
IDENTIFICAÇÃO E ANÁLISE DE REDE DE GENES … · FIGURA 7 – Mapa de interação dos genes...
131
FACULDADE INFÓRIUM Diego Vinícius de Castro Pereira IDENTIFICAÇÃO E ANÁLISE DE REDE DE GENES ENVOLVIDOS NO CÂNCER DE PELE DE MELANOMA ATRAVÉS DE MINERAÇÃO DE TEXTO Belo Horizonte Fevereiro de 2016
Transcript of IDENTIFICAÇÃO E ANÁLISE DE REDE DE GENES … · FIGURA 7 – Mapa de interação dos genes...

FACULDADE INFÓRIUM
Diego Vinícius de Castro Pereira
IDENTIFICAÇÃO E ANÁLISE DE REDE DE GENES ENVOLVIDOS NO CÂNCER
DE PELE DE MELANOMA ATRAVÉS DE MINERAÇÃO DE TEXTO
Belo Horizonte
Fevereiro de 2016

2
Diego Vinícius de Castro Pereira
IDENTIFICAÇÃO E ANÁLISE DE REDE DE GENES ENVOLVIDOS NO CÂNCER
DE PELE DE MELANOMA ATRAVÉS DE MINERAÇÃO DE TEXTO
Dissertação apresentada ao Programa de pós-
graduação da Faculdade Infórium de Belo horizonte,
como requisito parcial a obtenção do título de mestre
em Tecnologia da Informação Aplicada à Biologia
Computacional.
Orientador: Dr. Fabiano Sviatopolk-Mirsky Pais
Coorientador: André Luiz Sena Guimaraes
Belo Horizonte
Fevereiro de 2016

3
Diego Vinícius de Castro Pereira. Identificação e Análise de Rede de Genes Envolvidos no
Câncer de Pele de Melanoma Através de Mineração de Texto.
Dissertação apresentada ao Programa de pós-
graduação da Faculdade Infórium de Belo horizonte,
como requisito parcial a obtenção do título de mestre
em Tecnologia da Informação Aplicada à Biologia
Computacional.
Aprovada em ___/___/___ pela banca constituída dos seguintes professores:
______________________________________________________
Prof. Dr. Fabiano Sviatopolk-Mirsky Pais – Infórium
_____________________________________________________
Prof.
______________________________________________________
Prof.
Belo horizonte, fevereiro 2016.
Faculdade Infórium de Belo Horizonte
Rua dos Timbiras 1532 – 30.140-061 – Belo Horizonte, MG
Telefone (31) 2103-2103 – www.mestradoti.com.br

4
Dedico este trabalho ao meu pai Francisco de Assis
Cota Aguiar e à minha mãe Dalvemídia Pereira
Aguiar, meus amores, que apesar das dificuldades
soube transmitir toda sua sabedoria e apoio
constante.

5
AGRADECIMENTOS
A DEUS, que todos os dias de minha vida me deu forças para nunca desistir.
A faculdade Infórium, por ter me dado à oportunidade de aprimorar meus conhecimentos.
Aos meus colegas de mestrado, pelos bons momentos que passamos juntos.
Ao meu orientador. Professor Dr. Fabiano Sviatopolk-Mirsky Pais e coorientador André Luiz
Sena Guimarães, pelo apoio e amizade, além da dedicação, competência e especial atenção
nas revisões e sugestões, fatores fundamentais para a conclusão deste trabalho.
Aos professores que destinaram parte de seu precioso tempo para participarem desta pesquisa,
em especial Marcos Flávio Silveira Vasconcelos D’angelo e Eloá Santos.
A todos os professores do mestrado que de alguma forma contribuíram para a minha
formação.
Aos familiares e amigos que sempre me incentivaram e apoiaram nessa jornada.
A minha namorada Joyce (Bisnaguinha), pelo carinho, apoio, paciência e massagens.
Aos acadêmicos Tiago Caldeira e Raimundo pelo apoio no desenvolvimento da pesquisa.

6
Toda a sabedoria vem do Senhor Deus, ela sempre
esteve com ele. Ela existe antes de todos os séculos.
Quem pode contar os grãos de areia do mar, as
gotas de chuva, os dias do tempo? Quem pode medir
a altura do céu, a extensão da terra, a profundidade
do abismo?
Quem pode penetrar a sabedoria divina, anterior a
tudo?
A sabedoria foi criada antes de todas as coisas, a
inteligência prudente existe antes dos séculos!
O verbo de Deus nos céus é fonte de sabedoria, seus
caminhos são os mandamentos eternos
(ECLESIÁSTICO 1: 1-5).

7
RESUMO
O câncer é definido como uma enfermidade caracterizada pelo crescimento descontrolado das
células. É apontado como uma das principais causas de morte no mundo. Existem vários tipos
de cânceres, sendo os mais frequentes na população masculina próstata, pulmão, cólon e reto.
Entre as mulheres são mais comuns: mama, cólon, reto e pulmão. Contudo, o câncer de
melanoma cutâneo é um tipo de câncer que vem apresentando um dos maiores índices de
crescimento na última década. O melanoma inicia-se nos melanócitos, os quais são células
que produzem a melanina e que garantem a proteção contra os danos causados pela radiação
ultravioleta (UV). O desenvolvimento do melanoma é resultado de alterações no DNA. O
estudo sobre o funcionamento de diversos genes vem sendo auxiliado pelas ferramentas de
bioinformática. O objetivo deste trabalho foi identificar genes que podem estar envolvidos no
desenvolvimento e/ou manutenção do câncer de pele melanoma e realizar uma comparação
com os genes envolvidos na manutenção de uma pele saudável, através de pesquisa
bibliográfica, bancos de dados genômicos e, sobretudo, com a utilização de ferramentas de
bioinformática. Para a identificação dos genes envolvidos ou potencialmente envolvidos que
foram analisados quanto a sua relação com o melanoma, utilizou-se como referência artigos
científicos, livros, dissertações e teses da base de dados Pubmed. Após a identificação dos
genes, as interações entre eles foram mapeadas, e um escore referente a essa interação, foi
definida através do banco de dados STRING (Search Tool for the Retrieval of Interacting
Genes/Proteins). Com base nesse escore, os genes foram agrupados usando o algoritmo de
classificação K-means. Os genes do grupo que obteve o maior escore foram chamados genes
líderes. Os outros grupos foram classificados por “B”, “C”, “D” e assim sucessivamente, de
acordo com a força do escore. O estudo foi complementado por uma análise topológica dessa
interação com os softwares Cytoscape, enquanto uma análise ontológica foi realizada com o
software BinGO. A analise realizada permitiu sugerir genes com forte interação para o câncer
de pele melanoma. Os genes líderes sugeridos foram o TP53, porém os genes pertencentes aos
grupos B, C, e D (AKT1, JUN, STAT3 e MYC) também apresentaram escore relevante e
obtiveram destaque no trabalho. Para a pele saudável, os genes líderes sugeridos foram o
UBC, porém os genes dos grupos B, C, e D (TP53, JUN, AKT1, CREBBP, EP300 e SRC)
também apresentaram o escore relevante. Uma comparação entre os genes identificados para
câncer de pele melanoma e os genes da pele saudável, também foi realizada, sugerindo genes

8
que podem estar ativos, silenciados, ou apresentando variações em suas funções para este tipo
de câncer.
Palavras chave: bioinformática; câncer; melanoma; genes; pele; líder (res).

9
ABSTRACT
Cancer is defined as a illness characterized by the uncontrolled growing of cells. It is
appointed as the main cause of deaths in world. There are several types of cancers, being the
most frequents in the male population: prostate, lung, colon and rectum. Among the women
are more common: breast, colon, rectum and lung. However, cancer of cutaneous melanoma
is a type of cancer which is showing one of the highest factors of growing on the last decade.
Melanoma is started on melanocytes that are the cells that produce melanin and that staff the
protection against the damages caused by the UV radiation. The development of melanoma is
result of alterations on genes. The study of working of these genes has been helped by
bioinformatic's tools. The purpose of this work was identify genes that can make part of
development and maintenance of the skin cancer melanoma and make a comparison with the
genes that make part of maintenance of healthy skin, through bibliographic search, genomic
databases and mainly through the utilization of bioinformatic's tools. To identify the genes
that potentially make part of the development and maintenance of the skin cancer and that had
an analysis on their relations with the melanoma were utilized as reference: scientific articles,
books, compositions and theses of Pubmed database. After the genes identification, their
interactions were mapped, and a score (which was named WNL - Weighted Number of Links)
relative to this interaction, was defined through STRING database (Search Tool for the
Retrieval of Interacting Genes/Proteins). Based on this score, genes were grouped through the
K-means classification algorithm. Genes of the group with the highest WNL were named
leader genes. The others groups were classified as "B", "C", "D" etc. according to WNL. The
study was completed with an topologic analysis of this interaction with the software
Cytoscape, while an ontological analysis was made with the software BinGO. The analysis
allowed suggest genes with a strong interaction with the skin cancer melanoma. The
suggested leader genes were TP53's, but genes that belong to groups B, C, and D (AKT1,
JUN, STAT3 and MYC) have also shown relevant WNL and got emphasis in this work.
Suggested leader genes on healthy skin were UBC, but the genes of groups B, C and D (TP53,
JUN, AKT1, CREBBP, EP300 e SRC) have also shown relevant WNL. A comparison
between genes identified on skin cancer melanoma and genes of healthy skin was also made,
suggesting genes that can be actives, silent, or showing variations on their functions to this
type of cancer.
Keywords: bioinformatic, cancer, melanoma, genes, skin, leader (s).

10
LISTA DE ILUSTRAÇÕES
LISTA DE ILUSTRAÇÕES
FIGURA 1 - Estrutura da Pele Humana...................................................................21
FIGURA 2 - Subtipos de melanomas......................................................................28
FIGURA 3 – Sistema ABCD(E) para identificação de melanomas.......................31
FIGURA 4 – Resultados dos tipos de associações em uma análise na base de
dados STRING..........................................................................................................35
FIGURA 5 - Organização da Pesquisa....................................................................44
FIGURA 6 – Mapa de interação dos genes potencialmente envolvidos no
câncer de pele melanoma........................................................................................46
FIGURA 7 – Mapa de interação dos genes potencialmente envolvidos na pele
saudável....................................................................................................................47
FIGURA 8 – Tipos de associações entre genes e respectivas pontuações.......48
FIGURA 9 - Resultado da analise topológica da rede do câncer de pele
melanoma..................................................................................................................50
FIGURA 10 - Resultado da analise topológica da rede da pele saudável...........51
FIGURA 11 – Associações dos genes líderes, grupo B, C e D do Câncer de Pele
Melanoma..................................................................................................................56
FIGURA 12 – Associações dos genes líderes, grupo B, C e D na pele
saudável....................................................................................................................58
FIGURA 13 – Analise Ontológica (processos celulares/biológicos) dos genes
líderes, grupo B, C e D do Câncer de Pele Melanoma..........................................65
FIGURA 14 – Analise Ontológica (processos biológicos/celulares) dos genes
líderes, grupo B, C e D da Pele Saudável..............................................................69
FIGURA 15 – Analise Ontológica (função molecular) dos genes líderes, grupo
B, C e D do Câncer de Pele Melanoma...................................................................72
FIGURA 16 – Analise Ontológica (funções moleculares) dos genes líderes,
grupo B, C e D da Pele Saudável............................................................................73

11
FIGURA 17 – Analise Ontológica (Componentes celulares) dos genes líderes,
grupo B, C e D do Câncer de Pele Melanoma......................................................75
FIGURA 18 – Analise Ontológica (componentes celulares) dos genes líderes,
grupo B, C e D da Pele Saudável..........................................................................76
GRÁFICO 1 – Comportamento lei de potência da rede do câncer de pele
melanoma................................................................................................................50
GRÁFICO 2 – Comportamento lei de potência da rede da pele saudável.........51
GRÁFICO 3 – Distribuição dos genes do câncer de pele melanoma em relação
ao WNL.....................................................................................................................55
GRÁFICO 4 – Distribuição dos genes da pele saudável em relação ao WNL....57
GRÁFICO 5 – Porcentagem dos genes em comum ao CPM e PS nas 10 classes
com maior poder de interação...............................................................................81
LISTA DE TABELAS
TABELA 1 - Características das lesões benignas e malignas.............................26
TABELA 2 – Resultados da analise de variância (ANOVA) com o melanoma....49
TABELA 3 – Resultados da analise de variância (ANOVA) para a pele
saudável....................................................................................................................49
TABELA 4 – Variação entre classes de um gene do melanoma nas 10 classes
com maior poder de interação................................................................................78
TABELA 5 – Variação entre classes de um gene da pele saudável nas 10
classes com maior poder de interação..................................................................79
TABELA 6 – Genes identificados apenas no câncer de pele melanoma............82
TABELA 7 – Genes exclusivos da pele saudável.................................................84

12
LISTA DE ABREVIATURAS E SIGLAS
A.C. - Antes de CRISTO
AJCC - American Joint Committee on Cancer
AKT1 - v-akt Murine Thymoma Viral Oncogene Homolog 1
AP-1 - Activating Protein 1
APP - Amyloid beta (A4) precursor protein
AR - Receptor de Androgénio
ARN - Ribonucleic acid
BINGO - Biological Networks Gene Ontology Tool
BRAF - B-Raf Proto-oncogene, serine/threonine kinase
CAMP - Cathelicidin Antimicrobial Peptide
CBC - Carcinoma Basocelular
CCE - carcinoma de Células Escamosas
CDH1 - Cadherin 1, type 1, E-cadherin (epithelial)
CDK2 - Cyclin-dependent kinase 2
CPM - Câncer de Pele Melanoma
CREBBP - CREB Binding Protein
CTNNB1 - Catenin (cadherin-associated protein), beta 1
D.C. - Depois de CRISTO
DNA - Ácido desoxirribonucleico
EGF - Fator de Crescimento Epidérmico
EP300 - E1A Binding Protein p300
ER-a - Receptor de Estrogênio - alfa
ERBB2 - Erb-b2 receptor tyrosine kinase 2
ESR1 - Estrogen Receptor 1
FYN - FYN Proto-oncogene, Src family tyrosine kinase

13
GLI2 - GLI Family Zinc Finger 2
GO - Gene Ontology
HF - História Familiar
HPV - Vírus do Papiloma Humano
HSP90A - Heat Shock Protein 90 Alpha
HSP90AA1 - Heat Shock Protein 90 Alpha 1
HUGO - Human Genome Organisation
IGF - Insulin Like Growth Factor
INCA - Instituto Nacional do Câncer
lncRNAs - RNAs não-codificadores longos
INS - Insulin
JUN - Jun Proto-oncogene
MATLAB - Matrix Laboratory
MES - Melanoma Expansivo Superficial
MLA - Melanoma Lentiginoso Acral
MLM - Melanoma Lentigo Maligno
MMP-2 - Matrix Metallopeptidase 2
MN - Melanoma Nodular
mRNA - RNA mensageiro
MYC - v-myc Avian Myelocytomatosis Viral Oncogene Homolog
NFKB1 - Nuclear factor of kappa light polypeptide gene enhancer in B-cells 1
NOTCH1 - notch 1
NRAS - Neuroblastoma RAS Viral (v-ras) Oncogene Homolog
OH - Hidroxila
OMS - Organização Mundial da Saúde
PGH - Projeto Genoma Humano
polyA - Poliadenilação Rica em Adenina

14
PPARa - Peroxisome proliferator-activated receptor alfa
PPARg - Peroxisome proliferator-activated receptor gamma
PPARy - Peroxisome proliferator-activated receptor delta
PS - Pele Saudável
PTPN11 - Protein tyrosine phosphatase, non-receptor type 11
RAC1 - Ras-related C3 botulinum toxin substrate 1
RAS - Rat Sarcoma Viral Oncogene Homolog
RHOA - Ras Homolog Family Member A
RNA - Ácido Ribonucleico
rRNA - RNA ribossomal
RUV - Radiação Ultravioleta
SPSS - Statistical Package for Social Sciences
SRC - SRC Proto-oncogene, non-receptor tyrosine kinase
STAT3 - Transdutor de sinal e ativador da transcrição 3
STRING - Search Tool For The Retrieval Of Interacting Genes/Proteins
TP53 - Tumor Protein p53
tRNA - RNA transportador
UBC - Ubiquitin C
UV - Ultravioleta
UVA - Ultravioleta Alfa
UVB - Ultravioleta Beta
VEGF - Vascular Endothelial Growth Factor A
WNL - Weighted Number of Links
WNT - Wnt Oncogene Analog

15
SUMÁRIO
2. FUNDAMENTAÇÃO TEÓRICA ............................................................................................................. 17
2.1 FUNDAMENTOS BIOLÓGICOS...................................................................................................... 17
2.1.1 Ácidos Nucléicos ................................................................................................................... 17
2.1.2 Proteínas e Aminoácidos ...................................................................................................... 18
2.1.3 Genes .................................................................................................................................... 19
2.1.4 A Pele Humana ..................................................................................................................... 21
2.2 Câncer .......................................................................................................................................... 22
2.2.1 Câncer de Pele e Principais Subtipos Não Melanoma .......................................................... 24
2.3 Câncer de Pele Melanoma .......................................................................................................... 25
2.3.1 Subtipos de Melanoma Maligno Cutâneo – Características ................................................ 27
2.3.2 Fatores de Risco ................................................................................................................... 29
2.3.3 Exame ................................................................................................................................... 31
2.3.4 Prognóstico e Estadiamento ................................................................................................ 32
2.3.5 Tratamento ........................................................................................................................... 32
2.4 BIOINFORMÁTICA ........................................................................................................................ 32
2.4.1 Search Tool For The Retrieval Of Interacting Genes/Proteins (STRING) ............................... 34
2.4.2 Cytoscape ............................................................................................................................. 35
2.4.3 Classificação e Agrupamento De Dados ............................................................................... 36
2.4.4 Biological Networks Gene Ontology Tool (Bingo) ................................................................ 37
2.4.5 Genecards ............................................................................................................................. 38
3. OBJETIVOS ......................................................................................................................................... 38
3.1. Delimitação do Tema ................................................................................................................. 38
3.2. Objetivo Geral ............................................................................................................................ 38
3.3. Objetivos Específicos .................................................................................................................. 39
4. JUSTIFICATIVA ................................................................................................................................... 39
5. OBJETO .............................................................................................................................................. 40
5.1. Problema .................................................................................................................................... 40
5.2. Hipótese Básica .......................................................................................................................... 40
6. METODOLOGIA .................................................................................................................................. 41
6.1. Tipo de Pesquisa ......................................................................................................................... 41
6.2. Dados Utilizados ......................................................................................................................... 41

16
6.3. Análise dos Dados ...................................................................................................................... 41
6.4. Organização Desta Pesquisa....................................................................................................... 44
6.5. Contribuições da Pesquisa ......................................................................................................... 44
7. RESULTADOS E DISCUSSÃO ............................................................................................................... 45
7.1. Identificação dos genes .............................................................................................................. 45
7.2. Criação do Mapa de Interação Genômico através do software STRING .................................... 45
7.3 Cálculo do Weighted Number of Links (WNL) ............................................................................. 47
7.4. Agrupamento e Classificação dos Genes ................................................................................... 48
7.5. Analise Topológica Através do Software Cytoscape .................................................................. 49
7.7 Genes Líderes .............................................................................................................................. 54
7.7.1 Genes Líderes no Câncer de Pele Melanoma e Pele Saudável............................................. 59
7.7.2 Genes líderes somente no Câncer de Pele Melanoma ........................................................ 60
7.7.3 Genes líderes somente na Pele Saudável ............................................................................. 62
7.8 Ontologias dos Genes Líderes ..................................................................................................... 64
7.8.1 Processos Celulares/biológicos ............................................................................................ 64
7.8.1.1 Processos Biológicos dos Genes Líderes do Câncer de Pele Melanoma ........................... 65
7.8.1.2 Processos Biológicos dos Genes Líderes da Pele Saudável ............................................... 68
7.8.2 Função Molecular ................................................................................................................. 71
7.8.2.1 Função Molecular dos Genes Líderes do Câncer de Pele Melanoma ............................... 71
7.8.2.2 Função Molecular dos Genes Líderes da Pele Saudável ................................................... 72
7.8.3 Componentes Celulares ....................................................................................................... 74
7.8.3.1 Componentes Celulares dos Genes Líderes do Câncer de Pele Melanoma ...................... 74
7.8.3.1 Componentes Celulares dos Genes Líderes da Pele Saudável .......................................... 76
7.9 Comparações Entre os Resultados Identificados no Câncer de Pele Melanoma (CPM) e Genes
Da Pele Saudável (PS) ........................................................................................................................ 77
7.9.1 Relação de Genes Identificados Apenas no Câncer de Pele Melanoma .............................. 80
7.9.2 Relação de Genes Identificados Apenas na Manutenção da Pele Saudável ........................ 83
7.9.3 Principais Variações Entre Genes dos 10 grupos Com Maior WNL ...................................... 88
8. CONCLUSÃO ...................................................................................................................................... 90
9. REFERÊNCIAS ..................................................................................................................................... 91
10. ANEXOS ......................................................................................................................................... 114
Genes Órfãos ................................................................................................................................... 114
Genes e WNL do Câncer de Pele Melanoma ................................................................................... 115
Genes e WNL da Pele Saudável ....................................................................................................... 125

17
2. FUNDAMENTAÇÃO TEÓRICA
Este capítulo apresenta a fundamentação teórica relacionada aos tópicos
tratados nesta dissertação.
A seção 2.1 apresenta os fundamentos biológicos importantes para o
entendimento deste trabalho. A seção 2.2 aborda o conceito de câncer, câncer de
pele e seus principais subtipos não melanoma. A seção 2.3 trata do tema câncer de
pele melanoma. A seção 2.4 apresenta uma visão geral sobre bioinformática e
softwares utilizados no trabalho.
2.1 FUNDAMENTOS BIOLÓGICOS
2.1.1 Ácidos Nucléicos
2.1.1.1 Ácido desoxirribonucleico (DNA)
O DNA é um polímero, constituído de unidades que se repetem chamadas
nucleotídeos. Os diferentes nucleotídeos ligam-se entre si, formando uma cadeia
polinucleotídica. Cada um deles por sua vez contém um grupo de fosfato ligado a
um açúcar com cinco carbonos (pentose) e uma base orgânica nitrogenada (Watson
et al, 1953; MIR et al, 2004).
O açúcar é um derivado da ribose chamado desoxirribose porque nele falta o
agrupamento hidroxila (OH) no carbono 2, semelhante a hidroxila OH presente no
carbono 3. As estruturas da base nitrogenadas incluem duas purinas (adenina e
guanina) e duas pirimidinas (timina e citosina) (Watson et al, 1953; Burns et al,
1991).
A molécula de DNA é composta por duas destas cadeias de nucleotídeos que
se dispõem de forma helicoidal em sentido antiparalelo. As bases de uma cadeia se

18
prendem as de outras por “ligações de hidrogênio” (Watson et al, 1953; MIR et al,
2004).
Com sua replicação precisa, o ácido desoxirribonucleico presta-se para levar
a informação genética de célula a célula e de geração a geração (Watson et al,
1953; Burns et al, 1991).
2.1.1.2 Ácido Ribonucleico (RNA)
O ácido ribonucleico (RNA), é representado como uma fita única de ácidos
nucléicos, que diferente do DNA, possui o açúcar ribose e a base nitrogenada
uracila em substituição a timina. Essa fita é obtida através da transcrição de uma
das fitas do DNA. Os principais tipos de RNA são: O RNA mensageiro (mRNA), que
contém a informação para a codificação das proteínas, o RNA transportador (tRNA),
que é o responsável pelo transporte de aminoácidos, e o RNA ribossomal (rRNA),
que possui um papel estrutural (Mena-Chalco, 2005). Entretanto, diversos RNAs
não-codificadores, como os long-non-coding (lncRNAs) estão emergindo como
importante reguladores da fisiologia do tecido e processos de doenças, incluindo
Câncer (Iyer et al, 2015).
2.1.2 Proteínas e Aminoácidos
As proteínas são as macromoléculas mais abundantes nas células vivas. Elas
ocorrem em todas as células e em todas as partes destas. As proteínas também
ocorrem em grande variedade; milhares de diferentes tipos, desde peptídeos de
tamanho relativamente pequeno até enormes polímeros com pesos moleculares na
faixa de milhões de Daltons, podem ser encontrados em uma única célula. As
proteínas também exibem uma grande diversidade de funções biológicas.
(Lehninger, 2000).
Uma proteína corresponde a uma macromolécula composta de uma ou mais
cadeias polipeptídicas, cada uma com uma sequência característica de aminoácidos

19
ligados por ligações peptídicas, que são um tipo de ligação covalente (Lehninger,
2000).
O primeiro aminoácido descoberto nas proteínas foi a aspargina, em 1806. O
último dos 20, a treonina, somente foi identificado em 1938. Todos os 20
aminoácidos encontrados nas proteínas são α-aminoácidos. Eles têm um grupo
carboxila e um grupo amino, ligados ao mesmo átomo de carbono (o carbono α).
Diferem entre si por suas cadeias laterais ou grupos R, os quais variam em
estrutura, tamanho e carga elétrica e influenciam a solubilidade do aminoácido em
água (Lehninger, 2000).
2.1.3 Genes
Unidade física e funcional fundamental da hereditariedade. Um gene é uma
sequência ordenada de nucleotídeos localizada em uma posição particular em um
cromossomo específico que codifica um produto funcional específico isto é, uma
proteína ou mesmo uma molécula de RNA não codificante (JOAQUIM et al, 2010).
Por muito tempo as regiões não-codificadoras do genoma foram consideradas
“lixo genético”, devido à ausência de informação para síntese de proteínas nessas
sequências nucleotídicas. Atualmente, sabe-se que a propagação da informação
genética é muito mais complexa e não segue o antigo dogma central da biologia
baseado no simples fluxo DNA-RNA-Proteína. Inclusive, hoje se sabe, que a maior
porção do transcritoma produzido pelo genoma de eucariotos é composta por RNAs
não-codificadores de proteínas (Hui e Bindereif, 2005; Nagalakshmi et al., 2008;
Guttman et al., 2009; Nachtigall, 2012).
Um gene típico codificante pode ser constituído pelas seguintes regiões
características (Mena-Chalco, 2005):
Região de reconhecimento (região promotora);
Início de transcrição;
Região não-traduzida 5', que regula a transcrição gênica;
Início de tradução, ou códon de início (start codon);
Região para a codificação de proteínas.
Parada de tradução, ou códon de parada (stop codon);

20
Região não-traduzida 3';
Região de poliadenilação rica em Adenina (polyA) unicamente presente nos
eucariotos;
Parada de transcrição.
Existem classes de genes importantes na patogênese do cancro, sendo eles
os oncogenes e os supressores tumorais. Anormalidades tanto nos genes
estimuladores de divisão celular (chamados de oncogenes), como nos protetores ou
bloqueadores do ciclo celular (chamados de genes supressores tumorais) podem
levar à desregulação do ciclo celular (WARD, 2002).
De acordo com Rivoire et al (2006), os proto-oncogenes estimulam os
processos de divisão celular. São genes celulares normais que controlam o
crescimento celular e a diferenciação do organismo. Sob certas circunstâncias
podem transformar-se em oncogenes, que são genes dominantes no nível celular
que codificam proteínas estimuladoras do crescimento, que contribuem para o
descontrole da divisão celular e o fenótipo maligno da célula. A maioria dos
oncogenes se origina a partir de proto-oncogenes (Borges et al, 2001).
Originalmente os oncogêneses foram encontrados em vírus, e supôs-se que
seriam transferidos a células de vertebrados, causando assim o câncer. Contudo
soube-se mais tarde que esses genes já estavam presentes nas células na maioria
dos vertebrados (Burns et al, 1991).
Os proto-oncogenes são genes altamente preservados na evolução das
espécies. Há uma alta fidelidade de conservação da sua mensagem genética, ao
longo da evolução dos vertebrados, incluindo mesmo alguns invertebrados. Eles
parecem constituir uma família de genes altamente hierárquicos, desempenhando
funções críticas, e a mínima alteração causam graves transtornos nos processos de
divisão celular (Manso, 1986).
Em relação aos genes supressores, de acordo com Rivoire et al (2006),
inibem os processos de divisão celular. Os genes de supressão tumoral ou genes
supressores de tumor são genes recessivos no nível celular e tem a função de
reprimir a divisão, como uma maneira de controlar esta divisão. Mas a sua função
pode ser perdida ou alterada por deleção ou mutação pontual, contribuindo para o
desenvolvimento do câncer (Borges, 2001).

21
2.1.4 A Pele Humana
A pele (FIGURA 1) é um órgão complexo que protege o organismo do meio
ambiente e ao mesmo tempo interage com ele, sendo considerado o maior órgão do
corpo humano (CHU, 2008).
Desempenha um papel fundamental na proteção do organismo contra o
trauma físico, agentes patogénicos, alergenos, produtos químicos, radiação
ultravioleta (UV), a água em excesso e a perda de eletrólitos (WOLK, 2010).
A pele humana tem quatro funções principais: a homeostase endógena
(regulação da temperatura corporal e equilíbrio de fluidos), o metabolismo (a síntese
de vitamina D), a entrada sensorial, e para servir como uma barreira para ameaças
externas (infecção, lesão, luz ultravioleta) (SPELLBERG, 2000).
A pele pode ser dividida em três camadas principais: epiderme, derme e
hipoderme (SPELLBERG, 2000).
FIGURA 1 - Estrutura da Pele Humana (Wolk et al, 2010)

22
O sistema de pigmentação da pele humana está baseado em dois tipos
celulares: melanócitos e queratinócitos (Fitzpatrick et al, 1979).
Os melanócitos são células fenotipicamente importantes, responsáveis pela
pigmentação da pele e dos pelos, contribuindo para a tonalidade cutânea, conferindo
proteção direta aos danos causados pelos raios ultravioletas. Os melanócitos são
células produtoras de melanina (Miot et al, 2009).
A melanogênese é o processo responsável pela formação da cor da pele,
sendo constituídos pela interação de várias substâncias cromóforas presentes na
epiderme e derme, na qual se destaca a melanina (PETIT, 2003).
Nos mamíferos, a melanina é responsável pela coloração da pele, cabelos e
olhos, sendo que na espécie humana, possui papel fundamental na pigmentação da
pele e dos cabelos (Nerya et al, 2003).
2.2 Câncer
O câncer é uma patologia de etiologia multifatorial, resultante, principalmente,
de alterações genéticas, fatores ambientais e do estilo de vida (Popim et al, 2008;
Tsao, 2001).
De acordo com Silva (2005) e Chiattone (1996), a origem da palavra câncer
vem do grego Karkinos e do latim Câncer, ambos significando caranguejo, pela
semelhança entre as veias ao redor do tumor externo e as pernas do crustáceo,
embora alguns acreditassem que o nome teria relação com o fato de a doença
evoluir de modo semelhante ao movimento do animal.
O câncer é uma doença obviamente antiga. Escritos egípcios (1200 A.C.),
Hipócrates (460 A.C.) também faziam referências à enfermidade. O médico grego
Galeno (130 D.C.) foi o primeiro a classificar tumores de pele em malignos e
benignos (TRINCAUS, 2005).
Somente a partir do século XIX é que ocorreram avanços mais significativos
sobre o entendimento desta enfermidade (BIELEMANN, 1997).
De acordo com o Instituto Nacional do Câncer (INCA), câncer é o nome dado
a um conjunto de mais de 100 doenças que têm em comum o crescimento

23
desordenado de células, que invadem tecidos e órgãos. Dividindo-se rapidamente,
estas células tendem a ser muito agressivas e incontroláveis, determinando a
formação de tumores malignos, que podem espalhar-se para outras regiões do
corpo.
Jurberg et al (2006) afirma que o câncer é uma doença que pode ter cura,
porém para um bom prognóstico é necessário um diagnóstico precoce. Alguns
hábitos também têm estreita correlação com o aparecimento da doença: o fumo, a
vida sedentária, a dieta rica em alimentos gordurosos ou pobres em vegetais e frutas
que, associados a fatores genéticos, podem levar ao desenvolvimento do câncer.
Além disso, vírus como o da hepatite B e o vírus do Papiloma Humano (HPV)
também estão ligados ao aparecimento do câncer.
O câncer é considerado uma das maiores causas de morte no mundo
(DANTAS et al, 2009). Segundo o INCA, o câncer pode surgir em qualquer parte do
corpo, mas alguns órgãos são mais afetados do que outros. Entre os mais afetados
no Brasil estão pulmão, mama, colo do útero, próstata, cólon e reto (intestino
grosso), pele, estômago, esôfago, medula óssea (leucemias) e cavidade oral (boca).
Cada órgão, por sua vez, pode ser afetado por tipos diferenciados de tumor, menos
ou mais agressivos.
Estimativas mundiais do projeto Globocan (FERLAY et al, 2012), da Agência
Internacional para Pesquisa em Câncer da Organização Mundial da Saúde (OMS),
afirmam que houve 14,1 milhões de casos novos de câncer e um total de 8,2
milhões de mortes por câncer, em todo o mundo, em 2012. Em 2030, a carga global
será de 21,4 milhões de casos novos de câncer e 13,2 milhões de mortes por
câncer, em consequência do crescimento e do envelhecimento da população, bem
como da redução na mortalidade infantil e nas mortes por doenças infecciosas em
países em desenvolvimento.
Segundo informações do INCA, a estimativa para o ano de 2014 no Brasil,
que será válida também para o ano de 2015, aponta para a ocorrência de
aproximadamente 576 mil casos novos de câncer, incluindo os casos de pele não
melanoma, reforçando a magnitude do problema do câncer no país. O câncer de
pele do tipo não melanoma (182 mil casos novos) será o mais incidente na
população brasileira, seguido pelos tumores de próstata (69 mil), mama feminina (57
mil), cólon e reto (33 mil), pulmão (27 mil), estômago (20 mil) e colo do útero (15 mil).

24
De acordo com Ferreira et al (2008), muitos fatores contribuem para o
crescimento do câncer no Brasil, entre eles o envelhecimento da população,
decorrente das ações de saúde que evitam mortes prematuras por doenças
infecciosas ou parasitárias. O desenvolvimento socioeconômico, no entanto,
modifica os hábitos da população: não existe sociedade sem câncer, mas os tipos
de câncer mudam de acordo com o desenvolvimento do país e muitos deles podem
ser evitados pela conscientização.
2.2.1 Câncer de Pele e Principais Subtipos Não Melanoma
A ocorrência mais comum de câncer nos próximos anos será o de pele.
Cancro na pele é um processo de múltiplos passos, em que uma série de
acontecimentos genéticos e ambientais conduz ao aparecimento de células que se
multiplicam utilizando mecanismos anormais de controle do crescimento
(Ananthaswamy, 1990).
Os cânceres cutâneos não melanoma são as neoplasias malignas mais
comuns em humanos. O carcinoma basocelular e o carcinoma espinocelular,
representam aproximadamente 95% dos cânceres cutâneos não melanoma, o que
os torna um crescente problema para a saúde pública mundial devido a suas
prevalências cada vez maiores (Martinez et al, 2006).
2.2.1.1 Carcinoma Basocelular
O carcinoma basocelular (CBC) deriva de células basais da epiderme e do
aparelho folicular. É um tumor de baixo grau de malignidade, com capacidade de
invasão local, destruição tecidual, recidivante e com limitado poder de
metastatização. A taxa de mortalidade é baixa devido à precocidade do diagnóstico
do CBC nas áreas expostas e ao crescimento lento das lesões (Mantese et al,
2006).
O CBC é a neoplasia maligna mais comum em humanos, corresponde a
aproximadamente 75% do total dos tumores malignos cutâneos. Acometem

25
principalmente pacientes do sexo masculino, acima de 40 anos de idade, brancos,
com relato de exposição solar repetitiva e, frequentemente, localiza-se na face. É
facilmente tratável pela excisão cirúrgica, desde que diagnosticado precocemente
(Chinem et al, 2011; Mantese et al, 2006).
2.2.1.2 Carcinoma Espinocelular
O carcinoma de células escamosas (CCE), também denominado carcinoma
epidermóide, carcinoma escamocelular e carcinoma espinocelular, é uma neoplasia
maligna que se origina no epitélio de revestimento, sendo considerada a neoplasia
maligna mais comum nesta região (Brener et al, 2007).
É o segundo mais prevalente dentre todos os tipos de câncer de pele.
Manifesta-se nas células escamosas, que constituem a maior parte das camadas
superiores da pele. Pode se desenvolver em todas as partes do corpo, embora seja
mais comum nas áreas expostas ao sol, como orelhas, rosto, couro cabeludo,
pescoço, etc (SBD, 2015).
2.3 Câncer de Pele Melanoma
O melanoma origina-se dos melanócitos, células da pele produtoras de um
pigmento denominado melanina, que lhe proporciona proteção contra os danos
causados pela UV. O desenvolvimento do melanoma é consequência da perda dos
mecanismos genéticos de controle celular causados principalmente pelas radiações
UVA e UVB (Souza et al, 2009).
De acordo com Wainstein et al, 2004, a primeira descrição que se refere ao
melanoma surgiu durante o período 460 a 375 A.C. nas escrituras de Hipócrates,
mas o termo "melanoma" foi originalmente empregado em 1838 por Robert Carswell,
que o utilizou para descrever lesões malignas pigmentadas da pele. Já em 1858,
Pemberton realizava e advogava a excisão ampla e profunda como tratamento da
doença. Ainda de acordo com Wainstein et al (2004) pouco depois, em 1907,
Handley recomendava a ressecção em blocos com margens amplas. Nas décadas

26
de 50 e 60 vários pesquisadores como Allen, Spitz, Petersen e Bodenhan já
tentavam identificar os fatores prognósticos relacionados ao melanoma. Em 1969
Clark e colaboradores aprimoraram o sistema de micro estadiamento utilizando
como critério os níveis de invasão da pele, e em seguida, Breslow demonstrou a
importância da espessura do melanoma primário. Finalmente em 1992, Morton
introduziu o rastreamento linfático pré-operatório e a linfadenectomia seletiva do(s)
linfonodo(s) marcado(s) (linfonodo sentinela), técnica que atualmente foi incorporada
como procedimento de rotina na maioria dos grandes centros especializados em
câncer (Wainstein et al, 2004).
Nas últimas décadas, o melanoma tornou-se um assunto de grande interesse
devido ao aumento na sua incidência e à sua elevada letalidade. Embora represente
apenas 4% a 5% dos casos de câncer de pele, o melanoma provoca a maioria das
mortes por malignidades cutâneas, pelo seu alto potencial de enviar metástases a
órgãos distantes (Battisti et al, 2009).
De acordo com o INCA, o crescimento ou alteração da forma é progressivo e
se faz no sentido horizontal ou vertical. Na fase de crescimento horizontal
(superficial), a neoplasia invade a epiderme (camada mais superficial da pele),
podendo atingir ou não a derme papilar superior (camada intermediária da pele). No
sentido vertical, seu crescimento é acelerado através da espessura da pele,
formando nódulos visíveis e palpáveis.
Apesar de muitos fatores ambientais e genéticos contribuírem para o
desenvolvimento do câncer, uma das mais importantes é a exposição crônica à
radiação UV da luz solar (Grossman et al, 1997).
Vários estudos relacionam a exposição à radiação solar como uma das
principais causas de câncer de pele tipo melanoma e não melanoma (Szklo et al,
2007).
A tabela 1 apresenta as características identificadas nas lesões benignas e
malignas.
Tabela 1 Características das lesões benignas e malignas
Lesão Melanocítica
Critério Lesão benigna Lesão maligna

27
1. Tamanho Pequeno (<10mm) Grande (>10mm)
2. Simetria Simétrica Assimétrica
3. Delimitação lateral Bem delimitada Mal delimitada
4. Maturação Presente (focal ou
extensa)
Ausente
5. Disseminação
pagetóide
Ausente; se presente,
focal e delimitada.
Extensa
6. Necrose/ulceração Ausente Pode estar presente
7. Infiltrado
inflamatório
Ausente ou discreto Leve a intenso
8. Regressão Ausente Pode estar presente
9. Atipias celulares Ausentes ou peculiares Presentes/extensas
10.
Mitoses
Profundas Ausentes Presentes
Atípicas Ausentes Presentes
11. Melanização da
lesão
Regular e superficial Irregular, podendo ser
profunda
12. Proliferação de
células isoladas
Ausente ou discreta Presente e às vezes
predominante
TABELA 1 - Características das lesões benignas e malignas. Fonte:(Veronese et al, 2004)
2.3.1 Subtipos de Melanoma Maligno Cutâneo – Características
O melanoma cutâneo é classificado em quatro grupos clínico-histológicos:
melanoma em lentigo maligno, melanoma disseminativo superficial, melanoma
nodular, e melanoma lentiginoso acral, conforme apresentado na FIGURA 2 (Souza,
S.R.P. et al, 2004).

28
FIGURA 2 - Subtipos de melanomas. A. Melanoma Nodular. B. Melanoma sobre Lentigo
Maligno. C. Melanoma Nodular. D. Melanoma Nodular Ulcerado. E. Melanoma de Extensão
Superficial. F. Melanoma Subungueal. G. Melanoma de Extensão Superficial in situ. Fonte:
Neto (2008)
2.3.1.1 Melanoma Lentigo Maligno (MLM)
Corresponde a 5% dos melanomas em caucasianos, e é mais frequentemente
diagnosticado em mulheres, indivíduos com idade superior a 60 anos e em áreas
anatômicas mais intensamente expostas ao sol (Souza, S.R.P. et al, 2004).
2.3.1.2 Melanoma Expansivo Superficial (MES)
Melanoma expansivo superficial, ou extensivo superficial, ou de espalhamento
superficial (MES) é o mais frequente (70% dos casos). Aparece principalmente entre
a quarta e quinta décadas de vida, preferencialmente no tronco dos homens e
membros inferiores das mulheres, o que sugere que esta forma clínica esteja
associada à exposição solar sazonal (DIMATOS et al 2009).

29
2.3.1.3 Melanoma Nodular
Melanoma nodular (MN), é o segundo mais comum (15 a 30% dos casos),
ocorre mais frequentemente nas quinta e sexta décadas de vida, sexo masculino, na
proporção de 2:1. Apresenta-se como lesão papulosa ou nodular, elevada, de cor
castanha, negra ou azulada. São frequentes a ulceração e o sangramento; existe a
variante amelanótica, com superfície critematosa (Fernandes et al, 2005).
2.3.1.4 Melanoma Lentiginoso Acral
Melanoma Lentiginoso Acral (MLA) ocorre nas regiões palmoplantares,
extremidades digitais, mucosas e semimucosas. É mais frequente em não brancos.
Não tem predileção por sexo. Comum na sétima década de vida. Nas extremidades
digitais pode-se apresentar como lesão tumoral acastanhada subungueal,
melanoníquia estriada, fragmentação longitudinal da lâmina ungueal, além de
paroníquia crônica e persistente (Fernandes et al, 2005).
2.3.2 Fatores de Risco
Vários fatores podem ser responsáveis pela incidência de melanoma (Neto,
2008; Markovic et al, 2007). De acordo com Alves et al (2007) os fatores de riscos
mais relacionados ao melanoma são: histórico familiar e pessoal de nevos
displásicos ou melanoma, presença de nevos congênitos, nevos melanocíticos
adquiridos, superior a 100, o número de nevos atípicos superior a 50 e o diâmetro
dos mesmos (superior a 6 mm de diâmetro), pele tipo I ou II e radiação ultravioleta.
Além de queimaduras solares na infância e alta exposição ao sol. Molgó et al (2006);
Neto (2008) afirmam que o principal fator de risco constitucional ou endógeno para o
melanoma é o tipo de pele.
Segundo a classificação de Fitzpatrick et al (1997), existem seis fototipos
diferentes, os quais classificam a reação da pele quando exposta ao sol: tipo I
queima-se facilmente e nunca se bronzeia, tipo II queima-se facilmente e bronzeia-
se minimamente, tipo III queima-se pouco e bronzeia-se gradualmente, tipo IV

30
queima-se pouco e bronzeia-se bem, tipo V raramente se queima, bronzeia-se
muito e tipo VI nunca se queima, bronzeia-se profundamente. Salienta-se que o
risco de melanoma é maior nas pessoas com fototipos I, II e III.
Já em relação a U.V., está bem definido que a radiação ultravioleta provoca
depleção imunológica na pele, permitindo o desenvolvimento de tumores cutâneos
malignos. A maioria dos pacientes de cânceres da pele não melanomas são
considerados UVB-suscetíveis (Nasser, 2010).
Os dois mecanismos pelos quais a radiação ultravioleta pode danificar o DNA
são a excitação direta das moléculas, predominante na região do UVB, e a geração
de espécies altamente reativas de oxigênio, predominante na região do UVA. Os
danos oxidativos possivelmente são intermediados pela melanina. Células
previamente irradiadas com dose baixa de UVA e posteriormente com alta dose
apresentam duas vezes mais danos oxidativos do que aquelas sem a pré-irradiação.
Os danos causados ao DNA podem ser prontamente reparados pelo sistema de
excisão-reparo de nucleotídeo (Souza, S.R.P. et al, 2004).
Ainda de acordo com Nasser (2010), sabe-se que a capacidade de a radiação
UVB alterar o sistema imunitário cutâneo tem sido amplamente documentada, além
de ser um importante fator de risco para o desenvolvimento do câncer da pele
induzido pela luz solar (Nasser, 2010). Estima-se que até 65% dos casos de
melanoma possam estar relacionados à exposição solar (Carvalho et al, 2004).
Outro fator de risco é a hereditariedade. Análises genéticas foram realizadas
na década de 1980. Identificaram um padrão de herança autossômico dominante em
algumas famílias com múltiplos casos de melanoma. Atualmente, acredita-se que a
maioria dos casos dessa neoplasia resulte da interação de fatores de risco genéticos
e ambientais, constituindo um modelo de doença multifatorial (Carvalho et al, 2004).
Cerca de 8 a 14% dos pacientes que recebem o diagnóstico de melanoma
apresentam história familiar (HF) positiva para essa neoplasia. Além disso, estima-se
que cerca de 10% dos pacientes diagnosticados com mais de um melanoma
primário sejam portadores de uma mutação de predisposição hereditária a esse
tumor. Embora agrupamentos de casos de melanoma e/ou nevos displásicos
possam ocorrer em uma família devido a padrões comuns de exposição solar e não
a fatores genéticos, hoje estima-se que 10% de todos casos de melanoma sejam

31
causados por mutações germinativas em genes de suscetibilidade, sendo, portanto,
hereditários (Carvalho et al, 2004).
2.3.3 Exame
O diagnóstico de melanoma baseia-se na observação clínica, no
reconhecimento de sinais e sintomas, na presença de lesão suspeita, e aplicação de
algoritmos diagnósticos, sobretudo do sistema ABCD(E): A de assimetria da lesão, B
de características dos bordos (regular, irregular), C de cor (mudança de cor: preto,
castanho, cinzento, rosa, branco ou azul), D de diâmetro superior a 6 milímetros, E
de evolução (FIGURA 3) (Thomas et al, 2006).
FIGURA 3 – Sistema ABCD(E) para identificação de melanomas. A. Assimetria. B. Bordos.
C. Cor. D. Diâmetro. Adaptado de Manual de Dermatologia y Venerologia, Corell, J.J., 2008,
pq. 442. Adaptado por Neto, 2008.
As técnicas utilizadas para o diagnóstico são diversas como: a
dermatoscopia, biópsia de excisão, biópsia incisional, imunohistoquimica (Fitzpatrick
et al, 1979).

32
2.3.4 Prognóstico e Estadiamento
A espessura e a presença de ulceração do tumor são os principais fatores
prognósticos em pacientes com melanoma primário sem acometimento linfonodal. Já
naqueles com rastreamento e exérese eletiva do linfonodo sentinela, o fator mais
significativo é a característica histológica do linfonodo biopsiado (Wainstein et al,
2004).
Ainda de acordo Wainstein et al (2004), uma vez diagnosticadas metástases
linfonodais, os fatores prognósticos principais passam a ser o número de linfonodos
acometidos, a presença de macrometástases e ulceração da lesão.
O estadiamento da doença é feito de acordo com os protocolos do American
Joint Committee on Cancer (AJCC), publicados em 2002. Este estadiamento analisa
a espessura da lesão primária, a presença de ulceração, satelitose, acometimento
linfonodal e presença de metástases viscerais ou à distância (Ramos et al, 2009).
2.3.5 Tratamento
Em geral, as lesões são de fácil diagnóstico e possuem índices de cura
superiores a 95% quando tratados precoce e corretamente (Dimatos et al, 2009).
De acordo com o INCA, a cirurgia é o tratamento mais indicado. A
radioterapia e a quimioterapia também podem ser utilizadas dependendo do estágio
do câncer. Quando há metástase (o câncer já se espalhou para outros órgãos), o
melanoma é de difícil cura na maioria dos casos. A estratégia de tratamento para a
doença avançada deve ter então como objetivo aliviar os sintomas e melhorar a
qualidade de vida do paciente.
2.4 BIOINFORMÁTICA
A bioinformática é a ciência que utiliza a informática, a estatística e a
matemática na biologia molecular (Santos, 2010; Medini et al, 2008). Durante os

33
últimos anos, a bioinformática tornou-se um dos campos mais visíveis da ciência
moderna (OUZOUNIS et al, 2003).
O termo “Bioinformática” foi primeiramente usado por Pauline Hogeweg em
1979 para estudos de processos de informática aplicados na biologia de sistemas.
Desde então o seu principal uso tem sido nos ramos da genética e da genômica em
especial para auxiliar no manejo da grande quantidade de dados gerado no
sequenciamento de DNA, RNA e aminoácidos (Santos, 2010).
Atualmente, a bioinformática tem sido utilizada em diversas áreas como a
construção de banco de dados e a mineração de dados, análises de sequências,
identificação e analise de genes, relacionamento entre genes e proteínas, além de
prever a conformação tridimensional das proteínas, construir árvores filogenéticas e
modelos evolutivos, construir bibliotecas genômicas, estudar as funções biológicas,
design de drogas entre muitas outras coisas (Santos, 2010). Um tópico importante
na análise de sequências biológicas é a busca por genes, principalmente na
identificação de regiões codificantes de proteínas (Mena-Chalco, 2009).
Uma pergunta que se poderia fazer antes de sequenciar um genoma seria,
por exemplo, se esse organismo tem algum gene de potencial biotecnológico ou o
que há nos genes desse organismo que o faz conseguir viver nessa condição, ou
que gera uma doença. Vale observar que as ferramentas de bioinformática têm
permitido um grande avanço na identificação das funções de genes. A sequência de
um novo gene pode ser também comparada com aquelas armazenadas em um
banco de dados de genes de função conhecida, permitindo a rápida dedução da
possível função desse gene recém sequenciado. Testes experimentais para
descobrir a função de cada novo gene descoberto possivelmente exigiriam várias
décadas de pesquisa (Prosdocimi et al, 2004).
Nos últimos anos, o sequenciamento de DNA de diversos organismos tem
gerado uma grande quantidade de dados biológicos. Para possibilitar o
armazenamento, gerenciamento e disponibilização destes dados de forma a
potencializar ao máximo sua utilização, a bioinformática se desenvolveu
conjuntamente, e numa velocidade também impressionante (Formighieri, 2002).
A seguir, serão descritas algumas ferramentas de bioinformática utilizadas
neste trabalho.

34
2.4.1 Search Tool For The Retrieval Of Interacting Genes/Proteins (STRING)
A base de dados STRING tem como objetivo proporcionar uma avaliação da
integração nas interações gene-gene incluindo os efeitos diretos (físicos), bem como
associações indiretas (funcionais) (Franceschini et al, 2015).
Essas interações ajudam a descrever a função de um gene. O conhecimento
sobre os seus parceiros na interação é um requisito importante para entender
melhor a sua função (Franceschini et al 2015). Houve um progresso notável nos
últimos anos, tanto em termos de medidas experimentais e técnicas de previsão
computacionais (Szklarczyk et al, 2011).
A base de dados STRING abrange atualmente 2.031 organismos (STRING,
2015).
O STRING fornece uma base chamada primária experimental, originária a
partir de uma variedade de técnicas de bioquímicas, biofísicas e genéticas. As
interações também podem ser previstas computacionalmente, através de algoritmos
de previsão (Franceschini et al, 2015).
Szklarczyk et al, (2011) ressalta que todas as associações em STRING são
fornecidas com uma pontuação de confiança probabilística, utilizando o banco de
dados da Enciclopédia Kyoto Encyclopedia of Genes and Genomes (KEGG) e que
cada pontuação representa uma estimativa aproximada de uma determinada
associação.
O resultado é uma rede de associações previstas para um determinado grupo
de gene. Os nós da rede são os genes e as arestas representam as associações
funcionais previstas (STRING, 2015). Uma aresta pode ser representada com até
sete linhas de cores diferentes, sendo que estas linhas indicam a existência dos
tipos de provas utilizadas na previsão das associações. As provas podem indicar a
existência de conserved neighborhood, co-occurrence (coocorrência), fusion (fusão)
View, co-expression (coo expressão), experiments (experimentos), databases
(banco de dados), text mining (mineração de texto) e homology (homologia)
(STRING, 2015). A FIGURA 4 apresenta exemplos de resultados de cada tipo de
prova.

35
FIGURA 4 – Resultados dos tipos de associações em uma análise na base de dados
STRING. A: Conserved Neighborhood View, B: Co-occurrence, C: Fusion View, D: Co-
expression View; E: Experiments View, F: Databases View, G: Text Mining View. Fonte:
STRING, 2015.
2.4.2 Cytoscape
O Cytoscape é um projeto de software, de código aberto, desenvolvido para
visualizar e integrar interações de redes biomoleculares através de grandes bases
de dados de proteína-proteína, proteína-DNA e interações genéticas (Shannon,

36
2003). Utiliza os modelos computacionais baseados em grafos e redes, sendo que
um grafo é um conjunto não vazio de vértices (nós) e um conjunto finito de arestas
(arcos) tal que cada elemento do conjunto de arcos é representado por um par de
elementos de nós (Vaz, 2009). É importante ressaltar que topologia corresponde ao
modo pelo qual os vários elementos de uma rede são interconectados (Michaelis,
2015). Carvalho (2011) cita o termo topologia em seu trabalho e conceitua como
análise e correlação das propriedades da rede, ou seja, a interdependência entre
essa rede.
Existem disponíveis vários plugins para o Cytoscape que permitem interações
relacionadas com as mudanças na expressão gênica, analise de proteínas
envolvidas na recuperação celular voltadas a danos no DNA e entre outros
(Shannon, 2003). No presente trabalho foi utilizado o plugin network analyzer, o qual
realiza a analise de uma rede biológica com o objetivo de descobrir suas
propriedades topológicas (Cytoscape, 2015).
2.4.3 Classificação e Agrupamento De Dados
A técnica de agrupamento (ou clustering) procura identificar um conjunto de
categorias ou classes para descrever os dados (FAYYAD, 1996). O agrupamento de
dados em clusters (grupos) pode oferecer uma maneira de entender e extrair
informações relevantes de conjuntos de dados. A ideia é que dados de um mesmo
grupo tenham mais características em comum entre si do que com dados de
qualquer outro grupo (Vargas, 2010).
Existem algoritmos que realizam o agrupamento e classificação. O presente
trabalho utilizou o algoritmo fuzzy K-means. O K-means é um algoritmo muito
popular e bastante simples. O objetivo deste método é particionar os dados em uma
quantidade K de agrupamentos, minimizando a distância intraclusters e
maximizando a distância inter-clusters. Essa técnica utiliza um algoritmo de
agrupamento de dados com o objetivo de encontrar a melhor divisão de N dados em
K grupos. O centro do grupo inicial é formado em torno dos dados mais próximos e,
então, comparado com os pontos mais distantes, os outros grupos são então
formados. A partir daí, dentro de um processo de atualização contínua e de um

37
processo interativo encontram-se os centros dos grupos finais (EUZEBIO, 2012;
MONTEIRO E SILVA et al., 2001; TAN et al., 2006; SAMMA e SALAM, 2009).
2.4.4 Biological Networks Gene Ontology Tool (Bingo)
É uma ferramenta de código aberto, desenvolvida na linguagem de
programação Java. Tem como objetivo determinar a ontologia do Gene dentro de
um conjunto de genes (Maere et al, 2005; Liberman, 2004).
Ontologias oferecem um mecanismo pelo qual o conhecimento pode ser
representado numa forma capaz de ser processada por máquinas. O consórcio
“Gene Ontology” (GO) (“The Gene Ontology Consortium”, 2001) é um esforço
colaborativo para resolver a necessidade de descrições consistentes para produtos
de genes em diferentes bancos de dados, consistindo de um vocabulário padrão
para descrever a função de genes. No GO, foram desenvolvidas três classes
estruturadas (Liberman, 2004; The Gene Ontology Consortium”, 2001):
Função molecular: Palavras que descrevem as tarefas realizadas por
produtos gênicos individuais, como por exemplo, suas atividades enzimáticas,
catalíticas ou de ligação ao nível molecular.
Processos celulares/biológicos: Palavras que são usadas com objetivos
biológicos mais amplos, como a meiose, crescimento e manutenção celular
ou transdução de sinal.
Componentes celulares: Palavras que descrevem genes em termos das
estruturas subcelulares onde eles estão localizados, como organelas.
O Bingo mapeia as funções predominantes de cada gene, definindo, caso
necessário, hierarquias. Apresenta um visual intuitivo e personalizável dos
resultados (Maere et al, 2005).

38
2.4.5 Genecards
A base de dados GeneCards contém informações abrangentes sobre genes
humanos, incluindo dados sobre as funções celulares de seus produtos, e seu
envolvimento em doenças (Rebhan et al, 199).
Integra automaticamente mais de cem fontes web, apresentando informações
genômicas, transcritômicas, proteômicas, genéticas, clínicas e informações
funcionais (GeneCards, 2015).
3. OBJETIVOS
3.1. Delimitação do Tema
O tema apresentado utilizou como referências principais os artigos científicos,
livros, dissertações e teses das bases de dados eletrônicas Pubmed, Scielo e Web
of Science.
A metodologia aplicada neste trabalho utilizou como referência principal, o
artigo Bioinformatics and systems biology analysis of genes network involved in OLP
(Oral Lichen Planus) pathogenesis (Orlando et al, 2013).
As analises foram realizadas com os softwares STRING (Search Tool for the
Retrieval of Interacting Genes/Proteins), Matrix Laboratory (MATLAB), Statistical
Package for Social Sciences (SPSS), Cytoscape e a ferramenta Biológica Networks
Gene Ontology (Bingo)
3.2. Objetivo Geral
Identificar os genes potencialmente envolvidos no câncer de pele melanoma e
na manutenção da pele saudável através de pesquisas bibliográficas, bancos de
dados genômicos e ferramentas de bioinformática, bem como estudar as interações
realizadas por cada gene.

39
3.3. Objetivos Específicos
Para este objetivo geral, pretende-se de forma específica:
Identificar os genes possivelmente envolvidos no melanoma através de
pesquisa bibliográfica e os genes possivelmente envolvidos na manutenção
da pele saudável, através de pesquisa a base de dados GeneCards;
Criação do mapa de interação dos genes identificados utilizando o software
STRING 10;
Calcular o escore de cada gene, através da soma das pontuações referentes
às suas respectivas associações. Para esse escore foi definido o nome
Weighted Number of Links (WNL).
Análise topológica do mapa de interação através dos softwares Cytoscape;
Classificação dos genes através do algoritmo K-means;
Verificar a confiabilidade da rede, através de analises estatísticas utilizando o
software SPSS;
Realizar uma analise ontológica dos genes envolvidos através do software
Bingo;
Comparar os resultados das análises, considerando os dez grupos com maior
WNL, no câncer de pele melanoma e os genes potencialmente envolvidos na
pele saudável.
4. JUSTIFICATIVA
O câncer é uma doença genética. Inicia-se como consequência de alterações
múltiplas no ácido desoxirribonucleico (DNA) de uma única célula que muda a sua
constituição genômica (JR WAS, 2009). O dano genético ocorre em genes que
afetam a homeostase de vários processos biológicos como proliferação, crescimento
celular, apoptose, angiogênese, invasão e metástase (JR WAS, 2009).
Atualmente, ferramentas de bioinformática possibilitam que estudos sobre o
câncer apresentem resultados confiáveis a um baixo custo. Diante deste cenário, foi
proposto no presente trabalho um estudo dos genes possivelmente relacionados ao
câncer de pele melanoma.

40
Para uma melhor compreensão desta patogênese, é importante identificar os
genes envolvidos. Estes genes estão em diferentes processos biológicos, formando
assim uma complexa rede de interações. No entanto, apenas alguns deles têm um
elevado número de interações com outros genes na rede, portanto, eles podem ter
um papel mais importante. Os genes com o maior grau de interação serão
classificados como genes líderes, e poderão ser alvos de estudos para um
tratamento mais eficaz desta doença (ORLANDO et al, 2013).
A utilização de ferramentas de bioinformática e as analises realizadas podem
apresentar um melhor entendimento sobre essa doença, através de uma possível
identificação de genes importantes e a influência que estes genes exercem sobre
ela.
5. OBJETO
5.1. Problema
Com o objetivo de orientar a realização deste trabalho, o seguinte problema
foi formulado: Quais são os genes que possuem maior relevância no câncer de pele
melanoma?
5.2. Hipótese Básica
Genes que apresentam alto WNL e baixo TIS podem indicar maiores
relevância no câncer de pele melanoma, os quais poderão ser comprovados com
experimentos.

41
6. METODOLOGIA
6.1. Tipo de Pesquisa
Esta pesquisa será uma pesquisa exploratória. Pesquisas exploratórias são
desenvolvidas com o objetivo de proporcionar visão geral, de tipo aproximativo,
acerca de determinado fato. Ainda segundo GIL (2008), muitas vezes as pesquisas
exploratórias constituem a primeira etapa de uma investigação mais ampla.
6.2. Dados Utilizados
Para a identificação dos genes envolvidos ou potencialmente envolvidos no
câncer de pele melanoma que foram analisados neste trabalho utilizou-se, como
referência, apenas artigos científicos, livros, dissertações e teses da base de dados
Pubmed. Os descritos utilizados foram “Malignant melanoma and gene expression.”
para genes do câncer de pele melanoma. A nomenclatura oficial utilizada foi a
HUGO (Human Genome Organisation), sendo considerados apenas os genes
humanos. Os artigos pesquisados foram referentes aos últimos cinco anos,
apresentando como resultado 3.086 artigos. Foram considerados apenas os genes
identificados no resumo e conclusão dos artigos.
Em relação à manutenção da pele saudável, os genes potenciais também
foram identificados, apenas para fazer uma comparação com os resultados obtidos
no câncer de pele melanoma. Para determinar o principal conjunto de genes, foi
realizada uma pesquisa considerando apenas os genes humanos na base de dados
GeneCards, sendo os descritos “Skin Normal”. Também foi utilizado a nomenclatura
oficial HUGO (Human Genome Organisation), sendo considerados apenas os genes
humanos.
6.3. Análise dos Dados
Após a identificação dos genes, as interações entre eles foram mapeadas, e
uma pontuação, referente à interação, foi definida para cada um através do banco

42
de dados STRING versão 10.0 (Search Tool for the Retrieval of Interacting
Genes/Proteins) (Von, 2005; Jensen, 2009), permitindo assim a classificação destes
genes. Foram consideradas apenas as interações com um elevado grau de
confiança, acima de 0.9 (valor de confiança em STRING varia entre 0 e 0.99, sendo
0.99 a confiança mais alta). Foram mantidas as opções padrão para os outros
parâmetros.
Depois dessa etapa, foi gerada uma lista de genes candidatos potencialmente
relacionados com o melanoma e pele saudável. O software STRING foi usado para
definir cada interação e construir o mapa genômico com os genes identificados.
Para todos os genes identificados, foram somadas pontuações (escores)
referentes às suas respectivas associações. Somaram-se as pontuações dos
diferentes tipos de associações de cada gene, e este resultado foi então
multiplicados por 1000 (Poswar, 2015; Covani, 2008; Orlando, 2013; Giacomelli,
2006; Bragazzi, 2011), obtendo um escore único. Para esse escore foi definido o
nome Weighted Number of Links (WNL). Para o cálculo de WNL foram utilizados os
escores das associações encontrados nos arquivos Text Summary e Network
proteins description, disponibilizados após o mapeamento da rede de genes na base
de dados STRING.
Também foi realizado um escore global para os genes analisados neste
trabalho. Este escore foi chamado de TIS (significant global connectivity) (Barone,
2014; Poswar, 2015; Covani, 2008; Orlando, 2013; Giacomelli, 2006; Bragazzi,
2011). As pontuações, referentes ao TIS de cada gene, foi identificada utilizando a
mesma metodologia para a identificação do WNL. Entretanto o WNL utiliza as
pontuações apenas das associações da rede de genes aqui identificadas e o TIS
utiliza os escores de uma rede global de genes, disponível na base de dados
STRING. Para o cálculo do TIS foi utilizado os escores das associações encontradas
no arquivo Protein Network Data, disponível na base de dados STRING.
Com base na pontuação WNL, os genes foram agrupados através do
algoritmo de agrupamento K-means, para este agrupamento foi utilizado o software
SPSS PASW Statistics 18. A plataforma MatLab v.13 também foi utilizada apenas
para conferência dos resultados obtidos. Os parâmetros utilizados foram: Variável
WNL e para o Label case by (rótulo) foram utilizados os genes. O número de cluster

43
utilizado para o câncer de pele melanoma foi 20 e para a pele saudável foi utilizado
16 cluster. O método definido foi Iterate and classify (iteração e classificação). Em
Save (salvar), foi definida Cluster membership (participação do cluster). As opções
definidas foram: Initial cluster centers (Centros dos grupos iniciais), ANOVA table
(tabela ANOVA), Cluster information for each case (Informações sobre o cluster para
cada caso). Os outros parâmetros foram mantidos como padrão. A classificação foi
realizada de acordo com a média do WNL de cada grupo. Os genes líderes
obtiveram o posto mais alto (Poswar, 2015; Covani, 2008; Orlando, 2013;
Giacomelli, 2006; Bragazzi, 2011). Os outros genes foram denominados de acordo
com sua pontuação WNL. Genes sem interações, ou seja, com o WNL igual à zero
foram definidos como genes órfãos (Poswar, 2015; Orlando, 2013; Giacomelli, 2006;
Bragazzi, 2011).
Para avaliar as diferenças entre as classes em termos de WNL, foram
realizadas uma análise de variância (ANOVA) e TukeyKramer. Foram utilizados os
testes post hoc. A significância estatística foi fixada em um P valor <0,001. Em
Depedent List (lista dependente) foi utilizado a variável WNL e em Factor (fator) foi
utilizado o resultado obtido através da análise de cluster K-Means. Os outros
parâmetros foram mantidos como padrão.
Análise topológica foi realizada com Cytoscape, foi utilizado o arquivo Text
Summary para a visualização da rede, para as opções Source Interaction (fonte da
Interação) foi utilizado com campo Node1 (Nó1), para Target Interaction (Interação
alvo) foi utilizado com campo Node2 (Nó2) e para Interaction Type (Tipo de
Interação) foi utilizado com campo combined_score (Escore combinado). Foi
utilizado o plugin Network Analyzer e a opção Analyze Network (Topological
Coefficiente), foi selecionada a opção rede sem direção e todos os parâmetros foram
mantidos como padrão (Shannon, 2003).
A análise ontológica foi realizada com a ferramenta Biológica Networks Gene
Ontology (Bingo), para a visualização da rede foram utilizadas os mesmos
procedimentos adotados para a visualização da rede no Cytoscape. Somente foram
considerados os genes líderes, grupo B, C e D. O nível de significância foi fixado em
0,001, o testing correction definido foi Benamini & Hochberg False Discovery RATE
(FDR) correction, as ontologias utilizadas foram GO_Biological_Process,
GO_Molecular_Function, Go _Cellular_Component, todas as ontologias foram

44
relacionadas ao Homo sapiens. Os outros parâmetros não foram alterados, fazendo
uso das configurações padrão. Foram consideradas para analises as ontologias
mais frequentes no dataset (conjunto de dados) (Poswar, 2015; Covani, 2008;
Orlando, 2013; Giacomelli, 2006; Bragazzi, 2011; Maere, 2005).
6.4. Organização Desta Pesquisa
A organização dos conhecimentos envolvidos com o processo de
desenvolvimento do trabalho da dissertação para alcançar o objetivo geral está
esquematizada na FIGURA 5.
FIGURA 5 - Organização da Pesquisa. Fonte: Próprio autor.
6.5. Contribuições da Pesquisa
A principal contribuição desta pesquisa é apresentar um melhor entendimento
sobre o câncer de pele melanoma, sugerir genes que possam estar relacionados
com o desenvolvimento da doença.
Identificação dos possíveis
genes relacionados ao câncer de
pele melanoma e
na manutenção
da pele saudável
Criação do mapa de
interação dos genes
identificados utilizando o
software STRING 10
Cálculo do WNL de cada
gene identificado
Analise topológica da rede de genes
através dos softwares Cytoscape
Classificação dos genes através do
algoritmo K-means
Verificar a confiabilidade
da rede, através de
analises estatísticas utilizando o
software SPSS
Realizar uma analise
ontológica dos genes
envolvidos através do software
Bingo
Comparar os genes
potenciais do câncer de pele melanoma e pele saudável

45
7. RESULTADOS E DISCUSSÃO
Neste capítulo, serão apresentados os resultados da pesquisa que resultou
na presente dissertação. O objetivo foi verificar quais são os genes que
possivelmente possuem maior grau de interações no câncer de pele melanoma e as
principais diferenças em relação aos genes potenciais com maior interação na pele
saudável, no intuito de sugerir possíveis alvos de estudos e pesquisas.
7.1. Identificação dos genes
A análise dos artigos científicos, livros, dissertações e teses da base de dados
Pubmed permitiu a identificação de 820 genes para o câncer de pele melanoma. Em
relação à manutenção da pele saudável a lista inicial identificada no GeneCards
apresentou um total de 2.867 genes, sendo que todos apresentavam um escore de
confiança gerado pelo próprio GeneCards. Uma média de escores dos genes
identificados foi realizada e foram considerados somente os genes que possuíam
escores acima da média. Assim foram utilizados neste trabalho 503 genes para a
pele saudável.
7.2. Criação do Mapa de Interação Genômico através do software STRING
Após a identificação dos genes, as interações entre eles foram mapeadas,
através do banco de dados STRING versão 10.0 (Search Tool for the Retrieval of
Interacting Genes/Proteins) (Von, 2005; Jensen, 2009), Conforme descrito na seção
de metodologia do trabalho e cujo resultado está apresentado nas Figuras 6 e 7.
Para o mapeamento, foram consideradas apenas as interações com um
elevado grau de confiança, acima de 0.9 (valor de confiança em STRING varia entre
0 e 0.99, sendo 0.99 a confiança mais alta). É importante ressaltar que os genes são
representados como nós e as associações como arestas. Uma aresta pode ser
representada com até oito linhas de cores diferentes, sendo que estas linhas
indicam a existência dos tipos de provas utilizadas na previsão das associações
(STRING,2015).

46
FIGURA 6 – Mapa de interação dos genes potencialmente envolvidos no câncer de pele
melanoma. A associação vermelha indica fusion (fusão), a verde indica Conserved
Neighborhood (vizinhança conservada), a azul escuro indica co-occurrence (coocorrência),
roxa indica a existência de experiments (experimentos), a amarela indica textmining
(mineração de texto), a azul claro indica databases (banco de dados), a associação azul
indica homology (homologia) e finalmente, a preta indica co-expression (co-expressão)
(STRING, 2015). Fonte: Próprio autor.

47
FIGURA 7 – Mapa de interação dos genes potencialmente envolvidos na pele saudável.
Fonte: Próprio autor.
7.3 Cálculo do Weighted Number of Links (WNL)
A cada incidência do gene soma-se o escore encontrado (Poswar, 2015;
Covani, 2008; Orlando, 2013; Giacomelli, 2006; Bragazzi, 2011), obtendo um escore
único. Para esse escore foi definido o nome Weighted Number of Links (WNL)
(Poswar, 2015; Covani, 2008; Orlando, 2013; Giacomelli, 2006; Bragazzi, 2011).
A FIGURA 8 apresenta uma parcial de relatório obtido após o mapeamento no
banco de dados STRING, apresentando as pontuações das associações entre os
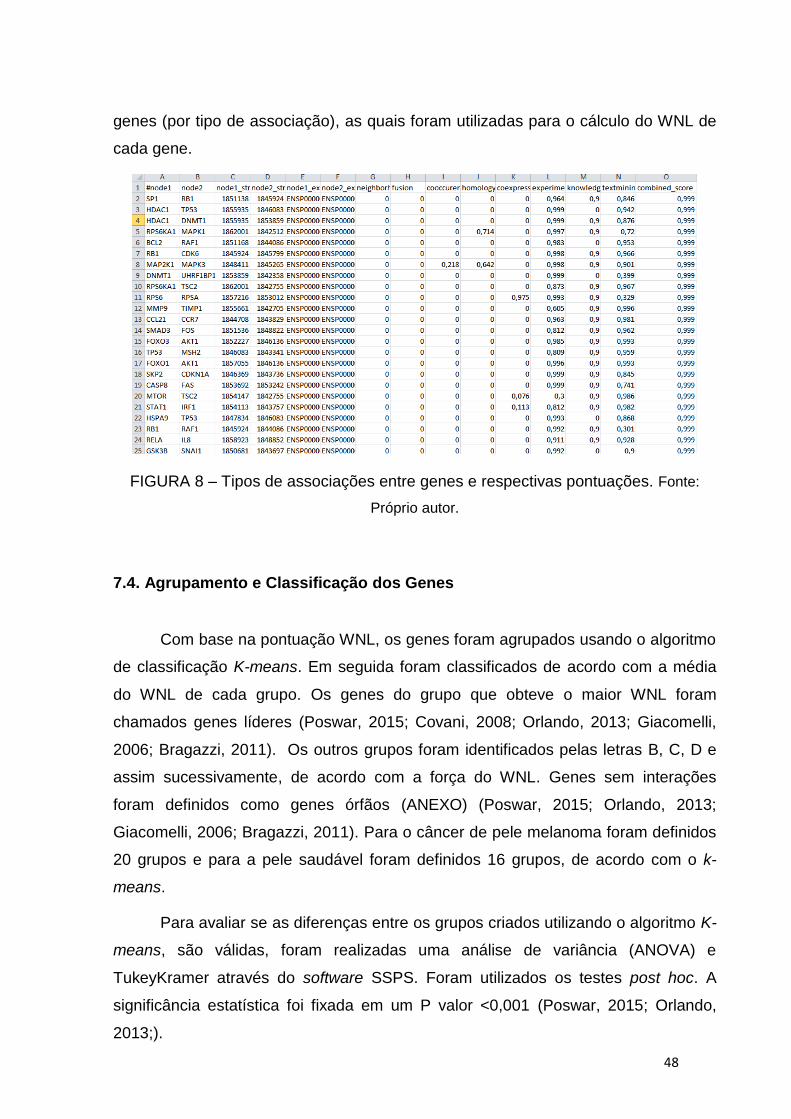
48
genes (por tipo de associação), as quais foram utilizadas para o cálculo do WNL de
cada gene.
FIGURA 8 – Tipos de associações entre genes e respectivas pontuações. Fonte:
Próprio autor.
7.4. Agrupamento e Classificação dos Genes
Com base na pontuação WNL, os genes foram agrupados usando o algoritmo
de classificação K-means. Em seguida foram classificados de acordo com a média
do WNL de cada grupo. Os genes do grupo que obteve o maior WNL foram
chamados genes líderes (Poswar, 2015; Covani, 2008; Orlando, 2013; Giacomelli,
2006; Bragazzi, 2011). Os outros grupos foram identificados pelas letras B, C, D e
assim sucessivamente, de acordo com a força do WNL. Genes sem interações
foram definidos como genes órfãos (ANEXO) (Poswar, 2015; Orlando, 2013;
Giacomelli, 2006; Bragazzi, 2011). Para o câncer de pele melanoma foram definidos
20 grupos e para a pele saudável foram definidos 16 grupos, de acordo com o k-
means.
Para avaliar se as diferenças entre os grupos criados utilizando o algoritmo K-
means, são válidas, foram realizadas uma análise de variância (ANOVA) e
TukeyKramer através do software SSPS. Foram utilizados os testes post hoc. A
significância estatística foi fixada em um P valor <0,001 (Poswar, 2015; Orlando,
2013;).

49
A diferença de pontuação WNL entre os grupos relacionados ao melanoma foi
confirmada pela ANOVA com o teste Tukey post hoc (P < 0,001), apresentando
significância igual à zero (TABELA 2).
TABELA 2 – Resultados da analise de variância (ANOVA) com o teste Tukey
post hoc utilizando o software SSPS.
A diferença de pontuação WNL entre os grupos relacionados à pele saudável
também foi confirmada pela ANOVA com o teste Tukey post hoc (P < 0,001),
(TABELA 3).
TABELA 3 – Resultados da analise de variância (ANOVA) com o teste Tukey
post hoc utilizando o software SSPS.
7.5. Analise Topológica Através do Software Cytoscape
Para verificar se a rede atende as propriedades da rede livres de escalas, a
análise topológica foi realizada com o software Cytoscape (Shannon, 2003), foi

50
utilizado o plugin Network Analyzer. Através desta analise foi possível confirmar que
a rede (mapeamento dos genes) do câncer melanoma exibe um comportamento lei
de potência (correlação = 0,914, R2 = 0,885) (FIGURA 9) de acordo com a teoria da
rede livre de escala (Poswar, 2015; Orlando, 2013).
FIGURA 9 - Resultado da analise topológica, obtida com o software cytoscape, da
rede de genes envolvidos no câncer de pele melanoma.
A analise topológica, realizada no software Cytoscape (plugin Network
Analyzer), também apresentou um comportamento compatível com lei de potência
da rede do câncer de pele melanoma, através do GRÁFICO 1.

51
GRÁFICO 1 – Comportamento lei de potência da rede do câncer de pele
melanoma. Resultado da analise topológica, obtida com o software Cytoscape.
Fonte: Próprio autor.
A rede de genes da pele saudável também exibiu um comportamento
compatível com a lei de potência (correlação = 0,934, R2 = 0,875) (FIGURA 10) de
acordo com a teoria da rede livre de escala (Poswar, 2015; Orlando, 2013).
FIGURA 10 - Resultado da analise topológica, obtida com o software
Cytoscape, da rede de genes envolvidos na manutenção da pele saudável.

52
GRÁFICO 2 – Comportamento lei de potência da rede da pele saudável.
Resultado da analise topológica, obtida com o software Cytoscape. Fonte: Próprio
autor.
A analise topológica realizada através software Cytoscape, permitiu observar
que os mapas de interação do câncer de pele melanoma e da pele saudável seguem
uma lei de potência, o que indica que a interação do mapa de genes possui
propriedades de rede livres de escala. Estas redes foram assim denominadas por
não apresentarem um número característico de ligações como acontece nas redes
de caráter aleatório que seguem uma distribuição de Poisson, onde a maioria dos
nós tem um número de ligações próximo da média, sendo pouco provável encontrar
nós com o número de ligações significativamente maior ou menor do que a média
(Barabasi et al, 1999).
Ainda de acordo com Barabasi et al (1999), uma das principais características
da rede livres de escalas é denominada de conexão preferencial, ou seja, existe
uma tendência de um novo vértice se conectar a um vértice da rede que tem um
grau elevado de conexões. Essa característica implica em redes com poucos
vértices altamente conectados, denominados hubs, e muitos vértices com poucas
conexões.
No presente trabalho, os nós com muitas ligações são os genes que possuem
maior interação, apresentando maior relevância neste trabalho. Os nós com poucas
ligações apresentam menor interação. Neste caso, quanto maior o número de
interações com outros genes, maior a possibilidade de interagir com novos genes.
As redes em escala-livre seguem uma distribuição em lei de potência, isto
quer dizer que a probabilidade de um nó estar conectado a k outros nós é
proporcional a k-y (Metz et al, 2007; Barabasi et al, 1999).
Umas das propriedades da lei de potência é a correlação. A correlação revela
se existe uma relação funcional entre uma variável e as variáveis restantes. A
correlação mostra o quão forte é a ligação entre as variáveis (UFPA, 2015). A
correlação nunca pode ser maior do que 1 ou menor do que menos 1. Uma
correlação próxima à zero indica que as duas variáveis não estão relacionadas. Uma
correlação positiva indica que as duas variáveis movem juntas, e a relação é forte

53
quanto mais à correlação se aproxima de um. Uma correlação negativa indica que
as duas variáveis movem-se em direções opostas. Duas variáveis que estão
perfeitamente correlacionadas positivamente movem-se essencialmente em perfeita
proporção na mesma direção (ME, 2016).
A correlação encontrada nas duas redes analisadas estão muito próximas de
1, o que quer dizer que é muito grande a possibilidade de que variações em um
gene serão acompanhadas por outros genes. Se um gene X aumentar sua força de
interação, outros genes que interagem com o gene X apresentarão altas
possibilidades de aumentar sua força de interação, o mesmo ocorrerá caso o gene X
diminua a sua força de interação.
Outra propriedade analisada foi o r-square (r quadrado). R quadrado é a
porcentagem da alteração da variável de resposta explicada pela relação com uma
ou mais variáveis preditoras. Normalmente, quanto maior o R quadrado, melhor o
modelo ajusta os dados. O valor de R quadrado está sempre entre 0 e 100%. O R
quadrado também é conhecido como coeficiente de determinação ou determinação
múltipla (em regressão linear múltipla) (MINITAB, 2016).
O r quadrado analisado foi de 88,5% para a rede do câncer de pele
melanoma e 87,5% para a pele saudável, o que representa uma alta porcentagem
de ajuste dos dados, caso aconteça alterações nas variáveis (genes) das redes
analisadas.
Os gráficos 1 e 2 apresentam um Power Law (comportamento lei de potência
da rede), mostrando a relação dos coeficientes topológicos em relação quantidade
de vizinhos de cada gene. É possível observar que quanto maior o número de
vizinhos, menor o coeficiente topológico. Os gráficos demonstram que os graus de
distribuição das redes analisadas satisfazem a lei de potência. O coeficiente
topológico foi calculado para cada gene da rede e representado graficamente em
função do número de ligações ( Tn = ( J(n,m) / Kn ) , em que J(n,m) indica o número
de nós a que ambos n e m estão ligados com o acréscimo de mais um, e Kn é o
número de ligações do nó n. Os nós que possuem menos que 2 vizinhos
apresentam um coeficiente topológico de zero (NA, 2016; Stelz et al, 2005).

54
7.7 Genes Líderes
Diante dos resultados da analise, para o melanoma os genes líderes
sugeridos foram o TP53, porém os genes pertencentes aos grupos B, C, e D (AKT1,
JUN, STAT3 e MYC) também apresentaram WNL relevante e foram considerados
líderes (FIGURA 11). O WNL de cada gene foi representado graficamente contra a
sua conectividade global, chamado de TIS (significant global connectivity). As
pontuações, referentes ao TIS de cada gene, foi identificada utilizando a mesma
metodologia para a identificação do WNL. Entretanto o WNL utiliza as pontuações
apenas das associações da rede de genes aqui identificadas e o TIS utiliza os
escores de uma rede global de genes, disponível na base de dados STRING. A
abordagem TIS foi descrita anteriormente (Barone, 2014; Poswar, 2015; Covani,
2008; Orlando, 2013; Giacomelli, 2006; Bragazzi, 2011).
Os gráficos 3 e 4 apresentam a dispersão entre os genes em relação ao WNL
e o TIS. Através desta análise é possível identificar que os genes líderes, grupo B, C
e D apresentam dispersão muito acima do valor esperado na regressão linear. O
objetivo deste gráfico é visualizar a relevância do gene na via analisada. Quanto
maior o WNL, maior a interação do gene na via analisada e mais relevante será o
gene neste trabalho. Porém quanto menor o TIS, mais interessante se torna o gene.
O TIS apresenta escores de uma rede global de genes, portanto se um determinado
gene apresenta um TIS muito elevado, este gene tende a ser comum em várias vias,
podendo apresentar funções mais gerais. Portanto, pode-se observar através destes
gráficos, que os genes líderes, grupo B, C e D se destacam em relação aos outros
genes.

55
GRÁFICO 3 – Distribuição dos genes do câncer de pele melanoma em relação ao
WNL
WNL

56
FIGURA 11 – Associações dos genes líderes, grupo B, C e D do Câncer de Pele Melanoma.
Fonte: Próprio autor.
Para a pele saudável, os genes líderes sugeridos foram o UBC, porém os
genes dos grupos B, C, e D (TP53, JUN, AKT1, CREBBP, EP300 e SRC) também
apresentaram o WNL relevante e foram considerados líderes (GRÁFICO 4)(FIGURA
12).

57
GRÁFICO 4 – Distribuição dos genes da pele saudável em relação ao WNL
WNL

58
FIGURA 12 – Associações dos genes líderes, grupo B, C e D na pele saudável. Fonte:
Próprio autor.
Em outras pesquisas, os genes citados neste tópico apresentou relação com
o câncer de pele melanoma. Entretanto, o gene UBC foi pouco relacionado a este
tipo de câncer, e também pode ser um interessante alvo para experimentos. Os
genes citados neste tópico serão discutidos logo abaixo.

59
7.7.1 Genes Líderes no Câncer de Pele Melanoma e Pele Saudável
TP53
Atua como um supressor tumoral em muitos tipos de tumores. Induz a
interrupção do crescimento ou apoptose, dependendo das circunstâncias fisiológicas
e tipo de célula. Está também envolvido na regulação do ciclo celular como um
ativador que atua para regular a divisão celular através do controle de um conjunto
de genes necessários para este processo (STRING, 2015).
Mutações em TP53 são as alterações genéticas mais comuns em
malignidades humanas. A super-expressão da proteína p53 tem sido relatada em
altas frequências em todos os tipos de câncer de pele. Cerca de 50% de todos os
cancros da pele apresentam mutações p53. (He, 2015; Weiss et al, 1995).
Avery-Kiejda et al (2011) apresenta como resultados de seu estudo
comprovações do envolvimento de TP53 no câncer de pele melanoma. Viros et al
(2014) acredita que este gene não tem um papel importante no melanoma.
AKT1
AKT é um componente de sinalização envolvido no desenvolvimento de
muitos cancros (Tang et al, 2015). Um aspecto importante da função AKT1 é o seu
envolvimento no metabolismo celular e na produção de energia celular. Sob algumas
circunstâncias, forte ativação de AKT1 aumenta o stress oxidativo, o que pode
causar a apoptose celular, como por exemplo, em células hematopoiéticas (Tang et
al, 2015).
Rebecca et al (2014) em seu trabalho cita que a inibição de AKT em
combinação com quimioterapia podem ter atividades clínica (tratamento) no
melanoma. Wan et al (2001) acredita que a ativação de PI 3-kinase / AKT pelo UV
pode aumentar a sobrevivência de células mutantes, promovendo, assim, o cancro
da pele, como foi encontrado em diversos outros tipos de cancro.

60
Shi et al (2014) apresenta uma ideia contrária, de acordo com o autor uma
minoria dos melanomas abrigam lesões genéticas, com mutações no AKT1.
JUN
JUN é a sigla de "JU-Nana", japonês para 17 (vírus do sarcoma aviário 17),
oncogene JUN (c-jun) codifica uma proteína nuclear que está envolvida no controle
transcricional relacionado com o crescimento celular (Carvalho C. H. P, 2010).
c-Jun (membro da família AP-1) foi inicialmente identificada como proto-
oncogenes e o seu papel central no desenvolvimento do cancro tem sido descrito
em muitos modelos. É frequentemente sobre-expressa em cancro. Entre outros
efeitos, c-Jun inibe p53 e promove à expressão da ciclina D. É altamente expresso e
ativo nas células com melanoma, e os mecanismos e as vias de sinalização que
regulam a proteína c-JUN são diversas (Kappelmann et al, 2014).
De acordo com Yang et al (2004) c-JUN pode oferecer um novo alvo para a
prevenção ou tratamento da progressão do melanoma humano.
7.7.2 Genes líderes somente no Câncer de Pele Melanoma
STAT3
Transdutor de sinal e ativador de transcrição que medeia às respostas
celulares a interleucinas, KITLG/SCF e outros fatores de crescimento (STRING,
2015).
Apresenta um papel crítico na sobrevivência, proliferação, angiogénese,
metástase e evasão de células de tumor (Kortylewski et al, 2005). Ainda de acordo
com Kortylewski et al (2005) contribui para o crescimento de células tumorais
através da regulação da expressão de genes que estão envolvidos na sobrevivência
e proliferação das células. Promove metástase e angiogénese através da indução da
expressão do gene metastático, MMP-2, e o gene angiogénico, VEGF (Kortylewski

61
et al, 2005). Participa na regulação da evasão do tumor através da inibição da
expressão de mediadores pró-inflamatórios, enquanto que promovem a expressão
de fatores imuno-supressores, que por sua vez ativa a sinalização STAT3 em
células dendríticas que conduzem a tolerância imunológica.
O gene STAT3 está ativo em diferentes tipos de cânceres humanos, incluindo
o melanoma. Muitos dos estudos que definem o papel da STAT3 na oncogênese
foram realizados em células de melanoma (Kortylewski et al, 2005; Emeagi et al,
2013).
MYC
A proteína codificada por este gene desempenha um papel na progressão do
ciclo celular, apoptose e transformação celular. Este gene funciona como um fator
de transcrição que regula a transcrição de genes alvos específicos. As mutações, a
sobre expressão, o rearranjo e translocação deste gene tem sido associado com
uma variedade de tumores hematopoiéticos, leucemias e linfomas (NCBI, 2016).
A sua expressão desregulada tem sido relatada para uma variedade de
tumores humanos e tem sido associada com o mau prognóstico no carcinoma do
pulmão, carcinoma de células escamosas da cabeça e pescoço, e carcinoma de
mama e colo do útero (Prins et al, 1993).
Existem três membros na família da oncoproteína MYC, são eles: c-myc, N-
myc, L-myc. Ambos apresentam papel na patogénese de muitas doenças
neoplásicas humanas (Chadd et al, 1999).
Para o melanoma humano a elevada expressão c-MYC é encontrada na fase
de crescimento vertical e relatada em metástases de tumores primários
(Schlagbauer-Wadl et al, 1999).
Zhuang et al (2008) em seus resultados sugerem que uma das principais
funções do C-MYC na progressão do melanoma é suprimir o gene BRAF ou NRAS,
já que cita em seu trabalho que melanomas malignos muitas vezes abrigam
mutações ativadoras em BRAF em NRAS.

62
7.7.3 Genes líderes somente na Pele Saudável
UBC
O gene UBC (Ubiquitin C) está associado com a degradação protéica, reparo
do DNA, regulação do ciclo celular, modificação quinase, endocitose e regulação de
outras vias de sinalização celular. Identificado em doenças como esclerose tuberosa
e nanismo de mulibrey (GeneCards, 2015).
CREBBP
O gene CREBBP codifica a proteína CREB. Este gene está envolvido em
muitos fatores de transcrição diferentes. Desempenha um papel crítico no
desenvolvimento embrionário e controle do crescimento. Mutações neste gene
podem causar síndrome de Rubinstein-Taybi. Também está associado com a
leucemia mielóide aguda (GeneCards, 2015). Além dessas doenças, um grande
número de estudos sugere que CREB está envolvida na progressão de melanoma
(Dobroff et al, 2009).
Jean et al (1998) em seu trabalho apresenta dados que indicam que CREB e
suas proteínas associadas atuam como fatores de sobrevivência para células de
melanoma humanos, e, por conseguinte, contribui para a aquisição do fenótipo
maligno. Melnikova et al (2010) e Dobroff et al (2009) afirmam que o aumento da
atividade de ligação ao elemento de cAMP (CREB) é uma característica da
progressão maligna do melanoma cutâneo.
EP300
EP300 é uma histona-acetiltransferase que regula a transcrição através da
remodelação da cromatina e é importante nos processos de proliferação e
diferenciação celular (Gayther et al, 2000). Além disso, medeia a ligação do gene

63
cAMP à proteína CREB. Os defeitos neste gene são uma das causas da síndrome
de Rubinstein-Taybi e podem também desempenhar um papel no cancro epitelial
(NCBI, 2015).
Gayther et al (2000) apresenta dados que demonstram que EP300 está
mutado em cancros epiteliais e fornece evidências de que ele se comporta como um
gene supressor de tumor.
Bhandaru et al (2014) combinou a sua proteína p300 e Braf no diagnóstico e
prognóstico de melanoma. Nos resultados de seu estudo, Bhandaru et al (2014)
demonstrou que a expressão Braf é inversamente correlacionada com p300 nuclear
e positivamente correlacionada com a expressão p300 citoplasmática. Braf e p300
citoplasmáticos foram associados com a progressão do melanoma, o tamanho do
tumor e estado de ulceração.
SRC
O SRC participa nas vias de sinalização que controlam um diversificado
espectro de atividades biológicas, incluindo a transcrição do gene, resposta imune, a
adesão celular, progressão do ciclo celular, apoptose, migração e transformação
(STRING, 2015).
Homsi et al (2009), Meierjohann et al (2006) comprovam um papel da família
src (proteína tirosina quinase citoplasmática) de Fyn no desenvolvimento e
progressão do melanoma. De acordo Homsi et al (2009), a sinalização Src implica
em várias doenças malignas, incluindo o melanoma. A prevalência da ativação de
Src no melanoma humano e o efeito dos inibidores de Src, como agentes isolados
ou em combinação, em células de melanoma não estão bem estabelecidos. Ainda
segundo Homsi et al (2009) a sua pesquisa comprovou que Src é expresso na
mucosa cutânea e melanoma metastático. Sugere ainda que inibidores de Src
podem ser uma terapia promissora no tratamento do melanoma.

64
7.8 Ontologias dos Genes Líderes
A análise das ontologias, a qual apresenta os processos biológicos/celulares,
funções moleculares e componentes celulares, para os genes líderes foi realizada
com a ferramenta Biológica Networks Gene Ontology (Bingo) (Poswar, 2015; Covani,
2008; Orlando, 2013; Giacomelli, 2006; Bragazzi, 2011; Maere, 2005).
Diante da analise de ontologias realizada no presente trabalho, é possível
perceber que variações em processos biológicos, funções moleculares e/ou em
componentes celulares podem influenciar o desenvolvimento da doença. Essas
alterações estão relacionadas com os genes líderes estudados e podem estar
ocorrendo por alterações das funções, inativação ou até mesmo ativação dos genes
analisados.
Entre as ontologias mais frequentes sugeridas no presente trabalho, existem
ocorrências em comum, ou seja, ontologias que são relacionadas com frequência
tanto no câncer de pele melanoma, quanto na manutenção da pele saudável.
Variações nestas ontologias podem de certa, forma influenciar no desenvolvimento
e/ou manutenção da doença. É importante destacar que existem ocorrências de
ontologias com alta frequência no dataset de uma via analisada que não apresentam
o mesmo nível de ocorrência na outra via aqui também analisada, ou até mesmo
ontologias que estão presentes em uma via e não estão presentes na outra via.
Podemos concluir que estas alterações podem ser significativas e devem ter
atenção especial no câncer de pele melanoma.
Este tópico apresentará as ontologias referentes aos genes líderes, grupo B,
C e D. Foram considerados para discussão as ontologias mais frequentes no dataset
(conjunto de dados).
7.8.1 Processos Celulares/biológicos
Este tópico apresenta as ontologias referentes aos processos biológicos para
os genes líderes, grupo B, C e D para o câncer de pele melanoma e pele saudável.
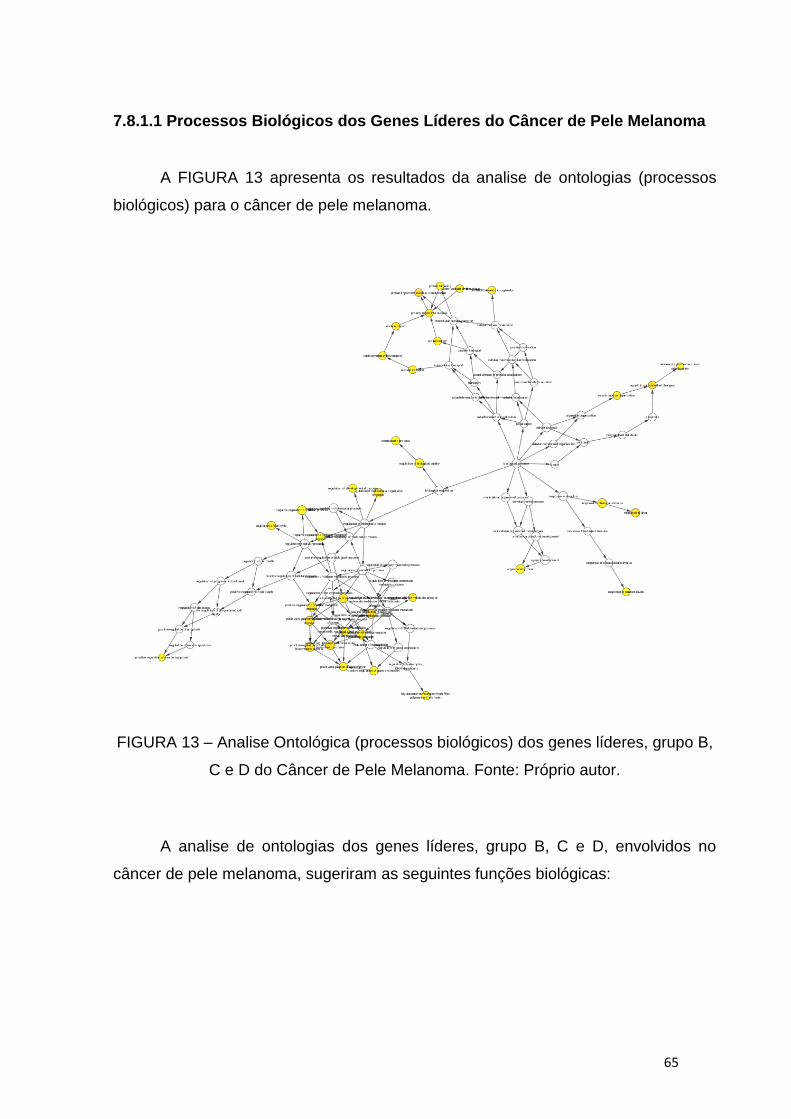
65
7.8.1.1 Processos Biológicos dos Genes Líderes do Câncer de Pele Melanoma
A FIGURA 13 apresenta os resultados da analise de ontologias (processos
biológicos) para o câncer de pele melanoma.
FIGURA 13 – Analise Ontológica (processos biológicos) dos genes líderes, grupo B,
C e D do Câncer de Pele Melanoma. Fonte: Próprio autor.
A analise de ontologias dos genes líderes, grupo B, C e D, envolvidos no
câncer de pele melanoma, sugeriram as seguintes funções biológicas:

66
Regulação Positiva do Processo de Biossíntese da Macromolécula
Qualquer processo que modula a velocidade, a frequência ou a extensão das
reações químicas, resultando na formação de uma macromolécula. Uma
macromolécula consiste em qualquer molécula de massa molecular relativamente
elevada, cuja estrutura compreende essencialmente a repetição múltipla de
unidades derivadas, efetiva, a partir de moléculas de massa molecular relativa baixa
(AmiGO2, 2016).
Regulação Positiva de Azoto Processo Metabólico Composto
Qualquer processo que ativa ou aumenta a frequência das reações químicas
e caminhos que envolvem compostos de azoto ou nitrogenados.
Ativação do metabolismo de nitrogênio, a regulação positiva do processo
metabólico de azoto, a regulação positiva do metabolismo do nitrogênio, a
estimulação do metabolismo de nitrogênio, a regulação do metabolismo de
nitrogênio, aumento da regulação do metabolismo de nitrogênio, regulação positiva
de processo metabólico de azoto (SGD, 2016).
Regulação Positiva do Processo de Biossíntese Celular
Qualquer processo que ativa ou aumenta a frequência das reações químicas,
resultando na formação de substâncias, realizadas por células individuais (AspGD,
2016).
Regulação Positiva do Processo de Biossíntese
Qualquer processo que ativa ou aumenta a frequência, a taxa ou extensão
das reações químicas, resultando na formação de substâncias (BioCyc, 2016).

67
Regulação Positiva do Processo Metabólico da Macromolécula
Qualquer processo que aumente a taxa, a frequência ou a extensão das
reações químicas, resultando na formação de uma macromolécula (AmiGO2, 2016).
Regulação Positiva do Processo Metabólico Celular
Qualquer processo que ativa ou aumenta a frequência das reações químicas
pelos quais as células individuais transformam substâncias químicas (BioCyc, 2016).
Regulação Positiva do Processo Metabólico
Qualquer processo que ativa ou aumenta a frequência das reações químicas
dentro de uma célula ou um organismo (BioCyc, 2016).
Regulamento de Processo do Organismo Multicelular
Qualquer processo que modula a frequência, a taxa de um processo do
organismo multicelular. Os processos relevantes para a função de um organismo
multicelular acima do nível celular; inclui os processos integrados de tecidos e
órgãos (AmiGO2, 2016).
Respostas a Estímulos Químicos
Qualquer processo que resulta em uma mudança de estado ou a atividade de
uma célula (em termos de movimento, secreção, produção de enzima, expressão de
gene, etc.) como um resultado de um estímulo químico (AmiGO2, 2016).

68
Regulamentação de Qualidade Biológica
Qualquer processo que modula uma característica qualitativa ou quantitativa
de uma qualidade biológica. Uma qualidade biológica é um atributo mensurável de
um organismo ou parte de um organismo, tais como o tamanho, massa, forma, cor,
etc (CGD, 2016).
Desenvolvimento de Órgãos
O processo cujo resultado específico é a progressão de órgãos ao longo do
tempo desde sua formação até a estrutura madura (AspGD, 2016).
Regulação Negativa de Processo Celular
Qualquer processo que interrompe, impede ou reduz a frequência, a taxa ou
extensão de um processo celular. São realizadas a nível celular, mas não estão
necessariamente limitados a uma única célula. Por exemplo, a comunicação entre
células ocorre em mais de uma célula, mas ocorre a nível celular (AmiGO2, 2016).
7.8.1.2 Processos Biológicos dos Genes Líderes da Pele Saudável
Para a pele saudável, também foi realizada a mesma analise com os genes
líderes, genes pertencentes aos grupos B, C e D. A FIGURA 14 apresenta os
resultados encontrados.

69
FIGURA 14 – Analise Ontológica (processos biológicos/celulares) dos genes líderes,
grupo B, C e D da Pele Saudável. Fonte: Próprio autor.
A analise ontológica destes genes líderes, grupo B, C e D, envolvidos na
manutenção da pele saudável, sugeriram os seguintes processos biológicos:
Respostas a Estímulos Químicos
Qualquer processo que resulta em uma mudança de estado ou a atividade de
uma célula (em termos de movimento, secreção, a produção da enzima, a
expressão do gene, etc.) como um resultado de um estímulo químico (AmiGO2,
2016).

70
Regulamentação de Qualidade Biológica
Qualquer processo que modula uma característica qualitativa ou quantitativa
de uma qualidade biológica. Uma qualidade biológica é um atributo mensurável de
um organismo ou parte de um organismo, tais como o tamanho, massa, forma, cor,
etc (CGD, 2016).
Transdução de Sinal
É o processo celular no qual um sinal é transmitido para provocar uma
alteração na atividade ou no estado de uma célula. A transdução do sinal começa
com um sinal de recepção. A Transdução de sinal termina com regulação de um
processo celular, por exemplo, regulação da transcrição ou regulamento de um
processo metabólico (AmiGO2, 2016).
Transmissão de Sinal
É o processo em que um sinal é liberado e/ou enviado de um local para outro
(AmiGO2, 2016).
Processo de Sinalização
A sinalização celular é um mecanismo de comunicação entre as células que
se encontram presente nas mais diversas formas de vida, desde organismos
unicelulares, como bactérias, fungos e protozoários, até seres multicelulares.
O mecanismo de sinalização celular envolve a participação de uma célula
sinalizadora, responsável pela produção e, na maioria dos casos, liberação de uma
molécula sinalizadora, denominada ligante, e uma célula alvo, que apresenta
receptores (proteínas que reconhecem especificamente o ligante) que serão

71
responsáveis pela propagação do sinal e consequente resposta celular (UFPB,
2016).
Regulação Positiva do Processo Biológico
Qualquer processo que ativa ou aumenta a frequência de um processo
biológico. Os processos biológicos são regulados por diversos meios, inclui o
controle da expressão do gene, modificação de proteínas ou interação com uma
molécula de proteína ou substrato (AmiGO2, 2016).
Organização do Componente Celular
Um processo que resulta na montagem, disposição das partes constituintes,
ou desmontagem de um componente celular (PANTHER, 2016).
7.8.2 Função Molecular
Este tópico apresenta as ontologias referentes à função molecular para os
genes líderes, grupo B, C e D para o câncer de pele melanoma e pele saudável.
7.8.2.1 Função Molecular dos Genes Líderes do Câncer de Pele Melanoma
A FIGURA 15 apresenta os resultados da analise ontológica (função
molecular) para o câncer de pele melanoma.

72
FIGURA 15 – Analise Ontológica (função molecular) dos genes líderes, grupo B, C e
D do Câncer de Pele Melanoma. Fonte: Próprio autor.
A analise ontológica destes genes líderes, grupo B, C e D, envolvidos no
câncer de pele melanoma, sugeriram as seguintes funções moleculares:
Atividade Ativadora de Transcrição
Qualquer atividade reguladora de transcrição necessária para a iniciação ou
regulação positiva da transcrição (AmiGO2, 2016).
Ligação de DNA com Sequência Específica
Interagir seletivamente e de forma não covalente com o DNA, através da
composição de nucleotídeos específicos (AmiGO2, 2016).
7.8.2.2 Função Molecular dos Genes Líderes da Pele Saudável

73
Para a pele saudável, também foi realizada a mesma analise com os genes
líderes, genes pertencentes aos grupos B, C e D. A FIGURA 16 apresenta os
resultados encontrados.
FIGURA 16 – Analise Ontológica (funções moleculares) dos genes líderes, grupo B,
C e D da Pele Saudável. Fonte: Próprio autor.
A analise ontológica destes genes líderes, grupo B, C e D, envolvidos na
manutenção da pele saudável, sugeriram as seguintes funções moleculares:
Atividade Ativadora de Transcrição
Qualquer atividade reguladora de transcrição necessária para a iniciação ou
regulação positiva da transcrição (AmiGO2, 2016).

74
Fator de Transcrição de Ligação
Interage seletivamente e de forma não covalente com um fator de transcrição
específico, o qual pode ser uma única proteína ou um complexo, de modo a modular
a transcrição. Um fator de transcrição de ligação à proteína pode ou não interagir
com o ácido nucleico (DNA ou RNA) (AmiGO2, 2016).
7.8.3 Componentes Celulares
Este tópico apresenta as ontologias referentes aos componentes celulares
para os genes líderes, grupo B, C e D para o câncer de pele melanoma e pele
saudável.
7.8.3.1 Componentes Celulares dos Genes Líderes do Câncer de Pele
Melanoma
A FIGURA 17 apresenta os resultados da analise ontológica (componentes
celulares) para o câncer de pele melanoma.

75
FIGURA 17 – Analise Ontológica (Componentes celulares) dos genes líderes, grupo
B, C e D do Câncer de Pele Melanoma. Fonte: Próprio autor.
A analise ontológica destes genes líderes, grupo B, C e D, envolvidos no
câncer de pele melanoma, sugeriram os seguintes componentes celulares:
Núcleo
Organela delimitada por membrana de células eucarióticas em que os
cromossomos são alojados e replicados (AmiGO2, 2016).

76
7.8.3.1 Componentes Celulares dos Genes Líderes da Pele Saudável
Para a pele saudável, também foi realizada a mesma analise com os genes
líderes, genes pertencentes aos grupos B, C e D. A FIGURA 20 apresenta os
resultados encontrados.
FIGURA 18 – Analise Ontológica (componentes celulares) dos genes líderes, grupo
B, C e D da Pele Saudável. Fonte: Próprio autor.
A analise ontológica destes genes líderes, grupo B, C e D, envolvidos na
manutenção da pele saudável, sugeriram os seguintes componentes celulares:

77
Organela Intracelular do Lúmen
Uma organela lúmen que faz parte de um organelo (pequenos órgãos)
intracelular (AmiGO2, 2016).
Organela do Lúmen
O volume interno fechado por membranas de um organelo particular, por
exemplo, retículo endoplasmático lúmen, lúmen nuclear (CATHACYC, 2016).
Membrana Fechada do Lúmen
O volume encerrado dentro de uma membrana selada entre duas
membranas. Engloba o volume de um organelo específico, por exemplo, retículo
endoplasmático lúmen (AmiGO2, 2016).
Parte Nuclear
Qualquer parte constituinte do núcleo, uma organela delimitada por
membrana de células eucarióticas em que os cromossomos são alojados e
replicados (AmiGO2, 2016).
7.9 Comparações Entre os Resultados Identificados no Câncer de Pele
Melanoma (CPM) e Genes Da Pele Saudável (PS)
Uma comparação entre os genes identificados para câncer de pele melanoma
e os genes da manutenção da pele saudável, foi realizada. Considerando, apenas,
os 10 grupos (identificados através do agrupamento realizado com o algoritmo k-

78
means) com maior WNL no CPM e na PS, variações que podem ser importantes na
patogênese foram sugeridas a seguir.
Os dez grupos com maior WNL no CPM apresentaram um total de 28 genes,
conforme apresentado na TABELA 4.
Gene Grupo no melanoma
TP53 A
AKT1 B
JUN C
MYC D
STAT3 D
EGFR E
MAPK1 E
HSP90AA1 F
MAPK14 F
MAPK8 F
RAC1 F
CCND1 G
FOS G
FYN G
PIK3CA G
SMAD3 G
HDAC1 H
HRAS H
RELA H
RHOA H
STAT1 H
BCL2 I
CDKN1A I
MAPK3 I
SMAD4 I
VEGFA I
ESR1 J
JAK2 J
TABELA 4 – Variação entre classes de um gene do melanoma nas 10 classes com maior
poder de interação. Fonte: Próprio Autor
Os dez grupos (identificados através do agrupamento realizado com o
algoritmo k-means) com maior WNL na PS apresentaram um total de 48 genes,
conforme apresentado na TABELA 5.

79
Gene Grupo na Pele Saudável
UBC A
TP53 B
JUN C
AKT1 D
CREBBP D
EP300 D
SRC D
FOS E
MYC E
STAT3 E
CTNNB1 F
EGFR F
ESR1 F
MAPK1 F
NFKB1 G
CCND1 G
PIK3CA G
RELA G
SMAD3 G
AR H
MAPK14 H
MAPK8 H
SMAD4 H
SP1 H
BCL2 I
INS I
PTPN11 I
CDKN1A I
HDAC1 I
HRAS I
IL6 I
JAK2 I
MAPK3 I
TGFB1 I
VEGFA I
APP J
CDK2 J
ERBB2 J
NOTCH1 J
PPARG J
CDH1 J
CREB1 J
EDN1 J
HIF1A J

80
IGF1 J
PPARA J
RB1 J
STAT1 J
TABELA 5 – Variação entre classes de um gene da pele saudável nas 10 classes com maior
poder de interação. Fonte: Próprio Autor
Considerando os 10 grupos citados, 24 genes estão presentes tanto no CPM
e na PS. Os genes em comum ao CPM representaram um total de 86%, enquanto
os genes em comum à pele saudável teve um total de 50%, o que indica que a
maioria dos genes com maior WNL no CPM também apresentam alto WNL na
manutenção da PS. Portanto é possível sugerir que pequenas variações em suas
funções possam estar associadas à manutenção e/ou desenvolvimento do câncer de
pele melanoma. Esta analise também indica que podem existir genes silenciados ou
ativados no câncer de pele melanoma. Estes valores estão representados no
GRÁFICO 5.
GRÁFICO 5 – Porcentagem dos genes em comum ao CPM e PS nas 10 classes com maior
poder de interação. Fonte: Próprio Autor
7.9.1 Relação de Genes Identificados Apenas no Câncer de Pele Melanoma
Considerando os 10 grupos com maior WNL no CPM e todos os grupos da
PS, foram identificados quatro genes incidentes apenas no CPM, o que sugere a
possibilidade de que estes genes possam estar ativados nesta doença (TABELA 6).
14%
86%
Genes Melanoma
Genes Melanoma Genes em comum
50% 50%
Pele Saudável
Pele Saudável Genes em comum

81
Genes Melanoma exclusivos
HSP90AA1
RAC1
FYN
RHOA
TABELA 6 – Genes identificados apenas no câncer de pele melanoma. Fonte: Próprio Autor
Estes genes serão discutidos abaixo.
HSP90AA1
O gene HSP90AA1 promove a maturação, manutenção estrutural e uma
regulamentação adequada de proteínas-alvo envolvidas, por exemplo, no controle
do ciclo celular e transdução de sinal. Além disso, codifica dois transcritos de mRNA,
sendo a proteína Hsp90A uma das proteínas codificadas (Zuehlke et al, 2015;
STRING, 2015).
McCarthy et al (2008), Becker et al (2004) sugerem que a proteína HSP90
seja um marcador de progressão no melanoma e que pode ter um efeito
estabilizador sobre as funções celulares nas células em proliferação de lesões
melanocíticas, podendo ser um pré-requisito para a progressão do tumor (McCarthy
et al, 2008).
RAC1
Em seu estado ativo, esta proteína conecta-se a uma variedade de proteínas
responsáveis pela regulação de respostas celulares, tais como os processos de
secreção, fagocitose de células apoptóticas, polarização celular epitelial (STRING,
2015).

82
Rac1 é um membro da superfamília GTPase de RAS, sendo que RAS foi
identificada como a terceira mutação recorrente mais comum em melanomas
(Halaban et al, 2015).
De acordo com Krauthammer et al (2012) as estruturas cristalinas, e estudos
bioquímicos e funcionais de Rac1 mostraram que ela pode promover a proliferação e
migração de melanócitos.
A mutação em RAC1 revelou-se através do sequenciamento de regiões
codificantes no melanoma e forneceram evidências de ser um novo "controlador" na
gênese da doença. A mutação é mais frequente em pacientes com melanomas
relacionados à exposição ao sol, que ocorre em cerca de 4-7% dos pacientes, sendo
altamente significativo (Halaban et al, 2015).
Krauthammer et al (2012) sugere a possibilidade de que RAC1 pode ter
benefício terapêutico da doença.
FYN
FYN é um proto-Oncogene, da família Src de tirosina-quinase (GeneCards,
2015). Desempenha um papel em muitos processos biológicos, incluindo a
regulação do crescimento e sobrevivência celular, a adesão celular, a sinalização
mediada pela integrina, remodelação do citoesqueleto, a motilidade celular, resposta
imune e orientação do axónio (STRING, 2015).
FYN contribui para o desenvolvimento e progressão de vários tipos de cancro,
através da sua participação no controle do crescimento celular, morte,
transformação morfogênica e motilidade celular. Estes cancros inclui o melanoma, o
carcinoma de células escamosas, da próstata e cancros da mama. Estudos recentes
demonstraram a importância de Fyn na resistência ou susceptibilidade das células
cancerosas em alguns tratamentos anticancerígenos (Elias et al, 2015).
RHOA

83
Membro da família RAS, regula uma via de transdução do sinal de ligação de
receptores da membrana plasmática para a montagem de adesões focais e fibras de
stress de actina. Está envolvido na formação do anel de miosina contrátil do ciclo
celular durante a citocinese. Também desempenha um papel essencial na formação
do sulco de clivagem e é necessário para a formação da junção apical de adesão
célula-célula de queratinócitos (STRING,2015).
Klein et al (2011) em seu estudo identifica papéis para o gene RHOA na
circulação das células de melanoma, mas não no crescimento ou sobrevivência das
células, quando tratadas com inibidores de BRAF.
7.9.2 Relação de Genes Identificados Apenas na Manutenção da Pele Saudável
Comparando os resultados dos 10 grupos com WNL mais forte na
manutenção da PS e todos os grupos do CPM, foram identificados treze genes
incidentes apenas na manutenção da PS, sugerindo a possibilidade de que alguns
destes genes possam estar silenciados nesta doença (TABELA 7).
Genes Pele Saudável exclusivos
UBC
CREBBP
EP300
SRC
CTNNB1
NFKB1
INS
PTPN11
APP
CDK2
ERBB2
NOTCH1
PPARG
TABELA 7 – Genes exclusivos da pele saudável. Fonte: Próprio Autor
Estes genes serão discutidos abaixo. Importante ressaltar que o gene
CTNNB1, PTPN11 e UBC menos relacionados a este tipo de câncer, estes genes

84
podem estar silenciados na CPM e podem ser interessantes para estudos
experimentais. Alguns destes genes já foram citados em tópicos anteriores.
APP
Mutações neste gene têm sido implicadas na doença de Alzheimer e
amiloidose cerebral arterial (NCBI, 2015).
Em relação à pele, Vega et al (1995) apresenta dados que demonstram que
APP beta está presente na pele humana glabra (regiões da pele sujeitas a maior
atrito, como palmas das mãos e pés).
Botelho et al (2010) em seu estudo fornece evidências de que APP é
expresso em níveis elevados em melanomas com estágio avançado. Ressalta ainda
que o bloqueio da expressão de APP pela interferência de RNA prejudica a
proliferação de células de melanoma metastático e leva à sua diferenciação terminal
e irreversível.
CDK2
O gene CDK2 é um membro da família da proteína-quinase (Chohan et al,
2015). Desempenha um papel importante na regulação de vários eventos do ciclo da
divisão celular eucariótica. Evidências indicam que a sobre-expressão de CDK2
pode causar a regulação anormal do ciclo celular, podendo estar diretamente
associada com a hiperproliferação de células cancerosas (Chohan et al, 2015).
Em relação ao câncer de pele, Macias et al (2008), Walker et al, 2001
afirmam que a ativação de CDK2 induz a proliferação de queratinócitos, mas não
afeta o desenvolvimento do tumor de pele. Contudo, de acordo com Du et al (2004)
os níveis da expressão de CDK2 estão fortemente correlacionados em melanomas.
Em seu estudo, Du et al (2004) afirma que o esgotamento de CDK2 suprimiu o
crescimento e progressão do ciclo celular em pacientes com melanoma, mas não em
outros tipos de câncer.

85
CTNNB1
CTNNB1 é o gene que codifica a proteína beta-catenina. É parte de um
complexo de proteínas que formam junções aderentes, que são importantes para a
criação e manutenção de camadas de células epiteliais, regulando o crescimento
celular e adesão entre as células adjacentes (Hartsock, 2008).
O nível de beta-catenina em células é rigorosamente controlado em um
complexo multiprotéico, e mutações no 3beta glicogénio-sintase-quinase (GSK-
3beta) resultam em um acumulo nuclear, transativação constitutiva do fator de
células T e fator intensificador de linfoides, um mecanismo que ocorre em muitos
tipos de cancro (Demunter et al, 2002).
Linhas celulares de melanoma podem abrigar mutações beta-catenina, no
entanto, o acúmulo celular de beta-catenina é raramente causado por mutações
CTNNB1 (Demunter et al, 2002; Omholt et al, 2001). No entanto, Castiglia et al
(2008) em seu estudo identificou mutações CTNNB1 em melanoma e sugeriu que a
desregulação da via Wnt/beta-catenina coopera no desenvolvimento e progressão
do melanoma.
ERBB2
Este gene codifica um membro da família de receptores do fator de
crescimento epidérmico (EGF). A sobre-expressão desse gene tem sido relatada em
numerosos cancros, incluindo cancro da mama e tumores do ovário (NCBI).
Justin et al (2006), Rao et al (2015) em seus estudos revelam a influência de
ErbB2 na resposta ao UV. A radiação UV ativa o receptor ErbB2/HER2. Sugerem
um papel para o ErbB2 em patologias induzidas por UV, tais como o cancro de pele,
já que apresentam dados que comprovam que a ativação de ErbB2 acelera a
progressão maligna do câncer de pele induzido por UV.

86
No melanoma ERBB2 dimeriza preferencialmente com ERBB3. A sinalização
NRG1-erbB3-ErbB2 promovem a sobrevivência celular, crescimento, a migração e
proliferação em células de melanoma (Rao et al, 2015; Zhang et al 2013).
INS
A insulina diminui a concentração de glicose no sangue. Aumenta a
permeabilidade da célula aos monossacáridos, aminoácidos e ácidos gordos. Além
disso, acelera a glicólise, o ciclo das pentoses fosfato, e síntese de glicogênio no
fígado (STRING, 2015).
A insulina e os fatores de crescimento semelhantes à insulina (IGFs) são
importantes reguladores do crescimento e metabolismo energético. Várias
descobertas têm delineado um papel importante desempenhado por esta família de
moléculas, tanto na manutenção e desenvolvimento do melanoma (Capoluongo,
2011). Apesar da contribuição estabelecida do sistema IGF na carcinogênese, foram
relatados poucos dados sobre as relações entre melanoma e esta família de
moléculas (Capoluongo, 2011).
Antoniadis et al (2011) no resultado de seu estudo aponta a resistência à
insulina como fator de risco potencial para o melanoma.
NFKB1
Também conhecido como: p50; KBF1; p105; A EBP-1; NF-KB1; NFKB-p50;
NFkappaB; NF-kappaB; NFKB-p105; NF-kappa-B
NF-kappa-B é um fator de transcrição presente em quase todos os tipos de
células e participa de uma série de eventos de transdução de sinal que são iniciados
por estímulos relacionados em muitos processos biológicos, tais como a inflamação,
a imunidade, diferenciação, de crescimento celular, tumorigénese e apoptose
(STRING, 2015).

87
O fator nuclear kappa B (NF-kB) aparece desregulada em muitos tipos de
tumores, resultando na expressão e/ ou ativação de complexos de transcrição de
NF-kappaB mutantes (Karst et al, 2009).
NFkB é uma via que os tumores de melanoma usam para alcançar a
sobrevivência, proliferação e resistência à apoptose (Madonna et al, 2012). Ainda de
acordo com Madonna et al (2012) a inibição da ativação do NF-kB parece ser uma
opção muito promissora para terapias anti-cancerígenas.
NOTCH1
Notch1 é um proto-oncogene em vários órgãos. Na pele, o silenciamento do
gene Notch1 conduz à formação de tumores, sugerindo que Notch1 é um supressor
de tumor (Demehri et al, 2009). Liu et al (2006) cita que a sinalização de Notch1
impulsiona a fase de crescimento vertical do melanoma primário para um fenótipo
mais agressivo.
Proteínas Notch são importantes nas decisões do destino celular e está
associada com tumorigénese. O papel da sinalização Notch na pele de mamíferos
não está bem caracterizado e é baseada principalmente em estudos in vitro (Nicolas
et al, 2003). Nicolas et al (2003) sugere que a sinalização Notch induz a
diferenciação na pele de mamíferos (Nicolas et al, 2003).
PPARg
Receptores ativados por proliferação de peroxissoma (PPARs) são membros
da superfamília de receptores nucleares que regulam lipídios, glicose, e o
metabolismo de aminoácidos. Na pele regulam as funções celulares, incluindo a
proliferação e diferenciação celular, assim como respostas inflamatórias (Sertznig et
al, 2008).
Kang et al (2004) identificou a expressão de todos os três subtipos de PPAR
em melanócitos humanos e Lee et al (2007) apresentou dados que comprovam que

88
a pigmentação em melanócitos foi acelerado com PPAR-alfa e PPAR-gama,
sugerindo um possível papel para PPAR-alfa e PPAR-gama na modulação da
proliferação e diferenciação dos melanócitos (pigmentação).
De acordo com Sertznig et al (2008), PPAR-delta têm um papel anti-
apoptótico e mantêm a sobrevivência e a diferenciação de células epiteliais. Já
PPAR-alfa e PPAR-gama induz a diferenciação e inibe a proliferação, além de
regular a apoptose. No melanoma, o efeito inibidor do crescimento através da
ativação de PPAR-gama é independente da apoptose e, aparentemente, ocorre
através da indução da paragem do ciclo celular na fase G1.
Ainda de acordo com Sertznig et al (2008), PPARs podem ser considerados
alvos potenciais para o tratamento de várias doenças de pele.
PTPN11
Atua no fluxo de vários receptores de tirosina-quinase citoplasmáticos e
participa na transdução do sinal a partir da superfície da célula para o núcleo
(STRING, 2015).
De acordo com Chan et al (2008) mutações germinativas em PTPN11
causam síndrome de Noonan e síndrome de leopardo, enquanto que mutações
somáticas em PTPN11 ocorrem em vários tipos de malignidades hematológicas,
principalmente leucemia mielomonocítica juvenil e, mais raramente, em tumores
sólidos.
Legius et al (2002) em seu estudo supõe que mutações PTPN11 são
associados com anomalias de pigmentação da pele, tais como várias lentigines ou
manchas café com leite.
7.9.3 Principais Variações Entre Genes dos 10 grupos Com Maior WNL
Considerando os 10 grupos com maior WNL, os genes ESR1 e PPARA
apresentaram uma variação significante entre grupos do CPM para a PS. No CPM, o
gene ESR1 se encontra no grupo F e na manutenção da pele saudável foi

89
identificado no grupo J. Já o gene PPARA, no CPM esta associado ao grupo J e na
pele saudável ao grupo O, o que sugere a possibilidade de perda ou ganho de
funções na doença.
Estes genes serão discutidos abaixo. Importante ressaltar que o gene ESR1
foi pouco relacionado a este tipo de câncer e também pode ser um valioso alvo para
estudos futuros. O gene PPARA foi discutido em outro tópico.
ESR1
Está diretamente envolvido na regulação da expressão de genes eucarióticos
e afetam a proliferação e diferenciação celular (STRING, 2015).
A ação do gene ESR1 é mais visível no câncer de mama. Mais de dois terços
dos casos expressam o receptor de estrogênio - alfa (ER-a, codificada por ESR1),
sendo que a maior parte destes casos são sensíveis à inibição ER-a. (Robinson et
al, 2013; Toy et al, 2013). O papel de ER-a em melanoma é desconhecido (Mori et
al, 2006).
PPARA (Proliferadores de Peroxisoma Alfa)
Peroxissomas são organelas subcelulares encontrados em plantas e animais
que contêm enzimas para a respiração e para o metabolismo do colesterol e lipídios.
A ação de proliferadores de peroxissomas é mediada através de receptores
específicos, denominados PPAR, que pertencem à superfamília dos receptores de
hormonios esteróides. PPARs afetam a expressão de genes envolvidos na
proliferação celular, diferenciação celular e nas respostas imunes e inflamatórias.
Este gene codifica o subtipo PPAR-alfa, que é um factor de transcrição nuclear
(NCBI, 2016).
De acordo com Sertznig et al (2008), PPARa induz a diferenciação e inibe a
proliferação, além de regular a apoptose.

90
Lee et al (2007) apresentou dados que comprovam que a pigmentação em
melanócitos foi acelerado com PPARa, sugerindo um possível papel para PPARa na
modulação da proliferação e diferenciação dos melanócitos (pigmentação).
Ainda de acordo com Sertznig et al (2008), PPARs podem ser considerados
alvos potenciais para o tratamento de várias doenças de pele.
8. CONCLUSÃO
O presente trabalho apresentou como objetivo identificar quais são os genes
que possuem maior grau de interações no câncer de pele melanoma e quais as
principais diferenças em relação aos genes que possivelmente interagem na
manutenção de uma pele saudável. Para isso foram realizadas analises utilizando
ferramentas de bioinformática e mineração de textos.
A analise realizada permitiu sugerir genes com forte interação para o câncer
de pele melanoma. Os genes líderes sugeridos foram o TP53, porém os genes
pertencentes aos grupos B, C, e D (AKT1, JUN, STAT3 e MYC) também
apresentaram WNL relevante e obtiveram destaque no trabalho. Estes genes líderes
possivelmente possuem uma forte influência do desenvolvimento e/ou evolução da
doença e são alvos importantes para estudos experimentais. Para a pele saudável,
os genes líderes sugeridos foram o UBC, porém os genes dos grupos B, C, e D
(TP53, JUN, AKT1, CREBBP, EP300 e SRC) também apresentaram o WNL
relevante.
Uma comparação entre os genes identificados para câncer de pele melanoma
e os genes da pele saudável, também foi realizada. Para esta comparação foram
analisados somente os genes alocados nos 10 grupos com maior WNL.
Através desta analise foi possível identificar que a maioria dos genes com
maior interação na manutenção da pele saudável, também possuem alta interação
no câncer de pele melanoma, e que pequenas variações na rede de genes podem
ter influências no desenvolvimento e/ ou evolução do câncer de pele melanoma.
Os genes HSP90AA1, RAC1, FYN e RHOA estiveram presentes apenas na
rede do Câncer de Pele Melanoma, o que sugere a possibilidade de que estes

91
genes possam estar ativados nesta doença. Os genes UBC, CREBBP, EP300, SRC,
CTNNB1, NFKB1, INS, PTPN11, APP, CDK2, ERBB2, NOTCH1 e PPARG foram
identificados apenas na rede de genes responsáveis pela manutenção da pele
saudável, sugerindo a possibilidade de que alguns destes genes possam estar
silenciados nesta doença. Os genes ESR1 e PPARA apresentaram variações
relevantes entre os grupos, identificados pelo K-means, da manutenção da pele
saudável para o câncer de pele melanoma, o que sugere a possibilidade de perda
ou ganho de funções na doença.
Os resultados, identificados no presente trabalho, reforçam que a
bioinformática e a mineração de textos podem ser um ponto de partida interessante
para aprimorar o conhecimento sobre interação gene-gene e proteína-proteína,
mecanismos moleculares de doenças, processos biológicos, entre outros. E que a
análise detalhada de mapas de interação genômicos e rankeamento de genes, de
acordo com seu número de confiança nas interações, pode apresentar resultados
relevantes e sugerir novos alvos para análises experimentais, além de sugerir
possíveis fatores de risco e novas terapias.
9. REFERÊNCIAS
Alves, R.; Esteves, T.; Marote, J.; CAldeira, C.; Neves, P. C.; Faria, A.; Malignant
Melanoma Review of years (1998 - 2004) at Hospital Central of Funchal. Skin
Cancer, vol. 22, pg. 181-189, 2007.
AmiGO2 - The Gene Ontology. Disponível em: <http://amigo.geneontology.org>.
Acessado em Janeiro de 2016.
Ananthaswamy HN, Pierceall WE. Molecular mechanisms of ultraviolet radiation carcinogenesis. Photochem Photo biol. 1990;52:1119-36.
Antoniadis AG, Petridou ET, Antonopoulos CN, Dessypris N, Panagopoulou P,
Chamberland JP, Adami HO, Gogas H, Mantzoros CS. Insulin resistance in
relation to melanoma risk. Melanoma Res. 2011 Dec;21(6):541-6. doi:
10.1097/CMR.0b013e32834b0eeb.

92
Aplin, Andrew E; Klein, R. Matthew. Rnd3 Regulation of the Actin Cytoskeleton
Promotes Melanoma Migration and Invasive Outgrowth in Three Dimensions.
Cancer Res March 15, 2009 69; 2224.
AspGD - Aspergillus Genome. Disponível em: <http://www.aspergillusgenome.org>
Acessado em Janeiro de 2016.
Avery-Kiejda, Kelly A ; Bowden, Nikola A; Croft, Amanda J.; Scurr, Lyndee L;
Kairupan, Carla F; Ashton, Katie A; Talseth-Palmer, Bente A; Rizos, Helen; Zhang,
Xu D; Scott, Rodney J.; Hersey, Peter. P53 in human melanoma fails to regulate
target genes associated with apoptosis and the cell cycle and may contribute
to proliferation. BMC Cancer 2011, 11:203
Barabasi, A. L. and Albert, R. Emergence of scaling in random networks. Science,
pages 286–509, 1999.
Barone, Antonio; Toti, Paolo; Giuca, Maria Rita; Derchi, Giacomo; Covani, Ugo. A
gene network bioinformatics analysis for pemphigoid autoimmune blistering
diseases. Springer-Verlag Berlin Heidelberg, 2014
Battisti, Rúbia ; Nunes, Daniel Holthausen ; Weber, Ariana Lebsa ; Schweitzer, Louise Cardoso ; Sgrot, Isadora. Avaliação do perfil epidemiológico e da mortalidade dos pacientes com diagnóstico de melanoma cutâneo primário no município de Florianópolis - SC, Brasi. An. Bras. Dermatol. vol.84 no.4 Rio de Janeiro July/Aug. 2009
Bayer-Garner IB, Smoller B. Androgen receptors: a marker to increase sensitivity
for identifying breast cancer in skin metastasis of unknown primary site. Mod
Pathol. 2000 Feb;13(2):119-22.
Becker B; Multhoff G; Farkas B; Wild PJ; Landthaler M; Stolz W; Vogt T. Induction
of Hsp90 protein expression in malignant melanomas and melanoma
metastases. Experimental Dermatology. 2004 Jan;13(1):27-32.
Bhandaru M; Ardekani GS; Zhang G; Martinka M; McElwee KJ; Li G; Rotte A. A
combination of p300 and Braf expression in the diagnosis and prognosis of
melanoma. BMC Cancer. 2014 Jun 3;14:398. doi: 10.1186/1471-2407-14-398.

93
Bielemann, V. L. M. O ser com câncer: uma experiência em família. Florianópolis: UFSC, 1997. 170p. Dissertação de Mestrado - Universidade Federal de Santa Catarina. bioinf.pdf>. Acessado em: 11 de julho de 2008.
BioCyc Database Collection. Disponível em: <http://biocyc.org>. Acessado em Janeiro de 2016.
Bonamigo, Renan Rangel;Carvalho, André Vicente Esteves de; Sebastiani, Vanessa Raquel Zaleski; Silva, Cristina Martino da; Pinto, Angela Caroline de Zorzi. HLA e câncer de pele. An. Bras. Dermatol. [online]. 2012, vol.87, n.1, pp. 9-18. ISSN 0365-0596.
BORGES-OSÓRIO, M. R., ROBINSON, W. M. Genética Humana. 2. ed. Porto
Alegre: Artmed, 2001. p. 278 – 299.
Botelho, Michelle G; Wang, Xiaolei; Arndt-Jovin, Donna J; Becker, Dorothea; Jovin,
Thomas M. Induction of Terminal Differentiation in Melanoma Cells on
Downregulation of ß-Amyloid Precursor Protein. Journal of Investigative
Dermatology (2010) 130, 1400–1410; doi:10.1038/jid.2009.296;
Bragazzi NL, Sivozhelezov V, Nicolini C. LeaderGene: a fast data-mining tool for
molecular genomics. J Proteomics Bioinform 2011;04:83–6.
Brener, Sylvie; Jeunon, Franca Arenare; Barbosa, Alvimar Afonso; Grandinetti, Helenice de Andrade Marigo. Carcinoma de células escamosas bucal: uma revisão de literatura entre o perfil do paciente, estadiamento clínico e tratamento proposto. RevistaBrasileira de Cancerologia 2007; 53(1): 63-69.
Burns, George W.; Bottino, Paul. Genética. Guanabara Koogan, 6ª ed. Rio de
janeiro, 1991;
Cao J; Dai X; Wan L; Wang H; Zhang J; Goff PS; Sviderskaya EV; Xuan Z; Xu Z; Xu
X; Hinds P; Flaherty KT; Faller DV; Goding CR; Wang Y; Wei W; Cui R. The E3
ligase APC/C(Cdh1) promotes ubiquitylation-mediated proteolysis of PAX3 to
suppress melanocyte proliferation and melanoma growth. Science Signaling.
2015 Sep 1;8(392):ra87. doi: 10.1126/scisignal.aab1995.

94
Capoluongo, Ettore. Insulin-Like Growth Factor System and Sporadic Malignant
Melanoma. The American Journal of Pathology 178.1 (2011): 26–31. PMC. Web. 20
Nov. 2015.
Carvalho, Charles André; Giugliani, Roberto; Ashton-Prolla, Patrícia; Cunha,
Mauricio Estrela da; Bakos, Lucio. Melanoma hereditário: prevalência de fatores
de risco em um grupo de pacientes no Sul do Brasil. An bras Dermatol, Rio de
Janeiro, 79(1):53-60, jan./fev. 2004.
Carvalho, Cyntia Helena Pereira. Relação da Imunoexpressão da BMP-2, BMPR-
IA e BMPR-II Com o Perfil Clínico-Patológico em Carcinoma Epidermóide de
Lábio Inferior. Dissertação, Universidade Federal do Rio Grande do Norte, 2010;
Carvalho, Lucas Miguel de. IIS - Integrated Interactome System : Uma plataforma
web para anotação, análise e visualização de interação de proteínas-genes-
drogas através da integração de diversas fontes de dados e ferramentas.
Instituto de Matemática, Estatística e Computação Científica (IMECC). Universidade
Estadual de Campinas ( Unicamp ), 2011;
Carvalho, Marcelo Sávio Revoredo Menezes de. A TRAJETÓRIA DA INTERNET
NO BRASIL: DO SURGIMENTO DAS REDES DE COMPUTADORES À
INSTITUIÇÃO DOS MECANISMOS DE GOVERNANÇA. RIO DE JANEIRO, RJ –
BRASIL, SETEMBRO DE 2006.
Castiglia D; Bernardini S; Alvino E; Pagani E; De Luca N; Falcinelli S; Pacchiarotti A;
Bonmassar E; Zambruno G; D'Atri S. Concomitant activation of Wnt pathway and
loss of mismatch repair function in human melanoma. Genes Chromosomes
Cancer. 2008 Jul;47(7):614-24. doi: 10.1002/gcc.20567.
CATHACYC - Metabolici Pathway Database of Catharanthus Roseus. Disponível
em: <http://www.cathacyc.org>, acessado em janeiro de 2016.
CGD - Candida Genome Database. Disponível em
<http://www.candidagenome.org>. Acessado em janeiro de 2016.

95
Chadd E Nesbit; Tersak, Jean M; Prochownik, Edward V. MYC oncogenes and
human neoplastic disease. Volume 18, Number 19, Pages 3004-3016, 1999.
Chan G; Kalaitzidis D; Neel BG. The tyrosine phosphatase Shp2 (PTPN11) in
cancer. Cancer Metastasis Rev. 2008 Jun;27(2):179-92. doi: 10.1007/s10555-008-
9126-y.
Chana Jagdeep S.; Grover, Rajiv; Tulley, Paul; Sanders, Roy; Grobbelaar, Adriaan
O.; Lohrer, Horst; Wilson, George D.. The c-myc oncogene: use of a biological
prognostic marker as a potential target for gene therapy in melanoma. British
Journal of Plastic Surgery, Volume 55, Issue 8, December 2002, Pages 623–627.
Chiattone, H. B. C. (1996). Uma vida para o câncer. Em: V. A. Angerami-Camon, (Org.). O doente, a psicologia e o hospital (pp. 75-89). São Paulo: Pioneira.
Chinem, Valquiria Pessoa; Miot, Hélio Amante. Epidemiologia do carcinoma
basocelular. AnbrasDermatol. 2011;86(2):292-305.
Chohan TA; Qian H; Pan Y; Chen JZ. Cyclin-dependent kinase-2 as a target for
cancer therapy: progress in the development of CDK2 inhibitors as anti-cancer
agents. Curr Med Chem. 2015;22(2):237-63.
Chu SH; Liu YW; Zhang L; Liu B; Li L; Shi JZ; Li L. Regulation of survival and
chemoresistance by HSP90AA1 in ovarian cancer SKOV3 cells. Mol Biol Rep.
2013 Jan;40(1):1-6. doi: 10.1007/s11033-012-1930-3. Epub 2012 Nov 8.
CHU, D.. Development and structure of skin. In: WOLF, K. et al. Fitzpatrick’s
Dermatology in General Medicine. New York: McGraw-Hill, 2008, p. 57-73.
CORRÊA, Marilena V..O Admirável Projeto Genoma Humano. PHYSIS: Rev. Saúde Coletiva, Rio de Janeiro, 12(2):277-299, 2002..
Covani U, Marconcini S, Giacomelli L, et al. Bioinformatic prediction of leader
genes in human periodontitis. J Periodontol 2008;79:1974–83.

96
Cytoscape - Network Data Integration, Analysis, and Visualization in a box.
Disponível em:< http://www.cytoscape.org/>.Acesso em: 20 set. 2015.
Dantas, Élida Lívia Rafael; Sá, Fernando Henrique de Lima; Carvalho, Sionara Melo de Figueiredo de; Arruda, Anderson Pontes;Ribeiro, Evelane Marques; Ribeiro, Erlane Marques. Genética do Câncer Hereditário. Revista Brasileira de Cancerologia 2009; 55(3): 263-269
Demehri, Shadmehr; Turkoz, Ahu; Kopan, Raphael. Epidermal Notch1 loss
promotes skin tumorigenesis by impacting the stromal microenvironment.
Cancer Cell. 2009 Jul 7; 16(1): 55–66.
Demunter A; Libbrecht L; Degreef H; De Wolf-Peeters C; van den Oord JJ. Loss of
membranous expression of beta-catenin is associated with tumor progression
in cutaneous melanoma and rarely caused by exon 3 mutations. Modern
Pathology. 2002 Apr;15(4):454-61.
Dimatos, Dimitri Cardoso; Duarte, Felipe Oliveira; Vieira, Vilberto José; Vasconcellos,
Zulmar Antonio Accioli De; Bins-Ely, Neves, Jorge Rodrigo D`Éça. Melanoma
Cutâneo No Brasil. Arquivos Catarinenses de Medicina - Volume 38 - Suplemento
01 - 2009.
Dobroff, Andrey S.; Wang,, Hua; Melnikova, Vladislava O.; Villares, Gabriel J.; Zigler,
Maya; Huang, Li; Bar-Eli, Menashe. Silencing cAMP-response Element-binding
Protein (CREB) Identifies CYR61 as a Tumor Suppressor Gene in Melanoma.
(2009) The Journal of Biological Chemistry, 284, 26194-26206.
doi:10.1093/bioinformatics/bti551
Du J; Widlund HR; Horstmann MA; Ramaswamy S; Ross K; Huber WE; Nishimura
EK; Golub TR; Fisher DE. Critical role of CDK2 for melanoma growth linked to its
melanocyte-specific transcriptional regulation by MITF. Cancer Cell. 2004
Dec;6(6):565-76
Elias D; Ditzel HJ. Fyn is an important molecule in cancer pathogenesis and
drug resistance. Pharmacol Res. 2015 Oct;100:250-4. doi:
10.1016/j.phrs.2015.08.010. Epub 2015 Aug 21

97
EUZEBIO, CARLOS DANILO GAIOLI. UMA CONTRIBUIÇÃO DA APLICAÇÃO DE
MODELOS FUZZY EMPREGADOS NA DETECÇÃO DA QUEIMA DE PEÇAS NA
RETIFICAÇÃO PLANA. Dissertação apresentada na Faculdade de Engenharia da
UNESP – campus de Bauru, São Paulo, 2012.
Evan, GI & Littlewood, TD: O papel do c-myc no crescimento celular. Curr Opin
Genet Dev, 1993 3, 44-49.
FAYYAD, U. Advances in Knowledge Discovery and Data Mining. [S.1.]:
AAAI/MIT Press, 1996.
FERLAY, J. et al. GLOBOCAN 2012 v1.0. Cancer incidence and mortality world wide.Lyon, France: IARC, 2013. Disponível em: <http://globocan.iarc.fr>. Acesso em: 10 jul. 2014.
Fernandes, Nurimar C.; Calmon, Roberto; Maceira, Juan P.; Cuzzi, Tullia; Silva,
Cesar S. Claudio da.. Melanoma cutâneo: estudo prospectivo de 65 casos. An.
Bras. Dermatol. vol.80 no.1 Rio de Janeiro Jan./Feb. 2005.
Ferreira, Flávia Regina; Nascimento, Luiz Fernando C.. Câncer cutâneo em Taubaté (SP) – Brasil, de 2001 a 2005: um estudo de prevalência. AnBrasDermatol. 2008;83(4):317-22.
Filho, Fernando Camargo; Prosdocime, Francisco; Cerqueira, Gustavo Coutinho; Binneck, Eliseu; Silva, Acácia Fernandes; Reis, Adriana Neves dos; Junqueira, Ana Carolina Martins; Santos, Ana Cecília Feio dos; Júnior, Antônio Nhani; Wust, Charles I.;Kessedjian, Jayme Lourenço; Petretski, Jorge H.; Camargo, Luiz Paulo; Ferreira, Ricardo de Godoi Mattos; Lima, Roceli P.; Pereira, Rodrigo Matheus; Jardim, Sílvia; Sampaio, Vanderson de Souza; Folgueras-Flatschart, Áurea V.. Bioinformática: Manual do Usuário. Biotecnologia Ciência & Desenvolvimento – nº 29, 2002. Disponível em: <http://www.biotecnologia.com.br/revista/bio29/
Fitzpatrick TB, Johnson RA, Wolff K. Color atlas and synopsis of clinical
dermatology. 1997:229-31.

98
Fitzpatrick, T.D.; Szabo, G.; Quevedo, W.C. Biology of the melanin pigmentarty
system. In. Biology of the melanocyte system., Fitzpatrick, T.D. editor. McGraw-Hill
Book Co., New York, 131-163, 1979.
Formighieri, Eduardo Fernandes. Bioinformática e anotação de genes em
Xanthomonas axonopodis pv. citri e Xylella fastidiosa: metabolismo de ferro e
biossíntese de pequenas moléculas. Piracicaba, 2002.
Franceschini A , Szklarczyk D, Wyder S, et al. STRING v10: protein–protein
interaction networks, integrated over the tree of life. Nucleic Acids Research.
2015;43(Database issue):D447-D452. doi:10.1093/nar/gku1003.
Gayther SA; Batley SJ; Linger L; Bannister A; Thorpe K; Chin SF; Daigo Y; Russell
P; Wilson A; Sowter HM; Delhanty JD; Ponder BA; Kouzarides T; Caldas C.
Mutations truncating the EP300 acetylase in human cancers. Nat Genet. 2000
Mar;24(3):300-3.
GeneCards - Human Gene Database. Disponível em: <http://www.genecards.org/>.
Acesso em: 15 de outubro de 2015;
Giacomelli L, Nicolini C. Gene expression of human T lymphocytes cell cycle:
experimental and bioinformatic analysis. J Cell Biochem 2006;99:1326–33.
Giglia-Mari, Giuseppina; Sarasin, Alain. TP53 Mutations in Human Skin
Cancers.P53 REVIEW ARTICLE. HUMAN MUTATION 21:217^228 (2003)
Gil, Antônio Carlos. Métodos e técnicas de pesquisa social. - 6. ed. - São Paulo : Atlas,2008.
Grossman D, Leffell DJ. The molecular basis of non melanoma skin cancer: new understanding. ArchDermatol. 1997;133:1263-70.

99
Guttman M et al. Chromatin signature reveals over a thousand highly conserved
large non-coding RNAs in mammals. Nature, 2009, 458(7235):223-7.
Halaban R. RAC1 and Melanoma. Clinical therapeutics. 2015;37(3):682-685.
doi:10.1016/j.clinthera.2014.10.027.
Hartsock A; Nelson WJ.. Adherens and tight junctions: structure, function and
connections to the actin cytoskeleton. Biochim Biophys Acta. 2008
Mar;1778(3):660-9.
He T, Wu J, Chen Y, Zhang J. TP53 polymorphisms and melanoma: A meta-
analysis. J Can Res Ther 2015;11:409-14
Homsi J; Cubitt CL; Zhang S, Munster PN; Yu H; Sullivan DM; Jove R; Messina JL;
Daud AI. Src activation in melanoma and Src inhibitors as therapeutic agents in
melanoma. Melanoma Res. 2009 Jun;19(3):167-75. doi:
10.1097/CMR.0b013e328304974c.
Hui J, Bindereif. Alternative pre-mRNA splicing in the human system:
Unexpected role of repetitive sequences as regulatory elements. Biol Chem,
2005, 386:1265-1271
Instituto Nacional de Câncer - INCA. Disponível em: http://www.inca.gov.br. Acesso em 3 de julho de 2014.
Iyer, Matthew K; Niknafs, Yashar S; Malik, Rohit; Singhal, Udit; Sahu, Anirban; Yasuyuki, Hosono; Barrette, Terrence R; Prensner, John R; Evans, Joseph R; Zhao, Shuang; Poliakov, Anton; Cao, Xuhong; Dhanasekaran, Saravana M; Wu, Yi-Mi; Robinson, Dan R; Beer, David G; Feng, Felix Y; Iyer, Hariharan K; Chinnaiyan, Arul M. The landscape of long noncoding RNAs in the human transcriptome. © 2015 Nature America, Inc. All rights reserved.
Jean, Didier; Harbison, Matthew; McConkey, David J.; Ronai, Zeèv; Bar-Eli,
Menashe. CREB and Its Associated Proteins Act as Survival Factors for Human
Melanoma Cells. (1998) The Journal of Biological Chemistry, 273, 24884-24890.

100
Jensen LJ, Kuhn M, Stark M, et al. STRING 8–a global view on proteins and their
functional interactions in 630 organisms. Nucleic Acids Res 2009;37(Database
issue):D412–6.
JOAQUIM, Leyla Mariane; EL-HANI, Charbel Niño. A genética em transformação:
crise e revisão do conceito de gene. Sci. stud. [online]. vol.8, n.1, 2010. [cited
2015-10-03], pp. 93-128.
Joaquim, Leyla Mariane; El-Hani, CharbelNiño. A genética em transformação: crise e revisão do conceito de gene.Sci. stud, São Paulo, v. 8, n. 1, p. 93-128, 2010.
Jr WAS. A importância dos estudos genéticos sobre câncer de pulmão. J BrasPneumol. 2009;35(8):721-722
Jurberg, Claudia; Gouveia, Maria Emmerick;Belisário, Camila. Na mira do câncer: o papel da mídia brasileira. Revista Brasileira de Cancerologia 2006; 52(2): 139-14
Justin G. Madson; David T. Lynch; Kelsey L. Tinkum; Sumanth K. Putta; Laura A.
Hansen. Erbb2 Regulates Inflammation and Proliferation in the Skin after
Ultraviolet Irradiation. Am J Pathol. 2006 Oct; 169(4): 1402–1414.
Kang HY, Chung E, Lee M, Y Cho, Kang WH. Expressão e função de receptores
activados pelo proliferador peroxissoma em melanócitos humanos. British
Journal of Dermatology. 2004; 150 (3): 462-468.
Kappelmann, Melanie; Bosserhoff, Anja; Kuphal, Silke. AP-1/c-Jun transcription
factors: Regulation and function in malignant melanoma. European Journal of
Cell Biology, Volume 93, Issues 1–2, January–February 2014, Pages 76–81.
Karst AM; Gao K; Nelson CC; Li G. Nuclear factor kappa B subunit p50 promotes
melanoma angiogenesis by upregulating interleukin-6 expression. Int J Cancer.
2009 Jan 15;124(2):494-501. doi: 10.1002/ijc.23973.
Klein, R Matthew, and Paul J Higgins. “A Switch in RND3-RHOA Signaling Is
Critical for Melanoma Cell Invasion Following Mutant-BRAF Inhibition.”
Molecular Cancer 10 (2011): 114. PMC. Web. 17 Nov. 2015.

101
Kortylewski, Marcin; Jove,, Richard; Yu, Hua. Targeting STAT3 affects melanoma
on multiple fronts. Cancer Metastasis Rev. 2005 Jun;24(2):315-27.
Krauthammer, Michael; Kong, Yong; Ha, Byung Hak; Evans, Perry; Bacchiocchi,
Antonella; McCusker, James P; Cheng, Elaine; Davis, Matthew J; Goh, Gerald;
Choi, Murim; Ariyan, Stephan; Narayan, Deepak; Dutton-Regester, Ken; Capatana,
Ana; Holman, Edna C; Bosenberg, Marcus; Sznol, Mario; Kluger, Harriet M; Brash,
Douglas E; Stern, David F; Materin, Miguel A; Lo, Roger S; Mane, Shrikant; Ma,
Shuangge; Kidd, Kenneth K; Hayward, Nicholas K; Lifton, Richard P; Schlessinger,
Joseph; Boggon, Titus J; Halaban, Ruth. Exome sequencing identifies recurrent
somatic RAC1 mutations in melanoma. Nature Genetics 44, 1006–1014 (2012)
doi:10.1038/ng.2359
Lai JJ; Chang P; Lai KP; Chen L; Chang C. The role of androgen and androgen
receptor in skin-related disorders. Arch Dermatol Res. 2012 Sep;304(7):499-510.
doi: 10.1007/s00403-012-1265-x. Epub 2012 Jul 25.
Lee JS, Choi YM, Kang HY. Agonista de PPAR-gama, ciglitazona, aumenta a
pigmentação e migração de melanócitos humanos. Experimental Dermatology.
2007; 16 (2):. 118-123
Legius E; Schrander-Stumpel C; Schollen E; Pulles-Heintzberger C; Gewillig M;
Fryns JP. Mutações no PTPN11 em síndrome Leopard. J Med Genet. 2002
agosto; 39 (8): 571-4.
Liberman, Felipe. Análise dos Fatores Determinantes Para a Qualidade da
Anotação Genômica Automática. Dissertação apresentada na Universidade
Católica de Brasília, 2004;
Liu ZJ; Xiao M; Balint K; Smalley KS; Brafford P; Qiu R; Pinnix CC; Li X; Herlyn M.
Notch1 signaling promotes primary melanoma progression by activating
mitogen-activated protein kinase/phosphatidylinositol 3-kinase-Akt pathways
and up-regulating N-cadherin expression. Cancer Res. 2006 Apr 15;66(8):4182-
90.
Macias, Everardo; Marval, Paula L. Miliani de; Siervi, Adriana De; Conti, Claudio J.;
Senderowicz, Adrian M.; Rodriguez-Puebla, Marcelo L.. CDK2 Activation in Mouse

102
Epidermis Induces Keratinocyte Proliferation but Does Not Affect Skin Tumor
Development. Am J Pathol. 2008 Aug; 173(2): 526–535.
Madonna, Gabriele; Ullman, Claudio Dansky; Gentilcore, Giusy; Palmieri, Giuseppe;
Ascierto, Antonio. NF-KB as potential target in the treatment of melanoma.
Journal of Translational Medicine 2012, 10:53 doi:10.1186/1479-5876-10-53
Maere S, Heymans K, Kuiper M. BiNGO: a Cytoscape plugin to assess
overrepresentation of gene ontology categories in biological networks.
Bioinformatics 2005;21: 3448–9
Maere, Steven; Heymans, Karel; Kuiper, Martin. BiNGO: a Cytoscape plugin to
assess overrepresentation of Gene Ontology categories in Biological
Networks. BIOINFORMATICS. Vol. 21 no. 16 Ghent, Belgium, 2005, pages 3448–
3449
Manso, Carlos. ONCOGENES. Faculdade de Medicina de Lisboa. Lisboa - 1986.
Mantese, Sônia Antunes Oliveira; Berbert, Alceu Luiz Camargo V.; Gomides, Mabel
Duarte Alves; Rocha, Ademir. Carcinoma basocelular - Análise de 300 casos
observados em Uberlândia - MG. AnBrasDermatol. 2006;81(2):136-42.
Markovic, S.N.; Erikson, L.A.; Rao, R.D.; Weenig, R H.; Pockaj, B.A.; Bardia, A.;
Vacon, C.M.; Schild, S.E.; Mcwilliams, R.R.; Hand, J.L.; Laman, S.D.;
Kottschade,L.A.; Maples, W.J.; Pittelkow, M.R.; Pulido, J.S.; Cameron, J.D.&
Creagan, E.T.; 2007. Malignant Melanoma in the 21 st Century, part 1:
epidemiology, risk factors, screening, prevention, and diagnosis. Mayo Clin
Proc., vol. 82, nº 3, pg. 364-380.
Martinez, Marcos Antônio Rodrigues; Francisco, Guilherme; Cabral, Luciana Sanches; Ruiz, Itamar Romano Garcia; Neto, Cyro Festa. Genética molecular aplicada ao câncer cutâneo não melanoma. An. Bras. Dermatol. vol.81 no.5 Rio de Janeiro Sept./Oct. 2006.

103
McCarthy MM; Pick E; Kluger Y; Gould-Rothberg B; Lazova R; Camp RL; Rimm DL;
Kluger HM. HSP90 as a marker of progression in melanoma. Ann Oncol. 2008
Mar;19(3):590-4. Epub 2007 Nov 23.
ME - Manual de Estatística. Disponível em
<http://www.bertolo.pro.br/AdminFin/StatFile/Manual_Estatistica.htm>, acessado em
janeiro de 2016.
MEDINI, D.; SERRUTO, D.; PARKHILL, J.; RELMAN, D.A.; DONATI, C.; MOXON,
R.; FALKOW, S.; RAPPUOLI, R. Microbiology in the post-genomic era. Nature
Reviews Microbiology, v.6, p. 419-430, 2008.
Meierjohann, Svenja; Wende, Elisabeth; Kraiss, Anita; Wellbrock, Claudia; Schartl,
Manfred. The Oncogenic Epidermal Growth Factor Receptor Variant
Xiphophorus Melanoma Receptor Kinase Induces Motility in Melanocytes by
Modulation of Focal Adhesions. Cancer Research. 2006 Mar 15;66(6):3145-52.
Melnikova, Vladislava O.; Dobroff, Andrey S.; Zigler, Maya; Villares, Gabriel J.;
Braeuer, Russell R.; Wang, Hua; Huang, Li; Bar-Eli, Menashe. CREB Inhibits AP-2a
Expression to Regulate the Malignant Phenotype of Melanoma. Chammas R, ed.
PLoS ONE. 2010;5(8):e12452. doi:10.1371/journal.pone.0012452.
Mena-Chalco, Jesús. Identificação de regiões codificantes de proteína através
da transformada modificada de Morlet. Instituto de Matemática e Estatística da
Universidade de São Paulo. São Paulo - Brasil, 2005.
Metz, Jean; Calvo, Rodrigo; Seno, Eloize Rossi Marques; Romero, Roseli A. F.;
Liang, Zhao. Redes Complexas: conceitos e aplicações. RELATÓRIOS
TÉCNICOS DO INSTITUTO DE CIÊNCIAS MATEMÁTICAS E COMPUTAÇÃO. N.
290, São Carlos, 2007. ISSN - 0103-2569
Michaelis - Moderno Dicionário da Língua Portuguesa. Disponível em:<
http://michaelis.uol.com.br/>.Acesso em: 31 out. 2015.
Minges JT, Su S, Grossman G, Blackwelder AJ, Pop EA, Mohler JL, Wilson EM.
Melanoma antigen-A11 (MAGE-A11) enhances transcriptional activity by linking
androgen receptor dimers. J Biol Chem. 2013 Jan 18;288(3):1939-52. doi:
10.1074/jbc.M112.428409. Epub 2012 Nov 21.

104
MINITAB. Disponível em <HTTPS://http://support.minitab.com/pt-br/minitab/17/topic-
library/modeling-statistics/regression-and-correlation/goodness-of-fit-statistics/r-
squared/> acessado em janeiro de 2016.
Miot, Luciane Donida Bartoli; Miot, Hélio Amante; Silva, Márcia Guimarães; Marques,
Mariângela Esther Alencar. Fisiopatologia do melasma. Na Bras Dermatol.
2009;84(6):623-35
MIR, Luiz; FILHO, Carlos Alberto Moreira; MENCK, Carlos Frederico Martins;
ABDELHAY, Eliana; RECH, Elíbio; NETO, Emmanuel Dias; ZALZANO, Francisco M.;
OLIVA, Glaucius; VANNUCHI, Helio; RAW, Isaias; HALIL, Jorge; OLIVEIRA, Jose
Eduardo Dutra; ZANETTINI, Maria Helena; VIEIRA, Maria Lucia Carneiro; ZATZ,
Mayana; NARDI, Nance; Velho, Natal; ARRUDA, Paulo; BISCH, Paulo Mascarello;
BOROJENIC, Rodovan; OLIVEIRA, Sergio Costa; PENA, Sergio. GENOMICA.
Conselhos de Informações sobre Biotecnologia. São Paulo: EDITORA Atheneu,
2004.
MMMP - Melanoma Molecular Map Project. Disponível em
<http://www.mmmp.org/>. Acesso em 13 de novembro de 2015;
Molgó, M., Santa Maria, M.L.S.; Lubiano, A.. Epidemiologia do Cancro Cutâneo. Piel,
Vol. 2, nº2, pg.41-45, 2007.
MONTEIRO & SILVA, A. B.; PORTUGAL, M. S.; CECHIN, A. L. Redes Neurais
Artificiais e Análise de Sensibilidade: Uma Aplicação à Demanda de
Importações Brasileira. Revecap v. 5 n. 4, 2001.
Mori, Takuji; Martinez, Steve R.; O’Day, Steven J.; Morton, Donald L.; Umetani,
Naoyuki; Kitago, Minoru; Tanemura, Atsushi; Nguyen, Sandy L.; Tran, Andy N.;
Wang, He-Jing; Hoon, Dave S.B. Estrogen Receptor-? Methylation Predicts
Melanoma Progression. Cancer research 66.13 (2006): 6692–6698. PMC. Web. 17
Nov. 2015.
Nachtigall, Pedro Gabriel. PERFIL DE EXPRESSÃO E ORGANIZAÇÃO
GENÔMICA DE MICRORNAS MÚSCULO-ESPECÍFICOS NA TILÁPIA DO NILO

105
Oreochromis niloticus. INSTITUTO DE BIOCIÊNCIAS - UNIVERSIDADE
ESTADUAL PAULISTA, Botucatu - SP, 2012.
Nagalakshmi U, Wang Z, Waern K, Shou C, Raha D, Gerstein M, Snyder M. The
transcriptional landscape of the yeast genome defined by RNA sequencing.
2008, Science, 320(5881):1344-9.
Nasser, Nilton. UVB: suscetibilidade no melanoma maligno. AnBrasDermatol.
2010; 85(6):843-8.
National Center for Biotechnoly Information - NCBI. Disponível em:
<http://www.ncbi.nlm.nih.gov/>. Acesso em 30 de abril de 2014.
Lehninger: Nelson, Danid L.; Cox, Michael M. Lehninger Principies of
Biochemistry. Third Edition, W.H. FREEMAN AND COMPANY, New York,
Copyright 2000
Nerya, O.; Vaya, J.; Musa, R.; Izrael, S.; Ben-aire, R.; Tamir, S.
Glabreneandisoquiritigenin as tyrosinase inhibitors from licorice roots. J. Agric.
Food. Chem. 51:1201-1207, 2003;
Neto, Ana Alexandra Ferreira. Análise Retrospectiva de Melanoma Maligno
Cutâneo Primário no Serviço de Dermatovenereologia do Hospital de Souza
Martins: 1995 - 2007. Faculdade de Ciências da Saúde - Universidade da Beira
Interior. Covilhã, Junho de 2008.
Neto, Ana Alexandra Ferreira. Análise Retrospectiva de Melanoma Maligno
Cutâneo Primário no Serviço de Dermatovenereologia do Hospital de Souza
Martins: 1995-2007. Universidade da Beira Interior - Faculdade de Ciências da
Saúde. Covilhã, 2008.

106
Nicolas M; Wolfer A; Raj K; Kummer JA; Mill P; Van Noort M; Hui CC; Clevers H;
Dotto GP; Radtke F. Notch1 functions as a tumor suppressor in mouse skin. Nat
Genet. 2003 Mar;33(3):416-21.
Nriagu J; Lin TS; Mazumder DG; Chatterjee D. E-cadherin polymorphisms and
susceptibility to arsenic-related skin lesions in West Bengal, India. Science of
The Total Environment. 2012 Mar 15;420:65-72. doi:
10.1016/j.scitotenv.2012.01.007. Epub 2012 Feb 11.
Omholt K; Platz A; Ringborg U; Hansson J. Cytoplasmic and nuclear
accumulation of beta-catenin is rarely caused by CTNNB1 exon 3 mutations in
cutaneous malignant melanoma.Int J Cancer. 2001 Jun 15;92(6):839-42.
Orlando B, Bragazzi N, Nicolini C. Bioinformatics and systems biology analysis
of genes network involved in OLP (Oral Lichen Planus) pathogenesis. Arch Oral
Biol 2013;58:664–73.
OUZOUNIS, C.A.; VALENCIA, A. Early bioinformatics: the birth of a discipline a
personal view. Bioinformatics, v. 19, p. 2176-2190, 2003.
P U Emeagi, S Maenhout, N Dang, C Heirman, K Thielemans et al. Downregulation
of Stat3 in melanoma: reprogramming the immune microenvironment as an
anticancer therapeutic strategy. Gene Therapy, Nature Publishing Group, Nov 1,
2013. Copyright © 2013, Rights Managed by Nature Publishing Group.
PANTHER - Geneontology Unifying Biology. Disponível em
<http://www.pantherdb.org>, acessado em janeiro de 2016.
Perrot CY; Gilbert C; Marsaud V; Postigo A; Javelaud D; Mauviel A. GLI2
cooperates with ZEB1 for transcriptional repression of CDH1 expression in
human melanoma cells. Pigment Cell Melanoma Res. 2013 Nov;26(6):861-73. doi:
10.1111/pcmr.12149. Epub 2013 Aug 12.
PETIT, L. PIERARD, G.E. Skin-lightening products revisited. Inter J Cosmetic Sci,
v.25, p. 169-181, 2003.

107
Popim, Regina Célia; Corrente, José Eduardo;Marino, Jaqueline Aparecida Geromel; Souza, Carolina Arantes de. Câncer de pele: uso de medidas preventivas e perfil demográfico de um grupo de risco na cidade de Botucatu. Ciência & Saúde Coletiva, 13(4):1331-1336, 2008
Poswar, Fabiano de Oliveira; Farias, Lucyana Conceição; Fraga, Carlos Alberto de
Carvalho; Bambirra, Wilson; Brito-Junior, Manoel; Sousa-Neto, Manoel Damiao,
Santos, Sergio Henrique Souza; de Paula, Alfredo Mauricio Batista; D’Angelo,
Marcos Flavio Silveira Vasconcelos; Guimaraes, Andre Luiz Sena. Bioinformatics,
Interaction Network Analysis, and Neural Networks to Characterize Gene
Expression of Radicular Cystand Periapical Granuloma. Journal of Endodontics,
Volume 41, Issue 6, June 2015, Pages 877–883, Copyright ª 2015 American
Association of Endodontists. http://dx.doi.org/10.1016/j.joen.2015.02.004
Prins, J, de Vries, Ege, Mulder, NH:. A família myc de oncogenes e sua presença
e importância no carcinoma do pulmão de células pequenas e outros tipos de
tumores. Anticancer Res, 1993 13, 1373-1386.
Prosdocimi, Francisco; Santos, Fabrício. Sobre bioinformática, genoma e ciência.
CIÊNCIA HOJE • vol. 35 • nº 209, Belo Horizonte, 2004.
Ramos, Cristiano D. Silveira; Gomes, Carlos Augusto; Lucinda, Antônio Fonseca;
Delgado, José Lindomar; Almeida, Marcelo Wilson Rocha; Aguiar, Leonardo
Fernandes de Souza; Pitanguy, Ivo. Tratamento cirúrgico do melanoma cutâneo
com o auxílio da cintilografia do linfonodo sentinela: estudo de 16 casos. Rev.
Bras. Cir. Plást. 2009; 24(1): 64-70.
Rao VH; Vogel K; Yanagida JK; Marwaha N; Kandel A; Trempus C; Repertinger SK;
Hansen LA1. Erbb2 up-regulation of ADAM12 expression accelerates skin
cancer progression. Mol Carcinog. 2015 Oct;54(10):1026-36. doi:
10.1002/mc.22171. Epub 2014 May 5.
Rebecca VW, Massaro RR, Fedorenko IV, Sondak VK, Anderson AR, Kim E, et al.
Inhibition of autophagy enhances the effects of the AKT inhibitor MK-2206

108
when combined with paclitaxel and carboplatin in BRAF wild-type melanoma.
Pigment Cell Melanoma Res. 2014;27:465–478.
Rebhan M, Chalifa-Caspi V, Prilusky J, Lancet D. GeneCards: integrating
information about genes, proteins and diseases. Trends Genet 1997;13:163.
Rivoire, Waldemar Augusto; Corleta, Helena Von; Brum, Ilma Simoni; Capp, Edison.
Molecular Biology of Cervical Cancer. Rev. Bras. Saúde Mater. Infant. Vol.6, nº.4
Recife, 2006.
Robinson, Dan R; Wu, Yi-Mi; Vats, Pankaj; Su, Fengyun; Lonigro, Robert J; Cao,
Xuhong; Kalyana-Sundaram, Shanker; Wang, Rui; Ning, Yu; Hodges, Lynda; Gursky,
Amy; Siddiqui, Javed; Tomlins, Scott A; Roychowdhury, Sameek; Pienta, Kenneth J;
Kim, Scott Y; Roberts, J Scott; Rae, James M; Poznak, Catherine H Van; Hayes,
Daniel F ; Chugh, Rashmi; Kunju, Lakshmi P; Talpaz, Moshe; Schott, Anne F;
Chinnaiyan, Arul M. Activating ESR1 mutations in hormone-resistant metastatic
breast cancer. Nature Genetics 45, 1446–1451 (2013) doi:10.1038/ng.2823
S Tiwary; M Preziosi; P G Rothberg; N Zeitouni; N Corson; L Xu. ERBB3 is required
for metastasis formation of melanoma cells. Oncogenesis (2014) 3, e110;
doi:10.1038/oncsis.2014.23
Saito YD; Jensen AR; Salgia R; Posadas EM. Fyn: a novel molecular target in
cancer. Cancer. 2010 Apr 1;116(7):1629-37. doi: 10.1002/cncr.24879
SAMMA, A. S. B. E SALAM, R. A. Adaptation of k-means algorithm for image
segmentation. In Proceedings of World Academy of Science, Engineering and
Technology, p 58–62, 2009.
Santos, Luciane Amorim. USO DE FERRAMENTAS DE BIOINFORMÁTICA PARA
ESTUDOS DE EPIDEMIOLOGIA MOLECULAR, FILOGEOGRAFIA E
FILODINÂMICA VIRAL. CENTRO DE PESQUISAS GONÇALO MONIZ -
FUNDAÇÃO OSWALDO CRUZ. Salvador - Brasil. 2010.

109
Sasaki, H., Okuda, K., Kawano, O., Yukiue, H., Yano, M., & Fujii, Y. (2008). AKT1
and AKT2 mutations in lung cancer in a Japanese population. Molecular
Medicine Reports, 1, 663-666. http://dx.doi.org/10.3892/mmr_00000009
SBD - Portal da Sociedade Brasileira de Dermatologia. Disponível em:
<http://www.sbd.org.br/>. Acesso em: 05 de novembro de 2015;
Scapin, Marcos Paulo. Um Algoritmo Genético Híbrido Aplicado à Predição da
Estrutura de Proteínas Utilizando o Modelo Hidrofóbico-Polar Bidimensional.
Dissertação apresentada ao Central Federal de Educação Tecnológica do Paraná,
2005.
Schlagbauer-Wadl, Hermine; Griffioen, Marieke; Elsas, Andrea van; Schrier, Peter I;
Pustelnik, Tom; Eichler, Hans-Georg; Wolff, Klaus; Pehamberger, Hubert; Jansen,
Burkhard. Influence of Increased c-Myc Expression on the Growth
Characteristics of Human Melanoma. Journal of Investigative Dermatology (1999)
112, 332–336; doi:10.1046/j.1523-1747.1999.00506.x
Sertznig P; Seifert M; Tilgen W; Reichrath J. Peroxisome proliferator-activated
receptors (PPARs) and the human skin: importance of PPARs in skin
physiology and dermatologic diseases. Am J Clin Dermatol. 2008;9(1):15-31.
SGD - Saccharomyces Genome Database. Disponível em:
<http://www.yeastgenome.org>. Acessado em Janeiro de 2016.
Shannon P, Markiel A, Ozier O, et al. Cytoscape: A Software Environment for
Integrated Models of Biomolecular Interaction Networks. Genome Research.
2003;13(11):2498-2504. doi:10.1101/gr.1239303.
Shi H, Hong A, Kong X, et al. A novel AKT1 mutant amplifies an adaptive
melanoma response to BRAF inhibition. Cancer discovery. 2014;4(1):69-79.
doi:10.1158/2159-8290.CD-13-0279.
Silva, V. C. E. (2005). O impacto da revelação do diagnóstico de câncer na percepção do paciente. Dissertação de Mestrado, Curso de Pós-Graduação em Enfermagem, Programa interinstitucional USP/ UEL/ UNOPAR, São Paulo.

110
Simanainen U; Ryan T; Li D; Suarez FG; Gao YR; Watson G; Wang Y; Handelsman
DJ. Androgen receptor actions modify skin structure and chemical carcinogen-
induced skin cancer susceptibility in mice. Horm Cancer. 2015 Feb;6(1):45-53.
doi: 10.1007/s12672-014-0210-1. Epub 2015 Jan 7.
Souza, Reynaldo José Sant’Anna Pereira de; Mattedi, Adriana Prest; Rezende,
Marcelo Lacerda; Corrêa, Marcelo de Paula; Duarte, Etiene Marques. Estimativa do
custo do tratamento de câncer de pele tipo melanoma no Estado de São Paulo
– Brasil. AnBrasDermatol. 2009;84(3):237-43.
Souza, Sonia R P de; Fischer, Souza, Frida M; José M P de.. Bronzeamento e
risco de melanoma cutâneo: revisão da literatura. Rev. Saúde Pública vol.38 no.4
São Paulo Aug. 2004.
SPELLBERG, B. The cutaneous citadel: a holistic view of skin and immunity.
Life Sciences, v. 67, p. 477-502, 2000.
STANSFIELD, W. D. Genética. 2ª ed. São Paulo: McGraw-Hill do Brasil, 1985.
STRING - Known and Predicted Protein-Protein Interactions. Disponível em:<
http://string-db.org/>.Acesso em: 16 set. 2015.
Szklarczyk D, Franceschini A, Kuhn M, et al. The STRING database in 2011:
functional interaction networks of proteins, globally integrated and scored.
Nucleic Acids Research. 2011;39(Database issue):D561-D568.
doi:10.1093/nar/gkq973.
Szklo, André Salem; Almeida, Liz Maria de; Figueiredo, Valeska; Lozana, José de Azevedo ; Mendonça, Guinar Azevedo e Silva ; Moura, Lenildo de; Szklo, Moysés. Comportamento relativo à exposição e proteção solar na população de 15 anos ou mais de 15 capitais brasileiras e Distrito Federal. Cad. Saúde Pública, Rio de Janeiro, 23(4):823-834, abr, 2007.

111
TAN, P. N.; STEINBACH, M.; V. KUMAR, Introduction to Data Mining. Pearson.
Addison-Wesley, Published: 05/02/2005, ISBN-10: 0321321367, ISBN-13:
9780321321367, 2006.
Tang, Yanjuan; Halvarsson, Camilla; Nordigården, Amanda; Kumar, Komal; Åhsberg,
Josefine; Rörby, Emma; Wong, Wan Man; Jönsson, Jan-Ingvar. Coexpression of
hyperactivated AKT1 with additional genes activated in leukemia drives
hematopoietic progenitor cells to cell cycle block and apoptosis. ISEH -
Experimental Hematology. Volume 43, Issue 7, July 2015, Pages 554–564.
The Gene Ontology Consortium. Creating the Gene Ontology resource. design
and implementation. Genome Res., v. 11, p. 1425-1433, 2001;
Thomas, J.M.; Giblin. V.. Cure of cutaneous melanoma. BMJ, vol. 332, pg. 987-
988, 2006;
Torres-Estay V; Carreño DV; San Francisco IF1; Sotomayor P1; Godoy AS; Smith
GJ. Androgen receptor in human endothelial cells. J Endocrinol. 2015
Mar;224(3):R131-7. doi: 10.1530/JOE-14-0611. Epub 2015 Jan 6.
Toy, Weiyi; Shen, Yang; Won, Helen; Green, Bradley; Sakr, Rita A.; Will, Marie; Li,
Zhiqiang; Gala, Kinisha; Fanning, Sean; King, Tari A.; Hudis, Clifford; Chen, David;
Taran, Tetiana; Hortobagyi, Gabriel; Greene, Geoffrey; Berger, Michael; Baselga,
Jose; Chandarlapaty, Sarat. “ESR1 Ligand Binding Domain Mutations in
Hormone-Resistant Breast Cancer.” Nature genetics 45.12 (2013): 1439–1445.
PMC. Web. 17 Nov. 2015.
Trincaus, Maria Regiane. A morte em seu mostrar-se ao paciente oncológico em situação de metástase. Dissertação de Mestrado. Programa de Pós – Graduação em Enfermagem Fundamental, Escola de Enfermagem de Ribeirão Preto da Universidade de São Paulo, São Paulo, 2005.
Tsao H. Genetics of nonmelanoma skin cancer. Arch Dermatol. 2001;137:1486-92.

112
UFPA - Universidade Federal do Pará. Disponível em: <http://www.ufpa.br/dicas/biome/bioreg.htm>. Acesso em 13 de novembro de 2015.
UFPB - Biologia e Fisiologia Celular - Sinalização Celular. Disponível em: <http://www.dbm.ufpb.br/~marques/Artigos/Biologia_e_Fisiologia_Celular_-_Unidade_7_-_Sinalizacao_Celular.pdf>, acessado em janeiro de 2016.
US National Library of Medicine - PUBMED. Disponível em http://www.ncbi.nlm.nih.gov/pubmed. Acesso em1 de julho de 2014.
Vargas, Rogério Rodrigues. Uma Nova Forma de Calcular os Centros dos Cluster
em Algoritmos de Agrupamento Tipo Fuzzy C-Means. Universidade Federal do
Rio Grande do Norte, Natal - RN, 2012.
Vaz, Glauber José. The construction of sociograms and the theory of graphs.
Rev. bras. psicodrama vol.17 no.2 São Paulo 2009.
Vega JA; Diaz-Trelles R; Haro JJ; del Valle ME; Naves FJ; Fernández-Sánchez MT.
Beta-amyloid precursor protein in human digital skin. Neurosci Lett. 1995 Jun
9;192(2):132-6.
Veronese, Luiz Alberto; Marques, Mariangela Esther Alencar. Critérios
anatomopatológicos para melanoma maligno cutâneo: análise qualitativa de
sua eficácia e revisão da literatura. J. Bras. Patol. Med. Lab. vol.40 no.2 Rio de
Janeiro Apr. 2004.
Vieira, Eduardo Alano; Nodari, Rubens Onofre; Carvalho, Fernando Irajá Félix; Fialho Josefino de Freitas. Mapeamento Genético de Caracteres Quantitativos e sua Interação com o Ambiente. EMBRAPA ISSN 1517-5111. Planaltina, 2006.
Viros A, Sanchez-Laorden B, Pedersen M, et al. Ultraviolet radiation accelerates
BRAF-driven melanomagenesis by targeting TP53. Nature. 2014;511(7510):478–
482
Vleugel, Marije M.; Greijer, Astrid E.; Bos, Reinhard; Wall, Elsken van der; Dies, Paul
J. van. c-Jun activation is associated with proliferation and angiogenesis in

113
invasive breast cancer. ELSEVIER. Human Pathology, Volume 37, Issue 6, June
2006, Pages 668–674.
Von Mering C, Jensen LJ, Snel B, et al. STRING: known and predicted protein-
protein associations, integrated and transferred across organisms. Nucleic
Acids Res 2005; 33(Database issue):D433–7.
WAINSTEIN, Alberto J. A.; BELFORT, Francisco A.. Conduta para o melanoma
cutâneo. Rev. Col. Bras. Cir., Rio de Janeiro , v. 31, n. 3, p. 204-214, June 2004 .
Available from <http://www.scielo.br/scielo.php?script=sci_arttext&pid=S0100-
69912004000300011&lng=en&nrm=iso>. access on 04 Nov. 2015.
http://dx.doi.org/10.1590/S0100-69912004000300011.
Walker G; Hayward N. No evidence of a role for activating CDK2 mutations in
melanoma. Melanoma Res. 2001 Aug;11(4):343-8.
Wan, Y.S., Wang, Z.Q., Shao, Y., Voorhees, J.J., & Fisher, G.J. (2001). Ultraviolet
irradiation activates PI 3-kinase/AKT survival pathway via EGF receptors in
human skin in vivo. International Journal of Oncology, 18, 461-466.
http://dx.doi.org/10.3892/ijo.18.3.461
Watson, J. D., & Crick, F. H. C. A structure for deoxyribose nucleic acid. Nature
171, 737–738 (1953)
Ward, Laura Sterian. Entendendo o Processo Molecular da Tumorigênese. Arq Bras Endocrinol Metab vol.46 nº.4 São Paulo Aug. 2002.
Weiss J, Heine M, Arden KC, Korner B, Pilch H, Herbst RA, et al. Mutation and
expression of TP53 in malignant melanomas. Recent Results Cancer Res.
1995;139:137–54.
WOLK, K.; WITTE, K., SABAT, R. Interleukin-28 and Interleukin-29: Novel
Regulators of Skin Biology. Journal of Interferon & Cytokine Research, v.30, nº 8,
p. 617 – 628, 2010.

114
Yang S; McNulty S; Meyskens FL Jr. During human melanoma progression AP-1
binding pairs are altered with loss of c-Jun in vitro. Pigment Cell Res. 2004
Feb;17(1):74-83.
Zhang K; Wong P; Duan J; Jacobs B; Borden EC; Bedogni B. An ERBB3/ERBB2
oncogenic unit plays a key role in NRG1 signaling and melanoma cell growth
and survival. Pigment Cell Melanoma Res. 2013 May;26(3):408-14. doi:
10.1111/pcmr.12089. Epub 2013 Apr 2.
Zhuang D, Mannava S, Grachtchouk V, et al. C-MYC overexpression is required
for continuous suppression of oncogene-induced senescence in melanoma
cells. Oncogene. 2008;27(52):10.1038/onc.2008.258. doi:10.1038/onc.2008.258.
Zuehlke, Abbey D.; Beebe, Kristin; Neckers, Len; Prince, Thomas. Regulation and
function of the human HSP90AA1 gene. Gene, Volume 570, Issue 1, 1 October
2015, Pages 8–16.
10. ANEXOS
Genes Órfãos
Para o câncer de pele melanoma foram identificados 196 genes órfãos (eles
serão apenas citados no presente trabalho), são eles: GPR52, STX17, GCA, COMP,
TH1L, SERPINB9, NUAK2, MCC, SDCBP, ALCAM, PCDHB11, RAP1GAP, PHGDH,
MIA2, DHODH, MAGEA4, PHIP, LILRB1, SELENBP1, MGRN1, MUM1, POU3F3,
MXI1, EPHB6, ARTN, HOXA1, SERPINA3, AQP9, orf, TMIGD2, CYGB, RNASE3,
BPTF, PHLDB1, SRPK1, IFNK, TRPM1, TUBB, TIPARP, VTCN1, RNASET2,
SP110, TCF21, MCAM, B1, CLEC2B, ESAM, AIM1, GOLPH3, IDH1, PLP1, STPG2,
RBMX, RWDD1, BRD3, NHS, AIM2, ROPN1B, ROS1, FAT2, CD53, MAGEC1,
SPARCL1, IL32, PAEP, DCTN6, MAGEA10, RXRB, IFI44L, DEAF1, ESF1,
POMGNT1, ABCF2, MAGEA6, SLN, CEACAM1, SSX2, CFH, DCSTAMP, SPHKAP,
CMIP, MAGEA12, FUK, TDG, TRPV2, MAGEA11, ANTXR1, SOX17, TRIM2,
CYP4A22, REG1A, NUAK1, NACC1, CD160, MAGEL2, PLEK2, TMEM47, MT3,
TOP1MT, GOLPH3L, PPM1E, PSAP, CSMD1, FABP3, VPS39, BAALC, RTEL1,
JHDM1D, CD68, CA2, MAGEA1, ROPN1, NAT8, HLA-DPB1, HEXA, PLCXD1,

115
ERP29, ZFP36L2, MEOX1, ME2, MIA, RAB28, FAT3, DDX11, CIC, ANXA5, MLX,
OLFM4, CRYAB, DIDO1, MGAT5, SULF1, TBX4, CDKL1, SLPI, DDX43, FAP, EBP,
MTAP, CTCFL, RHO, NLRP2, ABCB5, ACP5, CSTB, MAGEH1, ATP7B, IER5,
CYP27A1, MB, LOX, CTAG1B, MIA3, MOCOS, SFMBT1, NLRP6, D3, HDGF,
ALDH1A1, HHLA2, SERPINB2, MAGED2, EVPL, TTBK2, PRAME, FAT4,
RARRES1, SERPINB13, MIF, PGP, AMACR, CTHRC1, MAGEA3, SYN1, TFEB,
GCLC, OCA2, MAGEF1, ZHX3, FCRL1, IFI44, KLK8, MARCO, MAGEB18, HTRA1,
AQP3, KLK6, ATRAID, SULF2, HOXC13, HOXC11, ATL1, FXYD5, C17orf79, D4,
MYO1B.
Para a pele saudável foram identificados 46 genes órfãos, são eles: KLK8,
GJB3, ECM1, FLCN, TGM5, CASP14, ALOXE3, BMP6, SLC45A2, MAOA,
RARRES2, CYP21A2, GJB2, FZD6, EXT1, FMR1, CDSN, ABCC6, HERC2, ELL2,
SPINK5, CLDN1, TGM1, GAR1, SLC24A5, SLC6A4, AICDA, F8, GBA, OCA2, MICA,
CSTA, PLAG1, HFE, KRT6A, FERMT1, GJB6, PLAGL1, DSC3, ELN, C4B, NR3C2,
IL36RN, CRABP2, SLURP1, HR.
Genes e WNL do Câncer de Pele Melanoma
Gene WNL 431 RAB11A 7757
1 TP53 404905 432 SFRP1 7727
2 AKT1 319796 433 ATF1 7537
3 JUN 273673 434 CITED1 7508
4 STAT3 230391 435 CLTC 7454
5 MYC 227249 436 MGMT 7432
6 EGFR 205607 437 DDR1 7421
7 MAPK1 204434 438 FDFT1 7300
8 MAPK14 192814 439 CD82 7240
9 MAPK8 192702 440 HHEX 7190
10 RAC1 191658 441 ETV1 6899
11 HSP90AA1 185749 442 GRIN1 6840
12 FYN 182703 443 PDCD4 6835
13 SMAD3 175621 444 ONECUT1 6818
14 PIK3CA 173733 445 BAP1 6766
15 FOS 171902 446 CX3CL1 6689
16 CCND1 169581 447 TCF4 6642
17 HRAS 167064 448 KLRK1 6532
18 HDAC1 163430 449 ANGPTL4 6476

116
19 STAT1 163345 450 GDF15 6445
20 RHOA 162603 451 TBX3 6419
21 RELA 159470 452 NLRP1 6414
22 CDKN1A 153607 453 CYP24A1 6404
23 MAPK3 152061 454 TBX2 6395
24 SMAD4 152043 455 GHRH 6360
25 BCL2 150278 456 SPRY4 6340
26 VEGFA 144208 457 SRY 6216
27 ESR1 138194 458 IL24 6200
28 JAK2 137278 459 ADORA2B 6171
29 AR 123983 460 NOX4 6116
30 TGFB1 123719 461 IMP3 6111
31 IL2 120215 462 LIF 6046
32 CDH1 119944 463 FDPS 5958
33 SP1 119223 464 EIF2C2 5957
34 IL6 117217 465 IL21 5938
35 RB1 115490 466 NLRC5 5877
36 HIF1A 115371 467 TYRP1 5786
37 IL8 115129 468 CSPG4 5730
38 NRAS 113458 469 ZFP36 5677
39 CDKN1B 113077 470 CPEB1 5664
40 CDC42 112728 471 MST1R 5661
41 YES1 107436 472 TRPV6 5644
42 CXCL12 107419 473 CD34 5616
43 MMP9 103862 474 C1QB 5538
44 ATF2 103525 475 PDE1A 5486
45 RAF1 102572 476 PDE1B 5441
46 EGR1 101791 477 AXL 5416
47 CDK4 100539 478 FZD3 5399
48 SMAD2 99099 479 CAV3 5395
49 FGF2 97869 480 PRKAB1 5394
50 GSK3B 95308 481 VIM 5343
51 MMP2 95092 482 BNIP3 5248
52 IRF1 94470 483 DDB2 5222
53 EDN1 94305 484 MRFAP1 5207
54 PTK2 93846 485 TAX1BP1 5099
55 CREB1 93199 486 RABGEF1 5074
56 IGF1 92557 487 FLOT2 4918
57 MTOR 92336 488 NLRP3 4916
58 E2F1 89994 489 BRD7 4905
59 SIRT1 89624 490 CSE1L 4897
60 PTEN 89313 491 ZEB1 4845
61 CDKN2A 86717 492 BRMS1 4827
62 MAP3K1 86302 493 MBD4 4793
63 KIT 84796 494 RASSF2 4776
117
64 IRF3 83814 495 PLIN2 4726
65 BCL2L1 82864 496 NDN 4714
66 ICAM1 82671 497 KDM5B 4680
67 IRF7 81640 498 PREX2 4679
68 PDGFRB 81112 499 SATB1 4569
69 TRAF6 80699 500 UHRF1BP1 4555
70 CAV1 80008 501 RAG2 4545
71 FOXO1 78778 502 GP9 4491
72 MAP2K1 76715 503 LMO2 4367
73 NCK1 74203 504 FLI1 4276
74 PPBP 73524 505 DOCK3 4261
75 KRAS 71496 506 POU3F2 4244
76 SYK 71494 507 MAGED1 4228
77 EGF 70467 508 SLIT3 4186
78 CXCR4 69975 509 BAG6 4170
79 CCR1 69621 510 MTSS1 4111
80 STAT2 69523 511 ATP7A 4009
81 PPARA 69519 512 CTSK 4005
82 BMP4 69176 513 TGFBI 3891
83 KDR 67834 514 KIF5B 3889
84 IL1B 67372 515 RAB11FIP2 3884
85 IGF1R 66648 516 CAPN3 3870
86 CCL5 66319 517 RANBP1 3862
87 HDAC9 66113 518 PLEK 3836
88 BRCA1 65940 519 PDCD1 3781
89 MET 65563 520 ULBP3 3750
90 CDK6 65340 521 DLX2 3730
91 ITGAV 64595 522 TLR5 3713
92 IFNG 64471 523 TRPM8 3708
93 FOXP3 62735 524 IL15RA 3694
94 SOCS3 62479 525 TCF12 3671
95 PDGFB 62274 526 USP15 3647
96 TERT 62232 527 EPCAM 3636
97 FOXO3 62195 528 DARC 3606
98 RASA1 62194 529 S100A2 3573
99 TIMP1 61480 530 RFXANK 3561
100 CCR7 61397 531 DCT 3529
101 HLA-A 61189 532 CD27 3476
102 TLR4 60355 533 CD70 3476
103 MDM2 60203 534 CD55 3421
104 SOCS1 59854 535 CD97 3421
105 ETS1 59124 536 MERTK 3221
106 PLK1 59107 537 SET 3212
107 CXCR2 59022 538 RYK 3209
108 ISG15 58913 539 GFAP 3130

118
109 IL4 58344 540 FBLN1 3039
110 CXCL10 58149 541 PHF19 2983
111 MYD88 57067 542 LGALS3BP 2918
112 POMC 55370 543 CCL8 2898
113 CCND2 54967 544 PERP 2896
114 CDC25A 54601 545 EID1 2826
115 CXCL9 54590 546 ITGB8 2826
116 CD40 54262 547 NXF1 2794
117 TGFB2 53673 548 CHRDL1 2791
118 VAV2 53650 549 ADAMTS13 2788
119 TNF 52620 550 ULBP2 2782
120 MAP2K2 52617 551 TYRO3 2765
121 ANXA1 52545 552 DGCR8 2760
122 CCL2 52145 553 NDST1 2759
123 FN1 51855 554 MVD 2754
124 CASP8 51640 555 LCN2 2720
125 MMP1 50416 556 LRAT 2692
126 IFIT1 50400 557 RPE65 2692
127 BRAF 50376 558 BATF 2647
128 E2F3 50210 559 RAB22A 2645
129 SKP2 49881 560 IFI16 2612
130 CXCL11 49856 561 IDH2 2611
131 VWF 49524 562 PFN1 2593
132 CCR10 49151 563 CAPZA1 2559
133 LEF1 49138 564 SETDB1 2526
134 EZH2 49047 565 IL20 2494
135 CCL19 48634 566 EYA4 2487
136 IL10 48296 567 DACH2 2487
137 CCL21 48086 568 DEC1 2470
138 CSK 47922 569 PAXIP1 2439
139 MCL1 47838 570 BRD4 2437
140 RRAS 47765 571 REV1 2423
141 CXCL5 47603 572 EWSR1 2394
142 TEK 47311 573 TCF20 2394
143 FADD 47107 574 SOX4 2369
144 SNAI1 45851 575 CD63 2369
145 DNMT1 45381 576 MAGEA2 2369
146 HLA-C 44965 577 TFAP2C 2358
147 CD8A 44911 578 BCL2L12 2354
148 IFIT3 44763 579 DDX21 2350
149 CCL27 44269 580 MYBBP1A 2333
150 CHEK1 44246 581 DERL2 2322
151 CD44 44009 582 GSR 2316
152 IFIT2 43666 583 LIN28B 2302
153 EIF4E 42858 584 ROBO3 2288

119
154 CNR1 42742 585 STMN1 2287
155 CCNA1 42674 586 NEU3 2277
156 CCL11 42568 587 RND3 2250
157 EPHA2 42525 588 PTGDS 2189
158 NCK2 41798 589 WWP2 2146
159 HMOX1 41474 590 ALDH3A1 2138
160 HSPA4 41203 591 S100A13 2114
161 BAX 41177 592 RAB27A 2089
162 TSC2 40693 593 KLK7 2087
163 PDGFRA 40481 594 CADM1 2086
164 GRM3 40464 595 TAZ 2034
165 ERBB3 40440 596 TFAP2A 2012
166 RHOC 40091 597 KRT4 1998
167 ISG20 39918 598 ENPP2 1981
168 VEGFC 39829 599 CD33 1960
169 GAL 39701 600 F10 1955
170 HSPB1 38993 601 PIM3 1916
171 RHOB 38724 602 TRPV4 1914
172 HGF 38255 603 ISYNA1 1902
173 ATF3 37852 604 NPTN 1898
174 ALDOA 37774 605 NES 1886
175 MX2 37646 606 CD99 1882
176 IL12RB1 37634 607 SERPINF1 1874
177 MITF 37111 608 DDR2 1865
178 VCAM1 37092 609 TRPV3 1864
179 IFI6 36984 610 TUBA1B 1859
180 WNT5A 36925 611 LAG3 1856
181 GADD45A 36856 612 CLDN1 1852
182 AURKA 36732 613 CD81 1852
183 PTGS2 36523 614 CHST14 1848
184 THBS1 36521 615 FABP7 1842
185 RPS6KA1 36071 616 PTMA 1841
186 SMAD1 35778 617 BRD2 1822
187 IFI27 35749 618 GPNMB 1820
188 KLF4 35555 619 CAMP 1816
189 EPO 35521 620 TYR 1810
190 GAPDH 35421 621 MLANA 1810
191 E2F5 35256 622 GALNT7 1800
192 GAS6 35192 623 CPE 1800
193 ATF4 34899 624 CNTN1 1799
194 NANOG 34868 625 GPR52 0
195 XAF1 34863 626 STX17 0
196 DDX58 34674 627 GCA 0
197 VAV3 34659 628 COMP 0
198 TLR2 34256 629 TH1L 0

120
199 CD86 33767 630 SERPINB9 0
200 TRADD 33723 631 NUAK2 0
201 IFNB1 33674 632 MCC 0
202 INSR 33456 633 SDCBP 0
203 IFITM3 33184 634 ALCAM 0
204 EPOR 33003 635 PCDHB11 0
205 PAX6 32919 636 RAP1GAP 0
206 RAD51 32547 637 PHGDH 0
207 EPHB2 32414 638 MIA2 0
208 CD28 32300 639 DHODH 0
209 CTGF 32282 640 MAGEA4 0
210 MAVS 32173 641 PHIP 0
211 STAT4 32167 642 LILRB1 0
212 SNAI2 32130 643 SELENBP1 0
213 KLK3 32110 644 MGRN1 0
214 CTTN 32041 645 MUM1 0
215 RPS6 31880 646 POU3F3 0
216 MAP4K4 31749 647 MXI1 0
217 TGFBR2 31331 648 EPHB6 0
218 GNAQ 31274 649 ARTN 0
219 SUZ12 31174 650 HOXA1 0
220 CDKN2C 31001 651 SERPINA3 0
221 ANGPT2 30935 652 AQP9 0
222 CIITA 30681 653 orf 0
223 SERPING1 30562 654 TMIGD2 0
224 HSPA5 30464 655 CYGB 0
225 VDR 30017 656 RNASE3 0
226 RUNX1 29792 657 BPTF 0
227 CSNK1A1 29469 658 PHLDB1 0
228 SOX2 29347 659 SRPK1 0
229 RPSA 29327 660 IFNK 0
230 TNFSF10 28842 661 TRPM1 0
231 CD80 28796 662 TUBB 0
232 COL1A2 28122 663 TIPARP 0
233 HSPA6 27823 664 VTCN1 0
234 BRCA2 27687 665 RNASET2 0
235 F13A1 27644 666 SP110 0
236 CDKN2D 27527 667 TCF21 0
237 SMARCB1 27122 668 MCAM 0
238 DNAJA1 27056 669 B1 0
239 BMI1 26964 670 CLEC2B 0
240 TLR3 26464 671 ESAM 0
241 PLXNB1 26381 672 AIM1 0
242 GLI3 26380 673 GOLPH3 0
243 HSF1 26328 674 IDH1 0

121
244 HDAC10 26024 675 PLP1 0
245 APC 25866 676 STPG2 0
246 HSPH1 25786 677 RBMX 0
247 KSR1 25710 678 RWDD1 0
248 MSH2 25648 679 BRD3 0
249 FAS 25247 680 NHS 0
250 MORF4L1 25115 681 AIM2 0
251 GNA11 25099 682 ROPN1B 0
252 HDAC8 24970 683 ROS1 0
253 RPS15A 24906 684 FAT2 0
254 RUNX3 24409 685 CD53 0
255 TBK1 24300 686 MAGEC1 0
256 EIF4A2 24042 687 SPARCL1 0
257 CCL3 23914 688 IL32 0
258 EDNRB 23888 689 PAEP 0
259 HES1 23860 690 DCTN6 0
260 IL1A 23810 691 MAGEA10 0
261 GRM1 23783 692 RXRB 0
262 DNAJB6 23605 693 IFI44L 0
263 MLH1 23426 694 DEAF1 0
264 DDIT3 23335 695 ESF1 0
265 GATA2 23287 696 POMGNT1 0
266 WT1 23178 697 ABCF2 0
267 MDM4 22933 698 MAGEA6 0
268 MYBL2 22624 699 SLN 0
269 DAXX 22577 700 CEACAM1 0
270 HDAC11 22554 701 SSX2 0
271 TP73 22390 702 CFH 0
272 HDAC6 22259 703 DCSTAMP 0
273 BUB1 22067 704 SPHKAP 0
274 TEC 21915 705 CMIP 0
275 NF1 21800 706 MAGEA12 0
276 RPL18 21774 707 FUK 0
277 CTLA4 21638 708 TDG 0
278 EIF3F 21496 709 TRPV2 0
279 SOX9 21487 710 MAGEA11 0
280 AXIN2 21392 711 ANTXR1 0
281 SKI 21187 712 SOX17 0
282 CDK8 21167 713 TRIM2 0
283 CDX2 21156 714 CYP4A22 0
284 DNAJA4 21071 715 REG1A 0
285 KISS1 21005 716 NUAK1 0
286 FOXA1 20994 717 NACC1 0
287 ITGA3 20802 718 CD160 0
288 DNAJB4 20626 719 MAGEL2 0

122
289 GLI2 20575 720 PLEK2 0
290 CTCF 20430 721 TMEM47 0
291 NME1 20252 722 MT3 0
292 BCL3 20229 723 TOP1MT 0
293 PSMA3 20199 724 GOLPH3L 0
294 FOXA2 20107 725 PPM1E 0
295 CDC25C 20069 726 PSAP 0
296 IFNAR1 20023 727 CSMD1 0
297 TANK 20008 728 FABP3 0
298 SH3KBP1 19941 729 VPS39 0
299 RARA 19650 730 BAALC 0
300 TNFRSF10B 19582 731 RTEL1 0
301 PARP1 19531 732 JHDM1D 0
302 ENO1 19442 733 CD68 0
303 BMP7 19345 734 CA2 0
304 LAMA3 18452 735 MAGEA1 0
305 EIF3D 18359 736 ROPN1 0
306 ADAM10 18289 737 NAT8 0
307 MMP3 18027 738 HLA-DPB1 0
308 CYLD 18013 739 HEXA 0
309 HSPA1A 17770 740 PLCXD1 0
310 LAMB3 17689 741 ERP29 0
311 IL27 17636 742 ZFP36L2 0
312 NOX1 17591 743 MEOX1 0
313 RHOV 17508 744 ME2 0
314 IL18 17194 745 MIA 0
315 SEMA4D 17143 746 RAB28 0
316 CDKN1C 17106 747 FAT3 0
317 IL7 17010 748 DDX11 0
318 BMX 16813 749 CIC 0
319 PDGFA 16750 750 ANXA5 0
320 KLF5 16718 751 MLX 0
321 PARK7 16641 752 OLFM4 0
322 GPC3 16630 753 CRYAB 0
323 FRS2 16576 754 DIDO1 0
324 EBI3 16444 755 MGAT5 0
325 PAX3 16384 756 SULF1 0
326 BUB3 16232 757 TBX4 0
327 MAP3K8 16126 758 CDKL1 0
328 HMGA2 15947 759 SLPI 0
329 CCL22 15913 760 DDX43 0
330 PMAIP1 15898 761 FAP 0
331 DKK1 15698 762 EBP 0
332 HINT1 15677 763 MTAP 0
333 DNMT3B 15589 764 CTCFL 0

123
334 NCF1 15437 765 RHO 0
335 FOXD3 15415 766 NLRP2 0
336 TLR7 15353 767 ABCB5 0
337 BCL2A1 15242 768 ACP5 0
338 NOTCH3 15211 769 CSTB 0
339 TIMP2 15160 770 MAGEH1 0
340 SOD1 15147 771 ATP7B 0
341 CYR61 15115 772 IER5 0
342 PROK1 14990 773 CYP27A1 0
343 PAX2 14952 774 MB 0
344 LGALS3 14871 775 LOX 0
345 LPL 14770 776 CTAG1B 0
346 TCF3 14673 777 MIA3 0
347 LDHA 14601 778 MOCOS 0
348 GH1 14276 779 SFMBT1 0
349 STAG2 14005 780 NLRP6 0
350 IL10RA 13915 781 D3 0
351 AHR 13812 782 HDGF 0
352 ARPC2 13803 783 ALDH1A1 0
353 CENPE 13803 784 HHLA2 0
354 DPP4 13399 785 SERPINB2 0
355 UBE3A 13325 786 MAGED2 0
356 NOTCH2 13319 787 EVPL 0
357 DUSP6 13147 788 TTBK2 0
358 RAD51C 13090 789 PRAME 0
359 ADAM17 13090 790 FAT4 0
360 PTCH1 12963 791 RARRES1 0
361 NT5E 12957 792 SERPINB13 0
362 NEDD8 12812 793 MIF 0
363 ATG5 12727 794 PGP 0
364 EPHA3 12713 795 AMACR 0
365 PRKX 12675 796 CTHRC1 0
366 ABCB1 12590 797 MAGEA3 0
367 NEDD9 12412 798 SYN1 0
368 GRIN2A 12367 799 TFEB 0
369 IL22RA1 12335 800 GCLC 0
370 PDIA3 12273 801 OCA2 0
371 HSPA1L 12254 802 MAGEF1 0
372 PLD1 12244 803 ZHX3 0
373 PBX1 12122 804 FCRL1 0
374 FBXW7 12037 805 IFI44 0
375 HSPA9 11640 806 KLK8 0
376 APRT 11595 807 MARCO 0
377 JAG1 11415 808 MAGEB18 0
378 PTP4A3 11348 809 HTRA1 0
124
379 C1R 11299 810 AQP3 0
380 FGFR3 11279 811 KLK6 0
381 DDX41 11273 812 ATRAID 0
382 HPRT1 11252 813 SULF2 0
383 CAV2 11232 814 HOXC13 0
384 GPI 11159 815 HOXC11 0
385 ATF6 11091 816 ATL1 0
386 GSTP1 11050 817 FXYD5 0
387 SOX10 10784 818 C17orf79 0
388 S100B 10700 819 ID4 0
389 IL7R 10667 820 MYO1B 0
390 CD14 10574
391 TP63 10550
392 IFNA1 10448
393 ATIC 10193
394 TAOK1 10146
395 MC1R 9930
396 HRK 9877
397 CDK5R1 9850
398 TRPV1 9753
399 NID1 9738
400 UCHL1 9721
401 BHLHE40 9676
402 MUC1 9487
403 FAT1 9486
404 CD83 9412
405 C1S 9393
406 NOD2 9374
407 MORF4L2 9233
408 TNFRSF10D 9227
409 TRPC1 9159
410 MAPK8IP1 9038
411 CD4 8947
412 WNT6 8896
413 TFE3 8617
414 EXO1 8600
415 RRM2B 8574
416 TPI1 8529
417 FLT3 8484
418 IL15 8428
419 RASGRP3 8168
420 ERCC1 8154
421 PLA2G6 8144
422 APC2 8140
423 EGLN3 8115

125
424 RIN1 8080
425 MSX2 7970
426 MYH11 7945
427 MEIS1 7876
428 ETV4 7867
429 RECK 7802
430 CYP27B1 7782
Genes e WNL da Pele Saudável
Gene WNL 270 BRCA2 25139
1 UBC 387957 271 GH1 24910
2 TP53 365317 272 COL4A5 24715
3 JUN 305674 273 MTA1 24650
4 AKT1 281377 274 CD36 24648
5 EP300 273624 275 CLOCK 24417
6 CREBBP 269441 276 PTHLH 24342
7 SRC 261329 277 WT1 24109
8 STAT3 249155 278 CCL27 23819
9 FOS 245046 279 SOX2 23682
10 MYC 232503 280 PTCH1 23589
11 ESR1 214720 281 KITLG 23567
12 CTNNB1 212322 282 PROC 23209
13 EGFR 205468 283 MC1R 23145
14 MAPK1 201881 284 VIP 23039
15 RELA 193979 285 PER2 22935
16 NFKB1 188459 286 FAS 22746
17 SMAD3 185064 287 SPP1 22253
18 PIK3CA 180012 288 SOD1 21985
19 CCND1 177166 289 GNRHR 21899
20 MAPK8 170337 290 HLA-C 21706
21 SMAD4 169202 291 HLA-B 21565
22 MAPK14 166573 292 GNRH1 21486
23 AR 160409 293 AHR 21485
24 SP1 160211 294 BCR 21349
25 IL6 157085 295 TSHR 21332
26 CDKN1A 156160 296 IL4R 21317
27 JAK2 155560 297 GLI2 21247
28 TGFB1 155404 298 GSTP1 20625
29 INS 155146 299 HLA-DRB1 20570
30 VEGFA 154789 300 PMS2 20413
31 HDAC1 153600 301 PDX1 20390
32 PTPN11 152610 302 POU5F1 20247

126
33 MAPK3 147894 303 CYP17A1 19893
34 HRAS 145697 304 CSF1R 19873
35 BCL2 144120 305 ABCA1 19845
36 STAT1 136990 306 CFTR 19781
37 HIF1A 133841 307 NF1 19298
38 PPARG 131324 308 HTT 19289
39 NOTCH1 127395 309 SOD2 18435
40 RB1 125703 310 NR0B1 18288
41 CREB1 124300 311 FGFR2 18277
42 APP 123768 312 TNFSF11 18093
43 PPARA 123618 313 TSC1 17727
44 IGF1 122186 314 MUC1 17628
45 ERBB2 120122 315 PARK2 17449
46 CDH1 119443 316 CYP19A1 17254
47 CDK2 118905 317 NOTCH2 17159
48 EDN1 116786 318 RXRA 16918
49 SMAD2 109689 319 ENG 16814
50 SIRT1 108056 320 ABCB1 16741
51 ABL1 105458 321 PLEC 16725
52 FGF2 105424 322 PER1 16599
53 MMP9 105180 323 CYR61 16527
54 IL8 101672 324 XRCC1 16365
55 NRAS 99190 325 CYP1B1 15891
56 PTEN 97303 326 PRNP 15760
57 MMP2 97276 327 NPPA 15747
58 IL2 96074 328 PITX2 15255
59 ICAM1 95483 329 TP63 14948
60 RPS27A 95139 330 PDGFA 14929
61 CEBPB 94845 331 FGFR3 14869
62 CDK4 93838 332 LEPR 14869
63 FOXO1 93717 333 TNFSF10 14716
64 GSK3B 92952 334 DMD 14675
65 NFKBIA 91506 335 CYLD 14646
66 E2F1 90052 336 MAF 14262
67 CDKN1B 89841 337 TGFA 13927
68 IKBKB 88794 338 PTCH2 13901
69 AGT 87763 339 SIRT2 13461
70 LEP 87483 340 IVL 13439
71 CHUK 87189 341 MYOG 13287
72 CXCL12 87183 342 TH 13239
73 SMARCA4 85621 343 KRT1 13217
74 PRKCA 84878 344 ELANE 12810
75 NCOA3 84875 345 LDLR 12636
76 IL1B 84816 346 MMP3 12240
77 PCNA 84504 347 GSTM1 12017

127
78 NOS3 84065 348 ALOX12B 12009
79 CAV1 83639 349 ADAR 11995
80 RAF1 83224 350 IL1RN 11886
81 BCL2L1 83169 351 OGG1 11798
82 TERT 82107 352 SELE 11737
83 SERPINE1 80735 353 TSG101 11681
84 CSF2 80324 354 CAT 11616
85 EGR1 77295 355 RPS19 11352
86 KIT 77107 356 GSTT1 11194
87 MDM2 76888 357 UGT1A1 11181
88 RARA 76627 358 SERPINC1 10826
89 ITGB1 76214 359 POLH 10744
90 F2 75919 360 NKX2-1 10021
91 BRCA1 75765 361 THBD 10015
92 LEF1 75472 362 MSX2 9996
93 CDKN2A 75428 363 DHFR 9876
94 IGF1R 72901 364 KRT16 9524
95 PDGFRB 71991 365 NF2 9514
96 ESR2 71272 366 MMP13 9500
97 VDR 70867 367 ISL1 9335
98 PGR 70833 368 IFNA1 9224
99 EGF 69127 369 PKP1 8775
100 IL4 66658 370 FBN1 8694
101 MET 65312 371 CSF3 8646
102 BMP4 64965 372 LOR 8636
103 SOCS3 64542 373 SOX10 8628
104 NR3C1 64352 374 CTLA4 8483
105 KRAS 63630 375 ALOX12 8423
106 ALB 63604 376 HMGCR 8381
107 POMC 62649 377 RAG2 8297
108 NR5A1 62177 378 LIF 8106
109 IRF1 61700 379 SRY 8055
110 TGFB2 61226 380 PAX3 7806
111 NFATC1 61169 381 FLG 7786
112 MYD88 60824 382 MGMT 7432
113 TIMP1 60745 383 T 7415
114 ATM 60653 384 XBP1 7137
115 CDK6 59525 385 XRCC3 7136
116 RUNX2 58964 386 KRT5 7088
117 FOXP3 58237 387 KRT14 6848
118 CD44 57586 388 NR1I2 6833
119 KDR 57281 389 ATP2A2 6792
120 TLR4 57234 390 PTGS1 6672
121 KLF4 56167 391 SLC6A3 6587
122 CASP3 55928 392 CDH3 6524

128
123 PML 54821 393 NME1 6485
124 HSPA8 53627 394 S100A8 6155
125 TNF 53372 395 AREG 6153
126 IGF2 53292 396 FHIT 6118
127 CCR5 52715 397 ODC1 5987
128 PDGFB 52695 398 MPO 5927
129 CEBPA 52664 399 ACE 5679
130 PPARD 52078 400 NOD2 5679
131 NGFR 52048 401 CSF1 5582
132 MSH2 51620 402 F3 5037
133 RORA 51315 403 MME 4887
134 BDNF 50324 404 CRHR1 4865
135 CD40 49987 405 FLNA 4639
136 COL1A2 49554 406 HBA2 4601
137 SHH 49475 407 HBB 4601
138 NGF 49223 408 DSG3 4456
139 ERBB4 48786 409 PKD1 4401
140 NR1H2 48038 410 DSP 4365
141 IFNG 47670 411 TRPV4 4344
142 PTGS2 47568 412 LMNA 4339
143 TCF7L2 47562 413 DSG1 4308
144 IL10 47305 414 DICER1 4307
145 IL12B 46981 415 CBS 4294
146 FN1 46871 416 PMP22 4247
147 TGFBR1 46076 417 TPR 4242
148 EPAS1 45630 418 CRP 4202
149 HMOX1 45089 419 FECH 4186
150 MMP1 45070 420 KL 4170
151 HNF1A 44940 421 DES 4120
152 TSC2 44819 422 FBLN5 4085
153 GATA3 44710 423 KRT17 3970
154 BAX 44507 424 TFAP2A 3868
155 GATA4 42727 425 SIRT6 3803
156 IL2RA 42276 426 RAG1 3752
157 HSPA4 42112 427 ETV6 3746
158 CDKN2B 42004 428 S100A7 3724
159 RUNX1 41914 429 CYP27B1 3666
160 NPY 41825 430 CALCRL 3617
161 HSPB1 41770 431 CETP 3184
162 F5 41647 432 ASIP 3169
163 SST 41423 433 KRT10 2869
164 CASR 41283 434 ERRFI1 2857
165 PLAU 41264 435 DST 2788
166 NFKB2 41043 436 TYR 2516
167 GNAS 40627 437 MEIS1 2462

129
168 APOA1 40564 438 MPZ 2447
169 HBEGF 40145 439 CACNA1A 2379
170 NOS2 39836 440 RYR1 2379
171 ERCC2 39766 441 HPRT1 2360
172 PSEN1 39691 442 DEFB4A 2303
173 CCL2 39507 443 HCCS 2278
174 ADRB2 39474 444 TYRP1 2257
175 TLR2 39438 445 SMN1 2095
176 COL17A1 39377 446 KLK7 2087
177 ITGB4 39273 447 MIF 1988
178 TEK 38962 448 G6PD 1976
179 GHR 38808 449 EWSR1 1900
180 TGFBR2 38643 450 SLC4A1 1872
181 RARB 38578 451 MECP2 1841
182 HGF 38438 452 MTHFR 1822
183 RARG 38175 453 HLA-DQB1 1820
184 CTGF 38066 454 PI3 1816
185 EPO 37895 455 DTNBP1 1800
186 FGFR1 37709 456 KLK8 0
187 INSR 37509 457 GJB3 0
188 COL1A1 36989 458 ECM1 0
189 IL5 36886 459 FLCN 0
190 ADAM17 36674 460 TGM5 0
191 STK11 36428 461 CASP14 0
192 BIRC5 36082 462 ALOXE3 0
193 PARP1 35634 463 BMP6 0
194 MITF 35232 464 SLC45A2 0
195 TNFRSF1A 35195 465 MAOA 0
196 RET 35157 466 RARRES2 0
197 BRAF 34615 467 CYP21A2 0
198 PSEN2 34482 468 GJB2 0
199 DNMT1 34466 469 FZD6 0
200 COL7A1 34461 470 EXT1 0
201 CCL5 34324 471 FMR1 0
202 SNCA 34024 472 CDSN 0
203 CHEK2 33976 473 ABCC6 0
204 VWF 33933 474 HERC2 0
205 VHL 33827 475 ELL2 0
206 LPL 33667 476 SPINK5 0
207 RBPJ 33593 477 CLDN1 0
208 IL13 33341 478 TGM1 0
209 DCN 33274 479 GAR1 0
210 FGF7 33042 480 SLC24A5 0
211 XPC 32860 481 SLC6A4 0
130
212 EDNRB 32840 482 AICDA 0
213 FASLG 32778 483 F8 0
214 IGFBP3 32612 484 GBA 0
215 ERCC3 32528 485 OCA2 0
216 IKBKG 32507 486 MICA 0
217 ADIPOQ 32354 487 CSTA 0
218 EZH2 32287 488 PLAG1 0
219 MAPT 32137 489 HFE 0
220 CRH 32131 490 KRT6A 0
221 MLH1 32100 491 FERMT1 0
222 FGF10 32099 492 GJB6 0
223 NFKBIB 32012 493 PLAGL1 0
224 SRF 31678 494 DSC3 0
225 PDGFRA 31560 495 ELN 0
226 SOX9 31367 496 C4B 0
227 PAX6 31300 497 NR3C2 0
228 BCL6 31189 498 IL36RN 0
229 PRL 30837 499 CRABP2 0
230 AGTR1 30759 500 SLURP1 0
231 NPM1 30668 501 HR 0
232 TP73 30563
233 CYP1A1 30496
234 TAC1 30492
235 PRKAR1A 30323
236 GLI1 29997
237 NFE2L2 29421
238 ARNTL 29409
239 MSH6 29148
240 CALCA 29062
241 CIITA 29057
242 XPA 29036
243 COL3A1 28935
244 ERCC4 28705
245 NTRK1 28431
246 LAMB3 28308
247 CDX2 28291
248 DRD2 28272
249 APOE 28128
250 GLI3 27777
251 LAMA3 27660
252 COL2A1 27616
253 LAMC2 27607
254 IL1A 27597
255 COL4A1 27568
256 NOS1 27240

131
257 ERCC6 27094
258 GATA1 26968
259 NOTCH3 26573
260 SP3 26419
261 GJA1 26368
262 ERCC5 25980
263 IL18 25839
264 IL1R1 25828
265 OPRM1 25763
266 PRLR 25737
267 APC 25709
268 HTR2A 25680
269 COL4A3 25430


















