
INTERAÇÃO GENÓTIPOS POR AMBIENTES PARA … · experimentos, realização e interpretação das...
107
INSTITUTO AGRONÔMICO CURSO DE PÓS-GRADUAÇÃO EM AGRICULTURA TROPICAL E SUBTROPICAL “INTERAÇÃO GENÓTIPOS POR AMBIENTES PARA PRODUTIVIDADE DE GRÃOS, QUALIDADE TECNOLÓGICA E RESISTÊNCIA A PATÓGENOS EM FEIJOEIRO” Cleber Vinicius Giaretta Azevedo Orientador: Dr. Alisson Fernando Chiorato Dissertação submetida como requisito parcial para obtenção do grau de Mestre em Agricultura Tropical e Subtropical, Área de Concentração em Genética, Melhoramento Vegetal e Biotecnologia. Campinas, SP Fevereiro, 2014
-
Upload
phungtuong -
Category
Documents
-
view
215 -
download
0
Transcript of INTERAÇÃO GENÓTIPOS POR AMBIENTES PARA … · experimentos, realização e interpretação das...

INSTITUTO AGRONÔMICO
CURSO DE PÓS-GRADUAÇÃO EM AGRICULTURA
TROPICAL E SUBTROPICAL
“INTERAÇÃO GENÓTIPOS POR AMBIENTES PARA
PRODUTIVIDADE DE GRÃOS, QUALIDADE
TECNOLÓGICA E RESISTÊNCIA A PATÓGENOS EM
FEIJOEIRO”
Cleber Vinicius Giaretta Azevedo
Orientador: Dr. Alisson Fernando Chiorato
Dissertação submetida como requisito parcial para
obtenção do grau de Mestre em Agricultura
Tropical e Subtropical, Área de Concentração em
Genética, Melhoramento Vegetal e Biotecnologia.
Campinas, SP
Fevereiro, 2014

ii

iii
Aos meus pais Valéria e Cleber, que em
nenhum momento mediram esforços para
realização dos meus sonhos. A eles devo a
pessoa que me tornei, tenho muito orgulho
por chamá-los de pai e mãe.
DEDICO
A minha namorada Letícia, pelos
conselhos e companheirismo, estando
ao meu lado em todos os momentos,
me fazendo feliz e querer ser sempre
melhor.
OFEREÇO

iv
“Aprenda como se você fosse viver para
sempre. Viva como se você fosse morrer
amanhã”
Mahatma Gandhi

v
AGRADECIMENTOS
Agradeço primeiramente a Deus, pela vida, por estar sempre no meu caminho, iluminando e
guiando às escolhas certas.
A minha família, que sempre me apoiou em todos os momentos.
Ao Dr. Alisson Fernando Chiorato pela orientação, oportunidade, ensinamentos e tempo dedicado
para a conclusão deste trabalho.
Ao Dr. Sérgio Augusto Morais Carbonell pelo auxilio e sugestões durante o transcorrer do trabalho.
A Dra Maria Elisa Ayres Guidetti Zagatto Paterniani pelos ensinamentos, correções e sugestões em
minha dissertação.
Aos Dr. José Antônio de Fátima Esteves, Dr. João Guilherme Ribeiro Gonçalves, Dra Fátima
Bosseti, Msc. Daiana Alves da Silva e Msc. Vinicius Andrade pela ajuda na execução dos
experimentos, realização e interpretação das análises estatísticas.
Ao Instituto Agronômico – IAC pela oportunidade.
A FAPESP pelo financiamento do trabalho e concessão da bolsa de estudos.
Aos membros da banca examinadora pela contribuição no trabalho e pela disponibilidade de
participação.
Aos amigos que sempre estiveram ao meu lado na execução deste trabalho: Aurélio Teixeira, Fábio
Belo, Estela Reis de Andrade, Tamires Ribeiro, Rodrigo Lorenzetti Tunes e Graziele Sasseron que
também contribuíram durante este período.
A coordenadora da Pós Graduação do Instituto Agronômico, Adriana Parada Dias da Silveira, a
todos os professores, secretárias, colegas, estagiários do Programa de Melhoramento Genético do
Feijoeiro e técnicos de campo pelo apoio e compreensão.
A todos que contribuíam diretamente ou indiretamente para que este trabalho pudesse ser realizado.
Aos amigos que fiz durante o curso, pela verdadeira amizade que construímos em particular aqueles
que estavam sempre ao meu lado.
Aos meus amigos da área de Grãos e Fibras do Instituto Agronômico - IAC.
Enfim a todas as pessoas que me ajudaram, não poderia deixar de expressar à minha imensa
gratidão.

vi
SUMÁRIO
LISTA DE SIGLAS .......................................................................................................................... viii
LISTA DE TABELAS ........................................................................................................................ ix
LISTA DE FIGURAS ......................................................................................................................... xi
1. INTRODUÇÃO ........................................................................................................................ 1
2. REVISÃO DE LITERATURA ................................................................................................. 2
2.1 ASPECTOS GERAIS DA CULTURA DO FEIJOEIRO ........................................................................... 2
2.2 ENSAIOS DE VALOR DE CULTIVO E USO (VCU) .......................................................................... 2
2.3 RESISTÊNCIA A PATÓGENOS ......................................................................................................... 3
2.4 ANTRACNOSE (COLLETOTRICHUM LINDEMUTHIANUM) ................................................................. 4
2.5 MURCHA DE FUSARIUM (FUSARIUM OXYSPORUM F.SP. PHASEOLI) ................................................ 5
2.6 CRESTAMENTO BACTERIANO (XANTHOMONAS AXONOPODIS PV. PHASEOLI) .................................. 6
2.7 QUALIDADE TECNOLÓGICA DE GRÃOS ........................................................................................ 7
2.8 INTERAÇÃO GENÓTIPOS POR AMBIENTES (GXA) ......................................................................... 9
2.9 ESTRATIFICAÇÃO AMBIENTAL ................................................................................................... 10
2.10 ADAPTABILIDADE E ESTABILIDADE ........................................................................................... 11
3. MATERIAL E MÉTODOS .................................................................................................... 13
3.1 ENSAIOS DE VCU ...................................................................................................................... 13
3.2 AVALIAÇÃO DE DOENÇAS ......................................................................................................... 15
3.2.1 Antracnose (Colletotrichum lindemuthianum) ......................................................................... 15
3.2.2 Murcha de Fusarium (Fusarium. oxysporum f. sp. phaseoli) ................................................... 16
3.2.3 Crestamento Bacteriano (Xanthomonas axonopodis pv. phaseoli) .......................................... 17
3.3 AVALIAÇÃO DA QUALIDADE TECNOLÓGICA DOS GRÃOS .......................................................... 19
3.3.1 Análises do Tempo de Cocção dos Grãos ................................................................................ 19
3.3.2 Absorção de Água Antes do Cozimento (PEANC), Absorção de Água Após Cozimento e
Porcentagem de Grãos Inteiros (PGI) ................................................................................................ 20
3.3.3 Avaliação da Coloração do Tegumento .................................................................................... 20

vii
3.3.4 Avaliação da Porcentagem de Proteína nos Grãos ................................................................... 21
3.4 ANÁLISES ESTATÍSTICAS DOS DADOS ........................................................................................ 22
3.4.1 Análise de Variância Individual e Conjunta ............................................................................. 22
3.4.2 Estratificação Ambiental .......................................................................................................... 22
3.4.3 Adaptabilidade e Estabilidade .................................................................................................. 26
4. RESULTADOS E DISCUSSÃO ............................................................................................ 28
4.1 AVALIAÇÃO DE DOENÇAS ......................................................................................................... 28
4.1.1 Antracnose (Colletotrichum lindemuthianum) ......................................................................... 28
4.1.2 Murcha de Fusarium (Fusarium oxysporum f. sp, phaseoli) .................................................... 29
4.1.3 Crestamento Bacteriano (Xanthomonas axonopodis pv. phaseoli) .......................................... 30
4.2 ANÁLISES DOS DADOS ............................................................................................................... 31
4.3 QUALIDADE TECNOLÓGICA ....................................................................................................... 33
4.4 PRODUTIVIDADE DE GRÃOS ....................................................................................................... 40
4.5 ESTRATIFICAÇÃO AMBIENTAL ................................................................................................... 41
4.6 ADAPTABILIDADE E ESTABILIDADE ........................................................................................... 51
5. CONCLUSÕES ...................................................................................................................... 60
6. REFERÊNCIAS ...................................................................................................................... 61
7. ANEXOS ................................................................................................................................ 70

viii
LISTA DE SIGLAS
VCU – VALOR DE CULTIVO E USO
GxA – Genótipo x Ambiente
AF – Análise de Fatores
TC – Tempo de cozimento
PEANC – Porcentagem de embebição antes do cozimento
PEAPC – Porcentagem de embebição após o cozimento
PGI – Porcentagem de grãos inteiros
PROT – Porcentagem de proteína
PROD – Produtividade em kg.ha-1
CBA.11 – Capão Bonito – Safra das águas de 2011
MOA.11 – Mococa – Safra das águas de 2011
TQS.12 – Taquarituba – Safra da seca de 2012
AVS.12 – Avaré – Safra da seca de 2012
MOS.12 – Mococa – Safra da seca de 2012
VTI.12 – Votuporanga – Safra de inverno de 2012
RPI.12 – Ribeirão Preto – Safra de inverno de 2012
COI.12 – Colina – Safra de inverno de 2012
ANI.12 – Andradina – Safra de inverno de 2012
CBA.12 – Capão Bonito – Safra das águas de 2012
ITA.12 – Itararé – Safra das águas de 2012
TQA.12 – Taquarituba – Safra das águas de 2012
MOS.13 – Mococa – Safra da seca de 2013
ITS.13 – Itararé – Safra da seca de 2013
ARS.13 – Araras – Safra da seca de 2013
TQS.13 – Taquarituba – Safra da seca de 2013
ANI.13 – Andradina – Safra de inverno de 2013
VTI.13 – Votuporanda – Safra de inverno de 2013
RPI.13 – Ribeirão Preto – Safra de inverno de 2013

ix
LISTA DE TABELAS
Tabela 1. Relação dos 27 genótipos de feijoeiro avaliados nos ensaios de VCU para os anos
agrícolas 2011/2012 e 2013, no estado de São Paulo. .................................................... 14
Tabela 2. Municípios utilizados para condução dos ensaios de acordo com a safra e suas respectivas
latitudes e altitudes .......................................................................................................... 14
Tabela 3. Avaliação da reação de resistência dos genótipos de feijoeiro ao patógeno Colletotrichum
lindemuthianum. .............................................................................................................. 29
Tabela 4. Avaliação da reação de resistência dos genótipos de feijoeiro ao patógeno Fusarium
oxysporum f. sp, phaseoli. ............................................................................................... 30
Tabela 5. Avaliação da reação de resistência dos genótipos de feijoeiro ao patógeno Xanthomonas
axonopodis. pv, phaseoli. ................................................................................................ 31
Tabela 6. Resumo da análise de variância conjunta dos 27 genótipos de feijoeiro cultivados em 19
locais do estado de São Paulo por safra e geral. ............................................................. 32
Tabela 7. Avaliação de colorimetria (L*) para grãos de tegumento carioca. ................................... 34
Tabela 8. Avaliação de colorimetria (L*) para grãos de tegumento preto. ....................................... 34
Tabela 9. Estimativa de médias das avaliações de qualidade tecnológica tempo de cozimento,
porcentagem de embebição antes do cozimento (PEAPC), porcentagem de embebição
após o cozimento (PEAPC), porcentagem de grãos inteiros (PGI) e teor de proteína
bruta (PROT) para safra das águas. ................................................................................. 35
Tabela 10. Médias das avaliações para tempo de cozimento, porcentagem de embebição antes do
cozimento (PEAPC), porcentagem de embebição após o cozimento (PEAPC),
porcentagem de grãos inteiros (PGI) e teor de proteína bruta (PROT) para safra da seca.
......................................................................................................................................... 36
Tabela 11. Médias das avaliações para tempo de cozimento, porcentagem de embebição antes do
cozimento (PEAPC), porcentagem de embebição após o cozimento (PEAPC),
porcentagem de grãos inteiros (PGI) e teor de proteína bruta (PROT) para safra de
inverno ............................................................................................................................. 37
Tabela 12. Médias das avaliações de qualidade tecnológica tempo de cozimento, porcentagem de
embebição antes do cozimento (PEAPC), porcentagem de embebição após o cozimento
(PEAPC), porcentagem de grãos inteiros (PGI) e teor de proteína bruta (PROT). ........ 39
Tabela 13. Produtividade média dos genótipos em kg.ha-1
por safra e geral para todos ambientes. 41
Tabela 14. Porcentagem da parte simples da interação genótipo x ambiente para a variável
produção de grãos. .......................................................................................................... 42
Tabela 15. Estratificação ambiental para produtividade por meio da analise de fatores, com 27
genótipos de feijoeiro avaliados em 19 ambientes no estado de São Paulo. ................... 44

x
Tabela 16. Porcentagem da parte simples da interação genótipo x ambiente para a variável
coloração do tegumento dos grãos. ................................................................................. 46
Tabela 17. Estratificação ambiental para coloração do tegumento por meio da analise de fatores,
com 27 genótipos de feijoeiro avaliados em 19 ambientes no estado de São Paulo. ...... 47
Tabela 18. Porcentagem da parte simples da interação genótipo x ambiente para a variável
porcentagem de proteína nos grãos. ................................................................................ 48
Tabela 19. Estratificação ambiental para porcentagem de proteína nos grãos por meio da analise de
fatores, com 27 genótipos de feijoeiro avaliados em 19 ambientes no estado de São
Paulo. ............................................................................................................................... 49
Tabela 20. Porcentagem da parte simples da interação genótipo x ambiente para a variável tempo
médio de cozimento dos grãos. ....................................................................................... 50
Tabela 21. Estratificação ambiental para tempo de cozimento dos grãos por meio da analise de
fatores, com 27 genótipos de feijoeiro avaliados em 19 ambientes no estado de São
Paulo. ............................................................................................................................... 51
Tabela 23. Estimativas de parâmetros de estabilidade multivariada (Pi) de acordo com o método de
Lin e Binns (1988) modificada por Carneiro (1998) e média em kg.ha-1
para 27
genótipos de feijoeiro avaliados em 5 ambientes no estado de São Paulo na safra das
águas. ............................................................................................................................... 55
Tabela 24. Estimativas de parâmetros de estabilidade multivariada (Pi) de acordo com o método de
Lin e Binns (1988) modificada por Carneiro (1998) e média em kg.ha-1
para 27
genótipos de feijoeiro avaliados em 7 ambientes no estado de São Paulo na safra da
seca. ................................................................................................................................. 56
Tabela 25. Estimativas de parâmetros de estabilidade multivariada (Pi) de acordo com o método de
Lin e Binns (1988) modificada por Carneiro (1998) e média em kg.ha-1
para 27
genótipos de feijoeiro avaliados em 7 ambientes no estado de São Paulo na safra de
inverno. ............................................................................................................................ 57
Tabela 26. Estimativas de parâmetros de estabilidade multivariada de acordo com o método de Lin
e Binns (1988) modificada por Carneiro (1998) para 27 genótipos de feijoeiro avaliados
em 7 ambientes no estado de São Paulo considerando todas safras. .............................. 58

xi
LISTA DE FIGURAS
Figura 1. Procedimentos realizados para inoculação de C. lindemuthianum em 27 genótipos de
feijoeiro. .......................................................................................................................... 16
Figura 2. Inoculação de Fusarium oxysporum f. sp. phaseoli pelo método de imersão dipping
(COSTA et al., 1989). ..................................................................................................... 17
Figura 3. Plantas de feijoeiro resistentes (a esquerda), intermediarias (no centro) e suscetível (a
direita) ao Fusarium oxysporum f. sp. phaseoli. ............................................................. 17
Figura 4. Inoculação de Xanthomonas axonopodis pv. phaseoli pelo método agulhas múltiplas
(POMPEU et al. 1973). ................................................................................................... 18
Figura 5. Plantas de feijoeiro resistente (a esquerda), intermediaria (no centro) e suscetível (a
direita) a Xanthomonas axonopodis pv. phaseoli. .......................................................... 19
Figura 6. Análise de tempo de cozimento por meio do Cozedor de Mattson. ................................. 19
Figura 7. Colorímetro Minolta® - CR 410 ....................................................................................... 21
Figura 8. Análise gráfica da adaptabilidade de 27 genótipos de feijoeiro, por meio da análise de
fatores, considerando os ambientes contidos nos fatores 1 e 2. ...................................... 52
Figura 9. Análise gráfica da adaptabilidade de 27 genótipos de feijoeiro, por meio da análise de
fatores, considerando os ambientes contidos nos fatores 3 e 4. ...................................... 53

xii
LISTA DE ANEXOS
Anexo 1. Médias de PEANC, PEAPC, PGI, PROT, L* e PROD dos 27 genótipos avaliados em
Capão Bonito na safra das águas de 2011 ......................................................................... 70
Anexo 2. Médias de PEANC, PEAPC, PGI, PROT, L* e PROD dos 27 genótipos avaliados em
Mococa na safra das águas de 2011 .................................................................................. 71
Anexo 3. Médias de PEANC, PEAPC, PGI, PROT, L* e PROD dos 27 genótipos avaliados em
Capão Bonito na safra das águas de 2012. ........................................................................ 72
Anexo 4. Médias de PEANC, PEAPC, PGI, PROT, L* e PROD dos 27 genótipos avaliados em
Itararé na safra das águas de 2012. .................................................................................... 73
Anexo 5. Médias de PEANC, PEAPC, PGI, PROT, L* e PROD dos 27 genótipos avaliados em
Taquarituba na safra das águas de 2012. ........................................................................... 74
Anexo 6. Médias de PEANC, PEAPC, PGI, PROT, L* e PROD dos 27 genótipos avaliados em
Andradina na safra de inverno de 2012. ............................................................................ 75
Anexo 7. Médias de PEANC, PEAPC, PGI, PROT, L* e PROD dos 27 genótipos avaliados em
Colina na safra de inverno de 2012. .................................................................................. 76
Anexo 8. Médias de PEANC, PEAPC, PGI, PROT, L* e PROD dos 27 genótipos avaliados em
Ribeirão Preto na safra de inverno de 2012. ...................................................................... 77
Anexo 9. Médias de PEANC, PEAPC, PGI, PROT, L* e PROD dos 27 genótipos avaliados em
Votuporanga na safra de inverno de 2012. ........................................................................ 78
Anexo 10. Médias de PEANC, PEAPC, PGI, PROT, L* e PROD dos 27 genótipos avaliados em
Avaré na safra da seca de 2012. ........................................................................................ 79
Anexo 11. Médias de PEANC, PEAPC, PGI, PROT, L* e PROD dos 27 genótipos avaliados em
Mococa na safra da seca de 2012. ..................................................................................... 80
Anexo 12. Médias de PEANC, PEAPC, PGI, PROT, L* e PROD dos 27 genótipos avaliados em
Taquarituba na safra da seca de 2012. ............................................................................... 81
Anexo 13. Médias de PEANC, PEAPC, PGI, PROT, L* e PROD dos 27 genótipos avaliados em
Andradina na safra de inverno de 2013. ............................................................................ 82
Anexo 14. Médias de PEANC, PEAPC, PGI, PROT, L* e PROD dos 27 genótipos avaliados em
Ribeirão Preto na safra de inverno de 2013. ...................................................................... 83
Anexo 15. Médias de PEANC, PEAPC, PGI, PROT, L* e PROD dos 27 genótipos avaliados em
Votuporanga na safra de inverno de 2013. ........................................................................ 84
Anexo 16. Médias de PEANC, PEAPC, PGI, PROT, L* e PROD dos 27 genótipos avaliados em
Araras na safra da seca de 2013. ....................................................................................... 85

xiii
Anexo 17. Médias de PEANC, PEAPC, PGI, PROT, L* e PROD dos 27 genótipos avaliados em
Itararé na safra da seca de 2013. ........................................................................................ 86
Anexo 18. Médias de PEANC, PEAPC, PGI, PROT, L* e PROD dos 27 genótipos avaliados em
Mococa na safra da seca de 2013. ..................................................................................... 87
Anexo 19. Médias de PEANC, PEAPC, PGI, PROT, L* e PROD dos 27 genótipos avaliados em
Taquarituba na safra da seca de 2013. ............................................................................... 88
Anexo 20. Estimativas de correlação de Pearson entre as variáveis TC, PEANC, PEAPC, PGI,
PROT, COR e PROD ........................................................................................................ 88
Anexo 21. Estimativas de parâmetros da estabilidade de produção de grãosde acordo com o método
de Lin e Binns (1988) modificada por Carneiro (1998) para 27 genótipos de feijoeiro
avaliados na safra das águas no estado de São Paulo. ....................................................... 89
Anexo 22. Estimativas de parâmetros da estabilidade de produção de grãosde acordo com o método
de Lin e Binns (1988) modificada por Carneiro (1998) para 27 genótipos de feijoeiro
avaliados na safra da seca no estado de São Paulo. ........................................................... 90
Anexo 23. Estimativas de parâmetros da estabilidade de produção de grãosde acordo com o método
de Lin e Binns (1988) modificada por Carneiro (1998) para 27 genótipos de feijoeiro
avaliados na safra de inverno no estado de São Paulo. ..................................................... 91
Anexo 24. Estimativas de parâmetros da estabilidade de produção de grãosde acordo com o método
de Lin e Binns (1988) modificada por Carneiro (1998) para 27 genótipos de feijoeiro
avaliados no estado de São Paulo em três safras. .............................................................. 92

xiv
Interação genótipos por ambientes para produtividade de grãos, qualidade tecnológica e
resistência a patógenos em feijoeiro
RESUMO
O objetivo do trabalho foi avaliar 27 genótipos de feijoeiro nos ensaios de Valor de Cultivo e Uso -
VCU realizados pelo Instituto Agronômico – IAC em diferentes ambientes no estado de São Paulo.
Os genótipos foram avaliados para a produtividade de grãos, reação de resistência à patógenos e
qualidade tecnológica dos grãos, estimando-se também os efeitos da interação GxA para indicação
do genótipo mais adaptado as diferentes condições de cultivo. Por meio de inoculações artificiais,
os genótipos foram avaliados para os patógenos da antracnose, murcha de fusarium e crestamento
bacteriano. Para qualidade tecnológica foram realizados os testes de tempo de cozimento (TC),
porcentagem de embebição antes e após o cozimento (PEANC e PEAPC), porcentagem de grãos
inteiros após cozimento (PGI), teor de proteína bruta (PROT) e coloração do tegumento (COR). Os
efeitos da interação GxA foram estimados por meio da decomposição da parte simples da interação
GxA e estratificação de ambientes por meio da análise de fatores. Também foram realizadas
análises de adaptabilidade e estabilidade pelo método de LIN E BINNS (1988) modificado por
Carneiro (1998). Para as avaliações de reação de resistência aos patógenos foi observado que os
genótipos CNFC 10762, LP 09-40, MAII – 2, IAC – Una, Gen PR 14-2-10-1-2, Gen PR 14-2-2-1-1
e BRS Madrepérola foram resistentes à antracnose. Os genótipos Gen C 4-7-7-2-2, LP 09-40,
CNFC 10729, Gen PR 14-2-2-1-1, LP 08-90 e SM 1810 foram resistentes à murcha de fusarium e
os genótipos LP 07-80, LP 08-90, LP 09-192 e SM 1810 foram resistentes ao crestamento
bacteriano. Observou-se que a maior parte da interação GxA para produtividade, PROT e TC foi do
tipo complexa, sendo possível o agrupamento de alguns ambientes por meio da análise de fatores.
Para COR foi observado que a maior parte da interação GxA foi do tipo simples, sendo agrupado a
maioria dos ambientes em apenas um grupo por meio da análise de fatores. Nas análises de
adaptabilidade e estabilidade foi observado que a utilização apenas da produtividade de grãos
fornece resultados mais robustos sobre o genótipo de melhor desempenho quando comparado à
análise de adaptabilidade e estabilidade com diversas variáveis. Dentre os genótipos de tegumento
carioca os mais estáveis foram LP 09-40 e LP 07-80 e para os genótipos de tegumento preto foram
CNFP 10794 e LP 09-192. Considerando que para a correta indicação de um genótipo de feijoeiro
deve-se levar em consideração outras características além da produtividade de grãos, observou-se
que os genótipos superiores que reuniram estabilidade produtiva, alta qualidade tecnológica dos
grãos e resistência a doenças foram LP 09-40, CNFC 10762 e Gen C 4-7-7-2-2 para genótipos de
tegumento carioca e CNFP 10794, LP 09-192 para genótipos de tegumento preto.
Palavras-chave: Phaseolus vulgaris L., adaptabilidade e estabilidade, proteína bruta, tempo de
cozimento, proteína bruta

xv
Genotype by environment’s interaction of the production of grain, technological quality and
pathogens resistance of commom bean
ABSTRACT
The objective of this study was to evaluate 27 common bean genotypes in trials of Value
Cultivation and Use - VCU conducted by the Agronomic Institute - IAC in different environments
in the state of São Paulo. The genotypes were evaluated for the grain yield, reaction of resistance to
pathogens and technological quality of grain, also estimating the effects of GxE interaction for
indication of the most adapted genotype to different growing conditions. By artificial inoculation,
the genotypes were evaluated for the anthracnose pathogen, wilt fusarium and bacterial blight. For
the technological quality it was conducted cooking time test (TC), imbibition percentage before and
after cooking (PEANC and PEAPC), percentage of whole grains after cooking (PGI), row protein
level (PROT) and tegument coloration (COR). The effects of the GxE interaction were estimated
through the decomposition of the GxE´s simplest part interaction and environments stratification
through factor analysis. Adaptability and stability analysis by the method of Lin and Binns (1988)
modified by Carneiro (1998) were also performed. For evaluations of the resistance reactions to
pathogens it was observed that genotypes CNFC 10762 , LP 09-40 , Maii - 2 , IAC Una , Gen PR
14-2-10-1-2 , Gen PR 14-2-2-1-1 and BRS Madrepérola were resistant to anthracnose. The
genotypes Gen C 4-7-7-2-2 , LP 09-40 , CNFC 10729 , Gen PR 14-2-2-1-1 , SM 1810 LP 08-90
were resistant to fusarium wilt and genotypes LP 07-80 , LP 08-90 , LP 09-192 and SM 1810 were
resistant to bacterial blight. It was observed that most part of the GxE for productivity, PROT and
TC were the complex type, making possible the grouping of some environments through factors
analysis. For COR it was observed that the biggest part of the GxE interaction was considered
simple type, which was grouped to the majority of envirouments in only one group by fator
analysis. In the analyzes of adaptability and stability it has been observed that the use of only the
productivity parameters provide more robust results about the genotype of best performance when
compared to analysis of stability and adaptability to different variables. Among varieties of carioca
integument the most stable were LP 09-40 and LP 07-80 and for the black integument genotypes
were CNFP 10794 and LP 09-192. Considering that for the correct FEIJOEIRO genotype
indication, one should take into account some other characteristics besides grain yield, it was
observed that superior genotypes that put together yield stability , high technological grain quality
and disease resistance were LP 09-40 , CNFC 10762 and Gen C 4-7-7-2-2 for genotypes carioca
integument and CNFP 10794 , LP 09-192 genotypes for black coat .
Key words: Phaseolus vulgaris L., adaptability and stability, raw protein, cooking time

1
1. INTRODUÇÃO
Para uma nova cultivar de feijoeiro (Phaseolus vulgaris L.) ser aceita no mercado é
necessário que o genótipo indicado reúna características desejáveis para os produtores e para o
mercado consumidor. Neste sentido, os programas de melhoramento genético de feijoeiro visam à
seleção de genótipos que apresentem características agronômicas desejáveis, como por exemplo,
alta produtividade de grãos, resistência aos principais patógenos da cultura e arquitetura de plantas
para colheita mecanizada, além das características relativas à qualidade tecnológica dos grãos, como
maior teor de nutrientes, menor tempo de cozimento, coloração clara do tegumento para feijões
cariocas e grãos mais graúdos.
No melhoramento genético de plantas autógamas, as etapas iniciais de seleção dos genótipos
normalmente são realizadas em apenas um ambiente devido a baixa quantidade de sementes, onde
desta forma a manifestação fenotípica é o resultado da ação do genótipo sob influência deste meio.
Entretanto, quando se considera uma série de ambientes distintos, sendo esta, situação comum aos
produtores, observa-se além dos efeitos genéticos e ambientais, um efeito adicional, proporcionado
pela interação destes (CRUZ et al. 2004).
Segundo RESENDE (2007), o efeito da interação GxA é decorrente do comportamento
diferencial dos genótipos nos diferentes ambientes de cultivo e pode indicar que o melhor indivíduo
em um ambiente pode não sê-lo em outro. Assim, este pode ser um complicador na seleção e
indicação de novas cultivares, se não for considerado adequadamente.
Neste sentido, na etapa final do desenvolvimento de cultivares é necessário um estudo
detalhado do desempenho dos genótipos em diversos ambientes, para avaliar a magnitude e
significância da interação GxA, fornecendo subsídios que possibilitem adotar procedimentos para
sua minimização e, ou, seu aproveitamento (CRUZ et al. 2004).
Estas avaliações são denominadas “ensaios de VCU” e são indispensáveis para o registro
nacional de cultivares (RNC) junto ao Ministério da Agricultura Pecuária e Abastecimento (MAPA)
para testar os genótipos nas diferentes condições edafoclimáticas que serão recomendados, sendo
assim possível selecionar genótipos mais adaptados e que sofram menos com a interação GxA.
Do exposto, este trabalho teve o objetivo de avaliar 27 genótipos de feijoeiro pertencentes
aos ensaios de VCU realizados nos anos agrícolas de 2011, 2012 e 2013 em diferentes municípios
do estado de São Paulo para estimar a interação GxA para a produtividade de grãos e qualidade
tecnológica por meio de análises de estratificação ambiental, avaliar a eficiência dos métodos de
estabilidade fenotípica, e também a reação de resistência dos genótipos aos patógenos:
Colletotrichum lindemuthianum, Fusarium oxysporum e Xanthomonas axonopodis por meio de
inoculações artificiais em laboratório.

2
2. REVISÃO DE LITERATURA
2.1 ASPECTOS GERAIS DA CULTURA DO FEIJOEIRO
O feijão é uma espécie pertencente a família Leguminosae, gênero Phaseolus, e classificado
como Phaseolus vulgaris L. (SANTOS E GAVILANES, 1998). É uma planta predominantemente
autógama, de ciclo anual, originária do continente americano, porém sem centro de origem
específico, sendo considerado apenas dois locais de domesticação, Andino e Mesoamericano.
O Brasil se destaca entre os maiores produtores e consumidores mundiais de feijão, tendo
produzido no ano de 2013 cerca de 3,384 milhões de toneladas de grãos, onde este, é cultivado em
praticamente todo o território, em uma área total estimada de 3,129 milhões de ha-1
(CONAB,
2014).
Esta produção está concentrada principalmente nos estados do Paraná, Minas Gerais, Mato
Grosso, Goias e São Paulo os quais respondem por 70% da produção nacional. De acordo com o
zoneamento agrícola da cultura, no estado de São Paulo o cultivo concentra principalmente em três
safras, sendo: safra “das águas”, onde a semeadura é realizada predominantemente entre os meses
de agosto e setembro, safra “da seca”, realizada entre os meses de janeiro e fevereiro e o cultivo de
“inverno”, semeando-se entre os meses de abril e maio (CONAB, 2014).
Embora o Brasil se enquadre entre os maiores produtores mundiais de feijão, a
produtividade brasileira, estimada em 1.082 kg.ha-1
é considerada baixa frente ao potencial de
produção da cultura, sendo que em algumas regiões de cultivo nos estados de Minas Gerais, Paraná,
São Paulo e Goiás, têm alcançado produtividade acima de 3.000 kg.ha-1
(IBGE, 2012).
Segundo ZUCARELI et al. (2011), os principais fatores responsáveis por este baixo
rendimento são o uso de manejos culturais inadequados, incidência de pragas e doenças, uso de
sementes de baixa qualidade fisiológica e sanitária, pouco uso de insumos agrícolas como corretivos
e fertilizantes, além de problemas climáticos.
2.2 ENSAIOS DE VALOR DE CULTIVO E USO (VCU)
De acordo com a instrução normativa n° 25, de 23 de maio de 2006, do Ministério da
Agricultura, Pecuária e Abastecimento (MAPA), para serem lançadas novas cultivares de feijoeiro
no mercado, os genótipos devem ser testadas por um mínimo de dois anos, três locais por região e
safra representativas à cultura (MAPA, 2006). Estes ensaios são denominados como “ensaios de
VCU” (Valor de Cultivo e Uso) e propõem os métodos de campo e laboratório para avaliação destas
cultivares e linhagens.
A importância destes ensaios é para obtenção de informações biológicas sobre o

3
comportamento das cultivares nos ambientes em que estes serão recomendados, sendo também,
uma etapa indispensável para o registro nacional de cultivares (RNC) junto ao Ministério da
Agricultura Pecuária e Abastecimento (MAPA). Estas avaliações fazem parte da etapa final no
desenvolvimento de uma nova cultivar e visam proteger o agricultor da venda indiscriminada de
sementes e mudas de cultivares que não tenham sido testadas e validadas nas condições
edafoclimáticas de exploração agrícola no Brasil (MAPA, 2006).
Além das avaliações agronômicas referentes à produtividade de grãos e resistência a
patógenos, a partir de 1999, de acordo com a Portaria 294 de 14 de outubro de 1998, testes de
qualidade tecnológica também passaram a ser exigidos para o registro de novas cultivares de
feijoeiro junto ao MAPA, para sua inscrição no sistema de comercialização de sementes no Brasil.
Atualmente, os principais parâmetros de qualidade dos grãos exigidos são a determinação do tempo
médio de cozimento e do teor médio de proteína dos genótipos que estejam inseridos nos ensaios de
VCU.
Neste sentido, o Programa de Melhoramento Genético do Feijoeiro do Instituto Agronômico
– IAC é responsável pela realização dos ensaios de VCU no Estado de São Paulo, onde em parceria
com outros institutos de pesquisa é realizada a avaliação de linhagens e cultivares de diversas
regiões do país.
2.3 RESISTÊNCIA A PATÓGENOS
Dentre os fatores que contribuem para a instabilidade de produção e a baixa produtividade
do feijoeiro, MIKLAS et al. (2006) salientaram que o principal estresse biótico que afeta a
produtividade de grãos de feijoeiro no país, é a ocorrência de doenças causadas por vírus, fungos e
bactérias. Citam-se mais de 45 patógenos de maior ou menor importância incidindo sobre o
feijoeiro. Entre as mais comuns no Brasil, estão: antracnose (Colletotrichum lindemuthianum),
mancha-angular (Pseudocercospora griseola), ferrugem (Uromyces appendiculatus), mofo branco
(Sclerotinia sclerotiorum), oídio (Erysiphe polygoni) e murcha de fusarium (Fusarium oxysporum
f.sp. phaseoli), causadas por fungos, crestamento bacteriano (Xanthomonas axonopodis pv.
phaseoli), murcha de curtobacterium causada pela Curtobacterium flaccumfaciens pv.
flaccumfacienalém e mosaico dourado causado por vírus do grupo dos geminivírus (PAULA
JUNIOR E ZAMBOLIM, 2006).
De acordo com PIZA et. al. (1993), existem diversas medidas fitossanitárias que podem ser
adotadas para o manejo destas doenças, contudo a utilização de cultivares resistentes é a mais
eficiente e menos onerosa para o agricultor, sendo desta forma um dos principais objetivos dos
programas de melhoramento genético no Brasil e no mundo.

4
2.4 ANTRACNOSE (Colletotrichum lindemuthianum)
A antracnose é amplamente distribuída nos estados brasileiros, ocorrendo principalmente em
áreas serranas onde a temperatura moderada favorece o seu desenvolvimento, como é o caso dos
estados do Rio Grande do Sul, Paraná e Minas Gerais, sendo de menor importância em regiões mais
secas e quentes (VIEIRA et al. 2005).
A doença é favorecida por temperaturas entre 13 e 27 °C, com ótimo a 21 °C e umidade
relativa acima de 91%. Os conídios germinam entre seis e nove horas sob condições favoráveis,
penetrando mecanicamente na cutícula e epiderme do hospedeiro, sendo que os sintomas aparecem
a partir de seis dias após o inicio da infecção (PASTOR CORRALES E TU, 1994).
Os sintomas da antracnose podem ser observados em qualquer órgão da parte aérea da
planta e a disseminação do fungo ocorre principalmente por meio de respingos de água da chuva e
irrigação. Estudos relatam que a maior incidência deste patógeno ocorre na safra da seca, devido às
temperaturas amenas e ao uso da irrigação por aspersão (GARCIA, 1998; PINTO et al. 2001).
Para o controle da antracnose do feijoeiro devem ser utilizadas as práticas culturais como a
utilização de sementes sadias, rotação de culturas com plantas não hospedeiras, controle químico e
principalmente a utilização de cultivares resistentes.
Entretanto, a obtenção de genótipos resistentes ao C. lindemuthianum é dificultado pela
existência de grande número de raças, sendo mais de 50 raças foram identificadas no Brasil (SILVA
et al. 2007).
Segundo LANZA et al. (1997) a raça 81 é considerada de ocorrência frequente nas regiões
produtoras do estado de Minas Gerais e CARBONELL et al (1999) cita as raças 31, 65 e 81 sendo
as mais disseminadas do estado de São Paulo. CHIORATO et al. (2006), relataram a raça 65 como a
mais disseminada no Brasil, reforçando assim a importância da identificação e recomendação de
genótipos superiores e resistentes a estas raças. Melhor compreensão sobre a estrutura populacional
de C. lindemuthium é sugerida como estratégia fundamental para a incorporação de resistência à
antracnose do feijoeiro (BALARDIN E KELLY, 1998; MAHUKU et al. 2002).
De acordo com SINGH E SCHWARTZ (2010), o controle genético da resistência ao C.
lindemuthianum, tem sido estudado por diversos autores e embora existam duvidas a respeito do
número de genes envolvidos, sabe-se que ele é grande, sendo assim uma resistência complexa.
Uma relação de genes já descritos e a fonte de resistência são apresentadas nos trabalhos de
GONÇALVES-VIDIGAL (2011) e SINGH E SCHWARTZ (2010), onde os autores demonstraram
que existe variabilidade genética para resistência à antracnose, o que permite inferir que é possível
obter genótipos resistentes. Entretanto, o patógeno também apresenta variabilidade, assim, a

5
resistência obtida nem sempre é duradoura ou eficiente em todas as regiões de cultivo.
2.5 MURCHA DE FUSARIUM (FUSARIUM OXYSPORUM F.SP. PHASEOLI)
A murcha de fusarium causada pelo fungo Fusarium oxysporum f.sp. phaseoli destaca-se
como uma severa doença vascular, identificada em diversas regiões produtoras do mundo
(PASTOR-CORRALES E ABAWI, 1987; BURUCHARA E CAMACHO, 2000). No Brasil
existem relatos que esta doença é disseminada em diversas regiões produtoras por todo o país.
(COSTA et al. 1993; BALARDIN et al. 1990)
A infecção de F. oxysporum pode ocorrer em qualquer época do ciclo da planta e sua
incidência ocorre em reboleiras. (MOHAN et al. 1983). As plantas infectadas são identificadas pelo
sintoma de murcha, amarelecimento progressivo e senescência prematura (BIANCHINI et al.
2005).
Este fungo habita o solo e vive saprofiticamente sobre a matéria orgânica e restos culturais,
podendo sobreviver por vários anos na forma de clamidosporos (MOHAN et al 1983). A
temperatura ótima para o desenvolvimento da doença é de 20 °C, e sua disseminação ocorre
principalmente por meio de sementes contaminadas, pela água e pelo vento, que transportam
partículas de solo infestadas. Segundo SARTORATO E RAVA (1994), uma vez introduzido o
patógeno na área, seu controle por meio de práticas culturais é dificultado, sendo a prática mais
viável e eficaz o uso de cultivares resistentes.
Na literatura foram encontrados diversos trabalhos de avaliação da reação a este patógeno,
com diferenças entre os resultados obtidos entre eles, PEREIRA et al. (2011), em seus estudos com
349 genótipos de feijoeiro encontraram 134 genótipos resistentes (36%). A maior proporção foi
observada por RAVA et al. (1996) que encontraram 50% dos genótipos resistentes, entretanto seu
estudo contou com apenas 12 genótipos. Em contrapartida, ROCHA JUNIOR et al. (1998)
avaliando 169 genótipos constatou que 155 (92%) deles foram suscetíveis.
PEREIRA et al. (2011), ressaltaram que muitas das linhagens de feijoeiro recomendadas
para semeadura nos estados brasileiros apresentam alta suscetibilidade a este patógeno, entretanto
evidenciaram alta herdabilidade (h2=87%), indicando que a princípio, é esperado sucesso com a
seleção para este caráter. Há relatos de que o alelo dominante do gene Fop 1, confere resistência à
raça 2 de Fusarium, também denominada de “Brasileira” (RIBEIRO E HAGEDORN, 1979).
Também utilizando linhagens brasileiras, Pereira (2009), evidenciaram que o controle da resistência
é monogênico. Entretanto, a existência de outros genes de efeito menor não deve ser descartada, em
razão da ampla variação na expressão fenotípica (PEREIRA 2008).

6
2.6 CRESTAMENTO BACTERIANO (XANTHOMONAS AXONOPODIS PV. PHASEOLI)
O crestamento bacteriano comum, causado pela bactéria Xanthomonas axonopodis pv.
Phaseoli, é uma das principais doenças do feijoeiro em diversos países, sendo responsável por
prejuízos consideráveis à cultura, devido a agressividade e dificuldade de controle do patógeno
(SARTORATO E RAVA 1994; SAETTLER, 2005).
Esta doença é transmitida principalmente por sementes contaminadas, afetando toda a aérea
do feijoeiro dentro da lavoura, além disso, restos culturais contaminados como também respingos
de água e vento são importantes disseminadores do patógeno.
Segundo DIAZ et al. (2001), esta doença pode causar perdas superiores a 60% da produção,
sob condições favoráveis, devido a desfolha severa e formação de grãos chochos e de má qualidade.
Temperaturas altas e alta umidade são condições favoráveis a X. axonopodis, sendo assim,
normalmente de maior importância na safra das águas, principalmente em regiões de clima quente
(BIANCHINI et al. 2005).
A bactéria X. axonopodis, sobrevive em diferentes formas. Em sementes, pode resistir por
períodos variáveis de 2 a 15 anos, podendo estar localizada interna ou externamente, sem perder sua
patogenicidade. A sobrevivência em restos culturais infectados no solo é variável em função das
condições climáticas. Trabalhos desenvolvidos sob condições de campo, no estado do Paraná,
evidenciaram a sobrevivência desta bactéria por um período variável de 45 a 180 dias, em folhas
doentes sob a superfície do solo (BIANCHINI et al. 2005).
Os sintomas do crestamento bacteriano manifestam-se em toda a parte aérea da planta,
caracterizando-se inicialmente por pequenas áreas encharcadas, que crescem rapidamente de
tamanho e progridem para áreas necróticas (EMBRAPA, 2014). Temperatura e umidade elevadas
favorecem o desenvolvimento da doença, sendo que sob condições controladas, plantas incubadas a
28 °C desenvolvem os sintomas cerca de após nove dias após a inoculação (BIANCHINI et al.
2005).
O controle da doença é realizado por meio da adoção simultânea de várias medidas. O
emprego de sementes de boa qualidade sanitária é imprescindível, além disso, a rotação com
culturas não hospedeiras, a incorporação de restos culturais contaminados, o controle de insetos
disseminadores desta bactéria e a utilização de cultivares resistentes são algumas práticas que
podem ser adotadas no manejo desta doença (ZANATTA et al. 2007).
Diversos autores ressaltam a dificuldade na obtenção de genótipos resistentes a este
patógeno. DURSUN et al (2002), avaliando 22 genótipos de feijoeiro encontrou apenas 1 resistente,
RAVA et al (1990) obteve apenas 8 genótipos resistentes entre 60 genótipos avaliados. Contudo

7
SILVA (2009), classificou 21 genótipos com resistência de um total de 56 avaliados. Segundo
SANTOS et al. (2003), não há disponibilidade de cultivares comerciais de feijoeiro com alto nível
de resistência a este patógeno, enfatizando a necessidade de mais estudos relativos a esta doença.
2.7 QUALIDADE TECNOLÓGICA DE GRÃOS
As características denominadas de qualidade tecnológica de grãos são relativas às exigências
do mercado consumidor, aos que interessam apenas os aspectos relacionados com os grãos, como
cor, tamanho, forma, baixo tempo de cozimento, caldo espesso, sabor e nutrientes (BASSINELLO
et al. 2003).
Um aspecto importante da cultura do feijoeiro são suas características nutricionais, sendo
um alimento básico da população brasileira rico em carboidratos, fibras, minerais, vitaminas do
complexo B e principal fonte de proteína de origem vegetal consumida (MESQUITA et al. 2007).
Neste sentido, é crescente o interesse no melhoramento de feijoeiro para as características
relativas à qualidade tecnológica dos grãos, como por exemplo, o menor tempo de cozimento,
coloração adequada do tegumento e maiores teores de nutrientes.
No que diz respeito ao tempo de cozimento, encontra-se na literatura diversos trabalhos que
relatam a presença de variabilidade para esta característica, com valores entre 21 e 40 minutos
(RODRIGUES et al. 2005), de 15 a 25 minutos (LEMOS et al. 2004), de 35 a 45 minutos (RAMOS
JUNIOR et al. 2005) e 22 a 139 minutos (RIBEIRO et al. 2005), sendo assim possível a seleção de
genótipos superiores para esta característica.
COSTA et al. (2001), avaliaram a variabilidade genética para absorção de água em 100
genótipos do Banco de Germoplasma da Universidade Federal de Lavras (UFLA), como indicativo
do tempo de cozimento, cujas sementes permaneceram em embebição por quatro horas, sendo logo
após esse período estimada a porcentagem de água absorvida. Os autores constataram ampla
variação na capacidade de absorção de água dos genótipos, destacando-se os genótipos IAC - Aruã,
CI-107 e Ouro Negro.
Embora existam diversos trabalhos que sugerem a existência de variabilidade genética para
tempo de cozimento e absorção de água, poucos relacionam o tipo de controle genético desses
caracteres. Um dos trabalhos mais completos realizados a este respeito foi o de ELIA et al. (1996),
onde foram utilizaram 16 genótipos de feijoeiro, diferindo na capacidade de absorção de água e no
tempo de cozimento. Segundo os autores existe variabilidade genética para esta característica, sendo
que a variância genética observada foi predominantemente do tipo aditiva. Além disso, os valores

8
das estimativas de herdabilidade foram altos, indicando a possibilidade de sucesso para seleção de
genótipos superiores para estas características.
Em relação à variação do teor de proteínas, esta característica não depende apenas da
expressão genética que controla a síntese e o acúmulo de frações específicas de proteínas, mas
também de genes que controlam outros fatores, como aquisição de nutrientes, vigor da planta,
maturação, tamanho da semente, síntese e acúmulo de amido na semente (OSBORN, 1988). Há
evidências também, de que, além da própria cultivar, o conteúdo proteico dos feijões pode ser
influenciado por fatores climáticos e pelo local de cultivo (FARINELLI, 2006).
Segundo RIBEIRO et al. (2005), os teores de proteína encontrados em grãos de feijão
variam de 18 a 31%, dependendo do genótipo avaliado. O controle genético do teor de proteína nos
grãos é complexo, sendo que a variação do teor de proteína não depende somente do gene que
controla sua síntese e acumulação, mas também de genes que controlam outros fatores, tais como a
absorção de nutrientes, maturação, rendimento e da quantidade de adubação nitrogenada durante o
cultivo.
A composição de proteínas nos grãos, também pode ser alterada em função do genótipo,
ambiente e da interação genótipos por ambientes (BURATTO et al. 2009, RIBEIRO, 2010)
Consequentemente observa-se variação nos teores de minerais em cultivares e linhagens avaliadas
em diferentes locais, anos e safras. Contudo, os mesmos autores citam a existência de variabilidade
genética para características de qualidade nutricional em grãos de feijão, sendo possível assim a
obtenção de genótipos promissores por meio do melhoramento genético.
Outro problema relativo à seleção de genótipos com altos teores de proteína é a correlação
negativa deste caráter com o rendimento de grãos por planta (MELLO FILHO et al. 2004),
corroborado por LEMOS et al. (2004) citaram que genótipos de feijoeiro com produtividade de
grãos abaixo da media experimental apresentaram os maiores teores de proteína bruta.
Com relação à coloração de tegumento, o consumidor de feijão no Brasil tem diferentes
exigências de acordo com a região. O feijão de tegumento preto é consumido nos estados da região
sul,Rio de Janeiro, na região da Zona da Mata em Minas Gerais, bem como no Distrito Federal. No
nordeste a preferencia é por feijão do grupo mulatinho. Contudo, a preferencia por grãos tipo
carioca corresponde cerca de 70% do total consumido no Brasil (RAMALHO E ABREU, 2006).
Mais de 18 genes estão envolvidos com a cor do tegumento e nos padrões de listras e halo
no tegumento, o que implica em uma grande complexidade desta característica (GEPTS E
DEBOUCK, 1993).

9
2.8 INTERAÇÃO GENÓTIPOS POR AMBIENTES (GXA)
Na fase final do programa de melhoramento genético de plantas, os genótipos avançados
são avaliados em varias localidades, em diferentes condições de clima, fertilidade de solo e também
em diferentes safras (ACCIARESI E CHIDICHIMO, 1999), de modo a constituir as chamadas
redes de avaliações de cultivares ou ensaios de valor de cultivo e uso (VCU). Segundo FERREIRA
et al. (2006), a combinação das diferentes condições, definidas pelos locais e safras, pode ser
considerada como um único fator para as condições ambientais.
De acordo com RAMALHO et al. (2012), considerando um caráter como por exemplo a
produtividade de grãos, o valor observado para esse caráter, denominado fenótipo, é função do
genótipo (G), do ambiente (A) e da interação genótipos por ambientes (GxA). A ocorrência da
interação GxA é consequência do comportamento não coincidente dos genótipos nos diferentes
ambientes avaliados (CRUZ E CARNEIRO, 2003), sendo este, um fenômeno natural que faz parte
da evolução das espécies e seus efeitos permitem o aparecimento de genótipos adaptados, aptos a
um ambiente específico, assim como, de comportamento geral aptos a vários ambientes
(LAVORANTI, 2003).
Em termos genéticos, de acordo com KANG E GAUCH (1996) a interação ocorre quando a
contribuição dos alelos dos diferentes genes que controlam o caráter ou o nível de expressão dos
mesmos não é coincidente entre os ambientes. Isso significa que os efeitos genéticos e ambientais
não são independentes, isto porque a expressão dos genes é influenciada e ou regulada pelo
ambiente.
Segundo GARBUGLIO et al. (2007), nos programas de melhoramento genético de diversas
espécies, a interação GxA dificulta a seleção e indicação de cultivares, em razão da inconsistência
do desempenho dos genótipos em ambiente distintos. No entanto, CHAVES (2001) citou que a
utilização de ferramentas estatísticas, juntamente com o conhecimento do fenômeno no âmbito
biológico pode-se obter informações para entender a interação GxA e propor alternativas para
minimizá-la ou aproveitá-la em beneficio dos agricultores.
A interação GxA pode ser de dois tipos, simples ou complexa. A interação do tipo simples é
caracterizada pela não alteração da classificação relativa dos genótipos avaliados, dentro de um
conjunto de ambientes, tomados dois a dois, havendo porém diferença da magnitude das respostas.
As interações do tipo complexa ocorre quando a correlação entre o desempenho dos
genótipos ao longo dos ambientes é baixa, fazendo com que a posição relativa dos genótipos seja
alterada em virtude das diferentes respostas às variações ambientais (ROBERTSON, 1959).
GARBUGLIO et al. (2007), citaram que em situações reais o mais comum é que ocorram as

10
interações do tipo simples e complexa em conjunto, sendo esta última a que mais atrapalha o
melhorista, principalmente no momento da recomendação de cultivares.
Neste sentido, para a correta recomendação de novas cultivares, deve-se fazer estudos sobre
a interação GxA, realizando-se experimentos em diversos locais e anos, avaliando e quantificando
os efeitos desta interação, possibilitando assim adotar estratégias que possibilitem contornar estes
efeitos (RESENDE, 2002). Enfatiza-se que esta fase de avaliação é a mais etapa mais difícil e cara
no desenvolvimento de novas cultivares (RAMALHO et al., 2012)
Segundo CRUZ et al. (2004) do ponto de vista biométrico, dois procedimentos podem ser
empregados: estudos de adaptabilidade e estabilidade de genótipos, onde procura-se quantificar as
respostas de cada genótipo frente as variações ambientais, para identificar aqueles de
comportamento previsível e que sejam responsivos às variações ambientais, em condições
específicas ou amplas. E o relativo aos métodos de estratificação ambiental, que identifica entre os
ambientes disponíveis, padrões de similaridades de respostas dos genótipos, sendo possível avaliar
o grau de representatividade dos ensaios e tomar decisões com relação a descarte de ambientes
quando necessário identificando grupos de ambientes que a interação GxA possa ser não
significativa, ou com predominância de porção interação de natureza simples, ou seja, que não
venha a comprometer a recomendação de cultivares.
2.9 ESTRATIFICAÇÃO AMBIENTAL
DUARTE E ZIMERMANN (1991) ponderaram que em estudos para a determinação de
locais-chaves para realização de experimentos com genótipos, deve-se levar em consideração que as
avaliações devem ser realizadas em vários anos ou safras, devido as variações ambientais não
previstas que influenciam a interação de genótipos com anos e, com locais e anos.
Dentre os métodos de estratificação ambiental, citam-se os que procuram formar subgrupos
homogêneos, em que a interação GxA seja não significativa entre pares de ambientes (LIN, 1982)
ou, em outros casos significativa, mas de natureza predominantemente simples. O método definido
por CRUZ E CASTOLDI (1991), procura quantificar o porcentual de parte simples atuante (PS%)
da interação GxA, em que as posições relativas dos genótipos de um ambiente para o outro não
sofram alterações relevantes ou que comprometam as recomendações dos genótipos.
Outro método que vem sendo amplamente utilizado é a análise de fatores (AF), que segundo
FERREIRA (2008), consiste basicamente, em reduzir um número elevado de variáveis originais a
um pequeno número de variáveis abstratas, denominadas fatores. Cada fator irá agrupar um
conjunto de ambientes, altamente correlacionados entre si e fracamente correlacionados com os

11
ambientes agrupados nos demais fatores.
Entretanto, MURAKAMI E CRUZ (2004), citam que os estudos de estratificação ambiental
e adaptabilidade e estabilidade vêm sendo realizados de forma dissociada e propuseram um método
que contempla simultaneamente as duas análises, por meio do princípio da similaridade do
desempenho fenotípico, baseado na técnica de AF, descrita a seguir:
Na utilização da AF destinada à estratificação de ambientes, deve-se interpretar o
significado de cada fator e avaliar o interesse por ele. São analisados os elementos ou cargas
fatoriais, identificando, para cada variável (que neste caso representa um determinado ambiente), os
fatores com coeficientes de ponderação próximos da unidade.
Desta forma, o agrupamento de ambientes é realizado a partir das informações sobre a
magnitude das cargas fatoriais finais (obtidas após rotações), sendo que cargas fatoriais maiores ou
iguais a 0,70 indicam ambientes com altas correlações, devendo ser agrupados dentro de cada fator.
Cargas fatoriais com valores baixos indicam que o respectivo ambiente não deve pertencer ao
grupo.
Antes de serem obtidas as cargas fatoriais que permitirão identificar o melhor agrupamento
de ambientes, deve-se estabelecer o número de fatores a serem analisados. O número de fatores
finais pode ser admitido como igual ao número de autovalores maiores ou iguais a 1,0. No entanto,
nos casos em que a proporção da variabilidade explicada pelos autovalores maiores que 1,0 for
baixa, devem ser considerados mais fatores, até que se atinja uma proporção adequada de
explicação da variabilidade, geralmente próximo de 80% da variação total.
2.10 ADAPTABILIDADE E ESTABILIDADE
MARIOTTI et al. (1976), COSTA et al. (1999) E CRUZ et al. (2004), consideraram
estabilidade como sendo a capacidade dos genótipos mostrarem comportamento altamente
previsível em função do estímulo do ambiente enquanto que adaptabilidade refere-se à capacidade
dos genótipos aproveitarem vantajosamente o estímulo ambiental.
De acordo com PEREIRA et al. (2009), existem inúmeros métodos de análise de
adaptabilidade e estabilidade, baseados em diferentes princípios, entre eles: os baseados em
regressões lineares como de EBERHART E RUSSEL, 1966 e CRUZ et al. 2004, modelos não
paramétricos como os de LIN E BINNS (1988), LIN E BINNS modificado por CARNEIRO (1998)
e ANNICCHIARICO (1992), e os multivariados, como por exemplo o baseado em análises de
fatores (MURAKAMI E CRUZ, 2004), da análise da interação multiplicativa dos efeitos principais
aditivos (AMMI) e GGE-Biplot.

12
Devido à grande variedade de métodos para análise de adaptabilidade e estabilidade, cabe ao
melhorista analisar as vantagens e desvantagens de cada método e utilizar o que melhor se adequar
ao seu experimento e assim selecionar os genótipos efetivamente superiores, com menor distorção
ambiental possível, pois o que é passado para as próximas gerações é o genótipo (FONSECA
JUNIOR, 1996).
PEREIRA et al. (2009), relataram que um dos métodos mais utilizados na cultura do
feijoeiro é o não paramétrico proposto por LIN E BINNS (1988), modificado por CARNEIRO
(1998). Este método se baseia na avaliação da performance genotípica (Pi) dos cultivares avaliados,
que refere-se o estimador da performance genotípica (Pi) refere-se ao quadrado médio da
distância entre a média do cultivar e a resposta média máxima para todos os ambientes. Quanto
menor o valor de (Pi) maior a estabilidade do cultivar. A decomposição de Pi proposta por
CARNEIRO (1998) divide a estimativa Pi a ambientes favoráveis e desfavoráveis, em que a
classificação destes ambientes é feita com base nos índices ambientais, definidos como a diferença
entre a média dos cultivares avaliados em cada ambiente e a média geral.
Diversos autores relataram resultados satisfatórios em seus trabalhos utilizando esta
metodologia, sendo de fácil interpretação e possibilitando discriminar o desempenho dos genótipos
em ambientes favoráveis e desfavoráveis (MURAKAMI et al., 2004; ELIAS et al., 2005; RIBEIRO
et al., 2009 e PEREIRA et al., 2009).
Estudando adaptabilidade e estabilidade de soja por métodos paramétricos e não
paramétricos, BARROS (2008) concluiu que os métodos de LIN E BINNS (1988),
ANNICCHIARICO (1992) e Centroide de ROCHA et al. (2005) foram coerentes entre si e
permitiram identificar, entre os genótipos avaliados, aqueles de maior produtividade, adaptabilidade
e estabilidade.
A análise de adaptabilidade fenotípica, baseada na AF, é realizada graficamente por meio de
escores obtidos em relação aos fatores. De acordo com MURAKAMI E CRUZ (2004), nos
quadrantes II e IV, estarão os genótipos de adaptabilidade específicas aos ambientes agrupados em
cada um dos dois fatores utilizados na plotagem gráfica. No quadrante I, estarão os genótipos de
adaptabilidade ampla, ou seja, apresentarão escores altos para os ambientes agrupados nos dois
fatores, simultaneamente. No quadrante III estarão os genótipos de baixo desempenho, passíveis de
não indicação de cultivo para os ambientes em estudos.
GARBUGLIO et al. (2007), estudando a adaptabilidade de genótipos de milho e realizando
a estratificação em 22 ambientes no estado do Paraná, constataram que a adaptabilidade analisada
por meio de plotagens gráficas a partir dos escores fatoriais, apresentou respostas diferenciadas e

13
mais eficientes, quando comparada com os métodos de regressão bissegmentada.
MENDONÇA et al. (2007), avaliando a eficiência de diferentes métodos de estratificação
ambiental e a adaptabilidade e estabilidade de genótipos de soja nos estados do Paraná e Santa
Catarina, verificaram que a técnica de AF foi mais seletiva no processo de estratificação ambiental
do que o método tradicional de LIN (1982), e superior ao método de regressão linear simples de
EBERHART E RUSSELL (1966) para adaptabilidade e estabilidade fenotípica.
Nesse sentido, as análises de adaptabilidade e estabilidade fenotípica são procedimentos
estatísticos que permitem, de algum modo, identificar genótipos de comportamento mais estável e
que respondam previsivelmente às variações ambientais, sendo assim uma forma importante, entre
os melhorista de plantas, para avaliar novos genótipos antes de sua recomendação como cultivares
(ROCHA et al. 2005).
3. MATERIAL E MÉTODOS
3.1 ENSAIOS DE VCU
Foram utilizados 27 genótipos, compreendendo linhagens e cultivares de tegumento carioca
e preto, provenientes de diferentes instituições públicas que efetuam melhoramento genético do
feijoeiro no Brasil (Tabela 1). Do total de genótipos utilizados, quatro linhagens foram provenientes
da EMBRAPA Arroz e Feijão (Santo António do Goiás, GO), duas linhagens da EPAGRI (Empresa
de Pesquisa Agropecuária e Extensão Rural de Santa Catarina, SC), duas linhagens da FEPAGRO
(Fundação de Pesquisa Agropecuária, RS), quatro linhagens do IAPAR (Instituto Agronômico do
Paraná, PR), duas linhagens da UFLA (Universidade Estadual de Lavras, MG) e nove linhagens do
Instituto Agronômico - IAC.
Como testemunhas, para o grupo de grãos de tegumento carioca foram utilizadas as
cultivares IAC Alvorada (IAC), de alto potencial produtivo, porte semi-ereto, ciclo médio de 92
dias, com alta qualidade de grãos, resistência moderada à antracnose, suscetível à murcha de
fusarium e ao crestamento bacteriano e IPR 139 (IAPAR), de alto potencial produtivo, porte ereto,
ciclo médio de 89 dias, moderada resistência ao crestamento bacteriano e suscetível à antracnose e à
murcha de fusarium.
Para o grupo de grãos de tegumento preto foram utilizadas as cultivares IAC Diplomata, de
alto potencial produtivo, porte ereto, ciclo médio de 90 dias, resistência à antracnose e a murcha de
fusarium; e IAC Una, de alto potencial produtivo, porte ereto, ciclo médio de 94 dias, resistente a
antracnose e suscetível à murcha de fusarium e ao crestamento bacteriano.
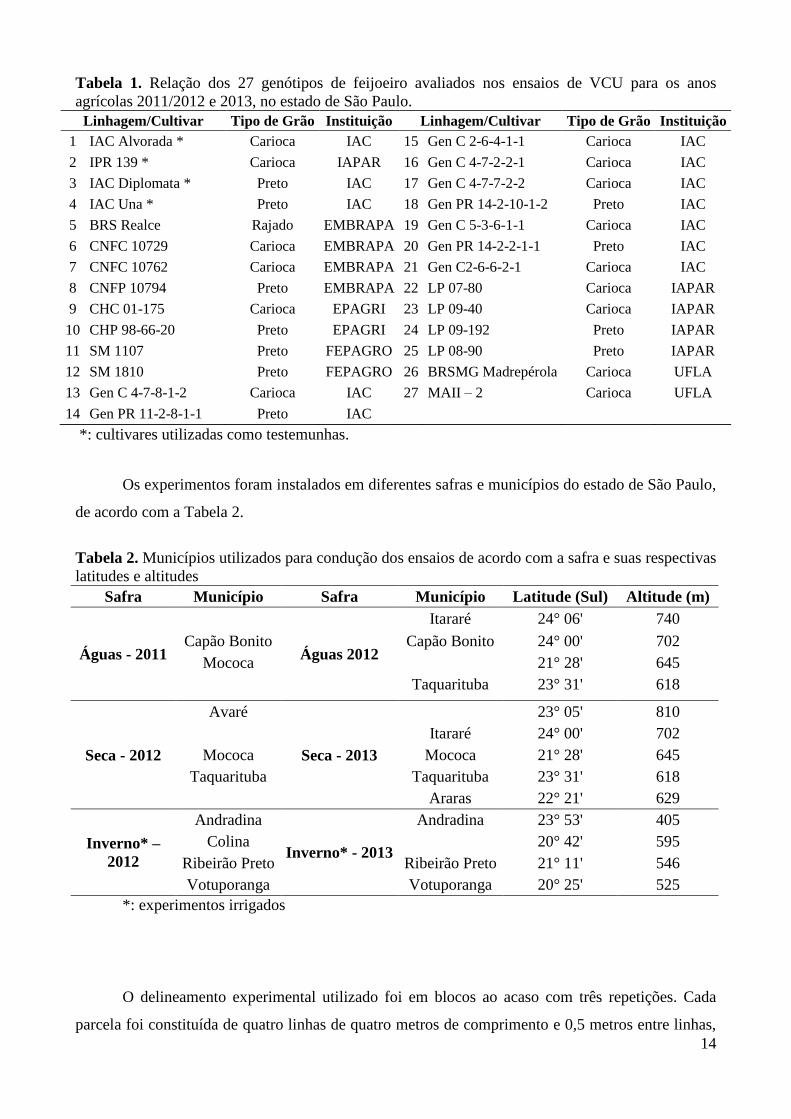
14
Tabela 1. Relação dos 27 genótipos de feijoeiro avaliados nos ensaios de VCU para os anos
agrícolas 2011/2012 e 2013, no estado de São Paulo.
Linhagem/Cultivar Tipo de Grão Instituição Linhagem/Cultivar Tipo de Grão Instituição
1 IAC Alvorada * Carioca IAC 15 Gen C 2-6-4-1-1 Carioca IAC
2 IPR 139 * Carioca IAPAR 16 Gen C 4-7-2-2-1 Carioca IAC
3 IAC Diplomata * Preto IAC 17 Gen C 4-7-7-2-2 Carioca IAC
4 IAC Una * Preto IAC 18 Gen PR 14-2-10-1-2 Preto IAC
5 BRS Realce Rajado EMBRAPA 19 Gen C 5-3-6-1-1 Carioca IAC
6 CNFC 10729 Carioca EMBRAPA 20 Gen PR 14-2-2-1-1 Preto IAC
7 CNFC 10762 Carioca EMBRAPA 21 Gen C2-6-6-2-1 Carioca IAC
8 CNFP 10794 Preto EMBRAPA 22 LP 07-80 Carioca IAPAR
9 CHC 01-175 Carioca EPAGRI 23 LP 09-40 Carioca IAPAR
10 CHP 98-66-20 Preto EPAGRI 24 LP 09-192 Preto IAPAR
11 SM 1107 Preto FEPAGRO 25 LP 08-90 Preto IAPAR
12 SM 1810 Preto FEPAGRO 26 BRSMG Madrepérola Carioca UFLA
13 Gen C 4-7-8-1-2 Carioca IAC 27 MAII – 2 Carioca UFLA
14 Gen PR 11-2-8-1-1 Preto IAC
*: cultivares utilizadas como testemunhas.
Os experimentos foram instalados em diferentes safras e municípios do estado de São Paulo,
de acordo com a Tabela 2.
Tabela 2. Municípios utilizados para condução dos ensaios de acordo com a safra e suas respectivas
latitudes e altitudes
Safra Município Safra Município Latitude (Sul) Altitude (m)
Águas - 2011
Águas 2012
Itararé 24° 06' 740
Capão Bonito Capão Bonito 24° 00' 702
Mococa
21° 28' 645
Taquarituba 23° 31' 618
Seca - 2012
Avaré
Seca - 2013
23° 05' 810
Itararé 24° 00' 702
Mococa Mococa 21° 28' 645
Taquarituba Taquarituba 23° 31' 618
Araras 22° 21' 629
Inverno* –
2012
Andradina
Inverno* - 2013
Andradina 23° 53' 405
Colina
20° 42' 595
Ribeirão Preto Ribeirão Preto 21° 11' 546
Votuporanga Votuporanga 20° 25' 525
*: experimentos irrigados
O delineamento experimental utilizado foi em blocos ao acaso com três repetições. Cada
parcela foi constituída de quatro linhas de quatro metros de comprimento e 0,5 metros entre linhas,

15
mantendo-se em média 10 plantas por metro linear. Para efeito de coleta de dados, as duas linhas
centrais foram consideradas como área útil (4 m2).
A adubação base foi feita de acordo com a análise de solo e necessidade da cultura em cada
local, e após 20 dias da emergência das plântulas foi realizada adubação nitrogenada de cobertura
com 200 kg.ha-1
de sulfato de amônio. Os demais tratos culturais realizados foram: irrigações,
capinas manuais e aplicação de herbicidas e inseticidas.
3.2 AVALIAÇÃO DE DOENÇAS
Neste trabalho o objetivo foi a identificação de genótipos com resistência aos patógenos da
Antracnose, Murcha de Fusarium e Crestamento Bacteriano para fins de recomendação e registro de
nova cultivar, portanto não foram realizados estudos detalhados sobre a natureza genética da
resistência dos genótipos aos patógenos.
3.2.1 Antracnose (Colletotrichum lindemuthianum)
Sementes dos 27 genótipos foram germinadas em papel germitest, com pH neutro, à
temperatura de 28 ºC, por um período de três dias. Para cada genótipo foram utilizadas 10 sementes.
Após esse período as plântulas foram transplantadas em linhas para bandejas com vermiculita
autoclavada e mantidas em casa de vegetação (Figura 1 A).
Em bandejas, as plântulas foram inoculadas para o C. lindemuthianum com as raças
fisiológicas 31, 65 e 81, conforme mencionado por CARBONELL et al. (1999) como as mais
disseminadas no estado de São Paulo (Figura 1 B). O inóculo foi obtido a partir da suspensão de
conídios filtrada em tecido de filó, com a concentração dos conídios ajustada para 1,2 x 106
conídios ml-1
.
A inoculação do patógeno foi feita por aspersão, em estádio de desenvolvimento V2. Após a
inoculação, as plântulas foram submetidas à câmara úmida por 48 horas, e transcorrido esse
período, foram retiradas da câmara e as plântulas permaneceram, em ambiente controlado por mais
cinco dias, quando foi realizada a avaliação quanto à severidade da doença (Figura 1 C e D).
A avaliação da virulência dos isolados foi feita com base na intensidade da doença
utilizando-se a escala descrita por BALARDIN E PASTOR CORRALES (1990), com notas de 1 a
9, sendo 1, planta completamente livre de sintomas visíveis da doença e 9, necrose, desfolhamento e
morte das plantas. Foram considerados resistentes genótipos que apresentaram notas de 1 a 3,
intermediários plântulas com notas 4 a 6 e suscetíveis as que apresentaram notas de 7 a 9.

16
Figura 1. Procedimentos realizados para
inoculação de C. lindemuthianum em 27 genótipos
de feijoeiro.
3.2.2 Murcha de Fusarium (Fusarium. oxysporum f. sp. phaseoli)
Sementes dos 27 genótipos foram desinfetadas com hipoclorito de sódio a 1,25% durante
cinco minutos, enxaguadas em água destilada e transferidas para folhas de papel de germinação
umedecidas com água destilada, para germinação em laboratório. As sementes pré-germinadas
foram transferidas para recipientes com vermiculita lavada e esterilizada.
Após dez dias da semeadura, as plantas foram retiradas da vermiculita esterilizada e foram
inoculadas pelo método de imersão proposto por COSTA (1989) (Figura 2), com inóculo ajustado a
concentrações de 1x106 conídios ml
-1. Plantas com dez dias de idade após o transplante foram
retiradas da vermiculita esterilizada, as raízes foram lavadas e as extremidades foram cortadas em
1/3 do seu comprimento e mergulhadas no inóculo durante 10 minutos, sendo transplantadas em
seguida duas plantas por vaso, contendo três repetições para cada linhagem testada (PASTOR-
CORRALES E ABAWI, 1987). Cada vaso foi constituído de 500g de substrato (composto orgânico
+ terra na proporção de 1:1) e acondicionado em casa de vegetação (Figura 2).
A avaliação dos sintomas foi efetuada 45 dias após a inoculação seguindo uma escala de
notas desenvolvida por NASCIMENTO (1995) onde, 0 e 1 = plantas resistentes 2 e 3 =
moderadamente resistentes, 4 = suscetíveis. As plantas com notas até 1 foram consideradas
resistentes e acima de 1 foram consideradas suscetíveis (Figura 3).

17
Figura 2. Inoculação de Fusarium oxysporum f. sp. phaseoli pelo método de imersão
dipping (COSTA et al., 1989).
Figura 3. Plantas de feijoeiro resistentes (a
esquerda), intermediarias (no centro) e
suscetível (a direita) ao Fusarium oxysporum f.
sp. phaseoli.
3.2.3 Crestamento Bacteriano (Xanthomonas axonopodis pv. phaseoli)
Foram germinadas sementes dos 27 genótipos em papel de germitest, com pH neutro, à
temperatura de 28 ºC, por um período de três dias. Para cada genótipo foram utilizadas em torno de
20 sementes. Após esse período as plântulas foram transplantadas para vasos contendo terra como
substrato e mantidas em casa de vegetação. Para cada genótipo de feijoeiro foram utilizados três
vasos com duas plântulas por vaso.
O isolado n° 11090 de X. axonopodis, registrado no Centro de Pesquisa e Desenvolvimento
de Fitossanidade - IAC foi repicado para placas de Petri, contendo o meio de cultura BDA para a

18
produção do inóculo. Em seguida, as placas foram incubadas em estufa à temperatura de 28 ºC, por
um período de 24 horas a 48 horas. O inóculo foi preparado pela adição de água destilada e
esterilizada à superfície da colônia bacteriana e posterior raspagem da superfície com uma lâmina
de vidro. A concentração do inóculo foi ajustada a 108 ufc/Ml (POMPEU et al. 1973).
A inoculação foi realizada pela técnica de agulhas múltiplas proposta por POMPEU et al.
(1973), com a perfuração das folhas primárias de plântulas de feijoeiro que apresentaram-se bem
expandidas. O inóculo foi colocado numa esponja e em seguida as folhas foram perfuradas sobre a
esponja com leve pressão entre a folha e a espuma, permitindo assim, a entrada do inóculo por meio
do ferimento. Após a aplicação do inóculo, os vasos foram mantidos em câmara úmida, por um
período de 48 horas (Figura 4).
Figura 4. Inoculação de Xanthomonas axonopodis pv. phaseoli pelo método agulhas múltiplas
(POMPEU et al. 1973).
A avaliação foi efetuada no sétimo dia e no décimo dia após inoculação (Figura 5) pela
escala de notas de 1 a 5 de acordo com SUGIMORI et al. (1989), sendo 1 = sem sintoma na área
inoculada (Resistente = R), 2 = encharcamento e/ou necrose da área inoculada (Resistente = R), 3 =
encharcamento de até 20% em torno dos pontos de inoculação (Moderadamente Resistente = MR),
4 = encharcamento e/ou necrose da área inoculada (Suscetível = S) e 5 = encharcamento, necrose da
folha e aparecimento de sintomas fora da área inoculada (Suscetível = S).

19
Figura 5. Plantas de feijoeiro resistente (a esquerda),
intermediaria (no centro) e suscetível (a direita) a
Xanthomonas axonopodis pv. phaseoli.
3.3 AVALIAÇÃO DA QUALIDADE TECNOLÓGICA DOS GRÃOS
As avaliações analíticas, da qualidade tecnológica dos 27 genótipos foram efetuadas em
triplicatas de sementes com até 60 dias após colhidas, uniformizadas por peneiras de classificação
(n° 13\64” pol.) e pré-selecionadas para a retirada de sementes visualmente danificadas por insetos
ou dano mecânico.
3.3.1 Análises do Tempo de Cocção dos Grãos
Foi determinado utilizando-se 30g de sementes uniformes e inteiras, embebidas em água
destilada por 16 horas a temperatura ambiente. Após este período, vinte e cinco grãos foram
colocados individualmente nos orifícios da base do cozedor de Mattson (Figura 6), sendo o aparelho
colocado dentro de um béquer contendo 1000 ml de água destilada em ebulição. Os tempos médios
de cozimento foram obtidos de acordo com o proposto por PROCTOR E WATTS (1987) e
SARTORI (1982), onde, o tempo é cronometrado em minutos até a queda da 13ª vareta, que
consiste em 50% dos grãos perfurados pelas hastes do aparelho.
Figura 6. Análise de tempo de cozimento por meio do Cozedor de Mattson.

20
3.3.2 Absorção de Água Antes do Cozimento (PEANC), Absorção de Água Após Cozimento
e Porcentagem de Grãos Inteiros (PGI)
Foram determinadas de acordo com os métodos modificados descritos por PLHAK et al.
(1989) e GARCIA-VELA E STANLEY (1989). De acordo com o método, uma amostra de 30g de
grãos de feijão foi colocada em béquer de 250 ml com 100 ml de água destilada por 16 horas à
temperatura ambiente. Após o período de embebição a água foi drenada e os grãos foram pesados e
a capacidade de absorção de água antes do cozimento (PEANC) foi calculada pela fórmula:
onde: ps = peso inicial; pu = peso dos grãos após maceração
Os grãos drenados foram colocados novamente no béquer com 100 ml de água destilada e
então foram aquecidos por 1 hora, utilizando-se de uma chapa aquecedora elétrica e iniciando a
contagem do tempo após o início da fervura da água. Em seguida, o caldo foi drenado e a
capacidade de absorção de água após o cozimento (PEAPC) foi calculada pela fórmula:
onde: pc = peso dos grãos após cozimento; ps = peso inicial dos grãos
Após o cozimento os grãos foram contados e separados em duas porções: inteiros e partidos.
Em seguida, o cálculo da porcentagem de grãos inteiros após o cozimento, foi efetuado pela
fórmula:
onde: ng = número total de grãos; ngi = número de grãos inteiros após cozimento
3.3.3 Avaliação da Coloração do Tegumento
Para e determinação da coloração do tegumento, foi utilizado o aparelho colorímetro marca
Minolta®, modelo CR-410 (Figura 7), seguindo o sistema CIELab, no qual obtém-se o parâmetro
L* que indica a luminosidade, variando do preto (L=0) ao branco (L=100).
% Peanc = pu − ps
ps x 100
PEAPC% = pc − ps
ps x 100

21
Figura 7. Colorímetro Minolta® - CR 410
3.3.4 Avaliação da Porcentagem de Proteína nos Grãos
A determinação do teor de proteína foi realizada pela técnica de espectroscopia na região do
infravermelho próximo (NIRS). Primeiramente os grãos de feijão foram moídos, utilizando moinho
IKA A11 Basic, em seguida o material foi armazenado em temperatura ambiente, em frascos
plásticos, permanecendo nessas condições até o momento da coleta dos espectros.
As coletas foram realizadas utilizando o espectrofotômetro FOSS NIRSystems, modelo
6500, equipado com detector PbS e lâmpada de filamento de tungstênio-halogênio para as medidas
de reflectância difusa dos grãos moídos. Por meio do software ISIscan™, versão 3.10
(INFRASOFT INTERNACIONAL, 2007), os espectros foram registrados de 1100 a 2498 nm, com
sinais expressos em log (1/R). As leituras foram realizadas em cela quarter cup, na temperatura de
20-22°C, com 64 varreduras e resolução espectral de 2nm. Foram registrados dois espectros por
genótipo e cada espectro correspondeu à média de duas leituras espectrofotométricas.
O teor de proteína foi determinado comparando-se os espectros gerados e corrigidos após
transformações matemáticas (normalização por área total, correção multiplicativa do sinal e
aplicação de primeira derivada com janela de 20 pontos), com uma curva de calibração multivariada
pré-estabelecida utilizando genótipos de feijão cultivados em anos anteriores.
Para a construção do modelo de regressão multivariado e quantificação do teor de proteína
nos grãos utilizou-se o método de regressão por Quadrados Mínimos Parciais (PLS, Parcial Least
Squares), por meio do software Pirouette 4.0, baseando-se no método oficial 989.03 da AOAC
(2005) e no método descrito por HACISALIHOGLU et al. (2010).

22
3.4 ANÁLISES ESTATÍSTICAS DOS DADOS
3.4.1 Análise de Variância Individual e Conjunta
Após a colheita e tabulação dos dados, foram realizadas análises de variância de cada local e
ano, utilizando-se o seguinte modelo matemático:
Yij= µ + ti + bj+ eij
onde: Yij= observação do tratamento i no bloco j; µ = média geral dos tratamentos no experimento;
ti= efeito do i-ésimo tratamento i (i = 1, 2, ..., I); bj= efeito do k-ésimo bloco j (j = 1, 2, ..., K); eij=
erro experimental associado à observação Yij.
Após verificadas as magnitudes dos quadrados médios residuais, foi realizada a análise de
variância conjunta de acordo com o seguinte modelo estatístico:
Yijk= µ + ti + lj + (tl)ij + (b/l) kj+ eijk
onde: Yijk= observação do tratamento i no bloco k no ambiente j; µ = média geral dos
tratamentos no experimento; ti= efeito do i-ésimo tratamento i (i = 1, 2, ..., I); lj = efeito do j-
ésimo ambiente j (j = 1, 2, ..., J); (tl) ij = efeito da interação do i-ésimo genótipo com o j-ésimo
ambiente; (b/l) kj= efeito do k-ésimo bloco (k = 1, 2, ..., K) dentro do j-ésimo ambiente; ijk= erro
experimental associado à observação Yijk.
As análises de variâncias individuais e conjuntas foram realizadas utilizando-se o modelo
PROC GLM de ANAVA do programa computacional SAS (Statistical Analysis System). Todos os
efeitos, exceto o erro, foram considerados como fixos. Para produtividade de grãos, as médias dos
genótipos avaliados foram comparadas em relação a melhor testemunha correspondente ao grupo de
tegumento carioca (IAC Alvorada e IPR 139) e de tegumento preto (IAC Una e IAC Diplomata)
pelo teste de Dunnett (5%). Para as variáveis relativas à qualidade tecnológica utilizou-se o teste de
comparação de médias de Scott-Knott (5%).
3.4.2 Estratificação Ambiental
Após comprovação da existência da interação entre os genótipos com ambientes, foi
realizada a análise de estratificação ambiental pelo método multivariado de análises de fatores,
proposto por MURAKAMI E CRUZ (2004). O modelo fatorial pressupõe que cada variável pode
ser expressa por uma combinação linear de fatores, além do erro, sendo dado por:

23
...
Ou
∑
Sendo: m < a; em que é a carga fatorial para a j-ésima variável associada ao k-ésimo
vetor; Fk é o k-ésimo fator comum e é o fator específico associado à j-ésima variável.
Para a utilização da análise de fatores em estudos de estratificação ambiental, deve-se
destacar o fato de que X1, X2 ... Xh representam uma única variável como, por exemplo,
produtividade de grãos, porém, avaliada em cada um dos “q” ambientes em que os genótipos foram
avaliados ou dos “p” genótipos referentes aos ambientes.
A carga fatorial do fator Fk é dada pela correlação entre o fator e a variável correspondente,
ou seja:
√ ( )
Na análise de fatores são impostas as seguintes restrições em relação aos fatores comuns: de
média nula, variância unitária e covariância entre fatores comuns também nula, de modo que eles
sejam ortogonais (CRUZ, 2006).
Também é calculada a comunalidade, que é uma medida da eficiência da representação de
uma variável xj por uma parte comum, também envolvida nas demais variáveis analisadas. De
acordo com CRUZ E CARNEIRO (2003), valores de comunalidade superiores a 0,64 são tidos
como razoáveis, por ser equivalente a um coeficiente de correlação entre xj e zj duperior a 0,80.
Portanto, a comunalidade é dada por:
∑
−

24
Onde: para cada variável xj tem-se que
, em que , denominado
comunalidade, representa a proporção da variância da variável padronizada “j” devida a fatores
comuns.
A forma funcional do modelo de análise de fatores é dada por:
em que:
X: é a matriz (GxA) de observações que apresenta valores obtidos para cada genótipo e
variável analisada, destacando-se o fato de que a variável é representada por uma mesma
característica medida em vários ambientes; Y: matriz (GxM) de fatores comuns; : matriz (MxA)
de cargas fatoriais; : matriz (GxA) de fatores específicos.
Objetiva-se na análise de fatores o estabelecimento de combinações lineares, que permitam a
interpretação quanto aos grupos de ambientes com mesmo padrão de discriminação genotípica,
capazes de reterem o máximo das associações originalmente disponíveis entre os ambientes
estudados. Como nesta análise procura-se relacionar o conjunto de variáveis observáveis,
representadas na matriz Y, e outra por fator específico, representadas na matriz , tem-se também
associação entre as respectivas matrizes de variâncias e covariâncias, tal como ilustrado a seguir:
em que:
R: matriz (a x a) de correlações entre o desempenho dos genótipos em cada par de ambientes
considerados, sendo:
−
A matriz Rc é denominada de matriz de correlações reduzidas do modelo de análise fatorial
e apresenta os mesmos elementos da matriz de correlação original, à exceção da sua diagonal, cujos
elementos são reduzidos de , sendo denominados comunalidade. Segundo Cruz e Carneiro
(2003), o problema fundamental da análise de fatores é o calculo da matriz de cargas e da matriz
diagonal de especificidades cujos elementos não nulos são , a partir de uma matriz R
conhecida.
Uma maneira de obter as cargas fatoriais é por meio da fatoração de Rc, usando o teorema da

25
decomposição espectral, em que qualquer matriz simétrica Rc pode ser descrita como:
− ∑
sendo: os autovalores de Rc; os autovetores correspondentes.
Para interpretação deste procedimento são analisados os elementos ou cargas fatoriais,
identificando, para cada variável os fatores com coeficientes de ponderação próximos da unidade. O
agrupamento é realizado portanto, a partir das informações sobre a magnitude das cargas fatoriais
finais obtidas após rotações. São adotados os seguintes critérios:
1) Cargas fatoriais maiores ou iguais a 0,70, em valor absoluto, indicam ambientes com
altas correlações e são agrupadas dentro de cada fator.
2) Cargas fatoriais de valores abaixo de 0,5 indicam que o respectivo ambiente não deve
pertencer ao grupo.
3) Cargas fatoriais com valores intermediários não garantem nenhuma definição de
agrupamento.
O número de fatores finais pode ser admitido como igual ao número de autovalores maiores
ou iguais a 1. No entanto, nos casos em que a proporção da variabilidade explicada pelos
autovalores maiores que 1 for baixa, pode-se considerar mais fatores, até que se consiga uma
proporção adequada da variabilidade, geralmente mais de 80% da variação total (CRUZ, 2006).
Foi estimada também a fração simples da interação GxA, de acordo com o método de CRUZ
E CASTOLDI (1991), em que a parte simples é expressa por:
em que: S=QMGjj’ – C, sendo C = 1-√ − , onde Qj e Qj’ correspondem aos
quadrados médios nos ambientes j e j’.
Esta decomposição pondera a contribuição da correlação e da diferença de variabilidade
genotípica nos ambientes, estimando assim a porcentagem da parte simples na interação.

26
3.4.3 Adaptabilidade e Estabilidade
Foram comparadas as análises de adaptabilidade e estabilidade por meio dos métodos da
análise de fatores, obtida em conjunto com a análise de estratificação ambiental e também pelo
modelo proposto por LIN E BINNS (1988), modificado por CARNEIRO (1998), considerando-se
para este método dois modelos, sendo, com apenas uma variável, neste caso a produtividade de
grãos e, o multivariado, onde considerou-se as variáveis de qualidade tecnológica em conjunto para
estimar os valores de Pi.
A análise de adaptabilidade baseada na análise de fatores foi realizada como descrita
anteriormente. Sendo que sua interpretação é realizada graficamente por meio dos escores em
relação aos fatores. Para isso, são traçados eixos paralelos, tomando-se a média dos escores, de
modo a estabelecer quatro quadrantes. Nos quadrantes II e IV ficam situados os genótipos com
adaptabilidade específica ao grupo de ambientes determinado pelo fator. No quadrante I ficam
situados os genótipos de adaptabilidade ampla e, no quadrante III os genótipos de baixo
desempenho, não adaptados aos ambientes de cultivo dos determinados fatores.
Na metodologia de LIN E BINNS o desempenho geral dos genótipos é definido como sendo
o quadrado médio da distância entre a média da cultivar e a resposta média máxima para todos os
locais, de acordo com o modelo:
onde: Pig = estimativa do parâmetro de estabilidade do cultivar i; Yij = produtividade de grãos
(kg.ha-1
) do i-ésimo cultivar no j-ésimo local; Mj = resposta máxima observada entre todos os
genótipos no j-ésimo ambiente; n = número de ambientes.
Nesta metodologia, os genótipos com menores valores de Pi correspondem aos de melhor
desempenho (MURAKAMI E CRUZ. 2004).
Para atender à identificação de genótipos superiores nos grupos de ambientes favoráveis e
desfavoráveis, CARNEIRO (1998) decompôs a estatística Pi utilizando a mesma metodologia de
classificação de ambientes de EBERHART E RUSSELL (1966). A classificação de ambiente em
favorável ou desfavorável, é feita com base nos índices ambientais (I), definidos como a diferença
entre a média dos genótipos avaliados em cada local e a média geral. Os ambientes que apresentam
maior média em relação à média geral constituem os ambientes favoráveis (Pif) e aqueles com
média menor que a média geral são classificados como ambientes desfavoráveis (Pid).
Pi = (Yij − Mj)
2nj=0
2n

27
Nesta decomposição, os seguintes estimadores são utilizados:
onde: Pif = estimador de Pig para ambientes favoráveis; Pid = estimador de Pid para ambientes
desfavoráveis; f = número de ambientes favoráveis; d = numero de ambientes desfavoráveis; Yij
e Mj como definidos anteriormente.
Considerando a recomendação de uma cultivar de feijoeiro não deve ser realizada baseada
em apenas uma variável, foi considerada também a decomposição multivariada do método LIN E
BINNS (1988), proposta por CARNEIRO (1998), alertando que este procedimento pode ser
realizado desde que seja possível somar os valores dos Pi’s do genótipo. Para isso, é necessária a
padronização destes Pi’s conforme:
Onde: Pmi = estimador do parâmetro de medida de adaptabilidade e estabilidade de
comportamento (MAEC), multiplicador para a cultivar i; Pik = estimador do parâmetro MAEC para
o cultivar i relativo à k-ésima variável; σpk = desvio padrão dos Pi’s para a k-ésima variável.
Além disso, CARNEIRO (1998) considerou um fator de multiplicação dado pelo inverso
dos pesos atribuídos a cada variável a fim de balancear a importância de cada variável. Assim, o
estimador do parâmetro MAEC é dado pela equação:
Os pesos atribuídos para cada variável foram : 1 PROD; 2 para COR e PROT; 3 para TC e 4
para PEANC e PEAPC.
Pif = (Yij
fj=l − Mj)
2 2f
e Pid = (Yij
dj=l − Mj)
2 2d
P𝑚𝑖 = ∑ Pik
1
pk
v
k=1
P𝑚𝑖 = ∑ Pik
1
pk
v
k=1
x 1
Pk

28
4. RESULTADOS E DISCUSSÃO
4.1 AVALIAÇÃO DE DOENÇAS
4.1.1 Antracnose (Colletotrichum lindemuthianum)
Na Tabela 3 encontram-se dispostas as notas dos genótipos avaliados para reação de
resistência às raças 31, 65 e 81 de C. lindemuthuanum, citadas por CARBONELL et al. (1999)
como as mais frequentes do estado de São Paulo. Observa-se que dos 27 genótipos avaliados, sete
(30%) genótipos foram considerados resistentes obtendo nota um, sendo, três genótipos de
tegumento carioca, CNFC 10762, LP 09-40 e MAII – 2 e quatro genótipos de tegumento preto, IAC
Una, Gen PR 14-2-10-1-2 e Gen PR 14-2-2-1-1 ressaltando-se neste caso o bom desempenho para
os genótipos de tegumentos preto provenientes do Instituto Agronômico - IAC, na qual foram os
únicos resistentes ao patógeno. Outro genótipo considerado resistente nesta avaliação foi o de grão
especial BRS-Realce.
Durante as observações de campo em diversos municípios houve a ocorrência da doença em
diferentes níveis de severidade, sendo observado em algumas ocasiões, que o genótipo Gen C 4-7-
7-2-2 embora considerado moderadamente resistente nas avaliações de inoculação, demonstrou
resistência ao patógeno em situações que outros genótipos foram suscetíveis. Uma hipótese para
isto pode ser a possível resistência horizontal deste genótipo ao C. lindemuthianum, que segundo
CAMARGO E BERGAMIM (1995), é conferida por um conjunto de genes de efeitos secundários,
aferindo um efeito quantitativo sobre a severidade da doença. Deste modo, embora o genótipo não
apresente resistência total ao patógeno, este, consegue completar seu ciclo sem grandes perdas de
produção. Além disso, a resistência horizontal é efetiva contra um numero maior de raças e mais
difícil de ser quebrada, se comparada à resistência vertical (BORÉM E MIRANDA, 2005).
Observa-se também que nenhuma das testemunhas comerciais utilizadas foram consideradas
resistentes ao patógeno. Entretanto, nas avaliações de campo foi observado que a cultivar IAC
Alvorada apresenta resistência à antracnose, assim como o genótipo Gen C 4-7-7-2-2, ressaltando
que mais estudos devem ser realizados para determinar a natureza desta resistência.
De acordo com ABREU et al. (2003), a ocorrência desta doença causa grandes prejuízos à
cultura, e a seleção de genótipos resistentes afeta positivamente a produtividade de grãos e a
estabilidade final do genótipo, sendo desta forma necessária a indicação de novas cultivares que
possuam resistência ao C. lindemuthuanum.
A existência de inúmeras raças do patógeno dificulta a seleção de genótipos de ampla
resistência, sendo necessária a inclusão de diferentes genes para a obtenção de um genótipo com

29
resistência horizontal às diferentes raças do patógeno. Na literatura há relatos que ocorrem mais de
13 genes de resistência a diferentes raças de antracnose (SINGH E SCHWARTZ, 2010).
Tabela 3. Avaliação da reação de resistência dos genótipos de
feijoeiro ao patógeno Colletotrichum lindemuthianum.
Resistentes Med. Resistentes Suscetível
CNFC 10762 LP 08-90 IAC Alvorada
IAC Diplomata SM 1107 IPR 139
IAC Una C 4-7-7-2-2 Gen C 2-6-4-1-1
Gen PR 14-2-10-1-2 C 4-7-8-1-2 Gen C 2-6-6-2-1
Gen PR 14-2-2-1-1 LP 07-80 Gen C 4-7-2-2-1
LP 09-40 BRSMG Madrepérola Gen C 5-3-6-1-1
MAII - 2
CHC 01-175
BRS Realce
CHP 98-66-20
CNFC 10729
CNFP 10794
LP 09-192
Gen PR 11-2-8-1-1
SM 1810
*: R: resistente; MR: medianamente resistente; S: suscetível.
4.1.2 Murcha de Fusarium (Fusarium oxysporum f. sp, phaseoli)
Na Tabela 4 seguem os resultados obtidos pelos 27 genótipos avaliados para reação de
resistência ao F. oxysporum. Dentre os genótipos avaliados sete foram considerados resistentes
(26%). Para os genótipos de tegumento carioca foram considerados resistentes os genótipos CNFC
10729, Gen C 4-7-7-2-2 e LP 09-40. Considerando os genótipos de tegumento preto, foram
resistentes Gen PR 14-2-2-1-1, LP 08-90, SM 1810 e a testemunha IAC Diplomata. Destacaram-se
os genótipos Gen PR 14-2-2-1-1 e LP 09-40 por apresentarem resistência mutua ao C.
lindemuthianum e F. oxysporum.
A murcha de fusarium, encontra-se amplamente disseminada nas principais regiões
produtivas do país, sendo citada entre as mais importantes doenças para as regiões sudeste, centro-
oeste e nordeste do país (THUNG E SARTORATO, 2002), ocasionando perdas de até 80% no
rendimento de grãos da cultura (COSTA et al., 2007). Como a maioria das cultivares de feijoeiro
recomendadas para semeadura nos estados brasileiros apresentam alta suscetibilidade a este
patógeno, é interessante que os programas de melhoramento genético de feijoeiro indiquem novas
cultivares de feijoeiro que sejam resistentes a este patógeno.
No presente trabalho foram observados genótipos resistentes a três raças citadas entre as
mais disseminadas e agressivas deste patógeno (CARBONELL et al. 1999, CHIORATO, 2004).

30
Estes genótipos devem ser candidatos a novas cultivares, desde que apresentarem também, boas
características agronômicas e de qualidade tecnológica dos grãos.
Tabela 4. Avaliação da reação de resistência dos genótipos de
feijoeiro ao patógeno Fusarium oxysporum f. sp, phaseoli.
Resistentes Med. Resistentes Suscetível
Gen C 4-7-7-2-2 Gen C 4-7-8-1-2 IAC Alvorada
CNFC 10729 CHP 98-66-20 IPR 139
LP 09-40 LP 09-192 Gen C 2-6-4-1-1
IAC Diplomata
Gen C 2-6-6-2-1
LP 08-90
Gen C 4-7-2-2-1
Gen PR 14-2-2-1-1
Gen C 5-3-6-1-1
SM 1810
CHC 01-175
CNFC 10762
LP 07-80
BRSMG Madrepérola
MAII - 2
IAC Una
CNFP 10794
Gen PR 11-2-8-1-1
Gen PR 14-2-10-1-2
SM 1107
BRS Realce
4.1.3 Crestamento Bacteriano (Xanthomonas axonopodis pv. phaseoli)
Na Tabela 5, estão dispostas as notas dos 27 genótipos , sendo identificados apenas quatro
genótipos resistentes ao patógeno (14%), sendo um genótipo de tegumento carioca LP 07-80 e três
genótipos de tegumento preto LP 08-90, LP 09-192 e SM 1810. Isso demonstra a dificuldade de se
encontrar genótipos com resistência o crestamento bacteriano, corroborando com os resultados de
DURSUN et al. (2002) e SANTOS et al. (2003), que também relatam o baixo número de genótipos
resistentes identificados, sendo importante assim a identificação destes para utilização em
cruzamentos e introdução de genes em programas de melhoramento genético de feijoeiro.

31
Tabela 5. Avaliação da reação de resistência dos genótipos de
feijoeiro ao patógeno Xanthomonas axonopodis. pv, phaseoli.
Resistentes Med. Resistentes Suscetível
Gen C 4-7-7-2-2 Gen C 4-7-8-1-2 IAC Alvorada
CNFC 10729 CHP 98-66-20 IPR 139
LP 09-40 LP 09-192 Gen C 2-6-4-1-1
IAC Diplomata
Gen C 2-6-6-2-1
LP 08-90
Gen C 4-7-2-2-1
Gen PR 14-2-2-1-1
Gen C 5-3-6-1-1
SM 1810
CHC 01-175
CNFC 10762
LP 07-80
BRSMG
Madrepérola
MAII - 2
IAC Una
CNFP 10794
Gen PR 11-2-8-1-1
Gen PR 14-2-10-1-2
SM 1107
BRS Realce
4.2 ANÁLISES DOS DADOS
Os resumos das análises de variância conjunta por safra e geral são apresentados na Tabela
6. Previamente a esta análise, foi verificado que a razão entre o maior e o menor valor para os
quadrados médios do erro das análises de variância individuais foram menores que 7, o que segundo
BANZATTO E KRONKA (1995), indica homogeneidade de variância, possibilitando assim a
realização da análise conjunta.
Pode-se observar que os coeficientes de variação ambiental (CV) obtidos foram aceitáveis
para todas as variáveis, sugerindo boa precisão nos experimentos. Para fins de recomendação de
cultivares, são aceitos experimentos com até 25% de CV (MAPA, 2006).
O teste F da análise de variância (Tabela 6) detectou diferenças significativas entre
tratamentos para a maioria das características avaliadas, e de acordo com SENNA et al. (2012), isto
indica a presença de diferença significativa entre os genótipos avaliados. Contudo, para a variável
cor do tegumento preto, os tratamentos não apresentaram diferenças significativas em nenhuma das
análises.
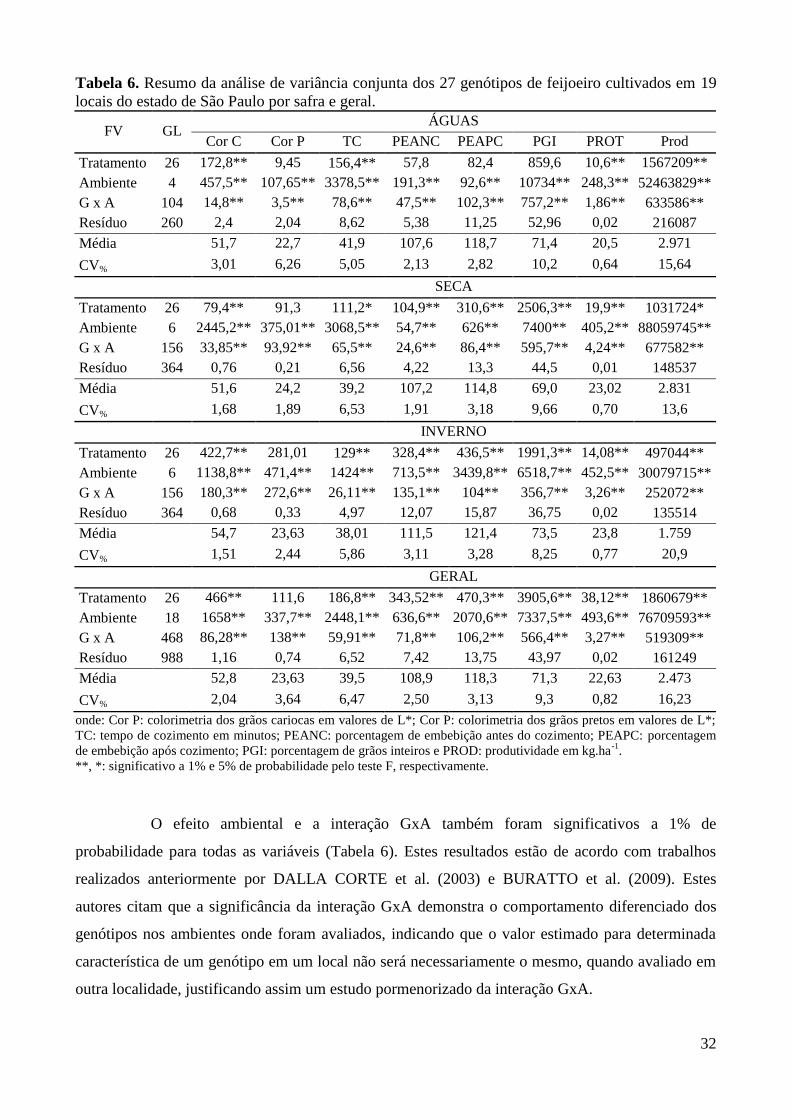
32
Tabela 6. Resumo da análise de variância conjunta dos 27 genótipos de feijoeiro cultivados em 19
locais do estado de São Paulo por safra e geral.
FV GL ÁGUAS
Cor C Cor P TC PEANC PEAPC PGI PROT Prod
Tratamento 26 172,8** 9,45 156,4** 57,8 82,4 859,6 10,6** 1567209**
Ambiente 4 457,5** 107,65** 3378,5** 191,3** 92,6** 10734** 248,3** 52463829**
G x A 104 14,8** 3,5** 78,6** 47,5** 102,3** 757,2** 1,86** 633586**
Resíduo 260 2,4 2,04 8,62 5,38 11,25 52,96 0,02 216087
Média
51,7 22,7 41,9 107,6 118,7 71,4 20,5 2.971
CV% 3,01 6,26 5,05 2,13 2,82 10,2 0,64 15,64
SECA
Tratamento 26 79,4** 91,3 111,2* 104,9** 310,6** 2506,3** 19,9** 1031724*
Ambiente 6 2445,2** 375,01** 3068,5** 54,7** 626** 7400** 405,2** 88059745**
G x A 156 33,85** 93,92** 65,5** 24,6** 86,4** 595,7** 4,24** 677582**
Resíduo 364 0,76 0,21 6,56 4,22 13,3 44,5 0,01 148537
Média
51,6 24,2 39,2 107,2 114,8 69,0 23,02 2.831
CV% 1,68 1,89 6,53 1,91 3,18 9,66 0,70 13,6
INVERNO
Tratamento 26 422,7** 281,01 129** 328,4** 436,5** 1991,3** 14,08** 497044**
Ambiente 6 1138,8** 471,4** 1424** 713,5** 3439,8** 6518,7** 452,5** 30079715**
G x A 156 180,3** 272,6** 26,11** 135,1** 104** 356,7** 3,26** 252072**
Resíduo 364 0,68 0,33 4,97 12,07 15,87 36,75 0,02 135514
Média
54,7 23,63 38,01 111,5 121,4 73,5 23,8 1.759
CV% 1,51 2,44 5,86 3,11 3,28 8,25 0,77 20,9
GERAL
Tratamento 26 466** 111,6 186,8** 343,52** 470,3** 3905,6** 38,12** 1860679**
Ambiente 18 1658** 337,7** 2448,1** 636,6** 2070,6** 7337,5** 493,6** 76709593**
G x A 468 86,28** 138** 59,91** 71,8** 106,2** 566,4** 3,27** 519309**
Resíduo 988 1,16 0,74 6,52 7,42 13,75 43,97 0,02 161249
Média
52,8 23,63 39,5 108,9 118,3 71,3 22,63 2.473
CV% 2,04 3,64 6,47 2,50 3,13 9,3 0,82 16,23
onde: Cor P: colorimetria dos grãos cariocas em valores de L*; Cor P: colorimetria dos grãos pretos em valores de L*;
TC: tempo de cozimento em minutos; PEANC: porcentagem de embebição antes do cozimento; PEAPC: porcentagem
de embebição após cozimento; PGI: porcentagem de grãos inteiros e PROD: produtividade em kg.ha-1
.
**, *: significativo a 1% e 5% de probabilidade pelo teste F, respectivamente.
O efeito ambiental e a interação GxA também foram significativos a 1% de
probabilidade para todas as variáveis (Tabela 6). Estes resultados estão de acordo com trabalhos
realizados anteriormente por DALLA CORTE et al. (2003) e BURATTO et al. (2009). Estes
autores citam que a significância da interação GxA demonstra o comportamento diferenciado dos
genótipos nos ambientes onde foram avaliados, indicando que o valor estimado para determinada
característica de um genótipo em um local não será necessariamente o mesmo, quando avaliado em
outra localidade, justificando assim um estudo pormenorizado da interação GxA.

33
4.3 QUALIDADE TECNOLÓGICA
Nas Tabelas 7 e 8, observam-se os valores médios de L* por safra para grãos de tegumento
carioca e preto respectivamente. De acordo com os dados obtidos, foi possível observar que os
maiores valores de L* para grãos de tegumento carioca (Tabela 7) foram obtidos durante a safra de
inverno, onde os genótipos foram divididos em sete grupos de acordo com o teste de Scott-Knott a
5% de probabilidade, destacando o genótipo BRSMG Madrepérola (58.56) como único no grupo
“a”.
Considerando todos os ambientes, a média de L* dos genótipos foi de 52.8, sendo
novamente o genótipo BRSMG Madrepérola (56,1) agrupado separado dos demais genótipos.
Outros trabalhos apontam esta cultivar como sendo de coloração bem clara e de tolerância ao
escurecimento por longo período de armazenamento. (ABREU et al 2011; ARAÚJO et al. 2012).
Entretanto, observa-se que os genótipos LP 09-40 (54.9), IAC Alvorada (54.2), Gen C 4-7-7-2-2
(53.2), Gen C 4-7-8-1-2 (53.9) e IPR 139 (53.6), pertencentes aos grupos “b” e “c” de acordo com o
teste de Scott-Knott a 5%, obtiveram médias de L* acima de 53, sendo assim genótipos que
apresentam grãos claros.
Para os genótipos de tegumento preto (Tabela 8) não foram observadas médias de L*
inferiores a 22 em nenhuma das safras. Na safra das águas, não houve diferenças significativas entre
os genótipos, porém isto era esperado tendo em vista a não significância obtida pelo teste F na
análise de variância.
Entre as médias da safra da seca, inverno e geral, foram obtidas diferenças significativas
entre os genótipos pelo teste de Scott-Knott e embora não tivessem sido detectadas diferenças
significativas pelo teste F na análise de variância, este fato, de acordo com PIMENTEL-GOMES
(1985) se deve por existirem divergências de natureza matemática do modelo, não sendo isto de
grande importância do ponto de vista estatístico, podendo isto ser comprovado observando a baixa
magnitude das diferenças entre as médias dos genótipos.
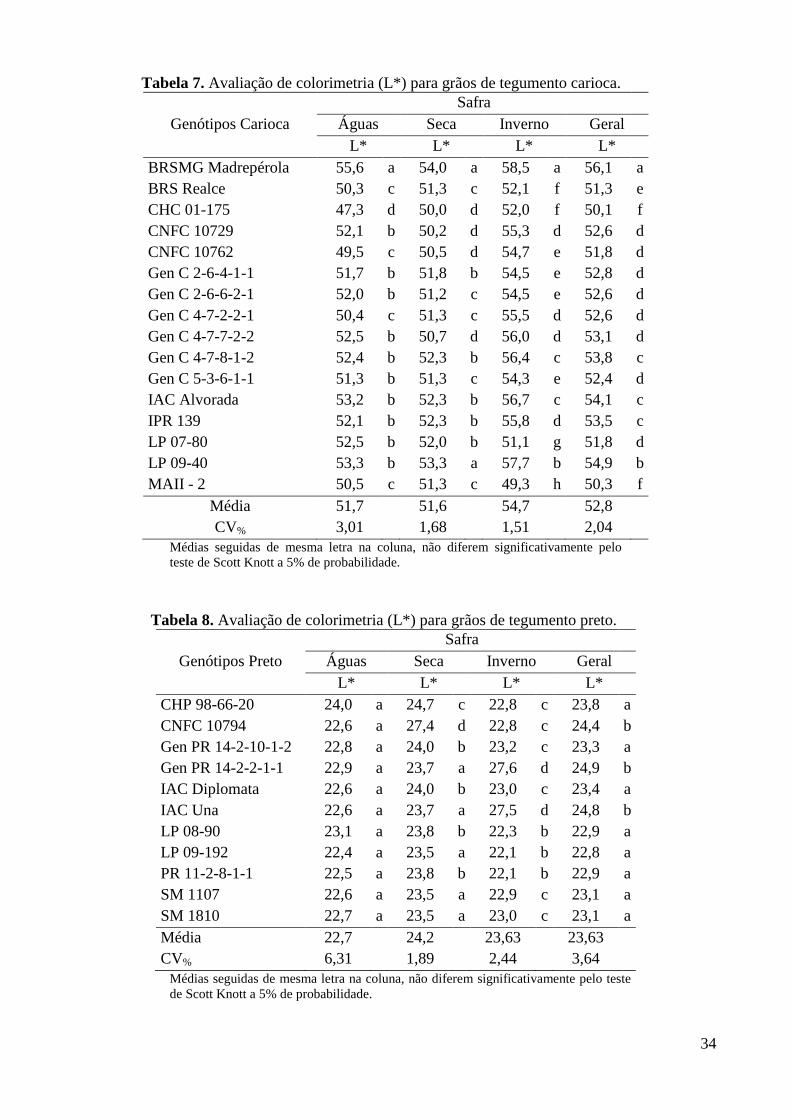
34
Tabela 7. Avaliação de colorimetria (L*) para grãos de tegumento carioca.
Genótipos Carioca
Safra
Águas Seca Inverno Geral
L* L* L* L*
BRSMG Madrepérola 55,6 a 54,0 a 58,5 a 56,1 a
BRS Realce 50,3 c 51,3 c 52,1 f 51,3 e
CHC 01-175 47,3 d 50,0 d 52,0 f 50,1 f
CNFC 10729 52,1 b 50,2 d 55,3 d 52,6 d
CNFC 10762 49,5 c 50,5 d 54,7 e 51,8 d
Gen C 2-6-4-1-1 51,7 b 51,8 b 54,5 e 52,8 d
Gen C 2-6-6-2-1 52,0 b 51,2 c 54,5 e 52,6 d
Gen C 4-7-2-2-1 50,4 c 51,3 c 55,5 d 52,6 d
Gen C 4-7-7-2-2 52,5 b 50,7 d 56,0 d 53,1 d
Gen C 4-7-8-1-2 52,4 b 52,3 b 56,4 c 53,8 c
Gen C 5-3-6-1-1 51,3 b 51,3 c 54,3 e 52,4 d
IAC Alvorada 53,2 b 52,3 b 56,7 c 54,1 c
IPR 139 52,1 b 52,3 b 55,8 d 53,5 c
LP 07-80 52,5 b 52,0 b 51,1 g 51,8 d
LP 09-40 53,3 b 53,3 a 57,7 b 54,9 b
MAII - 2 50,5 c 51,3 c 49,3 h 50,3 f
Média 51,7
51,6
54,7
52,8
CV% 3,01
1,68
1,51
2,04
Médias seguidas de mesma letra na coluna, não diferem significativamente pelo
teste de Scott Knott a 5% de probabilidade.
Tabela 8. Avaliação de colorimetria (L*) para grãos de tegumento preto.
Genótipos Preto
Safra
Águas Seca Inverno Geral
L* L* L* L*
CHP 98-66-20 24,0 a 24,7 c 22,8 c 23,8 a
CNFC 10794 22,6 a 27,4 d 22,8 c 24,4 b
Gen PR 14-2-10-1-2 22,8 a 24,0 b 23,2 c 23,3 a
Gen PR 14-2-2-1-1 22,9 a 23,7 a 27,6 d 24,9 b
IAC Diplomata 22,6 a 24,0 b 23,0 c 23,4 a
IAC Una 22,6 a 23,7 a 27,5 d 24,8 b
LP 08-90 23,1 a 23,8 b 22,3 b 22,9 a
LP 09-192 22,4 a 23,5 a 22,1 b 22,8 a
PR 11-2-8-1-1 22,5 a 23,8 b 22,1 b 22,9 a
SM 1107 22,6 a 23,5 a 22,9 c 23,1 a
SM 1810 22,7 a 23,5 a 23,0 c 23,1 a
Média 22,7 24,2 23,63 23,63
CV% 6,31 1,89 2,44 3,64
Médias seguidas de mesma letra na coluna, não diferem significativamente pelo teste
de Scott Knott a 5% de probabilidade.
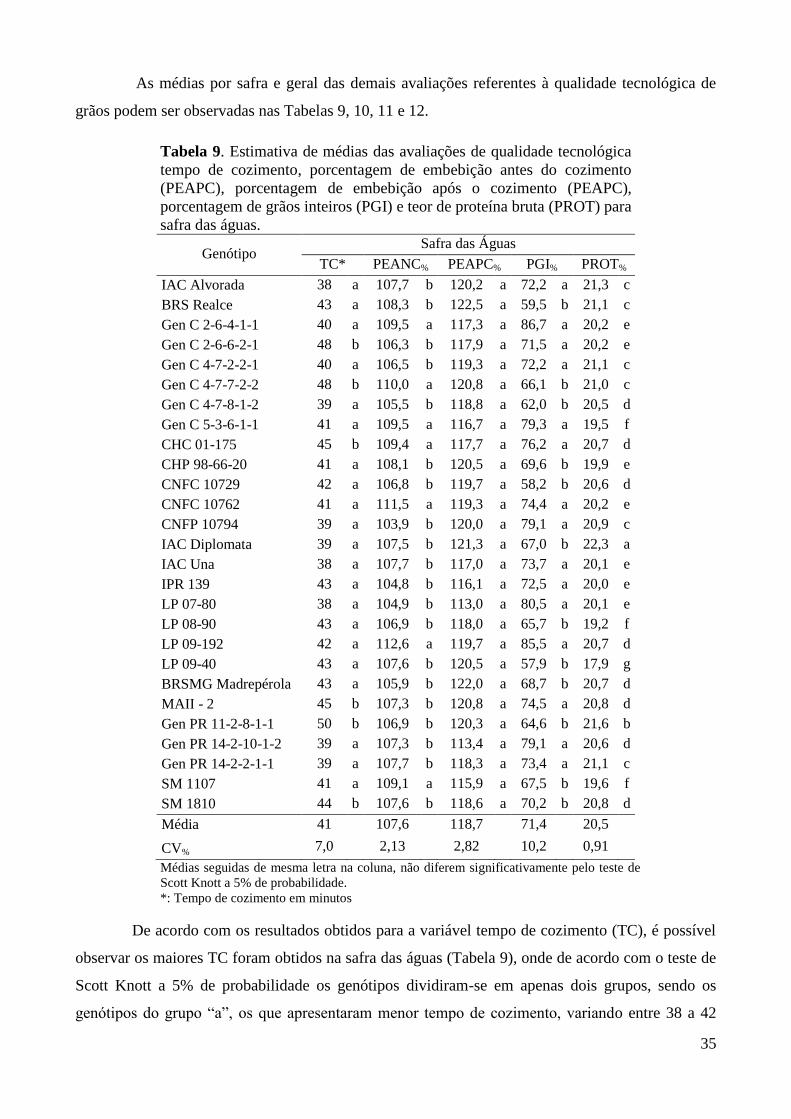
35
As médias por safra e geral das demais avaliações referentes à qualidade tecnológica de
grãos podem ser observadas nas Tabelas 9, 10, 11 e 12.
Tabela 9. Estimativa de médias das avaliações de qualidade tecnológica
tempo de cozimento, porcentagem de embebição antes do cozimento
(PEAPC), porcentagem de embebição após o cozimento (PEAPC),
porcentagem de grãos inteiros (PGI) e teor de proteína bruta (PROT) para
safra das águas.
Genótipo Safra das Águas
TC* PEANC% PEAPC% PGI% PROT%
IAC Alvorada 38 a 107,7 b 120,2 a 72,2 a 21,3 c
BRS Realce 43 a 108,3 b 122,5 a 59,5 b 21,1 c
Gen C 2-6-4-1-1 40 a 109,5 a 117,3 a 86,7 a 20,2 e
Gen C 2-6-6-2-1 48 b 106,3 b 117,9 a 71,5 a 20,2 e
Gen C 4-7-2-2-1 40 a 106,5 b 119,3 a 72,2 a 21,1 c
Gen C 4-7-7-2-2 48 b 110,0 a 120,8 a 66,1 b 21,0 c
Gen C 4-7-8-1-2 39 a 105,5 b 118,8 a 62,0 b 20,5 d
Gen C 5-3-6-1-1 41 a 109,5 a 116,7 a 79,3 a 19,5 f
CHC 01-175 45 b 109,4 a 117,7 a 76,2 a 20,7 d
CHP 98-66-20 41 a 108,1 b 120,5 a 69,6 b 19,9 e
CNFC 10729 42 a 106,8 b 119,7 a 58,2 b 20,6 d
CNFC 10762 41 a 111,5 a 119,3 a 74,4 a 20,2 e
CNFP 10794 39 a 103,9 b 120,0 a 79,1 a 20,9 c
IAC Diplomata 39 a 107,5 b 121,3 a 67,0 b 22,3 a
IAC Una 38 a 107,7 b 117,0 a 73,7 a 20,1 e
IPR 139 43 a 104,8 b 116,1 a 72,5 a 20,0 e
LP 07-80 38 a 104,9 b 113,0 a 80,5 a 20,1 e
LP 08-90 43 a 106,9 b 118,0 a 65,7 b 19,2 f
LP 09-192 42 a 112,6 a 119,7 a 85,5 a 20,7 d
LP 09-40 43 a 107,6 b 120,5 a 57,9 b 17,9 g
BRSMG Madrepérola 43 a 105,9 b 122,0 a 68,7 b 20,7 d
MAII - 2 45 b 107,3 b 120,8 a 74,5 a 20,8 d
Gen PR 11-2-8-1-1 50 b 106,9 b 120,3 a 64,6 b 21,6 b
Gen PR 14-2-10-1-2 39 a 107,3 b 113,4 a 79,1 a 20,6 d
Gen PR 14-2-2-1-1 39 a 107,7 b 118,3 a 73,4 a 21,1 c
SM 1107 41 a 109,1 a 115,9 a 67,5 b 19,6 f
SM 1810 44 b 107,6 b 118,6 a 70,2 b 20,8 d
Média 41 107,6 118,7 71,4 20,5
CV% 7,0 2,13 2,82 10,2 0,91
Médias seguidas de mesma letra na coluna, não diferem significativamente pelo teste de
Scott Knott a 5% de probabilidade.
*: Tempo de cozimento em minutos
De acordo com os resultados obtidos para a variável tempo de cozimento (TC), é possível
observar os maiores TC foram obtidos na safra das águas (Tabela 9), onde de acordo com o teste de
Scott Knott a 5% de probabilidade os genótipos dividiram-se em apenas dois grupos, sendo os
genótipos do grupo “a”, os que apresentaram menor tempo de cozimento, variando entre 38 a 42
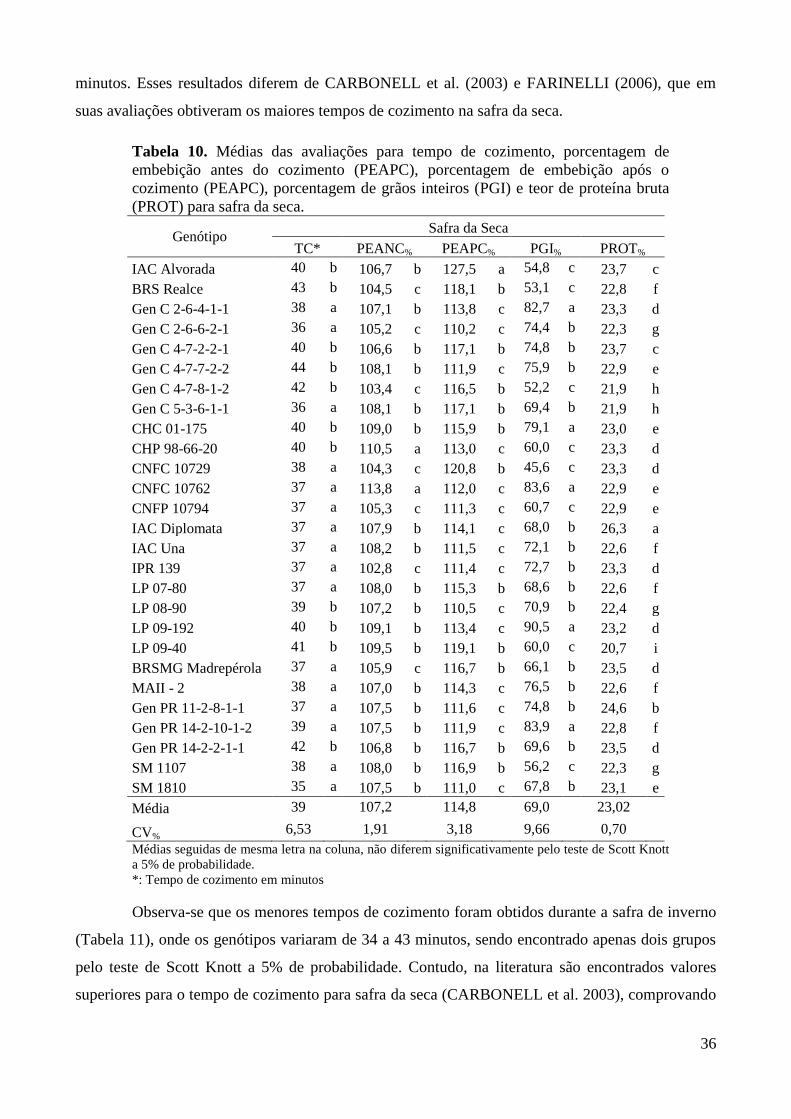
36
minutos. Esses resultados diferem de CARBONELL et al. (2003) e FARINELLI (2006), que em
suas avaliações obtiveram os maiores tempos de cozimento na safra da seca.
Tabela 10. Médias das avaliações para tempo de cozimento, porcentagem de
embebição antes do cozimento (PEAPC), porcentagem de embebição após o
cozimento (PEAPC), porcentagem de grãos inteiros (PGI) e teor de proteína bruta
(PROT) para safra da seca.
Genótipo Safra da Seca
TC* PEANC% PEAPC% PGI% PROT%
IAC Alvorada 40 b 106,7 b 127,5 a 54,8 c 23,7 c
BRS Realce 43 b 104,5 c 118,1 b 53,1 c 22,8 f
Gen C 2-6-4-1-1 38 a 107,1 b 113,8 c 82,7 a 23,3 d
Gen C 2-6-6-2-1 36 a 105,2 c 110,2 c 74,4 b 22,3 g
Gen C 4-7-2-2-1 40 b 106,6 b 117,1 b 74,8 b 23,7 c
Gen C 4-7-7-2-2 44 b 108,1 b 111,9 c 75,9 b 22,9 e
Gen C 4-7-8-1-2 42 b 103,4 c 116,5 b 52,2 c 21,9 h
Gen C 5-3-6-1-1 36 a 108,1 b 117,1 b 69,4 b 21,9 h
CHC 01-175 40 b 109,0 b 115,9 b 79,1 a 23,0 e
CHP 98-66-20 40 b 110,5 a 113,0 c 60,0 c 23,3 d
CNFC 10729 38 a 104,3 c 120,8 b 45,6 c 23,3 d
CNFC 10762 37 a 113,8 a 112,0 c 83,6 a 22,9 e
CNFP 10794 37 a 105,3 c 111,3 c 60,7 c 22,9 e
IAC Diplomata 37 a 107,9 b 114,1 c 68,0 b 26,3 a
IAC Una 37 a 108,2 b 111,5 c 72,1 b 22,6 f
IPR 139 37 a 102,8 c 111,4 c 72,7 b 23,3 d
LP 07-80 37 a 108,0 b 115,3 b 68,6 b 22,6 f
LP 08-90 39 b 107,2 b 110,5 c 70,9 b 22,4 g
LP 09-192 40 b 109,1 b 113,4 c 90,5 a 23,2 d
LP 09-40 41 b 109,5 b 119,1 b 60,0 c 20,7 i
BRSMG Madrepérola 37 a 105,9 c 116,7 b 66,1 b 23,5 d
MAII - 2 38 a 107,0 b 114,3 c 76,5 b 22,6 f
Gen PR 11-2-8-1-1 37 a 107,5 b 111,6 c 74,8 b 24,6 b
Gen PR 14-2-10-1-2 39 a 107,5 b 111,9 c 83,9 a 22,8 f
Gen PR 14-2-2-1-1 42 b 106,8 b 116,7 b 69,6 b 23,5 d
SM 1107 38 a 108,0 b 116,9 b 56,2 c 22,3 g
SM 1810 35 a 107,5 b 111,0 c 67,8 b 23,1 e
Média 39 107,2 114,8 69,0 23,02
CV% 6,53 1,91 3,18 9,66 0,70 Médias seguidas de mesma letra na coluna, não diferem significativamente pelo teste de Scott Knott
a 5% de probabilidade.
*: Tempo de cozimento em minutos
Observa-se que os menores tempos de cozimento foram obtidos durante a safra de inverno
(Tabela 11), onde os genótipos variaram de 34 a 43 minutos, sendo encontrado apenas dois grupos
pelo teste de Scott Knott a 5% de probabilidade. Contudo, na literatura são encontrados valores
superiores para o tempo de cozimento para safra da seca (CARBONELL et al. 2003), comprovando
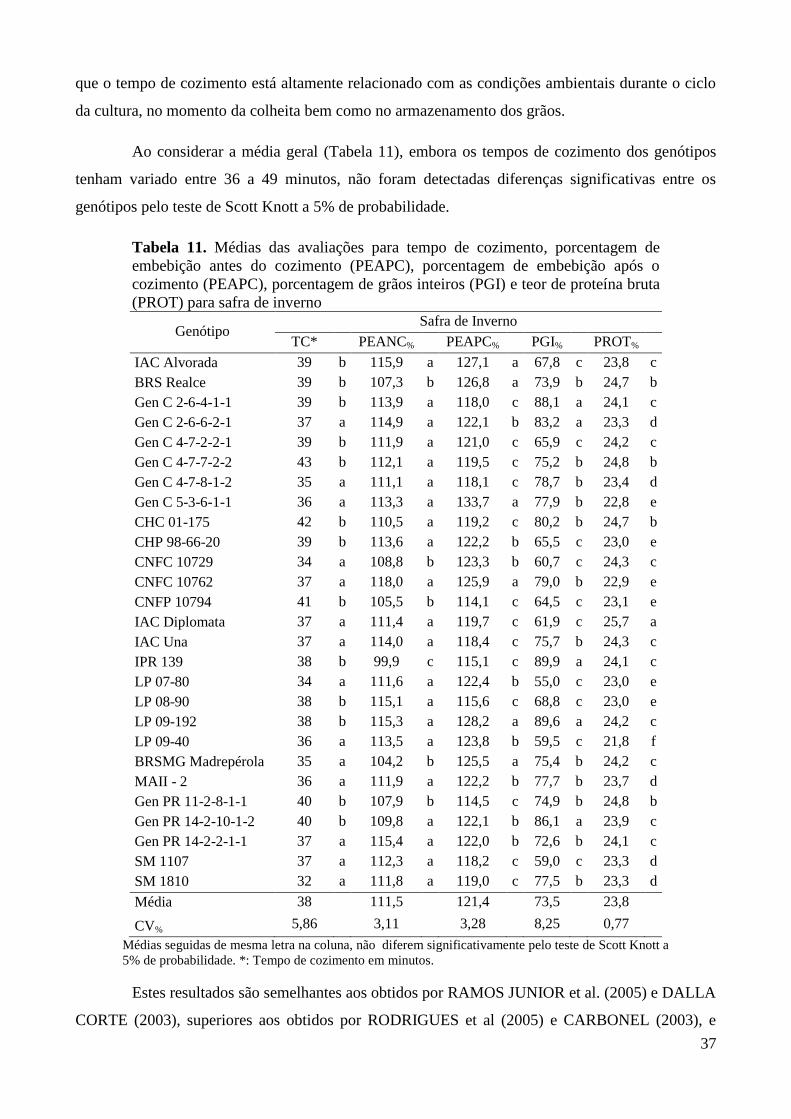
37
que o tempo de cozimento está altamente relacionado com as condições ambientais durante o ciclo
da cultura, no momento da colheita bem como no armazenamento dos grãos.
Ao considerar a média geral (Tabela 11), embora os tempos de cozimento dos genótipos
tenham variado entre 36 a 49 minutos, não foram detectadas diferenças significativas entre os
genótipos pelo teste de Scott Knott a 5% de probabilidade.
Tabela 11. Médias das avaliações para tempo de cozimento, porcentagem de
embebição antes do cozimento (PEAPC), porcentagem de embebição após o
cozimento (PEAPC), porcentagem de grãos inteiros (PGI) e teor de proteína bruta
(PROT) para safra de inverno
Genótipo Safra de Inverno
TC* PEANC% PEAPC% PGI% PROT%
IAC Alvorada 39 b 115,9 a 127,1 a 67,8 c 23,8 c
BRS Realce 39 b 107,3 b 126,8 a 73,9 b 24,7 b
Gen C 2-6-4-1-1 39 b 113,9 a 118,0 c 88,1 a 24,1 c
Gen C 2-6-6-2-1 37 a 114,9 a 122,1 b 83,2 a 23,3 d
Gen C 4-7-2-2-1 39 b 111,9 a 121,0 c 65,9 c 24,2 c
Gen C 4-7-7-2-2 43 b 112,1 a 119,5 c 75,2 b 24,8 b
Gen C 4-7-8-1-2 35 a 111,1 a 118,1 c 78,7 b 23,4 d
Gen C 5-3-6-1-1 36 a 113,3 a 133,7 a 77,9 b 22,8 e
CHC 01-175 42 b 110,5 a 119,2 c 80,2 b 24,7 b
CHP 98-66-20 39 b 113,6 a 122,2 b 65,5 c 23,0 e
CNFC 10729 34 a 108,8 b 123,3 b 60,7 c 24,3 c
CNFC 10762 37 a 118,0 a 125,9 a 79,0 b 22,9 e
CNFP 10794 41 b 105,5 b 114,1 c 64,5 c 23,1 e
IAC Diplomata 37 a 111,4 a 119,7 c 61,9 c 25,7 a
IAC Una 37 a 114,0 a 118,4 c 75,7 b 24,3 c
IPR 139 38 b 99,9 c 115,1 c 89,9 a 24,1 c
LP 07-80 34 a 111,6 a 122,4 b 55,0 c 23,0 e
LP 08-90 38 b 115,1 a 115,6 c 68,8 c 23,0 e
LP 09-192 38 b 115,3 a 128,2 a 89,6 a 24,2 c
LP 09-40 36 a 113,5 a 123,8 b 59,5 c 21,8 f
BRSMG Madrepérola 35 a 104,2 b 125,5 a 75,4 b 24,2 c
MAII - 2 36 a 111,9 a 122,2 b 77,7 b 23,7 d
Gen PR 11-2-8-1-1 40 b 107,9 b 114,5 c 74,9 b 24,8 b
Gen PR 14-2-10-1-2 40 b 109,8 a 122,1 b 86,1 a 23,9 c
Gen PR 14-2-2-1-1 37 a 115,4 a 122,0 b 72,6 b 24,1 c
SM 1107 37 a 112,3 a 118,2 c 59,0 c 23,3 d
SM 1810 32 a 111,8 a 119,0 c 77,5 b 23,3 d
Média 38 111,5 121,4 73,5 23,8
CV% 5,86 3,11 3,28 8,25 0,77
Médias seguidas de mesma letra na coluna, não diferem significativamente pelo teste de Scott Knott a
5% de probabilidade. *: Tempo de cozimento em minutos.
Estes resultados são semelhantes aos obtidos por RAMOS JUNIOR et al. (2005) e DALLA
CORTE (2003), superiores aos obtidos por RODRIGUES et al (2005) e CARBONEL (2003), e

38
inferiores aos obtidos por RIBEIRO et al. (2005). De acordo com RIBEIRO et al. (2007), estas
variações entre os tempos de cozimento obtidas podem ser explicadas devido aos diferentes
critérios utilizados para caracterizar o tempo de cozimento, à temperatura de aquecimento utilizada,
da qualidade, tempo de armazenamento, além da questão genotípica dos grãos avaliados.
Para os resultados PEANC e PEAPC, é possível observar que embora a magnitude das
diferenças seja baixa, a safra de inverno (Tabela 10) obteve as maiores porcentagens de embebição
e a safra da seca as menores porcentagens. Observa-se também que nas análises por safra e geral, os
genótipos foram divididos em apenas dois grupos pelo teste de Scott-Knott a 5% de probabilidade.
Na literatura existem relatos de correlação negativa entre absorção de água pelos grãos e o
tempo de cozimento SCHOLZ E FONSECA JUNIOR (1999), DALLA CORTE et al. (2003) e
RODRIGUES et al. (2005). Entretanto, neste trabalho não foram obtidas correlações significativas
entre estas características (Anexo 20), corroborando com a afirmação realizada por CARBONELL
et al. (2003) que em seleções de genótipos com base na utilização do teste de absorção de água
pelos grãos são pouco expressivas e nem sempre confiáveis, devido a inconstante correlação
encontrada entre essas características.
No que diz respeito à variável porcentagem de grãos inteiros (PGI), os aspectos
desejados pelo mercado consumidor são genótipos que apresentaram maior quantidade de grãos
inteiros após o cozimento, sendo desta forma de maior rendimento após cozimento. Neste caso, os
melhores resultados obtidos também foram na safra de inverno (Tabela 10) com uma média de
73,5%, em que os genótipos que se destacaram no grupo “a” pelo teste de Scott-Knott a 5% de
probabilidade foram IPR 139 (89,9%), Gen C 2-6-4-1-1 (88,1%), LP 09-192 (89,6%), Gen C 2-6-6-
2-1 (83,2%) e Gen PR 14-2-10-1-2 (86,1%).
Na safra da seca (Tabela 9) foram obtidos os menores valores para PGI, com uma média de
69%, destacando os genótipos Gen C 2-6-4-1-1 (82,7%), LP 09-192 (90,5%) e Gen PR 14-2-10-1-2
(83,9%), no grupo “a” pelo teste de Scott-Knott a 5%. Considerando a média geral (Tabela 11),
estes mesmos genótipos foram os únicos a diferirem estatisticamente dos demais, com 85,7%,
88,8% e 83,5% respectivamente.
Para a variável teor de proteína os valores médios obtidos variaram entre 17,9 e 26,3%,
confirmando os resultados obtidos por GUZMÁN-MALDONADO E PAREDE-LÓPEZ (1999) que
em seus trabalhos obtiveram teores entre 16 a 33%. BURATO et al. (2009), avaliando 18 genótipos
de tegumento preto em três locais do estado do Paraná relatam menor amplitude nos teores
proteicos obtidos, variando entre 23,2 a 26,3%.
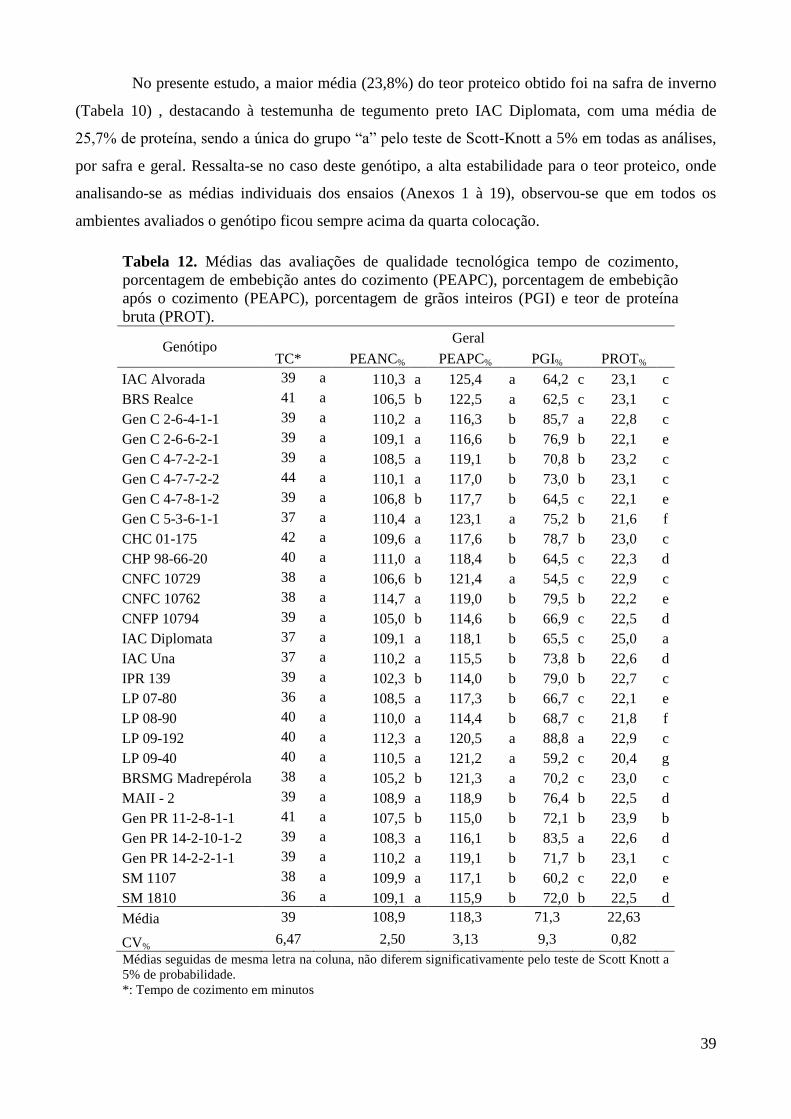
39
No presente estudo, a maior média (23,8%) do teor proteico obtido foi na safra de inverno
(Tabela 10) , destacando à testemunha de tegumento preto IAC Diplomata, com uma média de
25,7% de proteína, sendo a única do grupo “a” pelo teste de Scott-Knott a 5% em todas as análises,
por safra e geral. Ressalta-se no caso deste genótipo, a alta estabilidade para o teor proteico, onde
analisando-se as médias individuais dos ensaios (Anexos 1 à 19), observou-se que em todos os
ambientes avaliados o genótipo ficou sempre acima da quarta colocação.
Tabela 12. Médias das avaliações de qualidade tecnológica tempo de cozimento,
porcentagem de embebição antes do cozimento (PEAPC), porcentagem de embebição
após o cozimento (PEAPC), porcentagem de grãos inteiros (PGI) e teor de proteína
bruta (PROT).
Genótipo Geral
TC* PEANC% PEAPC% PGI% PROT%
IAC Alvorada 39 a 110,3 a 125,4 a 64,2 c 23,1 c
BRS Realce 41 a 106,5 b 122,5 a 62,5 c 23,1 c
Gen C 2-6-4-1-1 39 a 110,2 a 116,3 b 85,7 a 22,8 c
Gen C 2-6-6-2-1 39 a 109,1 a 116,6 b 76,9 b 22,1 e
Gen C 4-7-2-2-1 39 a 108,5 a 119,1 b 70,8 b 23,2 c
Gen C 4-7-7-2-2 44 a 110,1 a 117,0 b 73,0 b 23,1 c
Gen C 4-7-8-1-2 39 a 106,8 b 117,7 b 64,5 c 22,1 e
Gen C 5-3-6-1-1 37 a 110,4 a 123,1 a 75,2 b 21,6 f
CHC 01-175 42 a 109,6 a 117,6 b 78,7 b 23,0 c
CHP 98-66-20 40 a 111,0 a 118,4 b 64,5 c 22,3 d
CNFC 10729 38 a 106,6 b 121,4 a 54,5 c 22,9 c
CNFC 10762 38 a 114,7 a 119,0 b 79,5 b 22,2 e
CNFP 10794 39 a 105,0 b 114,6 b 66,9 c 22,5 d
IAC Diplomata 37 a 109,1 a 118,1 b 65,5 c 25,0 a
IAC Una 37 a 110,2 a 115,5 b 73,8 b 22,6 d
IPR 139 39 a 102,3 b 114,0 b 79,0 b 22,7 c
LP 07-80 36 a 108,5 a 117,3 b 66,7 c 22,1 e
LP 08-90 40 a 110,0 a 114,4 b 68,7 c 21,8 f
LP 09-192 40 a 112,3 a 120,5 a 88,8 a 22,9 c
LP 09-40 40 a 110,5 a 121,2 a 59,2 c 20,4 g
BRSMG Madrepérola 38 a 105,2 b 121,3 a 70,2 c 23,0 c
MAII - 2 39 a 108,9 a 118,9 b 76,4 b 22,5 d
Gen PR 11-2-8-1-1 41 a 107,5 b 115,0 b 72,1 b 23,9 b
Gen PR 14-2-10-1-2 39 a 108,3 a 116,1 b 83,5 a 22,6 d
Gen PR 14-2-2-1-1 39 a 110,2 a 119,1 b 71,7 b 23,1 c
SM 1107 38 a 109,9 a 117,1 b 60,2 c 22,0 e
SM 1810 36 a 109,1 a 115,9 b 72,0 b 22,5 d
Média 39 108,9 118,3 71,3 22,63
CV% 6,47 2,50 3,13 9,3 0,82 Médias seguidas de mesma letra na coluna, não diferem significativamente pelo teste de Scott Knott a
5% de probabilidade.
*: Tempo de cozimento em minutos

40
Entretanto, um problema na indicação de genótipos com alto teor de proteína é a
correlação negativa desta característica com a produtividade de grãos, citada por diversos autores
LEMOS et al. (2004), MELLO FILHO et al. (2004) e BASSO et al. (2011), sendo confirmada neste
trabalho (Anexo 20).
4.4 PRODUTIVIDADE DE GRÃOS
Na Tabela 13, são apresentados os dados de média de produtividade separados por safra e a
média geral dos genótipos em todos os ambientes avaliados, onde observa-se que a maior média de
produtividade foi obtida na safra das águas (2.971 kg.ha-1
) para a safra da seca a média foi de 2.831
kg.ha-1
e na safra de inverno foi obtida a menor média, 1.759 kg.ha-1
. Estes resultados são
semelhantes aos obtidos por PEREIRA et al (2010), que avaliando 16 genótipos de feijoeiro no
estado de Santa Catarina obtiveram a maior média de produção na safra das águas, com 2.977
kg.ha-1
, e de 2.177 kg.ha-1
na safra da seca.
Para safra das águas (Tabela 13), observou-se que para genótipos de tegumento preto,
apenas a linhagem LP 09-192 (3.455 kg.ha-1
) foi superior estatisticamente à testemunha de melhor
desempenho IAC Una (2.609 kg.ha-1
) pelo teste de Dunnet a 5% de probabilidade e considerando-se
genótipos de tegumento carioca nenhum genótipo diferiu estatisticamente da testemunha IAC
Alvorada (3.125 kg.ha-1
).
Considerando a safra da seca (Tabela 13), apenas o genótipo LP 09-40 (3.217 kg.ha-1
) de
tegumento carioca foi superior estatisticamente à testemunha de melhor desempenho IAC Alvorada
(2.753 kg.ha-1
) pelo teste de Dunnet a 5%. Entre os genótipos de tegumento preto, somente a
linhagem CNFP 10794 (3.323 kg.ha-1
) foi superior à testemunha de melhor desempenho IAC Una
(2.933 kg.ha-1
). Considerando-se a safra de inverno, nenhum genótipo obteve desempenho
estatisticamente superior às testemunhas de melhor desempenho.
Analisando a média geral dos genótipos (Tabela 13), quando considerado todos os
ambientes, nota-se que no caso dos genótipos de tegumento carioca, a testemunha de melhor
desempenho foi a cultivar IAC – Alvorada (2.543 kg.ha-1
), e que somente o genótipo LP 09-40 com
a produtividade média de 2.822 kg.ha-1
foi superior estatisticamente a esta testemunha pelo teste de
Dunnet a 5%. Para os genótipo de tegumento preto foi observado que somente o genótipo CNFP
10794 obteve produtividade média de 2.756 kg.ha-1
sendo estatisticamente superior à testemunha
IAC Una (2.477 kg.ha-1
).
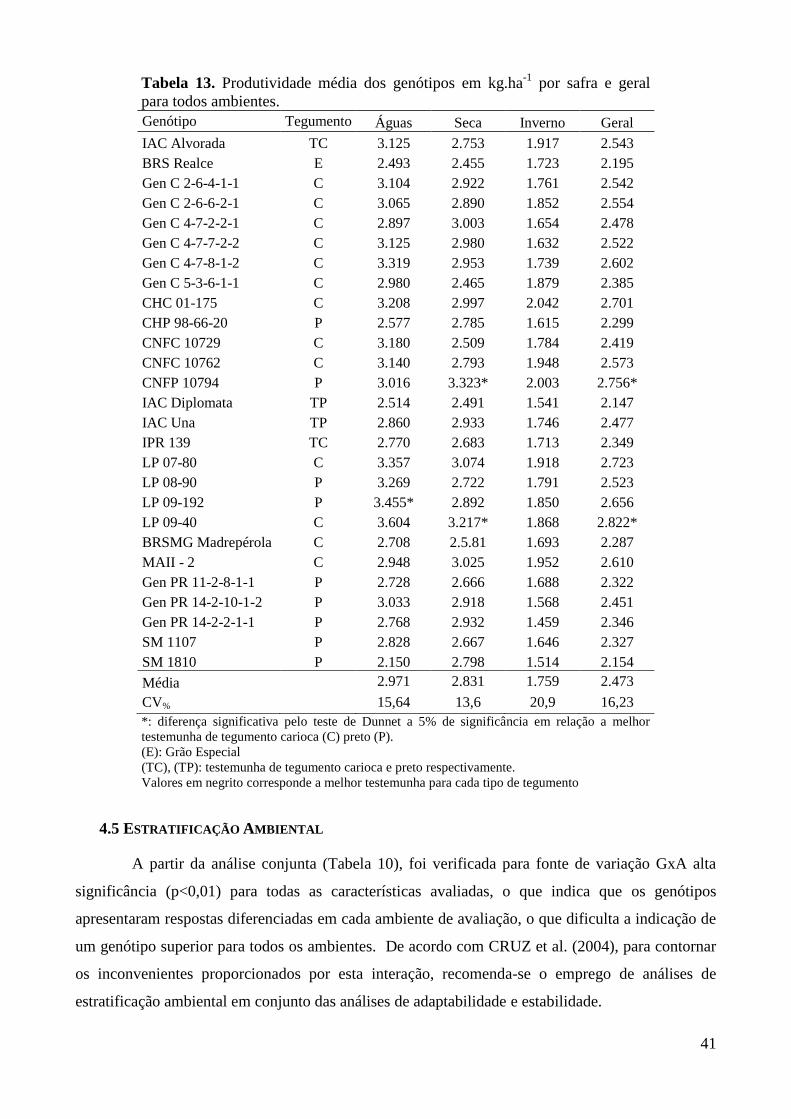
41
Tabela 13. Produtividade média dos genótipos em kg.ha-1
por safra e geral
para todos ambientes.
Genótipo Tegumento Águas Seca Inverno Geral
IAC Alvorada TC 3.125 2.753 1.917 2.543
BRS Realce E 2.493 2.455 1.723 2.195
Gen C 2-6-4-1-1 C 3.104 2.922 1.761 2.542
Gen C 2-6-6-2-1 C 3.065 2.890 1.852 2.554
Gen C 4-7-2-2-1 C 2.897 3.003 1.654 2.478
Gen C 4-7-7-2-2 C 3.125 2.980 1.632 2.522
Gen C 4-7-8-1-2 C 3.319 2.953 1.739 2.602
Gen C 5-3-6-1-1 C 2.980 2.465 1.879 2.385
CHC 01-175 C 3.208 2.997 2.042 2.701
CHP 98-66-20 P 2.577 2.785 1.615 2.299
CNFC 10729 C 3.180 2.509 1.784 2.419
CNFC 10762 C 3.140 2.793 1.948 2.573
CNFP 10794 P 3.016 3.323* 2.003 2.756*
IAC Diplomata TP 2.514 2.491 1.541 2.147
IAC Una TP 2.860 2.933 1.746 2.477
IPR 139 TC 2.770 2.683 1.713 2.349
LP 07-80 C 3.357 3.074 1.918 2.723
LP 08-90 P 3.269 2.722 1.791 2.523
LP 09-192 P 3.455* 2.892 1.850 2.656
LP 09-40 C 3.604 3.217* 1.868 2.822*
BRSMG Madrepérola C 2.708 2.5.81 1.693 2.287
MAII - 2 C 2.948 3.025 1.952 2.610
Gen PR 11-2-8-1-1 P 2.728 2.666 1.688 2.322
Gen PR 14-2-10-1-2 P 3.033 2.918 1.568 2.451
Gen PR 14-2-2-1-1 P 2.768 2.932 1.459 2.346
SM 1107 P 2.828 2.667 1.646 2.327
SM 1810 P 2.150 2.798 1.514 2.154
Média 2.971 2.831 1.759 2.473
CV% 15,64 13,6 20,9 16,23
*: diferença significativa pelo teste de Dunnet a 5% de significância em relação a melhor
testemunha de tegumento carioca (C) preto (P).
(E): Grão Especial
(TC), (TP): testemunha de tegumento carioca e preto respectivamente.
Valores em negrito corresponde a melhor testemunha para cada tipo de tegumento
4.5 ESTRATIFICAÇÃO AMBIENTAL
A partir da análise conjunta (Tabela 10), foi verificada para fonte de variação GxA alta
significância (p<0,01) para todas as características avaliadas, o que indica que os genótipos
apresentaram respostas diferenciadas em cada ambiente de avaliação, o que dificulta a indicação de
um genótipo superior para todos os ambientes. De acordo com CRUZ et al. (2004), para contornar
os inconvenientes proporcionados por esta interação, recomenda-se o emprego de análises de
estratificação ambiental em conjunto das análises de adaptabilidade e estabilidade.

42
O conceito básico da análise de estratificação ambiental se fundamenta na subdivisão de
regiões heterogêneas em grupos de sub-regiões mais uniformes, buscando reunir ambientes de
classificação genotípica similar, independentemente da qualidade.
CRUZ E CASTOLDI (1991) propuseram um método para formação de subgrupos com
pares de ambientes, onde a interação GxA apresente natureza predominantemente simples, de modo
que os genótipos apresentem comportamento coerente de um ambiente para o outro, ou que não
sofram alterações relevantes que comprometam as recomendações das cultivares.
Pelo método de CRUZ E CASTOLDI (1991), para os 175 pares combinações possíveis
entre os 19 ambientes de estudo (Tabela 14), foi observado que apenas 19 combinações
apresentaram interação GxA com predominância de parte simples (PS% > 50) para a produtividade
de grãos.
Tabela 14. Porcentagem da parte simples da interação genótipo x ambiente para a variável
produção de grãos. Pares PS(%) Pares PS(%) Pares PS(%) Pares PS(%)
CBA.11 x MOA.11 32,0
CBA.11 x TQS.12 22,09
VTI.12 x MOS.13 17,8
VTI.12 x ITS.13 42,6
CBA.11 x AVS.12 30,6
CBA.11 x MOS.12 47,82
VTI.12 x ARS.13 14,3
VTI.12 x TQS.13 18,0
CBA.11 x VTI.12 69,0
CBA.11 x RPI.12 60,87
VTI.12 x ANI.13 16,6
VTI.12 x VTI.13 10,6
CBA.11 x COI.12 26,1
CBA.11 x ANI.12 77,22
VTI.12 x RPI.13 6,3
RPI.12 x COI.12 23,5
CBA.11 x CBA.12 20,9
CBA.11 x ITA.12 36,21
RPI.12 x ANI.12 48,6
RPI.12 x CBA.12 32,6
CBA.11 x TQA.12 30,3
CBA.11 x MOS.13 23,92
RPI.12 x ITA.12 57,4
RPI.12 x TQA.12 15,9
CBA.11 x ITS.13 4,1
CBA.11 x ARS.13 41,74
RPI.12 x MOS.13 18,0
RPI.12 x ITS.13 50,1
CBA.11 x TQS.13 1,1
CBA.11 x ANI.13 45,44
RPI.12 x ARS.13 16,1
RPI.12 x TQS.13 22,7
CBA.11 x VTI.13 44,2
CBA.11 x RPI.13 7,21
RPI.12 x ANI.13 13,7
RPI.12 x VTI.13 17,2
MOA.11 x TQS.12 46,4
MOA.11 x AVS.12 21,99
RPI.12 x RPI.13 17,4
COI.12 x ANI.12 61,4
MOA.11 x MOS.12 20,6
MOA.11 x VTI.12 9,40
COI.12 x CBA.12 35,1
COI.12 x ITA.12 8,1
MOA.11 x RPI.12 9,6
MOA.11 x COI.12 20,31
COI.12 x TQA.12 16,7
COI.12 x MOS.13 2,5
MOA.11 x ANI.12 48,2
MOA.11 x CBA.12 20,42
COI.12 x ITS.13 3,2
COI.12 x ARS.13 17,4
MOA.11 x ITA.12 0,8
MOA.11 x TQA.12 6,47
COI.12 x TQS.13 9,3
COI.12 x ANI.13 31,7
MOA.11 x MOS.13 2,3
MOA.11 x ITS.13 1,27
COI.12 x VTI.13 18,9
COI.12 x RPI.13 0,4
MOA.11 x ARS.13 13,9
MOA.11 x TQS.13 15,21
ANI.12 x CBA.12 73,8
ANI.12 x ITA.12 69,4
MOA.11 x ANI.13 28,4
MOA.11 x VTI.13 30,58
ANI.12 x TQA.12 53,0
ANI.12 x MOS.13 59,7
MOA.11 x RPI.13 15,5
TQS.12 x AVS.12 24,70
ANI.12 x ITS.13 69,3
ANI.12 x ARS.13 26,2
TQS.12 x MOS.12 69,1
TQS.12 x VTI.12 36,84
ANI.12 x TQS.13 71,0
ANI.12 x ANI.13 19,5
TQS.12 x RPI.12 35,1
TQS.12 x COI.12 46,89
ANI.12 x VTI.13 48,7
ANI.12 x RPI.13 57,3
TQS.12 x ANI.12 68,2
TQS.12 x CBA.12 18,98
CBA.12 x ITA.12 13,6
CBA.12 x TQA.12 20,3
TQS.12 x ITA.12 6,8
TQS.12 x TQA.12 29,78
CBA.12 x MOS.13 44,7
CBA.12 x ITS.13 0,6
TQS.12 x MOS.13 33,1
TQS.12 x ITS.13 5,20
CBA.12 x ARS.13 41,6
CBA.12 x TQS.13 1,1
TQS.12 x ARS.13 48,0
TQS.12 x TQS.13 0,90
CBA.12 x ANI.13 56,9
CBA.12 x VTI.13 36,7
TQS.12 x ANI.13 40,9
TQS.12 x VTI.13 43,49
CBA.12 x RPI.13 12,1
ITA.12 x TQA.12 27,6
TQS.12 x RPI.13 4,2
AVS.12 x MOS.12 39,52
ITA.12 x MOS.13 12,5
ITA.12 x ITS.13 23,6
AVS.12 x VTI.12 28,0
AVS.12 x RPI.12 30,19
ITA.12 x ARS.13 30,8
ITA.12 x TQS.13 7,7
AVS.12 x COI.12 31,4
AVS.12 x ANI.12 55,51
ITA.12 x ANI.13 50,4
ITA.12 x VTI.13 10,6
AVS.12 x CBA.12 9,3
AVS.12 x ITA.12 13,89
ITA.12 x RPI.13 12,2
TQA.12 x MOS.13 10,5
AVS.12 x TQA.12 31,3
AVS.12 x MOS.13 2,44
TQA.12 x ITS.13 3,0
TQA.12 x ARS.13 18,1
AVS.12 x ITS.13 0,9
AVS.12 x ARS.13 30,01
TQA.12 x TQS.13 8,7
TQA.12 x ANI.13 27,3
AVS.12 x TQS.13 14,9
AVS.12 x ANI.13 44,32
TQA.12 x VTI.13 22,3
TQA.12 x RPI.13 8,6
AVS.12 x VTI.13 29,2
AVS.12 x RPI.13 2,54
MOS.13 x ITS.13 28,0
MOS.13 x ARS.13 23,3
MOS.12 x VTI.12 13,5
MOS.12 x RPI.12 3,34
MOS.13 x TQS.13 4,4
MOS.13 x ANI.13 13,8
MOS.12 x COI.12 35,0
MOS.12 x ANI.12 42,62
MOS.13 x VTI.13 15,1
MOS.13 x RPI.13 4,7

43
MOS.12 x CBA.12 44,3
MOS.12 x ITA.12 20,97
ITS.13 x ARS.13 35,4
ITS.13 x TQS.13 5,0
MOS.12 x TQA.12 24,4
MOS.12 x MOS.13 16,65
ITS.13 x ANI.13 39,7
ITS.13 x VTI.13 1,9
MOS.12 x ITS.13 20,9
MOS.12 x ARS.13 20,45
ITS.13 x RPI.13 15,7
ARS.13 x TQS.13 34,1
MOS.12 x TQS.13 11,7
MOS.12 x ANI.13 12,52
ARS.13 x ANI.13 3,5
ARS.13 x VTI.13 7,9
MOS.12 x VTI.13 25,8
MOS.12 x RPI.13 7,04
ARS.13 x RPI.13 49,0
TQS.13 x ANI.13 28,8
VTI.12 x RPI.12 30,8
VTI.12 x COI.12 7,42
TQS.13 x VTI.13 18,8
TQS.13 x RPI.13 2,3
VTI.12 x ANI.12 62,9
VTI.12 x CBA.12 41,23
ANI.13 x VTI.13 26,2
ANI.13 x RPI.13 30,4
VTI.12 x ITA.12 66,7 VTI.12 x TQA.12 18,80 VTI.13 x RPI.13 13,2
Valores em itálico correspondem aos pares de ambientes com % da parte simples da interação maior que 50%
Nota-se que houve um padrão entre pares de locais apenas para safra de inverno para os
locais VTI.12 x ANI.12, COI.12 x ANI.12 no mesmo ano agrícola e ANI.12 x RPI.13 em anos
diferentes na mesma safra, indicando que os genótipos não sofreram alterações relevantes em sua
classificação para esta safra, o que provavelmente se deve ao fato de que o cultivo de feijoeiro para
safra de inverno é recomendado apenas para municípios da região norte do estado de São Paulo,
desta maneira, as condições climáticas foram semelhantes entre os ambientes avaliados,
proporcionando assim, pouca alteração na classificação para produtividade de grãos dos genótipos
nesta safra.
Pela método da análise de fatores proposta por MURAKAMI E CRUZ (2004) permite-se a
formação de grupos de ambientes altamente correlacionados dentro de cada fator obtido. No
presente caso (Tabela 15), os sete autovalores acima de 1 absorveram 75,22% da variação total,
deste modo optou-se por utilizar mais 1 autovalor, fixando o número total de 8 fatores pare serem
utilizados nos agrupamentos passando assim a absorver 80,1% da variação total.
Com exceção dos ambientes RPI.12 e COI.12, as comunalidades (Tabela 15) apresentaram
valores acima de 0,70, o que indica alta eficiência do processo de fatoração para o agrupamento dos
ambientes (GARBUGLIO et al. 2007). Segundo CRUZ e CARNEIRO (2003), valores de
comunalidades superiores a 0,64 são aceitos como razoáveis, pois equivalem a uma correlação de
0,80 entre a variável padronizada (xj) e a parte comum que explica esta variável (zj).
O fator 1 (Tabela 15), permitiu o agrupamento dos ambientes TQS.12, AVS.12 e MOS.12,
que apresentaram cargas fatoriais acima de 0,70, sendo interessante, pois estes experimentos foram
realizados durante a safra da seca de 2012, levando a inferir que os genótipos tiveram
comportamentos similares durante esta safra nos locais avaliados. Desta forma, se estes resultados
se repetissem ao longo dos anos, poderia ser levado em consideração a exclusão de 2 destes locais
da rede experimental, o que representaria economia de recursos.

44
Tabela 15. Estratificação ambiental para produtividade por meio da analise de fatores, com 27
genótipos de feijoeiro avaliados em 19 ambientes no estado de São Paulo.
Autovalor %
Acumulada Ambiente
Cargas fatoriais após rotação Comunalidades
Fator 1 Fator 2 Fator 3 Fator 4 Fator 5 Fator 6 Fator 7 Fator 8
4,459 23,47 CBA.11 0,461 0,521 0,180 0,093 -0,005 0,022 -0,160 -0,179 0,833
2,782 38,11 MOA.11 0,306 0,091 -0,095 -0,132 0,038 0,169 -0,850 0,049 0,883
1,954 48,40 TQS.12 0,825 -0,064 0,001 0,136 -0,196 -0,198 -0,168 -0,129 0,826
1,569 56,65 AVS.12 0,715 0,148 0,375 -0,243 0,312 0,348 0,050 -0,078 0,826
1,327 63,64 MOS.12 0,798 0,146 0,053 0,205 -0,055 -0,001 -0,136 0,245 0,784
1,165 69,77 VTI.12 0,034 0,871 0,056 0,150 -0,039 0,032 -0,025 0,059 0,792
1,037 75,23 RPI.12 -0,059 0,769 -0,079 -0,027 0,055 0,181 -0,062 0,216 0,688
0,927 80,11 COI.12 0,660 0,003 0,157 0,116 0,220 0,191 -0,185 -0,095 0,602
0,886 84,77 ANI.12 -0,117 0,741 0,001 0,367 0,390 -0,278 -0,215 0,203 0,758
0,682 88,36 CBA.12 0,297 -0,013 0,103 0,751 -0,099 0,263 0,000 -0,206 0,784
0,604 91,54 ITA.12 0,086 0,732 0,215 0,005 -0,186 0,217 0,380 -0,319 0,917
0,461 93,97 TQA.12 0,252 0,228 0,829 0,074 0,061 0,062 -0,016 -0,009 0,816
0,369 95,91 MOS.13 0,083 0,190 -0,029 0,822 0,000 -0,068 0,034 0,159 0,750
0,292 97,45 ITS.13 -0,017 0,371 -0,533 0,137 0,171 0,158 0,557 -0,167 0,834
0,176 98,37 ARS.13 0,046 0,136 -0,032 -0,007 -0,041 -0,029 0,054 0,927 0,887
0,153 99,18 TQS.13 0,028 -0,049 0,019 -0,064 0,936 -0,011 -0,040 -0,056 0,890
0,065 99,52 ANI.13 0,028 0,211 0,042 0,096 -0,027 0,886 -0,183 0,038 0,878
0,051 99,79 VTI.13 0,289 0,163 0,265 0,259 0,144 0,162 -0,636 -0,132 0,716
0,040 100,00 RPI.13 -0,226 -0,249 0,711 0,130 -0,045 0,248 -0,325 0,548 0,757
Valores em itálico correspondem aos ambientes agrupados no fator
Entretanto, como mencionam PEREIRA et al. (2010), as variações ambientais variam ao
longo dos anos e a eliminação de um local de uma rede de ensaios por ser identificado como pouco
informativo em apenas uma safra, pode ser precipitada. Este resultado foi confirmado na safra da
seca do ano de 2013, onde embora não tenha sido realizado o experimento no município de Avaré,
os ambientes MOS.13 e TQS.13, não foram novamente alocados no mesmo grupo.
O fator 2 (Tabela 15) originou um subgrupo contendo os ambientes VTI.12, RPI.12,
ANI.12 e ITA.12, confirmando o resultado obtido por meio da metodologia de CRUZ E
CASTOLDI (1991), onde os genótipos apresentaram comportamentos similares para safra de
inverno. O fator 3 gerou um sub grupo contendo os ambientes TQA.12 e RPI.13, o fator 4 agrupou
os ambientes CBA.12 e MOS.13. Já os fatores 5, 6 e 7 contiveram apenas 1 ambiente cada, sendo o
TQS.13, ANI.13 e ARS.13 respectivamente. O fator 7 não agrupou nenhum ambiente.
Segundo CROSSA (1990), uma característica marcante dos modelos multivariados é que
eles buscam capitalizar grande parte do chamado “padrão” nos primeiros fatores principais,
relegando aos fatores subsequentes, cada vez menos “padrão” e mais “ruído”. Desta forma, quanto
mais variação for captada nos primeiros fatores, preferencialmente os dois primeiros, menos
“ruídos” associados haverá, melhorando assim a qualidade das estimativas.

45
É possível constatar que as metodologias foram eficientes em discriminar os conjuntos de
ambientes semelhantes, entretanto o método de análise de fatores proporciona uma melhor
interpretação dos resultados, agrupando um maior número de ambientes comuns, em comparação
com o método de CRUZ E CASTOLDI (1991) que apresenta a comparação apenas entre pares de
ambientes. Desta maneira, foi possível observar padrões na classificação dos genótipos avaliados na
safra de inverno e da seca, demonstrando que os genótipos sofreram maiores interações GxA entre
safras que dentro de safras, provavelmente devido a maior semelhança das condições climáticas.
Na literatura, o único trabalho de estratificação ambiental para a cultura do feijoeiro no
estado de São Paulo foi o realizado por CARBONELL E POMPEU (1997), que agruparam os
municípios de Ribeirão Preto, Votuporanga, Aguaí, Mococa e Riversul. Estes autores citaram a
importância do conhecimento da existência de ambientes contrastantes para avaliação de genótipos
quanto ao comportamento em relação aos agentes patogênicos e demais fatores estressantes. De
maneira geral, entre os ambientes deste estudo não foram detectados locais com alta correlação
entre a classificação dos genótipos para a variável produtividade de grãos, não sendo possível a
exclusão de ambientes da rede experimental.
Para coloração do tegumento (Tabela 16), de acordo com o método de CRUZ E
CASTOLDI (1991), foi possível observar que apenas 14 pares de ambientes não apresentaram
interação GxA com predominância do tipo simples (PS% > 50%), indicando que embora esta
característica sofra influência pela interação GxA, a interação é predominante do tipo simples, ou
seja, a classificação relativa dos genótipos não é alterada pelo estímulo ambiental. Desta maneira,
pode-se dizer que a seleção para cor de grão pode ser feita por meio da média de apenas um
ambiente.
Observa-se também, que a maioria das interações GxA do tipo complexa (Tabela 16)
ocorreram entre pares com o ambiente COI.12 , onde ocorreram fortes chuvas durante o período da
colheita o que proporcionou uma considerável perda na qualidade dos grãos.
Por meio da metodologia da análise de fatores (Tabela 17), os quatro autovalores acima de
1 absorveram 81,38% do total da variação, sendo assim fixado o número de 4 fatores para serem
utilizados nos agrupamentos. Com exceção dos ambientes ITS.13 e ARS.13, o restante das
comunalidades apresentaram valores acima de 0,70 indicando alta eficiência da representação das
variáveis (GARBUGLIO et al. 2007).

46
Tabela 16. Porcentagem da parte simples da interação genótipo x ambiente para a variável
coloração do tegumento dos grãos. Pares PS(%) Pares PS(%) Pares PS(%) Pares PS(%)
CBA.11 x MOA.11 93,5
CBA.11 x TQS.12 96,1
MOS.12 x VTI.12 98,5
MOS.12 x RPI.12 98,8
CBA.11 x AVS.12 74,3
CBA.11 x MOS.12 96,5
MOS.12 x COI.12 46,8
MOS.12 x ANI.12 98,4
CBA.11 x VTI.12 99,5
CBA.11 x RPI.12 99,4
MOS.12 x CBA.12 97,8
MOS.12 x ITA.12 95,0
CBA.11 x COI.12 47,4
CBA.11 x ANI.12 98,9
MOS.12 x TQA.12 94,2
MOS.12 x MOS.13 99,4
CBA.11 x CBA.12 96,2
CBA.11 x ITA.12 95,5
MOS.12 x AVS.12 91,7
MOS.12 x ARS.13 97,5
CBA.11 x TQA.12 99,0
CBA.11 x MOS.13 98,8
MOS.12 x TQS.13 90,5
MOS.12 x ANI.13 97,3
CBA.11 x AVS.12 90,6
CBA.11 x ARS.13 95,0
MOS.12 x VTI.13 96,3
MOS.12 x RPI.13 93,8
CBA.11 x TQS.13 89,3
CBA.11 x ANI.13 98,7
VTI.12 x RPI.12 96,3
VTI.12 x COI.12 45,8
CBA.11 x VTI.13 98,4
CBA.11 x RPI.13 91,8
VTI.12 x ANI.12 95,2
VTI.12 x CBA.12 99,2
MOA.11 x TQS.12 95,9
MOA.11 x AVS.12 71,7
VTI.12 x ITA.12 99,1
VTI.12 x TQA.12 99,4
MOA.11 x MOS.12 92,0
MOA.11 x VTI.12 96,8
COI.12 x CBA.12 48,0
COI.12 x ITA.12 46,8
MOA.11 x RPI.12 96,5
MOA.11 x COI.12 47,6
COI.12 x TQA.12 46,2
COI.12 x MOS.13 46,6
MOA.11 x ANI.12 97,1
MOA.11 x CBA.12 88,9
COI.12 x AVS.12 46,1
COI.12 x ARS.13 49,6
MOA.11 x ITA.12 92,6
MOA.11 x TQA.12 93,2
COI.12 x TQS.13 44,7
COI.12 x ANI.13 45,9
MOA.11 x MOS.13 95,7
MOA.11 x AVS.12 87,6
COI.12 x VTI.13 46,1
COI.12 x RPI.13 48,3
MOA.11 x ARS.13 90,7
MOA.11 x TQS.13 86,6
ANI.12 x CBA.12 98,6
ANI.12 x ITA.12 99,3
MOA.11 x ANI.13 94,7
MOA.11 x VTI.13 93,5
ANI.12 x TQA.12 98,0
ANI.12 x MOS.13 95,3
MOA.11 x RPI.13 86,1
TQS.12 x AVS.12 81,6
ANI.12 x AVS.12 97,5
ANI.12 x ARS.13 98,4
TQS.12 x MOS.12 95,1
TQS.12 x VTI.12 97,3
ANI.12 x TQS.13 95,9
ANI.12 x ANI.13 97,7
TQS.12 x RPI.12 96,7
TQS.12 x COI.12 64,2
ANI.12 x VTI.13 99,0
ANI.12 x RPI.13 97,1
TQS.12 x ANI.12 96,6
TQS.12 x CBA.12 94,4
CBA.12 x ITA.12 94,7
CBA.12 x TQA.12 98,4
TQS.12 x ITA.12 95,5
TQS.12 x TQA.12 96,6
CBA.12 x MOS.13 99,1
CBA.12 x AVS.12 91,7
TQS.12 x MOS.13 96,1
TQS.12 x AVS.12 93,9
CBA.12 x ARS.13 92,3
CBA.12 x TQS.13 90,3
TQS.12 x ARS.13 93,7
TQS.12 x TQS.13 92,9
CBA.12 x ANI.13 98,7
CBA.12 x VTI.13 98,9
TQS.12 x ANI.13 96,2
TQS.12 x VTI.13 95,8
CBA.12 x RPI.13 93,3
ITA.12 x TQA.12 95,4
TQS.12 x RPI.13 92,8
AVS.12 x MOS.12 76,6
ITA.12 x MOS.13 99,0
ITA.12 x AVS.12 92,3
AVS.12 x VTI.12 81,4
AVS.12 x RPI.12 80,5
ITA.12 x ARS.13 96,5
ITA.12 x TQS.13 90,2
AVS.12 x COI.12 43,4
AVS.12 x ANI.12 82,1
ITA.12 x ANI.13 97,8
ITA.12 x VTI.13 96,9
AVS.12 x CBA.12 74,0
AVS.12 x ITA.12 75,3
ITA.12 x RPI.13 90,3
TQA.12 x MOS.13 97,9
AVS.12 x TQA.12 76,5
AVS.12 x MOS.13 81,7
TQA.12 x AVS.12 90,8
TQA.12 x ARS.13 97,1
AVS.12 x AVS.12 74,6
AVS.12 x ARS.13 73,2
TQA.12 x TQS.13 89,4
TQA.12 x ANI.13 98,1
AVS.12 x TQS.13 73,3
AVS.12 x ANI.13 79,2
TQA.12 x VTI.13 97,2
TQA.12 x RPI.13 95,2
AVS.12 x VTI.13 78,2
AVS.12 x RPI.13 73,3
MOS.13 x AVS.12 97,3
MOS.13 x ARS.13 98,6
VTI.12 x MOS.13 94,7
VTI.12 x AVS.12 96,6
MOS.13 x TQS.13 96,3
MOS.13 x ANI.13 97,7
VTI.12 x ARS.13 99,3
VTI.12 x TQS.13 94,2
MOS.13 x VTI.13 98,1
MOS.13 x RPI.13 97,0
VTI.12 x ANI.13 97,2
VTI.12 x VTI.13 98,0
ITS.13 x ARS.13 94,5
ITS.13 x TQS.13 92,0
VTI.12 x RPI.13 97,3
RPI.12 x COI.12 46,5
ITS.13 x ANI.13 94,6
ITS.13 x VTI.13 92,8
RPI.12 x ANI.12 94,6
RPI.12 x CBA.12 99,3
ITS.13 x RPI.13 87,1
ARS.13 x TQS.13 93,0
RPI.12 x ITA.12 98,4
RPI.12 x TQA.12 98,9
ARS.13 x ANI.13 98,3
ARS.13 x VTI.13 97,2
RPI.12 x MOS.13 95,3
RPI.12 x AVS.12 96,5
ARS.13 x RPI.13 88,4
TQS.13 x ANI.13 94,0
RPI.12 x ARS.13 98,8
RPI.12 x TQS.13 95,3
TQS.13 x VTI.13 91,2
TQS.13 x RPI.13 86,0
RPI.12 x ANI.13 97,1
RPI.12 x VTI.13 97,5
ANI.13 x VTI.13 97,4
ANI.14 x RPI.13 96,2
RPI.12 x RPI.13 96,9 COI.12 x ANI.12 45,9 VTI.13 x RPI.13 97,7
Valores em itálico correspondem aos pares de ambientes com % da parte simples da interação maior que 50%
O primeiro fator (Tabela 17) agrupou 13 ambientes, sendo CBA11, MOA.11, AVS.12,
MOS.12, VTI.12, RPI.12, ANI.12, CBA.12, ITA.12, TQA.12, ANI.13, VTI.13 e RPI.13, ou seja,
74% dos ambientes foram agrupados no mesmo fatos, o que confirma os resultados obtidos pelo
método de CRUZ E CASTOLDI (1991). Isto demonstra que embora esta característica seja
considerada complexa, sendo controlada por mais de 18 genes (GEPTS E DEBOUCK, 1993), não
foram observadas diferenças na classificação dos genótipos na maioria dos ambientes avaliados.
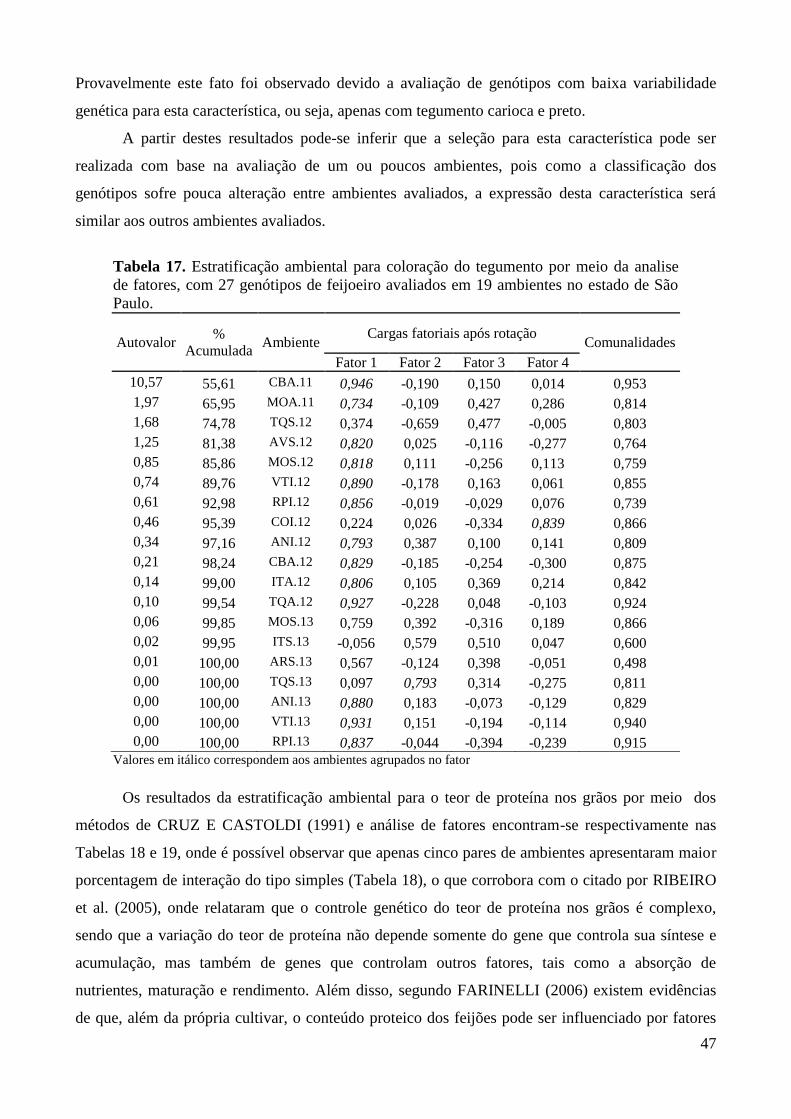
47
Provavelmente este fato foi observado devido a avaliação de genótipos com baixa variabilidade
genética para esta característica, ou seja, apenas com tegumento carioca e preto.
A partir destes resultados pode-se inferir que a seleção para esta característica pode ser
realizada com base na avaliação de um ou poucos ambientes, pois como a classificação dos
genótipos sofre pouca alteração entre ambientes avaliados, a expressão desta característica será
similar aos outros ambientes avaliados.
Tabela 17. Estratificação ambiental para coloração do tegumento por meio da analise
de fatores, com 27 genótipos de feijoeiro avaliados em 19 ambientes no estado de São
Paulo.
Autovalor %
Acumulada Ambiente
Cargas fatoriais após rotação Comunalidades
Fator 1 Fator 2 Fator 3 Fator 4
10,57 55,61 CBA.11 0,946 -0,190 0,150 0,014 0,953
1,97 65,95 MOA.11 0,734 -0,109 0,427 0,286 0,814
1,68 74,78 TQS.12 0,374 -0,659 0,477 -0,005 0,803
1,25 81,38 AVS.12 0,820 0,025 -0,116 -0,277 0,764
0,85 85,86 MOS.12 0,818 0,111 -0,256 0,113 0,759
0,74 89,76 VTI.12 0,890 -0,178 0,163 0,061 0,855
0,61 92,98 RPI.12 0,856 -0,019 -0,029 0,076 0,739
0,46 95,39 COI.12 0,224 0,026 -0,334 0,839 0,866
0,34 97,16 ANI.12 0,793 0,387 0,100 0,141 0,809
0,21 98,24 CBA.12 0,829 -0,185 -0,254 -0,300 0,875
0,14 99,00 ITA.12 0,806 0,105 0,369 0,214 0,842
0,10 99,54 TQA.12 0,927 -0,228 0,048 -0,103 0,924
0,06 99,85 MOS.13 0,759 0,392 -0,316 0,189 0,866
0,02 99,95 ITS.13 -0,056 0,579 0,510 0,047 0,600
0,01 100,00 ARS.13 0,567 -0,124 0,398 -0,051 0,498
0,00 100,00 TQS.13 0,097 0,793 0,314 -0,275 0,811
0,00 100,00 ANI.13 0,880 0,183 -0,073 -0,129 0,829
0,00 100,00 VTI.13 0,931 0,151 -0,194 -0,114 0,940
0,00 100,00 RPI.13 0,837 -0,044 -0,394 -0,239 0,915 Valores em itálico correspondem aos ambientes agrupados no fator
Os resultados da estratificação ambiental para o teor de proteína nos grãos por meio dos
métodos de CRUZ E CASTOLDI (1991) e análise de fatores encontram-se respectivamente nas
Tabelas 18 e 19, onde é possível observar que apenas cinco pares de ambientes apresentaram maior
porcentagem de interação do tipo simples (Tabela 18), o que corrobora com o citado por RIBEIRO
et al. (2005), onde relataram que o controle genético do teor de proteína nos grãos é complexo,
sendo que a variação do teor de proteína não depende somente do gene que controla sua síntese e
acumulação, mas também de genes que controlam outros fatores, tais como a absorção de
nutrientes, maturação e rendimento. Além disso, segundo FARINELLI (2006) existem evidências
de que, além da própria cultivar, o conteúdo proteico dos feijões pode ser influenciado por fatores
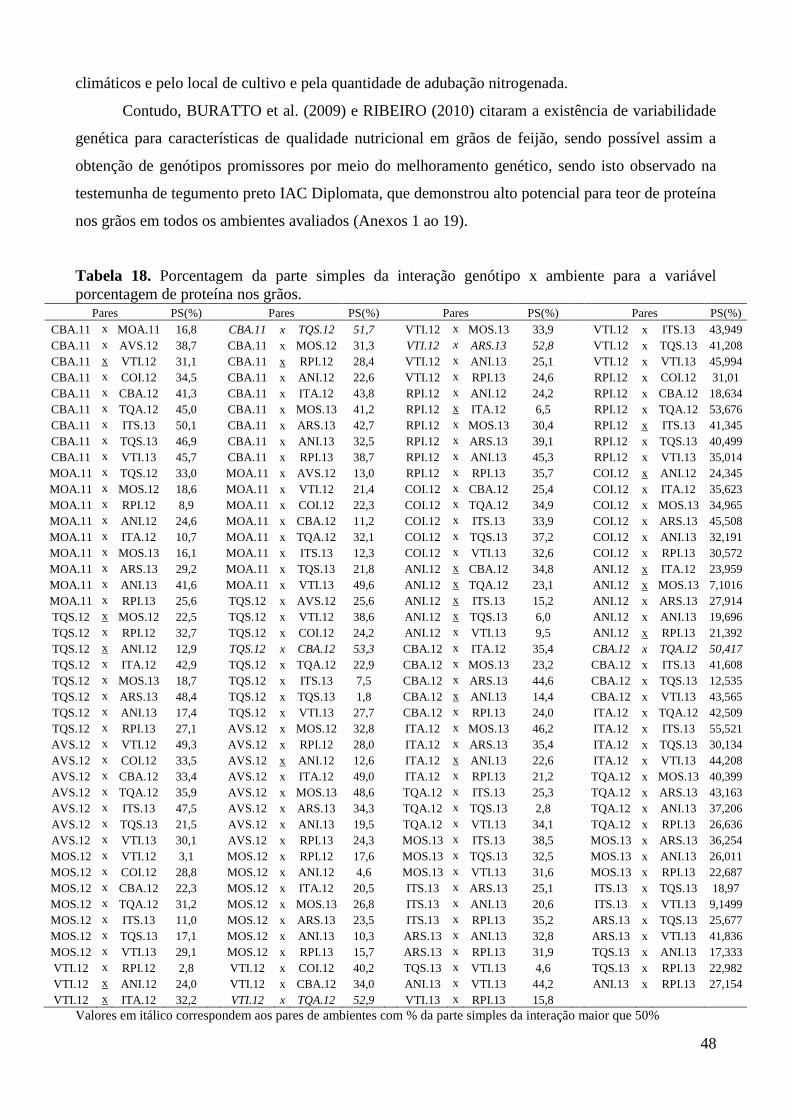
48
climáticos e pelo local de cultivo e pela quantidade de adubação nitrogenada.
Contudo, BURATTO et al. (2009) e RIBEIRO (2010) citaram a existência de variabilidade
genética para características de qualidade nutricional em grãos de feijão, sendo possível assim a
obtenção de genótipos promissores por meio do melhoramento genético, sendo isto observado na
testemunha de tegumento preto IAC Diplomata, que demonstrou alto potencial para teor de proteína
nos grãos em todos os ambientes avaliados (Anexos 1 ao 19).
Tabela 18. Porcentagem da parte simples da interação genótipo x ambiente para a variável
porcentagem de proteína nos grãos. Pares PS(%) Pares PS(%) Pares PS(%) Pares PS(%)
CBA.11 x MOA.11 16,8
CBA.11 x TQS.12 51,7
VTI.12 x MOS.13 33,9
VTI.12 x ITS.13 43,949
CBA.11 x AVS.12 38,7
CBA.11 x MOS.12 31,3
VTI.12 x ARS.13 52,8
VTI.12 x TQS.13 41,208
CBA.11 x VTI.12 31,1
CBA.11 x RPI.12 28,4
VTI.12 x ANI.13 25,1
VTI.12 x VTI.13 45,994
CBA.11 x COI.12 34,5
CBA.11 x ANI.12 22,6
VTI.12 x RPI.13 24,6
RPI.12 x COI.12 31,01
CBA.11 x CBA.12 41,3
CBA.11 x ITA.12 43,8
RPI.12 x ANI.12 24,2
RPI.12 x CBA.12 18,634
CBA.11 x TQA.12 45,0
CBA.11 x MOS.13 41,2
RPI.12 x ITA.12 6,5
RPI.12 x TQA.12 53,676
CBA.11 x ITS.13 50,1
CBA.11 x ARS.13 42,7
RPI.12 x MOS.13 30,4
RPI.12 x ITS.13 41,345
CBA.11 x TQS.13 46,9
CBA.11 x ANI.13 32,5
RPI.12 x ARS.13 39,1
RPI.12 x TQS.13 40,499
CBA.11 x VTI.13 45,7
CBA.11 x RPI.13 38,7
RPI.12 x ANI.13 45,3
RPI.12 x VTI.13 35,014
MOA.11 x TQS.12 33,0
MOA.11 x AVS.12 13,0
RPI.12 x RPI.13 35,7
COI.12 x ANI.12 24,345
MOA.11 x MOS.12 18,6
MOA.11 x VTI.12 21,4
COI.12 x CBA.12 25,4
COI.12 x ITA.12 35,623
MOA.11 x RPI.12 8,9
MOA.11 x COI.12 22,3
COI.12 x TQA.12 34,9
COI.12 x MOS.13 34,965
MOA.11 x ANI.12 24,6
MOA.11 x CBA.12 11,2
COI.12 x ITS.13 33,9
COI.12 x ARS.13 45,508
MOA.11 x ITA.12 10,7
MOA.11 x TQA.12 32,1
COI.12 x TQS.13 37,2
COI.12 x ANI.13 32,191
MOA.11 x MOS.13 16,1
MOA.11 x ITS.13 12,3
COI.12 x VTI.13 32,6
COI.12 x RPI.13 30,572
MOA.11 x ARS.13 29,2
MOA.11 x TQS.13 21,8
ANI.12 x CBA.12 34,8
ANI.12 x ITA.12 23,959
MOA.11 x ANI.13 41,6
MOA.11 x VTI.13 49,6
ANI.12 x TQA.12 23,1
ANI.12 x MOS.13 7,1016
MOA.11 x RPI.13 25,6
TQS.12 x AVS.12 25,6
ANI.12 x ITS.13 15,2
ANI.12 x ARS.13 27,914
TQS.12 x MOS.12 22,5
TQS.12 x VTI.12 38,6
ANI.12 x TQS.13 6,0
ANI.12 x ANI.13 19,696
TQS.12 x RPI.12 32,7
TQS.12 x COI.12 24,2
ANI.12 x VTI.13 9,5
ANI.12 x RPI.13 21,392
TQS.12 x ANI.12 12,9
TQS.12 x CBA.12 53,3
CBA.12 x ITA.12 35,4
CBA.12 x TQA.12 50,417
TQS.12 x ITA.12 42,9
TQS.12 x TQA.12 22,9
CBA.12 x MOS.13 23,2
CBA.12 x ITS.13 41,608
TQS.12 x MOS.13 18,7
TQS.12 x ITS.13 7,5
CBA.12 x ARS.13 44,6
CBA.12 x TQS.13 12,535
TQS.12 x ARS.13 48,4
TQS.12 x TQS.13 1,8
CBA.12 x ANI.13 14,4
CBA.12 x VTI.13 43,565
TQS.12 x ANI.13 17,4
TQS.12 x VTI.13 27,7
CBA.12 x RPI.13 24,0
ITA.12 x TQA.12 42,509
TQS.12 x RPI.13 27,1
AVS.12 x MOS.12 32,8
ITA.12 x MOS.13 46,2
ITA.12 x ITS.13 55,521
AVS.12 x VTI.12 49,3
AVS.12 x RPI.12 28,0
ITA.12 x ARS.13 35,4
ITA.12 x TQS.13 30,134
AVS.12 x COI.12 33,5
AVS.12 x ANI.12 12,6
ITA.12 x ANI.13 22,6
ITA.12 x VTI.13 44,208
AVS.12 x CBA.12 33,4
AVS.12 x ITA.12 49,0
ITA.12 x RPI.13 21,2
TQA.12 x MOS.13 40,399
AVS.12 x TQA.12 35,9
AVS.12 x MOS.13 48,6
TQA.12 x ITS.13 25,3
TQA.12 x ARS.13 43,163
AVS.12 x ITS.13 47,5
AVS.12 x ARS.13 34,3
TQA.12 x TQS.13 2,8
TQA.12 x ANI.13 37,206
AVS.12 x TQS.13 21,5
AVS.12 x ANI.13 19,5
TQA.12 x VTI.13 34,1
TQA.12 x RPI.13 26,636
AVS.12 x VTI.13 30,1
AVS.12 x RPI.13 24,3
MOS.13 x ITS.13 38,5
MOS.13 x ARS.13 36,254
MOS.12 x VTI.12 3,1
MOS.12 x RPI.12 17,6
MOS.13 x TQS.13 32,5
MOS.13 x ANI.13 26,011
MOS.12 x COI.12 28,8
MOS.12 x ANI.12 4,6
MOS.13 x VTI.13 31,6
MOS.13 x RPI.13 22,687
MOS.12 x CBA.12 22,3
MOS.12 x ITA.12 20,5
ITS.13 x ARS.13 25,1
ITS.13 x TQS.13 18,97
MOS.12 x TQA.12 31,2
MOS.12 x MOS.13 26,8
ITS.13 x ANI.13 20,6
ITS.13 x VTI.13 9,1499
MOS.12 x ITS.13 11,0
MOS.12 x ARS.13 23,5
ITS.13 x RPI.13 35,2
ARS.13 x TQS.13 25,677
MOS.12 x TQS.13 17,1
MOS.12 x ANI.13 10,3
ARS.13 x ANI.13 32,8
ARS.13 x VTI.13 41,836
MOS.12 x VTI.13 29,1
MOS.12 x RPI.13 15,7
ARS.13 x RPI.13 31,9
TQS.13 x ANI.13 17,333
VTI.12 x RPI.12 2,8
VTI.12 x COI.12 40,2
TQS.13 x VTI.13 4,6
TQS.13 x RPI.13 22,982
VTI.12 x ANI.12 24,0
VTI.12 x CBA.12 34,0
ANI.13 x VTI.13 44,2
ANI.13 x RPI.13 27,154
VTI.12 x ITA.12 32,2 VTI.12 x TQA.12 52,9 VTI.13 x RPI.13 15,8
Valores em itálico correspondem aos pares de ambientes com % da parte simples da interação maior que 50%
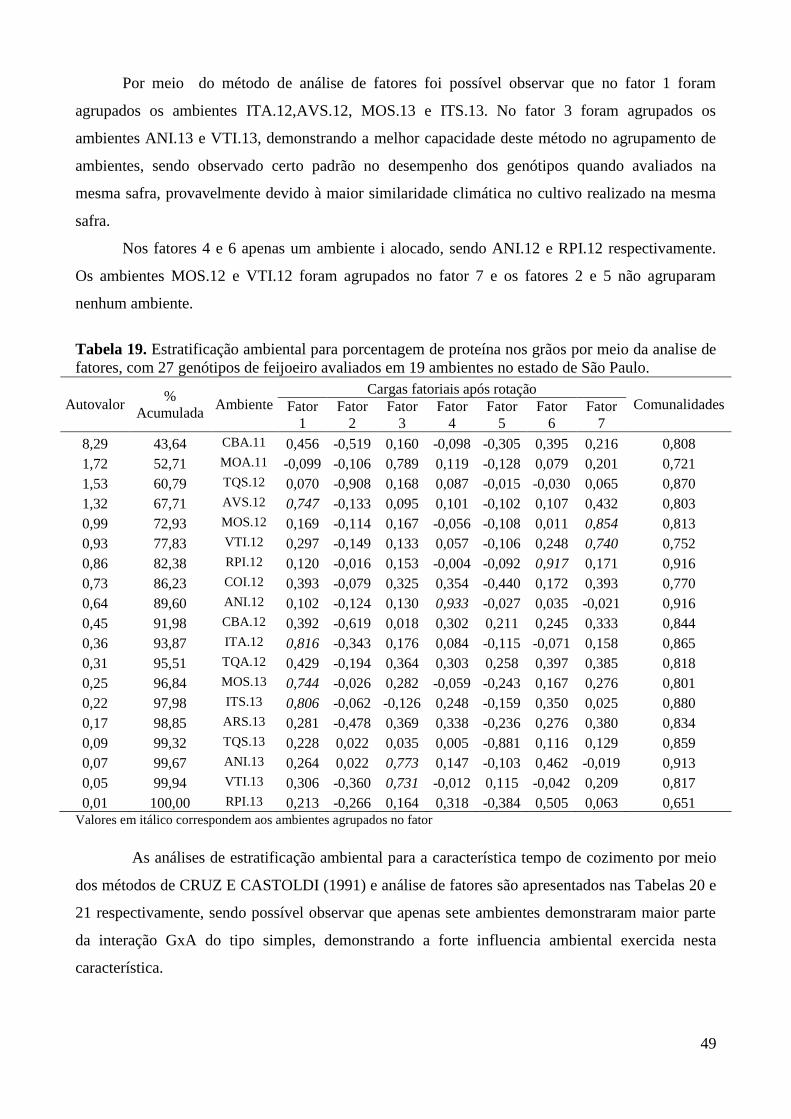
49
Por meio do método de análise de fatores foi possível observar que no fator 1 foram
agrupados os ambientes ITA.12,AVS.12, MOS.13 e ITS.13. No fator 3 foram agrupados os
ambientes ANI.13 e VTI.13, demonstrando a melhor capacidade deste método no agrupamento de
ambientes, sendo observado certo padrão no desempenho dos genótipos quando avaliados na
mesma safra, provavelmente devido à maior similaridade climática no cultivo realizado na mesma
safra.
Nos fatores 4 e 6 apenas um ambiente i alocado, sendo ANI.12 e RPI.12 respectivamente.
Os ambientes MOS.12 e VTI.12 foram agrupados no fator 7 e os fatores 2 e 5 não agruparam
nenhum ambiente.
Tabela 19. Estratificação ambiental para porcentagem de proteína nos grãos por meio da analise de
fatores, com 27 genótipos de feijoeiro avaliados em 19 ambientes no estado de São Paulo.
Autovalor %
Acumulada Ambiente
Cargas fatoriais após rotação Comunalidades Fator
1
Fator
2
Fator
3
Fator
4
Fator
5
Fator
6
Fator
7
8,29 43,64 CBA.11 0,456 -0,519 0,160 -0,098 -0,305 0,395 0,216 0,808
1,72 52,71 MOA.11 -0,099 -0,106 0,789 0,119 -0,128 0,079 0,201 0,721
1,53 60,79 TQS.12 0,070 -0,908 0,168 0,087 -0,015 -0,030 0,065 0,870
1,32 67,71 AVS.12 0,747 -0,133 0,095 0,101 -0,102 0,107 0,432 0,803
0,99 72,93 MOS.12 0,169 -0,114 0,167 -0,056 -0,108 0,011 0,854 0,813
0,93 77,83 VTI.12 0,297 -0,149 0,133 0,057 -0,106 0,248 0,740 0,752
0,86 82,38 RPI.12 0,120 -0,016 0,153 -0,004 -0,092 0,917 0,171 0,916
0,73 86,23 COI.12 0,393 -0,079 0,325 0,354 -0,440 0,172 0,393 0,770
0,64 89,60 ANI.12 0,102 -0,124 0,130 0,933 -0,027 0,035 -0,021 0,916
0,45 91,98 CBA.12 0,392 -0,619 0,018 0,302 0,211 0,245 0,333 0,844
0,36 93,87 ITA.12 0,816 -0,343 0,176 0,084 -0,115 -0,071 0,158 0,865
0,31 95,51 TQA.12 0,429 -0,194 0,364 0,303 0,258 0,397 0,385 0,818
0,25 96,84 MOS.13 0,744 -0,026 0,282 -0,059 -0,243 0,167 0,276 0,801
0,22 97,98 ITS.13 0,806 -0,062 -0,126 0,248 -0,159 0,350 0,025 0,880
0,17 98,85 ARS.13 0,281 -0,478 0,369 0,338 -0,236 0,276 0,380 0,834
0,09 99,32 TQS.13 0,228 0,022 0,035 0,005 -0,881 0,116 0,129 0,859
0,07 99,67 ANI.13 0,264 0,022 0,773 0,147 -0,103 0,462 -0,019 0,913
0,05 99,94 VTI.13 0,306 -0,360 0,731 -0,012 0,115 -0,042 0,209 0,817
0,01 100,00 RPI.13 0,213 -0,266 0,164 0,318 -0,384 0,505 0,063 0,651 Valores em itálico correspondem aos ambientes agrupados no fator
As análises de estratificação ambiental para a característica tempo de cozimento por meio
dos métodos de CRUZ E CASTOLDI (1991) e análise de fatores são apresentados nas Tabelas 20 e
21 respectivamente, sendo possível observar que apenas sete ambientes demonstraram maior parte
da interação GxA do tipo simples, demonstrando a forte influencia ambiental exercida nesta
característica.
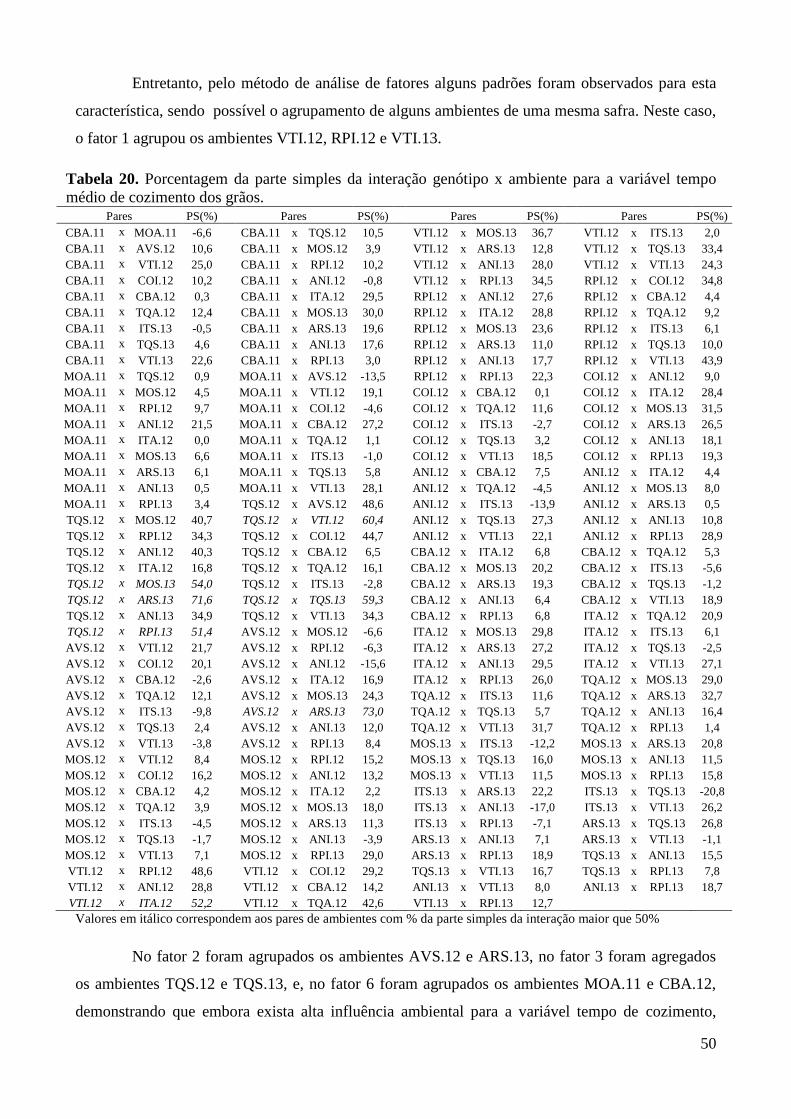
50
Entretanto, pelo método de análise de fatores alguns padrões foram observados para esta
característica, sendo possível o agrupamento de alguns ambientes de uma mesma safra. Neste caso,
o fator 1 agrupou os ambientes VTI.12, RPI.12 e VTI.13.
Tabela 20. Porcentagem da parte simples da interação genótipo x ambiente para a variável tempo
médio de cozimento dos grãos. Pares PS(%) Pares PS(%) Pares PS(%) Pares PS(%)
CBA.11 x MOA.11 -6,6
CBA.11 x TQS.12 10,5
VTI.12 x MOS.13 36,7
VTI.12 x ITS.13 2,0
CBA.11 x AVS.12 10,6
CBA.11 x MOS.12 3,9
VTI.12 x ARS.13 12,8
VTI.12 x TQS.13 33,4
CBA.11 x VTI.12 25,0
CBA.11 x RPI.12 10,2
VTI.12 x ANI.13 28,0
VTI.12 x VTI.13 24,3
CBA.11 x COI.12 10,2
CBA.11 x ANI.12 -0,8
VTI.12 x RPI.13 34,5
RPI.12 x COI.12 34,8
CBA.11 x CBA.12 0,3
CBA.11 x ITA.12 29,5
RPI.12 x ANI.12 27,6
RPI.12 x CBA.12 4,4
CBA.11 x TQA.12 12,4
CBA.11 x MOS.13 30,0
RPI.12 x ITA.12 28,8
RPI.12 x TQA.12 9,2
CBA.11 x ITS.13 -0,5
CBA.11 x ARS.13 19,6
RPI.12 x MOS.13 23,6
RPI.12 x ITS.13 6,1
CBA.11 x TQS.13 4,6
CBA.11 x ANI.13 17,6
RPI.12 x ARS.13 11,0
RPI.12 x TQS.13 10,0
CBA.11 x VTI.13 22,6
CBA.11 x RPI.13 3,0
RPI.12 x ANI.13 17,7
RPI.12 x VTI.13 43,9
MOA.11 x TQS.12 0,9
MOA.11 x AVS.12 -13,5
RPI.12 x RPI.13 22,3
COI.12 x ANI.12 9,0
MOA.11 x MOS.12 4,5
MOA.11 x VTI.12 19,1
COI.12 x CBA.12 0,1
COI.12 x ITA.12 28,4
MOA.11 x RPI.12 9,7
MOA.11 x COI.12 -4,6
COI.12 x TQA.12 11,6
COI.12 x MOS.13 31,5
MOA.11 x ANI.12 21,5
MOA.11 x CBA.12 27,2
COI.12 x ITS.13 -2,7
COI.12 x ARS.13 26,5
MOA.11 x ITA.12 0,0
MOA.11 x TQA.12 1,1
COI.12 x TQS.13 3,2
COI.12 x ANI.13 18,1
MOA.11 x MOS.13 6,6
MOA.11 x ITS.13 -1,0
COI.12 x VTI.13 18,5
COI.12 x RPI.13 19,3
MOA.11 x ARS.13 6,1
MOA.11 x TQS.13 5,8
ANI.12 x CBA.12 7,5
ANI.12 x ITA.12 4,4
MOA.11 x ANI.13 0,5
MOA.11 x VTI.13 28,1
ANI.12 x TQA.12 -4,5
ANI.12 x MOS.13 8,0
MOA.11 x RPI.13 3,4
TQS.12 x AVS.12 48,6
ANI.12 x ITS.13 -13,9
ANI.12 x ARS.13 0,5
TQS.12 x MOS.12 40,7
TQS.12 x VTI.12 60,4
ANI.12 x TQS.13 27,3
ANI.12 x ANI.13 10,8
TQS.12 x RPI.12 34,3
TQS.12 x COI.12 44,7
ANI.12 x VTI.13 22,1
ANI.12 x RPI.13 28,9
TQS.12 x ANI.12 40,3
TQS.12 x CBA.12 6,5
CBA.12 x ITA.12 6,8
CBA.12 x TQA.12 5,3
TQS.12 x ITA.12 16,8
TQS.12 x TQA.12 16,1
CBA.12 x MOS.13 20,2
CBA.12 x ITS.13 -5,6
TQS.12 x MOS.13 54,0
TQS.12 x ITS.13 -2,8
CBA.12 x ARS.13 19,3
CBA.12 x TQS.13 -1,2
TQS.12 x ARS.13 71,6
TQS.12 x TQS.13 59,3
CBA.12 x ANI.13 6,4
CBA.12 x VTI.13 18,9
TQS.12 x ANI.13 34,9
TQS.12 x VTI.13 34,3
CBA.12 x RPI.13 6,8
ITA.12 x TQA.12 20,9
TQS.12 x RPI.13 51,4
AVS.12 x MOS.12 -6,6
ITA.12 x MOS.13 29,8
ITA.12 x ITS.13 6,1
AVS.12 x VTI.12 21,7
AVS.12 x RPI.12 -6,3
ITA.12 x ARS.13 27,2
ITA.12 x TQS.13 -2,5
AVS.12 x COI.12 20,1
AVS.12 x ANI.12 -15,6
ITA.12 x ANI.13 29,5
ITA.12 x VTI.13 27,1
AVS.12 x CBA.12 -2,6
AVS.12 x ITA.12 16,9
ITA.12 x RPI.13 26,0
TQA.12 x MOS.13 29,0
AVS.12 x TQA.12 12,1
AVS.12 x MOS.13 24,3
TQA.12 x ITS.13 11,6
TQA.12 x ARS.13 32,7
AVS.12 x ITS.13 -9,8
AVS.12 x ARS.13 73,0
TQA.12 x TQS.13 5,7
TQA.12 x ANI.13 16,4
AVS.12 x TQS.13 2,4
AVS.12 x ANI.13 12,0
TQA.12 x VTI.13 31,7
TQA.12 x RPI.13 1,4
AVS.12 x VTI.13 -3,8
AVS.12 x RPI.13 8,4
MOS.13 x ITS.13 -12,2
MOS.13 x ARS.13 20,8
MOS.12 x VTI.12 8,4
MOS.12 x RPI.12 15,2
MOS.13 x TQS.13 16,0
MOS.13 x ANI.13 11,5
MOS.12 x COI.12 16,2
MOS.12 x ANI.12 13,2
MOS.13 x VTI.13 11,5
MOS.13 x RPI.13 15,8
MOS.12 x CBA.12 4,2
MOS.12 x ITA.12 2,2
ITS.13 x ARS.13 22,2
ITS.13 x TQS.13 -20,8
MOS.12 x TQA.12 3,9
MOS.12 x MOS.13 18,0
ITS.13 x ANI.13 -17,0
ITS.13 x VTI.13 26,2
MOS.12 x ITS.13 -4,5
MOS.12 x ARS.13 11,3
ITS.13 x RPI.13 -7,1
ARS.13 x TQS.13 26,8
MOS.12 x TQS.13 -1,7
MOS.12 x ANI.13 -3,9
ARS.13 x ANI.13 7,1
ARS.13 x VTI.13 -1,1
MOS.12 x VTI.13 7,1
MOS.12 x RPI.13 29,0
ARS.13 x RPI.13 18,9
TQS.13 x ANI.13 15,5
VTI.12 x RPI.12 48,6
VTI.12 x COI.12 29,2
TQS.13 x VTI.13 16,7
TQS.13 x RPI.13 7,8
VTI.12 x ANI.12 28,8
VTI.12 x CBA.12 14,2
ANI.13 x VTI.13 8,0
ANI.13 x RPI.13 18,7
VTI.12 x ITA.12 52,2 VTI.12 x TQA.12 42,6 VTI.13 x RPI.13 12,7
Valores em itálico correspondem aos pares de ambientes com % da parte simples da interação maior que 50%
No fator 2 foram agrupados os ambientes AVS.12 e ARS.13, no fator 3 foram agregados
os ambientes TQS.12 e TQS.13, e, no fator 6 foram agrupados os ambientes MOA.11 e CBA.12,
demonstrando que embora exista alta influência ambiental para a variável tempo de cozimento,
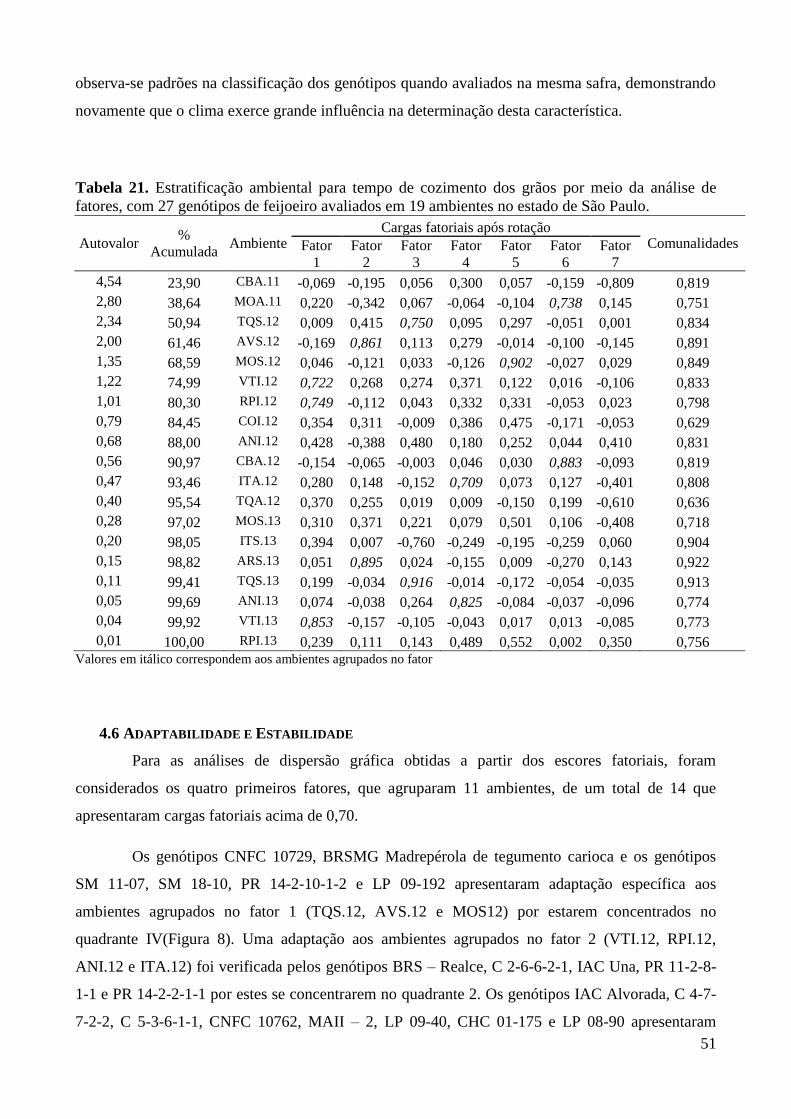
51
observa-se padrões na classificação dos genótipos quando avaliados na mesma safra, demonstrando
novamente que o clima exerce grande influência na determinação desta característica.
Tabela 21. Estratificação ambiental para tempo de cozimento dos grãos por meio da análise de
fatores, com 27 genótipos de feijoeiro avaliados em 19 ambientes no estado de São Paulo.
Autovalor %
Acumulada Ambiente
Cargas fatoriais após rotação Comunalidades Fator
1
Fator
2
Fator
3
Fator
4
Fator
5
Fator
6
Fator
7
4,54 23,90 CBA.11 -0,069 -0,195 0,056 0,300 0,057 -0,159 -0,809 0,819
2,80 38,64 MOA.11 0,220 -0,342 0,067 -0,064 -0,104 0,738 0,145 0,751
2,34 50,94 TQS.12 0,009 0,415 0,750 0,095 0,297 -0,051 0,001 0,834
2,00 61,46 AVS.12 -0,169 0,861 0,113 0,279 -0,014 -0,100 -0,145 0,891
1,35 68,59 MOS.12 0,046 -0,121 0,033 -0,126 0,902 -0,027 0,029 0,849
1,22 74,99 VTI.12 0,722 0,268 0,274 0,371 0,122 0,016 -0,106 0,833
1,01 80,30 RPI.12 0,749 -0,112 0,043 0,332 0,331 -0,053 0,023 0,798
0,79 84,45 COI.12 0,354 0,311 -0,009 0,386 0,475 -0,171 -0,053 0,629
0,68 88,00 ANI.12 0,428 -0,388 0,480 0,180 0,252 0,044 0,410 0,831
0,56 90,97 CBA.12 -0,154 -0,065 -0,003 0,046 0,030 0,883 -0,093 0,819
0,47 93,46 ITA.12 0,280 0,148 -0,152 0,709 0,073 0,127 -0,401 0,808
0,40 95,54 TQA.12 0,370 0,255 0,019 0,009 -0,150 0,199 -0,610 0,636
0,28 97,02 MOS.13 0,310 0,371 0,221 0,079 0,501 0,106 -0,408 0,718
0,20 98,05 ITS.13 0,394 0,007 -0,760 -0,249 -0,195 -0,259 0,060 0,904
0,15 98,82 ARS.13 0,051 0,895 0,024 -0,155 0,009 -0,270 0,143 0,922
0,11 99,41 TQS.13 0,199 -0,034 0,916 -0,014 -0,172 -0,054 -0,035 0,913
0,05 99,69 ANI.13 0,074 -0,038 0,264 0,825 -0,084 -0,037 -0,096 0,774
0,04 99,92 VTI.13 0,853 -0,157 -0,105 -0,043 0,017 0,013 -0,085 0,773
0,01 100,00 RPI.13 0,239 0,111 0,143 0,489 0,552 0,002 0,350 0,756 Valores em itálico correspondem aos ambientes agrupados no fator
4.6 ADAPTABILIDADE E ESTABILIDADE
Para as análises de dispersão gráfica obtidas a partir dos escores fatoriais, foram
considerados os quatro primeiros fatores, que agruparam 11 ambientes, de um total de 14 que
apresentaram cargas fatoriais acima de 0,70.
Os genótipos CNFC 10729, BRSMG Madrepérola de tegumento carioca e os genótipos
SM 11-07, SM 18-10, PR 14-2-10-1-2 e LP 09-192 apresentaram adaptação específica aos
ambientes agrupados no fator 1 (TQS.12, AVS.12 e MOS12) por estarem concentrados no
quadrante IV(Figura 8). Uma adaptação aos ambientes agrupados no fator 2 (VTI.12, RPI.12,
ANI.12 e ITA.12) foi verificada pelos genótipos BRS – Realce, C 2-6-6-2-1, IAC Una, PR 11-2-8-
1-1 e PR 14-2-2-1-1 por estes se concentrarem no quadrante 2. Os genótipos IAC Alvorada, C 4-7-
7-2-2, C 5-3-6-1-1, CNFC 10762, MAII – 2, LP 09-40, CHC 01-175 e LP 08-90 apresentaram
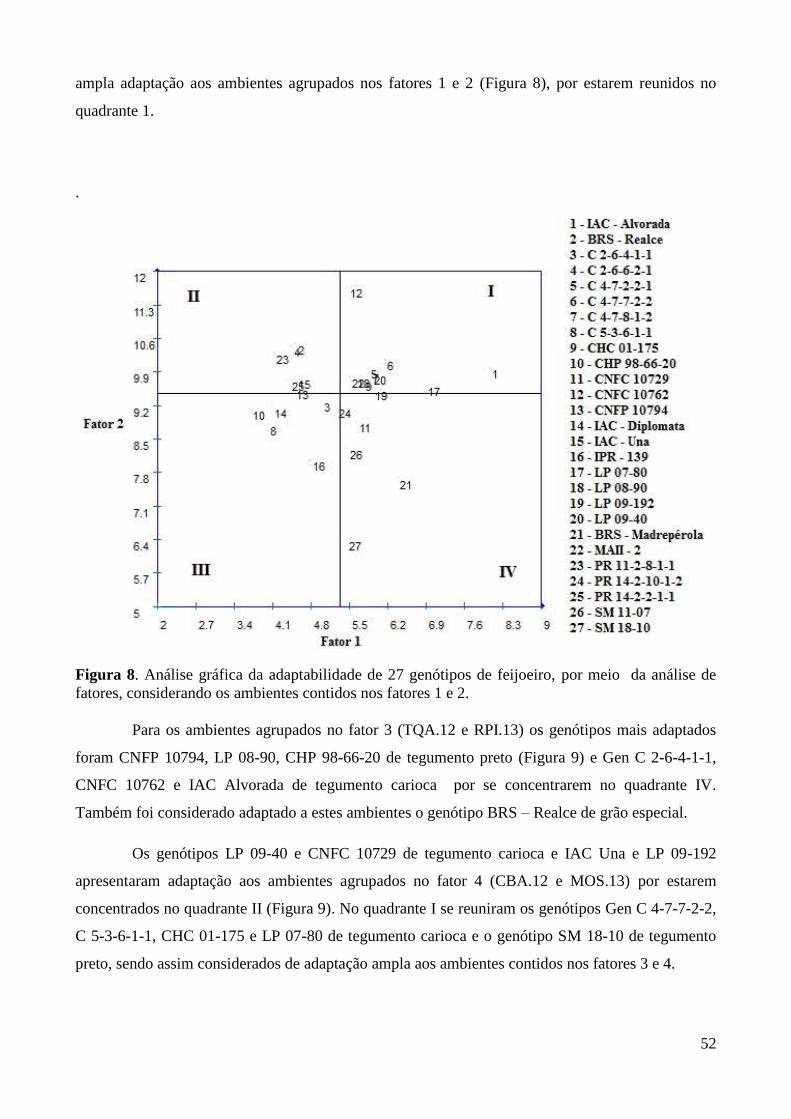
52
ampla adaptação aos ambientes agrupados nos fatores 1 e 2 (Figura 8), por estarem reunidos no
quadrante 1.
.
Figura 8. Análise gráfica da adaptabilidade de 27 genótipos de feijoeiro, por meio da análise de
fatores, considerando os ambientes contidos nos fatores 1 e 2.
Para os ambientes agrupados no fator 3 (TQA.12 e RPI.13) os genótipos mais adaptados
foram CNFP 10794, LP 08-90, CHP 98-66-20 de tegumento preto (Figura 9) e Gen C 2-6-4-1-1,
CNFC 10762 e IAC Alvorada de tegumento carioca por se concentrarem no quadrante IV.
Também foi considerado adaptado a estes ambientes o genótipo BRS – Realce de grão especial.
Os genótipos LP 09-40 e CNFC 10729 de tegumento carioca e IAC Una e LP 09-192
apresentaram adaptação aos ambientes agrupados no fator 4 (CBA.12 e MOS.13) por estarem
concentrados no quadrante II (Figura 9). No quadrante I se reuniram os genótipos Gen C 4-7-7-2-2,
C 5-3-6-1-1, CHC 01-175 e LP 07-80 de tegumento carioca e o genótipo SM 18-10 de tegumento
preto, sendo assim considerados de adaptação ampla aos ambientes contidos nos fatores 3 e 4.
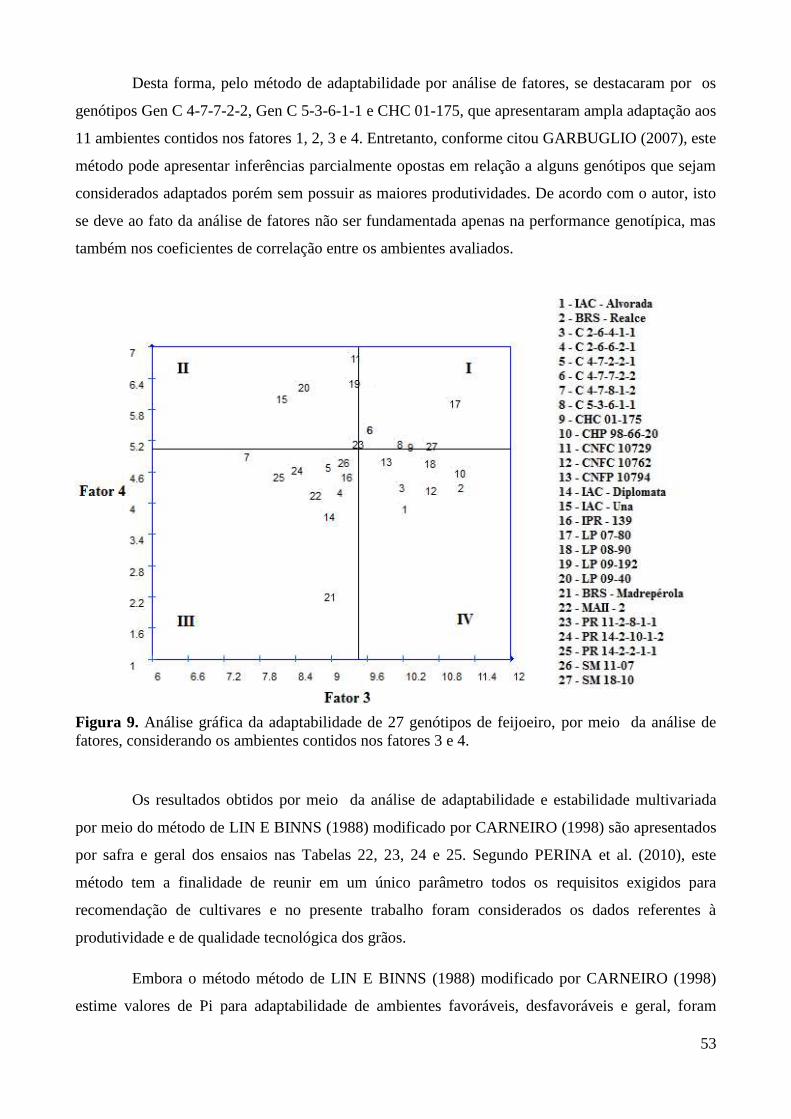
53
Desta forma, pelo método de adaptabilidade por análise de fatores, se destacaram por os
genótipos Gen C 4-7-7-2-2, Gen C 5-3-6-1-1 e CHC 01-175, que apresentaram ampla adaptação aos
11 ambientes contidos nos fatores 1, 2, 3 e 4. Entretanto, conforme citou GARBUGLIO (2007), este
método pode apresentar inferências parcialmente opostas em relação a alguns genótipos que sejam
considerados adaptados porém sem possuir as maiores produtividades. De acordo com o autor, isto
se deve ao fato da análise de fatores não ser fundamentada apenas na performance genotípica, mas
também nos coeficientes de correlação entre os ambientes avaliados.
Figura 9. Análise gráfica da adaptabilidade de 27 genótipos de feijoeiro, por meio da análise de
fatores, considerando os ambientes contidos nos fatores 3 e 4.
Os resultados obtidos por meio da análise de adaptabilidade e estabilidade multivariada
por meio do método de LIN E BINNS (1988) modificado por CARNEIRO (1998) são apresentados
por safra e geral dos ensaios nas Tabelas 22, 23, 24 e 25. Segundo PERINA et al. (2010), este
método tem a finalidade de reunir em um único parâmetro todos os requisitos exigidos para
recomendação de cultivares e no presente trabalho foram considerados os dados referentes à
produtividade e de qualidade tecnológica dos grãos.
Embora o método método de LIN E BINNS (1988) modificado por CARNEIRO (1998)
estime valores de Pi para adaptabilidade de ambientes favoráveis, desfavoráveis e geral, foram

54
considerados apenas os valores de Pi geral, pois um ambiente considerado favorável em
determinado ano, pode não sê-lo novamente em outro ano de cultivo, mesmo quando realizado na
mesma safra. Além disso, a classificação dos ambientes em favoráveis e desfavoráveis é realizada
com base no índice ambiental para cada variável analisada, não sendo possível a classificação de
ambientes em favorável e desfavorável quando utilizada a metodologia multivariada de
adaptabilidade e estabilidade.
Uma crítica que deve ser feita para este método é que neste trabalho observou-se que
ponderando diversos fatores, acaba-se penalizando o principal fator que é a produtividade de grãos
mesmo não havendo diferenças significativas entre os genótipos. Além disso, em ensaios finais de
VCU os genótipos avaliados já passaram por diversas etapas de seleção, e devido a isto, já possuem
o mínimo necessário para os padrões comerciais de qualidade tecnológica de grãos, sendo que
melhorias em outros caracteres somente se justificam quando associadas à alta produtividade de
grãos
Na Tabela 22 encontra-se as estimativas multivariadas de Pi para safra das águas, em que
para os genótipos de tegumento preto, observa-se que as menores estimativas foram obtidas pelo
genótipo LP 09-192, com uma média produtiva de 3.455 kg.ha-1
. Neste caso, de acordo com a
análise de adaptabilidade e estabilidade realizada apenas para produtividade (Anexo 21), nota-se
que este genótipo também obteve as menores estimativas de Pi entre os genótipos de tegumento
preto, demonstrando o bom potencial deste genótipo para produtividade e qualidade tecnológica dos
grãos para safra das águas, além disso este genótipo demonstrou-se resistente à antracnose e ao
crestamento bacteriano, sendo assim um possível genótipo a ser indicado como nova cultivar
Considerando os genótipos de tegumento carioca (Tabela 22) para safra das águas, a
testemunha IAC Alvorada foi considerada o genótipo de melhor desempenho com uma média
produtiva de 3.125 kg.ha-1
, apresentando a menor estimativa multivariada de Pi. Este resultado
confirma os obtidos por PERINA et al. (2010) que citaram a cultivar IAC Alvorada como genótipo
produtivo e estável, além de possuir boa qualidade tecnológica de grãos. Entretanto, considerando a
estabilidade apenas para produção de grãos (Anexo 21), observa-se que o genótipo com as menores
estimativas de Pi foi LP 09-40, com média de 3.604 kg.ha-1
. Nota-se também que não foram obtidas
diferenças significativas entre esses genótipos LP 09-40 e IAC Alvorada para os parâmetros de L* e
TC (Tabela 5), sendo observadas diferenças significativas entre estes genótipos apenas para o teor
de proteína.
Observa-se que a cultivar IAC Alvorada não apresentou resistência à nenhum dos
patógenos avaliados, sendo assim interessante a indicação de novas cultivares que apresentem

55
resistência. Neste caso destacam-se os genótipos Gen C 4-7-7-2-2 e LP 09-40 que não se diferiram
estatisticamente do genótipo IAC Alvorada, mas possuem reação de resistência à murcha de
fusarium e a antracnose do feijoeiro.
Tabela 22. Estimativas de parâmetros de estabilidade multivariada (Pi) de acordo com o método de
Lin e Binns (1988) modificada por Carneiro (1998) e média em kg.ha-1
para 27 genótipos de
feijoeiro avaliados em 5 ambientes no estado de São Paulo na safra das águas.
Gen Média Pi geral Gen Pi fav Gen Pi desf.
LP 09-192 3.455 0,254 LP 09-192 0,256 LP 09-192 0,243
IAC Alvorada 3.125 0,336 IAC Una 0,323 IAC Una 0,311
CNFP 10794 3.015 0,341 Gen PR 14-2-10-1-2 0,344 Gen PR 14-2-10-1-2 0,327
Gen PR 14-2-10-1-2 3.032 0,349 CNFP 10794 0,353 CNFP 10794 0,337
LP 07-80 3.356 0,357 SM 1107 0,362 SM 1107 0,348
LP 08-90 3.269 0,360 Gen PR 14-2-2-1-1 0,376 Gen C 2-6-4-1-1 0,353
Gen C 2-6-4-1-1 3.103 0,361 Gen C 4-7-8-1-2 0,393 Gen PR 14-2-2-1-1 0,357
Gen C 4-7-8-1-2 3.319 0,363 IAC Diplomata 0,393 Gen C 4-7-8-1-2 0,362
Gen PR 14-2-2-1-1 2.768 0,367 LP 09-40 0,405 LP 09-40 0,367
IAC Una 2.860 0,371 Gen C 2-6-4-1-1 0,412 MAII - 2 0,371
LP 09-40 3.603 0,387 IAC Alvorada 0,417 IAC Diplomata 0,375
IAC Diplomata 2.514 0,391 LP 08-90 0,417 LP 07-80 0,384
Gen C 4-7-7-2-2 3.124 0,396 Gen PR 11-2-8-1-1 0,427 IAC Alvorada 0,385
CNFC 10729 3.180 0,402 MAII - 2 0,434 Gen C 5-3-6-1-1 0,393
CNFC 10762 3.140 0,405 LP 07-80 0,436 Gen C 2-6-6-2-1 0,395
BRSMG Madrepérola 2.707 0,408 BRSMG Madrepérola 0,439 LP 08-90 0,402
Gen C 4-7-2-2-1 2.897 0,413 Gen C 5-3-6-1-1 0,443 Gen PR 11-2-8-1-1 0,415
SM 1107 2.827 0,420 Gen C 2-6-6-2-1 0,452 CNFC 10729 0,420
Gen PR 11-2-8-1-1 2.728 0,425 CHP 98-66-20 0,464 CNFC 10762 0,426
Gen C 5-3-6-1-1 2.980 0,426 CNFC 10729 0,465 BRSMG Madrepérola 0,430
Gen C 2-6-6-2-1 3.064 0,430 Gen C 4-7-7-2-2 0,478 Gen C 4-7-7-2-2 0,431
CHP 98-66-20 2.577 0,444 IPR 139 0,500 CHC 01-175 0,433
MAII – 2 2.947 0,447 CNFC 10762 0,502 Gen C 4-7-2-2-1 0,434
CHC 01-175 3.207 0,449 Gen C 4-7-2-2-1 0,509 CHP 98-66-20 0,444
IPR 139 2.770 0,465 BRS Realce 0,559 IPR 139 0,456
BRS Realce 2.493 0,522 SM 1810 0,565 BRS Realce 0,479
SM 1810 2.149 0,539 CHC 01-175 0,567 SM 1810 0,545
Na Tabela 23, estão dispostas as estimativas multivariadas de Pi obtidas para safra da seca,
onde observa-se que para os genótipos de tegumento preto, novamente LP 09-192 com uma média
de 2.892 kg.ha-1
obteve as menores estimativas de Pi. Entretanto, considerando a estabilidade da
produção de grãos (Anexo 22), observa-se que o genótipo superior foi CNFP 10794 que produziu
3.323 kg.ha-1
e que este genótipo, diferentemente do LP 09-192 se diferenciou estatisticamente da
testemunha IAC – Una (2.933 kg.ha-1
) pelo teste de Dunnet a 5% de probabilidade (Tabela 6). Para
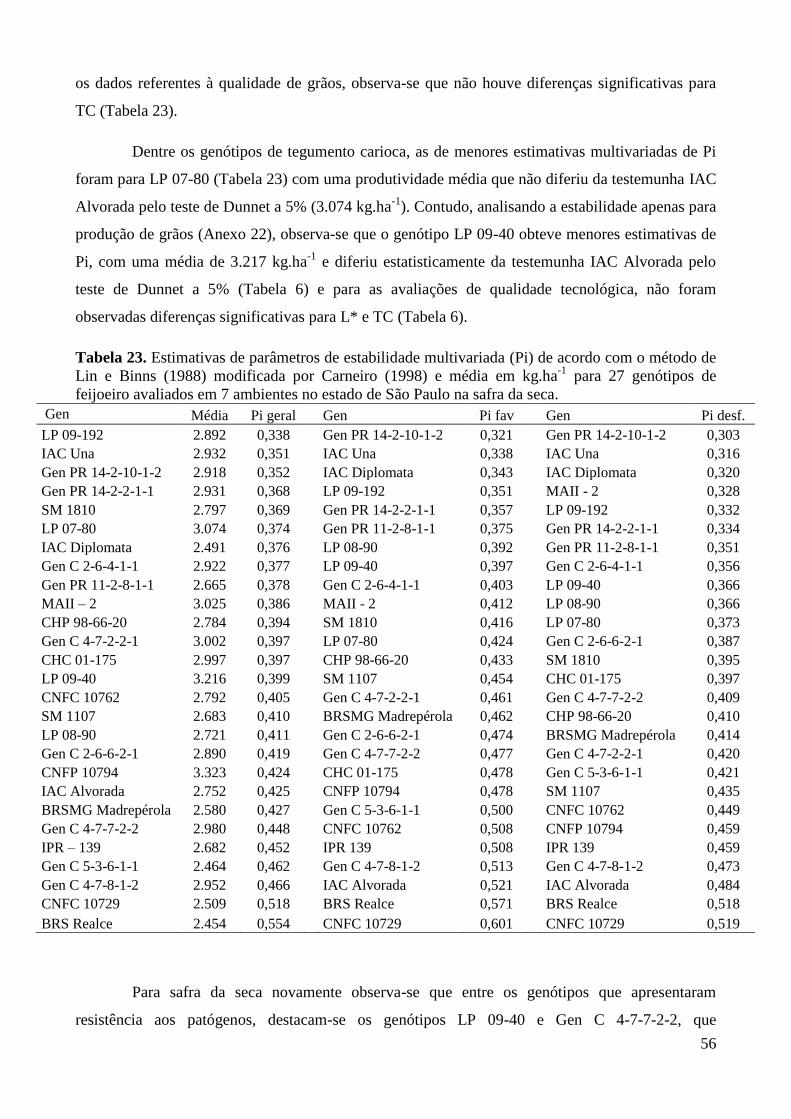
56
os dados referentes à qualidade de grãos, observa-se que não houve diferenças significativas para
TC (Tabela 23).
Dentre os genótipos de tegumento carioca, as de menores estimativas multivariadas de Pi
foram para LP 07-80 (Tabela 23) com uma produtividade média que não diferiu da testemunha IAC
Alvorada pelo teste de Dunnet a 5% (3.074 kg.ha-1
). Contudo, analisando a estabilidade apenas para
produção de grãos (Anexo 22), observa-se que o genótipo LP 09-40 obteve menores estimativas de
Pi, com uma média de 3.217 kg.ha-1
e diferiu estatisticamente da testemunha IAC Alvorada pelo
teste de Dunnet a 5% (Tabela 6) e para as avaliações de qualidade tecnológica, não foram
observadas diferenças significativas para L* e TC (Tabela 6).
Tabela 23. Estimativas de parâmetros de estabilidade multivariada (Pi) de acordo com o método de
Lin e Binns (1988) modificada por Carneiro (1998) e média em kg.ha-1
para 27 genótipos de
feijoeiro avaliados em 7 ambientes no estado de São Paulo na safra da seca.
Gen Média Pi geral Gen Pi fav Gen Pi desf.
LP 09-192 2.892 0,338 Gen PR 14-2-10-1-2 0,321 Gen PR 14-2-10-1-2 0,303
IAC Una 2.932 0,351 IAC Una 0,338 IAC Una 0,316
Gen PR 14-2-10-1-2 2.918 0,352 IAC Diplomata 0,343 IAC Diplomata 0,320
Gen PR 14-2-2-1-1 2.931 0,368 LP 09-192 0,351 MAII - 2 0,328
SM 1810 2.797 0,369 Gen PR 14-2-2-1-1 0,357 LP 09-192 0,332
LP 07-80 3.074 0,374 Gen PR 11-2-8-1-1 0,375 Gen PR 14-2-2-1-1 0,334
IAC Diplomata 2.491 0,376 LP 08-90 0,392 Gen PR 11-2-8-1-1 0,351
Gen C 2-6-4-1-1 2.922 0,377 LP 09-40 0,397 Gen C 2-6-4-1-1 0,356
Gen PR 11-2-8-1-1 2.665 0,378 Gen C 2-6-4-1-1 0,403 LP 09-40 0,366
MAII – 2 3.025 0,386 MAII - 2 0,412 LP 08-90 0,366
CHP 98-66-20 2.784 0,394 SM 1810 0,416 LP 07-80 0,373
Gen C 4-7-2-2-1 3.002 0,397 LP 07-80 0,424 Gen C 2-6-6-2-1 0,387
CHC 01-175 2.997 0,397 CHP 98-66-20 0,433 SM 1810 0,395
LP 09-40 3.216 0,399 SM 1107 0,454 CHC 01-175 0,397
CNFC 10762 2.792 0,405 Gen C 4-7-2-2-1 0,461 Gen C 4-7-7-2-2 0,409
SM 1107 2.683 0,410 BRSMG Madrepérola 0,462 CHP 98-66-20 0,410
LP 08-90 2.721 0,411 Gen C 2-6-6-2-1 0,474 BRSMG Madrepérola 0,414
Gen C 2-6-6-2-1 2.890 0,419 Gen C 4-7-7-2-2 0,477 Gen C 4-7-2-2-1 0,420
CNFP 10794 3.323 0,424 CHC 01-175 0,478 Gen C 5-3-6-1-1 0,421
IAC Alvorada 2.752 0,425 CNFP 10794 0,478 SM 1107 0,435
BRSMG Madrepérola 2.580 0,427 Gen C 5-3-6-1-1 0,500 CNFC 10762 0,449
Gen C 4-7-7-2-2 2.980 0,448 CNFC 10762 0,508 CNFP 10794 0,459
IPR – 139 2.682 0,452 IPR 139 0,508 IPR 139 0,459
Gen C 5-3-6-1-1 2.464 0,462 Gen C 4-7-8-1-2 0,513 Gen C 4-7-8-1-2 0,473
Gen C 4-7-8-1-2 2.952 0,466 IAC Alvorada 0,521 IAC Alvorada 0,484
CNFC 10729 2.509 0,518 BRS Realce 0,571 BRS Realce 0,518
BRS Realce 2.454 0,554 CNFC 10729 0,601 CNFC 10729 0,519
Para safra da seca novamente observa-se que entre os genótipos que apresentaram
resistência aos patógenos, destacam-se os genótipos LP 09-40 e Gen C 4-7-7-2-2, que
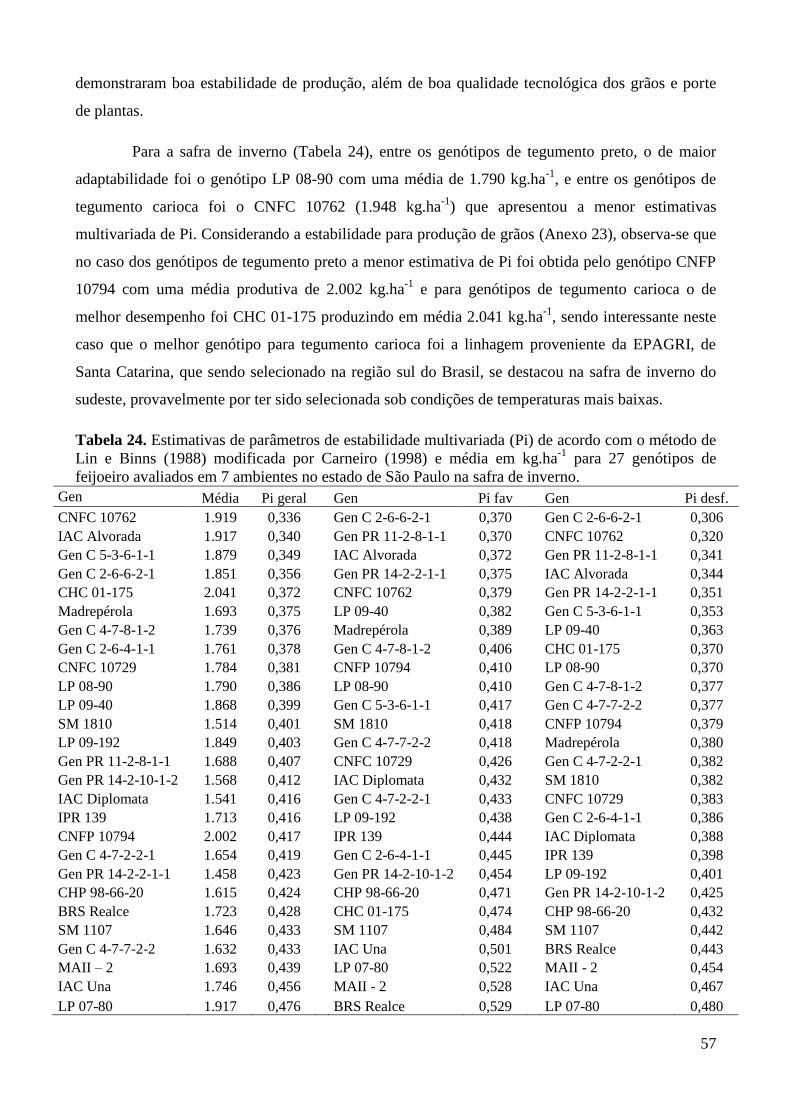
57
demonstraram boa estabilidade de produção, além de boa qualidade tecnológica dos grãos e porte
de plantas.
Para a safra de inverno (Tabela 24), entre os genótipos de tegumento preto, o de maior
adaptabilidade foi o genótipo LP 08-90 com uma média de 1.790 kg.ha-1
, e entre os genótipos de
tegumento carioca foi o CNFC 10762 (1.948 kg.ha-1
) que apresentou a menor estimativas
multivariada de Pi. Considerando a estabilidade para produção de grãos (Anexo 23), observa-se que
no caso dos genótipos de tegumento preto a menor estimativa de Pi foi obtida pelo genótipo CNFP
10794 com uma média produtiva de 2.002 kg.ha-1
e para genótipos de tegumento carioca o de
melhor desempenho foi CHC 01-175 produzindo em média 2.041 kg.ha-1
, sendo interessante neste
caso que o melhor genótipo para tegumento carioca foi a linhagem proveniente da EPAGRI, de
Santa Catarina, que sendo selecionado na região sul do Brasil, se destacou na safra de inverno do
sudeste, provavelmente por ter sido selecionada sob condições de temperaturas mais baixas.
Tabela 24. Estimativas de parâmetros de estabilidade multivariada (Pi) de acordo com o método de
Lin e Binns (1988) modificada por Carneiro (1998) e média em kg.ha-1
para 27 genótipos de
feijoeiro avaliados em 7 ambientes no estado de São Paulo na safra de inverno.
Gen Média Pi geral Gen Pi fav Gen Pi desf.
CNFC 10762 1.919 0,336 Gen C 2-6-6-2-1 0,370 Gen C 2-6-6-2-1 0,306
IAC Alvorada 1.917 0,340 Gen PR 11-2-8-1-1 0,370 CNFC 10762 0,320
Gen C 5-3-6-1-1 1.879 0,349 IAC Alvorada 0,372 Gen PR 11-2-8-1-1 0,341
Gen C 2-6-6-2-1 1.851 0,356 Gen PR 14-2-2-1-1 0,375 IAC Alvorada 0,344
CHC 01-175 2.041 0,372 CNFC 10762 0,379 Gen PR 14-2-2-1-1 0,351
Madrepérola 1.693 0,375 LP 09-40 0,382 Gen C 5-3-6-1-1 0,353
Gen C 4-7-8-1-2 1.739 0,376 Madrepérola 0,389 LP 09-40 0,363
Gen C 2-6-4-1-1 1.761 0,378 Gen C 4-7-8-1-2 0,406 CHC 01-175 0,370
CNFC 10729 1.784 0,381 CNFP 10794 0,410 LP 08-90 0,370
LP 08-90 1.790 0,386 LP 08-90 0,410 Gen C 4-7-8-1-2 0,377
LP 09-40 1.868 0,399 Gen C 5-3-6-1-1 0,417 Gen C 4-7-7-2-2 0,377
SM 1810 1.514 0,401 SM 1810 0,418 CNFP 10794 0,379
LP 09-192 1.849 0,403 Gen C 4-7-7-2-2 0,418 Madrepérola 0,380
Gen PR 11-2-8-1-1 1.688 0,407 CNFC 10729 0,426 Gen C 4-7-2-2-1 0,382
Gen PR 14-2-10-1-2 1.568 0,412 IAC Diplomata 0,432 SM 1810 0,382
IAC Diplomata 1.541 0,416 Gen C 4-7-2-2-1 0,433 CNFC 10729 0,383
IPR 139 1.713 0,416 LP 09-192 0,438 Gen C 2-6-4-1-1 0,386
CNFP 10794 2.002 0,417 IPR 139 0,444 IAC Diplomata 0,388
Gen C 4-7-2-2-1 1.654 0,419 Gen C 2-6-4-1-1 0,445 IPR 139 0,398
Gen PR 14-2-2-1-1 1.458 0,423 Gen PR 14-2-10-1-2 0,454 LP 09-192 0,401
CHP 98-66-20 1.615 0,424 CHP 98-66-20 0,471 Gen PR 14-2-10-1-2 0,425
BRS Realce 1.723 0,428 CHC 01-175 0,474 CHP 98-66-20 0,432
SM 1107 1.646 0,433 SM 1107 0,484 SM 1107 0,442
Gen C 4-7-7-2-2 1.632 0,433 IAC Una 0,501 BRS Realce 0,443
MAII – 2 1.693 0,439 LP 07-80 0,522 MAII - 2 0,454
IAC Una 1.746 0,456 MAII - 2 0,528 IAC Una 0,467
LP 07-80 1.917 0,476 BRS Realce 0,529 LP 07-80 0,480

58
Na Tabela 25, encontram-se as estimativas multivariadas de Pi obtidas considerando todas
as safras, onde para os genótipos de tegumento preto destacou-se o LP 09-192 que produziu em
média 2.656 kg.ha-1
e não diferiu estatisticamente da testemunha pelo teste de Dunnet a 5% (Tabela
8). Analisando as estimativas de Pi para produtividade (Anexo 24) observa-se que o melhor
genótipo neste caso foi CNFP 10794 (2.755 kg.ha-1
) que diferiu estatisticamente da testemunha IAC
Una com produtividade média de 2.477 kg.ha-1
(Tabela 8). Para as avaliações da qualidade
tecnológica dos grãos, não foram detectadas diferenças significativas entre estes genótipos para L*
e TC.
Tabela 25. Estimativas de parâmetros de estabilidade multivariada de acordo com o método de Lin e
Binns (1988) modificada por Carneiro (1998) para 27 genótipos de feijoeiro avaliados em 7 ambientes
no estado de São Paulo considerando todas safras.
Gen Média Pi geral Gen Pi fav Gen Pi desf.
LP 09-192 2.656 0,371
Gen PR 14-2-10-1-2 0,383
Gen PR 14-2-10-1-2 0,356
Gen PR 14-2-10-1-2 2.451 0,379
Gen PR 14-2-2-1-1 0,393
Gen PR 14-2-2-1-1 0,366
Gen C 2-6-6-2-1 2.553 0,388
IAC Diplomata 0,405
IAC Diplomata 0,369
IAC Alvorada 2.542 0,392
Gen PR 11-2-8-1-1 0,411
Gen PR 11-2-8-1-1 0,383
Gen PR 14-2-2-1-1 2.345 0,393
LP 09-192 0,417
Gen C 2-6-6-2-1 0,383
CNFC 10762 2.573 0,398
LP 08-90 0,425
LP 09-192 0,386
LP 08-90 2.522 0,402
LP 09-40 0,431
LP 08-90 0,391
IAC Diplomata 2.147 0,403
IAC Una 0,439
Gen C 4-7-2-2-1 0,391
CNFP 10794 2.755 0,409
CNFP 10794 0,439
LP 09-40 0,400
BRSMG Madrepérola 2.287 0,411
BRSMG Madrepérola 0,440
CNFC 10762 0,407
LP 09-40 2.821 0,412
Gen C 2-6-6-2-1 0,442
IAC Una 0,409
Gen C 4-7-2-2-1 2.478 0,415
IAC Alvorada 0,451
CHC 01-175 0,410
IAC Una 2.476 0,415
SM 1107 0,451
CNFP 10794 0,411
Gen PR 11-2-8-1-1 2.322 0,415
Gen C 5-3-6-1-1 0,456
BRSMG Madrepérola 0,412
Gen C 5-3-6-1-1 2.384 0,416
Gen C 4-7-2-2-1 0,462
IAC Alvorada 0,415
Gen C 4-7-7-2-2 2.521 0,419
SM 1810 0,464
Gen C 4-7-7-2-2 0,417
SM 1810 2.154 0,422
CHP 98-66-20 0,467
BRS Realce 0,417
BRS Realce 2.195 0,425
CNFC 10762 0,469
Gen C 5-3-6-1-1 0,417
CHP 98-66-20 2.299 0,426
Gen C 4-7-7-2-2 0,470
SM 1107 0,419
CHC 01-175 2.700 0,426
CHC 01-175 0,477
Gen C 4-7-8-1-2 0,426
SM 1107 2.339 0,429
IPR 139 0,479
IPR 139 0,430
LP 07-80 2.722 0,431
Gen C 4-7-8-1-2 0,481
SM 1810 0,432
Gen C 4-7-8-1-2 2.602 0,440
LP 07-80 0,501
CHP 98-66-20 0,434
IPR 139 2.348 0,442
CNFC 10729 0,510
MAII - 2 0,443
MAII - 2 2.609 0,448
MAII - 2 0,512
CNFC 10729 0,451
CNFC 10729 2.418 0,453
BRS Realce 0,512
LP 07-80 0,451
Gen C 2-6-4-1-1 2.542 0,508 Gen C 2-6-4-1-1 0,567 Gen C 2-6-4-1-1 0,496
Para os genótipos de tegumento carioca, observou-se que o genótipo Gen C 2-6-6-2-1
produziu em média 2.554 kg.ha-1
e obteve as menores estimativas multivariadas de Pi (Tabela 25),
entretanto, considerando a estabilidade para produção de grãos (Anexo 24), observa-se que o
genótipo de melhor desempenho foi LP 09-40 com uma produtividade média de 2.755 kg.ha-1
,

59
sendo o único a diferir estatisticamente da testemunha comercial IAC – Alvorada (2.543 kg.ha-1
)
pelo teste de Dunnet a 5% de probabilidade (Tabela 8). Contudo, não foram observadas diferenças
significativas para L* e TC.
Como destacado anteriormente, as metodologias de adaptabilidade e estabilidade não
consideram a resistência dos genótipos aos patógenos da cultura, sendo isto de grande importância
no lançamento de uma nova cultivar. Neste sentido, o melhorista deve considerar estas análises
como apoio na indicação do melhor genótipo em conjunto com outras análises realizadas. Dentre
os genótipos avaliados neste estudo, os que se destacaram por apresentar alta estabilidade produtiva,
aliado à boa qualidade tecnológica de grãos, arquitetura de planta e resistência aos patógenos foram
os genótipos LP 09-40 e Gen C 4-7-7-2-2 de tegumento carioca e o genótipo LP 09-192 de
tegumento preto, sendo desta forma candidatos a indicação como novas cultivares.
De acordo com os resultados acima, ressalta-se que não houve coerência entre os
resultados obtidos por meio da análise de estabilidade multivariada de LIN E BINNS (1988)
modificada por CARNEIRO (1998) e o desejado de um genótipo para ser recomendado como nova
cultivar, sendo que todos os genótipos avaliados possuem características mínimas requeridas para o
consumo, neste sentido, observa-se que a análise de estabilidade considerando apenas os parâmetros
de produtividade foi mais adequada na indicação do genótipo superior.
Entretanto, análises multivariadas podem ser de grande utilidade nas etapas iniciais de
seleção em programas de melhoramento genético, onde a utilização de índices de seleção, como por
exemplo o índice clássico de SMITH HAZEL (1943) e o índice de soma dos ranks (MULAMBA E
MOCK, 1978), podem sem úteis na seleção de genótipos que reúnam diversas características
desejáveis em populações segregantes, como por exemplo a qualidade tecnológica de grãos com a
produvidade, tal como relatado por MENDES et al. (2009), NEVES et al. (2011) e SANTOS et al.
(2007).
Comparando os métodos de adaptabilidade e estabilidade por análises de fatores e LIN E
BINNS (1988), modificado por CARNEIRO (1998), observa-se que estes métodos podem ser
complementares, pois com a análise gráfica obtida por meio da análise de fatores é possível
identificar os genótipos adaptados a regiões específicas ou amplamente adaptados, porém sem
quantificar a adaptabilidade, pois método pondera os genótipos que menos variaram entre
ambientes, fornecendo um indicativo dos genótipos mais estáveis. Já método de LINN E BINNS
(1988), modificado por CARNEIRO (1998), indica os genótipos que menos variaram em relação ao
melhor genótipo de cada ambiente, indicando genótipos mais adaptados e produtivos.

60
Desta forma, os genótipos considerados mais estáveis e adaptados aos ambientes avaliados
foram LP 09-40, Gen C 4-7-7-2-2 e CNFC 10762 para genótipos de tegumento carioca e CNFC
10794, LP 09-192 e LP 08-90 para genótipos de tegumento preto.
5. CONCLUSÕES
1) As análises de estratificação ambiental demonstraram a alta influência da interação GxA do
tipo complexa para produtividade de grãos, proteína e tempo de cozimento. Para coloração a
maior parte da interação GxA foi do tipo simples.
2) Os genótipos de tegumento preto mais produtivos e adaptados foram CNFP 10794, LP 09-192
com resistência ao crestamento bacteriano e o genótipo LP 08-90 com resistência à murcha de
fusarium e ao crestamento bacteriano.
3) Os genótipos de tegumento carioca superiores foram LP 09-40, com resistência à antracnose e
à murcha de fusarium, o genótipo Gen C 4-7-7-2-2, por apresentar adaptabilidade ampla e
resistência à murcha de fusarium e o genótipo CNFC 10762, com resistência à antracnose.
4) O método de LIN e BINNS modificado por CARNEIRO (1998) foi mais eficiente quando
utilizado apenas com a variável produtividade de grãos.
5) O método da análise de fatores é uma ferramenta eficiente para estudos de estratificação
ambiental e adaptabilidade fenotípica, podendo ser utilizado em conjunto com o método de
LINN e BINNS modificado por CARNEIRO (1998).

61
6. REFERÊNCIAS
ABREU, A. F. B.; CARNEIRO, J. E. S.; RAMALHO, M. A. P.; MELO, L. C.; PAULA JÚNIOR,
T. J.; PEREIRA FILHO, I. A.; MARTINS, M.; PEREIRA, H. S.; CARNEIRO, P. C. S.; DEL
GEÚDICE, M. P.; VIEIRA, R. F.; TEIXEIRA, H.; DEL PELOSO, M. J.; FARIA, L. C.; SANTOS,
J. B.; COSTA, J. G. C.; MOREIRA, J. A. A.; WENDLAND, A. BRSMG Madrepérola: cultivar de
feijão tipo carioca com escurecimento tardio dos grãos. Santo Antônio de Goiás: Embrapa, 2011.
ABREU, A. F. B.; RAMALHO, M. A. P.; GONÇALVES, F. M. A.; MENDONÇA, H. A.
Utilização da produtividade de grãos na seleção para resistência ao Colletotrichum lindemuthianum
no feijoeiro. Ciência e Agrotecnologia, p. 363-369, 2003.
ACCIARESI, H. A.; CHIDICHIMO, H. O. Genotype-environment interaction in Avena sativa L-
employing AMMI and factorial correspondence models. Pesquisa Agropecuária Brasileira, p. 1823-
1830, 1999.
ANNICCHIARICO, P. Cultivar adaptation and recommendation from alfafa trials in Northern
Italy. Journal of Genetics and Breeding, Rome, p. 269-278, 1992.
ARAÚJO, L. C. A.; RAMALHO, M. A. P.; ABREU, A. F. B. Estimates of genetic parameters of
late seed-coat darkening of carioca type dry beans. Ciência e Agrotecnologia, p. 156-162, 2012.
ASSOCIATION OF OFFICIAL AGRICULTURAL CHEMISTS - AOAC. Official methods of
analysis of AOAC International. 18. ed. Gaithersburg: AOAC International, p.32-33, 2005.
ARIOTTI, J. A.; OYARZABAL, E. S.; OSA, J. M.; BULACIO, A. N. R.; ALMADA, G. H.
Análisis de estabilidad y adaptabilidad de genótipos de caña-de-azúcar. I: interacciones dentro de
una localidad experimental. Revista Agronômica do Noroeste Argentino, p.105-127, 1976.
BALARDIN R. S.; KELLY J. D. Interaction between Colletotrichum lindemuthuanum races and
gene pool diversity in Phaseolus vulgaris. Journal of the American Society for Horticultural
Science. p. 1038-1047, 1998.
BALARDIN, R. S; PASTOR-CORRALES, M. A. e OTOYA, M. M. Variabilidade patogênica de
colletotrichum lindemuthianum no Estado de Santa Catarina. Fitopatologia Brasileira, p.243-245,
1990.
BANZATTO, D. A.; KRONKA, S. N. Experimentação agrícola. 3.ed. - FUNEP, p. 247, 1995.
BARROS, H. B.; SEDIYAMA, T.; TEIXEIRA, R. de C.; CRUZ, C.D. Análises paramétricas e não-
paramétricas para determinação da adaptabilidade e estabilidade de genótipos de soja. Scientia
Agraria, p.299-309, 2008.
BASSINELLO, P. Z.; COBUCCI, R. M. A.; ULHÔA, V. G.; MELO, L. C.; PELOSO, M. J.
D. Aceitabilidade de três cultivares de feijoeiro comum. Comunicado Técnico. EMBRAPA,
Santo Antônio de Goiás, p.5, 2003.
BASSO, F. C.; ANDREOTTI, M.; CARVALHO, M. P.; LODO, B.; MONTANARI, R. Correlação
linear e espacial entre a produtividade e o teor de proteína bruta do guandu anão e os atributos de
um Latossolo. Revista Brasileira de Ciências Agrárias, 2011.

62
BERTOLDO, J. G.; COIMBRA, J. L. M.; NODARI, B. O.; GUIDOLIN, A. F.; HEMP, S.;
BARILI, L. D.; VALE, N. M.; ROZZETO, D. S. Stratification of the state of Santa Catarina in
macro-environments for bean cultivation. Crop Breeding and Applied Biotechnology, p. 335-343,
2009.
BIANCHINI, A.; MARINGONI, A. C.; CARNEIRO, S. M. Doenças do feijoeiro. In: KIMATI, H.
et al. Manual de fitopatologia: doenças das plantas cultivadas 3. Ed São Paulo: Agronômica Ceres,
p. 376-399, 2005
BORÉM, A.; MIRANDA, G. V. Melhoramento de Plantas. 4. ed. Viçosa: UFV, p. 525, 2005
BURATTO, J. S.; CIRINO, V. M.; JÚNIOR, N. S. F; PRETE, C. E. C.; FARIA, R. T.
Adaptabilidade e estabilidade produtiva em genótipos precoces de feijão no estado do Paraná.
Semina: Ciências Agrárias, p. 373-380, 2008.
BURATTO, J. S.; MODA CIRINO, V.; SCHOLZ, M. B. S.; LANGAME, D.E.M.; FONSECA
JUNIOR, N. S.; PRETÉ, C. E. C. Variabilidade genética e efeito do ambiente para o teor de
proteína em grãos de feijão. Acta Scientiarum, p.593-597, 2009.
BURUCHARA R. A.; CAMACHO L. Common bean reaction to Fusarium oxysporum f.
sp. phaseoli, the cause of severe vascular wilt in Central Africa. Journal of Phytopathology. p. 39-
45, 2000.
CAMARGO, L. E. A.; BERGAMIN FILHO, A. Controle genético. In.: BERGAMIN FILHO, A.;
KIMATI, H.; AMORIM, L. Manual de Fitopatologia: Princípios e Conceitos. São Paulo:
Agronômica Ceres, 1995.
CARBONELL, S. A. M.; CARVALHO, C. R. L.; PEREIRA; V. R. Qualidade tecnológica de grãos
de genótipos de feijoeiro cultivados em diferentes ambientes. Bragantia, p.369-379, 2003.
CARBONELL, S.A.M.; POMPEU, A.S. Estratificação de ambientes em experimentos de feijoeiro
no Estado de São Paulo. Bragantia, Campinas, p.207-218, 1997.
CARBONELL, S. M.; ITO, M. F., POMPEU A. S.; FRANCISCO F. G.; RAVAGNANI S.;
ALMEIDA A. L. L. Raças fisiológicas de Colletotrichum lindemuthianum e reação de linhagens e
cultivares de feijoeiro no Estado de São Paulo. Fitopatologia Brasileira, 1999.
CARNEIRO, P. C. S. Novas metodologias de análise da adaptabilidade e estabilidade de
comportamento. Tese (Doutorado) - Universidade Federal de Viçosa, Viçosa, 1998.
CHAVES, L. J. Interação de cultivares com ambientes. In: NASS, L. L.; VALOIS, A. C. C.;
MELO, I. S.; VALADARES, M. C. Recursos Genéticos e Melhoramento: Plantas. Rondonópolis,
Fundação MT, p. 673-713, 2001.
CHIORATO, A. F. Análise da divergência genética de acessos de feijoeiro (Phaseolus vulgaris L.)
do Banco de germoplasma do Instituto Agronômico - IAC. Dissertação (Mestrado), Instituto
Agronômico - IAC, Campinas, S.P., p. 81, 2004.
CHIORATO, A. F.; CARBONELL, S. A. M.; MOURA, R. R. de; ITO, M. F.; COLOMBO, C. A.
Co‑evolução entre raças fisiológicas de Colletotrichum lindemuthianum em feijoeiro. Bragantia,
p.381-388, 2006

63
COIMBRA, R. R.; MIRANDA, G. V.; TEIXEIRA, F. F; DEITOS, A.; SOUZA, L.V. de; MELO,
A.V. de; DE MELLO, M.P. Estratificação ambiental e análise de adaptabilidade de genótipos de
milho baseada em análise de fatores. Revista Ciência Agroambiental, 2006.
CONAB – COMPANHIA NACIONAL DE ABASTECIMENTO, disponível em:
http//:www.conab.gov.br – acesso em 15-01-2014
COSTA, A. F.; FERRÃO, M.A.G; VENTURA, J.A; LIBERATO, J.R.; PACOVA, B.E.V. Murcha
de Fusarium do feijoeiro. Vitória EMCAPA, comunicado técnico, 1993.
COSTA, H.; ZAMBOLIM, L.; VENTURA, J. A. Doenças de hortaliças que se constituem em
desafio para o controle. In: Zambolim, L. et al. (Eds.). Manejo integrado de doenças e pragas:
hortaliças. Viçosa: Editora UFV, p.319–336, 2007.
COSTA, J. F. et al. Métodos de inoculação em plantas de feijoeiro. Fitopatologia Brasileira, p.125-
128, 1989.
COSTA, J.G.; MARINHO, J.T.S.; PEREIRA, R.C.A.; LEDO, F.J.S.; MORAES, R.N.S.
Adaptabilidade e estabilidade de produção de cultivares de milho recomendadas para o Estado do
Acre. Ciência e Agrotecnologia, p.7-11, 1999.
COSTA, R. G.; RAMALHO, P. A. M.; ABREU, B. F. A. Variabilidade para a absorção de água
nos grãos de feijão do germoplasma da UFLA. Ciência e Agrotecnologia, p.1017-1021, 2001
CROSSA, J. Statistical analysis of multilocations trials. Advances in agronomy, San Diego, p. 55-
85, 1990.
CRUZ, C. D.; CARNEIRO, P. C. S. Modelos biométricos aplicados ao melhoramento genético. 2.
ed. Viçosa: UFV, 2006.
CRUZ, C. D.; REGAZZI, A. J.; CARNEIRO, P.C.S. Modelos biométricos aplicados ao
melhoramento genético. Viçosa: UFV, 2004.
CRUZ, C. D.; CARNEIRO, P. C. S. Modelos biométricos aplicados ao melhoramento genético.
Viçosa: UFV, v.2. 2003.
CRUZ, C. D.; CASTOLDI, F. Decomposição da interação genotipos x ambientes em partes simples
e complexa. Revista Ceres, p. 35-39, 1991.
DALLA CORTE, A.; MODA-CIRINO, V.; SCHOLZ, M. B. S.; DESTRO, D. Environment effect
on grain quality in early common bean cultivars and lines. Crop Breeding and Applied
Biotechnology, 2003.
DIAZ, C. G.; BASSANEZI, R. B.; GODOY, C. V.; LOPES, D. B.; BERGAMIN FILHO, A.
Quantificação do efeito do crestamento bacteriano comum na eficiencia fotossintetica e na protução
do feijoeiro. Fitopatologia Brasileira, p. 71-76, 2001.
DUARTE, J. B.; ZIMMERMANN, M. J. Selection of location for common bean (Phaseolus
vulgaris L.) germplasm evaluation. Revista Brasileira de Genética, 1991.
DURSUN, A.; DONMEZ, F.; SAHIN, F. Identification of resistance to common bacterial blight
disease on bean genotypes grown in Turkey. European Journal of Plant Pathology, p.811-813, 2002.

64
EBERHART, S. A.; RUSSELL, W. A. Stability parameters for comparing varieties. Crop Science,
p. 36-40, 1966.
ELIA, F. M.; HOSFIELD, G. L.; KELLY, J. D.; UEBERSAX, M. A. Genetic analysis and
interrelationships between traits for cooking time, water absorption, and protein and tannin content
of Andean dry beans. Journal American Society Horticulture Science, p.512-518, 1996.
ELIAS, H. T.; SILMAR, H.; SCAPIM, C. A.; RODOVALHO, M. A.; ROYER, M. R.; MORA, F.;
BARRETO, R. R. Análise da estabilidade de genótipos de feijoeiro no Estado de Santa
Catarina. Acta Scientiarum Agronomy, p.623-628, 2005.
EMBRAPA. Doenças bacterianas: crestamento bacteriano comum. Disponivel em:
http://www.cnpaf.embrapa.br/feijao/pragasedoenças/crestamento.htm. Acesso em 24-01-2014
FARINELLI, R. Características agronômicas e tecnológicas em genótipos de feijoeiro. Tese
(Doutorado). Faculdade de Ciências Agronômicas, Universidade Estadual Paulista Júlio de
Mesquita Filho, Botucatu, 2006
FERREIRA, D. F. Estatística multivariada. Lavras: Ed UFLA, p. 662, 2008.
FERREIRA, D. F. et al. Statostocal models in agriculture: biometrical methods for evaluating
phenotypic stability in plant breeding. Cerne, p. 373-388, 2006.
FONSECA JÚNIOR, N. S.; ALVES, S. J.; OLIARI, L.; CIRINO, V. M. Análise de Estabilidade e
Estratificação de Rendimento da Cultivar Pérola no Paraná. In: REUNIÃO NACIONAL DE
PESQUISA DE FEIJÃO 5, Goiânia: Embrapa – CNPAF - APA. Anais.p.310 – 312, 1996.
GARBUGLIO, D. D.; GERAGE, A. C.; ARAÚJO, P. M.; FONSECA JUNIOR, N. S.; SHIOGA, P.
S. Análise de fatores e regressão bissegmentada em estudos de estratificação ambiental e
adaptabilidade em milho. Pesquisa Agropecuária Brasileira, p.183-191, 2007.
GARCIA, A. Análise temporal de doenças foliares do feijoeiro comum em Lavras, MG. 1998. 52 p.
Dissertação (Mestrado em Fitopatologia) – Universidade Federal de Lavras, Lavras, 1998.
GARCIA-VELA, L. A.; STANLEY, D. W. Water-holding capacity in hard-cook bean (P. vulgaris):
effect of pH and ionic strenght. Journal of Food Science, p. 1080-1081, 1989.
GEPTS, P.; DEBOUCK, D. Origin domestication and evotion of the common bean (Phaseolus
vulgaris L,) In: SHOONHOVENM, A,van; VOYSEST, O (Eds) Common beans. Reasarch for crop
improvement. Cali.CAB International. CIAT, p.7-53, 1993.
GONCALVES, J. G. R.; CHIORATO, A. F.; PERINA, E. F.; CARBONELL, S. A. M.
Estabilidade fenotípica em feijoeiro estimada por análise ammi com genótipo
suplementar. Bragantia, p. 863-871, 2009.
GONÇALVES-VIDIGAL M. C.; CRUZ A. S.; GARCIA A.; KAMI J.; VIDIGAL FILHO P. S.;
SOUSA L. L.; MCCLEAN P.; GEPTS P.; PASTOR-CORRALES A. Linkage mapping of the Phg-
1 and Co-14 genes for resistance to angular leaf spot and anthracnose in the common bean cultivar
AND 277. Theoretical and Applied Genetics 2011.
GUZMAN-MALDONADO, S. H.; PAREDES-LOPEZ, O. Biotechnology for the improvement of
nutritional quality of food crop plants. In PAREDES-LOPEZ, O. (Ed.). Molecular biotechnology
for plant food production. Lancaster: Technomic, p. 590-591, 1999.

65
HACISALIHOGLU, G.; LARBI, B.; SETTLES, A. M. Near-infrared reflectance spectroscopy
predicts protein, starch, and seed weigth in intact seeds of common bean (Phaseolus vulgaris L.).
Journal of Agricultural and Food Chemistry, p. 702-706, 2010.
IADEROZA, M., SALES, A. M., BALDINI, V. L. S. Atividade de polifenoloxidase e alterações da
cor e dos teores de taninos condensados em novas cultivares de feijão (Phaseolus vulgaris L.)
durante o armazenamento. Coletânea do Instituto de Tecnologia de Alimentos, Campinas, p.154-
164, 1989.
IBGE – Instituto Brasileiro de Geografia e Estatística. Levantamento sistemático da produção
agrícola. Rio de Janeiro: IBGE, 2012, disponível em:
http://www.ibge.gov.br/home/estatistica/indicadores/agropecuaria/lspa/ - acesso em 21-12-2013.
INFRASOFT INTERNATIONAL. Routine operation, calibration development and network system
management manual. Infrasoft International, Port Matilda, Pennsylvania, p. 335, 2007.
KANG, M. S.; GAUCH Júnior, H. G. Genotype by environment interaction. New York: CRC, p.
416, 1996.
LANZA, M.A.; PAULA J.R.; BARROS E.G.; MOREIRA, M.A. Avaliação da Resistencia de
cultivares de feijão recomendados para Minas Gerais à murcha de Fusarium. Fitopatologia
Brasileira, p. 22-27, 1997.
LAVORANTI, O. J. Estabilidade e adaptabilidade fenotípica por meio da reamostragem
“Bootstrap” no modelo AMMI. 2003. 166f. Tese (Doutorado em Estatística e Experimentação
Agronômica) – Escola Superior de Agricultura “Luiz de Queiroz”, Universidade de São Paulo,
Piracicaba, 2003.
LEMOS, L. B.; OLIVEIRA, R. S.; PALOMINO, E. C.; SILVA, T. R. B. Características
agronômicas e tecnológicas de genótipos de feijão do grupo comercial Carioca. Pesquisa
Agropecuária Brasileira, Brasília, p.319-326, 2004.
LIN, C. S.; BINNS, M. R. A method of analysing cultivars x location x year experiments : new
stability parameter. Theoretical Applied Genetics, p.425-430, 1988
LIN, C.S. Grouping genotypes by a cluster method directly relatedto genotype-environment
interaction mean square. Theoretical and Applied Genetics, p.277-280, 1982.
MAHUKU G. S.; JARA C.; CUASQUER J. B.; CASTELLANOS G. Genetc variability within
Phaeoisariopsis griseola from Central América and its implications for resistence breeding of
Common Bean. Plant Pathology, p. 594-604, 2002.
MAPA. Instrução normativa n° 25, de 23 de maio de 2006. Disponível em:
http://www.agricultura.gov.br/arq_editor/file/vegetal/Sementes_e_mudas/Registro_Nacional_de_C
ultivares.pdf
MARINGONI, A. C.; KIMATI, H.; E KUROZAWA, C. Variabilidade sorológica entre isolados
de Xanthomonas campestris pv. phaseoli. Summa Phytopathologica, p. 164-167, 1994.
MARIOTTI, J. A.; OYARZABAL, E. S.; OSA, J. M.; BULACIO, A. R. N.; ALMADA, G. H.
Analisis de estabilidad y adaptabilidad de genotipos de cana de azucar. i. interacciones dentro de
una localidad experimental. Revista Agronómica del Nordeste Argentino. p.104-118, 1976.

66
MELLO FILHO, O. L.; SEDIYAMA, C. S.; MOREIRA, M. A.; REIS, A.S.; MASSONI, G. A.;
PIOVESAN, N. D. Produtividade de grãos e qualidade de sementes de soja selecionada para alto
teor de proteína. Pesquisa Agropecuária Brasileira, p.445-450, 2004.
MENDES, F. F.; RAMALHO, M. A. P.; ABREU, Â. de F. B. Índice de seleção para escolha de
populações segregantes do feijoeiro-comum. Pesquisa Agropecuária Brasileira, p.1312-1318, 2009.
MESQUITA, R. F.; CORRÊA, D. A.; ABREU, P. M. C.; LIMA, Z. A. R.; ABREU, B. F. A.
Linhagens de feijão (Phaseolus vulgaris, L.): Composição química e digestibilidade protéica.
Ciência e Agrotecnologia, p.1114-1121, 2007.
MIKLAS P. N.; KELLY J. D.; BEEBE S. E.; BLAIR M. W. Common bean breeding for resistance
against biotic and abiotic stresses: From classical to MAS breeding. Euphytica, p.105-131, 2006.
MOHAN, S. K. BIANCHINI, A.; MENEZES, R. J. Doenças do feijoeiro no estado do Paraná: Guia
para identificação e controle. Londrina: IAPAR, p.26, 1983
MULAMBA, N. N.; MOCK, J. J. Improvement of yield potential of the Eto Blanco maize (Zea
mays L.) population by breeding for plant traits. Egyptian Journal of Genetics and Cytology, p. 40-
51, 1978.
MURAKAMI, D. M.; CRUZ, C. D. Proposal of methodologies for environment stratification and
analysis of genotype adaptability. Crop Breeding and Applied Biotechnology, p.7-11, 2004.
NASCIMENTO, S. R. C.; KUROZAWA, C.; MARINGONI, A. C. Avaliação de raças fisiológicas
de Fusarium oxysporum f. sp. phaseoli. Fitopatologia Brasileira, Brasília, p. 214-217, 1995.
NEVES, L. G.; BRUCKNER, C. H.; CRUZ, C. D.; VIANA, A. P.; CARELLI, M. A. A. Predição
de ganhos, com diferentes índices de seleção, para características de frutos do maracujazeiro-
amarelo. Revista Brasileira de Fruticultura, p. 1322-1330, 2011.
OSBORN, T. C. Genetic Control of Bean Seed Protein. Plant Science, p. 93-116, 1988.
PASTOR-CORRALES, M. A.; TU, J. C. Antracnosis. In: PASTOR-CORRALES, M. A.;
SCHWARTZ, H. F. (Ed). Problemas de produccion del frijol em los trópicos. 2. Ed. Cali: CIAT,
1994. p. 87-119.
PAULA JÚNIOR, T. J. de; ZAMBOLIM, L. Doenças. In VIEIRA, C.; PAULA JÚNIOR, T. J. de;
BORÉM, A. (org.). Feijão. 2. ed. Viçosa: UFV, 2006. p. 359-414.
PEREIRA H. S.; MELO L. C.; FÁRIA L. C.; DEL PELOSO M. J.; COSTA J. G. C., RAVA C. A.;
WENDLAND A. Adaptabilidade e Estabilidade de genótipos de feijoeiro-comum com grãos tipo
carioca na Região Central do Brasil. Pesquisa Agropecuária Brasileira, p.29-37, 2009.
PEREIRA, H. S.; MELO, L. C.; FARIA, L. C. de; DEL PELOSO, M. J.; WENDLAND, A.
Estratificação ambiental na avaliação de genótipos de feijoeiro-comum tipo Carioca em Goiás e no
Distrito Federal. Pesquisa Agropecuária Brasileira, p.554-562, 2010.
PEREIRA, M. J.; RAMALHO, M. A. P.; ABREU, A. F. Estratégias para eficiência da seleção de
feijoeiro para resistência à murcha-de-fusário. Pesquisa Agropecuária Brasileira, p.721-728, 2008.
PEREIRA, M. J. Z.; RAMALHO, M. A. P.; ABREU, A. F. B.. Reação de linhagens de feijoeiro ao
fungo Fusarium oxysporum f. sp. phaseoli em condições controladas. Ciência Agrotécnica, p.940-
947, 2011.

67
PIMENTEL GOMES, F. Curso de Estatística Experimental. São Paulo: Nobel, 1985.
PINTO, A. C. S. Análise padrão especial e do gradient da antracnose do feijoeiro em diferentes
safras de cultivo. Summa Phytopathologica, p. 392-398, 2001
PIZA, S. M. de T. Patogenicidade de Fusarium oxysporum f. sp. phaseoli e reação de germoplasma
de feijoeiro (Phaseolus vulgaris L.). Summa Phytopatologica, p. 156- 167, 1993.
PLHAK, L. C.; CALDWELL, K. B.; STANLEY, D. W. Comparison of methods used to
characterize water imbibition in hard-to-cook beans. Journal of Food Science, p. 326-336, 1989.
POMPEU, A. S.; CROWER, L. V. Methods of inoculation and bacterial concentrations of
Xanthomonas phaseoli Down. for the inheritance of disease reaction in Phaseolus vulgaris crosses
(dry bean), under growth chamber conditions. Ciência e Cultura, p. 1078-1081, 1973.
PROCTOR, J. R.; WATTS, B. M. Development of a Modified Mattson Bean Cooker Produre
Based on Sensory Panel Cookability Evaluation. Canadian Institute of Food Science and
Tecnology, Ottowa, p. 9-14, 1987.
RAMALHO, M. A. P.; ABREU, A. F. B. Cultivares. In: VIEIRA, C.; PAULA JÚNIOR, T. J.;
BORÉM, A. Feijão: aspectos gerais e cultura no Estado de Minas. 2. ed. Viçosa: UFV, p. 415-436.
2006.
RAMALHO, M. A. P.; ABREU A. F. B.; SANTOS J. B. NUNES, J. A. R. Aplicações da genética
quantitativa no melhoramento de plantas autógamas. Ciência Agrotécnica, p. 33-40, 2012.
RAMOS J. R.; LEMOS, L. B.; SILVA, T. R. B. Componentes da produção, produtividade de
grãos e características tecnológicas de cultivares de feijão. Bragantia, p.75-82, 2005.
RAVA, C. A.; SARTORATO, A.; COSTA, J. G. C. Reação de genótipos de feijoeiro comum ao
Fusarium oxysporum f. sp. phaseoli em casa de vegetação. Fitopatologia Brasileira, p.296-300,
1996.
RAVA, C.A.; SARTORATO, A.; ROMEIRO, R.S. Avaliação de cultivares de feijoeiro quanto a
resistência a Xanthomonas campestris pv. phaseoli em condições de campo e casa de vegetação.
Summa Phytopathologyca, p.83-91, 1990.
RESENDE, M. D. V. de. Matemática e estatística na análise de experimentos e no melhoramento
genético. Colombo: Embrapa Florestas, 2007.
RESENDE, M. D. V. Genética biométrica e estatística no melhoramento de plantas perenes.
Brasília: Embrapa Informação Tecnológica, 2002
RIBEIRO N. D., FILHO A. C., POERSCH, N. L.; ROSA, D. P. Critério de seleção indireta para a
produtividade de grãos em feijão. Ciencia. Rural, p. 4-9. 2010.
RIBEIRO, N. D.; LONDERO, P. M.G.; HOFFMANN JÚNIOR, L.; POERSCH, N. L.;
CARGNELUTTI FILHO, A. Dissimilaridade genética para teor de proteína e fibra em grãos de
feijão dos grupos preto e de cor. Revista Brasileira de Agrociência, p.167-173, 2005
RIBEIRO, N. D.; SOUZA, J. F. de; ANTUNES, I. F. e POERSCH, N. L. Estabilidade de
produção de cultivares de feijão de diferentes grupos comerciais no Estado do Rio Grande do
Sul. Bragantia, p. 339-346, 2009.

68
RIBEIRO, N. D. Potencial de aumento da qualidade nutricional do feijão por melhoramento
genético. Semina: Ciências Agrárias, p.1267-1376, 2010.
RIBEIRO, N. D.; RODRIGUES, J. A.; CARGNELUTTI FILHO, A.; POERSH, N. L.; TRENTIN,
M.; ROSA, S. S. Efeitos de períodos de semeadura e das condições de armazenamento sobre a
qualidade de grãos de feijão para o cozimento. Bragantia, p.157-163, 2007.
RIBEIRO, R. L. D. & HAGEDORN, D. J. Inheritance of resistance in beans to Fusarium
oxysporum f. sp. phaseoli. Phytopathology, p.859-61, 1979.
ROBERTSON, A. Experimental design on the measurement of heritabilities and genetic
correlations: biometrical genetics. Pergamon Press, p. 186, 1959.
ROCHA JÚNIOR, W. C.; SANTOS, J. B.; COSTA M. C. M. Reação de cultivares e linhagens de
feijão a Fusarium oxysporum f. sp. phaseoli. Fitopatologia Brasileira. p. 407-409, 1998.
ROCHA, R. B.; MURO-ABAD, J. I.; ARAUJO, E. F.; CRUZ, C. D. Avaliação do método
centroide para estudo de adaptabilidade ao ambiente de clones de Eucalyptus grandis. Ciência
Florestal, p.255-266, 2005.
RODRIGUES, J. A.; RIBEIRO, N. D.; CARGNELUTTI FILHO, A.; TRENTIN, M.;
LONDERO, P. M. G. Qualidade para o cozimento de grãos de feijão obtidos em diferentes
safras. Bragantia, p. 369-376, 2005.
SAETTLER, A.W. Common baxterial blight. In SCHWARTZ, H. F. et al (Ed.) Compendium of
bean diseases. 2nd
ed. Saint Paul: APS PRES, p. 46-47, 2005
SANTOS, A. S., BRESSAN-SMITH, R. E., PEREIRA, M. G., RODRIGUES, R., FERREIRA, C.
F. Genetic linkage map of Phaseolus vulgaris and identification of QTLs responsible for resistance
to Xanthomonas axonopodis pv. phaseoli. Fitopatologia. Brasileira 2003.
SARTORATO, A.; RAVA, C. A. Mancha angular. In: Sartorato, A. & Rava, C.A.(Eds.) Principais
Doenças do Feijoeiro Comum e seu Controle. Brasília. EMBRAPA-SPI. p. 41.68, 1994.
SANTOS, J. B.; GAVILANES, M. L. Botânica. In: VIEIRA, C.; PAULA JÚNIOR, T. J.; BORÉM,
A. (Eds.). Feijão.Aspectos gerais e cultura no Estado de Minas. Viçosa, MG: Universidade Federal
de Viçosa, 1998.
SARTORI, M. R. Technological quality of dry beans (Phaseolus vulgaris) stored under nitrogen.
PhD. Dissertation. Manhattan, Kansas, USA: Department of Grain Science and Industry, Kansas
State University, 1982.
SCHOLZ, S. B. M.; FONSECA JUNIOR, N. S. Influência ambiental, genotípica e sua interação na
qualidade tecnológica de feijão do grupo preto no Paraná. In: VI Reunião Nacional de Pesquisa do
Feijão, Goiânia, p.389.392, 1999.
SENNA S. N.; FREITAS M.L.M.; ZANATTO, A. C. S.; MORAIS, E.; ZANATA, M.; MORAES
M. L. T.; SEBENN A. M. Variação e parâmetros genéticos em teste de progenies de polinização
livre de Peltophorum dubium taubert em Luiz Antonio – SP. Scientia. Forestalis, p.345-352, 2012.
SILVA, K. J. D.; SOUZA, E. A.; ISHIKAWA, F. H. Characterization of Colletotrichum
lindemuthianum. Isolates from the state of Minas Gerais, Brazil. Journal of Phytopathology, p. 241-
247, 2007

69
SINGH, S. P.; SCHWARTZ, H. F. Breeding common bean for resistence to diseases: a review.
Crop Science, p. 2199-2223, 2010
SMITH, H. F. A discriminant function for plant selection. Annual Eugenics, [S.l.], p. 240-250,
1936.
SUGIMORI, M. H. Caracterização sorológica dos patovares brancos de Xanthomonas campestris e
Reação de Mangueiras (Mangifera indica L.) a Xanthomonas campestris pv. mangiferaeindicae.
Tese apresentada a escola superior de Agricultura Luiz de Queiroz, da Universidade de São Paulo,
1989.
THUNG, M.; SARTORATO, A. Alteração na ordem de importância das doenças do feijoeiro
comum (Phaseolus vulgaris L.) devido a mudanças nas práticas culturais e na preferência do tipo de
grão no Brasil. Série Documentos n° 132, CNPAF/EMBRAPA, 2002.
VIEIRA, C. Melhoramento do Feijão. In BOREM, A. Melhoramento de espécies cultivadas. 2. Ed
Viçosa, MG: UFV, p. 301-392, 2005.
ZANATTA Z. G. C.; MOURA A. B.; MAIA L. C.; SANTOS A.S. Bioassay for selection of
biocontroller bacteria against bean common blight (Xanthomonas axonopodis pv. phaseoli).
Brazilian Journal of Microbiology, p. 511–515, 2007
ZUCARELI, C; PRANDO, A. M.; RAMOS JUNIOR, E. U. e NAKAGAWA, J. Fósforo na
produtividade e qualidade de sementes de feijão Carioca Precoce cultivado no período das
águas. Ciência Agronomica. p. 32-38, 2011.
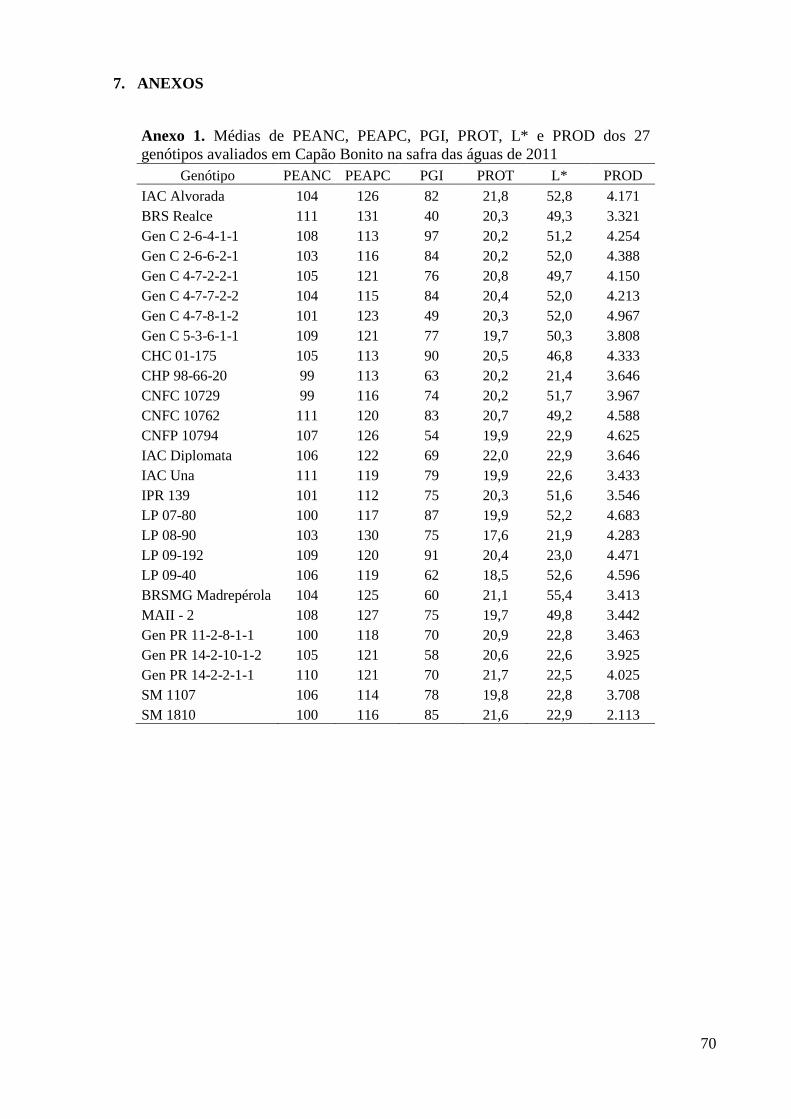
70
7. ANEXOS
Anexo 1. Médias de PEANC, PEAPC, PGI, PROT, L* e PROD dos 27
genótipos avaliados em Capão Bonito na safra das águas de 2011
Genótipo PEANC PEAPC PGI PROT L* PROD
IAC Alvorada 104 126 82 21,8 52,8 4.171
BRS Realce 111 131 40 20,3 49,3 3.321
Gen C 2-6-4-1-1 108 113 97 20,2 51,2 4.254
Gen C 2-6-6-2-1 103 116 84 20,2 52,0 4.388
Gen C 4-7-2-2-1 105 121 76 20,8 49,7 4.150
Gen C 4-7-7-2-2 104 115 84 20,4 52,0 4.213
Gen C 4-7-8-1-2 101 123 49 20,3 52,0 4.967
Gen C 5-3-6-1-1 109 121 77 19,7 50,3 3.808
CHC 01-175 105 113 90 20,5 46,8 4.333
CHP 98-66-20 99 113 63 20,2 21,4 3.646
CNFC 10729 99 116 74 20,2 51,7 3.967
CNFC 10762 111 120 83 20,7 49,2 4.588
CNFP 10794 107 126 54 19,9 22,9 4.625
IAC Diplomata 106 122 69 22,0 22,9 3.646
IAC Una 111 119 79 19,9 22,6 3.433
IPR 139 101 112 75 20,3 51,6 3.546
LP 07-80 100 117 87 19,9 52,2 4.683
LP 08-90 103 130 75 17,6 21,9 4.283
LP 09-192 109 120 91 20,4 23,0 4.471
LP 09-40 106 119 62 18,5 52,6 4.596
BRSMG Madrepérola 104 125 60 21,1 55,4 3.413
MAII - 2 108 127 75 19,7 49,8 3.442
Gen PR 11-2-8-1-1 100 118 70 20,9 22,8 3.463
Gen PR 14-2-10-1-2 105 121 58 20,6 22,6 3.925
Gen PR 14-2-2-1-1 110 121 70 21,7 22,5 4.025
SM 1107 106 114 78 19,8 22,8 3.708
SM 1810 100 116 85 21,6 22,9 2.113
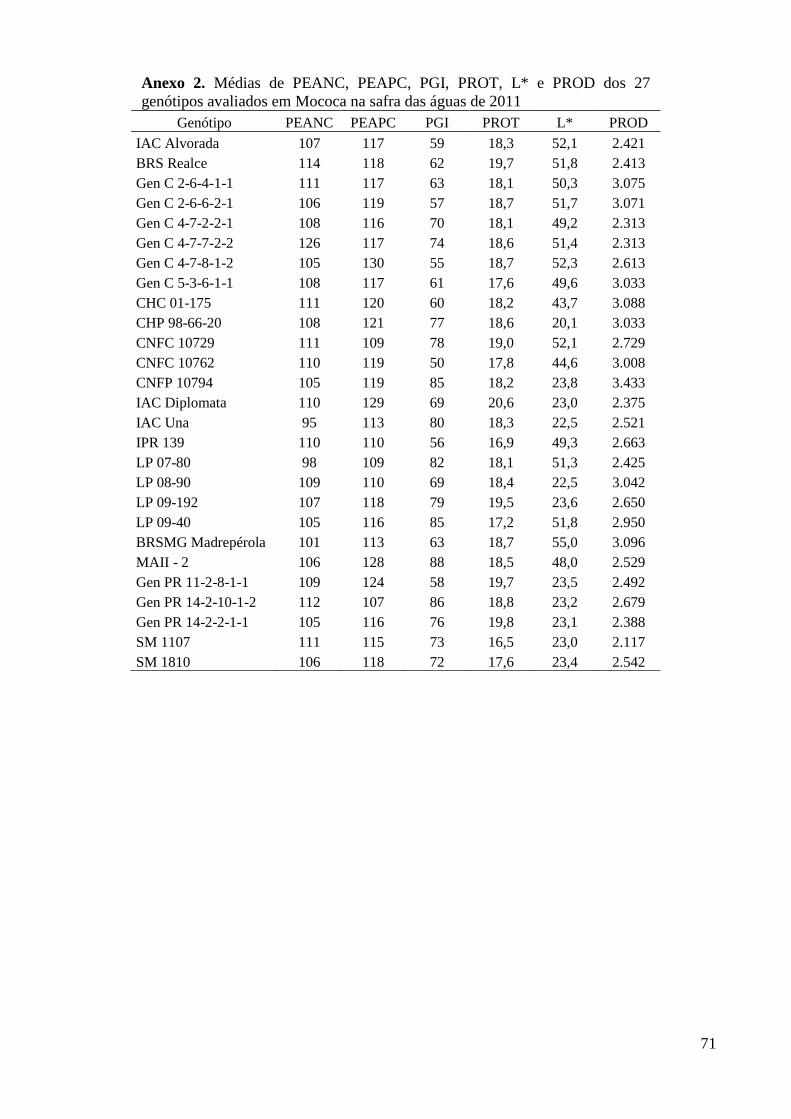
71
Anexo 2. Médias de PEANC, PEAPC, PGI, PROT, L* e PROD dos 27
genótipos avaliados em Mococa na safra das águas de 2011
Genótipo PEANC PEAPC PGI PROT L* PROD
IAC Alvorada 107 117 59 18,3 52,1 2.421
BRS Realce 114 118 62 19,7 51,8 2.413
Gen C 2-6-4-1-1 111 117 63 18,1 50,3 3.075
Gen C 2-6-6-2-1 106 119 57 18,7 51,7 3.071
Gen C 4-7-2-2-1 108 116 70 18,1 49,2 2.313
Gen C 4-7-7-2-2 126 117 74 18,6 51,4 2.313
Gen C 4-7-8-1-2 105 130 55 18,7 52,3 2.613
Gen C 5-3-6-1-1 108 117 61 17,6 49,6 3.033
CHC 01-175 111 120 60 18,2 43,7 3.088
CHP 98-66-20 108 121 77 18,6 20,1 3.033
CNFC 10729 111 109 78 19,0 52,1 2.729
CNFC 10762 110 119 50 17,8 44,6 3.008
CNFP 10794 105 119 85 18,2 23,8 3.433
IAC Diplomata 110 129 69 20,6 23,0 2.375
IAC Una 95 113 80 18,3 22,5 2.521
IPR 139 110 110 56 16,9 49,3 2.663
LP 07-80 98 109 82 18,1 51,3 2.425
LP 08-90 109 110 69 18,4 22,5 3.042
LP 09-192 107 118 79 19,5 23,6 2.650
LP 09-40 105 116 85 17,2 51,8 2.950
BRSMG Madrepérola 101 113 63 18,7 55,0 3.096
MAII - 2 106 128 88 18,5 48,0 2.529
Gen PR 11-2-8-1-1 109 124 58 19,7 23,5 2.492
Gen PR 14-2-10-1-2 112 107 86 18,8 23,2 2.679
Gen PR 14-2-2-1-1 105 116 76 19,8 23,1 2.388
SM 1107 111 115 73 16,5 23,0 2.117
SM 1810 106 118 72 17,6 23,4 2.542
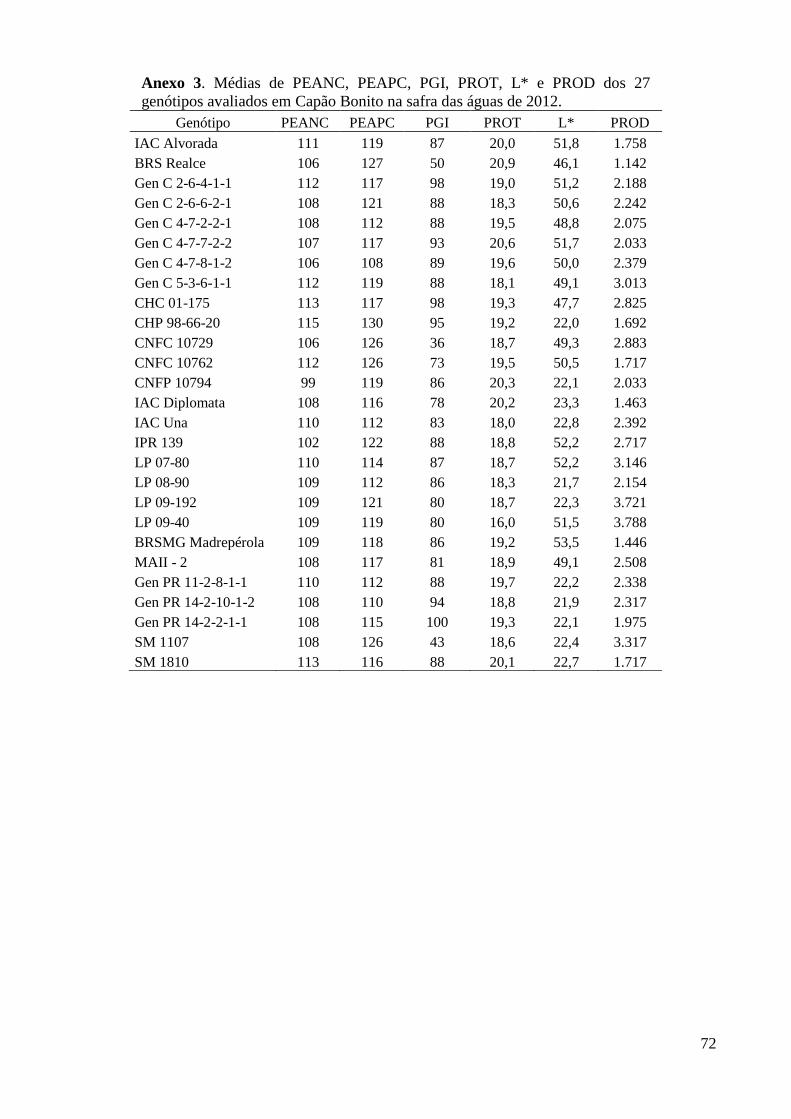
72
Anexo 3. Médias de PEANC, PEAPC, PGI, PROT, L* e PROD dos 27
genótipos avaliados em Capão Bonito na safra das águas de 2012.
Genótipo PEANC PEAPC PGI PROT L* PROD
IAC Alvorada 111 119 87 20,0 51,8 1.758
BRS Realce 106 127 50 20,9 46,1 1.142
Gen C 2-6-4-1-1 112 117 98 19,0 51,2 2.188
Gen C 2-6-6-2-1 108 121 88 18,3 50,6 2.242
Gen C 4-7-2-2-1 108 112 88 19,5 48,8 2.075
Gen C 4-7-7-2-2 107 117 93 20,6 51,7 2.033
Gen C 4-7-8-1-2 106 108 89 19,6 50,0 2.379
Gen C 5-3-6-1-1 112 119 88 18,1 49,1 3.013
CHC 01-175 113 117 98 19,3 47,7 2.825
CHP 98-66-20 115 130 95 19,2 22,0 1.692
CNFC 10729 106 126 36 18,7 49,3 2.883
CNFC 10762 112 126 73 19,5 50,5 1.717
CNFP 10794 99 119 86 20,3 22,1 2.033
IAC Diplomata 108 116 78 20,2 23,3 1.463
IAC Una 110 112 83 18,0 22,8 2.392
IPR 139 102 122 88 18,8 52,2 2.717
LP 07-80 110 114 87 18,7 52,2 3.146
LP 08-90 109 112 86 18,3 21,7 2.154
LP 09-192 109 121 80 18,7 22,3 3.721
LP 09-40 109 119 80 16,0 51,5 3.788
BRSMG Madrepérola 109 118 86 19,2 53,5 1.446
MAII - 2 108 117 81 18,9 49,1 2.508
Gen PR 11-2-8-1-1 110 112 88 19,7 22,2 2.338
Gen PR 14-2-10-1-2 108 110 94 18,8 21,9 2.317
Gen PR 14-2-2-1-1 108 115 100 19,3 22,1 1.975
SM 1107 108 126 43 18,6 22,4 3.317
SM 1810 113 116 88 20,1 22,7 1.717
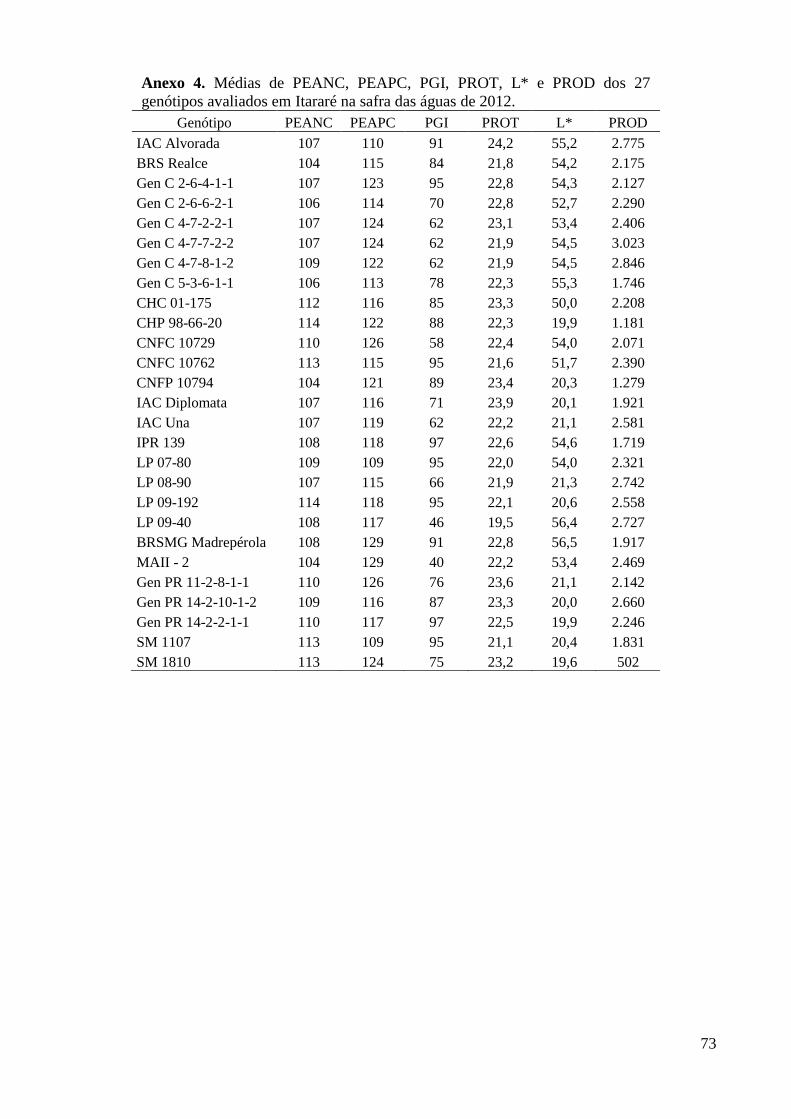
73
Anexo 4. Médias de PEANC, PEAPC, PGI, PROT, L* e PROD dos 27
genótipos avaliados em Itararé na safra das águas de 2012.
Genótipo PEANC PEAPC PGI PROT L* PROD
IAC Alvorada 107 110 91 24,2 55,2 2.775
BRS Realce 104 115 84 21,8 54,2 2.175
Gen C 2-6-4-1-1 107 123 95 22,8 54,3 2.127
Gen C 2-6-6-2-1 106 114 70 22,8 52,7 2.290
Gen C 4-7-2-2-1 107 124 62 23,1 53,4 2.406
Gen C 4-7-7-2-2 107 124 62 21,9 54,5 3.023
Gen C 4-7-8-1-2 109 122 62 21,9 54,5 2.846
Gen C 5-3-6-1-1 106 113 78 22,3 55,3 1.746
CHC 01-175 112 116 85 23,3 50,0 2.208
CHP 98-66-20 114 122 88 22,3 19,9 1.181
CNFC 10729 110 126 58 22,4 54,0 2.071
CNFC 10762 113 115 95 21,6 51,7 2.390
CNFP 10794 104 121 89 23,4 20,3 1.279
IAC Diplomata 107 116 71 23,9 20,1 1.921
IAC Una 107 119 62 22,2 21,1 2.581
IPR 139 108 118 97 22,6 54,6 1.719
LP 07-80 109 109 95 22,0 54,0 2.321
LP 08-90 107 115 66 21,9 21,3 2.742
LP 09-192 114 118 95 22,1 20,6 2.558
LP 09-40 108 117 46 19,5 56,4 2.727
BRSMG Madrepérola 108 129 91 22,8 56,5 1.917
MAII - 2 104 129 40 22,2 53,4 2.469
Gen PR 11-2-8-1-1 110 126 76 23,6 21,1 2.142
Gen PR 14-2-10-1-2 109 116 87 23,3 20,0 2.660
Gen PR 14-2-2-1-1 110 117 97 22,5 19,9 2.246
SM 1107 113 109 95 21,1 20,4 1.831
SM 1810 113 124 75 23,2 19,6 502
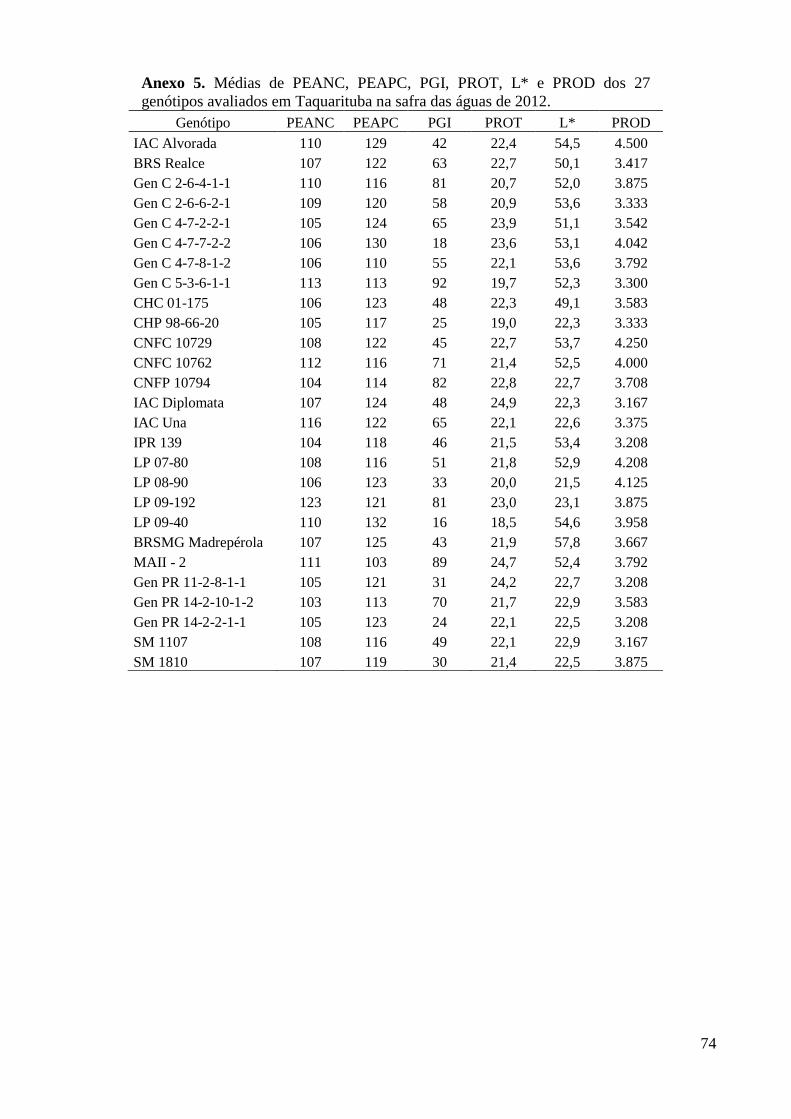
74
Anexo 5. Médias de PEANC, PEAPC, PGI, PROT, L* e PROD dos 27
genótipos avaliados em Taquarituba na safra das águas de 2012.
Genótipo PEANC PEAPC PGI PROT L* PROD
IAC Alvorada 110 129 42 22,4 54,5 4.500
BRS Realce 107 122 63 22,7 50,1 3.417
Gen C 2-6-4-1-1 110 116 81 20,7 52,0 3.875
Gen C 2-6-6-2-1 109 120 58 20,9 53,6 3.333
Gen C 4-7-2-2-1 105 124 65 23,9 51,1 3.542
Gen C 4-7-7-2-2 106 130 18 23,6 53,1 4.042
Gen C 4-7-8-1-2 106 110 55 22,1 53,6 3.792
Gen C 5-3-6-1-1 113 113 92 19,7 52,3 3.300
CHC 01-175 106 123 48 22,3 49,1 3.583
CHP 98-66-20 105 117 25 19,0 22,3 3.333
CNFC 10729 108 122 45 22,7 53,7 4.250
CNFC 10762 112 116 71 21,4 52,5 4.000
CNFP 10794 104 114 82 22,8 22,7 3.708
IAC Diplomata 107 124 48 24,9 22,3 3.167
IAC Una 116 122 65 22,1 22,6 3.375
IPR 139 104 118 46 21,5 53,4 3.208
LP 07-80 108 116 51 21,8 52,9 4.208
LP 08-90 106 123 33 20,0 21,5 4.125
LP 09-192 123 121 81 23,0 23,1 3.875
LP 09-40 110 132 16 18,5 54,6 3.958
BRSMG Madrepérola 107 125 43 21,9 57,8 3.667
MAII - 2 111 103 89 24,7 52,4 3.792
Gen PR 11-2-8-1-1 105 121 31 24,2 22,7 3.208
Gen PR 14-2-10-1-2 103 113 70 21,7 22,9 3.583
Gen PR 14-2-2-1-1 105 123 24 22,1 22,5 3.208
SM 1107 108 116 49 22,1 22,9 3.167
SM 1810 107 119 30 21,4 22,5 3.875
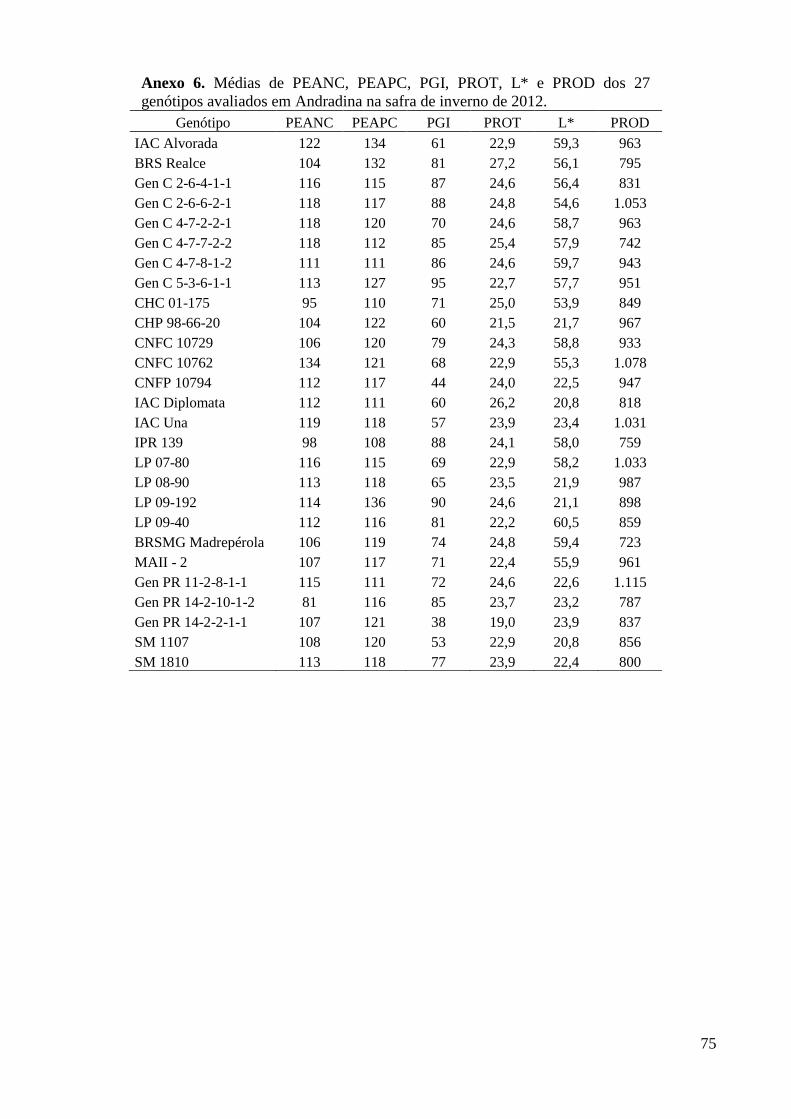
75
Anexo 6. Médias de PEANC, PEAPC, PGI, PROT, L* e PROD dos 27
genótipos avaliados em Andradina na safra de inverno de 2012.
Genótipo PEANC PEAPC PGI PROT L* PROD
IAC Alvorada 122 134 61 22,9 59,3 963
BRS Realce 104 132 81 27,2 56,1 795
Gen C 2-6-4-1-1 116 115 87 24,6 56,4 831
Gen C 2-6-6-2-1 118 117 88 24,8 54,6 1.053
Gen C 4-7-2-2-1 118 120 70 24,6 58,7 963
Gen C 4-7-7-2-2 118 112 85 25,4 57,9 742
Gen C 4-7-8-1-2 111 111 86 24,6 59,7 943
Gen C 5-3-6-1-1 113 127 95 22,7 57,7 951
CHC 01-175 95 110 71 25,0 53,9 849
CHP 98-66-20 104 122 60 21,5 21,7 967
CNFC 10729 106 120 79 24,3 58,8 933
CNFC 10762 134 121 68 22,9 55,3 1.078
CNFP 10794 112 117 44 24,0 22,5 947
IAC Diplomata 112 111 60 26,2 20,8 818
IAC Una 119 118 57 23,9 23,4 1.031
IPR 139 98 108 88 24,1 58,0 759
LP 07-80 116 115 69 22,9 58,2 1.033
LP 08-90 113 118 65 23,5 21,9 987
LP 09-192 114 136 90 24,6 21,1 898
LP 09-40 112 116 81 22,2 60,5 859
BRSMG Madrepérola 106 119 74 24,8 59,4 723
MAII - 2 107 117 71 22,4 55,9 961
Gen PR 11-2-8-1-1 115 111 72 24,6 22,6 1.115
Gen PR 14-2-10-1-2 81 116 85 23,7 23,2 787
Gen PR 14-2-2-1-1 107 121 38 19,0 23,9 837
SM 1107 108 120 53 22,9 20,8 856
SM 1810 113 118 77 23,9 22,4 800
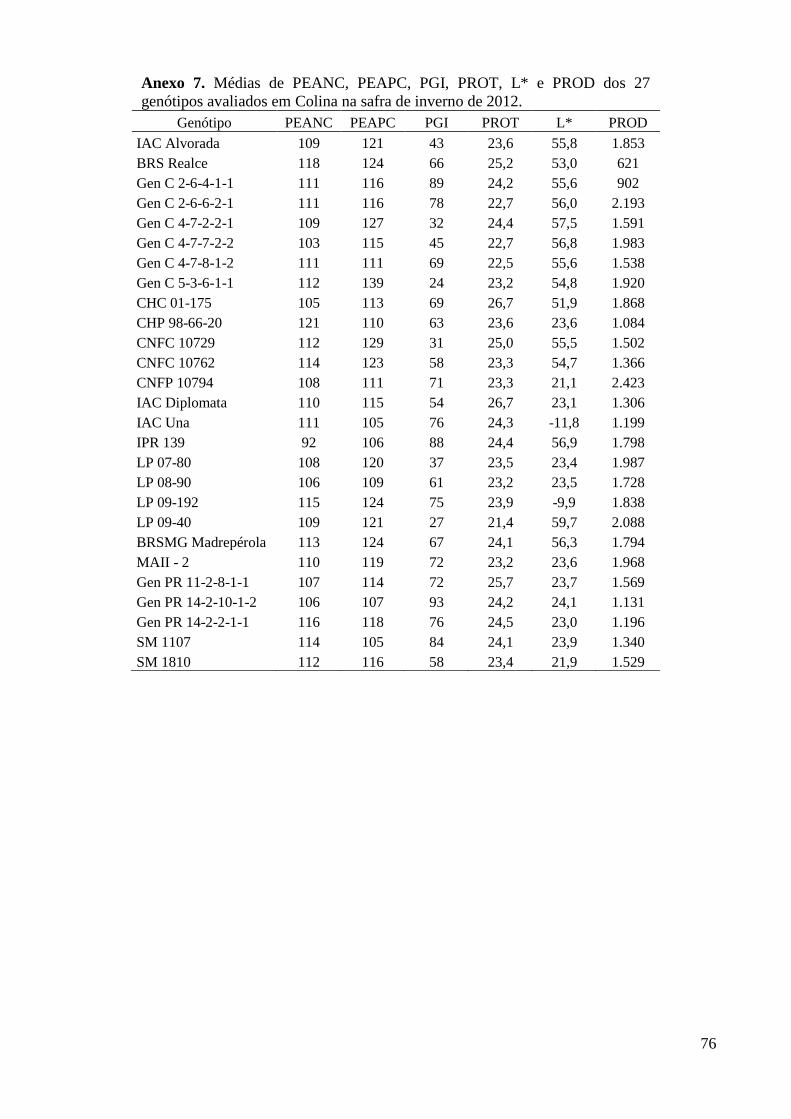
76
Anexo 7. Médias de PEANC, PEAPC, PGI, PROT, L* e PROD dos 27
genótipos avaliados em Colina na safra de inverno de 2012.
Genótipo PEANC PEAPC PGI PROT L* PROD
IAC Alvorada 109 121 43 23,6 55,8 1.853
BRS Realce 118 124 66 25,2 53,0 621
Gen C 2-6-4-1-1 111 116 89 24,2 55,6 902
Gen C 2-6-6-2-1 111 116 78 22,7 56,0 2.193
Gen C 4-7-2-2-1 109 127 32 24,4 57,5 1.591
Gen C 4-7-7-2-2 103 115 45 22,7 56,8 1.983
Gen C 4-7-8-1-2 111 111 69 22,5 55,6 1.538
Gen C 5-3-6-1-1 112 139 24 23,2 54,8 1.920
CHC 01-175 105 113 69 26,7 51,9 1.868
CHP 98-66-20 121 110 63 23,6 23,6 1.084
CNFC 10729 112 129 31 25,0 55,5 1.502
CNFC 10762 114 123 58 23,3 54,7 1.366
CNFP 10794 108 111 71 23,3 21,1 2.423
IAC Diplomata 110 115 54 26,7 23,1 1.306
IAC Una 111 105 76 24,3 -11,8 1.199
IPR 139 92 106 88 24,4 56,9 1.798
LP 07-80 108 120 37 23,5 23,4 1.987
LP 08-90 106 109 61 23,2 23,5 1.728
LP 09-192 115 124 75 23,9 -9,9 1.838
LP 09-40 109 121 27 21,4 59,7 2.088
BRSMG Madrepérola 113 124 67 24,1 56,3 1.794
MAII - 2 110 119 72 23,2 23,6 1.968
Gen PR 11-2-8-1-1 107 114 72 25,7 23,7 1.569
Gen PR 14-2-10-1-2 106 107 93 24,2 24,1 1.131
Gen PR 14-2-2-1-1 116 118 76 24,5 23,0 1.196
SM 1107 114 105 84 24,1 23,9 1.340
SM 1810 112 116 58 23,4 21,9 1.529
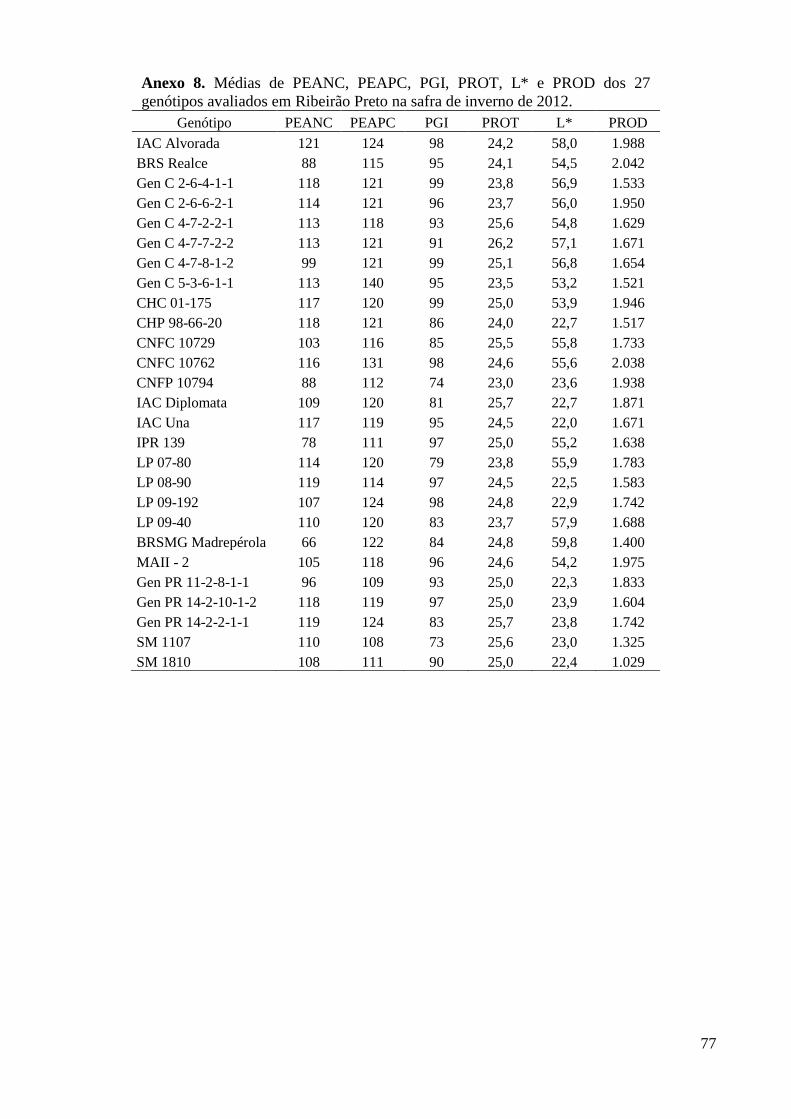
77
Anexo 8. Médias de PEANC, PEAPC, PGI, PROT, L* e PROD dos 27
genótipos avaliados em Ribeirão Preto na safra de inverno de 2012.
Genótipo PEANC PEAPC PGI PROT L* PROD
IAC Alvorada 121 124 98 24,2 58,0 1.988
BRS Realce 88 115 95 24,1 54,5 2.042
Gen C 2-6-4-1-1 118 121 99 23,8 56,9 1.533
Gen C 2-6-6-2-1 114 121 96 23,7 56,0 1.950
Gen C 4-7-2-2-1 113 118 93 25,6 54,8 1.629
Gen C 4-7-7-2-2 113 121 91 26,2 57,1 1.671
Gen C 4-7-8-1-2 99 121 99 25,1 56,8 1.654
Gen C 5-3-6-1-1 113 140 95 23,5 53,2 1.521
CHC 01-175 117 120 99 25,0 53,9 1.946
CHP 98-66-20 118 121 86 24,0 22,7 1.517
CNFC 10729 103 116 85 25,5 55,8 1.733
CNFC 10762 116 131 98 24,6 55,6 2.038
CNFP 10794 88 112 74 23,0 23,6 1.938
IAC Diplomata 109 120 81 25,7 22,7 1.871
IAC Una 117 119 95 24,5 22,0 1.671
IPR 139 78 111 97 25,0 55,2 1.638
LP 07-80 114 120 79 23,8 55,9 1.783
LP 08-90 119 114 97 24,5 22,5 1.583
LP 09-192 107 124 98 24,8 22,9 1.742
LP 09-40 110 120 83 23,7 57,9 1.688
BRSMG Madrepérola 66 122 84 24,8 59,8 1.400
MAII - 2 105 118 96 24,6 54,2 1.975
Gen PR 11-2-8-1-1 96 109 93 25,0 22,3 1.833
Gen PR 14-2-10-1-2 118 119 97 25,0 23,9 1.604
Gen PR 14-2-2-1-1 119 124 83 25,7 23,8 1.742
SM 1107 110 108 73 25,6 23,0 1.325
SM 1810 108 111 90 25,0 22,4 1.029
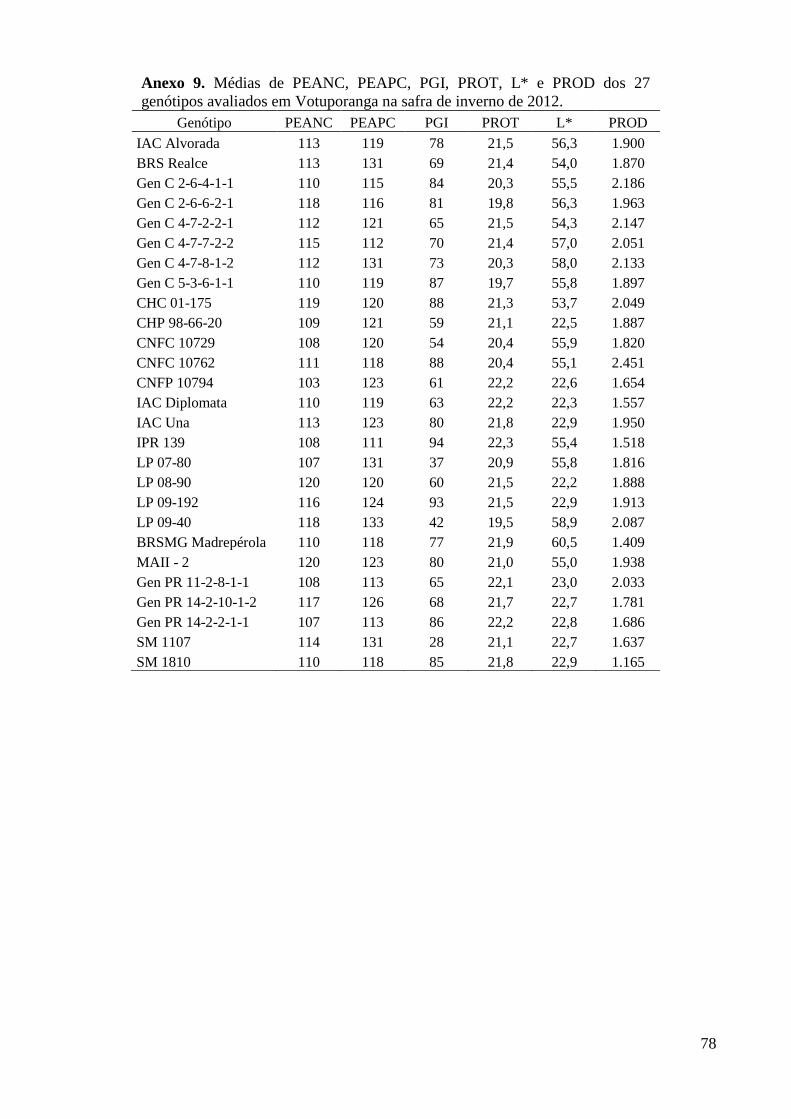
78
Anexo 9. Médias de PEANC, PEAPC, PGI, PROT, L* e PROD dos 27
genótipos avaliados em Votuporanga na safra de inverno de 2012.
Genótipo PEANC PEAPC PGI PROT L* PROD
IAC Alvorada 113 119 78 21,5 56,3 1.900
BRS Realce 113 131 69 21,4 54,0 1.870
Gen C 2-6-4-1-1 110 115 84 20,3 55,5 2.186
Gen C 2-6-6-2-1 118 116 81 19,8 56,3 1.963
Gen C 4-7-2-2-1 112 121 65 21,5 54,3 2.147
Gen C 4-7-7-2-2 115 112 70 21,4 57,0 2.051
Gen C 4-7-8-1-2 112 131 73 20,3 58,0 2.133
Gen C 5-3-6-1-1 110 119 87 19,7 55,8 1.897
CHC 01-175 119 120 88 21,3 53,7 2.049
CHP 98-66-20 109 121 59 21,1 22,5 1.887
CNFC 10729 108 120 54 20,4 55,9 1.820
CNFC 10762 111 118 88 20,4 55,1 2.451
CNFP 10794 103 123 61 22,2 22,6 1.654
IAC Diplomata 110 119 63 22,2 22,3 1.557
IAC Una 113 123 80 21,8 22,9 1.950
IPR 139 108 111 94 22,3 55,4 1.518
LP 07-80 107 131 37 20,9 55,8 1.816
LP 08-90 120 120 60 21,5 22,2 1.888
LP 09-192 116 124 93 21,5 22,9 1.913
LP 09-40 118 133 42 19,5 58,9 2.087
BRSMG Madrepérola 110 118 77 21,9 60,5 1.409
MAII - 2 120 123 80 21,0 55,0 1.938
Gen PR 11-2-8-1-1 108 113 65 22,1 23,0 2.033
Gen PR 14-2-10-1-2 117 126 68 21,7 22,7 1.781
Gen PR 14-2-2-1-1 107 113 86 22,2 22,8 1.686
SM 1107 114 131 28 21,1 22,7 1.637
SM 1810 110 118 85 21,8 22,9 1.165
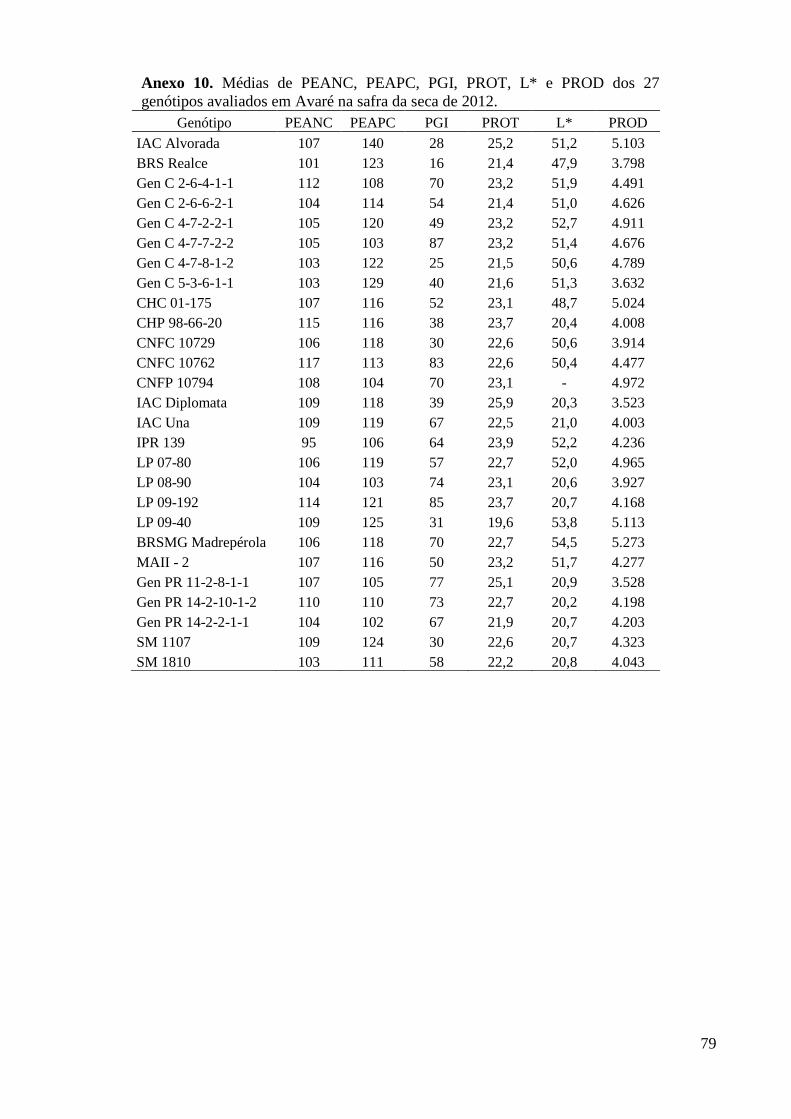
79
Anexo 10. Médias de PEANC, PEAPC, PGI, PROT, L* e PROD dos 27
genótipos avaliados em Avaré na safra da seca de 2012.
Genótipo PEANC PEAPC PGI PROT L* PROD
IAC Alvorada 107 140 28 25,2 51,2 5.103
BRS Realce 101 123 16 21,4 47,9 3.798
Gen C 2-6-4-1-1 112 108 70 23,2 51,9 4.491
Gen C 2-6-6-2-1 104 114 54 21,4 51,0 4.626
Gen C 4-7-2-2-1 105 120 49 23,2 52,7 4.911
Gen C 4-7-7-2-2 105 103 87 23,2 51,4 4.676
Gen C 4-7-8-1-2 103 122 25 21,5 50,6 4.789
Gen C 5-3-6-1-1 103 129 40 21,6 51,3 3.632
CHC 01-175 107 116 52 23,1 48,7 5.024
CHP 98-66-20 115 116 38 23,7 20,4 4.008
CNFC 10729 106 118 30 22,6 50,6 3.914
CNFC 10762 117 113 83 22,6 50,4 4.477
CNFP 10794 108 104 70 23,1 - 4.972
IAC Diplomata 109 118 39 25,9 20,3 3.523
IAC Una 109 119 67 22,5 21,0 4.003
IPR 139 95 106 64 23,9 52,2 4.236
LP 07-80 106 119 57 22,7 52,0 4.965
LP 08-90 104 103 74 23,1 20,6 3.927
LP 09-192 114 121 85 23,7 20,7 4.168
LP 09-40 109 125 31 19,6 53,8 5.113
BRSMG Madrepérola 106 118 70 22,7 54,5 5.273
MAII - 2 107 116 50 23,2 51,7 4.277
Gen PR 11-2-8-1-1 107 105 77 25,1 20,9 3.528
Gen PR 14-2-10-1-2 110 110 73 22,7 20,2 4.198
Gen PR 14-2-2-1-1 104 102 67 21,9 20,7 4.203
SM 1107 109 124 30 22,6 20,7 4.323
SM 1810 103 111 58 22,2 20,8 4.043
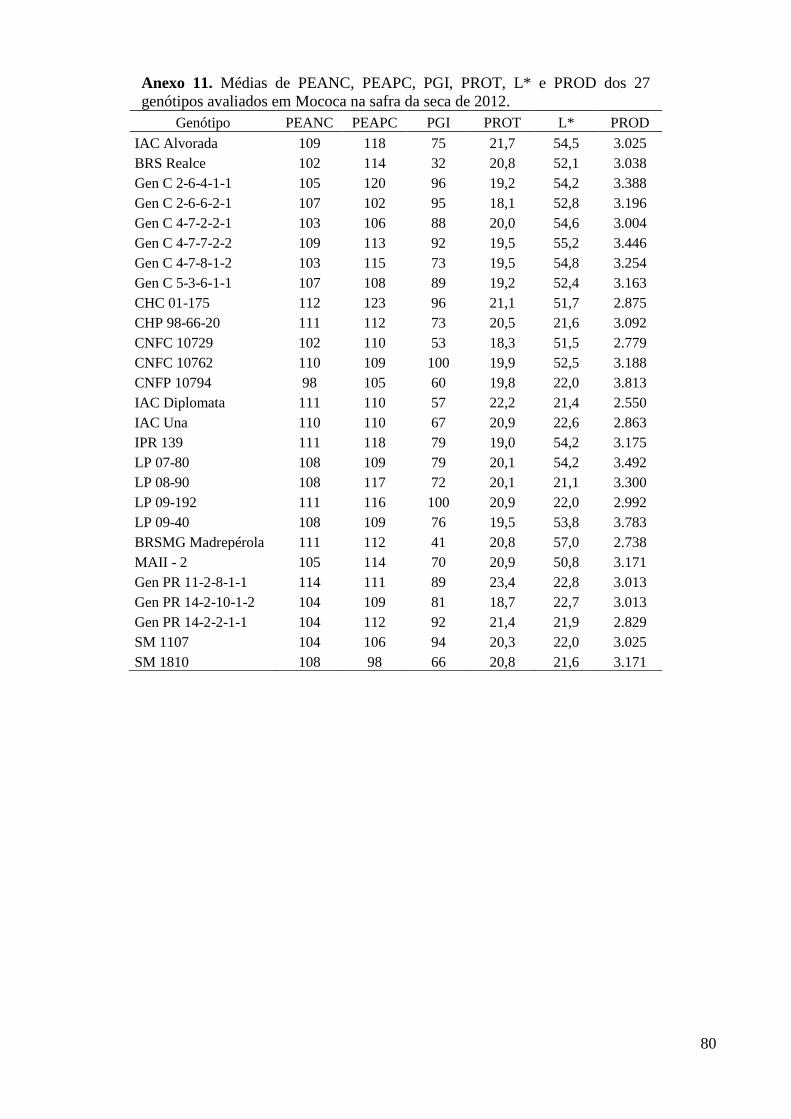
80
Anexo 11. Médias de PEANC, PEAPC, PGI, PROT, L* e PROD dos 27
genótipos avaliados em Mococa na safra da seca de 2012.
Genótipo PEANC PEAPC PGI PROT L* PROD
IAC Alvorada 109 118 75 21,7 54,5 3.025
BRS Realce 102 114 32 20,8 52,1 3.038
Gen C 2-6-4-1-1 105 120 96 19,2 54,2 3.388
Gen C 2-6-6-2-1 107 102 95 18,1 52,8 3.196
Gen C 4-7-2-2-1 103 106 88 20,0 54,6 3.004
Gen C 4-7-7-2-2 109 113 92 19,5 55,2 3.446
Gen C 4-7-8-1-2 103 115 73 19,5 54,8 3.254
Gen C 5-3-6-1-1 107 108 89 19,2 52,4 3.163
CHC 01-175 112 123 96 21,1 51,7 2.875
CHP 98-66-20 111 112 73 20,5 21,6 3.092
CNFC 10729 102 110 53 18,3 51,5 2.779
CNFC 10762 110 109 100 19,9 52,5 3.188
CNFP 10794 98 105 60 19,8 22,0 3.813
IAC Diplomata 111 110 57 22,2 21,4 2.550
IAC Una 110 110 67 20,9 22,6 2.863
IPR 139 111 118 79 19,0 54,2 3.175
LP 07-80 108 109 79 20,1 54,2 3.492
LP 08-90 108 117 72 20,1 21,1 3.300
LP 09-192 111 116 100 20,9 22,0 2.992
LP 09-40 108 109 76 19,5 53,8 3.783
BRSMG Madrepérola 111 112 41 20,8 57,0 2.738
MAII - 2 105 114 70 20,9 50,8 3.171
Gen PR 11-2-8-1-1 114 111 89 23,4 22,8 3.013
Gen PR 14-2-10-1-2 104 109 81 18,7 22,7 3.013
Gen PR 14-2-2-1-1 104 112 92 21,4 21,9 2.829
SM 1107 104 106 94 20,3 22,0 3.025
SM 1810 108 98 66 20,8 21,6 3.171
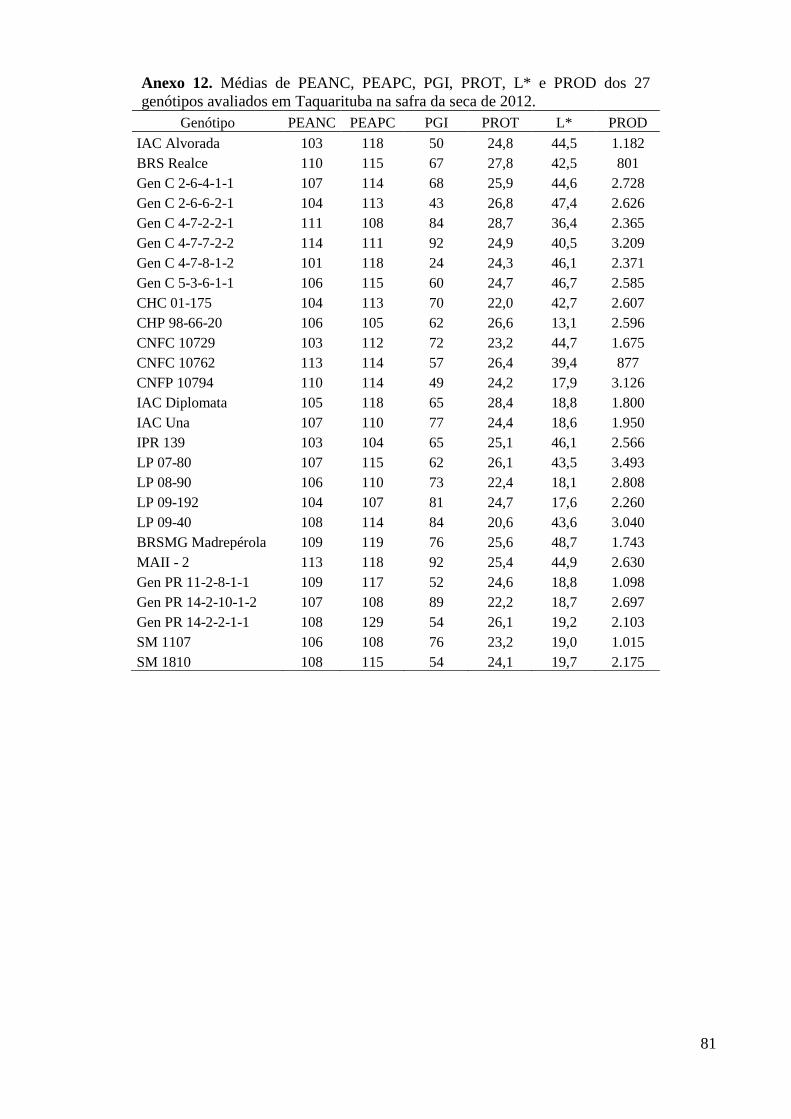
81
Anexo 12. Médias de PEANC, PEAPC, PGI, PROT, L* e PROD dos 27
genótipos avaliados em Taquarituba na safra da seca de 2012.
Genótipo PEANC PEAPC PGI PROT L* PROD
IAC Alvorada 103 118 50 24,8 44,5 1.182
BRS Realce 110 115 67 27,8 42,5 801
Gen C 2-6-4-1-1 107 114 68 25,9 44,6 2.728
Gen C 2-6-6-2-1 104 113 43 26,8 47,4 2.626
Gen C 4-7-2-2-1 111 108 84 28,7 36,4 2.365
Gen C 4-7-7-2-2 114 111 92 24,9 40,5 3.209
Gen C 4-7-8-1-2 101 118 24 24,3 46,1 2.371
Gen C 5-3-6-1-1 106 115 60 24,7 46,7 2.585
CHC 01-175 104 113 70 22,0 42,7 2.607
CHP 98-66-20 106 105 62 26,6 13,1 2.596
CNFC 10729 103 112 72 23,2 44,7 1.675
CNFC 10762 113 114 57 26,4 39,4 877
CNFP 10794 110 114 49 24,2 17,9 3.126
IAC Diplomata 105 118 65 28,4 18,8 1.800
IAC Una 107 110 77 24,4 18,6 1.950
IPR 139 103 104 65 25,1 46,1 2.566
LP 07-80 107 115 62 26,1 43,5 3.493
LP 08-90 106 110 73 22,4 18,1 2.808
LP 09-192 104 107 81 24,7 17,6 2.260
LP 09-40 108 114 84 20,6 43,6 3.040
BRSMG Madrepérola 109 119 76 25,6 48,7 1.743
MAII - 2 113 118 92 25,4 44,9 2.630
Gen PR 11-2-8-1-1 109 117 52 24,6 18,8 1.098
Gen PR 14-2-10-1-2 107 108 89 22,2 18,7 2.697
Gen PR 14-2-2-1-1 108 129 54 26,1 19,2 2.103
SM 1107 106 108 76 23,2 19,0 1.015
SM 1810 108 115 54 24,1 19,7 2.175
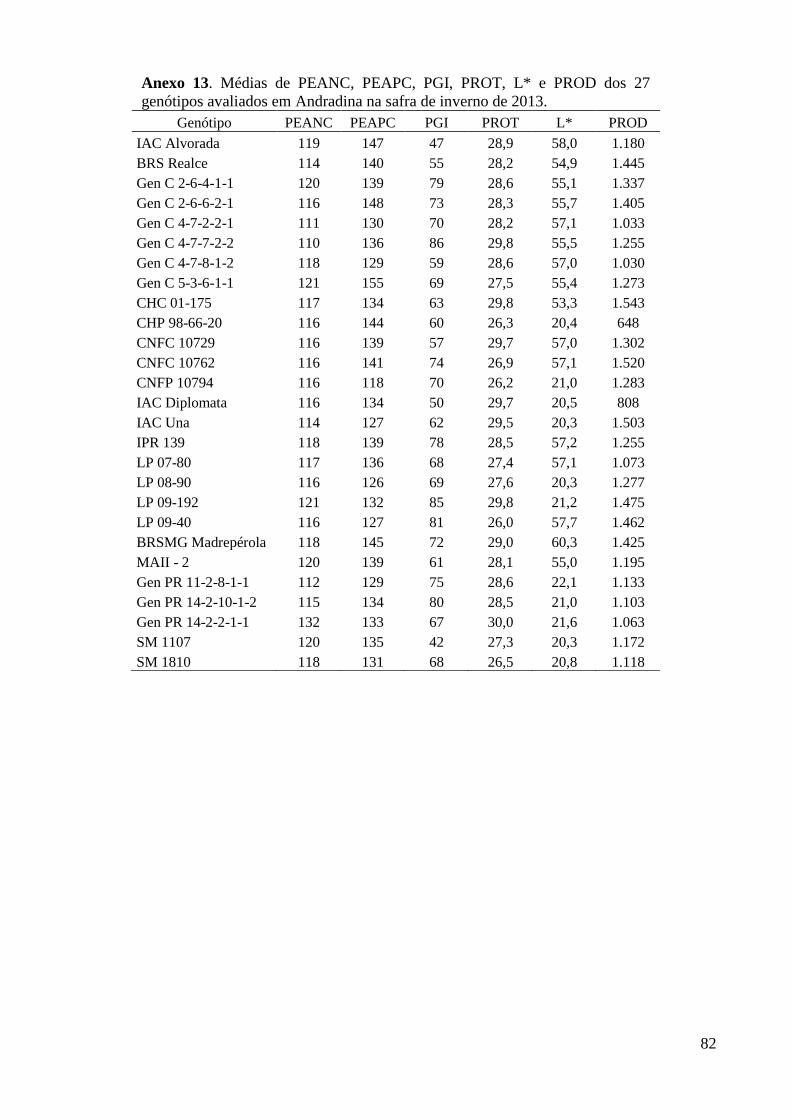
82
Anexo 13. Médias de PEANC, PEAPC, PGI, PROT, L* e PROD dos 27
genótipos avaliados em Andradina na safra de inverno de 2013.
Genótipo PEANC PEAPC PGI PROT L* PROD
IAC Alvorada 119 147 47 28,9 58,0 1.180
BRS Realce 114 140 55 28,2 54,9 1.445
Gen C 2-6-4-1-1 120 139 79 28,6 55,1 1.337
Gen C 2-6-6-2-1 116 148 73 28,3 55,7 1.405
Gen C 4-7-2-2-1 111 130 70 28,2 57,1 1.033
Gen C 4-7-7-2-2 110 136 86 29,8 55,5 1.255
Gen C 4-7-8-1-2 118 129 59 28,6 57,0 1.030
Gen C 5-3-6-1-1 121 155 69 27,5 55,4 1.273
CHC 01-175 117 134 63 29,8 53,3 1.543
CHP 98-66-20 116 144 60 26,3 20,4 648
CNFC 10729 116 139 57 29,7 57,0 1.302
CNFC 10762 116 141 74 26,9 57,1 1.520
CNFP 10794 116 118 70 26,2 21,0 1.283
IAC Diplomata 116 134 50 29,7 20,5 808
IAC Una 114 127 62 29,5 20,3 1.503
IPR 139 118 139 78 28,5 57,2 1.255
LP 07-80 117 136 68 27,4 57,1 1.073
LP 08-90 116 126 69 27,6 20,3 1.277
LP 09-192 121 132 85 29,8 21,2 1.475
LP 09-40 116 127 81 26,0 57,7 1.462
BRSMG Madrepérola 118 145 72 29,0 60,3 1.425
MAII - 2 120 139 61 28,1 55,0 1.195
Gen PR 11-2-8-1-1 112 129 75 28,6 22,1 1.133
Gen PR 14-2-10-1-2 115 134 80 28,5 21,0 1.103
Gen PR 14-2-2-1-1 132 133 67 30,0 21,6 1.063
SM 1107 120 135 42 27,3 20,3 1.172
SM 1810 118 131 68 26,5 20,8 1.118
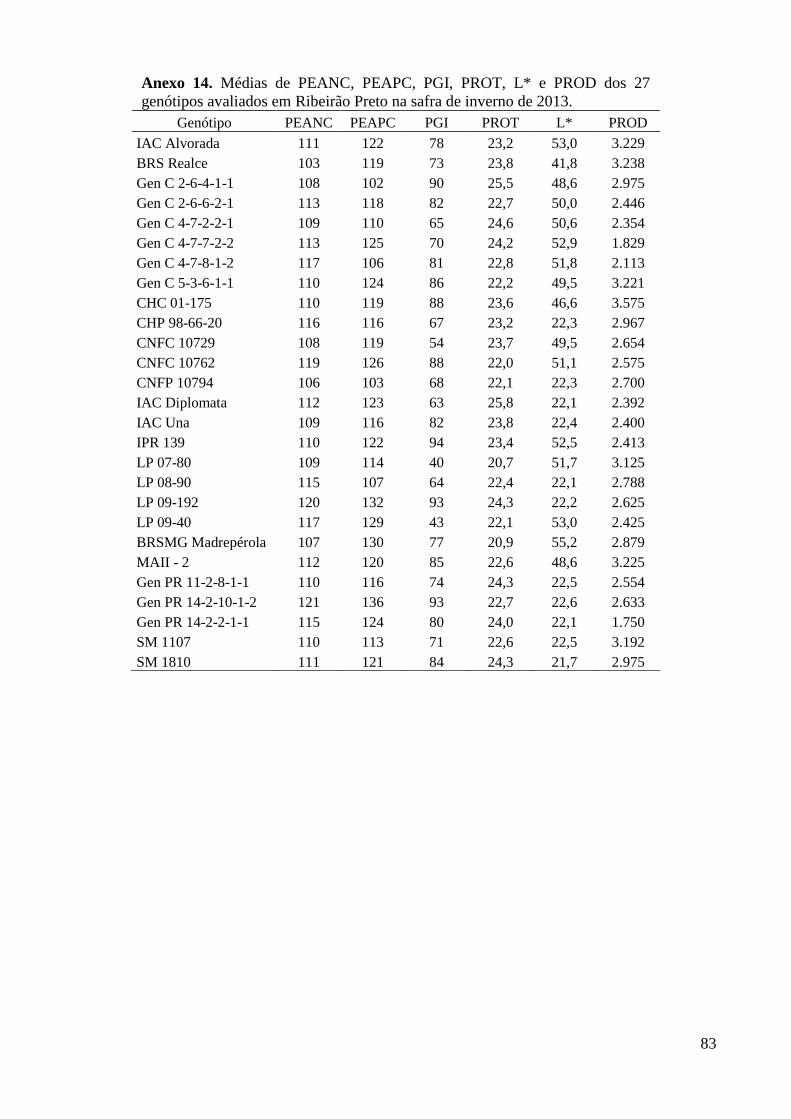
83
Anexo 14. Médias de PEANC, PEAPC, PGI, PROT, L* e PROD dos 27
genótipos avaliados em Ribeirão Preto na safra de inverno de 2013.
Genótipo PEANC PEAPC PGI PROT L* PROD
IAC Alvorada 111 122 78 23,2 53,0 3.229
BRS Realce 103 119 73 23,8 41,8 3.238
Gen C 2-6-4-1-1 108 102 90 25,5 48,6 2.975
Gen C 2-6-6-2-1 113 118 82 22,7 50,0 2.446
Gen C 4-7-2-2-1 109 110 65 24,6 50,6 2.354
Gen C 4-7-7-2-2 113 125 70 24,2 52,9 1.829
Gen C 4-7-8-1-2 117 106 81 22,8 51,8 2.113
Gen C 5-3-6-1-1 110 124 86 22,2 49,5 3.221
CHC 01-175 110 119 88 23,6 46,6 3.575
CHP 98-66-20 116 116 67 23,2 22,3 2.967
CNFC 10729 108 119 54 23,7 49,5 2.654
CNFC 10762 119 126 88 22,0 51,1 2.575
CNFP 10794 106 103 68 22,1 22,3 2.700
IAC Diplomata 112 123 63 25,8 22,1 2.392
IAC Una 109 116 82 23,8 22,4 2.400
IPR 139 110 122 94 23,4 52,5 2.413
LP 07-80 109 114 40 20,7 51,7 3.125
LP 08-90 115 107 64 22,4 22,1 2.788
LP 09-192 120 132 93 24,3 22,2 2.625
LP 09-40 117 129 43 22,1 53,0 2.425
BRSMG Madrepérola 107 130 77 20,9 55,2 2.879
MAII - 2 112 120 85 22,6 48,6 3.225
Gen PR 11-2-8-1-1 110 116 74 24,3 22,5 2.554
Gen PR 14-2-10-1-2 121 136 93 22,7 22,6 2.633
Gen PR 14-2-2-1-1 115 124 80 24,0 22,1 1.750
SM 1107 110 113 71 22,6 22,5 3.192
SM 1810 111 121 84 24,3 21,7 2.975
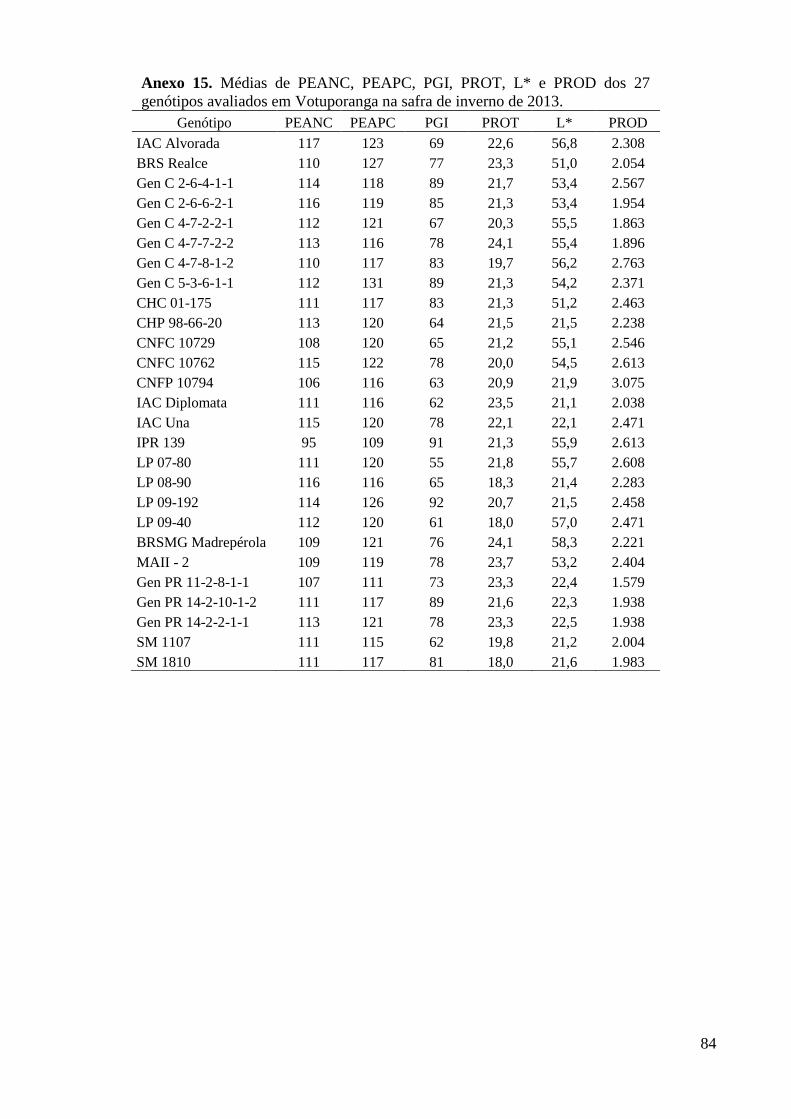
84
Anexo 15. Médias de PEANC, PEAPC, PGI, PROT, L* e PROD dos 27
genótipos avaliados em Votuporanga na safra de inverno de 2013.
Genótipo PEANC PEAPC PGI PROT L* PROD
IAC Alvorada 117 123 69 22,6 56,8 2.308
BRS Realce 110 127 77 23,3 51,0 2.054
Gen C 2-6-4-1-1 114 118 89 21,7 53,4 2.567
Gen C 2-6-6-2-1 116 119 85 21,3 53,4 1.954
Gen C 4-7-2-2-1 112 121 67 20,3 55,5 1.863
Gen C 4-7-7-2-2 113 116 78 24,1 55,4 1.896
Gen C 4-7-8-1-2 110 117 83 19,7 56,2 2.763
Gen C 5-3-6-1-1 112 131 89 21,3 54,2 2.371
CHC 01-175 111 117 83 21,3 51,2 2.463
CHP 98-66-20 113 120 64 21,5 21,5 2.238
CNFC 10729 108 120 65 21,2 55,1 2.546
CNFC 10762 115 122 78 20,0 54,5 2.613
CNFP 10794 106 116 63 20,9 21,9 3.075
IAC Diplomata 111 116 62 23,5 21,1 2.038
IAC Una 115 120 78 22,1 22,1 2.471
IPR 139 95 109 91 21,3 55,9 2.613
LP 07-80 111 120 55 21,8 55,7 2.608
LP 08-90 116 116 65 18,3 21,4 2.283
LP 09-192 114 126 92 20,7 21,5 2.458
LP 09-40 112 120 61 18,0 57,0 2.471
BRSMG Madrepérola 109 121 76 24,1 58,3 2.221
MAII - 2 109 119 78 23,7 53,2 2.404
Gen PR 11-2-8-1-1 107 111 73 23,3 22,4 1.579
Gen PR 14-2-10-1-2 111 117 89 21,6 22,3 1.938
Gen PR 14-2-2-1-1 113 121 78 23,3 22,5 1.938
SM 1107 111 115 62 19,8 21,2 2.004
SM 1810 111 117 81 18,0 21,6 1.983
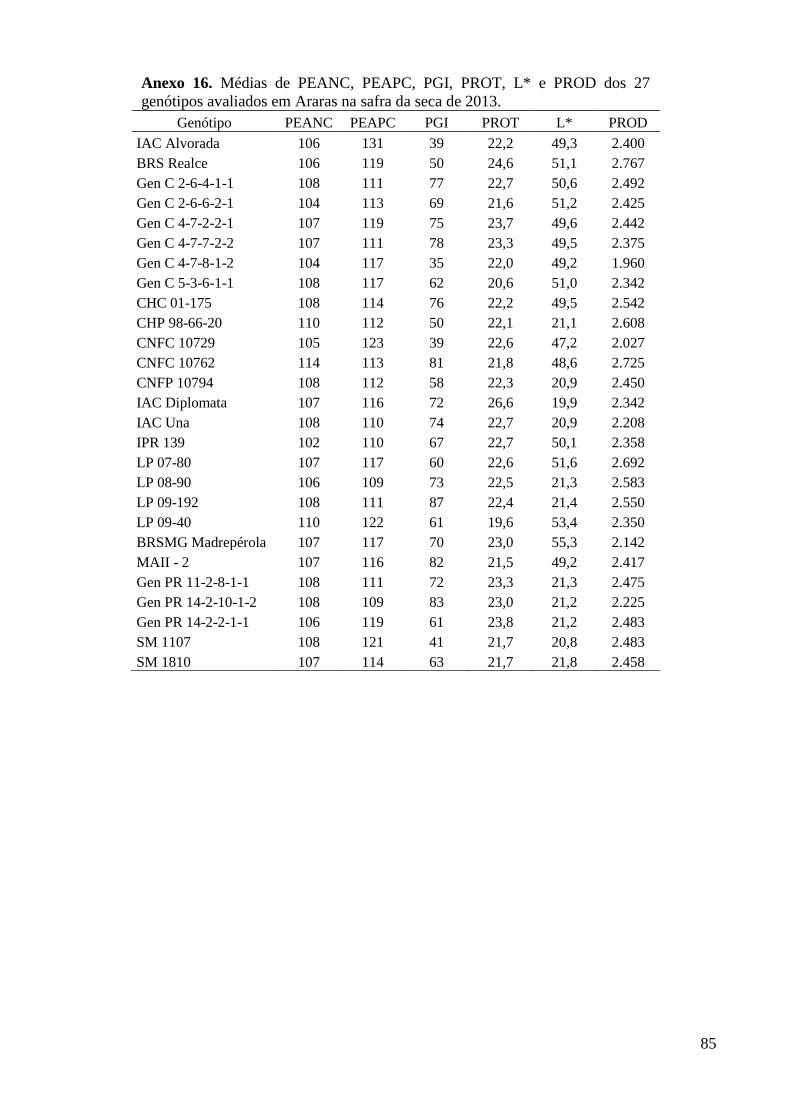
85
Anexo 16. Médias de PEANC, PEAPC, PGI, PROT, L* e PROD dos 27
genótipos avaliados em Araras na safra da seca de 2013.
Genótipo PEANC PEAPC PGI PROT L* PROD
IAC Alvorada 106 131 39 22,2 49,3 2.400
BRS Realce 106 119 50 24,6 51,1 2.767
Gen C 2-6-4-1-1 108 111 77 22,7 50,6 2.492
Gen C 2-6-6-2-1 104 113 69 21,6 51,2 2.425
Gen C 4-7-2-2-1 107 119 75 23,7 49,6 2.442
Gen C 4-7-7-2-2 107 111 78 23,3 49,5 2.375
Gen C 4-7-8-1-2 104 117 35 22,0 49,2 1.960
Gen C 5-3-6-1-1 108 117 62 20,6 51,0 2.342
CHC 01-175 108 114 76 22,2 49,5 2.542
CHP 98-66-20 110 112 50 22,1 21,1 2.608
CNFC 10729 105 123 39 22,6 47,2 2.027
CNFC 10762 114 113 81 21,8 48,6 2.725
CNFP 10794 108 112 58 22,3 20,9 2.450
IAC Diplomata 107 116 72 26,6 19,9 2.342
IAC Una 108 110 74 22,7 20,9 2.208
IPR 139 102 110 67 22,7 50,1 2.358
LP 07-80 107 117 60 22,6 51,6 2.692
LP 08-90 106 109 73 22,5 21,3 2.583
LP 09-192 108 111 87 22,4 21,4 2.550
LP 09-40 110 122 61 19,6 53,4 2.350
BRSMG Madrepérola 107 117 70 23,0 55,3 2.142
MAII - 2 107 116 82 21,5 49,2 2.417
Gen PR 11-2-8-1-1 108 111 72 23,3 21,3 2.475
Gen PR 14-2-10-1-2 108 109 83 23,0 21,2 2.225
Gen PR 14-2-2-1-1 106 119 61 23,8 21,2 2.483
SM 1107 108 121 41 21,7 20,8 2.483
SM 1810 107 114 63 21,7 21,8 2.458
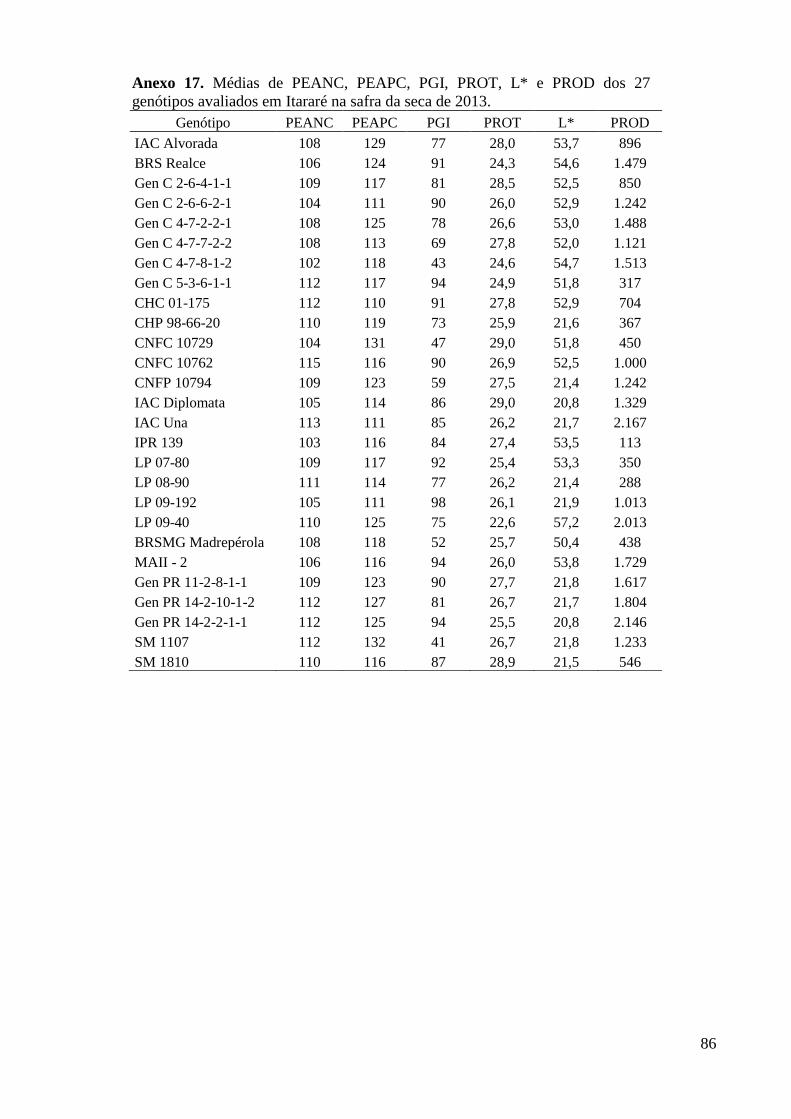
86
Anexo 17. Médias de PEANC, PEAPC, PGI, PROT, L* e PROD dos 27
genótipos avaliados em Itararé na safra da seca de 2013.
Genótipo PEANC PEAPC PGI PROT L* PROD
IAC Alvorada 108 129 77 28,0 53,7 896
BRS Realce 106 124 91 24,3 54,6 1.479
Gen C 2-6-4-1-1 109 117 81 28,5 52,5 850
Gen C 2-6-6-2-1 104 111 90 26,0 52,9 1.242
Gen C 4-7-2-2-1 108 125 78 26,6 53,0 1.488
Gen C 4-7-7-2-2 108 113 69 27,8 52,0 1.121
Gen C 4-7-8-1-2 102 118 43 24,6 54,7 1.513
Gen C 5-3-6-1-1 112 117 94 24,9 51,8 317
CHC 01-175 112 110 91 27,8 52,9 704
CHP 98-66-20 110 119 73 25,9 21,6 367
CNFC 10729 104 131 47 29,0 51,8 450
CNFC 10762 115 116 90 26,9 52,5 1.000
CNFP 10794 109 123 59 27,5 21,4 1.242
IAC Diplomata 105 114 86 29,0 20,8 1.329
IAC Una 113 111 85 26,2 21,7 2.167
IPR 139 103 116 84 27,4 53,5 113
LP 07-80 109 117 92 25,4 53,3 350
LP 08-90 111 114 77 26,2 21,4 288
LP 09-192 105 111 98 26,1 21,9 1.013
LP 09-40 110 125 75 22,6 57,2 2.013
BRSMG Madrepérola 108 118 52 25,7 50,4 438
MAII - 2 106 116 94 26,0 53,8 1.729
Gen PR 11-2-8-1-1 109 123 90 27,7 21,8 1.617
Gen PR 14-2-10-1-2 112 127 81 26,7 21,7 1.804
Gen PR 14-2-2-1-1 112 125 94 25,5 20,8 2.146
SM 1107 112 132 41 26,7 21,8 1.233
SM 1810 110 116 87 28,9 21,5 546
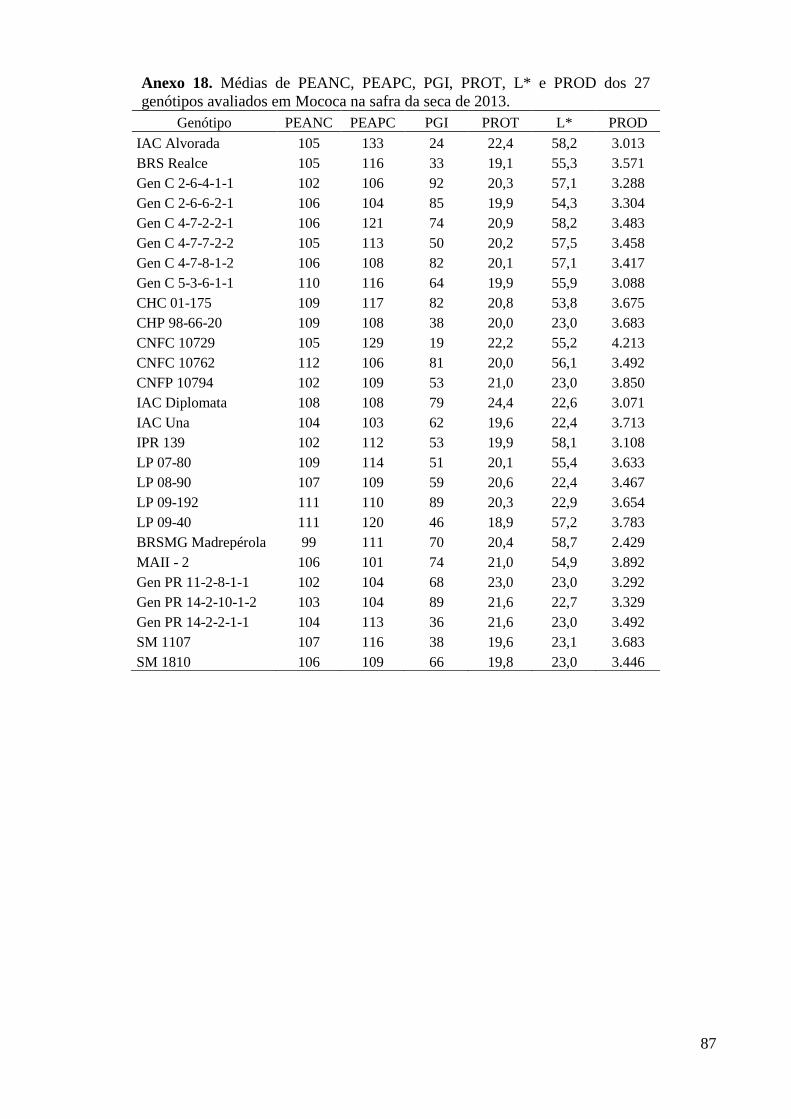
87
Anexo 18. Médias de PEANC, PEAPC, PGI, PROT, L* e PROD dos 27
genótipos avaliados em Mococa na safra da seca de 2013.
Genótipo PEANC PEAPC PGI PROT L* PROD
IAC Alvorada 105 133 24 22,4 58,2 3.013
BRS Realce 105 116 33 19,1 55,3 3.571
Gen C 2-6-4-1-1 102 106 92 20,3 57,1 3.288
Gen C 2-6-6-2-1 106 104 85 19,9 54,3 3.304
Gen C 4-7-2-2-1 106 121 74 20,9 58,2 3.483
Gen C 4-7-7-2-2 105 113 50 20,2 57,5 3.458
Gen C 4-7-8-1-2 106 108 82 20,1 57,1 3.417
Gen C 5-3-6-1-1 110 116 64 19,9 55,9 3.088
CHC 01-175 109 117 82 20,8 53,8 3.675
CHP 98-66-20 109 108 38 20,0 23,0 3.683
CNFC 10729 105 129 19 22,2 55,2 4.213
CNFC 10762 112 106 81 20,0 56,1 3.492
CNFP 10794 102 109 53 21,0 23,0 3.850
IAC Diplomata 108 108 79 24,4 22,6 3.071
IAC Una 104 103 62 19,6 22,4 3.713
IPR 139 102 112 53 19,9 58,1 3.108
LP 07-80 109 114 51 20,1 55,4 3.633
LP 08-90 107 109 59 20,6 22,4 3.467
LP 09-192 111 110 89 20,3 22,9 3.654
LP 09-40 111 120 46 18,9 57,2 3.783
BRSMG Madrepérola 99 111 70 20,4 58,7 2.429
MAII - 2 106 101 74 21,0 54,9 3.892
Gen PR 11-2-8-1-1 102 104 68 23,0 23,0 3.292
Gen PR 14-2-10-1-2 103 104 89 21,6 22,7 3.329
Gen PR 14-2-2-1-1 104 113 36 21,6 23,0 3.492
SM 1107 107 116 38 19,6 23,1 3.683
SM 1810 106 109 66 19,8 23,0 3.446
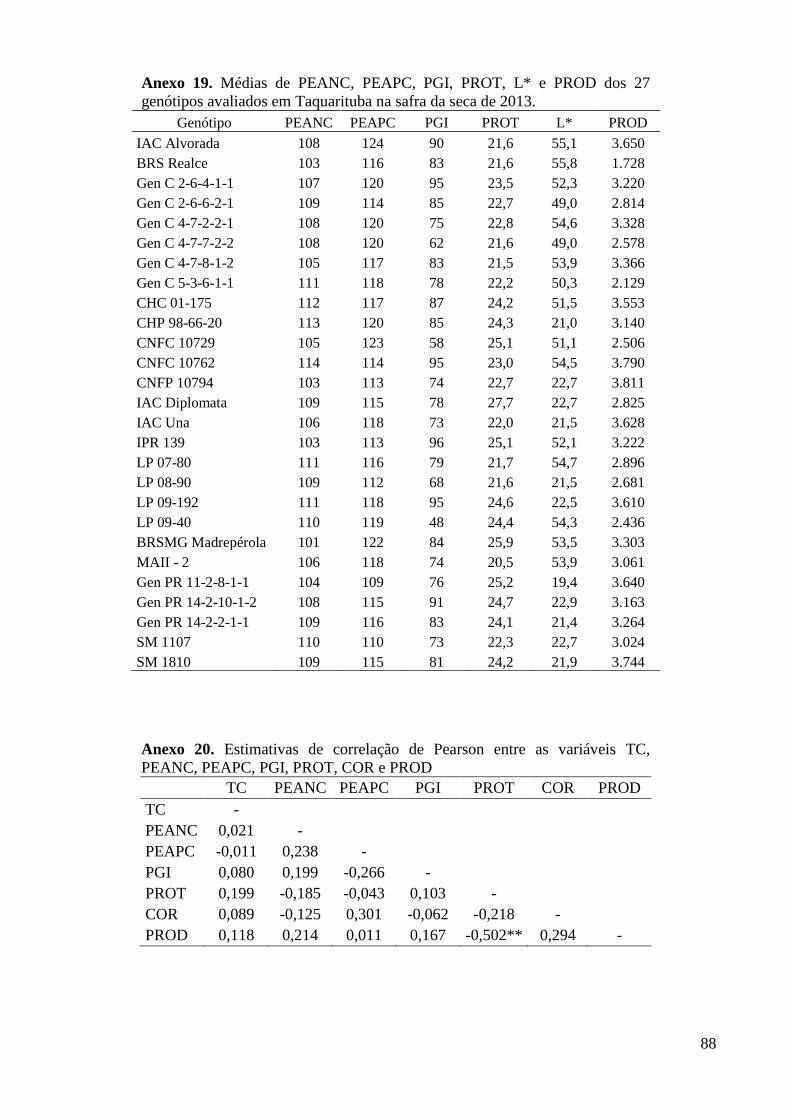
88
Anexo 19. Médias de PEANC, PEAPC, PGI, PROT, L* e PROD dos 27
genótipos avaliados em Taquarituba na safra da seca de 2013.
Genótipo PEANC PEAPC PGI PROT L* PROD
IAC Alvorada 108 124 90 21,6 55,1 3.650
BRS Realce 103 116 83 21,6 55,8 1.728
Gen C 2-6-4-1-1 107 120 95 23,5 52,3 3.220
Gen C 2-6-6-2-1 109 114 85 22,7 49,0 2.814
Gen C 4-7-2-2-1 108 120 75 22,8 54,6 3.328
Gen C 4-7-7-2-2 108 120 62 21,6 49,0 2.578
Gen C 4-7-8-1-2 105 117 83 21,5 53,9 3.366
Gen C 5-3-6-1-1 111 118 78 22,2 50,3 2.129
CHC 01-175 112 117 87 24,2 51,5 3.553
CHP 98-66-20 113 120 85 24,3 21,0 3.140
CNFC 10729 105 123 58 25,1 51,1 2.506
CNFC 10762 114 114 95 23,0 54,5 3.790
CNFP 10794 103 113 74 22,7 22,7 3.811
IAC Diplomata 109 115 78 27,7 22,7 2.825
IAC Una 106 118 73 22,0 21,5 3.628
IPR 139 103 113 96 25,1 52,1 3.222
LP 07-80 111 116 79 21,7 54,7 2.896
LP 08-90 109 112 68 21,6 21,5 2.681
LP 09-192 111 118 95 24,6 22,5 3.610
LP 09-40 110 119 48 24,4 54,3 2.436
BRSMG Madrepérola 101 122 84 25,9 53,5 3.303
MAII - 2 106 118 74 20,5 53,9 3.061
Gen PR 11-2-8-1-1 104 109 76 25,2 19,4 3.640
Gen PR 14-2-10-1-2 108 115 91 24,7 22,9 3.163
Gen PR 14-2-2-1-1 109 116 83 24,1 21,4 3.264
SM 1107 110 110 73 22,3 22,7 3.024
SM 1810 109 115 81 24,2 21,9 3.744
Anexo 20. Estimativas de correlação de Pearson entre as variáveis TC,
PEANC, PEAPC, PGI, PROT, COR e PROD
TC PEANC PEAPC PGI PROT COR PROD
TC -
PEANC 0,021 -
PEAPC -0,011 0,238 -
PGI 0,080 0,199 -0,266 -
PROT 0,199 -0,185 -0,043 0,103 -
COR 0,089 -0,125 0,301 -0,062 -0,218 -
PROD 0,118 0,214 0,011 0,167 -0,502** 0,294 -
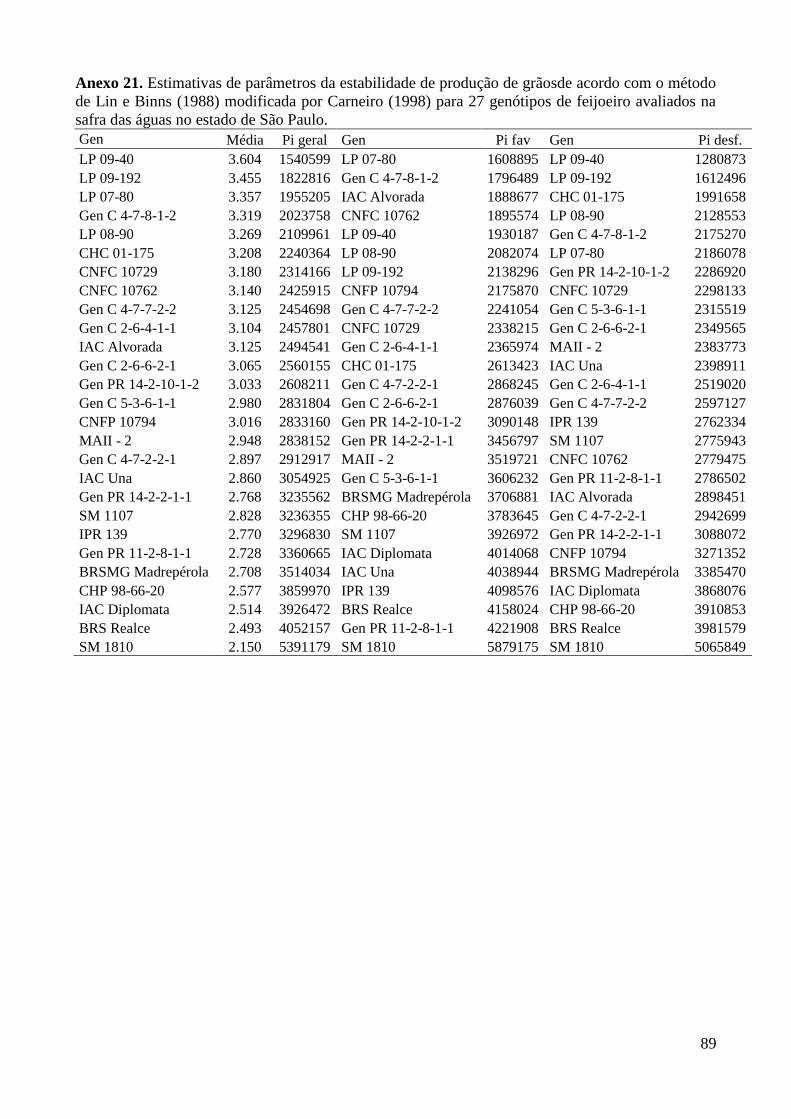
89
Anexo 21. Estimativas de parâmetros da estabilidade de produção de grãosde acordo com o método
de Lin e Binns (1988) modificada por Carneiro (1998) para 27 genótipos de feijoeiro avaliados na
safra das águas no estado de São Paulo.
Gen Média Pi geral Gen Pi fav Gen Pi desf.
LP 09-40 3.604 1540599 LP 07-80 1608895 LP 09-40 1280873
LP 09-192 3.455 1822816 Gen C 4-7-8-1-2 1796489 LP 09-192 1612496
LP 07-80 3.357 1955205 IAC Alvorada 1888677 CHC 01-175 1991658
Gen C 4-7-8-1-2 3.319 2023758 CNFC 10762 1895574 LP 08-90 2128553
LP 08-90 3.269 2109961 LP 09-40 1930187 Gen C 4-7-8-1-2 2175270
CHC 01-175 3.208 2240364 LP 08-90 2082074 LP 07-80 2186078
CNFC 10729 3.180 2314166 LP 09-192 2138296 Gen PR 14-2-10-1-2 2286920
CNFC 10762 3.140 2425915 CNFP 10794 2175870 CNFC 10729 2298133
Gen C 4-7-7-2-2 3.125 2454698 Gen C 4-7-7-2-2 2241054 Gen C 5-3-6-1-1 2315519
Gen C 2-6-4-1-1 3.104 2457801 CNFC 10729 2338215 Gen C 2-6-6-2-1 2349565
IAC Alvorada 3.125 2494541 Gen C 2-6-4-1-1 2365974 MAII - 2 2383773
Gen C 2-6-6-2-1 3.065 2560155 CHC 01-175 2613423 IAC Una 2398911
Gen PR 14-2-10-1-2 3.033 2608211 Gen C 4-7-2-2-1 2868245 Gen C 2-6-4-1-1 2519020
Gen C 5-3-6-1-1 2.980 2831804 Gen C 2-6-6-2-1 2876039 Gen C 4-7-7-2-2 2597127
CNFP 10794 3.016 2833160 Gen PR 14-2-10-1-2 3090148 IPR 139 2762334
MAII - 2 2.948 2838152 Gen PR 14-2-2-1-1 3456797 SM 1107 2775943
Gen C 4-7-2-2-1 2.897 2912917 MAII - 2 3519721 CNFC 10762 2779475
IAC Una 2.860 3054925 Gen C 5-3-6-1-1 3606232 Gen PR 11-2-8-1-1 2786502
Gen PR 14-2-2-1-1 2.768 3235562 BRSMG Madrepérola 3706881 IAC Alvorada 2898451
SM 1107 2.828 3236355 CHP 98-66-20 3783645 Gen C 4-7-2-2-1 2942699
IPR 139 2.770 3296830 SM 1107 3926972 Gen PR 14-2-2-1-1 3088072
Gen PR 11-2-8-1-1 2.728 3360665 IAC Diplomata 4014068 CNFP 10794 3271352
BRSMG Madrepérola 2.708 3514034 IAC Una 4038944 BRSMG Madrepérola 3385470
CHP 98-66-20 2.577 3859970 IPR 139 4098576 IAC Diplomata 3868076
IAC Diplomata 2.514 3926472 BRS Realce 4158024 CHP 98-66-20 3910853
BRS Realce 2.493 4052157 Gen PR 11-2-8-1-1 4221908 BRS Realce 3981579
SM 1810 2.150 5391179 SM 1810 5879175 SM 1810 5065849
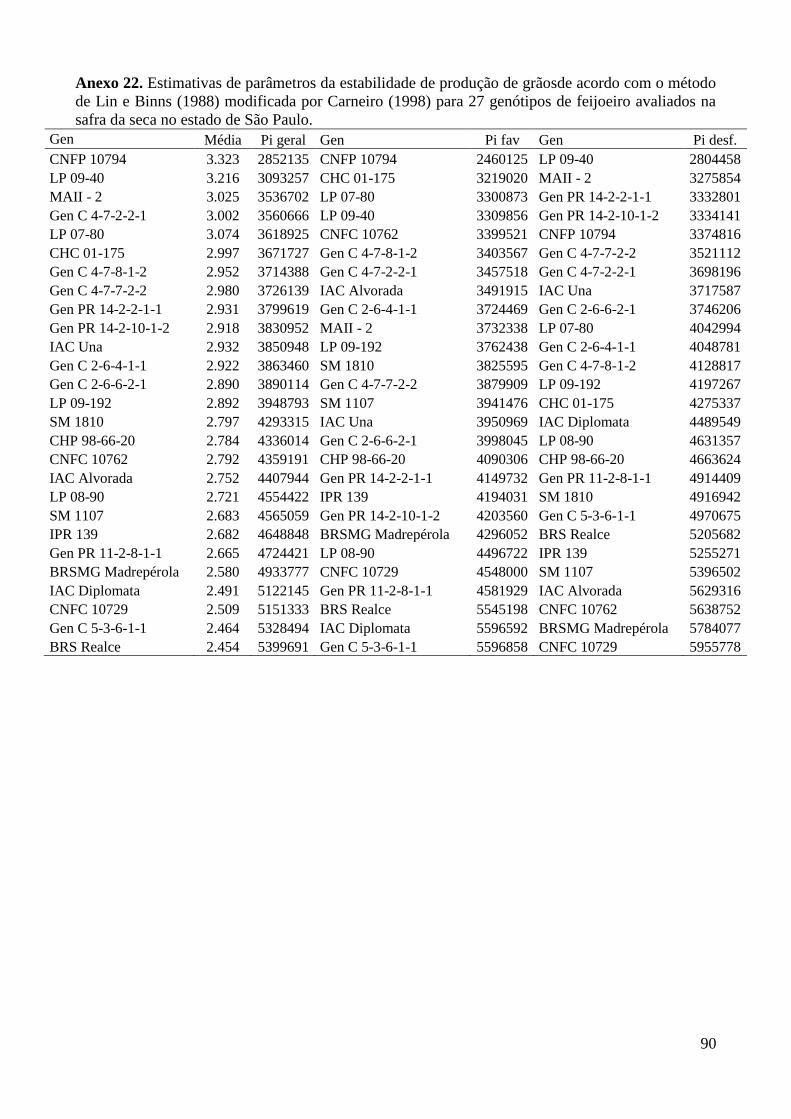
90
Anexo 22. Estimativas de parâmetros da estabilidade de produção de grãosde acordo com o método
de Lin e Binns (1988) modificada por Carneiro (1998) para 27 genótipos de feijoeiro avaliados na
safra da seca no estado de São Paulo.
Gen Média Pi geral Gen Pi fav Gen Pi desf.
CNFP 10794 3.323 2852135 CNFP 10794 2460125 LP 09-40 2804458
LP 09-40 3.216 3093257 CHC 01-175 3219020 MAII - 2 3275854
MAII - 2 3.025 3536702 LP 07-80 3300873 Gen PR 14-2-2-1-1 3332801
Gen C 4-7-2-2-1 3.002 3560666 LP 09-40 3309856 Gen PR 14-2-10-1-2 3334141
LP 07-80 3.074 3618925 CNFC 10762 3399521 CNFP 10794 3374816
CHC 01-175 2.997 3671727 Gen C 4-7-8-1-2 3403567 Gen C 4-7-7-2-2 3521112
Gen C 4-7-8-1-2 2.952 3714388 Gen C 4-7-2-2-1 3457518 Gen C 4-7-2-2-1 3698196
Gen C 4-7-7-2-2 2.980 3726139 IAC Alvorada 3491915 IAC Una 3717587
Gen PR 14-2-2-1-1 2.931 3799619 Gen C 2-6-4-1-1 3724469 Gen C 2-6-6-2-1 3746206
Gen PR 14-2-10-1-2 2.918 3830952 MAII - 2 3732338 LP 07-80 4042994
IAC Una 2.932 3850948 LP 09-192 3762438 Gen C 2-6-4-1-1 4048781
Gen C 2-6-4-1-1 2.922 3863460 SM 1810 3825595 Gen C 4-7-8-1-2 4128817
Gen C 2-6-6-2-1 2.890 3890114 Gen C 4-7-7-2-2 3879909 LP 09-192 4197267
LP 09-192 2.892 3948793 SM 1107 3941476 CHC 01-175 4275337
SM 1810 2.797 4293315 IAC Una 3950969 IAC Diplomata 4489549
CHP 98-66-20 2.784 4336014 Gen C 2-6-6-2-1 3998045 LP 08-90 4631357
CNFC 10762 2.792 4359191 CHP 98-66-20 4090306 CHP 98-66-20 4663624
IAC Alvorada 2.752 4407944 Gen PR 14-2-2-1-1 4149732 Gen PR 11-2-8-1-1 4914409
LP 08-90 2.721 4554422 IPR 139 4194031 SM 1810 4916942
SM 1107 2.683 4565059 Gen PR 14-2-10-1-2 4203560 Gen C 5-3-6-1-1 4970675
IPR 139 2.682 4648848 BRSMG Madrepérola 4296052 BRS Realce 5205682
Gen PR 11-2-8-1-1 2.665 4724421 LP 08-90 4496722 IPR 139 5255271
BRSMG Madrepérola 2.580 4933777 CNFC 10729 4548000 SM 1107 5396502
IAC Diplomata 2.491 5122145 Gen PR 11-2-8-1-1 4581929 IAC Alvorada 5629316
CNFC 10729 2.509 5151333 BRS Realce 5545198 CNFC 10762 5638752
Gen C 5-3-6-1-1 2.464 5328494 IAC Diplomata 5596592 BRSMG Madrepérola 5784077
BRS Realce 2.454 5399691 Gen C 5-3-6-1-1 5596858 CNFC 10729 5955778
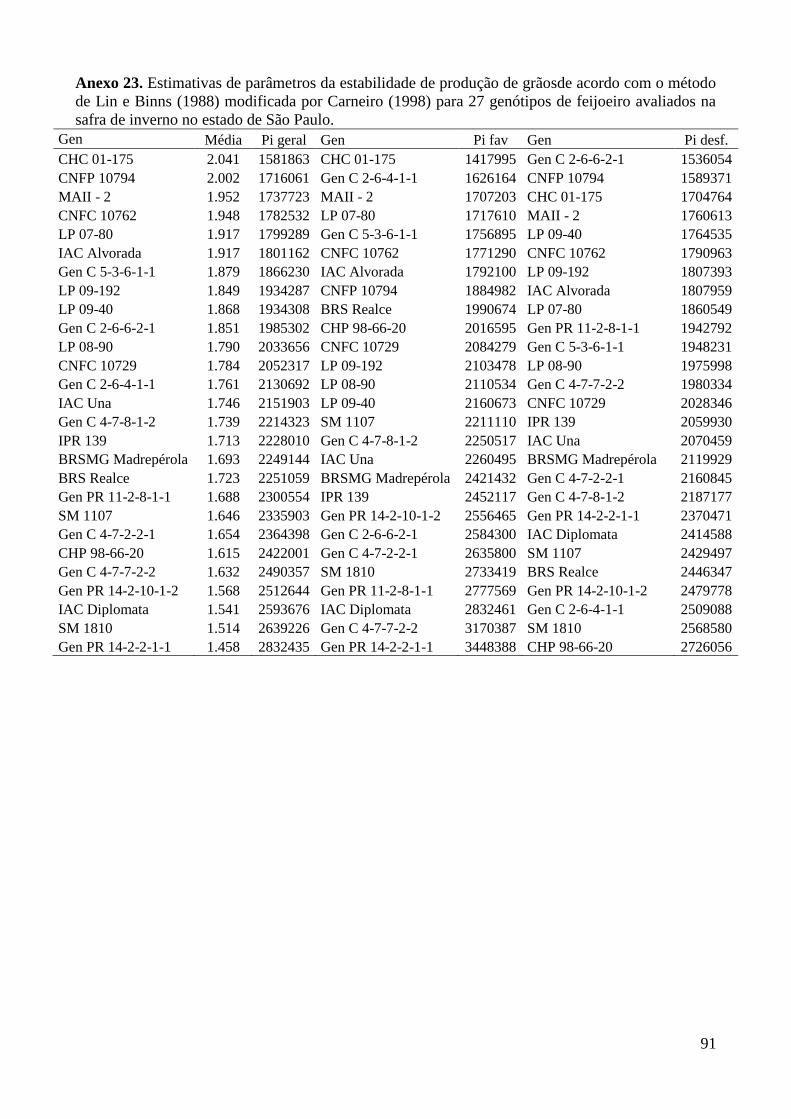
91
Anexo 23. Estimativas de parâmetros da estabilidade de produção de grãosde acordo com o método
de Lin e Binns (1988) modificada por Carneiro (1998) para 27 genótipos de feijoeiro avaliados na
safra de inverno no estado de São Paulo.
Gen Média Pi geral Gen Pi fav Gen Pi desf.
CHC 01-175 2.041 1581863 CHC 01-175 1417995 Gen C 2-6-6-2-1 1536054
CNFP 10794 2.002 1716061 Gen C 2-6-4-1-1 1626164 CNFP 10794 1589371
MAII - 2 1.952 1737723 MAII - 2 1707203 CHC 01-175 1704764
CNFC 10762 1.948 1782532 LP 07-80 1717610 MAII - 2 1760613
LP 07-80 1.917 1799289 Gen C 5-3-6-1-1 1756895 LP 09-40 1764535
IAC Alvorada 1.917 1801162 CNFC 10762 1771290 CNFC 10762 1790963
Gen C 5-3-6-1-1 1.879 1866230 IAC Alvorada 1792100 LP 09-192 1807393
LP 09-192 1.849 1934287 CNFP 10794 1884982 IAC Alvorada 1807959
LP 09-40 1.868 1934308 BRS Realce 1990674 LP 07-80 1860549
Gen C 2-6-6-2-1 1.851 1985302 CHP 98-66-20 2016595 Gen PR 11-2-8-1-1 1942792
LP 08-90 1.790 2033656 CNFC 10729 2084279 Gen C 5-3-6-1-1 1948231
CNFC 10729 1.784 2052317 LP 09-192 2103478 LP 08-90 1975998
Gen C 2-6-4-1-1 1.761 2130692 LP 08-90 2110534 Gen C 4-7-7-2-2 1980334
IAC Una 1.746 2151903 LP 09-40 2160673 CNFC 10729 2028346
Gen C 4-7-8-1-2 1.739 2214323 SM 1107 2211110 IPR 139 2059930
IPR 139 1.713 2228010 Gen C 4-7-8-1-2 2250517 IAC Una 2070459
BRSMG Madrepérola 1.693 2249144 IAC Una 2260495 BRSMG Madrepérola 2119929
BRS Realce 1.723 2251059 BRSMG Madrepérola 2421432 Gen C 4-7-2-2-1 2160845
Gen PR 11-2-8-1-1 1.688 2300554 IPR 139 2452117 Gen C 4-7-8-1-2 2187177
SM 1107 1.646 2335903 Gen PR 14-2-10-1-2 2556465 Gen PR 14-2-2-1-1 2370471
Gen C 4-7-2-2-1 1.654 2364398 Gen C 2-6-6-2-1 2584300 IAC Diplomata 2414588
CHP 98-66-20 1.615 2422001 Gen C 4-7-2-2-1 2635800 SM 1107 2429497
Gen C 4-7-7-2-2 1.632 2490357 SM 1810 2733419 BRS Realce 2446347
Gen PR 14-2-10-1-2 1.568 2512644 Gen PR 11-2-8-1-1 2777569 Gen PR 14-2-10-1-2 2479778
IAC Diplomata 1.541 2593676 IAC Diplomata 2832461 Gen C 2-6-4-1-1 2509088
SM 1810 1.514 2639226 Gen C 4-7-7-2-2 3170387 SM 1810 2568580
Gen PR 14-2-2-1-1 1.458 2832435 Gen PR 14-2-2-1-1 3448388 CHP 98-66-20 2726056
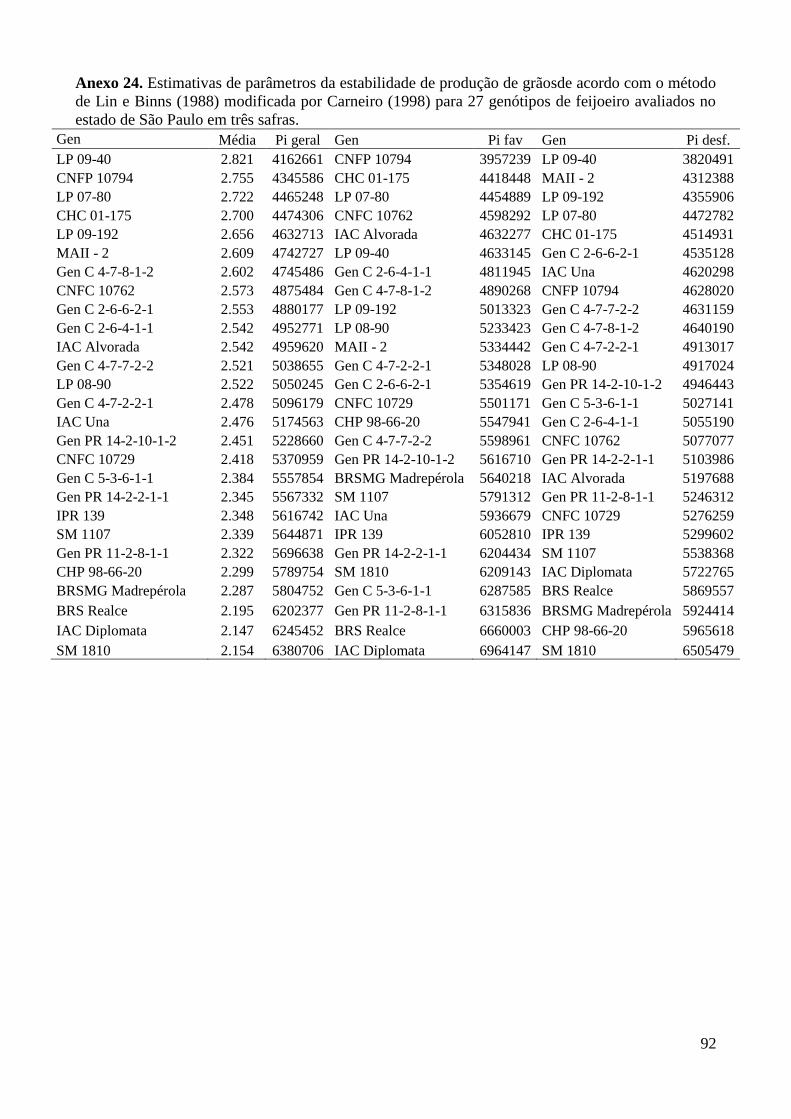
92
Anexo 24. Estimativas de parâmetros da estabilidade de produção de grãosde acordo com o método
de Lin e Binns (1988) modificada por Carneiro (1998) para 27 genótipos de feijoeiro avaliados no
estado de São Paulo em três safras.
Gen Média Pi geral Gen Pi fav Gen Pi desf.
LP 09-40 2.821 4162661 CNFP 10794 3957239 LP 09-40 3820491
CNFP 10794 2.755 4345586 CHC 01-175 4418448 MAII - 2 4312388
LP 07-80 2.722 4465248 LP 07-80 4454889 LP 09-192 4355906
CHC 01-175 2.700 4474306 CNFC 10762 4598292 LP 07-80 4472782
LP 09-192 2.656 4632713 IAC Alvorada 4632277 CHC 01-175 4514931
MAII - 2 2.609 4742727 LP 09-40 4633145 Gen C 2-6-6-2-1 4535128
Gen C 4-7-8-1-2 2.602 4745486 Gen C 2-6-4-1-1 4811945 IAC Una 4620298
CNFC 10762 2.573 4875484 Gen C 4-7-8-1-2 4890268 CNFP 10794 4628020
Gen C 2-6-6-2-1 2.553 4880177 LP 09-192 5013323 Gen C 4-7-7-2-2 4631159
Gen C 2-6-4-1-1 2.542 4952771 LP 08-90 5233423 Gen C 4-7-8-1-2 4640190
IAC Alvorada 2.542 4959620 MAII - 2 5334442 Gen C 4-7-2-2-1 4913017
Gen C 4-7-7-2-2 2.521 5038655 Gen C 4-7-2-2-1 5348028 LP 08-90 4917024
LP 08-90 2.522 5050245 Gen C 2-6-6-2-1 5354619 Gen PR 14-2-10-1-2 4946443
Gen C 4-7-2-2-1 2.478 5096179 CNFC 10729 5501171 Gen C 5-3-6-1-1 5027141
IAC Una 2.476 5174563 CHP 98-66-20 5547941 Gen C 2-6-4-1-1 5055190
Gen PR 14-2-10-1-2 2.451 5228660 Gen C 4-7-7-2-2 5598961 CNFC 10762 5077077
CNFC 10729 2.418 5370959 Gen PR 14-2-10-1-2 5616710 Gen PR 14-2-2-1-1 5103986
Gen C 5-3-6-1-1 2.384 5557854 BRSMG Madrepérola 5640218 IAC Alvorada 5197688
Gen PR 14-2-2-1-1 2.345 5567332 SM 1107 5791312 Gen PR 11-2-8-1-1 5246312
IPR 139 2.348 5616742 IAC Una 5936679 CNFC 10729 5276259
SM 1107 2.339 5644871 IPR 139 6052810 IPR 139 5299602
Gen PR 11-2-8-1-1 2.322 5696638 Gen PR 14-2-2-1-1 6204434 SM 1107 5538368
CHP 98-66-20 2.299 5789754 SM 1810 6209143 IAC Diplomata 5722765
BRSMG Madrepérola 2.287 5804752 Gen C 5-3-6-1-1 6287585 BRS Realce 5869557
BRS Realce 2.195 6202377 Gen PR 11-2-8-1-1 6315836 BRSMG Madrepérola 5924414
IAC Diplomata 2.147 6245452 BRS Realce 6660003 CHP 98-66-20 5965618
SM 1810 2.154 6380706 IAC Diplomata 6964147 SM 1810 6505479


















