extracts. This was necessary because very minor amounts of E. coli proteins inevitably 13
contaminated the recombinant LjGpxs employed to raise the antibodies. The resulting 14
antibodies specifically recognized recombinant LjGpx1 and LjGpx3 (Supplementary Fig. 15
S2A) and the respective proteins of nodule extracts (Supplementary Fig. S2B). The use of 16
these antibodies on immunoblots of yeast extracts revealed that LjGpx3 was expressed 17
during the 48 h of treatment with the peroxides and the other stress inducers, whereas 18
LjGpx1 was only detectable at 12 h and was probably degraded thereafter 19
(Supplementary Fig. S2C). This may explain the higher tolerance to t-butyl 20
hydroperoxide of yeast cells expressing LjGpx3 with respect to those expressing LjGpx1 21
(Fig. 3). 22
23
LjGpx1 and LjGpx3 mRNAs are abundant in the nodule infected zone and 24
the proteins are localized to various subcellular compartments 25 26 In situ hybridization of mature nodules of L. japonicus was used to localize the mRNAs 27
encoding the two LjGpx isoforms. The LjGpx1 (Fig. 4A, C) and LjGpx3 (Fig. 4E, G) 28
mRNAs were found to be preferentially localized to the infected zone. Besides, 29
significant amounts of LjGpx3 mRNA could be detected in the nodule cortex and in the 30
vascular bundles (Fig. 4E, G). However, in the case of LjGpx1, the control probe 31
produced a signal in the cortex (Fig. 4B, D) and hence some non-specific signal in this 32

14
nodule tissue cannot be ruled out. No background signal was seen for LjGpx3, confirming 1
genuine expression of this gene in the nodule cortex (Fig. 4F, H). 2
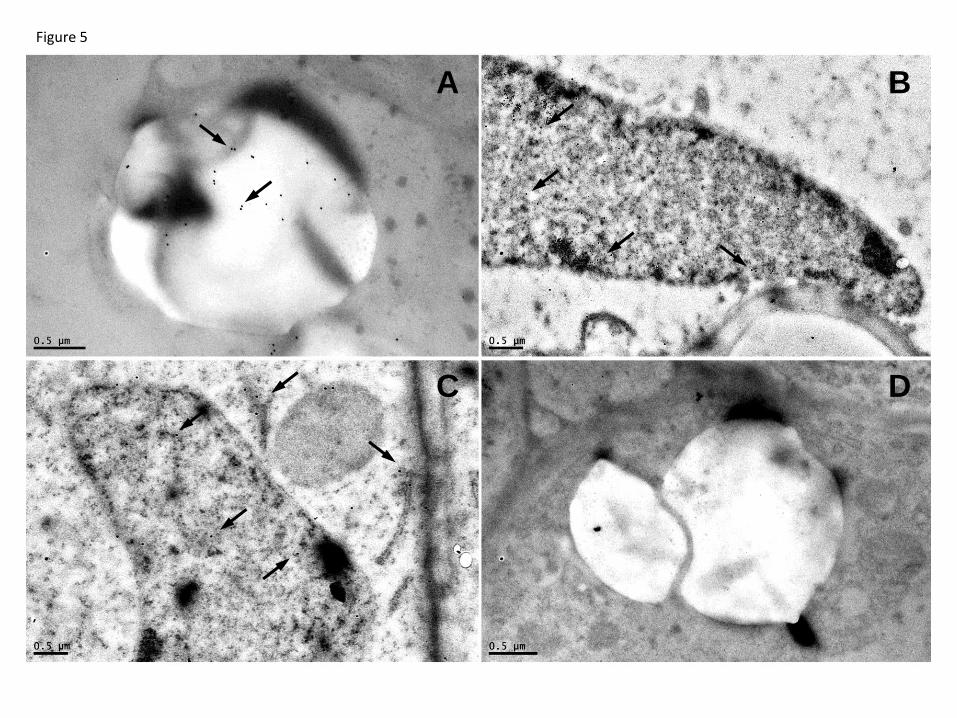
The LjGpx1 and LjGpx3 proteins were immunolocalized using our highly purified 3
antibodies (Fig. 5). For LjGpx1, gold labeling was evident in the amyloplasts (Fig. 5A) 4
and nuclei (Fig. 5B) of infected cells, cortical cells, and vascular bundle cells. For 5
LjGpx3, gold particles were mainly associated to the endoplasmic reticulum, cytosol, and 6
nuclei (Fig. 5C). A control in which preimmune serum replaced the primary antibodies 7
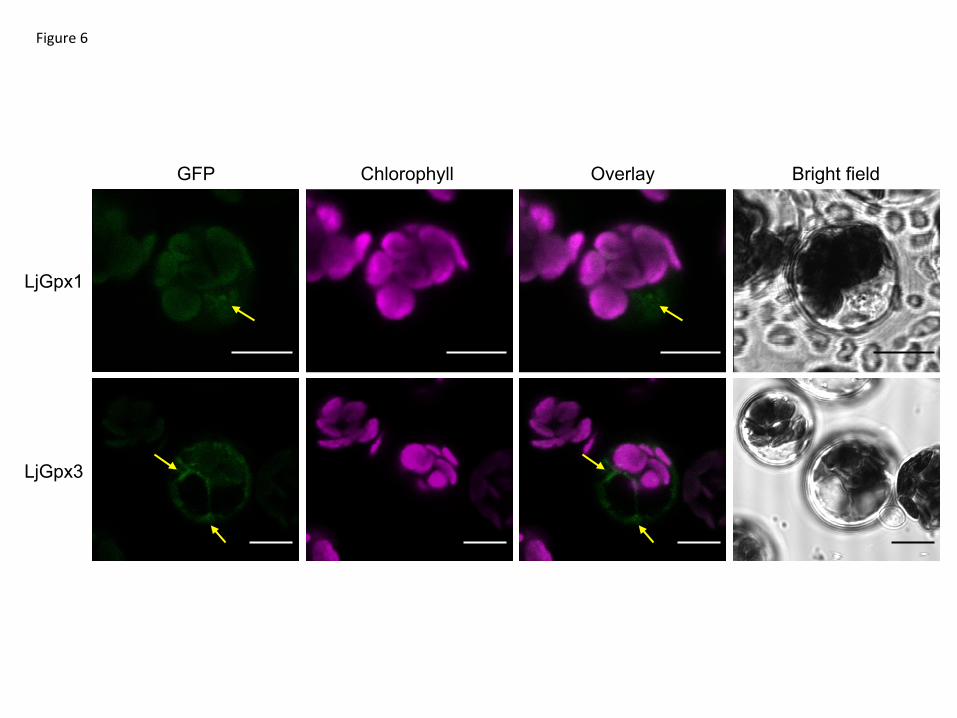
did not show any labeling in the amyloplasts or nuclei (Fig. 5D). The immunolocalization 8
study was complemented with fluorescence detection of the LjGpx-GFP fusion proteins 9
by confocal microscopy. The constructs were transfected into A. thaliana mesophyll 10
protoplasts (Fig. 6). GFP fluorescence was observed predominantly in the nuclei for 11
LjGpx1 (Fig. 6A) and in the cytosol for LjGpx3 (Fig. 6B). 12
13
LjGpx1 and LjGpx3 are nitrosylated in vitro and in vivo, which results in 14
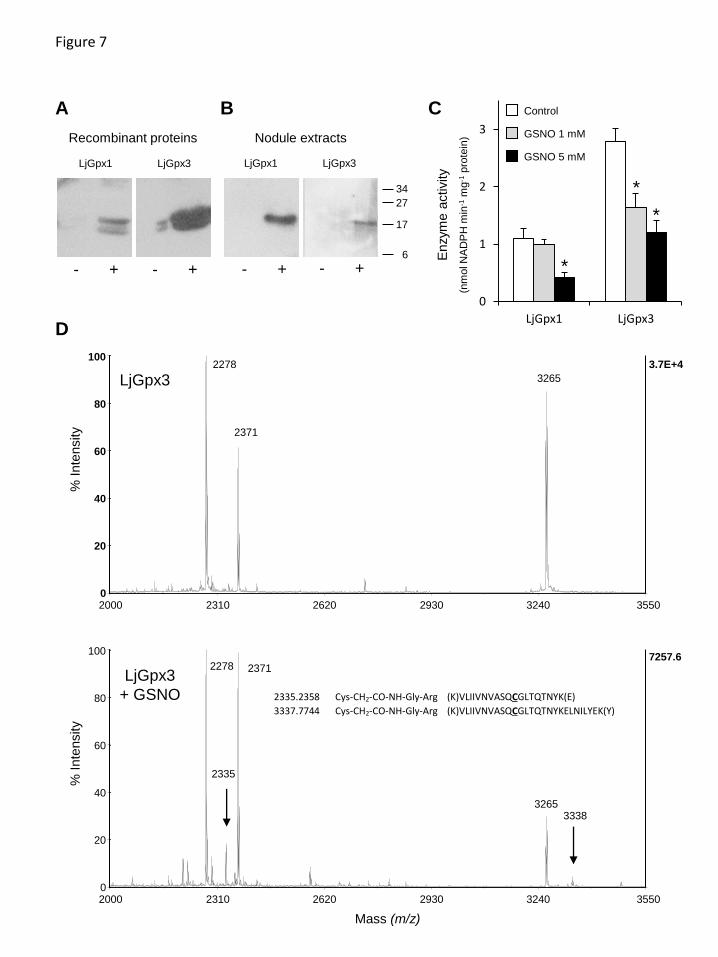
inhibition of enzyme activities 15 16 Protein S-nitrosylation is an important mechanism by which NO exerts regulatory 17
functions in all organisms (Astier et al., 2012). To elucidate whether the biological 18
activities of LjGpx1 and LjGpx3 could be modulated by NO, recombinant proteins were 19
treated with 1 mM GSNO and S-nitrosylation was evaluated with the biotin switch assay. 20
Immunoblots showed that both proteins can be nitrosylated in vitro to some extent (Fig. 21
7A). Also, this treatment caused a 40% reduction of LjGpx3 activity but had no effect on 22
LjGpx1 activity, whereas raising the GSNO concentration to 5 mM resulted in a 60% loss 23
of both LjGpx activities (Fig. 7C). Because the biotin switch does not permit the 24
identification of nitrosylated Cys, the His-tag switch was used. This method involves 25
derivatization of nitrosylated residues with a synthetic peptide. After trypsin digestion, 26
the dipeptide Gly-Arg remains bound to Cys and can be detected by MS (Camerini et al., 27
2007). The analysis demonstrated nitrosylation of Cys-85 in LjGpx3 (Fig. 7D), but could 28
not prove the equivalent nitrosylation in LjGpx1 (see Supplementary Fig. S1 for 29
numbering of Cys in the proteins). 30
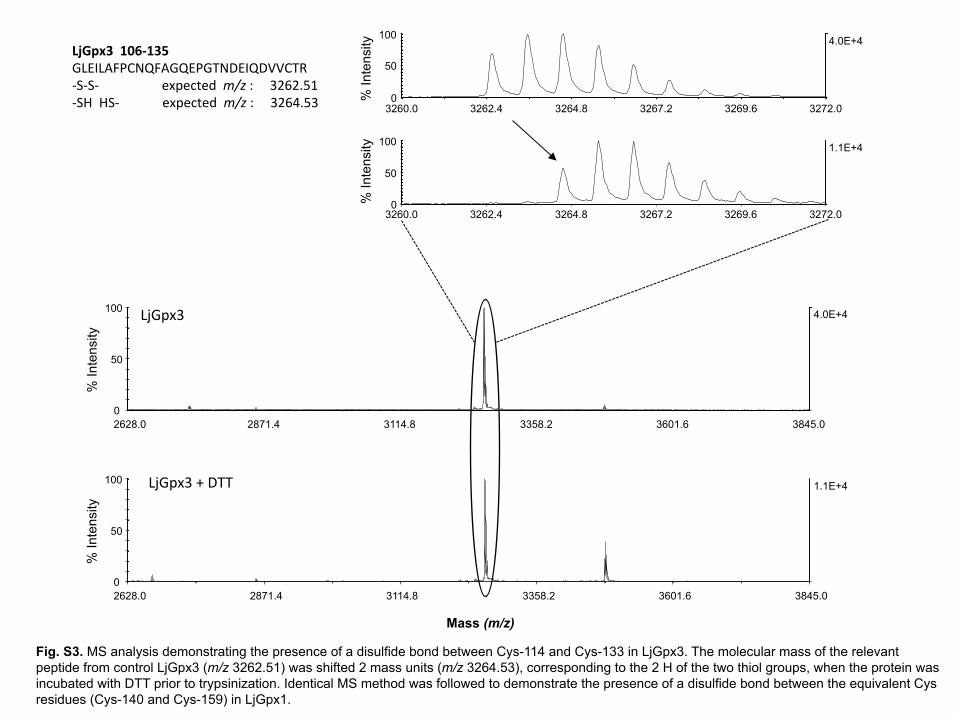
In this analysis, a disulfide bond was detected between Cys-140 and Cys-159 in 31
LjGpx1 and between Cys-114 and Cys-133 in LjGpx3. Addition of DTT before 32

15
trypsinization to reduce the disulfide increased the peptide molecular mass by 2 Da, 1
confirming the existence of the intramolecular bond (Supplementary Fig. S3). Further 2
controls were run by adding DTT to make all Cys accessible for nitrosylation and then 3
removing it prior to the biotin switch. This result further proved that nitrosylation was 4
restricted to Cys-85 of LjGpx3. The presence of disulfide bonds in LjGpx1 and LjGpx3 5
was already apparent on immunoblots of recombinant proteins or nodule extracts, in 6
which two immunoreactive bands (reduced and oxidized forms) were seen for each 7
protein (Fig. 7A). 8
Nodule extracts and intact plants were also incubated with 5 mM GSNO for 6 h and 9
nodule proteins were subjected to the biotin switch assay. Biotinylation of Cys residues 10
was observable after GSNO treatment of plants, but neither in nodule extracts nor in 11
nodules of plants treated with GSSG (control), indicating that LjGpx1 and LjGpx3 are 12
also amenable to nitrosylation in vivo (Fig. 7B). 13
Discussion 14
15 Legume nodules are endowed with major antioxidant defenses to keep ROS and NO 16
under control, thus allowing the onset and functioning of symbiosis. In this work, LjGpx1 17
and LjGpx3, two isoforms abundantly expressed in nodules, were found to catalyze the 18
efficient reduction of organic and lipid peroxides using Trx, but not GSH, as reductant 19
(Table 1). Both enzymes are, therefore, Trx-dependent phospholipid hydroperoxidases, 20
like other plant Gpxs for which kinetic parameters have been measured (Herbette et al., 21
2002; Jung et al., 2002; Navrot et al., 2006). By using an affinity binding assay with a 22
Cys-mutated Trx as a bait, it was shown that LjTrxh4 forms intermolecular disulfide 23
bonds with LjGpx1, LjGpx2, and LjGpx3 (Table 2), and hence that Trxs may act as in 24
vivo reductants of LjGpxs. Although the specificity of the Trx isoform was not examined, 25
LjGpxs may be targets of other LjTrxs because a high degree of interchangeability in the 26
affinity column procedure was observed for poplar Trxs (Balmer et al., 2004). 27
Further support for an in vivo role of LjGpxs as phospholipid hydroperoxidases is 28
lent by complementation of a S. cerevisiae gpx mutant. This microorganism expresses 29
three Gpxs, all of them identified as phospholipid hydroperoxidases (Avery and Avery, 30
2001). Accordingly, the triple-deletion mutant strain is hypersensitive to H2O2 and t-butyl 31
hydroperoxide. Expression of LjGpx1 or LjGpx3 in the mutant yeast conferred greater 32

16
tolerance to both peroxides (Fig. 3), indicating that the enzymes are functional. 1
Furthermore, it indicates that LjGpxs successfully recruit endogenous Trxs as reductants. 2
Because LjGpxs also afforded protection against linolenic acid, which sensitizes 3
membranes to lipid peroxidation (Avery and Avery, 2001), and against caffeine, which 4
also causes membrane lesions (Kuranda et al., 2006), it is concluded that these enzymes 5
protect against lipid peroxidation. Likewise, the beneficial effect of LjGpxs in yeast 6
treated with high NaCl concentrations (Fig. 3) may be indirect and attributable to the Gpx 7
capacity to offset oxidative stress, as proposed for the Gpx of salt-tolerant Citrus sinensis 8
(Gueta-Dahan et al., 1997; Avsian-Kretchmer et al., 2004). 9
Because LjGpx1 and LjGpx3 show similar kinetic properties and are unlikely to be 10
entirely redundant, the proteins may be differentially regulated by developmental and 11
environmental cues, and/or be localized in different tissues, cells, or organelles. All this 12
was found to be true. Thus, in nodules, LjGpx1 was induced by NO and LjGpx3 by Cd 13
and some hormones (Fig. 2). Earlier work in our laboratory showed that Gpx1 is down-14
regulated and Gpx3 is up-regulated in roots of non-nodulated L. corniculatus plants 15
treated with 20 M Cd in hydroponic cultures (Ramos et al., 2009). Although results are 16
difficult to compare due to differences in plant species and tissues and in growth and 17
treatment conditions, they show a consistent induction of the Gpx3 gene with Cd in the 18
two legumes. In A. thaliana, Gpx3 (At2g43350) is involved in the ABA response (Miao 19
et al., 2006) and expression of Gpx4 (At2g48150) and Gpx7 (At4g31870) is increased 20
upon auxin application (Passaia et al., 2014). In the present work, strong up-regulation of 21
LjGpx3 was seen with CK and less intense with ACC, but ABA had no effect. None of 22
the tested hormones altered LjGpx1 expression when applied at a high concentration of 23
50 M for 48 h (Fig. 2). A direct comparison of results obtained with A. thaliana and L. 24
japonicus is complicated because the functional equivalence of Gpx isoforms is uncertain 25
(Rodriguez Milla et al., 2003; Ramos et al., 2009). However, based on the observations 26
made with both model plants, it may be suggested that LjGpx1 and LjGpx3 isoforms 27
have functions beyond antioxidative defense (see also discussion below). In particular, 28
they might participate in signaling during plant development because their transcripts 29
accumulated in response to hormones in healthy, non-stressed plants, which do not 30
require an extra provision of antioxidants. 31

17
In a previous report, Gpx proteins were detected in root and nodule amyloplasts and 1
in leaf chloroplasts of L. japonicus and other legumes (Ramos et al., 2009) using an 2
antibody raised against poplar Gpx3.2 (Navrot et al., 2006). However, this antibody was 3
not isoform specific and probably recognized several LjGpx proteins. In the present 4
study, the tissue, cellular, and subcellular localizations of LjGpx1 and LjGpx3 were 5
examined using mRNA in situ hybridization (Fig. 4), immunoelectron microscopy (Fig. 6
5), and fluorescence detection of GFP-tagged proteins (Fig. 6). The mRNA and protein 7
levels of LjGpx1 and LjGpx3 are highest in the nodule infected zone. This pattern is in 8
line with their requirement for antioxidative protection in the host cells, which contain 9
copious amounts of symbiosomal membranes prone to peroxidation (Puppo et al., 1991). 10
Chloroplastic, cytosolic, and/or mitochondrial Gpxs have been reported in other vascular 11
plants (Mullineaux et al., 1998; Herbette et al., 2004; Navrot et al., 2006). In silico 12
analyses predict that LjGpx1 bears a transit peptide for possible targeting to the plastids 13
and mitochondria and that LjGpx3 is located to the cytosol and secretory pathway 14
(Supplementary Fig. S1). For LjGpx1, immunogold labeling was detected in the nodule 15
amyloplasts although GFP fluorescence was weak in A. thaliana chloroplasts. Neither 16
technique supported the presence of LjGpx1 in mitochondria. In contrast, both of them 17
indicated a nuclear localization. The immunolocalization study showed the presence of 18
LjGpx3 in nuclei but this could not be confirmed by GFP tagging. Until now, only 19
another plant Gpx, A. thaliana Gpx8 (At1g63460), was shown to be located to the 20
nucleus (Gaber et al., 2012). As for LjGpx3, immunogold labeling and GFP tagging were 21
consistent with in silico analysis, indicating that the protein is in the cytosol and 22
endoplasmic reticulum. 23
The differential regulation of LjGpx1 and LjGpx3 by the physiological NO donor 24
GSNO and by phytohormones, along with the localization of LjGpx1 in the nuclei, 25
provide indirect support for a role of LjGpxs beyond their antioxidant capacity. This 26
possibility was tested by a more direct approach aimed at determining whether LjGpxs 27
could be regulated by S-nitrosylation. The rationale for this set of experiments rests on 28
the observations that LjGpxs contain Cys residues essential for catalytic activity 29
(Supplementary Fig. S1) and that Cys nitrosylation is a major route for transmission of 30
NO bioactivity (Astier et al., 2012). In a first experiment, purified LjGpx1 and LjGpx3 31
were treated with GSNO and assayed for nitrosylation with the biotin switch (Fig. 7A). 32

18
The nitrosylation of LjGpx3 was confirmed with the His-tag switch followed by MS and 1
the target residue was identified as Cys-85 (Fig. 7D). However, LjGpx1 nitrosylation 2
could not be verified probably because MS was performed with proteins treated with 1 3
mM GSNO, a concentration at which LjGpx1 activity is not inhibited (Fig. 7C). Because 4
the biotin switch is a reliable and sensitive method (Astier et al., 2012), another likely 5
explanation is that the equivalent Cys residue of LjGpx1 is not readily accessible to the 6
peptide used for derivatization. In a second experiment, nodule extracts were treated with 7
GSNO or were made from plants treated with GSNO, and were assayed with the biotin 8
switch. The detection of nitrosylated LjGpxs indicates that both enzymes are targets of 9
nitrosylation in vitro and in vivo. An intriguing question is why nitrosylation of the 10
peroxidatic Cys (Cys-85) in LjGpx3 can take place while the resolving Cys (Cys-133) is 11
present. Maybe the conformational changes normally occurring to bring the resolving 12
Cys close to the peroxidatic Cys upon sulfenic acid formation cannot occur when Cys-85 13
is nitrosylated. The fact that nitrosylation of the peroxidatic Cys inhibits enzyme activity, 14
even in the presence of the Trx-reducing system, probably reflects the inability of Trx to 15
readily reduce the Cys-NO adduct. The MS analysis also pointed out the formation of a 16
disulfide bond between the second and third Cys in both LjGpxs (Supplementary Fig. 17
S3), as reported for a Chinese cabbage (Brassica rapa) Gpx (Jung et al., 2002). This 18
disulfide may regulate enzyme activity as it entails the third (resolving) Cys, required for 19
Trx-mediated regeneration (Navrot et al., 2006; Koh et al., 2007). None of the two other 20
possible internal disulfide bonds was detected. In particular, the disulfide bond between 21
the first and third Cys, essential for enzyme activity (Koh et al., 2007), may have been 22
missed because it involves two different tryptic peptides, which is often recalcitrant to 23
MS analysis. Overall, the observed GSNO-dependent inhibition of Gpxs may contribute 24
to the transient increase of the concentration of their targeted substrates, such as lipid 25
hydroperoxides, thus interconnecting NO and ROS signaling pathways, which are known, 26
for example, to play complementary roles during the plant immune response (Zaninotto et 27
al., 2006). 28
In summary, an extensive study of two Gpx isoforms abundant in legume nodules 29
has been conducted. LjGpx1 and LjGpx3 are phospholipid hydroperoxidases capable of 30
interacting in vitro with Trx endogenously present in nodules as Trxh4. The enzymes 31
protect from oxidative stress and membrane damage, are highly expressed in the nodule 32

19
infected cells, and are located to different cellular compartments. At least the LGpx1 1
isoform is present in the nucleus. Differential expression of LjGpx1 and LjGpx3 in 2
response to GSNO and hormones, localization in nuclei, and susceptibility to 3
nitrosylation of the catalytic Cys in vitro and probably in vivo provide strong support for 4
signaling roles in addition to their antioxidative properties. 5
6
Supplementary data
Supplementary data are available at JXB online
Figure S1. Amino acid sequence alignment of representative Gpxs mentioned in
this work.
Figure S2. Immunoblots showing the specificity of the LjGpx1 and LjGpx3
antibodies, and the expression of both proteins in nodules and in transformed yeast
cells.
Figure S3. MS analysis demonstrating the presence of a disulfide bond between
Cys-114 and Cys-133 in LjGpx3.
Table S1. Oligonucleotides used in this study.
Acknowledgements AS and PBS were the recipients of predoctoral (Formación de Personal Investigador) and
postdoctoral (Marie Curie) contracts, respectively. We thank Martin Crespi for help with
in situ RNA hybridization and Simon Avery for sharing the yeast mutant and for helpful
advice. This work was supported by Ministerio de Economía y Competitividad-Fondo
Europeo de Desarrollo Regional (AGL2011-24524 and AGL2014-53717-R). The
UMR1136 is supported by a grant overseen by the French National Research Agency
(ANR) as part of the "Investissements d'Avenir" program (ANR-11-LABX-0002-01, Lab
of Excellence ARBRE). MM and KJD acknowledge support within SPP1710. The

20
proteomic analysis was performed in the CSIC/UAB Proteomics Facility of IIBB-CSIC
that belongs to ProteoRed, PRB2-ISCIII, supported by grant PT13/0001.
References
Astier J, Kulik A, Koen E, Besson-Bard A, Bourque S, Jeandroz S, Lamotte O,
Wendehenne D. 2012. Protein S-nitrosylation: what’s going on in plants? Free Radical
Biology and Medicine 53, 1101–1110.
Avery AM, Avery SV. 2001. Saccharomyces cerevisiae expresses three phospholipid
hydroperoxide glutathione peroxidases. Journal of Biological Chemistry 276, 33730-33735.
Avsian-Kretchmer O, Gueta-Dahan Y, Lev-Yadun S, Ben-Hayyim G. 2004. The salt-stress
signal transduction pathway that activates the gpx1 promoter is mediated by intracellular
H2O2, different from the pathway induced by extracellular H2O2. Plant Physiology 135, 1685-
1696.
Balmer Y, Koller A, del Val, G., Manieri W, Schürmann P, Buchanan BB. 2003. Proteomics
gives insight into the regulatory function of chloroplast thioredoxins. Proceedings of the
National Academy of Sciences USA 100, 370-375.
Becana M, Matamoros MA, Udvardi M, Dalton DA. 2010. Recent insights into antioxidant
defenses of legume root nodules. New Phytologist 188, 960-976.
Brigelius-Flohé R, Maiorino M. 2013. Glutathione peroxidases. Biochimica Biophysica Acta
1830, 3289-3303.
Bright J, Desikan R, Hancock JT, Weir IS, Neill SJ. 2006. ABA-induced NO generation and
stomatal closure in Arabidopsis are dependent on H2O2 synthesis. The Plant Journal 45, 113–
122.
Broughton BJ, Dilworth MJ. 1971. Control of leghaemoglobin synthesis in snake beans.
Biochem Journal 125, 1075-1080.
Bustos-Sanmamed P, Laffont C, Frugier F, Lelandais-Brière C, Crespi M. 2013. Analyzing
small and long RNAs in plant development using non-radioactive in situ hybridization. In: De
Smet I, ed. Plant organogenesis: methods and protocols (Methods in Molecular Biology).
New York: Springer Science, vol. 959, 303-316.
Camerini S, Polci ML, Restuccia U, Usuelli V, Malgaroli A, Bachi A. 2007. A novel approach
to identify proteins modified by nitric oxide: the HIS-TAG switch method. Journal of
Proteome Research 6, 3224–3231.
Chang CCC, Slesak I, Jordá L, Sotnikov A, Melzer M, Miszalski Z, Mullineaux PM, Parker

21
JE, Karpinska B, Karpinski S. 2009. Arabidopsis chloroplastic glutathione peroxidases play
a role in cross talk between photooxidative stress and immune responses. Plant Physiology
150, 670-683.
Colebatch G, Kloska S, Trevakis B, Freund S, Altmann T, Udvardi MK. 2002. Novel aspects
of symbiotic nitrogen fixation uncovered by transcript profiling with cDNA arrays. Molecular
Plant-Microbe Interactions 15, 411-420.
Dalton DA. 1995. Antioxidant defenses of plants and fungi. In: Ahmad S, ed. Oxidative stress
and antioxidant defenses in biology. New York, USA: Chapman and Hall, 298-355.
Foyer CH, Noctor G. 2005. Oxidant and antioxidant signalling in plants: a re-evaluation of the
concept of oxidative stress in a physiological context. Plant Cell & Environment 28, 1056-
1071.
Fu L, Wang X, Eyal Y, She Y, Donald LJ, Standing KG, Ben-Hayyim G. 2002. A
selenoprotein in the plant kingdom. Journal of Biological Chemistry 277, 25983-25991.
Gaber A, Ogata T, Maruta T, Yoshimura K, Tamoi M, Shigeoka S. 2012. The involvement of
Arabidopsis glutathione peroxidase 8 in the suppression of oxidative damage in the nucleus
and cytosol. Plant Cell Physiology 53, 1596-1606.
Gietz, R.D., Woods, R.A. 2002. Transformation of yeast by lithium acetate/single-stranded
carrier DNA/polyethylene glycol method. Methods of Enzymology 350, 87-96.
Gueta-Dahan Y, Yaniv Z, Zilinskas BA, Ben-Hayyim G. 1997. Salt and oxidative stress:
similar and specific responses and their relation to salt tolerance in Citrus. Planta 203, 460-
469.
Guthrie C, Fink GR. 1991. Guide to yeast genetics and molecular and cell biology. New York:
Academic Press.
Herbette S, Lenne C, Leblanc N, Julien JL, Drevet JR, Roeckel-Drevet P. 2002. Two GPX-
like proteins from Lycopersicon esculentum and Helianthus annuus are antioxidant enzymes
with phospholipid hydroperoxide glutathione peroxidase and thioredoxin peroxidase activities.
European Journal of Biochemistry 269, 2414-2420.
Herbette S, Brunel N, Prensier G, Julien JL, Drevet JR, Roeckel-Drevet P. 2004.
Immunolocalization of a plant glutathione peroxidase-like protein. Planta 219, 784-789.
Herbette S, Roeckel-Drevet P, Drevet JR. 2007. Seleno-independent glutathione peroxidases.
More than simple antioxidant scavengers. FEBS Journal 274, 2163-2180.
Jaffrey SR, Erdjument-Bromage H, Ferris CD, Tempst P, Snyder SH. 2001. Protein S-
nitrosylation: a physiological signal for neuronal nitric oxide. Nature Cell Biology 3, 193–197.
Jung BG, Lee KO, Lee SS et al. 2002. A Chinese cabbage cDNA with high sequence identity to
phospholipid hydroperoxide glutathione peroxidases encodes a novel isoform of thioredoxin-

22
dependent peroxidase. Journal of Biological Chemistry 277, 12572-12578.
Koh CS, Didierjean C, Navrot N, Panjikar S, Mulliert G, Rouhier N, Jacquot JP, Aubry A,
Shawkataly O, Corbier C. 2007. Crystal structures of a poplar thioredoxin peroxidase that
exhibits the structure of glutathione peroxidases: insights int redox-driven conformational
changes. Journal of Molecular Biology 370, 512-529.
Kuranda K, Leberre V, Sokol S, Palamarczyk G, François J. 2006. Investigating the caffeine
effects in the yeast Saccharomyces cerevisiae brings new insights into the connection between
TOR, PKC and Ras/cAMP signalling pathways. Molecular Microbiology 61, 1147-1166.
Livak KJ, Schmittgen TD. 2001. Analysis of relative gene expression data using real-time
quantitative PCR and the 2-Ct method. Methods 25, 402-408.
Maiorino M, Gregolin C, Ursini F. 1990. Phospholipid hydroperoxide glutathione peroxidase.
Methods in Enzymology 186, 448–457.
Margis R, Dunand C, Teixeira FK, Margis-Pinheiro M. 2008. Glutathione peroxidase family -
an evolutionary overview. FEBS Journal 275, 3959-3970.
Miao Y, Lv D, Wang P, Wang X-C, Chen J, Miao C, Song C-P. 2006. An Arabidopsis
glutathione peroxidase functions as both a redox transducer and a scavenger in abscisic acid
and drought stress responses. The Plant Cell 18, 2749-2766.
Mullineaux PM, Karpinski S, Jiménez A, Cleary SP, Robinson C, Creissen GP. 1998.
Identification of cDNAs encoding plastid-targeted glutathione peroxidase. The Plant Journal
13, 375-379.
Nakagawa T, Kurose T, Hino T, Tanaka K, Kawamukai M, Niwa Y, Toyooka K, Matsuoka
K, Jinbo T, Kimura T. 2007. Development of series of gateway binary vectors, pGWBs, for
realizing effcient construction of fusion genes for plant transformation. Journal of Bioscience
and Bioengineering 104, 34-41.
Navrot N, Collin V, Gualberto J, Gelhaye E, Hirasawa M, Rey P, Knaff DB, Issakidis E,
Jacquot JP, Rouhier N. 2006. Plant glutathione peroxidases are functional peroxiredoxins
distributed in several subcellular compartments and regulated during biotic and abiotic
stresses. Plant Physiology 142, 1364-1379.
Passaia G, Queval G, Bai J, Margis-Pinheiro M, Foyer CH. 2014. The effects of redox
controls mediated by glutathione peroxidases on root architecture in Arabidopsis thaliana.
Journal of Experimental Botany 65, 1403-1413.
Perazzolli M, Dominici P, Romero-Puertas MC, Zago E, Zeier J, Sonoda M, Lamb C,
Delledonne M. 2004. Arabidopsis nonsymbiotic hemoglobin AHb1 modulates nitric oxide
bioactivity. The Plant Cell 16, 2785-2794.
Puppo A, Herrada G, Rigaud J. 1991. Lipid peroxidation in peribacteroid membranes from

23
French-bean nodules. Plant Physiology 96, 826-830.
Puppo A, Groten K, Bastian F, Carzaniga R, Soussi M, Lucas MM, de Felipe MR, Harrison
M, Vanacker H, Foyer CH. 2005. Legume nodule senescence: roles for redox and hormone
signalling in the orchestration of the natural aging process. New Phytologist 165, 683-701.
Puppo A, Pauly N, Boscari A, Mandon K, Brouquisse R. 2013. Hydrogen peroxide and nitric
oxide: key regulators of the legume-Rhizobium and mycorrhizal symbioses. Antioxidants and
Redox Signaling 18, 2202-2219.
Ramos J, Matamoros MA, Naya L, James EK, Rouhier N, Sato S, Tabata S, Becana M.
2009. The glutathione peroxidase gene family of Lotus japonicus: characterization of genomic
clones, expression analyses and immunolocalization in legumes. New Phytologist 181, 103-
114.
Rodríguez Milla MA, Maurer A, Rodríguez Huete A, Gustafson JP. 2003. Glutathione
peroxidase genes in Arabidopsis are ubiquitous and regulated by abiotic stresses through
diverse signaling pathways. The Plant Journal 36, 602-615.
Romero-Puertas MC, Rodríguez-Serrano M, Corpas FJ, Gómez M, del Río LA, Sandalio
LM. 2004. Cadmium-induced subcellular accumulation of O2- and H2O2 in pea leaves. Plant
Cell and Environment 27, 1122-1134.
Rubio MC, Becana M, Kanematsu S, Ushimaru T, James EK. 2009. Immunolocalization of
antioxidant enzymes in high-pressure frozen root and stem nodules of Sesbania rostrata. New
Phytologist 183, 395-407.
Sainz M, Pérez-Rontomé C, Ramos J, Mulet JM, James EK, Bhattacharjee U, Petrich JW,
Becana M. 2013. Plant hemoglobins may be maintained in functional form by reduced flavins
in the nuclei, and confer differential tolerance to nitro-oxidative stress. The Plant Journal 76,
875-887.
Sainz M, Calvo-Begueria L, Pérez-Rontomé C, Wienkoop S, Abián J, Staudinger C,
Bartesaghi S, Radi R, Becana M. 2015. Leghemoglobin is nitrated in functional legume
nodules in a tyrosine residue within the heme cavity by a nitrite/peroxide-dependent
mechanism. The Plant Journal (in press).
Seidel T, Kluge C, Hanitzsch M, Ross J, Sauer M, Dietz KJ, Golldack D. 2004.
Colocalization and FRET-analysis of subunits c and a of the vacuolar H+-ATPase in living
plant cells. Journal of Biotechnology 112, 165-175.
Serrano R, Mulet JM, Rios G et al. 1999. A glimpse of the mechanisms of ion homeostasis
during salt stress. Journal of Experimental Botany 50, 1023-1036.
Tovar-Méndez A, Matamoros MA, Bustos-Sanmamed P, Dietz K-J, Cejudo FJ, Rouhier N,
Sato S, Tabata S, Becana M. 2011. Peroxiredoxins and NADPH-dependent thioredoxin

24
systems in the model legume Lotus japonicus. Plant Physiology 156, 1535–1547.
Wolff SP. 1994. Ferrous ion oxidation of ferric ion indicator xylenol orange for measurement of
hydroperoxides. Methods in Enzymology 233, 182–189.
Zaninotto F, Camera SL, Polverari A, Delledonne M. 2006. Cross talk between reactive
nitrogen and oxygen species during the hypersensitive disease resistance response. Plant
Physiology 141, 379–383.
25
Table 1. Kinetic parameters of LjGpxs with various hydroperoxides as substrates and poplar thioredoxin (Trxh1) as the electron donor
Enzyme Peroxide Vmax (μmol min-1·mg-1) Km (μM) Vmax/Km
LjGpx1 H2O2 3.8 15.6 0.24 t-Butyl hydroperoxide 7.8 330.8 0.02 Cumene hydroperoxide 4.8 64.9 0.07 Phosphatidylcholine
hydroperoxide 3.2 1.6 2.00
LjGpx3 H2O2 4.0 20.5 0.20 t-Butyl hydroperoxide 3.6 166.6 0.02 Cumene hydroperoxide 14.7 213.5 0.07 Phosphatidylcholine
hydroperoxide 7.2 1.6 4.50

26
Table 2. Identification of LjGpxs that interact with LjTrxh4 Proteins from nodule extracts that interact with LjTrxh4 were trypsinized and the resulting peptides were analyzed by LC-MS/MS using the DDAM (data-dependent acquisition mode) or the TM (target mode). The mass to charge ratio (m/z) of the fragmented peptide ions are indicated. Three independent experiments were conducted, each one corresponding to a different nodule sample. +, ++, +++, positive identifications in one, two, or three experiments.
Proteins and peptides m/z DDAM TM
LjGpx1 FKAEFPVFDKVDVNGDSAAPLYK 853.10 ++ +++ AEFPVFDKVDVNGDSAAPLYK 761.38 ++ +++ VDVNGDSAAPLYK 674.84 ++ +++ GNDVNLGDYK 547.76 +++ +++ FLVDKEGNVVER 702.88 ++ +++
LjGpx2 FKSEFPIFDKIEVNGENSAPLYK 891.46 + +++ SEFPIFDKIEVNGENSAPLYK 799.73 + IEVNGENSAPLYK 717.37 ++ GSDVDLSTYK 1084.52 ++ +++ WGIFGDDIQWNFAK 848.90 +++ +++ FLVDKDGQVVDR 695.87 ++ +++
LjGpx3 SLYDFTVK 486.75 + +++ ELNILYEK 511.28 + +++

27
Figure Legends
Fig. 1. Procedure followed to demonstrate the interaction between the cytosolic thioredoxin
LjTrxh4 and LjGpxs. Two CNBr-Sepharose columns were prepared by covalently binding wild-
type (WT) and mutated (MUT) LjTrxh4. These columns were loaded with identical protein
amounts from soluble nodule extracts. After several washes with Tris-EDTA buffer (TE) without
and with NaCl, the proteins interacting with each of the LjTrxh4 proteins were eluted using a
DTT-containing buffer. Nodule proteins retained by the mutated LjTrxh4 but not by the wild-type
LjTrxh4 were considered as thioredoxin targets.
Fig. 2. Expression of LjGpx1 and LjGpx3 in nodules of L. japonicus plants exposed to nitro-
oxidative stress and hormones. Steady-state mRNA levels were normalized with respect to
ubiquitin and are expressed relative to those of untreated plants, which were given an arbitrary
value of 1. All data are means ± SE of 3-6 replicates. Asterisks denote significant up-regulation
(>2-fold).
Fig. 3. Functional complementation of LjGpx1 and LjGpx3 in yeast cells. The Gpx-
deficient mutant and transformed cells were grown on YPD medium for 48 h at 26°C with
inducers of oxidative stress [500 M H2O2 and 30 M t-butyl hydroperoxide (t-BuOOH)], of
salt stress (0.9 M NaCl), and of membrane damage (1.5 mM linolenic acid and 16 mM
caffeine). Serial dilutions (1:10, 1:100, and 1:1000) of saturated cultures (top to bottom), and
three replicates (left to right), are shown on the plates. The whole experiment was repeated
three times with similar results.
Fig. 4. In situ mRNA hybridization of (A-D) LjGpx1 and (E-H) LjGpx3 in mature nodules
(46-d-old plants). The figure shows nodule sections hybridized with (A, C, E, and G)
antisense probes and with (B, D, F, and H) sense probes (negative controls). Arrows mark
intense signal in the cortex, vascular bundles (for LjGpx3), and fixation zone. Bars, 75 μm
(A, B, E, and F); 300 μm (C, D, G, and H).

28
Fig. 5. Immunogold localization of LjGpx1 and LjGpx3 in nodules. Micrographs show
localization (arrows mark gold particles) of (A) LjGpx1 in amyloplast, (B) LjGpx1 in
nucleus, and (C) LjGpx3 in endoplasmic reticulum, cytosol, and nucleus. (D) Negative
control, in which non-immune serum substituted for antibodies against LjGpxs, shows the
absence of labeling in amyloplast. Bars, 0.5 µm.
Fig. 6. Subcellular localization of LjGpx1 and LjGpx3 using transient expression of GFP
fusions in A. thaliana protoplasts. GFP fluorescence is depicted in green and chlorophyll
autofluorescence in magenta. Arrows show localization of (A) LjGpx1 in nuclei and (B)
LjGpx3 in the cytosol. Bars, 10 m.
Fig. 7. Nitrosylation of LjGpx1 and LjGpx3. (A) Immunoblot showing nitrosylation of
purified LjGpx1 and LjGpx3. Recombinant proteins were treated with (-) 1 mM GSSG
(control) or with (+) 1 mM GSNO, subjected to the biotin-switch, and immunoblotted with
an anti-biotin antibody. (B) Immunoblot showing nitrosylation of LjGpx1 and LjGpx3 in
nodule extracts from plants treated (-) with 5 mM GSSG or (+) with 5 mM GSNO.
Biotinylated proteins were affinity purified using streptavidin-agarose and immunoblotted
with the LjGpx1 and LjGpx3 antibodies. (C) Effect of GSNO-mediated nitrosylation on
LjGpx activities measured using the NADPH-coupled assay with poplar Trxh1 and H2O2 as
substrates. Recombinant LjGpx1 and LjGpx3 were treated with 1 mM GSSG (control), 1
mM GSNO, or 5 mM GSNO for 1 h at 37ºC. Values are means ± SE of 6–8 replicates.
Means marked with an asterisk differ significantly from control at P<0.05 based on the
Student's t-test. (D) Mass spectra showing nitrosylation of Cys-85 using the His-tag switch.
Esentially, the nitrosyl group of Cys is released by ascorbate and the free thiol is then
alkylated by a synthetic peptide. During trypsinization, the synthetic peptide is cleaved,
producing a Gly-Arg dipeptide that remains bound to the Cys via an amide bond. Arrows
mark the presence of two peptides found in the tryptic digest of the nitrosylated protein
(LjGpx3 + GSNO; lower spectrum), which are not present in the control unmodified protein
(LjGpx3; upper spectrum). The molecular masses of these two peptides correspond to the
alkylation of the Cys residue by the Gly-Arg dipeptide, as indicated in the figure.
WT -‐SH
-‐SH
nodule extract
HS-‐ HS-‐
TE TE+NaCl wash
DTT elu:on
nodule extract
TE TE+NaCl wash
target proteins
nonspecific proteins
DTT elu:on
HS-‐
HS-‐
target + nonspecific proteins
-‐S -‐SH
S-‐ HS-‐ W
T -‐S -‐S
HS-‐ HS-‐ W
T -‐S -‐S W
T -‐SH -‐SH W
T
-‐SH -‐OH M
UT -‐S
-‐OH S-‐
HS-‐ MUT -‐S
-‐OH S-‐
HS-‐ MUT -‐SH
-‐OH HS-‐ HS-‐ M
UT
S-‐ S-‐
S-‐ S-‐
Figure 1
0
2
4
6
Cd GSNO ABA JA SA ACC CK
LjGpx1
LjGpx3 R
elat
ive
mR
NA
leve
l *
*
*
Treatment
*
0
2
4
6
Cd GSNO ABA JA SA ACC CK
GPX1
GPX3
Figure 2
Linolenic acid
Caffeine
t-‐BuOOH
Mutant LjGpx1 LjGpx3
NaCl
H2O2
Figure 3
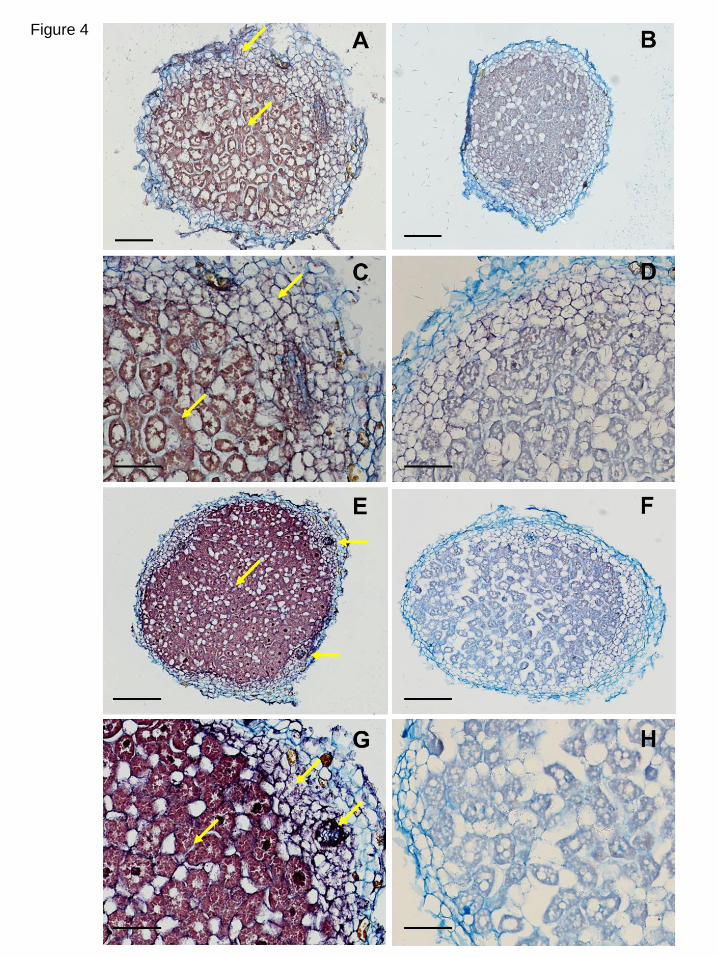
Figure 4
A B
C D
Figure 5
LjGpx1
LjGpx3
GFP Chlorophyll Overlay Bright field
Figure 6
2000 2310 2620 2930 3240 3550
Mass (m/z)
7257.6
0
20
40
60
80
100
% Inte
nsity
2000 2310 2620 2930 3240 3550 0
20
40
60
80
100
% Inte
nsity
3.7E+4
LjGpx3
LjGpx3
+ GSNO
0
1
2
3
LjGpx1 LjGpx3
6
17
27
34
- +
Control
GSNO 1 mM
Enzym
e a
ctivity
(nm
ol N
AD
PH
min
-1 m
g-1
pro
tein
)
GSNO 5 mM
Nodule extracts
- +
LjGpx3
- +
LjGpx1
2371
2278
3265
2371 2278
3265 3338
2335
2335.2358 Cys-CH2-CO-NH-Gly-Arg (K)VLIIVNVASQCGLTQTNYK(E)
3337.7744 Cys-CH2-CO-NH-Gly-Arg (K)VLIIVNVASQCGLTQTNYKELNILYEK(Y)
A C
D
*
*
Figure 7
*
B
- +
LjGpx3 LjGpx1
Recombinant proteins

PtGpx3 MLTSRSRILSQKYLNFASLSASFLLSKQSSFNSKQTLLPSLHNSPVSLYSQSIKAGVSRR AtGpx6 MLRSSIRLLYIRRTSPLLRSLSSSSSSSSSKRFDSAKPLFNSHRIISLPISTT LjGpx1 MLCTRILFFSRTIRFAAPLSSSSLHSFVFSNSPITLSRSYHSSLLTTTSFPIKSLVSTS LjGpx3 MQLLTFWNWISLVILAFAFFFFFFFFFFFFYSQ AtGpx3 MPRSSRWVNQRATSKIKKFILFLGVAFV BrGpx MASSSYAPFSAVFSGFAATKPNPPPTCSAFLVPKRRSNSRNLKNGVSLKSWNKHG * PtGpx3 LLGSVRFNHSMASQSSPQSAHDFTVKDAKGNDVDLSIYKGKVLLIVNVASQCGLTDSNYT AtGpx6 GAKLSRSEHSMAASSEPKSLYDFTVKDAKGNDVDLSIYKGKVLLIVNVASQCGLTNSNYT LjGpx1 TTPFSFTLRPDHTMAAPTSVYDFTVKDARGNDVNLGDYKGKVLLIVNVASQCGLTNSNYT LjGpx3 THPASPPSPSTMAEQTSKSLYDFTVKDIRGNDVSLSQYSGKVLIIVNVASQCGLTQTNYK AtGpx3 FYLYRYPSSPSTVEQSSTSIYNISVKDIEGKDVSLSKFTGKVLLIVNVASKCGLTHGNYK BrGpx FQFTSRNLSVYARATEEKTVHDFTVKDISGKDVSLDKFKGKPLLIVNVASKCGLTSSNYT * PtGpx3 ELTQLYAKYKDQGLEILAFPCNQFGSQEPGSSEEIVEFACTRFKAEYPIFDKVEVNGNNA AtGpx6 ELAQLYEKYKGHGFEILAFPCNQFGNQEPGTNEEIVQFACTRFKAEYPIFDKVDVNGDKA LjGpx1 ELSQLYEKYKSKGLEILGFPCNQFGAQEPGDNEQIQEFVCTRFKAEFPVFDKVDVNGDSA LjGpx3 ELNILYEKYKSKGLEILAFPCNQFAGQEPGTNDEIQDVVCTRFKSEFPVFDKVEVNGKNA AtGpx3 EMNILYAKYKTQGFEILAFPCNQFGSQEPGSNMEIKETVCNIFKAEFPIFDKIEVNGKNT BrGpx ELSQLYDKYRNQGFEILAFPCNQFGGQEPESNPDIKRFVCTRFKAEFPIFDKVDVNGPST * PtGpx3 APIYKYLKSSKGGLFGDNIKWNFSKFLVDKEGKVVDRYAPTTSPLSIEKEVKKLLGIA AtGpx6 APVYKFLKSSKGGLFGDGIKWNFAKFLVDKDGNVVDRFAPTTSPLSIEKDVKKLLGVTA LjGpx1 APLYKYLKSSKGGLFGDKIKWNFSKFLVDKEGNVVERYAPTTSPLSIEKDLVKLLGA LjGpx3 EPLFKFLKDQKGGIFGDGIKWNFTKFLVNKEGKVVERYAPTTSPMKIEKDLEKLLQSS AtGpx3 CPLYNFLKEQKGGLFGDAIKWNFAKFLVDRQGNVVDRYAPTTSPLEIEKDIVKLLASA BrGpx APIYQFLKSKSGGFLGDLIKWNFEKFLVDKKGNVVQRYPPTTSPLQIEKDIQKLLVA
Fig. S1. Amino acid sequences of representative Gpxs mentioned in this work. The
three Cys residues are indicated in red, the catalytic triad is marked with an asterisk, the
conserved domains are highlighted in yellow, and the putative transit peptides of LjGpxs
are highlighted in blue. At, Arabidopsis thaliana (Rodriguez Milla et al., 2003); Br,
Brassica rapa (Jung et al., 2002); Lj, Lotus japonicus (Ramos et al., 2009); Pt, Populus
trichocarpa (Navrot et al., 2006).
34.1
27.0
17.5
6.1
Ab LjGpx1 Ab LjGpx3 Ab LjGpx1 Ab LjGpx3
34.1
27.0
17.5
6.1
Nodule extract Recombinant proteins B A
10 µg 5 µg Gpx1 Gpx3 Gpx1 Gpx3 10 µg 5 µg
17.5
6.1
Ab LjGpx1 Ab LjGpx3
Yeast extract C
12 h 24 h
34.1
27.0
Fig. S2. Immunoblots showing the specificity of the LjGpx1 and LjGpx3 antibodies, and the expression of the two proteins in nodules and in transformed yeast cells. (A) Antibodies (Abs) recognize the respective purified recombinant proteins LjGpx1 and LjGpx3 (1 ng protein per lane). (B) Abs recognize the proteins in nodule extracts (5 or 10 µg protein per lane). (C) Abs recognize the proteins in transformed yeast cells (40 µg protein per lane). Note that LjGpx3 was visible after 12, 24, and 48 h of cell growth, whereas LjGpx1 was only visible at 12 h but not afterwards.
48 h 12 h 24 h 48 h
2628.0 2871.4 3114.8 3358.2 3601.6 3845.0
Mass (m/z)
1.1E+4
2871.4 3114.8 3358.2 3601.6 3845.0
4.0E+4
0
50
100
% In
tens
ity
LjGpx3 106-‐135 GLEILAFPCNQFAGQEPGTNDEIQDVVCTR -‐S-‐S-‐ expected m/z : 3262.51 -‐SH HS-‐ expected m/z : 3264.53
LjGpx3
LjGpx3 + DTT
0
50
100
% In
tens
ity
3260.0 3262.4 3264.8 3267.2 3269.6 3272.0
1.1E+4
0
50
100
3260.0 3262.4 3264.8 3267.2 3269.6 3272.0
4.0E+4
0
50
100
% In
tens
ity
% In
tens
ity
2628.0
Fig. S3. MS analysis demonstrating the presence of a disulfide bond between Cys-114 and Cys-133 in LjGpx3. The molecular mass of the relevant peptide from control LjGpx3 (m/z 3262.51) was shifted 2 mass units (m/z 3264.53), corresponding to the 2 H of the two thiol groups, when the protein was incubated with DTT prior to trypsinization. Identical MS method was followed to demonstrate the presence of a disulfide bond between the equivalent Cys residues (Cys-140 and Cys-159) in LjGpx1.
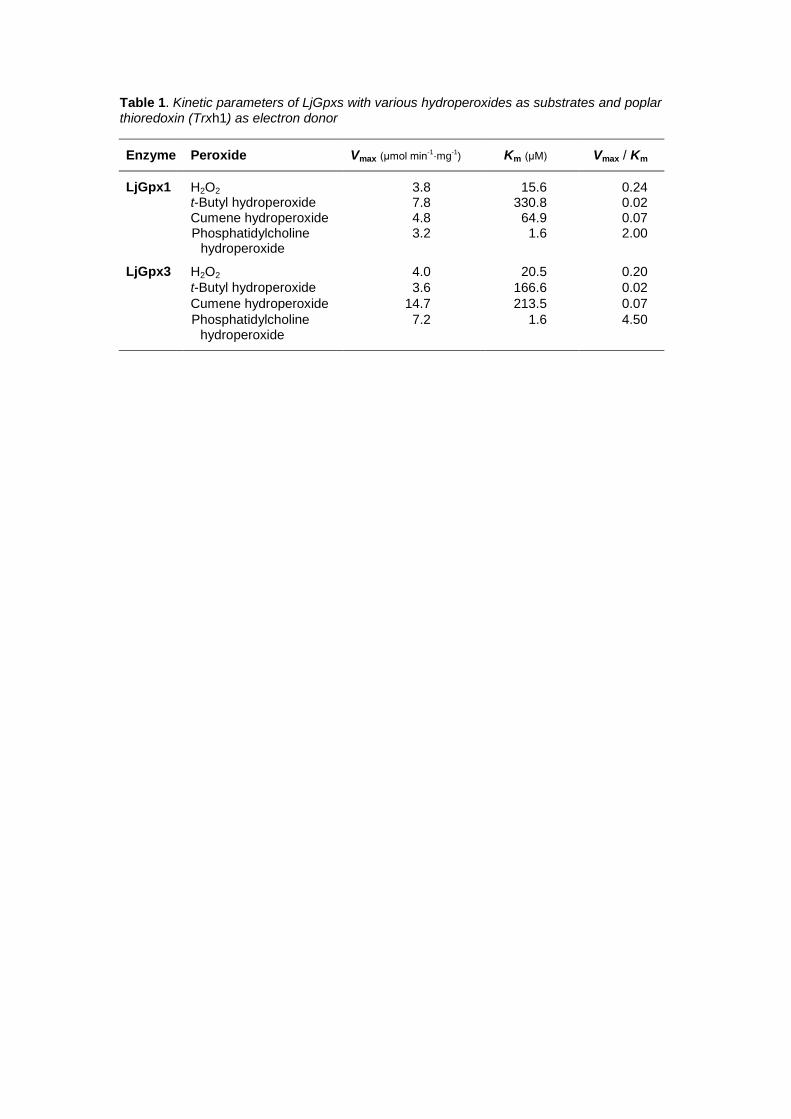
Table 1. Kinetic parameters of LjGpxs with various hydroperoxides as substrates and poplar
thioredoxin (Trxh1) as electron donor
Enzyme Peroxide Vmax (μmol min-1·mg
-1) Km (μM) Vmax / Km
LjGpx1 H2O2 3.8 15.6 0.24 t-Butyl hydroperoxide 7.8 330.8 0.02 Cumene hydroperoxide 4.8 64.9 0.07 Phosphatidylcholine
hydroperoxide 3.2 1.6 2.00
LjGpx3 H2O2 4.0 20.5 0.20
t-Butyl hydroperoxide 3.6 166.6 0.02
Cumene hydroperoxide 14.7 213.5 0.07
Phosphatidylcholine hydroperoxide
7.2 1.6 4.50
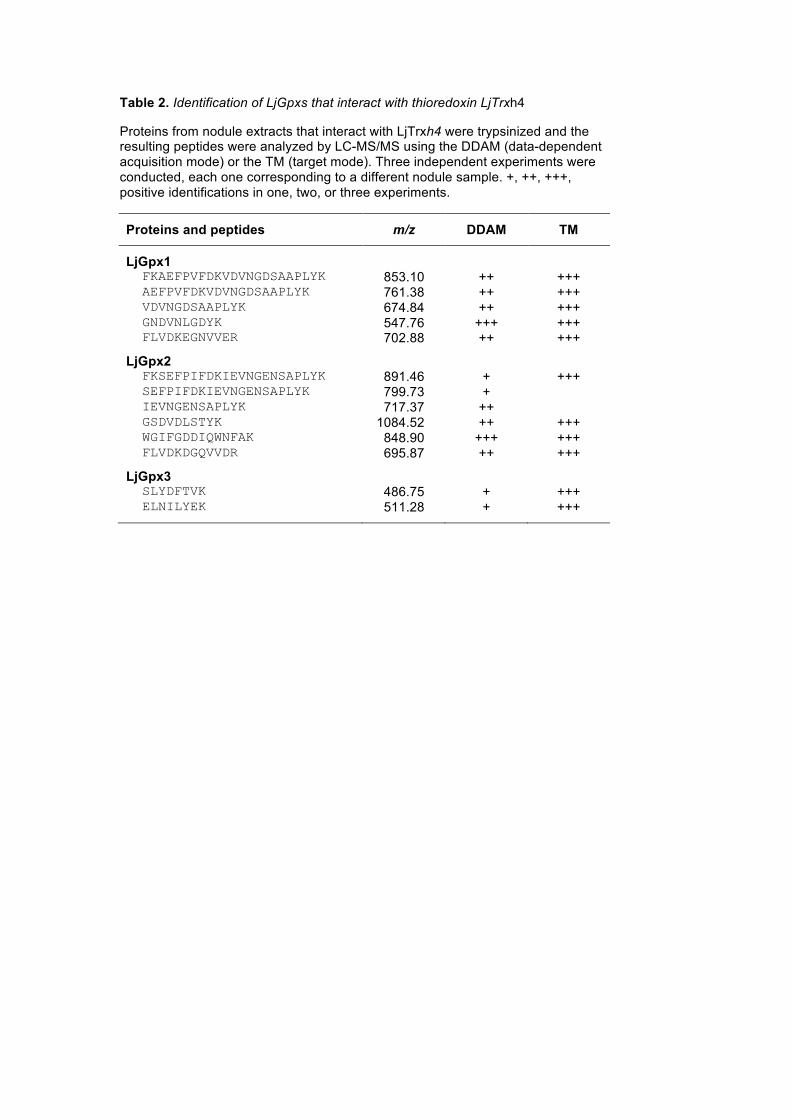
Table 2. Identification of LjGpxs that interact with thioredoxin LjTrxh4
Proteins from nodule extracts that interact with LjTrxh4 were trypsinized and the resulting peptides were analyzed by LC-MS/MS using the DDAM (data-dependent acquisition mode) or the TM (target mode). Three independent experiments were conducted, each one corresponding to a different nodule sample. +, ++, +++, positive identifications in one, two, or three experiments.
Proteins and peptides m/z DDAM TM
LjGpx1 FKAEFPVFDKVDVNGDSAAPLYK 853.10 ++ +++ AEFPVFDKVDVNGDSAAPLYK 761.38 ++ +++ VDVNGDSAAPLYK 674.84 ++ +++ GNDVNLGDYK 547.76 +++ +++ FLVDKEGNVVER 702.88 ++ +++
LjGpx2 FKSEFPIFDKIEVNGENSAPLYK 891.46 + +++ SEFPIFDKIEVNGENSAPLYK 799.73 + IEVNGENSAPLYK 717.37 ++ GSDVDLSTYK 1084.52 ++ +++ WGIFGDDIQWNFAK 848.90 +++ +++ FLVDKDGQVVDR 695.87 ++ +++
LjGpx3 SLYDFTVK 486.75 + +++ ELNILYEK 511.28 + +++

Table S1. Oligonucleotides used in this study
fw, forward primer (5' → 3'); rv, reverse primer (5' → 3'). Reference genes: LjUBQ, ubiquitin; LjeIF-‐4A, eukaryotic initiation factor 4A; LjPP2A, protein phosphatase 2A.
Production of recombinant proteins
LjGpx1 (fw) CAC CAT GGC TGC CCC CAC AT
LjGpx1 (rv) TCA TGC ACC CAA CAA TTT CAC
LjGpx3 (fw) CAC CAT GGC TGA ACA AAC CT
LjGpx3 (rv) TCA AGA AGA TTG TAA GAG CT
LjTrxh4 (fw) CAC CAT GGG CGG AGT CCT CTC T
LjTrxh4 (rv) CTA AGC TCG GAG CTG CTC A
Site-directed mutagenesis
LjTrxh4 (fw) TGG TGC GGG CCG AGC CGG TTC ATA G
LjTrxh4 (rv) CTA TGA ACC GGC TCG GCC CGC ACC A
In situ hybridization probes
LjGpx1 (fw) TCT GTA CTC GCA TCT TGT TCT TCT
LjGpx1 (rv) TGA TCT GGT CTG AGA GTG AAA GAG
LjGpx3 (fw) AGT TGT TGA CTT TCT GGA ATT GG
LjGpx3 (rv) ACT CAC ATC ATT TCC ACG GAT
LjGpx1 T7 (fw) ATT ATG CTG AGT GAT ATC CCT CTG TAC TCG CAT CTT GTT CTT CT
LjGpx1 T7 (rv) ATT ATG CTG AGT GAT ATC CCT GAT CTG GTC TGA GAG TGA AAG AG
LjGpx3 T7 (fw) ATT ATG CTG AGT GAT ATC CCA GTT GTT GAC TTT CTG GAA TTG G
LjGpx3 T7 (rv) ATT ATG CTG AGT GAT ATC CCA CTC ACA TCA TTT CCA CGG AT
Quantitative reverse-transcription PCR
LjGpx1 (fw) ACT ATG GCT GCC CCC ACA T
LjGpx1 (rv) TCT GGC ATC TTT GAC GGT GA
LjGpx3 (fw) TGG GAA GAA TGC AGA ACC ACT
LjGpx3 (rv) CCC CCT TTC TGA TCC TTC AAA
LjGpx1 (fw) ACT ATG GCT GCC CCC ACA T
LjUBQ (fw) TTC ACC TTG TGC TCC GTC TTC
LjUBQ (rv) AAC AAC AGC ACA CAC AGA CAA
LjeIF-4A (fw) AGA GGG TTT AAA GAT CAA AT
LjeIF-4A (rv) ATG TCA ATT CAT CAC GTT TT
LjPP2A (fw) TGA GCT ATG TGA AGC TGT TGG T
LjPP2A (rv) CAG CCT CAT TAT CAC GCA GTA G








