
INSTITUTO NACIONAL DE PESQUISAS DA AMAZÔNIA INPA PROGRAMA DE …‡ÃO_LUANA... · Dissertação...
105
INSTITUTO NACIONAL DE PESQUISAS DA AMAZÔNIA – INPA PROGRAMA DE PÓS-GRADUAÇÃO EM ENTOMOLOGIA BIONOMIA E MORFOLOGIA DOS ESTÁGIOS IMATUROS DE Hermetia illucens (LINNAEUS, 1758) (DIPTERA: STRATIOMYIDAE) DE INTERESSE FORENSE EM MANAUS, AMAZONAS, BRASIL LUANA MACHADO BARROS Manaus, Amazonas Maio, 2017
Transcript of INSTITUTO NACIONAL DE PESQUISAS DA AMAZÔNIA INPA PROGRAMA DE …‡ÃO_LUANA... · Dissertação...

INSTITUTO NACIONAL DE PESQUISAS DA AMAZÔNIA – INPA
PROGRAMA DE PÓS-GRADUAÇÃO EM ENTOMOLOGIA
BIONOMIA E MORFOLOGIA DOS ESTÁGIOS IMATUROS DE
Hermetia illucens (LINNAEUS, 1758) (DIPTERA: STRATIOMYIDAE) DE
INTERESSE FORENSE EM MANAUS, AMAZONAS, BRASIL
LUANA MACHADO BARROS
Manaus, Amazonas
Maio, 2017

LUANA MACHADO BARROS
BIONOMIA E MORFOLOGIA DOS ESTÁGIOS IMATUROS DE
Hermetia illucens (LINNAEUS, 1758) (DIPTERA: STRATIOMYIDAE) DE
INTERESSE FORENSE EM MANAUS, AMAZONAS, BRASIL
ORIENTADORA: DRA. RUTH LEILA FERREIRA-KEPPLER
Coorientadora: Dra. Ana Lúcia Nunes Gutjahr
Coorientador: Dr. Renato Tavares Martins
Dissertação apresentada ao conselho da
divisão do curso de Pós-Graduação em
Entomologia como parte dos requisitos
para a obtenção do título de Mestre em
Ciências Biológicas, área de
concentração em Entomologia.
Manaus, Amazonas
Maio, 2017

BANCA EXAMINADORA
Dra. Rosaly Ale Rocha– Titular (Presidente)
Instituto Nacional de Pesquisas da Amazônia
Dr. Rafael Augusto Pinheiro de Freitas Silva – Titular
Instituto Nacional de Pesquisas da Amazônia
Dra. Karime Rita de Souza Bentes – Titular
Universidade Federal do Amazonas
Dr. Alexandre Somavilla – Suplente
Instituto Nacional de Pesquisas da Amazônia
Dr. Vanderly Andrade Souza – Suplente
Instituto Nacional de Pesquisas da Amazônia

ii
Sinopse:
Estudaram-se dados bionômicos da fêmea adulta de Hermetia illucens e suas larvas
associadas à carcaça suína. Informações sobre comportamento de oviposição, tempo de
duração dos estágios de ovo, larva e pupa foram apresentados. O ovo e estágios imaturos
foram descritos morfologicamente e medidas corporais foram feitas em larvas para
estimar a biomassa seca e úmida de H. illucens.
Palavras-chave: Entomologia forense, tempo de desenvolvimento, descrição
morfológica.
B277 Barros , Luana Machado
Bionomia e morfologia dos estágios imaturos de Hermetia
Illucens (Linnaeus, 1758) (Diptera: Stratiomyidae) de interesse
forense em Manaus, Amazonas, Brasil /Luana Machado Barros. ---
Manaus: [s.n.], 2017.
xiv, 90 f.: il.
Dissertação (Mestrado) --- INPA, Manaus, 2017.
Orientador: Ruth Leila Ferreira-Keppler
Coorientador: Ana Lúcia Nunes Gutjahr e Renato Tavares M
Área de concentração: Entomologia
1. Hermetia illucens. 2.Mosca soldado negro. 3. Entomologia
forense. I. Título.
CDD 595.77

iii
Dedico esta dissertação aos meus pais Rose e Raimundo.

iv
AGRADECIMENTOS
Ao Instituto Nacional de Pesquisas da Amazônia (INPA) pelo apoio fornecido e
estrutura física para a realização desta dissertação.
À Coordenação de aperfeiçoamento de Pessoal de Nível Superior– CAPES pela
concessão da bolsa de Mestrado.
O apoio financeiro fornecido pelo projeto “Sistemática Integrada de Insetos Aquáticos,
com ênfase em Simuliidae (Diptera: Nematocera) na América do Sul” financiado pelo MCTI /
INPA e coordenado pela Dra. Neusa Hamada.
À Dra. Ruth Leila Ferreira-Keppler pela orientação, amizade e compreensão durante
estes dois anos. Ao Dr. Renato Tavares Martins pela coorientação, total apoio em minhas
dúvidas e por conselhos na vida acadêmica. À Dra. Ana Lúcia Nunes Gutjahr pela
coorientação, por despertar em minha pessoa o amor pelos insetos e incentivo para seguir nesta
área.
Ao Laboratório Temático de Microscopia Óptica e Eletrônica- LTMOE/
CPAAF/INPA e ao Laboratório de histologia e embriologia animal da Universidade Federal
Rural da Amazônia pela estrutura que possibilitaram a análise em microscopia de varredura.
À Dra. Karime Bentes pelo empréstimo da gaiola utilizada nos experimentos e ao Msc.
Fábio Godoi por compartilhar literatura utilizada nesta dissertação.
Ao Dr. Alexandre Ururahy pela experiência e troca de idéias responsáveis para o
amadurecimento desta pesquisa.
À Dra. Neusa Hamada por permitir a utilização de equipamentos necessários para a
execução desta dissertação.
À Dra. Vivian Campos por sempre ser solícita e me ajudar no agendamento de carros
pelo INPA.
A todos os professores que fizeram parte da minha formação profissional (graduação e
mestrado) e aos funcionários do Inpa que de forma direta ou indireta contribuíram para a
melhoria deste trabalho.
Aos membros da banca de qualificação Dr. Karime Bentes, Dra. Cínthia Barreto Chagas
e Dr. Rafael Freitas Silva pelas contribuições.
Aos amigos de Laboratório, Eduarda Denise, Eliane Solar, Irene Marques e demais
membros do Laboratório de Citotaxonomia e Insetos Aquáticos pelo apoio, conversas e
descontração.

v
Aos Msc. Tohnson Sales e Alex Barros, aos colegas Daniel de Paula, Gilderlânia e
Grazielle de Paula pelo apoio em campo e experiências compartilhadas.
A Léo Rossi por compartilhar seu conhecimento sobre o uso da lupa de automontagem
e por algumas das fotografias que compõem este trabalho.
Aos meus amigos do INPA e à turma de Mestrado em Entomologia 2015, especialmente
Renato Azevedo, Rafael Sobral, Ângelo Rêgo e Cris Utta pela convivência gratificante, por
muitas risadas, diversão e conhecimento. Sem dúvidas, vocês foram “minha família manauara”!
Aos amigos Msc. Márlon Breno, Nikolas Cipola, Gustavo Tavares, Marcus Bevilaqua,
Isis Sá e Dra Valéria Araújo pelo carinho, apoio, incentivo e conhecimento trocados.
Aos meus pais, Raimundo e Rose, que foram meu alicerce e incentivo em tudo que
executo em minha vida e a todos meus familiares, tios, primos, irmão, avós, que fizeram muita
falta nesse período de mestrado.
Ao meu companheiro Matheus Soares, o qual conheci neste mestrado e passou a fazer
parte de minha vida, sendo amigo, meu braço esquerdo, meu “mateiro” e compartilhando
sempre momentos de alegria e conhecimento e à família do Matheus que me acolheu como uma
filha e sempre desejam meu sucesso.
Aos amigos de Belém (Yan Corrêa, Taissa Miki, Wander Dias, Rafaela Alencar, Maysa
Oliveira, Railana Rodrigues), aos quais não posso citar todos, que fizeram parte do que sou e
se mantiveram companheiros mesmo à distância.
A todos que de alguma forma contribuíram para a realização desse trabalho e estiveram
presentes nessa etapa.
Muito obrigada!

vi
RESUMO
Hermetia possui 78 espécies conhecidas no mundo, sendo 54 registradas na região Neotropical
e 22 ocorrendo no Brasil. Dentre estas, Hermetia illucens é uma das espécies de Diptera de
interesse forense. No presente estudo buscou-se fornecer informações bionômicas desta espécie
durante o processo de decomposição cadavérica de suíno. Foi estudada a morfologia de seus
imaturos, sua biomassa foi estimada a partir da morfometria da larva e ajuste em diferentes
modelos de regressão. O estudo foi realizado em fragmento florestal urbano em Manaus,
Amazonas. Foi utilizado um suíno como isca atrativa e substrato para as observações
bionômicas. Fêmeas grávidas de H. illucens foram coletadas para a obtenção de imaturos a
partir de suas colônias de estoque. Para o estudo morfológico, utilizou-se microscopia ótica e
eletrônica de varredura. Para a estimativa de biomassa e obtenção do melhor ajuste, foram
mensuradas 280 larvas, sendo utilizadas quatro medidas morfométricas, testadas em três
modelos de regressão. Os resultados bionômicos mostram preferência da fêmea adulta por
ovipor em locais protegidos. As larvas se desenvolvem em locais úmidos, preferencialmente
abaixo do couro do suíno. As pupas completam seu desenvolvimento no substrato e não se
enterram. O tempo de desenvolvimento das larvas ocorreu no intervalo de 53 a 82 dias, com
seis estádios larvais e o de pupa. A fase de ovo durou 4-7 dias; primeiro estádio= 1-2 dias;
segundo= 2-3 dias; terceiro= 3-7 dias; quarto= 8-13; quinto= 9-15 dias; sexto= 11-15 dias e a
pupa= 15-20 dias. Morfologicamente, os ovos são ovoides alongados, com coloração branco-
leitoso à amarelado. Hermetia illucens pode ser identificada pela combinação das seguintes
características: cabeça comprida, onde sua largura não é superior à altura, quetotaxia dorsal e
ventral da cápsula cefálica, dos segmentos torácicos e abdominais, presença da mancha esternal
no sexto segmento e a morfologia dos espiráculos anteriores e posteriores. A pupa é adéctica e
coarctata, coloração castanho escuro com cerdas castanho-claras a douradas. As medidas com
melhor poder de predição da biomassa foram o comprimento total do corpo e largura do sexto
segmento abdominal. O melhor ajuste entre as dimensões corporais e biomassa ocorreu no
modelo exponencial. Estes resultados acrescentam informações sobre a bionomia, morfologia
e biomassa da espécie na região Amazônica, podendo ser aplicadas em áreas econômica e
forense.

vii
ABSTRACT
Hermetia is composed by 78 species worldwide, from which 54 occur in the neotropics, 22 in
Brazil. In particular, Hermetia illucens displays potential forensic importance and its biological
records may be useful in medico-legal circumstances and for economic activities. We recorded
bionomic data on the development of H. illucens on swine decomposition in an Amazonian
landscape. We described the immature morphology, and used larval body dimensions to
estimate biomass through different regression models. The study was conducted in an urban
forest fragment in Manaus City, State of Amazonas, Brazil. We used a swine carcass to attract
and function as substrate for the flies. We collected pregnant females of H. illucens for posterior
oviposition. For the morphological description, we used both stereomicroscopy and scanning
electronic microscopy. For the estimation of biomass and obtaining of the best fitting model,
we took four measurements of 20 individual larvae and tested three regression models. Our
results showed that females prefered to lay the eggs on sheltered areas. . Eggs tool four to seven
days to hatch. The larvae grew in humid zones, preferentially under the swine skin. The pupae
completed their development on the substrate, rather than buried underground. Immature stage
lasted from 53 to 82 days, and comprised six instars and the pupal stage. The eggs stage lasted
4-7 days. Duration of the immature stage: first instar = 1–2 days; second instar = 2–3 days; third
instar = 3–7 days; fourth instar = 8–13 days; fifth instar = 9–15 days; sixth instar = 11–15 days;
pupal stage = 15–20 days. Eggs are elliptical, elongated, color varying from white to yellowish.
Hermetia illucens is identified by the combination of the following set of characters: (1) head
elongated, its width never superior to its height; (2) dorsal and ventral chaetotaxy of the cephalic
capsule, thorax and abdomen; (3) sixth abdominal segment with a spot on the sternum; (4)
morphology of the anterior and posterior spiracles. The pupae is adectica and coarctata,
tegument dark brown, with setae varying from light brown to golden. The body measurements
that best predicted biomass were overall body length and width of the largest abdominal
segment. The exponential regression model best fitted body dimensions and biomass of larvae.
We provided information on bionomy, morphology and biomass of H. illucens in the Amazon
Region, which can be employed to economic and forensic activities.

viii
SUMÁRIO
LISTA DE TABELAS .............................................................................................................. ix
LISTA DE FIGURAS ................................................................................................................ x
INTRODUÇÃO GERAL ........................................................................................................... 1
Grupo Alvo: Hermetia illucens (Linnaeus, 1758) .................................................................. 2
Estudo da biologia e morfologia de Hermetia illucens ........................................................... 4
Estimativas de biomassa ......................................................................................................... 6
Organização dos resultados .................................................................................................... 7
OBJETIVO ................................................................................................................................. 8
Objetivo geral ......................................................................................................................... 8
Objetivos específicos .............................................................................................................. 8
Capítulo I: Bionomia de Hermetia illucens (Linnaeus, 1758) (Diptera:Stratiomyidae) em
suíno em decomposição em área urbana na cidade de Manaus, Amazonas, Brasil ................... 9
Capítulo II: Descrição morfológica do ovo e dos imaturos de Hermetia illucens (Linnaeus,
1758) (Diptera:Stratiomyidae) .................................................................................................. 33
Capítulo III: Estimativa de massa seca e úmida de larvas de Hermetia illucens (Linnaeus,
1758) (Diptera:Stratiomyidae) associadas a suíno em decomposição em ambiente urbano na
Amazônia Central ..................................................................................................................... 62
SÍNTESE .................................................................................................................................. 79
REFERÊNCIAS BIBLIOGRÁFICAS ..................................................................................... 80

ix
LISTA DE TABELAS
Capítulo I
Tabela A. Medidas de largura da cápsula cefálica (mm) em cada estádio larval encontradas por
diferentes autores.......................................................................................................................31
Tabela B. Períodos totais de desenvolvimento e para cada estádio larval encontradas por
diferentes autores.......................................................................................................................32
Capítulo III
Tabela 1- Amplitude, média, desvio padrão (DP), coeficiente de variação (CV = (DP/média)
*100, em %) e número de observações (N) para massa corporal e dimensões do corpo de larvas
de Hermetia illucens (Linnaeus, 1758) da Amazônia Central....................................................69
Tabela 2- Modelos linear, exponencial e de potência para a relação entre massa corporal (mg)
e dimensões do corpo (mm) de larvas de Hermetia illucens (Linnaeus, 1758) da Amazônia
Central. MU = Massa Úmida; MS = Massa Seca; CT = Comprimento Total; LC = Largura da
Cápsula Cefálica; CC = Comprimento da Cápsula Cefálica; LA =Largura do Maior Segmento
Abdominal. Para todos os modelos p < 0.001............................................................................70
Tabela 3- Poder de predição dos modelos de validação cruzada para estimar a biomassa de
Hermetia illucens (Linnaeus, 1758) da Amazônia Central. A diferença indica a porcentagem de
diferença entre dados previstos e observados. Diferença positiva indica que dados preditos
foram maiores que os dados observados. Os valores de F e P referem-se ao teste realizado entre
os valores observados e preditos. Os valores são referentes à média obtida para as predições e
testes repetidos por 100 vezes. CT = Comprimento Total; LC = Largura da Cápsula Cefálica;
CC = Comprimento da Cápsula Cefálica; LA =Largura do Maior Segmento
Abdominal.................................................................................................................................72

x
LISTA DE FIGURAS
Introdução geral
Figura 1- Asa de Hermetia illucens (Linnaeus, 1758) (Diptera: Stratiomyidae), com destaque
para a célula discal.......................................................................................................................1
Figura 2- Hermetia illucens (Linnaeus, 1758) ♀. A- Hábito lateral; B- Antena espatulada; C-
Escutelo, em destaque o espinho; D- Segundo segmento abdominal transparente. Escalas A, B
e D: 1 mm; C: 0,5 mm.................................................................................................................3
Capítulo I
Figura 1- Fêmea de Hermetia illucens (Linnaeus, 1758) ovipondo em tubo Falcon ® no
laboratório. Círculos destacam ovos depositados e a seta indica o ovipositor estendido da fêmea
no momento da oviposição........................................................................................................15
Figura 2- Posturas de ovos de Hermetia. illucens (Linnaeus, 1758): A- massa de ovos sob
estereomicroscópio; B- massa de ovo do crânio do suíno (forames palatinos maiores); C- massa
de ovo nas frestas da gaiola........................................................................................................16
Figura 3- Observações bionômicas de Hermetia illucens (Linnaeus, 1758): A- larvas de
diferentes estádios encontrados em ossos do côndilo femoral; B- larvas de sexto estádio
encontradas abaixo do couro do suíno (tecido conjuntivo) ........................................................16
Figura 4- Distribuição de frequência da largura da cápsula cefálica de Hermetia illucens
(Linnaeus, 1758) (Diptera: Stratiomyidae) evidenciando os seis estádios larvais: Cor verde=
Primeiro estádio; Cor azul escuro= Segundo estádio; Cor azul claro= Terceiro estádio; Cor
vermelha= Quarto estádio; Cor preta= Quinto estádio; Cor laranja= Sexto
estádio.......................................................................................................................................18
Figura 5- Análise de Componentes Principais (PCA) com as medidas corporais de Hermetia
illucens (Linnaeus, 1758). As cores dos pontos coloridos indicam os estádios larvais. O ápice
da seta indica o tipo de medida corporal. l.cap= Largura da cápsula cefálica, c.cap=
Comprimento da cápsula cefálica, l.seg= Largura do maior segmento abdominal e c.tot=
Comprimento total. Cor amarela= Primeiro estádio; Cor azul= Segundo estádio; Cor verde=
Terceiro estádio; Cor vermelha= Quarto estádio; Cor cinza= Quinto estádio; Cor laranja= Sexto
estádio.......................................................................................................................................19

xi
Figura 6- Tempo de desenvolvimento para cada estádio larval de Hermetia illucens (Linnaeus,
1758) em condições ambientais em Manaus-AM......................................................................20
Figura 7- Comprimento total (A) e massa úmida (B) para cada estádio larval de Hermetia
illucens (Linnaeus, 1758) em condições ambientais em Manaus-AM. .....................................21
Capítulo II
Figura 1- Morfologia dos ovos de Hermetia illucens (Linnaeus, 1758): A- Postura (massa de
ovos) sob estereomicroscópio ótico; B- Aspecto geral sob microscopia de varredura (MEV); C-
Superfície aproximada do córion. Escalas: A= 1 mm; B= 30 µm; C= 10 µm............................39
Figura 2- Morfologia da cápsula cefálica de uma larva do sexto estádio de Hermetia illucens
(Linnaeus, 1758) sob MEV: A- Vista antero-dorsal; Ecl= Esclerito lateral; Ecf= Esclerito
clípeo-frontal; L= Labro; O= Ocelo; An= Antena. B- Vista frontal da antena, em destaque uma
das sensilas campaniformes; Sc= Sensíla campaniforme. C- Vista ventral da cápsula cefálica;
Plv= Placa ventral; Prm= Premento; Am= Área molar. D- Aparelho bucal: Em 1= Escovas
mandibulares superiores; Em 2= Escovas mandibulares inferiores; Pm= Palpo maxilar; Cm=
Cerda maxilares; Lg= Lóbulos genais; Md= mandíbula. Escalas A= 100 µm; B= 20 µm; C=
500 µm; D= 200 µm...................................................................................................................41
Figura 3- Morfologia do tórax de uma larva do quinto estádio de Hermetia illucens (Linnaeus,
1758) sob MEV: A- Vista dorsal dos segmentos torácicos, em destaque o espiráculo anterior
esquerdo; B- Vestígio espiracular em destaque; C- Vista frontal do espiráculo anterior, em
destaque as duas aberturas espiraculares (AEsp) formando um V (indicado com seta vermelha)
e a cicatriz estigmática (CEst) acima (indicada com seta amarela); D- Vista ventral dos
segmentos torácicos, em destaque as sensilas (sen; indicada com seta amarela). Escalas A=2
mm; B= 100 µm, C= 30 µm e D= 2mm......................................................................................42
Figura 4- Morfologia do abdome de uma larva do sexto estádio de Hermetia illucens (Linnaeus,
1758) sob MEV: A- Sensilas presentes da região ventral do terceiro segmento abdominal; B-
Vestígio espiracular localizado em todos os segmentos abdominais em vista dorsal, com
exceção do último; C- Mancha esternal localizada no sexto segmento abdominal em vista
ventral; D- Abertura anal em destaque no último segmento abdominal, rodeada de espinhos,
em vista ventral; E- Abertura espiracular no último segmento abdominal em vista frontal.
Escalas A e B= 1 mm; C e E= 100 µm; D= 200 µm....................................................................43

xii
Figura 5- Larva de primeiro estádio de Hermetia illucens (Linnaeus, 1758) sob MEV: A- Vista
dorsal; B- Superfície corporal. Escalas A= 30 µm e B= 100 µm................................................44
Figura 6- Larva de segundo estádio de Hermetia illucens (Linnaeus, 1758) sob
estereomicroscópio ótico: A- Vista ventral; B- Vista dorsal. Escalas A e B= 1 mm...................45
Figura 7- Larvas de terceiro estádio de Hermetia illucens (Linnaeus, 1758) sob
estereomicroscópio ótico: A- Vista ventral; B- Vista dorsal. Escalas A e B= 1 mm...................46
Figura 8- Larvas de quarto estádio de Hermetia illucens (Linnaeus, 1758) sob
estereomicroscópio ótico: A- Vista ventral; B- Vista dorsal; C- Vista lateral. Escalas A, B e C=
1 mm..........................................................................................................................................47
Figura 9- Larvas de quinto estádio de Hermetia illucens (Linnaeus, 1758) sob
estereomicroscópio ótico: A- Vista ventral; B- Vista dorsal; C- Vista lateral. Escalas A, B e C=
2 mm..........................................................................................................................................48
Figura 10- Larvas de sexto estádio de Hermetia illucens (Linnaeus, 1758) sob
estereomicroscópio ótico: A- Vista ventral; B- Vista dorsal; C- Vista lateral. Escalas A, B e C=
5 mm..........................................................................................................................................49
Figura 11- Antena das larvas de Hermetia illucens (Linnaeus, 1758) evidenciando as diferenças
morfológicas entre os estádios: A- Primeiro estádio; B- Segundo estádio; C- Terceiro estádio;
D- Quarto estádio; E- Quinto estádio; F- Sexto estádio. Os números 1, 2, 3 e 4 indicam as
sensilas alongadas encontradas no segundo segmento da antena. Escalas A= 3 µm; B= 10 µm;
C= 5 µm; D= 100 µm; E= 200 µm e F= 20 µm...........................................................................50
Figura 12- Pupa de Hermetia illucens (Linnaeus, 1758) sob estereomicroscópio ótico: A- Vista
ventral; B- Vista dorsal; C- Vista lateral. Escalas A, B e C= 5 mm............................................51
Figura 13- Morfologia da cápsula cefálica da pupa de Hermetia illucens (Linnaeus, 1758) sob
MEV: A- Vista dorsal; Ecl= Esclerito lateral; Ecf= Esclerito clípeo-frontal; O= Ocelo; An=
Antena. B- Vista lateral da cápsula cefálica, evidenciando o olho direito (seta); L= Labro; C-
Vista dorsal da antena; D- Vista ventral da cápsula cefálica; Plv= Placa ventral; Prm= Premento;
Am= Área molar. Escalas A, B e D= 200 µm; C= 10 µm...........................................................52
Figura 14- Morfologia do tórax da pupa de Hermetia illucens (Linnaeus, 1758) sob MEV; A-
Vista lateral do primeiro segmento torácico, em destaque o espiráculo anterior direito; B- Vista

xiii
dorsal do espiráculo anterior; C- Vista dorsal do primeiro segmento do tórax; D- Vista ventral
dos segmentos torácicos; Escalas A= 500 µm; B= 100 µm; C e D= 1 mm.................................53
Figura 15- Morfologia do abdome da pupa de Hermetia illucens (Linnaeus, 1758) sob MEV:
A- Abertura espiracular no último segmento abdominal em vista dorsal; B- Abertura anal em
destaque no último segmento abdominal em vista ventral; C- Mancha esternal localizada no
sexto segmento abdominal em vista ventral; D- Visão aproximada da mancha esternal,
detalhando as células da superfície; E- Segmentos abdominais em vista dorsal; F- Superfície
corporal pruinosa da pupa. Escalas A e E= 1 mm; B= 200 µm; C, D e F = 100 µm....................55
Figura 16- Quetotaxia da cápsula cefálica de Hermetia illucens (Linnaeus, 1758): A- Vista
dorsal; B- Vista ventral..............................................................................................................56
Figura 17- Quetotaxia da larva de Hermetia illucens (Linnaeus, 1758) com ênfase nos
segmentos toráxicos e abdominais: A- Vista dorsal; B- Vista ventral. Os números 1-3
apresentados inicialmente indicam os segmentos torácicos. Os números de 1’ a 8’ indicam os
segmentos abdominais. Os Pontos pretos ( ) indicam as posições das cerdas nos segmentos
corporais da larva.......................................................................................................................57
Figura 18- Tipos de cerdas da superfície corporal de Hermetia illucens (Linnaeus, 1758): A-
Cerdas de primeiro estádio localizadas na região ventral; B- Cerdas de primeiro estádio
localizadas na região dorsal; C- Cerdas localizadas no segundo, terceiro, quarto, quinto, sexto
estádios e pupa. Escalas A= 10 µm; B= 10 µm e C= 100 µm.....................................................58
Capítulo III
Figura 1- Larva de quinto estádio de Hermetia illucens (Linnaeus, 1758), em vista ventral,
destacando as medidas realizadas. LC= Largura da cápsula cefálica; CC= Comprimento da
cápsula cefálica; CT= Comprimento total; LA= Largura do maior segmento abdominal..........68

1
INTRODUÇÃO GERAL
Os Diptera estão entre as quatro ordens megadiversas de insetos existentes. É composta
por 188 famílias e aproximadamente 159.000 espécies descritas, compreendendo de 10-12% de
toda a biodiversidade mundial (Carvalho et al., 2012; Lambkin et al., 2013). Muitos possuem
importância econômica, pois atuam como controladores de pragas (Carvalho et al., 2012).
Alguns são agentes polinizadores, outros possuem importância médico-veterinária, sendo
vetores de agentes infecciosos (Triplehorn e Johnson, 2011). Parte desses insetos tem papel
importante na decomposição da matéria orgânica, já que são os primeiros a colonizarem a
carcaça e a utilizam como fonte de alimento e substrato para oviposição, a exemplo:
Calliphoridae, Sarcophagidae, Muscidae, Stratiomyidae (Oliveira-Costa, 2011).
Stratiomyidae está incluído em Brachycera, infra-ordem Stratiomyomorpha (Pujol-Luz
e Pujol-Luz, 2014), onde encontra-se a espécie alvo do trabalho, Hermetia illucens (Linnaeus,
1758). Esta família possui uma grande variedade morfológica, sendo diferenciada dos outros
Diptera por possuírem a asa com a célula discal pequena (McFadden, 1967) (Figura 1). É
distribuída em grande parte das regiões biogeográficas, constituida 12 subfamílias, 375 gêneros
e aproximadamente 2.700 espécies. Na região Neotropical são conhecidas 987 espécies. Das 12
subfamílias, oito ocorrem na região Amazônica (Woodley, 2009).
Figura 1. Asa de Hermetia illucens (Linnaeus, 1758) (Diptera: Stratiomyidae), com destaque
para a célula discal.
Os adultos de Stratiomyidae podem ser encontrados descansando sobre folhas, frutos
maduros ou matéria em decomposição. Os imaturos possuem diferenças no formato, coloração
e quetotaxia, características importantes taxonomicamente. Além disso, suas larvas possuem

2
hábitos diferenciados em relação a outras famílias, podendo ser aquáticas, semiaquáticas ou
terrestres (James, 1981). Os imaturos aquáticos, por vezes, ocorrem em grande número em
lagoas, onde podem desempenhar papel importante nas cadeias tróficas, ao servir de alimento
para os peixes (Bondari e Sheppard, 1987). Os Stratiomyidae também podem ser encontrados
em matéria orgânica em decomposição, a exemplo, Beridinae, Clitellariinae, Chrysochlorinae,
Hermetiinae e Sargine (Woodley, 1989).
Grupo Alvo: Hermetia illucens (Linnaeus, 1758)
Hermetia possui 78 espécies conhecidas e destas, 54 ocorrem na região Neotropical,
incluindo 22 com distribuição no Brasil (McFadden, 1967; Woodley, 2011, Fachin DA, 2017).
Conhecidos como “Black soldier-fly”, apresentam o escutelo frequentemente adornado
com espinhos (Figura 2C), que em alusão ao armamento militar, justificaria seu nome comum
“mosca soldado” (Kovac e Rozkošný, 2004). O segundo segmento abdominal é transparente
(Figura 2D), as antenas possuem o segmento apical espatulado (Figura 2B; Üstuner et al.,
2003). A espécie está incluída à Hermetiinae, inserida em Hermetiini.
Os adultos de H. illucens (Figura 2A), em algumas ocasiões, exibem mimetismo com
certos grupos de Hymenoptera (Üstuner et al., 2003). A cópula ocorre aproximadamente cinco
dias após a emergência dos adultos (Gobbi, 2012). Não há consenso na literatura sobre o
procedimento de cópula em H. illucens. Tingle et al. (1975) observaram que as fêmeas que se
encontram em repouso atraem os machos em voo. Contrariamente, Copello (1926) observou
que a cópula se dava durante o voo e não em repouso. Tomberlin e Sheppard (2001) relatam
que o macho intercepta a fêmea no ar até encontrarem um lugar seguro, longe de outros machos
para realizar a cópula.
As larvas de H. illucens geralmente possuem seis estádios (Oliveira-Costa, 2011) e
podem medir até três centímetros de comprimento. Além disso, mudam de coloração no
decorrer de seu desenvolvimento, variando de cor esbranquiçada (estádios iniciais) à amarelado
(estádios finais). Possuem características na quetotaxia larval (dorsal e ventral), como posição
e tipo de cerdas, importantes para a taxonomia do grupo (Hall e Gerhardt, 2002). Têm hábito
detritívoro e são encontradas em microambientes como plantas, frutos e carne em
decomposição (James, 1981).

3
Figura 2. Hermetia illucens (Linnaeus, 1758) ♀. A- Hábito lateral esquerdo; B- Antena
espatulada; C- Escutelo, em destaque o espinho; D- Segundo segmento abdominal transparente.
Escalas A, B e D: 1 mm; C: 0,5 mm.
H. illucens vem despertando interesse na área médica e veterinária, pois foi associada a
miíases (Calderón-Arguedas et al., 2005). Na economia, elas são responsáveis por grande

4
consumo de matéria orgânica, sendo consideradas grandes conversoras de biomassa (Diener et
al., 2011). Desempenham papel de controle biológico, pois foram observadas inibindo e
controlando a oviposição e desenvolvimento de Musca domestica Linnaeus, 1758 (Sheppard,
1983). Além destas aplicações, H. illucens é alvo de estudos relacionados a entomologia forense
(Oliveira-Costa, 2011). Nesse contexto, a espécie foi considerada por Lord et al. (1994) como
oportunista ou necrófaga secundária, estando relacionada a estimativas de intervalo pós-morte
(IPM), ou seja, o tempo transcorrido desde a morte do cadáver até o momento em que o mesmo
é encontrado (Oliveira-Costa, 2011). Trabalhos recentes têm demonstrado a importância
forense desta espécie também na Europa (Turchetto et al., 2001; Martínez-Sanchez et al., 2011)
e na América do Sul (Pujol-Luz et al., 2008; Gobbi, 2012; Pujol-Luz e Pujol-Luz, 2014).
Estudo da biologia e morfologia de Hermetia illucens
Na literatura, há registros da biologia e morfologia de H. illucens. Copello (1926) gerou
informações sobre o comportamento de oviposição da espécie, afirmando que há preferência
por ser em fendas secas. Tingle et al. (1975) relataram que o acasalamento e oviposição foram
observados com frequência de três a seis vezes em gaiola instalada ao ar livre. Quando realizado
em estufa (temperatura de 29,3°C), o acasalamento obteve menor sucesso, sendo observado
apenas uma vez até a morte do espécime. Foi notado também que a luz do sol direta estimula o
acasalamento no campo.
Quanto ao ciclo de desenvolvimento, os autores divergem quanto a sua duração.
Furmand et al. (1959) relataram que larvas recém emergidas de H. illucens precisam de duas
semanas ou mais para alcançar seu desenvolvimento completo. May (1961) trabalhando com
criação na Nova Zelândia, em temperatura de 27,8 °C, contribuiu de maneira considerável para
os conhecimentos sobre o ciclo de vida com período de incubação dos ovos de cinco a 14 dias,
o período de larva de 31 dias e o de pupa de nove a dez dias. Tingle et al. (1975) obtiveram um
período total de 38 dias em temperatura de 29,3°C para a criação de ovos até adultos. Booth e
Shepard (1984) com base em criação determinaram somente o tempo em que a espécie
permanece na fase de ovo. Anos depois, Lord et al. (1994) produziram uma tabela com dados
do ciclo de desenvolvimento de H. illucens e concluíram que o desenvolvimento desta espécie
pode se estender até cinco meses sob diferentes condições. Kim et al. (2008) criaram H. illucens
e determinaram a existência de apenas cinco estádios para a espécie na Coreia do Sul. Ao
contrário do registrado por Kim et al. (2008), Hall e Gerhardt (2002) contabilizaram para a
América, seis estádios de desenvolvimento. Estudos mais recentes com H. illucens realizados

5
na Espanha trazem informações de biologia reprodutiva, avaliando a influência de diferentes
tipos de alimentação e temperaturas na biologia, além da determinação de fatores bióticos e
abióticos que afetam o desenvolvimento (Gobbi, 2012).
Em relação à morfologia, os imaturos de H. illucens foram retratados com desenhos,
bem como seu aparelho bucal, possibilitando um conhecimento inicial da morfologia das larvas
(Copello, 1926).
Além de Copello (1926), May (1961) contribuiu com considerável conhecimento sobre
o ciclo de vida de H. illucens na Nova Zelândia, descreveu características do tamanho e
coloração dos ovos e das larvas.
McFadden (1967) abordou características como: comprimento, coloração, presença ou
ausência de espiráculos na margem dos segmentos, apresentando figuras ilustrativas. No
entanto, não detalhou a morfologia em vistas dorsal, ventral e lateral e a disposição das cerdas
em cada estádio larval.
Teskey (1991) observou que as larvas possuem 11 segmentos no corpo com cerdas
conspícuas, podendo possuir ocelos laterais distintos, os quais tendem a se tornar cada vez mais
conspícuos nos estádios finais. Além disso, o autor pontuou algumas características da cabeça,
como o aparelho bucal.
Kim et al. (2010) contabilizaram a quantidade de mudas e a duração do desenvolvimento
da larva, peso, largura da cápsula cefálica e descrição da morfologia da boca com auxílio de
microscopia de varredura, sem descrever as cerdas nos estádios larvais.
Gobbi (2012) estudou características morfológicas de H. illucens foram em sua tese de
doutorado de. A autora realizou descrições sobre a estrutura corporal dos imaturos através de
microscopia de varredura e hidrocarbonetos cuticulares, abordando características como a
quetotaxia e aberturas espiraculares. O trabalho mais recente sobre a morfologia de H. illucens
foi realizado por Oliveira et al. (2016) com uma descrição sucinta e generalizada de todos os
estádios.
Barros-Cordeiro et al. (2014), além da morfologia externa das larvas, fez a descrição
com auxílio de microscopia de varredura de acontecimentos internos no processo de
desenvolvimento de um imaturo de H. illucens, no qual descreveram alguns processos, como a
duração e alterações morfológicas durante o desenvolvimento intra-pupal sob condições
controladas de laboratório.

6
Informações relacionadas ao ciclo de vida e ecologia desta espécie são essenciais para
a entomologia forense. Desta forma, o estudo de H. illucens justifica-se na medida em que são
necessárias complementações dos estudos bionômicos desta espécie, como o período que
adultos e imaturos podem ser encontrados na carcaça, o tempo de desenvolvimento das fases
imaturas, ferramentas para estimar a biomassa de larvas, as quais, são relevantes para auxiliar
em estimativas do IPM de cadáveres encontrados no Amazonas, devido diferenças climáticas
que influenciam no desenvolvimento da espécie e tornam importantes estudos locais. Estas
informações em toda a região Amazônica ainda são incipientes. Entretanto, a espécie é
frequente no processo de decomposição, e dados de sua biologia podem contribuir com a
criminalística. Quanto às descrições dos imaturos existentes, pode-se afirmar que estas, apesar
de possuírem abordagem na quetotaxia, não mostram o detalhamento das vistas do corpo de
uma larva (ventral e dorsal), sendo necessário mais estudos destas características, as quais
possuem importância significativa na taxonomia da espécie.
Estimativas de biomassa
Grande parte da biomassa animal é representada pelos insetos, os quais compõem
principalmente a base da cadeia alimentar (Gullan e Cranston, 1994). Estes são de extrema
importância, pois atuam em grande parte do ecossistema, seja como polinizador, controlador
biológico, vetor de doenças, decompositor, dentre outros (Triplehorn e Johnson, 2011). Desta
forma, a estimativa de biomassa é fundamental para entender processos ecológicos na natureza.
Entretanto, apesar de importante, esta não necessariamente é de fácil obtenção (Radtke e
Williamson, 2005).
Taxas de crescimento, comumente, são calculadas a partir da diferença entre as massas
do indivíduo (inicial e final), mas a destruição ou perda do material é a grande dificuldade
encontrada (Rogers et al., 1976). Assim, a estimativa da biomassa das larvas utilizando a
morfometria das mesmas é uma ferramenta importante em estudos longitudinais (Sample,
1993). A confiabilidade dos dados é maior quando alguns fatores são considerados, tais como:
modo de preservação das amostras, os quais influenciam a quantidade de água retida, e a
extrapolação de resultados dos modelos de regressão para indivíduos oriundos de diferentes
regiões geográficas, causadas por diferenças genéticas (Benke et al. 1999). Estimativas de
biomassa a partir de medidas morfométricas complementam as informações bionômicas da
espécie e podem servir de ferramenta para estudos aplicados. Portanto, uma vez que não há

7
equações de modelos de regressão para popupação de H. illucens da Amazônia central, é dada
importância deste estudo.
Organização dos resultados
Nesta dissertação foram estudados a bionomia, morfologia e estimativa de biomassa de
Hermetia illucens. Os resultados foram organizados em formato de artigos, distribuídos em três
capítulos. O primeiro capítulo aborda algumas características bionômicas da fêmea adulta em
carcaça suína, tais como: momento de chegada, preferência de lugares para a oviposição,
processo de postura de ovos e quantidade de ovos por fêmea adulta; assim como dos imaturos
associados ao processo de decomposição de suíno: permanência, abandono e pupação destes no
substrato.
O segundo capítulo refere-se ao estudo da morfologia do ovo e dos estágios imaturos de
H. illucens. Ovos, larvas e pupas foram obtidos a partir de fêmeas grávidas coletadas em suíno
em decomposição e descritos utilizando-se microscopia ótica e a técnica de microscopia
eletrônica de varredura.
No terceiro capítulo foi estimada a biomassa seca e úmida de H. illucens a partir de
medidas morfométricas. Foram mensuradas 280 larvas e utilizadas as seguintes medidas, para
determinar o melhor ajuste entre estas e a biomassa: largura da cápsula cefálica, comprimento
da cápsula cefálica, comprimento total do corpo e largura do maior segmento abdominal.

8
OBJETIVO
Objetivo geral
- Fornecer informações bionômicas, morfológicas e estimativa de biomassa de Hermetia
illucens associada com o processo de decomposição em carcaça suína em Manaus, Amazonas,
Brasil.
Objetivos específicos
- Apresentar aspectos bionômicos de H. illucens durante as fases de decomposição de suíno;
- Descrever estruturas morfológicas do ovo e dos imaturos de H. illucens participantes do
processo de decomposição em carcaça suína;
- Verificar o poder de predição de medidas morfométricas para o cálculo de biomassa seca e
úmida de H. illucens coletadas em carcaça suína.

9
Capítulo I
Barros, L. M.; Ferreira-Keppler, R. L.; Martins, R. T
& Gutjahr, A. L. N. 2017. Bionomia de Hermetia
illucens (Linnaeus, 1758) (Diptera: Stratiomyidade)
em suíno em decomposição em área urbana na cidade
de Manaus, Amazonas, Brasil. Manuscrito formatado
para Zoologischer Anzeiger.

10
Bionomia de Hermetia illucens (Linnaeus, 1758) (Diptera: Stratiomyidae) em suíno em
decomposição em área urbana na cidade de Manaus, Amazonas, Brasil
Luana Barros a*, Ruth Ferreira-Keppler a, Renato Martins a, Ana Lúcia Gutjahr b
a Programa de Pós-graduação em Entomologia, Instituto Nacional de Pesquisas da Amazônia -
INPA, Av. André Araújo, 2936, CEP 69067-375, Manaus, Amazonas, Brasil
b Departamento de Ciências Naturais da Universidade do Estado do Pará- UEPA, Tv. Djalma
Dutra, 156, Belém, PA, CEP 66050-540, Brasil, e-mail: [email protected]
* Autor correspondente: L. M. Barros, Programa de Pós-graduação em Entomologia,
Instituto Nacional de Pesquisas da Amazônia - INPA, Av. André Araújo, 2936, CEP 69067-
375, Manaus, Amazonas, Brasil. E-mail: [email protected]
Resumo
Estudos que abordam aspectos biológicos ou bionômicos de insetos podem resultar em
informações importantes para entomologia forense, devido a dados relevantes para o cálculo de
intervalo pós-morte, localização do óbito e tipo da morte auxiliando na solução de crimes. Desta
forma, o presente trabalho tem como objetivo fornecer informações bionômicas de Hermetia
illucens (Stratiomyidae), durante o processo de decomposição cadavérica de suíno Sus scrofa
Linnaeus, 1758, disposto em área urbana de Manaus-AM. Fêmeas grávidas e ovos de H.
illucens foram coletados sob carcaça para fornecer colônias de estoque a partir dos imaturos.
Após a oviposição e eclosão dos ovos, 6.215 larvas emergiram e foram distribuídas em 60 potes
de criação, observados diariamente e acompanhados até a emergência dos adultos. Os
recipientes foram mantidos em viveiro sob condições naturais. No total 1000 larvas de
diferentes estádios foram fixadas em álcool para determinar o período de cada estádio e realizar
as medidas de comprimento, largura e massa. A fêmea de H. illucens deposita de 620-700 ovos.
As larvas completaram seu desenvolvimento no intervalo de 53-82 dias, com seis estádios
larvais e o de pupa. A duração de cada estádio de desenvolvimento foi variável, sendo o período
de ovo 4-7 dias; primeiro estádio 1-2 dias; segundo estádio 2-3 dias; terceiro estádio 3-7 dias;

11
quarto estádio 8-13; quinto estádio 9-15 dias; sexto estádio 11-15 dias e a pupa 15-20 dias. A
morfometria evidenciou que as larvas de primeiro estádio possuem largura da cápsula cefálica
de 0,05-0,08 mm, segundo estádio 0,09-0,24 mm, terceiro estádio 0,25-0,45 mm, quarto estádio
0,46-0,57 mm, quinto estádio 0,58-1,04 mm e sexto estádio 1,05-1,14 mm, com razão média
do crescimento larval de 1,87. Estes resultados agregam informações sobre a espécie na região
Amazônica, auxiliando na sua aplicabilidade econômica e forense.
Palavras-chave: entomologia forense; observações bionômicas; estádios larvais.
1. Introdução
Hermetia illucens (Linnaeus, 1758) é uma das espécies de Diptera de interesse forense,
classificada em Stratiomyidae. É conhecida por “black soldier-fly” e possui distribuição
cosmopolita. Na área médica, as larvas podem eventualmente estar envolvidas em casos de
miíases (infestação de tecidos do corpo pelas larvas) em seres humanos e outros animais (Adler
e Brancato, 1995; Calderón-Arguedas et al., 2005). Economicamente os imaturos desta espécie
são utilizados na agricultura sustentável (Lalander et al., 2013), em ração de peixes (Barroso et
al., 2014) e de frangos (De Marco et al., 2015).
As larvas de H. illucens possuem coloração branco leitosa à amarelada, geralmente seis
estádios de desenvolvimento e, diferentemente dos adultos, possuem hábito detritívoro e
colonizam ambientes como plantas, frutos e carne em decomposição (Oliveira-Costa, 2011).
As larvas são vorazes e participam da ciclagem de matéria orgânica, sendo capazes de
transformar grandes quantidades de resíduos em biomassa animal (Diener et al., 2011). Além
disso, desempenham papel importante no controle biológico ou inibição da oviposição e
desenvolvimento de Musca domestica Linnaeus, 1758, através de feromônios (Sheppard,
1983). Na fase adulta, H. illucens possui pigmentação com predomínio de cor negra e a fêmea
geralmente possui tamanho superior ao macho (Üstuner et al., 2003). São comumente
encontrados próximos ao habitat das larvas (Woodley, 2009) e não se alimentam, mantendo-se
apenas com as reservas de energias adquiridas na fase larval (Tomberlin et al., 2002).
Assim como outras espécies de Diptera, H. illucens é um dos focos da entomologia
forense, com potencial aplicado à criminalística, devido possibilitar a determinação do intervalo
pós-morte (IPM), como já relatado por Lord et al. (1994), Oliveira-Costa (2011) e Pujol-Luz et
al. (2008). Portanto, dada sua importância e aplicabilidade, tornam-se necessárias
complementações dos estudos bionômicos desta espécie, visto que estes em toda a região

12
Amazônica ainda são incipientes. Desta forma, este estudo visa gerar informações bionômicas
de H. illucens associadas ao processo de decomposição em carcaça suína.
2. Material e métodos
O experimento foi executado de janeiro a março e junho a setembro de 2016, no Campus
II do Instituto Nacional de Pesquisas da Amazônia (03º5'47”S e 59º59'22”W), localizado na
região urbana da cidade de Manaus, Amazonas, Brasil. Uma área de aproximadamente 49.000
m2 (INPA, 2013), clima quente e úmido (INMET, 2017) com período mais chuvoso de
novembro a maio e menos chuvoso de junho a outubro. A precipitação média anual é de 2.286
mm3, umidade média de 80%, com temperatura média anual de 26°C (min = 23ºC, max = 31ºC;
Costa et al., 2013).
Para a atração das fêmeas grávidas de H. illucens foram utilizados dois suínos (Sus
scrofa Linnaeus, 1758) da raça Large White (45 kg cada). A eutanásia dos modelos foi realizada
no local do experimento com disparo de arma de fogo na região frontal da cabeça. Estes foram
deixados para decompor protegidos por uma gaiola com medida de 1 m² e estrutura metálica
galvanizada, revestida com tela aramada de 3 cm de malha de ferro, para impedir a influência
de vertebrados maiores. Ao redor da gaiola foi instalada uma armadilha Shannon (1,5 x 1,5 x
1,5 m) para captura de adultos.
As fêmeas de H. illucens utilizadas para a obtenção das colônias de estoque foram
coletadas no interior da Shannon com o auxílio de tubos Falcon® de 50 ml, onde permaneceram
até a oviposição. Nos tubos foi oferecido como substrato para oviposição 1 g de carne suína
crua. Cada postura de ovos obtidos foi transferida para placas de Petri forradas com papel filtro
umedecido e 1 g de carne suína crua. As larvas que emergiram foram transferidas para potes
plásticos cobertos por tecido do tipo voal com 100 g de carne suína.
Para o acompanhamento do desenvolvimento de H. illucens, foram montadas seis
réplicas para cada postura, com 100 larvas em cada pote, com exceção das posturas que
obtiveram um número superior a 600 larvas, sendo necessário em uma das réplicas adicionar a
quantidade superior a 100 indivíduos. Cada réplica recebeu identificação correspondente ao
número da criação e a data em que as larvas eclodiram. Destas seis réplicas, uma foi separada
para acompanhamento das fases de decomposição, da qual foi retirada uma larva por dia até
atingir o estágio de pupa. As outras cinco réplicas foram acompanhadas até a emergência dos
adultos. O alimento das larvas foi reposto uma vez por semana até o abandono da dieta, ou seja,
até atingir fase de pupa. Nesta fase, elas foram individualizadas em tubos Falcon® contendo

13
vermiculita, tampados com algodão hidrofóbico e observadas diariamente até a emergência dos
adultos. Os imaturos foram mantidos em viveiro próximo ao local do experimento, em ambiente
natural, com temperaturas variando de 24°C a 35°C e umidade de 66% a 88% (INMET, 2017).
2.1 Observações bionômicas na carcaça
As observações bionômicas de H. illucens foram feitas diariamente no cadáver entre os
horários de 8:00 às 12:00 e 14:00 às 18:00 horas, a partir da data de óbito do modelo. As fases
de decomposição observadas foram: fresca, enfisematosa, coliquativa, de esqueletização e de
restos, de acordo com a classificação de Oliveira-Costa (2011). As observações bionômicas de
H. illucens feitas durante o processo de decomposição foram: momento de chegada, preferência
de lugares para a oviposição, processo de postura de ovos, quantidade de ovos por fêmea adulta,
permanência, abandono e pupação dos imaturos no substrato.
2.2 Tempo de duração dos estádios e razão sexual
No ciclo de desenvolvimento da espécie foi observado: duração da fase de ovo, tempo
total de desenvolvimento (ovo a adulto), duração do período larval, determinação do número
de estádios larvais, duração da fase de pupa e a razão sexual dos adultos emergidos (número de
fêmeas dividido pelo número de adultos emergidos; Silveira-Neto et al., 1976). Entre os
caracteres utilizados para separar os sexos nos adultos de H. illucens, o principal refere-se a
terminália, que é diferenciada entre macho e fêmea, sendo que nesta última os escleritos finais
são mais largos do que os do macho (Gobbi, 2012).
Para a análise da morfometria, as larvas foram imersas em água aquecida
(aproximadamente 70°C), fixadas em solução KAAD (uma parte de querosene, 10 partes de
álcool (95%), duas partes de ácido acético e uma parte de dioxano) pelo período de 24 h, sendo
posteriormente acondicionadas em álcool 80% (Oliveira-Costa, 2011).
Foram mensurados o comprimento total do corpo, a largura e o comprimento da cápsula
cefálica e a largura do maior segmento abdominal dos seis estádios larvais em milímetro (mm)
e massa larval em miligrama (mg). As medidas de comprimento e largura foram realizadas sob
estereomicroscópio Leica M165C, com ocular micrométrica (acurácia = 0,001 mm), sendo o
comprimento total das larvas medido a partir do segmento cefálico até a extremidade distal do
último segmento (Lecheta, 2012). Além disso, foram realizadas pesagens para cada estádio,
utilizando uma balança de precisão (Mettler-Toledo; acurácia = 0,01 mg).
A taxa de crescimento foi calculada seguindo a regra de Dyar (1980), determinada a
partir da razão de crescimento (r), tendo como base a medida da cápsula cefálica. Desta forma,

14
r1 é a média de crescimento do segundo estádio dividida pela média de crescimento do
primeiro; r2 é a média de crescimento do terceiro estádio dividida pela média de crescimento
do segundo; r3 é a média de crescimento do quarto estádio dividida pela média de crescimento
do terceiro estádio; r4 é a média de crescimento do quinto estádio dividida pela média de
crescimento do quarto e r5 é a média de crescimento do sexto estádio dividida pela média de
crescimento do quinto.
As médias e desvios padrões de massa e comprimento de cada estádio foram calculados.
Os estádios foram caracterizados de acordo com os intervalos de medidas da largura da cápsula
cefálica, a qual é a utilizada pelos autores para H. illucens. Esta estrutura presente é a mais
esclerosada.Os dados obtidos diariamente das medidas foram agrupados em intervalos de classe
de 0,01 mm e plotados em gráficos para avaliação da distribuição da frequência. Para confirmar
a separação dos estádios através da largura da cápsula cefálica e das outras medidas avaliadas,
foi realizada uma Análise de Componentes Principais (PCA). Posteriormente, foi realizada uma
análise multivariada de variância baseada em distância (db-MANOVA; distancia euclidiana)
para verificar as diferenças dos dados morfométricos entre os estádios. Todas as análises foram
realizadas no software estatístico R (R Development Core Team (2011)).
Os adultos de H. illucens provenientes das criações foram montados em alfinetes
entomológicos e etiquetados e os imaturos conservados em álcool 80% para serem depositados
como material testemunho na Coleção de Invertebrados do INPA.
3. Resultados
3.1 Observações na carcaça
Durante o tempo de experimentação, não foi observado nenhum adulto macho de H.
illucens próximo à carcaça. Apenas fêmeas foram observadas e coletadas, atraídas para
oviposição. As fêmeas foram vistas em um intervalo de 24-72 horas após a morte do suíno,
quando este se encontrava na segunda fase de decomposição- enfisematosa. As fêmeas adultas
permaneceram visitando a carcaça até o 16º dia após o óbito do suíno. Estas pousavam próximo
ao cadáver, no interior da Shannon. Ao localizarem o recurso para a alimentação de seus
imaturos, estendem o ovipositor para fora do abdome (Figura 1) e depositam nas proximidades
da carcaça um por vez, formando uma massa de ovos (Figura 2A). As fêmeas ovipuseram
principalmente em locais onde outras fêmeas da mesma espécie já haviam realizado a
ovipostura. No experimento, os lugares escolhidos para oviposição foram escondidos e

15
protegidos, preferencialmente nos ossos do crânio do cadáver (Figura 2B) e em frestas da gaiola
(Figura 2C). Ovos foram visualizados a partir do 5º dia após a morte do suíno.
Figura 1. Fêmea de Hermetia illucens (Linnaeus, 1758) ovipondo em tubo Falcon ® no
laboratório. Círculos destacam ovos depositados e a seta indica o ovipositor estendido da fêmea
no momento da oviposição.
As larvas foram observadas na carcaça a partir do terceiro estádio (Figura 3A), devido
ao tamanho diminuto dos estádios iniciais. As larvas foram visualizadas na região abaixo do
couro do suíno em ambiente úmido (Figura 3B). Estas se movimentavam aleatoriamente e
continuamente no substrato, mantendo-se sempre aglomeradas. As larvas maiores, de último
estádio foram observadas na carcaça até aproximadamente 60 dias após o óbito.
Durante o processo de decomposição, após utilizar os recursos nutricionais do cadáver,
necessários para seu desenvolvimento, as larvas de H. illucens foram observadas se
movimentando lentamente. Ao contrário de outras espécies de Diptera, a mesma não procurou
ambientes mais secos para empupar, realizando este processo no próprio cadáver.
A empupação de H. illucens ocorreu na fase de restos, sendo as pupas adécticas e
exaradas, ou seja, não se alimentam e não se locomovem. Estas foram observadas a partir do
60º dia após o início da decomposição do suíno. Posteriormente ao estágio de pupa, o adulto
emergiu a partir da linha da ecdise. O adulto recém emergido, nas primeiras horas, possui o
corpo mole e locomove-se lentamente. As asas ficam atrofiadas por aproximadamente 24 horas
e os últimos segmentos abdominais na região ventral possuem coloração esverdeada.
Posteriormente, os segmentos abdominais passam a ter uma coloração escura e o segundo

16
segmento passa a ser hialino. Adultos emergidos na carcaça e nos potes de criação apresentaram
o mesmo padrão etológico e morfológico.
Figura 2. Posturas de ovos de Hermetia illucens (Linnaeus, 1758): A- massa de ovos sob
estereomicroscópio; B- massa de ovo no crânio do suíno (forames palatinos maiores); C- massa
de ovo nas frestas da gaiola.

17
Figura 3. Observações bionômicas de Hermetia illucens (Linnaeus, 1758): A- larvas de
diferentes estádios encontrados em ossos do côndilo femoral; B- larvas de sexto estádio
encontradas abaixo do couro do suíno (tecido conjuntivo).
3.2 Fases de desenvolvimento e razão sexual
Os potes de criação foram monitorados diariamente e foram obtidas 10 posturas de ovos
com seus respectivos ciclos de vida completos. Cada fêmea depositou de 620 a 700 ovos (em
média 660 ± 33 ovos) em uma única postura. Destes, aproximadamente 93% dos ovos
eclodiram e originaram larvas (6.215 larvas obtidas no experimento). Aproximadamente 72%
das larvas (n = 4.475) não atingiram o estágio de pupa e morreram em sua maioria nos primeiros
estádios larvais (1º a 3º). Além disso,17% das larvas chegaram à fase de pupa, mas não houve
emersão dos adultos (1.056 pupas). Apenas 11% das larvas (684 adultos emergidos)
conseguiram completar seu desenvolvimento e se tornar adultos. Em cada postura, a razão
sexual dos adultos emergidos foi de 0,49, correspondendo a aproximadamente, 50% machos e
50% fêmeas.
A partir da análise de frequência da largura da cápsula cefálica foi possível observar a
separação das larvas em seis estádios (Figura 4), embora seja evidente uma variação de tamanho
que foram expressos em intervalos de classes. Desta forma, o primeiro estádio possui largura
da cápsula cefálica variando de 0,05-0,08 mm; o segundo estádio de 0,09-0,24 mm; terceiro
estádio de 0,25-0,45 mm; quarto estádio de 0,46-0,57 mm; quinto estádio de 0,58-1,04 mm e
sexto estádio de 1,05-1,14 mm. Através da PCA, também foi possível observar a separação das
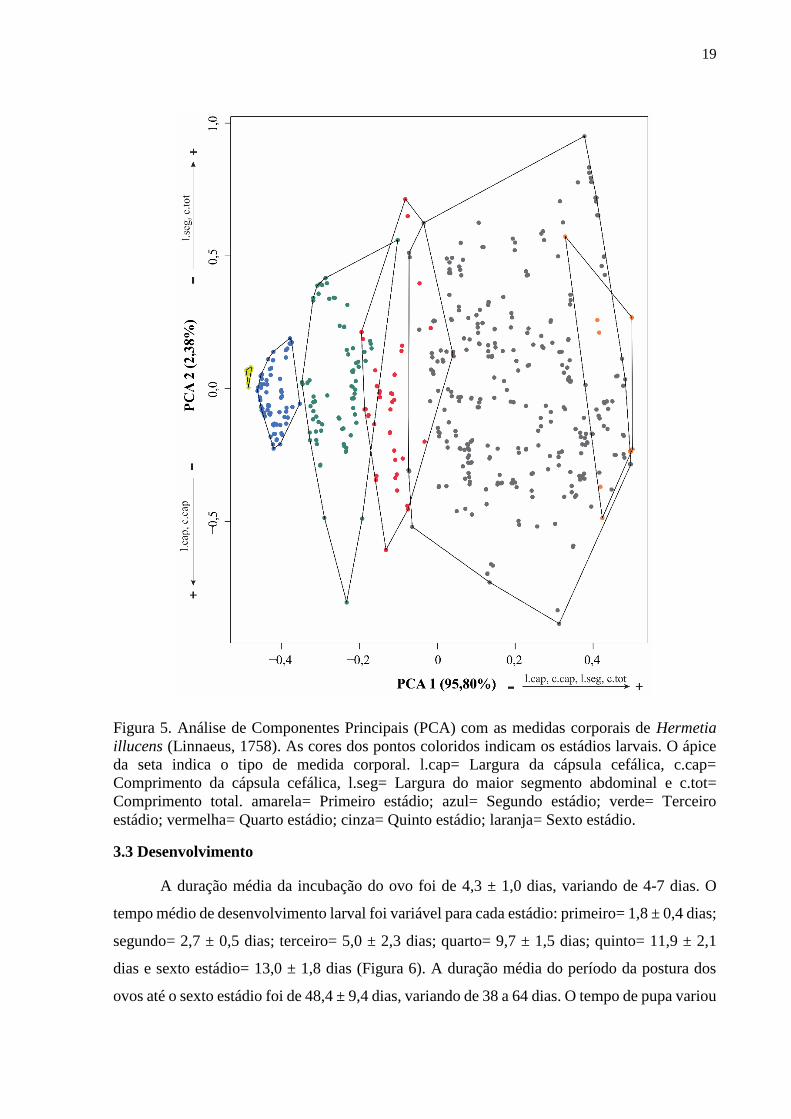
larvas em seis estádios (Figura 5). O primeiro eixo da PCA explicou ~96% da variação no
tamanho das larvas e esteve positivamente associado com o tamanho da larva (Largura da
cápsula cefálica, Comprimento da cápsula cefálica, Largura do maior segmento abdominal e
Comprimento total). Através da MANOVA foi possível confirmar que os estádios observados
na PCA são diferentes em tamanho (F5,562 = 1047,80; p = 0,001).

18
Figura 4. Distribuição de frequência da largura da cápsula cefálica de Hermetia illucens
(Linnaeus, 1758) (Diptera: Stratiomyidae) evidenciando os seis estádios larvais: verde=
Primeiro estádio; azul escuro= Segundo estádio; azul claro= Terceiro estádio; vermelha=
Quarto estádio; preta= Quinto estádio; laranja= Sexto estádio.
19
Figura 5. Análise de Componentes Principais (PCA) com as medidas corporais de Hermetia
illucens (Linnaeus, 1758). As cores dos pontos coloridos indicam os estádios larvais. O ápice
da seta indica o tipo de medida corporal. l.cap= Largura da cápsula cefálica, c.cap=
Comprimento da cápsula cefálica, l.seg= Largura do maior segmento abdominal e c.tot=
Comprimento total. amarela= Primeiro estádio; azul= Segundo estádio; verde= Terceiro
estádio; vermelha= Quarto estádio; cinza= Quinto estádio; laranja= Sexto estádio.
3.3 Desenvolvimento
A duração média da incubação do ovo foi de 4,3 ± 1,0 dias, variando de 4-7 dias. O
tempo médio de desenvolvimento larval foi variável para cada estádio: primeiro= 1,8 ± 0,4 dias;
segundo= 2,7 ± 0,5 dias; terceiro= 5,0 ± 2,3 dias; quarto= 9,7 ± 1,5 dias; quinto= 11,9 ± 2,1
dias e sexto estádio= 13,0 ± 1,8 dias (Figura 6). A duração média do período da postura dos
ovos até o sexto estádio foi de 48,4 ± 9,4 dias, variando de 38 a 64 dias. O tempo de pupa variou

20
de 15 a 20 dias. O tempo total de desenvolvimento, ou seja, de ovo até a emergência do adulto
variou de 53 a 82 dias.
Figura 6. Tempo de desenvolvimento para cada estádio larval de Hermetia illucens (Linnaeus,
1758) em condições ambientais em Manaus-AM.
A regra de Dyar indicou uma razão de crescimento para os estádios larvais com r = 1,87.
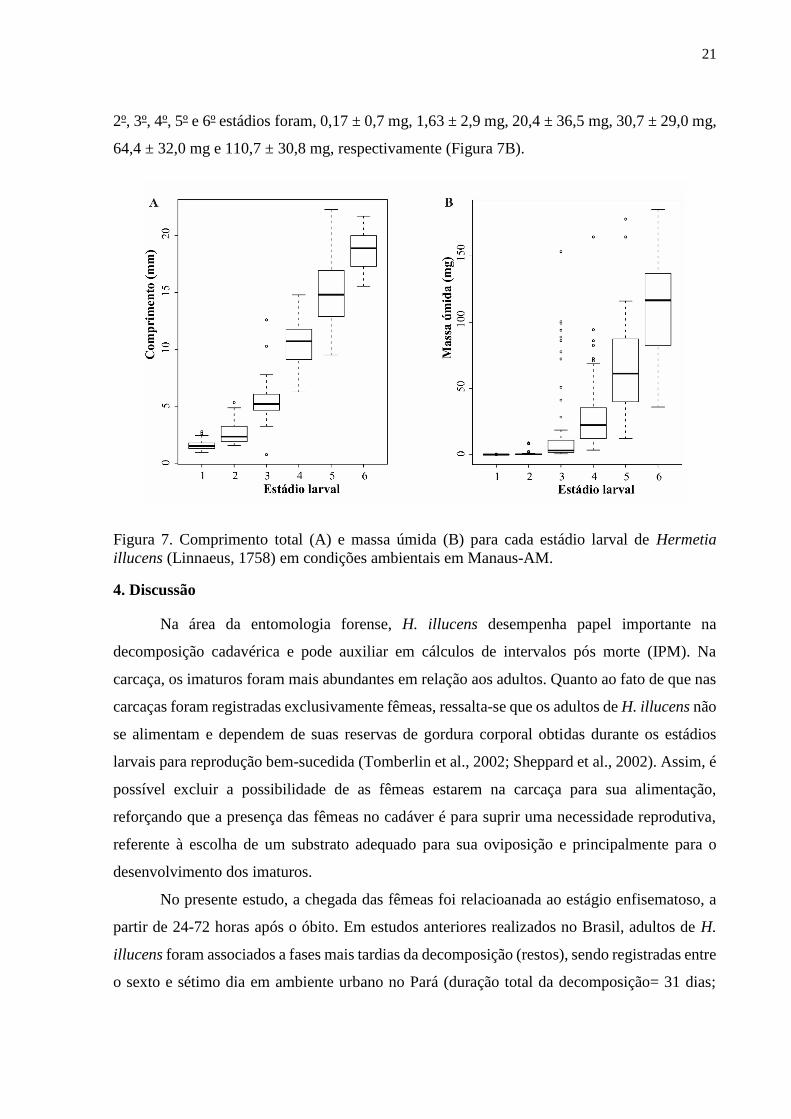
Durante o desenvolvimento de H. illucens, o comprimento larval aumentou aproximadamente
11,6 vezes em relação ao tamanho inicial. O comprimento total médio no primeiro estádio foi
de 1,6 ± 0,5 mm. Ao atingirem o sexto estádio a média foi de 18,6 ± 1,7 mm. Os valores do
comprimento médio no segundo, terceiro, quarto e quinto estádios foram 2,7 ± 1,0 mm, 5,5 ±
2,0 mm, 10,5 ± 2,2 mm e 14,7 ± 2,6 mm respectivamente (Figura 7A).
A massa das larvas de H. illucens aumentou gradativamente ao longo do
desenvolvimento. As pesagens do primeiro estádio foram realizadas, excluindo-se apenas as
que obtiveram insuficiência de massa na balança. As massas úmidas médias das larvas no 1º,
21
2º, 3º, 4º, 5º e 6º estádios foram, 0,17 ± 0,7 mg, 1,63 ± 2,9 mg, 20,4 ± 36,5 mg, 30,7 ± 29,0 mg,
64,4 ± 32,0 mg e 110,7 ± 30,8 mg, respectivamente (Figura 7B).
Figura 7. Comprimento total (A) e massa úmida (B) para cada estádio larval de Hermetia
illucens (Linnaeus, 1758) em condições ambientais em Manaus-AM.
4. Discussão
Na área da entomologia forense, H. illucens desempenha papel importante na
decomposição cadavérica e pode auxiliar em cálculos de intervalos pós morte (IPM). Na
carcaça, os imaturos foram mais abundantes em relação aos adultos. Quanto ao fato de que nas
carcaças foram registradas exclusivamente fêmeas, ressalta-se que os adultos de H. illucens não
se alimentam e dependem de suas reservas de gordura corporal obtidas durante os estádios
larvais para reprodução bem-sucedida (Tomberlin et al., 2002; Sheppard et al., 2002). Assim, é
possível excluir a possibilidade de as fêmeas estarem na carcaça para sua alimentação,
reforçando que a presença das fêmeas no cadáver é para suprir uma necessidade reprodutiva,
referente à escolha de um substrato adequado para sua oviposição e principalmente para o
desenvolvimento dos imaturos.
No presente estudo, a chegada das fêmeas foi relacioanada ao estágio enfisematoso, a
partir de 24-72 horas após o óbito. Em estudos anteriores realizados no Brasil, adultos de H.
illucens foram associados a fases mais tardias da decomposição (restos), sendo registradas entre
o sexto e sétimo dia em ambiente urbano no Pará (duração total da decomposição= 31 dias;

22
Anjos, 2001) e a partir do 18º dia em área urbana no Rio de Janeiro (duração total da
decomposição = 67 dias; Barbosa et al., 2006). Em estudos na região Temperada, a colonização
de H. illucens em carcaças foi registrada entre 20-30 dias após a morte do indivíduo (Tomberlin
et al., 2005). No entanto, no sul do estado da Geórgia (EUA), Tomberlin et al. (2005)
observaram o aparecimento da espécie na primeira semana após a morte. Tais diferenças na
colonização de H. illucens em carcaças em decomposição são devidas às diferentes localidades
e se extrapoladas podem resultar em erros nos cálculos de IPM, reforçando a importância e a
necessidade de estudos locais para sua aplicação médico-legal.
Fêmeas de H. illucens colocam ovos em massa, em média 660 ovos por postura. Para
Clark e Faeth (1998), a aglomeração dos ovos pode ser uma estratégia de oviposição da espécie,
para evitar o ressecamento dos ovos. Em estudos anteriores realizados em região temperada, o
número de ovos por postura variou de 320 a 600 ovos (Tomberlin et al., 2002; Sheppard, 1983).
O número de ovos pode ser variável e depende das condições onde se desenvolveram as larvas
que originaram as fêmeas (Blackmore e Lord, 2000). Também é importante afirmar que o
tamanho da fêmea pode influenciar a produção de ovos. Neste contexto, ressalta-se que estudos
realizados com mosquitos (Diptera: Culicidae) revelaram relação positiva entre o tamanho e
fecundidade (Clements, 1992; Renshaw et al., 1994). De acordo com Holmes et al. (2012),
fêmeas maiores de H. illucens foram mais férteis e apresentaram posturas maiores.
Independentemente do tamanho da prole, as fêmeas de H. illucens têm preferência por
ovipor em locais escondidos e protegidos. Além disso, na natureza ovipõem agrupamentos de
ovos em lugares úmidos, como exemplo, na parte inferior de uma folha úmida próxima à fonte
de alimento (Gobbi, 2012). Além disso, foram observadas também ovipondo em rachaduras
secas, fendas ao redor da matéria em decomposição, corroborando as observações de outros
estudos, como as de Copello (1926) e Gonzalez et al. (1963). Booth e Sheppard (1984)
encontraram fêmeas ovipondo rapidamente em aberturas diminutas das arestas de papelão
utilizadas em seu experimento. Este hábito das fêmeas, provavelmente, é uma forma de proteger
os ovos e as larvas da predação por outras espécies que também irão se alimentar da matéria
em decomposição. É importante destacar que as larvas de H. illucens demoram em média cinco
dias para eclodir e, sem esse comportamento protetor das fêmeas, provavelmente muitas larvas
não completariam seu desenvolvimento, devido a possível predação dos ovos.
As larvas de H. illucens foram relatadas por Lord et al. (1994), Oliveira-Costa (2011) e
Pujol-Luz et al. (2008) como ferramenta para determinar IPM, o que comprova sua grande
potencialidade na área forense, principalmente para o cálculo de intervalos de óbitos que

23
ultrapassam períodos de 15 dias. Neste período a maioria dos dípteros Calliphoridae e
Sarcophagidae completaram seu desenvolvimento e abandonaram o substrato. Durante esse
período as larvas de H. illucens foram observadas abaixo do couro, indicando que as mesmas
podem estar se alimentando dos restos da decomposição e de micro-organismos (e.g., fungos e
bactérias), pois a quantidade de carne em putrefação foi praticamente consumida pelas demais
espécies que estiveram presentes na carcaça.
Apesar da abundância de ovos depositados pelas fêmeas grávidas e de larvas eclodidas,
a maioria não alcançou a fase adulta. Estes resultados podem estar relacionados à
disponibilidade, condição ou ao tipo de recursos alimentares. Gobbi et al. (2013) observaram
que, dietas a base de carne bovina, frango ou a mistura destas duas, oferecidas para as larvas na
criação laboratorial influenciaram no sucesso de seu desenvolvimento e que a secagem rápida
do alimento e a textura deste também foram dados relevantes, visto que as larvas necessitam de
ambiente úmido. Quanto a isso, Souza e Kirst (2010) afirmam que é importante manter a
alimentação saudável para as larvas e recomendam o uso de carne bovina ou suína para a criação
de espécies de moscas de interesse forense. Outro aspecto relevante na criação de dípteros
refere-se aos fatores abióticos, os quais afetam o desenvolvimento e a sobrevivência desses
insetos. Por exemplo, é conhecida a existência de intervalos de temperaturas e umidade relativa
aceitáveis para o desenvolvimento de H. illucens (Booth e Sheppard, 1984; Sheppard et al.,
2002). Em geral, existem temperaturas ótimas para o desenvolvimento de insetos, sendo que
valores extremos, acima ou abaixo, resultam em mortalidade dos indivíduos (Byrd e Butler,
1996; Kingsolver e Huey, 2008). Quanto à umidade, este fator pode afetar a sobrevivência de
pupas para completar seu desenvolvimento e atingir a fase adulta (Sheppard et al., 1994).
No presente estudo foram registrados seis estádios larvais para H. illucens, resultados
semelhantes aos obtidos em estudos anteriores realizados na América do Norte por Hall e
Gerhardt (2002) e na Coréia do Sul por Kim et al. (2010). Entretanto, Kim et al. (2008)
determinaram a existência de apenas cinco estádios para a espécie na Coreia do Sul. Estes
resultados foram justificados por Kim et al. (2010) pela dificuldade de se distinguir as exúvias
do primeiro e segundo estádios, devido seu tamanho reduzido e similaridade das estruturas
larvais.
Os intervalos de medidas da largura da cápsula cefálica obtidas através de teste de
frequência neste experimento, em sua maioria, foram menores do que de outros trabalhos em
literatura (Tabela A). Ao mesmo tempo, o ciclo de vida observado variou de 53 a 82 dias, sendo
mais longo do que o registrado em outros locais para esta espécie. Kim et al. (2010) criaram

24
larvas de H. illucens na Coréia do Sul e contabilizaram um período larval menor (21-36 dias).
May (1961) criou os imaturos desta espécie na Nova Zelândia e o desenvolvimento larval durou
31 dias. As diferenças são ainda maiores quando levado em consideração o período total de
desenvolvimento (ovo à adulto). Furmand et al. (1959) afirmaram que H. illucens necessita de
14 dias ou meses para chegar à fase adulta. Por sua vez, as larvas criadas por May (1961)
demoraram 40-41 dias para completar seu ciclo. Nos Estados Unidos, Lord et al. (1994)
chegaram à conclusão que para H. illucens o período de desenvolvimento pode levar até cinco
meses para que ocorra a emersão dos adultos (Tabela B). As diferenças no tempo de
desenvolvimento e nas medidas corporais em relação a ouros estudos, provavelmente estão
relacionadas a fatores extrínsecos a elas, como temperatura, umidade e distribuição dos recursos
(Roubik 1989). Esses fatores podem resultar na aceleração ou retardamento da resposta do ciclo
de vida, ou seja, em todas as fases que vão de ovo a adulto (Souza e Kirst, 2010).
De acordo com Greenberg e Kunich (2002), características como tamanho e massa
corporal são influenciados principalmente pela temperatura e a dieta ao qual são submetidos.
Como H. illucens não se alimenta na fase adulta, precisa acumular energia necessária para seu
desenvolvimento. Desta forma, a qualidade dos alimentos disponíveis para as larvas tem
relevância para o sucesso dos adultos no ambiente (Parra, 1990). No que se refere à temperatura,
Atkinson (1994) propõe que seres ectotérmicos, como os insetos, que se desenvolvem em
temperaturas mais elevadas tendem a ser adultos menores do que indivíduos que crescem sob
temperaturas mais baixas. As larvas estudadas por Kim et al. (2010) possuíram mais massa
corporal do que as observadas neste experimento (primeiro e segundo estádios= não
mensurados, terceiro= 2,59-5,21 mg, quarto= 12,49-31,71 mg, quinto= 36,88- 95,72 mg, sexto=
151,51-219,29 mg).
Após a fase de imaturo, o adulto emerge em uma razão sexual de 0,49. Em Diptera essa
proporção é variável e depende da amplitude da fecundidade e sobrevivência (Serra et al.,
2007), e isso tem implicações diretas no crescimento populacional (Brown e Keller 2000). Por
exemplo, para Chrysomya albiceps (Wiedemann, 1819) (Diptera: Calliphoridae), dependendo
da estação do ano a prole pode ser toda de machos ou somente de fêmeas, isto ocorre pois,
diferentemente de H. illucens, trata-se de uma espécie monogênica (Serra et al., 2007). Em
Anastrepha Schiner, 1868 (Diptera: Tephritidae) foi encontrada uma razão sexual de
aproximadamente 0,50, semelhante aos resultados obtidos (Chiaradia et al., 2004).
Avaliando a dificuldade de determinação dos estádios larvais de H. illucens, bem como,
sua importância em diversos aspectos (econômico, médico, forense), estudos que possam

25
fornecer informações comportamentais e sobre o ciclo de desenvolvimento dessa espécie
poderão auxiliar na criminalística e minimizar possíveis erros na determinação de intervalos
pós morte. Além disso, informações bionômicas também são relevantes para a produção em
maior escala de larvas para estudos de conversão destas em biomassa, para a produção de ração,
entre outros.
Agradecimentos
Os autores agradecem a Tohnson Sales e Gilderlânia Oliveira pelo auxílio em campo.
A Léo Rossi e Daniel De Paula pela obtenção de algumas das imagens utilizadas. Ao
Laboratório de Citotaxonomia de insetos aquáticos pelo uso de equipamentos. À Coordenação
de aperfeiçoamento de Pessoal de Nível Superior – CAPES pela concessão da bolsa de
Mestrado. O apoio financeiro fornecido pelo projeto “Sistemática Integrada de Insetos
Aquáticos, com ênfase em Simuliidae (Diptera: Nematocera) na América do Sul” financiado
pelo MCTI / INPA.
Referências
Adler, A. I., Brancato, F.P., 1995. Human furuncular Myiasis caused by Hermetia illucens
(Diptera: Stratiomyidae). J. Med. Entomol. 32, 745-746.
Anjos, C. R. 2001. Entomofauna decompositora de carcaça de porcos na região de Belém, Pará,
Brasil, com ênfase na família Calliphoridae (Diptera). Museu Paraense Emilio Goeldi.
Belém, Pará.
Atkinson, D., 1994. Temperature and organism size – a biological law for ectotherms? Adv.
Ecol. Res. 3, 1–58.
Barbosa, R. R., Mello, R. P., Queiroz, M. M. C., 2006. Comportamento de Hermetia illucens
(Diptera: Stratiomyidae) como indicador forense na cidade do Rio de Janeiro, Brasil.
Resumo de congresso nacional. Recife, Pernambuco.
Barroso, F.G., De Haro, C., Sánchez-Muros, M.J., Venegas, E., Martínez-Sánchez, A., Pérez-
Bañón, C., 2014. The potencial of various insect species for use as food for fish.
Aquaculture. 422-423, 193-201.
Blackmore, M. S., Lord, C. C., 2000. The relationship between size and fecundity in Aedes
albopictus. J. of Vector Ecol. 25, 212-217.

26
Booth, D. C., Sheppard, C., 1984. Oviposition of the Black Soldier Fly, Hermetia illucens
(Diptera: Stratiomyidae): Eggs, Masses, Timing and Site Characteristics. Environ.
Entomol. 13, 421-423.
Brown, W.D., Keller, L., 2000. Colony sex ratios vary with queen number but note relatedness
asymmetry in the ant Formica exsecta. J. R. Soc. 267, 1751-1757.
Byrd , J . H., Butler, J. F., 1996. Effects of temperature on Cochliomyia macellaria. J. Med.
Entomol. 33, 901-905.
Calderón-Arguedas, O., Barrantes, M. J., Solano, M. E., 2005. Miíase entérica por Hermetia
illucens (Diptera: Stratiomyidae) en una paciente geriátrica de Costa Rica. Parasitol.
latinoam. 60 (3-4), 162-164.
Chiaradia, L. A., Milanez, J. M., Dittrich, R., 2004. Flutuação populacional de moscas-das-
frutas em pomares de citros no oeste de Santa Catarina, Brasil. Cienc. Rural. 34 (2), 337-
343.
Clark, B. R., Faeth, S. H., 1997. The consequences of larval aggregation in the butter ßy
Chlosyne lacinia. J. Ecol. Entomol. 22, 408-415.
Clements, A. N., 1992. The biology of Mosquitoes, development, nutricion, and reproduction.
Chapman e Hall, London. Vol. 1, 509 pp.
Copello, A. 1926. Biologia de Hermetia illucens Latr. Rev. Soc. Entomol. Argent. 1(2), 22-26.
Costa, A.C.L., Silva-Junior, J.A., Cunha, A.C., Feitosa, J.R.P., Portela, B.T.T., Silva, G.G.C.,
Costa, R.F., 2013. Índices de conforto térmico e suas variações sazonais em cidades de
diferentes dimensões na Região Amazônica. R. Bras. Geo. Fís. 6 (3), 478-487.
De Marco, M., Martínez, S., Hernandez, F., Madrid, J., Gai, F., Rotolo, L., Belforti, M.,
Bergero, D., Katz, H., Dadbou, S., Kovitvadhi, A., Zoccarato, I., Gasco, L., Schiavione,
A., 2015. Nutritional value of two insect latval meals (Tenebrio molitor and Hermetia
illucens) for broiler chickens: Apparent nutriente digestibility, apparent ileal amino acid
digestibility and apparent metabolizable energy. Anim. Feed Sci. Technol. 209, 211-218.

27
Diener, S., Solano, N. M. S., Gutiérrez, F. R., Zurbrügg, C., Tockner, K., 2011. Biological
treatment of municipal organic waste using black soldier fly larvae. Waste and Biomass
Valorization. 2 (4), 357-363.
Dyar, H. G., 1980. The number of molts Lepidopterous larvae. Psyche. 5, 420-422.
Furmand, P., Young, D., Catts, P. E., 1959. Hermetia illucens (Linnaeus) as a factor in the
natural control of Musca domestica (Linnaeus). J. of Econ. Entomol. 52 (5), 917-921.
Gobbi, F.P., 2012. Biología reproductiva y caracterización morfológica de los estadios larvarios
de Hermetia illucens (L., 1758) (Diptera: Stratiomyidae). Bases para su producción
masiva en Europa. Valência, Espanha.
Gobbi, P., Martínez-Sánchez, A., Rojo, S., 2013. The effects of larval diet n adult life-history
traits of the black soldier fly, Hermetia illucens (Diptera: Stratiomyidae). Eur. J. Entomol.
110 (3), 461–468.
Gonzalez, J. V., Young, W. R., Genel, M. R., 1963. Reducion de la problacion de mosca
domestica en gallinzaz por la mosca soldado en el Tropica. Agric. Tech. 2, 53-57.
Greenberg, B., Kunich, J.C., 2002. Entomology and the Law: Flies as Forensic Indicators.
Cambridge University Press, Cambridge.
Hall, D. C., Gerhardt, R. R., 2002. Flies (Diptera), in: Mullen, G., Durden, L. Academic Press.
Medical and veterinary entomology. San Diego. pp. 127- 161.
Holmes, L. A., Vanlaerhoven, S. L., Tomberlin, J. K., 2012. Relative Humidify effects on the
life history of Hermetia illucens (Diptera: Stratiomyidae). Environ. Entomol. 41 (4), 971-
978.
INMET – Instituto Nacional de Metereologia, 2017. Estações convencionais – gráficos.
http://www.inmet.gov.br/portal/index.php?r=home/page&page=rede_estacoes_conv_gr
af (accessed 24.01.2017).
INPA – Instituto Nacional de Pesquisas da Amazônia, 2013. Infraestrutura.
http://portal.inpa.gov.br/index.php/institucional/infraestrutura (accessed 09.01.17).

28
Kim, J. G., Choi, Y. C., Choi, J. Y., Kim, W. T., Jeong, G. S., Park, K. H., Hwang, S. J., 2008.
Ecology of the black soldier fly, Hermetia illucens (Diptera: Stratiomyidae) in Korea.
Korean J. Appl. Entomol. 47, 337-343.
Kim, W., Bae, S., Park, H., Park, K., Lee, S., Choi, Y., Han, S., Koh, Y., 2010. The larval age
and mouth morphology of the Black Soldier Fly, Hermetia illucens (Diptera:
Stratiomyidae). Int. J. Indust. Entomol. 21, 185-187.
Kingsolver, J. G., Huey, R. B., 2008. Size, temperature, and fitness: three rules. Evol. Ecol. Res.
10, 251–268.
Lalander, C, Diener, S, Magri, M. E., Zurbrügg, C., Lindström, A., Vinnerås, B., 2013. Faecal
sludge management with the larvae of the black soldier fly (Hermetia illucens) – From a
hygiene aspect. Sci. Total Environ. 458-460, 312-318.
Lecheta, M. C., 2012. Efeitos da temperatura no desenvolvimento de Sarconesia chlorogaster
(Wiedemam, 1830) (Diptera, Calliphoridae) e sua possível utilização na Entomologia
Forense. Curitiba, Paraná.
Lord, W. D., Goff, M. L., Adkins, T. R., Haskell, N. H., 1994. The black soldier fly Hermetia
illucens (Diptera: Stratiomyidae) as a potential measure of human postmortem interval:
observations and case histories. J. Forensic Sci. 39, 215–222.
May, B. M., 1961. The occurrence in New Zealand and the life-history of the soldier fly
Hermetia illucens (L.). New Zeal. J. Sci. 4, 55-65.
Oliveira-Costa, J., 2011. Entomologia Forense: Quando os insetos são vestígios. Millennium,
Campinas, São Paulo.
Parra, J.R.P., 1990. Consumo e utilização de alimentos por insetos, in: Panizzi, A.R., Parra,
J.R.P. (Manole): Ecologia Nutricional de Insetos e suas Implicações no Manejo de Pragas.
São Paulo, Brasil. pp. 9–57.
Pujol-Luz, J. R., Francez, P. A. C., Ururahy-Rodrigues, A., R. Constantino., 2008. The black
soldier-fly, Hermetia illucens (Díptera, Stratiomyidae), used to estimate the postmortem
interval in a case in Amapá state, Brazil. J. Forensic Sci. 53, 476–478.

29
Renshaw, M., Service, M. W., Birley, M. H., 1994. Size variation and reproductive success in
the mosquito Aedes cantans. Med. Vet. Entomol. 8, 179-186.
Roubik, D. W., 1989. Ecology and Natural History of Tropical Bees. Cambridge University.
Press, New York.
Serra, H., Godoy, W. A. C., Von-Zuben, F. J., Von-Zuben, C. J., Reis, S. F., 2007. Sex ratio
and dynamic behavior in populations of the exotic blowfly Chrysomya albiceps (Diptera,
Calliphoridae). Braz. J. Biol. 67(2), 347-353.
Sheppard, D. C., 1983. House fly and lesser fly control utilizing the black soldier fly in manure
management systems for caged laying hens. Environ. Entomol. 12, 1439- 1442.
Sheppard, D. C., Newton, G. L., Thompson, S. A., 1994. A value added manure management
system using the Black Soldier Fly. Bioresour. Technol. 50, 275-279.
Sheppard, D. C., Tomberlin, J. K., Joyce, J. A., Kiser, B. C., Sumner, S. M., 2002. Rearing and
colony maintenance of the black soldier fly, Hermetia illucens (L.) (Diptera:
Stratiomyidae). J. Med. Entomol. 39, 695-698.
Silveira-Neto, S., Nakano, O., Barbin, D., Villa-Nova, N. M., 1976. Manual de Ecologia de
Insetos. Editora Agronômica Ceres, São Paulo.
Souza, A. S. B., Kirst, F. D., 2010. Aspectos da bionomia e metodologia de criação de Dipteros
de interesse forense, In: Gomes, L. (Technical Books Editora), Entomologia Forense:
Novas tendências e tecnologias nas ciências criminais. Primeira edição. Rio de Janeiro.
pp. 169-182.
Tomberlin, J. K., Sheppard, D. C., Joyce, J. A., 2002. Selected life-history traits of Black
Soldier Flies (Diptera: Stratiomyidae) in a colony. J. Entomol. Sci. 37 (4): 345-352.
Tomberlin, J. K., Sheppard, D. C., Joyce, J. A., 2005. Black Soldier Fly (Diptera:
Stratiomyidae) colonization of pig carrion in South Georgia. J. Forensic Sci. 50 (1), 1-2.
Üstüner, T, Hasbenli, A., Rozkosny, R., 2003. The first record of Hermetia illucens (Linnaeus,
1758) (Diptera, Stratiomyidae) from the Near East. Stud. Dipterol. 10, 181-185.

30
Woodley, N. E., 2009. Family Stratiomyidae, In: Gerlach, J. (Pen soft Publishers) The Diptera
of the Seychelles islands. Sofia and Moscow. pp. 100-106.
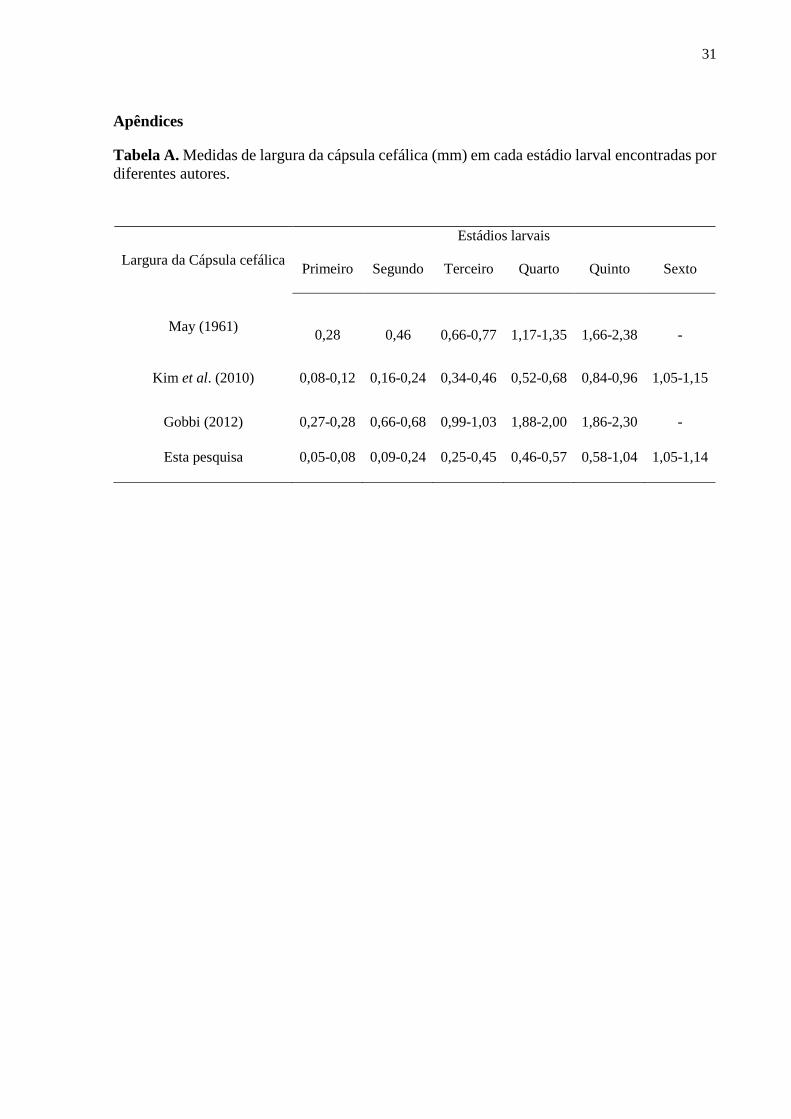
31
Apêndices
Tabela A. Medidas de largura da cápsula cefálica (mm) em cada estádio larval encontradas por
diferentes autores.
Largura da Cápsula cefálica
Estádios larvais
Primeiro Segundo Terceiro Quarto Quinto Sexto
May (1961)
0,28
0,46
0,66-0,77
1,17-1,35
1,66-2,38
-
Kim et al. (2010) 0,08-0,12 0,16-0,24 0,34-0,46 0,52-0,68 0,84-0,96 1,05-1,15
Gobbi (2012) 0,27-0,28 0,66-0,68 0,99-1,03 1,88-2,00 1,86-2,30 -
Esta pesquisa 0,05-0,08 0,09-0,24 0,25-0,45 0,46-0,57 0,58-1,04 1,05-1,14
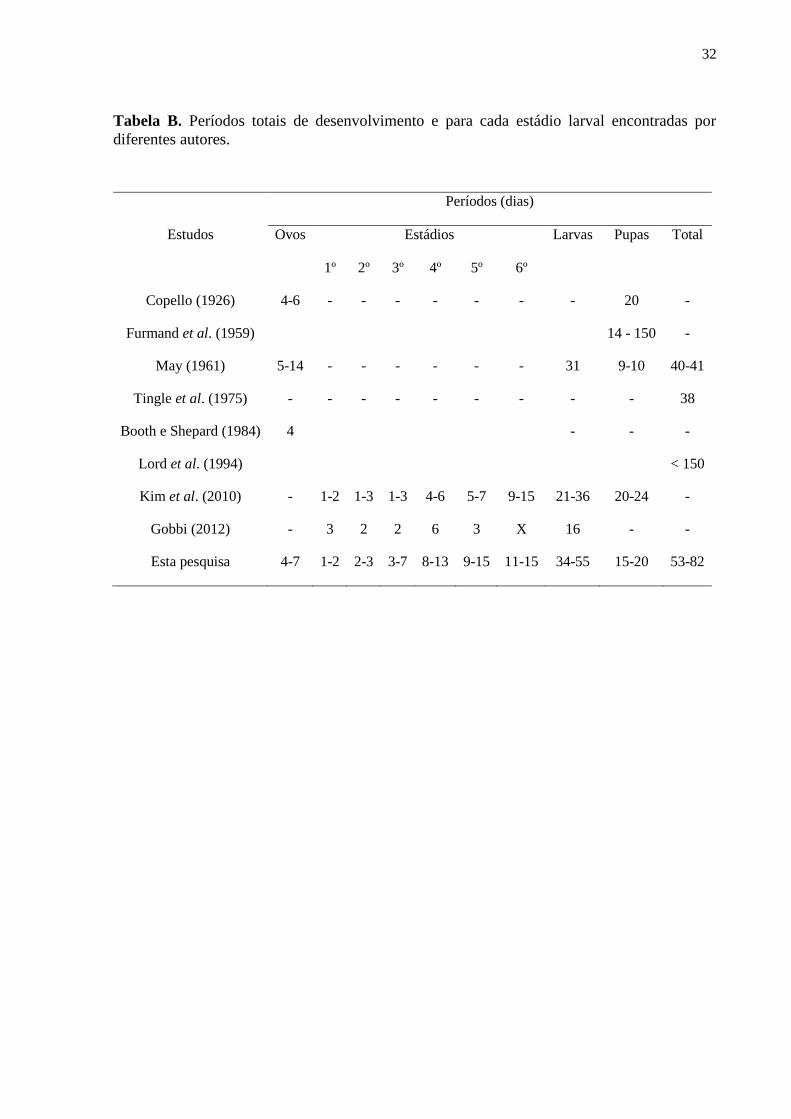
32
Tabela B. Períodos totais de desenvolvimento e para cada estádio larval encontradas por
diferentes autores.
Estudos
Períodos (dias)
Ovos Estádios Larvas Pupas Total
1º 2º 3º 4º 5º 6º
Copello (1926) 4-6 - - - - - - - 20 -
Furmand et al. (1959) 14 - 150 -
May (1961) 5-14 - - - - - - 31 9-10 40-41
Tingle et al. (1975) - - - - - - - - - 38
Booth e Shepard (1984) 4 - - -
Lord et al. (1994) < 150
Kim et al. (2010) - 1-2 1-3 1-3 4-6 5-7 9-15 21-36 20-24 -
Gobbi (2012) - 3 2 2 6 3 X 16 - -
Esta pesquisa 4-7 1-2 2-3 3-7 8-13 9-15 11-15 34-55 15-20 53-82

33
Capítulo II
Barros, L. M.; Gutjahr, A. L. N.; Ferreira-Keppler, R. L.;
& Martins, R. T. Descrição morfológica do ovo e dos
imaturos de Hermetia illucens (Linnaeus, 1758)
(Diptera: Stratiomyidae). Manuscrito formatado para
Neotropical Entomology.

34
Luana Machado Barros
Instituto Nacional de Pesquisas da Amazônia, INPA. Av. André Araújo 2936. Petrópolis.
69067-375 Manaus, AM, Brasil.
Descrição morfológica do ovo e dos imaturos de Hermetia illucens (Linnaeus) (Diptera:
Stratiomyidae)
L.M. BARROS¹, A.L.N. GUTJAHR², R.L. FERREIRA-KEPPLER¹, R.T. MARTINS¹
¹Programa de Pós-graduação em Entomologia, Instituto Nacional de Pesquisas da Amazônia -
INPA, Av. André Araújo, 2936, CEP 69067-375, Manaus, Amazonas, Brasil. ² Departamento
de Ciências Naturais da Universidade do Estado do Pará- UEPA, Tv. Djalma Dutra, 156,
Belém, PA, CEP 66050-540, Brasil.
Descrição dos imaturos de Hermetia illucens

35
Resumo - Dentro de Hermetia, há registros de descrição de imaturos para apenas seis espécies:
H. albitarsis Fabricius, 1805, H. aurata Bellardi, 1859, H. concinna Williston, 1900, H. illucens
(Linnaeus, 1758), H. panamensis Greene, 1940 e H. pulchra Wiedemann, 1830. Destas, H.
illucens, vem despertando interesse por sua aplicabilidade em grandes áreas da ciência (forense,
médica e econômica). Foi realizada a descrição morfológica de ovos e imaturos de H. illucens,
a fim de acrescentar informações sobre as modificações observadas entre os estádios larvais e
na fase de pupa, para auxiliar na identificação dos estádios larvais da espécie. No estudo
morfológico foi utilizada a microscopia ótica e a técnica de MEV. Os ovos têm formato ovoide
alongado retilíneo e possuem coloração branco leitosa à amarelada. As larvas são ápodes,
hemicéfalas e holopnêusticas, achatadas dorso-ventralmente e podem ser identificadas pela
combinação das seguintes características: cabeça longa, quetotaxia dorsal e ventral da cápsula
cefálica, dos segmentos torácicos e abdominais, presença da mancha esternal no sexto segmento
abdominal e morfologia dos espiráculos anteriores e posteriores. As pupas são ádécticas e
exaradas, castanho escuras com pruinosidade castanha à dourada. A morfologia dos estádios
larvais é semelhante, apresentando variação na forma dos artículos da antena. A quetotaxia das
larvas é semelhante nos estádios larvais e pupa, variando o formato das cerdas de primeiro
estádio com os demais. Os resultados obtidos ampliam o conhecimento da morfologia desse
díptero e podem ajudar peritos forenses na identificação e diferenciação dos estágios imaturos
em campo e, diminuir os erros no cálculo de intervalo pós morte.
Palavras-chave: Entomologia forense, Estádios larvais, morfologia, taxonomia.

36
Introdução
Stratiomyidae é formada por 12 subfamílias e por mais de 2.650 espécies, distribuídas
em 375 gêneros (Woodley 2009). Somente membros de Parhadestriinae e Raphiocerinae não
possuem registros de suas formas imaturas (Woodley 2001). As larvas da família geralmente
estão associadas à matéria orgânica vegetal ou animal em decomposição e podem habitar
ambientes terrestres, aquáticos ou semiaquáticos (McFadden 1967).
Em Stratiomyidae, Hermetiini é composta por cinco gêneros: Chaetohermetia (duas
espécies Neotropicais); Chaetosargus (quatro espécies Neotropicais); Hermetia (76 espécies
cosmopolitas), Nothohermetia (uma espécie Australiana) e Patagiomyia (uma espécie
Neotropical). Dentro de Hermetia existem 78 espécies conhecidas e destas, 54 ocorrem na
região Neotropical, incluindo 22 com distribuição no Brasil (McFadden 1967, Woodley 2011,
Fachin 2017). Dentre estas, há registros em literatura dos imaturos de apenas seis espécies:
Hermetia albitarsis Fabricius, 1805 (Brasil), H. aurata Bellardi, 1859 (México), H. concinna
Williston, 1900 (México), H. illucens (Linnaeus, 1758) (cosmopolita), H. panamensis Greene,
1940 (Panamá) e H. pulchra Wiedemann, 1830 (Neotropical) (McFadden 1967, Woodley
2011).
Hermetia illucens é conhecida popularmente como “The Black soldier-fly” e seus
representantes possuem hábito detritívoro. Diferentemente dos adultos que não se alimentam,
as larvas colonizam ambientes como plantas, frutos e carne em decomposição (Oliveira-Costa
2011). A espécie desperta interesse devido seus imaturos estarem relacionados à ciclagem de
matéria orgânica (Diener et al 2011) e por serem utilizados na produção de ração animal
(Barroso et al 2014, De Marco et al 2015). Além disso, estudos forenses utilizaram H. illucens
para determinar o intervalo pós morte (IPM), sendo esta espécie importante principalmente para
o cálculo de intervalos de óbitos que ultrapassam 15 dias (Lord et al 1994, Oliveira-Costa 2011,
Pujol-Luz et al 2008).
Portanto, o conhecimento sobre a morfologia desses imaturos faz-se necessário, na
medida que podem solucionar erros na identificação da espécie, do tempo de desenvolvimento
das larvas e, consequentemente, em estimativas de IPM, contribuindo de forma relevante com
a criminalística. Apesar da existência de dados em literatura sobre H. illucens, as descrições
dos imaturos ainda requerem detalhamento do corpo em vistas dorsal, ventral e lateral da
quetotaxia, sendo indispensável mais estudos destas características, as quais possuem
importância significativa na taxonomia do grupo. Desta forma, este artigo busca descrever e

37
detalhar a morfologia do ovo, dos estádios larvais e das pupas de H. illucens de Manaus,
Amazonas, Brasil.
Material e métodos
Os imaturos de H. illucens foram obtidos de fêmeas grávidas coletadas em suíno em
decomposição e criadas em condições naturais. O experimento foi realizado durante os meses
de janeiro a setembro de 2016 no Instituto Nacional de Pesquisas da Amazônia- INPA, campus
II, em Manaus, Amazonas, Brasil. Durante a coleta, uma postura de ovos, 45 larvas de cada
estádio e 45 pupas foram imersas em solução KAAD (1 parte de querosene, 10 partes de álcool
95%, 2 partes de ácido acético e 1 parte de dioxano) e posteriormente fixados em álcool 80%.
Este material foi utilizado para os registros fotográficos e descrição morfológica.
Os ovos e imaturos de H. illucens, coletados para análise em microscopia eletrônica de
varredura (MEV) foram lavados em soro fisiológico para retirar impurezas e depois fixados em
glutaraldeído 2%. Posteriormente, foram fixaçados em tetróxido de ósmio, lavados em solução
tampão de cacodilato de sódio 0,1 M em pH 7,2, sendo em seguida desidratados em solução de
etanol em diferentes concentrações (50%, 70%, 80%, 90%, 95% e 100%), por 20 minutos em
cada, com repetição na concentração de 100%. Os imaturos foram secos com gás carbônico
(CO2) pela técnica do ponto crítico, onde permaneceram por 60 minutos, e depois foram
montados em porta amostras (stubs) de alumínio e metalizados através de pulverização de sua
superfície em ouro. O processamento das amostras foi realizado no Laboratório Temático de
Microscopia Eletrônica do INPA. As visualizações e fotografias foram realizadas em
microscópio eletrônico de varredura VEGA 3 STEM (Tecscan) no Laboratório Temático de
Microscopia Eletrônica da Universidade Federal Rural da Amazônia (UFRA).
A determinação dos estádios larvais foi realizada através de medições da largura da base
da cápsula cefálica (LC) em vista ventral. As mensurações da LC e fotografias de ovos e
imaturos foram feitas sob estereomicroscópio Leica 165C com câmera fotográfica DFC420
acoplada. O programa de Processamento de Imagem Digital, Leica Application Suite V3. 4.1
foi utilizado para a digitalização das imagens. As descrições de estruturas menores dos ovos,
larvas e pupas foram feitas através das fotografias obtidas pela microscopia eletrônica de
varredura. As ilustrações foram realizadas através de software para vetorização de imagens e a
terminologia adotada para ovo e larva segue Rozkošný & Kovac (1998) e, para pupa, Costa &
Ide (2006). Todas as estruturas foram analisadas em vista dorsal, ventral e lateral.

38
Os imaturos foram armazenados em potes de vidro preenchidos com álcool 80% e
depositados como material testemunho na Coleção de Invertebrados do INPA.
Resultados
A descrição do ovo de H. illucens foi realizada com base na análise de 45 ovos. Esse
díptero depositada seus ovos em massa (Fig 1A), recobrindo sua superfície com muco, o que
proporciona a aderência do ovo ao substrato e entre eles. Os ovos ficam aglomerados em lugares
protegidos e escondidos, preferencialmente em orifícios pequenos, chegando uma postura a
totalizar entre 620 a 700 ovos (em média 660 ± 33 ovos).
Descrição do Ovo de Hermetia illucens
Dimensões variáveis, sendo o comprimento: 1,00-1,4 mm, diâmetro: 0,4-0,6 mm. Formato
ovóide alongado e retilíneo. Coloração variando de branco leitoso nas primeiras horas após a
oviposição, passando a amarelado com o tempo e o amadurecimento do embrião. Córion
brilhoso, com superfície irregular e ornamentações com aspecto enrugado (Fig 1B, C);
Micrópila não visualizada.
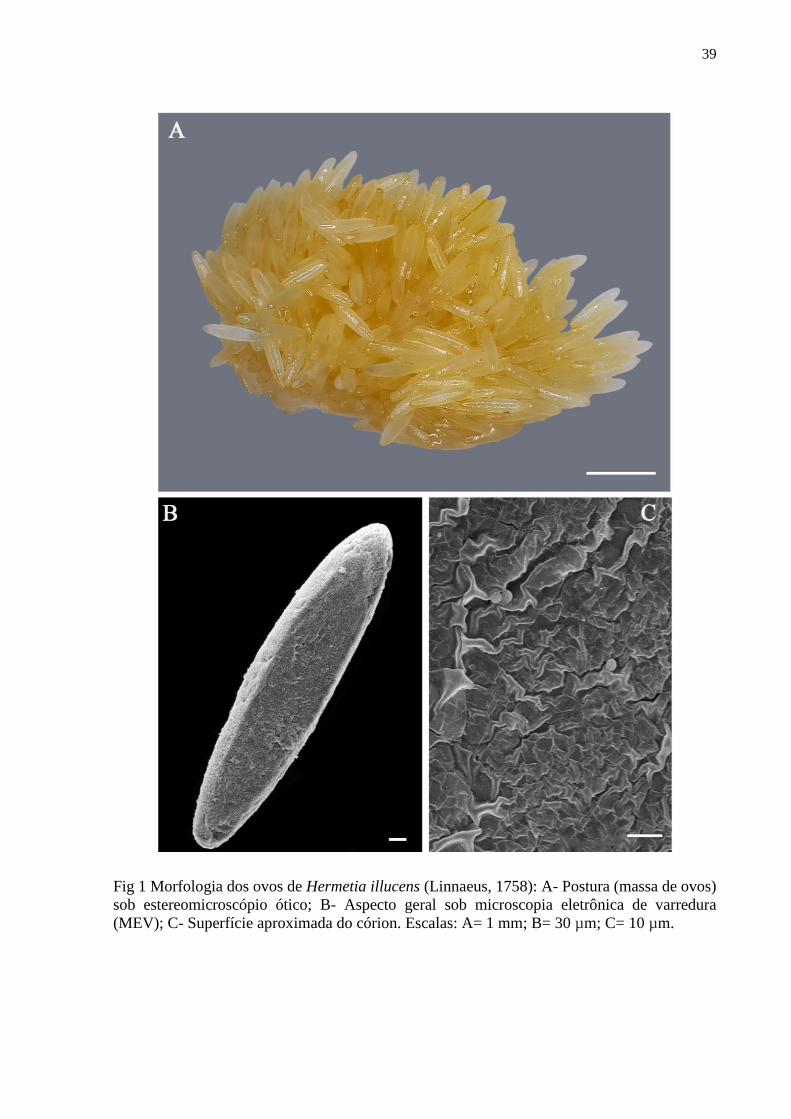
39
Fig 1 Morfologia dos ovos de Hermetia illucens (Linnaeus, 1758): A- Postura (massa de ovos)
sob estereomicroscópio ótico; B- Aspecto geral sob microscopia eletrônica de varredura
(MEV); C- Superfície aproximada do córion. Escalas: A= 1 mm; B= 30 µm; C= 10 µm.

40
Descrição da larva de Hermetia illucens
Larva ápode, hemicéfalica e holopnêustica. Corpo possui 12 segmentos (incluindo a cápsula
cefálica), alongado e achatado dorso-ventralmente (Figs 8A-C), evidenciando uma superfície
corpórea provida de cutícula ornamentada com aparência de mosaico ou formas circulares,
devido a deposição calcária (Fig 5B). Superfície corporal com cerdas e sensilas cuja coloração
varia em cada estádio.
Cápsula cefálica: castanha, tendendo a escurecer com o desenvolvimento larval, longa, mais
estreita que os segmentos seguintes e ligada ao protórax. Retrátil e esclerosada, provida de um
par de antena (An) (Fig 2B) situada lateralmente e formada por três segmentos. Segmento
antenal basal com sensilas campaniformes. Segundo segmento com sensilas longas e delgadas,
localizadas na região posterior (número varia dependendo do estádio larval). Terceiro segmento
oval, comprimento varia de acordo com o estádio (Fig 2B). Um par de ocelos ou olhos simples
(O) localizados lateralmente a cabeça. Dois escleritos laterais (Ecl) e um esclerito clípeo-frontal
(Ecf) alcançando a região apical. Labro (L) estreito, cônico e esclerosado (Fig 2A). Placa
ventral (Pv) ventrolateral, esclerosada e se estende na região basal. Premento (Prm) estreito na
base e alarga no ápice, localizado entre a placa ventral. Área molar (Am) bem desenvolvida e
localizada antero-ventralmente (Fig 2C). Lóbulos genais (Lg) estreitos e coriáceos. Complexo
mandíbulo- maxila, com mandíbula basal e maxila apical. Palpo maxilar (Pm) com cerdas
maxilares (Cm) ao redor da base pouco desenvolvidas. “Escovas mandibulares” (Em) na região
central, sendo uma superior (Em 1), localizada entre os palpos, e outra inferior (Em 2),
localizada entre os lóbulos genais (Fig 2D). Mandíbula (Md) estreita apicalmente com
pruinosidade concolor à cápsula cefálica.
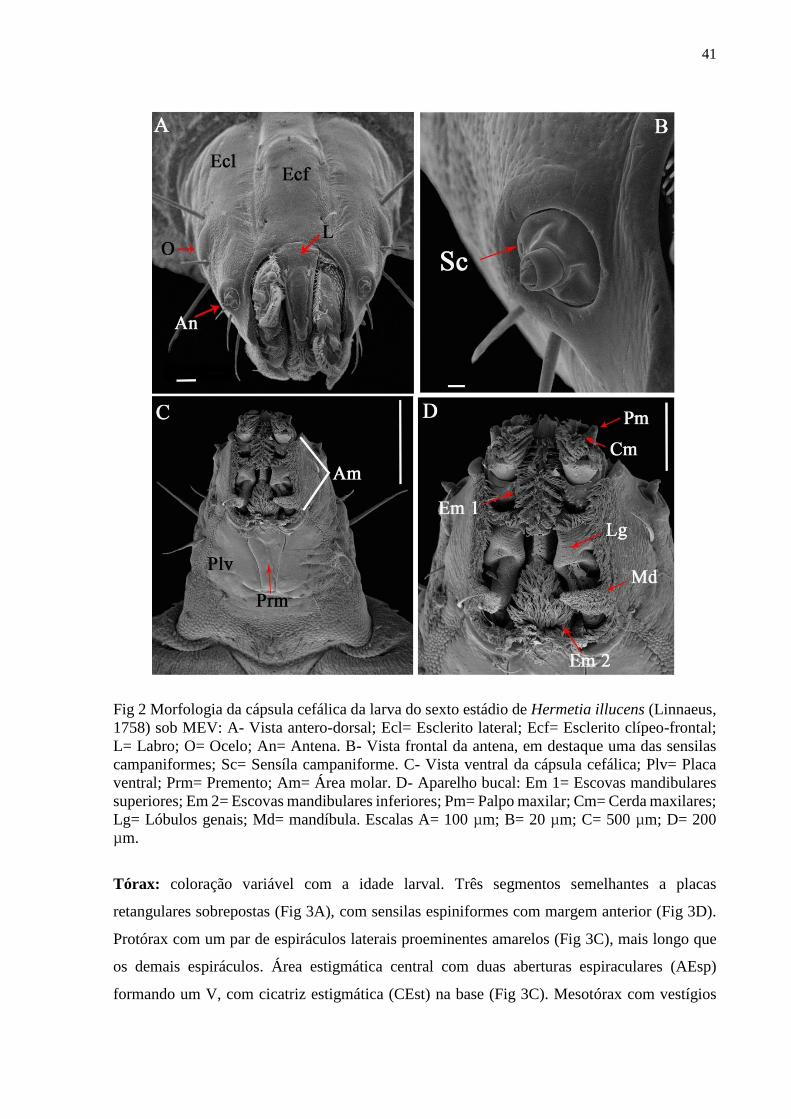
41
Fig 2 Morfologia da cápsula cefálica da larva do sexto estádio de Hermetia illucens (Linnaeus,
1758) sob MEV: A- Vista antero-dorsal; Ecl= Esclerito lateral; Ecf= Esclerito clípeo-frontal;
L= Labro; O= Ocelo; An= Antena. B- Vista frontal da antena, em destaque uma das sensilas
campaniformes; Sc= Sensíla campaniforme. C- Vista ventral da cápsula cefálica; Plv= Placa
ventral; Prm= Premento; Am= Área molar. D- Aparelho bucal: Em 1= Escovas mandibulares
superiores; Em 2= Escovas mandibulares inferiores; Pm= Palpo maxilar; Cm= Cerda maxilares;
Lg= Lóbulos genais; Md= mandíbula. Escalas A= 100 µm; B= 20 µm; C= 500 µm; D= 200
µm.
Tórax: coloração variável com a idade larval. Três segmentos semelhantes a placas
retangulares sobrepostas (Fig 3A), com sensilas espiniformes com margem anterior (Fig 3D).
Protórax com um par de espiráculos laterais proeminentes amarelos (Fig 3C), mais longo que
os demais espiráculos. Área estigmática central com duas aberturas espiraculares (AEsp)
formando um V, com cicatriz estigmática (CEst) na base (Fig 3C). Mesotórax com vestígios
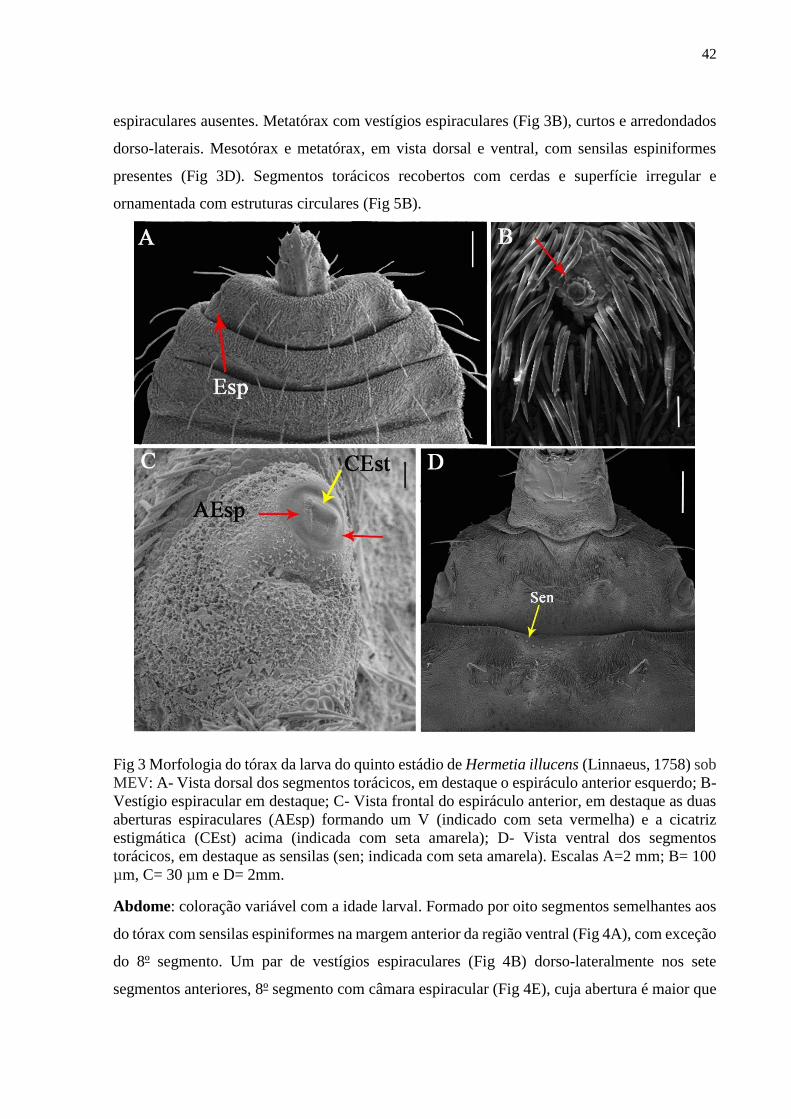
42
espiraculares ausentes. Metatórax com vestígios espiraculares (Fig 3B), curtos e arredondados
dorso-laterais. Mesotórax e metatórax, em vista dorsal e ventral, com sensilas espiniformes
presentes (Fig 3D). Segmentos torácicos recobertos com cerdas e superfície irregular e
ornamentada com estruturas circulares (Fig 5B).
Fig 3 Morfologia do tórax da larva do quinto estádio de Hermetia illucens (Linnaeus, 1758) sob
MEV: A- Vista dorsal dos segmentos torácicos, em destaque o espiráculo anterior esquerdo; B-
Vestígio espiracular em destaque; C- Vista frontal do espiráculo anterior, em destaque as duas
aberturas espiraculares (AEsp) formando um V (indicado com seta vermelha) e a cicatriz
estigmática (CEst) acima (indicada com seta amarela); D- Vista ventral dos segmentos
torácicos, em destaque as sensilas (sen; indicada com seta amarela). Escalas A=2 mm; B= 100
µm, C= 30 µm e D= 2mm.
Abdome: coloração variável com a idade larval. Formado por oito segmentos semelhantes aos
do tórax com sensilas espiniformes na margem anterior da região ventral (Fig 4A), com exceção
do 8º segmento. Um par de vestígios espiraculares (Fig 4B) dorso-lateralmente nos sete
segmentos anteriores, 8º segmento com câmara espiracular (Fig 4E), cuja abertura é maior que
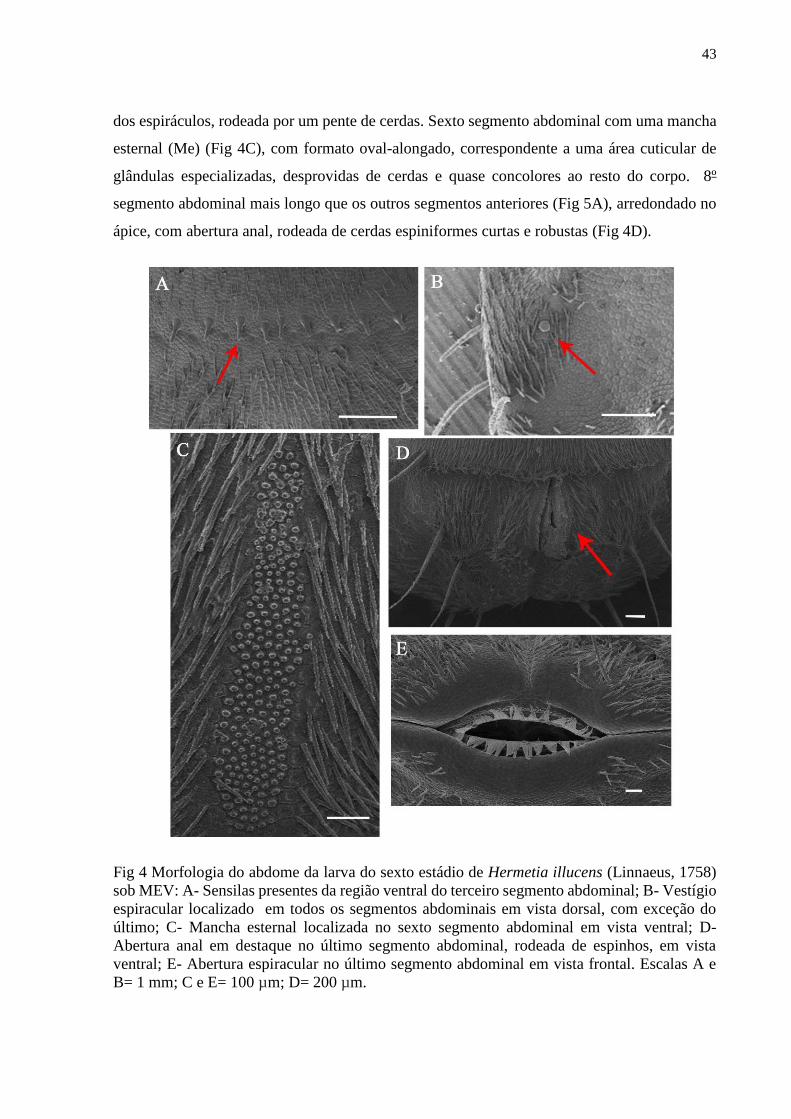
43
dos espiráculos, rodeada por um pente de cerdas. Sexto segmento abdominal com uma mancha
esternal (Me) (Fig 4C), com formato oval-alongado, correspondente a uma área cuticular de
glândulas especializadas, desprovidas de cerdas e quase concolores ao resto do corpo. 8º
segmento abdominal mais longo que os outros segmentos anteriores (Fig 5A), arredondado no
ápice, com abertura anal, rodeada de cerdas espiniformes curtas e robustas (Fig 4D).
Fig 4 Morfologia do abdome da larva do sexto estádio de Hermetia illucens (Linnaeus, 1758)
sob MEV: A- Sensilas presentes da região ventral do terceiro segmento abdominal; B- Vestígio
espiracular localizado em todos os segmentos abdominais em vista dorsal, com exceção do
último; C- Mancha esternal localizada no sexto segmento abdominal em vista ventral; D-
Abertura anal em destaque no último segmento abdominal, rodeada de espinhos, em vista
ventral; E- Abertura espiracular no último segmento abdominal em vista frontal. Escalas A e
B= 1 mm; C e E= 100 µm; D= 200 µm.
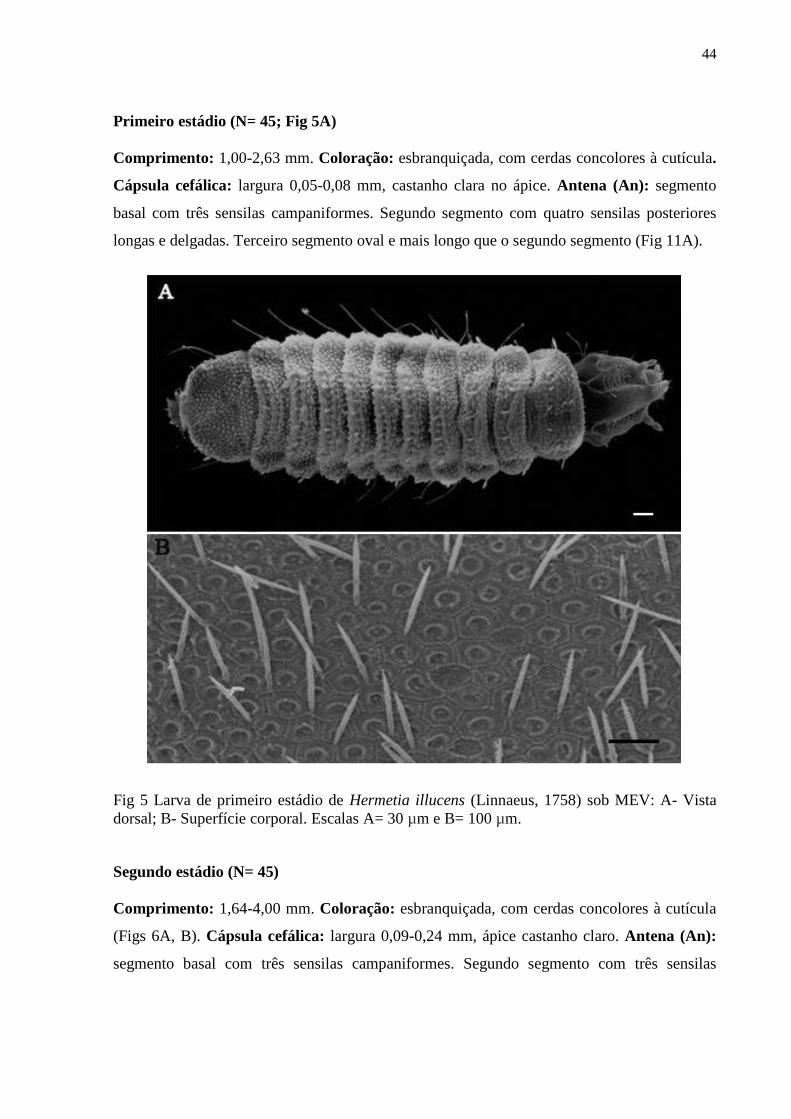
44
Primeiro estádio (N= 45; Fig 5A)
Comprimento: 1,00-2,63 mm. Coloração: esbranquiçada, com cerdas concolores à cutícula.
Cápsula cefálica: largura 0,05-0,08 mm, castanho clara no ápice. Antena (An): segmento
basal com três sensilas campaniformes. Segundo segmento com quatro sensilas posteriores
longas e delgadas. Terceiro segmento oval e mais longo que o segundo segmento (Fig 11A).
Fig 5 Larva de primeiro estádio de Hermetia illucens (Linnaeus, 1758) sob MEV: A- Vista
dorsal; B- Superfície corporal. Escalas A= 30 µm e B= 100 µm.
Segundo estádio (N= 45)
Comprimento: 1,64-4,00 mm. Coloração: esbranquiçada, com cerdas concolores à cutícula
(Figs 6A, B). Cápsula cefálica: largura 0,09-0,24 mm, ápice castanho claro. Antena (An):
segmento basal com três sensilas campaniformes. Segundo segmento com três sensilas
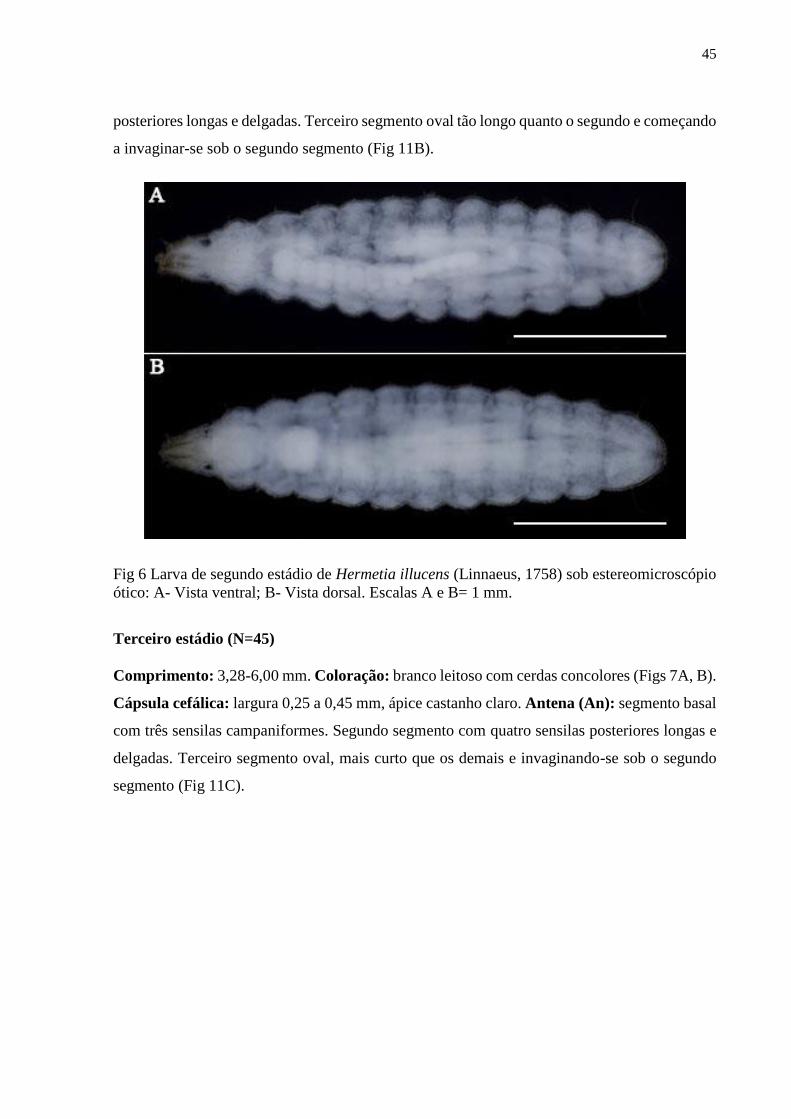
45
posteriores longas e delgadas. Terceiro segmento oval tão longo quanto o segundo e começando
a invaginar-se sob o segundo segmento (Fig 11B).
Fig 6 Larva de segundo estádio de Hermetia illucens (Linnaeus, 1758) sob estereomicroscópio
ótico: A- Vista ventral; B- Vista dorsal. Escalas A e B= 1 mm.
Terceiro estádio (N=45)
Comprimento: 3,28-6,00 mm. Coloração: branco leitoso com cerdas concolores (Figs 7A, B).
Cápsula cefálica: largura 0,25 a 0,45 mm, ápice castanho claro. Antena (An): segmento basal
com três sensilas campaniformes. Segundo segmento com quatro sensilas posteriores longas e
delgadas. Terceiro segmento oval, mais curto que os demais e invaginando-se sob o segundo
segmento (Fig 11C).
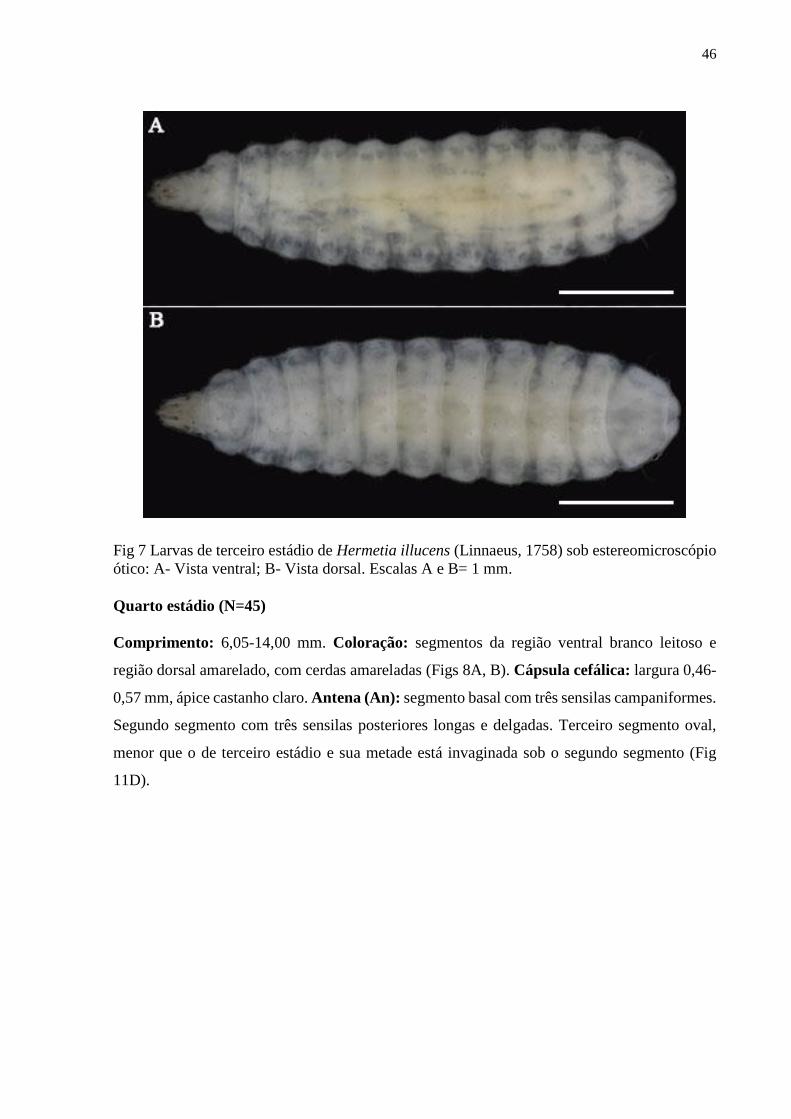
46
Fig 7 Larvas de terceiro estádio de Hermetia illucens (Linnaeus, 1758) sob estereomicroscópio
ótico: A- Vista ventral; B- Vista dorsal. Escalas A e B= 1 mm.
Quarto estádio (N=45)
Comprimento: 6,05-14,00 mm. Coloração: segmentos da região ventral branco leitoso e
região dorsal amarelado, com cerdas amareladas (Figs 8A, B). Cápsula cefálica: largura 0,46-
0,57 mm, ápice castanho claro. Antena (An): segmento basal com três sensilas campaniformes.
Segundo segmento com três sensilas posteriores longas e delgadas. Terceiro segmento oval,
menor que o de terceiro estádio e sua metade está invaginada sob o segundo segmento (Fig
11D).

47
Fig 8 Larvas de quarto estádio de Hermetia illucens (Linnaeus, 1758) sob estereomicroscópio
ótico: A- Vista ventral; B- Vista dorsal; C- Vista lateral. Escalas A, B e C= 1 mm.
Quinto estádio (N=45)
Comprimento: 9,53-20,29 mm. Coloração: branco leitoso, com cerdas amareladas (Figs 9A,
B). Cápsula cefálica: largura 0,58-1,04 mm, castanho claro e ápice mais escuro. Antena (An):
segmento basal com três sensilas campaniformes. Segundo segmento com três sensilas
posteriores longas e delgadas. Terceiro segmento oval, menor que o de quarto estádio e mais
de sua metade invaginou-se sob o segundo segmento (Fig 11E).

48
Fig 9 Larvas de quinto estádio de Hermetia illucens (Linnaeus, 1758) sob estereomicroscópio
ótico: A- Vista ventral; B- Vista dorsal; C- Vista lateral. Escalas A, B e C= 2 mm.
Sexto estádio (N= 45)
Comprimento: 9,53-20,29 mm. Coloração: branco leitoso, amarelada a castanho escuro, com
cerdas amareladas (Figs 10A, B). Cápsula cefálica: largura 1,05-1,14 mm, castanho clara e
ápice mais escuro. Antena (An): segmento basal com três sensilas campaniformes. Segundo
segmento com três sensilas posteriores longas e delgadas. Terceiro segmento oval, menor que
o de quinto estádio, ápice visível e o restante invaginado totalmente sob o segundo segmento
(Fig 11F).

49
Fig 10 Larvas de sexto estádio de Hermetia illucens (Linnaeus, 1758) sob estereomicroscópio
ótico: A- Vista ventral; B- Vista dorsal; C- Vista lateral. Escalas A, B e C= 5 mm.

50
Fig 11 Antena das larvas de Hermetia illucens (Linnaeus, 1758) evidenciando as diferenças
morfológicas entre os estádios: A- Primeiro estádio; B- Segundo estádio; C- Terceiro estádio;
D- Quarto estádio; E- Quinto estádio; F- Sexto estádio. Os números 1, 2, 3 e 4 indicam as
sensilas alongadas encontradas no segundo segmento da antena. Escalas A= 3 µm; B= 10 µm;
C= 5 µm; D= 100 µm; E= 200 µm e F= 20 µm.

51
Descrição da pupa de Hermetia illucens (n=45)
Comprimento: 15,00-22,29 mm. Pupa adéctica e coarctata. Corpo com 12 segmentos
(incluindo a cápsula cefálica), alongado e achatado dorso-ventralmente (Fig 12C). Superfície
áspera e rígida, retém estruturas cuticulares desenvolvidas na fase larval anterior. Cutícula com
aparência de mosaico, devido a deposição calcária. Cerdas mais rígidas do que nos estádios
larvais. Corpo coberto por pruinosidade densa, castanha a dourada. Coloração: castanho escura
tendendo ao marrom (Fig 12A).
Fig 12 Pupa de Hermetia illucens (Linnaeus, 1758) sob estereomicroscópio ótico: A- Vista
ventral; B- Vista dorsal; C- Vista lateral. Escalas A, B e C= 5 mm.
Cápsula cefálica: concolor ao restante do corpo. Curta e mais estreita que os segmentos
seguintes (Fig 12A). Largura: 0,73-0,86 mm. Totalmente esclerosada e provida de um par de
antenas (An) laterais, com segmento apical totalmente invaginado sob o segundo segmento
(Figs 13A, C). Um par de ocelos ou olho simples (O) laterais, correspondendo a ⅓ da altura da

52
cápsula cefálica, localizados próximo à base desta. Dois escleritos dorsais (Ecl), esclerito
clípeo-frontal (Ecf) atingindo a região apical (Fig 13A). Labro (L) estreito, cônico e esclerosado
(Fig 13B). Em posição ventro-lateral, encontra-se a placa ventral (Pv), que é esclerosada e se
estende na região basal (de cada lado da cabeça). O premento (Prm) é um esclerito estreito na
base que se alarga no ápice, localizado entre a placa ventral. Área molar apenas parcialmente
distinta. Complexo mandibulo-maxila seco e firmemente ligado aos lóbulos genais (Lg) da
cápsula cefálica. Aparelho bucal coberto por uma placa cuticular completa e esclerosada (Fig
13D).
Fig 13 Morfologia da cápsula cefálica da pupa de Hermetia illucens (Linnaeus, 1758) sob MEV:
A- Vista dorsal; Ecl= Esclerito lateral; Ecf= Esclerito clípeo-frontal; O= Ocelo; An= Antena.
B- Vista lateral da cápsula cefálica, evidenciando o olho direito (seta); L= Labro; C- Vista
dorsal da antena; D- Vista ventral da cápsula cefálica; Plv= Placa ventral; Prm= Premento; Am=
Área molar. Escalas A, B e D= 200 µm; C= 10 µm.
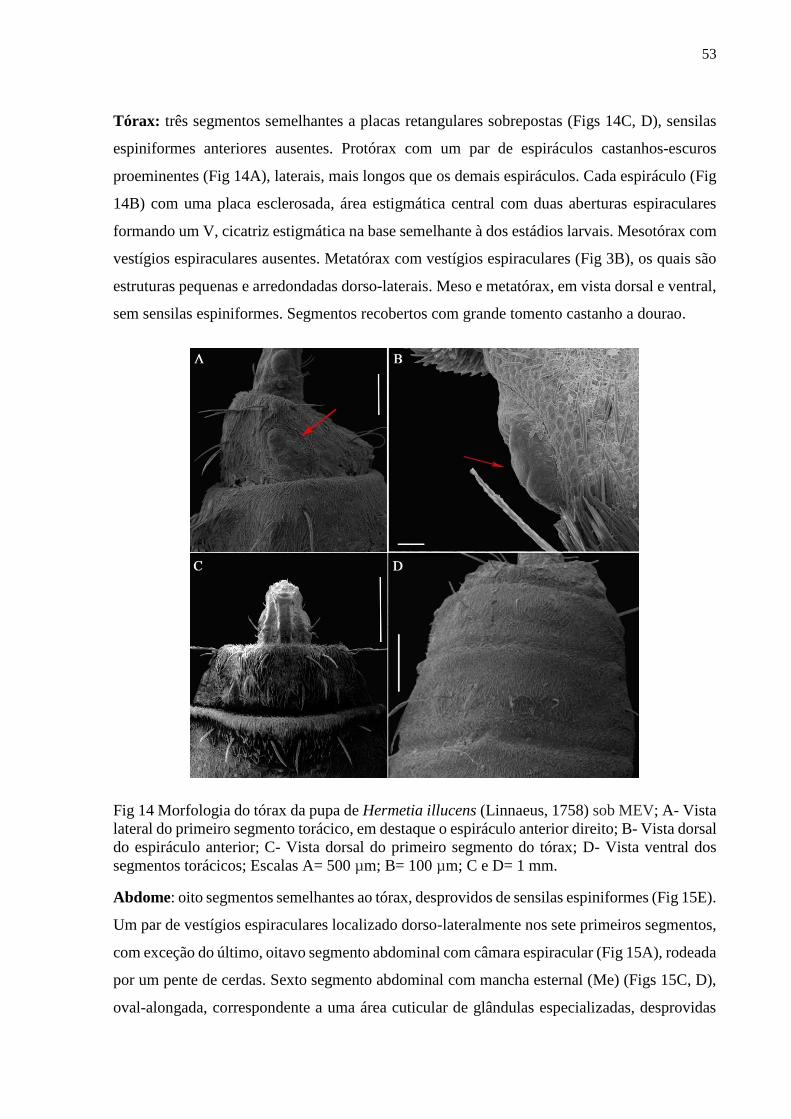
53
Tórax: três segmentos semelhantes a placas retangulares sobrepostas (Figs 14C, D), sensilas
espiniformes anteriores ausentes. Protórax com um par de espiráculos castanhos-escuros
proeminentes (Fig 14A), laterais, mais longos que os demais espiráculos. Cada espiráculo (Fig
14B) com uma placa esclerosada, área estigmática central com duas aberturas espiraculares
formando um V, cicatriz estigmática na base semelhante à dos estádios larvais. Mesotórax com
vestígios espiraculares ausentes. Metatórax com vestígios espiraculares (Fig 3B), os quais são
estruturas pequenas e arredondadas dorso-laterais. Meso e metatórax, em vista dorsal e ventral,
sem sensilas espiniformes. Segmentos recobertos com grande tomento castanho a dourao.
Fig 14 Morfologia do tórax da pupa de Hermetia illucens (Linnaeus, 1758) sob MEV; A- Vista
lateral do primeiro segmento torácico, em destaque o espiráculo anterior direito; B- Vista dorsal
do espiráculo anterior; C- Vista dorsal do primeiro segmento do tórax; D- Vista ventral dos
segmentos torácicos; Escalas A= 500 µm; B= 100 µm; C e D= 1 mm.
Abdome: oito segmentos semelhantes ao tórax, desprovidos de sensilas espiniformes (Fig 15E).
Um par de vestígios espiraculares localizado dorso-lateralmente nos sete primeiros segmentos,
com exceção do último, oitavo segmento abdominal com câmara espiracular (Fig 15A), rodeada
por um pente de cerdas. Sexto segmento abdominal com mancha esternal (Me) (Figs 15C, D),
oval-alongada, correspondente a uma área cuticular de glândulas especializadas, desprovidas
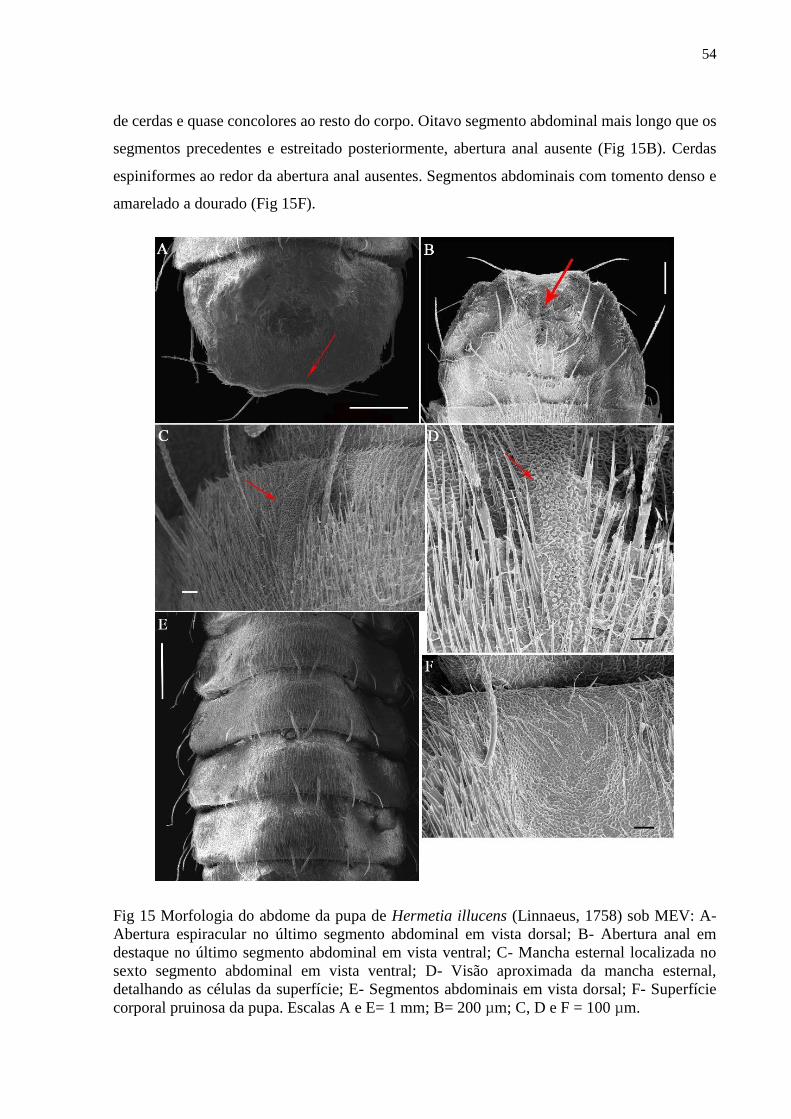
54
de cerdas e quase concolores ao resto do corpo. Oitavo segmento abdominal mais longo que os
segmentos precedentes e estreitado posteriormente, abertura anal ausente (Fig 15B). Cerdas
espiniformes ao redor da abertura anal ausentes. Segmentos abdominais com tomento denso e
amarelado a dourado (Fig 15F).
Fig 15 Morfologia do abdome da pupa de Hermetia illucens (Linnaeus, 1758) sob MEV: A-
Abertura espiracular no último segmento abdominal em vista dorsal; B- Abertura anal em
destaque no último segmento abdominal em vista ventral; C- Mancha esternal localizada no
sexto segmento abdominal em vista ventral; D- Visão aproximada da mancha esternal,
detalhando as células da superfície; E- Segmentos abdominais em vista dorsal; F- Superfície
corporal pruinosa da pupa. Escalas A e E= 1 mm; B= 200 µm; C, D e F = 100 µm.

55
Quetotaxia
Em todos os estádios larvais e na pupa de H. illucens, as posições das cerdas na cápsula
cefálica, região dorsal e ventral, são semelhantes (Figs 16 e 17). No entanto, foi observada
diferença no formato das cerdas. Larvas de primeiro estádio possuem dois tipos de cerdas: as
cerdas ventrais são lisas e subdivididas basalmente em dois filamentos, um menor e acuminado,
o outro longo e capitado distalmente (Fig 18A); e as cerdas dorsais são capitadas distalmente e
com numerosos dentículos (Fig 18B). Nos demais estádios, as cerdas são multiciliadas e
apicalmente acuminada e são distribuidas por todo corpo da larva (Fig 18C).
Cápsula cefálica: um par de cerdas dorso-laterais, dois pares de cerdas clípeo-frontais e dois
pares de cerdas labrais (Fig 16A). Ventralmente há três pares de cerdas ventrais e três pares
ventro-laterais (Fig 16B).
Fig 16 Quetotaxia da cápsula cefálica de Hermetia illucens (Linnaeus, 1758): A- Vista dorsal;
B- Vista ventral.

56
Tórax: protórax com dois pares de cerdas antero-dorsais, três pares de cerdas dorsais e um par
dorso-lateral. Meso e metatórax com três pares de cerdas dorsais e um par de cerdas dorso-
lateral (Fig 17A). Cada um dos três segmentos possue dois pares de cerdas ventrais e um par
de cerdas ventro-laterais (Fig 17B).
Abdome: primeiro ao sétimo segmento com um par de cerdas dorso-laterais e três pares de
cerdas dorsais cada. Oitavo segmento com dois pares de cerdas dorso-laterais e um par dorsal
(Fig 17A). Primeiro ao sétimo segmento com um par de cerdas ventro-lateral, três pares de
cerdas ventrais cada. Oitavo segmento com dois pares de cerdas póstero-ventrais, dois pares de
cerdas ventrais, um par de cerdas anais e dois pares de cerdas pré-anais (Fig 17B).
Fig 17 Quetotaxia da larva de Hermetia illucens (Linnaeus, 1758) com ênfase nos segmentos
toráxicos e abdominais: A- Vista dorsal; B- Vista ventral. Os números 1-3 apresentados
inicialmente indicam os segmentos torácicos. Os números de 1’ a 8’ indicam os segmentos
abdominais. Os pontos pretos ( ) indicam as posições das cerdas nos segmentos corporais da
larva.

57
Fig 18 Tipos de cerdas da superfície corporal de Hermetia illucens (Linnaeus, 1758): A- Cerdas
de primeiro estádio localizadas na região ventral; B- Cerdas de primeiro estádio localizadas na
região dorsal; C- Cerdas localizadas no segundo, terceiro, quarto, quinto, sexto estádios e pupa.
Escalas A= 10 µm; B= 10 µm e C= 100 µm.
Discussão
As larvas de H. illucens possuem a morfologia semelhante entre os estádios larvais. Este
provavelmente é o motivo pelo qual os estudos de morfologia da espécie são realizados apenas
com um único (Baez 1975, Kim et al 2010, Gobbi 2012, Oliveita et al 2015) ou dois estádios
(Oliveita et al 2016), sendo as descrições generalizadas para todos os estádios. Em literatura,
os estádios larvais desse diptero são definidos a partir de medidas da cápsula cefálica. Neste
artigo, além da morfometria, foram observadas diferenças na antena, a qual é modificada no

58
decorrer do desenvolvimento, sendo que o terceiro segmento invagina-se sob o segundo até não
ser mais visível no sexto estádio e pupa. Além disso, há uma grande modificação nas cerdas
das larvas de primeiro estádio em comparação com os demais.
O estudo da quetotaxia ainda não é tão utilizado nesta espécie. No entanto, Gobbi (2012)
destacou as posições das cerdas do quinto estádio larval, de maneira semelhante ao encontrado
neste artigo. O diferencial foi notar que há modificação no tipo e formato das cerdas e
caracterizá-las.
As cerdas presentes na cutícula dos imaturos de H. illucens, tanto na região dorsal
quanto ventral, proporcionaram melhor rendimento quanto a locomoção da larva,
principalmente para esta espécie que se desenvolve em matéria orgânica (Baez 1975).
Provavelmente, as larvas do primeiro estádio têm cerdas mais longas na região ventral para
facilitar sua aderência ao substrato e procurar ativaente locais mais favoráveis, pois são menores
e mais frágeis.
Hermetia illucens, assim como as outras espécies de Stratiomyidae e Xylomyidae fazem
parte de um grupo, no qual as pupas se originam dentro do tegumento do último estádio larval
(Smith 1989). Isto explica o fato da morfologia da pupa ser semelhante à da larva. Pupas de H.
illucens são incapazes de se locomover e se alimentar, por isso, provavelmente o aparelho bucal
vai se tornando mais esclerosado de acordo com a idade da larva até as peças se tornarem
totalmente fusionadas e fechadas no estágio de pupa.
Apesar das diferenças no formato das cerdas e da antena, a medida da largura da cápsula
cefálica é a característica mais eficiente para a separação dos estádios larvais e deve continuar
sendo utilizada para definir os estádios larvais de H. illucens, o que será muito importante para
determinar o calculo de intervalo pós morte, quando em casos forenses.
Informações relacionadas à morfologia de H. illucens, obtidas neste estudo, contribuiem
com a taxonomia do grupo e ampliação do conhecimento da espécie na Amazônia,
possibilitando mais segurança na identificação por parte de peritos que queiram usá-la na
criminalística.
Agradecimentos
Os autores agradecem ao Dr. Francisco Tiago Melo e aos técnicos Jackieline Veras,
Lucas Castanhola e Wilson Meirelles pelo auxílio no processamento das amostras em
microscopia eletrônica de varredura. Ao Laboratório Temático de Microscopia Óptica e

59
Eletrônica- LTMOE do INPA e ao Laboratório de histologia e embriologia animal da
Universidade Federal Rural da Amazônia – UFRA, pela estrutura que possibilitaram a
microscopia de varredura. A Léo Rossi por fotografar algumas das imagens utilizadas. Ao
Laboratório de Citotaxonomia de insetos aquáticos pelo uso do equipamento de
Automontagem. À Coordenação de aperfeiçoamento de Pessoal de Nível Superior – CAPES
pela concessão da bolsa de Mestrado. O apoio financeiro fornecido pelo projeto “Sistemática
Integrada de Insetos Aquáticos, com ênfase em Simuliidae (Diptera: Nematocera) na América
do Sul” financiado pelo MCTI / INPA, coordenado pela Dra. Neusa Hamada.
Referências
Baez M. (1975) Presencia de Hermetia illucens (L.) em las Islas Canarias (Diptera,
Stratiomyidae). Vieraea 4(1-2): 173-179.
Costa C., Ide S. (2006) Fases de desenvolvimento. p. 19-29. In: C. Costa, S. Ide e C.E. Simonka
(Holos editora). Insetos Imaturos: metamorfose e identificação. Ribeirão Preto, 249 p.
De Marco M, Martínez S, Hernandez F, Madrid J, Gai F, Rotolo L, Belforti M, Bergero D, Katz
H, Dadbou S, Kovitvadhi A, Zoccarato I, Gasco L, Schiavione A (2015) Nutritional value of
two insect latval meals (Tenebrio molitor and Hermetia illucens) for broiler chickens: Apparent
nutriente digestibility, apparent ileal amino acid digestibility and apparent metabolizable
energy. Anim. Feed Sci. Technol. 209: 211-218.
Diener S, Solano NMS, Gutiérrez FR, Zurbrügg C, Tockner K (2011) Biological treatment of
municipal organic waste using black soldier fly larvae. Waste and Biomass Valorization 2 (4):
357-363.
Fachin DA 2017. Stratiomyidae in Catálogo Taxonômico da Fauna do Brasil. PNUD.
(http://fauna.jbrj.gov.br/fauna/faunadobrasil/12536). Acesso: 24/05/2017.
Gobbi FP (2012) Biología reproductiva y caracterización morfológica de los estadios larvarios
de Hermetia illucens (L., 1758) (Diptera: Stratiomyidae). Bases para su producción masiva en
Europa. Tese de doutorado, Universidad de Alicante. 155 pp.

60
Kim W, Bae S, Park H, Park K, Lee S, Choi Y, Han S, Koh Y (2010) The larval age and mouth
morphology of the Black Soldier Fly, Hermetia illucens (Diptera: Stratiomyidae). Int. J. Indust.
Entomol. 21: 185-187.
Lord WD, Goff ML, Adkins TR, Haskell NH (1994) The black soldier fly Hermetia illucens
(Diptera: Stratiomyidae) as a potential measure of human postmortem interval: observations
and case histories. J. Forensic Sci. 39: 215–222.
McFaden MW (1967) Soldier fly larvae in America north of Mexico. Proceedings of the United
States National Museum 12:1-72.
Oliveira-Costa J (2011) Entomologia Forense: Quando os insetos são vestígios. Campinas, São
Paulo, Millennium, 257p.
Oliveira F, Doelle K, List R, O’Reilly J (2015) Assessment of Diptera: Stratiomyidae, genus
Hermetia illucens (L., 1758) using eléctron microscopy. J. Entomol. Zool. Stud. 3(5): 147-152.
Oliveira F, Doelle K, Smith RP (2016) External Mophology of Hermetia illucens
Stratiomyidae: Diptera (L. 1758) based on electron Microscopy. Annu. Res. Rev. Biol. 9(5): 1-
10.
Pujol-Luz JR, Francez PAC, Ururahy-Rodrigues A, Constantino R (2008) The black soldier-
fly, Hermetia illucens (Díptera, Stratiomyidae), used to estimate the postmortem interval in a
case in Amapá state, Brazil. J. Forensic Sci. 53: 476–478.
Rozkošný R, Kovac D (1998) Descriptions of bamboo-inhabiting larvae and puparia of oriental
soldier flies Ptecticus brunettii and P. flavife moratus (Diptera: Stratiomyidae: Sarginae) with
observations on their biology. Eur. J. Entomol. 95: 65-86.
Smith KGV (1989) An introduction to the immature stages of British flies. Handbook for
identification of British insects. Vol. 10, Part 14. Royan Entomological Society, London, 280
p.
Woodley NE (2001) A world catalogue of the Stratiornyidae (Insecta: Diptera). Myia, 11: 1-
473.

61
Woodley NE (2009) Family Stratiomyidae. p. 100-106. In: J. Gerlach (Pen soft Publishers).
The Diptera of the Seychelles islands. Sofia and Moscow, 431p.
Woodley, NE (2011) A World Catalog of the Stratiomyidae (Insecta: Diptera): A Supplement
with Revisionary Notes and Errata. Myia, 12: 379–415.

62
Capítulo III
Barros, L. M.; Martins, R. T; Ferreira-Keppler, R.
L. & Gutjahr, A. L. N. Estimativa de massa seca e
úmida de larvas de Hermetia illucens (Linnaeus,
1758) (Diptera: Stratiomyidae) associadas a suíno
em decomposição em ambiente urbano na
Amazônia Central. Manuscrito submetido para
Journal of Insect Science.

63
Estimativa de massa seca e úmida de larvas de Hermetia illucens (Linnaeus, 1758)
(Diptera: Stratiomyidae) associadas a suíno em decomposição em ambiente urbano na
Amazônia Central
Luana Machado Barros, 1,2 Renato Tavares Martins, ¹ Ruth Leila Ferreira-Keppler, ¹ e Ana
Lúcia Nunes Gutjahr³
1Programa de Pós-graduação em Entomologia, Instituto Nacional de Pesquisas da Amazônia -
INPA, Av. André Araújo, 2936, CEP 69067-375, Manaus, Amazonas, Brasil, 2 Autor
correspondente, e-mail: [email protected], 3Departamento de Ciências Naturais da
Universidade do Estado do Pará- UEPA, Tv. Djalma Dutra, 156, Belém, PA, CEP 66050-540,
Brasil.
Running head: Barros et al.: Estimativa de biomassa de Hermetia illucens da Amazônia
Central

64
Resumo
Dados de biomassa são relevantes para cálculo das taxas de crescimento e para compor
informações relacionadas com a aplicação econômica e médico-legal de Hermetia illucens
(Linnaeus, 1758). Apesar da biomassa ser importante para entender vários processos
ecológicos, ela não é necessariamente fácil de medir. A biomassa pode ser determinada através
de medição direta (pesagem) ou indiretamente por modelos de regressão entre dimensões do
corpo e massa seca e úmida do indivíduo. Neste artigo, analisou-se a relação entre a
morfometria e a biomassa de H. illucens, utilizando-se três modelos de regressão: linear,
exponencial e de potência. Para isso foram mensuradas 280 larvas, sendo utilizadas quatro
medidas morfométricas: largura da cápsula cefálica, comprimento da cápsula cefálica,
comprimento total do corpo e largura do maior segmento abdominal. As medidas que foram
melhores preditoras de biomassa foram o comprimento total do corpo e largura do maior
segmento abdominal. O modelo de regressão que obteve melhor ajuste entre as dimensões
corporais e biomassa (seca e úmida) foi o exponencial, seguido de potência e linear. Para todos
os modelos a biomassa úmida apresentou elevada relação com a biomassa seca (>75%). Não
houve diferença significativa entre os dados previstos pelos modelos e os valores observados,
e a capacidade de predição dos modelos variou de 27 a 79%. Os resultados demonstraram
grande ajuste entre as medidas corporais e a biomassa, podendo serem utilizados em estudos
aplicados envolvendo a espécie na Região Amazônica, para fins econômicos ou médico-legais.
Palavras-chave: modelos de regressão, morfometria, biomassa, insetos, relação alométrica.

65
Introdução
Os insetos representam grande parte da biomassa dos ecossistemas aquáticos e terrestres
(Stork 1988). Estes organismos são importantes fontes de alimento para vertebrados e
invertebrados (Price 1984, Huis et al. 2014) e fundamentais no processo de decomposição da
matéria orgânica animal (Deleport e Charrier 1996, Oliveira-Costa 2011, Martins et al. 2014).
Assim, a estimativa da biomassa é fundamental para entender o fluxo de energia (Ganihar
1997).
Embora a biomassa seja importante e necessária para entender vários processos
ecológicos, ela é um componente ambiental de difícil obtenção (Radtke e Williamson 2005). O
cálculo da biomassa de animais pequenos e numerosos requer tempo e é trabalhoso (Gowing e
Recher 1984, Ganihar 1997). Uma maneira de determiná-la de forma direta é pela secagem e
pesagem dos organismos, mas esta muitas vezes pode levar à destruição ou perda destes (Rogers
et al. 1976). Por outro lado, a biomassa pode ser obtida sem a destruição orgânica, através da
determinação do biovolume ou da relação entre medidas corporais e a massa do indivíduo
(Benke et al.1999). Este último método é considerado menos dispendioso e mais preciso do que
estimativas por biovolume e pesagens diretas, principalmente para invertebrados menores
(Brady e Noske 2006).
Diversos fatores devem ser levados em consideração ao utilizar modelos de regressão
para a determinação da biomassa. Os métodos de preservação e conservação das amostras
podem influenciar a quantidade de água e a conservação (e.g., quebra de partes do corpo) dos
indivíduos, afetando os valores da biomassa (Gruner 2003). A extrapolação de resultados dos
modelos de regressão para indivíduos oriundos de diferentes regiões geográficas pode resultar
estimativas errôneas de biomassa, devido às diferenças genéticas das populações e às condições
ambientais a que estão submetidas (Benke et al.1999). Além disso, o uso de dados de literatura
com regressões generalizadas (e.g., famílias e ordens) não devem substituir regressões
específicas do táxon (e.g., espécie) (Rogers et al.1976, Hódar 1996). Desta forma, é necessário
cautela ao extrapolar resultados de modelos de regressões para indivíduos de regiões e táxons
diferentes (Burgherr e Meyer 1997).
Para dípteros terrestres e aquáticos, alguns artigos que abordam estimativas de biomassa
de adultos por medidas corporais (e.g., Sabo et al. 2002, Gruner 2003, Brady e Noske 2006).
Em relação aos imaturos, muitos trabalhos estimam a densidade de larvas terrestres (Petersen e

66
Luxton 1982, Amorim et al. 2004, Beserra et al. 2009). No entanto, poucos tentaram quantificar
a biomassa desses indivíduos no ecossistema (Berg, 2000).
Larvas de Hermetia illucens (Linnaeus, 1758), espécie cosmopolita, têm seis estádios
(Oliveira-Costa 2011), e possuem hábito detritívoro, estando associadas à matéria orgânica
animal e vegetal em decomposição. Por isso, são conversoras muito eficientes de matéria
orgânica e podem transformar grandes quantidades de resíduos sólidos em biomassa rica em
proteínas (Diener et al. 2011). Possuem aplicação na entomologia forense, especificamente na
área médico-legal, auxiliando em cálculos de intervalo pós morte (IPM) (Lord et al. 1994,
Oliveira-Costa, 2011, Pujol-Luz et al. 2008). Além disso, despertam interesse economicamente,
pois são capazes de gerar biomassa rica em proteínas que podem substituir a farinha de peixe,
aplicadas à aquicultura sustentável (Bondari e Sheppard 1987).
O conhecimento da biomassa das larvas de H. illucens é relevante para o cálculo das
taxas de crescimento, gerando informações para a aplicabilidade desta espécie. Assim, pode ser
útil, especificamente na área forense, para peritos na obtenção de ganho de massa dos estádios
larvais, sendo aplicados na determinação de IPM. Apesar disso, há escassez de estudos que
contemplem estas estimativas na região Amazônica. Assim, avaliou-se a relação entre
dimensões do corpo e massa, a fim de verificar se as medidas corporais são boas preditoras da
biomassa seca e úmida.
Material e métodos
Foram coletadas 280 larvas de H. illucens de diferentes tamanhos (2º ao 6º estádios) em
um cadáver de suíno Sus scrofa Linnaeus, 1758. As coletas foram realizadas no período de
janeiro a setembro de 2016 (estação chuvosa) no Instituto Nacional de Pesquisas da Amazônia
(INPA; 03º05'47”S e 59º59'22”W), na região urbana da cidade de Manaus, Amazônia Central.
Após serem retiradas da carcaça, as larvas foram imersas e sacrificadas em água aquecida
(~70°C), fixadas em solução KAAD (1 parte de querosene, 10 partes de álcool 95%, 2 partes
de ácido acético e 1 parte de dioxano) por um período de 24 h. Posteriormente, as larvas foram
acondicionadas individualmente em microtúbulos contendo álcool 80%.
Para obtenção das imagens e mensuração das medidas corporais (largura e comprimento
da cápsula cefálica, comprimento total do corpo e largura do maior segmento abdominal) foi
utilizado o estereomicroscópio Leica M165C com câmera fotográfica DFC420, acoplada ao
equipamento e o programa de processamento de Imagem Digital, Leica Application Suite V3.
4.1. A largura de cápsula cefálica (LC) foi determinada como a maior largura desta estrutura.

67
O comprimento da cápsula cefálica (CC) foi medido pela distância entre o ápice da cápsula
cefálica e o primeiro segmento do tórax em vista ventral. O comprimento total do corpo (CT)
foi medido entre o ápice da cápsula cefálica e a parte posterior do último segmento abdominal.
A largura do maior segmento abdominal (LA) foi a distância entre as partes laterais deste (Fig.
1).
Fig. 1. Larva de quinto estádio de Hermetia illucens (Linnaeus, 1758), em vista ventral,
destacando as medidas realizadas. LC= Largura da cápsula cefálica; CC= Comprimento da
cápsula cefálica; CT= Comprimento total; LA= Largura do maior segmento abdominal.
Para as pesagens em meio úmido, as larvas foram retiradas dos microtúbulos, colocadas
em papel absorvente para retirada do excesso de umidade e pesadas individualmente. Para
determinar a massa seca, as larvas foram colocadas individualmente em estufa (60°C, 48 horas)
e posteriormente pesadas. Em ambas as pesagens foi utilizada uma balança de precisão
(Mettler-Toledo; acurácia = 0,01 mg).
Para determinar o melhor ajuste entre as medidas corporais e a biomassa (seca e úmida)
de H. illucens, foram usados três modelos de regressão: linear (y = b*x + a), exponencial (y =
a* e bx; na forma linear: ln y = ln a + b*x) e de potência (y = a* xb; na forma linear: ln y = ln a
+ b*ln x), onde y é o peso (mg) e x é a medida (mm). Para expressar os modelos exponencial e
de potência os dados foram transformados em base logarítmica (ln). Foram avaliados os ajustes
de modelos de regressão baseados em coeficientes de determinação “R2” (Zar 2010).
Afim de determinar o poder preditivo dos modelos obtidos com os dados do presente
estudo foi realizada uma análise de validação cruzada (Martins et al. 2014, Brito et al. 2015).
Para a realização desta análise, os dados foram divididos em dois subconjuntos: I) Treinamento:
para a obtenção dos modelos de regressão; e II) teste: para avaliar o poder de predição do
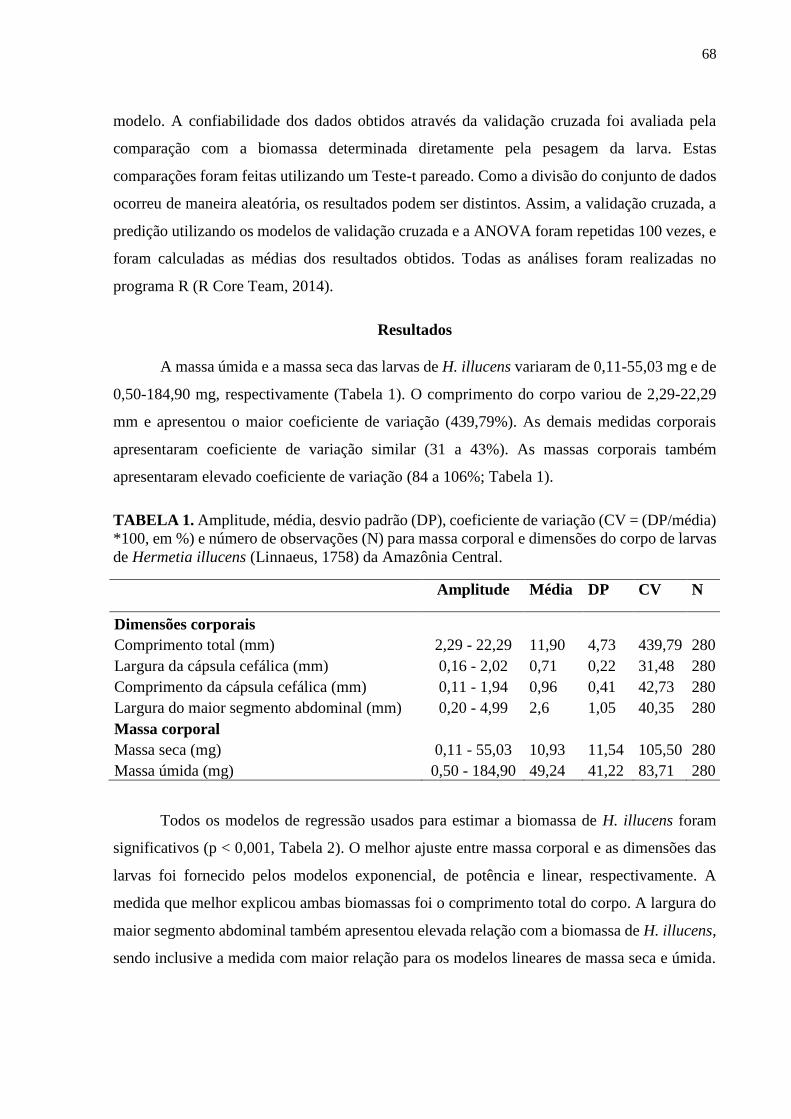
68
modelo. A confiabilidade dos dados obtidos através da validação cruzada foi avaliada pela
comparação com a biomassa determinada diretamente pela pesagem da larva. Estas
comparações foram feitas utilizando um Teste-t pareado. Como a divisão do conjunto de dados
ocorreu de maneira aleatória, os resultados podem ser distintos. Assim, a validação cruzada, a
predição utilizando os modelos de validação cruzada e a ANOVA foram repetidas 100 vezes, e
foram calculadas as médias dos resultados obtidos. Todas as análises foram realizadas no
programa R (R Core Team, 2014).
Resultados
A massa úmida e a massa seca das larvas de H. illucens variaram de 0,11-55,03 mg e de
0,50-184,90 mg, respectivamente (Tabela 1). O comprimento do corpo variou de 2,29-22,29
mm e apresentou o maior coeficiente de variação (439,79%). As demais medidas corporais
apresentaram coeficiente de variação similar (31 a 43%). As massas corporais também
apresentaram elevado coeficiente de variação (84 a 106%; Tabela 1).
TABELA 1. Amplitude, média, desvio padrão (DP), coeficiente de variação (CV = (DP/média)
*100, em %) e número de observações (N) para massa corporal e dimensões do corpo de larvas
de Hermetia illucens (Linnaeus, 1758) da Amazônia Central.
Amplitude Média DP CV N
Dimensões corporais
Comprimento total (mm) 2,29 - 22,29 11,90 4,73 439,79 280
Largura da cápsula cefálica (mm) 0,16 - 2,02 0,71 0,22 31,48 280
Comprimento da cápsula cefálica (mm) 0,11 - 1,94 0,96 0,41 42,73 280
Largura do maior segmento abdominal (mm) 0,20 - 4,99 2,6 1,05 40,35 280
Massa corporal
Massa seca (mg) 0,11 - 55,03 10,93 11,54 105,50 280
Massa úmida (mg) 0,50 - 184,90 49,24 41,22 83,71 280
Todos os modelos de regressão usados para estimar a biomassa de H. illucens foram
significativos (p < 0,001, Tabela 2). O melhor ajuste entre massa corporal e as dimensões das
larvas foi fornecido pelos modelos exponencial, de potência e linear, respectivamente. A
medida que melhor explicou ambas biomassas foi o comprimento total do corpo. A largura do
maior segmento abdominal também apresentou elevada relação com a biomassa de H. illucens,
sendo inclusive a medida com maior relação para os modelos lineares de massa seca e úmida.
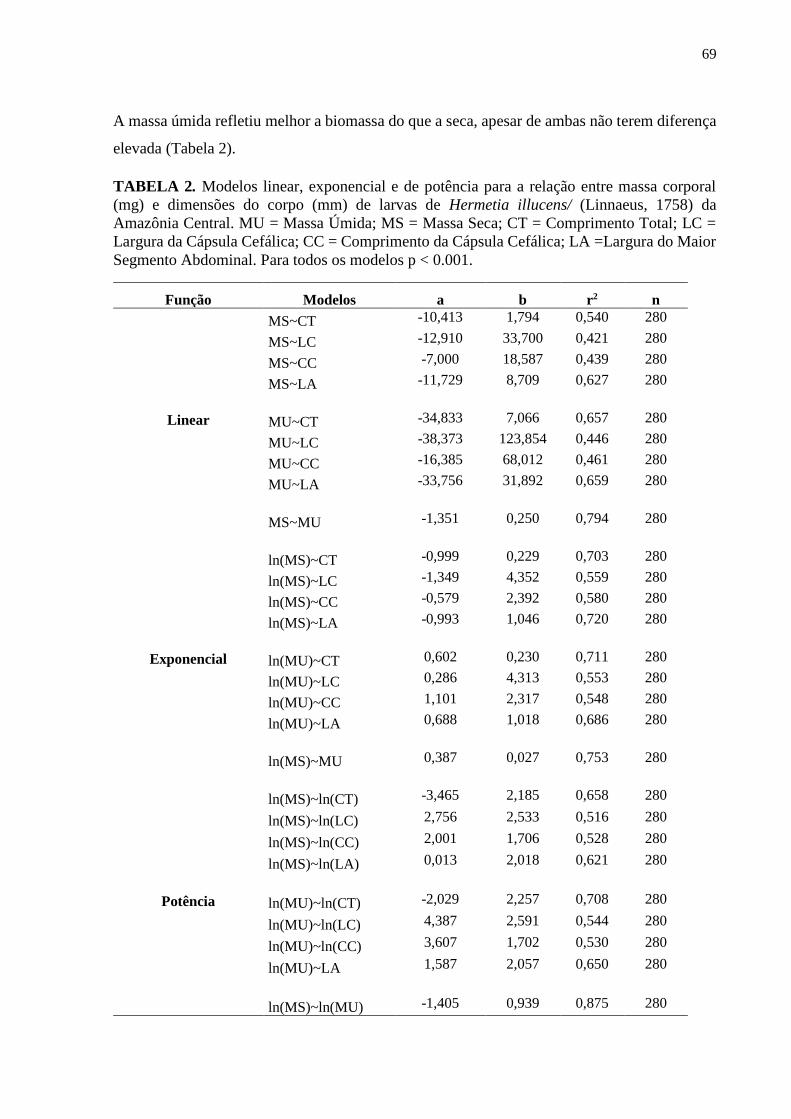
69
A massa úmida refletiu melhor a biomassa do que a seca, apesar de ambas não terem diferença
elevada (Tabela 2).
TABELA 2. Modelos linear, exponencial e de potência para a relação entre massa corporal
(mg) e dimensões do corpo (mm) de larvas de Hermetia illucens/ (Linnaeus, 1758) da
Amazônia Central. MU = Massa Úmida; MS = Massa Seca; CT = Comprimento Total; LC =
Largura da Cápsula Cefálica; CC = Comprimento da Cápsula Cefálica; LA =Largura do Maior
Segmento Abdominal. Para todos os modelos p < 0.001.
Função Modelos a b r2 n
Linear
MS~CT -10,413 1,794 0,540 280
MS~LC -12,910 33,700 0,421 280
MS~CC -7,000 18,587 0,439 280
MS~LA -11,729 8,709 0,627 280
MU~CT -34,833 7,066 0,657 280
MU~LC -38,373 123,854 0,446 280
MU~CC -16,385 68,012 0,461 280
MU~LA -33,756 31,892 0,659 280
MS~MU -1,351 0,250 0,794 280
Exponencial
ln(MS)~CT -0,999 0,229 0,703 280
ln(MS)~LC -1,349 4,352 0,559 280
ln(MS)~CC -0,579 2,392 0,580 280
ln(MS)~LA -0,993 1,046 0,720 280
ln(MU)~CT 0,602 0,230 0,711 280
ln(MU)~LC 0,286 4,313 0,553 280
ln(MU)~CC 1,101 2,317 0,548 280
ln(MU)~LA 0,688 1,018 0,686 280
ln(MS)~MU 0,387 0,027 0,753 280
Potência
ln(MS)~ln(CT) -3,465 2,185 0,658 280
ln(MS)~ln(LC) 2,756 2,533 0,516 280
ln(MS)~ln(CC) 2,001 1,706 0,528 280
ln(MS)~ln(LA) 0,013 2,018 0,621 280
ln(MU)~ln(CT) -2,029 2,257 0,708 280
ln(MU)~ln(LC) 4,387 2,591 0,544 280
ln(MU)~ln(CC) 3,607 1,702 0,530 280
ln(MU)~LA 1,587 2,057 0,650 280
ln(MS)~ln(MU) -1,405 0,939 0,875 280

70
A maioria dos coeficientes de determinação usando valores preditos (ex.: validação
cruzada) foram menores (Tabela 3) em relação aos valores estimados utilizando o mesmo
conjunto de dados (Tabela 2). Os coeficientes de determinação da validação cruzada para as
medidas corporais foram diferentes nos modelos linear, exponencial e de potência. Nos modelos
linear e de potência, as medidas corporais foram melhores preditoras de biomassa úmida do que
a seca. Os valores observados e estimados não foram diferentes entre os modelos de regressão.
Os valores dos modelos preditos têm tendência a superestimar os modelos observados (Tabela
3), evidenciados por valores de diferenças positivos.
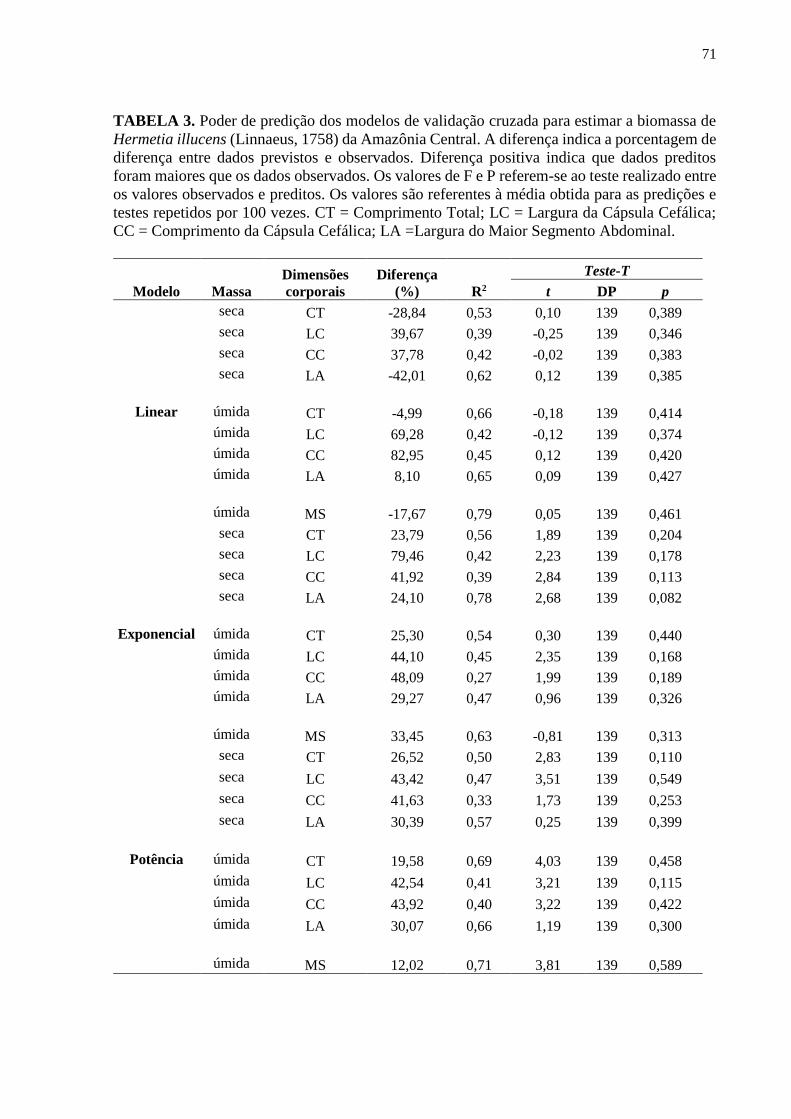
71
TABELA 3. Poder de predição dos modelos de validação cruzada para estimar a biomassa de
Hermetia illucens (Linnaeus, 1758) da Amazônia Central. A diferença indica a porcentagem de
diferença entre dados previstos e observados. Diferença positiva indica que dados preditos
foram maiores que os dados observados. Os valores de F e P referem-se ao teste realizado entre
os valores observados e preditos. Os valores são referentes à média obtida para as predições e
testes repetidos por 100 vezes. CT = Comprimento Total; LC = Largura da Cápsula Cefálica;
CC = Comprimento da Cápsula Cefálica; LA =Largura do Maior Segmento Abdominal.
Modelo Massa
Dimensões
corporais
Diferença
(%) R2
Teste-T
t DP p
Linear
seca CT -28,84 0,53 0,10 139 0,389
seca LC 39,67 0,39 -0,25 139 0,346
seca CC 37,78 0,42 -0,02 139 0,383
seca LA -42,01 0,62 0,12 139 0,385
úmida CT -4,99 0,66 -0,18 139 0,414
úmida LC 69,28 0,42 -0,12 139 0,374
úmida CC 82,95 0,45 0,12 139 0,420
úmida LA 8,10 0,65 0,09 139 0,427
úmida MS -17,67 0,79 0,05 139 0,461
Exponencial
seca CT 23,79 0,56 1,89 139 0,204
seca LC 79,46 0,42 2,23 139 0,178
seca CC 41,92 0,39 2,84 139 0,113
seca LA 24,10 0,78 2,68 139 0,082
úmida CT 25,30 0,54 0,30 139 0,440
úmida LC 44,10 0,45 2,35 139 0,168
úmida CC 48,09 0,27 1,99 139 0,189
úmida LA 29,27 0,47 0,96 139 0,326
úmida MS 33,45 0,63 -0,81 139 0,313
Potência
seca CT 26,52 0,50 2,83 139 0,110
seca LC 43,42 0,47 3,51 139 0,549
seca CC 41,63 0,33 1,73 139 0,253
seca LA 30,39 0,57 0,25 139 0,399
úmida CT 19,58 0,69 4,03 139 0,458
úmida LC 42,54 0,41 3,21 139 0,115
úmida CC 43,92 0,40 3,22 139 0,422
úmida LA 30,07 0,66 1,19 139 0,300
úmida MS 12,02 0,71 3,81 139 0,589

72
Discussão
O comprimento total do corpo e a largura do maior segmento abdominal foram os
melhores preditores da biomassa úmida e seca de H. illucens. Nestas larvas, o abdome é menos
esclerosado em relação à cápsula cefálica, o que permite o crescimento contínuo do indivíduo
ao longo do seu desenvolvimento, e possibilita um melhor ajuste de medidas corporais com a
biomassa (Benke et al. 1999, Johnston e Cunjak 1999). O comprimento do corpo é geralmente
um bom preditor da biomassa de larvas de insetos aquáticos (Becker 2009, Martins et al. 2014,
Brito et al. 2015) e larvas e adultos de insetos terrestres (Ganihar 1997, Johnson e Strong 2000,
Wardhaugh 2013). Além disso, assim como outras larvas de espécies de interesse forense, H.
illucens é detritívora e voraz em sua alimentação (Tomberlin et al. 2002), o que influencia
diretamente em suas taxas de crescimento (aumento do tamanho corporal) e mudanças nos
estádios.
A fixação de amostras em KAAD durante 24 horas e depois transferidas para álcool tem
a tendência a aumentar ligeiramente o comprimento corporal em uma semana de conservação
(Schiller e Solimini 2005). O álcool, na maioria das vezes, acaba sendo a escolha para o método
de preservação dos organismos, mas as concentrações utilizadas nem sempre são esclarecidas
(Lord e Rodriguez 1989, Anderson 1995, Benecke 2001). Entretanto, muitos estudos
demonstraram consideráveis reduções no tamanho de macroinvertebrados aquáticos após a
preservação química (Howmiller 1972, Landahl e Nagell 1978, Leuven et al. 1985). A
preservação em álcool é capaz de diminuir significativamente tanto o peso úmido quanto o seco
das amostras armazenadas (Knapp 2012), uma vez que substitui a água nos tecidos animais,
causando o encolhimento dos espécimes (Tucker e Chester 1984). Além disso, dissolve os
lipídios das larvas de insetos, causando perda de massa (Wetzel et al. 2005).
A diferença entre os ajustes dos valores de massa seca e úmida pode ser explicada pela
constituição do corpo do imaturo. Larvas de H. illucens possuem o corpo mole e apenas a
cápsula cefálica como uma estrutura esclerosada (Kim 2010). Desta forma, ao passar pelo
processo de secagem, a larva perde grande quantidade de líquido, por consequência perde sua
biomassa. Resultados semelhantes foram obtidos para larvas de Sciaridae (Diptera), onde as
regressões resultaram em melhores ajustes para massa úmida do que para a seca (Berg 2000).
Este resultado foi associado à alimentação das larvas (alimento com ~ 65% de líquido) e à
grande perda de biomassa durante o processo de secagem devido à evoporação dos líquidos
presentes no corpo (Berg 2000). Além disso, a forte relação (r2 > 0,75) entre a massa seca e

73
massa úmida pode ser uma importante ferramenta para estudos ecológicos, uma vez que a massa
seca é melhor indicativo de conteúdo orgânico do indivíduo (Dermott e Paterson 1974).
Os modelos exponenciais forneceram os melhores ajustes entre a biomassa de H.
illucens e as medidas corporais, apesar dos modelos de potência terem bons resultados. Brady
e Noske (2006) encontraram resultados semelhantes também para outros dípteros. Gowing e
Recher (1984) obtiveram melhores ajustes para as ordens Coleptera e Hymenoptera utilizando
modelos exponenciais, assim como Ganihar (1997) para Dermaptera e Lepidoptera. De acordo
com Becker et al. (2009), o modelo exponencial pode fornecer coeficientes de determinação
satisfatórios para verificar a relação entre massa seca e dimensões corporais. Em literatura,
observa-se que para invertebrados os modelos com melhores ajustes são geralmente os de
potência (Martins 2014). Para larvas de Simuliidae (Diptera), este modelo também foi o mais
ajustado (Belinic e Habdija 1992). Dados semelhantes, obtendo o modelo de potência com
melhores resultados também podem ser observados para Trichoptera (Becker 2009, Martins et
al. 2014, Brito et al. 2015), Gastropodas (Hawkins et al. 1997) e Arachnida (Brady e Noske
2006).
Neste estudo, os valores dos coeficientes de determinação dos modelos utilizando a
validação cruzada (conjunto teste) foram menores (Massa úmida: 7,8 ± 2,9%; Massa seca: 9,0
± 3,9%) quando comparados com os modelos estimados com os mesmos dados. Além disso
não foi registrada diferença significativa da biomassa observada e estimada. No entanto, deve-
se levar em consideração a superestimativa da biomassa (Massa úmida: 19,5 ± 14,6%; Massa
seca: 15,7 ± 15,9%) na utilização dos modelos obtidos. Isso pode ser resultado dos coeficientes
de determinação estimados a partir de uma amostra ser geralmente mais elevados do que os
produzidos utilizando a mesma regressão, mas aplicados a diferentes dados amostrados
(Martins et al. 2014). Por fim, com os dados deste estudo, afirma-se que a estimativa de
biomassa de H. illucens é confiável e pode servir de suporte para trabalhos posteriores
envolvendo aplicações desta espécie, principalmente na região onde foi realizado o presente
estudo, facilitando estimativas de biomassa a partir de medidas morfométricas. Esse resultado
é importante, pois pode auxiliar e diminuir o tempo de cálculos de taxas de crescimento na
criação de larvas para fins econômicos (produção de ração para peixes, compostagem). Além
disso, pode colaborar com a criminalística e peritos para obtenção de dados de biomassa mais
rápidos relacionados a espécie no local de crime, como: obtenção de ganho de massa durante
os estádios larvais, sem necessariamente a utilização de equipamentos de difícil acesso em
campo.

74
Agradecimentos
Os autores agradecem a Léo Rossi pela fotografia utilizada neste artigo. À Coordenação
de aperfeiçoamento de Pessoal de Nível Superior – CAPES pela concessão da bolsa de
Mestrado. O apoio financeiro fornecido pelo projeto “Sistemática Integrada de Insetos
Aquáticos, com ênfase em Simuliidae (Diptera: Nematocera) na América do Sul” financiado
pelo MCTI / INPA. RTM recebeu bolsa do Programa de Apoio à Fixação de Doutores no
Amazonas—FIXAM/AM. RLFK recebeu bolsa de produtividade (proc. 306081/2013-0) do
Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq). Aos projetos
“Sistemática Integrada de Insetos Aquáticos, com ênfase em Simuliidae (Diptera: Nematocera)
na América do Sul”, Pró-equipamentos Institucional/CAPES e INCT/ADAPTA II
(CNPq/FAPEAM), coordenado pela Dra. Neusa Hamada, pelo suporte na coleta de amostras e
análises laboratoriais.
Referências
Amorim, R. M., A. L. Henrique-Oliveira, and J. L. Nessimian. 2004. Distribuição espacial
e temporal das larvas de Chironomidae (Insecta: Diptera) na seção ritral do Rio Cascatinha,
Nova Friburgo, Rio de Janeiro, Brasil. Lundiana 5(2): 119-127.
Anderson, G. 1995. The use of insects in death investigations: an analysis of forensic
entomology cases in British Columbia over a five year period, Can. J. Forensic Sci. Soc. 28:
277–292.
Becker, B., M. S. Moretti, and M. Callisto. 2009. Length–dry mass relationships for a typical
shredder in Brazilian streams (Trichoptera: Calamoceratidae). Aquat. Insects 31(3): 227-234.
Belinic, I., and I. Habdija. 1992. Biomass of simuliid larvae as a function of body length.
Period. Biol. 94 (1): 47-52.
Benecke, M., and R. Lessig. 2001. Child neglect and forensic entomology. Forensic Sci. Int.
120: 155–159.
Benke, A., A. Huryn, L. Smock, and J. Wallace. 1999. Length–mass relationships for
freshwater macroinvertebrates in North America with particular reference to the southeastern
United States. J. N. Am. Benthol. Soc. 18 (3): 308-343.

75
Berg, M. P. 2000. Mass-length and mass-volume relationships of larvae of Bradysia paupera
(Diptera: Sciaridae) in laboratory cultures. Eur. J. Soil Biol. 36: 127-133.
Beserra, E. B., C. R. M. Fernandes, and P. S. Ribeiro. 2009. Relação entre densidade larval
e ciclo de vida, tamanho e fecundidade de Aedes (Stegomyia) aegypti (L.) (Diptera: Culicidae)
em Laboratório. Neotrop. Entomol. 38(6): 847-852.
Bondari, K., and D. C. Sheppard. 1987. Soldier fly, Hermetia illucens L., larvae as feed for
channel catfish, Ictalurus punctatus (Rafinesque), and blue tilapia, Oreochromis aureus
(Steindachner). Aquacult. Res. 18(3): 209-220.
Brady, C. J., and R. A. Noske. 2006. Generalised regressions provide good estimates of insect
and spider biomass in the monsoonal tropics of Australia. Aust. J. Entomol. 45(3): 187-191.
Brito, J. G., R. T. Martins, K. M. Soares, and N. Hamada. 2015. Biomass estimation of
Triplectides egleri Sattler (Trichoptera, Leptoceridae) in a stream at Ducke Reserve, Central
Amazonia. Rev. Bras. Entomol. 59: 332-336.
Burgherr, P., and E. I. Meyer. 1997. Regression analysis of linear body dimensions vs, dry
mass in stream macroinvertebrates. Arch. Hydrobiol. 139(1): 101-112.
Deleporte S., and M. Charrier. 1996. Comparison of digestive carbohydrases between two
forest sciarid (Diptera: Sciaridae) larvae in relation to their ecology. Pedobiologia. 40: 193–
200.
Dermott, R. M., and C. G. Paterson. 1974. Determining dry weight and percentage dry-matter
of chironomid larvae. Can. J. Zool. 52: 1243–1250.
Diener, S., N. M. S. Solano, F. R. Gutiérrez, C. Zurbrügg, and K. Tockner. 2011. Biological
treatment of municipal organic waste using black soldier fly larvae. Waste and Biomass
Valorization. 2(4): 357-363.
Ganihar, S. R. 1997. Biomass estimates of terrestrial arthropods based on body length. J.
Biosci. 22(2) 219-224.
Gowing, G., and H. F. Recher. 1984. Length- weight relationships for invertebrates from
forests in South- eastern New South Wales. Aust. J. Ecol. 9: 5-8.

76
Gruner, D. S. 2003. Regressions of length and width to predict Arthropod Biomass in the
Hawaiian Islands. Pac. Sci. 57(3): 325-336.
Hawkins, J. W., M. W. Lankester, R. A. Lautenschlager, and F. W. Bell. 1997. Length-
biomass and energy relationships of terrestrial gastropods in northern forest ecosystems. Can.
J. Zool. 75:501–505.
Hódar, J. A. 1996. The use of regression equations for estimation of arthropod biomass in
ecological studies. Acta Ecol. Sin. 17(5): 421-423.
Howmiller, R. P. 1972. Effects of preservatives on weights of some common macrobenthic
invertebrates. Trans. Am. Fish. Soc. 101 (4): 743-746.
Huis, A. V., H. V. Gurp, and M. Dicke. 2014. The insect cookbook, food for a sustainable
planet. Columbia University Press.
Johnston, T., and R. Cunjak. 1999. Dry mass-length relationships for benthic insects: a
review with new data from Catamaran Brook, New Brunswick. Canada. Freshwat. Biol. 41(4):
653-674.
Johnson, M. D., and A. M. Strong. 2000. Length- weight relationships of Jamaican
arthropods. Entomol. News. 111(4): 270-281.
Kim, W., S. Bae, H. Park, K. Park, S. Lee, Y. Choi, S. Han, and Y. Koh. 2010. The larval
age and mouth morphology of the Black Soldier Fly, Hermetia illucens (Diptera:
Stratiomyidae). Int. J. Indust. Entomol. 21: 185-187.
Knapp, M. 2012. Preservative fluid and storage conditions alter body mass estimation in a
terrestrial insect. Entomol. Exp. Appl. 143: 185-190.
Landahl, C. C., and B. Nagell. 1978. Influence of the season and of preservation methods on
wet- and dry weights of larvae of Chironomus plumosus L. Int. Rev. Hydrobiol. 63: 405–410.
Leuven, R. S. E. W., T. C. M. Brock, and H. A. M. Vandruten. 1985. Effects of preservation
on dry- and ash-free dry-weight biomass of some common aquatic macro- invertebrates.
Hydrobiologia 127: 151–159.

77
Lord, W. D., and W. C. Rodriguez. 1989. Forensic entomology: the use of insects in the
investigation of homicide and untimely death. Prosecutor. 22: 41–48.
Lord, W. D., M. L. Goff, T. R. Adkins, and N. H. Haskell. 1994. The black soldier fly
Hermetia illucens (Diptera: Stratiomyidae) as a potential measure of human postmortem
interval: observations and case histories. J. Forensic Sci. 39: 215–222.
Martins, R. T., A. S. Melo, J.F. Gonçalves Jr., and N. Hamada. 2014. Estimation of dry
massof caddisflies Phylloicus elektoros (Trichoptera: Calamoceratidae) in a Central Amazon
stream. Rev. Bras. Zool. 31: 337–342.
Oliveira-Costa, J. 2011. Entomologia forense: quando os insetos são vestígios. Millennium,
Campinas, SP.
Petersen, P., and M. Luxton. 1982. A comparative analysis of soil fauna populations and their
role in decomposition processes. Oikos. 39: 288–388.
Price, P. W. 1984. Insect ecology. John Wiley e Sons, NY.
Pujol-Luz, J. R., P. A. C. Francez, A. Ururahy-Rodrigues, and R. Constantino. 2008. The
black soldier-fly, Hermetia illucens (Díptera, Stratiomyidae), used to estimate the postmortem
interval in a case in Amapá state, Brazil. J. Forensic Sci. 53: 476–478.
Radtke, M. G., and G. B. Williamson. 2005. Volume and linear measurements as predictors
of dung beetle (Coleoptera: Scarabaeidae) biomass. Ann. Entomol. Soc. Am. 98(4): 548-551.
Rogers, L. E., R. L. Buschbom, and C. R. Watson. 1976. Length-Weight Relationships of
Shrub-Steppe Invertebrates. Ann. Entomol. Soc. Am. 70(1): 51-53.
Rogers, L. E., W. T. Hinds, and R. L. Buschbom. 1976. A general weight vs. length
relationship for insects. Ann. Entomol. Soc. Am. 69: 387-89.
Sabo, J. L., J. L. Bastow, and M. Power. 2002. Length- mass relationships for adult aquatic
and terrestrial invertebrates in a California Watershed. J. N. Am. Benthol. Soc. 21(2): 336- 343.

78
Schiller, D. V., and A. G. Solimini. 2005. Differential effects of preservation on the estimation
of biomass of two common mayfly species. Arch. Hydrobiol. J. 164(3): 325-334.
Stork, N. E. 1988. Insect diversity: facts, fiction and speculation. Biol. J. Linn. Soc. 35: 321–
337.
Tomberlin, J. K., D. C. Sheppard, and J. A. Joyce. 2002. A comparison of selected life
history traits of the black soldier ßy (Diptera: Stratiomyidae) whenreared on three diets. Ann.
Entomol. Soc. Am. 95: 379-387.
Tucker, J.W., and A. J. Chester. 1984. Effects of salinity, formalin concentration and
buffer on quality of preservation of Southern Flounder (Paralichthys lethostigma) larvae.
Copeia. 4: 981–988.
Wardhaugh, C. W. 2013. Estimation of biomass fron body length and width for tropical
rainforest canopy invertebrates. Aust. J. Entomol. 52: 291-298.
Wetzel, M. A., H. Leuchs, and J. H. E. Koop. 2005. Preservation effects on wet weight, dry
weight, and ash-free dry weight biomass estimates of four common estuarine macro-
invertebrates: no difference between ethanol and formalin. Helgol. Mar. Res. 59 (3): 206-213.
Zar, J. H. 2010. Biostatistical Analysis. Pearson Education, NJ.

79
SÍNTESE
Nesta dissertação foi realizado o estudo de Hermetia illucens, dividido em três capítulos
que se complementam e acrescentam informações sobre a espécie, formando um banco de
dados na região Amazônica, ainda inexistente. Com a abordagem bionômica, foi possível
observar o comportamento da fêmea adulta associada ao cadáver de suíno, bem como a
participação de suas larvas no processo de decomposição. Informações estas que podem ser
utilizadas por peritos na criminalística.
Além de dados bionômicos, a morfologia e definição dos estádios larvais são
importantes para a taxonomia da espécie, e também para peritos na identificação em campo
para utilização da mesma em cálculo do intervalo pós-morte. Quanto mais esclarecido é o
conhecimento da morfologia da larva, menos erros podem ser cometidos na identificação da
espécie e aplicabilidade na entomologia forense.
A relação de biomassa e medidas corporais torna-se interessante por ser uma ferramenta
para obtenção de ganho de massa durante os estádios larvais, com menor utilização de
equipamentos específicos. Pode contribuir a reduzir o esforço de cálculos de taxas de
crescimento na criação de larvas para fins econômicos. Além disso, destaca-se a importância
de trabalhos com essa abordagem que sejam específicos para a região Amazônica, pois dados
provenientes de outras regiões comumente não fornecem boas estimativas de biomassa.
Para melhor compreender a biologia e a importância de H. illucens para a entomologia
forense, estudos futuros devem avaliar:
1- A influência dos períodos mais e menos chuvoso no desenvolvimento de H. illucens na
região Amazônica.
2- O efeito da submersão em água na sobrevivência das pupas de H. illucens.
3- A variabilidade de hidrocarbonetos cuticulares na separação de estádios larvais.
4- Avaliar o poder de predição de modelos desenvolvidos para a biomassa de H. illucens de
outras regiões do Brasil.

80
REFERÊNCIAS BIBLIOGRÁFICAS
Adler, A.I.; Brancato, F.P. 1995. Human furuncular Myiasis caused by Hermetia illucens
(Diptera: Stratiomyidae). Journal Medical Entomology, 32: 745-746.
Amorim, R.M.; Henrique-Oliveira, A.L.; Nessimian, J.L. 2004. Distribuição espacial e
temporal das larvas de Chironomidae (Insecta: Diptera) na seção ritral do Rio Cascatinha, Nova
Friburgo, Rio de Janeiro, Brasil. Lundiana 5(2): 119-127.
Anderson, G. 1995. The use of insects in death investigations: an analysis of forensic
entomology cases in British Columbia over a five year period, Can. Journal of Forensic Science
Society, 28: 277–292.
Anjos, C.R. 2001. Entomofauna decompositora de carcaça de porcos na região de Belém,
Pará, Brasil, com ênfase na família Calliphoridae (Diptera). Dissertação de mestrado,
Universidade Federal do Pará, Belém, Pará. 122 pp.
Atkinson, D. 1994. Temperature and organism size – a biological law for ectotherms? Advances
in Ecological. Research, 3: 1–58.
Baez, M. 1975. Presencia de Hermetia illucens (L.) em las Islas Canarias (Diptera,
Stratiomyidae). Vieraea, 4(1-2): 173-179.
Barbosa, R.R.; Mello, R.P.; Queiroz, M.M.C. 2006. Comportamento de Hermetia illucens
(Diptera: Stratiomyidae) como indicador forense na cidade do Rio de Janeiro, Brasil. Recife,
Pernambuco.
Barros-Cordeiro, K.B.; Báo, S.N; Pujol-Luz, J.R. 2014. Intra-puparial development of the black
soldier-fly, Hermetia illucens. Journal of Insect Science, 14(83): 1-10.
Barroso, F.G.; De Haro, C.; Sánchez-Muros, M.J.; Venegas, E.; Martínez-Sánchez, A.; Pérez-
Bañón, C. 2014. The potencial of various insect species for use as food for fish. Aquaculture,
422-423: 193-201.
Becker, B.; Moretti, M.S.; Callisto, M. 2009. Length–dry mass relationships for a typical
shredder in Brazilian streams (Trichoptera: Calamoceratidae). Aquatic Insects, 31(3): 227-234.

81
Belinic, I.; Habdija, I. 1992. Biomass of simuliid larvae as a function of body length.
Periodicum Biologorum, 94 (1): 47-52.
Benecke, M.; Lessig, R. 2001. Child neglect and forensic entomology. Forensic Science
International, 120: 155–159.
Benke, A.; Huryn, A.; Smock, L.; Wallace, J. 1999. Length–mass relationships for freshwater
macroinvertebrates in North America with particular reference to the southeastern United
States. Journal of the North American Benthological Society, 18 (3): 308-343.
Berg, M.P. 2000. Mass-length and mass-volume relationships of larvae of Bradysia paupera
(Diptera: Sciaridae) in laboratory cultures. European Journal of Soil Biology, 36: 127-133.
Beserra, E.B.; Fernandes, C.R.M.; Ribeiro, P.S. 2009. Relação entre densidade larval e ciclo de
vida, tamanho e fecundidade de Aedes (Stegomyia) aegypti (L.) (Diptera: Culicidae) em
Laboratório. Neotropical Entomology, 38(6): 847-852.
Blackmore, M.S.; Lord, C.C. 2000. The relationship between size and fecundity in Aedes
albopictus. Journal of Vector Ecology, 25: 212-217.
Bondari, K.; Sheppard, D.C. 1987. Soldier fly, Hermetia illucens L., larvae as feed for channel
catfish, Ictalurus punctatus (Rafinesque), and blue tilapia, Oreochromis aureus
(Steindachner). Aquaculture Research, 18(3): 209-220.
Booth, D.C.; Sheppard, C. 1984. Oviposition of the Black Soldier Fly, Hermetia illucens
(Diptera: Stratiomyidae): Eggs, Masses, Timing and Site Characteristics. Environmental
Entomology, 13: 421-423.
Brady, C.J.; Noske, R.A. 2006. Generalised regressions provide good estimates of insect and
spider biomass in the monsoonal tropics of Australia. Australian Journal of Entomology, 45(3):
187-191.
Brito, J.G.; Martins, R.T.; Soares, K.M.; Hamada, N. 2015. Biomass estimation of Triplectides
egleri Sattler (Trichoptera, Leptoceridae) in a stream at Ducke Reserve, Central Amazonia.
Revista Brasileira de Entomologia, 59: 332-336.

82
Brown, W.D.; Keller, L. 2000. Colony sex ratios vary with queen number but note relatedness
asymmetry in the ant Formica exsecta. Journal of The Royal Society, 267:1751-1757.
Burgherr, P.; Meyer, E.I. 1997. Regression analysis of linear body dimensions vs, dry mass in
stream macroinvertebrates. Archiv fur Hydrobiologie, 139(1): 101-112.
Byrd, J.H.; Butler, J.F. 1996. Effects of temperature on Cochliomyia macellaria. Journal of
Medical Entomology 33, 901-905.
Calderón-Arguedas, O.; Murillo Barrantes, J.; Solano, M.E. 2005. Miíase entérica por Hermetia
illucens (Diptera: Stratiomyidae) en una paciente geriátrica de Costa Rica. Parasitología
latinoamericana, 60 (3-4), 162-164.
Carvalho, C.J.B.; Rafael, J.A.; Couri, M.S.; Silva, V.C. 2012. Capítulo 40: Diptera. In: Rafael,
J.A.; Melo, G.A.R.; Carvalho, C.J.B.; Casari, S.A.; Constantino, R. (Holos Editora). Insetos do
Brasil: Diversidade e Taxonomia. Ribeirão Preto, São Paulo. p. 701 -744.
Chiaradia, L.A.; Milanez, J.M.; Dittrich, R. 2004. Flutuação populacional de moscas-das-frutas
em pomares de citros no oeste de Santa Catarina, Brasil. Ciência Rural, 34 (2), 337-343.
Clark, B.R.; Faeth, S. H. 1997. The consequences of larval aggregation in the butter ßy
Chlosyne lacinia. Journal of Ecological Entomology, 22: 408-415.
Clements, A.N. 1992. The biology of Mosquitoes, development, nutricion, and reproduction.
Chapman e Hall, London. Vol. 1, 509 pp.
Copello, A. 1926. Biologia de Hermetia illucens Latr. Revista de la sociedade Entomológica
Argentina, 1(2): 22-26.
Costa C.; Ide S. 2006. Fases de desenvolvimento, In: C. Costa, S. Ide; Simonka, C.E. (Holos
editora). Insetos Imaturos: metamorfose e identificação. Ribeirão Preto. p. 19-29.
Costa, A.C.L.; Silva-Junior, J.A.; Cunha, A.C.; Feitosa, J.R.P.; Portela, B.T.T.; Silva, G.G.C.;
Costa, R.F. 2013. Índices de conforto térmico e suas variações sazonais em cidades de
diferentes dimensões na Região Amazônica. Revista Brasileira de Geografia Física, 6 (3): 478-
487.

83
Deleporte S.; Charrier, M. 1996. Comparison of digestive carbohydrases between two forest
sciarid (Diptera: Sciaridae) larvae in relation to their ecology. Pedobiologia, 40: 193–200.
De Marco, M.; Martínez, S.; Hernandez, F.; Madrid, J.; Gai, F.; Rotolo, L.; Belforti, M.;
Bergero, D.; Katz, H.; Dadbou, S.; Kovitvadhi, A.; Zoccarato, I.; Gasco, L.; Schiavione, A.
2015. Nutritional value of two insect latval meals (Tenebrio molitor and Hermetia illucens) for
broiler chickens: Apparent nutriente digestibility, apparent ileal amino acid digestibility and
apparent metabolizable energy. Animal Feed Science Technology, 209: 211-218.
Dermott, R.M.; Paterson, C.G. 1974. Determining dry weight and percentage dry-matter of
chironomid larvae. Canadian Journal of Zoology, 52: 1243–1250.
Diener, S.; Solano, N.M.S.; Gutiérrez, F.R.; Zurbrügg, C.; Tockner, K. 2011. Biological
treatment of municipal organic waste using black soldier fly larvae. Waste and Biomass
Valorization, 2 (4): 357-363.
Dyar, H.G. 1980. The number of molts Lepidopterous larvae. Psyche, 5: 420-422.
Fachin DA 2017. Stratiomyidae in Catálogo Taxonômico da Fauna do Brasil. PNUD.
(http://fauna.jbrj.gov.br/fauna/faunadobrasil/12536). Acesso: 24/05/2017.
Furmand, P.; Young, D.; Catts, P.E. 1959. Hermetia illucens (Linnaeus) as a factor in the
natural control of Musca domestica (Linnaeus). Journal of Economic Entomology, 52 (5): 917-
921.
Ganihar, S.R. 1997. Biomass estimates of terrestrial arthropods based on body length. Jounal
of Biosciences, 22(2) 219-224.
Gobbi, F.P. 2012. Biología reproductiva y caracterización morfológica de los estadios
larvarios de Hermetia illucens (L., 1758) (Diptera: Stratiomyidae). Bases para su producción
masiva en Europa. Tese de doutorado, Universidad de Alicante. 155 pp.
Gobbi, P.; Martínez-Sánchez, A.; Rojo, S. 2013. The effects of larval diet n adult life-history
traits of the black soldier fly, Hermetia illucens (Diptera: Stratiomyidae). European Jounal of
Entomology, 110 (3): 461–468.

84
Gonzalez, J.V., Young, W.R., Genel, M.R. 1963. Reducion de la problacion de mosca
domestica en gallinzaz por la mosca soldado en el Tropica. Agricultural Technology, 2: 53-57.
Gowing, G.; Recher, H.F. 1984. Length- weight relationships for invertebrates from forests in
South- eastern New South Wales. Australian Journal Ecology, 9: 5-8.
Greenberg, B.; Kunich, J.C. 2002. Entomology and the Law: Flies as Forensic Indicators.
Cambridge University Press, Cambridge. 306pp.
Gruner, D.S. 2003. Regressions of length and width to predict Arthropod Biomass in the
Hawaiian Islands. Pacific Science, 57(3): 325-336.
Gullan, P.J.; Cranston, P.S. 1994. The insects: an outline of entomology. London, Chapman e
Hall. 480pp.
Hall, D.C.; Gerhardt, R.R. 2002. Flies (Diptera). In: Mullen, G.; Durden, L. (Academic Press)
Medical and veterinary entomology. San Diego, Califórnia p. 127- 161.
Hawkins, J.W.; Lankester, M.W.; Lautenschlager, R.A.; Bell, F.W. 1997. Length-biomass and
energy relationships of terrestrial gastropods in northern forest ecosystems. Canadian Journal
of Zoology, 75:501–505.
Hódar, J.A. 1996. The use of regression equations for estimation of arthropod biomass in
ecological studies. Acta Ecologica Sinica, 17(5): 421-423.
Holmes, L.A.; Vanlaerhoven, S.L.; Tomberlin, J.K. 2012. Relative Humidify effects on the life
history of Hermetia illucens (Diptera: Stratiomyidae). Environmental Entomology, 41 (4): 971-
978.
Howmiller, R.P. 1972. Effects of preservatives on weights of some common macrobenthic
invertebrates. Transactions of the American Fisheries Society, 101 (4): 743-746.
Huis, A.V.; Gurp, H.V.; Dicke, M. 2014. The insect cookbook, food for a sustainable planet.
Columbia University Press. 216pp.

85
INMET – Instituto Nacional de Metereologia, 2017. Estações convencionais – gráficos.
(http://www.inmet.gov.br/portal/index.php?r=home/page&page=rede_estacoes_conv_graf).
Acesso: 24/01/2017.
INPA – Instituto Nacional de Pesquisas da Amazônia, 2013. Infraestrutura.
(http://portal.inpa.gov.br/index.php/institucional/infraestrutura). Acesso: 09/01/17.
Johnston, T.; Cunjak, R. 1999. Dry mass-length relationships for benthic insects: a review with
new data from Catamaran Brook, New Brunswick. Canada. Freshwater Biology, 41(4): 653-
674.
Johnson, M.D.; Strong, A.M. 2000. Length- weight relationships of Jamaican arthropods.
Entomological News. 111(4): 270-281.
Kim, J.G.; Choi, Y.C.; Choi, J.Y.; Kim, W.T.; Jeong, G.S.; Park, K.H.; Hwang, S.J. 2008.
Ecology of the black soldier fly, Hermetia illucens (Diptera: Stratiomyidae) in Korea. Korean
Journal of Applied Entomology, 47: 337-343.
Kim, W.; Bae, S.; Park, H.; Park, K.; Lee, S.; Choi, Y., Han, S.; Koh, Y. 2010. The larval age
and mouth morphology of the Black Soldier Fly, Hermetia illucens (Diptera: Stratiomyidae).
International Journal of Industrial Entomology, 21: 185-187.
Kingsolver, J. G.; Huey, R. B. 2008. Size, temperature, and fitness: three rules. Evololutionary
Ecology Research, 10, 251–268.
Knapp, M. 2012. Preservative fluid and storage conditions alter body mass estimation in a
terrestrial insect. Entomol. Experimental and Applied Acarology, 143: 185-190.
Kovac, D.; Rozkošný, R. 2004. Insecta: Diptera, Stratiomyidae. In: Yule, C.M.; Yong, H. S.
(Academy of Sciences) Freshwater Invertebrates of the Malaysian Region. Malaysia. p. 798-
804.
Lalander, C; Diener, S; Magri, M.E.; Zurbrügg, C.; Lindström, A.; Vinnerås, B. 2013. Faecal
sludge management with the larvae of the black soldier fly (Hermetia illucens) – From a
hygiene aspect. Science of the Total Environment, 458-460, 312-318.

86
Lambkin, C.L.; Sinclair, B.J.; Pape, T; Courtney, G.W.; Skevington, J.H; Meier, R.; Yeastes,
D.K.; Blagoderov, V.; Wiegmann, B.M. 2013. The phylogenetic relatioships among infraorders
and superfamilies of Diptera based on morphological evidence, Systematic Entomology, 38:
164-179.
Landahl, C. C.; Nagell, B. 1978. Influence of the season and of preservation methods on wet-
and dry weights of larvae of Chironomus plumosus L. International Review of Hydrobiology,
63: 405–410.
Lecheta, M.C. 2012. Efeitos da temperatura no desenvolvimento de Sarconesia chlorogaster
(Wiedemam, 1830) (Diptera, Calliphoridae) e sua possível utilização na Entomologia Forense.
Dissertação de Mestrado. Programa de pós-graduação em Ciências Biológicas, Universidade
Federal do Paraná. 77 pp.
Leuven, R.S.E.W.; Brock, T.C.M.; Vandruten, H. A. M. 1985. Effects of preservation on dry-
and ash-free dry-weight biomass of some common aquatic macro- invertebrates.
Hydrobiologia, 127: 151–159.
Lord, W.D.; Rodriguez, W.C. 1989. Forensic entomology: the use of insects in the investigation
of homicide and untimely death. Prosecutor, 22: 41–48.
Lord, W.D.; Goff, M.L.; Adkins, T.R.; Haskell, N.H. 1994. The black soldier fly Hermetia
illucens (Diptera: Stratiomyidae) as a potential measure of human postmortem interval:
observations and case histories. Journal of Forensic Science, 39: 215–222.
Martínez-Sánchez, A; Magaña, C; Saloña, M.; Rojo, S. 2011. First record of Hermetia illucens
(Diptera: Stratiomyidae) on humam corpses in Iberian Penísula. Forensic Science
International, 206:76-78.
Martins, R.T.; Melo, A.S.; Gonçalves, J.F.Jr.; Hamada, N. 2014. Estimation of dry massof
caddisflies Phylloicus elektoros (Trichoptera: Calamoceratidae) in a Central Amazon stream.
Revista Brasileira de Zoologia, 31: 337–342.
May, B.M. 1961. The occurrence in New Zealand and the life-history of the soldier fly
Hermetia illucens (L.). New Zealand Journal Science, 4: 55-65.

87
McFaden, M.W. 1967. Soldier fly larvae in America north of Mexico. Proceedings of the
United States National Museum, 12:1-72.
Oliveira-Costa, J. 2011. Entomologia Forense: Quando os insetos são vestígios. Millennium,
Campinas, São Paulo, BRA. 257pp.
Oliveira, F.; Doelle, K.; List, R.; O’Reilly, J. 2015. Assessment of Diptera: Stratiomyidae,
genus Hermetia illucens (L., 1758) using eléctron microscopy. Journal of Entomology and
Zoology Studies, 3(5): 147-152.
Oliveira, F.; Doelle, K.; Smith, R.P. 2016. External Mophology of Hermetia illucens
Stratiomyidae: Diptera (L. 1758) based on electron Microscopy. Annual Research e Review in
Biology, 9(5): 1-10.
Parra, J.R.P. 1990. Consumo e utilização de alimentos por insetos, In: Panizzi, A.R.; Parra,
J.R.P. (Manole): Ecologia Nutricional de Insetos e suas Implicações no Manejo de Pragas. São
Paulo, Brasil. p. 9–57.
Petersen, P.; Luxton, M. 1982. A comparative analysis of soil fauna populations and their role
in decomposition processes. Oikos, 39: 288–388.
Price, P.W. 1984. Insect ecology. John Wiley e Sons, NY.
Pujol-Luz, J.R.; Francez, P.A.C.; Ururahy-Rodrigues, A.; Constantino, R. 2008. The black
soldier-fly, Hermetia illucens (Díptera, Stratiomyidae), used to estimate the postmortem
interval in a case in Amapá state, Brazil. Journal of Forensic Science, 53: 476–478.
Pujol-Luz, J.R.; Pujol-Luz, C.V.A. 2014. Stratiomyidae. In: Roig-Juñent, S.; Claps, L.E.;
Morrone, J.J. (Editorial INSUE –UNT). Biodiversidad de Artrópodos Argentinos. Vol. 4. San
Miguel de Tucumán, Argentina. p. 399-406.
Radtke, M.G.; Williamson, G.B. 2005. Volume and linear measurements as predictors of dung
beetle (Coleoptera: Scarabaeidae) biomass. Annals of the Entomological Society of America,
98(4): 548-551.
Renshaw, M.; Service, M. W.; Birley, M.H. 1994. Size variation and reproductive success in
the mosquito Aedes cantans. Medical and Veterinary Entomology, 8: 179-186.

88
Rogers, L.E.; Buschbom, R.L.; Watson, C.R. 1976. Length-Weight Relationships of Shrub-
Steppe Invertebrates. Annals of the Entomological Society of America, 70(1): 51-53.
Rogers, L.E.; Hinds, W.T.; Buschbom, R.L. 1976. A general weight vs. length relationship for
insects. Annal of the Entomological Society of America Journal, 69: 387-89.
Roubik, D.W. 1989. Ecology and Natural History of Tropical Bees. Cambridge University.
Press, New York. 512pp.
Rozkošný, R.; Kovac, D. 1998. Descriptions of bamboo-inhabiting larvae and puparia of
oriental soldier flies Ptecticus brunettii and P. flavife moratus (Diptera: Stratiomyidae:
Sarginae) with observations on their biology. European Journal of Entomology, 95: 65-86.
Sabo, J.L.; Bastow, J.L.; Power, M. 2002. Length- mass relationships for adult aquatic and
terrestrial invertebrates in a California Watershed. Journal of the North American
Benthological Society, 21(2): 336- 343.
Sample, B.E.; Cooper, R.J.; Greer, R.D.; Whitmore, R.C. 1993. Estimation of Insect Biomass
by length and width. American Midland Naturalist, 129 (2): 234-240.
Schiller, D.V.; Solimini, A.G. 2005. Differential effects of preservation on the estimation of
biomass of two common mayfly species. Archiv fur Hydrobiologie Journal, 164(3): 325-334.
Serra, H.; Godoy, W.A.C.; Von-Zuben, F.J.; Von-Zuben, C.J.; Reis, S. F. 2007. Sex ratio and
dynamic behavior in populations of the exotic blowfly Chrysomya albiceps (Diptera,
Calliphoridae). Brazilian Journal of Biology, 67(2), 347-353.
Sheppard, D.C. 1983. House fly and lesser fly control utilizing the black soldier fly in manure
management systems for caged laying hens. Environmental Entomology, 12: 1439- 1442.
Sheppard, D.C.; Newton, G.L.; Thompson, S.A. 1994. A value added manure management
system using the Black Soldier Fly. Bioresource Technology, 50: 275-279.
Sheppard, D.C.; Tomberlin, J.K.; Joyce, J.A.; Kiser, B.C.; Sumner, S.M. 2002. Rearing and
colony maintenance of the black soldier fly, Hermetia illucens (L.) (Diptera: Stratiomyidae).
Journal of Medical Entomology, 39: 695-698.

89
Silveira-Neto, S.; Nakano, O.; Barbin, D.;Villa-Nova, N.M. 1976. Manual de Ecologia de
Insetos. Editora Agronômica Ceres, São Paulo. 420pp.
Smith, K.G.V. 1989. An introduction to the immature stages of British flies. Handbook for
identification of British insects. Vol. 10, Part 14. Royan Entomological Society, London, 280
pp.
Souza, A.S.B.; Kirst, F.D. 2010. Aspectos da bionomia e metodologia de criação de Dipteros
de interesse forense. In: Gomes, L. (Technical Books Editora), Entomologia Forense: Novas
tendências e tecnologias nas ciências criminais. Primeira edição. Rio de Janeiro. p. 169-182.
Stork, N.E. 1988. Insect diversity: facts, fiction and speculation. Biological Jounal Linnean
Society, 35: 321–337.
Teskey, H.J. 1991. Diptera. In: Stehr, F. W. (Kendall Hunt Publishing Company). Immature
insect. Vol. 2. Dubuque, Iowa. p. 690–706.
Tingle, F.C.; Mitchell, E.R; Copeland, W.W. 1975. The soldier fly, Hermetia illucens, in
poultry houses in north central Florida. Journal of the Georgia Entomological Society, 10: 179-
183.
Tomberlin, J.K.; Sheppard, D.C. 2001. Lekking behavior of the Black Soldier Fly (Diptera:
Stratiomyidae). Florida Entomologist, 84:729-730.
Tomberlin, J.K.; Sheppard, D.C.; Joyce, J.A. 2002. A comparison of selected life history traits
of the black soldier ßy (Diptera: Stratiomyidae) whenreared on three diets. Annals of the
Entomological Society of America, 95: 379-387.
Tomberlin, J.K.; Sheppard, D.C.; Joyce, J.A. 2005. Black Soldier Fly (Diptera: Stratiomyidae)
colonization of pig carrion in South Georgia. Journal of Forensic Sciences, 50 (1): 1-2.
Triplehorn, C.A.; Johnson, N.F. 2011. Estudo dos insetos. Tradução da 7ª edição de Borror and
Delong’s introduction to the study of insects. Cengage Learning, São Paulo, BR. 809pp.
Tucker, J.W.; Chester, A.J. 1984. Effects of salinity, formalin concentration and

90
buffer on quality of preservation of Southern Flounder (Paralichthys lethostigma) larvae.
Copeia. 4: 981–988.
Turchetto, M.; Lafiscam S.; Constantini, G. 2001. Postmortem interval (PMI) determined by
study sarcophagous biocenoses: three cases from the province of Venice (Italy). Forensic
Science International, 120:28-31.
Üstüner, T; Hasbenli, A.; Rozkosny, R. 2003. The first record of Hermetia illucens (Linnaeus,
1758) (Diptera, Stratiomyidae) from the Near East. Studia dipterologica, 10: 181-185.
Wardhaugh, C.W. 2013. Estimation of biomass fron body length and width for tropical
rainforest canopy invertebrates. Australian Journal of Entomology, 52: 291-298.
Wetzel, M.A.; Leuchs, H.; Koop, J. H. E. 2005. Preservation effects on wet weight, dry weight,
and ash-free dry weight biomass estimates of four common estuarine macro-invertebrates: no
difference between ethanol and formalin. Helgoland Marine Research, 59 (3): 206-213.
Woodley, N.E. 1989. Family Stratiomyidae. Catalog of the Diptera of the Australasian and
Oceanian Regions. Bishop Museum Special Publication, 86: 301-320.
Woodley, N.E. 2001. A world catalogue of the Stratiornyidae (Insecta: Diptera). Myia, 11: 1-
473.
Woodley, N.E. 2009. Family Stratiomyidae. In: J. Gerlach (Pen soft Publishers). The Diptera
of the Seychelles islands. Sofia and Moscow. p. 100-106.
Woodley, N.E. 2011. A World Catalog of the Stratiomyidae (Insecta: Diptera): A Supplement
with Revisionary Notes and Errata. Myia, 12: 379–415.
Zar, J.H. 2010. Biostatistical Analysis. Pearson Education, NJ. 944pp.


















![WEB VERSION Peppers [Pimentas] of Roraima Project – INPA ... · 2 Peppers [Pimentas] of Roraima Project – INPA/Embrapa/MIRR CAPSICUM (Solanaceae): varieties in Roraima, Brazil](https://static.fdocumentos.com/doc/165x107/5f5c2cd77b4fc02e9d73016c/web-version-peppers-pimentas-of-roraima-project-a-inpa-2-peppers-pimentas.jpg)