
INSTITUTO NACIONAL DE PESQUISAS DA...
56
INSTITUTO NACIONAL DE PESQUISAS DA AMAZÔNIA-INPA PROGRAMA DE PÓS-GRADUAÇÃO EM BOTÂNICA ATRIBUTOS FUNCIONAIS DA FLORA ARBÓREA E ARBUSTIVA DE CAMPINA E CAMPINARANA NA AMAZÔNIA CENTRAL LIANE LIMA Manaus, Amazonas Abril, 2015
Transcript of INSTITUTO NACIONAL DE PESQUISAS DA...

INSTITUTO NACIONAL DE PESQUISAS DA AMAZÔNIA-INPA
PROGRAMA DE PÓS-GRADUAÇÃO EM BOTÂNICA
ATRIBUTOS FUNCIONAIS DA FLORA ARBÓREA E ARBUSTIVA DE CAMPINA E
CAMPINARANA NA AMAZÔNIA CENTRAL
LIANE LIMA
Manaus, Amazonas
Abril, 2015

ii
LIANE LIMA
ATRIBUTOS FUNCIONAIS DA FLORA ARBÓREA E ARBUSTIVA DE CAMPINA E
CAMPINARANA NA AMAZÔNIA CENTRAL
Dr. Florian Wittmann
Dissertação apresentada ao Instituto
Nacional de Pesquisas da Amazônia
como parte dos requisitos para
obtenção do título de Mestre em
Botânica.
Manaus, Amazonas
Abril, 2015

iii
Relação Banca Julgadora
Qualificação:
Henrique Eduardo M. Nascimento
Juliana Schietti de Almeida
Michael John Gilbert Hopkins
Defesa:
Christopher Jon Baraloto
Jochen Schöngart
Juliana Schietti de Almeida

iv
L732 Lima, Liane
Atributos funcionais da flora arbórea e arbustiva de campina e
campinarana na Amazônia Central / Liane Lima. --- Manaus: [s.n.],
2015.
56 f. : il.
Dissertação (Mestrado) --- INPA, Manaus, 2015.
Orientador : Florian Karl Wittmann.
Área de concentração: Botânica.
1. Composição florística. 2. Campina. 3. Campinarana. I.
Título.
CDD 581.4
Sinopse:
Estudou-se os efeitos dos diferentes valores de atributos funcionais da flora arbórea e
arbustiva de campina e campinarana do Parque Nacional do Jaú e da Reserva de
Desenvolvimento Sustentável do Uatumã.
Palavras-chave: Atributos funcionais, composição florística, campina, campinarana.

v
AGRADECIMENTOS
Agradeço aos meus pais, minha irmã e meus avós por todo amor e por sempre me
apoiarem em todas as minhas escolhas. Agradeço também ao meu orientador, Florian
Wittmann, pela amizade e compreensão, que sempre disposto a ajudar, contribuiu muito para
minha formação.
A todos os participantes do grupo MAUA (“Ecologia, Monitoramente e Uso
Sustentável de Áreas Úmidas” - projeto INPA/Max Planck): pesquisadores, técnicos,
auxiliares de limpeza, alunos dos programas de pós-graduação e demais bolsitas do projeto, a
vocês o meu eterno carinho e gratidão.
Aos amigos queridos que ajudaram nos trabalhos de campo e na construção desse
projeto: Adriano Quaresma, Bianca Weiss, Boris Villa, Cyro Assahira, Deborah Castro, Ethan
Householder, Layon Demarchi, Rafael Assis, Thaiane Souza, Yuri Feitosa.
A minha família manauara: Maria Antonieta Dias, Mariana Dettmer e Thaiane Souza,
gratidão pela amizade e por tornarem a minha vida em Manaus muito mais feliz, sempre
amenizando a saudade de casa.
Ao Instituo Nacional de Pesquisas da Amazônia (INPA); ao Programa de pós-
graduação em Botânica, seus coordenadores e secretárias; à Coordenação de Aperfeiçoamento
de Pessoal de Nível Superior (CAPES), pela concessão da bolsa de estudos; ao Projeto
INPA/Max-Planck e a Fundação de Amparo à Pesquisa do Estado do Amazonas (FAPEAM),
pelo financiamento e suporte logístico para as atividades de campo e de laboratório; ao
Herbário do INPA, a todos os funcionários, em especial ao Sr. José Ramos.
Gratidão a todos.

vi
RESUMO
As fito-fisionomias encontradas sobre areia branca (podzol) na Amazônia Central ocorrem de
forma fragmentada ao longo do bioma e são nomeadas de campinas e campinaranas. Elas
formam um conjunto único de variações fisionômicas sobre componentes edáficos pobres em
nutrientes minerais e, em alguns casos, em condições de saturação hídrica. Para
compreendermos a variação fisionômica das campinas para as campinaranas é necessário
avaliar os atributos funcionais que respondem sobre variações ambientais, podendo ser eles
tanto morfológicos (p.ex.: densidade específica da madeira, área foliar específica, DAP-
diâmetro na altura do peito, altura) como fisiológicos (p.ex.: concentração de micronutrientes
nas folhas). Este presente trabalho teve os seguintes objetivos: (i) compreender o papel dos
fatores edáficos na distribuição dos valores dos atributos funcionais dos indivíduos, da flora
arbórea e arbustiva, em diferentes fisionomias de campinas e campinaranas na Amazônia
Central, (ii) determinar se há um padrão funcional entre as campinas e campinaranas de duas
diferentes localidades, independente de suas localizações geográficas e das composições
florísticas. Dois sítios amostrais foram selecionados para este estudo, o Parque Nacional do
Jaú (PARNA Jaú) e a Reserva de Desenvolvimento Sustentável Uatumã (RDS Uatumã),
ambos localizados na Amazônia Central. Análise de Componentes Principais (ACP) foi
aplicada e através dela foi possível observar a formação de dois grupos distintos, separando a
distribuição dos atributos funcionais dos indivíduos da campina dos indivíduos da
campinarana, em ambas as áreas de estudo, para a maioria dos atributos avaliados. Porém a
variação fisionômica não foi explicada pelos componentes edáficos da vegetação, pois estes
não variaram de acordo com as fisionomias. Sendo assim, o fator saturação hídrica do solo
arenoso, que é diferente nas campinas em relação às campinaranas, corresponde à provável
explicação da variação estrutural da vegetação entre essas duas fito-fisionomias.

vii
ABSTRACT
The phyto-physiognomies which are found on white sand (podzol) in the Central Amazon
occurr in a fragmented way along the biome and are named “campinas” and “campinaranas”.
They comprehend a unique set of physiognomic variations on edaphic components with low
contents of mineral nutrients and, in some cases, subjected to water saturation. To understand
the physiognomic variation of the “campinas” to the “campinaranas” is necessary to evaluate
the functional attributes that respond to environmental changes, being they either
morphological (eg., specific wood density, specific leaf area, DBH – diameter at breast
height, height) as physiological (eg., concentration of micronutrients in the leaves). This
present study had the following objectives: (i) understand the role the edaphic factors in the
distribution of values of the individual functional attributes of the tree flora and shrubs at
different physiognomies of “campinas” and campinaranas in Central Amazonia, (ii) determine
whether there is a functional pattern between the “campinas” and “campinaranas” from two
different localities, regardless of their geographical location and floral composition. Two sites
were selected for this study, the Jaú National Park (PARNA Jaú) and the Sustainable
Development Reserve Uatumã (RDS Uatumã), both located in the Central Amazon. Principal
Component Analysis (PCA) was applied and through it was possible to observe the formation
of two distinct groups, separating the distribution of functional attributes of the individuals
from the “campinas” from those of the “campinarana” in both study areas, for most of the
evaluated attributes. However, the physiognomic variation was not explained by edaphic
components of vegetation, since they did not vary between the different phyto-
physiognomies. Therefore, the factor water saturation of the sandy soils, which is different in
the “campinas” when compared to “campinaranas”, corresponds to the possible explanation of
the vegetational structural variation between these two phyto-physiognomies.

viii
Sumário
1. Introdução Geral ..................................................................................................................... 9
2. Objetivos ............................................................................................................................... 13
Capítulo 1
Atributos funcionais de flora arbórea e arbustiva de campina e campinarana na Amazônia
Central. ................................................................................................................................... 155
3. Conclusões ............................................................................................................................ 50
4. ANEXOS .............................................................................................................................. 51
5. Referências bibliográficas .................................................................................................... 52

9
1. Introdução Geral
1.1. Campinas e Campinaranas
As classes de vegetação que ocorrem ao longo de um gradiente edáfico sobre
areia branca (podzol), nomeadas de campina e campinarana, ocupam a depressão central
da região interfluvial Amazônica (Nascimento et al. 1997) de forma fragmentada, em
padrão insular, cercadas por uma matriz de floresta de terra-firme e totalizando mais de
480.000 km2
(Junk et al. 2011). Estas formações são divididas em três subgrupos cujas
estruturas variam e recebem as seguintes denominações: florestada, arborizada e
gramíneo-lenhosa, esta última também conhecida como campina (Veloso et al. 1991).
A variação fisionômica da vegetação em questão está diretamente relacionada à
variação sazonal na altura do lençol freático e ao tempo de alagamento anual (Daly &
Mitchell 2000), assim como à seca fisiológica (diminuição da altura do lençol freático
no período de seca), tais fatores ambientais favorecem ou limitam a dominância de
espécies, bem como seu porte e densidade de indivíduos (Daly & Mitchell 2000;
Silveira 2003; Vicentini 2004). A inundação é um fator limitante sobre crescimento,
sobrevivência e reprodução de ecossistemas inundáveis (Parolin & Wittmann 2010),
assim indivíduos localizados nas áreas sujeitas a longos períodos de inundação tem seu
crescimento limitado, quando comparado com indivíduos sujeitos a menores períodos
de inundação (Wittmann et al. 2002). Segundo Ferreira (1997), o lençol freático nos
habitats de campina está mais próximo da superfície do solo do que nos habitats de
campinarana, onde a água subjacente não é capaz de atingir a rizosfera, nem mesmo nos
períodos de maior nível de água.
A natureza oligotrófica do solo arenoso sob a vegetação deve-se principalmente
à sua origem, que inclui diversas causas, como o intemperismo local do material de
origem pobre, como arenitos, quartzitos e granitos; deposição aluvial de areia
proveniente dos escudos da Guiana ou da porção central do Brasil; e a podzolização,
devido à flutuação do lençol freático, que carrega a matéria orgânica e partículas
argilosas dos perfis superiores do solo (Anderson 1981). A principal diferença entre o
solo de campina e de campinarana é a presença de uma camada espessa de serrapilheira
e uma fina rede de raízes de 15 a 20 cm de espessura, presente nas campinaranas
(Nascimento et al. 1997; Worbes 1997; Silveira 2003). De modo geral o solo é

10
quimicamente ácido (pH 4.5 a 5.6), empobrecido nutricionalmente e com baixa soma de
bases (inferior a 0.5 cmolcdm-3
) (Mendonça et al. 2014).
Dentro dos subgrupos de variação estrutural da vegetação, classificam-se as
fisionomias abertas como campinas, com presença de herbáceas e arbustos isolados
variando entre 1 a 5 metros de altura, com alguns indivíduos atingindo 9 metros
(Ferreira 2009). Durante o período seco, quando as campinas estão sujeitas à escassez
hídrica, devido a rápida percolação da água pela areia branca, a presença de biomassa
seca e de herbáceas altamente inflamáveis torna esse habitat fortemente suscetível a
incêndios (Prance & Shubart 1978). Assim, muitas espécies que constituem esta
comunidade apresentam adaptações ao fogo, como folhas coriáceas, lenho duro,
cutículas espessas, tricomas bem desenvolvidos, estômatos crípticos (Mendonça 2011) e
colmo coberto por folhas velhas que protegem os meristemas (Vicentini 2004). As
campinaranas correspondem às fisionomias florestais, caracterizadas por apresentarem
sub-bosque relativamente aberto e escassez de cipós e lianas, com indivíduos arbóreos
podendo atingir altura de até 30 metros (Anderson 1981).
Campinas e campinaranas possuem espécies com alto potencial de dispersão a
longas distâncias e uma diversidade de tipos de mecanismos dispersores (vento, água),
característica de biotas isoladas sobre áreas empobrecidas em nutrientes minerais
(Macedo & Prance 1978), também como taxas lentas de crescimento e eficientes
mecanismos de defesa, divergindo em atributos que conferem defesa contra aqueles que
conferem crescimento (Fine et al. 2006). Muitas espécies possuem características que
permitem a sobrevivência sobre o solo arenoso oligotrófico, como por exemplo folhas
coriáceas, lenho duro, cutículas espessas, tricomas bem desenvolvidos, estômatos
crípticos, porém poucas possuem atributos que promovam seu estabelecimento em
níveis altos sobre uma área, tornando-as dominantes (Fine et al. 2004, 2006).
Dessa forma, verificamos que nas campinas há uma baixa diversidade florística
com predominância de espécies generalistas de áreas abertas, geralmente de ampla
distribuição como Landenbergia amazonensis, Ouratea spruceana, Clusia nemorosa,
Clusia insignis, Chanouchiton loranthoides e Platycarpum englei, espécies descritas
com endêmicas (Ducke & Black 1954), porém encontradas em demais trabalhos na
Amazônia Central (e.g. Anderson et al. 1975; Ferreira 1997; Silveira 2003; Vicentini
2004).
Formando um conjunto único de fisionomias adaptadas a recursos de pobreza
nutricional e ao regime de sazonalidade hídrico do solo, as campinas e campinaranas

11
representam um dos ambientes mais frágeis e vulneráveis ás atividades antrópicas
(Silveira 2003). Atividades extrativistas e agrícolas são contraproducentes
economicamente nas campinas e campinaranas, uma vez que a retirada da vegetação
local provoca a rápida lixiviação do solo arenoso, degradando sua fertilidade e o
reestabelecimento da flora culmina em gastos de recursos não recuperáveis pelas
atividades em questão (Fine et al. 2010).
Estas ilhas edaficamente secas são de extrema importância como refúgios
periódicos de organismos terrestres, contribuindo decisivamente para a manutenção dos
ciclos de biodiversidade, sendo assim consideradas como parte indispensável dos
grandes sistemas de áreas úmidas na região Amazônica (Junk 2013).
1.2. Diversidade Funcional
Para melhor compreender a variação fisionômica das campinas para as
campinaranas, é preciso avaliar atributos funcionais que respondam as variações
ambientais. Os atributos funcionais são quaisquer características que alterem a aptidão
do indivíduo em relação às condições ambientais e mostram como os indivíduos
utilizam os recursos disponíveis (Wright et al. 2004), exemplos de atributos funcionais
seriam a densidade específica da madeira, área foliar específica (AFE) e concentração
de micronutrientes nas folhas. Em geral, o investimento em densidade específica da
madeira é refletido nas taxas de crescimento, sendo uma importante adaptação em
ambientes de pobreza nutricional, onde há pouca energia disponível para eventual
rebrota ou substituição de tecido danificado (Worbes et al. 1992). As espécies
permanentemente ou temporariamente situadas em ambientes ricos em nutrientes
tendem a possuir uma maior área foliar específica, atributo relacionado positivamente a
taxa de crescimento relativo das espécies, quando comparada com espécies que ocupam
ambientes com recursos limitados (Perez-Hanguindeguy 2013). Já a concentração de
micronutrientes nas folhas, necessários em quantidades vestigiais, respondem de forma
abrupta quando não estão disponíveis em quantidades adequadas para a planta, afetando
sua fisiologia e refletindo no seu rendimento celular (Taiwo et al. 2001; Adediran et al.
2004).
Valores compartilhados de atributos funcionais em um determinado local
refletem em tolerâncias ecológicas compartilhadas, diminuindo a capacidade de
propagação de um atributo ao longo do gradiente estrutural da vegetação (Cornwell et
al. 2006). Pode ser pensado como a redução da gama de estratégias bem sucedidas entre

12
as espécies que coexistem (van der Valk 1981; Keddy 1992; Weiher et al. 1998; Weiher
& Keddy 1999), limitando o intervalo que um atributo pode ocupar no ambiente (van
der Valk 1981; Keddy 1992).
Distribuídos ao longo do gradiente estrutural da vegetação, que reflete um
gradiente subjacente de produtividade, fundamentado na mudança da fisionomia
florestal para arbustiva, ocorre uma forte troca evolutiva entre a necessidade de tolerar
ambientes improdutivos e a necessidade de crescer rapidamente em ambientes
produtivos (Grime 2006). Essa capacidade de dominar instalações produtivas ou alocar
recursos de alta qualidade contradiz-se com a capacidade de persistir com recursos de
baixa qualidade ou de tolerar condições adversas (McGill 2006). Isso corresponde a
uma moeda de adequação, que é quando uma mudança benéfica de uma característica
está associada a uma mudança prejudicial em outra (trade-offs) (Stearns 1989). A
diversidade funcional dos atributos e a convergência dos filtros ambientais podem
operar simultaneamente no recrutamento de um conjunto de espécies, impondo efeitos
sobre a similaridade de valores de atributos, exibidos pelas espécies coexistentes (Grime
2006).
O funcionamento do ecossistema é o resultado final da operação de vários filtros
ambientais em uma escala hierárquica. Esses filtros selecionam indivíduos com
respostas adequadas, resultando em assembleias com composições diversas de
características e limitando sucessivamente quais as espécies e as características que
persistem em um dado local (Keddy 1992; Díaz et al. 1999). Mudanças na composição
de espécies, resultante de mudanças ambientais, resultam em modificações no
funcionamento do ecossistema, através de mudanças na representação dos atributos das
espécies (Lavorel & Garnier 2002).
Medir diversidade funcional corresponde a medir a diversidade dos atributos
funcionais, componentes fenotípicos que influenciam nos processos ecossistêmicos da
vegetação e que podem inferir sobre predições das estratégias que os indivíduos tem
dentro de uma comunidade e ecossistema (Grime 2006). Com o intuito de descrever um
padrão de similaridade funcional entre os indivíduos que ocupam uma determinada
fisionomia, o presente trabalho teve os seguintes questionamentos:
(1) Os fatores edáficos são responsáveis pela variação fisionômica entre as
campinas e campinaranas? (2) Como é dada a distribuição dos valores dos atributos
funcionais de densidade específica da madeira, área foliar específica e concentração de
micronutrientes nas folhas, ao longo das campinas a campinaranas?

13
2. Objetivos
2.1. Objetivo geral
O trabalho tem como objetivo compreender o papel dos fatores edáficos na
distribuição dos valores dos atributos funcionais de indivíduos de flora arbórea e arbustiva nas
diferentes fisionomias de campina e campinarana na Amazônia Centra, para determinar se há
um padrão funcional em duas áreas distintas, Parque Nacional do Jaú (PARNA Jaú) e Reserva
de Desenvolvimento Sustentável Uatumã (RDS Uatumã).
2.2. Objetivos específicos
(1) Determinar se há similaridade funcional entre os indivíduos dentro da campina e
da campinarana, independente da localização geográfica e das diferenças na composição
florística em cada área de estudo.
(2) Determinar se diferenças nos valores dos atributos funcionais entre os indivíduos
na campina e campinarana, nas duas áreas de estudo, é refletida nos fatores edáficos,
restringindo a capacidade de propagação de um atributo funcional a um habitat específico.
(3) Determinar se ocorre uma variação abrupta entre os valores dos atributos
funcionais de densidade específica da madeira, área foliar específica e concentração de
micronutrientes nas folhas em cada fisionomia, nas duas áreas de estudo, justificando a
variação fisionômica da vegetação.

14
Capítulo 1
Lima, L.; Wittmann, F.; Householder, J.E.
Atributos funcionais da flora arbórea e arbustiva
de campina e campinarana na Amazônia Central.
Manuscrito formatado para Acta Amazonica.

15
Atributos funcionais da flora arbórea e arbustiva de campina e campinarana na
Amazônia Central.
Liane Lima1
Florian Wittmann2
John Ethan Householder1
1. INPA – Instituto Nacional de Pesquisas da Amazônia, Avenida André Araújo, 2936, Bairro Aleixo,
69067-375, Manaus - AM, Brasil.
2. Instituto Max-Planck de Química, Departamento de Bioquímica, Hahn-Meitner Weg 1, 55128, Mainz -
Alemanha.

16
Resumo
As fito-fisionomias alagáveis na Amazônia Central, nomeadas campinas e
campinaranas, ocupam de forma fragmentada em padrão insular uma área de
aproximadamente 480.000 km². Constituída sobre um solo de areia branca, ácido, com
elevado oligotrofismo e em alguns casos, sujeita a flutuações periódicas do lençol
freático, a variação fisionômica da vegetação é dada pelos distintos atributos funcionais,
que determinam como um indivíduo utiliza os recursos disponíveis no ambiente, e de
qual forma é dada a sua propagação em um local. Respondendo sobre a aptidão do
indivíduo em relação ao meio, densidade específica da madeira, área foliar específica e
concentração de micronutrientes nas folhas foram medidas em indivíduos da campina e
campinarana, no Parque Nacional do Jaú e na Reserva de Desenvolvimento Sustentável
do Uatumã. O trabalho tem como intuito avaliar se ocorre similaridade funcional entre
indivíduos que ocupam uma determinada fisionomia, e se ela é refletida nos
componentes edáficos, restringindo a capacidade de propagação de um atributo a um
habitat específico, independente da localização geográfica e da composição florística da
vegetação. Pela Análise de Componentes Principais foi possível observar a formação de
dois grupos distintos, separando a distribuição dos atributos funcionais dos indivíduos
da campina dos indivíduos da campinarana, em ambas as áreas de estudo. Porém a
variação fisionômica não foi explicada pelos componentes edáficos da vegetação, que
não variaram de acordo com as fisionomias. Sendo assim, a saturação hídrico do solo
arenoso, diferenciada pelas camadas de solo arejada acima do lençol freático, tendo a
água subjacente ascendo nas campinas e uma espessa camada de serrapilheira nas
campinaranas, corresponde a provável explicação da variação estrutural da vegetação.
Palavras-chaves: Campina, campinarana, atributos funcionais, composição florística.

17
Introdução
As fito-fisionomias adaptadas a recursos de pobreza nutricional, que ocorrem ao
longo do gradiente edáfico sobre areia branca (podzol), são nomeadas de campinas e
campinaranas. Ocupam a depressão central da região interfluvial Amazônica
(Nascimento et al. 1997) e totalizam uma área de 480.000 km2
(Junk et al. 2011). Tais
fisionomias ocorrem de forma fragmentada em grandes áreas ao longo do alto Rio
Negro, e também na Amazônia Central (Junk et al. 2011), e estão dispostas em um
padrão insular, cercada por uma matriz de floresta de terra-firme.
A vegetação, em geral, é caracterizada por elevada esclerofilia, baixa
diversidade, em relação às florestas de terra-firme (Fine et al. 2010; Stropp et al. 2011),
e alto grau de endemismo de espécies, gêneros e famílias (Silveira 2003; Vicentini
2004). O solo é ácido com elevado oligotrofismo (Luizão et al. 2007; Mendonça 2011)
e, em alguns casos, a vegetação pode ser superficialmente inundada, ou pela elevação
das águas subterrâneas, ou pela saturação do solo em períodos chuvosos (Bleackley &
Khan 1963; Franco & Dezzeo 1994; Adeney 2009). Esses fatores ambientais estão
relacionados com a variação fisionômica da vegetação (Daly & Mitchell 2000), podem
favorecer ou limitar a dominância de espécies, bem como seu porte e densidade nessas
áreas úmidas (Daly & Mitchell 2000; Silveira 2003; Vicentini 2004).
Diante da complexidade da variação gradual dessa vegetação, para melhor
compreender essa variação fisionômica, é importante avaliar atributos que respondam as
variações ambientais. Os atributos funcionais são quaisquer características que alterem a
aptidão do indivíduo em relação às condições ambientais e mostram como os indivíduos
utilizam os recursos disponíveis (Wright et al. 2004). Valores compartilhados de
atributos funcionais em um determinado local refletem em tolerâncias ecológicas
compartilhadas, reduzindo a sua propagação ao longo do gradiente estrutural da
vegetação (Cornwell et al. 2006).
Atributos funcionais morfológicos, como densidade específica da madeira e área
foliar específica, e atributos funcionais fisiológicos, como concentração de
micronutrientes nas folhas, respondem a aspectos ambientais através da variação dos
seus valores de forma abrupta ou gradual, diferenciando aspectos morfológicos e
fisiológicos dos indivíduos, diante da variação entre as fisionomias, reduzindo a gama
de estratégias bem sucedidas entre indivíduos que coexistem (van der Valk 1981, Keddy

18
1992, Weiher et al. 1998, Weiher & Keddy 1999), limitando o intervalo que um atributo
pode ocupar no habitat (van der Valk 1981; Keddy 1992).
Formando um conjunto único de fito-fisionomias adaptadas à pobreza
nutricional e em alguns casos, ao regime de sazonalidade hídrico nos solos, as campinas
e campinaranas representam um dos ambientes mais frágeis e vulneráveis às atividades
antrópicas (Silveira 2003; Ferreira 2009). O trabalho tem como objetivo determinar se
há similaridade funcional entre as fisionomias de campina e campinarana no Parque
Nacional do Jaú (PARNA Jaú) e na Reserva de Desenvolvimento Sustentável Uatumã
(RDS Uatumã) e avaliar se as variações dos valores dos atributos são refletidas nos
componentes edáficos, podendo assim, restringir a capacidade de propagação de um
atributo a um habitat específico, independente da sua localização geográfica e
composição florística.
Material e Métodos
Área de estudo
As duas áreas de estudo foram o Parque Nacional do Jaú (PARNA Jaú) e a
Reserva de Desenvolvimento Sustentável do Uatumã (RDS Uatumã), ambas banhadas
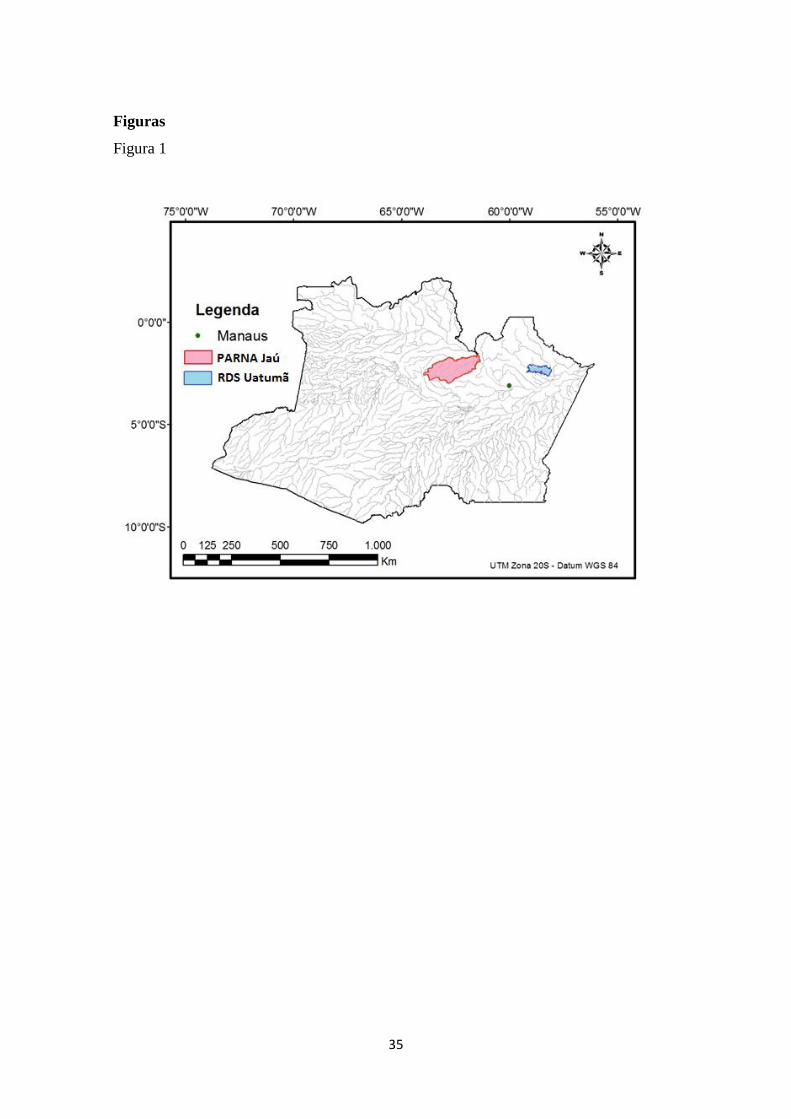
por tributários do rio Negro: rio Jaú e o rio Uatumã, ocupando uma área de 22.720 km²
e 4.244 km², respectivamente (Figura 1). O clima nas regiões é tropical úmido (Af –
classificação Köpper-Geiger) com temperatura média anual de 26.7° C e 28° C e
precipitação média anual de 2400 mm e 2376 mm, em cada área, respectivamente. No
PARNA Jaú o período de seca ocorre nos meses de junho a outubro, e o período de
chuva nos meses de dezembro a maio. Na RDS Uatumã o período de seca ocorre nos
meses se julho a outubro, e o período de chuva nos meses de fevereiro a abril.
Desenho amostral
O levantamento florístico para cada área de estudo, no PARNA Jaú nos meses
de outubro e novembro de 2013, e na RDS Uatumã nos meses de julho e agosto de
2014, foi feito através de dez transectos de 2 x 50 metros (0.01 ha) na campina e dez na
campinarana.
Variáveis ambientais
Foram coletadas três amostras de solo nas campinas e três amostras nas
campinaranas, nas duas áreas de estudo, ao longo dos transectos (0.01ha). As doze
amostras de solo foram coletadas entre 0 a 30 cm de profundidade, 200 metros entre si e

19
posteriormente, encaminhadas para o Laboratório de Análises de Solos e Plantas
(LASP) da EMBRAPA (Empresa Brasileira de Pesquisa Agropecuária) - Amazônia
Ocidental, para análise física (Areia Total, Silte, Argila, Classificação Textural) e
químicas (pH, P, K+, Na
+, Ca
2+, Mg
2+, Al
3+, soma de bases trocáveis (SB), capacidade
de troca catiônica (T), índice de saturação de bases (V), índice de saturação de alumínio
(m), Fe, Zn+, Mn
2+, Cu).
As campinas e campinaranas não tiveram a profundidade do lençol freático
medida, mas foi observado acúmulo de água com alguns centímetros de altura acima do
solo nas áreas do PARNA Jaú (fevereiro 2014) e na RDS Uatumã (abril 2014)
(observação pessoal).
Levantamento florístico
Todos os indivíduos com DAP > 0.5 cm (diâmetro na altura do peito) foram
inventariados nas campina e campinaranas, e cada indivíduo foi classificado segundo
DAP e altura: quando DAP < 10 cm e/ou não atingindo a altura do dossel, caracteriza-se
o indivíduo como arbustivo, quando DAP > 10 cm e/ou atingindo a altura do dossel,
caracteriza-se o indivíduo como arbóreo. Das espécies mais abundantes do
levantamento foram escolhidos um indivíduo de cada espécie, aleatoriamente dentro dos
transectos, e coletados dados de atributos funcionais morfológicos (densidade específica
da madeira e área foliar específica) e atributos funcionais fisiológicos (concentração de
micronutrientes nas folhas – Cu, Fe, Mn2+
, Zn+), altura e DAP. Os indivíduos do
levantamento foram identificados ou morfotipados em campo, com o auxílio de um
parataxônomo, e confirmados posteriormente, nível de espécie ou família, no Herbário
do INPA – Instituto Nacional de Pesquisas da Amazônia (Manaus – AM, Brasil) e os
espécimes férteis do levantamento florístico foram depositadas no Herbário do INPA.
Atributos funcionais
Com o auxílio de um scanner e do software Image J foi medida a área foliar de
10 folhas adultas, novas, expostas ao sol e totalmente estendidas, para cada indivíduo
escolhido de forma aleatória. As folhas foram armazenadas em sacos de papel e
secadas a 70°C por 72 horas, no laboratório de Ecofisiologia de Plantas do projeto
INPA/Max Planck, (Manaus - AM, Brasil), e pesadas após esse período para
determinação do peso seco. A razão área e peso seco determinaram a área foliar
específica (AFE) de cada indivíduo (Perez-Harguindeguy 2013). As amostras de

20
madeira (diâmetro interno de 5.15 mm, Haglöf Inc., Suécia) de 5 cm de comprimento
foram retiradas de diferentes secções da madeira, nas posições correspondentes ao DAP,
para cálculo da densidade específica da madeira. As amostras foram mantidas em
condições de umidade constante e medidas através do método de deslocamento de água,
permitindo uma medição confiável para amostras de forma irregular. No caso de
indivíduos arbustivos que não possuíam DAP suficiente para serem brocados, as
amostras retiradas correspondem a um corte transversal no caule central do indivíduo,
tendo a casca retirada. Com o auxílio de uma agulha fina e uma balança eletrônica de
precisão 0.01g, a amostra foi completamente submersa em um recipiente com água e o
peso da água deslocada corresponde ao volume da amostra (Chave et al. 2005). A
densidade da cada amostra foi calculada, após 72 horas de secagem em estufa (Heraeus
e Memmert) a temperatura de 105°C, como a razão entre o peso seco e o volume úmido
(Fearnside 1997; Nogueira et al. 2005). Também foram coletadas folhas para análise da
concentração de micronutrientes (Cu, Fe, Mn2+
, Zn+), realizados no Laboratório de
Análises de Solos e Plantas (LASP) da Embrapa Amazônia Ocidental.
Análise dos dados
As doze amostras de solo foram ordenadas segundo as 17 variáveis físico-
químicas analisadas pela EMBRAPA, pela Análise de Componentes Principais (ACP)
para caracterização máxima da variação do conjunto de dados, com o intuito de
descrever as duas áreas de estudo segundo suas fisionomias (campina e campinarana).
As distâncias florísticas entre os transectos foram analisados através do índice de
similaridade Bray-Curtis, indicando a probabilidade de amostrar ao acaso um táxon em
comum entre as duas áreas, na campina e na campinarana, comparando a fisionomia das
duas áreas com base na mínima abundância de cada espécie (Legendre & Legendre
1998). O índice varia de 0, quando não há coincidência entre as espécies / morfotipos, a
1, quando há total coincidência (Magurran 2004). Para os dados de atributos funcionais
morfológicos (DAP, altura, AFE, densidade específica da madeira) foi realizada uma
ACP para ordenação das quatro variáveis analisadas, posteriormente ANOVA (Análise
de Variância) em cada uma das variáveis para indicar diferenças significativas entre os
habitats de campina e campinarana (arbustivo e arbóreo). Para os dados de atributos
funcionais fisiológicos (concentração de micronutrientes nas folhas: Cu, Fe, Mn2+
, Zn+),
foi feita ANOVA para cada uma das quatro variáveis analisadas. Posteriormente esses

21
dados amostrados foram plotados em gráficos de dispersão para visualização das
diferenças entre as campinas e campinaranas (arbustivo e arbóreo). Os dados foram
analisados no programa R (The R Project for Statistical Computing).
Resultados
Composição do solo
A Análise de Componentes Principais (ACP) em seus dois primeiros eixos
sumarizaram as 17 variáveis físico-químicas do solo nas duas fisionomias, do PARNA
Jaú e da RDS Uatumã, e explicam, respectivamente, 63% e 31% da variação relativa
aos componentes nas duas áreas. As variáveis químicas e físicas do solo das duas áreas
são descritas nas Tabelas 6 e 7, onde é possível observar maiores valores de K
(potássio) e Na (sódio) na campina no PARNA Jaú, e Fe (ferro) e argila na campinarana
na RDS Uatumã, sendo esses valores os responsáveis pelos pontos com valores atípicos
na ACP (Figura 2). É possível observar na Figura 2 a formação de dois grupos distintos,
separando os pontos de coleta do PARNA Jaú da RDS Uatumã, porém não é obervado a
formação de grupos distintos segundo as fisionomias de campina e campinarana, nas
duas áreas.
Composição florística
Um total de 2358 indivíduos, com DAP > 0.5 cm, foram levantados nos 20
transectos de 50 x 2 metros, na campina e campinarana no PARNA Jaú. A campina
apresentou 836 indivíduos e a campinarana 1522 indivíduos. No levantamento
obtivemos 63 espécies ou morfotipos de 33 famílias botânicas. As dez famílias mais
abundantes correspondem a 60.09% dos indivíduos levantados (Tabela 1). As dez
espécies mais abundantes da área correspondem a 55.79% dos indivíduos do
levantamento (Tabela 2). Já na RDS Uatumã o total de indivíduos foi de 3016 (DAP >
0.5 cm), nas duas fisionomias, sendo 1779 indivíduos da campina e 1237 indivíduos da
campinarana. No levantamento obtivemos 73 espécies ou morfotipos de 31 famílias
botânicas. As dez famílias mais abundantes na área correspondem a 79.52% dos
indivíduos do levantamento (Tabela 1). As dez espécies mais abundantes correspondem
a 60.48% dos indivíduos do levantamento (Tabela 2).
A similaridade florística entre a campina e entre a campinarana, das duas áreas,
pode ser observada na Tabela 3. Das 10 famílias mais abundantes no PARNA Jaú,
somente 4 se encontram entre as 10 famílias mais abundantes na RDS Uatumã. As sete

22
espécies ou gêneros semelhantes no levantamento na campina correspondem a 38.04%
dos indivíduos no PARNA Jaú e a 49.52% dos indivíduos na RDS Uatumã (Tabela 4).
As dez espécies ou gêneros semelhantes no levantamento na campinarana correspondem
a 30.65% dos indivíduos no PARNA Jaú e a 11.48% dos indivíduos na RDS Uatumã
(Tabela 5).
Diversidade funcional
Um total de 118 indivíduos foram escolhidos ao acaso para coleta de atributos
funcionais dominantes, sendo esses indivíduos representados no PARNA Jaú por 43
espécies ou morfotipos de 30 famílias botânicas, essas espécies correspondendo a
86.05% dos indivíduos no levantamento da área, e na RDS Uatumã por 26 espécies ou
morfotipos de 15 famílias botânicas, essas espécies correspondendo a 80.24% dos
indivíduos levantados na área. Os indivíduos selecionados para coleta de atributos
funcionais na campina, do PARNA Jaú e da RDS Uatumã, são representados por 31
espécies ou morfotipos de 22 famílias botânicas, e na campinarana, também em ambas
as áreas, são representados por 43 espécies ou morfotipos de 24 famílias botânicas,
diferenciados entre indivíduos arbóreos e arbustivos (Tabela 8).
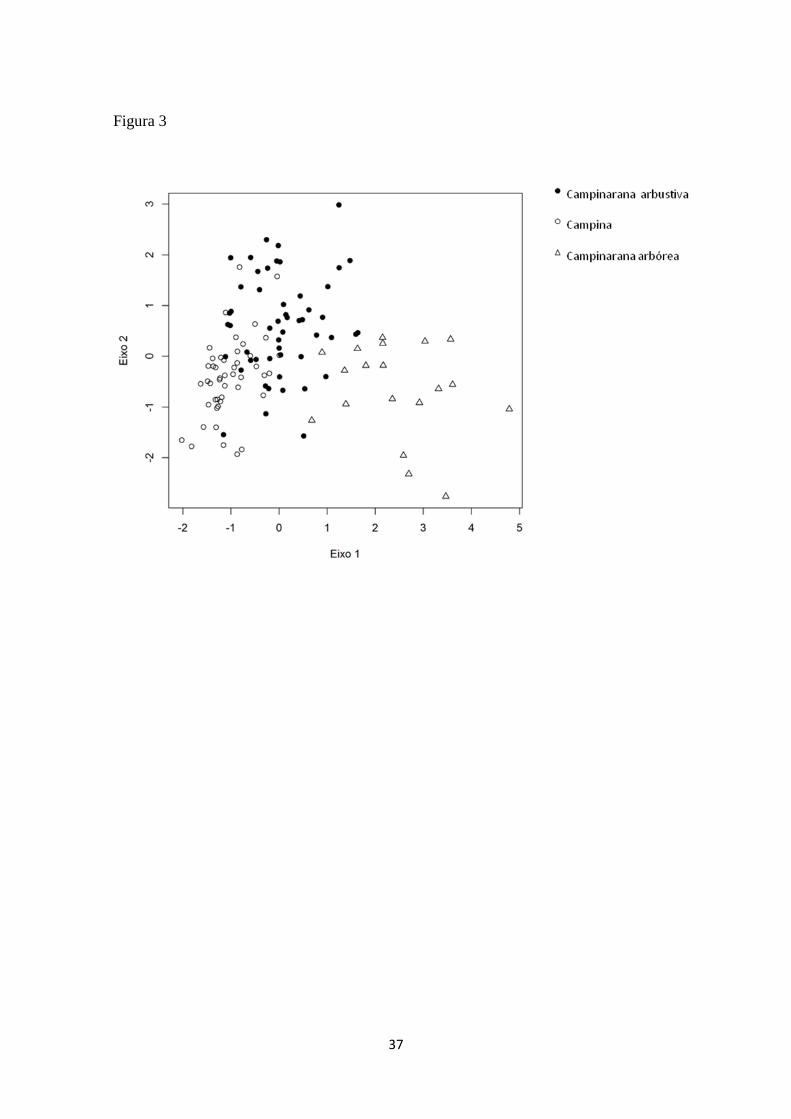
Os atributos funcionais, medidos em 118 indivíduos, segundo os parâmetros
morfológicos da vegetação (DAP, altura, AFE, densidade específica da madeira), pela
ANOVA apresentaram diferenças significativas nos seus valores de DAP, altura e AFE,
podendo assim agrupar os indivíduos segundo a sua distribuição entre as fisionomias de
campina e campinarana (arbórea e arbustiva), em ambas as áreas de estudo. Somente os
valores de densidade específica da madeira não apresentaram diferenças significativas
entre as fisionomias, não podendo agrupar os indivíduos das duas áreas (Tabela 9). O
agrupamento pode ser visualizado pela ACP em seus dois primeiros eixos, e sumarizam
as quatro variáveis morfológicas, explicando, respectivamente, 47% e 27% da variação
(Figura 3).
Os atributos funcionais, medidos em 64 indivíduos, segundo os parâmetros
fisiológicos da vegetação (concentração de micronutrientes nas folhas: Cu, Fe, Mn2+
,
Zn+), pela ANOVA também apresentaram diferenças significativas nos valores de
concentração de Cu, Fe e Mn2+
, permitindo o agrupamento dos indivíduos segundo a
sua distribuição entre as fisionomias, nas duas áreas de estudo. Somente os valores de
concentração de Zn+ não apresentaram diferenças significativas entre as fisionomias,

23
não podendo agrupar os indivíduos nas duas áreas (Tabela 9 e Figura 4). A ACP em
seus dois primeiros eixos sumarizaram as quatro variáveis fisiológicas, explicando,
respectivamente, 38% e 28% da variação.
Discussão
Composição do solo
Nas análises químicas do solo é possível caracterizá-lo como pobre
nutricionalmente e ácido, em ambas as áreas, pois a concentração hidrogeniônica das
fisionomias determina sua nutrição e distribuição, e também por apresentarem soma de
bases inferior a 1 cmol c /kg, pobreza na concentração de micronutrientes e capacidade
de reter cátions restrita à matéria orgânica pois são solos essencialmente arenosos
(Bravard & Righi 1990; Zech et al. 1997; Oliveira 2011).
O solo é descrito como importante na estruturação fisionômica de vegetação
inundável sobre areia branca (Damasco et al. 2013) , é um dos fatores determinantes dos
contrastes entre a vegetação de campinarana e as floretas de terra-firme circundantes
(Silveira 2003). Porém a variação das espécies ao longo do gradiente entre as
fisionomias se deu de acordo com as adaptações morfológicas e fisiológicas dos
indivíduos, permitindo que elas se desenvolvam e obtenham vantagens competitivas
sobre determinados fatores ambientais (Kubitzki 1987; Daly & Mitchell 2000; Silveira
2003).
As diferenças entre campina e campinarana já foram atribuídas aos teores de P e
K no solo (Vale 2011), porém nesse estudo a variação de K e P não se deu ao longo do
gradiente edáfico, mas sim entre os ambientes do PARNA Jaú e RDS Uatumã (Tabela
6).
Composição florística
Segundo Ferreira (2009), estudando campinaranas na Amazônia Central, as
famílias com maior dominância relativa são Humiriaceae, Rubiaceae, Myrtaceae,
Malpighiaceae e Clusiaceae, respectivamente, e no presente trabalho foram encontrada
com maior dominância relativa as famílias Malvaceae, Moraceae, Malpighiaceae,
Chrysobalanaceae e Myrtaceae no PARNA Jaú e Melastomataceae, Sapotaceae,
Chrysobalanaceae, Rubiaceae e Asteraceae na RDS Uatumã (Tabela 1) . Dentro do
levantamento ao longo da vegetação sobre areia branca, ocorre uma sobreposição de
espécies e esse padrão pode ser observado em áreas adjacentes ao longo do habitat (Fine

24
et al. 2010). Essa predominância de espécies é possivelmente o fator determinante da
baixa diversidade de espécies dentro dos levantamentos das duas áreas (Tabela 2). A
areia branca oferece um ambiente menos favorável para plantas não adaptadas à
escassez de nutrientes minerais e variação sazonal entre inundação e drenagem
excessiva do solo (Schubart 1975), e o número de espécies capazes de suportar os
fatores limitantes nestes substratos, é consideravelmente menor do que nos outros tipos
de solos Amazônicos, podendo ser muitas delas endêmicas à este tipo de solo (Vicentini
2004). Essas pequenas ilhas de areia branca dependem das espécies com potencial de
dispersão para longas distâncias, como pelo vento ou por pássaros, devida a sua
natureza isolada (Macedo 1975).
Devido às condições ambientais restritas nas campinas, há um pequeno número
de famílias registradas no levantamento, permitindo que cada família ganhe maior
dominância. O mesmo padrão é observado na campinarana nas duas áreas, porém ocorre
uma maior abundância de espécies nessas fisionomias (Tabela 10). Nas campinas, a
menor abundância de espécies pode ser dada por estarem sujeitas à maior período de
alagamento, devido a influencia da flutuação do lençol freático sobre o solo arenoso,
sem a presença de uma espessa camada de serrapilheira (Nascimento et al. 1997), o que
permite a rápida percolação da água subjacente. Essas poucas espécies que são
tolerantes aos longos períodos de alagamento se tornam dominantes, pois possuem
maior capacidade competitiva, obtendo maior sucesso na colonização dessas áreas
(Targhetta 2012).
Diversidade funcional
Dos atributos funcionais morfológicos (DAP, altura, AFE e densidade específica
da madeira), a elevada densidade específica da madeira (valor médio 0.68) apresentada
pelos indivíduos amostrados, parece indicar uma importante adaptação em ambientes de
pobreza nutricional, onde há pouca energia disponível para eventual rebrota ou
substituição de tecido danificado. O investimento em densidade específica da madeira,
assim como as baixas taxas fotossintéticas nos máximos dos períodos de seca e
alagamento, implicam em baixas taxas de crescimento (Worbes et al. 1992).
Os valores de densidade da madeira não diferiram nas fisionomias de campina e
campinarana. Isso demonstra que eventos extremos, no caso das campinas sujeitas a
maior penetração de luz no sub-bosque denso de arbustos (Anderson 1981) e saturação

25
hídrico do solo arenoso, pela flutuação do lençol freático, não interferem na forma de
alocação de recursos para esse atributo na vegetação, independente do seu habito de
vida (arbórea ou arbusto), como no caso da transição gradativa, ou às vezes abrupta,
entre fisionomias florestada para arbustiva.
A área foliar específica (AFE) é frequentemente usada em análises de
crescimento, por estar muitas vezes relacionada positivamente a taxa de crescimento
relativo das espécies (Perez-Hanguindeguy 2013). Em geral, as espécies
permanentemente ou temporariamente situadas em ambientes ricos em nutrientes,
tendem a possuir uma maior área foliar específica, quando comparada com espécies que
ocupam ambientes com recursos limitados (Perez-Hanguindeguy 2013). Os indivíduos
da campinarana apresentaram maior AFE, constituindo um investimento em folhas
“baratas” (Petchey & Gaston 2006) com curta duração e que cresçam rapidamente, pré-
condicionando recursos e competindo com outros indivíduos no ambiente. Já os
indivíduos da campina apresentaram uma baixa AFE, trocam crescimento rápido por
persistência, investindo em um rápido retorno, mas com segurança nos investimentos.
Essas espécies investem em folhas duras e bem protegidas, de maior penetração de luz
através do dossel, com baixos teores de nutrientes que podem diminuir herbivoria
(Coley 1983) e riscos físicos, levando a uma maior longevidade foliar e maior
persistência da planta (Poorter & Borgers 2006).
A diferença da altura dos indivíduos entre as fisionomias de campina e
campinarana, nas duas áreas de estudo, provavelmente se deve à predominância do
hábito de vida arbustivo na campina, principalmente devido às condições de radiação
extrema e flutuação do lençol freático no qual se desenvolve, fazendo com que haja
maior investimento em folhas em comparação ao caule, resultando em indivíduos de
baixa estatura (Foto 1) (Coomes & Grubb 1996). Isto se deve, em grande parte, ao fato
de espécies que habitam solos oligotróficos alocarem elevado conteúdo de seus recursos
na defesa contra herbivoria, em detrimento de seu crescimento (Fine et al. 2004; Fine et
al. 2006). Também pode ser dada pela diferença na profundidade da camada de solo
arejada acima do lençol freático (Bongers et al. 1985) caracterizado por uma camada de
15 a 20 cm de espessura formada por uma fina rede de raízes (Worbes 1997; Silveira
2003) e pela presença de uma espessa camada de serrapilheira na campinarana (Foto 2)
(Nascimento et al. 1997).

26
Dos atributos funcionais fisiológicos (concentração de micronutrientes nas
folhas: Cu, Fe, Mn2+
, Zn+), os elementos Fe, Mn
2+ e Zn
+ apresentam valores dentro dos
limites necessários dentro da planta e somente o elemento Cu apresenta valores abaixo
dos limites necessários na planta, caracterizando seu estado nutricional como deficiente
(Larcher 2000). Não há diferenciação do elemento Zn entre as campinas e campinaranas
(Tabela 5), isso pode se dever ao fato do elemento constituir um co-fator funcional,
estrutural e regulador de enzimas (Baker et al. 1982;. Clarkson & Hanson, 1980;
Marschner 1995; Tripathy & Mohanty 1980), podendo atuar e diversas funções
fisiológicos nos indivíduos. Já os elementos Cu, Mn+2
e Fe diferem nas campinas e
campinaranas (Tabela 5). Solos caracterizados como ácidos liberam íons Fe e Mn+2
em
excesso (Larcher 2000). Cu desempenha um papel na biossíntese de clorofila,
respiração, metabolismo de proteínas e carboidratos, lignificação das paredes celulares e
balanço hídrico (Clarkson & Hanson 1980; Droppa & Horvath 1990). Mn+2
atua
diretamente na fotossíntese e indiretamente na formação de carboidratos, os cloroplastos
são as organelas mais sensíveis ao micronutriente (Kirkby & Römheld 2007). A
interação de Zn-Fe é antagônica, porque o excesso de Zn ou Fe pode causar redução na
absorção de Fe ou Zn, respectivamente. Este efeito mútuo de Zn e Fe aumenta a
possibilidade da sua deficiência no crescimento das plantas (Kausar et al. 1976). Não
foi encontrada uma relação antagônica entre os micronutrientes, uma vez que o Zn não
apresentou variação significativa entre os habitats.
É comum ocorrer uma transição gradativa, ou às vezes abrupta, entre as
fisionomias de campina, campinarana e floresta de terra firme (Silveira 2003). Estas
transições são caracterizadas pela mudança na composição e estrutura da vegetação e
comumente são determinadas por variações edáficas, como alterações de textura e
fertilidade no solo (Worbes 1997; Silveira 2003; Mendonça 2011; Vale 2011). Porém
no presente trabalho, provavelmente o nível do lençol freático e tempo de alagamento
ocasionado pela elevação do lençol freático influenciando as mudanças nos padrões
florísticos e estruturais da vegetação (e.g. Coomes 1997; Daly & Mitchell 2000; Silveira
2003; Vicentini 2004; Junk et al. 2011).
Conclusão
Através dessa combinação de eventos abióticos é possível observar que uma
diversidade multivariada de atributos difere entre as fisionomias de campina e

27
campinarana, e é reduzida em habitats mais extremos, como a campina, potencialmente
porque a gama de estratégias ecológicas bem sucedidas é reduzido em habitats em
condições severas (maior penetração de luz no sub-bosque, saturação hídrica do solo).
Assim, apesar da estrutura fisionômica da vegetação ser obvia, mesmo retirando as
variações autoexplicativas dos atributos funcionais visualmente aparentes (DAP, altura),
ainda se encontra um padrão estrutural na vegetação (área foliar específica,
concentração de micronutriente nas folhas). Podendo ser devido ao nível do lençol
freático e pelo tempo de alagamento, que poderiam corresponder às mudanças nos
padrões florísticos e estruturais da vegetação, uma vez que as variáveis físico-químicas
do solo, nas duas áreas de estudo, não responderam as variações fisionômicas na
vegetação.
Bibliografia citada
Adeney, J.M.; Christensen, N.L.Jr.; Pimm, S.L. 2009. Reserves protect against
deforestation fires in the Amazon. PLoS ONE, 4(4):5014.
Anderson, A.B. 1981. White-sand vegetation of Brazilian Amazonia. Biotropica,
13:199-210.
Baker, N.R.; Fernyhough, P.; Meek, I.T. 1982. Light-dependent inhibition of
photosynthetic electron transport by zinc. Physiol. Plant, 56:217-222.
Baker, T.R.; Phillips, O.L.; Malhi, Y.; Almeida, S.; Arroyo, L.; Di Fiore, A.; Erwin, T.;
Higuchi, N.; Killeen, T.J.; et al. 2004. Increasing biomass in Amazonian forest plots.
Philosophical Transactions of the Royal Society London B: Biological Sciences, 359:
353 – 365.
Bleackley, D.; Khan, E.J.A. 1963. Observations on the white-sand areas of the Berbice
formation, British Guiana. Journal of Soil Science, 14(1):44-51.

28
Bongers, F.; Engelen, D.; Klinge, H. 1985. Phytomass structure of natural plant
communities on spodosols in southern Venezuela - the bana woodland. Vegetatio,
63:13–34
Chave, J.; Andalo, C.; Brown, S.; Cairns, M.A.; Chambers, J.Q.; Eamus, D.; Fölster, H.;
Fromard, F.; Higuchi, N.; Kira, T.; Lescure, J.P.; Nelson, B.W.; Ogawa, H.; Puig, H.;
Riera, B.; Yamakura, T. 2005. Tree allometry and improved estimation of carbon stocks
and balance in tropical forests. Oecologia, 145:87-99.
Clarkson, D.I.; Hanson, J.B. 1980. The mineral nutrition of higher plants. Annu. Rev.
Plant Physiol, 31:239-298.
Coley, P.D. 1983. Herbivory and defensive characteristics of tree species in a lowland
tropical forest. Ecological Monographs, 53:209-233.
Cornwell, W.K.; Schwilk, D.W.; Ackerly, D.D. 2006. A trait-based for habitat filtering:
convex hull volume. Ecology, 87:1465-1471.
Coomes, D. A.; Grubb, P. J. 1996. Amazonian caatinga and related communities at La
Esmeralda, Venezuela: forest structure, physiognomy and floristics, and control by soil
factors. Vegetatio, 122:167–191.
Coomes, D. A. 1997. Nutrient status of Amazonian caatinga forests in a seasonally dry
area: nutrient fluxes, litterfall and analyses of soils. Canadian Journal of Forest
Research, 27:831–839.
Daly, D.C.; Mitchaell, J.D. 2000. Lowland vegetation of Tropical South America – An
Overview. In: Lentz, D. (Ed.). Imperfect Balance: Landscape transformations in the pre-
Columbian Americas. Columbia University Press, p.391-454.

29
Damasco, G.; Vicentini, A.; Castilho, C.V.; Pimentel, T.P.; Nascimento, H.E.M. 2004.
Disentangling the role of edaphic variability, flooding regime and topography of
Amazonian white-sand vegetation. Journal of Vegetation Science, 24:384.394.
Droppa, M.; Horvath, G. 1990. The role of copper in photosynthesis. Crit. Rev. Plant
Sci., 9:111-123.
Fearnside, P.M. 1997. Wood density for estimating forest biomass in Brazilian
Amazonia. Forest Ecology and Management, 90:59–87.
Ferreira, A.C.A. 2009. Análise comparativa de vegetação lenhosa do ecossistema
campina na Amazônia Brasileira. Tese de Doutorado, Universidade Federal do
Amazonas, Manaus, Amazonas. 277p.
Ferreira, L.V.; Stohlgren, T.J. 1999. Effects of river level fluctuation on plant species
richness, diversity and distribution in a floodplain forest in Central Amazonia.
Oecologia, 120: 582-587.
Fine, P. V. A.; Mesones I.; Coley, P.D. 2004. Herbivores promote habitat specialization
by trees in Amazonian forests. Science, 305:663–665.
Fine, P.V.A.; Miller, Z.J.; Mesones I., Irazuzta S.; Appel H.M.; Stevens I.; Saaksjarvi I.;
Schultz J.C.; Coley, P.D. 2006. The growth–defense trade-off and habitat specialization
by plants in Amazonian forests. Ecology, 87:150–162.
Fine, P.V.A.; García-Villacorta, R.; Pitman, N.C.A.; Mesones, I.; Kembel, S.W. 2010.
A floristic study of the white-sand forest of Peru. Ann. Missouri Botanical Garden, 97:
283–305.
Franco, W.; Dezzeo, N. 1994. Soils and soil water regime in the terra firme-caatinga
forest complex near San Carlos de Rio Negro, state of Amazonas, Venezuela.
Interciencia 19(6):305–316.

30
IDESAM. 2009. Plano de Gestão da Reserva de Desenvolvimento Sustentável do
Uatumã. Versão para consulta pública. Instituto de Conservação e Desenvolvimento
Sustentável do Amazonas. Manaus, AM, Brasil.
Junk, W.J.; Piedade, M.T.F.; Schöngart, J.; Cohn-Haft, M.; Adeney, J.M.; Wittmann,F.
2011. A Classification of Major Naturally-Occurring Amazonian Lowland Wetlands.
Wetlands, 31:623-640.
Kausar, M. A.; Chaudhary, F. M.; Rashid, A.; Latif, A., Alam, S. M. 1976.
Micronutrient availability to cereals from calcareous soils. I. Comparative Zn and Cu
deficiency and their mutual interaction in rice and wheat. Plant Soil, 45:397–410..
Keddy, P.A. 1992. Assembly and response rules: two goals for predictive community
ecology. Jounal Vegetable Sciene, 3:157-164.
Kubitzki, K. 1987. The ecogeographical differentiation of Amazon inundation forests.
Plant Systematics and Evolution, 162:285–304.
Kirkby, E.A.; Römheld, V. 2007. Micronutriente na fisiologia de plantas: funções,
absorção e mobilidade. International Plant Nutrition Institute. Informações
Agronômicas, 118: 1-24.
Larcher, W. Ecofisiologia vegetal. São Carlos: Rima Artes e Textos, 2000. 531p.
Legendre, P. & Legendre, L. 1998. Numerical Ecology. Elsevier Science, B.V.,
Amsterdam.
Luizão, R.C.C.; Luizão, F.; Proctor, J. 2007. Fine root growth and nutrient release in
decomposing leaf litter in three contrasting vegetation types in central Amazonia. Plant
Ecol, 192:225-236.

31
Macedo, M. 1975. Dispersão de plantas lenhosas de uma campina Amazônica.
M.S.Thesis 1975.
Magurram, A.E. 2004. Measuring biological diversity. Blackwell, UK, 255p.
Marschner, H. 1995. Mineral Nutrition of Higher Plants. Academic Press, London.
Tripathy, B.C.; Mohanty, P. 1980. Zinc-inhibited electron transport of photosynthesis
in isolated barley chloroplasts. Plant Physiol, 66: 1174-1178
Mendonça, A.F.M. 2011. Campinaranas Amazônicas: pedogênese e relações solo-
vegetação. Tese de Doutorado, Universidade Federal de Viçosa, Viçosa, Minas Gerais.
110p.
Nascimento, N. R., R. Boulet, M. Lamotte e P. Magat. 1997. Relatório de campo da
equipe de solos em expedição à Campina do Patauá. Fundação Vitória Amazônica,
Manaus.
Nogueira, E. M.; Nelson, B.W.; Fearnside, P.M. 2005. Wood density in dense forest in
central Amazonia, Brazil. Forest Ecology and Management, 206:261–286.
Oliveira, J.B. Pedologia Aplicada. 4 ed. Piracicaba: Fealq, 2011. 592p.
Perez-Harguindeguy, N.; Díaz, S.; Garnier, E.; Lavorel, S.; Pooter, H.; Jaureguiberry,
P.; Bret-Harte, M.S.; Cornwell, W.K.; Craine, J.M.; Gurvich, D.E.; Urcelay, C.;
Veneklaas, E.J.; Reich, P.B.; Pooter, L.; Wright, I.J.; Ray, P. ; Enrico, L.; Pausas, J.G.;
de Vos, A.C.; Buchmann, N.; Funes, G.; Quetier, F.; Hodgson, J.G.; Thopson, K.;
Morgan, H.D.; ter Steege, H.; van der Heijden, M.G.A.; Sack, L.; Blonder, B.;
Poschilod, P.; Vaieretti, M.V.; Conti, G.; Staver, A.C.; Aquino, S.; Cornelissen, J.H.C.
2013. New handbook for standardised measurement of plant functional traits
worldwide. Australian Journal of Botany, 61:167-234.

32
Petchey, O.L.; Gaston, K.J. 2006. Functional diversity: back to basis and looking
forward. Ecology Letters, 9:741-758.
Pooter, L.; Bongers, F. 2006. Leaf traits are good predictors of plan performance across
53 rain forest species. Ecology, 87(7):1733–1743.
Stropp, J.; Van Der Sleen, P.; Assunção, P.A.; Silva, A.L.; Ter Steege, H. 2011. Tree
communities of white-sand and terra-firme forests of the upper Rio Negro. Acta
Amazônica, 41(4):521-544.
Silveira, M. 2003. Vegetação e Flora das Campinaranas do Sudoeste Amazônico (JU-
008). Relatório de Defesa Técnica, Associação S.O.S. Amazônia, Rio Branco. 28p.
Targhetta, N. 2012. Comparação florística e estrutural entre florestas de igapó e
campinarana ao longo de gradientes hidro-edáficos na Reserva de Desenvolvimento
Sustentável do Uatumã, Amazônia Central. Dissertação de Mestrado, Instituto Nacional
de Pesquisas da Amazônia, Manaus, Amazonas. 105p.
Tripathy, B.C.; Mohanty, P. 1980. Zinc-inhibited electron transport of photosynthesis in
isolated Barley chloroplasts. Plant Physiol, 66:1174-1178.
Vale, G.D. 2011. O efeito do solo, inundação e topografia sobre as campinaranas
inundáveis na Amazônia. Dissertação de Mestrado, Instituto de Pesquisas da Amazônia,
Manaus, Amazonas. 84p.
van der Valk, A.G. 1981. Succession in wetlands—a Gleasonian approach. Ecology,
62:688–696.
Vicentini, A. 2004. A Vegetação ao Longo de um Gradiente Edáfico no Parque
Nacional do Jaú. In: BORGES, S.H.; IWANAGA, S.; DURIGAN, C.C.; PINHEIRO,
M.R. (Eds.). Janelas para a Biodiversidade no Parque Nacional do Jaú: uma estratégia

33
para o estudo da biodiversidade na Amazônia. Fundação Vitória Amazônica
(FVA)/WWF/IBAMA, Manaus, Brasil, p.117-143.
Weiher, E.; Clarke, G.D.P.; Keddy, P.A. 1998. Community assembly rules,
morphological dispersion, and the coexistence of plant species. Oikos, 81:309–322.
Weiher, E.; Keddy, P. 1999. Ecological assembly rules: perspectives, advances,
retreats. Cambridge University Press, Cambridge, UK.
Worbes, M.; Klinge, H.; Revilla, J.D.; Martius, C. 1992. On the dynamics, floristic
subdivision and geographical distribution of várzea forests in Central Amazonia. J. Veg.
Science, 3: 553-564.
Worbes, M. 1997. The Forest Ecossystem of the Floodplains. In: JUNK, W.J. The
Central Amazon Floodplain: Ecology of a Pulsing System. Ecological Studies, vol. 126.
Springer, Berlim/Heidelberg/New York, p. 223-265.
Wright, I.J.; Reich, P.B,; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; et al.
2004. The worldwide leaf economics spectrum. Nature, 428:822-828.
Zech W, Senesi N, Guggenberger G, Kaiser K, Lehmann J, Miano T M, Miltner A,
Schroth G. 1997. Factors controlling humification and mineralization of soil organic
matter in the tropics. Geoderma, 79:117–161.

34
Legenda de figuras e tabelas
Figura 1: Mapa do Estado do Amazonas, em destque as aréas de estudo, o Parque
Nacional do Jaú (PARNA Jaú) e a Reserva de Desenvolvimento Sustentável (RDS
Uatumã).
Figura 2: ACP com os principais eixos das variáveis físico-químicas do solo no PARNA
Jaú e na RDS Uatumã.
Figura 3: ACP com os principais eixos dos atributos funcionais morfológicos (DAP.
Altura, AFE, densidade específica da madeira) entre a campina e campinarana
(arbustivo e arbóreo), em ambas as áreas.
Figura 4: Grágico de dispersão dos atributos funcionais fisiológicos (concentração de
micronutrientes nas folhas: Cu, Fe, Mn+2
, Zn+) entre a campina e campinarana
(arbustivo e arbóreo), em ambas as áreas.
Tabela 1: As dez famílias mais abundantes dentro do levantamento florístico na
campina e campinarana no PARNA Jaú e na RDS Uatumã.
Tabela 2: As dez espécies mais abundantes dentro do levantamento florístico na
campina e campinarana no PARNA Jaú e na RDS Uatumã.
Tabela 3: Índice de Similaridade Bray-Curtis para a composição florística entre as
campinas e campinaranas no PARNA Jaú e na RDS Uatumã.
Tabela 4: As sete espécies ou gêneros semelhantes na campina no PARNA Jaú e na
RDS Uatumã e a sua porcentagem de ocorrência no levantamento florístico.
Tabela 5: As dez espécies ou gêneros semelhantes na campinarana no PARNA Jaú e na
RDS Uatumã e a sua porcentagem de ocorrência no levantamento florístico.
Tabela 6: Variáveis químicas do solo no PARNA Jaú e RDS Uatumã.
Tabela 7: Variáveis físicas do solo no PARNA Jaú e RDS Uatumã.
Tabela 8: Espécies ou morfotipos selecionados para coleta de atributos funcionais na
campina e campinarana (arbórea e arbustiva), no PARNA Jaú e RDS Uatumã.
Tabela 9: Atributos funcionais medidos na campina e campinarana (arbórea e
arbustiva), no PARNA Jaú e RDS Uatumã.
Tabela 10: Famílias e espécies mais abundantes nos levantamentos na campina e
campinarana, no PARNA Jaú e na RDS Uatumã.
*a,b,c,d
: mesma letra representa similaridade significativa entre os valores.
*não foram excluídas as famílias/espécies/gêneros raros: < 0.10% de indivíduos nos levantamentos.
35
Figuras
Figura 1

36
Figura 2
37
Figura 3

38
Figura 4

39
Tabela 1
RDS Uatumã PARNA Jaú
Famílias % Famílias %
Melastomataceae 15.48 Malvaceae 12.64
Sapotaceae 13.79 Moraceae 7.76
Chrysobalanaceae 13.03 Malpighiaceae 7.12
Rubiaceae 10.94 Chrysobalanaceae 5.81
Asteraceae 8.42 Myrtaceae 5.30
Fabaceae 4.24 Asteraceae 4.71
Theaceae 4.14 Myrsinaceae 4.66
Ochnaceae 3.28 Burseraceae 4.50
Clusiaceae 3,18 Fabaceae 4.20
Rhabdodendraceae 3.02 Clusiaceae 3.39

40
Tabela 2 PARNA Jaú RDS Uatumã
Famílias Espécies % Famílias Espécies %
Malvaceae Pachira sordida (R.E.Schult.) W.S.Alverson
10.73 Melastomataceae Macairea theresiae Cogn.
11.31
Malphiguiaceae Byrsonimia eugeniifolia Sandwith.
7.12 Sapotaceae Pradosia schomburgkiana (A. DC.) Cronquist
9.15
Sapotaceae Elaeoluma schomburgkiana (Miq.) Baill.
6.28 Asteraceae Gongylolepis martiana (Baker) Steyrm. &
Cuatrec.
8.42
Moraceae Brosimum acutifolium Huber
6.15 Chrysobalanaceae Licania hypoleuca Benth.
8.09
Chrysobalanaceae Licania hypoleuca Benth.
5.81 Rubiaceae Alibertia rigida K.Schum.
6.20
Asteraceae Gongylolepis martiana (Baker) Steyrm. &
Cuatrec.
4.71 Theaceae Ternstroemia verticillata Klotzsch ex Wawra
4.14
Myrsinaceae Cybianthus fulvopulverulentus magnoliifolius
(Mez) Pipoly.
4.66 Sapotaceae Elaeoluma schomburgkiana (Miq.) Baill.
3.55
Myrtaceae Eugenea paniculiflora Steud.
4.58 Ochnaceae Ouratea spruceana Engl.
3.28
Theaceae Ternstroemia dentata (Aubl.) Sw.
2.88 Rubiaceae Pagamea coriacea (Spruce) Benth.
3.18
Ochnaceae Ouratea spruceana Engl. 2.84 Chrysobalanaceae Hirtella ulei Pilg.
3.15

41
Tabela 3 PARNA Jaú RDS Uatumã PARNA Jaú
Campinarana Campinarana Campina
RDS Uatumã Campinarana 0.0927 - -
PARNA Jaú Campina 0.2502 0.0376 -
RDS Uatumã Campina 0.2053 0.0988 0.1208
Tabela 4
Famílias Espécies
PARNA Jaú RDS Uatumã
Asteraceae Gongylolepis martiana (Baker) Steyrm.
& Cuatrec.
14.18 1.20
Chrysobalanecae
Licania hypoleuca Benth.
12.76 1.44
Humiraceae
Humiria balsamifera (Aubl.) J.St.-Hil.
0.22 3.55
Lauraceae
Ocotea Aubl.
0.11 2.87
Myrsinaceae
Cybianthus fulvopulverulentus
magnoliifolius (Mez) Pipoly.
0.84 13.16
Sapotaceae
Elaeoluma schomburgkiana (Miq.)
Baill.
5.96 14.95
Sapotaceae
Pradosia schomburgkiana (A. DC.)
Cronquist
15.35 1.08

42
Tabela 5
Famílias Espécies PARNA Jaú RDS Uatumã
Annonaceae
Xylopia spruceana Benth. Ex Spruce
0.16 1.91
Apocynaceae
Aspidosperma Mart 0.73 0.46
Bursenaceae
Protium sprucianum (Benth.) Engl.
1.37 3.48
Chrysobalanecae
Licania hypoleuca Benth.
1.37 8.21
Clusiaceae
Clusia grandifolia Engl.
0.24 3.39
Clusiaceae
Tovomita Aubl.
0.16 2.43
Myristicaceae
Iryanthera obovata Ducke
2.51 1.71
Ochnaceae
Ouratea spruceana Engl.
4.61 4.40
Sapotaceae
Elaeoluma schomburgkiana (Miq.) Baill.
0.08 1.51
Sapotaceae
Pradosia schomburgkiana (A. DC.)
Cronquist
0.24 3.15

43
Tabela 6
pH
P
K
Na Ca Mg Al H +Al
SB
t T V
m
Fe Zn Mn Cu
H20 mg/
dm3
mg/
dm3
mg/
dm3
cmolc/
dm3
cmolc/
dm3
cmolc/
dm3
cmolc/
dm3
cmolc/
dm3
cmolc/
dm3
cmolc/
dm3
% % mg/
dm3
mg/
dm3
mg/
dm3
mg/
dm3
PARNA Jaú
Campina 4.43
3.67 38.00 36.33 0.12 0.18 1.69 8.09 0.55 2.25 2.25 6.21 76.16 11.67 1.12 0.23 0.08
Campinarana 4.58 5.33 30.33 29.00 0.06 0.14 1.50 5.41 0.40 1.19 1.90 6.91 78.45 12.00 0.99 0.30 0.10
RDS Uatumã
Campina 4.19 2.67 10.33 2.67 0.03 0.07 0.97 2.72 0.14 1.10 1.10 7.19 87.09 8.00 0.50 0.59 0.18
Campinarana 4.45 2.33 11.33 6.67 0.03 0.08 1.19 3.75 0.17 1.35 1.35 4.34 87.56 25.67 0.54 0.44 0.15

44
Tabela 7
Areia Total
(g/kg)
Silte
(g/kg)
Argila
(g/kg)
Classificação
textural do solo
PARNA Jaú
Campina 877.71 105.79 16.50 Areia
Campinarana 884.08 103.09 12.83 Areia
RDS Uatumã
Campina 925.37 65.92 8.83 Areia
Campinarana 898.47 49.36 52.17 Areia

45
Tabela 8
PARNA Jaú
Campina – arbustivos
Família Espécies
Annonaceae Guateria maguirei R.E.Fr.
Aquifoliaceae Ilex L.
Asteraceae Gongylolepis martiana (Baker) Steyrm. & Cuatrec.
Chrysobalanaceae Licania hypoleuca Benth.
Clusiaceae Clusia grandifolia Engl.
Dilleniaceae Doliocrapus Rol.
Fabaceae Dimorphandra vernicosa Spreng. ex Benth.
Macrolobium campestre Huber.
Humiriaceae Humiria balsamifera (Aubl.) J.St.-Hil.
Lauraceae Ocotea Aubl.
Malphiguiaceae Byrsonimia eugeniifolia Sandwith.
Malvaceae Pachira sordida (R.E.Schult.) W.S.Alverson
Myrsinaceae Cybianthus fulvopulverulentus magnoliifolius (Mez) Pipoly.
Myrtaceae Myrcia clusifolia (Kunth) DC.
Ochnaceae Ouratea acuminata (A. DC.) Engl.
Olacaceae Chaunochiton angustifolium Sleumer
Phyllanthaceae Amanoa Aubl.
Rubiaceae Retiniphyllum schomburgkii (Benth.) Müll.Arg.
Sapotaceae Pradosia schomburgkiana (A. DC.) Cronquist
Elaeoluma schomburgkiana (Miq.) Baill.
Theaceae Ternstroemia dentata (Aubl.)Sw.
RDS Uatumã Campina – arbustivos Aquifoliaceae Ilex divaricata Mart. ex Reissek
Asteraceae Gongylolepis martiana (Baker) Steyrm. & Cuatrec.
Chrysobalanaceae Hirtella ulei Pilg.
Licania hypoleuca Benth.
Dilleniaceae Doliocarspus spatulifolius Kubitzki
Icacinaceae Emmotum acuminatum (Benth.) Miers
Lauraceae Ocotea boissieriana (Meisn.) Mez
Melastomataceae Macairea theresiae Cogn.
Ochnaceae Ouratea spruceana Engl.
Rubiaceae Alibertia rigida K.Schum.
Pagamea coriacea (Spruce) Benth.

46
Retiniphyllum schomburgkii (Benth.) Müll.Arg.
Sapotaceae Elaeoluma schomburgkiana (Miq.) Baill.
Pradosia schomburgkiana (A. DC.) Cronquist
Theaceae Ternstroemia verticillata Klotzsch ex Wawra
PARNA Jaú
Campinarana – arbóreos Annonaceae Xylopia spruceana Benth. Ex Spruce
Araliaceae Schefflera spruceana (Seem.) Maguire, Steyerm. & Frodin. Det:
Fiaschi, P.
Burseraceae Protium Burm.f.
Clusiaceae Tovomita Aubl.
Icacinaceae Emmotum nitens (Benth.) Miers
Malpighiaceae Byrsonimia eugeniifolia Sandwith.
Malvaceae Eritheca Schott & Endl.
Myrtaceae Eugenea paniculiflora Steud.
Ochnaceae Ouratea spruceana Engl.
Theaceae Ternstroemia dentata (Aubl.)Sw.
RDS Uatumã
Campinarana – arbóreos
Chrysobalanaceae Licania lata J.F.Macbr.
Licania gracilipes Taub.
Ochnaceae Ouratea spruceana Engl.
Rhabdodendraceae Rhabdodendron amazonicum (Spruce ex Benth.) Huber
Sapindaceae Matayba opaca Radlk.
Sapotaceae Chrysophyllum sanguinolentum (Pierre) Baehni
PARNA Jaú
Campinarana – arbustivos
Annonaceae
Xylopia spruceana Benth. Ex Spruce
Guateria maguirei R.E.Fr.
Apocynaceae Macoubea sprucei (Müll.Arg.) Markgr.
Asteraceae Gongylolepis martiana (Baker) Steyrm. & Cuatrec.
Burseraceae Protium Burm.f. (1)
Chrysobalanaceae Licania hypoleuca Benth.

47
Clusiaceae Clusia grandifolia Engl.
Tovomita Aubl.
Elaeocarpaceae Sloanea L.
Fabaceae Swartzia renticulata Ducke
Ormosia Jacks.
Linaceae Roucheria columbiana Hallier.
Melastomataceae Tococa Aubl.
Moraceae Brosimum acutifolium Huber
Brosimum guianense (Aubl.) Huber
Myristicaceae Iryanthera obovata Ducke
Myrtaceae Eugenea paniculiflora Steud.
Ochnaceae Ouratea spruceana Engl.
Primulaceae Stylogyne A.DC.
Sapindaceae Sapindacea sp.
Sapotaceae Pradosia schomburgkiana (A. DC.) Cronquist
Sapotaceae sp.
Theaceae Ternstroemia dentata (Aubl.)Sw.
RDS Uatumã
Campinarana – arbustivos
Apocynaceae Aspidosperma Mart.
Apocynaceae Lacmellea gracilis (Müll.Arg.) Markgr.
Chrysobalanaceae Licania lata J.F.Macbr.
Fabaceae Ormosia grossa Rudd.
Icacinaceae Emmotum acuminatum (Benth.) Miers
Melastomataceae Miconia argyrophylla DC.
Mouriri nervosa Pilg.
Ochnaceae Ouratea spruceana Engl.
Rhabdodendraceae Rhabdodendron amazonicum (Spruce ex Benth.) Huber
Rubiaceae Alibertia rigida K.Schum.
Duroia saccifera (Schult. & Schult.f.) K.Schum.
Sapindaceae Matayba opaca Radlk.
Sapotaceae Chrysophyllum sanguinolentum (Pierre) Baehni

48
Tabela 8 DAP
(cm)
Altura
(m)
AFE
(mm²/g)
Densidade
(g/cm³)
Cu
(mg/kg)
Fe
(mg/kg)
Mn
(mg/kg)
Zn
(mg/kg)
PARNA Jaú
Campina 0.5 – 5.3ª 1 – 3.5ª 35.390 – 105.160ª 0.44 – 0.86ª 0.84 – 2.84ª 14.47 – 109.67ª 2.9 – 93.49ª 2.49 – 23.53ª
Campinarana arbórea 9.5 – 19.7b 9 – 15
b 36.850 – 100.290
b 0.57 – 0.89ª 2.14 – 3.32
b 38.78 – 80.99
b 27.12 – 416.35
b 8.87 – 16.05ª
Campinarana arbustiva 0.5 – 4.5c 1.2 – 6
c 57.930 – 205.350
c 0.57 – 0.81ª 1.57 – 4.95
b 24.76 – 98.87ª
b 11.77 – 115.86ª
b 6.03 – 49.25ª
RDS Uatumã
Campina 0.5 – 4ª 1.1 – 5ª 42.495 – 90.885ª 0.63 – 0.89ª 1.37 – 6.35ª 45.84 – 90.71ª 5.97 – 96.32ª 3.72 – 13.68ª
Campinarana arbórea 6 – 11.4b 6 – 12
b 61.587 – 107.370
b 0.53 – 0.75ª 1.64 – 3.24
b 54.02 – 249.61
b 10.11 – 278.00
b 6.91 – 14.59ª
Campinarana arbustiva 1 – 6.1c 2.9 – 7.4
c 55.198 – 194.456
c 0.44 – 0.90ª 1.86 – 4.52
b 60.72 – 114.57ª
b 10.98 – 214.64ª
b 6.99 – 13.08ª

49
Tabela 9
PARNA Jaú % RDS Uatumã %
Campina Malvaceae 22.73 Sapotaceae 21.30
Sapotaceae 16.03 Asteraceae 14.28
Malpighiaceae 11.06 Rubiaceae 12.82
22 famílias botânicas,
28 espécies
18 famílias botânicas,
28 espécies
Campinarana Moraceae 12.02 Melastomataceae 37.75
Chrysobalanaceae 8.21 Rhabdodendraceae 7.36
Myrtaceae 7.36 Rubiaceae 8.25
24 famílias botânicas,
41 espécies
26 famílias botânicas,
59 espécies

50
3. Conclusões
A variação na composição de espécies entre as fisionomias pode estar
relacionada a fatores ambientais (Tuomisto & Ruokolainen 1994, Tuomisto et al. 1995).
Fatores edáficos têm sido descritos como importantes no papel da composição e
distribuição das espécies em vegetação inundável sobre areia branca (Ferreira 1991;
Worbes 1997; Damasco et al. 2013), porém a variação das espécies ao longo do
gradiente fisionômica pode ser dada também de acordo com as adaptações morfológicas
e fisiológicas dos indivíduos.
Tantos os atributos funcionais morfológicos (densidade da madeira e área foliar
específica), quanto os atributos funcionais fisiológicos (micronutrientes da folha: Cu,
Fe, Mn+2
, Zn+) apresentam um padrão de distribuição dos seus valores ao longo da
variação fisionômica da vegetação de campina a campinarana, nas duas áreas de estudo
(PARNA Jaú e RDS Uatumã). A diversidade multivariada de atributos difere entre as
fisionomias, provavelmente pelo fato das campinas atuarem como filtros ambientais,
reduzindo a gama de estratégias ecológicas que um atributo funcional ocupa em um
dado ambiente (Grime 2006).
Assim, apesar da estrutura fisionômica da vegetação ser obvia, mesmo retirando
as variações autoexplicativas dos atributos funcionais visualmente aparentes (atributos
morfológicos), ainda se encontra um padrão estrutural na vegetação que pode ser
explicado pelo nível do lençol freático e pelo tempo de alagamento, podendo ele
corresponder às mudanças nos padrões florísticos e estruturais da vegetação, uma vez
que as variáveis físico-químicas do solo, nas duas áreas de estudo, não se diferenciaram
segundo as variações fisionômicas na vegetação.

51
4. ANEXOS
Foto 1: Vegetação arbustiva da campina na RDS Uatumã.
Foto 2: Presença de serrapilheira na campinarana do PARNA Jaú.

52
5. Referências bibliográficas
Adediran, J.A.; Taiwo, L.B.; Akande, M.O.; Idowu, O.J.; Sobulo, R.A. 2004.
Application of organic and inorganic fertilizer for sustainable yield of maize and
cowpea in Nigeria. J. Plant Nut., 27: 1163-1181.
Anderson, A.B; Prance, G.T; Albuquerque, B.W.P. 1975. Estudos sobre as vegetações
de Campinas Amazônica III: a vegetação lenhosa da Campina da Reserva Biológica
INPA— SUFRAMA (Manaus- Caracaraí, km 62). Acta Amazonica 5(3):225 – 246.
Anderson, A.B. 1981. White-sand vegetation of Brazilian Amazonia. Biotropica,
13:199-210.
Bravard, S.; Righi, D. 1990. Podzols in Amazonia. Catena, 17:461-475.
Cornwell, W.K.; Schwilk, D.W.; Ackerly, D.D. 2006. A trait-based for habitat filtering:
convex hull volume. Ecology, 87:1465-1471.
Daly, D.C.; Mitchaell, J.D. 2000. Lowland vegetation of Tropical South America – An
Overview. In: Lentz, D. (Ed.). Imperfect Balance: Landscape transformations in the pre-
Columbian Americas. Columbia University Press, p.391-454.
Díaz, S. et al. 1999. Functional implications of trait-environment linkages in plant
communities. In: The Search for Assembly Rules in Ecological Communities (Weiher,
E. and Keddy, P.A., eds). Cambridge University Press, p.338-362.
Ducke, A; Black, G.A. 1954. Notas sobre a fitogeografia da Amazônia brasileira.
Boletim Técnico do Instituto Agronômico do Norte, Belém 29:1 – 62
Ferreira, C.A.C. 1997. Variação Florística e Fisionômica da Vegetação de Transição
Campina, Campinarana e Floresta de Terra Firme. Dissertação de Mestrado,
Universidade Federal Rural de Pernambuco.

53
Ferreira, C.A.C. 2009. Análise comparativa de vegetação lenhosa do ecossistema
campina na Amazônia Brasileira. Tese de Doutorado, Universidade Federal do
Amazonas, Manaus, Amazonas. 277p.
Fine, P. V. A.; Mesones I.; Coley, P.D. 2004. Herbivores promote habitat specialization
by trees in Amazonian forests. Science, 305:663–665.
Fine, P.V.A.; Z.J. Miller; I. Mesones, S.; Irazuzta H.M.; Appel, I. Stevens, I. Saaksjarvi,
J.C. Schultz, P.D. Coley. 2006. The growth–defense trade-off and habitat specialization
by plants in Amazonian forests. Ecology, 87:150–162.
Fine, P.V.A., R. Garcia-Villacorta, N. Pitman, I. Mesones, and S. W. Kembel. 2010. A
floristic study of the white sand forests of Peru. Annals of the Missouri Botanical
Garden, 97:283–305.
Grime, J.P. 2006. Trait convergence and trait divergence in herbaceous plant
communities: Mechanisms and consequences. Journal Vegetable Science, 17:255-260.
Junk, W.J.; Piedade, M.T.F.; Schöngart, J.; Cohn-Haft, M.; Adeney, J.M.; Wittmann,F.
2011. A Classification of Major Naturally-Occurring Amazonian Lowland Wetlands.
Wetlands, 31:623-640.
Junk, W.J. 2013. Current state of knowledge regarding South America wetlands and
their future under global climate change. Aquat Sci, 75:113–131.
Keddy, P.A. 1992. Assembly and response rules: two goals for predictive community
ecology. Jounal Vegetable Sciene, 3:157-164.
Lavorel, S.; Garnier, E. 2002. Predicting changes in community composition and
ecosystem functioning from plant traits: revisiting the Holy Grail. Funct Ecol, 16:545–
556.

54
Mendonça, A.F.M. 2011. Campinaranas Amazônicas: pedogênese e relações solo-
vegetação. Tese de Doutorado, Universidade Federal de Viçosa, Viçosa, Minas Gerais.
110p.
Mendonça, B. A. F; Filho, E. I. F.; Schaefer, C. E. G. R.; Carvalho, A.F. 2014. Use of
geophysical methods for the estudy of Sandy soils under Campinarana at the National
Park of Viruá, Roraima state, Brazilian Amazonia. J Soils Sediments, 14:525-537.
McBill, B.J. 2006. Rebuilding community ecology from functional traits. Trends in
Ecology and Evolution, 21:178–185.
Nascimento, N. R.; Boulet, R.; Lamotte, M.; Magat, P. 1997. Relatório de campo da
equipe de solos em expedição à Campina do Patauá. Fundação Vitória Amazônica,
Manaus.
Parolin, P.; Wittmann, F. 2010. Struggle in the flood: three responses to flooding stress
in four tropical floodplain systems. AoB Plants, doi: 10.1093/aobpla/plq003.
Prance, G.T.; Schubart, H.0.R. 1978. Notes of the vegetation of Amazonia. 1. A
preliminary note on the origin of the open white sand campinas of the lower Rio Negro.
Brittonia, 29:60-63.
Silveira, M. 2003. Vegetação e Flora das Campinaranas do Sudoeste Amazônico (JU-
008). Relatório de Defesa Técnica, Associação S.O.S. Amazônia, Rio Branco. 28p.
Stearns, S.C. 1989. Trade-offs in life-history evolution. Functional Ecology, 3: 259-268.
Taiwo, K.A.; Angersbach, A.; Adeomowaye, B.I.O.; Knorr, D. 2001. Effects of
pretreatments on the dissusion kinetics and some quality parameters of osmotically
dehydrated apple slices. Journal of Agricultural and Food Chemistry, 49:2804-2811.

55
van der Valk, A.G. 1981. Succession in wetlands—a Gleasonian approach. Ecology,
62:688–696.
Veloso, H.P.; Rangel-Filho, A.L.R.; Lima, J.C.A. 1991. Classificação da vegetação
brasileira, adaptada a um sistema universal. Rio de Janeiro: IBGE, 124p.
Vicentini, A. 2004. A Vegetação ao Longo de um Gradiente Edáfico no Parque
Nacional do Jaú. In: BORGES, S.H.; IWANAGA, S.; DURIGAN, C.C.; PINHEIRO,
M.R. (Eds.). Janelas para a Biodiversidade no Parque Nacional do Jaú: uma estratégia
para o estudo da biodiversidade na Amazônia. Fundação Vitória Amazônica
(FVA)/WWF/IBAMA, Manaus, Brasil, p.117-143.
Wittmann, F.; Anhuf, D.; Junk, W.J. (2002) Tree species distribution and community
structure of central Amazonian várzea forests by remote sensing techniques. Journal of
Tropical Ecology, 18, 805–820.
Weiher, E.; Clarke, G.D.P.; Keddy, P.A. 1998. Community assembly rules,
morphological dispersion, and the coexistence of plant species. Oikos, 81:309–322.
Weiher, E.; Keddy, P. 1999. Ecological assembly rules: perspectives, advances,
retreats. Cambridge University Press, Cambridge, UK.
Worbes, M.; Klinge, H.; Revilla, J.D.; Martius, C. 1992. On the dynamics, floristic
subdivision and geographical distribution of várzea forests in Central Amazonia. J. Veg.
Science, 3: 553-564.
Worbes, M. 1997. The Forest Ecossystem of the Floodplains. In: JUNK, W.J. The
Central Amazon Floodplain: Ecology of a Pulsing System. Ecological Studies, vol. 126.
Springer, Berlim/Heidelberg/New York, p. 223-265.
Wright, I.J.; Reich, P.B,; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; et al.
2004. The worldwide leaf economics spectrum. Nature. 428:822-828.

56


















