
Histocompatibilidade MHC
20
HISTOCOMPATIBILIDADE Victória Vendramini Müller, Victória Horn, Mariane Zanini Machado e Erico L. Silvestre.
-
Upload
vickmullerven -
Category
Health & Medicine
-
view
2.024 -
download
3
description
Histocompatibilidade
Transcript of Histocompatibilidade MHC

HISTOCOMPATIBILIDADE Victória Vendramini Müller, Victória
Horn, Mariane Zanini Machado e
Erico L. Silvestre.

DEFINIÇÃO
É uma região de genes altamente
polimórficos, cujos produtos são expressos na
superfície de diferentes tipos celulares.
PROPRIEDADES DAS MOLÉCULAS DO MHC
Cada molécula do MHC consiste em uma fenda, ou
sulco, extracelular que liga os peptídeos, seguida de
um par de domínios semelhantes a imunoglobulinas
(Igs), e está ancorada na membrana celular por
meio de domínios transmembrana e citoplasmático.
Os resíduos de aminoácidos polimórficos das
moléculas do MHC estão localizadas na fenda de
ligação de peptídeos e próximos a ela.

Produtos gênicos codificados no Complexo Principal de Histocompatibilidade foram inicialmente identificados como sendo importantes na rejeição a tecidos transplantados
Genes no MHC considerados altamente polimórficos
Genes no MHC também estavam envolvidos no controle tanto da respostas imune humoral como resposta na mediada por células.
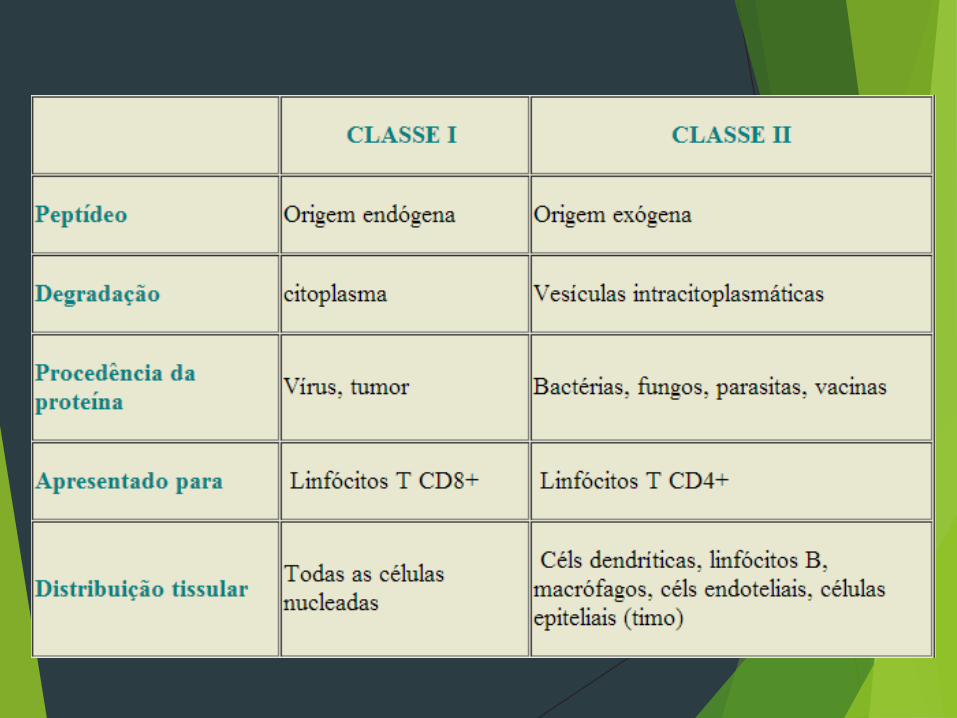
Moléculas de classe I e de classe II
-Moléculas de classe I são encontradas em todas a células nucleadas
-Moléculas de classe II são encontradas somente nas células apresentadoras de antígenos


ESTRUTURA GÊNICA DO MHC
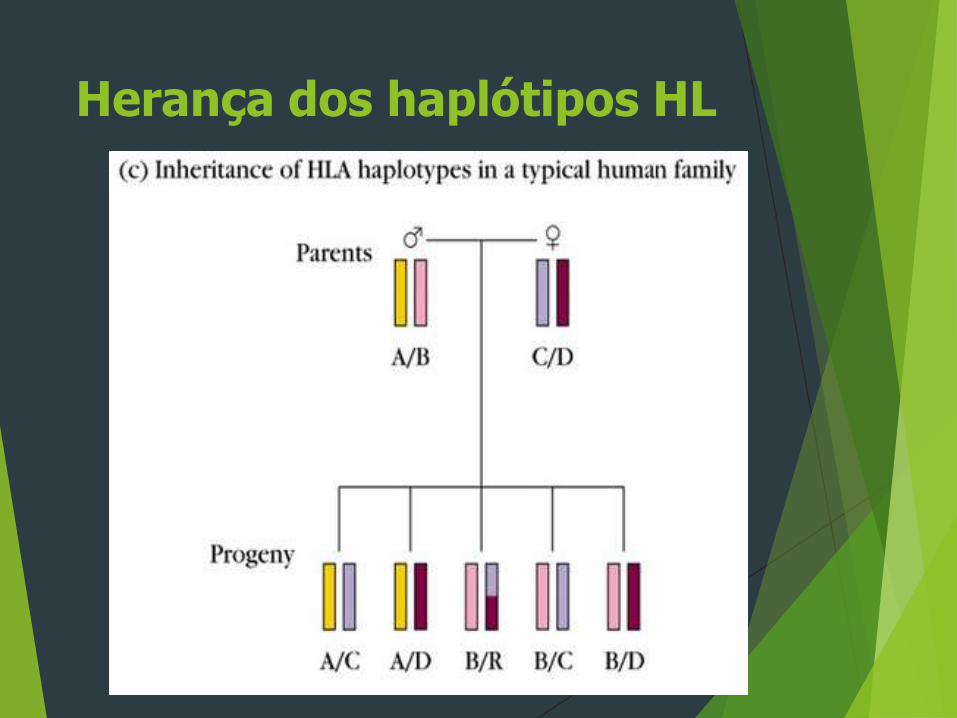
Herança dos haplótipos HL
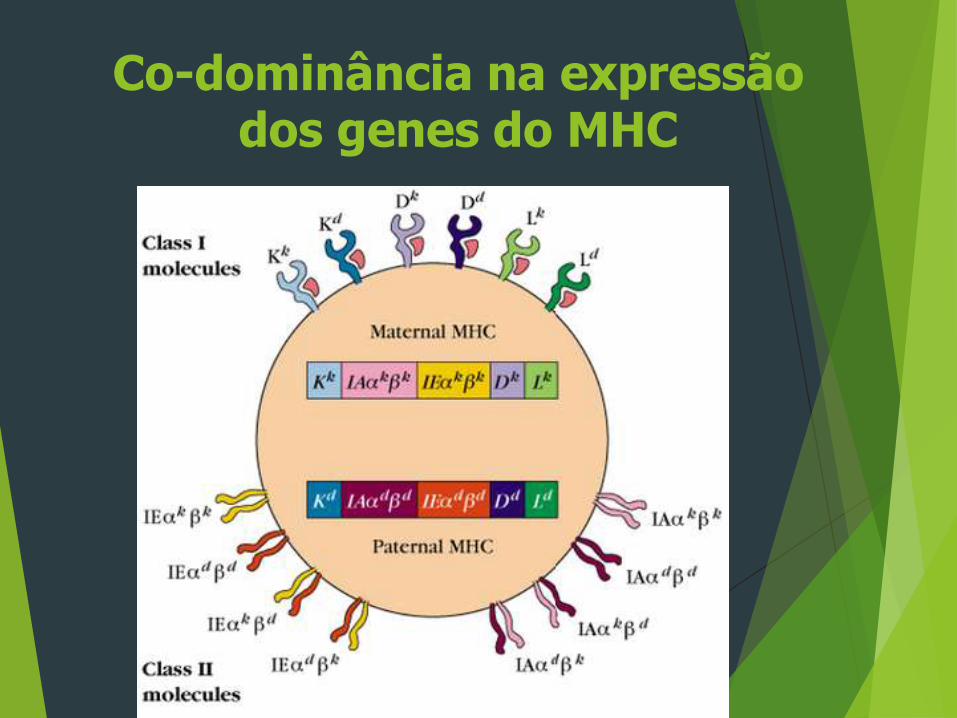
Co-dominância na expressão dos genes do MHC

CLASSES I e II
A função das moléculas de classes I e II codificadas pelo MHC é
se ligar a peptídeos antigênicos e os apresentar para serem
reconhecidos pelos linfócitos T específicos.
CLASSE I
As moléculas de classe I são compostas de uma cadeia α (ou
pesada). As extremidades da fenda de ligação de peptídeos
das moléculas da classe I são fechadas, impedindo a ligação
de peptídeos maiores. Assim sendo proteínas globulares
nativas precisam ser processadas para gerar fragmentos
pequenos o suficiente para se ligarem ás moléculas do MHC e
para serem reconhecidas pelas células T. Os resíduos
polimórficos das moléculas classe I estão confinados aos
domínios α1 e α2, onde contribuem para as variações na
ligação de peptídeos e no reconhecimento pelas células T dos
diversos alelos classe I.

CLASSE II
As moléculas do MHC classe II, são compostas de duas
cadeias polipeptídicas ligadas de forma não co-
valente. Ao contrário das moléculas do MHC classe
I, ambas as cadeias das moléculas classe II são
codificadas por genes do MHC polimórficos.
Alguns aminoácidos polimórficos determinam a
especificidade de ligação de peptídeos que formam
estruturas , chamadas de bolsa, as quais interagem
com aminoácidos complementares no peptídeo ligado.
Outros aminoácidos do MHC polimórficos e alguns dos
aminoácidos dos peptídeos não estão envolvidos na
ligação das moléculas do MHC, mas sim na formação
de estruturas que são reconhecidas pelas células T.
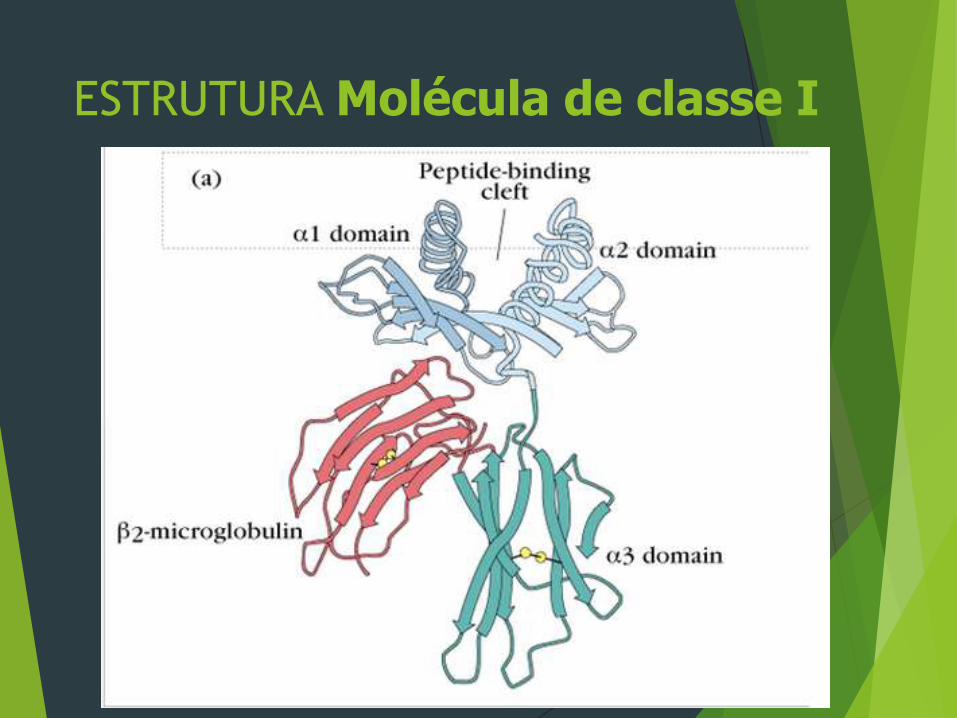
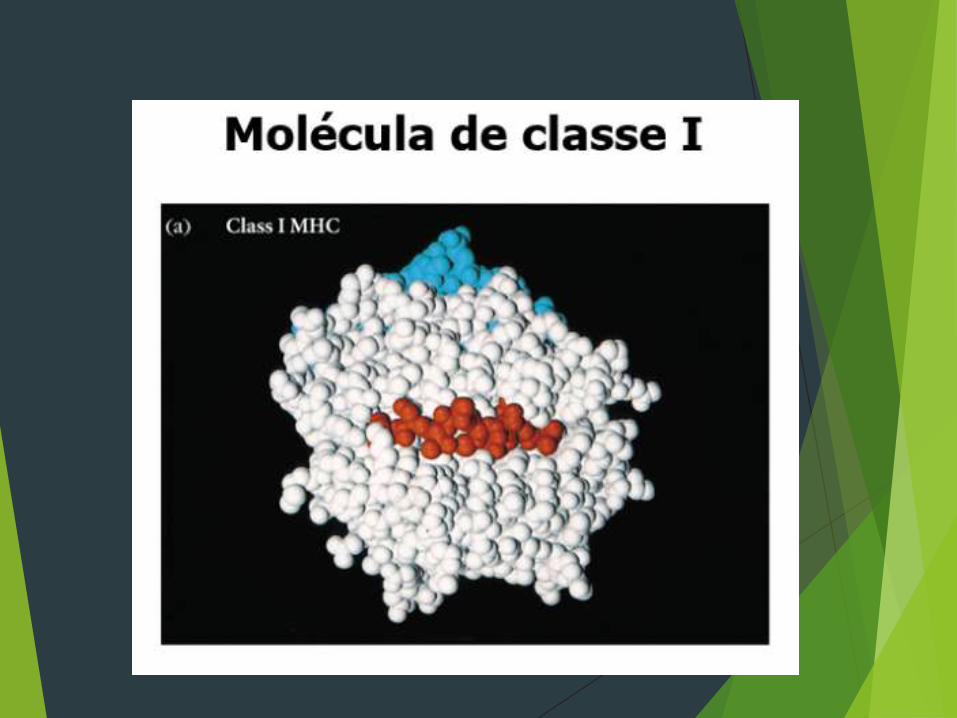
ESTRUTURA Molécula de classe I
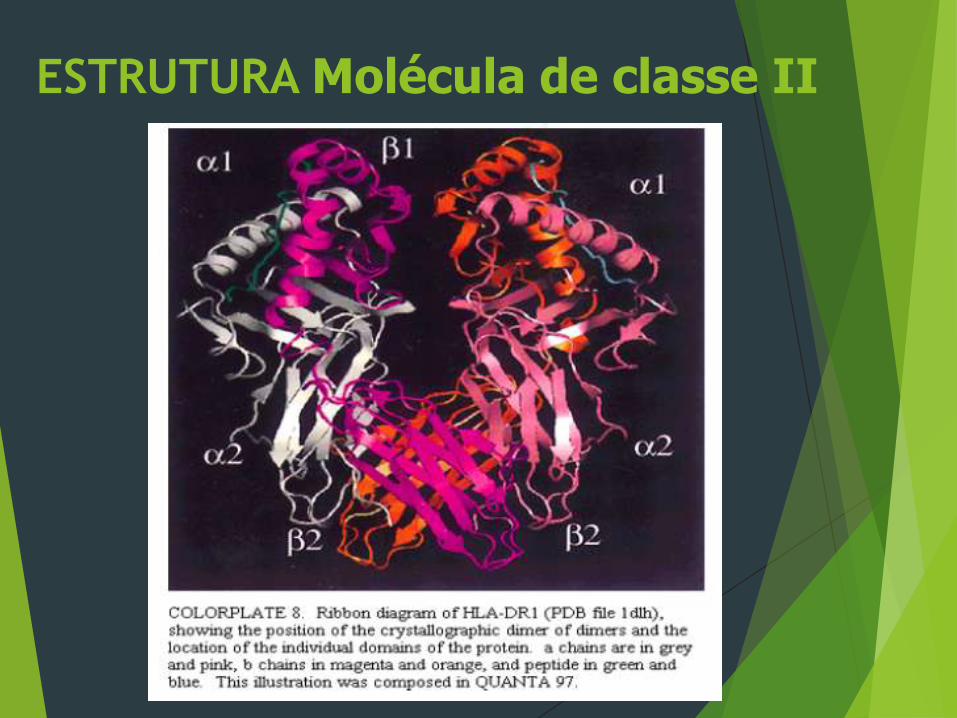
ESTRUTURA Molécula de classe II
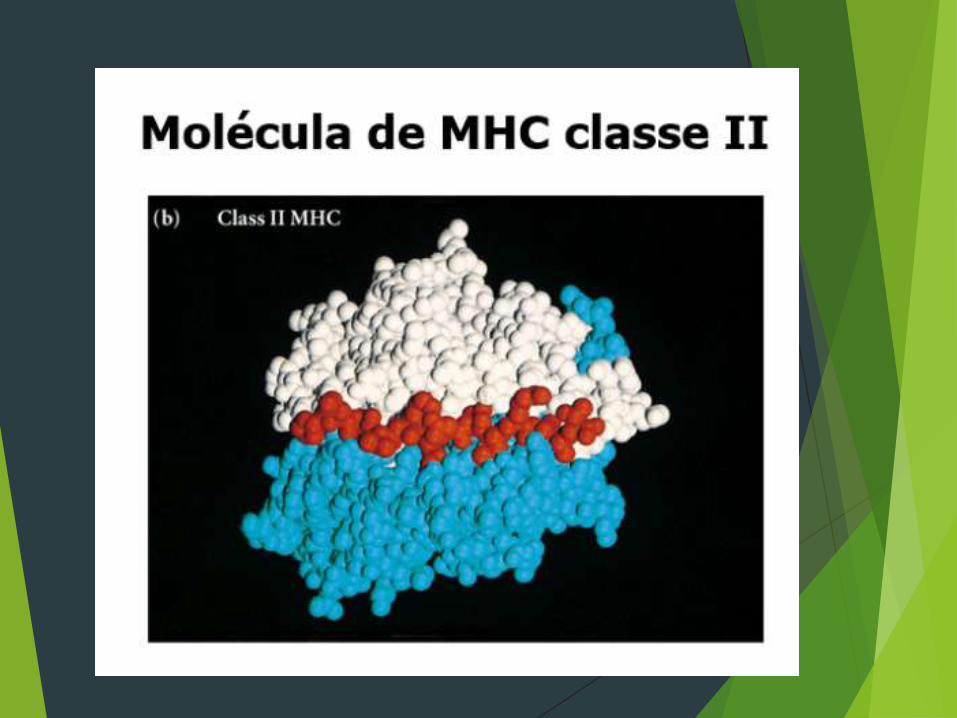

FUNÇÕES Interação entre CD4 e MHC II e CD8 E MHC I
Os genes MHC exercem um papel
central nas respostas imunes aos
antígenos protéicos. Isso é devido ao
fato de que os linfócitos T antígeno-
específicos não reconhecem antígenos
na forma livre ou solúvel, mas, ao
contrário, reconhecem porções de
antígenos protéicos ,isto é, peptídeos
não covalentemente ligados aos
produtos do gene MHC. Em outras
palavras, as moléculas do MHC
proporcionam um sistema para
apresentar peptídeos antigênicos às
células.
As moléculas do MHC são também
reconhecidas pelo seu papel em
desencadear respostas das células T
que causam rejeição de tecidos
transplantados.

Reconhecimento antigênico restrito pelo MHC

BIOSSÍNTESE
O processamento dos antígenos consiste na introdução de antígenos protéicos nas células apresentadoras de antígenos (APCs), na degradação proteolítica dessas proteínas em peptídeos, na ligação dos peptídeos as moléculas recém-montadas do MHC, e na exposição dos complexos peptídeo-MHC à superfície das APCs para o reconhecimento potencial pelas células T.
As vias de processamento do antígeno nas APCs utilizam os mecanismos proteolíticos celulares básicos, que também operam independentemente do sistema imune. Tanto as proteínas extracelulares como as intracelulares são selecionadas por essas vias de processamento do antígeno, e os peptídeos derivados tanto das proteínas próprias como das estranhas são exibidos às moléculas do MHC para a vigilância pelos linfócitos T.
As APCs especializadas, incluindo macrófagos, linfócitos B e células dendríticas, interiorizam as proteínas extracelulares nos endossomas para processamento pela via da classe II do MHC. Essas proteínas são clivadas proteoliticamente por enzimas que funcionam nas vesículas de pH ácido das vias endossômicas. Os heterodímeros recém-sintetizados da classe II do MHC associam-se à cadeia invariante e são dirigidos do RE para as vesículas endossômicas, onde a cadeia invariante é clivada proteoliticamente, e um pequeno remanescente peptídico da cadeia invariante é removido da fenda de ligação peptídica das moléculas do MHC.
Os peptídeos gerados pelas proteínas extracelulares ligam-se depois às moléculas da classe II do MHC, e o complexo peptídeo-MHC desloca-se para a superfície da célula.

COMPLEXO
As moléculas do MHC ligam apenas um peptídeo de cada vez, e todos os peptídeos que se ligam a uma forma alélica específica da molécula do MHC compartilham motivos estruturais comuns.
De um modo geral, a ligação peptídica é de baixa atividade (Kd = 10-6M), porém a velocidade de dissociação é muito lenta, de modo que os complexos, uma vez formados permanecem por um tempo suficientemente longo para ser reconhecido pelas células T. A fenda da molécula de classe I, onde ocorre a ligação dos peptídeos, é fechada nas suas extremidades, limitando a ligação dos peptídeos com resíduos com o comprimento de 11 aminoácidos; enquanto que a das moléculas da classe II é aberta, permitindo que maiores peptídeos (de até 30 resíduos aminoácidos ou mais) possam ligar-se.
Os resíduos polimórficos das moléculas do MHC são localizados no domínio de ligação dos peptídeos. Alguns resíduos de MHC polimórficos determinam as especificidades de ligação aos peptídeos pela formação de estruturas complementares (bolsos) que interagem com as peculiaridades conservadas da ligação peptídica.
Outros resíduos polimórficos do MHC em alguns resíduos dos peptídeos não são envolvidos na ligação peptídeo-molécula de MHC, mas, ao invés disso, formam um determinante antigênico reconhecido pelas células T.

GENÉTICA
O MHC humano é muito grande (cerca de 3.500 quilobases kb) e é organizado como se segue:
1. Genes da classe II (HLA-DP, HLA-DQ, HLA-DR)2. Genes do complemento3. Genes das proteínas do choque térmico e de algumas citocinas (TNF, LT e
LT-B)4. Genes da classe I (HLA-B, HLA-C e HLA-A)
Todos os genes do MHC têm estrutura semelhante, e a maioria das sequências reguladoras estão localizadas na região que flanqueia 5´. A expressão dos produtos do gene do MHC é, em grande parte, regulada ao nível da transcrição pelos fatores tipo celular-específicos como pelos estímulos inflamatórios e imunes incluindo citocinas como o IFN-gama. De modo geral, os genes da classe I são expressos mais largamente, isto é, em tipos celulares mais diversos do que os dos genes da classe II. Diferentes tipos celulares têm padrões distintos das expressão das moléculas da classe II do MHC. Algumas células, tais como os fagócitos mononucleares, podem ser induzidas por citocinas a expressar moléculas da classe II, especialmente IFN-gama. Outras células, como as dendríticas e os linfócitos B, expressam constitutivamente moléculas da classe II

REFERÊNCIAS
http://www.asbai.org.br/revistas/Vol292/aplicacoes.pdf
Http://biologia-
ap.no.comunidades.net/imagens/histocompatibilidade
Abbas; Lichtman; Pillai. Imunologia celular e molecular. 6ª
edição. Elsevier,2008.
Stites; Terr. Imunologia Básica. Prenttice-Hall, 1992.


















