
INFLUÊNCIA DA FRAGMENTAÇAO FLORESTAL E DA … Tatiana... · FÉ. Sonhei em realizar esse estudo...
143
Tatiana Bichara Dantas INFLUÊNCIA DA FRAGMENTAÇAO FLORESTAL E DA QUALIDADE DA MATRIZ DE MONOCULTURA DE EUCALIPTO SOBRE A COMPOSIÇÃO DAS COMUNIDADES DE VERTEBRADOS DE FOLHIÇO EM ÁREAS DE MATA ATLÂNTICA NO EXTREMO SUL DA BAHIA. Orientador: Dr. Pedro Luís Bernardo da Rocha Pós-Graduação IBIO/UFBA Dissertação apresentada ao Instituto de Biologia da Universidade Federal da Bahia, para obtenção do Título em Mestre em Ecologia e Biomonitoramento. SALVADOR 2004
Transcript of INFLUÊNCIA DA FRAGMENTAÇAO FLORESTAL E DA … Tatiana... · FÉ. Sonhei em realizar esse estudo...

Tatiana Bichara Dantas
INFLUÊNCIA DA FRAGMENTAÇAO FLORESTAL E DA QUALIDADE DA MATRIZ DE MONOCULTURA DE EUCALIPTO SOBRE A COMPOSIÇÃO DAS COMUNIDADES DE VERTEBRADOS DE FOLHIÇO EM ÁREAS DE MATA ATLÂNTICA NO EXTREMO SUL DA BAHIA.
Orientador: Dr. Pedro Luís Bernardo da Rocha
Pós-Graduação IBIO/UFBA
Dissertação apresentada ao Instituto de Biologia da Universidade Federal da Bahia, para obtenção do Título em Mestre em Ecologia e Biomonitoramento.
SALVADOR 2004

Biblioteca Central Reitor Macedo Costa - UFBA
D192 Dantas, Tatiana Bichara. Influência da fragmentação florestal e da qualidade da matriz de monocultura de
eucalipto sobre a composição das comunidades de vertebrados de folhiço em áreas de Mata Atlântica no Extremo Sul da Bahia / Tatiana Bichara Dantas. - 2004. 142 f. : il.
Orientador : Prof. Dr. Pedro Luís Bernardo da Rocha. Dissertação (mestrado) - Universidade Federal da Bahia, Instituto de Biologia, 2004.
Ecologia das florestas tropicais. 2.Eucalipto - Cultivo - Mata Atlântica (BA). 3. Comunidades animais - Conservação – Mata Atlântica (BA). 4. Lagarto - Mata Atlântica (BA). 5. Anfíbio – Mata Atlântica (BA). 7. Mata Atlântica - Conservação. I. Rocha, Pedro Luís Bernardo da. II. Universidade Federal da Bahia. Instituto de Biologia. III. Título.
CDD - 577.30913
CDU - 630*1
Comissão Julgadora:
Profa. Dra. Débora Faria
______________________Prof. Dr. Pedro Roch
________________________________
Profa. Dra. Blandina Viana
________________ a (Orientador)
2

3
Dedico essa dissertação aos meus pais, a André Luiz
e em especial ao meu “vô” Bichara,
que me proporcionaram o conhecimento de vida
e as alegrias, para que
eu pudesse seguir o meu caminho.

4
“Estamos aqui neste Planeta, por assim dizer, como turistas. Nenhum de nós pode
morar aqui para sempre. O maior tempo que podemos ficar são aproximadamente
cem anos. Sendo assim, enquanto estamos aqui, deveríamos ter um bom coração e
fazer de nossas vidas algo de positivo e útil. Quer vivamos poucos anos ou um
século inteiro, seria lamentável e triste passar esse tempo agravando os problemas
que afligem as outras pessoas, os animais e o ambiente. O mais importante de tudo
é ser uma boa pessoa.”
Dalai- Lama

5
Esta dissertação representa uma das maiores realizações profissionais e pessoais
da minha vida. Simboliza a superação de grandes dificuldades e desafios íntimos.
Porém, também representa amizade, família, companheirismo, amor, dedicação,
confiança, perseverança, espírito de luta, alegrias, e principalmente persistência na
FÉ. Sonhei em realizar esse estudo com intuito de compreender um pouco alguns
processos biológicos, e acima de tudo, tentar contribuir de alguma forma para a
conservação do nosso Planeta. Nessa jornada, diversas instituições, profissionais,
amigos e familiares foram fundamentais, e agora só tenho a explicitar os meus
agradecimentos. Gostaria muito de agradecer:
Ao Dr. Pedro Rocha, Coordenador Geral do Projeto Ecologia de Paisagem no
extremo Sul da Bahia, professor e coordenador do Mestrado em Ecologia e
Biomonitoramento da UFBA. Representa a minha maior fonte de inspiração
acadêmica e profissional: um exemplo de inteligência, ética, moral, conhecimento,
amizade, brilhantismo e excelência. Enfim, meu orientador querido, incansavelmente
presente em todos os momentos. Sem a sua existência a conclusão desse trabalho
nunca seria possível. Obrigada por tudo!!
Á Veracel Celulose S.A., especialmente na pessoa de Dr. Pedro Vaillant, que na
época ocupava a diretoria florestal, apostou no apoio logístico ao nosso projeto e
nos forneceu licença para realizarmos os estudos nas propriedades da empresa. Ao
engenheiro florestal Danilo Sette de Almeida, pela ajuda em muitos momentos, além
de ter sido o meu “primeiro mestre” sobre a Mata Atlântica. Á toda equipe da
Estação Veracruz, principalmente os meus queridos amigos: Valmir, Ivone, Hélio,
Luciano, Sandro, Jonas, Katão, Sara, Esperidião, Valdemir e Delgado, que me
ajudaram desde os tempos difíceis que passamos juntos, nos quatro anos de
trabalho na reserva.
Ás queridas Amada e Mison, pela atenção, desprendimento, ajuda e amizade em
todos os momentos dessa jornada. Aos colegas do LVT (Jorge, Érica, Roberta, Davi,
Alex e Luciano), pelo companheirismo e convivência nesses anos de luta! E também
a todos os pesquisadores e colaboradores do Projeto Ecologia da Paisagem.

6
Ao pessoal do Projeto Corredores Ecológicos, especialmente á Sidronio Bastos,
meu coordenador, que tanto suportou as minhas ausências, mau humor nesses 2
anos, e me deu forças para não desistir. À Andréia, que tanto me auxiliou com os
sapos e lagartos no laboratório e à Mel, pelo carinho e “segurada de barras” no
trabalho.
Aos doutores Marcelo Napoli, curador do Museu de Biologia da UFBA, e ao Dr.
Miguel Trefaut da USP, pelo auxílio na identificação dos anfíbios e répteis.
Á Eduardo Sarr, pelo milagroso apoio nas análises de Ecologia de Paisagem;
Silmara Magnobosco (analista de sistemas da Veracel Celulose), Daniela Blinder e
Eduardo Gabriel Palma (geógrafos SFC e CRA, respectivamente), Cristina Guerreiro
(arquiteta CRA) e Rômulo Morais (topógrafo Satélite) pelo auxílio com as bases
cartográficas. E à Annette, pela ajuda com os programas de geoprocessamento,
tradução dos resumos e pela amizade. À Marcos Franco, antigo gerente da Estação
Ecológica do Pau Brasil/CEPLAC, pelo acolhimento e auxílio logístico num momento
de desespero na última coleta de campo.
À Selma Ribeiro, engenheira florestal do IBAMA, amiga de muitas jornadas, que
mesmo á distância está sempre presente na minha vida. À Leo, meu amigo eterno.
À Rita Bastos, “Mary”, Dna Nadir e Simone também pela paciência, proteção e
amizade constantes nesse período. À Paty e Victor (ainda na barriga de Paty), pela
amizade e auxílio imprescindível com as figuras, tabelas e anexos dessa
dissertação.
À minha família, especialmente aos meus pais e avôs, que nunca hesitaram em
acreditar na importância do meu trabalho. Eles representam o meu alicerce e
segurança de vida, fonte maior de sabedoria e maturidade para o alcance de meu
crescimento interior. À Matheus, meu primo e Lú, minha irmã, que me socorriam em
finais de semana e madrugadas adentro com computadores e máquinas eletrônicas
desse caminho. E à Lúcia, pelo carinho e cuidados permanentes.
Á Deus, Meishu Sama, anjos protetores, espíritos de luz e guardiões, pela
oportunidade de concretizar mais esse sonho.

7
OBJETIVOS GERAIS
O presente estudo investigou o efeito da fragmentação e da qualidade da matriz de
monocultura de eucalipto sobre a composição das comunidades de vertebrados de
folhiço (anfíbios e répteis) em áreas de Mata Atlântica, no extremo Sul da Bahia.
Foram consideradas a abundância, biomassa e riqueza das espécies de anfíbios e
répteis em quatro fragmentos intermediários de sucessão (com áreas variando de 50
a 150 ha), quatro áreas de mata em excelente estado de conservação (considerada
Mata Contínua Primária) e quatro áreas com reflorestamento de eucalipto. Esta
pesquisa foi parte integrante de um projeto maior intitulado “Ecologia da Paisagem
no Extremo Sul da Bahia”, cujos objetivos principais seriam investigar,
comparativamente, diferentes grupos biológicos em relação à fragmentação e à
importância da matriz (eucaliptais), e a partir de dados empíricos, fornecer subsídios
para o estabelecimento de diretrizes de conservação para a região.
ORGANIZAÇÃO DA DISSERTAÇÃO Esta dissertação de Mestrado foi organizada em três capítulos. O primeiro traz uma
breve descrição sobre os principais aspectos teóricos relativos a fragmentação
florestal e seus processos associados; a Mata Atlântica brasileira e seu histórico de
destruição, incluindo uma breve abordagem sobre o Projeto Corredores Ecológicos,
uma nova perspectiva para manejo e conservação dos remanescentes existentes na
área do Corredor Central da Mata Atlântica. Este capítulo será publicado no livro -
texto do Seminário Brasil - Alemanha sobre Mata Atlântica e Biodiversidade.
O segundo capítulo visa a investigação do efeito da fragmentação, e a possível
viabilização da monocultura de eucalipto como uma matriz permeável à fauna de
répteis em relação à sua composição, o que permitiria a conectividade da paisagem
estudada. Além de abordar o desenho experimental básico do Projeto “Ecologia de
Paisagem no extremo Sul da Bahia”, sua região de estudo, as metodologias gerais
de coleta utilizadas para obtenção dos dados da Ecologia de Paisagem, micro-clima
e micro-habitat, os quais foram utilizados em comum para investigação dos diversos
grupos faunísticos.

8
O terceiro capítulo apresenta a mesma temática do anterior, só que do ponto de
vista da fauna de anfíbios e suas peculiaridades detectadas. Finalmente, será
apresentada uma discussão geral sobre as principais conclusões obtidas com esses
estudos, além de recomendações e propostas para manejo e conservação da região
em questão.

9
ÍNDICE
Capítulo 1: A Fragmentação Florestal e seus processos associados: o exemplo da Mata Atlântica no Brasil com suas perspectivas de manejo e conservação. 1.1 Resumo........................................................................................................... ..2 1.2 Abstract..............................................................................................................3 1.3 Introdução....................................................................................................... ..5 1.4 A Mata Atlântica no Brasil............................................................................. 11 1.5 Histórico de destruição da Mata Atlântica na Bahia................................... 15 1.6 Perspectivas de Manejo e Conservação para a Mata Atlântica da Bahia: o Projeto Corredores Ecológico e o Corredor Central da Mata Atl6antica na Bahia...................................................................................................................... 18 1.7 Modelo de Gestão do Projeto Corredores e suas ações para a formação do Corredor Central da Mata Atlântica na Bahia............................................... 24 1.8 Referências Bibliográficas............................................................................ 26 Capítulo 2: Influência da Fragmentação Florestal e da qualidade da matriz de monocultura de eucalipto sobre a composição das comunidades e da biomassa de lagartos de folhiço em áreas de Mata Atlântica no extremo Sul da Bahia. 2.1 Resumo........................................................................................................... 41 2.2 Introdução...................................................................................................... 42 2.3 Abstract............................................................................................................ 2.4. Material e Métodos.......................................................................... ............. 49
2.4.1 Área de Estudo: extremo Sul da Bahia................................................ 49 2.4.2 Configuração da paisagem regional e delineamento experimental do estudo ............................................................................................................. 55 2.4.3 Amostragem dos Lagartos de Folhiço................................................ 56 2.4.4 Variáveis da Fauna................................................................................ 58 2.4.5 Variáveis Ambientais Mensuradas....................................................... 59 2.4.6 Análise dos dados................................................................................. 65

10
2.5 Resultados..................................................................................................... 68 2.5.1 Descrição das Variáveis Ambientais.................................................. 75 2.5.2 Composição da comunidade de lagartos de folhiço entre os diversos componentes da paisagem........................................................... 81 2.5.3 Resultados da análise de ordenação em relação à comunidade de lagartos...................................................................................................... 85
2.6 Discussão....................................................................................................... 89 2.7 Referências Bibliográficas............................................................................ 96 Capítulo 3: Influência da Fragmentação Florestal e da qualidade da matriz de monocultura de eucalipto sobre a composição das comunidades e da biomassa de anfíbios de folhiço em áreas de Mata Atlântica no extremo Sul da Bahia. 3.1 Resumo......................................................................................................... 109 3.2 Introdução..................................................................................................... 110 3.3 Abstract.............................................................................................................. 3.4 Material e Métodos........................................................................... ........... 114 3.5 Resultados.................................................................................................... 114
3.5.1 Descrição das Variáveis Ambientais................................................. 121 3.5.2 Composição da comunidade de anfíbios de folhiço entre os diversos componentes da paisagem.......................................................... 121 3.5.3 Resultados da análise de ordenação em relação à comunidade de lagartos...................................................................................................... 85
3.6 Discussão....................................................................................................... 89 3.7 Referências Bibliográficas............................................................................ 96
Capítulo 4: Conclusões Gerais e considerações sobre propostas de conservação e manejo para a região de estudo....................................................................... 109
CAPÍTULO 1
A FRAGMENTAÇÃO FLORESTAL E SEUS PROCESSOS ASSOCIADOS: O EXEMPLO DA MATA ATLÂNTICA NO BRASIL COM SUAS PERSPECTIVAS DE MANEJO E CONSERVAÇÃO.
11

12
RESUMO
Mesmo abrigando mais da metade do número total de espécies no planeta, as
Florestas Tropicais correspondem aos ecossistemas mais ameaçados. A
fragmentação, que corresponde à conversão de habitats naturais em fragmentos de
diversos tamanhos, graus de conexão e níveis de perturbação, tem se tornado uma
das principais ameaças à biodiversidade em todo o mundo. Alterações promovidas
pela fragmentação, tais como extinções locais, alterações na composição e
abundância das espécies e redução do tamanho das populações são questões de
grande destaque que se fundamentam teoricamente em princípios gerais da
Ecologia. Predizem forte relação positiva entre o número de espécies e o tamanho
da área, devido à menor diversidade ou heterogeneidade de habitats associados a
áreas menores. Com o desenvolvimento de trabalhos sobre os efeitos da
fragmentação em florestas tropicais e o agravamento dos problemas ambientais
globais, ficou comprovado que as paisagens florestais fragmentadas são ambientes
extremamente complexos e variáveis, e que essa variabilidade está fortemente
relacionada aos habitats alterados, os quais envolvem os remanescentes
usualmente referidos como a matriz. A Mata Atlântica brasileira é um bom exemplo,
tanto em riqueza e grau de endemismo das Florestas Tropicais, como no tocante à
situação crítica de redução e alteração em que se encontram seus remanescentes
atuais, resumidos a menos de 8% de sua área original. Como o restante da Mata
Atlântica, o Sudeste baiano apresenta um histórico de degradação que se iniciou na
época do Descobrimento do Brasil e se mantém até os dias atuais, sob a forma de
desmatamentos e conversão das áreas naturais em ambientes antropizados. O
enfoque mais utilizado para minimizar os efeitos da fragmentação e isolamento de
florestas busca o estabelecimento de Unidades de Conservação em áreas
representativas dos habitats naturais, assegurando a preservação de amostras
protegidas da ação do homem. Os ramos da Biologia da Conservação e da Ecologia
de Paisagem indicam que essa estratégia, dissociada de abordagens que priorizem
a conservação de extensões mais abrangentes da paisagem natural e modificada,
não irá assegurar a manutenção de comunidades ecologicamente viáveis no longo
prazo. Dentre as várias abordagens possíveis, aquela dos “Corredores de
Biodiversidade” representa uma das mais promissoras para um planejamento
regional eficaz. Sendo assim, o Projeto Corredores Ecológicos representa um
componente do Ministério do Meio Ambiente, com objetivo principal de manter a

13
biodiversidade existente em grandes áreas de Floresta Tropical ("corredores" ou
"bio-regiões"), localizadas nas regiões de florestas da Amazônia e na Mata Atlântica,
através da gestão de uma paisagem dinâmica sustentada de larga escala, abrigando
comunidades e espécies com processos evolucionários e fluxos gênicos
assegurados, sob um regime de controle social local. ABSTRACT Even though tropical forests shelter more than half of the species on Planet Earth,
they are the most endangered ecosystems. Fragmentation, which results in the
conversion of natural habitats into fragments of diverse sizes and degrees of
connection and levels of disturbance, has become one of the main threats to
biodiversity around the world. Changes brought about by fragmentation, such as
local extinctions, alterations in the composition and abundance of species and
reduced populations, are outstanding questions that are theoretically fundamented
on general principles of ecology. A strong positive relationship between the number
of species and the size of an area is foreseen, as a result of less diversity or
heterogeneity of habitats associated with smaller areas. Studies on the effects of
fragmentation in tropical forests and increasing global environment problems have
proven that fragmented native forest landscapes are extremely complex and variable
environments, and that this variability is strongly related to altered habitats that
include forest remnants, usually referred to as matrix. The Brazilian Atlantic Forest is
a good example, because of the richness and degree of endemism of tropical forests
and the critical situation of its reduced and altered remnants, presently reduced to
less than 8% of its original area. Like the rest of the Atlantic Forest, the southeast of
Bahia presents a history of degradation that was initiated with the “Discovery” of
Brazil and continues to this day, by way of destruction of forests and conversion of
natural areas to human occupation. Aiming at minimizing the effects of fragmentation
and isolation of forests, focus has usually been on the establishment of protected
areas, representative of natural habitats, in the hope of ensuring the preservation of
samples, intending to protect them from encroaching human activities. Conservation
Biology and Landscape Ecology both indicate that this strategy, while disassociated
from methods that aim to secure the conservation of larger extensions of natural or
landscapes, will not ensure long-term maintenance of ecologically viable

14
communities. Among the possible alternatives, “Biodiversity Corridors” is one of the
most promising for efficient regional planning. The Ecology Corridor Project has as its
main goal, the sustainability of the biodiversity existent in large areas of tropical
forest (corridors or “bio-regions”), located in the native forested regions of the
Amazon and the Atlantic Forest, through the management of a large-scale sustained
dynamic landscape, sheltering communities and species, with evolutionary
processes and ensured genetic flux, under a controlled local social regime.

15
INTRODUÇÃO As Florestas Tropicais representam os ecossistemas mais ricos do mundo porque,
mesmo correspondendo apenas a 7% da superfície terrestre, elas abrigam mais da
metade do número total de espécies no planeta (Wilson, 1997; Wilson, 1988;
Withmore, 1997). Apesar de representarem grande relevância em termos de
diversidade e biologia das comunidades (Lewin, 1986), se apresentam como as
áreas mais ameaçadas do globo, em decorrência de intensa ação antrópica,
vinculadas principalmente ao desenvolvimento de atividades agrícolas, à extração
madeireira e à implantação da pecuária.
Dentre todas essas alterações desencadeadas pelo homem na natureza e
intensificadas neste último século, o desmatamento ou destruição das florestas
tropicais é considerado o principal processo responsável pela elevação das taxas de
extinção de espécies terrestres no mundo (Shafer, 1990, Henle et al.,1996; Pardini,
2001).
A redução dos ambientes naturais florestados teoricamente ocasiona a perda de
espécies, especialmente aquelas cuja distribuição é restrita aos trópicos, com altos
níveis de endemismos; e provoca a fragmentação florestal, acarretando no
isolamento dos habitats originais, circundados por ambientes alterados (Shafer,
1990).
A fragmentação, que corresponde à conversão de habitats naturais em fragmentos
de diversos tamanhos, graus de conexão e níveis de perturbação, tem se tornado
uma das principais ameaças à biodiversidade em todo o mundo (Shafer, 1990;
Debinski & Holt, 1999). Este processo teoricamente interfere no tamanho das
populações, na dispersão das espécies, na estrutura e quantidade de habitat
disponível e na probabilidade de invasões (Harrison & Bruna, 1999; Haila, 2002).
Tais aspectos podem acarretar alterações nas comunidades biológicas, não apenas
pela ação direta de cada um deles ou da sua interação, mas também por efeitos
indiretos, também conhecidos como efeitos cascata, através dos quais um grupo
biológico afeta outro, gerando uma dinâmica diferente daquela prevista para
sistemas florestais contínuos (Laurance & Bierregaard, 1997; Bierregaard, et al.
2001, Wilson, 1988; Lewinsohn & Prado, 2002; Faria, 2002).

16
Alterações promovidas pela fragmentação, tais como extinções locais, alterações na
composição e abundância das espécies e redução do tamanho das populações são
questões de grande destaque que se fundamentam teoricamente em princípios
gerais da Ecologia. Predizem forte relação positiva entre o número de espécies e o
tamanho da área devido à menor diversidade ou heterogeneidade de habitats
associados a áreas menores (Williams, 1943; Preston, 1962; Schoener, 1976;
Connor & MacCoy, 1979; Saunders et. al, 1991).
De acordo com a teoria de biogeografia de ilhas, postulada por MacArthur e Wilson
(1967), ilhas pequenas e isoladas apresentariam um menor número de espécies do
que aquelas maiores e próximas a outras ilhas. Essa teoria reuniu preceitos e fatos
já conhecidos e aceitos, baseando-se em duas variáveis - chaves: o tamanho das
ilhas e a distância ao continente. De modo análogo, fragmentos de um ecossistema
terrestre poderiam ser vistos como ilhas de vegetação distantes e isoladas entre si
por estarem circundados por um mar de ambientes modificados e inóspitos (Preston,
1962). Esta semelhança entre sistemas insulares e fragmentos florestais resultou na
condução de inúmeros estudos sobre o tema, e a teoria de biogeografia de ilhas
tornou-se, até recentemente, imprescindível na interpretação de dados empíricos e
no direcionamento de políticas de conservação em paisagens fragmentadas e ilhas
oceânicas.
Com o desenvolvimento de trabalhos sobre os efeitos da fragmentação em florestas
tropicais e o agravamento dos problemas ambientais globais ficou comprovado que
as paisagens florestais fragmentadas são ambientes extremamente complexos e
variáveis, e que essa variabilidade está fortemente relacionada aos habitats
alterados que envolvem os remanescentes, usualmente referidos como a matriz
(Laurance & Bierregaard, 1997; Bierregaard, et al. 2001, Pardini, 2001; Faria, 2002).
A influência da matriz ocorre em virtude da presença das espécies que nela habitam
e da alteração qualitativa do habitat original remanescente. Matrizes de diferentes
composições e natureza interagem com os fragmentos, de forma que a paisagem
como um todo exerça influência direta sobre a manutenção das comunidades nos
remanescentes (Gascon et al. 1999; Faria, 2002; Pardini, 2003).

17
A natureza e composição das matrizes e a tolerância de cada organismo a estes
ambientes geram respostas variáveis dos diferentes grupos taxonômicos à
fragmentação florestal, mostrando-se como um dos atributos-chave na determinação
da vulnerabilidade das espécies à fragmentação (Laurance, 1990; 1991a; Malcolm,
1997; Gascon et al. 1999; Joly et al.; 2000; Haila, 2002; Faria 2002 ) .
A proporção da paisagem ocupada pela matriz controla o fluxo de perturbações no
mosaico. Pode facilitar a propagação de distúrbios, como fogo ou espécies
invasoras ou generalistas ou ainda aumentar a heterogeneidade da paisagem,
podendo levar a uma convergência espacial de recursos e ao aumento do número
de espécies no mosaico (Metzger, 1999; Huenneke, 1992). Ou seja, as espécies
com maior capacidade de se dispersar e explorar a matriz podem se manter e até
são favorecidas nas áreas fragmentadas, enquanto espécies menos tolerantes
tendem a maior vulnerabilidade, mantendo-se isoladas e restringindo sua
distribuição no fragmento (Bierreggard et al. 1997; Gascon et al. 1999; Pardini,
2003.)
A capacidade de explorar as matrizes foi a característica ecológica mais importante
relacionada à vulnerabilidade de marsupiais numa região fragmentada na Austrália
(Laurance, 1991a), determinando também a vulnerabilidade de formigas, aves,
pequenos mamíferos e anuros em uma paisagem fragmentada na Amazônia
brasileira (Gascon et al. 1999; Perfecto & Vandermeer, 2002).
Janzen (1983) aponta que a influência das matrizes de habitats modificados, como
áreas secundárias, torna-se preocupante mesmo quando não acarreta redução
direta da riqueza total de espécies em um remanescente. Em alguns casos, deve
ocorrer uma profunda alteração na representatividade de determinadas espécies,
favorecendo aquelas tipicamente associadas a áreas mais perturbadas (Joly et al.
2000). Espécies pioneiras, generalistas e exóticas freqüentemente invadem ou são
favorecidas em remanescentes florestais, alterando a composição das comunidades
locais e aumentando a vulnerabilidade de espécies restritas às áreas da floresta
original (Faria, 2002; Rolim & Chiarello, 2003).

18
Outra conseqüência direta da fragmentação é a modificação do habitat
remanescentes através da influência dos habitats alterados criados ao seu redor,
denominados efeito de borda (Lovejoy et al. 1986). Definida como a zona mais
externa do remanescente, a borda sofre influência direta dos ambientes do entorno,
e nela ocorrem mudanças drásticas de caráter físico e biótico. Estas alterações são
particularmente mais acentuadas quando ocorre uma transição abrupta entre a
floresta e áreas abertas como pastagens e monoculturas (Mesquita et al. 1999;
Primack & Rodrigues, 2001).
Nas faixas de borda ocorre uma maior penetração de luz, redução na umidade
relativa, maior variação da temperatura e aumento da incidência de ventos, o que
resulta em um micro-clima distinto das regiões mais interiores da floresta (Lovejoy et
al. 1986). A penetração desse efeito de borda varia de acordo com a região e com o
tipo de parâmetro considerado. Características como o formato e tamanho do
fragmento e a natureza da matriz que o envolve podem ocasionar um drástico efeito
de borda, a ponto de todo o remanescente ser alterado, não permitindo a
sobrevivência de espécies mais sensíveis e geralmente restritas a áreas interiores
ou nucleares de um fragmento (Laurance, 1990, 1991)
Muitas evidências empíricas sugerem que, pelo menos no médio prazo, mudanças
no remanescente florestal decorrentes do efeito de borda acarretam alterações
determinísticas de suas comunidades biológicas, em muitos casos, consideradas
mais evidentes do que a redução do tamanho ou influência da matriz nos
remanescentes (Mesquita et al. 1999; Pardini, 2001; 2004).
Além da tolerância das espécies a habitats modificados, como matrizes e bordas,
sabe-se que características ecológicas e da história natural das espécies de
Florestas Tropicais as tornam particularmente vulneráveis aos efeitos da
fragmentação (Bierrgaard et al. 1997; Pardini, 2001; Dixo, 2001; Faria, 2002).

19
Conforme previsto na teoria de metapopulações, a destruição ou fragmentação do
habitat de uma população central pode resultar na extinção de numerosas
populações satélites que dependem da população central para sua colonização,
visto que a troca de indivíduos é bem mais freqüente entre grupos de fragmentos
próximos (Hanski, 1989; Lamberson et al., 1992; Harrison, 1994; Hokit et al., 2000;
Lopez & Pfister, 2001).
Fatores como baixa densidade populacional, pequena fecundidade, baixa taxa
reprodutiva, raridade natural, pouca plasticidade ecológica e a dinâmica da
estruturação natural das populações tornam certos componentes do ecossistema
mais sensíveis à redução e ao isolamento de áreas naturais do que outros
(Laurance, 1990).
Mais recentemente, os estudos sobre fragmentação começaram a mostrar que a
análise da paisagem como um todo - incluindo suas características gerais como
distribuição, forma e conectividade entre as porções fragmentadas, áreas
perturbadas e semi-naturais - aliada a pesquisas sobre distribuição e uso de habitat
pela biota, deveriam servir de unidade de manejo para permitir a conservação de
algumas espécies (Sauders et al 1991; Harrison, 1992; Tutin et al. 1997; Gascon et
al . 1999; Debinsk & Holt, 2000).
Diante da complexidade dos efeitos da fragmentação, da particularidade de cada
paisagem fragmentada e da conseqüente necessidade de incluir-se escalas
espaciais amplas, deve priorizar-se estudos sobre fragmentação de habitat na
determinação de seus efeitos, através da comparação de paisagens e dos fatores-
chave da variação na importância destes processos (Davies et al. 2001a; Laurance
et. al. 2001; Pardini, 2001).
Apesar dos avanços obtidos nos últimos anos em pesquisas sobre fragmentação de
habitat, ainda são raros os estudos que descrevam a resposta da biota tropical em
uma escala de paisagem, e estes englobam um número limitado de grupos
biológicos (Turner, 1996).

20
Com algumas exceções, a maior parte do conhecimento sobre as Florestas
Tropicais provêm de estudos na América Central (Estrada et al., 1993, 1994), do
Projeto de Dinâmica Biológica de Fragmentos Florestais na Amazônia brasileira
(Bierregaard et al., 1992; Laurance et al. 2001) e, mais recentemente, do Projeto
Resta Una, na Mata Atlântica no sul da Bahia (Mesquita et al. 1999; Pardini, 2001;
Dixo, 2001; Faria, 2002). A relativa escassez de dados tem limitado a capacidade de
prever e diagnosticar com segurança o impacto deste processo em áreas já
fragmentadas ou sujeitas à fragmentação (Gascon et al. 1993).
A Mata Atlântica brasileira é um bom exemplo, tanto em riqueza e grau de
endemismo das Florestas Tropicais, como em situação crítica de redução e
alteração em que se encontram seus remanescentes atuais (resumidos a menos de
8% de sua área original). Segundo comentado em Faria (2002), embora a maior
parte dos centros de pesquisa estejam localizados dentro do domínio da Mata
Atlântica, pouco conhecimento foi gerado sobre os efeitos do processo de
fragmentação nessas áreas, comparando-se fragmentos de diversos tamanhos,
grupos taxonômicos ou considerando os diversos elementos presentes nas
paisagens estudadas.

21
A MATA ATLÂNTICA NO BRASIL Dentre os países chamados de megadiversos em termos de riqueza de espécies, o
Brasil detém cerca de 1,4 milhão de organismos conhecidos pela Ciência (Wilson,
1988; Lewinsohn & Prado, 2002). Essa posição de destaque mundial está aliada à
presença do maior bloco de área verde do planeta - a Amazônia, e da presença de
outros dois biomas importantes, o Cerrado e a Mata Atlântica (CI, 1994; Mittermeyer
et al., 1992; Fonseca, 2004).
Para alguns autores, a distribuição original da Mata Atlântica restringia-se à floresta
densa ao longo do litoral brasileiro. Entretanto, outros estudiosos (Rizzini, 1979;
Eiten, 1983; Coimbra & Câmara, 1996 e Ab’saber, 2003), baseados em critérios
botânicos e fitofisionômicos, cruzados com considerações de natureza geológica e
geográfica, sugerem que o domínio da Mata Atlântica engloba um conjunto de
tipologias vegetais, incluindo a Floresta Ombrófila Densa, Floresta Estacional
Semidecidual, Floresta Estacional, ecossistemas associados como restingas,
manguezais, florestas costeiras, campos de altitude e encraves de campos, brejos de
altitude e cerrados, ocupando uma área aproximada de 1.300 mil quilômetros
quadrados, o equivalente a cerca de 15% do território brasileiro. Sua região de
ocorrência original abrangia integral ou parcialmente dezessete estados brasileiros,
do Rio Grande do Norte ao Rio Grande do Sul, sendo atualmente ocupada por 70%
da população brasileira (Figura 1).
Fatores como a grande variação latitudinal (de 5o a 25o de Latitude Sul), a variação
de altitudes (desde o nível do mar até mais de 1.000 metros acima deste), os
contatos temporários com a Floresta Amazônica durante períodos interglaciais,
contribuiram para que a Mata Atlântica seja um dos biomas de maior diversidade
biológica do planeta (Coimbra & Câmara, 1996; Almeida, 2000; Ab’saber, 2003). Tal
posição de destaque fundamenta-se na ocorrência de alta riqueza biológica aliada
aos significativos níveis de endemismo da sua fauna e flora (Fonseca, 1997) e no
elevado grau de fragmentação dos ecossistemas florestais remanescentes, reduzidos
a menos de 7% de sua extensão original (Fonseca, 1985; INPE, 2002; SOS Mata
Atlântica, 2004).

22

23
Em conjunto, os mamíferos, aves, répteis e anfíbios existentes na Mata Atlântica
somam em torno de 1.809 espécies, sendo 389 endêmicas. Além disso, a Mata
Atlântica abriga cerca de 7% das espécies conhecidas no mundo nesses grupos de
vertebrados (Ministério do Meio Ambiente, 2000).
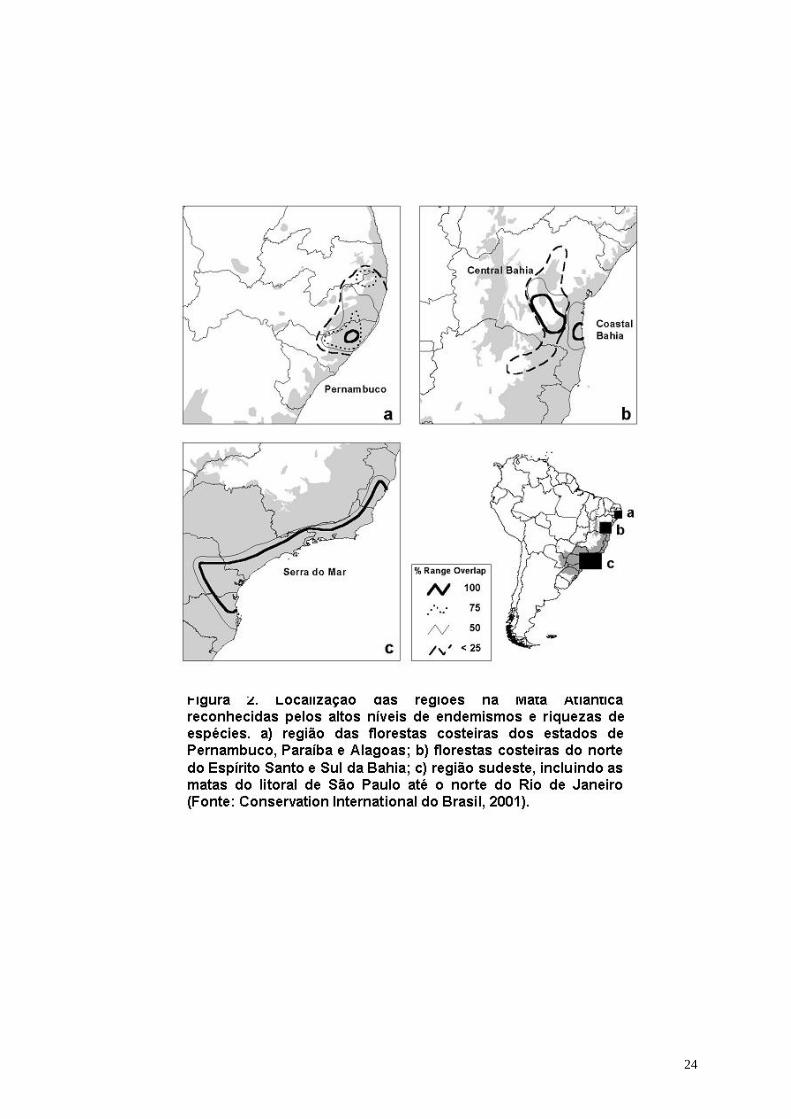
Espacialmente, pelo menos três regiões distintas são reconhecidas pelos altos níveis
de endemismos para vários grupos biológicos na Mata Atlântica (Thomas et al.
1998). Estas regiões foram delimitadas a partir da sobreposição de mapas em
relação a distribuição das espécies (Fonseca et al.). Um desses bolsões localiza-se
na região Sudeste, incluindo as matas do litoral de São Paulo ao Norte do Rio de
Janeiro e estendendo-se ainda em parte do extremo Sul do Espírito Santo. Outros
dois bolsões situam-se na região Nordeste: um, nas Florestas Costeiras dos estados
de Pernambuco, Paraíba e Alagoas, hoje restritos às áreas de brejos ou topos de
morros; e o outro localiza-se mais ao Sul, incluindo as Florestas Costeiras do Norte
do Espírito Santo e Sul da Bahia (Figura 2).
Este último bolsão de endemismo, localizado precisamente entre a Foz do Rio Doce
(ES) e o Sudeste da Bahia, forma uma faixa de Floresta Pluvial do complexo
atlântico com características florísticas e faunísticas peculiares, denominada Hiléia
Baiana (Coimbra & Câmara, 1996; Almeida, 2000; Ab’saber, 2003).
Esta região representa um dos principais centros de endemismo da Mata Atlântica
conforme descrição de plantas (Mori et al. 1981, Prance, 1982), borboletas (Brown,
1991), répteis (Jackson, 1978), aves (Haffer, 1974) e mamíferos (Rylands, 1982).
Detém ainda dois dos maiores recordes de diversidade botânica em todo o mundo: a
região serrana do Espírito Santo (Thomas & Monteiro, 1997) e a floresta próxima ao
Parque Estadual da Serra do Conduru, onde foi detectada a ocorrência de 458
espécies diferentes de plantas lenhosas por hectare (Carvalho & Thomas 1993;
Thomas et al., 1998).
Identificou-se mais de 50% espécies de aves endêmicas da Mata Atlântica, como o
gênero endêmico monotípico recentemente descrito - o graveteiro acrobata
(Acrobatornis fonsecai), e 60% das espécies endêmicas de primatas do bioma,
como é o caso do mico-leão-de-cara-dourada (Leontopithecus chrysomelas) e o
macaco-prego-do-peito-amarelo (Cebus xanthosternos).
24

25
HISTÓRICO DE DESTRUIÇÃO DA MATA ATLÂNTICA NA BAHIA Apesar de sua extraordinária biodiversidade, a Mata Atlântica presente da Bahia
sofreu desmatamento desde o período colonial, resumindo-se a poucos
remanescentes existentes. Dos seus iniciais 20.354.548 hectares (o equivalente a
quase 36% do território baiano), restam apenas 1.263.175 hectares, e isso
representa somente cerca de 6% de sua cobertura original (Ministério do Meio
Ambiente, 2000; Fundação SOS Mata Atlântica, 2004).
Como o restante da Mata Atlântica, o Sudeste baiano apresenta um histórico de
degradação que se iniciou na época do Descobrimento do Brasil, com a exploração
de toras de Pau-Brasil (Caesalphinia echinata) para abastecimento do mercado
europeu, cujo interesse comercial na indústria têxtil da espécie decaiu apenas por
volta de 1560, com o advento das anilinas (Dean, 1998).
A degradação teve continuidade na década de 50, com a exploração seletiva de
árvores nobres, dentre elas o Jacarandá da Bahia (Dalbergia nigra). Esta fase
caracterizou-se pela presença de diversas serrarias implantadas na região do
extremo Sul. No período de 1970 a 1980, ocorreu um outro grande ciclo de
destruição, iniciado a partir da construção da BR 101, o que propiciou a instalação de
pólos madeireiros em vários pontos às margens dessa rodovia (Coimbra-Filho &
Câmara, 1996; Dean, 1998). Foram instaladas diversas indústrias de madeiras e
serrarias, aumentando o desmatamento predatório. Após esta fase, houve
procedimentos de queimadas do solo e o seu subseqüente uso pela pecuária
extensiva agravou o estado de degradação. Em decorrência disso, as áreas
anteriormente florestadas foram substituídas por pastagens destinadas à criação de
gado de baixa produtividade (1 cabeça de gado / hectare) e, mais recentemente, por
florestas de plantação da monocultura de eucalipto (Figura 3).

Ao contrário do ocorrido no extremo Sul, até a década de 90 a fisionomia da
paisagem do Sul da Bahia ainda era a Floresta, em parte devido às peculiaridades do
cultivo do cacau (Theobroma cacao), principal produto agrícola local desde o século
XVIII. Este cultivo caracteriza-se pela cabruca, um sistema onde ocorre a substituição
dos estratos inferiores da floresta por cacau, com retenção de uma fração do dossel
para sombreamento da cultura. Apesar de causar uma profunda alteração na
composição florística, parte da estrutura da vegetação é mantida, e a plantação se
assemelha a uma floresta simplificada (Alves, 1990).
Com a decadência da lavoura cacaueira através da introdução e expansão do fungo
vassoura-de-bruxa (Crinipellis perniciosa), ocorreu uma considerável redução da
cobertura florestal nesta região. Endividados e sem perspectivas, os proprietários das
grandes fazendas começaram a explorar a madeira das cabrucas e fragmentos
florestais remanescentes, substituindo as lavouras por outros tipos de cultivos e
pastagens para garantir melhor retorno financeiro. Além disso, assentamentos
agrícolas e invasões em áreas de solos de menor produtividade impõem pressões
aos remanescentes florestais da região, acelerando assim o processo de
fragmentação das florestas.
26

27
Após sucessivos ciclos de exploração, evidencia-se que a derrubada da Mata
Atlântica é basicamente um evento da segunda metade do século XX, realizado, em
grande parte, nos últimos cinquenta anos. Na região Sudeste da Bahia,
especificamente, há estimativas da dizimação de 93% da Floresta original (Fundação
SOS Mata Atlântica, 2004), cujos remanescentes naturais apresentam-se espalhados
na paisagem regional como ilhas de florestas inseridas nesse contexto geográfico,
circundadas por grandes extensões de pastagens, áreas agrícolas ou florestas de
eucalipto.
Devido a essa atual situação de degradação e da importância ecológica dos
fragmentos florestais naturais ainda existentes, é evidente a necessidade do
desenvolvimento de estudos fundamentados na determinação de estratégias para
reverter essa fragmentação e conservar os remanescentes presentes.

PERSPECTIVAS DE MANEJO E CONSERVAÇÀO PARA A MATA ATLANTICA DA BAHIA: O PROJETO CORREDORES ECOLÓGICOS E O CORREDOR CENTRAL DA MATA ATLÂNTICA. O enfoque mais utilizado para minimizar os efeitos da fragmentação e isolamento de
florestas consiste no estabelecimento de Unidades de Conservação em áreas
representativas dos habitats naturais, assegurando a preservação de amostras
protegidas da ação do homem. Os ramos da Biologia da Conservação e da Ecologia
de Paisagem indicam que essa estratégia, dissociada de abordagens que visem
assegurar a conservação de extensões mais abrangentes da paisagem natural e
modificada, não irá assegurar a manutenção de comunidades ecologicamente viáveis
no longo prazo (Fonseca, 1997; Lindenmayer et. al., 2002). Dentre as várias
abordagens possíveis, aquela dos “Corredores de Biodiversidade” representa uma
das mais promissoras para um planejamento regional eficaz (Saunders et al., 1991;
Mittermeyer et al., 1999).
Um “Corredor de Biodiversidade” ou “Corredor Ecológico” corresponde a uma rede
de parques, reservas e outras áreas de uso menos intensivo, que são gerenciadas de
maneira integrada a garantir a sobrevivência do maior número possível de espécies
de uma região (Figura 4).
28

29
Segundo Simberloff et al. (1992) há 4 razões para se manter corredores: 1)
aumentar as taxas de imigração; 2) providenciar rotas de movimento para espécies
que apresentam ampla distribuição; 3) diminuir a depressão endogâmica; e 4)
reduzir a estocasticidade demográfica. Mesmo apresentando essas vantagens, tem
sido levantado que os corredores também podem mais rapidamente contribuir para
espalhar doenças e eventos catastróficos como fogos ou doenças exóticas, e se as
populações de uma espécie têm genótipos caracteristicamente diferentes, essas
diferenças podem ser rompidas (Simberloff & Cox, 1995).
A efetividade de um corredor está estreitamente ligada à escala, visto que as
espécies percebem a conectividade de formas distintas. Segundo as escalas
espaciais e temporais os corredores podem ser de 3 tipos (Noss, 1991): 1)
Corredores de faixas de habitat, que conectam partes pequenas de habitats e
próximas, efetivos para espécies pequenas e tolerantes, destinados principalmente
para a conservação da diversidade em escala local. 2) Corredores em mosaico
espacial, que são mais amplos e longos, e abrigam uma maior variedade de
paisagens. Estes contribuem tanto para espécies de borda como de interior,
resultando em um mosaico de reservas e permitem tanto o movimento diário e
sazonal de espécies. 3) Corredores em escala regional, que conectam grandes
extensões de terra, podendo conectar reservas naturais em um contexto de redes
regionais de unidades de conservação (Soulé & Terborgh, 1999).
A decisão pela implantação de corredores ao invés de outras alternativas de manejo
da paisagem, como “stepping stones” ou ilhas de passagem de biodiversidade,
representadas por pequenos bosques entre fragmentos de floresta nativa, utilizados
para possibilitar passagem de polinizadores, dispersores e outros animais, reduzindo
o efeito de isolamento entre os fragmentos, aumentando a heterogeneidade da
paisagem e estimulando movimentos saltitantes de dispersão para muitas espécies
(Valladares-Padua et. al., 2004) dependerá de alguns fatores como: a) influência
positiva no tamanho das populações e na persistência; b) os benefícios em termos
de custo econômico de sua manutenção; e c) na configuração e distribuição natural
das reservas. Os corredores devem ser implantados para reter ou garantir algum
nível de conectividade que naturalmente exista ou existia (Costa & Scariot, 2003).

30
O Projeto Corredores Ecológicos representa um dos componentes do Ministério do
Meio Ambiente. Tem como objetivo principal manter a biodiversidade existente em
grandes áreas de Floresta Tropical ("corredores" ou "bio-regiões"), localizadas nas
regiões de florestas da Amazônia e na Mata Atlântica, através da gestão de uma
paisagem dinâmica sustentada de larga escala, abrigando comunidades e espécies
com processos evolucionários e fluxos gênicos assegurados, sob um regime de
controle social local.
Para atingir esse objetivo são necessárias a criação de áreas protegidas adicionais,
a introdução de estratégias mais adequadas de uso da terra e a restauração de
trechos degradados em áreas-chave (Cracco & Guerrero, 2004).
Mosaicos com múltiplos usos da terra em uma paisagem manejada podem permitir
o movimento de populações por meio de “ligações” entre florestas próximas. Sob
uma perspectiva institucional, a estratégia do Corredor procura melhorar o manejo
de áreas protegidas, criar a capacidade de manejo na região e promover pesquisas
biológicas e sócio-econômicas que ajudem a reduzir a ameaça de extinção de
espécies. As aspirações das comunidades e lideranças locais devem ser levadas em
consideração como elementos-chave na equação da conservação, como garantia de
sustentabilidade em longo prazo de parques e reservas (Fonseca et al. 2004).
No Brasil, a abordagem dos Corredores de Biodiversidade vem sendo utilizada em
diferentes escalas de proteção ambiental, desde a local até a regional, utilizando-se
de métodos que assegurem a seleção criteriosa de porções suficientemente grandes
de ambientes naturais, buscando-se representar diferentes ecossistemas, além de
manter ou incrementar os níveis de conectividade entre as diferentes áreas.
Contudo, é essencial que a interferência humana nas Unidades de Conservação
(entendidas como de proteção integral) ou nos núcleos protegidos de outras
categorias de manejo, seja mantida em níveis mínimos, já que estes representam as
vértebras de sustentação do sistema (MMA, 2000).

31
O componente Corredor Central da Mata Atlântica do Projeto Corredores Ecológicos
é definido como segmentos do bioma Mata Atlântica biologicamente prioritários,
compostos por fragmentos florestais e de áreas naturais, inclusive ecossistemas
aquáticos, agrupados em padrões de dispersão e em áreas protegidas públicas e
privadas, que representam quase a totalidade dos remanescentes florestais
existentes, em geral, sob ameaça de alguma forma de exploração ou mesmo de
desflorestamento (MMA, 2000; Fonseca et al. 2004; Mores, 2004).
Na Bahia, foram propostos critérios biológicos específicos utilizados na seleção dos
corredores para serem incluídos no projeto, como: riqueza de espécies em relação a
biodiversidade regional, diversidade comunitária e de ecossistema, grau de
conectividade existente ao longo do corredor, integridade e tamanho das áreas de
apoio e espécies endêmicas. Posteriormente foram selecionadas áreas inseridas no
território delimitado pelo domínio legal da Floresta Ombrófila Densa, que vai desde a
bacia do rio Jequiriçá até o extremo Sul, no limite com o estado do Espírito Santo.
Essa extensa porção territorial abriga características locais que definem um conjunto
de três ecorregiões distintas, denominadas genericamente como: Baixo Sul, Sul e
Extremo Sul na Bahia (Mores, 2004) Figura 5.
Além da formação da Floresta Ombrófila Densa, também é relevante a diversidade
de ecossistemas presentes, dentre eles mosaicos de manguezais, restingas,
várzeas, brejos e recifes de corais, com extrema significância biológica e em rápido
processo de degradação (Myers et al., 2000). A implementação do Projeto
Corredores na Bahia propiciará a realização de ações para consolidação das
Unidades de Conservação existentes, controle e fiscalização na área do Corredor e
realização de programas de uso público. Todas essas ações serão realizadas com
apoio efetivo dos Órgãos Federais e Estaduais, priorizando assim a segurança
patrimonial de suas áreas naturais e possibilitando a sua consolidação física e a
efetividade na conservação dos ecossistemas presentes.

32
Considerando a área do Corredor Central da Mata Atlântica, uma análise da rede de
Unidades de Conservação dos estados de Bahia e Espírito Santo indica claramente
que o atual sistema não é geográficamente, nem ecologicamente bem distribuído.
No extremo Sul da Bahia encontra-se um dos blocos mais importantes de mata do
Corredor Central, o qual compreende quatro parques nacionais (Descobrimento,
Monte Pascoal, Pau-Brasil, e Abrolhos) e protege cerca de 50.000 hectares de mata
e 90.000 hectares de áreas marinhas (Ayres et al., 1997).
As pequenas bacias hidrográficas protegidas por estes parques nacionais são
extremamente importantes, não só para a biodiversidade da Mata Atlântica, como
também para os recifes de coral e outros ecossistemas marinhos no Banco de
Abrolhos e no Parque Nacional Marinho de Abrolhos, a zona mais rica em recifes de
coral no Atlântico Sul.
Na porção do Corredor no Espírito Santo encontra-se uma das principais seções de
Floresta Ombrófila Densa, na tipologia Mata de Tabuleiros, criada pela ligação da
Reserva Biológica de Sooretama à Reserva Florestal de Linhares, totalizando cerca
de 44.000 hectares (Ayres et al., 1997) IBAMA, 2004).

33

34
MODELO DE GESTÃO DO PROJETO CORREDORES E SUAS AÇÕES PARA A FORMAÇÃO DO CORREDOR CENTRAL DA MATA ATLÂNTICA NA BAHIA.
A nível de estrutura gerencial existe a Unidade de Coordenação Geral do Projeto,
sediada em Brasília, e no âmbito estadual, a Unidade de Coordenação Estadual e
Comitê de Gestão. Este último é formado por instâncias governamentais e não-
governamentais, responsável por deliberar sobre todas as ações do projeto em
instância estadual. Além disso, a execução das atividades previstas envolve
agências governamentais dos três níveis da administração pública (Federal,
Estadual e Municipal), ONGS e outros organismos públicos ou privados (Mores,
2004).
O Projeto é dividido em duas fases, cujos valores totais de investimentos estão na
ordem de vinte e dois milhões de dólares. Na primeira fase está previsto: apoio a
implementação de algumas Unidades de Conservação Estaduais e Federais,
realização de ações de fiscalização conjunta entre os Órgãos Estaduais de Meio
Ambiente, e a elaboração do Plano de Gestão do Corredor.
Quanto as ações de apoio a implementação de algumas Unidades de Conservação
Estaduais e Federais em 2004, conforme relatório de atividades realizadas pelo
Projeto (relatório não publicado) foi realizado: conclusão do diagnóstico que integra
uma das fases de elaboração do Plano de Manejo do Parque Estadual Serra do
Conduru; elaboração do Plano de Manejo da APA do Pratigi; diversos cursos de
capacitação; e oficinas para discussão de Planejamento Estratégico relativo à
Gestão Ambiental Municipal no Corredor Central da Mata Atlântica – Bahia,
Em relação as ações de fiscalização conjunta entre os Órgãos Estaduais de Meio
Ambiente, está previsto o aperfeiçoamento do sistema de fiscalização, com o
objetivo de aprimorar a eficiência e eficácia das ações de controle e fiscalização,
vigilância e monitoramento, com base no Plano Integrado de Fiscalização, Vigilância
e Monitoramento, e já foram realizadas operações de fiscalização nas três sub-
regiões do Corredor na Bahia, sendo que a última ocorreu na região do Baixo Sul.

35
A maior atividade prevista para a primeira fase do projeto e a elaboração do Plano
de Gestão do Corredor Central da Mata Atlântica. Este representará o documento
norteador das ações do Projeto, o qual definirá o zoneamento e as diretrizes para a
sua gestão. Sua elaboração ocorreria em 4 etapas: a) Fase Preparatória, que
consiste no planejamento prévio e consolidação do Plano; b) Fase Interpretativa,
composta pelos componentes “Caracterização Temática e Elaboração de mapas” e
“Elaboração do Cenário da Dinâmica Bioregional”. C) Fase Conjectural Propositiva, consiste na seleção e hierarquização de áreas prioritárias por unidades
de Paisagem e definição do desenho do Corredor; e d) Fase de Integração de Resultados, representa a formulação de instrumentos e mecanismos de
implementação do Corredor.
Segundo Mores (2004), o Projeto concentrou toda a responsabilidade do seu
processo de planejamento na elaboração do Plano de Gestão, envolvendo em uma
única atividade toda a responsabilidade de concepção e articulação dos critérios
biológicos, sociais, econômicos, culturais e políticos do Corredor. Assim, como a
segunda fase consiste na implementação do Corredor, caso o Plano e Gestão não
seja criteriosamente elaborado, os objetivos de formação do Corredor Central da
Mata Atlântica poderão estar totalmente comprometidos.
Outra questão a ser abordada, é que o Projeto não definiu desde o seu início o fluxo
de informações entre os membros do Conselho Gestor e os grupos por ele
representados, o que tem gerado deficiências de comunicação durante a concepção
do Projeto (Mores, 2004). Também é sugerida precaução para não criar expectativas
nas comunidades locais e parceiros com relação a implementação do Corredor.
Apesar das ações já realizadas, e do pouco conhecimento real espera-se a efetiva
implementação da abordagem proposta para a formação dos Corredores de
Biodiversidade, como teoricamente previsto para o Corredor Central da Mata
Atlântica. Para tal é necessário incorporar métodos que assegurem a seleção
criteriosa de porções suficientemente grandes de ambientes naturais, com a
intenção de assegurar diferentes ecossistemas, manter ou incrementar os níveis de
conectividade entre as diferentes áreas e não menosprezar que a “vontade política”
e o papel social são instrumentos fundamentais para a garantia de sucesso deste
processo.

36
REFERÊNCIAS BIBLIOGRÁFICAS ALMEIDA, DANILO SETTE DE. Recuperação Ambiental da Mata Atlântica. 2000. AB’SABER, AZIZ NACIB. 2003. Os Domínios de Natureza no Brasil: Potencialidades
Paisagísticas. ALVES, M. C.. 1990. The Role Cacao Plantations in the Conservation of the Atlantic
Forest of Southen Bahia, Brazil. Master Thesis, University of Florida. Gainesville, Flórida.
AYRES, J.M., FONSECA, G.A.B. DA, RYLANDS, A.B.,QUEIROZ, H.L., PINTO,
L.P.S., MASTERSON, D. & CAVALCANTI, R. (1997). Abordagens Inovadoras para Conservação da Biodiversidade no Brasil: Os Corredores das Florestas Neotropicais. Versão 3.0. PP/G7 - Programa Piloto para a Proteção das Florestas Neotropicais: Projeto Parques e Reservas. Ministério do Meio Ambiente, Recursos Hídricos e da Amazônia Legal (MMA), Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais Renováveis (Ibama), Brasília.
BIERREGAARD, R. O., JR., T. E. LOVEJOY, V. KAPOS, A. A. DOS SANTOS, and
R. W. HUTCHINGS. 1992. The biological dynamics of tropical rain forest fragments. Bioscinence 42: 859-66.
BIERREGAARD, R. O., JR., R.O., LAURANCE, W.F., SITESJR., J.W., et al. 1997.
Key priorities for the suty of fragmented tropical ecossystems. IN: Laurance, W.F. & Bierregaard Jr., R.O. [Eds]. Tropical forest remnants: ecology, management, and conservation of fragmented communities. Chicago, The University of Chicago Press. 515-525.
BIERREGAARD, R. O.,JR., R.O., GASCON, T.E. LOVEJOY, AND A. DOS
SANTOS. 2001. Lessons from Amazonia: The Ecology and Conservation of a Fragmented Forest. Yale University Press.
BROWN, K. S.. 1991. Conservation of Neotropical environments: Insects as
indicators. Em: Collins, N. M. e J. A. Thomas (eds.). Conservation of insects and their natural environments. R. E. S. Symposium Nr. 15. Academic Press, London, 350-404.
CARVALHO, A. M., e W. W. THOMAS. 1993. Projeto Mata Atântica Nordeste:
Estudo fitossociológico de Serra Grande, Uruçuca, Bahia – Brasil. Em: XLIV Congresso Nacional de Botânica, São Luís, Brasil.
COIMBRA-FILHO, A. F., e l. DE G. CÂMARA. 1996. Os limites originais do bioma
Mata Atlântica na região nordeste do Brasil. Fundação Brasileira para a Conservação da Natureza, Rio de Janeiro.
CONNOR, E.F. & E.D. MacCoy. 1979. The statistics and biology of the species-area
relationship. The American Naturalist, 113 (6):791-883.

37
CONSERVATION International do Brasil, Fundação Biodiversitas & Sociedade Nordestina de Ecologia (1994) Workshop “Áreas Prioritárias para Conservação da Mata Atlântica do Nordeste” Pernambuco, 1993. Mapa de Prioridades para Conservação da Mata Atlântica do Nordeste. Belo Horizonte.
COSTA, R. BRITO da. SCARIOT, A. 2003. A fragmentação florestal e seus recursos
genéticos. Fragmentação florestal e alternativas de desenvolvimento rural na Região Centro –Oeste. Campo Grande : UCDB. 3: 53 - 74.
CHUST, G., PRETUS, DUCROT, D., BEDOS, A., DEHARVENG, L., 2003.
Response of Soil Fauna to Landscap Heterogeneity: Determining Optimal Scales for Biodiversity Modeling. Conservation Biology, 17:1712-1723.
CRACCO, M. & GUERRERO, E. 2004. Aplicación del Enfoque Ecosistêmico a la
Gestión de Corredores en America del Sur. Memorias del Taller Regiona, 3 al 5 de junio, UICN. Quito, Ecuador.
CRUZ, C.A.G. & PIMENTA, B.V.S. – 2004 – New Species of Physalaemus Fitzinger,
1826 from Southern Bahia, Brazil (Anura, Leptodactylidae). Universidade Federal do Rio de Janeiro.
DAVIES, K.F., C. GASCON & C.R. MARGULES. 2001a. Habitat fragmentation:
consequences, management and future research priorities. In: Conservation Biology – Research Priorities for the Next Decade (Soulé, M.E. & G.H. Orians, eds.). Island Press, Washington.
DEAN, W.. 1998. A Ferro e Fogo. A História e a Devastação da mata Atlântica
Brasileira. Companhia das Letras, São Paulo, SP. DEBINSK, DIANE M., HOLT, ROBERT D. 1999. A Survey and Overview of Habitat
Fragmentation Experiments. Conservation Biology. 14:342-355. DIXO, M. B. de. 2001. Efeito da Fragmentação da Floresta sobre a comunidade de
sapos e lagartos de serrapilheira no sul da Bahia. Dissertação de mestrado, USP.
EITEN, G. Classificação da Vegetação do Brasil. Brasília: CNPq, 1983. ESTRADA, A., R. COATES-ESTRADA, e D. MERITT Jr.. 1993. Bat species richness
and abundance in tropical rain forest fragments and in agricultural habitats at Los Tuxtlas, mexico. Ecography 16(4): 309-318.
ESTRADA, A., R. COATES-ESTRADA, e D. MERITT Jr.. 1994. Non flyng mammals
and landscape changes in the tropical rain forest region of Los Tuxtlas, México. Ecography 17: 229-241.
FARIA, DEBORAH MARIA., 2002. Comunidade de Morcegos em uma paisagem
fragmentada da Mata Atlântica do Sul da Bahia, Brasil. Universidade Estadual de Campinas, SP. Tese de Doutorado.

38
FONSECA, G. A. B., HERRMANN, G., & LEITE, Y. L. R. no prelo. Macrogeography of Brazilian mammals. In: Eisenberg, J. F. (ed) Mammals of the Neotropics (Vol. III), University of Cicago. Press, Cicago.
FONSECA, G. A. B. 1985. The vanishing Brazilian Atlantic forest. Biological
Conservation 34: 17-34. FONSECA, G. A. B. 1997. Biodiversidade e impactos antrópicos. Em J. A. de Paula
(Org.) Biodiversidade, População e Economia: Uma região de Mata Atlântica. UFMG, Belo Horizonte, M. G.
FONSECA, GUSTAVO A. B., ALGER, K., PINTO, L. PAULO, ARAÚJO, M. E
CAVALCANTI, R. 2004. Corredores de Biodiversidade: O Corredor Central da Mata Atlântica. In: Corredores Ecológicos: uma abordagem integradora de ecossistemas no Brasil. Brasília; IBAMA. 2: 47-66.
GASCON, C. 1993. Breeding – habitat use by five Amazonian frogs at forest edges.
Biodiversity and Conservation, 2: 438-444. GASCON, C., T.E. LOVEJOY, R. O. BIERREGAARD, J. R. MALCOLM, P.C.
STTOUFER, H. L. VASCONCELOS, W. F. LAURANCE, B. ZIMMERMAN, M. TOCHER, E S. BORGES. 1999. Matrix habitat and species richness in tropical Forest remmants. Biological Conservation 91:223-229.
HAILA, Y. 2002. A conceptual genealogy of fragmentation research: from island
biogeography to landscape ecology. Ecological Applications 12: 321-334. HANSKI, I. 1989. Metapopulation dynamics: does it help to have more of the same?
Trends on Ecology and Evolution 4: 113-114. HARRISON, R. L. 1992. Towards theory of inter-refuge corridor design. Conservation
Biology 6: 293-95. HARRISON, S. 1994. Metapopulations and conservation. In P. J. Edwards, R.M. May
e N. R. Webb, Large-Scale Ecology and Conservation Biology, pp.111-128. Blackwell Scientific Publications, Oxford.
HARRISON, S. & BRUNA, E. 1999– Habitat fragmentation and large scale
conservation: what do we know for sure. Ecography 22: 225-232. HAFFER, J.. 1974. Avian speciation in tropical South America. Publication of the
Nuttall Ornithological Club 14: 1-390. HENLE, K.,P. POSCHLOD, C. MARGULES & J. SETTELE.1996. Species survival in
relation to habitat quality, size and isolation: summary conclusions and future directions. In: Species survival in fragment landscapes. (Settle, J.,C. Margules, & P. Poschlod, eds.).pp. 373-381. Kluwer Academic Publishers, Dordrecht.
HOKIT, D.G.; STITH, B.M. & BRANCH, L.C. – Comparison of two types of
metapopulation models in real and articifial landscapes – 2000 – University of Florida

39
HUENNEKE, LAURA F., HOBBS, RICHARD J. 1992. Disturbance, Diversity, and Invasion: Implications for Conservation. Conservation Biology 6: 324-333.
IBAMA. Corredores Ecológicos 2004. Uma Abordagem Integradora de Ecossistemas
no Brasil / Moacir Bueno Arruda, Luís Fernando S. Nogueira de Sá. INPE. 2002. Evolução dos Remanescentes Florestais e Ecossistemas Associados do
Domínio da Mata Atlântica – no período de 1985-1990. Ed. Fundação Mata Atlântica 46p.
JACKSON, J. F.. 1978. Differentiation in the genera Enyalius and Strobilurus
(Iguanidae): Implications for Pleistocene climatic changes in eastern Brazil. Arquivos de Zoologia 30: 1-79.
JANZEN, D. H.. 1983. No park is in an island: increase in interference from outside
as park size decreases. Oikos. 41: 402-410. JAX, K & ROZZI, R. Ecological theory and values in the determination of
conservation goals examples for temperate regions of Germany, United States of America, and Chile. 2004. Revista Chilena de Historia Natural 77: 349-366.
JOLY, PIERRE, MIAUD, CLAUDE, LEHMANN, ANTHONY, GROLET, ODILE. 2000.
Conservation Biology, 15:239-248. LAMBERSON, R. H., R. McElvey, B. R. Noon e C. Voss. 1992. A dynamic analysis of
Northern Spotted Owl viability in a fragmented forest landscape. Conservation Biology 6: 143-158.
LAURANCE, W. F..1990. Comparative response of five arboreal marsupials to
tropical forest fragmentation. Journal of Mammalogy 71(4): 641-653. LAURANCE, W.F. 1991. Ecological correlates of extinction proneness in Australian
tropical rain forest mammals. Conservation Biology, 5(1): 79-89. LAURANCE, W. F.. 1991a. Ecological correlates of extinction proneness in
Australian tropical rainforest mammals. Conservation Biology 5: 79-89. LAURANCE, WILLIAM F., LIVEJOY, THOMAS E., VASCONCELOS, HERALDO L.,
BRUNA, EMILIO M., DIDHAM, RAPHAEL K., STOUFFER, C. PHILIP, GASCON, CLAUDE, BIERREGAAD, O. RICHARD, LAURANCE, SUSAN G., SAMPAIO, ERICA. 2001. Ecosystem Decay of Amazonian Forest Fragments: a 22-Year Investigation. Conservation Biology 16:605-618.
LAURANCE, W. F.. 1997. Hyper – disturbed parks: edge effects and the ecology of
isolated rainforest reserves in tropical Australia. Em: Laurance, W. F. and Bierregaard Jr., R. O. (eds.), Tropical Forest Remnants: Ecology, Management, and Conservation of Fragmented Communities. University of Chicago Press, Chicago, 71-83.
LEWIN, R. 1986. A mass extintction withour asteroids. Science 234: 14-15.

40
LEWINSOHN, T. M. & PRADO, P.,I. 2002. Biodiversidade Brasileira: síntese do estado atual do conhecimento. Ministério do Meio Ambiente e Conservation International do Brasil. Contexto, São Paulo.176p.
LINDENMAYER, D.B. et al. – Tha Focal-Species Approach and Landscape
Restoration: a Critique. 2002 LOPES, J.E. & PFISTER, C. – Local Population Dynamics in Metapopulation Models:
Implications for Conservation – 2001 – University of`Chicago. LOVEJOY, T. E.; R. O. Bierregaard Jr.; A. B. RYLANDS; J. R. MALCOLM; C. E.
QUINTELA; L. H. HARPER; K. S. BROWN JR.; A. H. POWELL; H. O. R. SCHUBART, E M. B. HAYS. 1986. Edge and others effects of isolation on Amazonian forest fragments. Em: M.E. Soulé (editor). Conservation biology. The science of scarcity and diversity. Sinauer Associates, Inc. Sunderland, Massachusetts, 237-256.
MACARTHUR R.H. & Wilson E.O. 1967. The theory of island biogeography Princeton University Press, Princeton, New Jersey. MALCOLM, J. R.. 1997. Biomass and diversity of small mammals in Amazonian
forest fragments. Em: Laurence, W. F., Bierregaard Jr., R. O. (Eds), Tropical Forest Remmants: Ecology, Management, and Conservation of Fragmented Communities. University of Chicago Press, Chicago, 207-221.
MESQUITA, R. C. G., P. Delamonica, e W. F. LAURANCE. 1999. Effects of matrix
type on edge related tree mortality in Amazonian Forest fragments. Biological Conservation 91:129-134.
METZGER, J.P. 1999. Estrutura da paisagem e fragmentação: análise bibliográfica.
Anais da Academia Brasileira de Ciências, 71: 445-463. MILLER, J. R. & HOBBS, R. J. – Conservation where people live and work – 2002 -
University of Wisconsin, Madison, USA MINISTÉRIO DO MEIO AMBIENTE. BRASÍLIA 2000. Avaliação e Ações
Prioritárias Para a Conservação da Biodversidade da Mata Atlântica e Campos Sulinos.
MITTERMEIER, R. A., M. SCHWARZ, J. M. AYRES. 1992. A new species of
marmoset, genus Callithrix Erxleblen 1777 (Callitrichidae, primates), from the Rio Maués region, state of Amazonas, Central Brazilizm Amazonia. Goeldiana Zoologia (14): 1-17.
MITTERMEIER, R. A., P. ROBLES GIL, and C.G. MITTERMIER (eds). 1997.
Megadiversity: Earth´s Biologically Wealthiest nations CEMEX, México. MITTERMEIER, R. A., MYERS, P. ROBLES GIL, and C. G. MITTERMEIER. 1999.
Hotspots: Earth’s Biologically Rich-est and Most Endangered Terrestrial Ecoregions. CEMEX, México.

41
MORES, M. 2004. O Projeto Corredores Ecológicos e o desenvolvimento de novos cenários para a conservação da biodiversidade brasileira: estudo de caso para o Corredor Central da Mata Atlântica. In: Aplicación del Enfoque Ecosistêmico a la Gestión de Corredores en America del Sur. Memorias del Taller Regiona, 3 al 5 de junio, UICN. Quito, Ecuador. P. 26-30.
MORI, S. A.,B. M.BOOM, and G. T. PRANCE. 1981. Distribution patterns and
Conservation of eastern Brazilian Coastal Forest Species. Brittonia 33: 233-45. MYERS, N., MITTERMEIER, R.A.; MITTERMEIER,C.G.; FONSECA, G.A.B. &
KENT, J. (2000). Biodiversity hotspots for conservation priorities. Nature 403: 853-858.
NOSS, R. F. 1991. Landscape connectivity: Different functions at different scales. In:
HUDSON, W. E. (Ed.). Landscape linkages and biodiversity. Washington, D. C., Island Press.
PARDINI, RENATA. 2001. Pequenos mamíferos e a fragmentação da Mata Atlântica
de Una, Sul da Bahia – Processos e Conservação. PARDINI, RENATA 2003 – Effects of Forest fragmentation on small mammals in an
Atlantic Forest landscape – Universidade de São Paulo. PERFECTO, IVETTE, VANDERMEER, JOHN. 2002. Quality of Agroecological
Matrix in a Tropical Montane e landscape: Ants in Coffee Plantations in Southern Mexico. Conservation Biology 16: 174-182.
PHILPOTT, STACY M., DIETSCH, THOMAS. 2003. Coffee and Conservation: a
Global Context and the Value of Farmer Involvement. Conservation Biology 17:1844-1846.
PRANCE, G. T. 1982. Forest refuges: Evidence from woody angiosperms. Em G. T.
Prance (ed) Biological Diversification in the Tropics. Columbia University Press, New York.
PRESTON, F. W.. 1962. The canonical distribution of commonness and rarity: part I.
Ecology 48: 185 – 215; part II. 43:410-433. PRIMACK, RICHARD B. 2001. Biologia da Conservação / Richard B. Primack,
Efraim Rodrigues. – Londrina: E. Rodrigues. Vii, 328p. RICH, ADAM C., DOBKIN, DAVID S., NILES, LAWRENCE J. 1994. Defining Forest
Fragmentation by Corridor Width: The Influence of Narrow Forest-Dividing Corridors on Forest-Nesting Birds in Southern New Jersey. Conservation Biology 8: 1109-1121.
RYLANDS, A. B.. 1982. The behaviour and ecology of three species of marmosets
and tamarins (Callitrichdae, Primates) in Brasil. Tese de Doutorado. University of Cambridge, Cambridge, UK.
RIZZINI, C. T. 1979. Tratado de fitogeografia do Brasil: aspectos sociológicos e
florísticos. Editora da Universidade de São Paulo, São Paulo, Brasil. 374pp.

42
ROLIM, S.G. & CHIARELLO, A.G. Slow death of Atlantic forest trees in cocoa
agroforestry in southeastern Brazil Biodiversity and Conservation 13: 2679-2694-2004
SAUNDERS, D. A., R. J. HOBBS, C. R. MARGULES. 1991. Biological
consequences of ecosystem fragmentation: a review. Conservation Biology 7:18-32.
SCHOENER, T. W.. 1976. The species-area relation within archipelagos: models
and evidence from island land birds. Em: H. J. Frith and J. H. Calaby (eds.), Proceedings of the 61th international Ornithological Congress, Canberra, Australia, 12 – 17 August 1974 1974, pp. 629-642. Australian Academy of Science. Griffin Press Ltd., Netley.
SHAFER, C. L. 1990. Nature reserves: island theory and conservation practice.
Smithsonian Institution Press, Washington, DC. SIMBERLOFF, D.; J.COX. 1995. Consequences and costs of conservation corridors.
In: D. Eherenfeld (Ed.). The Landscape perspetive. The Society for Conservation Biology and Blackwell Science, Inc. p. 44-52.
SIMBERLOFF, D. J.A.FARR; J. COX; DW. MEHLMAN. 1992. Movement corridors:
conservation bargains or poor investments. Conservation biology, v.6, p. 493-504.
SOS MATA ATLÂNTICA E INPE. 2004. Atlas da Evolução dos Remanescentes
Florestais da Mata Atlântica. Fundação SOS Mata Atlântica e Instituto Nacional de Pesquisas Espaciais, São Paulo, SP.
SOULÉ, M. E.; J. TERBORGH. 1999. Continental Conservation: scientific
foundatiions of regional reserva networks. Washington, D. C., Island Press. THOMAS, W.M.W., A.M.V., Carvalho, A.M.A. AMORIM, J. GARRISON. & A.L.
ARBELÁEZ. 1998. Plant endemism in two forests in southern Bahia, Brasil. Biodversity and Conservation, 7:311-322.
THOMAZ, L.D. & MONTEIRO, R. (1997). Composição florística da Mata Atlântica de
encosta da Estação Biológica de Santa Lúcia, município de Santa Teresa,ES. Bol. Mus. Biol. Mello Leitao (N. Ser.), 7: 3-48.
TURNER, I. M.. 1996. Species loss in fragments of tropical rain Forest: a review
of the evidence. Journal of Applied Ecology 33:200-209. TUTIN, C. E. G., WHITE, L. J. T., MISSANDZOU, MACKANGA. 1997. The Use by
Rain Forest Mammals of Natural Forest Fragments in an Equatorial African Savanna. Conservation Biology, 11:1190-1203.

43
VALLADARES-PÁDUA, C.; CULLEN,L.JR. 2004. Combinando comunidade, conectividade e biodiversidade na restauração da paisagem do Pontal do Paranapanema como estratégia de Conservação do Corredor do Rio Paraná. In: Corredores Ecológicos: uma abordagem integradora de ecossistemas no Brasil. Brasília; IBAMA. 3: 67-80.
WILLIAMS, C. B.. 1943. Área and number of species. Nature 152:264-267. WILSON, E. O.,EDITOR; PETER, FRANCES M., 1997. Biodiversidade. WILSON, E. O.. 1988. The current state of biodiversity. Em E. O.Wilson (ed).
Biodiversity. Academic Press, Washington, DC WHITMORE, T. C. 1997. Tropical forest disturbance, disappearance and species
loss. In: Tropical forest remanants – ecology, management, and conservation of fragmented communities. (Laurance, w.f. & R.O. Biorregaard, eds.). pp. 3-12. The University of Chicago Press, Chicago.

CAPÍTULO 2
INFLUÊNCIA DA FRAGMENTAÇAO FLORESTAL E DA QUALIDADE DA MATRIZ DE MONOCULTURA DE EUCALIPTO SOBRE A COMPOSIÇÃO DAS COMUNIDADES DE LAGARTOS DE FOLHIÇO EM ÁREAS DE MATA ATLÂNTICA NO EXTREMO SUL DA BAHIA.
44

45
RESUMO
No extremo Sul da Bahia o processo histórico de destruição da Mata Atlântica
acarretou uma situação em que pastagens e a monocultura de eucalipto representam
os componentes dominantes da paisagem, onde os principais remanescentes
naturais existentes encontram-se isolados e imersos nessas matrizes. Ao contrário do
que acontece em outras áreas, os eucaliptais da região desenvolvem um sub-bosque
de espécies nativas que poderia ser permeável a estas espécies, promovendo a
conectividade entre os remanescentes florestais existentes. Este estudo objetivou
comparar a composição e biomassa total da comunidade de lagartos de serrapilheira
em três componentes florestados da paisagem regional, através da implantação de
432 armadilhas de queda pitfall traps. Foram realizadas campanhas amostrais na: a)
RPPN Estação Veracruz, mata de grande porte considerada como sistema de
referência; b) Remanescentes de pequeno porte em estágio intermediário de
regeneração (os mais comuns na paisagem); e c) Eucaliptais em estágio final de
crescimento (sete anos de idade), com o intuito de discutir os efeitos da fragmentação
e a qualidade da matriz de eucalipto na promoção de conectividade entre os
fragmentos. Avaliou-se também a existência de associação com variáveis ambientais
que expressam características de micro-habitat, micro-clima e Ecologia da Paisagem.
No total dos 40 dias de coleta, foram capturados 206 lagartos de folhiço,
correspondentes a doze espécies, pertencentes a sete famílias diferentes: Anguidae,
Gekkonidae, Gymnophthalmidae, Polychrotidae, Teiidae, Tropiduridae e Scincidae,
onde Gymnodactylus darwinii, Leposoma scincoides e Enyalius catenatus picus foram
as espécies mais dominantes. De acordo com os resultados obtidos, não foi possível
detectar a interferência negativa da fragmentação e de seus processos associados na
composição das comunidades de lagartos de folhiço nas áreas amostradas. Do ponto
de vista da biomassa total, foram identificadas diferenças no padrão de distribuição
entre os Eucaliptais e a Mata Contínua, associadas às variáveis ambientais como:
distância em relação as florestas nativas, aumento da temperatura e umidade do ar,
diminuição da altura de folhiço e densidade de folhagem nos estratos superiores da
floresta. A análise de ordenação com as variáveis ambientais apontou a existência de
gradientes entre a Mata Contínua, os Remanescentes e os Eucaliptais, porém não foi
identificada uma influência negativa da matriz sobre a comunidade de lagartos de
folhiço nas áreas estudadas.

46
ABSTRACT
In the extreme south of Bahia, the local historic process of destruction of the Atlantic
Forest has led to a current situation in which pastures and the monoculture of
eucalyptus represent the dominant components of the landscape, where the main
existing native forest remnants are found isolated and immersed in this matrix.
Contrary to what happens in eucalyptus plantations in other regions, an undergrowth
of native species is found here, which could be permeable to native species,
promoting a connectivity between the existing forest remnants. This study aims at
comparing the composition and total biomass of the litter lizard community in three
forested components of the regional landscape, through the setting up of 432 pitfall
traps. Sample campaigns were done at: a) the Natural Heritage Reserve - RPPN
Estação Veracruz, a well established native forest that is considered a reference
system; b) small native forest remnants in intermediary stage of regeneration (the
most common remnants landscape); and c) eucalyptus plantations, exotic forests in
final stage of growth before harvesting (7 years old); with the intention of discussing
the effects of fragmentation and a matrix quality of eucalyptus to promote connectivity
of the native forest fragments. The existence of an association amongst variable
environments that express micro-habitat, micro-climate and ecology landscape was
also evaluated. In a total of 40 days of sampled, 206 lizards were surveyed, of which
12 species were belonging to 7 different families: Anguidae, Gekkonidae,
Gymnophthalmidae, Polychrotidae, Teiidae, Tropiduridae and Scincidae.
Gymnodactylus darwinii, Leposoma scincoides and Enyalius catenatus picus were the
dominant species. According to the obtained results, it was not possible to detect that
fragmentation and its associated processes were interfering negatively on the
composition of the lizard communities in the sample areas. From the total biomass
point of view, there were differences in the distribution standard between eucalyptus
plantations and native continuous forests, associated with environmental variables,
such as: distance in relation to native forests, increase in temperature and air humidity
and diminution of the litter height and density of foliage in the superior stratus of the
forest. Ordination analysis with environmental variables pointed to existing
environmental gradients between continuous native forests, remnants of these and
eucalyptus plantations; however, a negative influence of the eucalyptus matrix, on the
studied community, was not identified.

47
INTRODUÇÃO
O bioma Mata Atlântica é reconhecido internacionalmente como prioritário para a
conservação da biodiversidade em todo o continente americano (CI, WWF, 1995),
cuja importância baseia-se na ocorrência de alta riqueza biológica, aliada aos
significativos níveis de endemismo da sua fauna e flora (Fonseca, 1997) e no
elevado grau de fragmentação dos ecossistemas florestais remanescentes,
reduzidos a menos de 7% de sua extensão original (Fonseca, 1985; SOS Mata
Atlântica e INPE, 2004).
No Brasil, a sub-região da Mata Atlântica no Sul da Bahia, em especial a do extremo
Sul, constitui um dos mais importantes centros de endemismos de todo o bioma, onde
atualmente, apesar de sua extraordinária biodiversidade, essa porção resume-se a
poucos remanescentes. Após ter sofrido sucessivos ciclos de destruição (Brown,
1992) as áreas anteriormente florestadas foram substituídas por pastagens
destinadas a criação de gado extensiva de baixa produtividade (1 cabeça de
gado/hectare) e, mais recentemente, em florestas plantadas com monocultura de
eucalipto.
De acordo com análises recentes de imagens de satélite sobre a região, (Veracel
Celulose 2002), existem apenas cerca de 4% de remanescentes florestais naturais
espalhados na paisagem regional, cujos principais fragmentos apresentam-se como
ilhas de florestas inseridas nesse contexto geográfico, circundadas por grandes
extensões de pastagens ou intercaladas em florestas de eucalipto, evidenciando uma
paisagem em mosaico.
Devido à atual situação de degradação e à importância ecológica dos fragmentos
florestais naturais ainda existentes, é evidente a necessidade de desenvolver-se
estudos fundamentados na determinação de estratégias que priorizem a reversão de
tal fragmentação e a conservação dos remanescentes presentes (Vide Capítulo 1).
O enfoque mais utilizado para minimizar esses efeitos em ecossistemas naturais
busca o estabelecimento de Unidades de Conservação em áreas representativas dos
habitats naturais, assegurando a preservação de amostras protegidas da ação do
homem. Por outro lado, os ramos da Biologia da Conservação e da Ecologia de

48
Paisagem evidenciam que essa estratégia, dissociada de abordagens que visem
compreender e assegurar a dinâmica dos principais processos biológicos existentes,
não proporcionará a garantia da manutenção de comunidades ecologicamente viáveis
a longo prazo (Fonseca, 1997).
A tendência à progressiva insularização dos ambientes naturais, particularmente em
ecossistemas florestais, indica que o cenário mais provável no médio prazo é a
formação de “ilhas” de matas, teoricamente preservadas (através de Unidades de
Conservação ou por determinação da Legislação Ambiental vigente), circundadas por
um “oceano” de ambientes inóspitos para os seus componentes florísticos e
faunísticos.
Os fragmentos a serem protegidos são invariavelmente reduzidos em tamanho –
devido à necessidade da manutenção de populações mínimas viáveis daquelas
espécies com um maior requerimento de área - além de sofrerem uma deterioração
progressiva de seus ambientes a partir das bordas, gerando uma reação em cadeia
que ocasiona a erosão da diversidade biológica original (Laurence et al., 1997a;
Laurence et al., 1997b).
Alguns estudos foram desenvolvidos visando elucidar os efeitos da insularização
sobre as comunidades ecológicas de área florestadas. Muitos, porém, não levaram
em consideração as matrizes em que estão inseridos os remanescentes e a
permeabilidade das mesmas às diferentes espécies de uma comunidade (Noss &
Csuti, 1994; Dixo, 2001).
Trabalhos recentes têm demonstrado que a capacidade de dispersão através da
matriz é um dos fatores determinantes na persistência de algumas espécies em áreas
fragmentadas (Laurance & Bierregaard, 1997; Gascon et al., 1999 e Dixo, 2001).
Porém grande parte das informações sobre os efeitos da fragmentação e do uso da
terra em florestas neotropicais é proveniente de estudos na América Central (Estrada
et al., 1993, 1994), na Região Amazônica (Bierregaard et al., 1992) e mais
recentemente, na região de UNA, Sul da Bahia (Dixo, 2001; Pardini, 2001; Faria,
2002).

49
No extremo Sul da Bahia, o processo histórico de destruição da Mata Atlântica levou
a uma situação em que pastagens e a monocultura de eucalipto representam os
componentes dominantes da paisagem. Porém, pouco conhecimento empírico foi
obtido no tocante à utilização das espécies locais quanto à tolerância às alterações
ambientais desta região. A silvicultura geralmente é reconhecida como um ambiente
inóspito para a fauna, mas os impactos gerados pelas práticas florestais intensivas
ainda causam controvérsias (Spellerberg & Sawyer, 1996). Sabe-se que a baixa
diversidade descrita em tais ambientes era atribuída à homogeneidade espacial e
estrutural, o que resulta, em última instância, em pequena disponibilidade de
recursos.
Ao contrário do que acontece em outras áreas, no extremo Sul da Bahia os
Eucaliptais desenvolvem um sub-bosque de espécies nativas, em função do
espaçamento entre mudas utilizado pelas empresas florestais, da alta disponibilidade
de luz, dos altos índices pluviométricos e da não - remoção do banco de sementes
naturais do solo. Essa situação poderia amenizar as condições ambientais da
monocultura, tornando-a mais permeável às espécies nativas de modo a promover a
conectividade entre os remanescentes florestais existentes. Contudo, não há,
evidências empíricas sobre a resposta da fauna local a essa situação.
O surgimento do sub-bosque revela-se importante pois, ampliando-se a diversidade
florística da área ocupada pela espécie florestal, há uma garantia de maior
estabilidade ecológica desses sítios, o que é teoricamente benéfico para a fauna
silvestre, devido ao acréscimo de disponibilidade de abrigo, refúgio e fonte de
alimento (Silva &Souza, 1994; Almeida 2000).
Estudos sobre a caracterização da avifauna realizados em propriedades da empresa
Veracel Celulose, no extremo Sul da Bahia, evidenciaram a presença de 23
espécies nos Eucaliptais e 16 espécies nas áreas com pastagem (Sampaio 2001),
sugerindo que Eucaliptais com sub-bosque denso apresentaram maior diversidade
de espécies vegetais e, consequentemente abrigavam um maior número de
espécies de aves.

50
A Veracel Celulose S.A instalou-se na região em 1992, com o propósito de
implantar uma unidade industrial para a fabricação de celulose branqueada de
eucalipto. Segundo a política ambiental desenvolvida pela empresa - através do
Programa Mata Atlântica Veracel - as florestas de eucaliptos ocupam áreas
degradadas e são direcionadas para a criação de condições que visem a melhoria
ambiental na região de influência do empreendimento, o que engloba 70.000 ha com
plantios de eucalipto e aproximadamente 60.000 ha de áreas destinadas à
recuperação e manejo de Mata Atlântica (Consórcio Mata Atlântica, 1992).
Com base em dados obtidos pela empresa a partir de análises georeferenciadas
(Veracel, 2001, dados não publicados), detectou-se que a maioria dos
Remanescentes florestais localizam-se nos vales e foram classificados em estágio
intermediário de sucessão, estando imersos em uma matriz formada
majoritariamente por plantações de eucalipto com a espécie híbrida Eucalyptus
urograndis (Koopmans, 1999).
A produtividade do eucalipto na região é uma das maiores do Globo (Veracel, 2000),
o que tem gerado pressão crescente para a expansão dessa cultura em substituição
a áreas de pasto e de floresta em estágios iniciais de sucessão. De fato, estão aí
instaladas as maiores empresas florestais do mundo (Aracruz Celulose, Veracel
Celulose e Bahia Sul Celulose). Sendo assim, urge a realização de estudos empíricos
que avaliem o estado de conservação dos remanescentes de floresta nativa da região
e a qualidade da matriz formada pelo eucalipto no quesito da promoção da
conectividade entre eles, com o objetivo de propor diretrizes de manejo e
conservação para a biota local (Koopmans, 1999).
Entre os vertebrados terrestres, os lagartos representam um modelo bastante
estudado, de fácil visualização, captura e manuseio, cuja taxonomia é relativamente
bem conhecida (Camp, 1923; Bellairs, 1969). São reconhecidos como um grupo de
grande interesse científico por apresentarem altas diversidades e abundâncias e por
serem mais generalizados que aves e mamíferos, em função de sua ectotermia
(Pianka, 1966; Vanzolini, 1972; Vanzolini, 1974; Vanzolini, 1976; Vanzolini, 1980).

51
Além disso, representam, junto com os anuros, um dos principais grupos taxonômicos
que convertem biomassa de artrópodes terrestres em biomassa disponível aos níveis
tróficos superiores.
Porém, grande parte da bibliografia ecológica de lagartos neotropicais refere-se a
estudos de auto-Ecologia de várias espécies de alguns gêneros, como: Ameiva na
Nicarágua (Vitt & Zanni 1996) e Cnemidophorus na Venezuela (Paulissen & Walker
1994) e na Nicarágua (Vitt et al. 1993). Alguns artigos de cunho faunístico
apresentam também informações sobre aspectos da Ecologia das espécies de
determinadas regiões (p.ex., Iquitos no Peru - Dixon & Soini 1975).
No Brasil, foram estudados, entre outros, Ameiva em caatinga (Vitt 1982a), cerrado
(Colli 1991) e em outras formações (Vitt & Colli 1994); Enyalius em floresta do Pará
(Vitt et al. 1996), Kentropyx em floresta (Vitt 1991) e cerrado (Vitt & Carvalho 1992) de
Rondônia; Mabuya em restinga no Espírito Santo (VRCibradic & Rocha 1996) e em
floresta de Rondônia (Vitt & Blackburn 1991), Polychrus (Vitt & Lacher 1981),
Tropidurus (Vitt & Golberg 1983, Van Sluys 1992, 1993), em restinga do Espírito
Santo (Figueira et al. 1994) e em afloramentos rochosos e cerrados em Roraima e
Pará (Vitt 1993; Vitt et al. 1996). Informações sobre aspectos da biologia de várias
espécies são também apresentadas para os répteis da caatinga em Vanzolini et al.,
1980 e para os lagartos da Amazônia em Avila-Pires, em 1995.
Assim, apesar da existência de inúmeros trabalhos sobre Ecologia de lagartos em
ambientes florestados (Kichener et al.,1980; Smith et al.,1996; Cosson et al., 1999;
Sarre & Meyers, 1995; Summer et al., 1999; Caldwell & Vitt, 1999; Schlaepfer &
Gavin, 2001; Renken et al.,2003), a resposta destes animais à fragmentação e
processos associados ainda é um assunto pouco explorado. Particularmente no
bioma Mata Atlântica, quase nenhum trabalho explorou a resposta desses animais à
redução dos ambientes florestados no contexto da paisagem, o papel das interações
ecológicas sobre a estrutura de suas comunidades e seus mecanismos de adaptação
frente às alterações ambientais (Duellman, 1990; Franco, 1998; Rodrigues, 1990).

52
Em 2001, Dixo realizou um estudo sobre a estrutura de comunidades de lagartos de
folhiço em áreas de Mata Atlântica no Sul da Bahia, na região da Reserva Biológica
de UNA e seu entorno. Foram comparadas abundância, riqueza, diversidade,
dominância e equitatividade de lagartos em seis habitats diferentes (interior e borda
de Mata Contínua; interior e borda de fragmento de mata; cabruca - plantação de
cacau sombreada; e capoeira - Mata Secundária). De acordo com os resultados
obtidos, a riqueza e a diversidade de lagartos não foram significativamente
diferentes entre os diversos habitats. Entretanto, a abundância, a equitatividade e a
dominância de lagartos diferiu entre os tratamentos. Os estudos indicaram que, no
contexto da paisagem estudada, as cabrucas e as capoeiras parecem funcionar
como extensões da floresta.
O presente estudo teve como objetivo comparar a composição e biomassa total da
comunidade de lagartos de serrapilheira entre os três principais componentes
florestados da paisagem do extremo Sul da Bahia: Remanescentes de grande porte
de mata em estágio avançado de regeneração (considerados sistemas de referência),
Remanescentes de pequeno porte em estágio intermediário de regeneração (os mais
comuns na paisagem) e Eucaliptais em estágio final de crescimento (7 anos de
idade). Além disso, foi avaliada a existência de associação entre as duas variáveis
indicadoras da fauna (composição e biomassa) e variáveis ambientais que expressam
características de micro-habitat, micro-clima e Ecologia da Paisagem. Com base nos
resultados, foram discutidos os efeitos da fragmentação sobre esse componente da
fauna e a qualidade da matriz de eucalipto na promoção de conectividade entre os
fragmentos.
Estudos com o mesmo delineamento amostral foram realizados sincronicamente na
região para anuros (Vide Capítulo 3 dessa Dissertação) e artrópodes de serrapilheira
(Tinoco, em preparação), abelhas (Melo, em preparação) e borboletas (Cardoso, em
preparação). Além disso, um estudo com delineamento semelhante foi realizado na
área com o objetivo de comparar a Mata de Referência com Eucaliptais que a
margeiam (Cavalcanti, em preparação).

53
De acordo com os objetivos propostos, pretendeu-se responder às seguintes
perguntas:
1. Quais as principais diferenças na composição das comunidades de lagartos
de folhiço presentes nos fragmentos do tipo mais comum na região
(remanescentes florestais em estágio intermediário de sucessão entre 100 e
250 ha) e a mata original (estimada com base na situação do fragmento de
6.000 ha)?
2. Quais variáveis ambientais e variáveis de paisagem mensuradas estão
correlacionadas a essas diferenças?
3. A monocultura de eucalipto pode ser considerada uma matriz adequada para
conectar pequenos fragmentos de floresta em estágios intermediários de
sucessão?

54
MATERIAL E MÉTODOS
ÁREA DE ESTUDO: EXTREMO SUL DA BAHIA
Este estudo é parte integrante de um projeto maior intitulado “Ecologia de Paisagem
no extremo Sul da Bahia”, realizado em propriedades da empresa Veracel Celulose,
localizadas na macro-região do extremo Sul da Bahia. Na década de 50, esta região
apresentava como principal tipologia a Floresta Ombrófila Densa, em quase 95 % de
toda sua extensão territorial (vide Capítulo 1).
Geograficamente, a macro-região do Extremo Sul da Bahia é limitada ao Norte pelo
Rio Jequitinhonha, ao Sul pela fronteira com o estado do Espírito Santo, ao Leste
pelo Oceano Atlântico e a Oeste pela fronteira com o estado de Minas Gerais. Esta
região possui uma área total de 30.000 Km2 e compreende vinte e um municípios
(Anexo 1).
Segundo levantamentos recentes (SOS Mata Atlântica, 2004), ainda existem na
região cerca de 7% de remanescentes florestais naturais, sendo os maiores núcleos
representados pelas seguintes Unidades de Conservação: Parque Nacional de
Monte Pascoal (14.000 ha); Parque Nacional do Pau Brasil (12.000 ha) e Parque
Nacional do Descobrimento (20.000 ha); RPPN Estação Veracruz/Estação Pau
Brasil (7.300ha); e as APAs de Santo Antônio, Coroa Vermelha e Caraíva /Trancoso
(Anexo 2).
Quanto às características climáticas, a região do Extremo Sul da Bahia se enquadra
no clima Af – chuvoso quente e úmido, característico do litoral e sem estação seca
(segundo a classificação de Köppen) e apresenta temperaturas elevadas, com baixa
amplitude de variação. A temperatura média é de 22,9ºC, sendo a temperatura
média máxima de 27,9ºC e média mínima de 18,9 ºC. A precipitação anual é
elevada, apresentando valores médios de 1.787,6 mm, e bem distribuída durante o
ano, sem déficit hídrico (Veracel Celulose, 1998).

Essa área apresenta topografia caracterizada por extensos platôs relativamente
planos recortados por vales em forma de “v”, associados a córregos, e inclui
remanescentes florestais imersos em uma matriz formada por pastos e amplas
áreas de Eucaliptais, estas últimas restritas aos platôs, formando uma paisagem em
mosaico. Os vales de mais difícil acesso apresentam alguns remanescentes
florestais em diferentes estágios sucessionais (Figura 6).
55

56
A RELEVÂNCIA BIOLÓGICA DA RPPN ESTAÇÃO VERACRUZ
A RPPN Estação Veracruz de propriedade da empresa Veracel Celulose, localizada
nos municípios de Porto Seguro e de Santa Cruz Cabrália, abrange um dos mais
significativos e complexos remanescentes do ecossistema de Floresta Atlântica
existentes no Nordeste do Brasil (Figura 7). Esta reserva foi considerada de alta
prioridade para a conservação da biodiversidade durante a realização do Workshop
sobre Prioridades para a Conservação da Biodiversidade da Mata Atlântica do
Nordeste Brasileiro (CI/FB/SNE. 1994; Ministério do Meio Ambiente, 2000). É
considerada uma das áreas que apresenta a maior concentração de espécies de
aves ameaçadas ou raras da região Neotropical (Collar et al.1992). Também figura
como uma região de especial interesse para a conservação de espécies ameaçadas
de borboletas Papilionideas da região Neotropical (Teixeira, 1995; 1996).
A riqueza de anfíbios e répteis é proverbial. Porto et al. (1995 a, b), identificaram 60
espécies de repteis e 40 espécies de anfíbios, confinados na área de 6.000 ha da
RPPN Estação Veracruz. Estes números são altamente significativos quando
comparados com a composição destes grupos em áreas maiores, intensamente
estudadas, apontando a reserva como uma das áreas de maior diversidade destes
grupos da região Neotropical (Franco,1998).
Devido a estas características singulares, a RPPN Estação Veracruz deve ser
tratada como uma região prioritária para o estabelecimento de estratégias de
conservação de sua biodiversidade e segundo o Plano de Manejo da Reserva
(Veracel Celulose, 2001), destaca-se pelos seguintes fatores:
• É considerada uma das regiões de maior diversidade de aves, répteis e anfíbios
da região Neotropical;
• É um dos dois sítios que protege o maior número de espécies de aves
ameaçadas de extinção da região Neotropical;
• Situa-se como rota de aves migratórias de ambientes florestais, originárias da
América do Norte e da América do Sul;

• Apresenta elevado número de espécies de interesse para a conservação, tais
como: espécies raras, endêmicas e numerosas espécies novas para a Ciência;
• É considerada a maior Reserva Particular de Mata Atlântica do Brasil e maior
Reserva de Mata Atlântica do Nordeste Brasileiro.
• É a reserva brasileira com maior número de espécies de vertebrados ameaçados
de extinção.
• É reconhecida como Sítio do Patrimônio Mundial Natural pela UNESCO.
57

58

59

60
CONFIGURAÇÃO DA PAISAGEM REGIONAL E DELINEAMENTO EXPERIMENTAL DO ESTUDO
A Veracel Celulose S.A, instalou-se no extremo Sul da Bahia em 1992, detendo mais
de 150.000 hectares (ha), os quais incluem florestas de eucaliptos e
aproximadamente 60.000 ha de áreas destinadas à recuperação e manejo de Mata
Atlântica (cerca de 43% de toda a Mata Atlântica da Região).
Com base em dados obtidos pela empresa a partir de análises georeferenciadas em
ortofotocartas de 1:10.000 e fotografias aéreas em escala de 1:32.500 (Veracel,
2002, dados não publicados), detectou-se que a maioria dos remanescentes
florestais estão distribuídos em fragmentos com área superficial variando entre 25 a
2588 ha, dos quais 7% representam florestas primárias - incluindo a RPPN Estação
Veracruz, maior reserva particular de Mata Atlântica do Brasil, com os seus 6069ha -
3% de florestas em estágio avançado de sucessão e 15% em estágio intermediário
(Figura 4). Destes últimos em estágio intermediário, 20 apresentam área maior que
100 ha (entre 102 e 229 ha), a maioria dos quais se encontram imersos em uma
matriz formada majoritariamente por plantações de eucalipto.
Tendo em vista a configuração e os diversos componentes da paisagem presentes
no extremo Sul da Bahia, a distribuição, o status de conservação e o número
reduzido dos principais remanescentes florestais naturais de propriedade da Veracel
Celulose; o reconhecimento da RPPN Estação Veracruz, com um dos últimos
remanescentes florestais de Mata Atlântica mais bem conservados do Brasil; e a
disponibilidade logística viável para a realização de experimentos de campo que
pudessem atender as premissas de replicabilidade e número satisfatório de
unidades amostrais necessárias, como estradas e vias de acesso, foram adotados
nesse estudo os seguintes componentes da paisagem (Anexo 3):
a) Mata de referência RPPN Estação Veracruz (M): em excelente estado de
conservação, trata-se de um dos maiores fragmentos de mata primária da região,
com superfície uma ordem de grandeza maior que os demais remanescentes
existentes. Foi considerada como área representativa de floresta contínua atual,
apresentando 35,4 km de perímetro, 16,9 dos quais em contato com áreas de Mata
Atlântica, 17,5 com áreas de pecuária e 1,0 com monocultura de eucalipto.

61
b) Sua topografia inclui amplos platôs recortados por alguns vales. Metade das
unidades amostrais foram realizadas no interior da reserva (M1 e M2); e as outras
duas foram realizadas próximo ao eucaliptal à margem da floresta contínua (M3 e
M4).
c) Remanescentes florestais (R): dentre remanescentes de Mata Atlântica da
região de propriedade da Veracel Celulose foram escolhidos os remanescentes mais
freqüentes na paisagem. Ou seja: aqueles em estado intermediário de sucessão,
que apresentassem áreas entre 100 e 250 ha, para os quais o perímetro em contato
com áreas de eucalipto representassem pelo menos 75% do perímetro total e que
estivessem localizados no platôs. Diante do seu número reduzido e localização
geográfica viável foram selecionados quatro remanescentes (R1, R2, R3 e R4),
próximos ao Projeto Sucupira II (Eucaliptal).
d) Eucaliptais (E): foram selecionadas quatro áreas de eucaliptais em estágio
avançado de crescimento (plantados há pelo menos sete anos, que já apresentam
sub-bosque desenvolvido, distando pelo menos 2 km da Estação Veracruz e 1 km
de fragmentos de mata com área superior a 100 ha e que apresentassem
diversidade de clones). Com essas escolhas pretendeu-se, por um lado, restringir a
diversidade de situações existentes (estágios de crescimento do eucalipto); e por
outro, caracterizar a variabilidade dos Eucaliptais suficientemente distantes das
fontes faunísticas originais, de modo que se pudesse considerar a fauna amostrada
como residente na monocultura. Os quatro eucaliptais selecionados (E1, E2, E3 e
E4) pertenciam ao Projeto Sucupira II.
Cada um dos três componentes da paisagem (M, R e E) foi caracterizado a partir de
quatro unidades amostrais (quatro réplicas espaciais). Estas foram distribuídas em
regiões de platôs nas áreas do universo de cada componente. Especificamente na
Mata Contínua (M) foram implantadas duas réplicas no interior da reserva (M1 e M2)
e duas na borda (M3 e M4).

62
Uma unidade amostral correspondeu a uma área interna ao componente da
paisagem, com dimensões de cerca de 60m x 100m (0,6ha), cuja área total
amostrada representou uma pequena fração da área total (no caso das áreas de
Mata Atlântica, M: 0,039%; R: 0,016%), indicando um impacto mínimo quanto à
fauna coletada.
Foram realizadas 4 campanhas de dez dias de coleta, percorrendo-se todos os
componentes da paisagem selecionados, entre os meses de fevereiro, março, junho,
julho, setembro, outubro e dezembro de 2003, e janeiro de 2004, períodos que
englobam maior e menor precipitação pluviométrica na região.

1

AMOSTRAGEM DOS LAGARTOS DE FOLHIÇO
Em cada unidade amostral, a captura da fauna de folhiço foi realizada por 36
armadilhas de queda do tipo pitfall traps. Uma armadilha corresponde a um balde de
40 litros (provido de tampa removível) enterrado no solo, associado a duas cercas
de direcionamento de 1,5 m de comprimento por 0,4 metros de altura, construídas
com lona plástica e fixada ao solo por ripas de madeira (Gotelli. et al., 2001; Dantas,
2003). Figura 8.
56

As 36 armadilhas foram distribuídas a cada dez metros em três linhas paralelas
espaçadas entre si por vinte metros. Tendo em vista o delineamento experimental
proposto nesse estudo, foram instaladas um total de 432 armadilhas (36
armadilhas/unidade amostral X 4 réplicas x 3 componentes da paisagem), as quais
eram vistoriadas diariamente durante os quarenta dias de coletas de campo. Após
tomadas as medidas de biomassa, os animais coletados foram devidamente
identificados, sofreram eutanásia com clorofórmio, foram fixados com formalina a
10% e mantidos em álcool a 70%. Todos os exemplares foram depositados no
Museu de História Natural da UFBA. (Figuras 9 e 10.)
57

VARIÁVEIS DA FAUNA
Foram calculadas com base no total dos dados de captura nas armadilhas de queda
de cada unidade amostral, somando-se as quatro excursões.
a) Abundância relativa da espécie – Abun rel (indivíduos . armadilha-1 . dia-1 :
índice): calculado para cada espécie (ou morfo-espécie) por réplica, através da
fórmula:
F_ABrel = (número de indivíduos capturados) . (esforço amostral)-1
b) Abundância relativa da comunidade – Abund com (indivíduos . armadilha-1 .
dia-1 : índice): calculado para cada comunidade por réplica de modo análogo ao
apresentado acima.
58

c) Riqueza de espécies – Riq (espécies): estimada para cada área a partir do
número de espécies amostradas para as comunidades em relação aos quarenta
dias de coletas de campo.
d) Biomassa total da comunidade– Biom (gramas: variável em escala
proporcional): mensurada em balança semi-analítica (precisão 0,01g) após coleta de
cada espécie. Foi realizada média da biomassa da comunidade por unidade
amostral.
VARIÁVEIS AMBIENTAIS MENSURADAS
Três grupos de variáveis foram quantificados nas unidades amostrais para auxiliar a
interpretação da composição das comunidades de lagartos de folhiço, nos três
componentes da paisagem estudada: variáveis de micro-habitat (a), micro-clima (b),
da paisagem (c).
A) VARIÁVEIS DE MICRO-HABITAT:
Foram medidas em apenas uma excursão e os valores utilizados nas análises
representam a média das 36 medidas de cada unidade amostral.
Distância média a plantas lenhosas – (Dist. árvores): estimada através do
método do quadrante (point-quarter method, Krebs 1999: 182), utilizando-se os
baldes como centro dos quadrantes e registrando-se todos os indivíduos.
Perímetro do tronco – (Per Tron): para cada indivíduo amostrado, o perímetro foi
medido a 1,5 metros do solo (altura do peito), perfazendo um total de 144 valores
por unidade amostral.
Densidade da folhagem – (Df): foi medida em 18 estações de coleta, em cada uma
das áreas amostradas, sendo avaliada para os seguintes estratos da vegetação: 0 a
5 metros de altura (Df05), 5 a 10 m (Df510), 10 a 15 (Df1015); 15 a 20 m (Df1520),
20 a 25 m (Df2025) e maior que 25 m (Dfm25). Foram realizados sorteios para
determinar as 18 estações amostradas, a direção (esquerda ou direita) e as
distâncias que determinariam cada local onde seriam feitas as medições. As
distâncias entre as estações variaram de 1 a 10 m, limite correspondente à largura
59

entre as linhas amostrais de cada área. Os limites superior e inferior de cada estrato
foram avaliados com o auxílio de uma vara metálica de três metros de comprimento
fincada no chão, em cada sítio. Esta funcionava como uma mira para o
estabelecimento de um retângulo vertical imaginário, de cerca de 15 cm de largura
(Figura 11).
Os valores por estrato foram atribuídos segundo a porcentagem de preenchimento
do retângulo por folhagem, de acordo com as categorias 0-25%, 26-50%, 51-75% e
76-100% (dezoito valores por estrato por unidade amostral). Para efeito de análise,
essas categorias foram substituídas pelos valores 1, 2, 3 e 4. Para cada unidade
amostral, a densidade de folhagem em um determinado estrato é a média da
metragem ocupada por folhagem naquele estrato, nos 18 pontos (Hubbell & Foster,
1986). (Figuras 12 e 13).
60

61

Densidade do estrato herbáceo – (Den Her) avaliada em um círculo de 1,5 m de
raio com centro do balde. Os valores foram atribuídos segundo as categorias 0-25%,
26-50%, 51-75%, 76-100% de área coberta por herbáceas (36 valores por unidade
amostral). Para efeito de análise, essas categorias também foram substituídas pelos
valores 1, 2, 3 e 4.
Densidade de serrapilheira – (Den Ser) avaliada em um círculo de 1,5 m de raio
com centro do balde. Os valores serão atribuídos segundo as categorias 0-25%, 26-
50%, 51-75%, 76-100% de área coberta por serrapilheira (36 valores por unidade
amostral). Para efeito de análise, essas categorias foram substituídas pelos valores
1, 2, 3 e 4.
Altura do folhiço – (Alt fol): avaliado à direita de cada balde, a uma distância de 3
metros do mesmo, através da penetração de uma barra de metal graduada na
camada de serrapilheira, até o contato com o solo compactado. (36 valores por
unidade amostral).
Número de troncos caídos – (N Tron): avaliada em um círculo de 5 m de raio com
centro no balde, através de contagem direta dos indivíduos observados, com CAP
superior a 10 centímetros).
Grau de decomposição de troncos caídos – (Dec Tron): avaliado para cada
tronco acima contado, de acordo com as seguintes categorias de estado de
decomposição: estágio recente, estágio intermediário, estágio avançado de
decomposição. Para efeito de análise, essas categorias foram substituídas pelos
valores 1, 2 e 3.
Percentual de troncos caídos – (Percent Tron): avaliado em um círculo de 1,5 m
de raio com centro do balde. Os valores foram atribuídos segundo a seguinte
percentagem de cobertura: 0-25%, <25-50%, <50-75%, <75-100% (36 valores por
unidade amostral), cujas categorias foram substituídas pelos valores 1, 2, 3 e 4.
Grau de compactação do solo – (Comp sol): avaliado à direita de cada balde, a
uma distância de 3 metros do mesmo, através da penetração de uma barra de metal
graduada no solo. Os valores foram atribuídos segundo as categorias “pouco
compacto”, “medianamente compacto” e “muito compacto” (36 valores por unidade
amostral). Para efeito de análise, essas categorias foram substituídas pelos valores
1, 2 e 3.
62

63
lativa do ar - (Um Ar) (unidade:%) avaliada na circunferência de 1,5 m
(unidade: lux): avaliada na circunferência de 1,5 m de raio a
centrados na unidade amostral, com as
b) Variáveis de micro-clima
Foram medidas em todas as excursões (40 valores) e os valores utilizados nas
análises representam a média das médias das 36 medidas de cada unidade
amostral.
Temperatura do ar - (Temp Ar) (unidade: ºC): avaliada na circunferência de 1,5 m
de raio com centro de cada balde, através da utilização de um termigrômetro de ar,
(36 valores por unidade amostral).
Umidade rede raio a partir do centro do balde, através da utilização de um termigrômetro de ar,
(36 valores por unidade amostral).
Luminosidade – (Lux) partir do centro do balde, através da utilização de um luxímetro.
c) Variáveis da paisagem
Foram medidas através do Programa Arcview 3.3, a partir da interpretação de
ortofotocartas de 1:10.000 e fotografias aéreas de 1:32.500 da região referentes ao
ano de 1998 (Veracel, 2002, dados não publicados).
Cobertura de vegetação por componente da paisagem- (“buff”): Foram gerados
três ‘buffers”, ou círculos diferenciados
seguintes medidas de raios: buff1= 250m2 ; buff2= 500m2, buff3= 1000m2, e
mensurado em m2, a quantidade de floresta (estágios inicial a avançado e floresta
primária) presente na área de cada “buffer”, calculando-se a média para cada
unidade amostral (12 valores/”buffer’) Figura 14.
Distância das unidades amostrais em relação ao centro de gravidade na Mata Contínua – (dist pto O): foram calculadas através do programa AUTOCAD 2000, as
distâncias (m) de cada unidade amostral, em relação ao ponto “O”, plotado
imaginariamente no centro da Mata Contínua.

FIG sen ers” os em ca o o da pai . a) “buff” m ro m2, rmelho. Lo iz o nto ar o “O”
URA
14. Repre250m
tação dos ‘buffsa; b) “buff”500
plotadem ve
da ccal
nente do po
sagembitrári2, e
, onde foram medidas as distâncias entre as diversas unida s amostrais.
O
mpaçãde

ANÁLISE DOS DADOS Para a realização das análises de dados foram elaboradas matrizes biomassa e de
abundância das espécies de lagartos por unidades amostrais, que representam as
variáveis dependentes das análises ou foram usadas para gerá-las. As matrizes de
abundância foram transformadas dividindo-se cada valor pelo somatório da linha
(= unidade amostral) de modo a que todas as unidades passassem a ter o m
peso na solução das análises (Magnusson, 2003).
Os números totais de indivíduos e de espécies capturados foram conside
índices comparativos de abundância e riqueza adequados, visto que o esfor
coleta foi idêntico em todas as unidades amostrais. Também foi elaborada
matriz de variáveis ambientais, que representam as variáveis independente
análises ou foram usadas para gerá-las.
Comparação entre os componentes da paisagem
Essa análise refere-se à detecção de relação entre a variável desc
“componente da paisagem” e as variáveis critério relacionadas tanto à
(composição e biomassa), como às variáveis ambientais. Assume-se que
componente de paisagem estabelecida representa uma unidade natura
paisagem, ou seja, que o valor esperado das variáveis não controladas pra tod
réplicas de cada componente de paisagem é o mesmo (Kachingan,
Magnusson, 2003).
Para testar a hipótese nula sobre a diferença na composição da comun
faunística e biomassa entre os componentes da paisagem (Mata Con
Remanescentes e Eucaliptais) utilizou-se o método de Procedimento de Permu
de Resposta Múltipla (MRPP) sobre a matriz de abundâncias transformada, us
como medida de distância a distância de Sorensen. Este teste não par
pressuposto de normalidade multivariada e homogeneidade de variâncias dos d
e, portanto, é o mais adequado para detectar diferenças entre comunid
biológicas (McCune & Grace, 2002). O mesmo procedimento foi adotado para
esmo
rados
ço de
uma
s das
ritora
fauna
cada
l na
as as
1986;
idade
tínua,
tação
ando
te do
ados
ades
testar
104

a hipótese nula da ausência de diferença entre as biomassas e as variáveis
Na segunda etapa das análises, as doze unidades amostrais foram consideradas
omo amostras independentes das formações florestais da região, utilizando-se
com base nos dados de composição da
omunidade com o objetivo de detectar variáveis ambientais associadas a
gradientes de abundância e biomassa das espécies. Nessa análise, assume-se que
os de abundância e
ma parte associada ao erro, que descreve o modo pelo qual a
perada.
ambientais nos diversos componentes da paisagem, diferenciando-se apenas pela
utilização na análise dos eixos de redução das variáveis ambientais (PC’s) e
distância euclidiana.
a) Análises de ordenação
c
modelos lineares para testar a hipótese de associação entre as variáveis
independentes (ambientais) e as dependentes (relativas às comunidades).
Esse procedimento organiza os sítios (unidades amostrais) envolvidos em
determinada análise ao longo de eixos
c
há uma estrutura subjacente (estrutura latente) aos dad
biomassa da fauna, ou seja, que as ocorrências de todas as espécies em análise
são determinadas por poucas variáveis ambientais desconhecidas (variáveis
latentes) de acordo com um modelo de resposta simples.
Tal modelo consiste em uma parte sistemática, uma equação de regressão, que
descreve o modo pelo qual a resposta esperada depende das variáveis
explanatórias e de u
resposta observada desvia da es
Para testar a hipótese nula de ausência de associação entre as principais
dimensões de variação ambiental e a principal variação da composição da
comunidade de lagartos, foram adotados os seguintes procedimentos: geração de
um vetor de ordenação das espécies (NMDS1) através do método de escalonamento
multidimensional não métrico (NMDS) sobre a matriz de abundâncias transformada,
utilizando-se a medida de distância de Sorensen.
105

Nesse procedimento, foram realizadas análises a partir de dez configurações iniciais
para os dados reais a fim de evitar o problema de mínimos locais, utilizando-se
como critério de estabilidade o valor de instabilidade de 0,0005, 20 iterações para
avaliar a estabilidade da solução e 500 como o número máximo de iterações.
For principais de variação da matriz de variáveis
mbientais através do método de Análise de Componentes Principais (PCA) com
múltipla
aseado no modelo: NMDS1 = constante + PC1*X1 + PC2*X2, representando os
calculada através de um
ste de correlação de Mantel entre a matriz original (usando a distância de
endente.
am extraídos dois vetores
a
base em matriz de correlação e rotação Varimax (Kashigan, 1986), calculando os
escores dos dois componentes sobre as unidades amostrais. Finalmente, foi testada
a hipótese de ausência de associação significativa entre os vetores principais (PC1 e
PC2) e o vetor de ordenação (NMDS1) através de um teste de regressão
b
gráficos de dispersão das regressões parciais (MacCune & Grace, 2002).
Também foi realizado um teste de Monte Carlo (40 randomizações) para avaliar a
força do padrão da matriz de abundâncias transformada, comparando o estresse
calculado na ordenação da matriz original a distribuição de estresses gerados na
ordenação das matrizes randomizadas. A porcentagem de explicação das medidas
de distância na matriz original pelas distâncias no vetor de ordenação foi avaliada
pelo quadrado da estatística de Mantel padronizada (r2),
te
Sorensen) e o vetor de ordenação (usando a distância euclidiana). O valor de
probabilidade da associação foi calculado através de um teste de Monte Carlo com
1000 randomizações. O mesmo procedimento foi repetido para a comunidade
faunistica e a biomassa como variável dep
A qualidade da ordenação indireta pelo NMDS foi apresentada através dos gráficos
de abundâncias das espécies (ordenadas pelos seus escores calculados pela média
recíproca) pelas unidades amostrais (ordenadas pelo vetor de ordenação da
comunidade NMSD1), e a qualidade da ordenação direta pelos PCs através dos
gráficos de abundâncias pelos vetores ambientais (PC1 ePC2) .
106

O nível de significância adotado foi de 0,05. As análises de PCA e regressão
múltipla foram realizadas pelo pacote estatístico SPSS for Windowns, versão 11.0, e
as análises de MRPP, NMDS, Monte Carlo e Correlação de Mantel foram realizadas
usando-se o pacote estatístico PC-ORD for Windows versão 4.25.
107

Resultados
No total dos 40 dias de coletas, foram capturados 206 lagartos de folhiço,
correspondentes a 12 espécies, pertencentes a 7 famílias diferentes: Anguidae,
Gekkonidae, Gymnophthalmidae, Polychrotidae, Teiidae, Tropiduridae e Scincidae.
A Tabela 1 apresenta o número total de indivíduos e o local de captura de cada
espécie.
A curva média do coletor para os lagartos capturados nos 3 componentes da
paisagem não atingiu um platô, evidenciando que não foram coletadas todas as
espécies existentes nas áreas estudadas (Figuras 15 a 17).
As espécies Gymnodactylus darwinii, Leposoma scincoides e Enyalius catenatus
picus foram as dominantes, correspondendo respectivamente a 51%, 24% e 12% do
total dos indivíduos coletados, presentes em todos os componentes da paisagem
amostrados. As demais 9 espécies representaram 13% do total coletado, sendo que
algumas foram exclusivas de algum componente da paisagem, como Mabuya
macrorhyncha e Polychrus marmoratus apenas coletadas na Mata Contínua e
Ophiodes striatus, cuja ocorrência foi exclusiva nos Eucaliptais. As abundâncias
individuais das espécies podem ser observadas detalhadamente na Figura 18.
De uma forma geral, a Mata de Referência foi o componente da paisagem que
apresentou maiores valores de riqueza (M2) e abundância (M3) de lagartos. Porém
em termos de riqueza (M1) apresentou valores similares ao Remanescente (R2). O
Remanescente (R4) obteve o menor valor de riqueza, enquanto que o Eucaliptal
(E1) o menor valor de abundância de lagartos. Embora a Mata Contínua
evidenciasse níveis de riqueza e abundância superiores, não se pode afirmar que tal
componente da paisagem apresente maior biodiversidade em relação aos
Remanescentes e aos Eucaliptais.
108

Como a riqueza total não evidencia a composição da comunidade de lagartos de
lhiços para cada unidade amostral, esse índice não foi utilizado nos cálculos
merianae apresentaram os maiores valores, enquanto a espécie Anolis
scoauratus obtiveram os menores valores se comparados às demais. (Tabela 2).
fo
estatísticos subsequentes (ex. M1 apresentou o mesmo valor de riqueza que R2,
porém composição de espécies diferenciadas). Figuras 19.
Quanto à biomassa, as espécies Enyalius catenatus picus, Gymnodactylus darwinii e
Tupinambis
fu
As Matas Contínuas representaram os componentes da paisagem com maiores
valores totais de biomassa, enquanto que o Remanescente (R4) obteve os menores
valores, seguido dos Eucaliptais (E2, E1 e E4). Figura abaixo
109
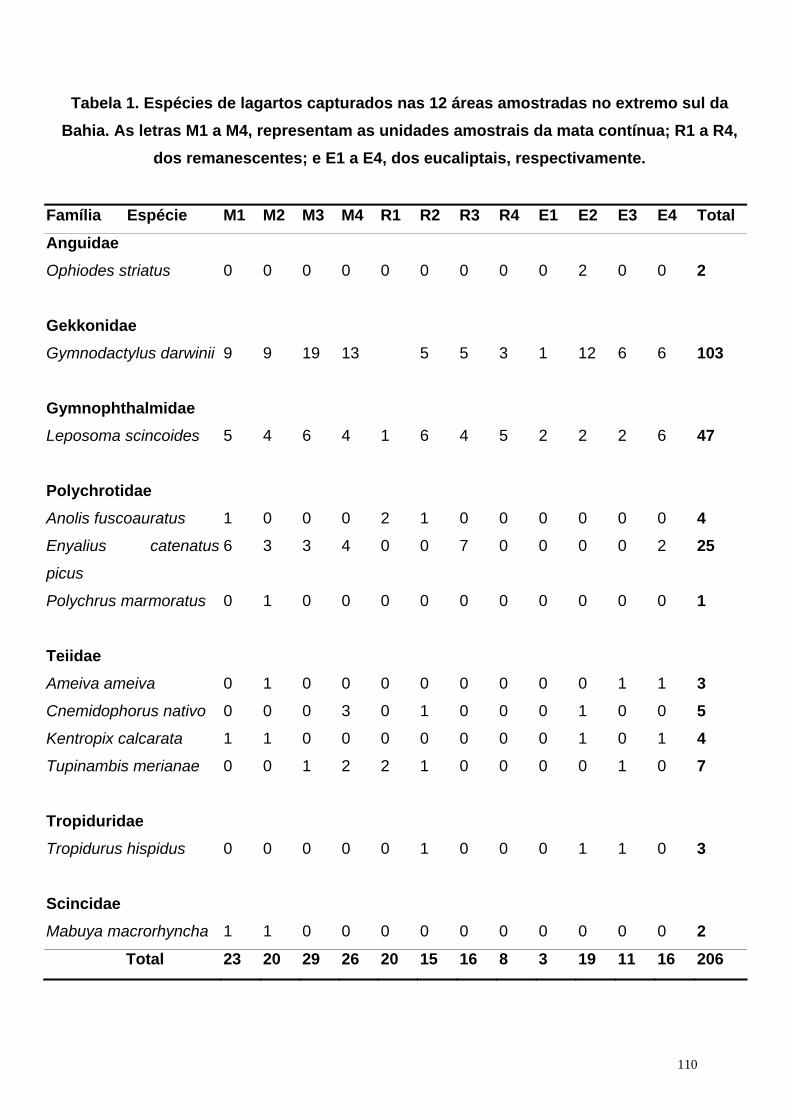
Ta da Bah R4,
Família Espécie M1 M2 M3 M4 R1 R2 R3 R4 E1 E2 E3 E4 Total
bela 1. Espécies de lagartos capturados nas 12 áreas amostradas no extremo sul ia. As letras M1 a M4, representam as unidades amostrais da mata contínua; R1 a
dos remanescentes; e E1 a E4, dos eucaliptais, respectivamente.
Anguidae Ophiod
GekkoGymno 03 Gymnophthalmidae Leposoma scincoides 5 4 6 4 1 6 4 5 2 2 2 6 47 Polychrotidae Anolis fuscoauratus 1 0 0 0 2 1 0 0 0 0 0 0 4 Enyalius catenatus
picus
6 3 3 4 0 0 7 0 0 0 0 2 25
Polychrus marmoratus 0 1 0 0 0 0 0 0 0 0 0 0 1 Teiidae Ameiva ameiva 0 1 0 0 0 0 0 0 0 0 1 1 3 Cnemidophorus nativo 0 0 0 3 0 1 0 0 0 1 0 0 5 Kentropix calcarata 1 1 0 0 0 0 0 0 0 1 0 1 4 Tupinambis merianae 0 0 1 2 2 1 0 0 0 0 1 0 7 Tropiduridae Tropidurus hispidus 0 0 0 0 0 1 0 0 0 1 1 0 3 Scincidae Mabuya macrorhyncha 1 1 0 0 0 0 0 0 0 0 0 0 2
Total 23 20 29 26 20 15 16 8 3 19 11 16 206
es striatus 0 0 0 0 0 0 0 0 0 2 0 0 2
nidae dactylus darwinii 9 9 19 13 5 5 3 1 12 6 6 1
110

Tab excontínua; R1 a nte.
M1 M2 M3 M4 R1 R2 R3 R4 E1 2 E3 E4
ela 2. Biomassa das espécies de lagartos capturados nas 12 áreas amostradas notremo sul da Bahia. As letras M1 a M4, representam as unidades amostrais da mata
R4, dos remanescentes; e E1 a E4, dos eucaliptais, respectivame
Família Espécie
E
Anguidae Ophiode 0 0 0 0 0 0 91 55 0 0
arwinii12 ,41 , 2 9 ,64 ,63 4,9 38,73 24,04 19,88
3 7 4 ,5 4, 1 6 2 2,5 2,02 1,93 6,1
0,25 0 2, 4,68 0 0 0 0
53,42 0 0 136,2
rus ratus
0 31,06 0 0 0 0 0 0 0 0 0 0
eiidae meiva ameiva 0 ,73
h 0 10, 0 4,32 0 0 5, 9 0 0
19,49 3 0 0 0 7, 0 15,04
,0 9, 62 67,0 0 0 7 6
ropidur s ispidus
0 0 0 5 0 0 5, 3 3,93
acrorhyncha 43 64 0 0 0 0 0
Total 160,9 1,3 53,8 162 11 ,4 1 2,1 66,3 16,1 98, 59 07 1 7,7 6,95
sstriatus
0 0 0 ,
Gekkonidae Gymnodactylus 34,54
24,
60
30
39 42,86
23
3
d 2,1 4
Gymnophthalmidae
Leposoma scincoides
15,6 3,9 9, 8 8 03 7,9 6,5 1 4 6,25
Polychrotidae Anolis
0
0
0
0
fuscoauratus 59
Enyalius catenatus picus
89,59
Polychmarmo
67,46 67,97 0 0 0 0 37,15
TA
65 0 0 0 0 0 0 0 0 35,24 18,63
Cnemidop nativo
orus 0 0 21 0 2
Kentropix calcarata Tupinambis 0
27, 0 0 0 0 99
merianae
0 16 5 8 ,9 0 0 8,4 0
Tropiduridae Th
u
0 0 ,9 0 1 0
Scincidae Mabuya 1,
11,
m0 0 0 0 0
23 1 ,4 2 2 1 7 51 , 4 9
111

112

113

114

DESCRIÇÃO DAS VARIÁVEIS AMBIENTAIS
eis de micro-habitat (a), micro-clima (b) e
amostrais estão expressos nas Figuras
Quanto ao micro-clima, foi observado um gradiente ambiental da Mata Contínua aos
Eucaliptais relacionado às variáveis de umidade, temperatura do ar e luminosidade
(Figura 20).
Em relação ao micro-habitat, apenas as variáveis grau de compactação do solo e
percentagem de troncos caídos apresentaram um gradiente ambiental da Mata
Contínua em relação aos Eucaliptais. Os Remanescentes apresentaram os maiores
valores em relação à variável número de troncos caídos, seguido da Mata Contínua
e por último, os Eucaliptais (Figura 21).
Quanto à densidade de herbáceas, os Remanescentes e os Eucaliptais
apresentaram médias similares e a Mata Contínua, os menores valores. Em relação
a altura de folhiço, foram encontrados maiores valores na Mata Contínua, enquanto
que os Eucaliptais obtiveram maiores médias relacionadas à espessura de troncos e
distâncias das árvores em relação ao centros dos pitfalls (Figura 22).
As variáveis relacionadas à estrutura vertical da vegetação evidenciaram que a Mata
Contínua apresenta os maiores valores médios de densidade de folhagem,
demonstrando um gradiente ambiental bem definido nos diversos estratos da
vegetação relativos às unidades amostrais estudadas. Os Remanescentes
presentam menor quantidade de folhagem que a Mata Contínua, principalmente em
lação aos estratos superiores a dez metros. Apesar de representarem formações
orestais homogêneas, foi detectado a presença de sub-bosque nos Eucaliptais
igura 23).
medição do percentual de vegetação presente nos “buffers”, ou círculos de raios
iferenciados nas unidades amostrais, apenas revelou a presença de formações
orestais nativas na Mata Contínua e nos Remanescentes em relação ao “buffer” de
50 m2. Aumentando-se o raio do “buffer” para 500 m2, evidencia-se um gradiente
mbiental da Mata Contínua aos Eucaliptais, que também foi detectado na avaliação
o “buffer” de 1000 m2. Quanto á distância dos pontos amostrais em relação a um
Os resultados obtidos em relação às variáv
da paisagem (c) quantificadas nas unidades
de 20 a 23.
a
re
fl
(F
A
d
fl
2
a
d
115

ponto arbitrário localizado no centro da Mata Contínua, percebeu-se que os
entes apresentaram as maiores variações, localizando-se mais dispersos
a 24). De um modo geral, a Mata Contínua
e os Remanescentes apresentaram maior percentual de florestais nativas próximas
Remanesc
na paisagem como todo, seguidos dos Eucaliptais, que apesar de mais distanciados
do ponto arbitrário em relação a Mata Contínua, suas unidades amostrais se
localizaram mais próximas entre si (Figur
em relação aos Eucaliptais (Figura 25).
116

117

118

119

120

121

122

COMPOSIÇÃO DA COMUNIDADE DE LAGARTOS DE FOLHIÇO ENTRE OS DIVERSOS COMPONENTES DA PAISAGEM.
Considerando que cada componente de paisagem estabelecida representa uma
unidade natural na paisagem, ou seja, que o valor esperado das variáveis não
controladas para todas as réplicas de cada componente de paisagem é o mesmo
(Kachingan 1986: 297), estatisticamente, baseado nos resultados da análise do
MRPP, não foi possível detectar diferenças na composição das comunidades de
lagartos de folhiço nos diversos componentes da paisagem avaliados (T = - 0,264;
p= 0,347).
Quanto á biomassa total de lagartos, a análise de MRPP indicou que há uma
diferença marginalmente significativa nos diversos componentes da paisagem
(T = - 1,66; p= 0,066), e que essa diferença ocorre entre a Mata Contínua e os
Eucaliptais (T = - 2,853; p= 0,021). Nos demais componentes da paisagem não foi
ossível detectar diferença na biomassa total de lagartos (Eucaliptais e
Remanescentes (T = 0,499; p= 0,665 e Mata Contínua e Remanescentes
= - 1,764;p= 0,061).
ESULTADOS DA ANÁLISE DE ORDENAÇÃO EM RELAÇÃO ÀS VARIÁVEIS AIS
a segunda etapa das análises, as 12 unidades amostrais foram consideradas como
mostras independentes das formações florestais da região.
A partir da análise de ordenação das variáveis ambientais foram extraídos 3 eixos
rincipais que explicaram 86% da variação dos dados, evidenciando gradientes
istintos entre os componentes da paisagem avaliados.
p
T
RAMBIENT
N
a
p
d
123

O primeiro eixo, que explica 44,4%, da variação original dos dados, representa um
gradiente ambiental direcionado da mata de referência aos Eucaliptais,
rrelacionado positivamente com as seguintes variáveis ambientais: proximidade
com florestas nativas (“buff” 250, 500, 1000m2), compactação do solo,
s dados originais, também representa um gradiente ambiental dos
sociado a uma específica unidade amostral, o primeiro
manescente (R1), em relação às variáveis: densidade de serrapilheira e densidade
análise de MRPP detectou que há diferenças entre as variáveis ambientais nos
0,006) entre a Mata
ontínua e os Eucaliptais E (T= -3,842, p= 0,007), mas não entre a Mata Contínua e
co
decomposição de troncos caídos, densidade de folhagem (df0-5/5-10/10-15m),
número e percentagem de troncos caídos.
As variáveis ambientais, perímetro de tronco, distância entre as árvores e
luminosidade também estavam associadas ao PC1, mas em direção contrária ao
gradiente. (Figuras 26 a, b). O segundo eixo de ordenação, que explica 29,8% da
variação do
Eucaliptais à Mata Contínua, relacionando-se positivamente com as variáveis:
distância em relação às florestas nativas, temperatura e umidade do ar. As variáveis
altura de folhiço e densidade de folhagem nos estratos superiores (df15-20/20-
25/>25m) também estiveram associadas a esse PC2, mas em sentido contrário
(Figura 27). O terceiro eixo principal extraído da análise (PC3), explicava 11,9% da
variação geral dos dados, porém não foi utilizado nas análises subseqüentes. Este
PC3 estava mais as
re
de herbáceas, não representando um gradiente ambiental.
A
diversos componentes da paisagem (T= -3,973, p= 0,003), o que reflete diferenças
entre os Remanescentes e os Eucaliptais (T= -3,916, p=
C
os Remanescentes (T= -0,251, p=0,320).
124

Figura 26 b) Representação do gradiente ambiental entre os Eucaliptais, a Mata Contínua e os Remanescentes, e das variáveis ambientais, com relação ao eixo principal PC1.
125

Figura 27. Representação do gradiente ambiental entre os Eucaliptais e os Remanescentes, e a Mata Contínua e das variáveis ambientais, com relação ao eixo principal PC2.
126
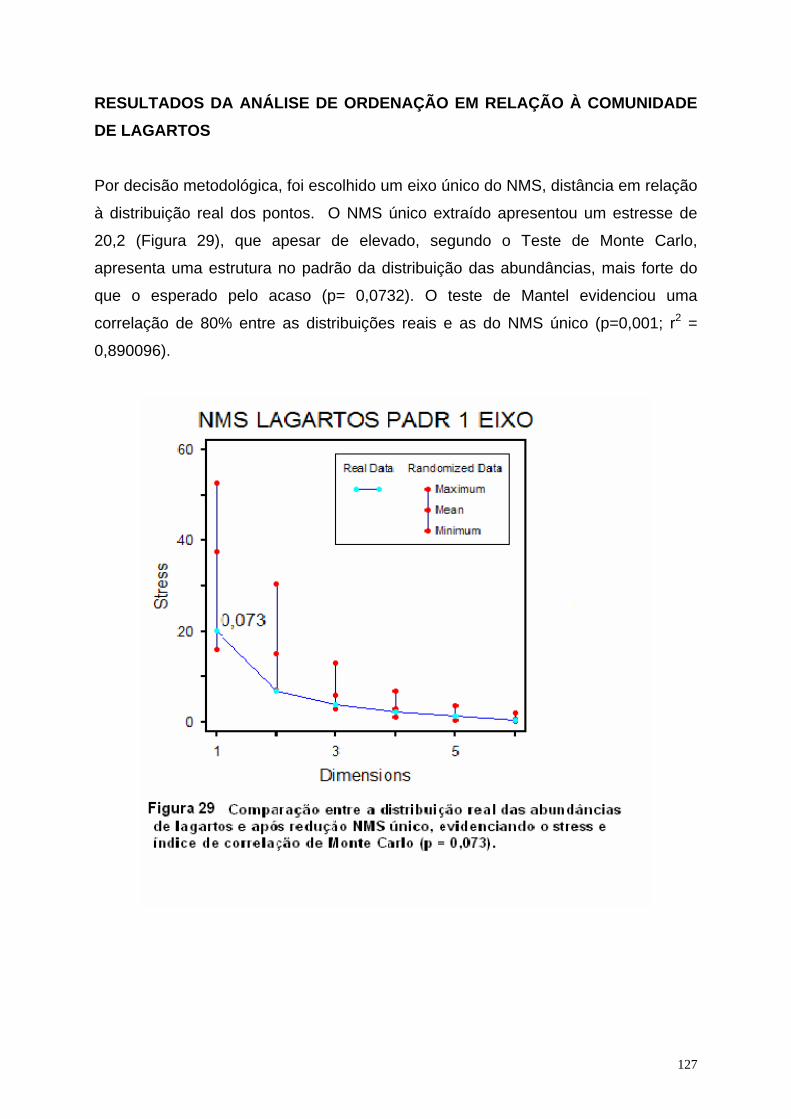
RESULTADOS DA ANÁLISE DE ORDENAÇÃO EM RELAÇÃO À COMUNIDADE DE LAGARTOS Por decisão metodológica, foi escolhido um eixo único do NMS, distância em relação
à distribuição real dos pontos. O NMS único extraído apresentou um estresse de
20,2 (Figura 29), que apesar de elevado, segundo o Teste de Monte Carlo,
apresenta uma estrutura no padrão da distribuição das abundâncias, mais forte do
que o esperado pelo acaso (p= 0,0732). O teste de Mantel evidenciou uma
correlação de 80% entre as distribuições reais e as do NMS único (p=0,001; r2 =
0,890096).
127

A análise de regressão linear entre o NMS (variável dependente) e os PC1 e PC2
(variáveis independentes) não evidenciou correlação entre o padrão de distribuição
as abundâncias dos lagartos e as variáveis ambientais mensuradas (r2 = 0,5; d
F= 0,744). Porém, foi estatisticamente significativo quando realizado para a
biomassa total de lagartos e o segundo eixo do componente principal da redução
das variáveis ambientais PC2 (r2 = 0,05; t = 3,654). Figura 30.
A qualidade da ordenação indireta pelo NMS foi apresentada através dos gráficos de
abundâncias das espécies, ordenadas pelos seus escores calculados pela média
recíproca, e pelas unidades amostrais, ordenadas pelo vetor de ordenação da
comunidade NMS (Figura 31). Esses gráficos evidenciam o eixo que melhor
presenta a variação ambiental na composição da comunidade de lagartos. re
128

Figura 31. Representação das distribuições de abundâncias das espécies de lagartos coletados nos diversos componentes da paisagem estudados no melhor eixo de ordenação.
129

Discussão Com a realização desse estudo, a partir dos 206 lagartos de folhiço capturados, foi
possível detectar a presença de 12 espécies nas áreas avaliadas, pertencentes a 7
famílias diferentes: Anguidae, Gekkonidae, Gymnophthalmidae, Polychrotidae,
Teiidae, Tropiduridae e Scincidae.
Embora a curva média do coletor não tenha atingido um platô para os lagartos
capturados nos três componentes da paisagem, a riqueza observada não se distancia
dos poucos trabalhos retratados sobre a fauna de lagartos de folhiço na Mata
Atlântica (Caldwell & Vitt, 1999; Schlaepfer & Gavin, 2001; Renken et al.,2002). Em
2001, Dixo realizou um estudo sobre a estrutur
folhiço no Sul da Bahia, através de armadilhas de
total. Franco (1988) realizou levantamentos na Es
estudo como área de Mata Contínua, em detrim
conservação florestal - e capturou 16 espécies de lagartos no
valendo-se dos mais diversos tipos de coleta, ativa e/ou passiva.
As espécies dominantes encontradas nos componentes da paisagem avaliadas
foram: Gymnodactylus darwinii, Leposoma scincoides, caracterizadas como espécies
de hábito generalistas, que utilizam divervos habitat. Tal resultado é similar àqueles
encontrados por autores como Vanzolini, 1980; Franco, 1988; Rodrigues & Borges,
1997; Freire, 1998; Caldwell & Vitt, 1999; Schlaepfer & Gavin, 2001; Dixo, 2001;
Rodrigues et. al., 2002; Renken et al.,2002 e Gerapue, 2004. No “Workshop de
“Avaliação e Ações Prioritárias para a Conservação da Biodiversidade da Mata
Atlântica e Campos Sulinos”, foram listadas 197 espécies de répteis, e ressaltou-se
que a fauna de lagartos da Mata Atlântica apresenta grande distribuição geográfica
ocorrendo, inclusive, em outras formações: na Amazônia, no Cerrado e na Caatinga
(MMA, 2000)
Mesmo ocorrendo no Eucaliptal (E2), a espécie Enyalius catenatus picus mostrou-se
mais abundante nos Remanescentes e na Mata Contínua. Segundo Etheridge, (1969)
esta espécie é arborícola, típica de ambientes florestados e sua distribuição
a de comunidades de lagartos de
queda, encontrando 17 espécies no
tação Veracruz -considerada nesse
ento ao seu excelente estado de
local durante 30 dias,
130

geográfica se restringe ao extremo Sul da Bahia. A espécie Anolis fuscoauratus
mbém apresenta hábito arborícola e esteve presente apenas na Mata Contínua e
s espécies Ameiva ameiva, Cnemidophorus nativo, Kentropix calcarata e
, com distribuição
eográfica em diversos biomas brasileiros (Freire, 1996; Rocha 2002; Rocha, 2003;
paisagem - Remanescentes) e Eucaliptais em estágio final de
rescimento: sete anos de idade.
ta
no Remanescente (R1), evidenciando que além de ambientes bem conservados,
pode ainda ocupar bordas de fragmentos, o que também foi descrito por Vitt, et al.,
2003, em estudos desenvolvidos na Amazônia.
A
Tupinambis merianae apresentaram baixas ocorrências nos diversos componentes da
paisagem estudados, entretanto mostraram-se abundantes na Mata Contínua e nos
Eucaliptais. Tais resultados estão em concordância com a literatura, uma vez que
essas espécies são forrageadoras, podendo ocupar diversos ambientes,
especialmente áreas abertas e bordas de florestas (Ruibal, 1952; Crump, 1971;
Vanzollini, 1974; Vitt, 1991; Vitt & Carvalho, 1992; Franco et al., 1998; Dias, et al.
2002; Rodrigues et.al 2002; Rocha, 2003; Marques & Sazima, 2004; Gerapue, 2004).
A espécie Tropidurus hispidus esteve presente apenas no Remanescente (R1) e nos
Eucaliptais (E1 e E2), sendo característica de aéreas abertas
g
Guerape, 2003). Mabuya Macrorhyncha ocorreu apenas na Mata Contínua, e apesar
de seu baixo índice de captura, tal espécie é capaz de ocupar diversos ambientes.
Em relação à composição das comunidades de lagartos de folhiço nas unidades
amostrais avaliadas, os dados indicaram que, apesar do intenso processo de
destruição florestal sofrido pela Mata Atlântica na região, em termos de composição,
não há diferenças entre os três principais componentes florestados da paisagem do
extremo Sul da Bahia: remanescentes de grande porte de mata em estágio avançado
de regeneração (considerados sistemas de referência – Mata Contínua),
remanescentes de pequeno porte em estágio intermediário de regeneração (os mais
comuns na
c
131

Sendo assim, não foi possível detectar se há interferência negativa da fragmentação
e seus processos associados na composição das comunidades de lagartos de folhiço
nessas áreas estudadas.
O padrão da distribuição das abundâncias de lagartos não parece estar associado
àquele evidenciado para as variáveis ambientais, indicando que a composição da
ariáveis ambientais como: distância em relação a florestas nativas, aumento da
m
iomassa disponível aos níveis tróficos superiores (Renken et.al.,2003). Sendo assim,
ram encontrados resultados concordantes a essa
uestão (Freire, 1996; Caldwell & Vitt, 1999; Schlaepfer & Gavin, 2001; Rocha et. al.
002; Rocha, 2002; Gerapue, 2003; Renken et al.,2003).
comunidade de lagartos pode estar vinculada a outros fatores além daqueles
estudados. Uma explicação para essa questão talvez esteja fundamentada no fato de
que a fauna de lagartos de folhiço estudada seja predominantemente generalista,
capaz de adaptar-se aos mais diversos tipos de ambientes, sem apresentar um
padrão específico de distribuição na paisagem (Leiden. et al., 1999; Vitt, 1998).
Porém, do ponto de vista da biomassa total, foram identificadas diferenças no padrão
de distribuição entre os Eucaliptais e a Mata Contínua, diferenças associadas às
v
temperatura e umidade do ar e diminuição da altura de folhiço e densidade de
folhagem nos estratos superiores da floresta.
Sabe-se que juntamente com os anfíbios anuros, os lagartos representam um dos
principais grupos taxonômicos que convertem biomassa de artrópodes terrestres e
b
os resultados indicam que, do ponto de vista da fauna de lagartos de folhiço, pode
existir maior disponibilidade de biomassa na Mata Contínua do que nos Eucaliptais, já
que as espécies arborícolas detectadas nesse estudo predominam nos ambientes
florestados mais conservados, a exemplo de Enyalius catenantus picus e Anolis
fuscoauratus.
Dos poucos estudos que investigaram as respostas de lagartos de folhiço perante a
fragmentação e seus processos associados, em especial no bioma Mata Atlântica, do
ponto de vista da paisagem não fo
q
2
132

No estudo de Dixo (2001), foram comparados fatores como abundância, riqueza,
diversidade, dominância e equitatividade de lagartos em seis habitats diferentes
(interior e borda de Mata Contínua; interior e borda de fragmento de mata; cabruca -
lantação de cacau sombreada; e capoeira - Mata Secundária). De acordo com os
ro de
spécies de lagartos de folhiço presentes nos diversos componentes da paisagem
esultados similares foram identificados em estudos sobre a estrutura da
onocultura de eucalipto com sub-bosque de espécies nativas pode
presentar um importante papel na conservação da diversidade regional em uma
de espécies generalistas
), levando a uma convergência espacial de recursos
e ao aumento do número de espécies no mosaico.
p
resultados obtidos, a riqueza e a diversidade de lagartos não foram significativamente
diferentes entre os diversos habitats. Por outro lado, fatores como abundância,
equitatividade e dominância de lagartos divergiram entre os tratamentos, o que indica
que, no contexto da paisagem, as cabrucas e as capoeiras parecem funcionar como
extensões da floresta.
Como não foram detectadas diferenças quanto à composição e ao núme
e
avaliados, tais resultados indicam que a qualidade da matriz de monocultura de
eucalipto pode ser considerada como adequada para a conexão de pequenos
fragmentos de floresta em estágios intermediários de sucessão do ponto de vista
dessa fauna.
R
comunidade de pequenos mamíferos em um fragmento de Mata Atlântica e em
monocultura de eucalipto sob prática de manejo que permite o desenvolvimento de
sub-bosque de espécies nativas, no Vale do Rio Doce, estado de Minas Gerais
(Fonseca, 1997). Foram utilizados os parâmetros de diversidade, riqueza e
abundância das espécies para a comparação das comunidades. Estas também não
apresentaram alterações que pudessem sugerir uma influência negativa da matriz
de monocultura de eucalipto. O estudo indicou que a riqueza e a diversidade de
espécies na m
re
região de grande vocação silvicultural (Fonseca, 1997).
De acordo com Metzger (1999), uma explicação para isso está fundamentada no
fato de que a proporção da paisagem ocupada pela matriz controla o fluxo de
perturbações no mosaico, podendo facilitar a propagação
(Vitt, 1991; Bull & Freake, 1999
133

Conforme comentário acima, a composição das comunidades de lagartos de folhiço
nos diversos componentes da paisagem avaliados está representada
majoritariamente por espécies generalistas. Esta fauna pode apresentar maior
capacidade de se dispersar e explorar a matriz formada pelos Eucaliptais, podendo
manter-se e até mesmo estar favorecida nas áreas fragmentadas (Janzen, 1983).
Em contrapartida, espécies menos tolerantes tendem a ser mais vulneráveis,
mantendo-se isoladas e restringindo sua distribuição a áreas mais conservadas,
como foi observado com a espécie Enyalius catenatus picus (Etheridge, 1969;
Bierreggard et al. 1997; Gascon et al. 1999).
za total de espécies de determinados segmentos da fauna, podendo
terferir na dinâmica de outros processos biológicos nas comunidades biológicas
A análise de ordenação com as variáveis ambientais apontou a existência de
gradientes entre a Mata Contínua, os Remanescentes e os Eucaliptais. Assim,
mesmo não sendo identificada uma influência negativa da matriz, a constante
redução das porções originais de habitat natural e conversão dessas áreas em
ambientes modificados, torna-se preocupante mesmo quando não acarreta redução
direta da rique
in
(Joly et al. 2000; Sauders. et al, 1991; Harison & Bruna, 1999; Debinski & Holt, 1999;
Faria, 2002).
134

R
A A
B
B . KAPOS, A. A. DOS SANTOS, and R.
amics of tropical rain forest fragments. Bioscience 42: 859-66.
B
B
B
sleep lizard, Tiliqua rugosa. CALDWELL, JANALEE P. VITT, LAURIE J. 1999. Dietary asymmetry in leaf litter
frogs and lizards in a transitional northern Amazonian rain forest. CAMP, C. L. 1923. Classification of the lizards. Bull Amer. Amer. Mus. Nat. Hist.,
48: 289-481.
CI/FB/SNE. 1994. Prioridades para a Conservação da Biodiversidade da Mata Atlântica do Nordeste. Mapa e texto. Conservation International, Fundação Biodiversitas e Sociedade Nordestina de Ecologia, Washington, D.C.
COLLI, G.R. 1991. Ecology of Ameiva ameiva (Sauria: Teiidae) in the cerrado of
central do Brazil. Copeia, n.4,p.1002-1012.
COLLAR, N. J., GONZAGA, L. P., KRABBE, N. 1992. Threatened birds of the Américas. Washington D. C.: Smthsonian Institution Press, 1150p.
CONSÓRCIO MATA ATLÂNTICA. 1992. Referências Bibliográficas. V.01.Ed.
UNICAMP.São Paulo. 101p.
EFERÊNCIAS BIBLIOGRÁFICAS
LMEIDA, DANILO SETTE DE. Recuperação Ambiental da Mata Atlântica. 2000.
VILA-PIRES, T.C.S. 1995. Lizard of Brazilian Amazonia (Reptilia: Squamata).Zoologische Verhandeligen, n.299.
ELLAIRS, A. D’A. 1969. The Life of Reptiles vols. 1-2 Wiedenfeld and Nicolson. London.
IERREGAARD, R. O., JR., T. E. LOVEJOY, VW. HUTCHINGS. 1992. The biological dyn
IERREGAARD JR., R.O., LAURANCE, W.F., SITESJR., J.W., et al. 1997. Key priorities for the suty of fragmented tropical ecossystems. IN: Laurance, W.F. & Bierregaard Jr., R.O. [Eds]. Tropical forest remnants: ecology, management, and conservation of fragmented communities. Chicago, The University of Chicago Press. 515-525.
ROWN, K. S. & G. G. BROWN. 1992. Habitat alteration and species loss in Brazilian forests. In: Tropical deforestation and species extinction (T. C. Whitmore & J. A. Sayer, eds). Pp. 119-142. Chapman & Hall, London.
ULL, C. MICHAEL, FREAKE, MICHAEL J. 1999. Home –range fidelity in the Australian
135

COSSON, J. F., RINGUET, S., CLAESSENS, O., DE MASSARY, J.C., DALECKY, A. , J. M. 1999. Ecological changes in recent
land-bridge island in French Guiana, with emphasis on vertebrate communities.
ursores de folhiço na Mata Atlântica do extremo sul da Bahia. Anais VI
DEAão Paulo, SP
DIAS
gartos do Gênero Cnemidophorus (Sáuria: Teiidae) das Dunas
DIXOsertação de mestrado, USP.
nians. Contrib. Biol. Geol., n.4, p. 1-58.
. [ Ed.] Four
ST OATES-ESTRADA, e D. MERITT Jr.. 1993. Bat species richness and abundance in tropical rain forest fragments and in agricultural habitats at Los
EST 994. Non flyng mammals
and landscape changes in the tropical rain forest region of Los Tuxtlas, México.
ETHERIDGE, RICHARD. 1969. A Review of the Iguanid Lizard Genus Enyalius.
FARIA, DEBORAH MARIA., 2002. Comunidade de Morcegos em uma paisagem
orado.
VILLIERS, J.F., GRANJON, L., PONS
Biological Conservation 91: 213-222. CRUMP, MARTHA L., 1971 – Quantitative Analysis of the Ecological Distribution of a
Tropical Herpetofauna. University of Kansas Lawrence, Kansas. DANTAS, T. B. 2003. Análise da efetividade das armadilhas de queda (pitfall traps)
com a utilização de diferentes números de cercas guias na amostragem de animais cCongresso Brasileiro de Ecologia. Pág 380.
N, W. 1996. À ferro e fogo,a história e devastação da Mata Atlântica brasileira. Companhia da Letras, S
DEBINSK, DIANE M., HOLT, ROBERT D. 1999. A Survey and Overview of Habitat
Fragmentation Experiments. 14:342-355.
, E. J. R., GOMES, A. S. CAMPOS, M. C.. 2002. Ecologia de Duas Espécies Simpátricas de laCosteiras do Parque Metropolitano do Abaeté – Salvador – Bahia – Brasil.
, M. B. de. 2001. Efeito da Fragmentação da Floresta sobre a comunidade de sapos e lagartos de serrapilheira no sul da Bahia. Dis
DIXON, J.R. & SOINI, P. 1975. The reptiles of the upper Amazon basin, Iquitos
Region, Peru: I. Lizards and amphisbae DUELLMAN, W.E. 1990. Herpetofaunas in neotropical forest: comparative
compositions, history, and resource use. IN: GENTRY, A.HNeotropical rain forests. New Haven, Yale University Press, p.455-505.
RADA, A., R. CE
Tuxtlas, mexico. Ecography 16(4): 309-318.
RADA, A., R. COATES-ESTRADA, e D. MERITT Jr.. 1
Ecography 17: 229-241.
18(8): 231-260.
fragmentada da Mata Atlântica do Sul da Bahia, Brasil. Universidade Estadual de Campinas, SP. Tese de Dout
136

FIGUEIRA, J.E.C.; VASCONCELOS-NETO, J.; GARCIA, M.A.; SOUZA, A.L.T. 1994. Saurocory in Melanocactus violaceus ( Cactacea). Biotropica v.26, n.3, p.295-301.
ONSECA, G. A. B. 1997. Biodiversidade e impactos antrópicos. Em J. A. de Paula (Org.)
FRA na Estação Vera Cruz / Franco,
Francisco Luís – Rio de janeiro: Veracel Celulose, 1998.
Zoogeográfico Sobre a Fauna de Lagartos (Sáuria) das Dunas de Natal, Rio Grande do Norte e da
REIRE, ELIZA MARIA XAVIER. 1998. Diferenciação Geográfica em Gmynodactylus
ASCON, C., T.E. LOVEJOY, R. O. BIERREGAARD, J. R. MALCOLM, P.C.
Forest remmants. Biological Conservation 91:223-229.
GOT ty: procedures and pitfalls in the measurement and comparison of species richness.
GER
UBBELL, S. P. &R. B. FOSTER. 1986. Canopy gaps and the dynamics of a
ANZEN, D. H.. 1983. No park is an island: increaseim interference from outside as
KICHENER, D. J., CHAPMAN, A. D. DELL . MUIR, B. G. 1980. Lizard assemblage
and reserve size and structure in the Western Australian wheatbelt – some implications for conservation. Biological Conservation 17: 25-62.
FBiodiversidade, População e Economia: Uma região de Mata Atlântica. UFMG, BeloHorizonte, M. G.
FONSECA, G. A. B. 1985. The vanishing Brazilian Atlantic forest. Biological
Conservation 34: 17-34.
NCO, FRANCISCO LUÍS. 1998. Répteis
FREIRE, ELIZA MARIA XAVIER. 1996. Estudo Ecológico e
Restinga de Ponta de Campina, Cabedelo, Paraíba, Brasil. 13(4): 903-921.
FDarwinii (Gray, 1845) (Sáuria, Gekkonidae). Museu de Zoologia da Universidade de São Paulo. 40(20): 311-322.
GSTTOUFER, H. L. VASCONCELOS, W. F. LAURANCE, B. ZIMMERMAN, M. TOCHER, E S. BORGES. 1999. Matrix habitat and species richness in tropical
ELLI, NICHOLAS J. COLWELL, ROBERT. 2001. Quantifylng biodiversi
APUE, EDUARDO AUGUSTO. 2004. As Ricas Caatingas. SciAm. Brasil. Ano n. 25.
HARRISON, SUSAN, BRUNA, EMILIO. 1999. Habitat fragmentation and large-scale
conservation: what do we know for sure?
HNeotropical forest. In: Plant Ecology (M. J. Crawley, ed.). pp. 77-95. Blackwell, Oxford.
Jpark size decreases. Oikos 41: 402-410.
JOLY, PIERRE,MIAUD, CLAUDE, LEHMANN, ANTHONY, GROLET, ODILE. 2000. Conservation Biology, 15:239-248.
, J
137

KOOOPMANS, Pe. JOSÉ. 1999. Além do eucalipto: o papel do extremo sul. Salvador: Momorial das Letras. 195 p.
KWET, A. & DI-BERNARDO, M. 1999. Pró-Mata – Anfíbios. Amphibians. EDIPUCRS,
LAURANCE, W. F., Bierregaard, R. O., Jr., Gascon, C., Didham, R. K., Smith, A. P.,
Lynam, A. J., Viana, V. M., Lovejoy, T. E., Sieving, K. E., Sites, J. W., Jr.,
tion: Synthesis of a diverse and dynamic discipline. Em W. F. Laurance e R. O. Bierregaard, Jr. (eds.) Tropical Forest Remnants: Ecology,
ment, and Conservation of Fragmented Communities. University of Chicago Press,
LAU Ferreira, L. V., Rankin-de Merona, J. M., Gascon,
C. e T. E. Lovejoy. (1997b).Biomass collapse in Amazonian forest fragments.
LEID
e Elisha Mitchel Scientific Society. 115(4):270-280. 1999.
MAG A ligação entre as questões e a análise.
MCCUNE, B. & GRACE, J.B. 2002 – Analysis of Ecological Communities
, 71: 445-463.
MITTERMEIER, R. A., MYERS, P. ROBLES GIL, and C. G. MITTERMEIER. 1999.
ULLER, P. 1973. The dispersal centers of terrestrial vertebrates in the Neotropical
OSS, R. F. e B. CSUTI. 1994. Habitat Fragmentation. Em: Meffe, K. G. & C. R.
PAR
Porto Alegre, Brasil. 107p., 138 figs.
Andersen, M., Tocher, M. D., Kramer, E. A., Restrepo, C. e C. Moritz (1997a). Tropical forest fragmenta
Management, and Conservation of Fragmented Communities. University of Chicago Press, Chicago.
LAURANCE, W. F.. 1997. Hyper – disturbed parks: edge effects and the ecology of
isolated rainforest reserves in tropical Australia. Em: Laurance, W. F. and Bierregaard Jr., R. O. (eds.), Tropical Forest Remnants: Ecology, Manage
Chicago, 71-83.
RANCE, W. F., Laurance, S. G.,
Science 278:1117-1118.
EN, Y.A., DORCAS, M.E. & GIBBONS, J. W. Herpetofaunal diversity in coastal plain communities of south carolina. The Journal of th
NUSSON, W.E. MOURÃO, GUILHERME. 2003. Estatística Sem Matemática:
METZGER, J.P. 1999. Estrutura da paisagem e fragmentação: análise bibliográfica.
Anais da Academia Brasileira de Ciências
MINISTÉRIO DO MEIO AMBIENTE. BRASÍLIA 2000. Avaliação e Ações Prioritárias Para a Conservação da Biodversidade da Mata Atlântica e Campos Sulinos.
Hotspots: Earth’s Biologically Rich-est and Most Endangered Terrestrial Ecoregions. CEMEX, México.
M
Realm. W. Junk Publishers, The Hague.
NCarroll (eds.) Principles of Conservation Biology. Sinauer Press. DINI, RENATA. Pequenos mamíferos e a fragmentação da Mata Atlântica de Una, Sul da Bahia – Processos e Conservação.
138

PAULISSEN, M.A. & WALKER, J.M. 1994. Diet of the insular lizard Cnemidophorus
nigricolor (Teiidae) from Grand Rocques Island, Venezuela. Journal of
IANKA, E.R. 1966. Convexity, desert lizards, and spatial heterogeneity.
PRA
REN
IMOTHY J., RICKE, KEVIN B., RUSSEL, BRADLEY, WANG, XIAOYIN. 2003. Effects of Forest Management on Amphibians and
ROD
ia Brasileira de Ciências, Rio de Janeiro, p. 305 - 315
OCHA, CARLOS FREDERICO DUARTE, et al. 2003. A Biodiversidade nos Grandes
OCHA, C. F. D., DUTRA, G. F., VRCIBRADIC, D., MENEZES, V.A. 2002. The
ents in South Brazil. Ph.D. Thesis, Harvard University Cambridge, Massachusetts.
ROD of Leposoma (Squamata: Gymnophthalmidae) From a relictual forest in semiarid northeasthern Brazil.
RODRIGUES, M. T. 1988. Distribution of lizards of the genus Ttopidurus in Brazil
Herpetology,v.28, n.4,p.524-526.
P
NCE, G. T. 1982. Forest refuges: Evidence from woody angiosperms. Em G. T. Prance (ed) Biological Diversification in the Tropics. Columbia University Press, New York.
KEN, ROCHELLE B., GRAM, WENDY K., FANTZ, DEBRA K., RICHTER, STEPHEN C., MILLER, T
Reptiles in Missouri Ozark Forests.
RIGUES, M.T. 1988 – Distribution of lizards of the genus Tropidurus in Brazil (Sauna Iguanidae) in Proceedings of a workshop on Neotropical distribution patterns. Academ
ROCHA, CARLOS FREDERICO DUARTE DA. 2002 Biogeografia de Répteis de
Restingas: Distribuição, Ocorrência e Endemismos. p. 99-113.
RRemanescentes Florestais do Estado do Rio de Janeiro e nas Restingas da Mata Atlântica / Carlos Frederico Duarte da Costa et al. – São Carlos: RiMa. P.133.
RTerrestrial Reptile Fauna of the Abrolhos Archipelago: Species List and Ecological Aspects. 62(2): 285-291.
RODRIGUES, E 1998 Edge effects on the regeneration of the forest fragm
RIGUES, M. T. & BORGES, M. D. 1997. A new species
Herpetologica. Vol 53. n01. pg 1-5.
(Sáuria, Iguanidae) in Procee-dings of a worksohop on Neotropical distribution patterns. W. R.Heyer & P. E. Vanzolini (eds.) Academia Brasileira de Ciências, Rio de Janeiro p. 305-315.
RODRIGUES, M. T.; DIXO, M. ; DANTE, P. & VERDADE, V.K. 2002. Papéis Avulsos de Zoologia. S. Paulo 42 (14): 335-350.
RUIBAL, RODOLFO. 1952. Revisionary Studies of Some South American Teiidae. 106(11): 477-529.
139

SAMPAIO,D. F. – Aviafauna Associada a Florestas de Eucalipto da Veracel Celulose S.
AUNDERS, D. A.,R. J. HOBBS, C. R. MARGULES. 1991. Biological
iology 7:18-32.
SCHpical Forest Fragments.
ARRE, S., SMITH, G. T. & MEYERS, J.A. 1995. Persistence of two species of Gecko
ILVA, E., SOUZA, A. L. – Perfil Ambiental das Florestas plantadas no Brasil. Viçosa,
MITH, G. T., ARNOLD, G. W., SARRE, S., ABENPERG – TRAUN, M., STEVEN, D. E.
Paulo, SP.
e ickly forest skink ( Gnypetoscincus queenslandiae ). Biological Conservation 91: 159-167.
TEIX/ Dante Martins Teixeira; colaboração de
marcovan Porto, Maria Lúcia Lourine, 51p. (Publicação técnico-científica / Estação
TEIX
(Aves, Cotingidae) / Dante Martins Teixeira, Antônio Cláudio Conceição de Almeida. – Eunápolis, BA: Veracruz Florestal. 60p.
VAN SLUYS, M. 1992. Aspectos da ecologia de lagarto Tropidurus itambere ( Sauria: Tropiduridae) , em uma área do Sudeste do Brasil. Rev. Brasil. Biol.v.52,
AN SLUYS, M. 1993. The reproductive cycle of Tropidurus itambere ( Sauria:
Tropiduridae) in southeastern Brazil. Journal of Herpetology, v.27, n.1,p.28-32.
A.,Eunápolis, Ba. Plano de Monitoramento de Sub-bosque de Eucalipto – Segunda Campanha, Eunapólis, Ba, 2001.
Sconsequences of ecosystem fragmentation: a review. Conservation B
LAEPEPFER, MARTIN A., GAVIN, THOMAS A. 2001. Edge Effcts on Lizards and Frogs in Tro
SPELLERBERG, I. F. & J. W. D. SAWYER. 1996. Standards for biodiversity: a proposal based on biodiversity standards for forest platantions. Biodiversity and Conservation 5: 447-459.
S(Oedura reticulata and Gehyra variegate) in remant habitat. Biological Conservatiion 71: 25-33.
SMG: SIF. 1994. 34P.
S1996. The effects of habitats fragmentation and livestock grazing on animal communities in remnants of gimlet Eucalypyus salubris woodland in the Western Australian wheatbelt. II. Lizards. Journal of Applied Ecology 33: 1302-1310.
SOS MATA ATLÂNTICA E INPE. 2004. Atlas da Evolução dos Remanescentes
Florestais da Mata Atlântica. Fundação SOS Mata Atlântica e Instituto Nacional de Pesquisas Espaciais, São
SUMMER, J., MORTIZ, C. & SHINE, R. 1999. shinking forest shrinks skink:
morphological change in response to rainforest fragmentation in th
EIRA, DANTE MARTINS. 1995. Animais vertebrados ameaçados de extinção presentes na Estação Veracruz
Veracruz; n. 1).
EIRA, DANTE MARTINS. 1996. Sobre a biologia da ‘’escarradeira’’, Xipholena atropurpurea (Wied, 1820)
n.1,p.181-185.
V
140

ZOLINI, P E. 1972. Miscellaneous notes on the ecology of some Brazilian lizards (Sauria). Pap. Av. Zoo 26(8): 83-115.
VAN
ANZOLINI. P. E. 1976. On the lizards of a Cerrado – Caatinga contact: evolutionary
ANZOLINI, P. E., RAMOS-COSTA. A M M & VITT. L. J. 1980. Répteis da
VERACEL CELULOSE S. A., 1998 – Plano de Manejo da Estação Veracruz, Eunápolis,
BA.. 64 p.
VER em 20 de setembro de 2000.
ITT, L. J. 1991. An introducion to the ecology of Cerrado lizards. J. Herpetol., 25(1):
VITT ae)
in Amazonian lowland rain forest. Can. J. Zool., v.71,p.2370-2390.
VITT
ITT, L.J. & CARVALHO, C. M. 1992. Life in the tress: the ecology and life histpry of
VANZOLINI. P. E. 1974. Ecological and geographical distribution of lizards in
Pernambuco, northeastern Bra (Sauria). Pap. Av. Zool., 28(4): 61-90.
Vand zoogeographical placations (Sauria). Pap. Av. Zool., 29(16) 111-119.
VCaatinga. Academia Brasileira Ciências, Rio de Janeiro.
ACEL CELULOSE S.A. Meio Ambiente. http://www.veracel.com.br/meio
/frame1.htm. Consultado
VERACEL CELULOSE S. A., 2001. Corredor Central da Mata Atlântica. CR-ROM, Ilhéus, IESB/CI/CRA.
VITT, LAURIE J., PIRES, TERESA CRISTINA S. AVILA, ZANI, PETER A., SARTORIUS, SHAWN S., EMPÓSITO, MARIA CRISTINA. 2003. Life above ground: ecology of Anolis fuscoauratus in the Amazon rain forest, and comparisons with its nearest relatives. 81: 142-156.
VITT, L.J. 1982a. Reproductive tactis of Ameiva ameiva ( Lacertilia: Teiidae) in a
seasonally fluctuating tropical habitat. Can.J. Zool.,v.60,p.3113-3120.
V79-90.
, L.J. 1993. Ecology of isolated open-formation Tropidurus ( Reptilia: Tropidurid
, L.J. & BLACKBURN, D.G. 1991. Ecology and life history of the viviparous lizard Mabuya bistriat ( Scindae) in the Brazilian Amazon. Copeia, v, 1991,n.4,p.916-927.
VKentropyx striatus ( Teiidae ) in the lavrado área of Roraima, Brazil, with comments on the life histories of tropical teiid lizards. Can. J. Zool., v.70, p. 1995-2006.
VITT, L.J. & COLLI, G.R. 1994. Geographical ecology of a Neotropical lizard: Ameiva ameiva ( Teiidae) in Brazil. Can. J. Ecol.,v.72,p.1986-2008.
VITT, L.J. & GOLDBERG, R. 1983. Reproductive ecology of two tropical iguanid lizards:
Tropidurus torquatus and Platynotus semitaeniatus. Copeia, 1983 (1): 131-141.
141

VITT, L.J. & LACHER, T. E. 1981. Behavior, habitat, diet, and reproduction of the
iguanid lizard Polychrus aucutirostris in the caatinga of nosteastern Brazil. Herpetologica v. 37, n.1.p.53-63.
VRC al sympatric skinks (Mabuya macrorhyncha and Mabuya agilis) in southeastern Brazil. J.
IBRADIC, D. & ROCHA, C.F.D. 1996. Ecological differences in tropic
Herpetology, v. 30, n.1, p.60-67.
142

CAPÍTULO 3
INFLUÊNCIA DA FRAGMENTAÇAO FLORESTAL E DA QUALIDADE DA MATRIZ DE MONOCULTURA DE EUCALIPTO SOBRE A COMPOSIÇÃO DAS COMUNIDADES DE ANFÍBIOS DE FOLHIÇO EM ÁREAS DE MATA ATLÂNTICA NO EXTREMO SUL DA BAHIA.
143

RESUMO
Os anfíbios apresentam grande relevância entre os habitantes da fauna de
florestas tropicais, porque várias espécies possuem ampla
servir como espécies-chave na
etecção de distúrbios do ambiente, pois a composição das suas comunidades pode
er afetada negativamente, em decorrência do efeito da fragmentação florestal. Este
studo objetivou comparar a composição e biomassa total da comunidade de anfíbios
e serrapilheira em três principais componentes florestados da paisagem do extremo
ul da Bahia: Mata Contínua, Remanescentes em estágio intermediário de
generação e Eucaliptais em estágio final de crescimento. Foi avaliada também a
smocleis
omponentes da paisagem amostrados. As espécies Stereocyclops incrassatus e
hiasmoclei schubarti apresentaram os maiores valores de biomassa total.
Estatisticamente, baseado no resultados da análise do MRPP, não foi possível
etectar diferenças na composição das comunidades (p= 0,4824; T = 0,1571), e na
biomassa total de anfíbios de folhiço nos diversos componentes da paisagem
avaliados de anfíbios (p= 0,1876; T= - 0,8132). A análise de regressão linear não
evidenciou correlação entre o padrão de distribuição das abundâncias dos anfíbios e
as variáveis ambientais mensuradas (r2 = 0,97; F= 0,028). Porém, foi marginalmente
significativo quando realizado para a biomassa total de anfíbios e o primeiro eixo do
componente principal da redução das variáveis ambientais PC1 (r2 = 0,05; T = 2,257).
Não foram detectadas diferenças significativas quanto à composição das
comunidades de anfíbios de folhiço nos componentes da paisagem estudados,
especialmente entre aqueles florestados (Mata Contínua e Remanescentes) e os
Eucaliptais. Tais resultados indicaram que a qualidade da matriz de monocultura de
eucalipto, com desenvolvimento de sub-bosque de nativas, pode ser considerada
como adequada para conectar pequenos fragmentos de floresta em estágios
termediários de sucessão e parece não estar interferindo negativamente na
composição dessa comunidade avaliada.
serrapilheira de
distribuição geográfica e potencialmente podem
d
s
e
d
S
re
existência de associação entre a composição e biomassa da fauna de anfíbios e as
variáveis ambientais que expressam características de micro-habitat, micro-clima e
geometria da paisagem. Foram capturados 523 anfíbios de folhiço, nos quarenta dias
de coletas, correspondentes a treze espécies distintas, das quais Chia
schubarti e Stereocyclops incrassatus foram as dominantes, presentes em todos os
c
C
d
in
144

ABSTRACT Amphibians are of great relevance amongst the inhabitants of the scrub vegetation
fauna of tropical forests, since various species possess ample geographic distribution,
potentially serving as key-species to detect environmental disturbances, whose
community composition can be negatively affected by the effect of forest
fragmentation. This study aimed at comparing the composition and total biomass of
the scrub vegetation amphibian community in three main forested landscapes of the
extreme south of Bahia: continuous native forest, forest remnants in intermediary
stage of regeneration and eucalyptus plantations in the final stage of growth. The
existence of an association between the composition and biomass of amphibians and
environmental variables that express micro-habitat, micro-climate and landscape
geometry, was also evaluated. 523 scrub vegetation amphibians were captured during
the 40 days of collection, corresponding to 13 distinct species. Chiasmocleis schubarti
and Stereocyclops incrassatus were dominant, present in all the landscape
components sampled. The species Stereocyclops incrassatus and Chiasmoclei
schubarti presented the largest value in total biomass. Statistically, based on the
results of the MRPP analysis, it was not possible to detect differences in the
composition of the communities (p= 0,4824; T = 0,1571) or in the total biomass of
scrub vegetation amphibians in the diverse components of the evaluated amphibian
landscape (p= 0,1876;T= - 0,8132). Linear regression analysis did not show a
correlation between the pattern of abundance distribution of the amphibians and the
measured environmental variables (r2 = 0,97; F= 0,028). However, it was marginally
significant when done for the total biomass of the amphibians and the first axle of the
main component of environmental variable PC1 (r2 = 0,05; T = 2,257). As for the
composition of the scrub vegetation amphibian community in the landscape
components studied, no significant differences were detected, especially between the
forested (continuous and remnant native forests) and the eucalyptus plantations,
indicating that the matrix quality of eucalyptus monoculture with development of native
undergrowth, can be considered adequate for connection of small forest fragments in
the intermediary stage of succession and seems not to be interfering negatively on the
composition of this community.
145

INTRODUÇÃO
scon, 1999; Vallan, 2000; Stallard, 2001; Toral et. al.,
001; Pineda & Halffter, 2004).
O conhecimento sobre a composição dos grupos de vertebrados de uma área é fator
de importância primordial em projetos para a sua conservação. Assim, a identificação
das espécies de anfíbios e o estudo de suas particularidades ecológicas revelam-se
decisivos para o sucesso de ações que busquem conservar a sua biodiversidade
(Heyer et al., 1994).
Os anfíbios apresentam grande relevância entre os habitantes da fauna de
serrapilheira de florestas tropicais, porque, segundo alguns autores, várias espécies
possuem ampla distribuição geográfica e podem servir potencialmente como
espécies-chave na detecção de distúrbios do ambiente (Jim, 1980; Watson. et al.,
1995; Silvano & Pimenta, 2003).
Fatores como a alteração da umidade, a profundidade do folhiço, a altitude e a
temperatura, podem influenciar no padrão da distribuição dos anfíbios de uma
determinada paisagem, ocasionando alterações na densidade, biomassa e riqueza
dessas comunidades, porque certas características fisiológicas (como a pele
permeável) e ecológicas (como clico de vida bifásico) tornam esses animais
extremamente dependentes da água e sensíveis a alterações de parâmetros físico-
químicos no ambiente.
Tais fatores evidenciam que a composição das comunidades de anfíbios mantém
íntima relação com o seu habitat natural (Bradford et. al., 1993; Blaustein et al., 1994;
Dixo, 2001) e podem ser negativamente afetadas quanto à sua riqueza e abundância
ao submeterem-se aos efeito da fragmentação florestal (Scott, 1976; Fauth et al,
1989; Wyman, 1990; Wake, 1991; Vallan, 2000).
Poucos estudos investigaram empiricamente o efeito da fragmentação e de seus
processos associados sobre as comunidades de anfíbios de regiões tropicais
(Lieberman, 1986; Heinen, 1992; Marsh & Pearman, 1997; Tocher et al, 1997;
Knutson et. al., 1999; Lima & Ga
2
146

A grande maioria dos trabalhos que abordaram esse tema no Brasil provém do
rojeto Dinâmica Biológica de Fragmentos Florestais na Amazônia (PDBFF) ou que
poeiras), ocasionando um aumento na riqueza dos fragmentos
pós o isolamento (Lima & Gascon, 1999).
levantamento em vinte e uma áreas inseridas no
orredor Central nos anos de 2000 e 2001, onde foram listadas noventa e duas
da paisagem em áreas de Mata Atlântica; ou o papel das
terações ecológicas sobre a estrutura de suas comunidades e seus mecanismos de
Mata Atlântica no Sul da Bahia.
oram comparadas abundância, riqueza, diversidade, dominância e equitatividade
ma fisionomia florestal nas cabrucas e capoeiras a
manutenção das comunidades.
P
retratam apenas a realidade da composição faunística deste grupo sobre a Região
Amazônica (Ramos & Gallatti, 2002). Após dez anos de investigação, foi detectado
que a riqueza de sapos estava positivamente relacionada com o tamanho da área,
em virtude da presença de espécies oportunistas associadas a matriz do habitat
alterado (pastos e ca
a
Na Mata Atlântica são conhecidas cerca de 340 espécies de anfíbios, das quais
aproximadamente oitenta são endêmicas do bioma (Kwet & Di-Bernardo, 1999; MMA,
2003; Silvano & Pimenta, 2003; Prado et al., 2003). Em relação à Mata Atlântica do
Sul da Bahia, foi realizado um
C
espécies de anfíbios anuros, das quais catorze foram consideradas como novas
descobertas para a Ciência (Silvano & Pimenta, 2003).
Apesar desse avanço sobre o conhecimento do grupo, a grande maioria dos
trabalhos sobre anfíbios de folhiço retratam levantamentos em área pontuais e quase
nenhum trabalho explorou a resposta desses animais à redução dos ambientes
florestados no contexto
in
adaptação diante das alterações ambientais.
O Projeto RESTA UNA contribuiu para o aprofundamento de tal abordagem por meio
dos estudos realizados por Dixo em 2001, o qual investigou a composição das
comunidades de anfíbios de folhiço em áreas de
F
dos anfíbios em seis habitats diferentes (interior e borda de Mata Contínua; interior e
borda de fragmento de mata; cabruca - plantação de cacau sombreada; e capoeira -
Mata Secundária). Os resultados obtidos não evidenciaram alterações no padrão da
composição das comunidades nas áreas amostradas e atribuíram aos fatores de
presença da serrapilheira e de u
147

Outro estudo relevante sobre a composição das comunidades de anfíbios anuros na
Mata da Bahia foi realizado por Silvano & Pimenta, em 2003, que investigaram
fragmentos florestais de diversos tamanhos, determinando sua riqueza, abundância,
freqüência relativa e similaridade entre as áreas amostradas, com objetivo de
fornecer indicadores úteis para as políticas de conservação da biodiversidade e a
efetiva implantação do Corredor Central.
Esses autores realizaram coletas amostrais nos anos de 2000 e 2001, através da
tilização de diversas metodologias (identificação de vocalizações, busca ativa e
dos sistemas de referência),
manescentes de pequeno porte em estágio intermediário de regeneração (os mais
a promoção de conectividade entre os
agmentos, conforme descrição detalhada contida no Capítulo 2 dessa Dissertação.
u
armadilhas de queda “pitfall traps”), onde identificaram a existência de 92 espécies de
anfíbios anuros nos 21 fragmentos estudados.
O presente estudo teve por objetivo comparar a composição e biomassa total da
comunidade de anfíbios de serrapilheira entre os três principais componentes
florestados da paisagem do extremo Sul da Bahia: remanescentes de grande porte de
mata em estágio avançado de regeneração (considera
re
comuns na paisagem) e eucaliptais em estágio final de crescimento (7 anos de
idade). Além disso, foi avaliada a existência de associação entre as duas variáveis
indicadoras da fauna (composição e biomassa) e variáveis ambientais que expressam
características de micro-habitat, micro-clima e Ecologia da Paisagem. Com base nos
resultados, foam discutidos os efeitos da fragmentação sobre esse componente da
fauna e a qualidade da matriz de eucalipto n
fr
148

De acordo com os objetivos propostos, pretendeu-se responder as seguintes
perguntas:
4. Quais as principais diferenças na composição das comunidades de anfíbios
de folhiço presentes nos fragmentos do tipo mais comum na região
(remanescentes florestais em estágio intermediário de sucessão entre 100 e
250 ha) e a mata original (estimada com base no fragmento de 6.000 ha)?
esta em estágios intermediários de
sucessão?
5. Quais variáveis ambientais e variáveis de paisagem mensuradas estão
correlacionadas a essas diferenças?
6. A monocultura de eucalipto pode ser considerada uma matriz adequada para
conectar pequenos fragmentos de flor
149

MATERIAL E MÉTODOS
Sim
dessa
Resultados Foram capturados 523 anfíbios de folhiço, nos quarenta dias de coletas realizadas,
cor
Leptod
número total de indivíduos e o local de captura de cada espécie.
A curva média do coletor para os anfíbios capturados nos três componentes da
paisagem não atingiu um platô, evidenciando que não foram coletadas todas as
espécies existentes nas áreas estudadas (Figura 32).
As espécies Chiasmocleis schubarti, Stereocyclops incrassatus foram as
dominantes, presentes em todos os componentes da paisagem amostrados,
correspondendo a 75% e 10,5% de todos os indivíduos coletados. As demais
espécies Eleutherodactylus binotatus, Leptodactylus gr. Fuscus, Proceratophrys
laticeps, Bufo crucifer, Scinax sp., Arasphenodon brunei e Dcsypops schirch, foram
exclusivas de algum componente da paisagem, com índice reduzido de captura. As
abundâncias individuais das espécies podem ser observadas detalhadamente na
Figura 33.
Quanto à biomassa, as espécies Stereocyclops incrassatus e Chiasmoclei schubarti
apresentaram os maiores valores, enquanto que as demais espécies obtiveram
valores muito inferiores quando comparados com as demais. (Tabela 4). Os
Remanescentes e a Mata de Referência, com destaque para (M4 e M2)
representaram os componentes da paisagem com maiores valores totais de
biomassa, enquanto os Eucaliptais apresentaram os menores valores. Figura 34.
ilares aos adotados no estudo de lagartos de folhiço, descritos no Capítulo 2
Dissertação.
respondentes a 13 espécies, pertencentes a 5 famílias diferentes: Microhylidae,
actylidae, Bufonidae, Hylidae e Dendrobatidae. A Tabela 3 apresenta o
150

De uma forma geral, todos os componentes da paisagem estudados apresentaram
alores similares de riqueza e abundância em relação aos anfíbios coletados.
Apenas a Mata Contínua (M1) e o Remanescente (R1) se destacaram dos demais,
v
apresentando os maiores valores, respectivamente (Figura 35).
151

Tabela 3. Espécies de anfíbios capturados nas 12 áreas amostradas no extremo sul etrarepresentam as unidades amostrais da mata contínua; R1 a R4, dos remanescentes; e E1 a E4, dorespectivamente.
Família Espécie M1 M2 M3 M4 R1 R2 R3 R4 E1 E2 E3
da Bahia. As l s M1 a M4, s eucaliptais,
E4 Total Microhylidae Chiasmoclei schubarti 4 182 3 2 179 11 7 5 Stereocyclops incrassatus 1 2 9 16 17 6 1 1 Leptodactylidae Eleutherodactylus binotatus 5 5 Leptodactylus gr. fuscus 1 Physalaemus gr. cuvieri
1 1 1 1
Physalaemus signifer 16 3 2 Proceratophyrys laticeps 1 Bufonidae Bufo crucifer 1 Bufo margaritifer 1 1 3 6 1 Hylidae Scinax sp. 1 1 Dendrobatidae Colostethus sp. 21 Aparasphenodon brunei 1 Dscypops schirchi 3 TOTAL 5 223 10 18 203 14 30 14 1 2 2 1
1 394 53 10 1 4
21 1 1 11 2 21 1 3
523

Tabela 4. Biomassa dos anfíbios capturados nas 12 áreas amostradas no extremo sul da Bahia. As letras M1 a M4, representam as unidades amostrais da mata contínua; R1 a R4, dos remanescentes; e E1 a E4, dos eucaliptais, respectivamente.
M1 M2 M3 M4
Família Espécie R1 R2 R3 R4 E1 E2 E3 E4 Total Microhylidae Chiasmoclei schubarti 8,Stereocyclops incrassatus
53 1 ,19 7,72 2,61 9,8 ,34 0923,23 257,062 269,55 472,69 136,51 23,64 24,11 1216,3
8,35 47,74 56,0
r. fuscus 9,33 9,33 i 0,1 4,87 1,2 4,87 11,04
42,1 3,54 52,0 5,85 5,22 5,22 2,48 2,48 35,88 123,0 1,09 0,5 1,59 1,45 1,45 18,03 18,03 ,14 36,14
Total 11,01 39,3 510,78 155,11 23,64 10,09 33,44 3,39
39 19 5 5,81 35,89 6 3,39 4 ,3 9,57
Leptodactylidae Eleutherodactylus binotatus
9
Leptodactylus g Physalaemus gr. cuvierPhysalaemus signifer 6,41 5 Proceratophyrys laticeps 5,85 Bufonidae
Bufo crucifer Bufo margaritifer 82,17 Hylidae
Scinax sp. Dendrobatidae
Colostethus sp. Aparasphenodon brunei Dscypops schirchi
330,3136
195,98 41,68 590,75 1945,49
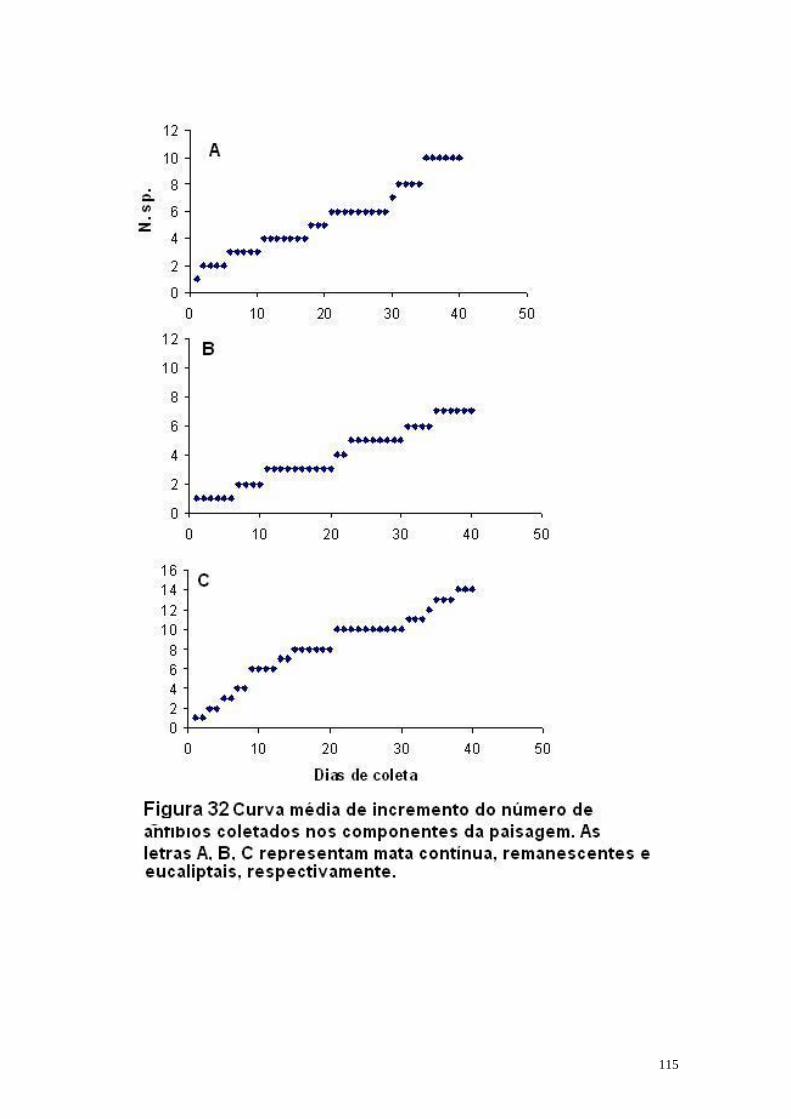
115

116

117

Descrição das Variáveis Ambientais Dados similares ao Capítulo 2.
Composição da comunidade de anfíbios de folhiço entre os diversos componentes da paisagem.
Considerando que cada componente de paisagem estabelecida representa
uma unidade natural na paisagem, ou seja: o valor esperado das variáveis não
controladas para todas as réplicas de cada componente de paisagem é o
mesmo (Kachingan 1986: 297), estatisticamente baseado no resultados da
análise do MRPP, tal como no caso dos lagartos, não foi possível detectar
iferenças na composição das comunidades de anfíbios de folhiço nos diversos
omponentes da paisagem avaliados (p= 0,4824; T = 0,1571). Resultados
omassa total de anfíbios, baseado na
nálise de (p= 0,1876; T= - 0,8132).
RESULTADOS ANÁLISE DE ORDENAÇÃO EM RELAÇÃO AS VARIÁVEIS AMBIENTAIS
Dados similares aos apresentados no Capítulo 2 desta Dissertação.
RESULTADOS DA ANÁLISE DE ORDENAÇÃO EM RELAÇÃO À COMUNIDADE DE ANFÍBIOS Tal como no estudo dos lagartos, foi escolhido um eixo único do NMS,
correspondendo ao eixo que melhor representa a distribuição de distâncias em
relação à distribuição real dos pontos.
d
c
similares aos encontrados quanto à bi
a
118

O NMS único extraído apresentou um estresse de 25 (Figura 36), que apesar
e elevado, segundo o Teste de Monte Carlo, demonstra existir uma estrutura
bundâncias, mais forte do que o esperado pelo
caso (p= 0,0976). O teste de Mantel evidenciou uma correlação de 74% entre
d
no padrão da distribuição das a
a
as distribuições reais e as do NMS único (p=0,001; r2 = 0,86).
A análise de regressão linear entre o NMS (variável dependente) e os PC1 e
PC2 (variáveis independentes) não evidenciou correlação entre o padrão de
distribuição das abundâncias dos anfíbios e as variáveis ambientais
ensuradas (r2 = 0,97; F= 0,028). Porém, foi marginalmente significativa
uando realizada para a biomassa total de anfíbios e o primeiro eixo do
omponente principal da redução das variáveis ambientais PC1 (r2 = 0,05; T =
2,257) Figura 37.
m
q
c
119

A qualidade da ordenação indireta pelo NMS foi apresentada através dos
gráficos de abundâncias das espécies, ordenadas pelos seus escores
calculados pela média recíproca, e pelas unidades amostrais, ordenadas pelo
etor de ordenação da comunidade NMS (Figura 38). Esses gráficos v
evidenciam o eixo que melhor representa a variação ambiental sobre a
distribuição das abundâncias dos anfíbios.
120

Figura 38. Representação das distribuições de abundâncias das espécies de sapos coletados nos diversos componentes da paisagem estudados no melhor eixo de ordenação.
121

DISCUSSÃO
Com a realização desse estudo, dos 523 anfíbios de folhiço capturados, foi
possível identificar 13 espécies diferenciadas nas áreas avaliadas, pertencentes
a 5 famílias diferentes: Microhylidae, Leptodactylidae, Bufonidae, Hylidae e
Dendrobatidae.
Trabalhos sobre a fauna de anfíbios de folhiço na Mata Atlântica citam a
presença de 340 espécies, correspondendo a 65% das espécies brasileiras
conhecidas, onde 80% são endêmicas. Na Bahia, são listadas a ocorrência de
aproximadamente 133 espécies distribuídas em todos os biomas do Estado,
onde foram citadas a presença de 115 espéc
Atlântica do Sul da Bahia, habitat da maioria do
(Bonino, 1987; Teixeira, 1957; Dixo, 2001; Silvano
Poucos são os trabalhos metódicos existentes d nga duração na
região, direcionados especificamente para a fauna de anfíbios anuros
representativos desse bioma, avaliando o efeito da fragmentação. As
informações disponíveis limitaram-se a descrições das espécies ocorrentes,
resultantes de coletas esporádicas realizadas principalmente por naturalistas
europeus no século passado e pelo extenso trabalho de Bokermann (1966a,
1966b, 1966c, 1968, 1973, 1974).
De acordo com levantamentos recentes, diversas espécies novas ainda estão
sendo descobertas na Mata Atlântica da Bahia, evidenciando incertezas quanto
à diversidade dessa fauna regional, e a necessidade de se obter mais dados
empíricos sobre a fauna de anfíbios desse bioma (Cardoso et al., 1989; Rossa-
Feres & Jim, 1994; Pombal –Jr, 1997; Haddad, 1998; .Silvano & Pimenta, 2003).
Segundo estudos realizados por Silvano & Pimenta em 2003, foi possível
identificar a existência de 92 espécies de anfíbios anuros em 21 fragmentos
florestais de diversos tamanhos na Mata Atlântica da Bahia. Esses autores
realizaram diversas metodologias de coleta (identificação de vocalizações,
usca ativa e utilização de armadilhas de queda “pitfall traps”). Porém, na RPPN
ies para o domínio da Mata
s anfíbios anuros conhecidos
& Pimenta 2003).
e média e lo
b
122

Estação Veracruz, foram detectadas 36 espécies, as quais cerca de 20 foram
entificadas pelas vocalizações ou buscas ativas. Cavalcanti, 2004 (em
ados obtidos nesse estudo (13 espécies) não se distanciam
aqueles realizados na mesma área de estudo, utilizando-se de metodologias
t al., 1997;
imenta & Silvano, 2003) e Stereocyclops incrassatus, que também apresenta
contradas nos Remanescentes e na Mata
ontínua, sendo que a espécie Physalaemus gr. signifer é considerada de
id
preparação) realizou estudo similar, utilizando semelhante metodologia de
coleta, na Mata Contínua (RPPN Estação Veracruz) e em Eucaliptais do
extremo Sul da Bahia, identificando a presença de 13 espécies de anfíbios.
Assim, os result
d
de captura através de coletas passivas para a captura dos anfíbios de folhiço
nessas áreas.
A maioria das espécies capturadas são consideradas como típicas de mata, e
ao mesmo tempo, foram as dominantes nos componentes da paisagem
avaliados neste estudo: Schiasmocleis schubarti - reconhecida como típica de
Mata Atlântica, recentemente registrada para a Bahia - (veja Cruz e
P
hábitos florestais, similares a Schiasmocleis, considerada como endêmica desse
bioma, presente nos estados do Espírito Santo, leste de Minas Gerais e Sul da
Bahia (Frost, 2002). Apesar de serem espécies típicas de ambientes florestais e
endêmicos do bioma, essas duas espécies também foram capturadas nos
Eucaliptais estudados.
As demais espécies foram mais en
C
hábito generalista, ocupando áreas abertas, mas a sua distribuição estava
restrita à REBIO UNA, no Parque Estadual da Serra do Conduru, em Itacaré,
Bahia. Esta espécie é filogeneticamente relacionada a Physalaemus signifer,
considerada como uma espécie típica de florestas (Izecksohn & Carvalho &
Silva, 2001; Weber & Carvalho & Silva, 2001; Pimenta & Silvano, 2003).
123

Bufo margaritifer também é considerada uma espécie de hábitos florestais,
endêmica do Sul da Bahia, podendo também ser encontrada nas bordas de
mata, enquanto que o status de Bufo crucifer ainda é duvidoso, podendo ser
encontrada em diferentes fisionomias do domínio do bioma, tanto em áreas
abertas quanto em matas (Heyer et. al. 1990; Izeckson & Carvalho & Silva,
2001; Weber & Carvalho & Silva, 2001; Pimenta & Silvano, 2003).
Eleutherodactylus binotatus, Scinax sp. podem ocupar áreas abertas e
florestadas, apresentando ampla distribuição geográfica na América do Sul. As
spécies da família Dendobratidae (Aparasphenodon bruenei, Colostethus sp. e
omponentes florestados da paisagem do extremo Sul da
ahia: remanescentes de grande porte de mata em estágio avançado de
mbora os anfíbios sejam reconhecidos pela grande fidelidade ao habitat natural
evido ao fato de apresentarem certas características fisiológicas (pele
ermeável) e ecológicas (ciclo de vida bifásico) (Toft, 1980; Bradford et. al.,
993; Blaustein et al., 1994; Dixo, 2001), o padrão da distribuição das
bundâncias de anfíbios não parece estar associado àquele evidenciado para as
e
Dcsypops schirchi) são predominantemente amazônicas e foram apenas
detectadas na Mata Continua. Todas essas espécies ocupam preferencialmente
ambientes florestados, consideradas como espécies bioindicadoras, vinculado
ao seu modo reprodutivo altamente especializado de colocar os seus ovos no
folhiço úmido (Rossa-Feres & Jim, 1994; Frost, 2002).
Em relação à composição das comunidades de anfíbios de folhiço avaliadas nas
unidades amostrais, os dados indicaram que, apesar do intenso processo de
destruição florestal sofrido pela Mata Atlântica na região, não há diferenças
entre os três principais c
B
regeneração (considerados sistemas de referência – Mata Contínua),
remanescentes de pequeno porte em estágio intermediário de regeneração (os
mais comuns na paisagem - Remanescentes) e Eucaliptais em estágio final de
crescimento, com 7 anos de idade. Não foi possível detectar se a fragmentação
e seus processos associados estão interferindo negativamente na composição
das comunidades de anfíbios de folhiço nas áreas amostradas.
E
d
p
1
a
124

variáveis ambientais, o que indica que a composição da comunidade pode estar
vinculada a outros fatores além daqueles estudados.
Do ponto de vista da biomassa total, foram identificadas diferenças no padrão de
distribuição entre a Mata Contínua e os Eucaliptais, diferenças associadas
positivamente com as variáveis ambientais: proximidade com florestas nativas,
ompactação do solo, decomposição de troncos caídos, densidade de folhagem,
nos Eucaliptais.
iço.
c
número e percentagem de troncos caídos; e negativamente com perímetro de
tronco, distância entre as árvores e luminosidade.
Sabe-se que, ao lado dos lagartos de folhiço, os anfíbios representam um dos
principais grupos taxonômicos que convertem biomassa de artrópodes terrestres
em biomassa disponível aos níveis tróficos superiores (Renken et.al.,2003),
indicando que, no tocante à fauna de anfíbios de folhiço, pode existir maior
disponibilidade de biomassa na Mata Contínua, do que
A grande maioria das espécies de anfíbios identificados nesse estudo são
típicas de ambientes florestais, ou endêmicas do Bioma Mata Atlântica. Como
não foram detectadas diferenças significativas quanto à composição e ao
número de espécies de anfíbios de folhiço presentes nos diversos componentes
da paisagem estudados, especialmente entre aqueles florestados (Mata
Contínua e Remanescentes) e os Eucaliptais. Esses resultados indicaram que a
qualidade da matriz de monocultura de eucalipto nas áreas avaliadas (com
desenvolvimento de sub-bosque de nativas, 7 anos de idade) pode ser
considerada como adequada para conectar pequenos fragmentos de floresta em
estágios intermediários de sucessão e não parece estar interferindo
negativamente na composição da comunidade de anfíbios de folh
Resultados similares também foram identificados por Demayanadier & Hunter
em 1998, os quais avaliaram o efeito de borda e o papel da matriz de
monocultura de eucalipto na parte central de Maine, USA, onde tanto espécies
generalistas como aquelas típicas de matas habitaram os diversos ambientes da
paisagem.
125

REFERÊNCIAS BIBLIOGRÁFICAS BONINO, A.R.L. 1987 - Levantamento da Faauna da Mata Atlântica do Sul da
gênero Phyllodites Wagler, 1830 (Anura, Hylidae). Anais da Academia Brasileira de Ciências, 38(2): 335-344.
1974). Três espécies novas de Eleutherodactylus do sudeste da Bahia, Brasil (Anura, leptodactylidae). Revista Brasileira de
ARDOSO, A. J.; ANDRADE, G. V.; HADDAD, C. F. B. 1989. Distribuição
Bahia. JOÃO PESSOA – FUNAPE BOKERMANN, W. C. A. (1966a). Duas novas espécies de Sphaenorhynchus
(Amphibia, Hylidae). Revista Brasileira de Biologia, 26(1): 15-21. BOKERMANN, W. C. A. (1966b). Notas sobre três espécies de Physalaemus
de Maracás, Bahia (Amphibia, Leptodactylidae). Revista Brasileira de Biologia, 26(3): 253-259.
BOKERMANN, W. C. A. (1966c). O
BOKERMANN, W. C. A. (1968). Three new Hyla from the Plateau of Maracás,
Central Bahia, Brazil. Journal of herpetology, 1(1-4): 25-31. BOKERMANN, W. C. A. (1973). Duas novas espécies de Sphaenorhynchus
da Bahia (Anura, Hylidae). Revista Brasileira de Biologia, 33(4): 589-594. BOKERMANN, W. C. A. (
Biologia, 34(1): 11-18. BRADFORD, D. F., TABATABAI, F. & GRABER, D. M. 1993. Isolation of the
native frog, Rana muscosa, by introduced fishes in Sequoia and Kings Canion National parks, California. Conservation Biology 7: 882-888.
BLAUSTEIN, A. R., WAKE, D. B., SOUSA, W. P. 1994. Amphibians declines:
Judging stability, persistence, and susceptibility of populations to local and global extinction. Conservation 2002.Biology 8, 60-71.
C., EDUARDO TORAL; FEINSINGER, PETER; CRUMP, MARTHA L. Frogs
and a Cloud-Forest Edge in Ecuador. Conservation Biology, v.16, n.3, p.735-744. 2002.
CARAMASCHI, U. ; CRUZ C.A.G.; FEIO, R.N. 2000 – Geography Distribution.
Phasmahyla jandaia. Herpetological Review 31(4):251 C
espacial em comunidades de anfíbios (Anura) no sudeste do Brasil. Revista Brasileira de Biologia, v.49, n.1, p.241-249.
CRUZ, C.A.G.; PIMENTA, B.V.S. e SILVANO, D.L.- The genus Chiasmocleis
Méhely, 1904 (Anura Microhylidae) in the Atlantic Rain Forest of Brazil, with description of three new species. Alytes, 15(2): 49 – 71
126

DEMAYNADIER PHILLIP G.; JR. MALCOSivicultural Edges on the Distribution and Abunda
LM L. HUNTER. 1998. Effects of nce of Anphibians in
Maine. Conservation Biology, 12(2), p.340-352.
DIXO tação da Floresta sobre a comunidade de sapos e lagartos de serrapilheira no sul da Bahia. Dissertação de
AUTH, J. E., CROTTHER, B.I. & SLOWINSKI, B. 1989. Elevational patterns
RAGA .L.; WEIDERHECKER, H. & SANTOS, P.S.- Anfíbios do
, M. B. de. 2001. Efeito da Fragmen
mestrado, USP.
Fof richness, evenness, and abundance of the Costa Rican leaf litter herpetofauna. Biotropica 21: 178-185.
FEIO, R.; B U.MParque Estadual do Rio Doce (Minas Gerais), Brasil, Revista Ceres, 42(239):53-61
FROST, W.D. 2002 – Amphibians species of the world. An online reference .
http://research.amnb.org/herpetology/amphibia/ HADDAD, C. F. B. (1998). Biodiversidade dos anfíbios no Estado de São
Paulo. In: Castro, R. M. C. (ed.). Biodiversidade do Estado de São Paulo, Brasil. São Paulo: FAPESP. P. 15-26. 71p. (Série Vertebrados).
HEINrimary rain forest in Costa Rica, some implications
for fauna restaurations. Biotropica 24, 431-438.
HEYersity.
Standard methods for Amphibians. Smithsonian Institution Press,
IZEC unicípio do Rio de
Janeiro: Editora UFRJ, 2001. 148p.
JIMPaulo (Amphibia, Anura). Dissertação de Doutorado,
Universidade de São Paulo, São Paulo.
KWE o. – Porto Alegre: EDIPUCRS,
KNUANNOO, MICHAEL J.
Conservation Biology, v.13, n.6, p.1437-1446. 1999
LIEB5 1-72
EN, J.T. 1992 Comparisons of the leaf litter herpetofauna in abandoned cacao plantations and p
ER, W. R.; DONNELLY, M. A. ; McDIARMID, R. W. ; HAYEK, L. C. e FOSTER, M. S. 1994. Measuring ands monitoring biological div
Washington.
KSOHN, E. & CARVALHO E SILVA, S.P.- Anfíbios do M
, J. 1980. Aspectos ecológicos dos anfíbios registrados na região de
Botucatu, São
T, AXEL. Anfíbios. / Axel Kwet, Marcos Di-Bernard
TSON, MELINDA G.; SAUER, JOHN R.; OLSEN, DOUGLAS A.; MOSSMAN, MICHAEL J.; HEMESATH, LISA M.; L
ERMAN, S.S. 1986 Ecology of the leaf litter herpetofauna of a neotropical rain forest: La Sela, Costa Rica. Acta 200L Mex. Nueva Ser 1
127

LIMA, M. G. & GASCON, C. 1999. The conservation value of linear forest remnants in central Amazonia. Biological Conservation 91: 241-247.
ARSH, D. M. & PEARMAN, P. B. 1997. Effects of habitat fragmentation on
MA 2003. Ministério do Meio Ambiente: Anexo à Instrução Normativa, n. 3, de
Fa
IMENTA, B.V.S. & SILVANO, D.L. 2002 – Geographic Distribution.
PINEDA, EDUARDO, HALFFTER, GONZALO.; Species diversity and habitat
fragmentation: frogs in a tropical montane landscape in Mexico. Biological
OMBAL Jr., J. P. (1997). Distribuição especial e temporal de anuros
d
RADO, P. L., LANDAU, E. C., MOURA, R. T., PINTO, L. P. S., FONSECA,
divCo
ENKEN, ROCHELLE B., GRAM, WENDY K., FANTZ, DEBRA K., RICHTER,
B 3. Effects of Forest Management on Amphibians and Reptiles in Missouri Ozark Forests.
ROStu, São Paulo. Revista Brasileira
de Biologia, 54(2): 323-334.
SCOTT, N. 1976. The abundance and diversity of the herpetofauna of tropical forest litter Biotropica 8, 41-58
SILV p.1-18.
De hives. Conservation Biology, v.15, n.4, p.943-953. 2001.
Mthe abundance of two species of Leptodactylidae frogs in the Andean montane forest. Conservation Biology 11: 1323-1328.
M27 de maio de 2003, do Ministério do Meio Ambiente – Lista das Espécies da
una Brasileira Ameaçadas de Extinção.
PChiasmocleis schubarti. Herpetological Review, 33(3).219.
Conservtion 117 (2004) 499-508.
P(Amphibia) em uma poça permanente na Serra de Paranapiacaba, sudeste o Brasil. Revista Brasileira de Biologia, 57(4): 583-594.
P
G. A. B., ALGER, K. (Orgs.) 2003. Corredor de Biodversidade da Mata Atlântica do SuL da Bahia. Publicação em CD-ROOM, Ilhéus, IESB/CI/CABS/UFMG/UNICAMP.
RAMOS, CLAUDIA AZEVEDO; GALATTI, ULISSES. Patterns of amphibian ersity in Brazilian Amazônia: conservation implications. Biological nservation, 103 (2002) 103-111.
RSTEPHEN C., MILLER, TIMOTHY J., RICKE, KEVIN B., RUSSEL,
RADLEY, WANG, XIAOYIN. 200
SA-FERES, D. C.; JIM, J. (1994). Distribuição sazonal em comunidades de anfíbios anuros na região de Botuca
ANO, LEITE DÉBORA; PIMENTA, BRUNO V. S.Diversidade e Distribuição de Anfíbios na Mata Atlântica do Sul da Bahia. 2003.
STALLARD, ROBERT. Possible Environmental Factors Underlying Amphibian
cline in Eastern Puerto Rico: analysis of U. S. Government Data Arc
128

TEIXEIRA, Dante Martins 1957 – Animais Vertebrados Ameaçados de Extinção Presentes na Mata Atlântica – Fundação Biblioteca Nacional
TOCHER, M. D., GASCON, C. & ZIMMERMAN, B. L. 1997. Fragmentation
effects on a Central Amazonian frog community: a ten year study, pp. 124-137. In: Laurance, W. F., Biergaard R. O. (Eds.). Tropical Forest Remnants: Ecology, Management, and Conservation of Fragmented
TO y of Thirteen Syntopic Species of
Anurans in a Seasonal Tropical Environment. Oecologia (Berl) 45, 131-
ALLAN, 2000 Influence of forest fragmentation on the amphibian diversity in
WAKE, D. B. 1991. Declining amphibian populations. Science 253: 860.
WE Lectodactylidae) e
informações sobrea reprodução e a distribuição geográfica da espécie.
WAT n temporary
ydrobiologia 299: 53-73.
Communities. University of Chicago Press, Chicago.
FT, CATHERINE A. 1980. Feeding Ecolog
141.
Vthe nature reserve of Ambohitantely. Highland Madagascar. Biological Conservation 96, 31-43
BER, L.N. & CARVALHO-E-SILVA, S.P. 2001. Descrição da larva de
Physalaemus signifer (Girard, 1853) (Amphibia, Anura,
Boletim do Museu Nacional, N.S., Zoologia (462):1-6
SON, G. F.; DAVIES, M.; TYLER, M. J. 1995. Observationsowaters in northwestern Australia. H
WYMAN, R. L. 1990. What is happening to the amphibians? Conservation Biology 4: 350-352.
129

ÍTULO 4 CAP
CON
ROPOSTAS DE CONSERVAÇÃO E MANEJO PARA A REGIÃO DE E
CLUSÕES GERAIS E CONSIDERAÇÕES SOBRE P
STUDO.
130

CONCLUSÕES FINAIS
ste estudo não pôde identificar se a fragmentação florestal e seus processos
ssociados interferem negativamente na composição das comunidades de
do Brasil (em especial, á da sub-região do extremo Sul da
ahia) é reconhecida como prioritária para a conservação no Planeta,
ncontrando-se reduzida a menos de 7% da sua cobertura original e ainda
presenta simultaneamente elevados índices de endemismos e de
iodiversidade local. Apesar desse estudo não ter detectado alterações
negativas na abundância e biomassa total nas comunidades investigadas, e ter
sido fundamentado a partir de um desenho amostral consistente e robusto,
tratando-se da primeira iniciativa empírica na região que avalia tal temática e
compara os diversos componentes da paisagem existentes, tais conclusões
ainda representam dados preliminares.
Diante da complexidade dos efeitos da fragmentação, da particularidade de
cada paisagem fragmentada e da conseqüente necessidade de incluir-se
escalas espaciais amplas, deve-se priorizar a realização de outros estudos que
possam investigar comparativamente os resultados obtidos nessa Dissertação,
com outros grupos biológicos que também habitam a região.
Apesar do padrão da distribuição das abundâncias de lagartos não estar
associado àquele evidenciado para as variáveis ambientais, esse estudo
propiciou detectar a existência de gradientes ambientais entre os Eucaliptais, os
Remanescentes e a Mata Contínua. A ordenação das variáveis ambientais a
partir dos dados mensurados de micro-clima, micro-habitat, estrutura da
vegetação e das variáveis da paisagem demonstrou que existem diferenças
concretas entre esses três ambientes, e que essas diferenças podem
eventualmente interferir na distribuição de biomassa total e na disponibilidade de
alimento para essas comunidades faunísticas habitantes dessas áreas.
E
a
lagartos e anfíbios de folhiço nas 12 unidades amostrais avaliadas no extremo
Sul da Bahia.
A Mata Atlântica
B
e
a
b
131

Quanto aos anfíbios, a grande maioria das espécies detectadas nesse estudo
e anfíbios realizadas até o momento demonstraram o pequeno
onhecimento existente sobre a fauna, visto o elevado número de espécies
ultados em relação à composição das
omunidades de anfíbios e répteis de folhiço para essa Unidade de
de ambientes
orestados e, principalmente em relação aos anfíbios), associadas a ambientes
são típicas do bioma Mata Atlântica, a qual, apesar da redução da sua cobertura
original, mantém sua importância, representada pelos poucos remanescentes
existentes, importantes centros de endemismo para vários grupos. As
amostragens d
c
ainda não identificadas ou recém descritas na Ciência.
Ao mesmo passo que não foi possível identificar efeito negativo da
fragmentação para as comunidades avaliadas, de acordo com os dados
evidenciados nesse estudo, a Mata Contínua localizada na RPPN Estação
Veracruz, reconhecida como uma das áreas mais conservadas de Mata
Atlântica primária no Brasil assemelha-se biologicamente, ao nível da paisagem
regional, a um remanescente em estado intermediário de conservação.
A expectativa inicial era de que os res
c
Conservação, indicassem maiores valores de abundância e riqueza para as
espécies típicas de matas conservadas. Porém, conforme apresentado ao longo
dessa Dissertação, as comunidades faunísticas estudadas parecem não
apresentar um padrão específico de distribuição na paisagem, comportando-se
como espécies generalistas (mesmo aquelas específicas
fl
mais antropizados.
Como não foram detectadas diferenças quanto à composição e ao número de
espécies de anfíbios e lagartos de folhiço presentes nos diversos componentes
da paisagem, os resultados indicaram que a qualidade da matriz de monocultura
de eucalipto, com desenvolvimento de sub-bosque de nativas e 7 anos de idade
- período que antecede a exploração florestal, pode não estar interferindo
negativamente na manutenção dessas comunidades na região.
132

Em contrapartida, a produtividade do eucalipto na região é uma das maiores do
A manutenção destes
istemas - e de pelo menos uma parcela de sua diversidade biológica - só se
izar os
cursos utilizados e o “ideal ambientalista”; na tentativa de que o
nos platôs, (plantados em curvas de nível, com
esenvolvimento de sub-bosque de plantas nativas) e os remanescentes
cuperar áreas
egradadas no seu entorno. Tais medidas devem ser adotadas para que sua
área superficial seja ampliada, seja reduzido o efeito de borda sobre a Unidade
globo, onde estão implantadas as maiores indústrias de celulose branqueada do
mundo, o que tem gerado pressão crescente para expansão dessa cultura em
substituição a áreas de pasto e de floresta em estágios iniciais de sucessão.
Numa região com relevante vocação para implantação de silvicultura, e
detentora de valiosa biodiversidade, conceitos como sustentabilidade e
conservação devem ser rigorosamente avaliadas.
s
concretizará se as práticas de manejo empregadas atuarem em uma escala
regional, levando-se em conta as plantações e os remanescentes de florestas
nativas.
Um dos principais anseios no desenvolvimento desta pesquisa era de subsidiar
a construção de propostas de manejo para a região, no sentido de otim
re
conhecimento biológico gerado pudesse auxiliar na formulação de estratégias
conservacionistas para a área estudada.
A partir desses princípios, a nível regional, urge garantir-se a conservação de
todos os remanescentes florestais existentes, independente de seus tamanhos
e formas, e a restauração de áreas –chave, de modo a permitir a conexão
entre eles, possibilitar a passagem de polinizadores, dispersores e outros
animais e reduzir o efeito de isolamento. A paisagem em mosaico formada
pelos eucaliptais
d
florestais localizados nos vales, também deve ser mantida, sendo de
fundamental importância para auxiliar na configuração de uma paisagem
balanceada e interligação entre os recursos hídricos existentes. Figura 39
Apesar do seu estado fisionômico de conservação, é imprescindível conectar a
RPPN Estação Veracruz a outros remanescentes próximos, e re
d
133

e minimize possíveis ocorrências de torná-la uma “floresta vazia”. Sua atual
condição de isolamento pode agir negativamente sobre a riqueza biologia
presente, diminuindo as taxas de imigração, o que pode levar populações
pequenas à extinção e favorecer espécies dominantes que conseguem se
anter em manchas isoladas. Desta forma, a diversidade diminui, não só pela m
redução da riqueza, mas também pela redução na equitabilidade.
Além de todas essas medidas citadas, deve-se priorizar e impleme
Figura 39. Apresentação visual da proposta de manejo para a região de estudo.
ntar
esquisas que determinem a viabilidade econômica da monocultura de p
eucalipto sob prática florestal que permitam o desenvolvimento de sub-bosque
de espécies nativas em todas as demais empresas implantadas na região do
extremo sul da Bahia.
134

É importante ressaltar que a diversidade biológica em sistemas de domínio
privado só será realmente preservada, se planos de conservação e manejo
forem estabelecidos conjuntamente com os de desenvolvimento econômico,
funcionando qual um sistema de integração entre os diferentes elementos da
paisagem.
Os fragmentos de floresta nativa devem ser tratados não exclusivamente como
mantenedor de uma fração da biodiversidade regional, mas como responsáveis
pela manutenção de um viável e representativo “pool” gênico de espécies
representantes de um ecossistema altamente ameaçado como a Mata
Atlântica. Por outro lado, se estes fragmentos forem tratados separadamente
da matriz de habitat no qual se insere, os tamanhos destes devem ser
necessariamente maiores para resguardar o mesmo potencial de conservação.
Diante da enorme extensão de áreas ocupadas por monocultura de eucalipto
esta atividade econômica pode ser utilizada como instrumento de auxílio para a
manutenção da diversidade biológica e, dessa forma, estabelecer uma aliança
entre os interesses econômico e os de conservação.
É importante ressaltar a necessidade de adoção emergencial de outras
alternativas de manejo, como a formação de Corredores de Biodiversidade, a
exemplo do Corredor Central da Mata Atlântica. Tais medidas não devem ser
meros instrumentos políticos e carecem implementação prioritária, sendo
fundamentais para a manutenção dos processos biológicos e sociais
xi
e stentes.
135


















