HERPESVÍRUS: O ATAQUE E A RESPOSTA DO ...portais.ufg.br/up/67/o/semi2011_Duane_Alves_1c.pdf1...
33
UNIVERSIDADE FEDERAL DE GOIÁS ESCOLA DE VETERINÁRIA E ZOOTECNIA PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIA ANIMAL Disciplina: SEMINÁRIOS APLICADOS HERPESVÍRUS: O ATAQUE E A RESPOSTA DO HOSPEDEIRO Duanne Alves da Silva Orientador (a): Profa. Dra. Ana Paula Junqueira Kipnis Goiânia 2011
Transcript of HERPESVÍRUS: O ATAQUE E A RESPOSTA DO ...portais.ufg.br/up/67/o/semi2011_Duane_Alves_1c.pdf1...

UNIVERSIDADE FEDERAL DE GOIÁS ESCOLA DE VETERINÁRIA E ZOOTECNIA
PROGRAMA DE PÓS-GRADUAÇÃO EM CIÊNCIA ANIMAL
Disciplina: SEMINÁRIOS APLICADOS
HERPESVÍRUS: O ATAQUE E A RESPOSTA DO
HOSPEDEIRO
Duanne Alves da Silva
Orientador (a): Profa. Dra. Ana Paula Junqueira Kipnis
Goiânia
2011

ii
DUANNE ALVES DA SILVA
HERPESVÍRUS: O ATAQUE E A RESPOSTA DO
HOSPEDEIRO
Seminário apresentado à disciplina de
Seminários Aplicados do Programa de
Pós-Graduação em Ciência Animal da
Escola de Veterinária e Zootecnia da
Universidade Federal de Goiás
Nível: Doutorado
Área de Concentração:
Sanidade Animal e Higiene e Tecnologia de Alimentos
Orientador (a):
Profa. Dra. Ana Paula Junqueira Kipnis
Comitê de Orientação:
Profa. Dra. Wilia Marta Elsner Diederichsen de Brito – EV/UFG
Profa. Dra Liliane Borges de Menezes – IPTSP/UFG
Goiânia
2011

iii
SUMÁRIO
1-INTRODUÇÃO ................................................................................................ 1
2-REVISÃO DE LITERATURA ........................................................................... 4
2.1- Entrada do vírus na célula ........................................................................... 4
2.1.1- Glicoproteínas .......................................................................................... 4
2.1.2- Diversidade de receptores de ligação e de entrada ................................. 5
2.1.3- Entrada de Alphaherpesvírus ................................................................... 7
2.2- Ataque; fusão; perfuração e penetração; desassociação.......................... 10
2.3- Resposta do hospedeiro ........................................................................... 15
3-CONSIDERAÇÕES FINAIS .......................................................................... 19
REFERÊNCIAS ................................................................................................ 20

iv
LISTA DE FIGURAS
FIGURA 1- Desenho esquemático e micrografia eletrônica da partícula viral de herpesvírus..........................................................................................................1 FIGURA 2- Participantes na entrada dos herpesvírus e indução da fusão celular..................................................................................................................7 FIGURA 3- Desenho esquemático dos eventos de fusão do envelope viral com a membrana plasmática.....................................................................................11 FIGURA 4- Observação por microscopia eletrônica da entrada do BoHV-1 em células MDBK....................................................................................................13 FIGURA 5- Observação por microscopia eletrônica da entrada do BoHV-5 em células MDBK....................................................................................................14 FIGURA 6- Ações biológicas dos interferons tipo I...........................................18

1
1-INTRODUÇÃO
Os herpesvírus são membros da ordem Herpesvirales (DAVISON et
al., 2009), família Herpesviridae estando classificados em três subfamílias, de
acordo com suas propriedades biológicas: Alphaherpesvirinae,
Betaherpesvirinae e Gammaherpesvirinae. Os membros das respectivas
subfamílias são agrupados em gêneros de acordo com a homologia das
sequências de DNA, similaridades na estrutura e organização genômica e
relação antigênica (FRANCO & ROEHE, 2007). Os vírus apresentam genoma
de DNA envolto em um capsídeo icosaédrico, que por sua vez, é revestido com
uma camada protéica chamada de tegumento e um envelope contendo de dez
a doze proteínas e glicoproteínas virais inseridas na bicamada lipídica que
formam essa estrutura (Figura 1). Pelo menos três, às vezes quatro, dessas
glicoproteínas do envelope são essenciais para a entrada do vírus na célula
(SPEAR & LONGNECKER, 2003).
FIGURA 1: desenho esquemático e micrografia eletrônica da partícula viral de herpesvírus.
Os herpesvírus são muito antigos, havendo indícios que eles co-
evoluiram com seus hospedeiros há quase um bilhão de anos. Estudos
genéticos apontam esse elevado nível de adaptação observado entre os vírus
e os seus hospedeiros naturais como uma possível explicação para essa
evolução paralela. Várias semelhanças podem ser observadas na estrutura de
diferentes herpesvírus, o que sugere que eles tenham surgido de um ancestral
comum, dando origem a duas linhagens: uma representada pelos herpesvírus

2
alfa, beta e gama, que infectam aves e mamíferos; e a outra representada
pelos herpesvírus de animais de sangue frio (FRANCO & ROEHE, 2007).
A infecção aguda causada pelos herpesvírus, tanto em animais
como em humanos, é capaz de gerar importantes quadros mórbidos podendo,
inclusive, levar a morte do hospedeiro. Em muitos casos a infecção pode ser
leve e muitas vezes inaparente. Entretanto, esses vírus têm como importante
característica a capacidade de estabelecer infecção por toda a vida do
hospedeiro, ou seja, uma vez infectado o indivíduo permanece portador do
vírus na forma latente. A presença do vírus em latência implica no
desenvolvimento ocasional de infecções recorrentes que acontecem na maioria
dos casos, de forma inaparente (FRANCO & ROEHE, 2007).
Estes agentes infecciosos estão amplamente distribuídos na
natureza e a maioria das espécies animais serve de hospedeiro natural de pelo
menos um membro da família. No entanto, até o momento, apenas um
pequeno número de herpesvírus foi identificado, compreendendo
aproximadamente 130 espécies. Destes, muitos são de importância veterinária,
visto que cada espécie doméstica alberga pelo menos um desses agentes.
Podem-se citar como exemplos: os herpesvírus bovino tipos 1, 2, 4 e 5 (BoHV-
1, BoHV-2, BoHV-4 e BoHV-5); os herpesvírus eqüino tipos 1, 3 e 4 (EHV-1,
EHV-3 e EHV-4); o vírus da doença de Aujeszky ou da pseudoraiva- PRV-
herpesvírus suíno tipo 1 (SuHV-1); o herpesvírus caprino tipo 1 (CpHV-1); o
herpesvírus canino tipo 1 (CaHV-1); o herpesvírus felino tipo 1 (FeHV-1); os
herpesvírus de galídeos tipo 1 e tipo 2 (GaHV-1 e 2) (FRANCO & ROEHE,
2007).
No que diz respeito a infecção em animais, os herpesvírus são
incriminados mundialmente como causadores de grandes prejuízos
econômicos, principalmente para a indústria bovina e suína (POMERANZ et al.,
2005) pois geram perdas produtivas e reprodutivas significativas. Estima-se
que os prejuízos causados por enfermidades relacionadas aos herpesvírus
bovinos alcancem um bilhão de dólares por ano em todo o mundo (DEL
MÉDICO ZAJAC, 2010).
Dentre os herpesvírus que infectam humanos, podem-se citar o
herpesvírus humano 1 (HHV-1 ou vírus do herpes simplex HSV-1), que é o
protótipo da família; o herpesvírus humano 2 (HHV-2 ou HSV-2); o agente da

3
varicela zoster (VZV) herpesvírus humano 3 (HHV-3); o vírus Epstein-Barr
(EBV ou HHV-4); o citomegalovírus humano (HCMV ou HHV-5) e os
herpesvírus humano 6A, 6B, 7 e 8 (HHV-6A, HHV-6B, HHV-7 e HHV-8)
(FRANCO & ROEHE, 2007).
Embora a infecção natural com a maioria dos herpesvírus seja
restrita a uma única espécie, alguns desses vírus podem infectar outras
espécies experimentalmente ou acidentalmente. Em geral, os alphaherpesvírus
são vistos como tendo a mais ampla gama de acometimento, com respeito a
espécies (com exceção do VZV) e tipo de célula. Os betaherpesvírus podem
infectar uma grande variedade de tipos de celulares, mas geralmente
restringem-se a infecções de seus hospedeiros naturais e, os gamaherpesvírus
são restritos em relação ao hospedeiro e tipo de célula (SPEAR &
LONGNECKER, 2003).
Por serem patógenos importantes, tanto para animais como para
humanos, os herpesvírus têm sido alvo de constantes estudos que visam
elucidar os mecanismos envolvidos no estabelecimento da infecção.
Entretanto, ainda existem muitas dúvidas acerca das estratégias utilizadas por
esses agentes para estabelecer uma infecção produtiva no organismo, desde a
sua entrada na célula-alvo até o estabelecimento de uma resposta eficiente
pelo hospedeiro que consiga combater o ataque. O modelo de estudo da
família Herpesviridae é o HHV-1, contudo, algumas diferenças entre ele e os
outros membros já foram observadas.
Assim, objetiva-se com este seminário, abordar de uma forma geral
os mecanismos de ataque dos herpesvírus e como o organismo do hospedeiro
reage a infecção, tendo como base os conhecimentos já estabelecidos para a
família Herpesviridae, apresentando os aspectos referentes aos herpesvírus de
bovinos.

4
2-REVISÃO DE LITERATURA
2.1- Entrada do vírus na célula
A entrada dos vírus na célula hospedeira envolve diversos
mecanismos, a saber: adsorção, fusão e penetração. Para entrarem na célula
os herpesvírus fazem uso de múltiplos receptores. A ligação inicial (adsorção)
pode ser reversível, servindo para concentrar os vírus na superfície celular,
entretanto não provoca as alterações necessárias para a fusão da membrana.
A maioria dos herpesvírus, provavelmente, reconhece múltiplos receptores
celulares, qualquer um dos quais pode ser suficiente para a entrada na célula
(SPEAR & LONGNECKER, 2003).
Acredita-se que os herpesvírus, geralmente, entram nas células por
meio da fusão do envelope viral com a membrana plasmática celular, onde o
nucleocapsídeo e as proteínas do tegumento ganham acesso ao citoplasma
(LYCKE et al., 1988; FULLER & LEE, 1992; GRANZOW et al., 1997; SODEIK
et al., 1997) ou por endocitose (GRANZOW et al., 1997).
2.1.1- Glicoproteínas
A entrada dos herpesvírus nas células pode ser mediada por
diferentes glicoproteínas do envelope (SPEAR, 1993; SCHWYZER &
ACKERMANN, 1996), no entanto, não se sabe ao certo o número de
glicoproteínas necessárias para mediar a adsorção (WUDUNN & SPEAR,
1989; LIANG et al., 1991; FLYNN & RYAN, 1995; LI et al., 1995; WHITBECK et
al., 1997), a fusão (PEETERS et al., 1992; TURNER et al., 1998) e a
penetração (FULLER et al., 1989; PEETERS et al., 1992).
Foi demonstrado que algumas glicoproteínas do HHV-1 são
necessárias e essenciais para fusão da membrana (TURNER et al., 1998). O
envolvimento de mais de duas glicoproteínas sugere um mecanismo
complicado de ligação, fusão e/ou penetração (WILD et al., 1998).

5
Três glicoproteínas parecem ser essenciais para a entrada de todos
os herpesvírus, designadas gB, gH e gL. Os genes codificadores destas
glicoproteínas são conservados, com a gB exibindo o maior grau de
similaridade de sequência entre os diferentes membros da família
Herpesviridade. Pelo menos para alguns herpesvírus, a gB é um homodímero
ou homotrímero exibido como um ponto de destaque. A heterodimerização de
gH e gL é uma característica conservada, com a adição de uma outra
subunidade protéica de alguns vírus (SPEAR & LONGNECKER, 2003).
2.1.2- Diversidade de receptores de ligação e de entrada
A maioria dos herpesvírus pode fazer o contato inicial com a célula
ligando-se a glicosaminoglicanos, (normalmente, heparan sulfato), e em
proteoglicanos de superfície celular (SHUKLA & SPEAR, 2001). A vinculação
ao heparan sulfato nem sempre é fundamental para a infecção, pois
glicoproteínas virais não essenciais para a entrada do vírus podem mediar esse
contato inicial (adsorção viral). No entanto, a presença do heparan sulfato na
superfície celular aumenta muito a eficiência da entrada viral, provavelmente
por concentrar os vírus na superfície celular para que os receptores de entrada
possam ser encontrados pelos ligantes virais apropriados (SPEAR &
LONGENECKER, 2003).
A ligação do HHV-1 e de outros alphaherpesvírus ao heparan sulfato
é reversível e os vírus eluídos permanecem infecciosos, indicando que a
atividade de fusão não foi desencadeada de forma irreversível. A interação com
um receptor de entrada parece ser irreversível, talvez, por levar imediatamente
a fusão da membrana. Essa definição operacional de receptores de ligação e
de entrada também pode ser aplicada a outros herpesvírus (SPEAR &
LONGNECKER, 2003).
Após a adsorção, a entrada dos herpesvírus na célula ocorre por
meio da fusão do envelope viral com a membrana plasmática (ou do
endossoma). Para a entrada de alguns herpesvírus como o HHV-1 em alguns
tipos celulares, é necessário ocorrer endocitose e acidificação do endossoma
(NICOLA et al., 2003; WANG et al., 2003).

6
A via de entrada e os mecanismos de fusão de outros vírus
envelopados, tais como o vírus da imunodeficiência humana (HIV), o vírus
Sendai e os vírus influenza já estão bem descritos (NIR et al., 1990; WHITE,
1992; HERNANDEZ et al., 1996; HUGHSON, 1997). O HIV e os vírus influenza
necessitam de apenas uma glicoproteína para mediar tanto a adsorção do vírus
quanto a fusão na célula hospedeira. Já o vírus Sendai requer duas
glicoproteínas para entrar na célula; uma para a ligação e outra para a fusão.
Os vírus influenza entram nas células por endocitose e são liberados no
citoplasma pela fusão do envelope viral com a membrana do vacúolo
endocítico. Os vírus Sendai e HIV entram nas células por fusão do envelope
viral com a membrana plasmática, assim como os Herpesvírus bovino tipos 1 e
5, que entram nas células por meio de uma fusão única do envelope com a
membrana celular (WILD et al., 1998).
As interações de uma ou mais glicoproteínas virais com os
receptores celulares pode provocar a fusão do envelope com a membrana ou a
fusão célula a célula. Presume-se que o mecanismo básico de fusão para a
entrada dos herpesvírus inclua as glicoproteínas gB, gH e gL, embora
receptores virais de ligação adicionais também possam ser necessários (Figura
2) (SPEAR & LONGNECKER, 2003).
O HHV-1, por exemplo, requer a gD como um ligante para os
receptores de entrada. Apenas os alphaherpesvirus (exceto VZV) codificam
genes dos membros da família da gD. O EBV requer a gp42, um componente
do complexo gH-gL-gp42, como um ligante para as moléculas de antígeno
leucocitário humano de classe II (HLA ou MHC) nos linfócitos B. O
citomegalovírus humano codifica a gO, que não está relacionada com a gp42,
mas também forma complexo com gH-gL (HUBER & COMPTON, 1998).
Similarmente, o HHV-6A codifica a gQ, que forma complexo com gH-gL (MORI
et al., 2003). Ainda não está claro se gO ou gQ são necessárias para a entrada
desses vírus. Os alphaherpesvirus não codificam homólogos de gp42, gO ou
gQ. Assim, parece que o mecanismo básico de fusão da membrana é
conservado entre os herpesvírus, enquanto que os ligantes virais que se
conectam aos receptores da superfície celular diferem entre as subfamílias, o
que explicaria, em partes, a diferença de tropismo por células e tecidos
(SPEAR & LONGNECKER, 2003).

7
Fonte: SPEAR & LONGNECKER, 2003.
2.1.3- Entrada de Alphaherpesvírus
Os alphaherpesvírus, incluindo HHV-1, HHV-2, PRV e BoHV-1
apresentam um número de glicoproteínas funcionalmente e estruturalmente
homólogas. As três glicoproteínas principais (gB, gC e gD) são parcialmente
homólogas as do HHV (CHOWDHURY, 1995).
As glicoproteínas gB e/ou gC podem mediar a ligação dos vírus ao
heparan sulfato das superfícies celulares. Apesar de a gC ser dispensável para
a infecção de culturas celulares, sua presença pode aumentar a eficiência da
ligação do HHV-1 em quase 10 vezes. Muitos autores já demonstraram que a
gC é uma importante glicoproteína no processo de ataque do BoHV-1, pois
FIGURA 2: Participantes na entrada dos herpesvírus e indução da fusão celular. A ligação dos alphaherpesvirus como dos gamaherpesvirus às células, pode ser mediada por glicoproteínas virais que não são essenciais para a entrada. O heparan sulfato é o receptor para a gC do HHV-1 e para a gpK8.1A do HHV-8. Para a gp350 do EBV o receptor é o CD21 (em caso de células B). A entrada requer a interação dos ligantes virais com outros receptores celulares. Para o HHV-1, a gD é o ligante para muitos receptores celulares de superfície como, HVEM, nectinas e 3-O-sulfated heparan sulfato e qualquer um destes pode mediar a entrada. Para o EBV entrar nas células B, a gp42 se liga às moléculas HLA de classe II. Para a entrada do HHV-8, a gB pode se ligar às integrinas. É pensado que qualquer uma dessas interações de um ligante viral com um receptor de entrada pode acionar a atividade de fusão de gB e gH-gL.

8
permite a adsorção do vírus na célula (COLLINS et al.,1984; VAN DRUNEN
LITTLE-VAN DEN HURK et al., 1984; VAN DRUNEN LITTLE-VAN DEN HURK
& BABIUK, 1986; MARSHALL et al., 1988).
Acredita-se que as glicoproteínas essenciais, gB, gD, gH e gL agem
em conjunto para induzir a fusão do envelope viral com a membrana.
Receptores celulares de entrada são demandados para provocar essa fusão e
o ligante viral para todos os receptores de HHV é a gD (COCCHI et al., 1998;
SPEAR et al., 2000; CAMPADELLI-FIUME et al., 2007). Esses requisitos para
a entrada dos vírus nas células se aplicam aos alphaherpesvírus de animais,
bem como o PRV e o BoHV-1 (WARNER et al., 1998).
Já foram identificadas três classes de receptores de entrada do
HHV-1 (SPEAR et al., 2000) que também são utilizados pelo BoHV-1 e -5.
Nestas classes encontram-se: o mediador de entrada de herpesvírus (HVEM),
que é um membro da família (HveA) de receptores do fator de necrose tumoral
(TNF); nectina-1 e nectina-2, que são dois membros da superfamília das
imunoglobulinas (MONTGOMERY et al., 1996; GERAGHTY et al., 1998;
WARNER et al., 1998); e sítios específicos no heparan sulfato. Qualquer uma
dessas moléculas de superfície pode ligar-se a gD, com cada sorotipo viral
apresentando diferenças na preferência pelo receptor (SPEAR et al., 2000;
CONNOLLY et al., 2001; SPEAR, 2004).
Além de a gD servir como glicoproteína de ligação aos receptores,
ela provoca a fusão por meio de um domínio especializado denominado
domínio de pró-fusão (PFD), localizado na região C-terminal da proteína
(CAMPADELLI-FIUME et al., 2007).
PERTEL et al. (2001), após transfectarem células deficientes de
receptores endógenos para HHV-1 com plasmídeos expressando receptores
de entrada, quantificaram a entrada viral usando vírus que expressavam genes
imediatamente após a entrada. Neste estudo foi possível verificar que,
enquanto o HVEM e a nectina-1 são excelentes receptores de entrada tanto
para o HHV-1 quanto o HHV-2, a nectina-2 é mais ativa para o HHV-2 do que
para o HHV-1. Já o heparan sulfato é, provavelmente, mais ativo para o HHV-1
do que para o HHV-2.
Ainda não se sabe como a interação da gD com um receptor de
entrada desencadeia a entrada viral ou a fusão celular. SPEAR &

9
LONGNECKER (2003) levantaram a hipótese de que, a ligação da gD com um
dos receptores resultaria numa mudança conformacional desta glicoproteína,
permitindo sua interação com gB ou gH-gL e o acionamento da atividade
fusiogênica. De acordo com estes autores, a gD não seria um componente
integral da maquinaria básica de fusão e, além disso, poderiam existir outros
receptores para gB e/ou gH-gL. Logo, ligações a esses outros receptores,
poderiam desencadear a atividade de fusão, não havendo necessidade da
presença da gD.
Essas idéias surgiram, em parte, de resultados obtidos com o vírus
da pseudoraiva. O PRV pode usar como receptores de entrada, vários
membros da família das nectinas de humanos e de animais (GERAGHTY et al.,
1998; MILNE et al., 2001). KLUPP et al. (2000) afirmaram que, apesar de todas
as quatro glicoproteínas do PRV, incluindo a gD, serem necessárias para a
entrada viral, gB, gH e gL são suficientes para a fusão em células de coelhos
(cujos receptores não foram identificados). Segundo os autores, este fato
sugere a existência de receptores de superfície celular para gB ou gH-gL.
Em 2007, CAMPADELLI-FIUME et al., confirmaram a hipótese
levantada em 2003, de que a gD em sua forma livre (não ligada a receptor)
apresentava a região C-terminal dobrada em torno da região N-terminal, de
modo que a glicoproteína adotava uma conformação fechada (auto inibida).
Quando ligada ao HVEM e a nectina-1, a região C-terminal é deslocada,
deixando a gD com uma conformação aberta.
Ainda segundo CAMPADELLI-FIUME et al. (2007), a estrutura
trimérica da gB, que é similar a de outra glicoproteína de fusão, faria desta
proteína uma ferramenta no mecanismo de fusão. Por outro lado, segundo os
mesmos autores, gH apresenta elementos moleculares típicos das
glicoproteínas de fusão, apresentando uma forte tendência a interagir com os
lipídios. Ainda não foi determinado se a fusão é executada por gB ou gH-gL,
complexadas ou em sequência.
GABEV et al. (2010) construíram um BoHV-1 mutante que carreava
a gD do BoHV-5 (gD5) e um BoHV-5 mutante que carreava a gD do BoHV-1
(gD1). Após infectarem camundongos com estes mutantes, verificaram que o
BoHV-1 com a gD5 foi quase tão virulento quanto o BoHV-5 selvagem. No
entanto, a mudança da gD5 pela gD1 no BoHV-5, não reduziu a habilidade

10
deste em invadir o cérebro, enquanto que a troca de gD1 por gD5 no BoHV-1
não desenvolveu a habilidade deste em infectar neurônios secundários.
Contudo, o sucesso na inibição da infectividade de mutantes de gD5 com
diferentes nectinas solúveis de origem humana, in vivo, sugeriu a hipótese
desta glicoproteína se ligar a uma ampla variedade de receptores, quando
comparada a gD de outros alphaherpesvírus, como o HHV-1 (GABEV et al.,
2010)
2.2- Ataque; fusão; perfuração e penetração; desassociação
Em 1998, WILD et al. realizaram um estudo com técnicas que
utilizam baixas temperaturas, onde puderam monitorar a incubação de células
com herpesvírus bovino tipos 1 e 5 na faixa de segundos, verificando o
processo de entrada dos vírus nas células em milésimos de segundos a
qualquer momento desejado. Após inocularem cepas de BoHV-1 e 5 em cultura
de células MDBK (―Madin Darby bovine kidney‖) à temperatura de 37°C, os
autores observaram que de 0 a 30 segundos após a incubação, algumas
partículas virais localizavam-se no meio extracelular próximas a membrana
citoplasmática das células. De 30 a 40 segundos após a incubação, o envelope
de alguns vírus estava parcialmente fusionado com a camada externa da
membrana; logo, apenas três camadas das duas bicamadas eram visíveis
sugerindo que, apenas a camada externa do envelope viral foi fusionada a
camada externa da membrana plasmática (Figura 3).
A natureza hidrofílica e hidrofóbica dos fosfolipídios exige que, os
que se encontram na camada exterior do envelope viral, sejam conduzidos
para a camada exterior de fosfolipídios da membrana plasmática, um processo
mediado por uma ou mais das glicoproteínas assumidas para atuarem como
proteínas de fusão (FULLER & LEE, 1992; PEETERS et al., 1992). A
separação das camadas de fosfolipídios para posterior fusão requer domínios
específicos em ambos os sítios (membrana plasmática e envelope viral) e um
mecanismo particular para desencadear a fusão. A fusão completa, ou seja, o
momento em que a camada externa do envelope viral se une a camada interna
da membrana plasmática e vice e versa, resultando no cruzamento (Figura 4D;
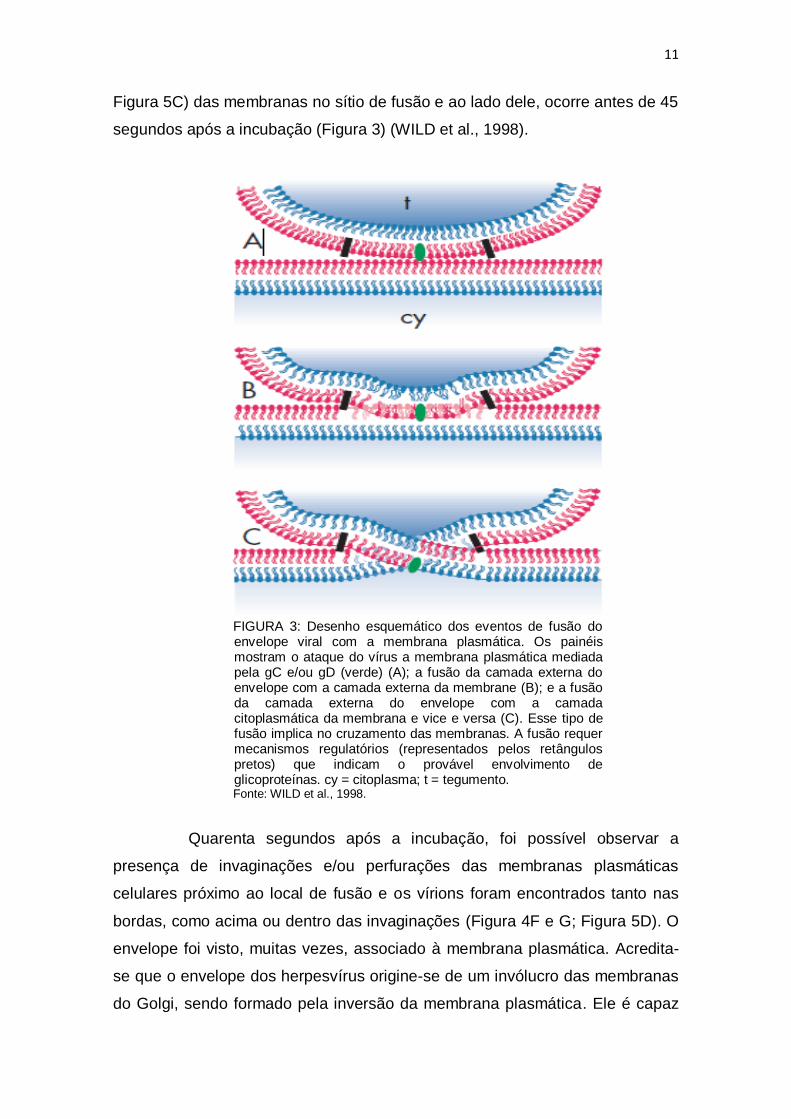
11
Figura 5C) das membranas no sítio de fusão e ao lado dele, ocorre antes de 45
segundos após a incubação (Figura 3) (WILD et al., 1998).
FIGURA 3: Desenho esquemático dos eventos de fusão do envelope viral com a membrana plasmática. Os painéis mostram o ataque do vírus a membrana plasmática mediada pela gC e/ou gD (verde) (A); a fusão da camada externa do envelope com a camada externa da membrane (B); e a fusão da camada externa do envelope com a camada citoplasmática da membrana e vice e versa (C). Esse tipo de fusão implica no cruzamento das membranas. A fusão requer mecanismos regulatórios (representados pelos retângulos pretos) que indicam o provável envolvimento de glicoproteínas. cy = citoplasma; t = tegumento. Fonte: WILD et al., 1998.
Quarenta segundos após a incubação, foi possível observar a
presença de invaginações e/ou perfurações das membranas plasmáticas
celulares próximo ao local de fusão e os vírions foram encontrados tanto nas
bordas, como acima ou dentro das invaginações (Figura 4F e G; Figura 5D). O
envelope foi visto, muitas vezes, associado à membrana plasmática. Acredita-
se que o envelope dos herpesvírus origine-se de um invólucro das membranas
do Golgi, sendo formado pela inversão da membrana plasmática. Ele é capaz

12
de fundir-se com a membrana plasmática mantendo a assimetria da
transmembrana (DEVAUX, 1992) como já foi demonstrado anteriormente para
a fusão do vírus da gripe com lipossomas (KLOTZ et al., 1996).
A fusão do envelope viral com a membrana plasmática resulta numa
inserção parcial do mesmo dentro da membrana. Por razões geométricas,
dobras na membrana plasmática devem surgir e/ou deve ser perdida a
integridade da membrana e do envelope viral em uma área circunscrita. O
significado deste tipo de fusão ainda não está claro. Certamente não permite
que o vírus penetre a membrana plasmática. Em vez disso, o vírus ganha
acesso ao citoplasma através de uma invaginação que se desenvolve nas
proximidades da zona de fusão e que se abre para o citoplasma (WILD et al.,
1998).
Partículas virais envelopadas (Figura 5F) são encontradas dentro do
citoplasma 50 segundos após a incubação. O envelope apresenta-se fusionado
ou associado à membrana plasmática. Em seguida, é possível observar os
vírions dentro da matriz citoplasmática podendo, ocasionalmente, estar
próximos ao núcleo em frente a um poro nuclear (Figura 4N) (WILD et al.,
1998).
Após a fusão, já no citoplasma celular, o envelope dos herpesvírus,
provavelmente neutraliza a partir da membrana plasmática, que por sua vez
deve ser restaurada para que a integridade celular seja mantida. Algumas
proteínas do tegumento se dissociam do nucleocapsídeo e permanecem no
citoplasma, enquanto outras são transportadas até o núcleo. O
nucleocapsídeo, ainda associado com algumas proteínas do tegumento, é
transportado ao longo dos microtúbulos até o centro de organização dos
mesmos (SODEIK et al., 1997), que está situado perto do núcleo. WILD et al.
(1998) observaram vírus envelopados próximos ao núcleo após 15 minutos do
início da incubação.
Os nucleocapsídeos associam-se aos complexos de poros nucleares
desintegrando-se e liberando o genoma no interior do núcleo para que ocorra a
replicação. Os restos do capsídeo ficam na região externa da membrana
nuclear (FRANCO & ROEHE, 2007).

13
FIGURA 4: observação por microscopia eletrônica da entrada do BoHV-1 em células MDBK. (A) ataque; (B) fusão da camada externa do envelope viral com a camada externa da membrana plasmática;(C a E) fusão da camada externa do envelope com a camada interna da membrana plasmática e vice e versa, resultando no cruzamento (setas) das membranas; (E a L) vários estágios da invaginação e perfuração da membrana plasmática, incluindo a perda da integridade da membrana, próximo ao local de fusão (E); partículas virais logo acima de uma pequena invaginação (F e G); envelope viral sendo conectado a membrana plasmática acima da invaginação (H), na borda da invaginação (I), acima de uma invaginação ampla complicada (K) e na entrada da invaginação (L); a integridade da membrana plasmática é perdida na ponta da invaginação, mas é atravessada (L- seta) ao lado da partícula viral. (M e N) os vírions estão dentro do citoplasma e o envelope está associado à membrana plasmática, onde mais de duas camadas são visíveis (seta), indicando dobras (M); uma localização perto de um poro nuclear é obervada (N) com as membranas nucleares, externa (o) e interna (i), e a membrana de Golgi (g) sendo indicadas. Fonte: WILD et al., 1998.

14
FIGURA 5: observação por microscopia eletrônica da entrada do BoHV-5 em células MDBK (A- direita) ataque das partículas virais a membrana plasmática, associadas a membrana (esquerda) e localizadas acima de uma pequena invaginação da membrana plasmática ao lado de uma microvilosidade (m). (B a D) fusão da camada externa do envelope viral com a camada externa da membrana plasmática (B); (C) fusão completa do envelope com a membrana formando um pequeno cruzamento (seta) e uma ―tricamada‖ (entre a flecha e a ponta da seta), perda da integridade da membrana e os primeiros sinais da perfuração na base da microvilosidade; (D) partículas virais acima de uma pequena invaginação e em associação com a ondulação da membrana plasmática (direita). (E) vírion no citoplasma; o envelope está fusionado (ponta da seta) com a membrana e, além do local de fusão, a matriz citoplasmática parece vazar através da membrana plasmática perfurada (setas). (F) partícula viral envelopada dentro do citoplasma. Fonte: WILD et al., 1998.

15
2.3- Resposta do hospedeiro
Até o momento, poucos estudos foram desenvolvidos buscando
elucidar os mecanismos de indução da resposta imune após a infecção pelos
herpesvírus, principalmente os de animais. Com relação aos herpesvírus
bovino tipos 1 e 5, devido as similaridades existentes entre eles, acredita-se
que ambos possam provocar mecanismos semelhantes no hospedeiro durante
a infecção (DEL MÉDICO ZAJAC et al., 2010).
Alguns estudos com HHV-1 já revelaram os mecanismos
imunológicos envolvidos na encefalite fatal causada por esse agente.
LUNDBERG et al. (2008), sugeriram que a variação da resposta inflamatória
desenvolvida pelos hospedeiros possa desempenhar um papel determinante
na fatalidade da doença. SERGERIE & BOIVIN (2007), observaram intensa
resposta inflamatória no sistema nervoso central de camundongos infectados
experimentalmente com HHV-1, com expressão proeminente do fator de
necrose tumoral.
Além disso, alguns estudos in vivo e in vitro já demonstraram que a
microglia de humanos e camundongos com infecção de HHV-1 não produtiva,
expressam uma variedade de quimiocinas e citocinas pró-inflamatórias de
acordo com o seu envolvimento no início da resposta inata contra o vírus.
Porém, não se sabe se essa resposta inflamatória inata é protetora ou deletéria
(LOKENSGARD et al., 2001; LOKENSGARD et al., 2002).
Outras ferramentas importantes na resposta imunológica aos
herpesvírus são as glicoproteínas, que além de mediarem a infecção das
células-alvo influenciando o tropismo celular e tecidual, são os principais
antígenos reconhecidos pelo sistema imune do hospedeiro infectado (LUPTON
& REED, 1980; LITTLE et al., 1981; GLORIOSO et al., 1984; BABIUK et al.,
1987; HEROLD et al., 1991). A gB é uma das principais glicoproteínas no
envelope viral e na membrana plasmática das células infectadas (MISRA et al.,
1981). Em bovinos infectados pelo BoHV-1, a resposta de anticorpos
específicos para gB é detectada antes da resposta a outros antígenos (VAN
DRUNEN LITTEL-VAN DEN HURK et al., 1990), e sua atividade neutralizante
já foi mapeada para, pelo menos, seis epítopos diferentes (FITZPATRICK et
al., 1990). Alguns autores já demonstraram a capacidade de uma vacina de

16
DNA de gB do BoHV-1 em induzir anticorpos e resposta de linfócitos T
citotóxicos (CTLs) em bovinos e camundongos (LOEHR et al., 2001; HUANG et
al., 2005).
Em infecções pelo BoHV-1, a primeira resposta do hospedeiro
constitui-se em inflamação e uma resposta imune inata. A resposta adaptativa
começa a partir do quinto dia após a infecção, com a indução de linfócitos
CD4+ e CD8+, apresentando sua maior atividade no décimo dia após a infecção
(DENIS et al., 1994; BABIUK et al., 1996). A resposta imune humoral é
verificada a partir do décimo dia após a infecção. Anticorpos contra o BoHV-1
participam na neutralização das partículas virais que se encontram no meio
extracelular, bem como na indução de células anticorpo-dependentes mediante
a citotoxicidade. A resposta de anticorpos é essencial na prevenção de
infecções secundárias e na limitação das conseqüências da reativação
(BABIUK et al., 1996).
Em 2007, DEL MÉDICO ZAJAC et al., após infectarem bovinos
experimentalmente com BoHV-5, verificaram a ocorrência de resposta
linfoproliferativa entre sete e 14 dias após a infecção. Os animais apresentaram
anticorpos totais no soro a partir do dia 12° após a infecção e IgA e IgG1 foram
detectadas na secreção nasal. Anticorpos neutralizantes foram detectáveis no
dia 14 após a infecção (MEYER et al., 2001; PEREZ et al., 2002; VOGEL et al.,
2003; DEL MÉDICO ZAJAC et al., 2006).
TIKOO et al. (1995) afirmam que os CTLs são componentes críticos
no processo de eliminação de herpesvírus, uma vez que a disseminação viral
célula a célula ocorre antes da disseminação hematógena. Nesse sentido, têm-
se buscado desenvolver vacinas recombinantes capazes de induzir CTLs
contra os herpesvírus, pois as vacinas inativadas disponíveis apresentam uma
boa atividade neutralizante, no entanto, não são boas indutoras da resposta
mediada por células, além de a resposta de anticorpos induzida por elas ser
curta (DESHPANDE et al., 2002).
Outra ferramenta importante no combate as infecções virais são os
sistemas interferon (IFN). Ao infectarem as células, os vírus induzem a
expressão de moléculas de IFN, que são uma grande família de citocinas
estruturalmente relacionadas. Os sistemas IFN-α e β influenciam a replicação
do vírus, diretamente por meio de sua atividade antiviral em nível celular

17
(MULLER et al., 1994; GROB et al., 1999) e, indiretamente, pela modulação da
resposta imune (Figura 6) (LANDOLFO, 1995; BOEHM et al., 1997; DE WAAL
MALEFTY, 1997; BIRON, 1998 e 1999).
Diferente dos humanos e camundongos, os bovinos apresentam três
genes para IFN- β com promotores distintos (WILSON et al., 1983;
VALARCHER et al., 2003). Todos os três isótipos de IFN- β apresentam
atividade antiviral mas, ainda não está claro se a infecção induz os três
subtipos de forma diferente. A regulação viral da resposta do IFN é crucial para
a sobrevivência do patógeno na natureza e para a patogênese (JONES &
SILVA, 2011). ABRIL et al. (2004) verificaram que camundongos que
apresentavam receptores para IFN tipos I e II combinados com deleções de
genes de ativação de recombinação (RAG-2) morreram em poucos dias após a
infecção pelo BoHV-1. Reciprocamente, camundongos selvagens (sem
deleções genéticas) infectados com BoHV-1 não desenvolveram sinais clínicos.
O BoHV-1 induz a resposta de IFN devido ao seu elevado nível de
CpG DNA não metilado. Assim como o HHV-1 ou HHV-2, o DNA do BoHV-1
contém elevados níveis de CpG DNA (LUNDBERG et al., 2003), que pode
desencadear a resposta imune inata (via receptores Toll-like 9 – TLR9, por
exemplo) (ADEREM & HUME, 2000; HACKER et al., 2000). Durante o curso da
infecção produtiva, proteínas virais são codificadas, incluindo a bICP0 (proteína
de células infectadas 0) interferindo na resposta imune inata. A bICP0 é uma
importante proteína regulatória que estimula a infecção produtiva e também
inibe a transcrição dependente de interferon, por meio da degradação do fator
estimulatório 3 de IFN (IRF-3) (JONES, 2009).
Embora pareça provável o envolvimento de outros produtos
codificados pelo herpesvírus bovino na indução da resposta imune inata, eles
ainda não foram identificados. Logo, o tipo de regulação da resposta imune
inata estabelecido pelo HHV-1 é aceito também para o BoHV-1. A infecção
pelo HHV-1 em cultura de células humanas leva a produção e secreção de
interferon. A gD induz a produção de IFN-α em células mononucleares (KATZE
et al., 2002), em parte, devido a ativação do IRF-3 em certos tipos celulares
(PRESTON et al., 2001). A entrada do envelope viral nas células também pode
induzir a resposta imune inata dependente de IFN (NOYCE et al., 2006).

18
FIGURA 6: ações biológicas dos interferons tipo I. Os IFNs tipo I (IFN-α e IFN-β) são produzidos por células infectadas por vírus em resposta a sinalização intracelular por TLR e outros sensores de RNA viral. Os IFNs tipo I se ligam a receptores nas células vizinhas não infectadas, e ativam vias de sinalização por Jak-STAT, as quais induzem expressão de genes cujos produtos interferem com a replicação viral. Os IFNs também se ligam a receptores nas células infectadas, e induzem a expressão de genes cujos produtos aumentam a suscetibilidade das células à destruição mediada por CTL. Fonte: ABBAS et al., 2008.

19
3-CONSIDERAÇÕES FINAIS
O Brasil está em uma posição de destaque como um dos maiores
produtores da indústria bovina mundial. Logo, para que a qualidade dos
produtos oriundos desta atividade seja garantida, o controle sanitário dos
rebanhos é imprescindível.
No entanto, patógenos causadores de enfermidades importantes em
bovinos, como os herpesvírus, ainda apresentam aspectos relacionados a sua
biologia não muito bem esclarecidos.
O grande prejuízo econômico gerado na pecuária mundial, devido as
perdas produtivas e reprodutivas causadas pelos herpesvírus, é um sinal de
alerta para produtores, pesquisadores e autoridades sanitárias que precisam
unir forças no combate a esta infecção.
Apesar de existirem muitos estudos sobre os herpesvírus de
animais, o conhecimento acerca da patogenia e da resposta imune
estabelecidas por eles ainda é incipiente. Os aspectos já estabelecidos sobre a
biologia do agente referem-se, em sua maioria, aos herpesvírus de humanos.
Assim, pesquisas que visam elucidar os mecanismos de indução da
resposta imunológica, bem como a patogênese do agente são extremamente
relevantes, pois podem auxiliar no desenvolvimento de tecnologias
diagnósticas e métodos de controle e erradicação eficazes.

20
REFERÊNCIAS
1. ABBAS, A. K.; LICHTMAN, A. H.; PILLAI, S. Imunologia celular e molecular. Ed.Elsevier Brasil, 2008. 576 p.
2. ABRIL, C.; ENGELS, M.; LIMMAN, A.; HILBE, M.; ALBINI, S.;
FRANCHINI, M.; SUTER, M.; ACKERMAN, M. Both viral and host factors contribute to neurovirulence of bovine herpesvirus 1 and 5 in interferon receptor-deficient mice. Journal of Virology, v. 78, n. 7, p. 3644–3653, 2004.
3. ADEREM, A.; HUME, D. A. How do you see CG? Cell, v.103, n. 7 p.
993-996, 2000.
4. BABIUK, L. A.; L'ITALIEN, J.; VAN DRUNEN LITTLE-VAN DEN HURK, S.; ZAMB, T.; LAWMAN, M. J. P.; HUGHES, G.; GIFFORD, G. A. Protection of cattle from bovine herpesvirus type 1 (BHV-1) infection by immunization with individual viral glycoproteins. Virology, v. 159, p. 57-
66, 1987.
5. BABIUK, L. A.; DRUNEN, H. L.; TIKOO, S. K. Imunology of bovine herpesvírus 1 infection. Veterinary Microbiology, Amsterdam, v.53, n.1-2, p.31-42, 1996.
6. BIRON, C. A. Role of early cytokines, including alpha and beta
interferons (IFN-alpha/beta), in innate and adaptive immune responses to viral infections. Seminars in Immunology, v. 10, p. 383–390, 1998.
7. BIRON, C. A. Initial and innate responses to viral infections—pattern
setting in immunity or disease. Current Opinion in Microbiology, v. 2, p. 374–381, 1999.
8. BOEHM, U.; KLAMP, T.; GROOT, M.; HOWARD, J. C. Cellular
responses to interferon-gamma. Annual Reviews of Immunology v. 5, p. 749–795, 1997
9. CAMPADELLI-FIUME, G.; AMASIO, M.; AVITABILE, E.; CERRETANI,
A.; FORGHIERI, C.; GIANNI, T.; MENOTTI, L. The multipartite system that mediates entry of herpes simplex virus into the cell. Reviews in Medical Virology, v. 17, p. 313–326, 2007.
10. CHOWDHURY, S. I. Molecular basics of antigenic variation between the
glycoprotein C of respiratory bovine herpesvírus 1 (BHV-1) and neurovirulent BHV-5. Virology, v. 213, p. 558-568, 1995.
11. COCCHI, F.; MENOTII, L.; MIRANDOLA, P.; LOPEZ, M.; CAMPADELLI-
FIUME, G. The ectodomain of a novel member of the immunoglobulin

21
subfamily related to the poliovirus receptor has the attributes of a bona fide receptor for herpes simplex virus types 1 and 2 in human cells. Journal of Virology, v. 72, p. 9992–10002, 1998.
12. COLLINS, J. K.; BUTCHER, A. C.; RIEGEL, C. A.; MCGRANE, V.;
BLAIR, C. D.; TARAMOTO, Y. A.; WINSTON, S. Neutralizing determinant defined by monoclonal antibodies on polypeptides specified by bovine herpesvirus 1. Journal of Virology, v. 52, p. 403-409, 1984.
13. CONNOLLY, S. A.; WHITBECK, J. J; RUX, A. H.; KRUMMENACHER, C.; VAN DRUNEN LITTLE-VAN DEN HURK, S.; COHEN, G. H.; EISENBERG, R. J. Glycoprotein D homologs in herpes simplex virus type 1, pseudorabies virus, and bovine herpes virus type 1 bind directly to human HveC (nectin-1) with different affinities. Virology, v. 280, p. 7–18, 2001.
14. DAVISON, A. J.; EBERLE, R.; EHLERS, B.; HAYWARD, G. S.;
McGEOCH, D. J.; MINSON, A. C.; PELLETT, P. E.; ROIZMAN, B.; STUDDERT, M.; THIRY, E. The order Herpesvirales. Archives of Virology, v. 154, p. 171-177, 2009.
15. DEL MÉDICO ZAJAC, M. P.; PUNTEL, M.; ZAMORANO, P.; SADIR, A.
M.; ROMERA, S. A. BHV-1 vaccine induces cross-protection against BHV-5 disease in cattle. Research in Veterinary Science, Edimburgo, v.81, p.327–334, 2006.
16. DEL MÉDICO ZAJAC, M. P. Herpesvírus bovino 5: estúdio de La infección, la respuesta inmune y los recombinantes com herpesvírus bovino 1 em El hospedador natural. 2007. 145 f. phD Tese. Buenos Aires.
17. DEL MÉDICO ZAJAC, M. P.; LADELFA, M. F.; KOTSIAS, F.;
MUYLKENS, B.; THIRY, J.; ROMERA, S. A. Biology of bovine herpesvirus 5. The Veterinary Journal, v. 184, p.138-145, 2010.
18. DENIS, M.; KAASHOEK, M. J.; VAN OIRSCHOT, J. T.; PASTORET, P. P.; THIRY, E. Quantitative assessment of the specific CD4+ T lymphocyte proliferative response in bovine herpesvirus 1 immune cattle. Veterinary Immunology and Immunopathology, v. 42, p. 275–286, 1994.
19. DESHPANDE, M. S.; AMBAGALA, C. T.; HEGDE, N. R.; HARIHARAN,
M. J.; NAVARATNAM, M.; SRIKUMARAN, S. Induction of cytotoxic T-lymphocytes specific for bovine herpesvírus-1 by DNA immunization. Vaccine, v. 20, p. 3744-3751, 2002.
20. DEVAUX, P. F. Protein involvement in transmembrane lipid asymmetry.
Annual Review of Biophysics and Biomolecular Structure, v. 21, p. 417–439, 1992.

22
21. DE WAAL MALEFYT, R. The role of type I interferons in the
differentiation and function of Th1 and Th2 cells. Seminars in Oncology, v. 24, p. 94–98, 1997.
22. ESKRA, L.; SPLITTER, G. A. Bovine herpesvirus-1 infects activated
CD4+ lymphocytes. Journal of General Virology, v. 78, p. 2159–66,
1997.
23. FITZPATRICK D. R.; REDMOND, M. J.; ATTAH-POKU, S. K.; VAN DRUNEN LITTELVAN DEN HURK, S.; BABIUK, L. A.; ZAMB,T. J. Mapping of 10 epitopes on bovine herpesvirus type 1 glycoproteins gI and gIII. Virology, v. 176, n. 1, p. 145–57, 1990.
24. FLYNN, S. J.; RYAN, P. A heterologous heparin-binding domain can
promote functional attachment of a pseudorabies virus gC mutant to cell surfaces. Journal of Virology, v. 69, p. 834–839, 1995.
25. FRANCO, A.; ROEHE, P. M. Herpesviridae. In: FLORES, E. F. Virologia
Veterinária. 1.ed. Santa Maria: Editora UFSM, 2007. cap. 17, p. 435-485.
26. FULLER, A. O.; SANTOS, R. E.; SPEAR, P. G. Neutralizing antibodies
specific for glycoprotein H of herpes simplex virus permit viral attachment to cells but prevent penetration. Journal of Virology, v. 63, p. 3435–
3443, 1989.
27. FULLER, A. O.; LEE, W. C. Herpes simplex virus type 1 entry through a cascade of virus-cell interactions requires different roles of gD and gH in penetration. Journal of Virology, v. 66, p. 5002–5012, 1992.
28. GABEV, E.; TOBLER, K.; ABRIL, C.; HILBE, M.; SENN, C.; FRANCHINI,
M.; CAMPADELLI-FIUME, G.; FRAEFEL, C.; ACKERMANN, M. Glycoprotein D of bovine herpesvirus 5 (BoHV-5) confers an extended host range to BoHV-1 but does not contribute to invasion of the brain. Journal of Virology, v. 84, n. 11, p. 5583-5593, 2010.
29. GERAGHTY, R. J.; KRUMMENACHER, C.; COHEN, G. H.;
EISENBERG, R. J.; SPEAR, P. G. Entry of alphaherpesviruses mediated by poliovirus receptor related protein 1 and poliovirus receptor. Science, v. 280, p. 1618–1620, 1998.
30. GLORIOSO, J.; SCHRODER, C. H.; KIMEL, G.; SZCZESIUL, M.; LEVINE, M. Immunogenicity of herpes simplex virus glycoproteins gC and gB and their role in protective immunity. Journal of Virology, v. 50, p. 805-812, 1984.
31. GRANZOW, H.; WEILAND, F.; JÖNS, A.; KLUPP, B. G.; KARGER, A.;
METTENLEITER, T. C. Ultrastructural analysis of the replication cycle of

23
pseudorabies virus in cell culture: a reassessment. Journal of Virology,
v. 71, p. 2072–2082, 1997.
32. GROB, P.; SCHIJNS, V. E.; VAN DEN BROEK, M. F.; COX, S. P.; ACKERMANN, M.; SUTER, M. Role of the individual interferon systems and specific immunity in mice in controlling systemic dissemination of attenuated pseudorabies virus infection. Journal of Virology, v. 73,
p.4748–4754, 1999.
33. HACKER, H.; VABULAS, R. M.; TAKEUCHI, O.; HOSHINO, K.; AKIRA, S.; WAGNER, H. Immune cell activation by bacterial CpG-DNA through myeloid differentiation marker 88 and tumor necrosis factor receptor-associated factor (TRAF) 6. Journal of Experimental Medicine, v. 192,
p. 595-600, 2000.
34. HARIHARAN, M. J.; NATARAJ, C.; SRIKUMARAN, S. Down regulation of murine MHC class I expression by bovine herpesvirus 1. Viral Immunology, v. 6, n. 4, p. 273–84, 1993.
35. HEROLD, B. C.; WUDUNN, D.; SOLTYS, N.; SPEAR, P. G. Glycoprotein C of herpes simplex virus type 1 plays a principal role in the adsorption of virus to cells and in infectivity. Journal of Virology, v. 65, p. 1090-1098,
1991.
36. HERNANDEZ, L. D.; HOFFMAN, L. R.; WOLFSBERG, T. G.; WHITE, J. M. Virus-cell and cell-cell fusion. Annual Review of Cell and Developmental Biology, v. 12, p. 627–661, 1996.
37. HUANG, Y.; BABIUK, L. A.; VAN DRUNEN LITTEL-VAN DEN HURK, S.
Immunization with a bovine herpesvirus 1 glycoprotein B DNA vaccine induces cytotoxic T-lymphocyte responses in mice and cattle. Journal of General Virology, v. 86, p. 887–98, 2005.
38. HUANG, Y.; BABIUK, L. A.; VAN DRUNEN LITTLE-VAN DEN HURK, S.
The cell-mediated immune response induced by plasmid encoding bovine herpesvírus 1 glycoprotein B is enhanced by plasmid encoding IL-12 when delivered intramuscularly or by gene gun, but not after intradermal injection. Vaccine, v. 24, p. 5349-5359, 2006.
39. HUBER, M. T.; COMPTON, T. The human cytomegalovirus UL74 gene
encodes the third component of the glycoprotein H-glycoprotein L containing envelope complex. Journal of Virology, v. 72, p.8191–8197,
1998.
40. HUGHSON, F. M. Enveloped viruses: a common mode of membrane fusion? Current Biology, v. 7, p. 565–569, 1997.
41. JONES, C. Regulation of Innate Immune Responses by Bovine
Herpesvirus 1 and Infected Cell Protein 0 (bICP0). Viruses, v. 1, p. 255-275, 2009.

24
42. JONES, C.; SILVA, L. F. Infection of cultured bovine cells with bovine
herpesvírus 1 (BHV-1) or Sendai virus induces different beta interferon subtypes. Virus Research, v. 157, p. 54-60, 2011.
43. KATZE, M. G.; HENG, Y.; GALE, M. Viruses and interferon: fight for
supremacy. Nature Reviews Immunology, v. 2, p. 675-686, 2002.
44. KLOTZ, K., H.; BARTOLDUS, I.; STEGMAN, T. Membrane asymmetry is
maintained during influenza-induced fusion. Journal of Biological Chemestry, v. 271, p. 2383–2386, 1996.
45. KLUPP, B. G.; NIXDORF, R.; METTENLEITER, T. C. Pseudorabies vírus
glycoprotein M inhibits membrane fusion. Journal of Virology, v. 74, p. 6760–6768., 2000.
46. KLUPP, B. G.; GRANZOW, H.; FUCHS, W.; KEIL, G. M.; FINKE, S.;
METTENLEITER, T. Vesicle formation from the nuclear membrane is induced by coexpression of two conserved herpesvirus proteins. Proceedings of the National Academy of Sciences of the United States of America, v. 104, n. 17, p. 72421-7246, 2007.
47. LANDOLFO, S.; GRIBAUDO, G.; ANGERETTI, A.; GARIGLIO, M.
Mechanisms of viral inhibition by interferons. Pharmacolology & Therapeutics, v. 65, p. 415–442, 1995.
48. LIANG, X.; BABIUK, L. A.; VAN DRUNEL LITTEL-VAN DEN HURK, S.
D. R. Fitzpatrick, and T. J. Zamb. Bovine herpesvirus 1 attachment to permissive cells is mediated by its major glycoproteins gI, gIII, and gIV. Journal of Virolology, v. 65, p. 1124–1132, 1991.
49. LI, Y.; VAN DRUNEN LITTEL-VAN DEN HURK, S.; BABIUK, L. A.;
LIANG, X. Characterization of cell-binding properties of bovine herpesvirus glycoproteins B, C, and D: identification of a dual cell-binding function of gB. Journal of Virology, v. 69, p. 4758–4768, 1995.
50. LITTLE, S. P.; JOFRE, J. T.; COURTNEY, R. J.; SCHAFFER, P. A. A virion-associated glycoprotein essential for infectivity of herpes simplex type 1. Virology, v. 115, p. 149-160, 1981.
51. LOEHR, B. I.; PONTAROLLO, R.; RANKIN, R.; LATIMER, L.;, WILLSON, P.; BABIUK, L. A. Priming by DNA immunization augments T-cell responses induced by modified live bovine herpesvirus vaccine. Journal of General Virology, v. 82, p. 3035–3043, 2001.
52. LOKENSGARD, J. R.; HU, S., SHENG, W.; VANOIJEN, M.; COX, D.; CHEERAN, M. C.; PETERSON, P. K. Robust expression of TNF-alpha, IL-1beta, RANTES, and IP-10 by human microglial cells during

25
nonproductive infection with herpes simplex virus. Journal for Neurovirology, v. 7, p. 208–219, 2001
53. LOKENSGARD, J. R. ; CHEERAN, M. C.; HU, S.; GEKKER, G. PETERSON, P. Glial cell responses to herpesvirus infections: role in defense and immunopathogenesis. Journal of Infectious Diseases, v.
186, p. 171–S179, 2002
54. LUNDBEG, P.; RAMAKRISHNA, C.; BROWN, J.; MICHAEL TYSZKA, J.; HAMAMURA, M.; HINTON, D. R.; KOVATS, S.; NALCIOGLU, O.; WEINBERG, K.; OPENSHAW, H.; CANTIN, E. M. The immune response to herpes simplex virus type 1 infection in susceptible mice is a major cause of central nervous system pathology resulting in fatal encephalitis. Journal of Virology, v. 82, n. 14, p. 7078-7088, 2008.
55. LUPTON, H. W.; REED, D. E. Evaluation of experimental subunit vaccines for infectious bovine rhinotracheitis. American Journal of Veterinary Research, v. 41, p. 383-390, 1980.
56. LYCKE, E.; HAMARK, B.; JOHANSSON, M.; KROTOCHWIL, A.; LYCKE, J.; SVENNERHOLM, B. Herpes simplex virus infection of the human sensory neuron. An electron microscopy study. Archives of Virology, v. 101, p. 87–104, 1988.
57. MARSHALL, R. L.; ISRAEL, B. A.; LETCHWORTH, G. J. Monoclonal
antibody analysis of bovine herpesvirusq glycoprotein antigenic areas relevant to natural infection. Virology, v. 165, p. 338 347, 1988.
58. MEYER, G.; LEMAIRE, M.; ROSS, C.; BELAK, K.; GABRIEL, A.;
CASSART, D.; COIGNOUL, F.; BELAK, K.; THIRY, E. Comparative pathogenesis of acute and latent infections of calves with bovine herpesvirus types 1 and 5. Archives of Virology, v. 146, p. 633-652, 2001
59. MILNE, R. S. B.; CONNOLLY, S. A.; KRUMMENACHER, C.;
EISENBERG, R. J.; COHEN, G. H. Porcine HveC, a member of the highly conserved HveC/ nectin 1 family, is a functional alphaherpesvirus receptor. Virology, v. 281, p. 315–328, 2001.
60. MISRA V.; BLUMENTHAL, R. M.; BABIUK, L. A. Proteins Specified by
bovine herpesvirus 1 (infectious bovine rhinotracheitis virus). Journal of Virology, v. 40, n. 2, p. 367–78, 1981
61. MONTGOMERY, R. I.; WARNER, M. S.; LUM, B. J.; SPEAR, P. G.
Herpes simplex virus-1 entry into cells mediated by a novel member of the TNF/NGF receptor family. Cell, v. 87, p. 427–436, 1996.

26
62. MORI, Y.; YANG, X.; AKKAPAIBOON, P.; OKUNO, T.; YAMANISHI, K. Human herpesvirus 6 variant A glycoprotein H-glycoprotein L-glycoprotein Q complex associates with human CD46. Journal of Virology, v. 77, p. 4992–4999, 2003.
63. MULLER, U.; STEINHOFF, U.; REIS, L. F.; HEMMI, S.; PAVLOVIC, J.;
ZINKERNAGEL, R. M.; AGUET, M. Functional role of type I and type II interferons in antiviral defense. Science, v. 264, p. 1918–1921, 1994.
64. NATARAJ, C.; EIDMANN, S.; HARIHARAN, M. J.; SUR, J. H.; PERRY, G. A.; SRIKUMARAN, S. Bovine herpesvirus 1 downregulates the expression of bovine MHC class I molecules. Viral Immunology, v. 10, n. 1, p. 21–34, 1997.
65. NICOLA, A. V.; MCEVOY, A. M.; STRAUS, S. E. Roles for endocytosis
and low pH in herpes simplex virus entry into HeLa and Chinese hamster ovary cells. Journal of Virology, v. 77, p. 5324–5332, 2003.
66. NIR, S.; DÜZGÜNES, N.; PEDROSO DE LIMA, M. C.; HOEKSTRA, D.
Fusion of enveloped viruses with cells and liposomes. Cell Biophysics, v. 17, p. 181-201, 1990.
67. NOYCE, R. S.; COLLINS, S. E.; MOSSMAN, K. L. Identification of a
novel pathway essential for the immediate-early, interferon-independent antiviral response to enveloped virions. Journal of Virology, v. 80, p.
226-235, 2006.
68. PEETERS, B.; DE WIND, N.; HOOISMA, M.; WAGENAAR, F.; GIELKENS, A. MOORMAN, R. Pseudorabies virus envelope glycoproteins gp50 and gII are essential for virus penetration, but only gII is involved in membrane fusion. Journal of Virology, v. 66, p. 894–905,
1992.
69. PERTEL, P.; FRIDBERG, A.; PARISH, M. L.; SPEAR, P. G. Cell fusion induced by herpes simplex virus glycoproteins gB, gD, and gH-gL requires a gD receptor but not necessarily heparan sulfate. Virology, v. 279, p. 313–324, 2001.
70. PEREZ, S. E.; BRETSCHNEIDER, G.; LEUNDA, M. R.; OSORIO, E. A.;
FLORES, E. F.; ODEON, A. C. Primary infection, latency and reactivation of bovine herpesvirus type 5 in the bovine nervous system. Veterinary Pathology, Baltimore, v. 39, p. 437- 444, 2002.
71. POMERANZ, L. E.; REYNOLDS, A. E.; HENGARTNERT, C. J. Molecular
biology of pseudorabies virus: impact on neurovirology and veterinary medicine. Microbiology and Molecular Biology Reviews, v. 69, n. 3, p.
462-500, 2005.
72. PRESTON, C. M.; HARMAN, A. N.; NICHOLL, M. J. Activation of interferon response factor-3 in human cells infected with herpes simplex

27
virus type 1 or human cytomegalovirus. Journal of Virology, v. 75, p.
8909-8916, 2001.
73. SCHWYZER, M.; ACKERMANN, M. Molecular Virology of ruminant herpesviruses. Veterinary Microbiology, v. 53, p. 17-29, 1996.
74. SERGERIE, Y.; RIVEST, S.; BOIVIN, G. Tumor necrosis factor-alpha
and interleukin 1-beta play a critical role in the resistance against lethal herpes simplex virus encephalitis. Journal of Infectious Diseases, v.
196, p. 853–860, 2007.
75. SHUKLA, D.; SPEAR, P. G. Herpesviruses and heparan sulfate: an intimate relationship in aid of viral entry. Journal of Clinical Investigation, v. 108, p. 503–510, 2001.
76. SODEIK, B.; EBERSOLD, M. W.; HELENIUS, A. Microtubule-mediated
transport of incoming herpes simplex virus 1 capsids to the nucleus. Journal of Cell Biology, v. 136, p. 1007–1021, 1997.
77. SPEAR, P. G. Entry of alphaherpesviruses into cells. Seminars in
Virology, v. 4, p. 167-180, 1993.
78. SPEAR, P. G.; EISENBERG, R. J.; COHEN, G. H. Three classes of cell
surface receptors for alphaherpesvirus entry. Virology, v. 275, p.1–8, 2000.
79. SPEAR, P. G.; LONGNECKER, R. Herpesvirus entry: an update.
Journal of Virology, v. 77, n. 19, p. 10179-10185, 2003.
80. SPEAR, P. G. Herpes simplex virus: receptors and ligands for cell entry. Cellular Microbiology, v. 6, p. 401–410, 2004.
81. SZABO, S. J.; SULLIVAN, B. M.; PENG, S. L.; GLIMCHER, L. H.
Molecular mechanisms regulating Th1 immune responses. Annual Reviews of Immunology, v. 21, p. 713–758, 2003.
82. TIKOO S. K,; CAMPOS, M.; BABIUK, L. A. Bovine herpesvirus-1 (BHV-
1): biology, pathogenesis, and control. Advances in Virus Research, v. 45, p.191–223, 1995.
83. TRINCHIERI, G. Interleukin-12 and the regulation of innate resistance
and adaptive immunity. Nature Reviews Immunology, v. 3, n. 2, p. 133–146, 2003.
84. TURNER, A.; BRUUN, B.; MINSON, T.; BROWNE, H. Glycoproteins gB,
gD, and gH.gL of herpes simplex virus type 1 are necessary and sufficient to mediate membrane fusion in a Cos cell transfection system. Journal of Virology, v. 72, p. 873 -875, 1998.

28
85. VAN DRUNEN LITTLE-VAN DEN HURK, S.; VAN DEN HURK, J. V.; GILCHRIST, J. E.; MISRA, V.; BABIUK, L. A. Interactions of monoclonal antibodies and bovine herpesvirus type 1 (BHV-1) glycoproteins. Virology, v. 135, p. 466-479, 1984.
86. VAN DRUNEN LITTLE-VAN DEN HURK, S.; BABIUK, L. A. Synthesis
and processing of bovine herpesvirus type 1 glycoproteins. Journal of Virolology, v. 59, p. 401-410, 1986.
87. VAN DRUNEN LITTEL-VAN DEN HURK, S.; GIFFORD, G. A; BABIUK,
L. A. Epitope specificity of the protective immune response induced by individual bovine herpesvirus-1 glycoproteins. Vaccine, v. 8, n. 4, p.358–68, 1990.
88. VALARCHER, J. F.; FURZE, J.; WYLD, S.; COOK, R.; CONZELMAN, K.
K.; TAYLOR, G. Role of alpha/beta interferons in the attenuation and immunogenecity of recombinant bovine respiratory syncitial viruses lacking NS proteins. Journal of Virology, v. 77, p. 8426–8439, 2003.
89. VOGEL, F. S.; CARON, L.; FLORES, E. F., WEIBLEN, R.;
WINKELMANN, E. R.; MAYER, S. V., BASTOS, R. G. Distribution of bovine herpesvirus type 5 DNA in the central nervous systems of latently, experimentally infected calves. Journal of Clinical Microbiology,
Washington, v. 41, p. 4512–4520, 2003.
90. WANG, F. Z.; AKULA, S. M.; SHARMA-WALIA, N.; ZENG, L.; CHANDRAN, B. Human herpesvirus 8 envelope glycoprotein B mediates cell adhesion via its RGD sequence. Journal of Virology, v. 77, p. 3131–3147, 2003.
91. WARNER, M. S.; GERAGHTY, R. J.; MARTINEZ, W. M.;
MONTGOMERY, R. I.; WHITBECK, J. C.; XU, R.; EISENBERG, R. J.; COHEN, G. H.; SPEAR, P. G. A cell surface protein with herpesvirus entry activity (HveB) confers susceptibility to infection by mutants of herpes simplex virus type 1, herpes simplex virus type 2, and pseudorabies virus. Virology, v. 246, p. 179–189, 1998.
92. WILD, P.; SCHRANER, E. M.; PETER, J.; LOEPFE, E.; ENGELS, M.
Novel entry pathway of bovine herpesvírus 1 and 5. Journal of Virology, v. 72, n. 12, p. 9561-9566, 1998.
93. WILSON, V.; JEFFREYS, A. J.; BARRIE, P. A. A comparison of
vertebrate interferon gene families deteced by hybridization with human interferon DNA. Journal of Molecular Biology, v. 166, p. 457–475,
1983.
94. WHITE, J. M. Membrane fusion. Science, v. 258, p. 917–924, 1992.
95. WHITBECK, J. C.; PENG, C.; LOU, H.; XU, R.; WILLIS, S. H.; PONCE DE LEON, M.; PENG, T.; NICOLA, A. V.; MONTGOMERY, R. I.;

29
WARNER, M. S.; SOULIKA, A. M.; SPRUCE, L. A.; MOORE, W. T.; LAMBRIS, J. D.; SPEAR, P. G.; COHEN, G. H.; EISENBERG, R. J. Glycoprotein D of herpes simplex virus (HSV) binds directlyto HVEM, a member of the tumor necrosis factor receptor superfamily and a mediator of HSV entry. Journal of Virology, v. 71, p. 6083–6093, 1997.
96. WUDUNN, D.; SPEAR, P. G. Initial interaction of herpes simplex virus
with cells is binding to heparan sulfate. Journal of Virolology, v. 63, p. 52–58, 1989.

![Aula esqueleto.ppt [Modo de Compatibilidade] - portais.ufg.br · Alterações da Coluna Vertebral Doença Disco Intervertebral Condição degenerativa Coluna. 18/11/2011 22 Borges,](https://static.fdocumentos.com/doc/165x107/5be7e1d509d3f2191b8d9085/aula-modo-de-compatibilidade-portaisufgbr-alteracoes-da-coluna-vertebral.jpg)