
INSTITUTO NACIONAL DE PESQUISAS DA AMAZÔNIA - INPA...
104
INSTITUTO NACIONAL DE PESQUISAS DA AMAZÔNIA - INPA PROGRAMA DE PÓS-GRADUAÇÃO DO INPA PROGRAMA DE PÓS-GRADUAÇÃO EM BIOLOGIA DE ÁGUA DOCE E PESCA INTERIOR COMPOSIÇÃO QUÍMICA DE OTÓLITOS DE TUCUNARÉ-AÇU (Cichla temensis HUMBOLT, 1821) E SEU POTENCIAL COMO MARCADOR AMBIENTAL EM POPULAÇÕES DE LAGOS DE VÁRZEA E IGAPÓ NA AMAZÔNIA-BRASILEIRA RANIERE GARCEZ COSTA SOUSA Manaus, Amazonas Fevereiro/2014
Transcript of INSTITUTO NACIONAL DE PESQUISAS DA AMAZÔNIA - INPA...

INSTITUTO NACIONAL DE PESQUISAS DA AMAZÔNIA - INPA
PROGRAMA DE PÓS-GRADUAÇÃO DO INPA
PROGRAMA DE PÓS-GRADUAÇÃO EM BIOLOGIA DE ÁGUA DOCE
E PESCA INTERIOR
COMPOSIÇÃO QUÍMICA DE OTÓLITOS DE TUCUNARÉ-AÇU
(Cichla temensis HUMBOLT, 1821) E SEU POTENCIAL COMO
MARCADOR AMBIENTAL EM POPULAÇÕES DE LAGOS DE
VÁRZEA E IGAPÓ NA AMAZÔNIA-BRASILEIRA
RANIERE GARCEZ COSTA SOUSA
Manaus, Amazonas
Fevereiro/2014

ii
RANIERE GARCEZ COSTA SOUSA
COMPOSIÇÃO QUÍMICA DE OTÓLITOS DE TUCUNARÉ-AÇU
(Cichla temensis HUMBOLT, 1821) E SEU POTENCIAL COMO
MARCADOR AMBIENTAL EM POPULAÇÕES DE LAGOS DE
VÁRZEA E IGAPÓ NA AMAZÔNIA-BRASILEIRA
Orientador: CARLOS EDWAR DE CARVALHO FREITAS, PhD.
Tese apresentada ao Programa de Pós-
Graduação em Biologia de Água Doce e Pesca Interior
do INPA, como parte dos requisitos para obtenção do
título de Doutor em Ciências Biológicas, área de
concentração Biologia de Água Doce e Pesca Interior.
Manaus, Amazonas
Fevereiro/2014
Fontes financiadoras: Projeto PRONEX, (Edital 023/2009) – FAPEAM; Programa Institucional de
Doutorado Sanduíche no Exterior – PDSE/CAPES; Instituto Nacional de Pesquisas na Amazônia –
INPA; Instituto de Inteligência Socioambiental Estratégica da Amazônia, I-PIATAM; Conselho
Nacional de Desenvolvimento Científico e Tecnológico – CNPq e Washington and Lee University –
WLU, U.S.A.

iii
Ficha catalográfica
S725 Sousa, Raniere Garcez Costa
Composição química de otólitos de tucunaré-açu (cichla
temensis humbolt, 1821) e seu potencial como marcador ambiental
em populações de lagos de várzea e igapó na Amazônia-brasileira /
Raniere Garcez Costa Sousa. --- Manaus: [s.n.], 2014.
xiii, 91 f. : il. color.
Tese (Doutorado) --- INPA, Manaus, 2014.
Orientador : Carlos Edwar de Carvalho Freitas.
Área de concentração : Biologia de Água Doce e Pesca Interior.
1. Microquímica de otólitos. 2. Tucunaré-açu. 3. Ecologia de
populações. I. Título.
CDD 597.58
Sinopse: Este estudo investiga o uso de isótopos de estrôncio e
elementos traços como marcadores ambientais em otólitos de
tucunarés-açu Cichla temensis, para a identificação dos locais de
nascimento de indivíduos jovens e adultos. Também, baseado na
microquímica dos otólitos e geologia, avalia a distribuição espacial,
padrões metapopulacionais e de filopatria de indivíduos adultos em
tributários da Bacia do Médio Rio Negro.
Palavras-chave: Microquímica de otólitos; isótopos e elementos
traços; movimentos de dispersão; ecologia de populações.

iv
Dedicatória
À Joyce Lara Garcez e meus filhotes, pelo amor
e carinho, fontes de motivação para continuar
sempre otimista na minha jornada.

v
AGRADECIMENTOS
A Deus pela oportunidade da vida.
Ao Professor Carlos Edwar de Carvalho Freitas, meu amigo e orientador nesta
tese, por sua integridade, paciência e companheirismo, os quais me incentivaram a
persistir e não fraquejar nos momentos difíceis, e, principalmente, pela imensa
dedicação na orientação dessa pesquisa, fato impossível de ser mensurado.
Ao Professor Robert Humston, meu supervisor durante o doutorado sanduiche
na Washington and Lee University, o qual me ajudou a desenvolver os modelos
estatísticos e de análises químicas de otólitos.
Ao Instituto Nacional de Pesquisas da Amazônia - INPA, através do Programa
de Pós Graduação em Biologia de Água doce e Pesca Interior - BADPI - pela ímpar
oportunidade de capacitação profissional que me foi concedida.
Aos órgãos de fomento: Conselho Nacional de Desenvolvimento Científico e
Tecnológico - CNPq e Programa Institucional de Doutorado Sanduíche no Exterior -
PDSE/CAPES, os quais, através de suas bolsas de estudos e auxílios foram de
fundamental importância para a provisão da minha subsistência durante este trabalho,
quando do meu estágio tanto aqui no Brasil como no exterior. Também ao Instituto
Brasileiro de Meio Ambiente – IBAMA - pelas licenças concedidas para a coleta e
transporte de materiais em campo.
A todos os integrantes do Instituto de Inteligência Socioambiental Estratégica da
Amazônia - I-PIATAM - especialmente aos amigos Caroline Campos, Antônio Oliveira,
Michel Catarino, Flávia Souza e Hostília Campos, os quais estiveram sempre dispostos
a ajudar-me em momentos pouco auspiciosos.
Ao amigo e compadre Júlio Alberto Dias Siqueira, por sua valiosa e rara
amizade, bem como pelo apoio concedido nas coletas de dados que tive que
empreender.
A todos os auxiliares de campo, mateiros e pescadores que foram parte
imprescindível deste trabalho, cujos nomes não foram anotados. Mas que anonimamente
deram suas parcelas de contribuição.

vi
Aos meus filhotes, Adrielly, Ranielly e Raniere, bênçãos do Criador em minha
vida, nos quais encontrei estímulo para lutar e motivação para perseverar na minha
caminhada.
À minha querida esposa Joyce Lara Araújo da Fonseca Garcez, que sempre
esteve ao meu lado como uma guerreira, principalmente quando, por meses, tive que
ausentar-me do lar em excursões ao exterior do país. Que Deus me conceda a graça de
tê-la por toda a vida.

vii
Salmos 37:5
Entrega o teu caminho ao SENHOR
confia Nele, e o mais Ele fará.
Salmos de Davi
Bíblia Sagrada

viii
RESUMO
Neste estudo foi avaliada a aplicação da geoquímica de otólitos como marcador
ambiental da origem de nascimento de tucunarés-açu Cichla temensis (Humboldt,
1833), jovens menores que um ano (YOY) e de indivíduos adultos, na bacia do Rio
Negro. Inicialmente, foram medidas as variações de isótopos de estrôncio e elementos
traços em otólitos de exemplares jovens em diferentes locais de coletas e comparados os
resultados coma geologia do local. Os resultados indicam que padrões na microquímica
dos otólitos dos peixes jovens, particularmente na razão isotópica 87Sr/86Sr,
correspondem aos padrões da base geológica do local de nascimento. Isto permitiu uma
classificação correta de 99% dos peixes juvenis com seus rios de origem, usando uma
tabela de correlação da análise de função discriminante linear (LDFA). Os resultados
mostram também que é possível usar inferências de mapas geológicos e assinatura
geoquímica dos otólitos para traçar movimentos de indivíduos adultos entre o Rio
Negro e seus tributários. A análise LDFA mostrou que a microquímica da parte central
dos otólitos dos tucunarés adultos classificou corretamente cada indivíduo com seu local
de nascimento, com 41,9% de acurácia. Ainda, a variação da razão isotópica 87Sr/86Sr da
linha transversal (do centro para a borda) da superfície dos otólitos dos tucunarés-açus
adultos, indicou a existência de populações isoladas e mistas de tucunarés-açu na área
do estudo. Os resultados também sugerem que os tucunarés adultos realizam
movimentos de dispersão entre os tributários estudados indicando um padrão
metapopulacional e filopátrico, percebidos com o retorno desses indivíduos aos seus
locais de nascimento, e pela permanência de outros nesses locais, mostrando fidelidade
ao sítio de origem. O presente estudo proporciona novas informações a respeito da
distribuição espacial do tucunaré-paca ou tucunaré-açu C. temensis em seu ambiente
natural, que poderiam ser usadas como apoio efetivo nas estratégias de gestão pesqueira
e fornecer informações básicas para trabalhos futuros em investigações sobre a história
inicial de vida e ecologia espacial de peixes na bacia Amazônica.
Palavras-chave: Manejo pesqueiro; geoquímica; assinatura digital química em otólitos;
isótopos de estrôncio; filopatria.

ix
ABSTRACT
This study examined the application of otolith geochemistry as a natural marker of natal
origins in peacock bass Cichla temensis (Humboldt, 1833) young-of-the-year (YOY)
and adults in the Negro River basin. Initially were assessed variations in strontium
isotopes and trace elements in otoliths of young individuals from different sampling
locations comparing the results with local geology. The results suggests that patterns in
otolith microchemistry of YOY, particularly 87Sr/86Sr isotopes, correspond to patterns in
origin bedrock geology. This approach allowed us to correctly classify 99% of juvenile
fish to their natal streams using a cross-validation table from the linear discriminant
function analysis (LDFA). The results also indicates that is possible to use inferences
from geologic maps to track movements of adults fish between Negro River and its
tributaries. The LDFA analyses shows that the microchemistry from the central part of
the adults peacock bass otolith, classified correctly each individual for its birth location
with 41.9% of accuracy. Also, variation in 87Sr/86Sr isotopic rate from the transect line
(from the core to the border) on the otolith surface of adults peacock bass, indicated the
existence of isolated and mixed populations in the study area. The results also suggests
that adult peacock bass ensure dispersal movements between studied tributaries showing
a metapopulational and philopatric pattern, noticed through the individuals returned to
their birth locations, and by the permanence of some individuals in those locations,
showing loyalty to their origin place. Therefore, the present report offers new
information about spatial distribution of C. temensis in their natural environment, which
could support effective strategies of fishery management, and provides the basic
information for future work to investigate the early life history and spatial ecology of
fresh water fish in the Amazon basin.
Keywords: Fish management; geochemistry; otolith chemical fingerprints; strontium
isotopes; philopatry.

x
SUMÁRIO
Introdução geral ............................................................................................................. 1
Biologia do tucunaré-açu Cichla temensis ....................................................................... 3
Mobilidade de dispersão de C. temensis ........................................................................... 4
Otólitos ............................................................................................................................. 5
Migração e microquímica de otólitos ............................................................................... 7
Distribuição da razão isotópica de 87Sr/86Sr no ambiente................................................. 7
Problema enfocado no estudo ........................................................................................ 9
Hipóteses ........................................................................................................................ 10
Objetivo geral ................................................................................................................ 11
Objetivos específicos ..................................................................................................... 12
Organização da Tese .................................................................................................... 12
Metodologia geral ......................................................................................................... 13
Área de estudo ................................................................................................................ 13
Coleta e análise dos dados .............................................................................................. 15
Capítulo I. Otolith geochemistry in young-of-the-year peacock bass Cichla temensis for
investigating natal dispersal in the Rio Negro (Amazon - Brazil) river system ............. 21
Abstract ......................................................................................................................... 23
Introduction .................................................................................................................. 24
Materials and Methods ................................................................................................ 25
Study Area ...................................................................................................................... 25
Sample collection ........................................................................................................... 29
Otolith preparation and analysis ..................................................................................... 29
Statistical analysis .......................................................................................................... 30
Geologic analyses ........................................................................................................... 31

xi
Results ............................................................................................................................ 32
Discussion ...................................................................................................................... 41
Acknowledgments ......................................................................................................... 42
Capítulo II. Philopatry and metapopulation patterns for the peacock bass Cichla
temensis of the middle Negro River Basin (Amazonas - Brazil): an otolith chemical
analysis. .......................................................................................................................... 43
Abstract ......................................................................................................................... 45
Introduction .................................................................................................................. 46
Material and Methods .................................................................................................. 48
Study area ....................................................................................................................... 48
Sample collections and preparations .............................................................................. 50
Strontium isotope and elemental analysis ...................................................................... 51
Water data collection and analysis ................................................................................. 51
Otoliths statistical analyses ............................................................................................. 52
Results ............................................................................................................................ 53
Young-of-year (YOY) and adult peacock bass otolith core microchemical analyses .... 53
Adult peacock bass otolith core chemical analysis ........................................................ 56
87Sr/86Sr isotope ratio analysis from otolith transects of adults peacock bass ................ 59
Water chemical analysis ................................................................................................. 62
Discussion ...................................................................................................................... 63
Cichla temensis movements, reproduction and phylopatric behaviour synchronism. ... 66
Hydrological barrier and C. temensis metapopulation structure .................................... 67
Peacock bass fisheries management proposition ............................................................ 72
Acknowledgments ......................................................................................................... 73
Conclusões ..................................................................................................................... 74
Referências Bibliográficas ........................................................................................... 75

xii
LISTA DE FIGURAS
Introdução geral
Figura 1. Exemplar de tucunaré-açu Cichla temensis, capturado na região do Rio
Padauari em 2012............................................................................................................17
Figura 1. Exemplar de tucunaré-açu Cichla temensis, capturado na região do Rio
Padauari em 2012...........................................................................................................19
Figura 3. Localização dos locais amostrais na região do Médio Rio Negro, município de
Barcelos-Amazonas…………………………………………………………………….27
Figura 4. Biometria e extração dos otólitos dos peixes adultos.......................................29
Figura 5. Material utilizado para a separação e montagem dos otólitos..........................30
Figura 6. Lixamento dos otólitos.....................................................................................30
Figura 7. Banheira ultrassônica para limpeza dos otólitos..............................................31
Figura 8. Acondicionamento dos otólitos em sacolas e caixas esterilizadas para
transporte……………………………………………………………………………….31
Figura 9. Coletor Múltiplo de Espectrometria de Massas por Plasma Indutivamente
Acoplado..........................................................................................................................32
Figura 10. Queima a laser da superfície do otólito..........................................................32
Capítulo I
Figure 1. Location and geological conditions of the study área......................................40
Figure 2. Mean isotopic and trace element concentration ratios in the otoliths of YOY
fish groups.......................................................................................................................49
Figure 3. Microchemical isotopic and trace element ratios from the otoliths of YOY C.
temensis distributed within groups and between areas....................................................52
Capítulo II
Figure 1. Study area and geographic distribution of sampling sites in the middle Negro
River, Barcelos Municipality...........................................................................................63
Figure 2. A scatter-plot displaying the distribution of Cichla temensis’ YOY and adult
otolith core chemical fingerprints....................................................................................68
Figure 3. Adult Cichla temensis peacock bass’ otolith core 87Sr/86Sr isotope ratio
distributed by sampling sites...........................................................................................71

xiii
Figure 4. Distribution of elemental compositions from adult Cichla temensis peacock
bass’ otolith core………………………………………………………………………..72
Figure 5. Cichla temensis peacock bass’ movement history from their birth to the
locations where they were caught as adult individuals, based on variation in 87Sr/86Sr
isotope ratio values on the otolith transect……………………………………………..73
Figure 6. Transect lines displaying Cichla temensis peacock bass’ movement histories
from their birth to catch locations………………………………………………………76
Figure 7. Organizational chart with C. temensis movements…………………………..79
Figure 8. The organogram illustrates C. temensis movement behaviour among different
water types in relation to differences in otoliths’ geochemical composition (87Sr/86Sr
isotopes)………………………………………………………………………………...83
Figure 9. Theoretical spatial movement distribution of Cichla temensis in the middle
Negro River basin………………………………………………………………………84

1
Introdução geral
A complexidade da pesca na Amazônia dificulta o pleno entendimento de alguns
pressupostos de modelos tradicionais para avaliação dos estoques pesqueiros, aumentando o
grau de incerteza dos resultados obtidos. Como regra geral, a aplicação dos modelos analíticos
é embasada em parâmetros de dinâmica populacional e ainda existem poucos estudos
conclusivos sobre a identificação das populações de peixes que habitam os ambientes
aquáticos amazônicos. Barthem e Petrere Jr. (1996) postularam, por meio de estudos de
crescimento, a existência de uma única população de piramutaba Brachyplatystoma vailantii
(Valenciennes, 1840) desde os trechos superiores dos tributários de águas brancas da calha do
rio Solimões/Amazonas até o estuário. No entanto, a maioria das espécies importantes para a
pesca comercial ainda são pouco estudadas, e não temos informações suficientes para afirmar
que estes indivíduos pertencem a uma única ou a várias populações (Bailey e Petrere, 1989;
Batista, 2001). Sendo assim, a falta de informações precisas sobre os comportamentos
migratórios das espécies se torna uma das principais limitações para a sua conservação, o que
precisa ser corrigido com mais pesquisas nessa área.
A descrição dos padrões de movimentos migratórios tem sido uma questão central na
biologia e manejo de peixes de água doce. As migrações entre os ambientes marinhos e de
água doce, realizadas por peixes anádromos e catádromos, são relativamente bem
conhecidas (Groot e Margolis, 1991, Walter e Thorrold, 2006), mas o mesmo não ocorre
com os movimentos migratórios de peixes de áreas continentais (Gowan et al., 1994;
Northcote, 1997).
Os processos de migração e dispersão dos peixes são considerados fatores
fundamentais na definição de populações e metapopulações (Hanski e Gilpin, 1997; Rieman
e Dunham, 2000). No entanto, para rastrear esses indivíduos entre ambientes distintos é
necessário o uso de alguns artifícios e metodologias. Dentre as metodologias mais utilizadas
se destacam a marcação e recaptura (Hoeinghaus et al., 2003), a telemetria (Semmens et al.,
2006) e, atualmente, o uso de marcadores ambientais (isótopos e elementos traços)
encontrados em otólitos e outras estruturas rígidas em peixes (Thorrold et al., 2001; Walther
et al., 2008).

2
Análises de isótopos e de elementos traços em otólitos têm apresentado resultados
mais acurados e completos na descrição retrospectiva da história dos movimentos de
dispersão de indivíduos e de grupos de peixes (Wells et al., 2003; Rich et al., 2004; Walther
et al., 2008; Walther e Thorrold, 2010), fornecendo informações robustas e confiáveis sobre
a dinâmica espacial de populações (Rieman et al. 1994; Humston & Harbor, 2006).
No entanto, a maioria dos estudos realizados com essa metodologia até o presente
momento, tem contemplado espécies de peixes que habitam ambientes marinhos ou
estuarinos (Gillanders 2002; Walther e Thorrold, 2010), onde é esperada uma grande
variação na concentração de solutos nos diferentes ambientes. Recentemente, alguns estudos
realizados em ambientes de águas interiores também têm mostrado bons resultados em
monitorar movimentos de peixes entre ambientes quimicamente distintos (Rich et al., 2004;
Humston et al., 2010; Johnson et al., 2012).
Na região amazônica, vários estudos sobre os processos migratórios de peixes foram
realizados (Ribeiro e Petrere Jr. 1990; Barthen et al., 1991; Freitas e Garcez, 2004).
Entretanto, devido às dificuldades em monitorar indivíduos pequenos (ou jovens) com os
modelos de marcação tradicionais, somados às adversidades encontradas em recapturar ou
acompanhar indivíduos em áreas remotas, a maioria desses estudos gerou resultados não
conclusivos. Assim, os isótopos de elementos traços detectados na composição de otólitos
de peixes, pode ser utilizados como marcadores ambientais naturais, para rastrear
movimentos migratórios de indivíduos que transitam entre ambientes quimicamente
diferentes (Johnson et al., 2012), e solucionar essa deficiência metodológica.
Nesse contexto, a análise química de otólitos de peixes pode contribuir para esclarecer
dúvidas relacionadas à existência de migração de indivíduos entre populações, melhorando o
entendimento dos parâmetros da dinâmica populacional de peixes. Isso poderá propiciar o
aprimoramento das estratégias atuais de manejo pesqueiro utilizadas na Região Amazônica,
principalmente nas áreas onde atuam várias modalidades de pesca sobre o mesmo recurso.
Na região do Rio Negro ocorrem quatro modalidades de pesca: a comercial, a
esportiva, a ornamental e a de subsistência (Sobreiro et al., 2010). Dentre estas, a que mais
vem crescendo nessa região é a pesca esportiva, com a modalidade pesque e solte (Holey et
al., 2008), a qual atua principalmente sobre as espécies de ciclídeos, com destaque para o

3
tucunaré-açu C. temensis, por apresentar maior tamanho e voracidade em relação aos seus
congêneres (Taphorn e Barbarino-Duque, 1993).
Alguns estudos sobre a distribuição espacial de espécies do gênero Cichla foram
realizadas na bacia do Rio Amazonas (Macrander 2010; Willis et al., 2007; 2010). No
entanto, os resultados apresentados por esses estudos não mostram claramente a existência
de movimentos de dispersão ou processos migratórios entre os diferentes sistemas aquáticos,
o que dificulta a compreensão adequada da dinâmica de movimentos dessas espécies na
região. Desse modo, também pouco se sabe sobre a distribuição espacial do tucunaré-açu C.
temensis na região do Médio Rio Negro, área onde a atividade da pesca esportiva ocorre
com maior intensidade.
Pelo exposto, e visando compreender a dinâmica de dispersão dos tucunarés-açu, e, ao
mesmo tempo, subsidiar informações que possam contribuir com o entendimento da
distribuição espacial dessa espécie na região do Médio Rio Negro, a presente pesquisa
analisou os isótopos de 87Sr/86Sr e a concentração dos elementos traços Sr/Ca e Ba/Ca em
otólitos de tucunaré-açu Cichla temensis, no intuito de identificar o local de nascimento de
juvenis menores de um ano e adultos. Este estudo verificou ainda, através das diferenças
químicas existentes nos otólitos dos tucunarés, variações indicando movimentos de
dispersão entre ambientes quimicamente distintos, o que, posteriormente, foi utilizado para
avaliar a existência ou não de uma estrutura de metapopulação de Cichla temensis na região.
Biologia do tucunaré-açu Cichla temensis
O tucunaré pertence à família Cichlidae, é peixe adaptado principalmente a ambientes
lênticos e ocorre naturalmente na América do Sul, em rios de água preta, clara ou branca
(Winemiller et al., 2008). Pertence ao gênero Cichla, que abriga 14 espécies válidas
(Kullander e Ferreira, 2006). Na região amazônica as espécies de tucunarés utilizadas na
pesca são: Cichla temensis Humbolt, 1821, C. ocellaris Bloch e Schneider, 1801, C.
orinocencis Humbolt, 1821, C. intermedia Machado-Allison, 1971 e C. monoculus Spix e
Agassiz, 1831.

4
O tucunaré-paca ou tucunaré-açu Cichla temensis (Figura 1) é um importante predador
de topo de cadeia alimentar em diversos ambientes aquáticos (Barros, 1980). Segundo Graef
(1995) o C. temensis atingem a idade adulta por volta dos dois anos, em ambiente natural. A
espécie possui desova parcelada, a qual ocorre durante todo o ano e se intensifica na época
da seca (Fontenele, 1950; Gomiero et al., 2009). É um peixe popular na pesca esportiva, mas
também tem alto valor na pesca comercial e de subsistência (Corrêa, 1998; Jepsen et al.,
1999). Por este motivo, conflitos são frequentes entre os diferentes usuários do recurso,
quando estes atuam nas mesmas áreas de exploração pesqueira (Sobreiro et al., 2010).
Figura 1. Exemplar de tucunaré-açu Cichla temensis, capturado na região do Rio Padauari em
2012.
Mobilidade de dispersão de C. temensis
Estudos indicam que o tucunaré Cichla temensis é uma espécie sedentária e
territorialista (Barros, 1980, Santos et al., 1984, Jepsen et al., 1999), por outro lado, alguns
estudos afirmam a existência de movimentos de dispersão desses indivíduos (Hoeinghaus et
al., 2003; Holley et al., 2008). Essa controvérsia nas informações causam incertezas quanto a
distribuição desses indivíduos dentro dos sistemas aquáticos amazônicos, e tampouco se estes
realizam movimentos territoriais entre populações distintas. Essa lacuna de informações
também se estende à maioria das espécies importantes para a pesca comercial no Amazonas
(Bailey e Petrere, 1989; Batista, 2001).

5
Pesquisas têm sido realizadas no intuito de compreender os mecanismos que regulam
o recrutamento e a ecologia espacial do tucunaré, particularmente em sistemas de rios em
áreas abertas de bacias hidrográficas brasileiras (Smith et al., 2005). A variabilidade de
recrutamento em estoques é geralmente alta, tornando-se visível a correlação com fatores
abióticos, particularmente descargas d'água e temperaturas (Smith et al., 2005).
Em recentes pesquisas realizadas na bacia amazônica sobre a dispersão gênica de
tucunarés, foi observada a separação de populações ao longo dos diversos tributários nessa
região (Macrander 2010; Willis et al., 2007; 2010). Estudos sobre marcação e recaptura
realizados com Cichla temensis, C. orinocensis e C. intermedia em rios da Venezuela
demonstraram que a maioria dos peixes recapturados estava dentro de um raio de 1 km, a
partir do ponto de sua marcação (Hoeinghaus et al., 2003). Por outro lado, indivíduos de C.
temensis marcados nesse mesmo estudo, foram recapturados com distâncias entre 17 a 21 km
(o tempo entre o período de marcação e a recaptura dos exemplares de C. temensis, não foi
divulgado no estudo). Holley et al. (2008) utilizaram a mesma metodologia de marcação e
recaptura com indivíduos de C. temensis na região do Médio rio Negro, e relataram que um
indivíduo dessa espécie foi recapturado a 40 km de distância de seu local de origem, após um
ano de sua marcação.
No entanto, estes esforços não foram suficientes para elucidar completamente o
processo de dispersão desses ciclídeos, o que torna necessária a realização de investigações
acerca dos padrões migratórios e/ou de movimentos de dispersão desses indivíduos entre
regiões distintas. Tais estudos poderão contribuir para um entendimento mais aprofundado
sobre a distribuição espacial de (sub)populações de tucunarés na região do Médio Rio Negro.
Otólitos
Os peixes teleósteos têm três pares de estruturas inorgânicas em seu sistema auditivo,
conhecidas como otólitos (lapillus, sagitta e asteriscus), que estão localizados atrás dos seus
globos oculares (Campana, 1999). Estas estruturas são formadas basicamente por carbonato
de cálcio (aragonita) que é sedimentado nos otólitos diretamente da química do ambiente
aquático onde vivem (Campana 1999; Bath et al., 2000; Campana e Thorrold, 2001; Walther
6
e Thorrold 2006). Os otólitos são metabolicamente inertes e crescem continuamente através
da sedimentação de finas camadas diárias (2 a 5 µm), que formam anéis concêntricos durante
toda a vida do peixe (Pannella, 1971; Gauldie, 1993; Lord et al., 2011).
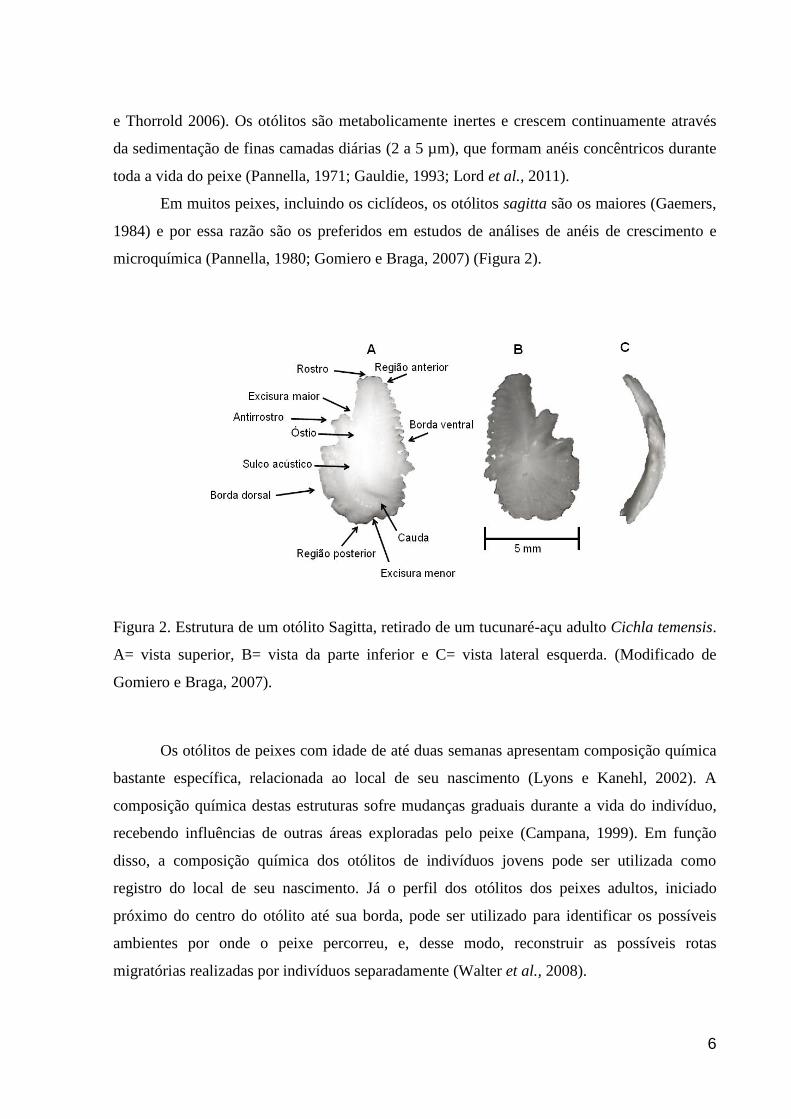
Em muitos peixes, incluindo os ciclídeos, os otólitos sagitta são os maiores (Gaemers,
1984) e por essa razão são os preferidos em estudos de análises de anéis de crescimento e
microquímica (Pannella, 1980; Gomiero e Braga, 2007) (Figura 2).
Figura 2. Estrutura de um otólito Sagitta, retirado de um tucunaré-açu adulto Cichla temensis.
A= vista superior, B= vista da parte inferior e C= vista lateral esquerda. (Modificado de
Gomiero e Braga, 2007).
Os otólitos de peixes com idade de até duas semanas apresentam composição química
bastante específica, relacionada ao local de seu nascimento (Lyons e Kanehl, 2002). A
composição química destas estruturas sofre mudanças graduais durante a vida do indivíduo,
recebendo influências de outras áreas exploradas pelo peixe (Campana, 1999). Em função
disso, a composição química dos otólitos de indivíduos jovens pode ser utilizada como
registro do local de seu nascimento. Já o perfil dos otólitos dos peixes adultos, iniciado
próximo do centro do otólito até sua borda, pode ser utilizado para identificar os possíveis
ambientes por onde o peixe percorreu, e, desse modo, reconstruir as possíveis rotas
migratórias realizadas por indivíduos separadamente (Walter et al., 2008).

7
Migração e microquímica de otólitos
Estudos recentes sugerem que a heterogeneidade na geologia do canal principal de um
rio pode apresentar diferenças detectáveis na química dos otólitos dos peixes, se estes
percorrerem diferentes seções de um único rio ao longo de seu crescimento e vida adulta
(Wells et al., 2003). Do mesmo modo, a variação química entre os sistemas aquáticos pode
contribuir para a distinção entre os locais de nascimento de peixes entre rios com diferentes
características químicas da água (Campana, 1999).
As análises microquímicas de otólitos mostram que é possível correlacionar tais
características com diferenças limnológicas entre ambientes aquáticos, através dos registros
dos isótopos e elementos traços sedimentados nessa estrutura ao longo da vida do peixe
(Campana e Thorrold, 2001). Essas análises podem ser utilizadas para gerenciamento
estratégico de uma determinada espécie que apresente padrões migratórios ou de dispersão,
como troca de indivíduos entre duas ou mais áreas ou populações, os quais podem afetar o
resultado do recrutamento de indivíduos de uma determinada região (Bilby et al., 2001) e sua
distribuição espacial (Ridgeway et al., 2002).
O presente estudo teve como objetivo analisar populações de tucunaré-açu Cichla
temensis da região do médio Rio Negro, por meio da aplicação da metodologia de análise de
isótopos e elementos traços ambientais encontrados em otólitos de peixes, procurando
contribuir com dados mais precisos sobre os movimentos inter-habitats para essa espécie. A
detecção desses movimentos é possível caso existam diferenças químicas significativas entre
os ambientes percorridos pelo peixe (Wells et al., 2003), uma vez que a química dos otólitos
registra com precisão os movimentos de saída e/ou de retorno desses indivíduos, desde o seu
local de nascimento/origem até o local de sua captura (Guido et al., 2006).
Distribuição da razão isotópica de 87Sr/86Sr no ambiente
Estrôncio (Sr) é um elemento traço encontrado na maioria das rochas ígneas,
metamórficas e sedimentares. Está distribuído em vários tipos de matérias e ambientes, como

8
na água, no solo, nas plantas e em animais (Slovak e Paytan, 2011). Naturalmente o Estrôncio
ocorre distribuído em quatro formas estáveis (84Sr, 86Sr, 87Sr, e 88Sr), das quais três não são
radioativas: 84Sr (0,56 %), 86Sr (9,87%) e 88Sr (82,53%). O isótopo 87Sr (7,04%) é radiogênico
(radioativo) e é formado pela queda da radioatividade do rubídio 87Rb (Faure e Mensing,
2005). Devido à seus valores proporcionais no ambiente terrestre o 87Sr e o 86Sr podem ser
correlacionados, para normalizar a distribuição do 87Sr em substituição ao rubídio,
representado na forma do isótopo de 87Sr/86Sr.
Faure (2001) relata que a proporção média do isótopo de 87Sr/86Sr na crosta
continental é de 0,70916. Entretanto, estudos recentes encontraram valores isotópicos de
87Sr/86Sr originários de duas medidas de referência: a Eimer e Amend (E&A) com valor de
87Sr/86Sr de 0,70800, e a U.S. National Bureau of Standards-NBS987, apresentando valor de
0,71025. Os mais altos valores de 87Sr/86Sr foram encontrados em rochas muitos antigas, tais
como granitos, que geralmente apresentam altos valores de Rb/Sr com valores de 87Sr/86Sr
acima de 0,710 (Faure 2001). Ao contrário, rochas jovens apresentam baixos valores de Rb/Sr
e também valores de 87Sr/86Sr menores que 0,704 (Faure 1977; Capo et al. 1998; Slovak e
Paytan 2011).
Uma vez que os valores de 87Sr/86Sr variam muito pouco quando passam das rochas
lixiviadas indo para o solo, de onde são absorvidos pela vegetação e posteriormente pela
cadeia alimentar (Graustein e Armstrong, 1983; Miller et al., 1993), seus valores não se
alteram significativamente nesse processo, e, consequentemente, preservam os valores
originais de suas rochas matrizes (Hurst e Davis 1981; Faure, 1986; Åberg et al., 1995;
Wakabayashi et al., 2007; Ruggeberg et al., 2008). Desse modo, o isótopo de 87Sr/86Sr pode
ser utilizado como marcador ambiental em estudos de movimentos de dispersão de animais
terrestres e/ou aquáticos que tenham incorporado esses elementos em suas estruturas rígidas
(Capo et al., 1998; Elsdon et al., 2008; Turner e Limburg, 2012).

9
Problema enfocado no estudo
Estudos como os de Hoeinghaus et al. (2003) e Holley et al. (2008), mostraram que os
ciclídeos, principalmente o C. temensis, realizam movimentos de dispersão consideráveis
entre os tributários de grandes rios, como o Rio Cinaruco (Venezuela) e o Rio Negro (Brasil).
Por outro lado, diversas pesquisas apontam o tununaré-açu como uma espécie sedentária
(Winemiller, 2001; Granado-Lorencio et al., 2005). Essa polêmica sobre o tucunaré-açu ser
ou não ser um peixe sedentário, é reforçada quando Macrander (2010) sugere a existência de
barreiras hidroquímicas que impedem o fluxo de indivíduos de C. temensis entre as bacias dos
rios Orinoco (Venezuela) e Rio Negro. Será que os indivíduos de tucunarés que não realizam
seus movimentos de dispersão entre os ambientes ondem residem é devido as barreiras
hidroquímicas ou porque são de fato sedentários? Ainda, se realizam movimentos de
dispersão entre ambientes distintos, formam estruturas metapopulacionais?
Procurando entender a dinâmica de dispersão do tucunaré-açu Cichla temensis, a
presente pesquisa, se propôs a estudar os movimentos de dispersão da espécie C. temensis na
bacia do Médio Rio Negro. Por haver nessa região vários tributários do Rio Negro, com tipos
distintos de água, onde habitam a espécie em questão. Alguns tributários desta região, foram
selecionados para a presente pesquisa por formarem pares de rios conectados entre si, mas
serem compostos por diferentes tipos de água. Por exemplo, o Rio Aracá (água preta) e o
Demeni (água branca), o Rio Preto (água preta) e o Padauari (água branca). A exceção foi o
Rio Cuiuni (água preta) que é diretamente conectado com o Rio Negro (água preta). No
entanto o Rio Cuiuni, apresenta sua bacia de drenagem situada em região geologicamente
diferenciada do leito do canal principal do Rio Negro. Outro fator importante dos tributários
dessa região é que eles drenam bacias com geologias distintas, o que esperamos ser visível na
microquímica dos otólitos dos exemplares de tucunaré-açu ali coletados.
Além disso, o presente estudo verifica se as diferenças microquímicas dos ambientes
registradas nos otólitos, podem indicar a formação de barreiras hidroquímicas entre os
tributários com diferentes tipos de água (água preta vs água branca), o que poderia indicar a
existências de populações isoladas localmente ou de metapopulações, quando conectadas pelo
fluxo de entrada e saída de indivíduos procedentes de outros tributários. Com esse panorama
ambiental surgiu a pergunta chave para o presente trabalho. Existem movimentos de dispersão

10
de tucunarés-açu Cichla temensis que mostram padrões de metapopulações entre o Rio Negro
e seus tributários (rios Cuiuni, Aracá, Demeni, Preto e Padauari)? É possível verificar esses
movimentos de dispersão comparando as diferenças microquímicas dos otólitos de tucunaré-
açu com os tipos de geologia das áreas por eles habitadas?
Sabe-se que os ciclídeos fazem pequenos deslocamentos entre ambientes, entretanto
ainda pouco se conhece sobre os padrões de movimentos de dispersão entre essas populações.
No entanto, estudos realizados sobre espécies de peixes que habitam ambientes de água doce
(Bataille & Bowen 2012; Hegg et al. 2013), mostram que é possível a verificação da origem
dessas populações de peixes através de registros químicos existentes em otólitos relacionados
aos ambientes em que estas espécies vivem ou viveram. Informações sobre a dinâmica de
dispersão de indivíduos de uma mesma população e implicações da existência de estrutura
metapopulacional para espécies de peixes de interesse múltiplos, como o C. temensis pode
contribuir para a criação de estratégias de manejo para essa espécie. Hoje em dia, existem
algumas tentativas de manejo participativo entre os usuários do recurso na região de Barcelos
no Amazonas, comunidades ribeirinhas, pescadores comerciais e pescadores esportivos. No
entanto, essas tentativas de acordo de pesca não tiveram êxito (Sobreiro et al., 2010). Nesse
sentido, se faz necessário produzir informações sobre a distribuição espacial do tucunaré-açu
que possa auxiliar no manejo efetivo das pescarias dessa espécie na região do Médio Rio
Negro.
Hipóteses
H0 = A composição química de otólitos não é um marcador natural do local de
nascimento de tucunaré-acú jovens e adultos;
H1 = A composição química de otólitos é um marcador natural do local de nascimento
de tucunaré-acú jovens e adultos;

11
H0 = As populações de tucunaré-açu Cichla temensis dos rios Cuiuni, Aracá, Demeni,
Preto, Padauari e Negro não são conectadas entre si através de movimentos de dispersão entre
diferentes ambientes;
H1 = As populações de tucunarés Cichla temensis dos rios Cuiuni, Aracá, Demeni,
Preto, Padauari e Negro, são conectadas entre si através de movimentos de dispersão entre
diferentes ambientes.
Objetivo geral
Verificar se a composição microquímica de otólitos atua como marcador ambiental do
local de nascimento de tucunaré-acú jovens e adultos e se existem populações diferentes dessa
espécie para os rios Cuiuni, Aracá, Demeni, Preto, Padauari e Negro.
Nesse contexto, esta pesquisa propõe:
1) Apresentar informações que indiquem a região de nascimento dos indivíduos de
tucunaré-açu e a distribuição espacial de suas populações, dentro do sistema aquático
estudado, utilizando isótopos e elementos traços acumulados temporalmente nos otólitos dos
peixes; e
2) Analisar a aplicação dos resultados dessa pesquisa para o estudo da dispersão e o
intercâmbio dos tucunaré-açu adultos entre os rios Cuiuni, Aracá, Demeni, Preto, Padauari e
Negro.

12
Objetivos específicos
a) Investigar a geoquímica de otólitos como potencial marcador natural do local
de nascimento de peixes jovens; ao longo dos rios Cuiuni, Aracá, Demeni, Preto, Padauari e
Negro na região do Médio Rio Negro;
b) Identificar o local de nascimento dos tucunarés com menos de um ano e
também de tucunarés adultos, coletados nesses diferentes locais, analisando a microquímica
da parte central dos otólitos;
c) Avaliar os padrões de movimentos de dispersão de indivíduos de tucunaré-açu
entre os ambientes de águas preta ou branca, através da variação microquímica do centro até a
borda dos otólitos, que possam indicar a ocorrência de metapopulação dessa espécie na
região; e
d) Sugerir estratégias de gestão do(s) estoque(s) de tucunaré-açu, incorporando
informações sobre a estrutura espacial e padrões de movimentos de dispersão entre os rios
acima citados.
Organização da Tese
O Capítulo I apresenta um estudo sobre a distribuição espacial dos berçários de
tucunarés-açu jovens, com menos de um ano, em diferentes tributários de águas brancas e
pretas na região do Médio Rio Negro. Esse estudo foi desenvolvido utilizando informações de
mapas geológicos relacionados aos tipos de sedimentos encontrados na região do Médio Rio
Negro. Os dados sobre a geologia da área do estudo foram posteriormente utilizados para
identificar a distribuição espacial dos isótopos de 87Sr/86Sr e dos elementos traços Sr/Ca e
Ba/Ca, encontrados na composição dos otólitos dos tucunarés-açu jovens.
No Capítulo II foram analisadas as razões isotópicas de 87Sr/86Sr e dos elementos
traços Sr/Ca e Ba/Ca, mensuradas na parte central dos otólitos de tucunarés adultos, para

13
determinar a origem de cada peixe. Foram comparadas as características microquímicas do
centro dos otólitos desses peixes adultos com a composição química dos otólitos dos peixes
jovens de origem conhecida. Ainda, foi examinado as diferenças microquímicas existentes na
parte central até a borda dos otólitos dos peixes adultos para verificar se esses indivíduos
realizavam movimentos de dispersão deixando os seus locais de nascimento e se retornavam
posteriormente aos seus locais de origem.
Uma seção de conclusões finais apresenta um sumário sobre os resultados obtidos em
ambos os capítulos. Os capítulos desta Tese encontram-se em formato de manuscritos,
conforme recomendação do Programa de Pós Graduação em Biologia de Água Doce e Pesca
Interior do INPA. Esses, por sua vez, estão formatados segundo as normas das revistas às
quais serão submetidos para publicação.
Metodologia geral
Área de estudo
A bacia do Rio Negro tem cerca de 715.000 km2 (Latrubesse e Franzinelli, 2005),
formando uma área que ultrapassa as fronteiras do nosso país (Franzinelli e Igreja, 2002). O
Rio Negro tem aproximadamente 1.700 km de extensão da nascente ao seu delta (Leon et al.,
2006), se tornando o maior rio de água preta do globo terrestre (Latrubesse e Franzinelli,
2005).
A bacia de drenagem desse rio banha regiões com rochas antigas e jovens (Horbe e
Santos, 2009), as quais, consequentemente, influenciam na coloração da água de seus
tributários (Sioli, 1984). A água preta é a que predomina na bacia do Rio Negro (Junk et
al.,1997; Latrubesse e Franzinelli, 2005), embora existam vários tributários de água branca ou
clara. Essa diversidade da geologia e os diferentes tipos de água de seus tributários foram de
crucial importância para o desenho amostral desta pesquisa (Figura 3).

14
Figura 3. Localização dos locais amostrais na região do Médio Rio Negro, município de Barcelos-Amazonas. Pt01 = Rio Cuiuni (água preta),
Pt02 = Confluência do Rio Demeni com o Rio Negro (água preta), Pt03 = confluência do Rio Aracá com o Rio Demeni (água mista), Pt04 = Rio
Aracá (água preta), Pt05 = Rio Demeni (água branca); Pt06 = confluência do Rio Padauari com o Rio Negro (água preta), Pt07 = confluência do
Rio Preto com o Rio Padauari (água mista), Pt08 = Rio Preto (água preta) e Pt09 = Rio Padauari (água branca).

15
Coleta e análise dos dados
Foram coletados 142 exemplares de Cichla temensis, sendo 99 de indivíduos jovens
com menos de um ano e 43 adultos. As coletas foram realizadas nos períodos de seca, em
janeiro de 2011 e de março 2012 (Tabela 1).
Tabela 1. Locais amostrais e comprimento padrão (mm) dos tucunarés-açu C. temensis
coletados na região do Médio Rio Negro – Amazonas.
Período das coletas
Locais amostrais
2011 2012
Juvenis
N (*Min-Máx)
Adultos
N (*Min-Máx)
Juvenis
N (*Min-Máx)
Adultos
N (*Min-Máx)
Pt 01 14 (10.5 – 180)
5 (245 -385)
Pt 02 - 4 (280-460) 15 (11-15) -
Pt 03 - - 15 (72-92) 6 (290-540)
Pt 04 - 1 (765) 11 (14-18) 5 (225-260)
Pt 05 - 2 (325-465) 15 (65-93) 4 (245-425)
Pt 06 - - 14 (40-53) 1 (240)
Pt 07 - 1 (570) - 4 (240-510)
Pt 08 - - 15 (13-22) 6 (465-625)
Pt 09 - 1 (500) - 5 (210-465)
Total 14 8 85 35
N = número de indivíduos (* Comprimento padrão em milímetros)
Os indivíduos foram submetidos a choque térmico em gelo logo após sua captura,
embalados, etiquetados e acondicionados em caixas térmicas. Os peixes adultos (maiores que
24 cm) foram submetidos a um processo de dissecação para a retirada dos otólitos ainda em
campo. Os otólitos dos indivíduos jovens foram extraídos em laboratório, com o auxílio de
equipamentos especializados (Figura 4).

16
Figura 4. A = Biometria dos peixes adultos, B = Extração dos otólitos dos peixes adultos, C =
Biometria dos peixes jovens e D = Extração dos otólitos dos peixes jovens.
Os otólitos extraídos foram limpos com água destilada, secos, identificados e
armazenados em tubos eppendorf, para posterior montagem. Em laboratório, cada otólito foi
colado sobre uma lamínula de vidro para microscopia com a sua superfície côncava voltada
para cima (Figura 5).
A B
C D

17
Figura 5. A = Material utilizado para a separação e montagem dos otólitos, B = Otólito fixado
em lamínulas.
Posteriormente cada otólito foi lixado com lixas diamantadas com textura entre 30 a
200 µm, até alcançar a visualização (sob microscópio óptico) dos anéis de crescimento com
distância aproximada de 30 micrometros (µm) do núcleo do otólito (Figura 6).
Figura 6. A = Lixamento dos otólitos, B = Microscópio óptico utilizado na visualização da
distância entre a superfície do otólito e sua parte central (µm).
Depois de lixados os otólitos foram submetidos a uma limpeza com água destilada em
banheira ultrassônica (Branson, 200), por dois minutos. Em seguida foram acomodados para
A B
A B

18
secagem. Todo o processo de limpeza e secagem foi realizado em uma câmara ultralimpa
(AirClean 600 PCR workstation) (Figura 7).
Figura 7. A = Banheira ultrassônica para limpeza dos otólitos, B = Câmara ultralimpa para
limpeza e secagem dos otólitos.
Após a limpeza e secagem dos otólitos, eles foram armazenados em caixas de
lamínulas limpas, e essas acondicionadas em sacolas plásticas do tipo zip lock (Figura 8).
Figura 8. A = Acondicionamento dos otólitos em sacolas e caixas esterilizadas, B =
Acondicionamento dos otólitos para transporte.
A B
A B

19
No Laboratório de Espectrometria de Massa do Instituto Oceanográfico Woods Hole
(WHOI) em Massachusetts nos Estados Unidos da América, os otólitos foram selecionados
para as análises isotópicas e de elementos traços, realizadas com queima a laser de parte da
superfície do otólito, com uso de equipamento óptico associado a um coletor múltiplo de
espectrometria de massa por plasma indutivamente acoplados (LA-MC-ICP-MS) (Figura 9).
Figura 9. A = Coletor Múltiplo de Espectrometria de Massas por Plasma Indutivamente
Acoplado, B = Imagem de um otólito com a região central mapeada (amarelo) e o transecto
estabelecido para queima (verde), prontos para a análise química a laser.
Cada otólito submetido à análise microquímica teve seus valores químicos
visualizados automaticamente através de uma tela de computador (Figura 10).
Figura 10. A = Queima a laser da superfície do otólito, B = Resultados das análises
microquímicas em computador.
A B
A B

20
Os dados coletados foram tabulados e posteriormente analisados estatisticamente
empregando-se os programas SPSS (SPSS, Inc) e Statistica 7.0 (StatSoft, Inc). Os resultados
obtidos com as análises foram utilizados na elaboração dos Capítulos I e II desta Tese.

21
Capítulo I
Garcez, R.C.S.; Humston, R.; Harbor, D. & Freitas, C.E.C.
Otolith geochemistry in young-of-the-year peacock bass
Cichla temensis for investigating natal dispersal in the Rio
Negro (Amazon - Brazil) river system. Submetido e aceito
na revista Ecology of Freshwater Fish, em 22 Março de
2014.

22
Otolith geochemistry in young-of-the-year peacock bass Cichla temensis for
investigating natal dispersal in the Rio Negro (Amazon - Brazil) river
system
R. C. S. Garcez1, R. Humston 2, D. Harbor3, and C. E. C. Freitas4
1 Programa de Pós-Graduação em Biologia de Água Doce e pesca Interior, Instituto Nacional
de pesquisas da Amazônia, Av. André Araújo, 2936. Aleixo, Manaus, AM 69.060-001,
Brazil.
2Department of Biology, Washington and Lee University, Lexington, VA 24450,
U.S.A.
3Department of Geology, Washington and Lee University, Lexington, VA 24450,
U.S.A.
4Departamento de Ciências Pesqueiras. Universidade Federal do Amazonas, Avenida General
Rodrigo Otávio Jordão Ramos, 3000. Manaus, AM 69.070-000, Brazil.
Running head: Otolith geochemistry of peacock bass C. temensis

23
Abstract
This study examined otolith geochemistry as a natural marker of natal origins in young-of-
the-year (YOY) C. temensis in the Negro River basin of Brazil. We analysed trace element
and isotopic composition of otoliths of YOY collected off spawning nests from the main stem
and major tributaries. These were compared to regional bedrock geologic composition to
explore underlying mechanisms of differences in otolith geochemistry. Our results suggest
that spatial differences in otolith geochemistry can be used to distinguish natal origins based
on 87Sr/86Sr, Sr/Ca, and Ba/Ca ratios. This approach allowed us to correctly classify 99% of
juvenile fish to their natal streams using cross-validation in a linear discriminant function
analysis (LDFA). Patterns of otolith isotopic composition correspond with patterns in regional
geology as expected based on previously demonstrated correlations, though some fine-scale
spatial differences cannot be accounted for by available geologic information. These results
demonstrate that otolith chemistry is valuable as a natural marker of natal origins in this
system and suggest that inferences from geologic maps may be useful for interpreting
movements based on otolith geochemical signatures. This information provides the basis for
future work to investigate the early life history and spatial ecology of this important cichlid.
Key words: Amazon Basin; Fish movement; Fresh water; Geochemistry; Strontium isotope;
Trace elements.

24
Introduction
The fish species of the Amazon Basin exhibit an incredible diversity of life histories,
and greater scientific understanding of fisheries ecology in the area is crucial to the
development of feasible and realistic strategies of conservation. Species of the genus Cichla
support economically important fisheries for small-scale commercial harvest as well as
recreational angling and associated ecotourism. The biology of Cichla species has been fairly
well documented, including population structure (Winemiller et al. 1997); age and growth
(Jepsen et al. 1997); abundance (Taphorn & Barbarino Duque 1993); feeding habits (Novaes
et al. 2004); spawning (Chellappa et al. 2003); and dispersal movement (Hoeinghaus et al.
2003). In general, reports about dispersal movements of Cichla spp. in the Amazon Basin
come from research using capture–mark–recapture technique (Taphorn & Barbarino Duque
1993; Hoeinghaus et al. 2003; Holley et al. 2008). However, fish must be large enough to bear
physical tags; therefore, these methods are less useful for tracking movement early in a fish’s
life history (Holley et al. 2008). Dispersal in the early life stages is important for
understanding spatial population dynamics and recruitment patterns across the river tributary
network (Humston et al. 2010) and therefore represents a considerable knowledge gap for
managing these fisheries.
Like many South American rivers, the Negro River near the equator in Brazil supports
a popular recreational sport fishery that focuses primarily on Cichla species (Hoeinghaus et
al. 2003; Holley et al. 2008). In this region, Cichla are abundant and colonise the main
channel of the Negro River and the majority of its white and black water tributaries (Jepsen et
al. 1997). Sport fishing occurs primarily during the receding water period between September
and April and brings approximately 1800 anglers to the Negro River Basin during this period
(Holley et al. 2008). Cichla reproduction takes place at the same time, following the receding
water period (Zaret 1980). These cichlids tend to prefer lentic habitats (Winemiller 2001),
make their nests on the river bottom (Jepsen et al. 1997) and demonstrate parental care
behaviour (Winemiller & Jepsen 1998). In dispersal following this period of parental care,
young-of-year could redistribute widely throughout the extensive network of rivers and
tributaries, particularly during the high flood season flows that follow in May–August.
Emigration and recruitment from spawning areas could therefore support distant fisheries,
obscuring the effects of harvest on local populations and/or creating source-sink dynamics.

25
Finding a way to effectively study movement across the full range of ontogeny is therefore
important for identifying such dynamics.
Isotopic and trace elemental composition of hard parts of animals has been
demonstrated to be useful as a natural fingerprint of past residency in studies of movement
ecology (Price et al. 2000; Walther et al. 2008; Walther & Limburg 2012; Wolff et al. 2012).
The chemistry of fish otoliths in particular has been demonstrated to be an effective tool for
tracking fish movement, as this structure readily incorporates the chemical signatures of
specific water bodies as influenced by their lithospheric geochemistry (Thorrold et al. 2001;
Humston & Harbor 2006; Walther et al. 2008; Nowling et al. 2011). The use of otolith
microchemistry as an alternative method of estimating natal origin and fish movement rates
could circumvent existing limitations to the use of physical tags in the study of movement by
Cichla; however, no study of Cichla species in the Amazon Basin has investigated this
possibility. In particular, isotopic chemistry (e.g., 87Sr/86Sr) holds potential as a distinct
marker in these rivers as it correlates with age and composition of underlying geology
(Barnett-Johnson et al. 2008; Bataille & Bowen 2012; Hegg et al. 2013), both of which vary
substantially across this system. In this study, we sought to determine whether otolith
geochemistry could provide a useful indicator of provenance of Cichla temensis in the Rio
Negro system. Correspondingly, we qualitatively explored whether regional differences in
otolith chemistry correspond with variation in underlying lithology based on available data.
Taken together, these sources of inference could provide a foundation for reconstructing
movement patterns across the full range of ontogeny in Cichla species in the Amazon Basin
based on retrospective analysis afforded by otolith chemical composition.
Materials and Methods
Study Area
The Negro River Basin (NRB) comprises over 715,000 km2 that extend from latitude
3°140S to 5°80N and from longitude 72°570W to 58°160W (Latrubesse & Franzinelli 2005)

26
and includes large, remote areas with minimal exploration or study. The NRB is a
transboundary basin, with its area divided among Brazil, Colombia, Venezuela and Guiana
(Franzinelli & Igreja 2002). The Negro River forms from the confluence of the Guiana River
and the Casiquiare Channel and flows roughly 1700 km to its terminus at the Solimões
(Amazon) River. It contributes a mean annual discharge of over 50,000 m3∙s-1 (Leon et al.
2006) to the system making it the second largest Amazon River tributary in this respect
(Latrubesse & Franzinelli 2005).
The Negro River catchment incorporates three distinct upper, middle and lower
regions: (i) the upper region beginning in the Guyana Shield, where the Negro River
headwaters are located, and ends in Santa Isabel do Rio Negro (Franzinelli 2011); (ii) the
middle region extends to the confluence with the Branco River and exhibits Cenozoic
sediments derived from Precambrian crystalline basement or Palaeozoic and Mesozoic
sedimentary rocks (Latrubesse & Franzinelli 2005); and (iii) the lower region begins at the
confluence with the Branco River, occurs nearly entirely in the Cenozoic floodplain, passing
across the Archipelago of Anavilhanas and ending at the confluence with the Solimões River
(Franzinelli & Igreja 2002; Fig. 1). The chemical composition of the water in the Negro River
and its tributaries is highly variable and strongly influenced by forest characteristics and
geological location (Horbe & Santos 2009). Within the NRB, black water with high acidity is
predominant (Stallard & Edmond 1987).

27
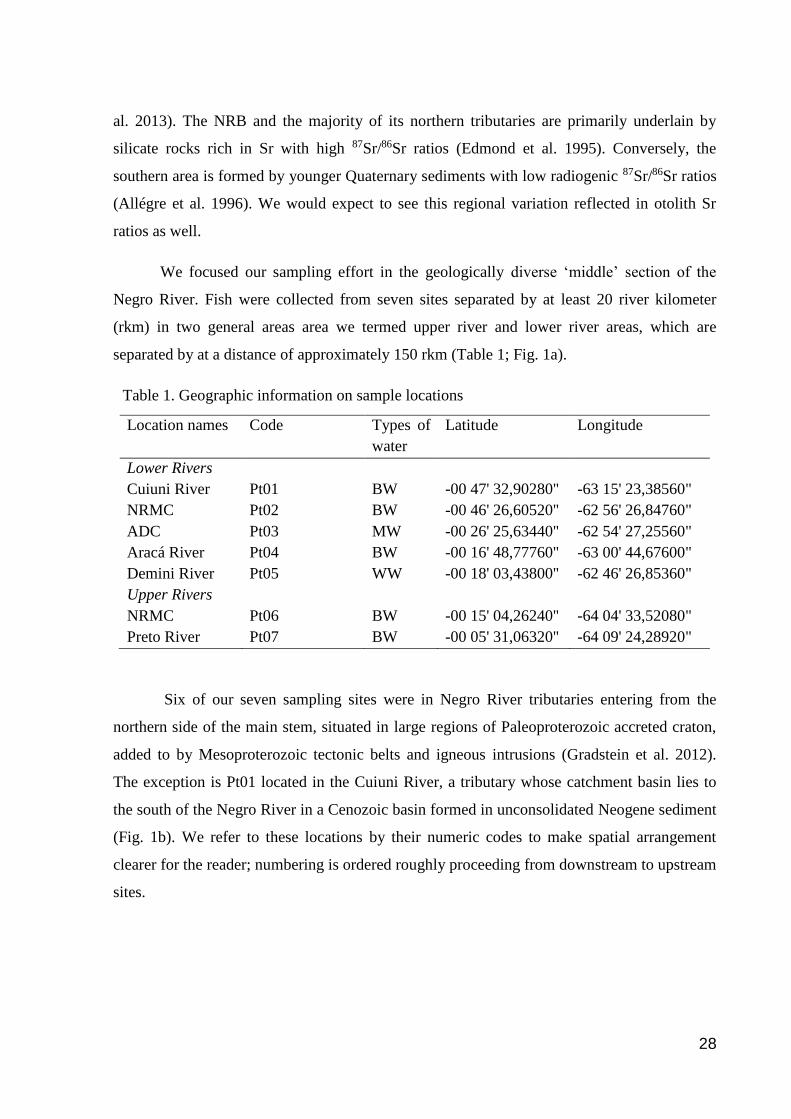
Fig.1. Location and geological conditions of the study area: (a) location of the sample
sites with respect to the major tributaries of the Negro River, (b) ages of the geological units
from Ferreira et al. (2005) and dashed outlines of the basins upstream of the sample sites, and
(c) geologic units as potential sources of isotopic differentiation.
Geologic age corresponds roughly with 87Sr/86Sr ratios due to radiogenic Sr
accumulation (Faure 2001), and geologic composition determines how geochemistry of
bedrock and corresponding weathering can contribute to Sr abundance and isotopic
composition in surface waters (Barnett-Johnson et al. 2008; Bataille & Bowen 2012; Hegg et
28
al. 2013). The NRB and the majority of its northern tributaries are primarily underlain by
silicate rocks rich in Sr with high 87Sr/86Sr ratios (Edmond et al. 1995). Conversely, the
southern area is formed by younger Quaternary sediments with low radiogenic 87Sr/86Sr ratios
(Allégre et al. 1996). We would expect to see this regional variation reflected in otolith Sr
ratios as well.
We focused our sampling effort in the geologically diverse ‘middle’ section of the
Negro River. Fish were collected from seven sites separated by at least 20 river kilometer
(rkm) in two general areas area we termed upper river and lower river areas, which are
separated by at a distance of approximately 150 rkm (Table 1; Fig. 1a).
Table 1. Geographic information on sample locations
Location names Code Types of
water
Latitude Longitude
Lower Rivers
Cuiuni River Pt01 BW -00 47' 32,90280'' -63 15' 23,38560"
NRMC Pt02 BW -00 46' 26,60520'' -62 56' 26,84760"
ADC Pt03 MW -00 26' 25,63440'' -62 54' 27,25560"
Aracá River Pt04 BW -00 16' 48,77760'' -63 00' 44,67600"
Demini River Pt05 WW -00 18' 03,43800'' -62 46' 26,85360"
Upper Rivers
NRMC Pt06 BW -00 15' 04,26240'' -64 04' 33,52080"
Preto River Pt07 BW -00 05' 31,06320'' -64 09' 24,28920"
Six of our seven sampling sites were in Negro River tributaries entering from the
northern side of the main stem, situated in large regions of Paleoproterozoic accreted craton,
added to by Mesoproterozoic tectonic belts and igneous intrusions (Gradstein et al. 2012).
The exception is Pt01 located in the Cuiuni River, a tributary whose catchment basin lies to
the south of the Negro River in a Cenozoic basin formed in unconsolidated Neogene sediment
(Fig. 1b). We refer to these locations by their numeric codes to make spatial arrangement
clearer for the reader; numbering is ordered roughly proceeding from downstream to upstream
sites.

29
Sample collection
Fish were collected at two separate times, with the first collection occurring in March
2011; only one site was sampled (Pt01) in this period due to poor conditions. A second
collection effort was organised in January 2012 to increase sample size and spatial coverage.
Collections were conducted during these months to correspond with spawning and so that
young-of-the-year (YOY) C. temensis could be collected near their natal nests in the shallow
floodplain streams and lake margins while still under parental care. Zaret (1980) reported that
YOY Cichla spp. Do not disperse from their nursery ground until they are at least 2 months in
age and 60 mm standard; therefore, we only collected YOY well below this size. Captured
fish were euthanized via thermal shock, covered with ice and stored in sealed plastic bags.
They were labelled and transported to the fish biology laboratory facility at the Federal
Amazon University. In the laboratory, the standard length (SL) of each YOY was measured to
the nearest 1 mm (±1), and the fish were weighed to the nearest 0.01 g before otolith
extraction.
Otolith preparation and analysis
Sagittal otoliths were removed under a dissecting microscope, triple-washed in
ultrapure deionised water to remove attached fleshy tissue, dried and stored in a 2 ml
Eppendorf tube for further analysis. Then, they were transported (IBAMA licence number
110752) to Washington and Lee University (WLU - Virginia, USA) for preparation. The right
otolith from each individual fish was selected for chemical analysis, and the left otolith was
saved as a potential replacement. Otoliths were mounted with the convex surface up (i.e.,
sulcus up) on microscope slides using gel-consistency cyanoacrylate. Otoliths were then
hand-polished on the saggital plane using 30 µm and 3 µm diamond lapping film to a distance
of approximately 50 µm above the core.
All mounted and polished otoliths were cleaned in ultrapure (Milli-Q, EMD Millipore,
Billerica, MA, USA) water for two minutes using an ultrasonic bath (Bransonic 200) under

30
class 100 clean conditions. Afterwards, the otoliths were dried in a laminar flow hood
(approximately 2 h) and stored for transport to the Woods Hole Oceanographic Institute
(WHOI).
Otoliths with a diameter of length and width of more than 250 µm were selected for
chemical analyses, to ensure enough area and mass for laser ablation. An exception was made
for the otoliths from Pt04, which were approximately 200 µm in length and 100 µm in width.
The analytical procedure was performed using a Neptune multicollector ICP-MS (Thermo
Scientific, Waltham, MA, USA) attached to a 193 nm laser ablation system (New Wave
Research, ESI Inc., Portland, OR, USA), which was used to ablate a raster centred on the core
of the otolith. The laser ablated a 100-µm-diameter spot per cycle while running at 100%
power, 10 Hz repetition rate and 6 µm∙s-1 scan speed. We were able to simultaneously
measure 87Sr/86Sr, Sr/Ca and Ba/Ca ratios from a single raster following procedures outlined
in Walther and Thorrold (2010). Blank samples and two different standard reference materials
were analysed between every 12 otolith samples. Reference materials included a strontium
carbonate isotopic standard (SRM 987) and a dissolved otolith certified reference (FEBS-01;
Sturgeon et al. 2005). Blank, mass bias and interference corrections were applied as outlined
in Walther et al. (2008) and Walther and Thorrold (2010). Mean 87Sr/86Sr in SRM 987
standard measurements (N = 17) was 0.7102502 with a standard deviation (SD) of 0.0000218,
which is within 1 SD of the known value of 0.71024 for SRM 987.
Statistical analysis
We initially employed analyses of variance – ANOVA (Zar 1999) to examine spatial
differences in otolith 87Sr/86Sr, Sr/Ca and Ba/Ca composition individually. Tukey’s HSD test
(Zar 1999) was employed to identify pairwise differences when significant differences were
identified among sampling locations. This allowed us to elucidate how each variable varied
among locations and corresponded to geologic variation. We then used linear discriminant
function analysis (LDFA) to determine the degree to which multivariate geochemical
signatures in otoliths could be used to distinguish origins of individuals from our sample.
Variables were entered into the model using a stepwise forward method, using Wilk’s

31
Lambda and F-statistic probabilities to determine sequence of variable addition and evaluate
model improvement, respectively (Huberty 1994). A serial deletion cross validation procedure
was used to quantify classification accuracy and error rates. All statistical analyses were
conducted using SPSS software (SPSS, Inc., IBM Corporation, Armonk, NY, USA).
Geologic analyses
Due to the sheer number of tributaries and the vast size and remoteness of this area, it
is not feasible to collect a ‘catalog’ of reference data on otolith chemical composition from all
possible natal sources in this system. Therefore, we investigated correspondence between
geologic composition of watersheds and chemical composition of otoliths collected from
sampling locations. We focused on 87Sr/86Sr in particular in this analysis, as previous
examples demonstrate that this can be correlated with geologic data (Barnett- Johnson et al.
2008; Bataille & Bowen 2012; Hegg et al. 2013) and that it is often less variable than trace
element chemistry in surface waters (with notable exceptions, e.g., Clow et al. 1997; Aubert et
al. 2002; Voss et al. 2014). Following Barnett-Johnson et al. (2008) and Bataille and Bowen
(2012), we used available data to examine the age of geologic features in watersheds as well
as their relative composition. Although limited data were available in this regard, we
attempted to infer the degree to which particular types of rocks in the study area would
contribute radiogenic strontium and followed data reported in Bataille and Bowen (2012) for
guidance. We divided the map units of middle Negro River region following Ferreira et al.
(2005) into the following broad units: (i) Paleoproterozoic basement of the Santa Izabel do
Rio Negro unit, which comprises metasedimentary, metamorphosed felsic volcanic and
plutonic rocks including granodiorite, quartz diorite, migmatites and monzogranites, and
gneiss in the northwest corner; (ii) Paleoproterozoic granite intrusions (syeno- and
monzogranite); (iii) Mesoproterozoic granite intrusions (syeno- and monzogranite, 93%) and
minor mafic and ultramafic intrusions (< 7%); (iv) Mesoproterozoic quartzite; (v) Cenozoic
sediments including river floodplains, Quaternary terraces and aeolian deposits. We then
quantified the relative composition of watersheds in these different geologic categories and
compared these with otolith 87Sr/86Sr composition from sampling locations in those
watersheds. Finally, we examined scatter plots of per cent rock type against mean 87Sr/86Sr

32
observed in collections and performed simple linear regressions to explore the degree to
which watershed geologic composition could be used to predict stream isotopic chemistry.
Results
We collected a total of 175 YOY C. temensis from seven experimental reaches in two
general areas, designated as upper river and lower river areas in this study (Table 2). The
sampled fish ranged in weight from 0.02 to 69.25 g (±25.30) and in standard length (SL) from
1.62 to 8.98 cm (±2.95). This size range suggests these fish were still in close proximity to
their spawning site at the time of collection. Their mean age in months was 2.41 ± 1.39. Of
these, a total of 99 otoliths were of sufficient size for laser ablation and chemical analyses.
Table 2. Strontium 87Sr/86Sr isotope and trace element ratio values (mean ± SD) from
the otoliths of young-of-the-year C. temensis (sampling site names are listed in Table 1).
Area Location
Code
N 87Sr/86Sr Sr/Ca (mmol/mol-1) Ba/Ca (mmol/mol-1)
Lower rivers
Pt01 14 0.71314 ± 0.00066 0.01458 ± 0.00303 0.00120 ± 0.00057
Pt02 15 0.74097 ± 0.00147 0.00624 ± 0.00098 0.00041 ± 0.00017
Pt03 15 0.78554 ± 0.00104 0.00945 ± 0.00155 0.00081 ± 0.00024
Pt04 11 0.78292 ± 0.00101 0.00621 ± 0.00130 0.00127 ± 0.00010
Pt05 15 0.73494 ± 0.00016 0.00480 ± 0.00034 0.00026 ± 0.00006
Upper rivers Pt06 14 0.74230 ± 0.00086 0.00847 ± 0.00051 0.00074 ± 0.00010
Pt07 15 0.74211 ± 0.00100 0.00552 ± 0.00236 0.00135 ± 0.00025
Results of univariate analyses (ANOVA) indicated significant differences among
sample sites in otolith 87Sr/86Sr (F6, 92 = 9919.871, P < 0.0001), Sr/Ca (F6, 92 = 55.323, P <
0.0001) and Ba/Ca ratios (F6, 92 = 35.250, P < 0.0001). Otolith strontium isotopic ratios
varied widely among locations, with location means ranging from 0.71314 (Pt01) to 0.78554
(Pt03) with low variance within sites. Tukey’s HSD tests revealed pairwise differences among
all locations except the two most upstream sampling sites (Pt06 and Pt07). Differences
between the two mainstem Negro River sites (Pt02 and Pt06) were subtle, with the
downstream location (Pt02) slightly lower than upstream. In the lower sampled section, the
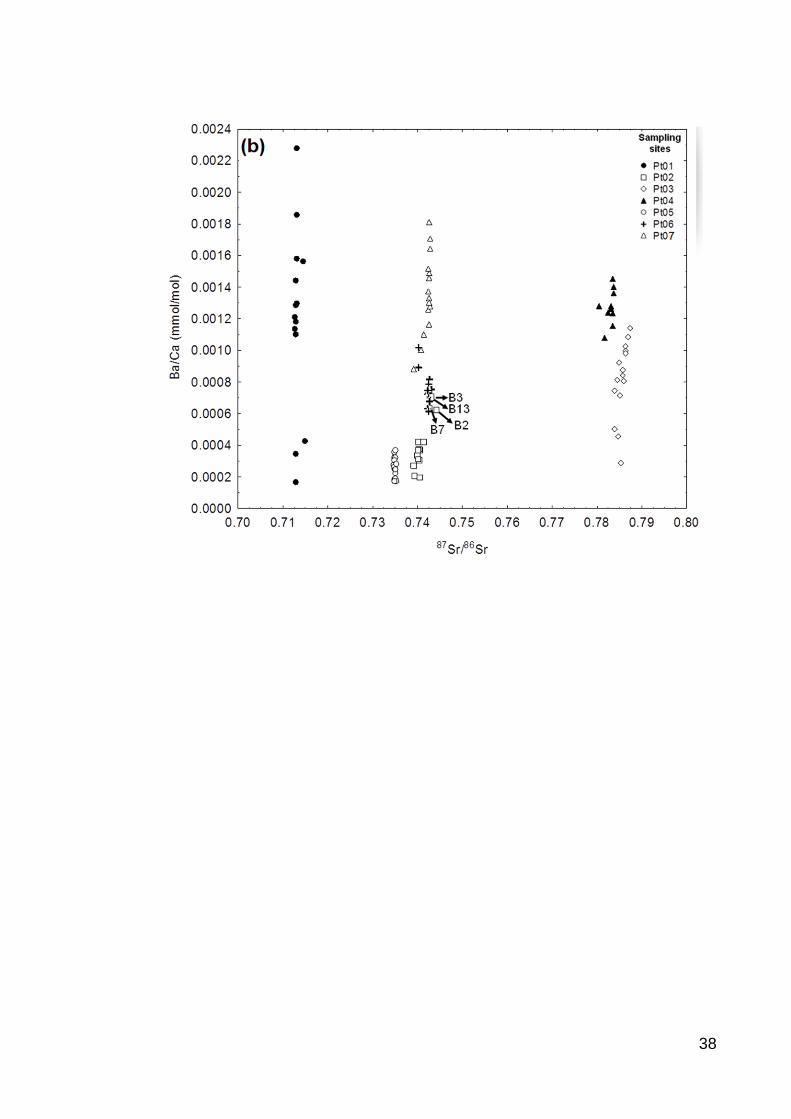
southern tributary had the lowest 87Sr/86Sr by far. An interesting pattern emerged among the

33
northern tributaries of the lower section, which includes three sites comprising two smaller
tributaries (Pt05 and Pt04) and one larger tributary (Pt03) formed by their confluence (Fig.
2a). Otoliths sampled from the western tributary (Pt04) were very high in radiogenic
strontium, while otoliths collected from the eastern tributary (Pt05) were much lower.
Although we would expect the area below the confluence of these rivers (Pt04) to produce
otoliths with 87Sr/86Sr somewhere between these two, we actually observed the highest overall
levels of radiogenic strontium in otoliths from this location.
Analyses suggested greater separation among sites based on Sr/Ca ratios. Locations
Pt02, Pt04 and Pt07 were indistinguishable with respect to Sr/Ca, but all other remaining
locations separated out individually (Fig. 2b). Uniquely high Sr/Ca was observed in the
southern tributary sampled at site Pt01. By contrast, univariate screening suggested that
otolith Ba/Ca appeared to hold the lowest potential as a discriminatory variable, with
sampling sites roughly separating into two groups: locations Pt02 and Pt05 form one group
with relatively low Ba/Ca, while all other locations had similarly higher Ba/Ca ratios (Fig.
2c). Variance within locations was high leading to substantial overlap in observed ranges
among sites.

34

35

36
Fig. 2. Mean isotopic and trace element concentration ratios in the otoliths of YOY
fish groups. (a) 87Sr/86Sr ratio, (b) Sr/Ca ratio and (c) Ba/Ca ratio. Means sharing a lowercase
letter are not significantly different (Tukey’s HSD pairwise mean comparisons, α = 0.05).
In LDFA, 99% of YOY C. temensis were classified accurately with respect to their
natal origin in cross-validation based on all three isotopic and trace element variables. As
expected from results of univariate analyses, 87Sr/86Sr was the most influential variable in
discriminant functions, with an eigenvalue of 894.476 (98.5% of variance) followed by Sr/Ca
with 11.835 (1.3% of variance) and by Ba/Ca 1.987 (0.2% of variance). All three variables
were retained in stepwise model construction (P < 0.001 for each variable addition). The only
error committed in crossvalidation was a single fish from location Pt02, which was predicted
to be from location Pt06. This individual fish is 12.0 mm and 0.0092 g, indicating that it was
still under parental care (Winemiller et al. 1997) and unlikely to have dispersed from its natal
site, especially over this great a distance (~150 rkm). Bivariate plots (Fig. 3a-c) demonstrate
overlap occurs between samples from these locations in all bivariate combinations, providing
a simpler, statistical explanation for this misclassification.

37
38

39
Fig. 3. Microchemical isotopic and trace element ratios from the otoliths of YOY C.
temensis distributed within groups and between areas. Sr/Ca versus 87Sr/86Sr ratios (a), Ba/Ca
versus 87Sr/86Sr ratios (b) and Ba/Ca versus Sr/Ca ratios (c). The sample location codes are
listed in Table 1. YOY, young-of-the-year.
Spatial patterns and correlations with geology
The most significant geological differentiation of the sample locations identified in our
analysis is the southern versus northern tributaries (Table 3). The southern watersheds
comprise depositional lowlands of the Solimões-Negro divide, which includes extraregional
sediments of the Amazon Basin derived in part from the Andean highlands. The northern
watersheds include Proterozoic igneous and metamorphic highlands that have shed sediments
into the basin during the Cenozoic. As would be predicted, 87Sr/86Sr was lowest in the
southern tributary sampled (Pt01), reflecting the dominance of unlithified Neogene sediments
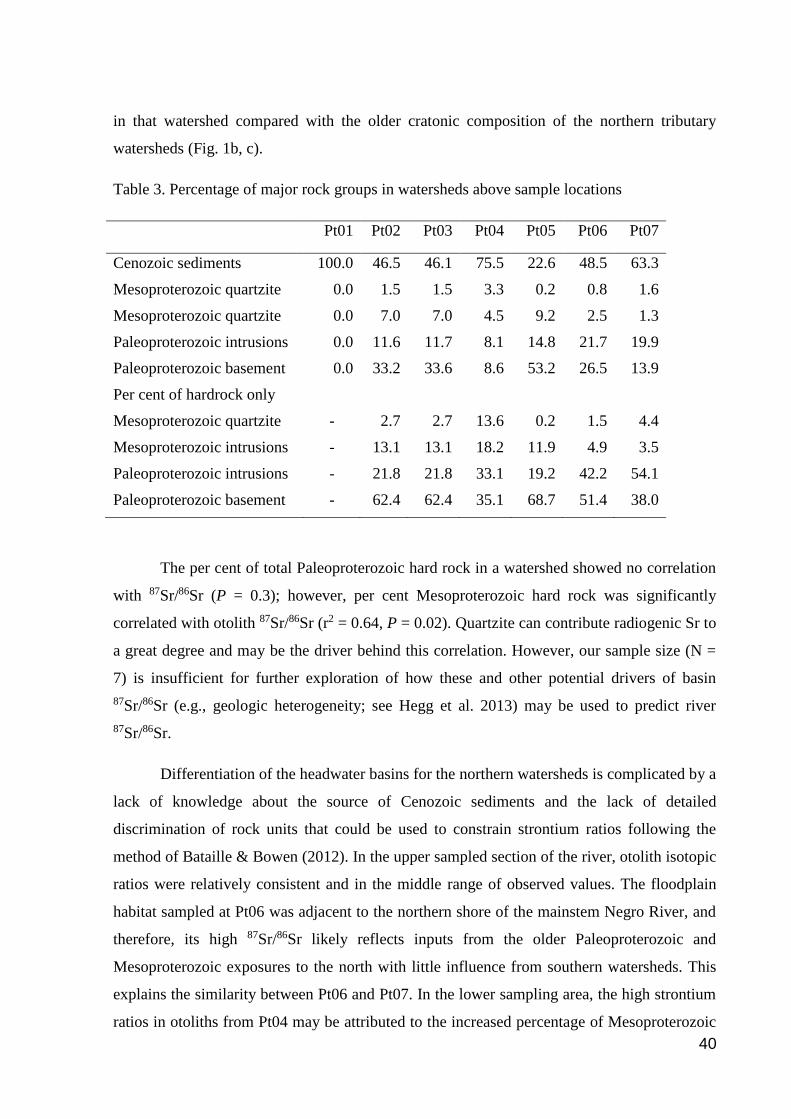
40
in that watershed compared with the older cratonic composition of the northern tributary
watersheds (Fig. 1b, c).
Table 3. Percentage of major rock groups in watersheds above sample locations
Pt01 Pt02 Pt03 Pt04 Pt05 Pt06 Pt07
Cenozoic sediments 100.0 46.5 46.1 75.5 22.6 48.5 63.3
Mesoproterozoic quartzite 0.0 1.5 1.5 3.3 0.2 0.8 1.6
Mesoproterozoic quartzite 0.0 7.0 7.0 4.5 9.2 2.5 1.3
Paleoproterozoic intrusions 0.0 11.6 11.7 8.1 14.8 21.7 19.9
Paleoproterozoic basement 0.0 33.2 33.6 8.6 53.2 26.5 13.9
Per cent of hardrock only
Mesoproterozoic quartzite - 2.7 2.7 13.6 0.2 1.5 4.4
Mesoproterozoic intrusions - 13.1 13.1 18.2 11.9 4.9 3.5
Paleoproterozoic intrusions - 21.8 21.8 33.1 19.2 42.2 54.1
Paleoproterozoic basement - 62.4 62.4 35.1 68.7 51.4 38.0
The per cent of total Paleoproterozoic hard rock in a watershed showed no correlation
with 87Sr/86Sr (P = 0.3); however, per cent Mesoproterozoic hard rock was significantly
correlated with otolith 87Sr/86Sr (r2 = 0.64, P = 0.02). Quartzite can contribute radiogenic Sr to
a great degree and may be the driver behind this correlation. However, our sample size (N =
7) is insufficient for further exploration of how these and other potential drivers of basin
87Sr/86Sr (e.g., geologic heterogeneity; see Hegg et al. 2013) may be used to predict river
87Sr/86Sr.
Differentiation of the headwater basins for the northern watersheds is complicated by a
lack of knowledge about the source of Cenozoic sediments and the lack of detailed
discrimination of rock units that could be used to constrain strontium ratios following the
method of Bataille & Bowen (2012). In the upper sampled section of the river, otolith isotopic
ratios were relatively consistent and in the middle range of observed values. The floodplain
habitat sampled at Pt06 was adjacent to the northern shore of the mainstem Negro River, and
therefore, its high 87Sr/86Sr likely reflects inputs from the older Paleoproterozoic and
Mesoproterozoic exposures to the north with little influence from southern watersheds. This
explains the similarity between Pt06 and Pt07. In the lower sampling area, the high strontium
ratios in otoliths from Pt04 may be attributed to the increased percentage of Mesoproterozoic

41
quartzite or the higher percentage of Mesoproterozoic granite intrusions in the exposed rocks
of the highlands (Fig. 1b,c and Table 3). In addition, aeolian sediments are only exposed in
basin 4, where they comprise over 5% of the basin (Fig. 1c). The source of these late
Quaternary sediments (Tatumi et al. 2002) is unknown but may include winnowing of
sediments from the igneous and metamorphic highlands and therefore could be a source of the
isotopic difference. Our geologic analyses do not shed light on why otoliths from fish
collected at Pt03 would be enriched in radiogenic strontium compared with its adjacent
tributary (Pt04).
Discussion
Chemical composition of YOY otoliths from C. temensis showed strong potential for
differentiating natal origins, particularly for distinguishing between fish emerging from
southern versus northern tributaries based on Sr isotopic and elemental composition. The
former is likely the most reliable indicator of provenance in this respect, as it is less
susceptible to temporal variation in stream water chemistry. Given the large differences in
87Sr/86Sr we observed between northern and southern tributaries and its strong correspondence
with regional geology, this marker is likely also useful for reconstructing movement patterns
between northern and southern tributaries over entire life histories from adult otoliths
(Walther & Limburg 2012). Differentiating movement between tributaries from upstream
versus downstream reaches of the Negro River (e.g., between upper and lower areas in our
study) based solely on isotopic ratios is less straightforward. As spawning takes place in
floodplain lakes adjacent to the rivers, it is possible that fine-scale variation in geology
influencing the water chemistry of lake inputs from groundwater or small feeder streams is
reflected in otolith chemistry of YOY from these habitats. Recent examples demonstrate
methods for characterising ‘isoscapes’ of spatial 87Sr/86Sr variation based on geologic
composition and weathering dynamics (Bataille & Bowen 2012; Hegg et al. 2013), and this
approach could be useful for explaining observed fine-scale variation and predicting tributary
87Sr/86Sr. However, the lack of sufficient data differentiating regional geology to the same
degree precludes applying these same methods in this study area.

42
Although temporal variation in trace element-to-calcium ratios in rivers can reduce
their reliability as a marker of provenance in a river tributary network, there is some
indication in the geology of the region that Sr/Ca could provide reasonable information for
retrospective analysis of movement from otoliths. Our limited sample was not sufficient to
explore this correlation at length; however, recent examples (Hegg et al. 2013) demonstrate
that this is a rich area for further consideration. Although geologic data can provide inference
on relative spatial variation in otolith trace element composition (Humston & Harbor 2006),
predictions of trace elemental composition of stream water (and hence otolith chemistry) from
geology are not possible to the same extent as for isotopic composition. Likewise, these
methods cannot account for differences in trace element incorporation in otoliths attributable
to changes in fish physiology. We therefore suggest that strontium isotopes provide the most
reliable ‘natural tag’ for reconstructing fish movement in this system from otolith chemistry
chronologies.
This research confirms that otolith geochemical signatures in YOY C. temensis are
distinct and specific to their river of origin. This method can therefore be applied to examine
the number of spawning sources contributing to a regional stock, and the relative contribution
of rivers differentiated by unique isotopic ‘signatures’ (e.g., northern vs. southern tributaries)
can be elucidated as well. Isotopic ratios show potential for retrospective analysis of adult
movement patterns; however, differentiating movement among northern tributaries may be
complicated by similarity of isotopic ratios among rivers. Applicability of these methods for
studying movement of Cichla and other fish species in the Negro River system can be
measurably enhanced by further characterization of bedrock and stream water geochemistry in
this region.
Acknowledgments
We thank B. A. Harris for his helpful review of previous drafts and Mr. J. A. D.
Siqueira for his assistance in the field collecting the samples. Financial support of this study
was provided by CAPES, CNPq, FAPEAM and INCT-Adapta. Laboratory, and logistic
support also was provided by UFAM, INPA, I-PIATAM and WLU.

43
Capítulo II
Garcez, R.C.S.; Humston, R. & Freitas, C.E.C. Philopatry
and metapopulation patterns for the peacock bass Cichla
temensis of the middle Negro River Basin (Amazonas -
Brazil): an otolith chemical analysis. Manuscrito
formatado para a Ecology of Freshwater Fish.

44
Philopatry and metapopulation patterns for the peacock bass Cichla
temensis of the middle Negro River Basin (Amazonas - Brazil): an otolith
chemical analysis.
Raniere C. S. Garcez1, R. Humston 2, and C. E. C. Freitas3
1 Programa de Pós-Graduação em Biologia de Água Doce e pesca Interior, Instituto Nacional
de pesquisas da Amazônia, Av. André Araújo, 2936. Aleixo, Manaus, AM 69.060-001,
Brazil.
2Department of Biology, Washington and Lee University, Lexington, VA 24450,
U.S.A.
3 Department of Fisheries Sciences. Federal University of the Amazonas, Avenida
General Rodrigo Otávio Jordão Ramos, 3000. Manaus, AM 69.070-000, Brazil.
Running head: Otolith geochemistry and philopatry of peacock bass C. temensis

45
Abstract
Strontium isotope and trace elements from otolith of peacock bass Cichla temensis
(Humboldt, 1833) indicated that it is possible to track movements of young-of-year (YOY)
and adult fish in the Negro River and its tributaries. A canonical discriminant function
analysis (CDFA) shows in a cross table that 93.9% of YOY and 41.9% of adult peacock bass
otolith core fingerprints matched correctly with their nursery areas. Variation in 87Sr/86Sr
isotopes ratio from a transect line on the otolith surface indicated the existence of isolated and
mixed populations for adult fish in the study area. Our results also confirm that adult peacock
bass present a meta-population structure and disperse up and down river with philopatric
behaviour. Therefore, the present report offers new information about spatial distribution of
C. temensis in their natural environment, which could be used to support effective strategies
for stocks and fisheries management and conservation.
Keywords: Fish management; geochemistry; homing, otolith fingerprints; trace elements.

46
Introduction
The Negro River basin is a vast and complex aquatic ecosystem that supports large
Cichla populations (Hoeinghaus et al., 2003). The main Cichla species in this huge aquatic
environment is the peacock bass Cichla temensis (Humboldt, 1833), which is the largest
representative of the family Cichlidae. It attains large sizes of approximately 81.5 cm
Standard Length (SL) (Taphorn and Barbarino-Duque, 1993) and more than 12kg (Montana et
al., 2006; Holley et al., 2008).
In this region three Cichla species, C. monoculus, C. temensis, and C. orinocensis,
(Kullander and Ferreira, 2006; Willis et al., 2010) co-exist, but among these species C.
temensis reportedly occurs in a greater variety of aquatic habitats than its congeners (Jepsen et
al., 1997; Hoeighaus et al., 2003). Other Cichla species naturally occupy black and white
waters in the Orinoco River (Colombia and Venezuela) and the Negro River in Brazil
(Winemiller et al., 1997; Brinn et al., 2004; Kullander and Ferreira, 2006).
The abundance and voracity of this large-sized predator (Winemiller 2001) gained the
attention of the catch-and-release sport-fishing industry (Macrander, 2010) that has emerged
in the Negro River, attracting over 2000 anglers to the region annually (Holley et al., 2008).
Despite the importance of the Cichla species, both as a food resource and sport fishing target
(Hoeinghaus et al., 2003; Willis et al., 2007), makes different users (subsistence, commercial
and sport fishing) competing for the same resource, that brought the necessity to increase
efforts to study the distribution of Cichla species populations in the Negro River Basin, in
order to develop an effective fisheries management strategy.
The knowledg about fish movements dispersiton among locations is the first step to
understand the fish population dynamics. For that is crucial to understand the concept of
metapopulations (Levins, 1970), that consicts in a conjunct of separated populations that are
conected by the emigration and immigrations of individuals in those aquatics system (Kritzer
and Sale, 2004).
The metapopulation is a potential approach to understand the spatial distribution of
animal populations (Levins, 1970) in a variety of aquatic and terrestrial systems (Hanski and
Simberloff, 1997; Kritzer and Sale, 2004). One way to better understand this idea for systems,
such as large rivers with adjacent floodplains, would be to assume that lakes and rivers are

47
patches of aquatic habitats for resident fish. The fish movements between the patches of these
open and connected systems could be interpreted in terms of metapopulational dynamics.
Mapping fish population connectivity through adults’ dispersal and their emigration or
immigration movements among different geological natal patches is the basis of
understanding this theory (Thorrold et al., 2001).
Adult peacock bass are presumed to be sedentary fish (Granado-Lorencio et al., 2005),
and have been classified as an equilibrium species (Winemiller, 2001). Meanwhile, some
studies have reported that the Cichla species realizes small-scale dispersal movements
between main channels and lakes (Hoeinghaus et al., 2003). On the other hand, this
supposition has basically been tested using conventional approaches such as field
observations (Lowe-McConnell, 1969), tagging (Taphorn and Barbarino-Duque 1993;
Hoeinghaus et al., 2003; Holley et al., 2008; Willis et al., 2010), radio telemetry (Thorstad et
al., 2001), and genetic procedures (Macrander, 2010). The information collected, however, is
inconclusive and leaves many lacunas, making it difficult to understand the movements of C.
temensis and its distribution pattern, which may be attributed to problems with these
methodologies.
Previous studies using fish’s otolith geochemical signatures (Brazner et al., 2004;
Walter and Thorrold, 2006; 2010; 2013) have shown accurate results tracking small and large
fish in marine (Humphreys Jr. et al., 2005) and fresh water habitats (Humston et al., 2010).
These findings confirm that this technique is useful as a natural marker approach (Thorrold et
al., 2001), and an effective apparatus to resolve the discrepancy regarding frequency of large
scale movements (between rivers with different geological composition) by drawing a
complete route history for each fish individually among river habitats.
It is possible to identify every different geochemical locations through which a fish
passes during its lifetime because fish otoliths are calcareous structures formed by the
sequential addition of inert layers of calcium carbonate (Campana 1997; 1999). The calcium
carbonate is derived from surrounding aquatic environment, and shapes a geochemical
fingerprint (Walther and Thorrold, 2006). Even if the chemical changes are minuscule, we can
track the fish movements across different geologic locations (Humston and Harbour, 2006).
Based on this premise, this study intends to verify the peacock bass spatial distribution
in the Negro River basin in Brazil, by investigating if the young-of-year C. temensis otoliths’

48
core microchemical signatures differ enough between different geological sampling sites to be
used as an effective reference for tracking back its birth place. If so, it seeks to verify the
otolith core isotope and elemental composition for the existence of similarities between YOY
and adult chemical fingerprints that could be used to predict adult fish origins (i.e. birth
place).
Also, in order to verify the existence of adult C. temensis’ spatial distribution and
movements down or upstream, this study analyses variation in 87Sr/86Sr isotopes in a laser
ablated transect from the otoliths surfaces of adults fish for the purpose of identifying some
pattern of philopatry or metapopulation structure. In this way, this study intends to test the
following null hypotheses:
i) There is no difference in the chemistry between otolith core of the YOY and adult
peacock bass; ii) Adult peacock bass’ otolith core isotope and elemental composition do not
vary among tributaries of the Negro River with different water types; iii) There are no
chemical differences in the values of 87Sr/86Sr isotopes from the adult otoliths expressed in a
transect line (from core to the edge) that could indicate fish movements among sampling sites;
and iv) Adult C. temensis do not return to their nursery areas after leaving it.
Material and Methods
Study area
The study area is located at the Middle Negro River region, about 500 kilometers
upstream from Manaus, the capital of Amazonas State, Brazil (Holley et al., 2008) (Figure 1).
The Negro River is the largest black water river in the globe (Latrubesse & Franzinelli, 2005),
and it is formed of a complex network of tributaries, which differ greatly in their geologic
basement and water types (Gaillardet et al., 1997; Queiroz et al., 2009).

49
Figure 1. Study area and geographic distribution of sampling sites in the middle Negro River, Barcelos Municipality. Different colors
in the basin rivers delineated its areas.
B
A

50
Sample collections and preparations
A total of 142 individuals of Cichla temensis were collected (43 adults and 99 young-
of-year) during two ocasions, March 2011 and January 2012 from nine sampling sites in the
Middle Negro River Basin. Data values from sampling sites Pt07 and Pt09 related to YOY
were absent. Consequently, to get data to represent adults birth from this two locations, the
otolith core chemical fingerprints from fish coded as G2 and ID1 were used to represent Pt07
and Pt09, respectively. The distance between samples location was about 25 rkm (river
kilometer), this procedure was made once C. temensis as reported by Hoeinghaus et al. (2003)
made movements dispersion from their targed locatins in a distance around 21 Km. The
sample locations were choose in trybutaries located in diferente geological formation and also
with diferente water types (black or white). Peacock bass were caught with rod and reel, dip
net, and line and hook. Standard length (SL) was measured for both adults and YOY to the
nearest centimeter and millimeter, respectively, and weight was recorded to the nearest gram.
To ensure that YOY individuals were collected from their natal rivers, they were sampled
while in their nests or close to them. Further, all specimens were subsequently euthanized
with ice, saved in plastic bags with identification labels, and transported in insulated boxes
with ice to the laboratory.
In the laboratory Sagittae otoliths were removed from each fish under dissecting
microscopes, rinsed in ultrapure deionizer water to facilitate the removal of remaining tissue,
and stored in numbered two-milliliter Eppendorf microcentrifuge tubes for subsequent
preparation. Afterward, one otolith of each pair was mounted with the convex surface up on
petrographic glass slides using ultra gel super glue. Otoliths were then hand-polished in the
horizontal plane using a 30μm and 3μm Al2O3 lapping film at an approximate 50µm distance
from the core (Jones and Chen, 2003). Once ground, the otolith was cleaned for two minutes
in ultraclean water in an ultrasonic bath machine (Bransonic 200®), and air-dried under a
laminar flow hood for two hours. All cleaning took place in an air-clean system (600 PCR
workstation). Next, the otoliths were saved in a slide box in Ziplock plastic bags.

51
Strontium isotope and elemental analysis
Strontium isotopes and elemental compositions of Ba/Ca and Sr/Ca ratios from
individual peacock bass otoliths were analyzed at the Wood Hole Oceanographic Institution
(WHOI), by means of a Thermo Finnigan Neptune multiple collector and an inductively
coupled plasma mass spectrometer (LA-MC-ICP-MS) in conjunction with UP-193 laser
ablation systems were used. Primarily, the otolith samples of YOY and adult peacock bass
were organized in a chamber. Subsequently the best annuli position in the YOY and adult fish
otoliths was determined using a microscope attached to a video camera and computer system.
Once the otolith annuli was found, a spot raster was drawn on the otolith surface of the YOY,
and a raster and transect line (from the core moving to the otolith edge) were drawn on the
otolith surface of adults. These procedures were programmed and saved for ablation.
This methodology was used to ablate a 250 µm x 250 µm spot raster centered on the
nucleus of each otolith in order to obtain the natal origin concentrations of trace elements and
87Sr/86Sr isotope ratios from adult and YOY peacock bass. A laser ablated a spot with a 100
µm diameter per blast on the adults otolith surface along the transect line while running at
100% power, or 10 Hz repetition rate and 6 µm·s-1scan speed. The methods used included
blank and mass bias corrections as outlined by Walther et al. (2008) and Walther and
Thorrold (2010). Thus, ratios of Sr/Ca, Ba/Ca and 87Sr/86Sr isotopes were quantified with a
single ablated spot raster or on the transect line through the microscope’s objective lens for
each YOY and adult peacock bass.
Water data collection and analysis
Water quality parameters were measured during the experimental fisheries, using a HI-
9828 Multi-Parameter Water Quality Portable Meter, in order to get data about hydrogenionic
potential (pH); temperature, TEMP (oC); conductivity, COND (µS·cm-¹); and Dissolved
oxygen, DO (mg·l-¹) from each selected tributaries in Negro River basin. To test water
parameters difference among sample locations were used analysis of variance (ANOVA)

52
applied separately for each parameter. When differences was confirmed were used a Tukey
HSD test to verify where the differences occurred within water locations.
Otoliths statistical analyses
To test the first hypothesis, the correlation between YOY and adult otolith core
chemical composition was analyzed by applying a Pearson's correlation analysis on the data
set for fish from the same place. Subsequently, a canonical discriminant function analysis
(CDFA) with a Wilks’ lambda (λ) test was performed to verify which variable (87Sr/86Sr,
Sr/Ca and Ba/Ca) better reflect the distribution of the data groups (Cruz-Castillo et al., 1994;
Nowling et al., 2011) through the difference of distance among canonical variables. A cross-
validation table procedure was also used to verify the probability of adult fish to be accurately
predicted in their nursery areas.
The second hypothesis was tested using an analysis of variance (ANOVA) applied
individually for each variable (87Sr/86Sr isotope, Sr/Ca and Ba/Ca) to verify significant
differences among groups of adults’ otolith core chemical fingerprint sampled in tributaries
with different water types. If this hypothesis was rejected, a post hoc Tukey’s HSD (honestly
significant difference) test was used to identify where these differences occur between
tributaries.
The third hypothesis was tested by direct observations of 87Sr/86Sr values from adult
fish’s otolith transects plotted in a scatter-plot in order to verify differences in 87Sr/86Sr values
that could imply fish movement among sampling sites.
Finally, the values of 87Sr/86Sr from adults fish's transect line (navigated route) were
plotted in a scatter-plot (x, y), to verify any correlation between the otolith microchemistry
from their nursery grounds to their caught location, in order to verify possible indications of
homing back movements.
The Pearson, ANOVA and Tukey tests were performed using Statistica 7.0 (StatSoft,
Inc), whereas CDFA analysis was executed with SPSS (SPSS, Inc). All data analysis were
carried out using probability values of 0.05 as the level of statistical significance.

53
Results
Young-of-year (YOY) and adult peacock bass otolith core microchemical analyses
A Pearson's analysis was applied in the Sr isotopes and elemental composition data
from YOY and adults’ otoliths core chemical compositions and showed positive correlation,
which were mainly explained by 87Sr/86Sr with r2= 0.8903 and by Sr/Ca ratios’ composition
with r2= 0.5512. These results indicated a strong correlation among the groups of variables
analyzed.
The CDFA analysis revealed function values of 87Sr/86Sr (Wilk's λ = 0.0001, χ2 =
971.26, df=18, p<0.0001), Sr/Ca (Wilk's λ = 0.026, χ2 = 339.11, df = 10, p<0.0001), and of
Ba/Ca (Wilk's λ = 0.335, χ2=101.76, df = 4, p<0.0001) presenting significant differences
among canonical variable groups. The distribution of canonical variables was largely driven
by variations in 87Sr/86Sr values emphasized in function 1, explained by the Eigenvalue of
98.5% and canonical correlation of 0.999.
The canonical scores of YOY and adult peacock bass were distributed according to
their influence to plot the data set in the canonical bidimentional space, resulting in similar
values for both YOYs and adults, which matched positively. This finding suggests that the
confidence with which adults were classified to a particular YOY birth area was based on the
homologous geochemical signatures. There were two adult fish that did not match properly
with YOY sampling sites: one fish from Pt03 and another from Pt04 (Figure 2).

54
Figure 2. A scatter-plot displaying the distribution of Cichla temensis’ YOY and adult
otolith core chemical fingerprints. The diamond symbols within the colored ellipses are the
YOY otolith fingerprints, and all of the other symbols correspond to the different adult fish
sampling sites. The same colors for the ellipses and for the solid symbols indicate possible
relationships between YOY and adult fish.
The CDFA joined the YOY and adults together according to similarities in their
microchemical signatures. This tendency was clearly observed in function 1, with the
distribution of adult peacock bass from sampling sites Pt01, Pt03, Pt04 and Pt05; the other
sampling sites were grouped together, which made interpretation more difficult.
These results were also confirmed using a cross-validation table procedure that shows
classification accuracies ranging from 14.3% to 100% and from 16.7% to 100% for both
YOY and adult fish, respectively. These chemical distribution signatures among fish groups
and sampling sites were predicted as follows:
Adult peacock bass caught at the Cuiuni (Pt01) and Demeni (Pt05) Rivers were
correctly matched to their birth locations with an accuracy of 100%. One fish from location
Pt06 was predicted to have originated from the Demeni River (Pt05) with 100% accuracy.
The fish caught at the Aracá River (Pt04) were classified as coming from the Pt03 with 83.3%
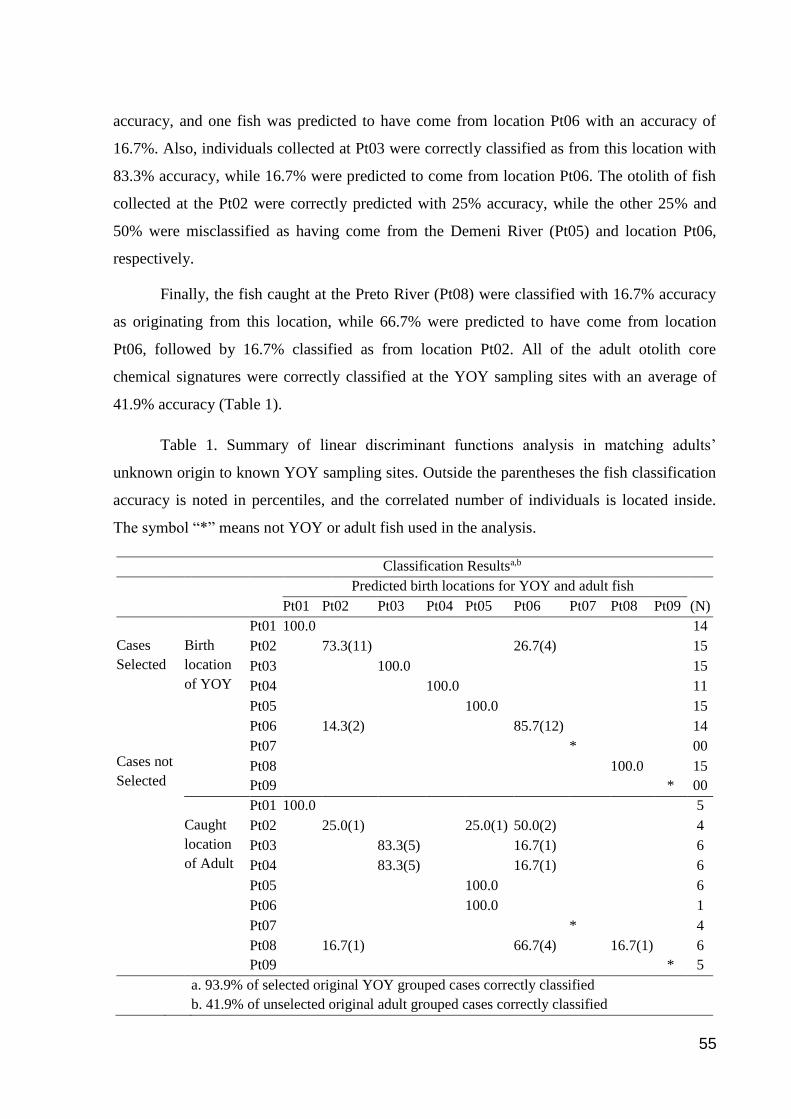
55
accuracy, and one fish was predicted to have come from location Pt06 with an accuracy of
16.7%. Also, individuals collected at Pt03 were correctly classified as from this location with
83.3% accuracy, while 16.7% were predicted to come from location Pt06. The otolith of fish
collected at the Pt02 were correctly predicted with 25% accuracy, while the other 25% and
50% were misclassified as having come from the Demeni River (Pt05) and location Pt06,
respectively.
Finally, the fish caught at the Preto River (Pt08) were classified with 16.7% accuracy
as originating from this location, while 66.7% were predicted to have come from location
Pt06, followed by 16.7% classified as from location Pt02. All of the adult otolith core
chemical signatures were correctly classified at the YOY sampling sites with an average of
41.9% accuracy (Table 1).
Table 1. Summary of linear discriminant functions analysis in matching adults’
unknown origin to known YOY sampling sites. Outside the parentheses the fish classification
accuracy is noted in percentiles, and the correlated number of individuals is located inside.
The symbol “*” means not YOY or adult fish used in the analysis.
Classification Resultsa,b
Predicted birth locations for YOY and adult fish
Pt01 Pt02 Pt03 Pt04 Pt05 Pt06 Pt07 Pt08 Pt09 (N)
Cases
Selected
Cases not
Selected
Birth
location
of YOY
Pt01 100.0 14
Pt02 73.3(11) 26.7(4) 15
Pt03 100.0 15
Pt04 100.0 11
Pt05 100.0 15
Pt06 14.3(2) 85.7(12) 14
Pt07 * 00
Pt08 100.0 15
Pt09 * 00
Caught
location
of Adult
Pt01 100.0 5
Pt02 25.0(1) 25.0(1) 50.0(2) 4
Pt03 83.3(5) 16.7(1) 6
Pt04 83.3(5) 16.7(1) 6
Pt05 100.0 6
Pt06 100.0 1
Pt07 * 4
Pt08 16.7(1) 66.7(4) 16.7(1) 6
Pt09 * 5
a. 93.9% of selected original YOY grouped cases correctly classified
b. 41.9% of unselected original adult grouped cases correctly classified

56
Adult peacock bass otolith core chemical analysis
Using 87Sr/86Sr ratio as response variable measured from the adults otoliths core, there
were differences between the otoliths chemistry of the adult peacock bass within sampling
sites (F8, 32 = 402.41, p < 0.0001). Also Sr/Ca presented values of F8, 32 = 12.97 with p <
0.0001, showing significant differences among groups of adult fish by sampling sites.
Contrary to Ba/Ca, ANOVA showed values of F8, 32 = 1.65 and p < 0.175, indicating that
there are no significant differences among fish groups. Tukey's HSD test for 87Sr/86Sr
presented no significant differences among sampling sites for Pt02 related to Pt06 (df = 32, p
= 0.936397) and to Pt08 (df = 32, p = 0.230759); Pt03 was correlated with Pt04 (df = 32, p =
0.994172); Pt05 was related to Pt06 (df = 32, p = 0.988656), Pt07 was related to Pt08 (df =
32, p = 0.121124) and to Pt09 (df = 32, p = 0.900394); Pt08 was related with Pt06 (df = 32, p
= 0.140686). Also for Sr/Ca no significant differences among sampling sites were found for
Pt02 and sampling sites Pt03 (df = 32, p = 1.000), Pt04 (df = 32, p = 0.999821), Pt06 (df = 32,
p = 1.000), Pt07 (df = 32, p = 0.999525), and Pt08 (df = 32, p = 0.998068); also between
sampling sites Pt05 and Pt09 (df = 32, p = 1.000), no significantly differences were found. As
well, Tukey's test for Ba/Ca were no significantly different among all sampling sites.
The evidence was apparent when looking at the values of 87Sr/86Sr isotopes from
sampling sites Pt03 and Pt04 (Aracá River) that presented the highest values of 87Sr/86Sr
isotopes. Intermediate values of 87Sr/86Sr were found from sampling sites Pt05 (Demeni
River), Pt07, Pt08 (Preto River) and Pt09 (Padauari River), also very similar to values of
87Sr/86Sr isotopes found in samplings Pt02 and Pt06 situated at the Negro River main channel.
On the other hand, the lowest radiogenic values of 87Sr/86Sr isotopes were found at the Cuiuni
River in sampling site Pt01 (Figure 3).

57
Figure 3. Adult Cichla temensis peacock bass’ otolith core 87Sr/86Sr isotope ratio distributed
by sampling sites. Identical letters mean homogeneous chemical values (ANOVA followed by
Tukey's test).
On the other hand, when comparing the values of Sr/Ca and Ba/Ca elemental
composition, only Sr/Ca presented notable differences in their chemical values with a
decreasing pattern occurring from sampling sites Pt01 (Cuiuni River) to Pt09 (Padauari
River). These variations occurred in three distinct situations: i) the samples collected at the
Cuiuni River (Pt01), situated at the southern margin of the Negro River, presented higher
elemental values than the others sampling sites, ii) all otolith cores sampled from black water
tributaries (Pt02 and Pt06), situated in the northern margin of the Negro River, presented
similar elemental composition values, iii) the otolith fingerprint from white water tributaries
(Pt05 and Pt09) exhibited homogeneous chemical values of Sr/Ca and Ba/Ca ratios, also
presenting lowest values of these elements/Ca when comparing its values with those of
nearby black water tributaries (Figure 4A and 4B).
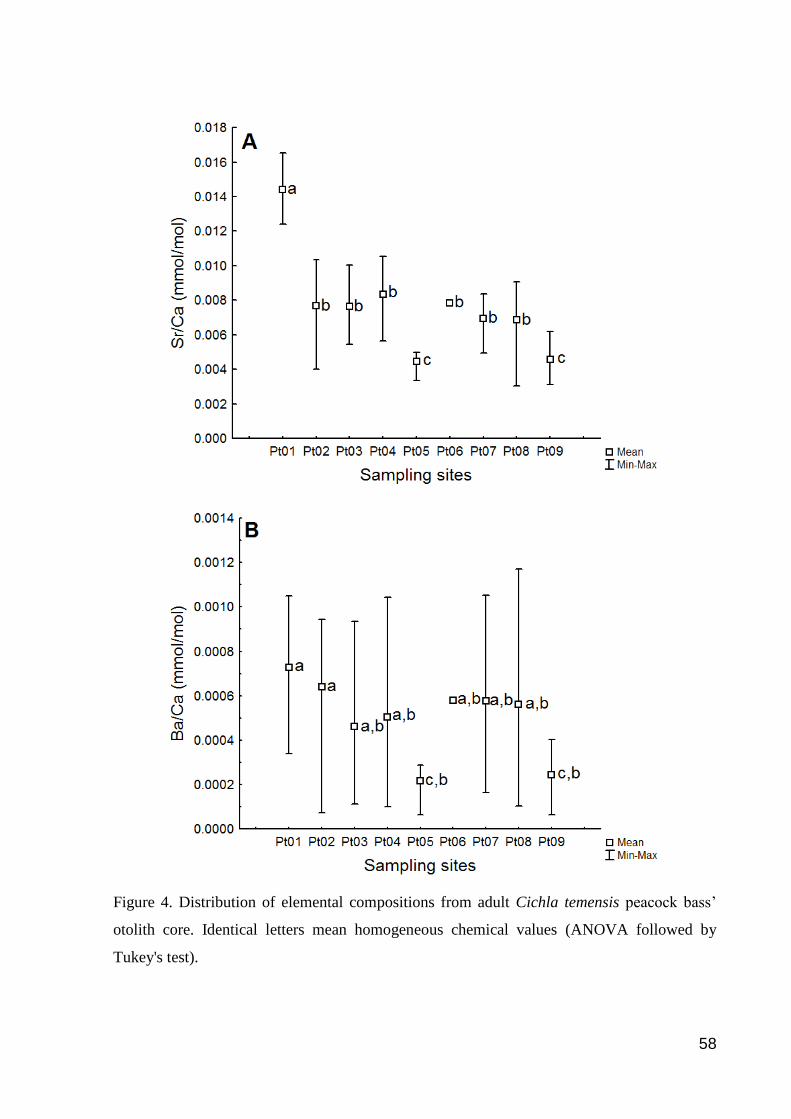
58
Figure 4. Distribution of elemental compositions from adult Cichla temensis peacock bass’
otolith core. Identical letters mean homogeneous chemical values (ANOVA followed by
Tukey's test).
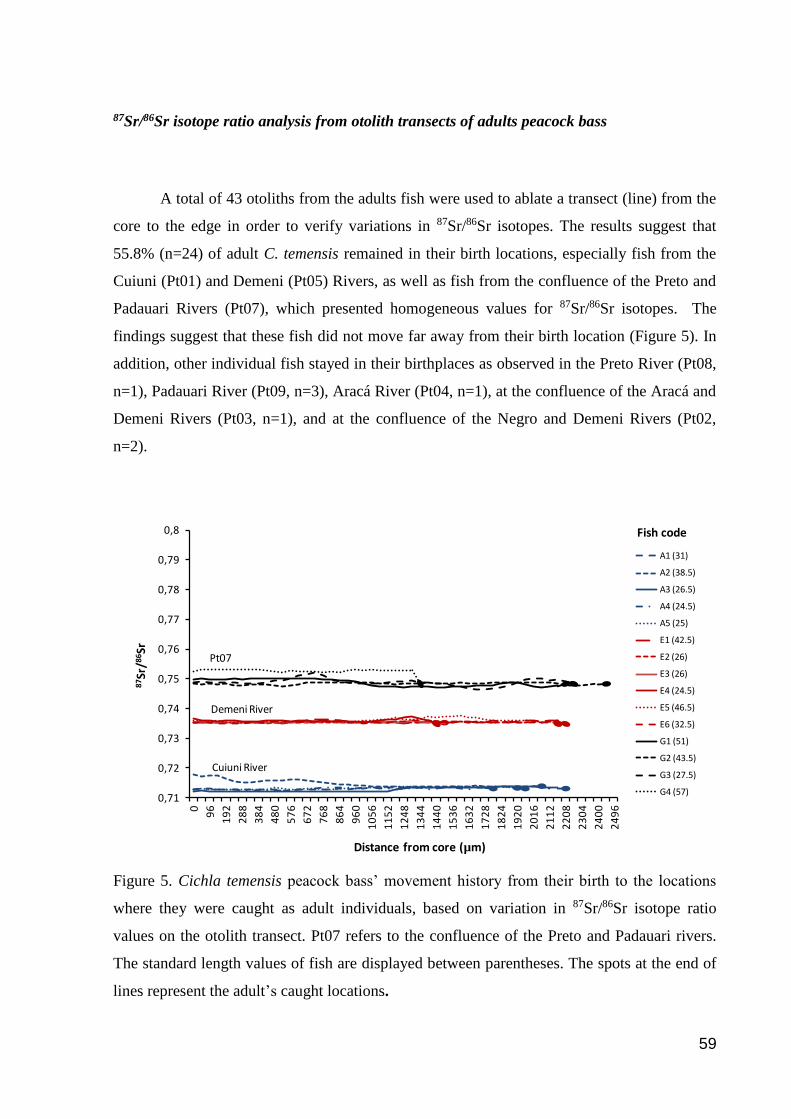
59
87Sr/86Sr isotope ratio analysis from otolith transects of adults peacock bass
A total of 43 otoliths from the adults fish were used to ablate a transect (line) from the
core to the edge in order to verify variations in 87Sr/86Sr isotopes. The results suggest that
55.8% (n=24) of adult C. temensis remained in their birth locations, especially fish from the
Cuiuni (Pt01) and Demeni (Pt05) Rivers, as well as fish from the confluence of the Preto and
Padauari Rivers (Pt07), which presented homogeneous values for 87Sr/86Sr isotopes. The
findings suggest that these fish did not move far away from their birth location (Figure 5). In
addition, other individual fish stayed in their birthplaces as observed in the Preto River (Pt08,
n=1), Padauari River (Pt09, n=3), Aracá River (Pt04, n=1), at the confluence of the Aracá and
Demeni Rivers (Pt03, n=1), and at the confluence of the Negro and Demeni Rivers (Pt02,
n=2).
Pt07
0,71
0,72
0,73
0,74
0,75
0,76
0,77
0,78
0,79
0,8
0
96
19
2
28
8
38
4
48
0
57
6
67
2
76
8
86
4
96
0
10
56
11
52
12
48
13
44
14
40
15
36
16
32
17
28
18
24
19
20
20
16
21
12
22
08
23
04
24
00
24
96
87Sr
/86Sr
Distance from core (µm)
A1 (31)
A2 (38.5)
A3 (26.5)
A4 (24.5)
A5 (25)
E1 (42.5)
E2 (26)
E3 (26)
E4 (24.5)
E5 (46.5)
E6 (32.5)
G1 (51)
G2 (43.5)
G3 (27.5)
G4 (57)
Fish code
Demeni River
Cuiuni River
Figure 5. Cichla temensis peacock bass’ movement history from their birth to the locations
where they were caught as adult individuals, based on variation in 87Sr/86Sr isotope ratio
values on the otolith transect. Pt07 refers to the confluence of the Preto and Padauari rivers.
The standard length values of fish are displayed between parentheses. The spots at the end of
lines represent the adult’s caught locations.
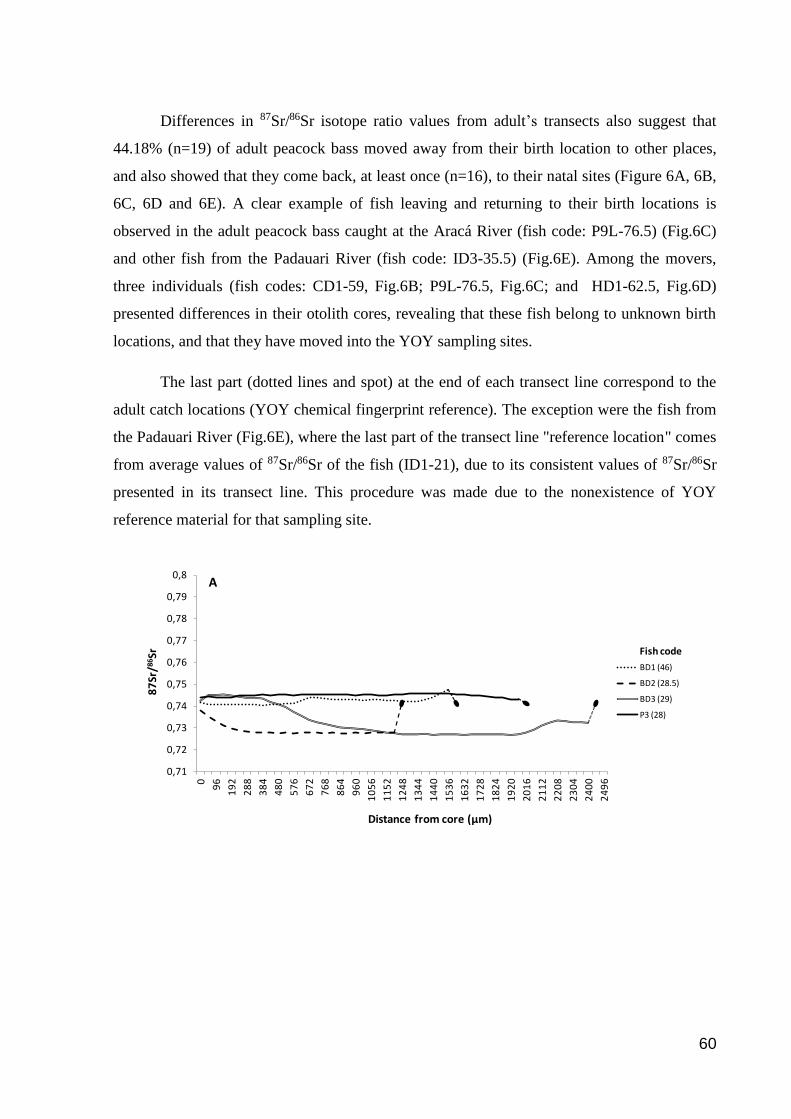
60
Differences in 87Sr/86Sr isotope ratio values from adult’s transects also suggest that
44.18% (n=19) of adult peacock bass moved away from their birth location to other places,
and also showed that they come back, at least once (n=16), to their natal sites (Figure 6A, 6B,
6C, 6D and 6E). A clear example of fish leaving and returning to their birth locations is
observed in the adult peacock bass caught at the Aracá River (fish code: P9L-76.5) (Fig.6C)
and other fish from the Padauari River (fish code: ID3-35.5) (Fig.6E). Among the movers,
three individuals (fish codes: CD1-59, Fig.6B; P9L-76.5, Fig.6C; and HD1-62.5, Fig.6D)
presented differences in their otolith cores, revealing that these fish belong to unknown birth
locations, and that they have moved into the YOY sampling sites.
The last part (dotted lines and spot) at the end of each transect line correspond to the
adult catch locations (YOY chemical fingerprint reference). The exception were the fish from
the Padauari River (Fig.6E), where the last part of the transect line "reference location" comes
from average values of 87Sr/86Sr of the fish (ID1-21), due to its consistent values of 87Sr/86Sr
presented in its transect line. This procedure was made due to the nonexistence of YOY
reference material for that sampling site.
0,71
0,72
0,73
0,74
0,75
0,76
0,77
0,78
0,79
0,8
0
96
19
2
28
8
38
4
48
0
57
6
67
2
76
8
86
4
96
0
10
56
11
52
12
48
13
44
14
40
15
36
16
32
17
28
18
24
19
20
20
16
21
12
22
08
23
04
24
00
24
96
87
Sr/86
Sr
Distance from core (µm)
BD1 (46)
BD2 (28.5)
BD3 (29)
P3 (28)
Fish code
A

61
0,71
0,72
0,73
0,74
0,75
0,76
0,77
0,78
0,79
0,8
0
14
4
28
8
43
2
57
6
72
0
86
4
10
08
11
52
12
96
14
40
15
84
17
28
18
72
20
16
21
60
23
04
24
48
25
92
27
36
28
80
30
24
87Sr
/86Sr
Distance from core (µm)
CD1 (59)
CD2 (28)
CD4 (27.5)
P4 (27.5)
P5 (26)
P6 (24)
Fish code
B
0,71
0,72
0,73
0,74
0,75
0,76
0,77
0,78
0,79
0,80
0
96
19
2
28
8
38
4
48
0
57
6
67
2
76
8
86
4
96
0
10
56
11
52
12
48
13
44
14
40
15
36
16
32
17
28
18
24
19
20
20
16
21
12
22
08
23
04
24
00
24
96
25
92
87Sr
/86Sr
Distance from core (µm)
DD1 (25.5)
DD2 (26)
DD3 (24.5)
P7 (24)
P8 (22.5)
P9L (76.5)
C
Fish code
0,71
0,72
0,73
0,74
0,75
0,76
0,77
0,78
0,79
0,8
0
96
19
2
28
8
38
4
48
0
57
6
67
2
76
8
86
4
96
0
10
56
11
52
12
48
13
44
14
40
15
36
16
32
17
28
18
24
19
20
20
16
21
12
22
08
23
04
24
00
24
96
87Sr
/86Sr
Distance from core (µm)
HD1 (62.5)
HD2 (56)
HD3 (51)
P14 (49)
P15 (46.5)
P16 (52.5)
Fish code
D
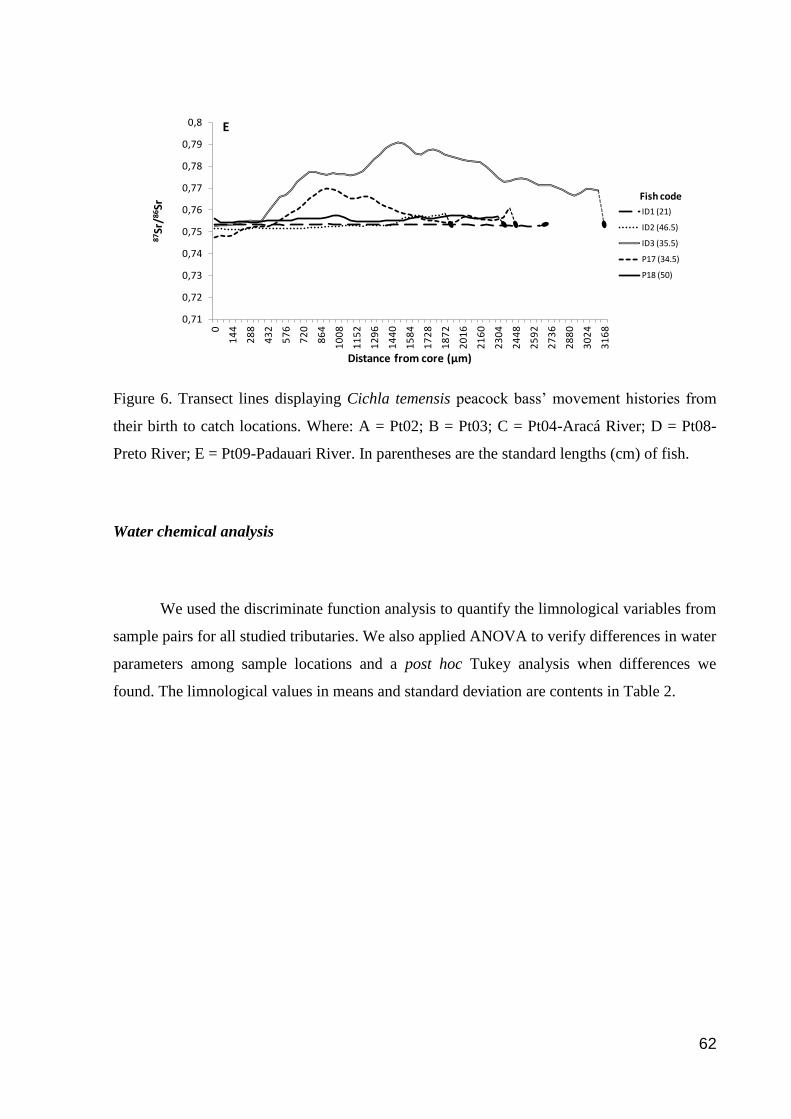
62
Figure 6. Transect lines displaying Cichla temensis peacock bass’ movement histories from
their birth to catch locations. Where: A = Pt02; B = Pt03; C = Pt04-Aracá River; D = Pt08-
Preto River; E = Pt09-Padauari River. In parentheses are the standard lengths (cm) of fish.
Water chemical analysis
We used the discriminate function analysis to quantify the limnological variables from
sample pairs for all studied tributaries. We also applied ANOVA to verify differences in water
parameters among sample locations and a post hoc Tukey analysis when differences we
found. The limnological values in means and standard deviation are contents in Table 2.
0,71
0,72
0,73
0,74
0,75
0,76
0,77
0,78
0,79
0,8
0
14
4
28
8
43
2
57
6
72
0
86
4
10
08
11
52
12
96
14
40
15
84
17
28
18
72
20
16
21
60
23
04
24
48
25
92
27
36
28
80
30
24
31
68
87Sr
/86Sr
Distance from core (µm)
ID1 (21)
ID2 (46.5)
ID3 (35.5)
P17 (34.5)
P18 (50)
Fish code
E

63
Table 2. Limnological variables from samples locations distributed at the Negro River
basin (Means ± standard deviation)
Sampling
sites pH TEMP DO COND
Pt01 4.68 ± 0.37 28.28 ± 1.62 1.86 ± 0.70 11.50 ± 2.12
Pt02 4.92 ± 0.22 27.86 ± 0.87 5.22 ± 2.05 11.50 ± 3.53
Pt03 4.69 ± 0.00 28.67 ± 1.03 1.97 ± 2.50 17.50 ± 0.70
Pt04 4.52 ± 0.00 28.79 ± 1.20 1.13 ± 0.89 17.00 ± 0.00
Pt05 5.38 ± 0.00 29.01 ± 2.26 1.02 ± 0.85 15.00 ± 5.65
Pt06 4.52 ± 0.11 26.42 ± 0,76 3.86 ± 0.55 18.50 ± 2.12
Pt07 4.34 ± 0.00 26.53 ± 1.56 3.23 ± 1.07 16.50 ± 4.94
Pt08 4.20 ± 0.14 26.22 ± 1.14 3.18 ± 0.45 26.00 ± 4.24
Pt09 4.70 ± 0.24 27.02 ± 2.19 3.35 ± 1.76 10.00 ± 2.82
ANOVA p = 0.02 p = 0.45 p = 0.16 p = 0.02
Tukey HSD test
pH: Pt05 ≠ Pt07 with p = 0.01 and Pt05 ≠ Pt08 with p = 0.03
CON: Pt08 ≠ Pt01, Pt02 with p = 0.03 and Pt08 ≠ Pt09, with p = 0.01
Where: DO, Dissolved oxygen (mg·l-¹); TEMP, temperature (oC); pH; COND,
conductivity (µS·cm-¹).
Discussion
The results confirm that otolith microchemistry presents spatial geochemistry
differentiation that is associated with stream beds’ geology, and can be used as a natural tag to
track individual fish movement among populations situated in habitats with different water
types (e.g. Humston et al., 2010; Walter and Limburg, 2012).
This method was effective and reveals the usual departure of Cichla temensis from
their nursery locations. It also shows that peacock bass may return to its birth location at
mature ages. These results are observed by analyzing 87Sr/86Sr isotope values from a transect

64
line ablated in the otoliths (from the core to the edge) of adult fish. Many studies using the
same technique have successfully tracked movements of small (Humston, 2004) and large fish
(Thorrold et al., 2001), proving that otoliths’ microchemistry really functions as an
environmental fingerprint (Walter et al., 2008) in terms of correctly identifying fish’s natal
places (Thorrold et al., 2001; Hamann and Kennedy, 2012), and inter-stream movements
(Gillanders, 2002; Muhlfeld et al., 2012; Wolff et al., 2012) thereby successfully surpassing
the conventional tagging techniques.
The peacock bass C. temensis and its congeners are included in the group of K-
strategist behaviour species (Pianka, 1970). Due to their particular parental care
characteristics, however, Winemiller (1991) classified them as equilibrium strategist species.
Thus far, Cichla spp. is recognized as a sedentary fish due to its short distance movements
between rivers and lakes (Hoeinghaus et al., 2003; Granado-Lorencio et al., 2005; Kehrig et
al., 2008; Olivares et al., 2013).
K-strategist fish commonly has a long life, reproduces on a seasonal basis, and
provides parental care (Pianka, 1970) meanwhile equilibrium strategists’ concept emphasizes
the elevated offspring survivorship as a result of the fathers’ rigorous parental care
(Winemiller and Rose, 1992). Our results also bring new information that could help to
improve C. temensis classification. We find that this fish species leaves their nursery grounds
and returns with a high level of geographic fidelity, once 55.8% of the fish stayed and 44.18%
returned at least once to their birth locations. Mayr (1963) theorizes this pattern as philopatric
behavior. This theoretical evolution of the movement of peacock bass C. temensis is
demonstrated in Figure 7, where i) fish did not move, ii) fish moved slightly and, iii) birth
place fidelity behavior where the fish left and returned to their birth places.

65
Figure 7. Organizational chart with C. temensis movements. Where: N.A = Nursery area.
References: a = Pianka (1970), b = Winemiller (1989), c = Holley et al. (2008), d =
Hoeinghaus et al. (2003), e = the present study. The narrow full line represents the fish
moving out of their nursery area; the dotted lines are the fish returning.
Recent studies also report that peacock bass moved out to their natal areas.
Hoeinghaus et al. (2003) studied C. temensis at the Cinaruco River (Venezuela), and collected
four tagged individuals approximately 21 kilometers from their initial marking places. Holley
et al. (2008), using the same mark-and-recapture (M-R) technique, caught one peacock bass
40 kilometers from the marking place in the Middle Negro River basin (Brazil). In addition,
other studies reported little information that had been collected on the movement of wild C.
temensis populations (Taphorn & Barbarino Duque, 1993; Thorstad et al., 2001).
However, these studies inconclusively indicate a movement pattern for C. temensis;
they only show dispersal movements from the fish’s tagged and catch locations. This situation
may be attributed to problems with M-R technique because traditional tagging methodology is
not appropriate for tracking small fish and neither for reconstructing adult fish’s movement
history (Thorrold et al., 2002; Hendry et al., 2004).

66
Cichla temensis movements, reproduction and phylopatric behaviour synchronism.
Our results showed a strong correlation between adult and YOY otoliths’ isotope and
elemental compositions from C. temensis, confirming that both fish groups belong to the same
nursery area. Further, by examining just otoliths’ 87Sr/86Sr isotope transect, variation in its
chemistry values were observed, showing that these fish left their nursery areas at some point
in their lives and returned later, at least once, to the same birth location. Previous studies with
introduced (Zaret, 1980) and native (Hoeinghaus et al. 2003) Cichla populations hypothesized
that most of fish remain close to their natal or capture locations (Macrander, 2010).
In contrast, our results showed that C. temensis move extensively to regions distant
from their nursery areas, but that they return with high levels of fidelity. This movement
pattern may be indicative of philopatry, which is a strong affection for the birth location that
sustains the individual fish permanently in its nursery ground or causes it to return repeatedly
(Mayr, 1963; Hueter et al., 2004).
This supposition is in agreement with Weatherhead et al. (1994), who reported that
philopatry is the return of a species to their birthplace in order to breed. Also, the philopatric
concept rests on the assumption that an individual moves out of its nursery area (known
location) to explore other environments and later navigates back to its original location with
high frequency. This behaviour was supposed to improve individuals’ familiarization with the
food and others resources in the local area, which would allow it to obtain greater advantages
than individual dispersers (Weatherhead and Forbes, 1994; Robertson and Cooke, 1999).
The degree of homecoming found in this study for C. temensis from otolith
fingerprints suggests a philopatric behaviour for this species in the Negro River basin. A
reasonable explanation to support this assumption for C. temensis is the possibility of these
individuals having to return to their nursery grounds in order to breed. This supposition is
supported by the premise that most of the C. temensis caught in this study occurred during the
dry season that corresponds with their reproduction period (Taphorn and Duque, 1996;
Winermiller et al., 1997; Jepsen et al., 1999; Hoeinghaus et al., 2003; Morales-Nin & Panfili,
2005; Montana et al., 2006; Gomiero et al., 2009). Another reason to sustain this assertion is
that all adult peacock bass were caught close to the YOY nests; and this could presumably be
associated with parental care, such as effectively protecting their nests (Taphorn and Duque,
1996), which is suggestive of recent breeding time.

67
Hydrological barrier and C. temensis metapopulation structure
The Negro River basin connects tributaries with significantly different water types,
including black-water and white-water. Many physical and chemical changes in aquatic
environments can interfer directly on fish movements between different locations, mainly
when such chemical variation are large enough to work as hydrochemical barriers
(Winemiller et al., 2008; Duncan and Fernandes, 2010; Willis et al., 2010).
Recent studies in the Amazon basin have hypothesized the existence of a biological
filter for gene flow between the Negro and Orinoco Rivers for C. temensis, and between the
Negro and Amazon rivers for C. monoculus (Macrander, 2010). Winemiller et al. (2008) and
Willis et al. (2010) also suggest that the gradient of environmental conditions along a river’s
course might create an ecological barrier for the dispersal of many fish species (see also
Torrente-Vilara et al., 2011).
The tributaries of the Negro River also differ greatly in 87Sr/86Sr isotopes ratio and
elemental compositions (Allègre et al., 1996), and these chemical differences in aquatic
habitats could work combined with others limnological parameters as a hydrochemical barrier
for fish groups in terms of spatial distribution (Winemiller et al., 2008; Macrander, 2010).
Corroborating with this statement, our results show that based on 87Sr/86Sr isotopes from
peacock bass otoliths’ transect, the occupancy of fish in tributaries with homogeneous values
of 87Sr/86Sr isotopes or with a soft increasing of Sr isotopes in gradients waters, permits C.
temensis to do movement dispersion among location. Contrary, when differences in values of
87Sr/86Sr isotopes occurs abruptly in the aquatic environment the fish did not leave their
nursery ground.
The chemical similarity between otoliths and ambient water is strongly correlated
when comparing the fish otolith environmental fingerprint from the Cuiuni River with its
87Sr/86Sr isotope values from sediment load, sampled by Allègre et al. (1996). The Cuiuni
River (Pt01) headwaters are located at the Solimões Formation where it receives sediments
from the Solimões River during the flood period, containing low 87Sr/86Sr isotope values,
around 0.71319 ± 0.00002 (Allègre et al., 1996), which closely resemble to the 87Sr/86Sr
isotopes found in the otolith core of both peacock bass YOY (0.71314 ± 0.00066) and adults
(0.71365 ± 0.00233) sampled in the same area. Also, analogous microchemistry from

68
surrounding habitats was found in samples from tributaries with catchment areas situated at
the Guyana Shield that presented high radiogenic 87Sr/86Sr isotopes with equivalent values
between YOY and adult peacock bass as observed in the Aracá River (Pt04) with 87Sr/86Sr
isotope values for YOY equal to 0.78292 ± 0.00101 and for adults equal to 0.78089 ±
0.00158. Samples from the Demini River (Pt05) presented values of 87Sr/86Sr isotopes from
YOY equal to 0.73494 ± 0.00016; these were very similar to those of adult fish with 0.73557
± 0.00055. Also, samples from the Preto River (Pt08) presented 87Sr/86Sr isotope values for
YOY equal to 0.74211 ± 0.00100, and these were very similar to values from adult fish with
0.74535 ± 0.00390.
According to otolith microchemistry, we can deduce that C. temensis from the Negro
River retained themselves away from aquatic environments with high geochemical
differentiation represented by the 87Sr/86Sr isotopes values, and it’s are not related directly
with water color, but straight with water geochemical and limnological composition. This
chemical discrepancy could be a indicative of local hydro-chemical barriers for C. temensis as
reported in several studies in the Amazon basin for Cichla spp. and for others fish species as
well (e.g. Winemiller et al., 2008; Macrander, 2010; Willis et al., 2010).
The main argument to support this assumption is that C. temensis move between
tributaries with similar geochemical compositions, but they do not leave their habitats if large
geochemical differences between tributaries exist. Accordingly, to Jepsen et al. (1997) and
Macrander (2010), Cichla species exhibit distinct habitat preferences and appear to be able to
disperse across habitats with relatively similar chemical compositions (Figure 8).
≈ ≈
≠
≈WW
BW
BW BW
Movements
Isolation Isolation

69
Figure 8. The organograme illustrates C. temensis movement behaviour among different water
types in relation to differences in otoliths’ geochemical composition (87Sr/86Sr isotopes). With
≈ (similar) or ≠ (dissimilar) 87Sr/86Sr isotopes values; BW = Black water and WW = white
water habitats.
According to Sr isotopes in this study, the peacock bass groups became isolated by the
local geochemical composition as presented from individuals caught at the Cuiuni (Pt01) and
Demeni (Pt05) rivers, which presented high microchemical differentiation from the
surrounding aquatic habitats, with the main Negro River channel and Aracá River,
respectively, that probably work as a hydro-chemical barrier for C. temensis. Contrastingly,
peacock bass movements occurred in other Negro River tributaries with similar watershed
chemical compositions but not with the same water types, such as the Preto River (black-
water) and Padauari River (white-water). This chemical homogeneity among the
environments, despite the differences in water type, allows C. temensis from the Preto River
to enter and leave the Padauari River. On the other hand, the fish group from the Padauari
River did not show a dispersal movement in the direction of the Preto River, but only moving
up Padauary River. (Figure 9).

70
22
31 11
1
21 1 11
1 4 1
3
1
1
?
?
Cuiuni River
b
c
a
d
e
f
g
h
Legend
i
Figure 9. Theoretical spatial movement distribution of Cichla temensis in the middle Negro
River basin. Where: ? = unknown locations; a = birth and catch place; b = catch place; c =
birth place; d = fish move out direction; e = homecoming direction; f = white water; g = black
water; h = water flow direction; and i = isolated fish group. Obs: the areas under the
confluences betweem Aracá and Demeni, and Padauari and Preto rivers are Mixed water.
The results also shows that the main factor that could acts as a hydrochemical barrier
for C. temensis, are both the local geological and limnological parameters, and not the water
types. One example that could corroborates with this assumption is the existence of an
isolated C. temensis population from location Pt07 that shows no movement dispersion, its
happened maybe due to the limnological parameter differences from surround locations as
Preto and Padauari Rivers, differing from the others isolated groups as Pt01 and Pt05 that
appears to be separated by the geochemical compositions as 87Sr/86Sr isotopes values. This
supposition is based on the premise that fish assemblages reacts when variations in water
physical and chemical parameters occurs (Garcez and Freitas, 2008), and can works as a
hydro-chemical barrier for many fish species (Winemiller et al., 2008; Duncan & Fernandes,
2010).

71
The small geochemical variation from the Preto River with low 87Sr/86Sr isotopes and
high values of Ba/Ca and Sr/Ca ratios in relation to the Padauari River (white water)
permitted the fish group from this area to engage in dispersal movements between both rivers.
According to variations in 87Sr/86Sr isotopes, the fish from the Padauari River moved up and
down in the main channel to unknown locations. Similar situations have been reported by
Winemiller et al. (2008) who inferred that only a few species are able to move from black
water in the Negro River to white water in the upper Orinoco River, this fish movements
limitations are linked directly with limnological differences in aquatic environments (Willis et
al. (2010).
Our results show that the presence of peacock bass populations established inside and
between tributaries with different water types and geochemical composition in the Negro
River micro-basins, predicts a possible homecoming behavior and spatial population
distribution with the existence of connectivity among groups that maybe works as a
metapopulation organization. To better understand the peacock bass as a metapopulation
structure, however, the aquatic environment needs to be considered as patches that are
seasonally linked by the Negro River through the flood pulse (Junk, 1997; Parolin et al.,
2004) and by the movements of fish between patches (aquatic habitats).
The fish’s ability and adaptability to navigate among habitats with different water
chemistry will determine which population will persist locally. An isolated population will be
more vulnerable to extinction than connected populations, especially when considering the
environmental conditions and fisheries activities in those areas (Ricklefs, 1996). In order to
support this affirmation, it is necessary to gather robust sampling data with sufficient
information about fish movement among sampling sites over a large time scale.
However, we have yet to address water microchemical analysis encompassing the
hydrological gradient in the sampling sites. The otoliths’ microchemical data set suggests that
water chemistry may be a factor that is influencing the dispersal movement of C. temensis and
the formation of isolated and connected populations, with changes in the water chemistry
directly affecting fish’s physiological conditions (Val & Almeida-Val, 1995) and its otolith
compounds as well (Walter and Thorrold, 2006). This affirmation is in agreement with many
studies realized in the Amazon basin, which suggest that some rivers with environmental
heterogeneity operate as a hydro-chemical barrier for fish (e.g. Willis et al., 2007; Toffoli et
al., 2008; Winemiller et al., 2008; Duncan & Fernandes, 2010).

72
Our results show much more spatial and temporal dynamics in the C. temensis
movements than reported in previous studies (e.g. Holley et al., 2008; Hoeinghaus et al.,
2003; Thorstad et al., 2001), and that this information may be useful when considering the
peacock bass population’s movement dynamics in the Negro River basin.
Peacock bass fisheries management proposition
The Negro River basin is composed of many tributaries, and its headwaters are located
on a different geologic formation where the water’s chemical composition differs from other
parts of the basin (Queiroz et al. 2009). As a result, if an individual fish passes across these
different geochemical environments its otolith will record this information as an
environmental fingerprint (Elsdon and Gillander, 2005). Thus, knowing the fish movements
within and between tributaries, we presume that it is possible to use this movement
information for fisheries management planning.
The fish’s movement dynamics are fundamental to understanding spatial distribution,
which is the first step in considering how to adequately manage fishery sectors. Our results
show that based on variation in the 87Sr/86Sr isotope, C. temensis exhibit dispersion
movements among Negro River tributaries, and that part of the individuals may return to their
birthplaces with a high level of fidelity. Also were observed in this study that some groups
and individuals of C. temensis did not leave their nursery grounds, and this may be a
consequence of a hydrochemical barrier between habitats for this fish species (e.g. Duncan
and Fernandes, 2010; Willis et al., 2010).
This scenario also, maybe interpreted as a metapopulation structure that rests on fish’s
ability to enter or leave an aquatic environment in a colonization process (Kritzer and Sale,
2004). Considering the existence of connected and isolated peacock bass populations, we can
view the possibility of managing the fisheries in those areas differently on a spatial and
temporal scale.
Spatially, fisheries management needs to consider the peacock bass’ dispersal
movements among network tributaries as fundamental information to the protection of the
host of habitats used by the fish, as a metapopulation space that is formed by a gathering of

73
fish populations (Levins, 1969). On the other hand, when a school of fish faces a
hydrochemical barrier between tributaries, this group of fish needs to be supervised
differently as an isolated population because these populations struggle to recover in a short
period, mainly when the fisheries effort occurs in those areas with high intensity and
frequency (Kritzer and Sale, 2004). On a temporal scale, the hydrological conditions also
need to be measured, mainly during the drought, period when peacock bass’ spawn
(Winermiller et al., 1997; Gomiero et al., 2009) that corresponds with the high season of sport
fishing in the Negro River basin (Freitas and Rivas, 2006).
For both, connected and isolated populations of peacock bass, the hydrological cycle
and fish physiological and biological conditions should be taken into consideration for
fisheries management procedures. This thinking is important to decide the areas where fishing
should be authorized, and during which times of the year, in order to avoid conflicts between
sport, commercial and subsistence fisheries (e.g. Sobreiro, 2007) and preserve the fish stock
as well.
We believe that mainly 87Sr/86Sr isotopes found in otoliths of peacock bass represent
an excellent, and yet underused record of the geochemical properties of aquatic environments
in the Negro River basin. The C. temensis otolith geochemical fingerprints also highlight the
potential to demarcate fisheries sectors and guide effective fisheries management plans in
freshwater environments.
Acknowledgments
We thank Mr. Julio. A. D. Siqueira for his assistance in the field collecting the
samples. Financial support of this study was provided by Coordination for the Improvement
of Higher Education Personnel (CAPES-PDSE 0909/12-2), National Council for Scientific
and Technological Development (CNPq), Amazonas Research Foundation (FAPEAM), and
INCT-Adapta. Laboratory and logistic support also was provided by Federal University of
Amazonas (UFAM), National Institute of Amazonian Research (INPA), Intelligence Social
Environment Strategic Petroleum Industry in the Amazon region (I-PIATAM), and
Washington and Lee University (WLU). Fish were collected under research license 25606-
1/2010 from Brazilian Institute of Environment and Renewable Natural Resources (IBAMA).

74
Conclusões
O uso da microquímica dos otólitos de tucunaré como marcadores ambientais, mostrou
ser eficiente para esclarecer os padrões de distribuição espacial da espécie C. temensis na
bacia do Médio Rio Negro. Com base nas informações dos isótopos de 87Sr/86Sr e da
concentração dos elementos traços Ba/Ca e Sr/Ca encontrados nos otólitos dos indivíduos
jovens e adultos, concluiu-se que:
i) É possível mapear a distribuição espacial de indivíduos jovens, menores que um
ano, através das diferenças microquímicas existentes em seus otólitos, que, por sua vez,
refletem a geoquímica da área de nascimento de cada grupo estudado;
ii) A microquímica da parte central dos otólitos dos tucunarés jovens foi similar à
encontrada nos otólitos de peixes adultos coletados nos mesmos locais, de forma que a
microquímica dos otólitos pode ser utilizada como impressão digital do local de nascimento
para prever a origem de indivíduos adultos capturados em diferentes locais da bacia do Rio
Negro;
iii) Os isótopos e elementos traços da parte central dos otólitos de tucunarés se
diferenciam nitidamente quando comparados à microquímica dos tributários de água preta
com os de água branca na bacia do Médio Rio Negro;
iv) A variação dos isótopos de 87Sr/86Sr em otólitos de tucunarés adultos, indicou a
existência de movimentos de dispersão de indivíduos entre os tributários do Rio Negro,
possivelmente formando metapopulações nessa região. Ainda, a microquímica dos otólitos
somadas as variações limnológicas ambientais, mostraram que as diferenças hidroquímicas
entre ambientes distintos podem atuar como barreiras hidroquímicas isolando algumas
populações;
v) Com o uso dos isótopos de 87Sr/86Sr, foi possível avaliar parte da história dos
movimentos de dispersão de cada tucunaré individualmente, onde foi verificado que parte dos
indivíduos estudados realizaram movimentos de dispersão entre os ambientes estudados, e
que outros permanecem em seus locais de origem. dos tucunarés adultos retornaram para seus
locais de nascimento com alto grau de filopatria.

75
Referências Bibliográficas
Allard, T., Weber, T., Bellot, C., Damblans, C., Bardy, M., Bueno, G., Nascimento, N., R.,
Fritsch, E., e Benedetti, M., F. 2011. Tracing source and evolution of suspended particles in
the Rio Negro Basin (Brazil) using chemical species of iron. Chemical Geology 280: 79-88.
Allègre C. J.; Dupré B.; Négrel P. e Gaillardet J. 1996. Sr-Nd-Pb isotopes systematics in
Amazon and Congo River systems. Constraints about erosion processes. Chemical Geology,
131: 93-112.
Aubert D.; Probst A.; Stille P. e Viville D. 2002. Evidence of hydrological control of Sr
behavior in stream water (Strengbach catchment, Vosges mountains, France). Applied
Geochemistry, 17: 285-300.
Bacon, C.R., Weber, P.K., Larsen, K.A., Reisenbichler, R., Fitzpatrick, J.A. e Wooden, J.L.
2004. Migration and rearing histories of chinook salmon (Oncorhynchus tshawytscha)
determined by ion microprobe Sr isotope and Sr/Ca transects of otoliths. Canadian Journal of
Fisheries and Aquatic Sciences, 61: 2425-2439.
Bailey, P.B. e Petrere Jr, M. 1989 - Amazon Fisheries: Assessment Methods, Current Status,
and Management Options. Dodge, D.P. Proceedings of the International Large River
Symposium: 385-398.
Bailey, S.W., Hornbeck, J.W., Driscoll, C.T. e Gaudette, H.E. 1996. Calcium inputs and
transport in a base-poor forest ecosystem as interpreted by Sr isotopes. Water Resourse
Research, 32: 707-719.
Barros, G.M.L. 1980. Os tucunarés (Actinopterygii, cichlidae) nos açudes públicos do
nordeste. Dissertação de Mestrado. Universidade Federal do Ceará - UFCE, Fortaleza, 64p.
Barthem R.B., Ribeiro M.C.L.B e Petrere Jr. M. 1991. Life stratategies of some long-distance
migratory catfish in relation to hydroelectric dams in the amazon basin. Biological
Conservation. 55(3): 339-345.

76
Barthem, R.B. e Petrere, M. 1996. Fisheries and population dynamics of Brachyplatystoma
vailantii (Pimelodidae) in the Amazon Estuary. In: Meyer, R.M.; Zhang, C.; Windsor, M.L.;
McCay, B.J.; Hushack, L.J. & Muth, R.M. [eds.] Fisheries resource utilization and policy.
Proceedings of the World Fisheries Congress, Theme 2. Oxford & IBH Publishing Co. 329-
340.
Bataille C. P. e Bowen G. J. 2012. Mapping 87Sr/86Sr variations in bedrock and water for large
scale provenance studies. Chemical Geology. 39-52.
Bath E.G.; Thorrold, S.R.; Jones, C.M.; Campana, S.E.; McLaren J.W. e Lam, J.W.H. 2000.
Strotium and barium uptake in aragonitic of marine fish. Geochimica et Cosmochimica Acta.
64 (10): 1705-1714.
Batista, V.S. 2001. Biologia e administração pesqueira de alguns caraciformes explotados na
Amazônia Central. Tese de professor titular. FUA. 131 p.
Bilby, R.E.; Fransen, B.R.; Walter, J.K.; Cederholm, C.J. e Scarlett, W.J. 2001. Preliminary
evaluation of the use of nitrogen stable isotope ratios to establish escapement levels for
Pacific salmon. Fisheries, 26: 6–13.
Blum J. D.; Taliaferro E. H., Weisse M. T. e Holmes R. T. 2000. Changes in Sr/Ca, Ba/Ca
and 87Sr/86Sr Ratios between Trophic Levels in Two Forest Ecosystems in the Northeastern
U.S.A. Biogeochemistry, 49(1): 87-101.
Bonin. B. 2004. Do coeval mafic and felsic magmas in post-collisional to within-plate
regimes necessarily imply two contrasting, mantle and crustal, sources? A review. Lithos 78:
1-24.
Bouchez, J., Gaillardet, J., France‐Lanord, C., Maurice, L. e Dutra‐Maia, P. 2011. Grain size
control of River suspended sediment geochemistry: Clues from Amazon River depth profiles.
Geochemistry, Geophysics, Geosystems, 12: 1-24.
Brazner, J.B., Campana, S.E., Tanner, D.K. e Schram, S.T. 2004. Reconstructing Habitat use
and wetland nursery origin of yellow perch from Lake Superior using otolith elemental
analysis. Journal of Great Lakes Research, 30: 492-507.

77
Brinn, M.N.A., Porto, J.I.R. e Feldberg, E. 2004. Karyological evidence for interspecific
hybridization between Cichla monoculus and C. temensis (Perciformes, Cichlidae) in the
Amazon. Hereditas, 141: 252-257.
Brown, R.J. e Severin, K.P. 2009. Otolith chemistry analyses indicate that water Sr:Ca is the
primary factor influencing otolith Sr:Ca for freshwater and diadromous fish but not for marine
fish. Canadian Journal of Fisheries and Aquatic Sciences, 66: 1790-1808.
Campana, S.E. 1999. Chemistry and composition of fish otoliths: pathways, mechanisms and
applications. Marine Ecology Progress Series, 188:263-297.
Capo, R.C., Stewart B.W. e Chadwick O.A. 1998. Strontium isotopes as tracers of ecosystem
processes: Theory and Methods, 82: 197-225.
Catella, A.C. 1992. Estrutura da comunidade e alimentação dos peixes da Baía da Onça, uma
Lagoa do Pantanal do Rio Aquidauana, MS. Dissertação de Mestrado, Universidade de
Campinas, UNICAMP. Campinas - SP.
Chellappa, S., Câmara, M.R., Chellappa, N.T., Beveridge, M.C.M. & Huntingford, F.A. 2003.
Reproductive ecology of a neotropical cichlid fish, Cichla monoculus (Osteichthyes:
Cichlidae). Brazilian Journal of Biology, 63: 17-26.
Chilton, D.E. e Beamish, R.J. 1982. Age determination methods for fishes studied by the
Groundfish Program at the Pacific Biological Station. Canadian Special Publishing in
Fisheries Aquatic Science, 60: 102 p.
Clow, D.W., Mast, M.A., Bullen, T.D. e Turk, J.T. 1997. Strontium 87/ strontium 86 as a
tracer of mineral weathering reactions and calcium sources in an alpine/subalpine watershed,
Loch Vale, Colorado. Water Resources Research, 33: 1335-1351.
Corrêa, M.F.M. e Vianna, M.S. 1992. Catálogo de otólitos de Sciaenidae (Osteichthyes-
Perciformes) do litoral do estado do Paraná, Brasil. Nerítica, Curitiba, 7: 13-41p.
Corrêa, R.O. 1998. Crescimento de tucunaré Cichla monoculus (Perciformes: Cichilidae) em
ambiente natural: seleção da melhor estrutura calcificada para a determinação da idade
Dissertação de Mestrado - Instituto Nacional de Pesquisas, Universidade Federal do
Amazonas, Manaus, Amazonas. 70 p.

78
Cruz-Castillo J. G., Ganeshanandam S., MacKay B.R., Lawes G.S., Lawoko C.R.O. e
Woolley D.J. 1994. Applications of Canonical Discriminant Analysis in Horticultural
Research. HortScience, 29(10): 1115-1119.
Dalai, T.K., Krishnaswami, S. e Sarin, M.M. 2002. Barium in the Yamuna River System in
the Himalaya: Sources, fluxes, and its behavior during weathering and transport.
Geochemistry, Geophysics, Geosystems, 12: 1076.
Devick, W.S. 1969. Life history study of the Tucunare, Cichla ocellaris. State of Hawaii Fed.
Aid Proj. F-4R-17, JCR 34 pp.
Do Nascimento, N.R., Fritsch, E., Bueno, G.T., Bardy, M., Grimaldi, C. e Melfi, A.J. 2008.
Podzolization as a deferralitization process: dynamics and chemistry of ground and surface
waters in an acrisol-podzol sequence of the upper Amazon Basin. European Journal of Soil
Science, 59: 911-924.
Dubroeucq, D. e Volkoff, B. 1998. From Oxisols to Spodosols and Histosols: evolution of the
soil mantles in the Rio Negro basin (Amazonia). Catena, 32: 245-280.
Duncan W. P., e Fernandes M. N. 2010. Physicochemical characterization of the white, black,
and clearwater rivers of the Amazon Basin and its implications on the distribution of
freshwater stingrays (Chondrichthyes, Potamotrygonidae). Pan-American Journal of Aquatic
Sciences, 5(3): 454-464.
Edmond, J.M., Palmer, M.R., Measures, I.C., Grant, B. e Stallard, R.F. 1995. The fluvial
geochemistry and denudation fate of Guayana Shield in Venezuela, Colombia, and Brazil.
Geochimical et Cosmochimica, 59: 3301-3325.
Elsdon, T.S. e Gillanders B.M. 2003. Relationship between water and otolith elemental
concentrations in juvenile black bream Acanthopagrus butcheri. Marine Ecology Progress
Series, 260: 263-272.
Elsdon T.S. e Gillanders B. M. 2005. Alternative life-history patterns of stuarine fish: barium
in otoliths elucidates freshwater residency. Canadian Journal of Fisheries and Aquatic
Sciences 62: 1143-1152.

79
FAO (Food and Agriculture Organization of the United Nations) 1995. Soil Map of the World,
Scale 1 : 5000,000, vol. I-X. United Nations Educational, Scientific, and Cultural
Organization: Paris.
Faure, G. 1986. Principles of isotope geology. 2nd edition, New York, John Wiley & Sons.
589 pp.
Faure, G. 2001. Origin of igneous rocks – the isotopic evidence. Springer, Berlin Heidelberg
New York. 496p.
Fittkau, E. J.; Irmler, U.; Junk, W. J.; Reiss, F. e Schmidt, G. W. 1975. Productivity, biomass,
and population dynamics in Amazonian water bodies. In Tropical Ecological Systems-Trends
in Terrestrial and Aquatic Research, Golley, F. B.; and Medina, E., (eds.); Springer Verlac
Inc., New York. 289-310.
Fontenele, O. 1950. Contribuição para o conhecimento da biologia dos tucunarés
(Actinopterygii, Cichlidae), em cativeiro. Aparelho de reprodução. Hábitos de desova e
incubação. Revista Brasileira de Biologia. 10(04): 503-519.
Fossum, P., Kalish, J. e Moksness, E. 2000. Foreword. Fisheries Research. 46: 1-2
Franzinelli, E. & Igreja, H. 2002. Modern sedimentation in the Lower Negro River,
Amazonas State, Brazil. Geomorphology 44: 259-271.
Franzinelli, E. 2011. Características morfológicas da confluência dos rios Negro e Solimões
(Amazonas, Brasil). Revista Brasileira de Geociências, 41 (4): 587-596.
Freitas, C.E.C. e Garcez, R.C.S. 2004. Fish communities of natural canals between floodplain
lakes and Solimões-Amazonas River. Acta Limnológica Brasiliensia, 16(3): 273-280.
Freitas, C.E.C. e Rivas, A.A.F. 2006. A pesca e os recursos pesqueiros na Amazônia
Ocidental. Ciência e Cultura, 58(3): 30-32.
Fritsch E.; Allard T. Benedetti M. F.; Bardy M.; do Nascimento N.R.; Li Y., e Calas G. 2009.
Organic complexation and translocation of ferric iron in podzols of the Negro River
watershed. Separation of secondary Fe species from Al species. Geochimica et Cosmochimica
Acta, 73:1813-1825.

80
Furch, K. 1984. Water chemistry of the Amazon Basin: The distribution of chemical elements
among freshwaters. In: Sioli, H. The Amazon Limnology and landscape ecology of a mighty
tropical river and its basin. Dordrecht: Dr. W. Junk. Publishers. 6: 167-199.
Gaillardet, J., Dupré, B. e Allégre, C.J. 1997. Chemical and physical denudation in the
Amazon River basin. Chemical Geology, 142: 141–173.
Gaillardet, J., Dupré, B. e Allégre, C.J. 1999. Geochemistry of large River suspended
sediments: Silicate weathering or recycling tracer? Geochimica et Cosmochimica Acta, 63:
4037-4051.
Galy A., France-Lanord C. e Derry L. 1999. The strontium isotopic budget of Himalayan
Rivers in Nepal and Bangladesh. Geochimica et Cosmochimica Acta. 63: 1905-1925.
Garcez, R.C.S. e Freitas, C.E.C. 2008. The influence of flood pulse on fish communities of
floodplain canals in the Middle Solimões River, Brazil. Neotropical Ichthyology. 6(2): 249-
255.
Garcez, R.C.S. e Freitas, C.E.C. 2011. Seasonal catch distribution of tambaqui (Colossoma
macropomum), Characidae in a central Amazon floodplain lake: implications for sustainable
fisheries management. Journal of Applied Ichthyology. 27(1): 118-121.
Garcez, R.C.S., Humston, R., Harbor, D. and Freitas, C.E.C. 2014, Otolith geochemistry in
young-of-the-year peacock bass Cichla temensis for investigating natal dispersal in the Rio
Negro (Amazon – Brazil) river system. Ecology of Freshwater Fish. doi: 10.1111/eff.12142
Gauldie, R.W. 1993. Polymorphic crystalline structure of fish otoliths. Journal of
Morphology, 218: 1-28.
Getirana, A.C.V., Bonnet, M.P, Rotunno Filho, O.C., Collischonn, W., Guyot, J.L., Seyler, F.
e Mansur, W.J. 2010. Hydrological modeling and water balance of the Negro River basin:
evaluation based on in situ and spatial altimetry data. Hydrological Processes, 24: 3219-
3236.
Gillanders, B.M. e Kingsford, M.J. 2000. Elemental fingerprints of otoliths of fish may
distinguish estuarine ‘‘nursery’’ habitats. Marine Ecology Progress Series, 201: 273-286.

81
Gillanders, B. M. 2002. Connectivity between juvenile and adult fish populations: do adults
remain near their recruitment estuaries? Marine Ecology Progress Series. 240: 215-223.
Gomiero, L.M. e Braga, F.M.S. 2007. Descrição dos otólitos de tucunarés (Cichla sp. E
Cichla monoculos) no reservatório da hidrelétrica de Volta Grande (SP-MG). Ciência Animal
Brasileira. 8(1): 119-126.
Gomiero, L.M.; Villares Junior, G.A. e Naous, F. 2009. Reproduction of Cichla kelberi
Kullander and Ferreira, 2006 introduced into an artificial lake in southeastern Brazilian.
Journal of Biology. 69(1): 175-183.
Goulding, M.; Carvalho, M. L. e Ferreira, E. G. 1988. Rio Negro, rich life poor water:
Amazonian diversity and floodchain ecology as seen through fish communities. The Hague,
SPB Academic Publishing, 200p.
Gowan, C.; Young, M.K.; Fausche, K.D. e Riley, S.C. 1994. Restricted movement in resident
stream salmonids: a paradigm lost? Canadian Journal of Fisheries and Aquatic Sciences,
51:2626–2637.
Garcez, R.C.S.; Freitas, C.E.C. 2008. The influence of flood pulse on fish communities of
floodplain canals in the Middle Solimões River, Brazil. Neotropical Ichthyology. 6(2): 249-
55.
Gradstein, F.M., Ogg, J., Schmitz, M.A. e Ogg, G. 2012. The Geologic Time Scale. 1st ed.
Elsevier Publishing Company, Oxford. 1176p.
Graef, E.W. 1995. As espécies de peixes com potencial para criação no Amazonas In:
Criando Peixes na Amazônia, Val, A. L. e Honczaryk, A. (eds). INPA. Manaus. 29-43.
Granado-Lorencio, C., Araujo Lima C. R. M. e Lobón-Cerviá J. 2005. Abundance distribution
relationships in fish assembly of the Amazonas floodplain lakes. Ecography, 28: 515-520.
Groot, C. e Margolis L. 1991. Pacific salmon life histories. University of British Columbia
Press, Vancouver.
Gross, M.L., Kapuscinski,A.R. e Faras, A.J. 1994. Nestspecific DNA fingerprints of
smallmouth bass in Lake Opeongo, Ontario . Transactions of the American Fisheries Society.
123: 449-459.

82
Guido, N.J.; Wang, X.; Adalsteinsson, D.; McMillen, D.; Hasty, J.; Cantor, C.R.; Elston, T.C.
e Collins, J.J. 2006. A bottom-up approach to gene regulation, Nature. 439: 856-860.
Hamann, E. J. e Kennedy, B. P. 2012. Juvenile dispersal affects straying behaviors of adults
in a migratory population. Ecology, 93(4):733-740.
Hanski, I. e Gaggiotti, O.E. 2004. Metapopulation biology: past, present, and future. In:
Hanski, I. e Gaggiotti, O.E. [eds.] Ecology, Genetics, and Evolution of Metapopulations.
Elsevier Academic Press. San Diego, 3-22.
Hanski, I. e Gilpin, M.E. 1997. Metapopulation biology: ecology, genetics, and evolution.
Academic Press, San Diego, California.
Hanski, I. e Simberloff, D. 1997. The metapopulation approach, its history, conceptual
domain, and application to conservation. pp. 5–26. In I. A. Hanski and M. E. Gilpin (eds.),
Metapopulation Biology. Academic Press, San Diego, California.
Hegg, J.C., Kennedy, B.P. e Fremier, A. K. 2013. Predicting strontium isotope variation and
fish location with bedrock geology: Understanding the effects of geologic heterogeneity.
Chemical Geology 361: 89-98.
Hoeinghaus, D. J.; Layman, C. A; Arrington D.A. e Winemiller, K. O. 2003, Movement of
Cichla species (Cichlidae) in a Venezuelan floondplain river. Neotropical Ichthyology, 1(2):
121-126.
Holley, M.H., Maceina, M.J., Thomé-Souza, M. e Forsberg, B.R. 2008. Analysis of the
trophy sport fishery for the speckled peacock bass in the Rio Negro River, Brazil. Fisheries
Management and Ecology, 15: 93-98.
Horbe, A.M.C., Gomes, I.L.F., Miranda, S.A.F. e Silva, M.S.R. 2005. Contribuição à
hidroquímica de drenagens no Município de Manaus – Amazonas. Acta Amazonica ,35: 119-
124.
Horbe, A.M.C. e Santos, A.G.S. 2009. Chemical Composition of Black-Watered Rivers in the
Western Amazon Region (Brazil). Journal of Brazilian Chemistry Society. 20(6): 1119-1126.

83
Hueter, R. E.; Heupel, M. R.; Heist, E. J. e Keeney, D. B. 2004. Evidence of Philopatry in
Sharks and Implications for the Management of Shark Fisheries. Journal of Northwest
Atlantic Fishery Science, 35: 239–247.
Humston, R. e D. Harbor. 2006. Geologic analyses for evaluating watershed heterogeneity:
implications for otolith microchemistry studies. Proceedings of the Southeastern Association
of Fish and Wildlife Agencies, 60: 132-139.
Humston R., Priest B. M., Hamilton W.C. e Bugas Jr. P. E. 2010. Dispersal between Tributary
and Main-Stem Rivers by Juvenile Smallmouth Bass Evaluated Using Otolith
Microchemistry, Transactions of the American Fisheries Society. 139(1): 171-184.
Jeffery, P.G . 1975. Chemical Methods of Rock Analysis, 2nd ed. Pergamon press, Oxford,
UK, vol 36, 525p.
Jepsen, D.B., Winemiller, K.O. e Taphorn, D.C. 1997. Temporal patterns of resource
partitioning among Cichla species in a Venezuelan blackwater River. Journal of Fish Biology,
51: 1085-1108.
Jepsen, D.B.; Winemiller, K.O. e Taphorn, D.C. 1999. Age structure and growth of peacock
cichlids from rivers and reservoirs of Venezuela. Journal of fish Biology, 55: 433-450.
Johnson R.C., Weber P.K., Wikert J. D., Workman M.L., MacFarlane R. B., Grove M. J., e
Schmitt A. K. 2012. Managed Metapopulations: Do Salmon Hatchery ‘Sources’ Lead to In-
River ‘Sinks’ in Conservation? PLoS ONE 7 (2).
Junk, W.J.; Bayley, P.B. e Sparks, R.E. 1989. The Flood Pulse Concept in River Floodplain
Systems. Canadian Special Publishing in Fisheries Aquatic Sciences. Canada, 106: 110-127.
Junk, W.J., 1997. The Central Amazon Floodplain: Ecology of a Pulsing System. Ecological
Studies, Vol. 126. Springer, Berlin.
Karim, A. & Veizer, J. 2000. Weathering processes in the Indus River Basin: implications
from Riverine carbon, sulfur, oxygen, and strontium isotopes. Chemical Geology, 170: 153–
177.

84
Kennedy M.J., Chadwick O.A., Vitousek P.M., Derry L.A. e Hendricks D.M. 1998. Changing
sources of base cations during ecosystem development, Hawaiian Islands. Geology 26: 1015-
1018
Kennedy, B.P., Klaue, A., Blum, J.D., Folt, C.L. e Nislow, K.H. 2002. Reconstructing the
lives of fish using Sr isotopes in otoliths. Canadian Journal of Fisheries and Aquatic Sciences
59(6): 925-929.
Kraus, R.T. e Secor, D. 2004. Incorporation of strontium into otoliths of an estuarine fish.
Journal of Experimental Marine Biology and Ecology, 302: 85-106.
Kritzer, J.P. e Sale, P.F. 2004. Metapopulation ecology in the sea: From Levins’ model to
marine ecology and fisheries science. Fish and Fisheries, 5: 131-140.
Küchler, I. L., Miekely, N. e Forsberg, B. R. 2000. A Contribution to the Chemical
Characterization of Rivers in the Rio Negro Basin, Brazil. Journal of Brazilian Chemical
Society, 11: 286-292.
Küchler, I.L., Miekeley, N. & Forsberg, B.R. 2000. A Contribution to the Chemical
Characterization of Rivers in the Rio Negro Basin, Brazil. Journal of the Brazilian Chemical
Society, 11: 286-292.
Kullander, S.O. e Ferreira, E.J.G. 2006. A review of the South American cichlid genus
Cichla, with descriptions of nine new species (Teleostei: Cichlidae). Ichthyological
Exploration Freshwaters, 17(4): 289-398.
Latrubesse, E.M. e Franzinelli E. 2005. The late Quaternary evolution of the Negro River,
Amazon, Brazil: Implications for island and floodplain formation in large anabranching
tropical systems. Geomorphology, 70: 372-397.
Layman, C.A., and Winemiller, K.O. 2004. Size-based response of prey to piscivore
exclusion in a species-rich Neotropical River. Ecology 85(5): 1311-1320.
Leon, J.G., Calmant, S., Seyler, F., Bonnet, M.P., Cauhope, M., Frappart, F., Filizola, N. e
Fraizy, P. 2006. Rating curves and estimation of average water depth at the upper Negro
River based on satellite altimeter data and modeled discharges. Journal of Hydrology, 328:
481-496.

85
Levins, R. 1969. Some demographic and genetic consequences of environmental
heterogeneity for biological control. Bulletin of the Entomological Society of America, 15:
237-240.
Levins, R. 1970. Extinction. Pages 75-107, in M. Gertenhaber, ed. Some mathematical
problems in biology. American Mathematical Society, Providence, Rhode Island, USA.
Lovejoy, N. R. e Araújo, M. Négrel, P., Fouillac, C., Brach, M. 1997. Occurrence of mineral
springs in the stream channel of the Allier River (Massif Central, France): chemical and Sr
isotope constraints. Journal of Hydrology. 203: 143-153.
Lowe-McConnell, R.H. 1969. The cichlid fishes of Guyana, South America, with notes on
their ecology and breeding behaviour. Zoological Journal of the Linnean Society, 48: 255-
302.
Lyons, J. e Kanehl, P. 2002. Seasonal movements of smallmouth bass in streams. In Philipp,
D.P. e Ridgway, M.S. [eds.] Black bass: Ecology, Conservation, and Management. American
Fisheries Society Symposium Proceedings. 31: 149-160.
Macrander J. C. 2010. Microsatellite development, population structure and demographic
histories for two species of amazonian peacock bass Cichla temensis and Cichla monoculus
(perciformes: cichlidae). Dissertations and Theses in Biological Sciences. Paper 20.
http://digitalcommons.unl.edu/bioscidiss/20.
Manly, B. F. J. 2005. Multivariate Statistical Methods, A Primer. 3rd ed. Chapman &
Hall/CRC, Boca Raton, 214p.
Mercier, L., Darnaude, A.M., Bruguier, O., Vasconcelos, R.P., Cabral, H.N., Costa, M.J.,
Lara, M., Jones, D. L. e Mouillo, T. D. 2011. Selecting statistical models and variable
combinations for optimal classification using otolith microchemistry. Ecological
Applications, 21: 1352-1364.
Muhlfeld, C. C.; Thorrold, S. R.; McMahon, T. E. e Marotz, B. 2012. Estimating westlope
cutthrout trout (Oncorhynchus clarkii lewisi) movements in river network using strontium
isocapes. Canadina Journal of Aquatic Science, 69: 906-915.

86
Négrel, P. e Dupré, B. 1993. Temporal variations of Sr isotopic ratios, major and trace
elements composition of the Oubangui River Basin: implications for the source of material.
Grands Bassins Fluviaux, Paris. 181-198.
Northcote, T. G. 1997. Potamodromy in salmonidae: living and moving in the fast lane. North
American Journal of Fisheries Management. 17: 1029-1045.
Novaes, J. L.C., Caramaschi, E.R.P. e Winemiller, K.O. 2004. Feeding of Cichla monoculus
Spix, 1829 (Teleostei: Cichlidae) during and after reservoir formation in the Tocantins River,
Central Brazil. Acta Limnologica Brasiliensia, 16: 41-49.
Nowling L., Gauldie R. W., Cowan Jr. J. H. e De Carlo E. 2011. Successful discrimination
using otolith microchemistry among samples of Red Snapper Lutjanus campechanus from
artificial reefs and samples of L. campechanus taken from Nearby oil and gas platforms. The
Open Fish Science Journal, 4: 1-9.
Olivares A.M., Hrbek T., Escobar M. D. e Caballero S. 2013. Population structure of the
black arowana (Osteoglossum ferreirai) in Brazil and Colombia: implications for its
management. Conservation Genetics. 14(3): 695-703.
Palmer, M.R. e Edmond J.M. 1989. The strontium isotope budget of the modern ocean. Earth
and Planetary Science Letters, 92: 11-26.
Parolin P., Simone O. D., Haase K., Waldhoff D., Rottenberger S., Kuhn U., Kesselmeier J.,
Kleiss B., Schmidt W., Piedade M. T. F. e Junk W. J. 2004. Central Amazonian floodplain
forests: Tree adaptations in a pulsing system. The Botanical Review, 70(3): 357-380
Pianka E.R. 1972. r and K Selection or b and d Selection? The American Naturalist,
106(951): 581-588.
Price, T.D., Manzanilla L. e Middleton W.D. 2000. Immigration and the Ancient City of
Teotihuacan in Mexico: a Study Using Strontium Isotope Ratios in Human Bone and Teeth.
Journal of Archaeological Science, 27: 903-913.
Queiroz, M. M. A., Horbe, A. M. C., Seyler, P. e Moura, C. A. V. 2009. Hidroquímica do rio
Solimões na região entre Manacapuru e Alvarães-Amazonas-Brasil. Acta Amazonica 39(4):
943-952.

87
Queiroz, M.M.A., Horbe, A.M.C., Seyler, P. e Moura, C. A. V. 2009. Hidroquímica do rio
Solimões na região entre Manacapuru e Alvarães-Amazonas-Brasil. Acta Amazonica. 39(4):
943-952.
Reis, N.J., Almeida, M. E., Riker, S. L. e Ferreira, A.L. 2006. Geologia e recursos minerais
do Estado do Amazonas. MME/CPRM/CIAMA, Programa de Geologia do Brasil (mapas
geológicos estaduais, escala 1:1.000.000), Manaus, AM. 125p.
Ribeiro, M.C.L. de B. e Petrere, M., Jr. 1990. Fisheries ecology and management of the
jaraqui (Semaprochilodus taeniurus, S. insignis) in central Amazonia. Regulated Rivers:
Research & Management. 5: 195-215.
Rich, T. D.; Beardmore, C. J.; Berlanga, H.; Blancher, P. B.; Bradstreet, M. S. W.; Butcher,
G. S.; Demarest, D.; Dunn, E. H.; Hunter, W. C.; Iñigo-Elias, E.; Kennedy, J. A.; Martell, A.;
Panjabi, A.; Pashley, D. N.; Rosenberg, K. V.; Rustay, C.; Wendt, S. e Will, T. 2004. Partners
in Flight North American landbird conservation plan. Cornell Laboratory of Ornithology,
Ithaca, New York, USA.
Ricklefs, R. E. e G. L. Miller. 2000. Ecology. 4th edition. W. H. Freeman and company,
New York, New York, USA.
Ridgeway, M.S.; Shuter, B.J.; Middel, T.A. e Gross, M.L. 2002. Spatial ecology and
density-dependent processes in smallmouth bass: the juvenile transition hypothesis. Pages 47-
60 in D.P. Philipp & M.S. Ridgeway (eds) Black bass: ecology, conservation, and
management. American Fisheries Society, Symposium 31, Bethesda, MD.
Rieman, B. E.; Myers, D. L. e Nielson, R. L. 1994. Use of otolith microchemistry to
discriminate Oncorhynchus nerka of resident and anadromous origin. Canadian Journal of
Fisheries and Aquatic Sciences. 51:68-77.
Rieman, B. E. e Dunham, J. B. 2000. Metapopulations and salmonids: a synthesis of life
history patterns and empirical observations. Ecology of Freshwater Fish. 9: 51-64.
Robertson, G.J. e Cooke, F. 1999. Winter philopatry in migratory waterfowl. The Auk, 116:
20-34.

88
Santos, G.M.; Jegu, M. e Merona, B. 1984. Catálogo de peixes comerciais do baixo rio
Tocantins. Eletronorte/CNPq/INPA, Manaus. 83p.
Scott, K.M., Vallance, J.W. e Pringle, P.T. 1995. Sedimentology, Behavior, and Hazards of
Debris Flows at Mount Rainier, Washington: U.S. Geological Survey Professional Paper.
1547, 56p.
Semmens, B., Luke, K.E., Bush, P.G. e Heppell, E. 2006. Investigating the reproductive
migration and spatial ecology of Nassau grouper (Epinephelus striatus) on Little Cayman
Island using acoustic tags – An Overview. FlSC Strategic Review in St. Petersburg, Florida,
USA.
Singh, S.K., Trivedi, J.R., Pande, K., Ramesh, R. e Krishnaswami, S. 1998. Chemical and
strontium, oxygen, and carbon isotopic compositions of carbonates from the Lesser Himalaya:
Implications to the strontium isotope composition of the source waters of the Ganga,
Ghaghara, and the Indus Rivers. Geochimica et Cosmochimica Acta, 62: 743-755.
Sioli, H. 1984. The Amazon and its main affluents: hydrography, morphology of the river
courses and river types. In: Sioli, H., ed. The Amazon (Monographiae biologicae), Dr. W.
Junk, The Hague, Netherlands, 65: 127-166.
Sioli H. 1991. Amazônia Fundamentos da ecologia da maior região de florestas tropicais.
3.ed., Petrópolis, Edit. Vozes, 72 p.
Smith, W.; Espíndola, E. L. G. e Rocha O. 2005. As introduções de espécies de peixes
exóticas e aloctones em bacias hidrográficas brasileiras. Pp. 25-44. In: Rocha, O. (Ed.).
Espécies invasoras em águas doces - estudo de caso e propostas de manejo. São Carlos,
Editora Universidade Federal de São Carlos, 416p.
Sobreiro, T. 2007. Territórios e Conflitos nas Pescarias do Médio Rio Negro (Barcelos,
Amazonas, Brasil). Dissertação de Mestrado - Instituto Nacional de Pesquisas na Amazônia-
INPA/UFAM. 172p.
Sobreiro T., Freitas C.E.C., Prado K.L., Nascimento F.A., Vicentini R. e Moraes A.M. 2010.
Na evaluation of fishery co-management experience in na Amazonian black-water river
(Unini River, Amazon, Brazil). Environment, Development and Sustainability. 12: 1013-
1024.

89
Stallard, R.F. e Edmond J.M. 1987. Geochemistry of the Amazon 3. Weathering Chemistry
and limits of dissolved inputs. Journal of Geophysical Research, 92: 8293-8302.
Sturgeon, R.E., Willie, S.N., Yang, L., Greenberg, R., Spatz, R.O., Chen, Z, Scriver, C.,
Clancy, V., Lam, J.W. e Thorrold, S. 2005. Certification of a fish otolith reference material in
support of quality assurance of trace element analysis. Journal of Analytical Atomic
Spectrometry, 20: 1067-1071.
Taphorn, D.C. e Barbarino Duque A. 1993. Evaluacion de la situacion actual de los pavones,
Cichla spp., en el Parque Nacional Capanaparo-Cinaruco, Estado Apure, Venezuela. Natura
96: 10-25.
Torrente-Vilara1, G.; Zuanon, J.; Leprieur, F.; Oberdorff, T.; Tedesco, P. A. 2011. Effects of
natural rapids and waterfalls on fish assemblage structure in the Madeira River (Amazon
Basin). Ecology of Freshwater Fish, 20: 588–597
Thorrold, S.R., Jones, C.M., Swart, P.K. e Targett, T.E. 1998. Accurate classification of
juvenile weakfish Cynoscion regalis to estuarine nursery areas based on chemical signatures
in otoliths. Marine Ecology Progress Series, 173: 253-265.
Thorrold, S. R., Latkoczy, C., Swart, P. K. e Jones C. B. M. 2001. Natal homing in a marine
fish metapopulation. Science 291, 297-299.
Thorstad, E. B.; Hay, C. J.; Næsje, T. F. e Økland, F. 2001. Movements and habitat utilization
of three cichlid species in the Zambezi River, Namibia. Ecology of Freshwater Fish, 10: 238-
246.
Tipper E. T.; Bickle M. J.; Galy A.; West A. J.; Pomie`s C. e Chapman H. J. 2006. The short
term climatic sensitivity of carbonate and silicate weathering fluxes: Insight from seasonal
variations in river chemistry. Geochimica et Cosmochimica Acta, 70: 2737-2754.
Vital, H. e Stattegger, K. 2000. Major and trace elements of stream sediments from the
lowermost Amazon River. Chemical Geology, 168: 151-168.
Walther, B. D. e Thorrold, S. R. 2010. Limited diversity in natal origins of immature
anadromous fish during ocean residency. Canadian Journal of Fisheries and Aquatic Science
67: 1699-1707.

90
Walther, B.D., Thorrold, S.R. e Olney J.E. 2008. Geochemical Signatures in Otoliths Record
Natal Origins of American Shad. Transactions of the American Fisheries Society, 137: 57-69.
Walther, B.D. e Thorrold, S.R. 2010. Limited diversity in natal origins of immature
anadromous fish during ocean residency. Canadian Journal of Fisheries and Aquatic
Sciences, 67: 1699-1707.
Walther, B.D. e Limburg, K.E. 2012. The use of otolith chemistry to characterize diadromous
migrations. Journal of Fish Biology, 81: 796-825.
Weatherhead, P.J.; Forbes, M.R.L. e Patrick J. 1994. Natal philopatry in passerine birds:
genetic or ecological influences?. International Society for Behavioural Ecology, 5(4): 426.
Wells, B.K., B.E. Rieman, J.L. Clayton, D.L. Horan e Jones, C.M. 2003. Relationships
between water, otolith, and scales chemistries of westlope cutthroat trout from the Coeur
d’Alene River, Idaho: the potential application of hard-part chemistry to describe movements
in freshwater. Transactions of the American Fisheries Society, 132: 409-424.
Willis S.C., Nunes M.S., Montana C.G., Farias I.P. e Lovejoy N.R. 2007. Systematics,
biogeography, and evolution of the neotropical peacock basses Cichla (Perciformes:
Cichlidae). Molecular Phylogenetics and Evolution, 44, 291-307.
Willis S. C., Nunes M. Montana C. G., Farias I. P. Orti G., e Lovejoy N. 2010. The
Casiquiare river acts as a corridor between the Amazonas and Orinoco river basins:
biogeographic analysis of the genus Cichla. Molecular Ecology, 19: 1014-1030.
Winemiller K. O. 1989. Patterns of variation in in life History among South American fishes
in seasonal environments. Oecologia,Spring-Verilag, 81: 225-241.
Winemiller, K.O., Taphorn, D.C. e Barbarino-Duque, A. 1997. The ecology of Cichla
(Cichlidae) in two blackwater Rivers of southern Venezuela. Copeia, 4: 690-696.
Winemiller, K.O. e Jepsen, D.B. 1998. Effects of seasonality and fish movement on tropical
River food webs. Journal of Fish Biology, 53: 267-296.
Winemiller, K.O. 2001. Ecology of peacock cichlids (Cichla spp.) in Venezuela. Journal of
Aquatic Sciences, 9: 93-112.

91
Winemiller, K.O.; López-Fernández, H.; Taphorn, D.C.; Nico, L.G. e Barbarino Duque A.
2008. Fish assemblages of the Casiquiare River, a corridor and zoogeographical filter for
dispersal between the Orinoco and Amazon basins. Journal of Biogeography. 35: 1551-1563.
Wolff, B.A.; Johnson, B.M.; Breton, A.R.; Martinez, P.J. e Winkelman, D.L. 2012. Origins of
invasive piscivores determined from the strotium isotope ratio (87Sr/86Sr) of otoliths.
Canadian Fish Science, 69: 724-739.
Zar, J.H. 1999. Biostatistical Analysis. Prentice Hall, New Jersey, 663 pp.
Zaret, T.M. 1980. Life history and growth relationships of Cichla ocellaris, a predatory South
American Cichlid. Biotropica, 12: 144-157.


















