
Interacção do vanádio com a miosina de músculo esquelético ... · A todos os meus colegas dos...
58
0 Relatório de Projecto de Bioquímica Interacção do vanádio com a miosina de músculo esquelético de coelho Joana Wilton Pereira n.º 20201 Licenciatura em Bioquímica Julho de 2009
Transcript of Interacção do vanádio com a miosina de músculo esquelético ... · A todos os meus colegas dos...

0
Relatoacuterio de Projecto de Bioquiacutemica
Interacccedilatildeo do vanaacutedio com a
miosina de muacutesculo
esqueleacutetico de coelho
Joana Wilton Pereira
nordm 20201
Licenciatura em Bioquiacutemica
Julho de 2009
1
Agradecimentos
Ao meu orientador Doutor Rui O Duarte pela oportunidade dada para estudar este sistema tatildeo
fascinante e pelos preciosos conselhos em relaccedilatildeo agrave espectroscopia de Ressonacircncia
Paramagneacutetica Electroacutenica
Agrave Susana Ramos pela orientaccedilatildeo equilibrada paciecircncia dedicaccedilatildeo conselhos preciosos humor e
amizade sem os quais natildeo teria sido possiacutevel realizar este trabalho Obrigada por me teres
desafiado a ser a melhor aprendiz de cientista que conseguisse ndash com planeamento e
determinaccedilatildeo tudo se consegue
Ao Professor Doutor Manuel Aureliano (supervisor) da Faculdade de Ciecircncias e Tecnologia da
Universidade do Algarve pela recepccedilatildeo calorosa disponibilidade das suas instalaccedilotildees para a
obtenccedilatildeo da actina e pela gentil cedecircncia do subfragmento-1 de miosina imprescindiacutevel para o
estudo que realizei
Ao Rui Almeida pelos preciosos conselhos e colaboraccedilatildeo que estimularam o meu fasciacutenio pela
ciecircncia A disponibilidade que mostraste para me esclarecer sempre com clareza e simpatia
tornaram o meu trabalho de todos os dias mais agradaacutevel
Ao Ameacuterico Duarte pela indispensaacutevel ajuda para a realizaccedilatildeo da electroforese em condiccedilotildees
desnaturantes e pela amizade demonstrada nos momentos em que o nervosismo tomou conta de
mim
Agrave Dra Maria do Rosaacuterio Caras-Altas pela disponibilidade concedida no uso do espectroacutemetro de
RMN equipamento crucial na realizaccedilatildeo deste trabalho
A todos os meus colegas dos laboratoacuterios 407 e 613 cujo bom-humor e entreajuda tornaram os
meus dias mais agradaacuteveis
2
Abreviaturas e Siacutembolos
Absx ndash Absorvecircncia ao comprimento de onda x (em nm)
ADP - Adenosina 5rsquo-difosfato
ATP ndash Adenosina 5rsquo-trifosfato
ATPase ndash Adenosina 5rsquo-trifosfatase
BSA ndash Albumina de soro bovino
-Me - -Mercaptoetanol
CNT ndash Aniatildeo 3-carboxilato-4-nitrotiofenolato
Cys ndash Cisteiacutena
DTNB ndash 55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico)
- Coeficiente de extinccedilatildeo (molar)
EDTA ndash Aacutecido etilenodiamino tetra-aceacutetico
ELC ndash Cadeia leve essencial de miosina
F-actina ndash Forma filamentosa da actina
g ndash Aceleraccedilatildeo da gravidade
G-actina ndash Forma monomeacuterica da actina
h ndash Horas
HEPES ndash Aacutecido 4-(2-hidroxiletil)-1-piperazineetanosulfoacutenico
HMM ndash Meromiosina pesada
IC50 ndash Concentraccedilatildeo inibitoacuteria 50
LDH ndash Lactato desidrogenase
Miosina-II ndash Miosina do tipo II
Met - Metionina
NAD+ ndash Nicotinamida adenina dinucleoacutetido (forma oxidada)
NADH ndash Nicotinamida adenina dinucleoacutetido (forma reduzida)
PDB ndash Protein Data Bank
Pi ndash Fosfato inorgacircnico
PEP ndash Fosfoenolpiruvato
PK ndash Piruvato cinase
PM ndash Peso Molecular
3
ppb ndash Partes por biliatildeo (1 gL)
ppm ndash Partes por milhatildeo (1 mgL)
PSA ndash persulfato de amoacutenio
Rf - Mobilidade relativa
RLC ndash Cadeia leve regulatoacuteria de miosina
RMN ndash Ressonacircncia Magneacutetica Nuclear
ROS ndash Espeacutecies reactivas de oxigeacutenio
RPE ndash Ressonacircncia Paramagneacutetica Electroacutenica
S1 ndash Subfragmento-1 da miosina II
SDS ndash Dodecilo sulfato de soacutedio
SDS-PAGE ndash Electroforese em gel de poliacrilamida com SDS (condiccedilotildees desnaturantes)
SH1 ndash Grupo tiol mais reactivo da miosina (resiacuteduo Cys-707)
SH2 ndash segundo grupo tiol mais reactivo da miosina (resiacuteduo Cys-697)
SNC ndash Sistema Nervoso Central
TCA ndash Aacutecido tricloroaceacutetico
TEMED ndash NNNrsquoNrsquo-tetrametiletilenodiamina
Tris ndash 2-amino-2-hidroximetil-13-propanodiol
Trp ndash triptofano
Tyr ndash tirosina
UV ndash gama ultravioleta do espectro electromagneacutetico
Vis ndash gama visiacutevel do espectro electromagneacutetico
4
Resumo
A exposiccedilatildeo a vanaacutedio conduz a uma diminuiccedilatildeo na actividade ATPaacutesica da miosina e provoca a
oxidaccedilatildeo dos grupos sulfidrilo desta proteiacutena No entanto natildeo estaacute integralmente esclarecido
quais os efeitos especiacuteficos que ocorrem na miosina quando exposta ao vanaacutedio Nesse sentido
interessou determinar in vitro quais as alteraccedilotildees estruturais funcionais e moleculares ocorridas
no Subfragmento-1 de miosina de muacutesculo esqueleacutetico de coelho apoacutes a exposiccedilatildeo croacutenica a
soluccedilotildees de metavanadato decavanadato e vanadilo Verificou-se que a exposiccedilatildeo de S1 (005
mgmL) a decavanadato inibe a sua actividade Mg2+
-ATPaacutesica sendo o valor de IC50 para esta
inibiccedilatildeo de 266 plusmn 07 microM decavanadato Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM
decavanadato A perda de funcionalidade proteica poderaacute dever-se agrave oxidaccedilatildeo de 2 resiacuteduos de
Cys de S1 natildeo expostos ao solvente apoacutes exposiccedilatildeo a 1000 microM de decavanadato Verificou-se
que as espeacutecies oligomeacutericas de vanaacutedio com maior afinidade para o S1 satildeo o decavanadato e o
tetravanadato Revelaram-se os possiacuteveis ligandos do vanaacutedio no S1 atraveacutes de espectroscopia de
RPE Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados
de oxidaccedilatildeo (IV) e (V) na estrutura e funccedilatildeo do S1 em concentraccedilotildees facilmente alcanccedilaacuteveis
daquele metal no ambiente terrestre
Summary
The exposure to vanadium leads to a decrease of the myosin ATPase activity and provokes the
oxidation of its sulfydryl groups However the specific effects of vanadium on myosin are not yet
fully understood Consequently the objectives of the present work were to establish the structural
funtional and molecular changes suffered by rabbit myosin subfragment-1 after cronic exposure to
metavanadate decavanadate and vanadyl solutions It was verified that the exposure of S1 (005
mgmL) to decavanadate results in the inhibition of its Mg2+
-ATPase activity resulting in the
IC50 value 266 plusmn 07 microM decavanadate In the presence of actin the IC50 value was 28 plusmn 1 microM
decavanadate It is suggested that the loss of protein function may be due to the oxidation of 2
core cysteine residues after exposure to 1000 microM decavanadate It was also observed that the
decavanadate and tetravanadate species show the highest affinity for S1 Lastly the possible S1
ligands to vanadium were revealed through EPR spectroscopy Globally the obtained results
point to significant effect of V(V) and V(IV) states on S1 structure and function for easily
reachable environmental vanadium concentrations
5
Iacutendice
1 Introduccedilatildeo
11 Muacutesculo esqueleacutetico
111 Miosina
112 Actina
113 Mecanismos moleculares da contracccedilatildeo muscular
1131 Regulaccedilatildeo da contracccedilatildeo muscular
12 Vanaacutedio
121 Propriedades quiacutemicas
1211 Vanaacutedio (IV)
1212 Vanaacutedio (V)
122 Metabolismo e efeito na actividade enzimaacutetica
13 Vanaacutedio e muacutesculo esqueleacutetico
14 Objectivos do projecto
2 Discussatildeo de Resultados
21 Caracterizaccedilatildeo das proteiacutenas isoladas
211 Anaacutelise do estado de pureza
212 Determinaccedilatildeo da concentraccedilatildeo proteica
2121 Subfragmento-1 de miosina
2122 Actina
22 Mediccedilatildeo da actividade enzimaacutetica de S1 de miosina
221 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de Fiske-
Subarow modificado
222 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
23 Preparaccedilatildeo e caracterizaccedilatildeo das soluccedilotildees de vanaacutedio
231 Anaacutelise da composiccedilatildeo das soluccedilotildees de vanadato por espectroscopia de 51V-RMN efeito da
concentraccedilatildeo da espeacutecie
232 Estabilidade das soluccedilotildees de decavanadato anaacutelise por espectroscopia de absorccedilatildeo UVVis
24 Interacccedilatildeo de S1 com vanaacutedio(V)
241 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
242 Actividade Mg2+
-ATPaacutesica de S1 de miosina na presenccedila de decanadato
243 Quantificaccedilatildeo de cisteiacutenas reduzidas pelo meacutetodo de Ellman
25 Interacccedilatildeo de S1 com vanaacutedio(IV)
251 Anaacutelise por espectroscopia de Ressonacircncia Paramagneacutetica Electroacutenica
3 Procedimento Experimental
31 Reagentes
32 Meacutetodos
321 Isolamento e purificaccedilatildeo das proteiacutenas de muacutesculo esqueleacutetico de coelho
3211 Purificaccedilatildeo da miosina preparaccedilatildeo de Subfragmento 1 por digestatildeo quimotriacutepsica da
miosina
32111 Purificaccedilatildeo da miosina
32112 Preparaccedilatildeo de Subfragmento-1 de miosina por digestatildeo quimotriacutepsica
3212 Isolamento e purificaccedilatildeo de actina
32121Obtenccedilatildeo do ldquopoacute de acetonardquo
32122 Purificaccedilatildeo da actina a partir de ldquopoacute de acetonardquo
331 Quantificaccedilatildeo das proteiacutenas isoladas
3311 Quantificaccedilatildeo pelo meacutetodo de Bradford
6
3312 Quantificaccedilatildeo pelo meacutetodo directo
332 Electroforese em condiccedilotildees desnaturantes (SDS-PAGE)
34 Mediccedilatildeo da actividade enzimaacutetica de S1
341 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de Fiske-
Subarow modificado
342 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
35 Preparaccedilatildeo e caracterizaccedilatildeo das soluccedilotildees de vanaacutedio por Espectroscopia UV-Visiacutevel e
Ressonacircncia Magneacutetica Nuclear
351 Preparaccedilatildeo das soluccedilotildees de vanadato
352 Anaacutelise por UV-Vis para determinaccedilatildeo da ordem da reacccedilatildeo
353 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
362 1 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
363 Interacccedilatildeo de S1 com V (V) por Ressonacircncia Magneacutetica Nuclear (RMN)
4 Conclusotildees gerais
5 Estudos futuros
6 Referecircncias bibliograacuteficas
7
1 Introduccedilatildeo
11 Muacutesculo esqueleacutetico
A capacidade de locomoccedilatildeo eacute uma das caracteriacutesticas fundamentais da maioria dos organismos
vivos A contracccedilatildeo muscular um dos fenoacutemenos que a torna possiacutevel apresenta grande
complexidade e tem vindo a ser utilizado como modelo para a compreensatildeo de sistemas menos
especializados de mobilidade celular [1]
No que diz respeito agrave sua fisiologia morfoloacutegica os muacutesculos dos vertebrados dividem-se em
lisos e estriados Estes uacuteltimos devem o seu nome agrave alternacircncia de zonas claras e escuras
observaacuteveis ao microscoacutepio electroacutenico de que resulta um aspecto estriado Os muacutesculos
estriados podem ainda ser divididos em duas subclasses muacutesculos esqueleacuteticos e cardiacuteacos [2]
consoante a forma celular (respectivamente longitudinal ou ramificada) ou tipo de contracccedilatildeo das
ceacutelulas que o compotildeem (respectivamente voluntaacuteria e involuntaacuteria) [3]
Os muacutesculos estriados esqueleacuteticos satildeo assim designados devido agrave sua localizaccedilatildeo no
organismo ndash ligados aos ossos da coluna vertical braccedilos e pernas Satildeo constituiacutedos por feixes de
ceacutelulas multinucleadas (miofibras) que contecircm miofibrilas paralelas de cerca de 1 m de
diacircmetro associadas agrave contracccedilatildeo voluntaacuteria raacutepida e forte em actividades de coordenaccedilatildeo
complexa [4] Cada feixe apresenta um diacircmetro compreendido entre 20 e 100 m e um
comprimento maacuteximo que pode atingir vaacuterios centiacutemetros (Fig 11)
Figura 11 ndash Organizaccedilatildeo hieraacuterquica geral do muacutesculo-esqueleacutetico (adaptado de Purves et al 2003 [3])
No muacutesculo-esqueleacutetico as zonas claras e escuras supramencionadas correspondem a zonas de
menor e maior densidade electroacutenica respectivamente As zonas claras constituem as bandas I (de
8
ldquoisotropiardquo ndash polarizaccedilatildeo uniforme em todas as direcccedilotildees) enquanto as zonas escuras constituem
as bandas A (de ldquoanisotropiardquo ndash polariza a luz visiacutevel) A banda I encontra-se dividida
transversalmente por uma linha escura ndash linha Z de densidade electroacutenica e refracccedilatildeo superiores
No interior de cada banda A existe uma zona transversal menos densa ndash zona H ndash rica em enzimas
importantes para o metabolismo celular No centro desta zona existe uma linha perpendicular
mais escura ndash linha M [2]
A unidade fundamental da contracccedilatildeo muscular eacute designada por sarcoacutemero (Fig 12) Encontra-se
repetida de forma regular ao longo das miofibrilas apresentando um comprimento de 25 a 3 m
aquando do relaxamento muscular diminuindo aquando da contracccedilatildeo muscular
Figura 12 ndash Representaccedilatildeo de fotomicrografia electroacutenica e esquemaacutetica do sarcoacutemero Os filamentos grossos e
finos satildeo maioritariamente constituiacutedos por miosina e actina respectivamente (Adaptado de Stryer 2002 [2])
As secccedilotildees de corte do sarcoacutemero datildeo conta da presenccedila de dois tipos de filamentos proteicos ndash os
filamentos grossos cujo diacircmetro eacute de 15 nm constituiacutedos sobretudo por miosina e os filamentos
finos com um diacircmetro de 9 nm constituiacutedos sobretudo por actina tropomiosina e pelo complexo
de troponina [2]
111 Miosina
Actualmente o termo miosina refere-se a uma superfamiacutelia diversa de motores moleculares
capazes de hidrolisar ATP e de se ligarem reversivelmente agrave actina A classificaccedilatildeo da
superfamiacutelia das miosinas nas 15 classes distintas faz-se atraveacutes de homologia entre as diferentes
sequecircncias [A] Eacute tambeacutem possiacutevel a classificaccedilatildeo por subclasses das quais se destaca a subclasse
4 proveniente do muacutesculo-esqueleacutetico e cardiacuteaco de vertebrados Este estudo focar-se-aacute apenas
nas miosinas de classe II (miosina-II) presente nos filamentos grossos do muacutesculo estriado
esqueleacutetico [B] e em ceacutelulas natildeo musculares A funccedilatildeo da miosina-II depende da sua localizaccedilatildeo
celular e tecidular
9
A miosina-II eacute constituiacuteda por duas cadeias polipeptiacutedicas idecircnticas denominadas cadeias pesadas
de miosina (HC) enroladas entre si e cujo peso molecular (PM) estaacute compreendido entre 171 e
241 kDa) Estruturalmente as HC dividem-se em 3 domiacutenios funcionais - cabeccedila pescoccedilo e
cauda como se pode observar na Fig 13 Agraves cabeccedilas ou zonas globulares estatildeo associados dois
pares de cadeias leves (peso molecular entre 15 e 30 kDa) que se dividem em dois tipos
essencial (ELC) e regulatoacuterio (RLC)
Figura 13 ndash Estrutura esquemaacutetica da moleacutecula de miosina-II Destacam-se as duas cabeccedilas globulares e a cauda
filamentosa ligadas entre si por uma zona flexiacutevel denominada pescoccedilo As cadeias leves (a verde e vermelho)
encontram-se associadas agraves cabeccedilas globulares
As cabeccedilas globulares contendo os locais de ligaccedilatildeo agrave actina e aos nucleoacutetidos mantecircm os seus
elementos estruturais baacutesicos conservados em todas as classes sendo estes constituiacutedos por 720-
80 aa A regiatildeo do pescoccedilo eacute constituiacuteda por uma longa heacutelice pertencente agrave cadeia pesada
que se encontra estabilizada atraveacutes da ligaccedilatildeo a subunidades de cadeia leve A estrutura do
pescoccedilo envolvida na ligaccedilatildeo agraves cadeias leves eacute denominada de motivo IQ cuja estrutura
consensual eacute IQXXXRGXXXR1 O pescoccedilo serve como extensatildeo do comprimento da cabeccedila de
modo a actuar como alavanca durante a contracccedilatildeo muscular [A]A regiatildeo da cauda ou acircncora
apresenta bastante divergecircncia no que respeita agrave sua sequecircncia estrutura e funccedilatildeo No entanto
pensa-se que a sua funccedilatildeo centrar-se-aacute na autoassociaccedilatildeo das vaacuterias moleacuteculas de miosina-II
atraveacutes da regiatildeo de coiled-coil Esta confere assimetria agrave moleacutecula e permite a formaccedilatildeo de
filamentos grossos capazes de translocar filamentos de actina [A]
Na regiatildeo da cabeccedila da miosina existem 2 regiotildees extremamente sensiacuteveis agrave proteoacutelise A
clivagem em ambos os locais resulta na produccedilatildeo de trecircs fragmentos um compreendendo o N-
terminal (fragmento 25 kDa) outro a regiatildeo central (50 kDa) e um terceiro o C-terminal (20 kDa)
(Fig 14) Estes fragmentos dissociam-se em condiccedilotildees desnaturantes e natildeo constituem domiacutenios
independentes da cabeccedila mas provavelmente loops flexiacuteveis uma vez que natildeo foram observadas
na estrutura cristalina Os vaacuterios fragmentos obtidos em diferentes condiccedilotildees foram cruciais para
a elucidaccedilatildeo das vaacuterias funccedilotildees da moleacutecula
A regiatildeo compreendida entre os fragmentos 25 e 50 kDa (loop 1) localiza-se directamente por
1 I = Isoleucina Q= Glutamina X = qualquer dos 20 aminoaacutecidos padratildeo R= Arginina G= Glicina
10
cima do local de ligaccedilatildeo ao nucleoacutetido A regiatildeo entre os fragmentos de 50 e 20 kDa (loop 2)
encontra-se perto da intersecccedilatildeo entre os domiacutenios da cabeccedila e do pescoccedilo e deveraacute interagir com
a actina uma vez que a ligaccedilatildeo desta a protege da proteoacutelise Na ausecircncia do loop 2 a miosina
liga mais fracamente agrave actina do que no caso da miosina intacta A sequecircncia primaacuteria de ambos
os loops apresenta conservaccedilatildeo reduzida sendo provaacutevel que sejam responsaacuteveis por muita da
diversidade das diferentes propriedades enzimaacuteticas presentes em diferentes isoformas
No fragmento de 20 kDa existem dois grupos sulfidrilo Cys-707 e -697 usualmente
denominados de SH1 e SH2 devido agrave ordem da sua reactividade quiacutemica Na presenccedila de
nucleoacutetidos pode ocorrer o ldquocross-linkingrdquo entre estes dois grupos incluindo a oxidaccedilatildeo a uma
ponte dissulfureto
Devido agrave insolubilidade da miosina-II nativa a estrateacutegia adoptada para os estudos com esta
proteiacutena eacute frequentemente a sua clivagem em fragmentos funcionais separaacuteveis Um dos locais de
clivagem ocorre tipicamente a 130 kDa do N-terminal produzindo a meromiosina pesada (HMM)
e a meromiosina leve (LMM) [C] A HMM eacute formada pelas 2 cabeccedilas e pescoccedilo e apresenta
propriedades semelhantes agrave miosina sem digestatildeo (ligaccedilatildeo e translocaccedilatildeo de actina e hidroacutelise de
MgATP)
A HMM pode ser novamente clivada atraveacutes de uma reacccedilatildeo de proteoacutelise produzindo os
subfragmentos 1 (S1) e 2 (S2)O S1 (90 kDa) eacute enzimaticamente activo sendo constituiacutedo pelos
domiacutenios da cabeccedila e do pescoccedilo e apresentando associaccedilatildeo agraves duas cadeias leves ou apenas agrave
ELC (Fig 14)Em alternativa o S1 pode ser produzido por clivagem directa da miosina intacta
formando-se tambeacutem um fragmento em forma de cilindro que consiste nas regiotildees S2 e LMM
(Ref)Utiliza-se extensivamente o S1 para estudos cineacuteticos e de ligaccedilatildeo agrave actina uma vez que a
sua solubilidade simplifica o planeamento da experiecircncia e a subsequente interpretaccedilatildeo dos dados
[A]
Figura 14- Estrutura do subfragmento-1 de miosina de muacutesculo esqueleacutetico com ecircnfase nos domiacutenios de 50 kDa
central (a vermelho) 25 kDa no N-terminal (a verde) e 20 kDa no C-terminal (a azul) As cadeias leves essenciais
encontram-se a amarelo e as cadeias leves regulatoacuterias a lilaacutes Os locais de ligaccedilatildeo agrave actina e ao ATP estatildeo
igualmente destacados (adaptado de Holmes e Geeves 1999 [D])
11
112 Actina
O termo ldquoactinardquo refere-se a uma famiacutelia extremamente conservada de proteiacutenas citoplasmaacuteticas
presente em todos os eucariotas Esta proteiacutena natildeo se encontra nos procariotas [E] A maioria das
actinas possui peso molecular proacuteximo de 42 kDa outra evidecircncia do seu grau elevado de
conservaccedilatildeo no que concerne agrave sequecircncia e ao nuacutemero de aminoaacutecidos Os eucariotas superiores
apresentam um grande nuacutemero de isoformas de actina que por sua vez se encontram codificados
por um ainda maior nuacutemero de genes (Ref) A expressatildeo das isoformas eacute diferencial consoante o
tecido e sua especializaccedilatildeo pelo que deveratildeo possuir funccedilotildees e mecanismos direccionais
diferentes (Ref) Foi ainda observado que os mRNA das diferentes isoformas apresentam uma
distribuiccedilatildeo heterogeacutenea nas ceacutelulas de forma semelhante aos seus produtos proteicos [F] Estes
dados suportam a hipoacutetese supramencionada do direccionamento das isoformas de actina para um
determinado local na ceacutelula de acordo com as suas funccedilotildees
A actina presente no citoesqueleto eacute responsaacutevel por processos como o mecanismo contraacutectil
fagocitose manutenccedilatildeo e alteraccedilatildeo da estrutura celular locomoccedilatildeo transporte de organelos
regulaccedilatildeo do transporte de solutos entre outras Nos eucariotas superiores estaacute ainda envolvida
no transporte de gracircnulos de pigmento retinoacuteide [G] transporte de gracircnulos no axoplasma [H] e
secreccedilatildeo entre outros [E] Actualmente a isoforma da actina (-actina) eacute a melhor estudada
encontrando-se envolvida no mecanismo contraacutectil [E]
Os filamentos de -actina tecircm cerca de 1m de comprimento e 80 Å Consistem em duas cadeias
de actina filamentosa (F-actina) organizados em heacutelice [I] (Holmes et al 1990) sobre os quais
se encontram outras proteiacutenas ndash a troponina (C I e T) e a tropomiosina (Azevedo 1999) (Figura
15) A troponina eacute filamentosa e localiza-se nas depressotildees da heacutelice de F-actina conferindo
rigidez aos filamentos finos Por sua vez a tropomiosina eacute essencial na regulaccedilatildeo do mecanismo
contraacutectil
Figura 15 ndash Modelo estrutural dos filamentos de F-actina contendo a tropomiosina e o complexo de troponina (I C
e T) (Adaptado de Susana Ramos 2005 [J])
12
A F-actina eacute formada atraveacutes da polimerizaccedilatildeo de subunidades globulares de actina (G-actina) A
G-actina eacute composta por 2 domiacutenios de tamanho semelhante separados pelos bolsos de ligaccedilatildeo(1)
de catiotildees divalentes e(2) de nucleoacutetidos [KLM] Estes por sua vez estatildeo subdivididos noutros
dois de menores dimensotildees (1-4) (Fig 16) cada um envolvido nas interacccedilotildees actina-actina ou
na formaccedilatildeo do topo de ligaccedilatildeo do nucleoacutetido As regiotildees N-terminal e C-terminal estatildeo proacuteximas
uma da outra no subdomiacutenio 1 [E]
Figura 16 ndash Estrutura do monoacutemero de actina Encontram-se representados os subdomiacutenios 1 a 4 e a vermelho a
moleacutecula de ATP junto do respectivo local de ligaccedilatildeo (Protein Data Bank ID 1j6z)
113 Mecanismos moleculares da contracccedilatildeo muscular
A contracccedilatildeo muscular ocorre atraveacutes do deslizamento dos filamentos finos (de actina) por entre
os filamentos grossos (de miosina) conseguida pela reacccedilatildeo de hidroacutelise de ATP e regulada pela
tropomiosina e pelo complexo de troponina em resposta a impulsos nervosos Este mecanismo foi
inicialmente proposto por Huxley-Niedergerke [N] e Huxley-Hanson [O] ambos em 1954 A base
experimental de ambos os estudos foi a inexistecircncia de variaccedilatildeo do tamanho dos filamentos
durante a contracccedilatildeo A banda A apresenta sempre as mesmas dimensotildees por outro lado a banda
I a zona H diminuem consideravelmente aquando da contracccedilatildeo (Figs 12 e 17)
Figura 17 ndash Modelo da contracccedilatildeo muscular ndash deslizamento dos filamentos finos (azuis) relativamente aos
filamentos grossos (rosa) (Adaptado de Stryer 2002 [2])
13
Na ausecircncia de ATP e supondo a ausecircncia de inibiccedilatildeo da interacccedilatildeo actina-miosina as proteiacutenas
encontram-se fortemente associadas formando o complexo actomiosina (AM) (Figura 15)
Nestas condiccedilotildees as cabeccedilas de miosina formam um acircngulo de 45ordm com o eixo dos filamentos
finos de actina [P]
Na presenccedila de ATP o complexo AM dissocia-se jaacute que a afinidade do complexo M-ATP para a
actina eacute 105 vezes inferior agrave afinidade da miosina para a actina O ATP liga-se agrave miosina
ocorrendo seguidamente a reacccedilatildeo de hidroacutelise de ATP Os produtos daiacute resultantes ndash ADP e Pi -
permanecem ligados agrave miosina formando o complexo M-ADP-Pi Com a ligaccedilatildeo subsequente agrave
actina forma-se o complexo AM-ADP-Pi no qual as cabeccedilas de miosina apresentam um acircngulo
recto com o eixo do muacutesculo [P] Neste ponto daacute-se a libertaccedilatildeo progressiva de Pi e ADP Eacute
provaacutevel que a energia proveniente da hidroacutelise do ATP provoque uma alteraccedilatildeo do acircngulo de
ligaccedilatildeo das cabeccedilas de miosina ao filamento de actina passando novamente de 90ordm para 45ordm [P]
Esta alteraccedilatildeo conformacional provoca o deslizamento do filamento de actina para o centro do
sarcoacutemero cujo encurtamento promove a contracccedilatildeo das miofibrilas O processo supracitado
ocorre em cerca de 1ms[Q] e encontra-se resumido na Fig 18
Figura 18 ndash Esquema cineacutetico da hidroacutelise de ATP pela miosina (Adaptado de Sellers 1999 [A])
1131 Regulaccedilatildeo da contracccedilatildeo muscular
O mecanismo pelo qual se processa a regulaccedilatildeo muscular foi inicialmente descrito por estudos
distintos de Haselgrove Huxley e Parrey-Squire todos em 1973 [RST] Aleacutem da implicaccedilatildeo
directa da tropomiosina e do complexo de troponina a concentraccedilatildeo de Ca2+
no muacutesculo em
relaxamento eacute bastante reduzida (10-7
M) aumentando notavelmente aquando da contracccedilatildeo (10-5
-
10-3
M)
Resumidamente quando ocorre a excitaccedilatildeo do muacutesculo esqueleacutetico a despolarizaccedilatildeo da
membrana plasmaacutetica eacute difundida ateacute ao interior do tecido atraveacutes dos tuacutebulos transversais
(invaginaccedilotildees da membrana plasmaacutetica da fibra muscular) A despolarizaccedilatildeo provoca a abertura
dos canais ioacutenicos de Ca2+
do retiacuteculo sarcoplasmaacutetico adjacente Como a concentraccedilatildeo de Ca2+
no interior do organelo eacute muito elevada o caacutelcio passa raacutepida e uniformemente para o citosol
14
Assim a regulaccedilatildeo da contracccedilatildeo ocorre com coordenaccedilatildeo e rapidez na totalidade das miofibrilas
do tecido muscular (Ref)
Em repouso a troponina I encontra-se posicionada de tal forma que a proteiacutena adjacente
troponina C impede estruturalmente a ligaccedilatildeo das cabeccedilas da miosina agrave actina (acircngulo =50ordm) O
aumento da concentraccedilatildeo intracelular de Ca2+
provoca um aumento da sua disponibilidade para
ligar agrave troponina C (Fig 19)
Figura 19ndash Regulaccedilatildeo do mecanismo contraacutectil no muacutesculo esqueleacutetico pelo complexo de troponina
Como consequecircncia a troponina C altera a sua conformaccedilatildeo (=70ordm) associando-se agrave troponina I
que deixa de inibir a ligaccedilatildeo das pontes transversais A alteraccedilatildeo anterior eacute transmitida agrave
tropomiosina atraveacutes da troponina T adjacente agrave C o que provoca o deslocamento da
tropomiosina na direcccedilatildeo do interior do sulco do filamento de actina Assim eacute possiacutevel a
interacccedilatildeo eficaz entre actina e miosina A Ca2+
-ATPase do retiacuteculo sarcoplasmaacutetico provoca a
reacumulaccedilatildeo do iatildeo caacutelcio no organelo impedindo o prolongamento desnecessaacuterio da contracccedilatildeo
(Refordf)
12 Vanaacutedio
O vanaacutedio eacute um metal de transiccedilatildeo pertencente ao grupo VB da tabela perioacutedica Foi
primeiramente descoberto em 1801 por Andreacutes Manuel del Riacuteo que concluiu erradamente que era
idecircntico ao croacutemio e definitivamente por Nils Gabriel Svenson em 1831 Recentemente foram
descobertas importantes funccedilotildees bioloacutegicas deste elemento provavelmente essencial para os
humanos de que se destaca no contexto do presente relatoacuterio a possibilidade de actuar como
anaacutelogo do grupo fosfato e a inibiccedilatildeo de enzimas in vitro como eacute o caso da actividade ATPaacutesica
da miosina
Eacute um metal soacutelido agrave temperatura ambiente que se encontra amplamente distribuiacutedo nos vaacuterios
ecossistemas em dois isoacutetopos naturais ndash 50
V e 51
V ndash raramente excedendo 01 partes por biliatildeo
15
(ppb) Estaacute associado a cerca de 70 minerais diferentes na natureza (dos quais se destacam o
uracircnio cobre chumbo zinco fosfato e titacircnio ferroso) especialmente na forma de vanadato ou
pentoacutexidos de vanaacutedio O vanaacutedio eacute um dos principais constituintes da mateacuteria orgacircnica foacutessil
com especial ecircnfase para o carvatildeo oacuteleo de crude e foacutesseis carbonaacuteceos [UV] bem como em
alimentos consumidos pelo Homem (salsa pimenta preta cogumelos marisco vegetais cereais
gorduras e oacuteleos) [W] Estes constituem a maior fonte de exposiccedilatildeo ao vanaacutedio da maioria da
populaccedilatildeo humana [X]
Normalmente a entrada do vanaacutedio no organismo ocorre por inalaccedilatildeo ou em menores
quantidades por absorccedilatildeo pelo tracto gastrointestinal ou pela pele O seu armazenamento centra-
se no fiacutegado rins ceacuterebro coraccedilatildeo muacutesculos e ossos No entanto a retenccedilatildeo de vanaacutedio no
organismo humano eacute reduzida ndash em condiccedilotildees normais cerca de 100 M (em muitos tecidos a
quantidade eacute inferior a 10 ngg de tecido) [Y]
Actualmente natildeo existem provas de deficiecircncia nutricional de vanaacutedio em humanos ou de
enzimas que contenham vanaacutedio em animais superiores No entanto vaacuterios estudos relacionam a
ocorrecircncia de deficiecircncia em galinhas e ratos devido a uma alimentaccedilatildeo pobre em vanaacutedio Os
sintomas relatados incluem retardamento do crescimento malformaccedilotildees oacutesseas perturbaccedilotildees no
metabolismo lipiacutedico e estagnaccedilatildeo reprodutiva entre outros [Z AA AB] Os efeitos fisioloacutegicos
do vanaacutedio podem depender do tipo de composto ou de oligoacutemero uma vez que consoante as
suas estabilidade labilidade e nuacutemero de coordenaccedilatildeo as respostas seratildeo diferenciadas ao niacutevel
dos organismos vivos
Por outro lado a toxicidade do vanaacutedio encontra-se bem caracterizada existindo sintomas
descritos em vaacuterios mamiacuteferos (inclusive o Homem) que incluem diarreias catildeibras irritaccedilatildeo da
pele e mucosas palpitaccedilotildees cardiacuteacas anemia pneumonia bronquite perturbaccedilotildees no Sistema
Nervoso Central (SNC) entre outros (Ref) A toxicidade varia geralmente com a natureza do
composto com aumento proporcional agrave sua valecircncia Verificou-se que a forma de entrada do
vanaacutedio tem tambeacutem efeito na sua toxicidade ndash os maiores niacuteveis verificaram-se por injecccedilatildeo
intravenosa da amostra sendo progressivamente inferiores por inalaccedilatildeo e finalmente por
ingestatildeo [X AA] A ampla utilizaccedilatildeo dos compostos de vanaacutedio na induacutestria quiacutemica bem como
as elevadas quantidades de vanaacutedio libertadas atraveacutes da combustatildeo dos oacuteleos de crude
contribuem consideravelmente para o aumento da biodisponibilidade do vanaacutedio Como tal o
interesse neste elemento e nos problemas ambientais por ele causados tem vindo a aumentar
significativamente [V]
121 Propriedades quiacutemicas
16
A quiacutemica do vanaacutedio eacute extraordinariamente complexa pelo que originou vaacuterias interpretaccedilotildees
incorrectas desde a sua descoberta No entanto as suas propriedades dependem em uacuteltima
anaacutelise da configuraccedilatildeo electroacutenica do elemento Assim a variabilidade encontrada no
comportamento quiacutemico do vanaacutedio pode dever-se agrave sua configuraccedilatildeo ndash [Ar] 3d2 4s
2 ndash que
permite a formaccedilatildeo de vaacuterios estados de oxidaccedilatildeo (de +2 a +5) Em soluccedilatildeo aquosa os estados de
oxidaccedilatildeo estaacuteveis vatildeo de V(V) a V(II) Destes apenas os +3 a +5 satildeo biologicamente importantes
jaacute que o V(II) apresenta demasiada capacidade redutora para existir nos organismos vivos
conhecidos [AC]
No fluido extracelular os compostos de vanaacutedio encontram-se predominantemente na forma de
monovanadatos (V(V)) livres ou ligados agrave transferina do soro ou agrave hemoglobina (Refordf) Os iotildees de
vanadato livres satildeo transportados para o meio intracelular atraveacutes de canais ioacutenicos inespeciacuteficos
Jaacute no ambiente intracelular os compostos sofreratildeo uma bio-reduccedilatildeo atraveacutes da acccedilatildeo da
glutationa ou outros compostos com grupos ndashOH como pentoses hexoses ou etanolaminas [V] A
reoxidaccedilatildeo de vanadilo (V(IV)) intracelular a vanadato (V(V)) provocaria a acumulaccedilatildeo toacutexica
destes uacuteltimos pelo que a complexaccedilatildeo da espeacutecie reduzida o impede O transporte de vanaacutedio
para o interior da ceacutelula encontra-se na Fig 110
Figura 110 ndash Transporte de vanaacutedio para a ceacuteluka e seu subsequente metabolismo (P=Proteiacutena) (Adaptado de
Zaporowska e Scibor 1998)
1211 Vanaacutedio(IV)
A espeacutecie de vanaacutedio (IV) mais bem conhecida eacute o iatildeo vanadilo (VO2+
) espeacutecie estaacutevel a pH
aacutecido sendo a maioritaacuteria a pH 3[AD] Apresenta uma cor azulada e forma complexos penta e
hexacoordenados Forma-se rapidamente a partir de V(V) na presenccedila de um agente redutor em
soluccedilatildeo aquosa No ambiente intracelular a presenccedila de glutationa reduzida aacutecido ascoacuterbico
NAD(P)H entre outros conduzem a que o principal estado de oxidaccedilatildeo nos sistemas bioloacutegicos
seja provavelmente o +4 encontrando-se o V(IV) fortemente complexado com proteiacutenas ou
aminoaacutecidos livres citrato ATP e pirofosfato (Refordfs) Em alternativa o V(IV) pode existir
17
livremente em ambientes fortemente aacutecidos como os lisossomas jaacute que a pH superior a 3 as
soluccedilotildees de vanadilo tendem a ser oxidadas pelo ar formando-se vanadato Esta grande
capacidade redutora justifica parcialmente a utilizaccedilatildeo pouco extensiva do iatildeo VO2+
como
inibidor [W]
1212 Vanaacutedio (V)
O vanaacutedio (V) eacute a forma mais estaacutevel num intervalo de pH amplo [W] O iatildeo anioacutenico VO3-tende
a constituir complexos polinucleares pelo que as soluccedilotildees aquosas de vanadato satildeo extremamente
complexas com ocorrecircncia simultacircnea de vaacuterios equiliacutebrios de protonaccedilatildeo (pKa(vanadato)= 35
78 e 125) e oligomerizaccedilatildeo As espeacutecies presentes (H3VO4 H2VO4- HVO4
2- e VO4
3-) dependem
do pH forccedila ioacutenica e temperatura da soluccedilatildeo e da concentraccedilatildeo total do elemento
[AEAFAGAHAI]
O VO43-
- monoacutemero ortovanadato V1 ndash estaacute associado a uma grande parte da importacircncia
bioloacutegica do vanaacutedio uma vez que se trata de um anaacutelogo electroacutenico e estrutural do ortofosfato
(PO43-
) A coordenaccedilatildeo do ortovanadato pode ser tetraeacutedrica ou bipiramidal trigonal (Refordfs)
A oligomerizaccedilatildeo do vanadato monomeacuterico daacute origem ao diacutemero (V2O72-
- V2) que apresenta
tambeacutem equiliacutebrios de protonaccedilatildeo Presume-se a existecircncia de analogia do vanadato dimeacuterico com
o pirofosfato embora natildeo haja evidecircncia estrutural (Refordf) A dimerizaccedilatildeo do V2 origina o
vanadato tetrameacuterico ou metavanadato (V4O124-
- V4) que constitui a espeacutecie maioritaacuteria em
soluccedilotildees aquosas concentradas (Refordf) As estruturas do metavanadato e a sua espeacutecie protonada
satildeo ciacuteclicas [AJ] Outra espeacutecie oligomeacuterica importante eacute o vanadato pentameacuterico (V5O153-
) cuja
estrutura presumida eacute ciacuteclica [AI]
O oligoacutemero com maior importacircncia bioquiacutemica eacute o vanadato decameacuterico (V10O286-
-V10)(Refordf) a
espeacutecie predominante em soluccedilotildees de pH 2-6 quando a concentraccedilatildeo total de vanadato eacute superior
a 100 M (Ref) A estrutura do V10 natildeo difere em soluccedilatildeo ou no estado soacutelidoOs 10 aacutetomos de
vanaacutedio da espeacutecie decameacuterica estatildeo divididos em trecircs grupos identificados na figura como VA-C
consoante o seu ambiente quiacutemico circundante (Refordf) (Fig 111)
A expressatildeo ldquodecavanadatordquo refere-se a uma soluccedilatildeo aquosa na qual a espeacutecie decameacuterica se
encontra em equiliacutebrio quiacutemico com as espeacutecies V1 V2 V4 e V5 sendo que a oligomerizaccedilatildeo
progressiva do vanaacutedio eacute proporcional agrave sua concentraccedilatildeo total (Ref) Nesta soluccedilatildeo a
concentraccedilatildeo de V10 corresponde a aproximadamente 10 da concentraccedilatildeo total de vanaacutedio [AK]
18
Figura 111 ndash Estruturas esquemaacuteticas das espeacutecies de vanadato (espeacutecie monomeacuterica (V1) dimeacuterica (V2)
tetrameacuterica (V4) pentameacuterica (V5) e decameacuterica (V10) (Adaptado de Aureliano e Crans 2008)
122 Metabolismo e efeito na actividade enzimaacutetica
Apesar de natildeo terem sido ainda encontradas as provas categoacutericas das suas funccedilotildees nos seres
vivos sabe-se que o iatildeo vanadato eacute capaz de inibir uma grande quantidade de enzimas in vitro
com especial ecircnfase na actividade ATPaacutesica da miosina [AL Teresa] Como referido
anteriormente o vanadato eacute capaz de actuar em alternativa ao fosfato nas reacccedilotildees bioquiacutemicas
incluindo a sua preferecircncia de ligaccedilatildeo agraves enzimas [W]
Eacute conhecido o papel do vanaacutedio no metabolismo do caacutelcio promovendo a mineralizaccedilatildeo dos
dentes e ossos na biossiacutentese de liacutepidos (trigliceacuteridos e colesterol) com posterior inibiccedilatildeo da
acumulaccedilatildeo destes nos vasos sanguiacuteneos e no SNC na reproduccedilatildeo e desenvolvimento por
mimetizaccedilatildeo e potenciaccedilatildeo de factores de crescimento na estimulaccedilatildeo do consumo de glucose e
da siacutentese de glicogeacutenio com actuaccedilatildeo equivalente aos efeitos da insulina ao niacutevel celular na
transduccedilatildeo de sinais celulares com regulaccedilatildeo da actividade de segundos mensageiros no
transporte de iotildees e aminoaacutecidos na transcriccedilatildeo geneacutetica e siacutentese de proteiacutenas e DNA entre
outros [W Y AB]
Assim o vanaacutedio cnseguiraacute provocar inuacutemeras mudanccedilas bioquiacutemicas nas ceacutelulas Alguns
exemplos da influecircncia bioquiacutemica do vanaacutedio incluem os seus efeitos nos processos de
fosforilaccedilatildeo e desfosforilaccedilatildeo de proteiacutenas e na participaccedilatildeo na formaccedilatildeo de espeacutecies reactivas de
oxigeacutenio devido ao seu efeito proacute-oxidante sobre agentes redutores a niacutevel celular [AM]
Quanto ao seu efeito na actividade enzimaacutetica pode dividir-se em dois tipos principais ndash inibitoacuterio
e estimulante - dependendo da enzima ou do tipo de reacccedilatildeo por esta catalisada No primeiro
grupo incluem-se enzimas como as ATPases (de miosina de canais ioacutenicos e fosfoenzimas
19
transportadoras) e as fosfatases (da tirosina aacutecida e alcalina) a fosfofrutocinase adenilato ciclase
e catalase entre outras [AA] O segundo grupo inclui a maioria das cinases a glucose-6-fosfato
desidrogenase a fosfoglicerato mutase e a fosfoglicerato fosfatase (Refordf)
O efeito do vanaacutedio (V) na actividade enzimaacutetica estaacute relacionado com o seu nuacutemero de
coordenaccedilatildeo podendo actuar como estado de transiccedilatildeo ou por mimetizaccedilatildeo de cofactores
substratos (especialmente o fosfato) e interacccedilotildees proteiacutena-proteiacutena (como no caso das
polimerases e transcriptases) Pode ainda estar envolvido na formaccedilatildeo de ligaccedilotildees de hidrogeacutenio
interacccedilotildees electrostaacuteticas esteacutereas e de oxidaccedilatildeo-reduccedilatildeo Uma evidecircncia desta uacuteltima eacute a sua
capacidade de alternacircncia entre os estados de oxidaccedilatildeo +5 a +3 o que torna faacutecil a sua interacccedilatildeo
redox com enzimas contendo resiacuteduos de cisteiacutenas (Cys) ou qualquer outro composto orgacircnico
com actividade redox [AN]
13 Vanaacutedio e muacutesculo esqueleacutetico
A primeira evidecircncia da presenccedila de vanaacutedio no muacutesculo esqueleacutetico proveio de uma amostra de
ATP isolado de muacutesculo de cavalo que continha uma impureza ndash vanaacutedio ndash cuja presenccedila
provocou a inibiccedilatildeo da actividade da bomba de Na+K
+-ATPase [AO] Desde entatildeo vaacuterios
estudos tecircm sido realizados com ecircnfase na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o
tecido muscular Destes destaca-se a descoberta da interacccedilatildeo forte da espeacutecie V10 com a miosina
fosforilada [AP] e agrave sua inibiccedilatildeo da actividade ATPaacutesica da miosina[AQ AR]
No quediz respeitoao iatildeo monovanadato destaca-se a sua inibiccedilatildeoda actividade Mg2+
-ATPaacutesica
da miosina [AS] que ocorre de duas formas distintas (1) na presenccedila de concentraccedilotildees reduzidas
de V1 a inibiccedilatildeo ocorre a velocidade reduzida dependente da concentraccedilatildeo da espeacutecie
monomeacuterica (2) na presenccedila de concentraccedilotildees elevadas de V1 o niacutevel de inibiccedilatildeo aproxima-se de
100 desde que [V1] seja superior agrave concentraccedilatildeo total de locais com actividade Mg2+
-ATPaacutesica
no ensaio Esta inibiccedilatildeo eacute irreversiacutevel tanto por diaacutelise ou adiccedilatildeo de agentes redutores do
vanadato [AL]Segundo Pate et al(1994)[AT] a espeacutecie monomeacuterica podeinibir a velocidade de
contracccedilatildeo a temperatura inferior a 20ordmC perdendogradual e rapidamente essa capacidade ateacute os
50ordmC temperatura agrave qual cessa a capacidade inibitoacuteria Os resultados obtidos sugeremainda uma
interacccedilatildeo altamente cooperativa entre o iatildeo vanadato e as proteiacutenas componentes do muacutesculo
esqueleacuteticoDestaca-se tambeacutem a descoberta da inibiccedilatildeo da geraccedilatildeo de forccedila contraacutectil nas fibras
musculares por parte da espeacutecie monomeacuterica[AT]Wilson et al(1995) [AU ]determinaram que a
afinidade da espeacutecie V1 para o S1 de miosina eacute significativamente mais elevada do que a do iatildeo
Pi seu anaacutelogo Descobriram ainda que que a ligaccedilatildeo do vanadato poderaacute envolver transiccedilotildees
entre estados energeacuteticos com energia livre independente dos impedimentos esteacutereos impostos
20
pela estrutura das miofibrilas Os estudos desenvolvidos por Smith et al (1990)[AV]
determinaram queo iatildeo monovanadato natildeo apresenta um efeito pronunciado na inibiccedilatildeo da
actividadeacto-S1-miosina-ATPaacutesica excepto em concentraccedilotildees superiores a 900microM a partir das
quais existe produccedilatildeo de espeacutecies oligomeacutericas para as quais o efeito inibitoacuterio seraacute mais
elevadoA espeacutecie monomeacuterica de vanadato consegue adoptar facilmente uma geometria do tipo
bipiramidal trigonal que mimetiza o intermediaacuterio formado (MgADP
Pi)aquando da reacccedilatildeo de
hidroacutelise de ATP no local activo da miosina [AW] De facto a interacccedilatildeo do vanaacutedio com a
miosina eacute inesperadamente estaacutevel na presenccedila de ADP Esta interacccedilatildeo resulta na formaccedilatildeo de
um complexo enzimaacutetico inactivo e estaacutevel com estequiometria de 11 ADPvanadato por siacutetio
activo da proteiacutena A formaccedilatildeo do complexo estaacutevel miosina-vanadato foi proposta por Goodnoet
al (1982) [AS] Nesta ocorre um preacute-equiliacutebrio raacutepido que induz a formaccedilatildeo de um complexo
ternaacuterio reversiacutevel (MADP
V) com formaccedilatildeo lenta do isoacutemero M
ADP
V A diferenccedila entre os
compostos de vanaacutedio e de fosfato centra-se no tempo de meia vida (t12) do Pi ou do V (no
primeiro t12=12 s enquanto no segundo t12=24 h) (Refordf) A diferenccedila dever-se-aacute agrave possiacutevel
complexaccedilatildeo do vanaacutedio com grupos nucleofiacutelicos no siacutetio activo o que provocaria uma
estabilizaccedilatildeo da miosina na conformaccedilatildeo MADP
V
No que respeita aos estudos realizados com a espeacutecie tetrameacuterica Cremo et al determinou que a
cadeia pesada do S1 de miosina pode ser clivada em dois locais distintos (designados V1 e V2)
pela espeacutecie V4 apoacutes irradiaccedilatildeo com luz UV na presenccedila de concentraccedilotildees de vanadato na ordem
dos milimolar e na ausecircncia de nucleoacutetidos ou metais divalentes
Jaacute no que concerne aos estudos realizados com ecircnfase na espeacutecie decameacuterica foi sugerido
porRamos et al (2006) [AK]que a interacccedilatildeo do decavanadato com a G-actina favorecida pela
polimerizaccedilatildeo desta uacuteltima estabiliza a espeacutecie V10 e induz a inibiccedilatildeo da formaccedilatildeo de F-actina A
estabilizaccedilatildeo da espeacutecie V10 por proteiacutenas tansmembranares e do citoesqueleto pode ser a razatildeo
pela qual ocorre a toxicidade do decavanadato reportada na avaliaccedilatildeo dos efeitos do V(V) nos
sistemas bioloacutegicosTiago et al (2007) [AR] concluiram que o V10 apresenta uma clara preferecircncia
de ligaccedilatildeo ao ldquoback-doorrdquo da S1-miosina-ATPase o que se verifica apenas na estrutura
conformacional desimpedida com acesso facilitado ao local de ligaccedilatildeo do Pi Sugere tambeacutem que
o V10 actua como um ldquoback-door stoprdquo bloqueando estaabertura do fragmento de 50-kDa
necessaacuteria para a hidroacutelise do ATP--fosfato o que constitui uma explicaccedilatildeo simples do
comportamento natildeo competitivo da espeacutecie decameacuterica estimulando a utilizaccedilatildeo deste oligoacutemero
como uma feramenta da elucidaccedilatildeo das mudanccedilas conformacionais ldquoback-doorrdquo da miosina no
21
processo da contracccedilatildeo muscular(ref)Destaca-se ainda a descoberta da capacidade inibitoacuteria do
V10 em relaccedilatildeo agrave fosforilaccedilatildeo oxidativa mitocondrial [AX]
Finalmente no que concerne aos estudos realizados com a presenccedila de vaacuterias espeacutecies
oligomeacutericas Aureliano e Madeira (1994) [AY]observaram que na presenccedila de Ca2+
as espeacutecies
V4 e V10se ligam agrave Ca2+
-ATPase do retiacuteculo sarcoplasmaacutetico enquanto a espeacutecie V1 se liga apenas
na presenccedila de ATP Aparentemente o aumento da concentraccedilatildeo de Ca2+
tem o efeito contraacuterio da
inibiccedilatildeo da ATPase do retiacuteculo sarcoplasmaacutetico pelo V1 natildeo tendo no entanto efeito significativo
sobre a inibiccedilatildeo por parte da espeacutecie V10 Tiago et al (2002) [AP] concluiacuteram que em condiccedilotildees
proacuteximas agraves fisioloacutegicas a espeacutecie V10 difere das espeacutecies presentes na soluccedilatildeo de metavanadato
devido agrave sua forte interacccedilatildeo com a miosina fosforilada e agrave sua inibiccedilatildeo da actividade ATPaacutesica
da miosina Para aleacutem disso o ATP diminui a afinidade da miosina para o V4 induzindo a
interacccedilatildeo com V1 e natildeo tendo qualquer efeito no que diz respeito ao V10 Foi tambeacutem proposto
[AQ] que a ligaccedilatildeo das espeacutecies oligomeacutericas de vanadato aoS1 de miosina nos locais onde o
fosfato coordenaeacute provavelmente a causa da inibiccedilatildeo da actividade ATPaacutesica da miosina
estimulada por actina atraveacutes da prevenccedilatildeo da troca entre ATPADP sendo esta favorecida para
aniotildees de vanadato com valecircncias superiores como eacute o caso do V10
Na presenccedila de actina as velocidades de dissociaccedilatildeo do Pi e do V aumentam consideravelmente
este uacuteltimo em menor quantidade Assim a ligaccedilatildeo da actina ao complexo MADP
V e
Pi eacute
bastante fraca pelo que se sugere que o ADPV teria um efeito de relaxamento semelhante ao
ATP No entanto a capacidade do vanaacutedio em inibir a actividade ATPaacutesica da miosina e da
actomiosina eacute diferenciada uma vez que na ausecircncia de actina a inibiccedilatildeo eacute total para
concentraccedilotildees na gama dos M mas na sua presenccedila a inibiccedilatildeo eacute totalmente conseguida apenas
em concentraccedilotildees na gama dos mM (Ref) No entanto eacute necessaacuterio considerar a possiacutevel
formaccedilatildeo de espeacutecies oligomeacutericas de vanadato cuja afinidade e efeito na enzima seratildeo
diferentes Assim pode concluir-se que o efeito de inibiccedilatildeo observado para a actividade ATPaacutesica
de miosina estimulada por actina se pode dever agrave presenccedila das diferentes espeacutecies oligomeacutericas o
que foi jaacute sugerido por Aureliano (2000) [AZ] ndash as espeacutecies tetrameacuterica e decameacuterica de vanadato
interactuariam com a miosina com alteraccedilatildeo da sua interacccedilatildeo com a actina No entanto natildeo eacute
conhecida a contribuiccedilatildeo absoluta das restantes espeacutecies oligomeacutericas para a inibiccedilatildeo da ATPase
da miosina estimulada por actina (Refs)
14 Objectivos do projecto
Pretende-se com este projecto o isolamento purificaccedilatildeo e caracterizaccedilatildeo da actina do muacutesculo
22
esqueleacutetico de coelho sendo esta uacuteltima tambeacutem realizada para o sufragmento-1 de miosina
Seguidamente realizar-se-aacute a quantificaccedilatildeo e caracterizaccedilatildeo das proteiacutenas por espectroscopia UV-
Vis e SDS-PAGE e a caracterizaccedilatildeo das soluccedilotildees de vanaacutedio por UV-Vis e ressonacircncia magneacutetica
nuclear (RMN) Por uacuteltimo a interacccedilatildeo do S1 de miosina com o vanaacutedio seraacute avaliada atraveacutes
das teacutecnicas de UVVis RMN e ressonacircncia paramagneacutetica electroacutenica (RPE)
2 Discussatildeo de Resultados
21 Caracterizaccedilatildeo das proteiacutenas isoladas
211Anaacutelise do estado de pureza
O estado de pureza das soluccedilotildees de actina e de S1 foi analisado por electroforese em gel de
poliacrilamida em condiccedilotildees desnaturantes (SDS-PAGE) (Fig 22) que separa as moleacuteculas de
acordo com o seu peso molecular (PM) A concentraccedilatildeo de acrilamida escolhida (10) permite a
detecccedilatildeo simultacircnea de componentes com peso molecular inferior a 20 kDa e outros com elevado
peso molecular
Figura 21 ndash Fotografia de gel de SDS-PAGE (10 acrilamida) Poccedilo (a)
10 L do marcador de elevado peso molecular () Poccedilo (b) 20 L de [G-
actina]=1 mgmL Poccedilo (c) 20 L de [S1]=1 mgmL Aplicou-se uma
voltagem de V durante h e min
A partir da recta de calibraccedilatildeo obtida log PM = -1177 Rf + 2247 (ver ponto 2 da secccedilatildeo
Anexos) pode verificar-se a presenccedila de actina na amostra do poccedilo (b) presente numa banda
bastante intensa (PMaparente=40 kDa) bem como a presenccedila minoritaacuteria de outros componentes
desconhecidos que natildeo teratildeo sido eliminados durante o processo de extracccedilatildeo e purificaccedilatildeo e natildeo
satildeo relevantes para o estudo em questatildeo Na amostra do poccedilo (c) nota-se a presenccedila de S1
(PMaparente=92 kDa) bem como a de outros componentes cuja banda eacute ligeiramente menos intensa
Estes poderatildeo ser fragmentos proteoliacuteticos de miosina e de S1 Estaacute ainda presente uma bada de
23
maior intensidade que poderaacute ser a proteiacutena-C (PM=135 kDa) uma vez que a sua remoccedilatildeo
completa da amostra necessitaria de um passo adicional de cromatografia de troca ioacutenica [BA]
No entanto natildeo podemos confirmar que se trata desta proteiacutena ou de qualquer outra uma vez que
se encontra fora do intervalo de migraccedilatildeo dos marcadores de peso molecular
212 Determinaccedilatildeo da concentraccedilatildeo proteica
2121Subfragmento-1 de miosina
A partir da recta de calibraccedilatildeo obtida pelo meacutetodo de Bradford [BB](ver secccedilatildeo 3311 do
Procedimento Experimental) A595nm = 00546[BSA] + 00209 (ver secccedilatildeo de Anexos)foi possiacutevel
determinar um intervalo de concentraccedilatildeo de S1 de miosina compreendido entre 226 e 117(plusmn
423M) (203 e 1056 (plusmn 381 mgmL)) (n=8) A concentraccedilatildeo meacutedia de S1 de miosina apoacutes
diaacutelise contra a soluccedilatildeo tampatildeo 10 mM HEPES pH 75 25 mM KCl determinada pela leitura da
Abs280 nmfoi de 20430M (1803 mgmL)(n=7)
As amostras a quantificar atraveacutes do meacutetodo de Bradford natildeo sofreram diaacutelise o que permite
explicar a concentraccedilatildeo superior de S1 de miosina em comparaccedilatildeo com o valor obtido atraveacutes do
meacutetodo directo Esta razatildeo aliada agrave sensibilidade e rapidez superiores deste uacuteltimo fez com que
se optasse pelo meacutetodo directo para a quantificaccedilatildeo da grande maioria dos ensaios
2122Actina
A concentraccedilatildeo de actina foi determinada quando esta se encontrava na forma monomeacuterica (G-
actina) por mediccedilatildeo da leitura directa da Abs290 nm tendo-se obtido uma concentraccedilatildeo meacutedia de
691481M (290202 mgmL) (n=3) em 2 mM Tris-HCl pH 80 02 mM CaCl2 02 mM Na-
ATP Ao comprimento de onda utilizado (290 nm) absorvem fortemente apenas os resiacuteduos de
Trp pelo que a mediccedilatildeo efectuada tem menor sensibilidade do que se tivesse sido efectuada a 280
nm uma vez que a actina apresenta apenas 4 resiacuteduos deste aminoaacutecido embora tenha 16
resiacuteduos de Tyr
22 Mediccedilatildeo da actividade enzimaacutetica de S1 de miosina
221 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
As actividades ATPaacutesicas de S1 de miosina dependentes de Ca2+
e K+foram analisadas pelo
caacutelculo das velocidades de consumo inicial do fosfato inorgacircnico (Pi) Definiu-se os tempos de 5
e 3 minutos para a mediccedilatildeo da Ca2+
e da K+EDTA-ATPase respectivamente Os valores obtidos
24
a partir do protocolo presente no ponto 341 da secccedilatildeo Procedimento experimental encontram-se
descritos na Tabela 2I
Tabela 2I ndash Acrividades natildeo-fisioloacutegicas de subfragmento-1 determinadas pelo meacutetodo de Fiske-Subarow
modificado (n=2)
Proteiacutena
Actividade
ATPaacutesica natildeo
fisioloacutegica
Actividade (nmol
Piminmg proteiacutena) Actividade (s-1)
S1
Ca2+
-ATPase 1400 plusmn 30 2100 plusmn 0045
K+EDTA-ATPase 2334 plusmn 50 3500 plusmn 0075
As actividades natildeo-fisioloacutegicas reflectem o estado de oxidaccedilatildeo dos resiacuteduos expostos de Cys 707
(SH1) e 697 (SH2) Se o grupo SH1 sofrer oxidaccedilatildeo a actividade ATPaacutesica dependente de Ca2+
aumenta e a actividade dependente de K+ diminui Por outro lado se o grupo SH2 sofrer
oxidaccedilatildeo regista-se a perda completa destas actividades
De acordo com Margossian e Lowey (1982) [BA] as actividades ATPaacutesica de S1 de miosina
dependente de Ca2+
e K+EDTA da miosina satildeo respectivamente de 24 e 112 s-1
Estaacute tambeacutem
descrito que a actividade ATPaacutesica dependente de K+ eacute em meacutedia cerca de 3 vezes superior agrave
dependente de Ca2+
reflectindo a quantidade das preparaccedilotildees A razatildeo obtida entre os valores das
actividades ATPaacutesicas dependentes de K+ e Ca
2+ eacute de 166 Os valores obtidos para as actividades
(Ca2+
-ATPase=21 s-1
K+EDTA-ATPase= 35 s
-1) satildeo inferiores agrave valores obtidos por
Margossian e Lowey [BA] e Tiago [BC] com forccedila ioacutenica inferior O valor obtido para a
actividade ATPaacutesica dependente de Ca2+
eacute superior ao obtido por Polosukhina et al (2000) [BD]
cujo meio de reacccedilatildeo continha MOPS e apresentava forccedila ioacutenica inferior Os dados
supramencionados comprovam a variaccedilatildeo da actividade a ATPaacutesica natildeo fisioloacutegica em relaccedilatildeo agrave
forccedila ioacutenica temperatura e pH
222 Mediccedilatildeo da actividadeMg2+
-ATPaacutesica de S1 pelo meacutetodo de enzimas acopladas
A actividade de Mg2+
-ATPase eacute a uacutenica que possui significado fisioloacutegico sendo as restantes
bastante uacuteteis apenas para indicaccedilatildeo da qualidade de preparaccedilatildeo do S1[BA] A 0575 M KCl a
actina e o S1 formam uma soluccedilatildeo viscosa e tuacuterbida Apoacutes a adiccedilatildeo de MgATP o complexo acto-
S1 dissocia-se e a turbidez da soluccedilatildeo diminui Se a forccedila ioacutenica for reduzida a dissociaccedilatildeo do
complexo actoS1 que se encontra na forma de suspensatildeo eacute induzida O MgATP eacute rapidamente
hidrolisado diminuindo a sua concentraccedilatildeo no meio reaccional Nestas condiccedilotildees haacute
reassociaccedilatildeo e subsequente contracccedilatildeo dos filamentos de actina e de S1 com formaccedilatildeo de
25
precipitado O meacutetodo descrito traduz de forma bastante elementar o fenoacutemeno da contracccedilatildeo in
vivo e elimina o problema anterior da solubilizaccedilatildeo ineficiente da miosina de modo a obter uma
mediccedilatildeo exacta da actividade [A] A utilizaccedilatildeo de forccedilas ioacutenicas reduzidas aumenta a afinidade da
actina para o S1 permitindo contornar a extrema dificuldade em obter a concentraccedilatildeo proteica dos
muacutesculos (cerca de 100 mgmL do complexo actomiosina) As actividades Mg2+
-ATPaacutesicas de S1
de miosina foram determinadas a partir do declive da variaccedilatildeo da Abs340 relativo agrave oxidaccedilatildeo de
NADH em funccedilatildeo do tempo como eacute descrito no ponto 342 do Procedimento Experimental Os
valores obtidos para a actividade Mg2+
-ATPaacutesica encontram-se apresentados na Tabela 2II
Tabela 2II ndash Actividade Mg2+
-ATPaacutesica de subfragmento-1 de miosina determinadas pelo meacutetodo das enzimas
acopladas
Actividade (nmol
ATPmin mg(S1)) Actividade (s
-1)
1097 plusmn 269 164 plusmn 04
Ramos (2005) [J] obteve o valor de 01 s-1
para a actividade Mg2+
-ATPaacutesica de S1 de miosina
com forccedila ioacutenica intermeacutedia Tiago (2005) [BC] obteve o valor de 011 s-1 em condiccedilotildees de forccedila
ioacutenica reduzida em relaccedilatildeo agrave utilizada no presente relatoacuterio
23 Preparaccedilatildeo e caracterizaccedilatildeo das soluccedilotildees de vanaacutedio
231 Anaacutelise da composiccedilatildeo das soluccedilotildees de vanadato por espectroscopia de 51
V-RMN
efeito da concentraccedilatildeo da espeacutecie
A diluiccedilatildeo das soluccedilotildees stock de meta- e decavanadato iraacute determinar fortemente a composiccedilatildeo
em soluccedilatildeo dos diferentes oligoacutemeros de vanadato (ref) No caso do decavanadato o periacuteodo de
tempo apoacutes a diluiccedilatildeo iraacute tambeacutem ser um factor importante a considerar devido agrave decomposiccedilatildeo
da espeacutecie V10(ref)Torna-se assim fundamental determinar a composiccedilatildeo quiacutemica das soluccedilotildees
no meio de ensaio utilizado (HEPES 10 mM KCl 25 mM) de modo a correlacionaacute-la
correctamente com os possiacuteveis efeitos promovidos pelas diferentes espeacutecies de vanadato no
sistema em estudo
Aleacutem disso deve ser dado ecircnfase agraves importantes interacccedilotildees que ocorrem entre o vanaacutedio e os
tampotildees escolhidos O tampatildeo com menor nuacutemero e magnitude de interacccedilatildeo eacute o HEPES [AD]
seguido do Tris sendo este uacuteltimo especialmente aconselhado em concentraccedilotildees baixas para casos
de incompatibilidade enzimaacutetica ou quiacutemica redox No caso especial da reacccedilatildeo redox
NAD(P)HNAD(P) o tampatildeo adequado eacute o imidazole de modo a natildeo haver reacccedilatildeo com o
26
vanadato Os tampotildees a evitar em qualquer situaccedilatildeo incluem a trietanolamina fosfato citrato e
glicina entre outros [AD] uma vez que todos interactuam fortemente com V(V) com formaccedilatildeo
de complexos estaacuteveis
Assim procedeu-se agrave anaacutelise da composiccedilatildeo das soluccedilotildees de meta- e decavanadato apoacutes diluiccedilatildeo
das soluccedilotildees stock (50 mM em vanadato total) no meio de reacccedilatildeo (tampatildeo D) atraveacutes da
espectroscopia de ressonacircncia magneacutetica nuclear de 51
V Nas figuras x1 e x2 apresentam-se os
espectros obtidos para as soluccedilotildees de deca- e metavanadato num intervalo de concentraccedilotildees de 1 a
10 mM A Tabela seguinte resume os paracircmetros espectroscoacutepicos obtidos para cada espeacutecie
presente nas soluccedilotildees analisadas
Figura 22ndash Espectros de RMN de 51
V de
concentraccedilotildees crescentes das espeacutecies oligomeacutericas
de vanadato obtidos apoacutes diluiccedilatildeo da soluccedilatildeo stock
de decavanadato em 10 mM HEPES pH 70 25 mM
KCl para concentraccedilotildees finais de A) 1 mM B)2
mM C) 4 mM D) 8 mM E) 10 mM de
decavanadato total V1refere-se agraves espeacutecies
monomeacutericade vanadato V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2IIIndash Desvios quiacutemicos meacutedios (part) das ressonacircncias das espeacutecies decameacuterica (V10) monomeacuterica (V1) e
tetrameacuterica (V4) presentes nos espectros da soluccedilatildeo de decavanadatoLargura a meia altura (∆12) da espeacutecie V10A
Paracircmetros V10A V10B V10C V1 V4
part (ppm) -5162 -5003 -4255 -5616 -5748
∆12 (Hz) 175 nd nd nd nd
Figura 23 ndash Distribuiccedilatildeo graacutefica das
espeacutecies oligomeacutericas de vanadato vs a
concentraccedilatildeo de decavanadato total As
concentraccedilotildees de cada espeacutecie foram
obtidas segundo a formula descrita na
secccedilatildeo 35 do Procedimento
Experimental (aacutereas dos sinais de 51
V-
RMN observadas na figura x1)V10 V1 e
V4 correspondem agraves espeacutecie monomeacuterica
dimeacuterica e tetrameacuterica respectivamente [Esp
eacuteci
e o
ligo
meacute
rica
de
van
aacutedio
] (m
M)
[Decavanadato]total (mM)
V10
V1
V4
27
Nas soluccedilotildees de decavanadato as espeacutecies monomeacuterica e decameacuterica de vanadato satildeo as uacutenicas
presentes para concentraccedilotildees de decavanadato inferiores a 8 mM Ambas as espeacutecies aumentam
linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado em [AY] e
em (Teresa) quando a concentraccedilatildeo de V1 eacute igual ou superior a 200 microM inicia-se a sua
oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica No entanto nas condiccedilotildees de realizaccedilatildeo dos
ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a espeacutecie tetrameacuterica neste
estudo em concentraccedilotildees inferiores a 8 mM tendo o mesmo sido observado por Ramoset al
(2006) [AK] Os resultados obtidos mostram claramente o equiliacutebrio entre a espeacutecie decameacuterica e
a monomeacuterica e que o aparecimento de outras espeacutecies oligomeacutericas se encontra dependente da
concentraccedilatildeo desta uacuteltima Apesar de natildeo ter sido realizado o ensaio com 5 mM de decavanadato
total no presente relatoacuterio estaacute reportado em Ramos et al (2006) [AK] que as concentraccedilotildees da
espeacutecie decameacuterica e monomeacuterica satildeo respectivamente de 500 microM e 200 microM Assim pode inferir-
se que para concentraccedilotildees inferiores a 5 mM e intervalos de tempo de cerca de 20 minutos (que
inclui o tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas
as espeacutecies decameacuterica e monomeacuterica de vanadato
Figura 24 ndash Espectros de RMN de 51
V de concentraccedilotildees
crescentes das espeacutecies oligomeacutericas de vanadato obtidos
apoacutes diluiccedilatildeo da soluccedilatildeo stock de metavanadato em 10 mM
HEPES pH 70 25 mM KCl para concentraccedilotildees finais de A)
1 mM B) 2 mM C) 4 mM D) 8 mM E) 10 mM de
metavanadato total V1 refere-se agraves espeacutecies monomeacuterica de
vanadatoV1 e V2 correspondem respectivamente agrave espeacutecie
monomeacuterica e dimeacuterica
Figura 25 ndash Distribuiccedilatildeo graacutefica das espeacutecies
oligomeacutericas de vanadato vs a concentraccedilatildeo de
metavanadato total As concentraccedilotildees de cada
espeacutecie foram obtidas segundo a formula descrita
na secccedilatildeo 35 do Procedimento Experimental
(aacutereas dos sinais de 51
V-RMN observadas na
figura x1)V10 V1 e V4 correspondem agraves espeacutecie
monomeacuterica dimeacuterica e tetrameacuterica
respectivamente
Esp
eacuteci
e o
ligo
meacute
rica
de
va
nad
ato
(m
M)
[Metavanadato]total (mM)
V1
V2
V4
V5
28
Tabela 2IVndash Desvios quiacutemicos meacutedios (part)das ressonacircncias das espeacutecies tetrameacuterica (V4) monomeacuterica (V1) e
dimeacuterica (V2) presentes no espectro da soluccedilatildeo de metavanadato (total) 10 mM
Paracircmetros V1 V2 V4 V5
part (ppm) -5615 -5748 -5789 -5869
Nas soluccedilotildees de metavanadato as espeacutecies monomeacuterica dimeacutericae tetrameacuterica de vanadato estatildeo
presentes para concentraccedilotildees de metavanadato inferiores a 4 mM Todas estas as espeacutecies
aumentam linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado
em (Aureliano e Madeira 1998) e em (Tiago et al 2004) quando a concentraccedilatildeo de V1 eacute igual ou
superior a 200 microM inicia-se a sua oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica Nas
condiccedilotildees de realizaccedilatildeo dos ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a
espeacutecie pentameacuterica neste estudo em concentraccedilotildees inferiores a 4 mM tendo o mesmo sido
observado por Ramos 2006 Os resultados obtidos mostram claramente o equiliacutebrio entre a
espeacutecie V4 e as espeacuteciesV1 e V2 no qual o aparecimento de outras espeacutecies oligomeacutericas se
encontra dependente da concentraccedilatildeo da espeacutecie V1 Por uacuteltimo pode inferir-se que para
concentraccedilotildees inferiores a 2 mM e intervalos de tempo de cerca de 20 minutos (que inclui o
tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas as
espeacutecies V1 V2 e V4 com concentraccedilotildees respectivas de 0796 0192 e 0196 mM
232Estabilidade das soluccedilotildees de decavanadato anaacutelise por espectroscopia de absorccedilatildeo
UVVis
Caracterizou-se a estabilidade da espeacutecie V10 numa soluccedilatildeo com 1 mM decavanadato total em
tampatildeo D atraveacutes da leitura da Abs400 Obteve-se uma cineacutetica de dissociaccedilatildeo de 1ordf ordem tal
como se pode verificar pela linearidade do graacutefico de ln(Abs400)vs o tempo de aquisiccedilatildeo espectral
(segundos) (Fig26 )
Figura 26ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do tempo
(em segundos) para uma soluccedilatildeo de decavanadato total de 1
mM em tampatildeo D a 25ordmC (n=2)
Sendo uma reacccedilatildeo de primeira ordem pode ser descrita pela equaccedilatildeo cujo
declive eacute ndashK (constante de dissociaccedilatildeo) e a intersecccedilatildeo no eixo das ordenadas corresponde
Ln
(Ab
s 400 n
m)
tempo (s)
29
aln(A0) Numa reacccedilatildeo de 1ordf ordem o tempo de meia-vida eacute independente da concentraccedilatildeo inicial
do reagente sendo descrito pela equaccedilatildeo [2] Deste modo a partir da recta
Ln(Abs400 nm) = -436E-5
K - 105 obteve-se um tempo de meia-vida de 4490 plusmn 0823 horas para
as condiccedilotildees supramencionadas
Apoacutes a adiccedilatildeo de S1 8 microM ao meio reaccional verificou-se que a reacccedilatildeo de desoligomerizaccedilatildeo
da espeacutecie decameacuterica de vanadato seguia igualmente uma cineacutetica de 1ordf ordem tal como se pode
verificar pela linearidade do graacutefico de ln(Abs400) versus o tempo de aquisiccedilatildeo espectral
(segundos) (Fig27)
Figura 27ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do
tempo (em segundos) para uma soluccedilatildeo de decavanadato
total de 1 mM em tampatildeo D a 25ordmC na presenccedila de 8
microM (n= 2)
A partir da equaccedilatildeo da recta Ln(Abs400 nm) = -7520E-5
K - 0613determinou-se o valor de K para
a espeacutecie decameacuterica de onde se obteve um tempo de meia-vida de 2430 plusmn 0191 horas para as
condiccedilotildees supramencionadas
Tanto este valor como o valor registado na ausecircncia de S1 satildeo substancialmente superiores agrave
duraccedilatildeo dos ensaios de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina descritos no presente
relatoacuterio
24 Interacccedilatildeo de S1 com vanaacutedio(V)
241Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
De modo a analisar qual o efeito da adiccedilatildeo de S1 de miosina (tampatildeo D) produzia nos sinais de
51V-NMR titulou-se uma soluccedilatildeo de 5mM de decavanadato total com concentraccedilotildees crescentes
de S1 (5 10 15 e 20 microM) Os espectros de 51
V-RMN encontram-se apresentados na Fig 2
No graacutefico da figura 2 Apresenta-se a concentraccedilatildeo de S1 em funccedilatildeo do factor de alargamento
corrigido dos espectros acima referidos
Ln
(Ab
s (4
00
nm
))
tempo (s)
30
Figura 28 ndash Graacutefico de
representaccedilatildeo da
concentraccedilatildeo de S1 em
funccedilatildeo do factor de
alargamento A
concentraccedilatildeo de
decavanadato total foi de 5
mM e as concentraccedilotildees
utilizadas de S1 foram de 0
5 10 e 15 microM
Verifica-se que o factor de alargamento aumenta linearmente em funccedilatildeo da concentraccedilatildeo de S1 no
ensaio o que indica a existecircncia de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina Observou-
se ainda que a adiccedilatildeo de S1 ao meio reaccional natildeo provocou qualquer alteraccedilatildeo dos desvios
quiacutemicos dos sinais de 51
V-RMN
Realizaram-se ensaios na presenccedila de ATP e de S1 com intuito de verificar o efeito do ATP na
interacccedilatildeo do S1 de miosina com o decavanadato
Figura 29 - Espectros de RMN de 51
V contendo 2
mM Decavanadato (A) (controlo) (B) na presenccedila
de 2 mM de ATP (C) na presenccedila de 2 mM de ATP
e 8 microM de S1 de miosina V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2Vndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies decameacuterica (V10) e monomeacuterica (V1) presentes nos
espectros da soluccedilatildeo de decavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V10A V10B V10C V1
part (ppm) -5161 -5002 -4254 -5614
Observa-se que a presenccedila de ATP (espectro B) natildeo provoca qualquer alteraccedilatildeo visiacutevel em
relaccedilatildeo ao espectro que contem apenas soluccedilatildeo de decavanadato Jaacute a presenccedila simultacircnea de ATP
e S1 provoca um alargamento da ∆12 do sinal do V10A de 1736 Hz (espectro (A)) para 2366 Hz
(espectro (C) Fig 29) Tendo em conta que a concentraccedilatildeo de S1 apresenta uma relaccedilatildeo linear
com o factor de alargamento converteu-se o valor da ∆12 e comparou-se com o valor esperado agrave
F = 00591[S1] - 00157 Rsup2 = 0998
Fact
or
de
ala
rgam
en
to
[S1] (microM)
31
concentraccedilatildeo de S1 utilizada O factor de alargamento obtido para 8 microM ndash 0376 ndash eacute inferior ao
valor esperado agrave mesma concentraccedilatildeo de S1 obtido atraveacutes da equaccedilatildeo da recta apresentada na
Fig 28 ndash 0457 ndash o que indica que o ATP poderaacute exercer um efeito de protecccedilatildeo do S1
diminuindo a sua interacccedilatildeo com a espeacutecieV10
Por uacuteltimo realizaram-se ensaios com metavanadato na presenccedila de 8 microM S1 de modo a verificar
se as espeacutecies oligomeacutericas contidas nesta soluccedilatildeo interagiam com a proteiacutena em estudo
Figura 210 - Espectros de RMN de 51
V contendo 2
mM metavanadato (A) (controlo) (B) na presenccedila
de 8 microM de S1 de miosina V1 V2 e V4
correspondem respectivamente aacutes espeacutecies
monomeacuterica dimeacuterica e tetrameacuterica
Tabela 2VIndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies monomeacuterica (V1) dimeacuterica (V2) e tetrameacuterica (V4)
presentes nos espectros da soluccedilatildeo de metavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V1 V2 V4
part (ppm) -5613 -5747 -5789
Como eacute possiacutevel observar na Fig 210 ocorre uma ligeiro alargamento dos sinal de V4 bem
como uma perda da intensidade pelo que se conclui quedas espeacutecies presentes na soluccedilatildeo de
metavanadato (V1 V2 e V4) apenas a espeacutecie tetrameacuterica interage com o S1 de miosina o que
estaacute de acordo com o reportado por Tiago et al 2002
242 Actividade Mg2+
-ATPaacutesica de S1 de miosina na presenccedila de decanadato
Com o intuito de avaliar o efeito do decavanadato na actividade Mg2+
-ATPaacutesica do S1 de
miosina monitorizou-se a reacccedilatildeo de oxidaccedilatildeo do NADH atraveacutes da leitura de Abs340 na
presenccedila de 005 mgmL de S1 de miosina e no caso da actividade estimulada 2microM de actina em
3125 mM Tris-HCl pH 71 e 375 mM MgCl2 pelo meacutetodo de enzimas acopladas (ver seccccedilatildeo xx
dos MM)Para tal realizaram-se ensaios com vaacuterias concentraccedilotildees de decavanadato total (10 50
100 250 500 e 1000) de modo a determinar qual o seu efeito na actividade MgATPaacutesica de S1
de miosina na ausecircncia e na presenccedila de um dos seus ligandos naturais Os ensaios foram
realizados agrave temperatura ambiente com incubaccedilatildeo preacutevia a 25ordm C durante 20 minutos
32
Figura 211 ndash Representaccedilatildeo
graacutefica da Actividade Mg2+
-
ATPaacutesica de S1 () (os ajustes
teoacutericos estatildeo representado a por
uma linha os resultados
experimentais estatildeo representados
por triacircngulos (Actividade
estimulada) e cruzes (Actividade
natildeo estimulada)) vs
[Decavanadato]total medidas pelo
meacutetodo das enzimas acopladas
(n=3) Os dados foram ajustados ao modelo de decaimento exponencial usando o programa GraphPad Prism para a
determinaccedilatildeo dos valores de IC50 (na ausecircncia de actina IC50= 266 plusmn 07 M decavanadato na presenccedila de actina
IC50= 28 plusmn 1 M decavanadato
A partir do graacutefico presente na Fig 29 verifica-se que o ajuste dos dados experimentais foi bem
sucedido A inibiccedilatildeo da actividade Mg2+
-ATPaacutesica do S1 de miosina eacute afectada em menor
amplitude na presenccedila de actina (60 de inibiccedilatildeo da actividade) do que na sua ausecircncia
(aproximadamente 80 de inibiccedilatildeo de actividade) Devido a esta diferenccedila pode sugerir-se que a
actina iraacute impedir parcialmente a ligaccedilatildeo do decavanadatoao S1 demiosina atraveacutes de
impedimentos esteacutereos ou interacccedilotildees electrostaacuteticas por exemplo A teacutecnica utilizada natildeo nos
permite concluir acerca de que regiatildeo(otildees) ou resiacuteduo(s) da actina participaratildeo na preservaccedilatildeo
parcial da actividade Mg2+
-ATPaacutesica
Por outro lado os valores de IC50 dos dois tipos de actividades natildeo apresentam diferenccedilas
significativas tendo-se obtido 266 plusmn 07 e 28 plusmn 1M decavanadato na ausecircncia e na presenccedila de
actina respectivamente o que corresponde a uma concentraccedilatildeo de cerca de 27 M de V10 O
meio de reacccedilatildeo utilizado nestes ensaios foi 10 mM HEPES e 25 mM KCl com 2 mM ATP Em
(Tiago et al 2004) o valor de IC50 para a actividade Mg2+-ATPaacutesica na ausecircncia de actina foi de
125 plusmn 025 microM enquanto o valor da actividade estimulada por actina foi de 136 plusmn 14 microM O
meio de reacccedilatildeo utilizado nos ensaios foi 20 mM HEPES 50 mM KCl e 2 mM ATP o que resulta
precisamente no dobro da forccedila ioacutenica a que foi realizado o ensaio do presente relatoacuterio
(exceptuando a contribuiccedilatildeo dos produtos da hidroacutelise de ATP uma vez que este meacutetodo
pressupotildee a ocorrecircncia de um ciclo de reacccedilotildees de hidroacutelise e siacutentese deste composto) Ora tendo
sido anteriormente referido que a afinidade do S1 de miosina para com a espeacutecie V10 diminui com
Act
ivid
ade
Mg
2+-A
TP
aacutesic
a d
e S
1 d
e
Mio
sin
a
[Decavanadato] (microM) Actividade Estimulada ()
Actividade Natildeo Estimulada ()
Ajuste Actividade Estimulada
Ajuste Actividade Natildeo Estimulada
33
o aumento da forccedila ioacutenica (Tiago et al 2004 tal poderaacute explicar a diferenccedila obtida entre estes
dois conjuntos de valores
243Quantificaccedilatildeo de cisteiacutenas reduzidas pelo meacutetodo de Ellman
A interacccedilatildeo do vanaacutedio com alguns tipos de aminoaacutecidos sensiacuteveis agrave oxidaccedilatildeo pode fornecer-nos
informaccedilatildeo bastante uacutetil em relaccedilatildeo agraves alteraccedilotildees estruturais que conduzem a uma diminuiccedilatildeo da
actividade fisioloacutegica do S1 de miosina Os estudos realizados por Chasteen 1983 concluiacuteram
que a reduccedilatildeo de vanadato (V) a vanadilo (IV) ocorre na presenccedila de agentes redutores comuns no
ambiente celular de que se destaca o aminoaacutecido cisteiacutena Esta eacute tambeacutem um alvo de
complexaccedilatildeo por parte do iatildeo VO2+
Jaacute Ramos et al (2009) demonstrou que a incubaccedilatildeo de
actina com a soluccedilatildeo de decavanadato induz a oxidaccedilatildeo de resiacuteduos de cisteiacutena e agrave formaccedilatildeo do
oxidovanaacutedio (IV) Este grupo de investigadores descobriu ainda que apenas a espeacutecie V10 induz a
oxidaccedilatildeo das cisteiacutenas ldquoraacutepidasrdquo da actina atraveacutes do processo proposto que eacute parcialmente
responsaacutevel pela inibiccedilatildeo jaacute descrita da actividade de ATPase do S1 de miosina estimulada por
actina quando exposta agrave soluccedilatildeo de decavanadato Esta inibiccedilatildeo apresenta-se afectada com a
presenccedila de ATP uma vez que os resiacuteduos de cisteiacutena permaneceram intactos nos ensaios
realizados com este ligando Jaacute Crans et al (2004) concluiu que o siacutetio activo de muitas fosfatases
(que conteacutem grupos tiol) pode formar um equiliacutebrio redox com o iatildeo vanadato (V) de forma natildeo
reversiacutevel
Os resultados obtidos por Sakurai e Shiromura (1980) sugeriram que o vanaacutedio (V) sofre reduccedilatildeo
a vanaacutedio (IV) sem dependecircncia do valor de pH do ensaio Registaram ainda uma estequiometria
de 31 na reacccedilatildeo do vanadato (V) com os resiacuteduos de Cys enquanto a estequiometria dos ensaios
com o vanadilo (IV) foi de 21
Berlett e Stadtman (1997) observaram que os tipos de aminoaacutecidos mais facilmente oxidaacuteveis
englobam entre outros os resiacuteduos de Cys Trp e MetAssim analisou-se o estado redox dos
grupos tiol dos resiacuteduos de Cys do S1 na presenccedila do decavanadato atraveacutes da monitorizaccedilatildeo da
Abs412 nm na presenccedila de 1 mgmL de S1 de miosina em tampatildeo 10 mM HEPES 25 mM KCl
variando a concentraccedilatildeo da soluccedilatildeo de decavanadato de modo a determinar que tipo de alteraccedilotildees
estruturais ocorrem na sua presenccedila no que diz respeito agrave diminuiccedilatildeo da actividade ATPaacutesica de
S1 na presenccedila de decavanadato
Os resultados referentes agrave quantificaccedilatildeo das cisteiacutenas reduzidas apoacutes o tratamento de S1 com
diferentes concentraccedilotildees de decavanadato em tampatildeo 10 mM HEPES pH 75 e 25 mM KCl a
25ordmC durante 20 minutos encontram-se descritos na Tabela 2VII
Tabela 2VII ndash Cisteiacutenas reduzidas totais e ldquoraacutepidas de S1 (1 mgmL) apoacutes tratamento com decavanadato
34
(010501002505001000 uM) em 10 mM HEPES 25 mM KCl pH 7 a 25ordmC durante 20 minutos A detecccedilatildeo
espectrofotomeacutetrica foi feita com DTNB (e 412=8400 uM-1 cm-1) (n variaacutevel entre 1 e 8)
Amostras Cys raacutepidas Cys totais
11 M S1 (controlo) 22 plusmn 04 106 plusmn 14
11 M S1 + 10 M Decavanadato 22 plusmn 05 95 plusmn 08
11 M S1 + 50 uM Decavanadato 20 plusmn 02 98 plusmn 14
11 M S1 + 100 uM Decavanadato 17 plusmn 03 97 plusmn14
11 M S1 + 250 uM Decavanadato 26 plusmn 02 98 plusmn 08
11 M S1 + 500 uM Decavanadato 21 plusmn 04 90 plusmn 06
11 M S1 + 1000 uM Decavanadato 24 plusmn 0 86 plusmn 08
A partir dos resultados obtidos pode sugerir-se que o S1 possui 11 resiacuteduos de Cys (controlo) o
que estaacute de acordo com o anteriormente descrito Verifica-se que o tratamento da proteiacutena com
concentraccedilotildees crescentes de decavanadato provoca a oxidaccedilatildeo de dois resiacuteduos de Cys do S1 de
miosina natildeo expostos ao solvente No entanto a teacutecnica utilizada natildeo nos permite identificar
quais
Tendo em consideraccedilatildeo que o valor de IC50 obtido para a reacccedilatildeo de inibiccedilatildeo daactividade Mg2+-
ATPaacutesica de S1 na presenccedila de decavanadato eacute de cerca de 27 microM pode sugerir-se que este seraacute o
valor ao qual ocorreraacute a oxidaccedilatildeo do primeiro resiacuteduo de Cys o que poderaacute contribuir para a
diminuiccedilatildeo da actividade de S1 observada
25Interacccedilatildeo de S1 com vanaacutedio(IV)
251Anaacutelise por espectroscopia de Ressonacircncia Paramagneacutetica Electroacutenica
A acumulaccedilatildeo intracelular de oxovanaacutedio (IV) in vivo constitui uma das principais formas de
apresentaccedilatildeo de vanaacutedio no organismo Deste modo torna-se bastante interessante analisar a
interacccedilatildeo entre este iatildeo e o S1 de miosina atraveacutes da teacutecnica de espectroscopia de Ressonacircncia
Paramagneacutetica Electroacutenica o meacutetodo de eleiccedilatildeo do estudo do iatildeo vanadilo e dos complexos por si
formados O electratildeo desemparelhado encontra-se fortemente acoplado ao nuacutecleo 51
V o que
35
resulta num espectro de RPE consistindo de 8 desdobramentos na direcccedilatildeo paralela e outros tantos
na direcccedilatildeo perpendicular (Chasteen 1983)
Os estudos realizados por Ahmed et al (1987) centraram-se na interacccedilatildeo do vanadilo com
proteiacutenas como eacute o caso da insulina transferrina entre outros obtendo um espectro caracterizado
por 8 linhas amplamente espaccediladas (na direcccedilatildeo paralela) Jaacute Ramos et al 2009 concluiu que a
exposiccedilatildeo da soluccedilatildeo de decavanadato agraves formas globular e filamentosa de actina produz um
espectro tiacutepico do iatildeo oxidovanaacutedio (IV) indicando a reduccedilatildeo do vanadato
Para aferir qual a concentraccedilatildeo de S1 que permitia obter uma boa razatildeo sinalruiacutedo titulou-se uma
soluccedilatildeo de sulfato de vanadilo 15 microM ([VOSO4]stock = 5 mM) com concentraccedilotildees crescentes de
proteiacutena (0 5 10 15 e 20 microM) em 10 mM HEPES 25 mM KCl A existecircncia de uma correlaccedilatildeo
linear entre a concentraccedilatildeo de proteiacutena e a intensidade dos sinais de RPE correspondente agrave banda
-12 perpendicular encontra-se representada na Fig 210
Figura 2 12ndash Representaccedilatildeo graacutefica da variaccedilatildeo da intensidade do sinal de RPE vs a concentraccedilatildeo de S1 utilizada
Apoacutes verificar que com uma concentraccedilatildeo de S1 igual a 20 microM se obtinham sinais com uma razatildeo
sinalruiacutedo adequada ao prosseguimento dos estudos sobre a interacccedilatildeo V(IV)-S1 de miosina
titulou-se uma soluccedilatildeo de 20 microM S1 com 0 25 5 10 15 25 50 75 100 150 200 500 e 1000
microM VOSO4
Traccedilou-se o graacutefico da intensidade da banda -12 perpendicular do sinal de EPR em funccedilatildeo da
concentraccedilatildeo de VOSO4 tendo-se determinado uma constante de dissociaccedilatildeo (KD) do complexo
S1-vanadilo de 8748 plusmn 2621 microM V(IV)
y = 016786x + 122435
Rsup2 = 092992759
Inte
nsi
dad
e d
o s
inal
de
RP
E
[S1] (microM)
36
Figura 213 ndash Representaccedilatildeo graacutefica da variaccedilatildeo do sinal de RPE vs a concentraccedilatildeo de vanadilo utilizada (os valores
experimentais estatildeo representados por losangos o seu ajuste estaacute representado pela linha)
A simulaccedilatildeo do espectro de EPR foi efectuada a partir do espectro obtido para uma concentraccedilatildeo
de 1 mM VOSO4 apoacutes adiccedilatildeo de 20 microM S1 de miosina tendo-se obtido os valores da constante de
acoplamento hiperfina (A=5016) e do factor g (g0=195)
Figura 214 ndash Representaccedilatildeo graacutefica do espectro de EPR obtido com 1000 microM de vanadilo e 20 microM de S1 (a cinzento
escuro) A simulaccedilatildeo do espectro encontra-se a cinzento claro
Por comparaccedilatildeo destes com os valores presentes na tabela de correspondecircncia xxx eacute possiacutevel
predizer quais os ligandos provaacuteveis do vanadilo na presenccedila de proteiacutena Assim concluiu-se que
o vanadilo estaacute coordenado com duas moleacuteculas de aacutegua um grupo hidroxilo e um grupo R=NR
O grupo hidroxilo pode indicar os resiacuteduos de aminoaacutecidos Tyr Treonina e Serina enquanto o
grupo R=NR poderaacute indicar a presenccedila de Arginina e Histidina como ligandos do vanadilo
Inte
nsi
dad
e d
os
sinai
s d
e R
PE
[Vanadilo] (microM)
Ajuste dos dados exp Dados experimentais
Campo magneacutetico (G)
Espectro Experimental Ajuste
37
3 Procedimento Experimental
31 Reagentes
Todos os reagentes utilizados satildeo de elevado grau de pureza (analiacutetico) Apresentam-se as suas
foacutermulas quiacutemicas ou abreviaturas quando necessaacuterio acrescentou-se a sua descriccedilatildeo Entre
parecircntesis colocou-se as empresas que os comercializam
Aacutecido aceacutetico glacial 100 (Riedel) Acrilamida 99 (Merck) Azul de bromofenol (Riedel-de-
Haumlen) -Mercaptoetanol (Sigma) Bissulfito de soacutedio (Sigma)Bradford (reagente de)
(BioRad)BSA (albumina de soro bovino Sigma-Aldrich) CaCl22H2O (cloreto de caacutelcio
dihidratado Fluka) CH3CH2OH 96 (etanol Panreac) Coomassie Blue R250 (Merck) D2O
(oacutexido de deuteacuterio SDS) DTNB (55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico) Aldrich) EDTA (aacutecido
etilenodiaminatetraaceacutetico Merck) Fosfoenolpiruvato 98 (Fluka) Glicerol (Sigma) Glicina
985 (Panreac) HCl 37 (Riedel-de Haumlen) H2SO4 96 (Pronalab) HEPES 99 (aacutecido 4-(2-
hidroxiletil)-1-piperazineetanosulfoacutenico Sigma) KH2PO4 99 (hidrogenofosfato de potaacutessio)
MgATP (Sigma) Na
ATP (Sigma) Na2HPO4 99 (hidrogenofosfato de soacutedio Riedel-de Haumlen)
NaH2PO4 (fosfato de soacutedio ) NaOH 986 (hidroacutexido de soacutedio Pronalab) KCl 99 (Cloreto de
potaacutessio Panreac) L-Cys 995 (enantioacutemero levoacutegiro do resiacuteduo de Cisteiacutena Fluka)Metanol
99 (Merck) Molibdato de amoacutenio (Sigma)NNrsquo-metil-bis-acrilamida 98 (Merck) NH4VO3
99 (metavanadato de amoacutenia Sigma) PSA 10 (persulfato de amoacutenio Merck) SDS
(dodecilo sulfato de soacutedio Sigma) Sulfito de soacutedio (Sigma) VOSO4 (sulfato de vanadilo
Aldrich) TEMED 99 (Tetrametiletilenodiamina Merck) Tris (2-amino-2-hidroximetil-13-
propanodiol Sigma)
32 Meacutetodos
321 Isolamento e purificaccedilatildeo das proteiacutenas de muacutesculo esqueleacutetico de coelho
3211 Purificaccedilatildeo da miosina preparaccedilatildeo de Subfragmento 1 por digestatildeo quimotriacutepsica
da miosina
O subfragmento-1 de miosina-II (S1) foi gentilmente cedido pelo Prof Doutor Manuel Aureliano
do Departamento de Quiacutemica Bioquiacutemica e Farmaacutecia da Faculdade de Ciecircncias e Tecnologia da
Universidade do Algarve
No entanto descreve-se abaixo de forma resumida o procedimento efectuado
38
32111 Purificaccedilatildeo da miosina
O protocolo de isolamento foi adaptado do meacutetodo previamente descrito por Margossian e Lowey
(1982) apresentando-se de forma esquemaacutetica na Fig 31
Sucintamente os tecidos musculares dorsais e das pernas foram extraiacutedos apoacutes o sacrifiacutecio do
animal (Fig31) De seguida foram adicionados 3 volumes da soluccedilatildeo de extracccedilatildeo A2
Centrifugou-se a soluccedilatildeo e filtrou-se o sobrenadante O sedimento foi entatildeo diluiacutedo em 14
volumes de aacutegua destilada de modo a que a forccedila ioacutenica atingisse 004 M Apoacutes adiccedilatildeo de szlig-
mercaptoetanol (szlig-me) deixou-se a precipitar a 4ordmC removendo o sobrenadante Centrifugou-se
novamente e ressuspendeu-se o sedimento na soluccedilatildeo de extracccedilatildeo A Adicionou-se a soluccedilatildeo
tampatildeo B3 para que a forccedila ioacutenica atingisse 05 M Homogeneizou-se e deixou-se repousar
durante 2h Homogeneizou-se novamente e centrifugou-se de modo a ocorrer precipitaccedilatildeo dos
complexos de actomiosina e proteiacutena desnaturada Apoacutes filtraccedilatildeo do sobrenadante em latilde de vidro
quantificou-se a soluccedilatildeo de miosina resultante pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental)
2 Soluccedilatildeo de extracccedilatildeo A 0012M K2CO3 004M KHCO3 06M KCl pH 65
3 Soluccedilatildeo-tampatildeo B Tris 100 mM pH 7 KCl 2M
39
Figura 31 ndash Representaccedilatildeo esquemaacutetica do protocolo de isolamento de miosina (Margossian e Lowey 1982) Todo o
procedimento foi realizado a 4ordmC (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
32112 Preparaccedilatildeo de Subfragmento-1 de miosina por digestatildeo com quimiotripsina
O S1 de miosina foi isolado apoacutes diaacutelise da miosina durante a noite contra a soluccedilatildeo C4 A
presenccedila do EDTA eacute essencial uma vez que a sua substituiccedilatildeo por MgCl2resulta na produccedilatildeo de
HMM em vez de S1 A suspensatildeo de miosina foi equilibrada e digerida com -quimotripsina
A digestatildeo foi interrompida pela adiccedilatildeo de PMSF Seguidamente ultracentrifugou-se o produto
da digestatildeo desprezando-se o sedimento (Fig 32) A soluccedilatildeo de sobrenadante maioritariamente
S1 foi quantificada pela leitura directa da Abs280 ou pelo meacutetodo de Bradford (ver as secccedilotildees
3311 e 3312 do Procedimento Experimental respectivamente) e armazenada em 10 sacarose
e 2 mM szlig-Me a -80ordmC
4 Soluccedilatildeo C 25mM Tris pH 70 012M KCl 1mM EDTA
40
Antes da realizaccedilatildeo dos ensaios descritos no presente relatoacuterio o S1 de miosina foi dialisado
contra a soluccedilatildeo D5 de modo a remover qualquer composto que pudesse complexar como
vanaacutedio Seguidamente quantificou-se a proteiacutena pelo meacutetodo de Bradford ou pelo meacutetodo
directo (ver secccedilotildees 3311 e 3312do Procedimento Experimtental)
Figura 3 2ndash Representaccedilatildeo esquemaacutetica do protocolo de preparaccedilatildeo do S1 de miosina por digestatildeo quimiotripsina de
acordo com o previamente descrito por Margossian e Lowey (1982) Todo o procedimento foi realizado a 4ordm C
excepto quando indicaccedilatildeo do contraacuterio (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
3212 Isolamento e purificaccedilatildeo de actina
Isolou-se e purificou-se a actina segundo o meacutetodo descrito por Pardee e Spudish que
compreende dois passos ndash a obtenccedilatildeo de um resiacuteduo seco denominado ldquopoacute de acetonardquo e a
extracccedilatildeo subsequente da actina a partir do ldquopoacute de acetonardquo na forma de G-Actina com a
posterior polimerizaccedilatildeo a F-Actina
32121 Obtenccedilatildeo do ldquopoacute de acetonardquo
Sacrificou-se o coelho com uma pancada atraacutes do pescoccedilo e sangrou-se o animal apoacutes o corte da
arteacuteria jugular Esfolou-se e colocou-se numa tina com gelo Retiraram-se as partes de tecido
conjuntivo que envolvem o tecido muscular com auxiacutelio de uma pinccedila e bisturi Seguidamente
retirou-se os muacutesculos dorsais e os muacutesculos brancos das pernas Passou-se os muacutesculos por aacutegua
destilada de modo a remover as impurezas e pesou-se num copo Trituraram-se os muacutesculos
numa maacutequina de triturar carne e extraiu-se o resultante com 3 volumes da soluccedilatildeo E6 durante 10
minutos com agitaccedilatildeo suave Centrifugou-se a suspensatildeo a 4000 g durante 10
5 Soluccedilatildeo D HEPES 10 mM pH 70 KCl 25 mM
6 Soluccedilatildeo E 01 M KCl 015 M K2HPO4 pH 65
41
minutosdesprezando-se o sobrenadante Ressuspendeu-se o sedimento em 6 volumes de soluccedilatildeo
F7durante 10 minutos com agitaccedilatildeo suave Verificou-se se o pH da soluccedilatildeo estava compreendido
entre 82 e 85 acertando-se estecom soluccedilatildeo G8 se necessaacuterio Centrifugou-se durante 10
minutos a 4000 g Desprezou-se o sobrenadante e ressuspendeu-se o sedimento em 3 volumes de
soluccedilatildeo H9 durante 10 minutos Centrifugou-se novamente a 4000 gdurante 10 minutos
desprezando-se o sobrenadante Ressuspendeu-se o sedimento duas vezes consecutivas em 6
volumes de aacutegua destilada durante 5 minutos Centrifugou-se a 4000 g durante 10 minutos apoacutes
cada adiccedilatildeo de H2O
Seguidamente ressuspendeu-se o sedimento 5 vezes consecutivas utilizando 3 volumes de
acetona de cada vez agitando suavemente durante 10 minutos agrave temperatura ambiente Colocou-
se o resiacuteduo filtrado numa tina de vidro sobre papel de filtro deixando-se evaporar o solvente
durante a noite num nicho Triturou-se o resiacuteduo seco numa maacutequina de triturar carne e deixou-se
novamente evaporar o solvente O ldquopoacute de acetonardquo obtido foi armazenado a -20 ordmC
32122 Purificaccedilatildeo da actina a partir de ldquopoacute de acetonardquo
Apoacutes a obtenccedilatildeo do poacute de acetona como acima indicado efectuou-se o protocolo seguidamente
descrito de modo a obter monoacutemeros de actina (G-Actina) e actina filamentosa (F-Actina)
Pesou-se 35 g de poacute de acetona e adicionaram-se 50 mL de soluccedilatildeo-tampatildeo I10
sendo a
extracccedilatildeo feita com agitaccedilatildeo suave durante 30 minutos Separou-se o extracto do poacute de acetona
hidratado filtrando a suspensatildeo com 4 camadas de gaze esterilizada tendo o cuidado de natildeo
espremer o resiacuteduo Voltou a extrair-se com 50 mL de soluccedilatildeo I durante 30 minutos mexendo
lentamente Adicionou-se este ao extracto obtido anteriormente Centrifugou-se o extracto total a
16 000 g durante 30 minutos Decantou-se para uma proveta sobre a qual se colocou gaze para
que o sedimento solto natildeo se juntasse ao sobrenadante e leu-se o volume De seguida adicionou-
se KCl ateacute uma concentraccedilatildeo final de 50 mM MgCl2 ateacute 2mM e ATP ateacute 1mM lentamente e com
agitaccedilatildeo contiacutenua de modo a promover a polimerizaccedilatildeo dos monoacutemeros de actina
Deixou-se polimerizar durante 2h a 25ordmC observando-se a presenccedila de pequenas bolhas de ar
indicativas de um aumento de viscosidade da soluccedilatildeo Arrefeceu-se em gelo durante 15 minutos e
adicionou-se KCl ateacute uma concentraccedilatildeo final de 06M Mexeu-se lentamente durante 30 minutos
apoacutes os quais se ultracentrifugou a 110 000g durante 1hora Desprezou-se o sobrenadante
ressuspendeu-se o sedimento com soluccedilatildeo tampatildeo E e transferiu-se para um potter
Dialisou-se durante a noite contra 1L de soluccedilatildeo-tampatildeo I a uma temperatura de 4ordmC com rotaccedilatildeo
7 Soluccedilatildeo F 005 M NaHCO3 pH 82-85
8 Soluccedilatildeo G 1M Na2CO3
9 Soluccedilatildeo H 1 mM EDTA pH 70
10 Soluccedilatildeo I 2 mM Tris-HCl pH 8 02 mM CaCl2 02 mM Na2ATP 05 mM szlig-Me 0005 NaN3
42
contiacutenua e mudanccedila de tampatildeo 2 a 3 vezes De seguida a actina foi centrifugada a 110
000gdurante 1hora Decantou-se a soluccedilatildeo de G-Actina (sobrenadante) e desprezou-se o
sedimento Determinou-se a concentraccedilatildeo de G-Actina pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental) A soluccedilatildeo de G-actina foi armazenada a uma temperatura de 4ordmC
apoacutes adiccedilatildeo de 002NaN3 mantendo-se estaacutevel durante 3 a 5 dias
331 Quantificaccedilatildeo das proteiacutenas isoladas
3311 Quantificaccedilatildeo pelo meacutetodo de Bradford
O meacutetodo de Bradford (Bradford 1976) baseia-se na utilizaccedilatildeo do reagente de Bradford
composto por entre outros aacutecido fosfoacuterico e o corante ldquoCoomassierdquo Este corante possui carga
negativa e a sua coloraccedilatildeo varia entre a vermelha (Absmax=465 nm) predominante em soluccedilatildeo e a
azul (Absmax=595 nm) presente aquando da ligaccedilatildeo aos grupos amina das proteiacutenas Em soluccedilotildees
aacutecidas diluiacutedas a coloraccedilatildeo varia proporcionalmente com a quantidade do corante ligado agrave
proteiacutena que por sua vez eacute directamente proporcional agrave concentraccedilatildeo da proteiacutena permitindo a
sua quantificaccedilatildeo ateacute a um limite de detecccedilatildeo de 20 gmL (Bradford 1976)
Deste modo prepararam-se amostras de BSA ([BSA]stock= 1mgmL) utilizando um intervalo de
concentraccedilotildees compreendido entre 0 e 15 gmL de acordo com a tabela 3I Apoacutes a adiccedilatildeo da
H2O MilliQ e do reagente de Bradford as soluccedilotildees foram sucessivamente homogeneizadas no
voacutertex
Tabela 3Indash Amostras utilizadas para a obtenccedilatildeo da recta de calibraccedilatildeo no meacutetodo de Bradford
BSA (L) BSA (g) H2O MilliQ (L) Reagente de Bradford (L)
0 0 800 200
125 125 79875 200
25 25 7975 200
5 5 795 200
10 10 790 200
15 15 785 200
Apoacutes incubaccedilatildeo a 25ordmC por 5 minutos leram-se os valores de Abs595 num
espectrofotoacutemetro (Shimadzu UV-160A) para todas as amostras Seguidamente traccedilou-se a recta
de calibraccedilatildeo (Abs595vs quantidade de BSA (g))
43
Para determinar a concentraccedilatildeo de proteiacutena apoacutes o processo de isolamento purificaccedilatildeo e
diaacutelise quando necessaacuterio colocou-se 5 L da amostra num tubo de Eppendorf e adicionou-se
795 L de H2O MilliQ homogeneizando a soluccedilatildeo no voacutertex Adicionou-se posteriormente 200
L de reagente de Bradford e repetiu-se a homogeneizaccedilatildeo Incubou-se a mistura a 25ordmC por 5
minutos apoacutes os quais se leu os valores de Abs595 nm no espectrofotoacutemetro Por interpolaccedilatildeo deste
valor a partir da recta de calibraccedilatildeo determinou-se a concentraccedilatildeo de proteiacutena na amostra
3312 Quantificaccedilatildeo pelo meacutetodo directo
A quantificaccedilatildeo das amostras proteicas pode tambeacutem ser efectuada atraveacutes da leitura directa da
absorvacircncia tendo por base a lei de Beer-Lambert onde Abs corresponde agrave
absorvacircncia ao comprimento de onda ao qual o ensaio foi realizado corresponde ao coeficiente
de extinccedilatildeo (substituiacutedo pela letra a se as unidades forem distintas da molaridade) b corresponde
ao percurso oacuteptico da radiaccedilatildeo na ceacutelula amostral e c corresponde agrave concentraccedilatildeo amostral
Assim no caso de S1 de miosina mediu-se a Abs280e calculou-se a concentraccedilatildeo da proteiacutena
sabendo que o coeficiente de extinccedilatildeo de S1 eacute (Ref)
No caso da actina mediu-se a Abs290ecalculou-se a concentraccedilatildeo da proteiacutena sabendo que o
coeficiente de extinccedilatildeo da actina eacute (Ref)
332Electroforese em condiccedilotildees desnaturantes (SDS-PAGE)
De modo a melhor caracterizar as proteiacutenas isoladas foi realizada uma electroforese em
condiccedilotildees desnaturantes (SDS-PAGE do inglecircs Sodium Dodecyl Sulfate Polyacrilamide Gel
Electrophoresis) O SDS (dodecilo sulfato de soacutedio) eacute um detergente anioacutenico utilizado neste
meacutetodo para desnaturar as proteiacutenas que se ligam ao SDS na proporccedilatildeo 14 g SDS g proteiacutena
(aproximadamente 1 moleacutecula de SDS por cada 2 resiacuteduos de aminoaacutecidos) (Stryer) O SDS induz
uma densidade de carga negativa significativa nas proteiacutenas a que se liga mascarando a sua carga
intriacutenseca pelo que as razotildees cargamassa satildeo tendencialmente idecircnticas Assim a SDS-PAGE
separa as proteiacutenas quando sujeitas a um campo eleacutectrico de acordo com o seu peso molecular
sendo a sua migraccedilatildeo directamente proporcional ao logaritmo deste uacuteltimo (Stryer)
O gel de resoluccedilatildeo com 10 de acrilamida foi obtido atraveacutes da adiccedilatildeo de 167 mL de soluccedilatildeo
stock de acrilamida11
075 mL de tampatildeo de resoluccedilatildeo12
251 mL de aacutegua destilada e 50 L de
SDS 10 Para promover a polimerizaccedilatildeo adicionou-se em seguida 38 L de PSA 10
11
soluccedilatildeo stock de acrilamida soluccedilatildeo de 30 (mv) de acrilamida e 08 (mv) de NNrsquo-metil-bis-acrilamida 12
tampatildeo de resoluccedilatildeo 15 M Tris-HCl pH 88
44
(persulfato de amoacutenia) 10 e 25 L de TEMED 99 (tetrametiletilenodiamina) Agitou-se
cuidadosamente de modo a natildeo formar bolhas e pipetou-se a soluccedilatildeo para a cassette de vidro
previamente montada no suporte De seguida adicionou-se uma peliacutecula de cerca de 2 mm de 2-
metil-2-butanol para verificar posteriormente a polimerizaccedilatildeo Apoacutes a polimerizaccedilatildeo que demora
cerca de 45 minutos verteu-se a agrave superfiacutecie e adicionou-se o gel de concentraccedilatildeo com 5 de
acrilamida constituiacutedo por 03 mL de soluccedilatildeo de stock acrilamida 045 mL de tampatildeo de
concentraccedilatildeo13
094 mL de aacutegua destilada 18 L de SDS 10 135 L de PSA 10 e 2L de
TEMED 99 Colocou-se o pente sobre o gel e deixou-se polimerizar durante cerca de 45
minutos
As amostras para aplicaccedilatildeo no gel de electroforese foram preparadas para uma concentraccedilatildeo de
proteiacutena de 1 mgmL num volume final de 100 L dos quais 25 L correspondem ao tampatildeo da
amostra14
As amostras foram aquecidas a 100ordmC durante 5 minutos Antes da aplicaccedilatildeo das
amostras transferiu-se a cassette para a tina de electroforese agrave qual se adicionou a soluccedilatildeo
tampatildeo de electroforese da tina superior15
e tampatildeo de electroforese da tina inferior16
Aplicaram-
se 10 L das soluccedilotildees de marcadores de peso molecular e 20 L das amostras proteicas
preparadas anteriormente A electroforese decorreu durante cerca de 3h30 min a uma voltagem
de 80 V De seguida transferiu-se o gel para uma tina corando as proteiacutenas durante 1h com
soluccedilatildeo corante17
Descorou-se o gel com soluccedilatildeo descorante18
tendo-se mudado esta soluccedilatildeo ateacute
as bandas se encontrarem bem definidas
Determinou-se a mobilidade relativa das bandas correspondentes aos marcadores de elevado peso
molecular e traccedilou-se a recta de calibraccedilatildeo ndash log(PM) vs log(Rf) onde Rf corresponde agrave
mobilidade relativa Por interpolaccedilatildeo determina-se o PM das proteiacutenas isoladas sabendo a sua
mobilidade relativa Por comparaccedilatildeo com os valores descritos na literatura pode concluir-se
acerca do grau de purificaccedilatildeo
34 Mediccedilatildeo da actividade enzimaacutetica de S1
341 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
13
tampatildeo de concentraccedilatildeo 05 M Tris-HCl pH 68 14
tampatildeo de amostra 320 mM Tris-HCl pH 68 04 M szlig-mercaptoetanol SDS 8 glicerol 15 (vv) e roxo de m-
cresol 0024 15
tampatildeo de electroforese da tina superior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS 16
tampatildeo de electroforese da tina inferior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS acetato de soacutedio
01 M 17
soluccedilatildeo corante ldquoCoomassie Blue R-250rdquo contendo metanol 45 (vv) e aacutecido aceacutetico glacial 10 (vv) 18
soluccedilatildeo descorante metanol 10 (vv) e aacutecido aceacutetico glacial 10 vv)
45
O meacutetodo de Fiske-Subarow modificado consiste na reacccedilatildeo do molibdato de amoacutenio com o
fosfato inorgacircnico em soluccedilatildeo aacutecida com formaccedilatildeo do aacutecido fosfomoliacutebdico Seguidamente daacute-
se a adiccedilatildeo de um agente redutor que provoca a reduccedilatildeo do composto a fosfomolibdato que forma
um complexo de cor azulada cuja absorvacircncia maacutexima ocorre para um comprimento de onda de
660 nm tornando possiacutevel a sua detecccedilatildeo por meacutetodos espectrofotomeacutetricos As actividades Ca2+
-
ATPaacutesica e K+EDTA-ATPaacutesica de S1 foram determinadas atraveacutes deste meacutetodo de acordo com
o descrito por Taussky e Shorr (1953) com mediccedilatildeo da quantidade de fosfato (Pi) libertado
atraveacutes da hidroacutelise de ATP
Para tal obteve-se primeiramente a recta de calibraccedilatildeo com as amostras presentes na Tabela 3II
utilizando uma soluccedilatildeo stock de 5 mM KH2PO4
Tabela 3II ndash Composiccedilatildeo das diferentes amostras utilizadas para determinar a recta de calibraccedilatildeo atraveacutes do meacutetodo
de Fiske-Subbarow modificado
KH2PO4 (L)
([KH2PO4]stock= 5mM)
PO43-
(nmole) H2O (uL)
0 0 1780
5 25 1775
10 50 1770
20 100 1760
40 200 1740
100 500 1680
A cada tubo de Eppendorf adicionaram-se seguidamente 200 L de soluccedilatildeo J19
e 20 L de agente
redutor20
homogeneizando no voacutertex apoacutes cada adiccedilatildeo Incubou-se durante 10 minutos e mediu-
se a Abs660 nm num espectrofotoacutemetro com intervalos de tempo iguais entre cada amostra Traccedilou-
se a recta de calibraccedilatildeo (Abs660 nmvs nmol PO43-
) a partir dos valores obtidos
Para determinar a actividade Ca2+
-ATPaacutesica de S1 prepararam-se tubos de Eppendorf com 400
L de meio de reacccedilatildeo21
25 L CaCl2 ([CaCl2]stock= 100 mM) 25 L ATP ([ATP]stock= 100 mM)
com [S1]ensaios=01 mgmL Perfez-se se necessaacuterio com H2O MilliQ ateacute perfazer um volume de 1
mL Para determinar a actividade K+EDTA-ATPaacutesica utilizou-se uma concentraccedilatildeo idecircntica de
19
Soluccedilatildeo J 5 N H2SO4 25 molibdato de amoacutenio 20
Agente redutor 5 mg aacutecido 1-amino-2-naftol-4-sulfoacutenico 30 mg bissulfito de soacutedio 30 mg sulfito de soacutedio em 26
mL H2O 21
meio de reacccedilatildeo 125 mM Tris-HCl pH 80 0575 M KCl
46
proteiacutena nos ensaios (01 mgmL) e o mesmo meio de reacccedilatildeo Aos tubos de Eppendorf
adicionou-se ainda 123 L de KCl ([KCl]stock = 3M) 25 L de ATP 10 L de EDTA
([EDTA]stock= 100 mM )e o volume de H2O MilliQ necessaacuterio para perfazer 1 mL Em ambas as
situaccedilotildees o uacuteltimo reagente adicionado foi o ATP uma vez que se trata do substrato da reacccedilatildeo
enzimaacutetica
A cineacutetica decorreu a 25ordmC durante 5 minutos para a actividade Ca2+
-ATPaacutesica e durante 3
minutos para a actividade K+EDTA-ATPaacutesica Apoacutes o periacuteodo de incubaccedilatildeo transferiu-se 150
L de amostra para tubos de Eppendorf contendo igual volume de TCA 10 Colocou-se no gelo
e homogeneizou-se no voacutertex Seguidamente centrifugou-se a 1500 g durante 10 minutos e
recolheu-se 200 L de sobrenadante para novos tubos de Eppendorf Adicionou-se a estes 158
mL de H2O MilliQ e 200 L de soluccedilatildeo J Homogeneizou-se no voacutertex e adicionou-se 20 L de
reagente redutor repetindo-se a homogeneizaccedilatildeo Incubou-se durante 10 minutos e mediu-se a
Abs660 nm com o mesmo intervalo de tempo de leitura para cada amostra Por intrapolaccedilatildeo a partir
da recta de calibraccedilatildeo obtida anteriormente determinou-se a quantidade de Pi libertado em cada
ensaio A partir destes resultados determinaram-se as actividades Ca2+
-ATPaacutesica e K+EDTA-
ATPaacutesica do S1 em nmol PO43-
minmg proteiacutena e em s-1
342 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
Da actividade Mg2+
-ATPaacutesica de S1 resulta a formaccedilatildeo dos produtos da reacccedilatildeo de
hidroacutelise do ATP - ADP e Pi Na presenccedila de ADP fosfoenolpiruvato (PEP) e da enzima piruvato
cinase (PK) os produtos resultantes satildeo ATP e piruvato Por acccedilatildeo da enzima lactato
desidrogenase (LDH) este uacuteltimo eacute convertido em lactato ocorrendo simultaneamente a oxidaccedilatildeo
do NADH a NAD+ monitorizada atraveacutes da Abs340 nm Quando o S1 de miosina se encontra no
meio reaccional o ATP eacute hidrolisado formando-se novamente ADP e Pi o que permite a
formaccedilatildeo de um ciclo (fig 21) Se as enzimas se encontrarem em excesso a velocidade de
conversatildeo do NADH eacute limitada pela produccedilatildeo de ADP e Pi Como o ATP eacute regenerado em cada
ciclo a sua concentraccedilatildeo inicial eacute sempre mantida (Bagshaw 1982)
Figura 33ndash Representaccedilatildeo esquemaacutetica do meacutetodo de enzimas acopladas
47
O protocolo utilizado e seguidamente descrito foi adaptado de Cuenda et al (1990 ndash Ver Refordf) A
uma cuvette de 1 mL adicionou-se S1 para uma concentraccedilatildeo final de 005 mgmL 800 L de
meio de reacccedilatildeo22
10 L de PEP ([PEP]stock = 42 mM) 10 L de PK+LDH 11 ([PK+LDH]stock =
1 mgmL) e 15 L de NADH ([NADH]stock=25 mM) Iniciou-se a monitorizaccedilatildeo da reacccedilatildeo
atraveacutes da leitura de Abs340 nm apoacutes a adiccedilatildeo de 20 L de ATP ([ATP]stock=100 mM) A
actividade Mg2+
-ATPaacutesica eacute obtida atraveacutes do declive correspondente agrave oxidaccedilatildeo do NADH Para
a mediccedilatildeo da Mg2+
-ATPase de S1 estimulada por actina adicionou-se F-actina para uma
concentraccedilatildeo final de 2 microM sendo a actividade obtida tambeacutem atraveacutes do declive correspondente
agrave oxidaccedilatildeo do NADH apoacutes a adiccedilatildeo da proteiacutena
35 Interaccedilatildeo entre vanaacutedio e subfragmento-1 de miosina
351 Preparaccedilatildeo das soluccedilotildees de vanaacutedio
As soluccedilotildees de vanadato foram preparadas em aacutegua MilliQ a partir de metavanadato de amoacutenio
(NH4VO3) numa concentraccedilatildeo stock de 50 mM com agitaccedilatildeo magneacutetica e aquecimento (~30 ordmC)
Esta soluccedilatildeo com um valor compreendido entre 6 e 7 designa-se por ldquometavanadatordquo e eacute
constituiacuteda pelas esspeacutecies V1 V2 V4 e V5 (Ref) Acidificando a soluccedilatildeo de metavanadato por
adiccedilatildeo de HCl ateacute um valor de pH proacuteximo de 4 observa-se uma mudanccedila de cor de amarelo
claro para laranja indicativa da formaccedilatildeo da espeacutecie decameacuterica de vanadato De seguida e
imediatamente antes da realizaccedilatildeo dos ensaios descritos o valor do pH da soluccedilatildeo de
decavanadato foi aumentado para cerca de 75 por adiccedilatildeo de NaOH A soluccedilatildeo (2) designada
por ldquometavanadatordquo natildeo foi posteriormente alterada ateacute agrave sua adiccedilatildeo para a realizaccedilatildeo dos ensaios
As soluccedilotildees foram armazenadas a 4ordmC e mantidas em gelo aquando da sua utilizaccedilatildeo
A soluccedilatildeo de V(IV) foi preparada em condiccedilotildees anaeroacutebias por diluiccedilatildeo de sulfato de vanadil
(VOSO4) para uma concentraccedilatildeo final de 5 mM em H2O MilliQ saturada com N2 gasoso
352Anaacutelise por espectroscopia de UV-Vis
A concentraccedilatildeo da espeacutecie decameacuterica em soluccedilatildeo depende do periacuteodo de tempo apoacutes a diluiccedilatildeo
no meio de reacccedilatildeo apropriado (Ref) A velocidade de desoligomerizaccedilatildeo da espeacutecie decameacuterica
pode ser analisada por espectroscopia de absorccedilatildeo na regiatildeo UVVis uma vez que esta espeacutecie
apresenta duas bandas caracteriacutesticas na regiatildeo do visiacutevel (a 358 e 400 nm) que natildeo se verifica
em quaisquer outras espeacutecies oligomeacutericas As bandas supramencionada sapresentam-se bem
22
Meio de reacccedilatildeo 3125 mM Tris-HCl pH 71 375 mM MgCl2
48
resolvidas atraveacutes da obtenccedilatildeo da derivada de 2ordf ordem do espectro apresentando a forma de 2
vales com miacutenimos centrados nos comprimentos de onda supracitados com dependecircncia linear
relativamente agrave concentraccedilatildeo de decavanadato total (Ramos et al 2006)
Assim a estabilidade da espeacutecie decameacuterica apoacutes diluiccedilatildeo da soluccedilatildeo de decavanadato no meio de
reacccedilatildeo dos ensaios (tampatildeo D) para uma concentraccedilatildeo final de 1 mM em vanadato total foi
caracterizada atraveacutes da leitura da Abs400 em funccedilatildeo do tempo tendo-se determinado qual a
ordem de reacccedilatildeo e o tempo de meia-vida da mesma Com base em estudos anteriores acerca da
estabilidade do V10 na presenccedila de diferentes proteiacutenas (Ramos et al 2006) determinou-se
igualmente o tempo de meia vida apoacutes a adiccedilatildeo de 20 microM de S1 de miosina ao meio reaccional
353 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
Os espectros de RMN do isoacutetopo 51
V foram realizados num espectroacutemetro Bruker AM-400 MHz
equipado com uma sonda multinuclear inversa de 5 mm Em tubos de RMN de 5 mm de diacircmetro
preparou-se soluccedilotildees de 500 L de deca- ou metavanadato correspondentes a um intervalo de
concentraccedilotildees de 1 a 10 mM suplementadas com 10 D2O Os espectros foram adquiridos agrave
temperatura ambiente com uma largura espectral de 23 923 Hz um tempo de aquisiccedilatildeo de 0086
s tempo de relaxaccedilatildeo de 001 s e uma sequecircncia de pulsos a 90ordm Utilizou-se a transformada de
Fourier com alargamento das bandas de 20Hz para o processamento dos espectros Os desvios
quiacutemicos das ressonacircncias de 51
V foram obtidos em relaccedilatildeo agrave espeacutecie VOCl3cuja frequecircncia de
ressonacircncia eacute considerada o padratildeo sendo todas as outras referenciadas a partir desta em ppm
As aacutereas dos sinais do RMN correspondentes agraves espeacutecies oligomeacutericas presentes foram obtidas
por integraccedilatildeo dos sinais As concentraccedilotildees relativas das espeacutecies foram calculadas com recurso agrave
equaccedilatildeo
onde A[Vx] corresponde agrave aacuterea referente agrave ressonacircncia da espeacutecie Vx com nordm de agregaccedilatildeo x
A[V]total agrave soma total das aacutereas medidas e [V] a concentraccedilatildeo total de vanaacutedio no ensaio
A teacutecnica de 51
V-RMN foi anteriormente aplicada em estudos de ligaccedilatildeo com S1 de miosina
(Ref) tendo sido demostrado que entre as espeacutecies V1 V2 V4 e V5 a espeacutecie tetrameacuterica possui a
maior afinidade para a miosina (Cremo et al) O desaparecimento ou alargamento dos sinais pode
implicar a ligaccedilatildeo de um dos oxovanadatos laacutebeis agrave miosina sem indicaccedilatildeo de qual uma vez que
49
se processa um equiliacutebrio entre eles (Crans 1990) Assim e tendo em conta as indicaccedilotildees de que
a espeacutecie V10 eacute distinta das restantes no que diz respeito agrave magnitude da interacccedilatildeo com a miosina
e inibiccedilatildeo da actividade ATPaacutesica estimulada por actina (Aureliano 2000 Tiago et al 2002)
torna-se fundamental investigar a interacccedilatildeo das diferentes espeacutecies de vanadato com o S1 de
miosina atraveacutes da teacutecnica de 51
V-RMN
Para tal obtiveram-se espectros de 51
V-RMN resultantes da titulaccedilatildeo de uma soluccedilatildeo 5 mM de
decavanadato total com concentraccedilotildees crescentes de S1 de miosina (0 5 10 15 e 20 microM) em
tampatildeo D Realizou-se ainda ensaios na presenccedila de 2 mM de decavanadato e 2 mM de ATP na
presenccedila e na ausecircncia de S1 de miosina de modo a verificar o efeito que estes ligandos naturais
da enzima teria na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o subfragmento-1
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
Os grupos sulfidrilo (-SH) das cisteiacutenas satildeo resiacuteduos de aminoaacutecidos extremamente sensiacuteveis agrave
oxidaccedilatildeo resultando na sua oxidaccedilatildeo (Dalle-Donne et al 1999) Logo a quantificaccedilatildeo destes
resiacuteduos eacute bastante importante nos estudos de interacccedilatildeo com vanaacutedio jaacute que os resiacuteduos de Cys
com ecircnfase para o SH1 e o SH2 satildeo bastante importantes na monitorizaccedilatildeo das modificaccedilotildees
funcionais da regiatildeo da cabeccedila
O procedimento seguidamente apresentado foi originalmente descrito por Gutieacuterrez-Merino
(1987)) Consiste na reacccedilatildeo entre os grupos ndashSH natildeo oxidados das proteiacutenas e o reagente de
Ellman (DTNB -55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico)) - formando-se o aniatildeo CNT (aniatildeo 3-
carboxilato-4-nitrotiofenolato) numa estequiometria de 11 Este apresenta um maacuteximo de
absorvecircncia a 412 nm (Robyt 1990) Assim apoacutes a incubaccedilatildeo da proteiacutena na presenccedila do metal
a quantidade de cisteiacutenas reduzidas por proteiacutena determina-se atraveacutes dos valores de Abs412
Deste modo colocou-se num tubo de Eppendorf o volume apropriado de S1 dialisado para que a
concentraccedilatildeo no ensaio fosse de 1 mgmL Adicionou-se o volume apropriado de decavanadato
para que a sua concentraccedilatildeo fosse de 0 10 50 100 250 500 ou 1000 M Perfez-se com soluccedilatildeo
D ateacute um volume final de 1 mL e incubou-se a 25ordmC por 20 minutos Apoacutes este periacuteodo colocou-
se 780 L da mistura reaccional numa cuvette de quartzo e adicionou-se 220 L de soluccedilatildeo K23
23
Tampatildeo K Tampatildeo fosfatohidrogenofosfato de soacutedio 025 M pH 72
50
Adicionou-se 15 L de DTNB (em soluccedilatildeo K [DTNB]stock=100 mM) ao meio reaccional e
registou-se o valor de absorvecircncia medido equivalente agraves cisteiacutenas ldquoraacutepidasrdquo (SH1 e SH2
expostos ao solvente) Adicionou-se 001 g de SDS que provoca a desnaturaccedilatildeo da proteiacutena e
consequentemente a exposiccedilatildeo de todos os resiacuteduos de cisteiacutenas que se encontravam ateacute aiacute
inacessiacuteveis no interior da proteiacutena (cisteiacutenas totais) Estes resiacuteduos podem assim reagir com o
DTNB
Estimou-se o nuacutemero de resiacuteduos de cisteiacutenas em todas as amostras com recurso ao coeficiente de
extinccedilatildeo molar () do DTNB (ver ponto 3 dos Anexos) que foi determinado atraveacutes da titulaccedilatildeo
de L-Cisteiacutena com intervalo de concentraccedilotildees compreendidos entre 0 e 50 microM na presenccedila de
25microM de DTNB em tampatildeo D Os valores de absorvecircncia foram obtidos para um comprimento de
onda de 412 nm
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
3621 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
A teacutecnica mais eficiente de anaacutelise da interacccedilatildeo entre o iatildeo vanadilo e proteiacutenas como a
calmodulina e a transferrina entre outras permite a obtenccedilatildeo raacutepida de informaccedilatildeo no que toca agrave
conformaccedilatildeo e amplitude da referida interacccedilatildeo
Os espectros de Ressonacircncia Paramagneacutetica Electroacutenica (RPE) da espeacutecie paramagneacutetica V(IV)
foram realizados a 70K num espectroacutemetro Bruker EMX-300 X-band Todas as soluccedilotildees
utilizadas e amostras foram preparadas em anaerobiose numa cacircmara de luvas de modo a evitar a
reduccedilatildeo de V(IV) a V(V) Em tubos capilares com 02 mL de amostra manteve-se a concentraccedilatildeo
de S1 a 20 M e variou-se a concentraccedilatildeo de vanadilo (0 25 5 10 15 25 50 75 100 150
200 500 e 1000 M [VOSO4]stock=5 mM) em tampatildeo D Apoacutes obtenccedilatildeo dos espectros estes
foram tratados atraveacutes do software SimFonia e WinEPR tendo-se obtido um graacutefico com a
variaccedilatildeo da intensidade do sinal de RPE em funccedilatildeo da concentraccedilatildeo de sulfato de vanadilo
presente Realizaram-se ainda simulaccedilotildees de espectros obtidos com 20 microM de S1 e 1000 microM de
V(IV) com vista agrave obtenccedilatildeo dos ligandos provaacuteveis do vanadilo atraveacutes da anaacutelise da constante
de acoplamento hiperfina (A) e do factor g (g)
4 Conclusotildees gerais
Apesar de jaacute ter sido demonstrado que o vanaacutedio inibe a Na+K
+-ATPase da miosina a funccedilatildeo
deste elemento no tecido muacutesculo esqueleacutetico continua largamente incompreendida A enorme
complexidade quiacutemica deste metal especialmente no que concerne agrave oligomerizaccedilatildeo e reacccedilotildees
51
redox constitui um obstaacuteculo agrave compreensatildeo do comportamento do vanaacutedio nos organismos
vivos Assim a caracterizaccedilatildeo da quiacutemica deste elemento eacute de grande utilidade antes de iniciar o
estudo da sua interacccedilatildeo com o presente sistema
A anaacutelise por espectroscopias de 51
V-RMN e de UV-Vis de soluccedilotildees de vanadato diluiacutedas nos
meios de reacccedilatildeo utilizados no presente relatoacuterio (10 mM HEPES pH 70 25 mM KCl) mostrou
claramente que a espeacutecie decameacuterica (V10) possui um tempo de vida suficientemente longo para a
determinaccedilatildeo do seu efeito isolado ao contraacuterio do que se verifica com as espeacutecies mono- di-
tetra- e pentameacutericas (V1 V2 V4 e V5) Por outro lado a anaacutelise verificou ainda que apoacutes diluiccedilatildeo
das soluccedilotildees de decavanadato no meio de reacccedilatildeo coexistem as espeacutecies V10 e V1 pelo que os
efeitos putativos desta uacuteltima deveratildeo ser subtraiacutedos aquando da monitorizaccedilatildeo dos efeitos da
espeacutecie decameacuterica
O vanaacutedio (V) (vanadato) inibe as enzimas que catalizam reacccedilotildees de transferecircncia de grupos
fosfato como a ATPase da miosina [Goodno 1979] devido agrave sua analogia estrutural com o iatildeo
fosfato Na sua utilizaccedilatildeo como inibidor enzimaacutetico natildeo eacute tida em conta a contribuiccedilatildeo individual
das espeacutecies oligomeacutericas considerando-se normalmente a espeacutecie V1 como inibidora efectiva
[Goodno e Taylor 1982] No presente relatoacuterio verificou-se por espectroscopia de 51
V-RMN que
outros oligoacutemeros de vanadato interagem com o S1 de miosina tendo as espeacutecies V10 e V4 os
maiores niacuteveis de afinidade A sua interacccedilatildeo envolve forccedilas electrostaacuteticas e eacute afectada
negativamente por ligandos naturais de enzima como eacute o caso do ATP
A exposiccedilatildeo do S1 de miosina (005 mgmL) a decavanadato conduz agrave inibiccedilatildeo do capacidade da
proteiacutena de estimular a actividade Mg2+
-ATPaacutesica com um valor de IC50 de 266 plusmn 07 M
decavanadato sendo o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM
decavanadato (80) Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM decavanadato sendo
o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM decavanadato (60)
exercendo um efeito de impedimento agrave ligaccedilatildeo entre o vanaacutedio e o S1
A uma concentraccedilatildeo de 27 microM decavanadato verifica-se a oxidaccedilatildeo de um dos resiacuteduos de Cys
de S1 de miosina seguida pela oxidaccedilatildeo gradual de outro resiacuteduo de Cys ateacute agrave concentraccedilatildeo de
1000 microM Ambos os resiacuteduos oxidados natildeo se encontravam expostos ao solvente utilizado A
oxidaccedilatildeo dos resiacuteduos permite sugerir que esta modificaccedilatildeo oxidativa sofrida pelo S1 contribui
para a diminuiccedilatildeo da sua actividade de ATPase
52
Por espectroscopia de RPE revelaram-se os possiacuteveis ligandos equatoriais do vanaacutedio aquando da
sua interacccedilatildeo com o S1 de miosina como sendo duas moleacuteculas de aacutegua um ligando contendo o
grupo R-OH e outro contendo o grupo R=NR
Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados de
oxidaccedilatildeo (IV) e (V) no S1 de miosina para concentraccedilotildees facilmente alcanccedilaacuteveis no ambiente
terrestre
53
5 Estudos futuros
No sentido do melhor esclarecimento das alteraccedilotildees moleculares e estruturais que ocorrem no
subfragmento 1 quando exposta ao vanaacutedio que conduzem a uma diminuiccedilatildeo da sua funccedilatildeo
sugere-se
- Quantificaccedilatildeo das cisteiacutenas reduzidas de S1 na presenccedila de metavanadato
- Determinaccedilatildeo das actividades Ca2+
- e K+-ATPaacutesica do S1 de miosina na presenccedila de
decavanadato
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina estimulada por actina
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina na presenccedila de
metavanadato
- Realizaccedilatildeo de RMN 2D com variaccedilatildeo das concentraccedilotildees das soluccedilotildees de decavanadato e
metavanadato na presenccedila de S1
-Mutageacutenese dos resiacuteduos de Cys doS1 de miosina de modo a verificar quais os resiacuteduos que
sofrem oxidaccedilatildeo na presenccedila de decavanadato
- Determinaccedilatildeo por calorimetria (DSC Differential Scanning Calorimetry) da variaccedilatildeo da
entalpia (∆H) e da temperatura de desnaturaccedilatildeo do S1 de miosina na presenccedila de vanaacutedio uma
vez que estas propriedades caracteriacutesticas de cada proteiacutena satildeo alteradas apoacutes ocorrecircncia de
modificaccedilotildees estruturais
- Anaacutelise por SDS-PAGE de S1 de miosina apoacutes tratamento com vanaacutedio de modo a determinar
quais os polipeacuteptidos eventualmente formados apoacutes clivagem da proteiacutena
54
6 Referecircncias bibliograacuteficas
- Sellers J Myosins 2nd Edition Oxford University Press 1999
- L Stryer inBiochemistry (Ed WH Freeman 2005 pp
Refordfas Materiais e Meacutetodos
[A] S S Margossian SLowey Methods Enzymol1982 85 55-71
[B] JD Pardue JA Spudich Methods Enzymol 1982 85 164-181
[C] M Bradford Anal Biochem1976 72 248-254
[D] H H Taussky E Shorr J Biol Chem 1953 202 675-685
[E] C R Bagshaw in Outline Studies in Biology ndash Muscle Contraction Chapman and Hall Ed
1982
[F] A Cuenda et al Eur J Biochem1990 194 663-670
55
Anexos
1 Determinaccedilatildeo da recta de calibraccedilatildeo meacutetodo de Bradford
Figura1ndashRepresentaccedilatildeo graacutefica da recta de calibraccedilatildeo (Abs595 nm vs [BSA]) utilizada para o meacutetodo de Bradford
2 Determinaccedilatildeo da recta calibraccedilatildeo para a electroforese em condiccedilotildees desnaturantes
Figura 2 ndash Representaccedilatildeo graacutefica da recta de calibraccedilatildeo (log(Peso Molecular) vs Rf dos marcadores de peso
molecular) onde Rf indica determinada atraveacutes do padratildeo de peso molecular
(poccedilo (a))
3Caacutelculo do coeficiente de extinccedilatildeo molar () para a quantificaccedilatildeo de cisteiacutenas reduzidas
atraveacutes do meacutetodo de Ellman
Traccedilou-se a recta de Abs412 nm meacutedia vs [L-Cisteiacutena] de modo a determinar o valor do coeficiente
de extinccedilatildeo molar definido como o declive daquela recta Assim =8400 M-1
cm-1
como
podemos verificar na Fig 3
A595 nm = 00546[BSA] + 00209 Rsup2 = 099051
Ab
s 59
5 n
m
[BSA] (mgmL)
log PM = -1177 Rf + 2247 Rsup2 = 098957
log
PM
Mobilidade relativa (Rf)
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero

1
Agradecimentos
Ao meu orientador Doutor Rui O Duarte pela oportunidade dada para estudar este sistema tatildeo
fascinante e pelos preciosos conselhos em relaccedilatildeo agrave espectroscopia de Ressonacircncia
Paramagneacutetica Electroacutenica
Agrave Susana Ramos pela orientaccedilatildeo equilibrada paciecircncia dedicaccedilatildeo conselhos preciosos humor e
amizade sem os quais natildeo teria sido possiacutevel realizar este trabalho Obrigada por me teres
desafiado a ser a melhor aprendiz de cientista que conseguisse ndash com planeamento e
determinaccedilatildeo tudo se consegue
Ao Professor Doutor Manuel Aureliano (supervisor) da Faculdade de Ciecircncias e Tecnologia da
Universidade do Algarve pela recepccedilatildeo calorosa disponibilidade das suas instalaccedilotildees para a
obtenccedilatildeo da actina e pela gentil cedecircncia do subfragmento-1 de miosina imprescindiacutevel para o
estudo que realizei
Ao Rui Almeida pelos preciosos conselhos e colaboraccedilatildeo que estimularam o meu fasciacutenio pela
ciecircncia A disponibilidade que mostraste para me esclarecer sempre com clareza e simpatia
tornaram o meu trabalho de todos os dias mais agradaacutevel
Ao Ameacuterico Duarte pela indispensaacutevel ajuda para a realizaccedilatildeo da electroforese em condiccedilotildees
desnaturantes e pela amizade demonstrada nos momentos em que o nervosismo tomou conta de
mim
Agrave Dra Maria do Rosaacuterio Caras-Altas pela disponibilidade concedida no uso do espectroacutemetro de
RMN equipamento crucial na realizaccedilatildeo deste trabalho
A todos os meus colegas dos laboratoacuterios 407 e 613 cujo bom-humor e entreajuda tornaram os
meus dias mais agradaacuteveis
2
Abreviaturas e Siacutembolos
Absx ndash Absorvecircncia ao comprimento de onda x (em nm)
ADP - Adenosina 5rsquo-difosfato
ATP ndash Adenosina 5rsquo-trifosfato
ATPase ndash Adenosina 5rsquo-trifosfatase
BSA ndash Albumina de soro bovino
-Me - -Mercaptoetanol
CNT ndash Aniatildeo 3-carboxilato-4-nitrotiofenolato
Cys ndash Cisteiacutena
DTNB ndash 55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico)
- Coeficiente de extinccedilatildeo (molar)
EDTA ndash Aacutecido etilenodiamino tetra-aceacutetico
ELC ndash Cadeia leve essencial de miosina
F-actina ndash Forma filamentosa da actina
g ndash Aceleraccedilatildeo da gravidade
G-actina ndash Forma monomeacuterica da actina
h ndash Horas
HEPES ndash Aacutecido 4-(2-hidroxiletil)-1-piperazineetanosulfoacutenico
HMM ndash Meromiosina pesada
IC50 ndash Concentraccedilatildeo inibitoacuteria 50
LDH ndash Lactato desidrogenase
Miosina-II ndash Miosina do tipo II
Met - Metionina
NAD+ ndash Nicotinamida adenina dinucleoacutetido (forma oxidada)
NADH ndash Nicotinamida adenina dinucleoacutetido (forma reduzida)
PDB ndash Protein Data Bank
Pi ndash Fosfato inorgacircnico
PEP ndash Fosfoenolpiruvato
PK ndash Piruvato cinase
PM ndash Peso Molecular
3
ppb ndash Partes por biliatildeo (1 gL)
ppm ndash Partes por milhatildeo (1 mgL)
PSA ndash persulfato de amoacutenio
Rf - Mobilidade relativa
RLC ndash Cadeia leve regulatoacuteria de miosina
RMN ndash Ressonacircncia Magneacutetica Nuclear
ROS ndash Espeacutecies reactivas de oxigeacutenio
RPE ndash Ressonacircncia Paramagneacutetica Electroacutenica
S1 ndash Subfragmento-1 da miosina II
SDS ndash Dodecilo sulfato de soacutedio
SDS-PAGE ndash Electroforese em gel de poliacrilamida com SDS (condiccedilotildees desnaturantes)
SH1 ndash Grupo tiol mais reactivo da miosina (resiacuteduo Cys-707)
SH2 ndash segundo grupo tiol mais reactivo da miosina (resiacuteduo Cys-697)
SNC ndash Sistema Nervoso Central
TCA ndash Aacutecido tricloroaceacutetico
TEMED ndash NNNrsquoNrsquo-tetrametiletilenodiamina
Tris ndash 2-amino-2-hidroximetil-13-propanodiol
Trp ndash triptofano
Tyr ndash tirosina
UV ndash gama ultravioleta do espectro electromagneacutetico
Vis ndash gama visiacutevel do espectro electromagneacutetico
4
Resumo
A exposiccedilatildeo a vanaacutedio conduz a uma diminuiccedilatildeo na actividade ATPaacutesica da miosina e provoca a
oxidaccedilatildeo dos grupos sulfidrilo desta proteiacutena No entanto natildeo estaacute integralmente esclarecido
quais os efeitos especiacuteficos que ocorrem na miosina quando exposta ao vanaacutedio Nesse sentido
interessou determinar in vitro quais as alteraccedilotildees estruturais funcionais e moleculares ocorridas
no Subfragmento-1 de miosina de muacutesculo esqueleacutetico de coelho apoacutes a exposiccedilatildeo croacutenica a
soluccedilotildees de metavanadato decavanadato e vanadilo Verificou-se que a exposiccedilatildeo de S1 (005
mgmL) a decavanadato inibe a sua actividade Mg2+
-ATPaacutesica sendo o valor de IC50 para esta
inibiccedilatildeo de 266 plusmn 07 microM decavanadato Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM
decavanadato A perda de funcionalidade proteica poderaacute dever-se agrave oxidaccedilatildeo de 2 resiacuteduos de
Cys de S1 natildeo expostos ao solvente apoacutes exposiccedilatildeo a 1000 microM de decavanadato Verificou-se
que as espeacutecies oligomeacutericas de vanaacutedio com maior afinidade para o S1 satildeo o decavanadato e o
tetravanadato Revelaram-se os possiacuteveis ligandos do vanaacutedio no S1 atraveacutes de espectroscopia de
RPE Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados
de oxidaccedilatildeo (IV) e (V) na estrutura e funccedilatildeo do S1 em concentraccedilotildees facilmente alcanccedilaacuteveis
daquele metal no ambiente terrestre
Summary
The exposure to vanadium leads to a decrease of the myosin ATPase activity and provokes the
oxidation of its sulfydryl groups However the specific effects of vanadium on myosin are not yet
fully understood Consequently the objectives of the present work were to establish the structural
funtional and molecular changes suffered by rabbit myosin subfragment-1 after cronic exposure to
metavanadate decavanadate and vanadyl solutions It was verified that the exposure of S1 (005
mgmL) to decavanadate results in the inhibition of its Mg2+
-ATPase activity resulting in the
IC50 value 266 plusmn 07 microM decavanadate In the presence of actin the IC50 value was 28 plusmn 1 microM
decavanadate It is suggested that the loss of protein function may be due to the oxidation of 2
core cysteine residues after exposure to 1000 microM decavanadate It was also observed that the
decavanadate and tetravanadate species show the highest affinity for S1 Lastly the possible S1
ligands to vanadium were revealed through EPR spectroscopy Globally the obtained results
point to significant effect of V(V) and V(IV) states on S1 structure and function for easily
reachable environmental vanadium concentrations
5
Iacutendice
1 Introduccedilatildeo
11 Muacutesculo esqueleacutetico
111 Miosina
112 Actina
113 Mecanismos moleculares da contracccedilatildeo muscular
1131 Regulaccedilatildeo da contracccedilatildeo muscular
12 Vanaacutedio
121 Propriedades quiacutemicas
1211 Vanaacutedio (IV)
1212 Vanaacutedio (V)
122 Metabolismo e efeito na actividade enzimaacutetica
13 Vanaacutedio e muacutesculo esqueleacutetico
14 Objectivos do projecto
2 Discussatildeo de Resultados
21 Caracterizaccedilatildeo das proteiacutenas isoladas
211 Anaacutelise do estado de pureza
212 Determinaccedilatildeo da concentraccedilatildeo proteica
2121 Subfragmento-1 de miosina
2122 Actina
22 Mediccedilatildeo da actividade enzimaacutetica de S1 de miosina
221 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de Fiske-
Subarow modificado
222 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
23 Preparaccedilatildeo e caracterizaccedilatildeo das soluccedilotildees de vanaacutedio
231 Anaacutelise da composiccedilatildeo das soluccedilotildees de vanadato por espectroscopia de 51V-RMN efeito da
concentraccedilatildeo da espeacutecie
232 Estabilidade das soluccedilotildees de decavanadato anaacutelise por espectroscopia de absorccedilatildeo UVVis
24 Interacccedilatildeo de S1 com vanaacutedio(V)
241 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
242 Actividade Mg2+
-ATPaacutesica de S1 de miosina na presenccedila de decanadato
243 Quantificaccedilatildeo de cisteiacutenas reduzidas pelo meacutetodo de Ellman
25 Interacccedilatildeo de S1 com vanaacutedio(IV)
251 Anaacutelise por espectroscopia de Ressonacircncia Paramagneacutetica Electroacutenica
3 Procedimento Experimental
31 Reagentes
32 Meacutetodos
321 Isolamento e purificaccedilatildeo das proteiacutenas de muacutesculo esqueleacutetico de coelho
3211 Purificaccedilatildeo da miosina preparaccedilatildeo de Subfragmento 1 por digestatildeo quimotriacutepsica da
miosina
32111 Purificaccedilatildeo da miosina
32112 Preparaccedilatildeo de Subfragmento-1 de miosina por digestatildeo quimotriacutepsica
3212 Isolamento e purificaccedilatildeo de actina
32121Obtenccedilatildeo do ldquopoacute de acetonardquo
32122 Purificaccedilatildeo da actina a partir de ldquopoacute de acetonardquo
331 Quantificaccedilatildeo das proteiacutenas isoladas
3311 Quantificaccedilatildeo pelo meacutetodo de Bradford
6
3312 Quantificaccedilatildeo pelo meacutetodo directo
332 Electroforese em condiccedilotildees desnaturantes (SDS-PAGE)
34 Mediccedilatildeo da actividade enzimaacutetica de S1
341 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de Fiske-
Subarow modificado
342 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
35 Preparaccedilatildeo e caracterizaccedilatildeo das soluccedilotildees de vanaacutedio por Espectroscopia UV-Visiacutevel e
Ressonacircncia Magneacutetica Nuclear
351 Preparaccedilatildeo das soluccedilotildees de vanadato
352 Anaacutelise por UV-Vis para determinaccedilatildeo da ordem da reacccedilatildeo
353 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
362 1 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
363 Interacccedilatildeo de S1 com V (V) por Ressonacircncia Magneacutetica Nuclear (RMN)
4 Conclusotildees gerais
5 Estudos futuros
6 Referecircncias bibliograacuteficas
7
1 Introduccedilatildeo
11 Muacutesculo esqueleacutetico
A capacidade de locomoccedilatildeo eacute uma das caracteriacutesticas fundamentais da maioria dos organismos
vivos A contracccedilatildeo muscular um dos fenoacutemenos que a torna possiacutevel apresenta grande
complexidade e tem vindo a ser utilizado como modelo para a compreensatildeo de sistemas menos
especializados de mobilidade celular [1]
No que diz respeito agrave sua fisiologia morfoloacutegica os muacutesculos dos vertebrados dividem-se em
lisos e estriados Estes uacuteltimos devem o seu nome agrave alternacircncia de zonas claras e escuras
observaacuteveis ao microscoacutepio electroacutenico de que resulta um aspecto estriado Os muacutesculos
estriados podem ainda ser divididos em duas subclasses muacutesculos esqueleacuteticos e cardiacuteacos [2]
consoante a forma celular (respectivamente longitudinal ou ramificada) ou tipo de contracccedilatildeo das
ceacutelulas que o compotildeem (respectivamente voluntaacuteria e involuntaacuteria) [3]
Os muacutesculos estriados esqueleacuteticos satildeo assim designados devido agrave sua localizaccedilatildeo no
organismo ndash ligados aos ossos da coluna vertical braccedilos e pernas Satildeo constituiacutedos por feixes de
ceacutelulas multinucleadas (miofibras) que contecircm miofibrilas paralelas de cerca de 1 m de
diacircmetro associadas agrave contracccedilatildeo voluntaacuteria raacutepida e forte em actividades de coordenaccedilatildeo
complexa [4] Cada feixe apresenta um diacircmetro compreendido entre 20 e 100 m e um
comprimento maacuteximo que pode atingir vaacuterios centiacutemetros (Fig 11)
Figura 11 ndash Organizaccedilatildeo hieraacuterquica geral do muacutesculo-esqueleacutetico (adaptado de Purves et al 2003 [3])
No muacutesculo-esqueleacutetico as zonas claras e escuras supramencionadas correspondem a zonas de
menor e maior densidade electroacutenica respectivamente As zonas claras constituem as bandas I (de
8
ldquoisotropiardquo ndash polarizaccedilatildeo uniforme em todas as direcccedilotildees) enquanto as zonas escuras constituem
as bandas A (de ldquoanisotropiardquo ndash polariza a luz visiacutevel) A banda I encontra-se dividida
transversalmente por uma linha escura ndash linha Z de densidade electroacutenica e refracccedilatildeo superiores
No interior de cada banda A existe uma zona transversal menos densa ndash zona H ndash rica em enzimas
importantes para o metabolismo celular No centro desta zona existe uma linha perpendicular
mais escura ndash linha M [2]
A unidade fundamental da contracccedilatildeo muscular eacute designada por sarcoacutemero (Fig 12) Encontra-se
repetida de forma regular ao longo das miofibrilas apresentando um comprimento de 25 a 3 m
aquando do relaxamento muscular diminuindo aquando da contracccedilatildeo muscular
Figura 12 ndash Representaccedilatildeo de fotomicrografia electroacutenica e esquemaacutetica do sarcoacutemero Os filamentos grossos e
finos satildeo maioritariamente constituiacutedos por miosina e actina respectivamente (Adaptado de Stryer 2002 [2])
As secccedilotildees de corte do sarcoacutemero datildeo conta da presenccedila de dois tipos de filamentos proteicos ndash os
filamentos grossos cujo diacircmetro eacute de 15 nm constituiacutedos sobretudo por miosina e os filamentos
finos com um diacircmetro de 9 nm constituiacutedos sobretudo por actina tropomiosina e pelo complexo
de troponina [2]
111 Miosina
Actualmente o termo miosina refere-se a uma superfamiacutelia diversa de motores moleculares
capazes de hidrolisar ATP e de se ligarem reversivelmente agrave actina A classificaccedilatildeo da
superfamiacutelia das miosinas nas 15 classes distintas faz-se atraveacutes de homologia entre as diferentes
sequecircncias [A] Eacute tambeacutem possiacutevel a classificaccedilatildeo por subclasses das quais se destaca a subclasse
4 proveniente do muacutesculo-esqueleacutetico e cardiacuteaco de vertebrados Este estudo focar-se-aacute apenas
nas miosinas de classe II (miosina-II) presente nos filamentos grossos do muacutesculo estriado
esqueleacutetico [B] e em ceacutelulas natildeo musculares A funccedilatildeo da miosina-II depende da sua localizaccedilatildeo
celular e tecidular
9
A miosina-II eacute constituiacuteda por duas cadeias polipeptiacutedicas idecircnticas denominadas cadeias pesadas
de miosina (HC) enroladas entre si e cujo peso molecular (PM) estaacute compreendido entre 171 e
241 kDa) Estruturalmente as HC dividem-se em 3 domiacutenios funcionais - cabeccedila pescoccedilo e
cauda como se pode observar na Fig 13 Agraves cabeccedilas ou zonas globulares estatildeo associados dois
pares de cadeias leves (peso molecular entre 15 e 30 kDa) que se dividem em dois tipos
essencial (ELC) e regulatoacuterio (RLC)
Figura 13 ndash Estrutura esquemaacutetica da moleacutecula de miosina-II Destacam-se as duas cabeccedilas globulares e a cauda
filamentosa ligadas entre si por uma zona flexiacutevel denominada pescoccedilo As cadeias leves (a verde e vermelho)
encontram-se associadas agraves cabeccedilas globulares
As cabeccedilas globulares contendo os locais de ligaccedilatildeo agrave actina e aos nucleoacutetidos mantecircm os seus
elementos estruturais baacutesicos conservados em todas as classes sendo estes constituiacutedos por 720-
80 aa A regiatildeo do pescoccedilo eacute constituiacuteda por uma longa heacutelice pertencente agrave cadeia pesada
que se encontra estabilizada atraveacutes da ligaccedilatildeo a subunidades de cadeia leve A estrutura do
pescoccedilo envolvida na ligaccedilatildeo agraves cadeias leves eacute denominada de motivo IQ cuja estrutura
consensual eacute IQXXXRGXXXR1 O pescoccedilo serve como extensatildeo do comprimento da cabeccedila de
modo a actuar como alavanca durante a contracccedilatildeo muscular [A]A regiatildeo da cauda ou acircncora
apresenta bastante divergecircncia no que respeita agrave sua sequecircncia estrutura e funccedilatildeo No entanto
pensa-se que a sua funccedilatildeo centrar-se-aacute na autoassociaccedilatildeo das vaacuterias moleacuteculas de miosina-II
atraveacutes da regiatildeo de coiled-coil Esta confere assimetria agrave moleacutecula e permite a formaccedilatildeo de
filamentos grossos capazes de translocar filamentos de actina [A]
Na regiatildeo da cabeccedila da miosina existem 2 regiotildees extremamente sensiacuteveis agrave proteoacutelise A
clivagem em ambos os locais resulta na produccedilatildeo de trecircs fragmentos um compreendendo o N-
terminal (fragmento 25 kDa) outro a regiatildeo central (50 kDa) e um terceiro o C-terminal (20 kDa)
(Fig 14) Estes fragmentos dissociam-se em condiccedilotildees desnaturantes e natildeo constituem domiacutenios
independentes da cabeccedila mas provavelmente loops flexiacuteveis uma vez que natildeo foram observadas
na estrutura cristalina Os vaacuterios fragmentos obtidos em diferentes condiccedilotildees foram cruciais para
a elucidaccedilatildeo das vaacuterias funccedilotildees da moleacutecula
A regiatildeo compreendida entre os fragmentos 25 e 50 kDa (loop 1) localiza-se directamente por
1 I = Isoleucina Q= Glutamina X = qualquer dos 20 aminoaacutecidos padratildeo R= Arginina G= Glicina
10
cima do local de ligaccedilatildeo ao nucleoacutetido A regiatildeo entre os fragmentos de 50 e 20 kDa (loop 2)
encontra-se perto da intersecccedilatildeo entre os domiacutenios da cabeccedila e do pescoccedilo e deveraacute interagir com
a actina uma vez que a ligaccedilatildeo desta a protege da proteoacutelise Na ausecircncia do loop 2 a miosina
liga mais fracamente agrave actina do que no caso da miosina intacta A sequecircncia primaacuteria de ambos
os loops apresenta conservaccedilatildeo reduzida sendo provaacutevel que sejam responsaacuteveis por muita da
diversidade das diferentes propriedades enzimaacuteticas presentes em diferentes isoformas
No fragmento de 20 kDa existem dois grupos sulfidrilo Cys-707 e -697 usualmente
denominados de SH1 e SH2 devido agrave ordem da sua reactividade quiacutemica Na presenccedila de
nucleoacutetidos pode ocorrer o ldquocross-linkingrdquo entre estes dois grupos incluindo a oxidaccedilatildeo a uma
ponte dissulfureto
Devido agrave insolubilidade da miosina-II nativa a estrateacutegia adoptada para os estudos com esta
proteiacutena eacute frequentemente a sua clivagem em fragmentos funcionais separaacuteveis Um dos locais de
clivagem ocorre tipicamente a 130 kDa do N-terminal produzindo a meromiosina pesada (HMM)
e a meromiosina leve (LMM) [C] A HMM eacute formada pelas 2 cabeccedilas e pescoccedilo e apresenta
propriedades semelhantes agrave miosina sem digestatildeo (ligaccedilatildeo e translocaccedilatildeo de actina e hidroacutelise de
MgATP)
A HMM pode ser novamente clivada atraveacutes de uma reacccedilatildeo de proteoacutelise produzindo os
subfragmentos 1 (S1) e 2 (S2)O S1 (90 kDa) eacute enzimaticamente activo sendo constituiacutedo pelos
domiacutenios da cabeccedila e do pescoccedilo e apresentando associaccedilatildeo agraves duas cadeias leves ou apenas agrave
ELC (Fig 14)Em alternativa o S1 pode ser produzido por clivagem directa da miosina intacta
formando-se tambeacutem um fragmento em forma de cilindro que consiste nas regiotildees S2 e LMM
(Ref)Utiliza-se extensivamente o S1 para estudos cineacuteticos e de ligaccedilatildeo agrave actina uma vez que a
sua solubilidade simplifica o planeamento da experiecircncia e a subsequente interpretaccedilatildeo dos dados
[A]
Figura 14- Estrutura do subfragmento-1 de miosina de muacutesculo esqueleacutetico com ecircnfase nos domiacutenios de 50 kDa
central (a vermelho) 25 kDa no N-terminal (a verde) e 20 kDa no C-terminal (a azul) As cadeias leves essenciais
encontram-se a amarelo e as cadeias leves regulatoacuterias a lilaacutes Os locais de ligaccedilatildeo agrave actina e ao ATP estatildeo
igualmente destacados (adaptado de Holmes e Geeves 1999 [D])
11
112 Actina
O termo ldquoactinardquo refere-se a uma famiacutelia extremamente conservada de proteiacutenas citoplasmaacuteticas
presente em todos os eucariotas Esta proteiacutena natildeo se encontra nos procariotas [E] A maioria das
actinas possui peso molecular proacuteximo de 42 kDa outra evidecircncia do seu grau elevado de
conservaccedilatildeo no que concerne agrave sequecircncia e ao nuacutemero de aminoaacutecidos Os eucariotas superiores
apresentam um grande nuacutemero de isoformas de actina que por sua vez se encontram codificados
por um ainda maior nuacutemero de genes (Ref) A expressatildeo das isoformas eacute diferencial consoante o
tecido e sua especializaccedilatildeo pelo que deveratildeo possuir funccedilotildees e mecanismos direccionais
diferentes (Ref) Foi ainda observado que os mRNA das diferentes isoformas apresentam uma
distribuiccedilatildeo heterogeacutenea nas ceacutelulas de forma semelhante aos seus produtos proteicos [F] Estes
dados suportam a hipoacutetese supramencionada do direccionamento das isoformas de actina para um
determinado local na ceacutelula de acordo com as suas funccedilotildees
A actina presente no citoesqueleto eacute responsaacutevel por processos como o mecanismo contraacutectil
fagocitose manutenccedilatildeo e alteraccedilatildeo da estrutura celular locomoccedilatildeo transporte de organelos
regulaccedilatildeo do transporte de solutos entre outras Nos eucariotas superiores estaacute ainda envolvida
no transporte de gracircnulos de pigmento retinoacuteide [G] transporte de gracircnulos no axoplasma [H] e
secreccedilatildeo entre outros [E] Actualmente a isoforma da actina (-actina) eacute a melhor estudada
encontrando-se envolvida no mecanismo contraacutectil [E]
Os filamentos de -actina tecircm cerca de 1m de comprimento e 80 Å Consistem em duas cadeias
de actina filamentosa (F-actina) organizados em heacutelice [I] (Holmes et al 1990) sobre os quais
se encontram outras proteiacutenas ndash a troponina (C I e T) e a tropomiosina (Azevedo 1999) (Figura
15) A troponina eacute filamentosa e localiza-se nas depressotildees da heacutelice de F-actina conferindo
rigidez aos filamentos finos Por sua vez a tropomiosina eacute essencial na regulaccedilatildeo do mecanismo
contraacutectil
Figura 15 ndash Modelo estrutural dos filamentos de F-actina contendo a tropomiosina e o complexo de troponina (I C
e T) (Adaptado de Susana Ramos 2005 [J])
12
A F-actina eacute formada atraveacutes da polimerizaccedilatildeo de subunidades globulares de actina (G-actina) A
G-actina eacute composta por 2 domiacutenios de tamanho semelhante separados pelos bolsos de ligaccedilatildeo(1)
de catiotildees divalentes e(2) de nucleoacutetidos [KLM] Estes por sua vez estatildeo subdivididos noutros
dois de menores dimensotildees (1-4) (Fig 16) cada um envolvido nas interacccedilotildees actina-actina ou
na formaccedilatildeo do topo de ligaccedilatildeo do nucleoacutetido As regiotildees N-terminal e C-terminal estatildeo proacuteximas
uma da outra no subdomiacutenio 1 [E]
Figura 16 ndash Estrutura do monoacutemero de actina Encontram-se representados os subdomiacutenios 1 a 4 e a vermelho a
moleacutecula de ATP junto do respectivo local de ligaccedilatildeo (Protein Data Bank ID 1j6z)
113 Mecanismos moleculares da contracccedilatildeo muscular
A contracccedilatildeo muscular ocorre atraveacutes do deslizamento dos filamentos finos (de actina) por entre
os filamentos grossos (de miosina) conseguida pela reacccedilatildeo de hidroacutelise de ATP e regulada pela
tropomiosina e pelo complexo de troponina em resposta a impulsos nervosos Este mecanismo foi
inicialmente proposto por Huxley-Niedergerke [N] e Huxley-Hanson [O] ambos em 1954 A base
experimental de ambos os estudos foi a inexistecircncia de variaccedilatildeo do tamanho dos filamentos
durante a contracccedilatildeo A banda A apresenta sempre as mesmas dimensotildees por outro lado a banda
I a zona H diminuem consideravelmente aquando da contracccedilatildeo (Figs 12 e 17)
Figura 17 ndash Modelo da contracccedilatildeo muscular ndash deslizamento dos filamentos finos (azuis) relativamente aos
filamentos grossos (rosa) (Adaptado de Stryer 2002 [2])
13
Na ausecircncia de ATP e supondo a ausecircncia de inibiccedilatildeo da interacccedilatildeo actina-miosina as proteiacutenas
encontram-se fortemente associadas formando o complexo actomiosina (AM) (Figura 15)
Nestas condiccedilotildees as cabeccedilas de miosina formam um acircngulo de 45ordm com o eixo dos filamentos
finos de actina [P]
Na presenccedila de ATP o complexo AM dissocia-se jaacute que a afinidade do complexo M-ATP para a
actina eacute 105 vezes inferior agrave afinidade da miosina para a actina O ATP liga-se agrave miosina
ocorrendo seguidamente a reacccedilatildeo de hidroacutelise de ATP Os produtos daiacute resultantes ndash ADP e Pi -
permanecem ligados agrave miosina formando o complexo M-ADP-Pi Com a ligaccedilatildeo subsequente agrave
actina forma-se o complexo AM-ADP-Pi no qual as cabeccedilas de miosina apresentam um acircngulo
recto com o eixo do muacutesculo [P] Neste ponto daacute-se a libertaccedilatildeo progressiva de Pi e ADP Eacute
provaacutevel que a energia proveniente da hidroacutelise do ATP provoque uma alteraccedilatildeo do acircngulo de
ligaccedilatildeo das cabeccedilas de miosina ao filamento de actina passando novamente de 90ordm para 45ordm [P]
Esta alteraccedilatildeo conformacional provoca o deslizamento do filamento de actina para o centro do
sarcoacutemero cujo encurtamento promove a contracccedilatildeo das miofibrilas O processo supracitado
ocorre em cerca de 1ms[Q] e encontra-se resumido na Fig 18
Figura 18 ndash Esquema cineacutetico da hidroacutelise de ATP pela miosina (Adaptado de Sellers 1999 [A])
1131 Regulaccedilatildeo da contracccedilatildeo muscular
O mecanismo pelo qual se processa a regulaccedilatildeo muscular foi inicialmente descrito por estudos
distintos de Haselgrove Huxley e Parrey-Squire todos em 1973 [RST] Aleacutem da implicaccedilatildeo
directa da tropomiosina e do complexo de troponina a concentraccedilatildeo de Ca2+
no muacutesculo em
relaxamento eacute bastante reduzida (10-7
M) aumentando notavelmente aquando da contracccedilatildeo (10-5
-
10-3
M)
Resumidamente quando ocorre a excitaccedilatildeo do muacutesculo esqueleacutetico a despolarizaccedilatildeo da
membrana plasmaacutetica eacute difundida ateacute ao interior do tecido atraveacutes dos tuacutebulos transversais
(invaginaccedilotildees da membrana plasmaacutetica da fibra muscular) A despolarizaccedilatildeo provoca a abertura
dos canais ioacutenicos de Ca2+
do retiacuteculo sarcoplasmaacutetico adjacente Como a concentraccedilatildeo de Ca2+
no interior do organelo eacute muito elevada o caacutelcio passa raacutepida e uniformemente para o citosol
14
Assim a regulaccedilatildeo da contracccedilatildeo ocorre com coordenaccedilatildeo e rapidez na totalidade das miofibrilas
do tecido muscular (Ref)
Em repouso a troponina I encontra-se posicionada de tal forma que a proteiacutena adjacente
troponina C impede estruturalmente a ligaccedilatildeo das cabeccedilas da miosina agrave actina (acircngulo =50ordm) O
aumento da concentraccedilatildeo intracelular de Ca2+
provoca um aumento da sua disponibilidade para
ligar agrave troponina C (Fig 19)
Figura 19ndash Regulaccedilatildeo do mecanismo contraacutectil no muacutesculo esqueleacutetico pelo complexo de troponina
Como consequecircncia a troponina C altera a sua conformaccedilatildeo (=70ordm) associando-se agrave troponina I
que deixa de inibir a ligaccedilatildeo das pontes transversais A alteraccedilatildeo anterior eacute transmitida agrave
tropomiosina atraveacutes da troponina T adjacente agrave C o que provoca o deslocamento da
tropomiosina na direcccedilatildeo do interior do sulco do filamento de actina Assim eacute possiacutevel a
interacccedilatildeo eficaz entre actina e miosina A Ca2+
-ATPase do retiacuteculo sarcoplasmaacutetico provoca a
reacumulaccedilatildeo do iatildeo caacutelcio no organelo impedindo o prolongamento desnecessaacuterio da contracccedilatildeo
(Refordf)
12 Vanaacutedio
O vanaacutedio eacute um metal de transiccedilatildeo pertencente ao grupo VB da tabela perioacutedica Foi
primeiramente descoberto em 1801 por Andreacutes Manuel del Riacuteo que concluiu erradamente que era
idecircntico ao croacutemio e definitivamente por Nils Gabriel Svenson em 1831 Recentemente foram
descobertas importantes funccedilotildees bioloacutegicas deste elemento provavelmente essencial para os
humanos de que se destaca no contexto do presente relatoacuterio a possibilidade de actuar como
anaacutelogo do grupo fosfato e a inibiccedilatildeo de enzimas in vitro como eacute o caso da actividade ATPaacutesica
da miosina
Eacute um metal soacutelido agrave temperatura ambiente que se encontra amplamente distribuiacutedo nos vaacuterios
ecossistemas em dois isoacutetopos naturais ndash 50
V e 51
V ndash raramente excedendo 01 partes por biliatildeo
15
(ppb) Estaacute associado a cerca de 70 minerais diferentes na natureza (dos quais se destacam o
uracircnio cobre chumbo zinco fosfato e titacircnio ferroso) especialmente na forma de vanadato ou
pentoacutexidos de vanaacutedio O vanaacutedio eacute um dos principais constituintes da mateacuteria orgacircnica foacutessil
com especial ecircnfase para o carvatildeo oacuteleo de crude e foacutesseis carbonaacuteceos [UV] bem como em
alimentos consumidos pelo Homem (salsa pimenta preta cogumelos marisco vegetais cereais
gorduras e oacuteleos) [W] Estes constituem a maior fonte de exposiccedilatildeo ao vanaacutedio da maioria da
populaccedilatildeo humana [X]
Normalmente a entrada do vanaacutedio no organismo ocorre por inalaccedilatildeo ou em menores
quantidades por absorccedilatildeo pelo tracto gastrointestinal ou pela pele O seu armazenamento centra-
se no fiacutegado rins ceacuterebro coraccedilatildeo muacutesculos e ossos No entanto a retenccedilatildeo de vanaacutedio no
organismo humano eacute reduzida ndash em condiccedilotildees normais cerca de 100 M (em muitos tecidos a
quantidade eacute inferior a 10 ngg de tecido) [Y]
Actualmente natildeo existem provas de deficiecircncia nutricional de vanaacutedio em humanos ou de
enzimas que contenham vanaacutedio em animais superiores No entanto vaacuterios estudos relacionam a
ocorrecircncia de deficiecircncia em galinhas e ratos devido a uma alimentaccedilatildeo pobre em vanaacutedio Os
sintomas relatados incluem retardamento do crescimento malformaccedilotildees oacutesseas perturbaccedilotildees no
metabolismo lipiacutedico e estagnaccedilatildeo reprodutiva entre outros [Z AA AB] Os efeitos fisioloacutegicos
do vanaacutedio podem depender do tipo de composto ou de oligoacutemero uma vez que consoante as
suas estabilidade labilidade e nuacutemero de coordenaccedilatildeo as respostas seratildeo diferenciadas ao niacutevel
dos organismos vivos
Por outro lado a toxicidade do vanaacutedio encontra-se bem caracterizada existindo sintomas
descritos em vaacuterios mamiacuteferos (inclusive o Homem) que incluem diarreias catildeibras irritaccedilatildeo da
pele e mucosas palpitaccedilotildees cardiacuteacas anemia pneumonia bronquite perturbaccedilotildees no Sistema
Nervoso Central (SNC) entre outros (Ref) A toxicidade varia geralmente com a natureza do
composto com aumento proporcional agrave sua valecircncia Verificou-se que a forma de entrada do
vanaacutedio tem tambeacutem efeito na sua toxicidade ndash os maiores niacuteveis verificaram-se por injecccedilatildeo
intravenosa da amostra sendo progressivamente inferiores por inalaccedilatildeo e finalmente por
ingestatildeo [X AA] A ampla utilizaccedilatildeo dos compostos de vanaacutedio na induacutestria quiacutemica bem como
as elevadas quantidades de vanaacutedio libertadas atraveacutes da combustatildeo dos oacuteleos de crude
contribuem consideravelmente para o aumento da biodisponibilidade do vanaacutedio Como tal o
interesse neste elemento e nos problemas ambientais por ele causados tem vindo a aumentar
significativamente [V]
121 Propriedades quiacutemicas
16
A quiacutemica do vanaacutedio eacute extraordinariamente complexa pelo que originou vaacuterias interpretaccedilotildees
incorrectas desde a sua descoberta No entanto as suas propriedades dependem em uacuteltima
anaacutelise da configuraccedilatildeo electroacutenica do elemento Assim a variabilidade encontrada no
comportamento quiacutemico do vanaacutedio pode dever-se agrave sua configuraccedilatildeo ndash [Ar] 3d2 4s
2 ndash que
permite a formaccedilatildeo de vaacuterios estados de oxidaccedilatildeo (de +2 a +5) Em soluccedilatildeo aquosa os estados de
oxidaccedilatildeo estaacuteveis vatildeo de V(V) a V(II) Destes apenas os +3 a +5 satildeo biologicamente importantes
jaacute que o V(II) apresenta demasiada capacidade redutora para existir nos organismos vivos
conhecidos [AC]
No fluido extracelular os compostos de vanaacutedio encontram-se predominantemente na forma de
monovanadatos (V(V)) livres ou ligados agrave transferina do soro ou agrave hemoglobina (Refordf) Os iotildees de
vanadato livres satildeo transportados para o meio intracelular atraveacutes de canais ioacutenicos inespeciacuteficos
Jaacute no ambiente intracelular os compostos sofreratildeo uma bio-reduccedilatildeo atraveacutes da acccedilatildeo da
glutationa ou outros compostos com grupos ndashOH como pentoses hexoses ou etanolaminas [V] A
reoxidaccedilatildeo de vanadilo (V(IV)) intracelular a vanadato (V(V)) provocaria a acumulaccedilatildeo toacutexica
destes uacuteltimos pelo que a complexaccedilatildeo da espeacutecie reduzida o impede O transporte de vanaacutedio
para o interior da ceacutelula encontra-se na Fig 110
Figura 110 ndash Transporte de vanaacutedio para a ceacuteluka e seu subsequente metabolismo (P=Proteiacutena) (Adaptado de
Zaporowska e Scibor 1998)
1211 Vanaacutedio(IV)
A espeacutecie de vanaacutedio (IV) mais bem conhecida eacute o iatildeo vanadilo (VO2+
) espeacutecie estaacutevel a pH
aacutecido sendo a maioritaacuteria a pH 3[AD] Apresenta uma cor azulada e forma complexos penta e
hexacoordenados Forma-se rapidamente a partir de V(V) na presenccedila de um agente redutor em
soluccedilatildeo aquosa No ambiente intracelular a presenccedila de glutationa reduzida aacutecido ascoacuterbico
NAD(P)H entre outros conduzem a que o principal estado de oxidaccedilatildeo nos sistemas bioloacutegicos
seja provavelmente o +4 encontrando-se o V(IV) fortemente complexado com proteiacutenas ou
aminoaacutecidos livres citrato ATP e pirofosfato (Refordfs) Em alternativa o V(IV) pode existir
17
livremente em ambientes fortemente aacutecidos como os lisossomas jaacute que a pH superior a 3 as
soluccedilotildees de vanadilo tendem a ser oxidadas pelo ar formando-se vanadato Esta grande
capacidade redutora justifica parcialmente a utilizaccedilatildeo pouco extensiva do iatildeo VO2+
como
inibidor [W]
1212 Vanaacutedio (V)
O vanaacutedio (V) eacute a forma mais estaacutevel num intervalo de pH amplo [W] O iatildeo anioacutenico VO3-tende
a constituir complexos polinucleares pelo que as soluccedilotildees aquosas de vanadato satildeo extremamente
complexas com ocorrecircncia simultacircnea de vaacuterios equiliacutebrios de protonaccedilatildeo (pKa(vanadato)= 35
78 e 125) e oligomerizaccedilatildeo As espeacutecies presentes (H3VO4 H2VO4- HVO4
2- e VO4
3-) dependem
do pH forccedila ioacutenica e temperatura da soluccedilatildeo e da concentraccedilatildeo total do elemento
[AEAFAGAHAI]
O VO43-
- monoacutemero ortovanadato V1 ndash estaacute associado a uma grande parte da importacircncia
bioloacutegica do vanaacutedio uma vez que se trata de um anaacutelogo electroacutenico e estrutural do ortofosfato
(PO43-
) A coordenaccedilatildeo do ortovanadato pode ser tetraeacutedrica ou bipiramidal trigonal (Refordfs)
A oligomerizaccedilatildeo do vanadato monomeacuterico daacute origem ao diacutemero (V2O72-
- V2) que apresenta
tambeacutem equiliacutebrios de protonaccedilatildeo Presume-se a existecircncia de analogia do vanadato dimeacuterico com
o pirofosfato embora natildeo haja evidecircncia estrutural (Refordf) A dimerizaccedilatildeo do V2 origina o
vanadato tetrameacuterico ou metavanadato (V4O124-
- V4) que constitui a espeacutecie maioritaacuteria em
soluccedilotildees aquosas concentradas (Refordf) As estruturas do metavanadato e a sua espeacutecie protonada
satildeo ciacuteclicas [AJ] Outra espeacutecie oligomeacuterica importante eacute o vanadato pentameacuterico (V5O153-
) cuja
estrutura presumida eacute ciacuteclica [AI]
O oligoacutemero com maior importacircncia bioquiacutemica eacute o vanadato decameacuterico (V10O286-
-V10)(Refordf) a
espeacutecie predominante em soluccedilotildees de pH 2-6 quando a concentraccedilatildeo total de vanadato eacute superior
a 100 M (Ref) A estrutura do V10 natildeo difere em soluccedilatildeo ou no estado soacutelidoOs 10 aacutetomos de
vanaacutedio da espeacutecie decameacuterica estatildeo divididos em trecircs grupos identificados na figura como VA-C
consoante o seu ambiente quiacutemico circundante (Refordf) (Fig 111)
A expressatildeo ldquodecavanadatordquo refere-se a uma soluccedilatildeo aquosa na qual a espeacutecie decameacuterica se
encontra em equiliacutebrio quiacutemico com as espeacutecies V1 V2 V4 e V5 sendo que a oligomerizaccedilatildeo
progressiva do vanaacutedio eacute proporcional agrave sua concentraccedilatildeo total (Ref) Nesta soluccedilatildeo a
concentraccedilatildeo de V10 corresponde a aproximadamente 10 da concentraccedilatildeo total de vanaacutedio [AK]
18
Figura 111 ndash Estruturas esquemaacuteticas das espeacutecies de vanadato (espeacutecie monomeacuterica (V1) dimeacuterica (V2)
tetrameacuterica (V4) pentameacuterica (V5) e decameacuterica (V10) (Adaptado de Aureliano e Crans 2008)
122 Metabolismo e efeito na actividade enzimaacutetica
Apesar de natildeo terem sido ainda encontradas as provas categoacutericas das suas funccedilotildees nos seres
vivos sabe-se que o iatildeo vanadato eacute capaz de inibir uma grande quantidade de enzimas in vitro
com especial ecircnfase na actividade ATPaacutesica da miosina [AL Teresa] Como referido
anteriormente o vanadato eacute capaz de actuar em alternativa ao fosfato nas reacccedilotildees bioquiacutemicas
incluindo a sua preferecircncia de ligaccedilatildeo agraves enzimas [W]
Eacute conhecido o papel do vanaacutedio no metabolismo do caacutelcio promovendo a mineralizaccedilatildeo dos
dentes e ossos na biossiacutentese de liacutepidos (trigliceacuteridos e colesterol) com posterior inibiccedilatildeo da
acumulaccedilatildeo destes nos vasos sanguiacuteneos e no SNC na reproduccedilatildeo e desenvolvimento por
mimetizaccedilatildeo e potenciaccedilatildeo de factores de crescimento na estimulaccedilatildeo do consumo de glucose e
da siacutentese de glicogeacutenio com actuaccedilatildeo equivalente aos efeitos da insulina ao niacutevel celular na
transduccedilatildeo de sinais celulares com regulaccedilatildeo da actividade de segundos mensageiros no
transporte de iotildees e aminoaacutecidos na transcriccedilatildeo geneacutetica e siacutentese de proteiacutenas e DNA entre
outros [W Y AB]
Assim o vanaacutedio cnseguiraacute provocar inuacutemeras mudanccedilas bioquiacutemicas nas ceacutelulas Alguns
exemplos da influecircncia bioquiacutemica do vanaacutedio incluem os seus efeitos nos processos de
fosforilaccedilatildeo e desfosforilaccedilatildeo de proteiacutenas e na participaccedilatildeo na formaccedilatildeo de espeacutecies reactivas de
oxigeacutenio devido ao seu efeito proacute-oxidante sobre agentes redutores a niacutevel celular [AM]
Quanto ao seu efeito na actividade enzimaacutetica pode dividir-se em dois tipos principais ndash inibitoacuterio
e estimulante - dependendo da enzima ou do tipo de reacccedilatildeo por esta catalisada No primeiro
grupo incluem-se enzimas como as ATPases (de miosina de canais ioacutenicos e fosfoenzimas
19
transportadoras) e as fosfatases (da tirosina aacutecida e alcalina) a fosfofrutocinase adenilato ciclase
e catalase entre outras [AA] O segundo grupo inclui a maioria das cinases a glucose-6-fosfato
desidrogenase a fosfoglicerato mutase e a fosfoglicerato fosfatase (Refordf)
O efeito do vanaacutedio (V) na actividade enzimaacutetica estaacute relacionado com o seu nuacutemero de
coordenaccedilatildeo podendo actuar como estado de transiccedilatildeo ou por mimetizaccedilatildeo de cofactores
substratos (especialmente o fosfato) e interacccedilotildees proteiacutena-proteiacutena (como no caso das
polimerases e transcriptases) Pode ainda estar envolvido na formaccedilatildeo de ligaccedilotildees de hidrogeacutenio
interacccedilotildees electrostaacuteticas esteacutereas e de oxidaccedilatildeo-reduccedilatildeo Uma evidecircncia desta uacuteltima eacute a sua
capacidade de alternacircncia entre os estados de oxidaccedilatildeo +5 a +3 o que torna faacutecil a sua interacccedilatildeo
redox com enzimas contendo resiacuteduos de cisteiacutenas (Cys) ou qualquer outro composto orgacircnico
com actividade redox [AN]
13 Vanaacutedio e muacutesculo esqueleacutetico
A primeira evidecircncia da presenccedila de vanaacutedio no muacutesculo esqueleacutetico proveio de uma amostra de
ATP isolado de muacutesculo de cavalo que continha uma impureza ndash vanaacutedio ndash cuja presenccedila
provocou a inibiccedilatildeo da actividade da bomba de Na+K
+-ATPase [AO] Desde entatildeo vaacuterios
estudos tecircm sido realizados com ecircnfase na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o
tecido muscular Destes destaca-se a descoberta da interacccedilatildeo forte da espeacutecie V10 com a miosina
fosforilada [AP] e agrave sua inibiccedilatildeo da actividade ATPaacutesica da miosina[AQ AR]
No quediz respeitoao iatildeo monovanadato destaca-se a sua inibiccedilatildeoda actividade Mg2+
-ATPaacutesica
da miosina [AS] que ocorre de duas formas distintas (1) na presenccedila de concentraccedilotildees reduzidas
de V1 a inibiccedilatildeo ocorre a velocidade reduzida dependente da concentraccedilatildeo da espeacutecie
monomeacuterica (2) na presenccedila de concentraccedilotildees elevadas de V1 o niacutevel de inibiccedilatildeo aproxima-se de
100 desde que [V1] seja superior agrave concentraccedilatildeo total de locais com actividade Mg2+
-ATPaacutesica
no ensaio Esta inibiccedilatildeo eacute irreversiacutevel tanto por diaacutelise ou adiccedilatildeo de agentes redutores do
vanadato [AL]Segundo Pate et al(1994)[AT] a espeacutecie monomeacuterica podeinibir a velocidade de
contracccedilatildeo a temperatura inferior a 20ordmC perdendogradual e rapidamente essa capacidade ateacute os
50ordmC temperatura agrave qual cessa a capacidade inibitoacuteria Os resultados obtidos sugeremainda uma
interacccedilatildeo altamente cooperativa entre o iatildeo vanadato e as proteiacutenas componentes do muacutesculo
esqueleacuteticoDestaca-se tambeacutem a descoberta da inibiccedilatildeo da geraccedilatildeo de forccedila contraacutectil nas fibras
musculares por parte da espeacutecie monomeacuterica[AT]Wilson et al(1995) [AU ]determinaram que a
afinidade da espeacutecie V1 para o S1 de miosina eacute significativamente mais elevada do que a do iatildeo
Pi seu anaacutelogo Descobriram ainda que que a ligaccedilatildeo do vanadato poderaacute envolver transiccedilotildees
entre estados energeacuteticos com energia livre independente dos impedimentos esteacutereos impostos
20
pela estrutura das miofibrilas Os estudos desenvolvidos por Smith et al (1990)[AV]
determinaram queo iatildeo monovanadato natildeo apresenta um efeito pronunciado na inibiccedilatildeo da
actividadeacto-S1-miosina-ATPaacutesica excepto em concentraccedilotildees superiores a 900microM a partir das
quais existe produccedilatildeo de espeacutecies oligomeacutericas para as quais o efeito inibitoacuterio seraacute mais
elevadoA espeacutecie monomeacuterica de vanadato consegue adoptar facilmente uma geometria do tipo
bipiramidal trigonal que mimetiza o intermediaacuterio formado (MgADP
Pi)aquando da reacccedilatildeo de
hidroacutelise de ATP no local activo da miosina [AW] De facto a interacccedilatildeo do vanaacutedio com a
miosina eacute inesperadamente estaacutevel na presenccedila de ADP Esta interacccedilatildeo resulta na formaccedilatildeo de
um complexo enzimaacutetico inactivo e estaacutevel com estequiometria de 11 ADPvanadato por siacutetio
activo da proteiacutena A formaccedilatildeo do complexo estaacutevel miosina-vanadato foi proposta por Goodnoet
al (1982) [AS] Nesta ocorre um preacute-equiliacutebrio raacutepido que induz a formaccedilatildeo de um complexo
ternaacuterio reversiacutevel (MADP
V) com formaccedilatildeo lenta do isoacutemero M
ADP
V A diferenccedila entre os
compostos de vanaacutedio e de fosfato centra-se no tempo de meia vida (t12) do Pi ou do V (no
primeiro t12=12 s enquanto no segundo t12=24 h) (Refordf) A diferenccedila dever-se-aacute agrave possiacutevel
complexaccedilatildeo do vanaacutedio com grupos nucleofiacutelicos no siacutetio activo o que provocaria uma
estabilizaccedilatildeo da miosina na conformaccedilatildeo MADP
V
No que respeita aos estudos realizados com a espeacutecie tetrameacuterica Cremo et al determinou que a
cadeia pesada do S1 de miosina pode ser clivada em dois locais distintos (designados V1 e V2)
pela espeacutecie V4 apoacutes irradiaccedilatildeo com luz UV na presenccedila de concentraccedilotildees de vanadato na ordem
dos milimolar e na ausecircncia de nucleoacutetidos ou metais divalentes
Jaacute no que concerne aos estudos realizados com ecircnfase na espeacutecie decameacuterica foi sugerido
porRamos et al (2006) [AK]que a interacccedilatildeo do decavanadato com a G-actina favorecida pela
polimerizaccedilatildeo desta uacuteltima estabiliza a espeacutecie V10 e induz a inibiccedilatildeo da formaccedilatildeo de F-actina A
estabilizaccedilatildeo da espeacutecie V10 por proteiacutenas tansmembranares e do citoesqueleto pode ser a razatildeo
pela qual ocorre a toxicidade do decavanadato reportada na avaliaccedilatildeo dos efeitos do V(V) nos
sistemas bioloacutegicosTiago et al (2007) [AR] concluiram que o V10 apresenta uma clara preferecircncia
de ligaccedilatildeo ao ldquoback-doorrdquo da S1-miosina-ATPase o que se verifica apenas na estrutura
conformacional desimpedida com acesso facilitado ao local de ligaccedilatildeo do Pi Sugere tambeacutem que
o V10 actua como um ldquoback-door stoprdquo bloqueando estaabertura do fragmento de 50-kDa
necessaacuteria para a hidroacutelise do ATP--fosfato o que constitui uma explicaccedilatildeo simples do
comportamento natildeo competitivo da espeacutecie decameacuterica estimulando a utilizaccedilatildeo deste oligoacutemero
como uma feramenta da elucidaccedilatildeo das mudanccedilas conformacionais ldquoback-doorrdquo da miosina no
21
processo da contracccedilatildeo muscular(ref)Destaca-se ainda a descoberta da capacidade inibitoacuteria do
V10 em relaccedilatildeo agrave fosforilaccedilatildeo oxidativa mitocondrial [AX]
Finalmente no que concerne aos estudos realizados com a presenccedila de vaacuterias espeacutecies
oligomeacutericas Aureliano e Madeira (1994) [AY]observaram que na presenccedila de Ca2+
as espeacutecies
V4 e V10se ligam agrave Ca2+
-ATPase do retiacuteculo sarcoplasmaacutetico enquanto a espeacutecie V1 se liga apenas
na presenccedila de ATP Aparentemente o aumento da concentraccedilatildeo de Ca2+
tem o efeito contraacuterio da
inibiccedilatildeo da ATPase do retiacuteculo sarcoplasmaacutetico pelo V1 natildeo tendo no entanto efeito significativo
sobre a inibiccedilatildeo por parte da espeacutecie V10 Tiago et al (2002) [AP] concluiacuteram que em condiccedilotildees
proacuteximas agraves fisioloacutegicas a espeacutecie V10 difere das espeacutecies presentes na soluccedilatildeo de metavanadato
devido agrave sua forte interacccedilatildeo com a miosina fosforilada e agrave sua inibiccedilatildeo da actividade ATPaacutesica
da miosina Para aleacutem disso o ATP diminui a afinidade da miosina para o V4 induzindo a
interacccedilatildeo com V1 e natildeo tendo qualquer efeito no que diz respeito ao V10 Foi tambeacutem proposto
[AQ] que a ligaccedilatildeo das espeacutecies oligomeacutericas de vanadato aoS1 de miosina nos locais onde o
fosfato coordenaeacute provavelmente a causa da inibiccedilatildeo da actividade ATPaacutesica da miosina
estimulada por actina atraveacutes da prevenccedilatildeo da troca entre ATPADP sendo esta favorecida para
aniotildees de vanadato com valecircncias superiores como eacute o caso do V10
Na presenccedila de actina as velocidades de dissociaccedilatildeo do Pi e do V aumentam consideravelmente
este uacuteltimo em menor quantidade Assim a ligaccedilatildeo da actina ao complexo MADP
V e
Pi eacute
bastante fraca pelo que se sugere que o ADPV teria um efeito de relaxamento semelhante ao
ATP No entanto a capacidade do vanaacutedio em inibir a actividade ATPaacutesica da miosina e da
actomiosina eacute diferenciada uma vez que na ausecircncia de actina a inibiccedilatildeo eacute total para
concentraccedilotildees na gama dos M mas na sua presenccedila a inibiccedilatildeo eacute totalmente conseguida apenas
em concentraccedilotildees na gama dos mM (Ref) No entanto eacute necessaacuterio considerar a possiacutevel
formaccedilatildeo de espeacutecies oligomeacutericas de vanadato cuja afinidade e efeito na enzima seratildeo
diferentes Assim pode concluir-se que o efeito de inibiccedilatildeo observado para a actividade ATPaacutesica
de miosina estimulada por actina se pode dever agrave presenccedila das diferentes espeacutecies oligomeacutericas o
que foi jaacute sugerido por Aureliano (2000) [AZ] ndash as espeacutecies tetrameacuterica e decameacuterica de vanadato
interactuariam com a miosina com alteraccedilatildeo da sua interacccedilatildeo com a actina No entanto natildeo eacute
conhecida a contribuiccedilatildeo absoluta das restantes espeacutecies oligomeacutericas para a inibiccedilatildeo da ATPase
da miosina estimulada por actina (Refs)
14 Objectivos do projecto
Pretende-se com este projecto o isolamento purificaccedilatildeo e caracterizaccedilatildeo da actina do muacutesculo
22
esqueleacutetico de coelho sendo esta uacuteltima tambeacutem realizada para o sufragmento-1 de miosina
Seguidamente realizar-se-aacute a quantificaccedilatildeo e caracterizaccedilatildeo das proteiacutenas por espectroscopia UV-
Vis e SDS-PAGE e a caracterizaccedilatildeo das soluccedilotildees de vanaacutedio por UV-Vis e ressonacircncia magneacutetica
nuclear (RMN) Por uacuteltimo a interacccedilatildeo do S1 de miosina com o vanaacutedio seraacute avaliada atraveacutes
das teacutecnicas de UVVis RMN e ressonacircncia paramagneacutetica electroacutenica (RPE)
2 Discussatildeo de Resultados
21 Caracterizaccedilatildeo das proteiacutenas isoladas
211Anaacutelise do estado de pureza
O estado de pureza das soluccedilotildees de actina e de S1 foi analisado por electroforese em gel de
poliacrilamida em condiccedilotildees desnaturantes (SDS-PAGE) (Fig 22) que separa as moleacuteculas de
acordo com o seu peso molecular (PM) A concentraccedilatildeo de acrilamida escolhida (10) permite a
detecccedilatildeo simultacircnea de componentes com peso molecular inferior a 20 kDa e outros com elevado
peso molecular
Figura 21 ndash Fotografia de gel de SDS-PAGE (10 acrilamida) Poccedilo (a)
10 L do marcador de elevado peso molecular () Poccedilo (b) 20 L de [G-
actina]=1 mgmL Poccedilo (c) 20 L de [S1]=1 mgmL Aplicou-se uma
voltagem de V durante h e min
A partir da recta de calibraccedilatildeo obtida log PM = -1177 Rf + 2247 (ver ponto 2 da secccedilatildeo
Anexos) pode verificar-se a presenccedila de actina na amostra do poccedilo (b) presente numa banda
bastante intensa (PMaparente=40 kDa) bem como a presenccedila minoritaacuteria de outros componentes
desconhecidos que natildeo teratildeo sido eliminados durante o processo de extracccedilatildeo e purificaccedilatildeo e natildeo
satildeo relevantes para o estudo em questatildeo Na amostra do poccedilo (c) nota-se a presenccedila de S1
(PMaparente=92 kDa) bem como a de outros componentes cuja banda eacute ligeiramente menos intensa
Estes poderatildeo ser fragmentos proteoliacuteticos de miosina e de S1 Estaacute ainda presente uma bada de
23
maior intensidade que poderaacute ser a proteiacutena-C (PM=135 kDa) uma vez que a sua remoccedilatildeo
completa da amostra necessitaria de um passo adicional de cromatografia de troca ioacutenica [BA]
No entanto natildeo podemos confirmar que se trata desta proteiacutena ou de qualquer outra uma vez que
se encontra fora do intervalo de migraccedilatildeo dos marcadores de peso molecular
212 Determinaccedilatildeo da concentraccedilatildeo proteica
2121Subfragmento-1 de miosina
A partir da recta de calibraccedilatildeo obtida pelo meacutetodo de Bradford [BB](ver secccedilatildeo 3311 do
Procedimento Experimental) A595nm = 00546[BSA] + 00209 (ver secccedilatildeo de Anexos)foi possiacutevel
determinar um intervalo de concentraccedilatildeo de S1 de miosina compreendido entre 226 e 117(plusmn
423M) (203 e 1056 (plusmn 381 mgmL)) (n=8) A concentraccedilatildeo meacutedia de S1 de miosina apoacutes
diaacutelise contra a soluccedilatildeo tampatildeo 10 mM HEPES pH 75 25 mM KCl determinada pela leitura da
Abs280 nmfoi de 20430M (1803 mgmL)(n=7)
As amostras a quantificar atraveacutes do meacutetodo de Bradford natildeo sofreram diaacutelise o que permite
explicar a concentraccedilatildeo superior de S1 de miosina em comparaccedilatildeo com o valor obtido atraveacutes do
meacutetodo directo Esta razatildeo aliada agrave sensibilidade e rapidez superiores deste uacuteltimo fez com que
se optasse pelo meacutetodo directo para a quantificaccedilatildeo da grande maioria dos ensaios
2122Actina
A concentraccedilatildeo de actina foi determinada quando esta se encontrava na forma monomeacuterica (G-
actina) por mediccedilatildeo da leitura directa da Abs290 nm tendo-se obtido uma concentraccedilatildeo meacutedia de
691481M (290202 mgmL) (n=3) em 2 mM Tris-HCl pH 80 02 mM CaCl2 02 mM Na-
ATP Ao comprimento de onda utilizado (290 nm) absorvem fortemente apenas os resiacuteduos de
Trp pelo que a mediccedilatildeo efectuada tem menor sensibilidade do que se tivesse sido efectuada a 280
nm uma vez que a actina apresenta apenas 4 resiacuteduos deste aminoaacutecido embora tenha 16
resiacuteduos de Tyr
22 Mediccedilatildeo da actividade enzimaacutetica de S1 de miosina
221 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
As actividades ATPaacutesicas de S1 de miosina dependentes de Ca2+
e K+foram analisadas pelo
caacutelculo das velocidades de consumo inicial do fosfato inorgacircnico (Pi) Definiu-se os tempos de 5
e 3 minutos para a mediccedilatildeo da Ca2+
e da K+EDTA-ATPase respectivamente Os valores obtidos
24
a partir do protocolo presente no ponto 341 da secccedilatildeo Procedimento experimental encontram-se
descritos na Tabela 2I
Tabela 2I ndash Acrividades natildeo-fisioloacutegicas de subfragmento-1 determinadas pelo meacutetodo de Fiske-Subarow
modificado (n=2)
Proteiacutena
Actividade
ATPaacutesica natildeo
fisioloacutegica
Actividade (nmol
Piminmg proteiacutena) Actividade (s-1)
S1
Ca2+
-ATPase 1400 plusmn 30 2100 plusmn 0045
K+EDTA-ATPase 2334 plusmn 50 3500 plusmn 0075
As actividades natildeo-fisioloacutegicas reflectem o estado de oxidaccedilatildeo dos resiacuteduos expostos de Cys 707
(SH1) e 697 (SH2) Se o grupo SH1 sofrer oxidaccedilatildeo a actividade ATPaacutesica dependente de Ca2+
aumenta e a actividade dependente de K+ diminui Por outro lado se o grupo SH2 sofrer
oxidaccedilatildeo regista-se a perda completa destas actividades
De acordo com Margossian e Lowey (1982) [BA] as actividades ATPaacutesica de S1 de miosina
dependente de Ca2+
e K+EDTA da miosina satildeo respectivamente de 24 e 112 s-1
Estaacute tambeacutem
descrito que a actividade ATPaacutesica dependente de K+ eacute em meacutedia cerca de 3 vezes superior agrave
dependente de Ca2+
reflectindo a quantidade das preparaccedilotildees A razatildeo obtida entre os valores das
actividades ATPaacutesicas dependentes de K+ e Ca
2+ eacute de 166 Os valores obtidos para as actividades
(Ca2+
-ATPase=21 s-1
K+EDTA-ATPase= 35 s
-1) satildeo inferiores agrave valores obtidos por
Margossian e Lowey [BA] e Tiago [BC] com forccedila ioacutenica inferior O valor obtido para a
actividade ATPaacutesica dependente de Ca2+
eacute superior ao obtido por Polosukhina et al (2000) [BD]
cujo meio de reacccedilatildeo continha MOPS e apresentava forccedila ioacutenica inferior Os dados
supramencionados comprovam a variaccedilatildeo da actividade a ATPaacutesica natildeo fisioloacutegica em relaccedilatildeo agrave
forccedila ioacutenica temperatura e pH
222 Mediccedilatildeo da actividadeMg2+
-ATPaacutesica de S1 pelo meacutetodo de enzimas acopladas
A actividade de Mg2+
-ATPase eacute a uacutenica que possui significado fisioloacutegico sendo as restantes
bastante uacuteteis apenas para indicaccedilatildeo da qualidade de preparaccedilatildeo do S1[BA] A 0575 M KCl a
actina e o S1 formam uma soluccedilatildeo viscosa e tuacuterbida Apoacutes a adiccedilatildeo de MgATP o complexo acto-
S1 dissocia-se e a turbidez da soluccedilatildeo diminui Se a forccedila ioacutenica for reduzida a dissociaccedilatildeo do
complexo actoS1 que se encontra na forma de suspensatildeo eacute induzida O MgATP eacute rapidamente
hidrolisado diminuindo a sua concentraccedilatildeo no meio reaccional Nestas condiccedilotildees haacute
reassociaccedilatildeo e subsequente contracccedilatildeo dos filamentos de actina e de S1 com formaccedilatildeo de
25
precipitado O meacutetodo descrito traduz de forma bastante elementar o fenoacutemeno da contracccedilatildeo in
vivo e elimina o problema anterior da solubilizaccedilatildeo ineficiente da miosina de modo a obter uma
mediccedilatildeo exacta da actividade [A] A utilizaccedilatildeo de forccedilas ioacutenicas reduzidas aumenta a afinidade da
actina para o S1 permitindo contornar a extrema dificuldade em obter a concentraccedilatildeo proteica dos
muacutesculos (cerca de 100 mgmL do complexo actomiosina) As actividades Mg2+
-ATPaacutesicas de S1
de miosina foram determinadas a partir do declive da variaccedilatildeo da Abs340 relativo agrave oxidaccedilatildeo de
NADH em funccedilatildeo do tempo como eacute descrito no ponto 342 do Procedimento Experimental Os
valores obtidos para a actividade Mg2+
-ATPaacutesica encontram-se apresentados na Tabela 2II
Tabela 2II ndash Actividade Mg2+
-ATPaacutesica de subfragmento-1 de miosina determinadas pelo meacutetodo das enzimas
acopladas
Actividade (nmol
ATPmin mg(S1)) Actividade (s
-1)
1097 plusmn 269 164 plusmn 04
Ramos (2005) [J] obteve o valor de 01 s-1
para a actividade Mg2+
-ATPaacutesica de S1 de miosina
com forccedila ioacutenica intermeacutedia Tiago (2005) [BC] obteve o valor de 011 s-1 em condiccedilotildees de forccedila
ioacutenica reduzida em relaccedilatildeo agrave utilizada no presente relatoacuterio
23 Preparaccedilatildeo e caracterizaccedilatildeo das soluccedilotildees de vanaacutedio
231 Anaacutelise da composiccedilatildeo das soluccedilotildees de vanadato por espectroscopia de 51
V-RMN
efeito da concentraccedilatildeo da espeacutecie
A diluiccedilatildeo das soluccedilotildees stock de meta- e decavanadato iraacute determinar fortemente a composiccedilatildeo
em soluccedilatildeo dos diferentes oligoacutemeros de vanadato (ref) No caso do decavanadato o periacuteodo de
tempo apoacutes a diluiccedilatildeo iraacute tambeacutem ser um factor importante a considerar devido agrave decomposiccedilatildeo
da espeacutecie V10(ref)Torna-se assim fundamental determinar a composiccedilatildeo quiacutemica das soluccedilotildees
no meio de ensaio utilizado (HEPES 10 mM KCl 25 mM) de modo a correlacionaacute-la
correctamente com os possiacuteveis efeitos promovidos pelas diferentes espeacutecies de vanadato no
sistema em estudo
Aleacutem disso deve ser dado ecircnfase agraves importantes interacccedilotildees que ocorrem entre o vanaacutedio e os
tampotildees escolhidos O tampatildeo com menor nuacutemero e magnitude de interacccedilatildeo eacute o HEPES [AD]
seguido do Tris sendo este uacuteltimo especialmente aconselhado em concentraccedilotildees baixas para casos
de incompatibilidade enzimaacutetica ou quiacutemica redox No caso especial da reacccedilatildeo redox
NAD(P)HNAD(P) o tampatildeo adequado eacute o imidazole de modo a natildeo haver reacccedilatildeo com o
26
vanadato Os tampotildees a evitar em qualquer situaccedilatildeo incluem a trietanolamina fosfato citrato e
glicina entre outros [AD] uma vez que todos interactuam fortemente com V(V) com formaccedilatildeo
de complexos estaacuteveis
Assim procedeu-se agrave anaacutelise da composiccedilatildeo das soluccedilotildees de meta- e decavanadato apoacutes diluiccedilatildeo
das soluccedilotildees stock (50 mM em vanadato total) no meio de reacccedilatildeo (tampatildeo D) atraveacutes da
espectroscopia de ressonacircncia magneacutetica nuclear de 51
V Nas figuras x1 e x2 apresentam-se os
espectros obtidos para as soluccedilotildees de deca- e metavanadato num intervalo de concentraccedilotildees de 1 a
10 mM A Tabela seguinte resume os paracircmetros espectroscoacutepicos obtidos para cada espeacutecie
presente nas soluccedilotildees analisadas
Figura 22ndash Espectros de RMN de 51
V de
concentraccedilotildees crescentes das espeacutecies oligomeacutericas
de vanadato obtidos apoacutes diluiccedilatildeo da soluccedilatildeo stock
de decavanadato em 10 mM HEPES pH 70 25 mM
KCl para concentraccedilotildees finais de A) 1 mM B)2
mM C) 4 mM D) 8 mM E) 10 mM de
decavanadato total V1refere-se agraves espeacutecies
monomeacutericade vanadato V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2IIIndash Desvios quiacutemicos meacutedios (part) das ressonacircncias das espeacutecies decameacuterica (V10) monomeacuterica (V1) e
tetrameacuterica (V4) presentes nos espectros da soluccedilatildeo de decavanadatoLargura a meia altura (∆12) da espeacutecie V10A
Paracircmetros V10A V10B V10C V1 V4
part (ppm) -5162 -5003 -4255 -5616 -5748
∆12 (Hz) 175 nd nd nd nd
Figura 23 ndash Distribuiccedilatildeo graacutefica das
espeacutecies oligomeacutericas de vanadato vs a
concentraccedilatildeo de decavanadato total As
concentraccedilotildees de cada espeacutecie foram
obtidas segundo a formula descrita na
secccedilatildeo 35 do Procedimento
Experimental (aacutereas dos sinais de 51
V-
RMN observadas na figura x1)V10 V1 e
V4 correspondem agraves espeacutecie monomeacuterica
dimeacuterica e tetrameacuterica respectivamente [Esp
eacuteci
e o
ligo
meacute
rica
de
van
aacutedio
] (m
M)
[Decavanadato]total (mM)
V10
V1
V4
27
Nas soluccedilotildees de decavanadato as espeacutecies monomeacuterica e decameacuterica de vanadato satildeo as uacutenicas
presentes para concentraccedilotildees de decavanadato inferiores a 8 mM Ambas as espeacutecies aumentam
linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado em [AY] e
em (Teresa) quando a concentraccedilatildeo de V1 eacute igual ou superior a 200 microM inicia-se a sua
oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica No entanto nas condiccedilotildees de realizaccedilatildeo dos
ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a espeacutecie tetrameacuterica neste
estudo em concentraccedilotildees inferiores a 8 mM tendo o mesmo sido observado por Ramoset al
(2006) [AK] Os resultados obtidos mostram claramente o equiliacutebrio entre a espeacutecie decameacuterica e
a monomeacuterica e que o aparecimento de outras espeacutecies oligomeacutericas se encontra dependente da
concentraccedilatildeo desta uacuteltima Apesar de natildeo ter sido realizado o ensaio com 5 mM de decavanadato
total no presente relatoacuterio estaacute reportado em Ramos et al (2006) [AK] que as concentraccedilotildees da
espeacutecie decameacuterica e monomeacuterica satildeo respectivamente de 500 microM e 200 microM Assim pode inferir-
se que para concentraccedilotildees inferiores a 5 mM e intervalos de tempo de cerca de 20 minutos (que
inclui o tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas
as espeacutecies decameacuterica e monomeacuterica de vanadato
Figura 24 ndash Espectros de RMN de 51
V de concentraccedilotildees
crescentes das espeacutecies oligomeacutericas de vanadato obtidos
apoacutes diluiccedilatildeo da soluccedilatildeo stock de metavanadato em 10 mM
HEPES pH 70 25 mM KCl para concentraccedilotildees finais de A)
1 mM B) 2 mM C) 4 mM D) 8 mM E) 10 mM de
metavanadato total V1 refere-se agraves espeacutecies monomeacuterica de
vanadatoV1 e V2 correspondem respectivamente agrave espeacutecie
monomeacuterica e dimeacuterica
Figura 25 ndash Distribuiccedilatildeo graacutefica das espeacutecies
oligomeacutericas de vanadato vs a concentraccedilatildeo de
metavanadato total As concentraccedilotildees de cada
espeacutecie foram obtidas segundo a formula descrita
na secccedilatildeo 35 do Procedimento Experimental
(aacutereas dos sinais de 51
V-RMN observadas na
figura x1)V10 V1 e V4 correspondem agraves espeacutecie
monomeacuterica dimeacuterica e tetrameacuterica
respectivamente
Esp
eacuteci
e o
ligo
meacute
rica
de
va
nad
ato
(m
M)
[Metavanadato]total (mM)
V1
V2
V4
V5
28
Tabela 2IVndash Desvios quiacutemicos meacutedios (part)das ressonacircncias das espeacutecies tetrameacuterica (V4) monomeacuterica (V1) e
dimeacuterica (V2) presentes no espectro da soluccedilatildeo de metavanadato (total) 10 mM
Paracircmetros V1 V2 V4 V5
part (ppm) -5615 -5748 -5789 -5869
Nas soluccedilotildees de metavanadato as espeacutecies monomeacuterica dimeacutericae tetrameacuterica de vanadato estatildeo
presentes para concentraccedilotildees de metavanadato inferiores a 4 mM Todas estas as espeacutecies
aumentam linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado
em (Aureliano e Madeira 1998) e em (Tiago et al 2004) quando a concentraccedilatildeo de V1 eacute igual ou
superior a 200 microM inicia-se a sua oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica Nas
condiccedilotildees de realizaccedilatildeo dos ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a
espeacutecie pentameacuterica neste estudo em concentraccedilotildees inferiores a 4 mM tendo o mesmo sido
observado por Ramos 2006 Os resultados obtidos mostram claramente o equiliacutebrio entre a
espeacutecie V4 e as espeacuteciesV1 e V2 no qual o aparecimento de outras espeacutecies oligomeacutericas se
encontra dependente da concentraccedilatildeo da espeacutecie V1 Por uacuteltimo pode inferir-se que para
concentraccedilotildees inferiores a 2 mM e intervalos de tempo de cerca de 20 minutos (que inclui o
tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas as
espeacutecies V1 V2 e V4 com concentraccedilotildees respectivas de 0796 0192 e 0196 mM
232Estabilidade das soluccedilotildees de decavanadato anaacutelise por espectroscopia de absorccedilatildeo
UVVis
Caracterizou-se a estabilidade da espeacutecie V10 numa soluccedilatildeo com 1 mM decavanadato total em
tampatildeo D atraveacutes da leitura da Abs400 Obteve-se uma cineacutetica de dissociaccedilatildeo de 1ordf ordem tal
como se pode verificar pela linearidade do graacutefico de ln(Abs400)vs o tempo de aquisiccedilatildeo espectral
(segundos) (Fig26 )
Figura 26ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do tempo
(em segundos) para uma soluccedilatildeo de decavanadato total de 1
mM em tampatildeo D a 25ordmC (n=2)
Sendo uma reacccedilatildeo de primeira ordem pode ser descrita pela equaccedilatildeo cujo
declive eacute ndashK (constante de dissociaccedilatildeo) e a intersecccedilatildeo no eixo das ordenadas corresponde
Ln
(Ab
s 400 n
m)
tempo (s)
29
aln(A0) Numa reacccedilatildeo de 1ordf ordem o tempo de meia-vida eacute independente da concentraccedilatildeo inicial
do reagente sendo descrito pela equaccedilatildeo [2] Deste modo a partir da recta
Ln(Abs400 nm) = -436E-5
K - 105 obteve-se um tempo de meia-vida de 4490 plusmn 0823 horas para
as condiccedilotildees supramencionadas
Apoacutes a adiccedilatildeo de S1 8 microM ao meio reaccional verificou-se que a reacccedilatildeo de desoligomerizaccedilatildeo
da espeacutecie decameacuterica de vanadato seguia igualmente uma cineacutetica de 1ordf ordem tal como se pode
verificar pela linearidade do graacutefico de ln(Abs400) versus o tempo de aquisiccedilatildeo espectral
(segundos) (Fig27)
Figura 27ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do
tempo (em segundos) para uma soluccedilatildeo de decavanadato
total de 1 mM em tampatildeo D a 25ordmC na presenccedila de 8
microM (n= 2)
A partir da equaccedilatildeo da recta Ln(Abs400 nm) = -7520E-5
K - 0613determinou-se o valor de K para
a espeacutecie decameacuterica de onde se obteve um tempo de meia-vida de 2430 plusmn 0191 horas para as
condiccedilotildees supramencionadas
Tanto este valor como o valor registado na ausecircncia de S1 satildeo substancialmente superiores agrave
duraccedilatildeo dos ensaios de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina descritos no presente
relatoacuterio
24 Interacccedilatildeo de S1 com vanaacutedio(V)
241Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
De modo a analisar qual o efeito da adiccedilatildeo de S1 de miosina (tampatildeo D) produzia nos sinais de
51V-NMR titulou-se uma soluccedilatildeo de 5mM de decavanadato total com concentraccedilotildees crescentes
de S1 (5 10 15 e 20 microM) Os espectros de 51
V-RMN encontram-se apresentados na Fig 2
No graacutefico da figura 2 Apresenta-se a concentraccedilatildeo de S1 em funccedilatildeo do factor de alargamento
corrigido dos espectros acima referidos
Ln
(Ab
s (4
00
nm
))
tempo (s)
30
Figura 28 ndash Graacutefico de
representaccedilatildeo da
concentraccedilatildeo de S1 em
funccedilatildeo do factor de
alargamento A
concentraccedilatildeo de
decavanadato total foi de 5
mM e as concentraccedilotildees
utilizadas de S1 foram de 0
5 10 e 15 microM
Verifica-se que o factor de alargamento aumenta linearmente em funccedilatildeo da concentraccedilatildeo de S1 no
ensaio o que indica a existecircncia de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina Observou-
se ainda que a adiccedilatildeo de S1 ao meio reaccional natildeo provocou qualquer alteraccedilatildeo dos desvios
quiacutemicos dos sinais de 51
V-RMN
Realizaram-se ensaios na presenccedila de ATP e de S1 com intuito de verificar o efeito do ATP na
interacccedilatildeo do S1 de miosina com o decavanadato
Figura 29 - Espectros de RMN de 51
V contendo 2
mM Decavanadato (A) (controlo) (B) na presenccedila
de 2 mM de ATP (C) na presenccedila de 2 mM de ATP
e 8 microM de S1 de miosina V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2Vndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies decameacuterica (V10) e monomeacuterica (V1) presentes nos
espectros da soluccedilatildeo de decavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V10A V10B V10C V1
part (ppm) -5161 -5002 -4254 -5614
Observa-se que a presenccedila de ATP (espectro B) natildeo provoca qualquer alteraccedilatildeo visiacutevel em
relaccedilatildeo ao espectro que contem apenas soluccedilatildeo de decavanadato Jaacute a presenccedila simultacircnea de ATP
e S1 provoca um alargamento da ∆12 do sinal do V10A de 1736 Hz (espectro (A)) para 2366 Hz
(espectro (C) Fig 29) Tendo em conta que a concentraccedilatildeo de S1 apresenta uma relaccedilatildeo linear
com o factor de alargamento converteu-se o valor da ∆12 e comparou-se com o valor esperado agrave
F = 00591[S1] - 00157 Rsup2 = 0998
Fact
or
de
ala
rgam
en
to
[S1] (microM)
31
concentraccedilatildeo de S1 utilizada O factor de alargamento obtido para 8 microM ndash 0376 ndash eacute inferior ao
valor esperado agrave mesma concentraccedilatildeo de S1 obtido atraveacutes da equaccedilatildeo da recta apresentada na
Fig 28 ndash 0457 ndash o que indica que o ATP poderaacute exercer um efeito de protecccedilatildeo do S1
diminuindo a sua interacccedilatildeo com a espeacutecieV10
Por uacuteltimo realizaram-se ensaios com metavanadato na presenccedila de 8 microM S1 de modo a verificar
se as espeacutecies oligomeacutericas contidas nesta soluccedilatildeo interagiam com a proteiacutena em estudo
Figura 210 - Espectros de RMN de 51
V contendo 2
mM metavanadato (A) (controlo) (B) na presenccedila
de 8 microM de S1 de miosina V1 V2 e V4
correspondem respectivamente aacutes espeacutecies
monomeacuterica dimeacuterica e tetrameacuterica
Tabela 2VIndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies monomeacuterica (V1) dimeacuterica (V2) e tetrameacuterica (V4)
presentes nos espectros da soluccedilatildeo de metavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V1 V2 V4
part (ppm) -5613 -5747 -5789
Como eacute possiacutevel observar na Fig 210 ocorre uma ligeiro alargamento dos sinal de V4 bem
como uma perda da intensidade pelo que se conclui quedas espeacutecies presentes na soluccedilatildeo de
metavanadato (V1 V2 e V4) apenas a espeacutecie tetrameacuterica interage com o S1 de miosina o que
estaacute de acordo com o reportado por Tiago et al 2002
242 Actividade Mg2+
-ATPaacutesica de S1 de miosina na presenccedila de decanadato
Com o intuito de avaliar o efeito do decavanadato na actividade Mg2+
-ATPaacutesica do S1 de
miosina monitorizou-se a reacccedilatildeo de oxidaccedilatildeo do NADH atraveacutes da leitura de Abs340 na
presenccedila de 005 mgmL de S1 de miosina e no caso da actividade estimulada 2microM de actina em
3125 mM Tris-HCl pH 71 e 375 mM MgCl2 pelo meacutetodo de enzimas acopladas (ver seccccedilatildeo xx
dos MM)Para tal realizaram-se ensaios com vaacuterias concentraccedilotildees de decavanadato total (10 50
100 250 500 e 1000) de modo a determinar qual o seu efeito na actividade MgATPaacutesica de S1
de miosina na ausecircncia e na presenccedila de um dos seus ligandos naturais Os ensaios foram
realizados agrave temperatura ambiente com incubaccedilatildeo preacutevia a 25ordm C durante 20 minutos
32
Figura 211 ndash Representaccedilatildeo
graacutefica da Actividade Mg2+
-
ATPaacutesica de S1 () (os ajustes
teoacutericos estatildeo representado a por
uma linha os resultados
experimentais estatildeo representados
por triacircngulos (Actividade
estimulada) e cruzes (Actividade
natildeo estimulada)) vs
[Decavanadato]total medidas pelo
meacutetodo das enzimas acopladas
(n=3) Os dados foram ajustados ao modelo de decaimento exponencial usando o programa GraphPad Prism para a
determinaccedilatildeo dos valores de IC50 (na ausecircncia de actina IC50= 266 plusmn 07 M decavanadato na presenccedila de actina
IC50= 28 plusmn 1 M decavanadato
A partir do graacutefico presente na Fig 29 verifica-se que o ajuste dos dados experimentais foi bem
sucedido A inibiccedilatildeo da actividade Mg2+
-ATPaacutesica do S1 de miosina eacute afectada em menor
amplitude na presenccedila de actina (60 de inibiccedilatildeo da actividade) do que na sua ausecircncia
(aproximadamente 80 de inibiccedilatildeo de actividade) Devido a esta diferenccedila pode sugerir-se que a
actina iraacute impedir parcialmente a ligaccedilatildeo do decavanadatoao S1 demiosina atraveacutes de
impedimentos esteacutereos ou interacccedilotildees electrostaacuteticas por exemplo A teacutecnica utilizada natildeo nos
permite concluir acerca de que regiatildeo(otildees) ou resiacuteduo(s) da actina participaratildeo na preservaccedilatildeo
parcial da actividade Mg2+
-ATPaacutesica
Por outro lado os valores de IC50 dos dois tipos de actividades natildeo apresentam diferenccedilas
significativas tendo-se obtido 266 plusmn 07 e 28 plusmn 1M decavanadato na ausecircncia e na presenccedila de
actina respectivamente o que corresponde a uma concentraccedilatildeo de cerca de 27 M de V10 O
meio de reacccedilatildeo utilizado nestes ensaios foi 10 mM HEPES e 25 mM KCl com 2 mM ATP Em
(Tiago et al 2004) o valor de IC50 para a actividade Mg2+-ATPaacutesica na ausecircncia de actina foi de
125 plusmn 025 microM enquanto o valor da actividade estimulada por actina foi de 136 plusmn 14 microM O
meio de reacccedilatildeo utilizado nos ensaios foi 20 mM HEPES 50 mM KCl e 2 mM ATP o que resulta
precisamente no dobro da forccedila ioacutenica a que foi realizado o ensaio do presente relatoacuterio
(exceptuando a contribuiccedilatildeo dos produtos da hidroacutelise de ATP uma vez que este meacutetodo
pressupotildee a ocorrecircncia de um ciclo de reacccedilotildees de hidroacutelise e siacutentese deste composto) Ora tendo
sido anteriormente referido que a afinidade do S1 de miosina para com a espeacutecie V10 diminui com
Act
ivid
ade
Mg
2+-A
TP
aacutesic
a d
e S
1 d
e
Mio
sin
a
[Decavanadato] (microM) Actividade Estimulada ()
Actividade Natildeo Estimulada ()
Ajuste Actividade Estimulada
Ajuste Actividade Natildeo Estimulada
33
o aumento da forccedila ioacutenica (Tiago et al 2004 tal poderaacute explicar a diferenccedila obtida entre estes
dois conjuntos de valores
243Quantificaccedilatildeo de cisteiacutenas reduzidas pelo meacutetodo de Ellman
A interacccedilatildeo do vanaacutedio com alguns tipos de aminoaacutecidos sensiacuteveis agrave oxidaccedilatildeo pode fornecer-nos
informaccedilatildeo bastante uacutetil em relaccedilatildeo agraves alteraccedilotildees estruturais que conduzem a uma diminuiccedilatildeo da
actividade fisioloacutegica do S1 de miosina Os estudos realizados por Chasteen 1983 concluiacuteram
que a reduccedilatildeo de vanadato (V) a vanadilo (IV) ocorre na presenccedila de agentes redutores comuns no
ambiente celular de que se destaca o aminoaacutecido cisteiacutena Esta eacute tambeacutem um alvo de
complexaccedilatildeo por parte do iatildeo VO2+
Jaacute Ramos et al (2009) demonstrou que a incubaccedilatildeo de
actina com a soluccedilatildeo de decavanadato induz a oxidaccedilatildeo de resiacuteduos de cisteiacutena e agrave formaccedilatildeo do
oxidovanaacutedio (IV) Este grupo de investigadores descobriu ainda que apenas a espeacutecie V10 induz a
oxidaccedilatildeo das cisteiacutenas ldquoraacutepidasrdquo da actina atraveacutes do processo proposto que eacute parcialmente
responsaacutevel pela inibiccedilatildeo jaacute descrita da actividade de ATPase do S1 de miosina estimulada por
actina quando exposta agrave soluccedilatildeo de decavanadato Esta inibiccedilatildeo apresenta-se afectada com a
presenccedila de ATP uma vez que os resiacuteduos de cisteiacutena permaneceram intactos nos ensaios
realizados com este ligando Jaacute Crans et al (2004) concluiu que o siacutetio activo de muitas fosfatases
(que conteacutem grupos tiol) pode formar um equiliacutebrio redox com o iatildeo vanadato (V) de forma natildeo
reversiacutevel
Os resultados obtidos por Sakurai e Shiromura (1980) sugeriram que o vanaacutedio (V) sofre reduccedilatildeo
a vanaacutedio (IV) sem dependecircncia do valor de pH do ensaio Registaram ainda uma estequiometria
de 31 na reacccedilatildeo do vanadato (V) com os resiacuteduos de Cys enquanto a estequiometria dos ensaios
com o vanadilo (IV) foi de 21
Berlett e Stadtman (1997) observaram que os tipos de aminoaacutecidos mais facilmente oxidaacuteveis
englobam entre outros os resiacuteduos de Cys Trp e MetAssim analisou-se o estado redox dos
grupos tiol dos resiacuteduos de Cys do S1 na presenccedila do decavanadato atraveacutes da monitorizaccedilatildeo da
Abs412 nm na presenccedila de 1 mgmL de S1 de miosina em tampatildeo 10 mM HEPES 25 mM KCl
variando a concentraccedilatildeo da soluccedilatildeo de decavanadato de modo a determinar que tipo de alteraccedilotildees
estruturais ocorrem na sua presenccedila no que diz respeito agrave diminuiccedilatildeo da actividade ATPaacutesica de
S1 na presenccedila de decavanadato
Os resultados referentes agrave quantificaccedilatildeo das cisteiacutenas reduzidas apoacutes o tratamento de S1 com
diferentes concentraccedilotildees de decavanadato em tampatildeo 10 mM HEPES pH 75 e 25 mM KCl a
25ordmC durante 20 minutos encontram-se descritos na Tabela 2VII
Tabela 2VII ndash Cisteiacutenas reduzidas totais e ldquoraacutepidas de S1 (1 mgmL) apoacutes tratamento com decavanadato
34
(010501002505001000 uM) em 10 mM HEPES 25 mM KCl pH 7 a 25ordmC durante 20 minutos A detecccedilatildeo
espectrofotomeacutetrica foi feita com DTNB (e 412=8400 uM-1 cm-1) (n variaacutevel entre 1 e 8)
Amostras Cys raacutepidas Cys totais
11 M S1 (controlo) 22 plusmn 04 106 plusmn 14
11 M S1 + 10 M Decavanadato 22 plusmn 05 95 plusmn 08
11 M S1 + 50 uM Decavanadato 20 plusmn 02 98 plusmn 14
11 M S1 + 100 uM Decavanadato 17 plusmn 03 97 plusmn14
11 M S1 + 250 uM Decavanadato 26 plusmn 02 98 plusmn 08
11 M S1 + 500 uM Decavanadato 21 plusmn 04 90 plusmn 06
11 M S1 + 1000 uM Decavanadato 24 plusmn 0 86 plusmn 08
A partir dos resultados obtidos pode sugerir-se que o S1 possui 11 resiacuteduos de Cys (controlo) o
que estaacute de acordo com o anteriormente descrito Verifica-se que o tratamento da proteiacutena com
concentraccedilotildees crescentes de decavanadato provoca a oxidaccedilatildeo de dois resiacuteduos de Cys do S1 de
miosina natildeo expostos ao solvente No entanto a teacutecnica utilizada natildeo nos permite identificar
quais
Tendo em consideraccedilatildeo que o valor de IC50 obtido para a reacccedilatildeo de inibiccedilatildeo daactividade Mg2+-
ATPaacutesica de S1 na presenccedila de decavanadato eacute de cerca de 27 microM pode sugerir-se que este seraacute o
valor ao qual ocorreraacute a oxidaccedilatildeo do primeiro resiacuteduo de Cys o que poderaacute contribuir para a
diminuiccedilatildeo da actividade de S1 observada
25Interacccedilatildeo de S1 com vanaacutedio(IV)
251Anaacutelise por espectroscopia de Ressonacircncia Paramagneacutetica Electroacutenica
A acumulaccedilatildeo intracelular de oxovanaacutedio (IV) in vivo constitui uma das principais formas de
apresentaccedilatildeo de vanaacutedio no organismo Deste modo torna-se bastante interessante analisar a
interacccedilatildeo entre este iatildeo e o S1 de miosina atraveacutes da teacutecnica de espectroscopia de Ressonacircncia
Paramagneacutetica Electroacutenica o meacutetodo de eleiccedilatildeo do estudo do iatildeo vanadilo e dos complexos por si
formados O electratildeo desemparelhado encontra-se fortemente acoplado ao nuacutecleo 51
V o que
35
resulta num espectro de RPE consistindo de 8 desdobramentos na direcccedilatildeo paralela e outros tantos
na direcccedilatildeo perpendicular (Chasteen 1983)
Os estudos realizados por Ahmed et al (1987) centraram-se na interacccedilatildeo do vanadilo com
proteiacutenas como eacute o caso da insulina transferrina entre outros obtendo um espectro caracterizado
por 8 linhas amplamente espaccediladas (na direcccedilatildeo paralela) Jaacute Ramos et al 2009 concluiu que a
exposiccedilatildeo da soluccedilatildeo de decavanadato agraves formas globular e filamentosa de actina produz um
espectro tiacutepico do iatildeo oxidovanaacutedio (IV) indicando a reduccedilatildeo do vanadato
Para aferir qual a concentraccedilatildeo de S1 que permitia obter uma boa razatildeo sinalruiacutedo titulou-se uma
soluccedilatildeo de sulfato de vanadilo 15 microM ([VOSO4]stock = 5 mM) com concentraccedilotildees crescentes de
proteiacutena (0 5 10 15 e 20 microM) em 10 mM HEPES 25 mM KCl A existecircncia de uma correlaccedilatildeo
linear entre a concentraccedilatildeo de proteiacutena e a intensidade dos sinais de RPE correspondente agrave banda
-12 perpendicular encontra-se representada na Fig 210
Figura 2 12ndash Representaccedilatildeo graacutefica da variaccedilatildeo da intensidade do sinal de RPE vs a concentraccedilatildeo de S1 utilizada
Apoacutes verificar que com uma concentraccedilatildeo de S1 igual a 20 microM se obtinham sinais com uma razatildeo
sinalruiacutedo adequada ao prosseguimento dos estudos sobre a interacccedilatildeo V(IV)-S1 de miosina
titulou-se uma soluccedilatildeo de 20 microM S1 com 0 25 5 10 15 25 50 75 100 150 200 500 e 1000
microM VOSO4
Traccedilou-se o graacutefico da intensidade da banda -12 perpendicular do sinal de EPR em funccedilatildeo da
concentraccedilatildeo de VOSO4 tendo-se determinado uma constante de dissociaccedilatildeo (KD) do complexo
S1-vanadilo de 8748 plusmn 2621 microM V(IV)
y = 016786x + 122435
Rsup2 = 092992759
Inte
nsi
dad
e d
o s
inal
de
RP
E
[S1] (microM)
36
Figura 213 ndash Representaccedilatildeo graacutefica da variaccedilatildeo do sinal de RPE vs a concentraccedilatildeo de vanadilo utilizada (os valores
experimentais estatildeo representados por losangos o seu ajuste estaacute representado pela linha)
A simulaccedilatildeo do espectro de EPR foi efectuada a partir do espectro obtido para uma concentraccedilatildeo
de 1 mM VOSO4 apoacutes adiccedilatildeo de 20 microM S1 de miosina tendo-se obtido os valores da constante de
acoplamento hiperfina (A=5016) e do factor g (g0=195)
Figura 214 ndash Representaccedilatildeo graacutefica do espectro de EPR obtido com 1000 microM de vanadilo e 20 microM de S1 (a cinzento
escuro) A simulaccedilatildeo do espectro encontra-se a cinzento claro
Por comparaccedilatildeo destes com os valores presentes na tabela de correspondecircncia xxx eacute possiacutevel
predizer quais os ligandos provaacuteveis do vanadilo na presenccedila de proteiacutena Assim concluiu-se que
o vanadilo estaacute coordenado com duas moleacuteculas de aacutegua um grupo hidroxilo e um grupo R=NR
O grupo hidroxilo pode indicar os resiacuteduos de aminoaacutecidos Tyr Treonina e Serina enquanto o
grupo R=NR poderaacute indicar a presenccedila de Arginina e Histidina como ligandos do vanadilo
Inte
nsi
dad
e d
os
sinai
s d
e R
PE
[Vanadilo] (microM)
Ajuste dos dados exp Dados experimentais
Campo magneacutetico (G)
Espectro Experimental Ajuste
37
3 Procedimento Experimental
31 Reagentes
Todos os reagentes utilizados satildeo de elevado grau de pureza (analiacutetico) Apresentam-se as suas
foacutermulas quiacutemicas ou abreviaturas quando necessaacuterio acrescentou-se a sua descriccedilatildeo Entre
parecircntesis colocou-se as empresas que os comercializam
Aacutecido aceacutetico glacial 100 (Riedel) Acrilamida 99 (Merck) Azul de bromofenol (Riedel-de-
Haumlen) -Mercaptoetanol (Sigma) Bissulfito de soacutedio (Sigma)Bradford (reagente de)
(BioRad)BSA (albumina de soro bovino Sigma-Aldrich) CaCl22H2O (cloreto de caacutelcio
dihidratado Fluka) CH3CH2OH 96 (etanol Panreac) Coomassie Blue R250 (Merck) D2O
(oacutexido de deuteacuterio SDS) DTNB (55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico) Aldrich) EDTA (aacutecido
etilenodiaminatetraaceacutetico Merck) Fosfoenolpiruvato 98 (Fluka) Glicerol (Sigma) Glicina
985 (Panreac) HCl 37 (Riedel-de Haumlen) H2SO4 96 (Pronalab) HEPES 99 (aacutecido 4-(2-
hidroxiletil)-1-piperazineetanosulfoacutenico Sigma) KH2PO4 99 (hidrogenofosfato de potaacutessio)
MgATP (Sigma) Na
ATP (Sigma) Na2HPO4 99 (hidrogenofosfato de soacutedio Riedel-de Haumlen)
NaH2PO4 (fosfato de soacutedio ) NaOH 986 (hidroacutexido de soacutedio Pronalab) KCl 99 (Cloreto de
potaacutessio Panreac) L-Cys 995 (enantioacutemero levoacutegiro do resiacuteduo de Cisteiacutena Fluka)Metanol
99 (Merck) Molibdato de amoacutenio (Sigma)NNrsquo-metil-bis-acrilamida 98 (Merck) NH4VO3
99 (metavanadato de amoacutenia Sigma) PSA 10 (persulfato de amoacutenio Merck) SDS
(dodecilo sulfato de soacutedio Sigma) Sulfito de soacutedio (Sigma) VOSO4 (sulfato de vanadilo
Aldrich) TEMED 99 (Tetrametiletilenodiamina Merck) Tris (2-amino-2-hidroximetil-13-
propanodiol Sigma)
32 Meacutetodos
321 Isolamento e purificaccedilatildeo das proteiacutenas de muacutesculo esqueleacutetico de coelho
3211 Purificaccedilatildeo da miosina preparaccedilatildeo de Subfragmento 1 por digestatildeo quimotriacutepsica
da miosina
O subfragmento-1 de miosina-II (S1) foi gentilmente cedido pelo Prof Doutor Manuel Aureliano
do Departamento de Quiacutemica Bioquiacutemica e Farmaacutecia da Faculdade de Ciecircncias e Tecnologia da
Universidade do Algarve
No entanto descreve-se abaixo de forma resumida o procedimento efectuado
38
32111 Purificaccedilatildeo da miosina
O protocolo de isolamento foi adaptado do meacutetodo previamente descrito por Margossian e Lowey
(1982) apresentando-se de forma esquemaacutetica na Fig 31
Sucintamente os tecidos musculares dorsais e das pernas foram extraiacutedos apoacutes o sacrifiacutecio do
animal (Fig31) De seguida foram adicionados 3 volumes da soluccedilatildeo de extracccedilatildeo A2
Centrifugou-se a soluccedilatildeo e filtrou-se o sobrenadante O sedimento foi entatildeo diluiacutedo em 14
volumes de aacutegua destilada de modo a que a forccedila ioacutenica atingisse 004 M Apoacutes adiccedilatildeo de szlig-
mercaptoetanol (szlig-me) deixou-se a precipitar a 4ordmC removendo o sobrenadante Centrifugou-se
novamente e ressuspendeu-se o sedimento na soluccedilatildeo de extracccedilatildeo A Adicionou-se a soluccedilatildeo
tampatildeo B3 para que a forccedila ioacutenica atingisse 05 M Homogeneizou-se e deixou-se repousar
durante 2h Homogeneizou-se novamente e centrifugou-se de modo a ocorrer precipitaccedilatildeo dos
complexos de actomiosina e proteiacutena desnaturada Apoacutes filtraccedilatildeo do sobrenadante em latilde de vidro
quantificou-se a soluccedilatildeo de miosina resultante pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental)
2 Soluccedilatildeo de extracccedilatildeo A 0012M K2CO3 004M KHCO3 06M KCl pH 65
3 Soluccedilatildeo-tampatildeo B Tris 100 mM pH 7 KCl 2M
39
Figura 31 ndash Representaccedilatildeo esquemaacutetica do protocolo de isolamento de miosina (Margossian e Lowey 1982) Todo o
procedimento foi realizado a 4ordmC (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
32112 Preparaccedilatildeo de Subfragmento-1 de miosina por digestatildeo com quimiotripsina
O S1 de miosina foi isolado apoacutes diaacutelise da miosina durante a noite contra a soluccedilatildeo C4 A
presenccedila do EDTA eacute essencial uma vez que a sua substituiccedilatildeo por MgCl2resulta na produccedilatildeo de
HMM em vez de S1 A suspensatildeo de miosina foi equilibrada e digerida com -quimotripsina
A digestatildeo foi interrompida pela adiccedilatildeo de PMSF Seguidamente ultracentrifugou-se o produto
da digestatildeo desprezando-se o sedimento (Fig 32) A soluccedilatildeo de sobrenadante maioritariamente
S1 foi quantificada pela leitura directa da Abs280 ou pelo meacutetodo de Bradford (ver as secccedilotildees
3311 e 3312 do Procedimento Experimental respectivamente) e armazenada em 10 sacarose
e 2 mM szlig-Me a -80ordmC
4 Soluccedilatildeo C 25mM Tris pH 70 012M KCl 1mM EDTA
40
Antes da realizaccedilatildeo dos ensaios descritos no presente relatoacuterio o S1 de miosina foi dialisado
contra a soluccedilatildeo D5 de modo a remover qualquer composto que pudesse complexar como
vanaacutedio Seguidamente quantificou-se a proteiacutena pelo meacutetodo de Bradford ou pelo meacutetodo
directo (ver secccedilotildees 3311 e 3312do Procedimento Experimtental)
Figura 3 2ndash Representaccedilatildeo esquemaacutetica do protocolo de preparaccedilatildeo do S1 de miosina por digestatildeo quimiotripsina de
acordo com o previamente descrito por Margossian e Lowey (1982) Todo o procedimento foi realizado a 4ordm C
excepto quando indicaccedilatildeo do contraacuterio (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
3212 Isolamento e purificaccedilatildeo de actina
Isolou-se e purificou-se a actina segundo o meacutetodo descrito por Pardee e Spudish que
compreende dois passos ndash a obtenccedilatildeo de um resiacuteduo seco denominado ldquopoacute de acetonardquo e a
extracccedilatildeo subsequente da actina a partir do ldquopoacute de acetonardquo na forma de G-Actina com a
posterior polimerizaccedilatildeo a F-Actina
32121 Obtenccedilatildeo do ldquopoacute de acetonardquo
Sacrificou-se o coelho com uma pancada atraacutes do pescoccedilo e sangrou-se o animal apoacutes o corte da
arteacuteria jugular Esfolou-se e colocou-se numa tina com gelo Retiraram-se as partes de tecido
conjuntivo que envolvem o tecido muscular com auxiacutelio de uma pinccedila e bisturi Seguidamente
retirou-se os muacutesculos dorsais e os muacutesculos brancos das pernas Passou-se os muacutesculos por aacutegua
destilada de modo a remover as impurezas e pesou-se num copo Trituraram-se os muacutesculos
numa maacutequina de triturar carne e extraiu-se o resultante com 3 volumes da soluccedilatildeo E6 durante 10
minutos com agitaccedilatildeo suave Centrifugou-se a suspensatildeo a 4000 g durante 10
5 Soluccedilatildeo D HEPES 10 mM pH 70 KCl 25 mM
6 Soluccedilatildeo E 01 M KCl 015 M K2HPO4 pH 65
41
minutosdesprezando-se o sobrenadante Ressuspendeu-se o sedimento em 6 volumes de soluccedilatildeo
F7durante 10 minutos com agitaccedilatildeo suave Verificou-se se o pH da soluccedilatildeo estava compreendido
entre 82 e 85 acertando-se estecom soluccedilatildeo G8 se necessaacuterio Centrifugou-se durante 10
minutos a 4000 g Desprezou-se o sobrenadante e ressuspendeu-se o sedimento em 3 volumes de
soluccedilatildeo H9 durante 10 minutos Centrifugou-se novamente a 4000 gdurante 10 minutos
desprezando-se o sobrenadante Ressuspendeu-se o sedimento duas vezes consecutivas em 6
volumes de aacutegua destilada durante 5 minutos Centrifugou-se a 4000 g durante 10 minutos apoacutes
cada adiccedilatildeo de H2O
Seguidamente ressuspendeu-se o sedimento 5 vezes consecutivas utilizando 3 volumes de
acetona de cada vez agitando suavemente durante 10 minutos agrave temperatura ambiente Colocou-
se o resiacuteduo filtrado numa tina de vidro sobre papel de filtro deixando-se evaporar o solvente
durante a noite num nicho Triturou-se o resiacuteduo seco numa maacutequina de triturar carne e deixou-se
novamente evaporar o solvente O ldquopoacute de acetonardquo obtido foi armazenado a -20 ordmC
32122 Purificaccedilatildeo da actina a partir de ldquopoacute de acetonardquo
Apoacutes a obtenccedilatildeo do poacute de acetona como acima indicado efectuou-se o protocolo seguidamente
descrito de modo a obter monoacutemeros de actina (G-Actina) e actina filamentosa (F-Actina)
Pesou-se 35 g de poacute de acetona e adicionaram-se 50 mL de soluccedilatildeo-tampatildeo I10
sendo a
extracccedilatildeo feita com agitaccedilatildeo suave durante 30 minutos Separou-se o extracto do poacute de acetona
hidratado filtrando a suspensatildeo com 4 camadas de gaze esterilizada tendo o cuidado de natildeo
espremer o resiacuteduo Voltou a extrair-se com 50 mL de soluccedilatildeo I durante 30 minutos mexendo
lentamente Adicionou-se este ao extracto obtido anteriormente Centrifugou-se o extracto total a
16 000 g durante 30 minutos Decantou-se para uma proveta sobre a qual se colocou gaze para
que o sedimento solto natildeo se juntasse ao sobrenadante e leu-se o volume De seguida adicionou-
se KCl ateacute uma concentraccedilatildeo final de 50 mM MgCl2 ateacute 2mM e ATP ateacute 1mM lentamente e com
agitaccedilatildeo contiacutenua de modo a promover a polimerizaccedilatildeo dos monoacutemeros de actina
Deixou-se polimerizar durante 2h a 25ordmC observando-se a presenccedila de pequenas bolhas de ar
indicativas de um aumento de viscosidade da soluccedilatildeo Arrefeceu-se em gelo durante 15 minutos e
adicionou-se KCl ateacute uma concentraccedilatildeo final de 06M Mexeu-se lentamente durante 30 minutos
apoacutes os quais se ultracentrifugou a 110 000g durante 1hora Desprezou-se o sobrenadante
ressuspendeu-se o sedimento com soluccedilatildeo tampatildeo E e transferiu-se para um potter
Dialisou-se durante a noite contra 1L de soluccedilatildeo-tampatildeo I a uma temperatura de 4ordmC com rotaccedilatildeo
7 Soluccedilatildeo F 005 M NaHCO3 pH 82-85
8 Soluccedilatildeo G 1M Na2CO3
9 Soluccedilatildeo H 1 mM EDTA pH 70
10 Soluccedilatildeo I 2 mM Tris-HCl pH 8 02 mM CaCl2 02 mM Na2ATP 05 mM szlig-Me 0005 NaN3
42
contiacutenua e mudanccedila de tampatildeo 2 a 3 vezes De seguida a actina foi centrifugada a 110
000gdurante 1hora Decantou-se a soluccedilatildeo de G-Actina (sobrenadante) e desprezou-se o
sedimento Determinou-se a concentraccedilatildeo de G-Actina pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental) A soluccedilatildeo de G-actina foi armazenada a uma temperatura de 4ordmC
apoacutes adiccedilatildeo de 002NaN3 mantendo-se estaacutevel durante 3 a 5 dias
331 Quantificaccedilatildeo das proteiacutenas isoladas
3311 Quantificaccedilatildeo pelo meacutetodo de Bradford
O meacutetodo de Bradford (Bradford 1976) baseia-se na utilizaccedilatildeo do reagente de Bradford
composto por entre outros aacutecido fosfoacuterico e o corante ldquoCoomassierdquo Este corante possui carga
negativa e a sua coloraccedilatildeo varia entre a vermelha (Absmax=465 nm) predominante em soluccedilatildeo e a
azul (Absmax=595 nm) presente aquando da ligaccedilatildeo aos grupos amina das proteiacutenas Em soluccedilotildees
aacutecidas diluiacutedas a coloraccedilatildeo varia proporcionalmente com a quantidade do corante ligado agrave
proteiacutena que por sua vez eacute directamente proporcional agrave concentraccedilatildeo da proteiacutena permitindo a
sua quantificaccedilatildeo ateacute a um limite de detecccedilatildeo de 20 gmL (Bradford 1976)
Deste modo prepararam-se amostras de BSA ([BSA]stock= 1mgmL) utilizando um intervalo de
concentraccedilotildees compreendido entre 0 e 15 gmL de acordo com a tabela 3I Apoacutes a adiccedilatildeo da
H2O MilliQ e do reagente de Bradford as soluccedilotildees foram sucessivamente homogeneizadas no
voacutertex
Tabela 3Indash Amostras utilizadas para a obtenccedilatildeo da recta de calibraccedilatildeo no meacutetodo de Bradford
BSA (L) BSA (g) H2O MilliQ (L) Reagente de Bradford (L)
0 0 800 200
125 125 79875 200
25 25 7975 200
5 5 795 200
10 10 790 200
15 15 785 200
Apoacutes incubaccedilatildeo a 25ordmC por 5 minutos leram-se os valores de Abs595 num
espectrofotoacutemetro (Shimadzu UV-160A) para todas as amostras Seguidamente traccedilou-se a recta
de calibraccedilatildeo (Abs595vs quantidade de BSA (g))
43
Para determinar a concentraccedilatildeo de proteiacutena apoacutes o processo de isolamento purificaccedilatildeo e
diaacutelise quando necessaacuterio colocou-se 5 L da amostra num tubo de Eppendorf e adicionou-se
795 L de H2O MilliQ homogeneizando a soluccedilatildeo no voacutertex Adicionou-se posteriormente 200
L de reagente de Bradford e repetiu-se a homogeneizaccedilatildeo Incubou-se a mistura a 25ordmC por 5
minutos apoacutes os quais se leu os valores de Abs595 nm no espectrofotoacutemetro Por interpolaccedilatildeo deste
valor a partir da recta de calibraccedilatildeo determinou-se a concentraccedilatildeo de proteiacutena na amostra
3312 Quantificaccedilatildeo pelo meacutetodo directo
A quantificaccedilatildeo das amostras proteicas pode tambeacutem ser efectuada atraveacutes da leitura directa da
absorvacircncia tendo por base a lei de Beer-Lambert onde Abs corresponde agrave
absorvacircncia ao comprimento de onda ao qual o ensaio foi realizado corresponde ao coeficiente
de extinccedilatildeo (substituiacutedo pela letra a se as unidades forem distintas da molaridade) b corresponde
ao percurso oacuteptico da radiaccedilatildeo na ceacutelula amostral e c corresponde agrave concentraccedilatildeo amostral
Assim no caso de S1 de miosina mediu-se a Abs280e calculou-se a concentraccedilatildeo da proteiacutena
sabendo que o coeficiente de extinccedilatildeo de S1 eacute (Ref)
No caso da actina mediu-se a Abs290ecalculou-se a concentraccedilatildeo da proteiacutena sabendo que o
coeficiente de extinccedilatildeo da actina eacute (Ref)
332Electroforese em condiccedilotildees desnaturantes (SDS-PAGE)
De modo a melhor caracterizar as proteiacutenas isoladas foi realizada uma electroforese em
condiccedilotildees desnaturantes (SDS-PAGE do inglecircs Sodium Dodecyl Sulfate Polyacrilamide Gel
Electrophoresis) O SDS (dodecilo sulfato de soacutedio) eacute um detergente anioacutenico utilizado neste
meacutetodo para desnaturar as proteiacutenas que se ligam ao SDS na proporccedilatildeo 14 g SDS g proteiacutena
(aproximadamente 1 moleacutecula de SDS por cada 2 resiacuteduos de aminoaacutecidos) (Stryer) O SDS induz
uma densidade de carga negativa significativa nas proteiacutenas a que se liga mascarando a sua carga
intriacutenseca pelo que as razotildees cargamassa satildeo tendencialmente idecircnticas Assim a SDS-PAGE
separa as proteiacutenas quando sujeitas a um campo eleacutectrico de acordo com o seu peso molecular
sendo a sua migraccedilatildeo directamente proporcional ao logaritmo deste uacuteltimo (Stryer)
O gel de resoluccedilatildeo com 10 de acrilamida foi obtido atraveacutes da adiccedilatildeo de 167 mL de soluccedilatildeo
stock de acrilamida11
075 mL de tampatildeo de resoluccedilatildeo12
251 mL de aacutegua destilada e 50 L de
SDS 10 Para promover a polimerizaccedilatildeo adicionou-se em seguida 38 L de PSA 10
11
soluccedilatildeo stock de acrilamida soluccedilatildeo de 30 (mv) de acrilamida e 08 (mv) de NNrsquo-metil-bis-acrilamida 12
tampatildeo de resoluccedilatildeo 15 M Tris-HCl pH 88
44
(persulfato de amoacutenia) 10 e 25 L de TEMED 99 (tetrametiletilenodiamina) Agitou-se
cuidadosamente de modo a natildeo formar bolhas e pipetou-se a soluccedilatildeo para a cassette de vidro
previamente montada no suporte De seguida adicionou-se uma peliacutecula de cerca de 2 mm de 2-
metil-2-butanol para verificar posteriormente a polimerizaccedilatildeo Apoacutes a polimerizaccedilatildeo que demora
cerca de 45 minutos verteu-se a agrave superfiacutecie e adicionou-se o gel de concentraccedilatildeo com 5 de
acrilamida constituiacutedo por 03 mL de soluccedilatildeo de stock acrilamida 045 mL de tampatildeo de
concentraccedilatildeo13
094 mL de aacutegua destilada 18 L de SDS 10 135 L de PSA 10 e 2L de
TEMED 99 Colocou-se o pente sobre o gel e deixou-se polimerizar durante cerca de 45
minutos
As amostras para aplicaccedilatildeo no gel de electroforese foram preparadas para uma concentraccedilatildeo de
proteiacutena de 1 mgmL num volume final de 100 L dos quais 25 L correspondem ao tampatildeo da
amostra14
As amostras foram aquecidas a 100ordmC durante 5 minutos Antes da aplicaccedilatildeo das
amostras transferiu-se a cassette para a tina de electroforese agrave qual se adicionou a soluccedilatildeo
tampatildeo de electroforese da tina superior15
e tampatildeo de electroforese da tina inferior16
Aplicaram-
se 10 L das soluccedilotildees de marcadores de peso molecular e 20 L das amostras proteicas
preparadas anteriormente A electroforese decorreu durante cerca de 3h30 min a uma voltagem
de 80 V De seguida transferiu-se o gel para uma tina corando as proteiacutenas durante 1h com
soluccedilatildeo corante17
Descorou-se o gel com soluccedilatildeo descorante18
tendo-se mudado esta soluccedilatildeo ateacute
as bandas se encontrarem bem definidas
Determinou-se a mobilidade relativa das bandas correspondentes aos marcadores de elevado peso
molecular e traccedilou-se a recta de calibraccedilatildeo ndash log(PM) vs log(Rf) onde Rf corresponde agrave
mobilidade relativa Por interpolaccedilatildeo determina-se o PM das proteiacutenas isoladas sabendo a sua
mobilidade relativa Por comparaccedilatildeo com os valores descritos na literatura pode concluir-se
acerca do grau de purificaccedilatildeo
34 Mediccedilatildeo da actividade enzimaacutetica de S1
341 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
13
tampatildeo de concentraccedilatildeo 05 M Tris-HCl pH 68 14
tampatildeo de amostra 320 mM Tris-HCl pH 68 04 M szlig-mercaptoetanol SDS 8 glicerol 15 (vv) e roxo de m-
cresol 0024 15
tampatildeo de electroforese da tina superior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS 16
tampatildeo de electroforese da tina inferior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS acetato de soacutedio
01 M 17
soluccedilatildeo corante ldquoCoomassie Blue R-250rdquo contendo metanol 45 (vv) e aacutecido aceacutetico glacial 10 (vv) 18
soluccedilatildeo descorante metanol 10 (vv) e aacutecido aceacutetico glacial 10 vv)
45
O meacutetodo de Fiske-Subarow modificado consiste na reacccedilatildeo do molibdato de amoacutenio com o
fosfato inorgacircnico em soluccedilatildeo aacutecida com formaccedilatildeo do aacutecido fosfomoliacutebdico Seguidamente daacute-
se a adiccedilatildeo de um agente redutor que provoca a reduccedilatildeo do composto a fosfomolibdato que forma
um complexo de cor azulada cuja absorvacircncia maacutexima ocorre para um comprimento de onda de
660 nm tornando possiacutevel a sua detecccedilatildeo por meacutetodos espectrofotomeacutetricos As actividades Ca2+
-
ATPaacutesica e K+EDTA-ATPaacutesica de S1 foram determinadas atraveacutes deste meacutetodo de acordo com
o descrito por Taussky e Shorr (1953) com mediccedilatildeo da quantidade de fosfato (Pi) libertado
atraveacutes da hidroacutelise de ATP
Para tal obteve-se primeiramente a recta de calibraccedilatildeo com as amostras presentes na Tabela 3II
utilizando uma soluccedilatildeo stock de 5 mM KH2PO4
Tabela 3II ndash Composiccedilatildeo das diferentes amostras utilizadas para determinar a recta de calibraccedilatildeo atraveacutes do meacutetodo
de Fiske-Subbarow modificado
KH2PO4 (L)
([KH2PO4]stock= 5mM)
PO43-
(nmole) H2O (uL)
0 0 1780
5 25 1775
10 50 1770
20 100 1760
40 200 1740
100 500 1680
A cada tubo de Eppendorf adicionaram-se seguidamente 200 L de soluccedilatildeo J19
e 20 L de agente
redutor20
homogeneizando no voacutertex apoacutes cada adiccedilatildeo Incubou-se durante 10 minutos e mediu-
se a Abs660 nm num espectrofotoacutemetro com intervalos de tempo iguais entre cada amostra Traccedilou-
se a recta de calibraccedilatildeo (Abs660 nmvs nmol PO43-
) a partir dos valores obtidos
Para determinar a actividade Ca2+
-ATPaacutesica de S1 prepararam-se tubos de Eppendorf com 400
L de meio de reacccedilatildeo21
25 L CaCl2 ([CaCl2]stock= 100 mM) 25 L ATP ([ATP]stock= 100 mM)
com [S1]ensaios=01 mgmL Perfez-se se necessaacuterio com H2O MilliQ ateacute perfazer um volume de 1
mL Para determinar a actividade K+EDTA-ATPaacutesica utilizou-se uma concentraccedilatildeo idecircntica de
19
Soluccedilatildeo J 5 N H2SO4 25 molibdato de amoacutenio 20
Agente redutor 5 mg aacutecido 1-amino-2-naftol-4-sulfoacutenico 30 mg bissulfito de soacutedio 30 mg sulfito de soacutedio em 26
mL H2O 21
meio de reacccedilatildeo 125 mM Tris-HCl pH 80 0575 M KCl
46
proteiacutena nos ensaios (01 mgmL) e o mesmo meio de reacccedilatildeo Aos tubos de Eppendorf
adicionou-se ainda 123 L de KCl ([KCl]stock = 3M) 25 L de ATP 10 L de EDTA
([EDTA]stock= 100 mM )e o volume de H2O MilliQ necessaacuterio para perfazer 1 mL Em ambas as
situaccedilotildees o uacuteltimo reagente adicionado foi o ATP uma vez que se trata do substrato da reacccedilatildeo
enzimaacutetica
A cineacutetica decorreu a 25ordmC durante 5 minutos para a actividade Ca2+
-ATPaacutesica e durante 3
minutos para a actividade K+EDTA-ATPaacutesica Apoacutes o periacuteodo de incubaccedilatildeo transferiu-se 150
L de amostra para tubos de Eppendorf contendo igual volume de TCA 10 Colocou-se no gelo
e homogeneizou-se no voacutertex Seguidamente centrifugou-se a 1500 g durante 10 minutos e
recolheu-se 200 L de sobrenadante para novos tubos de Eppendorf Adicionou-se a estes 158
mL de H2O MilliQ e 200 L de soluccedilatildeo J Homogeneizou-se no voacutertex e adicionou-se 20 L de
reagente redutor repetindo-se a homogeneizaccedilatildeo Incubou-se durante 10 minutos e mediu-se a
Abs660 nm com o mesmo intervalo de tempo de leitura para cada amostra Por intrapolaccedilatildeo a partir
da recta de calibraccedilatildeo obtida anteriormente determinou-se a quantidade de Pi libertado em cada
ensaio A partir destes resultados determinaram-se as actividades Ca2+
-ATPaacutesica e K+EDTA-
ATPaacutesica do S1 em nmol PO43-
minmg proteiacutena e em s-1
342 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
Da actividade Mg2+
-ATPaacutesica de S1 resulta a formaccedilatildeo dos produtos da reacccedilatildeo de
hidroacutelise do ATP - ADP e Pi Na presenccedila de ADP fosfoenolpiruvato (PEP) e da enzima piruvato
cinase (PK) os produtos resultantes satildeo ATP e piruvato Por acccedilatildeo da enzima lactato
desidrogenase (LDH) este uacuteltimo eacute convertido em lactato ocorrendo simultaneamente a oxidaccedilatildeo
do NADH a NAD+ monitorizada atraveacutes da Abs340 nm Quando o S1 de miosina se encontra no
meio reaccional o ATP eacute hidrolisado formando-se novamente ADP e Pi o que permite a
formaccedilatildeo de um ciclo (fig 21) Se as enzimas se encontrarem em excesso a velocidade de
conversatildeo do NADH eacute limitada pela produccedilatildeo de ADP e Pi Como o ATP eacute regenerado em cada
ciclo a sua concentraccedilatildeo inicial eacute sempre mantida (Bagshaw 1982)
Figura 33ndash Representaccedilatildeo esquemaacutetica do meacutetodo de enzimas acopladas
47
O protocolo utilizado e seguidamente descrito foi adaptado de Cuenda et al (1990 ndash Ver Refordf) A
uma cuvette de 1 mL adicionou-se S1 para uma concentraccedilatildeo final de 005 mgmL 800 L de
meio de reacccedilatildeo22
10 L de PEP ([PEP]stock = 42 mM) 10 L de PK+LDH 11 ([PK+LDH]stock =
1 mgmL) e 15 L de NADH ([NADH]stock=25 mM) Iniciou-se a monitorizaccedilatildeo da reacccedilatildeo
atraveacutes da leitura de Abs340 nm apoacutes a adiccedilatildeo de 20 L de ATP ([ATP]stock=100 mM) A
actividade Mg2+
-ATPaacutesica eacute obtida atraveacutes do declive correspondente agrave oxidaccedilatildeo do NADH Para
a mediccedilatildeo da Mg2+
-ATPase de S1 estimulada por actina adicionou-se F-actina para uma
concentraccedilatildeo final de 2 microM sendo a actividade obtida tambeacutem atraveacutes do declive correspondente
agrave oxidaccedilatildeo do NADH apoacutes a adiccedilatildeo da proteiacutena
35 Interaccedilatildeo entre vanaacutedio e subfragmento-1 de miosina
351 Preparaccedilatildeo das soluccedilotildees de vanaacutedio
As soluccedilotildees de vanadato foram preparadas em aacutegua MilliQ a partir de metavanadato de amoacutenio
(NH4VO3) numa concentraccedilatildeo stock de 50 mM com agitaccedilatildeo magneacutetica e aquecimento (~30 ordmC)
Esta soluccedilatildeo com um valor compreendido entre 6 e 7 designa-se por ldquometavanadatordquo e eacute
constituiacuteda pelas esspeacutecies V1 V2 V4 e V5 (Ref) Acidificando a soluccedilatildeo de metavanadato por
adiccedilatildeo de HCl ateacute um valor de pH proacuteximo de 4 observa-se uma mudanccedila de cor de amarelo
claro para laranja indicativa da formaccedilatildeo da espeacutecie decameacuterica de vanadato De seguida e
imediatamente antes da realizaccedilatildeo dos ensaios descritos o valor do pH da soluccedilatildeo de
decavanadato foi aumentado para cerca de 75 por adiccedilatildeo de NaOH A soluccedilatildeo (2) designada
por ldquometavanadatordquo natildeo foi posteriormente alterada ateacute agrave sua adiccedilatildeo para a realizaccedilatildeo dos ensaios
As soluccedilotildees foram armazenadas a 4ordmC e mantidas em gelo aquando da sua utilizaccedilatildeo
A soluccedilatildeo de V(IV) foi preparada em condiccedilotildees anaeroacutebias por diluiccedilatildeo de sulfato de vanadil
(VOSO4) para uma concentraccedilatildeo final de 5 mM em H2O MilliQ saturada com N2 gasoso
352Anaacutelise por espectroscopia de UV-Vis
A concentraccedilatildeo da espeacutecie decameacuterica em soluccedilatildeo depende do periacuteodo de tempo apoacutes a diluiccedilatildeo
no meio de reacccedilatildeo apropriado (Ref) A velocidade de desoligomerizaccedilatildeo da espeacutecie decameacuterica
pode ser analisada por espectroscopia de absorccedilatildeo na regiatildeo UVVis uma vez que esta espeacutecie
apresenta duas bandas caracteriacutesticas na regiatildeo do visiacutevel (a 358 e 400 nm) que natildeo se verifica
em quaisquer outras espeacutecies oligomeacutericas As bandas supramencionada sapresentam-se bem
22
Meio de reacccedilatildeo 3125 mM Tris-HCl pH 71 375 mM MgCl2
48
resolvidas atraveacutes da obtenccedilatildeo da derivada de 2ordf ordem do espectro apresentando a forma de 2
vales com miacutenimos centrados nos comprimentos de onda supracitados com dependecircncia linear
relativamente agrave concentraccedilatildeo de decavanadato total (Ramos et al 2006)
Assim a estabilidade da espeacutecie decameacuterica apoacutes diluiccedilatildeo da soluccedilatildeo de decavanadato no meio de
reacccedilatildeo dos ensaios (tampatildeo D) para uma concentraccedilatildeo final de 1 mM em vanadato total foi
caracterizada atraveacutes da leitura da Abs400 em funccedilatildeo do tempo tendo-se determinado qual a
ordem de reacccedilatildeo e o tempo de meia-vida da mesma Com base em estudos anteriores acerca da
estabilidade do V10 na presenccedila de diferentes proteiacutenas (Ramos et al 2006) determinou-se
igualmente o tempo de meia vida apoacutes a adiccedilatildeo de 20 microM de S1 de miosina ao meio reaccional
353 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
Os espectros de RMN do isoacutetopo 51
V foram realizados num espectroacutemetro Bruker AM-400 MHz
equipado com uma sonda multinuclear inversa de 5 mm Em tubos de RMN de 5 mm de diacircmetro
preparou-se soluccedilotildees de 500 L de deca- ou metavanadato correspondentes a um intervalo de
concentraccedilotildees de 1 a 10 mM suplementadas com 10 D2O Os espectros foram adquiridos agrave
temperatura ambiente com uma largura espectral de 23 923 Hz um tempo de aquisiccedilatildeo de 0086
s tempo de relaxaccedilatildeo de 001 s e uma sequecircncia de pulsos a 90ordm Utilizou-se a transformada de
Fourier com alargamento das bandas de 20Hz para o processamento dos espectros Os desvios
quiacutemicos das ressonacircncias de 51
V foram obtidos em relaccedilatildeo agrave espeacutecie VOCl3cuja frequecircncia de
ressonacircncia eacute considerada o padratildeo sendo todas as outras referenciadas a partir desta em ppm
As aacutereas dos sinais do RMN correspondentes agraves espeacutecies oligomeacutericas presentes foram obtidas
por integraccedilatildeo dos sinais As concentraccedilotildees relativas das espeacutecies foram calculadas com recurso agrave
equaccedilatildeo
onde A[Vx] corresponde agrave aacuterea referente agrave ressonacircncia da espeacutecie Vx com nordm de agregaccedilatildeo x
A[V]total agrave soma total das aacutereas medidas e [V] a concentraccedilatildeo total de vanaacutedio no ensaio
A teacutecnica de 51
V-RMN foi anteriormente aplicada em estudos de ligaccedilatildeo com S1 de miosina
(Ref) tendo sido demostrado que entre as espeacutecies V1 V2 V4 e V5 a espeacutecie tetrameacuterica possui a
maior afinidade para a miosina (Cremo et al) O desaparecimento ou alargamento dos sinais pode
implicar a ligaccedilatildeo de um dos oxovanadatos laacutebeis agrave miosina sem indicaccedilatildeo de qual uma vez que
49
se processa um equiliacutebrio entre eles (Crans 1990) Assim e tendo em conta as indicaccedilotildees de que
a espeacutecie V10 eacute distinta das restantes no que diz respeito agrave magnitude da interacccedilatildeo com a miosina
e inibiccedilatildeo da actividade ATPaacutesica estimulada por actina (Aureliano 2000 Tiago et al 2002)
torna-se fundamental investigar a interacccedilatildeo das diferentes espeacutecies de vanadato com o S1 de
miosina atraveacutes da teacutecnica de 51
V-RMN
Para tal obtiveram-se espectros de 51
V-RMN resultantes da titulaccedilatildeo de uma soluccedilatildeo 5 mM de
decavanadato total com concentraccedilotildees crescentes de S1 de miosina (0 5 10 15 e 20 microM) em
tampatildeo D Realizou-se ainda ensaios na presenccedila de 2 mM de decavanadato e 2 mM de ATP na
presenccedila e na ausecircncia de S1 de miosina de modo a verificar o efeito que estes ligandos naturais
da enzima teria na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o subfragmento-1
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
Os grupos sulfidrilo (-SH) das cisteiacutenas satildeo resiacuteduos de aminoaacutecidos extremamente sensiacuteveis agrave
oxidaccedilatildeo resultando na sua oxidaccedilatildeo (Dalle-Donne et al 1999) Logo a quantificaccedilatildeo destes
resiacuteduos eacute bastante importante nos estudos de interacccedilatildeo com vanaacutedio jaacute que os resiacuteduos de Cys
com ecircnfase para o SH1 e o SH2 satildeo bastante importantes na monitorizaccedilatildeo das modificaccedilotildees
funcionais da regiatildeo da cabeccedila
O procedimento seguidamente apresentado foi originalmente descrito por Gutieacuterrez-Merino
(1987)) Consiste na reacccedilatildeo entre os grupos ndashSH natildeo oxidados das proteiacutenas e o reagente de
Ellman (DTNB -55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico)) - formando-se o aniatildeo CNT (aniatildeo 3-
carboxilato-4-nitrotiofenolato) numa estequiometria de 11 Este apresenta um maacuteximo de
absorvecircncia a 412 nm (Robyt 1990) Assim apoacutes a incubaccedilatildeo da proteiacutena na presenccedila do metal
a quantidade de cisteiacutenas reduzidas por proteiacutena determina-se atraveacutes dos valores de Abs412
Deste modo colocou-se num tubo de Eppendorf o volume apropriado de S1 dialisado para que a
concentraccedilatildeo no ensaio fosse de 1 mgmL Adicionou-se o volume apropriado de decavanadato
para que a sua concentraccedilatildeo fosse de 0 10 50 100 250 500 ou 1000 M Perfez-se com soluccedilatildeo
D ateacute um volume final de 1 mL e incubou-se a 25ordmC por 20 minutos Apoacutes este periacuteodo colocou-
se 780 L da mistura reaccional numa cuvette de quartzo e adicionou-se 220 L de soluccedilatildeo K23
23
Tampatildeo K Tampatildeo fosfatohidrogenofosfato de soacutedio 025 M pH 72
50
Adicionou-se 15 L de DTNB (em soluccedilatildeo K [DTNB]stock=100 mM) ao meio reaccional e
registou-se o valor de absorvecircncia medido equivalente agraves cisteiacutenas ldquoraacutepidasrdquo (SH1 e SH2
expostos ao solvente) Adicionou-se 001 g de SDS que provoca a desnaturaccedilatildeo da proteiacutena e
consequentemente a exposiccedilatildeo de todos os resiacuteduos de cisteiacutenas que se encontravam ateacute aiacute
inacessiacuteveis no interior da proteiacutena (cisteiacutenas totais) Estes resiacuteduos podem assim reagir com o
DTNB
Estimou-se o nuacutemero de resiacuteduos de cisteiacutenas em todas as amostras com recurso ao coeficiente de
extinccedilatildeo molar () do DTNB (ver ponto 3 dos Anexos) que foi determinado atraveacutes da titulaccedilatildeo
de L-Cisteiacutena com intervalo de concentraccedilotildees compreendidos entre 0 e 50 microM na presenccedila de
25microM de DTNB em tampatildeo D Os valores de absorvecircncia foram obtidos para um comprimento de
onda de 412 nm
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
3621 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
A teacutecnica mais eficiente de anaacutelise da interacccedilatildeo entre o iatildeo vanadilo e proteiacutenas como a
calmodulina e a transferrina entre outras permite a obtenccedilatildeo raacutepida de informaccedilatildeo no que toca agrave
conformaccedilatildeo e amplitude da referida interacccedilatildeo
Os espectros de Ressonacircncia Paramagneacutetica Electroacutenica (RPE) da espeacutecie paramagneacutetica V(IV)
foram realizados a 70K num espectroacutemetro Bruker EMX-300 X-band Todas as soluccedilotildees
utilizadas e amostras foram preparadas em anaerobiose numa cacircmara de luvas de modo a evitar a
reduccedilatildeo de V(IV) a V(V) Em tubos capilares com 02 mL de amostra manteve-se a concentraccedilatildeo
de S1 a 20 M e variou-se a concentraccedilatildeo de vanadilo (0 25 5 10 15 25 50 75 100 150
200 500 e 1000 M [VOSO4]stock=5 mM) em tampatildeo D Apoacutes obtenccedilatildeo dos espectros estes
foram tratados atraveacutes do software SimFonia e WinEPR tendo-se obtido um graacutefico com a
variaccedilatildeo da intensidade do sinal de RPE em funccedilatildeo da concentraccedilatildeo de sulfato de vanadilo
presente Realizaram-se ainda simulaccedilotildees de espectros obtidos com 20 microM de S1 e 1000 microM de
V(IV) com vista agrave obtenccedilatildeo dos ligandos provaacuteveis do vanadilo atraveacutes da anaacutelise da constante
de acoplamento hiperfina (A) e do factor g (g)
4 Conclusotildees gerais
Apesar de jaacute ter sido demonstrado que o vanaacutedio inibe a Na+K
+-ATPase da miosina a funccedilatildeo
deste elemento no tecido muacutesculo esqueleacutetico continua largamente incompreendida A enorme
complexidade quiacutemica deste metal especialmente no que concerne agrave oligomerizaccedilatildeo e reacccedilotildees
51
redox constitui um obstaacuteculo agrave compreensatildeo do comportamento do vanaacutedio nos organismos
vivos Assim a caracterizaccedilatildeo da quiacutemica deste elemento eacute de grande utilidade antes de iniciar o
estudo da sua interacccedilatildeo com o presente sistema
A anaacutelise por espectroscopias de 51
V-RMN e de UV-Vis de soluccedilotildees de vanadato diluiacutedas nos
meios de reacccedilatildeo utilizados no presente relatoacuterio (10 mM HEPES pH 70 25 mM KCl) mostrou
claramente que a espeacutecie decameacuterica (V10) possui um tempo de vida suficientemente longo para a
determinaccedilatildeo do seu efeito isolado ao contraacuterio do que se verifica com as espeacutecies mono- di-
tetra- e pentameacutericas (V1 V2 V4 e V5) Por outro lado a anaacutelise verificou ainda que apoacutes diluiccedilatildeo
das soluccedilotildees de decavanadato no meio de reacccedilatildeo coexistem as espeacutecies V10 e V1 pelo que os
efeitos putativos desta uacuteltima deveratildeo ser subtraiacutedos aquando da monitorizaccedilatildeo dos efeitos da
espeacutecie decameacuterica
O vanaacutedio (V) (vanadato) inibe as enzimas que catalizam reacccedilotildees de transferecircncia de grupos
fosfato como a ATPase da miosina [Goodno 1979] devido agrave sua analogia estrutural com o iatildeo
fosfato Na sua utilizaccedilatildeo como inibidor enzimaacutetico natildeo eacute tida em conta a contribuiccedilatildeo individual
das espeacutecies oligomeacutericas considerando-se normalmente a espeacutecie V1 como inibidora efectiva
[Goodno e Taylor 1982] No presente relatoacuterio verificou-se por espectroscopia de 51
V-RMN que
outros oligoacutemeros de vanadato interagem com o S1 de miosina tendo as espeacutecies V10 e V4 os
maiores niacuteveis de afinidade A sua interacccedilatildeo envolve forccedilas electrostaacuteticas e eacute afectada
negativamente por ligandos naturais de enzima como eacute o caso do ATP
A exposiccedilatildeo do S1 de miosina (005 mgmL) a decavanadato conduz agrave inibiccedilatildeo do capacidade da
proteiacutena de estimular a actividade Mg2+
-ATPaacutesica com um valor de IC50 de 266 plusmn 07 M
decavanadato sendo o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM
decavanadato (80) Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM decavanadato sendo
o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM decavanadato (60)
exercendo um efeito de impedimento agrave ligaccedilatildeo entre o vanaacutedio e o S1
A uma concentraccedilatildeo de 27 microM decavanadato verifica-se a oxidaccedilatildeo de um dos resiacuteduos de Cys
de S1 de miosina seguida pela oxidaccedilatildeo gradual de outro resiacuteduo de Cys ateacute agrave concentraccedilatildeo de
1000 microM Ambos os resiacuteduos oxidados natildeo se encontravam expostos ao solvente utilizado A
oxidaccedilatildeo dos resiacuteduos permite sugerir que esta modificaccedilatildeo oxidativa sofrida pelo S1 contribui
para a diminuiccedilatildeo da sua actividade de ATPase
52
Por espectroscopia de RPE revelaram-se os possiacuteveis ligandos equatoriais do vanaacutedio aquando da
sua interacccedilatildeo com o S1 de miosina como sendo duas moleacuteculas de aacutegua um ligando contendo o
grupo R-OH e outro contendo o grupo R=NR
Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados de
oxidaccedilatildeo (IV) e (V) no S1 de miosina para concentraccedilotildees facilmente alcanccedilaacuteveis no ambiente
terrestre
53
5 Estudos futuros
No sentido do melhor esclarecimento das alteraccedilotildees moleculares e estruturais que ocorrem no
subfragmento 1 quando exposta ao vanaacutedio que conduzem a uma diminuiccedilatildeo da sua funccedilatildeo
sugere-se
- Quantificaccedilatildeo das cisteiacutenas reduzidas de S1 na presenccedila de metavanadato
- Determinaccedilatildeo das actividades Ca2+
- e K+-ATPaacutesica do S1 de miosina na presenccedila de
decavanadato
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina estimulada por actina
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina na presenccedila de
metavanadato
- Realizaccedilatildeo de RMN 2D com variaccedilatildeo das concentraccedilotildees das soluccedilotildees de decavanadato e
metavanadato na presenccedila de S1
-Mutageacutenese dos resiacuteduos de Cys doS1 de miosina de modo a verificar quais os resiacuteduos que
sofrem oxidaccedilatildeo na presenccedila de decavanadato
- Determinaccedilatildeo por calorimetria (DSC Differential Scanning Calorimetry) da variaccedilatildeo da
entalpia (∆H) e da temperatura de desnaturaccedilatildeo do S1 de miosina na presenccedila de vanaacutedio uma
vez que estas propriedades caracteriacutesticas de cada proteiacutena satildeo alteradas apoacutes ocorrecircncia de
modificaccedilotildees estruturais
- Anaacutelise por SDS-PAGE de S1 de miosina apoacutes tratamento com vanaacutedio de modo a determinar
quais os polipeacuteptidos eventualmente formados apoacutes clivagem da proteiacutena
54
6 Referecircncias bibliograacuteficas
- Sellers J Myosins 2nd Edition Oxford University Press 1999
- L Stryer inBiochemistry (Ed WH Freeman 2005 pp
Refordfas Materiais e Meacutetodos
[A] S S Margossian SLowey Methods Enzymol1982 85 55-71
[B] JD Pardue JA Spudich Methods Enzymol 1982 85 164-181
[C] M Bradford Anal Biochem1976 72 248-254
[D] H H Taussky E Shorr J Biol Chem 1953 202 675-685
[E] C R Bagshaw in Outline Studies in Biology ndash Muscle Contraction Chapman and Hall Ed
1982
[F] A Cuenda et al Eur J Biochem1990 194 663-670
55
Anexos
1 Determinaccedilatildeo da recta de calibraccedilatildeo meacutetodo de Bradford
Figura1ndashRepresentaccedilatildeo graacutefica da recta de calibraccedilatildeo (Abs595 nm vs [BSA]) utilizada para o meacutetodo de Bradford
2 Determinaccedilatildeo da recta calibraccedilatildeo para a electroforese em condiccedilotildees desnaturantes
Figura 2 ndash Representaccedilatildeo graacutefica da recta de calibraccedilatildeo (log(Peso Molecular) vs Rf dos marcadores de peso
molecular) onde Rf indica determinada atraveacutes do padratildeo de peso molecular
(poccedilo (a))
3Caacutelculo do coeficiente de extinccedilatildeo molar () para a quantificaccedilatildeo de cisteiacutenas reduzidas
atraveacutes do meacutetodo de Ellman
Traccedilou-se a recta de Abs412 nm meacutedia vs [L-Cisteiacutena] de modo a determinar o valor do coeficiente
de extinccedilatildeo molar definido como o declive daquela recta Assim =8400 M-1
cm-1
como
podemos verificar na Fig 3
A595 nm = 00546[BSA] + 00209 Rsup2 = 099051
Ab
s 59
5 n
m
[BSA] (mgmL)
log PM = -1177 Rf + 2247 Rsup2 = 098957
log
PM
Mobilidade relativa (Rf)
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero

2
Abreviaturas e Siacutembolos
Absx ndash Absorvecircncia ao comprimento de onda x (em nm)
ADP - Adenosina 5rsquo-difosfato
ATP ndash Adenosina 5rsquo-trifosfato
ATPase ndash Adenosina 5rsquo-trifosfatase
BSA ndash Albumina de soro bovino
-Me - -Mercaptoetanol
CNT ndash Aniatildeo 3-carboxilato-4-nitrotiofenolato
Cys ndash Cisteiacutena
DTNB ndash 55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico)
- Coeficiente de extinccedilatildeo (molar)
EDTA ndash Aacutecido etilenodiamino tetra-aceacutetico
ELC ndash Cadeia leve essencial de miosina
F-actina ndash Forma filamentosa da actina
g ndash Aceleraccedilatildeo da gravidade
G-actina ndash Forma monomeacuterica da actina
h ndash Horas
HEPES ndash Aacutecido 4-(2-hidroxiletil)-1-piperazineetanosulfoacutenico
HMM ndash Meromiosina pesada
IC50 ndash Concentraccedilatildeo inibitoacuteria 50
LDH ndash Lactato desidrogenase
Miosina-II ndash Miosina do tipo II
Met - Metionina
NAD+ ndash Nicotinamida adenina dinucleoacutetido (forma oxidada)
NADH ndash Nicotinamida adenina dinucleoacutetido (forma reduzida)
PDB ndash Protein Data Bank
Pi ndash Fosfato inorgacircnico
PEP ndash Fosfoenolpiruvato
PK ndash Piruvato cinase
PM ndash Peso Molecular
3
ppb ndash Partes por biliatildeo (1 gL)
ppm ndash Partes por milhatildeo (1 mgL)
PSA ndash persulfato de amoacutenio
Rf - Mobilidade relativa
RLC ndash Cadeia leve regulatoacuteria de miosina
RMN ndash Ressonacircncia Magneacutetica Nuclear
ROS ndash Espeacutecies reactivas de oxigeacutenio
RPE ndash Ressonacircncia Paramagneacutetica Electroacutenica
S1 ndash Subfragmento-1 da miosina II
SDS ndash Dodecilo sulfato de soacutedio
SDS-PAGE ndash Electroforese em gel de poliacrilamida com SDS (condiccedilotildees desnaturantes)
SH1 ndash Grupo tiol mais reactivo da miosina (resiacuteduo Cys-707)
SH2 ndash segundo grupo tiol mais reactivo da miosina (resiacuteduo Cys-697)
SNC ndash Sistema Nervoso Central
TCA ndash Aacutecido tricloroaceacutetico
TEMED ndash NNNrsquoNrsquo-tetrametiletilenodiamina
Tris ndash 2-amino-2-hidroximetil-13-propanodiol
Trp ndash triptofano
Tyr ndash tirosina
UV ndash gama ultravioleta do espectro electromagneacutetico
Vis ndash gama visiacutevel do espectro electromagneacutetico
4
Resumo
A exposiccedilatildeo a vanaacutedio conduz a uma diminuiccedilatildeo na actividade ATPaacutesica da miosina e provoca a
oxidaccedilatildeo dos grupos sulfidrilo desta proteiacutena No entanto natildeo estaacute integralmente esclarecido
quais os efeitos especiacuteficos que ocorrem na miosina quando exposta ao vanaacutedio Nesse sentido
interessou determinar in vitro quais as alteraccedilotildees estruturais funcionais e moleculares ocorridas
no Subfragmento-1 de miosina de muacutesculo esqueleacutetico de coelho apoacutes a exposiccedilatildeo croacutenica a
soluccedilotildees de metavanadato decavanadato e vanadilo Verificou-se que a exposiccedilatildeo de S1 (005
mgmL) a decavanadato inibe a sua actividade Mg2+
-ATPaacutesica sendo o valor de IC50 para esta
inibiccedilatildeo de 266 plusmn 07 microM decavanadato Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM
decavanadato A perda de funcionalidade proteica poderaacute dever-se agrave oxidaccedilatildeo de 2 resiacuteduos de
Cys de S1 natildeo expostos ao solvente apoacutes exposiccedilatildeo a 1000 microM de decavanadato Verificou-se
que as espeacutecies oligomeacutericas de vanaacutedio com maior afinidade para o S1 satildeo o decavanadato e o
tetravanadato Revelaram-se os possiacuteveis ligandos do vanaacutedio no S1 atraveacutes de espectroscopia de
RPE Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados
de oxidaccedilatildeo (IV) e (V) na estrutura e funccedilatildeo do S1 em concentraccedilotildees facilmente alcanccedilaacuteveis
daquele metal no ambiente terrestre
Summary
The exposure to vanadium leads to a decrease of the myosin ATPase activity and provokes the
oxidation of its sulfydryl groups However the specific effects of vanadium on myosin are not yet
fully understood Consequently the objectives of the present work were to establish the structural
funtional and molecular changes suffered by rabbit myosin subfragment-1 after cronic exposure to
metavanadate decavanadate and vanadyl solutions It was verified that the exposure of S1 (005
mgmL) to decavanadate results in the inhibition of its Mg2+
-ATPase activity resulting in the
IC50 value 266 plusmn 07 microM decavanadate In the presence of actin the IC50 value was 28 plusmn 1 microM
decavanadate It is suggested that the loss of protein function may be due to the oxidation of 2
core cysteine residues after exposure to 1000 microM decavanadate It was also observed that the
decavanadate and tetravanadate species show the highest affinity for S1 Lastly the possible S1
ligands to vanadium were revealed through EPR spectroscopy Globally the obtained results
point to significant effect of V(V) and V(IV) states on S1 structure and function for easily
reachable environmental vanadium concentrations
5
Iacutendice
1 Introduccedilatildeo
11 Muacutesculo esqueleacutetico
111 Miosina
112 Actina
113 Mecanismos moleculares da contracccedilatildeo muscular
1131 Regulaccedilatildeo da contracccedilatildeo muscular
12 Vanaacutedio
121 Propriedades quiacutemicas
1211 Vanaacutedio (IV)
1212 Vanaacutedio (V)
122 Metabolismo e efeito na actividade enzimaacutetica
13 Vanaacutedio e muacutesculo esqueleacutetico
14 Objectivos do projecto
2 Discussatildeo de Resultados
21 Caracterizaccedilatildeo das proteiacutenas isoladas
211 Anaacutelise do estado de pureza
212 Determinaccedilatildeo da concentraccedilatildeo proteica
2121 Subfragmento-1 de miosina
2122 Actina
22 Mediccedilatildeo da actividade enzimaacutetica de S1 de miosina
221 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de Fiske-
Subarow modificado
222 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
23 Preparaccedilatildeo e caracterizaccedilatildeo das soluccedilotildees de vanaacutedio
231 Anaacutelise da composiccedilatildeo das soluccedilotildees de vanadato por espectroscopia de 51V-RMN efeito da
concentraccedilatildeo da espeacutecie
232 Estabilidade das soluccedilotildees de decavanadato anaacutelise por espectroscopia de absorccedilatildeo UVVis
24 Interacccedilatildeo de S1 com vanaacutedio(V)
241 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
242 Actividade Mg2+
-ATPaacutesica de S1 de miosina na presenccedila de decanadato
243 Quantificaccedilatildeo de cisteiacutenas reduzidas pelo meacutetodo de Ellman
25 Interacccedilatildeo de S1 com vanaacutedio(IV)
251 Anaacutelise por espectroscopia de Ressonacircncia Paramagneacutetica Electroacutenica
3 Procedimento Experimental
31 Reagentes
32 Meacutetodos
321 Isolamento e purificaccedilatildeo das proteiacutenas de muacutesculo esqueleacutetico de coelho
3211 Purificaccedilatildeo da miosina preparaccedilatildeo de Subfragmento 1 por digestatildeo quimotriacutepsica da
miosina
32111 Purificaccedilatildeo da miosina
32112 Preparaccedilatildeo de Subfragmento-1 de miosina por digestatildeo quimotriacutepsica
3212 Isolamento e purificaccedilatildeo de actina
32121Obtenccedilatildeo do ldquopoacute de acetonardquo
32122 Purificaccedilatildeo da actina a partir de ldquopoacute de acetonardquo
331 Quantificaccedilatildeo das proteiacutenas isoladas
3311 Quantificaccedilatildeo pelo meacutetodo de Bradford
6
3312 Quantificaccedilatildeo pelo meacutetodo directo
332 Electroforese em condiccedilotildees desnaturantes (SDS-PAGE)
34 Mediccedilatildeo da actividade enzimaacutetica de S1
341 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de Fiske-
Subarow modificado
342 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
35 Preparaccedilatildeo e caracterizaccedilatildeo das soluccedilotildees de vanaacutedio por Espectroscopia UV-Visiacutevel e
Ressonacircncia Magneacutetica Nuclear
351 Preparaccedilatildeo das soluccedilotildees de vanadato
352 Anaacutelise por UV-Vis para determinaccedilatildeo da ordem da reacccedilatildeo
353 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
362 1 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
363 Interacccedilatildeo de S1 com V (V) por Ressonacircncia Magneacutetica Nuclear (RMN)
4 Conclusotildees gerais
5 Estudos futuros
6 Referecircncias bibliograacuteficas
7
1 Introduccedilatildeo
11 Muacutesculo esqueleacutetico
A capacidade de locomoccedilatildeo eacute uma das caracteriacutesticas fundamentais da maioria dos organismos
vivos A contracccedilatildeo muscular um dos fenoacutemenos que a torna possiacutevel apresenta grande
complexidade e tem vindo a ser utilizado como modelo para a compreensatildeo de sistemas menos
especializados de mobilidade celular [1]
No que diz respeito agrave sua fisiologia morfoloacutegica os muacutesculos dos vertebrados dividem-se em
lisos e estriados Estes uacuteltimos devem o seu nome agrave alternacircncia de zonas claras e escuras
observaacuteveis ao microscoacutepio electroacutenico de que resulta um aspecto estriado Os muacutesculos
estriados podem ainda ser divididos em duas subclasses muacutesculos esqueleacuteticos e cardiacuteacos [2]
consoante a forma celular (respectivamente longitudinal ou ramificada) ou tipo de contracccedilatildeo das
ceacutelulas que o compotildeem (respectivamente voluntaacuteria e involuntaacuteria) [3]
Os muacutesculos estriados esqueleacuteticos satildeo assim designados devido agrave sua localizaccedilatildeo no
organismo ndash ligados aos ossos da coluna vertical braccedilos e pernas Satildeo constituiacutedos por feixes de
ceacutelulas multinucleadas (miofibras) que contecircm miofibrilas paralelas de cerca de 1 m de
diacircmetro associadas agrave contracccedilatildeo voluntaacuteria raacutepida e forte em actividades de coordenaccedilatildeo
complexa [4] Cada feixe apresenta um diacircmetro compreendido entre 20 e 100 m e um
comprimento maacuteximo que pode atingir vaacuterios centiacutemetros (Fig 11)
Figura 11 ndash Organizaccedilatildeo hieraacuterquica geral do muacutesculo-esqueleacutetico (adaptado de Purves et al 2003 [3])
No muacutesculo-esqueleacutetico as zonas claras e escuras supramencionadas correspondem a zonas de
menor e maior densidade electroacutenica respectivamente As zonas claras constituem as bandas I (de
8
ldquoisotropiardquo ndash polarizaccedilatildeo uniforme em todas as direcccedilotildees) enquanto as zonas escuras constituem
as bandas A (de ldquoanisotropiardquo ndash polariza a luz visiacutevel) A banda I encontra-se dividida
transversalmente por uma linha escura ndash linha Z de densidade electroacutenica e refracccedilatildeo superiores
No interior de cada banda A existe uma zona transversal menos densa ndash zona H ndash rica em enzimas
importantes para o metabolismo celular No centro desta zona existe uma linha perpendicular
mais escura ndash linha M [2]
A unidade fundamental da contracccedilatildeo muscular eacute designada por sarcoacutemero (Fig 12) Encontra-se
repetida de forma regular ao longo das miofibrilas apresentando um comprimento de 25 a 3 m
aquando do relaxamento muscular diminuindo aquando da contracccedilatildeo muscular
Figura 12 ndash Representaccedilatildeo de fotomicrografia electroacutenica e esquemaacutetica do sarcoacutemero Os filamentos grossos e
finos satildeo maioritariamente constituiacutedos por miosina e actina respectivamente (Adaptado de Stryer 2002 [2])
As secccedilotildees de corte do sarcoacutemero datildeo conta da presenccedila de dois tipos de filamentos proteicos ndash os
filamentos grossos cujo diacircmetro eacute de 15 nm constituiacutedos sobretudo por miosina e os filamentos
finos com um diacircmetro de 9 nm constituiacutedos sobretudo por actina tropomiosina e pelo complexo
de troponina [2]
111 Miosina
Actualmente o termo miosina refere-se a uma superfamiacutelia diversa de motores moleculares
capazes de hidrolisar ATP e de se ligarem reversivelmente agrave actina A classificaccedilatildeo da
superfamiacutelia das miosinas nas 15 classes distintas faz-se atraveacutes de homologia entre as diferentes
sequecircncias [A] Eacute tambeacutem possiacutevel a classificaccedilatildeo por subclasses das quais se destaca a subclasse
4 proveniente do muacutesculo-esqueleacutetico e cardiacuteaco de vertebrados Este estudo focar-se-aacute apenas
nas miosinas de classe II (miosina-II) presente nos filamentos grossos do muacutesculo estriado
esqueleacutetico [B] e em ceacutelulas natildeo musculares A funccedilatildeo da miosina-II depende da sua localizaccedilatildeo
celular e tecidular
9
A miosina-II eacute constituiacuteda por duas cadeias polipeptiacutedicas idecircnticas denominadas cadeias pesadas
de miosina (HC) enroladas entre si e cujo peso molecular (PM) estaacute compreendido entre 171 e
241 kDa) Estruturalmente as HC dividem-se em 3 domiacutenios funcionais - cabeccedila pescoccedilo e
cauda como se pode observar na Fig 13 Agraves cabeccedilas ou zonas globulares estatildeo associados dois
pares de cadeias leves (peso molecular entre 15 e 30 kDa) que se dividem em dois tipos
essencial (ELC) e regulatoacuterio (RLC)
Figura 13 ndash Estrutura esquemaacutetica da moleacutecula de miosina-II Destacam-se as duas cabeccedilas globulares e a cauda
filamentosa ligadas entre si por uma zona flexiacutevel denominada pescoccedilo As cadeias leves (a verde e vermelho)
encontram-se associadas agraves cabeccedilas globulares
As cabeccedilas globulares contendo os locais de ligaccedilatildeo agrave actina e aos nucleoacutetidos mantecircm os seus
elementos estruturais baacutesicos conservados em todas as classes sendo estes constituiacutedos por 720-
80 aa A regiatildeo do pescoccedilo eacute constituiacuteda por uma longa heacutelice pertencente agrave cadeia pesada
que se encontra estabilizada atraveacutes da ligaccedilatildeo a subunidades de cadeia leve A estrutura do
pescoccedilo envolvida na ligaccedilatildeo agraves cadeias leves eacute denominada de motivo IQ cuja estrutura
consensual eacute IQXXXRGXXXR1 O pescoccedilo serve como extensatildeo do comprimento da cabeccedila de
modo a actuar como alavanca durante a contracccedilatildeo muscular [A]A regiatildeo da cauda ou acircncora
apresenta bastante divergecircncia no que respeita agrave sua sequecircncia estrutura e funccedilatildeo No entanto
pensa-se que a sua funccedilatildeo centrar-se-aacute na autoassociaccedilatildeo das vaacuterias moleacuteculas de miosina-II
atraveacutes da regiatildeo de coiled-coil Esta confere assimetria agrave moleacutecula e permite a formaccedilatildeo de
filamentos grossos capazes de translocar filamentos de actina [A]
Na regiatildeo da cabeccedila da miosina existem 2 regiotildees extremamente sensiacuteveis agrave proteoacutelise A
clivagem em ambos os locais resulta na produccedilatildeo de trecircs fragmentos um compreendendo o N-
terminal (fragmento 25 kDa) outro a regiatildeo central (50 kDa) e um terceiro o C-terminal (20 kDa)
(Fig 14) Estes fragmentos dissociam-se em condiccedilotildees desnaturantes e natildeo constituem domiacutenios
independentes da cabeccedila mas provavelmente loops flexiacuteveis uma vez que natildeo foram observadas
na estrutura cristalina Os vaacuterios fragmentos obtidos em diferentes condiccedilotildees foram cruciais para
a elucidaccedilatildeo das vaacuterias funccedilotildees da moleacutecula
A regiatildeo compreendida entre os fragmentos 25 e 50 kDa (loop 1) localiza-se directamente por
1 I = Isoleucina Q= Glutamina X = qualquer dos 20 aminoaacutecidos padratildeo R= Arginina G= Glicina
10
cima do local de ligaccedilatildeo ao nucleoacutetido A regiatildeo entre os fragmentos de 50 e 20 kDa (loop 2)
encontra-se perto da intersecccedilatildeo entre os domiacutenios da cabeccedila e do pescoccedilo e deveraacute interagir com
a actina uma vez que a ligaccedilatildeo desta a protege da proteoacutelise Na ausecircncia do loop 2 a miosina
liga mais fracamente agrave actina do que no caso da miosina intacta A sequecircncia primaacuteria de ambos
os loops apresenta conservaccedilatildeo reduzida sendo provaacutevel que sejam responsaacuteveis por muita da
diversidade das diferentes propriedades enzimaacuteticas presentes em diferentes isoformas
No fragmento de 20 kDa existem dois grupos sulfidrilo Cys-707 e -697 usualmente
denominados de SH1 e SH2 devido agrave ordem da sua reactividade quiacutemica Na presenccedila de
nucleoacutetidos pode ocorrer o ldquocross-linkingrdquo entre estes dois grupos incluindo a oxidaccedilatildeo a uma
ponte dissulfureto
Devido agrave insolubilidade da miosina-II nativa a estrateacutegia adoptada para os estudos com esta
proteiacutena eacute frequentemente a sua clivagem em fragmentos funcionais separaacuteveis Um dos locais de
clivagem ocorre tipicamente a 130 kDa do N-terminal produzindo a meromiosina pesada (HMM)
e a meromiosina leve (LMM) [C] A HMM eacute formada pelas 2 cabeccedilas e pescoccedilo e apresenta
propriedades semelhantes agrave miosina sem digestatildeo (ligaccedilatildeo e translocaccedilatildeo de actina e hidroacutelise de
MgATP)
A HMM pode ser novamente clivada atraveacutes de uma reacccedilatildeo de proteoacutelise produzindo os
subfragmentos 1 (S1) e 2 (S2)O S1 (90 kDa) eacute enzimaticamente activo sendo constituiacutedo pelos
domiacutenios da cabeccedila e do pescoccedilo e apresentando associaccedilatildeo agraves duas cadeias leves ou apenas agrave
ELC (Fig 14)Em alternativa o S1 pode ser produzido por clivagem directa da miosina intacta
formando-se tambeacutem um fragmento em forma de cilindro que consiste nas regiotildees S2 e LMM
(Ref)Utiliza-se extensivamente o S1 para estudos cineacuteticos e de ligaccedilatildeo agrave actina uma vez que a
sua solubilidade simplifica o planeamento da experiecircncia e a subsequente interpretaccedilatildeo dos dados
[A]
Figura 14- Estrutura do subfragmento-1 de miosina de muacutesculo esqueleacutetico com ecircnfase nos domiacutenios de 50 kDa
central (a vermelho) 25 kDa no N-terminal (a verde) e 20 kDa no C-terminal (a azul) As cadeias leves essenciais
encontram-se a amarelo e as cadeias leves regulatoacuterias a lilaacutes Os locais de ligaccedilatildeo agrave actina e ao ATP estatildeo
igualmente destacados (adaptado de Holmes e Geeves 1999 [D])
11
112 Actina
O termo ldquoactinardquo refere-se a uma famiacutelia extremamente conservada de proteiacutenas citoplasmaacuteticas
presente em todos os eucariotas Esta proteiacutena natildeo se encontra nos procariotas [E] A maioria das
actinas possui peso molecular proacuteximo de 42 kDa outra evidecircncia do seu grau elevado de
conservaccedilatildeo no que concerne agrave sequecircncia e ao nuacutemero de aminoaacutecidos Os eucariotas superiores
apresentam um grande nuacutemero de isoformas de actina que por sua vez se encontram codificados
por um ainda maior nuacutemero de genes (Ref) A expressatildeo das isoformas eacute diferencial consoante o
tecido e sua especializaccedilatildeo pelo que deveratildeo possuir funccedilotildees e mecanismos direccionais
diferentes (Ref) Foi ainda observado que os mRNA das diferentes isoformas apresentam uma
distribuiccedilatildeo heterogeacutenea nas ceacutelulas de forma semelhante aos seus produtos proteicos [F] Estes
dados suportam a hipoacutetese supramencionada do direccionamento das isoformas de actina para um
determinado local na ceacutelula de acordo com as suas funccedilotildees
A actina presente no citoesqueleto eacute responsaacutevel por processos como o mecanismo contraacutectil
fagocitose manutenccedilatildeo e alteraccedilatildeo da estrutura celular locomoccedilatildeo transporte de organelos
regulaccedilatildeo do transporte de solutos entre outras Nos eucariotas superiores estaacute ainda envolvida
no transporte de gracircnulos de pigmento retinoacuteide [G] transporte de gracircnulos no axoplasma [H] e
secreccedilatildeo entre outros [E] Actualmente a isoforma da actina (-actina) eacute a melhor estudada
encontrando-se envolvida no mecanismo contraacutectil [E]
Os filamentos de -actina tecircm cerca de 1m de comprimento e 80 Å Consistem em duas cadeias
de actina filamentosa (F-actina) organizados em heacutelice [I] (Holmes et al 1990) sobre os quais
se encontram outras proteiacutenas ndash a troponina (C I e T) e a tropomiosina (Azevedo 1999) (Figura
15) A troponina eacute filamentosa e localiza-se nas depressotildees da heacutelice de F-actina conferindo
rigidez aos filamentos finos Por sua vez a tropomiosina eacute essencial na regulaccedilatildeo do mecanismo
contraacutectil
Figura 15 ndash Modelo estrutural dos filamentos de F-actina contendo a tropomiosina e o complexo de troponina (I C
e T) (Adaptado de Susana Ramos 2005 [J])
12
A F-actina eacute formada atraveacutes da polimerizaccedilatildeo de subunidades globulares de actina (G-actina) A
G-actina eacute composta por 2 domiacutenios de tamanho semelhante separados pelos bolsos de ligaccedilatildeo(1)
de catiotildees divalentes e(2) de nucleoacutetidos [KLM] Estes por sua vez estatildeo subdivididos noutros
dois de menores dimensotildees (1-4) (Fig 16) cada um envolvido nas interacccedilotildees actina-actina ou
na formaccedilatildeo do topo de ligaccedilatildeo do nucleoacutetido As regiotildees N-terminal e C-terminal estatildeo proacuteximas
uma da outra no subdomiacutenio 1 [E]
Figura 16 ndash Estrutura do monoacutemero de actina Encontram-se representados os subdomiacutenios 1 a 4 e a vermelho a
moleacutecula de ATP junto do respectivo local de ligaccedilatildeo (Protein Data Bank ID 1j6z)
113 Mecanismos moleculares da contracccedilatildeo muscular
A contracccedilatildeo muscular ocorre atraveacutes do deslizamento dos filamentos finos (de actina) por entre
os filamentos grossos (de miosina) conseguida pela reacccedilatildeo de hidroacutelise de ATP e regulada pela
tropomiosina e pelo complexo de troponina em resposta a impulsos nervosos Este mecanismo foi
inicialmente proposto por Huxley-Niedergerke [N] e Huxley-Hanson [O] ambos em 1954 A base
experimental de ambos os estudos foi a inexistecircncia de variaccedilatildeo do tamanho dos filamentos
durante a contracccedilatildeo A banda A apresenta sempre as mesmas dimensotildees por outro lado a banda
I a zona H diminuem consideravelmente aquando da contracccedilatildeo (Figs 12 e 17)
Figura 17 ndash Modelo da contracccedilatildeo muscular ndash deslizamento dos filamentos finos (azuis) relativamente aos
filamentos grossos (rosa) (Adaptado de Stryer 2002 [2])
13
Na ausecircncia de ATP e supondo a ausecircncia de inibiccedilatildeo da interacccedilatildeo actina-miosina as proteiacutenas
encontram-se fortemente associadas formando o complexo actomiosina (AM) (Figura 15)
Nestas condiccedilotildees as cabeccedilas de miosina formam um acircngulo de 45ordm com o eixo dos filamentos
finos de actina [P]
Na presenccedila de ATP o complexo AM dissocia-se jaacute que a afinidade do complexo M-ATP para a
actina eacute 105 vezes inferior agrave afinidade da miosina para a actina O ATP liga-se agrave miosina
ocorrendo seguidamente a reacccedilatildeo de hidroacutelise de ATP Os produtos daiacute resultantes ndash ADP e Pi -
permanecem ligados agrave miosina formando o complexo M-ADP-Pi Com a ligaccedilatildeo subsequente agrave
actina forma-se o complexo AM-ADP-Pi no qual as cabeccedilas de miosina apresentam um acircngulo
recto com o eixo do muacutesculo [P] Neste ponto daacute-se a libertaccedilatildeo progressiva de Pi e ADP Eacute
provaacutevel que a energia proveniente da hidroacutelise do ATP provoque uma alteraccedilatildeo do acircngulo de
ligaccedilatildeo das cabeccedilas de miosina ao filamento de actina passando novamente de 90ordm para 45ordm [P]
Esta alteraccedilatildeo conformacional provoca o deslizamento do filamento de actina para o centro do
sarcoacutemero cujo encurtamento promove a contracccedilatildeo das miofibrilas O processo supracitado
ocorre em cerca de 1ms[Q] e encontra-se resumido na Fig 18
Figura 18 ndash Esquema cineacutetico da hidroacutelise de ATP pela miosina (Adaptado de Sellers 1999 [A])
1131 Regulaccedilatildeo da contracccedilatildeo muscular
O mecanismo pelo qual se processa a regulaccedilatildeo muscular foi inicialmente descrito por estudos
distintos de Haselgrove Huxley e Parrey-Squire todos em 1973 [RST] Aleacutem da implicaccedilatildeo
directa da tropomiosina e do complexo de troponina a concentraccedilatildeo de Ca2+
no muacutesculo em
relaxamento eacute bastante reduzida (10-7
M) aumentando notavelmente aquando da contracccedilatildeo (10-5
-
10-3
M)
Resumidamente quando ocorre a excitaccedilatildeo do muacutesculo esqueleacutetico a despolarizaccedilatildeo da
membrana plasmaacutetica eacute difundida ateacute ao interior do tecido atraveacutes dos tuacutebulos transversais
(invaginaccedilotildees da membrana plasmaacutetica da fibra muscular) A despolarizaccedilatildeo provoca a abertura
dos canais ioacutenicos de Ca2+
do retiacuteculo sarcoplasmaacutetico adjacente Como a concentraccedilatildeo de Ca2+
no interior do organelo eacute muito elevada o caacutelcio passa raacutepida e uniformemente para o citosol
14
Assim a regulaccedilatildeo da contracccedilatildeo ocorre com coordenaccedilatildeo e rapidez na totalidade das miofibrilas
do tecido muscular (Ref)
Em repouso a troponina I encontra-se posicionada de tal forma que a proteiacutena adjacente
troponina C impede estruturalmente a ligaccedilatildeo das cabeccedilas da miosina agrave actina (acircngulo =50ordm) O
aumento da concentraccedilatildeo intracelular de Ca2+
provoca um aumento da sua disponibilidade para
ligar agrave troponina C (Fig 19)
Figura 19ndash Regulaccedilatildeo do mecanismo contraacutectil no muacutesculo esqueleacutetico pelo complexo de troponina
Como consequecircncia a troponina C altera a sua conformaccedilatildeo (=70ordm) associando-se agrave troponina I
que deixa de inibir a ligaccedilatildeo das pontes transversais A alteraccedilatildeo anterior eacute transmitida agrave
tropomiosina atraveacutes da troponina T adjacente agrave C o que provoca o deslocamento da
tropomiosina na direcccedilatildeo do interior do sulco do filamento de actina Assim eacute possiacutevel a
interacccedilatildeo eficaz entre actina e miosina A Ca2+
-ATPase do retiacuteculo sarcoplasmaacutetico provoca a
reacumulaccedilatildeo do iatildeo caacutelcio no organelo impedindo o prolongamento desnecessaacuterio da contracccedilatildeo
(Refordf)
12 Vanaacutedio
O vanaacutedio eacute um metal de transiccedilatildeo pertencente ao grupo VB da tabela perioacutedica Foi
primeiramente descoberto em 1801 por Andreacutes Manuel del Riacuteo que concluiu erradamente que era
idecircntico ao croacutemio e definitivamente por Nils Gabriel Svenson em 1831 Recentemente foram
descobertas importantes funccedilotildees bioloacutegicas deste elemento provavelmente essencial para os
humanos de que se destaca no contexto do presente relatoacuterio a possibilidade de actuar como
anaacutelogo do grupo fosfato e a inibiccedilatildeo de enzimas in vitro como eacute o caso da actividade ATPaacutesica
da miosina
Eacute um metal soacutelido agrave temperatura ambiente que se encontra amplamente distribuiacutedo nos vaacuterios
ecossistemas em dois isoacutetopos naturais ndash 50
V e 51
V ndash raramente excedendo 01 partes por biliatildeo
15
(ppb) Estaacute associado a cerca de 70 minerais diferentes na natureza (dos quais se destacam o
uracircnio cobre chumbo zinco fosfato e titacircnio ferroso) especialmente na forma de vanadato ou
pentoacutexidos de vanaacutedio O vanaacutedio eacute um dos principais constituintes da mateacuteria orgacircnica foacutessil
com especial ecircnfase para o carvatildeo oacuteleo de crude e foacutesseis carbonaacuteceos [UV] bem como em
alimentos consumidos pelo Homem (salsa pimenta preta cogumelos marisco vegetais cereais
gorduras e oacuteleos) [W] Estes constituem a maior fonte de exposiccedilatildeo ao vanaacutedio da maioria da
populaccedilatildeo humana [X]
Normalmente a entrada do vanaacutedio no organismo ocorre por inalaccedilatildeo ou em menores
quantidades por absorccedilatildeo pelo tracto gastrointestinal ou pela pele O seu armazenamento centra-
se no fiacutegado rins ceacuterebro coraccedilatildeo muacutesculos e ossos No entanto a retenccedilatildeo de vanaacutedio no
organismo humano eacute reduzida ndash em condiccedilotildees normais cerca de 100 M (em muitos tecidos a
quantidade eacute inferior a 10 ngg de tecido) [Y]
Actualmente natildeo existem provas de deficiecircncia nutricional de vanaacutedio em humanos ou de
enzimas que contenham vanaacutedio em animais superiores No entanto vaacuterios estudos relacionam a
ocorrecircncia de deficiecircncia em galinhas e ratos devido a uma alimentaccedilatildeo pobre em vanaacutedio Os
sintomas relatados incluem retardamento do crescimento malformaccedilotildees oacutesseas perturbaccedilotildees no
metabolismo lipiacutedico e estagnaccedilatildeo reprodutiva entre outros [Z AA AB] Os efeitos fisioloacutegicos
do vanaacutedio podem depender do tipo de composto ou de oligoacutemero uma vez que consoante as
suas estabilidade labilidade e nuacutemero de coordenaccedilatildeo as respostas seratildeo diferenciadas ao niacutevel
dos organismos vivos
Por outro lado a toxicidade do vanaacutedio encontra-se bem caracterizada existindo sintomas
descritos em vaacuterios mamiacuteferos (inclusive o Homem) que incluem diarreias catildeibras irritaccedilatildeo da
pele e mucosas palpitaccedilotildees cardiacuteacas anemia pneumonia bronquite perturbaccedilotildees no Sistema
Nervoso Central (SNC) entre outros (Ref) A toxicidade varia geralmente com a natureza do
composto com aumento proporcional agrave sua valecircncia Verificou-se que a forma de entrada do
vanaacutedio tem tambeacutem efeito na sua toxicidade ndash os maiores niacuteveis verificaram-se por injecccedilatildeo
intravenosa da amostra sendo progressivamente inferiores por inalaccedilatildeo e finalmente por
ingestatildeo [X AA] A ampla utilizaccedilatildeo dos compostos de vanaacutedio na induacutestria quiacutemica bem como
as elevadas quantidades de vanaacutedio libertadas atraveacutes da combustatildeo dos oacuteleos de crude
contribuem consideravelmente para o aumento da biodisponibilidade do vanaacutedio Como tal o
interesse neste elemento e nos problemas ambientais por ele causados tem vindo a aumentar
significativamente [V]
121 Propriedades quiacutemicas
16
A quiacutemica do vanaacutedio eacute extraordinariamente complexa pelo que originou vaacuterias interpretaccedilotildees
incorrectas desde a sua descoberta No entanto as suas propriedades dependem em uacuteltima
anaacutelise da configuraccedilatildeo electroacutenica do elemento Assim a variabilidade encontrada no
comportamento quiacutemico do vanaacutedio pode dever-se agrave sua configuraccedilatildeo ndash [Ar] 3d2 4s
2 ndash que
permite a formaccedilatildeo de vaacuterios estados de oxidaccedilatildeo (de +2 a +5) Em soluccedilatildeo aquosa os estados de
oxidaccedilatildeo estaacuteveis vatildeo de V(V) a V(II) Destes apenas os +3 a +5 satildeo biologicamente importantes
jaacute que o V(II) apresenta demasiada capacidade redutora para existir nos organismos vivos
conhecidos [AC]
No fluido extracelular os compostos de vanaacutedio encontram-se predominantemente na forma de
monovanadatos (V(V)) livres ou ligados agrave transferina do soro ou agrave hemoglobina (Refordf) Os iotildees de
vanadato livres satildeo transportados para o meio intracelular atraveacutes de canais ioacutenicos inespeciacuteficos
Jaacute no ambiente intracelular os compostos sofreratildeo uma bio-reduccedilatildeo atraveacutes da acccedilatildeo da
glutationa ou outros compostos com grupos ndashOH como pentoses hexoses ou etanolaminas [V] A
reoxidaccedilatildeo de vanadilo (V(IV)) intracelular a vanadato (V(V)) provocaria a acumulaccedilatildeo toacutexica
destes uacuteltimos pelo que a complexaccedilatildeo da espeacutecie reduzida o impede O transporte de vanaacutedio
para o interior da ceacutelula encontra-se na Fig 110
Figura 110 ndash Transporte de vanaacutedio para a ceacuteluka e seu subsequente metabolismo (P=Proteiacutena) (Adaptado de
Zaporowska e Scibor 1998)
1211 Vanaacutedio(IV)
A espeacutecie de vanaacutedio (IV) mais bem conhecida eacute o iatildeo vanadilo (VO2+
) espeacutecie estaacutevel a pH
aacutecido sendo a maioritaacuteria a pH 3[AD] Apresenta uma cor azulada e forma complexos penta e
hexacoordenados Forma-se rapidamente a partir de V(V) na presenccedila de um agente redutor em
soluccedilatildeo aquosa No ambiente intracelular a presenccedila de glutationa reduzida aacutecido ascoacuterbico
NAD(P)H entre outros conduzem a que o principal estado de oxidaccedilatildeo nos sistemas bioloacutegicos
seja provavelmente o +4 encontrando-se o V(IV) fortemente complexado com proteiacutenas ou
aminoaacutecidos livres citrato ATP e pirofosfato (Refordfs) Em alternativa o V(IV) pode existir
17
livremente em ambientes fortemente aacutecidos como os lisossomas jaacute que a pH superior a 3 as
soluccedilotildees de vanadilo tendem a ser oxidadas pelo ar formando-se vanadato Esta grande
capacidade redutora justifica parcialmente a utilizaccedilatildeo pouco extensiva do iatildeo VO2+
como
inibidor [W]
1212 Vanaacutedio (V)
O vanaacutedio (V) eacute a forma mais estaacutevel num intervalo de pH amplo [W] O iatildeo anioacutenico VO3-tende
a constituir complexos polinucleares pelo que as soluccedilotildees aquosas de vanadato satildeo extremamente
complexas com ocorrecircncia simultacircnea de vaacuterios equiliacutebrios de protonaccedilatildeo (pKa(vanadato)= 35
78 e 125) e oligomerizaccedilatildeo As espeacutecies presentes (H3VO4 H2VO4- HVO4
2- e VO4
3-) dependem
do pH forccedila ioacutenica e temperatura da soluccedilatildeo e da concentraccedilatildeo total do elemento
[AEAFAGAHAI]
O VO43-
- monoacutemero ortovanadato V1 ndash estaacute associado a uma grande parte da importacircncia
bioloacutegica do vanaacutedio uma vez que se trata de um anaacutelogo electroacutenico e estrutural do ortofosfato
(PO43-
) A coordenaccedilatildeo do ortovanadato pode ser tetraeacutedrica ou bipiramidal trigonal (Refordfs)
A oligomerizaccedilatildeo do vanadato monomeacuterico daacute origem ao diacutemero (V2O72-
- V2) que apresenta
tambeacutem equiliacutebrios de protonaccedilatildeo Presume-se a existecircncia de analogia do vanadato dimeacuterico com
o pirofosfato embora natildeo haja evidecircncia estrutural (Refordf) A dimerizaccedilatildeo do V2 origina o
vanadato tetrameacuterico ou metavanadato (V4O124-
- V4) que constitui a espeacutecie maioritaacuteria em
soluccedilotildees aquosas concentradas (Refordf) As estruturas do metavanadato e a sua espeacutecie protonada
satildeo ciacuteclicas [AJ] Outra espeacutecie oligomeacuterica importante eacute o vanadato pentameacuterico (V5O153-
) cuja
estrutura presumida eacute ciacuteclica [AI]
O oligoacutemero com maior importacircncia bioquiacutemica eacute o vanadato decameacuterico (V10O286-
-V10)(Refordf) a
espeacutecie predominante em soluccedilotildees de pH 2-6 quando a concentraccedilatildeo total de vanadato eacute superior
a 100 M (Ref) A estrutura do V10 natildeo difere em soluccedilatildeo ou no estado soacutelidoOs 10 aacutetomos de
vanaacutedio da espeacutecie decameacuterica estatildeo divididos em trecircs grupos identificados na figura como VA-C
consoante o seu ambiente quiacutemico circundante (Refordf) (Fig 111)
A expressatildeo ldquodecavanadatordquo refere-se a uma soluccedilatildeo aquosa na qual a espeacutecie decameacuterica se
encontra em equiliacutebrio quiacutemico com as espeacutecies V1 V2 V4 e V5 sendo que a oligomerizaccedilatildeo
progressiva do vanaacutedio eacute proporcional agrave sua concentraccedilatildeo total (Ref) Nesta soluccedilatildeo a
concentraccedilatildeo de V10 corresponde a aproximadamente 10 da concentraccedilatildeo total de vanaacutedio [AK]
18
Figura 111 ndash Estruturas esquemaacuteticas das espeacutecies de vanadato (espeacutecie monomeacuterica (V1) dimeacuterica (V2)
tetrameacuterica (V4) pentameacuterica (V5) e decameacuterica (V10) (Adaptado de Aureliano e Crans 2008)
122 Metabolismo e efeito na actividade enzimaacutetica
Apesar de natildeo terem sido ainda encontradas as provas categoacutericas das suas funccedilotildees nos seres
vivos sabe-se que o iatildeo vanadato eacute capaz de inibir uma grande quantidade de enzimas in vitro
com especial ecircnfase na actividade ATPaacutesica da miosina [AL Teresa] Como referido
anteriormente o vanadato eacute capaz de actuar em alternativa ao fosfato nas reacccedilotildees bioquiacutemicas
incluindo a sua preferecircncia de ligaccedilatildeo agraves enzimas [W]
Eacute conhecido o papel do vanaacutedio no metabolismo do caacutelcio promovendo a mineralizaccedilatildeo dos
dentes e ossos na biossiacutentese de liacutepidos (trigliceacuteridos e colesterol) com posterior inibiccedilatildeo da
acumulaccedilatildeo destes nos vasos sanguiacuteneos e no SNC na reproduccedilatildeo e desenvolvimento por
mimetizaccedilatildeo e potenciaccedilatildeo de factores de crescimento na estimulaccedilatildeo do consumo de glucose e
da siacutentese de glicogeacutenio com actuaccedilatildeo equivalente aos efeitos da insulina ao niacutevel celular na
transduccedilatildeo de sinais celulares com regulaccedilatildeo da actividade de segundos mensageiros no
transporte de iotildees e aminoaacutecidos na transcriccedilatildeo geneacutetica e siacutentese de proteiacutenas e DNA entre
outros [W Y AB]
Assim o vanaacutedio cnseguiraacute provocar inuacutemeras mudanccedilas bioquiacutemicas nas ceacutelulas Alguns
exemplos da influecircncia bioquiacutemica do vanaacutedio incluem os seus efeitos nos processos de
fosforilaccedilatildeo e desfosforilaccedilatildeo de proteiacutenas e na participaccedilatildeo na formaccedilatildeo de espeacutecies reactivas de
oxigeacutenio devido ao seu efeito proacute-oxidante sobre agentes redutores a niacutevel celular [AM]
Quanto ao seu efeito na actividade enzimaacutetica pode dividir-se em dois tipos principais ndash inibitoacuterio
e estimulante - dependendo da enzima ou do tipo de reacccedilatildeo por esta catalisada No primeiro
grupo incluem-se enzimas como as ATPases (de miosina de canais ioacutenicos e fosfoenzimas
19
transportadoras) e as fosfatases (da tirosina aacutecida e alcalina) a fosfofrutocinase adenilato ciclase
e catalase entre outras [AA] O segundo grupo inclui a maioria das cinases a glucose-6-fosfato
desidrogenase a fosfoglicerato mutase e a fosfoglicerato fosfatase (Refordf)
O efeito do vanaacutedio (V) na actividade enzimaacutetica estaacute relacionado com o seu nuacutemero de
coordenaccedilatildeo podendo actuar como estado de transiccedilatildeo ou por mimetizaccedilatildeo de cofactores
substratos (especialmente o fosfato) e interacccedilotildees proteiacutena-proteiacutena (como no caso das
polimerases e transcriptases) Pode ainda estar envolvido na formaccedilatildeo de ligaccedilotildees de hidrogeacutenio
interacccedilotildees electrostaacuteticas esteacutereas e de oxidaccedilatildeo-reduccedilatildeo Uma evidecircncia desta uacuteltima eacute a sua
capacidade de alternacircncia entre os estados de oxidaccedilatildeo +5 a +3 o que torna faacutecil a sua interacccedilatildeo
redox com enzimas contendo resiacuteduos de cisteiacutenas (Cys) ou qualquer outro composto orgacircnico
com actividade redox [AN]
13 Vanaacutedio e muacutesculo esqueleacutetico
A primeira evidecircncia da presenccedila de vanaacutedio no muacutesculo esqueleacutetico proveio de uma amostra de
ATP isolado de muacutesculo de cavalo que continha uma impureza ndash vanaacutedio ndash cuja presenccedila
provocou a inibiccedilatildeo da actividade da bomba de Na+K
+-ATPase [AO] Desde entatildeo vaacuterios
estudos tecircm sido realizados com ecircnfase na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o
tecido muscular Destes destaca-se a descoberta da interacccedilatildeo forte da espeacutecie V10 com a miosina
fosforilada [AP] e agrave sua inibiccedilatildeo da actividade ATPaacutesica da miosina[AQ AR]
No quediz respeitoao iatildeo monovanadato destaca-se a sua inibiccedilatildeoda actividade Mg2+
-ATPaacutesica
da miosina [AS] que ocorre de duas formas distintas (1) na presenccedila de concentraccedilotildees reduzidas
de V1 a inibiccedilatildeo ocorre a velocidade reduzida dependente da concentraccedilatildeo da espeacutecie
monomeacuterica (2) na presenccedila de concentraccedilotildees elevadas de V1 o niacutevel de inibiccedilatildeo aproxima-se de
100 desde que [V1] seja superior agrave concentraccedilatildeo total de locais com actividade Mg2+
-ATPaacutesica
no ensaio Esta inibiccedilatildeo eacute irreversiacutevel tanto por diaacutelise ou adiccedilatildeo de agentes redutores do
vanadato [AL]Segundo Pate et al(1994)[AT] a espeacutecie monomeacuterica podeinibir a velocidade de
contracccedilatildeo a temperatura inferior a 20ordmC perdendogradual e rapidamente essa capacidade ateacute os
50ordmC temperatura agrave qual cessa a capacidade inibitoacuteria Os resultados obtidos sugeremainda uma
interacccedilatildeo altamente cooperativa entre o iatildeo vanadato e as proteiacutenas componentes do muacutesculo
esqueleacuteticoDestaca-se tambeacutem a descoberta da inibiccedilatildeo da geraccedilatildeo de forccedila contraacutectil nas fibras
musculares por parte da espeacutecie monomeacuterica[AT]Wilson et al(1995) [AU ]determinaram que a
afinidade da espeacutecie V1 para o S1 de miosina eacute significativamente mais elevada do que a do iatildeo
Pi seu anaacutelogo Descobriram ainda que que a ligaccedilatildeo do vanadato poderaacute envolver transiccedilotildees
entre estados energeacuteticos com energia livre independente dos impedimentos esteacutereos impostos
20
pela estrutura das miofibrilas Os estudos desenvolvidos por Smith et al (1990)[AV]
determinaram queo iatildeo monovanadato natildeo apresenta um efeito pronunciado na inibiccedilatildeo da
actividadeacto-S1-miosina-ATPaacutesica excepto em concentraccedilotildees superiores a 900microM a partir das
quais existe produccedilatildeo de espeacutecies oligomeacutericas para as quais o efeito inibitoacuterio seraacute mais
elevadoA espeacutecie monomeacuterica de vanadato consegue adoptar facilmente uma geometria do tipo
bipiramidal trigonal que mimetiza o intermediaacuterio formado (MgADP
Pi)aquando da reacccedilatildeo de
hidroacutelise de ATP no local activo da miosina [AW] De facto a interacccedilatildeo do vanaacutedio com a
miosina eacute inesperadamente estaacutevel na presenccedila de ADP Esta interacccedilatildeo resulta na formaccedilatildeo de
um complexo enzimaacutetico inactivo e estaacutevel com estequiometria de 11 ADPvanadato por siacutetio
activo da proteiacutena A formaccedilatildeo do complexo estaacutevel miosina-vanadato foi proposta por Goodnoet
al (1982) [AS] Nesta ocorre um preacute-equiliacutebrio raacutepido que induz a formaccedilatildeo de um complexo
ternaacuterio reversiacutevel (MADP
V) com formaccedilatildeo lenta do isoacutemero M
ADP
V A diferenccedila entre os
compostos de vanaacutedio e de fosfato centra-se no tempo de meia vida (t12) do Pi ou do V (no
primeiro t12=12 s enquanto no segundo t12=24 h) (Refordf) A diferenccedila dever-se-aacute agrave possiacutevel
complexaccedilatildeo do vanaacutedio com grupos nucleofiacutelicos no siacutetio activo o que provocaria uma
estabilizaccedilatildeo da miosina na conformaccedilatildeo MADP
V
No que respeita aos estudos realizados com a espeacutecie tetrameacuterica Cremo et al determinou que a
cadeia pesada do S1 de miosina pode ser clivada em dois locais distintos (designados V1 e V2)
pela espeacutecie V4 apoacutes irradiaccedilatildeo com luz UV na presenccedila de concentraccedilotildees de vanadato na ordem
dos milimolar e na ausecircncia de nucleoacutetidos ou metais divalentes
Jaacute no que concerne aos estudos realizados com ecircnfase na espeacutecie decameacuterica foi sugerido
porRamos et al (2006) [AK]que a interacccedilatildeo do decavanadato com a G-actina favorecida pela
polimerizaccedilatildeo desta uacuteltima estabiliza a espeacutecie V10 e induz a inibiccedilatildeo da formaccedilatildeo de F-actina A
estabilizaccedilatildeo da espeacutecie V10 por proteiacutenas tansmembranares e do citoesqueleto pode ser a razatildeo
pela qual ocorre a toxicidade do decavanadato reportada na avaliaccedilatildeo dos efeitos do V(V) nos
sistemas bioloacutegicosTiago et al (2007) [AR] concluiram que o V10 apresenta uma clara preferecircncia
de ligaccedilatildeo ao ldquoback-doorrdquo da S1-miosina-ATPase o que se verifica apenas na estrutura
conformacional desimpedida com acesso facilitado ao local de ligaccedilatildeo do Pi Sugere tambeacutem que
o V10 actua como um ldquoback-door stoprdquo bloqueando estaabertura do fragmento de 50-kDa
necessaacuteria para a hidroacutelise do ATP--fosfato o que constitui uma explicaccedilatildeo simples do
comportamento natildeo competitivo da espeacutecie decameacuterica estimulando a utilizaccedilatildeo deste oligoacutemero
como uma feramenta da elucidaccedilatildeo das mudanccedilas conformacionais ldquoback-doorrdquo da miosina no
21
processo da contracccedilatildeo muscular(ref)Destaca-se ainda a descoberta da capacidade inibitoacuteria do
V10 em relaccedilatildeo agrave fosforilaccedilatildeo oxidativa mitocondrial [AX]
Finalmente no que concerne aos estudos realizados com a presenccedila de vaacuterias espeacutecies
oligomeacutericas Aureliano e Madeira (1994) [AY]observaram que na presenccedila de Ca2+
as espeacutecies
V4 e V10se ligam agrave Ca2+
-ATPase do retiacuteculo sarcoplasmaacutetico enquanto a espeacutecie V1 se liga apenas
na presenccedila de ATP Aparentemente o aumento da concentraccedilatildeo de Ca2+
tem o efeito contraacuterio da
inibiccedilatildeo da ATPase do retiacuteculo sarcoplasmaacutetico pelo V1 natildeo tendo no entanto efeito significativo
sobre a inibiccedilatildeo por parte da espeacutecie V10 Tiago et al (2002) [AP] concluiacuteram que em condiccedilotildees
proacuteximas agraves fisioloacutegicas a espeacutecie V10 difere das espeacutecies presentes na soluccedilatildeo de metavanadato
devido agrave sua forte interacccedilatildeo com a miosina fosforilada e agrave sua inibiccedilatildeo da actividade ATPaacutesica
da miosina Para aleacutem disso o ATP diminui a afinidade da miosina para o V4 induzindo a
interacccedilatildeo com V1 e natildeo tendo qualquer efeito no que diz respeito ao V10 Foi tambeacutem proposto
[AQ] que a ligaccedilatildeo das espeacutecies oligomeacutericas de vanadato aoS1 de miosina nos locais onde o
fosfato coordenaeacute provavelmente a causa da inibiccedilatildeo da actividade ATPaacutesica da miosina
estimulada por actina atraveacutes da prevenccedilatildeo da troca entre ATPADP sendo esta favorecida para
aniotildees de vanadato com valecircncias superiores como eacute o caso do V10
Na presenccedila de actina as velocidades de dissociaccedilatildeo do Pi e do V aumentam consideravelmente
este uacuteltimo em menor quantidade Assim a ligaccedilatildeo da actina ao complexo MADP
V e
Pi eacute
bastante fraca pelo que se sugere que o ADPV teria um efeito de relaxamento semelhante ao
ATP No entanto a capacidade do vanaacutedio em inibir a actividade ATPaacutesica da miosina e da
actomiosina eacute diferenciada uma vez que na ausecircncia de actina a inibiccedilatildeo eacute total para
concentraccedilotildees na gama dos M mas na sua presenccedila a inibiccedilatildeo eacute totalmente conseguida apenas
em concentraccedilotildees na gama dos mM (Ref) No entanto eacute necessaacuterio considerar a possiacutevel
formaccedilatildeo de espeacutecies oligomeacutericas de vanadato cuja afinidade e efeito na enzima seratildeo
diferentes Assim pode concluir-se que o efeito de inibiccedilatildeo observado para a actividade ATPaacutesica
de miosina estimulada por actina se pode dever agrave presenccedila das diferentes espeacutecies oligomeacutericas o
que foi jaacute sugerido por Aureliano (2000) [AZ] ndash as espeacutecies tetrameacuterica e decameacuterica de vanadato
interactuariam com a miosina com alteraccedilatildeo da sua interacccedilatildeo com a actina No entanto natildeo eacute
conhecida a contribuiccedilatildeo absoluta das restantes espeacutecies oligomeacutericas para a inibiccedilatildeo da ATPase
da miosina estimulada por actina (Refs)
14 Objectivos do projecto
Pretende-se com este projecto o isolamento purificaccedilatildeo e caracterizaccedilatildeo da actina do muacutesculo
22
esqueleacutetico de coelho sendo esta uacuteltima tambeacutem realizada para o sufragmento-1 de miosina
Seguidamente realizar-se-aacute a quantificaccedilatildeo e caracterizaccedilatildeo das proteiacutenas por espectroscopia UV-
Vis e SDS-PAGE e a caracterizaccedilatildeo das soluccedilotildees de vanaacutedio por UV-Vis e ressonacircncia magneacutetica
nuclear (RMN) Por uacuteltimo a interacccedilatildeo do S1 de miosina com o vanaacutedio seraacute avaliada atraveacutes
das teacutecnicas de UVVis RMN e ressonacircncia paramagneacutetica electroacutenica (RPE)
2 Discussatildeo de Resultados
21 Caracterizaccedilatildeo das proteiacutenas isoladas
211Anaacutelise do estado de pureza
O estado de pureza das soluccedilotildees de actina e de S1 foi analisado por electroforese em gel de
poliacrilamida em condiccedilotildees desnaturantes (SDS-PAGE) (Fig 22) que separa as moleacuteculas de
acordo com o seu peso molecular (PM) A concentraccedilatildeo de acrilamida escolhida (10) permite a
detecccedilatildeo simultacircnea de componentes com peso molecular inferior a 20 kDa e outros com elevado
peso molecular
Figura 21 ndash Fotografia de gel de SDS-PAGE (10 acrilamida) Poccedilo (a)
10 L do marcador de elevado peso molecular () Poccedilo (b) 20 L de [G-
actina]=1 mgmL Poccedilo (c) 20 L de [S1]=1 mgmL Aplicou-se uma
voltagem de V durante h e min
A partir da recta de calibraccedilatildeo obtida log PM = -1177 Rf + 2247 (ver ponto 2 da secccedilatildeo
Anexos) pode verificar-se a presenccedila de actina na amostra do poccedilo (b) presente numa banda
bastante intensa (PMaparente=40 kDa) bem como a presenccedila minoritaacuteria de outros componentes
desconhecidos que natildeo teratildeo sido eliminados durante o processo de extracccedilatildeo e purificaccedilatildeo e natildeo
satildeo relevantes para o estudo em questatildeo Na amostra do poccedilo (c) nota-se a presenccedila de S1
(PMaparente=92 kDa) bem como a de outros componentes cuja banda eacute ligeiramente menos intensa
Estes poderatildeo ser fragmentos proteoliacuteticos de miosina e de S1 Estaacute ainda presente uma bada de
23
maior intensidade que poderaacute ser a proteiacutena-C (PM=135 kDa) uma vez que a sua remoccedilatildeo
completa da amostra necessitaria de um passo adicional de cromatografia de troca ioacutenica [BA]
No entanto natildeo podemos confirmar que se trata desta proteiacutena ou de qualquer outra uma vez que
se encontra fora do intervalo de migraccedilatildeo dos marcadores de peso molecular
212 Determinaccedilatildeo da concentraccedilatildeo proteica
2121Subfragmento-1 de miosina
A partir da recta de calibraccedilatildeo obtida pelo meacutetodo de Bradford [BB](ver secccedilatildeo 3311 do
Procedimento Experimental) A595nm = 00546[BSA] + 00209 (ver secccedilatildeo de Anexos)foi possiacutevel
determinar um intervalo de concentraccedilatildeo de S1 de miosina compreendido entre 226 e 117(plusmn
423M) (203 e 1056 (plusmn 381 mgmL)) (n=8) A concentraccedilatildeo meacutedia de S1 de miosina apoacutes
diaacutelise contra a soluccedilatildeo tampatildeo 10 mM HEPES pH 75 25 mM KCl determinada pela leitura da
Abs280 nmfoi de 20430M (1803 mgmL)(n=7)
As amostras a quantificar atraveacutes do meacutetodo de Bradford natildeo sofreram diaacutelise o que permite
explicar a concentraccedilatildeo superior de S1 de miosina em comparaccedilatildeo com o valor obtido atraveacutes do
meacutetodo directo Esta razatildeo aliada agrave sensibilidade e rapidez superiores deste uacuteltimo fez com que
se optasse pelo meacutetodo directo para a quantificaccedilatildeo da grande maioria dos ensaios
2122Actina
A concentraccedilatildeo de actina foi determinada quando esta se encontrava na forma monomeacuterica (G-
actina) por mediccedilatildeo da leitura directa da Abs290 nm tendo-se obtido uma concentraccedilatildeo meacutedia de
691481M (290202 mgmL) (n=3) em 2 mM Tris-HCl pH 80 02 mM CaCl2 02 mM Na-
ATP Ao comprimento de onda utilizado (290 nm) absorvem fortemente apenas os resiacuteduos de
Trp pelo que a mediccedilatildeo efectuada tem menor sensibilidade do que se tivesse sido efectuada a 280
nm uma vez que a actina apresenta apenas 4 resiacuteduos deste aminoaacutecido embora tenha 16
resiacuteduos de Tyr
22 Mediccedilatildeo da actividade enzimaacutetica de S1 de miosina
221 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
As actividades ATPaacutesicas de S1 de miosina dependentes de Ca2+
e K+foram analisadas pelo
caacutelculo das velocidades de consumo inicial do fosfato inorgacircnico (Pi) Definiu-se os tempos de 5
e 3 minutos para a mediccedilatildeo da Ca2+
e da K+EDTA-ATPase respectivamente Os valores obtidos
24
a partir do protocolo presente no ponto 341 da secccedilatildeo Procedimento experimental encontram-se
descritos na Tabela 2I
Tabela 2I ndash Acrividades natildeo-fisioloacutegicas de subfragmento-1 determinadas pelo meacutetodo de Fiske-Subarow
modificado (n=2)
Proteiacutena
Actividade
ATPaacutesica natildeo
fisioloacutegica
Actividade (nmol
Piminmg proteiacutena) Actividade (s-1)
S1
Ca2+
-ATPase 1400 plusmn 30 2100 plusmn 0045
K+EDTA-ATPase 2334 plusmn 50 3500 plusmn 0075
As actividades natildeo-fisioloacutegicas reflectem o estado de oxidaccedilatildeo dos resiacuteduos expostos de Cys 707
(SH1) e 697 (SH2) Se o grupo SH1 sofrer oxidaccedilatildeo a actividade ATPaacutesica dependente de Ca2+
aumenta e a actividade dependente de K+ diminui Por outro lado se o grupo SH2 sofrer
oxidaccedilatildeo regista-se a perda completa destas actividades
De acordo com Margossian e Lowey (1982) [BA] as actividades ATPaacutesica de S1 de miosina
dependente de Ca2+
e K+EDTA da miosina satildeo respectivamente de 24 e 112 s-1
Estaacute tambeacutem
descrito que a actividade ATPaacutesica dependente de K+ eacute em meacutedia cerca de 3 vezes superior agrave
dependente de Ca2+
reflectindo a quantidade das preparaccedilotildees A razatildeo obtida entre os valores das
actividades ATPaacutesicas dependentes de K+ e Ca
2+ eacute de 166 Os valores obtidos para as actividades
(Ca2+
-ATPase=21 s-1
K+EDTA-ATPase= 35 s
-1) satildeo inferiores agrave valores obtidos por
Margossian e Lowey [BA] e Tiago [BC] com forccedila ioacutenica inferior O valor obtido para a
actividade ATPaacutesica dependente de Ca2+
eacute superior ao obtido por Polosukhina et al (2000) [BD]
cujo meio de reacccedilatildeo continha MOPS e apresentava forccedila ioacutenica inferior Os dados
supramencionados comprovam a variaccedilatildeo da actividade a ATPaacutesica natildeo fisioloacutegica em relaccedilatildeo agrave
forccedila ioacutenica temperatura e pH
222 Mediccedilatildeo da actividadeMg2+
-ATPaacutesica de S1 pelo meacutetodo de enzimas acopladas
A actividade de Mg2+
-ATPase eacute a uacutenica que possui significado fisioloacutegico sendo as restantes
bastante uacuteteis apenas para indicaccedilatildeo da qualidade de preparaccedilatildeo do S1[BA] A 0575 M KCl a
actina e o S1 formam uma soluccedilatildeo viscosa e tuacuterbida Apoacutes a adiccedilatildeo de MgATP o complexo acto-
S1 dissocia-se e a turbidez da soluccedilatildeo diminui Se a forccedila ioacutenica for reduzida a dissociaccedilatildeo do
complexo actoS1 que se encontra na forma de suspensatildeo eacute induzida O MgATP eacute rapidamente
hidrolisado diminuindo a sua concentraccedilatildeo no meio reaccional Nestas condiccedilotildees haacute
reassociaccedilatildeo e subsequente contracccedilatildeo dos filamentos de actina e de S1 com formaccedilatildeo de
25
precipitado O meacutetodo descrito traduz de forma bastante elementar o fenoacutemeno da contracccedilatildeo in
vivo e elimina o problema anterior da solubilizaccedilatildeo ineficiente da miosina de modo a obter uma
mediccedilatildeo exacta da actividade [A] A utilizaccedilatildeo de forccedilas ioacutenicas reduzidas aumenta a afinidade da
actina para o S1 permitindo contornar a extrema dificuldade em obter a concentraccedilatildeo proteica dos
muacutesculos (cerca de 100 mgmL do complexo actomiosina) As actividades Mg2+
-ATPaacutesicas de S1
de miosina foram determinadas a partir do declive da variaccedilatildeo da Abs340 relativo agrave oxidaccedilatildeo de
NADH em funccedilatildeo do tempo como eacute descrito no ponto 342 do Procedimento Experimental Os
valores obtidos para a actividade Mg2+
-ATPaacutesica encontram-se apresentados na Tabela 2II
Tabela 2II ndash Actividade Mg2+
-ATPaacutesica de subfragmento-1 de miosina determinadas pelo meacutetodo das enzimas
acopladas
Actividade (nmol
ATPmin mg(S1)) Actividade (s
-1)
1097 plusmn 269 164 plusmn 04
Ramos (2005) [J] obteve o valor de 01 s-1
para a actividade Mg2+
-ATPaacutesica de S1 de miosina
com forccedila ioacutenica intermeacutedia Tiago (2005) [BC] obteve o valor de 011 s-1 em condiccedilotildees de forccedila
ioacutenica reduzida em relaccedilatildeo agrave utilizada no presente relatoacuterio
23 Preparaccedilatildeo e caracterizaccedilatildeo das soluccedilotildees de vanaacutedio
231 Anaacutelise da composiccedilatildeo das soluccedilotildees de vanadato por espectroscopia de 51
V-RMN
efeito da concentraccedilatildeo da espeacutecie
A diluiccedilatildeo das soluccedilotildees stock de meta- e decavanadato iraacute determinar fortemente a composiccedilatildeo
em soluccedilatildeo dos diferentes oligoacutemeros de vanadato (ref) No caso do decavanadato o periacuteodo de
tempo apoacutes a diluiccedilatildeo iraacute tambeacutem ser um factor importante a considerar devido agrave decomposiccedilatildeo
da espeacutecie V10(ref)Torna-se assim fundamental determinar a composiccedilatildeo quiacutemica das soluccedilotildees
no meio de ensaio utilizado (HEPES 10 mM KCl 25 mM) de modo a correlacionaacute-la
correctamente com os possiacuteveis efeitos promovidos pelas diferentes espeacutecies de vanadato no
sistema em estudo
Aleacutem disso deve ser dado ecircnfase agraves importantes interacccedilotildees que ocorrem entre o vanaacutedio e os
tampotildees escolhidos O tampatildeo com menor nuacutemero e magnitude de interacccedilatildeo eacute o HEPES [AD]
seguido do Tris sendo este uacuteltimo especialmente aconselhado em concentraccedilotildees baixas para casos
de incompatibilidade enzimaacutetica ou quiacutemica redox No caso especial da reacccedilatildeo redox
NAD(P)HNAD(P) o tampatildeo adequado eacute o imidazole de modo a natildeo haver reacccedilatildeo com o
26
vanadato Os tampotildees a evitar em qualquer situaccedilatildeo incluem a trietanolamina fosfato citrato e
glicina entre outros [AD] uma vez que todos interactuam fortemente com V(V) com formaccedilatildeo
de complexos estaacuteveis
Assim procedeu-se agrave anaacutelise da composiccedilatildeo das soluccedilotildees de meta- e decavanadato apoacutes diluiccedilatildeo
das soluccedilotildees stock (50 mM em vanadato total) no meio de reacccedilatildeo (tampatildeo D) atraveacutes da
espectroscopia de ressonacircncia magneacutetica nuclear de 51
V Nas figuras x1 e x2 apresentam-se os
espectros obtidos para as soluccedilotildees de deca- e metavanadato num intervalo de concentraccedilotildees de 1 a
10 mM A Tabela seguinte resume os paracircmetros espectroscoacutepicos obtidos para cada espeacutecie
presente nas soluccedilotildees analisadas
Figura 22ndash Espectros de RMN de 51
V de
concentraccedilotildees crescentes das espeacutecies oligomeacutericas
de vanadato obtidos apoacutes diluiccedilatildeo da soluccedilatildeo stock
de decavanadato em 10 mM HEPES pH 70 25 mM
KCl para concentraccedilotildees finais de A) 1 mM B)2
mM C) 4 mM D) 8 mM E) 10 mM de
decavanadato total V1refere-se agraves espeacutecies
monomeacutericade vanadato V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2IIIndash Desvios quiacutemicos meacutedios (part) das ressonacircncias das espeacutecies decameacuterica (V10) monomeacuterica (V1) e
tetrameacuterica (V4) presentes nos espectros da soluccedilatildeo de decavanadatoLargura a meia altura (∆12) da espeacutecie V10A
Paracircmetros V10A V10B V10C V1 V4
part (ppm) -5162 -5003 -4255 -5616 -5748
∆12 (Hz) 175 nd nd nd nd
Figura 23 ndash Distribuiccedilatildeo graacutefica das
espeacutecies oligomeacutericas de vanadato vs a
concentraccedilatildeo de decavanadato total As
concentraccedilotildees de cada espeacutecie foram
obtidas segundo a formula descrita na
secccedilatildeo 35 do Procedimento
Experimental (aacutereas dos sinais de 51
V-
RMN observadas na figura x1)V10 V1 e
V4 correspondem agraves espeacutecie monomeacuterica
dimeacuterica e tetrameacuterica respectivamente [Esp
eacuteci
e o
ligo
meacute
rica
de
van
aacutedio
] (m
M)
[Decavanadato]total (mM)
V10
V1
V4
27
Nas soluccedilotildees de decavanadato as espeacutecies monomeacuterica e decameacuterica de vanadato satildeo as uacutenicas
presentes para concentraccedilotildees de decavanadato inferiores a 8 mM Ambas as espeacutecies aumentam
linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado em [AY] e
em (Teresa) quando a concentraccedilatildeo de V1 eacute igual ou superior a 200 microM inicia-se a sua
oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica No entanto nas condiccedilotildees de realizaccedilatildeo dos
ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a espeacutecie tetrameacuterica neste
estudo em concentraccedilotildees inferiores a 8 mM tendo o mesmo sido observado por Ramoset al
(2006) [AK] Os resultados obtidos mostram claramente o equiliacutebrio entre a espeacutecie decameacuterica e
a monomeacuterica e que o aparecimento de outras espeacutecies oligomeacutericas se encontra dependente da
concentraccedilatildeo desta uacuteltima Apesar de natildeo ter sido realizado o ensaio com 5 mM de decavanadato
total no presente relatoacuterio estaacute reportado em Ramos et al (2006) [AK] que as concentraccedilotildees da
espeacutecie decameacuterica e monomeacuterica satildeo respectivamente de 500 microM e 200 microM Assim pode inferir-
se que para concentraccedilotildees inferiores a 5 mM e intervalos de tempo de cerca de 20 minutos (que
inclui o tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas
as espeacutecies decameacuterica e monomeacuterica de vanadato
Figura 24 ndash Espectros de RMN de 51
V de concentraccedilotildees
crescentes das espeacutecies oligomeacutericas de vanadato obtidos
apoacutes diluiccedilatildeo da soluccedilatildeo stock de metavanadato em 10 mM
HEPES pH 70 25 mM KCl para concentraccedilotildees finais de A)
1 mM B) 2 mM C) 4 mM D) 8 mM E) 10 mM de
metavanadato total V1 refere-se agraves espeacutecies monomeacuterica de
vanadatoV1 e V2 correspondem respectivamente agrave espeacutecie
monomeacuterica e dimeacuterica
Figura 25 ndash Distribuiccedilatildeo graacutefica das espeacutecies
oligomeacutericas de vanadato vs a concentraccedilatildeo de
metavanadato total As concentraccedilotildees de cada
espeacutecie foram obtidas segundo a formula descrita
na secccedilatildeo 35 do Procedimento Experimental
(aacutereas dos sinais de 51
V-RMN observadas na
figura x1)V10 V1 e V4 correspondem agraves espeacutecie
monomeacuterica dimeacuterica e tetrameacuterica
respectivamente
Esp
eacuteci
e o
ligo
meacute
rica
de
va
nad
ato
(m
M)
[Metavanadato]total (mM)
V1
V2
V4
V5
28
Tabela 2IVndash Desvios quiacutemicos meacutedios (part)das ressonacircncias das espeacutecies tetrameacuterica (V4) monomeacuterica (V1) e
dimeacuterica (V2) presentes no espectro da soluccedilatildeo de metavanadato (total) 10 mM
Paracircmetros V1 V2 V4 V5
part (ppm) -5615 -5748 -5789 -5869
Nas soluccedilotildees de metavanadato as espeacutecies monomeacuterica dimeacutericae tetrameacuterica de vanadato estatildeo
presentes para concentraccedilotildees de metavanadato inferiores a 4 mM Todas estas as espeacutecies
aumentam linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado
em (Aureliano e Madeira 1998) e em (Tiago et al 2004) quando a concentraccedilatildeo de V1 eacute igual ou
superior a 200 microM inicia-se a sua oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica Nas
condiccedilotildees de realizaccedilatildeo dos ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a
espeacutecie pentameacuterica neste estudo em concentraccedilotildees inferiores a 4 mM tendo o mesmo sido
observado por Ramos 2006 Os resultados obtidos mostram claramente o equiliacutebrio entre a
espeacutecie V4 e as espeacuteciesV1 e V2 no qual o aparecimento de outras espeacutecies oligomeacutericas se
encontra dependente da concentraccedilatildeo da espeacutecie V1 Por uacuteltimo pode inferir-se que para
concentraccedilotildees inferiores a 2 mM e intervalos de tempo de cerca de 20 minutos (que inclui o
tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas as
espeacutecies V1 V2 e V4 com concentraccedilotildees respectivas de 0796 0192 e 0196 mM
232Estabilidade das soluccedilotildees de decavanadato anaacutelise por espectroscopia de absorccedilatildeo
UVVis
Caracterizou-se a estabilidade da espeacutecie V10 numa soluccedilatildeo com 1 mM decavanadato total em
tampatildeo D atraveacutes da leitura da Abs400 Obteve-se uma cineacutetica de dissociaccedilatildeo de 1ordf ordem tal
como se pode verificar pela linearidade do graacutefico de ln(Abs400)vs o tempo de aquisiccedilatildeo espectral
(segundos) (Fig26 )
Figura 26ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do tempo
(em segundos) para uma soluccedilatildeo de decavanadato total de 1
mM em tampatildeo D a 25ordmC (n=2)
Sendo uma reacccedilatildeo de primeira ordem pode ser descrita pela equaccedilatildeo cujo
declive eacute ndashK (constante de dissociaccedilatildeo) e a intersecccedilatildeo no eixo das ordenadas corresponde
Ln
(Ab
s 400 n
m)
tempo (s)
29
aln(A0) Numa reacccedilatildeo de 1ordf ordem o tempo de meia-vida eacute independente da concentraccedilatildeo inicial
do reagente sendo descrito pela equaccedilatildeo [2] Deste modo a partir da recta
Ln(Abs400 nm) = -436E-5
K - 105 obteve-se um tempo de meia-vida de 4490 plusmn 0823 horas para
as condiccedilotildees supramencionadas
Apoacutes a adiccedilatildeo de S1 8 microM ao meio reaccional verificou-se que a reacccedilatildeo de desoligomerizaccedilatildeo
da espeacutecie decameacuterica de vanadato seguia igualmente uma cineacutetica de 1ordf ordem tal como se pode
verificar pela linearidade do graacutefico de ln(Abs400) versus o tempo de aquisiccedilatildeo espectral
(segundos) (Fig27)
Figura 27ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do
tempo (em segundos) para uma soluccedilatildeo de decavanadato
total de 1 mM em tampatildeo D a 25ordmC na presenccedila de 8
microM (n= 2)
A partir da equaccedilatildeo da recta Ln(Abs400 nm) = -7520E-5
K - 0613determinou-se o valor de K para
a espeacutecie decameacuterica de onde se obteve um tempo de meia-vida de 2430 plusmn 0191 horas para as
condiccedilotildees supramencionadas
Tanto este valor como o valor registado na ausecircncia de S1 satildeo substancialmente superiores agrave
duraccedilatildeo dos ensaios de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina descritos no presente
relatoacuterio
24 Interacccedilatildeo de S1 com vanaacutedio(V)
241Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
De modo a analisar qual o efeito da adiccedilatildeo de S1 de miosina (tampatildeo D) produzia nos sinais de
51V-NMR titulou-se uma soluccedilatildeo de 5mM de decavanadato total com concentraccedilotildees crescentes
de S1 (5 10 15 e 20 microM) Os espectros de 51
V-RMN encontram-se apresentados na Fig 2
No graacutefico da figura 2 Apresenta-se a concentraccedilatildeo de S1 em funccedilatildeo do factor de alargamento
corrigido dos espectros acima referidos
Ln
(Ab
s (4
00
nm
))
tempo (s)
30
Figura 28 ndash Graacutefico de
representaccedilatildeo da
concentraccedilatildeo de S1 em
funccedilatildeo do factor de
alargamento A
concentraccedilatildeo de
decavanadato total foi de 5
mM e as concentraccedilotildees
utilizadas de S1 foram de 0
5 10 e 15 microM
Verifica-se que o factor de alargamento aumenta linearmente em funccedilatildeo da concentraccedilatildeo de S1 no
ensaio o que indica a existecircncia de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina Observou-
se ainda que a adiccedilatildeo de S1 ao meio reaccional natildeo provocou qualquer alteraccedilatildeo dos desvios
quiacutemicos dos sinais de 51
V-RMN
Realizaram-se ensaios na presenccedila de ATP e de S1 com intuito de verificar o efeito do ATP na
interacccedilatildeo do S1 de miosina com o decavanadato
Figura 29 - Espectros de RMN de 51
V contendo 2
mM Decavanadato (A) (controlo) (B) na presenccedila
de 2 mM de ATP (C) na presenccedila de 2 mM de ATP
e 8 microM de S1 de miosina V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2Vndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies decameacuterica (V10) e monomeacuterica (V1) presentes nos
espectros da soluccedilatildeo de decavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V10A V10B V10C V1
part (ppm) -5161 -5002 -4254 -5614
Observa-se que a presenccedila de ATP (espectro B) natildeo provoca qualquer alteraccedilatildeo visiacutevel em
relaccedilatildeo ao espectro que contem apenas soluccedilatildeo de decavanadato Jaacute a presenccedila simultacircnea de ATP
e S1 provoca um alargamento da ∆12 do sinal do V10A de 1736 Hz (espectro (A)) para 2366 Hz
(espectro (C) Fig 29) Tendo em conta que a concentraccedilatildeo de S1 apresenta uma relaccedilatildeo linear
com o factor de alargamento converteu-se o valor da ∆12 e comparou-se com o valor esperado agrave
F = 00591[S1] - 00157 Rsup2 = 0998
Fact
or
de
ala
rgam
en
to
[S1] (microM)
31
concentraccedilatildeo de S1 utilizada O factor de alargamento obtido para 8 microM ndash 0376 ndash eacute inferior ao
valor esperado agrave mesma concentraccedilatildeo de S1 obtido atraveacutes da equaccedilatildeo da recta apresentada na
Fig 28 ndash 0457 ndash o que indica que o ATP poderaacute exercer um efeito de protecccedilatildeo do S1
diminuindo a sua interacccedilatildeo com a espeacutecieV10
Por uacuteltimo realizaram-se ensaios com metavanadato na presenccedila de 8 microM S1 de modo a verificar
se as espeacutecies oligomeacutericas contidas nesta soluccedilatildeo interagiam com a proteiacutena em estudo
Figura 210 - Espectros de RMN de 51
V contendo 2
mM metavanadato (A) (controlo) (B) na presenccedila
de 8 microM de S1 de miosina V1 V2 e V4
correspondem respectivamente aacutes espeacutecies
monomeacuterica dimeacuterica e tetrameacuterica
Tabela 2VIndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies monomeacuterica (V1) dimeacuterica (V2) e tetrameacuterica (V4)
presentes nos espectros da soluccedilatildeo de metavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V1 V2 V4
part (ppm) -5613 -5747 -5789
Como eacute possiacutevel observar na Fig 210 ocorre uma ligeiro alargamento dos sinal de V4 bem
como uma perda da intensidade pelo que se conclui quedas espeacutecies presentes na soluccedilatildeo de
metavanadato (V1 V2 e V4) apenas a espeacutecie tetrameacuterica interage com o S1 de miosina o que
estaacute de acordo com o reportado por Tiago et al 2002
242 Actividade Mg2+
-ATPaacutesica de S1 de miosina na presenccedila de decanadato
Com o intuito de avaliar o efeito do decavanadato na actividade Mg2+
-ATPaacutesica do S1 de
miosina monitorizou-se a reacccedilatildeo de oxidaccedilatildeo do NADH atraveacutes da leitura de Abs340 na
presenccedila de 005 mgmL de S1 de miosina e no caso da actividade estimulada 2microM de actina em
3125 mM Tris-HCl pH 71 e 375 mM MgCl2 pelo meacutetodo de enzimas acopladas (ver seccccedilatildeo xx
dos MM)Para tal realizaram-se ensaios com vaacuterias concentraccedilotildees de decavanadato total (10 50
100 250 500 e 1000) de modo a determinar qual o seu efeito na actividade MgATPaacutesica de S1
de miosina na ausecircncia e na presenccedila de um dos seus ligandos naturais Os ensaios foram
realizados agrave temperatura ambiente com incubaccedilatildeo preacutevia a 25ordm C durante 20 minutos
32
Figura 211 ndash Representaccedilatildeo
graacutefica da Actividade Mg2+
-
ATPaacutesica de S1 () (os ajustes
teoacutericos estatildeo representado a por
uma linha os resultados
experimentais estatildeo representados
por triacircngulos (Actividade
estimulada) e cruzes (Actividade
natildeo estimulada)) vs
[Decavanadato]total medidas pelo
meacutetodo das enzimas acopladas
(n=3) Os dados foram ajustados ao modelo de decaimento exponencial usando o programa GraphPad Prism para a
determinaccedilatildeo dos valores de IC50 (na ausecircncia de actina IC50= 266 plusmn 07 M decavanadato na presenccedila de actina
IC50= 28 plusmn 1 M decavanadato
A partir do graacutefico presente na Fig 29 verifica-se que o ajuste dos dados experimentais foi bem
sucedido A inibiccedilatildeo da actividade Mg2+
-ATPaacutesica do S1 de miosina eacute afectada em menor
amplitude na presenccedila de actina (60 de inibiccedilatildeo da actividade) do que na sua ausecircncia
(aproximadamente 80 de inibiccedilatildeo de actividade) Devido a esta diferenccedila pode sugerir-se que a
actina iraacute impedir parcialmente a ligaccedilatildeo do decavanadatoao S1 demiosina atraveacutes de
impedimentos esteacutereos ou interacccedilotildees electrostaacuteticas por exemplo A teacutecnica utilizada natildeo nos
permite concluir acerca de que regiatildeo(otildees) ou resiacuteduo(s) da actina participaratildeo na preservaccedilatildeo
parcial da actividade Mg2+
-ATPaacutesica
Por outro lado os valores de IC50 dos dois tipos de actividades natildeo apresentam diferenccedilas
significativas tendo-se obtido 266 plusmn 07 e 28 plusmn 1M decavanadato na ausecircncia e na presenccedila de
actina respectivamente o que corresponde a uma concentraccedilatildeo de cerca de 27 M de V10 O
meio de reacccedilatildeo utilizado nestes ensaios foi 10 mM HEPES e 25 mM KCl com 2 mM ATP Em
(Tiago et al 2004) o valor de IC50 para a actividade Mg2+-ATPaacutesica na ausecircncia de actina foi de
125 plusmn 025 microM enquanto o valor da actividade estimulada por actina foi de 136 plusmn 14 microM O
meio de reacccedilatildeo utilizado nos ensaios foi 20 mM HEPES 50 mM KCl e 2 mM ATP o que resulta
precisamente no dobro da forccedila ioacutenica a que foi realizado o ensaio do presente relatoacuterio
(exceptuando a contribuiccedilatildeo dos produtos da hidroacutelise de ATP uma vez que este meacutetodo
pressupotildee a ocorrecircncia de um ciclo de reacccedilotildees de hidroacutelise e siacutentese deste composto) Ora tendo
sido anteriormente referido que a afinidade do S1 de miosina para com a espeacutecie V10 diminui com
Act
ivid
ade
Mg
2+-A
TP
aacutesic
a d
e S
1 d
e
Mio
sin
a
[Decavanadato] (microM) Actividade Estimulada ()
Actividade Natildeo Estimulada ()
Ajuste Actividade Estimulada
Ajuste Actividade Natildeo Estimulada
33
o aumento da forccedila ioacutenica (Tiago et al 2004 tal poderaacute explicar a diferenccedila obtida entre estes
dois conjuntos de valores
243Quantificaccedilatildeo de cisteiacutenas reduzidas pelo meacutetodo de Ellman
A interacccedilatildeo do vanaacutedio com alguns tipos de aminoaacutecidos sensiacuteveis agrave oxidaccedilatildeo pode fornecer-nos
informaccedilatildeo bastante uacutetil em relaccedilatildeo agraves alteraccedilotildees estruturais que conduzem a uma diminuiccedilatildeo da
actividade fisioloacutegica do S1 de miosina Os estudos realizados por Chasteen 1983 concluiacuteram
que a reduccedilatildeo de vanadato (V) a vanadilo (IV) ocorre na presenccedila de agentes redutores comuns no
ambiente celular de que se destaca o aminoaacutecido cisteiacutena Esta eacute tambeacutem um alvo de
complexaccedilatildeo por parte do iatildeo VO2+
Jaacute Ramos et al (2009) demonstrou que a incubaccedilatildeo de
actina com a soluccedilatildeo de decavanadato induz a oxidaccedilatildeo de resiacuteduos de cisteiacutena e agrave formaccedilatildeo do
oxidovanaacutedio (IV) Este grupo de investigadores descobriu ainda que apenas a espeacutecie V10 induz a
oxidaccedilatildeo das cisteiacutenas ldquoraacutepidasrdquo da actina atraveacutes do processo proposto que eacute parcialmente
responsaacutevel pela inibiccedilatildeo jaacute descrita da actividade de ATPase do S1 de miosina estimulada por
actina quando exposta agrave soluccedilatildeo de decavanadato Esta inibiccedilatildeo apresenta-se afectada com a
presenccedila de ATP uma vez que os resiacuteduos de cisteiacutena permaneceram intactos nos ensaios
realizados com este ligando Jaacute Crans et al (2004) concluiu que o siacutetio activo de muitas fosfatases
(que conteacutem grupos tiol) pode formar um equiliacutebrio redox com o iatildeo vanadato (V) de forma natildeo
reversiacutevel
Os resultados obtidos por Sakurai e Shiromura (1980) sugeriram que o vanaacutedio (V) sofre reduccedilatildeo
a vanaacutedio (IV) sem dependecircncia do valor de pH do ensaio Registaram ainda uma estequiometria
de 31 na reacccedilatildeo do vanadato (V) com os resiacuteduos de Cys enquanto a estequiometria dos ensaios
com o vanadilo (IV) foi de 21
Berlett e Stadtman (1997) observaram que os tipos de aminoaacutecidos mais facilmente oxidaacuteveis
englobam entre outros os resiacuteduos de Cys Trp e MetAssim analisou-se o estado redox dos
grupos tiol dos resiacuteduos de Cys do S1 na presenccedila do decavanadato atraveacutes da monitorizaccedilatildeo da
Abs412 nm na presenccedila de 1 mgmL de S1 de miosina em tampatildeo 10 mM HEPES 25 mM KCl
variando a concentraccedilatildeo da soluccedilatildeo de decavanadato de modo a determinar que tipo de alteraccedilotildees
estruturais ocorrem na sua presenccedila no que diz respeito agrave diminuiccedilatildeo da actividade ATPaacutesica de
S1 na presenccedila de decavanadato
Os resultados referentes agrave quantificaccedilatildeo das cisteiacutenas reduzidas apoacutes o tratamento de S1 com
diferentes concentraccedilotildees de decavanadato em tampatildeo 10 mM HEPES pH 75 e 25 mM KCl a
25ordmC durante 20 minutos encontram-se descritos na Tabela 2VII
Tabela 2VII ndash Cisteiacutenas reduzidas totais e ldquoraacutepidas de S1 (1 mgmL) apoacutes tratamento com decavanadato
34
(010501002505001000 uM) em 10 mM HEPES 25 mM KCl pH 7 a 25ordmC durante 20 minutos A detecccedilatildeo
espectrofotomeacutetrica foi feita com DTNB (e 412=8400 uM-1 cm-1) (n variaacutevel entre 1 e 8)
Amostras Cys raacutepidas Cys totais
11 M S1 (controlo) 22 plusmn 04 106 plusmn 14
11 M S1 + 10 M Decavanadato 22 plusmn 05 95 plusmn 08
11 M S1 + 50 uM Decavanadato 20 plusmn 02 98 plusmn 14
11 M S1 + 100 uM Decavanadato 17 plusmn 03 97 plusmn14
11 M S1 + 250 uM Decavanadato 26 plusmn 02 98 plusmn 08
11 M S1 + 500 uM Decavanadato 21 plusmn 04 90 plusmn 06
11 M S1 + 1000 uM Decavanadato 24 plusmn 0 86 plusmn 08
A partir dos resultados obtidos pode sugerir-se que o S1 possui 11 resiacuteduos de Cys (controlo) o
que estaacute de acordo com o anteriormente descrito Verifica-se que o tratamento da proteiacutena com
concentraccedilotildees crescentes de decavanadato provoca a oxidaccedilatildeo de dois resiacuteduos de Cys do S1 de
miosina natildeo expostos ao solvente No entanto a teacutecnica utilizada natildeo nos permite identificar
quais
Tendo em consideraccedilatildeo que o valor de IC50 obtido para a reacccedilatildeo de inibiccedilatildeo daactividade Mg2+-
ATPaacutesica de S1 na presenccedila de decavanadato eacute de cerca de 27 microM pode sugerir-se que este seraacute o
valor ao qual ocorreraacute a oxidaccedilatildeo do primeiro resiacuteduo de Cys o que poderaacute contribuir para a
diminuiccedilatildeo da actividade de S1 observada
25Interacccedilatildeo de S1 com vanaacutedio(IV)
251Anaacutelise por espectroscopia de Ressonacircncia Paramagneacutetica Electroacutenica
A acumulaccedilatildeo intracelular de oxovanaacutedio (IV) in vivo constitui uma das principais formas de
apresentaccedilatildeo de vanaacutedio no organismo Deste modo torna-se bastante interessante analisar a
interacccedilatildeo entre este iatildeo e o S1 de miosina atraveacutes da teacutecnica de espectroscopia de Ressonacircncia
Paramagneacutetica Electroacutenica o meacutetodo de eleiccedilatildeo do estudo do iatildeo vanadilo e dos complexos por si
formados O electratildeo desemparelhado encontra-se fortemente acoplado ao nuacutecleo 51
V o que
35
resulta num espectro de RPE consistindo de 8 desdobramentos na direcccedilatildeo paralela e outros tantos
na direcccedilatildeo perpendicular (Chasteen 1983)
Os estudos realizados por Ahmed et al (1987) centraram-se na interacccedilatildeo do vanadilo com
proteiacutenas como eacute o caso da insulina transferrina entre outros obtendo um espectro caracterizado
por 8 linhas amplamente espaccediladas (na direcccedilatildeo paralela) Jaacute Ramos et al 2009 concluiu que a
exposiccedilatildeo da soluccedilatildeo de decavanadato agraves formas globular e filamentosa de actina produz um
espectro tiacutepico do iatildeo oxidovanaacutedio (IV) indicando a reduccedilatildeo do vanadato
Para aferir qual a concentraccedilatildeo de S1 que permitia obter uma boa razatildeo sinalruiacutedo titulou-se uma
soluccedilatildeo de sulfato de vanadilo 15 microM ([VOSO4]stock = 5 mM) com concentraccedilotildees crescentes de
proteiacutena (0 5 10 15 e 20 microM) em 10 mM HEPES 25 mM KCl A existecircncia de uma correlaccedilatildeo
linear entre a concentraccedilatildeo de proteiacutena e a intensidade dos sinais de RPE correspondente agrave banda
-12 perpendicular encontra-se representada na Fig 210
Figura 2 12ndash Representaccedilatildeo graacutefica da variaccedilatildeo da intensidade do sinal de RPE vs a concentraccedilatildeo de S1 utilizada
Apoacutes verificar que com uma concentraccedilatildeo de S1 igual a 20 microM se obtinham sinais com uma razatildeo
sinalruiacutedo adequada ao prosseguimento dos estudos sobre a interacccedilatildeo V(IV)-S1 de miosina
titulou-se uma soluccedilatildeo de 20 microM S1 com 0 25 5 10 15 25 50 75 100 150 200 500 e 1000
microM VOSO4
Traccedilou-se o graacutefico da intensidade da banda -12 perpendicular do sinal de EPR em funccedilatildeo da
concentraccedilatildeo de VOSO4 tendo-se determinado uma constante de dissociaccedilatildeo (KD) do complexo
S1-vanadilo de 8748 plusmn 2621 microM V(IV)
y = 016786x + 122435
Rsup2 = 092992759
Inte
nsi
dad
e d
o s
inal
de
RP
E
[S1] (microM)
36
Figura 213 ndash Representaccedilatildeo graacutefica da variaccedilatildeo do sinal de RPE vs a concentraccedilatildeo de vanadilo utilizada (os valores
experimentais estatildeo representados por losangos o seu ajuste estaacute representado pela linha)
A simulaccedilatildeo do espectro de EPR foi efectuada a partir do espectro obtido para uma concentraccedilatildeo
de 1 mM VOSO4 apoacutes adiccedilatildeo de 20 microM S1 de miosina tendo-se obtido os valores da constante de
acoplamento hiperfina (A=5016) e do factor g (g0=195)
Figura 214 ndash Representaccedilatildeo graacutefica do espectro de EPR obtido com 1000 microM de vanadilo e 20 microM de S1 (a cinzento
escuro) A simulaccedilatildeo do espectro encontra-se a cinzento claro
Por comparaccedilatildeo destes com os valores presentes na tabela de correspondecircncia xxx eacute possiacutevel
predizer quais os ligandos provaacuteveis do vanadilo na presenccedila de proteiacutena Assim concluiu-se que
o vanadilo estaacute coordenado com duas moleacuteculas de aacutegua um grupo hidroxilo e um grupo R=NR
O grupo hidroxilo pode indicar os resiacuteduos de aminoaacutecidos Tyr Treonina e Serina enquanto o
grupo R=NR poderaacute indicar a presenccedila de Arginina e Histidina como ligandos do vanadilo
Inte
nsi
dad
e d
os
sinai
s d
e R
PE
[Vanadilo] (microM)
Ajuste dos dados exp Dados experimentais
Campo magneacutetico (G)
Espectro Experimental Ajuste
37
3 Procedimento Experimental
31 Reagentes
Todos os reagentes utilizados satildeo de elevado grau de pureza (analiacutetico) Apresentam-se as suas
foacutermulas quiacutemicas ou abreviaturas quando necessaacuterio acrescentou-se a sua descriccedilatildeo Entre
parecircntesis colocou-se as empresas que os comercializam
Aacutecido aceacutetico glacial 100 (Riedel) Acrilamida 99 (Merck) Azul de bromofenol (Riedel-de-
Haumlen) -Mercaptoetanol (Sigma) Bissulfito de soacutedio (Sigma)Bradford (reagente de)
(BioRad)BSA (albumina de soro bovino Sigma-Aldrich) CaCl22H2O (cloreto de caacutelcio
dihidratado Fluka) CH3CH2OH 96 (etanol Panreac) Coomassie Blue R250 (Merck) D2O
(oacutexido de deuteacuterio SDS) DTNB (55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico) Aldrich) EDTA (aacutecido
etilenodiaminatetraaceacutetico Merck) Fosfoenolpiruvato 98 (Fluka) Glicerol (Sigma) Glicina
985 (Panreac) HCl 37 (Riedel-de Haumlen) H2SO4 96 (Pronalab) HEPES 99 (aacutecido 4-(2-
hidroxiletil)-1-piperazineetanosulfoacutenico Sigma) KH2PO4 99 (hidrogenofosfato de potaacutessio)
MgATP (Sigma) Na
ATP (Sigma) Na2HPO4 99 (hidrogenofosfato de soacutedio Riedel-de Haumlen)
NaH2PO4 (fosfato de soacutedio ) NaOH 986 (hidroacutexido de soacutedio Pronalab) KCl 99 (Cloreto de
potaacutessio Panreac) L-Cys 995 (enantioacutemero levoacutegiro do resiacuteduo de Cisteiacutena Fluka)Metanol
99 (Merck) Molibdato de amoacutenio (Sigma)NNrsquo-metil-bis-acrilamida 98 (Merck) NH4VO3
99 (metavanadato de amoacutenia Sigma) PSA 10 (persulfato de amoacutenio Merck) SDS
(dodecilo sulfato de soacutedio Sigma) Sulfito de soacutedio (Sigma) VOSO4 (sulfato de vanadilo
Aldrich) TEMED 99 (Tetrametiletilenodiamina Merck) Tris (2-amino-2-hidroximetil-13-
propanodiol Sigma)
32 Meacutetodos
321 Isolamento e purificaccedilatildeo das proteiacutenas de muacutesculo esqueleacutetico de coelho
3211 Purificaccedilatildeo da miosina preparaccedilatildeo de Subfragmento 1 por digestatildeo quimotriacutepsica
da miosina
O subfragmento-1 de miosina-II (S1) foi gentilmente cedido pelo Prof Doutor Manuel Aureliano
do Departamento de Quiacutemica Bioquiacutemica e Farmaacutecia da Faculdade de Ciecircncias e Tecnologia da
Universidade do Algarve
No entanto descreve-se abaixo de forma resumida o procedimento efectuado
38
32111 Purificaccedilatildeo da miosina
O protocolo de isolamento foi adaptado do meacutetodo previamente descrito por Margossian e Lowey
(1982) apresentando-se de forma esquemaacutetica na Fig 31
Sucintamente os tecidos musculares dorsais e das pernas foram extraiacutedos apoacutes o sacrifiacutecio do
animal (Fig31) De seguida foram adicionados 3 volumes da soluccedilatildeo de extracccedilatildeo A2
Centrifugou-se a soluccedilatildeo e filtrou-se o sobrenadante O sedimento foi entatildeo diluiacutedo em 14
volumes de aacutegua destilada de modo a que a forccedila ioacutenica atingisse 004 M Apoacutes adiccedilatildeo de szlig-
mercaptoetanol (szlig-me) deixou-se a precipitar a 4ordmC removendo o sobrenadante Centrifugou-se
novamente e ressuspendeu-se o sedimento na soluccedilatildeo de extracccedilatildeo A Adicionou-se a soluccedilatildeo
tampatildeo B3 para que a forccedila ioacutenica atingisse 05 M Homogeneizou-se e deixou-se repousar
durante 2h Homogeneizou-se novamente e centrifugou-se de modo a ocorrer precipitaccedilatildeo dos
complexos de actomiosina e proteiacutena desnaturada Apoacutes filtraccedilatildeo do sobrenadante em latilde de vidro
quantificou-se a soluccedilatildeo de miosina resultante pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental)
2 Soluccedilatildeo de extracccedilatildeo A 0012M K2CO3 004M KHCO3 06M KCl pH 65
3 Soluccedilatildeo-tampatildeo B Tris 100 mM pH 7 KCl 2M
39
Figura 31 ndash Representaccedilatildeo esquemaacutetica do protocolo de isolamento de miosina (Margossian e Lowey 1982) Todo o
procedimento foi realizado a 4ordmC (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
32112 Preparaccedilatildeo de Subfragmento-1 de miosina por digestatildeo com quimiotripsina
O S1 de miosina foi isolado apoacutes diaacutelise da miosina durante a noite contra a soluccedilatildeo C4 A
presenccedila do EDTA eacute essencial uma vez que a sua substituiccedilatildeo por MgCl2resulta na produccedilatildeo de
HMM em vez de S1 A suspensatildeo de miosina foi equilibrada e digerida com -quimotripsina
A digestatildeo foi interrompida pela adiccedilatildeo de PMSF Seguidamente ultracentrifugou-se o produto
da digestatildeo desprezando-se o sedimento (Fig 32) A soluccedilatildeo de sobrenadante maioritariamente
S1 foi quantificada pela leitura directa da Abs280 ou pelo meacutetodo de Bradford (ver as secccedilotildees
3311 e 3312 do Procedimento Experimental respectivamente) e armazenada em 10 sacarose
e 2 mM szlig-Me a -80ordmC
4 Soluccedilatildeo C 25mM Tris pH 70 012M KCl 1mM EDTA
40
Antes da realizaccedilatildeo dos ensaios descritos no presente relatoacuterio o S1 de miosina foi dialisado
contra a soluccedilatildeo D5 de modo a remover qualquer composto que pudesse complexar como
vanaacutedio Seguidamente quantificou-se a proteiacutena pelo meacutetodo de Bradford ou pelo meacutetodo
directo (ver secccedilotildees 3311 e 3312do Procedimento Experimtental)
Figura 3 2ndash Representaccedilatildeo esquemaacutetica do protocolo de preparaccedilatildeo do S1 de miosina por digestatildeo quimiotripsina de
acordo com o previamente descrito por Margossian e Lowey (1982) Todo o procedimento foi realizado a 4ordm C
excepto quando indicaccedilatildeo do contraacuterio (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
3212 Isolamento e purificaccedilatildeo de actina
Isolou-se e purificou-se a actina segundo o meacutetodo descrito por Pardee e Spudish que
compreende dois passos ndash a obtenccedilatildeo de um resiacuteduo seco denominado ldquopoacute de acetonardquo e a
extracccedilatildeo subsequente da actina a partir do ldquopoacute de acetonardquo na forma de G-Actina com a
posterior polimerizaccedilatildeo a F-Actina
32121 Obtenccedilatildeo do ldquopoacute de acetonardquo
Sacrificou-se o coelho com uma pancada atraacutes do pescoccedilo e sangrou-se o animal apoacutes o corte da
arteacuteria jugular Esfolou-se e colocou-se numa tina com gelo Retiraram-se as partes de tecido
conjuntivo que envolvem o tecido muscular com auxiacutelio de uma pinccedila e bisturi Seguidamente
retirou-se os muacutesculos dorsais e os muacutesculos brancos das pernas Passou-se os muacutesculos por aacutegua
destilada de modo a remover as impurezas e pesou-se num copo Trituraram-se os muacutesculos
numa maacutequina de triturar carne e extraiu-se o resultante com 3 volumes da soluccedilatildeo E6 durante 10
minutos com agitaccedilatildeo suave Centrifugou-se a suspensatildeo a 4000 g durante 10
5 Soluccedilatildeo D HEPES 10 mM pH 70 KCl 25 mM
6 Soluccedilatildeo E 01 M KCl 015 M K2HPO4 pH 65
41
minutosdesprezando-se o sobrenadante Ressuspendeu-se o sedimento em 6 volumes de soluccedilatildeo
F7durante 10 minutos com agitaccedilatildeo suave Verificou-se se o pH da soluccedilatildeo estava compreendido
entre 82 e 85 acertando-se estecom soluccedilatildeo G8 se necessaacuterio Centrifugou-se durante 10
minutos a 4000 g Desprezou-se o sobrenadante e ressuspendeu-se o sedimento em 3 volumes de
soluccedilatildeo H9 durante 10 minutos Centrifugou-se novamente a 4000 gdurante 10 minutos
desprezando-se o sobrenadante Ressuspendeu-se o sedimento duas vezes consecutivas em 6
volumes de aacutegua destilada durante 5 minutos Centrifugou-se a 4000 g durante 10 minutos apoacutes
cada adiccedilatildeo de H2O
Seguidamente ressuspendeu-se o sedimento 5 vezes consecutivas utilizando 3 volumes de
acetona de cada vez agitando suavemente durante 10 minutos agrave temperatura ambiente Colocou-
se o resiacuteduo filtrado numa tina de vidro sobre papel de filtro deixando-se evaporar o solvente
durante a noite num nicho Triturou-se o resiacuteduo seco numa maacutequina de triturar carne e deixou-se
novamente evaporar o solvente O ldquopoacute de acetonardquo obtido foi armazenado a -20 ordmC
32122 Purificaccedilatildeo da actina a partir de ldquopoacute de acetonardquo
Apoacutes a obtenccedilatildeo do poacute de acetona como acima indicado efectuou-se o protocolo seguidamente
descrito de modo a obter monoacutemeros de actina (G-Actina) e actina filamentosa (F-Actina)
Pesou-se 35 g de poacute de acetona e adicionaram-se 50 mL de soluccedilatildeo-tampatildeo I10
sendo a
extracccedilatildeo feita com agitaccedilatildeo suave durante 30 minutos Separou-se o extracto do poacute de acetona
hidratado filtrando a suspensatildeo com 4 camadas de gaze esterilizada tendo o cuidado de natildeo
espremer o resiacuteduo Voltou a extrair-se com 50 mL de soluccedilatildeo I durante 30 minutos mexendo
lentamente Adicionou-se este ao extracto obtido anteriormente Centrifugou-se o extracto total a
16 000 g durante 30 minutos Decantou-se para uma proveta sobre a qual se colocou gaze para
que o sedimento solto natildeo se juntasse ao sobrenadante e leu-se o volume De seguida adicionou-
se KCl ateacute uma concentraccedilatildeo final de 50 mM MgCl2 ateacute 2mM e ATP ateacute 1mM lentamente e com
agitaccedilatildeo contiacutenua de modo a promover a polimerizaccedilatildeo dos monoacutemeros de actina
Deixou-se polimerizar durante 2h a 25ordmC observando-se a presenccedila de pequenas bolhas de ar
indicativas de um aumento de viscosidade da soluccedilatildeo Arrefeceu-se em gelo durante 15 minutos e
adicionou-se KCl ateacute uma concentraccedilatildeo final de 06M Mexeu-se lentamente durante 30 minutos
apoacutes os quais se ultracentrifugou a 110 000g durante 1hora Desprezou-se o sobrenadante
ressuspendeu-se o sedimento com soluccedilatildeo tampatildeo E e transferiu-se para um potter
Dialisou-se durante a noite contra 1L de soluccedilatildeo-tampatildeo I a uma temperatura de 4ordmC com rotaccedilatildeo
7 Soluccedilatildeo F 005 M NaHCO3 pH 82-85
8 Soluccedilatildeo G 1M Na2CO3
9 Soluccedilatildeo H 1 mM EDTA pH 70
10 Soluccedilatildeo I 2 mM Tris-HCl pH 8 02 mM CaCl2 02 mM Na2ATP 05 mM szlig-Me 0005 NaN3
42
contiacutenua e mudanccedila de tampatildeo 2 a 3 vezes De seguida a actina foi centrifugada a 110
000gdurante 1hora Decantou-se a soluccedilatildeo de G-Actina (sobrenadante) e desprezou-se o
sedimento Determinou-se a concentraccedilatildeo de G-Actina pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental) A soluccedilatildeo de G-actina foi armazenada a uma temperatura de 4ordmC
apoacutes adiccedilatildeo de 002NaN3 mantendo-se estaacutevel durante 3 a 5 dias
331 Quantificaccedilatildeo das proteiacutenas isoladas
3311 Quantificaccedilatildeo pelo meacutetodo de Bradford
O meacutetodo de Bradford (Bradford 1976) baseia-se na utilizaccedilatildeo do reagente de Bradford
composto por entre outros aacutecido fosfoacuterico e o corante ldquoCoomassierdquo Este corante possui carga
negativa e a sua coloraccedilatildeo varia entre a vermelha (Absmax=465 nm) predominante em soluccedilatildeo e a
azul (Absmax=595 nm) presente aquando da ligaccedilatildeo aos grupos amina das proteiacutenas Em soluccedilotildees
aacutecidas diluiacutedas a coloraccedilatildeo varia proporcionalmente com a quantidade do corante ligado agrave
proteiacutena que por sua vez eacute directamente proporcional agrave concentraccedilatildeo da proteiacutena permitindo a
sua quantificaccedilatildeo ateacute a um limite de detecccedilatildeo de 20 gmL (Bradford 1976)
Deste modo prepararam-se amostras de BSA ([BSA]stock= 1mgmL) utilizando um intervalo de
concentraccedilotildees compreendido entre 0 e 15 gmL de acordo com a tabela 3I Apoacutes a adiccedilatildeo da
H2O MilliQ e do reagente de Bradford as soluccedilotildees foram sucessivamente homogeneizadas no
voacutertex
Tabela 3Indash Amostras utilizadas para a obtenccedilatildeo da recta de calibraccedilatildeo no meacutetodo de Bradford
BSA (L) BSA (g) H2O MilliQ (L) Reagente de Bradford (L)
0 0 800 200
125 125 79875 200
25 25 7975 200
5 5 795 200
10 10 790 200
15 15 785 200
Apoacutes incubaccedilatildeo a 25ordmC por 5 minutos leram-se os valores de Abs595 num
espectrofotoacutemetro (Shimadzu UV-160A) para todas as amostras Seguidamente traccedilou-se a recta
de calibraccedilatildeo (Abs595vs quantidade de BSA (g))
43
Para determinar a concentraccedilatildeo de proteiacutena apoacutes o processo de isolamento purificaccedilatildeo e
diaacutelise quando necessaacuterio colocou-se 5 L da amostra num tubo de Eppendorf e adicionou-se
795 L de H2O MilliQ homogeneizando a soluccedilatildeo no voacutertex Adicionou-se posteriormente 200
L de reagente de Bradford e repetiu-se a homogeneizaccedilatildeo Incubou-se a mistura a 25ordmC por 5
minutos apoacutes os quais se leu os valores de Abs595 nm no espectrofotoacutemetro Por interpolaccedilatildeo deste
valor a partir da recta de calibraccedilatildeo determinou-se a concentraccedilatildeo de proteiacutena na amostra
3312 Quantificaccedilatildeo pelo meacutetodo directo
A quantificaccedilatildeo das amostras proteicas pode tambeacutem ser efectuada atraveacutes da leitura directa da
absorvacircncia tendo por base a lei de Beer-Lambert onde Abs corresponde agrave
absorvacircncia ao comprimento de onda ao qual o ensaio foi realizado corresponde ao coeficiente
de extinccedilatildeo (substituiacutedo pela letra a se as unidades forem distintas da molaridade) b corresponde
ao percurso oacuteptico da radiaccedilatildeo na ceacutelula amostral e c corresponde agrave concentraccedilatildeo amostral
Assim no caso de S1 de miosina mediu-se a Abs280e calculou-se a concentraccedilatildeo da proteiacutena
sabendo que o coeficiente de extinccedilatildeo de S1 eacute (Ref)
No caso da actina mediu-se a Abs290ecalculou-se a concentraccedilatildeo da proteiacutena sabendo que o
coeficiente de extinccedilatildeo da actina eacute (Ref)
332Electroforese em condiccedilotildees desnaturantes (SDS-PAGE)
De modo a melhor caracterizar as proteiacutenas isoladas foi realizada uma electroforese em
condiccedilotildees desnaturantes (SDS-PAGE do inglecircs Sodium Dodecyl Sulfate Polyacrilamide Gel
Electrophoresis) O SDS (dodecilo sulfato de soacutedio) eacute um detergente anioacutenico utilizado neste
meacutetodo para desnaturar as proteiacutenas que se ligam ao SDS na proporccedilatildeo 14 g SDS g proteiacutena
(aproximadamente 1 moleacutecula de SDS por cada 2 resiacuteduos de aminoaacutecidos) (Stryer) O SDS induz
uma densidade de carga negativa significativa nas proteiacutenas a que se liga mascarando a sua carga
intriacutenseca pelo que as razotildees cargamassa satildeo tendencialmente idecircnticas Assim a SDS-PAGE
separa as proteiacutenas quando sujeitas a um campo eleacutectrico de acordo com o seu peso molecular
sendo a sua migraccedilatildeo directamente proporcional ao logaritmo deste uacuteltimo (Stryer)
O gel de resoluccedilatildeo com 10 de acrilamida foi obtido atraveacutes da adiccedilatildeo de 167 mL de soluccedilatildeo
stock de acrilamida11
075 mL de tampatildeo de resoluccedilatildeo12
251 mL de aacutegua destilada e 50 L de
SDS 10 Para promover a polimerizaccedilatildeo adicionou-se em seguida 38 L de PSA 10
11
soluccedilatildeo stock de acrilamida soluccedilatildeo de 30 (mv) de acrilamida e 08 (mv) de NNrsquo-metil-bis-acrilamida 12
tampatildeo de resoluccedilatildeo 15 M Tris-HCl pH 88
44
(persulfato de amoacutenia) 10 e 25 L de TEMED 99 (tetrametiletilenodiamina) Agitou-se
cuidadosamente de modo a natildeo formar bolhas e pipetou-se a soluccedilatildeo para a cassette de vidro
previamente montada no suporte De seguida adicionou-se uma peliacutecula de cerca de 2 mm de 2-
metil-2-butanol para verificar posteriormente a polimerizaccedilatildeo Apoacutes a polimerizaccedilatildeo que demora
cerca de 45 minutos verteu-se a agrave superfiacutecie e adicionou-se o gel de concentraccedilatildeo com 5 de
acrilamida constituiacutedo por 03 mL de soluccedilatildeo de stock acrilamida 045 mL de tampatildeo de
concentraccedilatildeo13
094 mL de aacutegua destilada 18 L de SDS 10 135 L de PSA 10 e 2L de
TEMED 99 Colocou-se o pente sobre o gel e deixou-se polimerizar durante cerca de 45
minutos
As amostras para aplicaccedilatildeo no gel de electroforese foram preparadas para uma concentraccedilatildeo de
proteiacutena de 1 mgmL num volume final de 100 L dos quais 25 L correspondem ao tampatildeo da
amostra14
As amostras foram aquecidas a 100ordmC durante 5 minutos Antes da aplicaccedilatildeo das
amostras transferiu-se a cassette para a tina de electroforese agrave qual se adicionou a soluccedilatildeo
tampatildeo de electroforese da tina superior15
e tampatildeo de electroforese da tina inferior16
Aplicaram-
se 10 L das soluccedilotildees de marcadores de peso molecular e 20 L das amostras proteicas
preparadas anteriormente A electroforese decorreu durante cerca de 3h30 min a uma voltagem
de 80 V De seguida transferiu-se o gel para uma tina corando as proteiacutenas durante 1h com
soluccedilatildeo corante17
Descorou-se o gel com soluccedilatildeo descorante18
tendo-se mudado esta soluccedilatildeo ateacute
as bandas se encontrarem bem definidas
Determinou-se a mobilidade relativa das bandas correspondentes aos marcadores de elevado peso
molecular e traccedilou-se a recta de calibraccedilatildeo ndash log(PM) vs log(Rf) onde Rf corresponde agrave
mobilidade relativa Por interpolaccedilatildeo determina-se o PM das proteiacutenas isoladas sabendo a sua
mobilidade relativa Por comparaccedilatildeo com os valores descritos na literatura pode concluir-se
acerca do grau de purificaccedilatildeo
34 Mediccedilatildeo da actividade enzimaacutetica de S1
341 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
13
tampatildeo de concentraccedilatildeo 05 M Tris-HCl pH 68 14
tampatildeo de amostra 320 mM Tris-HCl pH 68 04 M szlig-mercaptoetanol SDS 8 glicerol 15 (vv) e roxo de m-
cresol 0024 15
tampatildeo de electroforese da tina superior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS 16
tampatildeo de electroforese da tina inferior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS acetato de soacutedio
01 M 17
soluccedilatildeo corante ldquoCoomassie Blue R-250rdquo contendo metanol 45 (vv) e aacutecido aceacutetico glacial 10 (vv) 18
soluccedilatildeo descorante metanol 10 (vv) e aacutecido aceacutetico glacial 10 vv)
45
O meacutetodo de Fiske-Subarow modificado consiste na reacccedilatildeo do molibdato de amoacutenio com o
fosfato inorgacircnico em soluccedilatildeo aacutecida com formaccedilatildeo do aacutecido fosfomoliacutebdico Seguidamente daacute-
se a adiccedilatildeo de um agente redutor que provoca a reduccedilatildeo do composto a fosfomolibdato que forma
um complexo de cor azulada cuja absorvacircncia maacutexima ocorre para um comprimento de onda de
660 nm tornando possiacutevel a sua detecccedilatildeo por meacutetodos espectrofotomeacutetricos As actividades Ca2+
-
ATPaacutesica e K+EDTA-ATPaacutesica de S1 foram determinadas atraveacutes deste meacutetodo de acordo com
o descrito por Taussky e Shorr (1953) com mediccedilatildeo da quantidade de fosfato (Pi) libertado
atraveacutes da hidroacutelise de ATP
Para tal obteve-se primeiramente a recta de calibraccedilatildeo com as amostras presentes na Tabela 3II
utilizando uma soluccedilatildeo stock de 5 mM KH2PO4
Tabela 3II ndash Composiccedilatildeo das diferentes amostras utilizadas para determinar a recta de calibraccedilatildeo atraveacutes do meacutetodo
de Fiske-Subbarow modificado
KH2PO4 (L)
([KH2PO4]stock= 5mM)
PO43-
(nmole) H2O (uL)
0 0 1780
5 25 1775
10 50 1770
20 100 1760
40 200 1740
100 500 1680
A cada tubo de Eppendorf adicionaram-se seguidamente 200 L de soluccedilatildeo J19
e 20 L de agente
redutor20
homogeneizando no voacutertex apoacutes cada adiccedilatildeo Incubou-se durante 10 minutos e mediu-
se a Abs660 nm num espectrofotoacutemetro com intervalos de tempo iguais entre cada amostra Traccedilou-
se a recta de calibraccedilatildeo (Abs660 nmvs nmol PO43-
) a partir dos valores obtidos
Para determinar a actividade Ca2+
-ATPaacutesica de S1 prepararam-se tubos de Eppendorf com 400
L de meio de reacccedilatildeo21
25 L CaCl2 ([CaCl2]stock= 100 mM) 25 L ATP ([ATP]stock= 100 mM)
com [S1]ensaios=01 mgmL Perfez-se se necessaacuterio com H2O MilliQ ateacute perfazer um volume de 1
mL Para determinar a actividade K+EDTA-ATPaacutesica utilizou-se uma concentraccedilatildeo idecircntica de
19
Soluccedilatildeo J 5 N H2SO4 25 molibdato de amoacutenio 20
Agente redutor 5 mg aacutecido 1-amino-2-naftol-4-sulfoacutenico 30 mg bissulfito de soacutedio 30 mg sulfito de soacutedio em 26
mL H2O 21
meio de reacccedilatildeo 125 mM Tris-HCl pH 80 0575 M KCl
46
proteiacutena nos ensaios (01 mgmL) e o mesmo meio de reacccedilatildeo Aos tubos de Eppendorf
adicionou-se ainda 123 L de KCl ([KCl]stock = 3M) 25 L de ATP 10 L de EDTA
([EDTA]stock= 100 mM )e o volume de H2O MilliQ necessaacuterio para perfazer 1 mL Em ambas as
situaccedilotildees o uacuteltimo reagente adicionado foi o ATP uma vez que se trata do substrato da reacccedilatildeo
enzimaacutetica
A cineacutetica decorreu a 25ordmC durante 5 minutos para a actividade Ca2+
-ATPaacutesica e durante 3
minutos para a actividade K+EDTA-ATPaacutesica Apoacutes o periacuteodo de incubaccedilatildeo transferiu-se 150
L de amostra para tubos de Eppendorf contendo igual volume de TCA 10 Colocou-se no gelo
e homogeneizou-se no voacutertex Seguidamente centrifugou-se a 1500 g durante 10 minutos e
recolheu-se 200 L de sobrenadante para novos tubos de Eppendorf Adicionou-se a estes 158
mL de H2O MilliQ e 200 L de soluccedilatildeo J Homogeneizou-se no voacutertex e adicionou-se 20 L de
reagente redutor repetindo-se a homogeneizaccedilatildeo Incubou-se durante 10 minutos e mediu-se a
Abs660 nm com o mesmo intervalo de tempo de leitura para cada amostra Por intrapolaccedilatildeo a partir
da recta de calibraccedilatildeo obtida anteriormente determinou-se a quantidade de Pi libertado em cada
ensaio A partir destes resultados determinaram-se as actividades Ca2+
-ATPaacutesica e K+EDTA-
ATPaacutesica do S1 em nmol PO43-
minmg proteiacutena e em s-1
342 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
Da actividade Mg2+
-ATPaacutesica de S1 resulta a formaccedilatildeo dos produtos da reacccedilatildeo de
hidroacutelise do ATP - ADP e Pi Na presenccedila de ADP fosfoenolpiruvato (PEP) e da enzima piruvato
cinase (PK) os produtos resultantes satildeo ATP e piruvato Por acccedilatildeo da enzima lactato
desidrogenase (LDH) este uacuteltimo eacute convertido em lactato ocorrendo simultaneamente a oxidaccedilatildeo
do NADH a NAD+ monitorizada atraveacutes da Abs340 nm Quando o S1 de miosina se encontra no
meio reaccional o ATP eacute hidrolisado formando-se novamente ADP e Pi o que permite a
formaccedilatildeo de um ciclo (fig 21) Se as enzimas se encontrarem em excesso a velocidade de
conversatildeo do NADH eacute limitada pela produccedilatildeo de ADP e Pi Como o ATP eacute regenerado em cada
ciclo a sua concentraccedilatildeo inicial eacute sempre mantida (Bagshaw 1982)
Figura 33ndash Representaccedilatildeo esquemaacutetica do meacutetodo de enzimas acopladas
47
O protocolo utilizado e seguidamente descrito foi adaptado de Cuenda et al (1990 ndash Ver Refordf) A
uma cuvette de 1 mL adicionou-se S1 para uma concentraccedilatildeo final de 005 mgmL 800 L de
meio de reacccedilatildeo22
10 L de PEP ([PEP]stock = 42 mM) 10 L de PK+LDH 11 ([PK+LDH]stock =
1 mgmL) e 15 L de NADH ([NADH]stock=25 mM) Iniciou-se a monitorizaccedilatildeo da reacccedilatildeo
atraveacutes da leitura de Abs340 nm apoacutes a adiccedilatildeo de 20 L de ATP ([ATP]stock=100 mM) A
actividade Mg2+
-ATPaacutesica eacute obtida atraveacutes do declive correspondente agrave oxidaccedilatildeo do NADH Para
a mediccedilatildeo da Mg2+
-ATPase de S1 estimulada por actina adicionou-se F-actina para uma
concentraccedilatildeo final de 2 microM sendo a actividade obtida tambeacutem atraveacutes do declive correspondente
agrave oxidaccedilatildeo do NADH apoacutes a adiccedilatildeo da proteiacutena
35 Interaccedilatildeo entre vanaacutedio e subfragmento-1 de miosina
351 Preparaccedilatildeo das soluccedilotildees de vanaacutedio
As soluccedilotildees de vanadato foram preparadas em aacutegua MilliQ a partir de metavanadato de amoacutenio
(NH4VO3) numa concentraccedilatildeo stock de 50 mM com agitaccedilatildeo magneacutetica e aquecimento (~30 ordmC)
Esta soluccedilatildeo com um valor compreendido entre 6 e 7 designa-se por ldquometavanadatordquo e eacute
constituiacuteda pelas esspeacutecies V1 V2 V4 e V5 (Ref) Acidificando a soluccedilatildeo de metavanadato por
adiccedilatildeo de HCl ateacute um valor de pH proacuteximo de 4 observa-se uma mudanccedila de cor de amarelo
claro para laranja indicativa da formaccedilatildeo da espeacutecie decameacuterica de vanadato De seguida e
imediatamente antes da realizaccedilatildeo dos ensaios descritos o valor do pH da soluccedilatildeo de
decavanadato foi aumentado para cerca de 75 por adiccedilatildeo de NaOH A soluccedilatildeo (2) designada
por ldquometavanadatordquo natildeo foi posteriormente alterada ateacute agrave sua adiccedilatildeo para a realizaccedilatildeo dos ensaios
As soluccedilotildees foram armazenadas a 4ordmC e mantidas em gelo aquando da sua utilizaccedilatildeo
A soluccedilatildeo de V(IV) foi preparada em condiccedilotildees anaeroacutebias por diluiccedilatildeo de sulfato de vanadil
(VOSO4) para uma concentraccedilatildeo final de 5 mM em H2O MilliQ saturada com N2 gasoso
352Anaacutelise por espectroscopia de UV-Vis
A concentraccedilatildeo da espeacutecie decameacuterica em soluccedilatildeo depende do periacuteodo de tempo apoacutes a diluiccedilatildeo
no meio de reacccedilatildeo apropriado (Ref) A velocidade de desoligomerizaccedilatildeo da espeacutecie decameacuterica
pode ser analisada por espectroscopia de absorccedilatildeo na regiatildeo UVVis uma vez que esta espeacutecie
apresenta duas bandas caracteriacutesticas na regiatildeo do visiacutevel (a 358 e 400 nm) que natildeo se verifica
em quaisquer outras espeacutecies oligomeacutericas As bandas supramencionada sapresentam-se bem
22
Meio de reacccedilatildeo 3125 mM Tris-HCl pH 71 375 mM MgCl2
48
resolvidas atraveacutes da obtenccedilatildeo da derivada de 2ordf ordem do espectro apresentando a forma de 2
vales com miacutenimos centrados nos comprimentos de onda supracitados com dependecircncia linear
relativamente agrave concentraccedilatildeo de decavanadato total (Ramos et al 2006)
Assim a estabilidade da espeacutecie decameacuterica apoacutes diluiccedilatildeo da soluccedilatildeo de decavanadato no meio de
reacccedilatildeo dos ensaios (tampatildeo D) para uma concentraccedilatildeo final de 1 mM em vanadato total foi
caracterizada atraveacutes da leitura da Abs400 em funccedilatildeo do tempo tendo-se determinado qual a
ordem de reacccedilatildeo e o tempo de meia-vida da mesma Com base em estudos anteriores acerca da
estabilidade do V10 na presenccedila de diferentes proteiacutenas (Ramos et al 2006) determinou-se
igualmente o tempo de meia vida apoacutes a adiccedilatildeo de 20 microM de S1 de miosina ao meio reaccional
353 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
Os espectros de RMN do isoacutetopo 51
V foram realizados num espectroacutemetro Bruker AM-400 MHz
equipado com uma sonda multinuclear inversa de 5 mm Em tubos de RMN de 5 mm de diacircmetro
preparou-se soluccedilotildees de 500 L de deca- ou metavanadato correspondentes a um intervalo de
concentraccedilotildees de 1 a 10 mM suplementadas com 10 D2O Os espectros foram adquiridos agrave
temperatura ambiente com uma largura espectral de 23 923 Hz um tempo de aquisiccedilatildeo de 0086
s tempo de relaxaccedilatildeo de 001 s e uma sequecircncia de pulsos a 90ordm Utilizou-se a transformada de
Fourier com alargamento das bandas de 20Hz para o processamento dos espectros Os desvios
quiacutemicos das ressonacircncias de 51
V foram obtidos em relaccedilatildeo agrave espeacutecie VOCl3cuja frequecircncia de
ressonacircncia eacute considerada o padratildeo sendo todas as outras referenciadas a partir desta em ppm
As aacutereas dos sinais do RMN correspondentes agraves espeacutecies oligomeacutericas presentes foram obtidas
por integraccedilatildeo dos sinais As concentraccedilotildees relativas das espeacutecies foram calculadas com recurso agrave
equaccedilatildeo
onde A[Vx] corresponde agrave aacuterea referente agrave ressonacircncia da espeacutecie Vx com nordm de agregaccedilatildeo x
A[V]total agrave soma total das aacutereas medidas e [V] a concentraccedilatildeo total de vanaacutedio no ensaio
A teacutecnica de 51
V-RMN foi anteriormente aplicada em estudos de ligaccedilatildeo com S1 de miosina
(Ref) tendo sido demostrado que entre as espeacutecies V1 V2 V4 e V5 a espeacutecie tetrameacuterica possui a
maior afinidade para a miosina (Cremo et al) O desaparecimento ou alargamento dos sinais pode
implicar a ligaccedilatildeo de um dos oxovanadatos laacutebeis agrave miosina sem indicaccedilatildeo de qual uma vez que
49
se processa um equiliacutebrio entre eles (Crans 1990) Assim e tendo em conta as indicaccedilotildees de que
a espeacutecie V10 eacute distinta das restantes no que diz respeito agrave magnitude da interacccedilatildeo com a miosina
e inibiccedilatildeo da actividade ATPaacutesica estimulada por actina (Aureliano 2000 Tiago et al 2002)
torna-se fundamental investigar a interacccedilatildeo das diferentes espeacutecies de vanadato com o S1 de
miosina atraveacutes da teacutecnica de 51
V-RMN
Para tal obtiveram-se espectros de 51
V-RMN resultantes da titulaccedilatildeo de uma soluccedilatildeo 5 mM de
decavanadato total com concentraccedilotildees crescentes de S1 de miosina (0 5 10 15 e 20 microM) em
tampatildeo D Realizou-se ainda ensaios na presenccedila de 2 mM de decavanadato e 2 mM de ATP na
presenccedila e na ausecircncia de S1 de miosina de modo a verificar o efeito que estes ligandos naturais
da enzima teria na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o subfragmento-1
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
Os grupos sulfidrilo (-SH) das cisteiacutenas satildeo resiacuteduos de aminoaacutecidos extremamente sensiacuteveis agrave
oxidaccedilatildeo resultando na sua oxidaccedilatildeo (Dalle-Donne et al 1999) Logo a quantificaccedilatildeo destes
resiacuteduos eacute bastante importante nos estudos de interacccedilatildeo com vanaacutedio jaacute que os resiacuteduos de Cys
com ecircnfase para o SH1 e o SH2 satildeo bastante importantes na monitorizaccedilatildeo das modificaccedilotildees
funcionais da regiatildeo da cabeccedila
O procedimento seguidamente apresentado foi originalmente descrito por Gutieacuterrez-Merino
(1987)) Consiste na reacccedilatildeo entre os grupos ndashSH natildeo oxidados das proteiacutenas e o reagente de
Ellman (DTNB -55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico)) - formando-se o aniatildeo CNT (aniatildeo 3-
carboxilato-4-nitrotiofenolato) numa estequiometria de 11 Este apresenta um maacuteximo de
absorvecircncia a 412 nm (Robyt 1990) Assim apoacutes a incubaccedilatildeo da proteiacutena na presenccedila do metal
a quantidade de cisteiacutenas reduzidas por proteiacutena determina-se atraveacutes dos valores de Abs412
Deste modo colocou-se num tubo de Eppendorf o volume apropriado de S1 dialisado para que a
concentraccedilatildeo no ensaio fosse de 1 mgmL Adicionou-se o volume apropriado de decavanadato
para que a sua concentraccedilatildeo fosse de 0 10 50 100 250 500 ou 1000 M Perfez-se com soluccedilatildeo
D ateacute um volume final de 1 mL e incubou-se a 25ordmC por 20 minutos Apoacutes este periacuteodo colocou-
se 780 L da mistura reaccional numa cuvette de quartzo e adicionou-se 220 L de soluccedilatildeo K23
23
Tampatildeo K Tampatildeo fosfatohidrogenofosfato de soacutedio 025 M pH 72
50
Adicionou-se 15 L de DTNB (em soluccedilatildeo K [DTNB]stock=100 mM) ao meio reaccional e
registou-se o valor de absorvecircncia medido equivalente agraves cisteiacutenas ldquoraacutepidasrdquo (SH1 e SH2
expostos ao solvente) Adicionou-se 001 g de SDS que provoca a desnaturaccedilatildeo da proteiacutena e
consequentemente a exposiccedilatildeo de todos os resiacuteduos de cisteiacutenas que se encontravam ateacute aiacute
inacessiacuteveis no interior da proteiacutena (cisteiacutenas totais) Estes resiacuteduos podem assim reagir com o
DTNB
Estimou-se o nuacutemero de resiacuteduos de cisteiacutenas em todas as amostras com recurso ao coeficiente de
extinccedilatildeo molar () do DTNB (ver ponto 3 dos Anexos) que foi determinado atraveacutes da titulaccedilatildeo
de L-Cisteiacutena com intervalo de concentraccedilotildees compreendidos entre 0 e 50 microM na presenccedila de
25microM de DTNB em tampatildeo D Os valores de absorvecircncia foram obtidos para um comprimento de
onda de 412 nm
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
3621 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
A teacutecnica mais eficiente de anaacutelise da interacccedilatildeo entre o iatildeo vanadilo e proteiacutenas como a
calmodulina e a transferrina entre outras permite a obtenccedilatildeo raacutepida de informaccedilatildeo no que toca agrave
conformaccedilatildeo e amplitude da referida interacccedilatildeo
Os espectros de Ressonacircncia Paramagneacutetica Electroacutenica (RPE) da espeacutecie paramagneacutetica V(IV)
foram realizados a 70K num espectroacutemetro Bruker EMX-300 X-band Todas as soluccedilotildees
utilizadas e amostras foram preparadas em anaerobiose numa cacircmara de luvas de modo a evitar a
reduccedilatildeo de V(IV) a V(V) Em tubos capilares com 02 mL de amostra manteve-se a concentraccedilatildeo
de S1 a 20 M e variou-se a concentraccedilatildeo de vanadilo (0 25 5 10 15 25 50 75 100 150
200 500 e 1000 M [VOSO4]stock=5 mM) em tampatildeo D Apoacutes obtenccedilatildeo dos espectros estes
foram tratados atraveacutes do software SimFonia e WinEPR tendo-se obtido um graacutefico com a
variaccedilatildeo da intensidade do sinal de RPE em funccedilatildeo da concentraccedilatildeo de sulfato de vanadilo
presente Realizaram-se ainda simulaccedilotildees de espectros obtidos com 20 microM de S1 e 1000 microM de
V(IV) com vista agrave obtenccedilatildeo dos ligandos provaacuteveis do vanadilo atraveacutes da anaacutelise da constante
de acoplamento hiperfina (A) e do factor g (g)
4 Conclusotildees gerais
Apesar de jaacute ter sido demonstrado que o vanaacutedio inibe a Na+K
+-ATPase da miosina a funccedilatildeo
deste elemento no tecido muacutesculo esqueleacutetico continua largamente incompreendida A enorme
complexidade quiacutemica deste metal especialmente no que concerne agrave oligomerizaccedilatildeo e reacccedilotildees
51
redox constitui um obstaacuteculo agrave compreensatildeo do comportamento do vanaacutedio nos organismos
vivos Assim a caracterizaccedilatildeo da quiacutemica deste elemento eacute de grande utilidade antes de iniciar o
estudo da sua interacccedilatildeo com o presente sistema
A anaacutelise por espectroscopias de 51
V-RMN e de UV-Vis de soluccedilotildees de vanadato diluiacutedas nos
meios de reacccedilatildeo utilizados no presente relatoacuterio (10 mM HEPES pH 70 25 mM KCl) mostrou
claramente que a espeacutecie decameacuterica (V10) possui um tempo de vida suficientemente longo para a
determinaccedilatildeo do seu efeito isolado ao contraacuterio do que se verifica com as espeacutecies mono- di-
tetra- e pentameacutericas (V1 V2 V4 e V5) Por outro lado a anaacutelise verificou ainda que apoacutes diluiccedilatildeo
das soluccedilotildees de decavanadato no meio de reacccedilatildeo coexistem as espeacutecies V10 e V1 pelo que os
efeitos putativos desta uacuteltima deveratildeo ser subtraiacutedos aquando da monitorizaccedilatildeo dos efeitos da
espeacutecie decameacuterica
O vanaacutedio (V) (vanadato) inibe as enzimas que catalizam reacccedilotildees de transferecircncia de grupos
fosfato como a ATPase da miosina [Goodno 1979] devido agrave sua analogia estrutural com o iatildeo
fosfato Na sua utilizaccedilatildeo como inibidor enzimaacutetico natildeo eacute tida em conta a contribuiccedilatildeo individual
das espeacutecies oligomeacutericas considerando-se normalmente a espeacutecie V1 como inibidora efectiva
[Goodno e Taylor 1982] No presente relatoacuterio verificou-se por espectroscopia de 51
V-RMN que
outros oligoacutemeros de vanadato interagem com o S1 de miosina tendo as espeacutecies V10 e V4 os
maiores niacuteveis de afinidade A sua interacccedilatildeo envolve forccedilas electrostaacuteticas e eacute afectada
negativamente por ligandos naturais de enzima como eacute o caso do ATP
A exposiccedilatildeo do S1 de miosina (005 mgmL) a decavanadato conduz agrave inibiccedilatildeo do capacidade da
proteiacutena de estimular a actividade Mg2+
-ATPaacutesica com um valor de IC50 de 266 plusmn 07 M
decavanadato sendo o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM
decavanadato (80) Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM decavanadato sendo
o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM decavanadato (60)
exercendo um efeito de impedimento agrave ligaccedilatildeo entre o vanaacutedio e o S1
A uma concentraccedilatildeo de 27 microM decavanadato verifica-se a oxidaccedilatildeo de um dos resiacuteduos de Cys
de S1 de miosina seguida pela oxidaccedilatildeo gradual de outro resiacuteduo de Cys ateacute agrave concentraccedilatildeo de
1000 microM Ambos os resiacuteduos oxidados natildeo se encontravam expostos ao solvente utilizado A
oxidaccedilatildeo dos resiacuteduos permite sugerir que esta modificaccedilatildeo oxidativa sofrida pelo S1 contribui
para a diminuiccedilatildeo da sua actividade de ATPase
52
Por espectroscopia de RPE revelaram-se os possiacuteveis ligandos equatoriais do vanaacutedio aquando da
sua interacccedilatildeo com o S1 de miosina como sendo duas moleacuteculas de aacutegua um ligando contendo o
grupo R-OH e outro contendo o grupo R=NR
Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados de
oxidaccedilatildeo (IV) e (V) no S1 de miosina para concentraccedilotildees facilmente alcanccedilaacuteveis no ambiente
terrestre
53
5 Estudos futuros
No sentido do melhor esclarecimento das alteraccedilotildees moleculares e estruturais que ocorrem no
subfragmento 1 quando exposta ao vanaacutedio que conduzem a uma diminuiccedilatildeo da sua funccedilatildeo
sugere-se
- Quantificaccedilatildeo das cisteiacutenas reduzidas de S1 na presenccedila de metavanadato
- Determinaccedilatildeo das actividades Ca2+
- e K+-ATPaacutesica do S1 de miosina na presenccedila de
decavanadato
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina estimulada por actina
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina na presenccedila de
metavanadato
- Realizaccedilatildeo de RMN 2D com variaccedilatildeo das concentraccedilotildees das soluccedilotildees de decavanadato e
metavanadato na presenccedila de S1
-Mutageacutenese dos resiacuteduos de Cys doS1 de miosina de modo a verificar quais os resiacuteduos que
sofrem oxidaccedilatildeo na presenccedila de decavanadato
- Determinaccedilatildeo por calorimetria (DSC Differential Scanning Calorimetry) da variaccedilatildeo da
entalpia (∆H) e da temperatura de desnaturaccedilatildeo do S1 de miosina na presenccedila de vanaacutedio uma
vez que estas propriedades caracteriacutesticas de cada proteiacutena satildeo alteradas apoacutes ocorrecircncia de
modificaccedilotildees estruturais
- Anaacutelise por SDS-PAGE de S1 de miosina apoacutes tratamento com vanaacutedio de modo a determinar
quais os polipeacuteptidos eventualmente formados apoacutes clivagem da proteiacutena
54
6 Referecircncias bibliograacuteficas
- Sellers J Myosins 2nd Edition Oxford University Press 1999
- L Stryer inBiochemistry (Ed WH Freeman 2005 pp
Refordfas Materiais e Meacutetodos
[A] S S Margossian SLowey Methods Enzymol1982 85 55-71
[B] JD Pardue JA Spudich Methods Enzymol 1982 85 164-181
[C] M Bradford Anal Biochem1976 72 248-254
[D] H H Taussky E Shorr J Biol Chem 1953 202 675-685
[E] C R Bagshaw in Outline Studies in Biology ndash Muscle Contraction Chapman and Hall Ed
1982
[F] A Cuenda et al Eur J Biochem1990 194 663-670
55
Anexos
1 Determinaccedilatildeo da recta de calibraccedilatildeo meacutetodo de Bradford
Figura1ndashRepresentaccedilatildeo graacutefica da recta de calibraccedilatildeo (Abs595 nm vs [BSA]) utilizada para o meacutetodo de Bradford
2 Determinaccedilatildeo da recta calibraccedilatildeo para a electroforese em condiccedilotildees desnaturantes
Figura 2 ndash Representaccedilatildeo graacutefica da recta de calibraccedilatildeo (log(Peso Molecular) vs Rf dos marcadores de peso
molecular) onde Rf indica determinada atraveacutes do padratildeo de peso molecular
(poccedilo (a))
3Caacutelculo do coeficiente de extinccedilatildeo molar () para a quantificaccedilatildeo de cisteiacutenas reduzidas
atraveacutes do meacutetodo de Ellman
Traccedilou-se a recta de Abs412 nm meacutedia vs [L-Cisteiacutena] de modo a determinar o valor do coeficiente
de extinccedilatildeo molar definido como o declive daquela recta Assim =8400 M-1
cm-1
como
podemos verificar na Fig 3
A595 nm = 00546[BSA] + 00209 Rsup2 = 099051
Ab
s 59
5 n
m
[BSA] (mgmL)
log PM = -1177 Rf + 2247 Rsup2 = 098957
log
PM
Mobilidade relativa (Rf)
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero

3
ppb ndash Partes por biliatildeo (1 gL)
ppm ndash Partes por milhatildeo (1 mgL)
PSA ndash persulfato de amoacutenio
Rf - Mobilidade relativa
RLC ndash Cadeia leve regulatoacuteria de miosina
RMN ndash Ressonacircncia Magneacutetica Nuclear
ROS ndash Espeacutecies reactivas de oxigeacutenio
RPE ndash Ressonacircncia Paramagneacutetica Electroacutenica
S1 ndash Subfragmento-1 da miosina II
SDS ndash Dodecilo sulfato de soacutedio
SDS-PAGE ndash Electroforese em gel de poliacrilamida com SDS (condiccedilotildees desnaturantes)
SH1 ndash Grupo tiol mais reactivo da miosina (resiacuteduo Cys-707)
SH2 ndash segundo grupo tiol mais reactivo da miosina (resiacuteduo Cys-697)
SNC ndash Sistema Nervoso Central
TCA ndash Aacutecido tricloroaceacutetico
TEMED ndash NNNrsquoNrsquo-tetrametiletilenodiamina
Tris ndash 2-amino-2-hidroximetil-13-propanodiol
Trp ndash triptofano
Tyr ndash tirosina
UV ndash gama ultravioleta do espectro electromagneacutetico
Vis ndash gama visiacutevel do espectro electromagneacutetico
4
Resumo
A exposiccedilatildeo a vanaacutedio conduz a uma diminuiccedilatildeo na actividade ATPaacutesica da miosina e provoca a
oxidaccedilatildeo dos grupos sulfidrilo desta proteiacutena No entanto natildeo estaacute integralmente esclarecido
quais os efeitos especiacuteficos que ocorrem na miosina quando exposta ao vanaacutedio Nesse sentido
interessou determinar in vitro quais as alteraccedilotildees estruturais funcionais e moleculares ocorridas
no Subfragmento-1 de miosina de muacutesculo esqueleacutetico de coelho apoacutes a exposiccedilatildeo croacutenica a
soluccedilotildees de metavanadato decavanadato e vanadilo Verificou-se que a exposiccedilatildeo de S1 (005
mgmL) a decavanadato inibe a sua actividade Mg2+
-ATPaacutesica sendo o valor de IC50 para esta
inibiccedilatildeo de 266 plusmn 07 microM decavanadato Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM
decavanadato A perda de funcionalidade proteica poderaacute dever-se agrave oxidaccedilatildeo de 2 resiacuteduos de
Cys de S1 natildeo expostos ao solvente apoacutes exposiccedilatildeo a 1000 microM de decavanadato Verificou-se
que as espeacutecies oligomeacutericas de vanaacutedio com maior afinidade para o S1 satildeo o decavanadato e o
tetravanadato Revelaram-se os possiacuteveis ligandos do vanaacutedio no S1 atraveacutes de espectroscopia de
RPE Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados
de oxidaccedilatildeo (IV) e (V) na estrutura e funccedilatildeo do S1 em concentraccedilotildees facilmente alcanccedilaacuteveis
daquele metal no ambiente terrestre
Summary
The exposure to vanadium leads to a decrease of the myosin ATPase activity and provokes the
oxidation of its sulfydryl groups However the specific effects of vanadium on myosin are not yet
fully understood Consequently the objectives of the present work were to establish the structural
funtional and molecular changes suffered by rabbit myosin subfragment-1 after cronic exposure to
metavanadate decavanadate and vanadyl solutions It was verified that the exposure of S1 (005
mgmL) to decavanadate results in the inhibition of its Mg2+
-ATPase activity resulting in the
IC50 value 266 plusmn 07 microM decavanadate In the presence of actin the IC50 value was 28 plusmn 1 microM
decavanadate It is suggested that the loss of protein function may be due to the oxidation of 2
core cysteine residues after exposure to 1000 microM decavanadate It was also observed that the
decavanadate and tetravanadate species show the highest affinity for S1 Lastly the possible S1
ligands to vanadium were revealed through EPR spectroscopy Globally the obtained results
point to significant effect of V(V) and V(IV) states on S1 structure and function for easily
reachable environmental vanadium concentrations
5
Iacutendice
1 Introduccedilatildeo
11 Muacutesculo esqueleacutetico
111 Miosina
112 Actina
113 Mecanismos moleculares da contracccedilatildeo muscular
1131 Regulaccedilatildeo da contracccedilatildeo muscular
12 Vanaacutedio
121 Propriedades quiacutemicas
1211 Vanaacutedio (IV)
1212 Vanaacutedio (V)
122 Metabolismo e efeito na actividade enzimaacutetica
13 Vanaacutedio e muacutesculo esqueleacutetico
14 Objectivos do projecto
2 Discussatildeo de Resultados
21 Caracterizaccedilatildeo das proteiacutenas isoladas
211 Anaacutelise do estado de pureza
212 Determinaccedilatildeo da concentraccedilatildeo proteica
2121 Subfragmento-1 de miosina
2122 Actina
22 Mediccedilatildeo da actividade enzimaacutetica de S1 de miosina
221 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de Fiske-
Subarow modificado
222 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
23 Preparaccedilatildeo e caracterizaccedilatildeo das soluccedilotildees de vanaacutedio
231 Anaacutelise da composiccedilatildeo das soluccedilotildees de vanadato por espectroscopia de 51V-RMN efeito da
concentraccedilatildeo da espeacutecie
232 Estabilidade das soluccedilotildees de decavanadato anaacutelise por espectroscopia de absorccedilatildeo UVVis
24 Interacccedilatildeo de S1 com vanaacutedio(V)
241 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
242 Actividade Mg2+
-ATPaacutesica de S1 de miosina na presenccedila de decanadato
243 Quantificaccedilatildeo de cisteiacutenas reduzidas pelo meacutetodo de Ellman
25 Interacccedilatildeo de S1 com vanaacutedio(IV)
251 Anaacutelise por espectroscopia de Ressonacircncia Paramagneacutetica Electroacutenica
3 Procedimento Experimental
31 Reagentes
32 Meacutetodos
321 Isolamento e purificaccedilatildeo das proteiacutenas de muacutesculo esqueleacutetico de coelho
3211 Purificaccedilatildeo da miosina preparaccedilatildeo de Subfragmento 1 por digestatildeo quimotriacutepsica da
miosina
32111 Purificaccedilatildeo da miosina
32112 Preparaccedilatildeo de Subfragmento-1 de miosina por digestatildeo quimotriacutepsica
3212 Isolamento e purificaccedilatildeo de actina
32121Obtenccedilatildeo do ldquopoacute de acetonardquo
32122 Purificaccedilatildeo da actina a partir de ldquopoacute de acetonardquo
331 Quantificaccedilatildeo das proteiacutenas isoladas
3311 Quantificaccedilatildeo pelo meacutetodo de Bradford
6
3312 Quantificaccedilatildeo pelo meacutetodo directo
332 Electroforese em condiccedilotildees desnaturantes (SDS-PAGE)
34 Mediccedilatildeo da actividade enzimaacutetica de S1
341 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de Fiske-
Subarow modificado
342 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
35 Preparaccedilatildeo e caracterizaccedilatildeo das soluccedilotildees de vanaacutedio por Espectroscopia UV-Visiacutevel e
Ressonacircncia Magneacutetica Nuclear
351 Preparaccedilatildeo das soluccedilotildees de vanadato
352 Anaacutelise por UV-Vis para determinaccedilatildeo da ordem da reacccedilatildeo
353 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
362 1 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
363 Interacccedilatildeo de S1 com V (V) por Ressonacircncia Magneacutetica Nuclear (RMN)
4 Conclusotildees gerais
5 Estudos futuros
6 Referecircncias bibliograacuteficas
7
1 Introduccedilatildeo
11 Muacutesculo esqueleacutetico
A capacidade de locomoccedilatildeo eacute uma das caracteriacutesticas fundamentais da maioria dos organismos
vivos A contracccedilatildeo muscular um dos fenoacutemenos que a torna possiacutevel apresenta grande
complexidade e tem vindo a ser utilizado como modelo para a compreensatildeo de sistemas menos
especializados de mobilidade celular [1]
No que diz respeito agrave sua fisiologia morfoloacutegica os muacutesculos dos vertebrados dividem-se em
lisos e estriados Estes uacuteltimos devem o seu nome agrave alternacircncia de zonas claras e escuras
observaacuteveis ao microscoacutepio electroacutenico de que resulta um aspecto estriado Os muacutesculos
estriados podem ainda ser divididos em duas subclasses muacutesculos esqueleacuteticos e cardiacuteacos [2]
consoante a forma celular (respectivamente longitudinal ou ramificada) ou tipo de contracccedilatildeo das
ceacutelulas que o compotildeem (respectivamente voluntaacuteria e involuntaacuteria) [3]
Os muacutesculos estriados esqueleacuteticos satildeo assim designados devido agrave sua localizaccedilatildeo no
organismo ndash ligados aos ossos da coluna vertical braccedilos e pernas Satildeo constituiacutedos por feixes de
ceacutelulas multinucleadas (miofibras) que contecircm miofibrilas paralelas de cerca de 1 m de
diacircmetro associadas agrave contracccedilatildeo voluntaacuteria raacutepida e forte em actividades de coordenaccedilatildeo
complexa [4] Cada feixe apresenta um diacircmetro compreendido entre 20 e 100 m e um
comprimento maacuteximo que pode atingir vaacuterios centiacutemetros (Fig 11)
Figura 11 ndash Organizaccedilatildeo hieraacuterquica geral do muacutesculo-esqueleacutetico (adaptado de Purves et al 2003 [3])
No muacutesculo-esqueleacutetico as zonas claras e escuras supramencionadas correspondem a zonas de
menor e maior densidade electroacutenica respectivamente As zonas claras constituem as bandas I (de
8
ldquoisotropiardquo ndash polarizaccedilatildeo uniforme em todas as direcccedilotildees) enquanto as zonas escuras constituem
as bandas A (de ldquoanisotropiardquo ndash polariza a luz visiacutevel) A banda I encontra-se dividida
transversalmente por uma linha escura ndash linha Z de densidade electroacutenica e refracccedilatildeo superiores
No interior de cada banda A existe uma zona transversal menos densa ndash zona H ndash rica em enzimas
importantes para o metabolismo celular No centro desta zona existe uma linha perpendicular
mais escura ndash linha M [2]
A unidade fundamental da contracccedilatildeo muscular eacute designada por sarcoacutemero (Fig 12) Encontra-se
repetida de forma regular ao longo das miofibrilas apresentando um comprimento de 25 a 3 m
aquando do relaxamento muscular diminuindo aquando da contracccedilatildeo muscular
Figura 12 ndash Representaccedilatildeo de fotomicrografia electroacutenica e esquemaacutetica do sarcoacutemero Os filamentos grossos e
finos satildeo maioritariamente constituiacutedos por miosina e actina respectivamente (Adaptado de Stryer 2002 [2])
As secccedilotildees de corte do sarcoacutemero datildeo conta da presenccedila de dois tipos de filamentos proteicos ndash os
filamentos grossos cujo diacircmetro eacute de 15 nm constituiacutedos sobretudo por miosina e os filamentos
finos com um diacircmetro de 9 nm constituiacutedos sobretudo por actina tropomiosina e pelo complexo
de troponina [2]
111 Miosina
Actualmente o termo miosina refere-se a uma superfamiacutelia diversa de motores moleculares
capazes de hidrolisar ATP e de se ligarem reversivelmente agrave actina A classificaccedilatildeo da
superfamiacutelia das miosinas nas 15 classes distintas faz-se atraveacutes de homologia entre as diferentes
sequecircncias [A] Eacute tambeacutem possiacutevel a classificaccedilatildeo por subclasses das quais se destaca a subclasse
4 proveniente do muacutesculo-esqueleacutetico e cardiacuteaco de vertebrados Este estudo focar-se-aacute apenas
nas miosinas de classe II (miosina-II) presente nos filamentos grossos do muacutesculo estriado
esqueleacutetico [B] e em ceacutelulas natildeo musculares A funccedilatildeo da miosina-II depende da sua localizaccedilatildeo
celular e tecidular
9
A miosina-II eacute constituiacuteda por duas cadeias polipeptiacutedicas idecircnticas denominadas cadeias pesadas
de miosina (HC) enroladas entre si e cujo peso molecular (PM) estaacute compreendido entre 171 e
241 kDa) Estruturalmente as HC dividem-se em 3 domiacutenios funcionais - cabeccedila pescoccedilo e
cauda como se pode observar na Fig 13 Agraves cabeccedilas ou zonas globulares estatildeo associados dois
pares de cadeias leves (peso molecular entre 15 e 30 kDa) que se dividem em dois tipos
essencial (ELC) e regulatoacuterio (RLC)
Figura 13 ndash Estrutura esquemaacutetica da moleacutecula de miosina-II Destacam-se as duas cabeccedilas globulares e a cauda
filamentosa ligadas entre si por uma zona flexiacutevel denominada pescoccedilo As cadeias leves (a verde e vermelho)
encontram-se associadas agraves cabeccedilas globulares
As cabeccedilas globulares contendo os locais de ligaccedilatildeo agrave actina e aos nucleoacutetidos mantecircm os seus
elementos estruturais baacutesicos conservados em todas as classes sendo estes constituiacutedos por 720-
80 aa A regiatildeo do pescoccedilo eacute constituiacuteda por uma longa heacutelice pertencente agrave cadeia pesada
que se encontra estabilizada atraveacutes da ligaccedilatildeo a subunidades de cadeia leve A estrutura do
pescoccedilo envolvida na ligaccedilatildeo agraves cadeias leves eacute denominada de motivo IQ cuja estrutura
consensual eacute IQXXXRGXXXR1 O pescoccedilo serve como extensatildeo do comprimento da cabeccedila de
modo a actuar como alavanca durante a contracccedilatildeo muscular [A]A regiatildeo da cauda ou acircncora
apresenta bastante divergecircncia no que respeita agrave sua sequecircncia estrutura e funccedilatildeo No entanto
pensa-se que a sua funccedilatildeo centrar-se-aacute na autoassociaccedilatildeo das vaacuterias moleacuteculas de miosina-II
atraveacutes da regiatildeo de coiled-coil Esta confere assimetria agrave moleacutecula e permite a formaccedilatildeo de
filamentos grossos capazes de translocar filamentos de actina [A]
Na regiatildeo da cabeccedila da miosina existem 2 regiotildees extremamente sensiacuteveis agrave proteoacutelise A
clivagem em ambos os locais resulta na produccedilatildeo de trecircs fragmentos um compreendendo o N-
terminal (fragmento 25 kDa) outro a regiatildeo central (50 kDa) e um terceiro o C-terminal (20 kDa)
(Fig 14) Estes fragmentos dissociam-se em condiccedilotildees desnaturantes e natildeo constituem domiacutenios
independentes da cabeccedila mas provavelmente loops flexiacuteveis uma vez que natildeo foram observadas
na estrutura cristalina Os vaacuterios fragmentos obtidos em diferentes condiccedilotildees foram cruciais para
a elucidaccedilatildeo das vaacuterias funccedilotildees da moleacutecula
A regiatildeo compreendida entre os fragmentos 25 e 50 kDa (loop 1) localiza-se directamente por
1 I = Isoleucina Q= Glutamina X = qualquer dos 20 aminoaacutecidos padratildeo R= Arginina G= Glicina
10
cima do local de ligaccedilatildeo ao nucleoacutetido A regiatildeo entre os fragmentos de 50 e 20 kDa (loop 2)
encontra-se perto da intersecccedilatildeo entre os domiacutenios da cabeccedila e do pescoccedilo e deveraacute interagir com
a actina uma vez que a ligaccedilatildeo desta a protege da proteoacutelise Na ausecircncia do loop 2 a miosina
liga mais fracamente agrave actina do que no caso da miosina intacta A sequecircncia primaacuteria de ambos
os loops apresenta conservaccedilatildeo reduzida sendo provaacutevel que sejam responsaacuteveis por muita da
diversidade das diferentes propriedades enzimaacuteticas presentes em diferentes isoformas
No fragmento de 20 kDa existem dois grupos sulfidrilo Cys-707 e -697 usualmente
denominados de SH1 e SH2 devido agrave ordem da sua reactividade quiacutemica Na presenccedila de
nucleoacutetidos pode ocorrer o ldquocross-linkingrdquo entre estes dois grupos incluindo a oxidaccedilatildeo a uma
ponte dissulfureto
Devido agrave insolubilidade da miosina-II nativa a estrateacutegia adoptada para os estudos com esta
proteiacutena eacute frequentemente a sua clivagem em fragmentos funcionais separaacuteveis Um dos locais de
clivagem ocorre tipicamente a 130 kDa do N-terminal produzindo a meromiosina pesada (HMM)
e a meromiosina leve (LMM) [C] A HMM eacute formada pelas 2 cabeccedilas e pescoccedilo e apresenta
propriedades semelhantes agrave miosina sem digestatildeo (ligaccedilatildeo e translocaccedilatildeo de actina e hidroacutelise de
MgATP)
A HMM pode ser novamente clivada atraveacutes de uma reacccedilatildeo de proteoacutelise produzindo os
subfragmentos 1 (S1) e 2 (S2)O S1 (90 kDa) eacute enzimaticamente activo sendo constituiacutedo pelos
domiacutenios da cabeccedila e do pescoccedilo e apresentando associaccedilatildeo agraves duas cadeias leves ou apenas agrave
ELC (Fig 14)Em alternativa o S1 pode ser produzido por clivagem directa da miosina intacta
formando-se tambeacutem um fragmento em forma de cilindro que consiste nas regiotildees S2 e LMM
(Ref)Utiliza-se extensivamente o S1 para estudos cineacuteticos e de ligaccedilatildeo agrave actina uma vez que a
sua solubilidade simplifica o planeamento da experiecircncia e a subsequente interpretaccedilatildeo dos dados
[A]
Figura 14- Estrutura do subfragmento-1 de miosina de muacutesculo esqueleacutetico com ecircnfase nos domiacutenios de 50 kDa
central (a vermelho) 25 kDa no N-terminal (a verde) e 20 kDa no C-terminal (a azul) As cadeias leves essenciais
encontram-se a amarelo e as cadeias leves regulatoacuterias a lilaacutes Os locais de ligaccedilatildeo agrave actina e ao ATP estatildeo
igualmente destacados (adaptado de Holmes e Geeves 1999 [D])
11
112 Actina
O termo ldquoactinardquo refere-se a uma famiacutelia extremamente conservada de proteiacutenas citoplasmaacuteticas
presente em todos os eucariotas Esta proteiacutena natildeo se encontra nos procariotas [E] A maioria das
actinas possui peso molecular proacuteximo de 42 kDa outra evidecircncia do seu grau elevado de
conservaccedilatildeo no que concerne agrave sequecircncia e ao nuacutemero de aminoaacutecidos Os eucariotas superiores
apresentam um grande nuacutemero de isoformas de actina que por sua vez se encontram codificados
por um ainda maior nuacutemero de genes (Ref) A expressatildeo das isoformas eacute diferencial consoante o
tecido e sua especializaccedilatildeo pelo que deveratildeo possuir funccedilotildees e mecanismos direccionais
diferentes (Ref) Foi ainda observado que os mRNA das diferentes isoformas apresentam uma
distribuiccedilatildeo heterogeacutenea nas ceacutelulas de forma semelhante aos seus produtos proteicos [F] Estes
dados suportam a hipoacutetese supramencionada do direccionamento das isoformas de actina para um
determinado local na ceacutelula de acordo com as suas funccedilotildees
A actina presente no citoesqueleto eacute responsaacutevel por processos como o mecanismo contraacutectil
fagocitose manutenccedilatildeo e alteraccedilatildeo da estrutura celular locomoccedilatildeo transporte de organelos
regulaccedilatildeo do transporte de solutos entre outras Nos eucariotas superiores estaacute ainda envolvida
no transporte de gracircnulos de pigmento retinoacuteide [G] transporte de gracircnulos no axoplasma [H] e
secreccedilatildeo entre outros [E] Actualmente a isoforma da actina (-actina) eacute a melhor estudada
encontrando-se envolvida no mecanismo contraacutectil [E]
Os filamentos de -actina tecircm cerca de 1m de comprimento e 80 Å Consistem em duas cadeias
de actina filamentosa (F-actina) organizados em heacutelice [I] (Holmes et al 1990) sobre os quais
se encontram outras proteiacutenas ndash a troponina (C I e T) e a tropomiosina (Azevedo 1999) (Figura
15) A troponina eacute filamentosa e localiza-se nas depressotildees da heacutelice de F-actina conferindo
rigidez aos filamentos finos Por sua vez a tropomiosina eacute essencial na regulaccedilatildeo do mecanismo
contraacutectil
Figura 15 ndash Modelo estrutural dos filamentos de F-actina contendo a tropomiosina e o complexo de troponina (I C
e T) (Adaptado de Susana Ramos 2005 [J])
12
A F-actina eacute formada atraveacutes da polimerizaccedilatildeo de subunidades globulares de actina (G-actina) A
G-actina eacute composta por 2 domiacutenios de tamanho semelhante separados pelos bolsos de ligaccedilatildeo(1)
de catiotildees divalentes e(2) de nucleoacutetidos [KLM] Estes por sua vez estatildeo subdivididos noutros
dois de menores dimensotildees (1-4) (Fig 16) cada um envolvido nas interacccedilotildees actina-actina ou
na formaccedilatildeo do topo de ligaccedilatildeo do nucleoacutetido As regiotildees N-terminal e C-terminal estatildeo proacuteximas
uma da outra no subdomiacutenio 1 [E]
Figura 16 ndash Estrutura do monoacutemero de actina Encontram-se representados os subdomiacutenios 1 a 4 e a vermelho a
moleacutecula de ATP junto do respectivo local de ligaccedilatildeo (Protein Data Bank ID 1j6z)
113 Mecanismos moleculares da contracccedilatildeo muscular
A contracccedilatildeo muscular ocorre atraveacutes do deslizamento dos filamentos finos (de actina) por entre
os filamentos grossos (de miosina) conseguida pela reacccedilatildeo de hidroacutelise de ATP e regulada pela
tropomiosina e pelo complexo de troponina em resposta a impulsos nervosos Este mecanismo foi
inicialmente proposto por Huxley-Niedergerke [N] e Huxley-Hanson [O] ambos em 1954 A base
experimental de ambos os estudos foi a inexistecircncia de variaccedilatildeo do tamanho dos filamentos
durante a contracccedilatildeo A banda A apresenta sempre as mesmas dimensotildees por outro lado a banda
I a zona H diminuem consideravelmente aquando da contracccedilatildeo (Figs 12 e 17)
Figura 17 ndash Modelo da contracccedilatildeo muscular ndash deslizamento dos filamentos finos (azuis) relativamente aos
filamentos grossos (rosa) (Adaptado de Stryer 2002 [2])
13
Na ausecircncia de ATP e supondo a ausecircncia de inibiccedilatildeo da interacccedilatildeo actina-miosina as proteiacutenas
encontram-se fortemente associadas formando o complexo actomiosina (AM) (Figura 15)
Nestas condiccedilotildees as cabeccedilas de miosina formam um acircngulo de 45ordm com o eixo dos filamentos
finos de actina [P]
Na presenccedila de ATP o complexo AM dissocia-se jaacute que a afinidade do complexo M-ATP para a
actina eacute 105 vezes inferior agrave afinidade da miosina para a actina O ATP liga-se agrave miosina
ocorrendo seguidamente a reacccedilatildeo de hidroacutelise de ATP Os produtos daiacute resultantes ndash ADP e Pi -
permanecem ligados agrave miosina formando o complexo M-ADP-Pi Com a ligaccedilatildeo subsequente agrave
actina forma-se o complexo AM-ADP-Pi no qual as cabeccedilas de miosina apresentam um acircngulo
recto com o eixo do muacutesculo [P] Neste ponto daacute-se a libertaccedilatildeo progressiva de Pi e ADP Eacute
provaacutevel que a energia proveniente da hidroacutelise do ATP provoque uma alteraccedilatildeo do acircngulo de
ligaccedilatildeo das cabeccedilas de miosina ao filamento de actina passando novamente de 90ordm para 45ordm [P]
Esta alteraccedilatildeo conformacional provoca o deslizamento do filamento de actina para o centro do
sarcoacutemero cujo encurtamento promove a contracccedilatildeo das miofibrilas O processo supracitado
ocorre em cerca de 1ms[Q] e encontra-se resumido na Fig 18
Figura 18 ndash Esquema cineacutetico da hidroacutelise de ATP pela miosina (Adaptado de Sellers 1999 [A])
1131 Regulaccedilatildeo da contracccedilatildeo muscular
O mecanismo pelo qual se processa a regulaccedilatildeo muscular foi inicialmente descrito por estudos
distintos de Haselgrove Huxley e Parrey-Squire todos em 1973 [RST] Aleacutem da implicaccedilatildeo
directa da tropomiosina e do complexo de troponina a concentraccedilatildeo de Ca2+
no muacutesculo em
relaxamento eacute bastante reduzida (10-7
M) aumentando notavelmente aquando da contracccedilatildeo (10-5
-
10-3
M)
Resumidamente quando ocorre a excitaccedilatildeo do muacutesculo esqueleacutetico a despolarizaccedilatildeo da
membrana plasmaacutetica eacute difundida ateacute ao interior do tecido atraveacutes dos tuacutebulos transversais
(invaginaccedilotildees da membrana plasmaacutetica da fibra muscular) A despolarizaccedilatildeo provoca a abertura
dos canais ioacutenicos de Ca2+
do retiacuteculo sarcoplasmaacutetico adjacente Como a concentraccedilatildeo de Ca2+
no interior do organelo eacute muito elevada o caacutelcio passa raacutepida e uniformemente para o citosol
14
Assim a regulaccedilatildeo da contracccedilatildeo ocorre com coordenaccedilatildeo e rapidez na totalidade das miofibrilas
do tecido muscular (Ref)
Em repouso a troponina I encontra-se posicionada de tal forma que a proteiacutena adjacente
troponina C impede estruturalmente a ligaccedilatildeo das cabeccedilas da miosina agrave actina (acircngulo =50ordm) O
aumento da concentraccedilatildeo intracelular de Ca2+
provoca um aumento da sua disponibilidade para
ligar agrave troponina C (Fig 19)
Figura 19ndash Regulaccedilatildeo do mecanismo contraacutectil no muacutesculo esqueleacutetico pelo complexo de troponina
Como consequecircncia a troponina C altera a sua conformaccedilatildeo (=70ordm) associando-se agrave troponina I
que deixa de inibir a ligaccedilatildeo das pontes transversais A alteraccedilatildeo anterior eacute transmitida agrave
tropomiosina atraveacutes da troponina T adjacente agrave C o que provoca o deslocamento da
tropomiosina na direcccedilatildeo do interior do sulco do filamento de actina Assim eacute possiacutevel a
interacccedilatildeo eficaz entre actina e miosina A Ca2+
-ATPase do retiacuteculo sarcoplasmaacutetico provoca a
reacumulaccedilatildeo do iatildeo caacutelcio no organelo impedindo o prolongamento desnecessaacuterio da contracccedilatildeo
(Refordf)
12 Vanaacutedio
O vanaacutedio eacute um metal de transiccedilatildeo pertencente ao grupo VB da tabela perioacutedica Foi
primeiramente descoberto em 1801 por Andreacutes Manuel del Riacuteo que concluiu erradamente que era
idecircntico ao croacutemio e definitivamente por Nils Gabriel Svenson em 1831 Recentemente foram
descobertas importantes funccedilotildees bioloacutegicas deste elemento provavelmente essencial para os
humanos de que se destaca no contexto do presente relatoacuterio a possibilidade de actuar como
anaacutelogo do grupo fosfato e a inibiccedilatildeo de enzimas in vitro como eacute o caso da actividade ATPaacutesica
da miosina
Eacute um metal soacutelido agrave temperatura ambiente que se encontra amplamente distribuiacutedo nos vaacuterios
ecossistemas em dois isoacutetopos naturais ndash 50
V e 51
V ndash raramente excedendo 01 partes por biliatildeo
15
(ppb) Estaacute associado a cerca de 70 minerais diferentes na natureza (dos quais se destacam o
uracircnio cobre chumbo zinco fosfato e titacircnio ferroso) especialmente na forma de vanadato ou
pentoacutexidos de vanaacutedio O vanaacutedio eacute um dos principais constituintes da mateacuteria orgacircnica foacutessil
com especial ecircnfase para o carvatildeo oacuteleo de crude e foacutesseis carbonaacuteceos [UV] bem como em
alimentos consumidos pelo Homem (salsa pimenta preta cogumelos marisco vegetais cereais
gorduras e oacuteleos) [W] Estes constituem a maior fonte de exposiccedilatildeo ao vanaacutedio da maioria da
populaccedilatildeo humana [X]
Normalmente a entrada do vanaacutedio no organismo ocorre por inalaccedilatildeo ou em menores
quantidades por absorccedilatildeo pelo tracto gastrointestinal ou pela pele O seu armazenamento centra-
se no fiacutegado rins ceacuterebro coraccedilatildeo muacutesculos e ossos No entanto a retenccedilatildeo de vanaacutedio no
organismo humano eacute reduzida ndash em condiccedilotildees normais cerca de 100 M (em muitos tecidos a
quantidade eacute inferior a 10 ngg de tecido) [Y]
Actualmente natildeo existem provas de deficiecircncia nutricional de vanaacutedio em humanos ou de
enzimas que contenham vanaacutedio em animais superiores No entanto vaacuterios estudos relacionam a
ocorrecircncia de deficiecircncia em galinhas e ratos devido a uma alimentaccedilatildeo pobre em vanaacutedio Os
sintomas relatados incluem retardamento do crescimento malformaccedilotildees oacutesseas perturbaccedilotildees no
metabolismo lipiacutedico e estagnaccedilatildeo reprodutiva entre outros [Z AA AB] Os efeitos fisioloacutegicos
do vanaacutedio podem depender do tipo de composto ou de oligoacutemero uma vez que consoante as
suas estabilidade labilidade e nuacutemero de coordenaccedilatildeo as respostas seratildeo diferenciadas ao niacutevel
dos organismos vivos
Por outro lado a toxicidade do vanaacutedio encontra-se bem caracterizada existindo sintomas
descritos em vaacuterios mamiacuteferos (inclusive o Homem) que incluem diarreias catildeibras irritaccedilatildeo da
pele e mucosas palpitaccedilotildees cardiacuteacas anemia pneumonia bronquite perturbaccedilotildees no Sistema
Nervoso Central (SNC) entre outros (Ref) A toxicidade varia geralmente com a natureza do
composto com aumento proporcional agrave sua valecircncia Verificou-se que a forma de entrada do
vanaacutedio tem tambeacutem efeito na sua toxicidade ndash os maiores niacuteveis verificaram-se por injecccedilatildeo
intravenosa da amostra sendo progressivamente inferiores por inalaccedilatildeo e finalmente por
ingestatildeo [X AA] A ampla utilizaccedilatildeo dos compostos de vanaacutedio na induacutestria quiacutemica bem como
as elevadas quantidades de vanaacutedio libertadas atraveacutes da combustatildeo dos oacuteleos de crude
contribuem consideravelmente para o aumento da biodisponibilidade do vanaacutedio Como tal o
interesse neste elemento e nos problemas ambientais por ele causados tem vindo a aumentar
significativamente [V]
121 Propriedades quiacutemicas
16
A quiacutemica do vanaacutedio eacute extraordinariamente complexa pelo que originou vaacuterias interpretaccedilotildees
incorrectas desde a sua descoberta No entanto as suas propriedades dependem em uacuteltima
anaacutelise da configuraccedilatildeo electroacutenica do elemento Assim a variabilidade encontrada no
comportamento quiacutemico do vanaacutedio pode dever-se agrave sua configuraccedilatildeo ndash [Ar] 3d2 4s
2 ndash que
permite a formaccedilatildeo de vaacuterios estados de oxidaccedilatildeo (de +2 a +5) Em soluccedilatildeo aquosa os estados de
oxidaccedilatildeo estaacuteveis vatildeo de V(V) a V(II) Destes apenas os +3 a +5 satildeo biologicamente importantes
jaacute que o V(II) apresenta demasiada capacidade redutora para existir nos organismos vivos
conhecidos [AC]
No fluido extracelular os compostos de vanaacutedio encontram-se predominantemente na forma de
monovanadatos (V(V)) livres ou ligados agrave transferina do soro ou agrave hemoglobina (Refordf) Os iotildees de
vanadato livres satildeo transportados para o meio intracelular atraveacutes de canais ioacutenicos inespeciacuteficos
Jaacute no ambiente intracelular os compostos sofreratildeo uma bio-reduccedilatildeo atraveacutes da acccedilatildeo da
glutationa ou outros compostos com grupos ndashOH como pentoses hexoses ou etanolaminas [V] A
reoxidaccedilatildeo de vanadilo (V(IV)) intracelular a vanadato (V(V)) provocaria a acumulaccedilatildeo toacutexica
destes uacuteltimos pelo que a complexaccedilatildeo da espeacutecie reduzida o impede O transporte de vanaacutedio
para o interior da ceacutelula encontra-se na Fig 110
Figura 110 ndash Transporte de vanaacutedio para a ceacuteluka e seu subsequente metabolismo (P=Proteiacutena) (Adaptado de
Zaporowska e Scibor 1998)
1211 Vanaacutedio(IV)
A espeacutecie de vanaacutedio (IV) mais bem conhecida eacute o iatildeo vanadilo (VO2+
) espeacutecie estaacutevel a pH
aacutecido sendo a maioritaacuteria a pH 3[AD] Apresenta uma cor azulada e forma complexos penta e
hexacoordenados Forma-se rapidamente a partir de V(V) na presenccedila de um agente redutor em
soluccedilatildeo aquosa No ambiente intracelular a presenccedila de glutationa reduzida aacutecido ascoacuterbico
NAD(P)H entre outros conduzem a que o principal estado de oxidaccedilatildeo nos sistemas bioloacutegicos
seja provavelmente o +4 encontrando-se o V(IV) fortemente complexado com proteiacutenas ou
aminoaacutecidos livres citrato ATP e pirofosfato (Refordfs) Em alternativa o V(IV) pode existir
17
livremente em ambientes fortemente aacutecidos como os lisossomas jaacute que a pH superior a 3 as
soluccedilotildees de vanadilo tendem a ser oxidadas pelo ar formando-se vanadato Esta grande
capacidade redutora justifica parcialmente a utilizaccedilatildeo pouco extensiva do iatildeo VO2+
como
inibidor [W]
1212 Vanaacutedio (V)
O vanaacutedio (V) eacute a forma mais estaacutevel num intervalo de pH amplo [W] O iatildeo anioacutenico VO3-tende
a constituir complexos polinucleares pelo que as soluccedilotildees aquosas de vanadato satildeo extremamente
complexas com ocorrecircncia simultacircnea de vaacuterios equiliacutebrios de protonaccedilatildeo (pKa(vanadato)= 35
78 e 125) e oligomerizaccedilatildeo As espeacutecies presentes (H3VO4 H2VO4- HVO4
2- e VO4
3-) dependem
do pH forccedila ioacutenica e temperatura da soluccedilatildeo e da concentraccedilatildeo total do elemento
[AEAFAGAHAI]
O VO43-
- monoacutemero ortovanadato V1 ndash estaacute associado a uma grande parte da importacircncia
bioloacutegica do vanaacutedio uma vez que se trata de um anaacutelogo electroacutenico e estrutural do ortofosfato
(PO43-
) A coordenaccedilatildeo do ortovanadato pode ser tetraeacutedrica ou bipiramidal trigonal (Refordfs)
A oligomerizaccedilatildeo do vanadato monomeacuterico daacute origem ao diacutemero (V2O72-
- V2) que apresenta
tambeacutem equiliacutebrios de protonaccedilatildeo Presume-se a existecircncia de analogia do vanadato dimeacuterico com
o pirofosfato embora natildeo haja evidecircncia estrutural (Refordf) A dimerizaccedilatildeo do V2 origina o
vanadato tetrameacuterico ou metavanadato (V4O124-
- V4) que constitui a espeacutecie maioritaacuteria em
soluccedilotildees aquosas concentradas (Refordf) As estruturas do metavanadato e a sua espeacutecie protonada
satildeo ciacuteclicas [AJ] Outra espeacutecie oligomeacuterica importante eacute o vanadato pentameacuterico (V5O153-
) cuja
estrutura presumida eacute ciacuteclica [AI]
O oligoacutemero com maior importacircncia bioquiacutemica eacute o vanadato decameacuterico (V10O286-
-V10)(Refordf) a
espeacutecie predominante em soluccedilotildees de pH 2-6 quando a concentraccedilatildeo total de vanadato eacute superior
a 100 M (Ref) A estrutura do V10 natildeo difere em soluccedilatildeo ou no estado soacutelidoOs 10 aacutetomos de
vanaacutedio da espeacutecie decameacuterica estatildeo divididos em trecircs grupos identificados na figura como VA-C
consoante o seu ambiente quiacutemico circundante (Refordf) (Fig 111)
A expressatildeo ldquodecavanadatordquo refere-se a uma soluccedilatildeo aquosa na qual a espeacutecie decameacuterica se
encontra em equiliacutebrio quiacutemico com as espeacutecies V1 V2 V4 e V5 sendo que a oligomerizaccedilatildeo
progressiva do vanaacutedio eacute proporcional agrave sua concentraccedilatildeo total (Ref) Nesta soluccedilatildeo a
concentraccedilatildeo de V10 corresponde a aproximadamente 10 da concentraccedilatildeo total de vanaacutedio [AK]
18
Figura 111 ndash Estruturas esquemaacuteticas das espeacutecies de vanadato (espeacutecie monomeacuterica (V1) dimeacuterica (V2)
tetrameacuterica (V4) pentameacuterica (V5) e decameacuterica (V10) (Adaptado de Aureliano e Crans 2008)
122 Metabolismo e efeito na actividade enzimaacutetica
Apesar de natildeo terem sido ainda encontradas as provas categoacutericas das suas funccedilotildees nos seres
vivos sabe-se que o iatildeo vanadato eacute capaz de inibir uma grande quantidade de enzimas in vitro
com especial ecircnfase na actividade ATPaacutesica da miosina [AL Teresa] Como referido
anteriormente o vanadato eacute capaz de actuar em alternativa ao fosfato nas reacccedilotildees bioquiacutemicas
incluindo a sua preferecircncia de ligaccedilatildeo agraves enzimas [W]
Eacute conhecido o papel do vanaacutedio no metabolismo do caacutelcio promovendo a mineralizaccedilatildeo dos
dentes e ossos na biossiacutentese de liacutepidos (trigliceacuteridos e colesterol) com posterior inibiccedilatildeo da
acumulaccedilatildeo destes nos vasos sanguiacuteneos e no SNC na reproduccedilatildeo e desenvolvimento por
mimetizaccedilatildeo e potenciaccedilatildeo de factores de crescimento na estimulaccedilatildeo do consumo de glucose e
da siacutentese de glicogeacutenio com actuaccedilatildeo equivalente aos efeitos da insulina ao niacutevel celular na
transduccedilatildeo de sinais celulares com regulaccedilatildeo da actividade de segundos mensageiros no
transporte de iotildees e aminoaacutecidos na transcriccedilatildeo geneacutetica e siacutentese de proteiacutenas e DNA entre
outros [W Y AB]
Assim o vanaacutedio cnseguiraacute provocar inuacutemeras mudanccedilas bioquiacutemicas nas ceacutelulas Alguns
exemplos da influecircncia bioquiacutemica do vanaacutedio incluem os seus efeitos nos processos de
fosforilaccedilatildeo e desfosforilaccedilatildeo de proteiacutenas e na participaccedilatildeo na formaccedilatildeo de espeacutecies reactivas de
oxigeacutenio devido ao seu efeito proacute-oxidante sobre agentes redutores a niacutevel celular [AM]
Quanto ao seu efeito na actividade enzimaacutetica pode dividir-se em dois tipos principais ndash inibitoacuterio
e estimulante - dependendo da enzima ou do tipo de reacccedilatildeo por esta catalisada No primeiro
grupo incluem-se enzimas como as ATPases (de miosina de canais ioacutenicos e fosfoenzimas
19
transportadoras) e as fosfatases (da tirosina aacutecida e alcalina) a fosfofrutocinase adenilato ciclase
e catalase entre outras [AA] O segundo grupo inclui a maioria das cinases a glucose-6-fosfato
desidrogenase a fosfoglicerato mutase e a fosfoglicerato fosfatase (Refordf)
O efeito do vanaacutedio (V) na actividade enzimaacutetica estaacute relacionado com o seu nuacutemero de
coordenaccedilatildeo podendo actuar como estado de transiccedilatildeo ou por mimetizaccedilatildeo de cofactores
substratos (especialmente o fosfato) e interacccedilotildees proteiacutena-proteiacutena (como no caso das
polimerases e transcriptases) Pode ainda estar envolvido na formaccedilatildeo de ligaccedilotildees de hidrogeacutenio
interacccedilotildees electrostaacuteticas esteacutereas e de oxidaccedilatildeo-reduccedilatildeo Uma evidecircncia desta uacuteltima eacute a sua
capacidade de alternacircncia entre os estados de oxidaccedilatildeo +5 a +3 o que torna faacutecil a sua interacccedilatildeo
redox com enzimas contendo resiacuteduos de cisteiacutenas (Cys) ou qualquer outro composto orgacircnico
com actividade redox [AN]
13 Vanaacutedio e muacutesculo esqueleacutetico
A primeira evidecircncia da presenccedila de vanaacutedio no muacutesculo esqueleacutetico proveio de uma amostra de
ATP isolado de muacutesculo de cavalo que continha uma impureza ndash vanaacutedio ndash cuja presenccedila
provocou a inibiccedilatildeo da actividade da bomba de Na+K
+-ATPase [AO] Desde entatildeo vaacuterios
estudos tecircm sido realizados com ecircnfase na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o
tecido muscular Destes destaca-se a descoberta da interacccedilatildeo forte da espeacutecie V10 com a miosina
fosforilada [AP] e agrave sua inibiccedilatildeo da actividade ATPaacutesica da miosina[AQ AR]
No quediz respeitoao iatildeo monovanadato destaca-se a sua inibiccedilatildeoda actividade Mg2+
-ATPaacutesica
da miosina [AS] que ocorre de duas formas distintas (1) na presenccedila de concentraccedilotildees reduzidas
de V1 a inibiccedilatildeo ocorre a velocidade reduzida dependente da concentraccedilatildeo da espeacutecie
monomeacuterica (2) na presenccedila de concentraccedilotildees elevadas de V1 o niacutevel de inibiccedilatildeo aproxima-se de
100 desde que [V1] seja superior agrave concentraccedilatildeo total de locais com actividade Mg2+
-ATPaacutesica
no ensaio Esta inibiccedilatildeo eacute irreversiacutevel tanto por diaacutelise ou adiccedilatildeo de agentes redutores do
vanadato [AL]Segundo Pate et al(1994)[AT] a espeacutecie monomeacuterica podeinibir a velocidade de
contracccedilatildeo a temperatura inferior a 20ordmC perdendogradual e rapidamente essa capacidade ateacute os
50ordmC temperatura agrave qual cessa a capacidade inibitoacuteria Os resultados obtidos sugeremainda uma
interacccedilatildeo altamente cooperativa entre o iatildeo vanadato e as proteiacutenas componentes do muacutesculo
esqueleacuteticoDestaca-se tambeacutem a descoberta da inibiccedilatildeo da geraccedilatildeo de forccedila contraacutectil nas fibras
musculares por parte da espeacutecie monomeacuterica[AT]Wilson et al(1995) [AU ]determinaram que a
afinidade da espeacutecie V1 para o S1 de miosina eacute significativamente mais elevada do que a do iatildeo
Pi seu anaacutelogo Descobriram ainda que que a ligaccedilatildeo do vanadato poderaacute envolver transiccedilotildees
entre estados energeacuteticos com energia livre independente dos impedimentos esteacutereos impostos
20
pela estrutura das miofibrilas Os estudos desenvolvidos por Smith et al (1990)[AV]
determinaram queo iatildeo monovanadato natildeo apresenta um efeito pronunciado na inibiccedilatildeo da
actividadeacto-S1-miosina-ATPaacutesica excepto em concentraccedilotildees superiores a 900microM a partir das
quais existe produccedilatildeo de espeacutecies oligomeacutericas para as quais o efeito inibitoacuterio seraacute mais
elevadoA espeacutecie monomeacuterica de vanadato consegue adoptar facilmente uma geometria do tipo
bipiramidal trigonal que mimetiza o intermediaacuterio formado (MgADP
Pi)aquando da reacccedilatildeo de
hidroacutelise de ATP no local activo da miosina [AW] De facto a interacccedilatildeo do vanaacutedio com a
miosina eacute inesperadamente estaacutevel na presenccedila de ADP Esta interacccedilatildeo resulta na formaccedilatildeo de
um complexo enzimaacutetico inactivo e estaacutevel com estequiometria de 11 ADPvanadato por siacutetio
activo da proteiacutena A formaccedilatildeo do complexo estaacutevel miosina-vanadato foi proposta por Goodnoet
al (1982) [AS] Nesta ocorre um preacute-equiliacutebrio raacutepido que induz a formaccedilatildeo de um complexo
ternaacuterio reversiacutevel (MADP
V) com formaccedilatildeo lenta do isoacutemero M
ADP
V A diferenccedila entre os
compostos de vanaacutedio e de fosfato centra-se no tempo de meia vida (t12) do Pi ou do V (no
primeiro t12=12 s enquanto no segundo t12=24 h) (Refordf) A diferenccedila dever-se-aacute agrave possiacutevel
complexaccedilatildeo do vanaacutedio com grupos nucleofiacutelicos no siacutetio activo o que provocaria uma
estabilizaccedilatildeo da miosina na conformaccedilatildeo MADP
V
No que respeita aos estudos realizados com a espeacutecie tetrameacuterica Cremo et al determinou que a
cadeia pesada do S1 de miosina pode ser clivada em dois locais distintos (designados V1 e V2)
pela espeacutecie V4 apoacutes irradiaccedilatildeo com luz UV na presenccedila de concentraccedilotildees de vanadato na ordem
dos milimolar e na ausecircncia de nucleoacutetidos ou metais divalentes
Jaacute no que concerne aos estudos realizados com ecircnfase na espeacutecie decameacuterica foi sugerido
porRamos et al (2006) [AK]que a interacccedilatildeo do decavanadato com a G-actina favorecida pela
polimerizaccedilatildeo desta uacuteltima estabiliza a espeacutecie V10 e induz a inibiccedilatildeo da formaccedilatildeo de F-actina A
estabilizaccedilatildeo da espeacutecie V10 por proteiacutenas tansmembranares e do citoesqueleto pode ser a razatildeo
pela qual ocorre a toxicidade do decavanadato reportada na avaliaccedilatildeo dos efeitos do V(V) nos
sistemas bioloacutegicosTiago et al (2007) [AR] concluiram que o V10 apresenta uma clara preferecircncia
de ligaccedilatildeo ao ldquoback-doorrdquo da S1-miosina-ATPase o que se verifica apenas na estrutura
conformacional desimpedida com acesso facilitado ao local de ligaccedilatildeo do Pi Sugere tambeacutem que
o V10 actua como um ldquoback-door stoprdquo bloqueando estaabertura do fragmento de 50-kDa
necessaacuteria para a hidroacutelise do ATP--fosfato o que constitui uma explicaccedilatildeo simples do
comportamento natildeo competitivo da espeacutecie decameacuterica estimulando a utilizaccedilatildeo deste oligoacutemero
como uma feramenta da elucidaccedilatildeo das mudanccedilas conformacionais ldquoback-doorrdquo da miosina no
21
processo da contracccedilatildeo muscular(ref)Destaca-se ainda a descoberta da capacidade inibitoacuteria do
V10 em relaccedilatildeo agrave fosforilaccedilatildeo oxidativa mitocondrial [AX]
Finalmente no que concerne aos estudos realizados com a presenccedila de vaacuterias espeacutecies
oligomeacutericas Aureliano e Madeira (1994) [AY]observaram que na presenccedila de Ca2+
as espeacutecies
V4 e V10se ligam agrave Ca2+
-ATPase do retiacuteculo sarcoplasmaacutetico enquanto a espeacutecie V1 se liga apenas
na presenccedila de ATP Aparentemente o aumento da concentraccedilatildeo de Ca2+
tem o efeito contraacuterio da
inibiccedilatildeo da ATPase do retiacuteculo sarcoplasmaacutetico pelo V1 natildeo tendo no entanto efeito significativo
sobre a inibiccedilatildeo por parte da espeacutecie V10 Tiago et al (2002) [AP] concluiacuteram que em condiccedilotildees
proacuteximas agraves fisioloacutegicas a espeacutecie V10 difere das espeacutecies presentes na soluccedilatildeo de metavanadato
devido agrave sua forte interacccedilatildeo com a miosina fosforilada e agrave sua inibiccedilatildeo da actividade ATPaacutesica
da miosina Para aleacutem disso o ATP diminui a afinidade da miosina para o V4 induzindo a
interacccedilatildeo com V1 e natildeo tendo qualquer efeito no que diz respeito ao V10 Foi tambeacutem proposto
[AQ] que a ligaccedilatildeo das espeacutecies oligomeacutericas de vanadato aoS1 de miosina nos locais onde o
fosfato coordenaeacute provavelmente a causa da inibiccedilatildeo da actividade ATPaacutesica da miosina
estimulada por actina atraveacutes da prevenccedilatildeo da troca entre ATPADP sendo esta favorecida para
aniotildees de vanadato com valecircncias superiores como eacute o caso do V10
Na presenccedila de actina as velocidades de dissociaccedilatildeo do Pi e do V aumentam consideravelmente
este uacuteltimo em menor quantidade Assim a ligaccedilatildeo da actina ao complexo MADP
V e
Pi eacute
bastante fraca pelo que se sugere que o ADPV teria um efeito de relaxamento semelhante ao
ATP No entanto a capacidade do vanaacutedio em inibir a actividade ATPaacutesica da miosina e da
actomiosina eacute diferenciada uma vez que na ausecircncia de actina a inibiccedilatildeo eacute total para
concentraccedilotildees na gama dos M mas na sua presenccedila a inibiccedilatildeo eacute totalmente conseguida apenas
em concentraccedilotildees na gama dos mM (Ref) No entanto eacute necessaacuterio considerar a possiacutevel
formaccedilatildeo de espeacutecies oligomeacutericas de vanadato cuja afinidade e efeito na enzima seratildeo
diferentes Assim pode concluir-se que o efeito de inibiccedilatildeo observado para a actividade ATPaacutesica
de miosina estimulada por actina se pode dever agrave presenccedila das diferentes espeacutecies oligomeacutericas o
que foi jaacute sugerido por Aureliano (2000) [AZ] ndash as espeacutecies tetrameacuterica e decameacuterica de vanadato
interactuariam com a miosina com alteraccedilatildeo da sua interacccedilatildeo com a actina No entanto natildeo eacute
conhecida a contribuiccedilatildeo absoluta das restantes espeacutecies oligomeacutericas para a inibiccedilatildeo da ATPase
da miosina estimulada por actina (Refs)
14 Objectivos do projecto
Pretende-se com este projecto o isolamento purificaccedilatildeo e caracterizaccedilatildeo da actina do muacutesculo
22
esqueleacutetico de coelho sendo esta uacuteltima tambeacutem realizada para o sufragmento-1 de miosina
Seguidamente realizar-se-aacute a quantificaccedilatildeo e caracterizaccedilatildeo das proteiacutenas por espectroscopia UV-
Vis e SDS-PAGE e a caracterizaccedilatildeo das soluccedilotildees de vanaacutedio por UV-Vis e ressonacircncia magneacutetica
nuclear (RMN) Por uacuteltimo a interacccedilatildeo do S1 de miosina com o vanaacutedio seraacute avaliada atraveacutes
das teacutecnicas de UVVis RMN e ressonacircncia paramagneacutetica electroacutenica (RPE)
2 Discussatildeo de Resultados
21 Caracterizaccedilatildeo das proteiacutenas isoladas
211Anaacutelise do estado de pureza
O estado de pureza das soluccedilotildees de actina e de S1 foi analisado por electroforese em gel de
poliacrilamida em condiccedilotildees desnaturantes (SDS-PAGE) (Fig 22) que separa as moleacuteculas de
acordo com o seu peso molecular (PM) A concentraccedilatildeo de acrilamida escolhida (10) permite a
detecccedilatildeo simultacircnea de componentes com peso molecular inferior a 20 kDa e outros com elevado
peso molecular
Figura 21 ndash Fotografia de gel de SDS-PAGE (10 acrilamida) Poccedilo (a)
10 L do marcador de elevado peso molecular () Poccedilo (b) 20 L de [G-
actina]=1 mgmL Poccedilo (c) 20 L de [S1]=1 mgmL Aplicou-se uma
voltagem de V durante h e min
A partir da recta de calibraccedilatildeo obtida log PM = -1177 Rf + 2247 (ver ponto 2 da secccedilatildeo
Anexos) pode verificar-se a presenccedila de actina na amostra do poccedilo (b) presente numa banda
bastante intensa (PMaparente=40 kDa) bem como a presenccedila minoritaacuteria de outros componentes
desconhecidos que natildeo teratildeo sido eliminados durante o processo de extracccedilatildeo e purificaccedilatildeo e natildeo
satildeo relevantes para o estudo em questatildeo Na amostra do poccedilo (c) nota-se a presenccedila de S1
(PMaparente=92 kDa) bem como a de outros componentes cuja banda eacute ligeiramente menos intensa
Estes poderatildeo ser fragmentos proteoliacuteticos de miosina e de S1 Estaacute ainda presente uma bada de
23
maior intensidade que poderaacute ser a proteiacutena-C (PM=135 kDa) uma vez que a sua remoccedilatildeo
completa da amostra necessitaria de um passo adicional de cromatografia de troca ioacutenica [BA]
No entanto natildeo podemos confirmar que se trata desta proteiacutena ou de qualquer outra uma vez que
se encontra fora do intervalo de migraccedilatildeo dos marcadores de peso molecular
212 Determinaccedilatildeo da concentraccedilatildeo proteica
2121Subfragmento-1 de miosina
A partir da recta de calibraccedilatildeo obtida pelo meacutetodo de Bradford [BB](ver secccedilatildeo 3311 do
Procedimento Experimental) A595nm = 00546[BSA] + 00209 (ver secccedilatildeo de Anexos)foi possiacutevel
determinar um intervalo de concentraccedilatildeo de S1 de miosina compreendido entre 226 e 117(plusmn
423M) (203 e 1056 (plusmn 381 mgmL)) (n=8) A concentraccedilatildeo meacutedia de S1 de miosina apoacutes
diaacutelise contra a soluccedilatildeo tampatildeo 10 mM HEPES pH 75 25 mM KCl determinada pela leitura da
Abs280 nmfoi de 20430M (1803 mgmL)(n=7)
As amostras a quantificar atraveacutes do meacutetodo de Bradford natildeo sofreram diaacutelise o que permite
explicar a concentraccedilatildeo superior de S1 de miosina em comparaccedilatildeo com o valor obtido atraveacutes do
meacutetodo directo Esta razatildeo aliada agrave sensibilidade e rapidez superiores deste uacuteltimo fez com que
se optasse pelo meacutetodo directo para a quantificaccedilatildeo da grande maioria dos ensaios
2122Actina
A concentraccedilatildeo de actina foi determinada quando esta se encontrava na forma monomeacuterica (G-
actina) por mediccedilatildeo da leitura directa da Abs290 nm tendo-se obtido uma concentraccedilatildeo meacutedia de
691481M (290202 mgmL) (n=3) em 2 mM Tris-HCl pH 80 02 mM CaCl2 02 mM Na-
ATP Ao comprimento de onda utilizado (290 nm) absorvem fortemente apenas os resiacuteduos de
Trp pelo que a mediccedilatildeo efectuada tem menor sensibilidade do que se tivesse sido efectuada a 280
nm uma vez que a actina apresenta apenas 4 resiacuteduos deste aminoaacutecido embora tenha 16
resiacuteduos de Tyr
22 Mediccedilatildeo da actividade enzimaacutetica de S1 de miosina
221 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
As actividades ATPaacutesicas de S1 de miosina dependentes de Ca2+
e K+foram analisadas pelo
caacutelculo das velocidades de consumo inicial do fosfato inorgacircnico (Pi) Definiu-se os tempos de 5
e 3 minutos para a mediccedilatildeo da Ca2+
e da K+EDTA-ATPase respectivamente Os valores obtidos
24
a partir do protocolo presente no ponto 341 da secccedilatildeo Procedimento experimental encontram-se
descritos na Tabela 2I
Tabela 2I ndash Acrividades natildeo-fisioloacutegicas de subfragmento-1 determinadas pelo meacutetodo de Fiske-Subarow
modificado (n=2)
Proteiacutena
Actividade
ATPaacutesica natildeo
fisioloacutegica
Actividade (nmol
Piminmg proteiacutena) Actividade (s-1)
S1
Ca2+
-ATPase 1400 plusmn 30 2100 plusmn 0045
K+EDTA-ATPase 2334 plusmn 50 3500 plusmn 0075
As actividades natildeo-fisioloacutegicas reflectem o estado de oxidaccedilatildeo dos resiacuteduos expostos de Cys 707
(SH1) e 697 (SH2) Se o grupo SH1 sofrer oxidaccedilatildeo a actividade ATPaacutesica dependente de Ca2+
aumenta e a actividade dependente de K+ diminui Por outro lado se o grupo SH2 sofrer
oxidaccedilatildeo regista-se a perda completa destas actividades
De acordo com Margossian e Lowey (1982) [BA] as actividades ATPaacutesica de S1 de miosina
dependente de Ca2+
e K+EDTA da miosina satildeo respectivamente de 24 e 112 s-1
Estaacute tambeacutem
descrito que a actividade ATPaacutesica dependente de K+ eacute em meacutedia cerca de 3 vezes superior agrave
dependente de Ca2+
reflectindo a quantidade das preparaccedilotildees A razatildeo obtida entre os valores das
actividades ATPaacutesicas dependentes de K+ e Ca
2+ eacute de 166 Os valores obtidos para as actividades
(Ca2+
-ATPase=21 s-1
K+EDTA-ATPase= 35 s
-1) satildeo inferiores agrave valores obtidos por
Margossian e Lowey [BA] e Tiago [BC] com forccedila ioacutenica inferior O valor obtido para a
actividade ATPaacutesica dependente de Ca2+
eacute superior ao obtido por Polosukhina et al (2000) [BD]
cujo meio de reacccedilatildeo continha MOPS e apresentava forccedila ioacutenica inferior Os dados
supramencionados comprovam a variaccedilatildeo da actividade a ATPaacutesica natildeo fisioloacutegica em relaccedilatildeo agrave
forccedila ioacutenica temperatura e pH
222 Mediccedilatildeo da actividadeMg2+
-ATPaacutesica de S1 pelo meacutetodo de enzimas acopladas
A actividade de Mg2+
-ATPase eacute a uacutenica que possui significado fisioloacutegico sendo as restantes
bastante uacuteteis apenas para indicaccedilatildeo da qualidade de preparaccedilatildeo do S1[BA] A 0575 M KCl a
actina e o S1 formam uma soluccedilatildeo viscosa e tuacuterbida Apoacutes a adiccedilatildeo de MgATP o complexo acto-
S1 dissocia-se e a turbidez da soluccedilatildeo diminui Se a forccedila ioacutenica for reduzida a dissociaccedilatildeo do
complexo actoS1 que se encontra na forma de suspensatildeo eacute induzida O MgATP eacute rapidamente
hidrolisado diminuindo a sua concentraccedilatildeo no meio reaccional Nestas condiccedilotildees haacute
reassociaccedilatildeo e subsequente contracccedilatildeo dos filamentos de actina e de S1 com formaccedilatildeo de
25
precipitado O meacutetodo descrito traduz de forma bastante elementar o fenoacutemeno da contracccedilatildeo in
vivo e elimina o problema anterior da solubilizaccedilatildeo ineficiente da miosina de modo a obter uma
mediccedilatildeo exacta da actividade [A] A utilizaccedilatildeo de forccedilas ioacutenicas reduzidas aumenta a afinidade da
actina para o S1 permitindo contornar a extrema dificuldade em obter a concentraccedilatildeo proteica dos
muacutesculos (cerca de 100 mgmL do complexo actomiosina) As actividades Mg2+
-ATPaacutesicas de S1
de miosina foram determinadas a partir do declive da variaccedilatildeo da Abs340 relativo agrave oxidaccedilatildeo de
NADH em funccedilatildeo do tempo como eacute descrito no ponto 342 do Procedimento Experimental Os
valores obtidos para a actividade Mg2+
-ATPaacutesica encontram-se apresentados na Tabela 2II
Tabela 2II ndash Actividade Mg2+
-ATPaacutesica de subfragmento-1 de miosina determinadas pelo meacutetodo das enzimas
acopladas
Actividade (nmol
ATPmin mg(S1)) Actividade (s
-1)
1097 plusmn 269 164 plusmn 04
Ramos (2005) [J] obteve o valor de 01 s-1
para a actividade Mg2+
-ATPaacutesica de S1 de miosina
com forccedila ioacutenica intermeacutedia Tiago (2005) [BC] obteve o valor de 011 s-1 em condiccedilotildees de forccedila
ioacutenica reduzida em relaccedilatildeo agrave utilizada no presente relatoacuterio
23 Preparaccedilatildeo e caracterizaccedilatildeo das soluccedilotildees de vanaacutedio
231 Anaacutelise da composiccedilatildeo das soluccedilotildees de vanadato por espectroscopia de 51
V-RMN
efeito da concentraccedilatildeo da espeacutecie
A diluiccedilatildeo das soluccedilotildees stock de meta- e decavanadato iraacute determinar fortemente a composiccedilatildeo
em soluccedilatildeo dos diferentes oligoacutemeros de vanadato (ref) No caso do decavanadato o periacuteodo de
tempo apoacutes a diluiccedilatildeo iraacute tambeacutem ser um factor importante a considerar devido agrave decomposiccedilatildeo
da espeacutecie V10(ref)Torna-se assim fundamental determinar a composiccedilatildeo quiacutemica das soluccedilotildees
no meio de ensaio utilizado (HEPES 10 mM KCl 25 mM) de modo a correlacionaacute-la
correctamente com os possiacuteveis efeitos promovidos pelas diferentes espeacutecies de vanadato no
sistema em estudo
Aleacutem disso deve ser dado ecircnfase agraves importantes interacccedilotildees que ocorrem entre o vanaacutedio e os
tampotildees escolhidos O tampatildeo com menor nuacutemero e magnitude de interacccedilatildeo eacute o HEPES [AD]
seguido do Tris sendo este uacuteltimo especialmente aconselhado em concentraccedilotildees baixas para casos
de incompatibilidade enzimaacutetica ou quiacutemica redox No caso especial da reacccedilatildeo redox
NAD(P)HNAD(P) o tampatildeo adequado eacute o imidazole de modo a natildeo haver reacccedilatildeo com o
26
vanadato Os tampotildees a evitar em qualquer situaccedilatildeo incluem a trietanolamina fosfato citrato e
glicina entre outros [AD] uma vez que todos interactuam fortemente com V(V) com formaccedilatildeo
de complexos estaacuteveis
Assim procedeu-se agrave anaacutelise da composiccedilatildeo das soluccedilotildees de meta- e decavanadato apoacutes diluiccedilatildeo
das soluccedilotildees stock (50 mM em vanadato total) no meio de reacccedilatildeo (tampatildeo D) atraveacutes da
espectroscopia de ressonacircncia magneacutetica nuclear de 51
V Nas figuras x1 e x2 apresentam-se os
espectros obtidos para as soluccedilotildees de deca- e metavanadato num intervalo de concentraccedilotildees de 1 a
10 mM A Tabela seguinte resume os paracircmetros espectroscoacutepicos obtidos para cada espeacutecie
presente nas soluccedilotildees analisadas
Figura 22ndash Espectros de RMN de 51
V de
concentraccedilotildees crescentes das espeacutecies oligomeacutericas
de vanadato obtidos apoacutes diluiccedilatildeo da soluccedilatildeo stock
de decavanadato em 10 mM HEPES pH 70 25 mM
KCl para concentraccedilotildees finais de A) 1 mM B)2
mM C) 4 mM D) 8 mM E) 10 mM de
decavanadato total V1refere-se agraves espeacutecies
monomeacutericade vanadato V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2IIIndash Desvios quiacutemicos meacutedios (part) das ressonacircncias das espeacutecies decameacuterica (V10) monomeacuterica (V1) e
tetrameacuterica (V4) presentes nos espectros da soluccedilatildeo de decavanadatoLargura a meia altura (∆12) da espeacutecie V10A
Paracircmetros V10A V10B V10C V1 V4
part (ppm) -5162 -5003 -4255 -5616 -5748
∆12 (Hz) 175 nd nd nd nd
Figura 23 ndash Distribuiccedilatildeo graacutefica das
espeacutecies oligomeacutericas de vanadato vs a
concentraccedilatildeo de decavanadato total As
concentraccedilotildees de cada espeacutecie foram
obtidas segundo a formula descrita na
secccedilatildeo 35 do Procedimento
Experimental (aacutereas dos sinais de 51
V-
RMN observadas na figura x1)V10 V1 e
V4 correspondem agraves espeacutecie monomeacuterica
dimeacuterica e tetrameacuterica respectivamente [Esp
eacuteci
e o
ligo
meacute
rica
de
van
aacutedio
] (m
M)
[Decavanadato]total (mM)
V10
V1
V4
27
Nas soluccedilotildees de decavanadato as espeacutecies monomeacuterica e decameacuterica de vanadato satildeo as uacutenicas
presentes para concentraccedilotildees de decavanadato inferiores a 8 mM Ambas as espeacutecies aumentam
linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado em [AY] e
em (Teresa) quando a concentraccedilatildeo de V1 eacute igual ou superior a 200 microM inicia-se a sua
oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica No entanto nas condiccedilotildees de realizaccedilatildeo dos
ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a espeacutecie tetrameacuterica neste
estudo em concentraccedilotildees inferiores a 8 mM tendo o mesmo sido observado por Ramoset al
(2006) [AK] Os resultados obtidos mostram claramente o equiliacutebrio entre a espeacutecie decameacuterica e
a monomeacuterica e que o aparecimento de outras espeacutecies oligomeacutericas se encontra dependente da
concentraccedilatildeo desta uacuteltima Apesar de natildeo ter sido realizado o ensaio com 5 mM de decavanadato
total no presente relatoacuterio estaacute reportado em Ramos et al (2006) [AK] que as concentraccedilotildees da
espeacutecie decameacuterica e monomeacuterica satildeo respectivamente de 500 microM e 200 microM Assim pode inferir-
se que para concentraccedilotildees inferiores a 5 mM e intervalos de tempo de cerca de 20 minutos (que
inclui o tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas
as espeacutecies decameacuterica e monomeacuterica de vanadato
Figura 24 ndash Espectros de RMN de 51
V de concentraccedilotildees
crescentes das espeacutecies oligomeacutericas de vanadato obtidos
apoacutes diluiccedilatildeo da soluccedilatildeo stock de metavanadato em 10 mM
HEPES pH 70 25 mM KCl para concentraccedilotildees finais de A)
1 mM B) 2 mM C) 4 mM D) 8 mM E) 10 mM de
metavanadato total V1 refere-se agraves espeacutecies monomeacuterica de
vanadatoV1 e V2 correspondem respectivamente agrave espeacutecie
monomeacuterica e dimeacuterica
Figura 25 ndash Distribuiccedilatildeo graacutefica das espeacutecies
oligomeacutericas de vanadato vs a concentraccedilatildeo de
metavanadato total As concentraccedilotildees de cada
espeacutecie foram obtidas segundo a formula descrita
na secccedilatildeo 35 do Procedimento Experimental
(aacutereas dos sinais de 51
V-RMN observadas na
figura x1)V10 V1 e V4 correspondem agraves espeacutecie
monomeacuterica dimeacuterica e tetrameacuterica
respectivamente
Esp
eacuteci
e o
ligo
meacute
rica
de
va
nad
ato
(m
M)
[Metavanadato]total (mM)
V1
V2
V4
V5
28
Tabela 2IVndash Desvios quiacutemicos meacutedios (part)das ressonacircncias das espeacutecies tetrameacuterica (V4) monomeacuterica (V1) e
dimeacuterica (V2) presentes no espectro da soluccedilatildeo de metavanadato (total) 10 mM
Paracircmetros V1 V2 V4 V5
part (ppm) -5615 -5748 -5789 -5869
Nas soluccedilotildees de metavanadato as espeacutecies monomeacuterica dimeacutericae tetrameacuterica de vanadato estatildeo
presentes para concentraccedilotildees de metavanadato inferiores a 4 mM Todas estas as espeacutecies
aumentam linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado
em (Aureliano e Madeira 1998) e em (Tiago et al 2004) quando a concentraccedilatildeo de V1 eacute igual ou
superior a 200 microM inicia-se a sua oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica Nas
condiccedilotildees de realizaccedilatildeo dos ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a
espeacutecie pentameacuterica neste estudo em concentraccedilotildees inferiores a 4 mM tendo o mesmo sido
observado por Ramos 2006 Os resultados obtidos mostram claramente o equiliacutebrio entre a
espeacutecie V4 e as espeacuteciesV1 e V2 no qual o aparecimento de outras espeacutecies oligomeacutericas se
encontra dependente da concentraccedilatildeo da espeacutecie V1 Por uacuteltimo pode inferir-se que para
concentraccedilotildees inferiores a 2 mM e intervalos de tempo de cerca de 20 minutos (que inclui o
tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas as
espeacutecies V1 V2 e V4 com concentraccedilotildees respectivas de 0796 0192 e 0196 mM
232Estabilidade das soluccedilotildees de decavanadato anaacutelise por espectroscopia de absorccedilatildeo
UVVis
Caracterizou-se a estabilidade da espeacutecie V10 numa soluccedilatildeo com 1 mM decavanadato total em
tampatildeo D atraveacutes da leitura da Abs400 Obteve-se uma cineacutetica de dissociaccedilatildeo de 1ordf ordem tal
como se pode verificar pela linearidade do graacutefico de ln(Abs400)vs o tempo de aquisiccedilatildeo espectral
(segundos) (Fig26 )
Figura 26ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do tempo
(em segundos) para uma soluccedilatildeo de decavanadato total de 1
mM em tampatildeo D a 25ordmC (n=2)
Sendo uma reacccedilatildeo de primeira ordem pode ser descrita pela equaccedilatildeo cujo
declive eacute ndashK (constante de dissociaccedilatildeo) e a intersecccedilatildeo no eixo das ordenadas corresponde
Ln
(Ab
s 400 n
m)
tempo (s)
29
aln(A0) Numa reacccedilatildeo de 1ordf ordem o tempo de meia-vida eacute independente da concentraccedilatildeo inicial
do reagente sendo descrito pela equaccedilatildeo [2] Deste modo a partir da recta
Ln(Abs400 nm) = -436E-5
K - 105 obteve-se um tempo de meia-vida de 4490 plusmn 0823 horas para
as condiccedilotildees supramencionadas
Apoacutes a adiccedilatildeo de S1 8 microM ao meio reaccional verificou-se que a reacccedilatildeo de desoligomerizaccedilatildeo
da espeacutecie decameacuterica de vanadato seguia igualmente uma cineacutetica de 1ordf ordem tal como se pode
verificar pela linearidade do graacutefico de ln(Abs400) versus o tempo de aquisiccedilatildeo espectral
(segundos) (Fig27)
Figura 27ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do
tempo (em segundos) para uma soluccedilatildeo de decavanadato
total de 1 mM em tampatildeo D a 25ordmC na presenccedila de 8
microM (n= 2)
A partir da equaccedilatildeo da recta Ln(Abs400 nm) = -7520E-5
K - 0613determinou-se o valor de K para
a espeacutecie decameacuterica de onde se obteve um tempo de meia-vida de 2430 plusmn 0191 horas para as
condiccedilotildees supramencionadas
Tanto este valor como o valor registado na ausecircncia de S1 satildeo substancialmente superiores agrave
duraccedilatildeo dos ensaios de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina descritos no presente
relatoacuterio
24 Interacccedilatildeo de S1 com vanaacutedio(V)
241Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
De modo a analisar qual o efeito da adiccedilatildeo de S1 de miosina (tampatildeo D) produzia nos sinais de
51V-NMR titulou-se uma soluccedilatildeo de 5mM de decavanadato total com concentraccedilotildees crescentes
de S1 (5 10 15 e 20 microM) Os espectros de 51
V-RMN encontram-se apresentados na Fig 2
No graacutefico da figura 2 Apresenta-se a concentraccedilatildeo de S1 em funccedilatildeo do factor de alargamento
corrigido dos espectros acima referidos
Ln
(Ab
s (4
00
nm
))
tempo (s)
30
Figura 28 ndash Graacutefico de
representaccedilatildeo da
concentraccedilatildeo de S1 em
funccedilatildeo do factor de
alargamento A
concentraccedilatildeo de
decavanadato total foi de 5
mM e as concentraccedilotildees
utilizadas de S1 foram de 0
5 10 e 15 microM
Verifica-se que o factor de alargamento aumenta linearmente em funccedilatildeo da concentraccedilatildeo de S1 no
ensaio o que indica a existecircncia de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina Observou-
se ainda que a adiccedilatildeo de S1 ao meio reaccional natildeo provocou qualquer alteraccedilatildeo dos desvios
quiacutemicos dos sinais de 51
V-RMN
Realizaram-se ensaios na presenccedila de ATP e de S1 com intuito de verificar o efeito do ATP na
interacccedilatildeo do S1 de miosina com o decavanadato
Figura 29 - Espectros de RMN de 51
V contendo 2
mM Decavanadato (A) (controlo) (B) na presenccedila
de 2 mM de ATP (C) na presenccedila de 2 mM de ATP
e 8 microM de S1 de miosina V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2Vndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies decameacuterica (V10) e monomeacuterica (V1) presentes nos
espectros da soluccedilatildeo de decavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V10A V10B V10C V1
part (ppm) -5161 -5002 -4254 -5614
Observa-se que a presenccedila de ATP (espectro B) natildeo provoca qualquer alteraccedilatildeo visiacutevel em
relaccedilatildeo ao espectro que contem apenas soluccedilatildeo de decavanadato Jaacute a presenccedila simultacircnea de ATP
e S1 provoca um alargamento da ∆12 do sinal do V10A de 1736 Hz (espectro (A)) para 2366 Hz
(espectro (C) Fig 29) Tendo em conta que a concentraccedilatildeo de S1 apresenta uma relaccedilatildeo linear
com o factor de alargamento converteu-se o valor da ∆12 e comparou-se com o valor esperado agrave
F = 00591[S1] - 00157 Rsup2 = 0998
Fact
or
de
ala
rgam
en
to
[S1] (microM)
31
concentraccedilatildeo de S1 utilizada O factor de alargamento obtido para 8 microM ndash 0376 ndash eacute inferior ao
valor esperado agrave mesma concentraccedilatildeo de S1 obtido atraveacutes da equaccedilatildeo da recta apresentada na
Fig 28 ndash 0457 ndash o que indica que o ATP poderaacute exercer um efeito de protecccedilatildeo do S1
diminuindo a sua interacccedilatildeo com a espeacutecieV10
Por uacuteltimo realizaram-se ensaios com metavanadato na presenccedila de 8 microM S1 de modo a verificar
se as espeacutecies oligomeacutericas contidas nesta soluccedilatildeo interagiam com a proteiacutena em estudo
Figura 210 - Espectros de RMN de 51
V contendo 2
mM metavanadato (A) (controlo) (B) na presenccedila
de 8 microM de S1 de miosina V1 V2 e V4
correspondem respectivamente aacutes espeacutecies
monomeacuterica dimeacuterica e tetrameacuterica
Tabela 2VIndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies monomeacuterica (V1) dimeacuterica (V2) e tetrameacuterica (V4)
presentes nos espectros da soluccedilatildeo de metavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V1 V2 V4
part (ppm) -5613 -5747 -5789
Como eacute possiacutevel observar na Fig 210 ocorre uma ligeiro alargamento dos sinal de V4 bem
como uma perda da intensidade pelo que se conclui quedas espeacutecies presentes na soluccedilatildeo de
metavanadato (V1 V2 e V4) apenas a espeacutecie tetrameacuterica interage com o S1 de miosina o que
estaacute de acordo com o reportado por Tiago et al 2002
242 Actividade Mg2+
-ATPaacutesica de S1 de miosina na presenccedila de decanadato
Com o intuito de avaliar o efeito do decavanadato na actividade Mg2+
-ATPaacutesica do S1 de
miosina monitorizou-se a reacccedilatildeo de oxidaccedilatildeo do NADH atraveacutes da leitura de Abs340 na
presenccedila de 005 mgmL de S1 de miosina e no caso da actividade estimulada 2microM de actina em
3125 mM Tris-HCl pH 71 e 375 mM MgCl2 pelo meacutetodo de enzimas acopladas (ver seccccedilatildeo xx
dos MM)Para tal realizaram-se ensaios com vaacuterias concentraccedilotildees de decavanadato total (10 50
100 250 500 e 1000) de modo a determinar qual o seu efeito na actividade MgATPaacutesica de S1
de miosina na ausecircncia e na presenccedila de um dos seus ligandos naturais Os ensaios foram
realizados agrave temperatura ambiente com incubaccedilatildeo preacutevia a 25ordm C durante 20 minutos
32
Figura 211 ndash Representaccedilatildeo
graacutefica da Actividade Mg2+
-
ATPaacutesica de S1 () (os ajustes
teoacutericos estatildeo representado a por
uma linha os resultados
experimentais estatildeo representados
por triacircngulos (Actividade
estimulada) e cruzes (Actividade
natildeo estimulada)) vs
[Decavanadato]total medidas pelo
meacutetodo das enzimas acopladas
(n=3) Os dados foram ajustados ao modelo de decaimento exponencial usando o programa GraphPad Prism para a
determinaccedilatildeo dos valores de IC50 (na ausecircncia de actina IC50= 266 plusmn 07 M decavanadato na presenccedila de actina
IC50= 28 plusmn 1 M decavanadato
A partir do graacutefico presente na Fig 29 verifica-se que o ajuste dos dados experimentais foi bem
sucedido A inibiccedilatildeo da actividade Mg2+
-ATPaacutesica do S1 de miosina eacute afectada em menor
amplitude na presenccedila de actina (60 de inibiccedilatildeo da actividade) do que na sua ausecircncia
(aproximadamente 80 de inibiccedilatildeo de actividade) Devido a esta diferenccedila pode sugerir-se que a
actina iraacute impedir parcialmente a ligaccedilatildeo do decavanadatoao S1 demiosina atraveacutes de
impedimentos esteacutereos ou interacccedilotildees electrostaacuteticas por exemplo A teacutecnica utilizada natildeo nos
permite concluir acerca de que regiatildeo(otildees) ou resiacuteduo(s) da actina participaratildeo na preservaccedilatildeo
parcial da actividade Mg2+
-ATPaacutesica
Por outro lado os valores de IC50 dos dois tipos de actividades natildeo apresentam diferenccedilas
significativas tendo-se obtido 266 plusmn 07 e 28 plusmn 1M decavanadato na ausecircncia e na presenccedila de
actina respectivamente o que corresponde a uma concentraccedilatildeo de cerca de 27 M de V10 O
meio de reacccedilatildeo utilizado nestes ensaios foi 10 mM HEPES e 25 mM KCl com 2 mM ATP Em
(Tiago et al 2004) o valor de IC50 para a actividade Mg2+-ATPaacutesica na ausecircncia de actina foi de
125 plusmn 025 microM enquanto o valor da actividade estimulada por actina foi de 136 plusmn 14 microM O
meio de reacccedilatildeo utilizado nos ensaios foi 20 mM HEPES 50 mM KCl e 2 mM ATP o que resulta
precisamente no dobro da forccedila ioacutenica a que foi realizado o ensaio do presente relatoacuterio
(exceptuando a contribuiccedilatildeo dos produtos da hidroacutelise de ATP uma vez que este meacutetodo
pressupotildee a ocorrecircncia de um ciclo de reacccedilotildees de hidroacutelise e siacutentese deste composto) Ora tendo
sido anteriormente referido que a afinidade do S1 de miosina para com a espeacutecie V10 diminui com
Act
ivid
ade
Mg
2+-A
TP
aacutesic
a d
e S
1 d
e
Mio
sin
a
[Decavanadato] (microM) Actividade Estimulada ()
Actividade Natildeo Estimulada ()
Ajuste Actividade Estimulada
Ajuste Actividade Natildeo Estimulada
33
o aumento da forccedila ioacutenica (Tiago et al 2004 tal poderaacute explicar a diferenccedila obtida entre estes
dois conjuntos de valores
243Quantificaccedilatildeo de cisteiacutenas reduzidas pelo meacutetodo de Ellman
A interacccedilatildeo do vanaacutedio com alguns tipos de aminoaacutecidos sensiacuteveis agrave oxidaccedilatildeo pode fornecer-nos
informaccedilatildeo bastante uacutetil em relaccedilatildeo agraves alteraccedilotildees estruturais que conduzem a uma diminuiccedilatildeo da
actividade fisioloacutegica do S1 de miosina Os estudos realizados por Chasteen 1983 concluiacuteram
que a reduccedilatildeo de vanadato (V) a vanadilo (IV) ocorre na presenccedila de agentes redutores comuns no
ambiente celular de que se destaca o aminoaacutecido cisteiacutena Esta eacute tambeacutem um alvo de
complexaccedilatildeo por parte do iatildeo VO2+
Jaacute Ramos et al (2009) demonstrou que a incubaccedilatildeo de
actina com a soluccedilatildeo de decavanadato induz a oxidaccedilatildeo de resiacuteduos de cisteiacutena e agrave formaccedilatildeo do
oxidovanaacutedio (IV) Este grupo de investigadores descobriu ainda que apenas a espeacutecie V10 induz a
oxidaccedilatildeo das cisteiacutenas ldquoraacutepidasrdquo da actina atraveacutes do processo proposto que eacute parcialmente
responsaacutevel pela inibiccedilatildeo jaacute descrita da actividade de ATPase do S1 de miosina estimulada por
actina quando exposta agrave soluccedilatildeo de decavanadato Esta inibiccedilatildeo apresenta-se afectada com a
presenccedila de ATP uma vez que os resiacuteduos de cisteiacutena permaneceram intactos nos ensaios
realizados com este ligando Jaacute Crans et al (2004) concluiu que o siacutetio activo de muitas fosfatases
(que conteacutem grupos tiol) pode formar um equiliacutebrio redox com o iatildeo vanadato (V) de forma natildeo
reversiacutevel
Os resultados obtidos por Sakurai e Shiromura (1980) sugeriram que o vanaacutedio (V) sofre reduccedilatildeo
a vanaacutedio (IV) sem dependecircncia do valor de pH do ensaio Registaram ainda uma estequiometria
de 31 na reacccedilatildeo do vanadato (V) com os resiacuteduos de Cys enquanto a estequiometria dos ensaios
com o vanadilo (IV) foi de 21
Berlett e Stadtman (1997) observaram que os tipos de aminoaacutecidos mais facilmente oxidaacuteveis
englobam entre outros os resiacuteduos de Cys Trp e MetAssim analisou-se o estado redox dos
grupos tiol dos resiacuteduos de Cys do S1 na presenccedila do decavanadato atraveacutes da monitorizaccedilatildeo da
Abs412 nm na presenccedila de 1 mgmL de S1 de miosina em tampatildeo 10 mM HEPES 25 mM KCl
variando a concentraccedilatildeo da soluccedilatildeo de decavanadato de modo a determinar que tipo de alteraccedilotildees
estruturais ocorrem na sua presenccedila no que diz respeito agrave diminuiccedilatildeo da actividade ATPaacutesica de
S1 na presenccedila de decavanadato
Os resultados referentes agrave quantificaccedilatildeo das cisteiacutenas reduzidas apoacutes o tratamento de S1 com
diferentes concentraccedilotildees de decavanadato em tampatildeo 10 mM HEPES pH 75 e 25 mM KCl a
25ordmC durante 20 minutos encontram-se descritos na Tabela 2VII
Tabela 2VII ndash Cisteiacutenas reduzidas totais e ldquoraacutepidas de S1 (1 mgmL) apoacutes tratamento com decavanadato
34
(010501002505001000 uM) em 10 mM HEPES 25 mM KCl pH 7 a 25ordmC durante 20 minutos A detecccedilatildeo
espectrofotomeacutetrica foi feita com DTNB (e 412=8400 uM-1 cm-1) (n variaacutevel entre 1 e 8)
Amostras Cys raacutepidas Cys totais
11 M S1 (controlo) 22 plusmn 04 106 plusmn 14
11 M S1 + 10 M Decavanadato 22 plusmn 05 95 plusmn 08
11 M S1 + 50 uM Decavanadato 20 plusmn 02 98 plusmn 14
11 M S1 + 100 uM Decavanadato 17 plusmn 03 97 plusmn14
11 M S1 + 250 uM Decavanadato 26 plusmn 02 98 plusmn 08
11 M S1 + 500 uM Decavanadato 21 plusmn 04 90 plusmn 06
11 M S1 + 1000 uM Decavanadato 24 plusmn 0 86 plusmn 08
A partir dos resultados obtidos pode sugerir-se que o S1 possui 11 resiacuteduos de Cys (controlo) o
que estaacute de acordo com o anteriormente descrito Verifica-se que o tratamento da proteiacutena com
concentraccedilotildees crescentes de decavanadato provoca a oxidaccedilatildeo de dois resiacuteduos de Cys do S1 de
miosina natildeo expostos ao solvente No entanto a teacutecnica utilizada natildeo nos permite identificar
quais
Tendo em consideraccedilatildeo que o valor de IC50 obtido para a reacccedilatildeo de inibiccedilatildeo daactividade Mg2+-
ATPaacutesica de S1 na presenccedila de decavanadato eacute de cerca de 27 microM pode sugerir-se que este seraacute o
valor ao qual ocorreraacute a oxidaccedilatildeo do primeiro resiacuteduo de Cys o que poderaacute contribuir para a
diminuiccedilatildeo da actividade de S1 observada
25Interacccedilatildeo de S1 com vanaacutedio(IV)
251Anaacutelise por espectroscopia de Ressonacircncia Paramagneacutetica Electroacutenica
A acumulaccedilatildeo intracelular de oxovanaacutedio (IV) in vivo constitui uma das principais formas de
apresentaccedilatildeo de vanaacutedio no organismo Deste modo torna-se bastante interessante analisar a
interacccedilatildeo entre este iatildeo e o S1 de miosina atraveacutes da teacutecnica de espectroscopia de Ressonacircncia
Paramagneacutetica Electroacutenica o meacutetodo de eleiccedilatildeo do estudo do iatildeo vanadilo e dos complexos por si
formados O electratildeo desemparelhado encontra-se fortemente acoplado ao nuacutecleo 51
V o que
35
resulta num espectro de RPE consistindo de 8 desdobramentos na direcccedilatildeo paralela e outros tantos
na direcccedilatildeo perpendicular (Chasteen 1983)
Os estudos realizados por Ahmed et al (1987) centraram-se na interacccedilatildeo do vanadilo com
proteiacutenas como eacute o caso da insulina transferrina entre outros obtendo um espectro caracterizado
por 8 linhas amplamente espaccediladas (na direcccedilatildeo paralela) Jaacute Ramos et al 2009 concluiu que a
exposiccedilatildeo da soluccedilatildeo de decavanadato agraves formas globular e filamentosa de actina produz um
espectro tiacutepico do iatildeo oxidovanaacutedio (IV) indicando a reduccedilatildeo do vanadato
Para aferir qual a concentraccedilatildeo de S1 que permitia obter uma boa razatildeo sinalruiacutedo titulou-se uma
soluccedilatildeo de sulfato de vanadilo 15 microM ([VOSO4]stock = 5 mM) com concentraccedilotildees crescentes de
proteiacutena (0 5 10 15 e 20 microM) em 10 mM HEPES 25 mM KCl A existecircncia de uma correlaccedilatildeo
linear entre a concentraccedilatildeo de proteiacutena e a intensidade dos sinais de RPE correspondente agrave banda
-12 perpendicular encontra-se representada na Fig 210
Figura 2 12ndash Representaccedilatildeo graacutefica da variaccedilatildeo da intensidade do sinal de RPE vs a concentraccedilatildeo de S1 utilizada
Apoacutes verificar que com uma concentraccedilatildeo de S1 igual a 20 microM se obtinham sinais com uma razatildeo
sinalruiacutedo adequada ao prosseguimento dos estudos sobre a interacccedilatildeo V(IV)-S1 de miosina
titulou-se uma soluccedilatildeo de 20 microM S1 com 0 25 5 10 15 25 50 75 100 150 200 500 e 1000
microM VOSO4
Traccedilou-se o graacutefico da intensidade da banda -12 perpendicular do sinal de EPR em funccedilatildeo da
concentraccedilatildeo de VOSO4 tendo-se determinado uma constante de dissociaccedilatildeo (KD) do complexo
S1-vanadilo de 8748 plusmn 2621 microM V(IV)
y = 016786x + 122435
Rsup2 = 092992759
Inte
nsi
dad
e d
o s
inal
de
RP
E
[S1] (microM)
36
Figura 213 ndash Representaccedilatildeo graacutefica da variaccedilatildeo do sinal de RPE vs a concentraccedilatildeo de vanadilo utilizada (os valores
experimentais estatildeo representados por losangos o seu ajuste estaacute representado pela linha)
A simulaccedilatildeo do espectro de EPR foi efectuada a partir do espectro obtido para uma concentraccedilatildeo
de 1 mM VOSO4 apoacutes adiccedilatildeo de 20 microM S1 de miosina tendo-se obtido os valores da constante de
acoplamento hiperfina (A=5016) e do factor g (g0=195)
Figura 214 ndash Representaccedilatildeo graacutefica do espectro de EPR obtido com 1000 microM de vanadilo e 20 microM de S1 (a cinzento
escuro) A simulaccedilatildeo do espectro encontra-se a cinzento claro
Por comparaccedilatildeo destes com os valores presentes na tabela de correspondecircncia xxx eacute possiacutevel
predizer quais os ligandos provaacuteveis do vanadilo na presenccedila de proteiacutena Assim concluiu-se que
o vanadilo estaacute coordenado com duas moleacuteculas de aacutegua um grupo hidroxilo e um grupo R=NR
O grupo hidroxilo pode indicar os resiacuteduos de aminoaacutecidos Tyr Treonina e Serina enquanto o
grupo R=NR poderaacute indicar a presenccedila de Arginina e Histidina como ligandos do vanadilo
Inte
nsi
dad
e d
os
sinai
s d
e R
PE
[Vanadilo] (microM)
Ajuste dos dados exp Dados experimentais
Campo magneacutetico (G)
Espectro Experimental Ajuste
37
3 Procedimento Experimental
31 Reagentes
Todos os reagentes utilizados satildeo de elevado grau de pureza (analiacutetico) Apresentam-se as suas
foacutermulas quiacutemicas ou abreviaturas quando necessaacuterio acrescentou-se a sua descriccedilatildeo Entre
parecircntesis colocou-se as empresas que os comercializam
Aacutecido aceacutetico glacial 100 (Riedel) Acrilamida 99 (Merck) Azul de bromofenol (Riedel-de-
Haumlen) -Mercaptoetanol (Sigma) Bissulfito de soacutedio (Sigma)Bradford (reagente de)
(BioRad)BSA (albumina de soro bovino Sigma-Aldrich) CaCl22H2O (cloreto de caacutelcio
dihidratado Fluka) CH3CH2OH 96 (etanol Panreac) Coomassie Blue R250 (Merck) D2O
(oacutexido de deuteacuterio SDS) DTNB (55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico) Aldrich) EDTA (aacutecido
etilenodiaminatetraaceacutetico Merck) Fosfoenolpiruvato 98 (Fluka) Glicerol (Sigma) Glicina
985 (Panreac) HCl 37 (Riedel-de Haumlen) H2SO4 96 (Pronalab) HEPES 99 (aacutecido 4-(2-
hidroxiletil)-1-piperazineetanosulfoacutenico Sigma) KH2PO4 99 (hidrogenofosfato de potaacutessio)
MgATP (Sigma) Na
ATP (Sigma) Na2HPO4 99 (hidrogenofosfato de soacutedio Riedel-de Haumlen)
NaH2PO4 (fosfato de soacutedio ) NaOH 986 (hidroacutexido de soacutedio Pronalab) KCl 99 (Cloreto de
potaacutessio Panreac) L-Cys 995 (enantioacutemero levoacutegiro do resiacuteduo de Cisteiacutena Fluka)Metanol
99 (Merck) Molibdato de amoacutenio (Sigma)NNrsquo-metil-bis-acrilamida 98 (Merck) NH4VO3
99 (metavanadato de amoacutenia Sigma) PSA 10 (persulfato de amoacutenio Merck) SDS
(dodecilo sulfato de soacutedio Sigma) Sulfito de soacutedio (Sigma) VOSO4 (sulfato de vanadilo
Aldrich) TEMED 99 (Tetrametiletilenodiamina Merck) Tris (2-amino-2-hidroximetil-13-
propanodiol Sigma)
32 Meacutetodos
321 Isolamento e purificaccedilatildeo das proteiacutenas de muacutesculo esqueleacutetico de coelho
3211 Purificaccedilatildeo da miosina preparaccedilatildeo de Subfragmento 1 por digestatildeo quimotriacutepsica
da miosina
O subfragmento-1 de miosina-II (S1) foi gentilmente cedido pelo Prof Doutor Manuel Aureliano
do Departamento de Quiacutemica Bioquiacutemica e Farmaacutecia da Faculdade de Ciecircncias e Tecnologia da
Universidade do Algarve
No entanto descreve-se abaixo de forma resumida o procedimento efectuado
38
32111 Purificaccedilatildeo da miosina
O protocolo de isolamento foi adaptado do meacutetodo previamente descrito por Margossian e Lowey
(1982) apresentando-se de forma esquemaacutetica na Fig 31
Sucintamente os tecidos musculares dorsais e das pernas foram extraiacutedos apoacutes o sacrifiacutecio do
animal (Fig31) De seguida foram adicionados 3 volumes da soluccedilatildeo de extracccedilatildeo A2
Centrifugou-se a soluccedilatildeo e filtrou-se o sobrenadante O sedimento foi entatildeo diluiacutedo em 14
volumes de aacutegua destilada de modo a que a forccedila ioacutenica atingisse 004 M Apoacutes adiccedilatildeo de szlig-
mercaptoetanol (szlig-me) deixou-se a precipitar a 4ordmC removendo o sobrenadante Centrifugou-se
novamente e ressuspendeu-se o sedimento na soluccedilatildeo de extracccedilatildeo A Adicionou-se a soluccedilatildeo
tampatildeo B3 para que a forccedila ioacutenica atingisse 05 M Homogeneizou-se e deixou-se repousar
durante 2h Homogeneizou-se novamente e centrifugou-se de modo a ocorrer precipitaccedilatildeo dos
complexos de actomiosina e proteiacutena desnaturada Apoacutes filtraccedilatildeo do sobrenadante em latilde de vidro
quantificou-se a soluccedilatildeo de miosina resultante pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental)
2 Soluccedilatildeo de extracccedilatildeo A 0012M K2CO3 004M KHCO3 06M KCl pH 65
3 Soluccedilatildeo-tampatildeo B Tris 100 mM pH 7 KCl 2M
39
Figura 31 ndash Representaccedilatildeo esquemaacutetica do protocolo de isolamento de miosina (Margossian e Lowey 1982) Todo o
procedimento foi realizado a 4ordmC (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
32112 Preparaccedilatildeo de Subfragmento-1 de miosina por digestatildeo com quimiotripsina
O S1 de miosina foi isolado apoacutes diaacutelise da miosina durante a noite contra a soluccedilatildeo C4 A
presenccedila do EDTA eacute essencial uma vez que a sua substituiccedilatildeo por MgCl2resulta na produccedilatildeo de
HMM em vez de S1 A suspensatildeo de miosina foi equilibrada e digerida com -quimotripsina
A digestatildeo foi interrompida pela adiccedilatildeo de PMSF Seguidamente ultracentrifugou-se o produto
da digestatildeo desprezando-se o sedimento (Fig 32) A soluccedilatildeo de sobrenadante maioritariamente
S1 foi quantificada pela leitura directa da Abs280 ou pelo meacutetodo de Bradford (ver as secccedilotildees
3311 e 3312 do Procedimento Experimental respectivamente) e armazenada em 10 sacarose
e 2 mM szlig-Me a -80ordmC
4 Soluccedilatildeo C 25mM Tris pH 70 012M KCl 1mM EDTA
40
Antes da realizaccedilatildeo dos ensaios descritos no presente relatoacuterio o S1 de miosina foi dialisado
contra a soluccedilatildeo D5 de modo a remover qualquer composto que pudesse complexar como
vanaacutedio Seguidamente quantificou-se a proteiacutena pelo meacutetodo de Bradford ou pelo meacutetodo
directo (ver secccedilotildees 3311 e 3312do Procedimento Experimtental)
Figura 3 2ndash Representaccedilatildeo esquemaacutetica do protocolo de preparaccedilatildeo do S1 de miosina por digestatildeo quimiotripsina de
acordo com o previamente descrito por Margossian e Lowey (1982) Todo o procedimento foi realizado a 4ordm C
excepto quando indicaccedilatildeo do contraacuterio (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
3212 Isolamento e purificaccedilatildeo de actina
Isolou-se e purificou-se a actina segundo o meacutetodo descrito por Pardee e Spudish que
compreende dois passos ndash a obtenccedilatildeo de um resiacuteduo seco denominado ldquopoacute de acetonardquo e a
extracccedilatildeo subsequente da actina a partir do ldquopoacute de acetonardquo na forma de G-Actina com a
posterior polimerizaccedilatildeo a F-Actina
32121 Obtenccedilatildeo do ldquopoacute de acetonardquo
Sacrificou-se o coelho com uma pancada atraacutes do pescoccedilo e sangrou-se o animal apoacutes o corte da
arteacuteria jugular Esfolou-se e colocou-se numa tina com gelo Retiraram-se as partes de tecido
conjuntivo que envolvem o tecido muscular com auxiacutelio de uma pinccedila e bisturi Seguidamente
retirou-se os muacutesculos dorsais e os muacutesculos brancos das pernas Passou-se os muacutesculos por aacutegua
destilada de modo a remover as impurezas e pesou-se num copo Trituraram-se os muacutesculos
numa maacutequina de triturar carne e extraiu-se o resultante com 3 volumes da soluccedilatildeo E6 durante 10
minutos com agitaccedilatildeo suave Centrifugou-se a suspensatildeo a 4000 g durante 10
5 Soluccedilatildeo D HEPES 10 mM pH 70 KCl 25 mM
6 Soluccedilatildeo E 01 M KCl 015 M K2HPO4 pH 65
41
minutosdesprezando-se o sobrenadante Ressuspendeu-se o sedimento em 6 volumes de soluccedilatildeo
F7durante 10 minutos com agitaccedilatildeo suave Verificou-se se o pH da soluccedilatildeo estava compreendido
entre 82 e 85 acertando-se estecom soluccedilatildeo G8 se necessaacuterio Centrifugou-se durante 10
minutos a 4000 g Desprezou-se o sobrenadante e ressuspendeu-se o sedimento em 3 volumes de
soluccedilatildeo H9 durante 10 minutos Centrifugou-se novamente a 4000 gdurante 10 minutos
desprezando-se o sobrenadante Ressuspendeu-se o sedimento duas vezes consecutivas em 6
volumes de aacutegua destilada durante 5 minutos Centrifugou-se a 4000 g durante 10 minutos apoacutes
cada adiccedilatildeo de H2O
Seguidamente ressuspendeu-se o sedimento 5 vezes consecutivas utilizando 3 volumes de
acetona de cada vez agitando suavemente durante 10 minutos agrave temperatura ambiente Colocou-
se o resiacuteduo filtrado numa tina de vidro sobre papel de filtro deixando-se evaporar o solvente
durante a noite num nicho Triturou-se o resiacuteduo seco numa maacutequina de triturar carne e deixou-se
novamente evaporar o solvente O ldquopoacute de acetonardquo obtido foi armazenado a -20 ordmC
32122 Purificaccedilatildeo da actina a partir de ldquopoacute de acetonardquo
Apoacutes a obtenccedilatildeo do poacute de acetona como acima indicado efectuou-se o protocolo seguidamente
descrito de modo a obter monoacutemeros de actina (G-Actina) e actina filamentosa (F-Actina)
Pesou-se 35 g de poacute de acetona e adicionaram-se 50 mL de soluccedilatildeo-tampatildeo I10
sendo a
extracccedilatildeo feita com agitaccedilatildeo suave durante 30 minutos Separou-se o extracto do poacute de acetona
hidratado filtrando a suspensatildeo com 4 camadas de gaze esterilizada tendo o cuidado de natildeo
espremer o resiacuteduo Voltou a extrair-se com 50 mL de soluccedilatildeo I durante 30 minutos mexendo
lentamente Adicionou-se este ao extracto obtido anteriormente Centrifugou-se o extracto total a
16 000 g durante 30 minutos Decantou-se para uma proveta sobre a qual se colocou gaze para
que o sedimento solto natildeo se juntasse ao sobrenadante e leu-se o volume De seguida adicionou-
se KCl ateacute uma concentraccedilatildeo final de 50 mM MgCl2 ateacute 2mM e ATP ateacute 1mM lentamente e com
agitaccedilatildeo contiacutenua de modo a promover a polimerizaccedilatildeo dos monoacutemeros de actina
Deixou-se polimerizar durante 2h a 25ordmC observando-se a presenccedila de pequenas bolhas de ar
indicativas de um aumento de viscosidade da soluccedilatildeo Arrefeceu-se em gelo durante 15 minutos e
adicionou-se KCl ateacute uma concentraccedilatildeo final de 06M Mexeu-se lentamente durante 30 minutos
apoacutes os quais se ultracentrifugou a 110 000g durante 1hora Desprezou-se o sobrenadante
ressuspendeu-se o sedimento com soluccedilatildeo tampatildeo E e transferiu-se para um potter
Dialisou-se durante a noite contra 1L de soluccedilatildeo-tampatildeo I a uma temperatura de 4ordmC com rotaccedilatildeo
7 Soluccedilatildeo F 005 M NaHCO3 pH 82-85
8 Soluccedilatildeo G 1M Na2CO3
9 Soluccedilatildeo H 1 mM EDTA pH 70
10 Soluccedilatildeo I 2 mM Tris-HCl pH 8 02 mM CaCl2 02 mM Na2ATP 05 mM szlig-Me 0005 NaN3
42
contiacutenua e mudanccedila de tampatildeo 2 a 3 vezes De seguida a actina foi centrifugada a 110
000gdurante 1hora Decantou-se a soluccedilatildeo de G-Actina (sobrenadante) e desprezou-se o
sedimento Determinou-se a concentraccedilatildeo de G-Actina pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental) A soluccedilatildeo de G-actina foi armazenada a uma temperatura de 4ordmC
apoacutes adiccedilatildeo de 002NaN3 mantendo-se estaacutevel durante 3 a 5 dias
331 Quantificaccedilatildeo das proteiacutenas isoladas
3311 Quantificaccedilatildeo pelo meacutetodo de Bradford
O meacutetodo de Bradford (Bradford 1976) baseia-se na utilizaccedilatildeo do reagente de Bradford
composto por entre outros aacutecido fosfoacuterico e o corante ldquoCoomassierdquo Este corante possui carga
negativa e a sua coloraccedilatildeo varia entre a vermelha (Absmax=465 nm) predominante em soluccedilatildeo e a
azul (Absmax=595 nm) presente aquando da ligaccedilatildeo aos grupos amina das proteiacutenas Em soluccedilotildees
aacutecidas diluiacutedas a coloraccedilatildeo varia proporcionalmente com a quantidade do corante ligado agrave
proteiacutena que por sua vez eacute directamente proporcional agrave concentraccedilatildeo da proteiacutena permitindo a
sua quantificaccedilatildeo ateacute a um limite de detecccedilatildeo de 20 gmL (Bradford 1976)
Deste modo prepararam-se amostras de BSA ([BSA]stock= 1mgmL) utilizando um intervalo de
concentraccedilotildees compreendido entre 0 e 15 gmL de acordo com a tabela 3I Apoacutes a adiccedilatildeo da
H2O MilliQ e do reagente de Bradford as soluccedilotildees foram sucessivamente homogeneizadas no
voacutertex
Tabela 3Indash Amostras utilizadas para a obtenccedilatildeo da recta de calibraccedilatildeo no meacutetodo de Bradford
BSA (L) BSA (g) H2O MilliQ (L) Reagente de Bradford (L)
0 0 800 200
125 125 79875 200
25 25 7975 200
5 5 795 200
10 10 790 200
15 15 785 200
Apoacutes incubaccedilatildeo a 25ordmC por 5 minutos leram-se os valores de Abs595 num
espectrofotoacutemetro (Shimadzu UV-160A) para todas as amostras Seguidamente traccedilou-se a recta
de calibraccedilatildeo (Abs595vs quantidade de BSA (g))
43
Para determinar a concentraccedilatildeo de proteiacutena apoacutes o processo de isolamento purificaccedilatildeo e
diaacutelise quando necessaacuterio colocou-se 5 L da amostra num tubo de Eppendorf e adicionou-se
795 L de H2O MilliQ homogeneizando a soluccedilatildeo no voacutertex Adicionou-se posteriormente 200
L de reagente de Bradford e repetiu-se a homogeneizaccedilatildeo Incubou-se a mistura a 25ordmC por 5
minutos apoacutes os quais se leu os valores de Abs595 nm no espectrofotoacutemetro Por interpolaccedilatildeo deste
valor a partir da recta de calibraccedilatildeo determinou-se a concentraccedilatildeo de proteiacutena na amostra
3312 Quantificaccedilatildeo pelo meacutetodo directo
A quantificaccedilatildeo das amostras proteicas pode tambeacutem ser efectuada atraveacutes da leitura directa da
absorvacircncia tendo por base a lei de Beer-Lambert onde Abs corresponde agrave
absorvacircncia ao comprimento de onda ao qual o ensaio foi realizado corresponde ao coeficiente
de extinccedilatildeo (substituiacutedo pela letra a se as unidades forem distintas da molaridade) b corresponde
ao percurso oacuteptico da radiaccedilatildeo na ceacutelula amostral e c corresponde agrave concentraccedilatildeo amostral
Assim no caso de S1 de miosina mediu-se a Abs280e calculou-se a concentraccedilatildeo da proteiacutena
sabendo que o coeficiente de extinccedilatildeo de S1 eacute (Ref)
No caso da actina mediu-se a Abs290ecalculou-se a concentraccedilatildeo da proteiacutena sabendo que o
coeficiente de extinccedilatildeo da actina eacute (Ref)
332Electroforese em condiccedilotildees desnaturantes (SDS-PAGE)
De modo a melhor caracterizar as proteiacutenas isoladas foi realizada uma electroforese em
condiccedilotildees desnaturantes (SDS-PAGE do inglecircs Sodium Dodecyl Sulfate Polyacrilamide Gel
Electrophoresis) O SDS (dodecilo sulfato de soacutedio) eacute um detergente anioacutenico utilizado neste
meacutetodo para desnaturar as proteiacutenas que se ligam ao SDS na proporccedilatildeo 14 g SDS g proteiacutena
(aproximadamente 1 moleacutecula de SDS por cada 2 resiacuteduos de aminoaacutecidos) (Stryer) O SDS induz
uma densidade de carga negativa significativa nas proteiacutenas a que se liga mascarando a sua carga
intriacutenseca pelo que as razotildees cargamassa satildeo tendencialmente idecircnticas Assim a SDS-PAGE
separa as proteiacutenas quando sujeitas a um campo eleacutectrico de acordo com o seu peso molecular
sendo a sua migraccedilatildeo directamente proporcional ao logaritmo deste uacuteltimo (Stryer)
O gel de resoluccedilatildeo com 10 de acrilamida foi obtido atraveacutes da adiccedilatildeo de 167 mL de soluccedilatildeo
stock de acrilamida11
075 mL de tampatildeo de resoluccedilatildeo12
251 mL de aacutegua destilada e 50 L de
SDS 10 Para promover a polimerizaccedilatildeo adicionou-se em seguida 38 L de PSA 10
11
soluccedilatildeo stock de acrilamida soluccedilatildeo de 30 (mv) de acrilamida e 08 (mv) de NNrsquo-metil-bis-acrilamida 12
tampatildeo de resoluccedilatildeo 15 M Tris-HCl pH 88
44
(persulfato de amoacutenia) 10 e 25 L de TEMED 99 (tetrametiletilenodiamina) Agitou-se
cuidadosamente de modo a natildeo formar bolhas e pipetou-se a soluccedilatildeo para a cassette de vidro
previamente montada no suporte De seguida adicionou-se uma peliacutecula de cerca de 2 mm de 2-
metil-2-butanol para verificar posteriormente a polimerizaccedilatildeo Apoacutes a polimerizaccedilatildeo que demora
cerca de 45 minutos verteu-se a agrave superfiacutecie e adicionou-se o gel de concentraccedilatildeo com 5 de
acrilamida constituiacutedo por 03 mL de soluccedilatildeo de stock acrilamida 045 mL de tampatildeo de
concentraccedilatildeo13
094 mL de aacutegua destilada 18 L de SDS 10 135 L de PSA 10 e 2L de
TEMED 99 Colocou-se o pente sobre o gel e deixou-se polimerizar durante cerca de 45
minutos
As amostras para aplicaccedilatildeo no gel de electroforese foram preparadas para uma concentraccedilatildeo de
proteiacutena de 1 mgmL num volume final de 100 L dos quais 25 L correspondem ao tampatildeo da
amostra14
As amostras foram aquecidas a 100ordmC durante 5 minutos Antes da aplicaccedilatildeo das
amostras transferiu-se a cassette para a tina de electroforese agrave qual se adicionou a soluccedilatildeo
tampatildeo de electroforese da tina superior15
e tampatildeo de electroforese da tina inferior16
Aplicaram-
se 10 L das soluccedilotildees de marcadores de peso molecular e 20 L das amostras proteicas
preparadas anteriormente A electroforese decorreu durante cerca de 3h30 min a uma voltagem
de 80 V De seguida transferiu-se o gel para uma tina corando as proteiacutenas durante 1h com
soluccedilatildeo corante17
Descorou-se o gel com soluccedilatildeo descorante18
tendo-se mudado esta soluccedilatildeo ateacute
as bandas se encontrarem bem definidas
Determinou-se a mobilidade relativa das bandas correspondentes aos marcadores de elevado peso
molecular e traccedilou-se a recta de calibraccedilatildeo ndash log(PM) vs log(Rf) onde Rf corresponde agrave
mobilidade relativa Por interpolaccedilatildeo determina-se o PM das proteiacutenas isoladas sabendo a sua
mobilidade relativa Por comparaccedilatildeo com os valores descritos na literatura pode concluir-se
acerca do grau de purificaccedilatildeo
34 Mediccedilatildeo da actividade enzimaacutetica de S1
341 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
13
tampatildeo de concentraccedilatildeo 05 M Tris-HCl pH 68 14
tampatildeo de amostra 320 mM Tris-HCl pH 68 04 M szlig-mercaptoetanol SDS 8 glicerol 15 (vv) e roxo de m-
cresol 0024 15
tampatildeo de electroforese da tina superior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS 16
tampatildeo de electroforese da tina inferior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS acetato de soacutedio
01 M 17
soluccedilatildeo corante ldquoCoomassie Blue R-250rdquo contendo metanol 45 (vv) e aacutecido aceacutetico glacial 10 (vv) 18
soluccedilatildeo descorante metanol 10 (vv) e aacutecido aceacutetico glacial 10 vv)
45
O meacutetodo de Fiske-Subarow modificado consiste na reacccedilatildeo do molibdato de amoacutenio com o
fosfato inorgacircnico em soluccedilatildeo aacutecida com formaccedilatildeo do aacutecido fosfomoliacutebdico Seguidamente daacute-
se a adiccedilatildeo de um agente redutor que provoca a reduccedilatildeo do composto a fosfomolibdato que forma
um complexo de cor azulada cuja absorvacircncia maacutexima ocorre para um comprimento de onda de
660 nm tornando possiacutevel a sua detecccedilatildeo por meacutetodos espectrofotomeacutetricos As actividades Ca2+
-
ATPaacutesica e K+EDTA-ATPaacutesica de S1 foram determinadas atraveacutes deste meacutetodo de acordo com
o descrito por Taussky e Shorr (1953) com mediccedilatildeo da quantidade de fosfato (Pi) libertado
atraveacutes da hidroacutelise de ATP
Para tal obteve-se primeiramente a recta de calibraccedilatildeo com as amostras presentes na Tabela 3II
utilizando uma soluccedilatildeo stock de 5 mM KH2PO4
Tabela 3II ndash Composiccedilatildeo das diferentes amostras utilizadas para determinar a recta de calibraccedilatildeo atraveacutes do meacutetodo
de Fiske-Subbarow modificado
KH2PO4 (L)
([KH2PO4]stock= 5mM)
PO43-
(nmole) H2O (uL)
0 0 1780
5 25 1775
10 50 1770
20 100 1760
40 200 1740
100 500 1680
A cada tubo de Eppendorf adicionaram-se seguidamente 200 L de soluccedilatildeo J19
e 20 L de agente
redutor20
homogeneizando no voacutertex apoacutes cada adiccedilatildeo Incubou-se durante 10 minutos e mediu-
se a Abs660 nm num espectrofotoacutemetro com intervalos de tempo iguais entre cada amostra Traccedilou-
se a recta de calibraccedilatildeo (Abs660 nmvs nmol PO43-
) a partir dos valores obtidos
Para determinar a actividade Ca2+
-ATPaacutesica de S1 prepararam-se tubos de Eppendorf com 400
L de meio de reacccedilatildeo21
25 L CaCl2 ([CaCl2]stock= 100 mM) 25 L ATP ([ATP]stock= 100 mM)
com [S1]ensaios=01 mgmL Perfez-se se necessaacuterio com H2O MilliQ ateacute perfazer um volume de 1
mL Para determinar a actividade K+EDTA-ATPaacutesica utilizou-se uma concentraccedilatildeo idecircntica de
19
Soluccedilatildeo J 5 N H2SO4 25 molibdato de amoacutenio 20
Agente redutor 5 mg aacutecido 1-amino-2-naftol-4-sulfoacutenico 30 mg bissulfito de soacutedio 30 mg sulfito de soacutedio em 26
mL H2O 21
meio de reacccedilatildeo 125 mM Tris-HCl pH 80 0575 M KCl
46
proteiacutena nos ensaios (01 mgmL) e o mesmo meio de reacccedilatildeo Aos tubos de Eppendorf
adicionou-se ainda 123 L de KCl ([KCl]stock = 3M) 25 L de ATP 10 L de EDTA
([EDTA]stock= 100 mM )e o volume de H2O MilliQ necessaacuterio para perfazer 1 mL Em ambas as
situaccedilotildees o uacuteltimo reagente adicionado foi o ATP uma vez que se trata do substrato da reacccedilatildeo
enzimaacutetica
A cineacutetica decorreu a 25ordmC durante 5 minutos para a actividade Ca2+
-ATPaacutesica e durante 3
minutos para a actividade K+EDTA-ATPaacutesica Apoacutes o periacuteodo de incubaccedilatildeo transferiu-se 150
L de amostra para tubos de Eppendorf contendo igual volume de TCA 10 Colocou-se no gelo
e homogeneizou-se no voacutertex Seguidamente centrifugou-se a 1500 g durante 10 minutos e
recolheu-se 200 L de sobrenadante para novos tubos de Eppendorf Adicionou-se a estes 158
mL de H2O MilliQ e 200 L de soluccedilatildeo J Homogeneizou-se no voacutertex e adicionou-se 20 L de
reagente redutor repetindo-se a homogeneizaccedilatildeo Incubou-se durante 10 minutos e mediu-se a
Abs660 nm com o mesmo intervalo de tempo de leitura para cada amostra Por intrapolaccedilatildeo a partir
da recta de calibraccedilatildeo obtida anteriormente determinou-se a quantidade de Pi libertado em cada
ensaio A partir destes resultados determinaram-se as actividades Ca2+
-ATPaacutesica e K+EDTA-
ATPaacutesica do S1 em nmol PO43-
minmg proteiacutena e em s-1
342 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
Da actividade Mg2+
-ATPaacutesica de S1 resulta a formaccedilatildeo dos produtos da reacccedilatildeo de
hidroacutelise do ATP - ADP e Pi Na presenccedila de ADP fosfoenolpiruvato (PEP) e da enzima piruvato
cinase (PK) os produtos resultantes satildeo ATP e piruvato Por acccedilatildeo da enzima lactato
desidrogenase (LDH) este uacuteltimo eacute convertido em lactato ocorrendo simultaneamente a oxidaccedilatildeo
do NADH a NAD+ monitorizada atraveacutes da Abs340 nm Quando o S1 de miosina se encontra no
meio reaccional o ATP eacute hidrolisado formando-se novamente ADP e Pi o que permite a
formaccedilatildeo de um ciclo (fig 21) Se as enzimas se encontrarem em excesso a velocidade de
conversatildeo do NADH eacute limitada pela produccedilatildeo de ADP e Pi Como o ATP eacute regenerado em cada
ciclo a sua concentraccedilatildeo inicial eacute sempre mantida (Bagshaw 1982)
Figura 33ndash Representaccedilatildeo esquemaacutetica do meacutetodo de enzimas acopladas
47
O protocolo utilizado e seguidamente descrito foi adaptado de Cuenda et al (1990 ndash Ver Refordf) A
uma cuvette de 1 mL adicionou-se S1 para uma concentraccedilatildeo final de 005 mgmL 800 L de
meio de reacccedilatildeo22
10 L de PEP ([PEP]stock = 42 mM) 10 L de PK+LDH 11 ([PK+LDH]stock =
1 mgmL) e 15 L de NADH ([NADH]stock=25 mM) Iniciou-se a monitorizaccedilatildeo da reacccedilatildeo
atraveacutes da leitura de Abs340 nm apoacutes a adiccedilatildeo de 20 L de ATP ([ATP]stock=100 mM) A
actividade Mg2+
-ATPaacutesica eacute obtida atraveacutes do declive correspondente agrave oxidaccedilatildeo do NADH Para
a mediccedilatildeo da Mg2+
-ATPase de S1 estimulada por actina adicionou-se F-actina para uma
concentraccedilatildeo final de 2 microM sendo a actividade obtida tambeacutem atraveacutes do declive correspondente
agrave oxidaccedilatildeo do NADH apoacutes a adiccedilatildeo da proteiacutena
35 Interaccedilatildeo entre vanaacutedio e subfragmento-1 de miosina
351 Preparaccedilatildeo das soluccedilotildees de vanaacutedio
As soluccedilotildees de vanadato foram preparadas em aacutegua MilliQ a partir de metavanadato de amoacutenio
(NH4VO3) numa concentraccedilatildeo stock de 50 mM com agitaccedilatildeo magneacutetica e aquecimento (~30 ordmC)
Esta soluccedilatildeo com um valor compreendido entre 6 e 7 designa-se por ldquometavanadatordquo e eacute
constituiacuteda pelas esspeacutecies V1 V2 V4 e V5 (Ref) Acidificando a soluccedilatildeo de metavanadato por
adiccedilatildeo de HCl ateacute um valor de pH proacuteximo de 4 observa-se uma mudanccedila de cor de amarelo
claro para laranja indicativa da formaccedilatildeo da espeacutecie decameacuterica de vanadato De seguida e
imediatamente antes da realizaccedilatildeo dos ensaios descritos o valor do pH da soluccedilatildeo de
decavanadato foi aumentado para cerca de 75 por adiccedilatildeo de NaOH A soluccedilatildeo (2) designada
por ldquometavanadatordquo natildeo foi posteriormente alterada ateacute agrave sua adiccedilatildeo para a realizaccedilatildeo dos ensaios
As soluccedilotildees foram armazenadas a 4ordmC e mantidas em gelo aquando da sua utilizaccedilatildeo
A soluccedilatildeo de V(IV) foi preparada em condiccedilotildees anaeroacutebias por diluiccedilatildeo de sulfato de vanadil
(VOSO4) para uma concentraccedilatildeo final de 5 mM em H2O MilliQ saturada com N2 gasoso
352Anaacutelise por espectroscopia de UV-Vis
A concentraccedilatildeo da espeacutecie decameacuterica em soluccedilatildeo depende do periacuteodo de tempo apoacutes a diluiccedilatildeo
no meio de reacccedilatildeo apropriado (Ref) A velocidade de desoligomerizaccedilatildeo da espeacutecie decameacuterica
pode ser analisada por espectroscopia de absorccedilatildeo na regiatildeo UVVis uma vez que esta espeacutecie
apresenta duas bandas caracteriacutesticas na regiatildeo do visiacutevel (a 358 e 400 nm) que natildeo se verifica
em quaisquer outras espeacutecies oligomeacutericas As bandas supramencionada sapresentam-se bem
22
Meio de reacccedilatildeo 3125 mM Tris-HCl pH 71 375 mM MgCl2
48
resolvidas atraveacutes da obtenccedilatildeo da derivada de 2ordf ordem do espectro apresentando a forma de 2
vales com miacutenimos centrados nos comprimentos de onda supracitados com dependecircncia linear
relativamente agrave concentraccedilatildeo de decavanadato total (Ramos et al 2006)
Assim a estabilidade da espeacutecie decameacuterica apoacutes diluiccedilatildeo da soluccedilatildeo de decavanadato no meio de
reacccedilatildeo dos ensaios (tampatildeo D) para uma concentraccedilatildeo final de 1 mM em vanadato total foi
caracterizada atraveacutes da leitura da Abs400 em funccedilatildeo do tempo tendo-se determinado qual a
ordem de reacccedilatildeo e o tempo de meia-vida da mesma Com base em estudos anteriores acerca da
estabilidade do V10 na presenccedila de diferentes proteiacutenas (Ramos et al 2006) determinou-se
igualmente o tempo de meia vida apoacutes a adiccedilatildeo de 20 microM de S1 de miosina ao meio reaccional
353 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
Os espectros de RMN do isoacutetopo 51
V foram realizados num espectroacutemetro Bruker AM-400 MHz
equipado com uma sonda multinuclear inversa de 5 mm Em tubos de RMN de 5 mm de diacircmetro
preparou-se soluccedilotildees de 500 L de deca- ou metavanadato correspondentes a um intervalo de
concentraccedilotildees de 1 a 10 mM suplementadas com 10 D2O Os espectros foram adquiridos agrave
temperatura ambiente com uma largura espectral de 23 923 Hz um tempo de aquisiccedilatildeo de 0086
s tempo de relaxaccedilatildeo de 001 s e uma sequecircncia de pulsos a 90ordm Utilizou-se a transformada de
Fourier com alargamento das bandas de 20Hz para o processamento dos espectros Os desvios
quiacutemicos das ressonacircncias de 51
V foram obtidos em relaccedilatildeo agrave espeacutecie VOCl3cuja frequecircncia de
ressonacircncia eacute considerada o padratildeo sendo todas as outras referenciadas a partir desta em ppm
As aacutereas dos sinais do RMN correspondentes agraves espeacutecies oligomeacutericas presentes foram obtidas
por integraccedilatildeo dos sinais As concentraccedilotildees relativas das espeacutecies foram calculadas com recurso agrave
equaccedilatildeo
onde A[Vx] corresponde agrave aacuterea referente agrave ressonacircncia da espeacutecie Vx com nordm de agregaccedilatildeo x
A[V]total agrave soma total das aacutereas medidas e [V] a concentraccedilatildeo total de vanaacutedio no ensaio
A teacutecnica de 51
V-RMN foi anteriormente aplicada em estudos de ligaccedilatildeo com S1 de miosina
(Ref) tendo sido demostrado que entre as espeacutecies V1 V2 V4 e V5 a espeacutecie tetrameacuterica possui a
maior afinidade para a miosina (Cremo et al) O desaparecimento ou alargamento dos sinais pode
implicar a ligaccedilatildeo de um dos oxovanadatos laacutebeis agrave miosina sem indicaccedilatildeo de qual uma vez que
49
se processa um equiliacutebrio entre eles (Crans 1990) Assim e tendo em conta as indicaccedilotildees de que
a espeacutecie V10 eacute distinta das restantes no que diz respeito agrave magnitude da interacccedilatildeo com a miosina
e inibiccedilatildeo da actividade ATPaacutesica estimulada por actina (Aureliano 2000 Tiago et al 2002)
torna-se fundamental investigar a interacccedilatildeo das diferentes espeacutecies de vanadato com o S1 de
miosina atraveacutes da teacutecnica de 51
V-RMN
Para tal obtiveram-se espectros de 51
V-RMN resultantes da titulaccedilatildeo de uma soluccedilatildeo 5 mM de
decavanadato total com concentraccedilotildees crescentes de S1 de miosina (0 5 10 15 e 20 microM) em
tampatildeo D Realizou-se ainda ensaios na presenccedila de 2 mM de decavanadato e 2 mM de ATP na
presenccedila e na ausecircncia de S1 de miosina de modo a verificar o efeito que estes ligandos naturais
da enzima teria na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o subfragmento-1
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
Os grupos sulfidrilo (-SH) das cisteiacutenas satildeo resiacuteduos de aminoaacutecidos extremamente sensiacuteveis agrave
oxidaccedilatildeo resultando na sua oxidaccedilatildeo (Dalle-Donne et al 1999) Logo a quantificaccedilatildeo destes
resiacuteduos eacute bastante importante nos estudos de interacccedilatildeo com vanaacutedio jaacute que os resiacuteduos de Cys
com ecircnfase para o SH1 e o SH2 satildeo bastante importantes na monitorizaccedilatildeo das modificaccedilotildees
funcionais da regiatildeo da cabeccedila
O procedimento seguidamente apresentado foi originalmente descrito por Gutieacuterrez-Merino
(1987)) Consiste na reacccedilatildeo entre os grupos ndashSH natildeo oxidados das proteiacutenas e o reagente de
Ellman (DTNB -55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico)) - formando-se o aniatildeo CNT (aniatildeo 3-
carboxilato-4-nitrotiofenolato) numa estequiometria de 11 Este apresenta um maacuteximo de
absorvecircncia a 412 nm (Robyt 1990) Assim apoacutes a incubaccedilatildeo da proteiacutena na presenccedila do metal
a quantidade de cisteiacutenas reduzidas por proteiacutena determina-se atraveacutes dos valores de Abs412
Deste modo colocou-se num tubo de Eppendorf o volume apropriado de S1 dialisado para que a
concentraccedilatildeo no ensaio fosse de 1 mgmL Adicionou-se o volume apropriado de decavanadato
para que a sua concentraccedilatildeo fosse de 0 10 50 100 250 500 ou 1000 M Perfez-se com soluccedilatildeo
D ateacute um volume final de 1 mL e incubou-se a 25ordmC por 20 minutos Apoacutes este periacuteodo colocou-
se 780 L da mistura reaccional numa cuvette de quartzo e adicionou-se 220 L de soluccedilatildeo K23
23
Tampatildeo K Tampatildeo fosfatohidrogenofosfato de soacutedio 025 M pH 72
50
Adicionou-se 15 L de DTNB (em soluccedilatildeo K [DTNB]stock=100 mM) ao meio reaccional e
registou-se o valor de absorvecircncia medido equivalente agraves cisteiacutenas ldquoraacutepidasrdquo (SH1 e SH2
expostos ao solvente) Adicionou-se 001 g de SDS que provoca a desnaturaccedilatildeo da proteiacutena e
consequentemente a exposiccedilatildeo de todos os resiacuteduos de cisteiacutenas que se encontravam ateacute aiacute
inacessiacuteveis no interior da proteiacutena (cisteiacutenas totais) Estes resiacuteduos podem assim reagir com o
DTNB
Estimou-se o nuacutemero de resiacuteduos de cisteiacutenas em todas as amostras com recurso ao coeficiente de
extinccedilatildeo molar () do DTNB (ver ponto 3 dos Anexos) que foi determinado atraveacutes da titulaccedilatildeo
de L-Cisteiacutena com intervalo de concentraccedilotildees compreendidos entre 0 e 50 microM na presenccedila de
25microM de DTNB em tampatildeo D Os valores de absorvecircncia foram obtidos para um comprimento de
onda de 412 nm
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
3621 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
A teacutecnica mais eficiente de anaacutelise da interacccedilatildeo entre o iatildeo vanadilo e proteiacutenas como a
calmodulina e a transferrina entre outras permite a obtenccedilatildeo raacutepida de informaccedilatildeo no que toca agrave
conformaccedilatildeo e amplitude da referida interacccedilatildeo
Os espectros de Ressonacircncia Paramagneacutetica Electroacutenica (RPE) da espeacutecie paramagneacutetica V(IV)
foram realizados a 70K num espectroacutemetro Bruker EMX-300 X-band Todas as soluccedilotildees
utilizadas e amostras foram preparadas em anaerobiose numa cacircmara de luvas de modo a evitar a
reduccedilatildeo de V(IV) a V(V) Em tubos capilares com 02 mL de amostra manteve-se a concentraccedilatildeo
de S1 a 20 M e variou-se a concentraccedilatildeo de vanadilo (0 25 5 10 15 25 50 75 100 150
200 500 e 1000 M [VOSO4]stock=5 mM) em tampatildeo D Apoacutes obtenccedilatildeo dos espectros estes
foram tratados atraveacutes do software SimFonia e WinEPR tendo-se obtido um graacutefico com a
variaccedilatildeo da intensidade do sinal de RPE em funccedilatildeo da concentraccedilatildeo de sulfato de vanadilo
presente Realizaram-se ainda simulaccedilotildees de espectros obtidos com 20 microM de S1 e 1000 microM de
V(IV) com vista agrave obtenccedilatildeo dos ligandos provaacuteveis do vanadilo atraveacutes da anaacutelise da constante
de acoplamento hiperfina (A) e do factor g (g)
4 Conclusotildees gerais
Apesar de jaacute ter sido demonstrado que o vanaacutedio inibe a Na+K
+-ATPase da miosina a funccedilatildeo
deste elemento no tecido muacutesculo esqueleacutetico continua largamente incompreendida A enorme
complexidade quiacutemica deste metal especialmente no que concerne agrave oligomerizaccedilatildeo e reacccedilotildees
51
redox constitui um obstaacuteculo agrave compreensatildeo do comportamento do vanaacutedio nos organismos
vivos Assim a caracterizaccedilatildeo da quiacutemica deste elemento eacute de grande utilidade antes de iniciar o
estudo da sua interacccedilatildeo com o presente sistema
A anaacutelise por espectroscopias de 51
V-RMN e de UV-Vis de soluccedilotildees de vanadato diluiacutedas nos
meios de reacccedilatildeo utilizados no presente relatoacuterio (10 mM HEPES pH 70 25 mM KCl) mostrou
claramente que a espeacutecie decameacuterica (V10) possui um tempo de vida suficientemente longo para a
determinaccedilatildeo do seu efeito isolado ao contraacuterio do que se verifica com as espeacutecies mono- di-
tetra- e pentameacutericas (V1 V2 V4 e V5) Por outro lado a anaacutelise verificou ainda que apoacutes diluiccedilatildeo
das soluccedilotildees de decavanadato no meio de reacccedilatildeo coexistem as espeacutecies V10 e V1 pelo que os
efeitos putativos desta uacuteltima deveratildeo ser subtraiacutedos aquando da monitorizaccedilatildeo dos efeitos da
espeacutecie decameacuterica
O vanaacutedio (V) (vanadato) inibe as enzimas que catalizam reacccedilotildees de transferecircncia de grupos
fosfato como a ATPase da miosina [Goodno 1979] devido agrave sua analogia estrutural com o iatildeo
fosfato Na sua utilizaccedilatildeo como inibidor enzimaacutetico natildeo eacute tida em conta a contribuiccedilatildeo individual
das espeacutecies oligomeacutericas considerando-se normalmente a espeacutecie V1 como inibidora efectiva
[Goodno e Taylor 1982] No presente relatoacuterio verificou-se por espectroscopia de 51
V-RMN que
outros oligoacutemeros de vanadato interagem com o S1 de miosina tendo as espeacutecies V10 e V4 os
maiores niacuteveis de afinidade A sua interacccedilatildeo envolve forccedilas electrostaacuteticas e eacute afectada
negativamente por ligandos naturais de enzima como eacute o caso do ATP
A exposiccedilatildeo do S1 de miosina (005 mgmL) a decavanadato conduz agrave inibiccedilatildeo do capacidade da
proteiacutena de estimular a actividade Mg2+
-ATPaacutesica com um valor de IC50 de 266 plusmn 07 M
decavanadato sendo o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM
decavanadato (80) Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM decavanadato sendo
o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM decavanadato (60)
exercendo um efeito de impedimento agrave ligaccedilatildeo entre o vanaacutedio e o S1
A uma concentraccedilatildeo de 27 microM decavanadato verifica-se a oxidaccedilatildeo de um dos resiacuteduos de Cys
de S1 de miosina seguida pela oxidaccedilatildeo gradual de outro resiacuteduo de Cys ateacute agrave concentraccedilatildeo de
1000 microM Ambos os resiacuteduos oxidados natildeo se encontravam expostos ao solvente utilizado A
oxidaccedilatildeo dos resiacuteduos permite sugerir que esta modificaccedilatildeo oxidativa sofrida pelo S1 contribui
para a diminuiccedilatildeo da sua actividade de ATPase
52
Por espectroscopia de RPE revelaram-se os possiacuteveis ligandos equatoriais do vanaacutedio aquando da
sua interacccedilatildeo com o S1 de miosina como sendo duas moleacuteculas de aacutegua um ligando contendo o
grupo R-OH e outro contendo o grupo R=NR
Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados de
oxidaccedilatildeo (IV) e (V) no S1 de miosina para concentraccedilotildees facilmente alcanccedilaacuteveis no ambiente
terrestre
53
5 Estudos futuros
No sentido do melhor esclarecimento das alteraccedilotildees moleculares e estruturais que ocorrem no
subfragmento 1 quando exposta ao vanaacutedio que conduzem a uma diminuiccedilatildeo da sua funccedilatildeo
sugere-se
- Quantificaccedilatildeo das cisteiacutenas reduzidas de S1 na presenccedila de metavanadato
- Determinaccedilatildeo das actividades Ca2+
- e K+-ATPaacutesica do S1 de miosina na presenccedila de
decavanadato
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina estimulada por actina
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina na presenccedila de
metavanadato
- Realizaccedilatildeo de RMN 2D com variaccedilatildeo das concentraccedilotildees das soluccedilotildees de decavanadato e
metavanadato na presenccedila de S1
-Mutageacutenese dos resiacuteduos de Cys doS1 de miosina de modo a verificar quais os resiacuteduos que
sofrem oxidaccedilatildeo na presenccedila de decavanadato
- Determinaccedilatildeo por calorimetria (DSC Differential Scanning Calorimetry) da variaccedilatildeo da
entalpia (∆H) e da temperatura de desnaturaccedilatildeo do S1 de miosina na presenccedila de vanaacutedio uma
vez que estas propriedades caracteriacutesticas de cada proteiacutena satildeo alteradas apoacutes ocorrecircncia de
modificaccedilotildees estruturais
- Anaacutelise por SDS-PAGE de S1 de miosina apoacutes tratamento com vanaacutedio de modo a determinar
quais os polipeacuteptidos eventualmente formados apoacutes clivagem da proteiacutena
54
6 Referecircncias bibliograacuteficas
- Sellers J Myosins 2nd Edition Oxford University Press 1999
- L Stryer inBiochemistry (Ed WH Freeman 2005 pp
Refordfas Materiais e Meacutetodos
[A] S S Margossian SLowey Methods Enzymol1982 85 55-71
[B] JD Pardue JA Spudich Methods Enzymol 1982 85 164-181
[C] M Bradford Anal Biochem1976 72 248-254
[D] H H Taussky E Shorr J Biol Chem 1953 202 675-685
[E] C R Bagshaw in Outline Studies in Biology ndash Muscle Contraction Chapman and Hall Ed
1982
[F] A Cuenda et al Eur J Biochem1990 194 663-670
55
Anexos
1 Determinaccedilatildeo da recta de calibraccedilatildeo meacutetodo de Bradford
Figura1ndashRepresentaccedilatildeo graacutefica da recta de calibraccedilatildeo (Abs595 nm vs [BSA]) utilizada para o meacutetodo de Bradford
2 Determinaccedilatildeo da recta calibraccedilatildeo para a electroforese em condiccedilotildees desnaturantes
Figura 2 ndash Representaccedilatildeo graacutefica da recta de calibraccedilatildeo (log(Peso Molecular) vs Rf dos marcadores de peso
molecular) onde Rf indica determinada atraveacutes do padratildeo de peso molecular
(poccedilo (a))
3Caacutelculo do coeficiente de extinccedilatildeo molar () para a quantificaccedilatildeo de cisteiacutenas reduzidas
atraveacutes do meacutetodo de Ellman
Traccedilou-se a recta de Abs412 nm meacutedia vs [L-Cisteiacutena] de modo a determinar o valor do coeficiente
de extinccedilatildeo molar definido como o declive daquela recta Assim =8400 M-1
cm-1
como
podemos verificar na Fig 3
A595 nm = 00546[BSA] + 00209 Rsup2 = 099051
Ab
s 59
5 n
m
[BSA] (mgmL)
log PM = -1177 Rf + 2247 Rsup2 = 098957
log
PM
Mobilidade relativa (Rf)
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero

4
Resumo
A exposiccedilatildeo a vanaacutedio conduz a uma diminuiccedilatildeo na actividade ATPaacutesica da miosina e provoca a
oxidaccedilatildeo dos grupos sulfidrilo desta proteiacutena No entanto natildeo estaacute integralmente esclarecido
quais os efeitos especiacuteficos que ocorrem na miosina quando exposta ao vanaacutedio Nesse sentido
interessou determinar in vitro quais as alteraccedilotildees estruturais funcionais e moleculares ocorridas
no Subfragmento-1 de miosina de muacutesculo esqueleacutetico de coelho apoacutes a exposiccedilatildeo croacutenica a
soluccedilotildees de metavanadato decavanadato e vanadilo Verificou-se que a exposiccedilatildeo de S1 (005
mgmL) a decavanadato inibe a sua actividade Mg2+
-ATPaacutesica sendo o valor de IC50 para esta
inibiccedilatildeo de 266 plusmn 07 microM decavanadato Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM
decavanadato A perda de funcionalidade proteica poderaacute dever-se agrave oxidaccedilatildeo de 2 resiacuteduos de
Cys de S1 natildeo expostos ao solvente apoacutes exposiccedilatildeo a 1000 microM de decavanadato Verificou-se
que as espeacutecies oligomeacutericas de vanaacutedio com maior afinidade para o S1 satildeo o decavanadato e o
tetravanadato Revelaram-se os possiacuteveis ligandos do vanaacutedio no S1 atraveacutes de espectroscopia de
RPE Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados
de oxidaccedilatildeo (IV) e (V) na estrutura e funccedilatildeo do S1 em concentraccedilotildees facilmente alcanccedilaacuteveis
daquele metal no ambiente terrestre
Summary
The exposure to vanadium leads to a decrease of the myosin ATPase activity and provokes the
oxidation of its sulfydryl groups However the specific effects of vanadium on myosin are not yet
fully understood Consequently the objectives of the present work were to establish the structural
funtional and molecular changes suffered by rabbit myosin subfragment-1 after cronic exposure to
metavanadate decavanadate and vanadyl solutions It was verified that the exposure of S1 (005
mgmL) to decavanadate results in the inhibition of its Mg2+
-ATPase activity resulting in the
IC50 value 266 plusmn 07 microM decavanadate In the presence of actin the IC50 value was 28 plusmn 1 microM
decavanadate It is suggested that the loss of protein function may be due to the oxidation of 2
core cysteine residues after exposure to 1000 microM decavanadate It was also observed that the
decavanadate and tetravanadate species show the highest affinity for S1 Lastly the possible S1
ligands to vanadium were revealed through EPR spectroscopy Globally the obtained results
point to significant effect of V(V) and V(IV) states on S1 structure and function for easily
reachable environmental vanadium concentrations
5
Iacutendice
1 Introduccedilatildeo
11 Muacutesculo esqueleacutetico
111 Miosina
112 Actina
113 Mecanismos moleculares da contracccedilatildeo muscular
1131 Regulaccedilatildeo da contracccedilatildeo muscular
12 Vanaacutedio
121 Propriedades quiacutemicas
1211 Vanaacutedio (IV)
1212 Vanaacutedio (V)
122 Metabolismo e efeito na actividade enzimaacutetica
13 Vanaacutedio e muacutesculo esqueleacutetico
14 Objectivos do projecto
2 Discussatildeo de Resultados
21 Caracterizaccedilatildeo das proteiacutenas isoladas
211 Anaacutelise do estado de pureza
212 Determinaccedilatildeo da concentraccedilatildeo proteica
2121 Subfragmento-1 de miosina
2122 Actina
22 Mediccedilatildeo da actividade enzimaacutetica de S1 de miosina
221 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de Fiske-
Subarow modificado
222 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
23 Preparaccedilatildeo e caracterizaccedilatildeo das soluccedilotildees de vanaacutedio
231 Anaacutelise da composiccedilatildeo das soluccedilotildees de vanadato por espectroscopia de 51V-RMN efeito da
concentraccedilatildeo da espeacutecie
232 Estabilidade das soluccedilotildees de decavanadato anaacutelise por espectroscopia de absorccedilatildeo UVVis
24 Interacccedilatildeo de S1 com vanaacutedio(V)
241 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
242 Actividade Mg2+
-ATPaacutesica de S1 de miosina na presenccedila de decanadato
243 Quantificaccedilatildeo de cisteiacutenas reduzidas pelo meacutetodo de Ellman
25 Interacccedilatildeo de S1 com vanaacutedio(IV)
251 Anaacutelise por espectroscopia de Ressonacircncia Paramagneacutetica Electroacutenica
3 Procedimento Experimental
31 Reagentes
32 Meacutetodos
321 Isolamento e purificaccedilatildeo das proteiacutenas de muacutesculo esqueleacutetico de coelho
3211 Purificaccedilatildeo da miosina preparaccedilatildeo de Subfragmento 1 por digestatildeo quimotriacutepsica da
miosina
32111 Purificaccedilatildeo da miosina
32112 Preparaccedilatildeo de Subfragmento-1 de miosina por digestatildeo quimotriacutepsica
3212 Isolamento e purificaccedilatildeo de actina
32121Obtenccedilatildeo do ldquopoacute de acetonardquo
32122 Purificaccedilatildeo da actina a partir de ldquopoacute de acetonardquo
331 Quantificaccedilatildeo das proteiacutenas isoladas
3311 Quantificaccedilatildeo pelo meacutetodo de Bradford
6
3312 Quantificaccedilatildeo pelo meacutetodo directo
332 Electroforese em condiccedilotildees desnaturantes (SDS-PAGE)
34 Mediccedilatildeo da actividade enzimaacutetica de S1
341 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de Fiske-
Subarow modificado
342 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
35 Preparaccedilatildeo e caracterizaccedilatildeo das soluccedilotildees de vanaacutedio por Espectroscopia UV-Visiacutevel e
Ressonacircncia Magneacutetica Nuclear
351 Preparaccedilatildeo das soluccedilotildees de vanadato
352 Anaacutelise por UV-Vis para determinaccedilatildeo da ordem da reacccedilatildeo
353 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
362 1 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
363 Interacccedilatildeo de S1 com V (V) por Ressonacircncia Magneacutetica Nuclear (RMN)
4 Conclusotildees gerais
5 Estudos futuros
6 Referecircncias bibliograacuteficas
7
1 Introduccedilatildeo
11 Muacutesculo esqueleacutetico
A capacidade de locomoccedilatildeo eacute uma das caracteriacutesticas fundamentais da maioria dos organismos
vivos A contracccedilatildeo muscular um dos fenoacutemenos que a torna possiacutevel apresenta grande
complexidade e tem vindo a ser utilizado como modelo para a compreensatildeo de sistemas menos
especializados de mobilidade celular [1]
No que diz respeito agrave sua fisiologia morfoloacutegica os muacutesculos dos vertebrados dividem-se em
lisos e estriados Estes uacuteltimos devem o seu nome agrave alternacircncia de zonas claras e escuras
observaacuteveis ao microscoacutepio electroacutenico de que resulta um aspecto estriado Os muacutesculos
estriados podem ainda ser divididos em duas subclasses muacutesculos esqueleacuteticos e cardiacuteacos [2]
consoante a forma celular (respectivamente longitudinal ou ramificada) ou tipo de contracccedilatildeo das
ceacutelulas que o compotildeem (respectivamente voluntaacuteria e involuntaacuteria) [3]
Os muacutesculos estriados esqueleacuteticos satildeo assim designados devido agrave sua localizaccedilatildeo no
organismo ndash ligados aos ossos da coluna vertical braccedilos e pernas Satildeo constituiacutedos por feixes de
ceacutelulas multinucleadas (miofibras) que contecircm miofibrilas paralelas de cerca de 1 m de
diacircmetro associadas agrave contracccedilatildeo voluntaacuteria raacutepida e forte em actividades de coordenaccedilatildeo
complexa [4] Cada feixe apresenta um diacircmetro compreendido entre 20 e 100 m e um
comprimento maacuteximo que pode atingir vaacuterios centiacutemetros (Fig 11)
Figura 11 ndash Organizaccedilatildeo hieraacuterquica geral do muacutesculo-esqueleacutetico (adaptado de Purves et al 2003 [3])
No muacutesculo-esqueleacutetico as zonas claras e escuras supramencionadas correspondem a zonas de
menor e maior densidade electroacutenica respectivamente As zonas claras constituem as bandas I (de
8
ldquoisotropiardquo ndash polarizaccedilatildeo uniforme em todas as direcccedilotildees) enquanto as zonas escuras constituem
as bandas A (de ldquoanisotropiardquo ndash polariza a luz visiacutevel) A banda I encontra-se dividida
transversalmente por uma linha escura ndash linha Z de densidade electroacutenica e refracccedilatildeo superiores
No interior de cada banda A existe uma zona transversal menos densa ndash zona H ndash rica em enzimas
importantes para o metabolismo celular No centro desta zona existe uma linha perpendicular
mais escura ndash linha M [2]
A unidade fundamental da contracccedilatildeo muscular eacute designada por sarcoacutemero (Fig 12) Encontra-se
repetida de forma regular ao longo das miofibrilas apresentando um comprimento de 25 a 3 m
aquando do relaxamento muscular diminuindo aquando da contracccedilatildeo muscular
Figura 12 ndash Representaccedilatildeo de fotomicrografia electroacutenica e esquemaacutetica do sarcoacutemero Os filamentos grossos e
finos satildeo maioritariamente constituiacutedos por miosina e actina respectivamente (Adaptado de Stryer 2002 [2])
As secccedilotildees de corte do sarcoacutemero datildeo conta da presenccedila de dois tipos de filamentos proteicos ndash os
filamentos grossos cujo diacircmetro eacute de 15 nm constituiacutedos sobretudo por miosina e os filamentos
finos com um diacircmetro de 9 nm constituiacutedos sobretudo por actina tropomiosina e pelo complexo
de troponina [2]
111 Miosina
Actualmente o termo miosina refere-se a uma superfamiacutelia diversa de motores moleculares
capazes de hidrolisar ATP e de se ligarem reversivelmente agrave actina A classificaccedilatildeo da
superfamiacutelia das miosinas nas 15 classes distintas faz-se atraveacutes de homologia entre as diferentes
sequecircncias [A] Eacute tambeacutem possiacutevel a classificaccedilatildeo por subclasses das quais se destaca a subclasse
4 proveniente do muacutesculo-esqueleacutetico e cardiacuteaco de vertebrados Este estudo focar-se-aacute apenas
nas miosinas de classe II (miosina-II) presente nos filamentos grossos do muacutesculo estriado
esqueleacutetico [B] e em ceacutelulas natildeo musculares A funccedilatildeo da miosina-II depende da sua localizaccedilatildeo
celular e tecidular
9
A miosina-II eacute constituiacuteda por duas cadeias polipeptiacutedicas idecircnticas denominadas cadeias pesadas
de miosina (HC) enroladas entre si e cujo peso molecular (PM) estaacute compreendido entre 171 e
241 kDa) Estruturalmente as HC dividem-se em 3 domiacutenios funcionais - cabeccedila pescoccedilo e
cauda como se pode observar na Fig 13 Agraves cabeccedilas ou zonas globulares estatildeo associados dois
pares de cadeias leves (peso molecular entre 15 e 30 kDa) que se dividem em dois tipos
essencial (ELC) e regulatoacuterio (RLC)
Figura 13 ndash Estrutura esquemaacutetica da moleacutecula de miosina-II Destacam-se as duas cabeccedilas globulares e a cauda
filamentosa ligadas entre si por uma zona flexiacutevel denominada pescoccedilo As cadeias leves (a verde e vermelho)
encontram-se associadas agraves cabeccedilas globulares
As cabeccedilas globulares contendo os locais de ligaccedilatildeo agrave actina e aos nucleoacutetidos mantecircm os seus
elementos estruturais baacutesicos conservados em todas as classes sendo estes constituiacutedos por 720-
80 aa A regiatildeo do pescoccedilo eacute constituiacuteda por uma longa heacutelice pertencente agrave cadeia pesada
que se encontra estabilizada atraveacutes da ligaccedilatildeo a subunidades de cadeia leve A estrutura do
pescoccedilo envolvida na ligaccedilatildeo agraves cadeias leves eacute denominada de motivo IQ cuja estrutura
consensual eacute IQXXXRGXXXR1 O pescoccedilo serve como extensatildeo do comprimento da cabeccedila de
modo a actuar como alavanca durante a contracccedilatildeo muscular [A]A regiatildeo da cauda ou acircncora
apresenta bastante divergecircncia no que respeita agrave sua sequecircncia estrutura e funccedilatildeo No entanto
pensa-se que a sua funccedilatildeo centrar-se-aacute na autoassociaccedilatildeo das vaacuterias moleacuteculas de miosina-II
atraveacutes da regiatildeo de coiled-coil Esta confere assimetria agrave moleacutecula e permite a formaccedilatildeo de
filamentos grossos capazes de translocar filamentos de actina [A]
Na regiatildeo da cabeccedila da miosina existem 2 regiotildees extremamente sensiacuteveis agrave proteoacutelise A
clivagem em ambos os locais resulta na produccedilatildeo de trecircs fragmentos um compreendendo o N-
terminal (fragmento 25 kDa) outro a regiatildeo central (50 kDa) e um terceiro o C-terminal (20 kDa)
(Fig 14) Estes fragmentos dissociam-se em condiccedilotildees desnaturantes e natildeo constituem domiacutenios
independentes da cabeccedila mas provavelmente loops flexiacuteveis uma vez que natildeo foram observadas
na estrutura cristalina Os vaacuterios fragmentos obtidos em diferentes condiccedilotildees foram cruciais para
a elucidaccedilatildeo das vaacuterias funccedilotildees da moleacutecula
A regiatildeo compreendida entre os fragmentos 25 e 50 kDa (loop 1) localiza-se directamente por
1 I = Isoleucina Q= Glutamina X = qualquer dos 20 aminoaacutecidos padratildeo R= Arginina G= Glicina
10
cima do local de ligaccedilatildeo ao nucleoacutetido A regiatildeo entre os fragmentos de 50 e 20 kDa (loop 2)
encontra-se perto da intersecccedilatildeo entre os domiacutenios da cabeccedila e do pescoccedilo e deveraacute interagir com
a actina uma vez que a ligaccedilatildeo desta a protege da proteoacutelise Na ausecircncia do loop 2 a miosina
liga mais fracamente agrave actina do que no caso da miosina intacta A sequecircncia primaacuteria de ambos
os loops apresenta conservaccedilatildeo reduzida sendo provaacutevel que sejam responsaacuteveis por muita da
diversidade das diferentes propriedades enzimaacuteticas presentes em diferentes isoformas
No fragmento de 20 kDa existem dois grupos sulfidrilo Cys-707 e -697 usualmente
denominados de SH1 e SH2 devido agrave ordem da sua reactividade quiacutemica Na presenccedila de
nucleoacutetidos pode ocorrer o ldquocross-linkingrdquo entre estes dois grupos incluindo a oxidaccedilatildeo a uma
ponte dissulfureto
Devido agrave insolubilidade da miosina-II nativa a estrateacutegia adoptada para os estudos com esta
proteiacutena eacute frequentemente a sua clivagem em fragmentos funcionais separaacuteveis Um dos locais de
clivagem ocorre tipicamente a 130 kDa do N-terminal produzindo a meromiosina pesada (HMM)
e a meromiosina leve (LMM) [C] A HMM eacute formada pelas 2 cabeccedilas e pescoccedilo e apresenta
propriedades semelhantes agrave miosina sem digestatildeo (ligaccedilatildeo e translocaccedilatildeo de actina e hidroacutelise de
MgATP)
A HMM pode ser novamente clivada atraveacutes de uma reacccedilatildeo de proteoacutelise produzindo os
subfragmentos 1 (S1) e 2 (S2)O S1 (90 kDa) eacute enzimaticamente activo sendo constituiacutedo pelos
domiacutenios da cabeccedila e do pescoccedilo e apresentando associaccedilatildeo agraves duas cadeias leves ou apenas agrave
ELC (Fig 14)Em alternativa o S1 pode ser produzido por clivagem directa da miosina intacta
formando-se tambeacutem um fragmento em forma de cilindro que consiste nas regiotildees S2 e LMM
(Ref)Utiliza-se extensivamente o S1 para estudos cineacuteticos e de ligaccedilatildeo agrave actina uma vez que a
sua solubilidade simplifica o planeamento da experiecircncia e a subsequente interpretaccedilatildeo dos dados
[A]
Figura 14- Estrutura do subfragmento-1 de miosina de muacutesculo esqueleacutetico com ecircnfase nos domiacutenios de 50 kDa
central (a vermelho) 25 kDa no N-terminal (a verde) e 20 kDa no C-terminal (a azul) As cadeias leves essenciais
encontram-se a amarelo e as cadeias leves regulatoacuterias a lilaacutes Os locais de ligaccedilatildeo agrave actina e ao ATP estatildeo
igualmente destacados (adaptado de Holmes e Geeves 1999 [D])
11
112 Actina
O termo ldquoactinardquo refere-se a uma famiacutelia extremamente conservada de proteiacutenas citoplasmaacuteticas
presente em todos os eucariotas Esta proteiacutena natildeo se encontra nos procariotas [E] A maioria das
actinas possui peso molecular proacuteximo de 42 kDa outra evidecircncia do seu grau elevado de
conservaccedilatildeo no que concerne agrave sequecircncia e ao nuacutemero de aminoaacutecidos Os eucariotas superiores
apresentam um grande nuacutemero de isoformas de actina que por sua vez se encontram codificados
por um ainda maior nuacutemero de genes (Ref) A expressatildeo das isoformas eacute diferencial consoante o
tecido e sua especializaccedilatildeo pelo que deveratildeo possuir funccedilotildees e mecanismos direccionais
diferentes (Ref) Foi ainda observado que os mRNA das diferentes isoformas apresentam uma
distribuiccedilatildeo heterogeacutenea nas ceacutelulas de forma semelhante aos seus produtos proteicos [F] Estes
dados suportam a hipoacutetese supramencionada do direccionamento das isoformas de actina para um
determinado local na ceacutelula de acordo com as suas funccedilotildees
A actina presente no citoesqueleto eacute responsaacutevel por processos como o mecanismo contraacutectil
fagocitose manutenccedilatildeo e alteraccedilatildeo da estrutura celular locomoccedilatildeo transporte de organelos
regulaccedilatildeo do transporte de solutos entre outras Nos eucariotas superiores estaacute ainda envolvida
no transporte de gracircnulos de pigmento retinoacuteide [G] transporte de gracircnulos no axoplasma [H] e
secreccedilatildeo entre outros [E] Actualmente a isoforma da actina (-actina) eacute a melhor estudada
encontrando-se envolvida no mecanismo contraacutectil [E]
Os filamentos de -actina tecircm cerca de 1m de comprimento e 80 Å Consistem em duas cadeias
de actina filamentosa (F-actina) organizados em heacutelice [I] (Holmes et al 1990) sobre os quais
se encontram outras proteiacutenas ndash a troponina (C I e T) e a tropomiosina (Azevedo 1999) (Figura
15) A troponina eacute filamentosa e localiza-se nas depressotildees da heacutelice de F-actina conferindo
rigidez aos filamentos finos Por sua vez a tropomiosina eacute essencial na regulaccedilatildeo do mecanismo
contraacutectil
Figura 15 ndash Modelo estrutural dos filamentos de F-actina contendo a tropomiosina e o complexo de troponina (I C
e T) (Adaptado de Susana Ramos 2005 [J])
12
A F-actina eacute formada atraveacutes da polimerizaccedilatildeo de subunidades globulares de actina (G-actina) A
G-actina eacute composta por 2 domiacutenios de tamanho semelhante separados pelos bolsos de ligaccedilatildeo(1)
de catiotildees divalentes e(2) de nucleoacutetidos [KLM] Estes por sua vez estatildeo subdivididos noutros
dois de menores dimensotildees (1-4) (Fig 16) cada um envolvido nas interacccedilotildees actina-actina ou
na formaccedilatildeo do topo de ligaccedilatildeo do nucleoacutetido As regiotildees N-terminal e C-terminal estatildeo proacuteximas
uma da outra no subdomiacutenio 1 [E]
Figura 16 ndash Estrutura do monoacutemero de actina Encontram-se representados os subdomiacutenios 1 a 4 e a vermelho a
moleacutecula de ATP junto do respectivo local de ligaccedilatildeo (Protein Data Bank ID 1j6z)
113 Mecanismos moleculares da contracccedilatildeo muscular
A contracccedilatildeo muscular ocorre atraveacutes do deslizamento dos filamentos finos (de actina) por entre
os filamentos grossos (de miosina) conseguida pela reacccedilatildeo de hidroacutelise de ATP e regulada pela
tropomiosina e pelo complexo de troponina em resposta a impulsos nervosos Este mecanismo foi
inicialmente proposto por Huxley-Niedergerke [N] e Huxley-Hanson [O] ambos em 1954 A base
experimental de ambos os estudos foi a inexistecircncia de variaccedilatildeo do tamanho dos filamentos
durante a contracccedilatildeo A banda A apresenta sempre as mesmas dimensotildees por outro lado a banda
I a zona H diminuem consideravelmente aquando da contracccedilatildeo (Figs 12 e 17)
Figura 17 ndash Modelo da contracccedilatildeo muscular ndash deslizamento dos filamentos finos (azuis) relativamente aos
filamentos grossos (rosa) (Adaptado de Stryer 2002 [2])
13
Na ausecircncia de ATP e supondo a ausecircncia de inibiccedilatildeo da interacccedilatildeo actina-miosina as proteiacutenas
encontram-se fortemente associadas formando o complexo actomiosina (AM) (Figura 15)
Nestas condiccedilotildees as cabeccedilas de miosina formam um acircngulo de 45ordm com o eixo dos filamentos
finos de actina [P]
Na presenccedila de ATP o complexo AM dissocia-se jaacute que a afinidade do complexo M-ATP para a
actina eacute 105 vezes inferior agrave afinidade da miosina para a actina O ATP liga-se agrave miosina
ocorrendo seguidamente a reacccedilatildeo de hidroacutelise de ATP Os produtos daiacute resultantes ndash ADP e Pi -
permanecem ligados agrave miosina formando o complexo M-ADP-Pi Com a ligaccedilatildeo subsequente agrave
actina forma-se o complexo AM-ADP-Pi no qual as cabeccedilas de miosina apresentam um acircngulo
recto com o eixo do muacutesculo [P] Neste ponto daacute-se a libertaccedilatildeo progressiva de Pi e ADP Eacute
provaacutevel que a energia proveniente da hidroacutelise do ATP provoque uma alteraccedilatildeo do acircngulo de
ligaccedilatildeo das cabeccedilas de miosina ao filamento de actina passando novamente de 90ordm para 45ordm [P]
Esta alteraccedilatildeo conformacional provoca o deslizamento do filamento de actina para o centro do
sarcoacutemero cujo encurtamento promove a contracccedilatildeo das miofibrilas O processo supracitado
ocorre em cerca de 1ms[Q] e encontra-se resumido na Fig 18
Figura 18 ndash Esquema cineacutetico da hidroacutelise de ATP pela miosina (Adaptado de Sellers 1999 [A])
1131 Regulaccedilatildeo da contracccedilatildeo muscular
O mecanismo pelo qual se processa a regulaccedilatildeo muscular foi inicialmente descrito por estudos
distintos de Haselgrove Huxley e Parrey-Squire todos em 1973 [RST] Aleacutem da implicaccedilatildeo
directa da tropomiosina e do complexo de troponina a concentraccedilatildeo de Ca2+
no muacutesculo em
relaxamento eacute bastante reduzida (10-7
M) aumentando notavelmente aquando da contracccedilatildeo (10-5
-
10-3
M)
Resumidamente quando ocorre a excitaccedilatildeo do muacutesculo esqueleacutetico a despolarizaccedilatildeo da
membrana plasmaacutetica eacute difundida ateacute ao interior do tecido atraveacutes dos tuacutebulos transversais
(invaginaccedilotildees da membrana plasmaacutetica da fibra muscular) A despolarizaccedilatildeo provoca a abertura
dos canais ioacutenicos de Ca2+
do retiacuteculo sarcoplasmaacutetico adjacente Como a concentraccedilatildeo de Ca2+
no interior do organelo eacute muito elevada o caacutelcio passa raacutepida e uniformemente para o citosol
14
Assim a regulaccedilatildeo da contracccedilatildeo ocorre com coordenaccedilatildeo e rapidez na totalidade das miofibrilas
do tecido muscular (Ref)
Em repouso a troponina I encontra-se posicionada de tal forma que a proteiacutena adjacente
troponina C impede estruturalmente a ligaccedilatildeo das cabeccedilas da miosina agrave actina (acircngulo =50ordm) O
aumento da concentraccedilatildeo intracelular de Ca2+
provoca um aumento da sua disponibilidade para
ligar agrave troponina C (Fig 19)
Figura 19ndash Regulaccedilatildeo do mecanismo contraacutectil no muacutesculo esqueleacutetico pelo complexo de troponina
Como consequecircncia a troponina C altera a sua conformaccedilatildeo (=70ordm) associando-se agrave troponina I
que deixa de inibir a ligaccedilatildeo das pontes transversais A alteraccedilatildeo anterior eacute transmitida agrave
tropomiosina atraveacutes da troponina T adjacente agrave C o que provoca o deslocamento da
tropomiosina na direcccedilatildeo do interior do sulco do filamento de actina Assim eacute possiacutevel a
interacccedilatildeo eficaz entre actina e miosina A Ca2+
-ATPase do retiacuteculo sarcoplasmaacutetico provoca a
reacumulaccedilatildeo do iatildeo caacutelcio no organelo impedindo o prolongamento desnecessaacuterio da contracccedilatildeo
(Refordf)
12 Vanaacutedio
O vanaacutedio eacute um metal de transiccedilatildeo pertencente ao grupo VB da tabela perioacutedica Foi
primeiramente descoberto em 1801 por Andreacutes Manuel del Riacuteo que concluiu erradamente que era
idecircntico ao croacutemio e definitivamente por Nils Gabriel Svenson em 1831 Recentemente foram
descobertas importantes funccedilotildees bioloacutegicas deste elemento provavelmente essencial para os
humanos de que se destaca no contexto do presente relatoacuterio a possibilidade de actuar como
anaacutelogo do grupo fosfato e a inibiccedilatildeo de enzimas in vitro como eacute o caso da actividade ATPaacutesica
da miosina
Eacute um metal soacutelido agrave temperatura ambiente que se encontra amplamente distribuiacutedo nos vaacuterios
ecossistemas em dois isoacutetopos naturais ndash 50
V e 51
V ndash raramente excedendo 01 partes por biliatildeo
15
(ppb) Estaacute associado a cerca de 70 minerais diferentes na natureza (dos quais se destacam o
uracircnio cobre chumbo zinco fosfato e titacircnio ferroso) especialmente na forma de vanadato ou
pentoacutexidos de vanaacutedio O vanaacutedio eacute um dos principais constituintes da mateacuteria orgacircnica foacutessil
com especial ecircnfase para o carvatildeo oacuteleo de crude e foacutesseis carbonaacuteceos [UV] bem como em
alimentos consumidos pelo Homem (salsa pimenta preta cogumelos marisco vegetais cereais
gorduras e oacuteleos) [W] Estes constituem a maior fonte de exposiccedilatildeo ao vanaacutedio da maioria da
populaccedilatildeo humana [X]
Normalmente a entrada do vanaacutedio no organismo ocorre por inalaccedilatildeo ou em menores
quantidades por absorccedilatildeo pelo tracto gastrointestinal ou pela pele O seu armazenamento centra-
se no fiacutegado rins ceacuterebro coraccedilatildeo muacutesculos e ossos No entanto a retenccedilatildeo de vanaacutedio no
organismo humano eacute reduzida ndash em condiccedilotildees normais cerca de 100 M (em muitos tecidos a
quantidade eacute inferior a 10 ngg de tecido) [Y]
Actualmente natildeo existem provas de deficiecircncia nutricional de vanaacutedio em humanos ou de
enzimas que contenham vanaacutedio em animais superiores No entanto vaacuterios estudos relacionam a
ocorrecircncia de deficiecircncia em galinhas e ratos devido a uma alimentaccedilatildeo pobre em vanaacutedio Os
sintomas relatados incluem retardamento do crescimento malformaccedilotildees oacutesseas perturbaccedilotildees no
metabolismo lipiacutedico e estagnaccedilatildeo reprodutiva entre outros [Z AA AB] Os efeitos fisioloacutegicos
do vanaacutedio podem depender do tipo de composto ou de oligoacutemero uma vez que consoante as
suas estabilidade labilidade e nuacutemero de coordenaccedilatildeo as respostas seratildeo diferenciadas ao niacutevel
dos organismos vivos
Por outro lado a toxicidade do vanaacutedio encontra-se bem caracterizada existindo sintomas
descritos em vaacuterios mamiacuteferos (inclusive o Homem) que incluem diarreias catildeibras irritaccedilatildeo da
pele e mucosas palpitaccedilotildees cardiacuteacas anemia pneumonia bronquite perturbaccedilotildees no Sistema
Nervoso Central (SNC) entre outros (Ref) A toxicidade varia geralmente com a natureza do
composto com aumento proporcional agrave sua valecircncia Verificou-se que a forma de entrada do
vanaacutedio tem tambeacutem efeito na sua toxicidade ndash os maiores niacuteveis verificaram-se por injecccedilatildeo
intravenosa da amostra sendo progressivamente inferiores por inalaccedilatildeo e finalmente por
ingestatildeo [X AA] A ampla utilizaccedilatildeo dos compostos de vanaacutedio na induacutestria quiacutemica bem como
as elevadas quantidades de vanaacutedio libertadas atraveacutes da combustatildeo dos oacuteleos de crude
contribuem consideravelmente para o aumento da biodisponibilidade do vanaacutedio Como tal o
interesse neste elemento e nos problemas ambientais por ele causados tem vindo a aumentar
significativamente [V]
121 Propriedades quiacutemicas
16
A quiacutemica do vanaacutedio eacute extraordinariamente complexa pelo que originou vaacuterias interpretaccedilotildees
incorrectas desde a sua descoberta No entanto as suas propriedades dependem em uacuteltima
anaacutelise da configuraccedilatildeo electroacutenica do elemento Assim a variabilidade encontrada no
comportamento quiacutemico do vanaacutedio pode dever-se agrave sua configuraccedilatildeo ndash [Ar] 3d2 4s
2 ndash que
permite a formaccedilatildeo de vaacuterios estados de oxidaccedilatildeo (de +2 a +5) Em soluccedilatildeo aquosa os estados de
oxidaccedilatildeo estaacuteveis vatildeo de V(V) a V(II) Destes apenas os +3 a +5 satildeo biologicamente importantes
jaacute que o V(II) apresenta demasiada capacidade redutora para existir nos organismos vivos
conhecidos [AC]
No fluido extracelular os compostos de vanaacutedio encontram-se predominantemente na forma de
monovanadatos (V(V)) livres ou ligados agrave transferina do soro ou agrave hemoglobina (Refordf) Os iotildees de
vanadato livres satildeo transportados para o meio intracelular atraveacutes de canais ioacutenicos inespeciacuteficos
Jaacute no ambiente intracelular os compostos sofreratildeo uma bio-reduccedilatildeo atraveacutes da acccedilatildeo da
glutationa ou outros compostos com grupos ndashOH como pentoses hexoses ou etanolaminas [V] A
reoxidaccedilatildeo de vanadilo (V(IV)) intracelular a vanadato (V(V)) provocaria a acumulaccedilatildeo toacutexica
destes uacuteltimos pelo que a complexaccedilatildeo da espeacutecie reduzida o impede O transporte de vanaacutedio
para o interior da ceacutelula encontra-se na Fig 110
Figura 110 ndash Transporte de vanaacutedio para a ceacuteluka e seu subsequente metabolismo (P=Proteiacutena) (Adaptado de
Zaporowska e Scibor 1998)
1211 Vanaacutedio(IV)
A espeacutecie de vanaacutedio (IV) mais bem conhecida eacute o iatildeo vanadilo (VO2+
) espeacutecie estaacutevel a pH
aacutecido sendo a maioritaacuteria a pH 3[AD] Apresenta uma cor azulada e forma complexos penta e
hexacoordenados Forma-se rapidamente a partir de V(V) na presenccedila de um agente redutor em
soluccedilatildeo aquosa No ambiente intracelular a presenccedila de glutationa reduzida aacutecido ascoacuterbico
NAD(P)H entre outros conduzem a que o principal estado de oxidaccedilatildeo nos sistemas bioloacutegicos
seja provavelmente o +4 encontrando-se o V(IV) fortemente complexado com proteiacutenas ou
aminoaacutecidos livres citrato ATP e pirofosfato (Refordfs) Em alternativa o V(IV) pode existir
17
livremente em ambientes fortemente aacutecidos como os lisossomas jaacute que a pH superior a 3 as
soluccedilotildees de vanadilo tendem a ser oxidadas pelo ar formando-se vanadato Esta grande
capacidade redutora justifica parcialmente a utilizaccedilatildeo pouco extensiva do iatildeo VO2+
como
inibidor [W]
1212 Vanaacutedio (V)
O vanaacutedio (V) eacute a forma mais estaacutevel num intervalo de pH amplo [W] O iatildeo anioacutenico VO3-tende
a constituir complexos polinucleares pelo que as soluccedilotildees aquosas de vanadato satildeo extremamente
complexas com ocorrecircncia simultacircnea de vaacuterios equiliacutebrios de protonaccedilatildeo (pKa(vanadato)= 35
78 e 125) e oligomerizaccedilatildeo As espeacutecies presentes (H3VO4 H2VO4- HVO4
2- e VO4
3-) dependem
do pH forccedila ioacutenica e temperatura da soluccedilatildeo e da concentraccedilatildeo total do elemento
[AEAFAGAHAI]
O VO43-
- monoacutemero ortovanadato V1 ndash estaacute associado a uma grande parte da importacircncia
bioloacutegica do vanaacutedio uma vez que se trata de um anaacutelogo electroacutenico e estrutural do ortofosfato
(PO43-
) A coordenaccedilatildeo do ortovanadato pode ser tetraeacutedrica ou bipiramidal trigonal (Refordfs)
A oligomerizaccedilatildeo do vanadato monomeacuterico daacute origem ao diacutemero (V2O72-
- V2) que apresenta
tambeacutem equiliacutebrios de protonaccedilatildeo Presume-se a existecircncia de analogia do vanadato dimeacuterico com
o pirofosfato embora natildeo haja evidecircncia estrutural (Refordf) A dimerizaccedilatildeo do V2 origina o
vanadato tetrameacuterico ou metavanadato (V4O124-
- V4) que constitui a espeacutecie maioritaacuteria em
soluccedilotildees aquosas concentradas (Refordf) As estruturas do metavanadato e a sua espeacutecie protonada
satildeo ciacuteclicas [AJ] Outra espeacutecie oligomeacuterica importante eacute o vanadato pentameacuterico (V5O153-
) cuja
estrutura presumida eacute ciacuteclica [AI]
O oligoacutemero com maior importacircncia bioquiacutemica eacute o vanadato decameacuterico (V10O286-
-V10)(Refordf) a
espeacutecie predominante em soluccedilotildees de pH 2-6 quando a concentraccedilatildeo total de vanadato eacute superior
a 100 M (Ref) A estrutura do V10 natildeo difere em soluccedilatildeo ou no estado soacutelidoOs 10 aacutetomos de
vanaacutedio da espeacutecie decameacuterica estatildeo divididos em trecircs grupos identificados na figura como VA-C
consoante o seu ambiente quiacutemico circundante (Refordf) (Fig 111)
A expressatildeo ldquodecavanadatordquo refere-se a uma soluccedilatildeo aquosa na qual a espeacutecie decameacuterica se
encontra em equiliacutebrio quiacutemico com as espeacutecies V1 V2 V4 e V5 sendo que a oligomerizaccedilatildeo
progressiva do vanaacutedio eacute proporcional agrave sua concentraccedilatildeo total (Ref) Nesta soluccedilatildeo a
concentraccedilatildeo de V10 corresponde a aproximadamente 10 da concentraccedilatildeo total de vanaacutedio [AK]
18
Figura 111 ndash Estruturas esquemaacuteticas das espeacutecies de vanadato (espeacutecie monomeacuterica (V1) dimeacuterica (V2)
tetrameacuterica (V4) pentameacuterica (V5) e decameacuterica (V10) (Adaptado de Aureliano e Crans 2008)
122 Metabolismo e efeito na actividade enzimaacutetica
Apesar de natildeo terem sido ainda encontradas as provas categoacutericas das suas funccedilotildees nos seres
vivos sabe-se que o iatildeo vanadato eacute capaz de inibir uma grande quantidade de enzimas in vitro
com especial ecircnfase na actividade ATPaacutesica da miosina [AL Teresa] Como referido
anteriormente o vanadato eacute capaz de actuar em alternativa ao fosfato nas reacccedilotildees bioquiacutemicas
incluindo a sua preferecircncia de ligaccedilatildeo agraves enzimas [W]
Eacute conhecido o papel do vanaacutedio no metabolismo do caacutelcio promovendo a mineralizaccedilatildeo dos
dentes e ossos na biossiacutentese de liacutepidos (trigliceacuteridos e colesterol) com posterior inibiccedilatildeo da
acumulaccedilatildeo destes nos vasos sanguiacuteneos e no SNC na reproduccedilatildeo e desenvolvimento por
mimetizaccedilatildeo e potenciaccedilatildeo de factores de crescimento na estimulaccedilatildeo do consumo de glucose e
da siacutentese de glicogeacutenio com actuaccedilatildeo equivalente aos efeitos da insulina ao niacutevel celular na
transduccedilatildeo de sinais celulares com regulaccedilatildeo da actividade de segundos mensageiros no
transporte de iotildees e aminoaacutecidos na transcriccedilatildeo geneacutetica e siacutentese de proteiacutenas e DNA entre
outros [W Y AB]
Assim o vanaacutedio cnseguiraacute provocar inuacutemeras mudanccedilas bioquiacutemicas nas ceacutelulas Alguns
exemplos da influecircncia bioquiacutemica do vanaacutedio incluem os seus efeitos nos processos de
fosforilaccedilatildeo e desfosforilaccedilatildeo de proteiacutenas e na participaccedilatildeo na formaccedilatildeo de espeacutecies reactivas de
oxigeacutenio devido ao seu efeito proacute-oxidante sobre agentes redutores a niacutevel celular [AM]
Quanto ao seu efeito na actividade enzimaacutetica pode dividir-se em dois tipos principais ndash inibitoacuterio
e estimulante - dependendo da enzima ou do tipo de reacccedilatildeo por esta catalisada No primeiro
grupo incluem-se enzimas como as ATPases (de miosina de canais ioacutenicos e fosfoenzimas
19
transportadoras) e as fosfatases (da tirosina aacutecida e alcalina) a fosfofrutocinase adenilato ciclase
e catalase entre outras [AA] O segundo grupo inclui a maioria das cinases a glucose-6-fosfato
desidrogenase a fosfoglicerato mutase e a fosfoglicerato fosfatase (Refordf)
O efeito do vanaacutedio (V) na actividade enzimaacutetica estaacute relacionado com o seu nuacutemero de
coordenaccedilatildeo podendo actuar como estado de transiccedilatildeo ou por mimetizaccedilatildeo de cofactores
substratos (especialmente o fosfato) e interacccedilotildees proteiacutena-proteiacutena (como no caso das
polimerases e transcriptases) Pode ainda estar envolvido na formaccedilatildeo de ligaccedilotildees de hidrogeacutenio
interacccedilotildees electrostaacuteticas esteacutereas e de oxidaccedilatildeo-reduccedilatildeo Uma evidecircncia desta uacuteltima eacute a sua
capacidade de alternacircncia entre os estados de oxidaccedilatildeo +5 a +3 o que torna faacutecil a sua interacccedilatildeo
redox com enzimas contendo resiacuteduos de cisteiacutenas (Cys) ou qualquer outro composto orgacircnico
com actividade redox [AN]
13 Vanaacutedio e muacutesculo esqueleacutetico
A primeira evidecircncia da presenccedila de vanaacutedio no muacutesculo esqueleacutetico proveio de uma amostra de
ATP isolado de muacutesculo de cavalo que continha uma impureza ndash vanaacutedio ndash cuja presenccedila
provocou a inibiccedilatildeo da actividade da bomba de Na+K
+-ATPase [AO] Desde entatildeo vaacuterios
estudos tecircm sido realizados com ecircnfase na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o
tecido muscular Destes destaca-se a descoberta da interacccedilatildeo forte da espeacutecie V10 com a miosina
fosforilada [AP] e agrave sua inibiccedilatildeo da actividade ATPaacutesica da miosina[AQ AR]
No quediz respeitoao iatildeo monovanadato destaca-se a sua inibiccedilatildeoda actividade Mg2+
-ATPaacutesica
da miosina [AS] que ocorre de duas formas distintas (1) na presenccedila de concentraccedilotildees reduzidas
de V1 a inibiccedilatildeo ocorre a velocidade reduzida dependente da concentraccedilatildeo da espeacutecie
monomeacuterica (2) na presenccedila de concentraccedilotildees elevadas de V1 o niacutevel de inibiccedilatildeo aproxima-se de
100 desde que [V1] seja superior agrave concentraccedilatildeo total de locais com actividade Mg2+
-ATPaacutesica
no ensaio Esta inibiccedilatildeo eacute irreversiacutevel tanto por diaacutelise ou adiccedilatildeo de agentes redutores do
vanadato [AL]Segundo Pate et al(1994)[AT] a espeacutecie monomeacuterica podeinibir a velocidade de
contracccedilatildeo a temperatura inferior a 20ordmC perdendogradual e rapidamente essa capacidade ateacute os
50ordmC temperatura agrave qual cessa a capacidade inibitoacuteria Os resultados obtidos sugeremainda uma
interacccedilatildeo altamente cooperativa entre o iatildeo vanadato e as proteiacutenas componentes do muacutesculo
esqueleacuteticoDestaca-se tambeacutem a descoberta da inibiccedilatildeo da geraccedilatildeo de forccedila contraacutectil nas fibras
musculares por parte da espeacutecie monomeacuterica[AT]Wilson et al(1995) [AU ]determinaram que a
afinidade da espeacutecie V1 para o S1 de miosina eacute significativamente mais elevada do que a do iatildeo
Pi seu anaacutelogo Descobriram ainda que que a ligaccedilatildeo do vanadato poderaacute envolver transiccedilotildees
entre estados energeacuteticos com energia livre independente dos impedimentos esteacutereos impostos
20
pela estrutura das miofibrilas Os estudos desenvolvidos por Smith et al (1990)[AV]
determinaram queo iatildeo monovanadato natildeo apresenta um efeito pronunciado na inibiccedilatildeo da
actividadeacto-S1-miosina-ATPaacutesica excepto em concentraccedilotildees superiores a 900microM a partir das
quais existe produccedilatildeo de espeacutecies oligomeacutericas para as quais o efeito inibitoacuterio seraacute mais
elevadoA espeacutecie monomeacuterica de vanadato consegue adoptar facilmente uma geometria do tipo
bipiramidal trigonal que mimetiza o intermediaacuterio formado (MgADP
Pi)aquando da reacccedilatildeo de
hidroacutelise de ATP no local activo da miosina [AW] De facto a interacccedilatildeo do vanaacutedio com a
miosina eacute inesperadamente estaacutevel na presenccedila de ADP Esta interacccedilatildeo resulta na formaccedilatildeo de
um complexo enzimaacutetico inactivo e estaacutevel com estequiometria de 11 ADPvanadato por siacutetio
activo da proteiacutena A formaccedilatildeo do complexo estaacutevel miosina-vanadato foi proposta por Goodnoet
al (1982) [AS] Nesta ocorre um preacute-equiliacutebrio raacutepido que induz a formaccedilatildeo de um complexo
ternaacuterio reversiacutevel (MADP
V) com formaccedilatildeo lenta do isoacutemero M
ADP
V A diferenccedila entre os
compostos de vanaacutedio e de fosfato centra-se no tempo de meia vida (t12) do Pi ou do V (no
primeiro t12=12 s enquanto no segundo t12=24 h) (Refordf) A diferenccedila dever-se-aacute agrave possiacutevel
complexaccedilatildeo do vanaacutedio com grupos nucleofiacutelicos no siacutetio activo o que provocaria uma
estabilizaccedilatildeo da miosina na conformaccedilatildeo MADP
V
No que respeita aos estudos realizados com a espeacutecie tetrameacuterica Cremo et al determinou que a
cadeia pesada do S1 de miosina pode ser clivada em dois locais distintos (designados V1 e V2)
pela espeacutecie V4 apoacutes irradiaccedilatildeo com luz UV na presenccedila de concentraccedilotildees de vanadato na ordem
dos milimolar e na ausecircncia de nucleoacutetidos ou metais divalentes
Jaacute no que concerne aos estudos realizados com ecircnfase na espeacutecie decameacuterica foi sugerido
porRamos et al (2006) [AK]que a interacccedilatildeo do decavanadato com a G-actina favorecida pela
polimerizaccedilatildeo desta uacuteltima estabiliza a espeacutecie V10 e induz a inibiccedilatildeo da formaccedilatildeo de F-actina A
estabilizaccedilatildeo da espeacutecie V10 por proteiacutenas tansmembranares e do citoesqueleto pode ser a razatildeo
pela qual ocorre a toxicidade do decavanadato reportada na avaliaccedilatildeo dos efeitos do V(V) nos
sistemas bioloacutegicosTiago et al (2007) [AR] concluiram que o V10 apresenta uma clara preferecircncia
de ligaccedilatildeo ao ldquoback-doorrdquo da S1-miosina-ATPase o que se verifica apenas na estrutura
conformacional desimpedida com acesso facilitado ao local de ligaccedilatildeo do Pi Sugere tambeacutem que
o V10 actua como um ldquoback-door stoprdquo bloqueando estaabertura do fragmento de 50-kDa
necessaacuteria para a hidroacutelise do ATP--fosfato o que constitui uma explicaccedilatildeo simples do
comportamento natildeo competitivo da espeacutecie decameacuterica estimulando a utilizaccedilatildeo deste oligoacutemero
como uma feramenta da elucidaccedilatildeo das mudanccedilas conformacionais ldquoback-doorrdquo da miosina no
21
processo da contracccedilatildeo muscular(ref)Destaca-se ainda a descoberta da capacidade inibitoacuteria do
V10 em relaccedilatildeo agrave fosforilaccedilatildeo oxidativa mitocondrial [AX]
Finalmente no que concerne aos estudos realizados com a presenccedila de vaacuterias espeacutecies
oligomeacutericas Aureliano e Madeira (1994) [AY]observaram que na presenccedila de Ca2+
as espeacutecies
V4 e V10se ligam agrave Ca2+
-ATPase do retiacuteculo sarcoplasmaacutetico enquanto a espeacutecie V1 se liga apenas
na presenccedila de ATP Aparentemente o aumento da concentraccedilatildeo de Ca2+
tem o efeito contraacuterio da
inibiccedilatildeo da ATPase do retiacuteculo sarcoplasmaacutetico pelo V1 natildeo tendo no entanto efeito significativo
sobre a inibiccedilatildeo por parte da espeacutecie V10 Tiago et al (2002) [AP] concluiacuteram que em condiccedilotildees
proacuteximas agraves fisioloacutegicas a espeacutecie V10 difere das espeacutecies presentes na soluccedilatildeo de metavanadato
devido agrave sua forte interacccedilatildeo com a miosina fosforilada e agrave sua inibiccedilatildeo da actividade ATPaacutesica
da miosina Para aleacutem disso o ATP diminui a afinidade da miosina para o V4 induzindo a
interacccedilatildeo com V1 e natildeo tendo qualquer efeito no que diz respeito ao V10 Foi tambeacutem proposto
[AQ] que a ligaccedilatildeo das espeacutecies oligomeacutericas de vanadato aoS1 de miosina nos locais onde o
fosfato coordenaeacute provavelmente a causa da inibiccedilatildeo da actividade ATPaacutesica da miosina
estimulada por actina atraveacutes da prevenccedilatildeo da troca entre ATPADP sendo esta favorecida para
aniotildees de vanadato com valecircncias superiores como eacute o caso do V10
Na presenccedila de actina as velocidades de dissociaccedilatildeo do Pi e do V aumentam consideravelmente
este uacuteltimo em menor quantidade Assim a ligaccedilatildeo da actina ao complexo MADP
V e
Pi eacute
bastante fraca pelo que se sugere que o ADPV teria um efeito de relaxamento semelhante ao
ATP No entanto a capacidade do vanaacutedio em inibir a actividade ATPaacutesica da miosina e da
actomiosina eacute diferenciada uma vez que na ausecircncia de actina a inibiccedilatildeo eacute total para
concentraccedilotildees na gama dos M mas na sua presenccedila a inibiccedilatildeo eacute totalmente conseguida apenas
em concentraccedilotildees na gama dos mM (Ref) No entanto eacute necessaacuterio considerar a possiacutevel
formaccedilatildeo de espeacutecies oligomeacutericas de vanadato cuja afinidade e efeito na enzima seratildeo
diferentes Assim pode concluir-se que o efeito de inibiccedilatildeo observado para a actividade ATPaacutesica
de miosina estimulada por actina se pode dever agrave presenccedila das diferentes espeacutecies oligomeacutericas o
que foi jaacute sugerido por Aureliano (2000) [AZ] ndash as espeacutecies tetrameacuterica e decameacuterica de vanadato
interactuariam com a miosina com alteraccedilatildeo da sua interacccedilatildeo com a actina No entanto natildeo eacute
conhecida a contribuiccedilatildeo absoluta das restantes espeacutecies oligomeacutericas para a inibiccedilatildeo da ATPase
da miosina estimulada por actina (Refs)
14 Objectivos do projecto
Pretende-se com este projecto o isolamento purificaccedilatildeo e caracterizaccedilatildeo da actina do muacutesculo
22
esqueleacutetico de coelho sendo esta uacuteltima tambeacutem realizada para o sufragmento-1 de miosina
Seguidamente realizar-se-aacute a quantificaccedilatildeo e caracterizaccedilatildeo das proteiacutenas por espectroscopia UV-
Vis e SDS-PAGE e a caracterizaccedilatildeo das soluccedilotildees de vanaacutedio por UV-Vis e ressonacircncia magneacutetica
nuclear (RMN) Por uacuteltimo a interacccedilatildeo do S1 de miosina com o vanaacutedio seraacute avaliada atraveacutes
das teacutecnicas de UVVis RMN e ressonacircncia paramagneacutetica electroacutenica (RPE)
2 Discussatildeo de Resultados
21 Caracterizaccedilatildeo das proteiacutenas isoladas
211Anaacutelise do estado de pureza
O estado de pureza das soluccedilotildees de actina e de S1 foi analisado por electroforese em gel de
poliacrilamida em condiccedilotildees desnaturantes (SDS-PAGE) (Fig 22) que separa as moleacuteculas de
acordo com o seu peso molecular (PM) A concentraccedilatildeo de acrilamida escolhida (10) permite a
detecccedilatildeo simultacircnea de componentes com peso molecular inferior a 20 kDa e outros com elevado
peso molecular
Figura 21 ndash Fotografia de gel de SDS-PAGE (10 acrilamida) Poccedilo (a)
10 L do marcador de elevado peso molecular () Poccedilo (b) 20 L de [G-
actina]=1 mgmL Poccedilo (c) 20 L de [S1]=1 mgmL Aplicou-se uma
voltagem de V durante h e min
A partir da recta de calibraccedilatildeo obtida log PM = -1177 Rf + 2247 (ver ponto 2 da secccedilatildeo
Anexos) pode verificar-se a presenccedila de actina na amostra do poccedilo (b) presente numa banda
bastante intensa (PMaparente=40 kDa) bem como a presenccedila minoritaacuteria de outros componentes
desconhecidos que natildeo teratildeo sido eliminados durante o processo de extracccedilatildeo e purificaccedilatildeo e natildeo
satildeo relevantes para o estudo em questatildeo Na amostra do poccedilo (c) nota-se a presenccedila de S1
(PMaparente=92 kDa) bem como a de outros componentes cuja banda eacute ligeiramente menos intensa
Estes poderatildeo ser fragmentos proteoliacuteticos de miosina e de S1 Estaacute ainda presente uma bada de
23
maior intensidade que poderaacute ser a proteiacutena-C (PM=135 kDa) uma vez que a sua remoccedilatildeo
completa da amostra necessitaria de um passo adicional de cromatografia de troca ioacutenica [BA]
No entanto natildeo podemos confirmar que se trata desta proteiacutena ou de qualquer outra uma vez que
se encontra fora do intervalo de migraccedilatildeo dos marcadores de peso molecular
212 Determinaccedilatildeo da concentraccedilatildeo proteica
2121Subfragmento-1 de miosina
A partir da recta de calibraccedilatildeo obtida pelo meacutetodo de Bradford [BB](ver secccedilatildeo 3311 do
Procedimento Experimental) A595nm = 00546[BSA] + 00209 (ver secccedilatildeo de Anexos)foi possiacutevel
determinar um intervalo de concentraccedilatildeo de S1 de miosina compreendido entre 226 e 117(plusmn
423M) (203 e 1056 (plusmn 381 mgmL)) (n=8) A concentraccedilatildeo meacutedia de S1 de miosina apoacutes
diaacutelise contra a soluccedilatildeo tampatildeo 10 mM HEPES pH 75 25 mM KCl determinada pela leitura da
Abs280 nmfoi de 20430M (1803 mgmL)(n=7)
As amostras a quantificar atraveacutes do meacutetodo de Bradford natildeo sofreram diaacutelise o que permite
explicar a concentraccedilatildeo superior de S1 de miosina em comparaccedilatildeo com o valor obtido atraveacutes do
meacutetodo directo Esta razatildeo aliada agrave sensibilidade e rapidez superiores deste uacuteltimo fez com que
se optasse pelo meacutetodo directo para a quantificaccedilatildeo da grande maioria dos ensaios
2122Actina
A concentraccedilatildeo de actina foi determinada quando esta se encontrava na forma monomeacuterica (G-
actina) por mediccedilatildeo da leitura directa da Abs290 nm tendo-se obtido uma concentraccedilatildeo meacutedia de
691481M (290202 mgmL) (n=3) em 2 mM Tris-HCl pH 80 02 mM CaCl2 02 mM Na-
ATP Ao comprimento de onda utilizado (290 nm) absorvem fortemente apenas os resiacuteduos de
Trp pelo que a mediccedilatildeo efectuada tem menor sensibilidade do que se tivesse sido efectuada a 280
nm uma vez que a actina apresenta apenas 4 resiacuteduos deste aminoaacutecido embora tenha 16
resiacuteduos de Tyr
22 Mediccedilatildeo da actividade enzimaacutetica de S1 de miosina
221 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
As actividades ATPaacutesicas de S1 de miosina dependentes de Ca2+
e K+foram analisadas pelo
caacutelculo das velocidades de consumo inicial do fosfato inorgacircnico (Pi) Definiu-se os tempos de 5
e 3 minutos para a mediccedilatildeo da Ca2+
e da K+EDTA-ATPase respectivamente Os valores obtidos
24
a partir do protocolo presente no ponto 341 da secccedilatildeo Procedimento experimental encontram-se
descritos na Tabela 2I
Tabela 2I ndash Acrividades natildeo-fisioloacutegicas de subfragmento-1 determinadas pelo meacutetodo de Fiske-Subarow
modificado (n=2)
Proteiacutena
Actividade
ATPaacutesica natildeo
fisioloacutegica
Actividade (nmol
Piminmg proteiacutena) Actividade (s-1)
S1
Ca2+
-ATPase 1400 plusmn 30 2100 plusmn 0045
K+EDTA-ATPase 2334 plusmn 50 3500 plusmn 0075
As actividades natildeo-fisioloacutegicas reflectem o estado de oxidaccedilatildeo dos resiacuteduos expostos de Cys 707
(SH1) e 697 (SH2) Se o grupo SH1 sofrer oxidaccedilatildeo a actividade ATPaacutesica dependente de Ca2+
aumenta e a actividade dependente de K+ diminui Por outro lado se o grupo SH2 sofrer
oxidaccedilatildeo regista-se a perda completa destas actividades
De acordo com Margossian e Lowey (1982) [BA] as actividades ATPaacutesica de S1 de miosina
dependente de Ca2+
e K+EDTA da miosina satildeo respectivamente de 24 e 112 s-1
Estaacute tambeacutem
descrito que a actividade ATPaacutesica dependente de K+ eacute em meacutedia cerca de 3 vezes superior agrave
dependente de Ca2+
reflectindo a quantidade das preparaccedilotildees A razatildeo obtida entre os valores das
actividades ATPaacutesicas dependentes de K+ e Ca
2+ eacute de 166 Os valores obtidos para as actividades
(Ca2+
-ATPase=21 s-1
K+EDTA-ATPase= 35 s
-1) satildeo inferiores agrave valores obtidos por
Margossian e Lowey [BA] e Tiago [BC] com forccedila ioacutenica inferior O valor obtido para a
actividade ATPaacutesica dependente de Ca2+
eacute superior ao obtido por Polosukhina et al (2000) [BD]
cujo meio de reacccedilatildeo continha MOPS e apresentava forccedila ioacutenica inferior Os dados
supramencionados comprovam a variaccedilatildeo da actividade a ATPaacutesica natildeo fisioloacutegica em relaccedilatildeo agrave
forccedila ioacutenica temperatura e pH
222 Mediccedilatildeo da actividadeMg2+
-ATPaacutesica de S1 pelo meacutetodo de enzimas acopladas
A actividade de Mg2+
-ATPase eacute a uacutenica que possui significado fisioloacutegico sendo as restantes
bastante uacuteteis apenas para indicaccedilatildeo da qualidade de preparaccedilatildeo do S1[BA] A 0575 M KCl a
actina e o S1 formam uma soluccedilatildeo viscosa e tuacuterbida Apoacutes a adiccedilatildeo de MgATP o complexo acto-
S1 dissocia-se e a turbidez da soluccedilatildeo diminui Se a forccedila ioacutenica for reduzida a dissociaccedilatildeo do
complexo actoS1 que se encontra na forma de suspensatildeo eacute induzida O MgATP eacute rapidamente
hidrolisado diminuindo a sua concentraccedilatildeo no meio reaccional Nestas condiccedilotildees haacute
reassociaccedilatildeo e subsequente contracccedilatildeo dos filamentos de actina e de S1 com formaccedilatildeo de
25
precipitado O meacutetodo descrito traduz de forma bastante elementar o fenoacutemeno da contracccedilatildeo in
vivo e elimina o problema anterior da solubilizaccedilatildeo ineficiente da miosina de modo a obter uma
mediccedilatildeo exacta da actividade [A] A utilizaccedilatildeo de forccedilas ioacutenicas reduzidas aumenta a afinidade da
actina para o S1 permitindo contornar a extrema dificuldade em obter a concentraccedilatildeo proteica dos
muacutesculos (cerca de 100 mgmL do complexo actomiosina) As actividades Mg2+
-ATPaacutesicas de S1
de miosina foram determinadas a partir do declive da variaccedilatildeo da Abs340 relativo agrave oxidaccedilatildeo de
NADH em funccedilatildeo do tempo como eacute descrito no ponto 342 do Procedimento Experimental Os
valores obtidos para a actividade Mg2+
-ATPaacutesica encontram-se apresentados na Tabela 2II
Tabela 2II ndash Actividade Mg2+
-ATPaacutesica de subfragmento-1 de miosina determinadas pelo meacutetodo das enzimas
acopladas
Actividade (nmol
ATPmin mg(S1)) Actividade (s
-1)
1097 plusmn 269 164 plusmn 04
Ramos (2005) [J] obteve o valor de 01 s-1
para a actividade Mg2+
-ATPaacutesica de S1 de miosina
com forccedila ioacutenica intermeacutedia Tiago (2005) [BC] obteve o valor de 011 s-1 em condiccedilotildees de forccedila
ioacutenica reduzida em relaccedilatildeo agrave utilizada no presente relatoacuterio
23 Preparaccedilatildeo e caracterizaccedilatildeo das soluccedilotildees de vanaacutedio
231 Anaacutelise da composiccedilatildeo das soluccedilotildees de vanadato por espectroscopia de 51
V-RMN
efeito da concentraccedilatildeo da espeacutecie
A diluiccedilatildeo das soluccedilotildees stock de meta- e decavanadato iraacute determinar fortemente a composiccedilatildeo
em soluccedilatildeo dos diferentes oligoacutemeros de vanadato (ref) No caso do decavanadato o periacuteodo de
tempo apoacutes a diluiccedilatildeo iraacute tambeacutem ser um factor importante a considerar devido agrave decomposiccedilatildeo
da espeacutecie V10(ref)Torna-se assim fundamental determinar a composiccedilatildeo quiacutemica das soluccedilotildees
no meio de ensaio utilizado (HEPES 10 mM KCl 25 mM) de modo a correlacionaacute-la
correctamente com os possiacuteveis efeitos promovidos pelas diferentes espeacutecies de vanadato no
sistema em estudo
Aleacutem disso deve ser dado ecircnfase agraves importantes interacccedilotildees que ocorrem entre o vanaacutedio e os
tampotildees escolhidos O tampatildeo com menor nuacutemero e magnitude de interacccedilatildeo eacute o HEPES [AD]
seguido do Tris sendo este uacuteltimo especialmente aconselhado em concentraccedilotildees baixas para casos
de incompatibilidade enzimaacutetica ou quiacutemica redox No caso especial da reacccedilatildeo redox
NAD(P)HNAD(P) o tampatildeo adequado eacute o imidazole de modo a natildeo haver reacccedilatildeo com o
26
vanadato Os tampotildees a evitar em qualquer situaccedilatildeo incluem a trietanolamina fosfato citrato e
glicina entre outros [AD] uma vez que todos interactuam fortemente com V(V) com formaccedilatildeo
de complexos estaacuteveis
Assim procedeu-se agrave anaacutelise da composiccedilatildeo das soluccedilotildees de meta- e decavanadato apoacutes diluiccedilatildeo
das soluccedilotildees stock (50 mM em vanadato total) no meio de reacccedilatildeo (tampatildeo D) atraveacutes da
espectroscopia de ressonacircncia magneacutetica nuclear de 51
V Nas figuras x1 e x2 apresentam-se os
espectros obtidos para as soluccedilotildees de deca- e metavanadato num intervalo de concentraccedilotildees de 1 a
10 mM A Tabela seguinte resume os paracircmetros espectroscoacutepicos obtidos para cada espeacutecie
presente nas soluccedilotildees analisadas
Figura 22ndash Espectros de RMN de 51
V de
concentraccedilotildees crescentes das espeacutecies oligomeacutericas
de vanadato obtidos apoacutes diluiccedilatildeo da soluccedilatildeo stock
de decavanadato em 10 mM HEPES pH 70 25 mM
KCl para concentraccedilotildees finais de A) 1 mM B)2
mM C) 4 mM D) 8 mM E) 10 mM de
decavanadato total V1refere-se agraves espeacutecies
monomeacutericade vanadato V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2IIIndash Desvios quiacutemicos meacutedios (part) das ressonacircncias das espeacutecies decameacuterica (V10) monomeacuterica (V1) e
tetrameacuterica (V4) presentes nos espectros da soluccedilatildeo de decavanadatoLargura a meia altura (∆12) da espeacutecie V10A
Paracircmetros V10A V10B V10C V1 V4
part (ppm) -5162 -5003 -4255 -5616 -5748
∆12 (Hz) 175 nd nd nd nd
Figura 23 ndash Distribuiccedilatildeo graacutefica das
espeacutecies oligomeacutericas de vanadato vs a
concentraccedilatildeo de decavanadato total As
concentraccedilotildees de cada espeacutecie foram
obtidas segundo a formula descrita na
secccedilatildeo 35 do Procedimento
Experimental (aacutereas dos sinais de 51
V-
RMN observadas na figura x1)V10 V1 e
V4 correspondem agraves espeacutecie monomeacuterica
dimeacuterica e tetrameacuterica respectivamente [Esp
eacuteci
e o
ligo
meacute
rica
de
van
aacutedio
] (m
M)
[Decavanadato]total (mM)
V10
V1
V4
27
Nas soluccedilotildees de decavanadato as espeacutecies monomeacuterica e decameacuterica de vanadato satildeo as uacutenicas
presentes para concentraccedilotildees de decavanadato inferiores a 8 mM Ambas as espeacutecies aumentam
linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado em [AY] e
em (Teresa) quando a concentraccedilatildeo de V1 eacute igual ou superior a 200 microM inicia-se a sua
oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica No entanto nas condiccedilotildees de realizaccedilatildeo dos
ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a espeacutecie tetrameacuterica neste
estudo em concentraccedilotildees inferiores a 8 mM tendo o mesmo sido observado por Ramoset al
(2006) [AK] Os resultados obtidos mostram claramente o equiliacutebrio entre a espeacutecie decameacuterica e
a monomeacuterica e que o aparecimento de outras espeacutecies oligomeacutericas se encontra dependente da
concentraccedilatildeo desta uacuteltima Apesar de natildeo ter sido realizado o ensaio com 5 mM de decavanadato
total no presente relatoacuterio estaacute reportado em Ramos et al (2006) [AK] que as concentraccedilotildees da
espeacutecie decameacuterica e monomeacuterica satildeo respectivamente de 500 microM e 200 microM Assim pode inferir-
se que para concentraccedilotildees inferiores a 5 mM e intervalos de tempo de cerca de 20 minutos (que
inclui o tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas
as espeacutecies decameacuterica e monomeacuterica de vanadato
Figura 24 ndash Espectros de RMN de 51
V de concentraccedilotildees
crescentes das espeacutecies oligomeacutericas de vanadato obtidos
apoacutes diluiccedilatildeo da soluccedilatildeo stock de metavanadato em 10 mM
HEPES pH 70 25 mM KCl para concentraccedilotildees finais de A)
1 mM B) 2 mM C) 4 mM D) 8 mM E) 10 mM de
metavanadato total V1 refere-se agraves espeacutecies monomeacuterica de
vanadatoV1 e V2 correspondem respectivamente agrave espeacutecie
monomeacuterica e dimeacuterica
Figura 25 ndash Distribuiccedilatildeo graacutefica das espeacutecies
oligomeacutericas de vanadato vs a concentraccedilatildeo de
metavanadato total As concentraccedilotildees de cada
espeacutecie foram obtidas segundo a formula descrita
na secccedilatildeo 35 do Procedimento Experimental
(aacutereas dos sinais de 51
V-RMN observadas na
figura x1)V10 V1 e V4 correspondem agraves espeacutecie
monomeacuterica dimeacuterica e tetrameacuterica
respectivamente
Esp
eacuteci
e o
ligo
meacute
rica
de
va
nad
ato
(m
M)
[Metavanadato]total (mM)
V1
V2
V4
V5
28
Tabela 2IVndash Desvios quiacutemicos meacutedios (part)das ressonacircncias das espeacutecies tetrameacuterica (V4) monomeacuterica (V1) e
dimeacuterica (V2) presentes no espectro da soluccedilatildeo de metavanadato (total) 10 mM
Paracircmetros V1 V2 V4 V5
part (ppm) -5615 -5748 -5789 -5869
Nas soluccedilotildees de metavanadato as espeacutecies monomeacuterica dimeacutericae tetrameacuterica de vanadato estatildeo
presentes para concentraccedilotildees de metavanadato inferiores a 4 mM Todas estas as espeacutecies
aumentam linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado
em (Aureliano e Madeira 1998) e em (Tiago et al 2004) quando a concentraccedilatildeo de V1 eacute igual ou
superior a 200 microM inicia-se a sua oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica Nas
condiccedilotildees de realizaccedilatildeo dos ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a
espeacutecie pentameacuterica neste estudo em concentraccedilotildees inferiores a 4 mM tendo o mesmo sido
observado por Ramos 2006 Os resultados obtidos mostram claramente o equiliacutebrio entre a
espeacutecie V4 e as espeacuteciesV1 e V2 no qual o aparecimento de outras espeacutecies oligomeacutericas se
encontra dependente da concentraccedilatildeo da espeacutecie V1 Por uacuteltimo pode inferir-se que para
concentraccedilotildees inferiores a 2 mM e intervalos de tempo de cerca de 20 minutos (que inclui o
tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas as
espeacutecies V1 V2 e V4 com concentraccedilotildees respectivas de 0796 0192 e 0196 mM
232Estabilidade das soluccedilotildees de decavanadato anaacutelise por espectroscopia de absorccedilatildeo
UVVis
Caracterizou-se a estabilidade da espeacutecie V10 numa soluccedilatildeo com 1 mM decavanadato total em
tampatildeo D atraveacutes da leitura da Abs400 Obteve-se uma cineacutetica de dissociaccedilatildeo de 1ordf ordem tal
como se pode verificar pela linearidade do graacutefico de ln(Abs400)vs o tempo de aquisiccedilatildeo espectral
(segundos) (Fig26 )
Figura 26ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do tempo
(em segundos) para uma soluccedilatildeo de decavanadato total de 1
mM em tampatildeo D a 25ordmC (n=2)
Sendo uma reacccedilatildeo de primeira ordem pode ser descrita pela equaccedilatildeo cujo
declive eacute ndashK (constante de dissociaccedilatildeo) e a intersecccedilatildeo no eixo das ordenadas corresponde
Ln
(Ab
s 400 n
m)
tempo (s)
29
aln(A0) Numa reacccedilatildeo de 1ordf ordem o tempo de meia-vida eacute independente da concentraccedilatildeo inicial
do reagente sendo descrito pela equaccedilatildeo [2] Deste modo a partir da recta
Ln(Abs400 nm) = -436E-5
K - 105 obteve-se um tempo de meia-vida de 4490 plusmn 0823 horas para
as condiccedilotildees supramencionadas
Apoacutes a adiccedilatildeo de S1 8 microM ao meio reaccional verificou-se que a reacccedilatildeo de desoligomerizaccedilatildeo
da espeacutecie decameacuterica de vanadato seguia igualmente uma cineacutetica de 1ordf ordem tal como se pode
verificar pela linearidade do graacutefico de ln(Abs400) versus o tempo de aquisiccedilatildeo espectral
(segundos) (Fig27)
Figura 27ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do
tempo (em segundos) para uma soluccedilatildeo de decavanadato
total de 1 mM em tampatildeo D a 25ordmC na presenccedila de 8
microM (n= 2)
A partir da equaccedilatildeo da recta Ln(Abs400 nm) = -7520E-5
K - 0613determinou-se o valor de K para
a espeacutecie decameacuterica de onde se obteve um tempo de meia-vida de 2430 plusmn 0191 horas para as
condiccedilotildees supramencionadas
Tanto este valor como o valor registado na ausecircncia de S1 satildeo substancialmente superiores agrave
duraccedilatildeo dos ensaios de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina descritos no presente
relatoacuterio
24 Interacccedilatildeo de S1 com vanaacutedio(V)
241Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
De modo a analisar qual o efeito da adiccedilatildeo de S1 de miosina (tampatildeo D) produzia nos sinais de
51V-NMR titulou-se uma soluccedilatildeo de 5mM de decavanadato total com concentraccedilotildees crescentes
de S1 (5 10 15 e 20 microM) Os espectros de 51
V-RMN encontram-se apresentados na Fig 2
No graacutefico da figura 2 Apresenta-se a concentraccedilatildeo de S1 em funccedilatildeo do factor de alargamento
corrigido dos espectros acima referidos
Ln
(Ab
s (4
00
nm
))
tempo (s)
30
Figura 28 ndash Graacutefico de
representaccedilatildeo da
concentraccedilatildeo de S1 em
funccedilatildeo do factor de
alargamento A
concentraccedilatildeo de
decavanadato total foi de 5
mM e as concentraccedilotildees
utilizadas de S1 foram de 0
5 10 e 15 microM
Verifica-se que o factor de alargamento aumenta linearmente em funccedilatildeo da concentraccedilatildeo de S1 no
ensaio o que indica a existecircncia de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina Observou-
se ainda que a adiccedilatildeo de S1 ao meio reaccional natildeo provocou qualquer alteraccedilatildeo dos desvios
quiacutemicos dos sinais de 51
V-RMN
Realizaram-se ensaios na presenccedila de ATP e de S1 com intuito de verificar o efeito do ATP na
interacccedilatildeo do S1 de miosina com o decavanadato
Figura 29 - Espectros de RMN de 51
V contendo 2
mM Decavanadato (A) (controlo) (B) na presenccedila
de 2 mM de ATP (C) na presenccedila de 2 mM de ATP
e 8 microM de S1 de miosina V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2Vndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies decameacuterica (V10) e monomeacuterica (V1) presentes nos
espectros da soluccedilatildeo de decavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V10A V10B V10C V1
part (ppm) -5161 -5002 -4254 -5614
Observa-se que a presenccedila de ATP (espectro B) natildeo provoca qualquer alteraccedilatildeo visiacutevel em
relaccedilatildeo ao espectro que contem apenas soluccedilatildeo de decavanadato Jaacute a presenccedila simultacircnea de ATP
e S1 provoca um alargamento da ∆12 do sinal do V10A de 1736 Hz (espectro (A)) para 2366 Hz
(espectro (C) Fig 29) Tendo em conta que a concentraccedilatildeo de S1 apresenta uma relaccedilatildeo linear
com o factor de alargamento converteu-se o valor da ∆12 e comparou-se com o valor esperado agrave
F = 00591[S1] - 00157 Rsup2 = 0998
Fact
or
de
ala
rgam
en
to
[S1] (microM)
31
concentraccedilatildeo de S1 utilizada O factor de alargamento obtido para 8 microM ndash 0376 ndash eacute inferior ao
valor esperado agrave mesma concentraccedilatildeo de S1 obtido atraveacutes da equaccedilatildeo da recta apresentada na
Fig 28 ndash 0457 ndash o que indica que o ATP poderaacute exercer um efeito de protecccedilatildeo do S1
diminuindo a sua interacccedilatildeo com a espeacutecieV10
Por uacuteltimo realizaram-se ensaios com metavanadato na presenccedila de 8 microM S1 de modo a verificar
se as espeacutecies oligomeacutericas contidas nesta soluccedilatildeo interagiam com a proteiacutena em estudo
Figura 210 - Espectros de RMN de 51
V contendo 2
mM metavanadato (A) (controlo) (B) na presenccedila
de 8 microM de S1 de miosina V1 V2 e V4
correspondem respectivamente aacutes espeacutecies
monomeacuterica dimeacuterica e tetrameacuterica
Tabela 2VIndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies monomeacuterica (V1) dimeacuterica (V2) e tetrameacuterica (V4)
presentes nos espectros da soluccedilatildeo de metavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V1 V2 V4
part (ppm) -5613 -5747 -5789
Como eacute possiacutevel observar na Fig 210 ocorre uma ligeiro alargamento dos sinal de V4 bem
como uma perda da intensidade pelo que se conclui quedas espeacutecies presentes na soluccedilatildeo de
metavanadato (V1 V2 e V4) apenas a espeacutecie tetrameacuterica interage com o S1 de miosina o que
estaacute de acordo com o reportado por Tiago et al 2002
242 Actividade Mg2+
-ATPaacutesica de S1 de miosina na presenccedila de decanadato
Com o intuito de avaliar o efeito do decavanadato na actividade Mg2+
-ATPaacutesica do S1 de
miosina monitorizou-se a reacccedilatildeo de oxidaccedilatildeo do NADH atraveacutes da leitura de Abs340 na
presenccedila de 005 mgmL de S1 de miosina e no caso da actividade estimulada 2microM de actina em
3125 mM Tris-HCl pH 71 e 375 mM MgCl2 pelo meacutetodo de enzimas acopladas (ver seccccedilatildeo xx
dos MM)Para tal realizaram-se ensaios com vaacuterias concentraccedilotildees de decavanadato total (10 50
100 250 500 e 1000) de modo a determinar qual o seu efeito na actividade MgATPaacutesica de S1
de miosina na ausecircncia e na presenccedila de um dos seus ligandos naturais Os ensaios foram
realizados agrave temperatura ambiente com incubaccedilatildeo preacutevia a 25ordm C durante 20 minutos
32
Figura 211 ndash Representaccedilatildeo
graacutefica da Actividade Mg2+
-
ATPaacutesica de S1 () (os ajustes
teoacutericos estatildeo representado a por
uma linha os resultados
experimentais estatildeo representados
por triacircngulos (Actividade
estimulada) e cruzes (Actividade
natildeo estimulada)) vs
[Decavanadato]total medidas pelo
meacutetodo das enzimas acopladas
(n=3) Os dados foram ajustados ao modelo de decaimento exponencial usando o programa GraphPad Prism para a
determinaccedilatildeo dos valores de IC50 (na ausecircncia de actina IC50= 266 plusmn 07 M decavanadato na presenccedila de actina
IC50= 28 plusmn 1 M decavanadato
A partir do graacutefico presente na Fig 29 verifica-se que o ajuste dos dados experimentais foi bem
sucedido A inibiccedilatildeo da actividade Mg2+
-ATPaacutesica do S1 de miosina eacute afectada em menor
amplitude na presenccedila de actina (60 de inibiccedilatildeo da actividade) do que na sua ausecircncia
(aproximadamente 80 de inibiccedilatildeo de actividade) Devido a esta diferenccedila pode sugerir-se que a
actina iraacute impedir parcialmente a ligaccedilatildeo do decavanadatoao S1 demiosina atraveacutes de
impedimentos esteacutereos ou interacccedilotildees electrostaacuteticas por exemplo A teacutecnica utilizada natildeo nos
permite concluir acerca de que regiatildeo(otildees) ou resiacuteduo(s) da actina participaratildeo na preservaccedilatildeo
parcial da actividade Mg2+
-ATPaacutesica
Por outro lado os valores de IC50 dos dois tipos de actividades natildeo apresentam diferenccedilas
significativas tendo-se obtido 266 plusmn 07 e 28 plusmn 1M decavanadato na ausecircncia e na presenccedila de
actina respectivamente o que corresponde a uma concentraccedilatildeo de cerca de 27 M de V10 O
meio de reacccedilatildeo utilizado nestes ensaios foi 10 mM HEPES e 25 mM KCl com 2 mM ATP Em
(Tiago et al 2004) o valor de IC50 para a actividade Mg2+-ATPaacutesica na ausecircncia de actina foi de
125 plusmn 025 microM enquanto o valor da actividade estimulada por actina foi de 136 plusmn 14 microM O
meio de reacccedilatildeo utilizado nos ensaios foi 20 mM HEPES 50 mM KCl e 2 mM ATP o que resulta
precisamente no dobro da forccedila ioacutenica a que foi realizado o ensaio do presente relatoacuterio
(exceptuando a contribuiccedilatildeo dos produtos da hidroacutelise de ATP uma vez que este meacutetodo
pressupotildee a ocorrecircncia de um ciclo de reacccedilotildees de hidroacutelise e siacutentese deste composto) Ora tendo
sido anteriormente referido que a afinidade do S1 de miosina para com a espeacutecie V10 diminui com
Act
ivid
ade
Mg
2+-A
TP
aacutesic
a d
e S
1 d
e
Mio
sin
a
[Decavanadato] (microM) Actividade Estimulada ()
Actividade Natildeo Estimulada ()
Ajuste Actividade Estimulada
Ajuste Actividade Natildeo Estimulada
33
o aumento da forccedila ioacutenica (Tiago et al 2004 tal poderaacute explicar a diferenccedila obtida entre estes
dois conjuntos de valores
243Quantificaccedilatildeo de cisteiacutenas reduzidas pelo meacutetodo de Ellman
A interacccedilatildeo do vanaacutedio com alguns tipos de aminoaacutecidos sensiacuteveis agrave oxidaccedilatildeo pode fornecer-nos
informaccedilatildeo bastante uacutetil em relaccedilatildeo agraves alteraccedilotildees estruturais que conduzem a uma diminuiccedilatildeo da
actividade fisioloacutegica do S1 de miosina Os estudos realizados por Chasteen 1983 concluiacuteram
que a reduccedilatildeo de vanadato (V) a vanadilo (IV) ocorre na presenccedila de agentes redutores comuns no
ambiente celular de que se destaca o aminoaacutecido cisteiacutena Esta eacute tambeacutem um alvo de
complexaccedilatildeo por parte do iatildeo VO2+
Jaacute Ramos et al (2009) demonstrou que a incubaccedilatildeo de
actina com a soluccedilatildeo de decavanadato induz a oxidaccedilatildeo de resiacuteduos de cisteiacutena e agrave formaccedilatildeo do
oxidovanaacutedio (IV) Este grupo de investigadores descobriu ainda que apenas a espeacutecie V10 induz a
oxidaccedilatildeo das cisteiacutenas ldquoraacutepidasrdquo da actina atraveacutes do processo proposto que eacute parcialmente
responsaacutevel pela inibiccedilatildeo jaacute descrita da actividade de ATPase do S1 de miosina estimulada por
actina quando exposta agrave soluccedilatildeo de decavanadato Esta inibiccedilatildeo apresenta-se afectada com a
presenccedila de ATP uma vez que os resiacuteduos de cisteiacutena permaneceram intactos nos ensaios
realizados com este ligando Jaacute Crans et al (2004) concluiu que o siacutetio activo de muitas fosfatases
(que conteacutem grupos tiol) pode formar um equiliacutebrio redox com o iatildeo vanadato (V) de forma natildeo
reversiacutevel
Os resultados obtidos por Sakurai e Shiromura (1980) sugeriram que o vanaacutedio (V) sofre reduccedilatildeo
a vanaacutedio (IV) sem dependecircncia do valor de pH do ensaio Registaram ainda uma estequiometria
de 31 na reacccedilatildeo do vanadato (V) com os resiacuteduos de Cys enquanto a estequiometria dos ensaios
com o vanadilo (IV) foi de 21
Berlett e Stadtman (1997) observaram que os tipos de aminoaacutecidos mais facilmente oxidaacuteveis
englobam entre outros os resiacuteduos de Cys Trp e MetAssim analisou-se o estado redox dos
grupos tiol dos resiacuteduos de Cys do S1 na presenccedila do decavanadato atraveacutes da monitorizaccedilatildeo da
Abs412 nm na presenccedila de 1 mgmL de S1 de miosina em tampatildeo 10 mM HEPES 25 mM KCl
variando a concentraccedilatildeo da soluccedilatildeo de decavanadato de modo a determinar que tipo de alteraccedilotildees
estruturais ocorrem na sua presenccedila no que diz respeito agrave diminuiccedilatildeo da actividade ATPaacutesica de
S1 na presenccedila de decavanadato
Os resultados referentes agrave quantificaccedilatildeo das cisteiacutenas reduzidas apoacutes o tratamento de S1 com
diferentes concentraccedilotildees de decavanadato em tampatildeo 10 mM HEPES pH 75 e 25 mM KCl a
25ordmC durante 20 minutos encontram-se descritos na Tabela 2VII
Tabela 2VII ndash Cisteiacutenas reduzidas totais e ldquoraacutepidas de S1 (1 mgmL) apoacutes tratamento com decavanadato
34
(010501002505001000 uM) em 10 mM HEPES 25 mM KCl pH 7 a 25ordmC durante 20 minutos A detecccedilatildeo
espectrofotomeacutetrica foi feita com DTNB (e 412=8400 uM-1 cm-1) (n variaacutevel entre 1 e 8)
Amostras Cys raacutepidas Cys totais
11 M S1 (controlo) 22 plusmn 04 106 plusmn 14
11 M S1 + 10 M Decavanadato 22 plusmn 05 95 plusmn 08
11 M S1 + 50 uM Decavanadato 20 plusmn 02 98 plusmn 14
11 M S1 + 100 uM Decavanadato 17 plusmn 03 97 plusmn14
11 M S1 + 250 uM Decavanadato 26 plusmn 02 98 plusmn 08
11 M S1 + 500 uM Decavanadato 21 plusmn 04 90 plusmn 06
11 M S1 + 1000 uM Decavanadato 24 plusmn 0 86 plusmn 08
A partir dos resultados obtidos pode sugerir-se que o S1 possui 11 resiacuteduos de Cys (controlo) o
que estaacute de acordo com o anteriormente descrito Verifica-se que o tratamento da proteiacutena com
concentraccedilotildees crescentes de decavanadato provoca a oxidaccedilatildeo de dois resiacuteduos de Cys do S1 de
miosina natildeo expostos ao solvente No entanto a teacutecnica utilizada natildeo nos permite identificar
quais
Tendo em consideraccedilatildeo que o valor de IC50 obtido para a reacccedilatildeo de inibiccedilatildeo daactividade Mg2+-
ATPaacutesica de S1 na presenccedila de decavanadato eacute de cerca de 27 microM pode sugerir-se que este seraacute o
valor ao qual ocorreraacute a oxidaccedilatildeo do primeiro resiacuteduo de Cys o que poderaacute contribuir para a
diminuiccedilatildeo da actividade de S1 observada
25Interacccedilatildeo de S1 com vanaacutedio(IV)
251Anaacutelise por espectroscopia de Ressonacircncia Paramagneacutetica Electroacutenica
A acumulaccedilatildeo intracelular de oxovanaacutedio (IV) in vivo constitui uma das principais formas de
apresentaccedilatildeo de vanaacutedio no organismo Deste modo torna-se bastante interessante analisar a
interacccedilatildeo entre este iatildeo e o S1 de miosina atraveacutes da teacutecnica de espectroscopia de Ressonacircncia
Paramagneacutetica Electroacutenica o meacutetodo de eleiccedilatildeo do estudo do iatildeo vanadilo e dos complexos por si
formados O electratildeo desemparelhado encontra-se fortemente acoplado ao nuacutecleo 51
V o que
35
resulta num espectro de RPE consistindo de 8 desdobramentos na direcccedilatildeo paralela e outros tantos
na direcccedilatildeo perpendicular (Chasteen 1983)
Os estudos realizados por Ahmed et al (1987) centraram-se na interacccedilatildeo do vanadilo com
proteiacutenas como eacute o caso da insulina transferrina entre outros obtendo um espectro caracterizado
por 8 linhas amplamente espaccediladas (na direcccedilatildeo paralela) Jaacute Ramos et al 2009 concluiu que a
exposiccedilatildeo da soluccedilatildeo de decavanadato agraves formas globular e filamentosa de actina produz um
espectro tiacutepico do iatildeo oxidovanaacutedio (IV) indicando a reduccedilatildeo do vanadato
Para aferir qual a concentraccedilatildeo de S1 que permitia obter uma boa razatildeo sinalruiacutedo titulou-se uma
soluccedilatildeo de sulfato de vanadilo 15 microM ([VOSO4]stock = 5 mM) com concentraccedilotildees crescentes de
proteiacutena (0 5 10 15 e 20 microM) em 10 mM HEPES 25 mM KCl A existecircncia de uma correlaccedilatildeo
linear entre a concentraccedilatildeo de proteiacutena e a intensidade dos sinais de RPE correspondente agrave banda
-12 perpendicular encontra-se representada na Fig 210
Figura 2 12ndash Representaccedilatildeo graacutefica da variaccedilatildeo da intensidade do sinal de RPE vs a concentraccedilatildeo de S1 utilizada
Apoacutes verificar que com uma concentraccedilatildeo de S1 igual a 20 microM se obtinham sinais com uma razatildeo
sinalruiacutedo adequada ao prosseguimento dos estudos sobre a interacccedilatildeo V(IV)-S1 de miosina
titulou-se uma soluccedilatildeo de 20 microM S1 com 0 25 5 10 15 25 50 75 100 150 200 500 e 1000
microM VOSO4
Traccedilou-se o graacutefico da intensidade da banda -12 perpendicular do sinal de EPR em funccedilatildeo da
concentraccedilatildeo de VOSO4 tendo-se determinado uma constante de dissociaccedilatildeo (KD) do complexo
S1-vanadilo de 8748 plusmn 2621 microM V(IV)
y = 016786x + 122435
Rsup2 = 092992759
Inte
nsi
dad
e d
o s
inal
de
RP
E
[S1] (microM)
36
Figura 213 ndash Representaccedilatildeo graacutefica da variaccedilatildeo do sinal de RPE vs a concentraccedilatildeo de vanadilo utilizada (os valores
experimentais estatildeo representados por losangos o seu ajuste estaacute representado pela linha)
A simulaccedilatildeo do espectro de EPR foi efectuada a partir do espectro obtido para uma concentraccedilatildeo
de 1 mM VOSO4 apoacutes adiccedilatildeo de 20 microM S1 de miosina tendo-se obtido os valores da constante de
acoplamento hiperfina (A=5016) e do factor g (g0=195)
Figura 214 ndash Representaccedilatildeo graacutefica do espectro de EPR obtido com 1000 microM de vanadilo e 20 microM de S1 (a cinzento
escuro) A simulaccedilatildeo do espectro encontra-se a cinzento claro
Por comparaccedilatildeo destes com os valores presentes na tabela de correspondecircncia xxx eacute possiacutevel
predizer quais os ligandos provaacuteveis do vanadilo na presenccedila de proteiacutena Assim concluiu-se que
o vanadilo estaacute coordenado com duas moleacuteculas de aacutegua um grupo hidroxilo e um grupo R=NR
O grupo hidroxilo pode indicar os resiacuteduos de aminoaacutecidos Tyr Treonina e Serina enquanto o
grupo R=NR poderaacute indicar a presenccedila de Arginina e Histidina como ligandos do vanadilo
Inte
nsi
dad
e d
os
sinai
s d
e R
PE
[Vanadilo] (microM)
Ajuste dos dados exp Dados experimentais
Campo magneacutetico (G)
Espectro Experimental Ajuste
37
3 Procedimento Experimental
31 Reagentes
Todos os reagentes utilizados satildeo de elevado grau de pureza (analiacutetico) Apresentam-se as suas
foacutermulas quiacutemicas ou abreviaturas quando necessaacuterio acrescentou-se a sua descriccedilatildeo Entre
parecircntesis colocou-se as empresas que os comercializam
Aacutecido aceacutetico glacial 100 (Riedel) Acrilamida 99 (Merck) Azul de bromofenol (Riedel-de-
Haumlen) -Mercaptoetanol (Sigma) Bissulfito de soacutedio (Sigma)Bradford (reagente de)
(BioRad)BSA (albumina de soro bovino Sigma-Aldrich) CaCl22H2O (cloreto de caacutelcio
dihidratado Fluka) CH3CH2OH 96 (etanol Panreac) Coomassie Blue R250 (Merck) D2O
(oacutexido de deuteacuterio SDS) DTNB (55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico) Aldrich) EDTA (aacutecido
etilenodiaminatetraaceacutetico Merck) Fosfoenolpiruvato 98 (Fluka) Glicerol (Sigma) Glicina
985 (Panreac) HCl 37 (Riedel-de Haumlen) H2SO4 96 (Pronalab) HEPES 99 (aacutecido 4-(2-
hidroxiletil)-1-piperazineetanosulfoacutenico Sigma) KH2PO4 99 (hidrogenofosfato de potaacutessio)
MgATP (Sigma) Na
ATP (Sigma) Na2HPO4 99 (hidrogenofosfato de soacutedio Riedel-de Haumlen)
NaH2PO4 (fosfato de soacutedio ) NaOH 986 (hidroacutexido de soacutedio Pronalab) KCl 99 (Cloreto de
potaacutessio Panreac) L-Cys 995 (enantioacutemero levoacutegiro do resiacuteduo de Cisteiacutena Fluka)Metanol
99 (Merck) Molibdato de amoacutenio (Sigma)NNrsquo-metil-bis-acrilamida 98 (Merck) NH4VO3
99 (metavanadato de amoacutenia Sigma) PSA 10 (persulfato de amoacutenio Merck) SDS
(dodecilo sulfato de soacutedio Sigma) Sulfito de soacutedio (Sigma) VOSO4 (sulfato de vanadilo
Aldrich) TEMED 99 (Tetrametiletilenodiamina Merck) Tris (2-amino-2-hidroximetil-13-
propanodiol Sigma)
32 Meacutetodos
321 Isolamento e purificaccedilatildeo das proteiacutenas de muacutesculo esqueleacutetico de coelho
3211 Purificaccedilatildeo da miosina preparaccedilatildeo de Subfragmento 1 por digestatildeo quimotriacutepsica
da miosina
O subfragmento-1 de miosina-II (S1) foi gentilmente cedido pelo Prof Doutor Manuel Aureliano
do Departamento de Quiacutemica Bioquiacutemica e Farmaacutecia da Faculdade de Ciecircncias e Tecnologia da
Universidade do Algarve
No entanto descreve-se abaixo de forma resumida o procedimento efectuado
38
32111 Purificaccedilatildeo da miosina
O protocolo de isolamento foi adaptado do meacutetodo previamente descrito por Margossian e Lowey
(1982) apresentando-se de forma esquemaacutetica na Fig 31
Sucintamente os tecidos musculares dorsais e das pernas foram extraiacutedos apoacutes o sacrifiacutecio do
animal (Fig31) De seguida foram adicionados 3 volumes da soluccedilatildeo de extracccedilatildeo A2
Centrifugou-se a soluccedilatildeo e filtrou-se o sobrenadante O sedimento foi entatildeo diluiacutedo em 14
volumes de aacutegua destilada de modo a que a forccedila ioacutenica atingisse 004 M Apoacutes adiccedilatildeo de szlig-
mercaptoetanol (szlig-me) deixou-se a precipitar a 4ordmC removendo o sobrenadante Centrifugou-se
novamente e ressuspendeu-se o sedimento na soluccedilatildeo de extracccedilatildeo A Adicionou-se a soluccedilatildeo
tampatildeo B3 para que a forccedila ioacutenica atingisse 05 M Homogeneizou-se e deixou-se repousar
durante 2h Homogeneizou-se novamente e centrifugou-se de modo a ocorrer precipitaccedilatildeo dos
complexos de actomiosina e proteiacutena desnaturada Apoacutes filtraccedilatildeo do sobrenadante em latilde de vidro
quantificou-se a soluccedilatildeo de miosina resultante pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental)
2 Soluccedilatildeo de extracccedilatildeo A 0012M K2CO3 004M KHCO3 06M KCl pH 65
3 Soluccedilatildeo-tampatildeo B Tris 100 mM pH 7 KCl 2M
39
Figura 31 ndash Representaccedilatildeo esquemaacutetica do protocolo de isolamento de miosina (Margossian e Lowey 1982) Todo o
procedimento foi realizado a 4ordmC (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
32112 Preparaccedilatildeo de Subfragmento-1 de miosina por digestatildeo com quimiotripsina
O S1 de miosina foi isolado apoacutes diaacutelise da miosina durante a noite contra a soluccedilatildeo C4 A
presenccedila do EDTA eacute essencial uma vez que a sua substituiccedilatildeo por MgCl2resulta na produccedilatildeo de
HMM em vez de S1 A suspensatildeo de miosina foi equilibrada e digerida com -quimotripsina
A digestatildeo foi interrompida pela adiccedilatildeo de PMSF Seguidamente ultracentrifugou-se o produto
da digestatildeo desprezando-se o sedimento (Fig 32) A soluccedilatildeo de sobrenadante maioritariamente
S1 foi quantificada pela leitura directa da Abs280 ou pelo meacutetodo de Bradford (ver as secccedilotildees
3311 e 3312 do Procedimento Experimental respectivamente) e armazenada em 10 sacarose
e 2 mM szlig-Me a -80ordmC
4 Soluccedilatildeo C 25mM Tris pH 70 012M KCl 1mM EDTA
40
Antes da realizaccedilatildeo dos ensaios descritos no presente relatoacuterio o S1 de miosina foi dialisado
contra a soluccedilatildeo D5 de modo a remover qualquer composto que pudesse complexar como
vanaacutedio Seguidamente quantificou-se a proteiacutena pelo meacutetodo de Bradford ou pelo meacutetodo
directo (ver secccedilotildees 3311 e 3312do Procedimento Experimtental)
Figura 3 2ndash Representaccedilatildeo esquemaacutetica do protocolo de preparaccedilatildeo do S1 de miosina por digestatildeo quimiotripsina de
acordo com o previamente descrito por Margossian e Lowey (1982) Todo o procedimento foi realizado a 4ordm C
excepto quando indicaccedilatildeo do contraacuterio (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
3212 Isolamento e purificaccedilatildeo de actina
Isolou-se e purificou-se a actina segundo o meacutetodo descrito por Pardee e Spudish que
compreende dois passos ndash a obtenccedilatildeo de um resiacuteduo seco denominado ldquopoacute de acetonardquo e a
extracccedilatildeo subsequente da actina a partir do ldquopoacute de acetonardquo na forma de G-Actina com a
posterior polimerizaccedilatildeo a F-Actina
32121 Obtenccedilatildeo do ldquopoacute de acetonardquo
Sacrificou-se o coelho com uma pancada atraacutes do pescoccedilo e sangrou-se o animal apoacutes o corte da
arteacuteria jugular Esfolou-se e colocou-se numa tina com gelo Retiraram-se as partes de tecido
conjuntivo que envolvem o tecido muscular com auxiacutelio de uma pinccedila e bisturi Seguidamente
retirou-se os muacutesculos dorsais e os muacutesculos brancos das pernas Passou-se os muacutesculos por aacutegua
destilada de modo a remover as impurezas e pesou-se num copo Trituraram-se os muacutesculos
numa maacutequina de triturar carne e extraiu-se o resultante com 3 volumes da soluccedilatildeo E6 durante 10
minutos com agitaccedilatildeo suave Centrifugou-se a suspensatildeo a 4000 g durante 10
5 Soluccedilatildeo D HEPES 10 mM pH 70 KCl 25 mM
6 Soluccedilatildeo E 01 M KCl 015 M K2HPO4 pH 65
41
minutosdesprezando-se o sobrenadante Ressuspendeu-se o sedimento em 6 volumes de soluccedilatildeo
F7durante 10 minutos com agitaccedilatildeo suave Verificou-se se o pH da soluccedilatildeo estava compreendido
entre 82 e 85 acertando-se estecom soluccedilatildeo G8 se necessaacuterio Centrifugou-se durante 10
minutos a 4000 g Desprezou-se o sobrenadante e ressuspendeu-se o sedimento em 3 volumes de
soluccedilatildeo H9 durante 10 minutos Centrifugou-se novamente a 4000 gdurante 10 minutos
desprezando-se o sobrenadante Ressuspendeu-se o sedimento duas vezes consecutivas em 6
volumes de aacutegua destilada durante 5 minutos Centrifugou-se a 4000 g durante 10 minutos apoacutes
cada adiccedilatildeo de H2O
Seguidamente ressuspendeu-se o sedimento 5 vezes consecutivas utilizando 3 volumes de
acetona de cada vez agitando suavemente durante 10 minutos agrave temperatura ambiente Colocou-
se o resiacuteduo filtrado numa tina de vidro sobre papel de filtro deixando-se evaporar o solvente
durante a noite num nicho Triturou-se o resiacuteduo seco numa maacutequina de triturar carne e deixou-se
novamente evaporar o solvente O ldquopoacute de acetonardquo obtido foi armazenado a -20 ordmC
32122 Purificaccedilatildeo da actina a partir de ldquopoacute de acetonardquo
Apoacutes a obtenccedilatildeo do poacute de acetona como acima indicado efectuou-se o protocolo seguidamente
descrito de modo a obter monoacutemeros de actina (G-Actina) e actina filamentosa (F-Actina)
Pesou-se 35 g de poacute de acetona e adicionaram-se 50 mL de soluccedilatildeo-tampatildeo I10
sendo a
extracccedilatildeo feita com agitaccedilatildeo suave durante 30 minutos Separou-se o extracto do poacute de acetona
hidratado filtrando a suspensatildeo com 4 camadas de gaze esterilizada tendo o cuidado de natildeo
espremer o resiacuteduo Voltou a extrair-se com 50 mL de soluccedilatildeo I durante 30 minutos mexendo
lentamente Adicionou-se este ao extracto obtido anteriormente Centrifugou-se o extracto total a
16 000 g durante 30 minutos Decantou-se para uma proveta sobre a qual se colocou gaze para
que o sedimento solto natildeo se juntasse ao sobrenadante e leu-se o volume De seguida adicionou-
se KCl ateacute uma concentraccedilatildeo final de 50 mM MgCl2 ateacute 2mM e ATP ateacute 1mM lentamente e com
agitaccedilatildeo contiacutenua de modo a promover a polimerizaccedilatildeo dos monoacutemeros de actina
Deixou-se polimerizar durante 2h a 25ordmC observando-se a presenccedila de pequenas bolhas de ar
indicativas de um aumento de viscosidade da soluccedilatildeo Arrefeceu-se em gelo durante 15 minutos e
adicionou-se KCl ateacute uma concentraccedilatildeo final de 06M Mexeu-se lentamente durante 30 minutos
apoacutes os quais se ultracentrifugou a 110 000g durante 1hora Desprezou-se o sobrenadante
ressuspendeu-se o sedimento com soluccedilatildeo tampatildeo E e transferiu-se para um potter
Dialisou-se durante a noite contra 1L de soluccedilatildeo-tampatildeo I a uma temperatura de 4ordmC com rotaccedilatildeo
7 Soluccedilatildeo F 005 M NaHCO3 pH 82-85
8 Soluccedilatildeo G 1M Na2CO3
9 Soluccedilatildeo H 1 mM EDTA pH 70
10 Soluccedilatildeo I 2 mM Tris-HCl pH 8 02 mM CaCl2 02 mM Na2ATP 05 mM szlig-Me 0005 NaN3
42
contiacutenua e mudanccedila de tampatildeo 2 a 3 vezes De seguida a actina foi centrifugada a 110
000gdurante 1hora Decantou-se a soluccedilatildeo de G-Actina (sobrenadante) e desprezou-se o
sedimento Determinou-se a concentraccedilatildeo de G-Actina pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental) A soluccedilatildeo de G-actina foi armazenada a uma temperatura de 4ordmC
apoacutes adiccedilatildeo de 002NaN3 mantendo-se estaacutevel durante 3 a 5 dias
331 Quantificaccedilatildeo das proteiacutenas isoladas
3311 Quantificaccedilatildeo pelo meacutetodo de Bradford
O meacutetodo de Bradford (Bradford 1976) baseia-se na utilizaccedilatildeo do reagente de Bradford
composto por entre outros aacutecido fosfoacuterico e o corante ldquoCoomassierdquo Este corante possui carga
negativa e a sua coloraccedilatildeo varia entre a vermelha (Absmax=465 nm) predominante em soluccedilatildeo e a
azul (Absmax=595 nm) presente aquando da ligaccedilatildeo aos grupos amina das proteiacutenas Em soluccedilotildees
aacutecidas diluiacutedas a coloraccedilatildeo varia proporcionalmente com a quantidade do corante ligado agrave
proteiacutena que por sua vez eacute directamente proporcional agrave concentraccedilatildeo da proteiacutena permitindo a
sua quantificaccedilatildeo ateacute a um limite de detecccedilatildeo de 20 gmL (Bradford 1976)
Deste modo prepararam-se amostras de BSA ([BSA]stock= 1mgmL) utilizando um intervalo de
concentraccedilotildees compreendido entre 0 e 15 gmL de acordo com a tabela 3I Apoacutes a adiccedilatildeo da
H2O MilliQ e do reagente de Bradford as soluccedilotildees foram sucessivamente homogeneizadas no
voacutertex
Tabela 3Indash Amostras utilizadas para a obtenccedilatildeo da recta de calibraccedilatildeo no meacutetodo de Bradford
BSA (L) BSA (g) H2O MilliQ (L) Reagente de Bradford (L)
0 0 800 200
125 125 79875 200
25 25 7975 200
5 5 795 200
10 10 790 200
15 15 785 200
Apoacutes incubaccedilatildeo a 25ordmC por 5 minutos leram-se os valores de Abs595 num
espectrofotoacutemetro (Shimadzu UV-160A) para todas as amostras Seguidamente traccedilou-se a recta
de calibraccedilatildeo (Abs595vs quantidade de BSA (g))
43
Para determinar a concentraccedilatildeo de proteiacutena apoacutes o processo de isolamento purificaccedilatildeo e
diaacutelise quando necessaacuterio colocou-se 5 L da amostra num tubo de Eppendorf e adicionou-se
795 L de H2O MilliQ homogeneizando a soluccedilatildeo no voacutertex Adicionou-se posteriormente 200
L de reagente de Bradford e repetiu-se a homogeneizaccedilatildeo Incubou-se a mistura a 25ordmC por 5
minutos apoacutes os quais se leu os valores de Abs595 nm no espectrofotoacutemetro Por interpolaccedilatildeo deste
valor a partir da recta de calibraccedilatildeo determinou-se a concentraccedilatildeo de proteiacutena na amostra
3312 Quantificaccedilatildeo pelo meacutetodo directo
A quantificaccedilatildeo das amostras proteicas pode tambeacutem ser efectuada atraveacutes da leitura directa da
absorvacircncia tendo por base a lei de Beer-Lambert onde Abs corresponde agrave
absorvacircncia ao comprimento de onda ao qual o ensaio foi realizado corresponde ao coeficiente
de extinccedilatildeo (substituiacutedo pela letra a se as unidades forem distintas da molaridade) b corresponde
ao percurso oacuteptico da radiaccedilatildeo na ceacutelula amostral e c corresponde agrave concentraccedilatildeo amostral
Assim no caso de S1 de miosina mediu-se a Abs280e calculou-se a concentraccedilatildeo da proteiacutena
sabendo que o coeficiente de extinccedilatildeo de S1 eacute (Ref)
No caso da actina mediu-se a Abs290ecalculou-se a concentraccedilatildeo da proteiacutena sabendo que o
coeficiente de extinccedilatildeo da actina eacute (Ref)
332Electroforese em condiccedilotildees desnaturantes (SDS-PAGE)
De modo a melhor caracterizar as proteiacutenas isoladas foi realizada uma electroforese em
condiccedilotildees desnaturantes (SDS-PAGE do inglecircs Sodium Dodecyl Sulfate Polyacrilamide Gel
Electrophoresis) O SDS (dodecilo sulfato de soacutedio) eacute um detergente anioacutenico utilizado neste
meacutetodo para desnaturar as proteiacutenas que se ligam ao SDS na proporccedilatildeo 14 g SDS g proteiacutena
(aproximadamente 1 moleacutecula de SDS por cada 2 resiacuteduos de aminoaacutecidos) (Stryer) O SDS induz
uma densidade de carga negativa significativa nas proteiacutenas a que se liga mascarando a sua carga
intriacutenseca pelo que as razotildees cargamassa satildeo tendencialmente idecircnticas Assim a SDS-PAGE
separa as proteiacutenas quando sujeitas a um campo eleacutectrico de acordo com o seu peso molecular
sendo a sua migraccedilatildeo directamente proporcional ao logaritmo deste uacuteltimo (Stryer)
O gel de resoluccedilatildeo com 10 de acrilamida foi obtido atraveacutes da adiccedilatildeo de 167 mL de soluccedilatildeo
stock de acrilamida11
075 mL de tampatildeo de resoluccedilatildeo12
251 mL de aacutegua destilada e 50 L de
SDS 10 Para promover a polimerizaccedilatildeo adicionou-se em seguida 38 L de PSA 10
11
soluccedilatildeo stock de acrilamida soluccedilatildeo de 30 (mv) de acrilamida e 08 (mv) de NNrsquo-metil-bis-acrilamida 12
tampatildeo de resoluccedilatildeo 15 M Tris-HCl pH 88
44
(persulfato de amoacutenia) 10 e 25 L de TEMED 99 (tetrametiletilenodiamina) Agitou-se
cuidadosamente de modo a natildeo formar bolhas e pipetou-se a soluccedilatildeo para a cassette de vidro
previamente montada no suporte De seguida adicionou-se uma peliacutecula de cerca de 2 mm de 2-
metil-2-butanol para verificar posteriormente a polimerizaccedilatildeo Apoacutes a polimerizaccedilatildeo que demora
cerca de 45 minutos verteu-se a agrave superfiacutecie e adicionou-se o gel de concentraccedilatildeo com 5 de
acrilamida constituiacutedo por 03 mL de soluccedilatildeo de stock acrilamida 045 mL de tampatildeo de
concentraccedilatildeo13
094 mL de aacutegua destilada 18 L de SDS 10 135 L de PSA 10 e 2L de
TEMED 99 Colocou-se o pente sobre o gel e deixou-se polimerizar durante cerca de 45
minutos
As amostras para aplicaccedilatildeo no gel de electroforese foram preparadas para uma concentraccedilatildeo de
proteiacutena de 1 mgmL num volume final de 100 L dos quais 25 L correspondem ao tampatildeo da
amostra14
As amostras foram aquecidas a 100ordmC durante 5 minutos Antes da aplicaccedilatildeo das
amostras transferiu-se a cassette para a tina de electroforese agrave qual se adicionou a soluccedilatildeo
tampatildeo de electroforese da tina superior15
e tampatildeo de electroforese da tina inferior16
Aplicaram-
se 10 L das soluccedilotildees de marcadores de peso molecular e 20 L das amostras proteicas
preparadas anteriormente A electroforese decorreu durante cerca de 3h30 min a uma voltagem
de 80 V De seguida transferiu-se o gel para uma tina corando as proteiacutenas durante 1h com
soluccedilatildeo corante17
Descorou-se o gel com soluccedilatildeo descorante18
tendo-se mudado esta soluccedilatildeo ateacute
as bandas se encontrarem bem definidas
Determinou-se a mobilidade relativa das bandas correspondentes aos marcadores de elevado peso
molecular e traccedilou-se a recta de calibraccedilatildeo ndash log(PM) vs log(Rf) onde Rf corresponde agrave
mobilidade relativa Por interpolaccedilatildeo determina-se o PM das proteiacutenas isoladas sabendo a sua
mobilidade relativa Por comparaccedilatildeo com os valores descritos na literatura pode concluir-se
acerca do grau de purificaccedilatildeo
34 Mediccedilatildeo da actividade enzimaacutetica de S1
341 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
13
tampatildeo de concentraccedilatildeo 05 M Tris-HCl pH 68 14
tampatildeo de amostra 320 mM Tris-HCl pH 68 04 M szlig-mercaptoetanol SDS 8 glicerol 15 (vv) e roxo de m-
cresol 0024 15
tampatildeo de electroforese da tina superior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS 16
tampatildeo de electroforese da tina inferior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS acetato de soacutedio
01 M 17
soluccedilatildeo corante ldquoCoomassie Blue R-250rdquo contendo metanol 45 (vv) e aacutecido aceacutetico glacial 10 (vv) 18
soluccedilatildeo descorante metanol 10 (vv) e aacutecido aceacutetico glacial 10 vv)
45
O meacutetodo de Fiske-Subarow modificado consiste na reacccedilatildeo do molibdato de amoacutenio com o
fosfato inorgacircnico em soluccedilatildeo aacutecida com formaccedilatildeo do aacutecido fosfomoliacutebdico Seguidamente daacute-
se a adiccedilatildeo de um agente redutor que provoca a reduccedilatildeo do composto a fosfomolibdato que forma
um complexo de cor azulada cuja absorvacircncia maacutexima ocorre para um comprimento de onda de
660 nm tornando possiacutevel a sua detecccedilatildeo por meacutetodos espectrofotomeacutetricos As actividades Ca2+
-
ATPaacutesica e K+EDTA-ATPaacutesica de S1 foram determinadas atraveacutes deste meacutetodo de acordo com
o descrito por Taussky e Shorr (1953) com mediccedilatildeo da quantidade de fosfato (Pi) libertado
atraveacutes da hidroacutelise de ATP
Para tal obteve-se primeiramente a recta de calibraccedilatildeo com as amostras presentes na Tabela 3II
utilizando uma soluccedilatildeo stock de 5 mM KH2PO4
Tabela 3II ndash Composiccedilatildeo das diferentes amostras utilizadas para determinar a recta de calibraccedilatildeo atraveacutes do meacutetodo
de Fiske-Subbarow modificado
KH2PO4 (L)
([KH2PO4]stock= 5mM)
PO43-
(nmole) H2O (uL)
0 0 1780
5 25 1775
10 50 1770
20 100 1760
40 200 1740
100 500 1680
A cada tubo de Eppendorf adicionaram-se seguidamente 200 L de soluccedilatildeo J19
e 20 L de agente
redutor20
homogeneizando no voacutertex apoacutes cada adiccedilatildeo Incubou-se durante 10 minutos e mediu-
se a Abs660 nm num espectrofotoacutemetro com intervalos de tempo iguais entre cada amostra Traccedilou-
se a recta de calibraccedilatildeo (Abs660 nmvs nmol PO43-
) a partir dos valores obtidos
Para determinar a actividade Ca2+
-ATPaacutesica de S1 prepararam-se tubos de Eppendorf com 400
L de meio de reacccedilatildeo21
25 L CaCl2 ([CaCl2]stock= 100 mM) 25 L ATP ([ATP]stock= 100 mM)
com [S1]ensaios=01 mgmL Perfez-se se necessaacuterio com H2O MilliQ ateacute perfazer um volume de 1
mL Para determinar a actividade K+EDTA-ATPaacutesica utilizou-se uma concentraccedilatildeo idecircntica de
19
Soluccedilatildeo J 5 N H2SO4 25 molibdato de amoacutenio 20
Agente redutor 5 mg aacutecido 1-amino-2-naftol-4-sulfoacutenico 30 mg bissulfito de soacutedio 30 mg sulfito de soacutedio em 26
mL H2O 21
meio de reacccedilatildeo 125 mM Tris-HCl pH 80 0575 M KCl
46
proteiacutena nos ensaios (01 mgmL) e o mesmo meio de reacccedilatildeo Aos tubos de Eppendorf
adicionou-se ainda 123 L de KCl ([KCl]stock = 3M) 25 L de ATP 10 L de EDTA
([EDTA]stock= 100 mM )e o volume de H2O MilliQ necessaacuterio para perfazer 1 mL Em ambas as
situaccedilotildees o uacuteltimo reagente adicionado foi o ATP uma vez que se trata do substrato da reacccedilatildeo
enzimaacutetica
A cineacutetica decorreu a 25ordmC durante 5 minutos para a actividade Ca2+
-ATPaacutesica e durante 3
minutos para a actividade K+EDTA-ATPaacutesica Apoacutes o periacuteodo de incubaccedilatildeo transferiu-se 150
L de amostra para tubos de Eppendorf contendo igual volume de TCA 10 Colocou-se no gelo
e homogeneizou-se no voacutertex Seguidamente centrifugou-se a 1500 g durante 10 minutos e
recolheu-se 200 L de sobrenadante para novos tubos de Eppendorf Adicionou-se a estes 158
mL de H2O MilliQ e 200 L de soluccedilatildeo J Homogeneizou-se no voacutertex e adicionou-se 20 L de
reagente redutor repetindo-se a homogeneizaccedilatildeo Incubou-se durante 10 minutos e mediu-se a
Abs660 nm com o mesmo intervalo de tempo de leitura para cada amostra Por intrapolaccedilatildeo a partir
da recta de calibraccedilatildeo obtida anteriormente determinou-se a quantidade de Pi libertado em cada
ensaio A partir destes resultados determinaram-se as actividades Ca2+
-ATPaacutesica e K+EDTA-
ATPaacutesica do S1 em nmol PO43-
minmg proteiacutena e em s-1
342 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
Da actividade Mg2+
-ATPaacutesica de S1 resulta a formaccedilatildeo dos produtos da reacccedilatildeo de
hidroacutelise do ATP - ADP e Pi Na presenccedila de ADP fosfoenolpiruvato (PEP) e da enzima piruvato
cinase (PK) os produtos resultantes satildeo ATP e piruvato Por acccedilatildeo da enzima lactato
desidrogenase (LDH) este uacuteltimo eacute convertido em lactato ocorrendo simultaneamente a oxidaccedilatildeo
do NADH a NAD+ monitorizada atraveacutes da Abs340 nm Quando o S1 de miosina se encontra no
meio reaccional o ATP eacute hidrolisado formando-se novamente ADP e Pi o que permite a
formaccedilatildeo de um ciclo (fig 21) Se as enzimas se encontrarem em excesso a velocidade de
conversatildeo do NADH eacute limitada pela produccedilatildeo de ADP e Pi Como o ATP eacute regenerado em cada
ciclo a sua concentraccedilatildeo inicial eacute sempre mantida (Bagshaw 1982)
Figura 33ndash Representaccedilatildeo esquemaacutetica do meacutetodo de enzimas acopladas
47
O protocolo utilizado e seguidamente descrito foi adaptado de Cuenda et al (1990 ndash Ver Refordf) A
uma cuvette de 1 mL adicionou-se S1 para uma concentraccedilatildeo final de 005 mgmL 800 L de
meio de reacccedilatildeo22
10 L de PEP ([PEP]stock = 42 mM) 10 L de PK+LDH 11 ([PK+LDH]stock =
1 mgmL) e 15 L de NADH ([NADH]stock=25 mM) Iniciou-se a monitorizaccedilatildeo da reacccedilatildeo
atraveacutes da leitura de Abs340 nm apoacutes a adiccedilatildeo de 20 L de ATP ([ATP]stock=100 mM) A
actividade Mg2+
-ATPaacutesica eacute obtida atraveacutes do declive correspondente agrave oxidaccedilatildeo do NADH Para
a mediccedilatildeo da Mg2+
-ATPase de S1 estimulada por actina adicionou-se F-actina para uma
concentraccedilatildeo final de 2 microM sendo a actividade obtida tambeacutem atraveacutes do declive correspondente
agrave oxidaccedilatildeo do NADH apoacutes a adiccedilatildeo da proteiacutena
35 Interaccedilatildeo entre vanaacutedio e subfragmento-1 de miosina
351 Preparaccedilatildeo das soluccedilotildees de vanaacutedio
As soluccedilotildees de vanadato foram preparadas em aacutegua MilliQ a partir de metavanadato de amoacutenio
(NH4VO3) numa concentraccedilatildeo stock de 50 mM com agitaccedilatildeo magneacutetica e aquecimento (~30 ordmC)
Esta soluccedilatildeo com um valor compreendido entre 6 e 7 designa-se por ldquometavanadatordquo e eacute
constituiacuteda pelas esspeacutecies V1 V2 V4 e V5 (Ref) Acidificando a soluccedilatildeo de metavanadato por
adiccedilatildeo de HCl ateacute um valor de pH proacuteximo de 4 observa-se uma mudanccedila de cor de amarelo
claro para laranja indicativa da formaccedilatildeo da espeacutecie decameacuterica de vanadato De seguida e
imediatamente antes da realizaccedilatildeo dos ensaios descritos o valor do pH da soluccedilatildeo de
decavanadato foi aumentado para cerca de 75 por adiccedilatildeo de NaOH A soluccedilatildeo (2) designada
por ldquometavanadatordquo natildeo foi posteriormente alterada ateacute agrave sua adiccedilatildeo para a realizaccedilatildeo dos ensaios
As soluccedilotildees foram armazenadas a 4ordmC e mantidas em gelo aquando da sua utilizaccedilatildeo
A soluccedilatildeo de V(IV) foi preparada em condiccedilotildees anaeroacutebias por diluiccedilatildeo de sulfato de vanadil
(VOSO4) para uma concentraccedilatildeo final de 5 mM em H2O MilliQ saturada com N2 gasoso
352Anaacutelise por espectroscopia de UV-Vis
A concentraccedilatildeo da espeacutecie decameacuterica em soluccedilatildeo depende do periacuteodo de tempo apoacutes a diluiccedilatildeo
no meio de reacccedilatildeo apropriado (Ref) A velocidade de desoligomerizaccedilatildeo da espeacutecie decameacuterica
pode ser analisada por espectroscopia de absorccedilatildeo na regiatildeo UVVis uma vez que esta espeacutecie
apresenta duas bandas caracteriacutesticas na regiatildeo do visiacutevel (a 358 e 400 nm) que natildeo se verifica
em quaisquer outras espeacutecies oligomeacutericas As bandas supramencionada sapresentam-se bem
22
Meio de reacccedilatildeo 3125 mM Tris-HCl pH 71 375 mM MgCl2
48
resolvidas atraveacutes da obtenccedilatildeo da derivada de 2ordf ordem do espectro apresentando a forma de 2
vales com miacutenimos centrados nos comprimentos de onda supracitados com dependecircncia linear
relativamente agrave concentraccedilatildeo de decavanadato total (Ramos et al 2006)
Assim a estabilidade da espeacutecie decameacuterica apoacutes diluiccedilatildeo da soluccedilatildeo de decavanadato no meio de
reacccedilatildeo dos ensaios (tampatildeo D) para uma concentraccedilatildeo final de 1 mM em vanadato total foi
caracterizada atraveacutes da leitura da Abs400 em funccedilatildeo do tempo tendo-se determinado qual a
ordem de reacccedilatildeo e o tempo de meia-vida da mesma Com base em estudos anteriores acerca da
estabilidade do V10 na presenccedila de diferentes proteiacutenas (Ramos et al 2006) determinou-se
igualmente o tempo de meia vida apoacutes a adiccedilatildeo de 20 microM de S1 de miosina ao meio reaccional
353 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
Os espectros de RMN do isoacutetopo 51
V foram realizados num espectroacutemetro Bruker AM-400 MHz
equipado com uma sonda multinuclear inversa de 5 mm Em tubos de RMN de 5 mm de diacircmetro
preparou-se soluccedilotildees de 500 L de deca- ou metavanadato correspondentes a um intervalo de
concentraccedilotildees de 1 a 10 mM suplementadas com 10 D2O Os espectros foram adquiridos agrave
temperatura ambiente com uma largura espectral de 23 923 Hz um tempo de aquisiccedilatildeo de 0086
s tempo de relaxaccedilatildeo de 001 s e uma sequecircncia de pulsos a 90ordm Utilizou-se a transformada de
Fourier com alargamento das bandas de 20Hz para o processamento dos espectros Os desvios
quiacutemicos das ressonacircncias de 51
V foram obtidos em relaccedilatildeo agrave espeacutecie VOCl3cuja frequecircncia de
ressonacircncia eacute considerada o padratildeo sendo todas as outras referenciadas a partir desta em ppm
As aacutereas dos sinais do RMN correspondentes agraves espeacutecies oligomeacutericas presentes foram obtidas
por integraccedilatildeo dos sinais As concentraccedilotildees relativas das espeacutecies foram calculadas com recurso agrave
equaccedilatildeo
onde A[Vx] corresponde agrave aacuterea referente agrave ressonacircncia da espeacutecie Vx com nordm de agregaccedilatildeo x
A[V]total agrave soma total das aacutereas medidas e [V] a concentraccedilatildeo total de vanaacutedio no ensaio
A teacutecnica de 51
V-RMN foi anteriormente aplicada em estudos de ligaccedilatildeo com S1 de miosina
(Ref) tendo sido demostrado que entre as espeacutecies V1 V2 V4 e V5 a espeacutecie tetrameacuterica possui a
maior afinidade para a miosina (Cremo et al) O desaparecimento ou alargamento dos sinais pode
implicar a ligaccedilatildeo de um dos oxovanadatos laacutebeis agrave miosina sem indicaccedilatildeo de qual uma vez que
49
se processa um equiliacutebrio entre eles (Crans 1990) Assim e tendo em conta as indicaccedilotildees de que
a espeacutecie V10 eacute distinta das restantes no que diz respeito agrave magnitude da interacccedilatildeo com a miosina
e inibiccedilatildeo da actividade ATPaacutesica estimulada por actina (Aureliano 2000 Tiago et al 2002)
torna-se fundamental investigar a interacccedilatildeo das diferentes espeacutecies de vanadato com o S1 de
miosina atraveacutes da teacutecnica de 51
V-RMN
Para tal obtiveram-se espectros de 51
V-RMN resultantes da titulaccedilatildeo de uma soluccedilatildeo 5 mM de
decavanadato total com concentraccedilotildees crescentes de S1 de miosina (0 5 10 15 e 20 microM) em
tampatildeo D Realizou-se ainda ensaios na presenccedila de 2 mM de decavanadato e 2 mM de ATP na
presenccedila e na ausecircncia de S1 de miosina de modo a verificar o efeito que estes ligandos naturais
da enzima teria na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o subfragmento-1
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
Os grupos sulfidrilo (-SH) das cisteiacutenas satildeo resiacuteduos de aminoaacutecidos extremamente sensiacuteveis agrave
oxidaccedilatildeo resultando na sua oxidaccedilatildeo (Dalle-Donne et al 1999) Logo a quantificaccedilatildeo destes
resiacuteduos eacute bastante importante nos estudos de interacccedilatildeo com vanaacutedio jaacute que os resiacuteduos de Cys
com ecircnfase para o SH1 e o SH2 satildeo bastante importantes na monitorizaccedilatildeo das modificaccedilotildees
funcionais da regiatildeo da cabeccedila
O procedimento seguidamente apresentado foi originalmente descrito por Gutieacuterrez-Merino
(1987)) Consiste na reacccedilatildeo entre os grupos ndashSH natildeo oxidados das proteiacutenas e o reagente de
Ellman (DTNB -55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico)) - formando-se o aniatildeo CNT (aniatildeo 3-
carboxilato-4-nitrotiofenolato) numa estequiometria de 11 Este apresenta um maacuteximo de
absorvecircncia a 412 nm (Robyt 1990) Assim apoacutes a incubaccedilatildeo da proteiacutena na presenccedila do metal
a quantidade de cisteiacutenas reduzidas por proteiacutena determina-se atraveacutes dos valores de Abs412
Deste modo colocou-se num tubo de Eppendorf o volume apropriado de S1 dialisado para que a
concentraccedilatildeo no ensaio fosse de 1 mgmL Adicionou-se o volume apropriado de decavanadato
para que a sua concentraccedilatildeo fosse de 0 10 50 100 250 500 ou 1000 M Perfez-se com soluccedilatildeo
D ateacute um volume final de 1 mL e incubou-se a 25ordmC por 20 minutos Apoacutes este periacuteodo colocou-
se 780 L da mistura reaccional numa cuvette de quartzo e adicionou-se 220 L de soluccedilatildeo K23
23
Tampatildeo K Tampatildeo fosfatohidrogenofosfato de soacutedio 025 M pH 72
50
Adicionou-se 15 L de DTNB (em soluccedilatildeo K [DTNB]stock=100 mM) ao meio reaccional e
registou-se o valor de absorvecircncia medido equivalente agraves cisteiacutenas ldquoraacutepidasrdquo (SH1 e SH2
expostos ao solvente) Adicionou-se 001 g de SDS que provoca a desnaturaccedilatildeo da proteiacutena e
consequentemente a exposiccedilatildeo de todos os resiacuteduos de cisteiacutenas que se encontravam ateacute aiacute
inacessiacuteveis no interior da proteiacutena (cisteiacutenas totais) Estes resiacuteduos podem assim reagir com o
DTNB
Estimou-se o nuacutemero de resiacuteduos de cisteiacutenas em todas as amostras com recurso ao coeficiente de
extinccedilatildeo molar () do DTNB (ver ponto 3 dos Anexos) que foi determinado atraveacutes da titulaccedilatildeo
de L-Cisteiacutena com intervalo de concentraccedilotildees compreendidos entre 0 e 50 microM na presenccedila de
25microM de DTNB em tampatildeo D Os valores de absorvecircncia foram obtidos para um comprimento de
onda de 412 nm
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
3621 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
A teacutecnica mais eficiente de anaacutelise da interacccedilatildeo entre o iatildeo vanadilo e proteiacutenas como a
calmodulina e a transferrina entre outras permite a obtenccedilatildeo raacutepida de informaccedilatildeo no que toca agrave
conformaccedilatildeo e amplitude da referida interacccedilatildeo
Os espectros de Ressonacircncia Paramagneacutetica Electroacutenica (RPE) da espeacutecie paramagneacutetica V(IV)
foram realizados a 70K num espectroacutemetro Bruker EMX-300 X-band Todas as soluccedilotildees
utilizadas e amostras foram preparadas em anaerobiose numa cacircmara de luvas de modo a evitar a
reduccedilatildeo de V(IV) a V(V) Em tubos capilares com 02 mL de amostra manteve-se a concentraccedilatildeo
de S1 a 20 M e variou-se a concentraccedilatildeo de vanadilo (0 25 5 10 15 25 50 75 100 150
200 500 e 1000 M [VOSO4]stock=5 mM) em tampatildeo D Apoacutes obtenccedilatildeo dos espectros estes
foram tratados atraveacutes do software SimFonia e WinEPR tendo-se obtido um graacutefico com a
variaccedilatildeo da intensidade do sinal de RPE em funccedilatildeo da concentraccedilatildeo de sulfato de vanadilo
presente Realizaram-se ainda simulaccedilotildees de espectros obtidos com 20 microM de S1 e 1000 microM de
V(IV) com vista agrave obtenccedilatildeo dos ligandos provaacuteveis do vanadilo atraveacutes da anaacutelise da constante
de acoplamento hiperfina (A) e do factor g (g)
4 Conclusotildees gerais
Apesar de jaacute ter sido demonstrado que o vanaacutedio inibe a Na+K
+-ATPase da miosina a funccedilatildeo
deste elemento no tecido muacutesculo esqueleacutetico continua largamente incompreendida A enorme
complexidade quiacutemica deste metal especialmente no que concerne agrave oligomerizaccedilatildeo e reacccedilotildees
51
redox constitui um obstaacuteculo agrave compreensatildeo do comportamento do vanaacutedio nos organismos
vivos Assim a caracterizaccedilatildeo da quiacutemica deste elemento eacute de grande utilidade antes de iniciar o
estudo da sua interacccedilatildeo com o presente sistema
A anaacutelise por espectroscopias de 51
V-RMN e de UV-Vis de soluccedilotildees de vanadato diluiacutedas nos
meios de reacccedilatildeo utilizados no presente relatoacuterio (10 mM HEPES pH 70 25 mM KCl) mostrou
claramente que a espeacutecie decameacuterica (V10) possui um tempo de vida suficientemente longo para a
determinaccedilatildeo do seu efeito isolado ao contraacuterio do que se verifica com as espeacutecies mono- di-
tetra- e pentameacutericas (V1 V2 V4 e V5) Por outro lado a anaacutelise verificou ainda que apoacutes diluiccedilatildeo
das soluccedilotildees de decavanadato no meio de reacccedilatildeo coexistem as espeacutecies V10 e V1 pelo que os
efeitos putativos desta uacuteltima deveratildeo ser subtraiacutedos aquando da monitorizaccedilatildeo dos efeitos da
espeacutecie decameacuterica
O vanaacutedio (V) (vanadato) inibe as enzimas que catalizam reacccedilotildees de transferecircncia de grupos
fosfato como a ATPase da miosina [Goodno 1979] devido agrave sua analogia estrutural com o iatildeo
fosfato Na sua utilizaccedilatildeo como inibidor enzimaacutetico natildeo eacute tida em conta a contribuiccedilatildeo individual
das espeacutecies oligomeacutericas considerando-se normalmente a espeacutecie V1 como inibidora efectiva
[Goodno e Taylor 1982] No presente relatoacuterio verificou-se por espectroscopia de 51
V-RMN que
outros oligoacutemeros de vanadato interagem com o S1 de miosina tendo as espeacutecies V10 e V4 os
maiores niacuteveis de afinidade A sua interacccedilatildeo envolve forccedilas electrostaacuteticas e eacute afectada
negativamente por ligandos naturais de enzima como eacute o caso do ATP
A exposiccedilatildeo do S1 de miosina (005 mgmL) a decavanadato conduz agrave inibiccedilatildeo do capacidade da
proteiacutena de estimular a actividade Mg2+
-ATPaacutesica com um valor de IC50 de 266 plusmn 07 M
decavanadato sendo o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM
decavanadato (80) Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM decavanadato sendo
o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM decavanadato (60)
exercendo um efeito de impedimento agrave ligaccedilatildeo entre o vanaacutedio e o S1
A uma concentraccedilatildeo de 27 microM decavanadato verifica-se a oxidaccedilatildeo de um dos resiacuteduos de Cys
de S1 de miosina seguida pela oxidaccedilatildeo gradual de outro resiacuteduo de Cys ateacute agrave concentraccedilatildeo de
1000 microM Ambos os resiacuteduos oxidados natildeo se encontravam expostos ao solvente utilizado A
oxidaccedilatildeo dos resiacuteduos permite sugerir que esta modificaccedilatildeo oxidativa sofrida pelo S1 contribui
para a diminuiccedilatildeo da sua actividade de ATPase
52
Por espectroscopia de RPE revelaram-se os possiacuteveis ligandos equatoriais do vanaacutedio aquando da
sua interacccedilatildeo com o S1 de miosina como sendo duas moleacuteculas de aacutegua um ligando contendo o
grupo R-OH e outro contendo o grupo R=NR
Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados de
oxidaccedilatildeo (IV) e (V) no S1 de miosina para concentraccedilotildees facilmente alcanccedilaacuteveis no ambiente
terrestre
53
5 Estudos futuros
No sentido do melhor esclarecimento das alteraccedilotildees moleculares e estruturais que ocorrem no
subfragmento 1 quando exposta ao vanaacutedio que conduzem a uma diminuiccedilatildeo da sua funccedilatildeo
sugere-se
- Quantificaccedilatildeo das cisteiacutenas reduzidas de S1 na presenccedila de metavanadato
- Determinaccedilatildeo das actividades Ca2+
- e K+-ATPaacutesica do S1 de miosina na presenccedila de
decavanadato
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina estimulada por actina
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina na presenccedila de
metavanadato
- Realizaccedilatildeo de RMN 2D com variaccedilatildeo das concentraccedilotildees das soluccedilotildees de decavanadato e
metavanadato na presenccedila de S1
-Mutageacutenese dos resiacuteduos de Cys doS1 de miosina de modo a verificar quais os resiacuteduos que
sofrem oxidaccedilatildeo na presenccedila de decavanadato
- Determinaccedilatildeo por calorimetria (DSC Differential Scanning Calorimetry) da variaccedilatildeo da
entalpia (∆H) e da temperatura de desnaturaccedilatildeo do S1 de miosina na presenccedila de vanaacutedio uma
vez que estas propriedades caracteriacutesticas de cada proteiacutena satildeo alteradas apoacutes ocorrecircncia de
modificaccedilotildees estruturais
- Anaacutelise por SDS-PAGE de S1 de miosina apoacutes tratamento com vanaacutedio de modo a determinar
quais os polipeacuteptidos eventualmente formados apoacutes clivagem da proteiacutena
54
6 Referecircncias bibliograacuteficas
- Sellers J Myosins 2nd Edition Oxford University Press 1999
- L Stryer inBiochemistry (Ed WH Freeman 2005 pp
Refordfas Materiais e Meacutetodos
[A] S S Margossian SLowey Methods Enzymol1982 85 55-71
[B] JD Pardue JA Spudich Methods Enzymol 1982 85 164-181
[C] M Bradford Anal Biochem1976 72 248-254
[D] H H Taussky E Shorr J Biol Chem 1953 202 675-685
[E] C R Bagshaw in Outline Studies in Biology ndash Muscle Contraction Chapman and Hall Ed
1982
[F] A Cuenda et al Eur J Biochem1990 194 663-670
55
Anexos
1 Determinaccedilatildeo da recta de calibraccedilatildeo meacutetodo de Bradford
Figura1ndashRepresentaccedilatildeo graacutefica da recta de calibraccedilatildeo (Abs595 nm vs [BSA]) utilizada para o meacutetodo de Bradford
2 Determinaccedilatildeo da recta calibraccedilatildeo para a electroforese em condiccedilotildees desnaturantes
Figura 2 ndash Representaccedilatildeo graacutefica da recta de calibraccedilatildeo (log(Peso Molecular) vs Rf dos marcadores de peso
molecular) onde Rf indica determinada atraveacutes do padratildeo de peso molecular
(poccedilo (a))
3Caacutelculo do coeficiente de extinccedilatildeo molar () para a quantificaccedilatildeo de cisteiacutenas reduzidas
atraveacutes do meacutetodo de Ellman
Traccedilou-se a recta de Abs412 nm meacutedia vs [L-Cisteiacutena] de modo a determinar o valor do coeficiente
de extinccedilatildeo molar definido como o declive daquela recta Assim =8400 M-1
cm-1
como
podemos verificar na Fig 3
A595 nm = 00546[BSA] + 00209 Rsup2 = 099051
Ab
s 59
5 n
m
[BSA] (mgmL)
log PM = -1177 Rf + 2247 Rsup2 = 098957
log
PM
Mobilidade relativa (Rf)
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero

5
Iacutendice
1 Introduccedilatildeo
11 Muacutesculo esqueleacutetico
111 Miosina
112 Actina
113 Mecanismos moleculares da contracccedilatildeo muscular
1131 Regulaccedilatildeo da contracccedilatildeo muscular
12 Vanaacutedio
121 Propriedades quiacutemicas
1211 Vanaacutedio (IV)
1212 Vanaacutedio (V)
122 Metabolismo e efeito na actividade enzimaacutetica
13 Vanaacutedio e muacutesculo esqueleacutetico
14 Objectivos do projecto
2 Discussatildeo de Resultados
21 Caracterizaccedilatildeo das proteiacutenas isoladas
211 Anaacutelise do estado de pureza
212 Determinaccedilatildeo da concentraccedilatildeo proteica
2121 Subfragmento-1 de miosina
2122 Actina
22 Mediccedilatildeo da actividade enzimaacutetica de S1 de miosina
221 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de Fiske-
Subarow modificado
222 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
23 Preparaccedilatildeo e caracterizaccedilatildeo das soluccedilotildees de vanaacutedio
231 Anaacutelise da composiccedilatildeo das soluccedilotildees de vanadato por espectroscopia de 51V-RMN efeito da
concentraccedilatildeo da espeacutecie
232 Estabilidade das soluccedilotildees de decavanadato anaacutelise por espectroscopia de absorccedilatildeo UVVis
24 Interacccedilatildeo de S1 com vanaacutedio(V)
241 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
242 Actividade Mg2+
-ATPaacutesica de S1 de miosina na presenccedila de decanadato
243 Quantificaccedilatildeo de cisteiacutenas reduzidas pelo meacutetodo de Ellman
25 Interacccedilatildeo de S1 com vanaacutedio(IV)
251 Anaacutelise por espectroscopia de Ressonacircncia Paramagneacutetica Electroacutenica
3 Procedimento Experimental
31 Reagentes
32 Meacutetodos
321 Isolamento e purificaccedilatildeo das proteiacutenas de muacutesculo esqueleacutetico de coelho
3211 Purificaccedilatildeo da miosina preparaccedilatildeo de Subfragmento 1 por digestatildeo quimotriacutepsica da
miosina
32111 Purificaccedilatildeo da miosina
32112 Preparaccedilatildeo de Subfragmento-1 de miosina por digestatildeo quimotriacutepsica
3212 Isolamento e purificaccedilatildeo de actina
32121Obtenccedilatildeo do ldquopoacute de acetonardquo
32122 Purificaccedilatildeo da actina a partir de ldquopoacute de acetonardquo
331 Quantificaccedilatildeo das proteiacutenas isoladas
3311 Quantificaccedilatildeo pelo meacutetodo de Bradford
6
3312 Quantificaccedilatildeo pelo meacutetodo directo
332 Electroforese em condiccedilotildees desnaturantes (SDS-PAGE)
34 Mediccedilatildeo da actividade enzimaacutetica de S1
341 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de Fiske-
Subarow modificado
342 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
35 Preparaccedilatildeo e caracterizaccedilatildeo das soluccedilotildees de vanaacutedio por Espectroscopia UV-Visiacutevel e
Ressonacircncia Magneacutetica Nuclear
351 Preparaccedilatildeo das soluccedilotildees de vanadato
352 Anaacutelise por UV-Vis para determinaccedilatildeo da ordem da reacccedilatildeo
353 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
362 1 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
363 Interacccedilatildeo de S1 com V (V) por Ressonacircncia Magneacutetica Nuclear (RMN)
4 Conclusotildees gerais
5 Estudos futuros
6 Referecircncias bibliograacuteficas
7
1 Introduccedilatildeo
11 Muacutesculo esqueleacutetico
A capacidade de locomoccedilatildeo eacute uma das caracteriacutesticas fundamentais da maioria dos organismos
vivos A contracccedilatildeo muscular um dos fenoacutemenos que a torna possiacutevel apresenta grande
complexidade e tem vindo a ser utilizado como modelo para a compreensatildeo de sistemas menos
especializados de mobilidade celular [1]
No que diz respeito agrave sua fisiologia morfoloacutegica os muacutesculos dos vertebrados dividem-se em
lisos e estriados Estes uacuteltimos devem o seu nome agrave alternacircncia de zonas claras e escuras
observaacuteveis ao microscoacutepio electroacutenico de que resulta um aspecto estriado Os muacutesculos
estriados podem ainda ser divididos em duas subclasses muacutesculos esqueleacuteticos e cardiacuteacos [2]
consoante a forma celular (respectivamente longitudinal ou ramificada) ou tipo de contracccedilatildeo das
ceacutelulas que o compotildeem (respectivamente voluntaacuteria e involuntaacuteria) [3]
Os muacutesculos estriados esqueleacuteticos satildeo assim designados devido agrave sua localizaccedilatildeo no
organismo ndash ligados aos ossos da coluna vertical braccedilos e pernas Satildeo constituiacutedos por feixes de
ceacutelulas multinucleadas (miofibras) que contecircm miofibrilas paralelas de cerca de 1 m de
diacircmetro associadas agrave contracccedilatildeo voluntaacuteria raacutepida e forte em actividades de coordenaccedilatildeo
complexa [4] Cada feixe apresenta um diacircmetro compreendido entre 20 e 100 m e um
comprimento maacuteximo que pode atingir vaacuterios centiacutemetros (Fig 11)
Figura 11 ndash Organizaccedilatildeo hieraacuterquica geral do muacutesculo-esqueleacutetico (adaptado de Purves et al 2003 [3])
No muacutesculo-esqueleacutetico as zonas claras e escuras supramencionadas correspondem a zonas de
menor e maior densidade electroacutenica respectivamente As zonas claras constituem as bandas I (de
8
ldquoisotropiardquo ndash polarizaccedilatildeo uniforme em todas as direcccedilotildees) enquanto as zonas escuras constituem
as bandas A (de ldquoanisotropiardquo ndash polariza a luz visiacutevel) A banda I encontra-se dividida
transversalmente por uma linha escura ndash linha Z de densidade electroacutenica e refracccedilatildeo superiores
No interior de cada banda A existe uma zona transversal menos densa ndash zona H ndash rica em enzimas
importantes para o metabolismo celular No centro desta zona existe uma linha perpendicular
mais escura ndash linha M [2]
A unidade fundamental da contracccedilatildeo muscular eacute designada por sarcoacutemero (Fig 12) Encontra-se
repetida de forma regular ao longo das miofibrilas apresentando um comprimento de 25 a 3 m
aquando do relaxamento muscular diminuindo aquando da contracccedilatildeo muscular
Figura 12 ndash Representaccedilatildeo de fotomicrografia electroacutenica e esquemaacutetica do sarcoacutemero Os filamentos grossos e
finos satildeo maioritariamente constituiacutedos por miosina e actina respectivamente (Adaptado de Stryer 2002 [2])
As secccedilotildees de corte do sarcoacutemero datildeo conta da presenccedila de dois tipos de filamentos proteicos ndash os
filamentos grossos cujo diacircmetro eacute de 15 nm constituiacutedos sobretudo por miosina e os filamentos
finos com um diacircmetro de 9 nm constituiacutedos sobretudo por actina tropomiosina e pelo complexo
de troponina [2]
111 Miosina
Actualmente o termo miosina refere-se a uma superfamiacutelia diversa de motores moleculares
capazes de hidrolisar ATP e de se ligarem reversivelmente agrave actina A classificaccedilatildeo da
superfamiacutelia das miosinas nas 15 classes distintas faz-se atraveacutes de homologia entre as diferentes
sequecircncias [A] Eacute tambeacutem possiacutevel a classificaccedilatildeo por subclasses das quais se destaca a subclasse
4 proveniente do muacutesculo-esqueleacutetico e cardiacuteaco de vertebrados Este estudo focar-se-aacute apenas
nas miosinas de classe II (miosina-II) presente nos filamentos grossos do muacutesculo estriado
esqueleacutetico [B] e em ceacutelulas natildeo musculares A funccedilatildeo da miosina-II depende da sua localizaccedilatildeo
celular e tecidular
9
A miosina-II eacute constituiacuteda por duas cadeias polipeptiacutedicas idecircnticas denominadas cadeias pesadas
de miosina (HC) enroladas entre si e cujo peso molecular (PM) estaacute compreendido entre 171 e
241 kDa) Estruturalmente as HC dividem-se em 3 domiacutenios funcionais - cabeccedila pescoccedilo e
cauda como se pode observar na Fig 13 Agraves cabeccedilas ou zonas globulares estatildeo associados dois
pares de cadeias leves (peso molecular entre 15 e 30 kDa) que se dividem em dois tipos
essencial (ELC) e regulatoacuterio (RLC)
Figura 13 ndash Estrutura esquemaacutetica da moleacutecula de miosina-II Destacam-se as duas cabeccedilas globulares e a cauda
filamentosa ligadas entre si por uma zona flexiacutevel denominada pescoccedilo As cadeias leves (a verde e vermelho)
encontram-se associadas agraves cabeccedilas globulares
As cabeccedilas globulares contendo os locais de ligaccedilatildeo agrave actina e aos nucleoacutetidos mantecircm os seus
elementos estruturais baacutesicos conservados em todas as classes sendo estes constituiacutedos por 720-
80 aa A regiatildeo do pescoccedilo eacute constituiacuteda por uma longa heacutelice pertencente agrave cadeia pesada
que se encontra estabilizada atraveacutes da ligaccedilatildeo a subunidades de cadeia leve A estrutura do
pescoccedilo envolvida na ligaccedilatildeo agraves cadeias leves eacute denominada de motivo IQ cuja estrutura
consensual eacute IQXXXRGXXXR1 O pescoccedilo serve como extensatildeo do comprimento da cabeccedila de
modo a actuar como alavanca durante a contracccedilatildeo muscular [A]A regiatildeo da cauda ou acircncora
apresenta bastante divergecircncia no que respeita agrave sua sequecircncia estrutura e funccedilatildeo No entanto
pensa-se que a sua funccedilatildeo centrar-se-aacute na autoassociaccedilatildeo das vaacuterias moleacuteculas de miosina-II
atraveacutes da regiatildeo de coiled-coil Esta confere assimetria agrave moleacutecula e permite a formaccedilatildeo de
filamentos grossos capazes de translocar filamentos de actina [A]
Na regiatildeo da cabeccedila da miosina existem 2 regiotildees extremamente sensiacuteveis agrave proteoacutelise A
clivagem em ambos os locais resulta na produccedilatildeo de trecircs fragmentos um compreendendo o N-
terminal (fragmento 25 kDa) outro a regiatildeo central (50 kDa) e um terceiro o C-terminal (20 kDa)
(Fig 14) Estes fragmentos dissociam-se em condiccedilotildees desnaturantes e natildeo constituem domiacutenios
independentes da cabeccedila mas provavelmente loops flexiacuteveis uma vez que natildeo foram observadas
na estrutura cristalina Os vaacuterios fragmentos obtidos em diferentes condiccedilotildees foram cruciais para
a elucidaccedilatildeo das vaacuterias funccedilotildees da moleacutecula
A regiatildeo compreendida entre os fragmentos 25 e 50 kDa (loop 1) localiza-se directamente por
1 I = Isoleucina Q= Glutamina X = qualquer dos 20 aminoaacutecidos padratildeo R= Arginina G= Glicina
10
cima do local de ligaccedilatildeo ao nucleoacutetido A regiatildeo entre os fragmentos de 50 e 20 kDa (loop 2)
encontra-se perto da intersecccedilatildeo entre os domiacutenios da cabeccedila e do pescoccedilo e deveraacute interagir com
a actina uma vez que a ligaccedilatildeo desta a protege da proteoacutelise Na ausecircncia do loop 2 a miosina
liga mais fracamente agrave actina do que no caso da miosina intacta A sequecircncia primaacuteria de ambos
os loops apresenta conservaccedilatildeo reduzida sendo provaacutevel que sejam responsaacuteveis por muita da
diversidade das diferentes propriedades enzimaacuteticas presentes em diferentes isoformas
No fragmento de 20 kDa existem dois grupos sulfidrilo Cys-707 e -697 usualmente
denominados de SH1 e SH2 devido agrave ordem da sua reactividade quiacutemica Na presenccedila de
nucleoacutetidos pode ocorrer o ldquocross-linkingrdquo entre estes dois grupos incluindo a oxidaccedilatildeo a uma
ponte dissulfureto
Devido agrave insolubilidade da miosina-II nativa a estrateacutegia adoptada para os estudos com esta
proteiacutena eacute frequentemente a sua clivagem em fragmentos funcionais separaacuteveis Um dos locais de
clivagem ocorre tipicamente a 130 kDa do N-terminal produzindo a meromiosina pesada (HMM)
e a meromiosina leve (LMM) [C] A HMM eacute formada pelas 2 cabeccedilas e pescoccedilo e apresenta
propriedades semelhantes agrave miosina sem digestatildeo (ligaccedilatildeo e translocaccedilatildeo de actina e hidroacutelise de
MgATP)
A HMM pode ser novamente clivada atraveacutes de uma reacccedilatildeo de proteoacutelise produzindo os
subfragmentos 1 (S1) e 2 (S2)O S1 (90 kDa) eacute enzimaticamente activo sendo constituiacutedo pelos
domiacutenios da cabeccedila e do pescoccedilo e apresentando associaccedilatildeo agraves duas cadeias leves ou apenas agrave
ELC (Fig 14)Em alternativa o S1 pode ser produzido por clivagem directa da miosina intacta
formando-se tambeacutem um fragmento em forma de cilindro que consiste nas regiotildees S2 e LMM
(Ref)Utiliza-se extensivamente o S1 para estudos cineacuteticos e de ligaccedilatildeo agrave actina uma vez que a
sua solubilidade simplifica o planeamento da experiecircncia e a subsequente interpretaccedilatildeo dos dados
[A]
Figura 14- Estrutura do subfragmento-1 de miosina de muacutesculo esqueleacutetico com ecircnfase nos domiacutenios de 50 kDa
central (a vermelho) 25 kDa no N-terminal (a verde) e 20 kDa no C-terminal (a azul) As cadeias leves essenciais
encontram-se a amarelo e as cadeias leves regulatoacuterias a lilaacutes Os locais de ligaccedilatildeo agrave actina e ao ATP estatildeo
igualmente destacados (adaptado de Holmes e Geeves 1999 [D])
11
112 Actina
O termo ldquoactinardquo refere-se a uma famiacutelia extremamente conservada de proteiacutenas citoplasmaacuteticas
presente em todos os eucariotas Esta proteiacutena natildeo se encontra nos procariotas [E] A maioria das
actinas possui peso molecular proacuteximo de 42 kDa outra evidecircncia do seu grau elevado de
conservaccedilatildeo no que concerne agrave sequecircncia e ao nuacutemero de aminoaacutecidos Os eucariotas superiores
apresentam um grande nuacutemero de isoformas de actina que por sua vez se encontram codificados
por um ainda maior nuacutemero de genes (Ref) A expressatildeo das isoformas eacute diferencial consoante o
tecido e sua especializaccedilatildeo pelo que deveratildeo possuir funccedilotildees e mecanismos direccionais
diferentes (Ref) Foi ainda observado que os mRNA das diferentes isoformas apresentam uma
distribuiccedilatildeo heterogeacutenea nas ceacutelulas de forma semelhante aos seus produtos proteicos [F] Estes
dados suportam a hipoacutetese supramencionada do direccionamento das isoformas de actina para um
determinado local na ceacutelula de acordo com as suas funccedilotildees
A actina presente no citoesqueleto eacute responsaacutevel por processos como o mecanismo contraacutectil
fagocitose manutenccedilatildeo e alteraccedilatildeo da estrutura celular locomoccedilatildeo transporte de organelos
regulaccedilatildeo do transporte de solutos entre outras Nos eucariotas superiores estaacute ainda envolvida
no transporte de gracircnulos de pigmento retinoacuteide [G] transporte de gracircnulos no axoplasma [H] e
secreccedilatildeo entre outros [E] Actualmente a isoforma da actina (-actina) eacute a melhor estudada
encontrando-se envolvida no mecanismo contraacutectil [E]
Os filamentos de -actina tecircm cerca de 1m de comprimento e 80 Å Consistem em duas cadeias
de actina filamentosa (F-actina) organizados em heacutelice [I] (Holmes et al 1990) sobre os quais
se encontram outras proteiacutenas ndash a troponina (C I e T) e a tropomiosina (Azevedo 1999) (Figura
15) A troponina eacute filamentosa e localiza-se nas depressotildees da heacutelice de F-actina conferindo
rigidez aos filamentos finos Por sua vez a tropomiosina eacute essencial na regulaccedilatildeo do mecanismo
contraacutectil
Figura 15 ndash Modelo estrutural dos filamentos de F-actina contendo a tropomiosina e o complexo de troponina (I C
e T) (Adaptado de Susana Ramos 2005 [J])
12
A F-actina eacute formada atraveacutes da polimerizaccedilatildeo de subunidades globulares de actina (G-actina) A
G-actina eacute composta por 2 domiacutenios de tamanho semelhante separados pelos bolsos de ligaccedilatildeo(1)
de catiotildees divalentes e(2) de nucleoacutetidos [KLM] Estes por sua vez estatildeo subdivididos noutros
dois de menores dimensotildees (1-4) (Fig 16) cada um envolvido nas interacccedilotildees actina-actina ou
na formaccedilatildeo do topo de ligaccedilatildeo do nucleoacutetido As regiotildees N-terminal e C-terminal estatildeo proacuteximas
uma da outra no subdomiacutenio 1 [E]
Figura 16 ndash Estrutura do monoacutemero de actina Encontram-se representados os subdomiacutenios 1 a 4 e a vermelho a
moleacutecula de ATP junto do respectivo local de ligaccedilatildeo (Protein Data Bank ID 1j6z)
113 Mecanismos moleculares da contracccedilatildeo muscular
A contracccedilatildeo muscular ocorre atraveacutes do deslizamento dos filamentos finos (de actina) por entre
os filamentos grossos (de miosina) conseguida pela reacccedilatildeo de hidroacutelise de ATP e regulada pela
tropomiosina e pelo complexo de troponina em resposta a impulsos nervosos Este mecanismo foi
inicialmente proposto por Huxley-Niedergerke [N] e Huxley-Hanson [O] ambos em 1954 A base
experimental de ambos os estudos foi a inexistecircncia de variaccedilatildeo do tamanho dos filamentos
durante a contracccedilatildeo A banda A apresenta sempre as mesmas dimensotildees por outro lado a banda
I a zona H diminuem consideravelmente aquando da contracccedilatildeo (Figs 12 e 17)
Figura 17 ndash Modelo da contracccedilatildeo muscular ndash deslizamento dos filamentos finos (azuis) relativamente aos
filamentos grossos (rosa) (Adaptado de Stryer 2002 [2])
13
Na ausecircncia de ATP e supondo a ausecircncia de inibiccedilatildeo da interacccedilatildeo actina-miosina as proteiacutenas
encontram-se fortemente associadas formando o complexo actomiosina (AM) (Figura 15)
Nestas condiccedilotildees as cabeccedilas de miosina formam um acircngulo de 45ordm com o eixo dos filamentos
finos de actina [P]
Na presenccedila de ATP o complexo AM dissocia-se jaacute que a afinidade do complexo M-ATP para a
actina eacute 105 vezes inferior agrave afinidade da miosina para a actina O ATP liga-se agrave miosina
ocorrendo seguidamente a reacccedilatildeo de hidroacutelise de ATP Os produtos daiacute resultantes ndash ADP e Pi -
permanecem ligados agrave miosina formando o complexo M-ADP-Pi Com a ligaccedilatildeo subsequente agrave
actina forma-se o complexo AM-ADP-Pi no qual as cabeccedilas de miosina apresentam um acircngulo
recto com o eixo do muacutesculo [P] Neste ponto daacute-se a libertaccedilatildeo progressiva de Pi e ADP Eacute
provaacutevel que a energia proveniente da hidroacutelise do ATP provoque uma alteraccedilatildeo do acircngulo de
ligaccedilatildeo das cabeccedilas de miosina ao filamento de actina passando novamente de 90ordm para 45ordm [P]
Esta alteraccedilatildeo conformacional provoca o deslizamento do filamento de actina para o centro do
sarcoacutemero cujo encurtamento promove a contracccedilatildeo das miofibrilas O processo supracitado
ocorre em cerca de 1ms[Q] e encontra-se resumido na Fig 18
Figura 18 ndash Esquema cineacutetico da hidroacutelise de ATP pela miosina (Adaptado de Sellers 1999 [A])
1131 Regulaccedilatildeo da contracccedilatildeo muscular
O mecanismo pelo qual se processa a regulaccedilatildeo muscular foi inicialmente descrito por estudos
distintos de Haselgrove Huxley e Parrey-Squire todos em 1973 [RST] Aleacutem da implicaccedilatildeo
directa da tropomiosina e do complexo de troponina a concentraccedilatildeo de Ca2+
no muacutesculo em
relaxamento eacute bastante reduzida (10-7
M) aumentando notavelmente aquando da contracccedilatildeo (10-5
-
10-3
M)
Resumidamente quando ocorre a excitaccedilatildeo do muacutesculo esqueleacutetico a despolarizaccedilatildeo da
membrana plasmaacutetica eacute difundida ateacute ao interior do tecido atraveacutes dos tuacutebulos transversais
(invaginaccedilotildees da membrana plasmaacutetica da fibra muscular) A despolarizaccedilatildeo provoca a abertura
dos canais ioacutenicos de Ca2+
do retiacuteculo sarcoplasmaacutetico adjacente Como a concentraccedilatildeo de Ca2+
no interior do organelo eacute muito elevada o caacutelcio passa raacutepida e uniformemente para o citosol
14
Assim a regulaccedilatildeo da contracccedilatildeo ocorre com coordenaccedilatildeo e rapidez na totalidade das miofibrilas
do tecido muscular (Ref)
Em repouso a troponina I encontra-se posicionada de tal forma que a proteiacutena adjacente
troponina C impede estruturalmente a ligaccedilatildeo das cabeccedilas da miosina agrave actina (acircngulo =50ordm) O
aumento da concentraccedilatildeo intracelular de Ca2+
provoca um aumento da sua disponibilidade para
ligar agrave troponina C (Fig 19)
Figura 19ndash Regulaccedilatildeo do mecanismo contraacutectil no muacutesculo esqueleacutetico pelo complexo de troponina
Como consequecircncia a troponina C altera a sua conformaccedilatildeo (=70ordm) associando-se agrave troponina I
que deixa de inibir a ligaccedilatildeo das pontes transversais A alteraccedilatildeo anterior eacute transmitida agrave
tropomiosina atraveacutes da troponina T adjacente agrave C o que provoca o deslocamento da
tropomiosina na direcccedilatildeo do interior do sulco do filamento de actina Assim eacute possiacutevel a
interacccedilatildeo eficaz entre actina e miosina A Ca2+
-ATPase do retiacuteculo sarcoplasmaacutetico provoca a
reacumulaccedilatildeo do iatildeo caacutelcio no organelo impedindo o prolongamento desnecessaacuterio da contracccedilatildeo
(Refordf)
12 Vanaacutedio
O vanaacutedio eacute um metal de transiccedilatildeo pertencente ao grupo VB da tabela perioacutedica Foi
primeiramente descoberto em 1801 por Andreacutes Manuel del Riacuteo que concluiu erradamente que era
idecircntico ao croacutemio e definitivamente por Nils Gabriel Svenson em 1831 Recentemente foram
descobertas importantes funccedilotildees bioloacutegicas deste elemento provavelmente essencial para os
humanos de que se destaca no contexto do presente relatoacuterio a possibilidade de actuar como
anaacutelogo do grupo fosfato e a inibiccedilatildeo de enzimas in vitro como eacute o caso da actividade ATPaacutesica
da miosina
Eacute um metal soacutelido agrave temperatura ambiente que se encontra amplamente distribuiacutedo nos vaacuterios
ecossistemas em dois isoacutetopos naturais ndash 50
V e 51
V ndash raramente excedendo 01 partes por biliatildeo
15
(ppb) Estaacute associado a cerca de 70 minerais diferentes na natureza (dos quais se destacam o
uracircnio cobre chumbo zinco fosfato e titacircnio ferroso) especialmente na forma de vanadato ou
pentoacutexidos de vanaacutedio O vanaacutedio eacute um dos principais constituintes da mateacuteria orgacircnica foacutessil
com especial ecircnfase para o carvatildeo oacuteleo de crude e foacutesseis carbonaacuteceos [UV] bem como em
alimentos consumidos pelo Homem (salsa pimenta preta cogumelos marisco vegetais cereais
gorduras e oacuteleos) [W] Estes constituem a maior fonte de exposiccedilatildeo ao vanaacutedio da maioria da
populaccedilatildeo humana [X]
Normalmente a entrada do vanaacutedio no organismo ocorre por inalaccedilatildeo ou em menores
quantidades por absorccedilatildeo pelo tracto gastrointestinal ou pela pele O seu armazenamento centra-
se no fiacutegado rins ceacuterebro coraccedilatildeo muacutesculos e ossos No entanto a retenccedilatildeo de vanaacutedio no
organismo humano eacute reduzida ndash em condiccedilotildees normais cerca de 100 M (em muitos tecidos a
quantidade eacute inferior a 10 ngg de tecido) [Y]
Actualmente natildeo existem provas de deficiecircncia nutricional de vanaacutedio em humanos ou de
enzimas que contenham vanaacutedio em animais superiores No entanto vaacuterios estudos relacionam a
ocorrecircncia de deficiecircncia em galinhas e ratos devido a uma alimentaccedilatildeo pobre em vanaacutedio Os
sintomas relatados incluem retardamento do crescimento malformaccedilotildees oacutesseas perturbaccedilotildees no
metabolismo lipiacutedico e estagnaccedilatildeo reprodutiva entre outros [Z AA AB] Os efeitos fisioloacutegicos
do vanaacutedio podem depender do tipo de composto ou de oligoacutemero uma vez que consoante as
suas estabilidade labilidade e nuacutemero de coordenaccedilatildeo as respostas seratildeo diferenciadas ao niacutevel
dos organismos vivos
Por outro lado a toxicidade do vanaacutedio encontra-se bem caracterizada existindo sintomas
descritos em vaacuterios mamiacuteferos (inclusive o Homem) que incluem diarreias catildeibras irritaccedilatildeo da
pele e mucosas palpitaccedilotildees cardiacuteacas anemia pneumonia bronquite perturbaccedilotildees no Sistema
Nervoso Central (SNC) entre outros (Ref) A toxicidade varia geralmente com a natureza do
composto com aumento proporcional agrave sua valecircncia Verificou-se que a forma de entrada do
vanaacutedio tem tambeacutem efeito na sua toxicidade ndash os maiores niacuteveis verificaram-se por injecccedilatildeo
intravenosa da amostra sendo progressivamente inferiores por inalaccedilatildeo e finalmente por
ingestatildeo [X AA] A ampla utilizaccedilatildeo dos compostos de vanaacutedio na induacutestria quiacutemica bem como
as elevadas quantidades de vanaacutedio libertadas atraveacutes da combustatildeo dos oacuteleos de crude
contribuem consideravelmente para o aumento da biodisponibilidade do vanaacutedio Como tal o
interesse neste elemento e nos problemas ambientais por ele causados tem vindo a aumentar
significativamente [V]
121 Propriedades quiacutemicas
16
A quiacutemica do vanaacutedio eacute extraordinariamente complexa pelo que originou vaacuterias interpretaccedilotildees
incorrectas desde a sua descoberta No entanto as suas propriedades dependem em uacuteltima
anaacutelise da configuraccedilatildeo electroacutenica do elemento Assim a variabilidade encontrada no
comportamento quiacutemico do vanaacutedio pode dever-se agrave sua configuraccedilatildeo ndash [Ar] 3d2 4s
2 ndash que
permite a formaccedilatildeo de vaacuterios estados de oxidaccedilatildeo (de +2 a +5) Em soluccedilatildeo aquosa os estados de
oxidaccedilatildeo estaacuteveis vatildeo de V(V) a V(II) Destes apenas os +3 a +5 satildeo biologicamente importantes
jaacute que o V(II) apresenta demasiada capacidade redutora para existir nos organismos vivos
conhecidos [AC]
No fluido extracelular os compostos de vanaacutedio encontram-se predominantemente na forma de
monovanadatos (V(V)) livres ou ligados agrave transferina do soro ou agrave hemoglobina (Refordf) Os iotildees de
vanadato livres satildeo transportados para o meio intracelular atraveacutes de canais ioacutenicos inespeciacuteficos
Jaacute no ambiente intracelular os compostos sofreratildeo uma bio-reduccedilatildeo atraveacutes da acccedilatildeo da
glutationa ou outros compostos com grupos ndashOH como pentoses hexoses ou etanolaminas [V] A
reoxidaccedilatildeo de vanadilo (V(IV)) intracelular a vanadato (V(V)) provocaria a acumulaccedilatildeo toacutexica
destes uacuteltimos pelo que a complexaccedilatildeo da espeacutecie reduzida o impede O transporte de vanaacutedio
para o interior da ceacutelula encontra-se na Fig 110
Figura 110 ndash Transporte de vanaacutedio para a ceacuteluka e seu subsequente metabolismo (P=Proteiacutena) (Adaptado de
Zaporowska e Scibor 1998)
1211 Vanaacutedio(IV)
A espeacutecie de vanaacutedio (IV) mais bem conhecida eacute o iatildeo vanadilo (VO2+
) espeacutecie estaacutevel a pH
aacutecido sendo a maioritaacuteria a pH 3[AD] Apresenta uma cor azulada e forma complexos penta e
hexacoordenados Forma-se rapidamente a partir de V(V) na presenccedila de um agente redutor em
soluccedilatildeo aquosa No ambiente intracelular a presenccedila de glutationa reduzida aacutecido ascoacuterbico
NAD(P)H entre outros conduzem a que o principal estado de oxidaccedilatildeo nos sistemas bioloacutegicos
seja provavelmente o +4 encontrando-se o V(IV) fortemente complexado com proteiacutenas ou
aminoaacutecidos livres citrato ATP e pirofosfato (Refordfs) Em alternativa o V(IV) pode existir
17
livremente em ambientes fortemente aacutecidos como os lisossomas jaacute que a pH superior a 3 as
soluccedilotildees de vanadilo tendem a ser oxidadas pelo ar formando-se vanadato Esta grande
capacidade redutora justifica parcialmente a utilizaccedilatildeo pouco extensiva do iatildeo VO2+
como
inibidor [W]
1212 Vanaacutedio (V)
O vanaacutedio (V) eacute a forma mais estaacutevel num intervalo de pH amplo [W] O iatildeo anioacutenico VO3-tende
a constituir complexos polinucleares pelo que as soluccedilotildees aquosas de vanadato satildeo extremamente
complexas com ocorrecircncia simultacircnea de vaacuterios equiliacutebrios de protonaccedilatildeo (pKa(vanadato)= 35
78 e 125) e oligomerizaccedilatildeo As espeacutecies presentes (H3VO4 H2VO4- HVO4
2- e VO4
3-) dependem
do pH forccedila ioacutenica e temperatura da soluccedilatildeo e da concentraccedilatildeo total do elemento
[AEAFAGAHAI]
O VO43-
- monoacutemero ortovanadato V1 ndash estaacute associado a uma grande parte da importacircncia
bioloacutegica do vanaacutedio uma vez que se trata de um anaacutelogo electroacutenico e estrutural do ortofosfato
(PO43-
) A coordenaccedilatildeo do ortovanadato pode ser tetraeacutedrica ou bipiramidal trigonal (Refordfs)
A oligomerizaccedilatildeo do vanadato monomeacuterico daacute origem ao diacutemero (V2O72-
- V2) que apresenta
tambeacutem equiliacutebrios de protonaccedilatildeo Presume-se a existecircncia de analogia do vanadato dimeacuterico com
o pirofosfato embora natildeo haja evidecircncia estrutural (Refordf) A dimerizaccedilatildeo do V2 origina o
vanadato tetrameacuterico ou metavanadato (V4O124-
- V4) que constitui a espeacutecie maioritaacuteria em
soluccedilotildees aquosas concentradas (Refordf) As estruturas do metavanadato e a sua espeacutecie protonada
satildeo ciacuteclicas [AJ] Outra espeacutecie oligomeacuterica importante eacute o vanadato pentameacuterico (V5O153-
) cuja
estrutura presumida eacute ciacuteclica [AI]
O oligoacutemero com maior importacircncia bioquiacutemica eacute o vanadato decameacuterico (V10O286-
-V10)(Refordf) a
espeacutecie predominante em soluccedilotildees de pH 2-6 quando a concentraccedilatildeo total de vanadato eacute superior
a 100 M (Ref) A estrutura do V10 natildeo difere em soluccedilatildeo ou no estado soacutelidoOs 10 aacutetomos de
vanaacutedio da espeacutecie decameacuterica estatildeo divididos em trecircs grupos identificados na figura como VA-C
consoante o seu ambiente quiacutemico circundante (Refordf) (Fig 111)
A expressatildeo ldquodecavanadatordquo refere-se a uma soluccedilatildeo aquosa na qual a espeacutecie decameacuterica se
encontra em equiliacutebrio quiacutemico com as espeacutecies V1 V2 V4 e V5 sendo que a oligomerizaccedilatildeo
progressiva do vanaacutedio eacute proporcional agrave sua concentraccedilatildeo total (Ref) Nesta soluccedilatildeo a
concentraccedilatildeo de V10 corresponde a aproximadamente 10 da concentraccedilatildeo total de vanaacutedio [AK]
18
Figura 111 ndash Estruturas esquemaacuteticas das espeacutecies de vanadato (espeacutecie monomeacuterica (V1) dimeacuterica (V2)
tetrameacuterica (V4) pentameacuterica (V5) e decameacuterica (V10) (Adaptado de Aureliano e Crans 2008)
122 Metabolismo e efeito na actividade enzimaacutetica
Apesar de natildeo terem sido ainda encontradas as provas categoacutericas das suas funccedilotildees nos seres
vivos sabe-se que o iatildeo vanadato eacute capaz de inibir uma grande quantidade de enzimas in vitro
com especial ecircnfase na actividade ATPaacutesica da miosina [AL Teresa] Como referido
anteriormente o vanadato eacute capaz de actuar em alternativa ao fosfato nas reacccedilotildees bioquiacutemicas
incluindo a sua preferecircncia de ligaccedilatildeo agraves enzimas [W]
Eacute conhecido o papel do vanaacutedio no metabolismo do caacutelcio promovendo a mineralizaccedilatildeo dos
dentes e ossos na biossiacutentese de liacutepidos (trigliceacuteridos e colesterol) com posterior inibiccedilatildeo da
acumulaccedilatildeo destes nos vasos sanguiacuteneos e no SNC na reproduccedilatildeo e desenvolvimento por
mimetizaccedilatildeo e potenciaccedilatildeo de factores de crescimento na estimulaccedilatildeo do consumo de glucose e
da siacutentese de glicogeacutenio com actuaccedilatildeo equivalente aos efeitos da insulina ao niacutevel celular na
transduccedilatildeo de sinais celulares com regulaccedilatildeo da actividade de segundos mensageiros no
transporte de iotildees e aminoaacutecidos na transcriccedilatildeo geneacutetica e siacutentese de proteiacutenas e DNA entre
outros [W Y AB]
Assim o vanaacutedio cnseguiraacute provocar inuacutemeras mudanccedilas bioquiacutemicas nas ceacutelulas Alguns
exemplos da influecircncia bioquiacutemica do vanaacutedio incluem os seus efeitos nos processos de
fosforilaccedilatildeo e desfosforilaccedilatildeo de proteiacutenas e na participaccedilatildeo na formaccedilatildeo de espeacutecies reactivas de
oxigeacutenio devido ao seu efeito proacute-oxidante sobre agentes redutores a niacutevel celular [AM]
Quanto ao seu efeito na actividade enzimaacutetica pode dividir-se em dois tipos principais ndash inibitoacuterio
e estimulante - dependendo da enzima ou do tipo de reacccedilatildeo por esta catalisada No primeiro
grupo incluem-se enzimas como as ATPases (de miosina de canais ioacutenicos e fosfoenzimas
19
transportadoras) e as fosfatases (da tirosina aacutecida e alcalina) a fosfofrutocinase adenilato ciclase
e catalase entre outras [AA] O segundo grupo inclui a maioria das cinases a glucose-6-fosfato
desidrogenase a fosfoglicerato mutase e a fosfoglicerato fosfatase (Refordf)
O efeito do vanaacutedio (V) na actividade enzimaacutetica estaacute relacionado com o seu nuacutemero de
coordenaccedilatildeo podendo actuar como estado de transiccedilatildeo ou por mimetizaccedilatildeo de cofactores
substratos (especialmente o fosfato) e interacccedilotildees proteiacutena-proteiacutena (como no caso das
polimerases e transcriptases) Pode ainda estar envolvido na formaccedilatildeo de ligaccedilotildees de hidrogeacutenio
interacccedilotildees electrostaacuteticas esteacutereas e de oxidaccedilatildeo-reduccedilatildeo Uma evidecircncia desta uacuteltima eacute a sua
capacidade de alternacircncia entre os estados de oxidaccedilatildeo +5 a +3 o que torna faacutecil a sua interacccedilatildeo
redox com enzimas contendo resiacuteduos de cisteiacutenas (Cys) ou qualquer outro composto orgacircnico
com actividade redox [AN]
13 Vanaacutedio e muacutesculo esqueleacutetico
A primeira evidecircncia da presenccedila de vanaacutedio no muacutesculo esqueleacutetico proveio de uma amostra de
ATP isolado de muacutesculo de cavalo que continha uma impureza ndash vanaacutedio ndash cuja presenccedila
provocou a inibiccedilatildeo da actividade da bomba de Na+K
+-ATPase [AO] Desde entatildeo vaacuterios
estudos tecircm sido realizados com ecircnfase na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o
tecido muscular Destes destaca-se a descoberta da interacccedilatildeo forte da espeacutecie V10 com a miosina
fosforilada [AP] e agrave sua inibiccedilatildeo da actividade ATPaacutesica da miosina[AQ AR]
No quediz respeitoao iatildeo monovanadato destaca-se a sua inibiccedilatildeoda actividade Mg2+
-ATPaacutesica
da miosina [AS] que ocorre de duas formas distintas (1) na presenccedila de concentraccedilotildees reduzidas
de V1 a inibiccedilatildeo ocorre a velocidade reduzida dependente da concentraccedilatildeo da espeacutecie
monomeacuterica (2) na presenccedila de concentraccedilotildees elevadas de V1 o niacutevel de inibiccedilatildeo aproxima-se de
100 desde que [V1] seja superior agrave concentraccedilatildeo total de locais com actividade Mg2+
-ATPaacutesica
no ensaio Esta inibiccedilatildeo eacute irreversiacutevel tanto por diaacutelise ou adiccedilatildeo de agentes redutores do
vanadato [AL]Segundo Pate et al(1994)[AT] a espeacutecie monomeacuterica podeinibir a velocidade de
contracccedilatildeo a temperatura inferior a 20ordmC perdendogradual e rapidamente essa capacidade ateacute os
50ordmC temperatura agrave qual cessa a capacidade inibitoacuteria Os resultados obtidos sugeremainda uma
interacccedilatildeo altamente cooperativa entre o iatildeo vanadato e as proteiacutenas componentes do muacutesculo
esqueleacuteticoDestaca-se tambeacutem a descoberta da inibiccedilatildeo da geraccedilatildeo de forccedila contraacutectil nas fibras
musculares por parte da espeacutecie monomeacuterica[AT]Wilson et al(1995) [AU ]determinaram que a
afinidade da espeacutecie V1 para o S1 de miosina eacute significativamente mais elevada do que a do iatildeo
Pi seu anaacutelogo Descobriram ainda que que a ligaccedilatildeo do vanadato poderaacute envolver transiccedilotildees
entre estados energeacuteticos com energia livre independente dos impedimentos esteacutereos impostos
20
pela estrutura das miofibrilas Os estudos desenvolvidos por Smith et al (1990)[AV]
determinaram queo iatildeo monovanadato natildeo apresenta um efeito pronunciado na inibiccedilatildeo da
actividadeacto-S1-miosina-ATPaacutesica excepto em concentraccedilotildees superiores a 900microM a partir das
quais existe produccedilatildeo de espeacutecies oligomeacutericas para as quais o efeito inibitoacuterio seraacute mais
elevadoA espeacutecie monomeacuterica de vanadato consegue adoptar facilmente uma geometria do tipo
bipiramidal trigonal que mimetiza o intermediaacuterio formado (MgADP
Pi)aquando da reacccedilatildeo de
hidroacutelise de ATP no local activo da miosina [AW] De facto a interacccedilatildeo do vanaacutedio com a
miosina eacute inesperadamente estaacutevel na presenccedila de ADP Esta interacccedilatildeo resulta na formaccedilatildeo de
um complexo enzimaacutetico inactivo e estaacutevel com estequiometria de 11 ADPvanadato por siacutetio
activo da proteiacutena A formaccedilatildeo do complexo estaacutevel miosina-vanadato foi proposta por Goodnoet
al (1982) [AS] Nesta ocorre um preacute-equiliacutebrio raacutepido que induz a formaccedilatildeo de um complexo
ternaacuterio reversiacutevel (MADP
V) com formaccedilatildeo lenta do isoacutemero M
ADP
V A diferenccedila entre os
compostos de vanaacutedio e de fosfato centra-se no tempo de meia vida (t12) do Pi ou do V (no
primeiro t12=12 s enquanto no segundo t12=24 h) (Refordf) A diferenccedila dever-se-aacute agrave possiacutevel
complexaccedilatildeo do vanaacutedio com grupos nucleofiacutelicos no siacutetio activo o que provocaria uma
estabilizaccedilatildeo da miosina na conformaccedilatildeo MADP
V
No que respeita aos estudos realizados com a espeacutecie tetrameacuterica Cremo et al determinou que a
cadeia pesada do S1 de miosina pode ser clivada em dois locais distintos (designados V1 e V2)
pela espeacutecie V4 apoacutes irradiaccedilatildeo com luz UV na presenccedila de concentraccedilotildees de vanadato na ordem
dos milimolar e na ausecircncia de nucleoacutetidos ou metais divalentes
Jaacute no que concerne aos estudos realizados com ecircnfase na espeacutecie decameacuterica foi sugerido
porRamos et al (2006) [AK]que a interacccedilatildeo do decavanadato com a G-actina favorecida pela
polimerizaccedilatildeo desta uacuteltima estabiliza a espeacutecie V10 e induz a inibiccedilatildeo da formaccedilatildeo de F-actina A
estabilizaccedilatildeo da espeacutecie V10 por proteiacutenas tansmembranares e do citoesqueleto pode ser a razatildeo
pela qual ocorre a toxicidade do decavanadato reportada na avaliaccedilatildeo dos efeitos do V(V) nos
sistemas bioloacutegicosTiago et al (2007) [AR] concluiram que o V10 apresenta uma clara preferecircncia
de ligaccedilatildeo ao ldquoback-doorrdquo da S1-miosina-ATPase o que se verifica apenas na estrutura
conformacional desimpedida com acesso facilitado ao local de ligaccedilatildeo do Pi Sugere tambeacutem que
o V10 actua como um ldquoback-door stoprdquo bloqueando estaabertura do fragmento de 50-kDa
necessaacuteria para a hidroacutelise do ATP--fosfato o que constitui uma explicaccedilatildeo simples do
comportamento natildeo competitivo da espeacutecie decameacuterica estimulando a utilizaccedilatildeo deste oligoacutemero
como uma feramenta da elucidaccedilatildeo das mudanccedilas conformacionais ldquoback-doorrdquo da miosina no
21
processo da contracccedilatildeo muscular(ref)Destaca-se ainda a descoberta da capacidade inibitoacuteria do
V10 em relaccedilatildeo agrave fosforilaccedilatildeo oxidativa mitocondrial [AX]
Finalmente no que concerne aos estudos realizados com a presenccedila de vaacuterias espeacutecies
oligomeacutericas Aureliano e Madeira (1994) [AY]observaram que na presenccedila de Ca2+
as espeacutecies
V4 e V10se ligam agrave Ca2+
-ATPase do retiacuteculo sarcoplasmaacutetico enquanto a espeacutecie V1 se liga apenas
na presenccedila de ATP Aparentemente o aumento da concentraccedilatildeo de Ca2+
tem o efeito contraacuterio da
inibiccedilatildeo da ATPase do retiacuteculo sarcoplasmaacutetico pelo V1 natildeo tendo no entanto efeito significativo
sobre a inibiccedilatildeo por parte da espeacutecie V10 Tiago et al (2002) [AP] concluiacuteram que em condiccedilotildees
proacuteximas agraves fisioloacutegicas a espeacutecie V10 difere das espeacutecies presentes na soluccedilatildeo de metavanadato
devido agrave sua forte interacccedilatildeo com a miosina fosforilada e agrave sua inibiccedilatildeo da actividade ATPaacutesica
da miosina Para aleacutem disso o ATP diminui a afinidade da miosina para o V4 induzindo a
interacccedilatildeo com V1 e natildeo tendo qualquer efeito no que diz respeito ao V10 Foi tambeacutem proposto
[AQ] que a ligaccedilatildeo das espeacutecies oligomeacutericas de vanadato aoS1 de miosina nos locais onde o
fosfato coordenaeacute provavelmente a causa da inibiccedilatildeo da actividade ATPaacutesica da miosina
estimulada por actina atraveacutes da prevenccedilatildeo da troca entre ATPADP sendo esta favorecida para
aniotildees de vanadato com valecircncias superiores como eacute o caso do V10
Na presenccedila de actina as velocidades de dissociaccedilatildeo do Pi e do V aumentam consideravelmente
este uacuteltimo em menor quantidade Assim a ligaccedilatildeo da actina ao complexo MADP
V e
Pi eacute
bastante fraca pelo que se sugere que o ADPV teria um efeito de relaxamento semelhante ao
ATP No entanto a capacidade do vanaacutedio em inibir a actividade ATPaacutesica da miosina e da
actomiosina eacute diferenciada uma vez que na ausecircncia de actina a inibiccedilatildeo eacute total para
concentraccedilotildees na gama dos M mas na sua presenccedila a inibiccedilatildeo eacute totalmente conseguida apenas
em concentraccedilotildees na gama dos mM (Ref) No entanto eacute necessaacuterio considerar a possiacutevel
formaccedilatildeo de espeacutecies oligomeacutericas de vanadato cuja afinidade e efeito na enzima seratildeo
diferentes Assim pode concluir-se que o efeito de inibiccedilatildeo observado para a actividade ATPaacutesica
de miosina estimulada por actina se pode dever agrave presenccedila das diferentes espeacutecies oligomeacutericas o
que foi jaacute sugerido por Aureliano (2000) [AZ] ndash as espeacutecies tetrameacuterica e decameacuterica de vanadato
interactuariam com a miosina com alteraccedilatildeo da sua interacccedilatildeo com a actina No entanto natildeo eacute
conhecida a contribuiccedilatildeo absoluta das restantes espeacutecies oligomeacutericas para a inibiccedilatildeo da ATPase
da miosina estimulada por actina (Refs)
14 Objectivos do projecto
Pretende-se com este projecto o isolamento purificaccedilatildeo e caracterizaccedilatildeo da actina do muacutesculo
22
esqueleacutetico de coelho sendo esta uacuteltima tambeacutem realizada para o sufragmento-1 de miosina
Seguidamente realizar-se-aacute a quantificaccedilatildeo e caracterizaccedilatildeo das proteiacutenas por espectroscopia UV-
Vis e SDS-PAGE e a caracterizaccedilatildeo das soluccedilotildees de vanaacutedio por UV-Vis e ressonacircncia magneacutetica
nuclear (RMN) Por uacuteltimo a interacccedilatildeo do S1 de miosina com o vanaacutedio seraacute avaliada atraveacutes
das teacutecnicas de UVVis RMN e ressonacircncia paramagneacutetica electroacutenica (RPE)
2 Discussatildeo de Resultados
21 Caracterizaccedilatildeo das proteiacutenas isoladas
211Anaacutelise do estado de pureza
O estado de pureza das soluccedilotildees de actina e de S1 foi analisado por electroforese em gel de
poliacrilamida em condiccedilotildees desnaturantes (SDS-PAGE) (Fig 22) que separa as moleacuteculas de
acordo com o seu peso molecular (PM) A concentraccedilatildeo de acrilamida escolhida (10) permite a
detecccedilatildeo simultacircnea de componentes com peso molecular inferior a 20 kDa e outros com elevado
peso molecular
Figura 21 ndash Fotografia de gel de SDS-PAGE (10 acrilamida) Poccedilo (a)
10 L do marcador de elevado peso molecular () Poccedilo (b) 20 L de [G-
actina]=1 mgmL Poccedilo (c) 20 L de [S1]=1 mgmL Aplicou-se uma
voltagem de V durante h e min
A partir da recta de calibraccedilatildeo obtida log PM = -1177 Rf + 2247 (ver ponto 2 da secccedilatildeo
Anexos) pode verificar-se a presenccedila de actina na amostra do poccedilo (b) presente numa banda
bastante intensa (PMaparente=40 kDa) bem como a presenccedila minoritaacuteria de outros componentes
desconhecidos que natildeo teratildeo sido eliminados durante o processo de extracccedilatildeo e purificaccedilatildeo e natildeo
satildeo relevantes para o estudo em questatildeo Na amostra do poccedilo (c) nota-se a presenccedila de S1
(PMaparente=92 kDa) bem como a de outros componentes cuja banda eacute ligeiramente menos intensa
Estes poderatildeo ser fragmentos proteoliacuteticos de miosina e de S1 Estaacute ainda presente uma bada de
23
maior intensidade que poderaacute ser a proteiacutena-C (PM=135 kDa) uma vez que a sua remoccedilatildeo
completa da amostra necessitaria de um passo adicional de cromatografia de troca ioacutenica [BA]
No entanto natildeo podemos confirmar que se trata desta proteiacutena ou de qualquer outra uma vez que
se encontra fora do intervalo de migraccedilatildeo dos marcadores de peso molecular
212 Determinaccedilatildeo da concentraccedilatildeo proteica
2121Subfragmento-1 de miosina
A partir da recta de calibraccedilatildeo obtida pelo meacutetodo de Bradford [BB](ver secccedilatildeo 3311 do
Procedimento Experimental) A595nm = 00546[BSA] + 00209 (ver secccedilatildeo de Anexos)foi possiacutevel
determinar um intervalo de concentraccedilatildeo de S1 de miosina compreendido entre 226 e 117(plusmn
423M) (203 e 1056 (plusmn 381 mgmL)) (n=8) A concentraccedilatildeo meacutedia de S1 de miosina apoacutes
diaacutelise contra a soluccedilatildeo tampatildeo 10 mM HEPES pH 75 25 mM KCl determinada pela leitura da
Abs280 nmfoi de 20430M (1803 mgmL)(n=7)
As amostras a quantificar atraveacutes do meacutetodo de Bradford natildeo sofreram diaacutelise o que permite
explicar a concentraccedilatildeo superior de S1 de miosina em comparaccedilatildeo com o valor obtido atraveacutes do
meacutetodo directo Esta razatildeo aliada agrave sensibilidade e rapidez superiores deste uacuteltimo fez com que
se optasse pelo meacutetodo directo para a quantificaccedilatildeo da grande maioria dos ensaios
2122Actina
A concentraccedilatildeo de actina foi determinada quando esta se encontrava na forma monomeacuterica (G-
actina) por mediccedilatildeo da leitura directa da Abs290 nm tendo-se obtido uma concentraccedilatildeo meacutedia de
691481M (290202 mgmL) (n=3) em 2 mM Tris-HCl pH 80 02 mM CaCl2 02 mM Na-
ATP Ao comprimento de onda utilizado (290 nm) absorvem fortemente apenas os resiacuteduos de
Trp pelo que a mediccedilatildeo efectuada tem menor sensibilidade do que se tivesse sido efectuada a 280
nm uma vez que a actina apresenta apenas 4 resiacuteduos deste aminoaacutecido embora tenha 16
resiacuteduos de Tyr
22 Mediccedilatildeo da actividade enzimaacutetica de S1 de miosina
221 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
As actividades ATPaacutesicas de S1 de miosina dependentes de Ca2+
e K+foram analisadas pelo
caacutelculo das velocidades de consumo inicial do fosfato inorgacircnico (Pi) Definiu-se os tempos de 5
e 3 minutos para a mediccedilatildeo da Ca2+
e da K+EDTA-ATPase respectivamente Os valores obtidos
24
a partir do protocolo presente no ponto 341 da secccedilatildeo Procedimento experimental encontram-se
descritos na Tabela 2I
Tabela 2I ndash Acrividades natildeo-fisioloacutegicas de subfragmento-1 determinadas pelo meacutetodo de Fiske-Subarow
modificado (n=2)
Proteiacutena
Actividade
ATPaacutesica natildeo
fisioloacutegica
Actividade (nmol
Piminmg proteiacutena) Actividade (s-1)
S1
Ca2+
-ATPase 1400 plusmn 30 2100 plusmn 0045
K+EDTA-ATPase 2334 plusmn 50 3500 plusmn 0075
As actividades natildeo-fisioloacutegicas reflectem o estado de oxidaccedilatildeo dos resiacuteduos expostos de Cys 707
(SH1) e 697 (SH2) Se o grupo SH1 sofrer oxidaccedilatildeo a actividade ATPaacutesica dependente de Ca2+
aumenta e a actividade dependente de K+ diminui Por outro lado se o grupo SH2 sofrer
oxidaccedilatildeo regista-se a perda completa destas actividades
De acordo com Margossian e Lowey (1982) [BA] as actividades ATPaacutesica de S1 de miosina
dependente de Ca2+
e K+EDTA da miosina satildeo respectivamente de 24 e 112 s-1
Estaacute tambeacutem
descrito que a actividade ATPaacutesica dependente de K+ eacute em meacutedia cerca de 3 vezes superior agrave
dependente de Ca2+
reflectindo a quantidade das preparaccedilotildees A razatildeo obtida entre os valores das
actividades ATPaacutesicas dependentes de K+ e Ca
2+ eacute de 166 Os valores obtidos para as actividades
(Ca2+
-ATPase=21 s-1
K+EDTA-ATPase= 35 s
-1) satildeo inferiores agrave valores obtidos por
Margossian e Lowey [BA] e Tiago [BC] com forccedila ioacutenica inferior O valor obtido para a
actividade ATPaacutesica dependente de Ca2+
eacute superior ao obtido por Polosukhina et al (2000) [BD]
cujo meio de reacccedilatildeo continha MOPS e apresentava forccedila ioacutenica inferior Os dados
supramencionados comprovam a variaccedilatildeo da actividade a ATPaacutesica natildeo fisioloacutegica em relaccedilatildeo agrave
forccedila ioacutenica temperatura e pH
222 Mediccedilatildeo da actividadeMg2+
-ATPaacutesica de S1 pelo meacutetodo de enzimas acopladas
A actividade de Mg2+
-ATPase eacute a uacutenica que possui significado fisioloacutegico sendo as restantes
bastante uacuteteis apenas para indicaccedilatildeo da qualidade de preparaccedilatildeo do S1[BA] A 0575 M KCl a
actina e o S1 formam uma soluccedilatildeo viscosa e tuacuterbida Apoacutes a adiccedilatildeo de MgATP o complexo acto-
S1 dissocia-se e a turbidez da soluccedilatildeo diminui Se a forccedila ioacutenica for reduzida a dissociaccedilatildeo do
complexo actoS1 que se encontra na forma de suspensatildeo eacute induzida O MgATP eacute rapidamente
hidrolisado diminuindo a sua concentraccedilatildeo no meio reaccional Nestas condiccedilotildees haacute
reassociaccedilatildeo e subsequente contracccedilatildeo dos filamentos de actina e de S1 com formaccedilatildeo de
25
precipitado O meacutetodo descrito traduz de forma bastante elementar o fenoacutemeno da contracccedilatildeo in
vivo e elimina o problema anterior da solubilizaccedilatildeo ineficiente da miosina de modo a obter uma
mediccedilatildeo exacta da actividade [A] A utilizaccedilatildeo de forccedilas ioacutenicas reduzidas aumenta a afinidade da
actina para o S1 permitindo contornar a extrema dificuldade em obter a concentraccedilatildeo proteica dos
muacutesculos (cerca de 100 mgmL do complexo actomiosina) As actividades Mg2+
-ATPaacutesicas de S1
de miosina foram determinadas a partir do declive da variaccedilatildeo da Abs340 relativo agrave oxidaccedilatildeo de
NADH em funccedilatildeo do tempo como eacute descrito no ponto 342 do Procedimento Experimental Os
valores obtidos para a actividade Mg2+
-ATPaacutesica encontram-se apresentados na Tabela 2II
Tabela 2II ndash Actividade Mg2+
-ATPaacutesica de subfragmento-1 de miosina determinadas pelo meacutetodo das enzimas
acopladas
Actividade (nmol
ATPmin mg(S1)) Actividade (s
-1)
1097 plusmn 269 164 plusmn 04
Ramos (2005) [J] obteve o valor de 01 s-1
para a actividade Mg2+
-ATPaacutesica de S1 de miosina
com forccedila ioacutenica intermeacutedia Tiago (2005) [BC] obteve o valor de 011 s-1 em condiccedilotildees de forccedila
ioacutenica reduzida em relaccedilatildeo agrave utilizada no presente relatoacuterio
23 Preparaccedilatildeo e caracterizaccedilatildeo das soluccedilotildees de vanaacutedio
231 Anaacutelise da composiccedilatildeo das soluccedilotildees de vanadato por espectroscopia de 51
V-RMN
efeito da concentraccedilatildeo da espeacutecie
A diluiccedilatildeo das soluccedilotildees stock de meta- e decavanadato iraacute determinar fortemente a composiccedilatildeo
em soluccedilatildeo dos diferentes oligoacutemeros de vanadato (ref) No caso do decavanadato o periacuteodo de
tempo apoacutes a diluiccedilatildeo iraacute tambeacutem ser um factor importante a considerar devido agrave decomposiccedilatildeo
da espeacutecie V10(ref)Torna-se assim fundamental determinar a composiccedilatildeo quiacutemica das soluccedilotildees
no meio de ensaio utilizado (HEPES 10 mM KCl 25 mM) de modo a correlacionaacute-la
correctamente com os possiacuteveis efeitos promovidos pelas diferentes espeacutecies de vanadato no
sistema em estudo
Aleacutem disso deve ser dado ecircnfase agraves importantes interacccedilotildees que ocorrem entre o vanaacutedio e os
tampotildees escolhidos O tampatildeo com menor nuacutemero e magnitude de interacccedilatildeo eacute o HEPES [AD]
seguido do Tris sendo este uacuteltimo especialmente aconselhado em concentraccedilotildees baixas para casos
de incompatibilidade enzimaacutetica ou quiacutemica redox No caso especial da reacccedilatildeo redox
NAD(P)HNAD(P) o tampatildeo adequado eacute o imidazole de modo a natildeo haver reacccedilatildeo com o
26
vanadato Os tampotildees a evitar em qualquer situaccedilatildeo incluem a trietanolamina fosfato citrato e
glicina entre outros [AD] uma vez que todos interactuam fortemente com V(V) com formaccedilatildeo
de complexos estaacuteveis
Assim procedeu-se agrave anaacutelise da composiccedilatildeo das soluccedilotildees de meta- e decavanadato apoacutes diluiccedilatildeo
das soluccedilotildees stock (50 mM em vanadato total) no meio de reacccedilatildeo (tampatildeo D) atraveacutes da
espectroscopia de ressonacircncia magneacutetica nuclear de 51
V Nas figuras x1 e x2 apresentam-se os
espectros obtidos para as soluccedilotildees de deca- e metavanadato num intervalo de concentraccedilotildees de 1 a
10 mM A Tabela seguinte resume os paracircmetros espectroscoacutepicos obtidos para cada espeacutecie
presente nas soluccedilotildees analisadas
Figura 22ndash Espectros de RMN de 51
V de
concentraccedilotildees crescentes das espeacutecies oligomeacutericas
de vanadato obtidos apoacutes diluiccedilatildeo da soluccedilatildeo stock
de decavanadato em 10 mM HEPES pH 70 25 mM
KCl para concentraccedilotildees finais de A) 1 mM B)2
mM C) 4 mM D) 8 mM E) 10 mM de
decavanadato total V1refere-se agraves espeacutecies
monomeacutericade vanadato V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2IIIndash Desvios quiacutemicos meacutedios (part) das ressonacircncias das espeacutecies decameacuterica (V10) monomeacuterica (V1) e
tetrameacuterica (V4) presentes nos espectros da soluccedilatildeo de decavanadatoLargura a meia altura (∆12) da espeacutecie V10A
Paracircmetros V10A V10B V10C V1 V4
part (ppm) -5162 -5003 -4255 -5616 -5748
∆12 (Hz) 175 nd nd nd nd
Figura 23 ndash Distribuiccedilatildeo graacutefica das
espeacutecies oligomeacutericas de vanadato vs a
concentraccedilatildeo de decavanadato total As
concentraccedilotildees de cada espeacutecie foram
obtidas segundo a formula descrita na
secccedilatildeo 35 do Procedimento
Experimental (aacutereas dos sinais de 51
V-
RMN observadas na figura x1)V10 V1 e
V4 correspondem agraves espeacutecie monomeacuterica
dimeacuterica e tetrameacuterica respectivamente [Esp
eacuteci
e o
ligo
meacute
rica
de
van
aacutedio
] (m
M)
[Decavanadato]total (mM)
V10
V1
V4
27
Nas soluccedilotildees de decavanadato as espeacutecies monomeacuterica e decameacuterica de vanadato satildeo as uacutenicas
presentes para concentraccedilotildees de decavanadato inferiores a 8 mM Ambas as espeacutecies aumentam
linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado em [AY] e
em (Teresa) quando a concentraccedilatildeo de V1 eacute igual ou superior a 200 microM inicia-se a sua
oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica No entanto nas condiccedilotildees de realizaccedilatildeo dos
ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a espeacutecie tetrameacuterica neste
estudo em concentraccedilotildees inferiores a 8 mM tendo o mesmo sido observado por Ramoset al
(2006) [AK] Os resultados obtidos mostram claramente o equiliacutebrio entre a espeacutecie decameacuterica e
a monomeacuterica e que o aparecimento de outras espeacutecies oligomeacutericas se encontra dependente da
concentraccedilatildeo desta uacuteltima Apesar de natildeo ter sido realizado o ensaio com 5 mM de decavanadato
total no presente relatoacuterio estaacute reportado em Ramos et al (2006) [AK] que as concentraccedilotildees da
espeacutecie decameacuterica e monomeacuterica satildeo respectivamente de 500 microM e 200 microM Assim pode inferir-
se que para concentraccedilotildees inferiores a 5 mM e intervalos de tempo de cerca de 20 minutos (que
inclui o tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas
as espeacutecies decameacuterica e monomeacuterica de vanadato
Figura 24 ndash Espectros de RMN de 51
V de concentraccedilotildees
crescentes das espeacutecies oligomeacutericas de vanadato obtidos
apoacutes diluiccedilatildeo da soluccedilatildeo stock de metavanadato em 10 mM
HEPES pH 70 25 mM KCl para concentraccedilotildees finais de A)
1 mM B) 2 mM C) 4 mM D) 8 mM E) 10 mM de
metavanadato total V1 refere-se agraves espeacutecies monomeacuterica de
vanadatoV1 e V2 correspondem respectivamente agrave espeacutecie
monomeacuterica e dimeacuterica
Figura 25 ndash Distribuiccedilatildeo graacutefica das espeacutecies
oligomeacutericas de vanadato vs a concentraccedilatildeo de
metavanadato total As concentraccedilotildees de cada
espeacutecie foram obtidas segundo a formula descrita
na secccedilatildeo 35 do Procedimento Experimental
(aacutereas dos sinais de 51
V-RMN observadas na
figura x1)V10 V1 e V4 correspondem agraves espeacutecie
monomeacuterica dimeacuterica e tetrameacuterica
respectivamente
Esp
eacuteci
e o
ligo
meacute
rica
de
va
nad
ato
(m
M)
[Metavanadato]total (mM)
V1
V2
V4
V5
28
Tabela 2IVndash Desvios quiacutemicos meacutedios (part)das ressonacircncias das espeacutecies tetrameacuterica (V4) monomeacuterica (V1) e
dimeacuterica (V2) presentes no espectro da soluccedilatildeo de metavanadato (total) 10 mM
Paracircmetros V1 V2 V4 V5
part (ppm) -5615 -5748 -5789 -5869
Nas soluccedilotildees de metavanadato as espeacutecies monomeacuterica dimeacutericae tetrameacuterica de vanadato estatildeo
presentes para concentraccedilotildees de metavanadato inferiores a 4 mM Todas estas as espeacutecies
aumentam linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado
em (Aureliano e Madeira 1998) e em (Tiago et al 2004) quando a concentraccedilatildeo de V1 eacute igual ou
superior a 200 microM inicia-se a sua oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica Nas
condiccedilotildees de realizaccedilatildeo dos ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a
espeacutecie pentameacuterica neste estudo em concentraccedilotildees inferiores a 4 mM tendo o mesmo sido
observado por Ramos 2006 Os resultados obtidos mostram claramente o equiliacutebrio entre a
espeacutecie V4 e as espeacuteciesV1 e V2 no qual o aparecimento de outras espeacutecies oligomeacutericas se
encontra dependente da concentraccedilatildeo da espeacutecie V1 Por uacuteltimo pode inferir-se que para
concentraccedilotildees inferiores a 2 mM e intervalos de tempo de cerca de 20 minutos (que inclui o
tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas as
espeacutecies V1 V2 e V4 com concentraccedilotildees respectivas de 0796 0192 e 0196 mM
232Estabilidade das soluccedilotildees de decavanadato anaacutelise por espectroscopia de absorccedilatildeo
UVVis
Caracterizou-se a estabilidade da espeacutecie V10 numa soluccedilatildeo com 1 mM decavanadato total em
tampatildeo D atraveacutes da leitura da Abs400 Obteve-se uma cineacutetica de dissociaccedilatildeo de 1ordf ordem tal
como se pode verificar pela linearidade do graacutefico de ln(Abs400)vs o tempo de aquisiccedilatildeo espectral
(segundos) (Fig26 )
Figura 26ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do tempo
(em segundos) para uma soluccedilatildeo de decavanadato total de 1
mM em tampatildeo D a 25ordmC (n=2)
Sendo uma reacccedilatildeo de primeira ordem pode ser descrita pela equaccedilatildeo cujo
declive eacute ndashK (constante de dissociaccedilatildeo) e a intersecccedilatildeo no eixo das ordenadas corresponde
Ln
(Ab
s 400 n
m)
tempo (s)
29
aln(A0) Numa reacccedilatildeo de 1ordf ordem o tempo de meia-vida eacute independente da concentraccedilatildeo inicial
do reagente sendo descrito pela equaccedilatildeo [2] Deste modo a partir da recta
Ln(Abs400 nm) = -436E-5
K - 105 obteve-se um tempo de meia-vida de 4490 plusmn 0823 horas para
as condiccedilotildees supramencionadas
Apoacutes a adiccedilatildeo de S1 8 microM ao meio reaccional verificou-se que a reacccedilatildeo de desoligomerizaccedilatildeo
da espeacutecie decameacuterica de vanadato seguia igualmente uma cineacutetica de 1ordf ordem tal como se pode
verificar pela linearidade do graacutefico de ln(Abs400) versus o tempo de aquisiccedilatildeo espectral
(segundos) (Fig27)
Figura 27ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do
tempo (em segundos) para uma soluccedilatildeo de decavanadato
total de 1 mM em tampatildeo D a 25ordmC na presenccedila de 8
microM (n= 2)
A partir da equaccedilatildeo da recta Ln(Abs400 nm) = -7520E-5
K - 0613determinou-se o valor de K para
a espeacutecie decameacuterica de onde se obteve um tempo de meia-vida de 2430 plusmn 0191 horas para as
condiccedilotildees supramencionadas
Tanto este valor como o valor registado na ausecircncia de S1 satildeo substancialmente superiores agrave
duraccedilatildeo dos ensaios de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina descritos no presente
relatoacuterio
24 Interacccedilatildeo de S1 com vanaacutedio(V)
241Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
De modo a analisar qual o efeito da adiccedilatildeo de S1 de miosina (tampatildeo D) produzia nos sinais de
51V-NMR titulou-se uma soluccedilatildeo de 5mM de decavanadato total com concentraccedilotildees crescentes
de S1 (5 10 15 e 20 microM) Os espectros de 51
V-RMN encontram-se apresentados na Fig 2
No graacutefico da figura 2 Apresenta-se a concentraccedilatildeo de S1 em funccedilatildeo do factor de alargamento
corrigido dos espectros acima referidos
Ln
(Ab
s (4
00
nm
))
tempo (s)
30
Figura 28 ndash Graacutefico de
representaccedilatildeo da
concentraccedilatildeo de S1 em
funccedilatildeo do factor de
alargamento A
concentraccedilatildeo de
decavanadato total foi de 5
mM e as concentraccedilotildees
utilizadas de S1 foram de 0
5 10 e 15 microM
Verifica-se que o factor de alargamento aumenta linearmente em funccedilatildeo da concentraccedilatildeo de S1 no
ensaio o que indica a existecircncia de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina Observou-
se ainda que a adiccedilatildeo de S1 ao meio reaccional natildeo provocou qualquer alteraccedilatildeo dos desvios
quiacutemicos dos sinais de 51
V-RMN
Realizaram-se ensaios na presenccedila de ATP e de S1 com intuito de verificar o efeito do ATP na
interacccedilatildeo do S1 de miosina com o decavanadato
Figura 29 - Espectros de RMN de 51
V contendo 2
mM Decavanadato (A) (controlo) (B) na presenccedila
de 2 mM de ATP (C) na presenccedila de 2 mM de ATP
e 8 microM de S1 de miosina V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2Vndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies decameacuterica (V10) e monomeacuterica (V1) presentes nos
espectros da soluccedilatildeo de decavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V10A V10B V10C V1
part (ppm) -5161 -5002 -4254 -5614
Observa-se que a presenccedila de ATP (espectro B) natildeo provoca qualquer alteraccedilatildeo visiacutevel em
relaccedilatildeo ao espectro que contem apenas soluccedilatildeo de decavanadato Jaacute a presenccedila simultacircnea de ATP
e S1 provoca um alargamento da ∆12 do sinal do V10A de 1736 Hz (espectro (A)) para 2366 Hz
(espectro (C) Fig 29) Tendo em conta que a concentraccedilatildeo de S1 apresenta uma relaccedilatildeo linear
com o factor de alargamento converteu-se o valor da ∆12 e comparou-se com o valor esperado agrave
F = 00591[S1] - 00157 Rsup2 = 0998
Fact
or
de
ala
rgam
en
to
[S1] (microM)
31
concentraccedilatildeo de S1 utilizada O factor de alargamento obtido para 8 microM ndash 0376 ndash eacute inferior ao
valor esperado agrave mesma concentraccedilatildeo de S1 obtido atraveacutes da equaccedilatildeo da recta apresentada na
Fig 28 ndash 0457 ndash o que indica que o ATP poderaacute exercer um efeito de protecccedilatildeo do S1
diminuindo a sua interacccedilatildeo com a espeacutecieV10
Por uacuteltimo realizaram-se ensaios com metavanadato na presenccedila de 8 microM S1 de modo a verificar
se as espeacutecies oligomeacutericas contidas nesta soluccedilatildeo interagiam com a proteiacutena em estudo
Figura 210 - Espectros de RMN de 51
V contendo 2
mM metavanadato (A) (controlo) (B) na presenccedila
de 8 microM de S1 de miosina V1 V2 e V4
correspondem respectivamente aacutes espeacutecies
monomeacuterica dimeacuterica e tetrameacuterica
Tabela 2VIndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies monomeacuterica (V1) dimeacuterica (V2) e tetrameacuterica (V4)
presentes nos espectros da soluccedilatildeo de metavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V1 V2 V4
part (ppm) -5613 -5747 -5789
Como eacute possiacutevel observar na Fig 210 ocorre uma ligeiro alargamento dos sinal de V4 bem
como uma perda da intensidade pelo que se conclui quedas espeacutecies presentes na soluccedilatildeo de
metavanadato (V1 V2 e V4) apenas a espeacutecie tetrameacuterica interage com o S1 de miosina o que
estaacute de acordo com o reportado por Tiago et al 2002
242 Actividade Mg2+
-ATPaacutesica de S1 de miosina na presenccedila de decanadato
Com o intuito de avaliar o efeito do decavanadato na actividade Mg2+
-ATPaacutesica do S1 de
miosina monitorizou-se a reacccedilatildeo de oxidaccedilatildeo do NADH atraveacutes da leitura de Abs340 na
presenccedila de 005 mgmL de S1 de miosina e no caso da actividade estimulada 2microM de actina em
3125 mM Tris-HCl pH 71 e 375 mM MgCl2 pelo meacutetodo de enzimas acopladas (ver seccccedilatildeo xx
dos MM)Para tal realizaram-se ensaios com vaacuterias concentraccedilotildees de decavanadato total (10 50
100 250 500 e 1000) de modo a determinar qual o seu efeito na actividade MgATPaacutesica de S1
de miosina na ausecircncia e na presenccedila de um dos seus ligandos naturais Os ensaios foram
realizados agrave temperatura ambiente com incubaccedilatildeo preacutevia a 25ordm C durante 20 minutos
32
Figura 211 ndash Representaccedilatildeo
graacutefica da Actividade Mg2+
-
ATPaacutesica de S1 () (os ajustes
teoacutericos estatildeo representado a por
uma linha os resultados
experimentais estatildeo representados
por triacircngulos (Actividade
estimulada) e cruzes (Actividade
natildeo estimulada)) vs
[Decavanadato]total medidas pelo
meacutetodo das enzimas acopladas
(n=3) Os dados foram ajustados ao modelo de decaimento exponencial usando o programa GraphPad Prism para a
determinaccedilatildeo dos valores de IC50 (na ausecircncia de actina IC50= 266 plusmn 07 M decavanadato na presenccedila de actina
IC50= 28 plusmn 1 M decavanadato
A partir do graacutefico presente na Fig 29 verifica-se que o ajuste dos dados experimentais foi bem
sucedido A inibiccedilatildeo da actividade Mg2+
-ATPaacutesica do S1 de miosina eacute afectada em menor
amplitude na presenccedila de actina (60 de inibiccedilatildeo da actividade) do que na sua ausecircncia
(aproximadamente 80 de inibiccedilatildeo de actividade) Devido a esta diferenccedila pode sugerir-se que a
actina iraacute impedir parcialmente a ligaccedilatildeo do decavanadatoao S1 demiosina atraveacutes de
impedimentos esteacutereos ou interacccedilotildees electrostaacuteticas por exemplo A teacutecnica utilizada natildeo nos
permite concluir acerca de que regiatildeo(otildees) ou resiacuteduo(s) da actina participaratildeo na preservaccedilatildeo
parcial da actividade Mg2+
-ATPaacutesica
Por outro lado os valores de IC50 dos dois tipos de actividades natildeo apresentam diferenccedilas
significativas tendo-se obtido 266 plusmn 07 e 28 plusmn 1M decavanadato na ausecircncia e na presenccedila de
actina respectivamente o que corresponde a uma concentraccedilatildeo de cerca de 27 M de V10 O
meio de reacccedilatildeo utilizado nestes ensaios foi 10 mM HEPES e 25 mM KCl com 2 mM ATP Em
(Tiago et al 2004) o valor de IC50 para a actividade Mg2+-ATPaacutesica na ausecircncia de actina foi de
125 plusmn 025 microM enquanto o valor da actividade estimulada por actina foi de 136 plusmn 14 microM O
meio de reacccedilatildeo utilizado nos ensaios foi 20 mM HEPES 50 mM KCl e 2 mM ATP o que resulta
precisamente no dobro da forccedila ioacutenica a que foi realizado o ensaio do presente relatoacuterio
(exceptuando a contribuiccedilatildeo dos produtos da hidroacutelise de ATP uma vez que este meacutetodo
pressupotildee a ocorrecircncia de um ciclo de reacccedilotildees de hidroacutelise e siacutentese deste composto) Ora tendo
sido anteriormente referido que a afinidade do S1 de miosina para com a espeacutecie V10 diminui com
Act
ivid
ade
Mg
2+-A
TP
aacutesic
a d
e S
1 d
e
Mio
sin
a
[Decavanadato] (microM) Actividade Estimulada ()
Actividade Natildeo Estimulada ()
Ajuste Actividade Estimulada
Ajuste Actividade Natildeo Estimulada
33
o aumento da forccedila ioacutenica (Tiago et al 2004 tal poderaacute explicar a diferenccedila obtida entre estes
dois conjuntos de valores
243Quantificaccedilatildeo de cisteiacutenas reduzidas pelo meacutetodo de Ellman
A interacccedilatildeo do vanaacutedio com alguns tipos de aminoaacutecidos sensiacuteveis agrave oxidaccedilatildeo pode fornecer-nos
informaccedilatildeo bastante uacutetil em relaccedilatildeo agraves alteraccedilotildees estruturais que conduzem a uma diminuiccedilatildeo da
actividade fisioloacutegica do S1 de miosina Os estudos realizados por Chasteen 1983 concluiacuteram
que a reduccedilatildeo de vanadato (V) a vanadilo (IV) ocorre na presenccedila de agentes redutores comuns no
ambiente celular de que se destaca o aminoaacutecido cisteiacutena Esta eacute tambeacutem um alvo de
complexaccedilatildeo por parte do iatildeo VO2+
Jaacute Ramos et al (2009) demonstrou que a incubaccedilatildeo de
actina com a soluccedilatildeo de decavanadato induz a oxidaccedilatildeo de resiacuteduos de cisteiacutena e agrave formaccedilatildeo do
oxidovanaacutedio (IV) Este grupo de investigadores descobriu ainda que apenas a espeacutecie V10 induz a
oxidaccedilatildeo das cisteiacutenas ldquoraacutepidasrdquo da actina atraveacutes do processo proposto que eacute parcialmente
responsaacutevel pela inibiccedilatildeo jaacute descrita da actividade de ATPase do S1 de miosina estimulada por
actina quando exposta agrave soluccedilatildeo de decavanadato Esta inibiccedilatildeo apresenta-se afectada com a
presenccedila de ATP uma vez que os resiacuteduos de cisteiacutena permaneceram intactos nos ensaios
realizados com este ligando Jaacute Crans et al (2004) concluiu que o siacutetio activo de muitas fosfatases
(que conteacutem grupos tiol) pode formar um equiliacutebrio redox com o iatildeo vanadato (V) de forma natildeo
reversiacutevel
Os resultados obtidos por Sakurai e Shiromura (1980) sugeriram que o vanaacutedio (V) sofre reduccedilatildeo
a vanaacutedio (IV) sem dependecircncia do valor de pH do ensaio Registaram ainda uma estequiometria
de 31 na reacccedilatildeo do vanadato (V) com os resiacuteduos de Cys enquanto a estequiometria dos ensaios
com o vanadilo (IV) foi de 21
Berlett e Stadtman (1997) observaram que os tipos de aminoaacutecidos mais facilmente oxidaacuteveis
englobam entre outros os resiacuteduos de Cys Trp e MetAssim analisou-se o estado redox dos
grupos tiol dos resiacuteduos de Cys do S1 na presenccedila do decavanadato atraveacutes da monitorizaccedilatildeo da
Abs412 nm na presenccedila de 1 mgmL de S1 de miosina em tampatildeo 10 mM HEPES 25 mM KCl
variando a concentraccedilatildeo da soluccedilatildeo de decavanadato de modo a determinar que tipo de alteraccedilotildees
estruturais ocorrem na sua presenccedila no que diz respeito agrave diminuiccedilatildeo da actividade ATPaacutesica de
S1 na presenccedila de decavanadato
Os resultados referentes agrave quantificaccedilatildeo das cisteiacutenas reduzidas apoacutes o tratamento de S1 com
diferentes concentraccedilotildees de decavanadato em tampatildeo 10 mM HEPES pH 75 e 25 mM KCl a
25ordmC durante 20 minutos encontram-se descritos na Tabela 2VII
Tabela 2VII ndash Cisteiacutenas reduzidas totais e ldquoraacutepidas de S1 (1 mgmL) apoacutes tratamento com decavanadato
34
(010501002505001000 uM) em 10 mM HEPES 25 mM KCl pH 7 a 25ordmC durante 20 minutos A detecccedilatildeo
espectrofotomeacutetrica foi feita com DTNB (e 412=8400 uM-1 cm-1) (n variaacutevel entre 1 e 8)
Amostras Cys raacutepidas Cys totais
11 M S1 (controlo) 22 plusmn 04 106 plusmn 14
11 M S1 + 10 M Decavanadato 22 plusmn 05 95 plusmn 08
11 M S1 + 50 uM Decavanadato 20 plusmn 02 98 plusmn 14
11 M S1 + 100 uM Decavanadato 17 plusmn 03 97 plusmn14
11 M S1 + 250 uM Decavanadato 26 plusmn 02 98 plusmn 08
11 M S1 + 500 uM Decavanadato 21 plusmn 04 90 plusmn 06
11 M S1 + 1000 uM Decavanadato 24 plusmn 0 86 plusmn 08
A partir dos resultados obtidos pode sugerir-se que o S1 possui 11 resiacuteduos de Cys (controlo) o
que estaacute de acordo com o anteriormente descrito Verifica-se que o tratamento da proteiacutena com
concentraccedilotildees crescentes de decavanadato provoca a oxidaccedilatildeo de dois resiacuteduos de Cys do S1 de
miosina natildeo expostos ao solvente No entanto a teacutecnica utilizada natildeo nos permite identificar
quais
Tendo em consideraccedilatildeo que o valor de IC50 obtido para a reacccedilatildeo de inibiccedilatildeo daactividade Mg2+-
ATPaacutesica de S1 na presenccedila de decavanadato eacute de cerca de 27 microM pode sugerir-se que este seraacute o
valor ao qual ocorreraacute a oxidaccedilatildeo do primeiro resiacuteduo de Cys o que poderaacute contribuir para a
diminuiccedilatildeo da actividade de S1 observada
25Interacccedilatildeo de S1 com vanaacutedio(IV)
251Anaacutelise por espectroscopia de Ressonacircncia Paramagneacutetica Electroacutenica
A acumulaccedilatildeo intracelular de oxovanaacutedio (IV) in vivo constitui uma das principais formas de
apresentaccedilatildeo de vanaacutedio no organismo Deste modo torna-se bastante interessante analisar a
interacccedilatildeo entre este iatildeo e o S1 de miosina atraveacutes da teacutecnica de espectroscopia de Ressonacircncia
Paramagneacutetica Electroacutenica o meacutetodo de eleiccedilatildeo do estudo do iatildeo vanadilo e dos complexos por si
formados O electratildeo desemparelhado encontra-se fortemente acoplado ao nuacutecleo 51
V o que
35
resulta num espectro de RPE consistindo de 8 desdobramentos na direcccedilatildeo paralela e outros tantos
na direcccedilatildeo perpendicular (Chasteen 1983)
Os estudos realizados por Ahmed et al (1987) centraram-se na interacccedilatildeo do vanadilo com
proteiacutenas como eacute o caso da insulina transferrina entre outros obtendo um espectro caracterizado
por 8 linhas amplamente espaccediladas (na direcccedilatildeo paralela) Jaacute Ramos et al 2009 concluiu que a
exposiccedilatildeo da soluccedilatildeo de decavanadato agraves formas globular e filamentosa de actina produz um
espectro tiacutepico do iatildeo oxidovanaacutedio (IV) indicando a reduccedilatildeo do vanadato
Para aferir qual a concentraccedilatildeo de S1 que permitia obter uma boa razatildeo sinalruiacutedo titulou-se uma
soluccedilatildeo de sulfato de vanadilo 15 microM ([VOSO4]stock = 5 mM) com concentraccedilotildees crescentes de
proteiacutena (0 5 10 15 e 20 microM) em 10 mM HEPES 25 mM KCl A existecircncia de uma correlaccedilatildeo
linear entre a concentraccedilatildeo de proteiacutena e a intensidade dos sinais de RPE correspondente agrave banda
-12 perpendicular encontra-se representada na Fig 210
Figura 2 12ndash Representaccedilatildeo graacutefica da variaccedilatildeo da intensidade do sinal de RPE vs a concentraccedilatildeo de S1 utilizada
Apoacutes verificar que com uma concentraccedilatildeo de S1 igual a 20 microM se obtinham sinais com uma razatildeo
sinalruiacutedo adequada ao prosseguimento dos estudos sobre a interacccedilatildeo V(IV)-S1 de miosina
titulou-se uma soluccedilatildeo de 20 microM S1 com 0 25 5 10 15 25 50 75 100 150 200 500 e 1000
microM VOSO4
Traccedilou-se o graacutefico da intensidade da banda -12 perpendicular do sinal de EPR em funccedilatildeo da
concentraccedilatildeo de VOSO4 tendo-se determinado uma constante de dissociaccedilatildeo (KD) do complexo
S1-vanadilo de 8748 plusmn 2621 microM V(IV)
y = 016786x + 122435
Rsup2 = 092992759
Inte
nsi
dad
e d
o s
inal
de
RP
E
[S1] (microM)
36
Figura 213 ndash Representaccedilatildeo graacutefica da variaccedilatildeo do sinal de RPE vs a concentraccedilatildeo de vanadilo utilizada (os valores
experimentais estatildeo representados por losangos o seu ajuste estaacute representado pela linha)
A simulaccedilatildeo do espectro de EPR foi efectuada a partir do espectro obtido para uma concentraccedilatildeo
de 1 mM VOSO4 apoacutes adiccedilatildeo de 20 microM S1 de miosina tendo-se obtido os valores da constante de
acoplamento hiperfina (A=5016) e do factor g (g0=195)
Figura 214 ndash Representaccedilatildeo graacutefica do espectro de EPR obtido com 1000 microM de vanadilo e 20 microM de S1 (a cinzento
escuro) A simulaccedilatildeo do espectro encontra-se a cinzento claro
Por comparaccedilatildeo destes com os valores presentes na tabela de correspondecircncia xxx eacute possiacutevel
predizer quais os ligandos provaacuteveis do vanadilo na presenccedila de proteiacutena Assim concluiu-se que
o vanadilo estaacute coordenado com duas moleacuteculas de aacutegua um grupo hidroxilo e um grupo R=NR
O grupo hidroxilo pode indicar os resiacuteduos de aminoaacutecidos Tyr Treonina e Serina enquanto o
grupo R=NR poderaacute indicar a presenccedila de Arginina e Histidina como ligandos do vanadilo
Inte
nsi
dad
e d
os
sinai
s d
e R
PE
[Vanadilo] (microM)
Ajuste dos dados exp Dados experimentais
Campo magneacutetico (G)
Espectro Experimental Ajuste
37
3 Procedimento Experimental
31 Reagentes
Todos os reagentes utilizados satildeo de elevado grau de pureza (analiacutetico) Apresentam-se as suas
foacutermulas quiacutemicas ou abreviaturas quando necessaacuterio acrescentou-se a sua descriccedilatildeo Entre
parecircntesis colocou-se as empresas que os comercializam
Aacutecido aceacutetico glacial 100 (Riedel) Acrilamida 99 (Merck) Azul de bromofenol (Riedel-de-
Haumlen) -Mercaptoetanol (Sigma) Bissulfito de soacutedio (Sigma)Bradford (reagente de)
(BioRad)BSA (albumina de soro bovino Sigma-Aldrich) CaCl22H2O (cloreto de caacutelcio
dihidratado Fluka) CH3CH2OH 96 (etanol Panreac) Coomassie Blue R250 (Merck) D2O
(oacutexido de deuteacuterio SDS) DTNB (55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico) Aldrich) EDTA (aacutecido
etilenodiaminatetraaceacutetico Merck) Fosfoenolpiruvato 98 (Fluka) Glicerol (Sigma) Glicina
985 (Panreac) HCl 37 (Riedel-de Haumlen) H2SO4 96 (Pronalab) HEPES 99 (aacutecido 4-(2-
hidroxiletil)-1-piperazineetanosulfoacutenico Sigma) KH2PO4 99 (hidrogenofosfato de potaacutessio)
MgATP (Sigma) Na
ATP (Sigma) Na2HPO4 99 (hidrogenofosfato de soacutedio Riedel-de Haumlen)
NaH2PO4 (fosfato de soacutedio ) NaOH 986 (hidroacutexido de soacutedio Pronalab) KCl 99 (Cloreto de
potaacutessio Panreac) L-Cys 995 (enantioacutemero levoacutegiro do resiacuteduo de Cisteiacutena Fluka)Metanol
99 (Merck) Molibdato de amoacutenio (Sigma)NNrsquo-metil-bis-acrilamida 98 (Merck) NH4VO3
99 (metavanadato de amoacutenia Sigma) PSA 10 (persulfato de amoacutenio Merck) SDS
(dodecilo sulfato de soacutedio Sigma) Sulfito de soacutedio (Sigma) VOSO4 (sulfato de vanadilo
Aldrich) TEMED 99 (Tetrametiletilenodiamina Merck) Tris (2-amino-2-hidroximetil-13-
propanodiol Sigma)
32 Meacutetodos
321 Isolamento e purificaccedilatildeo das proteiacutenas de muacutesculo esqueleacutetico de coelho
3211 Purificaccedilatildeo da miosina preparaccedilatildeo de Subfragmento 1 por digestatildeo quimotriacutepsica
da miosina
O subfragmento-1 de miosina-II (S1) foi gentilmente cedido pelo Prof Doutor Manuel Aureliano
do Departamento de Quiacutemica Bioquiacutemica e Farmaacutecia da Faculdade de Ciecircncias e Tecnologia da
Universidade do Algarve
No entanto descreve-se abaixo de forma resumida o procedimento efectuado
38
32111 Purificaccedilatildeo da miosina
O protocolo de isolamento foi adaptado do meacutetodo previamente descrito por Margossian e Lowey
(1982) apresentando-se de forma esquemaacutetica na Fig 31
Sucintamente os tecidos musculares dorsais e das pernas foram extraiacutedos apoacutes o sacrifiacutecio do
animal (Fig31) De seguida foram adicionados 3 volumes da soluccedilatildeo de extracccedilatildeo A2
Centrifugou-se a soluccedilatildeo e filtrou-se o sobrenadante O sedimento foi entatildeo diluiacutedo em 14
volumes de aacutegua destilada de modo a que a forccedila ioacutenica atingisse 004 M Apoacutes adiccedilatildeo de szlig-
mercaptoetanol (szlig-me) deixou-se a precipitar a 4ordmC removendo o sobrenadante Centrifugou-se
novamente e ressuspendeu-se o sedimento na soluccedilatildeo de extracccedilatildeo A Adicionou-se a soluccedilatildeo
tampatildeo B3 para que a forccedila ioacutenica atingisse 05 M Homogeneizou-se e deixou-se repousar
durante 2h Homogeneizou-se novamente e centrifugou-se de modo a ocorrer precipitaccedilatildeo dos
complexos de actomiosina e proteiacutena desnaturada Apoacutes filtraccedilatildeo do sobrenadante em latilde de vidro
quantificou-se a soluccedilatildeo de miosina resultante pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental)
2 Soluccedilatildeo de extracccedilatildeo A 0012M K2CO3 004M KHCO3 06M KCl pH 65
3 Soluccedilatildeo-tampatildeo B Tris 100 mM pH 7 KCl 2M
39
Figura 31 ndash Representaccedilatildeo esquemaacutetica do protocolo de isolamento de miosina (Margossian e Lowey 1982) Todo o
procedimento foi realizado a 4ordmC (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
32112 Preparaccedilatildeo de Subfragmento-1 de miosina por digestatildeo com quimiotripsina
O S1 de miosina foi isolado apoacutes diaacutelise da miosina durante a noite contra a soluccedilatildeo C4 A
presenccedila do EDTA eacute essencial uma vez que a sua substituiccedilatildeo por MgCl2resulta na produccedilatildeo de
HMM em vez de S1 A suspensatildeo de miosina foi equilibrada e digerida com -quimotripsina
A digestatildeo foi interrompida pela adiccedilatildeo de PMSF Seguidamente ultracentrifugou-se o produto
da digestatildeo desprezando-se o sedimento (Fig 32) A soluccedilatildeo de sobrenadante maioritariamente
S1 foi quantificada pela leitura directa da Abs280 ou pelo meacutetodo de Bradford (ver as secccedilotildees
3311 e 3312 do Procedimento Experimental respectivamente) e armazenada em 10 sacarose
e 2 mM szlig-Me a -80ordmC
4 Soluccedilatildeo C 25mM Tris pH 70 012M KCl 1mM EDTA
40
Antes da realizaccedilatildeo dos ensaios descritos no presente relatoacuterio o S1 de miosina foi dialisado
contra a soluccedilatildeo D5 de modo a remover qualquer composto que pudesse complexar como
vanaacutedio Seguidamente quantificou-se a proteiacutena pelo meacutetodo de Bradford ou pelo meacutetodo
directo (ver secccedilotildees 3311 e 3312do Procedimento Experimtental)
Figura 3 2ndash Representaccedilatildeo esquemaacutetica do protocolo de preparaccedilatildeo do S1 de miosina por digestatildeo quimiotripsina de
acordo com o previamente descrito por Margossian e Lowey (1982) Todo o procedimento foi realizado a 4ordm C
excepto quando indicaccedilatildeo do contraacuterio (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
3212 Isolamento e purificaccedilatildeo de actina
Isolou-se e purificou-se a actina segundo o meacutetodo descrito por Pardee e Spudish que
compreende dois passos ndash a obtenccedilatildeo de um resiacuteduo seco denominado ldquopoacute de acetonardquo e a
extracccedilatildeo subsequente da actina a partir do ldquopoacute de acetonardquo na forma de G-Actina com a
posterior polimerizaccedilatildeo a F-Actina
32121 Obtenccedilatildeo do ldquopoacute de acetonardquo
Sacrificou-se o coelho com uma pancada atraacutes do pescoccedilo e sangrou-se o animal apoacutes o corte da
arteacuteria jugular Esfolou-se e colocou-se numa tina com gelo Retiraram-se as partes de tecido
conjuntivo que envolvem o tecido muscular com auxiacutelio de uma pinccedila e bisturi Seguidamente
retirou-se os muacutesculos dorsais e os muacutesculos brancos das pernas Passou-se os muacutesculos por aacutegua
destilada de modo a remover as impurezas e pesou-se num copo Trituraram-se os muacutesculos
numa maacutequina de triturar carne e extraiu-se o resultante com 3 volumes da soluccedilatildeo E6 durante 10
minutos com agitaccedilatildeo suave Centrifugou-se a suspensatildeo a 4000 g durante 10
5 Soluccedilatildeo D HEPES 10 mM pH 70 KCl 25 mM
6 Soluccedilatildeo E 01 M KCl 015 M K2HPO4 pH 65
41
minutosdesprezando-se o sobrenadante Ressuspendeu-se o sedimento em 6 volumes de soluccedilatildeo
F7durante 10 minutos com agitaccedilatildeo suave Verificou-se se o pH da soluccedilatildeo estava compreendido
entre 82 e 85 acertando-se estecom soluccedilatildeo G8 se necessaacuterio Centrifugou-se durante 10
minutos a 4000 g Desprezou-se o sobrenadante e ressuspendeu-se o sedimento em 3 volumes de
soluccedilatildeo H9 durante 10 minutos Centrifugou-se novamente a 4000 gdurante 10 minutos
desprezando-se o sobrenadante Ressuspendeu-se o sedimento duas vezes consecutivas em 6
volumes de aacutegua destilada durante 5 minutos Centrifugou-se a 4000 g durante 10 minutos apoacutes
cada adiccedilatildeo de H2O
Seguidamente ressuspendeu-se o sedimento 5 vezes consecutivas utilizando 3 volumes de
acetona de cada vez agitando suavemente durante 10 minutos agrave temperatura ambiente Colocou-
se o resiacuteduo filtrado numa tina de vidro sobre papel de filtro deixando-se evaporar o solvente
durante a noite num nicho Triturou-se o resiacuteduo seco numa maacutequina de triturar carne e deixou-se
novamente evaporar o solvente O ldquopoacute de acetonardquo obtido foi armazenado a -20 ordmC
32122 Purificaccedilatildeo da actina a partir de ldquopoacute de acetonardquo
Apoacutes a obtenccedilatildeo do poacute de acetona como acima indicado efectuou-se o protocolo seguidamente
descrito de modo a obter monoacutemeros de actina (G-Actina) e actina filamentosa (F-Actina)
Pesou-se 35 g de poacute de acetona e adicionaram-se 50 mL de soluccedilatildeo-tampatildeo I10
sendo a
extracccedilatildeo feita com agitaccedilatildeo suave durante 30 minutos Separou-se o extracto do poacute de acetona
hidratado filtrando a suspensatildeo com 4 camadas de gaze esterilizada tendo o cuidado de natildeo
espremer o resiacuteduo Voltou a extrair-se com 50 mL de soluccedilatildeo I durante 30 minutos mexendo
lentamente Adicionou-se este ao extracto obtido anteriormente Centrifugou-se o extracto total a
16 000 g durante 30 minutos Decantou-se para uma proveta sobre a qual se colocou gaze para
que o sedimento solto natildeo se juntasse ao sobrenadante e leu-se o volume De seguida adicionou-
se KCl ateacute uma concentraccedilatildeo final de 50 mM MgCl2 ateacute 2mM e ATP ateacute 1mM lentamente e com
agitaccedilatildeo contiacutenua de modo a promover a polimerizaccedilatildeo dos monoacutemeros de actina
Deixou-se polimerizar durante 2h a 25ordmC observando-se a presenccedila de pequenas bolhas de ar
indicativas de um aumento de viscosidade da soluccedilatildeo Arrefeceu-se em gelo durante 15 minutos e
adicionou-se KCl ateacute uma concentraccedilatildeo final de 06M Mexeu-se lentamente durante 30 minutos
apoacutes os quais se ultracentrifugou a 110 000g durante 1hora Desprezou-se o sobrenadante
ressuspendeu-se o sedimento com soluccedilatildeo tampatildeo E e transferiu-se para um potter
Dialisou-se durante a noite contra 1L de soluccedilatildeo-tampatildeo I a uma temperatura de 4ordmC com rotaccedilatildeo
7 Soluccedilatildeo F 005 M NaHCO3 pH 82-85
8 Soluccedilatildeo G 1M Na2CO3
9 Soluccedilatildeo H 1 mM EDTA pH 70
10 Soluccedilatildeo I 2 mM Tris-HCl pH 8 02 mM CaCl2 02 mM Na2ATP 05 mM szlig-Me 0005 NaN3
42
contiacutenua e mudanccedila de tampatildeo 2 a 3 vezes De seguida a actina foi centrifugada a 110
000gdurante 1hora Decantou-se a soluccedilatildeo de G-Actina (sobrenadante) e desprezou-se o
sedimento Determinou-se a concentraccedilatildeo de G-Actina pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental) A soluccedilatildeo de G-actina foi armazenada a uma temperatura de 4ordmC
apoacutes adiccedilatildeo de 002NaN3 mantendo-se estaacutevel durante 3 a 5 dias
331 Quantificaccedilatildeo das proteiacutenas isoladas
3311 Quantificaccedilatildeo pelo meacutetodo de Bradford
O meacutetodo de Bradford (Bradford 1976) baseia-se na utilizaccedilatildeo do reagente de Bradford
composto por entre outros aacutecido fosfoacuterico e o corante ldquoCoomassierdquo Este corante possui carga
negativa e a sua coloraccedilatildeo varia entre a vermelha (Absmax=465 nm) predominante em soluccedilatildeo e a
azul (Absmax=595 nm) presente aquando da ligaccedilatildeo aos grupos amina das proteiacutenas Em soluccedilotildees
aacutecidas diluiacutedas a coloraccedilatildeo varia proporcionalmente com a quantidade do corante ligado agrave
proteiacutena que por sua vez eacute directamente proporcional agrave concentraccedilatildeo da proteiacutena permitindo a
sua quantificaccedilatildeo ateacute a um limite de detecccedilatildeo de 20 gmL (Bradford 1976)
Deste modo prepararam-se amostras de BSA ([BSA]stock= 1mgmL) utilizando um intervalo de
concentraccedilotildees compreendido entre 0 e 15 gmL de acordo com a tabela 3I Apoacutes a adiccedilatildeo da
H2O MilliQ e do reagente de Bradford as soluccedilotildees foram sucessivamente homogeneizadas no
voacutertex
Tabela 3Indash Amostras utilizadas para a obtenccedilatildeo da recta de calibraccedilatildeo no meacutetodo de Bradford
BSA (L) BSA (g) H2O MilliQ (L) Reagente de Bradford (L)
0 0 800 200
125 125 79875 200
25 25 7975 200
5 5 795 200
10 10 790 200
15 15 785 200
Apoacutes incubaccedilatildeo a 25ordmC por 5 minutos leram-se os valores de Abs595 num
espectrofotoacutemetro (Shimadzu UV-160A) para todas as amostras Seguidamente traccedilou-se a recta
de calibraccedilatildeo (Abs595vs quantidade de BSA (g))
43
Para determinar a concentraccedilatildeo de proteiacutena apoacutes o processo de isolamento purificaccedilatildeo e
diaacutelise quando necessaacuterio colocou-se 5 L da amostra num tubo de Eppendorf e adicionou-se
795 L de H2O MilliQ homogeneizando a soluccedilatildeo no voacutertex Adicionou-se posteriormente 200
L de reagente de Bradford e repetiu-se a homogeneizaccedilatildeo Incubou-se a mistura a 25ordmC por 5
minutos apoacutes os quais se leu os valores de Abs595 nm no espectrofotoacutemetro Por interpolaccedilatildeo deste
valor a partir da recta de calibraccedilatildeo determinou-se a concentraccedilatildeo de proteiacutena na amostra
3312 Quantificaccedilatildeo pelo meacutetodo directo
A quantificaccedilatildeo das amostras proteicas pode tambeacutem ser efectuada atraveacutes da leitura directa da
absorvacircncia tendo por base a lei de Beer-Lambert onde Abs corresponde agrave
absorvacircncia ao comprimento de onda ao qual o ensaio foi realizado corresponde ao coeficiente
de extinccedilatildeo (substituiacutedo pela letra a se as unidades forem distintas da molaridade) b corresponde
ao percurso oacuteptico da radiaccedilatildeo na ceacutelula amostral e c corresponde agrave concentraccedilatildeo amostral
Assim no caso de S1 de miosina mediu-se a Abs280e calculou-se a concentraccedilatildeo da proteiacutena
sabendo que o coeficiente de extinccedilatildeo de S1 eacute (Ref)
No caso da actina mediu-se a Abs290ecalculou-se a concentraccedilatildeo da proteiacutena sabendo que o
coeficiente de extinccedilatildeo da actina eacute (Ref)
332Electroforese em condiccedilotildees desnaturantes (SDS-PAGE)
De modo a melhor caracterizar as proteiacutenas isoladas foi realizada uma electroforese em
condiccedilotildees desnaturantes (SDS-PAGE do inglecircs Sodium Dodecyl Sulfate Polyacrilamide Gel
Electrophoresis) O SDS (dodecilo sulfato de soacutedio) eacute um detergente anioacutenico utilizado neste
meacutetodo para desnaturar as proteiacutenas que se ligam ao SDS na proporccedilatildeo 14 g SDS g proteiacutena
(aproximadamente 1 moleacutecula de SDS por cada 2 resiacuteduos de aminoaacutecidos) (Stryer) O SDS induz
uma densidade de carga negativa significativa nas proteiacutenas a que se liga mascarando a sua carga
intriacutenseca pelo que as razotildees cargamassa satildeo tendencialmente idecircnticas Assim a SDS-PAGE
separa as proteiacutenas quando sujeitas a um campo eleacutectrico de acordo com o seu peso molecular
sendo a sua migraccedilatildeo directamente proporcional ao logaritmo deste uacuteltimo (Stryer)
O gel de resoluccedilatildeo com 10 de acrilamida foi obtido atraveacutes da adiccedilatildeo de 167 mL de soluccedilatildeo
stock de acrilamida11
075 mL de tampatildeo de resoluccedilatildeo12
251 mL de aacutegua destilada e 50 L de
SDS 10 Para promover a polimerizaccedilatildeo adicionou-se em seguida 38 L de PSA 10
11
soluccedilatildeo stock de acrilamida soluccedilatildeo de 30 (mv) de acrilamida e 08 (mv) de NNrsquo-metil-bis-acrilamida 12
tampatildeo de resoluccedilatildeo 15 M Tris-HCl pH 88
44
(persulfato de amoacutenia) 10 e 25 L de TEMED 99 (tetrametiletilenodiamina) Agitou-se
cuidadosamente de modo a natildeo formar bolhas e pipetou-se a soluccedilatildeo para a cassette de vidro
previamente montada no suporte De seguida adicionou-se uma peliacutecula de cerca de 2 mm de 2-
metil-2-butanol para verificar posteriormente a polimerizaccedilatildeo Apoacutes a polimerizaccedilatildeo que demora
cerca de 45 minutos verteu-se a agrave superfiacutecie e adicionou-se o gel de concentraccedilatildeo com 5 de
acrilamida constituiacutedo por 03 mL de soluccedilatildeo de stock acrilamida 045 mL de tampatildeo de
concentraccedilatildeo13
094 mL de aacutegua destilada 18 L de SDS 10 135 L de PSA 10 e 2L de
TEMED 99 Colocou-se o pente sobre o gel e deixou-se polimerizar durante cerca de 45
minutos
As amostras para aplicaccedilatildeo no gel de electroforese foram preparadas para uma concentraccedilatildeo de
proteiacutena de 1 mgmL num volume final de 100 L dos quais 25 L correspondem ao tampatildeo da
amostra14
As amostras foram aquecidas a 100ordmC durante 5 minutos Antes da aplicaccedilatildeo das
amostras transferiu-se a cassette para a tina de electroforese agrave qual se adicionou a soluccedilatildeo
tampatildeo de electroforese da tina superior15
e tampatildeo de electroforese da tina inferior16
Aplicaram-
se 10 L das soluccedilotildees de marcadores de peso molecular e 20 L das amostras proteicas
preparadas anteriormente A electroforese decorreu durante cerca de 3h30 min a uma voltagem
de 80 V De seguida transferiu-se o gel para uma tina corando as proteiacutenas durante 1h com
soluccedilatildeo corante17
Descorou-se o gel com soluccedilatildeo descorante18
tendo-se mudado esta soluccedilatildeo ateacute
as bandas se encontrarem bem definidas
Determinou-se a mobilidade relativa das bandas correspondentes aos marcadores de elevado peso
molecular e traccedilou-se a recta de calibraccedilatildeo ndash log(PM) vs log(Rf) onde Rf corresponde agrave
mobilidade relativa Por interpolaccedilatildeo determina-se o PM das proteiacutenas isoladas sabendo a sua
mobilidade relativa Por comparaccedilatildeo com os valores descritos na literatura pode concluir-se
acerca do grau de purificaccedilatildeo
34 Mediccedilatildeo da actividade enzimaacutetica de S1
341 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
13
tampatildeo de concentraccedilatildeo 05 M Tris-HCl pH 68 14
tampatildeo de amostra 320 mM Tris-HCl pH 68 04 M szlig-mercaptoetanol SDS 8 glicerol 15 (vv) e roxo de m-
cresol 0024 15
tampatildeo de electroforese da tina superior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS 16
tampatildeo de electroforese da tina inferior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS acetato de soacutedio
01 M 17
soluccedilatildeo corante ldquoCoomassie Blue R-250rdquo contendo metanol 45 (vv) e aacutecido aceacutetico glacial 10 (vv) 18
soluccedilatildeo descorante metanol 10 (vv) e aacutecido aceacutetico glacial 10 vv)
45
O meacutetodo de Fiske-Subarow modificado consiste na reacccedilatildeo do molibdato de amoacutenio com o
fosfato inorgacircnico em soluccedilatildeo aacutecida com formaccedilatildeo do aacutecido fosfomoliacutebdico Seguidamente daacute-
se a adiccedilatildeo de um agente redutor que provoca a reduccedilatildeo do composto a fosfomolibdato que forma
um complexo de cor azulada cuja absorvacircncia maacutexima ocorre para um comprimento de onda de
660 nm tornando possiacutevel a sua detecccedilatildeo por meacutetodos espectrofotomeacutetricos As actividades Ca2+
-
ATPaacutesica e K+EDTA-ATPaacutesica de S1 foram determinadas atraveacutes deste meacutetodo de acordo com
o descrito por Taussky e Shorr (1953) com mediccedilatildeo da quantidade de fosfato (Pi) libertado
atraveacutes da hidroacutelise de ATP
Para tal obteve-se primeiramente a recta de calibraccedilatildeo com as amostras presentes na Tabela 3II
utilizando uma soluccedilatildeo stock de 5 mM KH2PO4
Tabela 3II ndash Composiccedilatildeo das diferentes amostras utilizadas para determinar a recta de calibraccedilatildeo atraveacutes do meacutetodo
de Fiske-Subbarow modificado
KH2PO4 (L)
([KH2PO4]stock= 5mM)
PO43-
(nmole) H2O (uL)
0 0 1780
5 25 1775
10 50 1770
20 100 1760
40 200 1740
100 500 1680
A cada tubo de Eppendorf adicionaram-se seguidamente 200 L de soluccedilatildeo J19
e 20 L de agente
redutor20
homogeneizando no voacutertex apoacutes cada adiccedilatildeo Incubou-se durante 10 minutos e mediu-
se a Abs660 nm num espectrofotoacutemetro com intervalos de tempo iguais entre cada amostra Traccedilou-
se a recta de calibraccedilatildeo (Abs660 nmvs nmol PO43-
) a partir dos valores obtidos
Para determinar a actividade Ca2+
-ATPaacutesica de S1 prepararam-se tubos de Eppendorf com 400
L de meio de reacccedilatildeo21
25 L CaCl2 ([CaCl2]stock= 100 mM) 25 L ATP ([ATP]stock= 100 mM)
com [S1]ensaios=01 mgmL Perfez-se se necessaacuterio com H2O MilliQ ateacute perfazer um volume de 1
mL Para determinar a actividade K+EDTA-ATPaacutesica utilizou-se uma concentraccedilatildeo idecircntica de
19
Soluccedilatildeo J 5 N H2SO4 25 molibdato de amoacutenio 20
Agente redutor 5 mg aacutecido 1-amino-2-naftol-4-sulfoacutenico 30 mg bissulfito de soacutedio 30 mg sulfito de soacutedio em 26
mL H2O 21
meio de reacccedilatildeo 125 mM Tris-HCl pH 80 0575 M KCl
46
proteiacutena nos ensaios (01 mgmL) e o mesmo meio de reacccedilatildeo Aos tubos de Eppendorf
adicionou-se ainda 123 L de KCl ([KCl]stock = 3M) 25 L de ATP 10 L de EDTA
([EDTA]stock= 100 mM )e o volume de H2O MilliQ necessaacuterio para perfazer 1 mL Em ambas as
situaccedilotildees o uacuteltimo reagente adicionado foi o ATP uma vez que se trata do substrato da reacccedilatildeo
enzimaacutetica
A cineacutetica decorreu a 25ordmC durante 5 minutos para a actividade Ca2+
-ATPaacutesica e durante 3
minutos para a actividade K+EDTA-ATPaacutesica Apoacutes o periacuteodo de incubaccedilatildeo transferiu-se 150
L de amostra para tubos de Eppendorf contendo igual volume de TCA 10 Colocou-se no gelo
e homogeneizou-se no voacutertex Seguidamente centrifugou-se a 1500 g durante 10 minutos e
recolheu-se 200 L de sobrenadante para novos tubos de Eppendorf Adicionou-se a estes 158
mL de H2O MilliQ e 200 L de soluccedilatildeo J Homogeneizou-se no voacutertex e adicionou-se 20 L de
reagente redutor repetindo-se a homogeneizaccedilatildeo Incubou-se durante 10 minutos e mediu-se a
Abs660 nm com o mesmo intervalo de tempo de leitura para cada amostra Por intrapolaccedilatildeo a partir
da recta de calibraccedilatildeo obtida anteriormente determinou-se a quantidade de Pi libertado em cada
ensaio A partir destes resultados determinaram-se as actividades Ca2+
-ATPaacutesica e K+EDTA-
ATPaacutesica do S1 em nmol PO43-
minmg proteiacutena e em s-1
342 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
Da actividade Mg2+
-ATPaacutesica de S1 resulta a formaccedilatildeo dos produtos da reacccedilatildeo de
hidroacutelise do ATP - ADP e Pi Na presenccedila de ADP fosfoenolpiruvato (PEP) e da enzima piruvato
cinase (PK) os produtos resultantes satildeo ATP e piruvato Por acccedilatildeo da enzima lactato
desidrogenase (LDH) este uacuteltimo eacute convertido em lactato ocorrendo simultaneamente a oxidaccedilatildeo
do NADH a NAD+ monitorizada atraveacutes da Abs340 nm Quando o S1 de miosina se encontra no
meio reaccional o ATP eacute hidrolisado formando-se novamente ADP e Pi o que permite a
formaccedilatildeo de um ciclo (fig 21) Se as enzimas se encontrarem em excesso a velocidade de
conversatildeo do NADH eacute limitada pela produccedilatildeo de ADP e Pi Como o ATP eacute regenerado em cada
ciclo a sua concentraccedilatildeo inicial eacute sempre mantida (Bagshaw 1982)
Figura 33ndash Representaccedilatildeo esquemaacutetica do meacutetodo de enzimas acopladas
47
O protocolo utilizado e seguidamente descrito foi adaptado de Cuenda et al (1990 ndash Ver Refordf) A
uma cuvette de 1 mL adicionou-se S1 para uma concentraccedilatildeo final de 005 mgmL 800 L de
meio de reacccedilatildeo22
10 L de PEP ([PEP]stock = 42 mM) 10 L de PK+LDH 11 ([PK+LDH]stock =
1 mgmL) e 15 L de NADH ([NADH]stock=25 mM) Iniciou-se a monitorizaccedilatildeo da reacccedilatildeo
atraveacutes da leitura de Abs340 nm apoacutes a adiccedilatildeo de 20 L de ATP ([ATP]stock=100 mM) A
actividade Mg2+
-ATPaacutesica eacute obtida atraveacutes do declive correspondente agrave oxidaccedilatildeo do NADH Para
a mediccedilatildeo da Mg2+
-ATPase de S1 estimulada por actina adicionou-se F-actina para uma
concentraccedilatildeo final de 2 microM sendo a actividade obtida tambeacutem atraveacutes do declive correspondente
agrave oxidaccedilatildeo do NADH apoacutes a adiccedilatildeo da proteiacutena
35 Interaccedilatildeo entre vanaacutedio e subfragmento-1 de miosina
351 Preparaccedilatildeo das soluccedilotildees de vanaacutedio
As soluccedilotildees de vanadato foram preparadas em aacutegua MilliQ a partir de metavanadato de amoacutenio
(NH4VO3) numa concentraccedilatildeo stock de 50 mM com agitaccedilatildeo magneacutetica e aquecimento (~30 ordmC)
Esta soluccedilatildeo com um valor compreendido entre 6 e 7 designa-se por ldquometavanadatordquo e eacute
constituiacuteda pelas esspeacutecies V1 V2 V4 e V5 (Ref) Acidificando a soluccedilatildeo de metavanadato por
adiccedilatildeo de HCl ateacute um valor de pH proacuteximo de 4 observa-se uma mudanccedila de cor de amarelo
claro para laranja indicativa da formaccedilatildeo da espeacutecie decameacuterica de vanadato De seguida e
imediatamente antes da realizaccedilatildeo dos ensaios descritos o valor do pH da soluccedilatildeo de
decavanadato foi aumentado para cerca de 75 por adiccedilatildeo de NaOH A soluccedilatildeo (2) designada
por ldquometavanadatordquo natildeo foi posteriormente alterada ateacute agrave sua adiccedilatildeo para a realizaccedilatildeo dos ensaios
As soluccedilotildees foram armazenadas a 4ordmC e mantidas em gelo aquando da sua utilizaccedilatildeo
A soluccedilatildeo de V(IV) foi preparada em condiccedilotildees anaeroacutebias por diluiccedilatildeo de sulfato de vanadil
(VOSO4) para uma concentraccedilatildeo final de 5 mM em H2O MilliQ saturada com N2 gasoso
352Anaacutelise por espectroscopia de UV-Vis
A concentraccedilatildeo da espeacutecie decameacuterica em soluccedilatildeo depende do periacuteodo de tempo apoacutes a diluiccedilatildeo
no meio de reacccedilatildeo apropriado (Ref) A velocidade de desoligomerizaccedilatildeo da espeacutecie decameacuterica
pode ser analisada por espectroscopia de absorccedilatildeo na regiatildeo UVVis uma vez que esta espeacutecie
apresenta duas bandas caracteriacutesticas na regiatildeo do visiacutevel (a 358 e 400 nm) que natildeo se verifica
em quaisquer outras espeacutecies oligomeacutericas As bandas supramencionada sapresentam-se bem
22
Meio de reacccedilatildeo 3125 mM Tris-HCl pH 71 375 mM MgCl2
48
resolvidas atraveacutes da obtenccedilatildeo da derivada de 2ordf ordem do espectro apresentando a forma de 2
vales com miacutenimos centrados nos comprimentos de onda supracitados com dependecircncia linear
relativamente agrave concentraccedilatildeo de decavanadato total (Ramos et al 2006)
Assim a estabilidade da espeacutecie decameacuterica apoacutes diluiccedilatildeo da soluccedilatildeo de decavanadato no meio de
reacccedilatildeo dos ensaios (tampatildeo D) para uma concentraccedilatildeo final de 1 mM em vanadato total foi
caracterizada atraveacutes da leitura da Abs400 em funccedilatildeo do tempo tendo-se determinado qual a
ordem de reacccedilatildeo e o tempo de meia-vida da mesma Com base em estudos anteriores acerca da
estabilidade do V10 na presenccedila de diferentes proteiacutenas (Ramos et al 2006) determinou-se
igualmente o tempo de meia vida apoacutes a adiccedilatildeo de 20 microM de S1 de miosina ao meio reaccional
353 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
Os espectros de RMN do isoacutetopo 51
V foram realizados num espectroacutemetro Bruker AM-400 MHz
equipado com uma sonda multinuclear inversa de 5 mm Em tubos de RMN de 5 mm de diacircmetro
preparou-se soluccedilotildees de 500 L de deca- ou metavanadato correspondentes a um intervalo de
concentraccedilotildees de 1 a 10 mM suplementadas com 10 D2O Os espectros foram adquiridos agrave
temperatura ambiente com uma largura espectral de 23 923 Hz um tempo de aquisiccedilatildeo de 0086
s tempo de relaxaccedilatildeo de 001 s e uma sequecircncia de pulsos a 90ordm Utilizou-se a transformada de
Fourier com alargamento das bandas de 20Hz para o processamento dos espectros Os desvios
quiacutemicos das ressonacircncias de 51
V foram obtidos em relaccedilatildeo agrave espeacutecie VOCl3cuja frequecircncia de
ressonacircncia eacute considerada o padratildeo sendo todas as outras referenciadas a partir desta em ppm
As aacutereas dos sinais do RMN correspondentes agraves espeacutecies oligomeacutericas presentes foram obtidas
por integraccedilatildeo dos sinais As concentraccedilotildees relativas das espeacutecies foram calculadas com recurso agrave
equaccedilatildeo
onde A[Vx] corresponde agrave aacuterea referente agrave ressonacircncia da espeacutecie Vx com nordm de agregaccedilatildeo x
A[V]total agrave soma total das aacutereas medidas e [V] a concentraccedilatildeo total de vanaacutedio no ensaio
A teacutecnica de 51
V-RMN foi anteriormente aplicada em estudos de ligaccedilatildeo com S1 de miosina
(Ref) tendo sido demostrado que entre as espeacutecies V1 V2 V4 e V5 a espeacutecie tetrameacuterica possui a
maior afinidade para a miosina (Cremo et al) O desaparecimento ou alargamento dos sinais pode
implicar a ligaccedilatildeo de um dos oxovanadatos laacutebeis agrave miosina sem indicaccedilatildeo de qual uma vez que
49
se processa um equiliacutebrio entre eles (Crans 1990) Assim e tendo em conta as indicaccedilotildees de que
a espeacutecie V10 eacute distinta das restantes no que diz respeito agrave magnitude da interacccedilatildeo com a miosina
e inibiccedilatildeo da actividade ATPaacutesica estimulada por actina (Aureliano 2000 Tiago et al 2002)
torna-se fundamental investigar a interacccedilatildeo das diferentes espeacutecies de vanadato com o S1 de
miosina atraveacutes da teacutecnica de 51
V-RMN
Para tal obtiveram-se espectros de 51
V-RMN resultantes da titulaccedilatildeo de uma soluccedilatildeo 5 mM de
decavanadato total com concentraccedilotildees crescentes de S1 de miosina (0 5 10 15 e 20 microM) em
tampatildeo D Realizou-se ainda ensaios na presenccedila de 2 mM de decavanadato e 2 mM de ATP na
presenccedila e na ausecircncia de S1 de miosina de modo a verificar o efeito que estes ligandos naturais
da enzima teria na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o subfragmento-1
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
Os grupos sulfidrilo (-SH) das cisteiacutenas satildeo resiacuteduos de aminoaacutecidos extremamente sensiacuteveis agrave
oxidaccedilatildeo resultando na sua oxidaccedilatildeo (Dalle-Donne et al 1999) Logo a quantificaccedilatildeo destes
resiacuteduos eacute bastante importante nos estudos de interacccedilatildeo com vanaacutedio jaacute que os resiacuteduos de Cys
com ecircnfase para o SH1 e o SH2 satildeo bastante importantes na monitorizaccedilatildeo das modificaccedilotildees
funcionais da regiatildeo da cabeccedila
O procedimento seguidamente apresentado foi originalmente descrito por Gutieacuterrez-Merino
(1987)) Consiste na reacccedilatildeo entre os grupos ndashSH natildeo oxidados das proteiacutenas e o reagente de
Ellman (DTNB -55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico)) - formando-se o aniatildeo CNT (aniatildeo 3-
carboxilato-4-nitrotiofenolato) numa estequiometria de 11 Este apresenta um maacuteximo de
absorvecircncia a 412 nm (Robyt 1990) Assim apoacutes a incubaccedilatildeo da proteiacutena na presenccedila do metal
a quantidade de cisteiacutenas reduzidas por proteiacutena determina-se atraveacutes dos valores de Abs412
Deste modo colocou-se num tubo de Eppendorf o volume apropriado de S1 dialisado para que a
concentraccedilatildeo no ensaio fosse de 1 mgmL Adicionou-se o volume apropriado de decavanadato
para que a sua concentraccedilatildeo fosse de 0 10 50 100 250 500 ou 1000 M Perfez-se com soluccedilatildeo
D ateacute um volume final de 1 mL e incubou-se a 25ordmC por 20 minutos Apoacutes este periacuteodo colocou-
se 780 L da mistura reaccional numa cuvette de quartzo e adicionou-se 220 L de soluccedilatildeo K23
23
Tampatildeo K Tampatildeo fosfatohidrogenofosfato de soacutedio 025 M pH 72
50
Adicionou-se 15 L de DTNB (em soluccedilatildeo K [DTNB]stock=100 mM) ao meio reaccional e
registou-se o valor de absorvecircncia medido equivalente agraves cisteiacutenas ldquoraacutepidasrdquo (SH1 e SH2
expostos ao solvente) Adicionou-se 001 g de SDS que provoca a desnaturaccedilatildeo da proteiacutena e
consequentemente a exposiccedilatildeo de todos os resiacuteduos de cisteiacutenas que se encontravam ateacute aiacute
inacessiacuteveis no interior da proteiacutena (cisteiacutenas totais) Estes resiacuteduos podem assim reagir com o
DTNB
Estimou-se o nuacutemero de resiacuteduos de cisteiacutenas em todas as amostras com recurso ao coeficiente de
extinccedilatildeo molar () do DTNB (ver ponto 3 dos Anexos) que foi determinado atraveacutes da titulaccedilatildeo
de L-Cisteiacutena com intervalo de concentraccedilotildees compreendidos entre 0 e 50 microM na presenccedila de
25microM de DTNB em tampatildeo D Os valores de absorvecircncia foram obtidos para um comprimento de
onda de 412 nm
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
3621 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
A teacutecnica mais eficiente de anaacutelise da interacccedilatildeo entre o iatildeo vanadilo e proteiacutenas como a
calmodulina e a transferrina entre outras permite a obtenccedilatildeo raacutepida de informaccedilatildeo no que toca agrave
conformaccedilatildeo e amplitude da referida interacccedilatildeo
Os espectros de Ressonacircncia Paramagneacutetica Electroacutenica (RPE) da espeacutecie paramagneacutetica V(IV)
foram realizados a 70K num espectroacutemetro Bruker EMX-300 X-band Todas as soluccedilotildees
utilizadas e amostras foram preparadas em anaerobiose numa cacircmara de luvas de modo a evitar a
reduccedilatildeo de V(IV) a V(V) Em tubos capilares com 02 mL de amostra manteve-se a concentraccedilatildeo
de S1 a 20 M e variou-se a concentraccedilatildeo de vanadilo (0 25 5 10 15 25 50 75 100 150
200 500 e 1000 M [VOSO4]stock=5 mM) em tampatildeo D Apoacutes obtenccedilatildeo dos espectros estes
foram tratados atraveacutes do software SimFonia e WinEPR tendo-se obtido um graacutefico com a
variaccedilatildeo da intensidade do sinal de RPE em funccedilatildeo da concentraccedilatildeo de sulfato de vanadilo
presente Realizaram-se ainda simulaccedilotildees de espectros obtidos com 20 microM de S1 e 1000 microM de
V(IV) com vista agrave obtenccedilatildeo dos ligandos provaacuteveis do vanadilo atraveacutes da anaacutelise da constante
de acoplamento hiperfina (A) e do factor g (g)
4 Conclusotildees gerais
Apesar de jaacute ter sido demonstrado que o vanaacutedio inibe a Na+K
+-ATPase da miosina a funccedilatildeo
deste elemento no tecido muacutesculo esqueleacutetico continua largamente incompreendida A enorme
complexidade quiacutemica deste metal especialmente no que concerne agrave oligomerizaccedilatildeo e reacccedilotildees
51
redox constitui um obstaacuteculo agrave compreensatildeo do comportamento do vanaacutedio nos organismos
vivos Assim a caracterizaccedilatildeo da quiacutemica deste elemento eacute de grande utilidade antes de iniciar o
estudo da sua interacccedilatildeo com o presente sistema
A anaacutelise por espectroscopias de 51
V-RMN e de UV-Vis de soluccedilotildees de vanadato diluiacutedas nos
meios de reacccedilatildeo utilizados no presente relatoacuterio (10 mM HEPES pH 70 25 mM KCl) mostrou
claramente que a espeacutecie decameacuterica (V10) possui um tempo de vida suficientemente longo para a
determinaccedilatildeo do seu efeito isolado ao contraacuterio do que se verifica com as espeacutecies mono- di-
tetra- e pentameacutericas (V1 V2 V4 e V5) Por outro lado a anaacutelise verificou ainda que apoacutes diluiccedilatildeo
das soluccedilotildees de decavanadato no meio de reacccedilatildeo coexistem as espeacutecies V10 e V1 pelo que os
efeitos putativos desta uacuteltima deveratildeo ser subtraiacutedos aquando da monitorizaccedilatildeo dos efeitos da
espeacutecie decameacuterica
O vanaacutedio (V) (vanadato) inibe as enzimas que catalizam reacccedilotildees de transferecircncia de grupos
fosfato como a ATPase da miosina [Goodno 1979] devido agrave sua analogia estrutural com o iatildeo
fosfato Na sua utilizaccedilatildeo como inibidor enzimaacutetico natildeo eacute tida em conta a contribuiccedilatildeo individual
das espeacutecies oligomeacutericas considerando-se normalmente a espeacutecie V1 como inibidora efectiva
[Goodno e Taylor 1982] No presente relatoacuterio verificou-se por espectroscopia de 51
V-RMN que
outros oligoacutemeros de vanadato interagem com o S1 de miosina tendo as espeacutecies V10 e V4 os
maiores niacuteveis de afinidade A sua interacccedilatildeo envolve forccedilas electrostaacuteticas e eacute afectada
negativamente por ligandos naturais de enzima como eacute o caso do ATP
A exposiccedilatildeo do S1 de miosina (005 mgmL) a decavanadato conduz agrave inibiccedilatildeo do capacidade da
proteiacutena de estimular a actividade Mg2+
-ATPaacutesica com um valor de IC50 de 266 plusmn 07 M
decavanadato sendo o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM
decavanadato (80) Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM decavanadato sendo
o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM decavanadato (60)
exercendo um efeito de impedimento agrave ligaccedilatildeo entre o vanaacutedio e o S1
A uma concentraccedilatildeo de 27 microM decavanadato verifica-se a oxidaccedilatildeo de um dos resiacuteduos de Cys
de S1 de miosina seguida pela oxidaccedilatildeo gradual de outro resiacuteduo de Cys ateacute agrave concentraccedilatildeo de
1000 microM Ambos os resiacuteduos oxidados natildeo se encontravam expostos ao solvente utilizado A
oxidaccedilatildeo dos resiacuteduos permite sugerir que esta modificaccedilatildeo oxidativa sofrida pelo S1 contribui
para a diminuiccedilatildeo da sua actividade de ATPase
52
Por espectroscopia de RPE revelaram-se os possiacuteveis ligandos equatoriais do vanaacutedio aquando da
sua interacccedilatildeo com o S1 de miosina como sendo duas moleacuteculas de aacutegua um ligando contendo o
grupo R-OH e outro contendo o grupo R=NR
Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados de
oxidaccedilatildeo (IV) e (V) no S1 de miosina para concentraccedilotildees facilmente alcanccedilaacuteveis no ambiente
terrestre
53
5 Estudos futuros
No sentido do melhor esclarecimento das alteraccedilotildees moleculares e estruturais que ocorrem no
subfragmento 1 quando exposta ao vanaacutedio que conduzem a uma diminuiccedilatildeo da sua funccedilatildeo
sugere-se
- Quantificaccedilatildeo das cisteiacutenas reduzidas de S1 na presenccedila de metavanadato
- Determinaccedilatildeo das actividades Ca2+
- e K+-ATPaacutesica do S1 de miosina na presenccedila de
decavanadato
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina estimulada por actina
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina na presenccedila de
metavanadato
- Realizaccedilatildeo de RMN 2D com variaccedilatildeo das concentraccedilotildees das soluccedilotildees de decavanadato e
metavanadato na presenccedila de S1
-Mutageacutenese dos resiacuteduos de Cys doS1 de miosina de modo a verificar quais os resiacuteduos que
sofrem oxidaccedilatildeo na presenccedila de decavanadato
- Determinaccedilatildeo por calorimetria (DSC Differential Scanning Calorimetry) da variaccedilatildeo da
entalpia (∆H) e da temperatura de desnaturaccedilatildeo do S1 de miosina na presenccedila de vanaacutedio uma
vez que estas propriedades caracteriacutesticas de cada proteiacutena satildeo alteradas apoacutes ocorrecircncia de
modificaccedilotildees estruturais
- Anaacutelise por SDS-PAGE de S1 de miosina apoacutes tratamento com vanaacutedio de modo a determinar
quais os polipeacuteptidos eventualmente formados apoacutes clivagem da proteiacutena
54
6 Referecircncias bibliograacuteficas
- Sellers J Myosins 2nd Edition Oxford University Press 1999
- L Stryer inBiochemistry (Ed WH Freeman 2005 pp
Refordfas Materiais e Meacutetodos
[A] S S Margossian SLowey Methods Enzymol1982 85 55-71
[B] JD Pardue JA Spudich Methods Enzymol 1982 85 164-181
[C] M Bradford Anal Biochem1976 72 248-254
[D] H H Taussky E Shorr J Biol Chem 1953 202 675-685
[E] C R Bagshaw in Outline Studies in Biology ndash Muscle Contraction Chapman and Hall Ed
1982
[F] A Cuenda et al Eur J Biochem1990 194 663-670
55
Anexos
1 Determinaccedilatildeo da recta de calibraccedilatildeo meacutetodo de Bradford
Figura1ndashRepresentaccedilatildeo graacutefica da recta de calibraccedilatildeo (Abs595 nm vs [BSA]) utilizada para o meacutetodo de Bradford
2 Determinaccedilatildeo da recta calibraccedilatildeo para a electroforese em condiccedilotildees desnaturantes
Figura 2 ndash Representaccedilatildeo graacutefica da recta de calibraccedilatildeo (log(Peso Molecular) vs Rf dos marcadores de peso
molecular) onde Rf indica determinada atraveacutes do padratildeo de peso molecular
(poccedilo (a))
3Caacutelculo do coeficiente de extinccedilatildeo molar () para a quantificaccedilatildeo de cisteiacutenas reduzidas
atraveacutes do meacutetodo de Ellman
Traccedilou-se a recta de Abs412 nm meacutedia vs [L-Cisteiacutena] de modo a determinar o valor do coeficiente
de extinccedilatildeo molar definido como o declive daquela recta Assim =8400 M-1
cm-1
como
podemos verificar na Fig 3
A595 nm = 00546[BSA] + 00209 Rsup2 = 099051
Ab
s 59
5 n
m
[BSA] (mgmL)
log PM = -1177 Rf + 2247 Rsup2 = 098957
log
PM
Mobilidade relativa (Rf)
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero

6
3312 Quantificaccedilatildeo pelo meacutetodo directo
332 Electroforese em condiccedilotildees desnaturantes (SDS-PAGE)
34 Mediccedilatildeo da actividade enzimaacutetica de S1
341 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de Fiske-
Subarow modificado
342 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
35 Preparaccedilatildeo e caracterizaccedilatildeo das soluccedilotildees de vanaacutedio por Espectroscopia UV-Visiacutevel e
Ressonacircncia Magneacutetica Nuclear
351 Preparaccedilatildeo das soluccedilotildees de vanadato
352 Anaacutelise por UV-Vis para determinaccedilatildeo da ordem da reacccedilatildeo
353 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
362 1 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
363 Interacccedilatildeo de S1 com V (V) por Ressonacircncia Magneacutetica Nuclear (RMN)
4 Conclusotildees gerais
5 Estudos futuros
6 Referecircncias bibliograacuteficas
7
1 Introduccedilatildeo
11 Muacutesculo esqueleacutetico
A capacidade de locomoccedilatildeo eacute uma das caracteriacutesticas fundamentais da maioria dos organismos
vivos A contracccedilatildeo muscular um dos fenoacutemenos que a torna possiacutevel apresenta grande
complexidade e tem vindo a ser utilizado como modelo para a compreensatildeo de sistemas menos
especializados de mobilidade celular [1]
No que diz respeito agrave sua fisiologia morfoloacutegica os muacutesculos dos vertebrados dividem-se em
lisos e estriados Estes uacuteltimos devem o seu nome agrave alternacircncia de zonas claras e escuras
observaacuteveis ao microscoacutepio electroacutenico de que resulta um aspecto estriado Os muacutesculos
estriados podem ainda ser divididos em duas subclasses muacutesculos esqueleacuteticos e cardiacuteacos [2]
consoante a forma celular (respectivamente longitudinal ou ramificada) ou tipo de contracccedilatildeo das
ceacutelulas que o compotildeem (respectivamente voluntaacuteria e involuntaacuteria) [3]
Os muacutesculos estriados esqueleacuteticos satildeo assim designados devido agrave sua localizaccedilatildeo no
organismo ndash ligados aos ossos da coluna vertical braccedilos e pernas Satildeo constituiacutedos por feixes de
ceacutelulas multinucleadas (miofibras) que contecircm miofibrilas paralelas de cerca de 1 m de
diacircmetro associadas agrave contracccedilatildeo voluntaacuteria raacutepida e forte em actividades de coordenaccedilatildeo
complexa [4] Cada feixe apresenta um diacircmetro compreendido entre 20 e 100 m e um
comprimento maacuteximo que pode atingir vaacuterios centiacutemetros (Fig 11)
Figura 11 ndash Organizaccedilatildeo hieraacuterquica geral do muacutesculo-esqueleacutetico (adaptado de Purves et al 2003 [3])
No muacutesculo-esqueleacutetico as zonas claras e escuras supramencionadas correspondem a zonas de
menor e maior densidade electroacutenica respectivamente As zonas claras constituem as bandas I (de
8
ldquoisotropiardquo ndash polarizaccedilatildeo uniforme em todas as direcccedilotildees) enquanto as zonas escuras constituem
as bandas A (de ldquoanisotropiardquo ndash polariza a luz visiacutevel) A banda I encontra-se dividida
transversalmente por uma linha escura ndash linha Z de densidade electroacutenica e refracccedilatildeo superiores
No interior de cada banda A existe uma zona transversal menos densa ndash zona H ndash rica em enzimas
importantes para o metabolismo celular No centro desta zona existe uma linha perpendicular
mais escura ndash linha M [2]
A unidade fundamental da contracccedilatildeo muscular eacute designada por sarcoacutemero (Fig 12) Encontra-se
repetida de forma regular ao longo das miofibrilas apresentando um comprimento de 25 a 3 m
aquando do relaxamento muscular diminuindo aquando da contracccedilatildeo muscular
Figura 12 ndash Representaccedilatildeo de fotomicrografia electroacutenica e esquemaacutetica do sarcoacutemero Os filamentos grossos e
finos satildeo maioritariamente constituiacutedos por miosina e actina respectivamente (Adaptado de Stryer 2002 [2])
As secccedilotildees de corte do sarcoacutemero datildeo conta da presenccedila de dois tipos de filamentos proteicos ndash os
filamentos grossos cujo diacircmetro eacute de 15 nm constituiacutedos sobretudo por miosina e os filamentos
finos com um diacircmetro de 9 nm constituiacutedos sobretudo por actina tropomiosina e pelo complexo
de troponina [2]
111 Miosina
Actualmente o termo miosina refere-se a uma superfamiacutelia diversa de motores moleculares
capazes de hidrolisar ATP e de se ligarem reversivelmente agrave actina A classificaccedilatildeo da
superfamiacutelia das miosinas nas 15 classes distintas faz-se atraveacutes de homologia entre as diferentes
sequecircncias [A] Eacute tambeacutem possiacutevel a classificaccedilatildeo por subclasses das quais se destaca a subclasse
4 proveniente do muacutesculo-esqueleacutetico e cardiacuteaco de vertebrados Este estudo focar-se-aacute apenas
nas miosinas de classe II (miosina-II) presente nos filamentos grossos do muacutesculo estriado
esqueleacutetico [B] e em ceacutelulas natildeo musculares A funccedilatildeo da miosina-II depende da sua localizaccedilatildeo
celular e tecidular
9
A miosina-II eacute constituiacuteda por duas cadeias polipeptiacutedicas idecircnticas denominadas cadeias pesadas
de miosina (HC) enroladas entre si e cujo peso molecular (PM) estaacute compreendido entre 171 e
241 kDa) Estruturalmente as HC dividem-se em 3 domiacutenios funcionais - cabeccedila pescoccedilo e
cauda como se pode observar na Fig 13 Agraves cabeccedilas ou zonas globulares estatildeo associados dois
pares de cadeias leves (peso molecular entre 15 e 30 kDa) que se dividem em dois tipos
essencial (ELC) e regulatoacuterio (RLC)
Figura 13 ndash Estrutura esquemaacutetica da moleacutecula de miosina-II Destacam-se as duas cabeccedilas globulares e a cauda
filamentosa ligadas entre si por uma zona flexiacutevel denominada pescoccedilo As cadeias leves (a verde e vermelho)
encontram-se associadas agraves cabeccedilas globulares
As cabeccedilas globulares contendo os locais de ligaccedilatildeo agrave actina e aos nucleoacutetidos mantecircm os seus
elementos estruturais baacutesicos conservados em todas as classes sendo estes constituiacutedos por 720-
80 aa A regiatildeo do pescoccedilo eacute constituiacuteda por uma longa heacutelice pertencente agrave cadeia pesada
que se encontra estabilizada atraveacutes da ligaccedilatildeo a subunidades de cadeia leve A estrutura do
pescoccedilo envolvida na ligaccedilatildeo agraves cadeias leves eacute denominada de motivo IQ cuja estrutura
consensual eacute IQXXXRGXXXR1 O pescoccedilo serve como extensatildeo do comprimento da cabeccedila de
modo a actuar como alavanca durante a contracccedilatildeo muscular [A]A regiatildeo da cauda ou acircncora
apresenta bastante divergecircncia no que respeita agrave sua sequecircncia estrutura e funccedilatildeo No entanto
pensa-se que a sua funccedilatildeo centrar-se-aacute na autoassociaccedilatildeo das vaacuterias moleacuteculas de miosina-II
atraveacutes da regiatildeo de coiled-coil Esta confere assimetria agrave moleacutecula e permite a formaccedilatildeo de
filamentos grossos capazes de translocar filamentos de actina [A]
Na regiatildeo da cabeccedila da miosina existem 2 regiotildees extremamente sensiacuteveis agrave proteoacutelise A
clivagem em ambos os locais resulta na produccedilatildeo de trecircs fragmentos um compreendendo o N-
terminal (fragmento 25 kDa) outro a regiatildeo central (50 kDa) e um terceiro o C-terminal (20 kDa)
(Fig 14) Estes fragmentos dissociam-se em condiccedilotildees desnaturantes e natildeo constituem domiacutenios
independentes da cabeccedila mas provavelmente loops flexiacuteveis uma vez que natildeo foram observadas
na estrutura cristalina Os vaacuterios fragmentos obtidos em diferentes condiccedilotildees foram cruciais para
a elucidaccedilatildeo das vaacuterias funccedilotildees da moleacutecula
A regiatildeo compreendida entre os fragmentos 25 e 50 kDa (loop 1) localiza-se directamente por
1 I = Isoleucina Q= Glutamina X = qualquer dos 20 aminoaacutecidos padratildeo R= Arginina G= Glicina
10
cima do local de ligaccedilatildeo ao nucleoacutetido A regiatildeo entre os fragmentos de 50 e 20 kDa (loop 2)
encontra-se perto da intersecccedilatildeo entre os domiacutenios da cabeccedila e do pescoccedilo e deveraacute interagir com
a actina uma vez que a ligaccedilatildeo desta a protege da proteoacutelise Na ausecircncia do loop 2 a miosina
liga mais fracamente agrave actina do que no caso da miosina intacta A sequecircncia primaacuteria de ambos
os loops apresenta conservaccedilatildeo reduzida sendo provaacutevel que sejam responsaacuteveis por muita da
diversidade das diferentes propriedades enzimaacuteticas presentes em diferentes isoformas
No fragmento de 20 kDa existem dois grupos sulfidrilo Cys-707 e -697 usualmente
denominados de SH1 e SH2 devido agrave ordem da sua reactividade quiacutemica Na presenccedila de
nucleoacutetidos pode ocorrer o ldquocross-linkingrdquo entre estes dois grupos incluindo a oxidaccedilatildeo a uma
ponte dissulfureto
Devido agrave insolubilidade da miosina-II nativa a estrateacutegia adoptada para os estudos com esta
proteiacutena eacute frequentemente a sua clivagem em fragmentos funcionais separaacuteveis Um dos locais de
clivagem ocorre tipicamente a 130 kDa do N-terminal produzindo a meromiosina pesada (HMM)
e a meromiosina leve (LMM) [C] A HMM eacute formada pelas 2 cabeccedilas e pescoccedilo e apresenta
propriedades semelhantes agrave miosina sem digestatildeo (ligaccedilatildeo e translocaccedilatildeo de actina e hidroacutelise de
MgATP)
A HMM pode ser novamente clivada atraveacutes de uma reacccedilatildeo de proteoacutelise produzindo os
subfragmentos 1 (S1) e 2 (S2)O S1 (90 kDa) eacute enzimaticamente activo sendo constituiacutedo pelos
domiacutenios da cabeccedila e do pescoccedilo e apresentando associaccedilatildeo agraves duas cadeias leves ou apenas agrave
ELC (Fig 14)Em alternativa o S1 pode ser produzido por clivagem directa da miosina intacta
formando-se tambeacutem um fragmento em forma de cilindro que consiste nas regiotildees S2 e LMM
(Ref)Utiliza-se extensivamente o S1 para estudos cineacuteticos e de ligaccedilatildeo agrave actina uma vez que a
sua solubilidade simplifica o planeamento da experiecircncia e a subsequente interpretaccedilatildeo dos dados
[A]
Figura 14- Estrutura do subfragmento-1 de miosina de muacutesculo esqueleacutetico com ecircnfase nos domiacutenios de 50 kDa
central (a vermelho) 25 kDa no N-terminal (a verde) e 20 kDa no C-terminal (a azul) As cadeias leves essenciais
encontram-se a amarelo e as cadeias leves regulatoacuterias a lilaacutes Os locais de ligaccedilatildeo agrave actina e ao ATP estatildeo
igualmente destacados (adaptado de Holmes e Geeves 1999 [D])
11
112 Actina
O termo ldquoactinardquo refere-se a uma famiacutelia extremamente conservada de proteiacutenas citoplasmaacuteticas
presente em todos os eucariotas Esta proteiacutena natildeo se encontra nos procariotas [E] A maioria das
actinas possui peso molecular proacuteximo de 42 kDa outra evidecircncia do seu grau elevado de
conservaccedilatildeo no que concerne agrave sequecircncia e ao nuacutemero de aminoaacutecidos Os eucariotas superiores
apresentam um grande nuacutemero de isoformas de actina que por sua vez se encontram codificados
por um ainda maior nuacutemero de genes (Ref) A expressatildeo das isoformas eacute diferencial consoante o
tecido e sua especializaccedilatildeo pelo que deveratildeo possuir funccedilotildees e mecanismos direccionais
diferentes (Ref) Foi ainda observado que os mRNA das diferentes isoformas apresentam uma
distribuiccedilatildeo heterogeacutenea nas ceacutelulas de forma semelhante aos seus produtos proteicos [F] Estes
dados suportam a hipoacutetese supramencionada do direccionamento das isoformas de actina para um
determinado local na ceacutelula de acordo com as suas funccedilotildees
A actina presente no citoesqueleto eacute responsaacutevel por processos como o mecanismo contraacutectil
fagocitose manutenccedilatildeo e alteraccedilatildeo da estrutura celular locomoccedilatildeo transporte de organelos
regulaccedilatildeo do transporte de solutos entre outras Nos eucariotas superiores estaacute ainda envolvida
no transporte de gracircnulos de pigmento retinoacuteide [G] transporte de gracircnulos no axoplasma [H] e
secreccedilatildeo entre outros [E] Actualmente a isoforma da actina (-actina) eacute a melhor estudada
encontrando-se envolvida no mecanismo contraacutectil [E]
Os filamentos de -actina tecircm cerca de 1m de comprimento e 80 Å Consistem em duas cadeias
de actina filamentosa (F-actina) organizados em heacutelice [I] (Holmes et al 1990) sobre os quais
se encontram outras proteiacutenas ndash a troponina (C I e T) e a tropomiosina (Azevedo 1999) (Figura
15) A troponina eacute filamentosa e localiza-se nas depressotildees da heacutelice de F-actina conferindo
rigidez aos filamentos finos Por sua vez a tropomiosina eacute essencial na regulaccedilatildeo do mecanismo
contraacutectil
Figura 15 ndash Modelo estrutural dos filamentos de F-actina contendo a tropomiosina e o complexo de troponina (I C
e T) (Adaptado de Susana Ramos 2005 [J])
12
A F-actina eacute formada atraveacutes da polimerizaccedilatildeo de subunidades globulares de actina (G-actina) A
G-actina eacute composta por 2 domiacutenios de tamanho semelhante separados pelos bolsos de ligaccedilatildeo(1)
de catiotildees divalentes e(2) de nucleoacutetidos [KLM] Estes por sua vez estatildeo subdivididos noutros
dois de menores dimensotildees (1-4) (Fig 16) cada um envolvido nas interacccedilotildees actina-actina ou
na formaccedilatildeo do topo de ligaccedilatildeo do nucleoacutetido As regiotildees N-terminal e C-terminal estatildeo proacuteximas
uma da outra no subdomiacutenio 1 [E]
Figura 16 ndash Estrutura do monoacutemero de actina Encontram-se representados os subdomiacutenios 1 a 4 e a vermelho a
moleacutecula de ATP junto do respectivo local de ligaccedilatildeo (Protein Data Bank ID 1j6z)
113 Mecanismos moleculares da contracccedilatildeo muscular
A contracccedilatildeo muscular ocorre atraveacutes do deslizamento dos filamentos finos (de actina) por entre
os filamentos grossos (de miosina) conseguida pela reacccedilatildeo de hidroacutelise de ATP e regulada pela
tropomiosina e pelo complexo de troponina em resposta a impulsos nervosos Este mecanismo foi
inicialmente proposto por Huxley-Niedergerke [N] e Huxley-Hanson [O] ambos em 1954 A base
experimental de ambos os estudos foi a inexistecircncia de variaccedilatildeo do tamanho dos filamentos
durante a contracccedilatildeo A banda A apresenta sempre as mesmas dimensotildees por outro lado a banda
I a zona H diminuem consideravelmente aquando da contracccedilatildeo (Figs 12 e 17)
Figura 17 ndash Modelo da contracccedilatildeo muscular ndash deslizamento dos filamentos finos (azuis) relativamente aos
filamentos grossos (rosa) (Adaptado de Stryer 2002 [2])
13
Na ausecircncia de ATP e supondo a ausecircncia de inibiccedilatildeo da interacccedilatildeo actina-miosina as proteiacutenas
encontram-se fortemente associadas formando o complexo actomiosina (AM) (Figura 15)
Nestas condiccedilotildees as cabeccedilas de miosina formam um acircngulo de 45ordm com o eixo dos filamentos
finos de actina [P]
Na presenccedila de ATP o complexo AM dissocia-se jaacute que a afinidade do complexo M-ATP para a
actina eacute 105 vezes inferior agrave afinidade da miosina para a actina O ATP liga-se agrave miosina
ocorrendo seguidamente a reacccedilatildeo de hidroacutelise de ATP Os produtos daiacute resultantes ndash ADP e Pi -
permanecem ligados agrave miosina formando o complexo M-ADP-Pi Com a ligaccedilatildeo subsequente agrave
actina forma-se o complexo AM-ADP-Pi no qual as cabeccedilas de miosina apresentam um acircngulo
recto com o eixo do muacutesculo [P] Neste ponto daacute-se a libertaccedilatildeo progressiva de Pi e ADP Eacute
provaacutevel que a energia proveniente da hidroacutelise do ATP provoque uma alteraccedilatildeo do acircngulo de
ligaccedilatildeo das cabeccedilas de miosina ao filamento de actina passando novamente de 90ordm para 45ordm [P]
Esta alteraccedilatildeo conformacional provoca o deslizamento do filamento de actina para o centro do
sarcoacutemero cujo encurtamento promove a contracccedilatildeo das miofibrilas O processo supracitado
ocorre em cerca de 1ms[Q] e encontra-se resumido na Fig 18
Figura 18 ndash Esquema cineacutetico da hidroacutelise de ATP pela miosina (Adaptado de Sellers 1999 [A])
1131 Regulaccedilatildeo da contracccedilatildeo muscular
O mecanismo pelo qual se processa a regulaccedilatildeo muscular foi inicialmente descrito por estudos
distintos de Haselgrove Huxley e Parrey-Squire todos em 1973 [RST] Aleacutem da implicaccedilatildeo
directa da tropomiosina e do complexo de troponina a concentraccedilatildeo de Ca2+
no muacutesculo em
relaxamento eacute bastante reduzida (10-7
M) aumentando notavelmente aquando da contracccedilatildeo (10-5
-
10-3
M)
Resumidamente quando ocorre a excitaccedilatildeo do muacutesculo esqueleacutetico a despolarizaccedilatildeo da
membrana plasmaacutetica eacute difundida ateacute ao interior do tecido atraveacutes dos tuacutebulos transversais
(invaginaccedilotildees da membrana plasmaacutetica da fibra muscular) A despolarizaccedilatildeo provoca a abertura
dos canais ioacutenicos de Ca2+
do retiacuteculo sarcoplasmaacutetico adjacente Como a concentraccedilatildeo de Ca2+
no interior do organelo eacute muito elevada o caacutelcio passa raacutepida e uniformemente para o citosol
14
Assim a regulaccedilatildeo da contracccedilatildeo ocorre com coordenaccedilatildeo e rapidez na totalidade das miofibrilas
do tecido muscular (Ref)
Em repouso a troponina I encontra-se posicionada de tal forma que a proteiacutena adjacente
troponina C impede estruturalmente a ligaccedilatildeo das cabeccedilas da miosina agrave actina (acircngulo =50ordm) O
aumento da concentraccedilatildeo intracelular de Ca2+
provoca um aumento da sua disponibilidade para
ligar agrave troponina C (Fig 19)
Figura 19ndash Regulaccedilatildeo do mecanismo contraacutectil no muacutesculo esqueleacutetico pelo complexo de troponina
Como consequecircncia a troponina C altera a sua conformaccedilatildeo (=70ordm) associando-se agrave troponina I
que deixa de inibir a ligaccedilatildeo das pontes transversais A alteraccedilatildeo anterior eacute transmitida agrave
tropomiosina atraveacutes da troponina T adjacente agrave C o que provoca o deslocamento da
tropomiosina na direcccedilatildeo do interior do sulco do filamento de actina Assim eacute possiacutevel a
interacccedilatildeo eficaz entre actina e miosina A Ca2+
-ATPase do retiacuteculo sarcoplasmaacutetico provoca a
reacumulaccedilatildeo do iatildeo caacutelcio no organelo impedindo o prolongamento desnecessaacuterio da contracccedilatildeo
(Refordf)
12 Vanaacutedio
O vanaacutedio eacute um metal de transiccedilatildeo pertencente ao grupo VB da tabela perioacutedica Foi
primeiramente descoberto em 1801 por Andreacutes Manuel del Riacuteo que concluiu erradamente que era
idecircntico ao croacutemio e definitivamente por Nils Gabriel Svenson em 1831 Recentemente foram
descobertas importantes funccedilotildees bioloacutegicas deste elemento provavelmente essencial para os
humanos de que se destaca no contexto do presente relatoacuterio a possibilidade de actuar como
anaacutelogo do grupo fosfato e a inibiccedilatildeo de enzimas in vitro como eacute o caso da actividade ATPaacutesica
da miosina
Eacute um metal soacutelido agrave temperatura ambiente que se encontra amplamente distribuiacutedo nos vaacuterios
ecossistemas em dois isoacutetopos naturais ndash 50
V e 51
V ndash raramente excedendo 01 partes por biliatildeo
15
(ppb) Estaacute associado a cerca de 70 minerais diferentes na natureza (dos quais se destacam o
uracircnio cobre chumbo zinco fosfato e titacircnio ferroso) especialmente na forma de vanadato ou
pentoacutexidos de vanaacutedio O vanaacutedio eacute um dos principais constituintes da mateacuteria orgacircnica foacutessil
com especial ecircnfase para o carvatildeo oacuteleo de crude e foacutesseis carbonaacuteceos [UV] bem como em
alimentos consumidos pelo Homem (salsa pimenta preta cogumelos marisco vegetais cereais
gorduras e oacuteleos) [W] Estes constituem a maior fonte de exposiccedilatildeo ao vanaacutedio da maioria da
populaccedilatildeo humana [X]
Normalmente a entrada do vanaacutedio no organismo ocorre por inalaccedilatildeo ou em menores
quantidades por absorccedilatildeo pelo tracto gastrointestinal ou pela pele O seu armazenamento centra-
se no fiacutegado rins ceacuterebro coraccedilatildeo muacutesculos e ossos No entanto a retenccedilatildeo de vanaacutedio no
organismo humano eacute reduzida ndash em condiccedilotildees normais cerca de 100 M (em muitos tecidos a
quantidade eacute inferior a 10 ngg de tecido) [Y]
Actualmente natildeo existem provas de deficiecircncia nutricional de vanaacutedio em humanos ou de
enzimas que contenham vanaacutedio em animais superiores No entanto vaacuterios estudos relacionam a
ocorrecircncia de deficiecircncia em galinhas e ratos devido a uma alimentaccedilatildeo pobre em vanaacutedio Os
sintomas relatados incluem retardamento do crescimento malformaccedilotildees oacutesseas perturbaccedilotildees no
metabolismo lipiacutedico e estagnaccedilatildeo reprodutiva entre outros [Z AA AB] Os efeitos fisioloacutegicos
do vanaacutedio podem depender do tipo de composto ou de oligoacutemero uma vez que consoante as
suas estabilidade labilidade e nuacutemero de coordenaccedilatildeo as respostas seratildeo diferenciadas ao niacutevel
dos organismos vivos
Por outro lado a toxicidade do vanaacutedio encontra-se bem caracterizada existindo sintomas
descritos em vaacuterios mamiacuteferos (inclusive o Homem) que incluem diarreias catildeibras irritaccedilatildeo da
pele e mucosas palpitaccedilotildees cardiacuteacas anemia pneumonia bronquite perturbaccedilotildees no Sistema
Nervoso Central (SNC) entre outros (Ref) A toxicidade varia geralmente com a natureza do
composto com aumento proporcional agrave sua valecircncia Verificou-se que a forma de entrada do
vanaacutedio tem tambeacutem efeito na sua toxicidade ndash os maiores niacuteveis verificaram-se por injecccedilatildeo
intravenosa da amostra sendo progressivamente inferiores por inalaccedilatildeo e finalmente por
ingestatildeo [X AA] A ampla utilizaccedilatildeo dos compostos de vanaacutedio na induacutestria quiacutemica bem como
as elevadas quantidades de vanaacutedio libertadas atraveacutes da combustatildeo dos oacuteleos de crude
contribuem consideravelmente para o aumento da biodisponibilidade do vanaacutedio Como tal o
interesse neste elemento e nos problemas ambientais por ele causados tem vindo a aumentar
significativamente [V]
121 Propriedades quiacutemicas
16
A quiacutemica do vanaacutedio eacute extraordinariamente complexa pelo que originou vaacuterias interpretaccedilotildees
incorrectas desde a sua descoberta No entanto as suas propriedades dependem em uacuteltima
anaacutelise da configuraccedilatildeo electroacutenica do elemento Assim a variabilidade encontrada no
comportamento quiacutemico do vanaacutedio pode dever-se agrave sua configuraccedilatildeo ndash [Ar] 3d2 4s
2 ndash que
permite a formaccedilatildeo de vaacuterios estados de oxidaccedilatildeo (de +2 a +5) Em soluccedilatildeo aquosa os estados de
oxidaccedilatildeo estaacuteveis vatildeo de V(V) a V(II) Destes apenas os +3 a +5 satildeo biologicamente importantes
jaacute que o V(II) apresenta demasiada capacidade redutora para existir nos organismos vivos
conhecidos [AC]
No fluido extracelular os compostos de vanaacutedio encontram-se predominantemente na forma de
monovanadatos (V(V)) livres ou ligados agrave transferina do soro ou agrave hemoglobina (Refordf) Os iotildees de
vanadato livres satildeo transportados para o meio intracelular atraveacutes de canais ioacutenicos inespeciacuteficos
Jaacute no ambiente intracelular os compostos sofreratildeo uma bio-reduccedilatildeo atraveacutes da acccedilatildeo da
glutationa ou outros compostos com grupos ndashOH como pentoses hexoses ou etanolaminas [V] A
reoxidaccedilatildeo de vanadilo (V(IV)) intracelular a vanadato (V(V)) provocaria a acumulaccedilatildeo toacutexica
destes uacuteltimos pelo que a complexaccedilatildeo da espeacutecie reduzida o impede O transporte de vanaacutedio
para o interior da ceacutelula encontra-se na Fig 110
Figura 110 ndash Transporte de vanaacutedio para a ceacuteluka e seu subsequente metabolismo (P=Proteiacutena) (Adaptado de
Zaporowska e Scibor 1998)
1211 Vanaacutedio(IV)
A espeacutecie de vanaacutedio (IV) mais bem conhecida eacute o iatildeo vanadilo (VO2+
) espeacutecie estaacutevel a pH
aacutecido sendo a maioritaacuteria a pH 3[AD] Apresenta uma cor azulada e forma complexos penta e
hexacoordenados Forma-se rapidamente a partir de V(V) na presenccedila de um agente redutor em
soluccedilatildeo aquosa No ambiente intracelular a presenccedila de glutationa reduzida aacutecido ascoacuterbico
NAD(P)H entre outros conduzem a que o principal estado de oxidaccedilatildeo nos sistemas bioloacutegicos
seja provavelmente o +4 encontrando-se o V(IV) fortemente complexado com proteiacutenas ou
aminoaacutecidos livres citrato ATP e pirofosfato (Refordfs) Em alternativa o V(IV) pode existir
17
livremente em ambientes fortemente aacutecidos como os lisossomas jaacute que a pH superior a 3 as
soluccedilotildees de vanadilo tendem a ser oxidadas pelo ar formando-se vanadato Esta grande
capacidade redutora justifica parcialmente a utilizaccedilatildeo pouco extensiva do iatildeo VO2+
como
inibidor [W]
1212 Vanaacutedio (V)
O vanaacutedio (V) eacute a forma mais estaacutevel num intervalo de pH amplo [W] O iatildeo anioacutenico VO3-tende
a constituir complexos polinucleares pelo que as soluccedilotildees aquosas de vanadato satildeo extremamente
complexas com ocorrecircncia simultacircnea de vaacuterios equiliacutebrios de protonaccedilatildeo (pKa(vanadato)= 35
78 e 125) e oligomerizaccedilatildeo As espeacutecies presentes (H3VO4 H2VO4- HVO4
2- e VO4
3-) dependem
do pH forccedila ioacutenica e temperatura da soluccedilatildeo e da concentraccedilatildeo total do elemento
[AEAFAGAHAI]
O VO43-
- monoacutemero ortovanadato V1 ndash estaacute associado a uma grande parte da importacircncia
bioloacutegica do vanaacutedio uma vez que se trata de um anaacutelogo electroacutenico e estrutural do ortofosfato
(PO43-
) A coordenaccedilatildeo do ortovanadato pode ser tetraeacutedrica ou bipiramidal trigonal (Refordfs)
A oligomerizaccedilatildeo do vanadato monomeacuterico daacute origem ao diacutemero (V2O72-
- V2) que apresenta
tambeacutem equiliacutebrios de protonaccedilatildeo Presume-se a existecircncia de analogia do vanadato dimeacuterico com
o pirofosfato embora natildeo haja evidecircncia estrutural (Refordf) A dimerizaccedilatildeo do V2 origina o
vanadato tetrameacuterico ou metavanadato (V4O124-
- V4) que constitui a espeacutecie maioritaacuteria em
soluccedilotildees aquosas concentradas (Refordf) As estruturas do metavanadato e a sua espeacutecie protonada
satildeo ciacuteclicas [AJ] Outra espeacutecie oligomeacuterica importante eacute o vanadato pentameacuterico (V5O153-
) cuja
estrutura presumida eacute ciacuteclica [AI]
O oligoacutemero com maior importacircncia bioquiacutemica eacute o vanadato decameacuterico (V10O286-
-V10)(Refordf) a
espeacutecie predominante em soluccedilotildees de pH 2-6 quando a concentraccedilatildeo total de vanadato eacute superior
a 100 M (Ref) A estrutura do V10 natildeo difere em soluccedilatildeo ou no estado soacutelidoOs 10 aacutetomos de
vanaacutedio da espeacutecie decameacuterica estatildeo divididos em trecircs grupos identificados na figura como VA-C
consoante o seu ambiente quiacutemico circundante (Refordf) (Fig 111)
A expressatildeo ldquodecavanadatordquo refere-se a uma soluccedilatildeo aquosa na qual a espeacutecie decameacuterica se
encontra em equiliacutebrio quiacutemico com as espeacutecies V1 V2 V4 e V5 sendo que a oligomerizaccedilatildeo
progressiva do vanaacutedio eacute proporcional agrave sua concentraccedilatildeo total (Ref) Nesta soluccedilatildeo a
concentraccedilatildeo de V10 corresponde a aproximadamente 10 da concentraccedilatildeo total de vanaacutedio [AK]
18
Figura 111 ndash Estruturas esquemaacuteticas das espeacutecies de vanadato (espeacutecie monomeacuterica (V1) dimeacuterica (V2)
tetrameacuterica (V4) pentameacuterica (V5) e decameacuterica (V10) (Adaptado de Aureliano e Crans 2008)
122 Metabolismo e efeito na actividade enzimaacutetica
Apesar de natildeo terem sido ainda encontradas as provas categoacutericas das suas funccedilotildees nos seres
vivos sabe-se que o iatildeo vanadato eacute capaz de inibir uma grande quantidade de enzimas in vitro
com especial ecircnfase na actividade ATPaacutesica da miosina [AL Teresa] Como referido
anteriormente o vanadato eacute capaz de actuar em alternativa ao fosfato nas reacccedilotildees bioquiacutemicas
incluindo a sua preferecircncia de ligaccedilatildeo agraves enzimas [W]
Eacute conhecido o papel do vanaacutedio no metabolismo do caacutelcio promovendo a mineralizaccedilatildeo dos
dentes e ossos na biossiacutentese de liacutepidos (trigliceacuteridos e colesterol) com posterior inibiccedilatildeo da
acumulaccedilatildeo destes nos vasos sanguiacuteneos e no SNC na reproduccedilatildeo e desenvolvimento por
mimetizaccedilatildeo e potenciaccedilatildeo de factores de crescimento na estimulaccedilatildeo do consumo de glucose e
da siacutentese de glicogeacutenio com actuaccedilatildeo equivalente aos efeitos da insulina ao niacutevel celular na
transduccedilatildeo de sinais celulares com regulaccedilatildeo da actividade de segundos mensageiros no
transporte de iotildees e aminoaacutecidos na transcriccedilatildeo geneacutetica e siacutentese de proteiacutenas e DNA entre
outros [W Y AB]
Assim o vanaacutedio cnseguiraacute provocar inuacutemeras mudanccedilas bioquiacutemicas nas ceacutelulas Alguns
exemplos da influecircncia bioquiacutemica do vanaacutedio incluem os seus efeitos nos processos de
fosforilaccedilatildeo e desfosforilaccedilatildeo de proteiacutenas e na participaccedilatildeo na formaccedilatildeo de espeacutecies reactivas de
oxigeacutenio devido ao seu efeito proacute-oxidante sobre agentes redutores a niacutevel celular [AM]
Quanto ao seu efeito na actividade enzimaacutetica pode dividir-se em dois tipos principais ndash inibitoacuterio
e estimulante - dependendo da enzima ou do tipo de reacccedilatildeo por esta catalisada No primeiro
grupo incluem-se enzimas como as ATPases (de miosina de canais ioacutenicos e fosfoenzimas
19
transportadoras) e as fosfatases (da tirosina aacutecida e alcalina) a fosfofrutocinase adenilato ciclase
e catalase entre outras [AA] O segundo grupo inclui a maioria das cinases a glucose-6-fosfato
desidrogenase a fosfoglicerato mutase e a fosfoglicerato fosfatase (Refordf)
O efeito do vanaacutedio (V) na actividade enzimaacutetica estaacute relacionado com o seu nuacutemero de
coordenaccedilatildeo podendo actuar como estado de transiccedilatildeo ou por mimetizaccedilatildeo de cofactores
substratos (especialmente o fosfato) e interacccedilotildees proteiacutena-proteiacutena (como no caso das
polimerases e transcriptases) Pode ainda estar envolvido na formaccedilatildeo de ligaccedilotildees de hidrogeacutenio
interacccedilotildees electrostaacuteticas esteacutereas e de oxidaccedilatildeo-reduccedilatildeo Uma evidecircncia desta uacuteltima eacute a sua
capacidade de alternacircncia entre os estados de oxidaccedilatildeo +5 a +3 o que torna faacutecil a sua interacccedilatildeo
redox com enzimas contendo resiacuteduos de cisteiacutenas (Cys) ou qualquer outro composto orgacircnico
com actividade redox [AN]
13 Vanaacutedio e muacutesculo esqueleacutetico
A primeira evidecircncia da presenccedila de vanaacutedio no muacutesculo esqueleacutetico proveio de uma amostra de
ATP isolado de muacutesculo de cavalo que continha uma impureza ndash vanaacutedio ndash cuja presenccedila
provocou a inibiccedilatildeo da actividade da bomba de Na+K
+-ATPase [AO] Desde entatildeo vaacuterios
estudos tecircm sido realizados com ecircnfase na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o
tecido muscular Destes destaca-se a descoberta da interacccedilatildeo forte da espeacutecie V10 com a miosina
fosforilada [AP] e agrave sua inibiccedilatildeo da actividade ATPaacutesica da miosina[AQ AR]
No quediz respeitoao iatildeo monovanadato destaca-se a sua inibiccedilatildeoda actividade Mg2+
-ATPaacutesica
da miosina [AS] que ocorre de duas formas distintas (1) na presenccedila de concentraccedilotildees reduzidas
de V1 a inibiccedilatildeo ocorre a velocidade reduzida dependente da concentraccedilatildeo da espeacutecie
monomeacuterica (2) na presenccedila de concentraccedilotildees elevadas de V1 o niacutevel de inibiccedilatildeo aproxima-se de
100 desde que [V1] seja superior agrave concentraccedilatildeo total de locais com actividade Mg2+
-ATPaacutesica
no ensaio Esta inibiccedilatildeo eacute irreversiacutevel tanto por diaacutelise ou adiccedilatildeo de agentes redutores do
vanadato [AL]Segundo Pate et al(1994)[AT] a espeacutecie monomeacuterica podeinibir a velocidade de
contracccedilatildeo a temperatura inferior a 20ordmC perdendogradual e rapidamente essa capacidade ateacute os
50ordmC temperatura agrave qual cessa a capacidade inibitoacuteria Os resultados obtidos sugeremainda uma
interacccedilatildeo altamente cooperativa entre o iatildeo vanadato e as proteiacutenas componentes do muacutesculo
esqueleacuteticoDestaca-se tambeacutem a descoberta da inibiccedilatildeo da geraccedilatildeo de forccedila contraacutectil nas fibras
musculares por parte da espeacutecie monomeacuterica[AT]Wilson et al(1995) [AU ]determinaram que a
afinidade da espeacutecie V1 para o S1 de miosina eacute significativamente mais elevada do que a do iatildeo
Pi seu anaacutelogo Descobriram ainda que que a ligaccedilatildeo do vanadato poderaacute envolver transiccedilotildees
entre estados energeacuteticos com energia livre independente dos impedimentos esteacutereos impostos
20
pela estrutura das miofibrilas Os estudos desenvolvidos por Smith et al (1990)[AV]
determinaram queo iatildeo monovanadato natildeo apresenta um efeito pronunciado na inibiccedilatildeo da
actividadeacto-S1-miosina-ATPaacutesica excepto em concentraccedilotildees superiores a 900microM a partir das
quais existe produccedilatildeo de espeacutecies oligomeacutericas para as quais o efeito inibitoacuterio seraacute mais
elevadoA espeacutecie monomeacuterica de vanadato consegue adoptar facilmente uma geometria do tipo
bipiramidal trigonal que mimetiza o intermediaacuterio formado (MgADP
Pi)aquando da reacccedilatildeo de
hidroacutelise de ATP no local activo da miosina [AW] De facto a interacccedilatildeo do vanaacutedio com a
miosina eacute inesperadamente estaacutevel na presenccedila de ADP Esta interacccedilatildeo resulta na formaccedilatildeo de
um complexo enzimaacutetico inactivo e estaacutevel com estequiometria de 11 ADPvanadato por siacutetio
activo da proteiacutena A formaccedilatildeo do complexo estaacutevel miosina-vanadato foi proposta por Goodnoet
al (1982) [AS] Nesta ocorre um preacute-equiliacutebrio raacutepido que induz a formaccedilatildeo de um complexo
ternaacuterio reversiacutevel (MADP
V) com formaccedilatildeo lenta do isoacutemero M
ADP
V A diferenccedila entre os
compostos de vanaacutedio e de fosfato centra-se no tempo de meia vida (t12) do Pi ou do V (no
primeiro t12=12 s enquanto no segundo t12=24 h) (Refordf) A diferenccedila dever-se-aacute agrave possiacutevel
complexaccedilatildeo do vanaacutedio com grupos nucleofiacutelicos no siacutetio activo o que provocaria uma
estabilizaccedilatildeo da miosina na conformaccedilatildeo MADP
V
No que respeita aos estudos realizados com a espeacutecie tetrameacuterica Cremo et al determinou que a
cadeia pesada do S1 de miosina pode ser clivada em dois locais distintos (designados V1 e V2)
pela espeacutecie V4 apoacutes irradiaccedilatildeo com luz UV na presenccedila de concentraccedilotildees de vanadato na ordem
dos milimolar e na ausecircncia de nucleoacutetidos ou metais divalentes
Jaacute no que concerne aos estudos realizados com ecircnfase na espeacutecie decameacuterica foi sugerido
porRamos et al (2006) [AK]que a interacccedilatildeo do decavanadato com a G-actina favorecida pela
polimerizaccedilatildeo desta uacuteltima estabiliza a espeacutecie V10 e induz a inibiccedilatildeo da formaccedilatildeo de F-actina A
estabilizaccedilatildeo da espeacutecie V10 por proteiacutenas tansmembranares e do citoesqueleto pode ser a razatildeo
pela qual ocorre a toxicidade do decavanadato reportada na avaliaccedilatildeo dos efeitos do V(V) nos
sistemas bioloacutegicosTiago et al (2007) [AR] concluiram que o V10 apresenta uma clara preferecircncia
de ligaccedilatildeo ao ldquoback-doorrdquo da S1-miosina-ATPase o que se verifica apenas na estrutura
conformacional desimpedida com acesso facilitado ao local de ligaccedilatildeo do Pi Sugere tambeacutem que
o V10 actua como um ldquoback-door stoprdquo bloqueando estaabertura do fragmento de 50-kDa
necessaacuteria para a hidroacutelise do ATP--fosfato o que constitui uma explicaccedilatildeo simples do
comportamento natildeo competitivo da espeacutecie decameacuterica estimulando a utilizaccedilatildeo deste oligoacutemero
como uma feramenta da elucidaccedilatildeo das mudanccedilas conformacionais ldquoback-doorrdquo da miosina no
21
processo da contracccedilatildeo muscular(ref)Destaca-se ainda a descoberta da capacidade inibitoacuteria do
V10 em relaccedilatildeo agrave fosforilaccedilatildeo oxidativa mitocondrial [AX]
Finalmente no que concerne aos estudos realizados com a presenccedila de vaacuterias espeacutecies
oligomeacutericas Aureliano e Madeira (1994) [AY]observaram que na presenccedila de Ca2+
as espeacutecies
V4 e V10se ligam agrave Ca2+
-ATPase do retiacuteculo sarcoplasmaacutetico enquanto a espeacutecie V1 se liga apenas
na presenccedila de ATP Aparentemente o aumento da concentraccedilatildeo de Ca2+
tem o efeito contraacuterio da
inibiccedilatildeo da ATPase do retiacuteculo sarcoplasmaacutetico pelo V1 natildeo tendo no entanto efeito significativo
sobre a inibiccedilatildeo por parte da espeacutecie V10 Tiago et al (2002) [AP] concluiacuteram que em condiccedilotildees
proacuteximas agraves fisioloacutegicas a espeacutecie V10 difere das espeacutecies presentes na soluccedilatildeo de metavanadato
devido agrave sua forte interacccedilatildeo com a miosina fosforilada e agrave sua inibiccedilatildeo da actividade ATPaacutesica
da miosina Para aleacutem disso o ATP diminui a afinidade da miosina para o V4 induzindo a
interacccedilatildeo com V1 e natildeo tendo qualquer efeito no que diz respeito ao V10 Foi tambeacutem proposto
[AQ] que a ligaccedilatildeo das espeacutecies oligomeacutericas de vanadato aoS1 de miosina nos locais onde o
fosfato coordenaeacute provavelmente a causa da inibiccedilatildeo da actividade ATPaacutesica da miosina
estimulada por actina atraveacutes da prevenccedilatildeo da troca entre ATPADP sendo esta favorecida para
aniotildees de vanadato com valecircncias superiores como eacute o caso do V10
Na presenccedila de actina as velocidades de dissociaccedilatildeo do Pi e do V aumentam consideravelmente
este uacuteltimo em menor quantidade Assim a ligaccedilatildeo da actina ao complexo MADP
V e
Pi eacute
bastante fraca pelo que se sugere que o ADPV teria um efeito de relaxamento semelhante ao
ATP No entanto a capacidade do vanaacutedio em inibir a actividade ATPaacutesica da miosina e da
actomiosina eacute diferenciada uma vez que na ausecircncia de actina a inibiccedilatildeo eacute total para
concentraccedilotildees na gama dos M mas na sua presenccedila a inibiccedilatildeo eacute totalmente conseguida apenas
em concentraccedilotildees na gama dos mM (Ref) No entanto eacute necessaacuterio considerar a possiacutevel
formaccedilatildeo de espeacutecies oligomeacutericas de vanadato cuja afinidade e efeito na enzima seratildeo
diferentes Assim pode concluir-se que o efeito de inibiccedilatildeo observado para a actividade ATPaacutesica
de miosina estimulada por actina se pode dever agrave presenccedila das diferentes espeacutecies oligomeacutericas o
que foi jaacute sugerido por Aureliano (2000) [AZ] ndash as espeacutecies tetrameacuterica e decameacuterica de vanadato
interactuariam com a miosina com alteraccedilatildeo da sua interacccedilatildeo com a actina No entanto natildeo eacute
conhecida a contribuiccedilatildeo absoluta das restantes espeacutecies oligomeacutericas para a inibiccedilatildeo da ATPase
da miosina estimulada por actina (Refs)
14 Objectivos do projecto
Pretende-se com este projecto o isolamento purificaccedilatildeo e caracterizaccedilatildeo da actina do muacutesculo
22
esqueleacutetico de coelho sendo esta uacuteltima tambeacutem realizada para o sufragmento-1 de miosina
Seguidamente realizar-se-aacute a quantificaccedilatildeo e caracterizaccedilatildeo das proteiacutenas por espectroscopia UV-
Vis e SDS-PAGE e a caracterizaccedilatildeo das soluccedilotildees de vanaacutedio por UV-Vis e ressonacircncia magneacutetica
nuclear (RMN) Por uacuteltimo a interacccedilatildeo do S1 de miosina com o vanaacutedio seraacute avaliada atraveacutes
das teacutecnicas de UVVis RMN e ressonacircncia paramagneacutetica electroacutenica (RPE)
2 Discussatildeo de Resultados
21 Caracterizaccedilatildeo das proteiacutenas isoladas
211Anaacutelise do estado de pureza
O estado de pureza das soluccedilotildees de actina e de S1 foi analisado por electroforese em gel de
poliacrilamida em condiccedilotildees desnaturantes (SDS-PAGE) (Fig 22) que separa as moleacuteculas de
acordo com o seu peso molecular (PM) A concentraccedilatildeo de acrilamida escolhida (10) permite a
detecccedilatildeo simultacircnea de componentes com peso molecular inferior a 20 kDa e outros com elevado
peso molecular
Figura 21 ndash Fotografia de gel de SDS-PAGE (10 acrilamida) Poccedilo (a)
10 L do marcador de elevado peso molecular () Poccedilo (b) 20 L de [G-
actina]=1 mgmL Poccedilo (c) 20 L de [S1]=1 mgmL Aplicou-se uma
voltagem de V durante h e min
A partir da recta de calibraccedilatildeo obtida log PM = -1177 Rf + 2247 (ver ponto 2 da secccedilatildeo
Anexos) pode verificar-se a presenccedila de actina na amostra do poccedilo (b) presente numa banda
bastante intensa (PMaparente=40 kDa) bem como a presenccedila minoritaacuteria de outros componentes
desconhecidos que natildeo teratildeo sido eliminados durante o processo de extracccedilatildeo e purificaccedilatildeo e natildeo
satildeo relevantes para o estudo em questatildeo Na amostra do poccedilo (c) nota-se a presenccedila de S1
(PMaparente=92 kDa) bem como a de outros componentes cuja banda eacute ligeiramente menos intensa
Estes poderatildeo ser fragmentos proteoliacuteticos de miosina e de S1 Estaacute ainda presente uma bada de
23
maior intensidade que poderaacute ser a proteiacutena-C (PM=135 kDa) uma vez que a sua remoccedilatildeo
completa da amostra necessitaria de um passo adicional de cromatografia de troca ioacutenica [BA]
No entanto natildeo podemos confirmar que se trata desta proteiacutena ou de qualquer outra uma vez que
se encontra fora do intervalo de migraccedilatildeo dos marcadores de peso molecular
212 Determinaccedilatildeo da concentraccedilatildeo proteica
2121Subfragmento-1 de miosina
A partir da recta de calibraccedilatildeo obtida pelo meacutetodo de Bradford [BB](ver secccedilatildeo 3311 do
Procedimento Experimental) A595nm = 00546[BSA] + 00209 (ver secccedilatildeo de Anexos)foi possiacutevel
determinar um intervalo de concentraccedilatildeo de S1 de miosina compreendido entre 226 e 117(plusmn
423M) (203 e 1056 (plusmn 381 mgmL)) (n=8) A concentraccedilatildeo meacutedia de S1 de miosina apoacutes
diaacutelise contra a soluccedilatildeo tampatildeo 10 mM HEPES pH 75 25 mM KCl determinada pela leitura da
Abs280 nmfoi de 20430M (1803 mgmL)(n=7)
As amostras a quantificar atraveacutes do meacutetodo de Bradford natildeo sofreram diaacutelise o que permite
explicar a concentraccedilatildeo superior de S1 de miosina em comparaccedilatildeo com o valor obtido atraveacutes do
meacutetodo directo Esta razatildeo aliada agrave sensibilidade e rapidez superiores deste uacuteltimo fez com que
se optasse pelo meacutetodo directo para a quantificaccedilatildeo da grande maioria dos ensaios
2122Actina
A concentraccedilatildeo de actina foi determinada quando esta se encontrava na forma monomeacuterica (G-
actina) por mediccedilatildeo da leitura directa da Abs290 nm tendo-se obtido uma concentraccedilatildeo meacutedia de
691481M (290202 mgmL) (n=3) em 2 mM Tris-HCl pH 80 02 mM CaCl2 02 mM Na-
ATP Ao comprimento de onda utilizado (290 nm) absorvem fortemente apenas os resiacuteduos de
Trp pelo que a mediccedilatildeo efectuada tem menor sensibilidade do que se tivesse sido efectuada a 280
nm uma vez que a actina apresenta apenas 4 resiacuteduos deste aminoaacutecido embora tenha 16
resiacuteduos de Tyr
22 Mediccedilatildeo da actividade enzimaacutetica de S1 de miosina
221 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
As actividades ATPaacutesicas de S1 de miosina dependentes de Ca2+
e K+foram analisadas pelo
caacutelculo das velocidades de consumo inicial do fosfato inorgacircnico (Pi) Definiu-se os tempos de 5
e 3 minutos para a mediccedilatildeo da Ca2+
e da K+EDTA-ATPase respectivamente Os valores obtidos
24
a partir do protocolo presente no ponto 341 da secccedilatildeo Procedimento experimental encontram-se
descritos na Tabela 2I
Tabela 2I ndash Acrividades natildeo-fisioloacutegicas de subfragmento-1 determinadas pelo meacutetodo de Fiske-Subarow
modificado (n=2)
Proteiacutena
Actividade
ATPaacutesica natildeo
fisioloacutegica
Actividade (nmol
Piminmg proteiacutena) Actividade (s-1)
S1
Ca2+
-ATPase 1400 plusmn 30 2100 plusmn 0045
K+EDTA-ATPase 2334 plusmn 50 3500 plusmn 0075
As actividades natildeo-fisioloacutegicas reflectem o estado de oxidaccedilatildeo dos resiacuteduos expostos de Cys 707
(SH1) e 697 (SH2) Se o grupo SH1 sofrer oxidaccedilatildeo a actividade ATPaacutesica dependente de Ca2+
aumenta e a actividade dependente de K+ diminui Por outro lado se o grupo SH2 sofrer
oxidaccedilatildeo regista-se a perda completa destas actividades
De acordo com Margossian e Lowey (1982) [BA] as actividades ATPaacutesica de S1 de miosina
dependente de Ca2+
e K+EDTA da miosina satildeo respectivamente de 24 e 112 s-1
Estaacute tambeacutem
descrito que a actividade ATPaacutesica dependente de K+ eacute em meacutedia cerca de 3 vezes superior agrave
dependente de Ca2+
reflectindo a quantidade das preparaccedilotildees A razatildeo obtida entre os valores das
actividades ATPaacutesicas dependentes de K+ e Ca
2+ eacute de 166 Os valores obtidos para as actividades
(Ca2+
-ATPase=21 s-1
K+EDTA-ATPase= 35 s
-1) satildeo inferiores agrave valores obtidos por
Margossian e Lowey [BA] e Tiago [BC] com forccedila ioacutenica inferior O valor obtido para a
actividade ATPaacutesica dependente de Ca2+
eacute superior ao obtido por Polosukhina et al (2000) [BD]
cujo meio de reacccedilatildeo continha MOPS e apresentava forccedila ioacutenica inferior Os dados
supramencionados comprovam a variaccedilatildeo da actividade a ATPaacutesica natildeo fisioloacutegica em relaccedilatildeo agrave
forccedila ioacutenica temperatura e pH
222 Mediccedilatildeo da actividadeMg2+
-ATPaacutesica de S1 pelo meacutetodo de enzimas acopladas
A actividade de Mg2+
-ATPase eacute a uacutenica que possui significado fisioloacutegico sendo as restantes
bastante uacuteteis apenas para indicaccedilatildeo da qualidade de preparaccedilatildeo do S1[BA] A 0575 M KCl a
actina e o S1 formam uma soluccedilatildeo viscosa e tuacuterbida Apoacutes a adiccedilatildeo de MgATP o complexo acto-
S1 dissocia-se e a turbidez da soluccedilatildeo diminui Se a forccedila ioacutenica for reduzida a dissociaccedilatildeo do
complexo actoS1 que se encontra na forma de suspensatildeo eacute induzida O MgATP eacute rapidamente
hidrolisado diminuindo a sua concentraccedilatildeo no meio reaccional Nestas condiccedilotildees haacute
reassociaccedilatildeo e subsequente contracccedilatildeo dos filamentos de actina e de S1 com formaccedilatildeo de
25
precipitado O meacutetodo descrito traduz de forma bastante elementar o fenoacutemeno da contracccedilatildeo in
vivo e elimina o problema anterior da solubilizaccedilatildeo ineficiente da miosina de modo a obter uma
mediccedilatildeo exacta da actividade [A] A utilizaccedilatildeo de forccedilas ioacutenicas reduzidas aumenta a afinidade da
actina para o S1 permitindo contornar a extrema dificuldade em obter a concentraccedilatildeo proteica dos
muacutesculos (cerca de 100 mgmL do complexo actomiosina) As actividades Mg2+
-ATPaacutesicas de S1
de miosina foram determinadas a partir do declive da variaccedilatildeo da Abs340 relativo agrave oxidaccedilatildeo de
NADH em funccedilatildeo do tempo como eacute descrito no ponto 342 do Procedimento Experimental Os
valores obtidos para a actividade Mg2+
-ATPaacutesica encontram-se apresentados na Tabela 2II
Tabela 2II ndash Actividade Mg2+
-ATPaacutesica de subfragmento-1 de miosina determinadas pelo meacutetodo das enzimas
acopladas
Actividade (nmol
ATPmin mg(S1)) Actividade (s
-1)
1097 plusmn 269 164 plusmn 04
Ramos (2005) [J] obteve o valor de 01 s-1
para a actividade Mg2+
-ATPaacutesica de S1 de miosina
com forccedila ioacutenica intermeacutedia Tiago (2005) [BC] obteve o valor de 011 s-1 em condiccedilotildees de forccedila
ioacutenica reduzida em relaccedilatildeo agrave utilizada no presente relatoacuterio
23 Preparaccedilatildeo e caracterizaccedilatildeo das soluccedilotildees de vanaacutedio
231 Anaacutelise da composiccedilatildeo das soluccedilotildees de vanadato por espectroscopia de 51
V-RMN
efeito da concentraccedilatildeo da espeacutecie
A diluiccedilatildeo das soluccedilotildees stock de meta- e decavanadato iraacute determinar fortemente a composiccedilatildeo
em soluccedilatildeo dos diferentes oligoacutemeros de vanadato (ref) No caso do decavanadato o periacuteodo de
tempo apoacutes a diluiccedilatildeo iraacute tambeacutem ser um factor importante a considerar devido agrave decomposiccedilatildeo
da espeacutecie V10(ref)Torna-se assim fundamental determinar a composiccedilatildeo quiacutemica das soluccedilotildees
no meio de ensaio utilizado (HEPES 10 mM KCl 25 mM) de modo a correlacionaacute-la
correctamente com os possiacuteveis efeitos promovidos pelas diferentes espeacutecies de vanadato no
sistema em estudo
Aleacutem disso deve ser dado ecircnfase agraves importantes interacccedilotildees que ocorrem entre o vanaacutedio e os
tampotildees escolhidos O tampatildeo com menor nuacutemero e magnitude de interacccedilatildeo eacute o HEPES [AD]
seguido do Tris sendo este uacuteltimo especialmente aconselhado em concentraccedilotildees baixas para casos
de incompatibilidade enzimaacutetica ou quiacutemica redox No caso especial da reacccedilatildeo redox
NAD(P)HNAD(P) o tampatildeo adequado eacute o imidazole de modo a natildeo haver reacccedilatildeo com o
26
vanadato Os tampotildees a evitar em qualquer situaccedilatildeo incluem a trietanolamina fosfato citrato e
glicina entre outros [AD] uma vez que todos interactuam fortemente com V(V) com formaccedilatildeo
de complexos estaacuteveis
Assim procedeu-se agrave anaacutelise da composiccedilatildeo das soluccedilotildees de meta- e decavanadato apoacutes diluiccedilatildeo
das soluccedilotildees stock (50 mM em vanadato total) no meio de reacccedilatildeo (tampatildeo D) atraveacutes da
espectroscopia de ressonacircncia magneacutetica nuclear de 51
V Nas figuras x1 e x2 apresentam-se os
espectros obtidos para as soluccedilotildees de deca- e metavanadato num intervalo de concentraccedilotildees de 1 a
10 mM A Tabela seguinte resume os paracircmetros espectroscoacutepicos obtidos para cada espeacutecie
presente nas soluccedilotildees analisadas
Figura 22ndash Espectros de RMN de 51
V de
concentraccedilotildees crescentes das espeacutecies oligomeacutericas
de vanadato obtidos apoacutes diluiccedilatildeo da soluccedilatildeo stock
de decavanadato em 10 mM HEPES pH 70 25 mM
KCl para concentraccedilotildees finais de A) 1 mM B)2
mM C) 4 mM D) 8 mM E) 10 mM de
decavanadato total V1refere-se agraves espeacutecies
monomeacutericade vanadato V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2IIIndash Desvios quiacutemicos meacutedios (part) das ressonacircncias das espeacutecies decameacuterica (V10) monomeacuterica (V1) e
tetrameacuterica (V4) presentes nos espectros da soluccedilatildeo de decavanadatoLargura a meia altura (∆12) da espeacutecie V10A
Paracircmetros V10A V10B V10C V1 V4
part (ppm) -5162 -5003 -4255 -5616 -5748
∆12 (Hz) 175 nd nd nd nd
Figura 23 ndash Distribuiccedilatildeo graacutefica das
espeacutecies oligomeacutericas de vanadato vs a
concentraccedilatildeo de decavanadato total As
concentraccedilotildees de cada espeacutecie foram
obtidas segundo a formula descrita na
secccedilatildeo 35 do Procedimento
Experimental (aacutereas dos sinais de 51
V-
RMN observadas na figura x1)V10 V1 e
V4 correspondem agraves espeacutecie monomeacuterica
dimeacuterica e tetrameacuterica respectivamente [Esp
eacuteci
e o
ligo
meacute
rica
de
van
aacutedio
] (m
M)
[Decavanadato]total (mM)
V10
V1
V4
27
Nas soluccedilotildees de decavanadato as espeacutecies monomeacuterica e decameacuterica de vanadato satildeo as uacutenicas
presentes para concentraccedilotildees de decavanadato inferiores a 8 mM Ambas as espeacutecies aumentam
linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado em [AY] e
em (Teresa) quando a concentraccedilatildeo de V1 eacute igual ou superior a 200 microM inicia-se a sua
oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica No entanto nas condiccedilotildees de realizaccedilatildeo dos
ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a espeacutecie tetrameacuterica neste
estudo em concentraccedilotildees inferiores a 8 mM tendo o mesmo sido observado por Ramoset al
(2006) [AK] Os resultados obtidos mostram claramente o equiliacutebrio entre a espeacutecie decameacuterica e
a monomeacuterica e que o aparecimento de outras espeacutecies oligomeacutericas se encontra dependente da
concentraccedilatildeo desta uacuteltima Apesar de natildeo ter sido realizado o ensaio com 5 mM de decavanadato
total no presente relatoacuterio estaacute reportado em Ramos et al (2006) [AK] que as concentraccedilotildees da
espeacutecie decameacuterica e monomeacuterica satildeo respectivamente de 500 microM e 200 microM Assim pode inferir-
se que para concentraccedilotildees inferiores a 5 mM e intervalos de tempo de cerca de 20 minutos (que
inclui o tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas
as espeacutecies decameacuterica e monomeacuterica de vanadato
Figura 24 ndash Espectros de RMN de 51
V de concentraccedilotildees
crescentes das espeacutecies oligomeacutericas de vanadato obtidos
apoacutes diluiccedilatildeo da soluccedilatildeo stock de metavanadato em 10 mM
HEPES pH 70 25 mM KCl para concentraccedilotildees finais de A)
1 mM B) 2 mM C) 4 mM D) 8 mM E) 10 mM de
metavanadato total V1 refere-se agraves espeacutecies monomeacuterica de
vanadatoV1 e V2 correspondem respectivamente agrave espeacutecie
monomeacuterica e dimeacuterica
Figura 25 ndash Distribuiccedilatildeo graacutefica das espeacutecies
oligomeacutericas de vanadato vs a concentraccedilatildeo de
metavanadato total As concentraccedilotildees de cada
espeacutecie foram obtidas segundo a formula descrita
na secccedilatildeo 35 do Procedimento Experimental
(aacutereas dos sinais de 51
V-RMN observadas na
figura x1)V10 V1 e V4 correspondem agraves espeacutecie
monomeacuterica dimeacuterica e tetrameacuterica
respectivamente
Esp
eacuteci
e o
ligo
meacute
rica
de
va
nad
ato
(m
M)
[Metavanadato]total (mM)
V1
V2
V4
V5
28
Tabela 2IVndash Desvios quiacutemicos meacutedios (part)das ressonacircncias das espeacutecies tetrameacuterica (V4) monomeacuterica (V1) e
dimeacuterica (V2) presentes no espectro da soluccedilatildeo de metavanadato (total) 10 mM
Paracircmetros V1 V2 V4 V5
part (ppm) -5615 -5748 -5789 -5869
Nas soluccedilotildees de metavanadato as espeacutecies monomeacuterica dimeacutericae tetrameacuterica de vanadato estatildeo
presentes para concentraccedilotildees de metavanadato inferiores a 4 mM Todas estas as espeacutecies
aumentam linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado
em (Aureliano e Madeira 1998) e em (Tiago et al 2004) quando a concentraccedilatildeo de V1 eacute igual ou
superior a 200 microM inicia-se a sua oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica Nas
condiccedilotildees de realizaccedilatildeo dos ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a
espeacutecie pentameacuterica neste estudo em concentraccedilotildees inferiores a 4 mM tendo o mesmo sido
observado por Ramos 2006 Os resultados obtidos mostram claramente o equiliacutebrio entre a
espeacutecie V4 e as espeacuteciesV1 e V2 no qual o aparecimento de outras espeacutecies oligomeacutericas se
encontra dependente da concentraccedilatildeo da espeacutecie V1 Por uacuteltimo pode inferir-se que para
concentraccedilotildees inferiores a 2 mM e intervalos de tempo de cerca de 20 minutos (que inclui o
tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas as
espeacutecies V1 V2 e V4 com concentraccedilotildees respectivas de 0796 0192 e 0196 mM
232Estabilidade das soluccedilotildees de decavanadato anaacutelise por espectroscopia de absorccedilatildeo
UVVis
Caracterizou-se a estabilidade da espeacutecie V10 numa soluccedilatildeo com 1 mM decavanadato total em
tampatildeo D atraveacutes da leitura da Abs400 Obteve-se uma cineacutetica de dissociaccedilatildeo de 1ordf ordem tal
como se pode verificar pela linearidade do graacutefico de ln(Abs400)vs o tempo de aquisiccedilatildeo espectral
(segundos) (Fig26 )
Figura 26ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do tempo
(em segundos) para uma soluccedilatildeo de decavanadato total de 1
mM em tampatildeo D a 25ordmC (n=2)
Sendo uma reacccedilatildeo de primeira ordem pode ser descrita pela equaccedilatildeo cujo
declive eacute ndashK (constante de dissociaccedilatildeo) e a intersecccedilatildeo no eixo das ordenadas corresponde
Ln
(Ab
s 400 n
m)
tempo (s)
29
aln(A0) Numa reacccedilatildeo de 1ordf ordem o tempo de meia-vida eacute independente da concentraccedilatildeo inicial
do reagente sendo descrito pela equaccedilatildeo [2] Deste modo a partir da recta
Ln(Abs400 nm) = -436E-5
K - 105 obteve-se um tempo de meia-vida de 4490 plusmn 0823 horas para
as condiccedilotildees supramencionadas
Apoacutes a adiccedilatildeo de S1 8 microM ao meio reaccional verificou-se que a reacccedilatildeo de desoligomerizaccedilatildeo
da espeacutecie decameacuterica de vanadato seguia igualmente uma cineacutetica de 1ordf ordem tal como se pode
verificar pela linearidade do graacutefico de ln(Abs400) versus o tempo de aquisiccedilatildeo espectral
(segundos) (Fig27)
Figura 27ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do
tempo (em segundos) para uma soluccedilatildeo de decavanadato
total de 1 mM em tampatildeo D a 25ordmC na presenccedila de 8
microM (n= 2)
A partir da equaccedilatildeo da recta Ln(Abs400 nm) = -7520E-5
K - 0613determinou-se o valor de K para
a espeacutecie decameacuterica de onde se obteve um tempo de meia-vida de 2430 plusmn 0191 horas para as
condiccedilotildees supramencionadas
Tanto este valor como o valor registado na ausecircncia de S1 satildeo substancialmente superiores agrave
duraccedilatildeo dos ensaios de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina descritos no presente
relatoacuterio
24 Interacccedilatildeo de S1 com vanaacutedio(V)
241Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
De modo a analisar qual o efeito da adiccedilatildeo de S1 de miosina (tampatildeo D) produzia nos sinais de
51V-NMR titulou-se uma soluccedilatildeo de 5mM de decavanadato total com concentraccedilotildees crescentes
de S1 (5 10 15 e 20 microM) Os espectros de 51
V-RMN encontram-se apresentados na Fig 2
No graacutefico da figura 2 Apresenta-se a concentraccedilatildeo de S1 em funccedilatildeo do factor de alargamento
corrigido dos espectros acima referidos
Ln
(Ab
s (4
00
nm
))
tempo (s)
30
Figura 28 ndash Graacutefico de
representaccedilatildeo da
concentraccedilatildeo de S1 em
funccedilatildeo do factor de
alargamento A
concentraccedilatildeo de
decavanadato total foi de 5
mM e as concentraccedilotildees
utilizadas de S1 foram de 0
5 10 e 15 microM
Verifica-se que o factor de alargamento aumenta linearmente em funccedilatildeo da concentraccedilatildeo de S1 no
ensaio o que indica a existecircncia de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina Observou-
se ainda que a adiccedilatildeo de S1 ao meio reaccional natildeo provocou qualquer alteraccedilatildeo dos desvios
quiacutemicos dos sinais de 51
V-RMN
Realizaram-se ensaios na presenccedila de ATP e de S1 com intuito de verificar o efeito do ATP na
interacccedilatildeo do S1 de miosina com o decavanadato
Figura 29 - Espectros de RMN de 51
V contendo 2
mM Decavanadato (A) (controlo) (B) na presenccedila
de 2 mM de ATP (C) na presenccedila de 2 mM de ATP
e 8 microM de S1 de miosina V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2Vndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies decameacuterica (V10) e monomeacuterica (V1) presentes nos
espectros da soluccedilatildeo de decavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V10A V10B V10C V1
part (ppm) -5161 -5002 -4254 -5614
Observa-se que a presenccedila de ATP (espectro B) natildeo provoca qualquer alteraccedilatildeo visiacutevel em
relaccedilatildeo ao espectro que contem apenas soluccedilatildeo de decavanadato Jaacute a presenccedila simultacircnea de ATP
e S1 provoca um alargamento da ∆12 do sinal do V10A de 1736 Hz (espectro (A)) para 2366 Hz
(espectro (C) Fig 29) Tendo em conta que a concentraccedilatildeo de S1 apresenta uma relaccedilatildeo linear
com o factor de alargamento converteu-se o valor da ∆12 e comparou-se com o valor esperado agrave
F = 00591[S1] - 00157 Rsup2 = 0998
Fact
or
de
ala
rgam
en
to
[S1] (microM)
31
concentraccedilatildeo de S1 utilizada O factor de alargamento obtido para 8 microM ndash 0376 ndash eacute inferior ao
valor esperado agrave mesma concentraccedilatildeo de S1 obtido atraveacutes da equaccedilatildeo da recta apresentada na
Fig 28 ndash 0457 ndash o que indica que o ATP poderaacute exercer um efeito de protecccedilatildeo do S1
diminuindo a sua interacccedilatildeo com a espeacutecieV10
Por uacuteltimo realizaram-se ensaios com metavanadato na presenccedila de 8 microM S1 de modo a verificar
se as espeacutecies oligomeacutericas contidas nesta soluccedilatildeo interagiam com a proteiacutena em estudo
Figura 210 - Espectros de RMN de 51
V contendo 2
mM metavanadato (A) (controlo) (B) na presenccedila
de 8 microM de S1 de miosina V1 V2 e V4
correspondem respectivamente aacutes espeacutecies
monomeacuterica dimeacuterica e tetrameacuterica
Tabela 2VIndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies monomeacuterica (V1) dimeacuterica (V2) e tetrameacuterica (V4)
presentes nos espectros da soluccedilatildeo de metavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V1 V2 V4
part (ppm) -5613 -5747 -5789
Como eacute possiacutevel observar na Fig 210 ocorre uma ligeiro alargamento dos sinal de V4 bem
como uma perda da intensidade pelo que se conclui quedas espeacutecies presentes na soluccedilatildeo de
metavanadato (V1 V2 e V4) apenas a espeacutecie tetrameacuterica interage com o S1 de miosina o que
estaacute de acordo com o reportado por Tiago et al 2002
242 Actividade Mg2+
-ATPaacutesica de S1 de miosina na presenccedila de decanadato
Com o intuito de avaliar o efeito do decavanadato na actividade Mg2+
-ATPaacutesica do S1 de
miosina monitorizou-se a reacccedilatildeo de oxidaccedilatildeo do NADH atraveacutes da leitura de Abs340 na
presenccedila de 005 mgmL de S1 de miosina e no caso da actividade estimulada 2microM de actina em
3125 mM Tris-HCl pH 71 e 375 mM MgCl2 pelo meacutetodo de enzimas acopladas (ver seccccedilatildeo xx
dos MM)Para tal realizaram-se ensaios com vaacuterias concentraccedilotildees de decavanadato total (10 50
100 250 500 e 1000) de modo a determinar qual o seu efeito na actividade MgATPaacutesica de S1
de miosina na ausecircncia e na presenccedila de um dos seus ligandos naturais Os ensaios foram
realizados agrave temperatura ambiente com incubaccedilatildeo preacutevia a 25ordm C durante 20 minutos
32
Figura 211 ndash Representaccedilatildeo
graacutefica da Actividade Mg2+
-
ATPaacutesica de S1 () (os ajustes
teoacutericos estatildeo representado a por
uma linha os resultados
experimentais estatildeo representados
por triacircngulos (Actividade
estimulada) e cruzes (Actividade
natildeo estimulada)) vs
[Decavanadato]total medidas pelo
meacutetodo das enzimas acopladas
(n=3) Os dados foram ajustados ao modelo de decaimento exponencial usando o programa GraphPad Prism para a
determinaccedilatildeo dos valores de IC50 (na ausecircncia de actina IC50= 266 plusmn 07 M decavanadato na presenccedila de actina
IC50= 28 plusmn 1 M decavanadato
A partir do graacutefico presente na Fig 29 verifica-se que o ajuste dos dados experimentais foi bem
sucedido A inibiccedilatildeo da actividade Mg2+
-ATPaacutesica do S1 de miosina eacute afectada em menor
amplitude na presenccedila de actina (60 de inibiccedilatildeo da actividade) do que na sua ausecircncia
(aproximadamente 80 de inibiccedilatildeo de actividade) Devido a esta diferenccedila pode sugerir-se que a
actina iraacute impedir parcialmente a ligaccedilatildeo do decavanadatoao S1 demiosina atraveacutes de
impedimentos esteacutereos ou interacccedilotildees electrostaacuteticas por exemplo A teacutecnica utilizada natildeo nos
permite concluir acerca de que regiatildeo(otildees) ou resiacuteduo(s) da actina participaratildeo na preservaccedilatildeo
parcial da actividade Mg2+
-ATPaacutesica
Por outro lado os valores de IC50 dos dois tipos de actividades natildeo apresentam diferenccedilas
significativas tendo-se obtido 266 plusmn 07 e 28 plusmn 1M decavanadato na ausecircncia e na presenccedila de
actina respectivamente o que corresponde a uma concentraccedilatildeo de cerca de 27 M de V10 O
meio de reacccedilatildeo utilizado nestes ensaios foi 10 mM HEPES e 25 mM KCl com 2 mM ATP Em
(Tiago et al 2004) o valor de IC50 para a actividade Mg2+-ATPaacutesica na ausecircncia de actina foi de
125 plusmn 025 microM enquanto o valor da actividade estimulada por actina foi de 136 plusmn 14 microM O
meio de reacccedilatildeo utilizado nos ensaios foi 20 mM HEPES 50 mM KCl e 2 mM ATP o que resulta
precisamente no dobro da forccedila ioacutenica a que foi realizado o ensaio do presente relatoacuterio
(exceptuando a contribuiccedilatildeo dos produtos da hidroacutelise de ATP uma vez que este meacutetodo
pressupotildee a ocorrecircncia de um ciclo de reacccedilotildees de hidroacutelise e siacutentese deste composto) Ora tendo
sido anteriormente referido que a afinidade do S1 de miosina para com a espeacutecie V10 diminui com
Act
ivid
ade
Mg
2+-A
TP
aacutesic
a d
e S
1 d
e
Mio
sin
a
[Decavanadato] (microM) Actividade Estimulada ()
Actividade Natildeo Estimulada ()
Ajuste Actividade Estimulada
Ajuste Actividade Natildeo Estimulada
33
o aumento da forccedila ioacutenica (Tiago et al 2004 tal poderaacute explicar a diferenccedila obtida entre estes
dois conjuntos de valores
243Quantificaccedilatildeo de cisteiacutenas reduzidas pelo meacutetodo de Ellman
A interacccedilatildeo do vanaacutedio com alguns tipos de aminoaacutecidos sensiacuteveis agrave oxidaccedilatildeo pode fornecer-nos
informaccedilatildeo bastante uacutetil em relaccedilatildeo agraves alteraccedilotildees estruturais que conduzem a uma diminuiccedilatildeo da
actividade fisioloacutegica do S1 de miosina Os estudos realizados por Chasteen 1983 concluiacuteram
que a reduccedilatildeo de vanadato (V) a vanadilo (IV) ocorre na presenccedila de agentes redutores comuns no
ambiente celular de que se destaca o aminoaacutecido cisteiacutena Esta eacute tambeacutem um alvo de
complexaccedilatildeo por parte do iatildeo VO2+
Jaacute Ramos et al (2009) demonstrou que a incubaccedilatildeo de
actina com a soluccedilatildeo de decavanadato induz a oxidaccedilatildeo de resiacuteduos de cisteiacutena e agrave formaccedilatildeo do
oxidovanaacutedio (IV) Este grupo de investigadores descobriu ainda que apenas a espeacutecie V10 induz a
oxidaccedilatildeo das cisteiacutenas ldquoraacutepidasrdquo da actina atraveacutes do processo proposto que eacute parcialmente
responsaacutevel pela inibiccedilatildeo jaacute descrita da actividade de ATPase do S1 de miosina estimulada por
actina quando exposta agrave soluccedilatildeo de decavanadato Esta inibiccedilatildeo apresenta-se afectada com a
presenccedila de ATP uma vez que os resiacuteduos de cisteiacutena permaneceram intactos nos ensaios
realizados com este ligando Jaacute Crans et al (2004) concluiu que o siacutetio activo de muitas fosfatases
(que conteacutem grupos tiol) pode formar um equiliacutebrio redox com o iatildeo vanadato (V) de forma natildeo
reversiacutevel
Os resultados obtidos por Sakurai e Shiromura (1980) sugeriram que o vanaacutedio (V) sofre reduccedilatildeo
a vanaacutedio (IV) sem dependecircncia do valor de pH do ensaio Registaram ainda uma estequiometria
de 31 na reacccedilatildeo do vanadato (V) com os resiacuteduos de Cys enquanto a estequiometria dos ensaios
com o vanadilo (IV) foi de 21
Berlett e Stadtman (1997) observaram que os tipos de aminoaacutecidos mais facilmente oxidaacuteveis
englobam entre outros os resiacuteduos de Cys Trp e MetAssim analisou-se o estado redox dos
grupos tiol dos resiacuteduos de Cys do S1 na presenccedila do decavanadato atraveacutes da monitorizaccedilatildeo da
Abs412 nm na presenccedila de 1 mgmL de S1 de miosina em tampatildeo 10 mM HEPES 25 mM KCl
variando a concentraccedilatildeo da soluccedilatildeo de decavanadato de modo a determinar que tipo de alteraccedilotildees
estruturais ocorrem na sua presenccedila no que diz respeito agrave diminuiccedilatildeo da actividade ATPaacutesica de
S1 na presenccedila de decavanadato
Os resultados referentes agrave quantificaccedilatildeo das cisteiacutenas reduzidas apoacutes o tratamento de S1 com
diferentes concentraccedilotildees de decavanadato em tampatildeo 10 mM HEPES pH 75 e 25 mM KCl a
25ordmC durante 20 minutos encontram-se descritos na Tabela 2VII
Tabela 2VII ndash Cisteiacutenas reduzidas totais e ldquoraacutepidas de S1 (1 mgmL) apoacutes tratamento com decavanadato
34
(010501002505001000 uM) em 10 mM HEPES 25 mM KCl pH 7 a 25ordmC durante 20 minutos A detecccedilatildeo
espectrofotomeacutetrica foi feita com DTNB (e 412=8400 uM-1 cm-1) (n variaacutevel entre 1 e 8)
Amostras Cys raacutepidas Cys totais
11 M S1 (controlo) 22 plusmn 04 106 plusmn 14
11 M S1 + 10 M Decavanadato 22 plusmn 05 95 plusmn 08
11 M S1 + 50 uM Decavanadato 20 plusmn 02 98 plusmn 14
11 M S1 + 100 uM Decavanadato 17 plusmn 03 97 plusmn14
11 M S1 + 250 uM Decavanadato 26 plusmn 02 98 plusmn 08
11 M S1 + 500 uM Decavanadato 21 plusmn 04 90 plusmn 06
11 M S1 + 1000 uM Decavanadato 24 plusmn 0 86 plusmn 08
A partir dos resultados obtidos pode sugerir-se que o S1 possui 11 resiacuteduos de Cys (controlo) o
que estaacute de acordo com o anteriormente descrito Verifica-se que o tratamento da proteiacutena com
concentraccedilotildees crescentes de decavanadato provoca a oxidaccedilatildeo de dois resiacuteduos de Cys do S1 de
miosina natildeo expostos ao solvente No entanto a teacutecnica utilizada natildeo nos permite identificar
quais
Tendo em consideraccedilatildeo que o valor de IC50 obtido para a reacccedilatildeo de inibiccedilatildeo daactividade Mg2+-
ATPaacutesica de S1 na presenccedila de decavanadato eacute de cerca de 27 microM pode sugerir-se que este seraacute o
valor ao qual ocorreraacute a oxidaccedilatildeo do primeiro resiacuteduo de Cys o que poderaacute contribuir para a
diminuiccedilatildeo da actividade de S1 observada
25Interacccedilatildeo de S1 com vanaacutedio(IV)
251Anaacutelise por espectroscopia de Ressonacircncia Paramagneacutetica Electroacutenica
A acumulaccedilatildeo intracelular de oxovanaacutedio (IV) in vivo constitui uma das principais formas de
apresentaccedilatildeo de vanaacutedio no organismo Deste modo torna-se bastante interessante analisar a
interacccedilatildeo entre este iatildeo e o S1 de miosina atraveacutes da teacutecnica de espectroscopia de Ressonacircncia
Paramagneacutetica Electroacutenica o meacutetodo de eleiccedilatildeo do estudo do iatildeo vanadilo e dos complexos por si
formados O electratildeo desemparelhado encontra-se fortemente acoplado ao nuacutecleo 51
V o que
35
resulta num espectro de RPE consistindo de 8 desdobramentos na direcccedilatildeo paralela e outros tantos
na direcccedilatildeo perpendicular (Chasteen 1983)
Os estudos realizados por Ahmed et al (1987) centraram-se na interacccedilatildeo do vanadilo com
proteiacutenas como eacute o caso da insulina transferrina entre outros obtendo um espectro caracterizado
por 8 linhas amplamente espaccediladas (na direcccedilatildeo paralela) Jaacute Ramos et al 2009 concluiu que a
exposiccedilatildeo da soluccedilatildeo de decavanadato agraves formas globular e filamentosa de actina produz um
espectro tiacutepico do iatildeo oxidovanaacutedio (IV) indicando a reduccedilatildeo do vanadato
Para aferir qual a concentraccedilatildeo de S1 que permitia obter uma boa razatildeo sinalruiacutedo titulou-se uma
soluccedilatildeo de sulfato de vanadilo 15 microM ([VOSO4]stock = 5 mM) com concentraccedilotildees crescentes de
proteiacutena (0 5 10 15 e 20 microM) em 10 mM HEPES 25 mM KCl A existecircncia de uma correlaccedilatildeo
linear entre a concentraccedilatildeo de proteiacutena e a intensidade dos sinais de RPE correspondente agrave banda
-12 perpendicular encontra-se representada na Fig 210
Figura 2 12ndash Representaccedilatildeo graacutefica da variaccedilatildeo da intensidade do sinal de RPE vs a concentraccedilatildeo de S1 utilizada
Apoacutes verificar que com uma concentraccedilatildeo de S1 igual a 20 microM se obtinham sinais com uma razatildeo
sinalruiacutedo adequada ao prosseguimento dos estudos sobre a interacccedilatildeo V(IV)-S1 de miosina
titulou-se uma soluccedilatildeo de 20 microM S1 com 0 25 5 10 15 25 50 75 100 150 200 500 e 1000
microM VOSO4
Traccedilou-se o graacutefico da intensidade da banda -12 perpendicular do sinal de EPR em funccedilatildeo da
concentraccedilatildeo de VOSO4 tendo-se determinado uma constante de dissociaccedilatildeo (KD) do complexo
S1-vanadilo de 8748 plusmn 2621 microM V(IV)
y = 016786x + 122435
Rsup2 = 092992759
Inte
nsi
dad
e d
o s
inal
de
RP
E
[S1] (microM)
36
Figura 213 ndash Representaccedilatildeo graacutefica da variaccedilatildeo do sinal de RPE vs a concentraccedilatildeo de vanadilo utilizada (os valores
experimentais estatildeo representados por losangos o seu ajuste estaacute representado pela linha)
A simulaccedilatildeo do espectro de EPR foi efectuada a partir do espectro obtido para uma concentraccedilatildeo
de 1 mM VOSO4 apoacutes adiccedilatildeo de 20 microM S1 de miosina tendo-se obtido os valores da constante de
acoplamento hiperfina (A=5016) e do factor g (g0=195)
Figura 214 ndash Representaccedilatildeo graacutefica do espectro de EPR obtido com 1000 microM de vanadilo e 20 microM de S1 (a cinzento
escuro) A simulaccedilatildeo do espectro encontra-se a cinzento claro
Por comparaccedilatildeo destes com os valores presentes na tabela de correspondecircncia xxx eacute possiacutevel
predizer quais os ligandos provaacuteveis do vanadilo na presenccedila de proteiacutena Assim concluiu-se que
o vanadilo estaacute coordenado com duas moleacuteculas de aacutegua um grupo hidroxilo e um grupo R=NR
O grupo hidroxilo pode indicar os resiacuteduos de aminoaacutecidos Tyr Treonina e Serina enquanto o
grupo R=NR poderaacute indicar a presenccedila de Arginina e Histidina como ligandos do vanadilo
Inte
nsi
dad
e d
os
sinai
s d
e R
PE
[Vanadilo] (microM)
Ajuste dos dados exp Dados experimentais
Campo magneacutetico (G)
Espectro Experimental Ajuste
37
3 Procedimento Experimental
31 Reagentes
Todos os reagentes utilizados satildeo de elevado grau de pureza (analiacutetico) Apresentam-se as suas
foacutermulas quiacutemicas ou abreviaturas quando necessaacuterio acrescentou-se a sua descriccedilatildeo Entre
parecircntesis colocou-se as empresas que os comercializam
Aacutecido aceacutetico glacial 100 (Riedel) Acrilamida 99 (Merck) Azul de bromofenol (Riedel-de-
Haumlen) -Mercaptoetanol (Sigma) Bissulfito de soacutedio (Sigma)Bradford (reagente de)
(BioRad)BSA (albumina de soro bovino Sigma-Aldrich) CaCl22H2O (cloreto de caacutelcio
dihidratado Fluka) CH3CH2OH 96 (etanol Panreac) Coomassie Blue R250 (Merck) D2O
(oacutexido de deuteacuterio SDS) DTNB (55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico) Aldrich) EDTA (aacutecido
etilenodiaminatetraaceacutetico Merck) Fosfoenolpiruvato 98 (Fluka) Glicerol (Sigma) Glicina
985 (Panreac) HCl 37 (Riedel-de Haumlen) H2SO4 96 (Pronalab) HEPES 99 (aacutecido 4-(2-
hidroxiletil)-1-piperazineetanosulfoacutenico Sigma) KH2PO4 99 (hidrogenofosfato de potaacutessio)
MgATP (Sigma) Na
ATP (Sigma) Na2HPO4 99 (hidrogenofosfato de soacutedio Riedel-de Haumlen)
NaH2PO4 (fosfato de soacutedio ) NaOH 986 (hidroacutexido de soacutedio Pronalab) KCl 99 (Cloreto de
potaacutessio Panreac) L-Cys 995 (enantioacutemero levoacutegiro do resiacuteduo de Cisteiacutena Fluka)Metanol
99 (Merck) Molibdato de amoacutenio (Sigma)NNrsquo-metil-bis-acrilamida 98 (Merck) NH4VO3
99 (metavanadato de amoacutenia Sigma) PSA 10 (persulfato de amoacutenio Merck) SDS
(dodecilo sulfato de soacutedio Sigma) Sulfito de soacutedio (Sigma) VOSO4 (sulfato de vanadilo
Aldrich) TEMED 99 (Tetrametiletilenodiamina Merck) Tris (2-amino-2-hidroximetil-13-
propanodiol Sigma)
32 Meacutetodos
321 Isolamento e purificaccedilatildeo das proteiacutenas de muacutesculo esqueleacutetico de coelho
3211 Purificaccedilatildeo da miosina preparaccedilatildeo de Subfragmento 1 por digestatildeo quimotriacutepsica
da miosina
O subfragmento-1 de miosina-II (S1) foi gentilmente cedido pelo Prof Doutor Manuel Aureliano
do Departamento de Quiacutemica Bioquiacutemica e Farmaacutecia da Faculdade de Ciecircncias e Tecnologia da
Universidade do Algarve
No entanto descreve-se abaixo de forma resumida o procedimento efectuado
38
32111 Purificaccedilatildeo da miosina
O protocolo de isolamento foi adaptado do meacutetodo previamente descrito por Margossian e Lowey
(1982) apresentando-se de forma esquemaacutetica na Fig 31
Sucintamente os tecidos musculares dorsais e das pernas foram extraiacutedos apoacutes o sacrifiacutecio do
animal (Fig31) De seguida foram adicionados 3 volumes da soluccedilatildeo de extracccedilatildeo A2
Centrifugou-se a soluccedilatildeo e filtrou-se o sobrenadante O sedimento foi entatildeo diluiacutedo em 14
volumes de aacutegua destilada de modo a que a forccedila ioacutenica atingisse 004 M Apoacutes adiccedilatildeo de szlig-
mercaptoetanol (szlig-me) deixou-se a precipitar a 4ordmC removendo o sobrenadante Centrifugou-se
novamente e ressuspendeu-se o sedimento na soluccedilatildeo de extracccedilatildeo A Adicionou-se a soluccedilatildeo
tampatildeo B3 para que a forccedila ioacutenica atingisse 05 M Homogeneizou-se e deixou-se repousar
durante 2h Homogeneizou-se novamente e centrifugou-se de modo a ocorrer precipitaccedilatildeo dos
complexos de actomiosina e proteiacutena desnaturada Apoacutes filtraccedilatildeo do sobrenadante em latilde de vidro
quantificou-se a soluccedilatildeo de miosina resultante pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental)
2 Soluccedilatildeo de extracccedilatildeo A 0012M K2CO3 004M KHCO3 06M KCl pH 65
3 Soluccedilatildeo-tampatildeo B Tris 100 mM pH 7 KCl 2M
39
Figura 31 ndash Representaccedilatildeo esquemaacutetica do protocolo de isolamento de miosina (Margossian e Lowey 1982) Todo o
procedimento foi realizado a 4ordmC (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
32112 Preparaccedilatildeo de Subfragmento-1 de miosina por digestatildeo com quimiotripsina
O S1 de miosina foi isolado apoacutes diaacutelise da miosina durante a noite contra a soluccedilatildeo C4 A
presenccedila do EDTA eacute essencial uma vez que a sua substituiccedilatildeo por MgCl2resulta na produccedilatildeo de
HMM em vez de S1 A suspensatildeo de miosina foi equilibrada e digerida com -quimotripsina
A digestatildeo foi interrompida pela adiccedilatildeo de PMSF Seguidamente ultracentrifugou-se o produto
da digestatildeo desprezando-se o sedimento (Fig 32) A soluccedilatildeo de sobrenadante maioritariamente
S1 foi quantificada pela leitura directa da Abs280 ou pelo meacutetodo de Bradford (ver as secccedilotildees
3311 e 3312 do Procedimento Experimental respectivamente) e armazenada em 10 sacarose
e 2 mM szlig-Me a -80ordmC
4 Soluccedilatildeo C 25mM Tris pH 70 012M KCl 1mM EDTA
40
Antes da realizaccedilatildeo dos ensaios descritos no presente relatoacuterio o S1 de miosina foi dialisado
contra a soluccedilatildeo D5 de modo a remover qualquer composto que pudesse complexar como
vanaacutedio Seguidamente quantificou-se a proteiacutena pelo meacutetodo de Bradford ou pelo meacutetodo
directo (ver secccedilotildees 3311 e 3312do Procedimento Experimtental)
Figura 3 2ndash Representaccedilatildeo esquemaacutetica do protocolo de preparaccedilatildeo do S1 de miosina por digestatildeo quimiotripsina de
acordo com o previamente descrito por Margossian e Lowey (1982) Todo o procedimento foi realizado a 4ordm C
excepto quando indicaccedilatildeo do contraacuterio (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
3212 Isolamento e purificaccedilatildeo de actina
Isolou-se e purificou-se a actina segundo o meacutetodo descrito por Pardee e Spudish que
compreende dois passos ndash a obtenccedilatildeo de um resiacuteduo seco denominado ldquopoacute de acetonardquo e a
extracccedilatildeo subsequente da actina a partir do ldquopoacute de acetonardquo na forma de G-Actina com a
posterior polimerizaccedilatildeo a F-Actina
32121 Obtenccedilatildeo do ldquopoacute de acetonardquo
Sacrificou-se o coelho com uma pancada atraacutes do pescoccedilo e sangrou-se o animal apoacutes o corte da
arteacuteria jugular Esfolou-se e colocou-se numa tina com gelo Retiraram-se as partes de tecido
conjuntivo que envolvem o tecido muscular com auxiacutelio de uma pinccedila e bisturi Seguidamente
retirou-se os muacutesculos dorsais e os muacutesculos brancos das pernas Passou-se os muacutesculos por aacutegua
destilada de modo a remover as impurezas e pesou-se num copo Trituraram-se os muacutesculos
numa maacutequina de triturar carne e extraiu-se o resultante com 3 volumes da soluccedilatildeo E6 durante 10
minutos com agitaccedilatildeo suave Centrifugou-se a suspensatildeo a 4000 g durante 10
5 Soluccedilatildeo D HEPES 10 mM pH 70 KCl 25 mM
6 Soluccedilatildeo E 01 M KCl 015 M K2HPO4 pH 65
41
minutosdesprezando-se o sobrenadante Ressuspendeu-se o sedimento em 6 volumes de soluccedilatildeo
F7durante 10 minutos com agitaccedilatildeo suave Verificou-se se o pH da soluccedilatildeo estava compreendido
entre 82 e 85 acertando-se estecom soluccedilatildeo G8 se necessaacuterio Centrifugou-se durante 10
minutos a 4000 g Desprezou-se o sobrenadante e ressuspendeu-se o sedimento em 3 volumes de
soluccedilatildeo H9 durante 10 minutos Centrifugou-se novamente a 4000 gdurante 10 minutos
desprezando-se o sobrenadante Ressuspendeu-se o sedimento duas vezes consecutivas em 6
volumes de aacutegua destilada durante 5 minutos Centrifugou-se a 4000 g durante 10 minutos apoacutes
cada adiccedilatildeo de H2O
Seguidamente ressuspendeu-se o sedimento 5 vezes consecutivas utilizando 3 volumes de
acetona de cada vez agitando suavemente durante 10 minutos agrave temperatura ambiente Colocou-
se o resiacuteduo filtrado numa tina de vidro sobre papel de filtro deixando-se evaporar o solvente
durante a noite num nicho Triturou-se o resiacuteduo seco numa maacutequina de triturar carne e deixou-se
novamente evaporar o solvente O ldquopoacute de acetonardquo obtido foi armazenado a -20 ordmC
32122 Purificaccedilatildeo da actina a partir de ldquopoacute de acetonardquo
Apoacutes a obtenccedilatildeo do poacute de acetona como acima indicado efectuou-se o protocolo seguidamente
descrito de modo a obter monoacutemeros de actina (G-Actina) e actina filamentosa (F-Actina)
Pesou-se 35 g de poacute de acetona e adicionaram-se 50 mL de soluccedilatildeo-tampatildeo I10
sendo a
extracccedilatildeo feita com agitaccedilatildeo suave durante 30 minutos Separou-se o extracto do poacute de acetona
hidratado filtrando a suspensatildeo com 4 camadas de gaze esterilizada tendo o cuidado de natildeo
espremer o resiacuteduo Voltou a extrair-se com 50 mL de soluccedilatildeo I durante 30 minutos mexendo
lentamente Adicionou-se este ao extracto obtido anteriormente Centrifugou-se o extracto total a
16 000 g durante 30 minutos Decantou-se para uma proveta sobre a qual se colocou gaze para
que o sedimento solto natildeo se juntasse ao sobrenadante e leu-se o volume De seguida adicionou-
se KCl ateacute uma concentraccedilatildeo final de 50 mM MgCl2 ateacute 2mM e ATP ateacute 1mM lentamente e com
agitaccedilatildeo contiacutenua de modo a promover a polimerizaccedilatildeo dos monoacutemeros de actina
Deixou-se polimerizar durante 2h a 25ordmC observando-se a presenccedila de pequenas bolhas de ar
indicativas de um aumento de viscosidade da soluccedilatildeo Arrefeceu-se em gelo durante 15 minutos e
adicionou-se KCl ateacute uma concentraccedilatildeo final de 06M Mexeu-se lentamente durante 30 minutos
apoacutes os quais se ultracentrifugou a 110 000g durante 1hora Desprezou-se o sobrenadante
ressuspendeu-se o sedimento com soluccedilatildeo tampatildeo E e transferiu-se para um potter
Dialisou-se durante a noite contra 1L de soluccedilatildeo-tampatildeo I a uma temperatura de 4ordmC com rotaccedilatildeo
7 Soluccedilatildeo F 005 M NaHCO3 pH 82-85
8 Soluccedilatildeo G 1M Na2CO3
9 Soluccedilatildeo H 1 mM EDTA pH 70
10 Soluccedilatildeo I 2 mM Tris-HCl pH 8 02 mM CaCl2 02 mM Na2ATP 05 mM szlig-Me 0005 NaN3
42
contiacutenua e mudanccedila de tampatildeo 2 a 3 vezes De seguida a actina foi centrifugada a 110
000gdurante 1hora Decantou-se a soluccedilatildeo de G-Actina (sobrenadante) e desprezou-se o
sedimento Determinou-se a concentraccedilatildeo de G-Actina pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental) A soluccedilatildeo de G-actina foi armazenada a uma temperatura de 4ordmC
apoacutes adiccedilatildeo de 002NaN3 mantendo-se estaacutevel durante 3 a 5 dias
331 Quantificaccedilatildeo das proteiacutenas isoladas
3311 Quantificaccedilatildeo pelo meacutetodo de Bradford
O meacutetodo de Bradford (Bradford 1976) baseia-se na utilizaccedilatildeo do reagente de Bradford
composto por entre outros aacutecido fosfoacuterico e o corante ldquoCoomassierdquo Este corante possui carga
negativa e a sua coloraccedilatildeo varia entre a vermelha (Absmax=465 nm) predominante em soluccedilatildeo e a
azul (Absmax=595 nm) presente aquando da ligaccedilatildeo aos grupos amina das proteiacutenas Em soluccedilotildees
aacutecidas diluiacutedas a coloraccedilatildeo varia proporcionalmente com a quantidade do corante ligado agrave
proteiacutena que por sua vez eacute directamente proporcional agrave concentraccedilatildeo da proteiacutena permitindo a
sua quantificaccedilatildeo ateacute a um limite de detecccedilatildeo de 20 gmL (Bradford 1976)
Deste modo prepararam-se amostras de BSA ([BSA]stock= 1mgmL) utilizando um intervalo de
concentraccedilotildees compreendido entre 0 e 15 gmL de acordo com a tabela 3I Apoacutes a adiccedilatildeo da
H2O MilliQ e do reagente de Bradford as soluccedilotildees foram sucessivamente homogeneizadas no
voacutertex
Tabela 3Indash Amostras utilizadas para a obtenccedilatildeo da recta de calibraccedilatildeo no meacutetodo de Bradford
BSA (L) BSA (g) H2O MilliQ (L) Reagente de Bradford (L)
0 0 800 200
125 125 79875 200
25 25 7975 200
5 5 795 200
10 10 790 200
15 15 785 200
Apoacutes incubaccedilatildeo a 25ordmC por 5 minutos leram-se os valores de Abs595 num
espectrofotoacutemetro (Shimadzu UV-160A) para todas as amostras Seguidamente traccedilou-se a recta
de calibraccedilatildeo (Abs595vs quantidade de BSA (g))
43
Para determinar a concentraccedilatildeo de proteiacutena apoacutes o processo de isolamento purificaccedilatildeo e
diaacutelise quando necessaacuterio colocou-se 5 L da amostra num tubo de Eppendorf e adicionou-se
795 L de H2O MilliQ homogeneizando a soluccedilatildeo no voacutertex Adicionou-se posteriormente 200
L de reagente de Bradford e repetiu-se a homogeneizaccedilatildeo Incubou-se a mistura a 25ordmC por 5
minutos apoacutes os quais se leu os valores de Abs595 nm no espectrofotoacutemetro Por interpolaccedilatildeo deste
valor a partir da recta de calibraccedilatildeo determinou-se a concentraccedilatildeo de proteiacutena na amostra
3312 Quantificaccedilatildeo pelo meacutetodo directo
A quantificaccedilatildeo das amostras proteicas pode tambeacutem ser efectuada atraveacutes da leitura directa da
absorvacircncia tendo por base a lei de Beer-Lambert onde Abs corresponde agrave
absorvacircncia ao comprimento de onda ao qual o ensaio foi realizado corresponde ao coeficiente
de extinccedilatildeo (substituiacutedo pela letra a se as unidades forem distintas da molaridade) b corresponde
ao percurso oacuteptico da radiaccedilatildeo na ceacutelula amostral e c corresponde agrave concentraccedilatildeo amostral
Assim no caso de S1 de miosina mediu-se a Abs280e calculou-se a concentraccedilatildeo da proteiacutena
sabendo que o coeficiente de extinccedilatildeo de S1 eacute (Ref)
No caso da actina mediu-se a Abs290ecalculou-se a concentraccedilatildeo da proteiacutena sabendo que o
coeficiente de extinccedilatildeo da actina eacute (Ref)
332Electroforese em condiccedilotildees desnaturantes (SDS-PAGE)
De modo a melhor caracterizar as proteiacutenas isoladas foi realizada uma electroforese em
condiccedilotildees desnaturantes (SDS-PAGE do inglecircs Sodium Dodecyl Sulfate Polyacrilamide Gel
Electrophoresis) O SDS (dodecilo sulfato de soacutedio) eacute um detergente anioacutenico utilizado neste
meacutetodo para desnaturar as proteiacutenas que se ligam ao SDS na proporccedilatildeo 14 g SDS g proteiacutena
(aproximadamente 1 moleacutecula de SDS por cada 2 resiacuteduos de aminoaacutecidos) (Stryer) O SDS induz
uma densidade de carga negativa significativa nas proteiacutenas a que se liga mascarando a sua carga
intriacutenseca pelo que as razotildees cargamassa satildeo tendencialmente idecircnticas Assim a SDS-PAGE
separa as proteiacutenas quando sujeitas a um campo eleacutectrico de acordo com o seu peso molecular
sendo a sua migraccedilatildeo directamente proporcional ao logaritmo deste uacuteltimo (Stryer)
O gel de resoluccedilatildeo com 10 de acrilamida foi obtido atraveacutes da adiccedilatildeo de 167 mL de soluccedilatildeo
stock de acrilamida11
075 mL de tampatildeo de resoluccedilatildeo12
251 mL de aacutegua destilada e 50 L de
SDS 10 Para promover a polimerizaccedilatildeo adicionou-se em seguida 38 L de PSA 10
11
soluccedilatildeo stock de acrilamida soluccedilatildeo de 30 (mv) de acrilamida e 08 (mv) de NNrsquo-metil-bis-acrilamida 12
tampatildeo de resoluccedilatildeo 15 M Tris-HCl pH 88
44
(persulfato de amoacutenia) 10 e 25 L de TEMED 99 (tetrametiletilenodiamina) Agitou-se
cuidadosamente de modo a natildeo formar bolhas e pipetou-se a soluccedilatildeo para a cassette de vidro
previamente montada no suporte De seguida adicionou-se uma peliacutecula de cerca de 2 mm de 2-
metil-2-butanol para verificar posteriormente a polimerizaccedilatildeo Apoacutes a polimerizaccedilatildeo que demora
cerca de 45 minutos verteu-se a agrave superfiacutecie e adicionou-se o gel de concentraccedilatildeo com 5 de
acrilamida constituiacutedo por 03 mL de soluccedilatildeo de stock acrilamida 045 mL de tampatildeo de
concentraccedilatildeo13
094 mL de aacutegua destilada 18 L de SDS 10 135 L de PSA 10 e 2L de
TEMED 99 Colocou-se o pente sobre o gel e deixou-se polimerizar durante cerca de 45
minutos
As amostras para aplicaccedilatildeo no gel de electroforese foram preparadas para uma concentraccedilatildeo de
proteiacutena de 1 mgmL num volume final de 100 L dos quais 25 L correspondem ao tampatildeo da
amostra14
As amostras foram aquecidas a 100ordmC durante 5 minutos Antes da aplicaccedilatildeo das
amostras transferiu-se a cassette para a tina de electroforese agrave qual se adicionou a soluccedilatildeo
tampatildeo de electroforese da tina superior15
e tampatildeo de electroforese da tina inferior16
Aplicaram-
se 10 L das soluccedilotildees de marcadores de peso molecular e 20 L das amostras proteicas
preparadas anteriormente A electroforese decorreu durante cerca de 3h30 min a uma voltagem
de 80 V De seguida transferiu-se o gel para uma tina corando as proteiacutenas durante 1h com
soluccedilatildeo corante17
Descorou-se o gel com soluccedilatildeo descorante18
tendo-se mudado esta soluccedilatildeo ateacute
as bandas se encontrarem bem definidas
Determinou-se a mobilidade relativa das bandas correspondentes aos marcadores de elevado peso
molecular e traccedilou-se a recta de calibraccedilatildeo ndash log(PM) vs log(Rf) onde Rf corresponde agrave
mobilidade relativa Por interpolaccedilatildeo determina-se o PM das proteiacutenas isoladas sabendo a sua
mobilidade relativa Por comparaccedilatildeo com os valores descritos na literatura pode concluir-se
acerca do grau de purificaccedilatildeo
34 Mediccedilatildeo da actividade enzimaacutetica de S1
341 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
13
tampatildeo de concentraccedilatildeo 05 M Tris-HCl pH 68 14
tampatildeo de amostra 320 mM Tris-HCl pH 68 04 M szlig-mercaptoetanol SDS 8 glicerol 15 (vv) e roxo de m-
cresol 0024 15
tampatildeo de electroforese da tina superior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS 16
tampatildeo de electroforese da tina inferior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS acetato de soacutedio
01 M 17
soluccedilatildeo corante ldquoCoomassie Blue R-250rdquo contendo metanol 45 (vv) e aacutecido aceacutetico glacial 10 (vv) 18
soluccedilatildeo descorante metanol 10 (vv) e aacutecido aceacutetico glacial 10 vv)
45
O meacutetodo de Fiske-Subarow modificado consiste na reacccedilatildeo do molibdato de amoacutenio com o
fosfato inorgacircnico em soluccedilatildeo aacutecida com formaccedilatildeo do aacutecido fosfomoliacutebdico Seguidamente daacute-
se a adiccedilatildeo de um agente redutor que provoca a reduccedilatildeo do composto a fosfomolibdato que forma
um complexo de cor azulada cuja absorvacircncia maacutexima ocorre para um comprimento de onda de
660 nm tornando possiacutevel a sua detecccedilatildeo por meacutetodos espectrofotomeacutetricos As actividades Ca2+
-
ATPaacutesica e K+EDTA-ATPaacutesica de S1 foram determinadas atraveacutes deste meacutetodo de acordo com
o descrito por Taussky e Shorr (1953) com mediccedilatildeo da quantidade de fosfato (Pi) libertado
atraveacutes da hidroacutelise de ATP
Para tal obteve-se primeiramente a recta de calibraccedilatildeo com as amostras presentes na Tabela 3II
utilizando uma soluccedilatildeo stock de 5 mM KH2PO4
Tabela 3II ndash Composiccedilatildeo das diferentes amostras utilizadas para determinar a recta de calibraccedilatildeo atraveacutes do meacutetodo
de Fiske-Subbarow modificado
KH2PO4 (L)
([KH2PO4]stock= 5mM)
PO43-
(nmole) H2O (uL)
0 0 1780
5 25 1775
10 50 1770
20 100 1760
40 200 1740
100 500 1680
A cada tubo de Eppendorf adicionaram-se seguidamente 200 L de soluccedilatildeo J19
e 20 L de agente
redutor20
homogeneizando no voacutertex apoacutes cada adiccedilatildeo Incubou-se durante 10 minutos e mediu-
se a Abs660 nm num espectrofotoacutemetro com intervalos de tempo iguais entre cada amostra Traccedilou-
se a recta de calibraccedilatildeo (Abs660 nmvs nmol PO43-
) a partir dos valores obtidos
Para determinar a actividade Ca2+
-ATPaacutesica de S1 prepararam-se tubos de Eppendorf com 400
L de meio de reacccedilatildeo21
25 L CaCl2 ([CaCl2]stock= 100 mM) 25 L ATP ([ATP]stock= 100 mM)
com [S1]ensaios=01 mgmL Perfez-se se necessaacuterio com H2O MilliQ ateacute perfazer um volume de 1
mL Para determinar a actividade K+EDTA-ATPaacutesica utilizou-se uma concentraccedilatildeo idecircntica de
19
Soluccedilatildeo J 5 N H2SO4 25 molibdato de amoacutenio 20
Agente redutor 5 mg aacutecido 1-amino-2-naftol-4-sulfoacutenico 30 mg bissulfito de soacutedio 30 mg sulfito de soacutedio em 26
mL H2O 21
meio de reacccedilatildeo 125 mM Tris-HCl pH 80 0575 M KCl
46
proteiacutena nos ensaios (01 mgmL) e o mesmo meio de reacccedilatildeo Aos tubos de Eppendorf
adicionou-se ainda 123 L de KCl ([KCl]stock = 3M) 25 L de ATP 10 L de EDTA
([EDTA]stock= 100 mM )e o volume de H2O MilliQ necessaacuterio para perfazer 1 mL Em ambas as
situaccedilotildees o uacuteltimo reagente adicionado foi o ATP uma vez que se trata do substrato da reacccedilatildeo
enzimaacutetica
A cineacutetica decorreu a 25ordmC durante 5 minutos para a actividade Ca2+
-ATPaacutesica e durante 3
minutos para a actividade K+EDTA-ATPaacutesica Apoacutes o periacuteodo de incubaccedilatildeo transferiu-se 150
L de amostra para tubos de Eppendorf contendo igual volume de TCA 10 Colocou-se no gelo
e homogeneizou-se no voacutertex Seguidamente centrifugou-se a 1500 g durante 10 minutos e
recolheu-se 200 L de sobrenadante para novos tubos de Eppendorf Adicionou-se a estes 158
mL de H2O MilliQ e 200 L de soluccedilatildeo J Homogeneizou-se no voacutertex e adicionou-se 20 L de
reagente redutor repetindo-se a homogeneizaccedilatildeo Incubou-se durante 10 minutos e mediu-se a
Abs660 nm com o mesmo intervalo de tempo de leitura para cada amostra Por intrapolaccedilatildeo a partir
da recta de calibraccedilatildeo obtida anteriormente determinou-se a quantidade de Pi libertado em cada
ensaio A partir destes resultados determinaram-se as actividades Ca2+
-ATPaacutesica e K+EDTA-
ATPaacutesica do S1 em nmol PO43-
minmg proteiacutena e em s-1
342 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
Da actividade Mg2+
-ATPaacutesica de S1 resulta a formaccedilatildeo dos produtos da reacccedilatildeo de
hidroacutelise do ATP - ADP e Pi Na presenccedila de ADP fosfoenolpiruvato (PEP) e da enzima piruvato
cinase (PK) os produtos resultantes satildeo ATP e piruvato Por acccedilatildeo da enzima lactato
desidrogenase (LDH) este uacuteltimo eacute convertido em lactato ocorrendo simultaneamente a oxidaccedilatildeo
do NADH a NAD+ monitorizada atraveacutes da Abs340 nm Quando o S1 de miosina se encontra no
meio reaccional o ATP eacute hidrolisado formando-se novamente ADP e Pi o que permite a
formaccedilatildeo de um ciclo (fig 21) Se as enzimas se encontrarem em excesso a velocidade de
conversatildeo do NADH eacute limitada pela produccedilatildeo de ADP e Pi Como o ATP eacute regenerado em cada
ciclo a sua concentraccedilatildeo inicial eacute sempre mantida (Bagshaw 1982)
Figura 33ndash Representaccedilatildeo esquemaacutetica do meacutetodo de enzimas acopladas
47
O protocolo utilizado e seguidamente descrito foi adaptado de Cuenda et al (1990 ndash Ver Refordf) A
uma cuvette de 1 mL adicionou-se S1 para uma concentraccedilatildeo final de 005 mgmL 800 L de
meio de reacccedilatildeo22
10 L de PEP ([PEP]stock = 42 mM) 10 L de PK+LDH 11 ([PK+LDH]stock =
1 mgmL) e 15 L de NADH ([NADH]stock=25 mM) Iniciou-se a monitorizaccedilatildeo da reacccedilatildeo
atraveacutes da leitura de Abs340 nm apoacutes a adiccedilatildeo de 20 L de ATP ([ATP]stock=100 mM) A
actividade Mg2+
-ATPaacutesica eacute obtida atraveacutes do declive correspondente agrave oxidaccedilatildeo do NADH Para
a mediccedilatildeo da Mg2+
-ATPase de S1 estimulada por actina adicionou-se F-actina para uma
concentraccedilatildeo final de 2 microM sendo a actividade obtida tambeacutem atraveacutes do declive correspondente
agrave oxidaccedilatildeo do NADH apoacutes a adiccedilatildeo da proteiacutena
35 Interaccedilatildeo entre vanaacutedio e subfragmento-1 de miosina
351 Preparaccedilatildeo das soluccedilotildees de vanaacutedio
As soluccedilotildees de vanadato foram preparadas em aacutegua MilliQ a partir de metavanadato de amoacutenio
(NH4VO3) numa concentraccedilatildeo stock de 50 mM com agitaccedilatildeo magneacutetica e aquecimento (~30 ordmC)
Esta soluccedilatildeo com um valor compreendido entre 6 e 7 designa-se por ldquometavanadatordquo e eacute
constituiacuteda pelas esspeacutecies V1 V2 V4 e V5 (Ref) Acidificando a soluccedilatildeo de metavanadato por
adiccedilatildeo de HCl ateacute um valor de pH proacuteximo de 4 observa-se uma mudanccedila de cor de amarelo
claro para laranja indicativa da formaccedilatildeo da espeacutecie decameacuterica de vanadato De seguida e
imediatamente antes da realizaccedilatildeo dos ensaios descritos o valor do pH da soluccedilatildeo de
decavanadato foi aumentado para cerca de 75 por adiccedilatildeo de NaOH A soluccedilatildeo (2) designada
por ldquometavanadatordquo natildeo foi posteriormente alterada ateacute agrave sua adiccedilatildeo para a realizaccedilatildeo dos ensaios
As soluccedilotildees foram armazenadas a 4ordmC e mantidas em gelo aquando da sua utilizaccedilatildeo
A soluccedilatildeo de V(IV) foi preparada em condiccedilotildees anaeroacutebias por diluiccedilatildeo de sulfato de vanadil
(VOSO4) para uma concentraccedilatildeo final de 5 mM em H2O MilliQ saturada com N2 gasoso
352Anaacutelise por espectroscopia de UV-Vis
A concentraccedilatildeo da espeacutecie decameacuterica em soluccedilatildeo depende do periacuteodo de tempo apoacutes a diluiccedilatildeo
no meio de reacccedilatildeo apropriado (Ref) A velocidade de desoligomerizaccedilatildeo da espeacutecie decameacuterica
pode ser analisada por espectroscopia de absorccedilatildeo na regiatildeo UVVis uma vez que esta espeacutecie
apresenta duas bandas caracteriacutesticas na regiatildeo do visiacutevel (a 358 e 400 nm) que natildeo se verifica
em quaisquer outras espeacutecies oligomeacutericas As bandas supramencionada sapresentam-se bem
22
Meio de reacccedilatildeo 3125 mM Tris-HCl pH 71 375 mM MgCl2
48
resolvidas atraveacutes da obtenccedilatildeo da derivada de 2ordf ordem do espectro apresentando a forma de 2
vales com miacutenimos centrados nos comprimentos de onda supracitados com dependecircncia linear
relativamente agrave concentraccedilatildeo de decavanadato total (Ramos et al 2006)
Assim a estabilidade da espeacutecie decameacuterica apoacutes diluiccedilatildeo da soluccedilatildeo de decavanadato no meio de
reacccedilatildeo dos ensaios (tampatildeo D) para uma concentraccedilatildeo final de 1 mM em vanadato total foi
caracterizada atraveacutes da leitura da Abs400 em funccedilatildeo do tempo tendo-se determinado qual a
ordem de reacccedilatildeo e o tempo de meia-vida da mesma Com base em estudos anteriores acerca da
estabilidade do V10 na presenccedila de diferentes proteiacutenas (Ramos et al 2006) determinou-se
igualmente o tempo de meia vida apoacutes a adiccedilatildeo de 20 microM de S1 de miosina ao meio reaccional
353 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
Os espectros de RMN do isoacutetopo 51
V foram realizados num espectroacutemetro Bruker AM-400 MHz
equipado com uma sonda multinuclear inversa de 5 mm Em tubos de RMN de 5 mm de diacircmetro
preparou-se soluccedilotildees de 500 L de deca- ou metavanadato correspondentes a um intervalo de
concentraccedilotildees de 1 a 10 mM suplementadas com 10 D2O Os espectros foram adquiridos agrave
temperatura ambiente com uma largura espectral de 23 923 Hz um tempo de aquisiccedilatildeo de 0086
s tempo de relaxaccedilatildeo de 001 s e uma sequecircncia de pulsos a 90ordm Utilizou-se a transformada de
Fourier com alargamento das bandas de 20Hz para o processamento dos espectros Os desvios
quiacutemicos das ressonacircncias de 51
V foram obtidos em relaccedilatildeo agrave espeacutecie VOCl3cuja frequecircncia de
ressonacircncia eacute considerada o padratildeo sendo todas as outras referenciadas a partir desta em ppm
As aacutereas dos sinais do RMN correspondentes agraves espeacutecies oligomeacutericas presentes foram obtidas
por integraccedilatildeo dos sinais As concentraccedilotildees relativas das espeacutecies foram calculadas com recurso agrave
equaccedilatildeo
onde A[Vx] corresponde agrave aacuterea referente agrave ressonacircncia da espeacutecie Vx com nordm de agregaccedilatildeo x
A[V]total agrave soma total das aacutereas medidas e [V] a concentraccedilatildeo total de vanaacutedio no ensaio
A teacutecnica de 51
V-RMN foi anteriormente aplicada em estudos de ligaccedilatildeo com S1 de miosina
(Ref) tendo sido demostrado que entre as espeacutecies V1 V2 V4 e V5 a espeacutecie tetrameacuterica possui a
maior afinidade para a miosina (Cremo et al) O desaparecimento ou alargamento dos sinais pode
implicar a ligaccedilatildeo de um dos oxovanadatos laacutebeis agrave miosina sem indicaccedilatildeo de qual uma vez que
49
se processa um equiliacutebrio entre eles (Crans 1990) Assim e tendo em conta as indicaccedilotildees de que
a espeacutecie V10 eacute distinta das restantes no que diz respeito agrave magnitude da interacccedilatildeo com a miosina
e inibiccedilatildeo da actividade ATPaacutesica estimulada por actina (Aureliano 2000 Tiago et al 2002)
torna-se fundamental investigar a interacccedilatildeo das diferentes espeacutecies de vanadato com o S1 de
miosina atraveacutes da teacutecnica de 51
V-RMN
Para tal obtiveram-se espectros de 51
V-RMN resultantes da titulaccedilatildeo de uma soluccedilatildeo 5 mM de
decavanadato total com concentraccedilotildees crescentes de S1 de miosina (0 5 10 15 e 20 microM) em
tampatildeo D Realizou-se ainda ensaios na presenccedila de 2 mM de decavanadato e 2 mM de ATP na
presenccedila e na ausecircncia de S1 de miosina de modo a verificar o efeito que estes ligandos naturais
da enzima teria na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o subfragmento-1
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
Os grupos sulfidrilo (-SH) das cisteiacutenas satildeo resiacuteduos de aminoaacutecidos extremamente sensiacuteveis agrave
oxidaccedilatildeo resultando na sua oxidaccedilatildeo (Dalle-Donne et al 1999) Logo a quantificaccedilatildeo destes
resiacuteduos eacute bastante importante nos estudos de interacccedilatildeo com vanaacutedio jaacute que os resiacuteduos de Cys
com ecircnfase para o SH1 e o SH2 satildeo bastante importantes na monitorizaccedilatildeo das modificaccedilotildees
funcionais da regiatildeo da cabeccedila
O procedimento seguidamente apresentado foi originalmente descrito por Gutieacuterrez-Merino
(1987)) Consiste na reacccedilatildeo entre os grupos ndashSH natildeo oxidados das proteiacutenas e o reagente de
Ellman (DTNB -55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico)) - formando-se o aniatildeo CNT (aniatildeo 3-
carboxilato-4-nitrotiofenolato) numa estequiometria de 11 Este apresenta um maacuteximo de
absorvecircncia a 412 nm (Robyt 1990) Assim apoacutes a incubaccedilatildeo da proteiacutena na presenccedila do metal
a quantidade de cisteiacutenas reduzidas por proteiacutena determina-se atraveacutes dos valores de Abs412
Deste modo colocou-se num tubo de Eppendorf o volume apropriado de S1 dialisado para que a
concentraccedilatildeo no ensaio fosse de 1 mgmL Adicionou-se o volume apropriado de decavanadato
para que a sua concentraccedilatildeo fosse de 0 10 50 100 250 500 ou 1000 M Perfez-se com soluccedilatildeo
D ateacute um volume final de 1 mL e incubou-se a 25ordmC por 20 minutos Apoacutes este periacuteodo colocou-
se 780 L da mistura reaccional numa cuvette de quartzo e adicionou-se 220 L de soluccedilatildeo K23
23
Tampatildeo K Tampatildeo fosfatohidrogenofosfato de soacutedio 025 M pH 72
50
Adicionou-se 15 L de DTNB (em soluccedilatildeo K [DTNB]stock=100 mM) ao meio reaccional e
registou-se o valor de absorvecircncia medido equivalente agraves cisteiacutenas ldquoraacutepidasrdquo (SH1 e SH2
expostos ao solvente) Adicionou-se 001 g de SDS que provoca a desnaturaccedilatildeo da proteiacutena e
consequentemente a exposiccedilatildeo de todos os resiacuteduos de cisteiacutenas que se encontravam ateacute aiacute
inacessiacuteveis no interior da proteiacutena (cisteiacutenas totais) Estes resiacuteduos podem assim reagir com o
DTNB
Estimou-se o nuacutemero de resiacuteduos de cisteiacutenas em todas as amostras com recurso ao coeficiente de
extinccedilatildeo molar () do DTNB (ver ponto 3 dos Anexos) que foi determinado atraveacutes da titulaccedilatildeo
de L-Cisteiacutena com intervalo de concentraccedilotildees compreendidos entre 0 e 50 microM na presenccedila de
25microM de DTNB em tampatildeo D Os valores de absorvecircncia foram obtidos para um comprimento de
onda de 412 nm
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
3621 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
A teacutecnica mais eficiente de anaacutelise da interacccedilatildeo entre o iatildeo vanadilo e proteiacutenas como a
calmodulina e a transferrina entre outras permite a obtenccedilatildeo raacutepida de informaccedilatildeo no que toca agrave
conformaccedilatildeo e amplitude da referida interacccedilatildeo
Os espectros de Ressonacircncia Paramagneacutetica Electroacutenica (RPE) da espeacutecie paramagneacutetica V(IV)
foram realizados a 70K num espectroacutemetro Bruker EMX-300 X-band Todas as soluccedilotildees
utilizadas e amostras foram preparadas em anaerobiose numa cacircmara de luvas de modo a evitar a
reduccedilatildeo de V(IV) a V(V) Em tubos capilares com 02 mL de amostra manteve-se a concentraccedilatildeo
de S1 a 20 M e variou-se a concentraccedilatildeo de vanadilo (0 25 5 10 15 25 50 75 100 150
200 500 e 1000 M [VOSO4]stock=5 mM) em tampatildeo D Apoacutes obtenccedilatildeo dos espectros estes
foram tratados atraveacutes do software SimFonia e WinEPR tendo-se obtido um graacutefico com a
variaccedilatildeo da intensidade do sinal de RPE em funccedilatildeo da concentraccedilatildeo de sulfato de vanadilo
presente Realizaram-se ainda simulaccedilotildees de espectros obtidos com 20 microM de S1 e 1000 microM de
V(IV) com vista agrave obtenccedilatildeo dos ligandos provaacuteveis do vanadilo atraveacutes da anaacutelise da constante
de acoplamento hiperfina (A) e do factor g (g)
4 Conclusotildees gerais
Apesar de jaacute ter sido demonstrado que o vanaacutedio inibe a Na+K
+-ATPase da miosina a funccedilatildeo
deste elemento no tecido muacutesculo esqueleacutetico continua largamente incompreendida A enorme
complexidade quiacutemica deste metal especialmente no que concerne agrave oligomerizaccedilatildeo e reacccedilotildees
51
redox constitui um obstaacuteculo agrave compreensatildeo do comportamento do vanaacutedio nos organismos
vivos Assim a caracterizaccedilatildeo da quiacutemica deste elemento eacute de grande utilidade antes de iniciar o
estudo da sua interacccedilatildeo com o presente sistema
A anaacutelise por espectroscopias de 51
V-RMN e de UV-Vis de soluccedilotildees de vanadato diluiacutedas nos
meios de reacccedilatildeo utilizados no presente relatoacuterio (10 mM HEPES pH 70 25 mM KCl) mostrou
claramente que a espeacutecie decameacuterica (V10) possui um tempo de vida suficientemente longo para a
determinaccedilatildeo do seu efeito isolado ao contraacuterio do que se verifica com as espeacutecies mono- di-
tetra- e pentameacutericas (V1 V2 V4 e V5) Por outro lado a anaacutelise verificou ainda que apoacutes diluiccedilatildeo
das soluccedilotildees de decavanadato no meio de reacccedilatildeo coexistem as espeacutecies V10 e V1 pelo que os
efeitos putativos desta uacuteltima deveratildeo ser subtraiacutedos aquando da monitorizaccedilatildeo dos efeitos da
espeacutecie decameacuterica
O vanaacutedio (V) (vanadato) inibe as enzimas que catalizam reacccedilotildees de transferecircncia de grupos
fosfato como a ATPase da miosina [Goodno 1979] devido agrave sua analogia estrutural com o iatildeo
fosfato Na sua utilizaccedilatildeo como inibidor enzimaacutetico natildeo eacute tida em conta a contribuiccedilatildeo individual
das espeacutecies oligomeacutericas considerando-se normalmente a espeacutecie V1 como inibidora efectiva
[Goodno e Taylor 1982] No presente relatoacuterio verificou-se por espectroscopia de 51
V-RMN que
outros oligoacutemeros de vanadato interagem com o S1 de miosina tendo as espeacutecies V10 e V4 os
maiores niacuteveis de afinidade A sua interacccedilatildeo envolve forccedilas electrostaacuteticas e eacute afectada
negativamente por ligandos naturais de enzima como eacute o caso do ATP
A exposiccedilatildeo do S1 de miosina (005 mgmL) a decavanadato conduz agrave inibiccedilatildeo do capacidade da
proteiacutena de estimular a actividade Mg2+
-ATPaacutesica com um valor de IC50 de 266 plusmn 07 M
decavanadato sendo o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM
decavanadato (80) Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM decavanadato sendo
o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM decavanadato (60)
exercendo um efeito de impedimento agrave ligaccedilatildeo entre o vanaacutedio e o S1
A uma concentraccedilatildeo de 27 microM decavanadato verifica-se a oxidaccedilatildeo de um dos resiacuteduos de Cys
de S1 de miosina seguida pela oxidaccedilatildeo gradual de outro resiacuteduo de Cys ateacute agrave concentraccedilatildeo de
1000 microM Ambos os resiacuteduos oxidados natildeo se encontravam expostos ao solvente utilizado A
oxidaccedilatildeo dos resiacuteduos permite sugerir que esta modificaccedilatildeo oxidativa sofrida pelo S1 contribui
para a diminuiccedilatildeo da sua actividade de ATPase
52
Por espectroscopia de RPE revelaram-se os possiacuteveis ligandos equatoriais do vanaacutedio aquando da
sua interacccedilatildeo com o S1 de miosina como sendo duas moleacuteculas de aacutegua um ligando contendo o
grupo R-OH e outro contendo o grupo R=NR
Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados de
oxidaccedilatildeo (IV) e (V) no S1 de miosina para concentraccedilotildees facilmente alcanccedilaacuteveis no ambiente
terrestre
53
5 Estudos futuros
No sentido do melhor esclarecimento das alteraccedilotildees moleculares e estruturais que ocorrem no
subfragmento 1 quando exposta ao vanaacutedio que conduzem a uma diminuiccedilatildeo da sua funccedilatildeo
sugere-se
- Quantificaccedilatildeo das cisteiacutenas reduzidas de S1 na presenccedila de metavanadato
- Determinaccedilatildeo das actividades Ca2+
- e K+-ATPaacutesica do S1 de miosina na presenccedila de
decavanadato
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina estimulada por actina
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina na presenccedila de
metavanadato
- Realizaccedilatildeo de RMN 2D com variaccedilatildeo das concentraccedilotildees das soluccedilotildees de decavanadato e
metavanadato na presenccedila de S1
-Mutageacutenese dos resiacuteduos de Cys doS1 de miosina de modo a verificar quais os resiacuteduos que
sofrem oxidaccedilatildeo na presenccedila de decavanadato
- Determinaccedilatildeo por calorimetria (DSC Differential Scanning Calorimetry) da variaccedilatildeo da
entalpia (∆H) e da temperatura de desnaturaccedilatildeo do S1 de miosina na presenccedila de vanaacutedio uma
vez que estas propriedades caracteriacutesticas de cada proteiacutena satildeo alteradas apoacutes ocorrecircncia de
modificaccedilotildees estruturais
- Anaacutelise por SDS-PAGE de S1 de miosina apoacutes tratamento com vanaacutedio de modo a determinar
quais os polipeacuteptidos eventualmente formados apoacutes clivagem da proteiacutena
54
6 Referecircncias bibliograacuteficas
- Sellers J Myosins 2nd Edition Oxford University Press 1999
- L Stryer inBiochemistry (Ed WH Freeman 2005 pp
Refordfas Materiais e Meacutetodos
[A] S S Margossian SLowey Methods Enzymol1982 85 55-71
[B] JD Pardue JA Spudich Methods Enzymol 1982 85 164-181
[C] M Bradford Anal Biochem1976 72 248-254
[D] H H Taussky E Shorr J Biol Chem 1953 202 675-685
[E] C R Bagshaw in Outline Studies in Biology ndash Muscle Contraction Chapman and Hall Ed
1982
[F] A Cuenda et al Eur J Biochem1990 194 663-670
55
Anexos
1 Determinaccedilatildeo da recta de calibraccedilatildeo meacutetodo de Bradford
Figura1ndashRepresentaccedilatildeo graacutefica da recta de calibraccedilatildeo (Abs595 nm vs [BSA]) utilizada para o meacutetodo de Bradford
2 Determinaccedilatildeo da recta calibraccedilatildeo para a electroforese em condiccedilotildees desnaturantes
Figura 2 ndash Representaccedilatildeo graacutefica da recta de calibraccedilatildeo (log(Peso Molecular) vs Rf dos marcadores de peso
molecular) onde Rf indica determinada atraveacutes do padratildeo de peso molecular
(poccedilo (a))
3Caacutelculo do coeficiente de extinccedilatildeo molar () para a quantificaccedilatildeo de cisteiacutenas reduzidas
atraveacutes do meacutetodo de Ellman
Traccedilou-se a recta de Abs412 nm meacutedia vs [L-Cisteiacutena] de modo a determinar o valor do coeficiente
de extinccedilatildeo molar definido como o declive daquela recta Assim =8400 M-1
cm-1
como
podemos verificar na Fig 3
A595 nm = 00546[BSA] + 00209 Rsup2 = 099051
Ab
s 59
5 n
m
[BSA] (mgmL)
log PM = -1177 Rf + 2247 Rsup2 = 098957
log
PM
Mobilidade relativa (Rf)
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero

7
1 Introduccedilatildeo
11 Muacutesculo esqueleacutetico
A capacidade de locomoccedilatildeo eacute uma das caracteriacutesticas fundamentais da maioria dos organismos
vivos A contracccedilatildeo muscular um dos fenoacutemenos que a torna possiacutevel apresenta grande
complexidade e tem vindo a ser utilizado como modelo para a compreensatildeo de sistemas menos
especializados de mobilidade celular [1]
No que diz respeito agrave sua fisiologia morfoloacutegica os muacutesculos dos vertebrados dividem-se em
lisos e estriados Estes uacuteltimos devem o seu nome agrave alternacircncia de zonas claras e escuras
observaacuteveis ao microscoacutepio electroacutenico de que resulta um aspecto estriado Os muacutesculos
estriados podem ainda ser divididos em duas subclasses muacutesculos esqueleacuteticos e cardiacuteacos [2]
consoante a forma celular (respectivamente longitudinal ou ramificada) ou tipo de contracccedilatildeo das
ceacutelulas que o compotildeem (respectivamente voluntaacuteria e involuntaacuteria) [3]
Os muacutesculos estriados esqueleacuteticos satildeo assim designados devido agrave sua localizaccedilatildeo no
organismo ndash ligados aos ossos da coluna vertical braccedilos e pernas Satildeo constituiacutedos por feixes de
ceacutelulas multinucleadas (miofibras) que contecircm miofibrilas paralelas de cerca de 1 m de
diacircmetro associadas agrave contracccedilatildeo voluntaacuteria raacutepida e forte em actividades de coordenaccedilatildeo
complexa [4] Cada feixe apresenta um diacircmetro compreendido entre 20 e 100 m e um
comprimento maacuteximo que pode atingir vaacuterios centiacutemetros (Fig 11)
Figura 11 ndash Organizaccedilatildeo hieraacuterquica geral do muacutesculo-esqueleacutetico (adaptado de Purves et al 2003 [3])
No muacutesculo-esqueleacutetico as zonas claras e escuras supramencionadas correspondem a zonas de
menor e maior densidade electroacutenica respectivamente As zonas claras constituem as bandas I (de
8
ldquoisotropiardquo ndash polarizaccedilatildeo uniforme em todas as direcccedilotildees) enquanto as zonas escuras constituem
as bandas A (de ldquoanisotropiardquo ndash polariza a luz visiacutevel) A banda I encontra-se dividida
transversalmente por uma linha escura ndash linha Z de densidade electroacutenica e refracccedilatildeo superiores
No interior de cada banda A existe uma zona transversal menos densa ndash zona H ndash rica em enzimas
importantes para o metabolismo celular No centro desta zona existe uma linha perpendicular
mais escura ndash linha M [2]
A unidade fundamental da contracccedilatildeo muscular eacute designada por sarcoacutemero (Fig 12) Encontra-se
repetida de forma regular ao longo das miofibrilas apresentando um comprimento de 25 a 3 m
aquando do relaxamento muscular diminuindo aquando da contracccedilatildeo muscular
Figura 12 ndash Representaccedilatildeo de fotomicrografia electroacutenica e esquemaacutetica do sarcoacutemero Os filamentos grossos e
finos satildeo maioritariamente constituiacutedos por miosina e actina respectivamente (Adaptado de Stryer 2002 [2])
As secccedilotildees de corte do sarcoacutemero datildeo conta da presenccedila de dois tipos de filamentos proteicos ndash os
filamentos grossos cujo diacircmetro eacute de 15 nm constituiacutedos sobretudo por miosina e os filamentos
finos com um diacircmetro de 9 nm constituiacutedos sobretudo por actina tropomiosina e pelo complexo
de troponina [2]
111 Miosina
Actualmente o termo miosina refere-se a uma superfamiacutelia diversa de motores moleculares
capazes de hidrolisar ATP e de se ligarem reversivelmente agrave actina A classificaccedilatildeo da
superfamiacutelia das miosinas nas 15 classes distintas faz-se atraveacutes de homologia entre as diferentes
sequecircncias [A] Eacute tambeacutem possiacutevel a classificaccedilatildeo por subclasses das quais se destaca a subclasse
4 proveniente do muacutesculo-esqueleacutetico e cardiacuteaco de vertebrados Este estudo focar-se-aacute apenas
nas miosinas de classe II (miosina-II) presente nos filamentos grossos do muacutesculo estriado
esqueleacutetico [B] e em ceacutelulas natildeo musculares A funccedilatildeo da miosina-II depende da sua localizaccedilatildeo
celular e tecidular
9
A miosina-II eacute constituiacuteda por duas cadeias polipeptiacutedicas idecircnticas denominadas cadeias pesadas
de miosina (HC) enroladas entre si e cujo peso molecular (PM) estaacute compreendido entre 171 e
241 kDa) Estruturalmente as HC dividem-se em 3 domiacutenios funcionais - cabeccedila pescoccedilo e
cauda como se pode observar na Fig 13 Agraves cabeccedilas ou zonas globulares estatildeo associados dois
pares de cadeias leves (peso molecular entre 15 e 30 kDa) que se dividem em dois tipos
essencial (ELC) e regulatoacuterio (RLC)
Figura 13 ndash Estrutura esquemaacutetica da moleacutecula de miosina-II Destacam-se as duas cabeccedilas globulares e a cauda
filamentosa ligadas entre si por uma zona flexiacutevel denominada pescoccedilo As cadeias leves (a verde e vermelho)
encontram-se associadas agraves cabeccedilas globulares
As cabeccedilas globulares contendo os locais de ligaccedilatildeo agrave actina e aos nucleoacutetidos mantecircm os seus
elementos estruturais baacutesicos conservados em todas as classes sendo estes constituiacutedos por 720-
80 aa A regiatildeo do pescoccedilo eacute constituiacuteda por uma longa heacutelice pertencente agrave cadeia pesada
que se encontra estabilizada atraveacutes da ligaccedilatildeo a subunidades de cadeia leve A estrutura do
pescoccedilo envolvida na ligaccedilatildeo agraves cadeias leves eacute denominada de motivo IQ cuja estrutura
consensual eacute IQXXXRGXXXR1 O pescoccedilo serve como extensatildeo do comprimento da cabeccedila de
modo a actuar como alavanca durante a contracccedilatildeo muscular [A]A regiatildeo da cauda ou acircncora
apresenta bastante divergecircncia no que respeita agrave sua sequecircncia estrutura e funccedilatildeo No entanto
pensa-se que a sua funccedilatildeo centrar-se-aacute na autoassociaccedilatildeo das vaacuterias moleacuteculas de miosina-II
atraveacutes da regiatildeo de coiled-coil Esta confere assimetria agrave moleacutecula e permite a formaccedilatildeo de
filamentos grossos capazes de translocar filamentos de actina [A]
Na regiatildeo da cabeccedila da miosina existem 2 regiotildees extremamente sensiacuteveis agrave proteoacutelise A
clivagem em ambos os locais resulta na produccedilatildeo de trecircs fragmentos um compreendendo o N-
terminal (fragmento 25 kDa) outro a regiatildeo central (50 kDa) e um terceiro o C-terminal (20 kDa)
(Fig 14) Estes fragmentos dissociam-se em condiccedilotildees desnaturantes e natildeo constituem domiacutenios
independentes da cabeccedila mas provavelmente loops flexiacuteveis uma vez que natildeo foram observadas
na estrutura cristalina Os vaacuterios fragmentos obtidos em diferentes condiccedilotildees foram cruciais para
a elucidaccedilatildeo das vaacuterias funccedilotildees da moleacutecula
A regiatildeo compreendida entre os fragmentos 25 e 50 kDa (loop 1) localiza-se directamente por
1 I = Isoleucina Q= Glutamina X = qualquer dos 20 aminoaacutecidos padratildeo R= Arginina G= Glicina
10
cima do local de ligaccedilatildeo ao nucleoacutetido A regiatildeo entre os fragmentos de 50 e 20 kDa (loop 2)
encontra-se perto da intersecccedilatildeo entre os domiacutenios da cabeccedila e do pescoccedilo e deveraacute interagir com
a actina uma vez que a ligaccedilatildeo desta a protege da proteoacutelise Na ausecircncia do loop 2 a miosina
liga mais fracamente agrave actina do que no caso da miosina intacta A sequecircncia primaacuteria de ambos
os loops apresenta conservaccedilatildeo reduzida sendo provaacutevel que sejam responsaacuteveis por muita da
diversidade das diferentes propriedades enzimaacuteticas presentes em diferentes isoformas
No fragmento de 20 kDa existem dois grupos sulfidrilo Cys-707 e -697 usualmente
denominados de SH1 e SH2 devido agrave ordem da sua reactividade quiacutemica Na presenccedila de
nucleoacutetidos pode ocorrer o ldquocross-linkingrdquo entre estes dois grupos incluindo a oxidaccedilatildeo a uma
ponte dissulfureto
Devido agrave insolubilidade da miosina-II nativa a estrateacutegia adoptada para os estudos com esta
proteiacutena eacute frequentemente a sua clivagem em fragmentos funcionais separaacuteveis Um dos locais de
clivagem ocorre tipicamente a 130 kDa do N-terminal produzindo a meromiosina pesada (HMM)
e a meromiosina leve (LMM) [C] A HMM eacute formada pelas 2 cabeccedilas e pescoccedilo e apresenta
propriedades semelhantes agrave miosina sem digestatildeo (ligaccedilatildeo e translocaccedilatildeo de actina e hidroacutelise de
MgATP)
A HMM pode ser novamente clivada atraveacutes de uma reacccedilatildeo de proteoacutelise produzindo os
subfragmentos 1 (S1) e 2 (S2)O S1 (90 kDa) eacute enzimaticamente activo sendo constituiacutedo pelos
domiacutenios da cabeccedila e do pescoccedilo e apresentando associaccedilatildeo agraves duas cadeias leves ou apenas agrave
ELC (Fig 14)Em alternativa o S1 pode ser produzido por clivagem directa da miosina intacta
formando-se tambeacutem um fragmento em forma de cilindro que consiste nas regiotildees S2 e LMM
(Ref)Utiliza-se extensivamente o S1 para estudos cineacuteticos e de ligaccedilatildeo agrave actina uma vez que a
sua solubilidade simplifica o planeamento da experiecircncia e a subsequente interpretaccedilatildeo dos dados
[A]
Figura 14- Estrutura do subfragmento-1 de miosina de muacutesculo esqueleacutetico com ecircnfase nos domiacutenios de 50 kDa
central (a vermelho) 25 kDa no N-terminal (a verde) e 20 kDa no C-terminal (a azul) As cadeias leves essenciais
encontram-se a amarelo e as cadeias leves regulatoacuterias a lilaacutes Os locais de ligaccedilatildeo agrave actina e ao ATP estatildeo
igualmente destacados (adaptado de Holmes e Geeves 1999 [D])
11
112 Actina
O termo ldquoactinardquo refere-se a uma famiacutelia extremamente conservada de proteiacutenas citoplasmaacuteticas
presente em todos os eucariotas Esta proteiacutena natildeo se encontra nos procariotas [E] A maioria das
actinas possui peso molecular proacuteximo de 42 kDa outra evidecircncia do seu grau elevado de
conservaccedilatildeo no que concerne agrave sequecircncia e ao nuacutemero de aminoaacutecidos Os eucariotas superiores
apresentam um grande nuacutemero de isoformas de actina que por sua vez se encontram codificados
por um ainda maior nuacutemero de genes (Ref) A expressatildeo das isoformas eacute diferencial consoante o
tecido e sua especializaccedilatildeo pelo que deveratildeo possuir funccedilotildees e mecanismos direccionais
diferentes (Ref) Foi ainda observado que os mRNA das diferentes isoformas apresentam uma
distribuiccedilatildeo heterogeacutenea nas ceacutelulas de forma semelhante aos seus produtos proteicos [F] Estes
dados suportam a hipoacutetese supramencionada do direccionamento das isoformas de actina para um
determinado local na ceacutelula de acordo com as suas funccedilotildees
A actina presente no citoesqueleto eacute responsaacutevel por processos como o mecanismo contraacutectil
fagocitose manutenccedilatildeo e alteraccedilatildeo da estrutura celular locomoccedilatildeo transporte de organelos
regulaccedilatildeo do transporte de solutos entre outras Nos eucariotas superiores estaacute ainda envolvida
no transporte de gracircnulos de pigmento retinoacuteide [G] transporte de gracircnulos no axoplasma [H] e
secreccedilatildeo entre outros [E] Actualmente a isoforma da actina (-actina) eacute a melhor estudada
encontrando-se envolvida no mecanismo contraacutectil [E]
Os filamentos de -actina tecircm cerca de 1m de comprimento e 80 Å Consistem em duas cadeias
de actina filamentosa (F-actina) organizados em heacutelice [I] (Holmes et al 1990) sobre os quais
se encontram outras proteiacutenas ndash a troponina (C I e T) e a tropomiosina (Azevedo 1999) (Figura
15) A troponina eacute filamentosa e localiza-se nas depressotildees da heacutelice de F-actina conferindo
rigidez aos filamentos finos Por sua vez a tropomiosina eacute essencial na regulaccedilatildeo do mecanismo
contraacutectil
Figura 15 ndash Modelo estrutural dos filamentos de F-actina contendo a tropomiosina e o complexo de troponina (I C
e T) (Adaptado de Susana Ramos 2005 [J])
12
A F-actina eacute formada atraveacutes da polimerizaccedilatildeo de subunidades globulares de actina (G-actina) A
G-actina eacute composta por 2 domiacutenios de tamanho semelhante separados pelos bolsos de ligaccedilatildeo(1)
de catiotildees divalentes e(2) de nucleoacutetidos [KLM] Estes por sua vez estatildeo subdivididos noutros
dois de menores dimensotildees (1-4) (Fig 16) cada um envolvido nas interacccedilotildees actina-actina ou
na formaccedilatildeo do topo de ligaccedilatildeo do nucleoacutetido As regiotildees N-terminal e C-terminal estatildeo proacuteximas
uma da outra no subdomiacutenio 1 [E]
Figura 16 ndash Estrutura do monoacutemero de actina Encontram-se representados os subdomiacutenios 1 a 4 e a vermelho a
moleacutecula de ATP junto do respectivo local de ligaccedilatildeo (Protein Data Bank ID 1j6z)
113 Mecanismos moleculares da contracccedilatildeo muscular
A contracccedilatildeo muscular ocorre atraveacutes do deslizamento dos filamentos finos (de actina) por entre
os filamentos grossos (de miosina) conseguida pela reacccedilatildeo de hidroacutelise de ATP e regulada pela
tropomiosina e pelo complexo de troponina em resposta a impulsos nervosos Este mecanismo foi
inicialmente proposto por Huxley-Niedergerke [N] e Huxley-Hanson [O] ambos em 1954 A base
experimental de ambos os estudos foi a inexistecircncia de variaccedilatildeo do tamanho dos filamentos
durante a contracccedilatildeo A banda A apresenta sempre as mesmas dimensotildees por outro lado a banda
I a zona H diminuem consideravelmente aquando da contracccedilatildeo (Figs 12 e 17)
Figura 17 ndash Modelo da contracccedilatildeo muscular ndash deslizamento dos filamentos finos (azuis) relativamente aos
filamentos grossos (rosa) (Adaptado de Stryer 2002 [2])
13
Na ausecircncia de ATP e supondo a ausecircncia de inibiccedilatildeo da interacccedilatildeo actina-miosina as proteiacutenas
encontram-se fortemente associadas formando o complexo actomiosina (AM) (Figura 15)
Nestas condiccedilotildees as cabeccedilas de miosina formam um acircngulo de 45ordm com o eixo dos filamentos
finos de actina [P]
Na presenccedila de ATP o complexo AM dissocia-se jaacute que a afinidade do complexo M-ATP para a
actina eacute 105 vezes inferior agrave afinidade da miosina para a actina O ATP liga-se agrave miosina
ocorrendo seguidamente a reacccedilatildeo de hidroacutelise de ATP Os produtos daiacute resultantes ndash ADP e Pi -
permanecem ligados agrave miosina formando o complexo M-ADP-Pi Com a ligaccedilatildeo subsequente agrave
actina forma-se o complexo AM-ADP-Pi no qual as cabeccedilas de miosina apresentam um acircngulo
recto com o eixo do muacutesculo [P] Neste ponto daacute-se a libertaccedilatildeo progressiva de Pi e ADP Eacute
provaacutevel que a energia proveniente da hidroacutelise do ATP provoque uma alteraccedilatildeo do acircngulo de
ligaccedilatildeo das cabeccedilas de miosina ao filamento de actina passando novamente de 90ordm para 45ordm [P]
Esta alteraccedilatildeo conformacional provoca o deslizamento do filamento de actina para o centro do
sarcoacutemero cujo encurtamento promove a contracccedilatildeo das miofibrilas O processo supracitado
ocorre em cerca de 1ms[Q] e encontra-se resumido na Fig 18
Figura 18 ndash Esquema cineacutetico da hidroacutelise de ATP pela miosina (Adaptado de Sellers 1999 [A])
1131 Regulaccedilatildeo da contracccedilatildeo muscular
O mecanismo pelo qual se processa a regulaccedilatildeo muscular foi inicialmente descrito por estudos
distintos de Haselgrove Huxley e Parrey-Squire todos em 1973 [RST] Aleacutem da implicaccedilatildeo
directa da tropomiosina e do complexo de troponina a concentraccedilatildeo de Ca2+
no muacutesculo em
relaxamento eacute bastante reduzida (10-7
M) aumentando notavelmente aquando da contracccedilatildeo (10-5
-
10-3
M)
Resumidamente quando ocorre a excitaccedilatildeo do muacutesculo esqueleacutetico a despolarizaccedilatildeo da
membrana plasmaacutetica eacute difundida ateacute ao interior do tecido atraveacutes dos tuacutebulos transversais
(invaginaccedilotildees da membrana plasmaacutetica da fibra muscular) A despolarizaccedilatildeo provoca a abertura
dos canais ioacutenicos de Ca2+
do retiacuteculo sarcoplasmaacutetico adjacente Como a concentraccedilatildeo de Ca2+
no interior do organelo eacute muito elevada o caacutelcio passa raacutepida e uniformemente para o citosol
14
Assim a regulaccedilatildeo da contracccedilatildeo ocorre com coordenaccedilatildeo e rapidez na totalidade das miofibrilas
do tecido muscular (Ref)
Em repouso a troponina I encontra-se posicionada de tal forma que a proteiacutena adjacente
troponina C impede estruturalmente a ligaccedilatildeo das cabeccedilas da miosina agrave actina (acircngulo =50ordm) O
aumento da concentraccedilatildeo intracelular de Ca2+
provoca um aumento da sua disponibilidade para
ligar agrave troponina C (Fig 19)
Figura 19ndash Regulaccedilatildeo do mecanismo contraacutectil no muacutesculo esqueleacutetico pelo complexo de troponina
Como consequecircncia a troponina C altera a sua conformaccedilatildeo (=70ordm) associando-se agrave troponina I
que deixa de inibir a ligaccedilatildeo das pontes transversais A alteraccedilatildeo anterior eacute transmitida agrave
tropomiosina atraveacutes da troponina T adjacente agrave C o que provoca o deslocamento da
tropomiosina na direcccedilatildeo do interior do sulco do filamento de actina Assim eacute possiacutevel a
interacccedilatildeo eficaz entre actina e miosina A Ca2+
-ATPase do retiacuteculo sarcoplasmaacutetico provoca a
reacumulaccedilatildeo do iatildeo caacutelcio no organelo impedindo o prolongamento desnecessaacuterio da contracccedilatildeo
(Refordf)
12 Vanaacutedio
O vanaacutedio eacute um metal de transiccedilatildeo pertencente ao grupo VB da tabela perioacutedica Foi
primeiramente descoberto em 1801 por Andreacutes Manuel del Riacuteo que concluiu erradamente que era
idecircntico ao croacutemio e definitivamente por Nils Gabriel Svenson em 1831 Recentemente foram
descobertas importantes funccedilotildees bioloacutegicas deste elemento provavelmente essencial para os
humanos de que se destaca no contexto do presente relatoacuterio a possibilidade de actuar como
anaacutelogo do grupo fosfato e a inibiccedilatildeo de enzimas in vitro como eacute o caso da actividade ATPaacutesica
da miosina
Eacute um metal soacutelido agrave temperatura ambiente que se encontra amplamente distribuiacutedo nos vaacuterios
ecossistemas em dois isoacutetopos naturais ndash 50
V e 51
V ndash raramente excedendo 01 partes por biliatildeo
15
(ppb) Estaacute associado a cerca de 70 minerais diferentes na natureza (dos quais se destacam o
uracircnio cobre chumbo zinco fosfato e titacircnio ferroso) especialmente na forma de vanadato ou
pentoacutexidos de vanaacutedio O vanaacutedio eacute um dos principais constituintes da mateacuteria orgacircnica foacutessil
com especial ecircnfase para o carvatildeo oacuteleo de crude e foacutesseis carbonaacuteceos [UV] bem como em
alimentos consumidos pelo Homem (salsa pimenta preta cogumelos marisco vegetais cereais
gorduras e oacuteleos) [W] Estes constituem a maior fonte de exposiccedilatildeo ao vanaacutedio da maioria da
populaccedilatildeo humana [X]
Normalmente a entrada do vanaacutedio no organismo ocorre por inalaccedilatildeo ou em menores
quantidades por absorccedilatildeo pelo tracto gastrointestinal ou pela pele O seu armazenamento centra-
se no fiacutegado rins ceacuterebro coraccedilatildeo muacutesculos e ossos No entanto a retenccedilatildeo de vanaacutedio no
organismo humano eacute reduzida ndash em condiccedilotildees normais cerca de 100 M (em muitos tecidos a
quantidade eacute inferior a 10 ngg de tecido) [Y]
Actualmente natildeo existem provas de deficiecircncia nutricional de vanaacutedio em humanos ou de
enzimas que contenham vanaacutedio em animais superiores No entanto vaacuterios estudos relacionam a
ocorrecircncia de deficiecircncia em galinhas e ratos devido a uma alimentaccedilatildeo pobre em vanaacutedio Os
sintomas relatados incluem retardamento do crescimento malformaccedilotildees oacutesseas perturbaccedilotildees no
metabolismo lipiacutedico e estagnaccedilatildeo reprodutiva entre outros [Z AA AB] Os efeitos fisioloacutegicos
do vanaacutedio podem depender do tipo de composto ou de oligoacutemero uma vez que consoante as
suas estabilidade labilidade e nuacutemero de coordenaccedilatildeo as respostas seratildeo diferenciadas ao niacutevel
dos organismos vivos
Por outro lado a toxicidade do vanaacutedio encontra-se bem caracterizada existindo sintomas
descritos em vaacuterios mamiacuteferos (inclusive o Homem) que incluem diarreias catildeibras irritaccedilatildeo da
pele e mucosas palpitaccedilotildees cardiacuteacas anemia pneumonia bronquite perturbaccedilotildees no Sistema
Nervoso Central (SNC) entre outros (Ref) A toxicidade varia geralmente com a natureza do
composto com aumento proporcional agrave sua valecircncia Verificou-se que a forma de entrada do
vanaacutedio tem tambeacutem efeito na sua toxicidade ndash os maiores niacuteveis verificaram-se por injecccedilatildeo
intravenosa da amostra sendo progressivamente inferiores por inalaccedilatildeo e finalmente por
ingestatildeo [X AA] A ampla utilizaccedilatildeo dos compostos de vanaacutedio na induacutestria quiacutemica bem como
as elevadas quantidades de vanaacutedio libertadas atraveacutes da combustatildeo dos oacuteleos de crude
contribuem consideravelmente para o aumento da biodisponibilidade do vanaacutedio Como tal o
interesse neste elemento e nos problemas ambientais por ele causados tem vindo a aumentar
significativamente [V]
121 Propriedades quiacutemicas
16
A quiacutemica do vanaacutedio eacute extraordinariamente complexa pelo que originou vaacuterias interpretaccedilotildees
incorrectas desde a sua descoberta No entanto as suas propriedades dependem em uacuteltima
anaacutelise da configuraccedilatildeo electroacutenica do elemento Assim a variabilidade encontrada no
comportamento quiacutemico do vanaacutedio pode dever-se agrave sua configuraccedilatildeo ndash [Ar] 3d2 4s
2 ndash que
permite a formaccedilatildeo de vaacuterios estados de oxidaccedilatildeo (de +2 a +5) Em soluccedilatildeo aquosa os estados de
oxidaccedilatildeo estaacuteveis vatildeo de V(V) a V(II) Destes apenas os +3 a +5 satildeo biologicamente importantes
jaacute que o V(II) apresenta demasiada capacidade redutora para existir nos organismos vivos
conhecidos [AC]
No fluido extracelular os compostos de vanaacutedio encontram-se predominantemente na forma de
monovanadatos (V(V)) livres ou ligados agrave transferina do soro ou agrave hemoglobina (Refordf) Os iotildees de
vanadato livres satildeo transportados para o meio intracelular atraveacutes de canais ioacutenicos inespeciacuteficos
Jaacute no ambiente intracelular os compostos sofreratildeo uma bio-reduccedilatildeo atraveacutes da acccedilatildeo da
glutationa ou outros compostos com grupos ndashOH como pentoses hexoses ou etanolaminas [V] A
reoxidaccedilatildeo de vanadilo (V(IV)) intracelular a vanadato (V(V)) provocaria a acumulaccedilatildeo toacutexica
destes uacuteltimos pelo que a complexaccedilatildeo da espeacutecie reduzida o impede O transporte de vanaacutedio
para o interior da ceacutelula encontra-se na Fig 110
Figura 110 ndash Transporte de vanaacutedio para a ceacuteluka e seu subsequente metabolismo (P=Proteiacutena) (Adaptado de
Zaporowska e Scibor 1998)
1211 Vanaacutedio(IV)
A espeacutecie de vanaacutedio (IV) mais bem conhecida eacute o iatildeo vanadilo (VO2+
) espeacutecie estaacutevel a pH
aacutecido sendo a maioritaacuteria a pH 3[AD] Apresenta uma cor azulada e forma complexos penta e
hexacoordenados Forma-se rapidamente a partir de V(V) na presenccedila de um agente redutor em
soluccedilatildeo aquosa No ambiente intracelular a presenccedila de glutationa reduzida aacutecido ascoacuterbico
NAD(P)H entre outros conduzem a que o principal estado de oxidaccedilatildeo nos sistemas bioloacutegicos
seja provavelmente o +4 encontrando-se o V(IV) fortemente complexado com proteiacutenas ou
aminoaacutecidos livres citrato ATP e pirofosfato (Refordfs) Em alternativa o V(IV) pode existir
17
livremente em ambientes fortemente aacutecidos como os lisossomas jaacute que a pH superior a 3 as
soluccedilotildees de vanadilo tendem a ser oxidadas pelo ar formando-se vanadato Esta grande
capacidade redutora justifica parcialmente a utilizaccedilatildeo pouco extensiva do iatildeo VO2+
como
inibidor [W]
1212 Vanaacutedio (V)
O vanaacutedio (V) eacute a forma mais estaacutevel num intervalo de pH amplo [W] O iatildeo anioacutenico VO3-tende
a constituir complexos polinucleares pelo que as soluccedilotildees aquosas de vanadato satildeo extremamente
complexas com ocorrecircncia simultacircnea de vaacuterios equiliacutebrios de protonaccedilatildeo (pKa(vanadato)= 35
78 e 125) e oligomerizaccedilatildeo As espeacutecies presentes (H3VO4 H2VO4- HVO4
2- e VO4
3-) dependem
do pH forccedila ioacutenica e temperatura da soluccedilatildeo e da concentraccedilatildeo total do elemento
[AEAFAGAHAI]
O VO43-
- monoacutemero ortovanadato V1 ndash estaacute associado a uma grande parte da importacircncia
bioloacutegica do vanaacutedio uma vez que se trata de um anaacutelogo electroacutenico e estrutural do ortofosfato
(PO43-
) A coordenaccedilatildeo do ortovanadato pode ser tetraeacutedrica ou bipiramidal trigonal (Refordfs)
A oligomerizaccedilatildeo do vanadato monomeacuterico daacute origem ao diacutemero (V2O72-
- V2) que apresenta
tambeacutem equiliacutebrios de protonaccedilatildeo Presume-se a existecircncia de analogia do vanadato dimeacuterico com
o pirofosfato embora natildeo haja evidecircncia estrutural (Refordf) A dimerizaccedilatildeo do V2 origina o
vanadato tetrameacuterico ou metavanadato (V4O124-
- V4) que constitui a espeacutecie maioritaacuteria em
soluccedilotildees aquosas concentradas (Refordf) As estruturas do metavanadato e a sua espeacutecie protonada
satildeo ciacuteclicas [AJ] Outra espeacutecie oligomeacuterica importante eacute o vanadato pentameacuterico (V5O153-
) cuja
estrutura presumida eacute ciacuteclica [AI]
O oligoacutemero com maior importacircncia bioquiacutemica eacute o vanadato decameacuterico (V10O286-
-V10)(Refordf) a
espeacutecie predominante em soluccedilotildees de pH 2-6 quando a concentraccedilatildeo total de vanadato eacute superior
a 100 M (Ref) A estrutura do V10 natildeo difere em soluccedilatildeo ou no estado soacutelidoOs 10 aacutetomos de
vanaacutedio da espeacutecie decameacuterica estatildeo divididos em trecircs grupos identificados na figura como VA-C
consoante o seu ambiente quiacutemico circundante (Refordf) (Fig 111)
A expressatildeo ldquodecavanadatordquo refere-se a uma soluccedilatildeo aquosa na qual a espeacutecie decameacuterica se
encontra em equiliacutebrio quiacutemico com as espeacutecies V1 V2 V4 e V5 sendo que a oligomerizaccedilatildeo
progressiva do vanaacutedio eacute proporcional agrave sua concentraccedilatildeo total (Ref) Nesta soluccedilatildeo a
concentraccedilatildeo de V10 corresponde a aproximadamente 10 da concentraccedilatildeo total de vanaacutedio [AK]
18
Figura 111 ndash Estruturas esquemaacuteticas das espeacutecies de vanadato (espeacutecie monomeacuterica (V1) dimeacuterica (V2)
tetrameacuterica (V4) pentameacuterica (V5) e decameacuterica (V10) (Adaptado de Aureliano e Crans 2008)
122 Metabolismo e efeito na actividade enzimaacutetica
Apesar de natildeo terem sido ainda encontradas as provas categoacutericas das suas funccedilotildees nos seres
vivos sabe-se que o iatildeo vanadato eacute capaz de inibir uma grande quantidade de enzimas in vitro
com especial ecircnfase na actividade ATPaacutesica da miosina [AL Teresa] Como referido
anteriormente o vanadato eacute capaz de actuar em alternativa ao fosfato nas reacccedilotildees bioquiacutemicas
incluindo a sua preferecircncia de ligaccedilatildeo agraves enzimas [W]
Eacute conhecido o papel do vanaacutedio no metabolismo do caacutelcio promovendo a mineralizaccedilatildeo dos
dentes e ossos na biossiacutentese de liacutepidos (trigliceacuteridos e colesterol) com posterior inibiccedilatildeo da
acumulaccedilatildeo destes nos vasos sanguiacuteneos e no SNC na reproduccedilatildeo e desenvolvimento por
mimetizaccedilatildeo e potenciaccedilatildeo de factores de crescimento na estimulaccedilatildeo do consumo de glucose e
da siacutentese de glicogeacutenio com actuaccedilatildeo equivalente aos efeitos da insulina ao niacutevel celular na
transduccedilatildeo de sinais celulares com regulaccedilatildeo da actividade de segundos mensageiros no
transporte de iotildees e aminoaacutecidos na transcriccedilatildeo geneacutetica e siacutentese de proteiacutenas e DNA entre
outros [W Y AB]
Assim o vanaacutedio cnseguiraacute provocar inuacutemeras mudanccedilas bioquiacutemicas nas ceacutelulas Alguns
exemplos da influecircncia bioquiacutemica do vanaacutedio incluem os seus efeitos nos processos de
fosforilaccedilatildeo e desfosforilaccedilatildeo de proteiacutenas e na participaccedilatildeo na formaccedilatildeo de espeacutecies reactivas de
oxigeacutenio devido ao seu efeito proacute-oxidante sobre agentes redutores a niacutevel celular [AM]
Quanto ao seu efeito na actividade enzimaacutetica pode dividir-se em dois tipos principais ndash inibitoacuterio
e estimulante - dependendo da enzima ou do tipo de reacccedilatildeo por esta catalisada No primeiro
grupo incluem-se enzimas como as ATPases (de miosina de canais ioacutenicos e fosfoenzimas
19
transportadoras) e as fosfatases (da tirosina aacutecida e alcalina) a fosfofrutocinase adenilato ciclase
e catalase entre outras [AA] O segundo grupo inclui a maioria das cinases a glucose-6-fosfato
desidrogenase a fosfoglicerato mutase e a fosfoglicerato fosfatase (Refordf)
O efeito do vanaacutedio (V) na actividade enzimaacutetica estaacute relacionado com o seu nuacutemero de
coordenaccedilatildeo podendo actuar como estado de transiccedilatildeo ou por mimetizaccedilatildeo de cofactores
substratos (especialmente o fosfato) e interacccedilotildees proteiacutena-proteiacutena (como no caso das
polimerases e transcriptases) Pode ainda estar envolvido na formaccedilatildeo de ligaccedilotildees de hidrogeacutenio
interacccedilotildees electrostaacuteticas esteacutereas e de oxidaccedilatildeo-reduccedilatildeo Uma evidecircncia desta uacuteltima eacute a sua
capacidade de alternacircncia entre os estados de oxidaccedilatildeo +5 a +3 o que torna faacutecil a sua interacccedilatildeo
redox com enzimas contendo resiacuteduos de cisteiacutenas (Cys) ou qualquer outro composto orgacircnico
com actividade redox [AN]
13 Vanaacutedio e muacutesculo esqueleacutetico
A primeira evidecircncia da presenccedila de vanaacutedio no muacutesculo esqueleacutetico proveio de uma amostra de
ATP isolado de muacutesculo de cavalo que continha uma impureza ndash vanaacutedio ndash cuja presenccedila
provocou a inibiccedilatildeo da actividade da bomba de Na+K
+-ATPase [AO] Desde entatildeo vaacuterios
estudos tecircm sido realizados com ecircnfase na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o
tecido muscular Destes destaca-se a descoberta da interacccedilatildeo forte da espeacutecie V10 com a miosina
fosforilada [AP] e agrave sua inibiccedilatildeo da actividade ATPaacutesica da miosina[AQ AR]
No quediz respeitoao iatildeo monovanadato destaca-se a sua inibiccedilatildeoda actividade Mg2+
-ATPaacutesica
da miosina [AS] que ocorre de duas formas distintas (1) na presenccedila de concentraccedilotildees reduzidas
de V1 a inibiccedilatildeo ocorre a velocidade reduzida dependente da concentraccedilatildeo da espeacutecie
monomeacuterica (2) na presenccedila de concentraccedilotildees elevadas de V1 o niacutevel de inibiccedilatildeo aproxima-se de
100 desde que [V1] seja superior agrave concentraccedilatildeo total de locais com actividade Mg2+
-ATPaacutesica
no ensaio Esta inibiccedilatildeo eacute irreversiacutevel tanto por diaacutelise ou adiccedilatildeo de agentes redutores do
vanadato [AL]Segundo Pate et al(1994)[AT] a espeacutecie monomeacuterica podeinibir a velocidade de
contracccedilatildeo a temperatura inferior a 20ordmC perdendogradual e rapidamente essa capacidade ateacute os
50ordmC temperatura agrave qual cessa a capacidade inibitoacuteria Os resultados obtidos sugeremainda uma
interacccedilatildeo altamente cooperativa entre o iatildeo vanadato e as proteiacutenas componentes do muacutesculo
esqueleacuteticoDestaca-se tambeacutem a descoberta da inibiccedilatildeo da geraccedilatildeo de forccedila contraacutectil nas fibras
musculares por parte da espeacutecie monomeacuterica[AT]Wilson et al(1995) [AU ]determinaram que a
afinidade da espeacutecie V1 para o S1 de miosina eacute significativamente mais elevada do que a do iatildeo
Pi seu anaacutelogo Descobriram ainda que que a ligaccedilatildeo do vanadato poderaacute envolver transiccedilotildees
entre estados energeacuteticos com energia livre independente dos impedimentos esteacutereos impostos
20
pela estrutura das miofibrilas Os estudos desenvolvidos por Smith et al (1990)[AV]
determinaram queo iatildeo monovanadato natildeo apresenta um efeito pronunciado na inibiccedilatildeo da
actividadeacto-S1-miosina-ATPaacutesica excepto em concentraccedilotildees superiores a 900microM a partir das
quais existe produccedilatildeo de espeacutecies oligomeacutericas para as quais o efeito inibitoacuterio seraacute mais
elevadoA espeacutecie monomeacuterica de vanadato consegue adoptar facilmente uma geometria do tipo
bipiramidal trigonal que mimetiza o intermediaacuterio formado (MgADP
Pi)aquando da reacccedilatildeo de
hidroacutelise de ATP no local activo da miosina [AW] De facto a interacccedilatildeo do vanaacutedio com a
miosina eacute inesperadamente estaacutevel na presenccedila de ADP Esta interacccedilatildeo resulta na formaccedilatildeo de
um complexo enzimaacutetico inactivo e estaacutevel com estequiometria de 11 ADPvanadato por siacutetio
activo da proteiacutena A formaccedilatildeo do complexo estaacutevel miosina-vanadato foi proposta por Goodnoet
al (1982) [AS] Nesta ocorre um preacute-equiliacutebrio raacutepido que induz a formaccedilatildeo de um complexo
ternaacuterio reversiacutevel (MADP
V) com formaccedilatildeo lenta do isoacutemero M
ADP
V A diferenccedila entre os
compostos de vanaacutedio e de fosfato centra-se no tempo de meia vida (t12) do Pi ou do V (no
primeiro t12=12 s enquanto no segundo t12=24 h) (Refordf) A diferenccedila dever-se-aacute agrave possiacutevel
complexaccedilatildeo do vanaacutedio com grupos nucleofiacutelicos no siacutetio activo o que provocaria uma
estabilizaccedilatildeo da miosina na conformaccedilatildeo MADP
V
No que respeita aos estudos realizados com a espeacutecie tetrameacuterica Cremo et al determinou que a
cadeia pesada do S1 de miosina pode ser clivada em dois locais distintos (designados V1 e V2)
pela espeacutecie V4 apoacutes irradiaccedilatildeo com luz UV na presenccedila de concentraccedilotildees de vanadato na ordem
dos milimolar e na ausecircncia de nucleoacutetidos ou metais divalentes
Jaacute no que concerne aos estudos realizados com ecircnfase na espeacutecie decameacuterica foi sugerido
porRamos et al (2006) [AK]que a interacccedilatildeo do decavanadato com a G-actina favorecida pela
polimerizaccedilatildeo desta uacuteltima estabiliza a espeacutecie V10 e induz a inibiccedilatildeo da formaccedilatildeo de F-actina A
estabilizaccedilatildeo da espeacutecie V10 por proteiacutenas tansmembranares e do citoesqueleto pode ser a razatildeo
pela qual ocorre a toxicidade do decavanadato reportada na avaliaccedilatildeo dos efeitos do V(V) nos
sistemas bioloacutegicosTiago et al (2007) [AR] concluiram que o V10 apresenta uma clara preferecircncia
de ligaccedilatildeo ao ldquoback-doorrdquo da S1-miosina-ATPase o que se verifica apenas na estrutura
conformacional desimpedida com acesso facilitado ao local de ligaccedilatildeo do Pi Sugere tambeacutem que
o V10 actua como um ldquoback-door stoprdquo bloqueando estaabertura do fragmento de 50-kDa
necessaacuteria para a hidroacutelise do ATP--fosfato o que constitui uma explicaccedilatildeo simples do
comportamento natildeo competitivo da espeacutecie decameacuterica estimulando a utilizaccedilatildeo deste oligoacutemero
como uma feramenta da elucidaccedilatildeo das mudanccedilas conformacionais ldquoback-doorrdquo da miosina no
21
processo da contracccedilatildeo muscular(ref)Destaca-se ainda a descoberta da capacidade inibitoacuteria do
V10 em relaccedilatildeo agrave fosforilaccedilatildeo oxidativa mitocondrial [AX]
Finalmente no que concerne aos estudos realizados com a presenccedila de vaacuterias espeacutecies
oligomeacutericas Aureliano e Madeira (1994) [AY]observaram que na presenccedila de Ca2+
as espeacutecies
V4 e V10se ligam agrave Ca2+
-ATPase do retiacuteculo sarcoplasmaacutetico enquanto a espeacutecie V1 se liga apenas
na presenccedila de ATP Aparentemente o aumento da concentraccedilatildeo de Ca2+
tem o efeito contraacuterio da
inibiccedilatildeo da ATPase do retiacuteculo sarcoplasmaacutetico pelo V1 natildeo tendo no entanto efeito significativo
sobre a inibiccedilatildeo por parte da espeacutecie V10 Tiago et al (2002) [AP] concluiacuteram que em condiccedilotildees
proacuteximas agraves fisioloacutegicas a espeacutecie V10 difere das espeacutecies presentes na soluccedilatildeo de metavanadato
devido agrave sua forte interacccedilatildeo com a miosina fosforilada e agrave sua inibiccedilatildeo da actividade ATPaacutesica
da miosina Para aleacutem disso o ATP diminui a afinidade da miosina para o V4 induzindo a
interacccedilatildeo com V1 e natildeo tendo qualquer efeito no que diz respeito ao V10 Foi tambeacutem proposto
[AQ] que a ligaccedilatildeo das espeacutecies oligomeacutericas de vanadato aoS1 de miosina nos locais onde o
fosfato coordenaeacute provavelmente a causa da inibiccedilatildeo da actividade ATPaacutesica da miosina
estimulada por actina atraveacutes da prevenccedilatildeo da troca entre ATPADP sendo esta favorecida para
aniotildees de vanadato com valecircncias superiores como eacute o caso do V10
Na presenccedila de actina as velocidades de dissociaccedilatildeo do Pi e do V aumentam consideravelmente
este uacuteltimo em menor quantidade Assim a ligaccedilatildeo da actina ao complexo MADP
V e
Pi eacute
bastante fraca pelo que se sugere que o ADPV teria um efeito de relaxamento semelhante ao
ATP No entanto a capacidade do vanaacutedio em inibir a actividade ATPaacutesica da miosina e da
actomiosina eacute diferenciada uma vez que na ausecircncia de actina a inibiccedilatildeo eacute total para
concentraccedilotildees na gama dos M mas na sua presenccedila a inibiccedilatildeo eacute totalmente conseguida apenas
em concentraccedilotildees na gama dos mM (Ref) No entanto eacute necessaacuterio considerar a possiacutevel
formaccedilatildeo de espeacutecies oligomeacutericas de vanadato cuja afinidade e efeito na enzima seratildeo
diferentes Assim pode concluir-se que o efeito de inibiccedilatildeo observado para a actividade ATPaacutesica
de miosina estimulada por actina se pode dever agrave presenccedila das diferentes espeacutecies oligomeacutericas o
que foi jaacute sugerido por Aureliano (2000) [AZ] ndash as espeacutecies tetrameacuterica e decameacuterica de vanadato
interactuariam com a miosina com alteraccedilatildeo da sua interacccedilatildeo com a actina No entanto natildeo eacute
conhecida a contribuiccedilatildeo absoluta das restantes espeacutecies oligomeacutericas para a inibiccedilatildeo da ATPase
da miosina estimulada por actina (Refs)
14 Objectivos do projecto
Pretende-se com este projecto o isolamento purificaccedilatildeo e caracterizaccedilatildeo da actina do muacutesculo
22
esqueleacutetico de coelho sendo esta uacuteltima tambeacutem realizada para o sufragmento-1 de miosina
Seguidamente realizar-se-aacute a quantificaccedilatildeo e caracterizaccedilatildeo das proteiacutenas por espectroscopia UV-
Vis e SDS-PAGE e a caracterizaccedilatildeo das soluccedilotildees de vanaacutedio por UV-Vis e ressonacircncia magneacutetica
nuclear (RMN) Por uacuteltimo a interacccedilatildeo do S1 de miosina com o vanaacutedio seraacute avaliada atraveacutes
das teacutecnicas de UVVis RMN e ressonacircncia paramagneacutetica electroacutenica (RPE)
2 Discussatildeo de Resultados
21 Caracterizaccedilatildeo das proteiacutenas isoladas
211Anaacutelise do estado de pureza
O estado de pureza das soluccedilotildees de actina e de S1 foi analisado por electroforese em gel de
poliacrilamida em condiccedilotildees desnaturantes (SDS-PAGE) (Fig 22) que separa as moleacuteculas de
acordo com o seu peso molecular (PM) A concentraccedilatildeo de acrilamida escolhida (10) permite a
detecccedilatildeo simultacircnea de componentes com peso molecular inferior a 20 kDa e outros com elevado
peso molecular
Figura 21 ndash Fotografia de gel de SDS-PAGE (10 acrilamida) Poccedilo (a)
10 L do marcador de elevado peso molecular () Poccedilo (b) 20 L de [G-
actina]=1 mgmL Poccedilo (c) 20 L de [S1]=1 mgmL Aplicou-se uma
voltagem de V durante h e min
A partir da recta de calibraccedilatildeo obtida log PM = -1177 Rf + 2247 (ver ponto 2 da secccedilatildeo
Anexos) pode verificar-se a presenccedila de actina na amostra do poccedilo (b) presente numa banda
bastante intensa (PMaparente=40 kDa) bem como a presenccedila minoritaacuteria de outros componentes
desconhecidos que natildeo teratildeo sido eliminados durante o processo de extracccedilatildeo e purificaccedilatildeo e natildeo
satildeo relevantes para o estudo em questatildeo Na amostra do poccedilo (c) nota-se a presenccedila de S1
(PMaparente=92 kDa) bem como a de outros componentes cuja banda eacute ligeiramente menos intensa
Estes poderatildeo ser fragmentos proteoliacuteticos de miosina e de S1 Estaacute ainda presente uma bada de
23
maior intensidade que poderaacute ser a proteiacutena-C (PM=135 kDa) uma vez que a sua remoccedilatildeo
completa da amostra necessitaria de um passo adicional de cromatografia de troca ioacutenica [BA]
No entanto natildeo podemos confirmar que se trata desta proteiacutena ou de qualquer outra uma vez que
se encontra fora do intervalo de migraccedilatildeo dos marcadores de peso molecular
212 Determinaccedilatildeo da concentraccedilatildeo proteica
2121Subfragmento-1 de miosina
A partir da recta de calibraccedilatildeo obtida pelo meacutetodo de Bradford [BB](ver secccedilatildeo 3311 do
Procedimento Experimental) A595nm = 00546[BSA] + 00209 (ver secccedilatildeo de Anexos)foi possiacutevel
determinar um intervalo de concentraccedilatildeo de S1 de miosina compreendido entre 226 e 117(plusmn
423M) (203 e 1056 (plusmn 381 mgmL)) (n=8) A concentraccedilatildeo meacutedia de S1 de miosina apoacutes
diaacutelise contra a soluccedilatildeo tampatildeo 10 mM HEPES pH 75 25 mM KCl determinada pela leitura da
Abs280 nmfoi de 20430M (1803 mgmL)(n=7)
As amostras a quantificar atraveacutes do meacutetodo de Bradford natildeo sofreram diaacutelise o que permite
explicar a concentraccedilatildeo superior de S1 de miosina em comparaccedilatildeo com o valor obtido atraveacutes do
meacutetodo directo Esta razatildeo aliada agrave sensibilidade e rapidez superiores deste uacuteltimo fez com que
se optasse pelo meacutetodo directo para a quantificaccedilatildeo da grande maioria dos ensaios
2122Actina
A concentraccedilatildeo de actina foi determinada quando esta se encontrava na forma monomeacuterica (G-
actina) por mediccedilatildeo da leitura directa da Abs290 nm tendo-se obtido uma concentraccedilatildeo meacutedia de
691481M (290202 mgmL) (n=3) em 2 mM Tris-HCl pH 80 02 mM CaCl2 02 mM Na-
ATP Ao comprimento de onda utilizado (290 nm) absorvem fortemente apenas os resiacuteduos de
Trp pelo que a mediccedilatildeo efectuada tem menor sensibilidade do que se tivesse sido efectuada a 280
nm uma vez que a actina apresenta apenas 4 resiacuteduos deste aminoaacutecido embora tenha 16
resiacuteduos de Tyr
22 Mediccedilatildeo da actividade enzimaacutetica de S1 de miosina
221 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
As actividades ATPaacutesicas de S1 de miosina dependentes de Ca2+
e K+foram analisadas pelo
caacutelculo das velocidades de consumo inicial do fosfato inorgacircnico (Pi) Definiu-se os tempos de 5
e 3 minutos para a mediccedilatildeo da Ca2+
e da K+EDTA-ATPase respectivamente Os valores obtidos
24
a partir do protocolo presente no ponto 341 da secccedilatildeo Procedimento experimental encontram-se
descritos na Tabela 2I
Tabela 2I ndash Acrividades natildeo-fisioloacutegicas de subfragmento-1 determinadas pelo meacutetodo de Fiske-Subarow
modificado (n=2)
Proteiacutena
Actividade
ATPaacutesica natildeo
fisioloacutegica
Actividade (nmol
Piminmg proteiacutena) Actividade (s-1)
S1
Ca2+
-ATPase 1400 plusmn 30 2100 plusmn 0045
K+EDTA-ATPase 2334 plusmn 50 3500 plusmn 0075
As actividades natildeo-fisioloacutegicas reflectem o estado de oxidaccedilatildeo dos resiacuteduos expostos de Cys 707
(SH1) e 697 (SH2) Se o grupo SH1 sofrer oxidaccedilatildeo a actividade ATPaacutesica dependente de Ca2+
aumenta e a actividade dependente de K+ diminui Por outro lado se o grupo SH2 sofrer
oxidaccedilatildeo regista-se a perda completa destas actividades
De acordo com Margossian e Lowey (1982) [BA] as actividades ATPaacutesica de S1 de miosina
dependente de Ca2+
e K+EDTA da miosina satildeo respectivamente de 24 e 112 s-1
Estaacute tambeacutem
descrito que a actividade ATPaacutesica dependente de K+ eacute em meacutedia cerca de 3 vezes superior agrave
dependente de Ca2+
reflectindo a quantidade das preparaccedilotildees A razatildeo obtida entre os valores das
actividades ATPaacutesicas dependentes de K+ e Ca
2+ eacute de 166 Os valores obtidos para as actividades
(Ca2+
-ATPase=21 s-1
K+EDTA-ATPase= 35 s
-1) satildeo inferiores agrave valores obtidos por
Margossian e Lowey [BA] e Tiago [BC] com forccedila ioacutenica inferior O valor obtido para a
actividade ATPaacutesica dependente de Ca2+
eacute superior ao obtido por Polosukhina et al (2000) [BD]
cujo meio de reacccedilatildeo continha MOPS e apresentava forccedila ioacutenica inferior Os dados
supramencionados comprovam a variaccedilatildeo da actividade a ATPaacutesica natildeo fisioloacutegica em relaccedilatildeo agrave
forccedila ioacutenica temperatura e pH
222 Mediccedilatildeo da actividadeMg2+
-ATPaacutesica de S1 pelo meacutetodo de enzimas acopladas
A actividade de Mg2+
-ATPase eacute a uacutenica que possui significado fisioloacutegico sendo as restantes
bastante uacuteteis apenas para indicaccedilatildeo da qualidade de preparaccedilatildeo do S1[BA] A 0575 M KCl a
actina e o S1 formam uma soluccedilatildeo viscosa e tuacuterbida Apoacutes a adiccedilatildeo de MgATP o complexo acto-
S1 dissocia-se e a turbidez da soluccedilatildeo diminui Se a forccedila ioacutenica for reduzida a dissociaccedilatildeo do
complexo actoS1 que se encontra na forma de suspensatildeo eacute induzida O MgATP eacute rapidamente
hidrolisado diminuindo a sua concentraccedilatildeo no meio reaccional Nestas condiccedilotildees haacute
reassociaccedilatildeo e subsequente contracccedilatildeo dos filamentos de actina e de S1 com formaccedilatildeo de
25
precipitado O meacutetodo descrito traduz de forma bastante elementar o fenoacutemeno da contracccedilatildeo in
vivo e elimina o problema anterior da solubilizaccedilatildeo ineficiente da miosina de modo a obter uma
mediccedilatildeo exacta da actividade [A] A utilizaccedilatildeo de forccedilas ioacutenicas reduzidas aumenta a afinidade da
actina para o S1 permitindo contornar a extrema dificuldade em obter a concentraccedilatildeo proteica dos
muacutesculos (cerca de 100 mgmL do complexo actomiosina) As actividades Mg2+
-ATPaacutesicas de S1
de miosina foram determinadas a partir do declive da variaccedilatildeo da Abs340 relativo agrave oxidaccedilatildeo de
NADH em funccedilatildeo do tempo como eacute descrito no ponto 342 do Procedimento Experimental Os
valores obtidos para a actividade Mg2+
-ATPaacutesica encontram-se apresentados na Tabela 2II
Tabela 2II ndash Actividade Mg2+
-ATPaacutesica de subfragmento-1 de miosina determinadas pelo meacutetodo das enzimas
acopladas
Actividade (nmol
ATPmin mg(S1)) Actividade (s
-1)
1097 plusmn 269 164 plusmn 04
Ramos (2005) [J] obteve o valor de 01 s-1
para a actividade Mg2+
-ATPaacutesica de S1 de miosina
com forccedila ioacutenica intermeacutedia Tiago (2005) [BC] obteve o valor de 011 s-1 em condiccedilotildees de forccedila
ioacutenica reduzida em relaccedilatildeo agrave utilizada no presente relatoacuterio
23 Preparaccedilatildeo e caracterizaccedilatildeo das soluccedilotildees de vanaacutedio
231 Anaacutelise da composiccedilatildeo das soluccedilotildees de vanadato por espectroscopia de 51
V-RMN
efeito da concentraccedilatildeo da espeacutecie
A diluiccedilatildeo das soluccedilotildees stock de meta- e decavanadato iraacute determinar fortemente a composiccedilatildeo
em soluccedilatildeo dos diferentes oligoacutemeros de vanadato (ref) No caso do decavanadato o periacuteodo de
tempo apoacutes a diluiccedilatildeo iraacute tambeacutem ser um factor importante a considerar devido agrave decomposiccedilatildeo
da espeacutecie V10(ref)Torna-se assim fundamental determinar a composiccedilatildeo quiacutemica das soluccedilotildees
no meio de ensaio utilizado (HEPES 10 mM KCl 25 mM) de modo a correlacionaacute-la
correctamente com os possiacuteveis efeitos promovidos pelas diferentes espeacutecies de vanadato no
sistema em estudo
Aleacutem disso deve ser dado ecircnfase agraves importantes interacccedilotildees que ocorrem entre o vanaacutedio e os
tampotildees escolhidos O tampatildeo com menor nuacutemero e magnitude de interacccedilatildeo eacute o HEPES [AD]
seguido do Tris sendo este uacuteltimo especialmente aconselhado em concentraccedilotildees baixas para casos
de incompatibilidade enzimaacutetica ou quiacutemica redox No caso especial da reacccedilatildeo redox
NAD(P)HNAD(P) o tampatildeo adequado eacute o imidazole de modo a natildeo haver reacccedilatildeo com o
26
vanadato Os tampotildees a evitar em qualquer situaccedilatildeo incluem a trietanolamina fosfato citrato e
glicina entre outros [AD] uma vez que todos interactuam fortemente com V(V) com formaccedilatildeo
de complexos estaacuteveis
Assim procedeu-se agrave anaacutelise da composiccedilatildeo das soluccedilotildees de meta- e decavanadato apoacutes diluiccedilatildeo
das soluccedilotildees stock (50 mM em vanadato total) no meio de reacccedilatildeo (tampatildeo D) atraveacutes da
espectroscopia de ressonacircncia magneacutetica nuclear de 51
V Nas figuras x1 e x2 apresentam-se os
espectros obtidos para as soluccedilotildees de deca- e metavanadato num intervalo de concentraccedilotildees de 1 a
10 mM A Tabela seguinte resume os paracircmetros espectroscoacutepicos obtidos para cada espeacutecie
presente nas soluccedilotildees analisadas
Figura 22ndash Espectros de RMN de 51
V de
concentraccedilotildees crescentes das espeacutecies oligomeacutericas
de vanadato obtidos apoacutes diluiccedilatildeo da soluccedilatildeo stock
de decavanadato em 10 mM HEPES pH 70 25 mM
KCl para concentraccedilotildees finais de A) 1 mM B)2
mM C) 4 mM D) 8 mM E) 10 mM de
decavanadato total V1refere-se agraves espeacutecies
monomeacutericade vanadato V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2IIIndash Desvios quiacutemicos meacutedios (part) das ressonacircncias das espeacutecies decameacuterica (V10) monomeacuterica (V1) e
tetrameacuterica (V4) presentes nos espectros da soluccedilatildeo de decavanadatoLargura a meia altura (∆12) da espeacutecie V10A
Paracircmetros V10A V10B V10C V1 V4
part (ppm) -5162 -5003 -4255 -5616 -5748
∆12 (Hz) 175 nd nd nd nd
Figura 23 ndash Distribuiccedilatildeo graacutefica das
espeacutecies oligomeacutericas de vanadato vs a
concentraccedilatildeo de decavanadato total As
concentraccedilotildees de cada espeacutecie foram
obtidas segundo a formula descrita na
secccedilatildeo 35 do Procedimento
Experimental (aacutereas dos sinais de 51
V-
RMN observadas na figura x1)V10 V1 e
V4 correspondem agraves espeacutecie monomeacuterica
dimeacuterica e tetrameacuterica respectivamente [Esp
eacuteci
e o
ligo
meacute
rica
de
van
aacutedio
] (m
M)
[Decavanadato]total (mM)
V10
V1
V4
27
Nas soluccedilotildees de decavanadato as espeacutecies monomeacuterica e decameacuterica de vanadato satildeo as uacutenicas
presentes para concentraccedilotildees de decavanadato inferiores a 8 mM Ambas as espeacutecies aumentam
linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado em [AY] e
em (Teresa) quando a concentraccedilatildeo de V1 eacute igual ou superior a 200 microM inicia-se a sua
oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica No entanto nas condiccedilotildees de realizaccedilatildeo dos
ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a espeacutecie tetrameacuterica neste
estudo em concentraccedilotildees inferiores a 8 mM tendo o mesmo sido observado por Ramoset al
(2006) [AK] Os resultados obtidos mostram claramente o equiliacutebrio entre a espeacutecie decameacuterica e
a monomeacuterica e que o aparecimento de outras espeacutecies oligomeacutericas se encontra dependente da
concentraccedilatildeo desta uacuteltima Apesar de natildeo ter sido realizado o ensaio com 5 mM de decavanadato
total no presente relatoacuterio estaacute reportado em Ramos et al (2006) [AK] que as concentraccedilotildees da
espeacutecie decameacuterica e monomeacuterica satildeo respectivamente de 500 microM e 200 microM Assim pode inferir-
se que para concentraccedilotildees inferiores a 5 mM e intervalos de tempo de cerca de 20 minutos (que
inclui o tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas
as espeacutecies decameacuterica e monomeacuterica de vanadato
Figura 24 ndash Espectros de RMN de 51
V de concentraccedilotildees
crescentes das espeacutecies oligomeacutericas de vanadato obtidos
apoacutes diluiccedilatildeo da soluccedilatildeo stock de metavanadato em 10 mM
HEPES pH 70 25 mM KCl para concentraccedilotildees finais de A)
1 mM B) 2 mM C) 4 mM D) 8 mM E) 10 mM de
metavanadato total V1 refere-se agraves espeacutecies monomeacuterica de
vanadatoV1 e V2 correspondem respectivamente agrave espeacutecie
monomeacuterica e dimeacuterica
Figura 25 ndash Distribuiccedilatildeo graacutefica das espeacutecies
oligomeacutericas de vanadato vs a concentraccedilatildeo de
metavanadato total As concentraccedilotildees de cada
espeacutecie foram obtidas segundo a formula descrita
na secccedilatildeo 35 do Procedimento Experimental
(aacutereas dos sinais de 51
V-RMN observadas na
figura x1)V10 V1 e V4 correspondem agraves espeacutecie
monomeacuterica dimeacuterica e tetrameacuterica
respectivamente
Esp
eacuteci
e o
ligo
meacute
rica
de
va
nad
ato
(m
M)
[Metavanadato]total (mM)
V1
V2
V4
V5
28
Tabela 2IVndash Desvios quiacutemicos meacutedios (part)das ressonacircncias das espeacutecies tetrameacuterica (V4) monomeacuterica (V1) e
dimeacuterica (V2) presentes no espectro da soluccedilatildeo de metavanadato (total) 10 mM
Paracircmetros V1 V2 V4 V5
part (ppm) -5615 -5748 -5789 -5869
Nas soluccedilotildees de metavanadato as espeacutecies monomeacuterica dimeacutericae tetrameacuterica de vanadato estatildeo
presentes para concentraccedilotildees de metavanadato inferiores a 4 mM Todas estas as espeacutecies
aumentam linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado
em (Aureliano e Madeira 1998) e em (Tiago et al 2004) quando a concentraccedilatildeo de V1 eacute igual ou
superior a 200 microM inicia-se a sua oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica Nas
condiccedilotildees de realizaccedilatildeo dos ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a
espeacutecie pentameacuterica neste estudo em concentraccedilotildees inferiores a 4 mM tendo o mesmo sido
observado por Ramos 2006 Os resultados obtidos mostram claramente o equiliacutebrio entre a
espeacutecie V4 e as espeacuteciesV1 e V2 no qual o aparecimento de outras espeacutecies oligomeacutericas se
encontra dependente da concentraccedilatildeo da espeacutecie V1 Por uacuteltimo pode inferir-se que para
concentraccedilotildees inferiores a 2 mM e intervalos de tempo de cerca de 20 minutos (que inclui o
tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas as
espeacutecies V1 V2 e V4 com concentraccedilotildees respectivas de 0796 0192 e 0196 mM
232Estabilidade das soluccedilotildees de decavanadato anaacutelise por espectroscopia de absorccedilatildeo
UVVis
Caracterizou-se a estabilidade da espeacutecie V10 numa soluccedilatildeo com 1 mM decavanadato total em
tampatildeo D atraveacutes da leitura da Abs400 Obteve-se uma cineacutetica de dissociaccedilatildeo de 1ordf ordem tal
como se pode verificar pela linearidade do graacutefico de ln(Abs400)vs o tempo de aquisiccedilatildeo espectral
(segundos) (Fig26 )
Figura 26ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do tempo
(em segundos) para uma soluccedilatildeo de decavanadato total de 1
mM em tampatildeo D a 25ordmC (n=2)
Sendo uma reacccedilatildeo de primeira ordem pode ser descrita pela equaccedilatildeo cujo
declive eacute ndashK (constante de dissociaccedilatildeo) e a intersecccedilatildeo no eixo das ordenadas corresponde
Ln
(Ab
s 400 n
m)
tempo (s)
29
aln(A0) Numa reacccedilatildeo de 1ordf ordem o tempo de meia-vida eacute independente da concentraccedilatildeo inicial
do reagente sendo descrito pela equaccedilatildeo [2] Deste modo a partir da recta
Ln(Abs400 nm) = -436E-5
K - 105 obteve-se um tempo de meia-vida de 4490 plusmn 0823 horas para
as condiccedilotildees supramencionadas
Apoacutes a adiccedilatildeo de S1 8 microM ao meio reaccional verificou-se que a reacccedilatildeo de desoligomerizaccedilatildeo
da espeacutecie decameacuterica de vanadato seguia igualmente uma cineacutetica de 1ordf ordem tal como se pode
verificar pela linearidade do graacutefico de ln(Abs400) versus o tempo de aquisiccedilatildeo espectral
(segundos) (Fig27)
Figura 27ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do
tempo (em segundos) para uma soluccedilatildeo de decavanadato
total de 1 mM em tampatildeo D a 25ordmC na presenccedila de 8
microM (n= 2)
A partir da equaccedilatildeo da recta Ln(Abs400 nm) = -7520E-5
K - 0613determinou-se o valor de K para
a espeacutecie decameacuterica de onde se obteve um tempo de meia-vida de 2430 plusmn 0191 horas para as
condiccedilotildees supramencionadas
Tanto este valor como o valor registado na ausecircncia de S1 satildeo substancialmente superiores agrave
duraccedilatildeo dos ensaios de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina descritos no presente
relatoacuterio
24 Interacccedilatildeo de S1 com vanaacutedio(V)
241Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
De modo a analisar qual o efeito da adiccedilatildeo de S1 de miosina (tampatildeo D) produzia nos sinais de
51V-NMR titulou-se uma soluccedilatildeo de 5mM de decavanadato total com concentraccedilotildees crescentes
de S1 (5 10 15 e 20 microM) Os espectros de 51
V-RMN encontram-se apresentados na Fig 2
No graacutefico da figura 2 Apresenta-se a concentraccedilatildeo de S1 em funccedilatildeo do factor de alargamento
corrigido dos espectros acima referidos
Ln
(Ab
s (4
00
nm
))
tempo (s)
30
Figura 28 ndash Graacutefico de
representaccedilatildeo da
concentraccedilatildeo de S1 em
funccedilatildeo do factor de
alargamento A
concentraccedilatildeo de
decavanadato total foi de 5
mM e as concentraccedilotildees
utilizadas de S1 foram de 0
5 10 e 15 microM
Verifica-se que o factor de alargamento aumenta linearmente em funccedilatildeo da concentraccedilatildeo de S1 no
ensaio o que indica a existecircncia de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina Observou-
se ainda que a adiccedilatildeo de S1 ao meio reaccional natildeo provocou qualquer alteraccedilatildeo dos desvios
quiacutemicos dos sinais de 51
V-RMN
Realizaram-se ensaios na presenccedila de ATP e de S1 com intuito de verificar o efeito do ATP na
interacccedilatildeo do S1 de miosina com o decavanadato
Figura 29 - Espectros de RMN de 51
V contendo 2
mM Decavanadato (A) (controlo) (B) na presenccedila
de 2 mM de ATP (C) na presenccedila de 2 mM de ATP
e 8 microM de S1 de miosina V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2Vndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies decameacuterica (V10) e monomeacuterica (V1) presentes nos
espectros da soluccedilatildeo de decavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V10A V10B V10C V1
part (ppm) -5161 -5002 -4254 -5614
Observa-se que a presenccedila de ATP (espectro B) natildeo provoca qualquer alteraccedilatildeo visiacutevel em
relaccedilatildeo ao espectro que contem apenas soluccedilatildeo de decavanadato Jaacute a presenccedila simultacircnea de ATP
e S1 provoca um alargamento da ∆12 do sinal do V10A de 1736 Hz (espectro (A)) para 2366 Hz
(espectro (C) Fig 29) Tendo em conta que a concentraccedilatildeo de S1 apresenta uma relaccedilatildeo linear
com o factor de alargamento converteu-se o valor da ∆12 e comparou-se com o valor esperado agrave
F = 00591[S1] - 00157 Rsup2 = 0998
Fact
or
de
ala
rgam
en
to
[S1] (microM)
31
concentraccedilatildeo de S1 utilizada O factor de alargamento obtido para 8 microM ndash 0376 ndash eacute inferior ao
valor esperado agrave mesma concentraccedilatildeo de S1 obtido atraveacutes da equaccedilatildeo da recta apresentada na
Fig 28 ndash 0457 ndash o que indica que o ATP poderaacute exercer um efeito de protecccedilatildeo do S1
diminuindo a sua interacccedilatildeo com a espeacutecieV10
Por uacuteltimo realizaram-se ensaios com metavanadato na presenccedila de 8 microM S1 de modo a verificar
se as espeacutecies oligomeacutericas contidas nesta soluccedilatildeo interagiam com a proteiacutena em estudo
Figura 210 - Espectros de RMN de 51
V contendo 2
mM metavanadato (A) (controlo) (B) na presenccedila
de 8 microM de S1 de miosina V1 V2 e V4
correspondem respectivamente aacutes espeacutecies
monomeacuterica dimeacuterica e tetrameacuterica
Tabela 2VIndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies monomeacuterica (V1) dimeacuterica (V2) e tetrameacuterica (V4)
presentes nos espectros da soluccedilatildeo de metavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V1 V2 V4
part (ppm) -5613 -5747 -5789
Como eacute possiacutevel observar na Fig 210 ocorre uma ligeiro alargamento dos sinal de V4 bem
como uma perda da intensidade pelo que se conclui quedas espeacutecies presentes na soluccedilatildeo de
metavanadato (V1 V2 e V4) apenas a espeacutecie tetrameacuterica interage com o S1 de miosina o que
estaacute de acordo com o reportado por Tiago et al 2002
242 Actividade Mg2+
-ATPaacutesica de S1 de miosina na presenccedila de decanadato
Com o intuito de avaliar o efeito do decavanadato na actividade Mg2+
-ATPaacutesica do S1 de
miosina monitorizou-se a reacccedilatildeo de oxidaccedilatildeo do NADH atraveacutes da leitura de Abs340 na
presenccedila de 005 mgmL de S1 de miosina e no caso da actividade estimulada 2microM de actina em
3125 mM Tris-HCl pH 71 e 375 mM MgCl2 pelo meacutetodo de enzimas acopladas (ver seccccedilatildeo xx
dos MM)Para tal realizaram-se ensaios com vaacuterias concentraccedilotildees de decavanadato total (10 50
100 250 500 e 1000) de modo a determinar qual o seu efeito na actividade MgATPaacutesica de S1
de miosina na ausecircncia e na presenccedila de um dos seus ligandos naturais Os ensaios foram
realizados agrave temperatura ambiente com incubaccedilatildeo preacutevia a 25ordm C durante 20 minutos
32
Figura 211 ndash Representaccedilatildeo
graacutefica da Actividade Mg2+
-
ATPaacutesica de S1 () (os ajustes
teoacutericos estatildeo representado a por
uma linha os resultados
experimentais estatildeo representados
por triacircngulos (Actividade
estimulada) e cruzes (Actividade
natildeo estimulada)) vs
[Decavanadato]total medidas pelo
meacutetodo das enzimas acopladas
(n=3) Os dados foram ajustados ao modelo de decaimento exponencial usando o programa GraphPad Prism para a
determinaccedilatildeo dos valores de IC50 (na ausecircncia de actina IC50= 266 plusmn 07 M decavanadato na presenccedila de actina
IC50= 28 plusmn 1 M decavanadato
A partir do graacutefico presente na Fig 29 verifica-se que o ajuste dos dados experimentais foi bem
sucedido A inibiccedilatildeo da actividade Mg2+
-ATPaacutesica do S1 de miosina eacute afectada em menor
amplitude na presenccedila de actina (60 de inibiccedilatildeo da actividade) do que na sua ausecircncia
(aproximadamente 80 de inibiccedilatildeo de actividade) Devido a esta diferenccedila pode sugerir-se que a
actina iraacute impedir parcialmente a ligaccedilatildeo do decavanadatoao S1 demiosina atraveacutes de
impedimentos esteacutereos ou interacccedilotildees electrostaacuteticas por exemplo A teacutecnica utilizada natildeo nos
permite concluir acerca de que regiatildeo(otildees) ou resiacuteduo(s) da actina participaratildeo na preservaccedilatildeo
parcial da actividade Mg2+
-ATPaacutesica
Por outro lado os valores de IC50 dos dois tipos de actividades natildeo apresentam diferenccedilas
significativas tendo-se obtido 266 plusmn 07 e 28 plusmn 1M decavanadato na ausecircncia e na presenccedila de
actina respectivamente o que corresponde a uma concentraccedilatildeo de cerca de 27 M de V10 O
meio de reacccedilatildeo utilizado nestes ensaios foi 10 mM HEPES e 25 mM KCl com 2 mM ATP Em
(Tiago et al 2004) o valor de IC50 para a actividade Mg2+-ATPaacutesica na ausecircncia de actina foi de
125 plusmn 025 microM enquanto o valor da actividade estimulada por actina foi de 136 plusmn 14 microM O
meio de reacccedilatildeo utilizado nos ensaios foi 20 mM HEPES 50 mM KCl e 2 mM ATP o que resulta
precisamente no dobro da forccedila ioacutenica a que foi realizado o ensaio do presente relatoacuterio
(exceptuando a contribuiccedilatildeo dos produtos da hidroacutelise de ATP uma vez que este meacutetodo
pressupotildee a ocorrecircncia de um ciclo de reacccedilotildees de hidroacutelise e siacutentese deste composto) Ora tendo
sido anteriormente referido que a afinidade do S1 de miosina para com a espeacutecie V10 diminui com
Act
ivid
ade
Mg
2+-A
TP
aacutesic
a d
e S
1 d
e
Mio
sin
a
[Decavanadato] (microM) Actividade Estimulada ()
Actividade Natildeo Estimulada ()
Ajuste Actividade Estimulada
Ajuste Actividade Natildeo Estimulada
33
o aumento da forccedila ioacutenica (Tiago et al 2004 tal poderaacute explicar a diferenccedila obtida entre estes
dois conjuntos de valores
243Quantificaccedilatildeo de cisteiacutenas reduzidas pelo meacutetodo de Ellman
A interacccedilatildeo do vanaacutedio com alguns tipos de aminoaacutecidos sensiacuteveis agrave oxidaccedilatildeo pode fornecer-nos
informaccedilatildeo bastante uacutetil em relaccedilatildeo agraves alteraccedilotildees estruturais que conduzem a uma diminuiccedilatildeo da
actividade fisioloacutegica do S1 de miosina Os estudos realizados por Chasteen 1983 concluiacuteram
que a reduccedilatildeo de vanadato (V) a vanadilo (IV) ocorre na presenccedila de agentes redutores comuns no
ambiente celular de que se destaca o aminoaacutecido cisteiacutena Esta eacute tambeacutem um alvo de
complexaccedilatildeo por parte do iatildeo VO2+
Jaacute Ramos et al (2009) demonstrou que a incubaccedilatildeo de
actina com a soluccedilatildeo de decavanadato induz a oxidaccedilatildeo de resiacuteduos de cisteiacutena e agrave formaccedilatildeo do
oxidovanaacutedio (IV) Este grupo de investigadores descobriu ainda que apenas a espeacutecie V10 induz a
oxidaccedilatildeo das cisteiacutenas ldquoraacutepidasrdquo da actina atraveacutes do processo proposto que eacute parcialmente
responsaacutevel pela inibiccedilatildeo jaacute descrita da actividade de ATPase do S1 de miosina estimulada por
actina quando exposta agrave soluccedilatildeo de decavanadato Esta inibiccedilatildeo apresenta-se afectada com a
presenccedila de ATP uma vez que os resiacuteduos de cisteiacutena permaneceram intactos nos ensaios
realizados com este ligando Jaacute Crans et al (2004) concluiu que o siacutetio activo de muitas fosfatases
(que conteacutem grupos tiol) pode formar um equiliacutebrio redox com o iatildeo vanadato (V) de forma natildeo
reversiacutevel
Os resultados obtidos por Sakurai e Shiromura (1980) sugeriram que o vanaacutedio (V) sofre reduccedilatildeo
a vanaacutedio (IV) sem dependecircncia do valor de pH do ensaio Registaram ainda uma estequiometria
de 31 na reacccedilatildeo do vanadato (V) com os resiacuteduos de Cys enquanto a estequiometria dos ensaios
com o vanadilo (IV) foi de 21
Berlett e Stadtman (1997) observaram que os tipos de aminoaacutecidos mais facilmente oxidaacuteveis
englobam entre outros os resiacuteduos de Cys Trp e MetAssim analisou-se o estado redox dos
grupos tiol dos resiacuteduos de Cys do S1 na presenccedila do decavanadato atraveacutes da monitorizaccedilatildeo da
Abs412 nm na presenccedila de 1 mgmL de S1 de miosina em tampatildeo 10 mM HEPES 25 mM KCl
variando a concentraccedilatildeo da soluccedilatildeo de decavanadato de modo a determinar que tipo de alteraccedilotildees
estruturais ocorrem na sua presenccedila no que diz respeito agrave diminuiccedilatildeo da actividade ATPaacutesica de
S1 na presenccedila de decavanadato
Os resultados referentes agrave quantificaccedilatildeo das cisteiacutenas reduzidas apoacutes o tratamento de S1 com
diferentes concentraccedilotildees de decavanadato em tampatildeo 10 mM HEPES pH 75 e 25 mM KCl a
25ordmC durante 20 minutos encontram-se descritos na Tabela 2VII
Tabela 2VII ndash Cisteiacutenas reduzidas totais e ldquoraacutepidas de S1 (1 mgmL) apoacutes tratamento com decavanadato
34
(010501002505001000 uM) em 10 mM HEPES 25 mM KCl pH 7 a 25ordmC durante 20 minutos A detecccedilatildeo
espectrofotomeacutetrica foi feita com DTNB (e 412=8400 uM-1 cm-1) (n variaacutevel entre 1 e 8)
Amostras Cys raacutepidas Cys totais
11 M S1 (controlo) 22 plusmn 04 106 plusmn 14
11 M S1 + 10 M Decavanadato 22 plusmn 05 95 plusmn 08
11 M S1 + 50 uM Decavanadato 20 plusmn 02 98 plusmn 14
11 M S1 + 100 uM Decavanadato 17 plusmn 03 97 plusmn14
11 M S1 + 250 uM Decavanadato 26 plusmn 02 98 plusmn 08
11 M S1 + 500 uM Decavanadato 21 plusmn 04 90 plusmn 06
11 M S1 + 1000 uM Decavanadato 24 plusmn 0 86 plusmn 08
A partir dos resultados obtidos pode sugerir-se que o S1 possui 11 resiacuteduos de Cys (controlo) o
que estaacute de acordo com o anteriormente descrito Verifica-se que o tratamento da proteiacutena com
concentraccedilotildees crescentes de decavanadato provoca a oxidaccedilatildeo de dois resiacuteduos de Cys do S1 de
miosina natildeo expostos ao solvente No entanto a teacutecnica utilizada natildeo nos permite identificar
quais
Tendo em consideraccedilatildeo que o valor de IC50 obtido para a reacccedilatildeo de inibiccedilatildeo daactividade Mg2+-
ATPaacutesica de S1 na presenccedila de decavanadato eacute de cerca de 27 microM pode sugerir-se que este seraacute o
valor ao qual ocorreraacute a oxidaccedilatildeo do primeiro resiacuteduo de Cys o que poderaacute contribuir para a
diminuiccedilatildeo da actividade de S1 observada
25Interacccedilatildeo de S1 com vanaacutedio(IV)
251Anaacutelise por espectroscopia de Ressonacircncia Paramagneacutetica Electroacutenica
A acumulaccedilatildeo intracelular de oxovanaacutedio (IV) in vivo constitui uma das principais formas de
apresentaccedilatildeo de vanaacutedio no organismo Deste modo torna-se bastante interessante analisar a
interacccedilatildeo entre este iatildeo e o S1 de miosina atraveacutes da teacutecnica de espectroscopia de Ressonacircncia
Paramagneacutetica Electroacutenica o meacutetodo de eleiccedilatildeo do estudo do iatildeo vanadilo e dos complexos por si
formados O electratildeo desemparelhado encontra-se fortemente acoplado ao nuacutecleo 51
V o que
35
resulta num espectro de RPE consistindo de 8 desdobramentos na direcccedilatildeo paralela e outros tantos
na direcccedilatildeo perpendicular (Chasteen 1983)
Os estudos realizados por Ahmed et al (1987) centraram-se na interacccedilatildeo do vanadilo com
proteiacutenas como eacute o caso da insulina transferrina entre outros obtendo um espectro caracterizado
por 8 linhas amplamente espaccediladas (na direcccedilatildeo paralela) Jaacute Ramos et al 2009 concluiu que a
exposiccedilatildeo da soluccedilatildeo de decavanadato agraves formas globular e filamentosa de actina produz um
espectro tiacutepico do iatildeo oxidovanaacutedio (IV) indicando a reduccedilatildeo do vanadato
Para aferir qual a concentraccedilatildeo de S1 que permitia obter uma boa razatildeo sinalruiacutedo titulou-se uma
soluccedilatildeo de sulfato de vanadilo 15 microM ([VOSO4]stock = 5 mM) com concentraccedilotildees crescentes de
proteiacutena (0 5 10 15 e 20 microM) em 10 mM HEPES 25 mM KCl A existecircncia de uma correlaccedilatildeo
linear entre a concentraccedilatildeo de proteiacutena e a intensidade dos sinais de RPE correspondente agrave banda
-12 perpendicular encontra-se representada na Fig 210
Figura 2 12ndash Representaccedilatildeo graacutefica da variaccedilatildeo da intensidade do sinal de RPE vs a concentraccedilatildeo de S1 utilizada
Apoacutes verificar que com uma concentraccedilatildeo de S1 igual a 20 microM se obtinham sinais com uma razatildeo
sinalruiacutedo adequada ao prosseguimento dos estudos sobre a interacccedilatildeo V(IV)-S1 de miosina
titulou-se uma soluccedilatildeo de 20 microM S1 com 0 25 5 10 15 25 50 75 100 150 200 500 e 1000
microM VOSO4
Traccedilou-se o graacutefico da intensidade da banda -12 perpendicular do sinal de EPR em funccedilatildeo da
concentraccedilatildeo de VOSO4 tendo-se determinado uma constante de dissociaccedilatildeo (KD) do complexo
S1-vanadilo de 8748 plusmn 2621 microM V(IV)
y = 016786x + 122435
Rsup2 = 092992759
Inte
nsi
dad
e d
o s
inal
de
RP
E
[S1] (microM)
36
Figura 213 ndash Representaccedilatildeo graacutefica da variaccedilatildeo do sinal de RPE vs a concentraccedilatildeo de vanadilo utilizada (os valores
experimentais estatildeo representados por losangos o seu ajuste estaacute representado pela linha)
A simulaccedilatildeo do espectro de EPR foi efectuada a partir do espectro obtido para uma concentraccedilatildeo
de 1 mM VOSO4 apoacutes adiccedilatildeo de 20 microM S1 de miosina tendo-se obtido os valores da constante de
acoplamento hiperfina (A=5016) e do factor g (g0=195)
Figura 214 ndash Representaccedilatildeo graacutefica do espectro de EPR obtido com 1000 microM de vanadilo e 20 microM de S1 (a cinzento
escuro) A simulaccedilatildeo do espectro encontra-se a cinzento claro
Por comparaccedilatildeo destes com os valores presentes na tabela de correspondecircncia xxx eacute possiacutevel
predizer quais os ligandos provaacuteveis do vanadilo na presenccedila de proteiacutena Assim concluiu-se que
o vanadilo estaacute coordenado com duas moleacuteculas de aacutegua um grupo hidroxilo e um grupo R=NR
O grupo hidroxilo pode indicar os resiacuteduos de aminoaacutecidos Tyr Treonina e Serina enquanto o
grupo R=NR poderaacute indicar a presenccedila de Arginina e Histidina como ligandos do vanadilo
Inte
nsi
dad
e d
os
sinai
s d
e R
PE
[Vanadilo] (microM)
Ajuste dos dados exp Dados experimentais
Campo magneacutetico (G)
Espectro Experimental Ajuste
37
3 Procedimento Experimental
31 Reagentes
Todos os reagentes utilizados satildeo de elevado grau de pureza (analiacutetico) Apresentam-se as suas
foacutermulas quiacutemicas ou abreviaturas quando necessaacuterio acrescentou-se a sua descriccedilatildeo Entre
parecircntesis colocou-se as empresas que os comercializam
Aacutecido aceacutetico glacial 100 (Riedel) Acrilamida 99 (Merck) Azul de bromofenol (Riedel-de-
Haumlen) -Mercaptoetanol (Sigma) Bissulfito de soacutedio (Sigma)Bradford (reagente de)
(BioRad)BSA (albumina de soro bovino Sigma-Aldrich) CaCl22H2O (cloreto de caacutelcio
dihidratado Fluka) CH3CH2OH 96 (etanol Panreac) Coomassie Blue R250 (Merck) D2O
(oacutexido de deuteacuterio SDS) DTNB (55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico) Aldrich) EDTA (aacutecido
etilenodiaminatetraaceacutetico Merck) Fosfoenolpiruvato 98 (Fluka) Glicerol (Sigma) Glicina
985 (Panreac) HCl 37 (Riedel-de Haumlen) H2SO4 96 (Pronalab) HEPES 99 (aacutecido 4-(2-
hidroxiletil)-1-piperazineetanosulfoacutenico Sigma) KH2PO4 99 (hidrogenofosfato de potaacutessio)
MgATP (Sigma) Na
ATP (Sigma) Na2HPO4 99 (hidrogenofosfato de soacutedio Riedel-de Haumlen)
NaH2PO4 (fosfato de soacutedio ) NaOH 986 (hidroacutexido de soacutedio Pronalab) KCl 99 (Cloreto de
potaacutessio Panreac) L-Cys 995 (enantioacutemero levoacutegiro do resiacuteduo de Cisteiacutena Fluka)Metanol
99 (Merck) Molibdato de amoacutenio (Sigma)NNrsquo-metil-bis-acrilamida 98 (Merck) NH4VO3
99 (metavanadato de amoacutenia Sigma) PSA 10 (persulfato de amoacutenio Merck) SDS
(dodecilo sulfato de soacutedio Sigma) Sulfito de soacutedio (Sigma) VOSO4 (sulfato de vanadilo
Aldrich) TEMED 99 (Tetrametiletilenodiamina Merck) Tris (2-amino-2-hidroximetil-13-
propanodiol Sigma)
32 Meacutetodos
321 Isolamento e purificaccedilatildeo das proteiacutenas de muacutesculo esqueleacutetico de coelho
3211 Purificaccedilatildeo da miosina preparaccedilatildeo de Subfragmento 1 por digestatildeo quimotriacutepsica
da miosina
O subfragmento-1 de miosina-II (S1) foi gentilmente cedido pelo Prof Doutor Manuel Aureliano
do Departamento de Quiacutemica Bioquiacutemica e Farmaacutecia da Faculdade de Ciecircncias e Tecnologia da
Universidade do Algarve
No entanto descreve-se abaixo de forma resumida o procedimento efectuado
38
32111 Purificaccedilatildeo da miosina
O protocolo de isolamento foi adaptado do meacutetodo previamente descrito por Margossian e Lowey
(1982) apresentando-se de forma esquemaacutetica na Fig 31
Sucintamente os tecidos musculares dorsais e das pernas foram extraiacutedos apoacutes o sacrifiacutecio do
animal (Fig31) De seguida foram adicionados 3 volumes da soluccedilatildeo de extracccedilatildeo A2
Centrifugou-se a soluccedilatildeo e filtrou-se o sobrenadante O sedimento foi entatildeo diluiacutedo em 14
volumes de aacutegua destilada de modo a que a forccedila ioacutenica atingisse 004 M Apoacutes adiccedilatildeo de szlig-
mercaptoetanol (szlig-me) deixou-se a precipitar a 4ordmC removendo o sobrenadante Centrifugou-se
novamente e ressuspendeu-se o sedimento na soluccedilatildeo de extracccedilatildeo A Adicionou-se a soluccedilatildeo
tampatildeo B3 para que a forccedila ioacutenica atingisse 05 M Homogeneizou-se e deixou-se repousar
durante 2h Homogeneizou-se novamente e centrifugou-se de modo a ocorrer precipitaccedilatildeo dos
complexos de actomiosina e proteiacutena desnaturada Apoacutes filtraccedilatildeo do sobrenadante em latilde de vidro
quantificou-se a soluccedilatildeo de miosina resultante pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental)
2 Soluccedilatildeo de extracccedilatildeo A 0012M K2CO3 004M KHCO3 06M KCl pH 65
3 Soluccedilatildeo-tampatildeo B Tris 100 mM pH 7 KCl 2M
39
Figura 31 ndash Representaccedilatildeo esquemaacutetica do protocolo de isolamento de miosina (Margossian e Lowey 1982) Todo o
procedimento foi realizado a 4ordmC (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
32112 Preparaccedilatildeo de Subfragmento-1 de miosina por digestatildeo com quimiotripsina
O S1 de miosina foi isolado apoacutes diaacutelise da miosina durante a noite contra a soluccedilatildeo C4 A
presenccedila do EDTA eacute essencial uma vez que a sua substituiccedilatildeo por MgCl2resulta na produccedilatildeo de
HMM em vez de S1 A suspensatildeo de miosina foi equilibrada e digerida com -quimotripsina
A digestatildeo foi interrompida pela adiccedilatildeo de PMSF Seguidamente ultracentrifugou-se o produto
da digestatildeo desprezando-se o sedimento (Fig 32) A soluccedilatildeo de sobrenadante maioritariamente
S1 foi quantificada pela leitura directa da Abs280 ou pelo meacutetodo de Bradford (ver as secccedilotildees
3311 e 3312 do Procedimento Experimental respectivamente) e armazenada em 10 sacarose
e 2 mM szlig-Me a -80ordmC
4 Soluccedilatildeo C 25mM Tris pH 70 012M KCl 1mM EDTA
40
Antes da realizaccedilatildeo dos ensaios descritos no presente relatoacuterio o S1 de miosina foi dialisado
contra a soluccedilatildeo D5 de modo a remover qualquer composto que pudesse complexar como
vanaacutedio Seguidamente quantificou-se a proteiacutena pelo meacutetodo de Bradford ou pelo meacutetodo
directo (ver secccedilotildees 3311 e 3312do Procedimento Experimtental)
Figura 3 2ndash Representaccedilatildeo esquemaacutetica do protocolo de preparaccedilatildeo do S1 de miosina por digestatildeo quimiotripsina de
acordo com o previamente descrito por Margossian e Lowey (1982) Todo o procedimento foi realizado a 4ordm C
excepto quando indicaccedilatildeo do contraacuterio (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
3212 Isolamento e purificaccedilatildeo de actina
Isolou-se e purificou-se a actina segundo o meacutetodo descrito por Pardee e Spudish que
compreende dois passos ndash a obtenccedilatildeo de um resiacuteduo seco denominado ldquopoacute de acetonardquo e a
extracccedilatildeo subsequente da actina a partir do ldquopoacute de acetonardquo na forma de G-Actina com a
posterior polimerizaccedilatildeo a F-Actina
32121 Obtenccedilatildeo do ldquopoacute de acetonardquo
Sacrificou-se o coelho com uma pancada atraacutes do pescoccedilo e sangrou-se o animal apoacutes o corte da
arteacuteria jugular Esfolou-se e colocou-se numa tina com gelo Retiraram-se as partes de tecido
conjuntivo que envolvem o tecido muscular com auxiacutelio de uma pinccedila e bisturi Seguidamente
retirou-se os muacutesculos dorsais e os muacutesculos brancos das pernas Passou-se os muacutesculos por aacutegua
destilada de modo a remover as impurezas e pesou-se num copo Trituraram-se os muacutesculos
numa maacutequina de triturar carne e extraiu-se o resultante com 3 volumes da soluccedilatildeo E6 durante 10
minutos com agitaccedilatildeo suave Centrifugou-se a suspensatildeo a 4000 g durante 10
5 Soluccedilatildeo D HEPES 10 mM pH 70 KCl 25 mM
6 Soluccedilatildeo E 01 M KCl 015 M K2HPO4 pH 65
41
minutosdesprezando-se o sobrenadante Ressuspendeu-se o sedimento em 6 volumes de soluccedilatildeo
F7durante 10 minutos com agitaccedilatildeo suave Verificou-se se o pH da soluccedilatildeo estava compreendido
entre 82 e 85 acertando-se estecom soluccedilatildeo G8 se necessaacuterio Centrifugou-se durante 10
minutos a 4000 g Desprezou-se o sobrenadante e ressuspendeu-se o sedimento em 3 volumes de
soluccedilatildeo H9 durante 10 minutos Centrifugou-se novamente a 4000 gdurante 10 minutos
desprezando-se o sobrenadante Ressuspendeu-se o sedimento duas vezes consecutivas em 6
volumes de aacutegua destilada durante 5 minutos Centrifugou-se a 4000 g durante 10 minutos apoacutes
cada adiccedilatildeo de H2O
Seguidamente ressuspendeu-se o sedimento 5 vezes consecutivas utilizando 3 volumes de
acetona de cada vez agitando suavemente durante 10 minutos agrave temperatura ambiente Colocou-
se o resiacuteduo filtrado numa tina de vidro sobre papel de filtro deixando-se evaporar o solvente
durante a noite num nicho Triturou-se o resiacuteduo seco numa maacutequina de triturar carne e deixou-se
novamente evaporar o solvente O ldquopoacute de acetonardquo obtido foi armazenado a -20 ordmC
32122 Purificaccedilatildeo da actina a partir de ldquopoacute de acetonardquo
Apoacutes a obtenccedilatildeo do poacute de acetona como acima indicado efectuou-se o protocolo seguidamente
descrito de modo a obter monoacutemeros de actina (G-Actina) e actina filamentosa (F-Actina)
Pesou-se 35 g de poacute de acetona e adicionaram-se 50 mL de soluccedilatildeo-tampatildeo I10
sendo a
extracccedilatildeo feita com agitaccedilatildeo suave durante 30 minutos Separou-se o extracto do poacute de acetona
hidratado filtrando a suspensatildeo com 4 camadas de gaze esterilizada tendo o cuidado de natildeo
espremer o resiacuteduo Voltou a extrair-se com 50 mL de soluccedilatildeo I durante 30 minutos mexendo
lentamente Adicionou-se este ao extracto obtido anteriormente Centrifugou-se o extracto total a
16 000 g durante 30 minutos Decantou-se para uma proveta sobre a qual se colocou gaze para
que o sedimento solto natildeo se juntasse ao sobrenadante e leu-se o volume De seguida adicionou-
se KCl ateacute uma concentraccedilatildeo final de 50 mM MgCl2 ateacute 2mM e ATP ateacute 1mM lentamente e com
agitaccedilatildeo contiacutenua de modo a promover a polimerizaccedilatildeo dos monoacutemeros de actina
Deixou-se polimerizar durante 2h a 25ordmC observando-se a presenccedila de pequenas bolhas de ar
indicativas de um aumento de viscosidade da soluccedilatildeo Arrefeceu-se em gelo durante 15 minutos e
adicionou-se KCl ateacute uma concentraccedilatildeo final de 06M Mexeu-se lentamente durante 30 minutos
apoacutes os quais se ultracentrifugou a 110 000g durante 1hora Desprezou-se o sobrenadante
ressuspendeu-se o sedimento com soluccedilatildeo tampatildeo E e transferiu-se para um potter
Dialisou-se durante a noite contra 1L de soluccedilatildeo-tampatildeo I a uma temperatura de 4ordmC com rotaccedilatildeo
7 Soluccedilatildeo F 005 M NaHCO3 pH 82-85
8 Soluccedilatildeo G 1M Na2CO3
9 Soluccedilatildeo H 1 mM EDTA pH 70
10 Soluccedilatildeo I 2 mM Tris-HCl pH 8 02 mM CaCl2 02 mM Na2ATP 05 mM szlig-Me 0005 NaN3
42
contiacutenua e mudanccedila de tampatildeo 2 a 3 vezes De seguida a actina foi centrifugada a 110
000gdurante 1hora Decantou-se a soluccedilatildeo de G-Actina (sobrenadante) e desprezou-se o
sedimento Determinou-se a concentraccedilatildeo de G-Actina pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental) A soluccedilatildeo de G-actina foi armazenada a uma temperatura de 4ordmC
apoacutes adiccedilatildeo de 002NaN3 mantendo-se estaacutevel durante 3 a 5 dias
331 Quantificaccedilatildeo das proteiacutenas isoladas
3311 Quantificaccedilatildeo pelo meacutetodo de Bradford
O meacutetodo de Bradford (Bradford 1976) baseia-se na utilizaccedilatildeo do reagente de Bradford
composto por entre outros aacutecido fosfoacuterico e o corante ldquoCoomassierdquo Este corante possui carga
negativa e a sua coloraccedilatildeo varia entre a vermelha (Absmax=465 nm) predominante em soluccedilatildeo e a
azul (Absmax=595 nm) presente aquando da ligaccedilatildeo aos grupos amina das proteiacutenas Em soluccedilotildees
aacutecidas diluiacutedas a coloraccedilatildeo varia proporcionalmente com a quantidade do corante ligado agrave
proteiacutena que por sua vez eacute directamente proporcional agrave concentraccedilatildeo da proteiacutena permitindo a
sua quantificaccedilatildeo ateacute a um limite de detecccedilatildeo de 20 gmL (Bradford 1976)
Deste modo prepararam-se amostras de BSA ([BSA]stock= 1mgmL) utilizando um intervalo de
concentraccedilotildees compreendido entre 0 e 15 gmL de acordo com a tabela 3I Apoacutes a adiccedilatildeo da
H2O MilliQ e do reagente de Bradford as soluccedilotildees foram sucessivamente homogeneizadas no
voacutertex
Tabela 3Indash Amostras utilizadas para a obtenccedilatildeo da recta de calibraccedilatildeo no meacutetodo de Bradford
BSA (L) BSA (g) H2O MilliQ (L) Reagente de Bradford (L)
0 0 800 200
125 125 79875 200
25 25 7975 200
5 5 795 200
10 10 790 200
15 15 785 200
Apoacutes incubaccedilatildeo a 25ordmC por 5 minutos leram-se os valores de Abs595 num
espectrofotoacutemetro (Shimadzu UV-160A) para todas as amostras Seguidamente traccedilou-se a recta
de calibraccedilatildeo (Abs595vs quantidade de BSA (g))
43
Para determinar a concentraccedilatildeo de proteiacutena apoacutes o processo de isolamento purificaccedilatildeo e
diaacutelise quando necessaacuterio colocou-se 5 L da amostra num tubo de Eppendorf e adicionou-se
795 L de H2O MilliQ homogeneizando a soluccedilatildeo no voacutertex Adicionou-se posteriormente 200
L de reagente de Bradford e repetiu-se a homogeneizaccedilatildeo Incubou-se a mistura a 25ordmC por 5
minutos apoacutes os quais se leu os valores de Abs595 nm no espectrofotoacutemetro Por interpolaccedilatildeo deste
valor a partir da recta de calibraccedilatildeo determinou-se a concentraccedilatildeo de proteiacutena na amostra
3312 Quantificaccedilatildeo pelo meacutetodo directo
A quantificaccedilatildeo das amostras proteicas pode tambeacutem ser efectuada atraveacutes da leitura directa da
absorvacircncia tendo por base a lei de Beer-Lambert onde Abs corresponde agrave
absorvacircncia ao comprimento de onda ao qual o ensaio foi realizado corresponde ao coeficiente
de extinccedilatildeo (substituiacutedo pela letra a se as unidades forem distintas da molaridade) b corresponde
ao percurso oacuteptico da radiaccedilatildeo na ceacutelula amostral e c corresponde agrave concentraccedilatildeo amostral
Assim no caso de S1 de miosina mediu-se a Abs280e calculou-se a concentraccedilatildeo da proteiacutena
sabendo que o coeficiente de extinccedilatildeo de S1 eacute (Ref)
No caso da actina mediu-se a Abs290ecalculou-se a concentraccedilatildeo da proteiacutena sabendo que o
coeficiente de extinccedilatildeo da actina eacute (Ref)
332Electroforese em condiccedilotildees desnaturantes (SDS-PAGE)
De modo a melhor caracterizar as proteiacutenas isoladas foi realizada uma electroforese em
condiccedilotildees desnaturantes (SDS-PAGE do inglecircs Sodium Dodecyl Sulfate Polyacrilamide Gel
Electrophoresis) O SDS (dodecilo sulfato de soacutedio) eacute um detergente anioacutenico utilizado neste
meacutetodo para desnaturar as proteiacutenas que se ligam ao SDS na proporccedilatildeo 14 g SDS g proteiacutena
(aproximadamente 1 moleacutecula de SDS por cada 2 resiacuteduos de aminoaacutecidos) (Stryer) O SDS induz
uma densidade de carga negativa significativa nas proteiacutenas a que se liga mascarando a sua carga
intriacutenseca pelo que as razotildees cargamassa satildeo tendencialmente idecircnticas Assim a SDS-PAGE
separa as proteiacutenas quando sujeitas a um campo eleacutectrico de acordo com o seu peso molecular
sendo a sua migraccedilatildeo directamente proporcional ao logaritmo deste uacuteltimo (Stryer)
O gel de resoluccedilatildeo com 10 de acrilamida foi obtido atraveacutes da adiccedilatildeo de 167 mL de soluccedilatildeo
stock de acrilamida11
075 mL de tampatildeo de resoluccedilatildeo12
251 mL de aacutegua destilada e 50 L de
SDS 10 Para promover a polimerizaccedilatildeo adicionou-se em seguida 38 L de PSA 10
11
soluccedilatildeo stock de acrilamida soluccedilatildeo de 30 (mv) de acrilamida e 08 (mv) de NNrsquo-metil-bis-acrilamida 12
tampatildeo de resoluccedilatildeo 15 M Tris-HCl pH 88
44
(persulfato de amoacutenia) 10 e 25 L de TEMED 99 (tetrametiletilenodiamina) Agitou-se
cuidadosamente de modo a natildeo formar bolhas e pipetou-se a soluccedilatildeo para a cassette de vidro
previamente montada no suporte De seguida adicionou-se uma peliacutecula de cerca de 2 mm de 2-
metil-2-butanol para verificar posteriormente a polimerizaccedilatildeo Apoacutes a polimerizaccedilatildeo que demora
cerca de 45 minutos verteu-se a agrave superfiacutecie e adicionou-se o gel de concentraccedilatildeo com 5 de
acrilamida constituiacutedo por 03 mL de soluccedilatildeo de stock acrilamida 045 mL de tampatildeo de
concentraccedilatildeo13
094 mL de aacutegua destilada 18 L de SDS 10 135 L de PSA 10 e 2L de
TEMED 99 Colocou-se o pente sobre o gel e deixou-se polimerizar durante cerca de 45
minutos
As amostras para aplicaccedilatildeo no gel de electroforese foram preparadas para uma concentraccedilatildeo de
proteiacutena de 1 mgmL num volume final de 100 L dos quais 25 L correspondem ao tampatildeo da
amostra14
As amostras foram aquecidas a 100ordmC durante 5 minutos Antes da aplicaccedilatildeo das
amostras transferiu-se a cassette para a tina de electroforese agrave qual se adicionou a soluccedilatildeo
tampatildeo de electroforese da tina superior15
e tampatildeo de electroforese da tina inferior16
Aplicaram-
se 10 L das soluccedilotildees de marcadores de peso molecular e 20 L das amostras proteicas
preparadas anteriormente A electroforese decorreu durante cerca de 3h30 min a uma voltagem
de 80 V De seguida transferiu-se o gel para uma tina corando as proteiacutenas durante 1h com
soluccedilatildeo corante17
Descorou-se o gel com soluccedilatildeo descorante18
tendo-se mudado esta soluccedilatildeo ateacute
as bandas se encontrarem bem definidas
Determinou-se a mobilidade relativa das bandas correspondentes aos marcadores de elevado peso
molecular e traccedilou-se a recta de calibraccedilatildeo ndash log(PM) vs log(Rf) onde Rf corresponde agrave
mobilidade relativa Por interpolaccedilatildeo determina-se o PM das proteiacutenas isoladas sabendo a sua
mobilidade relativa Por comparaccedilatildeo com os valores descritos na literatura pode concluir-se
acerca do grau de purificaccedilatildeo
34 Mediccedilatildeo da actividade enzimaacutetica de S1
341 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
13
tampatildeo de concentraccedilatildeo 05 M Tris-HCl pH 68 14
tampatildeo de amostra 320 mM Tris-HCl pH 68 04 M szlig-mercaptoetanol SDS 8 glicerol 15 (vv) e roxo de m-
cresol 0024 15
tampatildeo de electroforese da tina superior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS 16
tampatildeo de electroforese da tina inferior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS acetato de soacutedio
01 M 17
soluccedilatildeo corante ldquoCoomassie Blue R-250rdquo contendo metanol 45 (vv) e aacutecido aceacutetico glacial 10 (vv) 18
soluccedilatildeo descorante metanol 10 (vv) e aacutecido aceacutetico glacial 10 vv)
45
O meacutetodo de Fiske-Subarow modificado consiste na reacccedilatildeo do molibdato de amoacutenio com o
fosfato inorgacircnico em soluccedilatildeo aacutecida com formaccedilatildeo do aacutecido fosfomoliacutebdico Seguidamente daacute-
se a adiccedilatildeo de um agente redutor que provoca a reduccedilatildeo do composto a fosfomolibdato que forma
um complexo de cor azulada cuja absorvacircncia maacutexima ocorre para um comprimento de onda de
660 nm tornando possiacutevel a sua detecccedilatildeo por meacutetodos espectrofotomeacutetricos As actividades Ca2+
-
ATPaacutesica e K+EDTA-ATPaacutesica de S1 foram determinadas atraveacutes deste meacutetodo de acordo com
o descrito por Taussky e Shorr (1953) com mediccedilatildeo da quantidade de fosfato (Pi) libertado
atraveacutes da hidroacutelise de ATP
Para tal obteve-se primeiramente a recta de calibraccedilatildeo com as amostras presentes na Tabela 3II
utilizando uma soluccedilatildeo stock de 5 mM KH2PO4
Tabela 3II ndash Composiccedilatildeo das diferentes amostras utilizadas para determinar a recta de calibraccedilatildeo atraveacutes do meacutetodo
de Fiske-Subbarow modificado
KH2PO4 (L)
([KH2PO4]stock= 5mM)
PO43-
(nmole) H2O (uL)
0 0 1780
5 25 1775
10 50 1770
20 100 1760
40 200 1740
100 500 1680
A cada tubo de Eppendorf adicionaram-se seguidamente 200 L de soluccedilatildeo J19
e 20 L de agente
redutor20
homogeneizando no voacutertex apoacutes cada adiccedilatildeo Incubou-se durante 10 minutos e mediu-
se a Abs660 nm num espectrofotoacutemetro com intervalos de tempo iguais entre cada amostra Traccedilou-
se a recta de calibraccedilatildeo (Abs660 nmvs nmol PO43-
) a partir dos valores obtidos
Para determinar a actividade Ca2+
-ATPaacutesica de S1 prepararam-se tubos de Eppendorf com 400
L de meio de reacccedilatildeo21
25 L CaCl2 ([CaCl2]stock= 100 mM) 25 L ATP ([ATP]stock= 100 mM)
com [S1]ensaios=01 mgmL Perfez-se se necessaacuterio com H2O MilliQ ateacute perfazer um volume de 1
mL Para determinar a actividade K+EDTA-ATPaacutesica utilizou-se uma concentraccedilatildeo idecircntica de
19
Soluccedilatildeo J 5 N H2SO4 25 molibdato de amoacutenio 20
Agente redutor 5 mg aacutecido 1-amino-2-naftol-4-sulfoacutenico 30 mg bissulfito de soacutedio 30 mg sulfito de soacutedio em 26
mL H2O 21
meio de reacccedilatildeo 125 mM Tris-HCl pH 80 0575 M KCl
46
proteiacutena nos ensaios (01 mgmL) e o mesmo meio de reacccedilatildeo Aos tubos de Eppendorf
adicionou-se ainda 123 L de KCl ([KCl]stock = 3M) 25 L de ATP 10 L de EDTA
([EDTA]stock= 100 mM )e o volume de H2O MilliQ necessaacuterio para perfazer 1 mL Em ambas as
situaccedilotildees o uacuteltimo reagente adicionado foi o ATP uma vez que se trata do substrato da reacccedilatildeo
enzimaacutetica
A cineacutetica decorreu a 25ordmC durante 5 minutos para a actividade Ca2+
-ATPaacutesica e durante 3
minutos para a actividade K+EDTA-ATPaacutesica Apoacutes o periacuteodo de incubaccedilatildeo transferiu-se 150
L de amostra para tubos de Eppendorf contendo igual volume de TCA 10 Colocou-se no gelo
e homogeneizou-se no voacutertex Seguidamente centrifugou-se a 1500 g durante 10 minutos e
recolheu-se 200 L de sobrenadante para novos tubos de Eppendorf Adicionou-se a estes 158
mL de H2O MilliQ e 200 L de soluccedilatildeo J Homogeneizou-se no voacutertex e adicionou-se 20 L de
reagente redutor repetindo-se a homogeneizaccedilatildeo Incubou-se durante 10 minutos e mediu-se a
Abs660 nm com o mesmo intervalo de tempo de leitura para cada amostra Por intrapolaccedilatildeo a partir
da recta de calibraccedilatildeo obtida anteriormente determinou-se a quantidade de Pi libertado em cada
ensaio A partir destes resultados determinaram-se as actividades Ca2+
-ATPaacutesica e K+EDTA-
ATPaacutesica do S1 em nmol PO43-
minmg proteiacutena e em s-1
342 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
Da actividade Mg2+
-ATPaacutesica de S1 resulta a formaccedilatildeo dos produtos da reacccedilatildeo de
hidroacutelise do ATP - ADP e Pi Na presenccedila de ADP fosfoenolpiruvato (PEP) e da enzima piruvato
cinase (PK) os produtos resultantes satildeo ATP e piruvato Por acccedilatildeo da enzima lactato
desidrogenase (LDH) este uacuteltimo eacute convertido em lactato ocorrendo simultaneamente a oxidaccedilatildeo
do NADH a NAD+ monitorizada atraveacutes da Abs340 nm Quando o S1 de miosina se encontra no
meio reaccional o ATP eacute hidrolisado formando-se novamente ADP e Pi o que permite a
formaccedilatildeo de um ciclo (fig 21) Se as enzimas se encontrarem em excesso a velocidade de
conversatildeo do NADH eacute limitada pela produccedilatildeo de ADP e Pi Como o ATP eacute regenerado em cada
ciclo a sua concentraccedilatildeo inicial eacute sempre mantida (Bagshaw 1982)
Figura 33ndash Representaccedilatildeo esquemaacutetica do meacutetodo de enzimas acopladas
47
O protocolo utilizado e seguidamente descrito foi adaptado de Cuenda et al (1990 ndash Ver Refordf) A
uma cuvette de 1 mL adicionou-se S1 para uma concentraccedilatildeo final de 005 mgmL 800 L de
meio de reacccedilatildeo22
10 L de PEP ([PEP]stock = 42 mM) 10 L de PK+LDH 11 ([PK+LDH]stock =
1 mgmL) e 15 L de NADH ([NADH]stock=25 mM) Iniciou-se a monitorizaccedilatildeo da reacccedilatildeo
atraveacutes da leitura de Abs340 nm apoacutes a adiccedilatildeo de 20 L de ATP ([ATP]stock=100 mM) A
actividade Mg2+
-ATPaacutesica eacute obtida atraveacutes do declive correspondente agrave oxidaccedilatildeo do NADH Para
a mediccedilatildeo da Mg2+
-ATPase de S1 estimulada por actina adicionou-se F-actina para uma
concentraccedilatildeo final de 2 microM sendo a actividade obtida tambeacutem atraveacutes do declive correspondente
agrave oxidaccedilatildeo do NADH apoacutes a adiccedilatildeo da proteiacutena
35 Interaccedilatildeo entre vanaacutedio e subfragmento-1 de miosina
351 Preparaccedilatildeo das soluccedilotildees de vanaacutedio
As soluccedilotildees de vanadato foram preparadas em aacutegua MilliQ a partir de metavanadato de amoacutenio
(NH4VO3) numa concentraccedilatildeo stock de 50 mM com agitaccedilatildeo magneacutetica e aquecimento (~30 ordmC)
Esta soluccedilatildeo com um valor compreendido entre 6 e 7 designa-se por ldquometavanadatordquo e eacute
constituiacuteda pelas esspeacutecies V1 V2 V4 e V5 (Ref) Acidificando a soluccedilatildeo de metavanadato por
adiccedilatildeo de HCl ateacute um valor de pH proacuteximo de 4 observa-se uma mudanccedila de cor de amarelo
claro para laranja indicativa da formaccedilatildeo da espeacutecie decameacuterica de vanadato De seguida e
imediatamente antes da realizaccedilatildeo dos ensaios descritos o valor do pH da soluccedilatildeo de
decavanadato foi aumentado para cerca de 75 por adiccedilatildeo de NaOH A soluccedilatildeo (2) designada
por ldquometavanadatordquo natildeo foi posteriormente alterada ateacute agrave sua adiccedilatildeo para a realizaccedilatildeo dos ensaios
As soluccedilotildees foram armazenadas a 4ordmC e mantidas em gelo aquando da sua utilizaccedilatildeo
A soluccedilatildeo de V(IV) foi preparada em condiccedilotildees anaeroacutebias por diluiccedilatildeo de sulfato de vanadil
(VOSO4) para uma concentraccedilatildeo final de 5 mM em H2O MilliQ saturada com N2 gasoso
352Anaacutelise por espectroscopia de UV-Vis
A concentraccedilatildeo da espeacutecie decameacuterica em soluccedilatildeo depende do periacuteodo de tempo apoacutes a diluiccedilatildeo
no meio de reacccedilatildeo apropriado (Ref) A velocidade de desoligomerizaccedilatildeo da espeacutecie decameacuterica
pode ser analisada por espectroscopia de absorccedilatildeo na regiatildeo UVVis uma vez que esta espeacutecie
apresenta duas bandas caracteriacutesticas na regiatildeo do visiacutevel (a 358 e 400 nm) que natildeo se verifica
em quaisquer outras espeacutecies oligomeacutericas As bandas supramencionada sapresentam-se bem
22
Meio de reacccedilatildeo 3125 mM Tris-HCl pH 71 375 mM MgCl2
48
resolvidas atraveacutes da obtenccedilatildeo da derivada de 2ordf ordem do espectro apresentando a forma de 2
vales com miacutenimos centrados nos comprimentos de onda supracitados com dependecircncia linear
relativamente agrave concentraccedilatildeo de decavanadato total (Ramos et al 2006)
Assim a estabilidade da espeacutecie decameacuterica apoacutes diluiccedilatildeo da soluccedilatildeo de decavanadato no meio de
reacccedilatildeo dos ensaios (tampatildeo D) para uma concentraccedilatildeo final de 1 mM em vanadato total foi
caracterizada atraveacutes da leitura da Abs400 em funccedilatildeo do tempo tendo-se determinado qual a
ordem de reacccedilatildeo e o tempo de meia-vida da mesma Com base em estudos anteriores acerca da
estabilidade do V10 na presenccedila de diferentes proteiacutenas (Ramos et al 2006) determinou-se
igualmente o tempo de meia vida apoacutes a adiccedilatildeo de 20 microM de S1 de miosina ao meio reaccional
353 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
Os espectros de RMN do isoacutetopo 51
V foram realizados num espectroacutemetro Bruker AM-400 MHz
equipado com uma sonda multinuclear inversa de 5 mm Em tubos de RMN de 5 mm de diacircmetro
preparou-se soluccedilotildees de 500 L de deca- ou metavanadato correspondentes a um intervalo de
concentraccedilotildees de 1 a 10 mM suplementadas com 10 D2O Os espectros foram adquiridos agrave
temperatura ambiente com uma largura espectral de 23 923 Hz um tempo de aquisiccedilatildeo de 0086
s tempo de relaxaccedilatildeo de 001 s e uma sequecircncia de pulsos a 90ordm Utilizou-se a transformada de
Fourier com alargamento das bandas de 20Hz para o processamento dos espectros Os desvios
quiacutemicos das ressonacircncias de 51
V foram obtidos em relaccedilatildeo agrave espeacutecie VOCl3cuja frequecircncia de
ressonacircncia eacute considerada o padratildeo sendo todas as outras referenciadas a partir desta em ppm
As aacutereas dos sinais do RMN correspondentes agraves espeacutecies oligomeacutericas presentes foram obtidas
por integraccedilatildeo dos sinais As concentraccedilotildees relativas das espeacutecies foram calculadas com recurso agrave
equaccedilatildeo
onde A[Vx] corresponde agrave aacuterea referente agrave ressonacircncia da espeacutecie Vx com nordm de agregaccedilatildeo x
A[V]total agrave soma total das aacutereas medidas e [V] a concentraccedilatildeo total de vanaacutedio no ensaio
A teacutecnica de 51
V-RMN foi anteriormente aplicada em estudos de ligaccedilatildeo com S1 de miosina
(Ref) tendo sido demostrado que entre as espeacutecies V1 V2 V4 e V5 a espeacutecie tetrameacuterica possui a
maior afinidade para a miosina (Cremo et al) O desaparecimento ou alargamento dos sinais pode
implicar a ligaccedilatildeo de um dos oxovanadatos laacutebeis agrave miosina sem indicaccedilatildeo de qual uma vez que
49
se processa um equiliacutebrio entre eles (Crans 1990) Assim e tendo em conta as indicaccedilotildees de que
a espeacutecie V10 eacute distinta das restantes no que diz respeito agrave magnitude da interacccedilatildeo com a miosina
e inibiccedilatildeo da actividade ATPaacutesica estimulada por actina (Aureliano 2000 Tiago et al 2002)
torna-se fundamental investigar a interacccedilatildeo das diferentes espeacutecies de vanadato com o S1 de
miosina atraveacutes da teacutecnica de 51
V-RMN
Para tal obtiveram-se espectros de 51
V-RMN resultantes da titulaccedilatildeo de uma soluccedilatildeo 5 mM de
decavanadato total com concentraccedilotildees crescentes de S1 de miosina (0 5 10 15 e 20 microM) em
tampatildeo D Realizou-se ainda ensaios na presenccedila de 2 mM de decavanadato e 2 mM de ATP na
presenccedila e na ausecircncia de S1 de miosina de modo a verificar o efeito que estes ligandos naturais
da enzima teria na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o subfragmento-1
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
Os grupos sulfidrilo (-SH) das cisteiacutenas satildeo resiacuteduos de aminoaacutecidos extremamente sensiacuteveis agrave
oxidaccedilatildeo resultando na sua oxidaccedilatildeo (Dalle-Donne et al 1999) Logo a quantificaccedilatildeo destes
resiacuteduos eacute bastante importante nos estudos de interacccedilatildeo com vanaacutedio jaacute que os resiacuteduos de Cys
com ecircnfase para o SH1 e o SH2 satildeo bastante importantes na monitorizaccedilatildeo das modificaccedilotildees
funcionais da regiatildeo da cabeccedila
O procedimento seguidamente apresentado foi originalmente descrito por Gutieacuterrez-Merino
(1987)) Consiste na reacccedilatildeo entre os grupos ndashSH natildeo oxidados das proteiacutenas e o reagente de
Ellman (DTNB -55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico)) - formando-se o aniatildeo CNT (aniatildeo 3-
carboxilato-4-nitrotiofenolato) numa estequiometria de 11 Este apresenta um maacuteximo de
absorvecircncia a 412 nm (Robyt 1990) Assim apoacutes a incubaccedilatildeo da proteiacutena na presenccedila do metal
a quantidade de cisteiacutenas reduzidas por proteiacutena determina-se atraveacutes dos valores de Abs412
Deste modo colocou-se num tubo de Eppendorf o volume apropriado de S1 dialisado para que a
concentraccedilatildeo no ensaio fosse de 1 mgmL Adicionou-se o volume apropriado de decavanadato
para que a sua concentraccedilatildeo fosse de 0 10 50 100 250 500 ou 1000 M Perfez-se com soluccedilatildeo
D ateacute um volume final de 1 mL e incubou-se a 25ordmC por 20 minutos Apoacutes este periacuteodo colocou-
se 780 L da mistura reaccional numa cuvette de quartzo e adicionou-se 220 L de soluccedilatildeo K23
23
Tampatildeo K Tampatildeo fosfatohidrogenofosfato de soacutedio 025 M pH 72
50
Adicionou-se 15 L de DTNB (em soluccedilatildeo K [DTNB]stock=100 mM) ao meio reaccional e
registou-se o valor de absorvecircncia medido equivalente agraves cisteiacutenas ldquoraacutepidasrdquo (SH1 e SH2
expostos ao solvente) Adicionou-se 001 g de SDS que provoca a desnaturaccedilatildeo da proteiacutena e
consequentemente a exposiccedilatildeo de todos os resiacuteduos de cisteiacutenas que se encontravam ateacute aiacute
inacessiacuteveis no interior da proteiacutena (cisteiacutenas totais) Estes resiacuteduos podem assim reagir com o
DTNB
Estimou-se o nuacutemero de resiacuteduos de cisteiacutenas em todas as amostras com recurso ao coeficiente de
extinccedilatildeo molar () do DTNB (ver ponto 3 dos Anexos) que foi determinado atraveacutes da titulaccedilatildeo
de L-Cisteiacutena com intervalo de concentraccedilotildees compreendidos entre 0 e 50 microM na presenccedila de
25microM de DTNB em tampatildeo D Os valores de absorvecircncia foram obtidos para um comprimento de
onda de 412 nm
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
3621 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
A teacutecnica mais eficiente de anaacutelise da interacccedilatildeo entre o iatildeo vanadilo e proteiacutenas como a
calmodulina e a transferrina entre outras permite a obtenccedilatildeo raacutepida de informaccedilatildeo no que toca agrave
conformaccedilatildeo e amplitude da referida interacccedilatildeo
Os espectros de Ressonacircncia Paramagneacutetica Electroacutenica (RPE) da espeacutecie paramagneacutetica V(IV)
foram realizados a 70K num espectroacutemetro Bruker EMX-300 X-band Todas as soluccedilotildees
utilizadas e amostras foram preparadas em anaerobiose numa cacircmara de luvas de modo a evitar a
reduccedilatildeo de V(IV) a V(V) Em tubos capilares com 02 mL de amostra manteve-se a concentraccedilatildeo
de S1 a 20 M e variou-se a concentraccedilatildeo de vanadilo (0 25 5 10 15 25 50 75 100 150
200 500 e 1000 M [VOSO4]stock=5 mM) em tampatildeo D Apoacutes obtenccedilatildeo dos espectros estes
foram tratados atraveacutes do software SimFonia e WinEPR tendo-se obtido um graacutefico com a
variaccedilatildeo da intensidade do sinal de RPE em funccedilatildeo da concentraccedilatildeo de sulfato de vanadilo
presente Realizaram-se ainda simulaccedilotildees de espectros obtidos com 20 microM de S1 e 1000 microM de
V(IV) com vista agrave obtenccedilatildeo dos ligandos provaacuteveis do vanadilo atraveacutes da anaacutelise da constante
de acoplamento hiperfina (A) e do factor g (g)
4 Conclusotildees gerais
Apesar de jaacute ter sido demonstrado que o vanaacutedio inibe a Na+K
+-ATPase da miosina a funccedilatildeo
deste elemento no tecido muacutesculo esqueleacutetico continua largamente incompreendida A enorme
complexidade quiacutemica deste metal especialmente no que concerne agrave oligomerizaccedilatildeo e reacccedilotildees
51
redox constitui um obstaacuteculo agrave compreensatildeo do comportamento do vanaacutedio nos organismos
vivos Assim a caracterizaccedilatildeo da quiacutemica deste elemento eacute de grande utilidade antes de iniciar o
estudo da sua interacccedilatildeo com o presente sistema
A anaacutelise por espectroscopias de 51
V-RMN e de UV-Vis de soluccedilotildees de vanadato diluiacutedas nos
meios de reacccedilatildeo utilizados no presente relatoacuterio (10 mM HEPES pH 70 25 mM KCl) mostrou
claramente que a espeacutecie decameacuterica (V10) possui um tempo de vida suficientemente longo para a
determinaccedilatildeo do seu efeito isolado ao contraacuterio do que se verifica com as espeacutecies mono- di-
tetra- e pentameacutericas (V1 V2 V4 e V5) Por outro lado a anaacutelise verificou ainda que apoacutes diluiccedilatildeo
das soluccedilotildees de decavanadato no meio de reacccedilatildeo coexistem as espeacutecies V10 e V1 pelo que os
efeitos putativos desta uacuteltima deveratildeo ser subtraiacutedos aquando da monitorizaccedilatildeo dos efeitos da
espeacutecie decameacuterica
O vanaacutedio (V) (vanadato) inibe as enzimas que catalizam reacccedilotildees de transferecircncia de grupos
fosfato como a ATPase da miosina [Goodno 1979] devido agrave sua analogia estrutural com o iatildeo
fosfato Na sua utilizaccedilatildeo como inibidor enzimaacutetico natildeo eacute tida em conta a contribuiccedilatildeo individual
das espeacutecies oligomeacutericas considerando-se normalmente a espeacutecie V1 como inibidora efectiva
[Goodno e Taylor 1982] No presente relatoacuterio verificou-se por espectroscopia de 51
V-RMN que
outros oligoacutemeros de vanadato interagem com o S1 de miosina tendo as espeacutecies V10 e V4 os
maiores niacuteveis de afinidade A sua interacccedilatildeo envolve forccedilas electrostaacuteticas e eacute afectada
negativamente por ligandos naturais de enzima como eacute o caso do ATP
A exposiccedilatildeo do S1 de miosina (005 mgmL) a decavanadato conduz agrave inibiccedilatildeo do capacidade da
proteiacutena de estimular a actividade Mg2+
-ATPaacutesica com um valor de IC50 de 266 plusmn 07 M
decavanadato sendo o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM
decavanadato (80) Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM decavanadato sendo
o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM decavanadato (60)
exercendo um efeito de impedimento agrave ligaccedilatildeo entre o vanaacutedio e o S1
A uma concentraccedilatildeo de 27 microM decavanadato verifica-se a oxidaccedilatildeo de um dos resiacuteduos de Cys
de S1 de miosina seguida pela oxidaccedilatildeo gradual de outro resiacuteduo de Cys ateacute agrave concentraccedilatildeo de
1000 microM Ambos os resiacuteduos oxidados natildeo se encontravam expostos ao solvente utilizado A
oxidaccedilatildeo dos resiacuteduos permite sugerir que esta modificaccedilatildeo oxidativa sofrida pelo S1 contribui
para a diminuiccedilatildeo da sua actividade de ATPase
52
Por espectroscopia de RPE revelaram-se os possiacuteveis ligandos equatoriais do vanaacutedio aquando da
sua interacccedilatildeo com o S1 de miosina como sendo duas moleacuteculas de aacutegua um ligando contendo o
grupo R-OH e outro contendo o grupo R=NR
Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados de
oxidaccedilatildeo (IV) e (V) no S1 de miosina para concentraccedilotildees facilmente alcanccedilaacuteveis no ambiente
terrestre
53
5 Estudos futuros
No sentido do melhor esclarecimento das alteraccedilotildees moleculares e estruturais que ocorrem no
subfragmento 1 quando exposta ao vanaacutedio que conduzem a uma diminuiccedilatildeo da sua funccedilatildeo
sugere-se
- Quantificaccedilatildeo das cisteiacutenas reduzidas de S1 na presenccedila de metavanadato
- Determinaccedilatildeo das actividades Ca2+
- e K+-ATPaacutesica do S1 de miosina na presenccedila de
decavanadato
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina estimulada por actina
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina na presenccedila de
metavanadato
- Realizaccedilatildeo de RMN 2D com variaccedilatildeo das concentraccedilotildees das soluccedilotildees de decavanadato e
metavanadato na presenccedila de S1
-Mutageacutenese dos resiacuteduos de Cys doS1 de miosina de modo a verificar quais os resiacuteduos que
sofrem oxidaccedilatildeo na presenccedila de decavanadato
- Determinaccedilatildeo por calorimetria (DSC Differential Scanning Calorimetry) da variaccedilatildeo da
entalpia (∆H) e da temperatura de desnaturaccedilatildeo do S1 de miosina na presenccedila de vanaacutedio uma
vez que estas propriedades caracteriacutesticas de cada proteiacutena satildeo alteradas apoacutes ocorrecircncia de
modificaccedilotildees estruturais
- Anaacutelise por SDS-PAGE de S1 de miosina apoacutes tratamento com vanaacutedio de modo a determinar
quais os polipeacuteptidos eventualmente formados apoacutes clivagem da proteiacutena
54
6 Referecircncias bibliograacuteficas
- Sellers J Myosins 2nd Edition Oxford University Press 1999
- L Stryer inBiochemistry (Ed WH Freeman 2005 pp
Refordfas Materiais e Meacutetodos
[A] S S Margossian SLowey Methods Enzymol1982 85 55-71
[B] JD Pardue JA Spudich Methods Enzymol 1982 85 164-181
[C] M Bradford Anal Biochem1976 72 248-254
[D] H H Taussky E Shorr J Biol Chem 1953 202 675-685
[E] C R Bagshaw in Outline Studies in Biology ndash Muscle Contraction Chapman and Hall Ed
1982
[F] A Cuenda et al Eur J Biochem1990 194 663-670
55
Anexos
1 Determinaccedilatildeo da recta de calibraccedilatildeo meacutetodo de Bradford
Figura1ndashRepresentaccedilatildeo graacutefica da recta de calibraccedilatildeo (Abs595 nm vs [BSA]) utilizada para o meacutetodo de Bradford
2 Determinaccedilatildeo da recta calibraccedilatildeo para a electroforese em condiccedilotildees desnaturantes
Figura 2 ndash Representaccedilatildeo graacutefica da recta de calibraccedilatildeo (log(Peso Molecular) vs Rf dos marcadores de peso
molecular) onde Rf indica determinada atraveacutes do padratildeo de peso molecular
(poccedilo (a))
3Caacutelculo do coeficiente de extinccedilatildeo molar () para a quantificaccedilatildeo de cisteiacutenas reduzidas
atraveacutes do meacutetodo de Ellman
Traccedilou-se a recta de Abs412 nm meacutedia vs [L-Cisteiacutena] de modo a determinar o valor do coeficiente
de extinccedilatildeo molar definido como o declive daquela recta Assim =8400 M-1
cm-1
como
podemos verificar na Fig 3
A595 nm = 00546[BSA] + 00209 Rsup2 = 099051
Ab
s 59
5 n
m
[BSA] (mgmL)
log PM = -1177 Rf + 2247 Rsup2 = 098957
log
PM
Mobilidade relativa (Rf)
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero

8
ldquoisotropiardquo ndash polarizaccedilatildeo uniforme em todas as direcccedilotildees) enquanto as zonas escuras constituem
as bandas A (de ldquoanisotropiardquo ndash polariza a luz visiacutevel) A banda I encontra-se dividida
transversalmente por uma linha escura ndash linha Z de densidade electroacutenica e refracccedilatildeo superiores
No interior de cada banda A existe uma zona transversal menos densa ndash zona H ndash rica em enzimas
importantes para o metabolismo celular No centro desta zona existe uma linha perpendicular
mais escura ndash linha M [2]
A unidade fundamental da contracccedilatildeo muscular eacute designada por sarcoacutemero (Fig 12) Encontra-se
repetida de forma regular ao longo das miofibrilas apresentando um comprimento de 25 a 3 m
aquando do relaxamento muscular diminuindo aquando da contracccedilatildeo muscular
Figura 12 ndash Representaccedilatildeo de fotomicrografia electroacutenica e esquemaacutetica do sarcoacutemero Os filamentos grossos e
finos satildeo maioritariamente constituiacutedos por miosina e actina respectivamente (Adaptado de Stryer 2002 [2])
As secccedilotildees de corte do sarcoacutemero datildeo conta da presenccedila de dois tipos de filamentos proteicos ndash os
filamentos grossos cujo diacircmetro eacute de 15 nm constituiacutedos sobretudo por miosina e os filamentos
finos com um diacircmetro de 9 nm constituiacutedos sobretudo por actina tropomiosina e pelo complexo
de troponina [2]
111 Miosina
Actualmente o termo miosina refere-se a uma superfamiacutelia diversa de motores moleculares
capazes de hidrolisar ATP e de se ligarem reversivelmente agrave actina A classificaccedilatildeo da
superfamiacutelia das miosinas nas 15 classes distintas faz-se atraveacutes de homologia entre as diferentes
sequecircncias [A] Eacute tambeacutem possiacutevel a classificaccedilatildeo por subclasses das quais se destaca a subclasse
4 proveniente do muacutesculo-esqueleacutetico e cardiacuteaco de vertebrados Este estudo focar-se-aacute apenas
nas miosinas de classe II (miosina-II) presente nos filamentos grossos do muacutesculo estriado
esqueleacutetico [B] e em ceacutelulas natildeo musculares A funccedilatildeo da miosina-II depende da sua localizaccedilatildeo
celular e tecidular
9
A miosina-II eacute constituiacuteda por duas cadeias polipeptiacutedicas idecircnticas denominadas cadeias pesadas
de miosina (HC) enroladas entre si e cujo peso molecular (PM) estaacute compreendido entre 171 e
241 kDa) Estruturalmente as HC dividem-se em 3 domiacutenios funcionais - cabeccedila pescoccedilo e
cauda como se pode observar na Fig 13 Agraves cabeccedilas ou zonas globulares estatildeo associados dois
pares de cadeias leves (peso molecular entre 15 e 30 kDa) que se dividem em dois tipos
essencial (ELC) e regulatoacuterio (RLC)
Figura 13 ndash Estrutura esquemaacutetica da moleacutecula de miosina-II Destacam-se as duas cabeccedilas globulares e a cauda
filamentosa ligadas entre si por uma zona flexiacutevel denominada pescoccedilo As cadeias leves (a verde e vermelho)
encontram-se associadas agraves cabeccedilas globulares
As cabeccedilas globulares contendo os locais de ligaccedilatildeo agrave actina e aos nucleoacutetidos mantecircm os seus
elementos estruturais baacutesicos conservados em todas as classes sendo estes constituiacutedos por 720-
80 aa A regiatildeo do pescoccedilo eacute constituiacuteda por uma longa heacutelice pertencente agrave cadeia pesada
que se encontra estabilizada atraveacutes da ligaccedilatildeo a subunidades de cadeia leve A estrutura do
pescoccedilo envolvida na ligaccedilatildeo agraves cadeias leves eacute denominada de motivo IQ cuja estrutura
consensual eacute IQXXXRGXXXR1 O pescoccedilo serve como extensatildeo do comprimento da cabeccedila de
modo a actuar como alavanca durante a contracccedilatildeo muscular [A]A regiatildeo da cauda ou acircncora
apresenta bastante divergecircncia no que respeita agrave sua sequecircncia estrutura e funccedilatildeo No entanto
pensa-se que a sua funccedilatildeo centrar-se-aacute na autoassociaccedilatildeo das vaacuterias moleacuteculas de miosina-II
atraveacutes da regiatildeo de coiled-coil Esta confere assimetria agrave moleacutecula e permite a formaccedilatildeo de
filamentos grossos capazes de translocar filamentos de actina [A]
Na regiatildeo da cabeccedila da miosina existem 2 regiotildees extremamente sensiacuteveis agrave proteoacutelise A
clivagem em ambos os locais resulta na produccedilatildeo de trecircs fragmentos um compreendendo o N-
terminal (fragmento 25 kDa) outro a regiatildeo central (50 kDa) e um terceiro o C-terminal (20 kDa)
(Fig 14) Estes fragmentos dissociam-se em condiccedilotildees desnaturantes e natildeo constituem domiacutenios
independentes da cabeccedila mas provavelmente loops flexiacuteveis uma vez que natildeo foram observadas
na estrutura cristalina Os vaacuterios fragmentos obtidos em diferentes condiccedilotildees foram cruciais para
a elucidaccedilatildeo das vaacuterias funccedilotildees da moleacutecula
A regiatildeo compreendida entre os fragmentos 25 e 50 kDa (loop 1) localiza-se directamente por
1 I = Isoleucina Q= Glutamina X = qualquer dos 20 aminoaacutecidos padratildeo R= Arginina G= Glicina
10
cima do local de ligaccedilatildeo ao nucleoacutetido A regiatildeo entre os fragmentos de 50 e 20 kDa (loop 2)
encontra-se perto da intersecccedilatildeo entre os domiacutenios da cabeccedila e do pescoccedilo e deveraacute interagir com
a actina uma vez que a ligaccedilatildeo desta a protege da proteoacutelise Na ausecircncia do loop 2 a miosina
liga mais fracamente agrave actina do que no caso da miosina intacta A sequecircncia primaacuteria de ambos
os loops apresenta conservaccedilatildeo reduzida sendo provaacutevel que sejam responsaacuteveis por muita da
diversidade das diferentes propriedades enzimaacuteticas presentes em diferentes isoformas
No fragmento de 20 kDa existem dois grupos sulfidrilo Cys-707 e -697 usualmente
denominados de SH1 e SH2 devido agrave ordem da sua reactividade quiacutemica Na presenccedila de
nucleoacutetidos pode ocorrer o ldquocross-linkingrdquo entre estes dois grupos incluindo a oxidaccedilatildeo a uma
ponte dissulfureto
Devido agrave insolubilidade da miosina-II nativa a estrateacutegia adoptada para os estudos com esta
proteiacutena eacute frequentemente a sua clivagem em fragmentos funcionais separaacuteveis Um dos locais de
clivagem ocorre tipicamente a 130 kDa do N-terminal produzindo a meromiosina pesada (HMM)
e a meromiosina leve (LMM) [C] A HMM eacute formada pelas 2 cabeccedilas e pescoccedilo e apresenta
propriedades semelhantes agrave miosina sem digestatildeo (ligaccedilatildeo e translocaccedilatildeo de actina e hidroacutelise de
MgATP)
A HMM pode ser novamente clivada atraveacutes de uma reacccedilatildeo de proteoacutelise produzindo os
subfragmentos 1 (S1) e 2 (S2)O S1 (90 kDa) eacute enzimaticamente activo sendo constituiacutedo pelos
domiacutenios da cabeccedila e do pescoccedilo e apresentando associaccedilatildeo agraves duas cadeias leves ou apenas agrave
ELC (Fig 14)Em alternativa o S1 pode ser produzido por clivagem directa da miosina intacta
formando-se tambeacutem um fragmento em forma de cilindro que consiste nas regiotildees S2 e LMM
(Ref)Utiliza-se extensivamente o S1 para estudos cineacuteticos e de ligaccedilatildeo agrave actina uma vez que a
sua solubilidade simplifica o planeamento da experiecircncia e a subsequente interpretaccedilatildeo dos dados
[A]
Figura 14- Estrutura do subfragmento-1 de miosina de muacutesculo esqueleacutetico com ecircnfase nos domiacutenios de 50 kDa
central (a vermelho) 25 kDa no N-terminal (a verde) e 20 kDa no C-terminal (a azul) As cadeias leves essenciais
encontram-se a amarelo e as cadeias leves regulatoacuterias a lilaacutes Os locais de ligaccedilatildeo agrave actina e ao ATP estatildeo
igualmente destacados (adaptado de Holmes e Geeves 1999 [D])
11
112 Actina
O termo ldquoactinardquo refere-se a uma famiacutelia extremamente conservada de proteiacutenas citoplasmaacuteticas
presente em todos os eucariotas Esta proteiacutena natildeo se encontra nos procariotas [E] A maioria das
actinas possui peso molecular proacuteximo de 42 kDa outra evidecircncia do seu grau elevado de
conservaccedilatildeo no que concerne agrave sequecircncia e ao nuacutemero de aminoaacutecidos Os eucariotas superiores
apresentam um grande nuacutemero de isoformas de actina que por sua vez se encontram codificados
por um ainda maior nuacutemero de genes (Ref) A expressatildeo das isoformas eacute diferencial consoante o
tecido e sua especializaccedilatildeo pelo que deveratildeo possuir funccedilotildees e mecanismos direccionais
diferentes (Ref) Foi ainda observado que os mRNA das diferentes isoformas apresentam uma
distribuiccedilatildeo heterogeacutenea nas ceacutelulas de forma semelhante aos seus produtos proteicos [F] Estes
dados suportam a hipoacutetese supramencionada do direccionamento das isoformas de actina para um
determinado local na ceacutelula de acordo com as suas funccedilotildees
A actina presente no citoesqueleto eacute responsaacutevel por processos como o mecanismo contraacutectil
fagocitose manutenccedilatildeo e alteraccedilatildeo da estrutura celular locomoccedilatildeo transporte de organelos
regulaccedilatildeo do transporte de solutos entre outras Nos eucariotas superiores estaacute ainda envolvida
no transporte de gracircnulos de pigmento retinoacuteide [G] transporte de gracircnulos no axoplasma [H] e
secreccedilatildeo entre outros [E] Actualmente a isoforma da actina (-actina) eacute a melhor estudada
encontrando-se envolvida no mecanismo contraacutectil [E]
Os filamentos de -actina tecircm cerca de 1m de comprimento e 80 Å Consistem em duas cadeias
de actina filamentosa (F-actina) organizados em heacutelice [I] (Holmes et al 1990) sobre os quais
se encontram outras proteiacutenas ndash a troponina (C I e T) e a tropomiosina (Azevedo 1999) (Figura
15) A troponina eacute filamentosa e localiza-se nas depressotildees da heacutelice de F-actina conferindo
rigidez aos filamentos finos Por sua vez a tropomiosina eacute essencial na regulaccedilatildeo do mecanismo
contraacutectil
Figura 15 ndash Modelo estrutural dos filamentos de F-actina contendo a tropomiosina e o complexo de troponina (I C
e T) (Adaptado de Susana Ramos 2005 [J])
12
A F-actina eacute formada atraveacutes da polimerizaccedilatildeo de subunidades globulares de actina (G-actina) A
G-actina eacute composta por 2 domiacutenios de tamanho semelhante separados pelos bolsos de ligaccedilatildeo(1)
de catiotildees divalentes e(2) de nucleoacutetidos [KLM] Estes por sua vez estatildeo subdivididos noutros
dois de menores dimensotildees (1-4) (Fig 16) cada um envolvido nas interacccedilotildees actina-actina ou
na formaccedilatildeo do topo de ligaccedilatildeo do nucleoacutetido As regiotildees N-terminal e C-terminal estatildeo proacuteximas
uma da outra no subdomiacutenio 1 [E]
Figura 16 ndash Estrutura do monoacutemero de actina Encontram-se representados os subdomiacutenios 1 a 4 e a vermelho a
moleacutecula de ATP junto do respectivo local de ligaccedilatildeo (Protein Data Bank ID 1j6z)
113 Mecanismos moleculares da contracccedilatildeo muscular
A contracccedilatildeo muscular ocorre atraveacutes do deslizamento dos filamentos finos (de actina) por entre
os filamentos grossos (de miosina) conseguida pela reacccedilatildeo de hidroacutelise de ATP e regulada pela
tropomiosina e pelo complexo de troponina em resposta a impulsos nervosos Este mecanismo foi
inicialmente proposto por Huxley-Niedergerke [N] e Huxley-Hanson [O] ambos em 1954 A base
experimental de ambos os estudos foi a inexistecircncia de variaccedilatildeo do tamanho dos filamentos
durante a contracccedilatildeo A banda A apresenta sempre as mesmas dimensotildees por outro lado a banda
I a zona H diminuem consideravelmente aquando da contracccedilatildeo (Figs 12 e 17)
Figura 17 ndash Modelo da contracccedilatildeo muscular ndash deslizamento dos filamentos finos (azuis) relativamente aos
filamentos grossos (rosa) (Adaptado de Stryer 2002 [2])
13
Na ausecircncia de ATP e supondo a ausecircncia de inibiccedilatildeo da interacccedilatildeo actina-miosina as proteiacutenas
encontram-se fortemente associadas formando o complexo actomiosina (AM) (Figura 15)
Nestas condiccedilotildees as cabeccedilas de miosina formam um acircngulo de 45ordm com o eixo dos filamentos
finos de actina [P]
Na presenccedila de ATP o complexo AM dissocia-se jaacute que a afinidade do complexo M-ATP para a
actina eacute 105 vezes inferior agrave afinidade da miosina para a actina O ATP liga-se agrave miosina
ocorrendo seguidamente a reacccedilatildeo de hidroacutelise de ATP Os produtos daiacute resultantes ndash ADP e Pi -
permanecem ligados agrave miosina formando o complexo M-ADP-Pi Com a ligaccedilatildeo subsequente agrave
actina forma-se o complexo AM-ADP-Pi no qual as cabeccedilas de miosina apresentam um acircngulo
recto com o eixo do muacutesculo [P] Neste ponto daacute-se a libertaccedilatildeo progressiva de Pi e ADP Eacute
provaacutevel que a energia proveniente da hidroacutelise do ATP provoque uma alteraccedilatildeo do acircngulo de
ligaccedilatildeo das cabeccedilas de miosina ao filamento de actina passando novamente de 90ordm para 45ordm [P]
Esta alteraccedilatildeo conformacional provoca o deslizamento do filamento de actina para o centro do
sarcoacutemero cujo encurtamento promove a contracccedilatildeo das miofibrilas O processo supracitado
ocorre em cerca de 1ms[Q] e encontra-se resumido na Fig 18
Figura 18 ndash Esquema cineacutetico da hidroacutelise de ATP pela miosina (Adaptado de Sellers 1999 [A])
1131 Regulaccedilatildeo da contracccedilatildeo muscular
O mecanismo pelo qual se processa a regulaccedilatildeo muscular foi inicialmente descrito por estudos
distintos de Haselgrove Huxley e Parrey-Squire todos em 1973 [RST] Aleacutem da implicaccedilatildeo
directa da tropomiosina e do complexo de troponina a concentraccedilatildeo de Ca2+
no muacutesculo em
relaxamento eacute bastante reduzida (10-7
M) aumentando notavelmente aquando da contracccedilatildeo (10-5
-
10-3
M)
Resumidamente quando ocorre a excitaccedilatildeo do muacutesculo esqueleacutetico a despolarizaccedilatildeo da
membrana plasmaacutetica eacute difundida ateacute ao interior do tecido atraveacutes dos tuacutebulos transversais
(invaginaccedilotildees da membrana plasmaacutetica da fibra muscular) A despolarizaccedilatildeo provoca a abertura
dos canais ioacutenicos de Ca2+
do retiacuteculo sarcoplasmaacutetico adjacente Como a concentraccedilatildeo de Ca2+
no interior do organelo eacute muito elevada o caacutelcio passa raacutepida e uniformemente para o citosol
14
Assim a regulaccedilatildeo da contracccedilatildeo ocorre com coordenaccedilatildeo e rapidez na totalidade das miofibrilas
do tecido muscular (Ref)
Em repouso a troponina I encontra-se posicionada de tal forma que a proteiacutena adjacente
troponina C impede estruturalmente a ligaccedilatildeo das cabeccedilas da miosina agrave actina (acircngulo =50ordm) O
aumento da concentraccedilatildeo intracelular de Ca2+
provoca um aumento da sua disponibilidade para
ligar agrave troponina C (Fig 19)
Figura 19ndash Regulaccedilatildeo do mecanismo contraacutectil no muacutesculo esqueleacutetico pelo complexo de troponina
Como consequecircncia a troponina C altera a sua conformaccedilatildeo (=70ordm) associando-se agrave troponina I
que deixa de inibir a ligaccedilatildeo das pontes transversais A alteraccedilatildeo anterior eacute transmitida agrave
tropomiosina atraveacutes da troponina T adjacente agrave C o que provoca o deslocamento da
tropomiosina na direcccedilatildeo do interior do sulco do filamento de actina Assim eacute possiacutevel a
interacccedilatildeo eficaz entre actina e miosina A Ca2+
-ATPase do retiacuteculo sarcoplasmaacutetico provoca a
reacumulaccedilatildeo do iatildeo caacutelcio no organelo impedindo o prolongamento desnecessaacuterio da contracccedilatildeo
(Refordf)
12 Vanaacutedio
O vanaacutedio eacute um metal de transiccedilatildeo pertencente ao grupo VB da tabela perioacutedica Foi
primeiramente descoberto em 1801 por Andreacutes Manuel del Riacuteo que concluiu erradamente que era
idecircntico ao croacutemio e definitivamente por Nils Gabriel Svenson em 1831 Recentemente foram
descobertas importantes funccedilotildees bioloacutegicas deste elemento provavelmente essencial para os
humanos de que se destaca no contexto do presente relatoacuterio a possibilidade de actuar como
anaacutelogo do grupo fosfato e a inibiccedilatildeo de enzimas in vitro como eacute o caso da actividade ATPaacutesica
da miosina
Eacute um metal soacutelido agrave temperatura ambiente que se encontra amplamente distribuiacutedo nos vaacuterios
ecossistemas em dois isoacutetopos naturais ndash 50
V e 51
V ndash raramente excedendo 01 partes por biliatildeo
15
(ppb) Estaacute associado a cerca de 70 minerais diferentes na natureza (dos quais se destacam o
uracircnio cobre chumbo zinco fosfato e titacircnio ferroso) especialmente na forma de vanadato ou
pentoacutexidos de vanaacutedio O vanaacutedio eacute um dos principais constituintes da mateacuteria orgacircnica foacutessil
com especial ecircnfase para o carvatildeo oacuteleo de crude e foacutesseis carbonaacuteceos [UV] bem como em
alimentos consumidos pelo Homem (salsa pimenta preta cogumelos marisco vegetais cereais
gorduras e oacuteleos) [W] Estes constituem a maior fonte de exposiccedilatildeo ao vanaacutedio da maioria da
populaccedilatildeo humana [X]
Normalmente a entrada do vanaacutedio no organismo ocorre por inalaccedilatildeo ou em menores
quantidades por absorccedilatildeo pelo tracto gastrointestinal ou pela pele O seu armazenamento centra-
se no fiacutegado rins ceacuterebro coraccedilatildeo muacutesculos e ossos No entanto a retenccedilatildeo de vanaacutedio no
organismo humano eacute reduzida ndash em condiccedilotildees normais cerca de 100 M (em muitos tecidos a
quantidade eacute inferior a 10 ngg de tecido) [Y]
Actualmente natildeo existem provas de deficiecircncia nutricional de vanaacutedio em humanos ou de
enzimas que contenham vanaacutedio em animais superiores No entanto vaacuterios estudos relacionam a
ocorrecircncia de deficiecircncia em galinhas e ratos devido a uma alimentaccedilatildeo pobre em vanaacutedio Os
sintomas relatados incluem retardamento do crescimento malformaccedilotildees oacutesseas perturbaccedilotildees no
metabolismo lipiacutedico e estagnaccedilatildeo reprodutiva entre outros [Z AA AB] Os efeitos fisioloacutegicos
do vanaacutedio podem depender do tipo de composto ou de oligoacutemero uma vez que consoante as
suas estabilidade labilidade e nuacutemero de coordenaccedilatildeo as respostas seratildeo diferenciadas ao niacutevel
dos organismos vivos
Por outro lado a toxicidade do vanaacutedio encontra-se bem caracterizada existindo sintomas
descritos em vaacuterios mamiacuteferos (inclusive o Homem) que incluem diarreias catildeibras irritaccedilatildeo da
pele e mucosas palpitaccedilotildees cardiacuteacas anemia pneumonia bronquite perturbaccedilotildees no Sistema
Nervoso Central (SNC) entre outros (Ref) A toxicidade varia geralmente com a natureza do
composto com aumento proporcional agrave sua valecircncia Verificou-se que a forma de entrada do
vanaacutedio tem tambeacutem efeito na sua toxicidade ndash os maiores niacuteveis verificaram-se por injecccedilatildeo
intravenosa da amostra sendo progressivamente inferiores por inalaccedilatildeo e finalmente por
ingestatildeo [X AA] A ampla utilizaccedilatildeo dos compostos de vanaacutedio na induacutestria quiacutemica bem como
as elevadas quantidades de vanaacutedio libertadas atraveacutes da combustatildeo dos oacuteleos de crude
contribuem consideravelmente para o aumento da biodisponibilidade do vanaacutedio Como tal o
interesse neste elemento e nos problemas ambientais por ele causados tem vindo a aumentar
significativamente [V]
121 Propriedades quiacutemicas
16
A quiacutemica do vanaacutedio eacute extraordinariamente complexa pelo que originou vaacuterias interpretaccedilotildees
incorrectas desde a sua descoberta No entanto as suas propriedades dependem em uacuteltima
anaacutelise da configuraccedilatildeo electroacutenica do elemento Assim a variabilidade encontrada no
comportamento quiacutemico do vanaacutedio pode dever-se agrave sua configuraccedilatildeo ndash [Ar] 3d2 4s
2 ndash que
permite a formaccedilatildeo de vaacuterios estados de oxidaccedilatildeo (de +2 a +5) Em soluccedilatildeo aquosa os estados de
oxidaccedilatildeo estaacuteveis vatildeo de V(V) a V(II) Destes apenas os +3 a +5 satildeo biologicamente importantes
jaacute que o V(II) apresenta demasiada capacidade redutora para existir nos organismos vivos
conhecidos [AC]
No fluido extracelular os compostos de vanaacutedio encontram-se predominantemente na forma de
monovanadatos (V(V)) livres ou ligados agrave transferina do soro ou agrave hemoglobina (Refordf) Os iotildees de
vanadato livres satildeo transportados para o meio intracelular atraveacutes de canais ioacutenicos inespeciacuteficos
Jaacute no ambiente intracelular os compostos sofreratildeo uma bio-reduccedilatildeo atraveacutes da acccedilatildeo da
glutationa ou outros compostos com grupos ndashOH como pentoses hexoses ou etanolaminas [V] A
reoxidaccedilatildeo de vanadilo (V(IV)) intracelular a vanadato (V(V)) provocaria a acumulaccedilatildeo toacutexica
destes uacuteltimos pelo que a complexaccedilatildeo da espeacutecie reduzida o impede O transporte de vanaacutedio
para o interior da ceacutelula encontra-se na Fig 110
Figura 110 ndash Transporte de vanaacutedio para a ceacuteluka e seu subsequente metabolismo (P=Proteiacutena) (Adaptado de
Zaporowska e Scibor 1998)
1211 Vanaacutedio(IV)
A espeacutecie de vanaacutedio (IV) mais bem conhecida eacute o iatildeo vanadilo (VO2+
) espeacutecie estaacutevel a pH
aacutecido sendo a maioritaacuteria a pH 3[AD] Apresenta uma cor azulada e forma complexos penta e
hexacoordenados Forma-se rapidamente a partir de V(V) na presenccedila de um agente redutor em
soluccedilatildeo aquosa No ambiente intracelular a presenccedila de glutationa reduzida aacutecido ascoacuterbico
NAD(P)H entre outros conduzem a que o principal estado de oxidaccedilatildeo nos sistemas bioloacutegicos
seja provavelmente o +4 encontrando-se o V(IV) fortemente complexado com proteiacutenas ou
aminoaacutecidos livres citrato ATP e pirofosfato (Refordfs) Em alternativa o V(IV) pode existir
17
livremente em ambientes fortemente aacutecidos como os lisossomas jaacute que a pH superior a 3 as
soluccedilotildees de vanadilo tendem a ser oxidadas pelo ar formando-se vanadato Esta grande
capacidade redutora justifica parcialmente a utilizaccedilatildeo pouco extensiva do iatildeo VO2+
como
inibidor [W]
1212 Vanaacutedio (V)
O vanaacutedio (V) eacute a forma mais estaacutevel num intervalo de pH amplo [W] O iatildeo anioacutenico VO3-tende
a constituir complexos polinucleares pelo que as soluccedilotildees aquosas de vanadato satildeo extremamente
complexas com ocorrecircncia simultacircnea de vaacuterios equiliacutebrios de protonaccedilatildeo (pKa(vanadato)= 35
78 e 125) e oligomerizaccedilatildeo As espeacutecies presentes (H3VO4 H2VO4- HVO4
2- e VO4
3-) dependem
do pH forccedila ioacutenica e temperatura da soluccedilatildeo e da concentraccedilatildeo total do elemento
[AEAFAGAHAI]
O VO43-
- monoacutemero ortovanadato V1 ndash estaacute associado a uma grande parte da importacircncia
bioloacutegica do vanaacutedio uma vez que se trata de um anaacutelogo electroacutenico e estrutural do ortofosfato
(PO43-
) A coordenaccedilatildeo do ortovanadato pode ser tetraeacutedrica ou bipiramidal trigonal (Refordfs)
A oligomerizaccedilatildeo do vanadato monomeacuterico daacute origem ao diacutemero (V2O72-
- V2) que apresenta
tambeacutem equiliacutebrios de protonaccedilatildeo Presume-se a existecircncia de analogia do vanadato dimeacuterico com
o pirofosfato embora natildeo haja evidecircncia estrutural (Refordf) A dimerizaccedilatildeo do V2 origina o
vanadato tetrameacuterico ou metavanadato (V4O124-
- V4) que constitui a espeacutecie maioritaacuteria em
soluccedilotildees aquosas concentradas (Refordf) As estruturas do metavanadato e a sua espeacutecie protonada
satildeo ciacuteclicas [AJ] Outra espeacutecie oligomeacuterica importante eacute o vanadato pentameacuterico (V5O153-
) cuja
estrutura presumida eacute ciacuteclica [AI]
O oligoacutemero com maior importacircncia bioquiacutemica eacute o vanadato decameacuterico (V10O286-
-V10)(Refordf) a
espeacutecie predominante em soluccedilotildees de pH 2-6 quando a concentraccedilatildeo total de vanadato eacute superior
a 100 M (Ref) A estrutura do V10 natildeo difere em soluccedilatildeo ou no estado soacutelidoOs 10 aacutetomos de
vanaacutedio da espeacutecie decameacuterica estatildeo divididos em trecircs grupos identificados na figura como VA-C
consoante o seu ambiente quiacutemico circundante (Refordf) (Fig 111)
A expressatildeo ldquodecavanadatordquo refere-se a uma soluccedilatildeo aquosa na qual a espeacutecie decameacuterica se
encontra em equiliacutebrio quiacutemico com as espeacutecies V1 V2 V4 e V5 sendo que a oligomerizaccedilatildeo
progressiva do vanaacutedio eacute proporcional agrave sua concentraccedilatildeo total (Ref) Nesta soluccedilatildeo a
concentraccedilatildeo de V10 corresponde a aproximadamente 10 da concentraccedilatildeo total de vanaacutedio [AK]
18
Figura 111 ndash Estruturas esquemaacuteticas das espeacutecies de vanadato (espeacutecie monomeacuterica (V1) dimeacuterica (V2)
tetrameacuterica (V4) pentameacuterica (V5) e decameacuterica (V10) (Adaptado de Aureliano e Crans 2008)
122 Metabolismo e efeito na actividade enzimaacutetica
Apesar de natildeo terem sido ainda encontradas as provas categoacutericas das suas funccedilotildees nos seres
vivos sabe-se que o iatildeo vanadato eacute capaz de inibir uma grande quantidade de enzimas in vitro
com especial ecircnfase na actividade ATPaacutesica da miosina [AL Teresa] Como referido
anteriormente o vanadato eacute capaz de actuar em alternativa ao fosfato nas reacccedilotildees bioquiacutemicas
incluindo a sua preferecircncia de ligaccedilatildeo agraves enzimas [W]
Eacute conhecido o papel do vanaacutedio no metabolismo do caacutelcio promovendo a mineralizaccedilatildeo dos
dentes e ossos na biossiacutentese de liacutepidos (trigliceacuteridos e colesterol) com posterior inibiccedilatildeo da
acumulaccedilatildeo destes nos vasos sanguiacuteneos e no SNC na reproduccedilatildeo e desenvolvimento por
mimetizaccedilatildeo e potenciaccedilatildeo de factores de crescimento na estimulaccedilatildeo do consumo de glucose e
da siacutentese de glicogeacutenio com actuaccedilatildeo equivalente aos efeitos da insulina ao niacutevel celular na
transduccedilatildeo de sinais celulares com regulaccedilatildeo da actividade de segundos mensageiros no
transporte de iotildees e aminoaacutecidos na transcriccedilatildeo geneacutetica e siacutentese de proteiacutenas e DNA entre
outros [W Y AB]
Assim o vanaacutedio cnseguiraacute provocar inuacutemeras mudanccedilas bioquiacutemicas nas ceacutelulas Alguns
exemplos da influecircncia bioquiacutemica do vanaacutedio incluem os seus efeitos nos processos de
fosforilaccedilatildeo e desfosforilaccedilatildeo de proteiacutenas e na participaccedilatildeo na formaccedilatildeo de espeacutecies reactivas de
oxigeacutenio devido ao seu efeito proacute-oxidante sobre agentes redutores a niacutevel celular [AM]
Quanto ao seu efeito na actividade enzimaacutetica pode dividir-se em dois tipos principais ndash inibitoacuterio
e estimulante - dependendo da enzima ou do tipo de reacccedilatildeo por esta catalisada No primeiro
grupo incluem-se enzimas como as ATPases (de miosina de canais ioacutenicos e fosfoenzimas
19
transportadoras) e as fosfatases (da tirosina aacutecida e alcalina) a fosfofrutocinase adenilato ciclase
e catalase entre outras [AA] O segundo grupo inclui a maioria das cinases a glucose-6-fosfato
desidrogenase a fosfoglicerato mutase e a fosfoglicerato fosfatase (Refordf)
O efeito do vanaacutedio (V) na actividade enzimaacutetica estaacute relacionado com o seu nuacutemero de
coordenaccedilatildeo podendo actuar como estado de transiccedilatildeo ou por mimetizaccedilatildeo de cofactores
substratos (especialmente o fosfato) e interacccedilotildees proteiacutena-proteiacutena (como no caso das
polimerases e transcriptases) Pode ainda estar envolvido na formaccedilatildeo de ligaccedilotildees de hidrogeacutenio
interacccedilotildees electrostaacuteticas esteacutereas e de oxidaccedilatildeo-reduccedilatildeo Uma evidecircncia desta uacuteltima eacute a sua
capacidade de alternacircncia entre os estados de oxidaccedilatildeo +5 a +3 o que torna faacutecil a sua interacccedilatildeo
redox com enzimas contendo resiacuteduos de cisteiacutenas (Cys) ou qualquer outro composto orgacircnico
com actividade redox [AN]
13 Vanaacutedio e muacutesculo esqueleacutetico
A primeira evidecircncia da presenccedila de vanaacutedio no muacutesculo esqueleacutetico proveio de uma amostra de
ATP isolado de muacutesculo de cavalo que continha uma impureza ndash vanaacutedio ndash cuja presenccedila
provocou a inibiccedilatildeo da actividade da bomba de Na+K
+-ATPase [AO] Desde entatildeo vaacuterios
estudos tecircm sido realizados com ecircnfase na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o
tecido muscular Destes destaca-se a descoberta da interacccedilatildeo forte da espeacutecie V10 com a miosina
fosforilada [AP] e agrave sua inibiccedilatildeo da actividade ATPaacutesica da miosina[AQ AR]
No quediz respeitoao iatildeo monovanadato destaca-se a sua inibiccedilatildeoda actividade Mg2+
-ATPaacutesica
da miosina [AS] que ocorre de duas formas distintas (1) na presenccedila de concentraccedilotildees reduzidas
de V1 a inibiccedilatildeo ocorre a velocidade reduzida dependente da concentraccedilatildeo da espeacutecie
monomeacuterica (2) na presenccedila de concentraccedilotildees elevadas de V1 o niacutevel de inibiccedilatildeo aproxima-se de
100 desde que [V1] seja superior agrave concentraccedilatildeo total de locais com actividade Mg2+
-ATPaacutesica
no ensaio Esta inibiccedilatildeo eacute irreversiacutevel tanto por diaacutelise ou adiccedilatildeo de agentes redutores do
vanadato [AL]Segundo Pate et al(1994)[AT] a espeacutecie monomeacuterica podeinibir a velocidade de
contracccedilatildeo a temperatura inferior a 20ordmC perdendogradual e rapidamente essa capacidade ateacute os
50ordmC temperatura agrave qual cessa a capacidade inibitoacuteria Os resultados obtidos sugeremainda uma
interacccedilatildeo altamente cooperativa entre o iatildeo vanadato e as proteiacutenas componentes do muacutesculo
esqueleacuteticoDestaca-se tambeacutem a descoberta da inibiccedilatildeo da geraccedilatildeo de forccedila contraacutectil nas fibras
musculares por parte da espeacutecie monomeacuterica[AT]Wilson et al(1995) [AU ]determinaram que a
afinidade da espeacutecie V1 para o S1 de miosina eacute significativamente mais elevada do que a do iatildeo
Pi seu anaacutelogo Descobriram ainda que que a ligaccedilatildeo do vanadato poderaacute envolver transiccedilotildees
entre estados energeacuteticos com energia livre independente dos impedimentos esteacutereos impostos
20
pela estrutura das miofibrilas Os estudos desenvolvidos por Smith et al (1990)[AV]
determinaram queo iatildeo monovanadato natildeo apresenta um efeito pronunciado na inibiccedilatildeo da
actividadeacto-S1-miosina-ATPaacutesica excepto em concentraccedilotildees superiores a 900microM a partir das
quais existe produccedilatildeo de espeacutecies oligomeacutericas para as quais o efeito inibitoacuterio seraacute mais
elevadoA espeacutecie monomeacuterica de vanadato consegue adoptar facilmente uma geometria do tipo
bipiramidal trigonal que mimetiza o intermediaacuterio formado (MgADP
Pi)aquando da reacccedilatildeo de
hidroacutelise de ATP no local activo da miosina [AW] De facto a interacccedilatildeo do vanaacutedio com a
miosina eacute inesperadamente estaacutevel na presenccedila de ADP Esta interacccedilatildeo resulta na formaccedilatildeo de
um complexo enzimaacutetico inactivo e estaacutevel com estequiometria de 11 ADPvanadato por siacutetio
activo da proteiacutena A formaccedilatildeo do complexo estaacutevel miosina-vanadato foi proposta por Goodnoet
al (1982) [AS] Nesta ocorre um preacute-equiliacutebrio raacutepido que induz a formaccedilatildeo de um complexo
ternaacuterio reversiacutevel (MADP
V) com formaccedilatildeo lenta do isoacutemero M
ADP
V A diferenccedila entre os
compostos de vanaacutedio e de fosfato centra-se no tempo de meia vida (t12) do Pi ou do V (no
primeiro t12=12 s enquanto no segundo t12=24 h) (Refordf) A diferenccedila dever-se-aacute agrave possiacutevel
complexaccedilatildeo do vanaacutedio com grupos nucleofiacutelicos no siacutetio activo o que provocaria uma
estabilizaccedilatildeo da miosina na conformaccedilatildeo MADP
V
No que respeita aos estudos realizados com a espeacutecie tetrameacuterica Cremo et al determinou que a
cadeia pesada do S1 de miosina pode ser clivada em dois locais distintos (designados V1 e V2)
pela espeacutecie V4 apoacutes irradiaccedilatildeo com luz UV na presenccedila de concentraccedilotildees de vanadato na ordem
dos milimolar e na ausecircncia de nucleoacutetidos ou metais divalentes
Jaacute no que concerne aos estudos realizados com ecircnfase na espeacutecie decameacuterica foi sugerido
porRamos et al (2006) [AK]que a interacccedilatildeo do decavanadato com a G-actina favorecida pela
polimerizaccedilatildeo desta uacuteltima estabiliza a espeacutecie V10 e induz a inibiccedilatildeo da formaccedilatildeo de F-actina A
estabilizaccedilatildeo da espeacutecie V10 por proteiacutenas tansmembranares e do citoesqueleto pode ser a razatildeo
pela qual ocorre a toxicidade do decavanadato reportada na avaliaccedilatildeo dos efeitos do V(V) nos
sistemas bioloacutegicosTiago et al (2007) [AR] concluiram que o V10 apresenta uma clara preferecircncia
de ligaccedilatildeo ao ldquoback-doorrdquo da S1-miosina-ATPase o que se verifica apenas na estrutura
conformacional desimpedida com acesso facilitado ao local de ligaccedilatildeo do Pi Sugere tambeacutem que
o V10 actua como um ldquoback-door stoprdquo bloqueando estaabertura do fragmento de 50-kDa
necessaacuteria para a hidroacutelise do ATP--fosfato o que constitui uma explicaccedilatildeo simples do
comportamento natildeo competitivo da espeacutecie decameacuterica estimulando a utilizaccedilatildeo deste oligoacutemero
como uma feramenta da elucidaccedilatildeo das mudanccedilas conformacionais ldquoback-doorrdquo da miosina no
21
processo da contracccedilatildeo muscular(ref)Destaca-se ainda a descoberta da capacidade inibitoacuteria do
V10 em relaccedilatildeo agrave fosforilaccedilatildeo oxidativa mitocondrial [AX]
Finalmente no que concerne aos estudos realizados com a presenccedila de vaacuterias espeacutecies
oligomeacutericas Aureliano e Madeira (1994) [AY]observaram que na presenccedila de Ca2+
as espeacutecies
V4 e V10se ligam agrave Ca2+
-ATPase do retiacuteculo sarcoplasmaacutetico enquanto a espeacutecie V1 se liga apenas
na presenccedila de ATP Aparentemente o aumento da concentraccedilatildeo de Ca2+
tem o efeito contraacuterio da
inibiccedilatildeo da ATPase do retiacuteculo sarcoplasmaacutetico pelo V1 natildeo tendo no entanto efeito significativo
sobre a inibiccedilatildeo por parte da espeacutecie V10 Tiago et al (2002) [AP] concluiacuteram que em condiccedilotildees
proacuteximas agraves fisioloacutegicas a espeacutecie V10 difere das espeacutecies presentes na soluccedilatildeo de metavanadato
devido agrave sua forte interacccedilatildeo com a miosina fosforilada e agrave sua inibiccedilatildeo da actividade ATPaacutesica
da miosina Para aleacutem disso o ATP diminui a afinidade da miosina para o V4 induzindo a
interacccedilatildeo com V1 e natildeo tendo qualquer efeito no que diz respeito ao V10 Foi tambeacutem proposto
[AQ] que a ligaccedilatildeo das espeacutecies oligomeacutericas de vanadato aoS1 de miosina nos locais onde o
fosfato coordenaeacute provavelmente a causa da inibiccedilatildeo da actividade ATPaacutesica da miosina
estimulada por actina atraveacutes da prevenccedilatildeo da troca entre ATPADP sendo esta favorecida para
aniotildees de vanadato com valecircncias superiores como eacute o caso do V10
Na presenccedila de actina as velocidades de dissociaccedilatildeo do Pi e do V aumentam consideravelmente
este uacuteltimo em menor quantidade Assim a ligaccedilatildeo da actina ao complexo MADP
V e
Pi eacute
bastante fraca pelo que se sugere que o ADPV teria um efeito de relaxamento semelhante ao
ATP No entanto a capacidade do vanaacutedio em inibir a actividade ATPaacutesica da miosina e da
actomiosina eacute diferenciada uma vez que na ausecircncia de actina a inibiccedilatildeo eacute total para
concentraccedilotildees na gama dos M mas na sua presenccedila a inibiccedilatildeo eacute totalmente conseguida apenas
em concentraccedilotildees na gama dos mM (Ref) No entanto eacute necessaacuterio considerar a possiacutevel
formaccedilatildeo de espeacutecies oligomeacutericas de vanadato cuja afinidade e efeito na enzima seratildeo
diferentes Assim pode concluir-se que o efeito de inibiccedilatildeo observado para a actividade ATPaacutesica
de miosina estimulada por actina se pode dever agrave presenccedila das diferentes espeacutecies oligomeacutericas o
que foi jaacute sugerido por Aureliano (2000) [AZ] ndash as espeacutecies tetrameacuterica e decameacuterica de vanadato
interactuariam com a miosina com alteraccedilatildeo da sua interacccedilatildeo com a actina No entanto natildeo eacute
conhecida a contribuiccedilatildeo absoluta das restantes espeacutecies oligomeacutericas para a inibiccedilatildeo da ATPase
da miosina estimulada por actina (Refs)
14 Objectivos do projecto
Pretende-se com este projecto o isolamento purificaccedilatildeo e caracterizaccedilatildeo da actina do muacutesculo
22
esqueleacutetico de coelho sendo esta uacuteltima tambeacutem realizada para o sufragmento-1 de miosina
Seguidamente realizar-se-aacute a quantificaccedilatildeo e caracterizaccedilatildeo das proteiacutenas por espectroscopia UV-
Vis e SDS-PAGE e a caracterizaccedilatildeo das soluccedilotildees de vanaacutedio por UV-Vis e ressonacircncia magneacutetica
nuclear (RMN) Por uacuteltimo a interacccedilatildeo do S1 de miosina com o vanaacutedio seraacute avaliada atraveacutes
das teacutecnicas de UVVis RMN e ressonacircncia paramagneacutetica electroacutenica (RPE)
2 Discussatildeo de Resultados
21 Caracterizaccedilatildeo das proteiacutenas isoladas
211Anaacutelise do estado de pureza
O estado de pureza das soluccedilotildees de actina e de S1 foi analisado por electroforese em gel de
poliacrilamida em condiccedilotildees desnaturantes (SDS-PAGE) (Fig 22) que separa as moleacuteculas de
acordo com o seu peso molecular (PM) A concentraccedilatildeo de acrilamida escolhida (10) permite a
detecccedilatildeo simultacircnea de componentes com peso molecular inferior a 20 kDa e outros com elevado
peso molecular
Figura 21 ndash Fotografia de gel de SDS-PAGE (10 acrilamida) Poccedilo (a)
10 L do marcador de elevado peso molecular () Poccedilo (b) 20 L de [G-
actina]=1 mgmL Poccedilo (c) 20 L de [S1]=1 mgmL Aplicou-se uma
voltagem de V durante h e min
A partir da recta de calibraccedilatildeo obtida log PM = -1177 Rf + 2247 (ver ponto 2 da secccedilatildeo
Anexos) pode verificar-se a presenccedila de actina na amostra do poccedilo (b) presente numa banda
bastante intensa (PMaparente=40 kDa) bem como a presenccedila minoritaacuteria de outros componentes
desconhecidos que natildeo teratildeo sido eliminados durante o processo de extracccedilatildeo e purificaccedilatildeo e natildeo
satildeo relevantes para o estudo em questatildeo Na amostra do poccedilo (c) nota-se a presenccedila de S1
(PMaparente=92 kDa) bem como a de outros componentes cuja banda eacute ligeiramente menos intensa
Estes poderatildeo ser fragmentos proteoliacuteticos de miosina e de S1 Estaacute ainda presente uma bada de
23
maior intensidade que poderaacute ser a proteiacutena-C (PM=135 kDa) uma vez que a sua remoccedilatildeo
completa da amostra necessitaria de um passo adicional de cromatografia de troca ioacutenica [BA]
No entanto natildeo podemos confirmar que se trata desta proteiacutena ou de qualquer outra uma vez que
se encontra fora do intervalo de migraccedilatildeo dos marcadores de peso molecular
212 Determinaccedilatildeo da concentraccedilatildeo proteica
2121Subfragmento-1 de miosina
A partir da recta de calibraccedilatildeo obtida pelo meacutetodo de Bradford [BB](ver secccedilatildeo 3311 do
Procedimento Experimental) A595nm = 00546[BSA] + 00209 (ver secccedilatildeo de Anexos)foi possiacutevel
determinar um intervalo de concentraccedilatildeo de S1 de miosina compreendido entre 226 e 117(plusmn
423M) (203 e 1056 (plusmn 381 mgmL)) (n=8) A concentraccedilatildeo meacutedia de S1 de miosina apoacutes
diaacutelise contra a soluccedilatildeo tampatildeo 10 mM HEPES pH 75 25 mM KCl determinada pela leitura da
Abs280 nmfoi de 20430M (1803 mgmL)(n=7)
As amostras a quantificar atraveacutes do meacutetodo de Bradford natildeo sofreram diaacutelise o que permite
explicar a concentraccedilatildeo superior de S1 de miosina em comparaccedilatildeo com o valor obtido atraveacutes do
meacutetodo directo Esta razatildeo aliada agrave sensibilidade e rapidez superiores deste uacuteltimo fez com que
se optasse pelo meacutetodo directo para a quantificaccedilatildeo da grande maioria dos ensaios
2122Actina
A concentraccedilatildeo de actina foi determinada quando esta se encontrava na forma monomeacuterica (G-
actina) por mediccedilatildeo da leitura directa da Abs290 nm tendo-se obtido uma concentraccedilatildeo meacutedia de
691481M (290202 mgmL) (n=3) em 2 mM Tris-HCl pH 80 02 mM CaCl2 02 mM Na-
ATP Ao comprimento de onda utilizado (290 nm) absorvem fortemente apenas os resiacuteduos de
Trp pelo que a mediccedilatildeo efectuada tem menor sensibilidade do que se tivesse sido efectuada a 280
nm uma vez que a actina apresenta apenas 4 resiacuteduos deste aminoaacutecido embora tenha 16
resiacuteduos de Tyr
22 Mediccedilatildeo da actividade enzimaacutetica de S1 de miosina
221 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
As actividades ATPaacutesicas de S1 de miosina dependentes de Ca2+
e K+foram analisadas pelo
caacutelculo das velocidades de consumo inicial do fosfato inorgacircnico (Pi) Definiu-se os tempos de 5
e 3 minutos para a mediccedilatildeo da Ca2+
e da K+EDTA-ATPase respectivamente Os valores obtidos
24
a partir do protocolo presente no ponto 341 da secccedilatildeo Procedimento experimental encontram-se
descritos na Tabela 2I
Tabela 2I ndash Acrividades natildeo-fisioloacutegicas de subfragmento-1 determinadas pelo meacutetodo de Fiske-Subarow
modificado (n=2)
Proteiacutena
Actividade
ATPaacutesica natildeo
fisioloacutegica
Actividade (nmol
Piminmg proteiacutena) Actividade (s-1)
S1
Ca2+
-ATPase 1400 plusmn 30 2100 plusmn 0045
K+EDTA-ATPase 2334 plusmn 50 3500 plusmn 0075
As actividades natildeo-fisioloacutegicas reflectem o estado de oxidaccedilatildeo dos resiacuteduos expostos de Cys 707
(SH1) e 697 (SH2) Se o grupo SH1 sofrer oxidaccedilatildeo a actividade ATPaacutesica dependente de Ca2+
aumenta e a actividade dependente de K+ diminui Por outro lado se o grupo SH2 sofrer
oxidaccedilatildeo regista-se a perda completa destas actividades
De acordo com Margossian e Lowey (1982) [BA] as actividades ATPaacutesica de S1 de miosina
dependente de Ca2+
e K+EDTA da miosina satildeo respectivamente de 24 e 112 s-1
Estaacute tambeacutem
descrito que a actividade ATPaacutesica dependente de K+ eacute em meacutedia cerca de 3 vezes superior agrave
dependente de Ca2+
reflectindo a quantidade das preparaccedilotildees A razatildeo obtida entre os valores das
actividades ATPaacutesicas dependentes de K+ e Ca
2+ eacute de 166 Os valores obtidos para as actividades
(Ca2+
-ATPase=21 s-1
K+EDTA-ATPase= 35 s
-1) satildeo inferiores agrave valores obtidos por
Margossian e Lowey [BA] e Tiago [BC] com forccedila ioacutenica inferior O valor obtido para a
actividade ATPaacutesica dependente de Ca2+
eacute superior ao obtido por Polosukhina et al (2000) [BD]
cujo meio de reacccedilatildeo continha MOPS e apresentava forccedila ioacutenica inferior Os dados
supramencionados comprovam a variaccedilatildeo da actividade a ATPaacutesica natildeo fisioloacutegica em relaccedilatildeo agrave
forccedila ioacutenica temperatura e pH
222 Mediccedilatildeo da actividadeMg2+
-ATPaacutesica de S1 pelo meacutetodo de enzimas acopladas
A actividade de Mg2+
-ATPase eacute a uacutenica que possui significado fisioloacutegico sendo as restantes
bastante uacuteteis apenas para indicaccedilatildeo da qualidade de preparaccedilatildeo do S1[BA] A 0575 M KCl a
actina e o S1 formam uma soluccedilatildeo viscosa e tuacuterbida Apoacutes a adiccedilatildeo de MgATP o complexo acto-
S1 dissocia-se e a turbidez da soluccedilatildeo diminui Se a forccedila ioacutenica for reduzida a dissociaccedilatildeo do
complexo actoS1 que se encontra na forma de suspensatildeo eacute induzida O MgATP eacute rapidamente
hidrolisado diminuindo a sua concentraccedilatildeo no meio reaccional Nestas condiccedilotildees haacute
reassociaccedilatildeo e subsequente contracccedilatildeo dos filamentos de actina e de S1 com formaccedilatildeo de
25
precipitado O meacutetodo descrito traduz de forma bastante elementar o fenoacutemeno da contracccedilatildeo in
vivo e elimina o problema anterior da solubilizaccedilatildeo ineficiente da miosina de modo a obter uma
mediccedilatildeo exacta da actividade [A] A utilizaccedilatildeo de forccedilas ioacutenicas reduzidas aumenta a afinidade da
actina para o S1 permitindo contornar a extrema dificuldade em obter a concentraccedilatildeo proteica dos
muacutesculos (cerca de 100 mgmL do complexo actomiosina) As actividades Mg2+
-ATPaacutesicas de S1
de miosina foram determinadas a partir do declive da variaccedilatildeo da Abs340 relativo agrave oxidaccedilatildeo de
NADH em funccedilatildeo do tempo como eacute descrito no ponto 342 do Procedimento Experimental Os
valores obtidos para a actividade Mg2+
-ATPaacutesica encontram-se apresentados na Tabela 2II
Tabela 2II ndash Actividade Mg2+
-ATPaacutesica de subfragmento-1 de miosina determinadas pelo meacutetodo das enzimas
acopladas
Actividade (nmol
ATPmin mg(S1)) Actividade (s
-1)
1097 plusmn 269 164 plusmn 04
Ramos (2005) [J] obteve o valor de 01 s-1
para a actividade Mg2+
-ATPaacutesica de S1 de miosina
com forccedila ioacutenica intermeacutedia Tiago (2005) [BC] obteve o valor de 011 s-1 em condiccedilotildees de forccedila
ioacutenica reduzida em relaccedilatildeo agrave utilizada no presente relatoacuterio
23 Preparaccedilatildeo e caracterizaccedilatildeo das soluccedilotildees de vanaacutedio
231 Anaacutelise da composiccedilatildeo das soluccedilotildees de vanadato por espectroscopia de 51
V-RMN
efeito da concentraccedilatildeo da espeacutecie
A diluiccedilatildeo das soluccedilotildees stock de meta- e decavanadato iraacute determinar fortemente a composiccedilatildeo
em soluccedilatildeo dos diferentes oligoacutemeros de vanadato (ref) No caso do decavanadato o periacuteodo de
tempo apoacutes a diluiccedilatildeo iraacute tambeacutem ser um factor importante a considerar devido agrave decomposiccedilatildeo
da espeacutecie V10(ref)Torna-se assim fundamental determinar a composiccedilatildeo quiacutemica das soluccedilotildees
no meio de ensaio utilizado (HEPES 10 mM KCl 25 mM) de modo a correlacionaacute-la
correctamente com os possiacuteveis efeitos promovidos pelas diferentes espeacutecies de vanadato no
sistema em estudo
Aleacutem disso deve ser dado ecircnfase agraves importantes interacccedilotildees que ocorrem entre o vanaacutedio e os
tampotildees escolhidos O tampatildeo com menor nuacutemero e magnitude de interacccedilatildeo eacute o HEPES [AD]
seguido do Tris sendo este uacuteltimo especialmente aconselhado em concentraccedilotildees baixas para casos
de incompatibilidade enzimaacutetica ou quiacutemica redox No caso especial da reacccedilatildeo redox
NAD(P)HNAD(P) o tampatildeo adequado eacute o imidazole de modo a natildeo haver reacccedilatildeo com o
26
vanadato Os tampotildees a evitar em qualquer situaccedilatildeo incluem a trietanolamina fosfato citrato e
glicina entre outros [AD] uma vez que todos interactuam fortemente com V(V) com formaccedilatildeo
de complexos estaacuteveis
Assim procedeu-se agrave anaacutelise da composiccedilatildeo das soluccedilotildees de meta- e decavanadato apoacutes diluiccedilatildeo
das soluccedilotildees stock (50 mM em vanadato total) no meio de reacccedilatildeo (tampatildeo D) atraveacutes da
espectroscopia de ressonacircncia magneacutetica nuclear de 51
V Nas figuras x1 e x2 apresentam-se os
espectros obtidos para as soluccedilotildees de deca- e metavanadato num intervalo de concentraccedilotildees de 1 a
10 mM A Tabela seguinte resume os paracircmetros espectroscoacutepicos obtidos para cada espeacutecie
presente nas soluccedilotildees analisadas
Figura 22ndash Espectros de RMN de 51
V de
concentraccedilotildees crescentes das espeacutecies oligomeacutericas
de vanadato obtidos apoacutes diluiccedilatildeo da soluccedilatildeo stock
de decavanadato em 10 mM HEPES pH 70 25 mM
KCl para concentraccedilotildees finais de A) 1 mM B)2
mM C) 4 mM D) 8 mM E) 10 mM de
decavanadato total V1refere-se agraves espeacutecies
monomeacutericade vanadato V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2IIIndash Desvios quiacutemicos meacutedios (part) das ressonacircncias das espeacutecies decameacuterica (V10) monomeacuterica (V1) e
tetrameacuterica (V4) presentes nos espectros da soluccedilatildeo de decavanadatoLargura a meia altura (∆12) da espeacutecie V10A
Paracircmetros V10A V10B V10C V1 V4
part (ppm) -5162 -5003 -4255 -5616 -5748
∆12 (Hz) 175 nd nd nd nd
Figura 23 ndash Distribuiccedilatildeo graacutefica das
espeacutecies oligomeacutericas de vanadato vs a
concentraccedilatildeo de decavanadato total As
concentraccedilotildees de cada espeacutecie foram
obtidas segundo a formula descrita na
secccedilatildeo 35 do Procedimento
Experimental (aacutereas dos sinais de 51
V-
RMN observadas na figura x1)V10 V1 e
V4 correspondem agraves espeacutecie monomeacuterica
dimeacuterica e tetrameacuterica respectivamente [Esp
eacuteci
e o
ligo
meacute
rica
de
van
aacutedio
] (m
M)
[Decavanadato]total (mM)
V10
V1
V4
27
Nas soluccedilotildees de decavanadato as espeacutecies monomeacuterica e decameacuterica de vanadato satildeo as uacutenicas
presentes para concentraccedilotildees de decavanadato inferiores a 8 mM Ambas as espeacutecies aumentam
linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado em [AY] e
em (Teresa) quando a concentraccedilatildeo de V1 eacute igual ou superior a 200 microM inicia-se a sua
oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica No entanto nas condiccedilotildees de realizaccedilatildeo dos
ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a espeacutecie tetrameacuterica neste
estudo em concentraccedilotildees inferiores a 8 mM tendo o mesmo sido observado por Ramoset al
(2006) [AK] Os resultados obtidos mostram claramente o equiliacutebrio entre a espeacutecie decameacuterica e
a monomeacuterica e que o aparecimento de outras espeacutecies oligomeacutericas se encontra dependente da
concentraccedilatildeo desta uacuteltima Apesar de natildeo ter sido realizado o ensaio com 5 mM de decavanadato
total no presente relatoacuterio estaacute reportado em Ramos et al (2006) [AK] que as concentraccedilotildees da
espeacutecie decameacuterica e monomeacuterica satildeo respectivamente de 500 microM e 200 microM Assim pode inferir-
se que para concentraccedilotildees inferiores a 5 mM e intervalos de tempo de cerca de 20 minutos (que
inclui o tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas
as espeacutecies decameacuterica e monomeacuterica de vanadato
Figura 24 ndash Espectros de RMN de 51
V de concentraccedilotildees
crescentes das espeacutecies oligomeacutericas de vanadato obtidos
apoacutes diluiccedilatildeo da soluccedilatildeo stock de metavanadato em 10 mM
HEPES pH 70 25 mM KCl para concentraccedilotildees finais de A)
1 mM B) 2 mM C) 4 mM D) 8 mM E) 10 mM de
metavanadato total V1 refere-se agraves espeacutecies monomeacuterica de
vanadatoV1 e V2 correspondem respectivamente agrave espeacutecie
monomeacuterica e dimeacuterica
Figura 25 ndash Distribuiccedilatildeo graacutefica das espeacutecies
oligomeacutericas de vanadato vs a concentraccedilatildeo de
metavanadato total As concentraccedilotildees de cada
espeacutecie foram obtidas segundo a formula descrita
na secccedilatildeo 35 do Procedimento Experimental
(aacutereas dos sinais de 51
V-RMN observadas na
figura x1)V10 V1 e V4 correspondem agraves espeacutecie
monomeacuterica dimeacuterica e tetrameacuterica
respectivamente
Esp
eacuteci
e o
ligo
meacute
rica
de
va
nad
ato
(m
M)
[Metavanadato]total (mM)
V1
V2
V4
V5
28
Tabela 2IVndash Desvios quiacutemicos meacutedios (part)das ressonacircncias das espeacutecies tetrameacuterica (V4) monomeacuterica (V1) e
dimeacuterica (V2) presentes no espectro da soluccedilatildeo de metavanadato (total) 10 mM
Paracircmetros V1 V2 V4 V5
part (ppm) -5615 -5748 -5789 -5869
Nas soluccedilotildees de metavanadato as espeacutecies monomeacuterica dimeacutericae tetrameacuterica de vanadato estatildeo
presentes para concentraccedilotildees de metavanadato inferiores a 4 mM Todas estas as espeacutecies
aumentam linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado
em (Aureliano e Madeira 1998) e em (Tiago et al 2004) quando a concentraccedilatildeo de V1 eacute igual ou
superior a 200 microM inicia-se a sua oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica Nas
condiccedilotildees de realizaccedilatildeo dos ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a
espeacutecie pentameacuterica neste estudo em concentraccedilotildees inferiores a 4 mM tendo o mesmo sido
observado por Ramos 2006 Os resultados obtidos mostram claramente o equiliacutebrio entre a
espeacutecie V4 e as espeacuteciesV1 e V2 no qual o aparecimento de outras espeacutecies oligomeacutericas se
encontra dependente da concentraccedilatildeo da espeacutecie V1 Por uacuteltimo pode inferir-se que para
concentraccedilotildees inferiores a 2 mM e intervalos de tempo de cerca de 20 minutos (que inclui o
tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas as
espeacutecies V1 V2 e V4 com concentraccedilotildees respectivas de 0796 0192 e 0196 mM
232Estabilidade das soluccedilotildees de decavanadato anaacutelise por espectroscopia de absorccedilatildeo
UVVis
Caracterizou-se a estabilidade da espeacutecie V10 numa soluccedilatildeo com 1 mM decavanadato total em
tampatildeo D atraveacutes da leitura da Abs400 Obteve-se uma cineacutetica de dissociaccedilatildeo de 1ordf ordem tal
como se pode verificar pela linearidade do graacutefico de ln(Abs400)vs o tempo de aquisiccedilatildeo espectral
(segundos) (Fig26 )
Figura 26ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do tempo
(em segundos) para uma soluccedilatildeo de decavanadato total de 1
mM em tampatildeo D a 25ordmC (n=2)
Sendo uma reacccedilatildeo de primeira ordem pode ser descrita pela equaccedilatildeo cujo
declive eacute ndashK (constante de dissociaccedilatildeo) e a intersecccedilatildeo no eixo das ordenadas corresponde
Ln
(Ab
s 400 n
m)
tempo (s)
29
aln(A0) Numa reacccedilatildeo de 1ordf ordem o tempo de meia-vida eacute independente da concentraccedilatildeo inicial
do reagente sendo descrito pela equaccedilatildeo [2] Deste modo a partir da recta
Ln(Abs400 nm) = -436E-5
K - 105 obteve-se um tempo de meia-vida de 4490 plusmn 0823 horas para
as condiccedilotildees supramencionadas
Apoacutes a adiccedilatildeo de S1 8 microM ao meio reaccional verificou-se que a reacccedilatildeo de desoligomerizaccedilatildeo
da espeacutecie decameacuterica de vanadato seguia igualmente uma cineacutetica de 1ordf ordem tal como se pode
verificar pela linearidade do graacutefico de ln(Abs400) versus o tempo de aquisiccedilatildeo espectral
(segundos) (Fig27)
Figura 27ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do
tempo (em segundos) para uma soluccedilatildeo de decavanadato
total de 1 mM em tampatildeo D a 25ordmC na presenccedila de 8
microM (n= 2)
A partir da equaccedilatildeo da recta Ln(Abs400 nm) = -7520E-5
K - 0613determinou-se o valor de K para
a espeacutecie decameacuterica de onde se obteve um tempo de meia-vida de 2430 plusmn 0191 horas para as
condiccedilotildees supramencionadas
Tanto este valor como o valor registado na ausecircncia de S1 satildeo substancialmente superiores agrave
duraccedilatildeo dos ensaios de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina descritos no presente
relatoacuterio
24 Interacccedilatildeo de S1 com vanaacutedio(V)
241Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
De modo a analisar qual o efeito da adiccedilatildeo de S1 de miosina (tampatildeo D) produzia nos sinais de
51V-NMR titulou-se uma soluccedilatildeo de 5mM de decavanadato total com concentraccedilotildees crescentes
de S1 (5 10 15 e 20 microM) Os espectros de 51
V-RMN encontram-se apresentados na Fig 2
No graacutefico da figura 2 Apresenta-se a concentraccedilatildeo de S1 em funccedilatildeo do factor de alargamento
corrigido dos espectros acima referidos
Ln
(Ab
s (4
00
nm
))
tempo (s)
30
Figura 28 ndash Graacutefico de
representaccedilatildeo da
concentraccedilatildeo de S1 em
funccedilatildeo do factor de
alargamento A
concentraccedilatildeo de
decavanadato total foi de 5
mM e as concentraccedilotildees
utilizadas de S1 foram de 0
5 10 e 15 microM
Verifica-se que o factor de alargamento aumenta linearmente em funccedilatildeo da concentraccedilatildeo de S1 no
ensaio o que indica a existecircncia de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina Observou-
se ainda que a adiccedilatildeo de S1 ao meio reaccional natildeo provocou qualquer alteraccedilatildeo dos desvios
quiacutemicos dos sinais de 51
V-RMN
Realizaram-se ensaios na presenccedila de ATP e de S1 com intuito de verificar o efeito do ATP na
interacccedilatildeo do S1 de miosina com o decavanadato
Figura 29 - Espectros de RMN de 51
V contendo 2
mM Decavanadato (A) (controlo) (B) na presenccedila
de 2 mM de ATP (C) na presenccedila de 2 mM de ATP
e 8 microM de S1 de miosina V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2Vndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies decameacuterica (V10) e monomeacuterica (V1) presentes nos
espectros da soluccedilatildeo de decavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V10A V10B V10C V1
part (ppm) -5161 -5002 -4254 -5614
Observa-se que a presenccedila de ATP (espectro B) natildeo provoca qualquer alteraccedilatildeo visiacutevel em
relaccedilatildeo ao espectro que contem apenas soluccedilatildeo de decavanadato Jaacute a presenccedila simultacircnea de ATP
e S1 provoca um alargamento da ∆12 do sinal do V10A de 1736 Hz (espectro (A)) para 2366 Hz
(espectro (C) Fig 29) Tendo em conta que a concentraccedilatildeo de S1 apresenta uma relaccedilatildeo linear
com o factor de alargamento converteu-se o valor da ∆12 e comparou-se com o valor esperado agrave
F = 00591[S1] - 00157 Rsup2 = 0998
Fact
or
de
ala
rgam
en
to
[S1] (microM)
31
concentraccedilatildeo de S1 utilizada O factor de alargamento obtido para 8 microM ndash 0376 ndash eacute inferior ao
valor esperado agrave mesma concentraccedilatildeo de S1 obtido atraveacutes da equaccedilatildeo da recta apresentada na
Fig 28 ndash 0457 ndash o que indica que o ATP poderaacute exercer um efeito de protecccedilatildeo do S1
diminuindo a sua interacccedilatildeo com a espeacutecieV10
Por uacuteltimo realizaram-se ensaios com metavanadato na presenccedila de 8 microM S1 de modo a verificar
se as espeacutecies oligomeacutericas contidas nesta soluccedilatildeo interagiam com a proteiacutena em estudo
Figura 210 - Espectros de RMN de 51
V contendo 2
mM metavanadato (A) (controlo) (B) na presenccedila
de 8 microM de S1 de miosina V1 V2 e V4
correspondem respectivamente aacutes espeacutecies
monomeacuterica dimeacuterica e tetrameacuterica
Tabela 2VIndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies monomeacuterica (V1) dimeacuterica (V2) e tetrameacuterica (V4)
presentes nos espectros da soluccedilatildeo de metavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V1 V2 V4
part (ppm) -5613 -5747 -5789
Como eacute possiacutevel observar na Fig 210 ocorre uma ligeiro alargamento dos sinal de V4 bem
como uma perda da intensidade pelo que se conclui quedas espeacutecies presentes na soluccedilatildeo de
metavanadato (V1 V2 e V4) apenas a espeacutecie tetrameacuterica interage com o S1 de miosina o que
estaacute de acordo com o reportado por Tiago et al 2002
242 Actividade Mg2+
-ATPaacutesica de S1 de miosina na presenccedila de decanadato
Com o intuito de avaliar o efeito do decavanadato na actividade Mg2+
-ATPaacutesica do S1 de
miosina monitorizou-se a reacccedilatildeo de oxidaccedilatildeo do NADH atraveacutes da leitura de Abs340 na
presenccedila de 005 mgmL de S1 de miosina e no caso da actividade estimulada 2microM de actina em
3125 mM Tris-HCl pH 71 e 375 mM MgCl2 pelo meacutetodo de enzimas acopladas (ver seccccedilatildeo xx
dos MM)Para tal realizaram-se ensaios com vaacuterias concentraccedilotildees de decavanadato total (10 50
100 250 500 e 1000) de modo a determinar qual o seu efeito na actividade MgATPaacutesica de S1
de miosina na ausecircncia e na presenccedila de um dos seus ligandos naturais Os ensaios foram
realizados agrave temperatura ambiente com incubaccedilatildeo preacutevia a 25ordm C durante 20 minutos
32
Figura 211 ndash Representaccedilatildeo
graacutefica da Actividade Mg2+
-
ATPaacutesica de S1 () (os ajustes
teoacutericos estatildeo representado a por
uma linha os resultados
experimentais estatildeo representados
por triacircngulos (Actividade
estimulada) e cruzes (Actividade
natildeo estimulada)) vs
[Decavanadato]total medidas pelo
meacutetodo das enzimas acopladas
(n=3) Os dados foram ajustados ao modelo de decaimento exponencial usando o programa GraphPad Prism para a
determinaccedilatildeo dos valores de IC50 (na ausecircncia de actina IC50= 266 plusmn 07 M decavanadato na presenccedila de actina
IC50= 28 plusmn 1 M decavanadato
A partir do graacutefico presente na Fig 29 verifica-se que o ajuste dos dados experimentais foi bem
sucedido A inibiccedilatildeo da actividade Mg2+
-ATPaacutesica do S1 de miosina eacute afectada em menor
amplitude na presenccedila de actina (60 de inibiccedilatildeo da actividade) do que na sua ausecircncia
(aproximadamente 80 de inibiccedilatildeo de actividade) Devido a esta diferenccedila pode sugerir-se que a
actina iraacute impedir parcialmente a ligaccedilatildeo do decavanadatoao S1 demiosina atraveacutes de
impedimentos esteacutereos ou interacccedilotildees electrostaacuteticas por exemplo A teacutecnica utilizada natildeo nos
permite concluir acerca de que regiatildeo(otildees) ou resiacuteduo(s) da actina participaratildeo na preservaccedilatildeo
parcial da actividade Mg2+
-ATPaacutesica
Por outro lado os valores de IC50 dos dois tipos de actividades natildeo apresentam diferenccedilas
significativas tendo-se obtido 266 plusmn 07 e 28 plusmn 1M decavanadato na ausecircncia e na presenccedila de
actina respectivamente o que corresponde a uma concentraccedilatildeo de cerca de 27 M de V10 O
meio de reacccedilatildeo utilizado nestes ensaios foi 10 mM HEPES e 25 mM KCl com 2 mM ATP Em
(Tiago et al 2004) o valor de IC50 para a actividade Mg2+-ATPaacutesica na ausecircncia de actina foi de
125 plusmn 025 microM enquanto o valor da actividade estimulada por actina foi de 136 plusmn 14 microM O
meio de reacccedilatildeo utilizado nos ensaios foi 20 mM HEPES 50 mM KCl e 2 mM ATP o que resulta
precisamente no dobro da forccedila ioacutenica a que foi realizado o ensaio do presente relatoacuterio
(exceptuando a contribuiccedilatildeo dos produtos da hidroacutelise de ATP uma vez que este meacutetodo
pressupotildee a ocorrecircncia de um ciclo de reacccedilotildees de hidroacutelise e siacutentese deste composto) Ora tendo
sido anteriormente referido que a afinidade do S1 de miosina para com a espeacutecie V10 diminui com
Act
ivid
ade
Mg
2+-A
TP
aacutesic
a d
e S
1 d
e
Mio
sin
a
[Decavanadato] (microM) Actividade Estimulada ()
Actividade Natildeo Estimulada ()
Ajuste Actividade Estimulada
Ajuste Actividade Natildeo Estimulada
33
o aumento da forccedila ioacutenica (Tiago et al 2004 tal poderaacute explicar a diferenccedila obtida entre estes
dois conjuntos de valores
243Quantificaccedilatildeo de cisteiacutenas reduzidas pelo meacutetodo de Ellman
A interacccedilatildeo do vanaacutedio com alguns tipos de aminoaacutecidos sensiacuteveis agrave oxidaccedilatildeo pode fornecer-nos
informaccedilatildeo bastante uacutetil em relaccedilatildeo agraves alteraccedilotildees estruturais que conduzem a uma diminuiccedilatildeo da
actividade fisioloacutegica do S1 de miosina Os estudos realizados por Chasteen 1983 concluiacuteram
que a reduccedilatildeo de vanadato (V) a vanadilo (IV) ocorre na presenccedila de agentes redutores comuns no
ambiente celular de que se destaca o aminoaacutecido cisteiacutena Esta eacute tambeacutem um alvo de
complexaccedilatildeo por parte do iatildeo VO2+
Jaacute Ramos et al (2009) demonstrou que a incubaccedilatildeo de
actina com a soluccedilatildeo de decavanadato induz a oxidaccedilatildeo de resiacuteduos de cisteiacutena e agrave formaccedilatildeo do
oxidovanaacutedio (IV) Este grupo de investigadores descobriu ainda que apenas a espeacutecie V10 induz a
oxidaccedilatildeo das cisteiacutenas ldquoraacutepidasrdquo da actina atraveacutes do processo proposto que eacute parcialmente
responsaacutevel pela inibiccedilatildeo jaacute descrita da actividade de ATPase do S1 de miosina estimulada por
actina quando exposta agrave soluccedilatildeo de decavanadato Esta inibiccedilatildeo apresenta-se afectada com a
presenccedila de ATP uma vez que os resiacuteduos de cisteiacutena permaneceram intactos nos ensaios
realizados com este ligando Jaacute Crans et al (2004) concluiu que o siacutetio activo de muitas fosfatases
(que conteacutem grupos tiol) pode formar um equiliacutebrio redox com o iatildeo vanadato (V) de forma natildeo
reversiacutevel
Os resultados obtidos por Sakurai e Shiromura (1980) sugeriram que o vanaacutedio (V) sofre reduccedilatildeo
a vanaacutedio (IV) sem dependecircncia do valor de pH do ensaio Registaram ainda uma estequiometria
de 31 na reacccedilatildeo do vanadato (V) com os resiacuteduos de Cys enquanto a estequiometria dos ensaios
com o vanadilo (IV) foi de 21
Berlett e Stadtman (1997) observaram que os tipos de aminoaacutecidos mais facilmente oxidaacuteveis
englobam entre outros os resiacuteduos de Cys Trp e MetAssim analisou-se o estado redox dos
grupos tiol dos resiacuteduos de Cys do S1 na presenccedila do decavanadato atraveacutes da monitorizaccedilatildeo da
Abs412 nm na presenccedila de 1 mgmL de S1 de miosina em tampatildeo 10 mM HEPES 25 mM KCl
variando a concentraccedilatildeo da soluccedilatildeo de decavanadato de modo a determinar que tipo de alteraccedilotildees
estruturais ocorrem na sua presenccedila no que diz respeito agrave diminuiccedilatildeo da actividade ATPaacutesica de
S1 na presenccedila de decavanadato
Os resultados referentes agrave quantificaccedilatildeo das cisteiacutenas reduzidas apoacutes o tratamento de S1 com
diferentes concentraccedilotildees de decavanadato em tampatildeo 10 mM HEPES pH 75 e 25 mM KCl a
25ordmC durante 20 minutos encontram-se descritos na Tabela 2VII
Tabela 2VII ndash Cisteiacutenas reduzidas totais e ldquoraacutepidas de S1 (1 mgmL) apoacutes tratamento com decavanadato
34
(010501002505001000 uM) em 10 mM HEPES 25 mM KCl pH 7 a 25ordmC durante 20 minutos A detecccedilatildeo
espectrofotomeacutetrica foi feita com DTNB (e 412=8400 uM-1 cm-1) (n variaacutevel entre 1 e 8)
Amostras Cys raacutepidas Cys totais
11 M S1 (controlo) 22 plusmn 04 106 plusmn 14
11 M S1 + 10 M Decavanadato 22 plusmn 05 95 plusmn 08
11 M S1 + 50 uM Decavanadato 20 plusmn 02 98 plusmn 14
11 M S1 + 100 uM Decavanadato 17 plusmn 03 97 plusmn14
11 M S1 + 250 uM Decavanadato 26 plusmn 02 98 plusmn 08
11 M S1 + 500 uM Decavanadato 21 plusmn 04 90 plusmn 06
11 M S1 + 1000 uM Decavanadato 24 plusmn 0 86 plusmn 08
A partir dos resultados obtidos pode sugerir-se que o S1 possui 11 resiacuteduos de Cys (controlo) o
que estaacute de acordo com o anteriormente descrito Verifica-se que o tratamento da proteiacutena com
concentraccedilotildees crescentes de decavanadato provoca a oxidaccedilatildeo de dois resiacuteduos de Cys do S1 de
miosina natildeo expostos ao solvente No entanto a teacutecnica utilizada natildeo nos permite identificar
quais
Tendo em consideraccedilatildeo que o valor de IC50 obtido para a reacccedilatildeo de inibiccedilatildeo daactividade Mg2+-
ATPaacutesica de S1 na presenccedila de decavanadato eacute de cerca de 27 microM pode sugerir-se que este seraacute o
valor ao qual ocorreraacute a oxidaccedilatildeo do primeiro resiacuteduo de Cys o que poderaacute contribuir para a
diminuiccedilatildeo da actividade de S1 observada
25Interacccedilatildeo de S1 com vanaacutedio(IV)
251Anaacutelise por espectroscopia de Ressonacircncia Paramagneacutetica Electroacutenica
A acumulaccedilatildeo intracelular de oxovanaacutedio (IV) in vivo constitui uma das principais formas de
apresentaccedilatildeo de vanaacutedio no organismo Deste modo torna-se bastante interessante analisar a
interacccedilatildeo entre este iatildeo e o S1 de miosina atraveacutes da teacutecnica de espectroscopia de Ressonacircncia
Paramagneacutetica Electroacutenica o meacutetodo de eleiccedilatildeo do estudo do iatildeo vanadilo e dos complexos por si
formados O electratildeo desemparelhado encontra-se fortemente acoplado ao nuacutecleo 51
V o que
35
resulta num espectro de RPE consistindo de 8 desdobramentos na direcccedilatildeo paralela e outros tantos
na direcccedilatildeo perpendicular (Chasteen 1983)
Os estudos realizados por Ahmed et al (1987) centraram-se na interacccedilatildeo do vanadilo com
proteiacutenas como eacute o caso da insulina transferrina entre outros obtendo um espectro caracterizado
por 8 linhas amplamente espaccediladas (na direcccedilatildeo paralela) Jaacute Ramos et al 2009 concluiu que a
exposiccedilatildeo da soluccedilatildeo de decavanadato agraves formas globular e filamentosa de actina produz um
espectro tiacutepico do iatildeo oxidovanaacutedio (IV) indicando a reduccedilatildeo do vanadato
Para aferir qual a concentraccedilatildeo de S1 que permitia obter uma boa razatildeo sinalruiacutedo titulou-se uma
soluccedilatildeo de sulfato de vanadilo 15 microM ([VOSO4]stock = 5 mM) com concentraccedilotildees crescentes de
proteiacutena (0 5 10 15 e 20 microM) em 10 mM HEPES 25 mM KCl A existecircncia de uma correlaccedilatildeo
linear entre a concentraccedilatildeo de proteiacutena e a intensidade dos sinais de RPE correspondente agrave banda
-12 perpendicular encontra-se representada na Fig 210
Figura 2 12ndash Representaccedilatildeo graacutefica da variaccedilatildeo da intensidade do sinal de RPE vs a concentraccedilatildeo de S1 utilizada
Apoacutes verificar que com uma concentraccedilatildeo de S1 igual a 20 microM se obtinham sinais com uma razatildeo
sinalruiacutedo adequada ao prosseguimento dos estudos sobre a interacccedilatildeo V(IV)-S1 de miosina
titulou-se uma soluccedilatildeo de 20 microM S1 com 0 25 5 10 15 25 50 75 100 150 200 500 e 1000
microM VOSO4
Traccedilou-se o graacutefico da intensidade da banda -12 perpendicular do sinal de EPR em funccedilatildeo da
concentraccedilatildeo de VOSO4 tendo-se determinado uma constante de dissociaccedilatildeo (KD) do complexo
S1-vanadilo de 8748 plusmn 2621 microM V(IV)
y = 016786x + 122435
Rsup2 = 092992759
Inte
nsi
dad
e d
o s
inal
de
RP
E
[S1] (microM)
36
Figura 213 ndash Representaccedilatildeo graacutefica da variaccedilatildeo do sinal de RPE vs a concentraccedilatildeo de vanadilo utilizada (os valores
experimentais estatildeo representados por losangos o seu ajuste estaacute representado pela linha)
A simulaccedilatildeo do espectro de EPR foi efectuada a partir do espectro obtido para uma concentraccedilatildeo
de 1 mM VOSO4 apoacutes adiccedilatildeo de 20 microM S1 de miosina tendo-se obtido os valores da constante de
acoplamento hiperfina (A=5016) e do factor g (g0=195)
Figura 214 ndash Representaccedilatildeo graacutefica do espectro de EPR obtido com 1000 microM de vanadilo e 20 microM de S1 (a cinzento
escuro) A simulaccedilatildeo do espectro encontra-se a cinzento claro
Por comparaccedilatildeo destes com os valores presentes na tabela de correspondecircncia xxx eacute possiacutevel
predizer quais os ligandos provaacuteveis do vanadilo na presenccedila de proteiacutena Assim concluiu-se que
o vanadilo estaacute coordenado com duas moleacuteculas de aacutegua um grupo hidroxilo e um grupo R=NR
O grupo hidroxilo pode indicar os resiacuteduos de aminoaacutecidos Tyr Treonina e Serina enquanto o
grupo R=NR poderaacute indicar a presenccedila de Arginina e Histidina como ligandos do vanadilo
Inte
nsi
dad
e d
os
sinai
s d
e R
PE
[Vanadilo] (microM)
Ajuste dos dados exp Dados experimentais
Campo magneacutetico (G)
Espectro Experimental Ajuste
37
3 Procedimento Experimental
31 Reagentes
Todos os reagentes utilizados satildeo de elevado grau de pureza (analiacutetico) Apresentam-se as suas
foacutermulas quiacutemicas ou abreviaturas quando necessaacuterio acrescentou-se a sua descriccedilatildeo Entre
parecircntesis colocou-se as empresas que os comercializam
Aacutecido aceacutetico glacial 100 (Riedel) Acrilamida 99 (Merck) Azul de bromofenol (Riedel-de-
Haumlen) -Mercaptoetanol (Sigma) Bissulfito de soacutedio (Sigma)Bradford (reagente de)
(BioRad)BSA (albumina de soro bovino Sigma-Aldrich) CaCl22H2O (cloreto de caacutelcio
dihidratado Fluka) CH3CH2OH 96 (etanol Panreac) Coomassie Blue R250 (Merck) D2O
(oacutexido de deuteacuterio SDS) DTNB (55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico) Aldrich) EDTA (aacutecido
etilenodiaminatetraaceacutetico Merck) Fosfoenolpiruvato 98 (Fluka) Glicerol (Sigma) Glicina
985 (Panreac) HCl 37 (Riedel-de Haumlen) H2SO4 96 (Pronalab) HEPES 99 (aacutecido 4-(2-
hidroxiletil)-1-piperazineetanosulfoacutenico Sigma) KH2PO4 99 (hidrogenofosfato de potaacutessio)
MgATP (Sigma) Na
ATP (Sigma) Na2HPO4 99 (hidrogenofosfato de soacutedio Riedel-de Haumlen)
NaH2PO4 (fosfato de soacutedio ) NaOH 986 (hidroacutexido de soacutedio Pronalab) KCl 99 (Cloreto de
potaacutessio Panreac) L-Cys 995 (enantioacutemero levoacutegiro do resiacuteduo de Cisteiacutena Fluka)Metanol
99 (Merck) Molibdato de amoacutenio (Sigma)NNrsquo-metil-bis-acrilamida 98 (Merck) NH4VO3
99 (metavanadato de amoacutenia Sigma) PSA 10 (persulfato de amoacutenio Merck) SDS
(dodecilo sulfato de soacutedio Sigma) Sulfito de soacutedio (Sigma) VOSO4 (sulfato de vanadilo
Aldrich) TEMED 99 (Tetrametiletilenodiamina Merck) Tris (2-amino-2-hidroximetil-13-
propanodiol Sigma)
32 Meacutetodos
321 Isolamento e purificaccedilatildeo das proteiacutenas de muacutesculo esqueleacutetico de coelho
3211 Purificaccedilatildeo da miosina preparaccedilatildeo de Subfragmento 1 por digestatildeo quimotriacutepsica
da miosina
O subfragmento-1 de miosina-II (S1) foi gentilmente cedido pelo Prof Doutor Manuel Aureliano
do Departamento de Quiacutemica Bioquiacutemica e Farmaacutecia da Faculdade de Ciecircncias e Tecnologia da
Universidade do Algarve
No entanto descreve-se abaixo de forma resumida o procedimento efectuado
38
32111 Purificaccedilatildeo da miosina
O protocolo de isolamento foi adaptado do meacutetodo previamente descrito por Margossian e Lowey
(1982) apresentando-se de forma esquemaacutetica na Fig 31
Sucintamente os tecidos musculares dorsais e das pernas foram extraiacutedos apoacutes o sacrifiacutecio do
animal (Fig31) De seguida foram adicionados 3 volumes da soluccedilatildeo de extracccedilatildeo A2
Centrifugou-se a soluccedilatildeo e filtrou-se o sobrenadante O sedimento foi entatildeo diluiacutedo em 14
volumes de aacutegua destilada de modo a que a forccedila ioacutenica atingisse 004 M Apoacutes adiccedilatildeo de szlig-
mercaptoetanol (szlig-me) deixou-se a precipitar a 4ordmC removendo o sobrenadante Centrifugou-se
novamente e ressuspendeu-se o sedimento na soluccedilatildeo de extracccedilatildeo A Adicionou-se a soluccedilatildeo
tampatildeo B3 para que a forccedila ioacutenica atingisse 05 M Homogeneizou-se e deixou-se repousar
durante 2h Homogeneizou-se novamente e centrifugou-se de modo a ocorrer precipitaccedilatildeo dos
complexos de actomiosina e proteiacutena desnaturada Apoacutes filtraccedilatildeo do sobrenadante em latilde de vidro
quantificou-se a soluccedilatildeo de miosina resultante pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental)
2 Soluccedilatildeo de extracccedilatildeo A 0012M K2CO3 004M KHCO3 06M KCl pH 65
3 Soluccedilatildeo-tampatildeo B Tris 100 mM pH 7 KCl 2M
39
Figura 31 ndash Representaccedilatildeo esquemaacutetica do protocolo de isolamento de miosina (Margossian e Lowey 1982) Todo o
procedimento foi realizado a 4ordmC (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
32112 Preparaccedilatildeo de Subfragmento-1 de miosina por digestatildeo com quimiotripsina
O S1 de miosina foi isolado apoacutes diaacutelise da miosina durante a noite contra a soluccedilatildeo C4 A
presenccedila do EDTA eacute essencial uma vez que a sua substituiccedilatildeo por MgCl2resulta na produccedilatildeo de
HMM em vez de S1 A suspensatildeo de miosina foi equilibrada e digerida com -quimotripsina
A digestatildeo foi interrompida pela adiccedilatildeo de PMSF Seguidamente ultracentrifugou-se o produto
da digestatildeo desprezando-se o sedimento (Fig 32) A soluccedilatildeo de sobrenadante maioritariamente
S1 foi quantificada pela leitura directa da Abs280 ou pelo meacutetodo de Bradford (ver as secccedilotildees
3311 e 3312 do Procedimento Experimental respectivamente) e armazenada em 10 sacarose
e 2 mM szlig-Me a -80ordmC
4 Soluccedilatildeo C 25mM Tris pH 70 012M KCl 1mM EDTA
40
Antes da realizaccedilatildeo dos ensaios descritos no presente relatoacuterio o S1 de miosina foi dialisado
contra a soluccedilatildeo D5 de modo a remover qualquer composto que pudesse complexar como
vanaacutedio Seguidamente quantificou-se a proteiacutena pelo meacutetodo de Bradford ou pelo meacutetodo
directo (ver secccedilotildees 3311 e 3312do Procedimento Experimtental)
Figura 3 2ndash Representaccedilatildeo esquemaacutetica do protocolo de preparaccedilatildeo do S1 de miosina por digestatildeo quimiotripsina de
acordo com o previamente descrito por Margossian e Lowey (1982) Todo o procedimento foi realizado a 4ordm C
excepto quando indicaccedilatildeo do contraacuterio (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
3212 Isolamento e purificaccedilatildeo de actina
Isolou-se e purificou-se a actina segundo o meacutetodo descrito por Pardee e Spudish que
compreende dois passos ndash a obtenccedilatildeo de um resiacuteduo seco denominado ldquopoacute de acetonardquo e a
extracccedilatildeo subsequente da actina a partir do ldquopoacute de acetonardquo na forma de G-Actina com a
posterior polimerizaccedilatildeo a F-Actina
32121 Obtenccedilatildeo do ldquopoacute de acetonardquo
Sacrificou-se o coelho com uma pancada atraacutes do pescoccedilo e sangrou-se o animal apoacutes o corte da
arteacuteria jugular Esfolou-se e colocou-se numa tina com gelo Retiraram-se as partes de tecido
conjuntivo que envolvem o tecido muscular com auxiacutelio de uma pinccedila e bisturi Seguidamente
retirou-se os muacutesculos dorsais e os muacutesculos brancos das pernas Passou-se os muacutesculos por aacutegua
destilada de modo a remover as impurezas e pesou-se num copo Trituraram-se os muacutesculos
numa maacutequina de triturar carne e extraiu-se o resultante com 3 volumes da soluccedilatildeo E6 durante 10
minutos com agitaccedilatildeo suave Centrifugou-se a suspensatildeo a 4000 g durante 10
5 Soluccedilatildeo D HEPES 10 mM pH 70 KCl 25 mM
6 Soluccedilatildeo E 01 M KCl 015 M K2HPO4 pH 65
41
minutosdesprezando-se o sobrenadante Ressuspendeu-se o sedimento em 6 volumes de soluccedilatildeo
F7durante 10 minutos com agitaccedilatildeo suave Verificou-se se o pH da soluccedilatildeo estava compreendido
entre 82 e 85 acertando-se estecom soluccedilatildeo G8 se necessaacuterio Centrifugou-se durante 10
minutos a 4000 g Desprezou-se o sobrenadante e ressuspendeu-se o sedimento em 3 volumes de
soluccedilatildeo H9 durante 10 minutos Centrifugou-se novamente a 4000 gdurante 10 minutos
desprezando-se o sobrenadante Ressuspendeu-se o sedimento duas vezes consecutivas em 6
volumes de aacutegua destilada durante 5 minutos Centrifugou-se a 4000 g durante 10 minutos apoacutes
cada adiccedilatildeo de H2O
Seguidamente ressuspendeu-se o sedimento 5 vezes consecutivas utilizando 3 volumes de
acetona de cada vez agitando suavemente durante 10 minutos agrave temperatura ambiente Colocou-
se o resiacuteduo filtrado numa tina de vidro sobre papel de filtro deixando-se evaporar o solvente
durante a noite num nicho Triturou-se o resiacuteduo seco numa maacutequina de triturar carne e deixou-se
novamente evaporar o solvente O ldquopoacute de acetonardquo obtido foi armazenado a -20 ordmC
32122 Purificaccedilatildeo da actina a partir de ldquopoacute de acetonardquo
Apoacutes a obtenccedilatildeo do poacute de acetona como acima indicado efectuou-se o protocolo seguidamente
descrito de modo a obter monoacutemeros de actina (G-Actina) e actina filamentosa (F-Actina)
Pesou-se 35 g de poacute de acetona e adicionaram-se 50 mL de soluccedilatildeo-tampatildeo I10
sendo a
extracccedilatildeo feita com agitaccedilatildeo suave durante 30 minutos Separou-se o extracto do poacute de acetona
hidratado filtrando a suspensatildeo com 4 camadas de gaze esterilizada tendo o cuidado de natildeo
espremer o resiacuteduo Voltou a extrair-se com 50 mL de soluccedilatildeo I durante 30 minutos mexendo
lentamente Adicionou-se este ao extracto obtido anteriormente Centrifugou-se o extracto total a
16 000 g durante 30 minutos Decantou-se para uma proveta sobre a qual se colocou gaze para
que o sedimento solto natildeo se juntasse ao sobrenadante e leu-se o volume De seguida adicionou-
se KCl ateacute uma concentraccedilatildeo final de 50 mM MgCl2 ateacute 2mM e ATP ateacute 1mM lentamente e com
agitaccedilatildeo contiacutenua de modo a promover a polimerizaccedilatildeo dos monoacutemeros de actina
Deixou-se polimerizar durante 2h a 25ordmC observando-se a presenccedila de pequenas bolhas de ar
indicativas de um aumento de viscosidade da soluccedilatildeo Arrefeceu-se em gelo durante 15 minutos e
adicionou-se KCl ateacute uma concentraccedilatildeo final de 06M Mexeu-se lentamente durante 30 minutos
apoacutes os quais se ultracentrifugou a 110 000g durante 1hora Desprezou-se o sobrenadante
ressuspendeu-se o sedimento com soluccedilatildeo tampatildeo E e transferiu-se para um potter
Dialisou-se durante a noite contra 1L de soluccedilatildeo-tampatildeo I a uma temperatura de 4ordmC com rotaccedilatildeo
7 Soluccedilatildeo F 005 M NaHCO3 pH 82-85
8 Soluccedilatildeo G 1M Na2CO3
9 Soluccedilatildeo H 1 mM EDTA pH 70
10 Soluccedilatildeo I 2 mM Tris-HCl pH 8 02 mM CaCl2 02 mM Na2ATP 05 mM szlig-Me 0005 NaN3
42
contiacutenua e mudanccedila de tampatildeo 2 a 3 vezes De seguida a actina foi centrifugada a 110
000gdurante 1hora Decantou-se a soluccedilatildeo de G-Actina (sobrenadante) e desprezou-se o
sedimento Determinou-se a concentraccedilatildeo de G-Actina pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental) A soluccedilatildeo de G-actina foi armazenada a uma temperatura de 4ordmC
apoacutes adiccedilatildeo de 002NaN3 mantendo-se estaacutevel durante 3 a 5 dias
331 Quantificaccedilatildeo das proteiacutenas isoladas
3311 Quantificaccedilatildeo pelo meacutetodo de Bradford
O meacutetodo de Bradford (Bradford 1976) baseia-se na utilizaccedilatildeo do reagente de Bradford
composto por entre outros aacutecido fosfoacuterico e o corante ldquoCoomassierdquo Este corante possui carga
negativa e a sua coloraccedilatildeo varia entre a vermelha (Absmax=465 nm) predominante em soluccedilatildeo e a
azul (Absmax=595 nm) presente aquando da ligaccedilatildeo aos grupos amina das proteiacutenas Em soluccedilotildees
aacutecidas diluiacutedas a coloraccedilatildeo varia proporcionalmente com a quantidade do corante ligado agrave
proteiacutena que por sua vez eacute directamente proporcional agrave concentraccedilatildeo da proteiacutena permitindo a
sua quantificaccedilatildeo ateacute a um limite de detecccedilatildeo de 20 gmL (Bradford 1976)
Deste modo prepararam-se amostras de BSA ([BSA]stock= 1mgmL) utilizando um intervalo de
concentraccedilotildees compreendido entre 0 e 15 gmL de acordo com a tabela 3I Apoacutes a adiccedilatildeo da
H2O MilliQ e do reagente de Bradford as soluccedilotildees foram sucessivamente homogeneizadas no
voacutertex
Tabela 3Indash Amostras utilizadas para a obtenccedilatildeo da recta de calibraccedilatildeo no meacutetodo de Bradford
BSA (L) BSA (g) H2O MilliQ (L) Reagente de Bradford (L)
0 0 800 200
125 125 79875 200
25 25 7975 200
5 5 795 200
10 10 790 200
15 15 785 200
Apoacutes incubaccedilatildeo a 25ordmC por 5 minutos leram-se os valores de Abs595 num
espectrofotoacutemetro (Shimadzu UV-160A) para todas as amostras Seguidamente traccedilou-se a recta
de calibraccedilatildeo (Abs595vs quantidade de BSA (g))
43
Para determinar a concentraccedilatildeo de proteiacutena apoacutes o processo de isolamento purificaccedilatildeo e
diaacutelise quando necessaacuterio colocou-se 5 L da amostra num tubo de Eppendorf e adicionou-se
795 L de H2O MilliQ homogeneizando a soluccedilatildeo no voacutertex Adicionou-se posteriormente 200
L de reagente de Bradford e repetiu-se a homogeneizaccedilatildeo Incubou-se a mistura a 25ordmC por 5
minutos apoacutes os quais se leu os valores de Abs595 nm no espectrofotoacutemetro Por interpolaccedilatildeo deste
valor a partir da recta de calibraccedilatildeo determinou-se a concentraccedilatildeo de proteiacutena na amostra
3312 Quantificaccedilatildeo pelo meacutetodo directo
A quantificaccedilatildeo das amostras proteicas pode tambeacutem ser efectuada atraveacutes da leitura directa da
absorvacircncia tendo por base a lei de Beer-Lambert onde Abs corresponde agrave
absorvacircncia ao comprimento de onda ao qual o ensaio foi realizado corresponde ao coeficiente
de extinccedilatildeo (substituiacutedo pela letra a se as unidades forem distintas da molaridade) b corresponde
ao percurso oacuteptico da radiaccedilatildeo na ceacutelula amostral e c corresponde agrave concentraccedilatildeo amostral
Assim no caso de S1 de miosina mediu-se a Abs280e calculou-se a concentraccedilatildeo da proteiacutena
sabendo que o coeficiente de extinccedilatildeo de S1 eacute (Ref)
No caso da actina mediu-se a Abs290ecalculou-se a concentraccedilatildeo da proteiacutena sabendo que o
coeficiente de extinccedilatildeo da actina eacute (Ref)
332Electroforese em condiccedilotildees desnaturantes (SDS-PAGE)
De modo a melhor caracterizar as proteiacutenas isoladas foi realizada uma electroforese em
condiccedilotildees desnaturantes (SDS-PAGE do inglecircs Sodium Dodecyl Sulfate Polyacrilamide Gel
Electrophoresis) O SDS (dodecilo sulfato de soacutedio) eacute um detergente anioacutenico utilizado neste
meacutetodo para desnaturar as proteiacutenas que se ligam ao SDS na proporccedilatildeo 14 g SDS g proteiacutena
(aproximadamente 1 moleacutecula de SDS por cada 2 resiacuteduos de aminoaacutecidos) (Stryer) O SDS induz
uma densidade de carga negativa significativa nas proteiacutenas a que se liga mascarando a sua carga
intriacutenseca pelo que as razotildees cargamassa satildeo tendencialmente idecircnticas Assim a SDS-PAGE
separa as proteiacutenas quando sujeitas a um campo eleacutectrico de acordo com o seu peso molecular
sendo a sua migraccedilatildeo directamente proporcional ao logaritmo deste uacuteltimo (Stryer)
O gel de resoluccedilatildeo com 10 de acrilamida foi obtido atraveacutes da adiccedilatildeo de 167 mL de soluccedilatildeo
stock de acrilamida11
075 mL de tampatildeo de resoluccedilatildeo12
251 mL de aacutegua destilada e 50 L de
SDS 10 Para promover a polimerizaccedilatildeo adicionou-se em seguida 38 L de PSA 10
11
soluccedilatildeo stock de acrilamida soluccedilatildeo de 30 (mv) de acrilamida e 08 (mv) de NNrsquo-metil-bis-acrilamida 12
tampatildeo de resoluccedilatildeo 15 M Tris-HCl pH 88
44
(persulfato de amoacutenia) 10 e 25 L de TEMED 99 (tetrametiletilenodiamina) Agitou-se
cuidadosamente de modo a natildeo formar bolhas e pipetou-se a soluccedilatildeo para a cassette de vidro
previamente montada no suporte De seguida adicionou-se uma peliacutecula de cerca de 2 mm de 2-
metil-2-butanol para verificar posteriormente a polimerizaccedilatildeo Apoacutes a polimerizaccedilatildeo que demora
cerca de 45 minutos verteu-se a agrave superfiacutecie e adicionou-se o gel de concentraccedilatildeo com 5 de
acrilamida constituiacutedo por 03 mL de soluccedilatildeo de stock acrilamida 045 mL de tampatildeo de
concentraccedilatildeo13
094 mL de aacutegua destilada 18 L de SDS 10 135 L de PSA 10 e 2L de
TEMED 99 Colocou-se o pente sobre o gel e deixou-se polimerizar durante cerca de 45
minutos
As amostras para aplicaccedilatildeo no gel de electroforese foram preparadas para uma concentraccedilatildeo de
proteiacutena de 1 mgmL num volume final de 100 L dos quais 25 L correspondem ao tampatildeo da
amostra14
As amostras foram aquecidas a 100ordmC durante 5 minutos Antes da aplicaccedilatildeo das
amostras transferiu-se a cassette para a tina de electroforese agrave qual se adicionou a soluccedilatildeo
tampatildeo de electroforese da tina superior15
e tampatildeo de electroforese da tina inferior16
Aplicaram-
se 10 L das soluccedilotildees de marcadores de peso molecular e 20 L das amostras proteicas
preparadas anteriormente A electroforese decorreu durante cerca de 3h30 min a uma voltagem
de 80 V De seguida transferiu-se o gel para uma tina corando as proteiacutenas durante 1h com
soluccedilatildeo corante17
Descorou-se o gel com soluccedilatildeo descorante18
tendo-se mudado esta soluccedilatildeo ateacute
as bandas se encontrarem bem definidas
Determinou-se a mobilidade relativa das bandas correspondentes aos marcadores de elevado peso
molecular e traccedilou-se a recta de calibraccedilatildeo ndash log(PM) vs log(Rf) onde Rf corresponde agrave
mobilidade relativa Por interpolaccedilatildeo determina-se o PM das proteiacutenas isoladas sabendo a sua
mobilidade relativa Por comparaccedilatildeo com os valores descritos na literatura pode concluir-se
acerca do grau de purificaccedilatildeo
34 Mediccedilatildeo da actividade enzimaacutetica de S1
341 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
13
tampatildeo de concentraccedilatildeo 05 M Tris-HCl pH 68 14
tampatildeo de amostra 320 mM Tris-HCl pH 68 04 M szlig-mercaptoetanol SDS 8 glicerol 15 (vv) e roxo de m-
cresol 0024 15
tampatildeo de electroforese da tina superior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS 16
tampatildeo de electroforese da tina inferior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS acetato de soacutedio
01 M 17
soluccedilatildeo corante ldquoCoomassie Blue R-250rdquo contendo metanol 45 (vv) e aacutecido aceacutetico glacial 10 (vv) 18
soluccedilatildeo descorante metanol 10 (vv) e aacutecido aceacutetico glacial 10 vv)
45
O meacutetodo de Fiske-Subarow modificado consiste na reacccedilatildeo do molibdato de amoacutenio com o
fosfato inorgacircnico em soluccedilatildeo aacutecida com formaccedilatildeo do aacutecido fosfomoliacutebdico Seguidamente daacute-
se a adiccedilatildeo de um agente redutor que provoca a reduccedilatildeo do composto a fosfomolibdato que forma
um complexo de cor azulada cuja absorvacircncia maacutexima ocorre para um comprimento de onda de
660 nm tornando possiacutevel a sua detecccedilatildeo por meacutetodos espectrofotomeacutetricos As actividades Ca2+
-
ATPaacutesica e K+EDTA-ATPaacutesica de S1 foram determinadas atraveacutes deste meacutetodo de acordo com
o descrito por Taussky e Shorr (1953) com mediccedilatildeo da quantidade de fosfato (Pi) libertado
atraveacutes da hidroacutelise de ATP
Para tal obteve-se primeiramente a recta de calibraccedilatildeo com as amostras presentes na Tabela 3II
utilizando uma soluccedilatildeo stock de 5 mM KH2PO4
Tabela 3II ndash Composiccedilatildeo das diferentes amostras utilizadas para determinar a recta de calibraccedilatildeo atraveacutes do meacutetodo
de Fiske-Subbarow modificado
KH2PO4 (L)
([KH2PO4]stock= 5mM)
PO43-
(nmole) H2O (uL)
0 0 1780
5 25 1775
10 50 1770
20 100 1760
40 200 1740
100 500 1680
A cada tubo de Eppendorf adicionaram-se seguidamente 200 L de soluccedilatildeo J19
e 20 L de agente
redutor20
homogeneizando no voacutertex apoacutes cada adiccedilatildeo Incubou-se durante 10 minutos e mediu-
se a Abs660 nm num espectrofotoacutemetro com intervalos de tempo iguais entre cada amostra Traccedilou-
se a recta de calibraccedilatildeo (Abs660 nmvs nmol PO43-
) a partir dos valores obtidos
Para determinar a actividade Ca2+
-ATPaacutesica de S1 prepararam-se tubos de Eppendorf com 400
L de meio de reacccedilatildeo21
25 L CaCl2 ([CaCl2]stock= 100 mM) 25 L ATP ([ATP]stock= 100 mM)
com [S1]ensaios=01 mgmL Perfez-se se necessaacuterio com H2O MilliQ ateacute perfazer um volume de 1
mL Para determinar a actividade K+EDTA-ATPaacutesica utilizou-se uma concentraccedilatildeo idecircntica de
19
Soluccedilatildeo J 5 N H2SO4 25 molibdato de amoacutenio 20
Agente redutor 5 mg aacutecido 1-amino-2-naftol-4-sulfoacutenico 30 mg bissulfito de soacutedio 30 mg sulfito de soacutedio em 26
mL H2O 21
meio de reacccedilatildeo 125 mM Tris-HCl pH 80 0575 M KCl
46
proteiacutena nos ensaios (01 mgmL) e o mesmo meio de reacccedilatildeo Aos tubos de Eppendorf
adicionou-se ainda 123 L de KCl ([KCl]stock = 3M) 25 L de ATP 10 L de EDTA
([EDTA]stock= 100 mM )e o volume de H2O MilliQ necessaacuterio para perfazer 1 mL Em ambas as
situaccedilotildees o uacuteltimo reagente adicionado foi o ATP uma vez que se trata do substrato da reacccedilatildeo
enzimaacutetica
A cineacutetica decorreu a 25ordmC durante 5 minutos para a actividade Ca2+
-ATPaacutesica e durante 3
minutos para a actividade K+EDTA-ATPaacutesica Apoacutes o periacuteodo de incubaccedilatildeo transferiu-se 150
L de amostra para tubos de Eppendorf contendo igual volume de TCA 10 Colocou-se no gelo
e homogeneizou-se no voacutertex Seguidamente centrifugou-se a 1500 g durante 10 minutos e
recolheu-se 200 L de sobrenadante para novos tubos de Eppendorf Adicionou-se a estes 158
mL de H2O MilliQ e 200 L de soluccedilatildeo J Homogeneizou-se no voacutertex e adicionou-se 20 L de
reagente redutor repetindo-se a homogeneizaccedilatildeo Incubou-se durante 10 minutos e mediu-se a
Abs660 nm com o mesmo intervalo de tempo de leitura para cada amostra Por intrapolaccedilatildeo a partir
da recta de calibraccedilatildeo obtida anteriormente determinou-se a quantidade de Pi libertado em cada
ensaio A partir destes resultados determinaram-se as actividades Ca2+
-ATPaacutesica e K+EDTA-
ATPaacutesica do S1 em nmol PO43-
minmg proteiacutena e em s-1
342 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
Da actividade Mg2+
-ATPaacutesica de S1 resulta a formaccedilatildeo dos produtos da reacccedilatildeo de
hidroacutelise do ATP - ADP e Pi Na presenccedila de ADP fosfoenolpiruvato (PEP) e da enzima piruvato
cinase (PK) os produtos resultantes satildeo ATP e piruvato Por acccedilatildeo da enzima lactato
desidrogenase (LDH) este uacuteltimo eacute convertido em lactato ocorrendo simultaneamente a oxidaccedilatildeo
do NADH a NAD+ monitorizada atraveacutes da Abs340 nm Quando o S1 de miosina se encontra no
meio reaccional o ATP eacute hidrolisado formando-se novamente ADP e Pi o que permite a
formaccedilatildeo de um ciclo (fig 21) Se as enzimas se encontrarem em excesso a velocidade de
conversatildeo do NADH eacute limitada pela produccedilatildeo de ADP e Pi Como o ATP eacute regenerado em cada
ciclo a sua concentraccedilatildeo inicial eacute sempre mantida (Bagshaw 1982)
Figura 33ndash Representaccedilatildeo esquemaacutetica do meacutetodo de enzimas acopladas
47
O protocolo utilizado e seguidamente descrito foi adaptado de Cuenda et al (1990 ndash Ver Refordf) A
uma cuvette de 1 mL adicionou-se S1 para uma concentraccedilatildeo final de 005 mgmL 800 L de
meio de reacccedilatildeo22
10 L de PEP ([PEP]stock = 42 mM) 10 L de PK+LDH 11 ([PK+LDH]stock =
1 mgmL) e 15 L de NADH ([NADH]stock=25 mM) Iniciou-se a monitorizaccedilatildeo da reacccedilatildeo
atraveacutes da leitura de Abs340 nm apoacutes a adiccedilatildeo de 20 L de ATP ([ATP]stock=100 mM) A
actividade Mg2+
-ATPaacutesica eacute obtida atraveacutes do declive correspondente agrave oxidaccedilatildeo do NADH Para
a mediccedilatildeo da Mg2+
-ATPase de S1 estimulada por actina adicionou-se F-actina para uma
concentraccedilatildeo final de 2 microM sendo a actividade obtida tambeacutem atraveacutes do declive correspondente
agrave oxidaccedilatildeo do NADH apoacutes a adiccedilatildeo da proteiacutena
35 Interaccedilatildeo entre vanaacutedio e subfragmento-1 de miosina
351 Preparaccedilatildeo das soluccedilotildees de vanaacutedio
As soluccedilotildees de vanadato foram preparadas em aacutegua MilliQ a partir de metavanadato de amoacutenio
(NH4VO3) numa concentraccedilatildeo stock de 50 mM com agitaccedilatildeo magneacutetica e aquecimento (~30 ordmC)
Esta soluccedilatildeo com um valor compreendido entre 6 e 7 designa-se por ldquometavanadatordquo e eacute
constituiacuteda pelas esspeacutecies V1 V2 V4 e V5 (Ref) Acidificando a soluccedilatildeo de metavanadato por
adiccedilatildeo de HCl ateacute um valor de pH proacuteximo de 4 observa-se uma mudanccedila de cor de amarelo
claro para laranja indicativa da formaccedilatildeo da espeacutecie decameacuterica de vanadato De seguida e
imediatamente antes da realizaccedilatildeo dos ensaios descritos o valor do pH da soluccedilatildeo de
decavanadato foi aumentado para cerca de 75 por adiccedilatildeo de NaOH A soluccedilatildeo (2) designada
por ldquometavanadatordquo natildeo foi posteriormente alterada ateacute agrave sua adiccedilatildeo para a realizaccedilatildeo dos ensaios
As soluccedilotildees foram armazenadas a 4ordmC e mantidas em gelo aquando da sua utilizaccedilatildeo
A soluccedilatildeo de V(IV) foi preparada em condiccedilotildees anaeroacutebias por diluiccedilatildeo de sulfato de vanadil
(VOSO4) para uma concentraccedilatildeo final de 5 mM em H2O MilliQ saturada com N2 gasoso
352Anaacutelise por espectroscopia de UV-Vis
A concentraccedilatildeo da espeacutecie decameacuterica em soluccedilatildeo depende do periacuteodo de tempo apoacutes a diluiccedilatildeo
no meio de reacccedilatildeo apropriado (Ref) A velocidade de desoligomerizaccedilatildeo da espeacutecie decameacuterica
pode ser analisada por espectroscopia de absorccedilatildeo na regiatildeo UVVis uma vez que esta espeacutecie
apresenta duas bandas caracteriacutesticas na regiatildeo do visiacutevel (a 358 e 400 nm) que natildeo se verifica
em quaisquer outras espeacutecies oligomeacutericas As bandas supramencionada sapresentam-se bem
22
Meio de reacccedilatildeo 3125 mM Tris-HCl pH 71 375 mM MgCl2
48
resolvidas atraveacutes da obtenccedilatildeo da derivada de 2ordf ordem do espectro apresentando a forma de 2
vales com miacutenimos centrados nos comprimentos de onda supracitados com dependecircncia linear
relativamente agrave concentraccedilatildeo de decavanadato total (Ramos et al 2006)
Assim a estabilidade da espeacutecie decameacuterica apoacutes diluiccedilatildeo da soluccedilatildeo de decavanadato no meio de
reacccedilatildeo dos ensaios (tampatildeo D) para uma concentraccedilatildeo final de 1 mM em vanadato total foi
caracterizada atraveacutes da leitura da Abs400 em funccedilatildeo do tempo tendo-se determinado qual a
ordem de reacccedilatildeo e o tempo de meia-vida da mesma Com base em estudos anteriores acerca da
estabilidade do V10 na presenccedila de diferentes proteiacutenas (Ramos et al 2006) determinou-se
igualmente o tempo de meia vida apoacutes a adiccedilatildeo de 20 microM de S1 de miosina ao meio reaccional
353 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
Os espectros de RMN do isoacutetopo 51
V foram realizados num espectroacutemetro Bruker AM-400 MHz
equipado com uma sonda multinuclear inversa de 5 mm Em tubos de RMN de 5 mm de diacircmetro
preparou-se soluccedilotildees de 500 L de deca- ou metavanadato correspondentes a um intervalo de
concentraccedilotildees de 1 a 10 mM suplementadas com 10 D2O Os espectros foram adquiridos agrave
temperatura ambiente com uma largura espectral de 23 923 Hz um tempo de aquisiccedilatildeo de 0086
s tempo de relaxaccedilatildeo de 001 s e uma sequecircncia de pulsos a 90ordm Utilizou-se a transformada de
Fourier com alargamento das bandas de 20Hz para o processamento dos espectros Os desvios
quiacutemicos das ressonacircncias de 51
V foram obtidos em relaccedilatildeo agrave espeacutecie VOCl3cuja frequecircncia de
ressonacircncia eacute considerada o padratildeo sendo todas as outras referenciadas a partir desta em ppm
As aacutereas dos sinais do RMN correspondentes agraves espeacutecies oligomeacutericas presentes foram obtidas
por integraccedilatildeo dos sinais As concentraccedilotildees relativas das espeacutecies foram calculadas com recurso agrave
equaccedilatildeo
onde A[Vx] corresponde agrave aacuterea referente agrave ressonacircncia da espeacutecie Vx com nordm de agregaccedilatildeo x
A[V]total agrave soma total das aacutereas medidas e [V] a concentraccedilatildeo total de vanaacutedio no ensaio
A teacutecnica de 51
V-RMN foi anteriormente aplicada em estudos de ligaccedilatildeo com S1 de miosina
(Ref) tendo sido demostrado que entre as espeacutecies V1 V2 V4 e V5 a espeacutecie tetrameacuterica possui a
maior afinidade para a miosina (Cremo et al) O desaparecimento ou alargamento dos sinais pode
implicar a ligaccedilatildeo de um dos oxovanadatos laacutebeis agrave miosina sem indicaccedilatildeo de qual uma vez que
49
se processa um equiliacutebrio entre eles (Crans 1990) Assim e tendo em conta as indicaccedilotildees de que
a espeacutecie V10 eacute distinta das restantes no que diz respeito agrave magnitude da interacccedilatildeo com a miosina
e inibiccedilatildeo da actividade ATPaacutesica estimulada por actina (Aureliano 2000 Tiago et al 2002)
torna-se fundamental investigar a interacccedilatildeo das diferentes espeacutecies de vanadato com o S1 de
miosina atraveacutes da teacutecnica de 51
V-RMN
Para tal obtiveram-se espectros de 51
V-RMN resultantes da titulaccedilatildeo de uma soluccedilatildeo 5 mM de
decavanadato total com concentraccedilotildees crescentes de S1 de miosina (0 5 10 15 e 20 microM) em
tampatildeo D Realizou-se ainda ensaios na presenccedila de 2 mM de decavanadato e 2 mM de ATP na
presenccedila e na ausecircncia de S1 de miosina de modo a verificar o efeito que estes ligandos naturais
da enzima teria na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o subfragmento-1
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
Os grupos sulfidrilo (-SH) das cisteiacutenas satildeo resiacuteduos de aminoaacutecidos extremamente sensiacuteveis agrave
oxidaccedilatildeo resultando na sua oxidaccedilatildeo (Dalle-Donne et al 1999) Logo a quantificaccedilatildeo destes
resiacuteduos eacute bastante importante nos estudos de interacccedilatildeo com vanaacutedio jaacute que os resiacuteduos de Cys
com ecircnfase para o SH1 e o SH2 satildeo bastante importantes na monitorizaccedilatildeo das modificaccedilotildees
funcionais da regiatildeo da cabeccedila
O procedimento seguidamente apresentado foi originalmente descrito por Gutieacuterrez-Merino
(1987)) Consiste na reacccedilatildeo entre os grupos ndashSH natildeo oxidados das proteiacutenas e o reagente de
Ellman (DTNB -55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico)) - formando-se o aniatildeo CNT (aniatildeo 3-
carboxilato-4-nitrotiofenolato) numa estequiometria de 11 Este apresenta um maacuteximo de
absorvecircncia a 412 nm (Robyt 1990) Assim apoacutes a incubaccedilatildeo da proteiacutena na presenccedila do metal
a quantidade de cisteiacutenas reduzidas por proteiacutena determina-se atraveacutes dos valores de Abs412
Deste modo colocou-se num tubo de Eppendorf o volume apropriado de S1 dialisado para que a
concentraccedilatildeo no ensaio fosse de 1 mgmL Adicionou-se o volume apropriado de decavanadato
para que a sua concentraccedilatildeo fosse de 0 10 50 100 250 500 ou 1000 M Perfez-se com soluccedilatildeo
D ateacute um volume final de 1 mL e incubou-se a 25ordmC por 20 minutos Apoacutes este periacuteodo colocou-
se 780 L da mistura reaccional numa cuvette de quartzo e adicionou-se 220 L de soluccedilatildeo K23
23
Tampatildeo K Tampatildeo fosfatohidrogenofosfato de soacutedio 025 M pH 72
50
Adicionou-se 15 L de DTNB (em soluccedilatildeo K [DTNB]stock=100 mM) ao meio reaccional e
registou-se o valor de absorvecircncia medido equivalente agraves cisteiacutenas ldquoraacutepidasrdquo (SH1 e SH2
expostos ao solvente) Adicionou-se 001 g de SDS que provoca a desnaturaccedilatildeo da proteiacutena e
consequentemente a exposiccedilatildeo de todos os resiacuteduos de cisteiacutenas que se encontravam ateacute aiacute
inacessiacuteveis no interior da proteiacutena (cisteiacutenas totais) Estes resiacuteduos podem assim reagir com o
DTNB
Estimou-se o nuacutemero de resiacuteduos de cisteiacutenas em todas as amostras com recurso ao coeficiente de
extinccedilatildeo molar () do DTNB (ver ponto 3 dos Anexos) que foi determinado atraveacutes da titulaccedilatildeo
de L-Cisteiacutena com intervalo de concentraccedilotildees compreendidos entre 0 e 50 microM na presenccedila de
25microM de DTNB em tampatildeo D Os valores de absorvecircncia foram obtidos para um comprimento de
onda de 412 nm
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
3621 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
A teacutecnica mais eficiente de anaacutelise da interacccedilatildeo entre o iatildeo vanadilo e proteiacutenas como a
calmodulina e a transferrina entre outras permite a obtenccedilatildeo raacutepida de informaccedilatildeo no que toca agrave
conformaccedilatildeo e amplitude da referida interacccedilatildeo
Os espectros de Ressonacircncia Paramagneacutetica Electroacutenica (RPE) da espeacutecie paramagneacutetica V(IV)
foram realizados a 70K num espectroacutemetro Bruker EMX-300 X-band Todas as soluccedilotildees
utilizadas e amostras foram preparadas em anaerobiose numa cacircmara de luvas de modo a evitar a
reduccedilatildeo de V(IV) a V(V) Em tubos capilares com 02 mL de amostra manteve-se a concentraccedilatildeo
de S1 a 20 M e variou-se a concentraccedilatildeo de vanadilo (0 25 5 10 15 25 50 75 100 150
200 500 e 1000 M [VOSO4]stock=5 mM) em tampatildeo D Apoacutes obtenccedilatildeo dos espectros estes
foram tratados atraveacutes do software SimFonia e WinEPR tendo-se obtido um graacutefico com a
variaccedilatildeo da intensidade do sinal de RPE em funccedilatildeo da concentraccedilatildeo de sulfato de vanadilo
presente Realizaram-se ainda simulaccedilotildees de espectros obtidos com 20 microM de S1 e 1000 microM de
V(IV) com vista agrave obtenccedilatildeo dos ligandos provaacuteveis do vanadilo atraveacutes da anaacutelise da constante
de acoplamento hiperfina (A) e do factor g (g)
4 Conclusotildees gerais
Apesar de jaacute ter sido demonstrado que o vanaacutedio inibe a Na+K
+-ATPase da miosina a funccedilatildeo
deste elemento no tecido muacutesculo esqueleacutetico continua largamente incompreendida A enorme
complexidade quiacutemica deste metal especialmente no que concerne agrave oligomerizaccedilatildeo e reacccedilotildees
51
redox constitui um obstaacuteculo agrave compreensatildeo do comportamento do vanaacutedio nos organismos
vivos Assim a caracterizaccedilatildeo da quiacutemica deste elemento eacute de grande utilidade antes de iniciar o
estudo da sua interacccedilatildeo com o presente sistema
A anaacutelise por espectroscopias de 51
V-RMN e de UV-Vis de soluccedilotildees de vanadato diluiacutedas nos
meios de reacccedilatildeo utilizados no presente relatoacuterio (10 mM HEPES pH 70 25 mM KCl) mostrou
claramente que a espeacutecie decameacuterica (V10) possui um tempo de vida suficientemente longo para a
determinaccedilatildeo do seu efeito isolado ao contraacuterio do que se verifica com as espeacutecies mono- di-
tetra- e pentameacutericas (V1 V2 V4 e V5) Por outro lado a anaacutelise verificou ainda que apoacutes diluiccedilatildeo
das soluccedilotildees de decavanadato no meio de reacccedilatildeo coexistem as espeacutecies V10 e V1 pelo que os
efeitos putativos desta uacuteltima deveratildeo ser subtraiacutedos aquando da monitorizaccedilatildeo dos efeitos da
espeacutecie decameacuterica
O vanaacutedio (V) (vanadato) inibe as enzimas que catalizam reacccedilotildees de transferecircncia de grupos
fosfato como a ATPase da miosina [Goodno 1979] devido agrave sua analogia estrutural com o iatildeo
fosfato Na sua utilizaccedilatildeo como inibidor enzimaacutetico natildeo eacute tida em conta a contribuiccedilatildeo individual
das espeacutecies oligomeacutericas considerando-se normalmente a espeacutecie V1 como inibidora efectiva
[Goodno e Taylor 1982] No presente relatoacuterio verificou-se por espectroscopia de 51
V-RMN que
outros oligoacutemeros de vanadato interagem com o S1 de miosina tendo as espeacutecies V10 e V4 os
maiores niacuteveis de afinidade A sua interacccedilatildeo envolve forccedilas electrostaacuteticas e eacute afectada
negativamente por ligandos naturais de enzima como eacute o caso do ATP
A exposiccedilatildeo do S1 de miosina (005 mgmL) a decavanadato conduz agrave inibiccedilatildeo do capacidade da
proteiacutena de estimular a actividade Mg2+
-ATPaacutesica com um valor de IC50 de 266 plusmn 07 M
decavanadato sendo o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM
decavanadato (80) Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM decavanadato sendo
o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM decavanadato (60)
exercendo um efeito de impedimento agrave ligaccedilatildeo entre o vanaacutedio e o S1
A uma concentraccedilatildeo de 27 microM decavanadato verifica-se a oxidaccedilatildeo de um dos resiacuteduos de Cys
de S1 de miosina seguida pela oxidaccedilatildeo gradual de outro resiacuteduo de Cys ateacute agrave concentraccedilatildeo de
1000 microM Ambos os resiacuteduos oxidados natildeo se encontravam expostos ao solvente utilizado A
oxidaccedilatildeo dos resiacuteduos permite sugerir que esta modificaccedilatildeo oxidativa sofrida pelo S1 contribui
para a diminuiccedilatildeo da sua actividade de ATPase
52
Por espectroscopia de RPE revelaram-se os possiacuteveis ligandos equatoriais do vanaacutedio aquando da
sua interacccedilatildeo com o S1 de miosina como sendo duas moleacuteculas de aacutegua um ligando contendo o
grupo R-OH e outro contendo o grupo R=NR
Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados de
oxidaccedilatildeo (IV) e (V) no S1 de miosina para concentraccedilotildees facilmente alcanccedilaacuteveis no ambiente
terrestre
53
5 Estudos futuros
No sentido do melhor esclarecimento das alteraccedilotildees moleculares e estruturais que ocorrem no
subfragmento 1 quando exposta ao vanaacutedio que conduzem a uma diminuiccedilatildeo da sua funccedilatildeo
sugere-se
- Quantificaccedilatildeo das cisteiacutenas reduzidas de S1 na presenccedila de metavanadato
- Determinaccedilatildeo das actividades Ca2+
- e K+-ATPaacutesica do S1 de miosina na presenccedila de
decavanadato
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina estimulada por actina
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina na presenccedila de
metavanadato
- Realizaccedilatildeo de RMN 2D com variaccedilatildeo das concentraccedilotildees das soluccedilotildees de decavanadato e
metavanadato na presenccedila de S1
-Mutageacutenese dos resiacuteduos de Cys doS1 de miosina de modo a verificar quais os resiacuteduos que
sofrem oxidaccedilatildeo na presenccedila de decavanadato
- Determinaccedilatildeo por calorimetria (DSC Differential Scanning Calorimetry) da variaccedilatildeo da
entalpia (∆H) e da temperatura de desnaturaccedilatildeo do S1 de miosina na presenccedila de vanaacutedio uma
vez que estas propriedades caracteriacutesticas de cada proteiacutena satildeo alteradas apoacutes ocorrecircncia de
modificaccedilotildees estruturais
- Anaacutelise por SDS-PAGE de S1 de miosina apoacutes tratamento com vanaacutedio de modo a determinar
quais os polipeacuteptidos eventualmente formados apoacutes clivagem da proteiacutena
54
6 Referecircncias bibliograacuteficas
- Sellers J Myosins 2nd Edition Oxford University Press 1999
- L Stryer inBiochemistry (Ed WH Freeman 2005 pp
Refordfas Materiais e Meacutetodos
[A] S S Margossian SLowey Methods Enzymol1982 85 55-71
[B] JD Pardue JA Spudich Methods Enzymol 1982 85 164-181
[C] M Bradford Anal Biochem1976 72 248-254
[D] H H Taussky E Shorr J Biol Chem 1953 202 675-685
[E] C R Bagshaw in Outline Studies in Biology ndash Muscle Contraction Chapman and Hall Ed
1982
[F] A Cuenda et al Eur J Biochem1990 194 663-670
55
Anexos
1 Determinaccedilatildeo da recta de calibraccedilatildeo meacutetodo de Bradford
Figura1ndashRepresentaccedilatildeo graacutefica da recta de calibraccedilatildeo (Abs595 nm vs [BSA]) utilizada para o meacutetodo de Bradford
2 Determinaccedilatildeo da recta calibraccedilatildeo para a electroforese em condiccedilotildees desnaturantes
Figura 2 ndash Representaccedilatildeo graacutefica da recta de calibraccedilatildeo (log(Peso Molecular) vs Rf dos marcadores de peso
molecular) onde Rf indica determinada atraveacutes do padratildeo de peso molecular
(poccedilo (a))
3Caacutelculo do coeficiente de extinccedilatildeo molar () para a quantificaccedilatildeo de cisteiacutenas reduzidas
atraveacutes do meacutetodo de Ellman
Traccedilou-se a recta de Abs412 nm meacutedia vs [L-Cisteiacutena] de modo a determinar o valor do coeficiente
de extinccedilatildeo molar definido como o declive daquela recta Assim =8400 M-1
cm-1
como
podemos verificar na Fig 3
A595 nm = 00546[BSA] + 00209 Rsup2 = 099051
Ab
s 59
5 n
m
[BSA] (mgmL)
log PM = -1177 Rf + 2247 Rsup2 = 098957
log
PM
Mobilidade relativa (Rf)
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero

9
A miosina-II eacute constituiacuteda por duas cadeias polipeptiacutedicas idecircnticas denominadas cadeias pesadas
de miosina (HC) enroladas entre si e cujo peso molecular (PM) estaacute compreendido entre 171 e
241 kDa) Estruturalmente as HC dividem-se em 3 domiacutenios funcionais - cabeccedila pescoccedilo e
cauda como se pode observar na Fig 13 Agraves cabeccedilas ou zonas globulares estatildeo associados dois
pares de cadeias leves (peso molecular entre 15 e 30 kDa) que se dividem em dois tipos
essencial (ELC) e regulatoacuterio (RLC)
Figura 13 ndash Estrutura esquemaacutetica da moleacutecula de miosina-II Destacam-se as duas cabeccedilas globulares e a cauda
filamentosa ligadas entre si por uma zona flexiacutevel denominada pescoccedilo As cadeias leves (a verde e vermelho)
encontram-se associadas agraves cabeccedilas globulares
As cabeccedilas globulares contendo os locais de ligaccedilatildeo agrave actina e aos nucleoacutetidos mantecircm os seus
elementos estruturais baacutesicos conservados em todas as classes sendo estes constituiacutedos por 720-
80 aa A regiatildeo do pescoccedilo eacute constituiacuteda por uma longa heacutelice pertencente agrave cadeia pesada
que se encontra estabilizada atraveacutes da ligaccedilatildeo a subunidades de cadeia leve A estrutura do
pescoccedilo envolvida na ligaccedilatildeo agraves cadeias leves eacute denominada de motivo IQ cuja estrutura
consensual eacute IQXXXRGXXXR1 O pescoccedilo serve como extensatildeo do comprimento da cabeccedila de
modo a actuar como alavanca durante a contracccedilatildeo muscular [A]A regiatildeo da cauda ou acircncora
apresenta bastante divergecircncia no que respeita agrave sua sequecircncia estrutura e funccedilatildeo No entanto
pensa-se que a sua funccedilatildeo centrar-se-aacute na autoassociaccedilatildeo das vaacuterias moleacuteculas de miosina-II
atraveacutes da regiatildeo de coiled-coil Esta confere assimetria agrave moleacutecula e permite a formaccedilatildeo de
filamentos grossos capazes de translocar filamentos de actina [A]
Na regiatildeo da cabeccedila da miosina existem 2 regiotildees extremamente sensiacuteveis agrave proteoacutelise A
clivagem em ambos os locais resulta na produccedilatildeo de trecircs fragmentos um compreendendo o N-
terminal (fragmento 25 kDa) outro a regiatildeo central (50 kDa) e um terceiro o C-terminal (20 kDa)
(Fig 14) Estes fragmentos dissociam-se em condiccedilotildees desnaturantes e natildeo constituem domiacutenios
independentes da cabeccedila mas provavelmente loops flexiacuteveis uma vez que natildeo foram observadas
na estrutura cristalina Os vaacuterios fragmentos obtidos em diferentes condiccedilotildees foram cruciais para
a elucidaccedilatildeo das vaacuterias funccedilotildees da moleacutecula
A regiatildeo compreendida entre os fragmentos 25 e 50 kDa (loop 1) localiza-se directamente por
1 I = Isoleucina Q= Glutamina X = qualquer dos 20 aminoaacutecidos padratildeo R= Arginina G= Glicina
10
cima do local de ligaccedilatildeo ao nucleoacutetido A regiatildeo entre os fragmentos de 50 e 20 kDa (loop 2)
encontra-se perto da intersecccedilatildeo entre os domiacutenios da cabeccedila e do pescoccedilo e deveraacute interagir com
a actina uma vez que a ligaccedilatildeo desta a protege da proteoacutelise Na ausecircncia do loop 2 a miosina
liga mais fracamente agrave actina do que no caso da miosina intacta A sequecircncia primaacuteria de ambos
os loops apresenta conservaccedilatildeo reduzida sendo provaacutevel que sejam responsaacuteveis por muita da
diversidade das diferentes propriedades enzimaacuteticas presentes em diferentes isoformas
No fragmento de 20 kDa existem dois grupos sulfidrilo Cys-707 e -697 usualmente
denominados de SH1 e SH2 devido agrave ordem da sua reactividade quiacutemica Na presenccedila de
nucleoacutetidos pode ocorrer o ldquocross-linkingrdquo entre estes dois grupos incluindo a oxidaccedilatildeo a uma
ponte dissulfureto
Devido agrave insolubilidade da miosina-II nativa a estrateacutegia adoptada para os estudos com esta
proteiacutena eacute frequentemente a sua clivagem em fragmentos funcionais separaacuteveis Um dos locais de
clivagem ocorre tipicamente a 130 kDa do N-terminal produzindo a meromiosina pesada (HMM)
e a meromiosina leve (LMM) [C] A HMM eacute formada pelas 2 cabeccedilas e pescoccedilo e apresenta
propriedades semelhantes agrave miosina sem digestatildeo (ligaccedilatildeo e translocaccedilatildeo de actina e hidroacutelise de
MgATP)
A HMM pode ser novamente clivada atraveacutes de uma reacccedilatildeo de proteoacutelise produzindo os
subfragmentos 1 (S1) e 2 (S2)O S1 (90 kDa) eacute enzimaticamente activo sendo constituiacutedo pelos
domiacutenios da cabeccedila e do pescoccedilo e apresentando associaccedilatildeo agraves duas cadeias leves ou apenas agrave
ELC (Fig 14)Em alternativa o S1 pode ser produzido por clivagem directa da miosina intacta
formando-se tambeacutem um fragmento em forma de cilindro que consiste nas regiotildees S2 e LMM
(Ref)Utiliza-se extensivamente o S1 para estudos cineacuteticos e de ligaccedilatildeo agrave actina uma vez que a
sua solubilidade simplifica o planeamento da experiecircncia e a subsequente interpretaccedilatildeo dos dados
[A]
Figura 14- Estrutura do subfragmento-1 de miosina de muacutesculo esqueleacutetico com ecircnfase nos domiacutenios de 50 kDa
central (a vermelho) 25 kDa no N-terminal (a verde) e 20 kDa no C-terminal (a azul) As cadeias leves essenciais
encontram-se a amarelo e as cadeias leves regulatoacuterias a lilaacutes Os locais de ligaccedilatildeo agrave actina e ao ATP estatildeo
igualmente destacados (adaptado de Holmes e Geeves 1999 [D])
11
112 Actina
O termo ldquoactinardquo refere-se a uma famiacutelia extremamente conservada de proteiacutenas citoplasmaacuteticas
presente em todos os eucariotas Esta proteiacutena natildeo se encontra nos procariotas [E] A maioria das
actinas possui peso molecular proacuteximo de 42 kDa outra evidecircncia do seu grau elevado de
conservaccedilatildeo no que concerne agrave sequecircncia e ao nuacutemero de aminoaacutecidos Os eucariotas superiores
apresentam um grande nuacutemero de isoformas de actina que por sua vez se encontram codificados
por um ainda maior nuacutemero de genes (Ref) A expressatildeo das isoformas eacute diferencial consoante o
tecido e sua especializaccedilatildeo pelo que deveratildeo possuir funccedilotildees e mecanismos direccionais
diferentes (Ref) Foi ainda observado que os mRNA das diferentes isoformas apresentam uma
distribuiccedilatildeo heterogeacutenea nas ceacutelulas de forma semelhante aos seus produtos proteicos [F] Estes
dados suportam a hipoacutetese supramencionada do direccionamento das isoformas de actina para um
determinado local na ceacutelula de acordo com as suas funccedilotildees
A actina presente no citoesqueleto eacute responsaacutevel por processos como o mecanismo contraacutectil
fagocitose manutenccedilatildeo e alteraccedilatildeo da estrutura celular locomoccedilatildeo transporte de organelos
regulaccedilatildeo do transporte de solutos entre outras Nos eucariotas superiores estaacute ainda envolvida
no transporte de gracircnulos de pigmento retinoacuteide [G] transporte de gracircnulos no axoplasma [H] e
secreccedilatildeo entre outros [E] Actualmente a isoforma da actina (-actina) eacute a melhor estudada
encontrando-se envolvida no mecanismo contraacutectil [E]
Os filamentos de -actina tecircm cerca de 1m de comprimento e 80 Å Consistem em duas cadeias
de actina filamentosa (F-actina) organizados em heacutelice [I] (Holmes et al 1990) sobre os quais
se encontram outras proteiacutenas ndash a troponina (C I e T) e a tropomiosina (Azevedo 1999) (Figura
15) A troponina eacute filamentosa e localiza-se nas depressotildees da heacutelice de F-actina conferindo
rigidez aos filamentos finos Por sua vez a tropomiosina eacute essencial na regulaccedilatildeo do mecanismo
contraacutectil
Figura 15 ndash Modelo estrutural dos filamentos de F-actina contendo a tropomiosina e o complexo de troponina (I C
e T) (Adaptado de Susana Ramos 2005 [J])
12
A F-actina eacute formada atraveacutes da polimerizaccedilatildeo de subunidades globulares de actina (G-actina) A
G-actina eacute composta por 2 domiacutenios de tamanho semelhante separados pelos bolsos de ligaccedilatildeo(1)
de catiotildees divalentes e(2) de nucleoacutetidos [KLM] Estes por sua vez estatildeo subdivididos noutros
dois de menores dimensotildees (1-4) (Fig 16) cada um envolvido nas interacccedilotildees actina-actina ou
na formaccedilatildeo do topo de ligaccedilatildeo do nucleoacutetido As regiotildees N-terminal e C-terminal estatildeo proacuteximas
uma da outra no subdomiacutenio 1 [E]
Figura 16 ndash Estrutura do monoacutemero de actina Encontram-se representados os subdomiacutenios 1 a 4 e a vermelho a
moleacutecula de ATP junto do respectivo local de ligaccedilatildeo (Protein Data Bank ID 1j6z)
113 Mecanismos moleculares da contracccedilatildeo muscular
A contracccedilatildeo muscular ocorre atraveacutes do deslizamento dos filamentos finos (de actina) por entre
os filamentos grossos (de miosina) conseguida pela reacccedilatildeo de hidroacutelise de ATP e regulada pela
tropomiosina e pelo complexo de troponina em resposta a impulsos nervosos Este mecanismo foi
inicialmente proposto por Huxley-Niedergerke [N] e Huxley-Hanson [O] ambos em 1954 A base
experimental de ambos os estudos foi a inexistecircncia de variaccedilatildeo do tamanho dos filamentos
durante a contracccedilatildeo A banda A apresenta sempre as mesmas dimensotildees por outro lado a banda
I a zona H diminuem consideravelmente aquando da contracccedilatildeo (Figs 12 e 17)
Figura 17 ndash Modelo da contracccedilatildeo muscular ndash deslizamento dos filamentos finos (azuis) relativamente aos
filamentos grossos (rosa) (Adaptado de Stryer 2002 [2])
13
Na ausecircncia de ATP e supondo a ausecircncia de inibiccedilatildeo da interacccedilatildeo actina-miosina as proteiacutenas
encontram-se fortemente associadas formando o complexo actomiosina (AM) (Figura 15)
Nestas condiccedilotildees as cabeccedilas de miosina formam um acircngulo de 45ordm com o eixo dos filamentos
finos de actina [P]
Na presenccedila de ATP o complexo AM dissocia-se jaacute que a afinidade do complexo M-ATP para a
actina eacute 105 vezes inferior agrave afinidade da miosina para a actina O ATP liga-se agrave miosina
ocorrendo seguidamente a reacccedilatildeo de hidroacutelise de ATP Os produtos daiacute resultantes ndash ADP e Pi -
permanecem ligados agrave miosina formando o complexo M-ADP-Pi Com a ligaccedilatildeo subsequente agrave
actina forma-se o complexo AM-ADP-Pi no qual as cabeccedilas de miosina apresentam um acircngulo
recto com o eixo do muacutesculo [P] Neste ponto daacute-se a libertaccedilatildeo progressiva de Pi e ADP Eacute
provaacutevel que a energia proveniente da hidroacutelise do ATP provoque uma alteraccedilatildeo do acircngulo de
ligaccedilatildeo das cabeccedilas de miosina ao filamento de actina passando novamente de 90ordm para 45ordm [P]
Esta alteraccedilatildeo conformacional provoca o deslizamento do filamento de actina para o centro do
sarcoacutemero cujo encurtamento promove a contracccedilatildeo das miofibrilas O processo supracitado
ocorre em cerca de 1ms[Q] e encontra-se resumido na Fig 18
Figura 18 ndash Esquema cineacutetico da hidroacutelise de ATP pela miosina (Adaptado de Sellers 1999 [A])
1131 Regulaccedilatildeo da contracccedilatildeo muscular
O mecanismo pelo qual se processa a regulaccedilatildeo muscular foi inicialmente descrito por estudos
distintos de Haselgrove Huxley e Parrey-Squire todos em 1973 [RST] Aleacutem da implicaccedilatildeo
directa da tropomiosina e do complexo de troponina a concentraccedilatildeo de Ca2+
no muacutesculo em
relaxamento eacute bastante reduzida (10-7
M) aumentando notavelmente aquando da contracccedilatildeo (10-5
-
10-3
M)
Resumidamente quando ocorre a excitaccedilatildeo do muacutesculo esqueleacutetico a despolarizaccedilatildeo da
membrana plasmaacutetica eacute difundida ateacute ao interior do tecido atraveacutes dos tuacutebulos transversais
(invaginaccedilotildees da membrana plasmaacutetica da fibra muscular) A despolarizaccedilatildeo provoca a abertura
dos canais ioacutenicos de Ca2+
do retiacuteculo sarcoplasmaacutetico adjacente Como a concentraccedilatildeo de Ca2+
no interior do organelo eacute muito elevada o caacutelcio passa raacutepida e uniformemente para o citosol
14
Assim a regulaccedilatildeo da contracccedilatildeo ocorre com coordenaccedilatildeo e rapidez na totalidade das miofibrilas
do tecido muscular (Ref)
Em repouso a troponina I encontra-se posicionada de tal forma que a proteiacutena adjacente
troponina C impede estruturalmente a ligaccedilatildeo das cabeccedilas da miosina agrave actina (acircngulo =50ordm) O
aumento da concentraccedilatildeo intracelular de Ca2+
provoca um aumento da sua disponibilidade para
ligar agrave troponina C (Fig 19)
Figura 19ndash Regulaccedilatildeo do mecanismo contraacutectil no muacutesculo esqueleacutetico pelo complexo de troponina
Como consequecircncia a troponina C altera a sua conformaccedilatildeo (=70ordm) associando-se agrave troponina I
que deixa de inibir a ligaccedilatildeo das pontes transversais A alteraccedilatildeo anterior eacute transmitida agrave
tropomiosina atraveacutes da troponina T adjacente agrave C o que provoca o deslocamento da
tropomiosina na direcccedilatildeo do interior do sulco do filamento de actina Assim eacute possiacutevel a
interacccedilatildeo eficaz entre actina e miosina A Ca2+
-ATPase do retiacuteculo sarcoplasmaacutetico provoca a
reacumulaccedilatildeo do iatildeo caacutelcio no organelo impedindo o prolongamento desnecessaacuterio da contracccedilatildeo
(Refordf)
12 Vanaacutedio
O vanaacutedio eacute um metal de transiccedilatildeo pertencente ao grupo VB da tabela perioacutedica Foi
primeiramente descoberto em 1801 por Andreacutes Manuel del Riacuteo que concluiu erradamente que era
idecircntico ao croacutemio e definitivamente por Nils Gabriel Svenson em 1831 Recentemente foram
descobertas importantes funccedilotildees bioloacutegicas deste elemento provavelmente essencial para os
humanos de que se destaca no contexto do presente relatoacuterio a possibilidade de actuar como
anaacutelogo do grupo fosfato e a inibiccedilatildeo de enzimas in vitro como eacute o caso da actividade ATPaacutesica
da miosina
Eacute um metal soacutelido agrave temperatura ambiente que se encontra amplamente distribuiacutedo nos vaacuterios
ecossistemas em dois isoacutetopos naturais ndash 50
V e 51
V ndash raramente excedendo 01 partes por biliatildeo
15
(ppb) Estaacute associado a cerca de 70 minerais diferentes na natureza (dos quais se destacam o
uracircnio cobre chumbo zinco fosfato e titacircnio ferroso) especialmente na forma de vanadato ou
pentoacutexidos de vanaacutedio O vanaacutedio eacute um dos principais constituintes da mateacuteria orgacircnica foacutessil
com especial ecircnfase para o carvatildeo oacuteleo de crude e foacutesseis carbonaacuteceos [UV] bem como em
alimentos consumidos pelo Homem (salsa pimenta preta cogumelos marisco vegetais cereais
gorduras e oacuteleos) [W] Estes constituem a maior fonte de exposiccedilatildeo ao vanaacutedio da maioria da
populaccedilatildeo humana [X]
Normalmente a entrada do vanaacutedio no organismo ocorre por inalaccedilatildeo ou em menores
quantidades por absorccedilatildeo pelo tracto gastrointestinal ou pela pele O seu armazenamento centra-
se no fiacutegado rins ceacuterebro coraccedilatildeo muacutesculos e ossos No entanto a retenccedilatildeo de vanaacutedio no
organismo humano eacute reduzida ndash em condiccedilotildees normais cerca de 100 M (em muitos tecidos a
quantidade eacute inferior a 10 ngg de tecido) [Y]
Actualmente natildeo existem provas de deficiecircncia nutricional de vanaacutedio em humanos ou de
enzimas que contenham vanaacutedio em animais superiores No entanto vaacuterios estudos relacionam a
ocorrecircncia de deficiecircncia em galinhas e ratos devido a uma alimentaccedilatildeo pobre em vanaacutedio Os
sintomas relatados incluem retardamento do crescimento malformaccedilotildees oacutesseas perturbaccedilotildees no
metabolismo lipiacutedico e estagnaccedilatildeo reprodutiva entre outros [Z AA AB] Os efeitos fisioloacutegicos
do vanaacutedio podem depender do tipo de composto ou de oligoacutemero uma vez que consoante as
suas estabilidade labilidade e nuacutemero de coordenaccedilatildeo as respostas seratildeo diferenciadas ao niacutevel
dos organismos vivos
Por outro lado a toxicidade do vanaacutedio encontra-se bem caracterizada existindo sintomas
descritos em vaacuterios mamiacuteferos (inclusive o Homem) que incluem diarreias catildeibras irritaccedilatildeo da
pele e mucosas palpitaccedilotildees cardiacuteacas anemia pneumonia bronquite perturbaccedilotildees no Sistema
Nervoso Central (SNC) entre outros (Ref) A toxicidade varia geralmente com a natureza do
composto com aumento proporcional agrave sua valecircncia Verificou-se que a forma de entrada do
vanaacutedio tem tambeacutem efeito na sua toxicidade ndash os maiores niacuteveis verificaram-se por injecccedilatildeo
intravenosa da amostra sendo progressivamente inferiores por inalaccedilatildeo e finalmente por
ingestatildeo [X AA] A ampla utilizaccedilatildeo dos compostos de vanaacutedio na induacutestria quiacutemica bem como
as elevadas quantidades de vanaacutedio libertadas atraveacutes da combustatildeo dos oacuteleos de crude
contribuem consideravelmente para o aumento da biodisponibilidade do vanaacutedio Como tal o
interesse neste elemento e nos problemas ambientais por ele causados tem vindo a aumentar
significativamente [V]
121 Propriedades quiacutemicas
16
A quiacutemica do vanaacutedio eacute extraordinariamente complexa pelo que originou vaacuterias interpretaccedilotildees
incorrectas desde a sua descoberta No entanto as suas propriedades dependem em uacuteltima
anaacutelise da configuraccedilatildeo electroacutenica do elemento Assim a variabilidade encontrada no
comportamento quiacutemico do vanaacutedio pode dever-se agrave sua configuraccedilatildeo ndash [Ar] 3d2 4s
2 ndash que
permite a formaccedilatildeo de vaacuterios estados de oxidaccedilatildeo (de +2 a +5) Em soluccedilatildeo aquosa os estados de
oxidaccedilatildeo estaacuteveis vatildeo de V(V) a V(II) Destes apenas os +3 a +5 satildeo biologicamente importantes
jaacute que o V(II) apresenta demasiada capacidade redutora para existir nos organismos vivos
conhecidos [AC]
No fluido extracelular os compostos de vanaacutedio encontram-se predominantemente na forma de
monovanadatos (V(V)) livres ou ligados agrave transferina do soro ou agrave hemoglobina (Refordf) Os iotildees de
vanadato livres satildeo transportados para o meio intracelular atraveacutes de canais ioacutenicos inespeciacuteficos
Jaacute no ambiente intracelular os compostos sofreratildeo uma bio-reduccedilatildeo atraveacutes da acccedilatildeo da
glutationa ou outros compostos com grupos ndashOH como pentoses hexoses ou etanolaminas [V] A
reoxidaccedilatildeo de vanadilo (V(IV)) intracelular a vanadato (V(V)) provocaria a acumulaccedilatildeo toacutexica
destes uacuteltimos pelo que a complexaccedilatildeo da espeacutecie reduzida o impede O transporte de vanaacutedio
para o interior da ceacutelula encontra-se na Fig 110
Figura 110 ndash Transporte de vanaacutedio para a ceacuteluka e seu subsequente metabolismo (P=Proteiacutena) (Adaptado de
Zaporowska e Scibor 1998)
1211 Vanaacutedio(IV)
A espeacutecie de vanaacutedio (IV) mais bem conhecida eacute o iatildeo vanadilo (VO2+
) espeacutecie estaacutevel a pH
aacutecido sendo a maioritaacuteria a pH 3[AD] Apresenta uma cor azulada e forma complexos penta e
hexacoordenados Forma-se rapidamente a partir de V(V) na presenccedila de um agente redutor em
soluccedilatildeo aquosa No ambiente intracelular a presenccedila de glutationa reduzida aacutecido ascoacuterbico
NAD(P)H entre outros conduzem a que o principal estado de oxidaccedilatildeo nos sistemas bioloacutegicos
seja provavelmente o +4 encontrando-se o V(IV) fortemente complexado com proteiacutenas ou
aminoaacutecidos livres citrato ATP e pirofosfato (Refordfs) Em alternativa o V(IV) pode existir
17
livremente em ambientes fortemente aacutecidos como os lisossomas jaacute que a pH superior a 3 as
soluccedilotildees de vanadilo tendem a ser oxidadas pelo ar formando-se vanadato Esta grande
capacidade redutora justifica parcialmente a utilizaccedilatildeo pouco extensiva do iatildeo VO2+
como
inibidor [W]
1212 Vanaacutedio (V)
O vanaacutedio (V) eacute a forma mais estaacutevel num intervalo de pH amplo [W] O iatildeo anioacutenico VO3-tende
a constituir complexos polinucleares pelo que as soluccedilotildees aquosas de vanadato satildeo extremamente
complexas com ocorrecircncia simultacircnea de vaacuterios equiliacutebrios de protonaccedilatildeo (pKa(vanadato)= 35
78 e 125) e oligomerizaccedilatildeo As espeacutecies presentes (H3VO4 H2VO4- HVO4
2- e VO4
3-) dependem
do pH forccedila ioacutenica e temperatura da soluccedilatildeo e da concentraccedilatildeo total do elemento
[AEAFAGAHAI]
O VO43-
- monoacutemero ortovanadato V1 ndash estaacute associado a uma grande parte da importacircncia
bioloacutegica do vanaacutedio uma vez que se trata de um anaacutelogo electroacutenico e estrutural do ortofosfato
(PO43-
) A coordenaccedilatildeo do ortovanadato pode ser tetraeacutedrica ou bipiramidal trigonal (Refordfs)
A oligomerizaccedilatildeo do vanadato monomeacuterico daacute origem ao diacutemero (V2O72-
- V2) que apresenta
tambeacutem equiliacutebrios de protonaccedilatildeo Presume-se a existecircncia de analogia do vanadato dimeacuterico com
o pirofosfato embora natildeo haja evidecircncia estrutural (Refordf) A dimerizaccedilatildeo do V2 origina o
vanadato tetrameacuterico ou metavanadato (V4O124-
- V4) que constitui a espeacutecie maioritaacuteria em
soluccedilotildees aquosas concentradas (Refordf) As estruturas do metavanadato e a sua espeacutecie protonada
satildeo ciacuteclicas [AJ] Outra espeacutecie oligomeacuterica importante eacute o vanadato pentameacuterico (V5O153-
) cuja
estrutura presumida eacute ciacuteclica [AI]
O oligoacutemero com maior importacircncia bioquiacutemica eacute o vanadato decameacuterico (V10O286-
-V10)(Refordf) a
espeacutecie predominante em soluccedilotildees de pH 2-6 quando a concentraccedilatildeo total de vanadato eacute superior
a 100 M (Ref) A estrutura do V10 natildeo difere em soluccedilatildeo ou no estado soacutelidoOs 10 aacutetomos de
vanaacutedio da espeacutecie decameacuterica estatildeo divididos em trecircs grupos identificados na figura como VA-C
consoante o seu ambiente quiacutemico circundante (Refordf) (Fig 111)
A expressatildeo ldquodecavanadatordquo refere-se a uma soluccedilatildeo aquosa na qual a espeacutecie decameacuterica se
encontra em equiliacutebrio quiacutemico com as espeacutecies V1 V2 V4 e V5 sendo que a oligomerizaccedilatildeo
progressiva do vanaacutedio eacute proporcional agrave sua concentraccedilatildeo total (Ref) Nesta soluccedilatildeo a
concentraccedilatildeo de V10 corresponde a aproximadamente 10 da concentraccedilatildeo total de vanaacutedio [AK]
18
Figura 111 ndash Estruturas esquemaacuteticas das espeacutecies de vanadato (espeacutecie monomeacuterica (V1) dimeacuterica (V2)
tetrameacuterica (V4) pentameacuterica (V5) e decameacuterica (V10) (Adaptado de Aureliano e Crans 2008)
122 Metabolismo e efeito na actividade enzimaacutetica
Apesar de natildeo terem sido ainda encontradas as provas categoacutericas das suas funccedilotildees nos seres
vivos sabe-se que o iatildeo vanadato eacute capaz de inibir uma grande quantidade de enzimas in vitro
com especial ecircnfase na actividade ATPaacutesica da miosina [AL Teresa] Como referido
anteriormente o vanadato eacute capaz de actuar em alternativa ao fosfato nas reacccedilotildees bioquiacutemicas
incluindo a sua preferecircncia de ligaccedilatildeo agraves enzimas [W]
Eacute conhecido o papel do vanaacutedio no metabolismo do caacutelcio promovendo a mineralizaccedilatildeo dos
dentes e ossos na biossiacutentese de liacutepidos (trigliceacuteridos e colesterol) com posterior inibiccedilatildeo da
acumulaccedilatildeo destes nos vasos sanguiacuteneos e no SNC na reproduccedilatildeo e desenvolvimento por
mimetizaccedilatildeo e potenciaccedilatildeo de factores de crescimento na estimulaccedilatildeo do consumo de glucose e
da siacutentese de glicogeacutenio com actuaccedilatildeo equivalente aos efeitos da insulina ao niacutevel celular na
transduccedilatildeo de sinais celulares com regulaccedilatildeo da actividade de segundos mensageiros no
transporte de iotildees e aminoaacutecidos na transcriccedilatildeo geneacutetica e siacutentese de proteiacutenas e DNA entre
outros [W Y AB]
Assim o vanaacutedio cnseguiraacute provocar inuacutemeras mudanccedilas bioquiacutemicas nas ceacutelulas Alguns
exemplos da influecircncia bioquiacutemica do vanaacutedio incluem os seus efeitos nos processos de
fosforilaccedilatildeo e desfosforilaccedilatildeo de proteiacutenas e na participaccedilatildeo na formaccedilatildeo de espeacutecies reactivas de
oxigeacutenio devido ao seu efeito proacute-oxidante sobre agentes redutores a niacutevel celular [AM]
Quanto ao seu efeito na actividade enzimaacutetica pode dividir-se em dois tipos principais ndash inibitoacuterio
e estimulante - dependendo da enzima ou do tipo de reacccedilatildeo por esta catalisada No primeiro
grupo incluem-se enzimas como as ATPases (de miosina de canais ioacutenicos e fosfoenzimas
19
transportadoras) e as fosfatases (da tirosina aacutecida e alcalina) a fosfofrutocinase adenilato ciclase
e catalase entre outras [AA] O segundo grupo inclui a maioria das cinases a glucose-6-fosfato
desidrogenase a fosfoglicerato mutase e a fosfoglicerato fosfatase (Refordf)
O efeito do vanaacutedio (V) na actividade enzimaacutetica estaacute relacionado com o seu nuacutemero de
coordenaccedilatildeo podendo actuar como estado de transiccedilatildeo ou por mimetizaccedilatildeo de cofactores
substratos (especialmente o fosfato) e interacccedilotildees proteiacutena-proteiacutena (como no caso das
polimerases e transcriptases) Pode ainda estar envolvido na formaccedilatildeo de ligaccedilotildees de hidrogeacutenio
interacccedilotildees electrostaacuteticas esteacutereas e de oxidaccedilatildeo-reduccedilatildeo Uma evidecircncia desta uacuteltima eacute a sua
capacidade de alternacircncia entre os estados de oxidaccedilatildeo +5 a +3 o que torna faacutecil a sua interacccedilatildeo
redox com enzimas contendo resiacuteduos de cisteiacutenas (Cys) ou qualquer outro composto orgacircnico
com actividade redox [AN]
13 Vanaacutedio e muacutesculo esqueleacutetico
A primeira evidecircncia da presenccedila de vanaacutedio no muacutesculo esqueleacutetico proveio de uma amostra de
ATP isolado de muacutesculo de cavalo que continha uma impureza ndash vanaacutedio ndash cuja presenccedila
provocou a inibiccedilatildeo da actividade da bomba de Na+K
+-ATPase [AO] Desde entatildeo vaacuterios
estudos tecircm sido realizados com ecircnfase na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o
tecido muscular Destes destaca-se a descoberta da interacccedilatildeo forte da espeacutecie V10 com a miosina
fosforilada [AP] e agrave sua inibiccedilatildeo da actividade ATPaacutesica da miosina[AQ AR]
No quediz respeitoao iatildeo monovanadato destaca-se a sua inibiccedilatildeoda actividade Mg2+
-ATPaacutesica
da miosina [AS] que ocorre de duas formas distintas (1) na presenccedila de concentraccedilotildees reduzidas
de V1 a inibiccedilatildeo ocorre a velocidade reduzida dependente da concentraccedilatildeo da espeacutecie
monomeacuterica (2) na presenccedila de concentraccedilotildees elevadas de V1 o niacutevel de inibiccedilatildeo aproxima-se de
100 desde que [V1] seja superior agrave concentraccedilatildeo total de locais com actividade Mg2+
-ATPaacutesica
no ensaio Esta inibiccedilatildeo eacute irreversiacutevel tanto por diaacutelise ou adiccedilatildeo de agentes redutores do
vanadato [AL]Segundo Pate et al(1994)[AT] a espeacutecie monomeacuterica podeinibir a velocidade de
contracccedilatildeo a temperatura inferior a 20ordmC perdendogradual e rapidamente essa capacidade ateacute os
50ordmC temperatura agrave qual cessa a capacidade inibitoacuteria Os resultados obtidos sugeremainda uma
interacccedilatildeo altamente cooperativa entre o iatildeo vanadato e as proteiacutenas componentes do muacutesculo
esqueleacuteticoDestaca-se tambeacutem a descoberta da inibiccedilatildeo da geraccedilatildeo de forccedila contraacutectil nas fibras
musculares por parte da espeacutecie monomeacuterica[AT]Wilson et al(1995) [AU ]determinaram que a
afinidade da espeacutecie V1 para o S1 de miosina eacute significativamente mais elevada do que a do iatildeo
Pi seu anaacutelogo Descobriram ainda que que a ligaccedilatildeo do vanadato poderaacute envolver transiccedilotildees
entre estados energeacuteticos com energia livre independente dos impedimentos esteacutereos impostos
20
pela estrutura das miofibrilas Os estudos desenvolvidos por Smith et al (1990)[AV]
determinaram queo iatildeo monovanadato natildeo apresenta um efeito pronunciado na inibiccedilatildeo da
actividadeacto-S1-miosina-ATPaacutesica excepto em concentraccedilotildees superiores a 900microM a partir das
quais existe produccedilatildeo de espeacutecies oligomeacutericas para as quais o efeito inibitoacuterio seraacute mais
elevadoA espeacutecie monomeacuterica de vanadato consegue adoptar facilmente uma geometria do tipo
bipiramidal trigonal que mimetiza o intermediaacuterio formado (MgADP
Pi)aquando da reacccedilatildeo de
hidroacutelise de ATP no local activo da miosina [AW] De facto a interacccedilatildeo do vanaacutedio com a
miosina eacute inesperadamente estaacutevel na presenccedila de ADP Esta interacccedilatildeo resulta na formaccedilatildeo de
um complexo enzimaacutetico inactivo e estaacutevel com estequiometria de 11 ADPvanadato por siacutetio
activo da proteiacutena A formaccedilatildeo do complexo estaacutevel miosina-vanadato foi proposta por Goodnoet
al (1982) [AS] Nesta ocorre um preacute-equiliacutebrio raacutepido que induz a formaccedilatildeo de um complexo
ternaacuterio reversiacutevel (MADP
V) com formaccedilatildeo lenta do isoacutemero M
ADP
V A diferenccedila entre os
compostos de vanaacutedio e de fosfato centra-se no tempo de meia vida (t12) do Pi ou do V (no
primeiro t12=12 s enquanto no segundo t12=24 h) (Refordf) A diferenccedila dever-se-aacute agrave possiacutevel
complexaccedilatildeo do vanaacutedio com grupos nucleofiacutelicos no siacutetio activo o que provocaria uma
estabilizaccedilatildeo da miosina na conformaccedilatildeo MADP
V
No que respeita aos estudos realizados com a espeacutecie tetrameacuterica Cremo et al determinou que a
cadeia pesada do S1 de miosina pode ser clivada em dois locais distintos (designados V1 e V2)
pela espeacutecie V4 apoacutes irradiaccedilatildeo com luz UV na presenccedila de concentraccedilotildees de vanadato na ordem
dos milimolar e na ausecircncia de nucleoacutetidos ou metais divalentes
Jaacute no que concerne aos estudos realizados com ecircnfase na espeacutecie decameacuterica foi sugerido
porRamos et al (2006) [AK]que a interacccedilatildeo do decavanadato com a G-actina favorecida pela
polimerizaccedilatildeo desta uacuteltima estabiliza a espeacutecie V10 e induz a inibiccedilatildeo da formaccedilatildeo de F-actina A
estabilizaccedilatildeo da espeacutecie V10 por proteiacutenas tansmembranares e do citoesqueleto pode ser a razatildeo
pela qual ocorre a toxicidade do decavanadato reportada na avaliaccedilatildeo dos efeitos do V(V) nos
sistemas bioloacutegicosTiago et al (2007) [AR] concluiram que o V10 apresenta uma clara preferecircncia
de ligaccedilatildeo ao ldquoback-doorrdquo da S1-miosina-ATPase o que se verifica apenas na estrutura
conformacional desimpedida com acesso facilitado ao local de ligaccedilatildeo do Pi Sugere tambeacutem que
o V10 actua como um ldquoback-door stoprdquo bloqueando estaabertura do fragmento de 50-kDa
necessaacuteria para a hidroacutelise do ATP--fosfato o que constitui uma explicaccedilatildeo simples do
comportamento natildeo competitivo da espeacutecie decameacuterica estimulando a utilizaccedilatildeo deste oligoacutemero
como uma feramenta da elucidaccedilatildeo das mudanccedilas conformacionais ldquoback-doorrdquo da miosina no
21
processo da contracccedilatildeo muscular(ref)Destaca-se ainda a descoberta da capacidade inibitoacuteria do
V10 em relaccedilatildeo agrave fosforilaccedilatildeo oxidativa mitocondrial [AX]
Finalmente no que concerne aos estudos realizados com a presenccedila de vaacuterias espeacutecies
oligomeacutericas Aureliano e Madeira (1994) [AY]observaram que na presenccedila de Ca2+
as espeacutecies
V4 e V10se ligam agrave Ca2+
-ATPase do retiacuteculo sarcoplasmaacutetico enquanto a espeacutecie V1 se liga apenas
na presenccedila de ATP Aparentemente o aumento da concentraccedilatildeo de Ca2+
tem o efeito contraacuterio da
inibiccedilatildeo da ATPase do retiacuteculo sarcoplasmaacutetico pelo V1 natildeo tendo no entanto efeito significativo
sobre a inibiccedilatildeo por parte da espeacutecie V10 Tiago et al (2002) [AP] concluiacuteram que em condiccedilotildees
proacuteximas agraves fisioloacutegicas a espeacutecie V10 difere das espeacutecies presentes na soluccedilatildeo de metavanadato
devido agrave sua forte interacccedilatildeo com a miosina fosforilada e agrave sua inibiccedilatildeo da actividade ATPaacutesica
da miosina Para aleacutem disso o ATP diminui a afinidade da miosina para o V4 induzindo a
interacccedilatildeo com V1 e natildeo tendo qualquer efeito no que diz respeito ao V10 Foi tambeacutem proposto
[AQ] que a ligaccedilatildeo das espeacutecies oligomeacutericas de vanadato aoS1 de miosina nos locais onde o
fosfato coordenaeacute provavelmente a causa da inibiccedilatildeo da actividade ATPaacutesica da miosina
estimulada por actina atraveacutes da prevenccedilatildeo da troca entre ATPADP sendo esta favorecida para
aniotildees de vanadato com valecircncias superiores como eacute o caso do V10
Na presenccedila de actina as velocidades de dissociaccedilatildeo do Pi e do V aumentam consideravelmente
este uacuteltimo em menor quantidade Assim a ligaccedilatildeo da actina ao complexo MADP
V e
Pi eacute
bastante fraca pelo que se sugere que o ADPV teria um efeito de relaxamento semelhante ao
ATP No entanto a capacidade do vanaacutedio em inibir a actividade ATPaacutesica da miosina e da
actomiosina eacute diferenciada uma vez que na ausecircncia de actina a inibiccedilatildeo eacute total para
concentraccedilotildees na gama dos M mas na sua presenccedila a inibiccedilatildeo eacute totalmente conseguida apenas
em concentraccedilotildees na gama dos mM (Ref) No entanto eacute necessaacuterio considerar a possiacutevel
formaccedilatildeo de espeacutecies oligomeacutericas de vanadato cuja afinidade e efeito na enzima seratildeo
diferentes Assim pode concluir-se que o efeito de inibiccedilatildeo observado para a actividade ATPaacutesica
de miosina estimulada por actina se pode dever agrave presenccedila das diferentes espeacutecies oligomeacutericas o
que foi jaacute sugerido por Aureliano (2000) [AZ] ndash as espeacutecies tetrameacuterica e decameacuterica de vanadato
interactuariam com a miosina com alteraccedilatildeo da sua interacccedilatildeo com a actina No entanto natildeo eacute
conhecida a contribuiccedilatildeo absoluta das restantes espeacutecies oligomeacutericas para a inibiccedilatildeo da ATPase
da miosina estimulada por actina (Refs)
14 Objectivos do projecto
Pretende-se com este projecto o isolamento purificaccedilatildeo e caracterizaccedilatildeo da actina do muacutesculo
22
esqueleacutetico de coelho sendo esta uacuteltima tambeacutem realizada para o sufragmento-1 de miosina
Seguidamente realizar-se-aacute a quantificaccedilatildeo e caracterizaccedilatildeo das proteiacutenas por espectroscopia UV-
Vis e SDS-PAGE e a caracterizaccedilatildeo das soluccedilotildees de vanaacutedio por UV-Vis e ressonacircncia magneacutetica
nuclear (RMN) Por uacuteltimo a interacccedilatildeo do S1 de miosina com o vanaacutedio seraacute avaliada atraveacutes
das teacutecnicas de UVVis RMN e ressonacircncia paramagneacutetica electroacutenica (RPE)
2 Discussatildeo de Resultados
21 Caracterizaccedilatildeo das proteiacutenas isoladas
211Anaacutelise do estado de pureza
O estado de pureza das soluccedilotildees de actina e de S1 foi analisado por electroforese em gel de
poliacrilamida em condiccedilotildees desnaturantes (SDS-PAGE) (Fig 22) que separa as moleacuteculas de
acordo com o seu peso molecular (PM) A concentraccedilatildeo de acrilamida escolhida (10) permite a
detecccedilatildeo simultacircnea de componentes com peso molecular inferior a 20 kDa e outros com elevado
peso molecular
Figura 21 ndash Fotografia de gel de SDS-PAGE (10 acrilamida) Poccedilo (a)
10 L do marcador de elevado peso molecular () Poccedilo (b) 20 L de [G-
actina]=1 mgmL Poccedilo (c) 20 L de [S1]=1 mgmL Aplicou-se uma
voltagem de V durante h e min
A partir da recta de calibraccedilatildeo obtida log PM = -1177 Rf + 2247 (ver ponto 2 da secccedilatildeo
Anexos) pode verificar-se a presenccedila de actina na amostra do poccedilo (b) presente numa banda
bastante intensa (PMaparente=40 kDa) bem como a presenccedila minoritaacuteria de outros componentes
desconhecidos que natildeo teratildeo sido eliminados durante o processo de extracccedilatildeo e purificaccedilatildeo e natildeo
satildeo relevantes para o estudo em questatildeo Na amostra do poccedilo (c) nota-se a presenccedila de S1
(PMaparente=92 kDa) bem como a de outros componentes cuja banda eacute ligeiramente menos intensa
Estes poderatildeo ser fragmentos proteoliacuteticos de miosina e de S1 Estaacute ainda presente uma bada de
23
maior intensidade que poderaacute ser a proteiacutena-C (PM=135 kDa) uma vez que a sua remoccedilatildeo
completa da amostra necessitaria de um passo adicional de cromatografia de troca ioacutenica [BA]
No entanto natildeo podemos confirmar que se trata desta proteiacutena ou de qualquer outra uma vez que
se encontra fora do intervalo de migraccedilatildeo dos marcadores de peso molecular
212 Determinaccedilatildeo da concentraccedilatildeo proteica
2121Subfragmento-1 de miosina
A partir da recta de calibraccedilatildeo obtida pelo meacutetodo de Bradford [BB](ver secccedilatildeo 3311 do
Procedimento Experimental) A595nm = 00546[BSA] + 00209 (ver secccedilatildeo de Anexos)foi possiacutevel
determinar um intervalo de concentraccedilatildeo de S1 de miosina compreendido entre 226 e 117(plusmn
423M) (203 e 1056 (plusmn 381 mgmL)) (n=8) A concentraccedilatildeo meacutedia de S1 de miosina apoacutes
diaacutelise contra a soluccedilatildeo tampatildeo 10 mM HEPES pH 75 25 mM KCl determinada pela leitura da
Abs280 nmfoi de 20430M (1803 mgmL)(n=7)
As amostras a quantificar atraveacutes do meacutetodo de Bradford natildeo sofreram diaacutelise o que permite
explicar a concentraccedilatildeo superior de S1 de miosina em comparaccedilatildeo com o valor obtido atraveacutes do
meacutetodo directo Esta razatildeo aliada agrave sensibilidade e rapidez superiores deste uacuteltimo fez com que
se optasse pelo meacutetodo directo para a quantificaccedilatildeo da grande maioria dos ensaios
2122Actina
A concentraccedilatildeo de actina foi determinada quando esta se encontrava na forma monomeacuterica (G-
actina) por mediccedilatildeo da leitura directa da Abs290 nm tendo-se obtido uma concentraccedilatildeo meacutedia de
691481M (290202 mgmL) (n=3) em 2 mM Tris-HCl pH 80 02 mM CaCl2 02 mM Na-
ATP Ao comprimento de onda utilizado (290 nm) absorvem fortemente apenas os resiacuteduos de
Trp pelo que a mediccedilatildeo efectuada tem menor sensibilidade do que se tivesse sido efectuada a 280
nm uma vez que a actina apresenta apenas 4 resiacuteduos deste aminoaacutecido embora tenha 16
resiacuteduos de Tyr
22 Mediccedilatildeo da actividade enzimaacutetica de S1 de miosina
221 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
As actividades ATPaacutesicas de S1 de miosina dependentes de Ca2+
e K+foram analisadas pelo
caacutelculo das velocidades de consumo inicial do fosfato inorgacircnico (Pi) Definiu-se os tempos de 5
e 3 minutos para a mediccedilatildeo da Ca2+
e da K+EDTA-ATPase respectivamente Os valores obtidos
24
a partir do protocolo presente no ponto 341 da secccedilatildeo Procedimento experimental encontram-se
descritos na Tabela 2I
Tabela 2I ndash Acrividades natildeo-fisioloacutegicas de subfragmento-1 determinadas pelo meacutetodo de Fiske-Subarow
modificado (n=2)
Proteiacutena
Actividade
ATPaacutesica natildeo
fisioloacutegica
Actividade (nmol
Piminmg proteiacutena) Actividade (s-1)
S1
Ca2+
-ATPase 1400 plusmn 30 2100 plusmn 0045
K+EDTA-ATPase 2334 plusmn 50 3500 plusmn 0075
As actividades natildeo-fisioloacutegicas reflectem o estado de oxidaccedilatildeo dos resiacuteduos expostos de Cys 707
(SH1) e 697 (SH2) Se o grupo SH1 sofrer oxidaccedilatildeo a actividade ATPaacutesica dependente de Ca2+
aumenta e a actividade dependente de K+ diminui Por outro lado se o grupo SH2 sofrer
oxidaccedilatildeo regista-se a perda completa destas actividades
De acordo com Margossian e Lowey (1982) [BA] as actividades ATPaacutesica de S1 de miosina
dependente de Ca2+
e K+EDTA da miosina satildeo respectivamente de 24 e 112 s-1
Estaacute tambeacutem
descrito que a actividade ATPaacutesica dependente de K+ eacute em meacutedia cerca de 3 vezes superior agrave
dependente de Ca2+
reflectindo a quantidade das preparaccedilotildees A razatildeo obtida entre os valores das
actividades ATPaacutesicas dependentes de K+ e Ca
2+ eacute de 166 Os valores obtidos para as actividades
(Ca2+
-ATPase=21 s-1
K+EDTA-ATPase= 35 s
-1) satildeo inferiores agrave valores obtidos por
Margossian e Lowey [BA] e Tiago [BC] com forccedila ioacutenica inferior O valor obtido para a
actividade ATPaacutesica dependente de Ca2+
eacute superior ao obtido por Polosukhina et al (2000) [BD]
cujo meio de reacccedilatildeo continha MOPS e apresentava forccedila ioacutenica inferior Os dados
supramencionados comprovam a variaccedilatildeo da actividade a ATPaacutesica natildeo fisioloacutegica em relaccedilatildeo agrave
forccedila ioacutenica temperatura e pH
222 Mediccedilatildeo da actividadeMg2+
-ATPaacutesica de S1 pelo meacutetodo de enzimas acopladas
A actividade de Mg2+
-ATPase eacute a uacutenica que possui significado fisioloacutegico sendo as restantes
bastante uacuteteis apenas para indicaccedilatildeo da qualidade de preparaccedilatildeo do S1[BA] A 0575 M KCl a
actina e o S1 formam uma soluccedilatildeo viscosa e tuacuterbida Apoacutes a adiccedilatildeo de MgATP o complexo acto-
S1 dissocia-se e a turbidez da soluccedilatildeo diminui Se a forccedila ioacutenica for reduzida a dissociaccedilatildeo do
complexo actoS1 que se encontra na forma de suspensatildeo eacute induzida O MgATP eacute rapidamente
hidrolisado diminuindo a sua concentraccedilatildeo no meio reaccional Nestas condiccedilotildees haacute
reassociaccedilatildeo e subsequente contracccedilatildeo dos filamentos de actina e de S1 com formaccedilatildeo de
25
precipitado O meacutetodo descrito traduz de forma bastante elementar o fenoacutemeno da contracccedilatildeo in
vivo e elimina o problema anterior da solubilizaccedilatildeo ineficiente da miosina de modo a obter uma
mediccedilatildeo exacta da actividade [A] A utilizaccedilatildeo de forccedilas ioacutenicas reduzidas aumenta a afinidade da
actina para o S1 permitindo contornar a extrema dificuldade em obter a concentraccedilatildeo proteica dos
muacutesculos (cerca de 100 mgmL do complexo actomiosina) As actividades Mg2+
-ATPaacutesicas de S1
de miosina foram determinadas a partir do declive da variaccedilatildeo da Abs340 relativo agrave oxidaccedilatildeo de
NADH em funccedilatildeo do tempo como eacute descrito no ponto 342 do Procedimento Experimental Os
valores obtidos para a actividade Mg2+
-ATPaacutesica encontram-se apresentados na Tabela 2II
Tabela 2II ndash Actividade Mg2+
-ATPaacutesica de subfragmento-1 de miosina determinadas pelo meacutetodo das enzimas
acopladas
Actividade (nmol
ATPmin mg(S1)) Actividade (s
-1)
1097 plusmn 269 164 plusmn 04
Ramos (2005) [J] obteve o valor de 01 s-1
para a actividade Mg2+
-ATPaacutesica de S1 de miosina
com forccedila ioacutenica intermeacutedia Tiago (2005) [BC] obteve o valor de 011 s-1 em condiccedilotildees de forccedila
ioacutenica reduzida em relaccedilatildeo agrave utilizada no presente relatoacuterio
23 Preparaccedilatildeo e caracterizaccedilatildeo das soluccedilotildees de vanaacutedio
231 Anaacutelise da composiccedilatildeo das soluccedilotildees de vanadato por espectroscopia de 51
V-RMN
efeito da concentraccedilatildeo da espeacutecie
A diluiccedilatildeo das soluccedilotildees stock de meta- e decavanadato iraacute determinar fortemente a composiccedilatildeo
em soluccedilatildeo dos diferentes oligoacutemeros de vanadato (ref) No caso do decavanadato o periacuteodo de
tempo apoacutes a diluiccedilatildeo iraacute tambeacutem ser um factor importante a considerar devido agrave decomposiccedilatildeo
da espeacutecie V10(ref)Torna-se assim fundamental determinar a composiccedilatildeo quiacutemica das soluccedilotildees
no meio de ensaio utilizado (HEPES 10 mM KCl 25 mM) de modo a correlacionaacute-la
correctamente com os possiacuteveis efeitos promovidos pelas diferentes espeacutecies de vanadato no
sistema em estudo
Aleacutem disso deve ser dado ecircnfase agraves importantes interacccedilotildees que ocorrem entre o vanaacutedio e os
tampotildees escolhidos O tampatildeo com menor nuacutemero e magnitude de interacccedilatildeo eacute o HEPES [AD]
seguido do Tris sendo este uacuteltimo especialmente aconselhado em concentraccedilotildees baixas para casos
de incompatibilidade enzimaacutetica ou quiacutemica redox No caso especial da reacccedilatildeo redox
NAD(P)HNAD(P) o tampatildeo adequado eacute o imidazole de modo a natildeo haver reacccedilatildeo com o
26
vanadato Os tampotildees a evitar em qualquer situaccedilatildeo incluem a trietanolamina fosfato citrato e
glicina entre outros [AD] uma vez que todos interactuam fortemente com V(V) com formaccedilatildeo
de complexos estaacuteveis
Assim procedeu-se agrave anaacutelise da composiccedilatildeo das soluccedilotildees de meta- e decavanadato apoacutes diluiccedilatildeo
das soluccedilotildees stock (50 mM em vanadato total) no meio de reacccedilatildeo (tampatildeo D) atraveacutes da
espectroscopia de ressonacircncia magneacutetica nuclear de 51
V Nas figuras x1 e x2 apresentam-se os
espectros obtidos para as soluccedilotildees de deca- e metavanadato num intervalo de concentraccedilotildees de 1 a
10 mM A Tabela seguinte resume os paracircmetros espectroscoacutepicos obtidos para cada espeacutecie
presente nas soluccedilotildees analisadas
Figura 22ndash Espectros de RMN de 51
V de
concentraccedilotildees crescentes das espeacutecies oligomeacutericas
de vanadato obtidos apoacutes diluiccedilatildeo da soluccedilatildeo stock
de decavanadato em 10 mM HEPES pH 70 25 mM
KCl para concentraccedilotildees finais de A) 1 mM B)2
mM C) 4 mM D) 8 mM E) 10 mM de
decavanadato total V1refere-se agraves espeacutecies
monomeacutericade vanadato V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2IIIndash Desvios quiacutemicos meacutedios (part) das ressonacircncias das espeacutecies decameacuterica (V10) monomeacuterica (V1) e
tetrameacuterica (V4) presentes nos espectros da soluccedilatildeo de decavanadatoLargura a meia altura (∆12) da espeacutecie V10A
Paracircmetros V10A V10B V10C V1 V4
part (ppm) -5162 -5003 -4255 -5616 -5748
∆12 (Hz) 175 nd nd nd nd
Figura 23 ndash Distribuiccedilatildeo graacutefica das
espeacutecies oligomeacutericas de vanadato vs a
concentraccedilatildeo de decavanadato total As
concentraccedilotildees de cada espeacutecie foram
obtidas segundo a formula descrita na
secccedilatildeo 35 do Procedimento
Experimental (aacutereas dos sinais de 51
V-
RMN observadas na figura x1)V10 V1 e
V4 correspondem agraves espeacutecie monomeacuterica
dimeacuterica e tetrameacuterica respectivamente [Esp
eacuteci
e o
ligo
meacute
rica
de
van
aacutedio
] (m
M)
[Decavanadato]total (mM)
V10
V1
V4
27
Nas soluccedilotildees de decavanadato as espeacutecies monomeacuterica e decameacuterica de vanadato satildeo as uacutenicas
presentes para concentraccedilotildees de decavanadato inferiores a 8 mM Ambas as espeacutecies aumentam
linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado em [AY] e
em (Teresa) quando a concentraccedilatildeo de V1 eacute igual ou superior a 200 microM inicia-se a sua
oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica No entanto nas condiccedilotildees de realizaccedilatildeo dos
ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a espeacutecie tetrameacuterica neste
estudo em concentraccedilotildees inferiores a 8 mM tendo o mesmo sido observado por Ramoset al
(2006) [AK] Os resultados obtidos mostram claramente o equiliacutebrio entre a espeacutecie decameacuterica e
a monomeacuterica e que o aparecimento de outras espeacutecies oligomeacutericas se encontra dependente da
concentraccedilatildeo desta uacuteltima Apesar de natildeo ter sido realizado o ensaio com 5 mM de decavanadato
total no presente relatoacuterio estaacute reportado em Ramos et al (2006) [AK] que as concentraccedilotildees da
espeacutecie decameacuterica e monomeacuterica satildeo respectivamente de 500 microM e 200 microM Assim pode inferir-
se que para concentraccedilotildees inferiores a 5 mM e intervalos de tempo de cerca de 20 minutos (que
inclui o tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas
as espeacutecies decameacuterica e monomeacuterica de vanadato
Figura 24 ndash Espectros de RMN de 51
V de concentraccedilotildees
crescentes das espeacutecies oligomeacutericas de vanadato obtidos
apoacutes diluiccedilatildeo da soluccedilatildeo stock de metavanadato em 10 mM
HEPES pH 70 25 mM KCl para concentraccedilotildees finais de A)
1 mM B) 2 mM C) 4 mM D) 8 mM E) 10 mM de
metavanadato total V1 refere-se agraves espeacutecies monomeacuterica de
vanadatoV1 e V2 correspondem respectivamente agrave espeacutecie
monomeacuterica e dimeacuterica
Figura 25 ndash Distribuiccedilatildeo graacutefica das espeacutecies
oligomeacutericas de vanadato vs a concentraccedilatildeo de
metavanadato total As concentraccedilotildees de cada
espeacutecie foram obtidas segundo a formula descrita
na secccedilatildeo 35 do Procedimento Experimental
(aacutereas dos sinais de 51
V-RMN observadas na
figura x1)V10 V1 e V4 correspondem agraves espeacutecie
monomeacuterica dimeacuterica e tetrameacuterica
respectivamente
Esp
eacuteci
e o
ligo
meacute
rica
de
va
nad
ato
(m
M)
[Metavanadato]total (mM)
V1
V2
V4
V5
28
Tabela 2IVndash Desvios quiacutemicos meacutedios (part)das ressonacircncias das espeacutecies tetrameacuterica (V4) monomeacuterica (V1) e
dimeacuterica (V2) presentes no espectro da soluccedilatildeo de metavanadato (total) 10 mM
Paracircmetros V1 V2 V4 V5
part (ppm) -5615 -5748 -5789 -5869
Nas soluccedilotildees de metavanadato as espeacutecies monomeacuterica dimeacutericae tetrameacuterica de vanadato estatildeo
presentes para concentraccedilotildees de metavanadato inferiores a 4 mM Todas estas as espeacutecies
aumentam linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado
em (Aureliano e Madeira 1998) e em (Tiago et al 2004) quando a concentraccedilatildeo de V1 eacute igual ou
superior a 200 microM inicia-se a sua oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica Nas
condiccedilotildees de realizaccedilatildeo dos ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a
espeacutecie pentameacuterica neste estudo em concentraccedilotildees inferiores a 4 mM tendo o mesmo sido
observado por Ramos 2006 Os resultados obtidos mostram claramente o equiliacutebrio entre a
espeacutecie V4 e as espeacuteciesV1 e V2 no qual o aparecimento de outras espeacutecies oligomeacutericas se
encontra dependente da concentraccedilatildeo da espeacutecie V1 Por uacuteltimo pode inferir-se que para
concentraccedilotildees inferiores a 2 mM e intervalos de tempo de cerca de 20 minutos (que inclui o
tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas as
espeacutecies V1 V2 e V4 com concentraccedilotildees respectivas de 0796 0192 e 0196 mM
232Estabilidade das soluccedilotildees de decavanadato anaacutelise por espectroscopia de absorccedilatildeo
UVVis
Caracterizou-se a estabilidade da espeacutecie V10 numa soluccedilatildeo com 1 mM decavanadato total em
tampatildeo D atraveacutes da leitura da Abs400 Obteve-se uma cineacutetica de dissociaccedilatildeo de 1ordf ordem tal
como se pode verificar pela linearidade do graacutefico de ln(Abs400)vs o tempo de aquisiccedilatildeo espectral
(segundos) (Fig26 )
Figura 26ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do tempo
(em segundos) para uma soluccedilatildeo de decavanadato total de 1
mM em tampatildeo D a 25ordmC (n=2)
Sendo uma reacccedilatildeo de primeira ordem pode ser descrita pela equaccedilatildeo cujo
declive eacute ndashK (constante de dissociaccedilatildeo) e a intersecccedilatildeo no eixo das ordenadas corresponde
Ln
(Ab
s 400 n
m)
tempo (s)
29
aln(A0) Numa reacccedilatildeo de 1ordf ordem o tempo de meia-vida eacute independente da concentraccedilatildeo inicial
do reagente sendo descrito pela equaccedilatildeo [2] Deste modo a partir da recta
Ln(Abs400 nm) = -436E-5
K - 105 obteve-se um tempo de meia-vida de 4490 plusmn 0823 horas para
as condiccedilotildees supramencionadas
Apoacutes a adiccedilatildeo de S1 8 microM ao meio reaccional verificou-se que a reacccedilatildeo de desoligomerizaccedilatildeo
da espeacutecie decameacuterica de vanadato seguia igualmente uma cineacutetica de 1ordf ordem tal como se pode
verificar pela linearidade do graacutefico de ln(Abs400) versus o tempo de aquisiccedilatildeo espectral
(segundos) (Fig27)
Figura 27ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do
tempo (em segundos) para uma soluccedilatildeo de decavanadato
total de 1 mM em tampatildeo D a 25ordmC na presenccedila de 8
microM (n= 2)
A partir da equaccedilatildeo da recta Ln(Abs400 nm) = -7520E-5
K - 0613determinou-se o valor de K para
a espeacutecie decameacuterica de onde se obteve um tempo de meia-vida de 2430 plusmn 0191 horas para as
condiccedilotildees supramencionadas
Tanto este valor como o valor registado na ausecircncia de S1 satildeo substancialmente superiores agrave
duraccedilatildeo dos ensaios de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina descritos no presente
relatoacuterio
24 Interacccedilatildeo de S1 com vanaacutedio(V)
241Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
De modo a analisar qual o efeito da adiccedilatildeo de S1 de miosina (tampatildeo D) produzia nos sinais de
51V-NMR titulou-se uma soluccedilatildeo de 5mM de decavanadato total com concentraccedilotildees crescentes
de S1 (5 10 15 e 20 microM) Os espectros de 51
V-RMN encontram-se apresentados na Fig 2
No graacutefico da figura 2 Apresenta-se a concentraccedilatildeo de S1 em funccedilatildeo do factor de alargamento
corrigido dos espectros acima referidos
Ln
(Ab
s (4
00
nm
))
tempo (s)
30
Figura 28 ndash Graacutefico de
representaccedilatildeo da
concentraccedilatildeo de S1 em
funccedilatildeo do factor de
alargamento A
concentraccedilatildeo de
decavanadato total foi de 5
mM e as concentraccedilotildees
utilizadas de S1 foram de 0
5 10 e 15 microM
Verifica-se que o factor de alargamento aumenta linearmente em funccedilatildeo da concentraccedilatildeo de S1 no
ensaio o que indica a existecircncia de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina Observou-
se ainda que a adiccedilatildeo de S1 ao meio reaccional natildeo provocou qualquer alteraccedilatildeo dos desvios
quiacutemicos dos sinais de 51
V-RMN
Realizaram-se ensaios na presenccedila de ATP e de S1 com intuito de verificar o efeito do ATP na
interacccedilatildeo do S1 de miosina com o decavanadato
Figura 29 - Espectros de RMN de 51
V contendo 2
mM Decavanadato (A) (controlo) (B) na presenccedila
de 2 mM de ATP (C) na presenccedila de 2 mM de ATP
e 8 microM de S1 de miosina V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2Vndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies decameacuterica (V10) e monomeacuterica (V1) presentes nos
espectros da soluccedilatildeo de decavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V10A V10B V10C V1
part (ppm) -5161 -5002 -4254 -5614
Observa-se que a presenccedila de ATP (espectro B) natildeo provoca qualquer alteraccedilatildeo visiacutevel em
relaccedilatildeo ao espectro que contem apenas soluccedilatildeo de decavanadato Jaacute a presenccedila simultacircnea de ATP
e S1 provoca um alargamento da ∆12 do sinal do V10A de 1736 Hz (espectro (A)) para 2366 Hz
(espectro (C) Fig 29) Tendo em conta que a concentraccedilatildeo de S1 apresenta uma relaccedilatildeo linear
com o factor de alargamento converteu-se o valor da ∆12 e comparou-se com o valor esperado agrave
F = 00591[S1] - 00157 Rsup2 = 0998
Fact
or
de
ala
rgam
en
to
[S1] (microM)
31
concentraccedilatildeo de S1 utilizada O factor de alargamento obtido para 8 microM ndash 0376 ndash eacute inferior ao
valor esperado agrave mesma concentraccedilatildeo de S1 obtido atraveacutes da equaccedilatildeo da recta apresentada na
Fig 28 ndash 0457 ndash o que indica que o ATP poderaacute exercer um efeito de protecccedilatildeo do S1
diminuindo a sua interacccedilatildeo com a espeacutecieV10
Por uacuteltimo realizaram-se ensaios com metavanadato na presenccedila de 8 microM S1 de modo a verificar
se as espeacutecies oligomeacutericas contidas nesta soluccedilatildeo interagiam com a proteiacutena em estudo
Figura 210 - Espectros de RMN de 51
V contendo 2
mM metavanadato (A) (controlo) (B) na presenccedila
de 8 microM de S1 de miosina V1 V2 e V4
correspondem respectivamente aacutes espeacutecies
monomeacuterica dimeacuterica e tetrameacuterica
Tabela 2VIndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies monomeacuterica (V1) dimeacuterica (V2) e tetrameacuterica (V4)
presentes nos espectros da soluccedilatildeo de metavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V1 V2 V4
part (ppm) -5613 -5747 -5789
Como eacute possiacutevel observar na Fig 210 ocorre uma ligeiro alargamento dos sinal de V4 bem
como uma perda da intensidade pelo que se conclui quedas espeacutecies presentes na soluccedilatildeo de
metavanadato (V1 V2 e V4) apenas a espeacutecie tetrameacuterica interage com o S1 de miosina o que
estaacute de acordo com o reportado por Tiago et al 2002
242 Actividade Mg2+
-ATPaacutesica de S1 de miosina na presenccedila de decanadato
Com o intuito de avaliar o efeito do decavanadato na actividade Mg2+
-ATPaacutesica do S1 de
miosina monitorizou-se a reacccedilatildeo de oxidaccedilatildeo do NADH atraveacutes da leitura de Abs340 na
presenccedila de 005 mgmL de S1 de miosina e no caso da actividade estimulada 2microM de actina em
3125 mM Tris-HCl pH 71 e 375 mM MgCl2 pelo meacutetodo de enzimas acopladas (ver seccccedilatildeo xx
dos MM)Para tal realizaram-se ensaios com vaacuterias concentraccedilotildees de decavanadato total (10 50
100 250 500 e 1000) de modo a determinar qual o seu efeito na actividade MgATPaacutesica de S1
de miosina na ausecircncia e na presenccedila de um dos seus ligandos naturais Os ensaios foram
realizados agrave temperatura ambiente com incubaccedilatildeo preacutevia a 25ordm C durante 20 minutos
32
Figura 211 ndash Representaccedilatildeo
graacutefica da Actividade Mg2+
-
ATPaacutesica de S1 () (os ajustes
teoacutericos estatildeo representado a por
uma linha os resultados
experimentais estatildeo representados
por triacircngulos (Actividade
estimulada) e cruzes (Actividade
natildeo estimulada)) vs
[Decavanadato]total medidas pelo
meacutetodo das enzimas acopladas
(n=3) Os dados foram ajustados ao modelo de decaimento exponencial usando o programa GraphPad Prism para a
determinaccedilatildeo dos valores de IC50 (na ausecircncia de actina IC50= 266 plusmn 07 M decavanadato na presenccedila de actina
IC50= 28 plusmn 1 M decavanadato
A partir do graacutefico presente na Fig 29 verifica-se que o ajuste dos dados experimentais foi bem
sucedido A inibiccedilatildeo da actividade Mg2+
-ATPaacutesica do S1 de miosina eacute afectada em menor
amplitude na presenccedila de actina (60 de inibiccedilatildeo da actividade) do que na sua ausecircncia
(aproximadamente 80 de inibiccedilatildeo de actividade) Devido a esta diferenccedila pode sugerir-se que a
actina iraacute impedir parcialmente a ligaccedilatildeo do decavanadatoao S1 demiosina atraveacutes de
impedimentos esteacutereos ou interacccedilotildees electrostaacuteticas por exemplo A teacutecnica utilizada natildeo nos
permite concluir acerca de que regiatildeo(otildees) ou resiacuteduo(s) da actina participaratildeo na preservaccedilatildeo
parcial da actividade Mg2+
-ATPaacutesica
Por outro lado os valores de IC50 dos dois tipos de actividades natildeo apresentam diferenccedilas
significativas tendo-se obtido 266 plusmn 07 e 28 plusmn 1M decavanadato na ausecircncia e na presenccedila de
actina respectivamente o que corresponde a uma concentraccedilatildeo de cerca de 27 M de V10 O
meio de reacccedilatildeo utilizado nestes ensaios foi 10 mM HEPES e 25 mM KCl com 2 mM ATP Em
(Tiago et al 2004) o valor de IC50 para a actividade Mg2+-ATPaacutesica na ausecircncia de actina foi de
125 plusmn 025 microM enquanto o valor da actividade estimulada por actina foi de 136 plusmn 14 microM O
meio de reacccedilatildeo utilizado nos ensaios foi 20 mM HEPES 50 mM KCl e 2 mM ATP o que resulta
precisamente no dobro da forccedila ioacutenica a que foi realizado o ensaio do presente relatoacuterio
(exceptuando a contribuiccedilatildeo dos produtos da hidroacutelise de ATP uma vez que este meacutetodo
pressupotildee a ocorrecircncia de um ciclo de reacccedilotildees de hidroacutelise e siacutentese deste composto) Ora tendo
sido anteriormente referido que a afinidade do S1 de miosina para com a espeacutecie V10 diminui com
Act
ivid
ade
Mg
2+-A
TP
aacutesic
a d
e S
1 d
e
Mio
sin
a
[Decavanadato] (microM) Actividade Estimulada ()
Actividade Natildeo Estimulada ()
Ajuste Actividade Estimulada
Ajuste Actividade Natildeo Estimulada
33
o aumento da forccedila ioacutenica (Tiago et al 2004 tal poderaacute explicar a diferenccedila obtida entre estes
dois conjuntos de valores
243Quantificaccedilatildeo de cisteiacutenas reduzidas pelo meacutetodo de Ellman
A interacccedilatildeo do vanaacutedio com alguns tipos de aminoaacutecidos sensiacuteveis agrave oxidaccedilatildeo pode fornecer-nos
informaccedilatildeo bastante uacutetil em relaccedilatildeo agraves alteraccedilotildees estruturais que conduzem a uma diminuiccedilatildeo da
actividade fisioloacutegica do S1 de miosina Os estudos realizados por Chasteen 1983 concluiacuteram
que a reduccedilatildeo de vanadato (V) a vanadilo (IV) ocorre na presenccedila de agentes redutores comuns no
ambiente celular de que se destaca o aminoaacutecido cisteiacutena Esta eacute tambeacutem um alvo de
complexaccedilatildeo por parte do iatildeo VO2+
Jaacute Ramos et al (2009) demonstrou que a incubaccedilatildeo de
actina com a soluccedilatildeo de decavanadato induz a oxidaccedilatildeo de resiacuteduos de cisteiacutena e agrave formaccedilatildeo do
oxidovanaacutedio (IV) Este grupo de investigadores descobriu ainda que apenas a espeacutecie V10 induz a
oxidaccedilatildeo das cisteiacutenas ldquoraacutepidasrdquo da actina atraveacutes do processo proposto que eacute parcialmente
responsaacutevel pela inibiccedilatildeo jaacute descrita da actividade de ATPase do S1 de miosina estimulada por
actina quando exposta agrave soluccedilatildeo de decavanadato Esta inibiccedilatildeo apresenta-se afectada com a
presenccedila de ATP uma vez que os resiacuteduos de cisteiacutena permaneceram intactos nos ensaios
realizados com este ligando Jaacute Crans et al (2004) concluiu que o siacutetio activo de muitas fosfatases
(que conteacutem grupos tiol) pode formar um equiliacutebrio redox com o iatildeo vanadato (V) de forma natildeo
reversiacutevel
Os resultados obtidos por Sakurai e Shiromura (1980) sugeriram que o vanaacutedio (V) sofre reduccedilatildeo
a vanaacutedio (IV) sem dependecircncia do valor de pH do ensaio Registaram ainda uma estequiometria
de 31 na reacccedilatildeo do vanadato (V) com os resiacuteduos de Cys enquanto a estequiometria dos ensaios
com o vanadilo (IV) foi de 21
Berlett e Stadtman (1997) observaram que os tipos de aminoaacutecidos mais facilmente oxidaacuteveis
englobam entre outros os resiacuteduos de Cys Trp e MetAssim analisou-se o estado redox dos
grupos tiol dos resiacuteduos de Cys do S1 na presenccedila do decavanadato atraveacutes da monitorizaccedilatildeo da
Abs412 nm na presenccedila de 1 mgmL de S1 de miosina em tampatildeo 10 mM HEPES 25 mM KCl
variando a concentraccedilatildeo da soluccedilatildeo de decavanadato de modo a determinar que tipo de alteraccedilotildees
estruturais ocorrem na sua presenccedila no que diz respeito agrave diminuiccedilatildeo da actividade ATPaacutesica de
S1 na presenccedila de decavanadato
Os resultados referentes agrave quantificaccedilatildeo das cisteiacutenas reduzidas apoacutes o tratamento de S1 com
diferentes concentraccedilotildees de decavanadato em tampatildeo 10 mM HEPES pH 75 e 25 mM KCl a
25ordmC durante 20 minutos encontram-se descritos na Tabela 2VII
Tabela 2VII ndash Cisteiacutenas reduzidas totais e ldquoraacutepidas de S1 (1 mgmL) apoacutes tratamento com decavanadato
34
(010501002505001000 uM) em 10 mM HEPES 25 mM KCl pH 7 a 25ordmC durante 20 minutos A detecccedilatildeo
espectrofotomeacutetrica foi feita com DTNB (e 412=8400 uM-1 cm-1) (n variaacutevel entre 1 e 8)
Amostras Cys raacutepidas Cys totais
11 M S1 (controlo) 22 plusmn 04 106 plusmn 14
11 M S1 + 10 M Decavanadato 22 plusmn 05 95 plusmn 08
11 M S1 + 50 uM Decavanadato 20 plusmn 02 98 plusmn 14
11 M S1 + 100 uM Decavanadato 17 plusmn 03 97 plusmn14
11 M S1 + 250 uM Decavanadato 26 plusmn 02 98 plusmn 08
11 M S1 + 500 uM Decavanadato 21 plusmn 04 90 plusmn 06
11 M S1 + 1000 uM Decavanadato 24 plusmn 0 86 plusmn 08
A partir dos resultados obtidos pode sugerir-se que o S1 possui 11 resiacuteduos de Cys (controlo) o
que estaacute de acordo com o anteriormente descrito Verifica-se que o tratamento da proteiacutena com
concentraccedilotildees crescentes de decavanadato provoca a oxidaccedilatildeo de dois resiacuteduos de Cys do S1 de
miosina natildeo expostos ao solvente No entanto a teacutecnica utilizada natildeo nos permite identificar
quais
Tendo em consideraccedilatildeo que o valor de IC50 obtido para a reacccedilatildeo de inibiccedilatildeo daactividade Mg2+-
ATPaacutesica de S1 na presenccedila de decavanadato eacute de cerca de 27 microM pode sugerir-se que este seraacute o
valor ao qual ocorreraacute a oxidaccedilatildeo do primeiro resiacuteduo de Cys o que poderaacute contribuir para a
diminuiccedilatildeo da actividade de S1 observada
25Interacccedilatildeo de S1 com vanaacutedio(IV)
251Anaacutelise por espectroscopia de Ressonacircncia Paramagneacutetica Electroacutenica
A acumulaccedilatildeo intracelular de oxovanaacutedio (IV) in vivo constitui uma das principais formas de
apresentaccedilatildeo de vanaacutedio no organismo Deste modo torna-se bastante interessante analisar a
interacccedilatildeo entre este iatildeo e o S1 de miosina atraveacutes da teacutecnica de espectroscopia de Ressonacircncia
Paramagneacutetica Electroacutenica o meacutetodo de eleiccedilatildeo do estudo do iatildeo vanadilo e dos complexos por si
formados O electratildeo desemparelhado encontra-se fortemente acoplado ao nuacutecleo 51
V o que
35
resulta num espectro de RPE consistindo de 8 desdobramentos na direcccedilatildeo paralela e outros tantos
na direcccedilatildeo perpendicular (Chasteen 1983)
Os estudos realizados por Ahmed et al (1987) centraram-se na interacccedilatildeo do vanadilo com
proteiacutenas como eacute o caso da insulina transferrina entre outros obtendo um espectro caracterizado
por 8 linhas amplamente espaccediladas (na direcccedilatildeo paralela) Jaacute Ramos et al 2009 concluiu que a
exposiccedilatildeo da soluccedilatildeo de decavanadato agraves formas globular e filamentosa de actina produz um
espectro tiacutepico do iatildeo oxidovanaacutedio (IV) indicando a reduccedilatildeo do vanadato
Para aferir qual a concentraccedilatildeo de S1 que permitia obter uma boa razatildeo sinalruiacutedo titulou-se uma
soluccedilatildeo de sulfato de vanadilo 15 microM ([VOSO4]stock = 5 mM) com concentraccedilotildees crescentes de
proteiacutena (0 5 10 15 e 20 microM) em 10 mM HEPES 25 mM KCl A existecircncia de uma correlaccedilatildeo
linear entre a concentraccedilatildeo de proteiacutena e a intensidade dos sinais de RPE correspondente agrave banda
-12 perpendicular encontra-se representada na Fig 210
Figura 2 12ndash Representaccedilatildeo graacutefica da variaccedilatildeo da intensidade do sinal de RPE vs a concentraccedilatildeo de S1 utilizada
Apoacutes verificar que com uma concentraccedilatildeo de S1 igual a 20 microM se obtinham sinais com uma razatildeo
sinalruiacutedo adequada ao prosseguimento dos estudos sobre a interacccedilatildeo V(IV)-S1 de miosina
titulou-se uma soluccedilatildeo de 20 microM S1 com 0 25 5 10 15 25 50 75 100 150 200 500 e 1000
microM VOSO4
Traccedilou-se o graacutefico da intensidade da banda -12 perpendicular do sinal de EPR em funccedilatildeo da
concentraccedilatildeo de VOSO4 tendo-se determinado uma constante de dissociaccedilatildeo (KD) do complexo
S1-vanadilo de 8748 plusmn 2621 microM V(IV)
y = 016786x + 122435
Rsup2 = 092992759
Inte
nsi
dad
e d
o s
inal
de
RP
E
[S1] (microM)
36
Figura 213 ndash Representaccedilatildeo graacutefica da variaccedilatildeo do sinal de RPE vs a concentraccedilatildeo de vanadilo utilizada (os valores
experimentais estatildeo representados por losangos o seu ajuste estaacute representado pela linha)
A simulaccedilatildeo do espectro de EPR foi efectuada a partir do espectro obtido para uma concentraccedilatildeo
de 1 mM VOSO4 apoacutes adiccedilatildeo de 20 microM S1 de miosina tendo-se obtido os valores da constante de
acoplamento hiperfina (A=5016) e do factor g (g0=195)
Figura 214 ndash Representaccedilatildeo graacutefica do espectro de EPR obtido com 1000 microM de vanadilo e 20 microM de S1 (a cinzento
escuro) A simulaccedilatildeo do espectro encontra-se a cinzento claro
Por comparaccedilatildeo destes com os valores presentes na tabela de correspondecircncia xxx eacute possiacutevel
predizer quais os ligandos provaacuteveis do vanadilo na presenccedila de proteiacutena Assim concluiu-se que
o vanadilo estaacute coordenado com duas moleacuteculas de aacutegua um grupo hidroxilo e um grupo R=NR
O grupo hidroxilo pode indicar os resiacuteduos de aminoaacutecidos Tyr Treonina e Serina enquanto o
grupo R=NR poderaacute indicar a presenccedila de Arginina e Histidina como ligandos do vanadilo
Inte
nsi
dad
e d
os
sinai
s d
e R
PE
[Vanadilo] (microM)
Ajuste dos dados exp Dados experimentais
Campo magneacutetico (G)
Espectro Experimental Ajuste
37
3 Procedimento Experimental
31 Reagentes
Todos os reagentes utilizados satildeo de elevado grau de pureza (analiacutetico) Apresentam-se as suas
foacutermulas quiacutemicas ou abreviaturas quando necessaacuterio acrescentou-se a sua descriccedilatildeo Entre
parecircntesis colocou-se as empresas que os comercializam
Aacutecido aceacutetico glacial 100 (Riedel) Acrilamida 99 (Merck) Azul de bromofenol (Riedel-de-
Haumlen) -Mercaptoetanol (Sigma) Bissulfito de soacutedio (Sigma)Bradford (reagente de)
(BioRad)BSA (albumina de soro bovino Sigma-Aldrich) CaCl22H2O (cloreto de caacutelcio
dihidratado Fluka) CH3CH2OH 96 (etanol Panreac) Coomassie Blue R250 (Merck) D2O
(oacutexido de deuteacuterio SDS) DTNB (55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico) Aldrich) EDTA (aacutecido
etilenodiaminatetraaceacutetico Merck) Fosfoenolpiruvato 98 (Fluka) Glicerol (Sigma) Glicina
985 (Panreac) HCl 37 (Riedel-de Haumlen) H2SO4 96 (Pronalab) HEPES 99 (aacutecido 4-(2-
hidroxiletil)-1-piperazineetanosulfoacutenico Sigma) KH2PO4 99 (hidrogenofosfato de potaacutessio)
MgATP (Sigma) Na
ATP (Sigma) Na2HPO4 99 (hidrogenofosfato de soacutedio Riedel-de Haumlen)
NaH2PO4 (fosfato de soacutedio ) NaOH 986 (hidroacutexido de soacutedio Pronalab) KCl 99 (Cloreto de
potaacutessio Panreac) L-Cys 995 (enantioacutemero levoacutegiro do resiacuteduo de Cisteiacutena Fluka)Metanol
99 (Merck) Molibdato de amoacutenio (Sigma)NNrsquo-metil-bis-acrilamida 98 (Merck) NH4VO3
99 (metavanadato de amoacutenia Sigma) PSA 10 (persulfato de amoacutenio Merck) SDS
(dodecilo sulfato de soacutedio Sigma) Sulfito de soacutedio (Sigma) VOSO4 (sulfato de vanadilo
Aldrich) TEMED 99 (Tetrametiletilenodiamina Merck) Tris (2-amino-2-hidroximetil-13-
propanodiol Sigma)
32 Meacutetodos
321 Isolamento e purificaccedilatildeo das proteiacutenas de muacutesculo esqueleacutetico de coelho
3211 Purificaccedilatildeo da miosina preparaccedilatildeo de Subfragmento 1 por digestatildeo quimotriacutepsica
da miosina
O subfragmento-1 de miosina-II (S1) foi gentilmente cedido pelo Prof Doutor Manuel Aureliano
do Departamento de Quiacutemica Bioquiacutemica e Farmaacutecia da Faculdade de Ciecircncias e Tecnologia da
Universidade do Algarve
No entanto descreve-se abaixo de forma resumida o procedimento efectuado
38
32111 Purificaccedilatildeo da miosina
O protocolo de isolamento foi adaptado do meacutetodo previamente descrito por Margossian e Lowey
(1982) apresentando-se de forma esquemaacutetica na Fig 31
Sucintamente os tecidos musculares dorsais e das pernas foram extraiacutedos apoacutes o sacrifiacutecio do
animal (Fig31) De seguida foram adicionados 3 volumes da soluccedilatildeo de extracccedilatildeo A2
Centrifugou-se a soluccedilatildeo e filtrou-se o sobrenadante O sedimento foi entatildeo diluiacutedo em 14
volumes de aacutegua destilada de modo a que a forccedila ioacutenica atingisse 004 M Apoacutes adiccedilatildeo de szlig-
mercaptoetanol (szlig-me) deixou-se a precipitar a 4ordmC removendo o sobrenadante Centrifugou-se
novamente e ressuspendeu-se o sedimento na soluccedilatildeo de extracccedilatildeo A Adicionou-se a soluccedilatildeo
tampatildeo B3 para que a forccedila ioacutenica atingisse 05 M Homogeneizou-se e deixou-se repousar
durante 2h Homogeneizou-se novamente e centrifugou-se de modo a ocorrer precipitaccedilatildeo dos
complexos de actomiosina e proteiacutena desnaturada Apoacutes filtraccedilatildeo do sobrenadante em latilde de vidro
quantificou-se a soluccedilatildeo de miosina resultante pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental)
2 Soluccedilatildeo de extracccedilatildeo A 0012M K2CO3 004M KHCO3 06M KCl pH 65
3 Soluccedilatildeo-tampatildeo B Tris 100 mM pH 7 KCl 2M
39
Figura 31 ndash Representaccedilatildeo esquemaacutetica do protocolo de isolamento de miosina (Margossian e Lowey 1982) Todo o
procedimento foi realizado a 4ordmC (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
32112 Preparaccedilatildeo de Subfragmento-1 de miosina por digestatildeo com quimiotripsina
O S1 de miosina foi isolado apoacutes diaacutelise da miosina durante a noite contra a soluccedilatildeo C4 A
presenccedila do EDTA eacute essencial uma vez que a sua substituiccedilatildeo por MgCl2resulta na produccedilatildeo de
HMM em vez de S1 A suspensatildeo de miosina foi equilibrada e digerida com -quimotripsina
A digestatildeo foi interrompida pela adiccedilatildeo de PMSF Seguidamente ultracentrifugou-se o produto
da digestatildeo desprezando-se o sedimento (Fig 32) A soluccedilatildeo de sobrenadante maioritariamente
S1 foi quantificada pela leitura directa da Abs280 ou pelo meacutetodo de Bradford (ver as secccedilotildees
3311 e 3312 do Procedimento Experimental respectivamente) e armazenada em 10 sacarose
e 2 mM szlig-Me a -80ordmC
4 Soluccedilatildeo C 25mM Tris pH 70 012M KCl 1mM EDTA
40
Antes da realizaccedilatildeo dos ensaios descritos no presente relatoacuterio o S1 de miosina foi dialisado
contra a soluccedilatildeo D5 de modo a remover qualquer composto que pudesse complexar como
vanaacutedio Seguidamente quantificou-se a proteiacutena pelo meacutetodo de Bradford ou pelo meacutetodo
directo (ver secccedilotildees 3311 e 3312do Procedimento Experimtental)
Figura 3 2ndash Representaccedilatildeo esquemaacutetica do protocolo de preparaccedilatildeo do S1 de miosina por digestatildeo quimiotripsina de
acordo com o previamente descrito por Margossian e Lowey (1982) Todo o procedimento foi realizado a 4ordm C
excepto quando indicaccedilatildeo do contraacuterio (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
3212 Isolamento e purificaccedilatildeo de actina
Isolou-se e purificou-se a actina segundo o meacutetodo descrito por Pardee e Spudish que
compreende dois passos ndash a obtenccedilatildeo de um resiacuteduo seco denominado ldquopoacute de acetonardquo e a
extracccedilatildeo subsequente da actina a partir do ldquopoacute de acetonardquo na forma de G-Actina com a
posterior polimerizaccedilatildeo a F-Actina
32121 Obtenccedilatildeo do ldquopoacute de acetonardquo
Sacrificou-se o coelho com uma pancada atraacutes do pescoccedilo e sangrou-se o animal apoacutes o corte da
arteacuteria jugular Esfolou-se e colocou-se numa tina com gelo Retiraram-se as partes de tecido
conjuntivo que envolvem o tecido muscular com auxiacutelio de uma pinccedila e bisturi Seguidamente
retirou-se os muacutesculos dorsais e os muacutesculos brancos das pernas Passou-se os muacutesculos por aacutegua
destilada de modo a remover as impurezas e pesou-se num copo Trituraram-se os muacutesculos
numa maacutequina de triturar carne e extraiu-se o resultante com 3 volumes da soluccedilatildeo E6 durante 10
minutos com agitaccedilatildeo suave Centrifugou-se a suspensatildeo a 4000 g durante 10
5 Soluccedilatildeo D HEPES 10 mM pH 70 KCl 25 mM
6 Soluccedilatildeo E 01 M KCl 015 M K2HPO4 pH 65
41
minutosdesprezando-se o sobrenadante Ressuspendeu-se o sedimento em 6 volumes de soluccedilatildeo
F7durante 10 minutos com agitaccedilatildeo suave Verificou-se se o pH da soluccedilatildeo estava compreendido
entre 82 e 85 acertando-se estecom soluccedilatildeo G8 se necessaacuterio Centrifugou-se durante 10
minutos a 4000 g Desprezou-se o sobrenadante e ressuspendeu-se o sedimento em 3 volumes de
soluccedilatildeo H9 durante 10 minutos Centrifugou-se novamente a 4000 gdurante 10 minutos
desprezando-se o sobrenadante Ressuspendeu-se o sedimento duas vezes consecutivas em 6
volumes de aacutegua destilada durante 5 minutos Centrifugou-se a 4000 g durante 10 minutos apoacutes
cada adiccedilatildeo de H2O
Seguidamente ressuspendeu-se o sedimento 5 vezes consecutivas utilizando 3 volumes de
acetona de cada vez agitando suavemente durante 10 minutos agrave temperatura ambiente Colocou-
se o resiacuteduo filtrado numa tina de vidro sobre papel de filtro deixando-se evaporar o solvente
durante a noite num nicho Triturou-se o resiacuteduo seco numa maacutequina de triturar carne e deixou-se
novamente evaporar o solvente O ldquopoacute de acetonardquo obtido foi armazenado a -20 ordmC
32122 Purificaccedilatildeo da actina a partir de ldquopoacute de acetonardquo
Apoacutes a obtenccedilatildeo do poacute de acetona como acima indicado efectuou-se o protocolo seguidamente
descrito de modo a obter monoacutemeros de actina (G-Actina) e actina filamentosa (F-Actina)
Pesou-se 35 g de poacute de acetona e adicionaram-se 50 mL de soluccedilatildeo-tampatildeo I10
sendo a
extracccedilatildeo feita com agitaccedilatildeo suave durante 30 minutos Separou-se o extracto do poacute de acetona
hidratado filtrando a suspensatildeo com 4 camadas de gaze esterilizada tendo o cuidado de natildeo
espremer o resiacuteduo Voltou a extrair-se com 50 mL de soluccedilatildeo I durante 30 minutos mexendo
lentamente Adicionou-se este ao extracto obtido anteriormente Centrifugou-se o extracto total a
16 000 g durante 30 minutos Decantou-se para uma proveta sobre a qual se colocou gaze para
que o sedimento solto natildeo se juntasse ao sobrenadante e leu-se o volume De seguida adicionou-
se KCl ateacute uma concentraccedilatildeo final de 50 mM MgCl2 ateacute 2mM e ATP ateacute 1mM lentamente e com
agitaccedilatildeo contiacutenua de modo a promover a polimerizaccedilatildeo dos monoacutemeros de actina
Deixou-se polimerizar durante 2h a 25ordmC observando-se a presenccedila de pequenas bolhas de ar
indicativas de um aumento de viscosidade da soluccedilatildeo Arrefeceu-se em gelo durante 15 minutos e
adicionou-se KCl ateacute uma concentraccedilatildeo final de 06M Mexeu-se lentamente durante 30 minutos
apoacutes os quais se ultracentrifugou a 110 000g durante 1hora Desprezou-se o sobrenadante
ressuspendeu-se o sedimento com soluccedilatildeo tampatildeo E e transferiu-se para um potter
Dialisou-se durante a noite contra 1L de soluccedilatildeo-tampatildeo I a uma temperatura de 4ordmC com rotaccedilatildeo
7 Soluccedilatildeo F 005 M NaHCO3 pH 82-85
8 Soluccedilatildeo G 1M Na2CO3
9 Soluccedilatildeo H 1 mM EDTA pH 70
10 Soluccedilatildeo I 2 mM Tris-HCl pH 8 02 mM CaCl2 02 mM Na2ATP 05 mM szlig-Me 0005 NaN3
42
contiacutenua e mudanccedila de tampatildeo 2 a 3 vezes De seguida a actina foi centrifugada a 110
000gdurante 1hora Decantou-se a soluccedilatildeo de G-Actina (sobrenadante) e desprezou-se o
sedimento Determinou-se a concentraccedilatildeo de G-Actina pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental) A soluccedilatildeo de G-actina foi armazenada a uma temperatura de 4ordmC
apoacutes adiccedilatildeo de 002NaN3 mantendo-se estaacutevel durante 3 a 5 dias
331 Quantificaccedilatildeo das proteiacutenas isoladas
3311 Quantificaccedilatildeo pelo meacutetodo de Bradford
O meacutetodo de Bradford (Bradford 1976) baseia-se na utilizaccedilatildeo do reagente de Bradford
composto por entre outros aacutecido fosfoacuterico e o corante ldquoCoomassierdquo Este corante possui carga
negativa e a sua coloraccedilatildeo varia entre a vermelha (Absmax=465 nm) predominante em soluccedilatildeo e a
azul (Absmax=595 nm) presente aquando da ligaccedilatildeo aos grupos amina das proteiacutenas Em soluccedilotildees
aacutecidas diluiacutedas a coloraccedilatildeo varia proporcionalmente com a quantidade do corante ligado agrave
proteiacutena que por sua vez eacute directamente proporcional agrave concentraccedilatildeo da proteiacutena permitindo a
sua quantificaccedilatildeo ateacute a um limite de detecccedilatildeo de 20 gmL (Bradford 1976)
Deste modo prepararam-se amostras de BSA ([BSA]stock= 1mgmL) utilizando um intervalo de
concentraccedilotildees compreendido entre 0 e 15 gmL de acordo com a tabela 3I Apoacutes a adiccedilatildeo da
H2O MilliQ e do reagente de Bradford as soluccedilotildees foram sucessivamente homogeneizadas no
voacutertex
Tabela 3Indash Amostras utilizadas para a obtenccedilatildeo da recta de calibraccedilatildeo no meacutetodo de Bradford
BSA (L) BSA (g) H2O MilliQ (L) Reagente de Bradford (L)
0 0 800 200
125 125 79875 200
25 25 7975 200
5 5 795 200
10 10 790 200
15 15 785 200
Apoacutes incubaccedilatildeo a 25ordmC por 5 minutos leram-se os valores de Abs595 num
espectrofotoacutemetro (Shimadzu UV-160A) para todas as amostras Seguidamente traccedilou-se a recta
de calibraccedilatildeo (Abs595vs quantidade de BSA (g))
43
Para determinar a concentraccedilatildeo de proteiacutena apoacutes o processo de isolamento purificaccedilatildeo e
diaacutelise quando necessaacuterio colocou-se 5 L da amostra num tubo de Eppendorf e adicionou-se
795 L de H2O MilliQ homogeneizando a soluccedilatildeo no voacutertex Adicionou-se posteriormente 200
L de reagente de Bradford e repetiu-se a homogeneizaccedilatildeo Incubou-se a mistura a 25ordmC por 5
minutos apoacutes os quais se leu os valores de Abs595 nm no espectrofotoacutemetro Por interpolaccedilatildeo deste
valor a partir da recta de calibraccedilatildeo determinou-se a concentraccedilatildeo de proteiacutena na amostra
3312 Quantificaccedilatildeo pelo meacutetodo directo
A quantificaccedilatildeo das amostras proteicas pode tambeacutem ser efectuada atraveacutes da leitura directa da
absorvacircncia tendo por base a lei de Beer-Lambert onde Abs corresponde agrave
absorvacircncia ao comprimento de onda ao qual o ensaio foi realizado corresponde ao coeficiente
de extinccedilatildeo (substituiacutedo pela letra a se as unidades forem distintas da molaridade) b corresponde
ao percurso oacuteptico da radiaccedilatildeo na ceacutelula amostral e c corresponde agrave concentraccedilatildeo amostral
Assim no caso de S1 de miosina mediu-se a Abs280e calculou-se a concentraccedilatildeo da proteiacutena
sabendo que o coeficiente de extinccedilatildeo de S1 eacute (Ref)
No caso da actina mediu-se a Abs290ecalculou-se a concentraccedilatildeo da proteiacutena sabendo que o
coeficiente de extinccedilatildeo da actina eacute (Ref)
332Electroforese em condiccedilotildees desnaturantes (SDS-PAGE)
De modo a melhor caracterizar as proteiacutenas isoladas foi realizada uma electroforese em
condiccedilotildees desnaturantes (SDS-PAGE do inglecircs Sodium Dodecyl Sulfate Polyacrilamide Gel
Electrophoresis) O SDS (dodecilo sulfato de soacutedio) eacute um detergente anioacutenico utilizado neste
meacutetodo para desnaturar as proteiacutenas que se ligam ao SDS na proporccedilatildeo 14 g SDS g proteiacutena
(aproximadamente 1 moleacutecula de SDS por cada 2 resiacuteduos de aminoaacutecidos) (Stryer) O SDS induz
uma densidade de carga negativa significativa nas proteiacutenas a que se liga mascarando a sua carga
intriacutenseca pelo que as razotildees cargamassa satildeo tendencialmente idecircnticas Assim a SDS-PAGE
separa as proteiacutenas quando sujeitas a um campo eleacutectrico de acordo com o seu peso molecular
sendo a sua migraccedilatildeo directamente proporcional ao logaritmo deste uacuteltimo (Stryer)
O gel de resoluccedilatildeo com 10 de acrilamida foi obtido atraveacutes da adiccedilatildeo de 167 mL de soluccedilatildeo
stock de acrilamida11
075 mL de tampatildeo de resoluccedilatildeo12
251 mL de aacutegua destilada e 50 L de
SDS 10 Para promover a polimerizaccedilatildeo adicionou-se em seguida 38 L de PSA 10
11
soluccedilatildeo stock de acrilamida soluccedilatildeo de 30 (mv) de acrilamida e 08 (mv) de NNrsquo-metil-bis-acrilamida 12
tampatildeo de resoluccedilatildeo 15 M Tris-HCl pH 88
44
(persulfato de amoacutenia) 10 e 25 L de TEMED 99 (tetrametiletilenodiamina) Agitou-se
cuidadosamente de modo a natildeo formar bolhas e pipetou-se a soluccedilatildeo para a cassette de vidro
previamente montada no suporte De seguida adicionou-se uma peliacutecula de cerca de 2 mm de 2-
metil-2-butanol para verificar posteriormente a polimerizaccedilatildeo Apoacutes a polimerizaccedilatildeo que demora
cerca de 45 minutos verteu-se a agrave superfiacutecie e adicionou-se o gel de concentraccedilatildeo com 5 de
acrilamida constituiacutedo por 03 mL de soluccedilatildeo de stock acrilamida 045 mL de tampatildeo de
concentraccedilatildeo13
094 mL de aacutegua destilada 18 L de SDS 10 135 L de PSA 10 e 2L de
TEMED 99 Colocou-se o pente sobre o gel e deixou-se polimerizar durante cerca de 45
minutos
As amostras para aplicaccedilatildeo no gel de electroforese foram preparadas para uma concentraccedilatildeo de
proteiacutena de 1 mgmL num volume final de 100 L dos quais 25 L correspondem ao tampatildeo da
amostra14
As amostras foram aquecidas a 100ordmC durante 5 minutos Antes da aplicaccedilatildeo das
amostras transferiu-se a cassette para a tina de electroforese agrave qual se adicionou a soluccedilatildeo
tampatildeo de electroforese da tina superior15
e tampatildeo de electroforese da tina inferior16
Aplicaram-
se 10 L das soluccedilotildees de marcadores de peso molecular e 20 L das amostras proteicas
preparadas anteriormente A electroforese decorreu durante cerca de 3h30 min a uma voltagem
de 80 V De seguida transferiu-se o gel para uma tina corando as proteiacutenas durante 1h com
soluccedilatildeo corante17
Descorou-se o gel com soluccedilatildeo descorante18
tendo-se mudado esta soluccedilatildeo ateacute
as bandas se encontrarem bem definidas
Determinou-se a mobilidade relativa das bandas correspondentes aos marcadores de elevado peso
molecular e traccedilou-se a recta de calibraccedilatildeo ndash log(PM) vs log(Rf) onde Rf corresponde agrave
mobilidade relativa Por interpolaccedilatildeo determina-se o PM das proteiacutenas isoladas sabendo a sua
mobilidade relativa Por comparaccedilatildeo com os valores descritos na literatura pode concluir-se
acerca do grau de purificaccedilatildeo
34 Mediccedilatildeo da actividade enzimaacutetica de S1
341 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
13
tampatildeo de concentraccedilatildeo 05 M Tris-HCl pH 68 14
tampatildeo de amostra 320 mM Tris-HCl pH 68 04 M szlig-mercaptoetanol SDS 8 glicerol 15 (vv) e roxo de m-
cresol 0024 15
tampatildeo de electroforese da tina superior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS 16
tampatildeo de electroforese da tina inferior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS acetato de soacutedio
01 M 17
soluccedilatildeo corante ldquoCoomassie Blue R-250rdquo contendo metanol 45 (vv) e aacutecido aceacutetico glacial 10 (vv) 18
soluccedilatildeo descorante metanol 10 (vv) e aacutecido aceacutetico glacial 10 vv)
45
O meacutetodo de Fiske-Subarow modificado consiste na reacccedilatildeo do molibdato de amoacutenio com o
fosfato inorgacircnico em soluccedilatildeo aacutecida com formaccedilatildeo do aacutecido fosfomoliacutebdico Seguidamente daacute-
se a adiccedilatildeo de um agente redutor que provoca a reduccedilatildeo do composto a fosfomolibdato que forma
um complexo de cor azulada cuja absorvacircncia maacutexima ocorre para um comprimento de onda de
660 nm tornando possiacutevel a sua detecccedilatildeo por meacutetodos espectrofotomeacutetricos As actividades Ca2+
-
ATPaacutesica e K+EDTA-ATPaacutesica de S1 foram determinadas atraveacutes deste meacutetodo de acordo com
o descrito por Taussky e Shorr (1953) com mediccedilatildeo da quantidade de fosfato (Pi) libertado
atraveacutes da hidroacutelise de ATP
Para tal obteve-se primeiramente a recta de calibraccedilatildeo com as amostras presentes na Tabela 3II
utilizando uma soluccedilatildeo stock de 5 mM KH2PO4
Tabela 3II ndash Composiccedilatildeo das diferentes amostras utilizadas para determinar a recta de calibraccedilatildeo atraveacutes do meacutetodo
de Fiske-Subbarow modificado
KH2PO4 (L)
([KH2PO4]stock= 5mM)
PO43-
(nmole) H2O (uL)
0 0 1780
5 25 1775
10 50 1770
20 100 1760
40 200 1740
100 500 1680
A cada tubo de Eppendorf adicionaram-se seguidamente 200 L de soluccedilatildeo J19
e 20 L de agente
redutor20
homogeneizando no voacutertex apoacutes cada adiccedilatildeo Incubou-se durante 10 minutos e mediu-
se a Abs660 nm num espectrofotoacutemetro com intervalos de tempo iguais entre cada amostra Traccedilou-
se a recta de calibraccedilatildeo (Abs660 nmvs nmol PO43-
) a partir dos valores obtidos
Para determinar a actividade Ca2+
-ATPaacutesica de S1 prepararam-se tubos de Eppendorf com 400
L de meio de reacccedilatildeo21
25 L CaCl2 ([CaCl2]stock= 100 mM) 25 L ATP ([ATP]stock= 100 mM)
com [S1]ensaios=01 mgmL Perfez-se se necessaacuterio com H2O MilliQ ateacute perfazer um volume de 1
mL Para determinar a actividade K+EDTA-ATPaacutesica utilizou-se uma concentraccedilatildeo idecircntica de
19
Soluccedilatildeo J 5 N H2SO4 25 molibdato de amoacutenio 20
Agente redutor 5 mg aacutecido 1-amino-2-naftol-4-sulfoacutenico 30 mg bissulfito de soacutedio 30 mg sulfito de soacutedio em 26
mL H2O 21
meio de reacccedilatildeo 125 mM Tris-HCl pH 80 0575 M KCl
46
proteiacutena nos ensaios (01 mgmL) e o mesmo meio de reacccedilatildeo Aos tubos de Eppendorf
adicionou-se ainda 123 L de KCl ([KCl]stock = 3M) 25 L de ATP 10 L de EDTA
([EDTA]stock= 100 mM )e o volume de H2O MilliQ necessaacuterio para perfazer 1 mL Em ambas as
situaccedilotildees o uacuteltimo reagente adicionado foi o ATP uma vez que se trata do substrato da reacccedilatildeo
enzimaacutetica
A cineacutetica decorreu a 25ordmC durante 5 minutos para a actividade Ca2+
-ATPaacutesica e durante 3
minutos para a actividade K+EDTA-ATPaacutesica Apoacutes o periacuteodo de incubaccedilatildeo transferiu-se 150
L de amostra para tubos de Eppendorf contendo igual volume de TCA 10 Colocou-se no gelo
e homogeneizou-se no voacutertex Seguidamente centrifugou-se a 1500 g durante 10 minutos e
recolheu-se 200 L de sobrenadante para novos tubos de Eppendorf Adicionou-se a estes 158
mL de H2O MilliQ e 200 L de soluccedilatildeo J Homogeneizou-se no voacutertex e adicionou-se 20 L de
reagente redutor repetindo-se a homogeneizaccedilatildeo Incubou-se durante 10 minutos e mediu-se a
Abs660 nm com o mesmo intervalo de tempo de leitura para cada amostra Por intrapolaccedilatildeo a partir
da recta de calibraccedilatildeo obtida anteriormente determinou-se a quantidade de Pi libertado em cada
ensaio A partir destes resultados determinaram-se as actividades Ca2+
-ATPaacutesica e K+EDTA-
ATPaacutesica do S1 em nmol PO43-
minmg proteiacutena e em s-1
342 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
Da actividade Mg2+
-ATPaacutesica de S1 resulta a formaccedilatildeo dos produtos da reacccedilatildeo de
hidroacutelise do ATP - ADP e Pi Na presenccedila de ADP fosfoenolpiruvato (PEP) e da enzima piruvato
cinase (PK) os produtos resultantes satildeo ATP e piruvato Por acccedilatildeo da enzima lactato
desidrogenase (LDH) este uacuteltimo eacute convertido em lactato ocorrendo simultaneamente a oxidaccedilatildeo
do NADH a NAD+ monitorizada atraveacutes da Abs340 nm Quando o S1 de miosina se encontra no
meio reaccional o ATP eacute hidrolisado formando-se novamente ADP e Pi o que permite a
formaccedilatildeo de um ciclo (fig 21) Se as enzimas se encontrarem em excesso a velocidade de
conversatildeo do NADH eacute limitada pela produccedilatildeo de ADP e Pi Como o ATP eacute regenerado em cada
ciclo a sua concentraccedilatildeo inicial eacute sempre mantida (Bagshaw 1982)
Figura 33ndash Representaccedilatildeo esquemaacutetica do meacutetodo de enzimas acopladas
47
O protocolo utilizado e seguidamente descrito foi adaptado de Cuenda et al (1990 ndash Ver Refordf) A
uma cuvette de 1 mL adicionou-se S1 para uma concentraccedilatildeo final de 005 mgmL 800 L de
meio de reacccedilatildeo22
10 L de PEP ([PEP]stock = 42 mM) 10 L de PK+LDH 11 ([PK+LDH]stock =
1 mgmL) e 15 L de NADH ([NADH]stock=25 mM) Iniciou-se a monitorizaccedilatildeo da reacccedilatildeo
atraveacutes da leitura de Abs340 nm apoacutes a adiccedilatildeo de 20 L de ATP ([ATP]stock=100 mM) A
actividade Mg2+
-ATPaacutesica eacute obtida atraveacutes do declive correspondente agrave oxidaccedilatildeo do NADH Para
a mediccedilatildeo da Mg2+
-ATPase de S1 estimulada por actina adicionou-se F-actina para uma
concentraccedilatildeo final de 2 microM sendo a actividade obtida tambeacutem atraveacutes do declive correspondente
agrave oxidaccedilatildeo do NADH apoacutes a adiccedilatildeo da proteiacutena
35 Interaccedilatildeo entre vanaacutedio e subfragmento-1 de miosina
351 Preparaccedilatildeo das soluccedilotildees de vanaacutedio
As soluccedilotildees de vanadato foram preparadas em aacutegua MilliQ a partir de metavanadato de amoacutenio
(NH4VO3) numa concentraccedilatildeo stock de 50 mM com agitaccedilatildeo magneacutetica e aquecimento (~30 ordmC)
Esta soluccedilatildeo com um valor compreendido entre 6 e 7 designa-se por ldquometavanadatordquo e eacute
constituiacuteda pelas esspeacutecies V1 V2 V4 e V5 (Ref) Acidificando a soluccedilatildeo de metavanadato por
adiccedilatildeo de HCl ateacute um valor de pH proacuteximo de 4 observa-se uma mudanccedila de cor de amarelo
claro para laranja indicativa da formaccedilatildeo da espeacutecie decameacuterica de vanadato De seguida e
imediatamente antes da realizaccedilatildeo dos ensaios descritos o valor do pH da soluccedilatildeo de
decavanadato foi aumentado para cerca de 75 por adiccedilatildeo de NaOH A soluccedilatildeo (2) designada
por ldquometavanadatordquo natildeo foi posteriormente alterada ateacute agrave sua adiccedilatildeo para a realizaccedilatildeo dos ensaios
As soluccedilotildees foram armazenadas a 4ordmC e mantidas em gelo aquando da sua utilizaccedilatildeo
A soluccedilatildeo de V(IV) foi preparada em condiccedilotildees anaeroacutebias por diluiccedilatildeo de sulfato de vanadil
(VOSO4) para uma concentraccedilatildeo final de 5 mM em H2O MilliQ saturada com N2 gasoso
352Anaacutelise por espectroscopia de UV-Vis
A concentraccedilatildeo da espeacutecie decameacuterica em soluccedilatildeo depende do periacuteodo de tempo apoacutes a diluiccedilatildeo
no meio de reacccedilatildeo apropriado (Ref) A velocidade de desoligomerizaccedilatildeo da espeacutecie decameacuterica
pode ser analisada por espectroscopia de absorccedilatildeo na regiatildeo UVVis uma vez que esta espeacutecie
apresenta duas bandas caracteriacutesticas na regiatildeo do visiacutevel (a 358 e 400 nm) que natildeo se verifica
em quaisquer outras espeacutecies oligomeacutericas As bandas supramencionada sapresentam-se bem
22
Meio de reacccedilatildeo 3125 mM Tris-HCl pH 71 375 mM MgCl2
48
resolvidas atraveacutes da obtenccedilatildeo da derivada de 2ordf ordem do espectro apresentando a forma de 2
vales com miacutenimos centrados nos comprimentos de onda supracitados com dependecircncia linear
relativamente agrave concentraccedilatildeo de decavanadato total (Ramos et al 2006)
Assim a estabilidade da espeacutecie decameacuterica apoacutes diluiccedilatildeo da soluccedilatildeo de decavanadato no meio de
reacccedilatildeo dos ensaios (tampatildeo D) para uma concentraccedilatildeo final de 1 mM em vanadato total foi
caracterizada atraveacutes da leitura da Abs400 em funccedilatildeo do tempo tendo-se determinado qual a
ordem de reacccedilatildeo e o tempo de meia-vida da mesma Com base em estudos anteriores acerca da
estabilidade do V10 na presenccedila de diferentes proteiacutenas (Ramos et al 2006) determinou-se
igualmente o tempo de meia vida apoacutes a adiccedilatildeo de 20 microM de S1 de miosina ao meio reaccional
353 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
Os espectros de RMN do isoacutetopo 51
V foram realizados num espectroacutemetro Bruker AM-400 MHz
equipado com uma sonda multinuclear inversa de 5 mm Em tubos de RMN de 5 mm de diacircmetro
preparou-se soluccedilotildees de 500 L de deca- ou metavanadato correspondentes a um intervalo de
concentraccedilotildees de 1 a 10 mM suplementadas com 10 D2O Os espectros foram adquiridos agrave
temperatura ambiente com uma largura espectral de 23 923 Hz um tempo de aquisiccedilatildeo de 0086
s tempo de relaxaccedilatildeo de 001 s e uma sequecircncia de pulsos a 90ordm Utilizou-se a transformada de
Fourier com alargamento das bandas de 20Hz para o processamento dos espectros Os desvios
quiacutemicos das ressonacircncias de 51
V foram obtidos em relaccedilatildeo agrave espeacutecie VOCl3cuja frequecircncia de
ressonacircncia eacute considerada o padratildeo sendo todas as outras referenciadas a partir desta em ppm
As aacutereas dos sinais do RMN correspondentes agraves espeacutecies oligomeacutericas presentes foram obtidas
por integraccedilatildeo dos sinais As concentraccedilotildees relativas das espeacutecies foram calculadas com recurso agrave
equaccedilatildeo
onde A[Vx] corresponde agrave aacuterea referente agrave ressonacircncia da espeacutecie Vx com nordm de agregaccedilatildeo x
A[V]total agrave soma total das aacutereas medidas e [V] a concentraccedilatildeo total de vanaacutedio no ensaio
A teacutecnica de 51
V-RMN foi anteriormente aplicada em estudos de ligaccedilatildeo com S1 de miosina
(Ref) tendo sido demostrado que entre as espeacutecies V1 V2 V4 e V5 a espeacutecie tetrameacuterica possui a
maior afinidade para a miosina (Cremo et al) O desaparecimento ou alargamento dos sinais pode
implicar a ligaccedilatildeo de um dos oxovanadatos laacutebeis agrave miosina sem indicaccedilatildeo de qual uma vez que
49
se processa um equiliacutebrio entre eles (Crans 1990) Assim e tendo em conta as indicaccedilotildees de que
a espeacutecie V10 eacute distinta das restantes no que diz respeito agrave magnitude da interacccedilatildeo com a miosina
e inibiccedilatildeo da actividade ATPaacutesica estimulada por actina (Aureliano 2000 Tiago et al 2002)
torna-se fundamental investigar a interacccedilatildeo das diferentes espeacutecies de vanadato com o S1 de
miosina atraveacutes da teacutecnica de 51
V-RMN
Para tal obtiveram-se espectros de 51
V-RMN resultantes da titulaccedilatildeo de uma soluccedilatildeo 5 mM de
decavanadato total com concentraccedilotildees crescentes de S1 de miosina (0 5 10 15 e 20 microM) em
tampatildeo D Realizou-se ainda ensaios na presenccedila de 2 mM de decavanadato e 2 mM de ATP na
presenccedila e na ausecircncia de S1 de miosina de modo a verificar o efeito que estes ligandos naturais
da enzima teria na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o subfragmento-1
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
Os grupos sulfidrilo (-SH) das cisteiacutenas satildeo resiacuteduos de aminoaacutecidos extremamente sensiacuteveis agrave
oxidaccedilatildeo resultando na sua oxidaccedilatildeo (Dalle-Donne et al 1999) Logo a quantificaccedilatildeo destes
resiacuteduos eacute bastante importante nos estudos de interacccedilatildeo com vanaacutedio jaacute que os resiacuteduos de Cys
com ecircnfase para o SH1 e o SH2 satildeo bastante importantes na monitorizaccedilatildeo das modificaccedilotildees
funcionais da regiatildeo da cabeccedila
O procedimento seguidamente apresentado foi originalmente descrito por Gutieacuterrez-Merino
(1987)) Consiste na reacccedilatildeo entre os grupos ndashSH natildeo oxidados das proteiacutenas e o reagente de
Ellman (DTNB -55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico)) - formando-se o aniatildeo CNT (aniatildeo 3-
carboxilato-4-nitrotiofenolato) numa estequiometria de 11 Este apresenta um maacuteximo de
absorvecircncia a 412 nm (Robyt 1990) Assim apoacutes a incubaccedilatildeo da proteiacutena na presenccedila do metal
a quantidade de cisteiacutenas reduzidas por proteiacutena determina-se atraveacutes dos valores de Abs412
Deste modo colocou-se num tubo de Eppendorf o volume apropriado de S1 dialisado para que a
concentraccedilatildeo no ensaio fosse de 1 mgmL Adicionou-se o volume apropriado de decavanadato
para que a sua concentraccedilatildeo fosse de 0 10 50 100 250 500 ou 1000 M Perfez-se com soluccedilatildeo
D ateacute um volume final de 1 mL e incubou-se a 25ordmC por 20 minutos Apoacutes este periacuteodo colocou-
se 780 L da mistura reaccional numa cuvette de quartzo e adicionou-se 220 L de soluccedilatildeo K23
23
Tampatildeo K Tampatildeo fosfatohidrogenofosfato de soacutedio 025 M pH 72
50
Adicionou-se 15 L de DTNB (em soluccedilatildeo K [DTNB]stock=100 mM) ao meio reaccional e
registou-se o valor de absorvecircncia medido equivalente agraves cisteiacutenas ldquoraacutepidasrdquo (SH1 e SH2
expostos ao solvente) Adicionou-se 001 g de SDS que provoca a desnaturaccedilatildeo da proteiacutena e
consequentemente a exposiccedilatildeo de todos os resiacuteduos de cisteiacutenas que se encontravam ateacute aiacute
inacessiacuteveis no interior da proteiacutena (cisteiacutenas totais) Estes resiacuteduos podem assim reagir com o
DTNB
Estimou-se o nuacutemero de resiacuteduos de cisteiacutenas em todas as amostras com recurso ao coeficiente de
extinccedilatildeo molar () do DTNB (ver ponto 3 dos Anexos) que foi determinado atraveacutes da titulaccedilatildeo
de L-Cisteiacutena com intervalo de concentraccedilotildees compreendidos entre 0 e 50 microM na presenccedila de
25microM de DTNB em tampatildeo D Os valores de absorvecircncia foram obtidos para um comprimento de
onda de 412 nm
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
3621 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
A teacutecnica mais eficiente de anaacutelise da interacccedilatildeo entre o iatildeo vanadilo e proteiacutenas como a
calmodulina e a transferrina entre outras permite a obtenccedilatildeo raacutepida de informaccedilatildeo no que toca agrave
conformaccedilatildeo e amplitude da referida interacccedilatildeo
Os espectros de Ressonacircncia Paramagneacutetica Electroacutenica (RPE) da espeacutecie paramagneacutetica V(IV)
foram realizados a 70K num espectroacutemetro Bruker EMX-300 X-band Todas as soluccedilotildees
utilizadas e amostras foram preparadas em anaerobiose numa cacircmara de luvas de modo a evitar a
reduccedilatildeo de V(IV) a V(V) Em tubos capilares com 02 mL de amostra manteve-se a concentraccedilatildeo
de S1 a 20 M e variou-se a concentraccedilatildeo de vanadilo (0 25 5 10 15 25 50 75 100 150
200 500 e 1000 M [VOSO4]stock=5 mM) em tampatildeo D Apoacutes obtenccedilatildeo dos espectros estes
foram tratados atraveacutes do software SimFonia e WinEPR tendo-se obtido um graacutefico com a
variaccedilatildeo da intensidade do sinal de RPE em funccedilatildeo da concentraccedilatildeo de sulfato de vanadilo
presente Realizaram-se ainda simulaccedilotildees de espectros obtidos com 20 microM de S1 e 1000 microM de
V(IV) com vista agrave obtenccedilatildeo dos ligandos provaacuteveis do vanadilo atraveacutes da anaacutelise da constante
de acoplamento hiperfina (A) e do factor g (g)
4 Conclusotildees gerais
Apesar de jaacute ter sido demonstrado que o vanaacutedio inibe a Na+K
+-ATPase da miosina a funccedilatildeo
deste elemento no tecido muacutesculo esqueleacutetico continua largamente incompreendida A enorme
complexidade quiacutemica deste metal especialmente no que concerne agrave oligomerizaccedilatildeo e reacccedilotildees
51
redox constitui um obstaacuteculo agrave compreensatildeo do comportamento do vanaacutedio nos organismos
vivos Assim a caracterizaccedilatildeo da quiacutemica deste elemento eacute de grande utilidade antes de iniciar o
estudo da sua interacccedilatildeo com o presente sistema
A anaacutelise por espectroscopias de 51
V-RMN e de UV-Vis de soluccedilotildees de vanadato diluiacutedas nos
meios de reacccedilatildeo utilizados no presente relatoacuterio (10 mM HEPES pH 70 25 mM KCl) mostrou
claramente que a espeacutecie decameacuterica (V10) possui um tempo de vida suficientemente longo para a
determinaccedilatildeo do seu efeito isolado ao contraacuterio do que se verifica com as espeacutecies mono- di-
tetra- e pentameacutericas (V1 V2 V4 e V5) Por outro lado a anaacutelise verificou ainda que apoacutes diluiccedilatildeo
das soluccedilotildees de decavanadato no meio de reacccedilatildeo coexistem as espeacutecies V10 e V1 pelo que os
efeitos putativos desta uacuteltima deveratildeo ser subtraiacutedos aquando da monitorizaccedilatildeo dos efeitos da
espeacutecie decameacuterica
O vanaacutedio (V) (vanadato) inibe as enzimas que catalizam reacccedilotildees de transferecircncia de grupos
fosfato como a ATPase da miosina [Goodno 1979] devido agrave sua analogia estrutural com o iatildeo
fosfato Na sua utilizaccedilatildeo como inibidor enzimaacutetico natildeo eacute tida em conta a contribuiccedilatildeo individual
das espeacutecies oligomeacutericas considerando-se normalmente a espeacutecie V1 como inibidora efectiva
[Goodno e Taylor 1982] No presente relatoacuterio verificou-se por espectroscopia de 51
V-RMN que
outros oligoacutemeros de vanadato interagem com o S1 de miosina tendo as espeacutecies V10 e V4 os
maiores niacuteveis de afinidade A sua interacccedilatildeo envolve forccedilas electrostaacuteticas e eacute afectada
negativamente por ligandos naturais de enzima como eacute o caso do ATP
A exposiccedilatildeo do S1 de miosina (005 mgmL) a decavanadato conduz agrave inibiccedilatildeo do capacidade da
proteiacutena de estimular a actividade Mg2+
-ATPaacutesica com um valor de IC50 de 266 plusmn 07 M
decavanadato sendo o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM
decavanadato (80) Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM decavanadato sendo
o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM decavanadato (60)
exercendo um efeito de impedimento agrave ligaccedilatildeo entre o vanaacutedio e o S1
A uma concentraccedilatildeo de 27 microM decavanadato verifica-se a oxidaccedilatildeo de um dos resiacuteduos de Cys
de S1 de miosina seguida pela oxidaccedilatildeo gradual de outro resiacuteduo de Cys ateacute agrave concentraccedilatildeo de
1000 microM Ambos os resiacuteduos oxidados natildeo se encontravam expostos ao solvente utilizado A
oxidaccedilatildeo dos resiacuteduos permite sugerir que esta modificaccedilatildeo oxidativa sofrida pelo S1 contribui
para a diminuiccedilatildeo da sua actividade de ATPase
52
Por espectroscopia de RPE revelaram-se os possiacuteveis ligandos equatoriais do vanaacutedio aquando da
sua interacccedilatildeo com o S1 de miosina como sendo duas moleacuteculas de aacutegua um ligando contendo o
grupo R-OH e outro contendo o grupo R=NR
Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados de
oxidaccedilatildeo (IV) e (V) no S1 de miosina para concentraccedilotildees facilmente alcanccedilaacuteveis no ambiente
terrestre
53
5 Estudos futuros
No sentido do melhor esclarecimento das alteraccedilotildees moleculares e estruturais que ocorrem no
subfragmento 1 quando exposta ao vanaacutedio que conduzem a uma diminuiccedilatildeo da sua funccedilatildeo
sugere-se
- Quantificaccedilatildeo das cisteiacutenas reduzidas de S1 na presenccedila de metavanadato
- Determinaccedilatildeo das actividades Ca2+
- e K+-ATPaacutesica do S1 de miosina na presenccedila de
decavanadato
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina estimulada por actina
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina na presenccedila de
metavanadato
- Realizaccedilatildeo de RMN 2D com variaccedilatildeo das concentraccedilotildees das soluccedilotildees de decavanadato e
metavanadato na presenccedila de S1
-Mutageacutenese dos resiacuteduos de Cys doS1 de miosina de modo a verificar quais os resiacuteduos que
sofrem oxidaccedilatildeo na presenccedila de decavanadato
- Determinaccedilatildeo por calorimetria (DSC Differential Scanning Calorimetry) da variaccedilatildeo da
entalpia (∆H) e da temperatura de desnaturaccedilatildeo do S1 de miosina na presenccedila de vanaacutedio uma
vez que estas propriedades caracteriacutesticas de cada proteiacutena satildeo alteradas apoacutes ocorrecircncia de
modificaccedilotildees estruturais
- Anaacutelise por SDS-PAGE de S1 de miosina apoacutes tratamento com vanaacutedio de modo a determinar
quais os polipeacuteptidos eventualmente formados apoacutes clivagem da proteiacutena
54
6 Referecircncias bibliograacuteficas
- Sellers J Myosins 2nd Edition Oxford University Press 1999
- L Stryer inBiochemistry (Ed WH Freeman 2005 pp
Refordfas Materiais e Meacutetodos
[A] S S Margossian SLowey Methods Enzymol1982 85 55-71
[B] JD Pardue JA Spudich Methods Enzymol 1982 85 164-181
[C] M Bradford Anal Biochem1976 72 248-254
[D] H H Taussky E Shorr J Biol Chem 1953 202 675-685
[E] C R Bagshaw in Outline Studies in Biology ndash Muscle Contraction Chapman and Hall Ed
1982
[F] A Cuenda et al Eur J Biochem1990 194 663-670
55
Anexos
1 Determinaccedilatildeo da recta de calibraccedilatildeo meacutetodo de Bradford
Figura1ndashRepresentaccedilatildeo graacutefica da recta de calibraccedilatildeo (Abs595 nm vs [BSA]) utilizada para o meacutetodo de Bradford
2 Determinaccedilatildeo da recta calibraccedilatildeo para a electroforese em condiccedilotildees desnaturantes
Figura 2 ndash Representaccedilatildeo graacutefica da recta de calibraccedilatildeo (log(Peso Molecular) vs Rf dos marcadores de peso
molecular) onde Rf indica determinada atraveacutes do padratildeo de peso molecular
(poccedilo (a))
3Caacutelculo do coeficiente de extinccedilatildeo molar () para a quantificaccedilatildeo de cisteiacutenas reduzidas
atraveacutes do meacutetodo de Ellman
Traccedilou-se a recta de Abs412 nm meacutedia vs [L-Cisteiacutena] de modo a determinar o valor do coeficiente
de extinccedilatildeo molar definido como o declive daquela recta Assim =8400 M-1
cm-1
como
podemos verificar na Fig 3
A595 nm = 00546[BSA] + 00209 Rsup2 = 099051
Ab
s 59
5 n
m
[BSA] (mgmL)
log PM = -1177 Rf + 2247 Rsup2 = 098957
log
PM
Mobilidade relativa (Rf)
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero

10
cima do local de ligaccedilatildeo ao nucleoacutetido A regiatildeo entre os fragmentos de 50 e 20 kDa (loop 2)
encontra-se perto da intersecccedilatildeo entre os domiacutenios da cabeccedila e do pescoccedilo e deveraacute interagir com
a actina uma vez que a ligaccedilatildeo desta a protege da proteoacutelise Na ausecircncia do loop 2 a miosina
liga mais fracamente agrave actina do que no caso da miosina intacta A sequecircncia primaacuteria de ambos
os loops apresenta conservaccedilatildeo reduzida sendo provaacutevel que sejam responsaacuteveis por muita da
diversidade das diferentes propriedades enzimaacuteticas presentes em diferentes isoformas
No fragmento de 20 kDa existem dois grupos sulfidrilo Cys-707 e -697 usualmente
denominados de SH1 e SH2 devido agrave ordem da sua reactividade quiacutemica Na presenccedila de
nucleoacutetidos pode ocorrer o ldquocross-linkingrdquo entre estes dois grupos incluindo a oxidaccedilatildeo a uma
ponte dissulfureto
Devido agrave insolubilidade da miosina-II nativa a estrateacutegia adoptada para os estudos com esta
proteiacutena eacute frequentemente a sua clivagem em fragmentos funcionais separaacuteveis Um dos locais de
clivagem ocorre tipicamente a 130 kDa do N-terminal produzindo a meromiosina pesada (HMM)
e a meromiosina leve (LMM) [C] A HMM eacute formada pelas 2 cabeccedilas e pescoccedilo e apresenta
propriedades semelhantes agrave miosina sem digestatildeo (ligaccedilatildeo e translocaccedilatildeo de actina e hidroacutelise de
MgATP)
A HMM pode ser novamente clivada atraveacutes de uma reacccedilatildeo de proteoacutelise produzindo os
subfragmentos 1 (S1) e 2 (S2)O S1 (90 kDa) eacute enzimaticamente activo sendo constituiacutedo pelos
domiacutenios da cabeccedila e do pescoccedilo e apresentando associaccedilatildeo agraves duas cadeias leves ou apenas agrave
ELC (Fig 14)Em alternativa o S1 pode ser produzido por clivagem directa da miosina intacta
formando-se tambeacutem um fragmento em forma de cilindro que consiste nas regiotildees S2 e LMM
(Ref)Utiliza-se extensivamente o S1 para estudos cineacuteticos e de ligaccedilatildeo agrave actina uma vez que a
sua solubilidade simplifica o planeamento da experiecircncia e a subsequente interpretaccedilatildeo dos dados
[A]
Figura 14- Estrutura do subfragmento-1 de miosina de muacutesculo esqueleacutetico com ecircnfase nos domiacutenios de 50 kDa
central (a vermelho) 25 kDa no N-terminal (a verde) e 20 kDa no C-terminal (a azul) As cadeias leves essenciais
encontram-se a amarelo e as cadeias leves regulatoacuterias a lilaacutes Os locais de ligaccedilatildeo agrave actina e ao ATP estatildeo
igualmente destacados (adaptado de Holmes e Geeves 1999 [D])
11
112 Actina
O termo ldquoactinardquo refere-se a uma famiacutelia extremamente conservada de proteiacutenas citoplasmaacuteticas
presente em todos os eucariotas Esta proteiacutena natildeo se encontra nos procariotas [E] A maioria das
actinas possui peso molecular proacuteximo de 42 kDa outra evidecircncia do seu grau elevado de
conservaccedilatildeo no que concerne agrave sequecircncia e ao nuacutemero de aminoaacutecidos Os eucariotas superiores
apresentam um grande nuacutemero de isoformas de actina que por sua vez se encontram codificados
por um ainda maior nuacutemero de genes (Ref) A expressatildeo das isoformas eacute diferencial consoante o
tecido e sua especializaccedilatildeo pelo que deveratildeo possuir funccedilotildees e mecanismos direccionais
diferentes (Ref) Foi ainda observado que os mRNA das diferentes isoformas apresentam uma
distribuiccedilatildeo heterogeacutenea nas ceacutelulas de forma semelhante aos seus produtos proteicos [F] Estes
dados suportam a hipoacutetese supramencionada do direccionamento das isoformas de actina para um
determinado local na ceacutelula de acordo com as suas funccedilotildees
A actina presente no citoesqueleto eacute responsaacutevel por processos como o mecanismo contraacutectil
fagocitose manutenccedilatildeo e alteraccedilatildeo da estrutura celular locomoccedilatildeo transporte de organelos
regulaccedilatildeo do transporte de solutos entre outras Nos eucariotas superiores estaacute ainda envolvida
no transporte de gracircnulos de pigmento retinoacuteide [G] transporte de gracircnulos no axoplasma [H] e
secreccedilatildeo entre outros [E] Actualmente a isoforma da actina (-actina) eacute a melhor estudada
encontrando-se envolvida no mecanismo contraacutectil [E]
Os filamentos de -actina tecircm cerca de 1m de comprimento e 80 Å Consistem em duas cadeias
de actina filamentosa (F-actina) organizados em heacutelice [I] (Holmes et al 1990) sobre os quais
se encontram outras proteiacutenas ndash a troponina (C I e T) e a tropomiosina (Azevedo 1999) (Figura
15) A troponina eacute filamentosa e localiza-se nas depressotildees da heacutelice de F-actina conferindo
rigidez aos filamentos finos Por sua vez a tropomiosina eacute essencial na regulaccedilatildeo do mecanismo
contraacutectil
Figura 15 ndash Modelo estrutural dos filamentos de F-actina contendo a tropomiosina e o complexo de troponina (I C
e T) (Adaptado de Susana Ramos 2005 [J])
12
A F-actina eacute formada atraveacutes da polimerizaccedilatildeo de subunidades globulares de actina (G-actina) A
G-actina eacute composta por 2 domiacutenios de tamanho semelhante separados pelos bolsos de ligaccedilatildeo(1)
de catiotildees divalentes e(2) de nucleoacutetidos [KLM] Estes por sua vez estatildeo subdivididos noutros
dois de menores dimensotildees (1-4) (Fig 16) cada um envolvido nas interacccedilotildees actina-actina ou
na formaccedilatildeo do topo de ligaccedilatildeo do nucleoacutetido As regiotildees N-terminal e C-terminal estatildeo proacuteximas
uma da outra no subdomiacutenio 1 [E]
Figura 16 ndash Estrutura do monoacutemero de actina Encontram-se representados os subdomiacutenios 1 a 4 e a vermelho a
moleacutecula de ATP junto do respectivo local de ligaccedilatildeo (Protein Data Bank ID 1j6z)
113 Mecanismos moleculares da contracccedilatildeo muscular
A contracccedilatildeo muscular ocorre atraveacutes do deslizamento dos filamentos finos (de actina) por entre
os filamentos grossos (de miosina) conseguida pela reacccedilatildeo de hidroacutelise de ATP e regulada pela
tropomiosina e pelo complexo de troponina em resposta a impulsos nervosos Este mecanismo foi
inicialmente proposto por Huxley-Niedergerke [N] e Huxley-Hanson [O] ambos em 1954 A base
experimental de ambos os estudos foi a inexistecircncia de variaccedilatildeo do tamanho dos filamentos
durante a contracccedilatildeo A banda A apresenta sempre as mesmas dimensotildees por outro lado a banda
I a zona H diminuem consideravelmente aquando da contracccedilatildeo (Figs 12 e 17)
Figura 17 ndash Modelo da contracccedilatildeo muscular ndash deslizamento dos filamentos finos (azuis) relativamente aos
filamentos grossos (rosa) (Adaptado de Stryer 2002 [2])
13
Na ausecircncia de ATP e supondo a ausecircncia de inibiccedilatildeo da interacccedilatildeo actina-miosina as proteiacutenas
encontram-se fortemente associadas formando o complexo actomiosina (AM) (Figura 15)
Nestas condiccedilotildees as cabeccedilas de miosina formam um acircngulo de 45ordm com o eixo dos filamentos
finos de actina [P]
Na presenccedila de ATP o complexo AM dissocia-se jaacute que a afinidade do complexo M-ATP para a
actina eacute 105 vezes inferior agrave afinidade da miosina para a actina O ATP liga-se agrave miosina
ocorrendo seguidamente a reacccedilatildeo de hidroacutelise de ATP Os produtos daiacute resultantes ndash ADP e Pi -
permanecem ligados agrave miosina formando o complexo M-ADP-Pi Com a ligaccedilatildeo subsequente agrave
actina forma-se o complexo AM-ADP-Pi no qual as cabeccedilas de miosina apresentam um acircngulo
recto com o eixo do muacutesculo [P] Neste ponto daacute-se a libertaccedilatildeo progressiva de Pi e ADP Eacute
provaacutevel que a energia proveniente da hidroacutelise do ATP provoque uma alteraccedilatildeo do acircngulo de
ligaccedilatildeo das cabeccedilas de miosina ao filamento de actina passando novamente de 90ordm para 45ordm [P]
Esta alteraccedilatildeo conformacional provoca o deslizamento do filamento de actina para o centro do
sarcoacutemero cujo encurtamento promove a contracccedilatildeo das miofibrilas O processo supracitado
ocorre em cerca de 1ms[Q] e encontra-se resumido na Fig 18
Figura 18 ndash Esquema cineacutetico da hidroacutelise de ATP pela miosina (Adaptado de Sellers 1999 [A])
1131 Regulaccedilatildeo da contracccedilatildeo muscular
O mecanismo pelo qual se processa a regulaccedilatildeo muscular foi inicialmente descrito por estudos
distintos de Haselgrove Huxley e Parrey-Squire todos em 1973 [RST] Aleacutem da implicaccedilatildeo
directa da tropomiosina e do complexo de troponina a concentraccedilatildeo de Ca2+
no muacutesculo em
relaxamento eacute bastante reduzida (10-7
M) aumentando notavelmente aquando da contracccedilatildeo (10-5
-
10-3
M)
Resumidamente quando ocorre a excitaccedilatildeo do muacutesculo esqueleacutetico a despolarizaccedilatildeo da
membrana plasmaacutetica eacute difundida ateacute ao interior do tecido atraveacutes dos tuacutebulos transversais
(invaginaccedilotildees da membrana plasmaacutetica da fibra muscular) A despolarizaccedilatildeo provoca a abertura
dos canais ioacutenicos de Ca2+
do retiacuteculo sarcoplasmaacutetico adjacente Como a concentraccedilatildeo de Ca2+
no interior do organelo eacute muito elevada o caacutelcio passa raacutepida e uniformemente para o citosol
14
Assim a regulaccedilatildeo da contracccedilatildeo ocorre com coordenaccedilatildeo e rapidez na totalidade das miofibrilas
do tecido muscular (Ref)
Em repouso a troponina I encontra-se posicionada de tal forma que a proteiacutena adjacente
troponina C impede estruturalmente a ligaccedilatildeo das cabeccedilas da miosina agrave actina (acircngulo =50ordm) O
aumento da concentraccedilatildeo intracelular de Ca2+
provoca um aumento da sua disponibilidade para
ligar agrave troponina C (Fig 19)
Figura 19ndash Regulaccedilatildeo do mecanismo contraacutectil no muacutesculo esqueleacutetico pelo complexo de troponina
Como consequecircncia a troponina C altera a sua conformaccedilatildeo (=70ordm) associando-se agrave troponina I
que deixa de inibir a ligaccedilatildeo das pontes transversais A alteraccedilatildeo anterior eacute transmitida agrave
tropomiosina atraveacutes da troponina T adjacente agrave C o que provoca o deslocamento da
tropomiosina na direcccedilatildeo do interior do sulco do filamento de actina Assim eacute possiacutevel a
interacccedilatildeo eficaz entre actina e miosina A Ca2+
-ATPase do retiacuteculo sarcoplasmaacutetico provoca a
reacumulaccedilatildeo do iatildeo caacutelcio no organelo impedindo o prolongamento desnecessaacuterio da contracccedilatildeo
(Refordf)
12 Vanaacutedio
O vanaacutedio eacute um metal de transiccedilatildeo pertencente ao grupo VB da tabela perioacutedica Foi
primeiramente descoberto em 1801 por Andreacutes Manuel del Riacuteo que concluiu erradamente que era
idecircntico ao croacutemio e definitivamente por Nils Gabriel Svenson em 1831 Recentemente foram
descobertas importantes funccedilotildees bioloacutegicas deste elemento provavelmente essencial para os
humanos de que se destaca no contexto do presente relatoacuterio a possibilidade de actuar como
anaacutelogo do grupo fosfato e a inibiccedilatildeo de enzimas in vitro como eacute o caso da actividade ATPaacutesica
da miosina
Eacute um metal soacutelido agrave temperatura ambiente que se encontra amplamente distribuiacutedo nos vaacuterios
ecossistemas em dois isoacutetopos naturais ndash 50
V e 51
V ndash raramente excedendo 01 partes por biliatildeo
15
(ppb) Estaacute associado a cerca de 70 minerais diferentes na natureza (dos quais se destacam o
uracircnio cobre chumbo zinco fosfato e titacircnio ferroso) especialmente na forma de vanadato ou
pentoacutexidos de vanaacutedio O vanaacutedio eacute um dos principais constituintes da mateacuteria orgacircnica foacutessil
com especial ecircnfase para o carvatildeo oacuteleo de crude e foacutesseis carbonaacuteceos [UV] bem como em
alimentos consumidos pelo Homem (salsa pimenta preta cogumelos marisco vegetais cereais
gorduras e oacuteleos) [W] Estes constituem a maior fonte de exposiccedilatildeo ao vanaacutedio da maioria da
populaccedilatildeo humana [X]
Normalmente a entrada do vanaacutedio no organismo ocorre por inalaccedilatildeo ou em menores
quantidades por absorccedilatildeo pelo tracto gastrointestinal ou pela pele O seu armazenamento centra-
se no fiacutegado rins ceacuterebro coraccedilatildeo muacutesculos e ossos No entanto a retenccedilatildeo de vanaacutedio no
organismo humano eacute reduzida ndash em condiccedilotildees normais cerca de 100 M (em muitos tecidos a
quantidade eacute inferior a 10 ngg de tecido) [Y]
Actualmente natildeo existem provas de deficiecircncia nutricional de vanaacutedio em humanos ou de
enzimas que contenham vanaacutedio em animais superiores No entanto vaacuterios estudos relacionam a
ocorrecircncia de deficiecircncia em galinhas e ratos devido a uma alimentaccedilatildeo pobre em vanaacutedio Os
sintomas relatados incluem retardamento do crescimento malformaccedilotildees oacutesseas perturbaccedilotildees no
metabolismo lipiacutedico e estagnaccedilatildeo reprodutiva entre outros [Z AA AB] Os efeitos fisioloacutegicos
do vanaacutedio podem depender do tipo de composto ou de oligoacutemero uma vez que consoante as
suas estabilidade labilidade e nuacutemero de coordenaccedilatildeo as respostas seratildeo diferenciadas ao niacutevel
dos organismos vivos
Por outro lado a toxicidade do vanaacutedio encontra-se bem caracterizada existindo sintomas
descritos em vaacuterios mamiacuteferos (inclusive o Homem) que incluem diarreias catildeibras irritaccedilatildeo da
pele e mucosas palpitaccedilotildees cardiacuteacas anemia pneumonia bronquite perturbaccedilotildees no Sistema
Nervoso Central (SNC) entre outros (Ref) A toxicidade varia geralmente com a natureza do
composto com aumento proporcional agrave sua valecircncia Verificou-se que a forma de entrada do
vanaacutedio tem tambeacutem efeito na sua toxicidade ndash os maiores niacuteveis verificaram-se por injecccedilatildeo
intravenosa da amostra sendo progressivamente inferiores por inalaccedilatildeo e finalmente por
ingestatildeo [X AA] A ampla utilizaccedilatildeo dos compostos de vanaacutedio na induacutestria quiacutemica bem como
as elevadas quantidades de vanaacutedio libertadas atraveacutes da combustatildeo dos oacuteleos de crude
contribuem consideravelmente para o aumento da biodisponibilidade do vanaacutedio Como tal o
interesse neste elemento e nos problemas ambientais por ele causados tem vindo a aumentar
significativamente [V]
121 Propriedades quiacutemicas
16
A quiacutemica do vanaacutedio eacute extraordinariamente complexa pelo que originou vaacuterias interpretaccedilotildees
incorrectas desde a sua descoberta No entanto as suas propriedades dependem em uacuteltima
anaacutelise da configuraccedilatildeo electroacutenica do elemento Assim a variabilidade encontrada no
comportamento quiacutemico do vanaacutedio pode dever-se agrave sua configuraccedilatildeo ndash [Ar] 3d2 4s
2 ndash que
permite a formaccedilatildeo de vaacuterios estados de oxidaccedilatildeo (de +2 a +5) Em soluccedilatildeo aquosa os estados de
oxidaccedilatildeo estaacuteveis vatildeo de V(V) a V(II) Destes apenas os +3 a +5 satildeo biologicamente importantes
jaacute que o V(II) apresenta demasiada capacidade redutora para existir nos organismos vivos
conhecidos [AC]
No fluido extracelular os compostos de vanaacutedio encontram-se predominantemente na forma de
monovanadatos (V(V)) livres ou ligados agrave transferina do soro ou agrave hemoglobina (Refordf) Os iotildees de
vanadato livres satildeo transportados para o meio intracelular atraveacutes de canais ioacutenicos inespeciacuteficos
Jaacute no ambiente intracelular os compostos sofreratildeo uma bio-reduccedilatildeo atraveacutes da acccedilatildeo da
glutationa ou outros compostos com grupos ndashOH como pentoses hexoses ou etanolaminas [V] A
reoxidaccedilatildeo de vanadilo (V(IV)) intracelular a vanadato (V(V)) provocaria a acumulaccedilatildeo toacutexica
destes uacuteltimos pelo que a complexaccedilatildeo da espeacutecie reduzida o impede O transporte de vanaacutedio
para o interior da ceacutelula encontra-se na Fig 110
Figura 110 ndash Transporte de vanaacutedio para a ceacuteluka e seu subsequente metabolismo (P=Proteiacutena) (Adaptado de
Zaporowska e Scibor 1998)
1211 Vanaacutedio(IV)
A espeacutecie de vanaacutedio (IV) mais bem conhecida eacute o iatildeo vanadilo (VO2+
) espeacutecie estaacutevel a pH
aacutecido sendo a maioritaacuteria a pH 3[AD] Apresenta uma cor azulada e forma complexos penta e
hexacoordenados Forma-se rapidamente a partir de V(V) na presenccedila de um agente redutor em
soluccedilatildeo aquosa No ambiente intracelular a presenccedila de glutationa reduzida aacutecido ascoacuterbico
NAD(P)H entre outros conduzem a que o principal estado de oxidaccedilatildeo nos sistemas bioloacutegicos
seja provavelmente o +4 encontrando-se o V(IV) fortemente complexado com proteiacutenas ou
aminoaacutecidos livres citrato ATP e pirofosfato (Refordfs) Em alternativa o V(IV) pode existir
17
livremente em ambientes fortemente aacutecidos como os lisossomas jaacute que a pH superior a 3 as
soluccedilotildees de vanadilo tendem a ser oxidadas pelo ar formando-se vanadato Esta grande
capacidade redutora justifica parcialmente a utilizaccedilatildeo pouco extensiva do iatildeo VO2+
como
inibidor [W]
1212 Vanaacutedio (V)
O vanaacutedio (V) eacute a forma mais estaacutevel num intervalo de pH amplo [W] O iatildeo anioacutenico VO3-tende
a constituir complexos polinucleares pelo que as soluccedilotildees aquosas de vanadato satildeo extremamente
complexas com ocorrecircncia simultacircnea de vaacuterios equiliacutebrios de protonaccedilatildeo (pKa(vanadato)= 35
78 e 125) e oligomerizaccedilatildeo As espeacutecies presentes (H3VO4 H2VO4- HVO4
2- e VO4
3-) dependem
do pH forccedila ioacutenica e temperatura da soluccedilatildeo e da concentraccedilatildeo total do elemento
[AEAFAGAHAI]
O VO43-
- monoacutemero ortovanadato V1 ndash estaacute associado a uma grande parte da importacircncia
bioloacutegica do vanaacutedio uma vez que se trata de um anaacutelogo electroacutenico e estrutural do ortofosfato
(PO43-
) A coordenaccedilatildeo do ortovanadato pode ser tetraeacutedrica ou bipiramidal trigonal (Refordfs)
A oligomerizaccedilatildeo do vanadato monomeacuterico daacute origem ao diacutemero (V2O72-
- V2) que apresenta
tambeacutem equiliacutebrios de protonaccedilatildeo Presume-se a existecircncia de analogia do vanadato dimeacuterico com
o pirofosfato embora natildeo haja evidecircncia estrutural (Refordf) A dimerizaccedilatildeo do V2 origina o
vanadato tetrameacuterico ou metavanadato (V4O124-
- V4) que constitui a espeacutecie maioritaacuteria em
soluccedilotildees aquosas concentradas (Refordf) As estruturas do metavanadato e a sua espeacutecie protonada
satildeo ciacuteclicas [AJ] Outra espeacutecie oligomeacuterica importante eacute o vanadato pentameacuterico (V5O153-
) cuja
estrutura presumida eacute ciacuteclica [AI]
O oligoacutemero com maior importacircncia bioquiacutemica eacute o vanadato decameacuterico (V10O286-
-V10)(Refordf) a
espeacutecie predominante em soluccedilotildees de pH 2-6 quando a concentraccedilatildeo total de vanadato eacute superior
a 100 M (Ref) A estrutura do V10 natildeo difere em soluccedilatildeo ou no estado soacutelidoOs 10 aacutetomos de
vanaacutedio da espeacutecie decameacuterica estatildeo divididos em trecircs grupos identificados na figura como VA-C
consoante o seu ambiente quiacutemico circundante (Refordf) (Fig 111)
A expressatildeo ldquodecavanadatordquo refere-se a uma soluccedilatildeo aquosa na qual a espeacutecie decameacuterica se
encontra em equiliacutebrio quiacutemico com as espeacutecies V1 V2 V4 e V5 sendo que a oligomerizaccedilatildeo
progressiva do vanaacutedio eacute proporcional agrave sua concentraccedilatildeo total (Ref) Nesta soluccedilatildeo a
concentraccedilatildeo de V10 corresponde a aproximadamente 10 da concentraccedilatildeo total de vanaacutedio [AK]
18
Figura 111 ndash Estruturas esquemaacuteticas das espeacutecies de vanadato (espeacutecie monomeacuterica (V1) dimeacuterica (V2)
tetrameacuterica (V4) pentameacuterica (V5) e decameacuterica (V10) (Adaptado de Aureliano e Crans 2008)
122 Metabolismo e efeito na actividade enzimaacutetica
Apesar de natildeo terem sido ainda encontradas as provas categoacutericas das suas funccedilotildees nos seres
vivos sabe-se que o iatildeo vanadato eacute capaz de inibir uma grande quantidade de enzimas in vitro
com especial ecircnfase na actividade ATPaacutesica da miosina [AL Teresa] Como referido
anteriormente o vanadato eacute capaz de actuar em alternativa ao fosfato nas reacccedilotildees bioquiacutemicas
incluindo a sua preferecircncia de ligaccedilatildeo agraves enzimas [W]
Eacute conhecido o papel do vanaacutedio no metabolismo do caacutelcio promovendo a mineralizaccedilatildeo dos
dentes e ossos na biossiacutentese de liacutepidos (trigliceacuteridos e colesterol) com posterior inibiccedilatildeo da
acumulaccedilatildeo destes nos vasos sanguiacuteneos e no SNC na reproduccedilatildeo e desenvolvimento por
mimetizaccedilatildeo e potenciaccedilatildeo de factores de crescimento na estimulaccedilatildeo do consumo de glucose e
da siacutentese de glicogeacutenio com actuaccedilatildeo equivalente aos efeitos da insulina ao niacutevel celular na
transduccedilatildeo de sinais celulares com regulaccedilatildeo da actividade de segundos mensageiros no
transporte de iotildees e aminoaacutecidos na transcriccedilatildeo geneacutetica e siacutentese de proteiacutenas e DNA entre
outros [W Y AB]
Assim o vanaacutedio cnseguiraacute provocar inuacutemeras mudanccedilas bioquiacutemicas nas ceacutelulas Alguns
exemplos da influecircncia bioquiacutemica do vanaacutedio incluem os seus efeitos nos processos de
fosforilaccedilatildeo e desfosforilaccedilatildeo de proteiacutenas e na participaccedilatildeo na formaccedilatildeo de espeacutecies reactivas de
oxigeacutenio devido ao seu efeito proacute-oxidante sobre agentes redutores a niacutevel celular [AM]
Quanto ao seu efeito na actividade enzimaacutetica pode dividir-se em dois tipos principais ndash inibitoacuterio
e estimulante - dependendo da enzima ou do tipo de reacccedilatildeo por esta catalisada No primeiro
grupo incluem-se enzimas como as ATPases (de miosina de canais ioacutenicos e fosfoenzimas
19
transportadoras) e as fosfatases (da tirosina aacutecida e alcalina) a fosfofrutocinase adenilato ciclase
e catalase entre outras [AA] O segundo grupo inclui a maioria das cinases a glucose-6-fosfato
desidrogenase a fosfoglicerato mutase e a fosfoglicerato fosfatase (Refordf)
O efeito do vanaacutedio (V) na actividade enzimaacutetica estaacute relacionado com o seu nuacutemero de
coordenaccedilatildeo podendo actuar como estado de transiccedilatildeo ou por mimetizaccedilatildeo de cofactores
substratos (especialmente o fosfato) e interacccedilotildees proteiacutena-proteiacutena (como no caso das
polimerases e transcriptases) Pode ainda estar envolvido na formaccedilatildeo de ligaccedilotildees de hidrogeacutenio
interacccedilotildees electrostaacuteticas esteacutereas e de oxidaccedilatildeo-reduccedilatildeo Uma evidecircncia desta uacuteltima eacute a sua
capacidade de alternacircncia entre os estados de oxidaccedilatildeo +5 a +3 o que torna faacutecil a sua interacccedilatildeo
redox com enzimas contendo resiacuteduos de cisteiacutenas (Cys) ou qualquer outro composto orgacircnico
com actividade redox [AN]
13 Vanaacutedio e muacutesculo esqueleacutetico
A primeira evidecircncia da presenccedila de vanaacutedio no muacutesculo esqueleacutetico proveio de uma amostra de
ATP isolado de muacutesculo de cavalo que continha uma impureza ndash vanaacutedio ndash cuja presenccedila
provocou a inibiccedilatildeo da actividade da bomba de Na+K
+-ATPase [AO] Desde entatildeo vaacuterios
estudos tecircm sido realizados com ecircnfase na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o
tecido muscular Destes destaca-se a descoberta da interacccedilatildeo forte da espeacutecie V10 com a miosina
fosforilada [AP] e agrave sua inibiccedilatildeo da actividade ATPaacutesica da miosina[AQ AR]
No quediz respeitoao iatildeo monovanadato destaca-se a sua inibiccedilatildeoda actividade Mg2+
-ATPaacutesica
da miosina [AS] que ocorre de duas formas distintas (1) na presenccedila de concentraccedilotildees reduzidas
de V1 a inibiccedilatildeo ocorre a velocidade reduzida dependente da concentraccedilatildeo da espeacutecie
monomeacuterica (2) na presenccedila de concentraccedilotildees elevadas de V1 o niacutevel de inibiccedilatildeo aproxima-se de
100 desde que [V1] seja superior agrave concentraccedilatildeo total de locais com actividade Mg2+
-ATPaacutesica
no ensaio Esta inibiccedilatildeo eacute irreversiacutevel tanto por diaacutelise ou adiccedilatildeo de agentes redutores do
vanadato [AL]Segundo Pate et al(1994)[AT] a espeacutecie monomeacuterica podeinibir a velocidade de
contracccedilatildeo a temperatura inferior a 20ordmC perdendogradual e rapidamente essa capacidade ateacute os
50ordmC temperatura agrave qual cessa a capacidade inibitoacuteria Os resultados obtidos sugeremainda uma
interacccedilatildeo altamente cooperativa entre o iatildeo vanadato e as proteiacutenas componentes do muacutesculo
esqueleacuteticoDestaca-se tambeacutem a descoberta da inibiccedilatildeo da geraccedilatildeo de forccedila contraacutectil nas fibras
musculares por parte da espeacutecie monomeacuterica[AT]Wilson et al(1995) [AU ]determinaram que a
afinidade da espeacutecie V1 para o S1 de miosina eacute significativamente mais elevada do que a do iatildeo
Pi seu anaacutelogo Descobriram ainda que que a ligaccedilatildeo do vanadato poderaacute envolver transiccedilotildees
entre estados energeacuteticos com energia livre independente dos impedimentos esteacutereos impostos
20
pela estrutura das miofibrilas Os estudos desenvolvidos por Smith et al (1990)[AV]
determinaram queo iatildeo monovanadato natildeo apresenta um efeito pronunciado na inibiccedilatildeo da
actividadeacto-S1-miosina-ATPaacutesica excepto em concentraccedilotildees superiores a 900microM a partir das
quais existe produccedilatildeo de espeacutecies oligomeacutericas para as quais o efeito inibitoacuterio seraacute mais
elevadoA espeacutecie monomeacuterica de vanadato consegue adoptar facilmente uma geometria do tipo
bipiramidal trigonal que mimetiza o intermediaacuterio formado (MgADP
Pi)aquando da reacccedilatildeo de
hidroacutelise de ATP no local activo da miosina [AW] De facto a interacccedilatildeo do vanaacutedio com a
miosina eacute inesperadamente estaacutevel na presenccedila de ADP Esta interacccedilatildeo resulta na formaccedilatildeo de
um complexo enzimaacutetico inactivo e estaacutevel com estequiometria de 11 ADPvanadato por siacutetio
activo da proteiacutena A formaccedilatildeo do complexo estaacutevel miosina-vanadato foi proposta por Goodnoet
al (1982) [AS] Nesta ocorre um preacute-equiliacutebrio raacutepido que induz a formaccedilatildeo de um complexo
ternaacuterio reversiacutevel (MADP
V) com formaccedilatildeo lenta do isoacutemero M
ADP
V A diferenccedila entre os
compostos de vanaacutedio e de fosfato centra-se no tempo de meia vida (t12) do Pi ou do V (no
primeiro t12=12 s enquanto no segundo t12=24 h) (Refordf) A diferenccedila dever-se-aacute agrave possiacutevel
complexaccedilatildeo do vanaacutedio com grupos nucleofiacutelicos no siacutetio activo o que provocaria uma
estabilizaccedilatildeo da miosina na conformaccedilatildeo MADP
V
No que respeita aos estudos realizados com a espeacutecie tetrameacuterica Cremo et al determinou que a
cadeia pesada do S1 de miosina pode ser clivada em dois locais distintos (designados V1 e V2)
pela espeacutecie V4 apoacutes irradiaccedilatildeo com luz UV na presenccedila de concentraccedilotildees de vanadato na ordem
dos milimolar e na ausecircncia de nucleoacutetidos ou metais divalentes
Jaacute no que concerne aos estudos realizados com ecircnfase na espeacutecie decameacuterica foi sugerido
porRamos et al (2006) [AK]que a interacccedilatildeo do decavanadato com a G-actina favorecida pela
polimerizaccedilatildeo desta uacuteltima estabiliza a espeacutecie V10 e induz a inibiccedilatildeo da formaccedilatildeo de F-actina A
estabilizaccedilatildeo da espeacutecie V10 por proteiacutenas tansmembranares e do citoesqueleto pode ser a razatildeo
pela qual ocorre a toxicidade do decavanadato reportada na avaliaccedilatildeo dos efeitos do V(V) nos
sistemas bioloacutegicosTiago et al (2007) [AR] concluiram que o V10 apresenta uma clara preferecircncia
de ligaccedilatildeo ao ldquoback-doorrdquo da S1-miosina-ATPase o que se verifica apenas na estrutura
conformacional desimpedida com acesso facilitado ao local de ligaccedilatildeo do Pi Sugere tambeacutem que
o V10 actua como um ldquoback-door stoprdquo bloqueando estaabertura do fragmento de 50-kDa
necessaacuteria para a hidroacutelise do ATP--fosfato o que constitui uma explicaccedilatildeo simples do
comportamento natildeo competitivo da espeacutecie decameacuterica estimulando a utilizaccedilatildeo deste oligoacutemero
como uma feramenta da elucidaccedilatildeo das mudanccedilas conformacionais ldquoback-doorrdquo da miosina no
21
processo da contracccedilatildeo muscular(ref)Destaca-se ainda a descoberta da capacidade inibitoacuteria do
V10 em relaccedilatildeo agrave fosforilaccedilatildeo oxidativa mitocondrial [AX]
Finalmente no que concerne aos estudos realizados com a presenccedila de vaacuterias espeacutecies
oligomeacutericas Aureliano e Madeira (1994) [AY]observaram que na presenccedila de Ca2+
as espeacutecies
V4 e V10se ligam agrave Ca2+
-ATPase do retiacuteculo sarcoplasmaacutetico enquanto a espeacutecie V1 se liga apenas
na presenccedila de ATP Aparentemente o aumento da concentraccedilatildeo de Ca2+
tem o efeito contraacuterio da
inibiccedilatildeo da ATPase do retiacuteculo sarcoplasmaacutetico pelo V1 natildeo tendo no entanto efeito significativo
sobre a inibiccedilatildeo por parte da espeacutecie V10 Tiago et al (2002) [AP] concluiacuteram que em condiccedilotildees
proacuteximas agraves fisioloacutegicas a espeacutecie V10 difere das espeacutecies presentes na soluccedilatildeo de metavanadato
devido agrave sua forte interacccedilatildeo com a miosina fosforilada e agrave sua inibiccedilatildeo da actividade ATPaacutesica
da miosina Para aleacutem disso o ATP diminui a afinidade da miosina para o V4 induzindo a
interacccedilatildeo com V1 e natildeo tendo qualquer efeito no que diz respeito ao V10 Foi tambeacutem proposto
[AQ] que a ligaccedilatildeo das espeacutecies oligomeacutericas de vanadato aoS1 de miosina nos locais onde o
fosfato coordenaeacute provavelmente a causa da inibiccedilatildeo da actividade ATPaacutesica da miosina
estimulada por actina atraveacutes da prevenccedilatildeo da troca entre ATPADP sendo esta favorecida para
aniotildees de vanadato com valecircncias superiores como eacute o caso do V10
Na presenccedila de actina as velocidades de dissociaccedilatildeo do Pi e do V aumentam consideravelmente
este uacuteltimo em menor quantidade Assim a ligaccedilatildeo da actina ao complexo MADP
V e
Pi eacute
bastante fraca pelo que se sugere que o ADPV teria um efeito de relaxamento semelhante ao
ATP No entanto a capacidade do vanaacutedio em inibir a actividade ATPaacutesica da miosina e da
actomiosina eacute diferenciada uma vez que na ausecircncia de actina a inibiccedilatildeo eacute total para
concentraccedilotildees na gama dos M mas na sua presenccedila a inibiccedilatildeo eacute totalmente conseguida apenas
em concentraccedilotildees na gama dos mM (Ref) No entanto eacute necessaacuterio considerar a possiacutevel
formaccedilatildeo de espeacutecies oligomeacutericas de vanadato cuja afinidade e efeito na enzima seratildeo
diferentes Assim pode concluir-se que o efeito de inibiccedilatildeo observado para a actividade ATPaacutesica
de miosina estimulada por actina se pode dever agrave presenccedila das diferentes espeacutecies oligomeacutericas o
que foi jaacute sugerido por Aureliano (2000) [AZ] ndash as espeacutecies tetrameacuterica e decameacuterica de vanadato
interactuariam com a miosina com alteraccedilatildeo da sua interacccedilatildeo com a actina No entanto natildeo eacute
conhecida a contribuiccedilatildeo absoluta das restantes espeacutecies oligomeacutericas para a inibiccedilatildeo da ATPase
da miosina estimulada por actina (Refs)
14 Objectivos do projecto
Pretende-se com este projecto o isolamento purificaccedilatildeo e caracterizaccedilatildeo da actina do muacutesculo
22
esqueleacutetico de coelho sendo esta uacuteltima tambeacutem realizada para o sufragmento-1 de miosina
Seguidamente realizar-se-aacute a quantificaccedilatildeo e caracterizaccedilatildeo das proteiacutenas por espectroscopia UV-
Vis e SDS-PAGE e a caracterizaccedilatildeo das soluccedilotildees de vanaacutedio por UV-Vis e ressonacircncia magneacutetica
nuclear (RMN) Por uacuteltimo a interacccedilatildeo do S1 de miosina com o vanaacutedio seraacute avaliada atraveacutes
das teacutecnicas de UVVis RMN e ressonacircncia paramagneacutetica electroacutenica (RPE)
2 Discussatildeo de Resultados
21 Caracterizaccedilatildeo das proteiacutenas isoladas
211Anaacutelise do estado de pureza
O estado de pureza das soluccedilotildees de actina e de S1 foi analisado por electroforese em gel de
poliacrilamida em condiccedilotildees desnaturantes (SDS-PAGE) (Fig 22) que separa as moleacuteculas de
acordo com o seu peso molecular (PM) A concentraccedilatildeo de acrilamida escolhida (10) permite a
detecccedilatildeo simultacircnea de componentes com peso molecular inferior a 20 kDa e outros com elevado
peso molecular
Figura 21 ndash Fotografia de gel de SDS-PAGE (10 acrilamida) Poccedilo (a)
10 L do marcador de elevado peso molecular () Poccedilo (b) 20 L de [G-
actina]=1 mgmL Poccedilo (c) 20 L de [S1]=1 mgmL Aplicou-se uma
voltagem de V durante h e min
A partir da recta de calibraccedilatildeo obtida log PM = -1177 Rf + 2247 (ver ponto 2 da secccedilatildeo
Anexos) pode verificar-se a presenccedila de actina na amostra do poccedilo (b) presente numa banda
bastante intensa (PMaparente=40 kDa) bem como a presenccedila minoritaacuteria de outros componentes
desconhecidos que natildeo teratildeo sido eliminados durante o processo de extracccedilatildeo e purificaccedilatildeo e natildeo
satildeo relevantes para o estudo em questatildeo Na amostra do poccedilo (c) nota-se a presenccedila de S1
(PMaparente=92 kDa) bem como a de outros componentes cuja banda eacute ligeiramente menos intensa
Estes poderatildeo ser fragmentos proteoliacuteticos de miosina e de S1 Estaacute ainda presente uma bada de
23
maior intensidade que poderaacute ser a proteiacutena-C (PM=135 kDa) uma vez que a sua remoccedilatildeo
completa da amostra necessitaria de um passo adicional de cromatografia de troca ioacutenica [BA]
No entanto natildeo podemos confirmar que se trata desta proteiacutena ou de qualquer outra uma vez que
se encontra fora do intervalo de migraccedilatildeo dos marcadores de peso molecular
212 Determinaccedilatildeo da concentraccedilatildeo proteica
2121Subfragmento-1 de miosina
A partir da recta de calibraccedilatildeo obtida pelo meacutetodo de Bradford [BB](ver secccedilatildeo 3311 do
Procedimento Experimental) A595nm = 00546[BSA] + 00209 (ver secccedilatildeo de Anexos)foi possiacutevel
determinar um intervalo de concentraccedilatildeo de S1 de miosina compreendido entre 226 e 117(plusmn
423M) (203 e 1056 (plusmn 381 mgmL)) (n=8) A concentraccedilatildeo meacutedia de S1 de miosina apoacutes
diaacutelise contra a soluccedilatildeo tampatildeo 10 mM HEPES pH 75 25 mM KCl determinada pela leitura da
Abs280 nmfoi de 20430M (1803 mgmL)(n=7)
As amostras a quantificar atraveacutes do meacutetodo de Bradford natildeo sofreram diaacutelise o que permite
explicar a concentraccedilatildeo superior de S1 de miosina em comparaccedilatildeo com o valor obtido atraveacutes do
meacutetodo directo Esta razatildeo aliada agrave sensibilidade e rapidez superiores deste uacuteltimo fez com que
se optasse pelo meacutetodo directo para a quantificaccedilatildeo da grande maioria dos ensaios
2122Actina
A concentraccedilatildeo de actina foi determinada quando esta se encontrava na forma monomeacuterica (G-
actina) por mediccedilatildeo da leitura directa da Abs290 nm tendo-se obtido uma concentraccedilatildeo meacutedia de
691481M (290202 mgmL) (n=3) em 2 mM Tris-HCl pH 80 02 mM CaCl2 02 mM Na-
ATP Ao comprimento de onda utilizado (290 nm) absorvem fortemente apenas os resiacuteduos de
Trp pelo que a mediccedilatildeo efectuada tem menor sensibilidade do que se tivesse sido efectuada a 280
nm uma vez que a actina apresenta apenas 4 resiacuteduos deste aminoaacutecido embora tenha 16
resiacuteduos de Tyr
22 Mediccedilatildeo da actividade enzimaacutetica de S1 de miosina
221 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
As actividades ATPaacutesicas de S1 de miosina dependentes de Ca2+
e K+foram analisadas pelo
caacutelculo das velocidades de consumo inicial do fosfato inorgacircnico (Pi) Definiu-se os tempos de 5
e 3 minutos para a mediccedilatildeo da Ca2+
e da K+EDTA-ATPase respectivamente Os valores obtidos
24
a partir do protocolo presente no ponto 341 da secccedilatildeo Procedimento experimental encontram-se
descritos na Tabela 2I
Tabela 2I ndash Acrividades natildeo-fisioloacutegicas de subfragmento-1 determinadas pelo meacutetodo de Fiske-Subarow
modificado (n=2)
Proteiacutena
Actividade
ATPaacutesica natildeo
fisioloacutegica
Actividade (nmol
Piminmg proteiacutena) Actividade (s-1)
S1
Ca2+
-ATPase 1400 plusmn 30 2100 plusmn 0045
K+EDTA-ATPase 2334 plusmn 50 3500 plusmn 0075
As actividades natildeo-fisioloacutegicas reflectem o estado de oxidaccedilatildeo dos resiacuteduos expostos de Cys 707
(SH1) e 697 (SH2) Se o grupo SH1 sofrer oxidaccedilatildeo a actividade ATPaacutesica dependente de Ca2+
aumenta e a actividade dependente de K+ diminui Por outro lado se o grupo SH2 sofrer
oxidaccedilatildeo regista-se a perda completa destas actividades
De acordo com Margossian e Lowey (1982) [BA] as actividades ATPaacutesica de S1 de miosina
dependente de Ca2+
e K+EDTA da miosina satildeo respectivamente de 24 e 112 s-1
Estaacute tambeacutem
descrito que a actividade ATPaacutesica dependente de K+ eacute em meacutedia cerca de 3 vezes superior agrave
dependente de Ca2+
reflectindo a quantidade das preparaccedilotildees A razatildeo obtida entre os valores das
actividades ATPaacutesicas dependentes de K+ e Ca
2+ eacute de 166 Os valores obtidos para as actividades
(Ca2+
-ATPase=21 s-1
K+EDTA-ATPase= 35 s
-1) satildeo inferiores agrave valores obtidos por
Margossian e Lowey [BA] e Tiago [BC] com forccedila ioacutenica inferior O valor obtido para a
actividade ATPaacutesica dependente de Ca2+
eacute superior ao obtido por Polosukhina et al (2000) [BD]
cujo meio de reacccedilatildeo continha MOPS e apresentava forccedila ioacutenica inferior Os dados
supramencionados comprovam a variaccedilatildeo da actividade a ATPaacutesica natildeo fisioloacutegica em relaccedilatildeo agrave
forccedila ioacutenica temperatura e pH
222 Mediccedilatildeo da actividadeMg2+
-ATPaacutesica de S1 pelo meacutetodo de enzimas acopladas
A actividade de Mg2+
-ATPase eacute a uacutenica que possui significado fisioloacutegico sendo as restantes
bastante uacuteteis apenas para indicaccedilatildeo da qualidade de preparaccedilatildeo do S1[BA] A 0575 M KCl a
actina e o S1 formam uma soluccedilatildeo viscosa e tuacuterbida Apoacutes a adiccedilatildeo de MgATP o complexo acto-
S1 dissocia-se e a turbidez da soluccedilatildeo diminui Se a forccedila ioacutenica for reduzida a dissociaccedilatildeo do
complexo actoS1 que se encontra na forma de suspensatildeo eacute induzida O MgATP eacute rapidamente
hidrolisado diminuindo a sua concentraccedilatildeo no meio reaccional Nestas condiccedilotildees haacute
reassociaccedilatildeo e subsequente contracccedilatildeo dos filamentos de actina e de S1 com formaccedilatildeo de
25
precipitado O meacutetodo descrito traduz de forma bastante elementar o fenoacutemeno da contracccedilatildeo in
vivo e elimina o problema anterior da solubilizaccedilatildeo ineficiente da miosina de modo a obter uma
mediccedilatildeo exacta da actividade [A] A utilizaccedilatildeo de forccedilas ioacutenicas reduzidas aumenta a afinidade da
actina para o S1 permitindo contornar a extrema dificuldade em obter a concentraccedilatildeo proteica dos
muacutesculos (cerca de 100 mgmL do complexo actomiosina) As actividades Mg2+
-ATPaacutesicas de S1
de miosina foram determinadas a partir do declive da variaccedilatildeo da Abs340 relativo agrave oxidaccedilatildeo de
NADH em funccedilatildeo do tempo como eacute descrito no ponto 342 do Procedimento Experimental Os
valores obtidos para a actividade Mg2+
-ATPaacutesica encontram-se apresentados na Tabela 2II
Tabela 2II ndash Actividade Mg2+
-ATPaacutesica de subfragmento-1 de miosina determinadas pelo meacutetodo das enzimas
acopladas
Actividade (nmol
ATPmin mg(S1)) Actividade (s
-1)
1097 plusmn 269 164 plusmn 04
Ramos (2005) [J] obteve o valor de 01 s-1
para a actividade Mg2+
-ATPaacutesica de S1 de miosina
com forccedila ioacutenica intermeacutedia Tiago (2005) [BC] obteve o valor de 011 s-1 em condiccedilotildees de forccedila
ioacutenica reduzida em relaccedilatildeo agrave utilizada no presente relatoacuterio
23 Preparaccedilatildeo e caracterizaccedilatildeo das soluccedilotildees de vanaacutedio
231 Anaacutelise da composiccedilatildeo das soluccedilotildees de vanadato por espectroscopia de 51
V-RMN
efeito da concentraccedilatildeo da espeacutecie
A diluiccedilatildeo das soluccedilotildees stock de meta- e decavanadato iraacute determinar fortemente a composiccedilatildeo
em soluccedilatildeo dos diferentes oligoacutemeros de vanadato (ref) No caso do decavanadato o periacuteodo de
tempo apoacutes a diluiccedilatildeo iraacute tambeacutem ser um factor importante a considerar devido agrave decomposiccedilatildeo
da espeacutecie V10(ref)Torna-se assim fundamental determinar a composiccedilatildeo quiacutemica das soluccedilotildees
no meio de ensaio utilizado (HEPES 10 mM KCl 25 mM) de modo a correlacionaacute-la
correctamente com os possiacuteveis efeitos promovidos pelas diferentes espeacutecies de vanadato no
sistema em estudo
Aleacutem disso deve ser dado ecircnfase agraves importantes interacccedilotildees que ocorrem entre o vanaacutedio e os
tampotildees escolhidos O tampatildeo com menor nuacutemero e magnitude de interacccedilatildeo eacute o HEPES [AD]
seguido do Tris sendo este uacuteltimo especialmente aconselhado em concentraccedilotildees baixas para casos
de incompatibilidade enzimaacutetica ou quiacutemica redox No caso especial da reacccedilatildeo redox
NAD(P)HNAD(P) o tampatildeo adequado eacute o imidazole de modo a natildeo haver reacccedilatildeo com o
26
vanadato Os tampotildees a evitar em qualquer situaccedilatildeo incluem a trietanolamina fosfato citrato e
glicina entre outros [AD] uma vez que todos interactuam fortemente com V(V) com formaccedilatildeo
de complexos estaacuteveis
Assim procedeu-se agrave anaacutelise da composiccedilatildeo das soluccedilotildees de meta- e decavanadato apoacutes diluiccedilatildeo
das soluccedilotildees stock (50 mM em vanadato total) no meio de reacccedilatildeo (tampatildeo D) atraveacutes da
espectroscopia de ressonacircncia magneacutetica nuclear de 51
V Nas figuras x1 e x2 apresentam-se os
espectros obtidos para as soluccedilotildees de deca- e metavanadato num intervalo de concentraccedilotildees de 1 a
10 mM A Tabela seguinte resume os paracircmetros espectroscoacutepicos obtidos para cada espeacutecie
presente nas soluccedilotildees analisadas
Figura 22ndash Espectros de RMN de 51
V de
concentraccedilotildees crescentes das espeacutecies oligomeacutericas
de vanadato obtidos apoacutes diluiccedilatildeo da soluccedilatildeo stock
de decavanadato em 10 mM HEPES pH 70 25 mM
KCl para concentraccedilotildees finais de A) 1 mM B)2
mM C) 4 mM D) 8 mM E) 10 mM de
decavanadato total V1refere-se agraves espeacutecies
monomeacutericade vanadato V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2IIIndash Desvios quiacutemicos meacutedios (part) das ressonacircncias das espeacutecies decameacuterica (V10) monomeacuterica (V1) e
tetrameacuterica (V4) presentes nos espectros da soluccedilatildeo de decavanadatoLargura a meia altura (∆12) da espeacutecie V10A
Paracircmetros V10A V10B V10C V1 V4
part (ppm) -5162 -5003 -4255 -5616 -5748
∆12 (Hz) 175 nd nd nd nd
Figura 23 ndash Distribuiccedilatildeo graacutefica das
espeacutecies oligomeacutericas de vanadato vs a
concentraccedilatildeo de decavanadato total As
concentraccedilotildees de cada espeacutecie foram
obtidas segundo a formula descrita na
secccedilatildeo 35 do Procedimento
Experimental (aacutereas dos sinais de 51
V-
RMN observadas na figura x1)V10 V1 e
V4 correspondem agraves espeacutecie monomeacuterica
dimeacuterica e tetrameacuterica respectivamente [Esp
eacuteci
e o
ligo
meacute
rica
de
van
aacutedio
] (m
M)
[Decavanadato]total (mM)
V10
V1
V4
27
Nas soluccedilotildees de decavanadato as espeacutecies monomeacuterica e decameacuterica de vanadato satildeo as uacutenicas
presentes para concentraccedilotildees de decavanadato inferiores a 8 mM Ambas as espeacutecies aumentam
linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado em [AY] e
em (Teresa) quando a concentraccedilatildeo de V1 eacute igual ou superior a 200 microM inicia-se a sua
oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica No entanto nas condiccedilotildees de realizaccedilatildeo dos
ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a espeacutecie tetrameacuterica neste
estudo em concentraccedilotildees inferiores a 8 mM tendo o mesmo sido observado por Ramoset al
(2006) [AK] Os resultados obtidos mostram claramente o equiliacutebrio entre a espeacutecie decameacuterica e
a monomeacuterica e que o aparecimento de outras espeacutecies oligomeacutericas se encontra dependente da
concentraccedilatildeo desta uacuteltima Apesar de natildeo ter sido realizado o ensaio com 5 mM de decavanadato
total no presente relatoacuterio estaacute reportado em Ramos et al (2006) [AK] que as concentraccedilotildees da
espeacutecie decameacuterica e monomeacuterica satildeo respectivamente de 500 microM e 200 microM Assim pode inferir-
se que para concentraccedilotildees inferiores a 5 mM e intervalos de tempo de cerca de 20 minutos (que
inclui o tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas
as espeacutecies decameacuterica e monomeacuterica de vanadato
Figura 24 ndash Espectros de RMN de 51
V de concentraccedilotildees
crescentes das espeacutecies oligomeacutericas de vanadato obtidos
apoacutes diluiccedilatildeo da soluccedilatildeo stock de metavanadato em 10 mM
HEPES pH 70 25 mM KCl para concentraccedilotildees finais de A)
1 mM B) 2 mM C) 4 mM D) 8 mM E) 10 mM de
metavanadato total V1 refere-se agraves espeacutecies monomeacuterica de
vanadatoV1 e V2 correspondem respectivamente agrave espeacutecie
monomeacuterica e dimeacuterica
Figura 25 ndash Distribuiccedilatildeo graacutefica das espeacutecies
oligomeacutericas de vanadato vs a concentraccedilatildeo de
metavanadato total As concentraccedilotildees de cada
espeacutecie foram obtidas segundo a formula descrita
na secccedilatildeo 35 do Procedimento Experimental
(aacutereas dos sinais de 51
V-RMN observadas na
figura x1)V10 V1 e V4 correspondem agraves espeacutecie
monomeacuterica dimeacuterica e tetrameacuterica
respectivamente
Esp
eacuteci
e o
ligo
meacute
rica
de
va
nad
ato
(m
M)
[Metavanadato]total (mM)
V1
V2
V4
V5
28
Tabela 2IVndash Desvios quiacutemicos meacutedios (part)das ressonacircncias das espeacutecies tetrameacuterica (V4) monomeacuterica (V1) e
dimeacuterica (V2) presentes no espectro da soluccedilatildeo de metavanadato (total) 10 mM
Paracircmetros V1 V2 V4 V5
part (ppm) -5615 -5748 -5789 -5869
Nas soluccedilotildees de metavanadato as espeacutecies monomeacuterica dimeacutericae tetrameacuterica de vanadato estatildeo
presentes para concentraccedilotildees de metavanadato inferiores a 4 mM Todas estas as espeacutecies
aumentam linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado
em (Aureliano e Madeira 1998) e em (Tiago et al 2004) quando a concentraccedilatildeo de V1 eacute igual ou
superior a 200 microM inicia-se a sua oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica Nas
condiccedilotildees de realizaccedilatildeo dos ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a
espeacutecie pentameacuterica neste estudo em concentraccedilotildees inferiores a 4 mM tendo o mesmo sido
observado por Ramos 2006 Os resultados obtidos mostram claramente o equiliacutebrio entre a
espeacutecie V4 e as espeacuteciesV1 e V2 no qual o aparecimento de outras espeacutecies oligomeacutericas se
encontra dependente da concentraccedilatildeo da espeacutecie V1 Por uacuteltimo pode inferir-se que para
concentraccedilotildees inferiores a 2 mM e intervalos de tempo de cerca de 20 minutos (que inclui o
tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas as
espeacutecies V1 V2 e V4 com concentraccedilotildees respectivas de 0796 0192 e 0196 mM
232Estabilidade das soluccedilotildees de decavanadato anaacutelise por espectroscopia de absorccedilatildeo
UVVis
Caracterizou-se a estabilidade da espeacutecie V10 numa soluccedilatildeo com 1 mM decavanadato total em
tampatildeo D atraveacutes da leitura da Abs400 Obteve-se uma cineacutetica de dissociaccedilatildeo de 1ordf ordem tal
como se pode verificar pela linearidade do graacutefico de ln(Abs400)vs o tempo de aquisiccedilatildeo espectral
(segundos) (Fig26 )
Figura 26ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do tempo
(em segundos) para uma soluccedilatildeo de decavanadato total de 1
mM em tampatildeo D a 25ordmC (n=2)
Sendo uma reacccedilatildeo de primeira ordem pode ser descrita pela equaccedilatildeo cujo
declive eacute ndashK (constante de dissociaccedilatildeo) e a intersecccedilatildeo no eixo das ordenadas corresponde
Ln
(Ab
s 400 n
m)
tempo (s)
29
aln(A0) Numa reacccedilatildeo de 1ordf ordem o tempo de meia-vida eacute independente da concentraccedilatildeo inicial
do reagente sendo descrito pela equaccedilatildeo [2] Deste modo a partir da recta
Ln(Abs400 nm) = -436E-5
K - 105 obteve-se um tempo de meia-vida de 4490 plusmn 0823 horas para
as condiccedilotildees supramencionadas
Apoacutes a adiccedilatildeo de S1 8 microM ao meio reaccional verificou-se que a reacccedilatildeo de desoligomerizaccedilatildeo
da espeacutecie decameacuterica de vanadato seguia igualmente uma cineacutetica de 1ordf ordem tal como se pode
verificar pela linearidade do graacutefico de ln(Abs400) versus o tempo de aquisiccedilatildeo espectral
(segundos) (Fig27)
Figura 27ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do
tempo (em segundos) para uma soluccedilatildeo de decavanadato
total de 1 mM em tampatildeo D a 25ordmC na presenccedila de 8
microM (n= 2)
A partir da equaccedilatildeo da recta Ln(Abs400 nm) = -7520E-5
K - 0613determinou-se o valor de K para
a espeacutecie decameacuterica de onde se obteve um tempo de meia-vida de 2430 plusmn 0191 horas para as
condiccedilotildees supramencionadas
Tanto este valor como o valor registado na ausecircncia de S1 satildeo substancialmente superiores agrave
duraccedilatildeo dos ensaios de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina descritos no presente
relatoacuterio
24 Interacccedilatildeo de S1 com vanaacutedio(V)
241Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
De modo a analisar qual o efeito da adiccedilatildeo de S1 de miosina (tampatildeo D) produzia nos sinais de
51V-NMR titulou-se uma soluccedilatildeo de 5mM de decavanadato total com concentraccedilotildees crescentes
de S1 (5 10 15 e 20 microM) Os espectros de 51
V-RMN encontram-se apresentados na Fig 2
No graacutefico da figura 2 Apresenta-se a concentraccedilatildeo de S1 em funccedilatildeo do factor de alargamento
corrigido dos espectros acima referidos
Ln
(Ab
s (4
00
nm
))
tempo (s)
30
Figura 28 ndash Graacutefico de
representaccedilatildeo da
concentraccedilatildeo de S1 em
funccedilatildeo do factor de
alargamento A
concentraccedilatildeo de
decavanadato total foi de 5
mM e as concentraccedilotildees
utilizadas de S1 foram de 0
5 10 e 15 microM
Verifica-se que o factor de alargamento aumenta linearmente em funccedilatildeo da concentraccedilatildeo de S1 no
ensaio o que indica a existecircncia de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina Observou-
se ainda que a adiccedilatildeo de S1 ao meio reaccional natildeo provocou qualquer alteraccedilatildeo dos desvios
quiacutemicos dos sinais de 51
V-RMN
Realizaram-se ensaios na presenccedila de ATP e de S1 com intuito de verificar o efeito do ATP na
interacccedilatildeo do S1 de miosina com o decavanadato
Figura 29 - Espectros de RMN de 51
V contendo 2
mM Decavanadato (A) (controlo) (B) na presenccedila
de 2 mM de ATP (C) na presenccedila de 2 mM de ATP
e 8 microM de S1 de miosina V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2Vndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies decameacuterica (V10) e monomeacuterica (V1) presentes nos
espectros da soluccedilatildeo de decavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V10A V10B V10C V1
part (ppm) -5161 -5002 -4254 -5614
Observa-se que a presenccedila de ATP (espectro B) natildeo provoca qualquer alteraccedilatildeo visiacutevel em
relaccedilatildeo ao espectro que contem apenas soluccedilatildeo de decavanadato Jaacute a presenccedila simultacircnea de ATP
e S1 provoca um alargamento da ∆12 do sinal do V10A de 1736 Hz (espectro (A)) para 2366 Hz
(espectro (C) Fig 29) Tendo em conta que a concentraccedilatildeo de S1 apresenta uma relaccedilatildeo linear
com o factor de alargamento converteu-se o valor da ∆12 e comparou-se com o valor esperado agrave
F = 00591[S1] - 00157 Rsup2 = 0998
Fact
or
de
ala
rgam
en
to
[S1] (microM)
31
concentraccedilatildeo de S1 utilizada O factor de alargamento obtido para 8 microM ndash 0376 ndash eacute inferior ao
valor esperado agrave mesma concentraccedilatildeo de S1 obtido atraveacutes da equaccedilatildeo da recta apresentada na
Fig 28 ndash 0457 ndash o que indica que o ATP poderaacute exercer um efeito de protecccedilatildeo do S1
diminuindo a sua interacccedilatildeo com a espeacutecieV10
Por uacuteltimo realizaram-se ensaios com metavanadato na presenccedila de 8 microM S1 de modo a verificar
se as espeacutecies oligomeacutericas contidas nesta soluccedilatildeo interagiam com a proteiacutena em estudo
Figura 210 - Espectros de RMN de 51
V contendo 2
mM metavanadato (A) (controlo) (B) na presenccedila
de 8 microM de S1 de miosina V1 V2 e V4
correspondem respectivamente aacutes espeacutecies
monomeacuterica dimeacuterica e tetrameacuterica
Tabela 2VIndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies monomeacuterica (V1) dimeacuterica (V2) e tetrameacuterica (V4)
presentes nos espectros da soluccedilatildeo de metavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V1 V2 V4
part (ppm) -5613 -5747 -5789
Como eacute possiacutevel observar na Fig 210 ocorre uma ligeiro alargamento dos sinal de V4 bem
como uma perda da intensidade pelo que se conclui quedas espeacutecies presentes na soluccedilatildeo de
metavanadato (V1 V2 e V4) apenas a espeacutecie tetrameacuterica interage com o S1 de miosina o que
estaacute de acordo com o reportado por Tiago et al 2002
242 Actividade Mg2+
-ATPaacutesica de S1 de miosina na presenccedila de decanadato
Com o intuito de avaliar o efeito do decavanadato na actividade Mg2+
-ATPaacutesica do S1 de
miosina monitorizou-se a reacccedilatildeo de oxidaccedilatildeo do NADH atraveacutes da leitura de Abs340 na
presenccedila de 005 mgmL de S1 de miosina e no caso da actividade estimulada 2microM de actina em
3125 mM Tris-HCl pH 71 e 375 mM MgCl2 pelo meacutetodo de enzimas acopladas (ver seccccedilatildeo xx
dos MM)Para tal realizaram-se ensaios com vaacuterias concentraccedilotildees de decavanadato total (10 50
100 250 500 e 1000) de modo a determinar qual o seu efeito na actividade MgATPaacutesica de S1
de miosina na ausecircncia e na presenccedila de um dos seus ligandos naturais Os ensaios foram
realizados agrave temperatura ambiente com incubaccedilatildeo preacutevia a 25ordm C durante 20 minutos
32
Figura 211 ndash Representaccedilatildeo
graacutefica da Actividade Mg2+
-
ATPaacutesica de S1 () (os ajustes
teoacutericos estatildeo representado a por
uma linha os resultados
experimentais estatildeo representados
por triacircngulos (Actividade
estimulada) e cruzes (Actividade
natildeo estimulada)) vs
[Decavanadato]total medidas pelo
meacutetodo das enzimas acopladas
(n=3) Os dados foram ajustados ao modelo de decaimento exponencial usando o programa GraphPad Prism para a
determinaccedilatildeo dos valores de IC50 (na ausecircncia de actina IC50= 266 plusmn 07 M decavanadato na presenccedila de actina
IC50= 28 plusmn 1 M decavanadato
A partir do graacutefico presente na Fig 29 verifica-se que o ajuste dos dados experimentais foi bem
sucedido A inibiccedilatildeo da actividade Mg2+
-ATPaacutesica do S1 de miosina eacute afectada em menor
amplitude na presenccedila de actina (60 de inibiccedilatildeo da actividade) do que na sua ausecircncia
(aproximadamente 80 de inibiccedilatildeo de actividade) Devido a esta diferenccedila pode sugerir-se que a
actina iraacute impedir parcialmente a ligaccedilatildeo do decavanadatoao S1 demiosina atraveacutes de
impedimentos esteacutereos ou interacccedilotildees electrostaacuteticas por exemplo A teacutecnica utilizada natildeo nos
permite concluir acerca de que regiatildeo(otildees) ou resiacuteduo(s) da actina participaratildeo na preservaccedilatildeo
parcial da actividade Mg2+
-ATPaacutesica
Por outro lado os valores de IC50 dos dois tipos de actividades natildeo apresentam diferenccedilas
significativas tendo-se obtido 266 plusmn 07 e 28 plusmn 1M decavanadato na ausecircncia e na presenccedila de
actina respectivamente o que corresponde a uma concentraccedilatildeo de cerca de 27 M de V10 O
meio de reacccedilatildeo utilizado nestes ensaios foi 10 mM HEPES e 25 mM KCl com 2 mM ATP Em
(Tiago et al 2004) o valor de IC50 para a actividade Mg2+-ATPaacutesica na ausecircncia de actina foi de
125 plusmn 025 microM enquanto o valor da actividade estimulada por actina foi de 136 plusmn 14 microM O
meio de reacccedilatildeo utilizado nos ensaios foi 20 mM HEPES 50 mM KCl e 2 mM ATP o que resulta
precisamente no dobro da forccedila ioacutenica a que foi realizado o ensaio do presente relatoacuterio
(exceptuando a contribuiccedilatildeo dos produtos da hidroacutelise de ATP uma vez que este meacutetodo
pressupotildee a ocorrecircncia de um ciclo de reacccedilotildees de hidroacutelise e siacutentese deste composto) Ora tendo
sido anteriormente referido que a afinidade do S1 de miosina para com a espeacutecie V10 diminui com
Act
ivid
ade
Mg
2+-A
TP
aacutesic
a d
e S
1 d
e
Mio
sin
a
[Decavanadato] (microM) Actividade Estimulada ()
Actividade Natildeo Estimulada ()
Ajuste Actividade Estimulada
Ajuste Actividade Natildeo Estimulada
33
o aumento da forccedila ioacutenica (Tiago et al 2004 tal poderaacute explicar a diferenccedila obtida entre estes
dois conjuntos de valores
243Quantificaccedilatildeo de cisteiacutenas reduzidas pelo meacutetodo de Ellman
A interacccedilatildeo do vanaacutedio com alguns tipos de aminoaacutecidos sensiacuteveis agrave oxidaccedilatildeo pode fornecer-nos
informaccedilatildeo bastante uacutetil em relaccedilatildeo agraves alteraccedilotildees estruturais que conduzem a uma diminuiccedilatildeo da
actividade fisioloacutegica do S1 de miosina Os estudos realizados por Chasteen 1983 concluiacuteram
que a reduccedilatildeo de vanadato (V) a vanadilo (IV) ocorre na presenccedila de agentes redutores comuns no
ambiente celular de que se destaca o aminoaacutecido cisteiacutena Esta eacute tambeacutem um alvo de
complexaccedilatildeo por parte do iatildeo VO2+
Jaacute Ramos et al (2009) demonstrou que a incubaccedilatildeo de
actina com a soluccedilatildeo de decavanadato induz a oxidaccedilatildeo de resiacuteduos de cisteiacutena e agrave formaccedilatildeo do
oxidovanaacutedio (IV) Este grupo de investigadores descobriu ainda que apenas a espeacutecie V10 induz a
oxidaccedilatildeo das cisteiacutenas ldquoraacutepidasrdquo da actina atraveacutes do processo proposto que eacute parcialmente
responsaacutevel pela inibiccedilatildeo jaacute descrita da actividade de ATPase do S1 de miosina estimulada por
actina quando exposta agrave soluccedilatildeo de decavanadato Esta inibiccedilatildeo apresenta-se afectada com a
presenccedila de ATP uma vez que os resiacuteduos de cisteiacutena permaneceram intactos nos ensaios
realizados com este ligando Jaacute Crans et al (2004) concluiu que o siacutetio activo de muitas fosfatases
(que conteacutem grupos tiol) pode formar um equiliacutebrio redox com o iatildeo vanadato (V) de forma natildeo
reversiacutevel
Os resultados obtidos por Sakurai e Shiromura (1980) sugeriram que o vanaacutedio (V) sofre reduccedilatildeo
a vanaacutedio (IV) sem dependecircncia do valor de pH do ensaio Registaram ainda uma estequiometria
de 31 na reacccedilatildeo do vanadato (V) com os resiacuteduos de Cys enquanto a estequiometria dos ensaios
com o vanadilo (IV) foi de 21
Berlett e Stadtman (1997) observaram que os tipos de aminoaacutecidos mais facilmente oxidaacuteveis
englobam entre outros os resiacuteduos de Cys Trp e MetAssim analisou-se o estado redox dos
grupos tiol dos resiacuteduos de Cys do S1 na presenccedila do decavanadato atraveacutes da monitorizaccedilatildeo da
Abs412 nm na presenccedila de 1 mgmL de S1 de miosina em tampatildeo 10 mM HEPES 25 mM KCl
variando a concentraccedilatildeo da soluccedilatildeo de decavanadato de modo a determinar que tipo de alteraccedilotildees
estruturais ocorrem na sua presenccedila no que diz respeito agrave diminuiccedilatildeo da actividade ATPaacutesica de
S1 na presenccedila de decavanadato
Os resultados referentes agrave quantificaccedilatildeo das cisteiacutenas reduzidas apoacutes o tratamento de S1 com
diferentes concentraccedilotildees de decavanadato em tampatildeo 10 mM HEPES pH 75 e 25 mM KCl a
25ordmC durante 20 minutos encontram-se descritos na Tabela 2VII
Tabela 2VII ndash Cisteiacutenas reduzidas totais e ldquoraacutepidas de S1 (1 mgmL) apoacutes tratamento com decavanadato
34
(010501002505001000 uM) em 10 mM HEPES 25 mM KCl pH 7 a 25ordmC durante 20 minutos A detecccedilatildeo
espectrofotomeacutetrica foi feita com DTNB (e 412=8400 uM-1 cm-1) (n variaacutevel entre 1 e 8)
Amostras Cys raacutepidas Cys totais
11 M S1 (controlo) 22 plusmn 04 106 plusmn 14
11 M S1 + 10 M Decavanadato 22 plusmn 05 95 plusmn 08
11 M S1 + 50 uM Decavanadato 20 plusmn 02 98 plusmn 14
11 M S1 + 100 uM Decavanadato 17 plusmn 03 97 plusmn14
11 M S1 + 250 uM Decavanadato 26 plusmn 02 98 plusmn 08
11 M S1 + 500 uM Decavanadato 21 plusmn 04 90 plusmn 06
11 M S1 + 1000 uM Decavanadato 24 plusmn 0 86 plusmn 08
A partir dos resultados obtidos pode sugerir-se que o S1 possui 11 resiacuteduos de Cys (controlo) o
que estaacute de acordo com o anteriormente descrito Verifica-se que o tratamento da proteiacutena com
concentraccedilotildees crescentes de decavanadato provoca a oxidaccedilatildeo de dois resiacuteduos de Cys do S1 de
miosina natildeo expostos ao solvente No entanto a teacutecnica utilizada natildeo nos permite identificar
quais
Tendo em consideraccedilatildeo que o valor de IC50 obtido para a reacccedilatildeo de inibiccedilatildeo daactividade Mg2+-
ATPaacutesica de S1 na presenccedila de decavanadato eacute de cerca de 27 microM pode sugerir-se que este seraacute o
valor ao qual ocorreraacute a oxidaccedilatildeo do primeiro resiacuteduo de Cys o que poderaacute contribuir para a
diminuiccedilatildeo da actividade de S1 observada
25Interacccedilatildeo de S1 com vanaacutedio(IV)
251Anaacutelise por espectroscopia de Ressonacircncia Paramagneacutetica Electroacutenica
A acumulaccedilatildeo intracelular de oxovanaacutedio (IV) in vivo constitui uma das principais formas de
apresentaccedilatildeo de vanaacutedio no organismo Deste modo torna-se bastante interessante analisar a
interacccedilatildeo entre este iatildeo e o S1 de miosina atraveacutes da teacutecnica de espectroscopia de Ressonacircncia
Paramagneacutetica Electroacutenica o meacutetodo de eleiccedilatildeo do estudo do iatildeo vanadilo e dos complexos por si
formados O electratildeo desemparelhado encontra-se fortemente acoplado ao nuacutecleo 51
V o que
35
resulta num espectro de RPE consistindo de 8 desdobramentos na direcccedilatildeo paralela e outros tantos
na direcccedilatildeo perpendicular (Chasteen 1983)
Os estudos realizados por Ahmed et al (1987) centraram-se na interacccedilatildeo do vanadilo com
proteiacutenas como eacute o caso da insulina transferrina entre outros obtendo um espectro caracterizado
por 8 linhas amplamente espaccediladas (na direcccedilatildeo paralela) Jaacute Ramos et al 2009 concluiu que a
exposiccedilatildeo da soluccedilatildeo de decavanadato agraves formas globular e filamentosa de actina produz um
espectro tiacutepico do iatildeo oxidovanaacutedio (IV) indicando a reduccedilatildeo do vanadato
Para aferir qual a concentraccedilatildeo de S1 que permitia obter uma boa razatildeo sinalruiacutedo titulou-se uma
soluccedilatildeo de sulfato de vanadilo 15 microM ([VOSO4]stock = 5 mM) com concentraccedilotildees crescentes de
proteiacutena (0 5 10 15 e 20 microM) em 10 mM HEPES 25 mM KCl A existecircncia de uma correlaccedilatildeo
linear entre a concentraccedilatildeo de proteiacutena e a intensidade dos sinais de RPE correspondente agrave banda
-12 perpendicular encontra-se representada na Fig 210
Figura 2 12ndash Representaccedilatildeo graacutefica da variaccedilatildeo da intensidade do sinal de RPE vs a concentraccedilatildeo de S1 utilizada
Apoacutes verificar que com uma concentraccedilatildeo de S1 igual a 20 microM se obtinham sinais com uma razatildeo
sinalruiacutedo adequada ao prosseguimento dos estudos sobre a interacccedilatildeo V(IV)-S1 de miosina
titulou-se uma soluccedilatildeo de 20 microM S1 com 0 25 5 10 15 25 50 75 100 150 200 500 e 1000
microM VOSO4
Traccedilou-se o graacutefico da intensidade da banda -12 perpendicular do sinal de EPR em funccedilatildeo da
concentraccedilatildeo de VOSO4 tendo-se determinado uma constante de dissociaccedilatildeo (KD) do complexo
S1-vanadilo de 8748 plusmn 2621 microM V(IV)
y = 016786x + 122435
Rsup2 = 092992759
Inte
nsi
dad
e d
o s
inal
de
RP
E
[S1] (microM)
36
Figura 213 ndash Representaccedilatildeo graacutefica da variaccedilatildeo do sinal de RPE vs a concentraccedilatildeo de vanadilo utilizada (os valores
experimentais estatildeo representados por losangos o seu ajuste estaacute representado pela linha)
A simulaccedilatildeo do espectro de EPR foi efectuada a partir do espectro obtido para uma concentraccedilatildeo
de 1 mM VOSO4 apoacutes adiccedilatildeo de 20 microM S1 de miosina tendo-se obtido os valores da constante de
acoplamento hiperfina (A=5016) e do factor g (g0=195)
Figura 214 ndash Representaccedilatildeo graacutefica do espectro de EPR obtido com 1000 microM de vanadilo e 20 microM de S1 (a cinzento
escuro) A simulaccedilatildeo do espectro encontra-se a cinzento claro
Por comparaccedilatildeo destes com os valores presentes na tabela de correspondecircncia xxx eacute possiacutevel
predizer quais os ligandos provaacuteveis do vanadilo na presenccedila de proteiacutena Assim concluiu-se que
o vanadilo estaacute coordenado com duas moleacuteculas de aacutegua um grupo hidroxilo e um grupo R=NR
O grupo hidroxilo pode indicar os resiacuteduos de aminoaacutecidos Tyr Treonina e Serina enquanto o
grupo R=NR poderaacute indicar a presenccedila de Arginina e Histidina como ligandos do vanadilo
Inte
nsi
dad
e d
os
sinai
s d
e R
PE
[Vanadilo] (microM)
Ajuste dos dados exp Dados experimentais
Campo magneacutetico (G)
Espectro Experimental Ajuste
37
3 Procedimento Experimental
31 Reagentes
Todos os reagentes utilizados satildeo de elevado grau de pureza (analiacutetico) Apresentam-se as suas
foacutermulas quiacutemicas ou abreviaturas quando necessaacuterio acrescentou-se a sua descriccedilatildeo Entre
parecircntesis colocou-se as empresas que os comercializam
Aacutecido aceacutetico glacial 100 (Riedel) Acrilamida 99 (Merck) Azul de bromofenol (Riedel-de-
Haumlen) -Mercaptoetanol (Sigma) Bissulfito de soacutedio (Sigma)Bradford (reagente de)
(BioRad)BSA (albumina de soro bovino Sigma-Aldrich) CaCl22H2O (cloreto de caacutelcio
dihidratado Fluka) CH3CH2OH 96 (etanol Panreac) Coomassie Blue R250 (Merck) D2O
(oacutexido de deuteacuterio SDS) DTNB (55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico) Aldrich) EDTA (aacutecido
etilenodiaminatetraaceacutetico Merck) Fosfoenolpiruvato 98 (Fluka) Glicerol (Sigma) Glicina
985 (Panreac) HCl 37 (Riedel-de Haumlen) H2SO4 96 (Pronalab) HEPES 99 (aacutecido 4-(2-
hidroxiletil)-1-piperazineetanosulfoacutenico Sigma) KH2PO4 99 (hidrogenofosfato de potaacutessio)
MgATP (Sigma) Na
ATP (Sigma) Na2HPO4 99 (hidrogenofosfato de soacutedio Riedel-de Haumlen)
NaH2PO4 (fosfato de soacutedio ) NaOH 986 (hidroacutexido de soacutedio Pronalab) KCl 99 (Cloreto de
potaacutessio Panreac) L-Cys 995 (enantioacutemero levoacutegiro do resiacuteduo de Cisteiacutena Fluka)Metanol
99 (Merck) Molibdato de amoacutenio (Sigma)NNrsquo-metil-bis-acrilamida 98 (Merck) NH4VO3
99 (metavanadato de amoacutenia Sigma) PSA 10 (persulfato de amoacutenio Merck) SDS
(dodecilo sulfato de soacutedio Sigma) Sulfito de soacutedio (Sigma) VOSO4 (sulfato de vanadilo
Aldrich) TEMED 99 (Tetrametiletilenodiamina Merck) Tris (2-amino-2-hidroximetil-13-
propanodiol Sigma)
32 Meacutetodos
321 Isolamento e purificaccedilatildeo das proteiacutenas de muacutesculo esqueleacutetico de coelho
3211 Purificaccedilatildeo da miosina preparaccedilatildeo de Subfragmento 1 por digestatildeo quimotriacutepsica
da miosina
O subfragmento-1 de miosina-II (S1) foi gentilmente cedido pelo Prof Doutor Manuel Aureliano
do Departamento de Quiacutemica Bioquiacutemica e Farmaacutecia da Faculdade de Ciecircncias e Tecnologia da
Universidade do Algarve
No entanto descreve-se abaixo de forma resumida o procedimento efectuado
38
32111 Purificaccedilatildeo da miosina
O protocolo de isolamento foi adaptado do meacutetodo previamente descrito por Margossian e Lowey
(1982) apresentando-se de forma esquemaacutetica na Fig 31
Sucintamente os tecidos musculares dorsais e das pernas foram extraiacutedos apoacutes o sacrifiacutecio do
animal (Fig31) De seguida foram adicionados 3 volumes da soluccedilatildeo de extracccedilatildeo A2
Centrifugou-se a soluccedilatildeo e filtrou-se o sobrenadante O sedimento foi entatildeo diluiacutedo em 14
volumes de aacutegua destilada de modo a que a forccedila ioacutenica atingisse 004 M Apoacutes adiccedilatildeo de szlig-
mercaptoetanol (szlig-me) deixou-se a precipitar a 4ordmC removendo o sobrenadante Centrifugou-se
novamente e ressuspendeu-se o sedimento na soluccedilatildeo de extracccedilatildeo A Adicionou-se a soluccedilatildeo
tampatildeo B3 para que a forccedila ioacutenica atingisse 05 M Homogeneizou-se e deixou-se repousar
durante 2h Homogeneizou-se novamente e centrifugou-se de modo a ocorrer precipitaccedilatildeo dos
complexos de actomiosina e proteiacutena desnaturada Apoacutes filtraccedilatildeo do sobrenadante em latilde de vidro
quantificou-se a soluccedilatildeo de miosina resultante pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental)
2 Soluccedilatildeo de extracccedilatildeo A 0012M K2CO3 004M KHCO3 06M KCl pH 65
3 Soluccedilatildeo-tampatildeo B Tris 100 mM pH 7 KCl 2M
39
Figura 31 ndash Representaccedilatildeo esquemaacutetica do protocolo de isolamento de miosina (Margossian e Lowey 1982) Todo o
procedimento foi realizado a 4ordmC (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
32112 Preparaccedilatildeo de Subfragmento-1 de miosina por digestatildeo com quimiotripsina
O S1 de miosina foi isolado apoacutes diaacutelise da miosina durante a noite contra a soluccedilatildeo C4 A
presenccedila do EDTA eacute essencial uma vez que a sua substituiccedilatildeo por MgCl2resulta na produccedilatildeo de
HMM em vez de S1 A suspensatildeo de miosina foi equilibrada e digerida com -quimotripsina
A digestatildeo foi interrompida pela adiccedilatildeo de PMSF Seguidamente ultracentrifugou-se o produto
da digestatildeo desprezando-se o sedimento (Fig 32) A soluccedilatildeo de sobrenadante maioritariamente
S1 foi quantificada pela leitura directa da Abs280 ou pelo meacutetodo de Bradford (ver as secccedilotildees
3311 e 3312 do Procedimento Experimental respectivamente) e armazenada em 10 sacarose
e 2 mM szlig-Me a -80ordmC
4 Soluccedilatildeo C 25mM Tris pH 70 012M KCl 1mM EDTA
40
Antes da realizaccedilatildeo dos ensaios descritos no presente relatoacuterio o S1 de miosina foi dialisado
contra a soluccedilatildeo D5 de modo a remover qualquer composto que pudesse complexar como
vanaacutedio Seguidamente quantificou-se a proteiacutena pelo meacutetodo de Bradford ou pelo meacutetodo
directo (ver secccedilotildees 3311 e 3312do Procedimento Experimtental)
Figura 3 2ndash Representaccedilatildeo esquemaacutetica do protocolo de preparaccedilatildeo do S1 de miosina por digestatildeo quimiotripsina de
acordo com o previamente descrito por Margossian e Lowey (1982) Todo o procedimento foi realizado a 4ordm C
excepto quando indicaccedilatildeo do contraacuterio (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
3212 Isolamento e purificaccedilatildeo de actina
Isolou-se e purificou-se a actina segundo o meacutetodo descrito por Pardee e Spudish que
compreende dois passos ndash a obtenccedilatildeo de um resiacuteduo seco denominado ldquopoacute de acetonardquo e a
extracccedilatildeo subsequente da actina a partir do ldquopoacute de acetonardquo na forma de G-Actina com a
posterior polimerizaccedilatildeo a F-Actina
32121 Obtenccedilatildeo do ldquopoacute de acetonardquo
Sacrificou-se o coelho com uma pancada atraacutes do pescoccedilo e sangrou-se o animal apoacutes o corte da
arteacuteria jugular Esfolou-se e colocou-se numa tina com gelo Retiraram-se as partes de tecido
conjuntivo que envolvem o tecido muscular com auxiacutelio de uma pinccedila e bisturi Seguidamente
retirou-se os muacutesculos dorsais e os muacutesculos brancos das pernas Passou-se os muacutesculos por aacutegua
destilada de modo a remover as impurezas e pesou-se num copo Trituraram-se os muacutesculos
numa maacutequina de triturar carne e extraiu-se o resultante com 3 volumes da soluccedilatildeo E6 durante 10
minutos com agitaccedilatildeo suave Centrifugou-se a suspensatildeo a 4000 g durante 10
5 Soluccedilatildeo D HEPES 10 mM pH 70 KCl 25 mM
6 Soluccedilatildeo E 01 M KCl 015 M K2HPO4 pH 65
41
minutosdesprezando-se o sobrenadante Ressuspendeu-se o sedimento em 6 volumes de soluccedilatildeo
F7durante 10 minutos com agitaccedilatildeo suave Verificou-se se o pH da soluccedilatildeo estava compreendido
entre 82 e 85 acertando-se estecom soluccedilatildeo G8 se necessaacuterio Centrifugou-se durante 10
minutos a 4000 g Desprezou-se o sobrenadante e ressuspendeu-se o sedimento em 3 volumes de
soluccedilatildeo H9 durante 10 minutos Centrifugou-se novamente a 4000 gdurante 10 minutos
desprezando-se o sobrenadante Ressuspendeu-se o sedimento duas vezes consecutivas em 6
volumes de aacutegua destilada durante 5 minutos Centrifugou-se a 4000 g durante 10 minutos apoacutes
cada adiccedilatildeo de H2O
Seguidamente ressuspendeu-se o sedimento 5 vezes consecutivas utilizando 3 volumes de
acetona de cada vez agitando suavemente durante 10 minutos agrave temperatura ambiente Colocou-
se o resiacuteduo filtrado numa tina de vidro sobre papel de filtro deixando-se evaporar o solvente
durante a noite num nicho Triturou-se o resiacuteduo seco numa maacutequina de triturar carne e deixou-se
novamente evaporar o solvente O ldquopoacute de acetonardquo obtido foi armazenado a -20 ordmC
32122 Purificaccedilatildeo da actina a partir de ldquopoacute de acetonardquo
Apoacutes a obtenccedilatildeo do poacute de acetona como acima indicado efectuou-se o protocolo seguidamente
descrito de modo a obter monoacutemeros de actina (G-Actina) e actina filamentosa (F-Actina)
Pesou-se 35 g de poacute de acetona e adicionaram-se 50 mL de soluccedilatildeo-tampatildeo I10
sendo a
extracccedilatildeo feita com agitaccedilatildeo suave durante 30 minutos Separou-se o extracto do poacute de acetona
hidratado filtrando a suspensatildeo com 4 camadas de gaze esterilizada tendo o cuidado de natildeo
espremer o resiacuteduo Voltou a extrair-se com 50 mL de soluccedilatildeo I durante 30 minutos mexendo
lentamente Adicionou-se este ao extracto obtido anteriormente Centrifugou-se o extracto total a
16 000 g durante 30 minutos Decantou-se para uma proveta sobre a qual se colocou gaze para
que o sedimento solto natildeo se juntasse ao sobrenadante e leu-se o volume De seguida adicionou-
se KCl ateacute uma concentraccedilatildeo final de 50 mM MgCl2 ateacute 2mM e ATP ateacute 1mM lentamente e com
agitaccedilatildeo contiacutenua de modo a promover a polimerizaccedilatildeo dos monoacutemeros de actina
Deixou-se polimerizar durante 2h a 25ordmC observando-se a presenccedila de pequenas bolhas de ar
indicativas de um aumento de viscosidade da soluccedilatildeo Arrefeceu-se em gelo durante 15 minutos e
adicionou-se KCl ateacute uma concentraccedilatildeo final de 06M Mexeu-se lentamente durante 30 minutos
apoacutes os quais se ultracentrifugou a 110 000g durante 1hora Desprezou-se o sobrenadante
ressuspendeu-se o sedimento com soluccedilatildeo tampatildeo E e transferiu-se para um potter
Dialisou-se durante a noite contra 1L de soluccedilatildeo-tampatildeo I a uma temperatura de 4ordmC com rotaccedilatildeo
7 Soluccedilatildeo F 005 M NaHCO3 pH 82-85
8 Soluccedilatildeo G 1M Na2CO3
9 Soluccedilatildeo H 1 mM EDTA pH 70
10 Soluccedilatildeo I 2 mM Tris-HCl pH 8 02 mM CaCl2 02 mM Na2ATP 05 mM szlig-Me 0005 NaN3
42
contiacutenua e mudanccedila de tampatildeo 2 a 3 vezes De seguida a actina foi centrifugada a 110
000gdurante 1hora Decantou-se a soluccedilatildeo de G-Actina (sobrenadante) e desprezou-se o
sedimento Determinou-se a concentraccedilatildeo de G-Actina pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental) A soluccedilatildeo de G-actina foi armazenada a uma temperatura de 4ordmC
apoacutes adiccedilatildeo de 002NaN3 mantendo-se estaacutevel durante 3 a 5 dias
331 Quantificaccedilatildeo das proteiacutenas isoladas
3311 Quantificaccedilatildeo pelo meacutetodo de Bradford
O meacutetodo de Bradford (Bradford 1976) baseia-se na utilizaccedilatildeo do reagente de Bradford
composto por entre outros aacutecido fosfoacuterico e o corante ldquoCoomassierdquo Este corante possui carga
negativa e a sua coloraccedilatildeo varia entre a vermelha (Absmax=465 nm) predominante em soluccedilatildeo e a
azul (Absmax=595 nm) presente aquando da ligaccedilatildeo aos grupos amina das proteiacutenas Em soluccedilotildees
aacutecidas diluiacutedas a coloraccedilatildeo varia proporcionalmente com a quantidade do corante ligado agrave
proteiacutena que por sua vez eacute directamente proporcional agrave concentraccedilatildeo da proteiacutena permitindo a
sua quantificaccedilatildeo ateacute a um limite de detecccedilatildeo de 20 gmL (Bradford 1976)
Deste modo prepararam-se amostras de BSA ([BSA]stock= 1mgmL) utilizando um intervalo de
concentraccedilotildees compreendido entre 0 e 15 gmL de acordo com a tabela 3I Apoacutes a adiccedilatildeo da
H2O MilliQ e do reagente de Bradford as soluccedilotildees foram sucessivamente homogeneizadas no
voacutertex
Tabela 3Indash Amostras utilizadas para a obtenccedilatildeo da recta de calibraccedilatildeo no meacutetodo de Bradford
BSA (L) BSA (g) H2O MilliQ (L) Reagente de Bradford (L)
0 0 800 200
125 125 79875 200
25 25 7975 200
5 5 795 200
10 10 790 200
15 15 785 200
Apoacutes incubaccedilatildeo a 25ordmC por 5 minutos leram-se os valores de Abs595 num
espectrofotoacutemetro (Shimadzu UV-160A) para todas as amostras Seguidamente traccedilou-se a recta
de calibraccedilatildeo (Abs595vs quantidade de BSA (g))
43
Para determinar a concentraccedilatildeo de proteiacutena apoacutes o processo de isolamento purificaccedilatildeo e
diaacutelise quando necessaacuterio colocou-se 5 L da amostra num tubo de Eppendorf e adicionou-se
795 L de H2O MilliQ homogeneizando a soluccedilatildeo no voacutertex Adicionou-se posteriormente 200
L de reagente de Bradford e repetiu-se a homogeneizaccedilatildeo Incubou-se a mistura a 25ordmC por 5
minutos apoacutes os quais se leu os valores de Abs595 nm no espectrofotoacutemetro Por interpolaccedilatildeo deste
valor a partir da recta de calibraccedilatildeo determinou-se a concentraccedilatildeo de proteiacutena na amostra
3312 Quantificaccedilatildeo pelo meacutetodo directo
A quantificaccedilatildeo das amostras proteicas pode tambeacutem ser efectuada atraveacutes da leitura directa da
absorvacircncia tendo por base a lei de Beer-Lambert onde Abs corresponde agrave
absorvacircncia ao comprimento de onda ao qual o ensaio foi realizado corresponde ao coeficiente
de extinccedilatildeo (substituiacutedo pela letra a se as unidades forem distintas da molaridade) b corresponde
ao percurso oacuteptico da radiaccedilatildeo na ceacutelula amostral e c corresponde agrave concentraccedilatildeo amostral
Assim no caso de S1 de miosina mediu-se a Abs280e calculou-se a concentraccedilatildeo da proteiacutena
sabendo que o coeficiente de extinccedilatildeo de S1 eacute (Ref)
No caso da actina mediu-se a Abs290ecalculou-se a concentraccedilatildeo da proteiacutena sabendo que o
coeficiente de extinccedilatildeo da actina eacute (Ref)
332Electroforese em condiccedilotildees desnaturantes (SDS-PAGE)
De modo a melhor caracterizar as proteiacutenas isoladas foi realizada uma electroforese em
condiccedilotildees desnaturantes (SDS-PAGE do inglecircs Sodium Dodecyl Sulfate Polyacrilamide Gel
Electrophoresis) O SDS (dodecilo sulfato de soacutedio) eacute um detergente anioacutenico utilizado neste
meacutetodo para desnaturar as proteiacutenas que se ligam ao SDS na proporccedilatildeo 14 g SDS g proteiacutena
(aproximadamente 1 moleacutecula de SDS por cada 2 resiacuteduos de aminoaacutecidos) (Stryer) O SDS induz
uma densidade de carga negativa significativa nas proteiacutenas a que se liga mascarando a sua carga
intriacutenseca pelo que as razotildees cargamassa satildeo tendencialmente idecircnticas Assim a SDS-PAGE
separa as proteiacutenas quando sujeitas a um campo eleacutectrico de acordo com o seu peso molecular
sendo a sua migraccedilatildeo directamente proporcional ao logaritmo deste uacuteltimo (Stryer)
O gel de resoluccedilatildeo com 10 de acrilamida foi obtido atraveacutes da adiccedilatildeo de 167 mL de soluccedilatildeo
stock de acrilamida11
075 mL de tampatildeo de resoluccedilatildeo12
251 mL de aacutegua destilada e 50 L de
SDS 10 Para promover a polimerizaccedilatildeo adicionou-se em seguida 38 L de PSA 10
11
soluccedilatildeo stock de acrilamida soluccedilatildeo de 30 (mv) de acrilamida e 08 (mv) de NNrsquo-metil-bis-acrilamida 12
tampatildeo de resoluccedilatildeo 15 M Tris-HCl pH 88
44
(persulfato de amoacutenia) 10 e 25 L de TEMED 99 (tetrametiletilenodiamina) Agitou-se
cuidadosamente de modo a natildeo formar bolhas e pipetou-se a soluccedilatildeo para a cassette de vidro
previamente montada no suporte De seguida adicionou-se uma peliacutecula de cerca de 2 mm de 2-
metil-2-butanol para verificar posteriormente a polimerizaccedilatildeo Apoacutes a polimerizaccedilatildeo que demora
cerca de 45 minutos verteu-se a agrave superfiacutecie e adicionou-se o gel de concentraccedilatildeo com 5 de
acrilamida constituiacutedo por 03 mL de soluccedilatildeo de stock acrilamida 045 mL de tampatildeo de
concentraccedilatildeo13
094 mL de aacutegua destilada 18 L de SDS 10 135 L de PSA 10 e 2L de
TEMED 99 Colocou-se o pente sobre o gel e deixou-se polimerizar durante cerca de 45
minutos
As amostras para aplicaccedilatildeo no gel de electroforese foram preparadas para uma concentraccedilatildeo de
proteiacutena de 1 mgmL num volume final de 100 L dos quais 25 L correspondem ao tampatildeo da
amostra14
As amostras foram aquecidas a 100ordmC durante 5 minutos Antes da aplicaccedilatildeo das
amostras transferiu-se a cassette para a tina de electroforese agrave qual se adicionou a soluccedilatildeo
tampatildeo de electroforese da tina superior15
e tampatildeo de electroforese da tina inferior16
Aplicaram-
se 10 L das soluccedilotildees de marcadores de peso molecular e 20 L das amostras proteicas
preparadas anteriormente A electroforese decorreu durante cerca de 3h30 min a uma voltagem
de 80 V De seguida transferiu-se o gel para uma tina corando as proteiacutenas durante 1h com
soluccedilatildeo corante17
Descorou-se o gel com soluccedilatildeo descorante18
tendo-se mudado esta soluccedilatildeo ateacute
as bandas se encontrarem bem definidas
Determinou-se a mobilidade relativa das bandas correspondentes aos marcadores de elevado peso
molecular e traccedilou-se a recta de calibraccedilatildeo ndash log(PM) vs log(Rf) onde Rf corresponde agrave
mobilidade relativa Por interpolaccedilatildeo determina-se o PM das proteiacutenas isoladas sabendo a sua
mobilidade relativa Por comparaccedilatildeo com os valores descritos na literatura pode concluir-se
acerca do grau de purificaccedilatildeo
34 Mediccedilatildeo da actividade enzimaacutetica de S1
341 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
13
tampatildeo de concentraccedilatildeo 05 M Tris-HCl pH 68 14
tampatildeo de amostra 320 mM Tris-HCl pH 68 04 M szlig-mercaptoetanol SDS 8 glicerol 15 (vv) e roxo de m-
cresol 0024 15
tampatildeo de electroforese da tina superior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS 16
tampatildeo de electroforese da tina inferior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS acetato de soacutedio
01 M 17
soluccedilatildeo corante ldquoCoomassie Blue R-250rdquo contendo metanol 45 (vv) e aacutecido aceacutetico glacial 10 (vv) 18
soluccedilatildeo descorante metanol 10 (vv) e aacutecido aceacutetico glacial 10 vv)
45
O meacutetodo de Fiske-Subarow modificado consiste na reacccedilatildeo do molibdato de amoacutenio com o
fosfato inorgacircnico em soluccedilatildeo aacutecida com formaccedilatildeo do aacutecido fosfomoliacutebdico Seguidamente daacute-
se a adiccedilatildeo de um agente redutor que provoca a reduccedilatildeo do composto a fosfomolibdato que forma
um complexo de cor azulada cuja absorvacircncia maacutexima ocorre para um comprimento de onda de
660 nm tornando possiacutevel a sua detecccedilatildeo por meacutetodos espectrofotomeacutetricos As actividades Ca2+
-
ATPaacutesica e K+EDTA-ATPaacutesica de S1 foram determinadas atraveacutes deste meacutetodo de acordo com
o descrito por Taussky e Shorr (1953) com mediccedilatildeo da quantidade de fosfato (Pi) libertado
atraveacutes da hidroacutelise de ATP
Para tal obteve-se primeiramente a recta de calibraccedilatildeo com as amostras presentes na Tabela 3II
utilizando uma soluccedilatildeo stock de 5 mM KH2PO4
Tabela 3II ndash Composiccedilatildeo das diferentes amostras utilizadas para determinar a recta de calibraccedilatildeo atraveacutes do meacutetodo
de Fiske-Subbarow modificado
KH2PO4 (L)
([KH2PO4]stock= 5mM)
PO43-
(nmole) H2O (uL)
0 0 1780
5 25 1775
10 50 1770
20 100 1760
40 200 1740
100 500 1680
A cada tubo de Eppendorf adicionaram-se seguidamente 200 L de soluccedilatildeo J19
e 20 L de agente
redutor20
homogeneizando no voacutertex apoacutes cada adiccedilatildeo Incubou-se durante 10 minutos e mediu-
se a Abs660 nm num espectrofotoacutemetro com intervalos de tempo iguais entre cada amostra Traccedilou-
se a recta de calibraccedilatildeo (Abs660 nmvs nmol PO43-
) a partir dos valores obtidos
Para determinar a actividade Ca2+
-ATPaacutesica de S1 prepararam-se tubos de Eppendorf com 400
L de meio de reacccedilatildeo21
25 L CaCl2 ([CaCl2]stock= 100 mM) 25 L ATP ([ATP]stock= 100 mM)
com [S1]ensaios=01 mgmL Perfez-se se necessaacuterio com H2O MilliQ ateacute perfazer um volume de 1
mL Para determinar a actividade K+EDTA-ATPaacutesica utilizou-se uma concentraccedilatildeo idecircntica de
19
Soluccedilatildeo J 5 N H2SO4 25 molibdato de amoacutenio 20
Agente redutor 5 mg aacutecido 1-amino-2-naftol-4-sulfoacutenico 30 mg bissulfito de soacutedio 30 mg sulfito de soacutedio em 26
mL H2O 21
meio de reacccedilatildeo 125 mM Tris-HCl pH 80 0575 M KCl
46
proteiacutena nos ensaios (01 mgmL) e o mesmo meio de reacccedilatildeo Aos tubos de Eppendorf
adicionou-se ainda 123 L de KCl ([KCl]stock = 3M) 25 L de ATP 10 L de EDTA
([EDTA]stock= 100 mM )e o volume de H2O MilliQ necessaacuterio para perfazer 1 mL Em ambas as
situaccedilotildees o uacuteltimo reagente adicionado foi o ATP uma vez que se trata do substrato da reacccedilatildeo
enzimaacutetica
A cineacutetica decorreu a 25ordmC durante 5 minutos para a actividade Ca2+
-ATPaacutesica e durante 3
minutos para a actividade K+EDTA-ATPaacutesica Apoacutes o periacuteodo de incubaccedilatildeo transferiu-se 150
L de amostra para tubos de Eppendorf contendo igual volume de TCA 10 Colocou-se no gelo
e homogeneizou-se no voacutertex Seguidamente centrifugou-se a 1500 g durante 10 minutos e
recolheu-se 200 L de sobrenadante para novos tubos de Eppendorf Adicionou-se a estes 158
mL de H2O MilliQ e 200 L de soluccedilatildeo J Homogeneizou-se no voacutertex e adicionou-se 20 L de
reagente redutor repetindo-se a homogeneizaccedilatildeo Incubou-se durante 10 minutos e mediu-se a
Abs660 nm com o mesmo intervalo de tempo de leitura para cada amostra Por intrapolaccedilatildeo a partir
da recta de calibraccedilatildeo obtida anteriormente determinou-se a quantidade de Pi libertado em cada
ensaio A partir destes resultados determinaram-se as actividades Ca2+
-ATPaacutesica e K+EDTA-
ATPaacutesica do S1 em nmol PO43-
minmg proteiacutena e em s-1
342 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
Da actividade Mg2+
-ATPaacutesica de S1 resulta a formaccedilatildeo dos produtos da reacccedilatildeo de
hidroacutelise do ATP - ADP e Pi Na presenccedila de ADP fosfoenolpiruvato (PEP) e da enzima piruvato
cinase (PK) os produtos resultantes satildeo ATP e piruvato Por acccedilatildeo da enzima lactato
desidrogenase (LDH) este uacuteltimo eacute convertido em lactato ocorrendo simultaneamente a oxidaccedilatildeo
do NADH a NAD+ monitorizada atraveacutes da Abs340 nm Quando o S1 de miosina se encontra no
meio reaccional o ATP eacute hidrolisado formando-se novamente ADP e Pi o que permite a
formaccedilatildeo de um ciclo (fig 21) Se as enzimas se encontrarem em excesso a velocidade de
conversatildeo do NADH eacute limitada pela produccedilatildeo de ADP e Pi Como o ATP eacute regenerado em cada
ciclo a sua concentraccedilatildeo inicial eacute sempre mantida (Bagshaw 1982)
Figura 33ndash Representaccedilatildeo esquemaacutetica do meacutetodo de enzimas acopladas
47
O protocolo utilizado e seguidamente descrito foi adaptado de Cuenda et al (1990 ndash Ver Refordf) A
uma cuvette de 1 mL adicionou-se S1 para uma concentraccedilatildeo final de 005 mgmL 800 L de
meio de reacccedilatildeo22
10 L de PEP ([PEP]stock = 42 mM) 10 L de PK+LDH 11 ([PK+LDH]stock =
1 mgmL) e 15 L de NADH ([NADH]stock=25 mM) Iniciou-se a monitorizaccedilatildeo da reacccedilatildeo
atraveacutes da leitura de Abs340 nm apoacutes a adiccedilatildeo de 20 L de ATP ([ATP]stock=100 mM) A
actividade Mg2+
-ATPaacutesica eacute obtida atraveacutes do declive correspondente agrave oxidaccedilatildeo do NADH Para
a mediccedilatildeo da Mg2+
-ATPase de S1 estimulada por actina adicionou-se F-actina para uma
concentraccedilatildeo final de 2 microM sendo a actividade obtida tambeacutem atraveacutes do declive correspondente
agrave oxidaccedilatildeo do NADH apoacutes a adiccedilatildeo da proteiacutena
35 Interaccedilatildeo entre vanaacutedio e subfragmento-1 de miosina
351 Preparaccedilatildeo das soluccedilotildees de vanaacutedio
As soluccedilotildees de vanadato foram preparadas em aacutegua MilliQ a partir de metavanadato de amoacutenio
(NH4VO3) numa concentraccedilatildeo stock de 50 mM com agitaccedilatildeo magneacutetica e aquecimento (~30 ordmC)
Esta soluccedilatildeo com um valor compreendido entre 6 e 7 designa-se por ldquometavanadatordquo e eacute
constituiacuteda pelas esspeacutecies V1 V2 V4 e V5 (Ref) Acidificando a soluccedilatildeo de metavanadato por
adiccedilatildeo de HCl ateacute um valor de pH proacuteximo de 4 observa-se uma mudanccedila de cor de amarelo
claro para laranja indicativa da formaccedilatildeo da espeacutecie decameacuterica de vanadato De seguida e
imediatamente antes da realizaccedilatildeo dos ensaios descritos o valor do pH da soluccedilatildeo de
decavanadato foi aumentado para cerca de 75 por adiccedilatildeo de NaOH A soluccedilatildeo (2) designada
por ldquometavanadatordquo natildeo foi posteriormente alterada ateacute agrave sua adiccedilatildeo para a realizaccedilatildeo dos ensaios
As soluccedilotildees foram armazenadas a 4ordmC e mantidas em gelo aquando da sua utilizaccedilatildeo
A soluccedilatildeo de V(IV) foi preparada em condiccedilotildees anaeroacutebias por diluiccedilatildeo de sulfato de vanadil
(VOSO4) para uma concentraccedilatildeo final de 5 mM em H2O MilliQ saturada com N2 gasoso
352Anaacutelise por espectroscopia de UV-Vis
A concentraccedilatildeo da espeacutecie decameacuterica em soluccedilatildeo depende do periacuteodo de tempo apoacutes a diluiccedilatildeo
no meio de reacccedilatildeo apropriado (Ref) A velocidade de desoligomerizaccedilatildeo da espeacutecie decameacuterica
pode ser analisada por espectroscopia de absorccedilatildeo na regiatildeo UVVis uma vez que esta espeacutecie
apresenta duas bandas caracteriacutesticas na regiatildeo do visiacutevel (a 358 e 400 nm) que natildeo se verifica
em quaisquer outras espeacutecies oligomeacutericas As bandas supramencionada sapresentam-se bem
22
Meio de reacccedilatildeo 3125 mM Tris-HCl pH 71 375 mM MgCl2
48
resolvidas atraveacutes da obtenccedilatildeo da derivada de 2ordf ordem do espectro apresentando a forma de 2
vales com miacutenimos centrados nos comprimentos de onda supracitados com dependecircncia linear
relativamente agrave concentraccedilatildeo de decavanadato total (Ramos et al 2006)
Assim a estabilidade da espeacutecie decameacuterica apoacutes diluiccedilatildeo da soluccedilatildeo de decavanadato no meio de
reacccedilatildeo dos ensaios (tampatildeo D) para uma concentraccedilatildeo final de 1 mM em vanadato total foi
caracterizada atraveacutes da leitura da Abs400 em funccedilatildeo do tempo tendo-se determinado qual a
ordem de reacccedilatildeo e o tempo de meia-vida da mesma Com base em estudos anteriores acerca da
estabilidade do V10 na presenccedila de diferentes proteiacutenas (Ramos et al 2006) determinou-se
igualmente o tempo de meia vida apoacutes a adiccedilatildeo de 20 microM de S1 de miosina ao meio reaccional
353 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
Os espectros de RMN do isoacutetopo 51
V foram realizados num espectroacutemetro Bruker AM-400 MHz
equipado com uma sonda multinuclear inversa de 5 mm Em tubos de RMN de 5 mm de diacircmetro
preparou-se soluccedilotildees de 500 L de deca- ou metavanadato correspondentes a um intervalo de
concentraccedilotildees de 1 a 10 mM suplementadas com 10 D2O Os espectros foram adquiridos agrave
temperatura ambiente com uma largura espectral de 23 923 Hz um tempo de aquisiccedilatildeo de 0086
s tempo de relaxaccedilatildeo de 001 s e uma sequecircncia de pulsos a 90ordm Utilizou-se a transformada de
Fourier com alargamento das bandas de 20Hz para o processamento dos espectros Os desvios
quiacutemicos das ressonacircncias de 51
V foram obtidos em relaccedilatildeo agrave espeacutecie VOCl3cuja frequecircncia de
ressonacircncia eacute considerada o padratildeo sendo todas as outras referenciadas a partir desta em ppm
As aacutereas dos sinais do RMN correspondentes agraves espeacutecies oligomeacutericas presentes foram obtidas
por integraccedilatildeo dos sinais As concentraccedilotildees relativas das espeacutecies foram calculadas com recurso agrave
equaccedilatildeo
onde A[Vx] corresponde agrave aacuterea referente agrave ressonacircncia da espeacutecie Vx com nordm de agregaccedilatildeo x
A[V]total agrave soma total das aacutereas medidas e [V] a concentraccedilatildeo total de vanaacutedio no ensaio
A teacutecnica de 51
V-RMN foi anteriormente aplicada em estudos de ligaccedilatildeo com S1 de miosina
(Ref) tendo sido demostrado que entre as espeacutecies V1 V2 V4 e V5 a espeacutecie tetrameacuterica possui a
maior afinidade para a miosina (Cremo et al) O desaparecimento ou alargamento dos sinais pode
implicar a ligaccedilatildeo de um dos oxovanadatos laacutebeis agrave miosina sem indicaccedilatildeo de qual uma vez que
49
se processa um equiliacutebrio entre eles (Crans 1990) Assim e tendo em conta as indicaccedilotildees de que
a espeacutecie V10 eacute distinta das restantes no que diz respeito agrave magnitude da interacccedilatildeo com a miosina
e inibiccedilatildeo da actividade ATPaacutesica estimulada por actina (Aureliano 2000 Tiago et al 2002)
torna-se fundamental investigar a interacccedilatildeo das diferentes espeacutecies de vanadato com o S1 de
miosina atraveacutes da teacutecnica de 51
V-RMN
Para tal obtiveram-se espectros de 51
V-RMN resultantes da titulaccedilatildeo de uma soluccedilatildeo 5 mM de
decavanadato total com concentraccedilotildees crescentes de S1 de miosina (0 5 10 15 e 20 microM) em
tampatildeo D Realizou-se ainda ensaios na presenccedila de 2 mM de decavanadato e 2 mM de ATP na
presenccedila e na ausecircncia de S1 de miosina de modo a verificar o efeito que estes ligandos naturais
da enzima teria na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o subfragmento-1
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
Os grupos sulfidrilo (-SH) das cisteiacutenas satildeo resiacuteduos de aminoaacutecidos extremamente sensiacuteveis agrave
oxidaccedilatildeo resultando na sua oxidaccedilatildeo (Dalle-Donne et al 1999) Logo a quantificaccedilatildeo destes
resiacuteduos eacute bastante importante nos estudos de interacccedilatildeo com vanaacutedio jaacute que os resiacuteduos de Cys
com ecircnfase para o SH1 e o SH2 satildeo bastante importantes na monitorizaccedilatildeo das modificaccedilotildees
funcionais da regiatildeo da cabeccedila
O procedimento seguidamente apresentado foi originalmente descrito por Gutieacuterrez-Merino
(1987)) Consiste na reacccedilatildeo entre os grupos ndashSH natildeo oxidados das proteiacutenas e o reagente de
Ellman (DTNB -55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico)) - formando-se o aniatildeo CNT (aniatildeo 3-
carboxilato-4-nitrotiofenolato) numa estequiometria de 11 Este apresenta um maacuteximo de
absorvecircncia a 412 nm (Robyt 1990) Assim apoacutes a incubaccedilatildeo da proteiacutena na presenccedila do metal
a quantidade de cisteiacutenas reduzidas por proteiacutena determina-se atraveacutes dos valores de Abs412
Deste modo colocou-se num tubo de Eppendorf o volume apropriado de S1 dialisado para que a
concentraccedilatildeo no ensaio fosse de 1 mgmL Adicionou-se o volume apropriado de decavanadato
para que a sua concentraccedilatildeo fosse de 0 10 50 100 250 500 ou 1000 M Perfez-se com soluccedilatildeo
D ateacute um volume final de 1 mL e incubou-se a 25ordmC por 20 minutos Apoacutes este periacuteodo colocou-
se 780 L da mistura reaccional numa cuvette de quartzo e adicionou-se 220 L de soluccedilatildeo K23
23
Tampatildeo K Tampatildeo fosfatohidrogenofosfato de soacutedio 025 M pH 72
50
Adicionou-se 15 L de DTNB (em soluccedilatildeo K [DTNB]stock=100 mM) ao meio reaccional e
registou-se o valor de absorvecircncia medido equivalente agraves cisteiacutenas ldquoraacutepidasrdquo (SH1 e SH2
expostos ao solvente) Adicionou-se 001 g de SDS que provoca a desnaturaccedilatildeo da proteiacutena e
consequentemente a exposiccedilatildeo de todos os resiacuteduos de cisteiacutenas que se encontravam ateacute aiacute
inacessiacuteveis no interior da proteiacutena (cisteiacutenas totais) Estes resiacuteduos podem assim reagir com o
DTNB
Estimou-se o nuacutemero de resiacuteduos de cisteiacutenas em todas as amostras com recurso ao coeficiente de
extinccedilatildeo molar () do DTNB (ver ponto 3 dos Anexos) que foi determinado atraveacutes da titulaccedilatildeo
de L-Cisteiacutena com intervalo de concentraccedilotildees compreendidos entre 0 e 50 microM na presenccedila de
25microM de DTNB em tampatildeo D Os valores de absorvecircncia foram obtidos para um comprimento de
onda de 412 nm
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
3621 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
A teacutecnica mais eficiente de anaacutelise da interacccedilatildeo entre o iatildeo vanadilo e proteiacutenas como a
calmodulina e a transferrina entre outras permite a obtenccedilatildeo raacutepida de informaccedilatildeo no que toca agrave
conformaccedilatildeo e amplitude da referida interacccedilatildeo
Os espectros de Ressonacircncia Paramagneacutetica Electroacutenica (RPE) da espeacutecie paramagneacutetica V(IV)
foram realizados a 70K num espectroacutemetro Bruker EMX-300 X-band Todas as soluccedilotildees
utilizadas e amostras foram preparadas em anaerobiose numa cacircmara de luvas de modo a evitar a
reduccedilatildeo de V(IV) a V(V) Em tubos capilares com 02 mL de amostra manteve-se a concentraccedilatildeo
de S1 a 20 M e variou-se a concentraccedilatildeo de vanadilo (0 25 5 10 15 25 50 75 100 150
200 500 e 1000 M [VOSO4]stock=5 mM) em tampatildeo D Apoacutes obtenccedilatildeo dos espectros estes
foram tratados atraveacutes do software SimFonia e WinEPR tendo-se obtido um graacutefico com a
variaccedilatildeo da intensidade do sinal de RPE em funccedilatildeo da concentraccedilatildeo de sulfato de vanadilo
presente Realizaram-se ainda simulaccedilotildees de espectros obtidos com 20 microM de S1 e 1000 microM de
V(IV) com vista agrave obtenccedilatildeo dos ligandos provaacuteveis do vanadilo atraveacutes da anaacutelise da constante
de acoplamento hiperfina (A) e do factor g (g)
4 Conclusotildees gerais
Apesar de jaacute ter sido demonstrado que o vanaacutedio inibe a Na+K
+-ATPase da miosina a funccedilatildeo
deste elemento no tecido muacutesculo esqueleacutetico continua largamente incompreendida A enorme
complexidade quiacutemica deste metal especialmente no que concerne agrave oligomerizaccedilatildeo e reacccedilotildees
51
redox constitui um obstaacuteculo agrave compreensatildeo do comportamento do vanaacutedio nos organismos
vivos Assim a caracterizaccedilatildeo da quiacutemica deste elemento eacute de grande utilidade antes de iniciar o
estudo da sua interacccedilatildeo com o presente sistema
A anaacutelise por espectroscopias de 51
V-RMN e de UV-Vis de soluccedilotildees de vanadato diluiacutedas nos
meios de reacccedilatildeo utilizados no presente relatoacuterio (10 mM HEPES pH 70 25 mM KCl) mostrou
claramente que a espeacutecie decameacuterica (V10) possui um tempo de vida suficientemente longo para a
determinaccedilatildeo do seu efeito isolado ao contraacuterio do que se verifica com as espeacutecies mono- di-
tetra- e pentameacutericas (V1 V2 V4 e V5) Por outro lado a anaacutelise verificou ainda que apoacutes diluiccedilatildeo
das soluccedilotildees de decavanadato no meio de reacccedilatildeo coexistem as espeacutecies V10 e V1 pelo que os
efeitos putativos desta uacuteltima deveratildeo ser subtraiacutedos aquando da monitorizaccedilatildeo dos efeitos da
espeacutecie decameacuterica
O vanaacutedio (V) (vanadato) inibe as enzimas que catalizam reacccedilotildees de transferecircncia de grupos
fosfato como a ATPase da miosina [Goodno 1979] devido agrave sua analogia estrutural com o iatildeo
fosfato Na sua utilizaccedilatildeo como inibidor enzimaacutetico natildeo eacute tida em conta a contribuiccedilatildeo individual
das espeacutecies oligomeacutericas considerando-se normalmente a espeacutecie V1 como inibidora efectiva
[Goodno e Taylor 1982] No presente relatoacuterio verificou-se por espectroscopia de 51
V-RMN que
outros oligoacutemeros de vanadato interagem com o S1 de miosina tendo as espeacutecies V10 e V4 os
maiores niacuteveis de afinidade A sua interacccedilatildeo envolve forccedilas electrostaacuteticas e eacute afectada
negativamente por ligandos naturais de enzima como eacute o caso do ATP
A exposiccedilatildeo do S1 de miosina (005 mgmL) a decavanadato conduz agrave inibiccedilatildeo do capacidade da
proteiacutena de estimular a actividade Mg2+
-ATPaacutesica com um valor de IC50 de 266 plusmn 07 M
decavanadato sendo o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM
decavanadato (80) Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM decavanadato sendo
o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM decavanadato (60)
exercendo um efeito de impedimento agrave ligaccedilatildeo entre o vanaacutedio e o S1
A uma concentraccedilatildeo de 27 microM decavanadato verifica-se a oxidaccedilatildeo de um dos resiacuteduos de Cys
de S1 de miosina seguida pela oxidaccedilatildeo gradual de outro resiacuteduo de Cys ateacute agrave concentraccedilatildeo de
1000 microM Ambos os resiacuteduos oxidados natildeo se encontravam expostos ao solvente utilizado A
oxidaccedilatildeo dos resiacuteduos permite sugerir que esta modificaccedilatildeo oxidativa sofrida pelo S1 contribui
para a diminuiccedilatildeo da sua actividade de ATPase
52
Por espectroscopia de RPE revelaram-se os possiacuteveis ligandos equatoriais do vanaacutedio aquando da
sua interacccedilatildeo com o S1 de miosina como sendo duas moleacuteculas de aacutegua um ligando contendo o
grupo R-OH e outro contendo o grupo R=NR
Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados de
oxidaccedilatildeo (IV) e (V) no S1 de miosina para concentraccedilotildees facilmente alcanccedilaacuteveis no ambiente
terrestre
53
5 Estudos futuros
No sentido do melhor esclarecimento das alteraccedilotildees moleculares e estruturais que ocorrem no
subfragmento 1 quando exposta ao vanaacutedio que conduzem a uma diminuiccedilatildeo da sua funccedilatildeo
sugere-se
- Quantificaccedilatildeo das cisteiacutenas reduzidas de S1 na presenccedila de metavanadato
- Determinaccedilatildeo das actividades Ca2+
- e K+-ATPaacutesica do S1 de miosina na presenccedila de
decavanadato
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina estimulada por actina
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina na presenccedila de
metavanadato
- Realizaccedilatildeo de RMN 2D com variaccedilatildeo das concentraccedilotildees das soluccedilotildees de decavanadato e
metavanadato na presenccedila de S1
-Mutageacutenese dos resiacuteduos de Cys doS1 de miosina de modo a verificar quais os resiacuteduos que
sofrem oxidaccedilatildeo na presenccedila de decavanadato
- Determinaccedilatildeo por calorimetria (DSC Differential Scanning Calorimetry) da variaccedilatildeo da
entalpia (∆H) e da temperatura de desnaturaccedilatildeo do S1 de miosina na presenccedila de vanaacutedio uma
vez que estas propriedades caracteriacutesticas de cada proteiacutena satildeo alteradas apoacutes ocorrecircncia de
modificaccedilotildees estruturais
- Anaacutelise por SDS-PAGE de S1 de miosina apoacutes tratamento com vanaacutedio de modo a determinar
quais os polipeacuteptidos eventualmente formados apoacutes clivagem da proteiacutena
54
6 Referecircncias bibliograacuteficas
- Sellers J Myosins 2nd Edition Oxford University Press 1999
- L Stryer inBiochemistry (Ed WH Freeman 2005 pp
Refordfas Materiais e Meacutetodos
[A] S S Margossian SLowey Methods Enzymol1982 85 55-71
[B] JD Pardue JA Spudich Methods Enzymol 1982 85 164-181
[C] M Bradford Anal Biochem1976 72 248-254
[D] H H Taussky E Shorr J Biol Chem 1953 202 675-685
[E] C R Bagshaw in Outline Studies in Biology ndash Muscle Contraction Chapman and Hall Ed
1982
[F] A Cuenda et al Eur J Biochem1990 194 663-670
55
Anexos
1 Determinaccedilatildeo da recta de calibraccedilatildeo meacutetodo de Bradford
Figura1ndashRepresentaccedilatildeo graacutefica da recta de calibraccedilatildeo (Abs595 nm vs [BSA]) utilizada para o meacutetodo de Bradford
2 Determinaccedilatildeo da recta calibraccedilatildeo para a electroforese em condiccedilotildees desnaturantes
Figura 2 ndash Representaccedilatildeo graacutefica da recta de calibraccedilatildeo (log(Peso Molecular) vs Rf dos marcadores de peso
molecular) onde Rf indica determinada atraveacutes do padratildeo de peso molecular
(poccedilo (a))
3Caacutelculo do coeficiente de extinccedilatildeo molar () para a quantificaccedilatildeo de cisteiacutenas reduzidas
atraveacutes do meacutetodo de Ellman
Traccedilou-se a recta de Abs412 nm meacutedia vs [L-Cisteiacutena] de modo a determinar o valor do coeficiente
de extinccedilatildeo molar definido como o declive daquela recta Assim =8400 M-1
cm-1
como
podemos verificar na Fig 3
A595 nm = 00546[BSA] + 00209 Rsup2 = 099051
Ab
s 59
5 n
m
[BSA] (mgmL)
log PM = -1177 Rf + 2247 Rsup2 = 098957
log
PM
Mobilidade relativa (Rf)
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero

11
112 Actina
O termo ldquoactinardquo refere-se a uma famiacutelia extremamente conservada de proteiacutenas citoplasmaacuteticas
presente em todos os eucariotas Esta proteiacutena natildeo se encontra nos procariotas [E] A maioria das
actinas possui peso molecular proacuteximo de 42 kDa outra evidecircncia do seu grau elevado de
conservaccedilatildeo no que concerne agrave sequecircncia e ao nuacutemero de aminoaacutecidos Os eucariotas superiores
apresentam um grande nuacutemero de isoformas de actina que por sua vez se encontram codificados
por um ainda maior nuacutemero de genes (Ref) A expressatildeo das isoformas eacute diferencial consoante o
tecido e sua especializaccedilatildeo pelo que deveratildeo possuir funccedilotildees e mecanismos direccionais
diferentes (Ref) Foi ainda observado que os mRNA das diferentes isoformas apresentam uma
distribuiccedilatildeo heterogeacutenea nas ceacutelulas de forma semelhante aos seus produtos proteicos [F] Estes
dados suportam a hipoacutetese supramencionada do direccionamento das isoformas de actina para um
determinado local na ceacutelula de acordo com as suas funccedilotildees
A actina presente no citoesqueleto eacute responsaacutevel por processos como o mecanismo contraacutectil
fagocitose manutenccedilatildeo e alteraccedilatildeo da estrutura celular locomoccedilatildeo transporte de organelos
regulaccedilatildeo do transporte de solutos entre outras Nos eucariotas superiores estaacute ainda envolvida
no transporte de gracircnulos de pigmento retinoacuteide [G] transporte de gracircnulos no axoplasma [H] e
secreccedilatildeo entre outros [E] Actualmente a isoforma da actina (-actina) eacute a melhor estudada
encontrando-se envolvida no mecanismo contraacutectil [E]
Os filamentos de -actina tecircm cerca de 1m de comprimento e 80 Å Consistem em duas cadeias
de actina filamentosa (F-actina) organizados em heacutelice [I] (Holmes et al 1990) sobre os quais
se encontram outras proteiacutenas ndash a troponina (C I e T) e a tropomiosina (Azevedo 1999) (Figura
15) A troponina eacute filamentosa e localiza-se nas depressotildees da heacutelice de F-actina conferindo
rigidez aos filamentos finos Por sua vez a tropomiosina eacute essencial na regulaccedilatildeo do mecanismo
contraacutectil
Figura 15 ndash Modelo estrutural dos filamentos de F-actina contendo a tropomiosina e o complexo de troponina (I C
e T) (Adaptado de Susana Ramos 2005 [J])
12
A F-actina eacute formada atraveacutes da polimerizaccedilatildeo de subunidades globulares de actina (G-actina) A
G-actina eacute composta por 2 domiacutenios de tamanho semelhante separados pelos bolsos de ligaccedilatildeo(1)
de catiotildees divalentes e(2) de nucleoacutetidos [KLM] Estes por sua vez estatildeo subdivididos noutros
dois de menores dimensotildees (1-4) (Fig 16) cada um envolvido nas interacccedilotildees actina-actina ou
na formaccedilatildeo do topo de ligaccedilatildeo do nucleoacutetido As regiotildees N-terminal e C-terminal estatildeo proacuteximas
uma da outra no subdomiacutenio 1 [E]
Figura 16 ndash Estrutura do monoacutemero de actina Encontram-se representados os subdomiacutenios 1 a 4 e a vermelho a
moleacutecula de ATP junto do respectivo local de ligaccedilatildeo (Protein Data Bank ID 1j6z)
113 Mecanismos moleculares da contracccedilatildeo muscular
A contracccedilatildeo muscular ocorre atraveacutes do deslizamento dos filamentos finos (de actina) por entre
os filamentos grossos (de miosina) conseguida pela reacccedilatildeo de hidroacutelise de ATP e regulada pela
tropomiosina e pelo complexo de troponina em resposta a impulsos nervosos Este mecanismo foi
inicialmente proposto por Huxley-Niedergerke [N] e Huxley-Hanson [O] ambos em 1954 A base
experimental de ambos os estudos foi a inexistecircncia de variaccedilatildeo do tamanho dos filamentos
durante a contracccedilatildeo A banda A apresenta sempre as mesmas dimensotildees por outro lado a banda
I a zona H diminuem consideravelmente aquando da contracccedilatildeo (Figs 12 e 17)
Figura 17 ndash Modelo da contracccedilatildeo muscular ndash deslizamento dos filamentos finos (azuis) relativamente aos
filamentos grossos (rosa) (Adaptado de Stryer 2002 [2])
13
Na ausecircncia de ATP e supondo a ausecircncia de inibiccedilatildeo da interacccedilatildeo actina-miosina as proteiacutenas
encontram-se fortemente associadas formando o complexo actomiosina (AM) (Figura 15)
Nestas condiccedilotildees as cabeccedilas de miosina formam um acircngulo de 45ordm com o eixo dos filamentos
finos de actina [P]
Na presenccedila de ATP o complexo AM dissocia-se jaacute que a afinidade do complexo M-ATP para a
actina eacute 105 vezes inferior agrave afinidade da miosina para a actina O ATP liga-se agrave miosina
ocorrendo seguidamente a reacccedilatildeo de hidroacutelise de ATP Os produtos daiacute resultantes ndash ADP e Pi -
permanecem ligados agrave miosina formando o complexo M-ADP-Pi Com a ligaccedilatildeo subsequente agrave
actina forma-se o complexo AM-ADP-Pi no qual as cabeccedilas de miosina apresentam um acircngulo
recto com o eixo do muacutesculo [P] Neste ponto daacute-se a libertaccedilatildeo progressiva de Pi e ADP Eacute
provaacutevel que a energia proveniente da hidroacutelise do ATP provoque uma alteraccedilatildeo do acircngulo de
ligaccedilatildeo das cabeccedilas de miosina ao filamento de actina passando novamente de 90ordm para 45ordm [P]
Esta alteraccedilatildeo conformacional provoca o deslizamento do filamento de actina para o centro do
sarcoacutemero cujo encurtamento promove a contracccedilatildeo das miofibrilas O processo supracitado
ocorre em cerca de 1ms[Q] e encontra-se resumido na Fig 18
Figura 18 ndash Esquema cineacutetico da hidroacutelise de ATP pela miosina (Adaptado de Sellers 1999 [A])
1131 Regulaccedilatildeo da contracccedilatildeo muscular
O mecanismo pelo qual se processa a regulaccedilatildeo muscular foi inicialmente descrito por estudos
distintos de Haselgrove Huxley e Parrey-Squire todos em 1973 [RST] Aleacutem da implicaccedilatildeo
directa da tropomiosina e do complexo de troponina a concentraccedilatildeo de Ca2+
no muacutesculo em
relaxamento eacute bastante reduzida (10-7
M) aumentando notavelmente aquando da contracccedilatildeo (10-5
-
10-3
M)
Resumidamente quando ocorre a excitaccedilatildeo do muacutesculo esqueleacutetico a despolarizaccedilatildeo da
membrana plasmaacutetica eacute difundida ateacute ao interior do tecido atraveacutes dos tuacutebulos transversais
(invaginaccedilotildees da membrana plasmaacutetica da fibra muscular) A despolarizaccedilatildeo provoca a abertura
dos canais ioacutenicos de Ca2+
do retiacuteculo sarcoplasmaacutetico adjacente Como a concentraccedilatildeo de Ca2+
no interior do organelo eacute muito elevada o caacutelcio passa raacutepida e uniformemente para o citosol
14
Assim a regulaccedilatildeo da contracccedilatildeo ocorre com coordenaccedilatildeo e rapidez na totalidade das miofibrilas
do tecido muscular (Ref)
Em repouso a troponina I encontra-se posicionada de tal forma que a proteiacutena adjacente
troponina C impede estruturalmente a ligaccedilatildeo das cabeccedilas da miosina agrave actina (acircngulo =50ordm) O
aumento da concentraccedilatildeo intracelular de Ca2+
provoca um aumento da sua disponibilidade para
ligar agrave troponina C (Fig 19)
Figura 19ndash Regulaccedilatildeo do mecanismo contraacutectil no muacutesculo esqueleacutetico pelo complexo de troponina
Como consequecircncia a troponina C altera a sua conformaccedilatildeo (=70ordm) associando-se agrave troponina I
que deixa de inibir a ligaccedilatildeo das pontes transversais A alteraccedilatildeo anterior eacute transmitida agrave
tropomiosina atraveacutes da troponina T adjacente agrave C o que provoca o deslocamento da
tropomiosina na direcccedilatildeo do interior do sulco do filamento de actina Assim eacute possiacutevel a
interacccedilatildeo eficaz entre actina e miosina A Ca2+
-ATPase do retiacuteculo sarcoplasmaacutetico provoca a
reacumulaccedilatildeo do iatildeo caacutelcio no organelo impedindo o prolongamento desnecessaacuterio da contracccedilatildeo
(Refordf)
12 Vanaacutedio
O vanaacutedio eacute um metal de transiccedilatildeo pertencente ao grupo VB da tabela perioacutedica Foi
primeiramente descoberto em 1801 por Andreacutes Manuel del Riacuteo que concluiu erradamente que era
idecircntico ao croacutemio e definitivamente por Nils Gabriel Svenson em 1831 Recentemente foram
descobertas importantes funccedilotildees bioloacutegicas deste elemento provavelmente essencial para os
humanos de que se destaca no contexto do presente relatoacuterio a possibilidade de actuar como
anaacutelogo do grupo fosfato e a inibiccedilatildeo de enzimas in vitro como eacute o caso da actividade ATPaacutesica
da miosina
Eacute um metal soacutelido agrave temperatura ambiente que se encontra amplamente distribuiacutedo nos vaacuterios
ecossistemas em dois isoacutetopos naturais ndash 50
V e 51
V ndash raramente excedendo 01 partes por biliatildeo
15
(ppb) Estaacute associado a cerca de 70 minerais diferentes na natureza (dos quais se destacam o
uracircnio cobre chumbo zinco fosfato e titacircnio ferroso) especialmente na forma de vanadato ou
pentoacutexidos de vanaacutedio O vanaacutedio eacute um dos principais constituintes da mateacuteria orgacircnica foacutessil
com especial ecircnfase para o carvatildeo oacuteleo de crude e foacutesseis carbonaacuteceos [UV] bem como em
alimentos consumidos pelo Homem (salsa pimenta preta cogumelos marisco vegetais cereais
gorduras e oacuteleos) [W] Estes constituem a maior fonte de exposiccedilatildeo ao vanaacutedio da maioria da
populaccedilatildeo humana [X]
Normalmente a entrada do vanaacutedio no organismo ocorre por inalaccedilatildeo ou em menores
quantidades por absorccedilatildeo pelo tracto gastrointestinal ou pela pele O seu armazenamento centra-
se no fiacutegado rins ceacuterebro coraccedilatildeo muacutesculos e ossos No entanto a retenccedilatildeo de vanaacutedio no
organismo humano eacute reduzida ndash em condiccedilotildees normais cerca de 100 M (em muitos tecidos a
quantidade eacute inferior a 10 ngg de tecido) [Y]
Actualmente natildeo existem provas de deficiecircncia nutricional de vanaacutedio em humanos ou de
enzimas que contenham vanaacutedio em animais superiores No entanto vaacuterios estudos relacionam a
ocorrecircncia de deficiecircncia em galinhas e ratos devido a uma alimentaccedilatildeo pobre em vanaacutedio Os
sintomas relatados incluem retardamento do crescimento malformaccedilotildees oacutesseas perturbaccedilotildees no
metabolismo lipiacutedico e estagnaccedilatildeo reprodutiva entre outros [Z AA AB] Os efeitos fisioloacutegicos
do vanaacutedio podem depender do tipo de composto ou de oligoacutemero uma vez que consoante as
suas estabilidade labilidade e nuacutemero de coordenaccedilatildeo as respostas seratildeo diferenciadas ao niacutevel
dos organismos vivos
Por outro lado a toxicidade do vanaacutedio encontra-se bem caracterizada existindo sintomas
descritos em vaacuterios mamiacuteferos (inclusive o Homem) que incluem diarreias catildeibras irritaccedilatildeo da
pele e mucosas palpitaccedilotildees cardiacuteacas anemia pneumonia bronquite perturbaccedilotildees no Sistema
Nervoso Central (SNC) entre outros (Ref) A toxicidade varia geralmente com a natureza do
composto com aumento proporcional agrave sua valecircncia Verificou-se que a forma de entrada do
vanaacutedio tem tambeacutem efeito na sua toxicidade ndash os maiores niacuteveis verificaram-se por injecccedilatildeo
intravenosa da amostra sendo progressivamente inferiores por inalaccedilatildeo e finalmente por
ingestatildeo [X AA] A ampla utilizaccedilatildeo dos compostos de vanaacutedio na induacutestria quiacutemica bem como
as elevadas quantidades de vanaacutedio libertadas atraveacutes da combustatildeo dos oacuteleos de crude
contribuem consideravelmente para o aumento da biodisponibilidade do vanaacutedio Como tal o
interesse neste elemento e nos problemas ambientais por ele causados tem vindo a aumentar
significativamente [V]
121 Propriedades quiacutemicas
16
A quiacutemica do vanaacutedio eacute extraordinariamente complexa pelo que originou vaacuterias interpretaccedilotildees
incorrectas desde a sua descoberta No entanto as suas propriedades dependem em uacuteltima
anaacutelise da configuraccedilatildeo electroacutenica do elemento Assim a variabilidade encontrada no
comportamento quiacutemico do vanaacutedio pode dever-se agrave sua configuraccedilatildeo ndash [Ar] 3d2 4s
2 ndash que
permite a formaccedilatildeo de vaacuterios estados de oxidaccedilatildeo (de +2 a +5) Em soluccedilatildeo aquosa os estados de
oxidaccedilatildeo estaacuteveis vatildeo de V(V) a V(II) Destes apenas os +3 a +5 satildeo biologicamente importantes
jaacute que o V(II) apresenta demasiada capacidade redutora para existir nos organismos vivos
conhecidos [AC]
No fluido extracelular os compostos de vanaacutedio encontram-se predominantemente na forma de
monovanadatos (V(V)) livres ou ligados agrave transferina do soro ou agrave hemoglobina (Refordf) Os iotildees de
vanadato livres satildeo transportados para o meio intracelular atraveacutes de canais ioacutenicos inespeciacuteficos
Jaacute no ambiente intracelular os compostos sofreratildeo uma bio-reduccedilatildeo atraveacutes da acccedilatildeo da
glutationa ou outros compostos com grupos ndashOH como pentoses hexoses ou etanolaminas [V] A
reoxidaccedilatildeo de vanadilo (V(IV)) intracelular a vanadato (V(V)) provocaria a acumulaccedilatildeo toacutexica
destes uacuteltimos pelo que a complexaccedilatildeo da espeacutecie reduzida o impede O transporte de vanaacutedio
para o interior da ceacutelula encontra-se na Fig 110
Figura 110 ndash Transporte de vanaacutedio para a ceacuteluka e seu subsequente metabolismo (P=Proteiacutena) (Adaptado de
Zaporowska e Scibor 1998)
1211 Vanaacutedio(IV)
A espeacutecie de vanaacutedio (IV) mais bem conhecida eacute o iatildeo vanadilo (VO2+
) espeacutecie estaacutevel a pH
aacutecido sendo a maioritaacuteria a pH 3[AD] Apresenta uma cor azulada e forma complexos penta e
hexacoordenados Forma-se rapidamente a partir de V(V) na presenccedila de um agente redutor em
soluccedilatildeo aquosa No ambiente intracelular a presenccedila de glutationa reduzida aacutecido ascoacuterbico
NAD(P)H entre outros conduzem a que o principal estado de oxidaccedilatildeo nos sistemas bioloacutegicos
seja provavelmente o +4 encontrando-se o V(IV) fortemente complexado com proteiacutenas ou
aminoaacutecidos livres citrato ATP e pirofosfato (Refordfs) Em alternativa o V(IV) pode existir
17
livremente em ambientes fortemente aacutecidos como os lisossomas jaacute que a pH superior a 3 as
soluccedilotildees de vanadilo tendem a ser oxidadas pelo ar formando-se vanadato Esta grande
capacidade redutora justifica parcialmente a utilizaccedilatildeo pouco extensiva do iatildeo VO2+
como
inibidor [W]
1212 Vanaacutedio (V)
O vanaacutedio (V) eacute a forma mais estaacutevel num intervalo de pH amplo [W] O iatildeo anioacutenico VO3-tende
a constituir complexos polinucleares pelo que as soluccedilotildees aquosas de vanadato satildeo extremamente
complexas com ocorrecircncia simultacircnea de vaacuterios equiliacutebrios de protonaccedilatildeo (pKa(vanadato)= 35
78 e 125) e oligomerizaccedilatildeo As espeacutecies presentes (H3VO4 H2VO4- HVO4
2- e VO4
3-) dependem
do pH forccedila ioacutenica e temperatura da soluccedilatildeo e da concentraccedilatildeo total do elemento
[AEAFAGAHAI]
O VO43-
- monoacutemero ortovanadato V1 ndash estaacute associado a uma grande parte da importacircncia
bioloacutegica do vanaacutedio uma vez que se trata de um anaacutelogo electroacutenico e estrutural do ortofosfato
(PO43-
) A coordenaccedilatildeo do ortovanadato pode ser tetraeacutedrica ou bipiramidal trigonal (Refordfs)
A oligomerizaccedilatildeo do vanadato monomeacuterico daacute origem ao diacutemero (V2O72-
- V2) que apresenta
tambeacutem equiliacutebrios de protonaccedilatildeo Presume-se a existecircncia de analogia do vanadato dimeacuterico com
o pirofosfato embora natildeo haja evidecircncia estrutural (Refordf) A dimerizaccedilatildeo do V2 origina o
vanadato tetrameacuterico ou metavanadato (V4O124-
- V4) que constitui a espeacutecie maioritaacuteria em
soluccedilotildees aquosas concentradas (Refordf) As estruturas do metavanadato e a sua espeacutecie protonada
satildeo ciacuteclicas [AJ] Outra espeacutecie oligomeacuterica importante eacute o vanadato pentameacuterico (V5O153-
) cuja
estrutura presumida eacute ciacuteclica [AI]
O oligoacutemero com maior importacircncia bioquiacutemica eacute o vanadato decameacuterico (V10O286-
-V10)(Refordf) a
espeacutecie predominante em soluccedilotildees de pH 2-6 quando a concentraccedilatildeo total de vanadato eacute superior
a 100 M (Ref) A estrutura do V10 natildeo difere em soluccedilatildeo ou no estado soacutelidoOs 10 aacutetomos de
vanaacutedio da espeacutecie decameacuterica estatildeo divididos em trecircs grupos identificados na figura como VA-C
consoante o seu ambiente quiacutemico circundante (Refordf) (Fig 111)
A expressatildeo ldquodecavanadatordquo refere-se a uma soluccedilatildeo aquosa na qual a espeacutecie decameacuterica se
encontra em equiliacutebrio quiacutemico com as espeacutecies V1 V2 V4 e V5 sendo que a oligomerizaccedilatildeo
progressiva do vanaacutedio eacute proporcional agrave sua concentraccedilatildeo total (Ref) Nesta soluccedilatildeo a
concentraccedilatildeo de V10 corresponde a aproximadamente 10 da concentraccedilatildeo total de vanaacutedio [AK]
18
Figura 111 ndash Estruturas esquemaacuteticas das espeacutecies de vanadato (espeacutecie monomeacuterica (V1) dimeacuterica (V2)
tetrameacuterica (V4) pentameacuterica (V5) e decameacuterica (V10) (Adaptado de Aureliano e Crans 2008)
122 Metabolismo e efeito na actividade enzimaacutetica
Apesar de natildeo terem sido ainda encontradas as provas categoacutericas das suas funccedilotildees nos seres
vivos sabe-se que o iatildeo vanadato eacute capaz de inibir uma grande quantidade de enzimas in vitro
com especial ecircnfase na actividade ATPaacutesica da miosina [AL Teresa] Como referido
anteriormente o vanadato eacute capaz de actuar em alternativa ao fosfato nas reacccedilotildees bioquiacutemicas
incluindo a sua preferecircncia de ligaccedilatildeo agraves enzimas [W]
Eacute conhecido o papel do vanaacutedio no metabolismo do caacutelcio promovendo a mineralizaccedilatildeo dos
dentes e ossos na biossiacutentese de liacutepidos (trigliceacuteridos e colesterol) com posterior inibiccedilatildeo da
acumulaccedilatildeo destes nos vasos sanguiacuteneos e no SNC na reproduccedilatildeo e desenvolvimento por
mimetizaccedilatildeo e potenciaccedilatildeo de factores de crescimento na estimulaccedilatildeo do consumo de glucose e
da siacutentese de glicogeacutenio com actuaccedilatildeo equivalente aos efeitos da insulina ao niacutevel celular na
transduccedilatildeo de sinais celulares com regulaccedilatildeo da actividade de segundos mensageiros no
transporte de iotildees e aminoaacutecidos na transcriccedilatildeo geneacutetica e siacutentese de proteiacutenas e DNA entre
outros [W Y AB]
Assim o vanaacutedio cnseguiraacute provocar inuacutemeras mudanccedilas bioquiacutemicas nas ceacutelulas Alguns
exemplos da influecircncia bioquiacutemica do vanaacutedio incluem os seus efeitos nos processos de
fosforilaccedilatildeo e desfosforilaccedilatildeo de proteiacutenas e na participaccedilatildeo na formaccedilatildeo de espeacutecies reactivas de
oxigeacutenio devido ao seu efeito proacute-oxidante sobre agentes redutores a niacutevel celular [AM]
Quanto ao seu efeito na actividade enzimaacutetica pode dividir-se em dois tipos principais ndash inibitoacuterio
e estimulante - dependendo da enzima ou do tipo de reacccedilatildeo por esta catalisada No primeiro
grupo incluem-se enzimas como as ATPases (de miosina de canais ioacutenicos e fosfoenzimas
19
transportadoras) e as fosfatases (da tirosina aacutecida e alcalina) a fosfofrutocinase adenilato ciclase
e catalase entre outras [AA] O segundo grupo inclui a maioria das cinases a glucose-6-fosfato
desidrogenase a fosfoglicerato mutase e a fosfoglicerato fosfatase (Refordf)
O efeito do vanaacutedio (V) na actividade enzimaacutetica estaacute relacionado com o seu nuacutemero de
coordenaccedilatildeo podendo actuar como estado de transiccedilatildeo ou por mimetizaccedilatildeo de cofactores
substratos (especialmente o fosfato) e interacccedilotildees proteiacutena-proteiacutena (como no caso das
polimerases e transcriptases) Pode ainda estar envolvido na formaccedilatildeo de ligaccedilotildees de hidrogeacutenio
interacccedilotildees electrostaacuteticas esteacutereas e de oxidaccedilatildeo-reduccedilatildeo Uma evidecircncia desta uacuteltima eacute a sua
capacidade de alternacircncia entre os estados de oxidaccedilatildeo +5 a +3 o que torna faacutecil a sua interacccedilatildeo
redox com enzimas contendo resiacuteduos de cisteiacutenas (Cys) ou qualquer outro composto orgacircnico
com actividade redox [AN]
13 Vanaacutedio e muacutesculo esqueleacutetico
A primeira evidecircncia da presenccedila de vanaacutedio no muacutesculo esqueleacutetico proveio de uma amostra de
ATP isolado de muacutesculo de cavalo que continha uma impureza ndash vanaacutedio ndash cuja presenccedila
provocou a inibiccedilatildeo da actividade da bomba de Na+K
+-ATPase [AO] Desde entatildeo vaacuterios
estudos tecircm sido realizados com ecircnfase na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o
tecido muscular Destes destaca-se a descoberta da interacccedilatildeo forte da espeacutecie V10 com a miosina
fosforilada [AP] e agrave sua inibiccedilatildeo da actividade ATPaacutesica da miosina[AQ AR]
No quediz respeitoao iatildeo monovanadato destaca-se a sua inibiccedilatildeoda actividade Mg2+
-ATPaacutesica
da miosina [AS] que ocorre de duas formas distintas (1) na presenccedila de concentraccedilotildees reduzidas
de V1 a inibiccedilatildeo ocorre a velocidade reduzida dependente da concentraccedilatildeo da espeacutecie
monomeacuterica (2) na presenccedila de concentraccedilotildees elevadas de V1 o niacutevel de inibiccedilatildeo aproxima-se de
100 desde que [V1] seja superior agrave concentraccedilatildeo total de locais com actividade Mg2+
-ATPaacutesica
no ensaio Esta inibiccedilatildeo eacute irreversiacutevel tanto por diaacutelise ou adiccedilatildeo de agentes redutores do
vanadato [AL]Segundo Pate et al(1994)[AT] a espeacutecie monomeacuterica podeinibir a velocidade de
contracccedilatildeo a temperatura inferior a 20ordmC perdendogradual e rapidamente essa capacidade ateacute os
50ordmC temperatura agrave qual cessa a capacidade inibitoacuteria Os resultados obtidos sugeremainda uma
interacccedilatildeo altamente cooperativa entre o iatildeo vanadato e as proteiacutenas componentes do muacutesculo
esqueleacuteticoDestaca-se tambeacutem a descoberta da inibiccedilatildeo da geraccedilatildeo de forccedila contraacutectil nas fibras
musculares por parte da espeacutecie monomeacuterica[AT]Wilson et al(1995) [AU ]determinaram que a
afinidade da espeacutecie V1 para o S1 de miosina eacute significativamente mais elevada do que a do iatildeo
Pi seu anaacutelogo Descobriram ainda que que a ligaccedilatildeo do vanadato poderaacute envolver transiccedilotildees
entre estados energeacuteticos com energia livre independente dos impedimentos esteacutereos impostos
20
pela estrutura das miofibrilas Os estudos desenvolvidos por Smith et al (1990)[AV]
determinaram queo iatildeo monovanadato natildeo apresenta um efeito pronunciado na inibiccedilatildeo da
actividadeacto-S1-miosina-ATPaacutesica excepto em concentraccedilotildees superiores a 900microM a partir das
quais existe produccedilatildeo de espeacutecies oligomeacutericas para as quais o efeito inibitoacuterio seraacute mais
elevadoA espeacutecie monomeacuterica de vanadato consegue adoptar facilmente uma geometria do tipo
bipiramidal trigonal que mimetiza o intermediaacuterio formado (MgADP
Pi)aquando da reacccedilatildeo de
hidroacutelise de ATP no local activo da miosina [AW] De facto a interacccedilatildeo do vanaacutedio com a
miosina eacute inesperadamente estaacutevel na presenccedila de ADP Esta interacccedilatildeo resulta na formaccedilatildeo de
um complexo enzimaacutetico inactivo e estaacutevel com estequiometria de 11 ADPvanadato por siacutetio
activo da proteiacutena A formaccedilatildeo do complexo estaacutevel miosina-vanadato foi proposta por Goodnoet
al (1982) [AS] Nesta ocorre um preacute-equiliacutebrio raacutepido que induz a formaccedilatildeo de um complexo
ternaacuterio reversiacutevel (MADP
V) com formaccedilatildeo lenta do isoacutemero M
ADP
V A diferenccedila entre os
compostos de vanaacutedio e de fosfato centra-se no tempo de meia vida (t12) do Pi ou do V (no
primeiro t12=12 s enquanto no segundo t12=24 h) (Refordf) A diferenccedila dever-se-aacute agrave possiacutevel
complexaccedilatildeo do vanaacutedio com grupos nucleofiacutelicos no siacutetio activo o que provocaria uma
estabilizaccedilatildeo da miosina na conformaccedilatildeo MADP
V
No que respeita aos estudos realizados com a espeacutecie tetrameacuterica Cremo et al determinou que a
cadeia pesada do S1 de miosina pode ser clivada em dois locais distintos (designados V1 e V2)
pela espeacutecie V4 apoacutes irradiaccedilatildeo com luz UV na presenccedila de concentraccedilotildees de vanadato na ordem
dos milimolar e na ausecircncia de nucleoacutetidos ou metais divalentes
Jaacute no que concerne aos estudos realizados com ecircnfase na espeacutecie decameacuterica foi sugerido
porRamos et al (2006) [AK]que a interacccedilatildeo do decavanadato com a G-actina favorecida pela
polimerizaccedilatildeo desta uacuteltima estabiliza a espeacutecie V10 e induz a inibiccedilatildeo da formaccedilatildeo de F-actina A
estabilizaccedilatildeo da espeacutecie V10 por proteiacutenas tansmembranares e do citoesqueleto pode ser a razatildeo
pela qual ocorre a toxicidade do decavanadato reportada na avaliaccedilatildeo dos efeitos do V(V) nos
sistemas bioloacutegicosTiago et al (2007) [AR] concluiram que o V10 apresenta uma clara preferecircncia
de ligaccedilatildeo ao ldquoback-doorrdquo da S1-miosina-ATPase o que se verifica apenas na estrutura
conformacional desimpedida com acesso facilitado ao local de ligaccedilatildeo do Pi Sugere tambeacutem que
o V10 actua como um ldquoback-door stoprdquo bloqueando estaabertura do fragmento de 50-kDa
necessaacuteria para a hidroacutelise do ATP--fosfato o que constitui uma explicaccedilatildeo simples do
comportamento natildeo competitivo da espeacutecie decameacuterica estimulando a utilizaccedilatildeo deste oligoacutemero
como uma feramenta da elucidaccedilatildeo das mudanccedilas conformacionais ldquoback-doorrdquo da miosina no
21
processo da contracccedilatildeo muscular(ref)Destaca-se ainda a descoberta da capacidade inibitoacuteria do
V10 em relaccedilatildeo agrave fosforilaccedilatildeo oxidativa mitocondrial [AX]
Finalmente no que concerne aos estudos realizados com a presenccedila de vaacuterias espeacutecies
oligomeacutericas Aureliano e Madeira (1994) [AY]observaram que na presenccedila de Ca2+
as espeacutecies
V4 e V10se ligam agrave Ca2+
-ATPase do retiacuteculo sarcoplasmaacutetico enquanto a espeacutecie V1 se liga apenas
na presenccedila de ATP Aparentemente o aumento da concentraccedilatildeo de Ca2+
tem o efeito contraacuterio da
inibiccedilatildeo da ATPase do retiacuteculo sarcoplasmaacutetico pelo V1 natildeo tendo no entanto efeito significativo
sobre a inibiccedilatildeo por parte da espeacutecie V10 Tiago et al (2002) [AP] concluiacuteram que em condiccedilotildees
proacuteximas agraves fisioloacutegicas a espeacutecie V10 difere das espeacutecies presentes na soluccedilatildeo de metavanadato
devido agrave sua forte interacccedilatildeo com a miosina fosforilada e agrave sua inibiccedilatildeo da actividade ATPaacutesica
da miosina Para aleacutem disso o ATP diminui a afinidade da miosina para o V4 induzindo a
interacccedilatildeo com V1 e natildeo tendo qualquer efeito no que diz respeito ao V10 Foi tambeacutem proposto
[AQ] que a ligaccedilatildeo das espeacutecies oligomeacutericas de vanadato aoS1 de miosina nos locais onde o
fosfato coordenaeacute provavelmente a causa da inibiccedilatildeo da actividade ATPaacutesica da miosina
estimulada por actina atraveacutes da prevenccedilatildeo da troca entre ATPADP sendo esta favorecida para
aniotildees de vanadato com valecircncias superiores como eacute o caso do V10
Na presenccedila de actina as velocidades de dissociaccedilatildeo do Pi e do V aumentam consideravelmente
este uacuteltimo em menor quantidade Assim a ligaccedilatildeo da actina ao complexo MADP
V e
Pi eacute
bastante fraca pelo que se sugere que o ADPV teria um efeito de relaxamento semelhante ao
ATP No entanto a capacidade do vanaacutedio em inibir a actividade ATPaacutesica da miosina e da
actomiosina eacute diferenciada uma vez que na ausecircncia de actina a inibiccedilatildeo eacute total para
concentraccedilotildees na gama dos M mas na sua presenccedila a inibiccedilatildeo eacute totalmente conseguida apenas
em concentraccedilotildees na gama dos mM (Ref) No entanto eacute necessaacuterio considerar a possiacutevel
formaccedilatildeo de espeacutecies oligomeacutericas de vanadato cuja afinidade e efeito na enzima seratildeo
diferentes Assim pode concluir-se que o efeito de inibiccedilatildeo observado para a actividade ATPaacutesica
de miosina estimulada por actina se pode dever agrave presenccedila das diferentes espeacutecies oligomeacutericas o
que foi jaacute sugerido por Aureliano (2000) [AZ] ndash as espeacutecies tetrameacuterica e decameacuterica de vanadato
interactuariam com a miosina com alteraccedilatildeo da sua interacccedilatildeo com a actina No entanto natildeo eacute
conhecida a contribuiccedilatildeo absoluta das restantes espeacutecies oligomeacutericas para a inibiccedilatildeo da ATPase
da miosina estimulada por actina (Refs)
14 Objectivos do projecto
Pretende-se com este projecto o isolamento purificaccedilatildeo e caracterizaccedilatildeo da actina do muacutesculo
22
esqueleacutetico de coelho sendo esta uacuteltima tambeacutem realizada para o sufragmento-1 de miosina
Seguidamente realizar-se-aacute a quantificaccedilatildeo e caracterizaccedilatildeo das proteiacutenas por espectroscopia UV-
Vis e SDS-PAGE e a caracterizaccedilatildeo das soluccedilotildees de vanaacutedio por UV-Vis e ressonacircncia magneacutetica
nuclear (RMN) Por uacuteltimo a interacccedilatildeo do S1 de miosina com o vanaacutedio seraacute avaliada atraveacutes
das teacutecnicas de UVVis RMN e ressonacircncia paramagneacutetica electroacutenica (RPE)
2 Discussatildeo de Resultados
21 Caracterizaccedilatildeo das proteiacutenas isoladas
211Anaacutelise do estado de pureza
O estado de pureza das soluccedilotildees de actina e de S1 foi analisado por electroforese em gel de
poliacrilamida em condiccedilotildees desnaturantes (SDS-PAGE) (Fig 22) que separa as moleacuteculas de
acordo com o seu peso molecular (PM) A concentraccedilatildeo de acrilamida escolhida (10) permite a
detecccedilatildeo simultacircnea de componentes com peso molecular inferior a 20 kDa e outros com elevado
peso molecular
Figura 21 ndash Fotografia de gel de SDS-PAGE (10 acrilamida) Poccedilo (a)
10 L do marcador de elevado peso molecular () Poccedilo (b) 20 L de [G-
actina]=1 mgmL Poccedilo (c) 20 L de [S1]=1 mgmL Aplicou-se uma
voltagem de V durante h e min
A partir da recta de calibraccedilatildeo obtida log PM = -1177 Rf + 2247 (ver ponto 2 da secccedilatildeo
Anexos) pode verificar-se a presenccedila de actina na amostra do poccedilo (b) presente numa banda
bastante intensa (PMaparente=40 kDa) bem como a presenccedila minoritaacuteria de outros componentes
desconhecidos que natildeo teratildeo sido eliminados durante o processo de extracccedilatildeo e purificaccedilatildeo e natildeo
satildeo relevantes para o estudo em questatildeo Na amostra do poccedilo (c) nota-se a presenccedila de S1
(PMaparente=92 kDa) bem como a de outros componentes cuja banda eacute ligeiramente menos intensa
Estes poderatildeo ser fragmentos proteoliacuteticos de miosina e de S1 Estaacute ainda presente uma bada de
23
maior intensidade que poderaacute ser a proteiacutena-C (PM=135 kDa) uma vez que a sua remoccedilatildeo
completa da amostra necessitaria de um passo adicional de cromatografia de troca ioacutenica [BA]
No entanto natildeo podemos confirmar que se trata desta proteiacutena ou de qualquer outra uma vez que
se encontra fora do intervalo de migraccedilatildeo dos marcadores de peso molecular
212 Determinaccedilatildeo da concentraccedilatildeo proteica
2121Subfragmento-1 de miosina
A partir da recta de calibraccedilatildeo obtida pelo meacutetodo de Bradford [BB](ver secccedilatildeo 3311 do
Procedimento Experimental) A595nm = 00546[BSA] + 00209 (ver secccedilatildeo de Anexos)foi possiacutevel
determinar um intervalo de concentraccedilatildeo de S1 de miosina compreendido entre 226 e 117(plusmn
423M) (203 e 1056 (plusmn 381 mgmL)) (n=8) A concentraccedilatildeo meacutedia de S1 de miosina apoacutes
diaacutelise contra a soluccedilatildeo tampatildeo 10 mM HEPES pH 75 25 mM KCl determinada pela leitura da
Abs280 nmfoi de 20430M (1803 mgmL)(n=7)
As amostras a quantificar atraveacutes do meacutetodo de Bradford natildeo sofreram diaacutelise o que permite
explicar a concentraccedilatildeo superior de S1 de miosina em comparaccedilatildeo com o valor obtido atraveacutes do
meacutetodo directo Esta razatildeo aliada agrave sensibilidade e rapidez superiores deste uacuteltimo fez com que
se optasse pelo meacutetodo directo para a quantificaccedilatildeo da grande maioria dos ensaios
2122Actina
A concentraccedilatildeo de actina foi determinada quando esta se encontrava na forma monomeacuterica (G-
actina) por mediccedilatildeo da leitura directa da Abs290 nm tendo-se obtido uma concentraccedilatildeo meacutedia de
691481M (290202 mgmL) (n=3) em 2 mM Tris-HCl pH 80 02 mM CaCl2 02 mM Na-
ATP Ao comprimento de onda utilizado (290 nm) absorvem fortemente apenas os resiacuteduos de
Trp pelo que a mediccedilatildeo efectuada tem menor sensibilidade do que se tivesse sido efectuada a 280
nm uma vez que a actina apresenta apenas 4 resiacuteduos deste aminoaacutecido embora tenha 16
resiacuteduos de Tyr
22 Mediccedilatildeo da actividade enzimaacutetica de S1 de miosina
221 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
As actividades ATPaacutesicas de S1 de miosina dependentes de Ca2+
e K+foram analisadas pelo
caacutelculo das velocidades de consumo inicial do fosfato inorgacircnico (Pi) Definiu-se os tempos de 5
e 3 minutos para a mediccedilatildeo da Ca2+
e da K+EDTA-ATPase respectivamente Os valores obtidos
24
a partir do protocolo presente no ponto 341 da secccedilatildeo Procedimento experimental encontram-se
descritos na Tabela 2I
Tabela 2I ndash Acrividades natildeo-fisioloacutegicas de subfragmento-1 determinadas pelo meacutetodo de Fiske-Subarow
modificado (n=2)
Proteiacutena
Actividade
ATPaacutesica natildeo
fisioloacutegica
Actividade (nmol
Piminmg proteiacutena) Actividade (s-1)
S1
Ca2+
-ATPase 1400 plusmn 30 2100 plusmn 0045
K+EDTA-ATPase 2334 plusmn 50 3500 plusmn 0075
As actividades natildeo-fisioloacutegicas reflectem o estado de oxidaccedilatildeo dos resiacuteduos expostos de Cys 707
(SH1) e 697 (SH2) Se o grupo SH1 sofrer oxidaccedilatildeo a actividade ATPaacutesica dependente de Ca2+
aumenta e a actividade dependente de K+ diminui Por outro lado se o grupo SH2 sofrer
oxidaccedilatildeo regista-se a perda completa destas actividades
De acordo com Margossian e Lowey (1982) [BA] as actividades ATPaacutesica de S1 de miosina
dependente de Ca2+
e K+EDTA da miosina satildeo respectivamente de 24 e 112 s-1
Estaacute tambeacutem
descrito que a actividade ATPaacutesica dependente de K+ eacute em meacutedia cerca de 3 vezes superior agrave
dependente de Ca2+
reflectindo a quantidade das preparaccedilotildees A razatildeo obtida entre os valores das
actividades ATPaacutesicas dependentes de K+ e Ca
2+ eacute de 166 Os valores obtidos para as actividades
(Ca2+
-ATPase=21 s-1
K+EDTA-ATPase= 35 s
-1) satildeo inferiores agrave valores obtidos por
Margossian e Lowey [BA] e Tiago [BC] com forccedila ioacutenica inferior O valor obtido para a
actividade ATPaacutesica dependente de Ca2+
eacute superior ao obtido por Polosukhina et al (2000) [BD]
cujo meio de reacccedilatildeo continha MOPS e apresentava forccedila ioacutenica inferior Os dados
supramencionados comprovam a variaccedilatildeo da actividade a ATPaacutesica natildeo fisioloacutegica em relaccedilatildeo agrave
forccedila ioacutenica temperatura e pH
222 Mediccedilatildeo da actividadeMg2+
-ATPaacutesica de S1 pelo meacutetodo de enzimas acopladas
A actividade de Mg2+
-ATPase eacute a uacutenica que possui significado fisioloacutegico sendo as restantes
bastante uacuteteis apenas para indicaccedilatildeo da qualidade de preparaccedilatildeo do S1[BA] A 0575 M KCl a
actina e o S1 formam uma soluccedilatildeo viscosa e tuacuterbida Apoacutes a adiccedilatildeo de MgATP o complexo acto-
S1 dissocia-se e a turbidez da soluccedilatildeo diminui Se a forccedila ioacutenica for reduzida a dissociaccedilatildeo do
complexo actoS1 que se encontra na forma de suspensatildeo eacute induzida O MgATP eacute rapidamente
hidrolisado diminuindo a sua concentraccedilatildeo no meio reaccional Nestas condiccedilotildees haacute
reassociaccedilatildeo e subsequente contracccedilatildeo dos filamentos de actina e de S1 com formaccedilatildeo de
25
precipitado O meacutetodo descrito traduz de forma bastante elementar o fenoacutemeno da contracccedilatildeo in
vivo e elimina o problema anterior da solubilizaccedilatildeo ineficiente da miosina de modo a obter uma
mediccedilatildeo exacta da actividade [A] A utilizaccedilatildeo de forccedilas ioacutenicas reduzidas aumenta a afinidade da
actina para o S1 permitindo contornar a extrema dificuldade em obter a concentraccedilatildeo proteica dos
muacutesculos (cerca de 100 mgmL do complexo actomiosina) As actividades Mg2+
-ATPaacutesicas de S1
de miosina foram determinadas a partir do declive da variaccedilatildeo da Abs340 relativo agrave oxidaccedilatildeo de
NADH em funccedilatildeo do tempo como eacute descrito no ponto 342 do Procedimento Experimental Os
valores obtidos para a actividade Mg2+
-ATPaacutesica encontram-se apresentados na Tabela 2II
Tabela 2II ndash Actividade Mg2+
-ATPaacutesica de subfragmento-1 de miosina determinadas pelo meacutetodo das enzimas
acopladas
Actividade (nmol
ATPmin mg(S1)) Actividade (s
-1)
1097 plusmn 269 164 plusmn 04
Ramos (2005) [J] obteve o valor de 01 s-1
para a actividade Mg2+
-ATPaacutesica de S1 de miosina
com forccedila ioacutenica intermeacutedia Tiago (2005) [BC] obteve o valor de 011 s-1 em condiccedilotildees de forccedila
ioacutenica reduzida em relaccedilatildeo agrave utilizada no presente relatoacuterio
23 Preparaccedilatildeo e caracterizaccedilatildeo das soluccedilotildees de vanaacutedio
231 Anaacutelise da composiccedilatildeo das soluccedilotildees de vanadato por espectroscopia de 51
V-RMN
efeito da concentraccedilatildeo da espeacutecie
A diluiccedilatildeo das soluccedilotildees stock de meta- e decavanadato iraacute determinar fortemente a composiccedilatildeo
em soluccedilatildeo dos diferentes oligoacutemeros de vanadato (ref) No caso do decavanadato o periacuteodo de
tempo apoacutes a diluiccedilatildeo iraacute tambeacutem ser um factor importante a considerar devido agrave decomposiccedilatildeo
da espeacutecie V10(ref)Torna-se assim fundamental determinar a composiccedilatildeo quiacutemica das soluccedilotildees
no meio de ensaio utilizado (HEPES 10 mM KCl 25 mM) de modo a correlacionaacute-la
correctamente com os possiacuteveis efeitos promovidos pelas diferentes espeacutecies de vanadato no
sistema em estudo
Aleacutem disso deve ser dado ecircnfase agraves importantes interacccedilotildees que ocorrem entre o vanaacutedio e os
tampotildees escolhidos O tampatildeo com menor nuacutemero e magnitude de interacccedilatildeo eacute o HEPES [AD]
seguido do Tris sendo este uacuteltimo especialmente aconselhado em concentraccedilotildees baixas para casos
de incompatibilidade enzimaacutetica ou quiacutemica redox No caso especial da reacccedilatildeo redox
NAD(P)HNAD(P) o tampatildeo adequado eacute o imidazole de modo a natildeo haver reacccedilatildeo com o
26
vanadato Os tampotildees a evitar em qualquer situaccedilatildeo incluem a trietanolamina fosfato citrato e
glicina entre outros [AD] uma vez que todos interactuam fortemente com V(V) com formaccedilatildeo
de complexos estaacuteveis
Assim procedeu-se agrave anaacutelise da composiccedilatildeo das soluccedilotildees de meta- e decavanadato apoacutes diluiccedilatildeo
das soluccedilotildees stock (50 mM em vanadato total) no meio de reacccedilatildeo (tampatildeo D) atraveacutes da
espectroscopia de ressonacircncia magneacutetica nuclear de 51
V Nas figuras x1 e x2 apresentam-se os
espectros obtidos para as soluccedilotildees de deca- e metavanadato num intervalo de concentraccedilotildees de 1 a
10 mM A Tabela seguinte resume os paracircmetros espectroscoacutepicos obtidos para cada espeacutecie
presente nas soluccedilotildees analisadas
Figura 22ndash Espectros de RMN de 51
V de
concentraccedilotildees crescentes das espeacutecies oligomeacutericas
de vanadato obtidos apoacutes diluiccedilatildeo da soluccedilatildeo stock
de decavanadato em 10 mM HEPES pH 70 25 mM
KCl para concentraccedilotildees finais de A) 1 mM B)2
mM C) 4 mM D) 8 mM E) 10 mM de
decavanadato total V1refere-se agraves espeacutecies
monomeacutericade vanadato V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2IIIndash Desvios quiacutemicos meacutedios (part) das ressonacircncias das espeacutecies decameacuterica (V10) monomeacuterica (V1) e
tetrameacuterica (V4) presentes nos espectros da soluccedilatildeo de decavanadatoLargura a meia altura (∆12) da espeacutecie V10A
Paracircmetros V10A V10B V10C V1 V4
part (ppm) -5162 -5003 -4255 -5616 -5748
∆12 (Hz) 175 nd nd nd nd
Figura 23 ndash Distribuiccedilatildeo graacutefica das
espeacutecies oligomeacutericas de vanadato vs a
concentraccedilatildeo de decavanadato total As
concentraccedilotildees de cada espeacutecie foram
obtidas segundo a formula descrita na
secccedilatildeo 35 do Procedimento
Experimental (aacutereas dos sinais de 51
V-
RMN observadas na figura x1)V10 V1 e
V4 correspondem agraves espeacutecie monomeacuterica
dimeacuterica e tetrameacuterica respectivamente [Esp
eacuteci
e o
ligo
meacute
rica
de
van
aacutedio
] (m
M)
[Decavanadato]total (mM)
V10
V1
V4
27
Nas soluccedilotildees de decavanadato as espeacutecies monomeacuterica e decameacuterica de vanadato satildeo as uacutenicas
presentes para concentraccedilotildees de decavanadato inferiores a 8 mM Ambas as espeacutecies aumentam
linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado em [AY] e
em (Teresa) quando a concentraccedilatildeo de V1 eacute igual ou superior a 200 microM inicia-se a sua
oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica No entanto nas condiccedilotildees de realizaccedilatildeo dos
ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a espeacutecie tetrameacuterica neste
estudo em concentraccedilotildees inferiores a 8 mM tendo o mesmo sido observado por Ramoset al
(2006) [AK] Os resultados obtidos mostram claramente o equiliacutebrio entre a espeacutecie decameacuterica e
a monomeacuterica e que o aparecimento de outras espeacutecies oligomeacutericas se encontra dependente da
concentraccedilatildeo desta uacuteltima Apesar de natildeo ter sido realizado o ensaio com 5 mM de decavanadato
total no presente relatoacuterio estaacute reportado em Ramos et al (2006) [AK] que as concentraccedilotildees da
espeacutecie decameacuterica e monomeacuterica satildeo respectivamente de 500 microM e 200 microM Assim pode inferir-
se que para concentraccedilotildees inferiores a 5 mM e intervalos de tempo de cerca de 20 minutos (que
inclui o tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas
as espeacutecies decameacuterica e monomeacuterica de vanadato
Figura 24 ndash Espectros de RMN de 51
V de concentraccedilotildees
crescentes das espeacutecies oligomeacutericas de vanadato obtidos
apoacutes diluiccedilatildeo da soluccedilatildeo stock de metavanadato em 10 mM
HEPES pH 70 25 mM KCl para concentraccedilotildees finais de A)
1 mM B) 2 mM C) 4 mM D) 8 mM E) 10 mM de
metavanadato total V1 refere-se agraves espeacutecies monomeacuterica de
vanadatoV1 e V2 correspondem respectivamente agrave espeacutecie
monomeacuterica e dimeacuterica
Figura 25 ndash Distribuiccedilatildeo graacutefica das espeacutecies
oligomeacutericas de vanadato vs a concentraccedilatildeo de
metavanadato total As concentraccedilotildees de cada
espeacutecie foram obtidas segundo a formula descrita
na secccedilatildeo 35 do Procedimento Experimental
(aacutereas dos sinais de 51
V-RMN observadas na
figura x1)V10 V1 e V4 correspondem agraves espeacutecie
monomeacuterica dimeacuterica e tetrameacuterica
respectivamente
Esp
eacuteci
e o
ligo
meacute
rica
de
va
nad
ato
(m
M)
[Metavanadato]total (mM)
V1
V2
V4
V5
28
Tabela 2IVndash Desvios quiacutemicos meacutedios (part)das ressonacircncias das espeacutecies tetrameacuterica (V4) monomeacuterica (V1) e
dimeacuterica (V2) presentes no espectro da soluccedilatildeo de metavanadato (total) 10 mM
Paracircmetros V1 V2 V4 V5
part (ppm) -5615 -5748 -5789 -5869
Nas soluccedilotildees de metavanadato as espeacutecies monomeacuterica dimeacutericae tetrameacuterica de vanadato estatildeo
presentes para concentraccedilotildees de metavanadato inferiores a 4 mM Todas estas as espeacutecies
aumentam linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado
em (Aureliano e Madeira 1998) e em (Tiago et al 2004) quando a concentraccedilatildeo de V1 eacute igual ou
superior a 200 microM inicia-se a sua oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica Nas
condiccedilotildees de realizaccedilatildeo dos ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a
espeacutecie pentameacuterica neste estudo em concentraccedilotildees inferiores a 4 mM tendo o mesmo sido
observado por Ramos 2006 Os resultados obtidos mostram claramente o equiliacutebrio entre a
espeacutecie V4 e as espeacuteciesV1 e V2 no qual o aparecimento de outras espeacutecies oligomeacutericas se
encontra dependente da concentraccedilatildeo da espeacutecie V1 Por uacuteltimo pode inferir-se que para
concentraccedilotildees inferiores a 2 mM e intervalos de tempo de cerca de 20 minutos (que inclui o
tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas as
espeacutecies V1 V2 e V4 com concentraccedilotildees respectivas de 0796 0192 e 0196 mM
232Estabilidade das soluccedilotildees de decavanadato anaacutelise por espectroscopia de absorccedilatildeo
UVVis
Caracterizou-se a estabilidade da espeacutecie V10 numa soluccedilatildeo com 1 mM decavanadato total em
tampatildeo D atraveacutes da leitura da Abs400 Obteve-se uma cineacutetica de dissociaccedilatildeo de 1ordf ordem tal
como se pode verificar pela linearidade do graacutefico de ln(Abs400)vs o tempo de aquisiccedilatildeo espectral
(segundos) (Fig26 )
Figura 26ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do tempo
(em segundos) para uma soluccedilatildeo de decavanadato total de 1
mM em tampatildeo D a 25ordmC (n=2)
Sendo uma reacccedilatildeo de primeira ordem pode ser descrita pela equaccedilatildeo cujo
declive eacute ndashK (constante de dissociaccedilatildeo) e a intersecccedilatildeo no eixo das ordenadas corresponde
Ln
(Ab
s 400 n
m)
tempo (s)
29
aln(A0) Numa reacccedilatildeo de 1ordf ordem o tempo de meia-vida eacute independente da concentraccedilatildeo inicial
do reagente sendo descrito pela equaccedilatildeo [2] Deste modo a partir da recta
Ln(Abs400 nm) = -436E-5
K - 105 obteve-se um tempo de meia-vida de 4490 plusmn 0823 horas para
as condiccedilotildees supramencionadas
Apoacutes a adiccedilatildeo de S1 8 microM ao meio reaccional verificou-se que a reacccedilatildeo de desoligomerizaccedilatildeo
da espeacutecie decameacuterica de vanadato seguia igualmente uma cineacutetica de 1ordf ordem tal como se pode
verificar pela linearidade do graacutefico de ln(Abs400) versus o tempo de aquisiccedilatildeo espectral
(segundos) (Fig27)
Figura 27ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do
tempo (em segundos) para uma soluccedilatildeo de decavanadato
total de 1 mM em tampatildeo D a 25ordmC na presenccedila de 8
microM (n= 2)
A partir da equaccedilatildeo da recta Ln(Abs400 nm) = -7520E-5
K - 0613determinou-se o valor de K para
a espeacutecie decameacuterica de onde se obteve um tempo de meia-vida de 2430 plusmn 0191 horas para as
condiccedilotildees supramencionadas
Tanto este valor como o valor registado na ausecircncia de S1 satildeo substancialmente superiores agrave
duraccedilatildeo dos ensaios de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina descritos no presente
relatoacuterio
24 Interacccedilatildeo de S1 com vanaacutedio(V)
241Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
De modo a analisar qual o efeito da adiccedilatildeo de S1 de miosina (tampatildeo D) produzia nos sinais de
51V-NMR titulou-se uma soluccedilatildeo de 5mM de decavanadato total com concentraccedilotildees crescentes
de S1 (5 10 15 e 20 microM) Os espectros de 51
V-RMN encontram-se apresentados na Fig 2
No graacutefico da figura 2 Apresenta-se a concentraccedilatildeo de S1 em funccedilatildeo do factor de alargamento
corrigido dos espectros acima referidos
Ln
(Ab
s (4
00
nm
))
tempo (s)
30
Figura 28 ndash Graacutefico de
representaccedilatildeo da
concentraccedilatildeo de S1 em
funccedilatildeo do factor de
alargamento A
concentraccedilatildeo de
decavanadato total foi de 5
mM e as concentraccedilotildees
utilizadas de S1 foram de 0
5 10 e 15 microM
Verifica-se que o factor de alargamento aumenta linearmente em funccedilatildeo da concentraccedilatildeo de S1 no
ensaio o que indica a existecircncia de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina Observou-
se ainda que a adiccedilatildeo de S1 ao meio reaccional natildeo provocou qualquer alteraccedilatildeo dos desvios
quiacutemicos dos sinais de 51
V-RMN
Realizaram-se ensaios na presenccedila de ATP e de S1 com intuito de verificar o efeito do ATP na
interacccedilatildeo do S1 de miosina com o decavanadato
Figura 29 - Espectros de RMN de 51
V contendo 2
mM Decavanadato (A) (controlo) (B) na presenccedila
de 2 mM de ATP (C) na presenccedila de 2 mM de ATP
e 8 microM de S1 de miosina V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2Vndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies decameacuterica (V10) e monomeacuterica (V1) presentes nos
espectros da soluccedilatildeo de decavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V10A V10B V10C V1
part (ppm) -5161 -5002 -4254 -5614
Observa-se que a presenccedila de ATP (espectro B) natildeo provoca qualquer alteraccedilatildeo visiacutevel em
relaccedilatildeo ao espectro que contem apenas soluccedilatildeo de decavanadato Jaacute a presenccedila simultacircnea de ATP
e S1 provoca um alargamento da ∆12 do sinal do V10A de 1736 Hz (espectro (A)) para 2366 Hz
(espectro (C) Fig 29) Tendo em conta que a concentraccedilatildeo de S1 apresenta uma relaccedilatildeo linear
com o factor de alargamento converteu-se o valor da ∆12 e comparou-se com o valor esperado agrave
F = 00591[S1] - 00157 Rsup2 = 0998
Fact
or
de
ala
rgam
en
to
[S1] (microM)
31
concentraccedilatildeo de S1 utilizada O factor de alargamento obtido para 8 microM ndash 0376 ndash eacute inferior ao
valor esperado agrave mesma concentraccedilatildeo de S1 obtido atraveacutes da equaccedilatildeo da recta apresentada na
Fig 28 ndash 0457 ndash o que indica que o ATP poderaacute exercer um efeito de protecccedilatildeo do S1
diminuindo a sua interacccedilatildeo com a espeacutecieV10
Por uacuteltimo realizaram-se ensaios com metavanadato na presenccedila de 8 microM S1 de modo a verificar
se as espeacutecies oligomeacutericas contidas nesta soluccedilatildeo interagiam com a proteiacutena em estudo
Figura 210 - Espectros de RMN de 51
V contendo 2
mM metavanadato (A) (controlo) (B) na presenccedila
de 8 microM de S1 de miosina V1 V2 e V4
correspondem respectivamente aacutes espeacutecies
monomeacuterica dimeacuterica e tetrameacuterica
Tabela 2VIndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies monomeacuterica (V1) dimeacuterica (V2) e tetrameacuterica (V4)
presentes nos espectros da soluccedilatildeo de metavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V1 V2 V4
part (ppm) -5613 -5747 -5789
Como eacute possiacutevel observar na Fig 210 ocorre uma ligeiro alargamento dos sinal de V4 bem
como uma perda da intensidade pelo que se conclui quedas espeacutecies presentes na soluccedilatildeo de
metavanadato (V1 V2 e V4) apenas a espeacutecie tetrameacuterica interage com o S1 de miosina o que
estaacute de acordo com o reportado por Tiago et al 2002
242 Actividade Mg2+
-ATPaacutesica de S1 de miosina na presenccedila de decanadato
Com o intuito de avaliar o efeito do decavanadato na actividade Mg2+
-ATPaacutesica do S1 de
miosina monitorizou-se a reacccedilatildeo de oxidaccedilatildeo do NADH atraveacutes da leitura de Abs340 na
presenccedila de 005 mgmL de S1 de miosina e no caso da actividade estimulada 2microM de actina em
3125 mM Tris-HCl pH 71 e 375 mM MgCl2 pelo meacutetodo de enzimas acopladas (ver seccccedilatildeo xx
dos MM)Para tal realizaram-se ensaios com vaacuterias concentraccedilotildees de decavanadato total (10 50
100 250 500 e 1000) de modo a determinar qual o seu efeito na actividade MgATPaacutesica de S1
de miosina na ausecircncia e na presenccedila de um dos seus ligandos naturais Os ensaios foram
realizados agrave temperatura ambiente com incubaccedilatildeo preacutevia a 25ordm C durante 20 minutos
32
Figura 211 ndash Representaccedilatildeo
graacutefica da Actividade Mg2+
-
ATPaacutesica de S1 () (os ajustes
teoacutericos estatildeo representado a por
uma linha os resultados
experimentais estatildeo representados
por triacircngulos (Actividade
estimulada) e cruzes (Actividade
natildeo estimulada)) vs
[Decavanadato]total medidas pelo
meacutetodo das enzimas acopladas
(n=3) Os dados foram ajustados ao modelo de decaimento exponencial usando o programa GraphPad Prism para a
determinaccedilatildeo dos valores de IC50 (na ausecircncia de actina IC50= 266 plusmn 07 M decavanadato na presenccedila de actina
IC50= 28 plusmn 1 M decavanadato
A partir do graacutefico presente na Fig 29 verifica-se que o ajuste dos dados experimentais foi bem
sucedido A inibiccedilatildeo da actividade Mg2+
-ATPaacutesica do S1 de miosina eacute afectada em menor
amplitude na presenccedila de actina (60 de inibiccedilatildeo da actividade) do que na sua ausecircncia
(aproximadamente 80 de inibiccedilatildeo de actividade) Devido a esta diferenccedila pode sugerir-se que a
actina iraacute impedir parcialmente a ligaccedilatildeo do decavanadatoao S1 demiosina atraveacutes de
impedimentos esteacutereos ou interacccedilotildees electrostaacuteticas por exemplo A teacutecnica utilizada natildeo nos
permite concluir acerca de que regiatildeo(otildees) ou resiacuteduo(s) da actina participaratildeo na preservaccedilatildeo
parcial da actividade Mg2+
-ATPaacutesica
Por outro lado os valores de IC50 dos dois tipos de actividades natildeo apresentam diferenccedilas
significativas tendo-se obtido 266 plusmn 07 e 28 plusmn 1M decavanadato na ausecircncia e na presenccedila de
actina respectivamente o que corresponde a uma concentraccedilatildeo de cerca de 27 M de V10 O
meio de reacccedilatildeo utilizado nestes ensaios foi 10 mM HEPES e 25 mM KCl com 2 mM ATP Em
(Tiago et al 2004) o valor de IC50 para a actividade Mg2+-ATPaacutesica na ausecircncia de actina foi de
125 plusmn 025 microM enquanto o valor da actividade estimulada por actina foi de 136 plusmn 14 microM O
meio de reacccedilatildeo utilizado nos ensaios foi 20 mM HEPES 50 mM KCl e 2 mM ATP o que resulta
precisamente no dobro da forccedila ioacutenica a que foi realizado o ensaio do presente relatoacuterio
(exceptuando a contribuiccedilatildeo dos produtos da hidroacutelise de ATP uma vez que este meacutetodo
pressupotildee a ocorrecircncia de um ciclo de reacccedilotildees de hidroacutelise e siacutentese deste composto) Ora tendo
sido anteriormente referido que a afinidade do S1 de miosina para com a espeacutecie V10 diminui com
Act
ivid
ade
Mg
2+-A
TP
aacutesic
a d
e S
1 d
e
Mio
sin
a
[Decavanadato] (microM) Actividade Estimulada ()
Actividade Natildeo Estimulada ()
Ajuste Actividade Estimulada
Ajuste Actividade Natildeo Estimulada
33
o aumento da forccedila ioacutenica (Tiago et al 2004 tal poderaacute explicar a diferenccedila obtida entre estes
dois conjuntos de valores
243Quantificaccedilatildeo de cisteiacutenas reduzidas pelo meacutetodo de Ellman
A interacccedilatildeo do vanaacutedio com alguns tipos de aminoaacutecidos sensiacuteveis agrave oxidaccedilatildeo pode fornecer-nos
informaccedilatildeo bastante uacutetil em relaccedilatildeo agraves alteraccedilotildees estruturais que conduzem a uma diminuiccedilatildeo da
actividade fisioloacutegica do S1 de miosina Os estudos realizados por Chasteen 1983 concluiacuteram
que a reduccedilatildeo de vanadato (V) a vanadilo (IV) ocorre na presenccedila de agentes redutores comuns no
ambiente celular de que se destaca o aminoaacutecido cisteiacutena Esta eacute tambeacutem um alvo de
complexaccedilatildeo por parte do iatildeo VO2+
Jaacute Ramos et al (2009) demonstrou que a incubaccedilatildeo de
actina com a soluccedilatildeo de decavanadato induz a oxidaccedilatildeo de resiacuteduos de cisteiacutena e agrave formaccedilatildeo do
oxidovanaacutedio (IV) Este grupo de investigadores descobriu ainda que apenas a espeacutecie V10 induz a
oxidaccedilatildeo das cisteiacutenas ldquoraacutepidasrdquo da actina atraveacutes do processo proposto que eacute parcialmente
responsaacutevel pela inibiccedilatildeo jaacute descrita da actividade de ATPase do S1 de miosina estimulada por
actina quando exposta agrave soluccedilatildeo de decavanadato Esta inibiccedilatildeo apresenta-se afectada com a
presenccedila de ATP uma vez que os resiacuteduos de cisteiacutena permaneceram intactos nos ensaios
realizados com este ligando Jaacute Crans et al (2004) concluiu que o siacutetio activo de muitas fosfatases
(que conteacutem grupos tiol) pode formar um equiliacutebrio redox com o iatildeo vanadato (V) de forma natildeo
reversiacutevel
Os resultados obtidos por Sakurai e Shiromura (1980) sugeriram que o vanaacutedio (V) sofre reduccedilatildeo
a vanaacutedio (IV) sem dependecircncia do valor de pH do ensaio Registaram ainda uma estequiometria
de 31 na reacccedilatildeo do vanadato (V) com os resiacuteduos de Cys enquanto a estequiometria dos ensaios
com o vanadilo (IV) foi de 21
Berlett e Stadtman (1997) observaram que os tipos de aminoaacutecidos mais facilmente oxidaacuteveis
englobam entre outros os resiacuteduos de Cys Trp e MetAssim analisou-se o estado redox dos
grupos tiol dos resiacuteduos de Cys do S1 na presenccedila do decavanadato atraveacutes da monitorizaccedilatildeo da
Abs412 nm na presenccedila de 1 mgmL de S1 de miosina em tampatildeo 10 mM HEPES 25 mM KCl
variando a concentraccedilatildeo da soluccedilatildeo de decavanadato de modo a determinar que tipo de alteraccedilotildees
estruturais ocorrem na sua presenccedila no que diz respeito agrave diminuiccedilatildeo da actividade ATPaacutesica de
S1 na presenccedila de decavanadato
Os resultados referentes agrave quantificaccedilatildeo das cisteiacutenas reduzidas apoacutes o tratamento de S1 com
diferentes concentraccedilotildees de decavanadato em tampatildeo 10 mM HEPES pH 75 e 25 mM KCl a
25ordmC durante 20 minutos encontram-se descritos na Tabela 2VII
Tabela 2VII ndash Cisteiacutenas reduzidas totais e ldquoraacutepidas de S1 (1 mgmL) apoacutes tratamento com decavanadato
34
(010501002505001000 uM) em 10 mM HEPES 25 mM KCl pH 7 a 25ordmC durante 20 minutos A detecccedilatildeo
espectrofotomeacutetrica foi feita com DTNB (e 412=8400 uM-1 cm-1) (n variaacutevel entre 1 e 8)
Amostras Cys raacutepidas Cys totais
11 M S1 (controlo) 22 plusmn 04 106 plusmn 14
11 M S1 + 10 M Decavanadato 22 plusmn 05 95 plusmn 08
11 M S1 + 50 uM Decavanadato 20 plusmn 02 98 plusmn 14
11 M S1 + 100 uM Decavanadato 17 plusmn 03 97 plusmn14
11 M S1 + 250 uM Decavanadato 26 plusmn 02 98 plusmn 08
11 M S1 + 500 uM Decavanadato 21 plusmn 04 90 plusmn 06
11 M S1 + 1000 uM Decavanadato 24 plusmn 0 86 plusmn 08
A partir dos resultados obtidos pode sugerir-se que o S1 possui 11 resiacuteduos de Cys (controlo) o
que estaacute de acordo com o anteriormente descrito Verifica-se que o tratamento da proteiacutena com
concentraccedilotildees crescentes de decavanadato provoca a oxidaccedilatildeo de dois resiacuteduos de Cys do S1 de
miosina natildeo expostos ao solvente No entanto a teacutecnica utilizada natildeo nos permite identificar
quais
Tendo em consideraccedilatildeo que o valor de IC50 obtido para a reacccedilatildeo de inibiccedilatildeo daactividade Mg2+-
ATPaacutesica de S1 na presenccedila de decavanadato eacute de cerca de 27 microM pode sugerir-se que este seraacute o
valor ao qual ocorreraacute a oxidaccedilatildeo do primeiro resiacuteduo de Cys o que poderaacute contribuir para a
diminuiccedilatildeo da actividade de S1 observada
25Interacccedilatildeo de S1 com vanaacutedio(IV)
251Anaacutelise por espectroscopia de Ressonacircncia Paramagneacutetica Electroacutenica
A acumulaccedilatildeo intracelular de oxovanaacutedio (IV) in vivo constitui uma das principais formas de
apresentaccedilatildeo de vanaacutedio no organismo Deste modo torna-se bastante interessante analisar a
interacccedilatildeo entre este iatildeo e o S1 de miosina atraveacutes da teacutecnica de espectroscopia de Ressonacircncia
Paramagneacutetica Electroacutenica o meacutetodo de eleiccedilatildeo do estudo do iatildeo vanadilo e dos complexos por si
formados O electratildeo desemparelhado encontra-se fortemente acoplado ao nuacutecleo 51
V o que
35
resulta num espectro de RPE consistindo de 8 desdobramentos na direcccedilatildeo paralela e outros tantos
na direcccedilatildeo perpendicular (Chasteen 1983)
Os estudos realizados por Ahmed et al (1987) centraram-se na interacccedilatildeo do vanadilo com
proteiacutenas como eacute o caso da insulina transferrina entre outros obtendo um espectro caracterizado
por 8 linhas amplamente espaccediladas (na direcccedilatildeo paralela) Jaacute Ramos et al 2009 concluiu que a
exposiccedilatildeo da soluccedilatildeo de decavanadato agraves formas globular e filamentosa de actina produz um
espectro tiacutepico do iatildeo oxidovanaacutedio (IV) indicando a reduccedilatildeo do vanadato
Para aferir qual a concentraccedilatildeo de S1 que permitia obter uma boa razatildeo sinalruiacutedo titulou-se uma
soluccedilatildeo de sulfato de vanadilo 15 microM ([VOSO4]stock = 5 mM) com concentraccedilotildees crescentes de
proteiacutena (0 5 10 15 e 20 microM) em 10 mM HEPES 25 mM KCl A existecircncia de uma correlaccedilatildeo
linear entre a concentraccedilatildeo de proteiacutena e a intensidade dos sinais de RPE correspondente agrave banda
-12 perpendicular encontra-se representada na Fig 210
Figura 2 12ndash Representaccedilatildeo graacutefica da variaccedilatildeo da intensidade do sinal de RPE vs a concentraccedilatildeo de S1 utilizada
Apoacutes verificar que com uma concentraccedilatildeo de S1 igual a 20 microM se obtinham sinais com uma razatildeo
sinalruiacutedo adequada ao prosseguimento dos estudos sobre a interacccedilatildeo V(IV)-S1 de miosina
titulou-se uma soluccedilatildeo de 20 microM S1 com 0 25 5 10 15 25 50 75 100 150 200 500 e 1000
microM VOSO4
Traccedilou-se o graacutefico da intensidade da banda -12 perpendicular do sinal de EPR em funccedilatildeo da
concentraccedilatildeo de VOSO4 tendo-se determinado uma constante de dissociaccedilatildeo (KD) do complexo
S1-vanadilo de 8748 plusmn 2621 microM V(IV)
y = 016786x + 122435
Rsup2 = 092992759
Inte
nsi
dad
e d
o s
inal
de
RP
E
[S1] (microM)
36
Figura 213 ndash Representaccedilatildeo graacutefica da variaccedilatildeo do sinal de RPE vs a concentraccedilatildeo de vanadilo utilizada (os valores
experimentais estatildeo representados por losangos o seu ajuste estaacute representado pela linha)
A simulaccedilatildeo do espectro de EPR foi efectuada a partir do espectro obtido para uma concentraccedilatildeo
de 1 mM VOSO4 apoacutes adiccedilatildeo de 20 microM S1 de miosina tendo-se obtido os valores da constante de
acoplamento hiperfina (A=5016) e do factor g (g0=195)
Figura 214 ndash Representaccedilatildeo graacutefica do espectro de EPR obtido com 1000 microM de vanadilo e 20 microM de S1 (a cinzento
escuro) A simulaccedilatildeo do espectro encontra-se a cinzento claro
Por comparaccedilatildeo destes com os valores presentes na tabela de correspondecircncia xxx eacute possiacutevel
predizer quais os ligandos provaacuteveis do vanadilo na presenccedila de proteiacutena Assim concluiu-se que
o vanadilo estaacute coordenado com duas moleacuteculas de aacutegua um grupo hidroxilo e um grupo R=NR
O grupo hidroxilo pode indicar os resiacuteduos de aminoaacutecidos Tyr Treonina e Serina enquanto o
grupo R=NR poderaacute indicar a presenccedila de Arginina e Histidina como ligandos do vanadilo
Inte
nsi
dad
e d
os
sinai
s d
e R
PE
[Vanadilo] (microM)
Ajuste dos dados exp Dados experimentais
Campo magneacutetico (G)
Espectro Experimental Ajuste
37
3 Procedimento Experimental
31 Reagentes
Todos os reagentes utilizados satildeo de elevado grau de pureza (analiacutetico) Apresentam-se as suas
foacutermulas quiacutemicas ou abreviaturas quando necessaacuterio acrescentou-se a sua descriccedilatildeo Entre
parecircntesis colocou-se as empresas que os comercializam
Aacutecido aceacutetico glacial 100 (Riedel) Acrilamida 99 (Merck) Azul de bromofenol (Riedel-de-
Haumlen) -Mercaptoetanol (Sigma) Bissulfito de soacutedio (Sigma)Bradford (reagente de)
(BioRad)BSA (albumina de soro bovino Sigma-Aldrich) CaCl22H2O (cloreto de caacutelcio
dihidratado Fluka) CH3CH2OH 96 (etanol Panreac) Coomassie Blue R250 (Merck) D2O
(oacutexido de deuteacuterio SDS) DTNB (55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico) Aldrich) EDTA (aacutecido
etilenodiaminatetraaceacutetico Merck) Fosfoenolpiruvato 98 (Fluka) Glicerol (Sigma) Glicina
985 (Panreac) HCl 37 (Riedel-de Haumlen) H2SO4 96 (Pronalab) HEPES 99 (aacutecido 4-(2-
hidroxiletil)-1-piperazineetanosulfoacutenico Sigma) KH2PO4 99 (hidrogenofosfato de potaacutessio)
MgATP (Sigma) Na
ATP (Sigma) Na2HPO4 99 (hidrogenofosfato de soacutedio Riedel-de Haumlen)
NaH2PO4 (fosfato de soacutedio ) NaOH 986 (hidroacutexido de soacutedio Pronalab) KCl 99 (Cloreto de
potaacutessio Panreac) L-Cys 995 (enantioacutemero levoacutegiro do resiacuteduo de Cisteiacutena Fluka)Metanol
99 (Merck) Molibdato de amoacutenio (Sigma)NNrsquo-metil-bis-acrilamida 98 (Merck) NH4VO3
99 (metavanadato de amoacutenia Sigma) PSA 10 (persulfato de amoacutenio Merck) SDS
(dodecilo sulfato de soacutedio Sigma) Sulfito de soacutedio (Sigma) VOSO4 (sulfato de vanadilo
Aldrich) TEMED 99 (Tetrametiletilenodiamina Merck) Tris (2-amino-2-hidroximetil-13-
propanodiol Sigma)
32 Meacutetodos
321 Isolamento e purificaccedilatildeo das proteiacutenas de muacutesculo esqueleacutetico de coelho
3211 Purificaccedilatildeo da miosina preparaccedilatildeo de Subfragmento 1 por digestatildeo quimotriacutepsica
da miosina
O subfragmento-1 de miosina-II (S1) foi gentilmente cedido pelo Prof Doutor Manuel Aureliano
do Departamento de Quiacutemica Bioquiacutemica e Farmaacutecia da Faculdade de Ciecircncias e Tecnologia da
Universidade do Algarve
No entanto descreve-se abaixo de forma resumida o procedimento efectuado
38
32111 Purificaccedilatildeo da miosina
O protocolo de isolamento foi adaptado do meacutetodo previamente descrito por Margossian e Lowey
(1982) apresentando-se de forma esquemaacutetica na Fig 31
Sucintamente os tecidos musculares dorsais e das pernas foram extraiacutedos apoacutes o sacrifiacutecio do
animal (Fig31) De seguida foram adicionados 3 volumes da soluccedilatildeo de extracccedilatildeo A2
Centrifugou-se a soluccedilatildeo e filtrou-se o sobrenadante O sedimento foi entatildeo diluiacutedo em 14
volumes de aacutegua destilada de modo a que a forccedila ioacutenica atingisse 004 M Apoacutes adiccedilatildeo de szlig-
mercaptoetanol (szlig-me) deixou-se a precipitar a 4ordmC removendo o sobrenadante Centrifugou-se
novamente e ressuspendeu-se o sedimento na soluccedilatildeo de extracccedilatildeo A Adicionou-se a soluccedilatildeo
tampatildeo B3 para que a forccedila ioacutenica atingisse 05 M Homogeneizou-se e deixou-se repousar
durante 2h Homogeneizou-se novamente e centrifugou-se de modo a ocorrer precipitaccedilatildeo dos
complexos de actomiosina e proteiacutena desnaturada Apoacutes filtraccedilatildeo do sobrenadante em latilde de vidro
quantificou-se a soluccedilatildeo de miosina resultante pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental)
2 Soluccedilatildeo de extracccedilatildeo A 0012M K2CO3 004M KHCO3 06M KCl pH 65
3 Soluccedilatildeo-tampatildeo B Tris 100 mM pH 7 KCl 2M
39
Figura 31 ndash Representaccedilatildeo esquemaacutetica do protocolo de isolamento de miosina (Margossian e Lowey 1982) Todo o
procedimento foi realizado a 4ordmC (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
32112 Preparaccedilatildeo de Subfragmento-1 de miosina por digestatildeo com quimiotripsina
O S1 de miosina foi isolado apoacutes diaacutelise da miosina durante a noite contra a soluccedilatildeo C4 A
presenccedila do EDTA eacute essencial uma vez que a sua substituiccedilatildeo por MgCl2resulta na produccedilatildeo de
HMM em vez de S1 A suspensatildeo de miosina foi equilibrada e digerida com -quimotripsina
A digestatildeo foi interrompida pela adiccedilatildeo de PMSF Seguidamente ultracentrifugou-se o produto
da digestatildeo desprezando-se o sedimento (Fig 32) A soluccedilatildeo de sobrenadante maioritariamente
S1 foi quantificada pela leitura directa da Abs280 ou pelo meacutetodo de Bradford (ver as secccedilotildees
3311 e 3312 do Procedimento Experimental respectivamente) e armazenada em 10 sacarose
e 2 mM szlig-Me a -80ordmC
4 Soluccedilatildeo C 25mM Tris pH 70 012M KCl 1mM EDTA
40
Antes da realizaccedilatildeo dos ensaios descritos no presente relatoacuterio o S1 de miosina foi dialisado
contra a soluccedilatildeo D5 de modo a remover qualquer composto que pudesse complexar como
vanaacutedio Seguidamente quantificou-se a proteiacutena pelo meacutetodo de Bradford ou pelo meacutetodo
directo (ver secccedilotildees 3311 e 3312do Procedimento Experimtental)
Figura 3 2ndash Representaccedilatildeo esquemaacutetica do protocolo de preparaccedilatildeo do S1 de miosina por digestatildeo quimiotripsina de
acordo com o previamente descrito por Margossian e Lowey (1982) Todo o procedimento foi realizado a 4ordm C
excepto quando indicaccedilatildeo do contraacuterio (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
3212 Isolamento e purificaccedilatildeo de actina
Isolou-se e purificou-se a actina segundo o meacutetodo descrito por Pardee e Spudish que
compreende dois passos ndash a obtenccedilatildeo de um resiacuteduo seco denominado ldquopoacute de acetonardquo e a
extracccedilatildeo subsequente da actina a partir do ldquopoacute de acetonardquo na forma de G-Actina com a
posterior polimerizaccedilatildeo a F-Actina
32121 Obtenccedilatildeo do ldquopoacute de acetonardquo
Sacrificou-se o coelho com uma pancada atraacutes do pescoccedilo e sangrou-se o animal apoacutes o corte da
arteacuteria jugular Esfolou-se e colocou-se numa tina com gelo Retiraram-se as partes de tecido
conjuntivo que envolvem o tecido muscular com auxiacutelio de uma pinccedila e bisturi Seguidamente
retirou-se os muacutesculos dorsais e os muacutesculos brancos das pernas Passou-se os muacutesculos por aacutegua
destilada de modo a remover as impurezas e pesou-se num copo Trituraram-se os muacutesculos
numa maacutequina de triturar carne e extraiu-se o resultante com 3 volumes da soluccedilatildeo E6 durante 10
minutos com agitaccedilatildeo suave Centrifugou-se a suspensatildeo a 4000 g durante 10
5 Soluccedilatildeo D HEPES 10 mM pH 70 KCl 25 mM
6 Soluccedilatildeo E 01 M KCl 015 M K2HPO4 pH 65
41
minutosdesprezando-se o sobrenadante Ressuspendeu-se o sedimento em 6 volumes de soluccedilatildeo
F7durante 10 minutos com agitaccedilatildeo suave Verificou-se se o pH da soluccedilatildeo estava compreendido
entre 82 e 85 acertando-se estecom soluccedilatildeo G8 se necessaacuterio Centrifugou-se durante 10
minutos a 4000 g Desprezou-se o sobrenadante e ressuspendeu-se o sedimento em 3 volumes de
soluccedilatildeo H9 durante 10 minutos Centrifugou-se novamente a 4000 gdurante 10 minutos
desprezando-se o sobrenadante Ressuspendeu-se o sedimento duas vezes consecutivas em 6
volumes de aacutegua destilada durante 5 minutos Centrifugou-se a 4000 g durante 10 minutos apoacutes
cada adiccedilatildeo de H2O
Seguidamente ressuspendeu-se o sedimento 5 vezes consecutivas utilizando 3 volumes de
acetona de cada vez agitando suavemente durante 10 minutos agrave temperatura ambiente Colocou-
se o resiacuteduo filtrado numa tina de vidro sobre papel de filtro deixando-se evaporar o solvente
durante a noite num nicho Triturou-se o resiacuteduo seco numa maacutequina de triturar carne e deixou-se
novamente evaporar o solvente O ldquopoacute de acetonardquo obtido foi armazenado a -20 ordmC
32122 Purificaccedilatildeo da actina a partir de ldquopoacute de acetonardquo
Apoacutes a obtenccedilatildeo do poacute de acetona como acima indicado efectuou-se o protocolo seguidamente
descrito de modo a obter monoacutemeros de actina (G-Actina) e actina filamentosa (F-Actina)
Pesou-se 35 g de poacute de acetona e adicionaram-se 50 mL de soluccedilatildeo-tampatildeo I10
sendo a
extracccedilatildeo feita com agitaccedilatildeo suave durante 30 minutos Separou-se o extracto do poacute de acetona
hidratado filtrando a suspensatildeo com 4 camadas de gaze esterilizada tendo o cuidado de natildeo
espremer o resiacuteduo Voltou a extrair-se com 50 mL de soluccedilatildeo I durante 30 minutos mexendo
lentamente Adicionou-se este ao extracto obtido anteriormente Centrifugou-se o extracto total a
16 000 g durante 30 minutos Decantou-se para uma proveta sobre a qual se colocou gaze para
que o sedimento solto natildeo se juntasse ao sobrenadante e leu-se o volume De seguida adicionou-
se KCl ateacute uma concentraccedilatildeo final de 50 mM MgCl2 ateacute 2mM e ATP ateacute 1mM lentamente e com
agitaccedilatildeo contiacutenua de modo a promover a polimerizaccedilatildeo dos monoacutemeros de actina
Deixou-se polimerizar durante 2h a 25ordmC observando-se a presenccedila de pequenas bolhas de ar
indicativas de um aumento de viscosidade da soluccedilatildeo Arrefeceu-se em gelo durante 15 minutos e
adicionou-se KCl ateacute uma concentraccedilatildeo final de 06M Mexeu-se lentamente durante 30 minutos
apoacutes os quais se ultracentrifugou a 110 000g durante 1hora Desprezou-se o sobrenadante
ressuspendeu-se o sedimento com soluccedilatildeo tampatildeo E e transferiu-se para um potter
Dialisou-se durante a noite contra 1L de soluccedilatildeo-tampatildeo I a uma temperatura de 4ordmC com rotaccedilatildeo
7 Soluccedilatildeo F 005 M NaHCO3 pH 82-85
8 Soluccedilatildeo G 1M Na2CO3
9 Soluccedilatildeo H 1 mM EDTA pH 70
10 Soluccedilatildeo I 2 mM Tris-HCl pH 8 02 mM CaCl2 02 mM Na2ATP 05 mM szlig-Me 0005 NaN3
42
contiacutenua e mudanccedila de tampatildeo 2 a 3 vezes De seguida a actina foi centrifugada a 110
000gdurante 1hora Decantou-se a soluccedilatildeo de G-Actina (sobrenadante) e desprezou-se o
sedimento Determinou-se a concentraccedilatildeo de G-Actina pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental) A soluccedilatildeo de G-actina foi armazenada a uma temperatura de 4ordmC
apoacutes adiccedilatildeo de 002NaN3 mantendo-se estaacutevel durante 3 a 5 dias
331 Quantificaccedilatildeo das proteiacutenas isoladas
3311 Quantificaccedilatildeo pelo meacutetodo de Bradford
O meacutetodo de Bradford (Bradford 1976) baseia-se na utilizaccedilatildeo do reagente de Bradford
composto por entre outros aacutecido fosfoacuterico e o corante ldquoCoomassierdquo Este corante possui carga
negativa e a sua coloraccedilatildeo varia entre a vermelha (Absmax=465 nm) predominante em soluccedilatildeo e a
azul (Absmax=595 nm) presente aquando da ligaccedilatildeo aos grupos amina das proteiacutenas Em soluccedilotildees
aacutecidas diluiacutedas a coloraccedilatildeo varia proporcionalmente com a quantidade do corante ligado agrave
proteiacutena que por sua vez eacute directamente proporcional agrave concentraccedilatildeo da proteiacutena permitindo a
sua quantificaccedilatildeo ateacute a um limite de detecccedilatildeo de 20 gmL (Bradford 1976)
Deste modo prepararam-se amostras de BSA ([BSA]stock= 1mgmL) utilizando um intervalo de
concentraccedilotildees compreendido entre 0 e 15 gmL de acordo com a tabela 3I Apoacutes a adiccedilatildeo da
H2O MilliQ e do reagente de Bradford as soluccedilotildees foram sucessivamente homogeneizadas no
voacutertex
Tabela 3Indash Amostras utilizadas para a obtenccedilatildeo da recta de calibraccedilatildeo no meacutetodo de Bradford
BSA (L) BSA (g) H2O MilliQ (L) Reagente de Bradford (L)
0 0 800 200
125 125 79875 200
25 25 7975 200
5 5 795 200
10 10 790 200
15 15 785 200
Apoacutes incubaccedilatildeo a 25ordmC por 5 minutos leram-se os valores de Abs595 num
espectrofotoacutemetro (Shimadzu UV-160A) para todas as amostras Seguidamente traccedilou-se a recta
de calibraccedilatildeo (Abs595vs quantidade de BSA (g))
43
Para determinar a concentraccedilatildeo de proteiacutena apoacutes o processo de isolamento purificaccedilatildeo e
diaacutelise quando necessaacuterio colocou-se 5 L da amostra num tubo de Eppendorf e adicionou-se
795 L de H2O MilliQ homogeneizando a soluccedilatildeo no voacutertex Adicionou-se posteriormente 200
L de reagente de Bradford e repetiu-se a homogeneizaccedilatildeo Incubou-se a mistura a 25ordmC por 5
minutos apoacutes os quais se leu os valores de Abs595 nm no espectrofotoacutemetro Por interpolaccedilatildeo deste
valor a partir da recta de calibraccedilatildeo determinou-se a concentraccedilatildeo de proteiacutena na amostra
3312 Quantificaccedilatildeo pelo meacutetodo directo
A quantificaccedilatildeo das amostras proteicas pode tambeacutem ser efectuada atraveacutes da leitura directa da
absorvacircncia tendo por base a lei de Beer-Lambert onde Abs corresponde agrave
absorvacircncia ao comprimento de onda ao qual o ensaio foi realizado corresponde ao coeficiente
de extinccedilatildeo (substituiacutedo pela letra a se as unidades forem distintas da molaridade) b corresponde
ao percurso oacuteptico da radiaccedilatildeo na ceacutelula amostral e c corresponde agrave concentraccedilatildeo amostral
Assim no caso de S1 de miosina mediu-se a Abs280e calculou-se a concentraccedilatildeo da proteiacutena
sabendo que o coeficiente de extinccedilatildeo de S1 eacute (Ref)
No caso da actina mediu-se a Abs290ecalculou-se a concentraccedilatildeo da proteiacutena sabendo que o
coeficiente de extinccedilatildeo da actina eacute (Ref)
332Electroforese em condiccedilotildees desnaturantes (SDS-PAGE)
De modo a melhor caracterizar as proteiacutenas isoladas foi realizada uma electroforese em
condiccedilotildees desnaturantes (SDS-PAGE do inglecircs Sodium Dodecyl Sulfate Polyacrilamide Gel
Electrophoresis) O SDS (dodecilo sulfato de soacutedio) eacute um detergente anioacutenico utilizado neste
meacutetodo para desnaturar as proteiacutenas que se ligam ao SDS na proporccedilatildeo 14 g SDS g proteiacutena
(aproximadamente 1 moleacutecula de SDS por cada 2 resiacuteduos de aminoaacutecidos) (Stryer) O SDS induz
uma densidade de carga negativa significativa nas proteiacutenas a que se liga mascarando a sua carga
intriacutenseca pelo que as razotildees cargamassa satildeo tendencialmente idecircnticas Assim a SDS-PAGE
separa as proteiacutenas quando sujeitas a um campo eleacutectrico de acordo com o seu peso molecular
sendo a sua migraccedilatildeo directamente proporcional ao logaritmo deste uacuteltimo (Stryer)
O gel de resoluccedilatildeo com 10 de acrilamida foi obtido atraveacutes da adiccedilatildeo de 167 mL de soluccedilatildeo
stock de acrilamida11
075 mL de tampatildeo de resoluccedilatildeo12
251 mL de aacutegua destilada e 50 L de
SDS 10 Para promover a polimerizaccedilatildeo adicionou-se em seguida 38 L de PSA 10
11
soluccedilatildeo stock de acrilamida soluccedilatildeo de 30 (mv) de acrilamida e 08 (mv) de NNrsquo-metil-bis-acrilamida 12
tampatildeo de resoluccedilatildeo 15 M Tris-HCl pH 88
44
(persulfato de amoacutenia) 10 e 25 L de TEMED 99 (tetrametiletilenodiamina) Agitou-se
cuidadosamente de modo a natildeo formar bolhas e pipetou-se a soluccedilatildeo para a cassette de vidro
previamente montada no suporte De seguida adicionou-se uma peliacutecula de cerca de 2 mm de 2-
metil-2-butanol para verificar posteriormente a polimerizaccedilatildeo Apoacutes a polimerizaccedilatildeo que demora
cerca de 45 minutos verteu-se a agrave superfiacutecie e adicionou-se o gel de concentraccedilatildeo com 5 de
acrilamida constituiacutedo por 03 mL de soluccedilatildeo de stock acrilamida 045 mL de tampatildeo de
concentraccedilatildeo13
094 mL de aacutegua destilada 18 L de SDS 10 135 L de PSA 10 e 2L de
TEMED 99 Colocou-se o pente sobre o gel e deixou-se polimerizar durante cerca de 45
minutos
As amostras para aplicaccedilatildeo no gel de electroforese foram preparadas para uma concentraccedilatildeo de
proteiacutena de 1 mgmL num volume final de 100 L dos quais 25 L correspondem ao tampatildeo da
amostra14
As amostras foram aquecidas a 100ordmC durante 5 minutos Antes da aplicaccedilatildeo das
amostras transferiu-se a cassette para a tina de electroforese agrave qual se adicionou a soluccedilatildeo
tampatildeo de electroforese da tina superior15
e tampatildeo de electroforese da tina inferior16
Aplicaram-
se 10 L das soluccedilotildees de marcadores de peso molecular e 20 L das amostras proteicas
preparadas anteriormente A electroforese decorreu durante cerca de 3h30 min a uma voltagem
de 80 V De seguida transferiu-se o gel para uma tina corando as proteiacutenas durante 1h com
soluccedilatildeo corante17
Descorou-se o gel com soluccedilatildeo descorante18
tendo-se mudado esta soluccedilatildeo ateacute
as bandas se encontrarem bem definidas
Determinou-se a mobilidade relativa das bandas correspondentes aos marcadores de elevado peso
molecular e traccedilou-se a recta de calibraccedilatildeo ndash log(PM) vs log(Rf) onde Rf corresponde agrave
mobilidade relativa Por interpolaccedilatildeo determina-se o PM das proteiacutenas isoladas sabendo a sua
mobilidade relativa Por comparaccedilatildeo com os valores descritos na literatura pode concluir-se
acerca do grau de purificaccedilatildeo
34 Mediccedilatildeo da actividade enzimaacutetica de S1
341 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
13
tampatildeo de concentraccedilatildeo 05 M Tris-HCl pH 68 14
tampatildeo de amostra 320 mM Tris-HCl pH 68 04 M szlig-mercaptoetanol SDS 8 glicerol 15 (vv) e roxo de m-
cresol 0024 15
tampatildeo de electroforese da tina superior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS 16
tampatildeo de electroforese da tina inferior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS acetato de soacutedio
01 M 17
soluccedilatildeo corante ldquoCoomassie Blue R-250rdquo contendo metanol 45 (vv) e aacutecido aceacutetico glacial 10 (vv) 18
soluccedilatildeo descorante metanol 10 (vv) e aacutecido aceacutetico glacial 10 vv)
45
O meacutetodo de Fiske-Subarow modificado consiste na reacccedilatildeo do molibdato de amoacutenio com o
fosfato inorgacircnico em soluccedilatildeo aacutecida com formaccedilatildeo do aacutecido fosfomoliacutebdico Seguidamente daacute-
se a adiccedilatildeo de um agente redutor que provoca a reduccedilatildeo do composto a fosfomolibdato que forma
um complexo de cor azulada cuja absorvacircncia maacutexima ocorre para um comprimento de onda de
660 nm tornando possiacutevel a sua detecccedilatildeo por meacutetodos espectrofotomeacutetricos As actividades Ca2+
-
ATPaacutesica e K+EDTA-ATPaacutesica de S1 foram determinadas atraveacutes deste meacutetodo de acordo com
o descrito por Taussky e Shorr (1953) com mediccedilatildeo da quantidade de fosfato (Pi) libertado
atraveacutes da hidroacutelise de ATP
Para tal obteve-se primeiramente a recta de calibraccedilatildeo com as amostras presentes na Tabela 3II
utilizando uma soluccedilatildeo stock de 5 mM KH2PO4
Tabela 3II ndash Composiccedilatildeo das diferentes amostras utilizadas para determinar a recta de calibraccedilatildeo atraveacutes do meacutetodo
de Fiske-Subbarow modificado
KH2PO4 (L)
([KH2PO4]stock= 5mM)
PO43-
(nmole) H2O (uL)
0 0 1780
5 25 1775
10 50 1770
20 100 1760
40 200 1740
100 500 1680
A cada tubo de Eppendorf adicionaram-se seguidamente 200 L de soluccedilatildeo J19
e 20 L de agente
redutor20
homogeneizando no voacutertex apoacutes cada adiccedilatildeo Incubou-se durante 10 minutos e mediu-
se a Abs660 nm num espectrofotoacutemetro com intervalos de tempo iguais entre cada amostra Traccedilou-
se a recta de calibraccedilatildeo (Abs660 nmvs nmol PO43-
) a partir dos valores obtidos
Para determinar a actividade Ca2+
-ATPaacutesica de S1 prepararam-se tubos de Eppendorf com 400
L de meio de reacccedilatildeo21
25 L CaCl2 ([CaCl2]stock= 100 mM) 25 L ATP ([ATP]stock= 100 mM)
com [S1]ensaios=01 mgmL Perfez-se se necessaacuterio com H2O MilliQ ateacute perfazer um volume de 1
mL Para determinar a actividade K+EDTA-ATPaacutesica utilizou-se uma concentraccedilatildeo idecircntica de
19
Soluccedilatildeo J 5 N H2SO4 25 molibdato de amoacutenio 20
Agente redutor 5 mg aacutecido 1-amino-2-naftol-4-sulfoacutenico 30 mg bissulfito de soacutedio 30 mg sulfito de soacutedio em 26
mL H2O 21
meio de reacccedilatildeo 125 mM Tris-HCl pH 80 0575 M KCl
46
proteiacutena nos ensaios (01 mgmL) e o mesmo meio de reacccedilatildeo Aos tubos de Eppendorf
adicionou-se ainda 123 L de KCl ([KCl]stock = 3M) 25 L de ATP 10 L de EDTA
([EDTA]stock= 100 mM )e o volume de H2O MilliQ necessaacuterio para perfazer 1 mL Em ambas as
situaccedilotildees o uacuteltimo reagente adicionado foi o ATP uma vez que se trata do substrato da reacccedilatildeo
enzimaacutetica
A cineacutetica decorreu a 25ordmC durante 5 minutos para a actividade Ca2+
-ATPaacutesica e durante 3
minutos para a actividade K+EDTA-ATPaacutesica Apoacutes o periacuteodo de incubaccedilatildeo transferiu-se 150
L de amostra para tubos de Eppendorf contendo igual volume de TCA 10 Colocou-se no gelo
e homogeneizou-se no voacutertex Seguidamente centrifugou-se a 1500 g durante 10 minutos e
recolheu-se 200 L de sobrenadante para novos tubos de Eppendorf Adicionou-se a estes 158
mL de H2O MilliQ e 200 L de soluccedilatildeo J Homogeneizou-se no voacutertex e adicionou-se 20 L de
reagente redutor repetindo-se a homogeneizaccedilatildeo Incubou-se durante 10 minutos e mediu-se a
Abs660 nm com o mesmo intervalo de tempo de leitura para cada amostra Por intrapolaccedilatildeo a partir
da recta de calibraccedilatildeo obtida anteriormente determinou-se a quantidade de Pi libertado em cada
ensaio A partir destes resultados determinaram-se as actividades Ca2+
-ATPaacutesica e K+EDTA-
ATPaacutesica do S1 em nmol PO43-
minmg proteiacutena e em s-1
342 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
Da actividade Mg2+
-ATPaacutesica de S1 resulta a formaccedilatildeo dos produtos da reacccedilatildeo de
hidroacutelise do ATP - ADP e Pi Na presenccedila de ADP fosfoenolpiruvato (PEP) e da enzima piruvato
cinase (PK) os produtos resultantes satildeo ATP e piruvato Por acccedilatildeo da enzima lactato
desidrogenase (LDH) este uacuteltimo eacute convertido em lactato ocorrendo simultaneamente a oxidaccedilatildeo
do NADH a NAD+ monitorizada atraveacutes da Abs340 nm Quando o S1 de miosina se encontra no
meio reaccional o ATP eacute hidrolisado formando-se novamente ADP e Pi o que permite a
formaccedilatildeo de um ciclo (fig 21) Se as enzimas se encontrarem em excesso a velocidade de
conversatildeo do NADH eacute limitada pela produccedilatildeo de ADP e Pi Como o ATP eacute regenerado em cada
ciclo a sua concentraccedilatildeo inicial eacute sempre mantida (Bagshaw 1982)
Figura 33ndash Representaccedilatildeo esquemaacutetica do meacutetodo de enzimas acopladas
47
O protocolo utilizado e seguidamente descrito foi adaptado de Cuenda et al (1990 ndash Ver Refordf) A
uma cuvette de 1 mL adicionou-se S1 para uma concentraccedilatildeo final de 005 mgmL 800 L de
meio de reacccedilatildeo22
10 L de PEP ([PEP]stock = 42 mM) 10 L de PK+LDH 11 ([PK+LDH]stock =
1 mgmL) e 15 L de NADH ([NADH]stock=25 mM) Iniciou-se a monitorizaccedilatildeo da reacccedilatildeo
atraveacutes da leitura de Abs340 nm apoacutes a adiccedilatildeo de 20 L de ATP ([ATP]stock=100 mM) A
actividade Mg2+
-ATPaacutesica eacute obtida atraveacutes do declive correspondente agrave oxidaccedilatildeo do NADH Para
a mediccedilatildeo da Mg2+
-ATPase de S1 estimulada por actina adicionou-se F-actina para uma
concentraccedilatildeo final de 2 microM sendo a actividade obtida tambeacutem atraveacutes do declive correspondente
agrave oxidaccedilatildeo do NADH apoacutes a adiccedilatildeo da proteiacutena
35 Interaccedilatildeo entre vanaacutedio e subfragmento-1 de miosina
351 Preparaccedilatildeo das soluccedilotildees de vanaacutedio
As soluccedilotildees de vanadato foram preparadas em aacutegua MilliQ a partir de metavanadato de amoacutenio
(NH4VO3) numa concentraccedilatildeo stock de 50 mM com agitaccedilatildeo magneacutetica e aquecimento (~30 ordmC)
Esta soluccedilatildeo com um valor compreendido entre 6 e 7 designa-se por ldquometavanadatordquo e eacute
constituiacuteda pelas esspeacutecies V1 V2 V4 e V5 (Ref) Acidificando a soluccedilatildeo de metavanadato por
adiccedilatildeo de HCl ateacute um valor de pH proacuteximo de 4 observa-se uma mudanccedila de cor de amarelo
claro para laranja indicativa da formaccedilatildeo da espeacutecie decameacuterica de vanadato De seguida e
imediatamente antes da realizaccedilatildeo dos ensaios descritos o valor do pH da soluccedilatildeo de
decavanadato foi aumentado para cerca de 75 por adiccedilatildeo de NaOH A soluccedilatildeo (2) designada
por ldquometavanadatordquo natildeo foi posteriormente alterada ateacute agrave sua adiccedilatildeo para a realizaccedilatildeo dos ensaios
As soluccedilotildees foram armazenadas a 4ordmC e mantidas em gelo aquando da sua utilizaccedilatildeo
A soluccedilatildeo de V(IV) foi preparada em condiccedilotildees anaeroacutebias por diluiccedilatildeo de sulfato de vanadil
(VOSO4) para uma concentraccedilatildeo final de 5 mM em H2O MilliQ saturada com N2 gasoso
352Anaacutelise por espectroscopia de UV-Vis
A concentraccedilatildeo da espeacutecie decameacuterica em soluccedilatildeo depende do periacuteodo de tempo apoacutes a diluiccedilatildeo
no meio de reacccedilatildeo apropriado (Ref) A velocidade de desoligomerizaccedilatildeo da espeacutecie decameacuterica
pode ser analisada por espectroscopia de absorccedilatildeo na regiatildeo UVVis uma vez que esta espeacutecie
apresenta duas bandas caracteriacutesticas na regiatildeo do visiacutevel (a 358 e 400 nm) que natildeo se verifica
em quaisquer outras espeacutecies oligomeacutericas As bandas supramencionada sapresentam-se bem
22
Meio de reacccedilatildeo 3125 mM Tris-HCl pH 71 375 mM MgCl2
48
resolvidas atraveacutes da obtenccedilatildeo da derivada de 2ordf ordem do espectro apresentando a forma de 2
vales com miacutenimos centrados nos comprimentos de onda supracitados com dependecircncia linear
relativamente agrave concentraccedilatildeo de decavanadato total (Ramos et al 2006)
Assim a estabilidade da espeacutecie decameacuterica apoacutes diluiccedilatildeo da soluccedilatildeo de decavanadato no meio de
reacccedilatildeo dos ensaios (tampatildeo D) para uma concentraccedilatildeo final de 1 mM em vanadato total foi
caracterizada atraveacutes da leitura da Abs400 em funccedilatildeo do tempo tendo-se determinado qual a
ordem de reacccedilatildeo e o tempo de meia-vida da mesma Com base em estudos anteriores acerca da
estabilidade do V10 na presenccedila de diferentes proteiacutenas (Ramos et al 2006) determinou-se
igualmente o tempo de meia vida apoacutes a adiccedilatildeo de 20 microM de S1 de miosina ao meio reaccional
353 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
Os espectros de RMN do isoacutetopo 51
V foram realizados num espectroacutemetro Bruker AM-400 MHz
equipado com uma sonda multinuclear inversa de 5 mm Em tubos de RMN de 5 mm de diacircmetro
preparou-se soluccedilotildees de 500 L de deca- ou metavanadato correspondentes a um intervalo de
concentraccedilotildees de 1 a 10 mM suplementadas com 10 D2O Os espectros foram adquiridos agrave
temperatura ambiente com uma largura espectral de 23 923 Hz um tempo de aquisiccedilatildeo de 0086
s tempo de relaxaccedilatildeo de 001 s e uma sequecircncia de pulsos a 90ordm Utilizou-se a transformada de
Fourier com alargamento das bandas de 20Hz para o processamento dos espectros Os desvios
quiacutemicos das ressonacircncias de 51
V foram obtidos em relaccedilatildeo agrave espeacutecie VOCl3cuja frequecircncia de
ressonacircncia eacute considerada o padratildeo sendo todas as outras referenciadas a partir desta em ppm
As aacutereas dos sinais do RMN correspondentes agraves espeacutecies oligomeacutericas presentes foram obtidas
por integraccedilatildeo dos sinais As concentraccedilotildees relativas das espeacutecies foram calculadas com recurso agrave
equaccedilatildeo
onde A[Vx] corresponde agrave aacuterea referente agrave ressonacircncia da espeacutecie Vx com nordm de agregaccedilatildeo x
A[V]total agrave soma total das aacutereas medidas e [V] a concentraccedilatildeo total de vanaacutedio no ensaio
A teacutecnica de 51
V-RMN foi anteriormente aplicada em estudos de ligaccedilatildeo com S1 de miosina
(Ref) tendo sido demostrado que entre as espeacutecies V1 V2 V4 e V5 a espeacutecie tetrameacuterica possui a
maior afinidade para a miosina (Cremo et al) O desaparecimento ou alargamento dos sinais pode
implicar a ligaccedilatildeo de um dos oxovanadatos laacutebeis agrave miosina sem indicaccedilatildeo de qual uma vez que
49
se processa um equiliacutebrio entre eles (Crans 1990) Assim e tendo em conta as indicaccedilotildees de que
a espeacutecie V10 eacute distinta das restantes no que diz respeito agrave magnitude da interacccedilatildeo com a miosina
e inibiccedilatildeo da actividade ATPaacutesica estimulada por actina (Aureliano 2000 Tiago et al 2002)
torna-se fundamental investigar a interacccedilatildeo das diferentes espeacutecies de vanadato com o S1 de
miosina atraveacutes da teacutecnica de 51
V-RMN
Para tal obtiveram-se espectros de 51
V-RMN resultantes da titulaccedilatildeo de uma soluccedilatildeo 5 mM de
decavanadato total com concentraccedilotildees crescentes de S1 de miosina (0 5 10 15 e 20 microM) em
tampatildeo D Realizou-se ainda ensaios na presenccedila de 2 mM de decavanadato e 2 mM de ATP na
presenccedila e na ausecircncia de S1 de miosina de modo a verificar o efeito que estes ligandos naturais
da enzima teria na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o subfragmento-1
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
Os grupos sulfidrilo (-SH) das cisteiacutenas satildeo resiacuteduos de aminoaacutecidos extremamente sensiacuteveis agrave
oxidaccedilatildeo resultando na sua oxidaccedilatildeo (Dalle-Donne et al 1999) Logo a quantificaccedilatildeo destes
resiacuteduos eacute bastante importante nos estudos de interacccedilatildeo com vanaacutedio jaacute que os resiacuteduos de Cys
com ecircnfase para o SH1 e o SH2 satildeo bastante importantes na monitorizaccedilatildeo das modificaccedilotildees
funcionais da regiatildeo da cabeccedila
O procedimento seguidamente apresentado foi originalmente descrito por Gutieacuterrez-Merino
(1987)) Consiste na reacccedilatildeo entre os grupos ndashSH natildeo oxidados das proteiacutenas e o reagente de
Ellman (DTNB -55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico)) - formando-se o aniatildeo CNT (aniatildeo 3-
carboxilato-4-nitrotiofenolato) numa estequiometria de 11 Este apresenta um maacuteximo de
absorvecircncia a 412 nm (Robyt 1990) Assim apoacutes a incubaccedilatildeo da proteiacutena na presenccedila do metal
a quantidade de cisteiacutenas reduzidas por proteiacutena determina-se atraveacutes dos valores de Abs412
Deste modo colocou-se num tubo de Eppendorf o volume apropriado de S1 dialisado para que a
concentraccedilatildeo no ensaio fosse de 1 mgmL Adicionou-se o volume apropriado de decavanadato
para que a sua concentraccedilatildeo fosse de 0 10 50 100 250 500 ou 1000 M Perfez-se com soluccedilatildeo
D ateacute um volume final de 1 mL e incubou-se a 25ordmC por 20 minutos Apoacutes este periacuteodo colocou-
se 780 L da mistura reaccional numa cuvette de quartzo e adicionou-se 220 L de soluccedilatildeo K23
23
Tampatildeo K Tampatildeo fosfatohidrogenofosfato de soacutedio 025 M pH 72
50
Adicionou-se 15 L de DTNB (em soluccedilatildeo K [DTNB]stock=100 mM) ao meio reaccional e
registou-se o valor de absorvecircncia medido equivalente agraves cisteiacutenas ldquoraacutepidasrdquo (SH1 e SH2
expostos ao solvente) Adicionou-se 001 g de SDS que provoca a desnaturaccedilatildeo da proteiacutena e
consequentemente a exposiccedilatildeo de todos os resiacuteduos de cisteiacutenas que se encontravam ateacute aiacute
inacessiacuteveis no interior da proteiacutena (cisteiacutenas totais) Estes resiacuteduos podem assim reagir com o
DTNB
Estimou-se o nuacutemero de resiacuteduos de cisteiacutenas em todas as amostras com recurso ao coeficiente de
extinccedilatildeo molar () do DTNB (ver ponto 3 dos Anexos) que foi determinado atraveacutes da titulaccedilatildeo
de L-Cisteiacutena com intervalo de concentraccedilotildees compreendidos entre 0 e 50 microM na presenccedila de
25microM de DTNB em tampatildeo D Os valores de absorvecircncia foram obtidos para um comprimento de
onda de 412 nm
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
3621 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
A teacutecnica mais eficiente de anaacutelise da interacccedilatildeo entre o iatildeo vanadilo e proteiacutenas como a
calmodulina e a transferrina entre outras permite a obtenccedilatildeo raacutepida de informaccedilatildeo no que toca agrave
conformaccedilatildeo e amplitude da referida interacccedilatildeo
Os espectros de Ressonacircncia Paramagneacutetica Electroacutenica (RPE) da espeacutecie paramagneacutetica V(IV)
foram realizados a 70K num espectroacutemetro Bruker EMX-300 X-band Todas as soluccedilotildees
utilizadas e amostras foram preparadas em anaerobiose numa cacircmara de luvas de modo a evitar a
reduccedilatildeo de V(IV) a V(V) Em tubos capilares com 02 mL de amostra manteve-se a concentraccedilatildeo
de S1 a 20 M e variou-se a concentraccedilatildeo de vanadilo (0 25 5 10 15 25 50 75 100 150
200 500 e 1000 M [VOSO4]stock=5 mM) em tampatildeo D Apoacutes obtenccedilatildeo dos espectros estes
foram tratados atraveacutes do software SimFonia e WinEPR tendo-se obtido um graacutefico com a
variaccedilatildeo da intensidade do sinal de RPE em funccedilatildeo da concentraccedilatildeo de sulfato de vanadilo
presente Realizaram-se ainda simulaccedilotildees de espectros obtidos com 20 microM de S1 e 1000 microM de
V(IV) com vista agrave obtenccedilatildeo dos ligandos provaacuteveis do vanadilo atraveacutes da anaacutelise da constante
de acoplamento hiperfina (A) e do factor g (g)
4 Conclusotildees gerais
Apesar de jaacute ter sido demonstrado que o vanaacutedio inibe a Na+K
+-ATPase da miosina a funccedilatildeo
deste elemento no tecido muacutesculo esqueleacutetico continua largamente incompreendida A enorme
complexidade quiacutemica deste metal especialmente no que concerne agrave oligomerizaccedilatildeo e reacccedilotildees
51
redox constitui um obstaacuteculo agrave compreensatildeo do comportamento do vanaacutedio nos organismos
vivos Assim a caracterizaccedilatildeo da quiacutemica deste elemento eacute de grande utilidade antes de iniciar o
estudo da sua interacccedilatildeo com o presente sistema
A anaacutelise por espectroscopias de 51
V-RMN e de UV-Vis de soluccedilotildees de vanadato diluiacutedas nos
meios de reacccedilatildeo utilizados no presente relatoacuterio (10 mM HEPES pH 70 25 mM KCl) mostrou
claramente que a espeacutecie decameacuterica (V10) possui um tempo de vida suficientemente longo para a
determinaccedilatildeo do seu efeito isolado ao contraacuterio do que se verifica com as espeacutecies mono- di-
tetra- e pentameacutericas (V1 V2 V4 e V5) Por outro lado a anaacutelise verificou ainda que apoacutes diluiccedilatildeo
das soluccedilotildees de decavanadato no meio de reacccedilatildeo coexistem as espeacutecies V10 e V1 pelo que os
efeitos putativos desta uacuteltima deveratildeo ser subtraiacutedos aquando da monitorizaccedilatildeo dos efeitos da
espeacutecie decameacuterica
O vanaacutedio (V) (vanadato) inibe as enzimas que catalizam reacccedilotildees de transferecircncia de grupos
fosfato como a ATPase da miosina [Goodno 1979] devido agrave sua analogia estrutural com o iatildeo
fosfato Na sua utilizaccedilatildeo como inibidor enzimaacutetico natildeo eacute tida em conta a contribuiccedilatildeo individual
das espeacutecies oligomeacutericas considerando-se normalmente a espeacutecie V1 como inibidora efectiva
[Goodno e Taylor 1982] No presente relatoacuterio verificou-se por espectroscopia de 51
V-RMN que
outros oligoacutemeros de vanadato interagem com o S1 de miosina tendo as espeacutecies V10 e V4 os
maiores niacuteveis de afinidade A sua interacccedilatildeo envolve forccedilas electrostaacuteticas e eacute afectada
negativamente por ligandos naturais de enzima como eacute o caso do ATP
A exposiccedilatildeo do S1 de miosina (005 mgmL) a decavanadato conduz agrave inibiccedilatildeo do capacidade da
proteiacutena de estimular a actividade Mg2+
-ATPaacutesica com um valor de IC50 de 266 plusmn 07 M
decavanadato sendo o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM
decavanadato (80) Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM decavanadato sendo
o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM decavanadato (60)
exercendo um efeito de impedimento agrave ligaccedilatildeo entre o vanaacutedio e o S1
A uma concentraccedilatildeo de 27 microM decavanadato verifica-se a oxidaccedilatildeo de um dos resiacuteduos de Cys
de S1 de miosina seguida pela oxidaccedilatildeo gradual de outro resiacuteduo de Cys ateacute agrave concentraccedilatildeo de
1000 microM Ambos os resiacuteduos oxidados natildeo se encontravam expostos ao solvente utilizado A
oxidaccedilatildeo dos resiacuteduos permite sugerir que esta modificaccedilatildeo oxidativa sofrida pelo S1 contribui
para a diminuiccedilatildeo da sua actividade de ATPase
52
Por espectroscopia de RPE revelaram-se os possiacuteveis ligandos equatoriais do vanaacutedio aquando da
sua interacccedilatildeo com o S1 de miosina como sendo duas moleacuteculas de aacutegua um ligando contendo o
grupo R-OH e outro contendo o grupo R=NR
Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados de
oxidaccedilatildeo (IV) e (V) no S1 de miosina para concentraccedilotildees facilmente alcanccedilaacuteveis no ambiente
terrestre
53
5 Estudos futuros
No sentido do melhor esclarecimento das alteraccedilotildees moleculares e estruturais que ocorrem no
subfragmento 1 quando exposta ao vanaacutedio que conduzem a uma diminuiccedilatildeo da sua funccedilatildeo
sugere-se
- Quantificaccedilatildeo das cisteiacutenas reduzidas de S1 na presenccedila de metavanadato
- Determinaccedilatildeo das actividades Ca2+
- e K+-ATPaacutesica do S1 de miosina na presenccedila de
decavanadato
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina estimulada por actina
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina na presenccedila de
metavanadato
- Realizaccedilatildeo de RMN 2D com variaccedilatildeo das concentraccedilotildees das soluccedilotildees de decavanadato e
metavanadato na presenccedila de S1
-Mutageacutenese dos resiacuteduos de Cys doS1 de miosina de modo a verificar quais os resiacuteduos que
sofrem oxidaccedilatildeo na presenccedila de decavanadato
- Determinaccedilatildeo por calorimetria (DSC Differential Scanning Calorimetry) da variaccedilatildeo da
entalpia (∆H) e da temperatura de desnaturaccedilatildeo do S1 de miosina na presenccedila de vanaacutedio uma
vez que estas propriedades caracteriacutesticas de cada proteiacutena satildeo alteradas apoacutes ocorrecircncia de
modificaccedilotildees estruturais
- Anaacutelise por SDS-PAGE de S1 de miosina apoacutes tratamento com vanaacutedio de modo a determinar
quais os polipeacuteptidos eventualmente formados apoacutes clivagem da proteiacutena
54
6 Referecircncias bibliograacuteficas
- Sellers J Myosins 2nd Edition Oxford University Press 1999
- L Stryer inBiochemistry (Ed WH Freeman 2005 pp
Refordfas Materiais e Meacutetodos
[A] S S Margossian SLowey Methods Enzymol1982 85 55-71
[B] JD Pardue JA Spudich Methods Enzymol 1982 85 164-181
[C] M Bradford Anal Biochem1976 72 248-254
[D] H H Taussky E Shorr J Biol Chem 1953 202 675-685
[E] C R Bagshaw in Outline Studies in Biology ndash Muscle Contraction Chapman and Hall Ed
1982
[F] A Cuenda et al Eur J Biochem1990 194 663-670
55
Anexos
1 Determinaccedilatildeo da recta de calibraccedilatildeo meacutetodo de Bradford
Figura1ndashRepresentaccedilatildeo graacutefica da recta de calibraccedilatildeo (Abs595 nm vs [BSA]) utilizada para o meacutetodo de Bradford
2 Determinaccedilatildeo da recta calibraccedilatildeo para a electroforese em condiccedilotildees desnaturantes
Figura 2 ndash Representaccedilatildeo graacutefica da recta de calibraccedilatildeo (log(Peso Molecular) vs Rf dos marcadores de peso
molecular) onde Rf indica determinada atraveacutes do padratildeo de peso molecular
(poccedilo (a))
3Caacutelculo do coeficiente de extinccedilatildeo molar () para a quantificaccedilatildeo de cisteiacutenas reduzidas
atraveacutes do meacutetodo de Ellman
Traccedilou-se a recta de Abs412 nm meacutedia vs [L-Cisteiacutena] de modo a determinar o valor do coeficiente
de extinccedilatildeo molar definido como o declive daquela recta Assim =8400 M-1
cm-1
como
podemos verificar na Fig 3
A595 nm = 00546[BSA] + 00209 Rsup2 = 099051
Ab
s 59
5 n
m
[BSA] (mgmL)
log PM = -1177 Rf + 2247 Rsup2 = 098957
log
PM
Mobilidade relativa (Rf)
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero

12
A F-actina eacute formada atraveacutes da polimerizaccedilatildeo de subunidades globulares de actina (G-actina) A
G-actina eacute composta por 2 domiacutenios de tamanho semelhante separados pelos bolsos de ligaccedilatildeo(1)
de catiotildees divalentes e(2) de nucleoacutetidos [KLM] Estes por sua vez estatildeo subdivididos noutros
dois de menores dimensotildees (1-4) (Fig 16) cada um envolvido nas interacccedilotildees actina-actina ou
na formaccedilatildeo do topo de ligaccedilatildeo do nucleoacutetido As regiotildees N-terminal e C-terminal estatildeo proacuteximas
uma da outra no subdomiacutenio 1 [E]
Figura 16 ndash Estrutura do monoacutemero de actina Encontram-se representados os subdomiacutenios 1 a 4 e a vermelho a
moleacutecula de ATP junto do respectivo local de ligaccedilatildeo (Protein Data Bank ID 1j6z)
113 Mecanismos moleculares da contracccedilatildeo muscular
A contracccedilatildeo muscular ocorre atraveacutes do deslizamento dos filamentos finos (de actina) por entre
os filamentos grossos (de miosina) conseguida pela reacccedilatildeo de hidroacutelise de ATP e regulada pela
tropomiosina e pelo complexo de troponina em resposta a impulsos nervosos Este mecanismo foi
inicialmente proposto por Huxley-Niedergerke [N] e Huxley-Hanson [O] ambos em 1954 A base
experimental de ambos os estudos foi a inexistecircncia de variaccedilatildeo do tamanho dos filamentos
durante a contracccedilatildeo A banda A apresenta sempre as mesmas dimensotildees por outro lado a banda
I a zona H diminuem consideravelmente aquando da contracccedilatildeo (Figs 12 e 17)
Figura 17 ndash Modelo da contracccedilatildeo muscular ndash deslizamento dos filamentos finos (azuis) relativamente aos
filamentos grossos (rosa) (Adaptado de Stryer 2002 [2])
13
Na ausecircncia de ATP e supondo a ausecircncia de inibiccedilatildeo da interacccedilatildeo actina-miosina as proteiacutenas
encontram-se fortemente associadas formando o complexo actomiosina (AM) (Figura 15)
Nestas condiccedilotildees as cabeccedilas de miosina formam um acircngulo de 45ordm com o eixo dos filamentos
finos de actina [P]
Na presenccedila de ATP o complexo AM dissocia-se jaacute que a afinidade do complexo M-ATP para a
actina eacute 105 vezes inferior agrave afinidade da miosina para a actina O ATP liga-se agrave miosina
ocorrendo seguidamente a reacccedilatildeo de hidroacutelise de ATP Os produtos daiacute resultantes ndash ADP e Pi -
permanecem ligados agrave miosina formando o complexo M-ADP-Pi Com a ligaccedilatildeo subsequente agrave
actina forma-se o complexo AM-ADP-Pi no qual as cabeccedilas de miosina apresentam um acircngulo
recto com o eixo do muacutesculo [P] Neste ponto daacute-se a libertaccedilatildeo progressiva de Pi e ADP Eacute
provaacutevel que a energia proveniente da hidroacutelise do ATP provoque uma alteraccedilatildeo do acircngulo de
ligaccedilatildeo das cabeccedilas de miosina ao filamento de actina passando novamente de 90ordm para 45ordm [P]
Esta alteraccedilatildeo conformacional provoca o deslizamento do filamento de actina para o centro do
sarcoacutemero cujo encurtamento promove a contracccedilatildeo das miofibrilas O processo supracitado
ocorre em cerca de 1ms[Q] e encontra-se resumido na Fig 18
Figura 18 ndash Esquema cineacutetico da hidroacutelise de ATP pela miosina (Adaptado de Sellers 1999 [A])
1131 Regulaccedilatildeo da contracccedilatildeo muscular
O mecanismo pelo qual se processa a regulaccedilatildeo muscular foi inicialmente descrito por estudos
distintos de Haselgrove Huxley e Parrey-Squire todos em 1973 [RST] Aleacutem da implicaccedilatildeo
directa da tropomiosina e do complexo de troponina a concentraccedilatildeo de Ca2+
no muacutesculo em
relaxamento eacute bastante reduzida (10-7
M) aumentando notavelmente aquando da contracccedilatildeo (10-5
-
10-3
M)
Resumidamente quando ocorre a excitaccedilatildeo do muacutesculo esqueleacutetico a despolarizaccedilatildeo da
membrana plasmaacutetica eacute difundida ateacute ao interior do tecido atraveacutes dos tuacutebulos transversais
(invaginaccedilotildees da membrana plasmaacutetica da fibra muscular) A despolarizaccedilatildeo provoca a abertura
dos canais ioacutenicos de Ca2+
do retiacuteculo sarcoplasmaacutetico adjacente Como a concentraccedilatildeo de Ca2+
no interior do organelo eacute muito elevada o caacutelcio passa raacutepida e uniformemente para o citosol
14
Assim a regulaccedilatildeo da contracccedilatildeo ocorre com coordenaccedilatildeo e rapidez na totalidade das miofibrilas
do tecido muscular (Ref)
Em repouso a troponina I encontra-se posicionada de tal forma que a proteiacutena adjacente
troponina C impede estruturalmente a ligaccedilatildeo das cabeccedilas da miosina agrave actina (acircngulo =50ordm) O
aumento da concentraccedilatildeo intracelular de Ca2+
provoca um aumento da sua disponibilidade para
ligar agrave troponina C (Fig 19)
Figura 19ndash Regulaccedilatildeo do mecanismo contraacutectil no muacutesculo esqueleacutetico pelo complexo de troponina
Como consequecircncia a troponina C altera a sua conformaccedilatildeo (=70ordm) associando-se agrave troponina I
que deixa de inibir a ligaccedilatildeo das pontes transversais A alteraccedilatildeo anterior eacute transmitida agrave
tropomiosina atraveacutes da troponina T adjacente agrave C o que provoca o deslocamento da
tropomiosina na direcccedilatildeo do interior do sulco do filamento de actina Assim eacute possiacutevel a
interacccedilatildeo eficaz entre actina e miosina A Ca2+
-ATPase do retiacuteculo sarcoplasmaacutetico provoca a
reacumulaccedilatildeo do iatildeo caacutelcio no organelo impedindo o prolongamento desnecessaacuterio da contracccedilatildeo
(Refordf)
12 Vanaacutedio
O vanaacutedio eacute um metal de transiccedilatildeo pertencente ao grupo VB da tabela perioacutedica Foi
primeiramente descoberto em 1801 por Andreacutes Manuel del Riacuteo que concluiu erradamente que era
idecircntico ao croacutemio e definitivamente por Nils Gabriel Svenson em 1831 Recentemente foram
descobertas importantes funccedilotildees bioloacutegicas deste elemento provavelmente essencial para os
humanos de que se destaca no contexto do presente relatoacuterio a possibilidade de actuar como
anaacutelogo do grupo fosfato e a inibiccedilatildeo de enzimas in vitro como eacute o caso da actividade ATPaacutesica
da miosina
Eacute um metal soacutelido agrave temperatura ambiente que se encontra amplamente distribuiacutedo nos vaacuterios
ecossistemas em dois isoacutetopos naturais ndash 50
V e 51
V ndash raramente excedendo 01 partes por biliatildeo
15
(ppb) Estaacute associado a cerca de 70 minerais diferentes na natureza (dos quais se destacam o
uracircnio cobre chumbo zinco fosfato e titacircnio ferroso) especialmente na forma de vanadato ou
pentoacutexidos de vanaacutedio O vanaacutedio eacute um dos principais constituintes da mateacuteria orgacircnica foacutessil
com especial ecircnfase para o carvatildeo oacuteleo de crude e foacutesseis carbonaacuteceos [UV] bem como em
alimentos consumidos pelo Homem (salsa pimenta preta cogumelos marisco vegetais cereais
gorduras e oacuteleos) [W] Estes constituem a maior fonte de exposiccedilatildeo ao vanaacutedio da maioria da
populaccedilatildeo humana [X]
Normalmente a entrada do vanaacutedio no organismo ocorre por inalaccedilatildeo ou em menores
quantidades por absorccedilatildeo pelo tracto gastrointestinal ou pela pele O seu armazenamento centra-
se no fiacutegado rins ceacuterebro coraccedilatildeo muacutesculos e ossos No entanto a retenccedilatildeo de vanaacutedio no
organismo humano eacute reduzida ndash em condiccedilotildees normais cerca de 100 M (em muitos tecidos a
quantidade eacute inferior a 10 ngg de tecido) [Y]
Actualmente natildeo existem provas de deficiecircncia nutricional de vanaacutedio em humanos ou de
enzimas que contenham vanaacutedio em animais superiores No entanto vaacuterios estudos relacionam a
ocorrecircncia de deficiecircncia em galinhas e ratos devido a uma alimentaccedilatildeo pobre em vanaacutedio Os
sintomas relatados incluem retardamento do crescimento malformaccedilotildees oacutesseas perturbaccedilotildees no
metabolismo lipiacutedico e estagnaccedilatildeo reprodutiva entre outros [Z AA AB] Os efeitos fisioloacutegicos
do vanaacutedio podem depender do tipo de composto ou de oligoacutemero uma vez que consoante as
suas estabilidade labilidade e nuacutemero de coordenaccedilatildeo as respostas seratildeo diferenciadas ao niacutevel
dos organismos vivos
Por outro lado a toxicidade do vanaacutedio encontra-se bem caracterizada existindo sintomas
descritos em vaacuterios mamiacuteferos (inclusive o Homem) que incluem diarreias catildeibras irritaccedilatildeo da
pele e mucosas palpitaccedilotildees cardiacuteacas anemia pneumonia bronquite perturbaccedilotildees no Sistema
Nervoso Central (SNC) entre outros (Ref) A toxicidade varia geralmente com a natureza do
composto com aumento proporcional agrave sua valecircncia Verificou-se que a forma de entrada do
vanaacutedio tem tambeacutem efeito na sua toxicidade ndash os maiores niacuteveis verificaram-se por injecccedilatildeo
intravenosa da amostra sendo progressivamente inferiores por inalaccedilatildeo e finalmente por
ingestatildeo [X AA] A ampla utilizaccedilatildeo dos compostos de vanaacutedio na induacutestria quiacutemica bem como
as elevadas quantidades de vanaacutedio libertadas atraveacutes da combustatildeo dos oacuteleos de crude
contribuem consideravelmente para o aumento da biodisponibilidade do vanaacutedio Como tal o
interesse neste elemento e nos problemas ambientais por ele causados tem vindo a aumentar
significativamente [V]
121 Propriedades quiacutemicas
16
A quiacutemica do vanaacutedio eacute extraordinariamente complexa pelo que originou vaacuterias interpretaccedilotildees
incorrectas desde a sua descoberta No entanto as suas propriedades dependem em uacuteltima
anaacutelise da configuraccedilatildeo electroacutenica do elemento Assim a variabilidade encontrada no
comportamento quiacutemico do vanaacutedio pode dever-se agrave sua configuraccedilatildeo ndash [Ar] 3d2 4s
2 ndash que
permite a formaccedilatildeo de vaacuterios estados de oxidaccedilatildeo (de +2 a +5) Em soluccedilatildeo aquosa os estados de
oxidaccedilatildeo estaacuteveis vatildeo de V(V) a V(II) Destes apenas os +3 a +5 satildeo biologicamente importantes
jaacute que o V(II) apresenta demasiada capacidade redutora para existir nos organismos vivos
conhecidos [AC]
No fluido extracelular os compostos de vanaacutedio encontram-se predominantemente na forma de
monovanadatos (V(V)) livres ou ligados agrave transferina do soro ou agrave hemoglobina (Refordf) Os iotildees de
vanadato livres satildeo transportados para o meio intracelular atraveacutes de canais ioacutenicos inespeciacuteficos
Jaacute no ambiente intracelular os compostos sofreratildeo uma bio-reduccedilatildeo atraveacutes da acccedilatildeo da
glutationa ou outros compostos com grupos ndashOH como pentoses hexoses ou etanolaminas [V] A
reoxidaccedilatildeo de vanadilo (V(IV)) intracelular a vanadato (V(V)) provocaria a acumulaccedilatildeo toacutexica
destes uacuteltimos pelo que a complexaccedilatildeo da espeacutecie reduzida o impede O transporte de vanaacutedio
para o interior da ceacutelula encontra-se na Fig 110
Figura 110 ndash Transporte de vanaacutedio para a ceacuteluka e seu subsequente metabolismo (P=Proteiacutena) (Adaptado de
Zaporowska e Scibor 1998)
1211 Vanaacutedio(IV)
A espeacutecie de vanaacutedio (IV) mais bem conhecida eacute o iatildeo vanadilo (VO2+
) espeacutecie estaacutevel a pH
aacutecido sendo a maioritaacuteria a pH 3[AD] Apresenta uma cor azulada e forma complexos penta e
hexacoordenados Forma-se rapidamente a partir de V(V) na presenccedila de um agente redutor em
soluccedilatildeo aquosa No ambiente intracelular a presenccedila de glutationa reduzida aacutecido ascoacuterbico
NAD(P)H entre outros conduzem a que o principal estado de oxidaccedilatildeo nos sistemas bioloacutegicos
seja provavelmente o +4 encontrando-se o V(IV) fortemente complexado com proteiacutenas ou
aminoaacutecidos livres citrato ATP e pirofosfato (Refordfs) Em alternativa o V(IV) pode existir
17
livremente em ambientes fortemente aacutecidos como os lisossomas jaacute que a pH superior a 3 as
soluccedilotildees de vanadilo tendem a ser oxidadas pelo ar formando-se vanadato Esta grande
capacidade redutora justifica parcialmente a utilizaccedilatildeo pouco extensiva do iatildeo VO2+
como
inibidor [W]
1212 Vanaacutedio (V)
O vanaacutedio (V) eacute a forma mais estaacutevel num intervalo de pH amplo [W] O iatildeo anioacutenico VO3-tende
a constituir complexos polinucleares pelo que as soluccedilotildees aquosas de vanadato satildeo extremamente
complexas com ocorrecircncia simultacircnea de vaacuterios equiliacutebrios de protonaccedilatildeo (pKa(vanadato)= 35
78 e 125) e oligomerizaccedilatildeo As espeacutecies presentes (H3VO4 H2VO4- HVO4
2- e VO4
3-) dependem
do pH forccedila ioacutenica e temperatura da soluccedilatildeo e da concentraccedilatildeo total do elemento
[AEAFAGAHAI]
O VO43-
- monoacutemero ortovanadato V1 ndash estaacute associado a uma grande parte da importacircncia
bioloacutegica do vanaacutedio uma vez que se trata de um anaacutelogo electroacutenico e estrutural do ortofosfato
(PO43-
) A coordenaccedilatildeo do ortovanadato pode ser tetraeacutedrica ou bipiramidal trigonal (Refordfs)
A oligomerizaccedilatildeo do vanadato monomeacuterico daacute origem ao diacutemero (V2O72-
- V2) que apresenta
tambeacutem equiliacutebrios de protonaccedilatildeo Presume-se a existecircncia de analogia do vanadato dimeacuterico com
o pirofosfato embora natildeo haja evidecircncia estrutural (Refordf) A dimerizaccedilatildeo do V2 origina o
vanadato tetrameacuterico ou metavanadato (V4O124-
- V4) que constitui a espeacutecie maioritaacuteria em
soluccedilotildees aquosas concentradas (Refordf) As estruturas do metavanadato e a sua espeacutecie protonada
satildeo ciacuteclicas [AJ] Outra espeacutecie oligomeacuterica importante eacute o vanadato pentameacuterico (V5O153-
) cuja
estrutura presumida eacute ciacuteclica [AI]
O oligoacutemero com maior importacircncia bioquiacutemica eacute o vanadato decameacuterico (V10O286-
-V10)(Refordf) a
espeacutecie predominante em soluccedilotildees de pH 2-6 quando a concentraccedilatildeo total de vanadato eacute superior
a 100 M (Ref) A estrutura do V10 natildeo difere em soluccedilatildeo ou no estado soacutelidoOs 10 aacutetomos de
vanaacutedio da espeacutecie decameacuterica estatildeo divididos em trecircs grupos identificados na figura como VA-C
consoante o seu ambiente quiacutemico circundante (Refordf) (Fig 111)
A expressatildeo ldquodecavanadatordquo refere-se a uma soluccedilatildeo aquosa na qual a espeacutecie decameacuterica se
encontra em equiliacutebrio quiacutemico com as espeacutecies V1 V2 V4 e V5 sendo que a oligomerizaccedilatildeo
progressiva do vanaacutedio eacute proporcional agrave sua concentraccedilatildeo total (Ref) Nesta soluccedilatildeo a
concentraccedilatildeo de V10 corresponde a aproximadamente 10 da concentraccedilatildeo total de vanaacutedio [AK]
18
Figura 111 ndash Estruturas esquemaacuteticas das espeacutecies de vanadato (espeacutecie monomeacuterica (V1) dimeacuterica (V2)
tetrameacuterica (V4) pentameacuterica (V5) e decameacuterica (V10) (Adaptado de Aureliano e Crans 2008)
122 Metabolismo e efeito na actividade enzimaacutetica
Apesar de natildeo terem sido ainda encontradas as provas categoacutericas das suas funccedilotildees nos seres
vivos sabe-se que o iatildeo vanadato eacute capaz de inibir uma grande quantidade de enzimas in vitro
com especial ecircnfase na actividade ATPaacutesica da miosina [AL Teresa] Como referido
anteriormente o vanadato eacute capaz de actuar em alternativa ao fosfato nas reacccedilotildees bioquiacutemicas
incluindo a sua preferecircncia de ligaccedilatildeo agraves enzimas [W]
Eacute conhecido o papel do vanaacutedio no metabolismo do caacutelcio promovendo a mineralizaccedilatildeo dos
dentes e ossos na biossiacutentese de liacutepidos (trigliceacuteridos e colesterol) com posterior inibiccedilatildeo da
acumulaccedilatildeo destes nos vasos sanguiacuteneos e no SNC na reproduccedilatildeo e desenvolvimento por
mimetizaccedilatildeo e potenciaccedilatildeo de factores de crescimento na estimulaccedilatildeo do consumo de glucose e
da siacutentese de glicogeacutenio com actuaccedilatildeo equivalente aos efeitos da insulina ao niacutevel celular na
transduccedilatildeo de sinais celulares com regulaccedilatildeo da actividade de segundos mensageiros no
transporte de iotildees e aminoaacutecidos na transcriccedilatildeo geneacutetica e siacutentese de proteiacutenas e DNA entre
outros [W Y AB]
Assim o vanaacutedio cnseguiraacute provocar inuacutemeras mudanccedilas bioquiacutemicas nas ceacutelulas Alguns
exemplos da influecircncia bioquiacutemica do vanaacutedio incluem os seus efeitos nos processos de
fosforilaccedilatildeo e desfosforilaccedilatildeo de proteiacutenas e na participaccedilatildeo na formaccedilatildeo de espeacutecies reactivas de
oxigeacutenio devido ao seu efeito proacute-oxidante sobre agentes redutores a niacutevel celular [AM]
Quanto ao seu efeito na actividade enzimaacutetica pode dividir-se em dois tipos principais ndash inibitoacuterio
e estimulante - dependendo da enzima ou do tipo de reacccedilatildeo por esta catalisada No primeiro
grupo incluem-se enzimas como as ATPases (de miosina de canais ioacutenicos e fosfoenzimas
19
transportadoras) e as fosfatases (da tirosina aacutecida e alcalina) a fosfofrutocinase adenilato ciclase
e catalase entre outras [AA] O segundo grupo inclui a maioria das cinases a glucose-6-fosfato
desidrogenase a fosfoglicerato mutase e a fosfoglicerato fosfatase (Refordf)
O efeito do vanaacutedio (V) na actividade enzimaacutetica estaacute relacionado com o seu nuacutemero de
coordenaccedilatildeo podendo actuar como estado de transiccedilatildeo ou por mimetizaccedilatildeo de cofactores
substratos (especialmente o fosfato) e interacccedilotildees proteiacutena-proteiacutena (como no caso das
polimerases e transcriptases) Pode ainda estar envolvido na formaccedilatildeo de ligaccedilotildees de hidrogeacutenio
interacccedilotildees electrostaacuteticas esteacutereas e de oxidaccedilatildeo-reduccedilatildeo Uma evidecircncia desta uacuteltima eacute a sua
capacidade de alternacircncia entre os estados de oxidaccedilatildeo +5 a +3 o que torna faacutecil a sua interacccedilatildeo
redox com enzimas contendo resiacuteduos de cisteiacutenas (Cys) ou qualquer outro composto orgacircnico
com actividade redox [AN]
13 Vanaacutedio e muacutesculo esqueleacutetico
A primeira evidecircncia da presenccedila de vanaacutedio no muacutesculo esqueleacutetico proveio de uma amostra de
ATP isolado de muacutesculo de cavalo que continha uma impureza ndash vanaacutedio ndash cuja presenccedila
provocou a inibiccedilatildeo da actividade da bomba de Na+K
+-ATPase [AO] Desde entatildeo vaacuterios
estudos tecircm sido realizados com ecircnfase na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o
tecido muscular Destes destaca-se a descoberta da interacccedilatildeo forte da espeacutecie V10 com a miosina
fosforilada [AP] e agrave sua inibiccedilatildeo da actividade ATPaacutesica da miosina[AQ AR]
No quediz respeitoao iatildeo monovanadato destaca-se a sua inibiccedilatildeoda actividade Mg2+
-ATPaacutesica
da miosina [AS] que ocorre de duas formas distintas (1) na presenccedila de concentraccedilotildees reduzidas
de V1 a inibiccedilatildeo ocorre a velocidade reduzida dependente da concentraccedilatildeo da espeacutecie
monomeacuterica (2) na presenccedila de concentraccedilotildees elevadas de V1 o niacutevel de inibiccedilatildeo aproxima-se de
100 desde que [V1] seja superior agrave concentraccedilatildeo total de locais com actividade Mg2+
-ATPaacutesica
no ensaio Esta inibiccedilatildeo eacute irreversiacutevel tanto por diaacutelise ou adiccedilatildeo de agentes redutores do
vanadato [AL]Segundo Pate et al(1994)[AT] a espeacutecie monomeacuterica podeinibir a velocidade de
contracccedilatildeo a temperatura inferior a 20ordmC perdendogradual e rapidamente essa capacidade ateacute os
50ordmC temperatura agrave qual cessa a capacidade inibitoacuteria Os resultados obtidos sugeremainda uma
interacccedilatildeo altamente cooperativa entre o iatildeo vanadato e as proteiacutenas componentes do muacutesculo
esqueleacuteticoDestaca-se tambeacutem a descoberta da inibiccedilatildeo da geraccedilatildeo de forccedila contraacutectil nas fibras
musculares por parte da espeacutecie monomeacuterica[AT]Wilson et al(1995) [AU ]determinaram que a
afinidade da espeacutecie V1 para o S1 de miosina eacute significativamente mais elevada do que a do iatildeo
Pi seu anaacutelogo Descobriram ainda que que a ligaccedilatildeo do vanadato poderaacute envolver transiccedilotildees
entre estados energeacuteticos com energia livre independente dos impedimentos esteacutereos impostos
20
pela estrutura das miofibrilas Os estudos desenvolvidos por Smith et al (1990)[AV]
determinaram queo iatildeo monovanadato natildeo apresenta um efeito pronunciado na inibiccedilatildeo da
actividadeacto-S1-miosina-ATPaacutesica excepto em concentraccedilotildees superiores a 900microM a partir das
quais existe produccedilatildeo de espeacutecies oligomeacutericas para as quais o efeito inibitoacuterio seraacute mais
elevadoA espeacutecie monomeacuterica de vanadato consegue adoptar facilmente uma geometria do tipo
bipiramidal trigonal que mimetiza o intermediaacuterio formado (MgADP
Pi)aquando da reacccedilatildeo de
hidroacutelise de ATP no local activo da miosina [AW] De facto a interacccedilatildeo do vanaacutedio com a
miosina eacute inesperadamente estaacutevel na presenccedila de ADP Esta interacccedilatildeo resulta na formaccedilatildeo de
um complexo enzimaacutetico inactivo e estaacutevel com estequiometria de 11 ADPvanadato por siacutetio
activo da proteiacutena A formaccedilatildeo do complexo estaacutevel miosina-vanadato foi proposta por Goodnoet
al (1982) [AS] Nesta ocorre um preacute-equiliacutebrio raacutepido que induz a formaccedilatildeo de um complexo
ternaacuterio reversiacutevel (MADP
V) com formaccedilatildeo lenta do isoacutemero M
ADP
V A diferenccedila entre os
compostos de vanaacutedio e de fosfato centra-se no tempo de meia vida (t12) do Pi ou do V (no
primeiro t12=12 s enquanto no segundo t12=24 h) (Refordf) A diferenccedila dever-se-aacute agrave possiacutevel
complexaccedilatildeo do vanaacutedio com grupos nucleofiacutelicos no siacutetio activo o que provocaria uma
estabilizaccedilatildeo da miosina na conformaccedilatildeo MADP
V
No que respeita aos estudos realizados com a espeacutecie tetrameacuterica Cremo et al determinou que a
cadeia pesada do S1 de miosina pode ser clivada em dois locais distintos (designados V1 e V2)
pela espeacutecie V4 apoacutes irradiaccedilatildeo com luz UV na presenccedila de concentraccedilotildees de vanadato na ordem
dos milimolar e na ausecircncia de nucleoacutetidos ou metais divalentes
Jaacute no que concerne aos estudos realizados com ecircnfase na espeacutecie decameacuterica foi sugerido
porRamos et al (2006) [AK]que a interacccedilatildeo do decavanadato com a G-actina favorecida pela
polimerizaccedilatildeo desta uacuteltima estabiliza a espeacutecie V10 e induz a inibiccedilatildeo da formaccedilatildeo de F-actina A
estabilizaccedilatildeo da espeacutecie V10 por proteiacutenas tansmembranares e do citoesqueleto pode ser a razatildeo
pela qual ocorre a toxicidade do decavanadato reportada na avaliaccedilatildeo dos efeitos do V(V) nos
sistemas bioloacutegicosTiago et al (2007) [AR] concluiram que o V10 apresenta uma clara preferecircncia
de ligaccedilatildeo ao ldquoback-doorrdquo da S1-miosina-ATPase o que se verifica apenas na estrutura
conformacional desimpedida com acesso facilitado ao local de ligaccedilatildeo do Pi Sugere tambeacutem que
o V10 actua como um ldquoback-door stoprdquo bloqueando estaabertura do fragmento de 50-kDa
necessaacuteria para a hidroacutelise do ATP--fosfato o que constitui uma explicaccedilatildeo simples do
comportamento natildeo competitivo da espeacutecie decameacuterica estimulando a utilizaccedilatildeo deste oligoacutemero
como uma feramenta da elucidaccedilatildeo das mudanccedilas conformacionais ldquoback-doorrdquo da miosina no
21
processo da contracccedilatildeo muscular(ref)Destaca-se ainda a descoberta da capacidade inibitoacuteria do
V10 em relaccedilatildeo agrave fosforilaccedilatildeo oxidativa mitocondrial [AX]
Finalmente no que concerne aos estudos realizados com a presenccedila de vaacuterias espeacutecies
oligomeacutericas Aureliano e Madeira (1994) [AY]observaram que na presenccedila de Ca2+
as espeacutecies
V4 e V10se ligam agrave Ca2+
-ATPase do retiacuteculo sarcoplasmaacutetico enquanto a espeacutecie V1 se liga apenas
na presenccedila de ATP Aparentemente o aumento da concentraccedilatildeo de Ca2+
tem o efeito contraacuterio da
inibiccedilatildeo da ATPase do retiacuteculo sarcoplasmaacutetico pelo V1 natildeo tendo no entanto efeito significativo
sobre a inibiccedilatildeo por parte da espeacutecie V10 Tiago et al (2002) [AP] concluiacuteram que em condiccedilotildees
proacuteximas agraves fisioloacutegicas a espeacutecie V10 difere das espeacutecies presentes na soluccedilatildeo de metavanadato
devido agrave sua forte interacccedilatildeo com a miosina fosforilada e agrave sua inibiccedilatildeo da actividade ATPaacutesica
da miosina Para aleacutem disso o ATP diminui a afinidade da miosina para o V4 induzindo a
interacccedilatildeo com V1 e natildeo tendo qualquer efeito no que diz respeito ao V10 Foi tambeacutem proposto
[AQ] que a ligaccedilatildeo das espeacutecies oligomeacutericas de vanadato aoS1 de miosina nos locais onde o
fosfato coordenaeacute provavelmente a causa da inibiccedilatildeo da actividade ATPaacutesica da miosina
estimulada por actina atraveacutes da prevenccedilatildeo da troca entre ATPADP sendo esta favorecida para
aniotildees de vanadato com valecircncias superiores como eacute o caso do V10
Na presenccedila de actina as velocidades de dissociaccedilatildeo do Pi e do V aumentam consideravelmente
este uacuteltimo em menor quantidade Assim a ligaccedilatildeo da actina ao complexo MADP
V e
Pi eacute
bastante fraca pelo que se sugere que o ADPV teria um efeito de relaxamento semelhante ao
ATP No entanto a capacidade do vanaacutedio em inibir a actividade ATPaacutesica da miosina e da
actomiosina eacute diferenciada uma vez que na ausecircncia de actina a inibiccedilatildeo eacute total para
concentraccedilotildees na gama dos M mas na sua presenccedila a inibiccedilatildeo eacute totalmente conseguida apenas
em concentraccedilotildees na gama dos mM (Ref) No entanto eacute necessaacuterio considerar a possiacutevel
formaccedilatildeo de espeacutecies oligomeacutericas de vanadato cuja afinidade e efeito na enzima seratildeo
diferentes Assim pode concluir-se que o efeito de inibiccedilatildeo observado para a actividade ATPaacutesica
de miosina estimulada por actina se pode dever agrave presenccedila das diferentes espeacutecies oligomeacutericas o
que foi jaacute sugerido por Aureliano (2000) [AZ] ndash as espeacutecies tetrameacuterica e decameacuterica de vanadato
interactuariam com a miosina com alteraccedilatildeo da sua interacccedilatildeo com a actina No entanto natildeo eacute
conhecida a contribuiccedilatildeo absoluta das restantes espeacutecies oligomeacutericas para a inibiccedilatildeo da ATPase
da miosina estimulada por actina (Refs)
14 Objectivos do projecto
Pretende-se com este projecto o isolamento purificaccedilatildeo e caracterizaccedilatildeo da actina do muacutesculo
22
esqueleacutetico de coelho sendo esta uacuteltima tambeacutem realizada para o sufragmento-1 de miosina
Seguidamente realizar-se-aacute a quantificaccedilatildeo e caracterizaccedilatildeo das proteiacutenas por espectroscopia UV-
Vis e SDS-PAGE e a caracterizaccedilatildeo das soluccedilotildees de vanaacutedio por UV-Vis e ressonacircncia magneacutetica
nuclear (RMN) Por uacuteltimo a interacccedilatildeo do S1 de miosina com o vanaacutedio seraacute avaliada atraveacutes
das teacutecnicas de UVVis RMN e ressonacircncia paramagneacutetica electroacutenica (RPE)
2 Discussatildeo de Resultados
21 Caracterizaccedilatildeo das proteiacutenas isoladas
211Anaacutelise do estado de pureza
O estado de pureza das soluccedilotildees de actina e de S1 foi analisado por electroforese em gel de
poliacrilamida em condiccedilotildees desnaturantes (SDS-PAGE) (Fig 22) que separa as moleacuteculas de
acordo com o seu peso molecular (PM) A concentraccedilatildeo de acrilamida escolhida (10) permite a
detecccedilatildeo simultacircnea de componentes com peso molecular inferior a 20 kDa e outros com elevado
peso molecular
Figura 21 ndash Fotografia de gel de SDS-PAGE (10 acrilamida) Poccedilo (a)
10 L do marcador de elevado peso molecular () Poccedilo (b) 20 L de [G-
actina]=1 mgmL Poccedilo (c) 20 L de [S1]=1 mgmL Aplicou-se uma
voltagem de V durante h e min
A partir da recta de calibraccedilatildeo obtida log PM = -1177 Rf + 2247 (ver ponto 2 da secccedilatildeo
Anexos) pode verificar-se a presenccedila de actina na amostra do poccedilo (b) presente numa banda
bastante intensa (PMaparente=40 kDa) bem como a presenccedila minoritaacuteria de outros componentes
desconhecidos que natildeo teratildeo sido eliminados durante o processo de extracccedilatildeo e purificaccedilatildeo e natildeo
satildeo relevantes para o estudo em questatildeo Na amostra do poccedilo (c) nota-se a presenccedila de S1
(PMaparente=92 kDa) bem como a de outros componentes cuja banda eacute ligeiramente menos intensa
Estes poderatildeo ser fragmentos proteoliacuteticos de miosina e de S1 Estaacute ainda presente uma bada de
23
maior intensidade que poderaacute ser a proteiacutena-C (PM=135 kDa) uma vez que a sua remoccedilatildeo
completa da amostra necessitaria de um passo adicional de cromatografia de troca ioacutenica [BA]
No entanto natildeo podemos confirmar que se trata desta proteiacutena ou de qualquer outra uma vez que
se encontra fora do intervalo de migraccedilatildeo dos marcadores de peso molecular
212 Determinaccedilatildeo da concentraccedilatildeo proteica
2121Subfragmento-1 de miosina
A partir da recta de calibraccedilatildeo obtida pelo meacutetodo de Bradford [BB](ver secccedilatildeo 3311 do
Procedimento Experimental) A595nm = 00546[BSA] + 00209 (ver secccedilatildeo de Anexos)foi possiacutevel
determinar um intervalo de concentraccedilatildeo de S1 de miosina compreendido entre 226 e 117(plusmn
423M) (203 e 1056 (plusmn 381 mgmL)) (n=8) A concentraccedilatildeo meacutedia de S1 de miosina apoacutes
diaacutelise contra a soluccedilatildeo tampatildeo 10 mM HEPES pH 75 25 mM KCl determinada pela leitura da
Abs280 nmfoi de 20430M (1803 mgmL)(n=7)
As amostras a quantificar atraveacutes do meacutetodo de Bradford natildeo sofreram diaacutelise o que permite
explicar a concentraccedilatildeo superior de S1 de miosina em comparaccedilatildeo com o valor obtido atraveacutes do
meacutetodo directo Esta razatildeo aliada agrave sensibilidade e rapidez superiores deste uacuteltimo fez com que
se optasse pelo meacutetodo directo para a quantificaccedilatildeo da grande maioria dos ensaios
2122Actina
A concentraccedilatildeo de actina foi determinada quando esta se encontrava na forma monomeacuterica (G-
actina) por mediccedilatildeo da leitura directa da Abs290 nm tendo-se obtido uma concentraccedilatildeo meacutedia de
691481M (290202 mgmL) (n=3) em 2 mM Tris-HCl pH 80 02 mM CaCl2 02 mM Na-
ATP Ao comprimento de onda utilizado (290 nm) absorvem fortemente apenas os resiacuteduos de
Trp pelo que a mediccedilatildeo efectuada tem menor sensibilidade do que se tivesse sido efectuada a 280
nm uma vez que a actina apresenta apenas 4 resiacuteduos deste aminoaacutecido embora tenha 16
resiacuteduos de Tyr
22 Mediccedilatildeo da actividade enzimaacutetica de S1 de miosina
221 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
As actividades ATPaacutesicas de S1 de miosina dependentes de Ca2+
e K+foram analisadas pelo
caacutelculo das velocidades de consumo inicial do fosfato inorgacircnico (Pi) Definiu-se os tempos de 5
e 3 minutos para a mediccedilatildeo da Ca2+
e da K+EDTA-ATPase respectivamente Os valores obtidos
24
a partir do protocolo presente no ponto 341 da secccedilatildeo Procedimento experimental encontram-se
descritos na Tabela 2I
Tabela 2I ndash Acrividades natildeo-fisioloacutegicas de subfragmento-1 determinadas pelo meacutetodo de Fiske-Subarow
modificado (n=2)
Proteiacutena
Actividade
ATPaacutesica natildeo
fisioloacutegica
Actividade (nmol
Piminmg proteiacutena) Actividade (s-1)
S1
Ca2+
-ATPase 1400 plusmn 30 2100 plusmn 0045
K+EDTA-ATPase 2334 plusmn 50 3500 plusmn 0075
As actividades natildeo-fisioloacutegicas reflectem o estado de oxidaccedilatildeo dos resiacuteduos expostos de Cys 707
(SH1) e 697 (SH2) Se o grupo SH1 sofrer oxidaccedilatildeo a actividade ATPaacutesica dependente de Ca2+
aumenta e a actividade dependente de K+ diminui Por outro lado se o grupo SH2 sofrer
oxidaccedilatildeo regista-se a perda completa destas actividades
De acordo com Margossian e Lowey (1982) [BA] as actividades ATPaacutesica de S1 de miosina
dependente de Ca2+
e K+EDTA da miosina satildeo respectivamente de 24 e 112 s-1
Estaacute tambeacutem
descrito que a actividade ATPaacutesica dependente de K+ eacute em meacutedia cerca de 3 vezes superior agrave
dependente de Ca2+
reflectindo a quantidade das preparaccedilotildees A razatildeo obtida entre os valores das
actividades ATPaacutesicas dependentes de K+ e Ca
2+ eacute de 166 Os valores obtidos para as actividades
(Ca2+
-ATPase=21 s-1
K+EDTA-ATPase= 35 s
-1) satildeo inferiores agrave valores obtidos por
Margossian e Lowey [BA] e Tiago [BC] com forccedila ioacutenica inferior O valor obtido para a
actividade ATPaacutesica dependente de Ca2+
eacute superior ao obtido por Polosukhina et al (2000) [BD]
cujo meio de reacccedilatildeo continha MOPS e apresentava forccedila ioacutenica inferior Os dados
supramencionados comprovam a variaccedilatildeo da actividade a ATPaacutesica natildeo fisioloacutegica em relaccedilatildeo agrave
forccedila ioacutenica temperatura e pH
222 Mediccedilatildeo da actividadeMg2+
-ATPaacutesica de S1 pelo meacutetodo de enzimas acopladas
A actividade de Mg2+
-ATPase eacute a uacutenica que possui significado fisioloacutegico sendo as restantes
bastante uacuteteis apenas para indicaccedilatildeo da qualidade de preparaccedilatildeo do S1[BA] A 0575 M KCl a
actina e o S1 formam uma soluccedilatildeo viscosa e tuacuterbida Apoacutes a adiccedilatildeo de MgATP o complexo acto-
S1 dissocia-se e a turbidez da soluccedilatildeo diminui Se a forccedila ioacutenica for reduzida a dissociaccedilatildeo do
complexo actoS1 que se encontra na forma de suspensatildeo eacute induzida O MgATP eacute rapidamente
hidrolisado diminuindo a sua concentraccedilatildeo no meio reaccional Nestas condiccedilotildees haacute
reassociaccedilatildeo e subsequente contracccedilatildeo dos filamentos de actina e de S1 com formaccedilatildeo de
25
precipitado O meacutetodo descrito traduz de forma bastante elementar o fenoacutemeno da contracccedilatildeo in
vivo e elimina o problema anterior da solubilizaccedilatildeo ineficiente da miosina de modo a obter uma
mediccedilatildeo exacta da actividade [A] A utilizaccedilatildeo de forccedilas ioacutenicas reduzidas aumenta a afinidade da
actina para o S1 permitindo contornar a extrema dificuldade em obter a concentraccedilatildeo proteica dos
muacutesculos (cerca de 100 mgmL do complexo actomiosina) As actividades Mg2+
-ATPaacutesicas de S1
de miosina foram determinadas a partir do declive da variaccedilatildeo da Abs340 relativo agrave oxidaccedilatildeo de
NADH em funccedilatildeo do tempo como eacute descrito no ponto 342 do Procedimento Experimental Os
valores obtidos para a actividade Mg2+
-ATPaacutesica encontram-se apresentados na Tabela 2II
Tabela 2II ndash Actividade Mg2+
-ATPaacutesica de subfragmento-1 de miosina determinadas pelo meacutetodo das enzimas
acopladas
Actividade (nmol
ATPmin mg(S1)) Actividade (s
-1)
1097 plusmn 269 164 plusmn 04
Ramos (2005) [J] obteve o valor de 01 s-1
para a actividade Mg2+
-ATPaacutesica de S1 de miosina
com forccedila ioacutenica intermeacutedia Tiago (2005) [BC] obteve o valor de 011 s-1 em condiccedilotildees de forccedila
ioacutenica reduzida em relaccedilatildeo agrave utilizada no presente relatoacuterio
23 Preparaccedilatildeo e caracterizaccedilatildeo das soluccedilotildees de vanaacutedio
231 Anaacutelise da composiccedilatildeo das soluccedilotildees de vanadato por espectroscopia de 51
V-RMN
efeito da concentraccedilatildeo da espeacutecie
A diluiccedilatildeo das soluccedilotildees stock de meta- e decavanadato iraacute determinar fortemente a composiccedilatildeo
em soluccedilatildeo dos diferentes oligoacutemeros de vanadato (ref) No caso do decavanadato o periacuteodo de
tempo apoacutes a diluiccedilatildeo iraacute tambeacutem ser um factor importante a considerar devido agrave decomposiccedilatildeo
da espeacutecie V10(ref)Torna-se assim fundamental determinar a composiccedilatildeo quiacutemica das soluccedilotildees
no meio de ensaio utilizado (HEPES 10 mM KCl 25 mM) de modo a correlacionaacute-la
correctamente com os possiacuteveis efeitos promovidos pelas diferentes espeacutecies de vanadato no
sistema em estudo
Aleacutem disso deve ser dado ecircnfase agraves importantes interacccedilotildees que ocorrem entre o vanaacutedio e os
tampotildees escolhidos O tampatildeo com menor nuacutemero e magnitude de interacccedilatildeo eacute o HEPES [AD]
seguido do Tris sendo este uacuteltimo especialmente aconselhado em concentraccedilotildees baixas para casos
de incompatibilidade enzimaacutetica ou quiacutemica redox No caso especial da reacccedilatildeo redox
NAD(P)HNAD(P) o tampatildeo adequado eacute o imidazole de modo a natildeo haver reacccedilatildeo com o
26
vanadato Os tampotildees a evitar em qualquer situaccedilatildeo incluem a trietanolamina fosfato citrato e
glicina entre outros [AD] uma vez que todos interactuam fortemente com V(V) com formaccedilatildeo
de complexos estaacuteveis
Assim procedeu-se agrave anaacutelise da composiccedilatildeo das soluccedilotildees de meta- e decavanadato apoacutes diluiccedilatildeo
das soluccedilotildees stock (50 mM em vanadato total) no meio de reacccedilatildeo (tampatildeo D) atraveacutes da
espectroscopia de ressonacircncia magneacutetica nuclear de 51
V Nas figuras x1 e x2 apresentam-se os
espectros obtidos para as soluccedilotildees de deca- e metavanadato num intervalo de concentraccedilotildees de 1 a
10 mM A Tabela seguinte resume os paracircmetros espectroscoacutepicos obtidos para cada espeacutecie
presente nas soluccedilotildees analisadas
Figura 22ndash Espectros de RMN de 51
V de
concentraccedilotildees crescentes das espeacutecies oligomeacutericas
de vanadato obtidos apoacutes diluiccedilatildeo da soluccedilatildeo stock
de decavanadato em 10 mM HEPES pH 70 25 mM
KCl para concentraccedilotildees finais de A) 1 mM B)2
mM C) 4 mM D) 8 mM E) 10 mM de
decavanadato total V1refere-se agraves espeacutecies
monomeacutericade vanadato V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2IIIndash Desvios quiacutemicos meacutedios (part) das ressonacircncias das espeacutecies decameacuterica (V10) monomeacuterica (V1) e
tetrameacuterica (V4) presentes nos espectros da soluccedilatildeo de decavanadatoLargura a meia altura (∆12) da espeacutecie V10A
Paracircmetros V10A V10B V10C V1 V4
part (ppm) -5162 -5003 -4255 -5616 -5748
∆12 (Hz) 175 nd nd nd nd
Figura 23 ndash Distribuiccedilatildeo graacutefica das
espeacutecies oligomeacutericas de vanadato vs a
concentraccedilatildeo de decavanadato total As
concentraccedilotildees de cada espeacutecie foram
obtidas segundo a formula descrita na
secccedilatildeo 35 do Procedimento
Experimental (aacutereas dos sinais de 51
V-
RMN observadas na figura x1)V10 V1 e
V4 correspondem agraves espeacutecie monomeacuterica
dimeacuterica e tetrameacuterica respectivamente [Esp
eacuteci
e o
ligo
meacute
rica
de
van
aacutedio
] (m
M)
[Decavanadato]total (mM)
V10
V1
V4
27
Nas soluccedilotildees de decavanadato as espeacutecies monomeacuterica e decameacuterica de vanadato satildeo as uacutenicas
presentes para concentraccedilotildees de decavanadato inferiores a 8 mM Ambas as espeacutecies aumentam
linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado em [AY] e
em (Teresa) quando a concentraccedilatildeo de V1 eacute igual ou superior a 200 microM inicia-se a sua
oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica No entanto nas condiccedilotildees de realizaccedilatildeo dos
ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a espeacutecie tetrameacuterica neste
estudo em concentraccedilotildees inferiores a 8 mM tendo o mesmo sido observado por Ramoset al
(2006) [AK] Os resultados obtidos mostram claramente o equiliacutebrio entre a espeacutecie decameacuterica e
a monomeacuterica e que o aparecimento de outras espeacutecies oligomeacutericas se encontra dependente da
concentraccedilatildeo desta uacuteltima Apesar de natildeo ter sido realizado o ensaio com 5 mM de decavanadato
total no presente relatoacuterio estaacute reportado em Ramos et al (2006) [AK] que as concentraccedilotildees da
espeacutecie decameacuterica e monomeacuterica satildeo respectivamente de 500 microM e 200 microM Assim pode inferir-
se que para concentraccedilotildees inferiores a 5 mM e intervalos de tempo de cerca de 20 minutos (que
inclui o tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas
as espeacutecies decameacuterica e monomeacuterica de vanadato
Figura 24 ndash Espectros de RMN de 51
V de concentraccedilotildees
crescentes das espeacutecies oligomeacutericas de vanadato obtidos
apoacutes diluiccedilatildeo da soluccedilatildeo stock de metavanadato em 10 mM
HEPES pH 70 25 mM KCl para concentraccedilotildees finais de A)
1 mM B) 2 mM C) 4 mM D) 8 mM E) 10 mM de
metavanadato total V1 refere-se agraves espeacutecies monomeacuterica de
vanadatoV1 e V2 correspondem respectivamente agrave espeacutecie
monomeacuterica e dimeacuterica
Figura 25 ndash Distribuiccedilatildeo graacutefica das espeacutecies
oligomeacutericas de vanadato vs a concentraccedilatildeo de
metavanadato total As concentraccedilotildees de cada
espeacutecie foram obtidas segundo a formula descrita
na secccedilatildeo 35 do Procedimento Experimental
(aacutereas dos sinais de 51
V-RMN observadas na
figura x1)V10 V1 e V4 correspondem agraves espeacutecie
monomeacuterica dimeacuterica e tetrameacuterica
respectivamente
Esp
eacuteci
e o
ligo
meacute
rica
de
va
nad
ato
(m
M)
[Metavanadato]total (mM)
V1
V2
V4
V5
28
Tabela 2IVndash Desvios quiacutemicos meacutedios (part)das ressonacircncias das espeacutecies tetrameacuterica (V4) monomeacuterica (V1) e
dimeacuterica (V2) presentes no espectro da soluccedilatildeo de metavanadato (total) 10 mM
Paracircmetros V1 V2 V4 V5
part (ppm) -5615 -5748 -5789 -5869
Nas soluccedilotildees de metavanadato as espeacutecies monomeacuterica dimeacutericae tetrameacuterica de vanadato estatildeo
presentes para concentraccedilotildees de metavanadato inferiores a 4 mM Todas estas as espeacutecies
aumentam linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado
em (Aureliano e Madeira 1998) e em (Tiago et al 2004) quando a concentraccedilatildeo de V1 eacute igual ou
superior a 200 microM inicia-se a sua oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica Nas
condiccedilotildees de realizaccedilatildeo dos ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a
espeacutecie pentameacuterica neste estudo em concentraccedilotildees inferiores a 4 mM tendo o mesmo sido
observado por Ramos 2006 Os resultados obtidos mostram claramente o equiliacutebrio entre a
espeacutecie V4 e as espeacuteciesV1 e V2 no qual o aparecimento de outras espeacutecies oligomeacutericas se
encontra dependente da concentraccedilatildeo da espeacutecie V1 Por uacuteltimo pode inferir-se que para
concentraccedilotildees inferiores a 2 mM e intervalos de tempo de cerca de 20 minutos (que inclui o
tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas as
espeacutecies V1 V2 e V4 com concentraccedilotildees respectivas de 0796 0192 e 0196 mM
232Estabilidade das soluccedilotildees de decavanadato anaacutelise por espectroscopia de absorccedilatildeo
UVVis
Caracterizou-se a estabilidade da espeacutecie V10 numa soluccedilatildeo com 1 mM decavanadato total em
tampatildeo D atraveacutes da leitura da Abs400 Obteve-se uma cineacutetica de dissociaccedilatildeo de 1ordf ordem tal
como se pode verificar pela linearidade do graacutefico de ln(Abs400)vs o tempo de aquisiccedilatildeo espectral
(segundos) (Fig26 )
Figura 26ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do tempo
(em segundos) para uma soluccedilatildeo de decavanadato total de 1
mM em tampatildeo D a 25ordmC (n=2)
Sendo uma reacccedilatildeo de primeira ordem pode ser descrita pela equaccedilatildeo cujo
declive eacute ndashK (constante de dissociaccedilatildeo) e a intersecccedilatildeo no eixo das ordenadas corresponde
Ln
(Ab
s 400 n
m)
tempo (s)
29
aln(A0) Numa reacccedilatildeo de 1ordf ordem o tempo de meia-vida eacute independente da concentraccedilatildeo inicial
do reagente sendo descrito pela equaccedilatildeo [2] Deste modo a partir da recta
Ln(Abs400 nm) = -436E-5
K - 105 obteve-se um tempo de meia-vida de 4490 plusmn 0823 horas para
as condiccedilotildees supramencionadas
Apoacutes a adiccedilatildeo de S1 8 microM ao meio reaccional verificou-se que a reacccedilatildeo de desoligomerizaccedilatildeo
da espeacutecie decameacuterica de vanadato seguia igualmente uma cineacutetica de 1ordf ordem tal como se pode
verificar pela linearidade do graacutefico de ln(Abs400) versus o tempo de aquisiccedilatildeo espectral
(segundos) (Fig27)
Figura 27ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do
tempo (em segundos) para uma soluccedilatildeo de decavanadato
total de 1 mM em tampatildeo D a 25ordmC na presenccedila de 8
microM (n= 2)
A partir da equaccedilatildeo da recta Ln(Abs400 nm) = -7520E-5
K - 0613determinou-se o valor de K para
a espeacutecie decameacuterica de onde se obteve um tempo de meia-vida de 2430 plusmn 0191 horas para as
condiccedilotildees supramencionadas
Tanto este valor como o valor registado na ausecircncia de S1 satildeo substancialmente superiores agrave
duraccedilatildeo dos ensaios de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina descritos no presente
relatoacuterio
24 Interacccedilatildeo de S1 com vanaacutedio(V)
241Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
De modo a analisar qual o efeito da adiccedilatildeo de S1 de miosina (tampatildeo D) produzia nos sinais de
51V-NMR titulou-se uma soluccedilatildeo de 5mM de decavanadato total com concentraccedilotildees crescentes
de S1 (5 10 15 e 20 microM) Os espectros de 51
V-RMN encontram-se apresentados na Fig 2
No graacutefico da figura 2 Apresenta-se a concentraccedilatildeo de S1 em funccedilatildeo do factor de alargamento
corrigido dos espectros acima referidos
Ln
(Ab
s (4
00
nm
))
tempo (s)
30
Figura 28 ndash Graacutefico de
representaccedilatildeo da
concentraccedilatildeo de S1 em
funccedilatildeo do factor de
alargamento A
concentraccedilatildeo de
decavanadato total foi de 5
mM e as concentraccedilotildees
utilizadas de S1 foram de 0
5 10 e 15 microM
Verifica-se que o factor de alargamento aumenta linearmente em funccedilatildeo da concentraccedilatildeo de S1 no
ensaio o que indica a existecircncia de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina Observou-
se ainda que a adiccedilatildeo de S1 ao meio reaccional natildeo provocou qualquer alteraccedilatildeo dos desvios
quiacutemicos dos sinais de 51
V-RMN
Realizaram-se ensaios na presenccedila de ATP e de S1 com intuito de verificar o efeito do ATP na
interacccedilatildeo do S1 de miosina com o decavanadato
Figura 29 - Espectros de RMN de 51
V contendo 2
mM Decavanadato (A) (controlo) (B) na presenccedila
de 2 mM de ATP (C) na presenccedila de 2 mM de ATP
e 8 microM de S1 de miosina V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2Vndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies decameacuterica (V10) e monomeacuterica (V1) presentes nos
espectros da soluccedilatildeo de decavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V10A V10B V10C V1
part (ppm) -5161 -5002 -4254 -5614
Observa-se que a presenccedila de ATP (espectro B) natildeo provoca qualquer alteraccedilatildeo visiacutevel em
relaccedilatildeo ao espectro que contem apenas soluccedilatildeo de decavanadato Jaacute a presenccedila simultacircnea de ATP
e S1 provoca um alargamento da ∆12 do sinal do V10A de 1736 Hz (espectro (A)) para 2366 Hz
(espectro (C) Fig 29) Tendo em conta que a concentraccedilatildeo de S1 apresenta uma relaccedilatildeo linear
com o factor de alargamento converteu-se o valor da ∆12 e comparou-se com o valor esperado agrave
F = 00591[S1] - 00157 Rsup2 = 0998
Fact
or
de
ala
rgam
en
to
[S1] (microM)
31
concentraccedilatildeo de S1 utilizada O factor de alargamento obtido para 8 microM ndash 0376 ndash eacute inferior ao
valor esperado agrave mesma concentraccedilatildeo de S1 obtido atraveacutes da equaccedilatildeo da recta apresentada na
Fig 28 ndash 0457 ndash o que indica que o ATP poderaacute exercer um efeito de protecccedilatildeo do S1
diminuindo a sua interacccedilatildeo com a espeacutecieV10
Por uacuteltimo realizaram-se ensaios com metavanadato na presenccedila de 8 microM S1 de modo a verificar
se as espeacutecies oligomeacutericas contidas nesta soluccedilatildeo interagiam com a proteiacutena em estudo
Figura 210 - Espectros de RMN de 51
V contendo 2
mM metavanadato (A) (controlo) (B) na presenccedila
de 8 microM de S1 de miosina V1 V2 e V4
correspondem respectivamente aacutes espeacutecies
monomeacuterica dimeacuterica e tetrameacuterica
Tabela 2VIndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies monomeacuterica (V1) dimeacuterica (V2) e tetrameacuterica (V4)
presentes nos espectros da soluccedilatildeo de metavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V1 V2 V4
part (ppm) -5613 -5747 -5789
Como eacute possiacutevel observar na Fig 210 ocorre uma ligeiro alargamento dos sinal de V4 bem
como uma perda da intensidade pelo que se conclui quedas espeacutecies presentes na soluccedilatildeo de
metavanadato (V1 V2 e V4) apenas a espeacutecie tetrameacuterica interage com o S1 de miosina o que
estaacute de acordo com o reportado por Tiago et al 2002
242 Actividade Mg2+
-ATPaacutesica de S1 de miosina na presenccedila de decanadato
Com o intuito de avaliar o efeito do decavanadato na actividade Mg2+
-ATPaacutesica do S1 de
miosina monitorizou-se a reacccedilatildeo de oxidaccedilatildeo do NADH atraveacutes da leitura de Abs340 na
presenccedila de 005 mgmL de S1 de miosina e no caso da actividade estimulada 2microM de actina em
3125 mM Tris-HCl pH 71 e 375 mM MgCl2 pelo meacutetodo de enzimas acopladas (ver seccccedilatildeo xx
dos MM)Para tal realizaram-se ensaios com vaacuterias concentraccedilotildees de decavanadato total (10 50
100 250 500 e 1000) de modo a determinar qual o seu efeito na actividade MgATPaacutesica de S1
de miosina na ausecircncia e na presenccedila de um dos seus ligandos naturais Os ensaios foram
realizados agrave temperatura ambiente com incubaccedilatildeo preacutevia a 25ordm C durante 20 minutos
32
Figura 211 ndash Representaccedilatildeo
graacutefica da Actividade Mg2+
-
ATPaacutesica de S1 () (os ajustes
teoacutericos estatildeo representado a por
uma linha os resultados
experimentais estatildeo representados
por triacircngulos (Actividade
estimulada) e cruzes (Actividade
natildeo estimulada)) vs
[Decavanadato]total medidas pelo
meacutetodo das enzimas acopladas
(n=3) Os dados foram ajustados ao modelo de decaimento exponencial usando o programa GraphPad Prism para a
determinaccedilatildeo dos valores de IC50 (na ausecircncia de actina IC50= 266 plusmn 07 M decavanadato na presenccedila de actina
IC50= 28 plusmn 1 M decavanadato
A partir do graacutefico presente na Fig 29 verifica-se que o ajuste dos dados experimentais foi bem
sucedido A inibiccedilatildeo da actividade Mg2+
-ATPaacutesica do S1 de miosina eacute afectada em menor
amplitude na presenccedila de actina (60 de inibiccedilatildeo da actividade) do que na sua ausecircncia
(aproximadamente 80 de inibiccedilatildeo de actividade) Devido a esta diferenccedila pode sugerir-se que a
actina iraacute impedir parcialmente a ligaccedilatildeo do decavanadatoao S1 demiosina atraveacutes de
impedimentos esteacutereos ou interacccedilotildees electrostaacuteticas por exemplo A teacutecnica utilizada natildeo nos
permite concluir acerca de que regiatildeo(otildees) ou resiacuteduo(s) da actina participaratildeo na preservaccedilatildeo
parcial da actividade Mg2+
-ATPaacutesica
Por outro lado os valores de IC50 dos dois tipos de actividades natildeo apresentam diferenccedilas
significativas tendo-se obtido 266 plusmn 07 e 28 plusmn 1M decavanadato na ausecircncia e na presenccedila de
actina respectivamente o que corresponde a uma concentraccedilatildeo de cerca de 27 M de V10 O
meio de reacccedilatildeo utilizado nestes ensaios foi 10 mM HEPES e 25 mM KCl com 2 mM ATP Em
(Tiago et al 2004) o valor de IC50 para a actividade Mg2+-ATPaacutesica na ausecircncia de actina foi de
125 plusmn 025 microM enquanto o valor da actividade estimulada por actina foi de 136 plusmn 14 microM O
meio de reacccedilatildeo utilizado nos ensaios foi 20 mM HEPES 50 mM KCl e 2 mM ATP o que resulta
precisamente no dobro da forccedila ioacutenica a que foi realizado o ensaio do presente relatoacuterio
(exceptuando a contribuiccedilatildeo dos produtos da hidroacutelise de ATP uma vez que este meacutetodo
pressupotildee a ocorrecircncia de um ciclo de reacccedilotildees de hidroacutelise e siacutentese deste composto) Ora tendo
sido anteriormente referido que a afinidade do S1 de miosina para com a espeacutecie V10 diminui com
Act
ivid
ade
Mg
2+-A
TP
aacutesic
a d
e S
1 d
e
Mio
sin
a
[Decavanadato] (microM) Actividade Estimulada ()
Actividade Natildeo Estimulada ()
Ajuste Actividade Estimulada
Ajuste Actividade Natildeo Estimulada
33
o aumento da forccedila ioacutenica (Tiago et al 2004 tal poderaacute explicar a diferenccedila obtida entre estes
dois conjuntos de valores
243Quantificaccedilatildeo de cisteiacutenas reduzidas pelo meacutetodo de Ellman
A interacccedilatildeo do vanaacutedio com alguns tipos de aminoaacutecidos sensiacuteveis agrave oxidaccedilatildeo pode fornecer-nos
informaccedilatildeo bastante uacutetil em relaccedilatildeo agraves alteraccedilotildees estruturais que conduzem a uma diminuiccedilatildeo da
actividade fisioloacutegica do S1 de miosina Os estudos realizados por Chasteen 1983 concluiacuteram
que a reduccedilatildeo de vanadato (V) a vanadilo (IV) ocorre na presenccedila de agentes redutores comuns no
ambiente celular de que se destaca o aminoaacutecido cisteiacutena Esta eacute tambeacutem um alvo de
complexaccedilatildeo por parte do iatildeo VO2+
Jaacute Ramos et al (2009) demonstrou que a incubaccedilatildeo de
actina com a soluccedilatildeo de decavanadato induz a oxidaccedilatildeo de resiacuteduos de cisteiacutena e agrave formaccedilatildeo do
oxidovanaacutedio (IV) Este grupo de investigadores descobriu ainda que apenas a espeacutecie V10 induz a
oxidaccedilatildeo das cisteiacutenas ldquoraacutepidasrdquo da actina atraveacutes do processo proposto que eacute parcialmente
responsaacutevel pela inibiccedilatildeo jaacute descrita da actividade de ATPase do S1 de miosina estimulada por
actina quando exposta agrave soluccedilatildeo de decavanadato Esta inibiccedilatildeo apresenta-se afectada com a
presenccedila de ATP uma vez que os resiacuteduos de cisteiacutena permaneceram intactos nos ensaios
realizados com este ligando Jaacute Crans et al (2004) concluiu que o siacutetio activo de muitas fosfatases
(que conteacutem grupos tiol) pode formar um equiliacutebrio redox com o iatildeo vanadato (V) de forma natildeo
reversiacutevel
Os resultados obtidos por Sakurai e Shiromura (1980) sugeriram que o vanaacutedio (V) sofre reduccedilatildeo
a vanaacutedio (IV) sem dependecircncia do valor de pH do ensaio Registaram ainda uma estequiometria
de 31 na reacccedilatildeo do vanadato (V) com os resiacuteduos de Cys enquanto a estequiometria dos ensaios
com o vanadilo (IV) foi de 21
Berlett e Stadtman (1997) observaram que os tipos de aminoaacutecidos mais facilmente oxidaacuteveis
englobam entre outros os resiacuteduos de Cys Trp e MetAssim analisou-se o estado redox dos
grupos tiol dos resiacuteduos de Cys do S1 na presenccedila do decavanadato atraveacutes da monitorizaccedilatildeo da
Abs412 nm na presenccedila de 1 mgmL de S1 de miosina em tampatildeo 10 mM HEPES 25 mM KCl
variando a concentraccedilatildeo da soluccedilatildeo de decavanadato de modo a determinar que tipo de alteraccedilotildees
estruturais ocorrem na sua presenccedila no que diz respeito agrave diminuiccedilatildeo da actividade ATPaacutesica de
S1 na presenccedila de decavanadato
Os resultados referentes agrave quantificaccedilatildeo das cisteiacutenas reduzidas apoacutes o tratamento de S1 com
diferentes concentraccedilotildees de decavanadato em tampatildeo 10 mM HEPES pH 75 e 25 mM KCl a
25ordmC durante 20 minutos encontram-se descritos na Tabela 2VII
Tabela 2VII ndash Cisteiacutenas reduzidas totais e ldquoraacutepidas de S1 (1 mgmL) apoacutes tratamento com decavanadato
34
(010501002505001000 uM) em 10 mM HEPES 25 mM KCl pH 7 a 25ordmC durante 20 minutos A detecccedilatildeo
espectrofotomeacutetrica foi feita com DTNB (e 412=8400 uM-1 cm-1) (n variaacutevel entre 1 e 8)
Amostras Cys raacutepidas Cys totais
11 M S1 (controlo) 22 plusmn 04 106 plusmn 14
11 M S1 + 10 M Decavanadato 22 plusmn 05 95 plusmn 08
11 M S1 + 50 uM Decavanadato 20 plusmn 02 98 plusmn 14
11 M S1 + 100 uM Decavanadato 17 plusmn 03 97 plusmn14
11 M S1 + 250 uM Decavanadato 26 plusmn 02 98 plusmn 08
11 M S1 + 500 uM Decavanadato 21 plusmn 04 90 plusmn 06
11 M S1 + 1000 uM Decavanadato 24 plusmn 0 86 plusmn 08
A partir dos resultados obtidos pode sugerir-se que o S1 possui 11 resiacuteduos de Cys (controlo) o
que estaacute de acordo com o anteriormente descrito Verifica-se que o tratamento da proteiacutena com
concentraccedilotildees crescentes de decavanadato provoca a oxidaccedilatildeo de dois resiacuteduos de Cys do S1 de
miosina natildeo expostos ao solvente No entanto a teacutecnica utilizada natildeo nos permite identificar
quais
Tendo em consideraccedilatildeo que o valor de IC50 obtido para a reacccedilatildeo de inibiccedilatildeo daactividade Mg2+-
ATPaacutesica de S1 na presenccedila de decavanadato eacute de cerca de 27 microM pode sugerir-se que este seraacute o
valor ao qual ocorreraacute a oxidaccedilatildeo do primeiro resiacuteduo de Cys o que poderaacute contribuir para a
diminuiccedilatildeo da actividade de S1 observada
25Interacccedilatildeo de S1 com vanaacutedio(IV)
251Anaacutelise por espectroscopia de Ressonacircncia Paramagneacutetica Electroacutenica
A acumulaccedilatildeo intracelular de oxovanaacutedio (IV) in vivo constitui uma das principais formas de
apresentaccedilatildeo de vanaacutedio no organismo Deste modo torna-se bastante interessante analisar a
interacccedilatildeo entre este iatildeo e o S1 de miosina atraveacutes da teacutecnica de espectroscopia de Ressonacircncia
Paramagneacutetica Electroacutenica o meacutetodo de eleiccedilatildeo do estudo do iatildeo vanadilo e dos complexos por si
formados O electratildeo desemparelhado encontra-se fortemente acoplado ao nuacutecleo 51
V o que
35
resulta num espectro de RPE consistindo de 8 desdobramentos na direcccedilatildeo paralela e outros tantos
na direcccedilatildeo perpendicular (Chasteen 1983)
Os estudos realizados por Ahmed et al (1987) centraram-se na interacccedilatildeo do vanadilo com
proteiacutenas como eacute o caso da insulina transferrina entre outros obtendo um espectro caracterizado
por 8 linhas amplamente espaccediladas (na direcccedilatildeo paralela) Jaacute Ramos et al 2009 concluiu que a
exposiccedilatildeo da soluccedilatildeo de decavanadato agraves formas globular e filamentosa de actina produz um
espectro tiacutepico do iatildeo oxidovanaacutedio (IV) indicando a reduccedilatildeo do vanadato
Para aferir qual a concentraccedilatildeo de S1 que permitia obter uma boa razatildeo sinalruiacutedo titulou-se uma
soluccedilatildeo de sulfato de vanadilo 15 microM ([VOSO4]stock = 5 mM) com concentraccedilotildees crescentes de
proteiacutena (0 5 10 15 e 20 microM) em 10 mM HEPES 25 mM KCl A existecircncia de uma correlaccedilatildeo
linear entre a concentraccedilatildeo de proteiacutena e a intensidade dos sinais de RPE correspondente agrave banda
-12 perpendicular encontra-se representada na Fig 210
Figura 2 12ndash Representaccedilatildeo graacutefica da variaccedilatildeo da intensidade do sinal de RPE vs a concentraccedilatildeo de S1 utilizada
Apoacutes verificar que com uma concentraccedilatildeo de S1 igual a 20 microM se obtinham sinais com uma razatildeo
sinalruiacutedo adequada ao prosseguimento dos estudos sobre a interacccedilatildeo V(IV)-S1 de miosina
titulou-se uma soluccedilatildeo de 20 microM S1 com 0 25 5 10 15 25 50 75 100 150 200 500 e 1000
microM VOSO4
Traccedilou-se o graacutefico da intensidade da banda -12 perpendicular do sinal de EPR em funccedilatildeo da
concentraccedilatildeo de VOSO4 tendo-se determinado uma constante de dissociaccedilatildeo (KD) do complexo
S1-vanadilo de 8748 plusmn 2621 microM V(IV)
y = 016786x + 122435
Rsup2 = 092992759
Inte
nsi
dad
e d
o s
inal
de
RP
E
[S1] (microM)
36
Figura 213 ndash Representaccedilatildeo graacutefica da variaccedilatildeo do sinal de RPE vs a concentraccedilatildeo de vanadilo utilizada (os valores
experimentais estatildeo representados por losangos o seu ajuste estaacute representado pela linha)
A simulaccedilatildeo do espectro de EPR foi efectuada a partir do espectro obtido para uma concentraccedilatildeo
de 1 mM VOSO4 apoacutes adiccedilatildeo de 20 microM S1 de miosina tendo-se obtido os valores da constante de
acoplamento hiperfina (A=5016) e do factor g (g0=195)
Figura 214 ndash Representaccedilatildeo graacutefica do espectro de EPR obtido com 1000 microM de vanadilo e 20 microM de S1 (a cinzento
escuro) A simulaccedilatildeo do espectro encontra-se a cinzento claro
Por comparaccedilatildeo destes com os valores presentes na tabela de correspondecircncia xxx eacute possiacutevel
predizer quais os ligandos provaacuteveis do vanadilo na presenccedila de proteiacutena Assim concluiu-se que
o vanadilo estaacute coordenado com duas moleacuteculas de aacutegua um grupo hidroxilo e um grupo R=NR
O grupo hidroxilo pode indicar os resiacuteduos de aminoaacutecidos Tyr Treonina e Serina enquanto o
grupo R=NR poderaacute indicar a presenccedila de Arginina e Histidina como ligandos do vanadilo
Inte
nsi
dad
e d
os
sinai
s d
e R
PE
[Vanadilo] (microM)
Ajuste dos dados exp Dados experimentais
Campo magneacutetico (G)
Espectro Experimental Ajuste
37
3 Procedimento Experimental
31 Reagentes
Todos os reagentes utilizados satildeo de elevado grau de pureza (analiacutetico) Apresentam-se as suas
foacutermulas quiacutemicas ou abreviaturas quando necessaacuterio acrescentou-se a sua descriccedilatildeo Entre
parecircntesis colocou-se as empresas que os comercializam
Aacutecido aceacutetico glacial 100 (Riedel) Acrilamida 99 (Merck) Azul de bromofenol (Riedel-de-
Haumlen) -Mercaptoetanol (Sigma) Bissulfito de soacutedio (Sigma)Bradford (reagente de)
(BioRad)BSA (albumina de soro bovino Sigma-Aldrich) CaCl22H2O (cloreto de caacutelcio
dihidratado Fluka) CH3CH2OH 96 (etanol Panreac) Coomassie Blue R250 (Merck) D2O
(oacutexido de deuteacuterio SDS) DTNB (55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico) Aldrich) EDTA (aacutecido
etilenodiaminatetraaceacutetico Merck) Fosfoenolpiruvato 98 (Fluka) Glicerol (Sigma) Glicina
985 (Panreac) HCl 37 (Riedel-de Haumlen) H2SO4 96 (Pronalab) HEPES 99 (aacutecido 4-(2-
hidroxiletil)-1-piperazineetanosulfoacutenico Sigma) KH2PO4 99 (hidrogenofosfato de potaacutessio)
MgATP (Sigma) Na
ATP (Sigma) Na2HPO4 99 (hidrogenofosfato de soacutedio Riedel-de Haumlen)
NaH2PO4 (fosfato de soacutedio ) NaOH 986 (hidroacutexido de soacutedio Pronalab) KCl 99 (Cloreto de
potaacutessio Panreac) L-Cys 995 (enantioacutemero levoacutegiro do resiacuteduo de Cisteiacutena Fluka)Metanol
99 (Merck) Molibdato de amoacutenio (Sigma)NNrsquo-metil-bis-acrilamida 98 (Merck) NH4VO3
99 (metavanadato de amoacutenia Sigma) PSA 10 (persulfato de amoacutenio Merck) SDS
(dodecilo sulfato de soacutedio Sigma) Sulfito de soacutedio (Sigma) VOSO4 (sulfato de vanadilo
Aldrich) TEMED 99 (Tetrametiletilenodiamina Merck) Tris (2-amino-2-hidroximetil-13-
propanodiol Sigma)
32 Meacutetodos
321 Isolamento e purificaccedilatildeo das proteiacutenas de muacutesculo esqueleacutetico de coelho
3211 Purificaccedilatildeo da miosina preparaccedilatildeo de Subfragmento 1 por digestatildeo quimotriacutepsica
da miosina
O subfragmento-1 de miosina-II (S1) foi gentilmente cedido pelo Prof Doutor Manuel Aureliano
do Departamento de Quiacutemica Bioquiacutemica e Farmaacutecia da Faculdade de Ciecircncias e Tecnologia da
Universidade do Algarve
No entanto descreve-se abaixo de forma resumida o procedimento efectuado
38
32111 Purificaccedilatildeo da miosina
O protocolo de isolamento foi adaptado do meacutetodo previamente descrito por Margossian e Lowey
(1982) apresentando-se de forma esquemaacutetica na Fig 31
Sucintamente os tecidos musculares dorsais e das pernas foram extraiacutedos apoacutes o sacrifiacutecio do
animal (Fig31) De seguida foram adicionados 3 volumes da soluccedilatildeo de extracccedilatildeo A2
Centrifugou-se a soluccedilatildeo e filtrou-se o sobrenadante O sedimento foi entatildeo diluiacutedo em 14
volumes de aacutegua destilada de modo a que a forccedila ioacutenica atingisse 004 M Apoacutes adiccedilatildeo de szlig-
mercaptoetanol (szlig-me) deixou-se a precipitar a 4ordmC removendo o sobrenadante Centrifugou-se
novamente e ressuspendeu-se o sedimento na soluccedilatildeo de extracccedilatildeo A Adicionou-se a soluccedilatildeo
tampatildeo B3 para que a forccedila ioacutenica atingisse 05 M Homogeneizou-se e deixou-se repousar
durante 2h Homogeneizou-se novamente e centrifugou-se de modo a ocorrer precipitaccedilatildeo dos
complexos de actomiosina e proteiacutena desnaturada Apoacutes filtraccedilatildeo do sobrenadante em latilde de vidro
quantificou-se a soluccedilatildeo de miosina resultante pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental)
2 Soluccedilatildeo de extracccedilatildeo A 0012M K2CO3 004M KHCO3 06M KCl pH 65
3 Soluccedilatildeo-tampatildeo B Tris 100 mM pH 7 KCl 2M
39
Figura 31 ndash Representaccedilatildeo esquemaacutetica do protocolo de isolamento de miosina (Margossian e Lowey 1982) Todo o
procedimento foi realizado a 4ordmC (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
32112 Preparaccedilatildeo de Subfragmento-1 de miosina por digestatildeo com quimiotripsina
O S1 de miosina foi isolado apoacutes diaacutelise da miosina durante a noite contra a soluccedilatildeo C4 A
presenccedila do EDTA eacute essencial uma vez que a sua substituiccedilatildeo por MgCl2resulta na produccedilatildeo de
HMM em vez de S1 A suspensatildeo de miosina foi equilibrada e digerida com -quimotripsina
A digestatildeo foi interrompida pela adiccedilatildeo de PMSF Seguidamente ultracentrifugou-se o produto
da digestatildeo desprezando-se o sedimento (Fig 32) A soluccedilatildeo de sobrenadante maioritariamente
S1 foi quantificada pela leitura directa da Abs280 ou pelo meacutetodo de Bradford (ver as secccedilotildees
3311 e 3312 do Procedimento Experimental respectivamente) e armazenada em 10 sacarose
e 2 mM szlig-Me a -80ordmC
4 Soluccedilatildeo C 25mM Tris pH 70 012M KCl 1mM EDTA
40
Antes da realizaccedilatildeo dos ensaios descritos no presente relatoacuterio o S1 de miosina foi dialisado
contra a soluccedilatildeo D5 de modo a remover qualquer composto que pudesse complexar como
vanaacutedio Seguidamente quantificou-se a proteiacutena pelo meacutetodo de Bradford ou pelo meacutetodo
directo (ver secccedilotildees 3311 e 3312do Procedimento Experimtental)
Figura 3 2ndash Representaccedilatildeo esquemaacutetica do protocolo de preparaccedilatildeo do S1 de miosina por digestatildeo quimiotripsina de
acordo com o previamente descrito por Margossian e Lowey (1982) Todo o procedimento foi realizado a 4ordm C
excepto quando indicaccedilatildeo do contraacuterio (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
3212 Isolamento e purificaccedilatildeo de actina
Isolou-se e purificou-se a actina segundo o meacutetodo descrito por Pardee e Spudish que
compreende dois passos ndash a obtenccedilatildeo de um resiacuteduo seco denominado ldquopoacute de acetonardquo e a
extracccedilatildeo subsequente da actina a partir do ldquopoacute de acetonardquo na forma de G-Actina com a
posterior polimerizaccedilatildeo a F-Actina
32121 Obtenccedilatildeo do ldquopoacute de acetonardquo
Sacrificou-se o coelho com uma pancada atraacutes do pescoccedilo e sangrou-se o animal apoacutes o corte da
arteacuteria jugular Esfolou-se e colocou-se numa tina com gelo Retiraram-se as partes de tecido
conjuntivo que envolvem o tecido muscular com auxiacutelio de uma pinccedila e bisturi Seguidamente
retirou-se os muacutesculos dorsais e os muacutesculos brancos das pernas Passou-se os muacutesculos por aacutegua
destilada de modo a remover as impurezas e pesou-se num copo Trituraram-se os muacutesculos
numa maacutequina de triturar carne e extraiu-se o resultante com 3 volumes da soluccedilatildeo E6 durante 10
minutos com agitaccedilatildeo suave Centrifugou-se a suspensatildeo a 4000 g durante 10
5 Soluccedilatildeo D HEPES 10 mM pH 70 KCl 25 mM
6 Soluccedilatildeo E 01 M KCl 015 M K2HPO4 pH 65
41
minutosdesprezando-se o sobrenadante Ressuspendeu-se o sedimento em 6 volumes de soluccedilatildeo
F7durante 10 minutos com agitaccedilatildeo suave Verificou-se se o pH da soluccedilatildeo estava compreendido
entre 82 e 85 acertando-se estecom soluccedilatildeo G8 se necessaacuterio Centrifugou-se durante 10
minutos a 4000 g Desprezou-se o sobrenadante e ressuspendeu-se o sedimento em 3 volumes de
soluccedilatildeo H9 durante 10 minutos Centrifugou-se novamente a 4000 gdurante 10 minutos
desprezando-se o sobrenadante Ressuspendeu-se o sedimento duas vezes consecutivas em 6
volumes de aacutegua destilada durante 5 minutos Centrifugou-se a 4000 g durante 10 minutos apoacutes
cada adiccedilatildeo de H2O
Seguidamente ressuspendeu-se o sedimento 5 vezes consecutivas utilizando 3 volumes de
acetona de cada vez agitando suavemente durante 10 minutos agrave temperatura ambiente Colocou-
se o resiacuteduo filtrado numa tina de vidro sobre papel de filtro deixando-se evaporar o solvente
durante a noite num nicho Triturou-se o resiacuteduo seco numa maacutequina de triturar carne e deixou-se
novamente evaporar o solvente O ldquopoacute de acetonardquo obtido foi armazenado a -20 ordmC
32122 Purificaccedilatildeo da actina a partir de ldquopoacute de acetonardquo
Apoacutes a obtenccedilatildeo do poacute de acetona como acima indicado efectuou-se o protocolo seguidamente
descrito de modo a obter monoacutemeros de actina (G-Actina) e actina filamentosa (F-Actina)
Pesou-se 35 g de poacute de acetona e adicionaram-se 50 mL de soluccedilatildeo-tampatildeo I10
sendo a
extracccedilatildeo feita com agitaccedilatildeo suave durante 30 minutos Separou-se o extracto do poacute de acetona
hidratado filtrando a suspensatildeo com 4 camadas de gaze esterilizada tendo o cuidado de natildeo
espremer o resiacuteduo Voltou a extrair-se com 50 mL de soluccedilatildeo I durante 30 minutos mexendo
lentamente Adicionou-se este ao extracto obtido anteriormente Centrifugou-se o extracto total a
16 000 g durante 30 minutos Decantou-se para uma proveta sobre a qual se colocou gaze para
que o sedimento solto natildeo se juntasse ao sobrenadante e leu-se o volume De seguida adicionou-
se KCl ateacute uma concentraccedilatildeo final de 50 mM MgCl2 ateacute 2mM e ATP ateacute 1mM lentamente e com
agitaccedilatildeo contiacutenua de modo a promover a polimerizaccedilatildeo dos monoacutemeros de actina
Deixou-se polimerizar durante 2h a 25ordmC observando-se a presenccedila de pequenas bolhas de ar
indicativas de um aumento de viscosidade da soluccedilatildeo Arrefeceu-se em gelo durante 15 minutos e
adicionou-se KCl ateacute uma concentraccedilatildeo final de 06M Mexeu-se lentamente durante 30 minutos
apoacutes os quais se ultracentrifugou a 110 000g durante 1hora Desprezou-se o sobrenadante
ressuspendeu-se o sedimento com soluccedilatildeo tampatildeo E e transferiu-se para um potter
Dialisou-se durante a noite contra 1L de soluccedilatildeo-tampatildeo I a uma temperatura de 4ordmC com rotaccedilatildeo
7 Soluccedilatildeo F 005 M NaHCO3 pH 82-85
8 Soluccedilatildeo G 1M Na2CO3
9 Soluccedilatildeo H 1 mM EDTA pH 70
10 Soluccedilatildeo I 2 mM Tris-HCl pH 8 02 mM CaCl2 02 mM Na2ATP 05 mM szlig-Me 0005 NaN3
42
contiacutenua e mudanccedila de tampatildeo 2 a 3 vezes De seguida a actina foi centrifugada a 110
000gdurante 1hora Decantou-se a soluccedilatildeo de G-Actina (sobrenadante) e desprezou-se o
sedimento Determinou-se a concentraccedilatildeo de G-Actina pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental) A soluccedilatildeo de G-actina foi armazenada a uma temperatura de 4ordmC
apoacutes adiccedilatildeo de 002NaN3 mantendo-se estaacutevel durante 3 a 5 dias
331 Quantificaccedilatildeo das proteiacutenas isoladas
3311 Quantificaccedilatildeo pelo meacutetodo de Bradford
O meacutetodo de Bradford (Bradford 1976) baseia-se na utilizaccedilatildeo do reagente de Bradford
composto por entre outros aacutecido fosfoacuterico e o corante ldquoCoomassierdquo Este corante possui carga
negativa e a sua coloraccedilatildeo varia entre a vermelha (Absmax=465 nm) predominante em soluccedilatildeo e a
azul (Absmax=595 nm) presente aquando da ligaccedilatildeo aos grupos amina das proteiacutenas Em soluccedilotildees
aacutecidas diluiacutedas a coloraccedilatildeo varia proporcionalmente com a quantidade do corante ligado agrave
proteiacutena que por sua vez eacute directamente proporcional agrave concentraccedilatildeo da proteiacutena permitindo a
sua quantificaccedilatildeo ateacute a um limite de detecccedilatildeo de 20 gmL (Bradford 1976)
Deste modo prepararam-se amostras de BSA ([BSA]stock= 1mgmL) utilizando um intervalo de
concentraccedilotildees compreendido entre 0 e 15 gmL de acordo com a tabela 3I Apoacutes a adiccedilatildeo da
H2O MilliQ e do reagente de Bradford as soluccedilotildees foram sucessivamente homogeneizadas no
voacutertex
Tabela 3Indash Amostras utilizadas para a obtenccedilatildeo da recta de calibraccedilatildeo no meacutetodo de Bradford
BSA (L) BSA (g) H2O MilliQ (L) Reagente de Bradford (L)
0 0 800 200
125 125 79875 200
25 25 7975 200
5 5 795 200
10 10 790 200
15 15 785 200
Apoacutes incubaccedilatildeo a 25ordmC por 5 minutos leram-se os valores de Abs595 num
espectrofotoacutemetro (Shimadzu UV-160A) para todas as amostras Seguidamente traccedilou-se a recta
de calibraccedilatildeo (Abs595vs quantidade de BSA (g))
43
Para determinar a concentraccedilatildeo de proteiacutena apoacutes o processo de isolamento purificaccedilatildeo e
diaacutelise quando necessaacuterio colocou-se 5 L da amostra num tubo de Eppendorf e adicionou-se
795 L de H2O MilliQ homogeneizando a soluccedilatildeo no voacutertex Adicionou-se posteriormente 200
L de reagente de Bradford e repetiu-se a homogeneizaccedilatildeo Incubou-se a mistura a 25ordmC por 5
minutos apoacutes os quais se leu os valores de Abs595 nm no espectrofotoacutemetro Por interpolaccedilatildeo deste
valor a partir da recta de calibraccedilatildeo determinou-se a concentraccedilatildeo de proteiacutena na amostra
3312 Quantificaccedilatildeo pelo meacutetodo directo
A quantificaccedilatildeo das amostras proteicas pode tambeacutem ser efectuada atraveacutes da leitura directa da
absorvacircncia tendo por base a lei de Beer-Lambert onde Abs corresponde agrave
absorvacircncia ao comprimento de onda ao qual o ensaio foi realizado corresponde ao coeficiente
de extinccedilatildeo (substituiacutedo pela letra a se as unidades forem distintas da molaridade) b corresponde
ao percurso oacuteptico da radiaccedilatildeo na ceacutelula amostral e c corresponde agrave concentraccedilatildeo amostral
Assim no caso de S1 de miosina mediu-se a Abs280e calculou-se a concentraccedilatildeo da proteiacutena
sabendo que o coeficiente de extinccedilatildeo de S1 eacute (Ref)
No caso da actina mediu-se a Abs290ecalculou-se a concentraccedilatildeo da proteiacutena sabendo que o
coeficiente de extinccedilatildeo da actina eacute (Ref)
332Electroforese em condiccedilotildees desnaturantes (SDS-PAGE)
De modo a melhor caracterizar as proteiacutenas isoladas foi realizada uma electroforese em
condiccedilotildees desnaturantes (SDS-PAGE do inglecircs Sodium Dodecyl Sulfate Polyacrilamide Gel
Electrophoresis) O SDS (dodecilo sulfato de soacutedio) eacute um detergente anioacutenico utilizado neste
meacutetodo para desnaturar as proteiacutenas que se ligam ao SDS na proporccedilatildeo 14 g SDS g proteiacutena
(aproximadamente 1 moleacutecula de SDS por cada 2 resiacuteduos de aminoaacutecidos) (Stryer) O SDS induz
uma densidade de carga negativa significativa nas proteiacutenas a que se liga mascarando a sua carga
intriacutenseca pelo que as razotildees cargamassa satildeo tendencialmente idecircnticas Assim a SDS-PAGE
separa as proteiacutenas quando sujeitas a um campo eleacutectrico de acordo com o seu peso molecular
sendo a sua migraccedilatildeo directamente proporcional ao logaritmo deste uacuteltimo (Stryer)
O gel de resoluccedilatildeo com 10 de acrilamida foi obtido atraveacutes da adiccedilatildeo de 167 mL de soluccedilatildeo
stock de acrilamida11
075 mL de tampatildeo de resoluccedilatildeo12
251 mL de aacutegua destilada e 50 L de
SDS 10 Para promover a polimerizaccedilatildeo adicionou-se em seguida 38 L de PSA 10
11
soluccedilatildeo stock de acrilamida soluccedilatildeo de 30 (mv) de acrilamida e 08 (mv) de NNrsquo-metil-bis-acrilamida 12
tampatildeo de resoluccedilatildeo 15 M Tris-HCl pH 88
44
(persulfato de amoacutenia) 10 e 25 L de TEMED 99 (tetrametiletilenodiamina) Agitou-se
cuidadosamente de modo a natildeo formar bolhas e pipetou-se a soluccedilatildeo para a cassette de vidro
previamente montada no suporte De seguida adicionou-se uma peliacutecula de cerca de 2 mm de 2-
metil-2-butanol para verificar posteriormente a polimerizaccedilatildeo Apoacutes a polimerizaccedilatildeo que demora
cerca de 45 minutos verteu-se a agrave superfiacutecie e adicionou-se o gel de concentraccedilatildeo com 5 de
acrilamida constituiacutedo por 03 mL de soluccedilatildeo de stock acrilamida 045 mL de tampatildeo de
concentraccedilatildeo13
094 mL de aacutegua destilada 18 L de SDS 10 135 L de PSA 10 e 2L de
TEMED 99 Colocou-se o pente sobre o gel e deixou-se polimerizar durante cerca de 45
minutos
As amostras para aplicaccedilatildeo no gel de electroforese foram preparadas para uma concentraccedilatildeo de
proteiacutena de 1 mgmL num volume final de 100 L dos quais 25 L correspondem ao tampatildeo da
amostra14
As amostras foram aquecidas a 100ordmC durante 5 minutos Antes da aplicaccedilatildeo das
amostras transferiu-se a cassette para a tina de electroforese agrave qual se adicionou a soluccedilatildeo
tampatildeo de electroforese da tina superior15
e tampatildeo de electroforese da tina inferior16
Aplicaram-
se 10 L das soluccedilotildees de marcadores de peso molecular e 20 L das amostras proteicas
preparadas anteriormente A electroforese decorreu durante cerca de 3h30 min a uma voltagem
de 80 V De seguida transferiu-se o gel para uma tina corando as proteiacutenas durante 1h com
soluccedilatildeo corante17
Descorou-se o gel com soluccedilatildeo descorante18
tendo-se mudado esta soluccedilatildeo ateacute
as bandas se encontrarem bem definidas
Determinou-se a mobilidade relativa das bandas correspondentes aos marcadores de elevado peso
molecular e traccedilou-se a recta de calibraccedilatildeo ndash log(PM) vs log(Rf) onde Rf corresponde agrave
mobilidade relativa Por interpolaccedilatildeo determina-se o PM das proteiacutenas isoladas sabendo a sua
mobilidade relativa Por comparaccedilatildeo com os valores descritos na literatura pode concluir-se
acerca do grau de purificaccedilatildeo
34 Mediccedilatildeo da actividade enzimaacutetica de S1
341 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
13
tampatildeo de concentraccedilatildeo 05 M Tris-HCl pH 68 14
tampatildeo de amostra 320 mM Tris-HCl pH 68 04 M szlig-mercaptoetanol SDS 8 glicerol 15 (vv) e roxo de m-
cresol 0024 15
tampatildeo de electroforese da tina superior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS 16
tampatildeo de electroforese da tina inferior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS acetato de soacutedio
01 M 17
soluccedilatildeo corante ldquoCoomassie Blue R-250rdquo contendo metanol 45 (vv) e aacutecido aceacutetico glacial 10 (vv) 18
soluccedilatildeo descorante metanol 10 (vv) e aacutecido aceacutetico glacial 10 vv)
45
O meacutetodo de Fiske-Subarow modificado consiste na reacccedilatildeo do molibdato de amoacutenio com o
fosfato inorgacircnico em soluccedilatildeo aacutecida com formaccedilatildeo do aacutecido fosfomoliacutebdico Seguidamente daacute-
se a adiccedilatildeo de um agente redutor que provoca a reduccedilatildeo do composto a fosfomolibdato que forma
um complexo de cor azulada cuja absorvacircncia maacutexima ocorre para um comprimento de onda de
660 nm tornando possiacutevel a sua detecccedilatildeo por meacutetodos espectrofotomeacutetricos As actividades Ca2+
-
ATPaacutesica e K+EDTA-ATPaacutesica de S1 foram determinadas atraveacutes deste meacutetodo de acordo com
o descrito por Taussky e Shorr (1953) com mediccedilatildeo da quantidade de fosfato (Pi) libertado
atraveacutes da hidroacutelise de ATP
Para tal obteve-se primeiramente a recta de calibraccedilatildeo com as amostras presentes na Tabela 3II
utilizando uma soluccedilatildeo stock de 5 mM KH2PO4
Tabela 3II ndash Composiccedilatildeo das diferentes amostras utilizadas para determinar a recta de calibraccedilatildeo atraveacutes do meacutetodo
de Fiske-Subbarow modificado
KH2PO4 (L)
([KH2PO4]stock= 5mM)
PO43-
(nmole) H2O (uL)
0 0 1780
5 25 1775
10 50 1770
20 100 1760
40 200 1740
100 500 1680
A cada tubo de Eppendorf adicionaram-se seguidamente 200 L de soluccedilatildeo J19
e 20 L de agente
redutor20
homogeneizando no voacutertex apoacutes cada adiccedilatildeo Incubou-se durante 10 minutos e mediu-
se a Abs660 nm num espectrofotoacutemetro com intervalos de tempo iguais entre cada amostra Traccedilou-
se a recta de calibraccedilatildeo (Abs660 nmvs nmol PO43-
) a partir dos valores obtidos
Para determinar a actividade Ca2+
-ATPaacutesica de S1 prepararam-se tubos de Eppendorf com 400
L de meio de reacccedilatildeo21
25 L CaCl2 ([CaCl2]stock= 100 mM) 25 L ATP ([ATP]stock= 100 mM)
com [S1]ensaios=01 mgmL Perfez-se se necessaacuterio com H2O MilliQ ateacute perfazer um volume de 1
mL Para determinar a actividade K+EDTA-ATPaacutesica utilizou-se uma concentraccedilatildeo idecircntica de
19
Soluccedilatildeo J 5 N H2SO4 25 molibdato de amoacutenio 20
Agente redutor 5 mg aacutecido 1-amino-2-naftol-4-sulfoacutenico 30 mg bissulfito de soacutedio 30 mg sulfito de soacutedio em 26
mL H2O 21
meio de reacccedilatildeo 125 mM Tris-HCl pH 80 0575 M KCl
46
proteiacutena nos ensaios (01 mgmL) e o mesmo meio de reacccedilatildeo Aos tubos de Eppendorf
adicionou-se ainda 123 L de KCl ([KCl]stock = 3M) 25 L de ATP 10 L de EDTA
([EDTA]stock= 100 mM )e o volume de H2O MilliQ necessaacuterio para perfazer 1 mL Em ambas as
situaccedilotildees o uacuteltimo reagente adicionado foi o ATP uma vez que se trata do substrato da reacccedilatildeo
enzimaacutetica
A cineacutetica decorreu a 25ordmC durante 5 minutos para a actividade Ca2+
-ATPaacutesica e durante 3
minutos para a actividade K+EDTA-ATPaacutesica Apoacutes o periacuteodo de incubaccedilatildeo transferiu-se 150
L de amostra para tubos de Eppendorf contendo igual volume de TCA 10 Colocou-se no gelo
e homogeneizou-se no voacutertex Seguidamente centrifugou-se a 1500 g durante 10 minutos e
recolheu-se 200 L de sobrenadante para novos tubos de Eppendorf Adicionou-se a estes 158
mL de H2O MilliQ e 200 L de soluccedilatildeo J Homogeneizou-se no voacutertex e adicionou-se 20 L de
reagente redutor repetindo-se a homogeneizaccedilatildeo Incubou-se durante 10 minutos e mediu-se a
Abs660 nm com o mesmo intervalo de tempo de leitura para cada amostra Por intrapolaccedilatildeo a partir
da recta de calibraccedilatildeo obtida anteriormente determinou-se a quantidade de Pi libertado em cada
ensaio A partir destes resultados determinaram-se as actividades Ca2+
-ATPaacutesica e K+EDTA-
ATPaacutesica do S1 em nmol PO43-
minmg proteiacutena e em s-1
342 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
Da actividade Mg2+
-ATPaacutesica de S1 resulta a formaccedilatildeo dos produtos da reacccedilatildeo de
hidroacutelise do ATP - ADP e Pi Na presenccedila de ADP fosfoenolpiruvato (PEP) e da enzima piruvato
cinase (PK) os produtos resultantes satildeo ATP e piruvato Por acccedilatildeo da enzima lactato
desidrogenase (LDH) este uacuteltimo eacute convertido em lactato ocorrendo simultaneamente a oxidaccedilatildeo
do NADH a NAD+ monitorizada atraveacutes da Abs340 nm Quando o S1 de miosina se encontra no
meio reaccional o ATP eacute hidrolisado formando-se novamente ADP e Pi o que permite a
formaccedilatildeo de um ciclo (fig 21) Se as enzimas se encontrarem em excesso a velocidade de
conversatildeo do NADH eacute limitada pela produccedilatildeo de ADP e Pi Como o ATP eacute regenerado em cada
ciclo a sua concentraccedilatildeo inicial eacute sempre mantida (Bagshaw 1982)
Figura 33ndash Representaccedilatildeo esquemaacutetica do meacutetodo de enzimas acopladas
47
O protocolo utilizado e seguidamente descrito foi adaptado de Cuenda et al (1990 ndash Ver Refordf) A
uma cuvette de 1 mL adicionou-se S1 para uma concentraccedilatildeo final de 005 mgmL 800 L de
meio de reacccedilatildeo22
10 L de PEP ([PEP]stock = 42 mM) 10 L de PK+LDH 11 ([PK+LDH]stock =
1 mgmL) e 15 L de NADH ([NADH]stock=25 mM) Iniciou-se a monitorizaccedilatildeo da reacccedilatildeo
atraveacutes da leitura de Abs340 nm apoacutes a adiccedilatildeo de 20 L de ATP ([ATP]stock=100 mM) A
actividade Mg2+
-ATPaacutesica eacute obtida atraveacutes do declive correspondente agrave oxidaccedilatildeo do NADH Para
a mediccedilatildeo da Mg2+
-ATPase de S1 estimulada por actina adicionou-se F-actina para uma
concentraccedilatildeo final de 2 microM sendo a actividade obtida tambeacutem atraveacutes do declive correspondente
agrave oxidaccedilatildeo do NADH apoacutes a adiccedilatildeo da proteiacutena
35 Interaccedilatildeo entre vanaacutedio e subfragmento-1 de miosina
351 Preparaccedilatildeo das soluccedilotildees de vanaacutedio
As soluccedilotildees de vanadato foram preparadas em aacutegua MilliQ a partir de metavanadato de amoacutenio
(NH4VO3) numa concentraccedilatildeo stock de 50 mM com agitaccedilatildeo magneacutetica e aquecimento (~30 ordmC)
Esta soluccedilatildeo com um valor compreendido entre 6 e 7 designa-se por ldquometavanadatordquo e eacute
constituiacuteda pelas esspeacutecies V1 V2 V4 e V5 (Ref) Acidificando a soluccedilatildeo de metavanadato por
adiccedilatildeo de HCl ateacute um valor de pH proacuteximo de 4 observa-se uma mudanccedila de cor de amarelo
claro para laranja indicativa da formaccedilatildeo da espeacutecie decameacuterica de vanadato De seguida e
imediatamente antes da realizaccedilatildeo dos ensaios descritos o valor do pH da soluccedilatildeo de
decavanadato foi aumentado para cerca de 75 por adiccedilatildeo de NaOH A soluccedilatildeo (2) designada
por ldquometavanadatordquo natildeo foi posteriormente alterada ateacute agrave sua adiccedilatildeo para a realizaccedilatildeo dos ensaios
As soluccedilotildees foram armazenadas a 4ordmC e mantidas em gelo aquando da sua utilizaccedilatildeo
A soluccedilatildeo de V(IV) foi preparada em condiccedilotildees anaeroacutebias por diluiccedilatildeo de sulfato de vanadil
(VOSO4) para uma concentraccedilatildeo final de 5 mM em H2O MilliQ saturada com N2 gasoso
352Anaacutelise por espectroscopia de UV-Vis
A concentraccedilatildeo da espeacutecie decameacuterica em soluccedilatildeo depende do periacuteodo de tempo apoacutes a diluiccedilatildeo
no meio de reacccedilatildeo apropriado (Ref) A velocidade de desoligomerizaccedilatildeo da espeacutecie decameacuterica
pode ser analisada por espectroscopia de absorccedilatildeo na regiatildeo UVVis uma vez que esta espeacutecie
apresenta duas bandas caracteriacutesticas na regiatildeo do visiacutevel (a 358 e 400 nm) que natildeo se verifica
em quaisquer outras espeacutecies oligomeacutericas As bandas supramencionada sapresentam-se bem
22
Meio de reacccedilatildeo 3125 mM Tris-HCl pH 71 375 mM MgCl2
48
resolvidas atraveacutes da obtenccedilatildeo da derivada de 2ordf ordem do espectro apresentando a forma de 2
vales com miacutenimos centrados nos comprimentos de onda supracitados com dependecircncia linear
relativamente agrave concentraccedilatildeo de decavanadato total (Ramos et al 2006)
Assim a estabilidade da espeacutecie decameacuterica apoacutes diluiccedilatildeo da soluccedilatildeo de decavanadato no meio de
reacccedilatildeo dos ensaios (tampatildeo D) para uma concentraccedilatildeo final de 1 mM em vanadato total foi
caracterizada atraveacutes da leitura da Abs400 em funccedilatildeo do tempo tendo-se determinado qual a
ordem de reacccedilatildeo e o tempo de meia-vida da mesma Com base em estudos anteriores acerca da
estabilidade do V10 na presenccedila de diferentes proteiacutenas (Ramos et al 2006) determinou-se
igualmente o tempo de meia vida apoacutes a adiccedilatildeo de 20 microM de S1 de miosina ao meio reaccional
353 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
Os espectros de RMN do isoacutetopo 51
V foram realizados num espectroacutemetro Bruker AM-400 MHz
equipado com uma sonda multinuclear inversa de 5 mm Em tubos de RMN de 5 mm de diacircmetro
preparou-se soluccedilotildees de 500 L de deca- ou metavanadato correspondentes a um intervalo de
concentraccedilotildees de 1 a 10 mM suplementadas com 10 D2O Os espectros foram adquiridos agrave
temperatura ambiente com uma largura espectral de 23 923 Hz um tempo de aquisiccedilatildeo de 0086
s tempo de relaxaccedilatildeo de 001 s e uma sequecircncia de pulsos a 90ordm Utilizou-se a transformada de
Fourier com alargamento das bandas de 20Hz para o processamento dos espectros Os desvios
quiacutemicos das ressonacircncias de 51
V foram obtidos em relaccedilatildeo agrave espeacutecie VOCl3cuja frequecircncia de
ressonacircncia eacute considerada o padratildeo sendo todas as outras referenciadas a partir desta em ppm
As aacutereas dos sinais do RMN correspondentes agraves espeacutecies oligomeacutericas presentes foram obtidas
por integraccedilatildeo dos sinais As concentraccedilotildees relativas das espeacutecies foram calculadas com recurso agrave
equaccedilatildeo
onde A[Vx] corresponde agrave aacuterea referente agrave ressonacircncia da espeacutecie Vx com nordm de agregaccedilatildeo x
A[V]total agrave soma total das aacutereas medidas e [V] a concentraccedilatildeo total de vanaacutedio no ensaio
A teacutecnica de 51
V-RMN foi anteriormente aplicada em estudos de ligaccedilatildeo com S1 de miosina
(Ref) tendo sido demostrado que entre as espeacutecies V1 V2 V4 e V5 a espeacutecie tetrameacuterica possui a
maior afinidade para a miosina (Cremo et al) O desaparecimento ou alargamento dos sinais pode
implicar a ligaccedilatildeo de um dos oxovanadatos laacutebeis agrave miosina sem indicaccedilatildeo de qual uma vez que
49
se processa um equiliacutebrio entre eles (Crans 1990) Assim e tendo em conta as indicaccedilotildees de que
a espeacutecie V10 eacute distinta das restantes no que diz respeito agrave magnitude da interacccedilatildeo com a miosina
e inibiccedilatildeo da actividade ATPaacutesica estimulada por actina (Aureliano 2000 Tiago et al 2002)
torna-se fundamental investigar a interacccedilatildeo das diferentes espeacutecies de vanadato com o S1 de
miosina atraveacutes da teacutecnica de 51
V-RMN
Para tal obtiveram-se espectros de 51
V-RMN resultantes da titulaccedilatildeo de uma soluccedilatildeo 5 mM de
decavanadato total com concentraccedilotildees crescentes de S1 de miosina (0 5 10 15 e 20 microM) em
tampatildeo D Realizou-se ainda ensaios na presenccedila de 2 mM de decavanadato e 2 mM de ATP na
presenccedila e na ausecircncia de S1 de miosina de modo a verificar o efeito que estes ligandos naturais
da enzima teria na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o subfragmento-1
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
Os grupos sulfidrilo (-SH) das cisteiacutenas satildeo resiacuteduos de aminoaacutecidos extremamente sensiacuteveis agrave
oxidaccedilatildeo resultando na sua oxidaccedilatildeo (Dalle-Donne et al 1999) Logo a quantificaccedilatildeo destes
resiacuteduos eacute bastante importante nos estudos de interacccedilatildeo com vanaacutedio jaacute que os resiacuteduos de Cys
com ecircnfase para o SH1 e o SH2 satildeo bastante importantes na monitorizaccedilatildeo das modificaccedilotildees
funcionais da regiatildeo da cabeccedila
O procedimento seguidamente apresentado foi originalmente descrito por Gutieacuterrez-Merino
(1987)) Consiste na reacccedilatildeo entre os grupos ndashSH natildeo oxidados das proteiacutenas e o reagente de
Ellman (DTNB -55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico)) - formando-se o aniatildeo CNT (aniatildeo 3-
carboxilato-4-nitrotiofenolato) numa estequiometria de 11 Este apresenta um maacuteximo de
absorvecircncia a 412 nm (Robyt 1990) Assim apoacutes a incubaccedilatildeo da proteiacutena na presenccedila do metal
a quantidade de cisteiacutenas reduzidas por proteiacutena determina-se atraveacutes dos valores de Abs412
Deste modo colocou-se num tubo de Eppendorf o volume apropriado de S1 dialisado para que a
concentraccedilatildeo no ensaio fosse de 1 mgmL Adicionou-se o volume apropriado de decavanadato
para que a sua concentraccedilatildeo fosse de 0 10 50 100 250 500 ou 1000 M Perfez-se com soluccedilatildeo
D ateacute um volume final de 1 mL e incubou-se a 25ordmC por 20 minutos Apoacutes este periacuteodo colocou-
se 780 L da mistura reaccional numa cuvette de quartzo e adicionou-se 220 L de soluccedilatildeo K23
23
Tampatildeo K Tampatildeo fosfatohidrogenofosfato de soacutedio 025 M pH 72
50
Adicionou-se 15 L de DTNB (em soluccedilatildeo K [DTNB]stock=100 mM) ao meio reaccional e
registou-se o valor de absorvecircncia medido equivalente agraves cisteiacutenas ldquoraacutepidasrdquo (SH1 e SH2
expostos ao solvente) Adicionou-se 001 g de SDS que provoca a desnaturaccedilatildeo da proteiacutena e
consequentemente a exposiccedilatildeo de todos os resiacuteduos de cisteiacutenas que se encontravam ateacute aiacute
inacessiacuteveis no interior da proteiacutena (cisteiacutenas totais) Estes resiacuteduos podem assim reagir com o
DTNB
Estimou-se o nuacutemero de resiacuteduos de cisteiacutenas em todas as amostras com recurso ao coeficiente de
extinccedilatildeo molar () do DTNB (ver ponto 3 dos Anexos) que foi determinado atraveacutes da titulaccedilatildeo
de L-Cisteiacutena com intervalo de concentraccedilotildees compreendidos entre 0 e 50 microM na presenccedila de
25microM de DTNB em tampatildeo D Os valores de absorvecircncia foram obtidos para um comprimento de
onda de 412 nm
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
3621 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
A teacutecnica mais eficiente de anaacutelise da interacccedilatildeo entre o iatildeo vanadilo e proteiacutenas como a
calmodulina e a transferrina entre outras permite a obtenccedilatildeo raacutepida de informaccedilatildeo no que toca agrave
conformaccedilatildeo e amplitude da referida interacccedilatildeo
Os espectros de Ressonacircncia Paramagneacutetica Electroacutenica (RPE) da espeacutecie paramagneacutetica V(IV)
foram realizados a 70K num espectroacutemetro Bruker EMX-300 X-band Todas as soluccedilotildees
utilizadas e amostras foram preparadas em anaerobiose numa cacircmara de luvas de modo a evitar a
reduccedilatildeo de V(IV) a V(V) Em tubos capilares com 02 mL de amostra manteve-se a concentraccedilatildeo
de S1 a 20 M e variou-se a concentraccedilatildeo de vanadilo (0 25 5 10 15 25 50 75 100 150
200 500 e 1000 M [VOSO4]stock=5 mM) em tampatildeo D Apoacutes obtenccedilatildeo dos espectros estes
foram tratados atraveacutes do software SimFonia e WinEPR tendo-se obtido um graacutefico com a
variaccedilatildeo da intensidade do sinal de RPE em funccedilatildeo da concentraccedilatildeo de sulfato de vanadilo
presente Realizaram-se ainda simulaccedilotildees de espectros obtidos com 20 microM de S1 e 1000 microM de
V(IV) com vista agrave obtenccedilatildeo dos ligandos provaacuteveis do vanadilo atraveacutes da anaacutelise da constante
de acoplamento hiperfina (A) e do factor g (g)
4 Conclusotildees gerais
Apesar de jaacute ter sido demonstrado que o vanaacutedio inibe a Na+K
+-ATPase da miosina a funccedilatildeo
deste elemento no tecido muacutesculo esqueleacutetico continua largamente incompreendida A enorme
complexidade quiacutemica deste metal especialmente no que concerne agrave oligomerizaccedilatildeo e reacccedilotildees
51
redox constitui um obstaacuteculo agrave compreensatildeo do comportamento do vanaacutedio nos organismos
vivos Assim a caracterizaccedilatildeo da quiacutemica deste elemento eacute de grande utilidade antes de iniciar o
estudo da sua interacccedilatildeo com o presente sistema
A anaacutelise por espectroscopias de 51
V-RMN e de UV-Vis de soluccedilotildees de vanadato diluiacutedas nos
meios de reacccedilatildeo utilizados no presente relatoacuterio (10 mM HEPES pH 70 25 mM KCl) mostrou
claramente que a espeacutecie decameacuterica (V10) possui um tempo de vida suficientemente longo para a
determinaccedilatildeo do seu efeito isolado ao contraacuterio do que se verifica com as espeacutecies mono- di-
tetra- e pentameacutericas (V1 V2 V4 e V5) Por outro lado a anaacutelise verificou ainda que apoacutes diluiccedilatildeo
das soluccedilotildees de decavanadato no meio de reacccedilatildeo coexistem as espeacutecies V10 e V1 pelo que os
efeitos putativos desta uacuteltima deveratildeo ser subtraiacutedos aquando da monitorizaccedilatildeo dos efeitos da
espeacutecie decameacuterica
O vanaacutedio (V) (vanadato) inibe as enzimas que catalizam reacccedilotildees de transferecircncia de grupos
fosfato como a ATPase da miosina [Goodno 1979] devido agrave sua analogia estrutural com o iatildeo
fosfato Na sua utilizaccedilatildeo como inibidor enzimaacutetico natildeo eacute tida em conta a contribuiccedilatildeo individual
das espeacutecies oligomeacutericas considerando-se normalmente a espeacutecie V1 como inibidora efectiva
[Goodno e Taylor 1982] No presente relatoacuterio verificou-se por espectroscopia de 51
V-RMN que
outros oligoacutemeros de vanadato interagem com o S1 de miosina tendo as espeacutecies V10 e V4 os
maiores niacuteveis de afinidade A sua interacccedilatildeo envolve forccedilas electrostaacuteticas e eacute afectada
negativamente por ligandos naturais de enzima como eacute o caso do ATP
A exposiccedilatildeo do S1 de miosina (005 mgmL) a decavanadato conduz agrave inibiccedilatildeo do capacidade da
proteiacutena de estimular a actividade Mg2+
-ATPaacutesica com um valor de IC50 de 266 plusmn 07 M
decavanadato sendo o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM
decavanadato (80) Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM decavanadato sendo
o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM decavanadato (60)
exercendo um efeito de impedimento agrave ligaccedilatildeo entre o vanaacutedio e o S1
A uma concentraccedilatildeo de 27 microM decavanadato verifica-se a oxidaccedilatildeo de um dos resiacuteduos de Cys
de S1 de miosina seguida pela oxidaccedilatildeo gradual de outro resiacuteduo de Cys ateacute agrave concentraccedilatildeo de
1000 microM Ambos os resiacuteduos oxidados natildeo se encontravam expostos ao solvente utilizado A
oxidaccedilatildeo dos resiacuteduos permite sugerir que esta modificaccedilatildeo oxidativa sofrida pelo S1 contribui
para a diminuiccedilatildeo da sua actividade de ATPase
52
Por espectroscopia de RPE revelaram-se os possiacuteveis ligandos equatoriais do vanaacutedio aquando da
sua interacccedilatildeo com o S1 de miosina como sendo duas moleacuteculas de aacutegua um ligando contendo o
grupo R-OH e outro contendo o grupo R=NR
Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados de
oxidaccedilatildeo (IV) e (V) no S1 de miosina para concentraccedilotildees facilmente alcanccedilaacuteveis no ambiente
terrestre
53
5 Estudos futuros
No sentido do melhor esclarecimento das alteraccedilotildees moleculares e estruturais que ocorrem no
subfragmento 1 quando exposta ao vanaacutedio que conduzem a uma diminuiccedilatildeo da sua funccedilatildeo
sugere-se
- Quantificaccedilatildeo das cisteiacutenas reduzidas de S1 na presenccedila de metavanadato
- Determinaccedilatildeo das actividades Ca2+
- e K+-ATPaacutesica do S1 de miosina na presenccedila de
decavanadato
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina estimulada por actina
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina na presenccedila de
metavanadato
- Realizaccedilatildeo de RMN 2D com variaccedilatildeo das concentraccedilotildees das soluccedilotildees de decavanadato e
metavanadato na presenccedila de S1
-Mutageacutenese dos resiacuteduos de Cys doS1 de miosina de modo a verificar quais os resiacuteduos que
sofrem oxidaccedilatildeo na presenccedila de decavanadato
- Determinaccedilatildeo por calorimetria (DSC Differential Scanning Calorimetry) da variaccedilatildeo da
entalpia (∆H) e da temperatura de desnaturaccedilatildeo do S1 de miosina na presenccedila de vanaacutedio uma
vez que estas propriedades caracteriacutesticas de cada proteiacutena satildeo alteradas apoacutes ocorrecircncia de
modificaccedilotildees estruturais
- Anaacutelise por SDS-PAGE de S1 de miosina apoacutes tratamento com vanaacutedio de modo a determinar
quais os polipeacuteptidos eventualmente formados apoacutes clivagem da proteiacutena
54
6 Referecircncias bibliograacuteficas
- Sellers J Myosins 2nd Edition Oxford University Press 1999
- L Stryer inBiochemistry (Ed WH Freeman 2005 pp
Refordfas Materiais e Meacutetodos
[A] S S Margossian SLowey Methods Enzymol1982 85 55-71
[B] JD Pardue JA Spudich Methods Enzymol 1982 85 164-181
[C] M Bradford Anal Biochem1976 72 248-254
[D] H H Taussky E Shorr J Biol Chem 1953 202 675-685
[E] C R Bagshaw in Outline Studies in Biology ndash Muscle Contraction Chapman and Hall Ed
1982
[F] A Cuenda et al Eur J Biochem1990 194 663-670
55
Anexos
1 Determinaccedilatildeo da recta de calibraccedilatildeo meacutetodo de Bradford
Figura1ndashRepresentaccedilatildeo graacutefica da recta de calibraccedilatildeo (Abs595 nm vs [BSA]) utilizada para o meacutetodo de Bradford
2 Determinaccedilatildeo da recta calibraccedilatildeo para a electroforese em condiccedilotildees desnaturantes
Figura 2 ndash Representaccedilatildeo graacutefica da recta de calibraccedilatildeo (log(Peso Molecular) vs Rf dos marcadores de peso
molecular) onde Rf indica determinada atraveacutes do padratildeo de peso molecular
(poccedilo (a))
3Caacutelculo do coeficiente de extinccedilatildeo molar () para a quantificaccedilatildeo de cisteiacutenas reduzidas
atraveacutes do meacutetodo de Ellman
Traccedilou-se a recta de Abs412 nm meacutedia vs [L-Cisteiacutena] de modo a determinar o valor do coeficiente
de extinccedilatildeo molar definido como o declive daquela recta Assim =8400 M-1
cm-1
como
podemos verificar na Fig 3
A595 nm = 00546[BSA] + 00209 Rsup2 = 099051
Ab
s 59
5 n
m
[BSA] (mgmL)
log PM = -1177 Rf + 2247 Rsup2 = 098957
log
PM
Mobilidade relativa (Rf)
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero

13
Na ausecircncia de ATP e supondo a ausecircncia de inibiccedilatildeo da interacccedilatildeo actina-miosina as proteiacutenas
encontram-se fortemente associadas formando o complexo actomiosina (AM) (Figura 15)
Nestas condiccedilotildees as cabeccedilas de miosina formam um acircngulo de 45ordm com o eixo dos filamentos
finos de actina [P]
Na presenccedila de ATP o complexo AM dissocia-se jaacute que a afinidade do complexo M-ATP para a
actina eacute 105 vezes inferior agrave afinidade da miosina para a actina O ATP liga-se agrave miosina
ocorrendo seguidamente a reacccedilatildeo de hidroacutelise de ATP Os produtos daiacute resultantes ndash ADP e Pi -
permanecem ligados agrave miosina formando o complexo M-ADP-Pi Com a ligaccedilatildeo subsequente agrave
actina forma-se o complexo AM-ADP-Pi no qual as cabeccedilas de miosina apresentam um acircngulo
recto com o eixo do muacutesculo [P] Neste ponto daacute-se a libertaccedilatildeo progressiva de Pi e ADP Eacute
provaacutevel que a energia proveniente da hidroacutelise do ATP provoque uma alteraccedilatildeo do acircngulo de
ligaccedilatildeo das cabeccedilas de miosina ao filamento de actina passando novamente de 90ordm para 45ordm [P]
Esta alteraccedilatildeo conformacional provoca o deslizamento do filamento de actina para o centro do
sarcoacutemero cujo encurtamento promove a contracccedilatildeo das miofibrilas O processo supracitado
ocorre em cerca de 1ms[Q] e encontra-se resumido na Fig 18
Figura 18 ndash Esquema cineacutetico da hidroacutelise de ATP pela miosina (Adaptado de Sellers 1999 [A])
1131 Regulaccedilatildeo da contracccedilatildeo muscular
O mecanismo pelo qual se processa a regulaccedilatildeo muscular foi inicialmente descrito por estudos
distintos de Haselgrove Huxley e Parrey-Squire todos em 1973 [RST] Aleacutem da implicaccedilatildeo
directa da tropomiosina e do complexo de troponina a concentraccedilatildeo de Ca2+
no muacutesculo em
relaxamento eacute bastante reduzida (10-7
M) aumentando notavelmente aquando da contracccedilatildeo (10-5
-
10-3
M)
Resumidamente quando ocorre a excitaccedilatildeo do muacutesculo esqueleacutetico a despolarizaccedilatildeo da
membrana plasmaacutetica eacute difundida ateacute ao interior do tecido atraveacutes dos tuacutebulos transversais
(invaginaccedilotildees da membrana plasmaacutetica da fibra muscular) A despolarizaccedilatildeo provoca a abertura
dos canais ioacutenicos de Ca2+
do retiacuteculo sarcoplasmaacutetico adjacente Como a concentraccedilatildeo de Ca2+
no interior do organelo eacute muito elevada o caacutelcio passa raacutepida e uniformemente para o citosol
14
Assim a regulaccedilatildeo da contracccedilatildeo ocorre com coordenaccedilatildeo e rapidez na totalidade das miofibrilas
do tecido muscular (Ref)
Em repouso a troponina I encontra-se posicionada de tal forma que a proteiacutena adjacente
troponina C impede estruturalmente a ligaccedilatildeo das cabeccedilas da miosina agrave actina (acircngulo =50ordm) O
aumento da concentraccedilatildeo intracelular de Ca2+
provoca um aumento da sua disponibilidade para
ligar agrave troponina C (Fig 19)
Figura 19ndash Regulaccedilatildeo do mecanismo contraacutectil no muacutesculo esqueleacutetico pelo complexo de troponina
Como consequecircncia a troponina C altera a sua conformaccedilatildeo (=70ordm) associando-se agrave troponina I
que deixa de inibir a ligaccedilatildeo das pontes transversais A alteraccedilatildeo anterior eacute transmitida agrave
tropomiosina atraveacutes da troponina T adjacente agrave C o que provoca o deslocamento da
tropomiosina na direcccedilatildeo do interior do sulco do filamento de actina Assim eacute possiacutevel a
interacccedilatildeo eficaz entre actina e miosina A Ca2+
-ATPase do retiacuteculo sarcoplasmaacutetico provoca a
reacumulaccedilatildeo do iatildeo caacutelcio no organelo impedindo o prolongamento desnecessaacuterio da contracccedilatildeo
(Refordf)
12 Vanaacutedio
O vanaacutedio eacute um metal de transiccedilatildeo pertencente ao grupo VB da tabela perioacutedica Foi
primeiramente descoberto em 1801 por Andreacutes Manuel del Riacuteo que concluiu erradamente que era
idecircntico ao croacutemio e definitivamente por Nils Gabriel Svenson em 1831 Recentemente foram
descobertas importantes funccedilotildees bioloacutegicas deste elemento provavelmente essencial para os
humanos de que se destaca no contexto do presente relatoacuterio a possibilidade de actuar como
anaacutelogo do grupo fosfato e a inibiccedilatildeo de enzimas in vitro como eacute o caso da actividade ATPaacutesica
da miosina
Eacute um metal soacutelido agrave temperatura ambiente que se encontra amplamente distribuiacutedo nos vaacuterios
ecossistemas em dois isoacutetopos naturais ndash 50
V e 51
V ndash raramente excedendo 01 partes por biliatildeo
15
(ppb) Estaacute associado a cerca de 70 minerais diferentes na natureza (dos quais se destacam o
uracircnio cobre chumbo zinco fosfato e titacircnio ferroso) especialmente na forma de vanadato ou
pentoacutexidos de vanaacutedio O vanaacutedio eacute um dos principais constituintes da mateacuteria orgacircnica foacutessil
com especial ecircnfase para o carvatildeo oacuteleo de crude e foacutesseis carbonaacuteceos [UV] bem como em
alimentos consumidos pelo Homem (salsa pimenta preta cogumelos marisco vegetais cereais
gorduras e oacuteleos) [W] Estes constituem a maior fonte de exposiccedilatildeo ao vanaacutedio da maioria da
populaccedilatildeo humana [X]
Normalmente a entrada do vanaacutedio no organismo ocorre por inalaccedilatildeo ou em menores
quantidades por absorccedilatildeo pelo tracto gastrointestinal ou pela pele O seu armazenamento centra-
se no fiacutegado rins ceacuterebro coraccedilatildeo muacutesculos e ossos No entanto a retenccedilatildeo de vanaacutedio no
organismo humano eacute reduzida ndash em condiccedilotildees normais cerca de 100 M (em muitos tecidos a
quantidade eacute inferior a 10 ngg de tecido) [Y]
Actualmente natildeo existem provas de deficiecircncia nutricional de vanaacutedio em humanos ou de
enzimas que contenham vanaacutedio em animais superiores No entanto vaacuterios estudos relacionam a
ocorrecircncia de deficiecircncia em galinhas e ratos devido a uma alimentaccedilatildeo pobre em vanaacutedio Os
sintomas relatados incluem retardamento do crescimento malformaccedilotildees oacutesseas perturbaccedilotildees no
metabolismo lipiacutedico e estagnaccedilatildeo reprodutiva entre outros [Z AA AB] Os efeitos fisioloacutegicos
do vanaacutedio podem depender do tipo de composto ou de oligoacutemero uma vez que consoante as
suas estabilidade labilidade e nuacutemero de coordenaccedilatildeo as respostas seratildeo diferenciadas ao niacutevel
dos organismos vivos
Por outro lado a toxicidade do vanaacutedio encontra-se bem caracterizada existindo sintomas
descritos em vaacuterios mamiacuteferos (inclusive o Homem) que incluem diarreias catildeibras irritaccedilatildeo da
pele e mucosas palpitaccedilotildees cardiacuteacas anemia pneumonia bronquite perturbaccedilotildees no Sistema
Nervoso Central (SNC) entre outros (Ref) A toxicidade varia geralmente com a natureza do
composto com aumento proporcional agrave sua valecircncia Verificou-se que a forma de entrada do
vanaacutedio tem tambeacutem efeito na sua toxicidade ndash os maiores niacuteveis verificaram-se por injecccedilatildeo
intravenosa da amostra sendo progressivamente inferiores por inalaccedilatildeo e finalmente por
ingestatildeo [X AA] A ampla utilizaccedilatildeo dos compostos de vanaacutedio na induacutestria quiacutemica bem como
as elevadas quantidades de vanaacutedio libertadas atraveacutes da combustatildeo dos oacuteleos de crude
contribuem consideravelmente para o aumento da biodisponibilidade do vanaacutedio Como tal o
interesse neste elemento e nos problemas ambientais por ele causados tem vindo a aumentar
significativamente [V]
121 Propriedades quiacutemicas
16
A quiacutemica do vanaacutedio eacute extraordinariamente complexa pelo que originou vaacuterias interpretaccedilotildees
incorrectas desde a sua descoberta No entanto as suas propriedades dependem em uacuteltima
anaacutelise da configuraccedilatildeo electroacutenica do elemento Assim a variabilidade encontrada no
comportamento quiacutemico do vanaacutedio pode dever-se agrave sua configuraccedilatildeo ndash [Ar] 3d2 4s
2 ndash que
permite a formaccedilatildeo de vaacuterios estados de oxidaccedilatildeo (de +2 a +5) Em soluccedilatildeo aquosa os estados de
oxidaccedilatildeo estaacuteveis vatildeo de V(V) a V(II) Destes apenas os +3 a +5 satildeo biologicamente importantes
jaacute que o V(II) apresenta demasiada capacidade redutora para existir nos organismos vivos
conhecidos [AC]
No fluido extracelular os compostos de vanaacutedio encontram-se predominantemente na forma de
monovanadatos (V(V)) livres ou ligados agrave transferina do soro ou agrave hemoglobina (Refordf) Os iotildees de
vanadato livres satildeo transportados para o meio intracelular atraveacutes de canais ioacutenicos inespeciacuteficos
Jaacute no ambiente intracelular os compostos sofreratildeo uma bio-reduccedilatildeo atraveacutes da acccedilatildeo da
glutationa ou outros compostos com grupos ndashOH como pentoses hexoses ou etanolaminas [V] A
reoxidaccedilatildeo de vanadilo (V(IV)) intracelular a vanadato (V(V)) provocaria a acumulaccedilatildeo toacutexica
destes uacuteltimos pelo que a complexaccedilatildeo da espeacutecie reduzida o impede O transporte de vanaacutedio
para o interior da ceacutelula encontra-se na Fig 110
Figura 110 ndash Transporte de vanaacutedio para a ceacuteluka e seu subsequente metabolismo (P=Proteiacutena) (Adaptado de
Zaporowska e Scibor 1998)
1211 Vanaacutedio(IV)
A espeacutecie de vanaacutedio (IV) mais bem conhecida eacute o iatildeo vanadilo (VO2+
) espeacutecie estaacutevel a pH
aacutecido sendo a maioritaacuteria a pH 3[AD] Apresenta uma cor azulada e forma complexos penta e
hexacoordenados Forma-se rapidamente a partir de V(V) na presenccedila de um agente redutor em
soluccedilatildeo aquosa No ambiente intracelular a presenccedila de glutationa reduzida aacutecido ascoacuterbico
NAD(P)H entre outros conduzem a que o principal estado de oxidaccedilatildeo nos sistemas bioloacutegicos
seja provavelmente o +4 encontrando-se o V(IV) fortemente complexado com proteiacutenas ou
aminoaacutecidos livres citrato ATP e pirofosfato (Refordfs) Em alternativa o V(IV) pode existir
17
livremente em ambientes fortemente aacutecidos como os lisossomas jaacute que a pH superior a 3 as
soluccedilotildees de vanadilo tendem a ser oxidadas pelo ar formando-se vanadato Esta grande
capacidade redutora justifica parcialmente a utilizaccedilatildeo pouco extensiva do iatildeo VO2+
como
inibidor [W]
1212 Vanaacutedio (V)
O vanaacutedio (V) eacute a forma mais estaacutevel num intervalo de pH amplo [W] O iatildeo anioacutenico VO3-tende
a constituir complexos polinucleares pelo que as soluccedilotildees aquosas de vanadato satildeo extremamente
complexas com ocorrecircncia simultacircnea de vaacuterios equiliacutebrios de protonaccedilatildeo (pKa(vanadato)= 35
78 e 125) e oligomerizaccedilatildeo As espeacutecies presentes (H3VO4 H2VO4- HVO4
2- e VO4
3-) dependem
do pH forccedila ioacutenica e temperatura da soluccedilatildeo e da concentraccedilatildeo total do elemento
[AEAFAGAHAI]
O VO43-
- monoacutemero ortovanadato V1 ndash estaacute associado a uma grande parte da importacircncia
bioloacutegica do vanaacutedio uma vez que se trata de um anaacutelogo electroacutenico e estrutural do ortofosfato
(PO43-
) A coordenaccedilatildeo do ortovanadato pode ser tetraeacutedrica ou bipiramidal trigonal (Refordfs)
A oligomerizaccedilatildeo do vanadato monomeacuterico daacute origem ao diacutemero (V2O72-
- V2) que apresenta
tambeacutem equiliacutebrios de protonaccedilatildeo Presume-se a existecircncia de analogia do vanadato dimeacuterico com
o pirofosfato embora natildeo haja evidecircncia estrutural (Refordf) A dimerizaccedilatildeo do V2 origina o
vanadato tetrameacuterico ou metavanadato (V4O124-
- V4) que constitui a espeacutecie maioritaacuteria em
soluccedilotildees aquosas concentradas (Refordf) As estruturas do metavanadato e a sua espeacutecie protonada
satildeo ciacuteclicas [AJ] Outra espeacutecie oligomeacuterica importante eacute o vanadato pentameacuterico (V5O153-
) cuja
estrutura presumida eacute ciacuteclica [AI]
O oligoacutemero com maior importacircncia bioquiacutemica eacute o vanadato decameacuterico (V10O286-
-V10)(Refordf) a
espeacutecie predominante em soluccedilotildees de pH 2-6 quando a concentraccedilatildeo total de vanadato eacute superior
a 100 M (Ref) A estrutura do V10 natildeo difere em soluccedilatildeo ou no estado soacutelidoOs 10 aacutetomos de
vanaacutedio da espeacutecie decameacuterica estatildeo divididos em trecircs grupos identificados na figura como VA-C
consoante o seu ambiente quiacutemico circundante (Refordf) (Fig 111)
A expressatildeo ldquodecavanadatordquo refere-se a uma soluccedilatildeo aquosa na qual a espeacutecie decameacuterica se
encontra em equiliacutebrio quiacutemico com as espeacutecies V1 V2 V4 e V5 sendo que a oligomerizaccedilatildeo
progressiva do vanaacutedio eacute proporcional agrave sua concentraccedilatildeo total (Ref) Nesta soluccedilatildeo a
concentraccedilatildeo de V10 corresponde a aproximadamente 10 da concentraccedilatildeo total de vanaacutedio [AK]
18
Figura 111 ndash Estruturas esquemaacuteticas das espeacutecies de vanadato (espeacutecie monomeacuterica (V1) dimeacuterica (V2)
tetrameacuterica (V4) pentameacuterica (V5) e decameacuterica (V10) (Adaptado de Aureliano e Crans 2008)
122 Metabolismo e efeito na actividade enzimaacutetica
Apesar de natildeo terem sido ainda encontradas as provas categoacutericas das suas funccedilotildees nos seres
vivos sabe-se que o iatildeo vanadato eacute capaz de inibir uma grande quantidade de enzimas in vitro
com especial ecircnfase na actividade ATPaacutesica da miosina [AL Teresa] Como referido
anteriormente o vanadato eacute capaz de actuar em alternativa ao fosfato nas reacccedilotildees bioquiacutemicas
incluindo a sua preferecircncia de ligaccedilatildeo agraves enzimas [W]
Eacute conhecido o papel do vanaacutedio no metabolismo do caacutelcio promovendo a mineralizaccedilatildeo dos
dentes e ossos na biossiacutentese de liacutepidos (trigliceacuteridos e colesterol) com posterior inibiccedilatildeo da
acumulaccedilatildeo destes nos vasos sanguiacuteneos e no SNC na reproduccedilatildeo e desenvolvimento por
mimetizaccedilatildeo e potenciaccedilatildeo de factores de crescimento na estimulaccedilatildeo do consumo de glucose e
da siacutentese de glicogeacutenio com actuaccedilatildeo equivalente aos efeitos da insulina ao niacutevel celular na
transduccedilatildeo de sinais celulares com regulaccedilatildeo da actividade de segundos mensageiros no
transporte de iotildees e aminoaacutecidos na transcriccedilatildeo geneacutetica e siacutentese de proteiacutenas e DNA entre
outros [W Y AB]
Assim o vanaacutedio cnseguiraacute provocar inuacutemeras mudanccedilas bioquiacutemicas nas ceacutelulas Alguns
exemplos da influecircncia bioquiacutemica do vanaacutedio incluem os seus efeitos nos processos de
fosforilaccedilatildeo e desfosforilaccedilatildeo de proteiacutenas e na participaccedilatildeo na formaccedilatildeo de espeacutecies reactivas de
oxigeacutenio devido ao seu efeito proacute-oxidante sobre agentes redutores a niacutevel celular [AM]
Quanto ao seu efeito na actividade enzimaacutetica pode dividir-se em dois tipos principais ndash inibitoacuterio
e estimulante - dependendo da enzima ou do tipo de reacccedilatildeo por esta catalisada No primeiro
grupo incluem-se enzimas como as ATPases (de miosina de canais ioacutenicos e fosfoenzimas
19
transportadoras) e as fosfatases (da tirosina aacutecida e alcalina) a fosfofrutocinase adenilato ciclase
e catalase entre outras [AA] O segundo grupo inclui a maioria das cinases a glucose-6-fosfato
desidrogenase a fosfoglicerato mutase e a fosfoglicerato fosfatase (Refordf)
O efeito do vanaacutedio (V) na actividade enzimaacutetica estaacute relacionado com o seu nuacutemero de
coordenaccedilatildeo podendo actuar como estado de transiccedilatildeo ou por mimetizaccedilatildeo de cofactores
substratos (especialmente o fosfato) e interacccedilotildees proteiacutena-proteiacutena (como no caso das
polimerases e transcriptases) Pode ainda estar envolvido na formaccedilatildeo de ligaccedilotildees de hidrogeacutenio
interacccedilotildees electrostaacuteticas esteacutereas e de oxidaccedilatildeo-reduccedilatildeo Uma evidecircncia desta uacuteltima eacute a sua
capacidade de alternacircncia entre os estados de oxidaccedilatildeo +5 a +3 o que torna faacutecil a sua interacccedilatildeo
redox com enzimas contendo resiacuteduos de cisteiacutenas (Cys) ou qualquer outro composto orgacircnico
com actividade redox [AN]
13 Vanaacutedio e muacutesculo esqueleacutetico
A primeira evidecircncia da presenccedila de vanaacutedio no muacutesculo esqueleacutetico proveio de uma amostra de
ATP isolado de muacutesculo de cavalo que continha uma impureza ndash vanaacutedio ndash cuja presenccedila
provocou a inibiccedilatildeo da actividade da bomba de Na+K
+-ATPase [AO] Desde entatildeo vaacuterios
estudos tecircm sido realizados com ecircnfase na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o
tecido muscular Destes destaca-se a descoberta da interacccedilatildeo forte da espeacutecie V10 com a miosina
fosforilada [AP] e agrave sua inibiccedilatildeo da actividade ATPaacutesica da miosina[AQ AR]
No quediz respeitoao iatildeo monovanadato destaca-se a sua inibiccedilatildeoda actividade Mg2+
-ATPaacutesica
da miosina [AS] que ocorre de duas formas distintas (1) na presenccedila de concentraccedilotildees reduzidas
de V1 a inibiccedilatildeo ocorre a velocidade reduzida dependente da concentraccedilatildeo da espeacutecie
monomeacuterica (2) na presenccedila de concentraccedilotildees elevadas de V1 o niacutevel de inibiccedilatildeo aproxima-se de
100 desde que [V1] seja superior agrave concentraccedilatildeo total de locais com actividade Mg2+
-ATPaacutesica
no ensaio Esta inibiccedilatildeo eacute irreversiacutevel tanto por diaacutelise ou adiccedilatildeo de agentes redutores do
vanadato [AL]Segundo Pate et al(1994)[AT] a espeacutecie monomeacuterica podeinibir a velocidade de
contracccedilatildeo a temperatura inferior a 20ordmC perdendogradual e rapidamente essa capacidade ateacute os
50ordmC temperatura agrave qual cessa a capacidade inibitoacuteria Os resultados obtidos sugeremainda uma
interacccedilatildeo altamente cooperativa entre o iatildeo vanadato e as proteiacutenas componentes do muacutesculo
esqueleacuteticoDestaca-se tambeacutem a descoberta da inibiccedilatildeo da geraccedilatildeo de forccedila contraacutectil nas fibras
musculares por parte da espeacutecie monomeacuterica[AT]Wilson et al(1995) [AU ]determinaram que a
afinidade da espeacutecie V1 para o S1 de miosina eacute significativamente mais elevada do que a do iatildeo
Pi seu anaacutelogo Descobriram ainda que que a ligaccedilatildeo do vanadato poderaacute envolver transiccedilotildees
entre estados energeacuteticos com energia livre independente dos impedimentos esteacutereos impostos
20
pela estrutura das miofibrilas Os estudos desenvolvidos por Smith et al (1990)[AV]
determinaram queo iatildeo monovanadato natildeo apresenta um efeito pronunciado na inibiccedilatildeo da
actividadeacto-S1-miosina-ATPaacutesica excepto em concentraccedilotildees superiores a 900microM a partir das
quais existe produccedilatildeo de espeacutecies oligomeacutericas para as quais o efeito inibitoacuterio seraacute mais
elevadoA espeacutecie monomeacuterica de vanadato consegue adoptar facilmente uma geometria do tipo
bipiramidal trigonal que mimetiza o intermediaacuterio formado (MgADP
Pi)aquando da reacccedilatildeo de
hidroacutelise de ATP no local activo da miosina [AW] De facto a interacccedilatildeo do vanaacutedio com a
miosina eacute inesperadamente estaacutevel na presenccedila de ADP Esta interacccedilatildeo resulta na formaccedilatildeo de
um complexo enzimaacutetico inactivo e estaacutevel com estequiometria de 11 ADPvanadato por siacutetio
activo da proteiacutena A formaccedilatildeo do complexo estaacutevel miosina-vanadato foi proposta por Goodnoet
al (1982) [AS] Nesta ocorre um preacute-equiliacutebrio raacutepido que induz a formaccedilatildeo de um complexo
ternaacuterio reversiacutevel (MADP
V) com formaccedilatildeo lenta do isoacutemero M
ADP
V A diferenccedila entre os
compostos de vanaacutedio e de fosfato centra-se no tempo de meia vida (t12) do Pi ou do V (no
primeiro t12=12 s enquanto no segundo t12=24 h) (Refordf) A diferenccedila dever-se-aacute agrave possiacutevel
complexaccedilatildeo do vanaacutedio com grupos nucleofiacutelicos no siacutetio activo o que provocaria uma
estabilizaccedilatildeo da miosina na conformaccedilatildeo MADP
V
No que respeita aos estudos realizados com a espeacutecie tetrameacuterica Cremo et al determinou que a
cadeia pesada do S1 de miosina pode ser clivada em dois locais distintos (designados V1 e V2)
pela espeacutecie V4 apoacutes irradiaccedilatildeo com luz UV na presenccedila de concentraccedilotildees de vanadato na ordem
dos milimolar e na ausecircncia de nucleoacutetidos ou metais divalentes
Jaacute no que concerne aos estudos realizados com ecircnfase na espeacutecie decameacuterica foi sugerido
porRamos et al (2006) [AK]que a interacccedilatildeo do decavanadato com a G-actina favorecida pela
polimerizaccedilatildeo desta uacuteltima estabiliza a espeacutecie V10 e induz a inibiccedilatildeo da formaccedilatildeo de F-actina A
estabilizaccedilatildeo da espeacutecie V10 por proteiacutenas tansmembranares e do citoesqueleto pode ser a razatildeo
pela qual ocorre a toxicidade do decavanadato reportada na avaliaccedilatildeo dos efeitos do V(V) nos
sistemas bioloacutegicosTiago et al (2007) [AR] concluiram que o V10 apresenta uma clara preferecircncia
de ligaccedilatildeo ao ldquoback-doorrdquo da S1-miosina-ATPase o que se verifica apenas na estrutura
conformacional desimpedida com acesso facilitado ao local de ligaccedilatildeo do Pi Sugere tambeacutem que
o V10 actua como um ldquoback-door stoprdquo bloqueando estaabertura do fragmento de 50-kDa
necessaacuteria para a hidroacutelise do ATP--fosfato o que constitui uma explicaccedilatildeo simples do
comportamento natildeo competitivo da espeacutecie decameacuterica estimulando a utilizaccedilatildeo deste oligoacutemero
como uma feramenta da elucidaccedilatildeo das mudanccedilas conformacionais ldquoback-doorrdquo da miosina no
21
processo da contracccedilatildeo muscular(ref)Destaca-se ainda a descoberta da capacidade inibitoacuteria do
V10 em relaccedilatildeo agrave fosforilaccedilatildeo oxidativa mitocondrial [AX]
Finalmente no que concerne aos estudos realizados com a presenccedila de vaacuterias espeacutecies
oligomeacutericas Aureliano e Madeira (1994) [AY]observaram que na presenccedila de Ca2+
as espeacutecies
V4 e V10se ligam agrave Ca2+
-ATPase do retiacuteculo sarcoplasmaacutetico enquanto a espeacutecie V1 se liga apenas
na presenccedila de ATP Aparentemente o aumento da concentraccedilatildeo de Ca2+
tem o efeito contraacuterio da
inibiccedilatildeo da ATPase do retiacuteculo sarcoplasmaacutetico pelo V1 natildeo tendo no entanto efeito significativo
sobre a inibiccedilatildeo por parte da espeacutecie V10 Tiago et al (2002) [AP] concluiacuteram que em condiccedilotildees
proacuteximas agraves fisioloacutegicas a espeacutecie V10 difere das espeacutecies presentes na soluccedilatildeo de metavanadato
devido agrave sua forte interacccedilatildeo com a miosina fosforilada e agrave sua inibiccedilatildeo da actividade ATPaacutesica
da miosina Para aleacutem disso o ATP diminui a afinidade da miosina para o V4 induzindo a
interacccedilatildeo com V1 e natildeo tendo qualquer efeito no que diz respeito ao V10 Foi tambeacutem proposto
[AQ] que a ligaccedilatildeo das espeacutecies oligomeacutericas de vanadato aoS1 de miosina nos locais onde o
fosfato coordenaeacute provavelmente a causa da inibiccedilatildeo da actividade ATPaacutesica da miosina
estimulada por actina atraveacutes da prevenccedilatildeo da troca entre ATPADP sendo esta favorecida para
aniotildees de vanadato com valecircncias superiores como eacute o caso do V10
Na presenccedila de actina as velocidades de dissociaccedilatildeo do Pi e do V aumentam consideravelmente
este uacuteltimo em menor quantidade Assim a ligaccedilatildeo da actina ao complexo MADP
V e
Pi eacute
bastante fraca pelo que se sugere que o ADPV teria um efeito de relaxamento semelhante ao
ATP No entanto a capacidade do vanaacutedio em inibir a actividade ATPaacutesica da miosina e da
actomiosina eacute diferenciada uma vez que na ausecircncia de actina a inibiccedilatildeo eacute total para
concentraccedilotildees na gama dos M mas na sua presenccedila a inibiccedilatildeo eacute totalmente conseguida apenas
em concentraccedilotildees na gama dos mM (Ref) No entanto eacute necessaacuterio considerar a possiacutevel
formaccedilatildeo de espeacutecies oligomeacutericas de vanadato cuja afinidade e efeito na enzima seratildeo
diferentes Assim pode concluir-se que o efeito de inibiccedilatildeo observado para a actividade ATPaacutesica
de miosina estimulada por actina se pode dever agrave presenccedila das diferentes espeacutecies oligomeacutericas o
que foi jaacute sugerido por Aureliano (2000) [AZ] ndash as espeacutecies tetrameacuterica e decameacuterica de vanadato
interactuariam com a miosina com alteraccedilatildeo da sua interacccedilatildeo com a actina No entanto natildeo eacute
conhecida a contribuiccedilatildeo absoluta das restantes espeacutecies oligomeacutericas para a inibiccedilatildeo da ATPase
da miosina estimulada por actina (Refs)
14 Objectivos do projecto
Pretende-se com este projecto o isolamento purificaccedilatildeo e caracterizaccedilatildeo da actina do muacutesculo
22
esqueleacutetico de coelho sendo esta uacuteltima tambeacutem realizada para o sufragmento-1 de miosina
Seguidamente realizar-se-aacute a quantificaccedilatildeo e caracterizaccedilatildeo das proteiacutenas por espectroscopia UV-
Vis e SDS-PAGE e a caracterizaccedilatildeo das soluccedilotildees de vanaacutedio por UV-Vis e ressonacircncia magneacutetica
nuclear (RMN) Por uacuteltimo a interacccedilatildeo do S1 de miosina com o vanaacutedio seraacute avaliada atraveacutes
das teacutecnicas de UVVis RMN e ressonacircncia paramagneacutetica electroacutenica (RPE)
2 Discussatildeo de Resultados
21 Caracterizaccedilatildeo das proteiacutenas isoladas
211Anaacutelise do estado de pureza
O estado de pureza das soluccedilotildees de actina e de S1 foi analisado por electroforese em gel de
poliacrilamida em condiccedilotildees desnaturantes (SDS-PAGE) (Fig 22) que separa as moleacuteculas de
acordo com o seu peso molecular (PM) A concentraccedilatildeo de acrilamida escolhida (10) permite a
detecccedilatildeo simultacircnea de componentes com peso molecular inferior a 20 kDa e outros com elevado
peso molecular
Figura 21 ndash Fotografia de gel de SDS-PAGE (10 acrilamida) Poccedilo (a)
10 L do marcador de elevado peso molecular () Poccedilo (b) 20 L de [G-
actina]=1 mgmL Poccedilo (c) 20 L de [S1]=1 mgmL Aplicou-se uma
voltagem de V durante h e min
A partir da recta de calibraccedilatildeo obtida log PM = -1177 Rf + 2247 (ver ponto 2 da secccedilatildeo
Anexos) pode verificar-se a presenccedila de actina na amostra do poccedilo (b) presente numa banda
bastante intensa (PMaparente=40 kDa) bem como a presenccedila minoritaacuteria de outros componentes
desconhecidos que natildeo teratildeo sido eliminados durante o processo de extracccedilatildeo e purificaccedilatildeo e natildeo
satildeo relevantes para o estudo em questatildeo Na amostra do poccedilo (c) nota-se a presenccedila de S1
(PMaparente=92 kDa) bem como a de outros componentes cuja banda eacute ligeiramente menos intensa
Estes poderatildeo ser fragmentos proteoliacuteticos de miosina e de S1 Estaacute ainda presente uma bada de
23
maior intensidade que poderaacute ser a proteiacutena-C (PM=135 kDa) uma vez que a sua remoccedilatildeo
completa da amostra necessitaria de um passo adicional de cromatografia de troca ioacutenica [BA]
No entanto natildeo podemos confirmar que se trata desta proteiacutena ou de qualquer outra uma vez que
se encontra fora do intervalo de migraccedilatildeo dos marcadores de peso molecular
212 Determinaccedilatildeo da concentraccedilatildeo proteica
2121Subfragmento-1 de miosina
A partir da recta de calibraccedilatildeo obtida pelo meacutetodo de Bradford [BB](ver secccedilatildeo 3311 do
Procedimento Experimental) A595nm = 00546[BSA] + 00209 (ver secccedilatildeo de Anexos)foi possiacutevel
determinar um intervalo de concentraccedilatildeo de S1 de miosina compreendido entre 226 e 117(plusmn
423M) (203 e 1056 (plusmn 381 mgmL)) (n=8) A concentraccedilatildeo meacutedia de S1 de miosina apoacutes
diaacutelise contra a soluccedilatildeo tampatildeo 10 mM HEPES pH 75 25 mM KCl determinada pela leitura da
Abs280 nmfoi de 20430M (1803 mgmL)(n=7)
As amostras a quantificar atraveacutes do meacutetodo de Bradford natildeo sofreram diaacutelise o que permite
explicar a concentraccedilatildeo superior de S1 de miosina em comparaccedilatildeo com o valor obtido atraveacutes do
meacutetodo directo Esta razatildeo aliada agrave sensibilidade e rapidez superiores deste uacuteltimo fez com que
se optasse pelo meacutetodo directo para a quantificaccedilatildeo da grande maioria dos ensaios
2122Actina
A concentraccedilatildeo de actina foi determinada quando esta se encontrava na forma monomeacuterica (G-
actina) por mediccedilatildeo da leitura directa da Abs290 nm tendo-se obtido uma concentraccedilatildeo meacutedia de
691481M (290202 mgmL) (n=3) em 2 mM Tris-HCl pH 80 02 mM CaCl2 02 mM Na-
ATP Ao comprimento de onda utilizado (290 nm) absorvem fortemente apenas os resiacuteduos de
Trp pelo que a mediccedilatildeo efectuada tem menor sensibilidade do que se tivesse sido efectuada a 280
nm uma vez que a actina apresenta apenas 4 resiacuteduos deste aminoaacutecido embora tenha 16
resiacuteduos de Tyr
22 Mediccedilatildeo da actividade enzimaacutetica de S1 de miosina
221 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
As actividades ATPaacutesicas de S1 de miosina dependentes de Ca2+
e K+foram analisadas pelo
caacutelculo das velocidades de consumo inicial do fosfato inorgacircnico (Pi) Definiu-se os tempos de 5
e 3 minutos para a mediccedilatildeo da Ca2+
e da K+EDTA-ATPase respectivamente Os valores obtidos
24
a partir do protocolo presente no ponto 341 da secccedilatildeo Procedimento experimental encontram-se
descritos na Tabela 2I
Tabela 2I ndash Acrividades natildeo-fisioloacutegicas de subfragmento-1 determinadas pelo meacutetodo de Fiske-Subarow
modificado (n=2)
Proteiacutena
Actividade
ATPaacutesica natildeo
fisioloacutegica
Actividade (nmol
Piminmg proteiacutena) Actividade (s-1)
S1
Ca2+
-ATPase 1400 plusmn 30 2100 plusmn 0045
K+EDTA-ATPase 2334 plusmn 50 3500 plusmn 0075
As actividades natildeo-fisioloacutegicas reflectem o estado de oxidaccedilatildeo dos resiacuteduos expostos de Cys 707
(SH1) e 697 (SH2) Se o grupo SH1 sofrer oxidaccedilatildeo a actividade ATPaacutesica dependente de Ca2+
aumenta e a actividade dependente de K+ diminui Por outro lado se o grupo SH2 sofrer
oxidaccedilatildeo regista-se a perda completa destas actividades
De acordo com Margossian e Lowey (1982) [BA] as actividades ATPaacutesica de S1 de miosina
dependente de Ca2+
e K+EDTA da miosina satildeo respectivamente de 24 e 112 s-1
Estaacute tambeacutem
descrito que a actividade ATPaacutesica dependente de K+ eacute em meacutedia cerca de 3 vezes superior agrave
dependente de Ca2+
reflectindo a quantidade das preparaccedilotildees A razatildeo obtida entre os valores das
actividades ATPaacutesicas dependentes de K+ e Ca
2+ eacute de 166 Os valores obtidos para as actividades
(Ca2+
-ATPase=21 s-1
K+EDTA-ATPase= 35 s
-1) satildeo inferiores agrave valores obtidos por
Margossian e Lowey [BA] e Tiago [BC] com forccedila ioacutenica inferior O valor obtido para a
actividade ATPaacutesica dependente de Ca2+
eacute superior ao obtido por Polosukhina et al (2000) [BD]
cujo meio de reacccedilatildeo continha MOPS e apresentava forccedila ioacutenica inferior Os dados
supramencionados comprovam a variaccedilatildeo da actividade a ATPaacutesica natildeo fisioloacutegica em relaccedilatildeo agrave
forccedila ioacutenica temperatura e pH
222 Mediccedilatildeo da actividadeMg2+
-ATPaacutesica de S1 pelo meacutetodo de enzimas acopladas
A actividade de Mg2+
-ATPase eacute a uacutenica que possui significado fisioloacutegico sendo as restantes
bastante uacuteteis apenas para indicaccedilatildeo da qualidade de preparaccedilatildeo do S1[BA] A 0575 M KCl a
actina e o S1 formam uma soluccedilatildeo viscosa e tuacuterbida Apoacutes a adiccedilatildeo de MgATP o complexo acto-
S1 dissocia-se e a turbidez da soluccedilatildeo diminui Se a forccedila ioacutenica for reduzida a dissociaccedilatildeo do
complexo actoS1 que se encontra na forma de suspensatildeo eacute induzida O MgATP eacute rapidamente
hidrolisado diminuindo a sua concentraccedilatildeo no meio reaccional Nestas condiccedilotildees haacute
reassociaccedilatildeo e subsequente contracccedilatildeo dos filamentos de actina e de S1 com formaccedilatildeo de
25
precipitado O meacutetodo descrito traduz de forma bastante elementar o fenoacutemeno da contracccedilatildeo in
vivo e elimina o problema anterior da solubilizaccedilatildeo ineficiente da miosina de modo a obter uma
mediccedilatildeo exacta da actividade [A] A utilizaccedilatildeo de forccedilas ioacutenicas reduzidas aumenta a afinidade da
actina para o S1 permitindo contornar a extrema dificuldade em obter a concentraccedilatildeo proteica dos
muacutesculos (cerca de 100 mgmL do complexo actomiosina) As actividades Mg2+
-ATPaacutesicas de S1
de miosina foram determinadas a partir do declive da variaccedilatildeo da Abs340 relativo agrave oxidaccedilatildeo de
NADH em funccedilatildeo do tempo como eacute descrito no ponto 342 do Procedimento Experimental Os
valores obtidos para a actividade Mg2+
-ATPaacutesica encontram-se apresentados na Tabela 2II
Tabela 2II ndash Actividade Mg2+
-ATPaacutesica de subfragmento-1 de miosina determinadas pelo meacutetodo das enzimas
acopladas
Actividade (nmol
ATPmin mg(S1)) Actividade (s
-1)
1097 plusmn 269 164 plusmn 04
Ramos (2005) [J] obteve o valor de 01 s-1
para a actividade Mg2+
-ATPaacutesica de S1 de miosina
com forccedila ioacutenica intermeacutedia Tiago (2005) [BC] obteve o valor de 011 s-1 em condiccedilotildees de forccedila
ioacutenica reduzida em relaccedilatildeo agrave utilizada no presente relatoacuterio
23 Preparaccedilatildeo e caracterizaccedilatildeo das soluccedilotildees de vanaacutedio
231 Anaacutelise da composiccedilatildeo das soluccedilotildees de vanadato por espectroscopia de 51
V-RMN
efeito da concentraccedilatildeo da espeacutecie
A diluiccedilatildeo das soluccedilotildees stock de meta- e decavanadato iraacute determinar fortemente a composiccedilatildeo
em soluccedilatildeo dos diferentes oligoacutemeros de vanadato (ref) No caso do decavanadato o periacuteodo de
tempo apoacutes a diluiccedilatildeo iraacute tambeacutem ser um factor importante a considerar devido agrave decomposiccedilatildeo
da espeacutecie V10(ref)Torna-se assim fundamental determinar a composiccedilatildeo quiacutemica das soluccedilotildees
no meio de ensaio utilizado (HEPES 10 mM KCl 25 mM) de modo a correlacionaacute-la
correctamente com os possiacuteveis efeitos promovidos pelas diferentes espeacutecies de vanadato no
sistema em estudo
Aleacutem disso deve ser dado ecircnfase agraves importantes interacccedilotildees que ocorrem entre o vanaacutedio e os
tampotildees escolhidos O tampatildeo com menor nuacutemero e magnitude de interacccedilatildeo eacute o HEPES [AD]
seguido do Tris sendo este uacuteltimo especialmente aconselhado em concentraccedilotildees baixas para casos
de incompatibilidade enzimaacutetica ou quiacutemica redox No caso especial da reacccedilatildeo redox
NAD(P)HNAD(P) o tampatildeo adequado eacute o imidazole de modo a natildeo haver reacccedilatildeo com o
26
vanadato Os tampotildees a evitar em qualquer situaccedilatildeo incluem a trietanolamina fosfato citrato e
glicina entre outros [AD] uma vez que todos interactuam fortemente com V(V) com formaccedilatildeo
de complexos estaacuteveis
Assim procedeu-se agrave anaacutelise da composiccedilatildeo das soluccedilotildees de meta- e decavanadato apoacutes diluiccedilatildeo
das soluccedilotildees stock (50 mM em vanadato total) no meio de reacccedilatildeo (tampatildeo D) atraveacutes da
espectroscopia de ressonacircncia magneacutetica nuclear de 51
V Nas figuras x1 e x2 apresentam-se os
espectros obtidos para as soluccedilotildees de deca- e metavanadato num intervalo de concentraccedilotildees de 1 a
10 mM A Tabela seguinte resume os paracircmetros espectroscoacutepicos obtidos para cada espeacutecie
presente nas soluccedilotildees analisadas
Figura 22ndash Espectros de RMN de 51
V de
concentraccedilotildees crescentes das espeacutecies oligomeacutericas
de vanadato obtidos apoacutes diluiccedilatildeo da soluccedilatildeo stock
de decavanadato em 10 mM HEPES pH 70 25 mM
KCl para concentraccedilotildees finais de A) 1 mM B)2
mM C) 4 mM D) 8 mM E) 10 mM de
decavanadato total V1refere-se agraves espeacutecies
monomeacutericade vanadato V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2IIIndash Desvios quiacutemicos meacutedios (part) das ressonacircncias das espeacutecies decameacuterica (V10) monomeacuterica (V1) e
tetrameacuterica (V4) presentes nos espectros da soluccedilatildeo de decavanadatoLargura a meia altura (∆12) da espeacutecie V10A
Paracircmetros V10A V10B V10C V1 V4
part (ppm) -5162 -5003 -4255 -5616 -5748
∆12 (Hz) 175 nd nd nd nd
Figura 23 ndash Distribuiccedilatildeo graacutefica das
espeacutecies oligomeacutericas de vanadato vs a
concentraccedilatildeo de decavanadato total As
concentraccedilotildees de cada espeacutecie foram
obtidas segundo a formula descrita na
secccedilatildeo 35 do Procedimento
Experimental (aacutereas dos sinais de 51
V-
RMN observadas na figura x1)V10 V1 e
V4 correspondem agraves espeacutecie monomeacuterica
dimeacuterica e tetrameacuterica respectivamente [Esp
eacuteci
e o
ligo
meacute
rica
de
van
aacutedio
] (m
M)
[Decavanadato]total (mM)
V10
V1
V4
27
Nas soluccedilotildees de decavanadato as espeacutecies monomeacuterica e decameacuterica de vanadato satildeo as uacutenicas
presentes para concentraccedilotildees de decavanadato inferiores a 8 mM Ambas as espeacutecies aumentam
linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado em [AY] e
em (Teresa) quando a concentraccedilatildeo de V1 eacute igual ou superior a 200 microM inicia-se a sua
oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica No entanto nas condiccedilotildees de realizaccedilatildeo dos
ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a espeacutecie tetrameacuterica neste
estudo em concentraccedilotildees inferiores a 8 mM tendo o mesmo sido observado por Ramoset al
(2006) [AK] Os resultados obtidos mostram claramente o equiliacutebrio entre a espeacutecie decameacuterica e
a monomeacuterica e que o aparecimento de outras espeacutecies oligomeacutericas se encontra dependente da
concentraccedilatildeo desta uacuteltima Apesar de natildeo ter sido realizado o ensaio com 5 mM de decavanadato
total no presente relatoacuterio estaacute reportado em Ramos et al (2006) [AK] que as concentraccedilotildees da
espeacutecie decameacuterica e monomeacuterica satildeo respectivamente de 500 microM e 200 microM Assim pode inferir-
se que para concentraccedilotildees inferiores a 5 mM e intervalos de tempo de cerca de 20 minutos (que
inclui o tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas
as espeacutecies decameacuterica e monomeacuterica de vanadato
Figura 24 ndash Espectros de RMN de 51
V de concentraccedilotildees
crescentes das espeacutecies oligomeacutericas de vanadato obtidos
apoacutes diluiccedilatildeo da soluccedilatildeo stock de metavanadato em 10 mM
HEPES pH 70 25 mM KCl para concentraccedilotildees finais de A)
1 mM B) 2 mM C) 4 mM D) 8 mM E) 10 mM de
metavanadato total V1 refere-se agraves espeacutecies monomeacuterica de
vanadatoV1 e V2 correspondem respectivamente agrave espeacutecie
monomeacuterica e dimeacuterica
Figura 25 ndash Distribuiccedilatildeo graacutefica das espeacutecies
oligomeacutericas de vanadato vs a concentraccedilatildeo de
metavanadato total As concentraccedilotildees de cada
espeacutecie foram obtidas segundo a formula descrita
na secccedilatildeo 35 do Procedimento Experimental
(aacutereas dos sinais de 51
V-RMN observadas na
figura x1)V10 V1 e V4 correspondem agraves espeacutecie
monomeacuterica dimeacuterica e tetrameacuterica
respectivamente
Esp
eacuteci
e o
ligo
meacute
rica
de
va
nad
ato
(m
M)
[Metavanadato]total (mM)
V1
V2
V4
V5
28
Tabela 2IVndash Desvios quiacutemicos meacutedios (part)das ressonacircncias das espeacutecies tetrameacuterica (V4) monomeacuterica (V1) e
dimeacuterica (V2) presentes no espectro da soluccedilatildeo de metavanadato (total) 10 mM
Paracircmetros V1 V2 V4 V5
part (ppm) -5615 -5748 -5789 -5869
Nas soluccedilotildees de metavanadato as espeacutecies monomeacuterica dimeacutericae tetrameacuterica de vanadato estatildeo
presentes para concentraccedilotildees de metavanadato inferiores a 4 mM Todas estas as espeacutecies
aumentam linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado
em (Aureliano e Madeira 1998) e em (Tiago et al 2004) quando a concentraccedilatildeo de V1 eacute igual ou
superior a 200 microM inicia-se a sua oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica Nas
condiccedilotildees de realizaccedilatildeo dos ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a
espeacutecie pentameacuterica neste estudo em concentraccedilotildees inferiores a 4 mM tendo o mesmo sido
observado por Ramos 2006 Os resultados obtidos mostram claramente o equiliacutebrio entre a
espeacutecie V4 e as espeacuteciesV1 e V2 no qual o aparecimento de outras espeacutecies oligomeacutericas se
encontra dependente da concentraccedilatildeo da espeacutecie V1 Por uacuteltimo pode inferir-se que para
concentraccedilotildees inferiores a 2 mM e intervalos de tempo de cerca de 20 minutos (que inclui o
tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas as
espeacutecies V1 V2 e V4 com concentraccedilotildees respectivas de 0796 0192 e 0196 mM
232Estabilidade das soluccedilotildees de decavanadato anaacutelise por espectroscopia de absorccedilatildeo
UVVis
Caracterizou-se a estabilidade da espeacutecie V10 numa soluccedilatildeo com 1 mM decavanadato total em
tampatildeo D atraveacutes da leitura da Abs400 Obteve-se uma cineacutetica de dissociaccedilatildeo de 1ordf ordem tal
como se pode verificar pela linearidade do graacutefico de ln(Abs400)vs o tempo de aquisiccedilatildeo espectral
(segundos) (Fig26 )
Figura 26ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do tempo
(em segundos) para uma soluccedilatildeo de decavanadato total de 1
mM em tampatildeo D a 25ordmC (n=2)
Sendo uma reacccedilatildeo de primeira ordem pode ser descrita pela equaccedilatildeo cujo
declive eacute ndashK (constante de dissociaccedilatildeo) e a intersecccedilatildeo no eixo das ordenadas corresponde
Ln
(Ab
s 400 n
m)
tempo (s)
29
aln(A0) Numa reacccedilatildeo de 1ordf ordem o tempo de meia-vida eacute independente da concentraccedilatildeo inicial
do reagente sendo descrito pela equaccedilatildeo [2] Deste modo a partir da recta
Ln(Abs400 nm) = -436E-5
K - 105 obteve-se um tempo de meia-vida de 4490 plusmn 0823 horas para
as condiccedilotildees supramencionadas
Apoacutes a adiccedilatildeo de S1 8 microM ao meio reaccional verificou-se que a reacccedilatildeo de desoligomerizaccedilatildeo
da espeacutecie decameacuterica de vanadato seguia igualmente uma cineacutetica de 1ordf ordem tal como se pode
verificar pela linearidade do graacutefico de ln(Abs400) versus o tempo de aquisiccedilatildeo espectral
(segundos) (Fig27)
Figura 27ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do
tempo (em segundos) para uma soluccedilatildeo de decavanadato
total de 1 mM em tampatildeo D a 25ordmC na presenccedila de 8
microM (n= 2)
A partir da equaccedilatildeo da recta Ln(Abs400 nm) = -7520E-5
K - 0613determinou-se o valor de K para
a espeacutecie decameacuterica de onde se obteve um tempo de meia-vida de 2430 plusmn 0191 horas para as
condiccedilotildees supramencionadas
Tanto este valor como o valor registado na ausecircncia de S1 satildeo substancialmente superiores agrave
duraccedilatildeo dos ensaios de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina descritos no presente
relatoacuterio
24 Interacccedilatildeo de S1 com vanaacutedio(V)
241Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
De modo a analisar qual o efeito da adiccedilatildeo de S1 de miosina (tampatildeo D) produzia nos sinais de
51V-NMR titulou-se uma soluccedilatildeo de 5mM de decavanadato total com concentraccedilotildees crescentes
de S1 (5 10 15 e 20 microM) Os espectros de 51
V-RMN encontram-se apresentados na Fig 2
No graacutefico da figura 2 Apresenta-se a concentraccedilatildeo de S1 em funccedilatildeo do factor de alargamento
corrigido dos espectros acima referidos
Ln
(Ab
s (4
00
nm
))
tempo (s)
30
Figura 28 ndash Graacutefico de
representaccedilatildeo da
concentraccedilatildeo de S1 em
funccedilatildeo do factor de
alargamento A
concentraccedilatildeo de
decavanadato total foi de 5
mM e as concentraccedilotildees
utilizadas de S1 foram de 0
5 10 e 15 microM
Verifica-se que o factor de alargamento aumenta linearmente em funccedilatildeo da concentraccedilatildeo de S1 no
ensaio o que indica a existecircncia de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina Observou-
se ainda que a adiccedilatildeo de S1 ao meio reaccional natildeo provocou qualquer alteraccedilatildeo dos desvios
quiacutemicos dos sinais de 51
V-RMN
Realizaram-se ensaios na presenccedila de ATP e de S1 com intuito de verificar o efeito do ATP na
interacccedilatildeo do S1 de miosina com o decavanadato
Figura 29 - Espectros de RMN de 51
V contendo 2
mM Decavanadato (A) (controlo) (B) na presenccedila
de 2 mM de ATP (C) na presenccedila de 2 mM de ATP
e 8 microM de S1 de miosina V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2Vndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies decameacuterica (V10) e monomeacuterica (V1) presentes nos
espectros da soluccedilatildeo de decavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V10A V10B V10C V1
part (ppm) -5161 -5002 -4254 -5614
Observa-se que a presenccedila de ATP (espectro B) natildeo provoca qualquer alteraccedilatildeo visiacutevel em
relaccedilatildeo ao espectro que contem apenas soluccedilatildeo de decavanadato Jaacute a presenccedila simultacircnea de ATP
e S1 provoca um alargamento da ∆12 do sinal do V10A de 1736 Hz (espectro (A)) para 2366 Hz
(espectro (C) Fig 29) Tendo em conta que a concentraccedilatildeo de S1 apresenta uma relaccedilatildeo linear
com o factor de alargamento converteu-se o valor da ∆12 e comparou-se com o valor esperado agrave
F = 00591[S1] - 00157 Rsup2 = 0998
Fact
or
de
ala
rgam
en
to
[S1] (microM)
31
concentraccedilatildeo de S1 utilizada O factor de alargamento obtido para 8 microM ndash 0376 ndash eacute inferior ao
valor esperado agrave mesma concentraccedilatildeo de S1 obtido atraveacutes da equaccedilatildeo da recta apresentada na
Fig 28 ndash 0457 ndash o que indica que o ATP poderaacute exercer um efeito de protecccedilatildeo do S1
diminuindo a sua interacccedilatildeo com a espeacutecieV10
Por uacuteltimo realizaram-se ensaios com metavanadato na presenccedila de 8 microM S1 de modo a verificar
se as espeacutecies oligomeacutericas contidas nesta soluccedilatildeo interagiam com a proteiacutena em estudo
Figura 210 - Espectros de RMN de 51
V contendo 2
mM metavanadato (A) (controlo) (B) na presenccedila
de 8 microM de S1 de miosina V1 V2 e V4
correspondem respectivamente aacutes espeacutecies
monomeacuterica dimeacuterica e tetrameacuterica
Tabela 2VIndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies monomeacuterica (V1) dimeacuterica (V2) e tetrameacuterica (V4)
presentes nos espectros da soluccedilatildeo de metavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V1 V2 V4
part (ppm) -5613 -5747 -5789
Como eacute possiacutevel observar na Fig 210 ocorre uma ligeiro alargamento dos sinal de V4 bem
como uma perda da intensidade pelo que se conclui quedas espeacutecies presentes na soluccedilatildeo de
metavanadato (V1 V2 e V4) apenas a espeacutecie tetrameacuterica interage com o S1 de miosina o que
estaacute de acordo com o reportado por Tiago et al 2002
242 Actividade Mg2+
-ATPaacutesica de S1 de miosina na presenccedila de decanadato
Com o intuito de avaliar o efeito do decavanadato na actividade Mg2+
-ATPaacutesica do S1 de
miosina monitorizou-se a reacccedilatildeo de oxidaccedilatildeo do NADH atraveacutes da leitura de Abs340 na
presenccedila de 005 mgmL de S1 de miosina e no caso da actividade estimulada 2microM de actina em
3125 mM Tris-HCl pH 71 e 375 mM MgCl2 pelo meacutetodo de enzimas acopladas (ver seccccedilatildeo xx
dos MM)Para tal realizaram-se ensaios com vaacuterias concentraccedilotildees de decavanadato total (10 50
100 250 500 e 1000) de modo a determinar qual o seu efeito na actividade MgATPaacutesica de S1
de miosina na ausecircncia e na presenccedila de um dos seus ligandos naturais Os ensaios foram
realizados agrave temperatura ambiente com incubaccedilatildeo preacutevia a 25ordm C durante 20 minutos
32
Figura 211 ndash Representaccedilatildeo
graacutefica da Actividade Mg2+
-
ATPaacutesica de S1 () (os ajustes
teoacutericos estatildeo representado a por
uma linha os resultados
experimentais estatildeo representados
por triacircngulos (Actividade
estimulada) e cruzes (Actividade
natildeo estimulada)) vs
[Decavanadato]total medidas pelo
meacutetodo das enzimas acopladas
(n=3) Os dados foram ajustados ao modelo de decaimento exponencial usando o programa GraphPad Prism para a
determinaccedilatildeo dos valores de IC50 (na ausecircncia de actina IC50= 266 plusmn 07 M decavanadato na presenccedila de actina
IC50= 28 plusmn 1 M decavanadato
A partir do graacutefico presente na Fig 29 verifica-se que o ajuste dos dados experimentais foi bem
sucedido A inibiccedilatildeo da actividade Mg2+
-ATPaacutesica do S1 de miosina eacute afectada em menor
amplitude na presenccedila de actina (60 de inibiccedilatildeo da actividade) do que na sua ausecircncia
(aproximadamente 80 de inibiccedilatildeo de actividade) Devido a esta diferenccedila pode sugerir-se que a
actina iraacute impedir parcialmente a ligaccedilatildeo do decavanadatoao S1 demiosina atraveacutes de
impedimentos esteacutereos ou interacccedilotildees electrostaacuteticas por exemplo A teacutecnica utilizada natildeo nos
permite concluir acerca de que regiatildeo(otildees) ou resiacuteduo(s) da actina participaratildeo na preservaccedilatildeo
parcial da actividade Mg2+
-ATPaacutesica
Por outro lado os valores de IC50 dos dois tipos de actividades natildeo apresentam diferenccedilas
significativas tendo-se obtido 266 plusmn 07 e 28 plusmn 1M decavanadato na ausecircncia e na presenccedila de
actina respectivamente o que corresponde a uma concentraccedilatildeo de cerca de 27 M de V10 O
meio de reacccedilatildeo utilizado nestes ensaios foi 10 mM HEPES e 25 mM KCl com 2 mM ATP Em
(Tiago et al 2004) o valor de IC50 para a actividade Mg2+-ATPaacutesica na ausecircncia de actina foi de
125 plusmn 025 microM enquanto o valor da actividade estimulada por actina foi de 136 plusmn 14 microM O
meio de reacccedilatildeo utilizado nos ensaios foi 20 mM HEPES 50 mM KCl e 2 mM ATP o que resulta
precisamente no dobro da forccedila ioacutenica a que foi realizado o ensaio do presente relatoacuterio
(exceptuando a contribuiccedilatildeo dos produtos da hidroacutelise de ATP uma vez que este meacutetodo
pressupotildee a ocorrecircncia de um ciclo de reacccedilotildees de hidroacutelise e siacutentese deste composto) Ora tendo
sido anteriormente referido que a afinidade do S1 de miosina para com a espeacutecie V10 diminui com
Act
ivid
ade
Mg
2+-A
TP
aacutesic
a d
e S
1 d
e
Mio
sin
a
[Decavanadato] (microM) Actividade Estimulada ()
Actividade Natildeo Estimulada ()
Ajuste Actividade Estimulada
Ajuste Actividade Natildeo Estimulada
33
o aumento da forccedila ioacutenica (Tiago et al 2004 tal poderaacute explicar a diferenccedila obtida entre estes
dois conjuntos de valores
243Quantificaccedilatildeo de cisteiacutenas reduzidas pelo meacutetodo de Ellman
A interacccedilatildeo do vanaacutedio com alguns tipos de aminoaacutecidos sensiacuteveis agrave oxidaccedilatildeo pode fornecer-nos
informaccedilatildeo bastante uacutetil em relaccedilatildeo agraves alteraccedilotildees estruturais que conduzem a uma diminuiccedilatildeo da
actividade fisioloacutegica do S1 de miosina Os estudos realizados por Chasteen 1983 concluiacuteram
que a reduccedilatildeo de vanadato (V) a vanadilo (IV) ocorre na presenccedila de agentes redutores comuns no
ambiente celular de que se destaca o aminoaacutecido cisteiacutena Esta eacute tambeacutem um alvo de
complexaccedilatildeo por parte do iatildeo VO2+
Jaacute Ramos et al (2009) demonstrou que a incubaccedilatildeo de
actina com a soluccedilatildeo de decavanadato induz a oxidaccedilatildeo de resiacuteduos de cisteiacutena e agrave formaccedilatildeo do
oxidovanaacutedio (IV) Este grupo de investigadores descobriu ainda que apenas a espeacutecie V10 induz a
oxidaccedilatildeo das cisteiacutenas ldquoraacutepidasrdquo da actina atraveacutes do processo proposto que eacute parcialmente
responsaacutevel pela inibiccedilatildeo jaacute descrita da actividade de ATPase do S1 de miosina estimulada por
actina quando exposta agrave soluccedilatildeo de decavanadato Esta inibiccedilatildeo apresenta-se afectada com a
presenccedila de ATP uma vez que os resiacuteduos de cisteiacutena permaneceram intactos nos ensaios
realizados com este ligando Jaacute Crans et al (2004) concluiu que o siacutetio activo de muitas fosfatases
(que conteacutem grupos tiol) pode formar um equiliacutebrio redox com o iatildeo vanadato (V) de forma natildeo
reversiacutevel
Os resultados obtidos por Sakurai e Shiromura (1980) sugeriram que o vanaacutedio (V) sofre reduccedilatildeo
a vanaacutedio (IV) sem dependecircncia do valor de pH do ensaio Registaram ainda uma estequiometria
de 31 na reacccedilatildeo do vanadato (V) com os resiacuteduos de Cys enquanto a estequiometria dos ensaios
com o vanadilo (IV) foi de 21
Berlett e Stadtman (1997) observaram que os tipos de aminoaacutecidos mais facilmente oxidaacuteveis
englobam entre outros os resiacuteduos de Cys Trp e MetAssim analisou-se o estado redox dos
grupos tiol dos resiacuteduos de Cys do S1 na presenccedila do decavanadato atraveacutes da monitorizaccedilatildeo da
Abs412 nm na presenccedila de 1 mgmL de S1 de miosina em tampatildeo 10 mM HEPES 25 mM KCl
variando a concentraccedilatildeo da soluccedilatildeo de decavanadato de modo a determinar que tipo de alteraccedilotildees
estruturais ocorrem na sua presenccedila no que diz respeito agrave diminuiccedilatildeo da actividade ATPaacutesica de
S1 na presenccedila de decavanadato
Os resultados referentes agrave quantificaccedilatildeo das cisteiacutenas reduzidas apoacutes o tratamento de S1 com
diferentes concentraccedilotildees de decavanadato em tampatildeo 10 mM HEPES pH 75 e 25 mM KCl a
25ordmC durante 20 minutos encontram-se descritos na Tabela 2VII
Tabela 2VII ndash Cisteiacutenas reduzidas totais e ldquoraacutepidas de S1 (1 mgmL) apoacutes tratamento com decavanadato
34
(010501002505001000 uM) em 10 mM HEPES 25 mM KCl pH 7 a 25ordmC durante 20 minutos A detecccedilatildeo
espectrofotomeacutetrica foi feita com DTNB (e 412=8400 uM-1 cm-1) (n variaacutevel entre 1 e 8)
Amostras Cys raacutepidas Cys totais
11 M S1 (controlo) 22 plusmn 04 106 plusmn 14
11 M S1 + 10 M Decavanadato 22 plusmn 05 95 plusmn 08
11 M S1 + 50 uM Decavanadato 20 plusmn 02 98 plusmn 14
11 M S1 + 100 uM Decavanadato 17 plusmn 03 97 plusmn14
11 M S1 + 250 uM Decavanadato 26 plusmn 02 98 plusmn 08
11 M S1 + 500 uM Decavanadato 21 plusmn 04 90 plusmn 06
11 M S1 + 1000 uM Decavanadato 24 plusmn 0 86 plusmn 08
A partir dos resultados obtidos pode sugerir-se que o S1 possui 11 resiacuteduos de Cys (controlo) o
que estaacute de acordo com o anteriormente descrito Verifica-se que o tratamento da proteiacutena com
concentraccedilotildees crescentes de decavanadato provoca a oxidaccedilatildeo de dois resiacuteduos de Cys do S1 de
miosina natildeo expostos ao solvente No entanto a teacutecnica utilizada natildeo nos permite identificar
quais
Tendo em consideraccedilatildeo que o valor de IC50 obtido para a reacccedilatildeo de inibiccedilatildeo daactividade Mg2+-
ATPaacutesica de S1 na presenccedila de decavanadato eacute de cerca de 27 microM pode sugerir-se que este seraacute o
valor ao qual ocorreraacute a oxidaccedilatildeo do primeiro resiacuteduo de Cys o que poderaacute contribuir para a
diminuiccedilatildeo da actividade de S1 observada
25Interacccedilatildeo de S1 com vanaacutedio(IV)
251Anaacutelise por espectroscopia de Ressonacircncia Paramagneacutetica Electroacutenica
A acumulaccedilatildeo intracelular de oxovanaacutedio (IV) in vivo constitui uma das principais formas de
apresentaccedilatildeo de vanaacutedio no organismo Deste modo torna-se bastante interessante analisar a
interacccedilatildeo entre este iatildeo e o S1 de miosina atraveacutes da teacutecnica de espectroscopia de Ressonacircncia
Paramagneacutetica Electroacutenica o meacutetodo de eleiccedilatildeo do estudo do iatildeo vanadilo e dos complexos por si
formados O electratildeo desemparelhado encontra-se fortemente acoplado ao nuacutecleo 51
V o que
35
resulta num espectro de RPE consistindo de 8 desdobramentos na direcccedilatildeo paralela e outros tantos
na direcccedilatildeo perpendicular (Chasteen 1983)
Os estudos realizados por Ahmed et al (1987) centraram-se na interacccedilatildeo do vanadilo com
proteiacutenas como eacute o caso da insulina transferrina entre outros obtendo um espectro caracterizado
por 8 linhas amplamente espaccediladas (na direcccedilatildeo paralela) Jaacute Ramos et al 2009 concluiu que a
exposiccedilatildeo da soluccedilatildeo de decavanadato agraves formas globular e filamentosa de actina produz um
espectro tiacutepico do iatildeo oxidovanaacutedio (IV) indicando a reduccedilatildeo do vanadato
Para aferir qual a concentraccedilatildeo de S1 que permitia obter uma boa razatildeo sinalruiacutedo titulou-se uma
soluccedilatildeo de sulfato de vanadilo 15 microM ([VOSO4]stock = 5 mM) com concentraccedilotildees crescentes de
proteiacutena (0 5 10 15 e 20 microM) em 10 mM HEPES 25 mM KCl A existecircncia de uma correlaccedilatildeo
linear entre a concentraccedilatildeo de proteiacutena e a intensidade dos sinais de RPE correspondente agrave banda
-12 perpendicular encontra-se representada na Fig 210
Figura 2 12ndash Representaccedilatildeo graacutefica da variaccedilatildeo da intensidade do sinal de RPE vs a concentraccedilatildeo de S1 utilizada
Apoacutes verificar que com uma concentraccedilatildeo de S1 igual a 20 microM se obtinham sinais com uma razatildeo
sinalruiacutedo adequada ao prosseguimento dos estudos sobre a interacccedilatildeo V(IV)-S1 de miosina
titulou-se uma soluccedilatildeo de 20 microM S1 com 0 25 5 10 15 25 50 75 100 150 200 500 e 1000
microM VOSO4
Traccedilou-se o graacutefico da intensidade da banda -12 perpendicular do sinal de EPR em funccedilatildeo da
concentraccedilatildeo de VOSO4 tendo-se determinado uma constante de dissociaccedilatildeo (KD) do complexo
S1-vanadilo de 8748 plusmn 2621 microM V(IV)
y = 016786x + 122435
Rsup2 = 092992759
Inte
nsi
dad
e d
o s
inal
de
RP
E
[S1] (microM)
36
Figura 213 ndash Representaccedilatildeo graacutefica da variaccedilatildeo do sinal de RPE vs a concentraccedilatildeo de vanadilo utilizada (os valores
experimentais estatildeo representados por losangos o seu ajuste estaacute representado pela linha)
A simulaccedilatildeo do espectro de EPR foi efectuada a partir do espectro obtido para uma concentraccedilatildeo
de 1 mM VOSO4 apoacutes adiccedilatildeo de 20 microM S1 de miosina tendo-se obtido os valores da constante de
acoplamento hiperfina (A=5016) e do factor g (g0=195)
Figura 214 ndash Representaccedilatildeo graacutefica do espectro de EPR obtido com 1000 microM de vanadilo e 20 microM de S1 (a cinzento
escuro) A simulaccedilatildeo do espectro encontra-se a cinzento claro
Por comparaccedilatildeo destes com os valores presentes na tabela de correspondecircncia xxx eacute possiacutevel
predizer quais os ligandos provaacuteveis do vanadilo na presenccedila de proteiacutena Assim concluiu-se que
o vanadilo estaacute coordenado com duas moleacuteculas de aacutegua um grupo hidroxilo e um grupo R=NR
O grupo hidroxilo pode indicar os resiacuteduos de aminoaacutecidos Tyr Treonina e Serina enquanto o
grupo R=NR poderaacute indicar a presenccedila de Arginina e Histidina como ligandos do vanadilo
Inte
nsi
dad
e d
os
sinai
s d
e R
PE
[Vanadilo] (microM)
Ajuste dos dados exp Dados experimentais
Campo magneacutetico (G)
Espectro Experimental Ajuste
37
3 Procedimento Experimental
31 Reagentes
Todos os reagentes utilizados satildeo de elevado grau de pureza (analiacutetico) Apresentam-se as suas
foacutermulas quiacutemicas ou abreviaturas quando necessaacuterio acrescentou-se a sua descriccedilatildeo Entre
parecircntesis colocou-se as empresas que os comercializam
Aacutecido aceacutetico glacial 100 (Riedel) Acrilamida 99 (Merck) Azul de bromofenol (Riedel-de-
Haumlen) -Mercaptoetanol (Sigma) Bissulfito de soacutedio (Sigma)Bradford (reagente de)
(BioRad)BSA (albumina de soro bovino Sigma-Aldrich) CaCl22H2O (cloreto de caacutelcio
dihidratado Fluka) CH3CH2OH 96 (etanol Panreac) Coomassie Blue R250 (Merck) D2O
(oacutexido de deuteacuterio SDS) DTNB (55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico) Aldrich) EDTA (aacutecido
etilenodiaminatetraaceacutetico Merck) Fosfoenolpiruvato 98 (Fluka) Glicerol (Sigma) Glicina
985 (Panreac) HCl 37 (Riedel-de Haumlen) H2SO4 96 (Pronalab) HEPES 99 (aacutecido 4-(2-
hidroxiletil)-1-piperazineetanosulfoacutenico Sigma) KH2PO4 99 (hidrogenofosfato de potaacutessio)
MgATP (Sigma) Na
ATP (Sigma) Na2HPO4 99 (hidrogenofosfato de soacutedio Riedel-de Haumlen)
NaH2PO4 (fosfato de soacutedio ) NaOH 986 (hidroacutexido de soacutedio Pronalab) KCl 99 (Cloreto de
potaacutessio Panreac) L-Cys 995 (enantioacutemero levoacutegiro do resiacuteduo de Cisteiacutena Fluka)Metanol
99 (Merck) Molibdato de amoacutenio (Sigma)NNrsquo-metil-bis-acrilamida 98 (Merck) NH4VO3
99 (metavanadato de amoacutenia Sigma) PSA 10 (persulfato de amoacutenio Merck) SDS
(dodecilo sulfato de soacutedio Sigma) Sulfito de soacutedio (Sigma) VOSO4 (sulfato de vanadilo
Aldrich) TEMED 99 (Tetrametiletilenodiamina Merck) Tris (2-amino-2-hidroximetil-13-
propanodiol Sigma)
32 Meacutetodos
321 Isolamento e purificaccedilatildeo das proteiacutenas de muacutesculo esqueleacutetico de coelho
3211 Purificaccedilatildeo da miosina preparaccedilatildeo de Subfragmento 1 por digestatildeo quimotriacutepsica
da miosina
O subfragmento-1 de miosina-II (S1) foi gentilmente cedido pelo Prof Doutor Manuel Aureliano
do Departamento de Quiacutemica Bioquiacutemica e Farmaacutecia da Faculdade de Ciecircncias e Tecnologia da
Universidade do Algarve
No entanto descreve-se abaixo de forma resumida o procedimento efectuado
38
32111 Purificaccedilatildeo da miosina
O protocolo de isolamento foi adaptado do meacutetodo previamente descrito por Margossian e Lowey
(1982) apresentando-se de forma esquemaacutetica na Fig 31
Sucintamente os tecidos musculares dorsais e das pernas foram extraiacutedos apoacutes o sacrifiacutecio do
animal (Fig31) De seguida foram adicionados 3 volumes da soluccedilatildeo de extracccedilatildeo A2
Centrifugou-se a soluccedilatildeo e filtrou-se o sobrenadante O sedimento foi entatildeo diluiacutedo em 14
volumes de aacutegua destilada de modo a que a forccedila ioacutenica atingisse 004 M Apoacutes adiccedilatildeo de szlig-
mercaptoetanol (szlig-me) deixou-se a precipitar a 4ordmC removendo o sobrenadante Centrifugou-se
novamente e ressuspendeu-se o sedimento na soluccedilatildeo de extracccedilatildeo A Adicionou-se a soluccedilatildeo
tampatildeo B3 para que a forccedila ioacutenica atingisse 05 M Homogeneizou-se e deixou-se repousar
durante 2h Homogeneizou-se novamente e centrifugou-se de modo a ocorrer precipitaccedilatildeo dos
complexos de actomiosina e proteiacutena desnaturada Apoacutes filtraccedilatildeo do sobrenadante em latilde de vidro
quantificou-se a soluccedilatildeo de miosina resultante pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental)
2 Soluccedilatildeo de extracccedilatildeo A 0012M K2CO3 004M KHCO3 06M KCl pH 65
3 Soluccedilatildeo-tampatildeo B Tris 100 mM pH 7 KCl 2M
39
Figura 31 ndash Representaccedilatildeo esquemaacutetica do protocolo de isolamento de miosina (Margossian e Lowey 1982) Todo o
procedimento foi realizado a 4ordmC (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
32112 Preparaccedilatildeo de Subfragmento-1 de miosina por digestatildeo com quimiotripsina
O S1 de miosina foi isolado apoacutes diaacutelise da miosina durante a noite contra a soluccedilatildeo C4 A
presenccedila do EDTA eacute essencial uma vez que a sua substituiccedilatildeo por MgCl2resulta na produccedilatildeo de
HMM em vez de S1 A suspensatildeo de miosina foi equilibrada e digerida com -quimotripsina
A digestatildeo foi interrompida pela adiccedilatildeo de PMSF Seguidamente ultracentrifugou-se o produto
da digestatildeo desprezando-se o sedimento (Fig 32) A soluccedilatildeo de sobrenadante maioritariamente
S1 foi quantificada pela leitura directa da Abs280 ou pelo meacutetodo de Bradford (ver as secccedilotildees
3311 e 3312 do Procedimento Experimental respectivamente) e armazenada em 10 sacarose
e 2 mM szlig-Me a -80ordmC
4 Soluccedilatildeo C 25mM Tris pH 70 012M KCl 1mM EDTA
40
Antes da realizaccedilatildeo dos ensaios descritos no presente relatoacuterio o S1 de miosina foi dialisado
contra a soluccedilatildeo D5 de modo a remover qualquer composto que pudesse complexar como
vanaacutedio Seguidamente quantificou-se a proteiacutena pelo meacutetodo de Bradford ou pelo meacutetodo
directo (ver secccedilotildees 3311 e 3312do Procedimento Experimtental)
Figura 3 2ndash Representaccedilatildeo esquemaacutetica do protocolo de preparaccedilatildeo do S1 de miosina por digestatildeo quimiotripsina de
acordo com o previamente descrito por Margossian e Lowey (1982) Todo o procedimento foi realizado a 4ordm C
excepto quando indicaccedilatildeo do contraacuterio (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
3212 Isolamento e purificaccedilatildeo de actina
Isolou-se e purificou-se a actina segundo o meacutetodo descrito por Pardee e Spudish que
compreende dois passos ndash a obtenccedilatildeo de um resiacuteduo seco denominado ldquopoacute de acetonardquo e a
extracccedilatildeo subsequente da actina a partir do ldquopoacute de acetonardquo na forma de G-Actina com a
posterior polimerizaccedilatildeo a F-Actina
32121 Obtenccedilatildeo do ldquopoacute de acetonardquo
Sacrificou-se o coelho com uma pancada atraacutes do pescoccedilo e sangrou-se o animal apoacutes o corte da
arteacuteria jugular Esfolou-se e colocou-se numa tina com gelo Retiraram-se as partes de tecido
conjuntivo que envolvem o tecido muscular com auxiacutelio de uma pinccedila e bisturi Seguidamente
retirou-se os muacutesculos dorsais e os muacutesculos brancos das pernas Passou-se os muacutesculos por aacutegua
destilada de modo a remover as impurezas e pesou-se num copo Trituraram-se os muacutesculos
numa maacutequina de triturar carne e extraiu-se o resultante com 3 volumes da soluccedilatildeo E6 durante 10
minutos com agitaccedilatildeo suave Centrifugou-se a suspensatildeo a 4000 g durante 10
5 Soluccedilatildeo D HEPES 10 mM pH 70 KCl 25 mM
6 Soluccedilatildeo E 01 M KCl 015 M K2HPO4 pH 65
41
minutosdesprezando-se o sobrenadante Ressuspendeu-se o sedimento em 6 volumes de soluccedilatildeo
F7durante 10 minutos com agitaccedilatildeo suave Verificou-se se o pH da soluccedilatildeo estava compreendido
entre 82 e 85 acertando-se estecom soluccedilatildeo G8 se necessaacuterio Centrifugou-se durante 10
minutos a 4000 g Desprezou-se o sobrenadante e ressuspendeu-se o sedimento em 3 volumes de
soluccedilatildeo H9 durante 10 minutos Centrifugou-se novamente a 4000 gdurante 10 minutos
desprezando-se o sobrenadante Ressuspendeu-se o sedimento duas vezes consecutivas em 6
volumes de aacutegua destilada durante 5 minutos Centrifugou-se a 4000 g durante 10 minutos apoacutes
cada adiccedilatildeo de H2O
Seguidamente ressuspendeu-se o sedimento 5 vezes consecutivas utilizando 3 volumes de
acetona de cada vez agitando suavemente durante 10 minutos agrave temperatura ambiente Colocou-
se o resiacuteduo filtrado numa tina de vidro sobre papel de filtro deixando-se evaporar o solvente
durante a noite num nicho Triturou-se o resiacuteduo seco numa maacutequina de triturar carne e deixou-se
novamente evaporar o solvente O ldquopoacute de acetonardquo obtido foi armazenado a -20 ordmC
32122 Purificaccedilatildeo da actina a partir de ldquopoacute de acetonardquo
Apoacutes a obtenccedilatildeo do poacute de acetona como acima indicado efectuou-se o protocolo seguidamente
descrito de modo a obter monoacutemeros de actina (G-Actina) e actina filamentosa (F-Actina)
Pesou-se 35 g de poacute de acetona e adicionaram-se 50 mL de soluccedilatildeo-tampatildeo I10
sendo a
extracccedilatildeo feita com agitaccedilatildeo suave durante 30 minutos Separou-se o extracto do poacute de acetona
hidratado filtrando a suspensatildeo com 4 camadas de gaze esterilizada tendo o cuidado de natildeo
espremer o resiacuteduo Voltou a extrair-se com 50 mL de soluccedilatildeo I durante 30 minutos mexendo
lentamente Adicionou-se este ao extracto obtido anteriormente Centrifugou-se o extracto total a
16 000 g durante 30 minutos Decantou-se para uma proveta sobre a qual se colocou gaze para
que o sedimento solto natildeo se juntasse ao sobrenadante e leu-se o volume De seguida adicionou-
se KCl ateacute uma concentraccedilatildeo final de 50 mM MgCl2 ateacute 2mM e ATP ateacute 1mM lentamente e com
agitaccedilatildeo contiacutenua de modo a promover a polimerizaccedilatildeo dos monoacutemeros de actina
Deixou-se polimerizar durante 2h a 25ordmC observando-se a presenccedila de pequenas bolhas de ar
indicativas de um aumento de viscosidade da soluccedilatildeo Arrefeceu-se em gelo durante 15 minutos e
adicionou-se KCl ateacute uma concentraccedilatildeo final de 06M Mexeu-se lentamente durante 30 minutos
apoacutes os quais se ultracentrifugou a 110 000g durante 1hora Desprezou-se o sobrenadante
ressuspendeu-se o sedimento com soluccedilatildeo tampatildeo E e transferiu-se para um potter
Dialisou-se durante a noite contra 1L de soluccedilatildeo-tampatildeo I a uma temperatura de 4ordmC com rotaccedilatildeo
7 Soluccedilatildeo F 005 M NaHCO3 pH 82-85
8 Soluccedilatildeo G 1M Na2CO3
9 Soluccedilatildeo H 1 mM EDTA pH 70
10 Soluccedilatildeo I 2 mM Tris-HCl pH 8 02 mM CaCl2 02 mM Na2ATP 05 mM szlig-Me 0005 NaN3
42
contiacutenua e mudanccedila de tampatildeo 2 a 3 vezes De seguida a actina foi centrifugada a 110
000gdurante 1hora Decantou-se a soluccedilatildeo de G-Actina (sobrenadante) e desprezou-se o
sedimento Determinou-se a concentraccedilatildeo de G-Actina pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental) A soluccedilatildeo de G-actina foi armazenada a uma temperatura de 4ordmC
apoacutes adiccedilatildeo de 002NaN3 mantendo-se estaacutevel durante 3 a 5 dias
331 Quantificaccedilatildeo das proteiacutenas isoladas
3311 Quantificaccedilatildeo pelo meacutetodo de Bradford
O meacutetodo de Bradford (Bradford 1976) baseia-se na utilizaccedilatildeo do reagente de Bradford
composto por entre outros aacutecido fosfoacuterico e o corante ldquoCoomassierdquo Este corante possui carga
negativa e a sua coloraccedilatildeo varia entre a vermelha (Absmax=465 nm) predominante em soluccedilatildeo e a
azul (Absmax=595 nm) presente aquando da ligaccedilatildeo aos grupos amina das proteiacutenas Em soluccedilotildees
aacutecidas diluiacutedas a coloraccedilatildeo varia proporcionalmente com a quantidade do corante ligado agrave
proteiacutena que por sua vez eacute directamente proporcional agrave concentraccedilatildeo da proteiacutena permitindo a
sua quantificaccedilatildeo ateacute a um limite de detecccedilatildeo de 20 gmL (Bradford 1976)
Deste modo prepararam-se amostras de BSA ([BSA]stock= 1mgmL) utilizando um intervalo de
concentraccedilotildees compreendido entre 0 e 15 gmL de acordo com a tabela 3I Apoacutes a adiccedilatildeo da
H2O MilliQ e do reagente de Bradford as soluccedilotildees foram sucessivamente homogeneizadas no
voacutertex
Tabela 3Indash Amostras utilizadas para a obtenccedilatildeo da recta de calibraccedilatildeo no meacutetodo de Bradford
BSA (L) BSA (g) H2O MilliQ (L) Reagente de Bradford (L)
0 0 800 200
125 125 79875 200
25 25 7975 200
5 5 795 200
10 10 790 200
15 15 785 200
Apoacutes incubaccedilatildeo a 25ordmC por 5 minutos leram-se os valores de Abs595 num
espectrofotoacutemetro (Shimadzu UV-160A) para todas as amostras Seguidamente traccedilou-se a recta
de calibraccedilatildeo (Abs595vs quantidade de BSA (g))
43
Para determinar a concentraccedilatildeo de proteiacutena apoacutes o processo de isolamento purificaccedilatildeo e
diaacutelise quando necessaacuterio colocou-se 5 L da amostra num tubo de Eppendorf e adicionou-se
795 L de H2O MilliQ homogeneizando a soluccedilatildeo no voacutertex Adicionou-se posteriormente 200
L de reagente de Bradford e repetiu-se a homogeneizaccedilatildeo Incubou-se a mistura a 25ordmC por 5
minutos apoacutes os quais se leu os valores de Abs595 nm no espectrofotoacutemetro Por interpolaccedilatildeo deste
valor a partir da recta de calibraccedilatildeo determinou-se a concentraccedilatildeo de proteiacutena na amostra
3312 Quantificaccedilatildeo pelo meacutetodo directo
A quantificaccedilatildeo das amostras proteicas pode tambeacutem ser efectuada atraveacutes da leitura directa da
absorvacircncia tendo por base a lei de Beer-Lambert onde Abs corresponde agrave
absorvacircncia ao comprimento de onda ao qual o ensaio foi realizado corresponde ao coeficiente
de extinccedilatildeo (substituiacutedo pela letra a se as unidades forem distintas da molaridade) b corresponde
ao percurso oacuteptico da radiaccedilatildeo na ceacutelula amostral e c corresponde agrave concentraccedilatildeo amostral
Assim no caso de S1 de miosina mediu-se a Abs280e calculou-se a concentraccedilatildeo da proteiacutena
sabendo que o coeficiente de extinccedilatildeo de S1 eacute (Ref)
No caso da actina mediu-se a Abs290ecalculou-se a concentraccedilatildeo da proteiacutena sabendo que o
coeficiente de extinccedilatildeo da actina eacute (Ref)
332Electroforese em condiccedilotildees desnaturantes (SDS-PAGE)
De modo a melhor caracterizar as proteiacutenas isoladas foi realizada uma electroforese em
condiccedilotildees desnaturantes (SDS-PAGE do inglecircs Sodium Dodecyl Sulfate Polyacrilamide Gel
Electrophoresis) O SDS (dodecilo sulfato de soacutedio) eacute um detergente anioacutenico utilizado neste
meacutetodo para desnaturar as proteiacutenas que se ligam ao SDS na proporccedilatildeo 14 g SDS g proteiacutena
(aproximadamente 1 moleacutecula de SDS por cada 2 resiacuteduos de aminoaacutecidos) (Stryer) O SDS induz
uma densidade de carga negativa significativa nas proteiacutenas a que se liga mascarando a sua carga
intriacutenseca pelo que as razotildees cargamassa satildeo tendencialmente idecircnticas Assim a SDS-PAGE
separa as proteiacutenas quando sujeitas a um campo eleacutectrico de acordo com o seu peso molecular
sendo a sua migraccedilatildeo directamente proporcional ao logaritmo deste uacuteltimo (Stryer)
O gel de resoluccedilatildeo com 10 de acrilamida foi obtido atraveacutes da adiccedilatildeo de 167 mL de soluccedilatildeo
stock de acrilamida11
075 mL de tampatildeo de resoluccedilatildeo12
251 mL de aacutegua destilada e 50 L de
SDS 10 Para promover a polimerizaccedilatildeo adicionou-se em seguida 38 L de PSA 10
11
soluccedilatildeo stock de acrilamida soluccedilatildeo de 30 (mv) de acrilamida e 08 (mv) de NNrsquo-metil-bis-acrilamida 12
tampatildeo de resoluccedilatildeo 15 M Tris-HCl pH 88
44
(persulfato de amoacutenia) 10 e 25 L de TEMED 99 (tetrametiletilenodiamina) Agitou-se
cuidadosamente de modo a natildeo formar bolhas e pipetou-se a soluccedilatildeo para a cassette de vidro
previamente montada no suporte De seguida adicionou-se uma peliacutecula de cerca de 2 mm de 2-
metil-2-butanol para verificar posteriormente a polimerizaccedilatildeo Apoacutes a polimerizaccedilatildeo que demora
cerca de 45 minutos verteu-se a agrave superfiacutecie e adicionou-se o gel de concentraccedilatildeo com 5 de
acrilamida constituiacutedo por 03 mL de soluccedilatildeo de stock acrilamida 045 mL de tampatildeo de
concentraccedilatildeo13
094 mL de aacutegua destilada 18 L de SDS 10 135 L de PSA 10 e 2L de
TEMED 99 Colocou-se o pente sobre o gel e deixou-se polimerizar durante cerca de 45
minutos
As amostras para aplicaccedilatildeo no gel de electroforese foram preparadas para uma concentraccedilatildeo de
proteiacutena de 1 mgmL num volume final de 100 L dos quais 25 L correspondem ao tampatildeo da
amostra14
As amostras foram aquecidas a 100ordmC durante 5 minutos Antes da aplicaccedilatildeo das
amostras transferiu-se a cassette para a tina de electroforese agrave qual se adicionou a soluccedilatildeo
tampatildeo de electroforese da tina superior15
e tampatildeo de electroforese da tina inferior16
Aplicaram-
se 10 L das soluccedilotildees de marcadores de peso molecular e 20 L das amostras proteicas
preparadas anteriormente A electroforese decorreu durante cerca de 3h30 min a uma voltagem
de 80 V De seguida transferiu-se o gel para uma tina corando as proteiacutenas durante 1h com
soluccedilatildeo corante17
Descorou-se o gel com soluccedilatildeo descorante18
tendo-se mudado esta soluccedilatildeo ateacute
as bandas se encontrarem bem definidas
Determinou-se a mobilidade relativa das bandas correspondentes aos marcadores de elevado peso
molecular e traccedilou-se a recta de calibraccedilatildeo ndash log(PM) vs log(Rf) onde Rf corresponde agrave
mobilidade relativa Por interpolaccedilatildeo determina-se o PM das proteiacutenas isoladas sabendo a sua
mobilidade relativa Por comparaccedilatildeo com os valores descritos na literatura pode concluir-se
acerca do grau de purificaccedilatildeo
34 Mediccedilatildeo da actividade enzimaacutetica de S1
341 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
13
tampatildeo de concentraccedilatildeo 05 M Tris-HCl pH 68 14
tampatildeo de amostra 320 mM Tris-HCl pH 68 04 M szlig-mercaptoetanol SDS 8 glicerol 15 (vv) e roxo de m-
cresol 0024 15
tampatildeo de electroforese da tina superior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS 16
tampatildeo de electroforese da tina inferior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS acetato de soacutedio
01 M 17
soluccedilatildeo corante ldquoCoomassie Blue R-250rdquo contendo metanol 45 (vv) e aacutecido aceacutetico glacial 10 (vv) 18
soluccedilatildeo descorante metanol 10 (vv) e aacutecido aceacutetico glacial 10 vv)
45
O meacutetodo de Fiske-Subarow modificado consiste na reacccedilatildeo do molibdato de amoacutenio com o
fosfato inorgacircnico em soluccedilatildeo aacutecida com formaccedilatildeo do aacutecido fosfomoliacutebdico Seguidamente daacute-
se a adiccedilatildeo de um agente redutor que provoca a reduccedilatildeo do composto a fosfomolibdato que forma
um complexo de cor azulada cuja absorvacircncia maacutexima ocorre para um comprimento de onda de
660 nm tornando possiacutevel a sua detecccedilatildeo por meacutetodos espectrofotomeacutetricos As actividades Ca2+
-
ATPaacutesica e K+EDTA-ATPaacutesica de S1 foram determinadas atraveacutes deste meacutetodo de acordo com
o descrito por Taussky e Shorr (1953) com mediccedilatildeo da quantidade de fosfato (Pi) libertado
atraveacutes da hidroacutelise de ATP
Para tal obteve-se primeiramente a recta de calibraccedilatildeo com as amostras presentes na Tabela 3II
utilizando uma soluccedilatildeo stock de 5 mM KH2PO4
Tabela 3II ndash Composiccedilatildeo das diferentes amostras utilizadas para determinar a recta de calibraccedilatildeo atraveacutes do meacutetodo
de Fiske-Subbarow modificado
KH2PO4 (L)
([KH2PO4]stock= 5mM)
PO43-
(nmole) H2O (uL)
0 0 1780
5 25 1775
10 50 1770
20 100 1760
40 200 1740
100 500 1680
A cada tubo de Eppendorf adicionaram-se seguidamente 200 L de soluccedilatildeo J19
e 20 L de agente
redutor20
homogeneizando no voacutertex apoacutes cada adiccedilatildeo Incubou-se durante 10 minutos e mediu-
se a Abs660 nm num espectrofotoacutemetro com intervalos de tempo iguais entre cada amostra Traccedilou-
se a recta de calibraccedilatildeo (Abs660 nmvs nmol PO43-
) a partir dos valores obtidos
Para determinar a actividade Ca2+
-ATPaacutesica de S1 prepararam-se tubos de Eppendorf com 400
L de meio de reacccedilatildeo21
25 L CaCl2 ([CaCl2]stock= 100 mM) 25 L ATP ([ATP]stock= 100 mM)
com [S1]ensaios=01 mgmL Perfez-se se necessaacuterio com H2O MilliQ ateacute perfazer um volume de 1
mL Para determinar a actividade K+EDTA-ATPaacutesica utilizou-se uma concentraccedilatildeo idecircntica de
19
Soluccedilatildeo J 5 N H2SO4 25 molibdato de amoacutenio 20
Agente redutor 5 mg aacutecido 1-amino-2-naftol-4-sulfoacutenico 30 mg bissulfito de soacutedio 30 mg sulfito de soacutedio em 26
mL H2O 21
meio de reacccedilatildeo 125 mM Tris-HCl pH 80 0575 M KCl
46
proteiacutena nos ensaios (01 mgmL) e o mesmo meio de reacccedilatildeo Aos tubos de Eppendorf
adicionou-se ainda 123 L de KCl ([KCl]stock = 3M) 25 L de ATP 10 L de EDTA
([EDTA]stock= 100 mM )e o volume de H2O MilliQ necessaacuterio para perfazer 1 mL Em ambas as
situaccedilotildees o uacuteltimo reagente adicionado foi o ATP uma vez que se trata do substrato da reacccedilatildeo
enzimaacutetica
A cineacutetica decorreu a 25ordmC durante 5 minutos para a actividade Ca2+
-ATPaacutesica e durante 3
minutos para a actividade K+EDTA-ATPaacutesica Apoacutes o periacuteodo de incubaccedilatildeo transferiu-se 150
L de amostra para tubos de Eppendorf contendo igual volume de TCA 10 Colocou-se no gelo
e homogeneizou-se no voacutertex Seguidamente centrifugou-se a 1500 g durante 10 minutos e
recolheu-se 200 L de sobrenadante para novos tubos de Eppendorf Adicionou-se a estes 158
mL de H2O MilliQ e 200 L de soluccedilatildeo J Homogeneizou-se no voacutertex e adicionou-se 20 L de
reagente redutor repetindo-se a homogeneizaccedilatildeo Incubou-se durante 10 minutos e mediu-se a
Abs660 nm com o mesmo intervalo de tempo de leitura para cada amostra Por intrapolaccedilatildeo a partir
da recta de calibraccedilatildeo obtida anteriormente determinou-se a quantidade de Pi libertado em cada
ensaio A partir destes resultados determinaram-se as actividades Ca2+
-ATPaacutesica e K+EDTA-
ATPaacutesica do S1 em nmol PO43-
minmg proteiacutena e em s-1
342 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
Da actividade Mg2+
-ATPaacutesica de S1 resulta a formaccedilatildeo dos produtos da reacccedilatildeo de
hidroacutelise do ATP - ADP e Pi Na presenccedila de ADP fosfoenolpiruvato (PEP) e da enzima piruvato
cinase (PK) os produtos resultantes satildeo ATP e piruvato Por acccedilatildeo da enzima lactato
desidrogenase (LDH) este uacuteltimo eacute convertido em lactato ocorrendo simultaneamente a oxidaccedilatildeo
do NADH a NAD+ monitorizada atraveacutes da Abs340 nm Quando o S1 de miosina se encontra no
meio reaccional o ATP eacute hidrolisado formando-se novamente ADP e Pi o que permite a
formaccedilatildeo de um ciclo (fig 21) Se as enzimas se encontrarem em excesso a velocidade de
conversatildeo do NADH eacute limitada pela produccedilatildeo de ADP e Pi Como o ATP eacute regenerado em cada
ciclo a sua concentraccedilatildeo inicial eacute sempre mantida (Bagshaw 1982)
Figura 33ndash Representaccedilatildeo esquemaacutetica do meacutetodo de enzimas acopladas
47
O protocolo utilizado e seguidamente descrito foi adaptado de Cuenda et al (1990 ndash Ver Refordf) A
uma cuvette de 1 mL adicionou-se S1 para uma concentraccedilatildeo final de 005 mgmL 800 L de
meio de reacccedilatildeo22
10 L de PEP ([PEP]stock = 42 mM) 10 L de PK+LDH 11 ([PK+LDH]stock =
1 mgmL) e 15 L de NADH ([NADH]stock=25 mM) Iniciou-se a monitorizaccedilatildeo da reacccedilatildeo
atraveacutes da leitura de Abs340 nm apoacutes a adiccedilatildeo de 20 L de ATP ([ATP]stock=100 mM) A
actividade Mg2+
-ATPaacutesica eacute obtida atraveacutes do declive correspondente agrave oxidaccedilatildeo do NADH Para
a mediccedilatildeo da Mg2+
-ATPase de S1 estimulada por actina adicionou-se F-actina para uma
concentraccedilatildeo final de 2 microM sendo a actividade obtida tambeacutem atraveacutes do declive correspondente
agrave oxidaccedilatildeo do NADH apoacutes a adiccedilatildeo da proteiacutena
35 Interaccedilatildeo entre vanaacutedio e subfragmento-1 de miosina
351 Preparaccedilatildeo das soluccedilotildees de vanaacutedio
As soluccedilotildees de vanadato foram preparadas em aacutegua MilliQ a partir de metavanadato de amoacutenio
(NH4VO3) numa concentraccedilatildeo stock de 50 mM com agitaccedilatildeo magneacutetica e aquecimento (~30 ordmC)
Esta soluccedilatildeo com um valor compreendido entre 6 e 7 designa-se por ldquometavanadatordquo e eacute
constituiacuteda pelas esspeacutecies V1 V2 V4 e V5 (Ref) Acidificando a soluccedilatildeo de metavanadato por
adiccedilatildeo de HCl ateacute um valor de pH proacuteximo de 4 observa-se uma mudanccedila de cor de amarelo
claro para laranja indicativa da formaccedilatildeo da espeacutecie decameacuterica de vanadato De seguida e
imediatamente antes da realizaccedilatildeo dos ensaios descritos o valor do pH da soluccedilatildeo de
decavanadato foi aumentado para cerca de 75 por adiccedilatildeo de NaOH A soluccedilatildeo (2) designada
por ldquometavanadatordquo natildeo foi posteriormente alterada ateacute agrave sua adiccedilatildeo para a realizaccedilatildeo dos ensaios
As soluccedilotildees foram armazenadas a 4ordmC e mantidas em gelo aquando da sua utilizaccedilatildeo
A soluccedilatildeo de V(IV) foi preparada em condiccedilotildees anaeroacutebias por diluiccedilatildeo de sulfato de vanadil
(VOSO4) para uma concentraccedilatildeo final de 5 mM em H2O MilliQ saturada com N2 gasoso
352Anaacutelise por espectroscopia de UV-Vis
A concentraccedilatildeo da espeacutecie decameacuterica em soluccedilatildeo depende do periacuteodo de tempo apoacutes a diluiccedilatildeo
no meio de reacccedilatildeo apropriado (Ref) A velocidade de desoligomerizaccedilatildeo da espeacutecie decameacuterica
pode ser analisada por espectroscopia de absorccedilatildeo na regiatildeo UVVis uma vez que esta espeacutecie
apresenta duas bandas caracteriacutesticas na regiatildeo do visiacutevel (a 358 e 400 nm) que natildeo se verifica
em quaisquer outras espeacutecies oligomeacutericas As bandas supramencionada sapresentam-se bem
22
Meio de reacccedilatildeo 3125 mM Tris-HCl pH 71 375 mM MgCl2
48
resolvidas atraveacutes da obtenccedilatildeo da derivada de 2ordf ordem do espectro apresentando a forma de 2
vales com miacutenimos centrados nos comprimentos de onda supracitados com dependecircncia linear
relativamente agrave concentraccedilatildeo de decavanadato total (Ramos et al 2006)
Assim a estabilidade da espeacutecie decameacuterica apoacutes diluiccedilatildeo da soluccedilatildeo de decavanadato no meio de
reacccedilatildeo dos ensaios (tampatildeo D) para uma concentraccedilatildeo final de 1 mM em vanadato total foi
caracterizada atraveacutes da leitura da Abs400 em funccedilatildeo do tempo tendo-se determinado qual a
ordem de reacccedilatildeo e o tempo de meia-vida da mesma Com base em estudos anteriores acerca da
estabilidade do V10 na presenccedila de diferentes proteiacutenas (Ramos et al 2006) determinou-se
igualmente o tempo de meia vida apoacutes a adiccedilatildeo de 20 microM de S1 de miosina ao meio reaccional
353 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
Os espectros de RMN do isoacutetopo 51
V foram realizados num espectroacutemetro Bruker AM-400 MHz
equipado com uma sonda multinuclear inversa de 5 mm Em tubos de RMN de 5 mm de diacircmetro
preparou-se soluccedilotildees de 500 L de deca- ou metavanadato correspondentes a um intervalo de
concentraccedilotildees de 1 a 10 mM suplementadas com 10 D2O Os espectros foram adquiridos agrave
temperatura ambiente com uma largura espectral de 23 923 Hz um tempo de aquisiccedilatildeo de 0086
s tempo de relaxaccedilatildeo de 001 s e uma sequecircncia de pulsos a 90ordm Utilizou-se a transformada de
Fourier com alargamento das bandas de 20Hz para o processamento dos espectros Os desvios
quiacutemicos das ressonacircncias de 51
V foram obtidos em relaccedilatildeo agrave espeacutecie VOCl3cuja frequecircncia de
ressonacircncia eacute considerada o padratildeo sendo todas as outras referenciadas a partir desta em ppm
As aacutereas dos sinais do RMN correspondentes agraves espeacutecies oligomeacutericas presentes foram obtidas
por integraccedilatildeo dos sinais As concentraccedilotildees relativas das espeacutecies foram calculadas com recurso agrave
equaccedilatildeo
onde A[Vx] corresponde agrave aacuterea referente agrave ressonacircncia da espeacutecie Vx com nordm de agregaccedilatildeo x
A[V]total agrave soma total das aacutereas medidas e [V] a concentraccedilatildeo total de vanaacutedio no ensaio
A teacutecnica de 51
V-RMN foi anteriormente aplicada em estudos de ligaccedilatildeo com S1 de miosina
(Ref) tendo sido demostrado que entre as espeacutecies V1 V2 V4 e V5 a espeacutecie tetrameacuterica possui a
maior afinidade para a miosina (Cremo et al) O desaparecimento ou alargamento dos sinais pode
implicar a ligaccedilatildeo de um dos oxovanadatos laacutebeis agrave miosina sem indicaccedilatildeo de qual uma vez que
49
se processa um equiliacutebrio entre eles (Crans 1990) Assim e tendo em conta as indicaccedilotildees de que
a espeacutecie V10 eacute distinta das restantes no que diz respeito agrave magnitude da interacccedilatildeo com a miosina
e inibiccedilatildeo da actividade ATPaacutesica estimulada por actina (Aureliano 2000 Tiago et al 2002)
torna-se fundamental investigar a interacccedilatildeo das diferentes espeacutecies de vanadato com o S1 de
miosina atraveacutes da teacutecnica de 51
V-RMN
Para tal obtiveram-se espectros de 51
V-RMN resultantes da titulaccedilatildeo de uma soluccedilatildeo 5 mM de
decavanadato total com concentraccedilotildees crescentes de S1 de miosina (0 5 10 15 e 20 microM) em
tampatildeo D Realizou-se ainda ensaios na presenccedila de 2 mM de decavanadato e 2 mM de ATP na
presenccedila e na ausecircncia de S1 de miosina de modo a verificar o efeito que estes ligandos naturais
da enzima teria na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o subfragmento-1
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
Os grupos sulfidrilo (-SH) das cisteiacutenas satildeo resiacuteduos de aminoaacutecidos extremamente sensiacuteveis agrave
oxidaccedilatildeo resultando na sua oxidaccedilatildeo (Dalle-Donne et al 1999) Logo a quantificaccedilatildeo destes
resiacuteduos eacute bastante importante nos estudos de interacccedilatildeo com vanaacutedio jaacute que os resiacuteduos de Cys
com ecircnfase para o SH1 e o SH2 satildeo bastante importantes na monitorizaccedilatildeo das modificaccedilotildees
funcionais da regiatildeo da cabeccedila
O procedimento seguidamente apresentado foi originalmente descrito por Gutieacuterrez-Merino
(1987)) Consiste na reacccedilatildeo entre os grupos ndashSH natildeo oxidados das proteiacutenas e o reagente de
Ellman (DTNB -55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico)) - formando-se o aniatildeo CNT (aniatildeo 3-
carboxilato-4-nitrotiofenolato) numa estequiometria de 11 Este apresenta um maacuteximo de
absorvecircncia a 412 nm (Robyt 1990) Assim apoacutes a incubaccedilatildeo da proteiacutena na presenccedila do metal
a quantidade de cisteiacutenas reduzidas por proteiacutena determina-se atraveacutes dos valores de Abs412
Deste modo colocou-se num tubo de Eppendorf o volume apropriado de S1 dialisado para que a
concentraccedilatildeo no ensaio fosse de 1 mgmL Adicionou-se o volume apropriado de decavanadato
para que a sua concentraccedilatildeo fosse de 0 10 50 100 250 500 ou 1000 M Perfez-se com soluccedilatildeo
D ateacute um volume final de 1 mL e incubou-se a 25ordmC por 20 minutos Apoacutes este periacuteodo colocou-
se 780 L da mistura reaccional numa cuvette de quartzo e adicionou-se 220 L de soluccedilatildeo K23
23
Tampatildeo K Tampatildeo fosfatohidrogenofosfato de soacutedio 025 M pH 72
50
Adicionou-se 15 L de DTNB (em soluccedilatildeo K [DTNB]stock=100 mM) ao meio reaccional e
registou-se o valor de absorvecircncia medido equivalente agraves cisteiacutenas ldquoraacutepidasrdquo (SH1 e SH2
expostos ao solvente) Adicionou-se 001 g de SDS que provoca a desnaturaccedilatildeo da proteiacutena e
consequentemente a exposiccedilatildeo de todos os resiacuteduos de cisteiacutenas que se encontravam ateacute aiacute
inacessiacuteveis no interior da proteiacutena (cisteiacutenas totais) Estes resiacuteduos podem assim reagir com o
DTNB
Estimou-se o nuacutemero de resiacuteduos de cisteiacutenas em todas as amostras com recurso ao coeficiente de
extinccedilatildeo molar () do DTNB (ver ponto 3 dos Anexos) que foi determinado atraveacutes da titulaccedilatildeo
de L-Cisteiacutena com intervalo de concentraccedilotildees compreendidos entre 0 e 50 microM na presenccedila de
25microM de DTNB em tampatildeo D Os valores de absorvecircncia foram obtidos para um comprimento de
onda de 412 nm
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
3621 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
A teacutecnica mais eficiente de anaacutelise da interacccedilatildeo entre o iatildeo vanadilo e proteiacutenas como a
calmodulina e a transferrina entre outras permite a obtenccedilatildeo raacutepida de informaccedilatildeo no que toca agrave
conformaccedilatildeo e amplitude da referida interacccedilatildeo
Os espectros de Ressonacircncia Paramagneacutetica Electroacutenica (RPE) da espeacutecie paramagneacutetica V(IV)
foram realizados a 70K num espectroacutemetro Bruker EMX-300 X-band Todas as soluccedilotildees
utilizadas e amostras foram preparadas em anaerobiose numa cacircmara de luvas de modo a evitar a
reduccedilatildeo de V(IV) a V(V) Em tubos capilares com 02 mL de amostra manteve-se a concentraccedilatildeo
de S1 a 20 M e variou-se a concentraccedilatildeo de vanadilo (0 25 5 10 15 25 50 75 100 150
200 500 e 1000 M [VOSO4]stock=5 mM) em tampatildeo D Apoacutes obtenccedilatildeo dos espectros estes
foram tratados atraveacutes do software SimFonia e WinEPR tendo-se obtido um graacutefico com a
variaccedilatildeo da intensidade do sinal de RPE em funccedilatildeo da concentraccedilatildeo de sulfato de vanadilo
presente Realizaram-se ainda simulaccedilotildees de espectros obtidos com 20 microM de S1 e 1000 microM de
V(IV) com vista agrave obtenccedilatildeo dos ligandos provaacuteveis do vanadilo atraveacutes da anaacutelise da constante
de acoplamento hiperfina (A) e do factor g (g)
4 Conclusotildees gerais
Apesar de jaacute ter sido demonstrado que o vanaacutedio inibe a Na+K
+-ATPase da miosina a funccedilatildeo
deste elemento no tecido muacutesculo esqueleacutetico continua largamente incompreendida A enorme
complexidade quiacutemica deste metal especialmente no que concerne agrave oligomerizaccedilatildeo e reacccedilotildees
51
redox constitui um obstaacuteculo agrave compreensatildeo do comportamento do vanaacutedio nos organismos
vivos Assim a caracterizaccedilatildeo da quiacutemica deste elemento eacute de grande utilidade antes de iniciar o
estudo da sua interacccedilatildeo com o presente sistema
A anaacutelise por espectroscopias de 51
V-RMN e de UV-Vis de soluccedilotildees de vanadato diluiacutedas nos
meios de reacccedilatildeo utilizados no presente relatoacuterio (10 mM HEPES pH 70 25 mM KCl) mostrou
claramente que a espeacutecie decameacuterica (V10) possui um tempo de vida suficientemente longo para a
determinaccedilatildeo do seu efeito isolado ao contraacuterio do que se verifica com as espeacutecies mono- di-
tetra- e pentameacutericas (V1 V2 V4 e V5) Por outro lado a anaacutelise verificou ainda que apoacutes diluiccedilatildeo
das soluccedilotildees de decavanadato no meio de reacccedilatildeo coexistem as espeacutecies V10 e V1 pelo que os
efeitos putativos desta uacuteltima deveratildeo ser subtraiacutedos aquando da monitorizaccedilatildeo dos efeitos da
espeacutecie decameacuterica
O vanaacutedio (V) (vanadato) inibe as enzimas que catalizam reacccedilotildees de transferecircncia de grupos
fosfato como a ATPase da miosina [Goodno 1979] devido agrave sua analogia estrutural com o iatildeo
fosfato Na sua utilizaccedilatildeo como inibidor enzimaacutetico natildeo eacute tida em conta a contribuiccedilatildeo individual
das espeacutecies oligomeacutericas considerando-se normalmente a espeacutecie V1 como inibidora efectiva
[Goodno e Taylor 1982] No presente relatoacuterio verificou-se por espectroscopia de 51
V-RMN que
outros oligoacutemeros de vanadato interagem com o S1 de miosina tendo as espeacutecies V10 e V4 os
maiores niacuteveis de afinidade A sua interacccedilatildeo envolve forccedilas electrostaacuteticas e eacute afectada
negativamente por ligandos naturais de enzima como eacute o caso do ATP
A exposiccedilatildeo do S1 de miosina (005 mgmL) a decavanadato conduz agrave inibiccedilatildeo do capacidade da
proteiacutena de estimular a actividade Mg2+
-ATPaacutesica com um valor de IC50 de 266 plusmn 07 M
decavanadato sendo o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM
decavanadato (80) Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM decavanadato sendo
o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM decavanadato (60)
exercendo um efeito de impedimento agrave ligaccedilatildeo entre o vanaacutedio e o S1
A uma concentraccedilatildeo de 27 microM decavanadato verifica-se a oxidaccedilatildeo de um dos resiacuteduos de Cys
de S1 de miosina seguida pela oxidaccedilatildeo gradual de outro resiacuteduo de Cys ateacute agrave concentraccedilatildeo de
1000 microM Ambos os resiacuteduos oxidados natildeo se encontravam expostos ao solvente utilizado A
oxidaccedilatildeo dos resiacuteduos permite sugerir que esta modificaccedilatildeo oxidativa sofrida pelo S1 contribui
para a diminuiccedilatildeo da sua actividade de ATPase
52
Por espectroscopia de RPE revelaram-se os possiacuteveis ligandos equatoriais do vanaacutedio aquando da
sua interacccedilatildeo com o S1 de miosina como sendo duas moleacuteculas de aacutegua um ligando contendo o
grupo R-OH e outro contendo o grupo R=NR
Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados de
oxidaccedilatildeo (IV) e (V) no S1 de miosina para concentraccedilotildees facilmente alcanccedilaacuteveis no ambiente
terrestre
53
5 Estudos futuros
No sentido do melhor esclarecimento das alteraccedilotildees moleculares e estruturais que ocorrem no
subfragmento 1 quando exposta ao vanaacutedio que conduzem a uma diminuiccedilatildeo da sua funccedilatildeo
sugere-se
- Quantificaccedilatildeo das cisteiacutenas reduzidas de S1 na presenccedila de metavanadato
- Determinaccedilatildeo das actividades Ca2+
- e K+-ATPaacutesica do S1 de miosina na presenccedila de
decavanadato
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina estimulada por actina
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina na presenccedila de
metavanadato
- Realizaccedilatildeo de RMN 2D com variaccedilatildeo das concentraccedilotildees das soluccedilotildees de decavanadato e
metavanadato na presenccedila de S1
-Mutageacutenese dos resiacuteduos de Cys doS1 de miosina de modo a verificar quais os resiacuteduos que
sofrem oxidaccedilatildeo na presenccedila de decavanadato
- Determinaccedilatildeo por calorimetria (DSC Differential Scanning Calorimetry) da variaccedilatildeo da
entalpia (∆H) e da temperatura de desnaturaccedilatildeo do S1 de miosina na presenccedila de vanaacutedio uma
vez que estas propriedades caracteriacutesticas de cada proteiacutena satildeo alteradas apoacutes ocorrecircncia de
modificaccedilotildees estruturais
- Anaacutelise por SDS-PAGE de S1 de miosina apoacutes tratamento com vanaacutedio de modo a determinar
quais os polipeacuteptidos eventualmente formados apoacutes clivagem da proteiacutena
54
6 Referecircncias bibliograacuteficas
- Sellers J Myosins 2nd Edition Oxford University Press 1999
- L Stryer inBiochemistry (Ed WH Freeman 2005 pp
Refordfas Materiais e Meacutetodos
[A] S S Margossian SLowey Methods Enzymol1982 85 55-71
[B] JD Pardue JA Spudich Methods Enzymol 1982 85 164-181
[C] M Bradford Anal Biochem1976 72 248-254
[D] H H Taussky E Shorr J Biol Chem 1953 202 675-685
[E] C R Bagshaw in Outline Studies in Biology ndash Muscle Contraction Chapman and Hall Ed
1982
[F] A Cuenda et al Eur J Biochem1990 194 663-670
55
Anexos
1 Determinaccedilatildeo da recta de calibraccedilatildeo meacutetodo de Bradford
Figura1ndashRepresentaccedilatildeo graacutefica da recta de calibraccedilatildeo (Abs595 nm vs [BSA]) utilizada para o meacutetodo de Bradford
2 Determinaccedilatildeo da recta calibraccedilatildeo para a electroforese em condiccedilotildees desnaturantes
Figura 2 ndash Representaccedilatildeo graacutefica da recta de calibraccedilatildeo (log(Peso Molecular) vs Rf dos marcadores de peso
molecular) onde Rf indica determinada atraveacutes do padratildeo de peso molecular
(poccedilo (a))
3Caacutelculo do coeficiente de extinccedilatildeo molar () para a quantificaccedilatildeo de cisteiacutenas reduzidas
atraveacutes do meacutetodo de Ellman
Traccedilou-se a recta de Abs412 nm meacutedia vs [L-Cisteiacutena] de modo a determinar o valor do coeficiente
de extinccedilatildeo molar definido como o declive daquela recta Assim =8400 M-1
cm-1
como
podemos verificar na Fig 3
A595 nm = 00546[BSA] + 00209 Rsup2 = 099051
Ab
s 59
5 n
m
[BSA] (mgmL)
log PM = -1177 Rf + 2247 Rsup2 = 098957
log
PM
Mobilidade relativa (Rf)
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero

14
Assim a regulaccedilatildeo da contracccedilatildeo ocorre com coordenaccedilatildeo e rapidez na totalidade das miofibrilas
do tecido muscular (Ref)
Em repouso a troponina I encontra-se posicionada de tal forma que a proteiacutena adjacente
troponina C impede estruturalmente a ligaccedilatildeo das cabeccedilas da miosina agrave actina (acircngulo =50ordm) O
aumento da concentraccedilatildeo intracelular de Ca2+
provoca um aumento da sua disponibilidade para
ligar agrave troponina C (Fig 19)
Figura 19ndash Regulaccedilatildeo do mecanismo contraacutectil no muacutesculo esqueleacutetico pelo complexo de troponina
Como consequecircncia a troponina C altera a sua conformaccedilatildeo (=70ordm) associando-se agrave troponina I
que deixa de inibir a ligaccedilatildeo das pontes transversais A alteraccedilatildeo anterior eacute transmitida agrave
tropomiosina atraveacutes da troponina T adjacente agrave C o que provoca o deslocamento da
tropomiosina na direcccedilatildeo do interior do sulco do filamento de actina Assim eacute possiacutevel a
interacccedilatildeo eficaz entre actina e miosina A Ca2+
-ATPase do retiacuteculo sarcoplasmaacutetico provoca a
reacumulaccedilatildeo do iatildeo caacutelcio no organelo impedindo o prolongamento desnecessaacuterio da contracccedilatildeo
(Refordf)
12 Vanaacutedio
O vanaacutedio eacute um metal de transiccedilatildeo pertencente ao grupo VB da tabela perioacutedica Foi
primeiramente descoberto em 1801 por Andreacutes Manuel del Riacuteo que concluiu erradamente que era
idecircntico ao croacutemio e definitivamente por Nils Gabriel Svenson em 1831 Recentemente foram
descobertas importantes funccedilotildees bioloacutegicas deste elemento provavelmente essencial para os
humanos de que se destaca no contexto do presente relatoacuterio a possibilidade de actuar como
anaacutelogo do grupo fosfato e a inibiccedilatildeo de enzimas in vitro como eacute o caso da actividade ATPaacutesica
da miosina
Eacute um metal soacutelido agrave temperatura ambiente que se encontra amplamente distribuiacutedo nos vaacuterios
ecossistemas em dois isoacutetopos naturais ndash 50
V e 51
V ndash raramente excedendo 01 partes por biliatildeo
15
(ppb) Estaacute associado a cerca de 70 minerais diferentes na natureza (dos quais se destacam o
uracircnio cobre chumbo zinco fosfato e titacircnio ferroso) especialmente na forma de vanadato ou
pentoacutexidos de vanaacutedio O vanaacutedio eacute um dos principais constituintes da mateacuteria orgacircnica foacutessil
com especial ecircnfase para o carvatildeo oacuteleo de crude e foacutesseis carbonaacuteceos [UV] bem como em
alimentos consumidos pelo Homem (salsa pimenta preta cogumelos marisco vegetais cereais
gorduras e oacuteleos) [W] Estes constituem a maior fonte de exposiccedilatildeo ao vanaacutedio da maioria da
populaccedilatildeo humana [X]
Normalmente a entrada do vanaacutedio no organismo ocorre por inalaccedilatildeo ou em menores
quantidades por absorccedilatildeo pelo tracto gastrointestinal ou pela pele O seu armazenamento centra-
se no fiacutegado rins ceacuterebro coraccedilatildeo muacutesculos e ossos No entanto a retenccedilatildeo de vanaacutedio no
organismo humano eacute reduzida ndash em condiccedilotildees normais cerca de 100 M (em muitos tecidos a
quantidade eacute inferior a 10 ngg de tecido) [Y]
Actualmente natildeo existem provas de deficiecircncia nutricional de vanaacutedio em humanos ou de
enzimas que contenham vanaacutedio em animais superiores No entanto vaacuterios estudos relacionam a
ocorrecircncia de deficiecircncia em galinhas e ratos devido a uma alimentaccedilatildeo pobre em vanaacutedio Os
sintomas relatados incluem retardamento do crescimento malformaccedilotildees oacutesseas perturbaccedilotildees no
metabolismo lipiacutedico e estagnaccedilatildeo reprodutiva entre outros [Z AA AB] Os efeitos fisioloacutegicos
do vanaacutedio podem depender do tipo de composto ou de oligoacutemero uma vez que consoante as
suas estabilidade labilidade e nuacutemero de coordenaccedilatildeo as respostas seratildeo diferenciadas ao niacutevel
dos organismos vivos
Por outro lado a toxicidade do vanaacutedio encontra-se bem caracterizada existindo sintomas
descritos em vaacuterios mamiacuteferos (inclusive o Homem) que incluem diarreias catildeibras irritaccedilatildeo da
pele e mucosas palpitaccedilotildees cardiacuteacas anemia pneumonia bronquite perturbaccedilotildees no Sistema
Nervoso Central (SNC) entre outros (Ref) A toxicidade varia geralmente com a natureza do
composto com aumento proporcional agrave sua valecircncia Verificou-se que a forma de entrada do
vanaacutedio tem tambeacutem efeito na sua toxicidade ndash os maiores niacuteveis verificaram-se por injecccedilatildeo
intravenosa da amostra sendo progressivamente inferiores por inalaccedilatildeo e finalmente por
ingestatildeo [X AA] A ampla utilizaccedilatildeo dos compostos de vanaacutedio na induacutestria quiacutemica bem como
as elevadas quantidades de vanaacutedio libertadas atraveacutes da combustatildeo dos oacuteleos de crude
contribuem consideravelmente para o aumento da biodisponibilidade do vanaacutedio Como tal o
interesse neste elemento e nos problemas ambientais por ele causados tem vindo a aumentar
significativamente [V]
121 Propriedades quiacutemicas
16
A quiacutemica do vanaacutedio eacute extraordinariamente complexa pelo que originou vaacuterias interpretaccedilotildees
incorrectas desde a sua descoberta No entanto as suas propriedades dependem em uacuteltima
anaacutelise da configuraccedilatildeo electroacutenica do elemento Assim a variabilidade encontrada no
comportamento quiacutemico do vanaacutedio pode dever-se agrave sua configuraccedilatildeo ndash [Ar] 3d2 4s
2 ndash que
permite a formaccedilatildeo de vaacuterios estados de oxidaccedilatildeo (de +2 a +5) Em soluccedilatildeo aquosa os estados de
oxidaccedilatildeo estaacuteveis vatildeo de V(V) a V(II) Destes apenas os +3 a +5 satildeo biologicamente importantes
jaacute que o V(II) apresenta demasiada capacidade redutora para existir nos organismos vivos
conhecidos [AC]
No fluido extracelular os compostos de vanaacutedio encontram-se predominantemente na forma de
monovanadatos (V(V)) livres ou ligados agrave transferina do soro ou agrave hemoglobina (Refordf) Os iotildees de
vanadato livres satildeo transportados para o meio intracelular atraveacutes de canais ioacutenicos inespeciacuteficos
Jaacute no ambiente intracelular os compostos sofreratildeo uma bio-reduccedilatildeo atraveacutes da acccedilatildeo da
glutationa ou outros compostos com grupos ndashOH como pentoses hexoses ou etanolaminas [V] A
reoxidaccedilatildeo de vanadilo (V(IV)) intracelular a vanadato (V(V)) provocaria a acumulaccedilatildeo toacutexica
destes uacuteltimos pelo que a complexaccedilatildeo da espeacutecie reduzida o impede O transporte de vanaacutedio
para o interior da ceacutelula encontra-se na Fig 110
Figura 110 ndash Transporte de vanaacutedio para a ceacuteluka e seu subsequente metabolismo (P=Proteiacutena) (Adaptado de
Zaporowska e Scibor 1998)
1211 Vanaacutedio(IV)
A espeacutecie de vanaacutedio (IV) mais bem conhecida eacute o iatildeo vanadilo (VO2+
) espeacutecie estaacutevel a pH
aacutecido sendo a maioritaacuteria a pH 3[AD] Apresenta uma cor azulada e forma complexos penta e
hexacoordenados Forma-se rapidamente a partir de V(V) na presenccedila de um agente redutor em
soluccedilatildeo aquosa No ambiente intracelular a presenccedila de glutationa reduzida aacutecido ascoacuterbico
NAD(P)H entre outros conduzem a que o principal estado de oxidaccedilatildeo nos sistemas bioloacutegicos
seja provavelmente o +4 encontrando-se o V(IV) fortemente complexado com proteiacutenas ou
aminoaacutecidos livres citrato ATP e pirofosfato (Refordfs) Em alternativa o V(IV) pode existir
17
livremente em ambientes fortemente aacutecidos como os lisossomas jaacute que a pH superior a 3 as
soluccedilotildees de vanadilo tendem a ser oxidadas pelo ar formando-se vanadato Esta grande
capacidade redutora justifica parcialmente a utilizaccedilatildeo pouco extensiva do iatildeo VO2+
como
inibidor [W]
1212 Vanaacutedio (V)
O vanaacutedio (V) eacute a forma mais estaacutevel num intervalo de pH amplo [W] O iatildeo anioacutenico VO3-tende
a constituir complexos polinucleares pelo que as soluccedilotildees aquosas de vanadato satildeo extremamente
complexas com ocorrecircncia simultacircnea de vaacuterios equiliacutebrios de protonaccedilatildeo (pKa(vanadato)= 35
78 e 125) e oligomerizaccedilatildeo As espeacutecies presentes (H3VO4 H2VO4- HVO4
2- e VO4
3-) dependem
do pH forccedila ioacutenica e temperatura da soluccedilatildeo e da concentraccedilatildeo total do elemento
[AEAFAGAHAI]
O VO43-
- monoacutemero ortovanadato V1 ndash estaacute associado a uma grande parte da importacircncia
bioloacutegica do vanaacutedio uma vez que se trata de um anaacutelogo electroacutenico e estrutural do ortofosfato
(PO43-
) A coordenaccedilatildeo do ortovanadato pode ser tetraeacutedrica ou bipiramidal trigonal (Refordfs)
A oligomerizaccedilatildeo do vanadato monomeacuterico daacute origem ao diacutemero (V2O72-
- V2) que apresenta
tambeacutem equiliacutebrios de protonaccedilatildeo Presume-se a existecircncia de analogia do vanadato dimeacuterico com
o pirofosfato embora natildeo haja evidecircncia estrutural (Refordf) A dimerizaccedilatildeo do V2 origina o
vanadato tetrameacuterico ou metavanadato (V4O124-
- V4) que constitui a espeacutecie maioritaacuteria em
soluccedilotildees aquosas concentradas (Refordf) As estruturas do metavanadato e a sua espeacutecie protonada
satildeo ciacuteclicas [AJ] Outra espeacutecie oligomeacuterica importante eacute o vanadato pentameacuterico (V5O153-
) cuja
estrutura presumida eacute ciacuteclica [AI]
O oligoacutemero com maior importacircncia bioquiacutemica eacute o vanadato decameacuterico (V10O286-
-V10)(Refordf) a
espeacutecie predominante em soluccedilotildees de pH 2-6 quando a concentraccedilatildeo total de vanadato eacute superior
a 100 M (Ref) A estrutura do V10 natildeo difere em soluccedilatildeo ou no estado soacutelidoOs 10 aacutetomos de
vanaacutedio da espeacutecie decameacuterica estatildeo divididos em trecircs grupos identificados na figura como VA-C
consoante o seu ambiente quiacutemico circundante (Refordf) (Fig 111)
A expressatildeo ldquodecavanadatordquo refere-se a uma soluccedilatildeo aquosa na qual a espeacutecie decameacuterica se
encontra em equiliacutebrio quiacutemico com as espeacutecies V1 V2 V4 e V5 sendo que a oligomerizaccedilatildeo
progressiva do vanaacutedio eacute proporcional agrave sua concentraccedilatildeo total (Ref) Nesta soluccedilatildeo a
concentraccedilatildeo de V10 corresponde a aproximadamente 10 da concentraccedilatildeo total de vanaacutedio [AK]
18
Figura 111 ndash Estruturas esquemaacuteticas das espeacutecies de vanadato (espeacutecie monomeacuterica (V1) dimeacuterica (V2)
tetrameacuterica (V4) pentameacuterica (V5) e decameacuterica (V10) (Adaptado de Aureliano e Crans 2008)
122 Metabolismo e efeito na actividade enzimaacutetica
Apesar de natildeo terem sido ainda encontradas as provas categoacutericas das suas funccedilotildees nos seres
vivos sabe-se que o iatildeo vanadato eacute capaz de inibir uma grande quantidade de enzimas in vitro
com especial ecircnfase na actividade ATPaacutesica da miosina [AL Teresa] Como referido
anteriormente o vanadato eacute capaz de actuar em alternativa ao fosfato nas reacccedilotildees bioquiacutemicas
incluindo a sua preferecircncia de ligaccedilatildeo agraves enzimas [W]
Eacute conhecido o papel do vanaacutedio no metabolismo do caacutelcio promovendo a mineralizaccedilatildeo dos
dentes e ossos na biossiacutentese de liacutepidos (trigliceacuteridos e colesterol) com posterior inibiccedilatildeo da
acumulaccedilatildeo destes nos vasos sanguiacuteneos e no SNC na reproduccedilatildeo e desenvolvimento por
mimetizaccedilatildeo e potenciaccedilatildeo de factores de crescimento na estimulaccedilatildeo do consumo de glucose e
da siacutentese de glicogeacutenio com actuaccedilatildeo equivalente aos efeitos da insulina ao niacutevel celular na
transduccedilatildeo de sinais celulares com regulaccedilatildeo da actividade de segundos mensageiros no
transporte de iotildees e aminoaacutecidos na transcriccedilatildeo geneacutetica e siacutentese de proteiacutenas e DNA entre
outros [W Y AB]
Assim o vanaacutedio cnseguiraacute provocar inuacutemeras mudanccedilas bioquiacutemicas nas ceacutelulas Alguns
exemplos da influecircncia bioquiacutemica do vanaacutedio incluem os seus efeitos nos processos de
fosforilaccedilatildeo e desfosforilaccedilatildeo de proteiacutenas e na participaccedilatildeo na formaccedilatildeo de espeacutecies reactivas de
oxigeacutenio devido ao seu efeito proacute-oxidante sobre agentes redutores a niacutevel celular [AM]
Quanto ao seu efeito na actividade enzimaacutetica pode dividir-se em dois tipos principais ndash inibitoacuterio
e estimulante - dependendo da enzima ou do tipo de reacccedilatildeo por esta catalisada No primeiro
grupo incluem-se enzimas como as ATPases (de miosina de canais ioacutenicos e fosfoenzimas
19
transportadoras) e as fosfatases (da tirosina aacutecida e alcalina) a fosfofrutocinase adenilato ciclase
e catalase entre outras [AA] O segundo grupo inclui a maioria das cinases a glucose-6-fosfato
desidrogenase a fosfoglicerato mutase e a fosfoglicerato fosfatase (Refordf)
O efeito do vanaacutedio (V) na actividade enzimaacutetica estaacute relacionado com o seu nuacutemero de
coordenaccedilatildeo podendo actuar como estado de transiccedilatildeo ou por mimetizaccedilatildeo de cofactores
substratos (especialmente o fosfato) e interacccedilotildees proteiacutena-proteiacutena (como no caso das
polimerases e transcriptases) Pode ainda estar envolvido na formaccedilatildeo de ligaccedilotildees de hidrogeacutenio
interacccedilotildees electrostaacuteticas esteacutereas e de oxidaccedilatildeo-reduccedilatildeo Uma evidecircncia desta uacuteltima eacute a sua
capacidade de alternacircncia entre os estados de oxidaccedilatildeo +5 a +3 o que torna faacutecil a sua interacccedilatildeo
redox com enzimas contendo resiacuteduos de cisteiacutenas (Cys) ou qualquer outro composto orgacircnico
com actividade redox [AN]
13 Vanaacutedio e muacutesculo esqueleacutetico
A primeira evidecircncia da presenccedila de vanaacutedio no muacutesculo esqueleacutetico proveio de uma amostra de
ATP isolado de muacutesculo de cavalo que continha uma impureza ndash vanaacutedio ndash cuja presenccedila
provocou a inibiccedilatildeo da actividade da bomba de Na+K
+-ATPase [AO] Desde entatildeo vaacuterios
estudos tecircm sido realizados com ecircnfase na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o
tecido muscular Destes destaca-se a descoberta da interacccedilatildeo forte da espeacutecie V10 com a miosina
fosforilada [AP] e agrave sua inibiccedilatildeo da actividade ATPaacutesica da miosina[AQ AR]
No quediz respeitoao iatildeo monovanadato destaca-se a sua inibiccedilatildeoda actividade Mg2+
-ATPaacutesica
da miosina [AS] que ocorre de duas formas distintas (1) na presenccedila de concentraccedilotildees reduzidas
de V1 a inibiccedilatildeo ocorre a velocidade reduzida dependente da concentraccedilatildeo da espeacutecie
monomeacuterica (2) na presenccedila de concentraccedilotildees elevadas de V1 o niacutevel de inibiccedilatildeo aproxima-se de
100 desde que [V1] seja superior agrave concentraccedilatildeo total de locais com actividade Mg2+
-ATPaacutesica
no ensaio Esta inibiccedilatildeo eacute irreversiacutevel tanto por diaacutelise ou adiccedilatildeo de agentes redutores do
vanadato [AL]Segundo Pate et al(1994)[AT] a espeacutecie monomeacuterica podeinibir a velocidade de
contracccedilatildeo a temperatura inferior a 20ordmC perdendogradual e rapidamente essa capacidade ateacute os
50ordmC temperatura agrave qual cessa a capacidade inibitoacuteria Os resultados obtidos sugeremainda uma
interacccedilatildeo altamente cooperativa entre o iatildeo vanadato e as proteiacutenas componentes do muacutesculo
esqueleacuteticoDestaca-se tambeacutem a descoberta da inibiccedilatildeo da geraccedilatildeo de forccedila contraacutectil nas fibras
musculares por parte da espeacutecie monomeacuterica[AT]Wilson et al(1995) [AU ]determinaram que a
afinidade da espeacutecie V1 para o S1 de miosina eacute significativamente mais elevada do que a do iatildeo
Pi seu anaacutelogo Descobriram ainda que que a ligaccedilatildeo do vanadato poderaacute envolver transiccedilotildees
entre estados energeacuteticos com energia livre independente dos impedimentos esteacutereos impostos
20
pela estrutura das miofibrilas Os estudos desenvolvidos por Smith et al (1990)[AV]
determinaram queo iatildeo monovanadato natildeo apresenta um efeito pronunciado na inibiccedilatildeo da
actividadeacto-S1-miosina-ATPaacutesica excepto em concentraccedilotildees superiores a 900microM a partir das
quais existe produccedilatildeo de espeacutecies oligomeacutericas para as quais o efeito inibitoacuterio seraacute mais
elevadoA espeacutecie monomeacuterica de vanadato consegue adoptar facilmente uma geometria do tipo
bipiramidal trigonal que mimetiza o intermediaacuterio formado (MgADP
Pi)aquando da reacccedilatildeo de
hidroacutelise de ATP no local activo da miosina [AW] De facto a interacccedilatildeo do vanaacutedio com a
miosina eacute inesperadamente estaacutevel na presenccedila de ADP Esta interacccedilatildeo resulta na formaccedilatildeo de
um complexo enzimaacutetico inactivo e estaacutevel com estequiometria de 11 ADPvanadato por siacutetio
activo da proteiacutena A formaccedilatildeo do complexo estaacutevel miosina-vanadato foi proposta por Goodnoet
al (1982) [AS] Nesta ocorre um preacute-equiliacutebrio raacutepido que induz a formaccedilatildeo de um complexo
ternaacuterio reversiacutevel (MADP
V) com formaccedilatildeo lenta do isoacutemero M
ADP
V A diferenccedila entre os
compostos de vanaacutedio e de fosfato centra-se no tempo de meia vida (t12) do Pi ou do V (no
primeiro t12=12 s enquanto no segundo t12=24 h) (Refordf) A diferenccedila dever-se-aacute agrave possiacutevel
complexaccedilatildeo do vanaacutedio com grupos nucleofiacutelicos no siacutetio activo o que provocaria uma
estabilizaccedilatildeo da miosina na conformaccedilatildeo MADP
V
No que respeita aos estudos realizados com a espeacutecie tetrameacuterica Cremo et al determinou que a
cadeia pesada do S1 de miosina pode ser clivada em dois locais distintos (designados V1 e V2)
pela espeacutecie V4 apoacutes irradiaccedilatildeo com luz UV na presenccedila de concentraccedilotildees de vanadato na ordem
dos milimolar e na ausecircncia de nucleoacutetidos ou metais divalentes
Jaacute no que concerne aos estudos realizados com ecircnfase na espeacutecie decameacuterica foi sugerido
porRamos et al (2006) [AK]que a interacccedilatildeo do decavanadato com a G-actina favorecida pela
polimerizaccedilatildeo desta uacuteltima estabiliza a espeacutecie V10 e induz a inibiccedilatildeo da formaccedilatildeo de F-actina A
estabilizaccedilatildeo da espeacutecie V10 por proteiacutenas tansmembranares e do citoesqueleto pode ser a razatildeo
pela qual ocorre a toxicidade do decavanadato reportada na avaliaccedilatildeo dos efeitos do V(V) nos
sistemas bioloacutegicosTiago et al (2007) [AR] concluiram que o V10 apresenta uma clara preferecircncia
de ligaccedilatildeo ao ldquoback-doorrdquo da S1-miosina-ATPase o que se verifica apenas na estrutura
conformacional desimpedida com acesso facilitado ao local de ligaccedilatildeo do Pi Sugere tambeacutem que
o V10 actua como um ldquoback-door stoprdquo bloqueando estaabertura do fragmento de 50-kDa
necessaacuteria para a hidroacutelise do ATP--fosfato o que constitui uma explicaccedilatildeo simples do
comportamento natildeo competitivo da espeacutecie decameacuterica estimulando a utilizaccedilatildeo deste oligoacutemero
como uma feramenta da elucidaccedilatildeo das mudanccedilas conformacionais ldquoback-doorrdquo da miosina no
21
processo da contracccedilatildeo muscular(ref)Destaca-se ainda a descoberta da capacidade inibitoacuteria do
V10 em relaccedilatildeo agrave fosforilaccedilatildeo oxidativa mitocondrial [AX]
Finalmente no que concerne aos estudos realizados com a presenccedila de vaacuterias espeacutecies
oligomeacutericas Aureliano e Madeira (1994) [AY]observaram que na presenccedila de Ca2+
as espeacutecies
V4 e V10se ligam agrave Ca2+
-ATPase do retiacuteculo sarcoplasmaacutetico enquanto a espeacutecie V1 se liga apenas
na presenccedila de ATP Aparentemente o aumento da concentraccedilatildeo de Ca2+
tem o efeito contraacuterio da
inibiccedilatildeo da ATPase do retiacuteculo sarcoplasmaacutetico pelo V1 natildeo tendo no entanto efeito significativo
sobre a inibiccedilatildeo por parte da espeacutecie V10 Tiago et al (2002) [AP] concluiacuteram que em condiccedilotildees
proacuteximas agraves fisioloacutegicas a espeacutecie V10 difere das espeacutecies presentes na soluccedilatildeo de metavanadato
devido agrave sua forte interacccedilatildeo com a miosina fosforilada e agrave sua inibiccedilatildeo da actividade ATPaacutesica
da miosina Para aleacutem disso o ATP diminui a afinidade da miosina para o V4 induzindo a
interacccedilatildeo com V1 e natildeo tendo qualquer efeito no que diz respeito ao V10 Foi tambeacutem proposto
[AQ] que a ligaccedilatildeo das espeacutecies oligomeacutericas de vanadato aoS1 de miosina nos locais onde o
fosfato coordenaeacute provavelmente a causa da inibiccedilatildeo da actividade ATPaacutesica da miosina
estimulada por actina atraveacutes da prevenccedilatildeo da troca entre ATPADP sendo esta favorecida para
aniotildees de vanadato com valecircncias superiores como eacute o caso do V10
Na presenccedila de actina as velocidades de dissociaccedilatildeo do Pi e do V aumentam consideravelmente
este uacuteltimo em menor quantidade Assim a ligaccedilatildeo da actina ao complexo MADP
V e
Pi eacute
bastante fraca pelo que se sugere que o ADPV teria um efeito de relaxamento semelhante ao
ATP No entanto a capacidade do vanaacutedio em inibir a actividade ATPaacutesica da miosina e da
actomiosina eacute diferenciada uma vez que na ausecircncia de actina a inibiccedilatildeo eacute total para
concentraccedilotildees na gama dos M mas na sua presenccedila a inibiccedilatildeo eacute totalmente conseguida apenas
em concentraccedilotildees na gama dos mM (Ref) No entanto eacute necessaacuterio considerar a possiacutevel
formaccedilatildeo de espeacutecies oligomeacutericas de vanadato cuja afinidade e efeito na enzima seratildeo
diferentes Assim pode concluir-se que o efeito de inibiccedilatildeo observado para a actividade ATPaacutesica
de miosina estimulada por actina se pode dever agrave presenccedila das diferentes espeacutecies oligomeacutericas o
que foi jaacute sugerido por Aureliano (2000) [AZ] ndash as espeacutecies tetrameacuterica e decameacuterica de vanadato
interactuariam com a miosina com alteraccedilatildeo da sua interacccedilatildeo com a actina No entanto natildeo eacute
conhecida a contribuiccedilatildeo absoluta das restantes espeacutecies oligomeacutericas para a inibiccedilatildeo da ATPase
da miosina estimulada por actina (Refs)
14 Objectivos do projecto
Pretende-se com este projecto o isolamento purificaccedilatildeo e caracterizaccedilatildeo da actina do muacutesculo
22
esqueleacutetico de coelho sendo esta uacuteltima tambeacutem realizada para o sufragmento-1 de miosina
Seguidamente realizar-se-aacute a quantificaccedilatildeo e caracterizaccedilatildeo das proteiacutenas por espectroscopia UV-
Vis e SDS-PAGE e a caracterizaccedilatildeo das soluccedilotildees de vanaacutedio por UV-Vis e ressonacircncia magneacutetica
nuclear (RMN) Por uacuteltimo a interacccedilatildeo do S1 de miosina com o vanaacutedio seraacute avaliada atraveacutes
das teacutecnicas de UVVis RMN e ressonacircncia paramagneacutetica electroacutenica (RPE)
2 Discussatildeo de Resultados
21 Caracterizaccedilatildeo das proteiacutenas isoladas
211Anaacutelise do estado de pureza
O estado de pureza das soluccedilotildees de actina e de S1 foi analisado por electroforese em gel de
poliacrilamida em condiccedilotildees desnaturantes (SDS-PAGE) (Fig 22) que separa as moleacuteculas de
acordo com o seu peso molecular (PM) A concentraccedilatildeo de acrilamida escolhida (10) permite a
detecccedilatildeo simultacircnea de componentes com peso molecular inferior a 20 kDa e outros com elevado
peso molecular
Figura 21 ndash Fotografia de gel de SDS-PAGE (10 acrilamida) Poccedilo (a)
10 L do marcador de elevado peso molecular () Poccedilo (b) 20 L de [G-
actina]=1 mgmL Poccedilo (c) 20 L de [S1]=1 mgmL Aplicou-se uma
voltagem de V durante h e min
A partir da recta de calibraccedilatildeo obtida log PM = -1177 Rf + 2247 (ver ponto 2 da secccedilatildeo
Anexos) pode verificar-se a presenccedila de actina na amostra do poccedilo (b) presente numa banda
bastante intensa (PMaparente=40 kDa) bem como a presenccedila minoritaacuteria de outros componentes
desconhecidos que natildeo teratildeo sido eliminados durante o processo de extracccedilatildeo e purificaccedilatildeo e natildeo
satildeo relevantes para o estudo em questatildeo Na amostra do poccedilo (c) nota-se a presenccedila de S1
(PMaparente=92 kDa) bem como a de outros componentes cuja banda eacute ligeiramente menos intensa
Estes poderatildeo ser fragmentos proteoliacuteticos de miosina e de S1 Estaacute ainda presente uma bada de
23
maior intensidade que poderaacute ser a proteiacutena-C (PM=135 kDa) uma vez que a sua remoccedilatildeo
completa da amostra necessitaria de um passo adicional de cromatografia de troca ioacutenica [BA]
No entanto natildeo podemos confirmar que se trata desta proteiacutena ou de qualquer outra uma vez que
se encontra fora do intervalo de migraccedilatildeo dos marcadores de peso molecular
212 Determinaccedilatildeo da concentraccedilatildeo proteica
2121Subfragmento-1 de miosina
A partir da recta de calibraccedilatildeo obtida pelo meacutetodo de Bradford [BB](ver secccedilatildeo 3311 do
Procedimento Experimental) A595nm = 00546[BSA] + 00209 (ver secccedilatildeo de Anexos)foi possiacutevel
determinar um intervalo de concentraccedilatildeo de S1 de miosina compreendido entre 226 e 117(plusmn
423M) (203 e 1056 (plusmn 381 mgmL)) (n=8) A concentraccedilatildeo meacutedia de S1 de miosina apoacutes
diaacutelise contra a soluccedilatildeo tampatildeo 10 mM HEPES pH 75 25 mM KCl determinada pela leitura da
Abs280 nmfoi de 20430M (1803 mgmL)(n=7)
As amostras a quantificar atraveacutes do meacutetodo de Bradford natildeo sofreram diaacutelise o que permite
explicar a concentraccedilatildeo superior de S1 de miosina em comparaccedilatildeo com o valor obtido atraveacutes do
meacutetodo directo Esta razatildeo aliada agrave sensibilidade e rapidez superiores deste uacuteltimo fez com que
se optasse pelo meacutetodo directo para a quantificaccedilatildeo da grande maioria dos ensaios
2122Actina
A concentraccedilatildeo de actina foi determinada quando esta se encontrava na forma monomeacuterica (G-
actina) por mediccedilatildeo da leitura directa da Abs290 nm tendo-se obtido uma concentraccedilatildeo meacutedia de
691481M (290202 mgmL) (n=3) em 2 mM Tris-HCl pH 80 02 mM CaCl2 02 mM Na-
ATP Ao comprimento de onda utilizado (290 nm) absorvem fortemente apenas os resiacuteduos de
Trp pelo que a mediccedilatildeo efectuada tem menor sensibilidade do que se tivesse sido efectuada a 280
nm uma vez que a actina apresenta apenas 4 resiacuteduos deste aminoaacutecido embora tenha 16
resiacuteduos de Tyr
22 Mediccedilatildeo da actividade enzimaacutetica de S1 de miosina
221 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
As actividades ATPaacutesicas de S1 de miosina dependentes de Ca2+
e K+foram analisadas pelo
caacutelculo das velocidades de consumo inicial do fosfato inorgacircnico (Pi) Definiu-se os tempos de 5
e 3 minutos para a mediccedilatildeo da Ca2+
e da K+EDTA-ATPase respectivamente Os valores obtidos
24
a partir do protocolo presente no ponto 341 da secccedilatildeo Procedimento experimental encontram-se
descritos na Tabela 2I
Tabela 2I ndash Acrividades natildeo-fisioloacutegicas de subfragmento-1 determinadas pelo meacutetodo de Fiske-Subarow
modificado (n=2)
Proteiacutena
Actividade
ATPaacutesica natildeo
fisioloacutegica
Actividade (nmol
Piminmg proteiacutena) Actividade (s-1)
S1
Ca2+
-ATPase 1400 plusmn 30 2100 plusmn 0045
K+EDTA-ATPase 2334 plusmn 50 3500 plusmn 0075
As actividades natildeo-fisioloacutegicas reflectem o estado de oxidaccedilatildeo dos resiacuteduos expostos de Cys 707
(SH1) e 697 (SH2) Se o grupo SH1 sofrer oxidaccedilatildeo a actividade ATPaacutesica dependente de Ca2+
aumenta e a actividade dependente de K+ diminui Por outro lado se o grupo SH2 sofrer
oxidaccedilatildeo regista-se a perda completa destas actividades
De acordo com Margossian e Lowey (1982) [BA] as actividades ATPaacutesica de S1 de miosina
dependente de Ca2+
e K+EDTA da miosina satildeo respectivamente de 24 e 112 s-1
Estaacute tambeacutem
descrito que a actividade ATPaacutesica dependente de K+ eacute em meacutedia cerca de 3 vezes superior agrave
dependente de Ca2+
reflectindo a quantidade das preparaccedilotildees A razatildeo obtida entre os valores das
actividades ATPaacutesicas dependentes de K+ e Ca
2+ eacute de 166 Os valores obtidos para as actividades
(Ca2+
-ATPase=21 s-1
K+EDTA-ATPase= 35 s
-1) satildeo inferiores agrave valores obtidos por
Margossian e Lowey [BA] e Tiago [BC] com forccedila ioacutenica inferior O valor obtido para a
actividade ATPaacutesica dependente de Ca2+
eacute superior ao obtido por Polosukhina et al (2000) [BD]
cujo meio de reacccedilatildeo continha MOPS e apresentava forccedila ioacutenica inferior Os dados
supramencionados comprovam a variaccedilatildeo da actividade a ATPaacutesica natildeo fisioloacutegica em relaccedilatildeo agrave
forccedila ioacutenica temperatura e pH
222 Mediccedilatildeo da actividadeMg2+
-ATPaacutesica de S1 pelo meacutetodo de enzimas acopladas
A actividade de Mg2+
-ATPase eacute a uacutenica que possui significado fisioloacutegico sendo as restantes
bastante uacuteteis apenas para indicaccedilatildeo da qualidade de preparaccedilatildeo do S1[BA] A 0575 M KCl a
actina e o S1 formam uma soluccedilatildeo viscosa e tuacuterbida Apoacutes a adiccedilatildeo de MgATP o complexo acto-
S1 dissocia-se e a turbidez da soluccedilatildeo diminui Se a forccedila ioacutenica for reduzida a dissociaccedilatildeo do
complexo actoS1 que se encontra na forma de suspensatildeo eacute induzida O MgATP eacute rapidamente
hidrolisado diminuindo a sua concentraccedilatildeo no meio reaccional Nestas condiccedilotildees haacute
reassociaccedilatildeo e subsequente contracccedilatildeo dos filamentos de actina e de S1 com formaccedilatildeo de
25
precipitado O meacutetodo descrito traduz de forma bastante elementar o fenoacutemeno da contracccedilatildeo in
vivo e elimina o problema anterior da solubilizaccedilatildeo ineficiente da miosina de modo a obter uma
mediccedilatildeo exacta da actividade [A] A utilizaccedilatildeo de forccedilas ioacutenicas reduzidas aumenta a afinidade da
actina para o S1 permitindo contornar a extrema dificuldade em obter a concentraccedilatildeo proteica dos
muacutesculos (cerca de 100 mgmL do complexo actomiosina) As actividades Mg2+
-ATPaacutesicas de S1
de miosina foram determinadas a partir do declive da variaccedilatildeo da Abs340 relativo agrave oxidaccedilatildeo de
NADH em funccedilatildeo do tempo como eacute descrito no ponto 342 do Procedimento Experimental Os
valores obtidos para a actividade Mg2+
-ATPaacutesica encontram-se apresentados na Tabela 2II
Tabela 2II ndash Actividade Mg2+
-ATPaacutesica de subfragmento-1 de miosina determinadas pelo meacutetodo das enzimas
acopladas
Actividade (nmol
ATPmin mg(S1)) Actividade (s
-1)
1097 plusmn 269 164 plusmn 04
Ramos (2005) [J] obteve o valor de 01 s-1
para a actividade Mg2+
-ATPaacutesica de S1 de miosina
com forccedila ioacutenica intermeacutedia Tiago (2005) [BC] obteve o valor de 011 s-1 em condiccedilotildees de forccedila
ioacutenica reduzida em relaccedilatildeo agrave utilizada no presente relatoacuterio
23 Preparaccedilatildeo e caracterizaccedilatildeo das soluccedilotildees de vanaacutedio
231 Anaacutelise da composiccedilatildeo das soluccedilotildees de vanadato por espectroscopia de 51
V-RMN
efeito da concentraccedilatildeo da espeacutecie
A diluiccedilatildeo das soluccedilotildees stock de meta- e decavanadato iraacute determinar fortemente a composiccedilatildeo
em soluccedilatildeo dos diferentes oligoacutemeros de vanadato (ref) No caso do decavanadato o periacuteodo de
tempo apoacutes a diluiccedilatildeo iraacute tambeacutem ser um factor importante a considerar devido agrave decomposiccedilatildeo
da espeacutecie V10(ref)Torna-se assim fundamental determinar a composiccedilatildeo quiacutemica das soluccedilotildees
no meio de ensaio utilizado (HEPES 10 mM KCl 25 mM) de modo a correlacionaacute-la
correctamente com os possiacuteveis efeitos promovidos pelas diferentes espeacutecies de vanadato no
sistema em estudo
Aleacutem disso deve ser dado ecircnfase agraves importantes interacccedilotildees que ocorrem entre o vanaacutedio e os
tampotildees escolhidos O tampatildeo com menor nuacutemero e magnitude de interacccedilatildeo eacute o HEPES [AD]
seguido do Tris sendo este uacuteltimo especialmente aconselhado em concentraccedilotildees baixas para casos
de incompatibilidade enzimaacutetica ou quiacutemica redox No caso especial da reacccedilatildeo redox
NAD(P)HNAD(P) o tampatildeo adequado eacute o imidazole de modo a natildeo haver reacccedilatildeo com o
26
vanadato Os tampotildees a evitar em qualquer situaccedilatildeo incluem a trietanolamina fosfato citrato e
glicina entre outros [AD] uma vez que todos interactuam fortemente com V(V) com formaccedilatildeo
de complexos estaacuteveis
Assim procedeu-se agrave anaacutelise da composiccedilatildeo das soluccedilotildees de meta- e decavanadato apoacutes diluiccedilatildeo
das soluccedilotildees stock (50 mM em vanadato total) no meio de reacccedilatildeo (tampatildeo D) atraveacutes da
espectroscopia de ressonacircncia magneacutetica nuclear de 51
V Nas figuras x1 e x2 apresentam-se os
espectros obtidos para as soluccedilotildees de deca- e metavanadato num intervalo de concentraccedilotildees de 1 a
10 mM A Tabela seguinte resume os paracircmetros espectroscoacutepicos obtidos para cada espeacutecie
presente nas soluccedilotildees analisadas
Figura 22ndash Espectros de RMN de 51
V de
concentraccedilotildees crescentes das espeacutecies oligomeacutericas
de vanadato obtidos apoacutes diluiccedilatildeo da soluccedilatildeo stock
de decavanadato em 10 mM HEPES pH 70 25 mM
KCl para concentraccedilotildees finais de A) 1 mM B)2
mM C) 4 mM D) 8 mM E) 10 mM de
decavanadato total V1refere-se agraves espeacutecies
monomeacutericade vanadato V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2IIIndash Desvios quiacutemicos meacutedios (part) das ressonacircncias das espeacutecies decameacuterica (V10) monomeacuterica (V1) e
tetrameacuterica (V4) presentes nos espectros da soluccedilatildeo de decavanadatoLargura a meia altura (∆12) da espeacutecie V10A
Paracircmetros V10A V10B V10C V1 V4
part (ppm) -5162 -5003 -4255 -5616 -5748
∆12 (Hz) 175 nd nd nd nd
Figura 23 ndash Distribuiccedilatildeo graacutefica das
espeacutecies oligomeacutericas de vanadato vs a
concentraccedilatildeo de decavanadato total As
concentraccedilotildees de cada espeacutecie foram
obtidas segundo a formula descrita na
secccedilatildeo 35 do Procedimento
Experimental (aacutereas dos sinais de 51
V-
RMN observadas na figura x1)V10 V1 e
V4 correspondem agraves espeacutecie monomeacuterica
dimeacuterica e tetrameacuterica respectivamente [Esp
eacuteci
e o
ligo
meacute
rica
de
van
aacutedio
] (m
M)
[Decavanadato]total (mM)
V10
V1
V4
27
Nas soluccedilotildees de decavanadato as espeacutecies monomeacuterica e decameacuterica de vanadato satildeo as uacutenicas
presentes para concentraccedilotildees de decavanadato inferiores a 8 mM Ambas as espeacutecies aumentam
linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado em [AY] e
em (Teresa) quando a concentraccedilatildeo de V1 eacute igual ou superior a 200 microM inicia-se a sua
oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica No entanto nas condiccedilotildees de realizaccedilatildeo dos
ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a espeacutecie tetrameacuterica neste
estudo em concentraccedilotildees inferiores a 8 mM tendo o mesmo sido observado por Ramoset al
(2006) [AK] Os resultados obtidos mostram claramente o equiliacutebrio entre a espeacutecie decameacuterica e
a monomeacuterica e que o aparecimento de outras espeacutecies oligomeacutericas se encontra dependente da
concentraccedilatildeo desta uacuteltima Apesar de natildeo ter sido realizado o ensaio com 5 mM de decavanadato
total no presente relatoacuterio estaacute reportado em Ramos et al (2006) [AK] que as concentraccedilotildees da
espeacutecie decameacuterica e monomeacuterica satildeo respectivamente de 500 microM e 200 microM Assim pode inferir-
se que para concentraccedilotildees inferiores a 5 mM e intervalos de tempo de cerca de 20 minutos (que
inclui o tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas
as espeacutecies decameacuterica e monomeacuterica de vanadato
Figura 24 ndash Espectros de RMN de 51
V de concentraccedilotildees
crescentes das espeacutecies oligomeacutericas de vanadato obtidos
apoacutes diluiccedilatildeo da soluccedilatildeo stock de metavanadato em 10 mM
HEPES pH 70 25 mM KCl para concentraccedilotildees finais de A)
1 mM B) 2 mM C) 4 mM D) 8 mM E) 10 mM de
metavanadato total V1 refere-se agraves espeacutecies monomeacuterica de
vanadatoV1 e V2 correspondem respectivamente agrave espeacutecie
monomeacuterica e dimeacuterica
Figura 25 ndash Distribuiccedilatildeo graacutefica das espeacutecies
oligomeacutericas de vanadato vs a concentraccedilatildeo de
metavanadato total As concentraccedilotildees de cada
espeacutecie foram obtidas segundo a formula descrita
na secccedilatildeo 35 do Procedimento Experimental
(aacutereas dos sinais de 51
V-RMN observadas na
figura x1)V10 V1 e V4 correspondem agraves espeacutecie
monomeacuterica dimeacuterica e tetrameacuterica
respectivamente
Esp
eacuteci
e o
ligo
meacute
rica
de
va
nad
ato
(m
M)
[Metavanadato]total (mM)
V1
V2
V4
V5
28
Tabela 2IVndash Desvios quiacutemicos meacutedios (part)das ressonacircncias das espeacutecies tetrameacuterica (V4) monomeacuterica (V1) e
dimeacuterica (V2) presentes no espectro da soluccedilatildeo de metavanadato (total) 10 mM
Paracircmetros V1 V2 V4 V5
part (ppm) -5615 -5748 -5789 -5869
Nas soluccedilotildees de metavanadato as espeacutecies monomeacuterica dimeacutericae tetrameacuterica de vanadato estatildeo
presentes para concentraccedilotildees de metavanadato inferiores a 4 mM Todas estas as espeacutecies
aumentam linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado
em (Aureliano e Madeira 1998) e em (Tiago et al 2004) quando a concentraccedilatildeo de V1 eacute igual ou
superior a 200 microM inicia-se a sua oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica Nas
condiccedilotildees de realizaccedilatildeo dos ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a
espeacutecie pentameacuterica neste estudo em concentraccedilotildees inferiores a 4 mM tendo o mesmo sido
observado por Ramos 2006 Os resultados obtidos mostram claramente o equiliacutebrio entre a
espeacutecie V4 e as espeacuteciesV1 e V2 no qual o aparecimento de outras espeacutecies oligomeacutericas se
encontra dependente da concentraccedilatildeo da espeacutecie V1 Por uacuteltimo pode inferir-se que para
concentraccedilotildees inferiores a 2 mM e intervalos de tempo de cerca de 20 minutos (que inclui o
tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas as
espeacutecies V1 V2 e V4 com concentraccedilotildees respectivas de 0796 0192 e 0196 mM
232Estabilidade das soluccedilotildees de decavanadato anaacutelise por espectroscopia de absorccedilatildeo
UVVis
Caracterizou-se a estabilidade da espeacutecie V10 numa soluccedilatildeo com 1 mM decavanadato total em
tampatildeo D atraveacutes da leitura da Abs400 Obteve-se uma cineacutetica de dissociaccedilatildeo de 1ordf ordem tal
como se pode verificar pela linearidade do graacutefico de ln(Abs400)vs o tempo de aquisiccedilatildeo espectral
(segundos) (Fig26 )
Figura 26ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do tempo
(em segundos) para uma soluccedilatildeo de decavanadato total de 1
mM em tampatildeo D a 25ordmC (n=2)
Sendo uma reacccedilatildeo de primeira ordem pode ser descrita pela equaccedilatildeo cujo
declive eacute ndashK (constante de dissociaccedilatildeo) e a intersecccedilatildeo no eixo das ordenadas corresponde
Ln
(Ab
s 400 n
m)
tempo (s)
29
aln(A0) Numa reacccedilatildeo de 1ordf ordem o tempo de meia-vida eacute independente da concentraccedilatildeo inicial
do reagente sendo descrito pela equaccedilatildeo [2] Deste modo a partir da recta
Ln(Abs400 nm) = -436E-5
K - 105 obteve-se um tempo de meia-vida de 4490 plusmn 0823 horas para
as condiccedilotildees supramencionadas
Apoacutes a adiccedilatildeo de S1 8 microM ao meio reaccional verificou-se que a reacccedilatildeo de desoligomerizaccedilatildeo
da espeacutecie decameacuterica de vanadato seguia igualmente uma cineacutetica de 1ordf ordem tal como se pode
verificar pela linearidade do graacutefico de ln(Abs400) versus o tempo de aquisiccedilatildeo espectral
(segundos) (Fig27)
Figura 27ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do
tempo (em segundos) para uma soluccedilatildeo de decavanadato
total de 1 mM em tampatildeo D a 25ordmC na presenccedila de 8
microM (n= 2)
A partir da equaccedilatildeo da recta Ln(Abs400 nm) = -7520E-5
K - 0613determinou-se o valor de K para
a espeacutecie decameacuterica de onde se obteve um tempo de meia-vida de 2430 plusmn 0191 horas para as
condiccedilotildees supramencionadas
Tanto este valor como o valor registado na ausecircncia de S1 satildeo substancialmente superiores agrave
duraccedilatildeo dos ensaios de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina descritos no presente
relatoacuterio
24 Interacccedilatildeo de S1 com vanaacutedio(V)
241Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
De modo a analisar qual o efeito da adiccedilatildeo de S1 de miosina (tampatildeo D) produzia nos sinais de
51V-NMR titulou-se uma soluccedilatildeo de 5mM de decavanadato total com concentraccedilotildees crescentes
de S1 (5 10 15 e 20 microM) Os espectros de 51
V-RMN encontram-se apresentados na Fig 2
No graacutefico da figura 2 Apresenta-se a concentraccedilatildeo de S1 em funccedilatildeo do factor de alargamento
corrigido dos espectros acima referidos
Ln
(Ab
s (4
00
nm
))
tempo (s)
30
Figura 28 ndash Graacutefico de
representaccedilatildeo da
concentraccedilatildeo de S1 em
funccedilatildeo do factor de
alargamento A
concentraccedilatildeo de
decavanadato total foi de 5
mM e as concentraccedilotildees
utilizadas de S1 foram de 0
5 10 e 15 microM
Verifica-se que o factor de alargamento aumenta linearmente em funccedilatildeo da concentraccedilatildeo de S1 no
ensaio o que indica a existecircncia de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina Observou-
se ainda que a adiccedilatildeo de S1 ao meio reaccional natildeo provocou qualquer alteraccedilatildeo dos desvios
quiacutemicos dos sinais de 51
V-RMN
Realizaram-se ensaios na presenccedila de ATP e de S1 com intuito de verificar o efeito do ATP na
interacccedilatildeo do S1 de miosina com o decavanadato
Figura 29 - Espectros de RMN de 51
V contendo 2
mM Decavanadato (A) (controlo) (B) na presenccedila
de 2 mM de ATP (C) na presenccedila de 2 mM de ATP
e 8 microM de S1 de miosina V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2Vndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies decameacuterica (V10) e monomeacuterica (V1) presentes nos
espectros da soluccedilatildeo de decavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V10A V10B V10C V1
part (ppm) -5161 -5002 -4254 -5614
Observa-se que a presenccedila de ATP (espectro B) natildeo provoca qualquer alteraccedilatildeo visiacutevel em
relaccedilatildeo ao espectro que contem apenas soluccedilatildeo de decavanadato Jaacute a presenccedila simultacircnea de ATP
e S1 provoca um alargamento da ∆12 do sinal do V10A de 1736 Hz (espectro (A)) para 2366 Hz
(espectro (C) Fig 29) Tendo em conta que a concentraccedilatildeo de S1 apresenta uma relaccedilatildeo linear
com o factor de alargamento converteu-se o valor da ∆12 e comparou-se com o valor esperado agrave
F = 00591[S1] - 00157 Rsup2 = 0998
Fact
or
de
ala
rgam
en
to
[S1] (microM)
31
concentraccedilatildeo de S1 utilizada O factor de alargamento obtido para 8 microM ndash 0376 ndash eacute inferior ao
valor esperado agrave mesma concentraccedilatildeo de S1 obtido atraveacutes da equaccedilatildeo da recta apresentada na
Fig 28 ndash 0457 ndash o que indica que o ATP poderaacute exercer um efeito de protecccedilatildeo do S1
diminuindo a sua interacccedilatildeo com a espeacutecieV10
Por uacuteltimo realizaram-se ensaios com metavanadato na presenccedila de 8 microM S1 de modo a verificar
se as espeacutecies oligomeacutericas contidas nesta soluccedilatildeo interagiam com a proteiacutena em estudo
Figura 210 - Espectros de RMN de 51
V contendo 2
mM metavanadato (A) (controlo) (B) na presenccedila
de 8 microM de S1 de miosina V1 V2 e V4
correspondem respectivamente aacutes espeacutecies
monomeacuterica dimeacuterica e tetrameacuterica
Tabela 2VIndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies monomeacuterica (V1) dimeacuterica (V2) e tetrameacuterica (V4)
presentes nos espectros da soluccedilatildeo de metavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V1 V2 V4
part (ppm) -5613 -5747 -5789
Como eacute possiacutevel observar na Fig 210 ocorre uma ligeiro alargamento dos sinal de V4 bem
como uma perda da intensidade pelo que se conclui quedas espeacutecies presentes na soluccedilatildeo de
metavanadato (V1 V2 e V4) apenas a espeacutecie tetrameacuterica interage com o S1 de miosina o que
estaacute de acordo com o reportado por Tiago et al 2002
242 Actividade Mg2+
-ATPaacutesica de S1 de miosina na presenccedila de decanadato
Com o intuito de avaliar o efeito do decavanadato na actividade Mg2+
-ATPaacutesica do S1 de
miosina monitorizou-se a reacccedilatildeo de oxidaccedilatildeo do NADH atraveacutes da leitura de Abs340 na
presenccedila de 005 mgmL de S1 de miosina e no caso da actividade estimulada 2microM de actina em
3125 mM Tris-HCl pH 71 e 375 mM MgCl2 pelo meacutetodo de enzimas acopladas (ver seccccedilatildeo xx
dos MM)Para tal realizaram-se ensaios com vaacuterias concentraccedilotildees de decavanadato total (10 50
100 250 500 e 1000) de modo a determinar qual o seu efeito na actividade MgATPaacutesica de S1
de miosina na ausecircncia e na presenccedila de um dos seus ligandos naturais Os ensaios foram
realizados agrave temperatura ambiente com incubaccedilatildeo preacutevia a 25ordm C durante 20 minutos
32
Figura 211 ndash Representaccedilatildeo
graacutefica da Actividade Mg2+
-
ATPaacutesica de S1 () (os ajustes
teoacutericos estatildeo representado a por
uma linha os resultados
experimentais estatildeo representados
por triacircngulos (Actividade
estimulada) e cruzes (Actividade
natildeo estimulada)) vs
[Decavanadato]total medidas pelo
meacutetodo das enzimas acopladas
(n=3) Os dados foram ajustados ao modelo de decaimento exponencial usando o programa GraphPad Prism para a
determinaccedilatildeo dos valores de IC50 (na ausecircncia de actina IC50= 266 plusmn 07 M decavanadato na presenccedila de actina
IC50= 28 plusmn 1 M decavanadato
A partir do graacutefico presente na Fig 29 verifica-se que o ajuste dos dados experimentais foi bem
sucedido A inibiccedilatildeo da actividade Mg2+
-ATPaacutesica do S1 de miosina eacute afectada em menor
amplitude na presenccedila de actina (60 de inibiccedilatildeo da actividade) do que na sua ausecircncia
(aproximadamente 80 de inibiccedilatildeo de actividade) Devido a esta diferenccedila pode sugerir-se que a
actina iraacute impedir parcialmente a ligaccedilatildeo do decavanadatoao S1 demiosina atraveacutes de
impedimentos esteacutereos ou interacccedilotildees electrostaacuteticas por exemplo A teacutecnica utilizada natildeo nos
permite concluir acerca de que regiatildeo(otildees) ou resiacuteduo(s) da actina participaratildeo na preservaccedilatildeo
parcial da actividade Mg2+
-ATPaacutesica
Por outro lado os valores de IC50 dos dois tipos de actividades natildeo apresentam diferenccedilas
significativas tendo-se obtido 266 plusmn 07 e 28 plusmn 1M decavanadato na ausecircncia e na presenccedila de
actina respectivamente o que corresponde a uma concentraccedilatildeo de cerca de 27 M de V10 O
meio de reacccedilatildeo utilizado nestes ensaios foi 10 mM HEPES e 25 mM KCl com 2 mM ATP Em
(Tiago et al 2004) o valor de IC50 para a actividade Mg2+-ATPaacutesica na ausecircncia de actina foi de
125 plusmn 025 microM enquanto o valor da actividade estimulada por actina foi de 136 plusmn 14 microM O
meio de reacccedilatildeo utilizado nos ensaios foi 20 mM HEPES 50 mM KCl e 2 mM ATP o que resulta
precisamente no dobro da forccedila ioacutenica a que foi realizado o ensaio do presente relatoacuterio
(exceptuando a contribuiccedilatildeo dos produtos da hidroacutelise de ATP uma vez que este meacutetodo
pressupotildee a ocorrecircncia de um ciclo de reacccedilotildees de hidroacutelise e siacutentese deste composto) Ora tendo
sido anteriormente referido que a afinidade do S1 de miosina para com a espeacutecie V10 diminui com
Act
ivid
ade
Mg
2+-A
TP
aacutesic
a d
e S
1 d
e
Mio
sin
a
[Decavanadato] (microM) Actividade Estimulada ()
Actividade Natildeo Estimulada ()
Ajuste Actividade Estimulada
Ajuste Actividade Natildeo Estimulada
33
o aumento da forccedila ioacutenica (Tiago et al 2004 tal poderaacute explicar a diferenccedila obtida entre estes
dois conjuntos de valores
243Quantificaccedilatildeo de cisteiacutenas reduzidas pelo meacutetodo de Ellman
A interacccedilatildeo do vanaacutedio com alguns tipos de aminoaacutecidos sensiacuteveis agrave oxidaccedilatildeo pode fornecer-nos
informaccedilatildeo bastante uacutetil em relaccedilatildeo agraves alteraccedilotildees estruturais que conduzem a uma diminuiccedilatildeo da
actividade fisioloacutegica do S1 de miosina Os estudos realizados por Chasteen 1983 concluiacuteram
que a reduccedilatildeo de vanadato (V) a vanadilo (IV) ocorre na presenccedila de agentes redutores comuns no
ambiente celular de que se destaca o aminoaacutecido cisteiacutena Esta eacute tambeacutem um alvo de
complexaccedilatildeo por parte do iatildeo VO2+
Jaacute Ramos et al (2009) demonstrou que a incubaccedilatildeo de
actina com a soluccedilatildeo de decavanadato induz a oxidaccedilatildeo de resiacuteduos de cisteiacutena e agrave formaccedilatildeo do
oxidovanaacutedio (IV) Este grupo de investigadores descobriu ainda que apenas a espeacutecie V10 induz a
oxidaccedilatildeo das cisteiacutenas ldquoraacutepidasrdquo da actina atraveacutes do processo proposto que eacute parcialmente
responsaacutevel pela inibiccedilatildeo jaacute descrita da actividade de ATPase do S1 de miosina estimulada por
actina quando exposta agrave soluccedilatildeo de decavanadato Esta inibiccedilatildeo apresenta-se afectada com a
presenccedila de ATP uma vez que os resiacuteduos de cisteiacutena permaneceram intactos nos ensaios
realizados com este ligando Jaacute Crans et al (2004) concluiu que o siacutetio activo de muitas fosfatases
(que conteacutem grupos tiol) pode formar um equiliacutebrio redox com o iatildeo vanadato (V) de forma natildeo
reversiacutevel
Os resultados obtidos por Sakurai e Shiromura (1980) sugeriram que o vanaacutedio (V) sofre reduccedilatildeo
a vanaacutedio (IV) sem dependecircncia do valor de pH do ensaio Registaram ainda uma estequiometria
de 31 na reacccedilatildeo do vanadato (V) com os resiacuteduos de Cys enquanto a estequiometria dos ensaios
com o vanadilo (IV) foi de 21
Berlett e Stadtman (1997) observaram que os tipos de aminoaacutecidos mais facilmente oxidaacuteveis
englobam entre outros os resiacuteduos de Cys Trp e MetAssim analisou-se o estado redox dos
grupos tiol dos resiacuteduos de Cys do S1 na presenccedila do decavanadato atraveacutes da monitorizaccedilatildeo da
Abs412 nm na presenccedila de 1 mgmL de S1 de miosina em tampatildeo 10 mM HEPES 25 mM KCl
variando a concentraccedilatildeo da soluccedilatildeo de decavanadato de modo a determinar que tipo de alteraccedilotildees
estruturais ocorrem na sua presenccedila no que diz respeito agrave diminuiccedilatildeo da actividade ATPaacutesica de
S1 na presenccedila de decavanadato
Os resultados referentes agrave quantificaccedilatildeo das cisteiacutenas reduzidas apoacutes o tratamento de S1 com
diferentes concentraccedilotildees de decavanadato em tampatildeo 10 mM HEPES pH 75 e 25 mM KCl a
25ordmC durante 20 minutos encontram-se descritos na Tabela 2VII
Tabela 2VII ndash Cisteiacutenas reduzidas totais e ldquoraacutepidas de S1 (1 mgmL) apoacutes tratamento com decavanadato
34
(010501002505001000 uM) em 10 mM HEPES 25 mM KCl pH 7 a 25ordmC durante 20 minutos A detecccedilatildeo
espectrofotomeacutetrica foi feita com DTNB (e 412=8400 uM-1 cm-1) (n variaacutevel entre 1 e 8)
Amostras Cys raacutepidas Cys totais
11 M S1 (controlo) 22 plusmn 04 106 plusmn 14
11 M S1 + 10 M Decavanadato 22 plusmn 05 95 plusmn 08
11 M S1 + 50 uM Decavanadato 20 plusmn 02 98 plusmn 14
11 M S1 + 100 uM Decavanadato 17 plusmn 03 97 plusmn14
11 M S1 + 250 uM Decavanadato 26 plusmn 02 98 plusmn 08
11 M S1 + 500 uM Decavanadato 21 plusmn 04 90 plusmn 06
11 M S1 + 1000 uM Decavanadato 24 plusmn 0 86 plusmn 08
A partir dos resultados obtidos pode sugerir-se que o S1 possui 11 resiacuteduos de Cys (controlo) o
que estaacute de acordo com o anteriormente descrito Verifica-se que o tratamento da proteiacutena com
concentraccedilotildees crescentes de decavanadato provoca a oxidaccedilatildeo de dois resiacuteduos de Cys do S1 de
miosina natildeo expostos ao solvente No entanto a teacutecnica utilizada natildeo nos permite identificar
quais
Tendo em consideraccedilatildeo que o valor de IC50 obtido para a reacccedilatildeo de inibiccedilatildeo daactividade Mg2+-
ATPaacutesica de S1 na presenccedila de decavanadato eacute de cerca de 27 microM pode sugerir-se que este seraacute o
valor ao qual ocorreraacute a oxidaccedilatildeo do primeiro resiacuteduo de Cys o que poderaacute contribuir para a
diminuiccedilatildeo da actividade de S1 observada
25Interacccedilatildeo de S1 com vanaacutedio(IV)
251Anaacutelise por espectroscopia de Ressonacircncia Paramagneacutetica Electroacutenica
A acumulaccedilatildeo intracelular de oxovanaacutedio (IV) in vivo constitui uma das principais formas de
apresentaccedilatildeo de vanaacutedio no organismo Deste modo torna-se bastante interessante analisar a
interacccedilatildeo entre este iatildeo e o S1 de miosina atraveacutes da teacutecnica de espectroscopia de Ressonacircncia
Paramagneacutetica Electroacutenica o meacutetodo de eleiccedilatildeo do estudo do iatildeo vanadilo e dos complexos por si
formados O electratildeo desemparelhado encontra-se fortemente acoplado ao nuacutecleo 51
V o que
35
resulta num espectro de RPE consistindo de 8 desdobramentos na direcccedilatildeo paralela e outros tantos
na direcccedilatildeo perpendicular (Chasteen 1983)
Os estudos realizados por Ahmed et al (1987) centraram-se na interacccedilatildeo do vanadilo com
proteiacutenas como eacute o caso da insulina transferrina entre outros obtendo um espectro caracterizado
por 8 linhas amplamente espaccediladas (na direcccedilatildeo paralela) Jaacute Ramos et al 2009 concluiu que a
exposiccedilatildeo da soluccedilatildeo de decavanadato agraves formas globular e filamentosa de actina produz um
espectro tiacutepico do iatildeo oxidovanaacutedio (IV) indicando a reduccedilatildeo do vanadato
Para aferir qual a concentraccedilatildeo de S1 que permitia obter uma boa razatildeo sinalruiacutedo titulou-se uma
soluccedilatildeo de sulfato de vanadilo 15 microM ([VOSO4]stock = 5 mM) com concentraccedilotildees crescentes de
proteiacutena (0 5 10 15 e 20 microM) em 10 mM HEPES 25 mM KCl A existecircncia de uma correlaccedilatildeo
linear entre a concentraccedilatildeo de proteiacutena e a intensidade dos sinais de RPE correspondente agrave banda
-12 perpendicular encontra-se representada na Fig 210
Figura 2 12ndash Representaccedilatildeo graacutefica da variaccedilatildeo da intensidade do sinal de RPE vs a concentraccedilatildeo de S1 utilizada
Apoacutes verificar que com uma concentraccedilatildeo de S1 igual a 20 microM se obtinham sinais com uma razatildeo
sinalruiacutedo adequada ao prosseguimento dos estudos sobre a interacccedilatildeo V(IV)-S1 de miosina
titulou-se uma soluccedilatildeo de 20 microM S1 com 0 25 5 10 15 25 50 75 100 150 200 500 e 1000
microM VOSO4
Traccedilou-se o graacutefico da intensidade da banda -12 perpendicular do sinal de EPR em funccedilatildeo da
concentraccedilatildeo de VOSO4 tendo-se determinado uma constante de dissociaccedilatildeo (KD) do complexo
S1-vanadilo de 8748 plusmn 2621 microM V(IV)
y = 016786x + 122435
Rsup2 = 092992759
Inte
nsi
dad
e d
o s
inal
de
RP
E
[S1] (microM)
36
Figura 213 ndash Representaccedilatildeo graacutefica da variaccedilatildeo do sinal de RPE vs a concentraccedilatildeo de vanadilo utilizada (os valores
experimentais estatildeo representados por losangos o seu ajuste estaacute representado pela linha)
A simulaccedilatildeo do espectro de EPR foi efectuada a partir do espectro obtido para uma concentraccedilatildeo
de 1 mM VOSO4 apoacutes adiccedilatildeo de 20 microM S1 de miosina tendo-se obtido os valores da constante de
acoplamento hiperfina (A=5016) e do factor g (g0=195)
Figura 214 ndash Representaccedilatildeo graacutefica do espectro de EPR obtido com 1000 microM de vanadilo e 20 microM de S1 (a cinzento
escuro) A simulaccedilatildeo do espectro encontra-se a cinzento claro
Por comparaccedilatildeo destes com os valores presentes na tabela de correspondecircncia xxx eacute possiacutevel
predizer quais os ligandos provaacuteveis do vanadilo na presenccedila de proteiacutena Assim concluiu-se que
o vanadilo estaacute coordenado com duas moleacuteculas de aacutegua um grupo hidroxilo e um grupo R=NR
O grupo hidroxilo pode indicar os resiacuteduos de aminoaacutecidos Tyr Treonina e Serina enquanto o
grupo R=NR poderaacute indicar a presenccedila de Arginina e Histidina como ligandos do vanadilo
Inte
nsi
dad
e d
os
sinai
s d
e R
PE
[Vanadilo] (microM)
Ajuste dos dados exp Dados experimentais
Campo magneacutetico (G)
Espectro Experimental Ajuste
37
3 Procedimento Experimental
31 Reagentes
Todos os reagentes utilizados satildeo de elevado grau de pureza (analiacutetico) Apresentam-se as suas
foacutermulas quiacutemicas ou abreviaturas quando necessaacuterio acrescentou-se a sua descriccedilatildeo Entre
parecircntesis colocou-se as empresas que os comercializam
Aacutecido aceacutetico glacial 100 (Riedel) Acrilamida 99 (Merck) Azul de bromofenol (Riedel-de-
Haumlen) -Mercaptoetanol (Sigma) Bissulfito de soacutedio (Sigma)Bradford (reagente de)
(BioRad)BSA (albumina de soro bovino Sigma-Aldrich) CaCl22H2O (cloreto de caacutelcio
dihidratado Fluka) CH3CH2OH 96 (etanol Panreac) Coomassie Blue R250 (Merck) D2O
(oacutexido de deuteacuterio SDS) DTNB (55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico) Aldrich) EDTA (aacutecido
etilenodiaminatetraaceacutetico Merck) Fosfoenolpiruvato 98 (Fluka) Glicerol (Sigma) Glicina
985 (Panreac) HCl 37 (Riedel-de Haumlen) H2SO4 96 (Pronalab) HEPES 99 (aacutecido 4-(2-
hidroxiletil)-1-piperazineetanosulfoacutenico Sigma) KH2PO4 99 (hidrogenofosfato de potaacutessio)
MgATP (Sigma) Na
ATP (Sigma) Na2HPO4 99 (hidrogenofosfato de soacutedio Riedel-de Haumlen)
NaH2PO4 (fosfato de soacutedio ) NaOH 986 (hidroacutexido de soacutedio Pronalab) KCl 99 (Cloreto de
potaacutessio Panreac) L-Cys 995 (enantioacutemero levoacutegiro do resiacuteduo de Cisteiacutena Fluka)Metanol
99 (Merck) Molibdato de amoacutenio (Sigma)NNrsquo-metil-bis-acrilamida 98 (Merck) NH4VO3
99 (metavanadato de amoacutenia Sigma) PSA 10 (persulfato de amoacutenio Merck) SDS
(dodecilo sulfato de soacutedio Sigma) Sulfito de soacutedio (Sigma) VOSO4 (sulfato de vanadilo
Aldrich) TEMED 99 (Tetrametiletilenodiamina Merck) Tris (2-amino-2-hidroximetil-13-
propanodiol Sigma)
32 Meacutetodos
321 Isolamento e purificaccedilatildeo das proteiacutenas de muacutesculo esqueleacutetico de coelho
3211 Purificaccedilatildeo da miosina preparaccedilatildeo de Subfragmento 1 por digestatildeo quimotriacutepsica
da miosina
O subfragmento-1 de miosina-II (S1) foi gentilmente cedido pelo Prof Doutor Manuel Aureliano
do Departamento de Quiacutemica Bioquiacutemica e Farmaacutecia da Faculdade de Ciecircncias e Tecnologia da
Universidade do Algarve
No entanto descreve-se abaixo de forma resumida o procedimento efectuado
38
32111 Purificaccedilatildeo da miosina
O protocolo de isolamento foi adaptado do meacutetodo previamente descrito por Margossian e Lowey
(1982) apresentando-se de forma esquemaacutetica na Fig 31
Sucintamente os tecidos musculares dorsais e das pernas foram extraiacutedos apoacutes o sacrifiacutecio do
animal (Fig31) De seguida foram adicionados 3 volumes da soluccedilatildeo de extracccedilatildeo A2
Centrifugou-se a soluccedilatildeo e filtrou-se o sobrenadante O sedimento foi entatildeo diluiacutedo em 14
volumes de aacutegua destilada de modo a que a forccedila ioacutenica atingisse 004 M Apoacutes adiccedilatildeo de szlig-
mercaptoetanol (szlig-me) deixou-se a precipitar a 4ordmC removendo o sobrenadante Centrifugou-se
novamente e ressuspendeu-se o sedimento na soluccedilatildeo de extracccedilatildeo A Adicionou-se a soluccedilatildeo
tampatildeo B3 para que a forccedila ioacutenica atingisse 05 M Homogeneizou-se e deixou-se repousar
durante 2h Homogeneizou-se novamente e centrifugou-se de modo a ocorrer precipitaccedilatildeo dos
complexos de actomiosina e proteiacutena desnaturada Apoacutes filtraccedilatildeo do sobrenadante em latilde de vidro
quantificou-se a soluccedilatildeo de miosina resultante pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental)
2 Soluccedilatildeo de extracccedilatildeo A 0012M K2CO3 004M KHCO3 06M KCl pH 65
3 Soluccedilatildeo-tampatildeo B Tris 100 mM pH 7 KCl 2M
39
Figura 31 ndash Representaccedilatildeo esquemaacutetica do protocolo de isolamento de miosina (Margossian e Lowey 1982) Todo o
procedimento foi realizado a 4ordmC (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
32112 Preparaccedilatildeo de Subfragmento-1 de miosina por digestatildeo com quimiotripsina
O S1 de miosina foi isolado apoacutes diaacutelise da miosina durante a noite contra a soluccedilatildeo C4 A
presenccedila do EDTA eacute essencial uma vez que a sua substituiccedilatildeo por MgCl2resulta na produccedilatildeo de
HMM em vez de S1 A suspensatildeo de miosina foi equilibrada e digerida com -quimotripsina
A digestatildeo foi interrompida pela adiccedilatildeo de PMSF Seguidamente ultracentrifugou-se o produto
da digestatildeo desprezando-se o sedimento (Fig 32) A soluccedilatildeo de sobrenadante maioritariamente
S1 foi quantificada pela leitura directa da Abs280 ou pelo meacutetodo de Bradford (ver as secccedilotildees
3311 e 3312 do Procedimento Experimental respectivamente) e armazenada em 10 sacarose
e 2 mM szlig-Me a -80ordmC
4 Soluccedilatildeo C 25mM Tris pH 70 012M KCl 1mM EDTA
40
Antes da realizaccedilatildeo dos ensaios descritos no presente relatoacuterio o S1 de miosina foi dialisado
contra a soluccedilatildeo D5 de modo a remover qualquer composto que pudesse complexar como
vanaacutedio Seguidamente quantificou-se a proteiacutena pelo meacutetodo de Bradford ou pelo meacutetodo
directo (ver secccedilotildees 3311 e 3312do Procedimento Experimtental)
Figura 3 2ndash Representaccedilatildeo esquemaacutetica do protocolo de preparaccedilatildeo do S1 de miosina por digestatildeo quimiotripsina de
acordo com o previamente descrito por Margossian e Lowey (1982) Todo o procedimento foi realizado a 4ordm C
excepto quando indicaccedilatildeo do contraacuterio (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
3212 Isolamento e purificaccedilatildeo de actina
Isolou-se e purificou-se a actina segundo o meacutetodo descrito por Pardee e Spudish que
compreende dois passos ndash a obtenccedilatildeo de um resiacuteduo seco denominado ldquopoacute de acetonardquo e a
extracccedilatildeo subsequente da actina a partir do ldquopoacute de acetonardquo na forma de G-Actina com a
posterior polimerizaccedilatildeo a F-Actina
32121 Obtenccedilatildeo do ldquopoacute de acetonardquo
Sacrificou-se o coelho com uma pancada atraacutes do pescoccedilo e sangrou-se o animal apoacutes o corte da
arteacuteria jugular Esfolou-se e colocou-se numa tina com gelo Retiraram-se as partes de tecido
conjuntivo que envolvem o tecido muscular com auxiacutelio de uma pinccedila e bisturi Seguidamente
retirou-se os muacutesculos dorsais e os muacutesculos brancos das pernas Passou-se os muacutesculos por aacutegua
destilada de modo a remover as impurezas e pesou-se num copo Trituraram-se os muacutesculos
numa maacutequina de triturar carne e extraiu-se o resultante com 3 volumes da soluccedilatildeo E6 durante 10
minutos com agitaccedilatildeo suave Centrifugou-se a suspensatildeo a 4000 g durante 10
5 Soluccedilatildeo D HEPES 10 mM pH 70 KCl 25 mM
6 Soluccedilatildeo E 01 M KCl 015 M K2HPO4 pH 65
41
minutosdesprezando-se o sobrenadante Ressuspendeu-se o sedimento em 6 volumes de soluccedilatildeo
F7durante 10 minutos com agitaccedilatildeo suave Verificou-se se o pH da soluccedilatildeo estava compreendido
entre 82 e 85 acertando-se estecom soluccedilatildeo G8 se necessaacuterio Centrifugou-se durante 10
minutos a 4000 g Desprezou-se o sobrenadante e ressuspendeu-se o sedimento em 3 volumes de
soluccedilatildeo H9 durante 10 minutos Centrifugou-se novamente a 4000 gdurante 10 minutos
desprezando-se o sobrenadante Ressuspendeu-se o sedimento duas vezes consecutivas em 6
volumes de aacutegua destilada durante 5 minutos Centrifugou-se a 4000 g durante 10 minutos apoacutes
cada adiccedilatildeo de H2O
Seguidamente ressuspendeu-se o sedimento 5 vezes consecutivas utilizando 3 volumes de
acetona de cada vez agitando suavemente durante 10 minutos agrave temperatura ambiente Colocou-
se o resiacuteduo filtrado numa tina de vidro sobre papel de filtro deixando-se evaporar o solvente
durante a noite num nicho Triturou-se o resiacuteduo seco numa maacutequina de triturar carne e deixou-se
novamente evaporar o solvente O ldquopoacute de acetonardquo obtido foi armazenado a -20 ordmC
32122 Purificaccedilatildeo da actina a partir de ldquopoacute de acetonardquo
Apoacutes a obtenccedilatildeo do poacute de acetona como acima indicado efectuou-se o protocolo seguidamente
descrito de modo a obter monoacutemeros de actina (G-Actina) e actina filamentosa (F-Actina)
Pesou-se 35 g de poacute de acetona e adicionaram-se 50 mL de soluccedilatildeo-tampatildeo I10
sendo a
extracccedilatildeo feita com agitaccedilatildeo suave durante 30 minutos Separou-se o extracto do poacute de acetona
hidratado filtrando a suspensatildeo com 4 camadas de gaze esterilizada tendo o cuidado de natildeo
espremer o resiacuteduo Voltou a extrair-se com 50 mL de soluccedilatildeo I durante 30 minutos mexendo
lentamente Adicionou-se este ao extracto obtido anteriormente Centrifugou-se o extracto total a
16 000 g durante 30 minutos Decantou-se para uma proveta sobre a qual se colocou gaze para
que o sedimento solto natildeo se juntasse ao sobrenadante e leu-se o volume De seguida adicionou-
se KCl ateacute uma concentraccedilatildeo final de 50 mM MgCl2 ateacute 2mM e ATP ateacute 1mM lentamente e com
agitaccedilatildeo contiacutenua de modo a promover a polimerizaccedilatildeo dos monoacutemeros de actina
Deixou-se polimerizar durante 2h a 25ordmC observando-se a presenccedila de pequenas bolhas de ar
indicativas de um aumento de viscosidade da soluccedilatildeo Arrefeceu-se em gelo durante 15 minutos e
adicionou-se KCl ateacute uma concentraccedilatildeo final de 06M Mexeu-se lentamente durante 30 minutos
apoacutes os quais se ultracentrifugou a 110 000g durante 1hora Desprezou-se o sobrenadante
ressuspendeu-se o sedimento com soluccedilatildeo tampatildeo E e transferiu-se para um potter
Dialisou-se durante a noite contra 1L de soluccedilatildeo-tampatildeo I a uma temperatura de 4ordmC com rotaccedilatildeo
7 Soluccedilatildeo F 005 M NaHCO3 pH 82-85
8 Soluccedilatildeo G 1M Na2CO3
9 Soluccedilatildeo H 1 mM EDTA pH 70
10 Soluccedilatildeo I 2 mM Tris-HCl pH 8 02 mM CaCl2 02 mM Na2ATP 05 mM szlig-Me 0005 NaN3
42
contiacutenua e mudanccedila de tampatildeo 2 a 3 vezes De seguida a actina foi centrifugada a 110
000gdurante 1hora Decantou-se a soluccedilatildeo de G-Actina (sobrenadante) e desprezou-se o
sedimento Determinou-se a concentraccedilatildeo de G-Actina pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental) A soluccedilatildeo de G-actina foi armazenada a uma temperatura de 4ordmC
apoacutes adiccedilatildeo de 002NaN3 mantendo-se estaacutevel durante 3 a 5 dias
331 Quantificaccedilatildeo das proteiacutenas isoladas
3311 Quantificaccedilatildeo pelo meacutetodo de Bradford
O meacutetodo de Bradford (Bradford 1976) baseia-se na utilizaccedilatildeo do reagente de Bradford
composto por entre outros aacutecido fosfoacuterico e o corante ldquoCoomassierdquo Este corante possui carga
negativa e a sua coloraccedilatildeo varia entre a vermelha (Absmax=465 nm) predominante em soluccedilatildeo e a
azul (Absmax=595 nm) presente aquando da ligaccedilatildeo aos grupos amina das proteiacutenas Em soluccedilotildees
aacutecidas diluiacutedas a coloraccedilatildeo varia proporcionalmente com a quantidade do corante ligado agrave
proteiacutena que por sua vez eacute directamente proporcional agrave concentraccedilatildeo da proteiacutena permitindo a
sua quantificaccedilatildeo ateacute a um limite de detecccedilatildeo de 20 gmL (Bradford 1976)
Deste modo prepararam-se amostras de BSA ([BSA]stock= 1mgmL) utilizando um intervalo de
concentraccedilotildees compreendido entre 0 e 15 gmL de acordo com a tabela 3I Apoacutes a adiccedilatildeo da
H2O MilliQ e do reagente de Bradford as soluccedilotildees foram sucessivamente homogeneizadas no
voacutertex
Tabela 3Indash Amostras utilizadas para a obtenccedilatildeo da recta de calibraccedilatildeo no meacutetodo de Bradford
BSA (L) BSA (g) H2O MilliQ (L) Reagente de Bradford (L)
0 0 800 200
125 125 79875 200
25 25 7975 200
5 5 795 200
10 10 790 200
15 15 785 200
Apoacutes incubaccedilatildeo a 25ordmC por 5 minutos leram-se os valores de Abs595 num
espectrofotoacutemetro (Shimadzu UV-160A) para todas as amostras Seguidamente traccedilou-se a recta
de calibraccedilatildeo (Abs595vs quantidade de BSA (g))
43
Para determinar a concentraccedilatildeo de proteiacutena apoacutes o processo de isolamento purificaccedilatildeo e
diaacutelise quando necessaacuterio colocou-se 5 L da amostra num tubo de Eppendorf e adicionou-se
795 L de H2O MilliQ homogeneizando a soluccedilatildeo no voacutertex Adicionou-se posteriormente 200
L de reagente de Bradford e repetiu-se a homogeneizaccedilatildeo Incubou-se a mistura a 25ordmC por 5
minutos apoacutes os quais se leu os valores de Abs595 nm no espectrofotoacutemetro Por interpolaccedilatildeo deste
valor a partir da recta de calibraccedilatildeo determinou-se a concentraccedilatildeo de proteiacutena na amostra
3312 Quantificaccedilatildeo pelo meacutetodo directo
A quantificaccedilatildeo das amostras proteicas pode tambeacutem ser efectuada atraveacutes da leitura directa da
absorvacircncia tendo por base a lei de Beer-Lambert onde Abs corresponde agrave
absorvacircncia ao comprimento de onda ao qual o ensaio foi realizado corresponde ao coeficiente
de extinccedilatildeo (substituiacutedo pela letra a se as unidades forem distintas da molaridade) b corresponde
ao percurso oacuteptico da radiaccedilatildeo na ceacutelula amostral e c corresponde agrave concentraccedilatildeo amostral
Assim no caso de S1 de miosina mediu-se a Abs280e calculou-se a concentraccedilatildeo da proteiacutena
sabendo que o coeficiente de extinccedilatildeo de S1 eacute (Ref)
No caso da actina mediu-se a Abs290ecalculou-se a concentraccedilatildeo da proteiacutena sabendo que o
coeficiente de extinccedilatildeo da actina eacute (Ref)
332Electroforese em condiccedilotildees desnaturantes (SDS-PAGE)
De modo a melhor caracterizar as proteiacutenas isoladas foi realizada uma electroforese em
condiccedilotildees desnaturantes (SDS-PAGE do inglecircs Sodium Dodecyl Sulfate Polyacrilamide Gel
Electrophoresis) O SDS (dodecilo sulfato de soacutedio) eacute um detergente anioacutenico utilizado neste
meacutetodo para desnaturar as proteiacutenas que se ligam ao SDS na proporccedilatildeo 14 g SDS g proteiacutena
(aproximadamente 1 moleacutecula de SDS por cada 2 resiacuteduos de aminoaacutecidos) (Stryer) O SDS induz
uma densidade de carga negativa significativa nas proteiacutenas a que se liga mascarando a sua carga
intriacutenseca pelo que as razotildees cargamassa satildeo tendencialmente idecircnticas Assim a SDS-PAGE
separa as proteiacutenas quando sujeitas a um campo eleacutectrico de acordo com o seu peso molecular
sendo a sua migraccedilatildeo directamente proporcional ao logaritmo deste uacuteltimo (Stryer)
O gel de resoluccedilatildeo com 10 de acrilamida foi obtido atraveacutes da adiccedilatildeo de 167 mL de soluccedilatildeo
stock de acrilamida11
075 mL de tampatildeo de resoluccedilatildeo12
251 mL de aacutegua destilada e 50 L de
SDS 10 Para promover a polimerizaccedilatildeo adicionou-se em seguida 38 L de PSA 10
11
soluccedilatildeo stock de acrilamida soluccedilatildeo de 30 (mv) de acrilamida e 08 (mv) de NNrsquo-metil-bis-acrilamida 12
tampatildeo de resoluccedilatildeo 15 M Tris-HCl pH 88
44
(persulfato de amoacutenia) 10 e 25 L de TEMED 99 (tetrametiletilenodiamina) Agitou-se
cuidadosamente de modo a natildeo formar bolhas e pipetou-se a soluccedilatildeo para a cassette de vidro
previamente montada no suporte De seguida adicionou-se uma peliacutecula de cerca de 2 mm de 2-
metil-2-butanol para verificar posteriormente a polimerizaccedilatildeo Apoacutes a polimerizaccedilatildeo que demora
cerca de 45 minutos verteu-se a agrave superfiacutecie e adicionou-se o gel de concentraccedilatildeo com 5 de
acrilamida constituiacutedo por 03 mL de soluccedilatildeo de stock acrilamida 045 mL de tampatildeo de
concentraccedilatildeo13
094 mL de aacutegua destilada 18 L de SDS 10 135 L de PSA 10 e 2L de
TEMED 99 Colocou-se o pente sobre o gel e deixou-se polimerizar durante cerca de 45
minutos
As amostras para aplicaccedilatildeo no gel de electroforese foram preparadas para uma concentraccedilatildeo de
proteiacutena de 1 mgmL num volume final de 100 L dos quais 25 L correspondem ao tampatildeo da
amostra14
As amostras foram aquecidas a 100ordmC durante 5 minutos Antes da aplicaccedilatildeo das
amostras transferiu-se a cassette para a tina de electroforese agrave qual se adicionou a soluccedilatildeo
tampatildeo de electroforese da tina superior15
e tampatildeo de electroforese da tina inferior16
Aplicaram-
se 10 L das soluccedilotildees de marcadores de peso molecular e 20 L das amostras proteicas
preparadas anteriormente A electroforese decorreu durante cerca de 3h30 min a uma voltagem
de 80 V De seguida transferiu-se o gel para uma tina corando as proteiacutenas durante 1h com
soluccedilatildeo corante17
Descorou-se o gel com soluccedilatildeo descorante18
tendo-se mudado esta soluccedilatildeo ateacute
as bandas se encontrarem bem definidas
Determinou-se a mobilidade relativa das bandas correspondentes aos marcadores de elevado peso
molecular e traccedilou-se a recta de calibraccedilatildeo ndash log(PM) vs log(Rf) onde Rf corresponde agrave
mobilidade relativa Por interpolaccedilatildeo determina-se o PM das proteiacutenas isoladas sabendo a sua
mobilidade relativa Por comparaccedilatildeo com os valores descritos na literatura pode concluir-se
acerca do grau de purificaccedilatildeo
34 Mediccedilatildeo da actividade enzimaacutetica de S1
341 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
13
tampatildeo de concentraccedilatildeo 05 M Tris-HCl pH 68 14
tampatildeo de amostra 320 mM Tris-HCl pH 68 04 M szlig-mercaptoetanol SDS 8 glicerol 15 (vv) e roxo de m-
cresol 0024 15
tampatildeo de electroforese da tina superior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS 16
tampatildeo de electroforese da tina inferior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS acetato de soacutedio
01 M 17
soluccedilatildeo corante ldquoCoomassie Blue R-250rdquo contendo metanol 45 (vv) e aacutecido aceacutetico glacial 10 (vv) 18
soluccedilatildeo descorante metanol 10 (vv) e aacutecido aceacutetico glacial 10 vv)
45
O meacutetodo de Fiske-Subarow modificado consiste na reacccedilatildeo do molibdato de amoacutenio com o
fosfato inorgacircnico em soluccedilatildeo aacutecida com formaccedilatildeo do aacutecido fosfomoliacutebdico Seguidamente daacute-
se a adiccedilatildeo de um agente redutor que provoca a reduccedilatildeo do composto a fosfomolibdato que forma
um complexo de cor azulada cuja absorvacircncia maacutexima ocorre para um comprimento de onda de
660 nm tornando possiacutevel a sua detecccedilatildeo por meacutetodos espectrofotomeacutetricos As actividades Ca2+
-
ATPaacutesica e K+EDTA-ATPaacutesica de S1 foram determinadas atraveacutes deste meacutetodo de acordo com
o descrito por Taussky e Shorr (1953) com mediccedilatildeo da quantidade de fosfato (Pi) libertado
atraveacutes da hidroacutelise de ATP
Para tal obteve-se primeiramente a recta de calibraccedilatildeo com as amostras presentes na Tabela 3II
utilizando uma soluccedilatildeo stock de 5 mM KH2PO4
Tabela 3II ndash Composiccedilatildeo das diferentes amostras utilizadas para determinar a recta de calibraccedilatildeo atraveacutes do meacutetodo
de Fiske-Subbarow modificado
KH2PO4 (L)
([KH2PO4]stock= 5mM)
PO43-
(nmole) H2O (uL)
0 0 1780
5 25 1775
10 50 1770
20 100 1760
40 200 1740
100 500 1680
A cada tubo de Eppendorf adicionaram-se seguidamente 200 L de soluccedilatildeo J19
e 20 L de agente
redutor20
homogeneizando no voacutertex apoacutes cada adiccedilatildeo Incubou-se durante 10 minutos e mediu-
se a Abs660 nm num espectrofotoacutemetro com intervalos de tempo iguais entre cada amostra Traccedilou-
se a recta de calibraccedilatildeo (Abs660 nmvs nmol PO43-
) a partir dos valores obtidos
Para determinar a actividade Ca2+
-ATPaacutesica de S1 prepararam-se tubos de Eppendorf com 400
L de meio de reacccedilatildeo21
25 L CaCl2 ([CaCl2]stock= 100 mM) 25 L ATP ([ATP]stock= 100 mM)
com [S1]ensaios=01 mgmL Perfez-se se necessaacuterio com H2O MilliQ ateacute perfazer um volume de 1
mL Para determinar a actividade K+EDTA-ATPaacutesica utilizou-se uma concentraccedilatildeo idecircntica de
19
Soluccedilatildeo J 5 N H2SO4 25 molibdato de amoacutenio 20
Agente redutor 5 mg aacutecido 1-amino-2-naftol-4-sulfoacutenico 30 mg bissulfito de soacutedio 30 mg sulfito de soacutedio em 26
mL H2O 21
meio de reacccedilatildeo 125 mM Tris-HCl pH 80 0575 M KCl
46
proteiacutena nos ensaios (01 mgmL) e o mesmo meio de reacccedilatildeo Aos tubos de Eppendorf
adicionou-se ainda 123 L de KCl ([KCl]stock = 3M) 25 L de ATP 10 L de EDTA
([EDTA]stock= 100 mM )e o volume de H2O MilliQ necessaacuterio para perfazer 1 mL Em ambas as
situaccedilotildees o uacuteltimo reagente adicionado foi o ATP uma vez que se trata do substrato da reacccedilatildeo
enzimaacutetica
A cineacutetica decorreu a 25ordmC durante 5 minutos para a actividade Ca2+
-ATPaacutesica e durante 3
minutos para a actividade K+EDTA-ATPaacutesica Apoacutes o periacuteodo de incubaccedilatildeo transferiu-se 150
L de amostra para tubos de Eppendorf contendo igual volume de TCA 10 Colocou-se no gelo
e homogeneizou-se no voacutertex Seguidamente centrifugou-se a 1500 g durante 10 minutos e
recolheu-se 200 L de sobrenadante para novos tubos de Eppendorf Adicionou-se a estes 158
mL de H2O MilliQ e 200 L de soluccedilatildeo J Homogeneizou-se no voacutertex e adicionou-se 20 L de
reagente redutor repetindo-se a homogeneizaccedilatildeo Incubou-se durante 10 minutos e mediu-se a
Abs660 nm com o mesmo intervalo de tempo de leitura para cada amostra Por intrapolaccedilatildeo a partir
da recta de calibraccedilatildeo obtida anteriormente determinou-se a quantidade de Pi libertado em cada
ensaio A partir destes resultados determinaram-se as actividades Ca2+
-ATPaacutesica e K+EDTA-
ATPaacutesica do S1 em nmol PO43-
minmg proteiacutena e em s-1
342 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
Da actividade Mg2+
-ATPaacutesica de S1 resulta a formaccedilatildeo dos produtos da reacccedilatildeo de
hidroacutelise do ATP - ADP e Pi Na presenccedila de ADP fosfoenolpiruvato (PEP) e da enzima piruvato
cinase (PK) os produtos resultantes satildeo ATP e piruvato Por acccedilatildeo da enzima lactato
desidrogenase (LDH) este uacuteltimo eacute convertido em lactato ocorrendo simultaneamente a oxidaccedilatildeo
do NADH a NAD+ monitorizada atraveacutes da Abs340 nm Quando o S1 de miosina se encontra no
meio reaccional o ATP eacute hidrolisado formando-se novamente ADP e Pi o que permite a
formaccedilatildeo de um ciclo (fig 21) Se as enzimas se encontrarem em excesso a velocidade de
conversatildeo do NADH eacute limitada pela produccedilatildeo de ADP e Pi Como o ATP eacute regenerado em cada
ciclo a sua concentraccedilatildeo inicial eacute sempre mantida (Bagshaw 1982)
Figura 33ndash Representaccedilatildeo esquemaacutetica do meacutetodo de enzimas acopladas
47
O protocolo utilizado e seguidamente descrito foi adaptado de Cuenda et al (1990 ndash Ver Refordf) A
uma cuvette de 1 mL adicionou-se S1 para uma concentraccedilatildeo final de 005 mgmL 800 L de
meio de reacccedilatildeo22
10 L de PEP ([PEP]stock = 42 mM) 10 L de PK+LDH 11 ([PK+LDH]stock =
1 mgmL) e 15 L de NADH ([NADH]stock=25 mM) Iniciou-se a monitorizaccedilatildeo da reacccedilatildeo
atraveacutes da leitura de Abs340 nm apoacutes a adiccedilatildeo de 20 L de ATP ([ATP]stock=100 mM) A
actividade Mg2+
-ATPaacutesica eacute obtida atraveacutes do declive correspondente agrave oxidaccedilatildeo do NADH Para
a mediccedilatildeo da Mg2+
-ATPase de S1 estimulada por actina adicionou-se F-actina para uma
concentraccedilatildeo final de 2 microM sendo a actividade obtida tambeacutem atraveacutes do declive correspondente
agrave oxidaccedilatildeo do NADH apoacutes a adiccedilatildeo da proteiacutena
35 Interaccedilatildeo entre vanaacutedio e subfragmento-1 de miosina
351 Preparaccedilatildeo das soluccedilotildees de vanaacutedio
As soluccedilotildees de vanadato foram preparadas em aacutegua MilliQ a partir de metavanadato de amoacutenio
(NH4VO3) numa concentraccedilatildeo stock de 50 mM com agitaccedilatildeo magneacutetica e aquecimento (~30 ordmC)
Esta soluccedilatildeo com um valor compreendido entre 6 e 7 designa-se por ldquometavanadatordquo e eacute
constituiacuteda pelas esspeacutecies V1 V2 V4 e V5 (Ref) Acidificando a soluccedilatildeo de metavanadato por
adiccedilatildeo de HCl ateacute um valor de pH proacuteximo de 4 observa-se uma mudanccedila de cor de amarelo
claro para laranja indicativa da formaccedilatildeo da espeacutecie decameacuterica de vanadato De seguida e
imediatamente antes da realizaccedilatildeo dos ensaios descritos o valor do pH da soluccedilatildeo de
decavanadato foi aumentado para cerca de 75 por adiccedilatildeo de NaOH A soluccedilatildeo (2) designada
por ldquometavanadatordquo natildeo foi posteriormente alterada ateacute agrave sua adiccedilatildeo para a realizaccedilatildeo dos ensaios
As soluccedilotildees foram armazenadas a 4ordmC e mantidas em gelo aquando da sua utilizaccedilatildeo
A soluccedilatildeo de V(IV) foi preparada em condiccedilotildees anaeroacutebias por diluiccedilatildeo de sulfato de vanadil
(VOSO4) para uma concentraccedilatildeo final de 5 mM em H2O MilliQ saturada com N2 gasoso
352Anaacutelise por espectroscopia de UV-Vis
A concentraccedilatildeo da espeacutecie decameacuterica em soluccedilatildeo depende do periacuteodo de tempo apoacutes a diluiccedilatildeo
no meio de reacccedilatildeo apropriado (Ref) A velocidade de desoligomerizaccedilatildeo da espeacutecie decameacuterica
pode ser analisada por espectroscopia de absorccedilatildeo na regiatildeo UVVis uma vez que esta espeacutecie
apresenta duas bandas caracteriacutesticas na regiatildeo do visiacutevel (a 358 e 400 nm) que natildeo se verifica
em quaisquer outras espeacutecies oligomeacutericas As bandas supramencionada sapresentam-se bem
22
Meio de reacccedilatildeo 3125 mM Tris-HCl pH 71 375 mM MgCl2
48
resolvidas atraveacutes da obtenccedilatildeo da derivada de 2ordf ordem do espectro apresentando a forma de 2
vales com miacutenimos centrados nos comprimentos de onda supracitados com dependecircncia linear
relativamente agrave concentraccedilatildeo de decavanadato total (Ramos et al 2006)
Assim a estabilidade da espeacutecie decameacuterica apoacutes diluiccedilatildeo da soluccedilatildeo de decavanadato no meio de
reacccedilatildeo dos ensaios (tampatildeo D) para uma concentraccedilatildeo final de 1 mM em vanadato total foi
caracterizada atraveacutes da leitura da Abs400 em funccedilatildeo do tempo tendo-se determinado qual a
ordem de reacccedilatildeo e o tempo de meia-vida da mesma Com base em estudos anteriores acerca da
estabilidade do V10 na presenccedila de diferentes proteiacutenas (Ramos et al 2006) determinou-se
igualmente o tempo de meia vida apoacutes a adiccedilatildeo de 20 microM de S1 de miosina ao meio reaccional
353 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
Os espectros de RMN do isoacutetopo 51
V foram realizados num espectroacutemetro Bruker AM-400 MHz
equipado com uma sonda multinuclear inversa de 5 mm Em tubos de RMN de 5 mm de diacircmetro
preparou-se soluccedilotildees de 500 L de deca- ou metavanadato correspondentes a um intervalo de
concentraccedilotildees de 1 a 10 mM suplementadas com 10 D2O Os espectros foram adquiridos agrave
temperatura ambiente com uma largura espectral de 23 923 Hz um tempo de aquisiccedilatildeo de 0086
s tempo de relaxaccedilatildeo de 001 s e uma sequecircncia de pulsos a 90ordm Utilizou-se a transformada de
Fourier com alargamento das bandas de 20Hz para o processamento dos espectros Os desvios
quiacutemicos das ressonacircncias de 51
V foram obtidos em relaccedilatildeo agrave espeacutecie VOCl3cuja frequecircncia de
ressonacircncia eacute considerada o padratildeo sendo todas as outras referenciadas a partir desta em ppm
As aacutereas dos sinais do RMN correspondentes agraves espeacutecies oligomeacutericas presentes foram obtidas
por integraccedilatildeo dos sinais As concentraccedilotildees relativas das espeacutecies foram calculadas com recurso agrave
equaccedilatildeo
onde A[Vx] corresponde agrave aacuterea referente agrave ressonacircncia da espeacutecie Vx com nordm de agregaccedilatildeo x
A[V]total agrave soma total das aacutereas medidas e [V] a concentraccedilatildeo total de vanaacutedio no ensaio
A teacutecnica de 51
V-RMN foi anteriormente aplicada em estudos de ligaccedilatildeo com S1 de miosina
(Ref) tendo sido demostrado que entre as espeacutecies V1 V2 V4 e V5 a espeacutecie tetrameacuterica possui a
maior afinidade para a miosina (Cremo et al) O desaparecimento ou alargamento dos sinais pode
implicar a ligaccedilatildeo de um dos oxovanadatos laacutebeis agrave miosina sem indicaccedilatildeo de qual uma vez que
49
se processa um equiliacutebrio entre eles (Crans 1990) Assim e tendo em conta as indicaccedilotildees de que
a espeacutecie V10 eacute distinta das restantes no que diz respeito agrave magnitude da interacccedilatildeo com a miosina
e inibiccedilatildeo da actividade ATPaacutesica estimulada por actina (Aureliano 2000 Tiago et al 2002)
torna-se fundamental investigar a interacccedilatildeo das diferentes espeacutecies de vanadato com o S1 de
miosina atraveacutes da teacutecnica de 51
V-RMN
Para tal obtiveram-se espectros de 51
V-RMN resultantes da titulaccedilatildeo de uma soluccedilatildeo 5 mM de
decavanadato total com concentraccedilotildees crescentes de S1 de miosina (0 5 10 15 e 20 microM) em
tampatildeo D Realizou-se ainda ensaios na presenccedila de 2 mM de decavanadato e 2 mM de ATP na
presenccedila e na ausecircncia de S1 de miosina de modo a verificar o efeito que estes ligandos naturais
da enzima teria na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o subfragmento-1
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
Os grupos sulfidrilo (-SH) das cisteiacutenas satildeo resiacuteduos de aminoaacutecidos extremamente sensiacuteveis agrave
oxidaccedilatildeo resultando na sua oxidaccedilatildeo (Dalle-Donne et al 1999) Logo a quantificaccedilatildeo destes
resiacuteduos eacute bastante importante nos estudos de interacccedilatildeo com vanaacutedio jaacute que os resiacuteduos de Cys
com ecircnfase para o SH1 e o SH2 satildeo bastante importantes na monitorizaccedilatildeo das modificaccedilotildees
funcionais da regiatildeo da cabeccedila
O procedimento seguidamente apresentado foi originalmente descrito por Gutieacuterrez-Merino
(1987)) Consiste na reacccedilatildeo entre os grupos ndashSH natildeo oxidados das proteiacutenas e o reagente de
Ellman (DTNB -55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico)) - formando-se o aniatildeo CNT (aniatildeo 3-
carboxilato-4-nitrotiofenolato) numa estequiometria de 11 Este apresenta um maacuteximo de
absorvecircncia a 412 nm (Robyt 1990) Assim apoacutes a incubaccedilatildeo da proteiacutena na presenccedila do metal
a quantidade de cisteiacutenas reduzidas por proteiacutena determina-se atraveacutes dos valores de Abs412
Deste modo colocou-se num tubo de Eppendorf o volume apropriado de S1 dialisado para que a
concentraccedilatildeo no ensaio fosse de 1 mgmL Adicionou-se o volume apropriado de decavanadato
para que a sua concentraccedilatildeo fosse de 0 10 50 100 250 500 ou 1000 M Perfez-se com soluccedilatildeo
D ateacute um volume final de 1 mL e incubou-se a 25ordmC por 20 minutos Apoacutes este periacuteodo colocou-
se 780 L da mistura reaccional numa cuvette de quartzo e adicionou-se 220 L de soluccedilatildeo K23
23
Tampatildeo K Tampatildeo fosfatohidrogenofosfato de soacutedio 025 M pH 72
50
Adicionou-se 15 L de DTNB (em soluccedilatildeo K [DTNB]stock=100 mM) ao meio reaccional e
registou-se o valor de absorvecircncia medido equivalente agraves cisteiacutenas ldquoraacutepidasrdquo (SH1 e SH2
expostos ao solvente) Adicionou-se 001 g de SDS que provoca a desnaturaccedilatildeo da proteiacutena e
consequentemente a exposiccedilatildeo de todos os resiacuteduos de cisteiacutenas que se encontravam ateacute aiacute
inacessiacuteveis no interior da proteiacutena (cisteiacutenas totais) Estes resiacuteduos podem assim reagir com o
DTNB
Estimou-se o nuacutemero de resiacuteduos de cisteiacutenas em todas as amostras com recurso ao coeficiente de
extinccedilatildeo molar () do DTNB (ver ponto 3 dos Anexos) que foi determinado atraveacutes da titulaccedilatildeo
de L-Cisteiacutena com intervalo de concentraccedilotildees compreendidos entre 0 e 50 microM na presenccedila de
25microM de DTNB em tampatildeo D Os valores de absorvecircncia foram obtidos para um comprimento de
onda de 412 nm
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
3621 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
A teacutecnica mais eficiente de anaacutelise da interacccedilatildeo entre o iatildeo vanadilo e proteiacutenas como a
calmodulina e a transferrina entre outras permite a obtenccedilatildeo raacutepida de informaccedilatildeo no que toca agrave
conformaccedilatildeo e amplitude da referida interacccedilatildeo
Os espectros de Ressonacircncia Paramagneacutetica Electroacutenica (RPE) da espeacutecie paramagneacutetica V(IV)
foram realizados a 70K num espectroacutemetro Bruker EMX-300 X-band Todas as soluccedilotildees
utilizadas e amostras foram preparadas em anaerobiose numa cacircmara de luvas de modo a evitar a
reduccedilatildeo de V(IV) a V(V) Em tubos capilares com 02 mL de amostra manteve-se a concentraccedilatildeo
de S1 a 20 M e variou-se a concentraccedilatildeo de vanadilo (0 25 5 10 15 25 50 75 100 150
200 500 e 1000 M [VOSO4]stock=5 mM) em tampatildeo D Apoacutes obtenccedilatildeo dos espectros estes
foram tratados atraveacutes do software SimFonia e WinEPR tendo-se obtido um graacutefico com a
variaccedilatildeo da intensidade do sinal de RPE em funccedilatildeo da concentraccedilatildeo de sulfato de vanadilo
presente Realizaram-se ainda simulaccedilotildees de espectros obtidos com 20 microM de S1 e 1000 microM de
V(IV) com vista agrave obtenccedilatildeo dos ligandos provaacuteveis do vanadilo atraveacutes da anaacutelise da constante
de acoplamento hiperfina (A) e do factor g (g)
4 Conclusotildees gerais
Apesar de jaacute ter sido demonstrado que o vanaacutedio inibe a Na+K
+-ATPase da miosina a funccedilatildeo
deste elemento no tecido muacutesculo esqueleacutetico continua largamente incompreendida A enorme
complexidade quiacutemica deste metal especialmente no que concerne agrave oligomerizaccedilatildeo e reacccedilotildees
51
redox constitui um obstaacuteculo agrave compreensatildeo do comportamento do vanaacutedio nos organismos
vivos Assim a caracterizaccedilatildeo da quiacutemica deste elemento eacute de grande utilidade antes de iniciar o
estudo da sua interacccedilatildeo com o presente sistema
A anaacutelise por espectroscopias de 51
V-RMN e de UV-Vis de soluccedilotildees de vanadato diluiacutedas nos
meios de reacccedilatildeo utilizados no presente relatoacuterio (10 mM HEPES pH 70 25 mM KCl) mostrou
claramente que a espeacutecie decameacuterica (V10) possui um tempo de vida suficientemente longo para a
determinaccedilatildeo do seu efeito isolado ao contraacuterio do que se verifica com as espeacutecies mono- di-
tetra- e pentameacutericas (V1 V2 V4 e V5) Por outro lado a anaacutelise verificou ainda que apoacutes diluiccedilatildeo
das soluccedilotildees de decavanadato no meio de reacccedilatildeo coexistem as espeacutecies V10 e V1 pelo que os
efeitos putativos desta uacuteltima deveratildeo ser subtraiacutedos aquando da monitorizaccedilatildeo dos efeitos da
espeacutecie decameacuterica
O vanaacutedio (V) (vanadato) inibe as enzimas que catalizam reacccedilotildees de transferecircncia de grupos
fosfato como a ATPase da miosina [Goodno 1979] devido agrave sua analogia estrutural com o iatildeo
fosfato Na sua utilizaccedilatildeo como inibidor enzimaacutetico natildeo eacute tida em conta a contribuiccedilatildeo individual
das espeacutecies oligomeacutericas considerando-se normalmente a espeacutecie V1 como inibidora efectiva
[Goodno e Taylor 1982] No presente relatoacuterio verificou-se por espectroscopia de 51
V-RMN que
outros oligoacutemeros de vanadato interagem com o S1 de miosina tendo as espeacutecies V10 e V4 os
maiores niacuteveis de afinidade A sua interacccedilatildeo envolve forccedilas electrostaacuteticas e eacute afectada
negativamente por ligandos naturais de enzima como eacute o caso do ATP
A exposiccedilatildeo do S1 de miosina (005 mgmL) a decavanadato conduz agrave inibiccedilatildeo do capacidade da
proteiacutena de estimular a actividade Mg2+
-ATPaacutesica com um valor de IC50 de 266 plusmn 07 M
decavanadato sendo o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM
decavanadato (80) Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM decavanadato sendo
o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM decavanadato (60)
exercendo um efeito de impedimento agrave ligaccedilatildeo entre o vanaacutedio e o S1
A uma concentraccedilatildeo de 27 microM decavanadato verifica-se a oxidaccedilatildeo de um dos resiacuteduos de Cys
de S1 de miosina seguida pela oxidaccedilatildeo gradual de outro resiacuteduo de Cys ateacute agrave concentraccedilatildeo de
1000 microM Ambos os resiacuteduos oxidados natildeo se encontravam expostos ao solvente utilizado A
oxidaccedilatildeo dos resiacuteduos permite sugerir que esta modificaccedilatildeo oxidativa sofrida pelo S1 contribui
para a diminuiccedilatildeo da sua actividade de ATPase
52
Por espectroscopia de RPE revelaram-se os possiacuteveis ligandos equatoriais do vanaacutedio aquando da
sua interacccedilatildeo com o S1 de miosina como sendo duas moleacuteculas de aacutegua um ligando contendo o
grupo R-OH e outro contendo o grupo R=NR
Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados de
oxidaccedilatildeo (IV) e (V) no S1 de miosina para concentraccedilotildees facilmente alcanccedilaacuteveis no ambiente
terrestre
53
5 Estudos futuros
No sentido do melhor esclarecimento das alteraccedilotildees moleculares e estruturais que ocorrem no
subfragmento 1 quando exposta ao vanaacutedio que conduzem a uma diminuiccedilatildeo da sua funccedilatildeo
sugere-se
- Quantificaccedilatildeo das cisteiacutenas reduzidas de S1 na presenccedila de metavanadato
- Determinaccedilatildeo das actividades Ca2+
- e K+-ATPaacutesica do S1 de miosina na presenccedila de
decavanadato
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina estimulada por actina
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina na presenccedila de
metavanadato
- Realizaccedilatildeo de RMN 2D com variaccedilatildeo das concentraccedilotildees das soluccedilotildees de decavanadato e
metavanadato na presenccedila de S1
-Mutageacutenese dos resiacuteduos de Cys doS1 de miosina de modo a verificar quais os resiacuteduos que
sofrem oxidaccedilatildeo na presenccedila de decavanadato
- Determinaccedilatildeo por calorimetria (DSC Differential Scanning Calorimetry) da variaccedilatildeo da
entalpia (∆H) e da temperatura de desnaturaccedilatildeo do S1 de miosina na presenccedila de vanaacutedio uma
vez que estas propriedades caracteriacutesticas de cada proteiacutena satildeo alteradas apoacutes ocorrecircncia de
modificaccedilotildees estruturais
- Anaacutelise por SDS-PAGE de S1 de miosina apoacutes tratamento com vanaacutedio de modo a determinar
quais os polipeacuteptidos eventualmente formados apoacutes clivagem da proteiacutena
54
6 Referecircncias bibliograacuteficas
- Sellers J Myosins 2nd Edition Oxford University Press 1999
- L Stryer inBiochemistry (Ed WH Freeman 2005 pp
Refordfas Materiais e Meacutetodos
[A] S S Margossian SLowey Methods Enzymol1982 85 55-71
[B] JD Pardue JA Spudich Methods Enzymol 1982 85 164-181
[C] M Bradford Anal Biochem1976 72 248-254
[D] H H Taussky E Shorr J Biol Chem 1953 202 675-685
[E] C R Bagshaw in Outline Studies in Biology ndash Muscle Contraction Chapman and Hall Ed
1982
[F] A Cuenda et al Eur J Biochem1990 194 663-670
55
Anexos
1 Determinaccedilatildeo da recta de calibraccedilatildeo meacutetodo de Bradford
Figura1ndashRepresentaccedilatildeo graacutefica da recta de calibraccedilatildeo (Abs595 nm vs [BSA]) utilizada para o meacutetodo de Bradford
2 Determinaccedilatildeo da recta calibraccedilatildeo para a electroforese em condiccedilotildees desnaturantes
Figura 2 ndash Representaccedilatildeo graacutefica da recta de calibraccedilatildeo (log(Peso Molecular) vs Rf dos marcadores de peso
molecular) onde Rf indica determinada atraveacutes do padratildeo de peso molecular
(poccedilo (a))
3Caacutelculo do coeficiente de extinccedilatildeo molar () para a quantificaccedilatildeo de cisteiacutenas reduzidas
atraveacutes do meacutetodo de Ellman
Traccedilou-se a recta de Abs412 nm meacutedia vs [L-Cisteiacutena] de modo a determinar o valor do coeficiente
de extinccedilatildeo molar definido como o declive daquela recta Assim =8400 M-1
cm-1
como
podemos verificar na Fig 3
A595 nm = 00546[BSA] + 00209 Rsup2 = 099051
Ab
s 59
5 n
m
[BSA] (mgmL)
log PM = -1177 Rf + 2247 Rsup2 = 098957
log
PM
Mobilidade relativa (Rf)
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero

15
(ppb) Estaacute associado a cerca de 70 minerais diferentes na natureza (dos quais se destacam o
uracircnio cobre chumbo zinco fosfato e titacircnio ferroso) especialmente na forma de vanadato ou
pentoacutexidos de vanaacutedio O vanaacutedio eacute um dos principais constituintes da mateacuteria orgacircnica foacutessil
com especial ecircnfase para o carvatildeo oacuteleo de crude e foacutesseis carbonaacuteceos [UV] bem como em
alimentos consumidos pelo Homem (salsa pimenta preta cogumelos marisco vegetais cereais
gorduras e oacuteleos) [W] Estes constituem a maior fonte de exposiccedilatildeo ao vanaacutedio da maioria da
populaccedilatildeo humana [X]
Normalmente a entrada do vanaacutedio no organismo ocorre por inalaccedilatildeo ou em menores
quantidades por absorccedilatildeo pelo tracto gastrointestinal ou pela pele O seu armazenamento centra-
se no fiacutegado rins ceacuterebro coraccedilatildeo muacutesculos e ossos No entanto a retenccedilatildeo de vanaacutedio no
organismo humano eacute reduzida ndash em condiccedilotildees normais cerca de 100 M (em muitos tecidos a
quantidade eacute inferior a 10 ngg de tecido) [Y]
Actualmente natildeo existem provas de deficiecircncia nutricional de vanaacutedio em humanos ou de
enzimas que contenham vanaacutedio em animais superiores No entanto vaacuterios estudos relacionam a
ocorrecircncia de deficiecircncia em galinhas e ratos devido a uma alimentaccedilatildeo pobre em vanaacutedio Os
sintomas relatados incluem retardamento do crescimento malformaccedilotildees oacutesseas perturbaccedilotildees no
metabolismo lipiacutedico e estagnaccedilatildeo reprodutiva entre outros [Z AA AB] Os efeitos fisioloacutegicos
do vanaacutedio podem depender do tipo de composto ou de oligoacutemero uma vez que consoante as
suas estabilidade labilidade e nuacutemero de coordenaccedilatildeo as respostas seratildeo diferenciadas ao niacutevel
dos organismos vivos
Por outro lado a toxicidade do vanaacutedio encontra-se bem caracterizada existindo sintomas
descritos em vaacuterios mamiacuteferos (inclusive o Homem) que incluem diarreias catildeibras irritaccedilatildeo da
pele e mucosas palpitaccedilotildees cardiacuteacas anemia pneumonia bronquite perturbaccedilotildees no Sistema
Nervoso Central (SNC) entre outros (Ref) A toxicidade varia geralmente com a natureza do
composto com aumento proporcional agrave sua valecircncia Verificou-se que a forma de entrada do
vanaacutedio tem tambeacutem efeito na sua toxicidade ndash os maiores niacuteveis verificaram-se por injecccedilatildeo
intravenosa da amostra sendo progressivamente inferiores por inalaccedilatildeo e finalmente por
ingestatildeo [X AA] A ampla utilizaccedilatildeo dos compostos de vanaacutedio na induacutestria quiacutemica bem como
as elevadas quantidades de vanaacutedio libertadas atraveacutes da combustatildeo dos oacuteleos de crude
contribuem consideravelmente para o aumento da biodisponibilidade do vanaacutedio Como tal o
interesse neste elemento e nos problemas ambientais por ele causados tem vindo a aumentar
significativamente [V]
121 Propriedades quiacutemicas
16
A quiacutemica do vanaacutedio eacute extraordinariamente complexa pelo que originou vaacuterias interpretaccedilotildees
incorrectas desde a sua descoberta No entanto as suas propriedades dependem em uacuteltima
anaacutelise da configuraccedilatildeo electroacutenica do elemento Assim a variabilidade encontrada no
comportamento quiacutemico do vanaacutedio pode dever-se agrave sua configuraccedilatildeo ndash [Ar] 3d2 4s
2 ndash que
permite a formaccedilatildeo de vaacuterios estados de oxidaccedilatildeo (de +2 a +5) Em soluccedilatildeo aquosa os estados de
oxidaccedilatildeo estaacuteveis vatildeo de V(V) a V(II) Destes apenas os +3 a +5 satildeo biologicamente importantes
jaacute que o V(II) apresenta demasiada capacidade redutora para existir nos organismos vivos
conhecidos [AC]
No fluido extracelular os compostos de vanaacutedio encontram-se predominantemente na forma de
monovanadatos (V(V)) livres ou ligados agrave transferina do soro ou agrave hemoglobina (Refordf) Os iotildees de
vanadato livres satildeo transportados para o meio intracelular atraveacutes de canais ioacutenicos inespeciacuteficos
Jaacute no ambiente intracelular os compostos sofreratildeo uma bio-reduccedilatildeo atraveacutes da acccedilatildeo da
glutationa ou outros compostos com grupos ndashOH como pentoses hexoses ou etanolaminas [V] A
reoxidaccedilatildeo de vanadilo (V(IV)) intracelular a vanadato (V(V)) provocaria a acumulaccedilatildeo toacutexica
destes uacuteltimos pelo que a complexaccedilatildeo da espeacutecie reduzida o impede O transporte de vanaacutedio
para o interior da ceacutelula encontra-se na Fig 110
Figura 110 ndash Transporte de vanaacutedio para a ceacuteluka e seu subsequente metabolismo (P=Proteiacutena) (Adaptado de
Zaporowska e Scibor 1998)
1211 Vanaacutedio(IV)
A espeacutecie de vanaacutedio (IV) mais bem conhecida eacute o iatildeo vanadilo (VO2+
) espeacutecie estaacutevel a pH
aacutecido sendo a maioritaacuteria a pH 3[AD] Apresenta uma cor azulada e forma complexos penta e
hexacoordenados Forma-se rapidamente a partir de V(V) na presenccedila de um agente redutor em
soluccedilatildeo aquosa No ambiente intracelular a presenccedila de glutationa reduzida aacutecido ascoacuterbico
NAD(P)H entre outros conduzem a que o principal estado de oxidaccedilatildeo nos sistemas bioloacutegicos
seja provavelmente o +4 encontrando-se o V(IV) fortemente complexado com proteiacutenas ou
aminoaacutecidos livres citrato ATP e pirofosfato (Refordfs) Em alternativa o V(IV) pode existir
17
livremente em ambientes fortemente aacutecidos como os lisossomas jaacute que a pH superior a 3 as
soluccedilotildees de vanadilo tendem a ser oxidadas pelo ar formando-se vanadato Esta grande
capacidade redutora justifica parcialmente a utilizaccedilatildeo pouco extensiva do iatildeo VO2+
como
inibidor [W]
1212 Vanaacutedio (V)
O vanaacutedio (V) eacute a forma mais estaacutevel num intervalo de pH amplo [W] O iatildeo anioacutenico VO3-tende
a constituir complexos polinucleares pelo que as soluccedilotildees aquosas de vanadato satildeo extremamente
complexas com ocorrecircncia simultacircnea de vaacuterios equiliacutebrios de protonaccedilatildeo (pKa(vanadato)= 35
78 e 125) e oligomerizaccedilatildeo As espeacutecies presentes (H3VO4 H2VO4- HVO4
2- e VO4
3-) dependem
do pH forccedila ioacutenica e temperatura da soluccedilatildeo e da concentraccedilatildeo total do elemento
[AEAFAGAHAI]
O VO43-
- monoacutemero ortovanadato V1 ndash estaacute associado a uma grande parte da importacircncia
bioloacutegica do vanaacutedio uma vez que se trata de um anaacutelogo electroacutenico e estrutural do ortofosfato
(PO43-
) A coordenaccedilatildeo do ortovanadato pode ser tetraeacutedrica ou bipiramidal trigonal (Refordfs)
A oligomerizaccedilatildeo do vanadato monomeacuterico daacute origem ao diacutemero (V2O72-
- V2) que apresenta
tambeacutem equiliacutebrios de protonaccedilatildeo Presume-se a existecircncia de analogia do vanadato dimeacuterico com
o pirofosfato embora natildeo haja evidecircncia estrutural (Refordf) A dimerizaccedilatildeo do V2 origina o
vanadato tetrameacuterico ou metavanadato (V4O124-
- V4) que constitui a espeacutecie maioritaacuteria em
soluccedilotildees aquosas concentradas (Refordf) As estruturas do metavanadato e a sua espeacutecie protonada
satildeo ciacuteclicas [AJ] Outra espeacutecie oligomeacuterica importante eacute o vanadato pentameacuterico (V5O153-
) cuja
estrutura presumida eacute ciacuteclica [AI]
O oligoacutemero com maior importacircncia bioquiacutemica eacute o vanadato decameacuterico (V10O286-
-V10)(Refordf) a
espeacutecie predominante em soluccedilotildees de pH 2-6 quando a concentraccedilatildeo total de vanadato eacute superior
a 100 M (Ref) A estrutura do V10 natildeo difere em soluccedilatildeo ou no estado soacutelidoOs 10 aacutetomos de
vanaacutedio da espeacutecie decameacuterica estatildeo divididos em trecircs grupos identificados na figura como VA-C
consoante o seu ambiente quiacutemico circundante (Refordf) (Fig 111)
A expressatildeo ldquodecavanadatordquo refere-se a uma soluccedilatildeo aquosa na qual a espeacutecie decameacuterica se
encontra em equiliacutebrio quiacutemico com as espeacutecies V1 V2 V4 e V5 sendo que a oligomerizaccedilatildeo
progressiva do vanaacutedio eacute proporcional agrave sua concentraccedilatildeo total (Ref) Nesta soluccedilatildeo a
concentraccedilatildeo de V10 corresponde a aproximadamente 10 da concentraccedilatildeo total de vanaacutedio [AK]
18
Figura 111 ndash Estruturas esquemaacuteticas das espeacutecies de vanadato (espeacutecie monomeacuterica (V1) dimeacuterica (V2)
tetrameacuterica (V4) pentameacuterica (V5) e decameacuterica (V10) (Adaptado de Aureliano e Crans 2008)
122 Metabolismo e efeito na actividade enzimaacutetica
Apesar de natildeo terem sido ainda encontradas as provas categoacutericas das suas funccedilotildees nos seres
vivos sabe-se que o iatildeo vanadato eacute capaz de inibir uma grande quantidade de enzimas in vitro
com especial ecircnfase na actividade ATPaacutesica da miosina [AL Teresa] Como referido
anteriormente o vanadato eacute capaz de actuar em alternativa ao fosfato nas reacccedilotildees bioquiacutemicas
incluindo a sua preferecircncia de ligaccedilatildeo agraves enzimas [W]
Eacute conhecido o papel do vanaacutedio no metabolismo do caacutelcio promovendo a mineralizaccedilatildeo dos
dentes e ossos na biossiacutentese de liacutepidos (trigliceacuteridos e colesterol) com posterior inibiccedilatildeo da
acumulaccedilatildeo destes nos vasos sanguiacuteneos e no SNC na reproduccedilatildeo e desenvolvimento por
mimetizaccedilatildeo e potenciaccedilatildeo de factores de crescimento na estimulaccedilatildeo do consumo de glucose e
da siacutentese de glicogeacutenio com actuaccedilatildeo equivalente aos efeitos da insulina ao niacutevel celular na
transduccedilatildeo de sinais celulares com regulaccedilatildeo da actividade de segundos mensageiros no
transporte de iotildees e aminoaacutecidos na transcriccedilatildeo geneacutetica e siacutentese de proteiacutenas e DNA entre
outros [W Y AB]
Assim o vanaacutedio cnseguiraacute provocar inuacutemeras mudanccedilas bioquiacutemicas nas ceacutelulas Alguns
exemplos da influecircncia bioquiacutemica do vanaacutedio incluem os seus efeitos nos processos de
fosforilaccedilatildeo e desfosforilaccedilatildeo de proteiacutenas e na participaccedilatildeo na formaccedilatildeo de espeacutecies reactivas de
oxigeacutenio devido ao seu efeito proacute-oxidante sobre agentes redutores a niacutevel celular [AM]
Quanto ao seu efeito na actividade enzimaacutetica pode dividir-se em dois tipos principais ndash inibitoacuterio
e estimulante - dependendo da enzima ou do tipo de reacccedilatildeo por esta catalisada No primeiro
grupo incluem-se enzimas como as ATPases (de miosina de canais ioacutenicos e fosfoenzimas
19
transportadoras) e as fosfatases (da tirosina aacutecida e alcalina) a fosfofrutocinase adenilato ciclase
e catalase entre outras [AA] O segundo grupo inclui a maioria das cinases a glucose-6-fosfato
desidrogenase a fosfoglicerato mutase e a fosfoglicerato fosfatase (Refordf)
O efeito do vanaacutedio (V) na actividade enzimaacutetica estaacute relacionado com o seu nuacutemero de
coordenaccedilatildeo podendo actuar como estado de transiccedilatildeo ou por mimetizaccedilatildeo de cofactores
substratos (especialmente o fosfato) e interacccedilotildees proteiacutena-proteiacutena (como no caso das
polimerases e transcriptases) Pode ainda estar envolvido na formaccedilatildeo de ligaccedilotildees de hidrogeacutenio
interacccedilotildees electrostaacuteticas esteacutereas e de oxidaccedilatildeo-reduccedilatildeo Uma evidecircncia desta uacuteltima eacute a sua
capacidade de alternacircncia entre os estados de oxidaccedilatildeo +5 a +3 o que torna faacutecil a sua interacccedilatildeo
redox com enzimas contendo resiacuteduos de cisteiacutenas (Cys) ou qualquer outro composto orgacircnico
com actividade redox [AN]
13 Vanaacutedio e muacutesculo esqueleacutetico
A primeira evidecircncia da presenccedila de vanaacutedio no muacutesculo esqueleacutetico proveio de uma amostra de
ATP isolado de muacutesculo de cavalo que continha uma impureza ndash vanaacutedio ndash cuja presenccedila
provocou a inibiccedilatildeo da actividade da bomba de Na+K
+-ATPase [AO] Desde entatildeo vaacuterios
estudos tecircm sido realizados com ecircnfase na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o
tecido muscular Destes destaca-se a descoberta da interacccedilatildeo forte da espeacutecie V10 com a miosina
fosforilada [AP] e agrave sua inibiccedilatildeo da actividade ATPaacutesica da miosina[AQ AR]
No quediz respeitoao iatildeo monovanadato destaca-se a sua inibiccedilatildeoda actividade Mg2+
-ATPaacutesica
da miosina [AS] que ocorre de duas formas distintas (1) na presenccedila de concentraccedilotildees reduzidas
de V1 a inibiccedilatildeo ocorre a velocidade reduzida dependente da concentraccedilatildeo da espeacutecie
monomeacuterica (2) na presenccedila de concentraccedilotildees elevadas de V1 o niacutevel de inibiccedilatildeo aproxima-se de
100 desde que [V1] seja superior agrave concentraccedilatildeo total de locais com actividade Mg2+
-ATPaacutesica
no ensaio Esta inibiccedilatildeo eacute irreversiacutevel tanto por diaacutelise ou adiccedilatildeo de agentes redutores do
vanadato [AL]Segundo Pate et al(1994)[AT] a espeacutecie monomeacuterica podeinibir a velocidade de
contracccedilatildeo a temperatura inferior a 20ordmC perdendogradual e rapidamente essa capacidade ateacute os
50ordmC temperatura agrave qual cessa a capacidade inibitoacuteria Os resultados obtidos sugeremainda uma
interacccedilatildeo altamente cooperativa entre o iatildeo vanadato e as proteiacutenas componentes do muacutesculo
esqueleacuteticoDestaca-se tambeacutem a descoberta da inibiccedilatildeo da geraccedilatildeo de forccedila contraacutectil nas fibras
musculares por parte da espeacutecie monomeacuterica[AT]Wilson et al(1995) [AU ]determinaram que a
afinidade da espeacutecie V1 para o S1 de miosina eacute significativamente mais elevada do que a do iatildeo
Pi seu anaacutelogo Descobriram ainda que que a ligaccedilatildeo do vanadato poderaacute envolver transiccedilotildees
entre estados energeacuteticos com energia livre independente dos impedimentos esteacutereos impostos
20
pela estrutura das miofibrilas Os estudos desenvolvidos por Smith et al (1990)[AV]
determinaram queo iatildeo monovanadato natildeo apresenta um efeito pronunciado na inibiccedilatildeo da
actividadeacto-S1-miosina-ATPaacutesica excepto em concentraccedilotildees superiores a 900microM a partir das
quais existe produccedilatildeo de espeacutecies oligomeacutericas para as quais o efeito inibitoacuterio seraacute mais
elevadoA espeacutecie monomeacuterica de vanadato consegue adoptar facilmente uma geometria do tipo
bipiramidal trigonal que mimetiza o intermediaacuterio formado (MgADP
Pi)aquando da reacccedilatildeo de
hidroacutelise de ATP no local activo da miosina [AW] De facto a interacccedilatildeo do vanaacutedio com a
miosina eacute inesperadamente estaacutevel na presenccedila de ADP Esta interacccedilatildeo resulta na formaccedilatildeo de
um complexo enzimaacutetico inactivo e estaacutevel com estequiometria de 11 ADPvanadato por siacutetio
activo da proteiacutena A formaccedilatildeo do complexo estaacutevel miosina-vanadato foi proposta por Goodnoet
al (1982) [AS] Nesta ocorre um preacute-equiliacutebrio raacutepido que induz a formaccedilatildeo de um complexo
ternaacuterio reversiacutevel (MADP
V) com formaccedilatildeo lenta do isoacutemero M
ADP
V A diferenccedila entre os
compostos de vanaacutedio e de fosfato centra-se no tempo de meia vida (t12) do Pi ou do V (no
primeiro t12=12 s enquanto no segundo t12=24 h) (Refordf) A diferenccedila dever-se-aacute agrave possiacutevel
complexaccedilatildeo do vanaacutedio com grupos nucleofiacutelicos no siacutetio activo o que provocaria uma
estabilizaccedilatildeo da miosina na conformaccedilatildeo MADP
V
No que respeita aos estudos realizados com a espeacutecie tetrameacuterica Cremo et al determinou que a
cadeia pesada do S1 de miosina pode ser clivada em dois locais distintos (designados V1 e V2)
pela espeacutecie V4 apoacutes irradiaccedilatildeo com luz UV na presenccedila de concentraccedilotildees de vanadato na ordem
dos milimolar e na ausecircncia de nucleoacutetidos ou metais divalentes
Jaacute no que concerne aos estudos realizados com ecircnfase na espeacutecie decameacuterica foi sugerido
porRamos et al (2006) [AK]que a interacccedilatildeo do decavanadato com a G-actina favorecida pela
polimerizaccedilatildeo desta uacuteltima estabiliza a espeacutecie V10 e induz a inibiccedilatildeo da formaccedilatildeo de F-actina A
estabilizaccedilatildeo da espeacutecie V10 por proteiacutenas tansmembranares e do citoesqueleto pode ser a razatildeo
pela qual ocorre a toxicidade do decavanadato reportada na avaliaccedilatildeo dos efeitos do V(V) nos
sistemas bioloacutegicosTiago et al (2007) [AR] concluiram que o V10 apresenta uma clara preferecircncia
de ligaccedilatildeo ao ldquoback-doorrdquo da S1-miosina-ATPase o que se verifica apenas na estrutura
conformacional desimpedida com acesso facilitado ao local de ligaccedilatildeo do Pi Sugere tambeacutem que
o V10 actua como um ldquoback-door stoprdquo bloqueando estaabertura do fragmento de 50-kDa
necessaacuteria para a hidroacutelise do ATP--fosfato o que constitui uma explicaccedilatildeo simples do
comportamento natildeo competitivo da espeacutecie decameacuterica estimulando a utilizaccedilatildeo deste oligoacutemero
como uma feramenta da elucidaccedilatildeo das mudanccedilas conformacionais ldquoback-doorrdquo da miosina no
21
processo da contracccedilatildeo muscular(ref)Destaca-se ainda a descoberta da capacidade inibitoacuteria do
V10 em relaccedilatildeo agrave fosforilaccedilatildeo oxidativa mitocondrial [AX]
Finalmente no que concerne aos estudos realizados com a presenccedila de vaacuterias espeacutecies
oligomeacutericas Aureliano e Madeira (1994) [AY]observaram que na presenccedila de Ca2+
as espeacutecies
V4 e V10se ligam agrave Ca2+
-ATPase do retiacuteculo sarcoplasmaacutetico enquanto a espeacutecie V1 se liga apenas
na presenccedila de ATP Aparentemente o aumento da concentraccedilatildeo de Ca2+
tem o efeito contraacuterio da
inibiccedilatildeo da ATPase do retiacuteculo sarcoplasmaacutetico pelo V1 natildeo tendo no entanto efeito significativo
sobre a inibiccedilatildeo por parte da espeacutecie V10 Tiago et al (2002) [AP] concluiacuteram que em condiccedilotildees
proacuteximas agraves fisioloacutegicas a espeacutecie V10 difere das espeacutecies presentes na soluccedilatildeo de metavanadato
devido agrave sua forte interacccedilatildeo com a miosina fosforilada e agrave sua inibiccedilatildeo da actividade ATPaacutesica
da miosina Para aleacutem disso o ATP diminui a afinidade da miosina para o V4 induzindo a
interacccedilatildeo com V1 e natildeo tendo qualquer efeito no que diz respeito ao V10 Foi tambeacutem proposto
[AQ] que a ligaccedilatildeo das espeacutecies oligomeacutericas de vanadato aoS1 de miosina nos locais onde o
fosfato coordenaeacute provavelmente a causa da inibiccedilatildeo da actividade ATPaacutesica da miosina
estimulada por actina atraveacutes da prevenccedilatildeo da troca entre ATPADP sendo esta favorecida para
aniotildees de vanadato com valecircncias superiores como eacute o caso do V10
Na presenccedila de actina as velocidades de dissociaccedilatildeo do Pi e do V aumentam consideravelmente
este uacuteltimo em menor quantidade Assim a ligaccedilatildeo da actina ao complexo MADP
V e
Pi eacute
bastante fraca pelo que se sugere que o ADPV teria um efeito de relaxamento semelhante ao
ATP No entanto a capacidade do vanaacutedio em inibir a actividade ATPaacutesica da miosina e da
actomiosina eacute diferenciada uma vez que na ausecircncia de actina a inibiccedilatildeo eacute total para
concentraccedilotildees na gama dos M mas na sua presenccedila a inibiccedilatildeo eacute totalmente conseguida apenas
em concentraccedilotildees na gama dos mM (Ref) No entanto eacute necessaacuterio considerar a possiacutevel
formaccedilatildeo de espeacutecies oligomeacutericas de vanadato cuja afinidade e efeito na enzima seratildeo
diferentes Assim pode concluir-se que o efeito de inibiccedilatildeo observado para a actividade ATPaacutesica
de miosina estimulada por actina se pode dever agrave presenccedila das diferentes espeacutecies oligomeacutericas o
que foi jaacute sugerido por Aureliano (2000) [AZ] ndash as espeacutecies tetrameacuterica e decameacuterica de vanadato
interactuariam com a miosina com alteraccedilatildeo da sua interacccedilatildeo com a actina No entanto natildeo eacute
conhecida a contribuiccedilatildeo absoluta das restantes espeacutecies oligomeacutericas para a inibiccedilatildeo da ATPase
da miosina estimulada por actina (Refs)
14 Objectivos do projecto
Pretende-se com este projecto o isolamento purificaccedilatildeo e caracterizaccedilatildeo da actina do muacutesculo
22
esqueleacutetico de coelho sendo esta uacuteltima tambeacutem realizada para o sufragmento-1 de miosina
Seguidamente realizar-se-aacute a quantificaccedilatildeo e caracterizaccedilatildeo das proteiacutenas por espectroscopia UV-
Vis e SDS-PAGE e a caracterizaccedilatildeo das soluccedilotildees de vanaacutedio por UV-Vis e ressonacircncia magneacutetica
nuclear (RMN) Por uacuteltimo a interacccedilatildeo do S1 de miosina com o vanaacutedio seraacute avaliada atraveacutes
das teacutecnicas de UVVis RMN e ressonacircncia paramagneacutetica electroacutenica (RPE)
2 Discussatildeo de Resultados
21 Caracterizaccedilatildeo das proteiacutenas isoladas
211Anaacutelise do estado de pureza
O estado de pureza das soluccedilotildees de actina e de S1 foi analisado por electroforese em gel de
poliacrilamida em condiccedilotildees desnaturantes (SDS-PAGE) (Fig 22) que separa as moleacuteculas de
acordo com o seu peso molecular (PM) A concentraccedilatildeo de acrilamida escolhida (10) permite a
detecccedilatildeo simultacircnea de componentes com peso molecular inferior a 20 kDa e outros com elevado
peso molecular
Figura 21 ndash Fotografia de gel de SDS-PAGE (10 acrilamida) Poccedilo (a)
10 L do marcador de elevado peso molecular () Poccedilo (b) 20 L de [G-
actina]=1 mgmL Poccedilo (c) 20 L de [S1]=1 mgmL Aplicou-se uma
voltagem de V durante h e min
A partir da recta de calibraccedilatildeo obtida log PM = -1177 Rf + 2247 (ver ponto 2 da secccedilatildeo
Anexos) pode verificar-se a presenccedila de actina na amostra do poccedilo (b) presente numa banda
bastante intensa (PMaparente=40 kDa) bem como a presenccedila minoritaacuteria de outros componentes
desconhecidos que natildeo teratildeo sido eliminados durante o processo de extracccedilatildeo e purificaccedilatildeo e natildeo
satildeo relevantes para o estudo em questatildeo Na amostra do poccedilo (c) nota-se a presenccedila de S1
(PMaparente=92 kDa) bem como a de outros componentes cuja banda eacute ligeiramente menos intensa
Estes poderatildeo ser fragmentos proteoliacuteticos de miosina e de S1 Estaacute ainda presente uma bada de
23
maior intensidade que poderaacute ser a proteiacutena-C (PM=135 kDa) uma vez que a sua remoccedilatildeo
completa da amostra necessitaria de um passo adicional de cromatografia de troca ioacutenica [BA]
No entanto natildeo podemos confirmar que se trata desta proteiacutena ou de qualquer outra uma vez que
se encontra fora do intervalo de migraccedilatildeo dos marcadores de peso molecular
212 Determinaccedilatildeo da concentraccedilatildeo proteica
2121Subfragmento-1 de miosina
A partir da recta de calibraccedilatildeo obtida pelo meacutetodo de Bradford [BB](ver secccedilatildeo 3311 do
Procedimento Experimental) A595nm = 00546[BSA] + 00209 (ver secccedilatildeo de Anexos)foi possiacutevel
determinar um intervalo de concentraccedilatildeo de S1 de miosina compreendido entre 226 e 117(plusmn
423M) (203 e 1056 (plusmn 381 mgmL)) (n=8) A concentraccedilatildeo meacutedia de S1 de miosina apoacutes
diaacutelise contra a soluccedilatildeo tampatildeo 10 mM HEPES pH 75 25 mM KCl determinada pela leitura da
Abs280 nmfoi de 20430M (1803 mgmL)(n=7)
As amostras a quantificar atraveacutes do meacutetodo de Bradford natildeo sofreram diaacutelise o que permite
explicar a concentraccedilatildeo superior de S1 de miosina em comparaccedilatildeo com o valor obtido atraveacutes do
meacutetodo directo Esta razatildeo aliada agrave sensibilidade e rapidez superiores deste uacuteltimo fez com que
se optasse pelo meacutetodo directo para a quantificaccedilatildeo da grande maioria dos ensaios
2122Actina
A concentraccedilatildeo de actina foi determinada quando esta se encontrava na forma monomeacuterica (G-
actina) por mediccedilatildeo da leitura directa da Abs290 nm tendo-se obtido uma concentraccedilatildeo meacutedia de
691481M (290202 mgmL) (n=3) em 2 mM Tris-HCl pH 80 02 mM CaCl2 02 mM Na-
ATP Ao comprimento de onda utilizado (290 nm) absorvem fortemente apenas os resiacuteduos de
Trp pelo que a mediccedilatildeo efectuada tem menor sensibilidade do que se tivesse sido efectuada a 280
nm uma vez que a actina apresenta apenas 4 resiacuteduos deste aminoaacutecido embora tenha 16
resiacuteduos de Tyr
22 Mediccedilatildeo da actividade enzimaacutetica de S1 de miosina
221 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
As actividades ATPaacutesicas de S1 de miosina dependentes de Ca2+
e K+foram analisadas pelo
caacutelculo das velocidades de consumo inicial do fosfato inorgacircnico (Pi) Definiu-se os tempos de 5
e 3 minutos para a mediccedilatildeo da Ca2+
e da K+EDTA-ATPase respectivamente Os valores obtidos
24
a partir do protocolo presente no ponto 341 da secccedilatildeo Procedimento experimental encontram-se
descritos na Tabela 2I
Tabela 2I ndash Acrividades natildeo-fisioloacutegicas de subfragmento-1 determinadas pelo meacutetodo de Fiske-Subarow
modificado (n=2)
Proteiacutena
Actividade
ATPaacutesica natildeo
fisioloacutegica
Actividade (nmol
Piminmg proteiacutena) Actividade (s-1)
S1
Ca2+
-ATPase 1400 plusmn 30 2100 plusmn 0045
K+EDTA-ATPase 2334 plusmn 50 3500 plusmn 0075
As actividades natildeo-fisioloacutegicas reflectem o estado de oxidaccedilatildeo dos resiacuteduos expostos de Cys 707
(SH1) e 697 (SH2) Se o grupo SH1 sofrer oxidaccedilatildeo a actividade ATPaacutesica dependente de Ca2+
aumenta e a actividade dependente de K+ diminui Por outro lado se o grupo SH2 sofrer
oxidaccedilatildeo regista-se a perda completa destas actividades
De acordo com Margossian e Lowey (1982) [BA] as actividades ATPaacutesica de S1 de miosina
dependente de Ca2+
e K+EDTA da miosina satildeo respectivamente de 24 e 112 s-1
Estaacute tambeacutem
descrito que a actividade ATPaacutesica dependente de K+ eacute em meacutedia cerca de 3 vezes superior agrave
dependente de Ca2+
reflectindo a quantidade das preparaccedilotildees A razatildeo obtida entre os valores das
actividades ATPaacutesicas dependentes de K+ e Ca
2+ eacute de 166 Os valores obtidos para as actividades
(Ca2+
-ATPase=21 s-1
K+EDTA-ATPase= 35 s
-1) satildeo inferiores agrave valores obtidos por
Margossian e Lowey [BA] e Tiago [BC] com forccedila ioacutenica inferior O valor obtido para a
actividade ATPaacutesica dependente de Ca2+
eacute superior ao obtido por Polosukhina et al (2000) [BD]
cujo meio de reacccedilatildeo continha MOPS e apresentava forccedila ioacutenica inferior Os dados
supramencionados comprovam a variaccedilatildeo da actividade a ATPaacutesica natildeo fisioloacutegica em relaccedilatildeo agrave
forccedila ioacutenica temperatura e pH
222 Mediccedilatildeo da actividadeMg2+
-ATPaacutesica de S1 pelo meacutetodo de enzimas acopladas
A actividade de Mg2+
-ATPase eacute a uacutenica que possui significado fisioloacutegico sendo as restantes
bastante uacuteteis apenas para indicaccedilatildeo da qualidade de preparaccedilatildeo do S1[BA] A 0575 M KCl a
actina e o S1 formam uma soluccedilatildeo viscosa e tuacuterbida Apoacutes a adiccedilatildeo de MgATP o complexo acto-
S1 dissocia-se e a turbidez da soluccedilatildeo diminui Se a forccedila ioacutenica for reduzida a dissociaccedilatildeo do
complexo actoS1 que se encontra na forma de suspensatildeo eacute induzida O MgATP eacute rapidamente
hidrolisado diminuindo a sua concentraccedilatildeo no meio reaccional Nestas condiccedilotildees haacute
reassociaccedilatildeo e subsequente contracccedilatildeo dos filamentos de actina e de S1 com formaccedilatildeo de
25
precipitado O meacutetodo descrito traduz de forma bastante elementar o fenoacutemeno da contracccedilatildeo in
vivo e elimina o problema anterior da solubilizaccedilatildeo ineficiente da miosina de modo a obter uma
mediccedilatildeo exacta da actividade [A] A utilizaccedilatildeo de forccedilas ioacutenicas reduzidas aumenta a afinidade da
actina para o S1 permitindo contornar a extrema dificuldade em obter a concentraccedilatildeo proteica dos
muacutesculos (cerca de 100 mgmL do complexo actomiosina) As actividades Mg2+
-ATPaacutesicas de S1
de miosina foram determinadas a partir do declive da variaccedilatildeo da Abs340 relativo agrave oxidaccedilatildeo de
NADH em funccedilatildeo do tempo como eacute descrito no ponto 342 do Procedimento Experimental Os
valores obtidos para a actividade Mg2+
-ATPaacutesica encontram-se apresentados na Tabela 2II
Tabela 2II ndash Actividade Mg2+
-ATPaacutesica de subfragmento-1 de miosina determinadas pelo meacutetodo das enzimas
acopladas
Actividade (nmol
ATPmin mg(S1)) Actividade (s
-1)
1097 plusmn 269 164 plusmn 04
Ramos (2005) [J] obteve o valor de 01 s-1
para a actividade Mg2+
-ATPaacutesica de S1 de miosina
com forccedila ioacutenica intermeacutedia Tiago (2005) [BC] obteve o valor de 011 s-1 em condiccedilotildees de forccedila
ioacutenica reduzida em relaccedilatildeo agrave utilizada no presente relatoacuterio
23 Preparaccedilatildeo e caracterizaccedilatildeo das soluccedilotildees de vanaacutedio
231 Anaacutelise da composiccedilatildeo das soluccedilotildees de vanadato por espectroscopia de 51
V-RMN
efeito da concentraccedilatildeo da espeacutecie
A diluiccedilatildeo das soluccedilotildees stock de meta- e decavanadato iraacute determinar fortemente a composiccedilatildeo
em soluccedilatildeo dos diferentes oligoacutemeros de vanadato (ref) No caso do decavanadato o periacuteodo de
tempo apoacutes a diluiccedilatildeo iraacute tambeacutem ser um factor importante a considerar devido agrave decomposiccedilatildeo
da espeacutecie V10(ref)Torna-se assim fundamental determinar a composiccedilatildeo quiacutemica das soluccedilotildees
no meio de ensaio utilizado (HEPES 10 mM KCl 25 mM) de modo a correlacionaacute-la
correctamente com os possiacuteveis efeitos promovidos pelas diferentes espeacutecies de vanadato no
sistema em estudo
Aleacutem disso deve ser dado ecircnfase agraves importantes interacccedilotildees que ocorrem entre o vanaacutedio e os
tampotildees escolhidos O tampatildeo com menor nuacutemero e magnitude de interacccedilatildeo eacute o HEPES [AD]
seguido do Tris sendo este uacuteltimo especialmente aconselhado em concentraccedilotildees baixas para casos
de incompatibilidade enzimaacutetica ou quiacutemica redox No caso especial da reacccedilatildeo redox
NAD(P)HNAD(P) o tampatildeo adequado eacute o imidazole de modo a natildeo haver reacccedilatildeo com o
26
vanadato Os tampotildees a evitar em qualquer situaccedilatildeo incluem a trietanolamina fosfato citrato e
glicina entre outros [AD] uma vez que todos interactuam fortemente com V(V) com formaccedilatildeo
de complexos estaacuteveis
Assim procedeu-se agrave anaacutelise da composiccedilatildeo das soluccedilotildees de meta- e decavanadato apoacutes diluiccedilatildeo
das soluccedilotildees stock (50 mM em vanadato total) no meio de reacccedilatildeo (tampatildeo D) atraveacutes da
espectroscopia de ressonacircncia magneacutetica nuclear de 51
V Nas figuras x1 e x2 apresentam-se os
espectros obtidos para as soluccedilotildees de deca- e metavanadato num intervalo de concentraccedilotildees de 1 a
10 mM A Tabela seguinte resume os paracircmetros espectroscoacutepicos obtidos para cada espeacutecie
presente nas soluccedilotildees analisadas
Figura 22ndash Espectros de RMN de 51
V de
concentraccedilotildees crescentes das espeacutecies oligomeacutericas
de vanadato obtidos apoacutes diluiccedilatildeo da soluccedilatildeo stock
de decavanadato em 10 mM HEPES pH 70 25 mM
KCl para concentraccedilotildees finais de A) 1 mM B)2
mM C) 4 mM D) 8 mM E) 10 mM de
decavanadato total V1refere-se agraves espeacutecies
monomeacutericade vanadato V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2IIIndash Desvios quiacutemicos meacutedios (part) das ressonacircncias das espeacutecies decameacuterica (V10) monomeacuterica (V1) e
tetrameacuterica (V4) presentes nos espectros da soluccedilatildeo de decavanadatoLargura a meia altura (∆12) da espeacutecie V10A
Paracircmetros V10A V10B V10C V1 V4
part (ppm) -5162 -5003 -4255 -5616 -5748
∆12 (Hz) 175 nd nd nd nd
Figura 23 ndash Distribuiccedilatildeo graacutefica das
espeacutecies oligomeacutericas de vanadato vs a
concentraccedilatildeo de decavanadato total As
concentraccedilotildees de cada espeacutecie foram
obtidas segundo a formula descrita na
secccedilatildeo 35 do Procedimento
Experimental (aacutereas dos sinais de 51
V-
RMN observadas na figura x1)V10 V1 e
V4 correspondem agraves espeacutecie monomeacuterica
dimeacuterica e tetrameacuterica respectivamente [Esp
eacuteci
e o
ligo
meacute
rica
de
van
aacutedio
] (m
M)
[Decavanadato]total (mM)
V10
V1
V4
27
Nas soluccedilotildees de decavanadato as espeacutecies monomeacuterica e decameacuterica de vanadato satildeo as uacutenicas
presentes para concentraccedilotildees de decavanadato inferiores a 8 mM Ambas as espeacutecies aumentam
linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado em [AY] e
em (Teresa) quando a concentraccedilatildeo de V1 eacute igual ou superior a 200 microM inicia-se a sua
oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica No entanto nas condiccedilotildees de realizaccedilatildeo dos
ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a espeacutecie tetrameacuterica neste
estudo em concentraccedilotildees inferiores a 8 mM tendo o mesmo sido observado por Ramoset al
(2006) [AK] Os resultados obtidos mostram claramente o equiliacutebrio entre a espeacutecie decameacuterica e
a monomeacuterica e que o aparecimento de outras espeacutecies oligomeacutericas se encontra dependente da
concentraccedilatildeo desta uacuteltima Apesar de natildeo ter sido realizado o ensaio com 5 mM de decavanadato
total no presente relatoacuterio estaacute reportado em Ramos et al (2006) [AK] que as concentraccedilotildees da
espeacutecie decameacuterica e monomeacuterica satildeo respectivamente de 500 microM e 200 microM Assim pode inferir-
se que para concentraccedilotildees inferiores a 5 mM e intervalos de tempo de cerca de 20 minutos (que
inclui o tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas
as espeacutecies decameacuterica e monomeacuterica de vanadato
Figura 24 ndash Espectros de RMN de 51
V de concentraccedilotildees
crescentes das espeacutecies oligomeacutericas de vanadato obtidos
apoacutes diluiccedilatildeo da soluccedilatildeo stock de metavanadato em 10 mM
HEPES pH 70 25 mM KCl para concentraccedilotildees finais de A)
1 mM B) 2 mM C) 4 mM D) 8 mM E) 10 mM de
metavanadato total V1 refere-se agraves espeacutecies monomeacuterica de
vanadatoV1 e V2 correspondem respectivamente agrave espeacutecie
monomeacuterica e dimeacuterica
Figura 25 ndash Distribuiccedilatildeo graacutefica das espeacutecies
oligomeacutericas de vanadato vs a concentraccedilatildeo de
metavanadato total As concentraccedilotildees de cada
espeacutecie foram obtidas segundo a formula descrita
na secccedilatildeo 35 do Procedimento Experimental
(aacutereas dos sinais de 51
V-RMN observadas na
figura x1)V10 V1 e V4 correspondem agraves espeacutecie
monomeacuterica dimeacuterica e tetrameacuterica
respectivamente
Esp
eacuteci
e o
ligo
meacute
rica
de
va
nad
ato
(m
M)
[Metavanadato]total (mM)
V1
V2
V4
V5
28
Tabela 2IVndash Desvios quiacutemicos meacutedios (part)das ressonacircncias das espeacutecies tetrameacuterica (V4) monomeacuterica (V1) e
dimeacuterica (V2) presentes no espectro da soluccedilatildeo de metavanadato (total) 10 mM
Paracircmetros V1 V2 V4 V5
part (ppm) -5615 -5748 -5789 -5869
Nas soluccedilotildees de metavanadato as espeacutecies monomeacuterica dimeacutericae tetrameacuterica de vanadato estatildeo
presentes para concentraccedilotildees de metavanadato inferiores a 4 mM Todas estas as espeacutecies
aumentam linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado
em (Aureliano e Madeira 1998) e em (Tiago et al 2004) quando a concentraccedilatildeo de V1 eacute igual ou
superior a 200 microM inicia-se a sua oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica Nas
condiccedilotildees de realizaccedilatildeo dos ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a
espeacutecie pentameacuterica neste estudo em concentraccedilotildees inferiores a 4 mM tendo o mesmo sido
observado por Ramos 2006 Os resultados obtidos mostram claramente o equiliacutebrio entre a
espeacutecie V4 e as espeacuteciesV1 e V2 no qual o aparecimento de outras espeacutecies oligomeacutericas se
encontra dependente da concentraccedilatildeo da espeacutecie V1 Por uacuteltimo pode inferir-se que para
concentraccedilotildees inferiores a 2 mM e intervalos de tempo de cerca de 20 minutos (que inclui o
tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas as
espeacutecies V1 V2 e V4 com concentraccedilotildees respectivas de 0796 0192 e 0196 mM
232Estabilidade das soluccedilotildees de decavanadato anaacutelise por espectroscopia de absorccedilatildeo
UVVis
Caracterizou-se a estabilidade da espeacutecie V10 numa soluccedilatildeo com 1 mM decavanadato total em
tampatildeo D atraveacutes da leitura da Abs400 Obteve-se uma cineacutetica de dissociaccedilatildeo de 1ordf ordem tal
como se pode verificar pela linearidade do graacutefico de ln(Abs400)vs o tempo de aquisiccedilatildeo espectral
(segundos) (Fig26 )
Figura 26ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do tempo
(em segundos) para uma soluccedilatildeo de decavanadato total de 1
mM em tampatildeo D a 25ordmC (n=2)
Sendo uma reacccedilatildeo de primeira ordem pode ser descrita pela equaccedilatildeo cujo
declive eacute ndashK (constante de dissociaccedilatildeo) e a intersecccedilatildeo no eixo das ordenadas corresponde
Ln
(Ab
s 400 n
m)
tempo (s)
29
aln(A0) Numa reacccedilatildeo de 1ordf ordem o tempo de meia-vida eacute independente da concentraccedilatildeo inicial
do reagente sendo descrito pela equaccedilatildeo [2] Deste modo a partir da recta
Ln(Abs400 nm) = -436E-5
K - 105 obteve-se um tempo de meia-vida de 4490 plusmn 0823 horas para
as condiccedilotildees supramencionadas
Apoacutes a adiccedilatildeo de S1 8 microM ao meio reaccional verificou-se que a reacccedilatildeo de desoligomerizaccedilatildeo
da espeacutecie decameacuterica de vanadato seguia igualmente uma cineacutetica de 1ordf ordem tal como se pode
verificar pela linearidade do graacutefico de ln(Abs400) versus o tempo de aquisiccedilatildeo espectral
(segundos) (Fig27)
Figura 27ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do
tempo (em segundos) para uma soluccedilatildeo de decavanadato
total de 1 mM em tampatildeo D a 25ordmC na presenccedila de 8
microM (n= 2)
A partir da equaccedilatildeo da recta Ln(Abs400 nm) = -7520E-5
K - 0613determinou-se o valor de K para
a espeacutecie decameacuterica de onde se obteve um tempo de meia-vida de 2430 plusmn 0191 horas para as
condiccedilotildees supramencionadas
Tanto este valor como o valor registado na ausecircncia de S1 satildeo substancialmente superiores agrave
duraccedilatildeo dos ensaios de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina descritos no presente
relatoacuterio
24 Interacccedilatildeo de S1 com vanaacutedio(V)
241Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
De modo a analisar qual o efeito da adiccedilatildeo de S1 de miosina (tampatildeo D) produzia nos sinais de
51V-NMR titulou-se uma soluccedilatildeo de 5mM de decavanadato total com concentraccedilotildees crescentes
de S1 (5 10 15 e 20 microM) Os espectros de 51
V-RMN encontram-se apresentados na Fig 2
No graacutefico da figura 2 Apresenta-se a concentraccedilatildeo de S1 em funccedilatildeo do factor de alargamento
corrigido dos espectros acima referidos
Ln
(Ab
s (4
00
nm
))
tempo (s)
30
Figura 28 ndash Graacutefico de
representaccedilatildeo da
concentraccedilatildeo de S1 em
funccedilatildeo do factor de
alargamento A
concentraccedilatildeo de
decavanadato total foi de 5
mM e as concentraccedilotildees
utilizadas de S1 foram de 0
5 10 e 15 microM
Verifica-se que o factor de alargamento aumenta linearmente em funccedilatildeo da concentraccedilatildeo de S1 no
ensaio o que indica a existecircncia de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina Observou-
se ainda que a adiccedilatildeo de S1 ao meio reaccional natildeo provocou qualquer alteraccedilatildeo dos desvios
quiacutemicos dos sinais de 51
V-RMN
Realizaram-se ensaios na presenccedila de ATP e de S1 com intuito de verificar o efeito do ATP na
interacccedilatildeo do S1 de miosina com o decavanadato
Figura 29 - Espectros de RMN de 51
V contendo 2
mM Decavanadato (A) (controlo) (B) na presenccedila
de 2 mM de ATP (C) na presenccedila de 2 mM de ATP
e 8 microM de S1 de miosina V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2Vndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies decameacuterica (V10) e monomeacuterica (V1) presentes nos
espectros da soluccedilatildeo de decavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V10A V10B V10C V1
part (ppm) -5161 -5002 -4254 -5614
Observa-se que a presenccedila de ATP (espectro B) natildeo provoca qualquer alteraccedilatildeo visiacutevel em
relaccedilatildeo ao espectro que contem apenas soluccedilatildeo de decavanadato Jaacute a presenccedila simultacircnea de ATP
e S1 provoca um alargamento da ∆12 do sinal do V10A de 1736 Hz (espectro (A)) para 2366 Hz
(espectro (C) Fig 29) Tendo em conta que a concentraccedilatildeo de S1 apresenta uma relaccedilatildeo linear
com o factor de alargamento converteu-se o valor da ∆12 e comparou-se com o valor esperado agrave
F = 00591[S1] - 00157 Rsup2 = 0998
Fact
or
de
ala
rgam
en
to
[S1] (microM)
31
concentraccedilatildeo de S1 utilizada O factor de alargamento obtido para 8 microM ndash 0376 ndash eacute inferior ao
valor esperado agrave mesma concentraccedilatildeo de S1 obtido atraveacutes da equaccedilatildeo da recta apresentada na
Fig 28 ndash 0457 ndash o que indica que o ATP poderaacute exercer um efeito de protecccedilatildeo do S1
diminuindo a sua interacccedilatildeo com a espeacutecieV10
Por uacuteltimo realizaram-se ensaios com metavanadato na presenccedila de 8 microM S1 de modo a verificar
se as espeacutecies oligomeacutericas contidas nesta soluccedilatildeo interagiam com a proteiacutena em estudo
Figura 210 - Espectros de RMN de 51
V contendo 2
mM metavanadato (A) (controlo) (B) na presenccedila
de 8 microM de S1 de miosina V1 V2 e V4
correspondem respectivamente aacutes espeacutecies
monomeacuterica dimeacuterica e tetrameacuterica
Tabela 2VIndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies monomeacuterica (V1) dimeacuterica (V2) e tetrameacuterica (V4)
presentes nos espectros da soluccedilatildeo de metavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V1 V2 V4
part (ppm) -5613 -5747 -5789
Como eacute possiacutevel observar na Fig 210 ocorre uma ligeiro alargamento dos sinal de V4 bem
como uma perda da intensidade pelo que se conclui quedas espeacutecies presentes na soluccedilatildeo de
metavanadato (V1 V2 e V4) apenas a espeacutecie tetrameacuterica interage com o S1 de miosina o que
estaacute de acordo com o reportado por Tiago et al 2002
242 Actividade Mg2+
-ATPaacutesica de S1 de miosina na presenccedila de decanadato
Com o intuito de avaliar o efeito do decavanadato na actividade Mg2+
-ATPaacutesica do S1 de
miosina monitorizou-se a reacccedilatildeo de oxidaccedilatildeo do NADH atraveacutes da leitura de Abs340 na
presenccedila de 005 mgmL de S1 de miosina e no caso da actividade estimulada 2microM de actina em
3125 mM Tris-HCl pH 71 e 375 mM MgCl2 pelo meacutetodo de enzimas acopladas (ver seccccedilatildeo xx
dos MM)Para tal realizaram-se ensaios com vaacuterias concentraccedilotildees de decavanadato total (10 50
100 250 500 e 1000) de modo a determinar qual o seu efeito na actividade MgATPaacutesica de S1
de miosina na ausecircncia e na presenccedila de um dos seus ligandos naturais Os ensaios foram
realizados agrave temperatura ambiente com incubaccedilatildeo preacutevia a 25ordm C durante 20 minutos
32
Figura 211 ndash Representaccedilatildeo
graacutefica da Actividade Mg2+
-
ATPaacutesica de S1 () (os ajustes
teoacutericos estatildeo representado a por
uma linha os resultados
experimentais estatildeo representados
por triacircngulos (Actividade
estimulada) e cruzes (Actividade
natildeo estimulada)) vs
[Decavanadato]total medidas pelo
meacutetodo das enzimas acopladas
(n=3) Os dados foram ajustados ao modelo de decaimento exponencial usando o programa GraphPad Prism para a
determinaccedilatildeo dos valores de IC50 (na ausecircncia de actina IC50= 266 plusmn 07 M decavanadato na presenccedila de actina
IC50= 28 plusmn 1 M decavanadato
A partir do graacutefico presente na Fig 29 verifica-se que o ajuste dos dados experimentais foi bem
sucedido A inibiccedilatildeo da actividade Mg2+
-ATPaacutesica do S1 de miosina eacute afectada em menor
amplitude na presenccedila de actina (60 de inibiccedilatildeo da actividade) do que na sua ausecircncia
(aproximadamente 80 de inibiccedilatildeo de actividade) Devido a esta diferenccedila pode sugerir-se que a
actina iraacute impedir parcialmente a ligaccedilatildeo do decavanadatoao S1 demiosina atraveacutes de
impedimentos esteacutereos ou interacccedilotildees electrostaacuteticas por exemplo A teacutecnica utilizada natildeo nos
permite concluir acerca de que regiatildeo(otildees) ou resiacuteduo(s) da actina participaratildeo na preservaccedilatildeo
parcial da actividade Mg2+
-ATPaacutesica
Por outro lado os valores de IC50 dos dois tipos de actividades natildeo apresentam diferenccedilas
significativas tendo-se obtido 266 plusmn 07 e 28 plusmn 1M decavanadato na ausecircncia e na presenccedila de
actina respectivamente o que corresponde a uma concentraccedilatildeo de cerca de 27 M de V10 O
meio de reacccedilatildeo utilizado nestes ensaios foi 10 mM HEPES e 25 mM KCl com 2 mM ATP Em
(Tiago et al 2004) o valor de IC50 para a actividade Mg2+-ATPaacutesica na ausecircncia de actina foi de
125 plusmn 025 microM enquanto o valor da actividade estimulada por actina foi de 136 plusmn 14 microM O
meio de reacccedilatildeo utilizado nos ensaios foi 20 mM HEPES 50 mM KCl e 2 mM ATP o que resulta
precisamente no dobro da forccedila ioacutenica a que foi realizado o ensaio do presente relatoacuterio
(exceptuando a contribuiccedilatildeo dos produtos da hidroacutelise de ATP uma vez que este meacutetodo
pressupotildee a ocorrecircncia de um ciclo de reacccedilotildees de hidroacutelise e siacutentese deste composto) Ora tendo
sido anteriormente referido que a afinidade do S1 de miosina para com a espeacutecie V10 diminui com
Act
ivid
ade
Mg
2+-A
TP
aacutesic
a d
e S
1 d
e
Mio
sin
a
[Decavanadato] (microM) Actividade Estimulada ()
Actividade Natildeo Estimulada ()
Ajuste Actividade Estimulada
Ajuste Actividade Natildeo Estimulada
33
o aumento da forccedila ioacutenica (Tiago et al 2004 tal poderaacute explicar a diferenccedila obtida entre estes
dois conjuntos de valores
243Quantificaccedilatildeo de cisteiacutenas reduzidas pelo meacutetodo de Ellman
A interacccedilatildeo do vanaacutedio com alguns tipos de aminoaacutecidos sensiacuteveis agrave oxidaccedilatildeo pode fornecer-nos
informaccedilatildeo bastante uacutetil em relaccedilatildeo agraves alteraccedilotildees estruturais que conduzem a uma diminuiccedilatildeo da
actividade fisioloacutegica do S1 de miosina Os estudos realizados por Chasteen 1983 concluiacuteram
que a reduccedilatildeo de vanadato (V) a vanadilo (IV) ocorre na presenccedila de agentes redutores comuns no
ambiente celular de que se destaca o aminoaacutecido cisteiacutena Esta eacute tambeacutem um alvo de
complexaccedilatildeo por parte do iatildeo VO2+
Jaacute Ramos et al (2009) demonstrou que a incubaccedilatildeo de
actina com a soluccedilatildeo de decavanadato induz a oxidaccedilatildeo de resiacuteduos de cisteiacutena e agrave formaccedilatildeo do
oxidovanaacutedio (IV) Este grupo de investigadores descobriu ainda que apenas a espeacutecie V10 induz a
oxidaccedilatildeo das cisteiacutenas ldquoraacutepidasrdquo da actina atraveacutes do processo proposto que eacute parcialmente
responsaacutevel pela inibiccedilatildeo jaacute descrita da actividade de ATPase do S1 de miosina estimulada por
actina quando exposta agrave soluccedilatildeo de decavanadato Esta inibiccedilatildeo apresenta-se afectada com a
presenccedila de ATP uma vez que os resiacuteduos de cisteiacutena permaneceram intactos nos ensaios
realizados com este ligando Jaacute Crans et al (2004) concluiu que o siacutetio activo de muitas fosfatases
(que conteacutem grupos tiol) pode formar um equiliacutebrio redox com o iatildeo vanadato (V) de forma natildeo
reversiacutevel
Os resultados obtidos por Sakurai e Shiromura (1980) sugeriram que o vanaacutedio (V) sofre reduccedilatildeo
a vanaacutedio (IV) sem dependecircncia do valor de pH do ensaio Registaram ainda uma estequiometria
de 31 na reacccedilatildeo do vanadato (V) com os resiacuteduos de Cys enquanto a estequiometria dos ensaios
com o vanadilo (IV) foi de 21
Berlett e Stadtman (1997) observaram que os tipos de aminoaacutecidos mais facilmente oxidaacuteveis
englobam entre outros os resiacuteduos de Cys Trp e MetAssim analisou-se o estado redox dos
grupos tiol dos resiacuteduos de Cys do S1 na presenccedila do decavanadato atraveacutes da monitorizaccedilatildeo da
Abs412 nm na presenccedila de 1 mgmL de S1 de miosina em tampatildeo 10 mM HEPES 25 mM KCl
variando a concentraccedilatildeo da soluccedilatildeo de decavanadato de modo a determinar que tipo de alteraccedilotildees
estruturais ocorrem na sua presenccedila no que diz respeito agrave diminuiccedilatildeo da actividade ATPaacutesica de
S1 na presenccedila de decavanadato
Os resultados referentes agrave quantificaccedilatildeo das cisteiacutenas reduzidas apoacutes o tratamento de S1 com
diferentes concentraccedilotildees de decavanadato em tampatildeo 10 mM HEPES pH 75 e 25 mM KCl a
25ordmC durante 20 minutos encontram-se descritos na Tabela 2VII
Tabela 2VII ndash Cisteiacutenas reduzidas totais e ldquoraacutepidas de S1 (1 mgmL) apoacutes tratamento com decavanadato
34
(010501002505001000 uM) em 10 mM HEPES 25 mM KCl pH 7 a 25ordmC durante 20 minutos A detecccedilatildeo
espectrofotomeacutetrica foi feita com DTNB (e 412=8400 uM-1 cm-1) (n variaacutevel entre 1 e 8)
Amostras Cys raacutepidas Cys totais
11 M S1 (controlo) 22 plusmn 04 106 plusmn 14
11 M S1 + 10 M Decavanadato 22 plusmn 05 95 plusmn 08
11 M S1 + 50 uM Decavanadato 20 plusmn 02 98 plusmn 14
11 M S1 + 100 uM Decavanadato 17 plusmn 03 97 plusmn14
11 M S1 + 250 uM Decavanadato 26 plusmn 02 98 plusmn 08
11 M S1 + 500 uM Decavanadato 21 plusmn 04 90 plusmn 06
11 M S1 + 1000 uM Decavanadato 24 plusmn 0 86 plusmn 08
A partir dos resultados obtidos pode sugerir-se que o S1 possui 11 resiacuteduos de Cys (controlo) o
que estaacute de acordo com o anteriormente descrito Verifica-se que o tratamento da proteiacutena com
concentraccedilotildees crescentes de decavanadato provoca a oxidaccedilatildeo de dois resiacuteduos de Cys do S1 de
miosina natildeo expostos ao solvente No entanto a teacutecnica utilizada natildeo nos permite identificar
quais
Tendo em consideraccedilatildeo que o valor de IC50 obtido para a reacccedilatildeo de inibiccedilatildeo daactividade Mg2+-
ATPaacutesica de S1 na presenccedila de decavanadato eacute de cerca de 27 microM pode sugerir-se que este seraacute o
valor ao qual ocorreraacute a oxidaccedilatildeo do primeiro resiacuteduo de Cys o que poderaacute contribuir para a
diminuiccedilatildeo da actividade de S1 observada
25Interacccedilatildeo de S1 com vanaacutedio(IV)
251Anaacutelise por espectroscopia de Ressonacircncia Paramagneacutetica Electroacutenica
A acumulaccedilatildeo intracelular de oxovanaacutedio (IV) in vivo constitui uma das principais formas de
apresentaccedilatildeo de vanaacutedio no organismo Deste modo torna-se bastante interessante analisar a
interacccedilatildeo entre este iatildeo e o S1 de miosina atraveacutes da teacutecnica de espectroscopia de Ressonacircncia
Paramagneacutetica Electroacutenica o meacutetodo de eleiccedilatildeo do estudo do iatildeo vanadilo e dos complexos por si
formados O electratildeo desemparelhado encontra-se fortemente acoplado ao nuacutecleo 51
V o que
35
resulta num espectro de RPE consistindo de 8 desdobramentos na direcccedilatildeo paralela e outros tantos
na direcccedilatildeo perpendicular (Chasteen 1983)
Os estudos realizados por Ahmed et al (1987) centraram-se na interacccedilatildeo do vanadilo com
proteiacutenas como eacute o caso da insulina transferrina entre outros obtendo um espectro caracterizado
por 8 linhas amplamente espaccediladas (na direcccedilatildeo paralela) Jaacute Ramos et al 2009 concluiu que a
exposiccedilatildeo da soluccedilatildeo de decavanadato agraves formas globular e filamentosa de actina produz um
espectro tiacutepico do iatildeo oxidovanaacutedio (IV) indicando a reduccedilatildeo do vanadato
Para aferir qual a concentraccedilatildeo de S1 que permitia obter uma boa razatildeo sinalruiacutedo titulou-se uma
soluccedilatildeo de sulfato de vanadilo 15 microM ([VOSO4]stock = 5 mM) com concentraccedilotildees crescentes de
proteiacutena (0 5 10 15 e 20 microM) em 10 mM HEPES 25 mM KCl A existecircncia de uma correlaccedilatildeo
linear entre a concentraccedilatildeo de proteiacutena e a intensidade dos sinais de RPE correspondente agrave banda
-12 perpendicular encontra-se representada na Fig 210
Figura 2 12ndash Representaccedilatildeo graacutefica da variaccedilatildeo da intensidade do sinal de RPE vs a concentraccedilatildeo de S1 utilizada
Apoacutes verificar que com uma concentraccedilatildeo de S1 igual a 20 microM se obtinham sinais com uma razatildeo
sinalruiacutedo adequada ao prosseguimento dos estudos sobre a interacccedilatildeo V(IV)-S1 de miosina
titulou-se uma soluccedilatildeo de 20 microM S1 com 0 25 5 10 15 25 50 75 100 150 200 500 e 1000
microM VOSO4
Traccedilou-se o graacutefico da intensidade da banda -12 perpendicular do sinal de EPR em funccedilatildeo da
concentraccedilatildeo de VOSO4 tendo-se determinado uma constante de dissociaccedilatildeo (KD) do complexo
S1-vanadilo de 8748 plusmn 2621 microM V(IV)
y = 016786x + 122435
Rsup2 = 092992759
Inte
nsi
dad
e d
o s
inal
de
RP
E
[S1] (microM)
36
Figura 213 ndash Representaccedilatildeo graacutefica da variaccedilatildeo do sinal de RPE vs a concentraccedilatildeo de vanadilo utilizada (os valores
experimentais estatildeo representados por losangos o seu ajuste estaacute representado pela linha)
A simulaccedilatildeo do espectro de EPR foi efectuada a partir do espectro obtido para uma concentraccedilatildeo
de 1 mM VOSO4 apoacutes adiccedilatildeo de 20 microM S1 de miosina tendo-se obtido os valores da constante de
acoplamento hiperfina (A=5016) e do factor g (g0=195)
Figura 214 ndash Representaccedilatildeo graacutefica do espectro de EPR obtido com 1000 microM de vanadilo e 20 microM de S1 (a cinzento
escuro) A simulaccedilatildeo do espectro encontra-se a cinzento claro
Por comparaccedilatildeo destes com os valores presentes na tabela de correspondecircncia xxx eacute possiacutevel
predizer quais os ligandos provaacuteveis do vanadilo na presenccedila de proteiacutena Assim concluiu-se que
o vanadilo estaacute coordenado com duas moleacuteculas de aacutegua um grupo hidroxilo e um grupo R=NR
O grupo hidroxilo pode indicar os resiacuteduos de aminoaacutecidos Tyr Treonina e Serina enquanto o
grupo R=NR poderaacute indicar a presenccedila de Arginina e Histidina como ligandos do vanadilo
Inte
nsi
dad
e d
os
sinai
s d
e R
PE
[Vanadilo] (microM)
Ajuste dos dados exp Dados experimentais
Campo magneacutetico (G)
Espectro Experimental Ajuste
37
3 Procedimento Experimental
31 Reagentes
Todos os reagentes utilizados satildeo de elevado grau de pureza (analiacutetico) Apresentam-se as suas
foacutermulas quiacutemicas ou abreviaturas quando necessaacuterio acrescentou-se a sua descriccedilatildeo Entre
parecircntesis colocou-se as empresas que os comercializam
Aacutecido aceacutetico glacial 100 (Riedel) Acrilamida 99 (Merck) Azul de bromofenol (Riedel-de-
Haumlen) -Mercaptoetanol (Sigma) Bissulfito de soacutedio (Sigma)Bradford (reagente de)
(BioRad)BSA (albumina de soro bovino Sigma-Aldrich) CaCl22H2O (cloreto de caacutelcio
dihidratado Fluka) CH3CH2OH 96 (etanol Panreac) Coomassie Blue R250 (Merck) D2O
(oacutexido de deuteacuterio SDS) DTNB (55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico) Aldrich) EDTA (aacutecido
etilenodiaminatetraaceacutetico Merck) Fosfoenolpiruvato 98 (Fluka) Glicerol (Sigma) Glicina
985 (Panreac) HCl 37 (Riedel-de Haumlen) H2SO4 96 (Pronalab) HEPES 99 (aacutecido 4-(2-
hidroxiletil)-1-piperazineetanosulfoacutenico Sigma) KH2PO4 99 (hidrogenofosfato de potaacutessio)
MgATP (Sigma) Na
ATP (Sigma) Na2HPO4 99 (hidrogenofosfato de soacutedio Riedel-de Haumlen)
NaH2PO4 (fosfato de soacutedio ) NaOH 986 (hidroacutexido de soacutedio Pronalab) KCl 99 (Cloreto de
potaacutessio Panreac) L-Cys 995 (enantioacutemero levoacutegiro do resiacuteduo de Cisteiacutena Fluka)Metanol
99 (Merck) Molibdato de amoacutenio (Sigma)NNrsquo-metil-bis-acrilamida 98 (Merck) NH4VO3
99 (metavanadato de amoacutenia Sigma) PSA 10 (persulfato de amoacutenio Merck) SDS
(dodecilo sulfato de soacutedio Sigma) Sulfito de soacutedio (Sigma) VOSO4 (sulfato de vanadilo
Aldrich) TEMED 99 (Tetrametiletilenodiamina Merck) Tris (2-amino-2-hidroximetil-13-
propanodiol Sigma)
32 Meacutetodos
321 Isolamento e purificaccedilatildeo das proteiacutenas de muacutesculo esqueleacutetico de coelho
3211 Purificaccedilatildeo da miosina preparaccedilatildeo de Subfragmento 1 por digestatildeo quimotriacutepsica
da miosina
O subfragmento-1 de miosina-II (S1) foi gentilmente cedido pelo Prof Doutor Manuel Aureliano
do Departamento de Quiacutemica Bioquiacutemica e Farmaacutecia da Faculdade de Ciecircncias e Tecnologia da
Universidade do Algarve
No entanto descreve-se abaixo de forma resumida o procedimento efectuado
38
32111 Purificaccedilatildeo da miosina
O protocolo de isolamento foi adaptado do meacutetodo previamente descrito por Margossian e Lowey
(1982) apresentando-se de forma esquemaacutetica na Fig 31
Sucintamente os tecidos musculares dorsais e das pernas foram extraiacutedos apoacutes o sacrifiacutecio do
animal (Fig31) De seguida foram adicionados 3 volumes da soluccedilatildeo de extracccedilatildeo A2
Centrifugou-se a soluccedilatildeo e filtrou-se o sobrenadante O sedimento foi entatildeo diluiacutedo em 14
volumes de aacutegua destilada de modo a que a forccedila ioacutenica atingisse 004 M Apoacutes adiccedilatildeo de szlig-
mercaptoetanol (szlig-me) deixou-se a precipitar a 4ordmC removendo o sobrenadante Centrifugou-se
novamente e ressuspendeu-se o sedimento na soluccedilatildeo de extracccedilatildeo A Adicionou-se a soluccedilatildeo
tampatildeo B3 para que a forccedila ioacutenica atingisse 05 M Homogeneizou-se e deixou-se repousar
durante 2h Homogeneizou-se novamente e centrifugou-se de modo a ocorrer precipitaccedilatildeo dos
complexos de actomiosina e proteiacutena desnaturada Apoacutes filtraccedilatildeo do sobrenadante em latilde de vidro
quantificou-se a soluccedilatildeo de miosina resultante pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental)
2 Soluccedilatildeo de extracccedilatildeo A 0012M K2CO3 004M KHCO3 06M KCl pH 65
3 Soluccedilatildeo-tampatildeo B Tris 100 mM pH 7 KCl 2M
39
Figura 31 ndash Representaccedilatildeo esquemaacutetica do protocolo de isolamento de miosina (Margossian e Lowey 1982) Todo o
procedimento foi realizado a 4ordmC (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
32112 Preparaccedilatildeo de Subfragmento-1 de miosina por digestatildeo com quimiotripsina
O S1 de miosina foi isolado apoacutes diaacutelise da miosina durante a noite contra a soluccedilatildeo C4 A
presenccedila do EDTA eacute essencial uma vez que a sua substituiccedilatildeo por MgCl2resulta na produccedilatildeo de
HMM em vez de S1 A suspensatildeo de miosina foi equilibrada e digerida com -quimotripsina
A digestatildeo foi interrompida pela adiccedilatildeo de PMSF Seguidamente ultracentrifugou-se o produto
da digestatildeo desprezando-se o sedimento (Fig 32) A soluccedilatildeo de sobrenadante maioritariamente
S1 foi quantificada pela leitura directa da Abs280 ou pelo meacutetodo de Bradford (ver as secccedilotildees
3311 e 3312 do Procedimento Experimental respectivamente) e armazenada em 10 sacarose
e 2 mM szlig-Me a -80ordmC
4 Soluccedilatildeo C 25mM Tris pH 70 012M KCl 1mM EDTA
40
Antes da realizaccedilatildeo dos ensaios descritos no presente relatoacuterio o S1 de miosina foi dialisado
contra a soluccedilatildeo D5 de modo a remover qualquer composto que pudesse complexar como
vanaacutedio Seguidamente quantificou-se a proteiacutena pelo meacutetodo de Bradford ou pelo meacutetodo
directo (ver secccedilotildees 3311 e 3312do Procedimento Experimtental)
Figura 3 2ndash Representaccedilatildeo esquemaacutetica do protocolo de preparaccedilatildeo do S1 de miosina por digestatildeo quimiotripsina de
acordo com o previamente descrito por Margossian e Lowey (1982) Todo o procedimento foi realizado a 4ordm C
excepto quando indicaccedilatildeo do contraacuterio (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
3212 Isolamento e purificaccedilatildeo de actina
Isolou-se e purificou-se a actina segundo o meacutetodo descrito por Pardee e Spudish que
compreende dois passos ndash a obtenccedilatildeo de um resiacuteduo seco denominado ldquopoacute de acetonardquo e a
extracccedilatildeo subsequente da actina a partir do ldquopoacute de acetonardquo na forma de G-Actina com a
posterior polimerizaccedilatildeo a F-Actina
32121 Obtenccedilatildeo do ldquopoacute de acetonardquo
Sacrificou-se o coelho com uma pancada atraacutes do pescoccedilo e sangrou-se o animal apoacutes o corte da
arteacuteria jugular Esfolou-se e colocou-se numa tina com gelo Retiraram-se as partes de tecido
conjuntivo que envolvem o tecido muscular com auxiacutelio de uma pinccedila e bisturi Seguidamente
retirou-se os muacutesculos dorsais e os muacutesculos brancos das pernas Passou-se os muacutesculos por aacutegua
destilada de modo a remover as impurezas e pesou-se num copo Trituraram-se os muacutesculos
numa maacutequina de triturar carne e extraiu-se o resultante com 3 volumes da soluccedilatildeo E6 durante 10
minutos com agitaccedilatildeo suave Centrifugou-se a suspensatildeo a 4000 g durante 10
5 Soluccedilatildeo D HEPES 10 mM pH 70 KCl 25 mM
6 Soluccedilatildeo E 01 M KCl 015 M K2HPO4 pH 65
41
minutosdesprezando-se o sobrenadante Ressuspendeu-se o sedimento em 6 volumes de soluccedilatildeo
F7durante 10 minutos com agitaccedilatildeo suave Verificou-se se o pH da soluccedilatildeo estava compreendido
entre 82 e 85 acertando-se estecom soluccedilatildeo G8 se necessaacuterio Centrifugou-se durante 10
minutos a 4000 g Desprezou-se o sobrenadante e ressuspendeu-se o sedimento em 3 volumes de
soluccedilatildeo H9 durante 10 minutos Centrifugou-se novamente a 4000 gdurante 10 minutos
desprezando-se o sobrenadante Ressuspendeu-se o sedimento duas vezes consecutivas em 6
volumes de aacutegua destilada durante 5 minutos Centrifugou-se a 4000 g durante 10 minutos apoacutes
cada adiccedilatildeo de H2O
Seguidamente ressuspendeu-se o sedimento 5 vezes consecutivas utilizando 3 volumes de
acetona de cada vez agitando suavemente durante 10 minutos agrave temperatura ambiente Colocou-
se o resiacuteduo filtrado numa tina de vidro sobre papel de filtro deixando-se evaporar o solvente
durante a noite num nicho Triturou-se o resiacuteduo seco numa maacutequina de triturar carne e deixou-se
novamente evaporar o solvente O ldquopoacute de acetonardquo obtido foi armazenado a -20 ordmC
32122 Purificaccedilatildeo da actina a partir de ldquopoacute de acetonardquo
Apoacutes a obtenccedilatildeo do poacute de acetona como acima indicado efectuou-se o protocolo seguidamente
descrito de modo a obter monoacutemeros de actina (G-Actina) e actina filamentosa (F-Actina)
Pesou-se 35 g de poacute de acetona e adicionaram-se 50 mL de soluccedilatildeo-tampatildeo I10
sendo a
extracccedilatildeo feita com agitaccedilatildeo suave durante 30 minutos Separou-se o extracto do poacute de acetona
hidratado filtrando a suspensatildeo com 4 camadas de gaze esterilizada tendo o cuidado de natildeo
espremer o resiacuteduo Voltou a extrair-se com 50 mL de soluccedilatildeo I durante 30 minutos mexendo
lentamente Adicionou-se este ao extracto obtido anteriormente Centrifugou-se o extracto total a
16 000 g durante 30 minutos Decantou-se para uma proveta sobre a qual se colocou gaze para
que o sedimento solto natildeo se juntasse ao sobrenadante e leu-se o volume De seguida adicionou-
se KCl ateacute uma concentraccedilatildeo final de 50 mM MgCl2 ateacute 2mM e ATP ateacute 1mM lentamente e com
agitaccedilatildeo contiacutenua de modo a promover a polimerizaccedilatildeo dos monoacutemeros de actina
Deixou-se polimerizar durante 2h a 25ordmC observando-se a presenccedila de pequenas bolhas de ar
indicativas de um aumento de viscosidade da soluccedilatildeo Arrefeceu-se em gelo durante 15 minutos e
adicionou-se KCl ateacute uma concentraccedilatildeo final de 06M Mexeu-se lentamente durante 30 minutos
apoacutes os quais se ultracentrifugou a 110 000g durante 1hora Desprezou-se o sobrenadante
ressuspendeu-se o sedimento com soluccedilatildeo tampatildeo E e transferiu-se para um potter
Dialisou-se durante a noite contra 1L de soluccedilatildeo-tampatildeo I a uma temperatura de 4ordmC com rotaccedilatildeo
7 Soluccedilatildeo F 005 M NaHCO3 pH 82-85
8 Soluccedilatildeo G 1M Na2CO3
9 Soluccedilatildeo H 1 mM EDTA pH 70
10 Soluccedilatildeo I 2 mM Tris-HCl pH 8 02 mM CaCl2 02 mM Na2ATP 05 mM szlig-Me 0005 NaN3
42
contiacutenua e mudanccedila de tampatildeo 2 a 3 vezes De seguida a actina foi centrifugada a 110
000gdurante 1hora Decantou-se a soluccedilatildeo de G-Actina (sobrenadante) e desprezou-se o
sedimento Determinou-se a concentraccedilatildeo de G-Actina pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental) A soluccedilatildeo de G-actina foi armazenada a uma temperatura de 4ordmC
apoacutes adiccedilatildeo de 002NaN3 mantendo-se estaacutevel durante 3 a 5 dias
331 Quantificaccedilatildeo das proteiacutenas isoladas
3311 Quantificaccedilatildeo pelo meacutetodo de Bradford
O meacutetodo de Bradford (Bradford 1976) baseia-se na utilizaccedilatildeo do reagente de Bradford
composto por entre outros aacutecido fosfoacuterico e o corante ldquoCoomassierdquo Este corante possui carga
negativa e a sua coloraccedilatildeo varia entre a vermelha (Absmax=465 nm) predominante em soluccedilatildeo e a
azul (Absmax=595 nm) presente aquando da ligaccedilatildeo aos grupos amina das proteiacutenas Em soluccedilotildees
aacutecidas diluiacutedas a coloraccedilatildeo varia proporcionalmente com a quantidade do corante ligado agrave
proteiacutena que por sua vez eacute directamente proporcional agrave concentraccedilatildeo da proteiacutena permitindo a
sua quantificaccedilatildeo ateacute a um limite de detecccedilatildeo de 20 gmL (Bradford 1976)
Deste modo prepararam-se amostras de BSA ([BSA]stock= 1mgmL) utilizando um intervalo de
concentraccedilotildees compreendido entre 0 e 15 gmL de acordo com a tabela 3I Apoacutes a adiccedilatildeo da
H2O MilliQ e do reagente de Bradford as soluccedilotildees foram sucessivamente homogeneizadas no
voacutertex
Tabela 3Indash Amostras utilizadas para a obtenccedilatildeo da recta de calibraccedilatildeo no meacutetodo de Bradford
BSA (L) BSA (g) H2O MilliQ (L) Reagente de Bradford (L)
0 0 800 200
125 125 79875 200
25 25 7975 200
5 5 795 200
10 10 790 200
15 15 785 200
Apoacutes incubaccedilatildeo a 25ordmC por 5 minutos leram-se os valores de Abs595 num
espectrofotoacutemetro (Shimadzu UV-160A) para todas as amostras Seguidamente traccedilou-se a recta
de calibraccedilatildeo (Abs595vs quantidade de BSA (g))
43
Para determinar a concentraccedilatildeo de proteiacutena apoacutes o processo de isolamento purificaccedilatildeo e
diaacutelise quando necessaacuterio colocou-se 5 L da amostra num tubo de Eppendorf e adicionou-se
795 L de H2O MilliQ homogeneizando a soluccedilatildeo no voacutertex Adicionou-se posteriormente 200
L de reagente de Bradford e repetiu-se a homogeneizaccedilatildeo Incubou-se a mistura a 25ordmC por 5
minutos apoacutes os quais se leu os valores de Abs595 nm no espectrofotoacutemetro Por interpolaccedilatildeo deste
valor a partir da recta de calibraccedilatildeo determinou-se a concentraccedilatildeo de proteiacutena na amostra
3312 Quantificaccedilatildeo pelo meacutetodo directo
A quantificaccedilatildeo das amostras proteicas pode tambeacutem ser efectuada atraveacutes da leitura directa da
absorvacircncia tendo por base a lei de Beer-Lambert onde Abs corresponde agrave
absorvacircncia ao comprimento de onda ao qual o ensaio foi realizado corresponde ao coeficiente
de extinccedilatildeo (substituiacutedo pela letra a se as unidades forem distintas da molaridade) b corresponde
ao percurso oacuteptico da radiaccedilatildeo na ceacutelula amostral e c corresponde agrave concentraccedilatildeo amostral
Assim no caso de S1 de miosina mediu-se a Abs280e calculou-se a concentraccedilatildeo da proteiacutena
sabendo que o coeficiente de extinccedilatildeo de S1 eacute (Ref)
No caso da actina mediu-se a Abs290ecalculou-se a concentraccedilatildeo da proteiacutena sabendo que o
coeficiente de extinccedilatildeo da actina eacute (Ref)
332Electroforese em condiccedilotildees desnaturantes (SDS-PAGE)
De modo a melhor caracterizar as proteiacutenas isoladas foi realizada uma electroforese em
condiccedilotildees desnaturantes (SDS-PAGE do inglecircs Sodium Dodecyl Sulfate Polyacrilamide Gel
Electrophoresis) O SDS (dodecilo sulfato de soacutedio) eacute um detergente anioacutenico utilizado neste
meacutetodo para desnaturar as proteiacutenas que se ligam ao SDS na proporccedilatildeo 14 g SDS g proteiacutena
(aproximadamente 1 moleacutecula de SDS por cada 2 resiacuteduos de aminoaacutecidos) (Stryer) O SDS induz
uma densidade de carga negativa significativa nas proteiacutenas a que se liga mascarando a sua carga
intriacutenseca pelo que as razotildees cargamassa satildeo tendencialmente idecircnticas Assim a SDS-PAGE
separa as proteiacutenas quando sujeitas a um campo eleacutectrico de acordo com o seu peso molecular
sendo a sua migraccedilatildeo directamente proporcional ao logaritmo deste uacuteltimo (Stryer)
O gel de resoluccedilatildeo com 10 de acrilamida foi obtido atraveacutes da adiccedilatildeo de 167 mL de soluccedilatildeo
stock de acrilamida11
075 mL de tampatildeo de resoluccedilatildeo12
251 mL de aacutegua destilada e 50 L de
SDS 10 Para promover a polimerizaccedilatildeo adicionou-se em seguida 38 L de PSA 10
11
soluccedilatildeo stock de acrilamida soluccedilatildeo de 30 (mv) de acrilamida e 08 (mv) de NNrsquo-metil-bis-acrilamida 12
tampatildeo de resoluccedilatildeo 15 M Tris-HCl pH 88
44
(persulfato de amoacutenia) 10 e 25 L de TEMED 99 (tetrametiletilenodiamina) Agitou-se
cuidadosamente de modo a natildeo formar bolhas e pipetou-se a soluccedilatildeo para a cassette de vidro
previamente montada no suporte De seguida adicionou-se uma peliacutecula de cerca de 2 mm de 2-
metil-2-butanol para verificar posteriormente a polimerizaccedilatildeo Apoacutes a polimerizaccedilatildeo que demora
cerca de 45 minutos verteu-se a agrave superfiacutecie e adicionou-se o gel de concentraccedilatildeo com 5 de
acrilamida constituiacutedo por 03 mL de soluccedilatildeo de stock acrilamida 045 mL de tampatildeo de
concentraccedilatildeo13
094 mL de aacutegua destilada 18 L de SDS 10 135 L de PSA 10 e 2L de
TEMED 99 Colocou-se o pente sobre o gel e deixou-se polimerizar durante cerca de 45
minutos
As amostras para aplicaccedilatildeo no gel de electroforese foram preparadas para uma concentraccedilatildeo de
proteiacutena de 1 mgmL num volume final de 100 L dos quais 25 L correspondem ao tampatildeo da
amostra14
As amostras foram aquecidas a 100ordmC durante 5 minutos Antes da aplicaccedilatildeo das
amostras transferiu-se a cassette para a tina de electroforese agrave qual se adicionou a soluccedilatildeo
tampatildeo de electroforese da tina superior15
e tampatildeo de electroforese da tina inferior16
Aplicaram-
se 10 L das soluccedilotildees de marcadores de peso molecular e 20 L das amostras proteicas
preparadas anteriormente A electroforese decorreu durante cerca de 3h30 min a uma voltagem
de 80 V De seguida transferiu-se o gel para uma tina corando as proteiacutenas durante 1h com
soluccedilatildeo corante17
Descorou-se o gel com soluccedilatildeo descorante18
tendo-se mudado esta soluccedilatildeo ateacute
as bandas se encontrarem bem definidas
Determinou-se a mobilidade relativa das bandas correspondentes aos marcadores de elevado peso
molecular e traccedilou-se a recta de calibraccedilatildeo ndash log(PM) vs log(Rf) onde Rf corresponde agrave
mobilidade relativa Por interpolaccedilatildeo determina-se o PM das proteiacutenas isoladas sabendo a sua
mobilidade relativa Por comparaccedilatildeo com os valores descritos na literatura pode concluir-se
acerca do grau de purificaccedilatildeo
34 Mediccedilatildeo da actividade enzimaacutetica de S1
341 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
13
tampatildeo de concentraccedilatildeo 05 M Tris-HCl pH 68 14
tampatildeo de amostra 320 mM Tris-HCl pH 68 04 M szlig-mercaptoetanol SDS 8 glicerol 15 (vv) e roxo de m-
cresol 0024 15
tampatildeo de electroforese da tina superior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS 16
tampatildeo de electroforese da tina inferior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS acetato de soacutedio
01 M 17
soluccedilatildeo corante ldquoCoomassie Blue R-250rdquo contendo metanol 45 (vv) e aacutecido aceacutetico glacial 10 (vv) 18
soluccedilatildeo descorante metanol 10 (vv) e aacutecido aceacutetico glacial 10 vv)
45
O meacutetodo de Fiske-Subarow modificado consiste na reacccedilatildeo do molibdato de amoacutenio com o
fosfato inorgacircnico em soluccedilatildeo aacutecida com formaccedilatildeo do aacutecido fosfomoliacutebdico Seguidamente daacute-
se a adiccedilatildeo de um agente redutor que provoca a reduccedilatildeo do composto a fosfomolibdato que forma
um complexo de cor azulada cuja absorvacircncia maacutexima ocorre para um comprimento de onda de
660 nm tornando possiacutevel a sua detecccedilatildeo por meacutetodos espectrofotomeacutetricos As actividades Ca2+
-
ATPaacutesica e K+EDTA-ATPaacutesica de S1 foram determinadas atraveacutes deste meacutetodo de acordo com
o descrito por Taussky e Shorr (1953) com mediccedilatildeo da quantidade de fosfato (Pi) libertado
atraveacutes da hidroacutelise de ATP
Para tal obteve-se primeiramente a recta de calibraccedilatildeo com as amostras presentes na Tabela 3II
utilizando uma soluccedilatildeo stock de 5 mM KH2PO4
Tabela 3II ndash Composiccedilatildeo das diferentes amostras utilizadas para determinar a recta de calibraccedilatildeo atraveacutes do meacutetodo
de Fiske-Subbarow modificado
KH2PO4 (L)
([KH2PO4]stock= 5mM)
PO43-
(nmole) H2O (uL)
0 0 1780
5 25 1775
10 50 1770
20 100 1760
40 200 1740
100 500 1680
A cada tubo de Eppendorf adicionaram-se seguidamente 200 L de soluccedilatildeo J19
e 20 L de agente
redutor20
homogeneizando no voacutertex apoacutes cada adiccedilatildeo Incubou-se durante 10 minutos e mediu-
se a Abs660 nm num espectrofotoacutemetro com intervalos de tempo iguais entre cada amostra Traccedilou-
se a recta de calibraccedilatildeo (Abs660 nmvs nmol PO43-
) a partir dos valores obtidos
Para determinar a actividade Ca2+
-ATPaacutesica de S1 prepararam-se tubos de Eppendorf com 400
L de meio de reacccedilatildeo21
25 L CaCl2 ([CaCl2]stock= 100 mM) 25 L ATP ([ATP]stock= 100 mM)
com [S1]ensaios=01 mgmL Perfez-se se necessaacuterio com H2O MilliQ ateacute perfazer um volume de 1
mL Para determinar a actividade K+EDTA-ATPaacutesica utilizou-se uma concentraccedilatildeo idecircntica de
19
Soluccedilatildeo J 5 N H2SO4 25 molibdato de amoacutenio 20
Agente redutor 5 mg aacutecido 1-amino-2-naftol-4-sulfoacutenico 30 mg bissulfito de soacutedio 30 mg sulfito de soacutedio em 26
mL H2O 21
meio de reacccedilatildeo 125 mM Tris-HCl pH 80 0575 M KCl
46
proteiacutena nos ensaios (01 mgmL) e o mesmo meio de reacccedilatildeo Aos tubos de Eppendorf
adicionou-se ainda 123 L de KCl ([KCl]stock = 3M) 25 L de ATP 10 L de EDTA
([EDTA]stock= 100 mM )e o volume de H2O MilliQ necessaacuterio para perfazer 1 mL Em ambas as
situaccedilotildees o uacuteltimo reagente adicionado foi o ATP uma vez que se trata do substrato da reacccedilatildeo
enzimaacutetica
A cineacutetica decorreu a 25ordmC durante 5 minutos para a actividade Ca2+
-ATPaacutesica e durante 3
minutos para a actividade K+EDTA-ATPaacutesica Apoacutes o periacuteodo de incubaccedilatildeo transferiu-se 150
L de amostra para tubos de Eppendorf contendo igual volume de TCA 10 Colocou-se no gelo
e homogeneizou-se no voacutertex Seguidamente centrifugou-se a 1500 g durante 10 minutos e
recolheu-se 200 L de sobrenadante para novos tubos de Eppendorf Adicionou-se a estes 158
mL de H2O MilliQ e 200 L de soluccedilatildeo J Homogeneizou-se no voacutertex e adicionou-se 20 L de
reagente redutor repetindo-se a homogeneizaccedilatildeo Incubou-se durante 10 minutos e mediu-se a
Abs660 nm com o mesmo intervalo de tempo de leitura para cada amostra Por intrapolaccedilatildeo a partir
da recta de calibraccedilatildeo obtida anteriormente determinou-se a quantidade de Pi libertado em cada
ensaio A partir destes resultados determinaram-se as actividades Ca2+
-ATPaacutesica e K+EDTA-
ATPaacutesica do S1 em nmol PO43-
minmg proteiacutena e em s-1
342 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
Da actividade Mg2+
-ATPaacutesica de S1 resulta a formaccedilatildeo dos produtos da reacccedilatildeo de
hidroacutelise do ATP - ADP e Pi Na presenccedila de ADP fosfoenolpiruvato (PEP) e da enzima piruvato
cinase (PK) os produtos resultantes satildeo ATP e piruvato Por acccedilatildeo da enzima lactato
desidrogenase (LDH) este uacuteltimo eacute convertido em lactato ocorrendo simultaneamente a oxidaccedilatildeo
do NADH a NAD+ monitorizada atraveacutes da Abs340 nm Quando o S1 de miosina se encontra no
meio reaccional o ATP eacute hidrolisado formando-se novamente ADP e Pi o que permite a
formaccedilatildeo de um ciclo (fig 21) Se as enzimas se encontrarem em excesso a velocidade de
conversatildeo do NADH eacute limitada pela produccedilatildeo de ADP e Pi Como o ATP eacute regenerado em cada
ciclo a sua concentraccedilatildeo inicial eacute sempre mantida (Bagshaw 1982)
Figura 33ndash Representaccedilatildeo esquemaacutetica do meacutetodo de enzimas acopladas
47
O protocolo utilizado e seguidamente descrito foi adaptado de Cuenda et al (1990 ndash Ver Refordf) A
uma cuvette de 1 mL adicionou-se S1 para uma concentraccedilatildeo final de 005 mgmL 800 L de
meio de reacccedilatildeo22
10 L de PEP ([PEP]stock = 42 mM) 10 L de PK+LDH 11 ([PK+LDH]stock =
1 mgmL) e 15 L de NADH ([NADH]stock=25 mM) Iniciou-se a monitorizaccedilatildeo da reacccedilatildeo
atraveacutes da leitura de Abs340 nm apoacutes a adiccedilatildeo de 20 L de ATP ([ATP]stock=100 mM) A
actividade Mg2+
-ATPaacutesica eacute obtida atraveacutes do declive correspondente agrave oxidaccedilatildeo do NADH Para
a mediccedilatildeo da Mg2+
-ATPase de S1 estimulada por actina adicionou-se F-actina para uma
concentraccedilatildeo final de 2 microM sendo a actividade obtida tambeacutem atraveacutes do declive correspondente
agrave oxidaccedilatildeo do NADH apoacutes a adiccedilatildeo da proteiacutena
35 Interaccedilatildeo entre vanaacutedio e subfragmento-1 de miosina
351 Preparaccedilatildeo das soluccedilotildees de vanaacutedio
As soluccedilotildees de vanadato foram preparadas em aacutegua MilliQ a partir de metavanadato de amoacutenio
(NH4VO3) numa concentraccedilatildeo stock de 50 mM com agitaccedilatildeo magneacutetica e aquecimento (~30 ordmC)
Esta soluccedilatildeo com um valor compreendido entre 6 e 7 designa-se por ldquometavanadatordquo e eacute
constituiacuteda pelas esspeacutecies V1 V2 V4 e V5 (Ref) Acidificando a soluccedilatildeo de metavanadato por
adiccedilatildeo de HCl ateacute um valor de pH proacuteximo de 4 observa-se uma mudanccedila de cor de amarelo
claro para laranja indicativa da formaccedilatildeo da espeacutecie decameacuterica de vanadato De seguida e
imediatamente antes da realizaccedilatildeo dos ensaios descritos o valor do pH da soluccedilatildeo de
decavanadato foi aumentado para cerca de 75 por adiccedilatildeo de NaOH A soluccedilatildeo (2) designada
por ldquometavanadatordquo natildeo foi posteriormente alterada ateacute agrave sua adiccedilatildeo para a realizaccedilatildeo dos ensaios
As soluccedilotildees foram armazenadas a 4ordmC e mantidas em gelo aquando da sua utilizaccedilatildeo
A soluccedilatildeo de V(IV) foi preparada em condiccedilotildees anaeroacutebias por diluiccedilatildeo de sulfato de vanadil
(VOSO4) para uma concentraccedilatildeo final de 5 mM em H2O MilliQ saturada com N2 gasoso
352Anaacutelise por espectroscopia de UV-Vis
A concentraccedilatildeo da espeacutecie decameacuterica em soluccedilatildeo depende do periacuteodo de tempo apoacutes a diluiccedilatildeo
no meio de reacccedilatildeo apropriado (Ref) A velocidade de desoligomerizaccedilatildeo da espeacutecie decameacuterica
pode ser analisada por espectroscopia de absorccedilatildeo na regiatildeo UVVis uma vez que esta espeacutecie
apresenta duas bandas caracteriacutesticas na regiatildeo do visiacutevel (a 358 e 400 nm) que natildeo se verifica
em quaisquer outras espeacutecies oligomeacutericas As bandas supramencionada sapresentam-se bem
22
Meio de reacccedilatildeo 3125 mM Tris-HCl pH 71 375 mM MgCl2
48
resolvidas atraveacutes da obtenccedilatildeo da derivada de 2ordf ordem do espectro apresentando a forma de 2
vales com miacutenimos centrados nos comprimentos de onda supracitados com dependecircncia linear
relativamente agrave concentraccedilatildeo de decavanadato total (Ramos et al 2006)
Assim a estabilidade da espeacutecie decameacuterica apoacutes diluiccedilatildeo da soluccedilatildeo de decavanadato no meio de
reacccedilatildeo dos ensaios (tampatildeo D) para uma concentraccedilatildeo final de 1 mM em vanadato total foi
caracterizada atraveacutes da leitura da Abs400 em funccedilatildeo do tempo tendo-se determinado qual a
ordem de reacccedilatildeo e o tempo de meia-vida da mesma Com base em estudos anteriores acerca da
estabilidade do V10 na presenccedila de diferentes proteiacutenas (Ramos et al 2006) determinou-se
igualmente o tempo de meia vida apoacutes a adiccedilatildeo de 20 microM de S1 de miosina ao meio reaccional
353 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
Os espectros de RMN do isoacutetopo 51
V foram realizados num espectroacutemetro Bruker AM-400 MHz
equipado com uma sonda multinuclear inversa de 5 mm Em tubos de RMN de 5 mm de diacircmetro
preparou-se soluccedilotildees de 500 L de deca- ou metavanadato correspondentes a um intervalo de
concentraccedilotildees de 1 a 10 mM suplementadas com 10 D2O Os espectros foram adquiridos agrave
temperatura ambiente com uma largura espectral de 23 923 Hz um tempo de aquisiccedilatildeo de 0086
s tempo de relaxaccedilatildeo de 001 s e uma sequecircncia de pulsos a 90ordm Utilizou-se a transformada de
Fourier com alargamento das bandas de 20Hz para o processamento dos espectros Os desvios
quiacutemicos das ressonacircncias de 51
V foram obtidos em relaccedilatildeo agrave espeacutecie VOCl3cuja frequecircncia de
ressonacircncia eacute considerada o padratildeo sendo todas as outras referenciadas a partir desta em ppm
As aacutereas dos sinais do RMN correspondentes agraves espeacutecies oligomeacutericas presentes foram obtidas
por integraccedilatildeo dos sinais As concentraccedilotildees relativas das espeacutecies foram calculadas com recurso agrave
equaccedilatildeo
onde A[Vx] corresponde agrave aacuterea referente agrave ressonacircncia da espeacutecie Vx com nordm de agregaccedilatildeo x
A[V]total agrave soma total das aacutereas medidas e [V] a concentraccedilatildeo total de vanaacutedio no ensaio
A teacutecnica de 51
V-RMN foi anteriormente aplicada em estudos de ligaccedilatildeo com S1 de miosina
(Ref) tendo sido demostrado que entre as espeacutecies V1 V2 V4 e V5 a espeacutecie tetrameacuterica possui a
maior afinidade para a miosina (Cremo et al) O desaparecimento ou alargamento dos sinais pode
implicar a ligaccedilatildeo de um dos oxovanadatos laacutebeis agrave miosina sem indicaccedilatildeo de qual uma vez que
49
se processa um equiliacutebrio entre eles (Crans 1990) Assim e tendo em conta as indicaccedilotildees de que
a espeacutecie V10 eacute distinta das restantes no que diz respeito agrave magnitude da interacccedilatildeo com a miosina
e inibiccedilatildeo da actividade ATPaacutesica estimulada por actina (Aureliano 2000 Tiago et al 2002)
torna-se fundamental investigar a interacccedilatildeo das diferentes espeacutecies de vanadato com o S1 de
miosina atraveacutes da teacutecnica de 51
V-RMN
Para tal obtiveram-se espectros de 51
V-RMN resultantes da titulaccedilatildeo de uma soluccedilatildeo 5 mM de
decavanadato total com concentraccedilotildees crescentes de S1 de miosina (0 5 10 15 e 20 microM) em
tampatildeo D Realizou-se ainda ensaios na presenccedila de 2 mM de decavanadato e 2 mM de ATP na
presenccedila e na ausecircncia de S1 de miosina de modo a verificar o efeito que estes ligandos naturais
da enzima teria na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o subfragmento-1
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
Os grupos sulfidrilo (-SH) das cisteiacutenas satildeo resiacuteduos de aminoaacutecidos extremamente sensiacuteveis agrave
oxidaccedilatildeo resultando na sua oxidaccedilatildeo (Dalle-Donne et al 1999) Logo a quantificaccedilatildeo destes
resiacuteduos eacute bastante importante nos estudos de interacccedilatildeo com vanaacutedio jaacute que os resiacuteduos de Cys
com ecircnfase para o SH1 e o SH2 satildeo bastante importantes na monitorizaccedilatildeo das modificaccedilotildees
funcionais da regiatildeo da cabeccedila
O procedimento seguidamente apresentado foi originalmente descrito por Gutieacuterrez-Merino
(1987)) Consiste na reacccedilatildeo entre os grupos ndashSH natildeo oxidados das proteiacutenas e o reagente de
Ellman (DTNB -55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico)) - formando-se o aniatildeo CNT (aniatildeo 3-
carboxilato-4-nitrotiofenolato) numa estequiometria de 11 Este apresenta um maacuteximo de
absorvecircncia a 412 nm (Robyt 1990) Assim apoacutes a incubaccedilatildeo da proteiacutena na presenccedila do metal
a quantidade de cisteiacutenas reduzidas por proteiacutena determina-se atraveacutes dos valores de Abs412
Deste modo colocou-se num tubo de Eppendorf o volume apropriado de S1 dialisado para que a
concentraccedilatildeo no ensaio fosse de 1 mgmL Adicionou-se o volume apropriado de decavanadato
para que a sua concentraccedilatildeo fosse de 0 10 50 100 250 500 ou 1000 M Perfez-se com soluccedilatildeo
D ateacute um volume final de 1 mL e incubou-se a 25ordmC por 20 minutos Apoacutes este periacuteodo colocou-
se 780 L da mistura reaccional numa cuvette de quartzo e adicionou-se 220 L de soluccedilatildeo K23
23
Tampatildeo K Tampatildeo fosfatohidrogenofosfato de soacutedio 025 M pH 72
50
Adicionou-se 15 L de DTNB (em soluccedilatildeo K [DTNB]stock=100 mM) ao meio reaccional e
registou-se o valor de absorvecircncia medido equivalente agraves cisteiacutenas ldquoraacutepidasrdquo (SH1 e SH2
expostos ao solvente) Adicionou-se 001 g de SDS que provoca a desnaturaccedilatildeo da proteiacutena e
consequentemente a exposiccedilatildeo de todos os resiacuteduos de cisteiacutenas que se encontravam ateacute aiacute
inacessiacuteveis no interior da proteiacutena (cisteiacutenas totais) Estes resiacuteduos podem assim reagir com o
DTNB
Estimou-se o nuacutemero de resiacuteduos de cisteiacutenas em todas as amostras com recurso ao coeficiente de
extinccedilatildeo molar () do DTNB (ver ponto 3 dos Anexos) que foi determinado atraveacutes da titulaccedilatildeo
de L-Cisteiacutena com intervalo de concentraccedilotildees compreendidos entre 0 e 50 microM na presenccedila de
25microM de DTNB em tampatildeo D Os valores de absorvecircncia foram obtidos para um comprimento de
onda de 412 nm
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
3621 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
A teacutecnica mais eficiente de anaacutelise da interacccedilatildeo entre o iatildeo vanadilo e proteiacutenas como a
calmodulina e a transferrina entre outras permite a obtenccedilatildeo raacutepida de informaccedilatildeo no que toca agrave
conformaccedilatildeo e amplitude da referida interacccedilatildeo
Os espectros de Ressonacircncia Paramagneacutetica Electroacutenica (RPE) da espeacutecie paramagneacutetica V(IV)
foram realizados a 70K num espectroacutemetro Bruker EMX-300 X-band Todas as soluccedilotildees
utilizadas e amostras foram preparadas em anaerobiose numa cacircmara de luvas de modo a evitar a
reduccedilatildeo de V(IV) a V(V) Em tubos capilares com 02 mL de amostra manteve-se a concentraccedilatildeo
de S1 a 20 M e variou-se a concentraccedilatildeo de vanadilo (0 25 5 10 15 25 50 75 100 150
200 500 e 1000 M [VOSO4]stock=5 mM) em tampatildeo D Apoacutes obtenccedilatildeo dos espectros estes
foram tratados atraveacutes do software SimFonia e WinEPR tendo-se obtido um graacutefico com a
variaccedilatildeo da intensidade do sinal de RPE em funccedilatildeo da concentraccedilatildeo de sulfato de vanadilo
presente Realizaram-se ainda simulaccedilotildees de espectros obtidos com 20 microM de S1 e 1000 microM de
V(IV) com vista agrave obtenccedilatildeo dos ligandos provaacuteveis do vanadilo atraveacutes da anaacutelise da constante
de acoplamento hiperfina (A) e do factor g (g)
4 Conclusotildees gerais
Apesar de jaacute ter sido demonstrado que o vanaacutedio inibe a Na+K
+-ATPase da miosina a funccedilatildeo
deste elemento no tecido muacutesculo esqueleacutetico continua largamente incompreendida A enorme
complexidade quiacutemica deste metal especialmente no que concerne agrave oligomerizaccedilatildeo e reacccedilotildees
51
redox constitui um obstaacuteculo agrave compreensatildeo do comportamento do vanaacutedio nos organismos
vivos Assim a caracterizaccedilatildeo da quiacutemica deste elemento eacute de grande utilidade antes de iniciar o
estudo da sua interacccedilatildeo com o presente sistema
A anaacutelise por espectroscopias de 51
V-RMN e de UV-Vis de soluccedilotildees de vanadato diluiacutedas nos
meios de reacccedilatildeo utilizados no presente relatoacuterio (10 mM HEPES pH 70 25 mM KCl) mostrou
claramente que a espeacutecie decameacuterica (V10) possui um tempo de vida suficientemente longo para a
determinaccedilatildeo do seu efeito isolado ao contraacuterio do que se verifica com as espeacutecies mono- di-
tetra- e pentameacutericas (V1 V2 V4 e V5) Por outro lado a anaacutelise verificou ainda que apoacutes diluiccedilatildeo
das soluccedilotildees de decavanadato no meio de reacccedilatildeo coexistem as espeacutecies V10 e V1 pelo que os
efeitos putativos desta uacuteltima deveratildeo ser subtraiacutedos aquando da monitorizaccedilatildeo dos efeitos da
espeacutecie decameacuterica
O vanaacutedio (V) (vanadato) inibe as enzimas que catalizam reacccedilotildees de transferecircncia de grupos
fosfato como a ATPase da miosina [Goodno 1979] devido agrave sua analogia estrutural com o iatildeo
fosfato Na sua utilizaccedilatildeo como inibidor enzimaacutetico natildeo eacute tida em conta a contribuiccedilatildeo individual
das espeacutecies oligomeacutericas considerando-se normalmente a espeacutecie V1 como inibidora efectiva
[Goodno e Taylor 1982] No presente relatoacuterio verificou-se por espectroscopia de 51
V-RMN que
outros oligoacutemeros de vanadato interagem com o S1 de miosina tendo as espeacutecies V10 e V4 os
maiores niacuteveis de afinidade A sua interacccedilatildeo envolve forccedilas electrostaacuteticas e eacute afectada
negativamente por ligandos naturais de enzima como eacute o caso do ATP
A exposiccedilatildeo do S1 de miosina (005 mgmL) a decavanadato conduz agrave inibiccedilatildeo do capacidade da
proteiacutena de estimular a actividade Mg2+
-ATPaacutesica com um valor de IC50 de 266 plusmn 07 M
decavanadato sendo o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM
decavanadato (80) Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM decavanadato sendo
o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM decavanadato (60)
exercendo um efeito de impedimento agrave ligaccedilatildeo entre o vanaacutedio e o S1
A uma concentraccedilatildeo de 27 microM decavanadato verifica-se a oxidaccedilatildeo de um dos resiacuteduos de Cys
de S1 de miosina seguida pela oxidaccedilatildeo gradual de outro resiacuteduo de Cys ateacute agrave concentraccedilatildeo de
1000 microM Ambos os resiacuteduos oxidados natildeo se encontravam expostos ao solvente utilizado A
oxidaccedilatildeo dos resiacuteduos permite sugerir que esta modificaccedilatildeo oxidativa sofrida pelo S1 contribui
para a diminuiccedilatildeo da sua actividade de ATPase
52
Por espectroscopia de RPE revelaram-se os possiacuteveis ligandos equatoriais do vanaacutedio aquando da
sua interacccedilatildeo com o S1 de miosina como sendo duas moleacuteculas de aacutegua um ligando contendo o
grupo R-OH e outro contendo o grupo R=NR
Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados de
oxidaccedilatildeo (IV) e (V) no S1 de miosina para concentraccedilotildees facilmente alcanccedilaacuteveis no ambiente
terrestre
53
5 Estudos futuros
No sentido do melhor esclarecimento das alteraccedilotildees moleculares e estruturais que ocorrem no
subfragmento 1 quando exposta ao vanaacutedio que conduzem a uma diminuiccedilatildeo da sua funccedilatildeo
sugere-se
- Quantificaccedilatildeo das cisteiacutenas reduzidas de S1 na presenccedila de metavanadato
- Determinaccedilatildeo das actividades Ca2+
- e K+-ATPaacutesica do S1 de miosina na presenccedila de
decavanadato
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina estimulada por actina
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina na presenccedila de
metavanadato
- Realizaccedilatildeo de RMN 2D com variaccedilatildeo das concentraccedilotildees das soluccedilotildees de decavanadato e
metavanadato na presenccedila de S1
-Mutageacutenese dos resiacuteduos de Cys doS1 de miosina de modo a verificar quais os resiacuteduos que
sofrem oxidaccedilatildeo na presenccedila de decavanadato
- Determinaccedilatildeo por calorimetria (DSC Differential Scanning Calorimetry) da variaccedilatildeo da
entalpia (∆H) e da temperatura de desnaturaccedilatildeo do S1 de miosina na presenccedila de vanaacutedio uma
vez que estas propriedades caracteriacutesticas de cada proteiacutena satildeo alteradas apoacutes ocorrecircncia de
modificaccedilotildees estruturais
- Anaacutelise por SDS-PAGE de S1 de miosina apoacutes tratamento com vanaacutedio de modo a determinar
quais os polipeacuteptidos eventualmente formados apoacutes clivagem da proteiacutena
54
6 Referecircncias bibliograacuteficas
- Sellers J Myosins 2nd Edition Oxford University Press 1999
- L Stryer inBiochemistry (Ed WH Freeman 2005 pp
Refordfas Materiais e Meacutetodos
[A] S S Margossian SLowey Methods Enzymol1982 85 55-71
[B] JD Pardue JA Spudich Methods Enzymol 1982 85 164-181
[C] M Bradford Anal Biochem1976 72 248-254
[D] H H Taussky E Shorr J Biol Chem 1953 202 675-685
[E] C R Bagshaw in Outline Studies in Biology ndash Muscle Contraction Chapman and Hall Ed
1982
[F] A Cuenda et al Eur J Biochem1990 194 663-670
55
Anexos
1 Determinaccedilatildeo da recta de calibraccedilatildeo meacutetodo de Bradford
Figura1ndashRepresentaccedilatildeo graacutefica da recta de calibraccedilatildeo (Abs595 nm vs [BSA]) utilizada para o meacutetodo de Bradford
2 Determinaccedilatildeo da recta calibraccedilatildeo para a electroforese em condiccedilotildees desnaturantes
Figura 2 ndash Representaccedilatildeo graacutefica da recta de calibraccedilatildeo (log(Peso Molecular) vs Rf dos marcadores de peso
molecular) onde Rf indica determinada atraveacutes do padratildeo de peso molecular
(poccedilo (a))
3Caacutelculo do coeficiente de extinccedilatildeo molar () para a quantificaccedilatildeo de cisteiacutenas reduzidas
atraveacutes do meacutetodo de Ellman
Traccedilou-se a recta de Abs412 nm meacutedia vs [L-Cisteiacutena] de modo a determinar o valor do coeficiente
de extinccedilatildeo molar definido como o declive daquela recta Assim =8400 M-1
cm-1
como
podemos verificar na Fig 3
A595 nm = 00546[BSA] + 00209 Rsup2 = 099051
Ab
s 59
5 n
m
[BSA] (mgmL)
log PM = -1177 Rf + 2247 Rsup2 = 098957
log
PM
Mobilidade relativa (Rf)
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero

16
A quiacutemica do vanaacutedio eacute extraordinariamente complexa pelo que originou vaacuterias interpretaccedilotildees
incorrectas desde a sua descoberta No entanto as suas propriedades dependem em uacuteltima
anaacutelise da configuraccedilatildeo electroacutenica do elemento Assim a variabilidade encontrada no
comportamento quiacutemico do vanaacutedio pode dever-se agrave sua configuraccedilatildeo ndash [Ar] 3d2 4s
2 ndash que
permite a formaccedilatildeo de vaacuterios estados de oxidaccedilatildeo (de +2 a +5) Em soluccedilatildeo aquosa os estados de
oxidaccedilatildeo estaacuteveis vatildeo de V(V) a V(II) Destes apenas os +3 a +5 satildeo biologicamente importantes
jaacute que o V(II) apresenta demasiada capacidade redutora para existir nos organismos vivos
conhecidos [AC]
No fluido extracelular os compostos de vanaacutedio encontram-se predominantemente na forma de
monovanadatos (V(V)) livres ou ligados agrave transferina do soro ou agrave hemoglobina (Refordf) Os iotildees de
vanadato livres satildeo transportados para o meio intracelular atraveacutes de canais ioacutenicos inespeciacuteficos
Jaacute no ambiente intracelular os compostos sofreratildeo uma bio-reduccedilatildeo atraveacutes da acccedilatildeo da
glutationa ou outros compostos com grupos ndashOH como pentoses hexoses ou etanolaminas [V] A
reoxidaccedilatildeo de vanadilo (V(IV)) intracelular a vanadato (V(V)) provocaria a acumulaccedilatildeo toacutexica
destes uacuteltimos pelo que a complexaccedilatildeo da espeacutecie reduzida o impede O transporte de vanaacutedio
para o interior da ceacutelula encontra-se na Fig 110
Figura 110 ndash Transporte de vanaacutedio para a ceacuteluka e seu subsequente metabolismo (P=Proteiacutena) (Adaptado de
Zaporowska e Scibor 1998)
1211 Vanaacutedio(IV)
A espeacutecie de vanaacutedio (IV) mais bem conhecida eacute o iatildeo vanadilo (VO2+
) espeacutecie estaacutevel a pH
aacutecido sendo a maioritaacuteria a pH 3[AD] Apresenta uma cor azulada e forma complexos penta e
hexacoordenados Forma-se rapidamente a partir de V(V) na presenccedila de um agente redutor em
soluccedilatildeo aquosa No ambiente intracelular a presenccedila de glutationa reduzida aacutecido ascoacuterbico
NAD(P)H entre outros conduzem a que o principal estado de oxidaccedilatildeo nos sistemas bioloacutegicos
seja provavelmente o +4 encontrando-se o V(IV) fortemente complexado com proteiacutenas ou
aminoaacutecidos livres citrato ATP e pirofosfato (Refordfs) Em alternativa o V(IV) pode existir
17
livremente em ambientes fortemente aacutecidos como os lisossomas jaacute que a pH superior a 3 as
soluccedilotildees de vanadilo tendem a ser oxidadas pelo ar formando-se vanadato Esta grande
capacidade redutora justifica parcialmente a utilizaccedilatildeo pouco extensiva do iatildeo VO2+
como
inibidor [W]
1212 Vanaacutedio (V)
O vanaacutedio (V) eacute a forma mais estaacutevel num intervalo de pH amplo [W] O iatildeo anioacutenico VO3-tende
a constituir complexos polinucleares pelo que as soluccedilotildees aquosas de vanadato satildeo extremamente
complexas com ocorrecircncia simultacircnea de vaacuterios equiliacutebrios de protonaccedilatildeo (pKa(vanadato)= 35
78 e 125) e oligomerizaccedilatildeo As espeacutecies presentes (H3VO4 H2VO4- HVO4
2- e VO4
3-) dependem
do pH forccedila ioacutenica e temperatura da soluccedilatildeo e da concentraccedilatildeo total do elemento
[AEAFAGAHAI]
O VO43-
- monoacutemero ortovanadato V1 ndash estaacute associado a uma grande parte da importacircncia
bioloacutegica do vanaacutedio uma vez que se trata de um anaacutelogo electroacutenico e estrutural do ortofosfato
(PO43-
) A coordenaccedilatildeo do ortovanadato pode ser tetraeacutedrica ou bipiramidal trigonal (Refordfs)
A oligomerizaccedilatildeo do vanadato monomeacuterico daacute origem ao diacutemero (V2O72-
- V2) que apresenta
tambeacutem equiliacutebrios de protonaccedilatildeo Presume-se a existecircncia de analogia do vanadato dimeacuterico com
o pirofosfato embora natildeo haja evidecircncia estrutural (Refordf) A dimerizaccedilatildeo do V2 origina o
vanadato tetrameacuterico ou metavanadato (V4O124-
- V4) que constitui a espeacutecie maioritaacuteria em
soluccedilotildees aquosas concentradas (Refordf) As estruturas do metavanadato e a sua espeacutecie protonada
satildeo ciacuteclicas [AJ] Outra espeacutecie oligomeacuterica importante eacute o vanadato pentameacuterico (V5O153-
) cuja
estrutura presumida eacute ciacuteclica [AI]
O oligoacutemero com maior importacircncia bioquiacutemica eacute o vanadato decameacuterico (V10O286-
-V10)(Refordf) a
espeacutecie predominante em soluccedilotildees de pH 2-6 quando a concentraccedilatildeo total de vanadato eacute superior
a 100 M (Ref) A estrutura do V10 natildeo difere em soluccedilatildeo ou no estado soacutelidoOs 10 aacutetomos de
vanaacutedio da espeacutecie decameacuterica estatildeo divididos em trecircs grupos identificados na figura como VA-C
consoante o seu ambiente quiacutemico circundante (Refordf) (Fig 111)
A expressatildeo ldquodecavanadatordquo refere-se a uma soluccedilatildeo aquosa na qual a espeacutecie decameacuterica se
encontra em equiliacutebrio quiacutemico com as espeacutecies V1 V2 V4 e V5 sendo que a oligomerizaccedilatildeo
progressiva do vanaacutedio eacute proporcional agrave sua concentraccedilatildeo total (Ref) Nesta soluccedilatildeo a
concentraccedilatildeo de V10 corresponde a aproximadamente 10 da concentraccedilatildeo total de vanaacutedio [AK]
18
Figura 111 ndash Estruturas esquemaacuteticas das espeacutecies de vanadato (espeacutecie monomeacuterica (V1) dimeacuterica (V2)
tetrameacuterica (V4) pentameacuterica (V5) e decameacuterica (V10) (Adaptado de Aureliano e Crans 2008)
122 Metabolismo e efeito na actividade enzimaacutetica
Apesar de natildeo terem sido ainda encontradas as provas categoacutericas das suas funccedilotildees nos seres
vivos sabe-se que o iatildeo vanadato eacute capaz de inibir uma grande quantidade de enzimas in vitro
com especial ecircnfase na actividade ATPaacutesica da miosina [AL Teresa] Como referido
anteriormente o vanadato eacute capaz de actuar em alternativa ao fosfato nas reacccedilotildees bioquiacutemicas
incluindo a sua preferecircncia de ligaccedilatildeo agraves enzimas [W]
Eacute conhecido o papel do vanaacutedio no metabolismo do caacutelcio promovendo a mineralizaccedilatildeo dos
dentes e ossos na biossiacutentese de liacutepidos (trigliceacuteridos e colesterol) com posterior inibiccedilatildeo da
acumulaccedilatildeo destes nos vasos sanguiacuteneos e no SNC na reproduccedilatildeo e desenvolvimento por
mimetizaccedilatildeo e potenciaccedilatildeo de factores de crescimento na estimulaccedilatildeo do consumo de glucose e
da siacutentese de glicogeacutenio com actuaccedilatildeo equivalente aos efeitos da insulina ao niacutevel celular na
transduccedilatildeo de sinais celulares com regulaccedilatildeo da actividade de segundos mensageiros no
transporte de iotildees e aminoaacutecidos na transcriccedilatildeo geneacutetica e siacutentese de proteiacutenas e DNA entre
outros [W Y AB]
Assim o vanaacutedio cnseguiraacute provocar inuacutemeras mudanccedilas bioquiacutemicas nas ceacutelulas Alguns
exemplos da influecircncia bioquiacutemica do vanaacutedio incluem os seus efeitos nos processos de
fosforilaccedilatildeo e desfosforilaccedilatildeo de proteiacutenas e na participaccedilatildeo na formaccedilatildeo de espeacutecies reactivas de
oxigeacutenio devido ao seu efeito proacute-oxidante sobre agentes redutores a niacutevel celular [AM]
Quanto ao seu efeito na actividade enzimaacutetica pode dividir-se em dois tipos principais ndash inibitoacuterio
e estimulante - dependendo da enzima ou do tipo de reacccedilatildeo por esta catalisada No primeiro
grupo incluem-se enzimas como as ATPases (de miosina de canais ioacutenicos e fosfoenzimas
19
transportadoras) e as fosfatases (da tirosina aacutecida e alcalina) a fosfofrutocinase adenilato ciclase
e catalase entre outras [AA] O segundo grupo inclui a maioria das cinases a glucose-6-fosfato
desidrogenase a fosfoglicerato mutase e a fosfoglicerato fosfatase (Refordf)
O efeito do vanaacutedio (V) na actividade enzimaacutetica estaacute relacionado com o seu nuacutemero de
coordenaccedilatildeo podendo actuar como estado de transiccedilatildeo ou por mimetizaccedilatildeo de cofactores
substratos (especialmente o fosfato) e interacccedilotildees proteiacutena-proteiacutena (como no caso das
polimerases e transcriptases) Pode ainda estar envolvido na formaccedilatildeo de ligaccedilotildees de hidrogeacutenio
interacccedilotildees electrostaacuteticas esteacutereas e de oxidaccedilatildeo-reduccedilatildeo Uma evidecircncia desta uacuteltima eacute a sua
capacidade de alternacircncia entre os estados de oxidaccedilatildeo +5 a +3 o que torna faacutecil a sua interacccedilatildeo
redox com enzimas contendo resiacuteduos de cisteiacutenas (Cys) ou qualquer outro composto orgacircnico
com actividade redox [AN]
13 Vanaacutedio e muacutesculo esqueleacutetico
A primeira evidecircncia da presenccedila de vanaacutedio no muacutesculo esqueleacutetico proveio de uma amostra de
ATP isolado de muacutesculo de cavalo que continha uma impureza ndash vanaacutedio ndash cuja presenccedila
provocou a inibiccedilatildeo da actividade da bomba de Na+K
+-ATPase [AO] Desde entatildeo vaacuterios
estudos tecircm sido realizados com ecircnfase na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o
tecido muscular Destes destaca-se a descoberta da interacccedilatildeo forte da espeacutecie V10 com a miosina
fosforilada [AP] e agrave sua inibiccedilatildeo da actividade ATPaacutesica da miosina[AQ AR]
No quediz respeitoao iatildeo monovanadato destaca-se a sua inibiccedilatildeoda actividade Mg2+
-ATPaacutesica
da miosina [AS] que ocorre de duas formas distintas (1) na presenccedila de concentraccedilotildees reduzidas
de V1 a inibiccedilatildeo ocorre a velocidade reduzida dependente da concentraccedilatildeo da espeacutecie
monomeacuterica (2) na presenccedila de concentraccedilotildees elevadas de V1 o niacutevel de inibiccedilatildeo aproxima-se de
100 desde que [V1] seja superior agrave concentraccedilatildeo total de locais com actividade Mg2+
-ATPaacutesica
no ensaio Esta inibiccedilatildeo eacute irreversiacutevel tanto por diaacutelise ou adiccedilatildeo de agentes redutores do
vanadato [AL]Segundo Pate et al(1994)[AT] a espeacutecie monomeacuterica podeinibir a velocidade de
contracccedilatildeo a temperatura inferior a 20ordmC perdendogradual e rapidamente essa capacidade ateacute os
50ordmC temperatura agrave qual cessa a capacidade inibitoacuteria Os resultados obtidos sugeremainda uma
interacccedilatildeo altamente cooperativa entre o iatildeo vanadato e as proteiacutenas componentes do muacutesculo
esqueleacuteticoDestaca-se tambeacutem a descoberta da inibiccedilatildeo da geraccedilatildeo de forccedila contraacutectil nas fibras
musculares por parte da espeacutecie monomeacuterica[AT]Wilson et al(1995) [AU ]determinaram que a
afinidade da espeacutecie V1 para o S1 de miosina eacute significativamente mais elevada do que a do iatildeo
Pi seu anaacutelogo Descobriram ainda que que a ligaccedilatildeo do vanadato poderaacute envolver transiccedilotildees
entre estados energeacuteticos com energia livre independente dos impedimentos esteacutereos impostos
20
pela estrutura das miofibrilas Os estudos desenvolvidos por Smith et al (1990)[AV]
determinaram queo iatildeo monovanadato natildeo apresenta um efeito pronunciado na inibiccedilatildeo da
actividadeacto-S1-miosina-ATPaacutesica excepto em concentraccedilotildees superiores a 900microM a partir das
quais existe produccedilatildeo de espeacutecies oligomeacutericas para as quais o efeito inibitoacuterio seraacute mais
elevadoA espeacutecie monomeacuterica de vanadato consegue adoptar facilmente uma geometria do tipo
bipiramidal trigonal que mimetiza o intermediaacuterio formado (MgADP
Pi)aquando da reacccedilatildeo de
hidroacutelise de ATP no local activo da miosina [AW] De facto a interacccedilatildeo do vanaacutedio com a
miosina eacute inesperadamente estaacutevel na presenccedila de ADP Esta interacccedilatildeo resulta na formaccedilatildeo de
um complexo enzimaacutetico inactivo e estaacutevel com estequiometria de 11 ADPvanadato por siacutetio
activo da proteiacutena A formaccedilatildeo do complexo estaacutevel miosina-vanadato foi proposta por Goodnoet
al (1982) [AS] Nesta ocorre um preacute-equiliacutebrio raacutepido que induz a formaccedilatildeo de um complexo
ternaacuterio reversiacutevel (MADP
V) com formaccedilatildeo lenta do isoacutemero M
ADP
V A diferenccedila entre os
compostos de vanaacutedio e de fosfato centra-se no tempo de meia vida (t12) do Pi ou do V (no
primeiro t12=12 s enquanto no segundo t12=24 h) (Refordf) A diferenccedila dever-se-aacute agrave possiacutevel
complexaccedilatildeo do vanaacutedio com grupos nucleofiacutelicos no siacutetio activo o que provocaria uma
estabilizaccedilatildeo da miosina na conformaccedilatildeo MADP
V
No que respeita aos estudos realizados com a espeacutecie tetrameacuterica Cremo et al determinou que a
cadeia pesada do S1 de miosina pode ser clivada em dois locais distintos (designados V1 e V2)
pela espeacutecie V4 apoacutes irradiaccedilatildeo com luz UV na presenccedila de concentraccedilotildees de vanadato na ordem
dos milimolar e na ausecircncia de nucleoacutetidos ou metais divalentes
Jaacute no que concerne aos estudos realizados com ecircnfase na espeacutecie decameacuterica foi sugerido
porRamos et al (2006) [AK]que a interacccedilatildeo do decavanadato com a G-actina favorecida pela
polimerizaccedilatildeo desta uacuteltima estabiliza a espeacutecie V10 e induz a inibiccedilatildeo da formaccedilatildeo de F-actina A
estabilizaccedilatildeo da espeacutecie V10 por proteiacutenas tansmembranares e do citoesqueleto pode ser a razatildeo
pela qual ocorre a toxicidade do decavanadato reportada na avaliaccedilatildeo dos efeitos do V(V) nos
sistemas bioloacutegicosTiago et al (2007) [AR] concluiram que o V10 apresenta uma clara preferecircncia
de ligaccedilatildeo ao ldquoback-doorrdquo da S1-miosina-ATPase o que se verifica apenas na estrutura
conformacional desimpedida com acesso facilitado ao local de ligaccedilatildeo do Pi Sugere tambeacutem que
o V10 actua como um ldquoback-door stoprdquo bloqueando estaabertura do fragmento de 50-kDa
necessaacuteria para a hidroacutelise do ATP--fosfato o que constitui uma explicaccedilatildeo simples do
comportamento natildeo competitivo da espeacutecie decameacuterica estimulando a utilizaccedilatildeo deste oligoacutemero
como uma feramenta da elucidaccedilatildeo das mudanccedilas conformacionais ldquoback-doorrdquo da miosina no
21
processo da contracccedilatildeo muscular(ref)Destaca-se ainda a descoberta da capacidade inibitoacuteria do
V10 em relaccedilatildeo agrave fosforilaccedilatildeo oxidativa mitocondrial [AX]
Finalmente no que concerne aos estudos realizados com a presenccedila de vaacuterias espeacutecies
oligomeacutericas Aureliano e Madeira (1994) [AY]observaram que na presenccedila de Ca2+
as espeacutecies
V4 e V10se ligam agrave Ca2+
-ATPase do retiacuteculo sarcoplasmaacutetico enquanto a espeacutecie V1 se liga apenas
na presenccedila de ATP Aparentemente o aumento da concentraccedilatildeo de Ca2+
tem o efeito contraacuterio da
inibiccedilatildeo da ATPase do retiacuteculo sarcoplasmaacutetico pelo V1 natildeo tendo no entanto efeito significativo
sobre a inibiccedilatildeo por parte da espeacutecie V10 Tiago et al (2002) [AP] concluiacuteram que em condiccedilotildees
proacuteximas agraves fisioloacutegicas a espeacutecie V10 difere das espeacutecies presentes na soluccedilatildeo de metavanadato
devido agrave sua forte interacccedilatildeo com a miosina fosforilada e agrave sua inibiccedilatildeo da actividade ATPaacutesica
da miosina Para aleacutem disso o ATP diminui a afinidade da miosina para o V4 induzindo a
interacccedilatildeo com V1 e natildeo tendo qualquer efeito no que diz respeito ao V10 Foi tambeacutem proposto
[AQ] que a ligaccedilatildeo das espeacutecies oligomeacutericas de vanadato aoS1 de miosina nos locais onde o
fosfato coordenaeacute provavelmente a causa da inibiccedilatildeo da actividade ATPaacutesica da miosina
estimulada por actina atraveacutes da prevenccedilatildeo da troca entre ATPADP sendo esta favorecida para
aniotildees de vanadato com valecircncias superiores como eacute o caso do V10
Na presenccedila de actina as velocidades de dissociaccedilatildeo do Pi e do V aumentam consideravelmente
este uacuteltimo em menor quantidade Assim a ligaccedilatildeo da actina ao complexo MADP
V e
Pi eacute
bastante fraca pelo que se sugere que o ADPV teria um efeito de relaxamento semelhante ao
ATP No entanto a capacidade do vanaacutedio em inibir a actividade ATPaacutesica da miosina e da
actomiosina eacute diferenciada uma vez que na ausecircncia de actina a inibiccedilatildeo eacute total para
concentraccedilotildees na gama dos M mas na sua presenccedila a inibiccedilatildeo eacute totalmente conseguida apenas
em concentraccedilotildees na gama dos mM (Ref) No entanto eacute necessaacuterio considerar a possiacutevel
formaccedilatildeo de espeacutecies oligomeacutericas de vanadato cuja afinidade e efeito na enzima seratildeo
diferentes Assim pode concluir-se que o efeito de inibiccedilatildeo observado para a actividade ATPaacutesica
de miosina estimulada por actina se pode dever agrave presenccedila das diferentes espeacutecies oligomeacutericas o
que foi jaacute sugerido por Aureliano (2000) [AZ] ndash as espeacutecies tetrameacuterica e decameacuterica de vanadato
interactuariam com a miosina com alteraccedilatildeo da sua interacccedilatildeo com a actina No entanto natildeo eacute
conhecida a contribuiccedilatildeo absoluta das restantes espeacutecies oligomeacutericas para a inibiccedilatildeo da ATPase
da miosina estimulada por actina (Refs)
14 Objectivos do projecto
Pretende-se com este projecto o isolamento purificaccedilatildeo e caracterizaccedilatildeo da actina do muacutesculo
22
esqueleacutetico de coelho sendo esta uacuteltima tambeacutem realizada para o sufragmento-1 de miosina
Seguidamente realizar-se-aacute a quantificaccedilatildeo e caracterizaccedilatildeo das proteiacutenas por espectroscopia UV-
Vis e SDS-PAGE e a caracterizaccedilatildeo das soluccedilotildees de vanaacutedio por UV-Vis e ressonacircncia magneacutetica
nuclear (RMN) Por uacuteltimo a interacccedilatildeo do S1 de miosina com o vanaacutedio seraacute avaliada atraveacutes
das teacutecnicas de UVVis RMN e ressonacircncia paramagneacutetica electroacutenica (RPE)
2 Discussatildeo de Resultados
21 Caracterizaccedilatildeo das proteiacutenas isoladas
211Anaacutelise do estado de pureza
O estado de pureza das soluccedilotildees de actina e de S1 foi analisado por electroforese em gel de
poliacrilamida em condiccedilotildees desnaturantes (SDS-PAGE) (Fig 22) que separa as moleacuteculas de
acordo com o seu peso molecular (PM) A concentraccedilatildeo de acrilamida escolhida (10) permite a
detecccedilatildeo simultacircnea de componentes com peso molecular inferior a 20 kDa e outros com elevado
peso molecular
Figura 21 ndash Fotografia de gel de SDS-PAGE (10 acrilamida) Poccedilo (a)
10 L do marcador de elevado peso molecular () Poccedilo (b) 20 L de [G-
actina]=1 mgmL Poccedilo (c) 20 L de [S1]=1 mgmL Aplicou-se uma
voltagem de V durante h e min
A partir da recta de calibraccedilatildeo obtida log PM = -1177 Rf + 2247 (ver ponto 2 da secccedilatildeo
Anexos) pode verificar-se a presenccedila de actina na amostra do poccedilo (b) presente numa banda
bastante intensa (PMaparente=40 kDa) bem como a presenccedila minoritaacuteria de outros componentes
desconhecidos que natildeo teratildeo sido eliminados durante o processo de extracccedilatildeo e purificaccedilatildeo e natildeo
satildeo relevantes para o estudo em questatildeo Na amostra do poccedilo (c) nota-se a presenccedila de S1
(PMaparente=92 kDa) bem como a de outros componentes cuja banda eacute ligeiramente menos intensa
Estes poderatildeo ser fragmentos proteoliacuteticos de miosina e de S1 Estaacute ainda presente uma bada de
23
maior intensidade que poderaacute ser a proteiacutena-C (PM=135 kDa) uma vez que a sua remoccedilatildeo
completa da amostra necessitaria de um passo adicional de cromatografia de troca ioacutenica [BA]
No entanto natildeo podemos confirmar que se trata desta proteiacutena ou de qualquer outra uma vez que
se encontra fora do intervalo de migraccedilatildeo dos marcadores de peso molecular
212 Determinaccedilatildeo da concentraccedilatildeo proteica
2121Subfragmento-1 de miosina
A partir da recta de calibraccedilatildeo obtida pelo meacutetodo de Bradford [BB](ver secccedilatildeo 3311 do
Procedimento Experimental) A595nm = 00546[BSA] + 00209 (ver secccedilatildeo de Anexos)foi possiacutevel
determinar um intervalo de concentraccedilatildeo de S1 de miosina compreendido entre 226 e 117(plusmn
423M) (203 e 1056 (plusmn 381 mgmL)) (n=8) A concentraccedilatildeo meacutedia de S1 de miosina apoacutes
diaacutelise contra a soluccedilatildeo tampatildeo 10 mM HEPES pH 75 25 mM KCl determinada pela leitura da
Abs280 nmfoi de 20430M (1803 mgmL)(n=7)
As amostras a quantificar atraveacutes do meacutetodo de Bradford natildeo sofreram diaacutelise o que permite
explicar a concentraccedilatildeo superior de S1 de miosina em comparaccedilatildeo com o valor obtido atraveacutes do
meacutetodo directo Esta razatildeo aliada agrave sensibilidade e rapidez superiores deste uacuteltimo fez com que
se optasse pelo meacutetodo directo para a quantificaccedilatildeo da grande maioria dos ensaios
2122Actina
A concentraccedilatildeo de actina foi determinada quando esta se encontrava na forma monomeacuterica (G-
actina) por mediccedilatildeo da leitura directa da Abs290 nm tendo-se obtido uma concentraccedilatildeo meacutedia de
691481M (290202 mgmL) (n=3) em 2 mM Tris-HCl pH 80 02 mM CaCl2 02 mM Na-
ATP Ao comprimento de onda utilizado (290 nm) absorvem fortemente apenas os resiacuteduos de
Trp pelo que a mediccedilatildeo efectuada tem menor sensibilidade do que se tivesse sido efectuada a 280
nm uma vez que a actina apresenta apenas 4 resiacuteduos deste aminoaacutecido embora tenha 16
resiacuteduos de Tyr
22 Mediccedilatildeo da actividade enzimaacutetica de S1 de miosina
221 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
As actividades ATPaacutesicas de S1 de miosina dependentes de Ca2+
e K+foram analisadas pelo
caacutelculo das velocidades de consumo inicial do fosfato inorgacircnico (Pi) Definiu-se os tempos de 5
e 3 minutos para a mediccedilatildeo da Ca2+
e da K+EDTA-ATPase respectivamente Os valores obtidos
24
a partir do protocolo presente no ponto 341 da secccedilatildeo Procedimento experimental encontram-se
descritos na Tabela 2I
Tabela 2I ndash Acrividades natildeo-fisioloacutegicas de subfragmento-1 determinadas pelo meacutetodo de Fiske-Subarow
modificado (n=2)
Proteiacutena
Actividade
ATPaacutesica natildeo
fisioloacutegica
Actividade (nmol
Piminmg proteiacutena) Actividade (s-1)
S1
Ca2+
-ATPase 1400 plusmn 30 2100 plusmn 0045
K+EDTA-ATPase 2334 plusmn 50 3500 plusmn 0075
As actividades natildeo-fisioloacutegicas reflectem o estado de oxidaccedilatildeo dos resiacuteduos expostos de Cys 707
(SH1) e 697 (SH2) Se o grupo SH1 sofrer oxidaccedilatildeo a actividade ATPaacutesica dependente de Ca2+
aumenta e a actividade dependente de K+ diminui Por outro lado se o grupo SH2 sofrer
oxidaccedilatildeo regista-se a perda completa destas actividades
De acordo com Margossian e Lowey (1982) [BA] as actividades ATPaacutesica de S1 de miosina
dependente de Ca2+
e K+EDTA da miosina satildeo respectivamente de 24 e 112 s-1
Estaacute tambeacutem
descrito que a actividade ATPaacutesica dependente de K+ eacute em meacutedia cerca de 3 vezes superior agrave
dependente de Ca2+
reflectindo a quantidade das preparaccedilotildees A razatildeo obtida entre os valores das
actividades ATPaacutesicas dependentes de K+ e Ca
2+ eacute de 166 Os valores obtidos para as actividades
(Ca2+
-ATPase=21 s-1
K+EDTA-ATPase= 35 s
-1) satildeo inferiores agrave valores obtidos por
Margossian e Lowey [BA] e Tiago [BC] com forccedila ioacutenica inferior O valor obtido para a
actividade ATPaacutesica dependente de Ca2+
eacute superior ao obtido por Polosukhina et al (2000) [BD]
cujo meio de reacccedilatildeo continha MOPS e apresentava forccedila ioacutenica inferior Os dados
supramencionados comprovam a variaccedilatildeo da actividade a ATPaacutesica natildeo fisioloacutegica em relaccedilatildeo agrave
forccedila ioacutenica temperatura e pH
222 Mediccedilatildeo da actividadeMg2+
-ATPaacutesica de S1 pelo meacutetodo de enzimas acopladas
A actividade de Mg2+
-ATPase eacute a uacutenica que possui significado fisioloacutegico sendo as restantes
bastante uacuteteis apenas para indicaccedilatildeo da qualidade de preparaccedilatildeo do S1[BA] A 0575 M KCl a
actina e o S1 formam uma soluccedilatildeo viscosa e tuacuterbida Apoacutes a adiccedilatildeo de MgATP o complexo acto-
S1 dissocia-se e a turbidez da soluccedilatildeo diminui Se a forccedila ioacutenica for reduzida a dissociaccedilatildeo do
complexo actoS1 que se encontra na forma de suspensatildeo eacute induzida O MgATP eacute rapidamente
hidrolisado diminuindo a sua concentraccedilatildeo no meio reaccional Nestas condiccedilotildees haacute
reassociaccedilatildeo e subsequente contracccedilatildeo dos filamentos de actina e de S1 com formaccedilatildeo de
25
precipitado O meacutetodo descrito traduz de forma bastante elementar o fenoacutemeno da contracccedilatildeo in
vivo e elimina o problema anterior da solubilizaccedilatildeo ineficiente da miosina de modo a obter uma
mediccedilatildeo exacta da actividade [A] A utilizaccedilatildeo de forccedilas ioacutenicas reduzidas aumenta a afinidade da
actina para o S1 permitindo contornar a extrema dificuldade em obter a concentraccedilatildeo proteica dos
muacutesculos (cerca de 100 mgmL do complexo actomiosina) As actividades Mg2+
-ATPaacutesicas de S1
de miosina foram determinadas a partir do declive da variaccedilatildeo da Abs340 relativo agrave oxidaccedilatildeo de
NADH em funccedilatildeo do tempo como eacute descrito no ponto 342 do Procedimento Experimental Os
valores obtidos para a actividade Mg2+
-ATPaacutesica encontram-se apresentados na Tabela 2II
Tabela 2II ndash Actividade Mg2+
-ATPaacutesica de subfragmento-1 de miosina determinadas pelo meacutetodo das enzimas
acopladas
Actividade (nmol
ATPmin mg(S1)) Actividade (s
-1)
1097 plusmn 269 164 plusmn 04
Ramos (2005) [J] obteve o valor de 01 s-1
para a actividade Mg2+
-ATPaacutesica de S1 de miosina
com forccedila ioacutenica intermeacutedia Tiago (2005) [BC] obteve o valor de 011 s-1 em condiccedilotildees de forccedila
ioacutenica reduzida em relaccedilatildeo agrave utilizada no presente relatoacuterio
23 Preparaccedilatildeo e caracterizaccedilatildeo das soluccedilotildees de vanaacutedio
231 Anaacutelise da composiccedilatildeo das soluccedilotildees de vanadato por espectroscopia de 51
V-RMN
efeito da concentraccedilatildeo da espeacutecie
A diluiccedilatildeo das soluccedilotildees stock de meta- e decavanadato iraacute determinar fortemente a composiccedilatildeo
em soluccedilatildeo dos diferentes oligoacutemeros de vanadato (ref) No caso do decavanadato o periacuteodo de
tempo apoacutes a diluiccedilatildeo iraacute tambeacutem ser um factor importante a considerar devido agrave decomposiccedilatildeo
da espeacutecie V10(ref)Torna-se assim fundamental determinar a composiccedilatildeo quiacutemica das soluccedilotildees
no meio de ensaio utilizado (HEPES 10 mM KCl 25 mM) de modo a correlacionaacute-la
correctamente com os possiacuteveis efeitos promovidos pelas diferentes espeacutecies de vanadato no
sistema em estudo
Aleacutem disso deve ser dado ecircnfase agraves importantes interacccedilotildees que ocorrem entre o vanaacutedio e os
tampotildees escolhidos O tampatildeo com menor nuacutemero e magnitude de interacccedilatildeo eacute o HEPES [AD]
seguido do Tris sendo este uacuteltimo especialmente aconselhado em concentraccedilotildees baixas para casos
de incompatibilidade enzimaacutetica ou quiacutemica redox No caso especial da reacccedilatildeo redox
NAD(P)HNAD(P) o tampatildeo adequado eacute o imidazole de modo a natildeo haver reacccedilatildeo com o
26
vanadato Os tampotildees a evitar em qualquer situaccedilatildeo incluem a trietanolamina fosfato citrato e
glicina entre outros [AD] uma vez que todos interactuam fortemente com V(V) com formaccedilatildeo
de complexos estaacuteveis
Assim procedeu-se agrave anaacutelise da composiccedilatildeo das soluccedilotildees de meta- e decavanadato apoacutes diluiccedilatildeo
das soluccedilotildees stock (50 mM em vanadato total) no meio de reacccedilatildeo (tampatildeo D) atraveacutes da
espectroscopia de ressonacircncia magneacutetica nuclear de 51
V Nas figuras x1 e x2 apresentam-se os
espectros obtidos para as soluccedilotildees de deca- e metavanadato num intervalo de concentraccedilotildees de 1 a
10 mM A Tabela seguinte resume os paracircmetros espectroscoacutepicos obtidos para cada espeacutecie
presente nas soluccedilotildees analisadas
Figura 22ndash Espectros de RMN de 51
V de
concentraccedilotildees crescentes das espeacutecies oligomeacutericas
de vanadato obtidos apoacutes diluiccedilatildeo da soluccedilatildeo stock
de decavanadato em 10 mM HEPES pH 70 25 mM
KCl para concentraccedilotildees finais de A) 1 mM B)2
mM C) 4 mM D) 8 mM E) 10 mM de
decavanadato total V1refere-se agraves espeacutecies
monomeacutericade vanadato V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2IIIndash Desvios quiacutemicos meacutedios (part) das ressonacircncias das espeacutecies decameacuterica (V10) monomeacuterica (V1) e
tetrameacuterica (V4) presentes nos espectros da soluccedilatildeo de decavanadatoLargura a meia altura (∆12) da espeacutecie V10A
Paracircmetros V10A V10B V10C V1 V4
part (ppm) -5162 -5003 -4255 -5616 -5748
∆12 (Hz) 175 nd nd nd nd
Figura 23 ndash Distribuiccedilatildeo graacutefica das
espeacutecies oligomeacutericas de vanadato vs a
concentraccedilatildeo de decavanadato total As
concentraccedilotildees de cada espeacutecie foram
obtidas segundo a formula descrita na
secccedilatildeo 35 do Procedimento
Experimental (aacutereas dos sinais de 51
V-
RMN observadas na figura x1)V10 V1 e
V4 correspondem agraves espeacutecie monomeacuterica
dimeacuterica e tetrameacuterica respectivamente [Esp
eacuteci
e o
ligo
meacute
rica
de
van
aacutedio
] (m
M)
[Decavanadato]total (mM)
V10
V1
V4
27
Nas soluccedilotildees de decavanadato as espeacutecies monomeacuterica e decameacuterica de vanadato satildeo as uacutenicas
presentes para concentraccedilotildees de decavanadato inferiores a 8 mM Ambas as espeacutecies aumentam
linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado em [AY] e
em (Teresa) quando a concentraccedilatildeo de V1 eacute igual ou superior a 200 microM inicia-se a sua
oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica No entanto nas condiccedilotildees de realizaccedilatildeo dos
ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a espeacutecie tetrameacuterica neste
estudo em concentraccedilotildees inferiores a 8 mM tendo o mesmo sido observado por Ramoset al
(2006) [AK] Os resultados obtidos mostram claramente o equiliacutebrio entre a espeacutecie decameacuterica e
a monomeacuterica e que o aparecimento de outras espeacutecies oligomeacutericas se encontra dependente da
concentraccedilatildeo desta uacuteltima Apesar de natildeo ter sido realizado o ensaio com 5 mM de decavanadato
total no presente relatoacuterio estaacute reportado em Ramos et al (2006) [AK] que as concentraccedilotildees da
espeacutecie decameacuterica e monomeacuterica satildeo respectivamente de 500 microM e 200 microM Assim pode inferir-
se que para concentraccedilotildees inferiores a 5 mM e intervalos de tempo de cerca de 20 minutos (que
inclui o tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas
as espeacutecies decameacuterica e monomeacuterica de vanadato
Figura 24 ndash Espectros de RMN de 51
V de concentraccedilotildees
crescentes das espeacutecies oligomeacutericas de vanadato obtidos
apoacutes diluiccedilatildeo da soluccedilatildeo stock de metavanadato em 10 mM
HEPES pH 70 25 mM KCl para concentraccedilotildees finais de A)
1 mM B) 2 mM C) 4 mM D) 8 mM E) 10 mM de
metavanadato total V1 refere-se agraves espeacutecies monomeacuterica de
vanadatoV1 e V2 correspondem respectivamente agrave espeacutecie
monomeacuterica e dimeacuterica
Figura 25 ndash Distribuiccedilatildeo graacutefica das espeacutecies
oligomeacutericas de vanadato vs a concentraccedilatildeo de
metavanadato total As concentraccedilotildees de cada
espeacutecie foram obtidas segundo a formula descrita
na secccedilatildeo 35 do Procedimento Experimental
(aacutereas dos sinais de 51
V-RMN observadas na
figura x1)V10 V1 e V4 correspondem agraves espeacutecie
monomeacuterica dimeacuterica e tetrameacuterica
respectivamente
Esp
eacuteci
e o
ligo
meacute
rica
de
va
nad
ato
(m
M)
[Metavanadato]total (mM)
V1
V2
V4
V5
28
Tabela 2IVndash Desvios quiacutemicos meacutedios (part)das ressonacircncias das espeacutecies tetrameacuterica (V4) monomeacuterica (V1) e
dimeacuterica (V2) presentes no espectro da soluccedilatildeo de metavanadato (total) 10 mM
Paracircmetros V1 V2 V4 V5
part (ppm) -5615 -5748 -5789 -5869
Nas soluccedilotildees de metavanadato as espeacutecies monomeacuterica dimeacutericae tetrameacuterica de vanadato estatildeo
presentes para concentraccedilotildees de metavanadato inferiores a 4 mM Todas estas as espeacutecies
aumentam linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado
em (Aureliano e Madeira 1998) e em (Tiago et al 2004) quando a concentraccedilatildeo de V1 eacute igual ou
superior a 200 microM inicia-se a sua oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica Nas
condiccedilotildees de realizaccedilatildeo dos ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a
espeacutecie pentameacuterica neste estudo em concentraccedilotildees inferiores a 4 mM tendo o mesmo sido
observado por Ramos 2006 Os resultados obtidos mostram claramente o equiliacutebrio entre a
espeacutecie V4 e as espeacuteciesV1 e V2 no qual o aparecimento de outras espeacutecies oligomeacutericas se
encontra dependente da concentraccedilatildeo da espeacutecie V1 Por uacuteltimo pode inferir-se que para
concentraccedilotildees inferiores a 2 mM e intervalos de tempo de cerca de 20 minutos (que inclui o
tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas as
espeacutecies V1 V2 e V4 com concentraccedilotildees respectivas de 0796 0192 e 0196 mM
232Estabilidade das soluccedilotildees de decavanadato anaacutelise por espectroscopia de absorccedilatildeo
UVVis
Caracterizou-se a estabilidade da espeacutecie V10 numa soluccedilatildeo com 1 mM decavanadato total em
tampatildeo D atraveacutes da leitura da Abs400 Obteve-se uma cineacutetica de dissociaccedilatildeo de 1ordf ordem tal
como se pode verificar pela linearidade do graacutefico de ln(Abs400)vs o tempo de aquisiccedilatildeo espectral
(segundos) (Fig26 )
Figura 26ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do tempo
(em segundos) para uma soluccedilatildeo de decavanadato total de 1
mM em tampatildeo D a 25ordmC (n=2)
Sendo uma reacccedilatildeo de primeira ordem pode ser descrita pela equaccedilatildeo cujo
declive eacute ndashK (constante de dissociaccedilatildeo) e a intersecccedilatildeo no eixo das ordenadas corresponde
Ln
(Ab
s 400 n
m)
tempo (s)
29
aln(A0) Numa reacccedilatildeo de 1ordf ordem o tempo de meia-vida eacute independente da concentraccedilatildeo inicial
do reagente sendo descrito pela equaccedilatildeo [2] Deste modo a partir da recta
Ln(Abs400 nm) = -436E-5
K - 105 obteve-se um tempo de meia-vida de 4490 plusmn 0823 horas para
as condiccedilotildees supramencionadas
Apoacutes a adiccedilatildeo de S1 8 microM ao meio reaccional verificou-se que a reacccedilatildeo de desoligomerizaccedilatildeo
da espeacutecie decameacuterica de vanadato seguia igualmente uma cineacutetica de 1ordf ordem tal como se pode
verificar pela linearidade do graacutefico de ln(Abs400) versus o tempo de aquisiccedilatildeo espectral
(segundos) (Fig27)
Figura 27ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do
tempo (em segundos) para uma soluccedilatildeo de decavanadato
total de 1 mM em tampatildeo D a 25ordmC na presenccedila de 8
microM (n= 2)
A partir da equaccedilatildeo da recta Ln(Abs400 nm) = -7520E-5
K - 0613determinou-se o valor de K para
a espeacutecie decameacuterica de onde se obteve um tempo de meia-vida de 2430 plusmn 0191 horas para as
condiccedilotildees supramencionadas
Tanto este valor como o valor registado na ausecircncia de S1 satildeo substancialmente superiores agrave
duraccedilatildeo dos ensaios de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina descritos no presente
relatoacuterio
24 Interacccedilatildeo de S1 com vanaacutedio(V)
241Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
De modo a analisar qual o efeito da adiccedilatildeo de S1 de miosina (tampatildeo D) produzia nos sinais de
51V-NMR titulou-se uma soluccedilatildeo de 5mM de decavanadato total com concentraccedilotildees crescentes
de S1 (5 10 15 e 20 microM) Os espectros de 51
V-RMN encontram-se apresentados na Fig 2
No graacutefico da figura 2 Apresenta-se a concentraccedilatildeo de S1 em funccedilatildeo do factor de alargamento
corrigido dos espectros acima referidos
Ln
(Ab
s (4
00
nm
))
tempo (s)
30
Figura 28 ndash Graacutefico de
representaccedilatildeo da
concentraccedilatildeo de S1 em
funccedilatildeo do factor de
alargamento A
concentraccedilatildeo de
decavanadato total foi de 5
mM e as concentraccedilotildees
utilizadas de S1 foram de 0
5 10 e 15 microM
Verifica-se que o factor de alargamento aumenta linearmente em funccedilatildeo da concentraccedilatildeo de S1 no
ensaio o que indica a existecircncia de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina Observou-
se ainda que a adiccedilatildeo de S1 ao meio reaccional natildeo provocou qualquer alteraccedilatildeo dos desvios
quiacutemicos dos sinais de 51
V-RMN
Realizaram-se ensaios na presenccedila de ATP e de S1 com intuito de verificar o efeito do ATP na
interacccedilatildeo do S1 de miosina com o decavanadato
Figura 29 - Espectros de RMN de 51
V contendo 2
mM Decavanadato (A) (controlo) (B) na presenccedila
de 2 mM de ATP (C) na presenccedila de 2 mM de ATP
e 8 microM de S1 de miosina V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2Vndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies decameacuterica (V10) e monomeacuterica (V1) presentes nos
espectros da soluccedilatildeo de decavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V10A V10B V10C V1
part (ppm) -5161 -5002 -4254 -5614
Observa-se que a presenccedila de ATP (espectro B) natildeo provoca qualquer alteraccedilatildeo visiacutevel em
relaccedilatildeo ao espectro que contem apenas soluccedilatildeo de decavanadato Jaacute a presenccedila simultacircnea de ATP
e S1 provoca um alargamento da ∆12 do sinal do V10A de 1736 Hz (espectro (A)) para 2366 Hz
(espectro (C) Fig 29) Tendo em conta que a concentraccedilatildeo de S1 apresenta uma relaccedilatildeo linear
com o factor de alargamento converteu-se o valor da ∆12 e comparou-se com o valor esperado agrave
F = 00591[S1] - 00157 Rsup2 = 0998
Fact
or
de
ala
rgam
en
to
[S1] (microM)
31
concentraccedilatildeo de S1 utilizada O factor de alargamento obtido para 8 microM ndash 0376 ndash eacute inferior ao
valor esperado agrave mesma concentraccedilatildeo de S1 obtido atraveacutes da equaccedilatildeo da recta apresentada na
Fig 28 ndash 0457 ndash o que indica que o ATP poderaacute exercer um efeito de protecccedilatildeo do S1
diminuindo a sua interacccedilatildeo com a espeacutecieV10
Por uacuteltimo realizaram-se ensaios com metavanadato na presenccedila de 8 microM S1 de modo a verificar
se as espeacutecies oligomeacutericas contidas nesta soluccedilatildeo interagiam com a proteiacutena em estudo
Figura 210 - Espectros de RMN de 51
V contendo 2
mM metavanadato (A) (controlo) (B) na presenccedila
de 8 microM de S1 de miosina V1 V2 e V4
correspondem respectivamente aacutes espeacutecies
monomeacuterica dimeacuterica e tetrameacuterica
Tabela 2VIndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies monomeacuterica (V1) dimeacuterica (V2) e tetrameacuterica (V4)
presentes nos espectros da soluccedilatildeo de metavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V1 V2 V4
part (ppm) -5613 -5747 -5789
Como eacute possiacutevel observar na Fig 210 ocorre uma ligeiro alargamento dos sinal de V4 bem
como uma perda da intensidade pelo que se conclui quedas espeacutecies presentes na soluccedilatildeo de
metavanadato (V1 V2 e V4) apenas a espeacutecie tetrameacuterica interage com o S1 de miosina o que
estaacute de acordo com o reportado por Tiago et al 2002
242 Actividade Mg2+
-ATPaacutesica de S1 de miosina na presenccedila de decanadato
Com o intuito de avaliar o efeito do decavanadato na actividade Mg2+
-ATPaacutesica do S1 de
miosina monitorizou-se a reacccedilatildeo de oxidaccedilatildeo do NADH atraveacutes da leitura de Abs340 na
presenccedila de 005 mgmL de S1 de miosina e no caso da actividade estimulada 2microM de actina em
3125 mM Tris-HCl pH 71 e 375 mM MgCl2 pelo meacutetodo de enzimas acopladas (ver seccccedilatildeo xx
dos MM)Para tal realizaram-se ensaios com vaacuterias concentraccedilotildees de decavanadato total (10 50
100 250 500 e 1000) de modo a determinar qual o seu efeito na actividade MgATPaacutesica de S1
de miosina na ausecircncia e na presenccedila de um dos seus ligandos naturais Os ensaios foram
realizados agrave temperatura ambiente com incubaccedilatildeo preacutevia a 25ordm C durante 20 minutos
32
Figura 211 ndash Representaccedilatildeo
graacutefica da Actividade Mg2+
-
ATPaacutesica de S1 () (os ajustes
teoacutericos estatildeo representado a por
uma linha os resultados
experimentais estatildeo representados
por triacircngulos (Actividade
estimulada) e cruzes (Actividade
natildeo estimulada)) vs
[Decavanadato]total medidas pelo
meacutetodo das enzimas acopladas
(n=3) Os dados foram ajustados ao modelo de decaimento exponencial usando o programa GraphPad Prism para a
determinaccedilatildeo dos valores de IC50 (na ausecircncia de actina IC50= 266 plusmn 07 M decavanadato na presenccedila de actina
IC50= 28 plusmn 1 M decavanadato
A partir do graacutefico presente na Fig 29 verifica-se que o ajuste dos dados experimentais foi bem
sucedido A inibiccedilatildeo da actividade Mg2+
-ATPaacutesica do S1 de miosina eacute afectada em menor
amplitude na presenccedila de actina (60 de inibiccedilatildeo da actividade) do que na sua ausecircncia
(aproximadamente 80 de inibiccedilatildeo de actividade) Devido a esta diferenccedila pode sugerir-se que a
actina iraacute impedir parcialmente a ligaccedilatildeo do decavanadatoao S1 demiosina atraveacutes de
impedimentos esteacutereos ou interacccedilotildees electrostaacuteticas por exemplo A teacutecnica utilizada natildeo nos
permite concluir acerca de que regiatildeo(otildees) ou resiacuteduo(s) da actina participaratildeo na preservaccedilatildeo
parcial da actividade Mg2+
-ATPaacutesica
Por outro lado os valores de IC50 dos dois tipos de actividades natildeo apresentam diferenccedilas
significativas tendo-se obtido 266 plusmn 07 e 28 plusmn 1M decavanadato na ausecircncia e na presenccedila de
actina respectivamente o que corresponde a uma concentraccedilatildeo de cerca de 27 M de V10 O
meio de reacccedilatildeo utilizado nestes ensaios foi 10 mM HEPES e 25 mM KCl com 2 mM ATP Em
(Tiago et al 2004) o valor de IC50 para a actividade Mg2+-ATPaacutesica na ausecircncia de actina foi de
125 plusmn 025 microM enquanto o valor da actividade estimulada por actina foi de 136 plusmn 14 microM O
meio de reacccedilatildeo utilizado nos ensaios foi 20 mM HEPES 50 mM KCl e 2 mM ATP o que resulta
precisamente no dobro da forccedila ioacutenica a que foi realizado o ensaio do presente relatoacuterio
(exceptuando a contribuiccedilatildeo dos produtos da hidroacutelise de ATP uma vez que este meacutetodo
pressupotildee a ocorrecircncia de um ciclo de reacccedilotildees de hidroacutelise e siacutentese deste composto) Ora tendo
sido anteriormente referido que a afinidade do S1 de miosina para com a espeacutecie V10 diminui com
Act
ivid
ade
Mg
2+-A
TP
aacutesic
a d
e S
1 d
e
Mio
sin
a
[Decavanadato] (microM) Actividade Estimulada ()
Actividade Natildeo Estimulada ()
Ajuste Actividade Estimulada
Ajuste Actividade Natildeo Estimulada
33
o aumento da forccedila ioacutenica (Tiago et al 2004 tal poderaacute explicar a diferenccedila obtida entre estes
dois conjuntos de valores
243Quantificaccedilatildeo de cisteiacutenas reduzidas pelo meacutetodo de Ellman
A interacccedilatildeo do vanaacutedio com alguns tipos de aminoaacutecidos sensiacuteveis agrave oxidaccedilatildeo pode fornecer-nos
informaccedilatildeo bastante uacutetil em relaccedilatildeo agraves alteraccedilotildees estruturais que conduzem a uma diminuiccedilatildeo da
actividade fisioloacutegica do S1 de miosina Os estudos realizados por Chasteen 1983 concluiacuteram
que a reduccedilatildeo de vanadato (V) a vanadilo (IV) ocorre na presenccedila de agentes redutores comuns no
ambiente celular de que se destaca o aminoaacutecido cisteiacutena Esta eacute tambeacutem um alvo de
complexaccedilatildeo por parte do iatildeo VO2+
Jaacute Ramos et al (2009) demonstrou que a incubaccedilatildeo de
actina com a soluccedilatildeo de decavanadato induz a oxidaccedilatildeo de resiacuteduos de cisteiacutena e agrave formaccedilatildeo do
oxidovanaacutedio (IV) Este grupo de investigadores descobriu ainda que apenas a espeacutecie V10 induz a
oxidaccedilatildeo das cisteiacutenas ldquoraacutepidasrdquo da actina atraveacutes do processo proposto que eacute parcialmente
responsaacutevel pela inibiccedilatildeo jaacute descrita da actividade de ATPase do S1 de miosina estimulada por
actina quando exposta agrave soluccedilatildeo de decavanadato Esta inibiccedilatildeo apresenta-se afectada com a
presenccedila de ATP uma vez que os resiacuteduos de cisteiacutena permaneceram intactos nos ensaios
realizados com este ligando Jaacute Crans et al (2004) concluiu que o siacutetio activo de muitas fosfatases
(que conteacutem grupos tiol) pode formar um equiliacutebrio redox com o iatildeo vanadato (V) de forma natildeo
reversiacutevel
Os resultados obtidos por Sakurai e Shiromura (1980) sugeriram que o vanaacutedio (V) sofre reduccedilatildeo
a vanaacutedio (IV) sem dependecircncia do valor de pH do ensaio Registaram ainda uma estequiometria
de 31 na reacccedilatildeo do vanadato (V) com os resiacuteduos de Cys enquanto a estequiometria dos ensaios
com o vanadilo (IV) foi de 21
Berlett e Stadtman (1997) observaram que os tipos de aminoaacutecidos mais facilmente oxidaacuteveis
englobam entre outros os resiacuteduos de Cys Trp e MetAssim analisou-se o estado redox dos
grupos tiol dos resiacuteduos de Cys do S1 na presenccedila do decavanadato atraveacutes da monitorizaccedilatildeo da
Abs412 nm na presenccedila de 1 mgmL de S1 de miosina em tampatildeo 10 mM HEPES 25 mM KCl
variando a concentraccedilatildeo da soluccedilatildeo de decavanadato de modo a determinar que tipo de alteraccedilotildees
estruturais ocorrem na sua presenccedila no que diz respeito agrave diminuiccedilatildeo da actividade ATPaacutesica de
S1 na presenccedila de decavanadato
Os resultados referentes agrave quantificaccedilatildeo das cisteiacutenas reduzidas apoacutes o tratamento de S1 com
diferentes concentraccedilotildees de decavanadato em tampatildeo 10 mM HEPES pH 75 e 25 mM KCl a
25ordmC durante 20 minutos encontram-se descritos na Tabela 2VII
Tabela 2VII ndash Cisteiacutenas reduzidas totais e ldquoraacutepidas de S1 (1 mgmL) apoacutes tratamento com decavanadato
34
(010501002505001000 uM) em 10 mM HEPES 25 mM KCl pH 7 a 25ordmC durante 20 minutos A detecccedilatildeo
espectrofotomeacutetrica foi feita com DTNB (e 412=8400 uM-1 cm-1) (n variaacutevel entre 1 e 8)
Amostras Cys raacutepidas Cys totais
11 M S1 (controlo) 22 plusmn 04 106 plusmn 14
11 M S1 + 10 M Decavanadato 22 plusmn 05 95 plusmn 08
11 M S1 + 50 uM Decavanadato 20 plusmn 02 98 plusmn 14
11 M S1 + 100 uM Decavanadato 17 plusmn 03 97 plusmn14
11 M S1 + 250 uM Decavanadato 26 plusmn 02 98 plusmn 08
11 M S1 + 500 uM Decavanadato 21 plusmn 04 90 plusmn 06
11 M S1 + 1000 uM Decavanadato 24 plusmn 0 86 plusmn 08
A partir dos resultados obtidos pode sugerir-se que o S1 possui 11 resiacuteduos de Cys (controlo) o
que estaacute de acordo com o anteriormente descrito Verifica-se que o tratamento da proteiacutena com
concentraccedilotildees crescentes de decavanadato provoca a oxidaccedilatildeo de dois resiacuteduos de Cys do S1 de
miosina natildeo expostos ao solvente No entanto a teacutecnica utilizada natildeo nos permite identificar
quais
Tendo em consideraccedilatildeo que o valor de IC50 obtido para a reacccedilatildeo de inibiccedilatildeo daactividade Mg2+-
ATPaacutesica de S1 na presenccedila de decavanadato eacute de cerca de 27 microM pode sugerir-se que este seraacute o
valor ao qual ocorreraacute a oxidaccedilatildeo do primeiro resiacuteduo de Cys o que poderaacute contribuir para a
diminuiccedilatildeo da actividade de S1 observada
25Interacccedilatildeo de S1 com vanaacutedio(IV)
251Anaacutelise por espectroscopia de Ressonacircncia Paramagneacutetica Electroacutenica
A acumulaccedilatildeo intracelular de oxovanaacutedio (IV) in vivo constitui uma das principais formas de
apresentaccedilatildeo de vanaacutedio no organismo Deste modo torna-se bastante interessante analisar a
interacccedilatildeo entre este iatildeo e o S1 de miosina atraveacutes da teacutecnica de espectroscopia de Ressonacircncia
Paramagneacutetica Electroacutenica o meacutetodo de eleiccedilatildeo do estudo do iatildeo vanadilo e dos complexos por si
formados O electratildeo desemparelhado encontra-se fortemente acoplado ao nuacutecleo 51
V o que
35
resulta num espectro de RPE consistindo de 8 desdobramentos na direcccedilatildeo paralela e outros tantos
na direcccedilatildeo perpendicular (Chasteen 1983)
Os estudos realizados por Ahmed et al (1987) centraram-se na interacccedilatildeo do vanadilo com
proteiacutenas como eacute o caso da insulina transferrina entre outros obtendo um espectro caracterizado
por 8 linhas amplamente espaccediladas (na direcccedilatildeo paralela) Jaacute Ramos et al 2009 concluiu que a
exposiccedilatildeo da soluccedilatildeo de decavanadato agraves formas globular e filamentosa de actina produz um
espectro tiacutepico do iatildeo oxidovanaacutedio (IV) indicando a reduccedilatildeo do vanadato
Para aferir qual a concentraccedilatildeo de S1 que permitia obter uma boa razatildeo sinalruiacutedo titulou-se uma
soluccedilatildeo de sulfato de vanadilo 15 microM ([VOSO4]stock = 5 mM) com concentraccedilotildees crescentes de
proteiacutena (0 5 10 15 e 20 microM) em 10 mM HEPES 25 mM KCl A existecircncia de uma correlaccedilatildeo
linear entre a concentraccedilatildeo de proteiacutena e a intensidade dos sinais de RPE correspondente agrave banda
-12 perpendicular encontra-se representada na Fig 210
Figura 2 12ndash Representaccedilatildeo graacutefica da variaccedilatildeo da intensidade do sinal de RPE vs a concentraccedilatildeo de S1 utilizada
Apoacutes verificar que com uma concentraccedilatildeo de S1 igual a 20 microM se obtinham sinais com uma razatildeo
sinalruiacutedo adequada ao prosseguimento dos estudos sobre a interacccedilatildeo V(IV)-S1 de miosina
titulou-se uma soluccedilatildeo de 20 microM S1 com 0 25 5 10 15 25 50 75 100 150 200 500 e 1000
microM VOSO4
Traccedilou-se o graacutefico da intensidade da banda -12 perpendicular do sinal de EPR em funccedilatildeo da
concentraccedilatildeo de VOSO4 tendo-se determinado uma constante de dissociaccedilatildeo (KD) do complexo
S1-vanadilo de 8748 plusmn 2621 microM V(IV)
y = 016786x + 122435
Rsup2 = 092992759
Inte
nsi
dad
e d
o s
inal
de
RP
E
[S1] (microM)
36
Figura 213 ndash Representaccedilatildeo graacutefica da variaccedilatildeo do sinal de RPE vs a concentraccedilatildeo de vanadilo utilizada (os valores
experimentais estatildeo representados por losangos o seu ajuste estaacute representado pela linha)
A simulaccedilatildeo do espectro de EPR foi efectuada a partir do espectro obtido para uma concentraccedilatildeo
de 1 mM VOSO4 apoacutes adiccedilatildeo de 20 microM S1 de miosina tendo-se obtido os valores da constante de
acoplamento hiperfina (A=5016) e do factor g (g0=195)
Figura 214 ndash Representaccedilatildeo graacutefica do espectro de EPR obtido com 1000 microM de vanadilo e 20 microM de S1 (a cinzento
escuro) A simulaccedilatildeo do espectro encontra-se a cinzento claro
Por comparaccedilatildeo destes com os valores presentes na tabela de correspondecircncia xxx eacute possiacutevel
predizer quais os ligandos provaacuteveis do vanadilo na presenccedila de proteiacutena Assim concluiu-se que
o vanadilo estaacute coordenado com duas moleacuteculas de aacutegua um grupo hidroxilo e um grupo R=NR
O grupo hidroxilo pode indicar os resiacuteduos de aminoaacutecidos Tyr Treonina e Serina enquanto o
grupo R=NR poderaacute indicar a presenccedila de Arginina e Histidina como ligandos do vanadilo
Inte
nsi
dad
e d
os
sinai
s d
e R
PE
[Vanadilo] (microM)
Ajuste dos dados exp Dados experimentais
Campo magneacutetico (G)
Espectro Experimental Ajuste
37
3 Procedimento Experimental
31 Reagentes
Todos os reagentes utilizados satildeo de elevado grau de pureza (analiacutetico) Apresentam-se as suas
foacutermulas quiacutemicas ou abreviaturas quando necessaacuterio acrescentou-se a sua descriccedilatildeo Entre
parecircntesis colocou-se as empresas que os comercializam
Aacutecido aceacutetico glacial 100 (Riedel) Acrilamida 99 (Merck) Azul de bromofenol (Riedel-de-
Haumlen) -Mercaptoetanol (Sigma) Bissulfito de soacutedio (Sigma)Bradford (reagente de)
(BioRad)BSA (albumina de soro bovino Sigma-Aldrich) CaCl22H2O (cloreto de caacutelcio
dihidratado Fluka) CH3CH2OH 96 (etanol Panreac) Coomassie Blue R250 (Merck) D2O
(oacutexido de deuteacuterio SDS) DTNB (55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico) Aldrich) EDTA (aacutecido
etilenodiaminatetraaceacutetico Merck) Fosfoenolpiruvato 98 (Fluka) Glicerol (Sigma) Glicina
985 (Panreac) HCl 37 (Riedel-de Haumlen) H2SO4 96 (Pronalab) HEPES 99 (aacutecido 4-(2-
hidroxiletil)-1-piperazineetanosulfoacutenico Sigma) KH2PO4 99 (hidrogenofosfato de potaacutessio)
MgATP (Sigma) Na
ATP (Sigma) Na2HPO4 99 (hidrogenofosfato de soacutedio Riedel-de Haumlen)
NaH2PO4 (fosfato de soacutedio ) NaOH 986 (hidroacutexido de soacutedio Pronalab) KCl 99 (Cloreto de
potaacutessio Panreac) L-Cys 995 (enantioacutemero levoacutegiro do resiacuteduo de Cisteiacutena Fluka)Metanol
99 (Merck) Molibdato de amoacutenio (Sigma)NNrsquo-metil-bis-acrilamida 98 (Merck) NH4VO3
99 (metavanadato de amoacutenia Sigma) PSA 10 (persulfato de amoacutenio Merck) SDS
(dodecilo sulfato de soacutedio Sigma) Sulfito de soacutedio (Sigma) VOSO4 (sulfato de vanadilo
Aldrich) TEMED 99 (Tetrametiletilenodiamina Merck) Tris (2-amino-2-hidroximetil-13-
propanodiol Sigma)
32 Meacutetodos
321 Isolamento e purificaccedilatildeo das proteiacutenas de muacutesculo esqueleacutetico de coelho
3211 Purificaccedilatildeo da miosina preparaccedilatildeo de Subfragmento 1 por digestatildeo quimotriacutepsica
da miosina
O subfragmento-1 de miosina-II (S1) foi gentilmente cedido pelo Prof Doutor Manuel Aureliano
do Departamento de Quiacutemica Bioquiacutemica e Farmaacutecia da Faculdade de Ciecircncias e Tecnologia da
Universidade do Algarve
No entanto descreve-se abaixo de forma resumida o procedimento efectuado
38
32111 Purificaccedilatildeo da miosina
O protocolo de isolamento foi adaptado do meacutetodo previamente descrito por Margossian e Lowey
(1982) apresentando-se de forma esquemaacutetica na Fig 31
Sucintamente os tecidos musculares dorsais e das pernas foram extraiacutedos apoacutes o sacrifiacutecio do
animal (Fig31) De seguida foram adicionados 3 volumes da soluccedilatildeo de extracccedilatildeo A2
Centrifugou-se a soluccedilatildeo e filtrou-se o sobrenadante O sedimento foi entatildeo diluiacutedo em 14
volumes de aacutegua destilada de modo a que a forccedila ioacutenica atingisse 004 M Apoacutes adiccedilatildeo de szlig-
mercaptoetanol (szlig-me) deixou-se a precipitar a 4ordmC removendo o sobrenadante Centrifugou-se
novamente e ressuspendeu-se o sedimento na soluccedilatildeo de extracccedilatildeo A Adicionou-se a soluccedilatildeo
tampatildeo B3 para que a forccedila ioacutenica atingisse 05 M Homogeneizou-se e deixou-se repousar
durante 2h Homogeneizou-se novamente e centrifugou-se de modo a ocorrer precipitaccedilatildeo dos
complexos de actomiosina e proteiacutena desnaturada Apoacutes filtraccedilatildeo do sobrenadante em latilde de vidro
quantificou-se a soluccedilatildeo de miosina resultante pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental)
2 Soluccedilatildeo de extracccedilatildeo A 0012M K2CO3 004M KHCO3 06M KCl pH 65
3 Soluccedilatildeo-tampatildeo B Tris 100 mM pH 7 KCl 2M
39
Figura 31 ndash Representaccedilatildeo esquemaacutetica do protocolo de isolamento de miosina (Margossian e Lowey 1982) Todo o
procedimento foi realizado a 4ordmC (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
32112 Preparaccedilatildeo de Subfragmento-1 de miosina por digestatildeo com quimiotripsina
O S1 de miosina foi isolado apoacutes diaacutelise da miosina durante a noite contra a soluccedilatildeo C4 A
presenccedila do EDTA eacute essencial uma vez que a sua substituiccedilatildeo por MgCl2resulta na produccedilatildeo de
HMM em vez de S1 A suspensatildeo de miosina foi equilibrada e digerida com -quimotripsina
A digestatildeo foi interrompida pela adiccedilatildeo de PMSF Seguidamente ultracentrifugou-se o produto
da digestatildeo desprezando-se o sedimento (Fig 32) A soluccedilatildeo de sobrenadante maioritariamente
S1 foi quantificada pela leitura directa da Abs280 ou pelo meacutetodo de Bradford (ver as secccedilotildees
3311 e 3312 do Procedimento Experimental respectivamente) e armazenada em 10 sacarose
e 2 mM szlig-Me a -80ordmC
4 Soluccedilatildeo C 25mM Tris pH 70 012M KCl 1mM EDTA
40
Antes da realizaccedilatildeo dos ensaios descritos no presente relatoacuterio o S1 de miosina foi dialisado
contra a soluccedilatildeo D5 de modo a remover qualquer composto que pudesse complexar como
vanaacutedio Seguidamente quantificou-se a proteiacutena pelo meacutetodo de Bradford ou pelo meacutetodo
directo (ver secccedilotildees 3311 e 3312do Procedimento Experimtental)
Figura 3 2ndash Representaccedilatildeo esquemaacutetica do protocolo de preparaccedilatildeo do S1 de miosina por digestatildeo quimiotripsina de
acordo com o previamente descrito por Margossian e Lowey (1982) Todo o procedimento foi realizado a 4ordm C
excepto quando indicaccedilatildeo do contraacuterio (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
3212 Isolamento e purificaccedilatildeo de actina
Isolou-se e purificou-se a actina segundo o meacutetodo descrito por Pardee e Spudish que
compreende dois passos ndash a obtenccedilatildeo de um resiacuteduo seco denominado ldquopoacute de acetonardquo e a
extracccedilatildeo subsequente da actina a partir do ldquopoacute de acetonardquo na forma de G-Actina com a
posterior polimerizaccedilatildeo a F-Actina
32121 Obtenccedilatildeo do ldquopoacute de acetonardquo
Sacrificou-se o coelho com uma pancada atraacutes do pescoccedilo e sangrou-se o animal apoacutes o corte da
arteacuteria jugular Esfolou-se e colocou-se numa tina com gelo Retiraram-se as partes de tecido
conjuntivo que envolvem o tecido muscular com auxiacutelio de uma pinccedila e bisturi Seguidamente
retirou-se os muacutesculos dorsais e os muacutesculos brancos das pernas Passou-se os muacutesculos por aacutegua
destilada de modo a remover as impurezas e pesou-se num copo Trituraram-se os muacutesculos
numa maacutequina de triturar carne e extraiu-se o resultante com 3 volumes da soluccedilatildeo E6 durante 10
minutos com agitaccedilatildeo suave Centrifugou-se a suspensatildeo a 4000 g durante 10
5 Soluccedilatildeo D HEPES 10 mM pH 70 KCl 25 mM
6 Soluccedilatildeo E 01 M KCl 015 M K2HPO4 pH 65
41
minutosdesprezando-se o sobrenadante Ressuspendeu-se o sedimento em 6 volumes de soluccedilatildeo
F7durante 10 minutos com agitaccedilatildeo suave Verificou-se se o pH da soluccedilatildeo estava compreendido
entre 82 e 85 acertando-se estecom soluccedilatildeo G8 se necessaacuterio Centrifugou-se durante 10
minutos a 4000 g Desprezou-se o sobrenadante e ressuspendeu-se o sedimento em 3 volumes de
soluccedilatildeo H9 durante 10 minutos Centrifugou-se novamente a 4000 gdurante 10 minutos
desprezando-se o sobrenadante Ressuspendeu-se o sedimento duas vezes consecutivas em 6
volumes de aacutegua destilada durante 5 minutos Centrifugou-se a 4000 g durante 10 minutos apoacutes
cada adiccedilatildeo de H2O
Seguidamente ressuspendeu-se o sedimento 5 vezes consecutivas utilizando 3 volumes de
acetona de cada vez agitando suavemente durante 10 minutos agrave temperatura ambiente Colocou-
se o resiacuteduo filtrado numa tina de vidro sobre papel de filtro deixando-se evaporar o solvente
durante a noite num nicho Triturou-se o resiacuteduo seco numa maacutequina de triturar carne e deixou-se
novamente evaporar o solvente O ldquopoacute de acetonardquo obtido foi armazenado a -20 ordmC
32122 Purificaccedilatildeo da actina a partir de ldquopoacute de acetonardquo
Apoacutes a obtenccedilatildeo do poacute de acetona como acima indicado efectuou-se o protocolo seguidamente
descrito de modo a obter monoacutemeros de actina (G-Actina) e actina filamentosa (F-Actina)
Pesou-se 35 g de poacute de acetona e adicionaram-se 50 mL de soluccedilatildeo-tampatildeo I10
sendo a
extracccedilatildeo feita com agitaccedilatildeo suave durante 30 minutos Separou-se o extracto do poacute de acetona
hidratado filtrando a suspensatildeo com 4 camadas de gaze esterilizada tendo o cuidado de natildeo
espremer o resiacuteduo Voltou a extrair-se com 50 mL de soluccedilatildeo I durante 30 minutos mexendo
lentamente Adicionou-se este ao extracto obtido anteriormente Centrifugou-se o extracto total a
16 000 g durante 30 minutos Decantou-se para uma proveta sobre a qual se colocou gaze para
que o sedimento solto natildeo se juntasse ao sobrenadante e leu-se o volume De seguida adicionou-
se KCl ateacute uma concentraccedilatildeo final de 50 mM MgCl2 ateacute 2mM e ATP ateacute 1mM lentamente e com
agitaccedilatildeo contiacutenua de modo a promover a polimerizaccedilatildeo dos monoacutemeros de actina
Deixou-se polimerizar durante 2h a 25ordmC observando-se a presenccedila de pequenas bolhas de ar
indicativas de um aumento de viscosidade da soluccedilatildeo Arrefeceu-se em gelo durante 15 minutos e
adicionou-se KCl ateacute uma concentraccedilatildeo final de 06M Mexeu-se lentamente durante 30 minutos
apoacutes os quais se ultracentrifugou a 110 000g durante 1hora Desprezou-se o sobrenadante
ressuspendeu-se o sedimento com soluccedilatildeo tampatildeo E e transferiu-se para um potter
Dialisou-se durante a noite contra 1L de soluccedilatildeo-tampatildeo I a uma temperatura de 4ordmC com rotaccedilatildeo
7 Soluccedilatildeo F 005 M NaHCO3 pH 82-85
8 Soluccedilatildeo G 1M Na2CO3
9 Soluccedilatildeo H 1 mM EDTA pH 70
10 Soluccedilatildeo I 2 mM Tris-HCl pH 8 02 mM CaCl2 02 mM Na2ATP 05 mM szlig-Me 0005 NaN3
42
contiacutenua e mudanccedila de tampatildeo 2 a 3 vezes De seguida a actina foi centrifugada a 110
000gdurante 1hora Decantou-se a soluccedilatildeo de G-Actina (sobrenadante) e desprezou-se o
sedimento Determinou-se a concentraccedilatildeo de G-Actina pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental) A soluccedilatildeo de G-actina foi armazenada a uma temperatura de 4ordmC
apoacutes adiccedilatildeo de 002NaN3 mantendo-se estaacutevel durante 3 a 5 dias
331 Quantificaccedilatildeo das proteiacutenas isoladas
3311 Quantificaccedilatildeo pelo meacutetodo de Bradford
O meacutetodo de Bradford (Bradford 1976) baseia-se na utilizaccedilatildeo do reagente de Bradford
composto por entre outros aacutecido fosfoacuterico e o corante ldquoCoomassierdquo Este corante possui carga
negativa e a sua coloraccedilatildeo varia entre a vermelha (Absmax=465 nm) predominante em soluccedilatildeo e a
azul (Absmax=595 nm) presente aquando da ligaccedilatildeo aos grupos amina das proteiacutenas Em soluccedilotildees
aacutecidas diluiacutedas a coloraccedilatildeo varia proporcionalmente com a quantidade do corante ligado agrave
proteiacutena que por sua vez eacute directamente proporcional agrave concentraccedilatildeo da proteiacutena permitindo a
sua quantificaccedilatildeo ateacute a um limite de detecccedilatildeo de 20 gmL (Bradford 1976)
Deste modo prepararam-se amostras de BSA ([BSA]stock= 1mgmL) utilizando um intervalo de
concentraccedilotildees compreendido entre 0 e 15 gmL de acordo com a tabela 3I Apoacutes a adiccedilatildeo da
H2O MilliQ e do reagente de Bradford as soluccedilotildees foram sucessivamente homogeneizadas no
voacutertex
Tabela 3Indash Amostras utilizadas para a obtenccedilatildeo da recta de calibraccedilatildeo no meacutetodo de Bradford
BSA (L) BSA (g) H2O MilliQ (L) Reagente de Bradford (L)
0 0 800 200
125 125 79875 200
25 25 7975 200
5 5 795 200
10 10 790 200
15 15 785 200
Apoacutes incubaccedilatildeo a 25ordmC por 5 minutos leram-se os valores de Abs595 num
espectrofotoacutemetro (Shimadzu UV-160A) para todas as amostras Seguidamente traccedilou-se a recta
de calibraccedilatildeo (Abs595vs quantidade de BSA (g))
43
Para determinar a concentraccedilatildeo de proteiacutena apoacutes o processo de isolamento purificaccedilatildeo e
diaacutelise quando necessaacuterio colocou-se 5 L da amostra num tubo de Eppendorf e adicionou-se
795 L de H2O MilliQ homogeneizando a soluccedilatildeo no voacutertex Adicionou-se posteriormente 200
L de reagente de Bradford e repetiu-se a homogeneizaccedilatildeo Incubou-se a mistura a 25ordmC por 5
minutos apoacutes os quais se leu os valores de Abs595 nm no espectrofotoacutemetro Por interpolaccedilatildeo deste
valor a partir da recta de calibraccedilatildeo determinou-se a concentraccedilatildeo de proteiacutena na amostra
3312 Quantificaccedilatildeo pelo meacutetodo directo
A quantificaccedilatildeo das amostras proteicas pode tambeacutem ser efectuada atraveacutes da leitura directa da
absorvacircncia tendo por base a lei de Beer-Lambert onde Abs corresponde agrave
absorvacircncia ao comprimento de onda ao qual o ensaio foi realizado corresponde ao coeficiente
de extinccedilatildeo (substituiacutedo pela letra a se as unidades forem distintas da molaridade) b corresponde
ao percurso oacuteptico da radiaccedilatildeo na ceacutelula amostral e c corresponde agrave concentraccedilatildeo amostral
Assim no caso de S1 de miosina mediu-se a Abs280e calculou-se a concentraccedilatildeo da proteiacutena
sabendo que o coeficiente de extinccedilatildeo de S1 eacute (Ref)
No caso da actina mediu-se a Abs290ecalculou-se a concentraccedilatildeo da proteiacutena sabendo que o
coeficiente de extinccedilatildeo da actina eacute (Ref)
332Electroforese em condiccedilotildees desnaturantes (SDS-PAGE)
De modo a melhor caracterizar as proteiacutenas isoladas foi realizada uma electroforese em
condiccedilotildees desnaturantes (SDS-PAGE do inglecircs Sodium Dodecyl Sulfate Polyacrilamide Gel
Electrophoresis) O SDS (dodecilo sulfato de soacutedio) eacute um detergente anioacutenico utilizado neste
meacutetodo para desnaturar as proteiacutenas que se ligam ao SDS na proporccedilatildeo 14 g SDS g proteiacutena
(aproximadamente 1 moleacutecula de SDS por cada 2 resiacuteduos de aminoaacutecidos) (Stryer) O SDS induz
uma densidade de carga negativa significativa nas proteiacutenas a que se liga mascarando a sua carga
intriacutenseca pelo que as razotildees cargamassa satildeo tendencialmente idecircnticas Assim a SDS-PAGE
separa as proteiacutenas quando sujeitas a um campo eleacutectrico de acordo com o seu peso molecular
sendo a sua migraccedilatildeo directamente proporcional ao logaritmo deste uacuteltimo (Stryer)
O gel de resoluccedilatildeo com 10 de acrilamida foi obtido atraveacutes da adiccedilatildeo de 167 mL de soluccedilatildeo
stock de acrilamida11
075 mL de tampatildeo de resoluccedilatildeo12
251 mL de aacutegua destilada e 50 L de
SDS 10 Para promover a polimerizaccedilatildeo adicionou-se em seguida 38 L de PSA 10
11
soluccedilatildeo stock de acrilamida soluccedilatildeo de 30 (mv) de acrilamida e 08 (mv) de NNrsquo-metil-bis-acrilamida 12
tampatildeo de resoluccedilatildeo 15 M Tris-HCl pH 88
44
(persulfato de amoacutenia) 10 e 25 L de TEMED 99 (tetrametiletilenodiamina) Agitou-se
cuidadosamente de modo a natildeo formar bolhas e pipetou-se a soluccedilatildeo para a cassette de vidro
previamente montada no suporte De seguida adicionou-se uma peliacutecula de cerca de 2 mm de 2-
metil-2-butanol para verificar posteriormente a polimerizaccedilatildeo Apoacutes a polimerizaccedilatildeo que demora
cerca de 45 minutos verteu-se a agrave superfiacutecie e adicionou-se o gel de concentraccedilatildeo com 5 de
acrilamida constituiacutedo por 03 mL de soluccedilatildeo de stock acrilamida 045 mL de tampatildeo de
concentraccedilatildeo13
094 mL de aacutegua destilada 18 L de SDS 10 135 L de PSA 10 e 2L de
TEMED 99 Colocou-se o pente sobre o gel e deixou-se polimerizar durante cerca de 45
minutos
As amostras para aplicaccedilatildeo no gel de electroforese foram preparadas para uma concentraccedilatildeo de
proteiacutena de 1 mgmL num volume final de 100 L dos quais 25 L correspondem ao tampatildeo da
amostra14
As amostras foram aquecidas a 100ordmC durante 5 minutos Antes da aplicaccedilatildeo das
amostras transferiu-se a cassette para a tina de electroforese agrave qual se adicionou a soluccedilatildeo
tampatildeo de electroforese da tina superior15
e tampatildeo de electroforese da tina inferior16
Aplicaram-
se 10 L das soluccedilotildees de marcadores de peso molecular e 20 L das amostras proteicas
preparadas anteriormente A electroforese decorreu durante cerca de 3h30 min a uma voltagem
de 80 V De seguida transferiu-se o gel para uma tina corando as proteiacutenas durante 1h com
soluccedilatildeo corante17
Descorou-se o gel com soluccedilatildeo descorante18
tendo-se mudado esta soluccedilatildeo ateacute
as bandas se encontrarem bem definidas
Determinou-se a mobilidade relativa das bandas correspondentes aos marcadores de elevado peso
molecular e traccedilou-se a recta de calibraccedilatildeo ndash log(PM) vs log(Rf) onde Rf corresponde agrave
mobilidade relativa Por interpolaccedilatildeo determina-se o PM das proteiacutenas isoladas sabendo a sua
mobilidade relativa Por comparaccedilatildeo com os valores descritos na literatura pode concluir-se
acerca do grau de purificaccedilatildeo
34 Mediccedilatildeo da actividade enzimaacutetica de S1
341 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
13
tampatildeo de concentraccedilatildeo 05 M Tris-HCl pH 68 14
tampatildeo de amostra 320 mM Tris-HCl pH 68 04 M szlig-mercaptoetanol SDS 8 glicerol 15 (vv) e roxo de m-
cresol 0024 15
tampatildeo de electroforese da tina superior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS 16
tampatildeo de electroforese da tina inferior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS acetato de soacutedio
01 M 17
soluccedilatildeo corante ldquoCoomassie Blue R-250rdquo contendo metanol 45 (vv) e aacutecido aceacutetico glacial 10 (vv) 18
soluccedilatildeo descorante metanol 10 (vv) e aacutecido aceacutetico glacial 10 vv)
45
O meacutetodo de Fiske-Subarow modificado consiste na reacccedilatildeo do molibdato de amoacutenio com o
fosfato inorgacircnico em soluccedilatildeo aacutecida com formaccedilatildeo do aacutecido fosfomoliacutebdico Seguidamente daacute-
se a adiccedilatildeo de um agente redutor que provoca a reduccedilatildeo do composto a fosfomolibdato que forma
um complexo de cor azulada cuja absorvacircncia maacutexima ocorre para um comprimento de onda de
660 nm tornando possiacutevel a sua detecccedilatildeo por meacutetodos espectrofotomeacutetricos As actividades Ca2+
-
ATPaacutesica e K+EDTA-ATPaacutesica de S1 foram determinadas atraveacutes deste meacutetodo de acordo com
o descrito por Taussky e Shorr (1953) com mediccedilatildeo da quantidade de fosfato (Pi) libertado
atraveacutes da hidroacutelise de ATP
Para tal obteve-se primeiramente a recta de calibraccedilatildeo com as amostras presentes na Tabela 3II
utilizando uma soluccedilatildeo stock de 5 mM KH2PO4
Tabela 3II ndash Composiccedilatildeo das diferentes amostras utilizadas para determinar a recta de calibraccedilatildeo atraveacutes do meacutetodo
de Fiske-Subbarow modificado
KH2PO4 (L)
([KH2PO4]stock= 5mM)
PO43-
(nmole) H2O (uL)
0 0 1780
5 25 1775
10 50 1770
20 100 1760
40 200 1740
100 500 1680
A cada tubo de Eppendorf adicionaram-se seguidamente 200 L de soluccedilatildeo J19
e 20 L de agente
redutor20
homogeneizando no voacutertex apoacutes cada adiccedilatildeo Incubou-se durante 10 minutos e mediu-
se a Abs660 nm num espectrofotoacutemetro com intervalos de tempo iguais entre cada amostra Traccedilou-
se a recta de calibraccedilatildeo (Abs660 nmvs nmol PO43-
) a partir dos valores obtidos
Para determinar a actividade Ca2+
-ATPaacutesica de S1 prepararam-se tubos de Eppendorf com 400
L de meio de reacccedilatildeo21
25 L CaCl2 ([CaCl2]stock= 100 mM) 25 L ATP ([ATP]stock= 100 mM)
com [S1]ensaios=01 mgmL Perfez-se se necessaacuterio com H2O MilliQ ateacute perfazer um volume de 1
mL Para determinar a actividade K+EDTA-ATPaacutesica utilizou-se uma concentraccedilatildeo idecircntica de
19
Soluccedilatildeo J 5 N H2SO4 25 molibdato de amoacutenio 20
Agente redutor 5 mg aacutecido 1-amino-2-naftol-4-sulfoacutenico 30 mg bissulfito de soacutedio 30 mg sulfito de soacutedio em 26
mL H2O 21
meio de reacccedilatildeo 125 mM Tris-HCl pH 80 0575 M KCl
46
proteiacutena nos ensaios (01 mgmL) e o mesmo meio de reacccedilatildeo Aos tubos de Eppendorf
adicionou-se ainda 123 L de KCl ([KCl]stock = 3M) 25 L de ATP 10 L de EDTA
([EDTA]stock= 100 mM )e o volume de H2O MilliQ necessaacuterio para perfazer 1 mL Em ambas as
situaccedilotildees o uacuteltimo reagente adicionado foi o ATP uma vez que se trata do substrato da reacccedilatildeo
enzimaacutetica
A cineacutetica decorreu a 25ordmC durante 5 minutos para a actividade Ca2+
-ATPaacutesica e durante 3
minutos para a actividade K+EDTA-ATPaacutesica Apoacutes o periacuteodo de incubaccedilatildeo transferiu-se 150
L de amostra para tubos de Eppendorf contendo igual volume de TCA 10 Colocou-se no gelo
e homogeneizou-se no voacutertex Seguidamente centrifugou-se a 1500 g durante 10 minutos e
recolheu-se 200 L de sobrenadante para novos tubos de Eppendorf Adicionou-se a estes 158
mL de H2O MilliQ e 200 L de soluccedilatildeo J Homogeneizou-se no voacutertex e adicionou-se 20 L de
reagente redutor repetindo-se a homogeneizaccedilatildeo Incubou-se durante 10 minutos e mediu-se a
Abs660 nm com o mesmo intervalo de tempo de leitura para cada amostra Por intrapolaccedilatildeo a partir
da recta de calibraccedilatildeo obtida anteriormente determinou-se a quantidade de Pi libertado em cada
ensaio A partir destes resultados determinaram-se as actividades Ca2+
-ATPaacutesica e K+EDTA-
ATPaacutesica do S1 em nmol PO43-
minmg proteiacutena e em s-1
342 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
Da actividade Mg2+
-ATPaacutesica de S1 resulta a formaccedilatildeo dos produtos da reacccedilatildeo de
hidroacutelise do ATP - ADP e Pi Na presenccedila de ADP fosfoenolpiruvato (PEP) e da enzima piruvato
cinase (PK) os produtos resultantes satildeo ATP e piruvato Por acccedilatildeo da enzima lactato
desidrogenase (LDH) este uacuteltimo eacute convertido em lactato ocorrendo simultaneamente a oxidaccedilatildeo
do NADH a NAD+ monitorizada atraveacutes da Abs340 nm Quando o S1 de miosina se encontra no
meio reaccional o ATP eacute hidrolisado formando-se novamente ADP e Pi o que permite a
formaccedilatildeo de um ciclo (fig 21) Se as enzimas se encontrarem em excesso a velocidade de
conversatildeo do NADH eacute limitada pela produccedilatildeo de ADP e Pi Como o ATP eacute regenerado em cada
ciclo a sua concentraccedilatildeo inicial eacute sempre mantida (Bagshaw 1982)
Figura 33ndash Representaccedilatildeo esquemaacutetica do meacutetodo de enzimas acopladas
47
O protocolo utilizado e seguidamente descrito foi adaptado de Cuenda et al (1990 ndash Ver Refordf) A
uma cuvette de 1 mL adicionou-se S1 para uma concentraccedilatildeo final de 005 mgmL 800 L de
meio de reacccedilatildeo22
10 L de PEP ([PEP]stock = 42 mM) 10 L de PK+LDH 11 ([PK+LDH]stock =
1 mgmL) e 15 L de NADH ([NADH]stock=25 mM) Iniciou-se a monitorizaccedilatildeo da reacccedilatildeo
atraveacutes da leitura de Abs340 nm apoacutes a adiccedilatildeo de 20 L de ATP ([ATP]stock=100 mM) A
actividade Mg2+
-ATPaacutesica eacute obtida atraveacutes do declive correspondente agrave oxidaccedilatildeo do NADH Para
a mediccedilatildeo da Mg2+
-ATPase de S1 estimulada por actina adicionou-se F-actina para uma
concentraccedilatildeo final de 2 microM sendo a actividade obtida tambeacutem atraveacutes do declive correspondente
agrave oxidaccedilatildeo do NADH apoacutes a adiccedilatildeo da proteiacutena
35 Interaccedilatildeo entre vanaacutedio e subfragmento-1 de miosina
351 Preparaccedilatildeo das soluccedilotildees de vanaacutedio
As soluccedilotildees de vanadato foram preparadas em aacutegua MilliQ a partir de metavanadato de amoacutenio
(NH4VO3) numa concentraccedilatildeo stock de 50 mM com agitaccedilatildeo magneacutetica e aquecimento (~30 ordmC)
Esta soluccedilatildeo com um valor compreendido entre 6 e 7 designa-se por ldquometavanadatordquo e eacute
constituiacuteda pelas esspeacutecies V1 V2 V4 e V5 (Ref) Acidificando a soluccedilatildeo de metavanadato por
adiccedilatildeo de HCl ateacute um valor de pH proacuteximo de 4 observa-se uma mudanccedila de cor de amarelo
claro para laranja indicativa da formaccedilatildeo da espeacutecie decameacuterica de vanadato De seguida e
imediatamente antes da realizaccedilatildeo dos ensaios descritos o valor do pH da soluccedilatildeo de
decavanadato foi aumentado para cerca de 75 por adiccedilatildeo de NaOH A soluccedilatildeo (2) designada
por ldquometavanadatordquo natildeo foi posteriormente alterada ateacute agrave sua adiccedilatildeo para a realizaccedilatildeo dos ensaios
As soluccedilotildees foram armazenadas a 4ordmC e mantidas em gelo aquando da sua utilizaccedilatildeo
A soluccedilatildeo de V(IV) foi preparada em condiccedilotildees anaeroacutebias por diluiccedilatildeo de sulfato de vanadil
(VOSO4) para uma concentraccedilatildeo final de 5 mM em H2O MilliQ saturada com N2 gasoso
352Anaacutelise por espectroscopia de UV-Vis
A concentraccedilatildeo da espeacutecie decameacuterica em soluccedilatildeo depende do periacuteodo de tempo apoacutes a diluiccedilatildeo
no meio de reacccedilatildeo apropriado (Ref) A velocidade de desoligomerizaccedilatildeo da espeacutecie decameacuterica
pode ser analisada por espectroscopia de absorccedilatildeo na regiatildeo UVVis uma vez que esta espeacutecie
apresenta duas bandas caracteriacutesticas na regiatildeo do visiacutevel (a 358 e 400 nm) que natildeo se verifica
em quaisquer outras espeacutecies oligomeacutericas As bandas supramencionada sapresentam-se bem
22
Meio de reacccedilatildeo 3125 mM Tris-HCl pH 71 375 mM MgCl2
48
resolvidas atraveacutes da obtenccedilatildeo da derivada de 2ordf ordem do espectro apresentando a forma de 2
vales com miacutenimos centrados nos comprimentos de onda supracitados com dependecircncia linear
relativamente agrave concentraccedilatildeo de decavanadato total (Ramos et al 2006)
Assim a estabilidade da espeacutecie decameacuterica apoacutes diluiccedilatildeo da soluccedilatildeo de decavanadato no meio de
reacccedilatildeo dos ensaios (tampatildeo D) para uma concentraccedilatildeo final de 1 mM em vanadato total foi
caracterizada atraveacutes da leitura da Abs400 em funccedilatildeo do tempo tendo-se determinado qual a
ordem de reacccedilatildeo e o tempo de meia-vida da mesma Com base em estudos anteriores acerca da
estabilidade do V10 na presenccedila de diferentes proteiacutenas (Ramos et al 2006) determinou-se
igualmente o tempo de meia vida apoacutes a adiccedilatildeo de 20 microM de S1 de miosina ao meio reaccional
353 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
Os espectros de RMN do isoacutetopo 51
V foram realizados num espectroacutemetro Bruker AM-400 MHz
equipado com uma sonda multinuclear inversa de 5 mm Em tubos de RMN de 5 mm de diacircmetro
preparou-se soluccedilotildees de 500 L de deca- ou metavanadato correspondentes a um intervalo de
concentraccedilotildees de 1 a 10 mM suplementadas com 10 D2O Os espectros foram adquiridos agrave
temperatura ambiente com uma largura espectral de 23 923 Hz um tempo de aquisiccedilatildeo de 0086
s tempo de relaxaccedilatildeo de 001 s e uma sequecircncia de pulsos a 90ordm Utilizou-se a transformada de
Fourier com alargamento das bandas de 20Hz para o processamento dos espectros Os desvios
quiacutemicos das ressonacircncias de 51
V foram obtidos em relaccedilatildeo agrave espeacutecie VOCl3cuja frequecircncia de
ressonacircncia eacute considerada o padratildeo sendo todas as outras referenciadas a partir desta em ppm
As aacutereas dos sinais do RMN correspondentes agraves espeacutecies oligomeacutericas presentes foram obtidas
por integraccedilatildeo dos sinais As concentraccedilotildees relativas das espeacutecies foram calculadas com recurso agrave
equaccedilatildeo
onde A[Vx] corresponde agrave aacuterea referente agrave ressonacircncia da espeacutecie Vx com nordm de agregaccedilatildeo x
A[V]total agrave soma total das aacutereas medidas e [V] a concentraccedilatildeo total de vanaacutedio no ensaio
A teacutecnica de 51
V-RMN foi anteriormente aplicada em estudos de ligaccedilatildeo com S1 de miosina
(Ref) tendo sido demostrado que entre as espeacutecies V1 V2 V4 e V5 a espeacutecie tetrameacuterica possui a
maior afinidade para a miosina (Cremo et al) O desaparecimento ou alargamento dos sinais pode
implicar a ligaccedilatildeo de um dos oxovanadatos laacutebeis agrave miosina sem indicaccedilatildeo de qual uma vez que
49
se processa um equiliacutebrio entre eles (Crans 1990) Assim e tendo em conta as indicaccedilotildees de que
a espeacutecie V10 eacute distinta das restantes no que diz respeito agrave magnitude da interacccedilatildeo com a miosina
e inibiccedilatildeo da actividade ATPaacutesica estimulada por actina (Aureliano 2000 Tiago et al 2002)
torna-se fundamental investigar a interacccedilatildeo das diferentes espeacutecies de vanadato com o S1 de
miosina atraveacutes da teacutecnica de 51
V-RMN
Para tal obtiveram-se espectros de 51
V-RMN resultantes da titulaccedilatildeo de uma soluccedilatildeo 5 mM de
decavanadato total com concentraccedilotildees crescentes de S1 de miosina (0 5 10 15 e 20 microM) em
tampatildeo D Realizou-se ainda ensaios na presenccedila de 2 mM de decavanadato e 2 mM de ATP na
presenccedila e na ausecircncia de S1 de miosina de modo a verificar o efeito que estes ligandos naturais
da enzima teria na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o subfragmento-1
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
Os grupos sulfidrilo (-SH) das cisteiacutenas satildeo resiacuteduos de aminoaacutecidos extremamente sensiacuteveis agrave
oxidaccedilatildeo resultando na sua oxidaccedilatildeo (Dalle-Donne et al 1999) Logo a quantificaccedilatildeo destes
resiacuteduos eacute bastante importante nos estudos de interacccedilatildeo com vanaacutedio jaacute que os resiacuteduos de Cys
com ecircnfase para o SH1 e o SH2 satildeo bastante importantes na monitorizaccedilatildeo das modificaccedilotildees
funcionais da regiatildeo da cabeccedila
O procedimento seguidamente apresentado foi originalmente descrito por Gutieacuterrez-Merino
(1987)) Consiste na reacccedilatildeo entre os grupos ndashSH natildeo oxidados das proteiacutenas e o reagente de
Ellman (DTNB -55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico)) - formando-se o aniatildeo CNT (aniatildeo 3-
carboxilato-4-nitrotiofenolato) numa estequiometria de 11 Este apresenta um maacuteximo de
absorvecircncia a 412 nm (Robyt 1990) Assim apoacutes a incubaccedilatildeo da proteiacutena na presenccedila do metal
a quantidade de cisteiacutenas reduzidas por proteiacutena determina-se atraveacutes dos valores de Abs412
Deste modo colocou-se num tubo de Eppendorf o volume apropriado de S1 dialisado para que a
concentraccedilatildeo no ensaio fosse de 1 mgmL Adicionou-se o volume apropriado de decavanadato
para que a sua concentraccedilatildeo fosse de 0 10 50 100 250 500 ou 1000 M Perfez-se com soluccedilatildeo
D ateacute um volume final de 1 mL e incubou-se a 25ordmC por 20 minutos Apoacutes este periacuteodo colocou-
se 780 L da mistura reaccional numa cuvette de quartzo e adicionou-se 220 L de soluccedilatildeo K23
23
Tampatildeo K Tampatildeo fosfatohidrogenofosfato de soacutedio 025 M pH 72
50
Adicionou-se 15 L de DTNB (em soluccedilatildeo K [DTNB]stock=100 mM) ao meio reaccional e
registou-se o valor de absorvecircncia medido equivalente agraves cisteiacutenas ldquoraacutepidasrdquo (SH1 e SH2
expostos ao solvente) Adicionou-se 001 g de SDS que provoca a desnaturaccedilatildeo da proteiacutena e
consequentemente a exposiccedilatildeo de todos os resiacuteduos de cisteiacutenas que se encontravam ateacute aiacute
inacessiacuteveis no interior da proteiacutena (cisteiacutenas totais) Estes resiacuteduos podem assim reagir com o
DTNB
Estimou-se o nuacutemero de resiacuteduos de cisteiacutenas em todas as amostras com recurso ao coeficiente de
extinccedilatildeo molar () do DTNB (ver ponto 3 dos Anexos) que foi determinado atraveacutes da titulaccedilatildeo
de L-Cisteiacutena com intervalo de concentraccedilotildees compreendidos entre 0 e 50 microM na presenccedila de
25microM de DTNB em tampatildeo D Os valores de absorvecircncia foram obtidos para um comprimento de
onda de 412 nm
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
3621 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
A teacutecnica mais eficiente de anaacutelise da interacccedilatildeo entre o iatildeo vanadilo e proteiacutenas como a
calmodulina e a transferrina entre outras permite a obtenccedilatildeo raacutepida de informaccedilatildeo no que toca agrave
conformaccedilatildeo e amplitude da referida interacccedilatildeo
Os espectros de Ressonacircncia Paramagneacutetica Electroacutenica (RPE) da espeacutecie paramagneacutetica V(IV)
foram realizados a 70K num espectroacutemetro Bruker EMX-300 X-band Todas as soluccedilotildees
utilizadas e amostras foram preparadas em anaerobiose numa cacircmara de luvas de modo a evitar a
reduccedilatildeo de V(IV) a V(V) Em tubos capilares com 02 mL de amostra manteve-se a concentraccedilatildeo
de S1 a 20 M e variou-se a concentraccedilatildeo de vanadilo (0 25 5 10 15 25 50 75 100 150
200 500 e 1000 M [VOSO4]stock=5 mM) em tampatildeo D Apoacutes obtenccedilatildeo dos espectros estes
foram tratados atraveacutes do software SimFonia e WinEPR tendo-se obtido um graacutefico com a
variaccedilatildeo da intensidade do sinal de RPE em funccedilatildeo da concentraccedilatildeo de sulfato de vanadilo
presente Realizaram-se ainda simulaccedilotildees de espectros obtidos com 20 microM de S1 e 1000 microM de
V(IV) com vista agrave obtenccedilatildeo dos ligandos provaacuteveis do vanadilo atraveacutes da anaacutelise da constante
de acoplamento hiperfina (A) e do factor g (g)
4 Conclusotildees gerais
Apesar de jaacute ter sido demonstrado que o vanaacutedio inibe a Na+K
+-ATPase da miosina a funccedilatildeo
deste elemento no tecido muacutesculo esqueleacutetico continua largamente incompreendida A enorme
complexidade quiacutemica deste metal especialmente no que concerne agrave oligomerizaccedilatildeo e reacccedilotildees
51
redox constitui um obstaacuteculo agrave compreensatildeo do comportamento do vanaacutedio nos organismos
vivos Assim a caracterizaccedilatildeo da quiacutemica deste elemento eacute de grande utilidade antes de iniciar o
estudo da sua interacccedilatildeo com o presente sistema
A anaacutelise por espectroscopias de 51
V-RMN e de UV-Vis de soluccedilotildees de vanadato diluiacutedas nos
meios de reacccedilatildeo utilizados no presente relatoacuterio (10 mM HEPES pH 70 25 mM KCl) mostrou
claramente que a espeacutecie decameacuterica (V10) possui um tempo de vida suficientemente longo para a
determinaccedilatildeo do seu efeito isolado ao contraacuterio do que se verifica com as espeacutecies mono- di-
tetra- e pentameacutericas (V1 V2 V4 e V5) Por outro lado a anaacutelise verificou ainda que apoacutes diluiccedilatildeo
das soluccedilotildees de decavanadato no meio de reacccedilatildeo coexistem as espeacutecies V10 e V1 pelo que os
efeitos putativos desta uacuteltima deveratildeo ser subtraiacutedos aquando da monitorizaccedilatildeo dos efeitos da
espeacutecie decameacuterica
O vanaacutedio (V) (vanadato) inibe as enzimas que catalizam reacccedilotildees de transferecircncia de grupos
fosfato como a ATPase da miosina [Goodno 1979] devido agrave sua analogia estrutural com o iatildeo
fosfato Na sua utilizaccedilatildeo como inibidor enzimaacutetico natildeo eacute tida em conta a contribuiccedilatildeo individual
das espeacutecies oligomeacutericas considerando-se normalmente a espeacutecie V1 como inibidora efectiva
[Goodno e Taylor 1982] No presente relatoacuterio verificou-se por espectroscopia de 51
V-RMN que
outros oligoacutemeros de vanadato interagem com o S1 de miosina tendo as espeacutecies V10 e V4 os
maiores niacuteveis de afinidade A sua interacccedilatildeo envolve forccedilas electrostaacuteticas e eacute afectada
negativamente por ligandos naturais de enzima como eacute o caso do ATP
A exposiccedilatildeo do S1 de miosina (005 mgmL) a decavanadato conduz agrave inibiccedilatildeo do capacidade da
proteiacutena de estimular a actividade Mg2+
-ATPaacutesica com um valor de IC50 de 266 plusmn 07 M
decavanadato sendo o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM
decavanadato (80) Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM decavanadato sendo
o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM decavanadato (60)
exercendo um efeito de impedimento agrave ligaccedilatildeo entre o vanaacutedio e o S1
A uma concentraccedilatildeo de 27 microM decavanadato verifica-se a oxidaccedilatildeo de um dos resiacuteduos de Cys
de S1 de miosina seguida pela oxidaccedilatildeo gradual de outro resiacuteduo de Cys ateacute agrave concentraccedilatildeo de
1000 microM Ambos os resiacuteduos oxidados natildeo se encontravam expostos ao solvente utilizado A
oxidaccedilatildeo dos resiacuteduos permite sugerir que esta modificaccedilatildeo oxidativa sofrida pelo S1 contribui
para a diminuiccedilatildeo da sua actividade de ATPase
52
Por espectroscopia de RPE revelaram-se os possiacuteveis ligandos equatoriais do vanaacutedio aquando da
sua interacccedilatildeo com o S1 de miosina como sendo duas moleacuteculas de aacutegua um ligando contendo o
grupo R-OH e outro contendo o grupo R=NR
Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados de
oxidaccedilatildeo (IV) e (V) no S1 de miosina para concentraccedilotildees facilmente alcanccedilaacuteveis no ambiente
terrestre
53
5 Estudos futuros
No sentido do melhor esclarecimento das alteraccedilotildees moleculares e estruturais que ocorrem no
subfragmento 1 quando exposta ao vanaacutedio que conduzem a uma diminuiccedilatildeo da sua funccedilatildeo
sugere-se
- Quantificaccedilatildeo das cisteiacutenas reduzidas de S1 na presenccedila de metavanadato
- Determinaccedilatildeo das actividades Ca2+
- e K+-ATPaacutesica do S1 de miosina na presenccedila de
decavanadato
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina estimulada por actina
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina na presenccedila de
metavanadato
- Realizaccedilatildeo de RMN 2D com variaccedilatildeo das concentraccedilotildees das soluccedilotildees de decavanadato e
metavanadato na presenccedila de S1
-Mutageacutenese dos resiacuteduos de Cys doS1 de miosina de modo a verificar quais os resiacuteduos que
sofrem oxidaccedilatildeo na presenccedila de decavanadato
- Determinaccedilatildeo por calorimetria (DSC Differential Scanning Calorimetry) da variaccedilatildeo da
entalpia (∆H) e da temperatura de desnaturaccedilatildeo do S1 de miosina na presenccedila de vanaacutedio uma
vez que estas propriedades caracteriacutesticas de cada proteiacutena satildeo alteradas apoacutes ocorrecircncia de
modificaccedilotildees estruturais
- Anaacutelise por SDS-PAGE de S1 de miosina apoacutes tratamento com vanaacutedio de modo a determinar
quais os polipeacuteptidos eventualmente formados apoacutes clivagem da proteiacutena
54
6 Referecircncias bibliograacuteficas
- Sellers J Myosins 2nd Edition Oxford University Press 1999
- L Stryer inBiochemistry (Ed WH Freeman 2005 pp
Refordfas Materiais e Meacutetodos
[A] S S Margossian SLowey Methods Enzymol1982 85 55-71
[B] JD Pardue JA Spudich Methods Enzymol 1982 85 164-181
[C] M Bradford Anal Biochem1976 72 248-254
[D] H H Taussky E Shorr J Biol Chem 1953 202 675-685
[E] C R Bagshaw in Outline Studies in Biology ndash Muscle Contraction Chapman and Hall Ed
1982
[F] A Cuenda et al Eur J Biochem1990 194 663-670
55
Anexos
1 Determinaccedilatildeo da recta de calibraccedilatildeo meacutetodo de Bradford
Figura1ndashRepresentaccedilatildeo graacutefica da recta de calibraccedilatildeo (Abs595 nm vs [BSA]) utilizada para o meacutetodo de Bradford
2 Determinaccedilatildeo da recta calibraccedilatildeo para a electroforese em condiccedilotildees desnaturantes
Figura 2 ndash Representaccedilatildeo graacutefica da recta de calibraccedilatildeo (log(Peso Molecular) vs Rf dos marcadores de peso
molecular) onde Rf indica determinada atraveacutes do padratildeo de peso molecular
(poccedilo (a))
3Caacutelculo do coeficiente de extinccedilatildeo molar () para a quantificaccedilatildeo de cisteiacutenas reduzidas
atraveacutes do meacutetodo de Ellman
Traccedilou-se a recta de Abs412 nm meacutedia vs [L-Cisteiacutena] de modo a determinar o valor do coeficiente
de extinccedilatildeo molar definido como o declive daquela recta Assim =8400 M-1
cm-1
como
podemos verificar na Fig 3
A595 nm = 00546[BSA] + 00209 Rsup2 = 099051
Ab
s 59
5 n
m
[BSA] (mgmL)
log PM = -1177 Rf + 2247 Rsup2 = 098957
log
PM
Mobilidade relativa (Rf)
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero

17
livremente em ambientes fortemente aacutecidos como os lisossomas jaacute que a pH superior a 3 as
soluccedilotildees de vanadilo tendem a ser oxidadas pelo ar formando-se vanadato Esta grande
capacidade redutora justifica parcialmente a utilizaccedilatildeo pouco extensiva do iatildeo VO2+
como
inibidor [W]
1212 Vanaacutedio (V)
O vanaacutedio (V) eacute a forma mais estaacutevel num intervalo de pH amplo [W] O iatildeo anioacutenico VO3-tende
a constituir complexos polinucleares pelo que as soluccedilotildees aquosas de vanadato satildeo extremamente
complexas com ocorrecircncia simultacircnea de vaacuterios equiliacutebrios de protonaccedilatildeo (pKa(vanadato)= 35
78 e 125) e oligomerizaccedilatildeo As espeacutecies presentes (H3VO4 H2VO4- HVO4
2- e VO4
3-) dependem
do pH forccedila ioacutenica e temperatura da soluccedilatildeo e da concentraccedilatildeo total do elemento
[AEAFAGAHAI]
O VO43-
- monoacutemero ortovanadato V1 ndash estaacute associado a uma grande parte da importacircncia
bioloacutegica do vanaacutedio uma vez que se trata de um anaacutelogo electroacutenico e estrutural do ortofosfato
(PO43-
) A coordenaccedilatildeo do ortovanadato pode ser tetraeacutedrica ou bipiramidal trigonal (Refordfs)
A oligomerizaccedilatildeo do vanadato monomeacuterico daacute origem ao diacutemero (V2O72-
- V2) que apresenta
tambeacutem equiliacutebrios de protonaccedilatildeo Presume-se a existecircncia de analogia do vanadato dimeacuterico com
o pirofosfato embora natildeo haja evidecircncia estrutural (Refordf) A dimerizaccedilatildeo do V2 origina o
vanadato tetrameacuterico ou metavanadato (V4O124-
- V4) que constitui a espeacutecie maioritaacuteria em
soluccedilotildees aquosas concentradas (Refordf) As estruturas do metavanadato e a sua espeacutecie protonada
satildeo ciacuteclicas [AJ] Outra espeacutecie oligomeacuterica importante eacute o vanadato pentameacuterico (V5O153-
) cuja
estrutura presumida eacute ciacuteclica [AI]
O oligoacutemero com maior importacircncia bioquiacutemica eacute o vanadato decameacuterico (V10O286-
-V10)(Refordf) a
espeacutecie predominante em soluccedilotildees de pH 2-6 quando a concentraccedilatildeo total de vanadato eacute superior
a 100 M (Ref) A estrutura do V10 natildeo difere em soluccedilatildeo ou no estado soacutelidoOs 10 aacutetomos de
vanaacutedio da espeacutecie decameacuterica estatildeo divididos em trecircs grupos identificados na figura como VA-C
consoante o seu ambiente quiacutemico circundante (Refordf) (Fig 111)
A expressatildeo ldquodecavanadatordquo refere-se a uma soluccedilatildeo aquosa na qual a espeacutecie decameacuterica se
encontra em equiliacutebrio quiacutemico com as espeacutecies V1 V2 V4 e V5 sendo que a oligomerizaccedilatildeo
progressiva do vanaacutedio eacute proporcional agrave sua concentraccedilatildeo total (Ref) Nesta soluccedilatildeo a
concentraccedilatildeo de V10 corresponde a aproximadamente 10 da concentraccedilatildeo total de vanaacutedio [AK]
18
Figura 111 ndash Estruturas esquemaacuteticas das espeacutecies de vanadato (espeacutecie monomeacuterica (V1) dimeacuterica (V2)
tetrameacuterica (V4) pentameacuterica (V5) e decameacuterica (V10) (Adaptado de Aureliano e Crans 2008)
122 Metabolismo e efeito na actividade enzimaacutetica
Apesar de natildeo terem sido ainda encontradas as provas categoacutericas das suas funccedilotildees nos seres
vivos sabe-se que o iatildeo vanadato eacute capaz de inibir uma grande quantidade de enzimas in vitro
com especial ecircnfase na actividade ATPaacutesica da miosina [AL Teresa] Como referido
anteriormente o vanadato eacute capaz de actuar em alternativa ao fosfato nas reacccedilotildees bioquiacutemicas
incluindo a sua preferecircncia de ligaccedilatildeo agraves enzimas [W]
Eacute conhecido o papel do vanaacutedio no metabolismo do caacutelcio promovendo a mineralizaccedilatildeo dos
dentes e ossos na biossiacutentese de liacutepidos (trigliceacuteridos e colesterol) com posterior inibiccedilatildeo da
acumulaccedilatildeo destes nos vasos sanguiacuteneos e no SNC na reproduccedilatildeo e desenvolvimento por
mimetizaccedilatildeo e potenciaccedilatildeo de factores de crescimento na estimulaccedilatildeo do consumo de glucose e
da siacutentese de glicogeacutenio com actuaccedilatildeo equivalente aos efeitos da insulina ao niacutevel celular na
transduccedilatildeo de sinais celulares com regulaccedilatildeo da actividade de segundos mensageiros no
transporte de iotildees e aminoaacutecidos na transcriccedilatildeo geneacutetica e siacutentese de proteiacutenas e DNA entre
outros [W Y AB]
Assim o vanaacutedio cnseguiraacute provocar inuacutemeras mudanccedilas bioquiacutemicas nas ceacutelulas Alguns
exemplos da influecircncia bioquiacutemica do vanaacutedio incluem os seus efeitos nos processos de
fosforilaccedilatildeo e desfosforilaccedilatildeo de proteiacutenas e na participaccedilatildeo na formaccedilatildeo de espeacutecies reactivas de
oxigeacutenio devido ao seu efeito proacute-oxidante sobre agentes redutores a niacutevel celular [AM]
Quanto ao seu efeito na actividade enzimaacutetica pode dividir-se em dois tipos principais ndash inibitoacuterio
e estimulante - dependendo da enzima ou do tipo de reacccedilatildeo por esta catalisada No primeiro
grupo incluem-se enzimas como as ATPases (de miosina de canais ioacutenicos e fosfoenzimas
19
transportadoras) e as fosfatases (da tirosina aacutecida e alcalina) a fosfofrutocinase adenilato ciclase
e catalase entre outras [AA] O segundo grupo inclui a maioria das cinases a glucose-6-fosfato
desidrogenase a fosfoglicerato mutase e a fosfoglicerato fosfatase (Refordf)
O efeito do vanaacutedio (V) na actividade enzimaacutetica estaacute relacionado com o seu nuacutemero de
coordenaccedilatildeo podendo actuar como estado de transiccedilatildeo ou por mimetizaccedilatildeo de cofactores
substratos (especialmente o fosfato) e interacccedilotildees proteiacutena-proteiacutena (como no caso das
polimerases e transcriptases) Pode ainda estar envolvido na formaccedilatildeo de ligaccedilotildees de hidrogeacutenio
interacccedilotildees electrostaacuteticas esteacutereas e de oxidaccedilatildeo-reduccedilatildeo Uma evidecircncia desta uacuteltima eacute a sua
capacidade de alternacircncia entre os estados de oxidaccedilatildeo +5 a +3 o que torna faacutecil a sua interacccedilatildeo
redox com enzimas contendo resiacuteduos de cisteiacutenas (Cys) ou qualquer outro composto orgacircnico
com actividade redox [AN]
13 Vanaacutedio e muacutesculo esqueleacutetico
A primeira evidecircncia da presenccedila de vanaacutedio no muacutesculo esqueleacutetico proveio de uma amostra de
ATP isolado de muacutesculo de cavalo que continha uma impureza ndash vanaacutedio ndash cuja presenccedila
provocou a inibiccedilatildeo da actividade da bomba de Na+K
+-ATPase [AO] Desde entatildeo vaacuterios
estudos tecircm sido realizados com ecircnfase na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o
tecido muscular Destes destaca-se a descoberta da interacccedilatildeo forte da espeacutecie V10 com a miosina
fosforilada [AP] e agrave sua inibiccedilatildeo da actividade ATPaacutesica da miosina[AQ AR]
No quediz respeitoao iatildeo monovanadato destaca-se a sua inibiccedilatildeoda actividade Mg2+
-ATPaacutesica
da miosina [AS] que ocorre de duas formas distintas (1) na presenccedila de concentraccedilotildees reduzidas
de V1 a inibiccedilatildeo ocorre a velocidade reduzida dependente da concentraccedilatildeo da espeacutecie
monomeacuterica (2) na presenccedila de concentraccedilotildees elevadas de V1 o niacutevel de inibiccedilatildeo aproxima-se de
100 desde que [V1] seja superior agrave concentraccedilatildeo total de locais com actividade Mg2+
-ATPaacutesica
no ensaio Esta inibiccedilatildeo eacute irreversiacutevel tanto por diaacutelise ou adiccedilatildeo de agentes redutores do
vanadato [AL]Segundo Pate et al(1994)[AT] a espeacutecie monomeacuterica podeinibir a velocidade de
contracccedilatildeo a temperatura inferior a 20ordmC perdendogradual e rapidamente essa capacidade ateacute os
50ordmC temperatura agrave qual cessa a capacidade inibitoacuteria Os resultados obtidos sugeremainda uma
interacccedilatildeo altamente cooperativa entre o iatildeo vanadato e as proteiacutenas componentes do muacutesculo
esqueleacuteticoDestaca-se tambeacutem a descoberta da inibiccedilatildeo da geraccedilatildeo de forccedila contraacutectil nas fibras
musculares por parte da espeacutecie monomeacuterica[AT]Wilson et al(1995) [AU ]determinaram que a
afinidade da espeacutecie V1 para o S1 de miosina eacute significativamente mais elevada do que a do iatildeo
Pi seu anaacutelogo Descobriram ainda que que a ligaccedilatildeo do vanadato poderaacute envolver transiccedilotildees
entre estados energeacuteticos com energia livre independente dos impedimentos esteacutereos impostos
20
pela estrutura das miofibrilas Os estudos desenvolvidos por Smith et al (1990)[AV]
determinaram queo iatildeo monovanadato natildeo apresenta um efeito pronunciado na inibiccedilatildeo da
actividadeacto-S1-miosina-ATPaacutesica excepto em concentraccedilotildees superiores a 900microM a partir das
quais existe produccedilatildeo de espeacutecies oligomeacutericas para as quais o efeito inibitoacuterio seraacute mais
elevadoA espeacutecie monomeacuterica de vanadato consegue adoptar facilmente uma geometria do tipo
bipiramidal trigonal que mimetiza o intermediaacuterio formado (MgADP
Pi)aquando da reacccedilatildeo de
hidroacutelise de ATP no local activo da miosina [AW] De facto a interacccedilatildeo do vanaacutedio com a
miosina eacute inesperadamente estaacutevel na presenccedila de ADP Esta interacccedilatildeo resulta na formaccedilatildeo de
um complexo enzimaacutetico inactivo e estaacutevel com estequiometria de 11 ADPvanadato por siacutetio
activo da proteiacutena A formaccedilatildeo do complexo estaacutevel miosina-vanadato foi proposta por Goodnoet
al (1982) [AS] Nesta ocorre um preacute-equiliacutebrio raacutepido que induz a formaccedilatildeo de um complexo
ternaacuterio reversiacutevel (MADP
V) com formaccedilatildeo lenta do isoacutemero M
ADP
V A diferenccedila entre os
compostos de vanaacutedio e de fosfato centra-se no tempo de meia vida (t12) do Pi ou do V (no
primeiro t12=12 s enquanto no segundo t12=24 h) (Refordf) A diferenccedila dever-se-aacute agrave possiacutevel
complexaccedilatildeo do vanaacutedio com grupos nucleofiacutelicos no siacutetio activo o que provocaria uma
estabilizaccedilatildeo da miosina na conformaccedilatildeo MADP
V
No que respeita aos estudos realizados com a espeacutecie tetrameacuterica Cremo et al determinou que a
cadeia pesada do S1 de miosina pode ser clivada em dois locais distintos (designados V1 e V2)
pela espeacutecie V4 apoacutes irradiaccedilatildeo com luz UV na presenccedila de concentraccedilotildees de vanadato na ordem
dos milimolar e na ausecircncia de nucleoacutetidos ou metais divalentes
Jaacute no que concerne aos estudos realizados com ecircnfase na espeacutecie decameacuterica foi sugerido
porRamos et al (2006) [AK]que a interacccedilatildeo do decavanadato com a G-actina favorecida pela
polimerizaccedilatildeo desta uacuteltima estabiliza a espeacutecie V10 e induz a inibiccedilatildeo da formaccedilatildeo de F-actina A
estabilizaccedilatildeo da espeacutecie V10 por proteiacutenas tansmembranares e do citoesqueleto pode ser a razatildeo
pela qual ocorre a toxicidade do decavanadato reportada na avaliaccedilatildeo dos efeitos do V(V) nos
sistemas bioloacutegicosTiago et al (2007) [AR] concluiram que o V10 apresenta uma clara preferecircncia
de ligaccedilatildeo ao ldquoback-doorrdquo da S1-miosina-ATPase o que se verifica apenas na estrutura
conformacional desimpedida com acesso facilitado ao local de ligaccedilatildeo do Pi Sugere tambeacutem que
o V10 actua como um ldquoback-door stoprdquo bloqueando estaabertura do fragmento de 50-kDa
necessaacuteria para a hidroacutelise do ATP--fosfato o que constitui uma explicaccedilatildeo simples do
comportamento natildeo competitivo da espeacutecie decameacuterica estimulando a utilizaccedilatildeo deste oligoacutemero
como uma feramenta da elucidaccedilatildeo das mudanccedilas conformacionais ldquoback-doorrdquo da miosina no
21
processo da contracccedilatildeo muscular(ref)Destaca-se ainda a descoberta da capacidade inibitoacuteria do
V10 em relaccedilatildeo agrave fosforilaccedilatildeo oxidativa mitocondrial [AX]
Finalmente no que concerne aos estudos realizados com a presenccedila de vaacuterias espeacutecies
oligomeacutericas Aureliano e Madeira (1994) [AY]observaram que na presenccedila de Ca2+
as espeacutecies
V4 e V10se ligam agrave Ca2+
-ATPase do retiacuteculo sarcoplasmaacutetico enquanto a espeacutecie V1 se liga apenas
na presenccedila de ATP Aparentemente o aumento da concentraccedilatildeo de Ca2+
tem o efeito contraacuterio da
inibiccedilatildeo da ATPase do retiacuteculo sarcoplasmaacutetico pelo V1 natildeo tendo no entanto efeito significativo
sobre a inibiccedilatildeo por parte da espeacutecie V10 Tiago et al (2002) [AP] concluiacuteram que em condiccedilotildees
proacuteximas agraves fisioloacutegicas a espeacutecie V10 difere das espeacutecies presentes na soluccedilatildeo de metavanadato
devido agrave sua forte interacccedilatildeo com a miosina fosforilada e agrave sua inibiccedilatildeo da actividade ATPaacutesica
da miosina Para aleacutem disso o ATP diminui a afinidade da miosina para o V4 induzindo a
interacccedilatildeo com V1 e natildeo tendo qualquer efeito no que diz respeito ao V10 Foi tambeacutem proposto
[AQ] que a ligaccedilatildeo das espeacutecies oligomeacutericas de vanadato aoS1 de miosina nos locais onde o
fosfato coordenaeacute provavelmente a causa da inibiccedilatildeo da actividade ATPaacutesica da miosina
estimulada por actina atraveacutes da prevenccedilatildeo da troca entre ATPADP sendo esta favorecida para
aniotildees de vanadato com valecircncias superiores como eacute o caso do V10
Na presenccedila de actina as velocidades de dissociaccedilatildeo do Pi e do V aumentam consideravelmente
este uacuteltimo em menor quantidade Assim a ligaccedilatildeo da actina ao complexo MADP
V e
Pi eacute
bastante fraca pelo que se sugere que o ADPV teria um efeito de relaxamento semelhante ao
ATP No entanto a capacidade do vanaacutedio em inibir a actividade ATPaacutesica da miosina e da
actomiosina eacute diferenciada uma vez que na ausecircncia de actina a inibiccedilatildeo eacute total para
concentraccedilotildees na gama dos M mas na sua presenccedila a inibiccedilatildeo eacute totalmente conseguida apenas
em concentraccedilotildees na gama dos mM (Ref) No entanto eacute necessaacuterio considerar a possiacutevel
formaccedilatildeo de espeacutecies oligomeacutericas de vanadato cuja afinidade e efeito na enzima seratildeo
diferentes Assim pode concluir-se que o efeito de inibiccedilatildeo observado para a actividade ATPaacutesica
de miosina estimulada por actina se pode dever agrave presenccedila das diferentes espeacutecies oligomeacutericas o
que foi jaacute sugerido por Aureliano (2000) [AZ] ndash as espeacutecies tetrameacuterica e decameacuterica de vanadato
interactuariam com a miosina com alteraccedilatildeo da sua interacccedilatildeo com a actina No entanto natildeo eacute
conhecida a contribuiccedilatildeo absoluta das restantes espeacutecies oligomeacutericas para a inibiccedilatildeo da ATPase
da miosina estimulada por actina (Refs)
14 Objectivos do projecto
Pretende-se com este projecto o isolamento purificaccedilatildeo e caracterizaccedilatildeo da actina do muacutesculo
22
esqueleacutetico de coelho sendo esta uacuteltima tambeacutem realizada para o sufragmento-1 de miosina
Seguidamente realizar-se-aacute a quantificaccedilatildeo e caracterizaccedilatildeo das proteiacutenas por espectroscopia UV-
Vis e SDS-PAGE e a caracterizaccedilatildeo das soluccedilotildees de vanaacutedio por UV-Vis e ressonacircncia magneacutetica
nuclear (RMN) Por uacuteltimo a interacccedilatildeo do S1 de miosina com o vanaacutedio seraacute avaliada atraveacutes
das teacutecnicas de UVVis RMN e ressonacircncia paramagneacutetica electroacutenica (RPE)
2 Discussatildeo de Resultados
21 Caracterizaccedilatildeo das proteiacutenas isoladas
211Anaacutelise do estado de pureza
O estado de pureza das soluccedilotildees de actina e de S1 foi analisado por electroforese em gel de
poliacrilamida em condiccedilotildees desnaturantes (SDS-PAGE) (Fig 22) que separa as moleacuteculas de
acordo com o seu peso molecular (PM) A concentraccedilatildeo de acrilamida escolhida (10) permite a
detecccedilatildeo simultacircnea de componentes com peso molecular inferior a 20 kDa e outros com elevado
peso molecular
Figura 21 ndash Fotografia de gel de SDS-PAGE (10 acrilamida) Poccedilo (a)
10 L do marcador de elevado peso molecular () Poccedilo (b) 20 L de [G-
actina]=1 mgmL Poccedilo (c) 20 L de [S1]=1 mgmL Aplicou-se uma
voltagem de V durante h e min
A partir da recta de calibraccedilatildeo obtida log PM = -1177 Rf + 2247 (ver ponto 2 da secccedilatildeo
Anexos) pode verificar-se a presenccedila de actina na amostra do poccedilo (b) presente numa banda
bastante intensa (PMaparente=40 kDa) bem como a presenccedila minoritaacuteria de outros componentes
desconhecidos que natildeo teratildeo sido eliminados durante o processo de extracccedilatildeo e purificaccedilatildeo e natildeo
satildeo relevantes para o estudo em questatildeo Na amostra do poccedilo (c) nota-se a presenccedila de S1
(PMaparente=92 kDa) bem como a de outros componentes cuja banda eacute ligeiramente menos intensa
Estes poderatildeo ser fragmentos proteoliacuteticos de miosina e de S1 Estaacute ainda presente uma bada de
23
maior intensidade que poderaacute ser a proteiacutena-C (PM=135 kDa) uma vez que a sua remoccedilatildeo
completa da amostra necessitaria de um passo adicional de cromatografia de troca ioacutenica [BA]
No entanto natildeo podemos confirmar que se trata desta proteiacutena ou de qualquer outra uma vez que
se encontra fora do intervalo de migraccedilatildeo dos marcadores de peso molecular
212 Determinaccedilatildeo da concentraccedilatildeo proteica
2121Subfragmento-1 de miosina
A partir da recta de calibraccedilatildeo obtida pelo meacutetodo de Bradford [BB](ver secccedilatildeo 3311 do
Procedimento Experimental) A595nm = 00546[BSA] + 00209 (ver secccedilatildeo de Anexos)foi possiacutevel
determinar um intervalo de concentraccedilatildeo de S1 de miosina compreendido entre 226 e 117(plusmn
423M) (203 e 1056 (plusmn 381 mgmL)) (n=8) A concentraccedilatildeo meacutedia de S1 de miosina apoacutes
diaacutelise contra a soluccedilatildeo tampatildeo 10 mM HEPES pH 75 25 mM KCl determinada pela leitura da
Abs280 nmfoi de 20430M (1803 mgmL)(n=7)
As amostras a quantificar atraveacutes do meacutetodo de Bradford natildeo sofreram diaacutelise o que permite
explicar a concentraccedilatildeo superior de S1 de miosina em comparaccedilatildeo com o valor obtido atraveacutes do
meacutetodo directo Esta razatildeo aliada agrave sensibilidade e rapidez superiores deste uacuteltimo fez com que
se optasse pelo meacutetodo directo para a quantificaccedilatildeo da grande maioria dos ensaios
2122Actina
A concentraccedilatildeo de actina foi determinada quando esta se encontrava na forma monomeacuterica (G-
actina) por mediccedilatildeo da leitura directa da Abs290 nm tendo-se obtido uma concentraccedilatildeo meacutedia de
691481M (290202 mgmL) (n=3) em 2 mM Tris-HCl pH 80 02 mM CaCl2 02 mM Na-
ATP Ao comprimento de onda utilizado (290 nm) absorvem fortemente apenas os resiacuteduos de
Trp pelo que a mediccedilatildeo efectuada tem menor sensibilidade do que se tivesse sido efectuada a 280
nm uma vez que a actina apresenta apenas 4 resiacuteduos deste aminoaacutecido embora tenha 16
resiacuteduos de Tyr
22 Mediccedilatildeo da actividade enzimaacutetica de S1 de miosina
221 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
As actividades ATPaacutesicas de S1 de miosina dependentes de Ca2+
e K+foram analisadas pelo
caacutelculo das velocidades de consumo inicial do fosfato inorgacircnico (Pi) Definiu-se os tempos de 5
e 3 minutos para a mediccedilatildeo da Ca2+
e da K+EDTA-ATPase respectivamente Os valores obtidos
24
a partir do protocolo presente no ponto 341 da secccedilatildeo Procedimento experimental encontram-se
descritos na Tabela 2I
Tabela 2I ndash Acrividades natildeo-fisioloacutegicas de subfragmento-1 determinadas pelo meacutetodo de Fiske-Subarow
modificado (n=2)
Proteiacutena
Actividade
ATPaacutesica natildeo
fisioloacutegica
Actividade (nmol
Piminmg proteiacutena) Actividade (s-1)
S1
Ca2+
-ATPase 1400 plusmn 30 2100 plusmn 0045
K+EDTA-ATPase 2334 plusmn 50 3500 plusmn 0075
As actividades natildeo-fisioloacutegicas reflectem o estado de oxidaccedilatildeo dos resiacuteduos expostos de Cys 707
(SH1) e 697 (SH2) Se o grupo SH1 sofrer oxidaccedilatildeo a actividade ATPaacutesica dependente de Ca2+
aumenta e a actividade dependente de K+ diminui Por outro lado se o grupo SH2 sofrer
oxidaccedilatildeo regista-se a perda completa destas actividades
De acordo com Margossian e Lowey (1982) [BA] as actividades ATPaacutesica de S1 de miosina
dependente de Ca2+
e K+EDTA da miosina satildeo respectivamente de 24 e 112 s-1
Estaacute tambeacutem
descrito que a actividade ATPaacutesica dependente de K+ eacute em meacutedia cerca de 3 vezes superior agrave
dependente de Ca2+
reflectindo a quantidade das preparaccedilotildees A razatildeo obtida entre os valores das
actividades ATPaacutesicas dependentes de K+ e Ca
2+ eacute de 166 Os valores obtidos para as actividades
(Ca2+
-ATPase=21 s-1
K+EDTA-ATPase= 35 s
-1) satildeo inferiores agrave valores obtidos por
Margossian e Lowey [BA] e Tiago [BC] com forccedila ioacutenica inferior O valor obtido para a
actividade ATPaacutesica dependente de Ca2+
eacute superior ao obtido por Polosukhina et al (2000) [BD]
cujo meio de reacccedilatildeo continha MOPS e apresentava forccedila ioacutenica inferior Os dados
supramencionados comprovam a variaccedilatildeo da actividade a ATPaacutesica natildeo fisioloacutegica em relaccedilatildeo agrave
forccedila ioacutenica temperatura e pH
222 Mediccedilatildeo da actividadeMg2+
-ATPaacutesica de S1 pelo meacutetodo de enzimas acopladas
A actividade de Mg2+
-ATPase eacute a uacutenica que possui significado fisioloacutegico sendo as restantes
bastante uacuteteis apenas para indicaccedilatildeo da qualidade de preparaccedilatildeo do S1[BA] A 0575 M KCl a
actina e o S1 formam uma soluccedilatildeo viscosa e tuacuterbida Apoacutes a adiccedilatildeo de MgATP o complexo acto-
S1 dissocia-se e a turbidez da soluccedilatildeo diminui Se a forccedila ioacutenica for reduzida a dissociaccedilatildeo do
complexo actoS1 que se encontra na forma de suspensatildeo eacute induzida O MgATP eacute rapidamente
hidrolisado diminuindo a sua concentraccedilatildeo no meio reaccional Nestas condiccedilotildees haacute
reassociaccedilatildeo e subsequente contracccedilatildeo dos filamentos de actina e de S1 com formaccedilatildeo de
25
precipitado O meacutetodo descrito traduz de forma bastante elementar o fenoacutemeno da contracccedilatildeo in
vivo e elimina o problema anterior da solubilizaccedilatildeo ineficiente da miosina de modo a obter uma
mediccedilatildeo exacta da actividade [A] A utilizaccedilatildeo de forccedilas ioacutenicas reduzidas aumenta a afinidade da
actina para o S1 permitindo contornar a extrema dificuldade em obter a concentraccedilatildeo proteica dos
muacutesculos (cerca de 100 mgmL do complexo actomiosina) As actividades Mg2+
-ATPaacutesicas de S1
de miosina foram determinadas a partir do declive da variaccedilatildeo da Abs340 relativo agrave oxidaccedilatildeo de
NADH em funccedilatildeo do tempo como eacute descrito no ponto 342 do Procedimento Experimental Os
valores obtidos para a actividade Mg2+
-ATPaacutesica encontram-se apresentados na Tabela 2II
Tabela 2II ndash Actividade Mg2+
-ATPaacutesica de subfragmento-1 de miosina determinadas pelo meacutetodo das enzimas
acopladas
Actividade (nmol
ATPmin mg(S1)) Actividade (s
-1)
1097 plusmn 269 164 plusmn 04
Ramos (2005) [J] obteve o valor de 01 s-1
para a actividade Mg2+
-ATPaacutesica de S1 de miosina
com forccedila ioacutenica intermeacutedia Tiago (2005) [BC] obteve o valor de 011 s-1 em condiccedilotildees de forccedila
ioacutenica reduzida em relaccedilatildeo agrave utilizada no presente relatoacuterio
23 Preparaccedilatildeo e caracterizaccedilatildeo das soluccedilotildees de vanaacutedio
231 Anaacutelise da composiccedilatildeo das soluccedilotildees de vanadato por espectroscopia de 51
V-RMN
efeito da concentraccedilatildeo da espeacutecie
A diluiccedilatildeo das soluccedilotildees stock de meta- e decavanadato iraacute determinar fortemente a composiccedilatildeo
em soluccedilatildeo dos diferentes oligoacutemeros de vanadato (ref) No caso do decavanadato o periacuteodo de
tempo apoacutes a diluiccedilatildeo iraacute tambeacutem ser um factor importante a considerar devido agrave decomposiccedilatildeo
da espeacutecie V10(ref)Torna-se assim fundamental determinar a composiccedilatildeo quiacutemica das soluccedilotildees
no meio de ensaio utilizado (HEPES 10 mM KCl 25 mM) de modo a correlacionaacute-la
correctamente com os possiacuteveis efeitos promovidos pelas diferentes espeacutecies de vanadato no
sistema em estudo
Aleacutem disso deve ser dado ecircnfase agraves importantes interacccedilotildees que ocorrem entre o vanaacutedio e os
tampotildees escolhidos O tampatildeo com menor nuacutemero e magnitude de interacccedilatildeo eacute o HEPES [AD]
seguido do Tris sendo este uacuteltimo especialmente aconselhado em concentraccedilotildees baixas para casos
de incompatibilidade enzimaacutetica ou quiacutemica redox No caso especial da reacccedilatildeo redox
NAD(P)HNAD(P) o tampatildeo adequado eacute o imidazole de modo a natildeo haver reacccedilatildeo com o
26
vanadato Os tampotildees a evitar em qualquer situaccedilatildeo incluem a trietanolamina fosfato citrato e
glicina entre outros [AD] uma vez que todos interactuam fortemente com V(V) com formaccedilatildeo
de complexos estaacuteveis
Assim procedeu-se agrave anaacutelise da composiccedilatildeo das soluccedilotildees de meta- e decavanadato apoacutes diluiccedilatildeo
das soluccedilotildees stock (50 mM em vanadato total) no meio de reacccedilatildeo (tampatildeo D) atraveacutes da
espectroscopia de ressonacircncia magneacutetica nuclear de 51
V Nas figuras x1 e x2 apresentam-se os
espectros obtidos para as soluccedilotildees de deca- e metavanadato num intervalo de concentraccedilotildees de 1 a
10 mM A Tabela seguinte resume os paracircmetros espectroscoacutepicos obtidos para cada espeacutecie
presente nas soluccedilotildees analisadas
Figura 22ndash Espectros de RMN de 51
V de
concentraccedilotildees crescentes das espeacutecies oligomeacutericas
de vanadato obtidos apoacutes diluiccedilatildeo da soluccedilatildeo stock
de decavanadato em 10 mM HEPES pH 70 25 mM
KCl para concentraccedilotildees finais de A) 1 mM B)2
mM C) 4 mM D) 8 mM E) 10 mM de
decavanadato total V1refere-se agraves espeacutecies
monomeacutericade vanadato V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2IIIndash Desvios quiacutemicos meacutedios (part) das ressonacircncias das espeacutecies decameacuterica (V10) monomeacuterica (V1) e
tetrameacuterica (V4) presentes nos espectros da soluccedilatildeo de decavanadatoLargura a meia altura (∆12) da espeacutecie V10A
Paracircmetros V10A V10B V10C V1 V4
part (ppm) -5162 -5003 -4255 -5616 -5748
∆12 (Hz) 175 nd nd nd nd
Figura 23 ndash Distribuiccedilatildeo graacutefica das
espeacutecies oligomeacutericas de vanadato vs a
concentraccedilatildeo de decavanadato total As
concentraccedilotildees de cada espeacutecie foram
obtidas segundo a formula descrita na
secccedilatildeo 35 do Procedimento
Experimental (aacutereas dos sinais de 51
V-
RMN observadas na figura x1)V10 V1 e
V4 correspondem agraves espeacutecie monomeacuterica
dimeacuterica e tetrameacuterica respectivamente [Esp
eacuteci
e o
ligo
meacute
rica
de
van
aacutedio
] (m
M)
[Decavanadato]total (mM)
V10
V1
V4
27
Nas soluccedilotildees de decavanadato as espeacutecies monomeacuterica e decameacuterica de vanadato satildeo as uacutenicas
presentes para concentraccedilotildees de decavanadato inferiores a 8 mM Ambas as espeacutecies aumentam
linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado em [AY] e
em (Teresa) quando a concentraccedilatildeo de V1 eacute igual ou superior a 200 microM inicia-se a sua
oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica No entanto nas condiccedilotildees de realizaccedilatildeo dos
ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a espeacutecie tetrameacuterica neste
estudo em concentraccedilotildees inferiores a 8 mM tendo o mesmo sido observado por Ramoset al
(2006) [AK] Os resultados obtidos mostram claramente o equiliacutebrio entre a espeacutecie decameacuterica e
a monomeacuterica e que o aparecimento de outras espeacutecies oligomeacutericas se encontra dependente da
concentraccedilatildeo desta uacuteltima Apesar de natildeo ter sido realizado o ensaio com 5 mM de decavanadato
total no presente relatoacuterio estaacute reportado em Ramos et al (2006) [AK] que as concentraccedilotildees da
espeacutecie decameacuterica e monomeacuterica satildeo respectivamente de 500 microM e 200 microM Assim pode inferir-
se que para concentraccedilotildees inferiores a 5 mM e intervalos de tempo de cerca de 20 minutos (que
inclui o tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas
as espeacutecies decameacuterica e monomeacuterica de vanadato
Figura 24 ndash Espectros de RMN de 51
V de concentraccedilotildees
crescentes das espeacutecies oligomeacutericas de vanadato obtidos
apoacutes diluiccedilatildeo da soluccedilatildeo stock de metavanadato em 10 mM
HEPES pH 70 25 mM KCl para concentraccedilotildees finais de A)
1 mM B) 2 mM C) 4 mM D) 8 mM E) 10 mM de
metavanadato total V1 refere-se agraves espeacutecies monomeacuterica de
vanadatoV1 e V2 correspondem respectivamente agrave espeacutecie
monomeacuterica e dimeacuterica
Figura 25 ndash Distribuiccedilatildeo graacutefica das espeacutecies
oligomeacutericas de vanadato vs a concentraccedilatildeo de
metavanadato total As concentraccedilotildees de cada
espeacutecie foram obtidas segundo a formula descrita
na secccedilatildeo 35 do Procedimento Experimental
(aacutereas dos sinais de 51
V-RMN observadas na
figura x1)V10 V1 e V4 correspondem agraves espeacutecie
monomeacuterica dimeacuterica e tetrameacuterica
respectivamente
Esp
eacuteci
e o
ligo
meacute
rica
de
va
nad
ato
(m
M)
[Metavanadato]total (mM)
V1
V2
V4
V5
28
Tabela 2IVndash Desvios quiacutemicos meacutedios (part)das ressonacircncias das espeacutecies tetrameacuterica (V4) monomeacuterica (V1) e
dimeacuterica (V2) presentes no espectro da soluccedilatildeo de metavanadato (total) 10 mM
Paracircmetros V1 V2 V4 V5
part (ppm) -5615 -5748 -5789 -5869
Nas soluccedilotildees de metavanadato as espeacutecies monomeacuterica dimeacutericae tetrameacuterica de vanadato estatildeo
presentes para concentraccedilotildees de metavanadato inferiores a 4 mM Todas estas as espeacutecies
aumentam linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado
em (Aureliano e Madeira 1998) e em (Tiago et al 2004) quando a concentraccedilatildeo de V1 eacute igual ou
superior a 200 microM inicia-se a sua oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica Nas
condiccedilotildees de realizaccedilatildeo dos ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a
espeacutecie pentameacuterica neste estudo em concentraccedilotildees inferiores a 4 mM tendo o mesmo sido
observado por Ramos 2006 Os resultados obtidos mostram claramente o equiliacutebrio entre a
espeacutecie V4 e as espeacuteciesV1 e V2 no qual o aparecimento de outras espeacutecies oligomeacutericas se
encontra dependente da concentraccedilatildeo da espeacutecie V1 Por uacuteltimo pode inferir-se que para
concentraccedilotildees inferiores a 2 mM e intervalos de tempo de cerca de 20 minutos (que inclui o
tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas as
espeacutecies V1 V2 e V4 com concentraccedilotildees respectivas de 0796 0192 e 0196 mM
232Estabilidade das soluccedilotildees de decavanadato anaacutelise por espectroscopia de absorccedilatildeo
UVVis
Caracterizou-se a estabilidade da espeacutecie V10 numa soluccedilatildeo com 1 mM decavanadato total em
tampatildeo D atraveacutes da leitura da Abs400 Obteve-se uma cineacutetica de dissociaccedilatildeo de 1ordf ordem tal
como se pode verificar pela linearidade do graacutefico de ln(Abs400)vs o tempo de aquisiccedilatildeo espectral
(segundos) (Fig26 )
Figura 26ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do tempo
(em segundos) para uma soluccedilatildeo de decavanadato total de 1
mM em tampatildeo D a 25ordmC (n=2)
Sendo uma reacccedilatildeo de primeira ordem pode ser descrita pela equaccedilatildeo cujo
declive eacute ndashK (constante de dissociaccedilatildeo) e a intersecccedilatildeo no eixo das ordenadas corresponde
Ln
(Ab
s 400 n
m)
tempo (s)
29
aln(A0) Numa reacccedilatildeo de 1ordf ordem o tempo de meia-vida eacute independente da concentraccedilatildeo inicial
do reagente sendo descrito pela equaccedilatildeo [2] Deste modo a partir da recta
Ln(Abs400 nm) = -436E-5
K - 105 obteve-se um tempo de meia-vida de 4490 plusmn 0823 horas para
as condiccedilotildees supramencionadas
Apoacutes a adiccedilatildeo de S1 8 microM ao meio reaccional verificou-se que a reacccedilatildeo de desoligomerizaccedilatildeo
da espeacutecie decameacuterica de vanadato seguia igualmente uma cineacutetica de 1ordf ordem tal como se pode
verificar pela linearidade do graacutefico de ln(Abs400) versus o tempo de aquisiccedilatildeo espectral
(segundos) (Fig27)
Figura 27ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do
tempo (em segundos) para uma soluccedilatildeo de decavanadato
total de 1 mM em tampatildeo D a 25ordmC na presenccedila de 8
microM (n= 2)
A partir da equaccedilatildeo da recta Ln(Abs400 nm) = -7520E-5
K - 0613determinou-se o valor de K para
a espeacutecie decameacuterica de onde se obteve um tempo de meia-vida de 2430 plusmn 0191 horas para as
condiccedilotildees supramencionadas
Tanto este valor como o valor registado na ausecircncia de S1 satildeo substancialmente superiores agrave
duraccedilatildeo dos ensaios de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina descritos no presente
relatoacuterio
24 Interacccedilatildeo de S1 com vanaacutedio(V)
241Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
De modo a analisar qual o efeito da adiccedilatildeo de S1 de miosina (tampatildeo D) produzia nos sinais de
51V-NMR titulou-se uma soluccedilatildeo de 5mM de decavanadato total com concentraccedilotildees crescentes
de S1 (5 10 15 e 20 microM) Os espectros de 51
V-RMN encontram-se apresentados na Fig 2
No graacutefico da figura 2 Apresenta-se a concentraccedilatildeo de S1 em funccedilatildeo do factor de alargamento
corrigido dos espectros acima referidos
Ln
(Ab
s (4
00
nm
))
tempo (s)
30
Figura 28 ndash Graacutefico de
representaccedilatildeo da
concentraccedilatildeo de S1 em
funccedilatildeo do factor de
alargamento A
concentraccedilatildeo de
decavanadato total foi de 5
mM e as concentraccedilotildees
utilizadas de S1 foram de 0
5 10 e 15 microM
Verifica-se que o factor de alargamento aumenta linearmente em funccedilatildeo da concentraccedilatildeo de S1 no
ensaio o que indica a existecircncia de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina Observou-
se ainda que a adiccedilatildeo de S1 ao meio reaccional natildeo provocou qualquer alteraccedilatildeo dos desvios
quiacutemicos dos sinais de 51
V-RMN
Realizaram-se ensaios na presenccedila de ATP e de S1 com intuito de verificar o efeito do ATP na
interacccedilatildeo do S1 de miosina com o decavanadato
Figura 29 - Espectros de RMN de 51
V contendo 2
mM Decavanadato (A) (controlo) (B) na presenccedila
de 2 mM de ATP (C) na presenccedila de 2 mM de ATP
e 8 microM de S1 de miosina V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2Vndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies decameacuterica (V10) e monomeacuterica (V1) presentes nos
espectros da soluccedilatildeo de decavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V10A V10B V10C V1
part (ppm) -5161 -5002 -4254 -5614
Observa-se que a presenccedila de ATP (espectro B) natildeo provoca qualquer alteraccedilatildeo visiacutevel em
relaccedilatildeo ao espectro que contem apenas soluccedilatildeo de decavanadato Jaacute a presenccedila simultacircnea de ATP
e S1 provoca um alargamento da ∆12 do sinal do V10A de 1736 Hz (espectro (A)) para 2366 Hz
(espectro (C) Fig 29) Tendo em conta que a concentraccedilatildeo de S1 apresenta uma relaccedilatildeo linear
com o factor de alargamento converteu-se o valor da ∆12 e comparou-se com o valor esperado agrave
F = 00591[S1] - 00157 Rsup2 = 0998
Fact
or
de
ala
rgam
en
to
[S1] (microM)
31
concentraccedilatildeo de S1 utilizada O factor de alargamento obtido para 8 microM ndash 0376 ndash eacute inferior ao
valor esperado agrave mesma concentraccedilatildeo de S1 obtido atraveacutes da equaccedilatildeo da recta apresentada na
Fig 28 ndash 0457 ndash o que indica que o ATP poderaacute exercer um efeito de protecccedilatildeo do S1
diminuindo a sua interacccedilatildeo com a espeacutecieV10
Por uacuteltimo realizaram-se ensaios com metavanadato na presenccedila de 8 microM S1 de modo a verificar
se as espeacutecies oligomeacutericas contidas nesta soluccedilatildeo interagiam com a proteiacutena em estudo
Figura 210 - Espectros de RMN de 51
V contendo 2
mM metavanadato (A) (controlo) (B) na presenccedila
de 8 microM de S1 de miosina V1 V2 e V4
correspondem respectivamente aacutes espeacutecies
monomeacuterica dimeacuterica e tetrameacuterica
Tabela 2VIndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies monomeacuterica (V1) dimeacuterica (V2) e tetrameacuterica (V4)
presentes nos espectros da soluccedilatildeo de metavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V1 V2 V4
part (ppm) -5613 -5747 -5789
Como eacute possiacutevel observar na Fig 210 ocorre uma ligeiro alargamento dos sinal de V4 bem
como uma perda da intensidade pelo que se conclui quedas espeacutecies presentes na soluccedilatildeo de
metavanadato (V1 V2 e V4) apenas a espeacutecie tetrameacuterica interage com o S1 de miosina o que
estaacute de acordo com o reportado por Tiago et al 2002
242 Actividade Mg2+
-ATPaacutesica de S1 de miosina na presenccedila de decanadato
Com o intuito de avaliar o efeito do decavanadato na actividade Mg2+
-ATPaacutesica do S1 de
miosina monitorizou-se a reacccedilatildeo de oxidaccedilatildeo do NADH atraveacutes da leitura de Abs340 na
presenccedila de 005 mgmL de S1 de miosina e no caso da actividade estimulada 2microM de actina em
3125 mM Tris-HCl pH 71 e 375 mM MgCl2 pelo meacutetodo de enzimas acopladas (ver seccccedilatildeo xx
dos MM)Para tal realizaram-se ensaios com vaacuterias concentraccedilotildees de decavanadato total (10 50
100 250 500 e 1000) de modo a determinar qual o seu efeito na actividade MgATPaacutesica de S1
de miosina na ausecircncia e na presenccedila de um dos seus ligandos naturais Os ensaios foram
realizados agrave temperatura ambiente com incubaccedilatildeo preacutevia a 25ordm C durante 20 minutos
32
Figura 211 ndash Representaccedilatildeo
graacutefica da Actividade Mg2+
-
ATPaacutesica de S1 () (os ajustes
teoacutericos estatildeo representado a por
uma linha os resultados
experimentais estatildeo representados
por triacircngulos (Actividade
estimulada) e cruzes (Actividade
natildeo estimulada)) vs
[Decavanadato]total medidas pelo
meacutetodo das enzimas acopladas
(n=3) Os dados foram ajustados ao modelo de decaimento exponencial usando o programa GraphPad Prism para a
determinaccedilatildeo dos valores de IC50 (na ausecircncia de actina IC50= 266 plusmn 07 M decavanadato na presenccedila de actina
IC50= 28 plusmn 1 M decavanadato
A partir do graacutefico presente na Fig 29 verifica-se que o ajuste dos dados experimentais foi bem
sucedido A inibiccedilatildeo da actividade Mg2+
-ATPaacutesica do S1 de miosina eacute afectada em menor
amplitude na presenccedila de actina (60 de inibiccedilatildeo da actividade) do que na sua ausecircncia
(aproximadamente 80 de inibiccedilatildeo de actividade) Devido a esta diferenccedila pode sugerir-se que a
actina iraacute impedir parcialmente a ligaccedilatildeo do decavanadatoao S1 demiosina atraveacutes de
impedimentos esteacutereos ou interacccedilotildees electrostaacuteticas por exemplo A teacutecnica utilizada natildeo nos
permite concluir acerca de que regiatildeo(otildees) ou resiacuteduo(s) da actina participaratildeo na preservaccedilatildeo
parcial da actividade Mg2+
-ATPaacutesica
Por outro lado os valores de IC50 dos dois tipos de actividades natildeo apresentam diferenccedilas
significativas tendo-se obtido 266 plusmn 07 e 28 plusmn 1M decavanadato na ausecircncia e na presenccedila de
actina respectivamente o que corresponde a uma concentraccedilatildeo de cerca de 27 M de V10 O
meio de reacccedilatildeo utilizado nestes ensaios foi 10 mM HEPES e 25 mM KCl com 2 mM ATP Em
(Tiago et al 2004) o valor de IC50 para a actividade Mg2+-ATPaacutesica na ausecircncia de actina foi de
125 plusmn 025 microM enquanto o valor da actividade estimulada por actina foi de 136 plusmn 14 microM O
meio de reacccedilatildeo utilizado nos ensaios foi 20 mM HEPES 50 mM KCl e 2 mM ATP o que resulta
precisamente no dobro da forccedila ioacutenica a que foi realizado o ensaio do presente relatoacuterio
(exceptuando a contribuiccedilatildeo dos produtos da hidroacutelise de ATP uma vez que este meacutetodo
pressupotildee a ocorrecircncia de um ciclo de reacccedilotildees de hidroacutelise e siacutentese deste composto) Ora tendo
sido anteriormente referido que a afinidade do S1 de miosina para com a espeacutecie V10 diminui com
Act
ivid
ade
Mg
2+-A
TP
aacutesic
a d
e S
1 d
e
Mio
sin
a
[Decavanadato] (microM) Actividade Estimulada ()
Actividade Natildeo Estimulada ()
Ajuste Actividade Estimulada
Ajuste Actividade Natildeo Estimulada
33
o aumento da forccedila ioacutenica (Tiago et al 2004 tal poderaacute explicar a diferenccedila obtida entre estes
dois conjuntos de valores
243Quantificaccedilatildeo de cisteiacutenas reduzidas pelo meacutetodo de Ellman
A interacccedilatildeo do vanaacutedio com alguns tipos de aminoaacutecidos sensiacuteveis agrave oxidaccedilatildeo pode fornecer-nos
informaccedilatildeo bastante uacutetil em relaccedilatildeo agraves alteraccedilotildees estruturais que conduzem a uma diminuiccedilatildeo da
actividade fisioloacutegica do S1 de miosina Os estudos realizados por Chasteen 1983 concluiacuteram
que a reduccedilatildeo de vanadato (V) a vanadilo (IV) ocorre na presenccedila de agentes redutores comuns no
ambiente celular de que se destaca o aminoaacutecido cisteiacutena Esta eacute tambeacutem um alvo de
complexaccedilatildeo por parte do iatildeo VO2+
Jaacute Ramos et al (2009) demonstrou que a incubaccedilatildeo de
actina com a soluccedilatildeo de decavanadato induz a oxidaccedilatildeo de resiacuteduos de cisteiacutena e agrave formaccedilatildeo do
oxidovanaacutedio (IV) Este grupo de investigadores descobriu ainda que apenas a espeacutecie V10 induz a
oxidaccedilatildeo das cisteiacutenas ldquoraacutepidasrdquo da actina atraveacutes do processo proposto que eacute parcialmente
responsaacutevel pela inibiccedilatildeo jaacute descrita da actividade de ATPase do S1 de miosina estimulada por
actina quando exposta agrave soluccedilatildeo de decavanadato Esta inibiccedilatildeo apresenta-se afectada com a
presenccedila de ATP uma vez que os resiacuteduos de cisteiacutena permaneceram intactos nos ensaios
realizados com este ligando Jaacute Crans et al (2004) concluiu que o siacutetio activo de muitas fosfatases
(que conteacutem grupos tiol) pode formar um equiliacutebrio redox com o iatildeo vanadato (V) de forma natildeo
reversiacutevel
Os resultados obtidos por Sakurai e Shiromura (1980) sugeriram que o vanaacutedio (V) sofre reduccedilatildeo
a vanaacutedio (IV) sem dependecircncia do valor de pH do ensaio Registaram ainda uma estequiometria
de 31 na reacccedilatildeo do vanadato (V) com os resiacuteduos de Cys enquanto a estequiometria dos ensaios
com o vanadilo (IV) foi de 21
Berlett e Stadtman (1997) observaram que os tipos de aminoaacutecidos mais facilmente oxidaacuteveis
englobam entre outros os resiacuteduos de Cys Trp e MetAssim analisou-se o estado redox dos
grupos tiol dos resiacuteduos de Cys do S1 na presenccedila do decavanadato atraveacutes da monitorizaccedilatildeo da
Abs412 nm na presenccedila de 1 mgmL de S1 de miosina em tampatildeo 10 mM HEPES 25 mM KCl
variando a concentraccedilatildeo da soluccedilatildeo de decavanadato de modo a determinar que tipo de alteraccedilotildees
estruturais ocorrem na sua presenccedila no que diz respeito agrave diminuiccedilatildeo da actividade ATPaacutesica de
S1 na presenccedila de decavanadato
Os resultados referentes agrave quantificaccedilatildeo das cisteiacutenas reduzidas apoacutes o tratamento de S1 com
diferentes concentraccedilotildees de decavanadato em tampatildeo 10 mM HEPES pH 75 e 25 mM KCl a
25ordmC durante 20 minutos encontram-se descritos na Tabela 2VII
Tabela 2VII ndash Cisteiacutenas reduzidas totais e ldquoraacutepidas de S1 (1 mgmL) apoacutes tratamento com decavanadato
34
(010501002505001000 uM) em 10 mM HEPES 25 mM KCl pH 7 a 25ordmC durante 20 minutos A detecccedilatildeo
espectrofotomeacutetrica foi feita com DTNB (e 412=8400 uM-1 cm-1) (n variaacutevel entre 1 e 8)
Amostras Cys raacutepidas Cys totais
11 M S1 (controlo) 22 plusmn 04 106 plusmn 14
11 M S1 + 10 M Decavanadato 22 plusmn 05 95 plusmn 08
11 M S1 + 50 uM Decavanadato 20 plusmn 02 98 plusmn 14
11 M S1 + 100 uM Decavanadato 17 plusmn 03 97 plusmn14
11 M S1 + 250 uM Decavanadato 26 plusmn 02 98 plusmn 08
11 M S1 + 500 uM Decavanadato 21 plusmn 04 90 plusmn 06
11 M S1 + 1000 uM Decavanadato 24 plusmn 0 86 plusmn 08
A partir dos resultados obtidos pode sugerir-se que o S1 possui 11 resiacuteduos de Cys (controlo) o
que estaacute de acordo com o anteriormente descrito Verifica-se que o tratamento da proteiacutena com
concentraccedilotildees crescentes de decavanadato provoca a oxidaccedilatildeo de dois resiacuteduos de Cys do S1 de
miosina natildeo expostos ao solvente No entanto a teacutecnica utilizada natildeo nos permite identificar
quais
Tendo em consideraccedilatildeo que o valor de IC50 obtido para a reacccedilatildeo de inibiccedilatildeo daactividade Mg2+-
ATPaacutesica de S1 na presenccedila de decavanadato eacute de cerca de 27 microM pode sugerir-se que este seraacute o
valor ao qual ocorreraacute a oxidaccedilatildeo do primeiro resiacuteduo de Cys o que poderaacute contribuir para a
diminuiccedilatildeo da actividade de S1 observada
25Interacccedilatildeo de S1 com vanaacutedio(IV)
251Anaacutelise por espectroscopia de Ressonacircncia Paramagneacutetica Electroacutenica
A acumulaccedilatildeo intracelular de oxovanaacutedio (IV) in vivo constitui uma das principais formas de
apresentaccedilatildeo de vanaacutedio no organismo Deste modo torna-se bastante interessante analisar a
interacccedilatildeo entre este iatildeo e o S1 de miosina atraveacutes da teacutecnica de espectroscopia de Ressonacircncia
Paramagneacutetica Electroacutenica o meacutetodo de eleiccedilatildeo do estudo do iatildeo vanadilo e dos complexos por si
formados O electratildeo desemparelhado encontra-se fortemente acoplado ao nuacutecleo 51
V o que
35
resulta num espectro de RPE consistindo de 8 desdobramentos na direcccedilatildeo paralela e outros tantos
na direcccedilatildeo perpendicular (Chasteen 1983)
Os estudos realizados por Ahmed et al (1987) centraram-se na interacccedilatildeo do vanadilo com
proteiacutenas como eacute o caso da insulina transferrina entre outros obtendo um espectro caracterizado
por 8 linhas amplamente espaccediladas (na direcccedilatildeo paralela) Jaacute Ramos et al 2009 concluiu que a
exposiccedilatildeo da soluccedilatildeo de decavanadato agraves formas globular e filamentosa de actina produz um
espectro tiacutepico do iatildeo oxidovanaacutedio (IV) indicando a reduccedilatildeo do vanadato
Para aferir qual a concentraccedilatildeo de S1 que permitia obter uma boa razatildeo sinalruiacutedo titulou-se uma
soluccedilatildeo de sulfato de vanadilo 15 microM ([VOSO4]stock = 5 mM) com concentraccedilotildees crescentes de
proteiacutena (0 5 10 15 e 20 microM) em 10 mM HEPES 25 mM KCl A existecircncia de uma correlaccedilatildeo
linear entre a concentraccedilatildeo de proteiacutena e a intensidade dos sinais de RPE correspondente agrave banda
-12 perpendicular encontra-se representada na Fig 210
Figura 2 12ndash Representaccedilatildeo graacutefica da variaccedilatildeo da intensidade do sinal de RPE vs a concentraccedilatildeo de S1 utilizada
Apoacutes verificar que com uma concentraccedilatildeo de S1 igual a 20 microM se obtinham sinais com uma razatildeo
sinalruiacutedo adequada ao prosseguimento dos estudos sobre a interacccedilatildeo V(IV)-S1 de miosina
titulou-se uma soluccedilatildeo de 20 microM S1 com 0 25 5 10 15 25 50 75 100 150 200 500 e 1000
microM VOSO4
Traccedilou-se o graacutefico da intensidade da banda -12 perpendicular do sinal de EPR em funccedilatildeo da
concentraccedilatildeo de VOSO4 tendo-se determinado uma constante de dissociaccedilatildeo (KD) do complexo
S1-vanadilo de 8748 plusmn 2621 microM V(IV)
y = 016786x + 122435
Rsup2 = 092992759
Inte
nsi
dad
e d
o s
inal
de
RP
E
[S1] (microM)
36
Figura 213 ndash Representaccedilatildeo graacutefica da variaccedilatildeo do sinal de RPE vs a concentraccedilatildeo de vanadilo utilizada (os valores
experimentais estatildeo representados por losangos o seu ajuste estaacute representado pela linha)
A simulaccedilatildeo do espectro de EPR foi efectuada a partir do espectro obtido para uma concentraccedilatildeo
de 1 mM VOSO4 apoacutes adiccedilatildeo de 20 microM S1 de miosina tendo-se obtido os valores da constante de
acoplamento hiperfina (A=5016) e do factor g (g0=195)
Figura 214 ndash Representaccedilatildeo graacutefica do espectro de EPR obtido com 1000 microM de vanadilo e 20 microM de S1 (a cinzento
escuro) A simulaccedilatildeo do espectro encontra-se a cinzento claro
Por comparaccedilatildeo destes com os valores presentes na tabela de correspondecircncia xxx eacute possiacutevel
predizer quais os ligandos provaacuteveis do vanadilo na presenccedila de proteiacutena Assim concluiu-se que
o vanadilo estaacute coordenado com duas moleacuteculas de aacutegua um grupo hidroxilo e um grupo R=NR
O grupo hidroxilo pode indicar os resiacuteduos de aminoaacutecidos Tyr Treonina e Serina enquanto o
grupo R=NR poderaacute indicar a presenccedila de Arginina e Histidina como ligandos do vanadilo
Inte
nsi
dad
e d
os
sinai
s d
e R
PE
[Vanadilo] (microM)
Ajuste dos dados exp Dados experimentais
Campo magneacutetico (G)
Espectro Experimental Ajuste
37
3 Procedimento Experimental
31 Reagentes
Todos os reagentes utilizados satildeo de elevado grau de pureza (analiacutetico) Apresentam-se as suas
foacutermulas quiacutemicas ou abreviaturas quando necessaacuterio acrescentou-se a sua descriccedilatildeo Entre
parecircntesis colocou-se as empresas que os comercializam
Aacutecido aceacutetico glacial 100 (Riedel) Acrilamida 99 (Merck) Azul de bromofenol (Riedel-de-
Haumlen) -Mercaptoetanol (Sigma) Bissulfito de soacutedio (Sigma)Bradford (reagente de)
(BioRad)BSA (albumina de soro bovino Sigma-Aldrich) CaCl22H2O (cloreto de caacutelcio
dihidratado Fluka) CH3CH2OH 96 (etanol Panreac) Coomassie Blue R250 (Merck) D2O
(oacutexido de deuteacuterio SDS) DTNB (55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico) Aldrich) EDTA (aacutecido
etilenodiaminatetraaceacutetico Merck) Fosfoenolpiruvato 98 (Fluka) Glicerol (Sigma) Glicina
985 (Panreac) HCl 37 (Riedel-de Haumlen) H2SO4 96 (Pronalab) HEPES 99 (aacutecido 4-(2-
hidroxiletil)-1-piperazineetanosulfoacutenico Sigma) KH2PO4 99 (hidrogenofosfato de potaacutessio)
MgATP (Sigma) Na
ATP (Sigma) Na2HPO4 99 (hidrogenofosfato de soacutedio Riedel-de Haumlen)
NaH2PO4 (fosfato de soacutedio ) NaOH 986 (hidroacutexido de soacutedio Pronalab) KCl 99 (Cloreto de
potaacutessio Panreac) L-Cys 995 (enantioacutemero levoacutegiro do resiacuteduo de Cisteiacutena Fluka)Metanol
99 (Merck) Molibdato de amoacutenio (Sigma)NNrsquo-metil-bis-acrilamida 98 (Merck) NH4VO3
99 (metavanadato de amoacutenia Sigma) PSA 10 (persulfato de amoacutenio Merck) SDS
(dodecilo sulfato de soacutedio Sigma) Sulfito de soacutedio (Sigma) VOSO4 (sulfato de vanadilo
Aldrich) TEMED 99 (Tetrametiletilenodiamina Merck) Tris (2-amino-2-hidroximetil-13-
propanodiol Sigma)
32 Meacutetodos
321 Isolamento e purificaccedilatildeo das proteiacutenas de muacutesculo esqueleacutetico de coelho
3211 Purificaccedilatildeo da miosina preparaccedilatildeo de Subfragmento 1 por digestatildeo quimotriacutepsica
da miosina
O subfragmento-1 de miosina-II (S1) foi gentilmente cedido pelo Prof Doutor Manuel Aureliano
do Departamento de Quiacutemica Bioquiacutemica e Farmaacutecia da Faculdade de Ciecircncias e Tecnologia da
Universidade do Algarve
No entanto descreve-se abaixo de forma resumida o procedimento efectuado
38
32111 Purificaccedilatildeo da miosina
O protocolo de isolamento foi adaptado do meacutetodo previamente descrito por Margossian e Lowey
(1982) apresentando-se de forma esquemaacutetica na Fig 31
Sucintamente os tecidos musculares dorsais e das pernas foram extraiacutedos apoacutes o sacrifiacutecio do
animal (Fig31) De seguida foram adicionados 3 volumes da soluccedilatildeo de extracccedilatildeo A2
Centrifugou-se a soluccedilatildeo e filtrou-se o sobrenadante O sedimento foi entatildeo diluiacutedo em 14
volumes de aacutegua destilada de modo a que a forccedila ioacutenica atingisse 004 M Apoacutes adiccedilatildeo de szlig-
mercaptoetanol (szlig-me) deixou-se a precipitar a 4ordmC removendo o sobrenadante Centrifugou-se
novamente e ressuspendeu-se o sedimento na soluccedilatildeo de extracccedilatildeo A Adicionou-se a soluccedilatildeo
tampatildeo B3 para que a forccedila ioacutenica atingisse 05 M Homogeneizou-se e deixou-se repousar
durante 2h Homogeneizou-se novamente e centrifugou-se de modo a ocorrer precipitaccedilatildeo dos
complexos de actomiosina e proteiacutena desnaturada Apoacutes filtraccedilatildeo do sobrenadante em latilde de vidro
quantificou-se a soluccedilatildeo de miosina resultante pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental)
2 Soluccedilatildeo de extracccedilatildeo A 0012M K2CO3 004M KHCO3 06M KCl pH 65
3 Soluccedilatildeo-tampatildeo B Tris 100 mM pH 7 KCl 2M
39
Figura 31 ndash Representaccedilatildeo esquemaacutetica do protocolo de isolamento de miosina (Margossian e Lowey 1982) Todo o
procedimento foi realizado a 4ordmC (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
32112 Preparaccedilatildeo de Subfragmento-1 de miosina por digestatildeo com quimiotripsina
O S1 de miosina foi isolado apoacutes diaacutelise da miosina durante a noite contra a soluccedilatildeo C4 A
presenccedila do EDTA eacute essencial uma vez que a sua substituiccedilatildeo por MgCl2resulta na produccedilatildeo de
HMM em vez de S1 A suspensatildeo de miosina foi equilibrada e digerida com -quimotripsina
A digestatildeo foi interrompida pela adiccedilatildeo de PMSF Seguidamente ultracentrifugou-se o produto
da digestatildeo desprezando-se o sedimento (Fig 32) A soluccedilatildeo de sobrenadante maioritariamente
S1 foi quantificada pela leitura directa da Abs280 ou pelo meacutetodo de Bradford (ver as secccedilotildees
3311 e 3312 do Procedimento Experimental respectivamente) e armazenada em 10 sacarose
e 2 mM szlig-Me a -80ordmC
4 Soluccedilatildeo C 25mM Tris pH 70 012M KCl 1mM EDTA
40
Antes da realizaccedilatildeo dos ensaios descritos no presente relatoacuterio o S1 de miosina foi dialisado
contra a soluccedilatildeo D5 de modo a remover qualquer composto que pudesse complexar como
vanaacutedio Seguidamente quantificou-se a proteiacutena pelo meacutetodo de Bradford ou pelo meacutetodo
directo (ver secccedilotildees 3311 e 3312do Procedimento Experimtental)
Figura 3 2ndash Representaccedilatildeo esquemaacutetica do protocolo de preparaccedilatildeo do S1 de miosina por digestatildeo quimiotripsina de
acordo com o previamente descrito por Margossian e Lowey (1982) Todo o procedimento foi realizado a 4ordm C
excepto quando indicaccedilatildeo do contraacuterio (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
3212 Isolamento e purificaccedilatildeo de actina
Isolou-se e purificou-se a actina segundo o meacutetodo descrito por Pardee e Spudish que
compreende dois passos ndash a obtenccedilatildeo de um resiacuteduo seco denominado ldquopoacute de acetonardquo e a
extracccedilatildeo subsequente da actina a partir do ldquopoacute de acetonardquo na forma de G-Actina com a
posterior polimerizaccedilatildeo a F-Actina
32121 Obtenccedilatildeo do ldquopoacute de acetonardquo
Sacrificou-se o coelho com uma pancada atraacutes do pescoccedilo e sangrou-se o animal apoacutes o corte da
arteacuteria jugular Esfolou-se e colocou-se numa tina com gelo Retiraram-se as partes de tecido
conjuntivo que envolvem o tecido muscular com auxiacutelio de uma pinccedila e bisturi Seguidamente
retirou-se os muacutesculos dorsais e os muacutesculos brancos das pernas Passou-se os muacutesculos por aacutegua
destilada de modo a remover as impurezas e pesou-se num copo Trituraram-se os muacutesculos
numa maacutequina de triturar carne e extraiu-se o resultante com 3 volumes da soluccedilatildeo E6 durante 10
minutos com agitaccedilatildeo suave Centrifugou-se a suspensatildeo a 4000 g durante 10
5 Soluccedilatildeo D HEPES 10 mM pH 70 KCl 25 mM
6 Soluccedilatildeo E 01 M KCl 015 M K2HPO4 pH 65
41
minutosdesprezando-se o sobrenadante Ressuspendeu-se o sedimento em 6 volumes de soluccedilatildeo
F7durante 10 minutos com agitaccedilatildeo suave Verificou-se se o pH da soluccedilatildeo estava compreendido
entre 82 e 85 acertando-se estecom soluccedilatildeo G8 se necessaacuterio Centrifugou-se durante 10
minutos a 4000 g Desprezou-se o sobrenadante e ressuspendeu-se o sedimento em 3 volumes de
soluccedilatildeo H9 durante 10 minutos Centrifugou-se novamente a 4000 gdurante 10 minutos
desprezando-se o sobrenadante Ressuspendeu-se o sedimento duas vezes consecutivas em 6
volumes de aacutegua destilada durante 5 minutos Centrifugou-se a 4000 g durante 10 minutos apoacutes
cada adiccedilatildeo de H2O
Seguidamente ressuspendeu-se o sedimento 5 vezes consecutivas utilizando 3 volumes de
acetona de cada vez agitando suavemente durante 10 minutos agrave temperatura ambiente Colocou-
se o resiacuteduo filtrado numa tina de vidro sobre papel de filtro deixando-se evaporar o solvente
durante a noite num nicho Triturou-se o resiacuteduo seco numa maacutequina de triturar carne e deixou-se
novamente evaporar o solvente O ldquopoacute de acetonardquo obtido foi armazenado a -20 ordmC
32122 Purificaccedilatildeo da actina a partir de ldquopoacute de acetonardquo
Apoacutes a obtenccedilatildeo do poacute de acetona como acima indicado efectuou-se o protocolo seguidamente
descrito de modo a obter monoacutemeros de actina (G-Actina) e actina filamentosa (F-Actina)
Pesou-se 35 g de poacute de acetona e adicionaram-se 50 mL de soluccedilatildeo-tampatildeo I10
sendo a
extracccedilatildeo feita com agitaccedilatildeo suave durante 30 minutos Separou-se o extracto do poacute de acetona
hidratado filtrando a suspensatildeo com 4 camadas de gaze esterilizada tendo o cuidado de natildeo
espremer o resiacuteduo Voltou a extrair-se com 50 mL de soluccedilatildeo I durante 30 minutos mexendo
lentamente Adicionou-se este ao extracto obtido anteriormente Centrifugou-se o extracto total a
16 000 g durante 30 minutos Decantou-se para uma proveta sobre a qual se colocou gaze para
que o sedimento solto natildeo se juntasse ao sobrenadante e leu-se o volume De seguida adicionou-
se KCl ateacute uma concentraccedilatildeo final de 50 mM MgCl2 ateacute 2mM e ATP ateacute 1mM lentamente e com
agitaccedilatildeo contiacutenua de modo a promover a polimerizaccedilatildeo dos monoacutemeros de actina
Deixou-se polimerizar durante 2h a 25ordmC observando-se a presenccedila de pequenas bolhas de ar
indicativas de um aumento de viscosidade da soluccedilatildeo Arrefeceu-se em gelo durante 15 minutos e
adicionou-se KCl ateacute uma concentraccedilatildeo final de 06M Mexeu-se lentamente durante 30 minutos
apoacutes os quais se ultracentrifugou a 110 000g durante 1hora Desprezou-se o sobrenadante
ressuspendeu-se o sedimento com soluccedilatildeo tampatildeo E e transferiu-se para um potter
Dialisou-se durante a noite contra 1L de soluccedilatildeo-tampatildeo I a uma temperatura de 4ordmC com rotaccedilatildeo
7 Soluccedilatildeo F 005 M NaHCO3 pH 82-85
8 Soluccedilatildeo G 1M Na2CO3
9 Soluccedilatildeo H 1 mM EDTA pH 70
10 Soluccedilatildeo I 2 mM Tris-HCl pH 8 02 mM CaCl2 02 mM Na2ATP 05 mM szlig-Me 0005 NaN3
42
contiacutenua e mudanccedila de tampatildeo 2 a 3 vezes De seguida a actina foi centrifugada a 110
000gdurante 1hora Decantou-se a soluccedilatildeo de G-Actina (sobrenadante) e desprezou-se o
sedimento Determinou-se a concentraccedilatildeo de G-Actina pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental) A soluccedilatildeo de G-actina foi armazenada a uma temperatura de 4ordmC
apoacutes adiccedilatildeo de 002NaN3 mantendo-se estaacutevel durante 3 a 5 dias
331 Quantificaccedilatildeo das proteiacutenas isoladas
3311 Quantificaccedilatildeo pelo meacutetodo de Bradford
O meacutetodo de Bradford (Bradford 1976) baseia-se na utilizaccedilatildeo do reagente de Bradford
composto por entre outros aacutecido fosfoacuterico e o corante ldquoCoomassierdquo Este corante possui carga
negativa e a sua coloraccedilatildeo varia entre a vermelha (Absmax=465 nm) predominante em soluccedilatildeo e a
azul (Absmax=595 nm) presente aquando da ligaccedilatildeo aos grupos amina das proteiacutenas Em soluccedilotildees
aacutecidas diluiacutedas a coloraccedilatildeo varia proporcionalmente com a quantidade do corante ligado agrave
proteiacutena que por sua vez eacute directamente proporcional agrave concentraccedilatildeo da proteiacutena permitindo a
sua quantificaccedilatildeo ateacute a um limite de detecccedilatildeo de 20 gmL (Bradford 1976)
Deste modo prepararam-se amostras de BSA ([BSA]stock= 1mgmL) utilizando um intervalo de
concentraccedilotildees compreendido entre 0 e 15 gmL de acordo com a tabela 3I Apoacutes a adiccedilatildeo da
H2O MilliQ e do reagente de Bradford as soluccedilotildees foram sucessivamente homogeneizadas no
voacutertex
Tabela 3Indash Amostras utilizadas para a obtenccedilatildeo da recta de calibraccedilatildeo no meacutetodo de Bradford
BSA (L) BSA (g) H2O MilliQ (L) Reagente de Bradford (L)
0 0 800 200
125 125 79875 200
25 25 7975 200
5 5 795 200
10 10 790 200
15 15 785 200
Apoacutes incubaccedilatildeo a 25ordmC por 5 minutos leram-se os valores de Abs595 num
espectrofotoacutemetro (Shimadzu UV-160A) para todas as amostras Seguidamente traccedilou-se a recta
de calibraccedilatildeo (Abs595vs quantidade de BSA (g))
43
Para determinar a concentraccedilatildeo de proteiacutena apoacutes o processo de isolamento purificaccedilatildeo e
diaacutelise quando necessaacuterio colocou-se 5 L da amostra num tubo de Eppendorf e adicionou-se
795 L de H2O MilliQ homogeneizando a soluccedilatildeo no voacutertex Adicionou-se posteriormente 200
L de reagente de Bradford e repetiu-se a homogeneizaccedilatildeo Incubou-se a mistura a 25ordmC por 5
minutos apoacutes os quais se leu os valores de Abs595 nm no espectrofotoacutemetro Por interpolaccedilatildeo deste
valor a partir da recta de calibraccedilatildeo determinou-se a concentraccedilatildeo de proteiacutena na amostra
3312 Quantificaccedilatildeo pelo meacutetodo directo
A quantificaccedilatildeo das amostras proteicas pode tambeacutem ser efectuada atraveacutes da leitura directa da
absorvacircncia tendo por base a lei de Beer-Lambert onde Abs corresponde agrave
absorvacircncia ao comprimento de onda ao qual o ensaio foi realizado corresponde ao coeficiente
de extinccedilatildeo (substituiacutedo pela letra a se as unidades forem distintas da molaridade) b corresponde
ao percurso oacuteptico da radiaccedilatildeo na ceacutelula amostral e c corresponde agrave concentraccedilatildeo amostral
Assim no caso de S1 de miosina mediu-se a Abs280e calculou-se a concentraccedilatildeo da proteiacutena
sabendo que o coeficiente de extinccedilatildeo de S1 eacute (Ref)
No caso da actina mediu-se a Abs290ecalculou-se a concentraccedilatildeo da proteiacutena sabendo que o
coeficiente de extinccedilatildeo da actina eacute (Ref)
332Electroforese em condiccedilotildees desnaturantes (SDS-PAGE)
De modo a melhor caracterizar as proteiacutenas isoladas foi realizada uma electroforese em
condiccedilotildees desnaturantes (SDS-PAGE do inglecircs Sodium Dodecyl Sulfate Polyacrilamide Gel
Electrophoresis) O SDS (dodecilo sulfato de soacutedio) eacute um detergente anioacutenico utilizado neste
meacutetodo para desnaturar as proteiacutenas que se ligam ao SDS na proporccedilatildeo 14 g SDS g proteiacutena
(aproximadamente 1 moleacutecula de SDS por cada 2 resiacuteduos de aminoaacutecidos) (Stryer) O SDS induz
uma densidade de carga negativa significativa nas proteiacutenas a que se liga mascarando a sua carga
intriacutenseca pelo que as razotildees cargamassa satildeo tendencialmente idecircnticas Assim a SDS-PAGE
separa as proteiacutenas quando sujeitas a um campo eleacutectrico de acordo com o seu peso molecular
sendo a sua migraccedilatildeo directamente proporcional ao logaritmo deste uacuteltimo (Stryer)
O gel de resoluccedilatildeo com 10 de acrilamida foi obtido atraveacutes da adiccedilatildeo de 167 mL de soluccedilatildeo
stock de acrilamida11
075 mL de tampatildeo de resoluccedilatildeo12
251 mL de aacutegua destilada e 50 L de
SDS 10 Para promover a polimerizaccedilatildeo adicionou-se em seguida 38 L de PSA 10
11
soluccedilatildeo stock de acrilamida soluccedilatildeo de 30 (mv) de acrilamida e 08 (mv) de NNrsquo-metil-bis-acrilamida 12
tampatildeo de resoluccedilatildeo 15 M Tris-HCl pH 88
44
(persulfato de amoacutenia) 10 e 25 L de TEMED 99 (tetrametiletilenodiamina) Agitou-se
cuidadosamente de modo a natildeo formar bolhas e pipetou-se a soluccedilatildeo para a cassette de vidro
previamente montada no suporte De seguida adicionou-se uma peliacutecula de cerca de 2 mm de 2-
metil-2-butanol para verificar posteriormente a polimerizaccedilatildeo Apoacutes a polimerizaccedilatildeo que demora
cerca de 45 minutos verteu-se a agrave superfiacutecie e adicionou-se o gel de concentraccedilatildeo com 5 de
acrilamida constituiacutedo por 03 mL de soluccedilatildeo de stock acrilamida 045 mL de tampatildeo de
concentraccedilatildeo13
094 mL de aacutegua destilada 18 L de SDS 10 135 L de PSA 10 e 2L de
TEMED 99 Colocou-se o pente sobre o gel e deixou-se polimerizar durante cerca de 45
minutos
As amostras para aplicaccedilatildeo no gel de electroforese foram preparadas para uma concentraccedilatildeo de
proteiacutena de 1 mgmL num volume final de 100 L dos quais 25 L correspondem ao tampatildeo da
amostra14
As amostras foram aquecidas a 100ordmC durante 5 minutos Antes da aplicaccedilatildeo das
amostras transferiu-se a cassette para a tina de electroforese agrave qual se adicionou a soluccedilatildeo
tampatildeo de electroforese da tina superior15
e tampatildeo de electroforese da tina inferior16
Aplicaram-
se 10 L das soluccedilotildees de marcadores de peso molecular e 20 L das amostras proteicas
preparadas anteriormente A electroforese decorreu durante cerca de 3h30 min a uma voltagem
de 80 V De seguida transferiu-se o gel para uma tina corando as proteiacutenas durante 1h com
soluccedilatildeo corante17
Descorou-se o gel com soluccedilatildeo descorante18
tendo-se mudado esta soluccedilatildeo ateacute
as bandas se encontrarem bem definidas
Determinou-se a mobilidade relativa das bandas correspondentes aos marcadores de elevado peso
molecular e traccedilou-se a recta de calibraccedilatildeo ndash log(PM) vs log(Rf) onde Rf corresponde agrave
mobilidade relativa Por interpolaccedilatildeo determina-se o PM das proteiacutenas isoladas sabendo a sua
mobilidade relativa Por comparaccedilatildeo com os valores descritos na literatura pode concluir-se
acerca do grau de purificaccedilatildeo
34 Mediccedilatildeo da actividade enzimaacutetica de S1
341 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
13
tampatildeo de concentraccedilatildeo 05 M Tris-HCl pH 68 14
tampatildeo de amostra 320 mM Tris-HCl pH 68 04 M szlig-mercaptoetanol SDS 8 glicerol 15 (vv) e roxo de m-
cresol 0024 15
tampatildeo de electroforese da tina superior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS 16
tampatildeo de electroforese da tina inferior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS acetato de soacutedio
01 M 17
soluccedilatildeo corante ldquoCoomassie Blue R-250rdquo contendo metanol 45 (vv) e aacutecido aceacutetico glacial 10 (vv) 18
soluccedilatildeo descorante metanol 10 (vv) e aacutecido aceacutetico glacial 10 vv)
45
O meacutetodo de Fiske-Subarow modificado consiste na reacccedilatildeo do molibdato de amoacutenio com o
fosfato inorgacircnico em soluccedilatildeo aacutecida com formaccedilatildeo do aacutecido fosfomoliacutebdico Seguidamente daacute-
se a adiccedilatildeo de um agente redutor que provoca a reduccedilatildeo do composto a fosfomolibdato que forma
um complexo de cor azulada cuja absorvacircncia maacutexima ocorre para um comprimento de onda de
660 nm tornando possiacutevel a sua detecccedilatildeo por meacutetodos espectrofotomeacutetricos As actividades Ca2+
-
ATPaacutesica e K+EDTA-ATPaacutesica de S1 foram determinadas atraveacutes deste meacutetodo de acordo com
o descrito por Taussky e Shorr (1953) com mediccedilatildeo da quantidade de fosfato (Pi) libertado
atraveacutes da hidroacutelise de ATP
Para tal obteve-se primeiramente a recta de calibraccedilatildeo com as amostras presentes na Tabela 3II
utilizando uma soluccedilatildeo stock de 5 mM KH2PO4
Tabela 3II ndash Composiccedilatildeo das diferentes amostras utilizadas para determinar a recta de calibraccedilatildeo atraveacutes do meacutetodo
de Fiske-Subbarow modificado
KH2PO4 (L)
([KH2PO4]stock= 5mM)
PO43-
(nmole) H2O (uL)
0 0 1780
5 25 1775
10 50 1770
20 100 1760
40 200 1740
100 500 1680
A cada tubo de Eppendorf adicionaram-se seguidamente 200 L de soluccedilatildeo J19
e 20 L de agente
redutor20
homogeneizando no voacutertex apoacutes cada adiccedilatildeo Incubou-se durante 10 minutos e mediu-
se a Abs660 nm num espectrofotoacutemetro com intervalos de tempo iguais entre cada amostra Traccedilou-
se a recta de calibraccedilatildeo (Abs660 nmvs nmol PO43-
) a partir dos valores obtidos
Para determinar a actividade Ca2+
-ATPaacutesica de S1 prepararam-se tubos de Eppendorf com 400
L de meio de reacccedilatildeo21
25 L CaCl2 ([CaCl2]stock= 100 mM) 25 L ATP ([ATP]stock= 100 mM)
com [S1]ensaios=01 mgmL Perfez-se se necessaacuterio com H2O MilliQ ateacute perfazer um volume de 1
mL Para determinar a actividade K+EDTA-ATPaacutesica utilizou-se uma concentraccedilatildeo idecircntica de
19
Soluccedilatildeo J 5 N H2SO4 25 molibdato de amoacutenio 20
Agente redutor 5 mg aacutecido 1-amino-2-naftol-4-sulfoacutenico 30 mg bissulfito de soacutedio 30 mg sulfito de soacutedio em 26
mL H2O 21
meio de reacccedilatildeo 125 mM Tris-HCl pH 80 0575 M KCl
46
proteiacutena nos ensaios (01 mgmL) e o mesmo meio de reacccedilatildeo Aos tubos de Eppendorf
adicionou-se ainda 123 L de KCl ([KCl]stock = 3M) 25 L de ATP 10 L de EDTA
([EDTA]stock= 100 mM )e o volume de H2O MilliQ necessaacuterio para perfazer 1 mL Em ambas as
situaccedilotildees o uacuteltimo reagente adicionado foi o ATP uma vez que se trata do substrato da reacccedilatildeo
enzimaacutetica
A cineacutetica decorreu a 25ordmC durante 5 minutos para a actividade Ca2+
-ATPaacutesica e durante 3
minutos para a actividade K+EDTA-ATPaacutesica Apoacutes o periacuteodo de incubaccedilatildeo transferiu-se 150
L de amostra para tubos de Eppendorf contendo igual volume de TCA 10 Colocou-se no gelo
e homogeneizou-se no voacutertex Seguidamente centrifugou-se a 1500 g durante 10 minutos e
recolheu-se 200 L de sobrenadante para novos tubos de Eppendorf Adicionou-se a estes 158
mL de H2O MilliQ e 200 L de soluccedilatildeo J Homogeneizou-se no voacutertex e adicionou-se 20 L de
reagente redutor repetindo-se a homogeneizaccedilatildeo Incubou-se durante 10 minutos e mediu-se a
Abs660 nm com o mesmo intervalo de tempo de leitura para cada amostra Por intrapolaccedilatildeo a partir
da recta de calibraccedilatildeo obtida anteriormente determinou-se a quantidade de Pi libertado em cada
ensaio A partir destes resultados determinaram-se as actividades Ca2+
-ATPaacutesica e K+EDTA-
ATPaacutesica do S1 em nmol PO43-
minmg proteiacutena e em s-1
342 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
Da actividade Mg2+
-ATPaacutesica de S1 resulta a formaccedilatildeo dos produtos da reacccedilatildeo de
hidroacutelise do ATP - ADP e Pi Na presenccedila de ADP fosfoenolpiruvato (PEP) e da enzima piruvato
cinase (PK) os produtos resultantes satildeo ATP e piruvato Por acccedilatildeo da enzima lactato
desidrogenase (LDH) este uacuteltimo eacute convertido em lactato ocorrendo simultaneamente a oxidaccedilatildeo
do NADH a NAD+ monitorizada atraveacutes da Abs340 nm Quando o S1 de miosina se encontra no
meio reaccional o ATP eacute hidrolisado formando-se novamente ADP e Pi o que permite a
formaccedilatildeo de um ciclo (fig 21) Se as enzimas se encontrarem em excesso a velocidade de
conversatildeo do NADH eacute limitada pela produccedilatildeo de ADP e Pi Como o ATP eacute regenerado em cada
ciclo a sua concentraccedilatildeo inicial eacute sempre mantida (Bagshaw 1982)
Figura 33ndash Representaccedilatildeo esquemaacutetica do meacutetodo de enzimas acopladas
47
O protocolo utilizado e seguidamente descrito foi adaptado de Cuenda et al (1990 ndash Ver Refordf) A
uma cuvette de 1 mL adicionou-se S1 para uma concentraccedilatildeo final de 005 mgmL 800 L de
meio de reacccedilatildeo22
10 L de PEP ([PEP]stock = 42 mM) 10 L de PK+LDH 11 ([PK+LDH]stock =
1 mgmL) e 15 L de NADH ([NADH]stock=25 mM) Iniciou-se a monitorizaccedilatildeo da reacccedilatildeo
atraveacutes da leitura de Abs340 nm apoacutes a adiccedilatildeo de 20 L de ATP ([ATP]stock=100 mM) A
actividade Mg2+
-ATPaacutesica eacute obtida atraveacutes do declive correspondente agrave oxidaccedilatildeo do NADH Para
a mediccedilatildeo da Mg2+
-ATPase de S1 estimulada por actina adicionou-se F-actina para uma
concentraccedilatildeo final de 2 microM sendo a actividade obtida tambeacutem atraveacutes do declive correspondente
agrave oxidaccedilatildeo do NADH apoacutes a adiccedilatildeo da proteiacutena
35 Interaccedilatildeo entre vanaacutedio e subfragmento-1 de miosina
351 Preparaccedilatildeo das soluccedilotildees de vanaacutedio
As soluccedilotildees de vanadato foram preparadas em aacutegua MilliQ a partir de metavanadato de amoacutenio
(NH4VO3) numa concentraccedilatildeo stock de 50 mM com agitaccedilatildeo magneacutetica e aquecimento (~30 ordmC)
Esta soluccedilatildeo com um valor compreendido entre 6 e 7 designa-se por ldquometavanadatordquo e eacute
constituiacuteda pelas esspeacutecies V1 V2 V4 e V5 (Ref) Acidificando a soluccedilatildeo de metavanadato por
adiccedilatildeo de HCl ateacute um valor de pH proacuteximo de 4 observa-se uma mudanccedila de cor de amarelo
claro para laranja indicativa da formaccedilatildeo da espeacutecie decameacuterica de vanadato De seguida e
imediatamente antes da realizaccedilatildeo dos ensaios descritos o valor do pH da soluccedilatildeo de
decavanadato foi aumentado para cerca de 75 por adiccedilatildeo de NaOH A soluccedilatildeo (2) designada
por ldquometavanadatordquo natildeo foi posteriormente alterada ateacute agrave sua adiccedilatildeo para a realizaccedilatildeo dos ensaios
As soluccedilotildees foram armazenadas a 4ordmC e mantidas em gelo aquando da sua utilizaccedilatildeo
A soluccedilatildeo de V(IV) foi preparada em condiccedilotildees anaeroacutebias por diluiccedilatildeo de sulfato de vanadil
(VOSO4) para uma concentraccedilatildeo final de 5 mM em H2O MilliQ saturada com N2 gasoso
352Anaacutelise por espectroscopia de UV-Vis
A concentraccedilatildeo da espeacutecie decameacuterica em soluccedilatildeo depende do periacuteodo de tempo apoacutes a diluiccedilatildeo
no meio de reacccedilatildeo apropriado (Ref) A velocidade de desoligomerizaccedilatildeo da espeacutecie decameacuterica
pode ser analisada por espectroscopia de absorccedilatildeo na regiatildeo UVVis uma vez que esta espeacutecie
apresenta duas bandas caracteriacutesticas na regiatildeo do visiacutevel (a 358 e 400 nm) que natildeo se verifica
em quaisquer outras espeacutecies oligomeacutericas As bandas supramencionada sapresentam-se bem
22
Meio de reacccedilatildeo 3125 mM Tris-HCl pH 71 375 mM MgCl2
48
resolvidas atraveacutes da obtenccedilatildeo da derivada de 2ordf ordem do espectro apresentando a forma de 2
vales com miacutenimos centrados nos comprimentos de onda supracitados com dependecircncia linear
relativamente agrave concentraccedilatildeo de decavanadato total (Ramos et al 2006)
Assim a estabilidade da espeacutecie decameacuterica apoacutes diluiccedilatildeo da soluccedilatildeo de decavanadato no meio de
reacccedilatildeo dos ensaios (tampatildeo D) para uma concentraccedilatildeo final de 1 mM em vanadato total foi
caracterizada atraveacutes da leitura da Abs400 em funccedilatildeo do tempo tendo-se determinado qual a
ordem de reacccedilatildeo e o tempo de meia-vida da mesma Com base em estudos anteriores acerca da
estabilidade do V10 na presenccedila de diferentes proteiacutenas (Ramos et al 2006) determinou-se
igualmente o tempo de meia vida apoacutes a adiccedilatildeo de 20 microM de S1 de miosina ao meio reaccional
353 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
Os espectros de RMN do isoacutetopo 51
V foram realizados num espectroacutemetro Bruker AM-400 MHz
equipado com uma sonda multinuclear inversa de 5 mm Em tubos de RMN de 5 mm de diacircmetro
preparou-se soluccedilotildees de 500 L de deca- ou metavanadato correspondentes a um intervalo de
concentraccedilotildees de 1 a 10 mM suplementadas com 10 D2O Os espectros foram adquiridos agrave
temperatura ambiente com uma largura espectral de 23 923 Hz um tempo de aquisiccedilatildeo de 0086
s tempo de relaxaccedilatildeo de 001 s e uma sequecircncia de pulsos a 90ordm Utilizou-se a transformada de
Fourier com alargamento das bandas de 20Hz para o processamento dos espectros Os desvios
quiacutemicos das ressonacircncias de 51
V foram obtidos em relaccedilatildeo agrave espeacutecie VOCl3cuja frequecircncia de
ressonacircncia eacute considerada o padratildeo sendo todas as outras referenciadas a partir desta em ppm
As aacutereas dos sinais do RMN correspondentes agraves espeacutecies oligomeacutericas presentes foram obtidas
por integraccedilatildeo dos sinais As concentraccedilotildees relativas das espeacutecies foram calculadas com recurso agrave
equaccedilatildeo
onde A[Vx] corresponde agrave aacuterea referente agrave ressonacircncia da espeacutecie Vx com nordm de agregaccedilatildeo x
A[V]total agrave soma total das aacutereas medidas e [V] a concentraccedilatildeo total de vanaacutedio no ensaio
A teacutecnica de 51
V-RMN foi anteriormente aplicada em estudos de ligaccedilatildeo com S1 de miosina
(Ref) tendo sido demostrado que entre as espeacutecies V1 V2 V4 e V5 a espeacutecie tetrameacuterica possui a
maior afinidade para a miosina (Cremo et al) O desaparecimento ou alargamento dos sinais pode
implicar a ligaccedilatildeo de um dos oxovanadatos laacutebeis agrave miosina sem indicaccedilatildeo de qual uma vez que
49
se processa um equiliacutebrio entre eles (Crans 1990) Assim e tendo em conta as indicaccedilotildees de que
a espeacutecie V10 eacute distinta das restantes no que diz respeito agrave magnitude da interacccedilatildeo com a miosina
e inibiccedilatildeo da actividade ATPaacutesica estimulada por actina (Aureliano 2000 Tiago et al 2002)
torna-se fundamental investigar a interacccedilatildeo das diferentes espeacutecies de vanadato com o S1 de
miosina atraveacutes da teacutecnica de 51
V-RMN
Para tal obtiveram-se espectros de 51
V-RMN resultantes da titulaccedilatildeo de uma soluccedilatildeo 5 mM de
decavanadato total com concentraccedilotildees crescentes de S1 de miosina (0 5 10 15 e 20 microM) em
tampatildeo D Realizou-se ainda ensaios na presenccedila de 2 mM de decavanadato e 2 mM de ATP na
presenccedila e na ausecircncia de S1 de miosina de modo a verificar o efeito que estes ligandos naturais
da enzima teria na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o subfragmento-1
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
Os grupos sulfidrilo (-SH) das cisteiacutenas satildeo resiacuteduos de aminoaacutecidos extremamente sensiacuteveis agrave
oxidaccedilatildeo resultando na sua oxidaccedilatildeo (Dalle-Donne et al 1999) Logo a quantificaccedilatildeo destes
resiacuteduos eacute bastante importante nos estudos de interacccedilatildeo com vanaacutedio jaacute que os resiacuteduos de Cys
com ecircnfase para o SH1 e o SH2 satildeo bastante importantes na monitorizaccedilatildeo das modificaccedilotildees
funcionais da regiatildeo da cabeccedila
O procedimento seguidamente apresentado foi originalmente descrito por Gutieacuterrez-Merino
(1987)) Consiste na reacccedilatildeo entre os grupos ndashSH natildeo oxidados das proteiacutenas e o reagente de
Ellman (DTNB -55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico)) - formando-se o aniatildeo CNT (aniatildeo 3-
carboxilato-4-nitrotiofenolato) numa estequiometria de 11 Este apresenta um maacuteximo de
absorvecircncia a 412 nm (Robyt 1990) Assim apoacutes a incubaccedilatildeo da proteiacutena na presenccedila do metal
a quantidade de cisteiacutenas reduzidas por proteiacutena determina-se atraveacutes dos valores de Abs412
Deste modo colocou-se num tubo de Eppendorf o volume apropriado de S1 dialisado para que a
concentraccedilatildeo no ensaio fosse de 1 mgmL Adicionou-se o volume apropriado de decavanadato
para que a sua concentraccedilatildeo fosse de 0 10 50 100 250 500 ou 1000 M Perfez-se com soluccedilatildeo
D ateacute um volume final de 1 mL e incubou-se a 25ordmC por 20 minutos Apoacutes este periacuteodo colocou-
se 780 L da mistura reaccional numa cuvette de quartzo e adicionou-se 220 L de soluccedilatildeo K23
23
Tampatildeo K Tampatildeo fosfatohidrogenofosfato de soacutedio 025 M pH 72
50
Adicionou-se 15 L de DTNB (em soluccedilatildeo K [DTNB]stock=100 mM) ao meio reaccional e
registou-se o valor de absorvecircncia medido equivalente agraves cisteiacutenas ldquoraacutepidasrdquo (SH1 e SH2
expostos ao solvente) Adicionou-se 001 g de SDS que provoca a desnaturaccedilatildeo da proteiacutena e
consequentemente a exposiccedilatildeo de todos os resiacuteduos de cisteiacutenas que se encontravam ateacute aiacute
inacessiacuteveis no interior da proteiacutena (cisteiacutenas totais) Estes resiacuteduos podem assim reagir com o
DTNB
Estimou-se o nuacutemero de resiacuteduos de cisteiacutenas em todas as amostras com recurso ao coeficiente de
extinccedilatildeo molar () do DTNB (ver ponto 3 dos Anexos) que foi determinado atraveacutes da titulaccedilatildeo
de L-Cisteiacutena com intervalo de concentraccedilotildees compreendidos entre 0 e 50 microM na presenccedila de
25microM de DTNB em tampatildeo D Os valores de absorvecircncia foram obtidos para um comprimento de
onda de 412 nm
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
3621 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
A teacutecnica mais eficiente de anaacutelise da interacccedilatildeo entre o iatildeo vanadilo e proteiacutenas como a
calmodulina e a transferrina entre outras permite a obtenccedilatildeo raacutepida de informaccedilatildeo no que toca agrave
conformaccedilatildeo e amplitude da referida interacccedilatildeo
Os espectros de Ressonacircncia Paramagneacutetica Electroacutenica (RPE) da espeacutecie paramagneacutetica V(IV)
foram realizados a 70K num espectroacutemetro Bruker EMX-300 X-band Todas as soluccedilotildees
utilizadas e amostras foram preparadas em anaerobiose numa cacircmara de luvas de modo a evitar a
reduccedilatildeo de V(IV) a V(V) Em tubos capilares com 02 mL de amostra manteve-se a concentraccedilatildeo
de S1 a 20 M e variou-se a concentraccedilatildeo de vanadilo (0 25 5 10 15 25 50 75 100 150
200 500 e 1000 M [VOSO4]stock=5 mM) em tampatildeo D Apoacutes obtenccedilatildeo dos espectros estes
foram tratados atraveacutes do software SimFonia e WinEPR tendo-se obtido um graacutefico com a
variaccedilatildeo da intensidade do sinal de RPE em funccedilatildeo da concentraccedilatildeo de sulfato de vanadilo
presente Realizaram-se ainda simulaccedilotildees de espectros obtidos com 20 microM de S1 e 1000 microM de
V(IV) com vista agrave obtenccedilatildeo dos ligandos provaacuteveis do vanadilo atraveacutes da anaacutelise da constante
de acoplamento hiperfina (A) e do factor g (g)
4 Conclusotildees gerais
Apesar de jaacute ter sido demonstrado que o vanaacutedio inibe a Na+K
+-ATPase da miosina a funccedilatildeo
deste elemento no tecido muacutesculo esqueleacutetico continua largamente incompreendida A enorme
complexidade quiacutemica deste metal especialmente no que concerne agrave oligomerizaccedilatildeo e reacccedilotildees
51
redox constitui um obstaacuteculo agrave compreensatildeo do comportamento do vanaacutedio nos organismos
vivos Assim a caracterizaccedilatildeo da quiacutemica deste elemento eacute de grande utilidade antes de iniciar o
estudo da sua interacccedilatildeo com o presente sistema
A anaacutelise por espectroscopias de 51
V-RMN e de UV-Vis de soluccedilotildees de vanadato diluiacutedas nos
meios de reacccedilatildeo utilizados no presente relatoacuterio (10 mM HEPES pH 70 25 mM KCl) mostrou
claramente que a espeacutecie decameacuterica (V10) possui um tempo de vida suficientemente longo para a
determinaccedilatildeo do seu efeito isolado ao contraacuterio do que se verifica com as espeacutecies mono- di-
tetra- e pentameacutericas (V1 V2 V4 e V5) Por outro lado a anaacutelise verificou ainda que apoacutes diluiccedilatildeo
das soluccedilotildees de decavanadato no meio de reacccedilatildeo coexistem as espeacutecies V10 e V1 pelo que os
efeitos putativos desta uacuteltima deveratildeo ser subtraiacutedos aquando da monitorizaccedilatildeo dos efeitos da
espeacutecie decameacuterica
O vanaacutedio (V) (vanadato) inibe as enzimas que catalizam reacccedilotildees de transferecircncia de grupos
fosfato como a ATPase da miosina [Goodno 1979] devido agrave sua analogia estrutural com o iatildeo
fosfato Na sua utilizaccedilatildeo como inibidor enzimaacutetico natildeo eacute tida em conta a contribuiccedilatildeo individual
das espeacutecies oligomeacutericas considerando-se normalmente a espeacutecie V1 como inibidora efectiva
[Goodno e Taylor 1982] No presente relatoacuterio verificou-se por espectroscopia de 51
V-RMN que
outros oligoacutemeros de vanadato interagem com o S1 de miosina tendo as espeacutecies V10 e V4 os
maiores niacuteveis de afinidade A sua interacccedilatildeo envolve forccedilas electrostaacuteticas e eacute afectada
negativamente por ligandos naturais de enzima como eacute o caso do ATP
A exposiccedilatildeo do S1 de miosina (005 mgmL) a decavanadato conduz agrave inibiccedilatildeo do capacidade da
proteiacutena de estimular a actividade Mg2+
-ATPaacutesica com um valor de IC50 de 266 plusmn 07 M
decavanadato sendo o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM
decavanadato (80) Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM decavanadato sendo
o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM decavanadato (60)
exercendo um efeito de impedimento agrave ligaccedilatildeo entre o vanaacutedio e o S1
A uma concentraccedilatildeo de 27 microM decavanadato verifica-se a oxidaccedilatildeo de um dos resiacuteduos de Cys
de S1 de miosina seguida pela oxidaccedilatildeo gradual de outro resiacuteduo de Cys ateacute agrave concentraccedilatildeo de
1000 microM Ambos os resiacuteduos oxidados natildeo se encontravam expostos ao solvente utilizado A
oxidaccedilatildeo dos resiacuteduos permite sugerir que esta modificaccedilatildeo oxidativa sofrida pelo S1 contribui
para a diminuiccedilatildeo da sua actividade de ATPase
52
Por espectroscopia de RPE revelaram-se os possiacuteveis ligandos equatoriais do vanaacutedio aquando da
sua interacccedilatildeo com o S1 de miosina como sendo duas moleacuteculas de aacutegua um ligando contendo o
grupo R-OH e outro contendo o grupo R=NR
Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados de
oxidaccedilatildeo (IV) e (V) no S1 de miosina para concentraccedilotildees facilmente alcanccedilaacuteveis no ambiente
terrestre
53
5 Estudos futuros
No sentido do melhor esclarecimento das alteraccedilotildees moleculares e estruturais que ocorrem no
subfragmento 1 quando exposta ao vanaacutedio que conduzem a uma diminuiccedilatildeo da sua funccedilatildeo
sugere-se
- Quantificaccedilatildeo das cisteiacutenas reduzidas de S1 na presenccedila de metavanadato
- Determinaccedilatildeo das actividades Ca2+
- e K+-ATPaacutesica do S1 de miosina na presenccedila de
decavanadato
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina estimulada por actina
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina na presenccedila de
metavanadato
- Realizaccedilatildeo de RMN 2D com variaccedilatildeo das concentraccedilotildees das soluccedilotildees de decavanadato e
metavanadato na presenccedila de S1
-Mutageacutenese dos resiacuteduos de Cys doS1 de miosina de modo a verificar quais os resiacuteduos que
sofrem oxidaccedilatildeo na presenccedila de decavanadato
- Determinaccedilatildeo por calorimetria (DSC Differential Scanning Calorimetry) da variaccedilatildeo da
entalpia (∆H) e da temperatura de desnaturaccedilatildeo do S1 de miosina na presenccedila de vanaacutedio uma
vez que estas propriedades caracteriacutesticas de cada proteiacutena satildeo alteradas apoacutes ocorrecircncia de
modificaccedilotildees estruturais
- Anaacutelise por SDS-PAGE de S1 de miosina apoacutes tratamento com vanaacutedio de modo a determinar
quais os polipeacuteptidos eventualmente formados apoacutes clivagem da proteiacutena
54
6 Referecircncias bibliograacuteficas
- Sellers J Myosins 2nd Edition Oxford University Press 1999
- L Stryer inBiochemistry (Ed WH Freeman 2005 pp
Refordfas Materiais e Meacutetodos
[A] S S Margossian SLowey Methods Enzymol1982 85 55-71
[B] JD Pardue JA Spudich Methods Enzymol 1982 85 164-181
[C] M Bradford Anal Biochem1976 72 248-254
[D] H H Taussky E Shorr J Biol Chem 1953 202 675-685
[E] C R Bagshaw in Outline Studies in Biology ndash Muscle Contraction Chapman and Hall Ed
1982
[F] A Cuenda et al Eur J Biochem1990 194 663-670
55
Anexos
1 Determinaccedilatildeo da recta de calibraccedilatildeo meacutetodo de Bradford
Figura1ndashRepresentaccedilatildeo graacutefica da recta de calibraccedilatildeo (Abs595 nm vs [BSA]) utilizada para o meacutetodo de Bradford
2 Determinaccedilatildeo da recta calibraccedilatildeo para a electroforese em condiccedilotildees desnaturantes
Figura 2 ndash Representaccedilatildeo graacutefica da recta de calibraccedilatildeo (log(Peso Molecular) vs Rf dos marcadores de peso
molecular) onde Rf indica determinada atraveacutes do padratildeo de peso molecular
(poccedilo (a))
3Caacutelculo do coeficiente de extinccedilatildeo molar () para a quantificaccedilatildeo de cisteiacutenas reduzidas
atraveacutes do meacutetodo de Ellman
Traccedilou-se a recta de Abs412 nm meacutedia vs [L-Cisteiacutena] de modo a determinar o valor do coeficiente
de extinccedilatildeo molar definido como o declive daquela recta Assim =8400 M-1
cm-1
como
podemos verificar na Fig 3
A595 nm = 00546[BSA] + 00209 Rsup2 = 099051
Ab
s 59
5 n
m
[BSA] (mgmL)
log PM = -1177 Rf + 2247 Rsup2 = 098957
log
PM
Mobilidade relativa (Rf)
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero

18
Figura 111 ndash Estruturas esquemaacuteticas das espeacutecies de vanadato (espeacutecie monomeacuterica (V1) dimeacuterica (V2)
tetrameacuterica (V4) pentameacuterica (V5) e decameacuterica (V10) (Adaptado de Aureliano e Crans 2008)
122 Metabolismo e efeito na actividade enzimaacutetica
Apesar de natildeo terem sido ainda encontradas as provas categoacutericas das suas funccedilotildees nos seres
vivos sabe-se que o iatildeo vanadato eacute capaz de inibir uma grande quantidade de enzimas in vitro
com especial ecircnfase na actividade ATPaacutesica da miosina [AL Teresa] Como referido
anteriormente o vanadato eacute capaz de actuar em alternativa ao fosfato nas reacccedilotildees bioquiacutemicas
incluindo a sua preferecircncia de ligaccedilatildeo agraves enzimas [W]
Eacute conhecido o papel do vanaacutedio no metabolismo do caacutelcio promovendo a mineralizaccedilatildeo dos
dentes e ossos na biossiacutentese de liacutepidos (trigliceacuteridos e colesterol) com posterior inibiccedilatildeo da
acumulaccedilatildeo destes nos vasos sanguiacuteneos e no SNC na reproduccedilatildeo e desenvolvimento por
mimetizaccedilatildeo e potenciaccedilatildeo de factores de crescimento na estimulaccedilatildeo do consumo de glucose e
da siacutentese de glicogeacutenio com actuaccedilatildeo equivalente aos efeitos da insulina ao niacutevel celular na
transduccedilatildeo de sinais celulares com regulaccedilatildeo da actividade de segundos mensageiros no
transporte de iotildees e aminoaacutecidos na transcriccedilatildeo geneacutetica e siacutentese de proteiacutenas e DNA entre
outros [W Y AB]
Assim o vanaacutedio cnseguiraacute provocar inuacutemeras mudanccedilas bioquiacutemicas nas ceacutelulas Alguns
exemplos da influecircncia bioquiacutemica do vanaacutedio incluem os seus efeitos nos processos de
fosforilaccedilatildeo e desfosforilaccedilatildeo de proteiacutenas e na participaccedilatildeo na formaccedilatildeo de espeacutecies reactivas de
oxigeacutenio devido ao seu efeito proacute-oxidante sobre agentes redutores a niacutevel celular [AM]
Quanto ao seu efeito na actividade enzimaacutetica pode dividir-se em dois tipos principais ndash inibitoacuterio
e estimulante - dependendo da enzima ou do tipo de reacccedilatildeo por esta catalisada No primeiro
grupo incluem-se enzimas como as ATPases (de miosina de canais ioacutenicos e fosfoenzimas
19
transportadoras) e as fosfatases (da tirosina aacutecida e alcalina) a fosfofrutocinase adenilato ciclase
e catalase entre outras [AA] O segundo grupo inclui a maioria das cinases a glucose-6-fosfato
desidrogenase a fosfoglicerato mutase e a fosfoglicerato fosfatase (Refordf)
O efeito do vanaacutedio (V) na actividade enzimaacutetica estaacute relacionado com o seu nuacutemero de
coordenaccedilatildeo podendo actuar como estado de transiccedilatildeo ou por mimetizaccedilatildeo de cofactores
substratos (especialmente o fosfato) e interacccedilotildees proteiacutena-proteiacutena (como no caso das
polimerases e transcriptases) Pode ainda estar envolvido na formaccedilatildeo de ligaccedilotildees de hidrogeacutenio
interacccedilotildees electrostaacuteticas esteacutereas e de oxidaccedilatildeo-reduccedilatildeo Uma evidecircncia desta uacuteltima eacute a sua
capacidade de alternacircncia entre os estados de oxidaccedilatildeo +5 a +3 o que torna faacutecil a sua interacccedilatildeo
redox com enzimas contendo resiacuteduos de cisteiacutenas (Cys) ou qualquer outro composto orgacircnico
com actividade redox [AN]
13 Vanaacutedio e muacutesculo esqueleacutetico
A primeira evidecircncia da presenccedila de vanaacutedio no muacutesculo esqueleacutetico proveio de uma amostra de
ATP isolado de muacutesculo de cavalo que continha uma impureza ndash vanaacutedio ndash cuja presenccedila
provocou a inibiccedilatildeo da actividade da bomba de Na+K
+-ATPase [AO] Desde entatildeo vaacuterios
estudos tecircm sido realizados com ecircnfase na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o
tecido muscular Destes destaca-se a descoberta da interacccedilatildeo forte da espeacutecie V10 com a miosina
fosforilada [AP] e agrave sua inibiccedilatildeo da actividade ATPaacutesica da miosina[AQ AR]
No quediz respeitoao iatildeo monovanadato destaca-se a sua inibiccedilatildeoda actividade Mg2+
-ATPaacutesica
da miosina [AS] que ocorre de duas formas distintas (1) na presenccedila de concentraccedilotildees reduzidas
de V1 a inibiccedilatildeo ocorre a velocidade reduzida dependente da concentraccedilatildeo da espeacutecie
monomeacuterica (2) na presenccedila de concentraccedilotildees elevadas de V1 o niacutevel de inibiccedilatildeo aproxima-se de
100 desde que [V1] seja superior agrave concentraccedilatildeo total de locais com actividade Mg2+
-ATPaacutesica
no ensaio Esta inibiccedilatildeo eacute irreversiacutevel tanto por diaacutelise ou adiccedilatildeo de agentes redutores do
vanadato [AL]Segundo Pate et al(1994)[AT] a espeacutecie monomeacuterica podeinibir a velocidade de
contracccedilatildeo a temperatura inferior a 20ordmC perdendogradual e rapidamente essa capacidade ateacute os
50ordmC temperatura agrave qual cessa a capacidade inibitoacuteria Os resultados obtidos sugeremainda uma
interacccedilatildeo altamente cooperativa entre o iatildeo vanadato e as proteiacutenas componentes do muacutesculo
esqueleacuteticoDestaca-se tambeacutem a descoberta da inibiccedilatildeo da geraccedilatildeo de forccedila contraacutectil nas fibras
musculares por parte da espeacutecie monomeacuterica[AT]Wilson et al(1995) [AU ]determinaram que a
afinidade da espeacutecie V1 para o S1 de miosina eacute significativamente mais elevada do que a do iatildeo
Pi seu anaacutelogo Descobriram ainda que que a ligaccedilatildeo do vanadato poderaacute envolver transiccedilotildees
entre estados energeacuteticos com energia livre independente dos impedimentos esteacutereos impostos
20
pela estrutura das miofibrilas Os estudos desenvolvidos por Smith et al (1990)[AV]
determinaram queo iatildeo monovanadato natildeo apresenta um efeito pronunciado na inibiccedilatildeo da
actividadeacto-S1-miosina-ATPaacutesica excepto em concentraccedilotildees superiores a 900microM a partir das
quais existe produccedilatildeo de espeacutecies oligomeacutericas para as quais o efeito inibitoacuterio seraacute mais
elevadoA espeacutecie monomeacuterica de vanadato consegue adoptar facilmente uma geometria do tipo
bipiramidal trigonal que mimetiza o intermediaacuterio formado (MgADP
Pi)aquando da reacccedilatildeo de
hidroacutelise de ATP no local activo da miosina [AW] De facto a interacccedilatildeo do vanaacutedio com a
miosina eacute inesperadamente estaacutevel na presenccedila de ADP Esta interacccedilatildeo resulta na formaccedilatildeo de
um complexo enzimaacutetico inactivo e estaacutevel com estequiometria de 11 ADPvanadato por siacutetio
activo da proteiacutena A formaccedilatildeo do complexo estaacutevel miosina-vanadato foi proposta por Goodnoet
al (1982) [AS] Nesta ocorre um preacute-equiliacutebrio raacutepido que induz a formaccedilatildeo de um complexo
ternaacuterio reversiacutevel (MADP
V) com formaccedilatildeo lenta do isoacutemero M
ADP
V A diferenccedila entre os
compostos de vanaacutedio e de fosfato centra-se no tempo de meia vida (t12) do Pi ou do V (no
primeiro t12=12 s enquanto no segundo t12=24 h) (Refordf) A diferenccedila dever-se-aacute agrave possiacutevel
complexaccedilatildeo do vanaacutedio com grupos nucleofiacutelicos no siacutetio activo o que provocaria uma
estabilizaccedilatildeo da miosina na conformaccedilatildeo MADP
V
No que respeita aos estudos realizados com a espeacutecie tetrameacuterica Cremo et al determinou que a
cadeia pesada do S1 de miosina pode ser clivada em dois locais distintos (designados V1 e V2)
pela espeacutecie V4 apoacutes irradiaccedilatildeo com luz UV na presenccedila de concentraccedilotildees de vanadato na ordem
dos milimolar e na ausecircncia de nucleoacutetidos ou metais divalentes
Jaacute no que concerne aos estudos realizados com ecircnfase na espeacutecie decameacuterica foi sugerido
porRamos et al (2006) [AK]que a interacccedilatildeo do decavanadato com a G-actina favorecida pela
polimerizaccedilatildeo desta uacuteltima estabiliza a espeacutecie V10 e induz a inibiccedilatildeo da formaccedilatildeo de F-actina A
estabilizaccedilatildeo da espeacutecie V10 por proteiacutenas tansmembranares e do citoesqueleto pode ser a razatildeo
pela qual ocorre a toxicidade do decavanadato reportada na avaliaccedilatildeo dos efeitos do V(V) nos
sistemas bioloacutegicosTiago et al (2007) [AR] concluiram que o V10 apresenta uma clara preferecircncia
de ligaccedilatildeo ao ldquoback-doorrdquo da S1-miosina-ATPase o que se verifica apenas na estrutura
conformacional desimpedida com acesso facilitado ao local de ligaccedilatildeo do Pi Sugere tambeacutem que
o V10 actua como um ldquoback-door stoprdquo bloqueando estaabertura do fragmento de 50-kDa
necessaacuteria para a hidroacutelise do ATP--fosfato o que constitui uma explicaccedilatildeo simples do
comportamento natildeo competitivo da espeacutecie decameacuterica estimulando a utilizaccedilatildeo deste oligoacutemero
como uma feramenta da elucidaccedilatildeo das mudanccedilas conformacionais ldquoback-doorrdquo da miosina no
21
processo da contracccedilatildeo muscular(ref)Destaca-se ainda a descoberta da capacidade inibitoacuteria do
V10 em relaccedilatildeo agrave fosforilaccedilatildeo oxidativa mitocondrial [AX]
Finalmente no que concerne aos estudos realizados com a presenccedila de vaacuterias espeacutecies
oligomeacutericas Aureliano e Madeira (1994) [AY]observaram que na presenccedila de Ca2+
as espeacutecies
V4 e V10se ligam agrave Ca2+
-ATPase do retiacuteculo sarcoplasmaacutetico enquanto a espeacutecie V1 se liga apenas
na presenccedila de ATP Aparentemente o aumento da concentraccedilatildeo de Ca2+
tem o efeito contraacuterio da
inibiccedilatildeo da ATPase do retiacuteculo sarcoplasmaacutetico pelo V1 natildeo tendo no entanto efeito significativo
sobre a inibiccedilatildeo por parte da espeacutecie V10 Tiago et al (2002) [AP] concluiacuteram que em condiccedilotildees
proacuteximas agraves fisioloacutegicas a espeacutecie V10 difere das espeacutecies presentes na soluccedilatildeo de metavanadato
devido agrave sua forte interacccedilatildeo com a miosina fosforilada e agrave sua inibiccedilatildeo da actividade ATPaacutesica
da miosina Para aleacutem disso o ATP diminui a afinidade da miosina para o V4 induzindo a
interacccedilatildeo com V1 e natildeo tendo qualquer efeito no que diz respeito ao V10 Foi tambeacutem proposto
[AQ] que a ligaccedilatildeo das espeacutecies oligomeacutericas de vanadato aoS1 de miosina nos locais onde o
fosfato coordenaeacute provavelmente a causa da inibiccedilatildeo da actividade ATPaacutesica da miosina
estimulada por actina atraveacutes da prevenccedilatildeo da troca entre ATPADP sendo esta favorecida para
aniotildees de vanadato com valecircncias superiores como eacute o caso do V10
Na presenccedila de actina as velocidades de dissociaccedilatildeo do Pi e do V aumentam consideravelmente
este uacuteltimo em menor quantidade Assim a ligaccedilatildeo da actina ao complexo MADP
V e
Pi eacute
bastante fraca pelo que se sugere que o ADPV teria um efeito de relaxamento semelhante ao
ATP No entanto a capacidade do vanaacutedio em inibir a actividade ATPaacutesica da miosina e da
actomiosina eacute diferenciada uma vez que na ausecircncia de actina a inibiccedilatildeo eacute total para
concentraccedilotildees na gama dos M mas na sua presenccedila a inibiccedilatildeo eacute totalmente conseguida apenas
em concentraccedilotildees na gama dos mM (Ref) No entanto eacute necessaacuterio considerar a possiacutevel
formaccedilatildeo de espeacutecies oligomeacutericas de vanadato cuja afinidade e efeito na enzima seratildeo
diferentes Assim pode concluir-se que o efeito de inibiccedilatildeo observado para a actividade ATPaacutesica
de miosina estimulada por actina se pode dever agrave presenccedila das diferentes espeacutecies oligomeacutericas o
que foi jaacute sugerido por Aureliano (2000) [AZ] ndash as espeacutecies tetrameacuterica e decameacuterica de vanadato
interactuariam com a miosina com alteraccedilatildeo da sua interacccedilatildeo com a actina No entanto natildeo eacute
conhecida a contribuiccedilatildeo absoluta das restantes espeacutecies oligomeacutericas para a inibiccedilatildeo da ATPase
da miosina estimulada por actina (Refs)
14 Objectivos do projecto
Pretende-se com este projecto o isolamento purificaccedilatildeo e caracterizaccedilatildeo da actina do muacutesculo
22
esqueleacutetico de coelho sendo esta uacuteltima tambeacutem realizada para o sufragmento-1 de miosina
Seguidamente realizar-se-aacute a quantificaccedilatildeo e caracterizaccedilatildeo das proteiacutenas por espectroscopia UV-
Vis e SDS-PAGE e a caracterizaccedilatildeo das soluccedilotildees de vanaacutedio por UV-Vis e ressonacircncia magneacutetica
nuclear (RMN) Por uacuteltimo a interacccedilatildeo do S1 de miosina com o vanaacutedio seraacute avaliada atraveacutes
das teacutecnicas de UVVis RMN e ressonacircncia paramagneacutetica electroacutenica (RPE)
2 Discussatildeo de Resultados
21 Caracterizaccedilatildeo das proteiacutenas isoladas
211Anaacutelise do estado de pureza
O estado de pureza das soluccedilotildees de actina e de S1 foi analisado por electroforese em gel de
poliacrilamida em condiccedilotildees desnaturantes (SDS-PAGE) (Fig 22) que separa as moleacuteculas de
acordo com o seu peso molecular (PM) A concentraccedilatildeo de acrilamida escolhida (10) permite a
detecccedilatildeo simultacircnea de componentes com peso molecular inferior a 20 kDa e outros com elevado
peso molecular
Figura 21 ndash Fotografia de gel de SDS-PAGE (10 acrilamida) Poccedilo (a)
10 L do marcador de elevado peso molecular () Poccedilo (b) 20 L de [G-
actina]=1 mgmL Poccedilo (c) 20 L de [S1]=1 mgmL Aplicou-se uma
voltagem de V durante h e min
A partir da recta de calibraccedilatildeo obtida log PM = -1177 Rf + 2247 (ver ponto 2 da secccedilatildeo
Anexos) pode verificar-se a presenccedila de actina na amostra do poccedilo (b) presente numa banda
bastante intensa (PMaparente=40 kDa) bem como a presenccedila minoritaacuteria de outros componentes
desconhecidos que natildeo teratildeo sido eliminados durante o processo de extracccedilatildeo e purificaccedilatildeo e natildeo
satildeo relevantes para o estudo em questatildeo Na amostra do poccedilo (c) nota-se a presenccedila de S1
(PMaparente=92 kDa) bem como a de outros componentes cuja banda eacute ligeiramente menos intensa
Estes poderatildeo ser fragmentos proteoliacuteticos de miosina e de S1 Estaacute ainda presente uma bada de
23
maior intensidade que poderaacute ser a proteiacutena-C (PM=135 kDa) uma vez que a sua remoccedilatildeo
completa da amostra necessitaria de um passo adicional de cromatografia de troca ioacutenica [BA]
No entanto natildeo podemos confirmar que se trata desta proteiacutena ou de qualquer outra uma vez que
se encontra fora do intervalo de migraccedilatildeo dos marcadores de peso molecular
212 Determinaccedilatildeo da concentraccedilatildeo proteica
2121Subfragmento-1 de miosina
A partir da recta de calibraccedilatildeo obtida pelo meacutetodo de Bradford [BB](ver secccedilatildeo 3311 do
Procedimento Experimental) A595nm = 00546[BSA] + 00209 (ver secccedilatildeo de Anexos)foi possiacutevel
determinar um intervalo de concentraccedilatildeo de S1 de miosina compreendido entre 226 e 117(plusmn
423M) (203 e 1056 (plusmn 381 mgmL)) (n=8) A concentraccedilatildeo meacutedia de S1 de miosina apoacutes
diaacutelise contra a soluccedilatildeo tampatildeo 10 mM HEPES pH 75 25 mM KCl determinada pela leitura da
Abs280 nmfoi de 20430M (1803 mgmL)(n=7)
As amostras a quantificar atraveacutes do meacutetodo de Bradford natildeo sofreram diaacutelise o que permite
explicar a concentraccedilatildeo superior de S1 de miosina em comparaccedilatildeo com o valor obtido atraveacutes do
meacutetodo directo Esta razatildeo aliada agrave sensibilidade e rapidez superiores deste uacuteltimo fez com que
se optasse pelo meacutetodo directo para a quantificaccedilatildeo da grande maioria dos ensaios
2122Actina
A concentraccedilatildeo de actina foi determinada quando esta se encontrava na forma monomeacuterica (G-
actina) por mediccedilatildeo da leitura directa da Abs290 nm tendo-se obtido uma concentraccedilatildeo meacutedia de
691481M (290202 mgmL) (n=3) em 2 mM Tris-HCl pH 80 02 mM CaCl2 02 mM Na-
ATP Ao comprimento de onda utilizado (290 nm) absorvem fortemente apenas os resiacuteduos de
Trp pelo que a mediccedilatildeo efectuada tem menor sensibilidade do que se tivesse sido efectuada a 280
nm uma vez que a actina apresenta apenas 4 resiacuteduos deste aminoaacutecido embora tenha 16
resiacuteduos de Tyr
22 Mediccedilatildeo da actividade enzimaacutetica de S1 de miosina
221 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
As actividades ATPaacutesicas de S1 de miosina dependentes de Ca2+
e K+foram analisadas pelo
caacutelculo das velocidades de consumo inicial do fosfato inorgacircnico (Pi) Definiu-se os tempos de 5
e 3 minutos para a mediccedilatildeo da Ca2+
e da K+EDTA-ATPase respectivamente Os valores obtidos
24
a partir do protocolo presente no ponto 341 da secccedilatildeo Procedimento experimental encontram-se
descritos na Tabela 2I
Tabela 2I ndash Acrividades natildeo-fisioloacutegicas de subfragmento-1 determinadas pelo meacutetodo de Fiske-Subarow
modificado (n=2)
Proteiacutena
Actividade
ATPaacutesica natildeo
fisioloacutegica
Actividade (nmol
Piminmg proteiacutena) Actividade (s-1)
S1
Ca2+
-ATPase 1400 plusmn 30 2100 plusmn 0045
K+EDTA-ATPase 2334 plusmn 50 3500 plusmn 0075
As actividades natildeo-fisioloacutegicas reflectem o estado de oxidaccedilatildeo dos resiacuteduos expostos de Cys 707
(SH1) e 697 (SH2) Se o grupo SH1 sofrer oxidaccedilatildeo a actividade ATPaacutesica dependente de Ca2+
aumenta e a actividade dependente de K+ diminui Por outro lado se o grupo SH2 sofrer
oxidaccedilatildeo regista-se a perda completa destas actividades
De acordo com Margossian e Lowey (1982) [BA] as actividades ATPaacutesica de S1 de miosina
dependente de Ca2+
e K+EDTA da miosina satildeo respectivamente de 24 e 112 s-1
Estaacute tambeacutem
descrito que a actividade ATPaacutesica dependente de K+ eacute em meacutedia cerca de 3 vezes superior agrave
dependente de Ca2+
reflectindo a quantidade das preparaccedilotildees A razatildeo obtida entre os valores das
actividades ATPaacutesicas dependentes de K+ e Ca
2+ eacute de 166 Os valores obtidos para as actividades
(Ca2+
-ATPase=21 s-1
K+EDTA-ATPase= 35 s
-1) satildeo inferiores agrave valores obtidos por
Margossian e Lowey [BA] e Tiago [BC] com forccedila ioacutenica inferior O valor obtido para a
actividade ATPaacutesica dependente de Ca2+
eacute superior ao obtido por Polosukhina et al (2000) [BD]
cujo meio de reacccedilatildeo continha MOPS e apresentava forccedila ioacutenica inferior Os dados
supramencionados comprovam a variaccedilatildeo da actividade a ATPaacutesica natildeo fisioloacutegica em relaccedilatildeo agrave
forccedila ioacutenica temperatura e pH
222 Mediccedilatildeo da actividadeMg2+
-ATPaacutesica de S1 pelo meacutetodo de enzimas acopladas
A actividade de Mg2+
-ATPase eacute a uacutenica que possui significado fisioloacutegico sendo as restantes
bastante uacuteteis apenas para indicaccedilatildeo da qualidade de preparaccedilatildeo do S1[BA] A 0575 M KCl a
actina e o S1 formam uma soluccedilatildeo viscosa e tuacuterbida Apoacutes a adiccedilatildeo de MgATP o complexo acto-
S1 dissocia-se e a turbidez da soluccedilatildeo diminui Se a forccedila ioacutenica for reduzida a dissociaccedilatildeo do
complexo actoS1 que se encontra na forma de suspensatildeo eacute induzida O MgATP eacute rapidamente
hidrolisado diminuindo a sua concentraccedilatildeo no meio reaccional Nestas condiccedilotildees haacute
reassociaccedilatildeo e subsequente contracccedilatildeo dos filamentos de actina e de S1 com formaccedilatildeo de
25
precipitado O meacutetodo descrito traduz de forma bastante elementar o fenoacutemeno da contracccedilatildeo in
vivo e elimina o problema anterior da solubilizaccedilatildeo ineficiente da miosina de modo a obter uma
mediccedilatildeo exacta da actividade [A] A utilizaccedilatildeo de forccedilas ioacutenicas reduzidas aumenta a afinidade da
actina para o S1 permitindo contornar a extrema dificuldade em obter a concentraccedilatildeo proteica dos
muacutesculos (cerca de 100 mgmL do complexo actomiosina) As actividades Mg2+
-ATPaacutesicas de S1
de miosina foram determinadas a partir do declive da variaccedilatildeo da Abs340 relativo agrave oxidaccedilatildeo de
NADH em funccedilatildeo do tempo como eacute descrito no ponto 342 do Procedimento Experimental Os
valores obtidos para a actividade Mg2+
-ATPaacutesica encontram-se apresentados na Tabela 2II
Tabela 2II ndash Actividade Mg2+
-ATPaacutesica de subfragmento-1 de miosina determinadas pelo meacutetodo das enzimas
acopladas
Actividade (nmol
ATPmin mg(S1)) Actividade (s
-1)
1097 plusmn 269 164 plusmn 04
Ramos (2005) [J] obteve o valor de 01 s-1
para a actividade Mg2+
-ATPaacutesica de S1 de miosina
com forccedila ioacutenica intermeacutedia Tiago (2005) [BC] obteve o valor de 011 s-1 em condiccedilotildees de forccedila
ioacutenica reduzida em relaccedilatildeo agrave utilizada no presente relatoacuterio
23 Preparaccedilatildeo e caracterizaccedilatildeo das soluccedilotildees de vanaacutedio
231 Anaacutelise da composiccedilatildeo das soluccedilotildees de vanadato por espectroscopia de 51
V-RMN
efeito da concentraccedilatildeo da espeacutecie
A diluiccedilatildeo das soluccedilotildees stock de meta- e decavanadato iraacute determinar fortemente a composiccedilatildeo
em soluccedilatildeo dos diferentes oligoacutemeros de vanadato (ref) No caso do decavanadato o periacuteodo de
tempo apoacutes a diluiccedilatildeo iraacute tambeacutem ser um factor importante a considerar devido agrave decomposiccedilatildeo
da espeacutecie V10(ref)Torna-se assim fundamental determinar a composiccedilatildeo quiacutemica das soluccedilotildees
no meio de ensaio utilizado (HEPES 10 mM KCl 25 mM) de modo a correlacionaacute-la
correctamente com os possiacuteveis efeitos promovidos pelas diferentes espeacutecies de vanadato no
sistema em estudo
Aleacutem disso deve ser dado ecircnfase agraves importantes interacccedilotildees que ocorrem entre o vanaacutedio e os
tampotildees escolhidos O tampatildeo com menor nuacutemero e magnitude de interacccedilatildeo eacute o HEPES [AD]
seguido do Tris sendo este uacuteltimo especialmente aconselhado em concentraccedilotildees baixas para casos
de incompatibilidade enzimaacutetica ou quiacutemica redox No caso especial da reacccedilatildeo redox
NAD(P)HNAD(P) o tampatildeo adequado eacute o imidazole de modo a natildeo haver reacccedilatildeo com o
26
vanadato Os tampotildees a evitar em qualquer situaccedilatildeo incluem a trietanolamina fosfato citrato e
glicina entre outros [AD] uma vez que todos interactuam fortemente com V(V) com formaccedilatildeo
de complexos estaacuteveis
Assim procedeu-se agrave anaacutelise da composiccedilatildeo das soluccedilotildees de meta- e decavanadato apoacutes diluiccedilatildeo
das soluccedilotildees stock (50 mM em vanadato total) no meio de reacccedilatildeo (tampatildeo D) atraveacutes da
espectroscopia de ressonacircncia magneacutetica nuclear de 51
V Nas figuras x1 e x2 apresentam-se os
espectros obtidos para as soluccedilotildees de deca- e metavanadato num intervalo de concentraccedilotildees de 1 a
10 mM A Tabela seguinte resume os paracircmetros espectroscoacutepicos obtidos para cada espeacutecie
presente nas soluccedilotildees analisadas
Figura 22ndash Espectros de RMN de 51
V de
concentraccedilotildees crescentes das espeacutecies oligomeacutericas
de vanadato obtidos apoacutes diluiccedilatildeo da soluccedilatildeo stock
de decavanadato em 10 mM HEPES pH 70 25 mM
KCl para concentraccedilotildees finais de A) 1 mM B)2
mM C) 4 mM D) 8 mM E) 10 mM de
decavanadato total V1refere-se agraves espeacutecies
monomeacutericade vanadato V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2IIIndash Desvios quiacutemicos meacutedios (part) das ressonacircncias das espeacutecies decameacuterica (V10) monomeacuterica (V1) e
tetrameacuterica (V4) presentes nos espectros da soluccedilatildeo de decavanadatoLargura a meia altura (∆12) da espeacutecie V10A
Paracircmetros V10A V10B V10C V1 V4
part (ppm) -5162 -5003 -4255 -5616 -5748
∆12 (Hz) 175 nd nd nd nd
Figura 23 ndash Distribuiccedilatildeo graacutefica das
espeacutecies oligomeacutericas de vanadato vs a
concentraccedilatildeo de decavanadato total As
concentraccedilotildees de cada espeacutecie foram
obtidas segundo a formula descrita na
secccedilatildeo 35 do Procedimento
Experimental (aacutereas dos sinais de 51
V-
RMN observadas na figura x1)V10 V1 e
V4 correspondem agraves espeacutecie monomeacuterica
dimeacuterica e tetrameacuterica respectivamente [Esp
eacuteci
e o
ligo
meacute
rica
de
van
aacutedio
] (m
M)
[Decavanadato]total (mM)
V10
V1
V4
27
Nas soluccedilotildees de decavanadato as espeacutecies monomeacuterica e decameacuterica de vanadato satildeo as uacutenicas
presentes para concentraccedilotildees de decavanadato inferiores a 8 mM Ambas as espeacutecies aumentam
linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado em [AY] e
em (Teresa) quando a concentraccedilatildeo de V1 eacute igual ou superior a 200 microM inicia-se a sua
oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica No entanto nas condiccedilotildees de realizaccedilatildeo dos
ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a espeacutecie tetrameacuterica neste
estudo em concentraccedilotildees inferiores a 8 mM tendo o mesmo sido observado por Ramoset al
(2006) [AK] Os resultados obtidos mostram claramente o equiliacutebrio entre a espeacutecie decameacuterica e
a monomeacuterica e que o aparecimento de outras espeacutecies oligomeacutericas se encontra dependente da
concentraccedilatildeo desta uacuteltima Apesar de natildeo ter sido realizado o ensaio com 5 mM de decavanadato
total no presente relatoacuterio estaacute reportado em Ramos et al (2006) [AK] que as concentraccedilotildees da
espeacutecie decameacuterica e monomeacuterica satildeo respectivamente de 500 microM e 200 microM Assim pode inferir-
se que para concentraccedilotildees inferiores a 5 mM e intervalos de tempo de cerca de 20 minutos (que
inclui o tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas
as espeacutecies decameacuterica e monomeacuterica de vanadato
Figura 24 ndash Espectros de RMN de 51
V de concentraccedilotildees
crescentes das espeacutecies oligomeacutericas de vanadato obtidos
apoacutes diluiccedilatildeo da soluccedilatildeo stock de metavanadato em 10 mM
HEPES pH 70 25 mM KCl para concentraccedilotildees finais de A)
1 mM B) 2 mM C) 4 mM D) 8 mM E) 10 mM de
metavanadato total V1 refere-se agraves espeacutecies monomeacuterica de
vanadatoV1 e V2 correspondem respectivamente agrave espeacutecie
monomeacuterica e dimeacuterica
Figura 25 ndash Distribuiccedilatildeo graacutefica das espeacutecies
oligomeacutericas de vanadato vs a concentraccedilatildeo de
metavanadato total As concentraccedilotildees de cada
espeacutecie foram obtidas segundo a formula descrita
na secccedilatildeo 35 do Procedimento Experimental
(aacutereas dos sinais de 51
V-RMN observadas na
figura x1)V10 V1 e V4 correspondem agraves espeacutecie
monomeacuterica dimeacuterica e tetrameacuterica
respectivamente
Esp
eacuteci
e o
ligo
meacute
rica
de
va
nad
ato
(m
M)
[Metavanadato]total (mM)
V1
V2
V4
V5
28
Tabela 2IVndash Desvios quiacutemicos meacutedios (part)das ressonacircncias das espeacutecies tetrameacuterica (V4) monomeacuterica (V1) e
dimeacuterica (V2) presentes no espectro da soluccedilatildeo de metavanadato (total) 10 mM
Paracircmetros V1 V2 V4 V5
part (ppm) -5615 -5748 -5789 -5869
Nas soluccedilotildees de metavanadato as espeacutecies monomeacuterica dimeacutericae tetrameacuterica de vanadato estatildeo
presentes para concentraccedilotildees de metavanadato inferiores a 4 mM Todas estas as espeacutecies
aumentam linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado
em (Aureliano e Madeira 1998) e em (Tiago et al 2004) quando a concentraccedilatildeo de V1 eacute igual ou
superior a 200 microM inicia-se a sua oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica Nas
condiccedilotildees de realizaccedilatildeo dos ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a
espeacutecie pentameacuterica neste estudo em concentraccedilotildees inferiores a 4 mM tendo o mesmo sido
observado por Ramos 2006 Os resultados obtidos mostram claramente o equiliacutebrio entre a
espeacutecie V4 e as espeacuteciesV1 e V2 no qual o aparecimento de outras espeacutecies oligomeacutericas se
encontra dependente da concentraccedilatildeo da espeacutecie V1 Por uacuteltimo pode inferir-se que para
concentraccedilotildees inferiores a 2 mM e intervalos de tempo de cerca de 20 minutos (que inclui o
tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas as
espeacutecies V1 V2 e V4 com concentraccedilotildees respectivas de 0796 0192 e 0196 mM
232Estabilidade das soluccedilotildees de decavanadato anaacutelise por espectroscopia de absorccedilatildeo
UVVis
Caracterizou-se a estabilidade da espeacutecie V10 numa soluccedilatildeo com 1 mM decavanadato total em
tampatildeo D atraveacutes da leitura da Abs400 Obteve-se uma cineacutetica de dissociaccedilatildeo de 1ordf ordem tal
como se pode verificar pela linearidade do graacutefico de ln(Abs400)vs o tempo de aquisiccedilatildeo espectral
(segundos) (Fig26 )
Figura 26ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do tempo
(em segundos) para uma soluccedilatildeo de decavanadato total de 1
mM em tampatildeo D a 25ordmC (n=2)
Sendo uma reacccedilatildeo de primeira ordem pode ser descrita pela equaccedilatildeo cujo
declive eacute ndashK (constante de dissociaccedilatildeo) e a intersecccedilatildeo no eixo das ordenadas corresponde
Ln
(Ab
s 400 n
m)
tempo (s)
29
aln(A0) Numa reacccedilatildeo de 1ordf ordem o tempo de meia-vida eacute independente da concentraccedilatildeo inicial
do reagente sendo descrito pela equaccedilatildeo [2] Deste modo a partir da recta
Ln(Abs400 nm) = -436E-5
K - 105 obteve-se um tempo de meia-vida de 4490 plusmn 0823 horas para
as condiccedilotildees supramencionadas
Apoacutes a adiccedilatildeo de S1 8 microM ao meio reaccional verificou-se que a reacccedilatildeo de desoligomerizaccedilatildeo
da espeacutecie decameacuterica de vanadato seguia igualmente uma cineacutetica de 1ordf ordem tal como se pode
verificar pela linearidade do graacutefico de ln(Abs400) versus o tempo de aquisiccedilatildeo espectral
(segundos) (Fig27)
Figura 27ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do
tempo (em segundos) para uma soluccedilatildeo de decavanadato
total de 1 mM em tampatildeo D a 25ordmC na presenccedila de 8
microM (n= 2)
A partir da equaccedilatildeo da recta Ln(Abs400 nm) = -7520E-5
K - 0613determinou-se o valor de K para
a espeacutecie decameacuterica de onde se obteve um tempo de meia-vida de 2430 plusmn 0191 horas para as
condiccedilotildees supramencionadas
Tanto este valor como o valor registado na ausecircncia de S1 satildeo substancialmente superiores agrave
duraccedilatildeo dos ensaios de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina descritos no presente
relatoacuterio
24 Interacccedilatildeo de S1 com vanaacutedio(V)
241Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
De modo a analisar qual o efeito da adiccedilatildeo de S1 de miosina (tampatildeo D) produzia nos sinais de
51V-NMR titulou-se uma soluccedilatildeo de 5mM de decavanadato total com concentraccedilotildees crescentes
de S1 (5 10 15 e 20 microM) Os espectros de 51
V-RMN encontram-se apresentados na Fig 2
No graacutefico da figura 2 Apresenta-se a concentraccedilatildeo de S1 em funccedilatildeo do factor de alargamento
corrigido dos espectros acima referidos
Ln
(Ab
s (4
00
nm
))
tempo (s)
30
Figura 28 ndash Graacutefico de
representaccedilatildeo da
concentraccedilatildeo de S1 em
funccedilatildeo do factor de
alargamento A
concentraccedilatildeo de
decavanadato total foi de 5
mM e as concentraccedilotildees
utilizadas de S1 foram de 0
5 10 e 15 microM
Verifica-se que o factor de alargamento aumenta linearmente em funccedilatildeo da concentraccedilatildeo de S1 no
ensaio o que indica a existecircncia de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina Observou-
se ainda que a adiccedilatildeo de S1 ao meio reaccional natildeo provocou qualquer alteraccedilatildeo dos desvios
quiacutemicos dos sinais de 51
V-RMN
Realizaram-se ensaios na presenccedila de ATP e de S1 com intuito de verificar o efeito do ATP na
interacccedilatildeo do S1 de miosina com o decavanadato
Figura 29 - Espectros de RMN de 51
V contendo 2
mM Decavanadato (A) (controlo) (B) na presenccedila
de 2 mM de ATP (C) na presenccedila de 2 mM de ATP
e 8 microM de S1 de miosina V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2Vndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies decameacuterica (V10) e monomeacuterica (V1) presentes nos
espectros da soluccedilatildeo de decavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V10A V10B V10C V1
part (ppm) -5161 -5002 -4254 -5614
Observa-se que a presenccedila de ATP (espectro B) natildeo provoca qualquer alteraccedilatildeo visiacutevel em
relaccedilatildeo ao espectro que contem apenas soluccedilatildeo de decavanadato Jaacute a presenccedila simultacircnea de ATP
e S1 provoca um alargamento da ∆12 do sinal do V10A de 1736 Hz (espectro (A)) para 2366 Hz
(espectro (C) Fig 29) Tendo em conta que a concentraccedilatildeo de S1 apresenta uma relaccedilatildeo linear
com o factor de alargamento converteu-se o valor da ∆12 e comparou-se com o valor esperado agrave
F = 00591[S1] - 00157 Rsup2 = 0998
Fact
or
de
ala
rgam
en
to
[S1] (microM)
31
concentraccedilatildeo de S1 utilizada O factor de alargamento obtido para 8 microM ndash 0376 ndash eacute inferior ao
valor esperado agrave mesma concentraccedilatildeo de S1 obtido atraveacutes da equaccedilatildeo da recta apresentada na
Fig 28 ndash 0457 ndash o que indica que o ATP poderaacute exercer um efeito de protecccedilatildeo do S1
diminuindo a sua interacccedilatildeo com a espeacutecieV10
Por uacuteltimo realizaram-se ensaios com metavanadato na presenccedila de 8 microM S1 de modo a verificar
se as espeacutecies oligomeacutericas contidas nesta soluccedilatildeo interagiam com a proteiacutena em estudo
Figura 210 - Espectros de RMN de 51
V contendo 2
mM metavanadato (A) (controlo) (B) na presenccedila
de 8 microM de S1 de miosina V1 V2 e V4
correspondem respectivamente aacutes espeacutecies
monomeacuterica dimeacuterica e tetrameacuterica
Tabela 2VIndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies monomeacuterica (V1) dimeacuterica (V2) e tetrameacuterica (V4)
presentes nos espectros da soluccedilatildeo de metavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V1 V2 V4
part (ppm) -5613 -5747 -5789
Como eacute possiacutevel observar na Fig 210 ocorre uma ligeiro alargamento dos sinal de V4 bem
como uma perda da intensidade pelo que se conclui quedas espeacutecies presentes na soluccedilatildeo de
metavanadato (V1 V2 e V4) apenas a espeacutecie tetrameacuterica interage com o S1 de miosina o que
estaacute de acordo com o reportado por Tiago et al 2002
242 Actividade Mg2+
-ATPaacutesica de S1 de miosina na presenccedila de decanadato
Com o intuito de avaliar o efeito do decavanadato na actividade Mg2+
-ATPaacutesica do S1 de
miosina monitorizou-se a reacccedilatildeo de oxidaccedilatildeo do NADH atraveacutes da leitura de Abs340 na
presenccedila de 005 mgmL de S1 de miosina e no caso da actividade estimulada 2microM de actina em
3125 mM Tris-HCl pH 71 e 375 mM MgCl2 pelo meacutetodo de enzimas acopladas (ver seccccedilatildeo xx
dos MM)Para tal realizaram-se ensaios com vaacuterias concentraccedilotildees de decavanadato total (10 50
100 250 500 e 1000) de modo a determinar qual o seu efeito na actividade MgATPaacutesica de S1
de miosina na ausecircncia e na presenccedila de um dos seus ligandos naturais Os ensaios foram
realizados agrave temperatura ambiente com incubaccedilatildeo preacutevia a 25ordm C durante 20 minutos
32
Figura 211 ndash Representaccedilatildeo
graacutefica da Actividade Mg2+
-
ATPaacutesica de S1 () (os ajustes
teoacutericos estatildeo representado a por
uma linha os resultados
experimentais estatildeo representados
por triacircngulos (Actividade
estimulada) e cruzes (Actividade
natildeo estimulada)) vs
[Decavanadato]total medidas pelo
meacutetodo das enzimas acopladas
(n=3) Os dados foram ajustados ao modelo de decaimento exponencial usando o programa GraphPad Prism para a
determinaccedilatildeo dos valores de IC50 (na ausecircncia de actina IC50= 266 plusmn 07 M decavanadato na presenccedila de actina
IC50= 28 plusmn 1 M decavanadato
A partir do graacutefico presente na Fig 29 verifica-se que o ajuste dos dados experimentais foi bem
sucedido A inibiccedilatildeo da actividade Mg2+
-ATPaacutesica do S1 de miosina eacute afectada em menor
amplitude na presenccedila de actina (60 de inibiccedilatildeo da actividade) do que na sua ausecircncia
(aproximadamente 80 de inibiccedilatildeo de actividade) Devido a esta diferenccedila pode sugerir-se que a
actina iraacute impedir parcialmente a ligaccedilatildeo do decavanadatoao S1 demiosina atraveacutes de
impedimentos esteacutereos ou interacccedilotildees electrostaacuteticas por exemplo A teacutecnica utilizada natildeo nos
permite concluir acerca de que regiatildeo(otildees) ou resiacuteduo(s) da actina participaratildeo na preservaccedilatildeo
parcial da actividade Mg2+
-ATPaacutesica
Por outro lado os valores de IC50 dos dois tipos de actividades natildeo apresentam diferenccedilas
significativas tendo-se obtido 266 plusmn 07 e 28 plusmn 1M decavanadato na ausecircncia e na presenccedila de
actina respectivamente o que corresponde a uma concentraccedilatildeo de cerca de 27 M de V10 O
meio de reacccedilatildeo utilizado nestes ensaios foi 10 mM HEPES e 25 mM KCl com 2 mM ATP Em
(Tiago et al 2004) o valor de IC50 para a actividade Mg2+-ATPaacutesica na ausecircncia de actina foi de
125 plusmn 025 microM enquanto o valor da actividade estimulada por actina foi de 136 plusmn 14 microM O
meio de reacccedilatildeo utilizado nos ensaios foi 20 mM HEPES 50 mM KCl e 2 mM ATP o que resulta
precisamente no dobro da forccedila ioacutenica a que foi realizado o ensaio do presente relatoacuterio
(exceptuando a contribuiccedilatildeo dos produtos da hidroacutelise de ATP uma vez que este meacutetodo
pressupotildee a ocorrecircncia de um ciclo de reacccedilotildees de hidroacutelise e siacutentese deste composto) Ora tendo
sido anteriormente referido que a afinidade do S1 de miosina para com a espeacutecie V10 diminui com
Act
ivid
ade
Mg
2+-A
TP
aacutesic
a d
e S
1 d
e
Mio
sin
a
[Decavanadato] (microM) Actividade Estimulada ()
Actividade Natildeo Estimulada ()
Ajuste Actividade Estimulada
Ajuste Actividade Natildeo Estimulada
33
o aumento da forccedila ioacutenica (Tiago et al 2004 tal poderaacute explicar a diferenccedila obtida entre estes
dois conjuntos de valores
243Quantificaccedilatildeo de cisteiacutenas reduzidas pelo meacutetodo de Ellman
A interacccedilatildeo do vanaacutedio com alguns tipos de aminoaacutecidos sensiacuteveis agrave oxidaccedilatildeo pode fornecer-nos
informaccedilatildeo bastante uacutetil em relaccedilatildeo agraves alteraccedilotildees estruturais que conduzem a uma diminuiccedilatildeo da
actividade fisioloacutegica do S1 de miosina Os estudos realizados por Chasteen 1983 concluiacuteram
que a reduccedilatildeo de vanadato (V) a vanadilo (IV) ocorre na presenccedila de agentes redutores comuns no
ambiente celular de que se destaca o aminoaacutecido cisteiacutena Esta eacute tambeacutem um alvo de
complexaccedilatildeo por parte do iatildeo VO2+
Jaacute Ramos et al (2009) demonstrou que a incubaccedilatildeo de
actina com a soluccedilatildeo de decavanadato induz a oxidaccedilatildeo de resiacuteduos de cisteiacutena e agrave formaccedilatildeo do
oxidovanaacutedio (IV) Este grupo de investigadores descobriu ainda que apenas a espeacutecie V10 induz a
oxidaccedilatildeo das cisteiacutenas ldquoraacutepidasrdquo da actina atraveacutes do processo proposto que eacute parcialmente
responsaacutevel pela inibiccedilatildeo jaacute descrita da actividade de ATPase do S1 de miosina estimulada por
actina quando exposta agrave soluccedilatildeo de decavanadato Esta inibiccedilatildeo apresenta-se afectada com a
presenccedila de ATP uma vez que os resiacuteduos de cisteiacutena permaneceram intactos nos ensaios
realizados com este ligando Jaacute Crans et al (2004) concluiu que o siacutetio activo de muitas fosfatases
(que conteacutem grupos tiol) pode formar um equiliacutebrio redox com o iatildeo vanadato (V) de forma natildeo
reversiacutevel
Os resultados obtidos por Sakurai e Shiromura (1980) sugeriram que o vanaacutedio (V) sofre reduccedilatildeo
a vanaacutedio (IV) sem dependecircncia do valor de pH do ensaio Registaram ainda uma estequiometria
de 31 na reacccedilatildeo do vanadato (V) com os resiacuteduos de Cys enquanto a estequiometria dos ensaios
com o vanadilo (IV) foi de 21
Berlett e Stadtman (1997) observaram que os tipos de aminoaacutecidos mais facilmente oxidaacuteveis
englobam entre outros os resiacuteduos de Cys Trp e MetAssim analisou-se o estado redox dos
grupos tiol dos resiacuteduos de Cys do S1 na presenccedila do decavanadato atraveacutes da monitorizaccedilatildeo da
Abs412 nm na presenccedila de 1 mgmL de S1 de miosina em tampatildeo 10 mM HEPES 25 mM KCl
variando a concentraccedilatildeo da soluccedilatildeo de decavanadato de modo a determinar que tipo de alteraccedilotildees
estruturais ocorrem na sua presenccedila no que diz respeito agrave diminuiccedilatildeo da actividade ATPaacutesica de
S1 na presenccedila de decavanadato
Os resultados referentes agrave quantificaccedilatildeo das cisteiacutenas reduzidas apoacutes o tratamento de S1 com
diferentes concentraccedilotildees de decavanadato em tampatildeo 10 mM HEPES pH 75 e 25 mM KCl a
25ordmC durante 20 minutos encontram-se descritos na Tabela 2VII
Tabela 2VII ndash Cisteiacutenas reduzidas totais e ldquoraacutepidas de S1 (1 mgmL) apoacutes tratamento com decavanadato
34
(010501002505001000 uM) em 10 mM HEPES 25 mM KCl pH 7 a 25ordmC durante 20 minutos A detecccedilatildeo
espectrofotomeacutetrica foi feita com DTNB (e 412=8400 uM-1 cm-1) (n variaacutevel entre 1 e 8)
Amostras Cys raacutepidas Cys totais
11 M S1 (controlo) 22 plusmn 04 106 plusmn 14
11 M S1 + 10 M Decavanadato 22 plusmn 05 95 plusmn 08
11 M S1 + 50 uM Decavanadato 20 plusmn 02 98 plusmn 14
11 M S1 + 100 uM Decavanadato 17 plusmn 03 97 plusmn14
11 M S1 + 250 uM Decavanadato 26 plusmn 02 98 plusmn 08
11 M S1 + 500 uM Decavanadato 21 plusmn 04 90 plusmn 06
11 M S1 + 1000 uM Decavanadato 24 plusmn 0 86 plusmn 08
A partir dos resultados obtidos pode sugerir-se que o S1 possui 11 resiacuteduos de Cys (controlo) o
que estaacute de acordo com o anteriormente descrito Verifica-se que o tratamento da proteiacutena com
concentraccedilotildees crescentes de decavanadato provoca a oxidaccedilatildeo de dois resiacuteduos de Cys do S1 de
miosina natildeo expostos ao solvente No entanto a teacutecnica utilizada natildeo nos permite identificar
quais
Tendo em consideraccedilatildeo que o valor de IC50 obtido para a reacccedilatildeo de inibiccedilatildeo daactividade Mg2+-
ATPaacutesica de S1 na presenccedila de decavanadato eacute de cerca de 27 microM pode sugerir-se que este seraacute o
valor ao qual ocorreraacute a oxidaccedilatildeo do primeiro resiacuteduo de Cys o que poderaacute contribuir para a
diminuiccedilatildeo da actividade de S1 observada
25Interacccedilatildeo de S1 com vanaacutedio(IV)
251Anaacutelise por espectroscopia de Ressonacircncia Paramagneacutetica Electroacutenica
A acumulaccedilatildeo intracelular de oxovanaacutedio (IV) in vivo constitui uma das principais formas de
apresentaccedilatildeo de vanaacutedio no organismo Deste modo torna-se bastante interessante analisar a
interacccedilatildeo entre este iatildeo e o S1 de miosina atraveacutes da teacutecnica de espectroscopia de Ressonacircncia
Paramagneacutetica Electroacutenica o meacutetodo de eleiccedilatildeo do estudo do iatildeo vanadilo e dos complexos por si
formados O electratildeo desemparelhado encontra-se fortemente acoplado ao nuacutecleo 51
V o que
35
resulta num espectro de RPE consistindo de 8 desdobramentos na direcccedilatildeo paralela e outros tantos
na direcccedilatildeo perpendicular (Chasteen 1983)
Os estudos realizados por Ahmed et al (1987) centraram-se na interacccedilatildeo do vanadilo com
proteiacutenas como eacute o caso da insulina transferrina entre outros obtendo um espectro caracterizado
por 8 linhas amplamente espaccediladas (na direcccedilatildeo paralela) Jaacute Ramos et al 2009 concluiu que a
exposiccedilatildeo da soluccedilatildeo de decavanadato agraves formas globular e filamentosa de actina produz um
espectro tiacutepico do iatildeo oxidovanaacutedio (IV) indicando a reduccedilatildeo do vanadato
Para aferir qual a concentraccedilatildeo de S1 que permitia obter uma boa razatildeo sinalruiacutedo titulou-se uma
soluccedilatildeo de sulfato de vanadilo 15 microM ([VOSO4]stock = 5 mM) com concentraccedilotildees crescentes de
proteiacutena (0 5 10 15 e 20 microM) em 10 mM HEPES 25 mM KCl A existecircncia de uma correlaccedilatildeo
linear entre a concentraccedilatildeo de proteiacutena e a intensidade dos sinais de RPE correspondente agrave banda
-12 perpendicular encontra-se representada na Fig 210
Figura 2 12ndash Representaccedilatildeo graacutefica da variaccedilatildeo da intensidade do sinal de RPE vs a concentraccedilatildeo de S1 utilizada
Apoacutes verificar que com uma concentraccedilatildeo de S1 igual a 20 microM se obtinham sinais com uma razatildeo
sinalruiacutedo adequada ao prosseguimento dos estudos sobre a interacccedilatildeo V(IV)-S1 de miosina
titulou-se uma soluccedilatildeo de 20 microM S1 com 0 25 5 10 15 25 50 75 100 150 200 500 e 1000
microM VOSO4
Traccedilou-se o graacutefico da intensidade da banda -12 perpendicular do sinal de EPR em funccedilatildeo da
concentraccedilatildeo de VOSO4 tendo-se determinado uma constante de dissociaccedilatildeo (KD) do complexo
S1-vanadilo de 8748 plusmn 2621 microM V(IV)
y = 016786x + 122435
Rsup2 = 092992759
Inte
nsi
dad
e d
o s
inal
de
RP
E
[S1] (microM)
36
Figura 213 ndash Representaccedilatildeo graacutefica da variaccedilatildeo do sinal de RPE vs a concentraccedilatildeo de vanadilo utilizada (os valores
experimentais estatildeo representados por losangos o seu ajuste estaacute representado pela linha)
A simulaccedilatildeo do espectro de EPR foi efectuada a partir do espectro obtido para uma concentraccedilatildeo
de 1 mM VOSO4 apoacutes adiccedilatildeo de 20 microM S1 de miosina tendo-se obtido os valores da constante de
acoplamento hiperfina (A=5016) e do factor g (g0=195)
Figura 214 ndash Representaccedilatildeo graacutefica do espectro de EPR obtido com 1000 microM de vanadilo e 20 microM de S1 (a cinzento
escuro) A simulaccedilatildeo do espectro encontra-se a cinzento claro
Por comparaccedilatildeo destes com os valores presentes na tabela de correspondecircncia xxx eacute possiacutevel
predizer quais os ligandos provaacuteveis do vanadilo na presenccedila de proteiacutena Assim concluiu-se que
o vanadilo estaacute coordenado com duas moleacuteculas de aacutegua um grupo hidroxilo e um grupo R=NR
O grupo hidroxilo pode indicar os resiacuteduos de aminoaacutecidos Tyr Treonina e Serina enquanto o
grupo R=NR poderaacute indicar a presenccedila de Arginina e Histidina como ligandos do vanadilo
Inte
nsi
dad
e d
os
sinai
s d
e R
PE
[Vanadilo] (microM)
Ajuste dos dados exp Dados experimentais
Campo magneacutetico (G)
Espectro Experimental Ajuste
37
3 Procedimento Experimental
31 Reagentes
Todos os reagentes utilizados satildeo de elevado grau de pureza (analiacutetico) Apresentam-se as suas
foacutermulas quiacutemicas ou abreviaturas quando necessaacuterio acrescentou-se a sua descriccedilatildeo Entre
parecircntesis colocou-se as empresas que os comercializam
Aacutecido aceacutetico glacial 100 (Riedel) Acrilamida 99 (Merck) Azul de bromofenol (Riedel-de-
Haumlen) -Mercaptoetanol (Sigma) Bissulfito de soacutedio (Sigma)Bradford (reagente de)
(BioRad)BSA (albumina de soro bovino Sigma-Aldrich) CaCl22H2O (cloreto de caacutelcio
dihidratado Fluka) CH3CH2OH 96 (etanol Panreac) Coomassie Blue R250 (Merck) D2O
(oacutexido de deuteacuterio SDS) DTNB (55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico) Aldrich) EDTA (aacutecido
etilenodiaminatetraaceacutetico Merck) Fosfoenolpiruvato 98 (Fluka) Glicerol (Sigma) Glicina
985 (Panreac) HCl 37 (Riedel-de Haumlen) H2SO4 96 (Pronalab) HEPES 99 (aacutecido 4-(2-
hidroxiletil)-1-piperazineetanosulfoacutenico Sigma) KH2PO4 99 (hidrogenofosfato de potaacutessio)
MgATP (Sigma) Na
ATP (Sigma) Na2HPO4 99 (hidrogenofosfato de soacutedio Riedel-de Haumlen)
NaH2PO4 (fosfato de soacutedio ) NaOH 986 (hidroacutexido de soacutedio Pronalab) KCl 99 (Cloreto de
potaacutessio Panreac) L-Cys 995 (enantioacutemero levoacutegiro do resiacuteduo de Cisteiacutena Fluka)Metanol
99 (Merck) Molibdato de amoacutenio (Sigma)NNrsquo-metil-bis-acrilamida 98 (Merck) NH4VO3
99 (metavanadato de amoacutenia Sigma) PSA 10 (persulfato de amoacutenio Merck) SDS
(dodecilo sulfato de soacutedio Sigma) Sulfito de soacutedio (Sigma) VOSO4 (sulfato de vanadilo
Aldrich) TEMED 99 (Tetrametiletilenodiamina Merck) Tris (2-amino-2-hidroximetil-13-
propanodiol Sigma)
32 Meacutetodos
321 Isolamento e purificaccedilatildeo das proteiacutenas de muacutesculo esqueleacutetico de coelho
3211 Purificaccedilatildeo da miosina preparaccedilatildeo de Subfragmento 1 por digestatildeo quimotriacutepsica
da miosina
O subfragmento-1 de miosina-II (S1) foi gentilmente cedido pelo Prof Doutor Manuel Aureliano
do Departamento de Quiacutemica Bioquiacutemica e Farmaacutecia da Faculdade de Ciecircncias e Tecnologia da
Universidade do Algarve
No entanto descreve-se abaixo de forma resumida o procedimento efectuado
38
32111 Purificaccedilatildeo da miosina
O protocolo de isolamento foi adaptado do meacutetodo previamente descrito por Margossian e Lowey
(1982) apresentando-se de forma esquemaacutetica na Fig 31
Sucintamente os tecidos musculares dorsais e das pernas foram extraiacutedos apoacutes o sacrifiacutecio do
animal (Fig31) De seguida foram adicionados 3 volumes da soluccedilatildeo de extracccedilatildeo A2
Centrifugou-se a soluccedilatildeo e filtrou-se o sobrenadante O sedimento foi entatildeo diluiacutedo em 14
volumes de aacutegua destilada de modo a que a forccedila ioacutenica atingisse 004 M Apoacutes adiccedilatildeo de szlig-
mercaptoetanol (szlig-me) deixou-se a precipitar a 4ordmC removendo o sobrenadante Centrifugou-se
novamente e ressuspendeu-se o sedimento na soluccedilatildeo de extracccedilatildeo A Adicionou-se a soluccedilatildeo
tampatildeo B3 para que a forccedila ioacutenica atingisse 05 M Homogeneizou-se e deixou-se repousar
durante 2h Homogeneizou-se novamente e centrifugou-se de modo a ocorrer precipitaccedilatildeo dos
complexos de actomiosina e proteiacutena desnaturada Apoacutes filtraccedilatildeo do sobrenadante em latilde de vidro
quantificou-se a soluccedilatildeo de miosina resultante pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental)
2 Soluccedilatildeo de extracccedilatildeo A 0012M K2CO3 004M KHCO3 06M KCl pH 65
3 Soluccedilatildeo-tampatildeo B Tris 100 mM pH 7 KCl 2M
39
Figura 31 ndash Representaccedilatildeo esquemaacutetica do protocolo de isolamento de miosina (Margossian e Lowey 1982) Todo o
procedimento foi realizado a 4ordmC (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
32112 Preparaccedilatildeo de Subfragmento-1 de miosina por digestatildeo com quimiotripsina
O S1 de miosina foi isolado apoacutes diaacutelise da miosina durante a noite contra a soluccedilatildeo C4 A
presenccedila do EDTA eacute essencial uma vez que a sua substituiccedilatildeo por MgCl2resulta na produccedilatildeo de
HMM em vez de S1 A suspensatildeo de miosina foi equilibrada e digerida com -quimotripsina
A digestatildeo foi interrompida pela adiccedilatildeo de PMSF Seguidamente ultracentrifugou-se o produto
da digestatildeo desprezando-se o sedimento (Fig 32) A soluccedilatildeo de sobrenadante maioritariamente
S1 foi quantificada pela leitura directa da Abs280 ou pelo meacutetodo de Bradford (ver as secccedilotildees
3311 e 3312 do Procedimento Experimental respectivamente) e armazenada em 10 sacarose
e 2 mM szlig-Me a -80ordmC
4 Soluccedilatildeo C 25mM Tris pH 70 012M KCl 1mM EDTA
40
Antes da realizaccedilatildeo dos ensaios descritos no presente relatoacuterio o S1 de miosina foi dialisado
contra a soluccedilatildeo D5 de modo a remover qualquer composto que pudesse complexar como
vanaacutedio Seguidamente quantificou-se a proteiacutena pelo meacutetodo de Bradford ou pelo meacutetodo
directo (ver secccedilotildees 3311 e 3312do Procedimento Experimtental)
Figura 3 2ndash Representaccedilatildeo esquemaacutetica do protocolo de preparaccedilatildeo do S1 de miosina por digestatildeo quimiotripsina de
acordo com o previamente descrito por Margossian e Lowey (1982) Todo o procedimento foi realizado a 4ordm C
excepto quando indicaccedilatildeo do contraacuterio (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
3212 Isolamento e purificaccedilatildeo de actina
Isolou-se e purificou-se a actina segundo o meacutetodo descrito por Pardee e Spudish que
compreende dois passos ndash a obtenccedilatildeo de um resiacuteduo seco denominado ldquopoacute de acetonardquo e a
extracccedilatildeo subsequente da actina a partir do ldquopoacute de acetonardquo na forma de G-Actina com a
posterior polimerizaccedilatildeo a F-Actina
32121 Obtenccedilatildeo do ldquopoacute de acetonardquo
Sacrificou-se o coelho com uma pancada atraacutes do pescoccedilo e sangrou-se o animal apoacutes o corte da
arteacuteria jugular Esfolou-se e colocou-se numa tina com gelo Retiraram-se as partes de tecido
conjuntivo que envolvem o tecido muscular com auxiacutelio de uma pinccedila e bisturi Seguidamente
retirou-se os muacutesculos dorsais e os muacutesculos brancos das pernas Passou-se os muacutesculos por aacutegua
destilada de modo a remover as impurezas e pesou-se num copo Trituraram-se os muacutesculos
numa maacutequina de triturar carne e extraiu-se o resultante com 3 volumes da soluccedilatildeo E6 durante 10
minutos com agitaccedilatildeo suave Centrifugou-se a suspensatildeo a 4000 g durante 10
5 Soluccedilatildeo D HEPES 10 mM pH 70 KCl 25 mM
6 Soluccedilatildeo E 01 M KCl 015 M K2HPO4 pH 65
41
minutosdesprezando-se o sobrenadante Ressuspendeu-se o sedimento em 6 volumes de soluccedilatildeo
F7durante 10 minutos com agitaccedilatildeo suave Verificou-se se o pH da soluccedilatildeo estava compreendido
entre 82 e 85 acertando-se estecom soluccedilatildeo G8 se necessaacuterio Centrifugou-se durante 10
minutos a 4000 g Desprezou-se o sobrenadante e ressuspendeu-se o sedimento em 3 volumes de
soluccedilatildeo H9 durante 10 minutos Centrifugou-se novamente a 4000 gdurante 10 minutos
desprezando-se o sobrenadante Ressuspendeu-se o sedimento duas vezes consecutivas em 6
volumes de aacutegua destilada durante 5 minutos Centrifugou-se a 4000 g durante 10 minutos apoacutes
cada adiccedilatildeo de H2O
Seguidamente ressuspendeu-se o sedimento 5 vezes consecutivas utilizando 3 volumes de
acetona de cada vez agitando suavemente durante 10 minutos agrave temperatura ambiente Colocou-
se o resiacuteduo filtrado numa tina de vidro sobre papel de filtro deixando-se evaporar o solvente
durante a noite num nicho Triturou-se o resiacuteduo seco numa maacutequina de triturar carne e deixou-se
novamente evaporar o solvente O ldquopoacute de acetonardquo obtido foi armazenado a -20 ordmC
32122 Purificaccedilatildeo da actina a partir de ldquopoacute de acetonardquo
Apoacutes a obtenccedilatildeo do poacute de acetona como acima indicado efectuou-se o protocolo seguidamente
descrito de modo a obter monoacutemeros de actina (G-Actina) e actina filamentosa (F-Actina)
Pesou-se 35 g de poacute de acetona e adicionaram-se 50 mL de soluccedilatildeo-tampatildeo I10
sendo a
extracccedilatildeo feita com agitaccedilatildeo suave durante 30 minutos Separou-se o extracto do poacute de acetona
hidratado filtrando a suspensatildeo com 4 camadas de gaze esterilizada tendo o cuidado de natildeo
espremer o resiacuteduo Voltou a extrair-se com 50 mL de soluccedilatildeo I durante 30 minutos mexendo
lentamente Adicionou-se este ao extracto obtido anteriormente Centrifugou-se o extracto total a
16 000 g durante 30 minutos Decantou-se para uma proveta sobre a qual se colocou gaze para
que o sedimento solto natildeo se juntasse ao sobrenadante e leu-se o volume De seguida adicionou-
se KCl ateacute uma concentraccedilatildeo final de 50 mM MgCl2 ateacute 2mM e ATP ateacute 1mM lentamente e com
agitaccedilatildeo contiacutenua de modo a promover a polimerizaccedilatildeo dos monoacutemeros de actina
Deixou-se polimerizar durante 2h a 25ordmC observando-se a presenccedila de pequenas bolhas de ar
indicativas de um aumento de viscosidade da soluccedilatildeo Arrefeceu-se em gelo durante 15 minutos e
adicionou-se KCl ateacute uma concentraccedilatildeo final de 06M Mexeu-se lentamente durante 30 minutos
apoacutes os quais se ultracentrifugou a 110 000g durante 1hora Desprezou-se o sobrenadante
ressuspendeu-se o sedimento com soluccedilatildeo tampatildeo E e transferiu-se para um potter
Dialisou-se durante a noite contra 1L de soluccedilatildeo-tampatildeo I a uma temperatura de 4ordmC com rotaccedilatildeo
7 Soluccedilatildeo F 005 M NaHCO3 pH 82-85
8 Soluccedilatildeo G 1M Na2CO3
9 Soluccedilatildeo H 1 mM EDTA pH 70
10 Soluccedilatildeo I 2 mM Tris-HCl pH 8 02 mM CaCl2 02 mM Na2ATP 05 mM szlig-Me 0005 NaN3
42
contiacutenua e mudanccedila de tampatildeo 2 a 3 vezes De seguida a actina foi centrifugada a 110
000gdurante 1hora Decantou-se a soluccedilatildeo de G-Actina (sobrenadante) e desprezou-se o
sedimento Determinou-se a concentraccedilatildeo de G-Actina pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental) A soluccedilatildeo de G-actina foi armazenada a uma temperatura de 4ordmC
apoacutes adiccedilatildeo de 002NaN3 mantendo-se estaacutevel durante 3 a 5 dias
331 Quantificaccedilatildeo das proteiacutenas isoladas
3311 Quantificaccedilatildeo pelo meacutetodo de Bradford
O meacutetodo de Bradford (Bradford 1976) baseia-se na utilizaccedilatildeo do reagente de Bradford
composto por entre outros aacutecido fosfoacuterico e o corante ldquoCoomassierdquo Este corante possui carga
negativa e a sua coloraccedilatildeo varia entre a vermelha (Absmax=465 nm) predominante em soluccedilatildeo e a
azul (Absmax=595 nm) presente aquando da ligaccedilatildeo aos grupos amina das proteiacutenas Em soluccedilotildees
aacutecidas diluiacutedas a coloraccedilatildeo varia proporcionalmente com a quantidade do corante ligado agrave
proteiacutena que por sua vez eacute directamente proporcional agrave concentraccedilatildeo da proteiacutena permitindo a
sua quantificaccedilatildeo ateacute a um limite de detecccedilatildeo de 20 gmL (Bradford 1976)
Deste modo prepararam-se amostras de BSA ([BSA]stock= 1mgmL) utilizando um intervalo de
concentraccedilotildees compreendido entre 0 e 15 gmL de acordo com a tabela 3I Apoacutes a adiccedilatildeo da
H2O MilliQ e do reagente de Bradford as soluccedilotildees foram sucessivamente homogeneizadas no
voacutertex
Tabela 3Indash Amostras utilizadas para a obtenccedilatildeo da recta de calibraccedilatildeo no meacutetodo de Bradford
BSA (L) BSA (g) H2O MilliQ (L) Reagente de Bradford (L)
0 0 800 200
125 125 79875 200
25 25 7975 200
5 5 795 200
10 10 790 200
15 15 785 200
Apoacutes incubaccedilatildeo a 25ordmC por 5 minutos leram-se os valores de Abs595 num
espectrofotoacutemetro (Shimadzu UV-160A) para todas as amostras Seguidamente traccedilou-se a recta
de calibraccedilatildeo (Abs595vs quantidade de BSA (g))
43
Para determinar a concentraccedilatildeo de proteiacutena apoacutes o processo de isolamento purificaccedilatildeo e
diaacutelise quando necessaacuterio colocou-se 5 L da amostra num tubo de Eppendorf e adicionou-se
795 L de H2O MilliQ homogeneizando a soluccedilatildeo no voacutertex Adicionou-se posteriormente 200
L de reagente de Bradford e repetiu-se a homogeneizaccedilatildeo Incubou-se a mistura a 25ordmC por 5
minutos apoacutes os quais se leu os valores de Abs595 nm no espectrofotoacutemetro Por interpolaccedilatildeo deste
valor a partir da recta de calibraccedilatildeo determinou-se a concentraccedilatildeo de proteiacutena na amostra
3312 Quantificaccedilatildeo pelo meacutetodo directo
A quantificaccedilatildeo das amostras proteicas pode tambeacutem ser efectuada atraveacutes da leitura directa da
absorvacircncia tendo por base a lei de Beer-Lambert onde Abs corresponde agrave
absorvacircncia ao comprimento de onda ao qual o ensaio foi realizado corresponde ao coeficiente
de extinccedilatildeo (substituiacutedo pela letra a se as unidades forem distintas da molaridade) b corresponde
ao percurso oacuteptico da radiaccedilatildeo na ceacutelula amostral e c corresponde agrave concentraccedilatildeo amostral
Assim no caso de S1 de miosina mediu-se a Abs280e calculou-se a concentraccedilatildeo da proteiacutena
sabendo que o coeficiente de extinccedilatildeo de S1 eacute (Ref)
No caso da actina mediu-se a Abs290ecalculou-se a concentraccedilatildeo da proteiacutena sabendo que o
coeficiente de extinccedilatildeo da actina eacute (Ref)
332Electroforese em condiccedilotildees desnaturantes (SDS-PAGE)
De modo a melhor caracterizar as proteiacutenas isoladas foi realizada uma electroforese em
condiccedilotildees desnaturantes (SDS-PAGE do inglecircs Sodium Dodecyl Sulfate Polyacrilamide Gel
Electrophoresis) O SDS (dodecilo sulfato de soacutedio) eacute um detergente anioacutenico utilizado neste
meacutetodo para desnaturar as proteiacutenas que se ligam ao SDS na proporccedilatildeo 14 g SDS g proteiacutena
(aproximadamente 1 moleacutecula de SDS por cada 2 resiacuteduos de aminoaacutecidos) (Stryer) O SDS induz
uma densidade de carga negativa significativa nas proteiacutenas a que se liga mascarando a sua carga
intriacutenseca pelo que as razotildees cargamassa satildeo tendencialmente idecircnticas Assim a SDS-PAGE
separa as proteiacutenas quando sujeitas a um campo eleacutectrico de acordo com o seu peso molecular
sendo a sua migraccedilatildeo directamente proporcional ao logaritmo deste uacuteltimo (Stryer)
O gel de resoluccedilatildeo com 10 de acrilamida foi obtido atraveacutes da adiccedilatildeo de 167 mL de soluccedilatildeo
stock de acrilamida11
075 mL de tampatildeo de resoluccedilatildeo12
251 mL de aacutegua destilada e 50 L de
SDS 10 Para promover a polimerizaccedilatildeo adicionou-se em seguida 38 L de PSA 10
11
soluccedilatildeo stock de acrilamida soluccedilatildeo de 30 (mv) de acrilamida e 08 (mv) de NNrsquo-metil-bis-acrilamida 12
tampatildeo de resoluccedilatildeo 15 M Tris-HCl pH 88
44
(persulfato de amoacutenia) 10 e 25 L de TEMED 99 (tetrametiletilenodiamina) Agitou-se
cuidadosamente de modo a natildeo formar bolhas e pipetou-se a soluccedilatildeo para a cassette de vidro
previamente montada no suporte De seguida adicionou-se uma peliacutecula de cerca de 2 mm de 2-
metil-2-butanol para verificar posteriormente a polimerizaccedilatildeo Apoacutes a polimerizaccedilatildeo que demora
cerca de 45 minutos verteu-se a agrave superfiacutecie e adicionou-se o gel de concentraccedilatildeo com 5 de
acrilamida constituiacutedo por 03 mL de soluccedilatildeo de stock acrilamida 045 mL de tampatildeo de
concentraccedilatildeo13
094 mL de aacutegua destilada 18 L de SDS 10 135 L de PSA 10 e 2L de
TEMED 99 Colocou-se o pente sobre o gel e deixou-se polimerizar durante cerca de 45
minutos
As amostras para aplicaccedilatildeo no gel de electroforese foram preparadas para uma concentraccedilatildeo de
proteiacutena de 1 mgmL num volume final de 100 L dos quais 25 L correspondem ao tampatildeo da
amostra14
As amostras foram aquecidas a 100ordmC durante 5 minutos Antes da aplicaccedilatildeo das
amostras transferiu-se a cassette para a tina de electroforese agrave qual se adicionou a soluccedilatildeo
tampatildeo de electroforese da tina superior15
e tampatildeo de electroforese da tina inferior16
Aplicaram-
se 10 L das soluccedilotildees de marcadores de peso molecular e 20 L das amostras proteicas
preparadas anteriormente A electroforese decorreu durante cerca de 3h30 min a uma voltagem
de 80 V De seguida transferiu-se o gel para uma tina corando as proteiacutenas durante 1h com
soluccedilatildeo corante17
Descorou-se o gel com soluccedilatildeo descorante18
tendo-se mudado esta soluccedilatildeo ateacute
as bandas se encontrarem bem definidas
Determinou-se a mobilidade relativa das bandas correspondentes aos marcadores de elevado peso
molecular e traccedilou-se a recta de calibraccedilatildeo ndash log(PM) vs log(Rf) onde Rf corresponde agrave
mobilidade relativa Por interpolaccedilatildeo determina-se o PM das proteiacutenas isoladas sabendo a sua
mobilidade relativa Por comparaccedilatildeo com os valores descritos na literatura pode concluir-se
acerca do grau de purificaccedilatildeo
34 Mediccedilatildeo da actividade enzimaacutetica de S1
341 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
13
tampatildeo de concentraccedilatildeo 05 M Tris-HCl pH 68 14
tampatildeo de amostra 320 mM Tris-HCl pH 68 04 M szlig-mercaptoetanol SDS 8 glicerol 15 (vv) e roxo de m-
cresol 0024 15
tampatildeo de electroforese da tina superior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS 16
tampatildeo de electroforese da tina inferior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS acetato de soacutedio
01 M 17
soluccedilatildeo corante ldquoCoomassie Blue R-250rdquo contendo metanol 45 (vv) e aacutecido aceacutetico glacial 10 (vv) 18
soluccedilatildeo descorante metanol 10 (vv) e aacutecido aceacutetico glacial 10 vv)
45
O meacutetodo de Fiske-Subarow modificado consiste na reacccedilatildeo do molibdato de amoacutenio com o
fosfato inorgacircnico em soluccedilatildeo aacutecida com formaccedilatildeo do aacutecido fosfomoliacutebdico Seguidamente daacute-
se a adiccedilatildeo de um agente redutor que provoca a reduccedilatildeo do composto a fosfomolibdato que forma
um complexo de cor azulada cuja absorvacircncia maacutexima ocorre para um comprimento de onda de
660 nm tornando possiacutevel a sua detecccedilatildeo por meacutetodos espectrofotomeacutetricos As actividades Ca2+
-
ATPaacutesica e K+EDTA-ATPaacutesica de S1 foram determinadas atraveacutes deste meacutetodo de acordo com
o descrito por Taussky e Shorr (1953) com mediccedilatildeo da quantidade de fosfato (Pi) libertado
atraveacutes da hidroacutelise de ATP
Para tal obteve-se primeiramente a recta de calibraccedilatildeo com as amostras presentes na Tabela 3II
utilizando uma soluccedilatildeo stock de 5 mM KH2PO4
Tabela 3II ndash Composiccedilatildeo das diferentes amostras utilizadas para determinar a recta de calibraccedilatildeo atraveacutes do meacutetodo
de Fiske-Subbarow modificado
KH2PO4 (L)
([KH2PO4]stock= 5mM)
PO43-
(nmole) H2O (uL)
0 0 1780
5 25 1775
10 50 1770
20 100 1760
40 200 1740
100 500 1680
A cada tubo de Eppendorf adicionaram-se seguidamente 200 L de soluccedilatildeo J19
e 20 L de agente
redutor20
homogeneizando no voacutertex apoacutes cada adiccedilatildeo Incubou-se durante 10 minutos e mediu-
se a Abs660 nm num espectrofotoacutemetro com intervalos de tempo iguais entre cada amostra Traccedilou-
se a recta de calibraccedilatildeo (Abs660 nmvs nmol PO43-
) a partir dos valores obtidos
Para determinar a actividade Ca2+
-ATPaacutesica de S1 prepararam-se tubos de Eppendorf com 400
L de meio de reacccedilatildeo21
25 L CaCl2 ([CaCl2]stock= 100 mM) 25 L ATP ([ATP]stock= 100 mM)
com [S1]ensaios=01 mgmL Perfez-se se necessaacuterio com H2O MilliQ ateacute perfazer um volume de 1
mL Para determinar a actividade K+EDTA-ATPaacutesica utilizou-se uma concentraccedilatildeo idecircntica de
19
Soluccedilatildeo J 5 N H2SO4 25 molibdato de amoacutenio 20
Agente redutor 5 mg aacutecido 1-amino-2-naftol-4-sulfoacutenico 30 mg bissulfito de soacutedio 30 mg sulfito de soacutedio em 26
mL H2O 21
meio de reacccedilatildeo 125 mM Tris-HCl pH 80 0575 M KCl
46
proteiacutena nos ensaios (01 mgmL) e o mesmo meio de reacccedilatildeo Aos tubos de Eppendorf
adicionou-se ainda 123 L de KCl ([KCl]stock = 3M) 25 L de ATP 10 L de EDTA
([EDTA]stock= 100 mM )e o volume de H2O MilliQ necessaacuterio para perfazer 1 mL Em ambas as
situaccedilotildees o uacuteltimo reagente adicionado foi o ATP uma vez que se trata do substrato da reacccedilatildeo
enzimaacutetica
A cineacutetica decorreu a 25ordmC durante 5 minutos para a actividade Ca2+
-ATPaacutesica e durante 3
minutos para a actividade K+EDTA-ATPaacutesica Apoacutes o periacuteodo de incubaccedilatildeo transferiu-se 150
L de amostra para tubos de Eppendorf contendo igual volume de TCA 10 Colocou-se no gelo
e homogeneizou-se no voacutertex Seguidamente centrifugou-se a 1500 g durante 10 minutos e
recolheu-se 200 L de sobrenadante para novos tubos de Eppendorf Adicionou-se a estes 158
mL de H2O MilliQ e 200 L de soluccedilatildeo J Homogeneizou-se no voacutertex e adicionou-se 20 L de
reagente redutor repetindo-se a homogeneizaccedilatildeo Incubou-se durante 10 minutos e mediu-se a
Abs660 nm com o mesmo intervalo de tempo de leitura para cada amostra Por intrapolaccedilatildeo a partir
da recta de calibraccedilatildeo obtida anteriormente determinou-se a quantidade de Pi libertado em cada
ensaio A partir destes resultados determinaram-se as actividades Ca2+
-ATPaacutesica e K+EDTA-
ATPaacutesica do S1 em nmol PO43-
minmg proteiacutena e em s-1
342 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
Da actividade Mg2+
-ATPaacutesica de S1 resulta a formaccedilatildeo dos produtos da reacccedilatildeo de
hidroacutelise do ATP - ADP e Pi Na presenccedila de ADP fosfoenolpiruvato (PEP) e da enzima piruvato
cinase (PK) os produtos resultantes satildeo ATP e piruvato Por acccedilatildeo da enzima lactato
desidrogenase (LDH) este uacuteltimo eacute convertido em lactato ocorrendo simultaneamente a oxidaccedilatildeo
do NADH a NAD+ monitorizada atraveacutes da Abs340 nm Quando o S1 de miosina se encontra no
meio reaccional o ATP eacute hidrolisado formando-se novamente ADP e Pi o que permite a
formaccedilatildeo de um ciclo (fig 21) Se as enzimas se encontrarem em excesso a velocidade de
conversatildeo do NADH eacute limitada pela produccedilatildeo de ADP e Pi Como o ATP eacute regenerado em cada
ciclo a sua concentraccedilatildeo inicial eacute sempre mantida (Bagshaw 1982)
Figura 33ndash Representaccedilatildeo esquemaacutetica do meacutetodo de enzimas acopladas
47
O protocolo utilizado e seguidamente descrito foi adaptado de Cuenda et al (1990 ndash Ver Refordf) A
uma cuvette de 1 mL adicionou-se S1 para uma concentraccedilatildeo final de 005 mgmL 800 L de
meio de reacccedilatildeo22
10 L de PEP ([PEP]stock = 42 mM) 10 L de PK+LDH 11 ([PK+LDH]stock =
1 mgmL) e 15 L de NADH ([NADH]stock=25 mM) Iniciou-se a monitorizaccedilatildeo da reacccedilatildeo
atraveacutes da leitura de Abs340 nm apoacutes a adiccedilatildeo de 20 L de ATP ([ATP]stock=100 mM) A
actividade Mg2+
-ATPaacutesica eacute obtida atraveacutes do declive correspondente agrave oxidaccedilatildeo do NADH Para
a mediccedilatildeo da Mg2+
-ATPase de S1 estimulada por actina adicionou-se F-actina para uma
concentraccedilatildeo final de 2 microM sendo a actividade obtida tambeacutem atraveacutes do declive correspondente
agrave oxidaccedilatildeo do NADH apoacutes a adiccedilatildeo da proteiacutena
35 Interaccedilatildeo entre vanaacutedio e subfragmento-1 de miosina
351 Preparaccedilatildeo das soluccedilotildees de vanaacutedio
As soluccedilotildees de vanadato foram preparadas em aacutegua MilliQ a partir de metavanadato de amoacutenio
(NH4VO3) numa concentraccedilatildeo stock de 50 mM com agitaccedilatildeo magneacutetica e aquecimento (~30 ordmC)
Esta soluccedilatildeo com um valor compreendido entre 6 e 7 designa-se por ldquometavanadatordquo e eacute
constituiacuteda pelas esspeacutecies V1 V2 V4 e V5 (Ref) Acidificando a soluccedilatildeo de metavanadato por
adiccedilatildeo de HCl ateacute um valor de pH proacuteximo de 4 observa-se uma mudanccedila de cor de amarelo
claro para laranja indicativa da formaccedilatildeo da espeacutecie decameacuterica de vanadato De seguida e
imediatamente antes da realizaccedilatildeo dos ensaios descritos o valor do pH da soluccedilatildeo de
decavanadato foi aumentado para cerca de 75 por adiccedilatildeo de NaOH A soluccedilatildeo (2) designada
por ldquometavanadatordquo natildeo foi posteriormente alterada ateacute agrave sua adiccedilatildeo para a realizaccedilatildeo dos ensaios
As soluccedilotildees foram armazenadas a 4ordmC e mantidas em gelo aquando da sua utilizaccedilatildeo
A soluccedilatildeo de V(IV) foi preparada em condiccedilotildees anaeroacutebias por diluiccedilatildeo de sulfato de vanadil
(VOSO4) para uma concentraccedilatildeo final de 5 mM em H2O MilliQ saturada com N2 gasoso
352Anaacutelise por espectroscopia de UV-Vis
A concentraccedilatildeo da espeacutecie decameacuterica em soluccedilatildeo depende do periacuteodo de tempo apoacutes a diluiccedilatildeo
no meio de reacccedilatildeo apropriado (Ref) A velocidade de desoligomerizaccedilatildeo da espeacutecie decameacuterica
pode ser analisada por espectroscopia de absorccedilatildeo na regiatildeo UVVis uma vez que esta espeacutecie
apresenta duas bandas caracteriacutesticas na regiatildeo do visiacutevel (a 358 e 400 nm) que natildeo se verifica
em quaisquer outras espeacutecies oligomeacutericas As bandas supramencionada sapresentam-se bem
22
Meio de reacccedilatildeo 3125 mM Tris-HCl pH 71 375 mM MgCl2
48
resolvidas atraveacutes da obtenccedilatildeo da derivada de 2ordf ordem do espectro apresentando a forma de 2
vales com miacutenimos centrados nos comprimentos de onda supracitados com dependecircncia linear
relativamente agrave concentraccedilatildeo de decavanadato total (Ramos et al 2006)
Assim a estabilidade da espeacutecie decameacuterica apoacutes diluiccedilatildeo da soluccedilatildeo de decavanadato no meio de
reacccedilatildeo dos ensaios (tampatildeo D) para uma concentraccedilatildeo final de 1 mM em vanadato total foi
caracterizada atraveacutes da leitura da Abs400 em funccedilatildeo do tempo tendo-se determinado qual a
ordem de reacccedilatildeo e o tempo de meia-vida da mesma Com base em estudos anteriores acerca da
estabilidade do V10 na presenccedila de diferentes proteiacutenas (Ramos et al 2006) determinou-se
igualmente o tempo de meia vida apoacutes a adiccedilatildeo de 20 microM de S1 de miosina ao meio reaccional
353 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
Os espectros de RMN do isoacutetopo 51
V foram realizados num espectroacutemetro Bruker AM-400 MHz
equipado com uma sonda multinuclear inversa de 5 mm Em tubos de RMN de 5 mm de diacircmetro
preparou-se soluccedilotildees de 500 L de deca- ou metavanadato correspondentes a um intervalo de
concentraccedilotildees de 1 a 10 mM suplementadas com 10 D2O Os espectros foram adquiridos agrave
temperatura ambiente com uma largura espectral de 23 923 Hz um tempo de aquisiccedilatildeo de 0086
s tempo de relaxaccedilatildeo de 001 s e uma sequecircncia de pulsos a 90ordm Utilizou-se a transformada de
Fourier com alargamento das bandas de 20Hz para o processamento dos espectros Os desvios
quiacutemicos das ressonacircncias de 51
V foram obtidos em relaccedilatildeo agrave espeacutecie VOCl3cuja frequecircncia de
ressonacircncia eacute considerada o padratildeo sendo todas as outras referenciadas a partir desta em ppm
As aacutereas dos sinais do RMN correspondentes agraves espeacutecies oligomeacutericas presentes foram obtidas
por integraccedilatildeo dos sinais As concentraccedilotildees relativas das espeacutecies foram calculadas com recurso agrave
equaccedilatildeo
onde A[Vx] corresponde agrave aacuterea referente agrave ressonacircncia da espeacutecie Vx com nordm de agregaccedilatildeo x
A[V]total agrave soma total das aacutereas medidas e [V] a concentraccedilatildeo total de vanaacutedio no ensaio
A teacutecnica de 51
V-RMN foi anteriormente aplicada em estudos de ligaccedilatildeo com S1 de miosina
(Ref) tendo sido demostrado que entre as espeacutecies V1 V2 V4 e V5 a espeacutecie tetrameacuterica possui a
maior afinidade para a miosina (Cremo et al) O desaparecimento ou alargamento dos sinais pode
implicar a ligaccedilatildeo de um dos oxovanadatos laacutebeis agrave miosina sem indicaccedilatildeo de qual uma vez que
49
se processa um equiliacutebrio entre eles (Crans 1990) Assim e tendo em conta as indicaccedilotildees de que
a espeacutecie V10 eacute distinta das restantes no que diz respeito agrave magnitude da interacccedilatildeo com a miosina
e inibiccedilatildeo da actividade ATPaacutesica estimulada por actina (Aureliano 2000 Tiago et al 2002)
torna-se fundamental investigar a interacccedilatildeo das diferentes espeacutecies de vanadato com o S1 de
miosina atraveacutes da teacutecnica de 51
V-RMN
Para tal obtiveram-se espectros de 51
V-RMN resultantes da titulaccedilatildeo de uma soluccedilatildeo 5 mM de
decavanadato total com concentraccedilotildees crescentes de S1 de miosina (0 5 10 15 e 20 microM) em
tampatildeo D Realizou-se ainda ensaios na presenccedila de 2 mM de decavanadato e 2 mM de ATP na
presenccedila e na ausecircncia de S1 de miosina de modo a verificar o efeito que estes ligandos naturais
da enzima teria na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o subfragmento-1
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
Os grupos sulfidrilo (-SH) das cisteiacutenas satildeo resiacuteduos de aminoaacutecidos extremamente sensiacuteveis agrave
oxidaccedilatildeo resultando na sua oxidaccedilatildeo (Dalle-Donne et al 1999) Logo a quantificaccedilatildeo destes
resiacuteduos eacute bastante importante nos estudos de interacccedilatildeo com vanaacutedio jaacute que os resiacuteduos de Cys
com ecircnfase para o SH1 e o SH2 satildeo bastante importantes na monitorizaccedilatildeo das modificaccedilotildees
funcionais da regiatildeo da cabeccedila
O procedimento seguidamente apresentado foi originalmente descrito por Gutieacuterrez-Merino
(1987)) Consiste na reacccedilatildeo entre os grupos ndashSH natildeo oxidados das proteiacutenas e o reagente de
Ellman (DTNB -55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico)) - formando-se o aniatildeo CNT (aniatildeo 3-
carboxilato-4-nitrotiofenolato) numa estequiometria de 11 Este apresenta um maacuteximo de
absorvecircncia a 412 nm (Robyt 1990) Assim apoacutes a incubaccedilatildeo da proteiacutena na presenccedila do metal
a quantidade de cisteiacutenas reduzidas por proteiacutena determina-se atraveacutes dos valores de Abs412
Deste modo colocou-se num tubo de Eppendorf o volume apropriado de S1 dialisado para que a
concentraccedilatildeo no ensaio fosse de 1 mgmL Adicionou-se o volume apropriado de decavanadato
para que a sua concentraccedilatildeo fosse de 0 10 50 100 250 500 ou 1000 M Perfez-se com soluccedilatildeo
D ateacute um volume final de 1 mL e incubou-se a 25ordmC por 20 minutos Apoacutes este periacuteodo colocou-
se 780 L da mistura reaccional numa cuvette de quartzo e adicionou-se 220 L de soluccedilatildeo K23
23
Tampatildeo K Tampatildeo fosfatohidrogenofosfato de soacutedio 025 M pH 72
50
Adicionou-se 15 L de DTNB (em soluccedilatildeo K [DTNB]stock=100 mM) ao meio reaccional e
registou-se o valor de absorvecircncia medido equivalente agraves cisteiacutenas ldquoraacutepidasrdquo (SH1 e SH2
expostos ao solvente) Adicionou-se 001 g de SDS que provoca a desnaturaccedilatildeo da proteiacutena e
consequentemente a exposiccedilatildeo de todos os resiacuteduos de cisteiacutenas que se encontravam ateacute aiacute
inacessiacuteveis no interior da proteiacutena (cisteiacutenas totais) Estes resiacuteduos podem assim reagir com o
DTNB
Estimou-se o nuacutemero de resiacuteduos de cisteiacutenas em todas as amostras com recurso ao coeficiente de
extinccedilatildeo molar () do DTNB (ver ponto 3 dos Anexos) que foi determinado atraveacutes da titulaccedilatildeo
de L-Cisteiacutena com intervalo de concentraccedilotildees compreendidos entre 0 e 50 microM na presenccedila de
25microM de DTNB em tampatildeo D Os valores de absorvecircncia foram obtidos para um comprimento de
onda de 412 nm
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
3621 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
A teacutecnica mais eficiente de anaacutelise da interacccedilatildeo entre o iatildeo vanadilo e proteiacutenas como a
calmodulina e a transferrina entre outras permite a obtenccedilatildeo raacutepida de informaccedilatildeo no que toca agrave
conformaccedilatildeo e amplitude da referida interacccedilatildeo
Os espectros de Ressonacircncia Paramagneacutetica Electroacutenica (RPE) da espeacutecie paramagneacutetica V(IV)
foram realizados a 70K num espectroacutemetro Bruker EMX-300 X-band Todas as soluccedilotildees
utilizadas e amostras foram preparadas em anaerobiose numa cacircmara de luvas de modo a evitar a
reduccedilatildeo de V(IV) a V(V) Em tubos capilares com 02 mL de amostra manteve-se a concentraccedilatildeo
de S1 a 20 M e variou-se a concentraccedilatildeo de vanadilo (0 25 5 10 15 25 50 75 100 150
200 500 e 1000 M [VOSO4]stock=5 mM) em tampatildeo D Apoacutes obtenccedilatildeo dos espectros estes
foram tratados atraveacutes do software SimFonia e WinEPR tendo-se obtido um graacutefico com a
variaccedilatildeo da intensidade do sinal de RPE em funccedilatildeo da concentraccedilatildeo de sulfato de vanadilo
presente Realizaram-se ainda simulaccedilotildees de espectros obtidos com 20 microM de S1 e 1000 microM de
V(IV) com vista agrave obtenccedilatildeo dos ligandos provaacuteveis do vanadilo atraveacutes da anaacutelise da constante
de acoplamento hiperfina (A) e do factor g (g)
4 Conclusotildees gerais
Apesar de jaacute ter sido demonstrado que o vanaacutedio inibe a Na+K
+-ATPase da miosina a funccedilatildeo
deste elemento no tecido muacutesculo esqueleacutetico continua largamente incompreendida A enorme
complexidade quiacutemica deste metal especialmente no que concerne agrave oligomerizaccedilatildeo e reacccedilotildees
51
redox constitui um obstaacuteculo agrave compreensatildeo do comportamento do vanaacutedio nos organismos
vivos Assim a caracterizaccedilatildeo da quiacutemica deste elemento eacute de grande utilidade antes de iniciar o
estudo da sua interacccedilatildeo com o presente sistema
A anaacutelise por espectroscopias de 51
V-RMN e de UV-Vis de soluccedilotildees de vanadato diluiacutedas nos
meios de reacccedilatildeo utilizados no presente relatoacuterio (10 mM HEPES pH 70 25 mM KCl) mostrou
claramente que a espeacutecie decameacuterica (V10) possui um tempo de vida suficientemente longo para a
determinaccedilatildeo do seu efeito isolado ao contraacuterio do que se verifica com as espeacutecies mono- di-
tetra- e pentameacutericas (V1 V2 V4 e V5) Por outro lado a anaacutelise verificou ainda que apoacutes diluiccedilatildeo
das soluccedilotildees de decavanadato no meio de reacccedilatildeo coexistem as espeacutecies V10 e V1 pelo que os
efeitos putativos desta uacuteltima deveratildeo ser subtraiacutedos aquando da monitorizaccedilatildeo dos efeitos da
espeacutecie decameacuterica
O vanaacutedio (V) (vanadato) inibe as enzimas que catalizam reacccedilotildees de transferecircncia de grupos
fosfato como a ATPase da miosina [Goodno 1979] devido agrave sua analogia estrutural com o iatildeo
fosfato Na sua utilizaccedilatildeo como inibidor enzimaacutetico natildeo eacute tida em conta a contribuiccedilatildeo individual
das espeacutecies oligomeacutericas considerando-se normalmente a espeacutecie V1 como inibidora efectiva
[Goodno e Taylor 1982] No presente relatoacuterio verificou-se por espectroscopia de 51
V-RMN que
outros oligoacutemeros de vanadato interagem com o S1 de miosina tendo as espeacutecies V10 e V4 os
maiores niacuteveis de afinidade A sua interacccedilatildeo envolve forccedilas electrostaacuteticas e eacute afectada
negativamente por ligandos naturais de enzima como eacute o caso do ATP
A exposiccedilatildeo do S1 de miosina (005 mgmL) a decavanadato conduz agrave inibiccedilatildeo do capacidade da
proteiacutena de estimular a actividade Mg2+
-ATPaacutesica com um valor de IC50 de 266 plusmn 07 M
decavanadato sendo o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM
decavanadato (80) Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM decavanadato sendo
o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM decavanadato (60)
exercendo um efeito de impedimento agrave ligaccedilatildeo entre o vanaacutedio e o S1
A uma concentraccedilatildeo de 27 microM decavanadato verifica-se a oxidaccedilatildeo de um dos resiacuteduos de Cys
de S1 de miosina seguida pela oxidaccedilatildeo gradual de outro resiacuteduo de Cys ateacute agrave concentraccedilatildeo de
1000 microM Ambos os resiacuteduos oxidados natildeo se encontravam expostos ao solvente utilizado A
oxidaccedilatildeo dos resiacuteduos permite sugerir que esta modificaccedilatildeo oxidativa sofrida pelo S1 contribui
para a diminuiccedilatildeo da sua actividade de ATPase
52
Por espectroscopia de RPE revelaram-se os possiacuteveis ligandos equatoriais do vanaacutedio aquando da
sua interacccedilatildeo com o S1 de miosina como sendo duas moleacuteculas de aacutegua um ligando contendo o
grupo R-OH e outro contendo o grupo R=NR
Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados de
oxidaccedilatildeo (IV) e (V) no S1 de miosina para concentraccedilotildees facilmente alcanccedilaacuteveis no ambiente
terrestre
53
5 Estudos futuros
No sentido do melhor esclarecimento das alteraccedilotildees moleculares e estruturais que ocorrem no
subfragmento 1 quando exposta ao vanaacutedio que conduzem a uma diminuiccedilatildeo da sua funccedilatildeo
sugere-se
- Quantificaccedilatildeo das cisteiacutenas reduzidas de S1 na presenccedila de metavanadato
- Determinaccedilatildeo das actividades Ca2+
- e K+-ATPaacutesica do S1 de miosina na presenccedila de
decavanadato
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina estimulada por actina
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina na presenccedila de
metavanadato
- Realizaccedilatildeo de RMN 2D com variaccedilatildeo das concentraccedilotildees das soluccedilotildees de decavanadato e
metavanadato na presenccedila de S1
-Mutageacutenese dos resiacuteduos de Cys doS1 de miosina de modo a verificar quais os resiacuteduos que
sofrem oxidaccedilatildeo na presenccedila de decavanadato
- Determinaccedilatildeo por calorimetria (DSC Differential Scanning Calorimetry) da variaccedilatildeo da
entalpia (∆H) e da temperatura de desnaturaccedilatildeo do S1 de miosina na presenccedila de vanaacutedio uma
vez que estas propriedades caracteriacutesticas de cada proteiacutena satildeo alteradas apoacutes ocorrecircncia de
modificaccedilotildees estruturais
- Anaacutelise por SDS-PAGE de S1 de miosina apoacutes tratamento com vanaacutedio de modo a determinar
quais os polipeacuteptidos eventualmente formados apoacutes clivagem da proteiacutena
54
6 Referecircncias bibliograacuteficas
- Sellers J Myosins 2nd Edition Oxford University Press 1999
- L Stryer inBiochemistry (Ed WH Freeman 2005 pp
Refordfas Materiais e Meacutetodos
[A] S S Margossian SLowey Methods Enzymol1982 85 55-71
[B] JD Pardue JA Spudich Methods Enzymol 1982 85 164-181
[C] M Bradford Anal Biochem1976 72 248-254
[D] H H Taussky E Shorr J Biol Chem 1953 202 675-685
[E] C R Bagshaw in Outline Studies in Biology ndash Muscle Contraction Chapman and Hall Ed
1982
[F] A Cuenda et al Eur J Biochem1990 194 663-670
55
Anexos
1 Determinaccedilatildeo da recta de calibraccedilatildeo meacutetodo de Bradford
Figura1ndashRepresentaccedilatildeo graacutefica da recta de calibraccedilatildeo (Abs595 nm vs [BSA]) utilizada para o meacutetodo de Bradford
2 Determinaccedilatildeo da recta calibraccedilatildeo para a electroforese em condiccedilotildees desnaturantes
Figura 2 ndash Representaccedilatildeo graacutefica da recta de calibraccedilatildeo (log(Peso Molecular) vs Rf dos marcadores de peso
molecular) onde Rf indica determinada atraveacutes do padratildeo de peso molecular
(poccedilo (a))
3Caacutelculo do coeficiente de extinccedilatildeo molar () para a quantificaccedilatildeo de cisteiacutenas reduzidas
atraveacutes do meacutetodo de Ellman
Traccedilou-se a recta de Abs412 nm meacutedia vs [L-Cisteiacutena] de modo a determinar o valor do coeficiente
de extinccedilatildeo molar definido como o declive daquela recta Assim =8400 M-1
cm-1
como
podemos verificar na Fig 3
A595 nm = 00546[BSA] + 00209 Rsup2 = 099051
Ab
s 59
5 n
m
[BSA] (mgmL)
log PM = -1177 Rf + 2247 Rsup2 = 098957
log
PM
Mobilidade relativa (Rf)
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero

19
transportadoras) e as fosfatases (da tirosina aacutecida e alcalina) a fosfofrutocinase adenilato ciclase
e catalase entre outras [AA] O segundo grupo inclui a maioria das cinases a glucose-6-fosfato
desidrogenase a fosfoglicerato mutase e a fosfoglicerato fosfatase (Refordf)
O efeito do vanaacutedio (V) na actividade enzimaacutetica estaacute relacionado com o seu nuacutemero de
coordenaccedilatildeo podendo actuar como estado de transiccedilatildeo ou por mimetizaccedilatildeo de cofactores
substratos (especialmente o fosfato) e interacccedilotildees proteiacutena-proteiacutena (como no caso das
polimerases e transcriptases) Pode ainda estar envolvido na formaccedilatildeo de ligaccedilotildees de hidrogeacutenio
interacccedilotildees electrostaacuteticas esteacutereas e de oxidaccedilatildeo-reduccedilatildeo Uma evidecircncia desta uacuteltima eacute a sua
capacidade de alternacircncia entre os estados de oxidaccedilatildeo +5 a +3 o que torna faacutecil a sua interacccedilatildeo
redox com enzimas contendo resiacuteduos de cisteiacutenas (Cys) ou qualquer outro composto orgacircnico
com actividade redox [AN]
13 Vanaacutedio e muacutesculo esqueleacutetico
A primeira evidecircncia da presenccedila de vanaacutedio no muacutesculo esqueleacutetico proveio de uma amostra de
ATP isolado de muacutesculo de cavalo que continha uma impureza ndash vanaacutedio ndash cuja presenccedila
provocou a inibiccedilatildeo da actividade da bomba de Na+K
+-ATPase [AO] Desde entatildeo vaacuterios
estudos tecircm sido realizados com ecircnfase na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o
tecido muscular Destes destaca-se a descoberta da interacccedilatildeo forte da espeacutecie V10 com a miosina
fosforilada [AP] e agrave sua inibiccedilatildeo da actividade ATPaacutesica da miosina[AQ AR]
No quediz respeitoao iatildeo monovanadato destaca-se a sua inibiccedilatildeoda actividade Mg2+
-ATPaacutesica
da miosina [AS] que ocorre de duas formas distintas (1) na presenccedila de concentraccedilotildees reduzidas
de V1 a inibiccedilatildeo ocorre a velocidade reduzida dependente da concentraccedilatildeo da espeacutecie
monomeacuterica (2) na presenccedila de concentraccedilotildees elevadas de V1 o niacutevel de inibiccedilatildeo aproxima-se de
100 desde que [V1] seja superior agrave concentraccedilatildeo total de locais com actividade Mg2+
-ATPaacutesica
no ensaio Esta inibiccedilatildeo eacute irreversiacutevel tanto por diaacutelise ou adiccedilatildeo de agentes redutores do
vanadato [AL]Segundo Pate et al(1994)[AT] a espeacutecie monomeacuterica podeinibir a velocidade de
contracccedilatildeo a temperatura inferior a 20ordmC perdendogradual e rapidamente essa capacidade ateacute os
50ordmC temperatura agrave qual cessa a capacidade inibitoacuteria Os resultados obtidos sugeremainda uma
interacccedilatildeo altamente cooperativa entre o iatildeo vanadato e as proteiacutenas componentes do muacutesculo
esqueleacuteticoDestaca-se tambeacutem a descoberta da inibiccedilatildeo da geraccedilatildeo de forccedila contraacutectil nas fibras
musculares por parte da espeacutecie monomeacuterica[AT]Wilson et al(1995) [AU ]determinaram que a
afinidade da espeacutecie V1 para o S1 de miosina eacute significativamente mais elevada do que a do iatildeo
Pi seu anaacutelogo Descobriram ainda que que a ligaccedilatildeo do vanadato poderaacute envolver transiccedilotildees
entre estados energeacuteticos com energia livre independente dos impedimentos esteacutereos impostos
20
pela estrutura das miofibrilas Os estudos desenvolvidos por Smith et al (1990)[AV]
determinaram queo iatildeo monovanadato natildeo apresenta um efeito pronunciado na inibiccedilatildeo da
actividadeacto-S1-miosina-ATPaacutesica excepto em concentraccedilotildees superiores a 900microM a partir das
quais existe produccedilatildeo de espeacutecies oligomeacutericas para as quais o efeito inibitoacuterio seraacute mais
elevadoA espeacutecie monomeacuterica de vanadato consegue adoptar facilmente uma geometria do tipo
bipiramidal trigonal que mimetiza o intermediaacuterio formado (MgADP
Pi)aquando da reacccedilatildeo de
hidroacutelise de ATP no local activo da miosina [AW] De facto a interacccedilatildeo do vanaacutedio com a
miosina eacute inesperadamente estaacutevel na presenccedila de ADP Esta interacccedilatildeo resulta na formaccedilatildeo de
um complexo enzimaacutetico inactivo e estaacutevel com estequiometria de 11 ADPvanadato por siacutetio
activo da proteiacutena A formaccedilatildeo do complexo estaacutevel miosina-vanadato foi proposta por Goodnoet
al (1982) [AS] Nesta ocorre um preacute-equiliacutebrio raacutepido que induz a formaccedilatildeo de um complexo
ternaacuterio reversiacutevel (MADP
V) com formaccedilatildeo lenta do isoacutemero M
ADP
V A diferenccedila entre os
compostos de vanaacutedio e de fosfato centra-se no tempo de meia vida (t12) do Pi ou do V (no
primeiro t12=12 s enquanto no segundo t12=24 h) (Refordf) A diferenccedila dever-se-aacute agrave possiacutevel
complexaccedilatildeo do vanaacutedio com grupos nucleofiacutelicos no siacutetio activo o que provocaria uma
estabilizaccedilatildeo da miosina na conformaccedilatildeo MADP
V
No que respeita aos estudos realizados com a espeacutecie tetrameacuterica Cremo et al determinou que a
cadeia pesada do S1 de miosina pode ser clivada em dois locais distintos (designados V1 e V2)
pela espeacutecie V4 apoacutes irradiaccedilatildeo com luz UV na presenccedila de concentraccedilotildees de vanadato na ordem
dos milimolar e na ausecircncia de nucleoacutetidos ou metais divalentes
Jaacute no que concerne aos estudos realizados com ecircnfase na espeacutecie decameacuterica foi sugerido
porRamos et al (2006) [AK]que a interacccedilatildeo do decavanadato com a G-actina favorecida pela
polimerizaccedilatildeo desta uacuteltima estabiliza a espeacutecie V10 e induz a inibiccedilatildeo da formaccedilatildeo de F-actina A
estabilizaccedilatildeo da espeacutecie V10 por proteiacutenas tansmembranares e do citoesqueleto pode ser a razatildeo
pela qual ocorre a toxicidade do decavanadato reportada na avaliaccedilatildeo dos efeitos do V(V) nos
sistemas bioloacutegicosTiago et al (2007) [AR] concluiram que o V10 apresenta uma clara preferecircncia
de ligaccedilatildeo ao ldquoback-doorrdquo da S1-miosina-ATPase o que se verifica apenas na estrutura
conformacional desimpedida com acesso facilitado ao local de ligaccedilatildeo do Pi Sugere tambeacutem que
o V10 actua como um ldquoback-door stoprdquo bloqueando estaabertura do fragmento de 50-kDa
necessaacuteria para a hidroacutelise do ATP--fosfato o que constitui uma explicaccedilatildeo simples do
comportamento natildeo competitivo da espeacutecie decameacuterica estimulando a utilizaccedilatildeo deste oligoacutemero
como uma feramenta da elucidaccedilatildeo das mudanccedilas conformacionais ldquoback-doorrdquo da miosina no
21
processo da contracccedilatildeo muscular(ref)Destaca-se ainda a descoberta da capacidade inibitoacuteria do
V10 em relaccedilatildeo agrave fosforilaccedilatildeo oxidativa mitocondrial [AX]
Finalmente no que concerne aos estudos realizados com a presenccedila de vaacuterias espeacutecies
oligomeacutericas Aureliano e Madeira (1994) [AY]observaram que na presenccedila de Ca2+
as espeacutecies
V4 e V10se ligam agrave Ca2+
-ATPase do retiacuteculo sarcoplasmaacutetico enquanto a espeacutecie V1 se liga apenas
na presenccedila de ATP Aparentemente o aumento da concentraccedilatildeo de Ca2+
tem o efeito contraacuterio da
inibiccedilatildeo da ATPase do retiacuteculo sarcoplasmaacutetico pelo V1 natildeo tendo no entanto efeito significativo
sobre a inibiccedilatildeo por parte da espeacutecie V10 Tiago et al (2002) [AP] concluiacuteram que em condiccedilotildees
proacuteximas agraves fisioloacutegicas a espeacutecie V10 difere das espeacutecies presentes na soluccedilatildeo de metavanadato
devido agrave sua forte interacccedilatildeo com a miosina fosforilada e agrave sua inibiccedilatildeo da actividade ATPaacutesica
da miosina Para aleacutem disso o ATP diminui a afinidade da miosina para o V4 induzindo a
interacccedilatildeo com V1 e natildeo tendo qualquer efeito no que diz respeito ao V10 Foi tambeacutem proposto
[AQ] que a ligaccedilatildeo das espeacutecies oligomeacutericas de vanadato aoS1 de miosina nos locais onde o
fosfato coordenaeacute provavelmente a causa da inibiccedilatildeo da actividade ATPaacutesica da miosina
estimulada por actina atraveacutes da prevenccedilatildeo da troca entre ATPADP sendo esta favorecida para
aniotildees de vanadato com valecircncias superiores como eacute o caso do V10
Na presenccedila de actina as velocidades de dissociaccedilatildeo do Pi e do V aumentam consideravelmente
este uacuteltimo em menor quantidade Assim a ligaccedilatildeo da actina ao complexo MADP
V e
Pi eacute
bastante fraca pelo que se sugere que o ADPV teria um efeito de relaxamento semelhante ao
ATP No entanto a capacidade do vanaacutedio em inibir a actividade ATPaacutesica da miosina e da
actomiosina eacute diferenciada uma vez que na ausecircncia de actina a inibiccedilatildeo eacute total para
concentraccedilotildees na gama dos M mas na sua presenccedila a inibiccedilatildeo eacute totalmente conseguida apenas
em concentraccedilotildees na gama dos mM (Ref) No entanto eacute necessaacuterio considerar a possiacutevel
formaccedilatildeo de espeacutecies oligomeacutericas de vanadato cuja afinidade e efeito na enzima seratildeo
diferentes Assim pode concluir-se que o efeito de inibiccedilatildeo observado para a actividade ATPaacutesica
de miosina estimulada por actina se pode dever agrave presenccedila das diferentes espeacutecies oligomeacutericas o
que foi jaacute sugerido por Aureliano (2000) [AZ] ndash as espeacutecies tetrameacuterica e decameacuterica de vanadato
interactuariam com a miosina com alteraccedilatildeo da sua interacccedilatildeo com a actina No entanto natildeo eacute
conhecida a contribuiccedilatildeo absoluta das restantes espeacutecies oligomeacutericas para a inibiccedilatildeo da ATPase
da miosina estimulada por actina (Refs)
14 Objectivos do projecto
Pretende-se com este projecto o isolamento purificaccedilatildeo e caracterizaccedilatildeo da actina do muacutesculo
22
esqueleacutetico de coelho sendo esta uacuteltima tambeacutem realizada para o sufragmento-1 de miosina
Seguidamente realizar-se-aacute a quantificaccedilatildeo e caracterizaccedilatildeo das proteiacutenas por espectroscopia UV-
Vis e SDS-PAGE e a caracterizaccedilatildeo das soluccedilotildees de vanaacutedio por UV-Vis e ressonacircncia magneacutetica
nuclear (RMN) Por uacuteltimo a interacccedilatildeo do S1 de miosina com o vanaacutedio seraacute avaliada atraveacutes
das teacutecnicas de UVVis RMN e ressonacircncia paramagneacutetica electroacutenica (RPE)
2 Discussatildeo de Resultados
21 Caracterizaccedilatildeo das proteiacutenas isoladas
211Anaacutelise do estado de pureza
O estado de pureza das soluccedilotildees de actina e de S1 foi analisado por electroforese em gel de
poliacrilamida em condiccedilotildees desnaturantes (SDS-PAGE) (Fig 22) que separa as moleacuteculas de
acordo com o seu peso molecular (PM) A concentraccedilatildeo de acrilamida escolhida (10) permite a
detecccedilatildeo simultacircnea de componentes com peso molecular inferior a 20 kDa e outros com elevado
peso molecular
Figura 21 ndash Fotografia de gel de SDS-PAGE (10 acrilamida) Poccedilo (a)
10 L do marcador de elevado peso molecular () Poccedilo (b) 20 L de [G-
actina]=1 mgmL Poccedilo (c) 20 L de [S1]=1 mgmL Aplicou-se uma
voltagem de V durante h e min
A partir da recta de calibraccedilatildeo obtida log PM = -1177 Rf + 2247 (ver ponto 2 da secccedilatildeo
Anexos) pode verificar-se a presenccedila de actina na amostra do poccedilo (b) presente numa banda
bastante intensa (PMaparente=40 kDa) bem como a presenccedila minoritaacuteria de outros componentes
desconhecidos que natildeo teratildeo sido eliminados durante o processo de extracccedilatildeo e purificaccedilatildeo e natildeo
satildeo relevantes para o estudo em questatildeo Na amostra do poccedilo (c) nota-se a presenccedila de S1
(PMaparente=92 kDa) bem como a de outros componentes cuja banda eacute ligeiramente menos intensa
Estes poderatildeo ser fragmentos proteoliacuteticos de miosina e de S1 Estaacute ainda presente uma bada de
23
maior intensidade que poderaacute ser a proteiacutena-C (PM=135 kDa) uma vez que a sua remoccedilatildeo
completa da amostra necessitaria de um passo adicional de cromatografia de troca ioacutenica [BA]
No entanto natildeo podemos confirmar que se trata desta proteiacutena ou de qualquer outra uma vez que
se encontra fora do intervalo de migraccedilatildeo dos marcadores de peso molecular
212 Determinaccedilatildeo da concentraccedilatildeo proteica
2121Subfragmento-1 de miosina
A partir da recta de calibraccedilatildeo obtida pelo meacutetodo de Bradford [BB](ver secccedilatildeo 3311 do
Procedimento Experimental) A595nm = 00546[BSA] + 00209 (ver secccedilatildeo de Anexos)foi possiacutevel
determinar um intervalo de concentraccedilatildeo de S1 de miosina compreendido entre 226 e 117(plusmn
423M) (203 e 1056 (plusmn 381 mgmL)) (n=8) A concentraccedilatildeo meacutedia de S1 de miosina apoacutes
diaacutelise contra a soluccedilatildeo tampatildeo 10 mM HEPES pH 75 25 mM KCl determinada pela leitura da
Abs280 nmfoi de 20430M (1803 mgmL)(n=7)
As amostras a quantificar atraveacutes do meacutetodo de Bradford natildeo sofreram diaacutelise o que permite
explicar a concentraccedilatildeo superior de S1 de miosina em comparaccedilatildeo com o valor obtido atraveacutes do
meacutetodo directo Esta razatildeo aliada agrave sensibilidade e rapidez superiores deste uacuteltimo fez com que
se optasse pelo meacutetodo directo para a quantificaccedilatildeo da grande maioria dos ensaios
2122Actina
A concentraccedilatildeo de actina foi determinada quando esta se encontrava na forma monomeacuterica (G-
actina) por mediccedilatildeo da leitura directa da Abs290 nm tendo-se obtido uma concentraccedilatildeo meacutedia de
691481M (290202 mgmL) (n=3) em 2 mM Tris-HCl pH 80 02 mM CaCl2 02 mM Na-
ATP Ao comprimento de onda utilizado (290 nm) absorvem fortemente apenas os resiacuteduos de
Trp pelo que a mediccedilatildeo efectuada tem menor sensibilidade do que se tivesse sido efectuada a 280
nm uma vez que a actina apresenta apenas 4 resiacuteduos deste aminoaacutecido embora tenha 16
resiacuteduos de Tyr
22 Mediccedilatildeo da actividade enzimaacutetica de S1 de miosina
221 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
As actividades ATPaacutesicas de S1 de miosina dependentes de Ca2+
e K+foram analisadas pelo
caacutelculo das velocidades de consumo inicial do fosfato inorgacircnico (Pi) Definiu-se os tempos de 5
e 3 minutos para a mediccedilatildeo da Ca2+
e da K+EDTA-ATPase respectivamente Os valores obtidos
24
a partir do protocolo presente no ponto 341 da secccedilatildeo Procedimento experimental encontram-se
descritos na Tabela 2I
Tabela 2I ndash Acrividades natildeo-fisioloacutegicas de subfragmento-1 determinadas pelo meacutetodo de Fiske-Subarow
modificado (n=2)
Proteiacutena
Actividade
ATPaacutesica natildeo
fisioloacutegica
Actividade (nmol
Piminmg proteiacutena) Actividade (s-1)
S1
Ca2+
-ATPase 1400 plusmn 30 2100 plusmn 0045
K+EDTA-ATPase 2334 plusmn 50 3500 plusmn 0075
As actividades natildeo-fisioloacutegicas reflectem o estado de oxidaccedilatildeo dos resiacuteduos expostos de Cys 707
(SH1) e 697 (SH2) Se o grupo SH1 sofrer oxidaccedilatildeo a actividade ATPaacutesica dependente de Ca2+
aumenta e a actividade dependente de K+ diminui Por outro lado se o grupo SH2 sofrer
oxidaccedilatildeo regista-se a perda completa destas actividades
De acordo com Margossian e Lowey (1982) [BA] as actividades ATPaacutesica de S1 de miosina
dependente de Ca2+
e K+EDTA da miosina satildeo respectivamente de 24 e 112 s-1
Estaacute tambeacutem
descrito que a actividade ATPaacutesica dependente de K+ eacute em meacutedia cerca de 3 vezes superior agrave
dependente de Ca2+
reflectindo a quantidade das preparaccedilotildees A razatildeo obtida entre os valores das
actividades ATPaacutesicas dependentes de K+ e Ca
2+ eacute de 166 Os valores obtidos para as actividades
(Ca2+
-ATPase=21 s-1
K+EDTA-ATPase= 35 s
-1) satildeo inferiores agrave valores obtidos por
Margossian e Lowey [BA] e Tiago [BC] com forccedila ioacutenica inferior O valor obtido para a
actividade ATPaacutesica dependente de Ca2+
eacute superior ao obtido por Polosukhina et al (2000) [BD]
cujo meio de reacccedilatildeo continha MOPS e apresentava forccedila ioacutenica inferior Os dados
supramencionados comprovam a variaccedilatildeo da actividade a ATPaacutesica natildeo fisioloacutegica em relaccedilatildeo agrave
forccedila ioacutenica temperatura e pH
222 Mediccedilatildeo da actividadeMg2+
-ATPaacutesica de S1 pelo meacutetodo de enzimas acopladas
A actividade de Mg2+
-ATPase eacute a uacutenica que possui significado fisioloacutegico sendo as restantes
bastante uacuteteis apenas para indicaccedilatildeo da qualidade de preparaccedilatildeo do S1[BA] A 0575 M KCl a
actina e o S1 formam uma soluccedilatildeo viscosa e tuacuterbida Apoacutes a adiccedilatildeo de MgATP o complexo acto-
S1 dissocia-se e a turbidez da soluccedilatildeo diminui Se a forccedila ioacutenica for reduzida a dissociaccedilatildeo do
complexo actoS1 que se encontra na forma de suspensatildeo eacute induzida O MgATP eacute rapidamente
hidrolisado diminuindo a sua concentraccedilatildeo no meio reaccional Nestas condiccedilotildees haacute
reassociaccedilatildeo e subsequente contracccedilatildeo dos filamentos de actina e de S1 com formaccedilatildeo de
25
precipitado O meacutetodo descrito traduz de forma bastante elementar o fenoacutemeno da contracccedilatildeo in
vivo e elimina o problema anterior da solubilizaccedilatildeo ineficiente da miosina de modo a obter uma
mediccedilatildeo exacta da actividade [A] A utilizaccedilatildeo de forccedilas ioacutenicas reduzidas aumenta a afinidade da
actina para o S1 permitindo contornar a extrema dificuldade em obter a concentraccedilatildeo proteica dos
muacutesculos (cerca de 100 mgmL do complexo actomiosina) As actividades Mg2+
-ATPaacutesicas de S1
de miosina foram determinadas a partir do declive da variaccedilatildeo da Abs340 relativo agrave oxidaccedilatildeo de
NADH em funccedilatildeo do tempo como eacute descrito no ponto 342 do Procedimento Experimental Os
valores obtidos para a actividade Mg2+
-ATPaacutesica encontram-se apresentados na Tabela 2II
Tabela 2II ndash Actividade Mg2+
-ATPaacutesica de subfragmento-1 de miosina determinadas pelo meacutetodo das enzimas
acopladas
Actividade (nmol
ATPmin mg(S1)) Actividade (s
-1)
1097 plusmn 269 164 plusmn 04
Ramos (2005) [J] obteve o valor de 01 s-1
para a actividade Mg2+
-ATPaacutesica de S1 de miosina
com forccedila ioacutenica intermeacutedia Tiago (2005) [BC] obteve o valor de 011 s-1 em condiccedilotildees de forccedila
ioacutenica reduzida em relaccedilatildeo agrave utilizada no presente relatoacuterio
23 Preparaccedilatildeo e caracterizaccedilatildeo das soluccedilotildees de vanaacutedio
231 Anaacutelise da composiccedilatildeo das soluccedilotildees de vanadato por espectroscopia de 51
V-RMN
efeito da concentraccedilatildeo da espeacutecie
A diluiccedilatildeo das soluccedilotildees stock de meta- e decavanadato iraacute determinar fortemente a composiccedilatildeo
em soluccedilatildeo dos diferentes oligoacutemeros de vanadato (ref) No caso do decavanadato o periacuteodo de
tempo apoacutes a diluiccedilatildeo iraacute tambeacutem ser um factor importante a considerar devido agrave decomposiccedilatildeo
da espeacutecie V10(ref)Torna-se assim fundamental determinar a composiccedilatildeo quiacutemica das soluccedilotildees
no meio de ensaio utilizado (HEPES 10 mM KCl 25 mM) de modo a correlacionaacute-la
correctamente com os possiacuteveis efeitos promovidos pelas diferentes espeacutecies de vanadato no
sistema em estudo
Aleacutem disso deve ser dado ecircnfase agraves importantes interacccedilotildees que ocorrem entre o vanaacutedio e os
tampotildees escolhidos O tampatildeo com menor nuacutemero e magnitude de interacccedilatildeo eacute o HEPES [AD]
seguido do Tris sendo este uacuteltimo especialmente aconselhado em concentraccedilotildees baixas para casos
de incompatibilidade enzimaacutetica ou quiacutemica redox No caso especial da reacccedilatildeo redox
NAD(P)HNAD(P) o tampatildeo adequado eacute o imidazole de modo a natildeo haver reacccedilatildeo com o
26
vanadato Os tampotildees a evitar em qualquer situaccedilatildeo incluem a trietanolamina fosfato citrato e
glicina entre outros [AD] uma vez que todos interactuam fortemente com V(V) com formaccedilatildeo
de complexos estaacuteveis
Assim procedeu-se agrave anaacutelise da composiccedilatildeo das soluccedilotildees de meta- e decavanadato apoacutes diluiccedilatildeo
das soluccedilotildees stock (50 mM em vanadato total) no meio de reacccedilatildeo (tampatildeo D) atraveacutes da
espectroscopia de ressonacircncia magneacutetica nuclear de 51
V Nas figuras x1 e x2 apresentam-se os
espectros obtidos para as soluccedilotildees de deca- e metavanadato num intervalo de concentraccedilotildees de 1 a
10 mM A Tabela seguinte resume os paracircmetros espectroscoacutepicos obtidos para cada espeacutecie
presente nas soluccedilotildees analisadas
Figura 22ndash Espectros de RMN de 51
V de
concentraccedilotildees crescentes das espeacutecies oligomeacutericas
de vanadato obtidos apoacutes diluiccedilatildeo da soluccedilatildeo stock
de decavanadato em 10 mM HEPES pH 70 25 mM
KCl para concentraccedilotildees finais de A) 1 mM B)2
mM C) 4 mM D) 8 mM E) 10 mM de
decavanadato total V1refere-se agraves espeacutecies
monomeacutericade vanadato V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2IIIndash Desvios quiacutemicos meacutedios (part) das ressonacircncias das espeacutecies decameacuterica (V10) monomeacuterica (V1) e
tetrameacuterica (V4) presentes nos espectros da soluccedilatildeo de decavanadatoLargura a meia altura (∆12) da espeacutecie V10A
Paracircmetros V10A V10B V10C V1 V4
part (ppm) -5162 -5003 -4255 -5616 -5748
∆12 (Hz) 175 nd nd nd nd
Figura 23 ndash Distribuiccedilatildeo graacutefica das
espeacutecies oligomeacutericas de vanadato vs a
concentraccedilatildeo de decavanadato total As
concentraccedilotildees de cada espeacutecie foram
obtidas segundo a formula descrita na
secccedilatildeo 35 do Procedimento
Experimental (aacutereas dos sinais de 51
V-
RMN observadas na figura x1)V10 V1 e
V4 correspondem agraves espeacutecie monomeacuterica
dimeacuterica e tetrameacuterica respectivamente [Esp
eacuteci
e o
ligo
meacute
rica
de
van
aacutedio
] (m
M)
[Decavanadato]total (mM)
V10
V1
V4
27
Nas soluccedilotildees de decavanadato as espeacutecies monomeacuterica e decameacuterica de vanadato satildeo as uacutenicas
presentes para concentraccedilotildees de decavanadato inferiores a 8 mM Ambas as espeacutecies aumentam
linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado em [AY] e
em (Teresa) quando a concentraccedilatildeo de V1 eacute igual ou superior a 200 microM inicia-se a sua
oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica No entanto nas condiccedilotildees de realizaccedilatildeo dos
ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a espeacutecie tetrameacuterica neste
estudo em concentraccedilotildees inferiores a 8 mM tendo o mesmo sido observado por Ramoset al
(2006) [AK] Os resultados obtidos mostram claramente o equiliacutebrio entre a espeacutecie decameacuterica e
a monomeacuterica e que o aparecimento de outras espeacutecies oligomeacutericas se encontra dependente da
concentraccedilatildeo desta uacuteltima Apesar de natildeo ter sido realizado o ensaio com 5 mM de decavanadato
total no presente relatoacuterio estaacute reportado em Ramos et al (2006) [AK] que as concentraccedilotildees da
espeacutecie decameacuterica e monomeacuterica satildeo respectivamente de 500 microM e 200 microM Assim pode inferir-
se que para concentraccedilotildees inferiores a 5 mM e intervalos de tempo de cerca de 20 minutos (que
inclui o tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas
as espeacutecies decameacuterica e monomeacuterica de vanadato
Figura 24 ndash Espectros de RMN de 51
V de concentraccedilotildees
crescentes das espeacutecies oligomeacutericas de vanadato obtidos
apoacutes diluiccedilatildeo da soluccedilatildeo stock de metavanadato em 10 mM
HEPES pH 70 25 mM KCl para concentraccedilotildees finais de A)
1 mM B) 2 mM C) 4 mM D) 8 mM E) 10 mM de
metavanadato total V1 refere-se agraves espeacutecies monomeacuterica de
vanadatoV1 e V2 correspondem respectivamente agrave espeacutecie
monomeacuterica e dimeacuterica
Figura 25 ndash Distribuiccedilatildeo graacutefica das espeacutecies
oligomeacutericas de vanadato vs a concentraccedilatildeo de
metavanadato total As concentraccedilotildees de cada
espeacutecie foram obtidas segundo a formula descrita
na secccedilatildeo 35 do Procedimento Experimental
(aacutereas dos sinais de 51
V-RMN observadas na
figura x1)V10 V1 e V4 correspondem agraves espeacutecie
monomeacuterica dimeacuterica e tetrameacuterica
respectivamente
Esp
eacuteci
e o
ligo
meacute
rica
de
va
nad
ato
(m
M)
[Metavanadato]total (mM)
V1
V2
V4
V5
28
Tabela 2IVndash Desvios quiacutemicos meacutedios (part)das ressonacircncias das espeacutecies tetrameacuterica (V4) monomeacuterica (V1) e
dimeacuterica (V2) presentes no espectro da soluccedilatildeo de metavanadato (total) 10 mM
Paracircmetros V1 V2 V4 V5
part (ppm) -5615 -5748 -5789 -5869
Nas soluccedilotildees de metavanadato as espeacutecies monomeacuterica dimeacutericae tetrameacuterica de vanadato estatildeo
presentes para concentraccedilotildees de metavanadato inferiores a 4 mM Todas estas as espeacutecies
aumentam linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado
em (Aureliano e Madeira 1998) e em (Tiago et al 2004) quando a concentraccedilatildeo de V1 eacute igual ou
superior a 200 microM inicia-se a sua oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica Nas
condiccedilotildees de realizaccedilatildeo dos ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a
espeacutecie pentameacuterica neste estudo em concentraccedilotildees inferiores a 4 mM tendo o mesmo sido
observado por Ramos 2006 Os resultados obtidos mostram claramente o equiliacutebrio entre a
espeacutecie V4 e as espeacuteciesV1 e V2 no qual o aparecimento de outras espeacutecies oligomeacutericas se
encontra dependente da concentraccedilatildeo da espeacutecie V1 Por uacuteltimo pode inferir-se que para
concentraccedilotildees inferiores a 2 mM e intervalos de tempo de cerca de 20 minutos (que inclui o
tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas as
espeacutecies V1 V2 e V4 com concentraccedilotildees respectivas de 0796 0192 e 0196 mM
232Estabilidade das soluccedilotildees de decavanadato anaacutelise por espectroscopia de absorccedilatildeo
UVVis
Caracterizou-se a estabilidade da espeacutecie V10 numa soluccedilatildeo com 1 mM decavanadato total em
tampatildeo D atraveacutes da leitura da Abs400 Obteve-se uma cineacutetica de dissociaccedilatildeo de 1ordf ordem tal
como se pode verificar pela linearidade do graacutefico de ln(Abs400)vs o tempo de aquisiccedilatildeo espectral
(segundos) (Fig26 )
Figura 26ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do tempo
(em segundos) para uma soluccedilatildeo de decavanadato total de 1
mM em tampatildeo D a 25ordmC (n=2)
Sendo uma reacccedilatildeo de primeira ordem pode ser descrita pela equaccedilatildeo cujo
declive eacute ndashK (constante de dissociaccedilatildeo) e a intersecccedilatildeo no eixo das ordenadas corresponde
Ln
(Ab
s 400 n
m)
tempo (s)
29
aln(A0) Numa reacccedilatildeo de 1ordf ordem o tempo de meia-vida eacute independente da concentraccedilatildeo inicial
do reagente sendo descrito pela equaccedilatildeo [2] Deste modo a partir da recta
Ln(Abs400 nm) = -436E-5
K - 105 obteve-se um tempo de meia-vida de 4490 plusmn 0823 horas para
as condiccedilotildees supramencionadas
Apoacutes a adiccedilatildeo de S1 8 microM ao meio reaccional verificou-se que a reacccedilatildeo de desoligomerizaccedilatildeo
da espeacutecie decameacuterica de vanadato seguia igualmente uma cineacutetica de 1ordf ordem tal como se pode
verificar pela linearidade do graacutefico de ln(Abs400) versus o tempo de aquisiccedilatildeo espectral
(segundos) (Fig27)
Figura 27ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do
tempo (em segundos) para uma soluccedilatildeo de decavanadato
total de 1 mM em tampatildeo D a 25ordmC na presenccedila de 8
microM (n= 2)
A partir da equaccedilatildeo da recta Ln(Abs400 nm) = -7520E-5
K - 0613determinou-se o valor de K para
a espeacutecie decameacuterica de onde se obteve um tempo de meia-vida de 2430 plusmn 0191 horas para as
condiccedilotildees supramencionadas
Tanto este valor como o valor registado na ausecircncia de S1 satildeo substancialmente superiores agrave
duraccedilatildeo dos ensaios de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina descritos no presente
relatoacuterio
24 Interacccedilatildeo de S1 com vanaacutedio(V)
241Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
De modo a analisar qual o efeito da adiccedilatildeo de S1 de miosina (tampatildeo D) produzia nos sinais de
51V-NMR titulou-se uma soluccedilatildeo de 5mM de decavanadato total com concentraccedilotildees crescentes
de S1 (5 10 15 e 20 microM) Os espectros de 51
V-RMN encontram-se apresentados na Fig 2
No graacutefico da figura 2 Apresenta-se a concentraccedilatildeo de S1 em funccedilatildeo do factor de alargamento
corrigido dos espectros acima referidos
Ln
(Ab
s (4
00
nm
))
tempo (s)
30
Figura 28 ndash Graacutefico de
representaccedilatildeo da
concentraccedilatildeo de S1 em
funccedilatildeo do factor de
alargamento A
concentraccedilatildeo de
decavanadato total foi de 5
mM e as concentraccedilotildees
utilizadas de S1 foram de 0
5 10 e 15 microM
Verifica-se que o factor de alargamento aumenta linearmente em funccedilatildeo da concentraccedilatildeo de S1 no
ensaio o que indica a existecircncia de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina Observou-
se ainda que a adiccedilatildeo de S1 ao meio reaccional natildeo provocou qualquer alteraccedilatildeo dos desvios
quiacutemicos dos sinais de 51
V-RMN
Realizaram-se ensaios na presenccedila de ATP e de S1 com intuito de verificar o efeito do ATP na
interacccedilatildeo do S1 de miosina com o decavanadato
Figura 29 - Espectros de RMN de 51
V contendo 2
mM Decavanadato (A) (controlo) (B) na presenccedila
de 2 mM de ATP (C) na presenccedila de 2 mM de ATP
e 8 microM de S1 de miosina V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2Vndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies decameacuterica (V10) e monomeacuterica (V1) presentes nos
espectros da soluccedilatildeo de decavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V10A V10B V10C V1
part (ppm) -5161 -5002 -4254 -5614
Observa-se que a presenccedila de ATP (espectro B) natildeo provoca qualquer alteraccedilatildeo visiacutevel em
relaccedilatildeo ao espectro que contem apenas soluccedilatildeo de decavanadato Jaacute a presenccedila simultacircnea de ATP
e S1 provoca um alargamento da ∆12 do sinal do V10A de 1736 Hz (espectro (A)) para 2366 Hz
(espectro (C) Fig 29) Tendo em conta que a concentraccedilatildeo de S1 apresenta uma relaccedilatildeo linear
com o factor de alargamento converteu-se o valor da ∆12 e comparou-se com o valor esperado agrave
F = 00591[S1] - 00157 Rsup2 = 0998
Fact
or
de
ala
rgam
en
to
[S1] (microM)
31
concentraccedilatildeo de S1 utilizada O factor de alargamento obtido para 8 microM ndash 0376 ndash eacute inferior ao
valor esperado agrave mesma concentraccedilatildeo de S1 obtido atraveacutes da equaccedilatildeo da recta apresentada na
Fig 28 ndash 0457 ndash o que indica que o ATP poderaacute exercer um efeito de protecccedilatildeo do S1
diminuindo a sua interacccedilatildeo com a espeacutecieV10
Por uacuteltimo realizaram-se ensaios com metavanadato na presenccedila de 8 microM S1 de modo a verificar
se as espeacutecies oligomeacutericas contidas nesta soluccedilatildeo interagiam com a proteiacutena em estudo
Figura 210 - Espectros de RMN de 51
V contendo 2
mM metavanadato (A) (controlo) (B) na presenccedila
de 8 microM de S1 de miosina V1 V2 e V4
correspondem respectivamente aacutes espeacutecies
monomeacuterica dimeacuterica e tetrameacuterica
Tabela 2VIndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies monomeacuterica (V1) dimeacuterica (V2) e tetrameacuterica (V4)
presentes nos espectros da soluccedilatildeo de metavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V1 V2 V4
part (ppm) -5613 -5747 -5789
Como eacute possiacutevel observar na Fig 210 ocorre uma ligeiro alargamento dos sinal de V4 bem
como uma perda da intensidade pelo que se conclui quedas espeacutecies presentes na soluccedilatildeo de
metavanadato (V1 V2 e V4) apenas a espeacutecie tetrameacuterica interage com o S1 de miosina o que
estaacute de acordo com o reportado por Tiago et al 2002
242 Actividade Mg2+
-ATPaacutesica de S1 de miosina na presenccedila de decanadato
Com o intuito de avaliar o efeito do decavanadato na actividade Mg2+
-ATPaacutesica do S1 de
miosina monitorizou-se a reacccedilatildeo de oxidaccedilatildeo do NADH atraveacutes da leitura de Abs340 na
presenccedila de 005 mgmL de S1 de miosina e no caso da actividade estimulada 2microM de actina em
3125 mM Tris-HCl pH 71 e 375 mM MgCl2 pelo meacutetodo de enzimas acopladas (ver seccccedilatildeo xx
dos MM)Para tal realizaram-se ensaios com vaacuterias concentraccedilotildees de decavanadato total (10 50
100 250 500 e 1000) de modo a determinar qual o seu efeito na actividade MgATPaacutesica de S1
de miosina na ausecircncia e na presenccedila de um dos seus ligandos naturais Os ensaios foram
realizados agrave temperatura ambiente com incubaccedilatildeo preacutevia a 25ordm C durante 20 minutos
32
Figura 211 ndash Representaccedilatildeo
graacutefica da Actividade Mg2+
-
ATPaacutesica de S1 () (os ajustes
teoacutericos estatildeo representado a por
uma linha os resultados
experimentais estatildeo representados
por triacircngulos (Actividade
estimulada) e cruzes (Actividade
natildeo estimulada)) vs
[Decavanadato]total medidas pelo
meacutetodo das enzimas acopladas
(n=3) Os dados foram ajustados ao modelo de decaimento exponencial usando o programa GraphPad Prism para a
determinaccedilatildeo dos valores de IC50 (na ausecircncia de actina IC50= 266 plusmn 07 M decavanadato na presenccedila de actina
IC50= 28 plusmn 1 M decavanadato
A partir do graacutefico presente na Fig 29 verifica-se que o ajuste dos dados experimentais foi bem
sucedido A inibiccedilatildeo da actividade Mg2+
-ATPaacutesica do S1 de miosina eacute afectada em menor
amplitude na presenccedila de actina (60 de inibiccedilatildeo da actividade) do que na sua ausecircncia
(aproximadamente 80 de inibiccedilatildeo de actividade) Devido a esta diferenccedila pode sugerir-se que a
actina iraacute impedir parcialmente a ligaccedilatildeo do decavanadatoao S1 demiosina atraveacutes de
impedimentos esteacutereos ou interacccedilotildees electrostaacuteticas por exemplo A teacutecnica utilizada natildeo nos
permite concluir acerca de que regiatildeo(otildees) ou resiacuteduo(s) da actina participaratildeo na preservaccedilatildeo
parcial da actividade Mg2+
-ATPaacutesica
Por outro lado os valores de IC50 dos dois tipos de actividades natildeo apresentam diferenccedilas
significativas tendo-se obtido 266 plusmn 07 e 28 plusmn 1M decavanadato na ausecircncia e na presenccedila de
actina respectivamente o que corresponde a uma concentraccedilatildeo de cerca de 27 M de V10 O
meio de reacccedilatildeo utilizado nestes ensaios foi 10 mM HEPES e 25 mM KCl com 2 mM ATP Em
(Tiago et al 2004) o valor de IC50 para a actividade Mg2+-ATPaacutesica na ausecircncia de actina foi de
125 plusmn 025 microM enquanto o valor da actividade estimulada por actina foi de 136 plusmn 14 microM O
meio de reacccedilatildeo utilizado nos ensaios foi 20 mM HEPES 50 mM KCl e 2 mM ATP o que resulta
precisamente no dobro da forccedila ioacutenica a que foi realizado o ensaio do presente relatoacuterio
(exceptuando a contribuiccedilatildeo dos produtos da hidroacutelise de ATP uma vez que este meacutetodo
pressupotildee a ocorrecircncia de um ciclo de reacccedilotildees de hidroacutelise e siacutentese deste composto) Ora tendo
sido anteriormente referido que a afinidade do S1 de miosina para com a espeacutecie V10 diminui com
Act
ivid
ade
Mg
2+-A
TP
aacutesic
a d
e S
1 d
e
Mio
sin
a
[Decavanadato] (microM) Actividade Estimulada ()
Actividade Natildeo Estimulada ()
Ajuste Actividade Estimulada
Ajuste Actividade Natildeo Estimulada
33
o aumento da forccedila ioacutenica (Tiago et al 2004 tal poderaacute explicar a diferenccedila obtida entre estes
dois conjuntos de valores
243Quantificaccedilatildeo de cisteiacutenas reduzidas pelo meacutetodo de Ellman
A interacccedilatildeo do vanaacutedio com alguns tipos de aminoaacutecidos sensiacuteveis agrave oxidaccedilatildeo pode fornecer-nos
informaccedilatildeo bastante uacutetil em relaccedilatildeo agraves alteraccedilotildees estruturais que conduzem a uma diminuiccedilatildeo da
actividade fisioloacutegica do S1 de miosina Os estudos realizados por Chasteen 1983 concluiacuteram
que a reduccedilatildeo de vanadato (V) a vanadilo (IV) ocorre na presenccedila de agentes redutores comuns no
ambiente celular de que se destaca o aminoaacutecido cisteiacutena Esta eacute tambeacutem um alvo de
complexaccedilatildeo por parte do iatildeo VO2+
Jaacute Ramos et al (2009) demonstrou que a incubaccedilatildeo de
actina com a soluccedilatildeo de decavanadato induz a oxidaccedilatildeo de resiacuteduos de cisteiacutena e agrave formaccedilatildeo do
oxidovanaacutedio (IV) Este grupo de investigadores descobriu ainda que apenas a espeacutecie V10 induz a
oxidaccedilatildeo das cisteiacutenas ldquoraacutepidasrdquo da actina atraveacutes do processo proposto que eacute parcialmente
responsaacutevel pela inibiccedilatildeo jaacute descrita da actividade de ATPase do S1 de miosina estimulada por
actina quando exposta agrave soluccedilatildeo de decavanadato Esta inibiccedilatildeo apresenta-se afectada com a
presenccedila de ATP uma vez que os resiacuteduos de cisteiacutena permaneceram intactos nos ensaios
realizados com este ligando Jaacute Crans et al (2004) concluiu que o siacutetio activo de muitas fosfatases
(que conteacutem grupos tiol) pode formar um equiliacutebrio redox com o iatildeo vanadato (V) de forma natildeo
reversiacutevel
Os resultados obtidos por Sakurai e Shiromura (1980) sugeriram que o vanaacutedio (V) sofre reduccedilatildeo
a vanaacutedio (IV) sem dependecircncia do valor de pH do ensaio Registaram ainda uma estequiometria
de 31 na reacccedilatildeo do vanadato (V) com os resiacuteduos de Cys enquanto a estequiometria dos ensaios
com o vanadilo (IV) foi de 21
Berlett e Stadtman (1997) observaram que os tipos de aminoaacutecidos mais facilmente oxidaacuteveis
englobam entre outros os resiacuteduos de Cys Trp e MetAssim analisou-se o estado redox dos
grupos tiol dos resiacuteduos de Cys do S1 na presenccedila do decavanadato atraveacutes da monitorizaccedilatildeo da
Abs412 nm na presenccedila de 1 mgmL de S1 de miosina em tampatildeo 10 mM HEPES 25 mM KCl
variando a concentraccedilatildeo da soluccedilatildeo de decavanadato de modo a determinar que tipo de alteraccedilotildees
estruturais ocorrem na sua presenccedila no que diz respeito agrave diminuiccedilatildeo da actividade ATPaacutesica de
S1 na presenccedila de decavanadato
Os resultados referentes agrave quantificaccedilatildeo das cisteiacutenas reduzidas apoacutes o tratamento de S1 com
diferentes concentraccedilotildees de decavanadato em tampatildeo 10 mM HEPES pH 75 e 25 mM KCl a
25ordmC durante 20 minutos encontram-se descritos na Tabela 2VII
Tabela 2VII ndash Cisteiacutenas reduzidas totais e ldquoraacutepidas de S1 (1 mgmL) apoacutes tratamento com decavanadato
34
(010501002505001000 uM) em 10 mM HEPES 25 mM KCl pH 7 a 25ordmC durante 20 minutos A detecccedilatildeo
espectrofotomeacutetrica foi feita com DTNB (e 412=8400 uM-1 cm-1) (n variaacutevel entre 1 e 8)
Amostras Cys raacutepidas Cys totais
11 M S1 (controlo) 22 plusmn 04 106 plusmn 14
11 M S1 + 10 M Decavanadato 22 plusmn 05 95 plusmn 08
11 M S1 + 50 uM Decavanadato 20 plusmn 02 98 plusmn 14
11 M S1 + 100 uM Decavanadato 17 plusmn 03 97 plusmn14
11 M S1 + 250 uM Decavanadato 26 plusmn 02 98 plusmn 08
11 M S1 + 500 uM Decavanadato 21 plusmn 04 90 plusmn 06
11 M S1 + 1000 uM Decavanadato 24 plusmn 0 86 plusmn 08
A partir dos resultados obtidos pode sugerir-se que o S1 possui 11 resiacuteduos de Cys (controlo) o
que estaacute de acordo com o anteriormente descrito Verifica-se que o tratamento da proteiacutena com
concentraccedilotildees crescentes de decavanadato provoca a oxidaccedilatildeo de dois resiacuteduos de Cys do S1 de
miosina natildeo expostos ao solvente No entanto a teacutecnica utilizada natildeo nos permite identificar
quais
Tendo em consideraccedilatildeo que o valor de IC50 obtido para a reacccedilatildeo de inibiccedilatildeo daactividade Mg2+-
ATPaacutesica de S1 na presenccedila de decavanadato eacute de cerca de 27 microM pode sugerir-se que este seraacute o
valor ao qual ocorreraacute a oxidaccedilatildeo do primeiro resiacuteduo de Cys o que poderaacute contribuir para a
diminuiccedilatildeo da actividade de S1 observada
25Interacccedilatildeo de S1 com vanaacutedio(IV)
251Anaacutelise por espectroscopia de Ressonacircncia Paramagneacutetica Electroacutenica
A acumulaccedilatildeo intracelular de oxovanaacutedio (IV) in vivo constitui uma das principais formas de
apresentaccedilatildeo de vanaacutedio no organismo Deste modo torna-se bastante interessante analisar a
interacccedilatildeo entre este iatildeo e o S1 de miosina atraveacutes da teacutecnica de espectroscopia de Ressonacircncia
Paramagneacutetica Electroacutenica o meacutetodo de eleiccedilatildeo do estudo do iatildeo vanadilo e dos complexos por si
formados O electratildeo desemparelhado encontra-se fortemente acoplado ao nuacutecleo 51
V o que
35
resulta num espectro de RPE consistindo de 8 desdobramentos na direcccedilatildeo paralela e outros tantos
na direcccedilatildeo perpendicular (Chasteen 1983)
Os estudos realizados por Ahmed et al (1987) centraram-se na interacccedilatildeo do vanadilo com
proteiacutenas como eacute o caso da insulina transferrina entre outros obtendo um espectro caracterizado
por 8 linhas amplamente espaccediladas (na direcccedilatildeo paralela) Jaacute Ramos et al 2009 concluiu que a
exposiccedilatildeo da soluccedilatildeo de decavanadato agraves formas globular e filamentosa de actina produz um
espectro tiacutepico do iatildeo oxidovanaacutedio (IV) indicando a reduccedilatildeo do vanadato
Para aferir qual a concentraccedilatildeo de S1 que permitia obter uma boa razatildeo sinalruiacutedo titulou-se uma
soluccedilatildeo de sulfato de vanadilo 15 microM ([VOSO4]stock = 5 mM) com concentraccedilotildees crescentes de
proteiacutena (0 5 10 15 e 20 microM) em 10 mM HEPES 25 mM KCl A existecircncia de uma correlaccedilatildeo
linear entre a concentraccedilatildeo de proteiacutena e a intensidade dos sinais de RPE correspondente agrave banda
-12 perpendicular encontra-se representada na Fig 210
Figura 2 12ndash Representaccedilatildeo graacutefica da variaccedilatildeo da intensidade do sinal de RPE vs a concentraccedilatildeo de S1 utilizada
Apoacutes verificar que com uma concentraccedilatildeo de S1 igual a 20 microM se obtinham sinais com uma razatildeo
sinalruiacutedo adequada ao prosseguimento dos estudos sobre a interacccedilatildeo V(IV)-S1 de miosina
titulou-se uma soluccedilatildeo de 20 microM S1 com 0 25 5 10 15 25 50 75 100 150 200 500 e 1000
microM VOSO4
Traccedilou-se o graacutefico da intensidade da banda -12 perpendicular do sinal de EPR em funccedilatildeo da
concentraccedilatildeo de VOSO4 tendo-se determinado uma constante de dissociaccedilatildeo (KD) do complexo
S1-vanadilo de 8748 plusmn 2621 microM V(IV)
y = 016786x + 122435
Rsup2 = 092992759
Inte
nsi
dad
e d
o s
inal
de
RP
E
[S1] (microM)
36
Figura 213 ndash Representaccedilatildeo graacutefica da variaccedilatildeo do sinal de RPE vs a concentraccedilatildeo de vanadilo utilizada (os valores
experimentais estatildeo representados por losangos o seu ajuste estaacute representado pela linha)
A simulaccedilatildeo do espectro de EPR foi efectuada a partir do espectro obtido para uma concentraccedilatildeo
de 1 mM VOSO4 apoacutes adiccedilatildeo de 20 microM S1 de miosina tendo-se obtido os valores da constante de
acoplamento hiperfina (A=5016) e do factor g (g0=195)
Figura 214 ndash Representaccedilatildeo graacutefica do espectro de EPR obtido com 1000 microM de vanadilo e 20 microM de S1 (a cinzento
escuro) A simulaccedilatildeo do espectro encontra-se a cinzento claro
Por comparaccedilatildeo destes com os valores presentes na tabela de correspondecircncia xxx eacute possiacutevel
predizer quais os ligandos provaacuteveis do vanadilo na presenccedila de proteiacutena Assim concluiu-se que
o vanadilo estaacute coordenado com duas moleacuteculas de aacutegua um grupo hidroxilo e um grupo R=NR
O grupo hidroxilo pode indicar os resiacuteduos de aminoaacutecidos Tyr Treonina e Serina enquanto o
grupo R=NR poderaacute indicar a presenccedila de Arginina e Histidina como ligandos do vanadilo
Inte
nsi
dad
e d
os
sinai
s d
e R
PE
[Vanadilo] (microM)
Ajuste dos dados exp Dados experimentais
Campo magneacutetico (G)
Espectro Experimental Ajuste
37
3 Procedimento Experimental
31 Reagentes
Todos os reagentes utilizados satildeo de elevado grau de pureza (analiacutetico) Apresentam-se as suas
foacutermulas quiacutemicas ou abreviaturas quando necessaacuterio acrescentou-se a sua descriccedilatildeo Entre
parecircntesis colocou-se as empresas que os comercializam
Aacutecido aceacutetico glacial 100 (Riedel) Acrilamida 99 (Merck) Azul de bromofenol (Riedel-de-
Haumlen) -Mercaptoetanol (Sigma) Bissulfito de soacutedio (Sigma)Bradford (reagente de)
(BioRad)BSA (albumina de soro bovino Sigma-Aldrich) CaCl22H2O (cloreto de caacutelcio
dihidratado Fluka) CH3CH2OH 96 (etanol Panreac) Coomassie Blue R250 (Merck) D2O
(oacutexido de deuteacuterio SDS) DTNB (55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico) Aldrich) EDTA (aacutecido
etilenodiaminatetraaceacutetico Merck) Fosfoenolpiruvato 98 (Fluka) Glicerol (Sigma) Glicina
985 (Panreac) HCl 37 (Riedel-de Haumlen) H2SO4 96 (Pronalab) HEPES 99 (aacutecido 4-(2-
hidroxiletil)-1-piperazineetanosulfoacutenico Sigma) KH2PO4 99 (hidrogenofosfato de potaacutessio)
MgATP (Sigma) Na
ATP (Sigma) Na2HPO4 99 (hidrogenofosfato de soacutedio Riedel-de Haumlen)
NaH2PO4 (fosfato de soacutedio ) NaOH 986 (hidroacutexido de soacutedio Pronalab) KCl 99 (Cloreto de
potaacutessio Panreac) L-Cys 995 (enantioacutemero levoacutegiro do resiacuteduo de Cisteiacutena Fluka)Metanol
99 (Merck) Molibdato de amoacutenio (Sigma)NNrsquo-metil-bis-acrilamida 98 (Merck) NH4VO3
99 (metavanadato de amoacutenia Sigma) PSA 10 (persulfato de amoacutenio Merck) SDS
(dodecilo sulfato de soacutedio Sigma) Sulfito de soacutedio (Sigma) VOSO4 (sulfato de vanadilo
Aldrich) TEMED 99 (Tetrametiletilenodiamina Merck) Tris (2-amino-2-hidroximetil-13-
propanodiol Sigma)
32 Meacutetodos
321 Isolamento e purificaccedilatildeo das proteiacutenas de muacutesculo esqueleacutetico de coelho
3211 Purificaccedilatildeo da miosina preparaccedilatildeo de Subfragmento 1 por digestatildeo quimotriacutepsica
da miosina
O subfragmento-1 de miosina-II (S1) foi gentilmente cedido pelo Prof Doutor Manuel Aureliano
do Departamento de Quiacutemica Bioquiacutemica e Farmaacutecia da Faculdade de Ciecircncias e Tecnologia da
Universidade do Algarve
No entanto descreve-se abaixo de forma resumida o procedimento efectuado
38
32111 Purificaccedilatildeo da miosina
O protocolo de isolamento foi adaptado do meacutetodo previamente descrito por Margossian e Lowey
(1982) apresentando-se de forma esquemaacutetica na Fig 31
Sucintamente os tecidos musculares dorsais e das pernas foram extraiacutedos apoacutes o sacrifiacutecio do
animal (Fig31) De seguida foram adicionados 3 volumes da soluccedilatildeo de extracccedilatildeo A2
Centrifugou-se a soluccedilatildeo e filtrou-se o sobrenadante O sedimento foi entatildeo diluiacutedo em 14
volumes de aacutegua destilada de modo a que a forccedila ioacutenica atingisse 004 M Apoacutes adiccedilatildeo de szlig-
mercaptoetanol (szlig-me) deixou-se a precipitar a 4ordmC removendo o sobrenadante Centrifugou-se
novamente e ressuspendeu-se o sedimento na soluccedilatildeo de extracccedilatildeo A Adicionou-se a soluccedilatildeo
tampatildeo B3 para que a forccedila ioacutenica atingisse 05 M Homogeneizou-se e deixou-se repousar
durante 2h Homogeneizou-se novamente e centrifugou-se de modo a ocorrer precipitaccedilatildeo dos
complexos de actomiosina e proteiacutena desnaturada Apoacutes filtraccedilatildeo do sobrenadante em latilde de vidro
quantificou-se a soluccedilatildeo de miosina resultante pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental)
2 Soluccedilatildeo de extracccedilatildeo A 0012M K2CO3 004M KHCO3 06M KCl pH 65
3 Soluccedilatildeo-tampatildeo B Tris 100 mM pH 7 KCl 2M
39
Figura 31 ndash Representaccedilatildeo esquemaacutetica do protocolo de isolamento de miosina (Margossian e Lowey 1982) Todo o
procedimento foi realizado a 4ordmC (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
32112 Preparaccedilatildeo de Subfragmento-1 de miosina por digestatildeo com quimiotripsina
O S1 de miosina foi isolado apoacutes diaacutelise da miosina durante a noite contra a soluccedilatildeo C4 A
presenccedila do EDTA eacute essencial uma vez que a sua substituiccedilatildeo por MgCl2resulta na produccedilatildeo de
HMM em vez de S1 A suspensatildeo de miosina foi equilibrada e digerida com -quimotripsina
A digestatildeo foi interrompida pela adiccedilatildeo de PMSF Seguidamente ultracentrifugou-se o produto
da digestatildeo desprezando-se o sedimento (Fig 32) A soluccedilatildeo de sobrenadante maioritariamente
S1 foi quantificada pela leitura directa da Abs280 ou pelo meacutetodo de Bradford (ver as secccedilotildees
3311 e 3312 do Procedimento Experimental respectivamente) e armazenada em 10 sacarose
e 2 mM szlig-Me a -80ordmC
4 Soluccedilatildeo C 25mM Tris pH 70 012M KCl 1mM EDTA
40
Antes da realizaccedilatildeo dos ensaios descritos no presente relatoacuterio o S1 de miosina foi dialisado
contra a soluccedilatildeo D5 de modo a remover qualquer composto que pudesse complexar como
vanaacutedio Seguidamente quantificou-se a proteiacutena pelo meacutetodo de Bradford ou pelo meacutetodo
directo (ver secccedilotildees 3311 e 3312do Procedimento Experimtental)
Figura 3 2ndash Representaccedilatildeo esquemaacutetica do protocolo de preparaccedilatildeo do S1 de miosina por digestatildeo quimiotripsina de
acordo com o previamente descrito por Margossian e Lowey (1982) Todo o procedimento foi realizado a 4ordm C
excepto quando indicaccedilatildeo do contraacuterio (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
3212 Isolamento e purificaccedilatildeo de actina
Isolou-se e purificou-se a actina segundo o meacutetodo descrito por Pardee e Spudish que
compreende dois passos ndash a obtenccedilatildeo de um resiacuteduo seco denominado ldquopoacute de acetonardquo e a
extracccedilatildeo subsequente da actina a partir do ldquopoacute de acetonardquo na forma de G-Actina com a
posterior polimerizaccedilatildeo a F-Actina
32121 Obtenccedilatildeo do ldquopoacute de acetonardquo
Sacrificou-se o coelho com uma pancada atraacutes do pescoccedilo e sangrou-se o animal apoacutes o corte da
arteacuteria jugular Esfolou-se e colocou-se numa tina com gelo Retiraram-se as partes de tecido
conjuntivo que envolvem o tecido muscular com auxiacutelio de uma pinccedila e bisturi Seguidamente
retirou-se os muacutesculos dorsais e os muacutesculos brancos das pernas Passou-se os muacutesculos por aacutegua
destilada de modo a remover as impurezas e pesou-se num copo Trituraram-se os muacutesculos
numa maacutequina de triturar carne e extraiu-se o resultante com 3 volumes da soluccedilatildeo E6 durante 10
minutos com agitaccedilatildeo suave Centrifugou-se a suspensatildeo a 4000 g durante 10
5 Soluccedilatildeo D HEPES 10 mM pH 70 KCl 25 mM
6 Soluccedilatildeo E 01 M KCl 015 M K2HPO4 pH 65
41
minutosdesprezando-se o sobrenadante Ressuspendeu-se o sedimento em 6 volumes de soluccedilatildeo
F7durante 10 minutos com agitaccedilatildeo suave Verificou-se se o pH da soluccedilatildeo estava compreendido
entre 82 e 85 acertando-se estecom soluccedilatildeo G8 se necessaacuterio Centrifugou-se durante 10
minutos a 4000 g Desprezou-se o sobrenadante e ressuspendeu-se o sedimento em 3 volumes de
soluccedilatildeo H9 durante 10 minutos Centrifugou-se novamente a 4000 gdurante 10 minutos
desprezando-se o sobrenadante Ressuspendeu-se o sedimento duas vezes consecutivas em 6
volumes de aacutegua destilada durante 5 minutos Centrifugou-se a 4000 g durante 10 minutos apoacutes
cada adiccedilatildeo de H2O
Seguidamente ressuspendeu-se o sedimento 5 vezes consecutivas utilizando 3 volumes de
acetona de cada vez agitando suavemente durante 10 minutos agrave temperatura ambiente Colocou-
se o resiacuteduo filtrado numa tina de vidro sobre papel de filtro deixando-se evaporar o solvente
durante a noite num nicho Triturou-se o resiacuteduo seco numa maacutequina de triturar carne e deixou-se
novamente evaporar o solvente O ldquopoacute de acetonardquo obtido foi armazenado a -20 ordmC
32122 Purificaccedilatildeo da actina a partir de ldquopoacute de acetonardquo
Apoacutes a obtenccedilatildeo do poacute de acetona como acima indicado efectuou-se o protocolo seguidamente
descrito de modo a obter monoacutemeros de actina (G-Actina) e actina filamentosa (F-Actina)
Pesou-se 35 g de poacute de acetona e adicionaram-se 50 mL de soluccedilatildeo-tampatildeo I10
sendo a
extracccedilatildeo feita com agitaccedilatildeo suave durante 30 minutos Separou-se o extracto do poacute de acetona
hidratado filtrando a suspensatildeo com 4 camadas de gaze esterilizada tendo o cuidado de natildeo
espremer o resiacuteduo Voltou a extrair-se com 50 mL de soluccedilatildeo I durante 30 minutos mexendo
lentamente Adicionou-se este ao extracto obtido anteriormente Centrifugou-se o extracto total a
16 000 g durante 30 minutos Decantou-se para uma proveta sobre a qual se colocou gaze para
que o sedimento solto natildeo se juntasse ao sobrenadante e leu-se o volume De seguida adicionou-
se KCl ateacute uma concentraccedilatildeo final de 50 mM MgCl2 ateacute 2mM e ATP ateacute 1mM lentamente e com
agitaccedilatildeo contiacutenua de modo a promover a polimerizaccedilatildeo dos monoacutemeros de actina
Deixou-se polimerizar durante 2h a 25ordmC observando-se a presenccedila de pequenas bolhas de ar
indicativas de um aumento de viscosidade da soluccedilatildeo Arrefeceu-se em gelo durante 15 minutos e
adicionou-se KCl ateacute uma concentraccedilatildeo final de 06M Mexeu-se lentamente durante 30 minutos
apoacutes os quais se ultracentrifugou a 110 000g durante 1hora Desprezou-se o sobrenadante
ressuspendeu-se o sedimento com soluccedilatildeo tampatildeo E e transferiu-se para um potter
Dialisou-se durante a noite contra 1L de soluccedilatildeo-tampatildeo I a uma temperatura de 4ordmC com rotaccedilatildeo
7 Soluccedilatildeo F 005 M NaHCO3 pH 82-85
8 Soluccedilatildeo G 1M Na2CO3
9 Soluccedilatildeo H 1 mM EDTA pH 70
10 Soluccedilatildeo I 2 mM Tris-HCl pH 8 02 mM CaCl2 02 mM Na2ATP 05 mM szlig-Me 0005 NaN3
42
contiacutenua e mudanccedila de tampatildeo 2 a 3 vezes De seguida a actina foi centrifugada a 110
000gdurante 1hora Decantou-se a soluccedilatildeo de G-Actina (sobrenadante) e desprezou-se o
sedimento Determinou-se a concentraccedilatildeo de G-Actina pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental) A soluccedilatildeo de G-actina foi armazenada a uma temperatura de 4ordmC
apoacutes adiccedilatildeo de 002NaN3 mantendo-se estaacutevel durante 3 a 5 dias
331 Quantificaccedilatildeo das proteiacutenas isoladas
3311 Quantificaccedilatildeo pelo meacutetodo de Bradford
O meacutetodo de Bradford (Bradford 1976) baseia-se na utilizaccedilatildeo do reagente de Bradford
composto por entre outros aacutecido fosfoacuterico e o corante ldquoCoomassierdquo Este corante possui carga
negativa e a sua coloraccedilatildeo varia entre a vermelha (Absmax=465 nm) predominante em soluccedilatildeo e a
azul (Absmax=595 nm) presente aquando da ligaccedilatildeo aos grupos amina das proteiacutenas Em soluccedilotildees
aacutecidas diluiacutedas a coloraccedilatildeo varia proporcionalmente com a quantidade do corante ligado agrave
proteiacutena que por sua vez eacute directamente proporcional agrave concentraccedilatildeo da proteiacutena permitindo a
sua quantificaccedilatildeo ateacute a um limite de detecccedilatildeo de 20 gmL (Bradford 1976)
Deste modo prepararam-se amostras de BSA ([BSA]stock= 1mgmL) utilizando um intervalo de
concentraccedilotildees compreendido entre 0 e 15 gmL de acordo com a tabela 3I Apoacutes a adiccedilatildeo da
H2O MilliQ e do reagente de Bradford as soluccedilotildees foram sucessivamente homogeneizadas no
voacutertex
Tabela 3Indash Amostras utilizadas para a obtenccedilatildeo da recta de calibraccedilatildeo no meacutetodo de Bradford
BSA (L) BSA (g) H2O MilliQ (L) Reagente de Bradford (L)
0 0 800 200
125 125 79875 200
25 25 7975 200
5 5 795 200
10 10 790 200
15 15 785 200
Apoacutes incubaccedilatildeo a 25ordmC por 5 minutos leram-se os valores de Abs595 num
espectrofotoacutemetro (Shimadzu UV-160A) para todas as amostras Seguidamente traccedilou-se a recta
de calibraccedilatildeo (Abs595vs quantidade de BSA (g))
43
Para determinar a concentraccedilatildeo de proteiacutena apoacutes o processo de isolamento purificaccedilatildeo e
diaacutelise quando necessaacuterio colocou-se 5 L da amostra num tubo de Eppendorf e adicionou-se
795 L de H2O MilliQ homogeneizando a soluccedilatildeo no voacutertex Adicionou-se posteriormente 200
L de reagente de Bradford e repetiu-se a homogeneizaccedilatildeo Incubou-se a mistura a 25ordmC por 5
minutos apoacutes os quais se leu os valores de Abs595 nm no espectrofotoacutemetro Por interpolaccedilatildeo deste
valor a partir da recta de calibraccedilatildeo determinou-se a concentraccedilatildeo de proteiacutena na amostra
3312 Quantificaccedilatildeo pelo meacutetodo directo
A quantificaccedilatildeo das amostras proteicas pode tambeacutem ser efectuada atraveacutes da leitura directa da
absorvacircncia tendo por base a lei de Beer-Lambert onde Abs corresponde agrave
absorvacircncia ao comprimento de onda ao qual o ensaio foi realizado corresponde ao coeficiente
de extinccedilatildeo (substituiacutedo pela letra a se as unidades forem distintas da molaridade) b corresponde
ao percurso oacuteptico da radiaccedilatildeo na ceacutelula amostral e c corresponde agrave concentraccedilatildeo amostral
Assim no caso de S1 de miosina mediu-se a Abs280e calculou-se a concentraccedilatildeo da proteiacutena
sabendo que o coeficiente de extinccedilatildeo de S1 eacute (Ref)
No caso da actina mediu-se a Abs290ecalculou-se a concentraccedilatildeo da proteiacutena sabendo que o
coeficiente de extinccedilatildeo da actina eacute (Ref)
332Electroforese em condiccedilotildees desnaturantes (SDS-PAGE)
De modo a melhor caracterizar as proteiacutenas isoladas foi realizada uma electroforese em
condiccedilotildees desnaturantes (SDS-PAGE do inglecircs Sodium Dodecyl Sulfate Polyacrilamide Gel
Electrophoresis) O SDS (dodecilo sulfato de soacutedio) eacute um detergente anioacutenico utilizado neste
meacutetodo para desnaturar as proteiacutenas que se ligam ao SDS na proporccedilatildeo 14 g SDS g proteiacutena
(aproximadamente 1 moleacutecula de SDS por cada 2 resiacuteduos de aminoaacutecidos) (Stryer) O SDS induz
uma densidade de carga negativa significativa nas proteiacutenas a que se liga mascarando a sua carga
intriacutenseca pelo que as razotildees cargamassa satildeo tendencialmente idecircnticas Assim a SDS-PAGE
separa as proteiacutenas quando sujeitas a um campo eleacutectrico de acordo com o seu peso molecular
sendo a sua migraccedilatildeo directamente proporcional ao logaritmo deste uacuteltimo (Stryer)
O gel de resoluccedilatildeo com 10 de acrilamida foi obtido atraveacutes da adiccedilatildeo de 167 mL de soluccedilatildeo
stock de acrilamida11
075 mL de tampatildeo de resoluccedilatildeo12
251 mL de aacutegua destilada e 50 L de
SDS 10 Para promover a polimerizaccedilatildeo adicionou-se em seguida 38 L de PSA 10
11
soluccedilatildeo stock de acrilamida soluccedilatildeo de 30 (mv) de acrilamida e 08 (mv) de NNrsquo-metil-bis-acrilamida 12
tampatildeo de resoluccedilatildeo 15 M Tris-HCl pH 88
44
(persulfato de amoacutenia) 10 e 25 L de TEMED 99 (tetrametiletilenodiamina) Agitou-se
cuidadosamente de modo a natildeo formar bolhas e pipetou-se a soluccedilatildeo para a cassette de vidro
previamente montada no suporte De seguida adicionou-se uma peliacutecula de cerca de 2 mm de 2-
metil-2-butanol para verificar posteriormente a polimerizaccedilatildeo Apoacutes a polimerizaccedilatildeo que demora
cerca de 45 minutos verteu-se a agrave superfiacutecie e adicionou-se o gel de concentraccedilatildeo com 5 de
acrilamida constituiacutedo por 03 mL de soluccedilatildeo de stock acrilamida 045 mL de tampatildeo de
concentraccedilatildeo13
094 mL de aacutegua destilada 18 L de SDS 10 135 L de PSA 10 e 2L de
TEMED 99 Colocou-se o pente sobre o gel e deixou-se polimerizar durante cerca de 45
minutos
As amostras para aplicaccedilatildeo no gel de electroforese foram preparadas para uma concentraccedilatildeo de
proteiacutena de 1 mgmL num volume final de 100 L dos quais 25 L correspondem ao tampatildeo da
amostra14
As amostras foram aquecidas a 100ordmC durante 5 minutos Antes da aplicaccedilatildeo das
amostras transferiu-se a cassette para a tina de electroforese agrave qual se adicionou a soluccedilatildeo
tampatildeo de electroforese da tina superior15
e tampatildeo de electroforese da tina inferior16
Aplicaram-
se 10 L das soluccedilotildees de marcadores de peso molecular e 20 L das amostras proteicas
preparadas anteriormente A electroforese decorreu durante cerca de 3h30 min a uma voltagem
de 80 V De seguida transferiu-se o gel para uma tina corando as proteiacutenas durante 1h com
soluccedilatildeo corante17
Descorou-se o gel com soluccedilatildeo descorante18
tendo-se mudado esta soluccedilatildeo ateacute
as bandas se encontrarem bem definidas
Determinou-se a mobilidade relativa das bandas correspondentes aos marcadores de elevado peso
molecular e traccedilou-se a recta de calibraccedilatildeo ndash log(PM) vs log(Rf) onde Rf corresponde agrave
mobilidade relativa Por interpolaccedilatildeo determina-se o PM das proteiacutenas isoladas sabendo a sua
mobilidade relativa Por comparaccedilatildeo com os valores descritos na literatura pode concluir-se
acerca do grau de purificaccedilatildeo
34 Mediccedilatildeo da actividade enzimaacutetica de S1
341 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
13
tampatildeo de concentraccedilatildeo 05 M Tris-HCl pH 68 14
tampatildeo de amostra 320 mM Tris-HCl pH 68 04 M szlig-mercaptoetanol SDS 8 glicerol 15 (vv) e roxo de m-
cresol 0024 15
tampatildeo de electroforese da tina superior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS 16
tampatildeo de electroforese da tina inferior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS acetato de soacutedio
01 M 17
soluccedilatildeo corante ldquoCoomassie Blue R-250rdquo contendo metanol 45 (vv) e aacutecido aceacutetico glacial 10 (vv) 18
soluccedilatildeo descorante metanol 10 (vv) e aacutecido aceacutetico glacial 10 vv)
45
O meacutetodo de Fiske-Subarow modificado consiste na reacccedilatildeo do molibdato de amoacutenio com o
fosfato inorgacircnico em soluccedilatildeo aacutecida com formaccedilatildeo do aacutecido fosfomoliacutebdico Seguidamente daacute-
se a adiccedilatildeo de um agente redutor que provoca a reduccedilatildeo do composto a fosfomolibdato que forma
um complexo de cor azulada cuja absorvacircncia maacutexima ocorre para um comprimento de onda de
660 nm tornando possiacutevel a sua detecccedilatildeo por meacutetodos espectrofotomeacutetricos As actividades Ca2+
-
ATPaacutesica e K+EDTA-ATPaacutesica de S1 foram determinadas atraveacutes deste meacutetodo de acordo com
o descrito por Taussky e Shorr (1953) com mediccedilatildeo da quantidade de fosfato (Pi) libertado
atraveacutes da hidroacutelise de ATP
Para tal obteve-se primeiramente a recta de calibraccedilatildeo com as amostras presentes na Tabela 3II
utilizando uma soluccedilatildeo stock de 5 mM KH2PO4
Tabela 3II ndash Composiccedilatildeo das diferentes amostras utilizadas para determinar a recta de calibraccedilatildeo atraveacutes do meacutetodo
de Fiske-Subbarow modificado
KH2PO4 (L)
([KH2PO4]stock= 5mM)
PO43-
(nmole) H2O (uL)
0 0 1780
5 25 1775
10 50 1770
20 100 1760
40 200 1740
100 500 1680
A cada tubo de Eppendorf adicionaram-se seguidamente 200 L de soluccedilatildeo J19
e 20 L de agente
redutor20
homogeneizando no voacutertex apoacutes cada adiccedilatildeo Incubou-se durante 10 minutos e mediu-
se a Abs660 nm num espectrofotoacutemetro com intervalos de tempo iguais entre cada amostra Traccedilou-
se a recta de calibraccedilatildeo (Abs660 nmvs nmol PO43-
) a partir dos valores obtidos
Para determinar a actividade Ca2+
-ATPaacutesica de S1 prepararam-se tubos de Eppendorf com 400
L de meio de reacccedilatildeo21
25 L CaCl2 ([CaCl2]stock= 100 mM) 25 L ATP ([ATP]stock= 100 mM)
com [S1]ensaios=01 mgmL Perfez-se se necessaacuterio com H2O MilliQ ateacute perfazer um volume de 1
mL Para determinar a actividade K+EDTA-ATPaacutesica utilizou-se uma concentraccedilatildeo idecircntica de
19
Soluccedilatildeo J 5 N H2SO4 25 molibdato de amoacutenio 20
Agente redutor 5 mg aacutecido 1-amino-2-naftol-4-sulfoacutenico 30 mg bissulfito de soacutedio 30 mg sulfito de soacutedio em 26
mL H2O 21
meio de reacccedilatildeo 125 mM Tris-HCl pH 80 0575 M KCl
46
proteiacutena nos ensaios (01 mgmL) e o mesmo meio de reacccedilatildeo Aos tubos de Eppendorf
adicionou-se ainda 123 L de KCl ([KCl]stock = 3M) 25 L de ATP 10 L de EDTA
([EDTA]stock= 100 mM )e o volume de H2O MilliQ necessaacuterio para perfazer 1 mL Em ambas as
situaccedilotildees o uacuteltimo reagente adicionado foi o ATP uma vez que se trata do substrato da reacccedilatildeo
enzimaacutetica
A cineacutetica decorreu a 25ordmC durante 5 minutos para a actividade Ca2+
-ATPaacutesica e durante 3
minutos para a actividade K+EDTA-ATPaacutesica Apoacutes o periacuteodo de incubaccedilatildeo transferiu-se 150
L de amostra para tubos de Eppendorf contendo igual volume de TCA 10 Colocou-se no gelo
e homogeneizou-se no voacutertex Seguidamente centrifugou-se a 1500 g durante 10 minutos e
recolheu-se 200 L de sobrenadante para novos tubos de Eppendorf Adicionou-se a estes 158
mL de H2O MilliQ e 200 L de soluccedilatildeo J Homogeneizou-se no voacutertex e adicionou-se 20 L de
reagente redutor repetindo-se a homogeneizaccedilatildeo Incubou-se durante 10 minutos e mediu-se a
Abs660 nm com o mesmo intervalo de tempo de leitura para cada amostra Por intrapolaccedilatildeo a partir
da recta de calibraccedilatildeo obtida anteriormente determinou-se a quantidade de Pi libertado em cada
ensaio A partir destes resultados determinaram-se as actividades Ca2+
-ATPaacutesica e K+EDTA-
ATPaacutesica do S1 em nmol PO43-
minmg proteiacutena e em s-1
342 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
Da actividade Mg2+
-ATPaacutesica de S1 resulta a formaccedilatildeo dos produtos da reacccedilatildeo de
hidroacutelise do ATP - ADP e Pi Na presenccedila de ADP fosfoenolpiruvato (PEP) e da enzima piruvato
cinase (PK) os produtos resultantes satildeo ATP e piruvato Por acccedilatildeo da enzima lactato
desidrogenase (LDH) este uacuteltimo eacute convertido em lactato ocorrendo simultaneamente a oxidaccedilatildeo
do NADH a NAD+ monitorizada atraveacutes da Abs340 nm Quando o S1 de miosina se encontra no
meio reaccional o ATP eacute hidrolisado formando-se novamente ADP e Pi o que permite a
formaccedilatildeo de um ciclo (fig 21) Se as enzimas se encontrarem em excesso a velocidade de
conversatildeo do NADH eacute limitada pela produccedilatildeo de ADP e Pi Como o ATP eacute regenerado em cada
ciclo a sua concentraccedilatildeo inicial eacute sempre mantida (Bagshaw 1982)
Figura 33ndash Representaccedilatildeo esquemaacutetica do meacutetodo de enzimas acopladas
47
O protocolo utilizado e seguidamente descrito foi adaptado de Cuenda et al (1990 ndash Ver Refordf) A
uma cuvette de 1 mL adicionou-se S1 para uma concentraccedilatildeo final de 005 mgmL 800 L de
meio de reacccedilatildeo22
10 L de PEP ([PEP]stock = 42 mM) 10 L de PK+LDH 11 ([PK+LDH]stock =
1 mgmL) e 15 L de NADH ([NADH]stock=25 mM) Iniciou-se a monitorizaccedilatildeo da reacccedilatildeo
atraveacutes da leitura de Abs340 nm apoacutes a adiccedilatildeo de 20 L de ATP ([ATP]stock=100 mM) A
actividade Mg2+
-ATPaacutesica eacute obtida atraveacutes do declive correspondente agrave oxidaccedilatildeo do NADH Para
a mediccedilatildeo da Mg2+
-ATPase de S1 estimulada por actina adicionou-se F-actina para uma
concentraccedilatildeo final de 2 microM sendo a actividade obtida tambeacutem atraveacutes do declive correspondente
agrave oxidaccedilatildeo do NADH apoacutes a adiccedilatildeo da proteiacutena
35 Interaccedilatildeo entre vanaacutedio e subfragmento-1 de miosina
351 Preparaccedilatildeo das soluccedilotildees de vanaacutedio
As soluccedilotildees de vanadato foram preparadas em aacutegua MilliQ a partir de metavanadato de amoacutenio
(NH4VO3) numa concentraccedilatildeo stock de 50 mM com agitaccedilatildeo magneacutetica e aquecimento (~30 ordmC)
Esta soluccedilatildeo com um valor compreendido entre 6 e 7 designa-se por ldquometavanadatordquo e eacute
constituiacuteda pelas esspeacutecies V1 V2 V4 e V5 (Ref) Acidificando a soluccedilatildeo de metavanadato por
adiccedilatildeo de HCl ateacute um valor de pH proacuteximo de 4 observa-se uma mudanccedila de cor de amarelo
claro para laranja indicativa da formaccedilatildeo da espeacutecie decameacuterica de vanadato De seguida e
imediatamente antes da realizaccedilatildeo dos ensaios descritos o valor do pH da soluccedilatildeo de
decavanadato foi aumentado para cerca de 75 por adiccedilatildeo de NaOH A soluccedilatildeo (2) designada
por ldquometavanadatordquo natildeo foi posteriormente alterada ateacute agrave sua adiccedilatildeo para a realizaccedilatildeo dos ensaios
As soluccedilotildees foram armazenadas a 4ordmC e mantidas em gelo aquando da sua utilizaccedilatildeo
A soluccedilatildeo de V(IV) foi preparada em condiccedilotildees anaeroacutebias por diluiccedilatildeo de sulfato de vanadil
(VOSO4) para uma concentraccedilatildeo final de 5 mM em H2O MilliQ saturada com N2 gasoso
352Anaacutelise por espectroscopia de UV-Vis
A concentraccedilatildeo da espeacutecie decameacuterica em soluccedilatildeo depende do periacuteodo de tempo apoacutes a diluiccedilatildeo
no meio de reacccedilatildeo apropriado (Ref) A velocidade de desoligomerizaccedilatildeo da espeacutecie decameacuterica
pode ser analisada por espectroscopia de absorccedilatildeo na regiatildeo UVVis uma vez que esta espeacutecie
apresenta duas bandas caracteriacutesticas na regiatildeo do visiacutevel (a 358 e 400 nm) que natildeo se verifica
em quaisquer outras espeacutecies oligomeacutericas As bandas supramencionada sapresentam-se bem
22
Meio de reacccedilatildeo 3125 mM Tris-HCl pH 71 375 mM MgCl2
48
resolvidas atraveacutes da obtenccedilatildeo da derivada de 2ordf ordem do espectro apresentando a forma de 2
vales com miacutenimos centrados nos comprimentos de onda supracitados com dependecircncia linear
relativamente agrave concentraccedilatildeo de decavanadato total (Ramos et al 2006)
Assim a estabilidade da espeacutecie decameacuterica apoacutes diluiccedilatildeo da soluccedilatildeo de decavanadato no meio de
reacccedilatildeo dos ensaios (tampatildeo D) para uma concentraccedilatildeo final de 1 mM em vanadato total foi
caracterizada atraveacutes da leitura da Abs400 em funccedilatildeo do tempo tendo-se determinado qual a
ordem de reacccedilatildeo e o tempo de meia-vida da mesma Com base em estudos anteriores acerca da
estabilidade do V10 na presenccedila de diferentes proteiacutenas (Ramos et al 2006) determinou-se
igualmente o tempo de meia vida apoacutes a adiccedilatildeo de 20 microM de S1 de miosina ao meio reaccional
353 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
Os espectros de RMN do isoacutetopo 51
V foram realizados num espectroacutemetro Bruker AM-400 MHz
equipado com uma sonda multinuclear inversa de 5 mm Em tubos de RMN de 5 mm de diacircmetro
preparou-se soluccedilotildees de 500 L de deca- ou metavanadato correspondentes a um intervalo de
concentraccedilotildees de 1 a 10 mM suplementadas com 10 D2O Os espectros foram adquiridos agrave
temperatura ambiente com uma largura espectral de 23 923 Hz um tempo de aquisiccedilatildeo de 0086
s tempo de relaxaccedilatildeo de 001 s e uma sequecircncia de pulsos a 90ordm Utilizou-se a transformada de
Fourier com alargamento das bandas de 20Hz para o processamento dos espectros Os desvios
quiacutemicos das ressonacircncias de 51
V foram obtidos em relaccedilatildeo agrave espeacutecie VOCl3cuja frequecircncia de
ressonacircncia eacute considerada o padratildeo sendo todas as outras referenciadas a partir desta em ppm
As aacutereas dos sinais do RMN correspondentes agraves espeacutecies oligomeacutericas presentes foram obtidas
por integraccedilatildeo dos sinais As concentraccedilotildees relativas das espeacutecies foram calculadas com recurso agrave
equaccedilatildeo
onde A[Vx] corresponde agrave aacuterea referente agrave ressonacircncia da espeacutecie Vx com nordm de agregaccedilatildeo x
A[V]total agrave soma total das aacutereas medidas e [V] a concentraccedilatildeo total de vanaacutedio no ensaio
A teacutecnica de 51
V-RMN foi anteriormente aplicada em estudos de ligaccedilatildeo com S1 de miosina
(Ref) tendo sido demostrado que entre as espeacutecies V1 V2 V4 e V5 a espeacutecie tetrameacuterica possui a
maior afinidade para a miosina (Cremo et al) O desaparecimento ou alargamento dos sinais pode
implicar a ligaccedilatildeo de um dos oxovanadatos laacutebeis agrave miosina sem indicaccedilatildeo de qual uma vez que
49
se processa um equiliacutebrio entre eles (Crans 1990) Assim e tendo em conta as indicaccedilotildees de que
a espeacutecie V10 eacute distinta das restantes no que diz respeito agrave magnitude da interacccedilatildeo com a miosina
e inibiccedilatildeo da actividade ATPaacutesica estimulada por actina (Aureliano 2000 Tiago et al 2002)
torna-se fundamental investigar a interacccedilatildeo das diferentes espeacutecies de vanadato com o S1 de
miosina atraveacutes da teacutecnica de 51
V-RMN
Para tal obtiveram-se espectros de 51
V-RMN resultantes da titulaccedilatildeo de uma soluccedilatildeo 5 mM de
decavanadato total com concentraccedilotildees crescentes de S1 de miosina (0 5 10 15 e 20 microM) em
tampatildeo D Realizou-se ainda ensaios na presenccedila de 2 mM de decavanadato e 2 mM de ATP na
presenccedila e na ausecircncia de S1 de miosina de modo a verificar o efeito que estes ligandos naturais
da enzima teria na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o subfragmento-1
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
Os grupos sulfidrilo (-SH) das cisteiacutenas satildeo resiacuteduos de aminoaacutecidos extremamente sensiacuteveis agrave
oxidaccedilatildeo resultando na sua oxidaccedilatildeo (Dalle-Donne et al 1999) Logo a quantificaccedilatildeo destes
resiacuteduos eacute bastante importante nos estudos de interacccedilatildeo com vanaacutedio jaacute que os resiacuteduos de Cys
com ecircnfase para o SH1 e o SH2 satildeo bastante importantes na monitorizaccedilatildeo das modificaccedilotildees
funcionais da regiatildeo da cabeccedila
O procedimento seguidamente apresentado foi originalmente descrito por Gutieacuterrez-Merino
(1987)) Consiste na reacccedilatildeo entre os grupos ndashSH natildeo oxidados das proteiacutenas e o reagente de
Ellman (DTNB -55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico)) - formando-se o aniatildeo CNT (aniatildeo 3-
carboxilato-4-nitrotiofenolato) numa estequiometria de 11 Este apresenta um maacuteximo de
absorvecircncia a 412 nm (Robyt 1990) Assim apoacutes a incubaccedilatildeo da proteiacutena na presenccedila do metal
a quantidade de cisteiacutenas reduzidas por proteiacutena determina-se atraveacutes dos valores de Abs412
Deste modo colocou-se num tubo de Eppendorf o volume apropriado de S1 dialisado para que a
concentraccedilatildeo no ensaio fosse de 1 mgmL Adicionou-se o volume apropriado de decavanadato
para que a sua concentraccedilatildeo fosse de 0 10 50 100 250 500 ou 1000 M Perfez-se com soluccedilatildeo
D ateacute um volume final de 1 mL e incubou-se a 25ordmC por 20 minutos Apoacutes este periacuteodo colocou-
se 780 L da mistura reaccional numa cuvette de quartzo e adicionou-se 220 L de soluccedilatildeo K23
23
Tampatildeo K Tampatildeo fosfatohidrogenofosfato de soacutedio 025 M pH 72
50
Adicionou-se 15 L de DTNB (em soluccedilatildeo K [DTNB]stock=100 mM) ao meio reaccional e
registou-se o valor de absorvecircncia medido equivalente agraves cisteiacutenas ldquoraacutepidasrdquo (SH1 e SH2
expostos ao solvente) Adicionou-se 001 g de SDS que provoca a desnaturaccedilatildeo da proteiacutena e
consequentemente a exposiccedilatildeo de todos os resiacuteduos de cisteiacutenas que se encontravam ateacute aiacute
inacessiacuteveis no interior da proteiacutena (cisteiacutenas totais) Estes resiacuteduos podem assim reagir com o
DTNB
Estimou-se o nuacutemero de resiacuteduos de cisteiacutenas em todas as amostras com recurso ao coeficiente de
extinccedilatildeo molar () do DTNB (ver ponto 3 dos Anexos) que foi determinado atraveacutes da titulaccedilatildeo
de L-Cisteiacutena com intervalo de concentraccedilotildees compreendidos entre 0 e 50 microM na presenccedila de
25microM de DTNB em tampatildeo D Os valores de absorvecircncia foram obtidos para um comprimento de
onda de 412 nm
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
3621 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
A teacutecnica mais eficiente de anaacutelise da interacccedilatildeo entre o iatildeo vanadilo e proteiacutenas como a
calmodulina e a transferrina entre outras permite a obtenccedilatildeo raacutepida de informaccedilatildeo no que toca agrave
conformaccedilatildeo e amplitude da referida interacccedilatildeo
Os espectros de Ressonacircncia Paramagneacutetica Electroacutenica (RPE) da espeacutecie paramagneacutetica V(IV)
foram realizados a 70K num espectroacutemetro Bruker EMX-300 X-band Todas as soluccedilotildees
utilizadas e amostras foram preparadas em anaerobiose numa cacircmara de luvas de modo a evitar a
reduccedilatildeo de V(IV) a V(V) Em tubos capilares com 02 mL de amostra manteve-se a concentraccedilatildeo
de S1 a 20 M e variou-se a concentraccedilatildeo de vanadilo (0 25 5 10 15 25 50 75 100 150
200 500 e 1000 M [VOSO4]stock=5 mM) em tampatildeo D Apoacutes obtenccedilatildeo dos espectros estes
foram tratados atraveacutes do software SimFonia e WinEPR tendo-se obtido um graacutefico com a
variaccedilatildeo da intensidade do sinal de RPE em funccedilatildeo da concentraccedilatildeo de sulfato de vanadilo
presente Realizaram-se ainda simulaccedilotildees de espectros obtidos com 20 microM de S1 e 1000 microM de
V(IV) com vista agrave obtenccedilatildeo dos ligandos provaacuteveis do vanadilo atraveacutes da anaacutelise da constante
de acoplamento hiperfina (A) e do factor g (g)
4 Conclusotildees gerais
Apesar de jaacute ter sido demonstrado que o vanaacutedio inibe a Na+K
+-ATPase da miosina a funccedilatildeo
deste elemento no tecido muacutesculo esqueleacutetico continua largamente incompreendida A enorme
complexidade quiacutemica deste metal especialmente no que concerne agrave oligomerizaccedilatildeo e reacccedilotildees
51
redox constitui um obstaacuteculo agrave compreensatildeo do comportamento do vanaacutedio nos organismos
vivos Assim a caracterizaccedilatildeo da quiacutemica deste elemento eacute de grande utilidade antes de iniciar o
estudo da sua interacccedilatildeo com o presente sistema
A anaacutelise por espectroscopias de 51
V-RMN e de UV-Vis de soluccedilotildees de vanadato diluiacutedas nos
meios de reacccedilatildeo utilizados no presente relatoacuterio (10 mM HEPES pH 70 25 mM KCl) mostrou
claramente que a espeacutecie decameacuterica (V10) possui um tempo de vida suficientemente longo para a
determinaccedilatildeo do seu efeito isolado ao contraacuterio do que se verifica com as espeacutecies mono- di-
tetra- e pentameacutericas (V1 V2 V4 e V5) Por outro lado a anaacutelise verificou ainda que apoacutes diluiccedilatildeo
das soluccedilotildees de decavanadato no meio de reacccedilatildeo coexistem as espeacutecies V10 e V1 pelo que os
efeitos putativos desta uacuteltima deveratildeo ser subtraiacutedos aquando da monitorizaccedilatildeo dos efeitos da
espeacutecie decameacuterica
O vanaacutedio (V) (vanadato) inibe as enzimas que catalizam reacccedilotildees de transferecircncia de grupos
fosfato como a ATPase da miosina [Goodno 1979] devido agrave sua analogia estrutural com o iatildeo
fosfato Na sua utilizaccedilatildeo como inibidor enzimaacutetico natildeo eacute tida em conta a contribuiccedilatildeo individual
das espeacutecies oligomeacutericas considerando-se normalmente a espeacutecie V1 como inibidora efectiva
[Goodno e Taylor 1982] No presente relatoacuterio verificou-se por espectroscopia de 51
V-RMN que
outros oligoacutemeros de vanadato interagem com o S1 de miosina tendo as espeacutecies V10 e V4 os
maiores niacuteveis de afinidade A sua interacccedilatildeo envolve forccedilas electrostaacuteticas e eacute afectada
negativamente por ligandos naturais de enzima como eacute o caso do ATP
A exposiccedilatildeo do S1 de miosina (005 mgmL) a decavanadato conduz agrave inibiccedilatildeo do capacidade da
proteiacutena de estimular a actividade Mg2+
-ATPaacutesica com um valor de IC50 de 266 plusmn 07 M
decavanadato sendo o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM
decavanadato (80) Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM decavanadato sendo
o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM decavanadato (60)
exercendo um efeito de impedimento agrave ligaccedilatildeo entre o vanaacutedio e o S1
A uma concentraccedilatildeo de 27 microM decavanadato verifica-se a oxidaccedilatildeo de um dos resiacuteduos de Cys
de S1 de miosina seguida pela oxidaccedilatildeo gradual de outro resiacuteduo de Cys ateacute agrave concentraccedilatildeo de
1000 microM Ambos os resiacuteduos oxidados natildeo se encontravam expostos ao solvente utilizado A
oxidaccedilatildeo dos resiacuteduos permite sugerir que esta modificaccedilatildeo oxidativa sofrida pelo S1 contribui
para a diminuiccedilatildeo da sua actividade de ATPase
52
Por espectroscopia de RPE revelaram-se os possiacuteveis ligandos equatoriais do vanaacutedio aquando da
sua interacccedilatildeo com o S1 de miosina como sendo duas moleacuteculas de aacutegua um ligando contendo o
grupo R-OH e outro contendo o grupo R=NR
Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados de
oxidaccedilatildeo (IV) e (V) no S1 de miosina para concentraccedilotildees facilmente alcanccedilaacuteveis no ambiente
terrestre
53
5 Estudos futuros
No sentido do melhor esclarecimento das alteraccedilotildees moleculares e estruturais que ocorrem no
subfragmento 1 quando exposta ao vanaacutedio que conduzem a uma diminuiccedilatildeo da sua funccedilatildeo
sugere-se
- Quantificaccedilatildeo das cisteiacutenas reduzidas de S1 na presenccedila de metavanadato
- Determinaccedilatildeo das actividades Ca2+
- e K+-ATPaacutesica do S1 de miosina na presenccedila de
decavanadato
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina estimulada por actina
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina na presenccedila de
metavanadato
- Realizaccedilatildeo de RMN 2D com variaccedilatildeo das concentraccedilotildees das soluccedilotildees de decavanadato e
metavanadato na presenccedila de S1
-Mutageacutenese dos resiacuteduos de Cys doS1 de miosina de modo a verificar quais os resiacuteduos que
sofrem oxidaccedilatildeo na presenccedila de decavanadato
- Determinaccedilatildeo por calorimetria (DSC Differential Scanning Calorimetry) da variaccedilatildeo da
entalpia (∆H) e da temperatura de desnaturaccedilatildeo do S1 de miosina na presenccedila de vanaacutedio uma
vez que estas propriedades caracteriacutesticas de cada proteiacutena satildeo alteradas apoacutes ocorrecircncia de
modificaccedilotildees estruturais
- Anaacutelise por SDS-PAGE de S1 de miosina apoacutes tratamento com vanaacutedio de modo a determinar
quais os polipeacuteptidos eventualmente formados apoacutes clivagem da proteiacutena
54
6 Referecircncias bibliograacuteficas
- Sellers J Myosins 2nd Edition Oxford University Press 1999
- L Stryer inBiochemistry (Ed WH Freeman 2005 pp
Refordfas Materiais e Meacutetodos
[A] S S Margossian SLowey Methods Enzymol1982 85 55-71
[B] JD Pardue JA Spudich Methods Enzymol 1982 85 164-181
[C] M Bradford Anal Biochem1976 72 248-254
[D] H H Taussky E Shorr J Biol Chem 1953 202 675-685
[E] C R Bagshaw in Outline Studies in Biology ndash Muscle Contraction Chapman and Hall Ed
1982
[F] A Cuenda et al Eur J Biochem1990 194 663-670
55
Anexos
1 Determinaccedilatildeo da recta de calibraccedilatildeo meacutetodo de Bradford
Figura1ndashRepresentaccedilatildeo graacutefica da recta de calibraccedilatildeo (Abs595 nm vs [BSA]) utilizada para o meacutetodo de Bradford
2 Determinaccedilatildeo da recta calibraccedilatildeo para a electroforese em condiccedilotildees desnaturantes
Figura 2 ndash Representaccedilatildeo graacutefica da recta de calibraccedilatildeo (log(Peso Molecular) vs Rf dos marcadores de peso
molecular) onde Rf indica determinada atraveacutes do padratildeo de peso molecular
(poccedilo (a))
3Caacutelculo do coeficiente de extinccedilatildeo molar () para a quantificaccedilatildeo de cisteiacutenas reduzidas
atraveacutes do meacutetodo de Ellman
Traccedilou-se a recta de Abs412 nm meacutedia vs [L-Cisteiacutena] de modo a determinar o valor do coeficiente
de extinccedilatildeo molar definido como o declive daquela recta Assim =8400 M-1
cm-1
como
podemos verificar na Fig 3
A595 nm = 00546[BSA] + 00209 Rsup2 = 099051
Ab
s 59
5 n
m
[BSA] (mgmL)
log PM = -1177 Rf + 2247 Rsup2 = 098957
log
PM
Mobilidade relativa (Rf)
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero

20
pela estrutura das miofibrilas Os estudos desenvolvidos por Smith et al (1990)[AV]
determinaram queo iatildeo monovanadato natildeo apresenta um efeito pronunciado na inibiccedilatildeo da
actividadeacto-S1-miosina-ATPaacutesica excepto em concentraccedilotildees superiores a 900microM a partir das
quais existe produccedilatildeo de espeacutecies oligomeacutericas para as quais o efeito inibitoacuterio seraacute mais
elevadoA espeacutecie monomeacuterica de vanadato consegue adoptar facilmente uma geometria do tipo
bipiramidal trigonal que mimetiza o intermediaacuterio formado (MgADP
Pi)aquando da reacccedilatildeo de
hidroacutelise de ATP no local activo da miosina [AW] De facto a interacccedilatildeo do vanaacutedio com a
miosina eacute inesperadamente estaacutevel na presenccedila de ADP Esta interacccedilatildeo resulta na formaccedilatildeo de
um complexo enzimaacutetico inactivo e estaacutevel com estequiometria de 11 ADPvanadato por siacutetio
activo da proteiacutena A formaccedilatildeo do complexo estaacutevel miosina-vanadato foi proposta por Goodnoet
al (1982) [AS] Nesta ocorre um preacute-equiliacutebrio raacutepido que induz a formaccedilatildeo de um complexo
ternaacuterio reversiacutevel (MADP
V) com formaccedilatildeo lenta do isoacutemero M
ADP
V A diferenccedila entre os
compostos de vanaacutedio e de fosfato centra-se no tempo de meia vida (t12) do Pi ou do V (no
primeiro t12=12 s enquanto no segundo t12=24 h) (Refordf) A diferenccedila dever-se-aacute agrave possiacutevel
complexaccedilatildeo do vanaacutedio com grupos nucleofiacutelicos no siacutetio activo o que provocaria uma
estabilizaccedilatildeo da miosina na conformaccedilatildeo MADP
V
No que respeita aos estudos realizados com a espeacutecie tetrameacuterica Cremo et al determinou que a
cadeia pesada do S1 de miosina pode ser clivada em dois locais distintos (designados V1 e V2)
pela espeacutecie V4 apoacutes irradiaccedilatildeo com luz UV na presenccedila de concentraccedilotildees de vanadato na ordem
dos milimolar e na ausecircncia de nucleoacutetidos ou metais divalentes
Jaacute no que concerne aos estudos realizados com ecircnfase na espeacutecie decameacuterica foi sugerido
porRamos et al (2006) [AK]que a interacccedilatildeo do decavanadato com a G-actina favorecida pela
polimerizaccedilatildeo desta uacuteltima estabiliza a espeacutecie V10 e induz a inibiccedilatildeo da formaccedilatildeo de F-actina A
estabilizaccedilatildeo da espeacutecie V10 por proteiacutenas tansmembranares e do citoesqueleto pode ser a razatildeo
pela qual ocorre a toxicidade do decavanadato reportada na avaliaccedilatildeo dos efeitos do V(V) nos
sistemas bioloacutegicosTiago et al (2007) [AR] concluiram que o V10 apresenta uma clara preferecircncia
de ligaccedilatildeo ao ldquoback-doorrdquo da S1-miosina-ATPase o que se verifica apenas na estrutura
conformacional desimpedida com acesso facilitado ao local de ligaccedilatildeo do Pi Sugere tambeacutem que
o V10 actua como um ldquoback-door stoprdquo bloqueando estaabertura do fragmento de 50-kDa
necessaacuteria para a hidroacutelise do ATP--fosfato o que constitui uma explicaccedilatildeo simples do
comportamento natildeo competitivo da espeacutecie decameacuterica estimulando a utilizaccedilatildeo deste oligoacutemero
como uma feramenta da elucidaccedilatildeo das mudanccedilas conformacionais ldquoback-doorrdquo da miosina no
21
processo da contracccedilatildeo muscular(ref)Destaca-se ainda a descoberta da capacidade inibitoacuteria do
V10 em relaccedilatildeo agrave fosforilaccedilatildeo oxidativa mitocondrial [AX]
Finalmente no que concerne aos estudos realizados com a presenccedila de vaacuterias espeacutecies
oligomeacutericas Aureliano e Madeira (1994) [AY]observaram que na presenccedila de Ca2+
as espeacutecies
V4 e V10se ligam agrave Ca2+
-ATPase do retiacuteculo sarcoplasmaacutetico enquanto a espeacutecie V1 se liga apenas
na presenccedila de ATP Aparentemente o aumento da concentraccedilatildeo de Ca2+
tem o efeito contraacuterio da
inibiccedilatildeo da ATPase do retiacuteculo sarcoplasmaacutetico pelo V1 natildeo tendo no entanto efeito significativo
sobre a inibiccedilatildeo por parte da espeacutecie V10 Tiago et al (2002) [AP] concluiacuteram que em condiccedilotildees
proacuteximas agraves fisioloacutegicas a espeacutecie V10 difere das espeacutecies presentes na soluccedilatildeo de metavanadato
devido agrave sua forte interacccedilatildeo com a miosina fosforilada e agrave sua inibiccedilatildeo da actividade ATPaacutesica
da miosina Para aleacutem disso o ATP diminui a afinidade da miosina para o V4 induzindo a
interacccedilatildeo com V1 e natildeo tendo qualquer efeito no que diz respeito ao V10 Foi tambeacutem proposto
[AQ] que a ligaccedilatildeo das espeacutecies oligomeacutericas de vanadato aoS1 de miosina nos locais onde o
fosfato coordenaeacute provavelmente a causa da inibiccedilatildeo da actividade ATPaacutesica da miosina
estimulada por actina atraveacutes da prevenccedilatildeo da troca entre ATPADP sendo esta favorecida para
aniotildees de vanadato com valecircncias superiores como eacute o caso do V10
Na presenccedila de actina as velocidades de dissociaccedilatildeo do Pi e do V aumentam consideravelmente
este uacuteltimo em menor quantidade Assim a ligaccedilatildeo da actina ao complexo MADP
V e
Pi eacute
bastante fraca pelo que se sugere que o ADPV teria um efeito de relaxamento semelhante ao
ATP No entanto a capacidade do vanaacutedio em inibir a actividade ATPaacutesica da miosina e da
actomiosina eacute diferenciada uma vez que na ausecircncia de actina a inibiccedilatildeo eacute total para
concentraccedilotildees na gama dos M mas na sua presenccedila a inibiccedilatildeo eacute totalmente conseguida apenas
em concentraccedilotildees na gama dos mM (Ref) No entanto eacute necessaacuterio considerar a possiacutevel
formaccedilatildeo de espeacutecies oligomeacutericas de vanadato cuja afinidade e efeito na enzima seratildeo
diferentes Assim pode concluir-se que o efeito de inibiccedilatildeo observado para a actividade ATPaacutesica
de miosina estimulada por actina se pode dever agrave presenccedila das diferentes espeacutecies oligomeacutericas o
que foi jaacute sugerido por Aureliano (2000) [AZ] ndash as espeacutecies tetrameacuterica e decameacuterica de vanadato
interactuariam com a miosina com alteraccedilatildeo da sua interacccedilatildeo com a actina No entanto natildeo eacute
conhecida a contribuiccedilatildeo absoluta das restantes espeacutecies oligomeacutericas para a inibiccedilatildeo da ATPase
da miosina estimulada por actina (Refs)
14 Objectivos do projecto
Pretende-se com este projecto o isolamento purificaccedilatildeo e caracterizaccedilatildeo da actina do muacutesculo
22
esqueleacutetico de coelho sendo esta uacuteltima tambeacutem realizada para o sufragmento-1 de miosina
Seguidamente realizar-se-aacute a quantificaccedilatildeo e caracterizaccedilatildeo das proteiacutenas por espectroscopia UV-
Vis e SDS-PAGE e a caracterizaccedilatildeo das soluccedilotildees de vanaacutedio por UV-Vis e ressonacircncia magneacutetica
nuclear (RMN) Por uacuteltimo a interacccedilatildeo do S1 de miosina com o vanaacutedio seraacute avaliada atraveacutes
das teacutecnicas de UVVis RMN e ressonacircncia paramagneacutetica electroacutenica (RPE)
2 Discussatildeo de Resultados
21 Caracterizaccedilatildeo das proteiacutenas isoladas
211Anaacutelise do estado de pureza
O estado de pureza das soluccedilotildees de actina e de S1 foi analisado por electroforese em gel de
poliacrilamida em condiccedilotildees desnaturantes (SDS-PAGE) (Fig 22) que separa as moleacuteculas de
acordo com o seu peso molecular (PM) A concentraccedilatildeo de acrilamida escolhida (10) permite a
detecccedilatildeo simultacircnea de componentes com peso molecular inferior a 20 kDa e outros com elevado
peso molecular
Figura 21 ndash Fotografia de gel de SDS-PAGE (10 acrilamida) Poccedilo (a)
10 L do marcador de elevado peso molecular () Poccedilo (b) 20 L de [G-
actina]=1 mgmL Poccedilo (c) 20 L de [S1]=1 mgmL Aplicou-se uma
voltagem de V durante h e min
A partir da recta de calibraccedilatildeo obtida log PM = -1177 Rf + 2247 (ver ponto 2 da secccedilatildeo
Anexos) pode verificar-se a presenccedila de actina na amostra do poccedilo (b) presente numa banda
bastante intensa (PMaparente=40 kDa) bem como a presenccedila minoritaacuteria de outros componentes
desconhecidos que natildeo teratildeo sido eliminados durante o processo de extracccedilatildeo e purificaccedilatildeo e natildeo
satildeo relevantes para o estudo em questatildeo Na amostra do poccedilo (c) nota-se a presenccedila de S1
(PMaparente=92 kDa) bem como a de outros componentes cuja banda eacute ligeiramente menos intensa
Estes poderatildeo ser fragmentos proteoliacuteticos de miosina e de S1 Estaacute ainda presente uma bada de
23
maior intensidade que poderaacute ser a proteiacutena-C (PM=135 kDa) uma vez que a sua remoccedilatildeo
completa da amostra necessitaria de um passo adicional de cromatografia de troca ioacutenica [BA]
No entanto natildeo podemos confirmar que se trata desta proteiacutena ou de qualquer outra uma vez que
se encontra fora do intervalo de migraccedilatildeo dos marcadores de peso molecular
212 Determinaccedilatildeo da concentraccedilatildeo proteica
2121Subfragmento-1 de miosina
A partir da recta de calibraccedilatildeo obtida pelo meacutetodo de Bradford [BB](ver secccedilatildeo 3311 do
Procedimento Experimental) A595nm = 00546[BSA] + 00209 (ver secccedilatildeo de Anexos)foi possiacutevel
determinar um intervalo de concentraccedilatildeo de S1 de miosina compreendido entre 226 e 117(plusmn
423M) (203 e 1056 (plusmn 381 mgmL)) (n=8) A concentraccedilatildeo meacutedia de S1 de miosina apoacutes
diaacutelise contra a soluccedilatildeo tampatildeo 10 mM HEPES pH 75 25 mM KCl determinada pela leitura da
Abs280 nmfoi de 20430M (1803 mgmL)(n=7)
As amostras a quantificar atraveacutes do meacutetodo de Bradford natildeo sofreram diaacutelise o que permite
explicar a concentraccedilatildeo superior de S1 de miosina em comparaccedilatildeo com o valor obtido atraveacutes do
meacutetodo directo Esta razatildeo aliada agrave sensibilidade e rapidez superiores deste uacuteltimo fez com que
se optasse pelo meacutetodo directo para a quantificaccedilatildeo da grande maioria dos ensaios
2122Actina
A concentraccedilatildeo de actina foi determinada quando esta se encontrava na forma monomeacuterica (G-
actina) por mediccedilatildeo da leitura directa da Abs290 nm tendo-se obtido uma concentraccedilatildeo meacutedia de
691481M (290202 mgmL) (n=3) em 2 mM Tris-HCl pH 80 02 mM CaCl2 02 mM Na-
ATP Ao comprimento de onda utilizado (290 nm) absorvem fortemente apenas os resiacuteduos de
Trp pelo que a mediccedilatildeo efectuada tem menor sensibilidade do que se tivesse sido efectuada a 280
nm uma vez que a actina apresenta apenas 4 resiacuteduos deste aminoaacutecido embora tenha 16
resiacuteduos de Tyr
22 Mediccedilatildeo da actividade enzimaacutetica de S1 de miosina
221 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
As actividades ATPaacutesicas de S1 de miosina dependentes de Ca2+
e K+foram analisadas pelo
caacutelculo das velocidades de consumo inicial do fosfato inorgacircnico (Pi) Definiu-se os tempos de 5
e 3 minutos para a mediccedilatildeo da Ca2+
e da K+EDTA-ATPase respectivamente Os valores obtidos
24
a partir do protocolo presente no ponto 341 da secccedilatildeo Procedimento experimental encontram-se
descritos na Tabela 2I
Tabela 2I ndash Acrividades natildeo-fisioloacutegicas de subfragmento-1 determinadas pelo meacutetodo de Fiske-Subarow
modificado (n=2)
Proteiacutena
Actividade
ATPaacutesica natildeo
fisioloacutegica
Actividade (nmol
Piminmg proteiacutena) Actividade (s-1)
S1
Ca2+
-ATPase 1400 plusmn 30 2100 plusmn 0045
K+EDTA-ATPase 2334 plusmn 50 3500 plusmn 0075
As actividades natildeo-fisioloacutegicas reflectem o estado de oxidaccedilatildeo dos resiacuteduos expostos de Cys 707
(SH1) e 697 (SH2) Se o grupo SH1 sofrer oxidaccedilatildeo a actividade ATPaacutesica dependente de Ca2+
aumenta e a actividade dependente de K+ diminui Por outro lado se o grupo SH2 sofrer
oxidaccedilatildeo regista-se a perda completa destas actividades
De acordo com Margossian e Lowey (1982) [BA] as actividades ATPaacutesica de S1 de miosina
dependente de Ca2+
e K+EDTA da miosina satildeo respectivamente de 24 e 112 s-1
Estaacute tambeacutem
descrito que a actividade ATPaacutesica dependente de K+ eacute em meacutedia cerca de 3 vezes superior agrave
dependente de Ca2+
reflectindo a quantidade das preparaccedilotildees A razatildeo obtida entre os valores das
actividades ATPaacutesicas dependentes de K+ e Ca
2+ eacute de 166 Os valores obtidos para as actividades
(Ca2+
-ATPase=21 s-1
K+EDTA-ATPase= 35 s
-1) satildeo inferiores agrave valores obtidos por
Margossian e Lowey [BA] e Tiago [BC] com forccedila ioacutenica inferior O valor obtido para a
actividade ATPaacutesica dependente de Ca2+
eacute superior ao obtido por Polosukhina et al (2000) [BD]
cujo meio de reacccedilatildeo continha MOPS e apresentava forccedila ioacutenica inferior Os dados
supramencionados comprovam a variaccedilatildeo da actividade a ATPaacutesica natildeo fisioloacutegica em relaccedilatildeo agrave
forccedila ioacutenica temperatura e pH
222 Mediccedilatildeo da actividadeMg2+
-ATPaacutesica de S1 pelo meacutetodo de enzimas acopladas
A actividade de Mg2+
-ATPase eacute a uacutenica que possui significado fisioloacutegico sendo as restantes
bastante uacuteteis apenas para indicaccedilatildeo da qualidade de preparaccedilatildeo do S1[BA] A 0575 M KCl a
actina e o S1 formam uma soluccedilatildeo viscosa e tuacuterbida Apoacutes a adiccedilatildeo de MgATP o complexo acto-
S1 dissocia-se e a turbidez da soluccedilatildeo diminui Se a forccedila ioacutenica for reduzida a dissociaccedilatildeo do
complexo actoS1 que se encontra na forma de suspensatildeo eacute induzida O MgATP eacute rapidamente
hidrolisado diminuindo a sua concentraccedilatildeo no meio reaccional Nestas condiccedilotildees haacute
reassociaccedilatildeo e subsequente contracccedilatildeo dos filamentos de actina e de S1 com formaccedilatildeo de
25
precipitado O meacutetodo descrito traduz de forma bastante elementar o fenoacutemeno da contracccedilatildeo in
vivo e elimina o problema anterior da solubilizaccedilatildeo ineficiente da miosina de modo a obter uma
mediccedilatildeo exacta da actividade [A] A utilizaccedilatildeo de forccedilas ioacutenicas reduzidas aumenta a afinidade da
actina para o S1 permitindo contornar a extrema dificuldade em obter a concentraccedilatildeo proteica dos
muacutesculos (cerca de 100 mgmL do complexo actomiosina) As actividades Mg2+
-ATPaacutesicas de S1
de miosina foram determinadas a partir do declive da variaccedilatildeo da Abs340 relativo agrave oxidaccedilatildeo de
NADH em funccedilatildeo do tempo como eacute descrito no ponto 342 do Procedimento Experimental Os
valores obtidos para a actividade Mg2+
-ATPaacutesica encontram-se apresentados na Tabela 2II
Tabela 2II ndash Actividade Mg2+
-ATPaacutesica de subfragmento-1 de miosina determinadas pelo meacutetodo das enzimas
acopladas
Actividade (nmol
ATPmin mg(S1)) Actividade (s
-1)
1097 plusmn 269 164 plusmn 04
Ramos (2005) [J] obteve o valor de 01 s-1
para a actividade Mg2+
-ATPaacutesica de S1 de miosina
com forccedila ioacutenica intermeacutedia Tiago (2005) [BC] obteve o valor de 011 s-1 em condiccedilotildees de forccedila
ioacutenica reduzida em relaccedilatildeo agrave utilizada no presente relatoacuterio
23 Preparaccedilatildeo e caracterizaccedilatildeo das soluccedilotildees de vanaacutedio
231 Anaacutelise da composiccedilatildeo das soluccedilotildees de vanadato por espectroscopia de 51
V-RMN
efeito da concentraccedilatildeo da espeacutecie
A diluiccedilatildeo das soluccedilotildees stock de meta- e decavanadato iraacute determinar fortemente a composiccedilatildeo
em soluccedilatildeo dos diferentes oligoacutemeros de vanadato (ref) No caso do decavanadato o periacuteodo de
tempo apoacutes a diluiccedilatildeo iraacute tambeacutem ser um factor importante a considerar devido agrave decomposiccedilatildeo
da espeacutecie V10(ref)Torna-se assim fundamental determinar a composiccedilatildeo quiacutemica das soluccedilotildees
no meio de ensaio utilizado (HEPES 10 mM KCl 25 mM) de modo a correlacionaacute-la
correctamente com os possiacuteveis efeitos promovidos pelas diferentes espeacutecies de vanadato no
sistema em estudo
Aleacutem disso deve ser dado ecircnfase agraves importantes interacccedilotildees que ocorrem entre o vanaacutedio e os
tampotildees escolhidos O tampatildeo com menor nuacutemero e magnitude de interacccedilatildeo eacute o HEPES [AD]
seguido do Tris sendo este uacuteltimo especialmente aconselhado em concentraccedilotildees baixas para casos
de incompatibilidade enzimaacutetica ou quiacutemica redox No caso especial da reacccedilatildeo redox
NAD(P)HNAD(P) o tampatildeo adequado eacute o imidazole de modo a natildeo haver reacccedilatildeo com o
26
vanadato Os tampotildees a evitar em qualquer situaccedilatildeo incluem a trietanolamina fosfato citrato e
glicina entre outros [AD] uma vez que todos interactuam fortemente com V(V) com formaccedilatildeo
de complexos estaacuteveis
Assim procedeu-se agrave anaacutelise da composiccedilatildeo das soluccedilotildees de meta- e decavanadato apoacutes diluiccedilatildeo
das soluccedilotildees stock (50 mM em vanadato total) no meio de reacccedilatildeo (tampatildeo D) atraveacutes da
espectroscopia de ressonacircncia magneacutetica nuclear de 51
V Nas figuras x1 e x2 apresentam-se os
espectros obtidos para as soluccedilotildees de deca- e metavanadato num intervalo de concentraccedilotildees de 1 a
10 mM A Tabela seguinte resume os paracircmetros espectroscoacutepicos obtidos para cada espeacutecie
presente nas soluccedilotildees analisadas
Figura 22ndash Espectros de RMN de 51
V de
concentraccedilotildees crescentes das espeacutecies oligomeacutericas
de vanadato obtidos apoacutes diluiccedilatildeo da soluccedilatildeo stock
de decavanadato em 10 mM HEPES pH 70 25 mM
KCl para concentraccedilotildees finais de A) 1 mM B)2
mM C) 4 mM D) 8 mM E) 10 mM de
decavanadato total V1refere-se agraves espeacutecies
monomeacutericade vanadato V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2IIIndash Desvios quiacutemicos meacutedios (part) das ressonacircncias das espeacutecies decameacuterica (V10) monomeacuterica (V1) e
tetrameacuterica (V4) presentes nos espectros da soluccedilatildeo de decavanadatoLargura a meia altura (∆12) da espeacutecie V10A
Paracircmetros V10A V10B V10C V1 V4
part (ppm) -5162 -5003 -4255 -5616 -5748
∆12 (Hz) 175 nd nd nd nd
Figura 23 ndash Distribuiccedilatildeo graacutefica das
espeacutecies oligomeacutericas de vanadato vs a
concentraccedilatildeo de decavanadato total As
concentraccedilotildees de cada espeacutecie foram
obtidas segundo a formula descrita na
secccedilatildeo 35 do Procedimento
Experimental (aacutereas dos sinais de 51
V-
RMN observadas na figura x1)V10 V1 e
V4 correspondem agraves espeacutecie monomeacuterica
dimeacuterica e tetrameacuterica respectivamente [Esp
eacuteci
e o
ligo
meacute
rica
de
van
aacutedio
] (m
M)
[Decavanadato]total (mM)
V10
V1
V4
27
Nas soluccedilotildees de decavanadato as espeacutecies monomeacuterica e decameacuterica de vanadato satildeo as uacutenicas
presentes para concentraccedilotildees de decavanadato inferiores a 8 mM Ambas as espeacutecies aumentam
linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado em [AY] e
em (Teresa) quando a concentraccedilatildeo de V1 eacute igual ou superior a 200 microM inicia-se a sua
oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica No entanto nas condiccedilotildees de realizaccedilatildeo dos
ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a espeacutecie tetrameacuterica neste
estudo em concentraccedilotildees inferiores a 8 mM tendo o mesmo sido observado por Ramoset al
(2006) [AK] Os resultados obtidos mostram claramente o equiliacutebrio entre a espeacutecie decameacuterica e
a monomeacuterica e que o aparecimento de outras espeacutecies oligomeacutericas se encontra dependente da
concentraccedilatildeo desta uacuteltima Apesar de natildeo ter sido realizado o ensaio com 5 mM de decavanadato
total no presente relatoacuterio estaacute reportado em Ramos et al (2006) [AK] que as concentraccedilotildees da
espeacutecie decameacuterica e monomeacuterica satildeo respectivamente de 500 microM e 200 microM Assim pode inferir-
se que para concentraccedilotildees inferiores a 5 mM e intervalos de tempo de cerca de 20 minutos (que
inclui o tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas
as espeacutecies decameacuterica e monomeacuterica de vanadato
Figura 24 ndash Espectros de RMN de 51
V de concentraccedilotildees
crescentes das espeacutecies oligomeacutericas de vanadato obtidos
apoacutes diluiccedilatildeo da soluccedilatildeo stock de metavanadato em 10 mM
HEPES pH 70 25 mM KCl para concentraccedilotildees finais de A)
1 mM B) 2 mM C) 4 mM D) 8 mM E) 10 mM de
metavanadato total V1 refere-se agraves espeacutecies monomeacuterica de
vanadatoV1 e V2 correspondem respectivamente agrave espeacutecie
monomeacuterica e dimeacuterica
Figura 25 ndash Distribuiccedilatildeo graacutefica das espeacutecies
oligomeacutericas de vanadato vs a concentraccedilatildeo de
metavanadato total As concentraccedilotildees de cada
espeacutecie foram obtidas segundo a formula descrita
na secccedilatildeo 35 do Procedimento Experimental
(aacutereas dos sinais de 51
V-RMN observadas na
figura x1)V10 V1 e V4 correspondem agraves espeacutecie
monomeacuterica dimeacuterica e tetrameacuterica
respectivamente
Esp
eacuteci
e o
ligo
meacute
rica
de
va
nad
ato
(m
M)
[Metavanadato]total (mM)
V1
V2
V4
V5
28
Tabela 2IVndash Desvios quiacutemicos meacutedios (part)das ressonacircncias das espeacutecies tetrameacuterica (V4) monomeacuterica (V1) e
dimeacuterica (V2) presentes no espectro da soluccedilatildeo de metavanadato (total) 10 mM
Paracircmetros V1 V2 V4 V5
part (ppm) -5615 -5748 -5789 -5869
Nas soluccedilotildees de metavanadato as espeacutecies monomeacuterica dimeacutericae tetrameacuterica de vanadato estatildeo
presentes para concentraccedilotildees de metavanadato inferiores a 4 mM Todas estas as espeacutecies
aumentam linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado
em (Aureliano e Madeira 1998) e em (Tiago et al 2004) quando a concentraccedilatildeo de V1 eacute igual ou
superior a 200 microM inicia-se a sua oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica Nas
condiccedilotildees de realizaccedilatildeo dos ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a
espeacutecie pentameacuterica neste estudo em concentraccedilotildees inferiores a 4 mM tendo o mesmo sido
observado por Ramos 2006 Os resultados obtidos mostram claramente o equiliacutebrio entre a
espeacutecie V4 e as espeacuteciesV1 e V2 no qual o aparecimento de outras espeacutecies oligomeacutericas se
encontra dependente da concentraccedilatildeo da espeacutecie V1 Por uacuteltimo pode inferir-se que para
concentraccedilotildees inferiores a 2 mM e intervalos de tempo de cerca de 20 minutos (que inclui o
tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas as
espeacutecies V1 V2 e V4 com concentraccedilotildees respectivas de 0796 0192 e 0196 mM
232Estabilidade das soluccedilotildees de decavanadato anaacutelise por espectroscopia de absorccedilatildeo
UVVis
Caracterizou-se a estabilidade da espeacutecie V10 numa soluccedilatildeo com 1 mM decavanadato total em
tampatildeo D atraveacutes da leitura da Abs400 Obteve-se uma cineacutetica de dissociaccedilatildeo de 1ordf ordem tal
como se pode verificar pela linearidade do graacutefico de ln(Abs400)vs o tempo de aquisiccedilatildeo espectral
(segundos) (Fig26 )
Figura 26ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do tempo
(em segundos) para uma soluccedilatildeo de decavanadato total de 1
mM em tampatildeo D a 25ordmC (n=2)
Sendo uma reacccedilatildeo de primeira ordem pode ser descrita pela equaccedilatildeo cujo
declive eacute ndashK (constante de dissociaccedilatildeo) e a intersecccedilatildeo no eixo das ordenadas corresponde
Ln
(Ab
s 400 n
m)
tempo (s)
29
aln(A0) Numa reacccedilatildeo de 1ordf ordem o tempo de meia-vida eacute independente da concentraccedilatildeo inicial
do reagente sendo descrito pela equaccedilatildeo [2] Deste modo a partir da recta
Ln(Abs400 nm) = -436E-5
K - 105 obteve-se um tempo de meia-vida de 4490 plusmn 0823 horas para
as condiccedilotildees supramencionadas
Apoacutes a adiccedilatildeo de S1 8 microM ao meio reaccional verificou-se que a reacccedilatildeo de desoligomerizaccedilatildeo
da espeacutecie decameacuterica de vanadato seguia igualmente uma cineacutetica de 1ordf ordem tal como se pode
verificar pela linearidade do graacutefico de ln(Abs400) versus o tempo de aquisiccedilatildeo espectral
(segundos) (Fig27)
Figura 27ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do
tempo (em segundos) para uma soluccedilatildeo de decavanadato
total de 1 mM em tampatildeo D a 25ordmC na presenccedila de 8
microM (n= 2)
A partir da equaccedilatildeo da recta Ln(Abs400 nm) = -7520E-5
K - 0613determinou-se o valor de K para
a espeacutecie decameacuterica de onde se obteve um tempo de meia-vida de 2430 plusmn 0191 horas para as
condiccedilotildees supramencionadas
Tanto este valor como o valor registado na ausecircncia de S1 satildeo substancialmente superiores agrave
duraccedilatildeo dos ensaios de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina descritos no presente
relatoacuterio
24 Interacccedilatildeo de S1 com vanaacutedio(V)
241Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
De modo a analisar qual o efeito da adiccedilatildeo de S1 de miosina (tampatildeo D) produzia nos sinais de
51V-NMR titulou-se uma soluccedilatildeo de 5mM de decavanadato total com concentraccedilotildees crescentes
de S1 (5 10 15 e 20 microM) Os espectros de 51
V-RMN encontram-se apresentados na Fig 2
No graacutefico da figura 2 Apresenta-se a concentraccedilatildeo de S1 em funccedilatildeo do factor de alargamento
corrigido dos espectros acima referidos
Ln
(Ab
s (4
00
nm
))
tempo (s)
30
Figura 28 ndash Graacutefico de
representaccedilatildeo da
concentraccedilatildeo de S1 em
funccedilatildeo do factor de
alargamento A
concentraccedilatildeo de
decavanadato total foi de 5
mM e as concentraccedilotildees
utilizadas de S1 foram de 0
5 10 e 15 microM
Verifica-se que o factor de alargamento aumenta linearmente em funccedilatildeo da concentraccedilatildeo de S1 no
ensaio o que indica a existecircncia de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina Observou-
se ainda que a adiccedilatildeo de S1 ao meio reaccional natildeo provocou qualquer alteraccedilatildeo dos desvios
quiacutemicos dos sinais de 51
V-RMN
Realizaram-se ensaios na presenccedila de ATP e de S1 com intuito de verificar o efeito do ATP na
interacccedilatildeo do S1 de miosina com o decavanadato
Figura 29 - Espectros de RMN de 51
V contendo 2
mM Decavanadato (A) (controlo) (B) na presenccedila
de 2 mM de ATP (C) na presenccedila de 2 mM de ATP
e 8 microM de S1 de miosina V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2Vndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies decameacuterica (V10) e monomeacuterica (V1) presentes nos
espectros da soluccedilatildeo de decavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V10A V10B V10C V1
part (ppm) -5161 -5002 -4254 -5614
Observa-se que a presenccedila de ATP (espectro B) natildeo provoca qualquer alteraccedilatildeo visiacutevel em
relaccedilatildeo ao espectro que contem apenas soluccedilatildeo de decavanadato Jaacute a presenccedila simultacircnea de ATP
e S1 provoca um alargamento da ∆12 do sinal do V10A de 1736 Hz (espectro (A)) para 2366 Hz
(espectro (C) Fig 29) Tendo em conta que a concentraccedilatildeo de S1 apresenta uma relaccedilatildeo linear
com o factor de alargamento converteu-se o valor da ∆12 e comparou-se com o valor esperado agrave
F = 00591[S1] - 00157 Rsup2 = 0998
Fact
or
de
ala
rgam
en
to
[S1] (microM)
31
concentraccedilatildeo de S1 utilizada O factor de alargamento obtido para 8 microM ndash 0376 ndash eacute inferior ao
valor esperado agrave mesma concentraccedilatildeo de S1 obtido atraveacutes da equaccedilatildeo da recta apresentada na
Fig 28 ndash 0457 ndash o que indica que o ATP poderaacute exercer um efeito de protecccedilatildeo do S1
diminuindo a sua interacccedilatildeo com a espeacutecieV10
Por uacuteltimo realizaram-se ensaios com metavanadato na presenccedila de 8 microM S1 de modo a verificar
se as espeacutecies oligomeacutericas contidas nesta soluccedilatildeo interagiam com a proteiacutena em estudo
Figura 210 - Espectros de RMN de 51
V contendo 2
mM metavanadato (A) (controlo) (B) na presenccedila
de 8 microM de S1 de miosina V1 V2 e V4
correspondem respectivamente aacutes espeacutecies
monomeacuterica dimeacuterica e tetrameacuterica
Tabela 2VIndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies monomeacuterica (V1) dimeacuterica (V2) e tetrameacuterica (V4)
presentes nos espectros da soluccedilatildeo de metavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V1 V2 V4
part (ppm) -5613 -5747 -5789
Como eacute possiacutevel observar na Fig 210 ocorre uma ligeiro alargamento dos sinal de V4 bem
como uma perda da intensidade pelo que se conclui quedas espeacutecies presentes na soluccedilatildeo de
metavanadato (V1 V2 e V4) apenas a espeacutecie tetrameacuterica interage com o S1 de miosina o que
estaacute de acordo com o reportado por Tiago et al 2002
242 Actividade Mg2+
-ATPaacutesica de S1 de miosina na presenccedila de decanadato
Com o intuito de avaliar o efeito do decavanadato na actividade Mg2+
-ATPaacutesica do S1 de
miosina monitorizou-se a reacccedilatildeo de oxidaccedilatildeo do NADH atraveacutes da leitura de Abs340 na
presenccedila de 005 mgmL de S1 de miosina e no caso da actividade estimulada 2microM de actina em
3125 mM Tris-HCl pH 71 e 375 mM MgCl2 pelo meacutetodo de enzimas acopladas (ver seccccedilatildeo xx
dos MM)Para tal realizaram-se ensaios com vaacuterias concentraccedilotildees de decavanadato total (10 50
100 250 500 e 1000) de modo a determinar qual o seu efeito na actividade MgATPaacutesica de S1
de miosina na ausecircncia e na presenccedila de um dos seus ligandos naturais Os ensaios foram
realizados agrave temperatura ambiente com incubaccedilatildeo preacutevia a 25ordm C durante 20 minutos
32
Figura 211 ndash Representaccedilatildeo
graacutefica da Actividade Mg2+
-
ATPaacutesica de S1 () (os ajustes
teoacutericos estatildeo representado a por
uma linha os resultados
experimentais estatildeo representados
por triacircngulos (Actividade
estimulada) e cruzes (Actividade
natildeo estimulada)) vs
[Decavanadato]total medidas pelo
meacutetodo das enzimas acopladas
(n=3) Os dados foram ajustados ao modelo de decaimento exponencial usando o programa GraphPad Prism para a
determinaccedilatildeo dos valores de IC50 (na ausecircncia de actina IC50= 266 plusmn 07 M decavanadato na presenccedila de actina
IC50= 28 plusmn 1 M decavanadato
A partir do graacutefico presente na Fig 29 verifica-se que o ajuste dos dados experimentais foi bem
sucedido A inibiccedilatildeo da actividade Mg2+
-ATPaacutesica do S1 de miosina eacute afectada em menor
amplitude na presenccedila de actina (60 de inibiccedilatildeo da actividade) do que na sua ausecircncia
(aproximadamente 80 de inibiccedilatildeo de actividade) Devido a esta diferenccedila pode sugerir-se que a
actina iraacute impedir parcialmente a ligaccedilatildeo do decavanadatoao S1 demiosina atraveacutes de
impedimentos esteacutereos ou interacccedilotildees electrostaacuteticas por exemplo A teacutecnica utilizada natildeo nos
permite concluir acerca de que regiatildeo(otildees) ou resiacuteduo(s) da actina participaratildeo na preservaccedilatildeo
parcial da actividade Mg2+
-ATPaacutesica
Por outro lado os valores de IC50 dos dois tipos de actividades natildeo apresentam diferenccedilas
significativas tendo-se obtido 266 plusmn 07 e 28 plusmn 1M decavanadato na ausecircncia e na presenccedila de
actina respectivamente o que corresponde a uma concentraccedilatildeo de cerca de 27 M de V10 O
meio de reacccedilatildeo utilizado nestes ensaios foi 10 mM HEPES e 25 mM KCl com 2 mM ATP Em
(Tiago et al 2004) o valor de IC50 para a actividade Mg2+-ATPaacutesica na ausecircncia de actina foi de
125 plusmn 025 microM enquanto o valor da actividade estimulada por actina foi de 136 plusmn 14 microM O
meio de reacccedilatildeo utilizado nos ensaios foi 20 mM HEPES 50 mM KCl e 2 mM ATP o que resulta
precisamente no dobro da forccedila ioacutenica a que foi realizado o ensaio do presente relatoacuterio
(exceptuando a contribuiccedilatildeo dos produtos da hidroacutelise de ATP uma vez que este meacutetodo
pressupotildee a ocorrecircncia de um ciclo de reacccedilotildees de hidroacutelise e siacutentese deste composto) Ora tendo
sido anteriormente referido que a afinidade do S1 de miosina para com a espeacutecie V10 diminui com
Act
ivid
ade
Mg
2+-A
TP
aacutesic
a d
e S
1 d
e
Mio
sin
a
[Decavanadato] (microM) Actividade Estimulada ()
Actividade Natildeo Estimulada ()
Ajuste Actividade Estimulada
Ajuste Actividade Natildeo Estimulada
33
o aumento da forccedila ioacutenica (Tiago et al 2004 tal poderaacute explicar a diferenccedila obtida entre estes
dois conjuntos de valores
243Quantificaccedilatildeo de cisteiacutenas reduzidas pelo meacutetodo de Ellman
A interacccedilatildeo do vanaacutedio com alguns tipos de aminoaacutecidos sensiacuteveis agrave oxidaccedilatildeo pode fornecer-nos
informaccedilatildeo bastante uacutetil em relaccedilatildeo agraves alteraccedilotildees estruturais que conduzem a uma diminuiccedilatildeo da
actividade fisioloacutegica do S1 de miosina Os estudos realizados por Chasteen 1983 concluiacuteram
que a reduccedilatildeo de vanadato (V) a vanadilo (IV) ocorre na presenccedila de agentes redutores comuns no
ambiente celular de que se destaca o aminoaacutecido cisteiacutena Esta eacute tambeacutem um alvo de
complexaccedilatildeo por parte do iatildeo VO2+
Jaacute Ramos et al (2009) demonstrou que a incubaccedilatildeo de
actina com a soluccedilatildeo de decavanadato induz a oxidaccedilatildeo de resiacuteduos de cisteiacutena e agrave formaccedilatildeo do
oxidovanaacutedio (IV) Este grupo de investigadores descobriu ainda que apenas a espeacutecie V10 induz a
oxidaccedilatildeo das cisteiacutenas ldquoraacutepidasrdquo da actina atraveacutes do processo proposto que eacute parcialmente
responsaacutevel pela inibiccedilatildeo jaacute descrita da actividade de ATPase do S1 de miosina estimulada por
actina quando exposta agrave soluccedilatildeo de decavanadato Esta inibiccedilatildeo apresenta-se afectada com a
presenccedila de ATP uma vez que os resiacuteduos de cisteiacutena permaneceram intactos nos ensaios
realizados com este ligando Jaacute Crans et al (2004) concluiu que o siacutetio activo de muitas fosfatases
(que conteacutem grupos tiol) pode formar um equiliacutebrio redox com o iatildeo vanadato (V) de forma natildeo
reversiacutevel
Os resultados obtidos por Sakurai e Shiromura (1980) sugeriram que o vanaacutedio (V) sofre reduccedilatildeo
a vanaacutedio (IV) sem dependecircncia do valor de pH do ensaio Registaram ainda uma estequiometria
de 31 na reacccedilatildeo do vanadato (V) com os resiacuteduos de Cys enquanto a estequiometria dos ensaios
com o vanadilo (IV) foi de 21
Berlett e Stadtman (1997) observaram que os tipos de aminoaacutecidos mais facilmente oxidaacuteveis
englobam entre outros os resiacuteduos de Cys Trp e MetAssim analisou-se o estado redox dos
grupos tiol dos resiacuteduos de Cys do S1 na presenccedila do decavanadato atraveacutes da monitorizaccedilatildeo da
Abs412 nm na presenccedila de 1 mgmL de S1 de miosina em tampatildeo 10 mM HEPES 25 mM KCl
variando a concentraccedilatildeo da soluccedilatildeo de decavanadato de modo a determinar que tipo de alteraccedilotildees
estruturais ocorrem na sua presenccedila no que diz respeito agrave diminuiccedilatildeo da actividade ATPaacutesica de
S1 na presenccedila de decavanadato
Os resultados referentes agrave quantificaccedilatildeo das cisteiacutenas reduzidas apoacutes o tratamento de S1 com
diferentes concentraccedilotildees de decavanadato em tampatildeo 10 mM HEPES pH 75 e 25 mM KCl a
25ordmC durante 20 minutos encontram-se descritos na Tabela 2VII
Tabela 2VII ndash Cisteiacutenas reduzidas totais e ldquoraacutepidas de S1 (1 mgmL) apoacutes tratamento com decavanadato
34
(010501002505001000 uM) em 10 mM HEPES 25 mM KCl pH 7 a 25ordmC durante 20 minutos A detecccedilatildeo
espectrofotomeacutetrica foi feita com DTNB (e 412=8400 uM-1 cm-1) (n variaacutevel entre 1 e 8)
Amostras Cys raacutepidas Cys totais
11 M S1 (controlo) 22 plusmn 04 106 plusmn 14
11 M S1 + 10 M Decavanadato 22 plusmn 05 95 plusmn 08
11 M S1 + 50 uM Decavanadato 20 plusmn 02 98 plusmn 14
11 M S1 + 100 uM Decavanadato 17 plusmn 03 97 plusmn14
11 M S1 + 250 uM Decavanadato 26 plusmn 02 98 plusmn 08
11 M S1 + 500 uM Decavanadato 21 plusmn 04 90 plusmn 06
11 M S1 + 1000 uM Decavanadato 24 plusmn 0 86 plusmn 08
A partir dos resultados obtidos pode sugerir-se que o S1 possui 11 resiacuteduos de Cys (controlo) o
que estaacute de acordo com o anteriormente descrito Verifica-se que o tratamento da proteiacutena com
concentraccedilotildees crescentes de decavanadato provoca a oxidaccedilatildeo de dois resiacuteduos de Cys do S1 de
miosina natildeo expostos ao solvente No entanto a teacutecnica utilizada natildeo nos permite identificar
quais
Tendo em consideraccedilatildeo que o valor de IC50 obtido para a reacccedilatildeo de inibiccedilatildeo daactividade Mg2+-
ATPaacutesica de S1 na presenccedila de decavanadato eacute de cerca de 27 microM pode sugerir-se que este seraacute o
valor ao qual ocorreraacute a oxidaccedilatildeo do primeiro resiacuteduo de Cys o que poderaacute contribuir para a
diminuiccedilatildeo da actividade de S1 observada
25Interacccedilatildeo de S1 com vanaacutedio(IV)
251Anaacutelise por espectroscopia de Ressonacircncia Paramagneacutetica Electroacutenica
A acumulaccedilatildeo intracelular de oxovanaacutedio (IV) in vivo constitui uma das principais formas de
apresentaccedilatildeo de vanaacutedio no organismo Deste modo torna-se bastante interessante analisar a
interacccedilatildeo entre este iatildeo e o S1 de miosina atraveacutes da teacutecnica de espectroscopia de Ressonacircncia
Paramagneacutetica Electroacutenica o meacutetodo de eleiccedilatildeo do estudo do iatildeo vanadilo e dos complexos por si
formados O electratildeo desemparelhado encontra-se fortemente acoplado ao nuacutecleo 51
V o que
35
resulta num espectro de RPE consistindo de 8 desdobramentos na direcccedilatildeo paralela e outros tantos
na direcccedilatildeo perpendicular (Chasteen 1983)
Os estudos realizados por Ahmed et al (1987) centraram-se na interacccedilatildeo do vanadilo com
proteiacutenas como eacute o caso da insulina transferrina entre outros obtendo um espectro caracterizado
por 8 linhas amplamente espaccediladas (na direcccedilatildeo paralela) Jaacute Ramos et al 2009 concluiu que a
exposiccedilatildeo da soluccedilatildeo de decavanadato agraves formas globular e filamentosa de actina produz um
espectro tiacutepico do iatildeo oxidovanaacutedio (IV) indicando a reduccedilatildeo do vanadato
Para aferir qual a concentraccedilatildeo de S1 que permitia obter uma boa razatildeo sinalruiacutedo titulou-se uma
soluccedilatildeo de sulfato de vanadilo 15 microM ([VOSO4]stock = 5 mM) com concentraccedilotildees crescentes de
proteiacutena (0 5 10 15 e 20 microM) em 10 mM HEPES 25 mM KCl A existecircncia de uma correlaccedilatildeo
linear entre a concentraccedilatildeo de proteiacutena e a intensidade dos sinais de RPE correspondente agrave banda
-12 perpendicular encontra-se representada na Fig 210
Figura 2 12ndash Representaccedilatildeo graacutefica da variaccedilatildeo da intensidade do sinal de RPE vs a concentraccedilatildeo de S1 utilizada
Apoacutes verificar que com uma concentraccedilatildeo de S1 igual a 20 microM se obtinham sinais com uma razatildeo
sinalruiacutedo adequada ao prosseguimento dos estudos sobre a interacccedilatildeo V(IV)-S1 de miosina
titulou-se uma soluccedilatildeo de 20 microM S1 com 0 25 5 10 15 25 50 75 100 150 200 500 e 1000
microM VOSO4
Traccedilou-se o graacutefico da intensidade da banda -12 perpendicular do sinal de EPR em funccedilatildeo da
concentraccedilatildeo de VOSO4 tendo-se determinado uma constante de dissociaccedilatildeo (KD) do complexo
S1-vanadilo de 8748 plusmn 2621 microM V(IV)
y = 016786x + 122435
Rsup2 = 092992759
Inte
nsi
dad
e d
o s
inal
de
RP
E
[S1] (microM)
36
Figura 213 ndash Representaccedilatildeo graacutefica da variaccedilatildeo do sinal de RPE vs a concentraccedilatildeo de vanadilo utilizada (os valores
experimentais estatildeo representados por losangos o seu ajuste estaacute representado pela linha)
A simulaccedilatildeo do espectro de EPR foi efectuada a partir do espectro obtido para uma concentraccedilatildeo
de 1 mM VOSO4 apoacutes adiccedilatildeo de 20 microM S1 de miosina tendo-se obtido os valores da constante de
acoplamento hiperfina (A=5016) e do factor g (g0=195)
Figura 214 ndash Representaccedilatildeo graacutefica do espectro de EPR obtido com 1000 microM de vanadilo e 20 microM de S1 (a cinzento
escuro) A simulaccedilatildeo do espectro encontra-se a cinzento claro
Por comparaccedilatildeo destes com os valores presentes na tabela de correspondecircncia xxx eacute possiacutevel
predizer quais os ligandos provaacuteveis do vanadilo na presenccedila de proteiacutena Assim concluiu-se que
o vanadilo estaacute coordenado com duas moleacuteculas de aacutegua um grupo hidroxilo e um grupo R=NR
O grupo hidroxilo pode indicar os resiacuteduos de aminoaacutecidos Tyr Treonina e Serina enquanto o
grupo R=NR poderaacute indicar a presenccedila de Arginina e Histidina como ligandos do vanadilo
Inte
nsi
dad
e d
os
sinai
s d
e R
PE
[Vanadilo] (microM)
Ajuste dos dados exp Dados experimentais
Campo magneacutetico (G)
Espectro Experimental Ajuste
37
3 Procedimento Experimental
31 Reagentes
Todos os reagentes utilizados satildeo de elevado grau de pureza (analiacutetico) Apresentam-se as suas
foacutermulas quiacutemicas ou abreviaturas quando necessaacuterio acrescentou-se a sua descriccedilatildeo Entre
parecircntesis colocou-se as empresas que os comercializam
Aacutecido aceacutetico glacial 100 (Riedel) Acrilamida 99 (Merck) Azul de bromofenol (Riedel-de-
Haumlen) -Mercaptoetanol (Sigma) Bissulfito de soacutedio (Sigma)Bradford (reagente de)
(BioRad)BSA (albumina de soro bovino Sigma-Aldrich) CaCl22H2O (cloreto de caacutelcio
dihidratado Fluka) CH3CH2OH 96 (etanol Panreac) Coomassie Blue R250 (Merck) D2O
(oacutexido de deuteacuterio SDS) DTNB (55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico) Aldrich) EDTA (aacutecido
etilenodiaminatetraaceacutetico Merck) Fosfoenolpiruvato 98 (Fluka) Glicerol (Sigma) Glicina
985 (Panreac) HCl 37 (Riedel-de Haumlen) H2SO4 96 (Pronalab) HEPES 99 (aacutecido 4-(2-
hidroxiletil)-1-piperazineetanosulfoacutenico Sigma) KH2PO4 99 (hidrogenofosfato de potaacutessio)
MgATP (Sigma) Na
ATP (Sigma) Na2HPO4 99 (hidrogenofosfato de soacutedio Riedel-de Haumlen)
NaH2PO4 (fosfato de soacutedio ) NaOH 986 (hidroacutexido de soacutedio Pronalab) KCl 99 (Cloreto de
potaacutessio Panreac) L-Cys 995 (enantioacutemero levoacutegiro do resiacuteduo de Cisteiacutena Fluka)Metanol
99 (Merck) Molibdato de amoacutenio (Sigma)NNrsquo-metil-bis-acrilamida 98 (Merck) NH4VO3
99 (metavanadato de amoacutenia Sigma) PSA 10 (persulfato de amoacutenio Merck) SDS
(dodecilo sulfato de soacutedio Sigma) Sulfito de soacutedio (Sigma) VOSO4 (sulfato de vanadilo
Aldrich) TEMED 99 (Tetrametiletilenodiamina Merck) Tris (2-amino-2-hidroximetil-13-
propanodiol Sigma)
32 Meacutetodos
321 Isolamento e purificaccedilatildeo das proteiacutenas de muacutesculo esqueleacutetico de coelho
3211 Purificaccedilatildeo da miosina preparaccedilatildeo de Subfragmento 1 por digestatildeo quimotriacutepsica
da miosina
O subfragmento-1 de miosina-II (S1) foi gentilmente cedido pelo Prof Doutor Manuel Aureliano
do Departamento de Quiacutemica Bioquiacutemica e Farmaacutecia da Faculdade de Ciecircncias e Tecnologia da
Universidade do Algarve
No entanto descreve-se abaixo de forma resumida o procedimento efectuado
38
32111 Purificaccedilatildeo da miosina
O protocolo de isolamento foi adaptado do meacutetodo previamente descrito por Margossian e Lowey
(1982) apresentando-se de forma esquemaacutetica na Fig 31
Sucintamente os tecidos musculares dorsais e das pernas foram extraiacutedos apoacutes o sacrifiacutecio do
animal (Fig31) De seguida foram adicionados 3 volumes da soluccedilatildeo de extracccedilatildeo A2
Centrifugou-se a soluccedilatildeo e filtrou-se o sobrenadante O sedimento foi entatildeo diluiacutedo em 14
volumes de aacutegua destilada de modo a que a forccedila ioacutenica atingisse 004 M Apoacutes adiccedilatildeo de szlig-
mercaptoetanol (szlig-me) deixou-se a precipitar a 4ordmC removendo o sobrenadante Centrifugou-se
novamente e ressuspendeu-se o sedimento na soluccedilatildeo de extracccedilatildeo A Adicionou-se a soluccedilatildeo
tampatildeo B3 para que a forccedila ioacutenica atingisse 05 M Homogeneizou-se e deixou-se repousar
durante 2h Homogeneizou-se novamente e centrifugou-se de modo a ocorrer precipitaccedilatildeo dos
complexos de actomiosina e proteiacutena desnaturada Apoacutes filtraccedilatildeo do sobrenadante em latilde de vidro
quantificou-se a soluccedilatildeo de miosina resultante pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental)
2 Soluccedilatildeo de extracccedilatildeo A 0012M K2CO3 004M KHCO3 06M KCl pH 65
3 Soluccedilatildeo-tampatildeo B Tris 100 mM pH 7 KCl 2M
39
Figura 31 ndash Representaccedilatildeo esquemaacutetica do protocolo de isolamento de miosina (Margossian e Lowey 1982) Todo o
procedimento foi realizado a 4ordmC (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
32112 Preparaccedilatildeo de Subfragmento-1 de miosina por digestatildeo com quimiotripsina
O S1 de miosina foi isolado apoacutes diaacutelise da miosina durante a noite contra a soluccedilatildeo C4 A
presenccedila do EDTA eacute essencial uma vez que a sua substituiccedilatildeo por MgCl2resulta na produccedilatildeo de
HMM em vez de S1 A suspensatildeo de miosina foi equilibrada e digerida com -quimotripsina
A digestatildeo foi interrompida pela adiccedilatildeo de PMSF Seguidamente ultracentrifugou-se o produto
da digestatildeo desprezando-se o sedimento (Fig 32) A soluccedilatildeo de sobrenadante maioritariamente
S1 foi quantificada pela leitura directa da Abs280 ou pelo meacutetodo de Bradford (ver as secccedilotildees
3311 e 3312 do Procedimento Experimental respectivamente) e armazenada em 10 sacarose
e 2 mM szlig-Me a -80ordmC
4 Soluccedilatildeo C 25mM Tris pH 70 012M KCl 1mM EDTA
40
Antes da realizaccedilatildeo dos ensaios descritos no presente relatoacuterio o S1 de miosina foi dialisado
contra a soluccedilatildeo D5 de modo a remover qualquer composto que pudesse complexar como
vanaacutedio Seguidamente quantificou-se a proteiacutena pelo meacutetodo de Bradford ou pelo meacutetodo
directo (ver secccedilotildees 3311 e 3312do Procedimento Experimtental)
Figura 3 2ndash Representaccedilatildeo esquemaacutetica do protocolo de preparaccedilatildeo do S1 de miosina por digestatildeo quimiotripsina de
acordo com o previamente descrito por Margossian e Lowey (1982) Todo o procedimento foi realizado a 4ordm C
excepto quando indicaccedilatildeo do contraacuterio (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
3212 Isolamento e purificaccedilatildeo de actina
Isolou-se e purificou-se a actina segundo o meacutetodo descrito por Pardee e Spudish que
compreende dois passos ndash a obtenccedilatildeo de um resiacuteduo seco denominado ldquopoacute de acetonardquo e a
extracccedilatildeo subsequente da actina a partir do ldquopoacute de acetonardquo na forma de G-Actina com a
posterior polimerizaccedilatildeo a F-Actina
32121 Obtenccedilatildeo do ldquopoacute de acetonardquo
Sacrificou-se o coelho com uma pancada atraacutes do pescoccedilo e sangrou-se o animal apoacutes o corte da
arteacuteria jugular Esfolou-se e colocou-se numa tina com gelo Retiraram-se as partes de tecido
conjuntivo que envolvem o tecido muscular com auxiacutelio de uma pinccedila e bisturi Seguidamente
retirou-se os muacutesculos dorsais e os muacutesculos brancos das pernas Passou-se os muacutesculos por aacutegua
destilada de modo a remover as impurezas e pesou-se num copo Trituraram-se os muacutesculos
numa maacutequina de triturar carne e extraiu-se o resultante com 3 volumes da soluccedilatildeo E6 durante 10
minutos com agitaccedilatildeo suave Centrifugou-se a suspensatildeo a 4000 g durante 10
5 Soluccedilatildeo D HEPES 10 mM pH 70 KCl 25 mM
6 Soluccedilatildeo E 01 M KCl 015 M K2HPO4 pH 65
41
minutosdesprezando-se o sobrenadante Ressuspendeu-se o sedimento em 6 volumes de soluccedilatildeo
F7durante 10 minutos com agitaccedilatildeo suave Verificou-se se o pH da soluccedilatildeo estava compreendido
entre 82 e 85 acertando-se estecom soluccedilatildeo G8 se necessaacuterio Centrifugou-se durante 10
minutos a 4000 g Desprezou-se o sobrenadante e ressuspendeu-se o sedimento em 3 volumes de
soluccedilatildeo H9 durante 10 minutos Centrifugou-se novamente a 4000 gdurante 10 minutos
desprezando-se o sobrenadante Ressuspendeu-se o sedimento duas vezes consecutivas em 6
volumes de aacutegua destilada durante 5 minutos Centrifugou-se a 4000 g durante 10 minutos apoacutes
cada adiccedilatildeo de H2O
Seguidamente ressuspendeu-se o sedimento 5 vezes consecutivas utilizando 3 volumes de
acetona de cada vez agitando suavemente durante 10 minutos agrave temperatura ambiente Colocou-
se o resiacuteduo filtrado numa tina de vidro sobre papel de filtro deixando-se evaporar o solvente
durante a noite num nicho Triturou-se o resiacuteduo seco numa maacutequina de triturar carne e deixou-se
novamente evaporar o solvente O ldquopoacute de acetonardquo obtido foi armazenado a -20 ordmC
32122 Purificaccedilatildeo da actina a partir de ldquopoacute de acetonardquo
Apoacutes a obtenccedilatildeo do poacute de acetona como acima indicado efectuou-se o protocolo seguidamente
descrito de modo a obter monoacutemeros de actina (G-Actina) e actina filamentosa (F-Actina)
Pesou-se 35 g de poacute de acetona e adicionaram-se 50 mL de soluccedilatildeo-tampatildeo I10
sendo a
extracccedilatildeo feita com agitaccedilatildeo suave durante 30 minutos Separou-se o extracto do poacute de acetona
hidratado filtrando a suspensatildeo com 4 camadas de gaze esterilizada tendo o cuidado de natildeo
espremer o resiacuteduo Voltou a extrair-se com 50 mL de soluccedilatildeo I durante 30 minutos mexendo
lentamente Adicionou-se este ao extracto obtido anteriormente Centrifugou-se o extracto total a
16 000 g durante 30 minutos Decantou-se para uma proveta sobre a qual se colocou gaze para
que o sedimento solto natildeo se juntasse ao sobrenadante e leu-se o volume De seguida adicionou-
se KCl ateacute uma concentraccedilatildeo final de 50 mM MgCl2 ateacute 2mM e ATP ateacute 1mM lentamente e com
agitaccedilatildeo contiacutenua de modo a promover a polimerizaccedilatildeo dos monoacutemeros de actina
Deixou-se polimerizar durante 2h a 25ordmC observando-se a presenccedila de pequenas bolhas de ar
indicativas de um aumento de viscosidade da soluccedilatildeo Arrefeceu-se em gelo durante 15 minutos e
adicionou-se KCl ateacute uma concentraccedilatildeo final de 06M Mexeu-se lentamente durante 30 minutos
apoacutes os quais se ultracentrifugou a 110 000g durante 1hora Desprezou-se o sobrenadante
ressuspendeu-se o sedimento com soluccedilatildeo tampatildeo E e transferiu-se para um potter
Dialisou-se durante a noite contra 1L de soluccedilatildeo-tampatildeo I a uma temperatura de 4ordmC com rotaccedilatildeo
7 Soluccedilatildeo F 005 M NaHCO3 pH 82-85
8 Soluccedilatildeo G 1M Na2CO3
9 Soluccedilatildeo H 1 mM EDTA pH 70
10 Soluccedilatildeo I 2 mM Tris-HCl pH 8 02 mM CaCl2 02 mM Na2ATP 05 mM szlig-Me 0005 NaN3
42
contiacutenua e mudanccedila de tampatildeo 2 a 3 vezes De seguida a actina foi centrifugada a 110
000gdurante 1hora Decantou-se a soluccedilatildeo de G-Actina (sobrenadante) e desprezou-se o
sedimento Determinou-se a concentraccedilatildeo de G-Actina pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental) A soluccedilatildeo de G-actina foi armazenada a uma temperatura de 4ordmC
apoacutes adiccedilatildeo de 002NaN3 mantendo-se estaacutevel durante 3 a 5 dias
331 Quantificaccedilatildeo das proteiacutenas isoladas
3311 Quantificaccedilatildeo pelo meacutetodo de Bradford
O meacutetodo de Bradford (Bradford 1976) baseia-se na utilizaccedilatildeo do reagente de Bradford
composto por entre outros aacutecido fosfoacuterico e o corante ldquoCoomassierdquo Este corante possui carga
negativa e a sua coloraccedilatildeo varia entre a vermelha (Absmax=465 nm) predominante em soluccedilatildeo e a
azul (Absmax=595 nm) presente aquando da ligaccedilatildeo aos grupos amina das proteiacutenas Em soluccedilotildees
aacutecidas diluiacutedas a coloraccedilatildeo varia proporcionalmente com a quantidade do corante ligado agrave
proteiacutena que por sua vez eacute directamente proporcional agrave concentraccedilatildeo da proteiacutena permitindo a
sua quantificaccedilatildeo ateacute a um limite de detecccedilatildeo de 20 gmL (Bradford 1976)
Deste modo prepararam-se amostras de BSA ([BSA]stock= 1mgmL) utilizando um intervalo de
concentraccedilotildees compreendido entre 0 e 15 gmL de acordo com a tabela 3I Apoacutes a adiccedilatildeo da
H2O MilliQ e do reagente de Bradford as soluccedilotildees foram sucessivamente homogeneizadas no
voacutertex
Tabela 3Indash Amostras utilizadas para a obtenccedilatildeo da recta de calibraccedilatildeo no meacutetodo de Bradford
BSA (L) BSA (g) H2O MilliQ (L) Reagente de Bradford (L)
0 0 800 200
125 125 79875 200
25 25 7975 200
5 5 795 200
10 10 790 200
15 15 785 200
Apoacutes incubaccedilatildeo a 25ordmC por 5 minutos leram-se os valores de Abs595 num
espectrofotoacutemetro (Shimadzu UV-160A) para todas as amostras Seguidamente traccedilou-se a recta
de calibraccedilatildeo (Abs595vs quantidade de BSA (g))
43
Para determinar a concentraccedilatildeo de proteiacutena apoacutes o processo de isolamento purificaccedilatildeo e
diaacutelise quando necessaacuterio colocou-se 5 L da amostra num tubo de Eppendorf e adicionou-se
795 L de H2O MilliQ homogeneizando a soluccedilatildeo no voacutertex Adicionou-se posteriormente 200
L de reagente de Bradford e repetiu-se a homogeneizaccedilatildeo Incubou-se a mistura a 25ordmC por 5
minutos apoacutes os quais se leu os valores de Abs595 nm no espectrofotoacutemetro Por interpolaccedilatildeo deste
valor a partir da recta de calibraccedilatildeo determinou-se a concentraccedilatildeo de proteiacutena na amostra
3312 Quantificaccedilatildeo pelo meacutetodo directo
A quantificaccedilatildeo das amostras proteicas pode tambeacutem ser efectuada atraveacutes da leitura directa da
absorvacircncia tendo por base a lei de Beer-Lambert onde Abs corresponde agrave
absorvacircncia ao comprimento de onda ao qual o ensaio foi realizado corresponde ao coeficiente
de extinccedilatildeo (substituiacutedo pela letra a se as unidades forem distintas da molaridade) b corresponde
ao percurso oacuteptico da radiaccedilatildeo na ceacutelula amostral e c corresponde agrave concentraccedilatildeo amostral
Assim no caso de S1 de miosina mediu-se a Abs280e calculou-se a concentraccedilatildeo da proteiacutena
sabendo que o coeficiente de extinccedilatildeo de S1 eacute (Ref)
No caso da actina mediu-se a Abs290ecalculou-se a concentraccedilatildeo da proteiacutena sabendo que o
coeficiente de extinccedilatildeo da actina eacute (Ref)
332Electroforese em condiccedilotildees desnaturantes (SDS-PAGE)
De modo a melhor caracterizar as proteiacutenas isoladas foi realizada uma electroforese em
condiccedilotildees desnaturantes (SDS-PAGE do inglecircs Sodium Dodecyl Sulfate Polyacrilamide Gel
Electrophoresis) O SDS (dodecilo sulfato de soacutedio) eacute um detergente anioacutenico utilizado neste
meacutetodo para desnaturar as proteiacutenas que se ligam ao SDS na proporccedilatildeo 14 g SDS g proteiacutena
(aproximadamente 1 moleacutecula de SDS por cada 2 resiacuteduos de aminoaacutecidos) (Stryer) O SDS induz
uma densidade de carga negativa significativa nas proteiacutenas a que se liga mascarando a sua carga
intriacutenseca pelo que as razotildees cargamassa satildeo tendencialmente idecircnticas Assim a SDS-PAGE
separa as proteiacutenas quando sujeitas a um campo eleacutectrico de acordo com o seu peso molecular
sendo a sua migraccedilatildeo directamente proporcional ao logaritmo deste uacuteltimo (Stryer)
O gel de resoluccedilatildeo com 10 de acrilamida foi obtido atraveacutes da adiccedilatildeo de 167 mL de soluccedilatildeo
stock de acrilamida11
075 mL de tampatildeo de resoluccedilatildeo12
251 mL de aacutegua destilada e 50 L de
SDS 10 Para promover a polimerizaccedilatildeo adicionou-se em seguida 38 L de PSA 10
11
soluccedilatildeo stock de acrilamida soluccedilatildeo de 30 (mv) de acrilamida e 08 (mv) de NNrsquo-metil-bis-acrilamida 12
tampatildeo de resoluccedilatildeo 15 M Tris-HCl pH 88
44
(persulfato de amoacutenia) 10 e 25 L de TEMED 99 (tetrametiletilenodiamina) Agitou-se
cuidadosamente de modo a natildeo formar bolhas e pipetou-se a soluccedilatildeo para a cassette de vidro
previamente montada no suporte De seguida adicionou-se uma peliacutecula de cerca de 2 mm de 2-
metil-2-butanol para verificar posteriormente a polimerizaccedilatildeo Apoacutes a polimerizaccedilatildeo que demora
cerca de 45 minutos verteu-se a agrave superfiacutecie e adicionou-se o gel de concentraccedilatildeo com 5 de
acrilamida constituiacutedo por 03 mL de soluccedilatildeo de stock acrilamida 045 mL de tampatildeo de
concentraccedilatildeo13
094 mL de aacutegua destilada 18 L de SDS 10 135 L de PSA 10 e 2L de
TEMED 99 Colocou-se o pente sobre o gel e deixou-se polimerizar durante cerca de 45
minutos
As amostras para aplicaccedilatildeo no gel de electroforese foram preparadas para uma concentraccedilatildeo de
proteiacutena de 1 mgmL num volume final de 100 L dos quais 25 L correspondem ao tampatildeo da
amostra14
As amostras foram aquecidas a 100ordmC durante 5 minutos Antes da aplicaccedilatildeo das
amostras transferiu-se a cassette para a tina de electroforese agrave qual se adicionou a soluccedilatildeo
tampatildeo de electroforese da tina superior15
e tampatildeo de electroforese da tina inferior16
Aplicaram-
se 10 L das soluccedilotildees de marcadores de peso molecular e 20 L das amostras proteicas
preparadas anteriormente A electroforese decorreu durante cerca de 3h30 min a uma voltagem
de 80 V De seguida transferiu-se o gel para uma tina corando as proteiacutenas durante 1h com
soluccedilatildeo corante17
Descorou-se o gel com soluccedilatildeo descorante18
tendo-se mudado esta soluccedilatildeo ateacute
as bandas se encontrarem bem definidas
Determinou-se a mobilidade relativa das bandas correspondentes aos marcadores de elevado peso
molecular e traccedilou-se a recta de calibraccedilatildeo ndash log(PM) vs log(Rf) onde Rf corresponde agrave
mobilidade relativa Por interpolaccedilatildeo determina-se o PM das proteiacutenas isoladas sabendo a sua
mobilidade relativa Por comparaccedilatildeo com os valores descritos na literatura pode concluir-se
acerca do grau de purificaccedilatildeo
34 Mediccedilatildeo da actividade enzimaacutetica de S1
341 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
13
tampatildeo de concentraccedilatildeo 05 M Tris-HCl pH 68 14
tampatildeo de amostra 320 mM Tris-HCl pH 68 04 M szlig-mercaptoetanol SDS 8 glicerol 15 (vv) e roxo de m-
cresol 0024 15
tampatildeo de electroforese da tina superior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS 16
tampatildeo de electroforese da tina inferior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS acetato de soacutedio
01 M 17
soluccedilatildeo corante ldquoCoomassie Blue R-250rdquo contendo metanol 45 (vv) e aacutecido aceacutetico glacial 10 (vv) 18
soluccedilatildeo descorante metanol 10 (vv) e aacutecido aceacutetico glacial 10 vv)
45
O meacutetodo de Fiske-Subarow modificado consiste na reacccedilatildeo do molibdato de amoacutenio com o
fosfato inorgacircnico em soluccedilatildeo aacutecida com formaccedilatildeo do aacutecido fosfomoliacutebdico Seguidamente daacute-
se a adiccedilatildeo de um agente redutor que provoca a reduccedilatildeo do composto a fosfomolibdato que forma
um complexo de cor azulada cuja absorvacircncia maacutexima ocorre para um comprimento de onda de
660 nm tornando possiacutevel a sua detecccedilatildeo por meacutetodos espectrofotomeacutetricos As actividades Ca2+
-
ATPaacutesica e K+EDTA-ATPaacutesica de S1 foram determinadas atraveacutes deste meacutetodo de acordo com
o descrito por Taussky e Shorr (1953) com mediccedilatildeo da quantidade de fosfato (Pi) libertado
atraveacutes da hidroacutelise de ATP
Para tal obteve-se primeiramente a recta de calibraccedilatildeo com as amostras presentes na Tabela 3II
utilizando uma soluccedilatildeo stock de 5 mM KH2PO4
Tabela 3II ndash Composiccedilatildeo das diferentes amostras utilizadas para determinar a recta de calibraccedilatildeo atraveacutes do meacutetodo
de Fiske-Subbarow modificado
KH2PO4 (L)
([KH2PO4]stock= 5mM)
PO43-
(nmole) H2O (uL)
0 0 1780
5 25 1775
10 50 1770
20 100 1760
40 200 1740
100 500 1680
A cada tubo de Eppendorf adicionaram-se seguidamente 200 L de soluccedilatildeo J19
e 20 L de agente
redutor20
homogeneizando no voacutertex apoacutes cada adiccedilatildeo Incubou-se durante 10 minutos e mediu-
se a Abs660 nm num espectrofotoacutemetro com intervalos de tempo iguais entre cada amostra Traccedilou-
se a recta de calibraccedilatildeo (Abs660 nmvs nmol PO43-
) a partir dos valores obtidos
Para determinar a actividade Ca2+
-ATPaacutesica de S1 prepararam-se tubos de Eppendorf com 400
L de meio de reacccedilatildeo21
25 L CaCl2 ([CaCl2]stock= 100 mM) 25 L ATP ([ATP]stock= 100 mM)
com [S1]ensaios=01 mgmL Perfez-se se necessaacuterio com H2O MilliQ ateacute perfazer um volume de 1
mL Para determinar a actividade K+EDTA-ATPaacutesica utilizou-se uma concentraccedilatildeo idecircntica de
19
Soluccedilatildeo J 5 N H2SO4 25 molibdato de amoacutenio 20
Agente redutor 5 mg aacutecido 1-amino-2-naftol-4-sulfoacutenico 30 mg bissulfito de soacutedio 30 mg sulfito de soacutedio em 26
mL H2O 21
meio de reacccedilatildeo 125 mM Tris-HCl pH 80 0575 M KCl
46
proteiacutena nos ensaios (01 mgmL) e o mesmo meio de reacccedilatildeo Aos tubos de Eppendorf
adicionou-se ainda 123 L de KCl ([KCl]stock = 3M) 25 L de ATP 10 L de EDTA
([EDTA]stock= 100 mM )e o volume de H2O MilliQ necessaacuterio para perfazer 1 mL Em ambas as
situaccedilotildees o uacuteltimo reagente adicionado foi o ATP uma vez que se trata do substrato da reacccedilatildeo
enzimaacutetica
A cineacutetica decorreu a 25ordmC durante 5 minutos para a actividade Ca2+
-ATPaacutesica e durante 3
minutos para a actividade K+EDTA-ATPaacutesica Apoacutes o periacuteodo de incubaccedilatildeo transferiu-se 150
L de amostra para tubos de Eppendorf contendo igual volume de TCA 10 Colocou-se no gelo
e homogeneizou-se no voacutertex Seguidamente centrifugou-se a 1500 g durante 10 minutos e
recolheu-se 200 L de sobrenadante para novos tubos de Eppendorf Adicionou-se a estes 158
mL de H2O MilliQ e 200 L de soluccedilatildeo J Homogeneizou-se no voacutertex e adicionou-se 20 L de
reagente redutor repetindo-se a homogeneizaccedilatildeo Incubou-se durante 10 minutos e mediu-se a
Abs660 nm com o mesmo intervalo de tempo de leitura para cada amostra Por intrapolaccedilatildeo a partir
da recta de calibraccedilatildeo obtida anteriormente determinou-se a quantidade de Pi libertado em cada
ensaio A partir destes resultados determinaram-se as actividades Ca2+
-ATPaacutesica e K+EDTA-
ATPaacutesica do S1 em nmol PO43-
minmg proteiacutena e em s-1
342 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
Da actividade Mg2+
-ATPaacutesica de S1 resulta a formaccedilatildeo dos produtos da reacccedilatildeo de
hidroacutelise do ATP - ADP e Pi Na presenccedila de ADP fosfoenolpiruvato (PEP) e da enzima piruvato
cinase (PK) os produtos resultantes satildeo ATP e piruvato Por acccedilatildeo da enzima lactato
desidrogenase (LDH) este uacuteltimo eacute convertido em lactato ocorrendo simultaneamente a oxidaccedilatildeo
do NADH a NAD+ monitorizada atraveacutes da Abs340 nm Quando o S1 de miosina se encontra no
meio reaccional o ATP eacute hidrolisado formando-se novamente ADP e Pi o que permite a
formaccedilatildeo de um ciclo (fig 21) Se as enzimas se encontrarem em excesso a velocidade de
conversatildeo do NADH eacute limitada pela produccedilatildeo de ADP e Pi Como o ATP eacute regenerado em cada
ciclo a sua concentraccedilatildeo inicial eacute sempre mantida (Bagshaw 1982)
Figura 33ndash Representaccedilatildeo esquemaacutetica do meacutetodo de enzimas acopladas
47
O protocolo utilizado e seguidamente descrito foi adaptado de Cuenda et al (1990 ndash Ver Refordf) A
uma cuvette de 1 mL adicionou-se S1 para uma concentraccedilatildeo final de 005 mgmL 800 L de
meio de reacccedilatildeo22
10 L de PEP ([PEP]stock = 42 mM) 10 L de PK+LDH 11 ([PK+LDH]stock =
1 mgmL) e 15 L de NADH ([NADH]stock=25 mM) Iniciou-se a monitorizaccedilatildeo da reacccedilatildeo
atraveacutes da leitura de Abs340 nm apoacutes a adiccedilatildeo de 20 L de ATP ([ATP]stock=100 mM) A
actividade Mg2+
-ATPaacutesica eacute obtida atraveacutes do declive correspondente agrave oxidaccedilatildeo do NADH Para
a mediccedilatildeo da Mg2+
-ATPase de S1 estimulada por actina adicionou-se F-actina para uma
concentraccedilatildeo final de 2 microM sendo a actividade obtida tambeacutem atraveacutes do declive correspondente
agrave oxidaccedilatildeo do NADH apoacutes a adiccedilatildeo da proteiacutena
35 Interaccedilatildeo entre vanaacutedio e subfragmento-1 de miosina
351 Preparaccedilatildeo das soluccedilotildees de vanaacutedio
As soluccedilotildees de vanadato foram preparadas em aacutegua MilliQ a partir de metavanadato de amoacutenio
(NH4VO3) numa concentraccedilatildeo stock de 50 mM com agitaccedilatildeo magneacutetica e aquecimento (~30 ordmC)
Esta soluccedilatildeo com um valor compreendido entre 6 e 7 designa-se por ldquometavanadatordquo e eacute
constituiacuteda pelas esspeacutecies V1 V2 V4 e V5 (Ref) Acidificando a soluccedilatildeo de metavanadato por
adiccedilatildeo de HCl ateacute um valor de pH proacuteximo de 4 observa-se uma mudanccedila de cor de amarelo
claro para laranja indicativa da formaccedilatildeo da espeacutecie decameacuterica de vanadato De seguida e
imediatamente antes da realizaccedilatildeo dos ensaios descritos o valor do pH da soluccedilatildeo de
decavanadato foi aumentado para cerca de 75 por adiccedilatildeo de NaOH A soluccedilatildeo (2) designada
por ldquometavanadatordquo natildeo foi posteriormente alterada ateacute agrave sua adiccedilatildeo para a realizaccedilatildeo dos ensaios
As soluccedilotildees foram armazenadas a 4ordmC e mantidas em gelo aquando da sua utilizaccedilatildeo
A soluccedilatildeo de V(IV) foi preparada em condiccedilotildees anaeroacutebias por diluiccedilatildeo de sulfato de vanadil
(VOSO4) para uma concentraccedilatildeo final de 5 mM em H2O MilliQ saturada com N2 gasoso
352Anaacutelise por espectroscopia de UV-Vis
A concentraccedilatildeo da espeacutecie decameacuterica em soluccedilatildeo depende do periacuteodo de tempo apoacutes a diluiccedilatildeo
no meio de reacccedilatildeo apropriado (Ref) A velocidade de desoligomerizaccedilatildeo da espeacutecie decameacuterica
pode ser analisada por espectroscopia de absorccedilatildeo na regiatildeo UVVis uma vez que esta espeacutecie
apresenta duas bandas caracteriacutesticas na regiatildeo do visiacutevel (a 358 e 400 nm) que natildeo se verifica
em quaisquer outras espeacutecies oligomeacutericas As bandas supramencionada sapresentam-se bem
22
Meio de reacccedilatildeo 3125 mM Tris-HCl pH 71 375 mM MgCl2
48
resolvidas atraveacutes da obtenccedilatildeo da derivada de 2ordf ordem do espectro apresentando a forma de 2
vales com miacutenimos centrados nos comprimentos de onda supracitados com dependecircncia linear
relativamente agrave concentraccedilatildeo de decavanadato total (Ramos et al 2006)
Assim a estabilidade da espeacutecie decameacuterica apoacutes diluiccedilatildeo da soluccedilatildeo de decavanadato no meio de
reacccedilatildeo dos ensaios (tampatildeo D) para uma concentraccedilatildeo final de 1 mM em vanadato total foi
caracterizada atraveacutes da leitura da Abs400 em funccedilatildeo do tempo tendo-se determinado qual a
ordem de reacccedilatildeo e o tempo de meia-vida da mesma Com base em estudos anteriores acerca da
estabilidade do V10 na presenccedila de diferentes proteiacutenas (Ramos et al 2006) determinou-se
igualmente o tempo de meia vida apoacutes a adiccedilatildeo de 20 microM de S1 de miosina ao meio reaccional
353 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
Os espectros de RMN do isoacutetopo 51
V foram realizados num espectroacutemetro Bruker AM-400 MHz
equipado com uma sonda multinuclear inversa de 5 mm Em tubos de RMN de 5 mm de diacircmetro
preparou-se soluccedilotildees de 500 L de deca- ou metavanadato correspondentes a um intervalo de
concentraccedilotildees de 1 a 10 mM suplementadas com 10 D2O Os espectros foram adquiridos agrave
temperatura ambiente com uma largura espectral de 23 923 Hz um tempo de aquisiccedilatildeo de 0086
s tempo de relaxaccedilatildeo de 001 s e uma sequecircncia de pulsos a 90ordm Utilizou-se a transformada de
Fourier com alargamento das bandas de 20Hz para o processamento dos espectros Os desvios
quiacutemicos das ressonacircncias de 51
V foram obtidos em relaccedilatildeo agrave espeacutecie VOCl3cuja frequecircncia de
ressonacircncia eacute considerada o padratildeo sendo todas as outras referenciadas a partir desta em ppm
As aacutereas dos sinais do RMN correspondentes agraves espeacutecies oligomeacutericas presentes foram obtidas
por integraccedilatildeo dos sinais As concentraccedilotildees relativas das espeacutecies foram calculadas com recurso agrave
equaccedilatildeo
onde A[Vx] corresponde agrave aacuterea referente agrave ressonacircncia da espeacutecie Vx com nordm de agregaccedilatildeo x
A[V]total agrave soma total das aacutereas medidas e [V] a concentraccedilatildeo total de vanaacutedio no ensaio
A teacutecnica de 51
V-RMN foi anteriormente aplicada em estudos de ligaccedilatildeo com S1 de miosina
(Ref) tendo sido demostrado que entre as espeacutecies V1 V2 V4 e V5 a espeacutecie tetrameacuterica possui a
maior afinidade para a miosina (Cremo et al) O desaparecimento ou alargamento dos sinais pode
implicar a ligaccedilatildeo de um dos oxovanadatos laacutebeis agrave miosina sem indicaccedilatildeo de qual uma vez que
49
se processa um equiliacutebrio entre eles (Crans 1990) Assim e tendo em conta as indicaccedilotildees de que
a espeacutecie V10 eacute distinta das restantes no que diz respeito agrave magnitude da interacccedilatildeo com a miosina
e inibiccedilatildeo da actividade ATPaacutesica estimulada por actina (Aureliano 2000 Tiago et al 2002)
torna-se fundamental investigar a interacccedilatildeo das diferentes espeacutecies de vanadato com o S1 de
miosina atraveacutes da teacutecnica de 51
V-RMN
Para tal obtiveram-se espectros de 51
V-RMN resultantes da titulaccedilatildeo de uma soluccedilatildeo 5 mM de
decavanadato total com concentraccedilotildees crescentes de S1 de miosina (0 5 10 15 e 20 microM) em
tampatildeo D Realizou-se ainda ensaios na presenccedila de 2 mM de decavanadato e 2 mM de ATP na
presenccedila e na ausecircncia de S1 de miosina de modo a verificar o efeito que estes ligandos naturais
da enzima teria na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o subfragmento-1
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
Os grupos sulfidrilo (-SH) das cisteiacutenas satildeo resiacuteduos de aminoaacutecidos extremamente sensiacuteveis agrave
oxidaccedilatildeo resultando na sua oxidaccedilatildeo (Dalle-Donne et al 1999) Logo a quantificaccedilatildeo destes
resiacuteduos eacute bastante importante nos estudos de interacccedilatildeo com vanaacutedio jaacute que os resiacuteduos de Cys
com ecircnfase para o SH1 e o SH2 satildeo bastante importantes na monitorizaccedilatildeo das modificaccedilotildees
funcionais da regiatildeo da cabeccedila
O procedimento seguidamente apresentado foi originalmente descrito por Gutieacuterrez-Merino
(1987)) Consiste na reacccedilatildeo entre os grupos ndashSH natildeo oxidados das proteiacutenas e o reagente de
Ellman (DTNB -55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico)) - formando-se o aniatildeo CNT (aniatildeo 3-
carboxilato-4-nitrotiofenolato) numa estequiometria de 11 Este apresenta um maacuteximo de
absorvecircncia a 412 nm (Robyt 1990) Assim apoacutes a incubaccedilatildeo da proteiacutena na presenccedila do metal
a quantidade de cisteiacutenas reduzidas por proteiacutena determina-se atraveacutes dos valores de Abs412
Deste modo colocou-se num tubo de Eppendorf o volume apropriado de S1 dialisado para que a
concentraccedilatildeo no ensaio fosse de 1 mgmL Adicionou-se o volume apropriado de decavanadato
para que a sua concentraccedilatildeo fosse de 0 10 50 100 250 500 ou 1000 M Perfez-se com soluccedilatildeo
D ateacute um volume final de 1 mL e incubou-se a 25ordmC por 20 minutos Apoacutes este periacuteodo colocou-
se 780 L da mistura reaccional numa cuvette de quartzo e adicionou-se 220 L de soluccedilatildeo K23
23
Tampatildeo K Tampatildeo fosfatohidrogenofosfato de soacutedio 025 M pH 72
50
Adicionou-se 15 L de DTNB (em soluccedilatildeo K [DTNB]stock=100 mM) ao meio reaccional e
registou-se o valor de absorvecircncia medido equivalente agraves cisteiacutenas ldquoraacutepidasrdquo (SH1 e SH2
expostos ao solvente) Adicionou-se 001 g de SDS que provoca a desnaturaccedilatildeo da proteiacutena e
consequentemente a exposiccedilatildeo de todos os resiacuteduos de cisteiacutenas que se encontravam ateacute aiacute
inacessiacuteveis no interior da proteiacutena (cisteiacutenas totais) Estes resiacuteduos podem assim reagir com o
DTNB
Estimou-se o nuacutemero de resiacuteduos de cisteiacutenas em todas as amostras com recurso ao coeficiente de
extinccedilatildeo molar () do DTNB (ver ponto 3 dos Anexos) que foi determinado atraveacutes da titulaccedilatildeo
de L-Cisteiacutena com intervalo de concentraccedilotildees compreendidos entre 0 e 50 microM na presenccedila de
25microM de DTNB em tampatildeo D Os valores de absorvecircncia foram obtidos para um comprimento de
onda de 412 nm
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
3621 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
A teacutecnica mais eficiente de anaacutelise da interacccedilatildeo entre o iatildeo vanadilo e proteiacutenas como a
calmodulina e a transferrina entre outras permite a obtenccedilatildeo raacutepida de informaccedilatildeo no que toca agrave
conformaccedilatildeo e amplitude da referida interacccedilatildeo
Os espectros de Ressonacircncia Paramagneacutetica Electroacutenica (RPE) da espeacutecie paramagneacutetica V(IV)
foram realizados a 70K num espectroacutemetro Bruker EMX-300 X-band Todas as soluccedilotildees
utilizadas e amostras foram preparadas em anaerobiose numa cacircmara de luvas de modo a evitar a
reduccedilatildeo de V(IV) a V(V) Em tubos capilares com 02 mL de amostra manteve-se a concentraccedilatildeo
de S1 a 20 M e variou-se a concentraccedilatildeo de vanadilo (0 25 5 10 15 25 50 75 100 150
200 500 e 1000 M [VOSO4]stock=5 mM) em tampatildeo D Apoacutes obtenccedilatildeo dos espectros estes
foram tratados atraveacutes do software SimFonia e WinEPR tendo-se obtido um graacutefico com a
variaccedilatildeo da intensidade do sinal de RPE em funccedilatildeo da concentraccedilatildeo de sulfato de vanadilo
presente Realizaram-se ainda simulaccedilotildees de espectros obtidos com 20 microM de S1 e 1000 microM de
V(IV) com vista agrave obtenccedilatildeo dos ligandos provaacuteveis do vanadilo atraveacutes da anaacutelise da constante
de acoplamento hiperfina (A) e do factor g (g)
4 Conclusotildees gerais
Apesar de jaacute ter sido demonstrado que o vanaacutedio inibe a Na+K
+-ATPase da miosina a funccedilatildeo
deste elemento no tecido muacutesculo esqueleacutetico continua largamente incompreendida A enorme
complexidade quiacutemica deste metal especialmente no que concerne agrave oligomerizaccedilatildeo e reacccedilotildees
51
redox constitui um obstaacuteculo agrave compreensatildeo do comportamento do vanaacutedio nos organismos
vivos Assim a caracterizaccedilatildeo da quiacutemica deste elemento eacute de grande utilidade antes de iniciar o
estudo da sua interacccedilatildeo com o presente sistema
A anaacutelise por espectroscopias de 51
V-RMN e de UV-Vis de soluccedilotildees de vanadato diluiacutedas nos
meios de reacccedilatildeo utilizados no presente relatoacuterio (10 mM HEPES pH 70 25 mM KCl) mostrou
claramente que a espeacutecie decameacuterica (V10) possui um tempo de vida suficientemente longo para a
determinaccedilatildeo do seu efeito isolado ao contraacuterio do que se verifica com as espeacutecies mono- di-
tetra- e pentameacutericas (V1 V2 V4 e V5) Por outro lado a anaacutelise verificou ainda que apoacutes diluiccedilatildeo
das soluccedilotildees de decavanadato no meio de reacccedilatildeo coexistem as espeacutecies V10 e V1 pelo que os
efeitos putativos desta uacuteltima deveratildeo ser subtraiacutedos aquando da monitorizaccedilatildeo dos efeitos da
espeacutecie decameacuterica
O vanaacutedio (V) (vanadato) inibe as enzimas que catalizam reacccedilotildees de transferecircncia de grupos
fosfato como a ATPase da miosina [Goodno 1979] devido agrave sua analogia estrutural com o iatildeo
fosfato Na sua utilizaccedilatildeo como inibidor enzimaacutetico natildeo eacute tida em conta a contribuiccedilatildeo individual
das espeacutecies oligomeacutericas considerando-se normalmente a espeacutecie V1 como inibidora efectiva
[Goodno e Taylor 1982] No presente relatoacuterio verificou-se por espectroscopia de 51
V-RMN que
outros oligoacutemeros de vanadato interagem com o S1 de miosina tendo as espeacutecies V10 e V4 os
maiores niacuteveis de afinidade A sua interacccedilatildeo envolve forccedilas electrostaacuteticas e eacute afectada
negativamente por ligandos naturais de enzima como eacute o caso do ATP
A exposiccedilatildeo do S1 de miosina (005 mgmL) a decavanadato conduz agrave inibiccedilatildeo do capacidade da
proteiacutena de estimular a actividade Mg2+
-ATPaacutesica com um valor de IC50 de 266 plusmn 07 M
decavanadato sendo o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM
decavanadato (80) Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM decavanadato sendo
o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM decavanadato (60)
exercendo um efeito de impedimento agrave ligaccedilatildeo entre o vanaacutedio e o S1
A uma concentraccedilatildeo de 27 microM decavanadato verifica-se a oxidaccedilatildeo de um dos resiacuteduos de Cys
de S1 de miosina seguida pela oxidaccedilatildeo gradual de outro resiacuteduo de Cys ateacute agrave concentraccedilatildeo de
1000 microM Ambos os resiacuteduos oxidados natildeo se encontravam expostos ao solvente utilizado A
oxidaccedilatildeo dos resiacuteduos permite sugerir que esta modificaccedilatildeo oxidativa sofrida pelo S1 contribui
para a diminuiccedilatildeo da sua actividade de ATPase
52
Por espectroscopia de RPE revelaram-se os possiacuteveis ligandos equatoriais do vanaacutedio aquando da
sua interacccedilatildeo com o S1 de miosina como sendo duas moleacuteculas de aacutegua um ligando contendo o
grupo R-OH e outro contendo o grupo R=NR
Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados de
oxidaccedilatildeo (IV) e (V) no S1 de miosina para concentraccedilotildees facilmente alcanccedilaacuteveis no ambiente
terrestre
53
5 Estudos futuros
No sentido do melhor esclarecimento das alteraccedilotildees moleculares e estruturais que ocorrem no
subfragmento 1 quando exposta ao vanaacutedio que conduzem a uma diminuiccedilatildeo da sua funccedilatildeo
sugere-se
- Quantificaccedilatildeo das cisteiacutenas reduzidas de S1 na presenccedila de metavanadato
- Determinaccedilatildeo das actividades Ca2+
- e K+-ATPaacutesica do S1 de miosina na presenccedila de
decavanadato
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina estimulada por actina
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina na presenccedila de
metavanadato
- Realizaccedilatildeo de RMN 2D com variaccedilatildeo das concentraccedilotildees das soluccedilotildees de decavanadato e
metavanadato na presenccedila de S1
-Mutageacutenese dos resiacuteduos de Cys doS1 de miosina de modo a verificar quais os resiacuteduos que
sofrem oxidaccedilatildeo na presenccedila de decavanadato
- Determinaccedilatildeo por calorimetria (DSC Differential Scanning Calorimetry) da variaccedilatildeo da
entalpia (∆H) e da temperatura de desnaturaccedilatildeo do S1 de miosina na presenccedila de vanaacutedio uma
vez que estas propriedades caracteriacutesticas de cada proteiacutena satildeo alteradas apoacutes ocorrecircncia de
modificaccedilotildees estruturais
- Anaacutelise por SDS-PAGE de S1 de miosina apoacutes tratamento com vanaacutedio de modo a determinar
quais os polipeacuteptidos eventualmente formados apoacutes clivagem da proteiacutena
54
6 Referecircncias bibliograacuteficas
- Sellers J Myosins 2nd Edition Oxford University Press 1999
- L Stryer inBiochemistry (Ed WH Freeman 2005 pp
Refordfas Materiais e Meacutetodos
[A] S S Margossian SLowey Methods Enzymol1982 85 55-71
[B] JD Pardue JA Spudich Methods Enzymol 1982 85 164-181
[C] M Bradford Anal Biochem1976 72 248-254
[D] H H Taussky E Shorr J Biol Chem 1953 202 675-685
[E] C R Bagshaw in Outline Studies in Biology ndash Muscle Contraction Chapman and Hall Ed
1982
[F] A Cuenda et al Eur J Biochem1990 194 663-670
55
Anexos
1 Determinaccedilatildeo da recta de calibraccedilatildeo meacutetodo de Bradford
Figura1ndashRepresentaccedilatildeo graacutefica da recta de calibraccedilatildeo (Abs595 nm vs [BSA]) utilizada para o meacutetodo de Bradford
2 Determinaccedilatildeo da recta calibraccedilatildeo para a electroforese em condiccedilotildees desnaturantes
Figura 2 ndash Representaccedilatildeo graacutefica da recta de calibraccedilatildeo (log(Peso Molecular) vs Rf dos marcadores de peso
molecular) onde Rf indica determinada atraveacutes do padratildeo de peso molecular
(poccedilo (a))
3Caacutelculo do coeficiente de extinccedilatildeo molar () para a quantificaccedilatildeo de cisteiacutenas reduzidas
atraveacutes do meacutetodo de Ellman
Traccedilou-se a recta de Abs412 nm meacutedia vs [L-Cisteiacutena] de modo a determinar o valor do coeficiente
de extinccedilatildeo molar definido como o declive daquela recta Assim =8400 M-1
cm-1
como
podemos verificar na Fig 3
A595 nm = 00546[BSA] + 00209 Rsup2 = 099051
Ab
s 59
5 n
m
[BSA] (mgmL)
log PM = -1177 Rf + 2247 Rsup2 = 098957
log
PM
Mobilidade relativa (Rf)
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero

21
processo da contracccedilatildeo muscular(ref)Destaca-se ainda a descoberta da capacidade inibitoacuteria do
V10 em relaccedilatildeo agrave fosforilaccedilatildeo oxidativa mitocondrial [AX]
Finalmente no que concerne aos estudos realizados com a presenccedila de vaacuterias espeacutecies
oligomeacutericas Aureliano e Madeira (1994) [AY]observaram que na presenccedila de Ca2+
as espeacutecies
V4 e V10se ligam agrave Ca2+
-ATPase do retiacuteculo sarcoplasmaacutetico enquanto a espeacutecie V1 se liga apenas
na presenccedila de ATP Aparentemente o aumento da concentraccedilatildeo de Ca2+
tem o efeito contraacuterio da
inibiccedilatildeo da ATPase do retiacuteculo sarcoplasmaacutetico pelo V1 natildeo tendo no entanto efeito significativo
sobre a inibiccedilatildeo por parte da espeacutecie V10 Tiago et al (2002) [AP] concluiacuteram que em condiccedilotildees
proacuteximas agraves fisioloacutegicas a espeacutecie V10 difere das espeacutecies presentes na soluccedilatildeo de metavanadato
devido agrave sua forte interacccedilatildeo com a miosina fosforilada e agrave sua inibiccedilatildeo da actividade ATPaacutesica
da miosina Para aleacutem disso o ATP diminui a afinidade da miosina para o V4 induzindo a
interacccedilatildeo com V1 e natildeo tendo qualquer efeito no que diz respeito ao V10 Foi tambeacutem proposto
[AQ] que a ligaccedilatildeo das espeacutecies oligomeacutericas de vanadato aoS1 de miosina nos locais onde o
fosfato coordenaeacute provavelmente a causa da inibiccedilatildeo da actividade ATPaacutesica da miosina
estimulada por actina atraveacutes da prevenccedilatildeo da troca entre ATPADP sendo esta favorecida para
aniotildees de vanadato com valecircncias superiores como eacute o caso do V10
Na presenccedila de actina as velocidades de dissociaccedilatildeo do Pi e do V aumentam consideravelmente
este uacuteltimo em menor quantidade Assim a ligaccedilatildeo da actina ao complexo MADP
V e
Pi eacute
bastante fraca pelo que se sugere que o ADPV teria um efeito de relaxamento semelhante ao
ATP No entanto a capacidade do vanaacutedio em inibir a actividade ATPaacutesica da miosina e da
actomiosina eacute diferenciada uma vez que na ausecircncia de actina a inibiccedilatildeo eacute total para
concentraccedilotildees na gama dos M mas na sua presenccedila a inibiccedilatildeo eacute totalmente conseguida apenas
em concentraccedilotildees na gama dos mM (Ref) No entanto eacute necessaacuterio considerar a possiacutevel
formaccedilatildeo de espeacutecies oligomeacutericas de vanadato cuja afinidade e efeito na enzima seratildeo
diferentes Assim pode concluir-se que o efeito de inibiccedilatildeo observado para a actividade ATPaacutesica
de miosina estimulada por actina se pode dever agrave presenccedila das diferentes espeacutecies oligomeacutericas o
que foi jaacute sugerido por Aureliano (2000) [AZ] ndash as espeacutecies tetrameacuterica e decameacuterica de vanadato
interactuariam com a miosina com alteraccedilatildeo da sua interacccedilatildeo com a actina No entanto natildeo eacute
conhecida a contribuiccedilatildeo absoluta das restantes espeacutecies oligomeacutericas para a inibiccedilatildeo da ATPase
da miosina estimulada por actina (Refs)
14 Objectivos do projecto
Pretende-se com este projecto o isolamento purificaccedilatildeo e caracterizaccedilatildeo da actina do muacutesculo
22
esqueleacutetico de coelho sendo esta uacuteltima tambeacutem realizada para o sufragmento-1 de miosina
Seguidamente realizar-se-aacute a quantificaccedilatildeo e caracterizaccedilatildeo das proteiacutenas por espectroscopia UV-
Vis e SDS-PAGE e a caracterizaccedilatildeo das soluccedilotildees de vanaacutedio por UV-Vis e ressonacircncia magneacutetica
nuclear (RMN) Por uacuteltimo a interacccedilatildeo do S1 de miosina com o vanaacutedio seraacute avaliada atraveacutes
das teacutecnicas de UVVis RMN e ressonacircncia paramagneacutetica electroacutenica (RPE)
2 Discussatildeo de Resultados
21 Caracterizaccedilatildeo das proteiacutenas isoladas
211Anaacutelise do estado de pureza
O estado de pureza das soluccedilotildees de actina e de S1 foi analisado por electroforese em gel de
poliacrilamida em condiccedilotildees desnaturantes (SDS-PAGE) (Fig 22) que separa as moleacuteculas de
acordo com o seu peso molecular (PM) A concentraccedilatildeo de acrilamida escolhida (10) permite a
detecccedilatildeo simultacircnea de componentes com peso molecular inferior a 20 kDa e outros com elevado
peso molecular
Figura 21 ndash Fotografia de gel de SDS-PAGE (10 acrilamida) Poccedilo (a)
10 L do marcador de elevado peso molecular () Poccedilo (b) 20 L de [G-
actina]=1 mgmL Poccedilo (c) 20 L de [S1]=1 mgmL Aplicou-se uma
voltagem de V durante h e min
A partir da recta de calibraccedilatildeo obtida log PM = -1177 Rf + 2247 (ver ponto 2 da secccedilatildeo
Anexos) pode verificar-se a presenccedila de actina na amostra do poccedilo (b) presente numa banda
bastante intensa (PMaparente=40 kDa) bem como a presenccedila minoritaacuteria de outros componentes
desconhecidos que natildeo teratildeo sido eliminados durante o processo de extracccedilatildeo e purificaccedilatildeo e natildeo
satildeo relevantes para o estudo em questatildeo Na amostra do poccedilo (c) nota-se a presenccedila de S1
(PMaparente=92 kDa) bem como a de outros componentes cuja banda eacute ligeiramente menos intensa
Estes poderatildeo ser fragmentos proteoliacuteticos de miosina e de S1 Estaacute ainda presente uma bada de
23
maior intensidade que poderaacute ser a proteiacutena-C (PM=135 kDa) uma vez que a sua remoccedilatildeo
completa da amostra necessitaria de um passo adicional de cromatografia de troca ioacutenica [BA]
No entanto natildeo podemos confirmar que se trata desta proteiacutena ou de qualquer outra uma vez que
se encontra fora do intervalo de migraccedilatildeo dos marcadores de peso molecular
212 Determinaccedilatildeo da concentraccedilatildeo proteica
2121Subfragmento-1 de miosina
A partir da recta de calibraccedilatildeo obtida pelo meacutetodo de Bradford [BB](ver secccedilatildeo 3311 do
Procedimento Experimental) A595nm = 00546[BSA] + 00209 (ver secccedilatildeo de Anexos)foi possiacutevel
determinar um intervalo de concentraccedilatildeo de S1 de miosina compreendido entre 226 e 117(plusmn
423M) (203 e 1056 (plusmn 381 mgmL)) (n=8) A concentraccedilatildeo meacutedia de S1 de miosina apoacutes
diaacutelise contra a soluccedilatildeo tampatildeo 10 mM HEPES pH 75 25 mM KCl determinada pela leitura da
Abs280 nmfoi de 20430M (1803 mgmL)(n=7)
As amostras a quantificar atraveacutes do meacutetodo de Bradford natildeo sofreram diaacutelise o que permite
explicar a concentraccedilatildeo superior de S1 de miosina em comparaccedilatildeo com o valor obtido atraveacutes do
meacutetodo directo Esta razatildeo aliada agrave sensibilidade e rapidez superiores deste uacuteltimo fez com que
se optasse pelo meacutetodo directo para a quantificaccedilatildeo da grande maioria dos ensaios
2122Actina
A concentraccedilatildeo de actina foi determinada quando esta se encontrava na forma monomeacuterica (G-
actina) por mediccedilatildeo da leitura directa da Abs290 nm tendo-se obtido uma concentraccedilatildeo meacutedia de
691481M (290202 mgmL) (n=3) em 2 mM Tris-HCl pH 80 02 mM CaCl2 02 mM Na-
ATP Ao comprimento de onda utilizado (290 nm) absorvem fortemente apenas os resiacuteduos de
Trp pelo que a mediccedilatildeo efectuada tem menor sensibilidade do que se tivesse sido efectuada a 280
nm uma vez que a actina apresenta apenas 4 resiacuteduos deste aminoaacutecido embora tenha 16
resiacuteduos de Tyr
22 Mediccedilatildeo da actividade enzimaacutetica de S1 de miosina
221 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
As actividades ATPaacutesicas de S1 de miosina dependentes de Ca2+
e K+foram analisadas pelo
caacutelculo das velocidades de consumo inicial do fosfato inorgacircnico (Pi) Definiu-se os tempos de 5
e 3 minutos para a mediccedilatildeo da Ca2+
e da K+EDTA-ATPase respectivamente Os valores obtidos
24
a partir do protocolo presente no ponto 341 da secccedilatildeo Procedimento experimental encontram-se
descritos na Tabela 2I
Tabela 2I ndash Acrividades natildeo-fisioloacutegicas de subfragmento-1 determinadas pelo meacutetodo de Fiske-Subarow
modificado (n=2)
Proteiacutena
Actividade
ATPaacutesica natildeo
fisioloacutegica
Actividade (nmol
Piminmg proteiacutena) Actividade (s-1)
S1
Ca2+
-ATPase 1400 plusmn 30 2100 plusmn 0045
K+EDTA-ATPase 2334 plusmn 50 3500 plusmn 0075
As actividades natildeo-fisioloacutegicas reflectem o estado de oxidaccedilatildeo dos resiacuteduos expostos de Cys 707
(SH1) e 697 (SH2) Se o grupo SH1 sofrer oxidaccedilatildeo a actividade ATPaacutesica dependente de Ca2+
aumenta e a actividade dependente de K+ diminui Por outro lado se o grupo SH2 sofrer
oxidaccedilatildeo regista-se a perda completa destas actividades
De acordo com Margossian e Lowey (1982) [BA] as actividades ATPaacutesica de S1 de miosina
dependente de Ca2+
e K+EDTA da miosina satildeo respectivamente de 24 e 112 s-1
Estaacute tambeacutem
descrito que a actividade ATPaacutesica dependente de K+ eacute em meacutedia cerca de 3 vezes superior agrave
dependente de Ca2+
reflectindo a quantidade das preparaccedilotildees A razatildeo obtida entre os valores das
actividades ATPaacutesicas dependentes de K+ e Ca
2+ eacute de 166 Os valores obtidos para as actividades
(Ca2+
-ATPase=21 s-1
K+EDTA-ATPase= 35 s
-1) satildeo inferiores agrave valores obtidos por
Margossian e Lowey [BA] e Tiago [BC] com forccedila ioacutenica inferior O valor obtido para a
actividade ATPaacutesica dependente de Ca2+
eacute superior ao obtido por Polosukhina et al (2000) [BD]
cujo meio de reacccedilatildeo continha MOPS e apresentava forccedila ioacutenica inferior Os dados
supramencionados comprovam a variaccedilatildeo da actividade a ATPaacutesica natildeo fisioloacutegica em relaccedilatildeo agrave
forccedila ioacutenica temperatura e pH
222 Mediccedilatildeo da actividadeMg2+
-ATPaacutesica de S1 pelo meacutetodo de enzimas acopladas
A actividade de Mg2+
-ATPase eacute a uacutenica que possui significado fisioloacutegico sendo as restantes
bastante uacuteteis apenas para indicaccedilatildeo da qualidade de preparaccedilatildeo do S1[BA] A 0575 M KCl a
actina e o S1 formam uma soluccedilatildeo viscosa e tuacuterbida Apoacutes a adiccedilatildeo de MgATP o complexo acto-
S1 dissocia-se e a turbidez da soluccedilatildeo diminui Se a forccedila ioacutenica for reduzida a dissociaccedilatildeo do
complexo actoS1 que se encontra na forma de suspensatildeo eacute induzida O MgATP eacute rapidamente
hidrolisado diminuindo a sua concentraccedilatildeo no meio reaccional Nestas condiccedilotildees haacute
reassociaccedilatildeo e subsequente contracccedilatildeo dos filamentos de actina e de S1 com formaccedilatildeo de
25
precipitado O meacutetodo descrito traduz de forma bastante elementar o fenoacutemeno da contracccedilatildeo in
vivo e elimina o problema anterior da solubilizaccedilatildeo ineficiente da miosina de modo a obter uma
mediccedilatildeo exacta da actividade [A] A utilizaccedilatildeo de forccedilas ioacutenicas reduzidas aumenta a afinidade da
actina para o S1 permitindo contornar a extrema dificuldade em obter a concentraccedilatildeo proteica dos
muacutesculos (cerca de 100 mgmL do complexo actomiosina) As actividades Mg2+
-ATPaacutesicas de S1
de miosina foram determinadas a partir do declive da variaccedilatildeo da Abs340 relativo agrave oxidaccedilatildeo de
NADH em funccedilatildeo do tempo como eacute descrito no ponto 342 do Procedimento Experimental Os
valores obtidos para a actividade Mg2+
-ATPaacutesica encontram-se apresentados na Tabela 2II
Tabela 2II ndash Actividade Mg2+
-ATPaacutesica de subfragmento-1 de miosina determinadas pelo meacutetodo das enzimas
acopladas
Actividade (nmol
ATPmin mg(S1)) Actividade (s
-1)
1097 plusmn 269 164 plusmn 04
Ramos (2005) [J] obteve o valor de 01 s-1
para a actividade Mg2+
-ATPaacutesica de S1 de miosina
com forccedila ioacutenica intermeacutedia Tiago (2005) [BC] obteve o valor de 011 s-1 em condiccedilotildees de forccedila
ioacutenica reduzida em relaccedilatildeo agrave utilizada no presente relatoacuterio
23 Preparaccedilatildeo e caracterizaccedilatildeo das soluccedilotildees de vanaacutedio
231 Anaacutelise da composiccedilatildeo das soluccedilotildees de vanadato por espectroscopia de 51
V-RMN
efeito da concentraccedilatildeo da espeacutecie
A diluiccedilatildeo das soluccedilotildees stock de meta- e decavanadato iraacute determinar fortemente a composiccedilatildeo
em soluccedilatildeo dos diferentes oligoacutemeros de vanadato (ref) No caso do decavanadato o periacuteodo de
tempo apoacutes a diluiccedilatildeo iraacute tambeacutem ser um factor importante a considerar devido agrave decomposiccedilatildeo
da espeacutecie V10(ref)Torna-se assim fundamental determinar a composiccedilatildeo quiacutemica das soluccedilotildees
no meio de ensaio utilizado (HEPES 10 mM KCl 25 mM) de modo a correlacionaacute-la
correctamente com os possiacuteveis efeitos promovidos pelas diferentes espeacutecies de vanadato no
sistema em estudo
Aleacutem disso deve ser dado ecircnfase agraves importantes interacccedilotildees que ocorrem entre o vanaacutedio e os
tampotildees escolhidos O tampatildeo com menor nuacutemero e magnitude de interacccedilatildeo eacute o HEPES [AD]
seguido do Tris sendo este uacuteltimo especialmente aconselhado em concentraccedilotildees baixas para casos
de incompatibilidade enzimaacutetica ou quiacutemica redox No caso especial da reacccedilatildeo redox
NAD(P)HNAD(P) o tampatildeo adequado eacute o imidazole de modo a natildeo haver reacccedilatildeo com o
26
vanadato Os tampotildees a evitar em qualquer situaccedilatildeo incluem a trietanolamina fosfato citrato e
glicina entre outros [AD] uma vez que todos interactuam fortemente com V(V) com formaccedilatildeo
de complexos estaacuteveis
Assim procedeu-se agrave anaacutelise da composiccedilatildeo das soluccedilotildees de meta- e decavanadato apoacutes diluiccedilatildeo
das soluccedilotildees stock (50 mM em vanadato total) no meio de reacccedilatildeo (tampatildeo D) atraveacutes da
espectroscopia de ressonacircncia magneacutetica nuclear de 51
V Nas figuras x1 e x2 apresentam-se os
espectros obtidos para as soluccedilotildees de deca- e metavanadato num intervalo de concentraccedilotildees de 1 a
10 mM A Tabela seguinte resume os paracircmetros espectroscoacutepicos obtidos para cada espeacutecie
presente nas soluccedilotildees analisadas
Figura 22ndash Espectros de RMN de 51
V de
concentraccedilotildees crescentes das espeacutecies oligomeacutericas
de vanadato obtidos apoacutes diluiccedilatildeo da soluccedilatildeo stock
de decavanadato em 10 mM HEPES pH 70 25 mM
KCl para concentraccedilotildees finais de A) 1 mM B)2
mM C) 4 mM D) 8 mM E) 10 mM de
decavanadato total V1refere-se agraves espeacutecies
monomeacutericade vanadato V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2IIIndash Desvios quiacutemicos meacutedios (part) das ressonacircncias das espeacutecies decameacuterica (V10) monomeacuterica (V1) e
tetrameacuterica (V4) presentes nos espectros da soluccedilatildeo de decavanadatoLargura a meia altura (∆12) da espeacutecie V10A
Paracircmetros V10A V10B V10C V1 V4
part (ppm) -5162 -5003 -4255 -5616 -5748
∆12 (Hz) 175 nd nd nd nd
Figura 23 ndash Distribuiccedilatildeo graacutefica das
espeacutecies oligomeacutericas de vanadato vs a
concentraccedilatildeo de decavanadato total As
concentraccedilotildees de cada espeacutecie foram
obtidas segundo a formula descrita na
secccedilatildeo 35 do Procedimento
Experimental (aacutereas dos sinais de 51
V-
RMN observadas na figura x1)V10 V1 e
V4 correspondem agraves espeacutecie monomeacuterica
dimeacuterica e tetrameacuterica respectivamente [Esp
eacuteci
e o
ligo
meacute
rica
de
van
aacutedio
] (m
M)
[Decavanadato]total (mM)
V10
V1
V4
27
Nas soluccedilotildees de decavanadato as espeacutecies monomeacuterica e decameacuterica de vanadato satildeo as uacutenicas
presentes para concentraccedilotildees de decavanadato inferiores a 8 mM Ambas as espeacutecies aumentam
linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado em [AY] e
em (Teresa) quando a concentraccedilatildeo de V1 eacute igual ou superior a 200 microM inicia-se a sua
oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica No entanto nas condiccedilotildees de realizaccedilatildeo dos
ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a espeacutecie tetrameacuterica neste
estudo em concentraccedilotildees inferiores a 8 mM tendo o mesmo sido observado por Ramoset al
(2006) [AK] Os resultados obtidos mostram claramente o equiliacutebrio entre a espeacutecie decameacuterica e
a monomeacuterica e que o aparecimento de outras espeacutecies oligomeacutericas se encontra dependente da
concentraccedilatildeo desta uacuteltima Apesar de natildeo ter sido realizado o ensaio com 5 mM de decavanadato
total no presente relatoacuterio estaacute reportado em Ramos et al (2006) [AK] que as concentraccedilotildees da
espeacutecie decameacuterica e monomeacuterica satildeo respectivamente de 500 microM e 200 microM Assim pode inferir-
se que para concentraccedilotildees inferiores a 5 mM e intervalos de tempo de cerca de 20 minutos (que
inclui o tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas
as espeacutecies decameacuterica e monomeacuterica de vanadato
Figura 24 ndash Espectros de RMN de 51
V de concentraccedilotildees
crescentes das espeacutecies oligomeacutericas de vanadato obtidos
apoacutes diluiccedilatildeo da soluccedilatildeo stock de metavanadato em 10 mM
HEPES pH 70 25 mM KCl para concentraccedilotildees finais de A)
1 mM B) 2 mM C) 4 mM D) 8 mM E) 10 mM de
metavanadato total V1 refere-se agraves espeacutecies monomeacuterica de
vanadatoV1 e V2 correspondem respectivamente agrave espeacutecie
monomeacuterica e dimeacuterica
Figura 25 ndash Distribuiccedilatildeo graacutefica das espeacutecies
oligomeacutericas de vanadato vs a concentraccedilatildeo de
metavanadato total As concentraccedilotildees de cada
espeacutecie foram obtidas segundo a formula descrita
na secccedilatildeo 35 do Procedimento Experimental
(aacutereas dos sinais de 51
V-RMN observadas na
figura x1)V10 V1 e V4 correspondem agraves espeacutecie
monomeacuterica dimeacuterica e tetrameacuterica
respectivamente
Esp
eacuteci
e o
ligo
meacute
rica
de
va
nad
ato
(m
M)
[Metavanadato]total (mM)
V1
V2
V4
V5
28
Tabela 2IVndash Desvios quiacutemicos meacutedios (part)das ressonacircncias das espeacutecies tetrameacuterica (V4) monomeacuterica (V1) e
dimeacuterica (V2) presentes no espectro da soluccedilatildeo de metavanadato (total) 10 mM
Paracircmetros V1 V2 V4 V5
part (ppm) -5615 -5748 -5789 -5869
Nas soluccedilotildees de metavanadato as espeacutecies monomeacuterica dimeacutericae tetrameacuterica de vanadato estatildeo
presentes para concentraccedilotildees de metavanadato inferiores a 4 mM Todas estas as espeacutecies
aumentam linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado
em (Aureliano e Madeira 1998) e em (Tiago et al 2004) quando a concentraccedilatildeo de V1 eacute igual ou
superior a 200 microM inicia-se a sua oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica Nas
condiccedilotildees de realizaccedilatildeo dos ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a
espeacutecie pentameacuterica neste estudo em concentraccedilotildees inferiores a 4 mM tendo o mesmo sido
observado por Ramos 2006 Os resultados obtidos mostram claramente o equiliacutebrio entre a
espeacutecie V4 e as espeacuteciesV1 e V2 no qual o aparecimento de outras espeacutecies oligomeacutericas se
encontra dependente da concentraccedilatildeo da espeacutecie V1 Por uacuteltimo pode inferir-se que para
concentraccedilotildees inferiores a 2 mM e intervalos de tempo de cerca de 20 minutos (que inclui o
tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas as
espeacutecies V1 V2 e V4 com concentraccedilotildees respectivas de 0796 0192 e 0196 mM
232Estabilidade das soluccedilotildees de decavanadato anaacutelise por espectroscopia de absorccedilatildeo
UVVis
Caracterizou-se a estabilidade da espeacutecie V10 numa soluccedilatildeo com 1 mM decavanadato total em
tampatildeo D atraveacutes da leitura da Abs400 Obteve-se uma cineacutetica de dissociaccedilatildeo de 1ordf ordem tal
como se pode verificar pela linearidade do graacutefico de ln(Abs400)vs o tempo de aquisiccedilatildeo espectral
(segundos) (Fig26 )
Figura 26ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do tempo
(em segundos) para uma soluccedilatildeo de decavanadato total de 1
mM em tampatildeo D a 25ordmC (n=2)
Sendo uma reacccedilatildeo de primeira ordem pode ser descrita pela equaccedilatildeo cujo
declive eacute ndashK (constante de dissociaccedilatildeo) e a intersecccedilatildeo no eixo das ordenadas corresponde
Ln
(Ab
s 400 n
m)
tempo (s)
29
aln(A0) Numa reacccedilatildeo de 1ordf ordem o tempo de meia-vida eacute independente da concentraccedilatildeo inicial
do reagente sendo descrito pela equaccedilatildeo [2] Deste modo a partir da recta
Ln(Abs400 nm) = -436E-5
K - 105 obteve-se um tempo de meia-vida de 4490 plusmn 0823 horas para
as condiccedilotildees supramencionadas
Apoacutes a adiccedilatildeo de S1 8 microM ao meio reaccional verificou-se que a reacccedilatildeo de desoligomerizaccedilatildeo
da espeacutecie decameacuterica de vanadato seguia igualmente uma cineacutetica de 1ordf ordem tal como se pode
verificar pela linearidade do graacutefico de ln(Abs400) versus o tempo de aquisiccedilatildeo espectral
(segundos) (Fig27)
Figura 27ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do
tempo (em segundos) para uma soluccedilatildeo de decavanadato
total de 1 mM em tampatildeo D a 25ordmC na presenccedila de 8
microM (n= 2)
A partir da equaccedilatildeo da recta Ln(Abs400 nm) = -7520E-5
K - 0613determinou-se o valor de K para
a espeacutecie decameacuterica de onde se obteve um tempo de meia-vida de 2430 plusmn 0191 horas para as
condiccedilotildees supramencionadas
Tanto este valor como o valor registado na ausecircncia de S1 satildeo substancialmente superiores agrave
duraccedilatildeo dos ensaios de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina descritos no presente
relatoacuterio
24 Interacccedilatildeo de S1 com vanaacutedio(V)
241Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
De modo a analisar qual o efeito da adiccedilatildeo de S1 de miosina (tampatildeo D) produzia nos sinais de
51V-NMR titulou-se uma soluccedilatildeo de 5mM de decavanadato total com concentraccedilotildees crescentes
de S1 (5 10 15 e 20 microM) Os espectros de 51
V-RMN encontram-se apresentados na Fig 2
No graacutefico da figura 2 Apresenta-se a concentraccedilatildeo de S1 em funccedilatildeo do factor de alargamento
corrigido dos espectros acima referidos
Ln
(Ab
s (4
00
nm
))
tempo (s)
30
Figura 28 ndash Graacutefico de
representaccedilatildeo da
concentraccedilatildeo de S1 em
funccedilatildeo do factor de
alargamento A
concentraccedilatildeo de
decavanadato total foi de 5
mM e as concentraccedilotildees
utilizadas de S1 foram de 0
5 10 e 15 microM
Verifica-se que o factor de alargamento aumenta linearmente em funccedilatildeo da concentraccedilatildeo de S1 no
ensaio o que indica a existecircncia de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina Observou-
se ainda que a adiccedilatildeo de S1 ao meio reaccional natildeo provocou qualquer alteraccedilatildeo dos desvios
quiacutemicos dos sinais de 51
V-RMN
Realizaram-se ensaios na presenccedila de ATP e de S1 com intuito de verificar o efeito do ATP na
interacccedilatildeo do S1 de miosina com o decavanadato
Figura 29 - Espectros de RMN de 51
V contendo 2
mM Decavanadato (A) (controlo) (B) na presenccedila
de 2 mM de ATP (C) na presenccedila de 2 mM de ATP
e 8 microM de S1 de miosina V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2Vndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies decameacuterica (V10) e monomeacuterica (V1) presentes nos
espectros da soluccedilatildeo de decavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V10A V10B V10C V1
part (ppm) -5161 -5002 -4254 -5614
Observa-se que a presenccedila de ATP (espectro B) natildeo provoca qualquer alteraccedilatildeo visiacutevel em
relaccedilatildeo ao espectro que contem apenas soluccedilatildeo de decavanadato Jaacute a presenccedila simultacircnea de ATP
e S1 provoca um alargamento da ∆12 do sinal do V10A de 1736 Hz (espectro (A)) para 2366 Hz
(espectro (C) Fig 29) Tendo em conta que a concentraccedilatildeo de S1 apresenta uma relaccedilatildeo linear
com o factor de alargamento converteu-se o valor da ∆12 e comparou-se com o valor esperado agrave
F = 00591[S1] - 00157 Rsup2 = 0998
Fact
or
de
ala
rgam
en
to
[S1] (microM)
31
concentraccedilatildeo de S1 utilizada O factor de alargamento obtido para 8 microM ndash 0376 ndash eacute inferior ao
valor esperado agrave mesma concentraccedilatildeo de S1 obtido atraveacutes da equaccedilatildeo da recta apresentada na
Fig 28 ndash 0457 ndash o que indica que o ATP poderaacute exercer um efeito de protecccedilatildeo do S1
diminuindo a sua interacccedilatildeo com a espeacutecieV10
Por uacuteltimo realizaram-se ensaios com metavanadato na presenccedila de 8 microM S1 de modo a verificar
se as espeacutecies oligomeacutericas contidas nesta soluccedilatildeo interagiam com a proteiacutena em estudo
Figura 210 - Espectros de RMN de 51
V contendo 2
mM metavanadato (A) (controlo) (B) na presenccedila
de 8 microM de S1 de miosina V1 V2 e V4
correspondem respectivamente aacutes espeacutecies
monomeacuterica dimeacuterica e tetrameacuterica
Tabela 2VIndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies monomeacuterica (V1) dimeacuterica (V2) e tetrameacuterica (V4)
presentes nos espectros da soluccedilatildeo de metavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V1 V2 V4
part (ppm) -5613 -5747 -5789
Como eacute possiacutevel observar na Fig 210 ocorre uma ligeiro alargamento dos sinal de V4 bem
como uma perda da intensidade pelo que se conclui quedas espeacutecies presentes na soluccedilatildeo de
metavanadato (V1 V2 e V4) apenas a espeacutecie tetrameacuterica interage com o S1 de miosina o que
estaacute de acordo com o reportado por Tiago et al 2002
242 Actividade Mg2+
-ATPaacutesica de S1 de miosina na presenccedila de decanadato
Com o intuito de avaliar o efeito do decavanadato na actividade Mg2+
-ATPaacutesica do S1 de
miosina monitorizou-se a reacccedilatildeo de oxidaccedilatildeo do NADH atraveacutes da leitura de Abs340 na
presenccedila de 005 mgmL de S1 de miosina e no caso da actividade estimulada 2microM de actina em
3125 mM Tris-HCl pH 71 e 375 mM MgCl2 pelo meacutetodo de enzimas acopladas (ver seccccedilatildeo xx
dos MM)Para tal realizaram-se ensaios com vaacuterias concentraccedilotildees de decavanadato total (10 50
100 250 500 e 1000) de modo a determinar qual o seu efeito na actividade MgATPaacutesica de S1
de miosina na ausecircncia e na presenccedila de um dos seus ligandos naturais Os ensaios foram
realizados agrave temperatura ambiente com incubaccedilatildeo preacutevia a 25ordm C durante 20 minutos
32
Figura 211 ndash Representaccedilatildeo
graacutefica da Actividade Mg2+
-
ATPaacutesica de S1 () (os ajustes
teoacutericos estatildeo representado a por
uma linha os resultados
experimentais estatildeo representados
por triacircngulos (Actividade
estimulada) e cruzes (Actividade
natildeo estimulada)) vs
[Decavanadato]total medidas pelo
meacutetodo das enzimas acopladas
(n=3) Os dados foram ajustados ao modelo de decaimento exponencial usando o programa GraphPad Prism para a
determinaccedilatildeo dos valores de IC50 (na ausecircncia de actina IC50= 266 plusmn 07 M decavanadato na presenccedila de actina
IC50= 28 plusmn 1 M decavanadato
A partir do graacutefico presente na Fig 29 verifica-se que o ajuste dos dados experimentais foi bem
sucedido A inibiccedilatildeo da actividade Mg2+
-ATPaacutesica do S1 de miosina eacute afectada em menor
amplitude na presenccedila de actina (60 de inibiccedilatildeo da actividade) do que na sua ausecircncia
(aproximadamente 80 de inibiccedilatildeo de actividade) Devido a esta diferenccedila pode sugerir-se que a
actina iraacute impedir parcialmente a ligaccedilatildeo do decavanadatoao S1 demiosina atraveacutes de
impedimentos esteacutereos ou interacccedilotildees electrostaacuteticas por exemplo A teacutecnica utilizada natildeo nos
permite concluir acerca de que regiatildeo(otildees) ou resiacuteduo(s) da actina participaratildeo na preservaccedilatildeo
parcial da actividade Mg2+
-ATPaacutesica
Por outro lado os valores de IC50 dos dois tipos de actividades natildeo apresentam diferenccedilas
significativas tendo-se obtido 266 plusmn 07 e 28 plusmn 1M decavanadato na ausecircncia e na presenccedila de
actina respectivamente o que corresponde a uma concentraccedilatildeo de cerca de 27 M de V10 O
meio de reacccedilatildeo utilizado nestes ensaios foi 10 mM HEPES e 25 mM KCl com 2 mM ATP Em
(Tiago et al 2004) o valor de IC50 para a actividade Mg2+-ATPaacutesica na ausecircncia de actina foi de
125 plusmn 025 microM enquanto o valor da actividade estimulada por actina foi de 136 plusmn 14 microM O
meio de reacccedilatildeo utilizado nos ensaios foi 20 mM HEPES 50 mM KCl e 2 mM ATP o que resulta
precisamente no dobro da forccedila ioacutenica a que foi realizado o ensaio do presente relatoacuterio
(exceptuando a contribuiccedilatildeo dos produtos da hidroacutelise de ATP uma vez que este meacutetodo
pressupotildee a ocorrecircncia de um ciclo de reacccedilotildees de hidroacutelise e siacutentese deste composto) Ora tendo
sido anteriormente referido que a afinidade do S1 de miosina para com a espeacutecie V10 diminui com
Act
ivid
ade
Mg
2+-A
TP
aacutesic
a d
e S
1 d
e
Mio
sin
a
[Decavanadato] (microM) Actividade Estimulada ()
Actividade Natildeo Estimulada ()
Ajuste Actividade Estimulada
Ajuste Actividade Natildeo Estimulada
33
o aumento da forccedila ioacutenica (Tiago et al 2004 tal poderaacute explicar a diferenccedila obtida entre estes
dois conjuntos de valores
243Quantificaccedilatildeo de cisteiacutenas reduzidas pelo meacutetodo de Ellman
A interacccedilatildeo do vanaacutedio com alguns tipos de aminoaacutecidos sensiacuteveis agrave oxidaccedilatildeo pode fornecer-nos
informaccedilatildeo bastante uacutetil em relaccedilatildeo agraves alteraccedilotildees estruturais que conduzem a uma diminuiccedilatildeo da
actividade fisioloacutegica do S1 de miosina Os estudos realizados por Chasteen 1983 concluiacuteram
que a reduccedilatildeo de vanadato (V) a vanadilo (IV) ocorre na presenccedila de agentes redutores comuns no
ambiente celular de que se destaca o aminoaacutecido cisteiacutena Esta eacute tambeacutem um alvo de
complexaccedilatildeo por parte do iatildeo VO2+
Jaacute Ramos et al (2009) demonstrou que a incubaccedilatildeo de
actina com a soluccedilatildeo de decavanadato induz a oxidaccedilatildeo de resiacuteduos de cisteiacutena e agrave formaccedilatildeo do
oxidovanaacutedio (IV) Este grupo de investigadores descobriu ainda que apenas a espeacutecie V10 induz a
oxidaccedilatildeo das cisteiacutenas ldquoraacutepidasrdquo da actina atraveacutes do processo proposto que eacute parcialmente
responsaacutevel pela inibiccedilatildeo jaacute descrita da actividade de ATPase do S1 de miosina estimulada por
actina quando exposta agrave soluccedilatildeo de decavanadato Esta inibiccedilatildeo apresenta-se afectada com a
presenccedila de ATP uma vez que os resiacuteduos de cisteiacutena permaneceram intactos nos ensaios
realizados com este ligando Jaacute Crans et al (2004) concluiu que o siacutetio activo de muitas fosfatases
(que conteacutem grupos tiol) pode formar um equiliacutebrio redox com o iatildeo vanadato (V) de forma natildeo
reversiacutevel
Os resultados obtidos por Sakurai e Shiromura (1980) sugeriram que o vanaacutedio (V) sofre reduccedilatildeo
a vanaacutedio (IV) sem dependecircncia do valor de pH do ensaio Registaram ainda uma estequiometria
de 31 na reacccedilatildeo do vanadato (V) com os resiacuteduos de Cys enquanto a estequiometria dos ensaios
com o vanadilo (IV) foi de 21
Berlett e Stadtman (1997) observaram que os tipos de aminoaacutecidos mais facilmente oxidaacuteveis
englobam entre outros os resiacuteduos de Cys Trp e MetAssim analisou-se o estado redox dos
grupos tiol dos resiacuteduos de Cys do S1 na presenccedila do decavanadato atraveacutes da monitorizaccedilatildeo da
Abs412 nm na presenccedila de 1 mgmL de S1 de miosina em tampatildeo 10 mM HEPES 25 mM KCl
variando a concentraccedilatildeo da soluccedilatildeo de decavanadato de modo a determinar que tipo de alteraccedilotildees
estruturais ocorrem na sua presenccedila no que diz respeito agrave diminuiccedilatildeo da actividade ATPaacutesica de
S1 na presenccedila de decavanadato
Os resultados referentes agrave quantificaccedilatildeo das cisteiacutenas reduzidas apoacutes o tratamento de S1 com
diferentes concentraccedilotildees de decavanadato em tampatildeo 10 mM HEPES pH 75 e 25 mM KCl a
25ordmC durante 20 minutos encontram-se descritos na Tabela 2VII
Tabela 2VII ndash Cisteiacutenas reduzidas totais e ldquoraacutepidas de S1 (1 mgmL) apoacutes tratamento com decavanadato
34
(010501002505001000 uM) em 10 mM HEPES 25 mM KCl pH 7 a 25ordmC durante 20 minutos A detecccedilatildeo
espectrofotomeacutetrica foi feita com DTNB (e 412=8400 uM-1 cm-1) (n variaacutevel entre 1 e 8)
Amostras Cys raacutepidas Cys totais
11 M S1 (controlo) 22 plusmn 04 106 plusmn 14
11 M S1 + 10 M Decavanadato 22 plusmn 05 95 plusmn 08
11 M S1 + 50 uM Decavanadato 20 plusmn 02 98 plusmn 14
11 M S1 + 100 uM Decavanadato 17 plusmn 03 97 plusmn14
11 M S1 + 250 uM Decavanadato 26 plusmn 02 98 plusmn 08
11 M S1 + 500 uM Decavanadato 21 plusmn 04 90 plusmn 06
11 M S1 + 1000 uM Decavanadato 24 plusmn 0 86 plusmn 08
A partir dos resultados obtidos pode sugerir-se que o S1 possui 11 resiacuteduos de Cys (controlo) o
que estaacute de acordo com o anteriormente descrito Verifica-se que o tratamento da proteiacutena com
concentraccedilotildees crescentes de decavanadato provoca a oxidaccedilatildeo de dois resiacuteduos de Cys do S1 de
miosina natildeo expostos ao solvente No entanto a teacutecnica utilizada natildeo nos permite identificar
quais
Tendo em consideraccedilatildeo que o valor de IC50 obtido para a reacccedilatildeo de inibiccedilatildeo daactividade Mg2+-
ATPaacutesica de S1 na presenccedila de decavanadato eacute de cerca de 27 microM pode sugerir-se que este seraacute o
valor ao qual ocorreraacute a oxidaccedilatildeo do primeiro resiacuteduo de Cys o que poderaacute contribuir para a
diminuiccedilatildeo da actividade de S1 observada
25Interacccedilatildeo de S1 com vanaacutedio(IV)
251Anaacutelise por espectroscopia de Ressonacircncia Paramagneacutetica Electroacutenica
A acumulaccedilatildeo intracelular de oxovanaacutedio (IV) in vivo constitui uma das principais formas de
apresentaccedilatildeo de vanaacutedio no organismo Deste modo torna-se bastante interessante analisar a
interacccedilatildeo entre este iatildeo e o S1 de miosina atraveacutes da teacutecnica de espectroscopia de Ressonacircncia
Paramagneacutetica Electroacutenica o meacutetodo de eleiccedilatildeo do estudo do iatildeo vanadilo e dos complexos por si
formados O electratildeo desemparelhado encontra-se fortemente acoplado ao nuacutecleo 51
V o que
35
resulta num espectro de RPE consistindo de 8 desdobramentos na direcccedilatildeo paralela e outros tantos
na direcccedilatildeo perpendicular (Chasteen 1983)
Os estudos realizados por Ahmed et al (1987) centraram-se na interacccedilatildeo do vanadilo com
proteiacutenas como eacute o caso da insulina transferrina entre outros obtendo um espectro caracterizado
por 8 linhas amplamente espaccediladas (na direcccedilatildeo paralela) Jaacute Ramos et al 2009 concluiu que a
exposiccedilatildeo da soluccedilatildeo de decavanadato agraves formas globular e filamentosa de actina produz um
espectro tiacutepico do iatildeo oxidovanaacutedio (IV) indicando a reduccedilatildeo do vanadato
Para aferir qual a concentraccedilatildeo de S1 que permitia obter uma boa razatildeo sinalruiacutedo titulou-se uma
soluccedilatildeo de sulfato de vanadilo 15 microM ([VOSO4]stock = 5 mM) com concentraccedilotildees crescentes de
proteiacutena (0 5 10 15 e 20 microM) em 10 mM HEPES 25 mM KCl A existecircncia de uma correlaccedilatildeo
linear entre a concentraccedilatildeo de proteiacutena e a intensidade dos sinais de RPE correspondente agrave banda
-12 perpendicular encontra-se representada na Fig 210
Figura 2 12ndash Representaccedilatildeo graacutefica da variaccedilatildeo da intensidade do sinal de RPE vs a concentraccedilatildeo de S1 utilizada
Apoacutes verificar que com uma concentraccedilatildeo de S1 igual a 20 microM se obtinham sinais com uma razatildeo
sinalruiacutedo adequada ao prosseguimento dos estudos sobre a interacccedilatildeo V(IV)-S1 de miosina
titulou-se uma soluccedilatildeo de 20 microM S1 com 0 25 5 10 15 25 50 75 100 150 200 500 e 1000
microM VOSO4
Traccedilou-se o graacutefico da intensidade da banda -12 perpendicular do sinal de EPR em funccedilatildeo da
concentraccedilatildeo de VOSO4 tendo-se determinado uma constante de dissociaccedilatildeo (KD) do complexo
S1-vanadilo de 8748 plusmn 2621 microM V(IV)
y = 016786x + 122435
Rsup2 = 092992759
Inte
nsi
dad
e d
o s
inal
de
RP
E
[S1] (microM)
36
Figura 213 ndash Representaccedilatildeo graacutefica da variaccedilatildeo do sinal de RPE vs a concentraccedilatildeo de vanadilo utilizada (os valores
experimentais estatildeo representados por losangos o seu ajuste estaacute representado pela linha)
A simulaccedilatildeo do espectro de EPR foi efectuada a partir do espectro obtido para uma concentraccedilatildeo
de 1 mM VOSO4 apoacutes adiccedilatildeo de 20 microM S1 de miosina tendo-se obtido os valores da constante de
acoplamento hiperfina (A=5016) e do factor g (g0=195)
Figura 214 ndash Representaccedilatildeo graacutefica do espectro de EPR obtido com 1000 microM de vanadilo e 20 microM de S1 (a cinzento
escuro) A simulaccedilatildeo do espectro encontra-se a cinzento claro
Por comparaccedilatildeo destes com os valores presentes na tabela de correspondecircncia xxx eacute possiacutevel
predizer quais os ligandos provaacuteveis do vanadilo na presenccedila de proteiacutena Assim concluiu-se que
o vanadilo estaacute coordenado com duas moleacuteculas de aacutegua um grupo hidroxilo e um grupo R=NR
O grupo hidroxilo pode indicar os resiacuteduos de aminoaacutecidos Tyr Treonina e Serina enquanto o
grupo R=NR poderaacute indicar a presenccedila de Arginina e Histidina como ligandos do vanadilo
Inte
nsi
dad
e d
os
sinai
s d
e R
PE
[Vanadilo] (microM)
Ajuste dos dados exp Dados experimentais
Campo magneacutetico (G)
Espectro Experimental Ajuste
37
3 Procedimento Experimental
31 Reagentes
Todos os reagentes utilizados satildeo de elevado grau de pureza (analiacutetico) Apresentam-se as suas
foacutermulas quiacutemicas ou abreviaturas quando necessaacuterio acrescentou-se a sua descriccedilatildeo Entre
parecircntesis colocou-se as empresas que os comercializam
Aacutecido aceacutetico glacial 100 (Riedel) Acrilamida 99 (Merck) Azul de bromofenol (Riedel-de-
Haumlen) -Mercaptoetanol (Sigma) Bissulfito de soacutedio (Sigma)Bradford (reagente de)
(BioRad)BSA (albumina de soro bovino Sigma-Aldrich) CaCl22H2O (cloreto de caacutelcio
dihidratado Fluka) CH3CH2OH 96 (etanol Panreac) Coomassie Blue R250 (Merck) D2O
(oacutexido de deuteacuterio SDS) DTNB (55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico) Aldrich) EDTA (aacutecido
etilenodiaminatetraaceacutetico Merck) Fosfoenolpiruvato 98 (Fluka) Glicerol (Sigma) Glicina
985 (Panreac) HCl 37 (Riedel-de Haumlen) H2SO4 96 (Pronalab) HEPES 99 (aacutecido 4-(2-
hidroxiletil)-1-piperazineetanosulfoacutenico Sigma) KH2PO4 99 (hidrogenofosfato de potaacutessio)
MgATP (Sigma) Na
ATP (Sigma) Na2HPO4 99 (hidrogenofosfato de soacutedio Riedel-de Haumlen)
NaH2PO4 (fosfato de soacutedio ) NaOH 986 (hidroacutexido de soacutedio Pronalab) KCl 99 (Cloreto de
potaacutessio Panreac) L-Cys 995 (enantioacutemero levoacutegiro do resiacuteduo de Cisteiacutena Fluka)Metanol
99 (Merck) Molibdato de amoacutenio (Sigma)NNrsquo-metil-bis-acrilamida 98 (Merck) NH4VO3
99 (metavanadato de amoacutenia Sigma) PSA 10 (persulfato de amoacutenio Merck) SDS
(dodecilo sulfato de soacutedio Sigma) Sulfito de soacutedio (Sigma) VOSO4 (sulfato de vanadilo
Aldrich) TEMED 99 (Tetrametiletilenodiamina Merck) Tris (2-amino-2-hidroximetil-13-
propanodiol Sigma)
32 Meacutetodos
321 Isolamento e purificaccedilatildeo das proteiacutenas de muacutesculo esqueleacutetico de coelho
3211 Purificaccedilatildeo da miosina preparaccedilatildeo de Subfragmento 1 por digestatildeo quimotriacutepsica
da miosina
O subfragmento-1 de miosina-II (S1) foi gentilmente cedido pelo Prof Doutor Manuel Aureliano
do Departamento de Quiacutemica Bioquiacutemica e Farmaacutecia da Faculdade de Ciecircncias e Tecnologia da
Universidade do Algarve
No entanto descreve-se abaixo de forma resumida o procedimento efectuado
38
32111 Purificaccedilatildeo da miosina
O protocolo de isolamento foi adaptado do meacutetodo previamente descrito por Margossian e Lowey
(1982) apresentando-se de forma esquemaacutetica na Fig 31
Sucintamente os tecidos musculares dorsais e das pernas foram extraiacutedos apoacutes o sacrifiacutecio do
animal (Fig31) De seguida foram adicionados 3 volumes da soluccedilatildeo de extracccedilatildeo A2
Centrifugou-se a soluccedilatildeo e filtrou-se o sobrenadante O sedimento foi entatildeo diluiacutedo em 14
volumes de aacutegua destilada de modo a que a forccedila ioacutenica atingisse 004 M Apoacutes adiccedilatildeo de szlig-
mercaptoetanol (szlig-me) deixou-se a precipitar a 4ordmC removendo o sobrenadante Centrifugou-se
novamente e ressuspendeu-se o sedimento na soluccedilatildeo de extracccedilatildeo A Adicionou-se a soluccedilatildeo
tampatildeo B3 para que a forccedila ioacutenica atingisse 05 M Homogeneizou-se e deixou-se repousar
durante 2h Homogeneizou-se novamente e centrifugou-se de modo a ocorrer precipitaccedilatildeo dos
complexos de actomiosina e proteiacutena desnaturada Apoacutes filtraccedilatildeo do sobrenadante em latilde de vidro
quantificou-se a soluccedilatildeo de miosina resultante pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental)
2 Soluccedilatildeo de extracccedilatildeo A 0012M K2CO3 004M KHCO3 06M KCl pH 65
3 Soluccedilatildeo-tampatildeo B Tris 100 mM pH 7 KCl 2M
39
Figura 31 ndash Representaccedilatildeo esquemaacutetica do protocolo de isolamento de miosina (Margossian e Lowey 1982) Todo o
procedimento foi realizado a 4ordmC (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
32112 Preparaccedilatildeo de Subfragmento-1 de miosina por digestatildeo com quimiotripsina
O S1 de miosina foi isolado apoacutes diaacutelise da miosina durante a noite contra a soluccedilatildeo C4 A
presenccedila do EDTA eacute essencial uma vez que a sua substituiccedilatildeo por MgCl2resulta na produccedilatildeo de
HMM em vez de S1 A suspensatildeo de miosina foi equilibrada e digerida com -quimotripsina
A digestatildeo foi interrompida pela adiccedilatildeo de PMSF Seguidamente ultracentrifugou-se o produto
da digestatildeo desprezando-se o sedimento (Fig 32) A soluccedilatildeo de sobrenadante maioritariamente
S1 foi quantificada pela leitura directa da Abs280 ou pelo meacutetodo de Bradford (ver as secccedilotildees
3311 e 3312 do Procedimento Experimental respectivamente) e armazenada em 10 sacarose
e 2 mM szlig-Me a -80ordmC
4 Soluccedilatildeo C 25mM Tris pH 70 012M KCl 1mM EDTA
40
Antes da realizaccedilatildeo dos ensaios descritos no presente relatoacuterio o S1 de miosina foi dialisado
contra a soluccedilatildeo D5 de modo a remover qualquer composto que pudesse complexar como
vanaacutedio Seguidamente quantificou-se a proteiacutena pelo meacutetodo de Bradford ou pelo meacutetodo
directo (ver secccedilotildees 3311 e 3312do Procedimento Experimtental)
Figura 3 2ndash Representaccedilatildeo esquemaacutetica do protocolo de preparaccedilatildeo do S1 de miosina por digestatildeo quimiotripsina de
acordo com o previamente descrito por Margossian e Lowey (1982) Todo o procedimento foi realizado a 4ordm C
excepto quando indicaccedilatildeo do contraacuterio (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
3212 Isolamento e purificaccedilatildeo de actina
Isolou-se e purificou-se a actina segundo o meacutetodo descrito por Pardee e Spudish que
compreende dois passos ndash a obtenccedilatildeo de um resiacuteduo seco denominado ldquopoacute de acetonardquo e a
extracccedilatildeo subsequente da actina a partir do ldquopoacute de acetonardquo na forma de G-Actina com a
posterior polimerizaccedilatildeo a F-Actina
32121 Obtenccedilatildeo do ldquopoacute de acetonardquo
Sacrificou-se o coelho com uma pancada atraacutes do pescoccedilo e sangrou-se o animal apoacutes o corte da
arteacuteria jugular Esfolou-se e colocou-se numa tina com gelo Retiraram-se as partes de tecido
conjuntivo que envolvem o tecido muscular com auxiacutelio de uma pinccedila e bisturi Seguidamente
retirou-se os muacutesculos dorsais e os muacutesculos brancos das pernas Passou-se os muacutesculos por aacutegua
destilada de modo a remover as impurezas e pesou-se num copo Trituraram-se os muacutesculos
numa maacutequina de triturar carne e extraiu-se o resultante com 3 volumes da soluccedilatildeo E6 durante 10
minutos com agitaccedilatildeo suave Centrifugou-se a suspensatildeo a 4000 g durante 10
5 Soluccedilatildeo D HEPES 10 mM pH 70 KCl 25 mM
6 Soluccedilatildeo E 01 M KCl 015 M K2HPO4 pH 65
41
minutosdesprezando-se o sobrenadante Ressuspendeu-se o sedimento em 6 volumes de soluccedilatildeo
F7durante 10 minutos com agitaccedilatildeo suave Verificou-se se o pH da soluccedilatildeo estava compreendido
entre 82 e 85 acertando-se estecom soluccedilatildeo G8 se necessaacuterio Centrifugou-se durante 10
minutos a 4000 g Desprezou-se o sobrenadante e ressuspendeu-se o sedimento em 3 volumes de
soluccedilatildeo H9 durante 10 minutos Centrifugou-se novamente a 4000 gdurante 10 minutos
desprezando-se o sobrenadante Ressuspendeu-se o sedimento duas vezes consecutivas em 6
volumes de aacutegua destilada durante 5 minutos Centrifugou-se a 4000 g durante 10 minutos apoacutes
cada adiccedilatildeo de H2O
Seguidamente ressuspendeu-se o sedimento 5 vezes consecutivas utilizando 3 volumes de
acetona de cada vez agitando suavemente durante 10 minutos agrave temperatura ambiente Colocou-
se o resiacuteduo filtrado numa tina de vidro sobre papel de filtro deixando-se evaporar o solvente
durante a noite num nicho Triturou-se o resiacuteduo seco numa maacutequina de triturar carne e deixou-se
novamente evaporar o solvente O ldquopoacute de acetonardquo obtido foi armazenado a -20 ordmC
32122 Purificaccedilatildeo da actina a partir de ldquopoacute de acetonardquo
Apoacutes a obtenccedilatildeo do poacute de acetona como acima indicado efectuou-se o protocolo seguidamente
descrito de modo a obter monoacutemeros de actina (G-Actina) e actina filamentosa (F-Actina)
Pesou-se 35 g de poacute de acetona e adicionaram-se 50 mL de soluccedilatildeo-tampatildeo I10
sendo a
extracccedilatildeo feita com agitaccedilatildeo suave durante 30 minutos Separou-se o extracto do poacute de acetona
hidratado filtrando a suspensatildeo com 4 camadas de gaze esterilizada tendo o cuidado de natildeo
espremer o resiacuteduo Voltou a extrair-se com 50 mL de soluccedilatildeo I durante 30 minutos mexendo
lentamente Adicionou-se este ao extracto obtido anteriormente Centrifugou-se o extracto total a
16 000 g durante 30 minutos Decantou-se para uma proveta sobre a qual se colocou gaze para
que o sedimento solto natildeo se juntasse ao sobrenadante e leu-se o volume De seguida adicionou-
se KCl ateacute uma concentraccedilatildeo final de 50 mM MgCl2 ateacute 2mM e ATP ateacute 1mM lentamente e com
agitaccedilatildeo contiacutenua de modo a promover a polimerizaccedilatildeo dos monoacutemeros de actina
Deixou-se polimerizar durante 2h a 25ordmC observando-se a presenccedila de pequenas bolhas de ar
indicativas de um aumento de viscosidade da soluccedilatildeo Arrefeceu-se em gelo durante 15 minutos e
adicionou-se KCl ateacute uma concentraccedilatildeo final de 06M Mexeu-se lentamente durante 30 minutos
apoacutes os quais se ultracentrifugou a 110 000g durante 1hora Desprezou-se o sobrenadante
ressuspendeu-se o sedimento com soluccedilatildeo tampatildeo E e transferiu-se para um potter
Dialisou-se durante a noite contra 1L de soluccedilatildeo-tampatildeo I a uma temperatura de 4ordmC com rotaccedilatildeo
7 Soluccedilatildeo F 005 M NaHCO3 pH 82-85
8 Soluccedilatildeo G 1M Na2CO3
9 Soluccedilatildeo H 1 mM EDTA pH 70
10 Soluccedilatildeo I 2 mM Tris-HCl pH 8 02 mM CaCl2 02 mM Na2ATP 05 mM szlig-Me 0005 NaN3
42
contiacutenua e mudanccedila de tampatildeo 2 a 3 vezes De seguida a actina foi centrifugada a 110
000gdurante 1hora Decantou-se a soluccedilatildeo de G-Actina (sobrenadante) e desprezou-se o
sedimento Determinou-se a concentraccedilatildeo de G-Actina pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental) A soluccedilatildeo de G-actina foi armazenada a uma temperatura de 4ordmC
apoacutes adiccedilatildeo de 002NaN3 mantendo-se estaacutevel durante 3 a 5 dias
331 Quantificaccedilatildeo das proteiacutenas isoladas
3311 Quantificaccedilatildeo pelo meacutetodo de Bradford
O meacutetodo de Bradford (Bradford 1976) baseia-se na utilizaccedilatildeo do reagente de Bradford
composto por entre outros aacutecido fosfoacuterico e o corante ldquoCoomassierdquo Este corante possui carga
negativa e a sua coloraccedilatildeo varia entre a vermelha (Absmax=465 nm) predominante em soluccedilatildeo e a
azul (Absmax=595 nm) presente aquando da ligaccedilatildeo aos grupos amina das proteiacutenas Em soluccedilotildees
aacutecidas diluiacutedas a coloraccedilatildeo varia proporcionalmente com a quantidade do corante ligado agrave
proteiacutena que por sua vez eacute directamente proporcional agrave concentraccedilatildeo da proteiacutena permitindo a
sua quantificaccedilatildeo ateacute a um limite de detecccedilatildeo de 20 gmL (Bradford 1976)
Deste modo prepararam-se amostras de BSA ([BSA]stock= 1mgmL) utilizando um intervalo de
concentraccedilotildees compreendido entre 0 e 15 gmL de acordo com a tabela 3I Apoacutes a adiccedilatildeo da
H2O MilliQ e do reagente de Bradford as soluccedilotildees foram sucessivamente homogeneizadas no
voacutertex
Tabela 3Indash Amostras utilizadas para a obtenccedilatildeo da recta de calibraccedilatildeo no meacutetodo de Bradford
BSA (L) BSA (g) H2O MilliQ (L) Reagente de Bradford (L)
0 0 800 200
125 125 79875 200
25 25 7975 200
5 5 795 200
10 10 790 200
15 15 785 200
Apoacutes incubaccedilatildeo a 25ordmC por 5 minutos leram-se os valores de Abs595 num
espectrofotoacutemetro (Shimadzu UV-160A) para todas as amostras Seguidamente traccedilou-se a recta
de calibraccedilatildeo (Abs595vs quantidade de BSA (g))
43
Para determinar a concentraccedilatildeo de proteiacutena apoacutes o processo de isolamento purificaccedilatildeo e
diaacutelise quando necessaacuterio colocou-se 5 L da amostra num tubo de Eppendorf e adicionou-se
795 L de H2O MilliQ homogeneizando a soluccedilatildeo no voacutertex Adicionou-se posteriormente 200
L de reagente de Bradford e repetiu-se a homogeneizaccedilatildeo Incubou-se a mistura a 25ordmC por 5
minutos apoacutes os quais se leu os valores de Abs595 nm no espectrofotoacutemetro Por interpolaccedilatildeo deste
valor a partir da recta de calibraccedilatildeo determinou-se a concentraccedilatildeo de proteiacutena na amostra
3312 Quantificaccedilatildeo pelo meacutetodo directo
A quantificaccedilatildeo das amostras proteicas pode tambeacutem ser efectuada atraveacutes da leitura directa da
absorvacircncia tendo por base a lei de Beer-Lambert onde Abs corresponde agrave
absorvacircncia ao comprimento de onda ao qual o ensaio foi realizado corresponde ao coeficiente
de extinccedilatildeo (substituiacutedo pela letra a se as unidades forem distintas da molaridade) b corresponde
ao percurso oacuteptico da radiaccedilatildeo na ceacutelula amostral e c corresponde agrave concentraccedilatildeo amostral
Assim no caso de S1 de miosina mediu-se a Abs280e calculou-se a concentraccedilatildeo da proteiacutena
sabendo que o coeficiente de extinccedilatildeo de S1 eacute (Ref)
No caso da actina mediu-se a Abs290ecalculou-se a concentraccedilatildeo da proteiacutena sabendo que o
coeficiente de extinccedilatildeo da actina eacute (Ref)
332Electroforese em condiccedilotildees desnaturantes (SDS-PAGE)
De modo a melhor caracterizar as proteiacutenas isoladas foi realizada uma electroforese em
condiccedilotildees desnaturantes (SDS-PAGE do inglecircs Sodium Dodecyl Sulfate Polyacrilamide Gel
Electrophoresis) O SDS (dodecilo sulfato de soacutedio) eacute um detergente anioacutenico utilizado neste
meacutetodo para desnaturar as proteiacutenas que se ligam ao SDS na proporccedilatildeo 14 g SDS g proteiacutena
(aproximadamente 1 moleacutecula de SDS por cada 2 resiacuteduos de aminoaacutecidos) (Stryer) O SDS induz
uma densidade de carga negativa significativa nas proteiacutenas a que se liga mascarando a sua carga
intriacutenseca pelo que as razotildees cargamassa satildeo tendencialmente idecircnticas Assim a SDS-PAGE
separa as proteiacutenas quando sujeitas a um campo eleacutectrico de acordo com o seu peso molecular
sendo a sua migraccedilatildeo directamente proporcional ao logaritmo deste uacuteltimo (Stryer)
O gel de resoluccedilatildeo com 10 de acrilamida foi obtido atraveacutes da adiccedilatildeo de 167 mL de soluccedilatildeo
stock de acrilamida11
075 mL de tampatildeo de resoluccedilatildeo12
251 mL de aacutegua destilada e 50 L de
SDS 10 Para promover a polimerizaccedilatildeo adicionou-se em seguida 38 L de PSA 10
11
soluccedilatildeo stock de acrilamida soluccedilatildeo de 30 (mv) de acrilamida e 08 (mv) de NNrsquo-metil-bis-acrilamida 12
tampatildeo de resoluccedilatildeo 15 M Tris-HCl pH 88
44
(persulfato de amoacutenia) 10 e 25 L de TEMED 99 (tetrametiletilenodiamina) Agitou-se
cuidadosamente de modo a natildeo formar bolhas e pipetou-se a soluccedilatildeo para a cassette de vidro
previamente montada no suporte De seguida adicionou-se uma peliacutecula de cerca de 2 mm de 2-
metil-2-butanol para verificar posteriormente a polimerizaccedilatildeo Apoacutes a polimerizaccedilatildeo que demora
cerca de 45 minutos verteu-se a agrave superfiacutecie e adicionou-se o gel de concentraccedilatildeo com 5 de
acrilamida constituiacutedo por 03 mL de soluccedilatildeo de stock acrilamida 045 mL de tampatildeo de
concentraccedilatildeo13
094 mL de aacutegua destilada 18 L de SDS 10 135 L de PSA 10 e 2L de
TEMED 99 Colocou-se o pente sobre o gel e deixou-se polimerizar durante cerca de 45
minutos
As amostras para aplicaccedilatildeo no gel de electroforese foram preparadas para uma concentraccedilatildeo de
proteiacutena de 1 mgmL num volume final de 100 L dos quais 25 L correspondem ao tampatildeo da
amostra14
As amostras foram aquecidas a 100ordmC durante 5 minutos Antes da aplicaccedilatildeo das
amostras transferiu-se a cassette para a tina de electroforese agrave qual se adicionou a soluccedilatildeo
tampatildeo de electroforese da tina superior15
e tampatildeo de electroforese da tina inferior16
Aplicaram-
se 10 L das soluccedilotildees de marcadores de peso molecular e 20 L das amostras proteicas
preparadas anteriormente A electroforese decorreu durante cerca de 3h30 min a uma voltagem
de 80 V De seguida transferiu-se o gel para uma tina corando as proteiacutenas durante 1h com
soluccedilatildeo corante17
Descorou-se o gel com soluccedilatildeo descorante18
tendo-se mudado esta soluccedilatildeo ateacute
as bandas se encontrarem bem definidas
Determinou-se a mobilidade relativa das bandas correspondentes aos marcadores de elevado peso
molecular e traccedilou-se a recta de calibraccedilatildeo ndash log(PM) vs log(Rf) onde Rf corresponde agrave
mobilidade relativa Por interpolaccedilatildeo determina-se o PM das proteiacutenas isoladas sabendo a sua
mobilidade relativa Por comparaccedilatildeo com os valores descritos na literatura pode concluir-se
acerca do grau de purificaccedilatildeo
34 Mediccedilatildeo da actividade enzimaacutetica de S1
341 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
13
tampatildeo de concentraccedilatildeo 05 M Tris-HCl pH 68 14
tampatildeo de amostra 320 mM Tris-HCl pH 68 04 M szlig-mercaptoetanol SDS 8 glicerol 15 (vv) e roxo de m-
cresol 0024 15
tampatildeo de electroforese da tina superior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS 16
tampatildeo de electroforese da tina inferior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS acetato de soacutedio
01 M 17
soluccedilatildeo corante ldquoCoomassie Blue R-250rdquo contendo metanol 45 (vv) e aacutecido aceacutetico glacial 10 (vv) 18
soluccedilatildeo descorante metanol 10 (vv) e aacutecido aceacutetico glacial 10 vv)
45
O meacutetodo de Fiske-Subarow modificado consiste na reacccedilatildeo do molibdato de amoacutenio com o
fosfato inorgacircnico em soluccedilatildeo aacutecida com formaccedilatildeo do aacutecido fosfomoliacutebdico Seguidamente daacute-
se a adiccedilatildeo de um agente redutor que provoca a reduccedilatildeo do composto a fosfomolibdato que forma
um complexo de cor azulada cuja absorvacircncia maacutexima ocorre para um comprimento de onda de
660 nm tornando possiacutevel a sua detecccedilatildeo por meacutetodos espectrofotomeacutetricos As actividades Ca2+
-
ATPaacutesica e K+EDTA-ATPaacutesica de S1 foram determinadas atraveacutes deste meacutetodo de acordo com
o descrito por Taussky e Shorr (1953) com mediccedilatildeo da quantidade de fosfato (Pi) libertado
atraveacutes da hidroacutelise de ATP
Para tal obteve-se primeiramente a recta de calibraccedilatildeo com as amostras presentes na Tabela 3II
utilizando uma soluccedilatildeo stock de 5 mM KH2PO4
Tabela 3II ndash Composiccedilatildeo das diferentes amostras utilizadas para determinar a recta de calibraccedilatildeo atraveacutes do meacutetodo
de Fiske-Subbarow modificado
KH2PO4 (L)
([KH2PO4]stock= 5mM)
PO43-
(nmole) H2O (uL)
0 0 1780
5 25 1775
10 50 1770
20 100 1760
40 200 1740
100 500 1680
A cada tubo de Eppendorf adicionaram-se seguidamente 200 L de soluccedilatildeo J19
e 20 L de agente
redutor20
homogeneizando no voacutertex apoacutes cada adiccedilatildeo Incubou-se durante 10 minutos e mediu-
se a Abs660 nm num espectrofotoacutemetro com intervalos de tempo iguais entre cada amostra Traccedilou-
se a recta de calibraccedilatildeo (Abs660 nmvs nmol PO43-
) a partir dos valores obtidos
Para determinar a actividade Ca2+
-ATPaacutesica de S1 prepararam-se tubos de Eppendorf com 400
L de meio de reacccedilatildeo21
25 L CaCl2 ([CaCl2]stock= 100 mM) 25 L ATP ([ATP]stock= 100 mM)
com [S1]ensaios=01 mgmL Perfez-se se necessaacuterio com H2O MilliQ ateacute perfazer um volume de 1
mL Para determinar a actividade K+EDTA-ATPaacutesica utilizou-se uma concentraccedilatildeo idecircntica de
19
Soluccedilatildeo J 5 N H2SO4 25 molibdato de amoacutenio 20
Agente redutor 5 mg aacutecido 1-amino-2-naftol-4-sulfoacutenico 30 mg bissulfito de soacutedio 30 mg sulfito de soacutedio em 26
mL H2O 21
meio de reacccedilatildeo 125 mM Tris-HCl pH 80 0575 M KCl
46
proteiacutena nos ensaios (01 mgmL) e o mesmo meio de reacccedilatildeo Aos tubos de Eppendorf
adicionou-se ainda 123 L de KCl ([KCl]stock = 3M) 25 L de ATP 10 L de EDTA
([EDTA]stock= 100 mM )e o volume de H2O MilliQ necessaacuterio para perfazer 1 mL Em ambas as
situaccedilotildees o uacuteltimo reagente adicionado foi o ATP uma vez que se trata do substrato da reacccedilatildeo
enzimaacutetica
A cineacutetica decorreu a 25ordmC durante 5 minutos para a actividade Ca2+
-ATPaacutesica e durante 3
minutos para a actividade K+EDTA-ATPaacutesica Apoacutes o periacuteodo de incubaccedilatildeo transferiu-se 150
L de amostra para tubos de Eppendorf contendo igual volume de TCA 10 Colocou-se no gelo
e homogeneizou-se no voacutertex Seguidamente centrifugou-se a 1500 g durante 10 minutos e
recolheu-se 200 L de sobrenadante para novos tubos de Eppendorf Adicionou-se a estes 158
mL de H2O MilliQ e 200 L de soluccedilatildeo J Homogeneizou-se no voacutertex e adicionou-se 20 L de
reagente redutor repetindo-se a homogeneizaccedilatildeo Incubou-se durante 10 minutos e mediu-se a
Abs660 nm com o mesmo intervalo de tempo de leitura para cada amostra Por intrapolaccedilatildeo a partir
da recta de calibraccedilatildeo obtida anteriormente determinou-se a quantidade de Pi libertado em cada
ensaio A partir destes resultados determinaram-se as actividades Ca2+
-ATPaacutesica e K+EDTA-
ATPaacutesica do S1 em nmol PO43-
minmg proteiacutena e em s-1
342 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
Da actividade Mg2+
-ATPaacutesica de S1 resulta a formaccedilatildeo dos produtos da reacccedilatildeo de
hidroacutelise do ATP - ADP e Pi Na presenccedila de ADP fosfoenolpiruvato (PEP) e da enzima piruvato
cinase (PK) os produtos resultantes satildeo ATP e piruvato Por acccedilatildeo da enzima lactato
desidrogenase (LDH) este uacuteltimo eacute convertido em lactato ocorrendo simultaneamente a oxidaccedilatildeo
do NADH a NAD+ monitorizada atraveacutes da Abs340 nm Quando o S1 de miosina se encontra no
meio reaccional o ATP eacute hidrolisado formando-se novamente ADP e Pi o que permite a
formaccedilatildeo de um ciclo (fig 21) Se as enzimas se encontrarem em excesso a velocidade de
conversatildeo do NADH eacute limitada pela produccedilatildeo de ADP e Pi Como o ATP eacute regenerado em cada
ciclo a sua concentraccedilatildeo inicial eacute sempre mantida (Bagshaw 1982)
Figura 33ndash Representaccedilatildeo esquemaacutetica do meacutetodo de enzimas acopladas
47
O protocolo utilizado e seguidamente descrito foi adaptado de Cuenda et al (1990 ndash Ver Refordf) A
uma cuvette de 1 mL adicionou-se S1 para uma concentraccedilatildeo final de 005 mgmL 800 L de
meio de reacccedilatildeo22
10 L de PEP ([PEP]stock = 42 mM) 10 L de PK+LDH 11 ([PK+LDH]stock =
1 mgmL) e 15 L de NADH ([NADH]stock=25 mM) Iniciou-se a monitorizaccedilatildeo da reacccedilatildeo
atraveacutes da leitura de Abs340 nm apoacutes a adiccedilatildeo de 20 L de ATP ([ATP]stock=100 mM) A
actividade Mg2+
-ATPaacutesica eacute obtida atraveacutes do declive correspondente agrave oxidaccedilatildeo do NADH Para
a mediccedilatildeo da Mg2+
-ATPase de S1 estimulada por actina adicionou-se F-actina para uma
concentraccedilatildeo final de 2 microM sendo a actividade obtida tambeacutem atraveacutes do declive correspondente
agrave oxidaccedilatildeo do NADH apoacutes a adiccedilatildeo da proteiacutena
35 Interaccedilatildeo entre vanaacutedio e subfragmento-1 de miosina
351 Preparaccedilatildeo das soluccedilotildees de vanaacutedio
As soluccedilotildees de vanadato foram preparadas em aacutegua MilliQ a partir de metavanadato de amoacutenio
(NH4VO3) numa concentraccedilatildeo stock de 50 mM com agitaccedilatildeo magneacutetica e aquecimento (~30 ordmC)
Esta soluccedilatildeo com um valor compreendido entre 6 e 7 designa-se por ldquometavanadatordquo e eacute
constituiacuteda pelas esspeacutecies V1 V2 V4 e V5 (Ref) Acidificando a soluccedilatildeo de metavanadato por
adiccedilatildeo de HCl ateacute um valor de pH proacuteximo de 4 observa-se uma mudanccedila de cor de amarelo
claro para laranja indicativa da formaccedilatildeo da espeacutecie decameacuterica de vanadato De seguida e
imediatamente antes da realizaccedilatildeo dos ensaios descritos o valor do pH da soluccedilatildeo de
decavanadato foi aumentado para cerca de 75 por adiccedilatildeo de NaOH A soluccedilatildeo (2) designada
por ldquometavanadatordquo natildeo foi posteriormente alterada ateacute agrave sua adiccedilatildeo para a realizaccedilatildeo dos ensaios
As soluccedilotildees foram armazenadas a 4ordmC e mantidas em gelo aquando da sua utilizaccedilatildeo
A soluccedilatildeo de V(IV) foi preparada em condiccedilotildees anaeroacutebias por diluiccedilatildeo de sulfato de vanadil
(VOSO4) para uma concentraccedilatildeo final de 5 mM em H2O MilliQ saturada com N2 gasoso
352Anaacutelise por espectroscopia de UV-Vis
A concentraccedilatildeo da espeacutecie decameacuterica em soluccedilatildeo depende do periacuteodo de tempo apoacutes a diluiccedilatildeo
no meio de reacccedilatildeo apropriado (Ref) A velocidade de desoligomerizaccedilatildeo da espeacutecie decameacuterica
pode ser analisada por espectroscopia de absorccedilatildeo na regiatildeo UVVis uma vez que esta espeacutecie
apresenta duas bandas caracteriacutesticas na regiatildeo do visiacutevel (a 358 e 400 nm) que natildeo se verifica
em quaisquer outras espeacutecies oligomeacutericas As bandas supramencionada sapresentam-se bem
22
Meio de reacccedilatildeo 3125 mM Tris-HCl pH 71 375 mM MgCl2
48
resolvidas atraveacutes da obtenccedilatildeo da derivada de 2ordf ordem do espectro apresentando a forma de 2
vales com miacutenimos centrados nos comprimentos de onda supracitados com dependecircncia linear
relativamente agrave concentraccedilatildeo de decavanadato total (Ramos et al 2006)
Assim a estabilidade da espeacutecie decameacuterica apoacutes diluiccedilatildeo da soluccedilatildeo de decavanadato no meio de
reacccedilatildeo dos ensaios (tampatildeo D) para uma concentraccedilatildeo final de 1 mM em vanadato total foi
caracterizada atraveacutes da leitura da Abs400 em funccedilatildeo do tempo tendo-se determinado qual a
ordem de reacccedilatildeo e o tempo de meia-vida da mesma Com base em estudos anteriores acerca da
estabilidade do V10 na presenccedila de diferentes proteiacutenas (Ramos et al 2006) determinou-se
igualmente o tempo de meia vida apoacutes a adiccedilatildeo de 20 microM de S1 de miosina ao meio reaccional
353 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
Os espectros de RMN do isoacutetopo 51
V foram realizados num espectroacutemetro Bruker AM-400 MHz
equipado com uma sonda multinuclear inversa de 5 mm Em tubos de RMN de 5 mm de diacircmetro
preparou-se soluccedilotildees de 500 L de deca- ou metavanadato correspondentes a um intervalo de
concentraccedilotildees de 1 a 10 mM suplementadas com 10 D2O Os espectros foram adquiridos agrave
temperatura ambiente com uma largura espectral de 23 923 Hz um tempo de aquisiccedilatildeo de 0086
s tempo de relaxaccedilatildeo de 001 s e uma sequecircncia de pulsos a 90ordm Utilizou-se a transformada de
Fourier com alargamento das bandas de 20Hz para o processamento dos espectros Os desvios
quiacutemicos das ressonacircncias de 51
V foram obtidos em relaccedilatildeo agrave espeacutecie VOCl3cuja frequecircncia de
ressonacircncia eacute considerada o padratildeo sendo todas as outras referenciadas a partir desta em ppm
As aacutereas dos sinais do RMN correspondentes agraves espeacutecies oligomeacutericas presentes foram obtidas
por integraccedilatildeo dos sinais As concentraccedilotildees relativas das espeacutecies foram calculadas com recurso agrave
equaccedilatildeo
onde A[Vx] corresponde agrave aacuterea referente agrave ressonacircncia da espeacutecie Vx com nordm de agregaccedilatildeo x
A[V]total agrave soma total das aacutereas medidas e [V] a concentraccedilatildeo total de vanaacutedio no ensaio
A teacutecnica de 51
V-RMN foi anteriormente aplicada em estudos de ligaccedilatildeo com S1 de miosina
(Ref) tendo sido demostrado que entre as espeacutecies V1 V2 V4 e V5 a espeacutecie tetrameacuterica possui a
maior afinidade para a miosina (Cremo et al) O desaparecimento ou alargamento dos sinais pode
implicar a ligaccedilatildeo de um dos oxovanadatos laacutebeis agrave miosina sem indicaccedilatildeo de qual uma vez que
49
se processa um equiliacutebrio entre eles (Crans 1990) Assim e tendo em conta as indicaccedilotildees de que
a espeacutecie V10 eacute distinta das restantes no que diz respeito agrave magnitude da interacccedilatildeo com a miosina
e inibiccedilatildeo da actividade ATPaacutesica estimulada por actina (Aureliano 2000 Tiago et al 2002)
torna-se fundamental investigar a interacccedilatildeo das diferentes espeacutecies de vanadato com o S1 de
miosina atraveacutes da teacutecnica de 51
V-RMN
Para tal obtiveram-se espectros de 51
V-RMN resultantes da titulaccedilatildeo de uma soluccedilatildeo 5 mM de
decavanadato total com concentraccedilotildees crescentes de S1 de miosina (0 5 10 15 e 20 microM) em
tampatildeo D Realizou-se ainda ensaios na presenccedila de 2 mM de decavanadato e 2 mM de ATP na
presenccedila e na ausecircncia de S1 de miosina de modo a verificar o efeito que estes ligandos naturais
da enzima teria na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o subfragmento-1
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
Os grupos sulfidrilo (-SH) das cisteiacutenas satildeo resiacuteduos de aminoaacutecidos extremamente sensiacuteveis agrave
oxidaccedilatildeo resultando na sua oxidaccedilatildeo (Dalle-Donne et al 1999) Logo a quantificaccedilatildeo destes
resiacuteduos eacute bastante importante nos estudos de interacccedilatildeo com vanaacutedio jaacute que os resiacuteduos de Cys
com ecircnfase para o SH1 e o SH2 satildeo bastante importantes na monitorizaccedilatildeo das modificaccedilotildees
funcionais da regiatildeo da cabeccedila
O procedimento seguidamente apresentado foi originalmente descrito por Gutieacuterrez-Merino
(1987)) Consiste na reacccedilatildeo entre os grupos ndashSH natildeo oxidados das proteiacutenas e o reagente de
Ellman (DTNB -55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico)) - formando-se o aniatildeo CNT (aniatildeo 3-
carboxilato-4-nitrotiofenolato) numa estequiometria de 11 Este apresenta um maacuteximo de
absorvecircncia a 412 nm (Robyt 1990) Assim apoacutes a incubaccedilatildeo da proteiacutena na presenccedila do metal
a quantidade de cisteiacutenas reduzidas por proteiacutena determina-se atraveacutes dos valores de Abs412
Deste modo colocou-se num tubo de Eppendorf o volume apropriado de S1 dialisado para que a
concentraccedilatildeo no ensaio fosse de 1 mgmL Adicionou-se o volume apropriado de decavanadato
para que a sua concentraccedilatildeo fosse de 0 10 50 100 250 500 ou 1000 M Perfez-se com soluccedilatildeo
D ateacute um volume final de 1 mL e incubou-se a 25ordmC por 20 minutos Apoacutes este periacuteodo colocou-
se 780 L da mistura reaccional numa cuvette de quartzo e adicionou-se 220 L de soluccedilatildeo K23
23
Tampatildeo K Tampatildeo fosfatohidrogenofosfato de soacutedio 025 M pH 72
50
Adicionou-se 15 L de DTNB (em soluccedilatildeo K [DTNB]stock=100 mM) ao meio reaccional e
registou-se o valor de absorvecircncia medido equivalente agraves cisteiacutenas ldquoraacutepidasrdquo (SH1 e SH2
expostos ao solvente) Adicionou-se 001 g de SDS que provoca a desnaturaccedilatildeo da proteiacutena e
consequentemente a exposiccedilatildeo de todos os resiacuteduos de cisteiacutenas que se encontravam ateacute aiacute
inacessiacuteveis no interior da proteiacutena (cisteiacutenas totais) Estes resiacuteduos podem assim reagir com o
DTNB
Estimou-se o nuacutemero de resiacuteduos de cisteiacutenas em todas as amostras com recurso ao coeficiente de
extinccedilatildeo molar () do DTNB (ver ponto 3 dos Anexos) que foi determinado atraveacutes da titulaccedilatildeo
de L-Cisteiacutena com intervalo de concentraccedilotildees compreendidos entre 0 e 50 microM na presenccedila de
25microM de DTNB em tampatildeo D Os valores de absorvecircncia foram obtidos para um comprimento de
onda de 412 nm
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
3621 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
A teacutecnica mais eficiente de anaacutelise da interacccedilatildeo entre o iatildeo vanadilo e proteiacutenas como a
calmodulina e a transferrina entre outras permite a obtenccedilatildeo raacutepida de informaccedilatildeo no que toca agrave
conformaccedilatildeo e amplitude da referida interacccedilatildeo
Os espectros de Ressonacircncia Paramagneacutetica Electroacutenica (RPE) da espeacutecie paramagneacutetica V(IV)
foram realizados a 70K num espectroacutemetro Bruker EMX-300 X-band Todas as soluccedilotildees
utilizadas e amostras foram preparadas em anaerobiose numa cacircmara de luvas de modo a evitar a
reduccedilatildeo de V(IV) a V(V) Em tubos capilares com 02 mL de amostra manteve-se a concentraccedilatildeo
de S1 a 20 M e variou-se a concentraccedilatildeo de vanadilo (0 25 5 10 15 25 50 75 100 150
200 500 e 1000 M [VOSO4]stock=5 mM) em tampatildeo D Apoacutes obtenccedilatildeo dos espectros estes
foram tratados atraveacutes do software SimFonia e WinEPR tendo-se obtido um graacutefico com a
variaccedilatildeo da intensidade do sinal de RPE em funccedilatildeo da concentraccedilatildeo de sulfato de vanadilo
presente Realizaram-se ainda simulaccedilotildees de espectros obtidos com 20 microM de S1 e 1000 microM de
V(IV) com vista agrave obtenccedilatildeo dos ligandos provaacuteveis do vanadilo atraveacutes da anaacutelise da constante
de acoplamento hiperfina (A) e do factor g (g)
4 Conclusotildees gerais
Apesar de jaacute ter sido demonstrado que o vanaacutedio inibe a Na+K
+-ATPase da miosina a funccedilatildeo
deste elemento no tecido muacutesculo esqueleacutetico continua largamente incompreendida A enorme
complexidade quiacutemica deste metal especialmente no que concerne agrave oligomerizaccedilatildeo e reacccedilotildees
51
redox constitui um obstaacuteculo agrave compreensatildeo do comportamento do vanaacutedio nos organismos
vivos Assim a caracterizaccedilatildeo da quiacutemica deste elemento eacute de grande utilidade antes de iniciar o
estudo da sua interacccedilatildeo com o presente sistema
A anaacutelise por espectroscopias de 51
V-RMN e de UV-Vis de soluccedilotildees de vanadato diluiacutedas nos
meios de reacccedilatildeo utilizados no presente relatoacuterio (10 mM HEPES pH 70 25 mM KCl) mostrou
claramente que a espeacutecie decameacuterica (V10) possui um tempo de vida suficientemente longo para a
determinaccedilatildeo do seu efeito isolado ao contraacuterio do que se verifica com as espeacutecies mono- di-
tetra- e pentameacutericas (V1 V2 V4 e V5) Por outro lado a anaacutelise verificou ainda que apoacutes diluiccedilatildeo
das soluccedilotildees de decavanadato no meio de reacccedilatildeo coexistem as espeacutecies V10 e V1 pelo que os
efeitos putativos desta uacuteltima deveratildeo ser subtraiacutedos aquando da monitorizaccedilatildeo dos efeitos da
espeacutecie decameacuterica
O vanaacutedio (V) (vanadato) inibe as enzimas que catalizam reacccedilotildees de transferecircncia de grupos
fosfato como a ATPase da miosina [Goodno 1979] devido agrave sua analogia estrutural com o iatildeo
fosfato Na sua utilizaccedilatildeo como inibidor enzimaacutetico natildeo eacute tida em conta a contribuiccedilatildeo individual
das espeacutecies oligomeacutericas considerando-se normalmente a espeacutecie V1 como inibidora efectiva
[Goodno e Taylor 1982] No presente relatoacuterio verificou-se por espectroscopia de 51
V-RMN que
outros oligoacutemeros de vanadato interagem com o S1 de miosina tendo as espeacutecies V10 e V4 os
maiores niacuteveis de afinidade A sua interacccedilatildeo envolve forccedilas electrostaacuteticas e eacute afectada
negativamente por ligandos naturais de enzima como eacute o caso do ATP
A exposiccedilatildeo do S1 de miosina (005 mgmL) a decavanadato conduz agrave inibiccedilatildeo do capacidade da
proteiacutena de estimular a actividade Mg2+
-ATPaacutesica com um valor de IC50 de 266 plusmn 07 M
decavanadato sendo o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM
decavanadato (80) Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM decavanadato sendo
o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM decavanadato (60)
exercendo um efeito de impedimento agrave ligaccedilatildeo entre o vanaacutedio e o S1
A uma concentraccedilatildeo de 27 microM decavanadato verifica-se a oxidaccedilatildeo de um dos resiacuteduos de Cys
de S1 de miosina seguida pela oxidaccedilatildeo gradual de outro resiacuteduo de Cys ateacute agrave concentraccedilatildeo de
1000 microM Ambos os resiacuteduos oxidados natildeo se encontravam expostos ao solvente utilizado A
oxidaccedilatildeo dos resiacuteduos permite sugerir que esta modificaccedilatildeo oxidativa sofrida pelo S1 contribui
para a diminuiccedilatildeo da sua actividade de ATPase
52
Por espectroscopia de RPE revelaram-se os possiacuteveis ligandos equatoriais do vanaacutedio aquando da
sua interacccedilatildeo com o S1 de miosina como sendo duas moleacuteculas de aacutegua um ligando contendo o
grupo R-OH e outro contendo o grupo R=NR
Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados de
oxidaccedilatildeo (IV) e (V) no S1 de miosina para concentraccedilotildees facilmente alcanccedilaacuteveis no ambiente
terrestre
53
5 Estudos futuros
No sentido do melhor esclarecimento das alteraccedilotildees moleculares e estruturais que ocorrem no
subfragmento 1 quando exposta ao vanaacutedio que conduzem a uma diminuiccedilatildeo da sua funccedilatildeo
sugere-se
- Quantificaccedilatildeo das cisteiacutenas reduzidas de S1 na presenccedila de metavanadato
- Determinaccedilatildeo das actividades Ca2+
- e K+-ATPaacutesica do S1 de miosina na presenccedila de
decavanadato
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina estimulada por actina
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina na presenccedila de
metavanadato
- Realizaccedilatildeo de RMN 2D com variaccedilatildeo das concentraccedilotildees das soluccedilotildees de decavanadato e
metavanadato na presenccedila de S1
-Mutageacutenese dos resiacuteduos de Cys doS1 de miosina de modo a verificar quais os resiacuteduos que
sofrem oxidaccedilatildeo na presenccedila de decavanadato
- Determinaccedilatildeo por calorimetria (DSC Differential Scanning Calorimetry) da variaccedilatildeo da
entalpia (∆H) e da temperatura de desnaturaccedilatildeo do S1 de miosina na presenccedila de vanaacutedio uma
vez que estas propriedades caracteriacutesticas de cada proteiacutena satildeo alteradas apoacutes ocorrecircncia de
modificaccedilotildees estruturais
- Anaacutelise por SDS-PAGE de S1 de miosina apoacutes tratamento com vanaacutedio de modo a determinar
quais os polipeacuteptidos eventualmente formados apoacutes clivagem da proteiacutena
54
6 Referecircncias bibliograacuteficas
- Sellers J Myosins 2nd Edition Oxford University Press 1999
- L Stryer inBiochemistry (Ed WH Freeman 2005 pp
Refordfas Materiais e Meacutetodos
[A] S S Margossian SLowey Methods Enzymol1982 85 55-71
[B] JD Pardue JA Spudich Methods Enzymol 1982 85 164-181
[C] M Bradford Anal Biochem1976 72 248-254
[D] H H Taussky E Shorr J Biol Chem 1953 202 675-685
[E] C R Bagshaw in Outline Studies in Biology ndash Muscle Contraction Chapman and Hall Ed
1982
[F] A Cuenda et al Eur J Biochem1990 194 663-670
55
Anexos
1 Determinaccedilatildeo da recta de calibraccedilatildeo meacutetodo de Bradford
Figura1ndashRepresentaccedilatildeo graacutefica da recta de calibraccedilatildeo (Abs595 nm vs [BSA]) utilizada para o meacutetodo de Bradford
2 Determinaccedilatildeo da recta calibraccedilatildeo para a electroforese em condiccedilotildees desnaturantes
Figura 2 ndash Representaccedilatildeo graacutefica da recta de calibraccedilatildeo (log(Peso Molecular) vs Rf dos marcadores de peso
molecular) onde Rf indica determinada atraveacutes do padratildeo de peso molecular
(poccedilo (a))
3Caacutelculo do coeficiente de extinccedilatildeo molar () para a quantificaccedilatildeo de cisteiacutenas reduzidas
atraveacutes do meacutetodo de Ellman
Traccedilou-se a recta de Abs412 nm meacutedia vs [L-Cisteiacutena] de modo a determinar o valor do coeficiente
de extinccedilatildeo molar definido como o declive daquela recta Assim =8400 M-1
cm-1
como
podemos verificar na Fig 3
A595 nm = 00546[BSA] + 00209 Rsup2 = 099051
Ab
s 59
5 n
m
[BSA] (mgmL)
log PM = -1177 Rf + 2247 Rsup2 = 098957
log
PM
Mobilidade relativa (Rf)
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero

22
esqueleacutetico de coelho sendo esta uacuteltima tambeacutem realizada para o sufragmento-1 de miosina
Seguidamente realizar-se-aacute a quantificaccedilatildeo e caracterizaccedilatildeo das proteiacutenas por espectroscopia UV-
Vis e SDS-PAGE e a caracterizaccedilatildeo das soluccedilotildees de vanaacutedio por UV-Vis e ressonacircncia magneacutetica
nuclear (RMN) Por uacuteltimo a interacccedilatildeo do S1 de miosina com o vanaacutedio seraacute avaliada atraveacutes
das teacutecnicas de UVVis RMN e ressonacircncia paramagneacutetica electroacutenica (RPE)
2 Discussatildeo de Resultados
21 Caracterizaccedilatildeo das proteiacutenas isoladas
211Anaacutelise do estado de pureza
O estado de pureza das soluccedilotildees de actina e de S1 foi analisado por electroforese em gel de
poliacrilamida em condiccedilotildees desnaturantes (SDS-PAGE) (Fig 22) que separa as moleacuteculas de
acordo com o seu peso molecular (PM) A concentraccedilatildeo de acrilamida escolhida (10) permite a
detecccedilatildeo simultacircnea de componentes com peso molecular inferior a 20 kDa e outros com elevado
peso molecular
Figura 21 ndash Fotografia de gel de SDS-PAGE (10 acrilamida) Poccedilo (a)
10 L do marcador de elevado peso molecular () Poccedilo (b) 20 L de [G-
actina]=1 mgmL Poccedilo (c) 20 L de [S1]=1 mgmL Aplicou-se uma
voltagem de V durante h e min
A partir da recta de calibraccedilatildeo obtida log PM = -1177 Rf + 2247 (ver ponto 2 da secccedilatildeo
Anexos) pode verificar-se a presenccedila de actina na amostra do poccedilo (b) presente numa banda
bastante intensa (PMaparente=40 kDa) bem como a presenccedila minoritaacuteria de outros componentes
desconhecidos que natildeo teratildeo sido eliminados durante o processo de extracccedilatildeo e purificaccedilatildeo e natildeo
satildeo relevantes para o estudo em questatildeo Na amostra do poccedilo (c) nota-se a presenccedila de S1
(PMaparente=92 kDa) bem como a de outros componentes cuja banda eacute ligeiramente menos intensa
Estes poderatildeo ser fragmentos proteoliacuteticos de miosina e de S1 Estaacute ainda presente uma bada de
23
maior intensidade que poderaacute ser a proteiacutena-C (PM=135 kDa) uma vez que a sua remoccedilatildeo
completa da amostra necessitaria de um passo adicional de cromatografia de troca ioacutenica [BA]
No entanto natildeo podemos confirmar que se trata desta proteiacutena ou de qualquer outra uma vez que
se encontra fora do intervalo de migraccedilatildeo dos marcadores de peso molecular
212 Determinaccedilatildeo da concentraccedilatildeo proteica
2121Subfragmento-1 de miosina
A partir da recta de calibraccedilatildeo obtida pelo meacutetodo de Bradford [BB](ver secccedilatildeo 3311 do
Procedimento Experimental) A595nm = 00546[BSA] + 00209 (ver secccedilatildeo de Anexos)foi possiacutevel
determinar um intervalo de concentraccedilatildeo de S1 de miosina compreendido entre 226 e 117(plusmn
423M) (203 e 1056 (plusmn 381 mgmL)) (n=8) A concentraccedilatildeo meacutedia de S1 de miosina apoacutes
diaacutelise contra a soluccedilatildeo tampatildeo 10 mM HEPES pH 75 25 mM KCl determinada pela leitura da
Abs280 nmfoi de 20430M (1803 mgmL)(n=7)
As amostras a quantificar atraveacutes do meacutetodo de Bradford natildeo sofreram diaacutelise o que permite
explicar a concentraccedilatildeo superior de S1 de miosina em comparaccedilatildeo com o valor obtido atraveacutes do
meacutetodo directo Esta razatildeo aliada agrave sensibilidade e rapidez superiores deste uacuteltimo fez com que
se optasse pelo meacutetodo directo para a quantificaccedilatildeo da grande maioria dos ensaios
2122Actina
A concentraccedilatildeo de actina foi determinada quando esta se encontrava na forma monomeacuterica (G-
actina) por mediccedilatildeo da leitura directa da Abs290 nm tendo-se obtido uma concentraccedilatildeo meacutedia de
691481M (290202 mgmL) (n=3) em 2 mM Tris-HCl pH 80 02 mM CaCl2 02 mM Na-
ATP Ao comprimento de onda utilizado (290 nm) absorvem fortemente apenas os resiacuteduos de
Trp pelo que a mediccedilatildeo efectuada tem menor sensibilidade do que se tivesse sido efectuada a 280
nm uma vez que a actina apresenta apenas 4 resiacuteduos deste aminoaacutecido embora tenha 16
resiacuteduos de Tyr
22 Mediccedilatildeo da actividade enzimaacutetica de S1 de miosina
221 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
As actividades ATPaacutesicas de S1 de miosina dependentes de Ca2+
e K+foram analisadas pelo
caacutelculo das velocidades de consumo inicial do fosfato inorgacircnico (Pi) Definiu-se os tempos de 5
e 3 minutos para a mediccedilatildeo da Ca2+
e da K+EDTA-ATPase respectivamente Os valores obtidos
24
a partir do protocolo presente no ponto 341 da secccedilatildeo Procedimento experimental encontram-se
descritos na Tabela 2I
Tabela 2I ndash Acrividades natildeo-fisioloacutegicas de subfragmento-1 determinadas pelo meacutetodo de Fiske-Subarow
modificado (n=2)
Proteiacutena
Actividade
ATPaacutesica natildeo
fisioloacutegica
Actividade (nmol
Piminmg proteiacutena) Actividade (s-1)
S1
Ca2+
-ATPase 1400 plusmn 30 2100 plusmn 0045
K+EDTA-ATPase 2334 plusmn 50 3500 plusmn 0075
As actividades natildeo-fisioloacutegicas reflectem o estado de oxidaccedilatildeo dos resiacuteduos expostos de Cys 707
(SH1) e 697 (SH2) Se o grupo SH1 sofrer oxidaccedilatildeo a actividade ATPaacutesica dependente de Ca2+
aumenta e a actividade dependente de K+ diminui Por outro lado se o grupo SH2 sofrer
oxidaccedilatildeo regista-se a perda completa destas actividades
De acordo com Margossian e Lowey (1982) [BA] as actividades ATPaacutesica de S1 de miosina
dependente de Ca2+
e K+EDTA da miosina satildeo respectivamente de 24 e 112 s-1
Estaacute tambeacutem
descrito que a actividade ATPaacutesica dependente de K+ eacute em meacutedia cerca de 3 vezes superior agrave
dependente de Ca2+
reflectindo a quantidade das preparaccedilotildees A razatildeo obtida entre os valores das
actividades ATPaacutesicas dependentes de K+ e Ca
2+ eacute de 166 Os valores obtidos para as actividades
(Ca2+
-ATPase=21 s-1
K+EDTA-ATPase= 35 s
-1) satildeo inferiores agrave valores obtidos por
Margossian e Lowey [BA] e Tiago [BC] com forccedila ioacutenica inferior O valor obtido para a
actividade ATPaacutesica dependente de Ca2+
eacute superior ao obtido por Polosukhina et al (2000) [BD]
cujo meio de reacccedilatildeo continha MOPS e apresentava forccedila ioacutenica inferior Os dados
supramencionados comprovam a variaccedilatildeo da actividade a ATPaacutesica natildeo fisioloacutegica em relaccedilatildeo agrave
forccedila ioacutenica temperatura e pH
222 Mediccedilatildeo da actividadeMg2+
-ATPaacutesica de S1 pelo meacutetodo de enzimas acopladas
A actividade de Mg2+
-ATPase eacute a uacutenica que possui significado fisioloacutegico sendo as restantes
bastante uacuteteis apenas para indicaccedilatildeo da qualidade de preparaccedilatildeo do S1[BA] A 0575 M KCl a
actina e o S1 formam uma soluccedilatildeo viscosa e tuacuterbida Apoacutes a adiccedilatildeo de MgATP o complexo acto-
S1 dissocia-se e a turbidez da soluccedilatildeo diminui Se a forccedila ioacutenica for reduzida a dissociaccedilatildeo do
complexo actoS1 que se encontra na forma de suspensatildeo eacute induzida O MgATP eacute rapidamente
hidrolisado diminuindo a sua concentraccedilatildeo no meio reaccional Nestas condiccedilotildees haacute
reassociaccedilatildeo e subsequente contracccedilatildeo dos filamentos de actina e de S1 com formaccedilatildeo de
25
precipitado O meacutetodo descrito traduz de forma bastante elementar o fenoacutemeno da contracccedilatildeo in
vivo e elimina o problema anterior da solubilizaccedilatildeo ineficiente da miosina de modo a obter uma
mediccedilatildeo exacta da actividade [A] A utilizaccedilatildeo de forccedilas ioacutenicas reduzidas aumenta a afinidade da
actina para o S1 permitindo contornar a extrema dificuldade em obter a concentraccedilatildeo proteica dos
muacutesculos (cerca de 100 mgmL do complexo actomiosina) As actividades Mg2+
-ATPaacutesicas de S1
de miosina foram determinadas a partir do declive da variaccedilatildeo da Abs340 relativo agrave oxidaccedilatildeo de
NADH em funccedilatildeo do tempo como eacute descrito no ponto 342 do Procedimento Experimental Os
valores obtidos para a actividade Mg2+
-ATPaacutesica encontram-se apresentados na Tabela 2II
Tabela 2II ndash Actividade Mg2+
-ATPaacutesica de subfragmento-1 de miosina determinadas pelo meacutetodo das enzimas
acopladas
Actividade (nmol
ATPmin mg(S1)) Actividade (s
-1)
1097 plusmn 269 164 plusmn 04
Ramos (2005) [J] obteve o valor de 01 s-1
para a actividade Mg2+
-ATPaacutesica de S1 de miosina
com forccedila ioacutenica intermeacutedia Tiago (2005) [BC] obteve o valor de 011 s-1 em condiccedilotildees de forccedila
ioacutenica reduzida em relaccedilatildeo agrave utilizada no presente relatoacuterio
23 Preparaccedilatildeo e caracterizaccedilatildeo das soluccedilotildees de vanaacutedio
231 Anaacutelise da composiccedilatildeo das soluccedilotildees de vanadato por espectroscopia de 51
V-RMN
efeito da concentraccedilatildeo da espeacutecie
A diluiccedilatildeo das soluccedilotildees stock de meta- e decavanadato iraacute determinar fortemente a composiccedilatildeo
em soluccedilatildeo dos diferentes oligoacutemeros de vanadato (ref) No caso do decavanadato o periacuteodo de
tempo apoacutes a diluiccedilatildeo iraacute tambeacutem ser um factor importante a considerar devido agrave decomposiccedilatildeo
da espeacutecie V10(ref)Torna-se assim fundamental determinar a composiccedilatildeo quiacutemica das soluccedilotildees
no meio de ensaio utilizado (HEPES 10 mM KCl 25 mM) de modo a correlacionaacute-la
correctamente com os possiacuteveis efeitos promovidos pelas diferentes espeacutecies de vanadato no
sistema em estudo
Aleacutem disso deve ser dado ecircnfase agraves importantes interacccedilotildees que ocorrem entre o vanaacutedio e os
tampotildees escolhidos O tampatildeo com menor nuacutemero e magnitude de interacccedilatildeo eacute o HEPES [AD]
seguido do Tris sendo este uacuteltimo especialmente aconselhado em concentraccedilotildees baixas para casos
de incompatibilidade enzimaacutetica ou quiacutemica redox No caso especial da reacccedilatildeo redox
NAD(P)HNAD(P) o tampatildeo adequado eacute o imidazole de modo a natildeo haver reacccedilatildeo com o
26
vanadato Os tampotildees a evitar em qualquer situaccedilatildeo incluem a trietanolamina fosfato citrato e
glicina entre outros [AD] uma vez que todos interactuam fortemente com V(V) com formaccedilatildeo
de complexos estaacuteveis
Assim procedeu-se agrave anaacutelise da composiccedilatildeo das soluccedilotildees de meta- e decavanadato apoacutes diluiccedilatildeo
das soluccedilotildees stock (50 mM em vanadato total) no meio de reacccedilatildeo (tampatildeo D) atraveacutes da
espectroscopia de ressonacircncia magneacutetica nuclear de 51
V Nas figuras x1 e x2 apresentam-se os
espectros obtidos para as soluccedilotildees de deca- e metavanadato num intervalo de concentraccedilotildees de 1 a
10 mM A Tabela seguinte resume os paracircmetros espectroscoacutepicos obtidos para cada espeacutecie
presente nas soluccedilotildees analisadas
Figura 22ndash Espectros de RMN de 51
V de
concentraccedilotildees crescentes das espeacutecies oligomeacutericas
de vanadato obtidos apoacutes diluiccedilatildeo da soluccedilatildeo stock
de decavanadato em 10 mM HEPES pH 70 25 mM
KCl para concentraccedilotildees finais de A) 1 mM B)2
mM C) 4 mM D) 8 mM E) 10 mM de
decavanadato total V1refere-se agraves espeacutecies
monomeacutericade vanadato V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2IIIndash Desvios quiacutemicos meacutedios (part) das ressonacircncias das espeacutecies decameacuterica (V10) monomeacuterica (V1) e
tetrameacuterica (V4) presentes nos espectros da soluccedilatildeo de decavanadatoLargura a meia altura (∆12) da espeacutecie V10A
Paracircmetros V10A V10B V10C V1 V4
part (ppm) -5162 -5003 -4255 -5616 -5748
∆12 (Hz) 175 nd nd nd nd
Figura 23 ndash Distribuiccedilatildeo graacutefica das
espeacutecies oligomeacutericas de vanadato vs a
concentraccedilatildeo de decavanadato total As
concentraccedilotildees de cada espeacutecie foram
obtidas segundo a formula descrita na
secccedilatildeo 35 do Procedimento
Experimental (aacutereas dos sinais de 51
V-
RMN observadas na figura x1)V10 V1 e
V4 correspondem agraves espeacutecie monomeacuterica
dimeacuterica e tetrameacuterica respectivamente [Esp
eacuteci
e o
ligo
meacute
rica
de
van
aacutedio
] (m
M)
[Decavanadato]total (mM)
V10
V1
V4
27
Nas soluccedilotildees de decavanadato as espeacutecies monomeacuterica e decameacuterica de vanadato satildeo as uacutenicas
presentes para concentraccedilotildees de decavanadato inferiores a 8 mM Ambas as espeacutecies aumentam
linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado em [AY] e
em (Teresa) quando a concentraccedilatildeo de V1 eacute igual ou superior a 200 microM inicia-se a sua
oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica No entanto nas condiccedilotildees de realizaccedilatildeo dos
ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a espeacutecie tetrameacuterica neste
estudo em concentraccedilotildees inferiores a 8 mM tendo o mesmo sido observado por Ramoset al
(2006) [AK] Os resultados obtidos mostram claramente o equiliacutebrio entre a espeacutecie decameacuterica e
a monomeacuterica e que o aparecimento de outras espeacutecies oligomeacutericas se encontra dependente da
concentraccedilatildeo desta uacuteltima Apesar de natildeo ter sido realizado o ensaio com 5 mM de decavanadato
total no presente relatoacuterio estaacute reportado em Ramos et al (2006) [AK] que as concentraccedilotildees da
espeacutecie decameacuterica e monomeacuterica satildeo respectivamente de 500 microM e 200 microM Assim pode inferir-
se que para concentraccedilotildees inferiores a 5 mM e intervalos de tempo de cerca de 20 minutos (que
inclui o tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas
as espeacutecies decameacuterica e monomeacuterica de vanadato
Figura 24 ndash Espectros de RMN de 51
V de concentraccedilotildees
crescentes das espeacutecies oligomeacutericas de vanadato obtidos
apoacutes diluiccedilatildeo da soluccedilatildeo stock de metavanadato em 10 mM
HEPES pH 70 25 mM KCl para concentraccedilotildees finais de A)
1 mM B) 2 mM C) 4 mM D) 8 mM E) 10 mM de
metavanadato total V1 refere-se agraves espeacutecies monomeacuterica de
vanadatoV1 e V2 correspondem respectivamente agrave espeacutecie
monomeacuterica e dimeacuterica
Figura 25 ndash Distribuiccedilatildeo graacutefica das espeacutecies
oligomeacutericas de vanadato vs a concentraccedilatildeo de
metavanadato total As concentraccedilotildees de cada
espeacutecie foram obtidas segundo a formula descrita
na secccedilatildeo 35 do Procedimento Experimental
(aacutereas dos sinais de 51
V-RMN observadas na
figura x1)V10 V1 e V4 correspondem agraves espeacutecie
monomeacuterica dimeacuterica e tetrameacuterica
respectivamente
Esp
eacuteci
e o
ligo
meacute
rica
de
va
nad
ato
(m
M)
[Metavanadato]total (mM)
V1
V2
V4
V5
28
Tabela 2IVndash Desvios quiacutemicos meacutedios (part)das ressonacircncias das espeacutecies tetrameacuterica (V4) monomeacuterica (V1) e
dimeacuterica (V2) presentes no espectro da soluccedilatildeo de metavanadato (total) 10 mM
Paracircmetros V1 V2 V4 V5
part (ppm) -5615 -5748 -5789 -5869
Nas soluccedilotildees de metavanadato as espeacutecies monomeacuterica dimeacutericae tetrameacuterica de vanadato estatildeo
presentes para concentraccedilotildees de metavanadato inferiores a 4 mM Todas estas as espeacutecies
aumentam linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado
em (Aureliano e Madeira 1998) e em (Tiago et al 2004) quando a concentraccedilatildeo de V1 eacute igual ou
superior a 200 microM inicia-se a sua oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica Nas
condiccedilotildees de realizaccedilatildeo dos ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a
espeacutecie pentameacuterica neste estudo em concentraccedilotildees inferiores a 4 mM tendo o mesmo sido
observado por Ramos 2006 Os resultados obtidos mostram claramente o equiliacutebrio entre a
espeacutecie V4 e as espeacuteciesV1 e V2 no qual o aparecimento de outras espeacutecies oligomeacutericas se
encontra dependente da concentraccedilatildeo da espeacutecie V1 Por uacuteltimo pode inferir-se que para
concentraccedilotildees inferiores a 2 mM e intervalos de tempo de cerca de 20 minutos (que inclui o
tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas as
espeacutecies V1 V2 e V4 com concentraccedilotildees respectivas de 0796 0192 e 0196 mM
232Estabilidade das soluccedilotildees de decavanadato anaacutelise por espectroscopia de absorccedilatildeo
UVVis
Caracterizou-se a estabilidade da espeacutecie V10 numa soluccedilatildeo com 1 mM decavanadato total em
tampatildeo D atraveacutes da leitura da Abs400 Obteve-se uma cineacutetica de dissociaccedilatildeo de 1ordf ordem tal
como se pode verificar pela linearidade do graacutefico de ln(Abs400)vs o tempo de aquisiccedilatildeo espectral
(segundos) (Fig26 )
Figura 26ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do tempo
(em segundos) para uma soluccedilatildeo de decavanadato total de 1
mM em tampatildeo D a 25ordmC (n=2)
Sendo uma reacccedilatildeo de primeira ordem pode ser descrita pela equaccedilatildeo cujo
declive eacute ndashK (constante de dissociaccedilatildeo) e a intersecccedilatildeo no eixo das ordenadas corresponde
Ln
(Ab
s 400 n
m)
tempo (s)
29
aln(A0) Numa reacccedilatildeo de 1ordf ordem o tempo de meia-vida eacute independente da concentraccedilatildeo inicial
do reagente sendo descrito pela equaccedilatildeo [2] Deste modo a partir da recta
Ln(Abs400 nm) = -436E-5
K - 105 obteve-se um tempo de meia-vida de 4490 plusmn 0823 horas para
as condiccedilotildees supramencionadas
Apoacutes a adiccedilatildeo de S1 8 microM ao meio reaccional verificou-se que a reacccedilatildeo de desoligomerizaccedilatildeo
da espeacutecie decameacuterica de vanadato seguia igualmente uma cineacutetica de 1ordf ordem tal como se pode
verificar pela linearidade do graacutefico de ln(Abs400) versus o tempo de aquisiccedilatildeo espectral
(segundos) (Fig27)
Figura 27ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do
tempo (em segundos) para uma soluccedilatildeo de decavanadato
total de 1 mM em tampatildeo D a 25ordmC na presenccedila de 8
microM (n= 2)
A partir da equaccedilatildeo da recta Ln(Abs400 nm) = -7520E-5
K - 0613determinou-se o valor de K para
a espeacutecie decameacuterica de onde se obteve um tempo de meia-vida de 2430 plusmn 0191 horas para as
condiccedilotildees supramencionadas
Tanto este valor como o valor registado na ausecircncia de S1 satildeo substancialmente superiores agrave
duraccedilatildeo dos ensaios de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina descritos no presente
relatoacuterio
24 Interacccedilatildeo de S1 com vanaacutedio(V)
241Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
De modo a analisar qual o efeito da adiccedilatildeo de S1 de miosina (tampatildeo D) produzia nos sinais de
51V-NMR titulou-se uma soluccedilatildeo de 5mM de decavanadato total com concentraccedilotildees crescentes
de S1 (5 10 15 e 20 microM) Os espectros de 51
V-RMN encontram-se apresentados na Fig 2
No graacutefico da figura 2 Apresenta-se a concentraccedilatildeo de S1 em funccedilatildeo do factor de alargamento
corrigido dos espectros acima referidos
Ln
(Ab
s (4
00
nm
))
tempo (s)
30
Figura 28 ndash Graacutefico de
representaccedilatildeo da
concentraccedilatildeo de S1 em
funccedilatildeo do factor de
alargamento A
concentraccedilatildeo de
decavanadato total foi de 5
mM e as concentraccedilotildees
utilizadas de S1 foram de 0
5 10 e 15 microM
Verifica-se que o factor de alargamento aumenta linearmente em funccedilatildeo da concentraccedilatildeo de S1 no
ensaio o que indica a existecircncia de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina Observou-
se ainda que a adiccedilatildeo de S1 ao meio reaccional natildeo provocou qualquer alteraccedilatildeo dos desvios
quiacutemicos dos sinais de 51
V-RMN
Realizaram-se ensaios na presenccedila de ATP e de S1 com intuito de verificar o efeito do ATP na
interacccedilatildeo do S1 de miosina com o decavanadato
Figura 29 - Espectros de RMN de 51
V contendo 2
mM Decavanadato (A) (controlo) (B) na presenccedila
de 2 mM de ATP (C) na presenccedila de 2 mM de ATP
e 8 microM de S1 de miosina V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2Vndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies decameacuterica (V10) e monomeacuterica (V1) presentes nos
espectros da soluccedilatildeo de decavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V10A V10B V10C V1
part (ppm) -5161 -5002 -4254 -5614
Observa-se que a presenccedila de ATP (espectro B) natildeo provoca qualquer alteraccedilatildeo visiacutevel em
relaccedilatildeo ao espectro que contem apenas soluccedilatildeo de decavanadato Jaacute a presenccedila simultacircnea de ATP
e S1 provoca um alargamento da ∆12 do sinal do V10A de 1736 Hz (espectro (A)) para 2366 Hz
(espectro (C) Fig 29) Tendo em conta que a concentraccedilatildeo de S1 apresenta uma relaccedilatildeo linear
com o factor de alargamento converteu-se o valor da ∆12 e comparou-se com o valor esperado agrave
F = 00591[S1] - 00157 Rsup2 = 0998
Fact
or
de
ala
rgam
en
to
[S1] (microM)
31
concentraccedilatildeo de S1 utilizada O factor de alargamento obtido para 8 microM ndash 0376 ndash eacute inferior ao
valor esperado agrave mesma concentraccedilatildeo de S1 obtido atraveacutes da equaccedilatildeo da recta apresentada na
Fig 28 ndash 0457 ndash o que indica que o ATP poderaacute exercer um efeito de protecccedilatildeo do S1
diminuindo a sua interacccedilatildeo com a espeacutecieV10
Por uacuteltimo realizaram-se ensaios com metavanadato na presenccedila de 8 microM S1 de modo a verificar
se as espeacutecies oligomeacutericas contidas nesta soluccedilatildeo interagiam com a proteiacutena em estudo
Figura 210 - Espectros de RMN de 51
V contendo 2
mM metavanadato (A) (controlo) (B) na presenccedila
de 8 microM de S1 de miosina V1 V2 e V4
correspondem respectivamente aacutes espeacutecies
monomeacuterica dimeacuterica e tetrameacuterica
Tabela 2VIndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies monomeacuterica (V1) dimeacuterica (V2) e tetrameacuterica (V4)
presentes nos espectros da soluccedilatildeo de metavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V1 V2 V4
part (ppm) -5613 -5747 -5789
Como eacute possiacutevel observar na Fig 210 ocorre uma ligeiro alargamento dos sinal de V4 bem
como uma perda da intensidade pelo que se conclui quedas espeacutecies presentes na soluccedilatildeo de
metavanadato (V1 V2 e V4) apenas a espeacutecie tetrameacuterica interage com o S1 de miosina o que
estaacute de acordo com o reportado por Tiago et al 2002
242 Actividade Mg2+
-ATPaacutesica de S1 de miosina na presenccedila de decanadato
Com o intuito de avaliar o efeito do decavanadato na actividade Mg2+
-ATPaacutesica do S1 de
miosina monitorizou-se a reacccedilatildeo de oxidaccedilatildeo do NADH atraveacutes da leitura de Abs340 na
presenccedila de 005 mgmL de S1 de miosina e no caso da actividade estimulada 2microM de actina em
3125 mM Tris-HCl pH 71 e 375 mM MgCl2 pelo meacutetodo de enzimas acopladas (ver seccccedilatildeo xx
dos MM)Para tal realizaram-se ensaios com vaacuterias concentraccedilotildees de decavanadato total (10 50
100 250 500 e 1000) de modo a determinar qual o seu efeito na actividade MgATPaacutesica de S1
de miosina na ausecircncia e na presenccedila de um dos seus ligandos naturais Os ensaios foram
realizados agrave temperatura ambiente com incubaccedilatildeo preacutevia a 25ordm C durante 20 minutos
32
Figura 211 ndash Representaccedilatildeo
graacutefica da Actividade Mg2+
-
ATPaacutesica de S1 () (os ajustes
teoacutericos estatildeo representado a por
uma linha os resultados
experimentais estatildeo representados
por triacircngulos (Actividade
estimulada) e cruzes (Actividade
natildeo estimulada)) vs
[Decavanadato]total medidas pelo
meacutetodo das enzimas acopladas
(n=3) Os dados foram ajustados ao modelo de decaimento exponencial usando o programa GraphPad Prism para a
determinaccedilatildeo dos valores de IC50 (na ausecircncia de actina IC50= 266 plusmn 07 M decavanadato na presenccedila de actina
IC50= 28 plusmn 1 M decavanadato
A partir do graacutefico presente na Fig 29 verifica-se que o ajuste dos dados experimentais foi bem
sucedido A inibiccedilatildeo da actividade Mg2+
-ATPaacutesica do S1 de miosina eacute afectada em menor
amplitude na presenccedila de actina (60 de inibiccedilatildeo da actividade) do que na sua ausecircncia
(aproximadamente 80 de inibiccedilatildeo de actividade) Devido a esta diferenccedila pode sugerir-se que a
actina iraacute impedir parcialmente a ligaccedilatildeo do decavanadatoao S1 demiosina atraveacutes de
impedimentos esteacutereos ou interacccedilotildees electrostaacuteticas por exemplo A teacutecnica utilizada natildeo nos
permite concluir acerca de que regiatildeo(otildees) ou resiacuteduo(s) da actina participaratildeo na preservaccedilatildeo
parcial da actividade Mg2+
-ATPaacutesica
Por outro lado os valores de IC50 dos dois tipos de actividades natildeo apresentam diferenccedilas
significativas tendo-se obtido 266 plusmn 07 e 28 plusmn 1M decavanadato na ausecircncia e na presenccedila de
actina respectivamente o que corresponde a uma concentraccedilatildeo de cerca de 27 M de V10 O
meio de reacccedilatildeo utilizado nestes ensaios foi 10 mM HEPES e 25 mM KCl com 2 mM ATP Em
(Tiago et al 2004) o valor de IC50 para a actividade Mg2+-ATPaacutesica na ausecircncia de actina foi de
125 plusmn 025 microM enquanto o valor da actividade estimulada por actina foi de 136 plusmn 14 microM O
meio de reacccedilatildeo utilizado nos ensaios foi 20 mM HEPES 50 mM KCl e 2 mM ATP o que resulta
precisamente no dobro da forccedila ioacutenica a que foi realizado o ensaio do presente relatoacuterio
(exceptuando a contribuiccedilatildeo dos produtos da hidroacutelise de ATP uma vez que este meacutetodo
pressupotildee a ocorrecircncia de um ciclo de reacccedilotildees de hidroacutelise e siacutentese deste composto) Ora tendo
sido anteriormente referido que a afinidade do S1 de miosina para com a espeacutecie V10 diminui com
Act
ivid
ade
Mg
2+-A
TP
aacutesic
a d
e S
1 d
e
Mio
sin
a
[Decavanadato] (microM) Actividade Estimulada ()
Actividade Natildeo Estimulada ()
Ajuste Actividade Estimulada
Ajuste Actividade Natildeo Estimulada
33
o aumento da forccedila ioacutenica (Tiago et al 2004 tal poderaacute explicar a diferenccedila obtida entre estes
dois conjuntos de valores
243Quantificaccedilatildeo de cisteiacutenas reduzidas pelo meacutetodo de Ellman
A interacccedilatildeo do vanaacutedio com alguns tipos de aminoaacutecidos sensiacuteveis agrave oxidaccedilatildeo pode fornecer-nos
informaccedilatildeo bastante uacutetil em relaccedilatildeo agraves alteraccedilotildees estruturais que conduzem a uma diminuiccedilatildeo da
actividade fisioloacutegica do S1 de miosina Os estudos realizados por Chasteen 1983 concluiacuteram
que a reduccedilatildeo de vanadato (V) a vanadilo (IV) ocorre na presenccedila de agentes redutores comuns no
ambiente celular de que se destaca o aminoaacutecido cisteiacutena Esta eacute tambeacutem um alvo de
complexaccedilatildeo por parte do iatildeo VO2+
Jaacute Ramos et al (2009) demonstrou que a incubaccedilatildeo de
actina com a soluccedilatildeo de decavanadato induz a oxidaccedilatildeo de resiacuteduos de cisteiacutena e agrave formaccedilatildeo do
oxidovanaacutedio (IV) Este grupo de investigadores descobriu ainda que apenas a espeacutecie V10 induz a
oxidaccedilatildeo das cisteiacutenas ldquoraacutepidasrdquo da actina atraveacutes do processo proposto que eacute parcialmente
responsaacutevel pela inibiccedilatildeo jaacute descrita da actividade de ATPase do S1 de miosina estimulada por
actina quando exposta agrave soluccedilatildeo de decavanadato Esta inibiccedilatildeo apresenta-se afectada com a
presenccedila de ATP uma vez que os resiacuteduos de cisteiacutena permaneceram intactos nos ensaios
realizados com este ligando Jaacute Crans et al (2004) concluiu que o siacutetio activo de muitas fosfatases
(que conteacutem grupos tiol) pode formar um equiliacutebrio redox com o iatildeo vanadato (V) de forma natildeo
reversiacutevel
Os resultados obtidos por Sakurai e Shiromura (1980) sugeriram que o vanaacutedio (V) sofre reduccedilatildeo
a vanaacutedio (IV) sem dependecircncia do valor de pH do ensaio Registaram ainda uma estequiometria
de 31 na reacccedilatildeo do vanadato (V) com os resiacuteduos de Cys enquanto a estequiometria dos ensaios
com o vanadilo (IV) foi de 21
Berlett e Stadtman (1997) observaram que os tipos de aminoaacutecidos mais facilmente oxidaacuteveis
englobam entre outros os resiacuteduos de Cys Trp e MetAssim analisou-se o estado redox dos
grupos tiol dos resiacuteduos de Cys do S1 na presenccedila do decavanadato atraveacutes da monitorizaccedilatildeo da
Abs412 nm na presenccedila de 1 mgmL de S1 de miosina em tampatildeo 10 mM HEPES 25 mM KCl
variando a concentraccedilatildeo da soluccedilatildeo de decavanadato de modo a determinar que tipo de alteraccedilotildees
estruturais ocorrem na sua presenccedila no que diz respeito agrave diminuiccedilatildeo da actividade ATPaacutesica de
S1 na presenccedila de decavanadato
Os resultados referentes agrave quantificaccedilatildeo das cisteiacutenas reduzidas apoacutes o tratamento de S1 com
diferentes concentraccedilotildees de decavanadato em tampatildeo 10 mM HEPES pH 75 e 25 mM KCl a
25ordmC durante 20 minutos encontram-se descritos na Tabela 2VII
Tabela 2VII ndash Cisteiacutenas reduzidas totais e ldquoraacutepidas de S1 (1 mgmL) apoacutes tratamento com decavanadato
34
(010501002505001000 uM) em 10 mM HEPES 25 mM KCl pH 7 a 25ordmC durante 20 minutos A detecccedilatildeo
espectrofotomeacutetrica foi feita com DTNB (e 412=8400 uM-1 cm-1) (n variaacutevel entre 1 e 8)
Amostras Cys raacutepidas Cys totais
11 M S1 (controlo) 22 plusmn 04 106 plusmn 14
11 M S1 + 10 M Decavanadato 22 plusmn 05 95 plusmn 08
11 M S1 + 50 uM Decavanadato 20 plusmn 02 98 plusmn 14
11 M S1 + 100 uM Decavanadato 17 plusmn 03 97 plusmn14
11 M S1 + 250 uM Decavanadato 26 plusmn 02 98 plusmn 08
11 M S1 + 500 uM Decavanadato 21 plusmn 04 90 plusmn 06
11 M S1 + 1000 uM Decavanadato 24 plusmn 0 86 plusmn 08
A partir dos resultados obtidos pode sugerir-se que o S1 possui 11 resiacuteduos de Cys (controlo) o
que estaacute de acordo com o anteriormente descrito Verifica-se que o tratamento da proteiacutena com
concentraccedilotildees crescentes de decavanadato provoca a oxidaccedilatildeo de dois resiacuteduos de Cys do S1 de
miosina natildeo expostos ao solvente No entanto a teacutecnica utilizada natildeo nos permite identificar
quais
Tendo em consideraccedilatildeo que o valor de IC50 obtido para a reacccedilatildeo de inibiccedilatildeo daactividade Mg2+-
ATPaacutesica de S1 na presenccedila de decavanadato eacute de cerca de 27 microM pode sugerir-se que este seraacute o
valor ao qual ocorreraacute a oxidaccedilatildeo do primeiro resiacuteduo de Cys o que poderaacute contribuir para a
diminuiccedilatildeo da actividade de S1 observada
25Interacccedilatildeo de S1 com vanaacutedio(IV)
251Anaacutelise por espectroscopia de Ressonacircncia Paramagneacutetica Electroacutenica
A acumulaccedilatildeo intracelular de oxovanaacutedio (IV) in vivo constitui uma das principais formas de
apresentaccedilatildeo de vanaacutedio no organismo Deste modo torna-se bastante interessante analisar a
interacccedilatildeo entre este iatildeo e o S1 de miosina atraveacutes da teacutecnica de espectroscopia de Ressonacircncia
Paramagneacutetica Electroacutenica o meacutetodo de eleiccedilatildeo do estudo do iatildeo vanadilo e dos complexos por si
formados O electratildeo desemparelhado encontra-se fortemente acoplado ao nuacutecleo 51
V o que
35
resulta num espectro de RPE consistindo de 8 desdobramentos na direcccedilatildeo paralela e outros tantos
na direcccedilatildeo perpendicular (Chasteen 1983)
Os estudos realizados por Ahmed et al (1987) centraram-se na interacccedilatildeo do vanadilo com
proteiacutenas como eacute o caso da insulina transferrina entre outros obtendo um espectro caracterizado
por 8 linhas amplamente espaccediladas (na direcccedilatildeo paralela) Jaacute Ramos et al 2009 concluiu que a
exposiccedilatildeo da soluccedilatildeo de decavanadato agraves formas globular e filamentosa de actina produz um
espectro tiacutepico do iatildeo oxidovanaacutedio (IV) indicando a reduccedilatildeo do vanadato
Para aferir qual a concentraccedilatildeo de S1 que permitia obter uma boa razatildeo sinalruiacutedo titulou-se uma
soluccedilatildeo de sulfato de vanadilo 15 microM ([VOSO4]stock = 5 mM) com concentraccedilotildees crescentes de
proteiacutena (0 5 10 15 e 20 microM) em 10 mM HEPES 25 mM KCl A existecircncia de uma correlaccedilatildeo
linear entre a concentraccedilatildeo de proteiacutena e a intensidade dos sinais de RPE correspondente agrave banda
-12 perpendicular encontra-se representada na Fig 210
Figura 2 12ndash Representaccedilatildeo graacutefica da variaccedilatildeo da intensidade do sinal de RPE vs a concentraccedilatildeo de S1 utilizada
Apoacutes verificar que com uma concentraccedilatildeo de S1 igual a 20 microM se obtinham sinais com uma razatildeo
sinalruiacutedo adequada ao prosseguimento dos estudos sobre a interacccedilatildeo V(IV)-S1 de miosina
titulou-se uma soluccedilatildeo de 20 microM S1 com 0 25 5 10 15 25 50 75 100 150 200 500 e 1000
microM VOSO4
Traccedilou-se o graacutefico da intensidade da banda -12 perpendicular do sinal de EPR em funccedilatildeo da
concentraccedilatildeo de VOSO4 tendo-se determinado uma constante de dissociaccedilatildeo (KD) do complexo
S1-vanadilo de 8748 plusmn 2621 microM V(IV)
y = 016786x + 122435
Rsup2 = 092992759
Inte
nsi
dad
e d
o s
inal
de
RP
E
[S1] (microM)
36
Figura 213 ndash Representaccedilatildeo graacutefica da variaccedilatildeo do sinal de RPE vs a concentraccedilatildeo de vanadilo utilizada (os valores
experimentais estatildeo representados por losangos o seu ajuste estaacute representado pela linha)
A simulaccedilatildeo do espectro de EPR foi efectuada a partir do espectro obtido para uma concentraccedilatildeo
de 1 mM VOSO4 apoacutes adiccedilatildeo de 20 microM S1 de miosina tendo-se obtido os valores da constante de
acoplamento hiperfina (A=5016) e do factor g (g0=195)
Figura 214 ndash Representaccedilatildeo graacutefica do espectro de EPR obtido com 1000 microM de vanadilo e 20 microM de S1 (a cinzento
escuro) A simulaccedilatildeo do espectro encontra-se a cinzento claro
Por comparaccedilatildeo destes com os valores presentes na tabela de correspondecircncia xxx eacute possiacutevel
predizer quais os ligandos provaacuteveis do vanadilo na presenccedila de proteiacutena Assim concluiu-se que
o vanadilo estaacute coordenado com duas moleacuteculas de aacutegua um grupo hidroxilo e um grupo R=NR
O grupo hidroxilo pode indicar os resiacuteduos de aminoaacutecidos Tyr Treonina e Serina enquanto o
grupo R=NR poderaacute indicar a presenccedila de Arginina e Histidina como ligandos do vanadilo
Inte
nsi
dad
e d
os
sinai
s d
e R
PE
[Vanadilo] (microM)
Ajuste dos dados exp Dados experimentais
Campo magneacutetico (G)
Espectro Experimental Ajuste
37
3 Procedimento Experimental
31 Reagentes
Todos os reagentes utilizados satildeo de elevado grau de pureza (analiacutetico) Apresentam-se as suas
foacutermulas quiacutemicas ou abreviaturas quando necessaacuterio acrescentou-se a sua descriccedilatildeo Entre
parecircntesis colocou-se as empresas que os comercializam
Aacutecido aceacutetico glacial 100 (Riedel) Acrilamida 99 (Merck) Azul de bromofenol (Riedel-de-
Haumlen) -Mercaptoetanol (Sigma) Bissulfito de soacutedio (Sigma)Bradford (reagente de)
(BioRad)BSA (albumina de soro bovino Sigma-Aldrich) CaCl22H2O (cloreto de caacutelcio
dihidratado Fluka) CH3CH2OH 96 (etanol Panreac) Coomassie Blue R250 (Merck) D2O
(oacutexido de deuteacuterio SDS) DTNB (55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico) Aldrich) EDTA (aacutecido
etilenodiaminatetraaceacutetico Merck) Fosfoenolpiruvato 98 (Fluka) Glicerol (Sigma) Glicina
985 (Panreac) HCl 37 (Riedel-de Haumlen) H2SO4 96 (Pronalab) HEPES 99 (aacutecido 4-(2-
hidroxiletil)-1-piperazineetanosulfoacutenico Sigma) KH2PO4 99 (hidrogenofosfato de potaacutessio)
MgATP (Sigma) Na
ATP (Sigma) Na2HPO4 99 (hidrogenofosfato de soacutedio Riedel-de Haumlen)
NaH2PO4 (fosfato de soacutedio ) NaOH 986 (hidroacutexido de soacutedio Pronalab) KCl 99 (Cloreto de
potaacutessio Panreac) L-Cys 995 (enantioacutemero levoacutegiro do resiacuteduo de Cisteiacutena Fluka)Metanol
99 (Merck) Molibdato de amoacutenio (Sigma)NNrsquo-metil-bis-acrilamida 98 (Merck) NH4VO3
99 (metavanadato de amoacutenia Sigma) PSA 10 (persulfato de amoacutenio Merck) SDS
(dodecilo sulfato de soacutedio Sigma) Sulfito de soacutedio (Sigma) VOSO4 (sulfato de vanadilo
Aldrich) TEMED 99 (Tetrametiletilenodiamina Merck) Tris (2-amino-2-hidroximetil-13-
propanodiol Sigma)
32 Meacutetodos
321 Isolamento e purificaccedilatildeo das proteiacutenas de muacutesculo esqueleacutetico de coelho
3211 Purificaccedilatildeo da miosina preparaccedilatildeo de Subfragmento 1 por digestatildeo quimotriacutepsica
da miosina
O subfragmento-1 de miosina-II (S1) foi gentilmente cedido pelo Prof Doutor Manuel Aureliano
do Departamento de Quiacutemica Bioquiacutemica e Farmaacutecia da Faculdade de Ciecircncias e Tecnologia da
Universidade do Algarve
No entanto descreve-se abaixo de forma resumida o procedimento efectuado
38
32111 Purificaccedilatildeo da miosina
O protocolo de isolamento foi adaptado do meacutetodo previamente descrito por Margossian e Lowey
(1982) apresentando-se de forma esquemaacutetica na Fig 31
Sucintamente os tecidos musculares dorsais e das pernas foram extraiacutedos apoacutes o sacrifiacutecio do
animal (Fig31) De seguida foram adicionados 3 volumes da soluccedilatildeo de extracccedilatildeo A2
Centrifugou-se a soluccedilatildeo e filtrou-se o sobrenadante O sedimento foi entatildeo diluiacutedo em 14
volumes de aacutegua destilada de modo a que a forccedila ioacutenica atingisse 004 M Apoacutes adiccedilatildeo de szlig-
mercaptoetanol (szlig-me) deixou-se a precipitar a 4ordmC removendo o sobrenadante Centrifugou-se
novamente e ressuspendeu-se o sedimento na soluccedilatildeo de extracccedilatildeo A Adicionou-se a soluccedilatildeo
tampatildeo B3 para que a forccedila ioacutenica atingisse 05 M Homogeneizou-se e deixou-se repousar
durante 2h Homogeneizou-se novamente e centrifugou-se de modo a ocorrer precipitaccedilatildeo dos
complexos de actomiosina e proteiacutena desnaturada Apoacutes filtraccedilatildeo do sobrenadante em latilde de vidro
quantificou-se a soluccedilatildeo de miosina resultante pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental)
2 Soluccedilatildeo de extracccedilatildeo A 0012M K2CO3 004M KHCO3 06M KCl pH 65
3 Soluccedilatildeo-tampatildeo B Tris 100 mM pH 7 KCl 2M
39
Figura 31 ndash Representaccedilatildeo esquemaacutetica do protocolo de isolamento de miosina (Margossian e Lowey 1982) Todo o
procedimento foi realizado a 4ordmC (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
32112 Preparaccedilatildeo de Subfragmento-1 de miosina por digestatildeo com quimiotripsina
O S1 de miosina foi isolado apoacutes diaacutelise da miosina durante a noite contra a soluccedilatildeo C4 A
presenccedila do EDTA eacute essencial uma vez que a sua substituiccedilatildeo por MgCl2resulta na produccedilatildeo de
HMM em vez de S1 A suspensatildeo de miosina foi equilibrada e digerida com -quimotripsina
A digestatildeo foi interrompida pela adiccedilatildeo de PMSF Seguidamente ultracentrifugou-se o produto
da digestatildeo desprezando-se o sedimento (Fig 32) A soluccedilatildeo de sobrenadante maioritariamente
S1 foi quantificada pela leitura directa da Abs280 ou pelo meacutetodo de Bradford (ver as secccedilotildees
3311 e 3312 do Procedimento Experimental respectivamente) e armazenada em 10 sacarose
e 2 mM szlig-Me a -80ordmC
4 Soluccedilatildeo C 25mM Tris pH 70 012M KCl 1mM EDTA
40
Antes da realizaccedilatildeo dos ensaios descritos no presente relatoacuterio o S1 de miosina foi dialisado
contra a soluccedilatildeo D5 de modo a remover qualquer composto que pudesse complexar como
vanaacutedio Seguidamente quantificou-se a proteiacutena pelo meacutetodo de Bradford ou pelo meacutetodo
directo (ver secccedilotildees 3311 e 3312do Procedimento Experimtental)
Figura 3 2ndash Representaccedilatildeo esquemaacutetica do protocolo de preparaccedilatildeo do S1 de miosina por digestatildeo quimiotripsina de
acordo com o previamente descrito por Margossian e Lowey (1982) Todo o procedimento foi realizado a 4ordm C
excepto quando indicaccedilatildeo do contraacuterio (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
3212 Isolamento e purificaccedilatildeo de actina
Isolou-se e purificou-se a actina segundo o meacutetodo descrito por Pardee e Spudish que
compreende dois passos ndash a obtenccedilatildeo de um resiacuteduo seco denominado ldquopoacute de acetonardquo e a
extracccedilatildeo subsequente da actina a partir do ldquopoacute de acetonardquo na forma de G-Actina com a
posterior polimerizaccedilatildeo a F-Actina
32121 Obtenccedilatildeo do ldquopoacute de acetonardquo
Sacrificou-se o coelho com uma pancada atraacutes do pescoccedilo e sangrou-se o animal apoacutes o corte da
arteacuteria jugular Esfolou-se e colocou-se numa tina com gelo Retiraram-se as partes de tecido
conjuntivo que envolvem o tecido muscular com auxiacutelio de uma pinccedila e bisturi Seguidamente
retirou-se os muacutesculos dorsais e os muacutesculos brancos das pernas Passou-se os muacutesculos por aacutegua
destilada de modo a remover as impurezas e pesou-se num copo Trituraram-se os muacutesculos
numa maacutequina de triturar carne e extraiu-se o resultante com 3 volumes da soluccedilatildeo E6 durante 10
minutos com agitaccedilatildeo suave Centrifugou-se a suspensatildeo a 4000 g durante 10
5 Soluccedilatildeo D HEPES 10 mM pH 70 KCl 25 mM
6 Soluccedilatildeo E 01 M KCl 015 M K2HPO4 pH 65
41
minutosdesprezando-se o sobrenadante Ressuspendeu-se o sedimento em 6 volumes de soluccedilatildeo
F7durante 10 minutos com agitaccedilatildeo suave Verificou-se se o pH da soluccedilatildeo estava compreendido
entre 82 e 85 acertando-se estecom soluccedilatildeo G8 se necessaacuterio Centrifugou-se durante 10
minutos a 4000 g Desprezou-se o sobrenadante e ressuspendeu-se o sedimento em 3 volumes de
soluccedilatildeo H9 durante 10 minutos Centrifugou-se novamente a 4000 gdurante 10 minutos
desprezando-se o sobrenadante Ressuspendeu-se o sedimento duas vezes consecutivas em 6
volumes de aacutegua destilada durante 5 minutos Centrifugou-se a 4000 g durante 10 minutos apoacutes
cada adiccedilatildeo de H2O
Seguidamente ressuspendeu-se o sedimento 5 vezes consecutivas utilizando 3 volumes de
acetona de cada vez agitando suavemente durante 10 minutos agrave temperatura ambiente Colocou-
se o resiacuteduo filtrado numa tina de vidro sobre papel de filtro deixando-se evaporar o solvente
durante a noite num nicho Triturou-se o resiacuteduo seco numa maacutequina de triturar carne e deixou-se
novamente evaporar o solvente O ldquopoacute de acetonardquo obtido foi armazenado a -20 ordmC
32122 Purificaccedilatildeo da actina a partir de ldquopoacute de acetonardquo
Apoacutes a obtenccedilatildeo do poacute de acetona como acima indicado efectuou-se o protocolo seguidamente
descrito de modo a obter monoacutemeros de actina (G-Actina) e actina filamentosa (F-Actina)
Pesou-se 35 g de poacute de acetona e adicionaram-se 50 mL de soluccedilatildeo-tampatildeo I10
sendo a
extracccedilatildeo feita com agitaccedilatildeo suave durante 30 minutos Separou-se o extracto do poacute de acetona
hidratado filtrando a suspensatildeo com 4 camadas de gaze esterilizada tendo o cuidado de natildeo
espremer o resiacuteduo Voltou a extrair-se com 50 mL de soluccedilatildeo I durante 30 minutos mexendo
lentamente Adicionou-se este ao extracto obtido anteriormente Centrifugou-se o extracto total a
16 000 g durante 30 minutos Decantou-se para uma proveta sobre a qual se colocou gaze para
que o sedimento solto natildeo se juntasse ao sobrenadante e leu-se o volume De seguida adicionou-
se KCl ateacute uma concentraccedilatildeo final de 50 mM MgCl2 ateacute 2mM e ATP ateacute 1mM lentamente e com
agitaccedilatildeo contiacutenua de modo a promover a polimerizaccedilatildeo dos monoacutemeros de actina
Deixou-se polimerizar durante 2h a 25ordmC observando-se a presenccedila de pequenas bolhas de ar
indicativas de um aumento de viscosidade da soluccedilatildeo Arrefeceu-se em gelo durante 15 minutos e
adicionou-se KCl ateacute uma concentraccedilatildeo final de 06M Mexeu-se lentamente durante 30 minutos
apoacutes os quais se ultracentrifugou a 110 000g durante 1hora Desprezou-se o sobrenadante
ressuspendeu-se o sedimento com soluccedilatildeo tampatildeo E e transferiu-se para um potter
Dialisou-se durante a noite contra 1L de soluccedilatildeo-tampatildeo I a uma temperatura de 4ordmC com rotaccedilatildeo
7 Soluccedilatildeo F 005 M NaHCO3 pH 82-85
8 Soluccedilatildeo G 1M Na2CO3
9 Soluccedilatildeo H 1 mM EDTA pH 70
10 Soluccedilatildeo I 2 mM Tris-HCl pH 8 02 mM CaCl2 02 mM Na2ATP 05 mM szlig-Me 0005 NaN3
42
contiacutenua e mudanccedila de tampatildeo 2 a 3 vezes De seguida a actina foi centrifugada a 110
000gdurante 1hora Decantou-se a soluccedilatildeo de G-Actina (sobrenadante) e desprezou-se o
sedimento Determinou-se a concentraccedilatildeo de G-Actina pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental) A soluccedilatildeo de G-actina foi armazenada a uma temperatura de 4ordmC
apoacutes adiccedilatildeo de 002NaN3 mantendo-se estaacutevel durante 3 a 5 dias
331 Quantificaccedilatildeo das proteiacutenas isoladas
3311 Quantificaccedilatildeo pelo meacutetodo de Bradford
O meacutetodo de Bradford (Bradford 1976) baseia-se na utilizaccedilatildeo do reagente de Bradford
composto por entre outros aacutecido fosfoacuterico e o corante ldquoCoomassierdquo Este corante possui carga
negativa e a sua coloraccedilatildeo varia entre a vermelha (Absmax=465 nm) predominante em soluccedilatildeo e a
azul (Absmax=595 nm) presente aquando da ligaccedilatildeo aos grupos amina das proteiacutenas Em soluccedilotildees
aacutecidas diluiacutedas a coloraccedilatildeo varia proporcionalmente com a quantidade do corante ligado agrave
proteiacutena que por sua vez eacute directamente proporcional agrave concentraccedilatildeo da proteiacutena permitindo a
sua quantificaccedilatildeo ateacute a um limite de detecccedilatildeo de 20 gmL (Bradford 1976)
Deste modo prepararam-se amostras de BSA ([BSA]stock= 1mgmL) utilizando um intervalo de
concentraccedilotildees compreendido entre 0 e 15 gmL de acordo com a tabela 3I Apoacutes a adiccedilatildeo da
H2O MilliQ e do reagente de Bradford as soluccedilotildees foram sucessivamente homogeneizadas no
voacutertex
Tabela 3Indash Amostras utilizadas para a obtenccedilatildeo da recta de calibraccedilatildeo no meacutetodo de Bradford
BSA (L) BSA (g) H2O MilliQ (L) Reagente de Bradford (L)
0 0 800 200
125 125 79875 200
25 25 7975 200
5 5 795 200
10 10 790 200
15 15 785 200
Apoacutes incubaccedilatildeo a 25ordmC por 5 minutos leram-se os valores de Abs595 num
espectrofotoacutemetro (Shimadzu UV-160A) para todas as amostras Seguidamente traccedilou-se a recta
de calibraccedilatildeo (Abs595vs quantidade de BSA (g))
43
Para determinar a concentraccedilatildeo de proteiacutena apoacutes o processo de isolamento purificaccedilatildeo e
diaacutelise quando necessaacuterio colocou-se 5 L da amostra num tubo de Eppendorf e adicionou-se
795 L de H2O MilliQ homogeneizando a soluccedilatildeo no voacutertex Adicionou-se posteriormente 200
L de reagente de Bradford e repetiu-se a homogeneizaccedilatildeo Incubou-se a mistura a 25ordmC por 5
minutos apoacutes os quais se leu os valores de Abs595 nm no espectrofotoacutemetro Por interpolaccedilatildeo deste
valor a partir da recta de calibraccedilatildeo determinou-se a concentraccedilatildeo de proteiacutena na amostra
3312 Quantificaccedilatildeo pelo meacutetodo directo
A quantificaccedilatildeo das amostras proteicas pode tambeacutem ser efectuada atraveacutes da leitura directa da
absorvacircncia tendo por base a lei de Beer-Lambert onde Abs corresponde agrave
absorvacircncia ao comprimento de onda ao qual o ensaio foi realizado corresponde ao coeficiente
de extinccedilatildeo (substituiacutedo pela letra a se as unidades forem distintas da molaridade) b corresponde
ao percurso oacuteptico da radiaccedilatildeo na ceacutelula amostral e c corresponde agrave concentraccedilatildeo amostral
Assim no caso de S1 de miosina mediu-se a Abs280e calculou-se a concentraccedilatildeo da proteiacutena
sabendo que o coeficiente de extinccedilatildeo de S1 eacute (Ref)
No caso da actina mediu-se a Abs290ecalculou-se a concentraccedilatildeo da proteiacutena sabendo que o
coeficiente de extinccedilatildeo da actina eacute (Ref)
332Electroforese em condiccedilotildees desnaturantes (SDS-PAGE)
De modo a melhor caracterizar as proteiacutenas isoladas foi realizada uma electroforese em
condiccedilotildees desnaturantes (SDS-PAGE do inglecircs Sodium Dodecyl Sulfate Polyacrilamide Gel
Electrophoresis) O SDS (dodecilo sulfato de soacutedio) eacute um detergente anioacutenico utilizado neste
meacutetodo para desnaturar as proteiacutenas que se ligam ao SDS na proporccedilatildeo 14 g SDS g proteiacutena
(aproximadamente 1 moleacutecula de SDS por cada 2 resiacuteduos de aminoaacutecidos) (Stryer) O SDS induz
uma densidade de carga negativa significativa nas proteiacutenas a que se liga mascarando a sua carga
intriacutenseca pelo que as razotildees cargamassa satildeo tendencialmente idecircnticas Assim a SDS-PAGE
separa as proteiacutenas quando sujeitas a um campo eleacutectrico de acordo com o seu peso molecular
sendo a sua migraccedilatildeo directamente proporcional ao logaritmo deste uacuteltimo (Stryer)
O gel de resoluccedilatildeo com 10 de acrilamida foi obtido atraveacutes da adiccedilatildeo de 167 mL de soluccedilatildeo
stock de acrilamida11
075 mL de tampatildeo de resoluccedilatildeo12
251 mL de aacutegua destilada e 50 L de
SDS 10 Para promover a polimerizaccedilatildeo adicionou-se em seguida 38 L de PSA 10
11
soluccedilatildeo stock de acrilamida soluccedilatildeo de 30 (mv) de acrilamida e 08 (mv) de NNrsquo-metil-bis-acrilamida 12
tampatildeo de resoluccedilatildeo 15 M Tris-HCl pH 88
44
(persulfato de amoacutenia) 10 e 25 L de TEMED 99 (tetrametiletilenodiamina) Agitou-se
cuidadosamente de modo a natildeo formar bolhas e pipetou-se a soluccedilatildeo para a cassette de vidro
previamente montada no suporte De seguida adicionou-se uma peliacutecula de cerca de 2 mm de 2-
metil-2-butanol para verificar posteriormente a polimerizaccedilatildeo Apoacutes a polimerizaccedilatildeo que demora
cerca de 45 minutos verteu-se a agrave superfiacutecie e adicionou-se o gel de concentraccedilatildeo com 5 de
acrilamida constituiacutedo por 03 mL de soluccedilatildeo de stock acrilamida 045 mL de tampatildeo de
concentraccedilatildeo13
094 mL de aacutegua destilada 18 L de SDS 10 135 L de PSA 10 e 2L de
TEMED 99 Colocou-se o pente sobre o gel e deixou-se polimerizar durante cerca de 45
minutos
As amostras para aplicaccedilatildeo no gel de electroforese foram preparadas para uma concentraccedilatildeo de
proteiacutena de 1 mgmL num volume final de 100 L dos quais 25 L correspondem ao tampatildeo da
amostra14
As amostras foram aquecidas a 100ordmC durante 5 minutos Antes da aplicaccedilatildeo das
amostras transferiu-se a cassette para a tina de electroforese agrave qual se adicionou a soluccedilatildeo
tampatildeo de electroforese da tina superior15
e tampatildeo de electroforese da tina inferior16
Aplicaram-
se 10 L das soluccedilotildees de marcadores de peso molecular e 20 L das amostras proteicas
preparadas anteriormente A electroforese decorreu durante cerca de 3h30 min a uma voltagem
de 80 V De seguida transferiu-se o gel para uma tina corando as proteiacutenas durante 1h com
soluccedilatildeo corante17
Descorou-se o gel com soluccedilatildeo descorante18
tendo-se mudado esta soluccedilatildeo ateacute
as bandas se encontrarem bem definidas
Determinou-se a mobilidade relativa das bandas correspondentes aos marcadores de elevado peso
molecular e traccedilou-se a recta de calibraccedilatildeo ndash log(PM) vs log(Rf) onde Rf corresponde agrave
mobilidade relativa Por interpolaccedilatildeo determina-se o PM das proteiacutenas isoladas sabendo a sua
mobilidade relativa Por comparaccedilatildeo com os valores descritos na literatura pode concluir-se
acerca do grau de purificaccedilatildeo
34 Mediccedilatildeo da actividade enzimaacutetica de S1
341 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
13
tampatildeo de concentraccedilatildeo 05 M Tris-HCl pH 68 14
tampatildeo de amostra 320 mM Tris-HCl pH 68 04 M szlig-mercaptoetanol SDS 8 glicerol 15 (vv) e roxo de m-
cresol 0024 15
tampatildeo de electroforese da tina superior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS 16
tampatildeo de electroforese da tina inferior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS acetato de soacutedio
01 M 17
soluccedilatildeo corante ldquoCoomassie Blue R-250rdquo contendo metanol 45 (vv) e aacutecido aceacutetico glacial 10 (vv) 18
soluccedilatildeo descorante metanol 10 (vv) e aacutecido aceacutetico glacial 10 vv)
45
O meacutetodo de Fiske-Subarow modificado consiste na reacccedilatildeo do molibdato de amoacutenio com o
fosfato inorgacircnico em soluccedilatildeo aacutecida com formaccedilatildeo do aacutecido fosfomoliacutebdico Seguidamente daacute-
se a adiccedilatildeo de um agente redutor que provoca a reduccedilatildeo do composto a fosfomolibdato que forma
um complexo de cor azulada cuja absorvacircncia maacutexima ocorre para um comprimento de onda de
660 nm tornando possiacutevel a sua detecccedilatildeo por meacutetodos espectrofotomeacutetricos As actividades Ca2+
-
ATPaacutesica e K+EDTA-ATPaacutesica de S1 foram determinadas atraveacutes deste meacutetodo de acordo com
o descrito por Taussky e Shorr (1953) com mediccedilatildeo da quantidade de fosfato (Pi) libertado
atraveacutes da hidroacutelise de ATP
Para tal obteve-se primeiramente a recta de calibraccedilatildeo com as amostras presentes na Tabela 3II
utilizando uma soluccedilatildeo stock de 5 mM KH2PO4
Tabela 3II ndash Composiccedilatildeo das diferentes amostras utilizadas para determinar a recta de calibraccedilatildeo atraveacutes do meacutetodo
de Fiske-Subbarow modificado
KH2PO4 (L)
([KH2PO4]stock= 5mM)
PO43-
(nmole) H2O (uL)
0 0 1780
5 25 1775
10 50 1770
20 100 1760
40 200 1740
100 500 1680
A cada tubo de Eppendorf adicionaram-se seguidamente 200 L de soluccedilatildeo J19
e 20 L de agente
redutor20
homogeneizando no voacutertex apoacutes cada adiccedilatildeo Incubou-se durante 10 minutos e mediu-
se a Abs660 nm num espectrofotoacutemetro com intervalos de tempo iguais entre cada amostra Traccedilou-
se a recta de calibraccedilatildeo (Abs660 nmvs nmol PO43-
) a partir dos valores obtidos
Para determinar a actividade Ca2+
-ATPaacutesica de S1 prepararam-se tubos de Eppendorf com 400
L de meio de reacccedilatildeo21
25 L CaCl2 ([CaCl2]stock= 100 mM) 25 L ATP ([ATP]stock= 100 mM)
com [S1]ensaios=01 mgmL Perfez-se se necessaacuterio com H2O MilliQ ateacute perfazer um volume de 1
mL Para determinar a actividade K+EDTA-ATPaacutesica utilizou-se uma concentraccedilatildeo idecircntica de
19
Soluccedilatildeo J 5 N H2SO4 25 molibdato de amoacutenio 20
Agente redutor 5 mg aacutecido 1-amino-2-naftol-4-sulfoacutenico 30 mg bissulfito de soacutedio 30 mg sulfito de soacutedio em 26
mL H2O 21
meio de reacccedilatildeo 125 mM Tris-HCl pH 80 0575 M KCl
46
proteiacutena nos ensaios (01 mgmL) e o mesmo meio de reacccedilatildeo Aos tubos de Eppendorf
adicionou-se ainda 123 L de KCl ([KCl]stock = 3M) 25 L de ATP 10 L de EDTA
([EDTA]stock= 100 mM )e o volume de H2O MilliQ necessaacuterio para perfazer 1 mL Em ambas as
situaccedilotildees o uacuteltimo reagente adicionado foi o ATP uma vez que se trata do substrato da reacccedilatildeo
enzimaacutetica
A cineacutetica decorreu a 25ordmC durante 5 minutos para a actividade Ca2+
-ATPaacutesica e durante 3
minutos para a actividade K+EDTA-ATPaacutesica Apoacutes o periacuteodo de incubaccedilatildeo transferiu-se 150
L de amostra para tubos de Eppendorf contendo igual volume de TCA 10 Colocou-se no gelo
e homogeneizou-se no voacutertex Seguidamente centrifugou-se a 1500 g durante 10 minutos e
recolheu-se 200 L de sobrenadante para novos tubos de Eppendorf Adicionou-se a estes 158
mL de H2O MilliQ e 200 L de soluccedilatildeo J Homogeneizou-se no voacutertex e adicionou-se 20 L de
reagente redutor repetindo-se a homogeneizaccedilatildeo Incubou-se durante 10 minutos e mediu-se a
Abs660 nm com o mesmo intervalo de tempo de leitura para cada amostra Por intrapolaccedilatildeo a partir
da recta de calibraccedilatildeo obtida anteriormente determinou-se a quantidade de Pi libertado em cada
ensaio A partir destes resultados determinaram-se as actividades Ca2+
-ATPaacutesica e K+EDTA-
ATPaacutesica do S1 em nmol PO43-
minmg proteiacutena e em s-1
342 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
Da actividade Mg2+
-ATPaacutesica de S1 resulta a formaccedilatildeo dos produtos da reacccedilatildeo de
hidroacutelise do ATP - ADP e Pi Na presenccedila de ADP fosfoenolpiruvato (PEP) e da enzima piruvato
cinase (PK) os produtos resultantes satildeo ATP e piruvato Por acccedilatildeo da enzima lactato
desidrogenase (LDH) este uacuteltimo eacute convertido em lactato ocorrendo simultaneamente a oxidaccedilatildeo
do NADH a NAD+ monitorizada atraveacutes da Abs340 nm Quando o S1 de miosina se encontra no
meio reaccional o ATP eacute hidrolisado formando-se novamente ADP e Pi o que permite a
formaccedilatildeo de um ciclo (fig 21) Se as enzimas se encontrarem em excesso a velocidade de
conversatildeo do NADH eacute limitada pela produccedilatildeo de ADP e Pi Como o ATP eacute regenerado em cada
ciclo a sua concentraccedilatildeo inicial eacute sempre mantida (Bagshaw 1982)
Figura 33ndash Representaccedilatildeo esquemaacutetica do meacutetodo de enzimas acopladas
47
O protocolo utilizado e seguidamente descrito foi adaptado de Cuenda et al (1990 ndash Ver Refordf) A
uma cuvette de 1 mL adicionou-se S1 para uma concentraccedilatildeo final de 005 mgmL 800 L de
meio de reacccedilatildeo22
10 L de PEP ([PEP]stock = 42 mM) 10 L de PK+LDH 11 ([PK+LDH]stock =
1 mgmL) e 15 L de NADH ([NADH]stock=25 mM) Iniciou-se a monitorizaccedilatildeo da reacccedilatildeo
atraveacutes da leitura de Abs340 nm apoacutes a adiccedilatildeo de 20 L de ATP ([ATP]stock=100 mM) A
actividade Mg2+
-ATPaacutesica eacute obtida atraveacutes do declive correspondente agrave oxidaccedilatildeo do NADH Para
a mediccedilatildeo da Mg2+
-ATPase de S1 estimulada por actina adicionou-se F-actina para uma
concentraccedilatildeo final de 2 microM sendo a actividade obtida tambeacutem atraveacutes do declive correspondente
agrave oxidaccedilatildeo do NADH apoacutes a adiccedilatildeo da proteiacutena
35 Interaccedilatildeo entre vanaacutedio e subfragmento-1 de miosina
351 Preparaccedilatildeo das soluccedilotildees de vanaacutedio
As soluccedilotildees de vanadato foram preparadas em aacutegua MilliQ a partir de metavanadato de amoacutenio
(NH4VO3) numa concentraccedilatildeo stock de 50 mM com agitaccedilatildeo magneacutetica e aquecimento (~30 ordmC)
Esta soluccedilatildeo com um valor compreendido entre 6 e 7 designa-se por ldquometavanadatordquo e eacute
constituiacuteda pelas esspeacutecies V1 V2 V4 e V5 (Ref) Acidificando a soluccedilatildeo de metavanadato por
adiccedilatildeo de HCl ateacute um valor de pH proacuteximo de 4 observa-se uma mudanccedila de cor de amarelo
claro para laranja indicativa da formaccedilatildeo da espeacutecie decameacuterica de vanadato De seguida e
imediatamente antes da realizaccedilatildeo dos ensaios descritos o valor do pH da soluccedilatildeo de
decavanadato foi aumentado para cerca de 75 por adiccedilatildeo de NaOH A soluccedilatildeo (2) designada
por ldquometavanadatordquo natildeo foi posteriormente alterada ateacute agrave sua adiccedilatildeo para a realizaccedilatildeo dos ensaios
As soluccedilotildees foram armazenadas a 4ordmC e mantidas em gelo aquando da sua utilizaccedilatildeo
A soluccedilatildeo de V(IV) foi preparada em condiccedilotildees anaeroacutebias por diluiccedilatildeo de sulfato de vanadil
(VOSO4) para uma concentraccedilatildeo final de 5 mM em H2O MilliQ saturada com N2 gasoso
352Anaacutelise por espectroscopia de UV-Vis
A concentraccedilatildeo da espeacutecie decameacuterica em soluccedilatildeo depende do periacuteodo de tempo apoacutes a diluiccedilatildeo
no meio de reacccedilatildeo apropriado (Ref) A velocidade de desoligomerizaccedilatildeo da espeacutecie decameacuterica
pode ser analisada por espectroscopia de absorccedilatildeo na regiatildeo UVVis uma vez que esta espeacutecie
apresenta duas bandas caracteriacutesticas na regiatildeo do visiacutevel (a 358 e 400 nm) que natildeo se verifica
em quaisquer outras espeacutecies oligomeacutericas As bandas supramencionada sapresentam-se bem
22
Meio de reacccedilatildeo 3125 mM Tris-HCl pH 71 375 mM MgCl2
48
resolvidas atraveacutes da obtenccedilatildeo da derivada de 2ordf ordem do espectro apresentando a forma de 2
vales com miacutenimos centrados nos comprimentos de onda supracitados com dependecircncia linear
relativamente agrave concentraccedilatildeo de decavanadato total (Ramos et al 2006)
Assim a estabilidade da espeacutecie decameacuterica apoacutes diluiccedilatildeo da soluccedilatildeo de decavanadato no meio de
reacccedilatildeo dos ensaios (tampatildeo D) para uma concentraccedilatildeo final de 1 mM em vanadato total foi
caracterizada atraveacutes da leitura da Abs400 em funccedilatildeo do tempo tendo-se determinado qual a
ordem de reacccedilatildeo e o tempo de meia-vida da mesma Com base em estudos anteriores acerca da
estabilidade do V10 na presenccedila de diferentes proteiacutenas (Ramos et al 2006) determinou-se
igualmente o tempo de meia vida apoacutes a adiccedilatildeo de 20 microM de S1 de miosina ao meio reaccional
353 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
Os espectros de RMN do isoacutetopo 51
V foram realizados num espectroacutemetro Bruker AM-400 MHz
equipado com uma sonda multinuclear inversa de 5 mm Em tubos de RMN de 5 mm de diacircmetro
preparou-se soluccedilotildees de 500 L de deca- ou metavanadato correspondentes a um intervalo de
concentraccedilotildees de 1 a 10 mM suplementadas com 10 D2O Os espectros foram adquiridos agrave
temperatura ambiente com uma largura espectral de 23 923 Hz um tempo de aquisiccedilatildeo de 0086
s tempo de relaxaccedilatildeo de 001 s e uma sequecircncia de pulsos a 90ordm Utilizou-se a transformada de
Fourier com alargamento das bandas de 20Hz para o processamento dos espectros Os desvios
quiacutemicos das ressonacircncias de 51
V foram obtidos em relaccedilatildeo agrave espeacutecie VOCl3cuja frequecircncia de
ressonacircncia eacute considerada o padratildeo sendo todas as outras referenciadas a partir desta em ppm
As aacutereas dos sinais do RMN correspondentes agraves espeacutecies oligomeacutericas presentes foram obtidas
por integraccedilatildeo dos sinais As concentraccedilotildees relativas das espeacutecies foram calculadas com recurso agrave
equaccedilatildeo
onde A[Vx] corresponde agrave aacuterea referente agrave ressonacircncia da espeacutecie Vx com nordm de agregaccedilatildeo x
A[V]total agrave soma total das aacutereas medidas e [V] a concentraccedilatildeo total de vanaacutedio no ensaio
A teacutecnica de 51
V-RMN foi anteriormente aplicada em estudos de ligaccedilatildeo com S1 de miosina
(Ref) tendo sido demostrado que entre as espeacutecies V1 V2 V4 e V5 a espeacutecie tetrameacuterica possui a
maior afinidade para a miosina (Cremo et al) O desaparecimento ou alargamento dos sinais pode
implicar a ligaccedilatildeo de um dos oxovanadatos laacutebeis agrave miosina sem indicaccedilatildeo de qual uma vez que
49
se processa um equiliacutebrio entre eles (Crans 1990) Assim e tendo em conta as indicaccedilotildees de que
a espeacutecie V10 eacute distinta das restantes no que diz respeito agrave magnitude da interacccedilatildeo com a miosina
e inibiccedilatildeo da actividade ATPaacutesica estimulada por actina (Aureliano 2000 Tiago et al 2002)
torna-se fundamental investigar a interacccedilatildeo das diferentes espeacutecies de vanadato com o S1 de
miosina atraveacutes da teacutecnica de 51
V-RMN
Para tal obtiveram-se espectros de 51
V-RMN resultantes da titulaccedilatildeo de uma soluccedilatildeo 5 mM de
decavanadato total com concentraccedilotildees crescentes de S1 de miosina (0 5 10 15 e 20 microM) em
tampatildeo D Realizou-se ainda ensaios na presenccedila de 2 mM de decavanadato e 2 mM de ATP na
presenccedila e na ausecircncia de S1 de miosina de modo a verificar o efeito que estes ligandos naturais
da enzima teria na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o subfragmento-1
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
Os grupos sulfidrilo (-SH) das cisteiacutenas satildeo resiacuteduos de aminoaacutecidos extremamente sensiacuteveis agrave
oxidaccedilatildeo resultando na sua oxidaccedilatildeo (Dalle-Donne et al 1999) Logo a quantificaccedilatildeo destes
resiacuteduos eacute bastante importante nos estudos de interacccedilatildeo com vanaacutedio jaacute que os resiacuteduos de Cys
com ecircnfase para o SH1 e o SH2 satildeo bastante importantes na monitorizaccedilatildeo das modificaccedilotildees
funcionais da regiatildeo da cabeccedila
O procedimento seguidamente apresentado foi originalmente descrito por Gutieacuterrez-Merino
(1987)) Consiste na reacccedilatildeo entre os grupos ndashSH natildeo oxidados das proteiacutenas e o reagente de
Ellman (DTNB -55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico)) - formando-se o aniatildeo CNT (aniatildeo 3-
carboxilato-4-nitrotiofenolato) numa estequiometria de 11 Este apresenta um maacuteximo de
absorvecircncia a 412 nm (Robyt 1990) Assim apoacutes a incubaccedilatildeo da proteiacutena na presenccedila do metal
a quantidade de cisteiacutenas reduzidas por proteiacutena determina-se atraveacutes dos valores de Abs412
Deste modo colocou-se num tubo de Eppendorf o volume apropriado de S1 dialisado para que a
concentraccedilatildeo no ensaio fosse de 1 mgmL Adicionou-se o volume apropriado de decavanadato
para que a sua concentraccedilatildeo fosse de 0 10 50 100 250 500 ou 1000 M Perfez-se com soluccedilatildeo
D ateacute um volume final de 1 mL e incubou-se a 25ordmC por 20 minutos Apoacutes este periacuteodo colocou-
se 780 L da mistura reaccional numa cuvette de quartzo e adicionou-se 220 L de soluccedilatildeo K23
23
Tampatildeo K Tampatildeo fosfatohidrogenofosfato de soacutedio 025 M pH 72
50
Adicionou-se 15 L de DTNB (em soluccedilatildeo K [DTNB]stock=100 mM) ao meio reaccional e
registou-se o valor de absorvecircncia medido equivalente agraves cisteiacutenas ldquoraacutepidasrdquo (SH1 e SH2
expostos ao solvente) Adicionou-se 001 g de SDS que provoca a desnaturaccedilatildeo da proteiacutena e
consequentemente a exposiccedilatildeo de todos os resiacuteduos de cisteiacutenas que se encontravam ateacute aiacute
inacessiacuteveis no interior da proteiacutena (cisteiacutenas totais) Estes resiacuteduos podem assim reagir com o
DTNB
Estimou-se o nuacutemero de resiacuteduos de cisteiacutenas em todas as amostras com recurso ao coeficiente de
extinccedilatildeo molar () do DTNB (ver ponto 3 dos Anexos) que foi determinado atraveacutes da titulaccedilatildeo
de L-Cisteiacutena com intervalo de concentraccedilotildees compreendidos entre 0 e 50 microM na presenccedila de
25microM de DTNB em tampatildeo D Os valores de absorvecircncia foram obtidos para um comprimento de
onda de 412 nm
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
3621 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
A teacutecnica mais eficiente de anaacutelise da interacccedilatildeo entre o iatildeo vanadilo e proteiacutenas como a
calmodulina e a transferrina entre outras permite a obtenccedilatildeo raacutepida de informaccedilatildeo no que toca agrave
conformaccedilatildeo e amplitude da referida interacccedilatildeo
Os espectros de Ressonacircncia Paramagneacutetica Electroacutenica (RPE) da espeacutecie paramagneacutetica V(IV)
foram realizados a 70K num espectroacutemetro Bruker EMX-300 X-band Todas as soluccedilotildees
utilizadas e amostras foram preparadas em anaerobiose numa cacircmara de luvas de modo a evitar a
reduccedilatildeo de V(IV) a V(V) Em tubos capilares com 02 mL de amostra manteve-se a concentraccedilatildeo
de S1 a 20 M e variou-se a concentraccedilatildeo de vanadilo (0 25 5 10 15 25 50 75 100 150
200 500 e 1000 M [VOSO4]stock=5 mM) em tampatildeo D Apoacutes obtenccedilatildeo dos espectros estes
foram tratados atraveacutes do software SimFonia e WinEPR tendo-se obtido um graacutefico com a
variaccedilatildeo da intensidade do sinal de RPE em funccedilatildeo da concentraccedilatildeo de sulfato de vanadilo
presente Realizaram-se ainda simulaccedilotildees de espectros obtidos com 20 microM de S1 e 1000 microM de
V(IV) com vista agrave obtenccedilatildeo dos ligandos provaacuteveis do vanadilo atraveacutes da anaacutelise da constante
de acoplamento hiperfina (A) e do factor g (g)
4 Conclusotildees gerais
Apesar de jaacute ter sido demonstrado que o vanaacutedio inibe a Na+K
+-ATPase da miosina a funccedilatildeo
deste elemento no tecido muacutesculo esqueleacutetico continua largamente incompreendida A enorme
complexidade quiacutemica deste metal especialmente no que concerne agrave oligomerizaccedilatildeo e reacccedilotildees
51
redox constitui um obstaacuteculo agrave compreensatildeo do comportamento do vanaacutedio nos organismos
vivos Assim a caracterizaccedilatildeo da quiacutemica deste elemento eacute de grande utilidade antes de iniciar o
estudo da sua interacccedilatildeo com o presente sistema
A anaacutelise por espectroscopias de 51
V-RMN e de UV-Vis de soluccedilotildees de vanadato diluiacutedas nos
meios de reacccedilatildeo utilizados no presente relatoacuterio (10 mM HEPES pH 70 25 mM KCl) mostrou
claramente que a espeacutecie decameacuterica (V10) possui um tempo de vida suficientemente longo para a
determinaccedilatildeo do seu efeito isolado ao contraacuterio do que se verifica com as espeacutecies mono- di-
tetra- e pentameacutericas (V1 V2 V4 e V5) Por outro lado a anaacutelise verificou ainda que apoacutes diluiccedilatildeo
das soluccedilotildees de decavanadato no meio de reacccedilatildeo coexistem as espeacutecies V10 e V1 pelo que os
efeitos putativos desta uacuteltima deveratildeo ser subtraiacutedos aquando da monitorizaccedilatildeo dos efeitos da
espeacutecie decameacuterica
O vanaacutedio (V) (vanadato) inibe as enzimas que catalizam reacccedilotildees de transferecircncia de grupos
fosfato como a ATPase da miosina [Goodno 1979] devido agrave sua analogia estrutural com o iatildeo
fosfato Na sua utilizaccedilatildeo como inibidor enzimaacutetico natildeo eacute tida em conta a contribuiccedilatildeo individual
das espeacutecies oligomeacutericas considerando-se normalmente a espeacutecie V1 como inibidora efectiva
[Goodno e Taylor 1982] No presente relatoacuterio verificou-se por espectroscopia de 51
V-RMN que
outros oligoacutemeros de vanadato interagem com o S1 de miosina tendo as espeacutecies V10 e V4 os
maiores niacuteveis de afinidade A sua interacccedilatildeo envolve forccedilas electrostaacuteticas e eacute afectada
negativamente por ligandos naturais de enzima como eacute o caso do ATP
A exposiccedilatildeo do S1 de miosina (005 mgmL) a decavanadato conduz agrave inibiccedilatildeo do capacidade da
proteiacutena de estimular a actividade Mg2+
-ATPaacutesica com um valor de IC50 de 266 plusmn 07 M
decavanadato sendo o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM
decavanadato (80) Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM decavanadato sendo
o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM decavanadato (60)
exercendo um efeito de impedimento agrave ligaccedilatildeo entre o vanaacutedio e o S1
A uma concentraccedilatildeo de 27 microM decavanadato verifica-se a oxidaccedilatildeo de um dos resiacuteduos de Cys
de S1 de miosina seguida pela oxidaccedilatildeo gradual de outro resiacuteduo de Cys ateacute agrave concentraccedilatildeo de
1000 microM Ambos os resiacuteduos oxidados natildeo se encontravam expostos ao solvente utilizado A
oxidaccedilatildeo dos resiacuteduos permite sugerir que esta modificaccedilatildeo oxidativa sofrida pelo S1 contribui
para a diminuiccedilatildeo da sua actividade de ATPase
52
Por espectroscopia de RPE revelaram-se os possiacuteveis ligandos equatoriais do vanaacutedio aquando da
sua interacccedilatildeo com o S1 de miosina como sendo duas moleacuteculas de aacutegua um ligando contendo o
grupo R-OH e outro contendo o grupo R=NR
Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados de
oxidaccedilatildeo (IV) e (V) no S1 de miosina para concentraccedilotildees facilmente alcanccedilaacuteveis no ambiente
terrestre
53
5 Estudos futuros
No sentido do melhor esclarecimento das alteraccedilotildees moleculares e estruturais que ocorrem no
subfragmento 1 quando exposta ao vanaacutedio que conduzem a uma diminuiccedilatildeo da sua funccedilatildeo
sugere-se
- Quantificaccedilatildeo das cisteiacutenas reduzidas de S1 na presenccedila de metavanadato
- Determinaccedilatildeo das actividades Ca2+
- e K+-ATPaacutesica do S1 de miosina na presenccedila de
decavanadato
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina estimulada por actina
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina na presenccedila de
metavanadato
- Realizaccedilatildeo de RMN 2D com variaccedilatildeo das concentraccedilotildees das soluccedilotildees de decavanadato e
metavanadato na presenccedila de S1
-Mutageacutenese dos resiacuteduos de Cys doS1 de miosina de modo a verificar quais os resiacuteduos que
sofrem oxidaccedilatildeo na presenccedila de decavanadato
- Determinaccedilatildeo por calorimetria (DSC Differential Scanning Calorimetry) da variaccedilatildeo da
entalpia (∆H) e da temperatura de desnaturaccedilatildeo do S1 de miosina na presenccedila de vanaacutedio uma
vez que estas propriedades caracteriacutesticas de cada proteiacutena satildeo alteradas apoacutes ocorrecircncia de
modificaccedilotildees estruturais
- Anaacutelise por SDS-PAGE de S1 de miosina apoacutes tratamento com vanaacutedio de modo a determinar
quais os polipeacuteptidos eventualmente formados apoacutes clivagem da proteiacutena
54
6 Referecircncias bibliograacuteficas
- Sellers J Myosins 2nd Edition Oxford University Press 1999
- L Stryer inBiochemistry (Ed WH Freeman 2005 pp
Refordfas Materiais e Meacutetodos
[A] S S Margossian SLowey Methods Enzymol1982 85 55-71
[B] JD Pardue JA Spudich Methods Enzymol 1982 85 164-181
[C] M Bradford Anal Biochem1976 72 248-254
[D] H H Taussky E Shorr J Biol Chem 1953 202 675-685
[E] C R Bagshaw in Outline Studies in Biology ndash Muscle Contraction Chapman and Hall Ed
1982
[F] A Cuenda et al Eur J Biochem1990 194 663-670
55
Anexos
1 Determinaccedilatildeo da recta de calibraccedilatildeo meacutetodo de Bradford
Figura1ndashRepresentaccedilatildeo graacutefica da recta de calibraccedilatildeo (Abs595 nm vs [BSA]) utilizada para o meacutetodo de Bradford
2 Determinaccedilatildeo da recta calibraccedilatildeo para a electroforese em condiccedilotildees desnaturantes
Figura 2 ndash Representaccedilatildeo graacutefica da recta de calibraccedilatildeo (log(Peso Molecular) vs Rf dos marcadores de peso
molecular) onde Rf indica determinada atraveacutes do padratildeo de peso molecular
(poccedilo (a))
3Caacutelculo do coeficiente de extinccedilatildeo molar () para a quantificaccedilatildeo de cisteiacutenas reduzidas
atraveacutes do meacutetodo de Ellman
Traccedilou-se a recta de Abs412 nm meacutedia vs [L-Cisteiacutena] de modo a determinar o valor do coeficiente
de extinccedilatildeo molar definido como o declive daquela recta Assim =8400 M-1
cm-1
como
podemos verificar na Fig 3
A595 nm = 00546[BSA] + 00209 Rsup2 = 099051
Ab
s 59
5 n
m
[BSA] (mgmL)
log PM = -1177 Rf + 2247 Rsup2 = 098957
log
PM
Mobilidade relativa (Rf)
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero

23
maior intensidade que poderaacute ser a proteiacutena-C (PM=135 kDa) uma vez que a sua remoccedilatildeo
completa da amostra necessitaria de um passo adicional de cromatografia de troca ioacutenica [BA]
No entanto natildeo podemos confirmar que se trata desta proteiacutena ou de qualquer outra uma vez que
se encontra fora do intervalo de migraccedilatildeo dos marcadores de peso molecular
212 Determinaccedilatildeo da concentraccedilatildeo proteica
2121Subfragmento-1 de miosina
A partir da recta de calibraccedilatildeo obtida pelo meacutetodo de Bradford [BB](ver secccedilatildeo 3311 do
Procedimento Experimental) A595nm = 00546[BSA] + 00209 (ver secccedilatildeo de Anexos)foi possiacutevel
determinar um intervalo de concentraccedilatildeo de S1 de miosina compreendido entre 226 e 117(plusmn
423M) (203 e 1056 (plusmn 381 mgmL)) (n=8) A concentraccedilatildeo meacutedia de S1 de miosina apoacutes
diaacutelise contra a soluccedilatildeo tampatildeo 10 mM HEPES pH 75 25 mM KCl determinada pela leitura da
Abs280 nmfoi de 20430M (1803 mgmL)(n=7)
As amostras a quantificar atraveacutes do meacutetodo de Bradford natildeo sofreram diaacutelise o que permite
explicar a concentraccedilatildeo superior de S1 de miosina em comparaccedilatildeo com o valor obtido atraveacutes do
meacutetodo directo Esta razatildeo aliada agrave sensibilidade e rapidez superiores deste uacuteltimo fez com que
se optasse pelo meacutetodo directo para a quantificaccedilatildeo da grande maioria dos ensaios
2122Actina
A concentraccedilatildeo de actina foi determinada quando esta se encontrava na forma monomeacuterica (G-
actina) por mediccedilatildeo da leitura directa da Abs290 nm tendo-se obtido uma concentraccedilatildeo meacutedia de
691481M (290202 mgmL) (n=3) em 2 mM Tris-HCl pH 80 02 mM CaCl2 02 mM Na-
ATP Ao comprimento de onda utilizado (290 nm) absorvem fortemente apenas os resiacuteduos de
Trp pelo que a mediccedilatildeo efectuada tem menor sensibilidade do que se tivesse sido efectuada a 280
nm uma vez que a actina apresenta apenas 4 resiacuteduos deste aminoaacutecido embora tenha 16
resiacuteduos de Tyr
22 Mediccedilatildeo da actividade enzimaacutetica de S1 de miosina
221 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
As actividades ATPaacutesicas de S1 de miosina dependentes de Ca2+
e K+foram analisadas pelo
caacutelculo das velocidades de consumo inicial do fosfato inorgacircnico (Pi) Definiu-se os tempos de 5
e 3 minutos para a mediccedilatildeo da Ca2+
e da K+EDTA-ATPase respectivamente Os valores obtidos
24
a partir do protocolo presente no ponto 341 da secccedilatildeo Procedimento experimental encontram-se
descritos na Tabela 2I
Tabela 2I ndash Acrividades natildeo-fisioloacutegicas de subfragmento-1 determinadas pelo meacutetodo de Fiske-Subarow
modificado (n=2)
Proteiacutena
Actividade
ATPaacutesica natildeo
fisioloacutegica
Actividade (nmol
Piminmg proteiacutena) Actividade (s-1)
S1
Ca2+
-ATPase 1400 plusmn 30 2100 plusmn 0045
K+EDTA-ATPase 2334 plusmn 50 3500 plusmn 0075
As actividades natildeo-fisioloacutegicas reflectem o estado de oxidaccedilatildeo dos resiacuteduos expostos de Cys 707
(SH1) e 697 (SH2) Se o grupo SH1 sofrer oxidaccedilatildeo a actividade ATPaacutesica dependente de Ca2+
aumenta e a actividade dependente de K+ diminui Por outro lado se o grupo SH2 sofrer
oxidaccedilatildeo regista-se a perda completa destas actividades
De acordo com Margossian e Lowey (1982) [BA] as actividades ATPaacutesica de S1 de miosina
dependente de Ca2+
e K+EDTA da miosina satildeo respectivamente de 24 e 112 s-1
Estaacute tambeacutem
descrito que a actividade ATPaacutesica dependente de K+ eacute em meacutedia cerca de 3 vezes superior agrave
dependente de Ca2+
reflectindo a quantidade das preparaccedilotildees A razatildeo obtida entre os valores das
actividades ATPaacutesicas dependentes de K+ e Ca
2+ eacute de 166 Os valores obtidos para as actividades
(Ca2+
-ATPase=21 s-1
K+EDTA-ATPase= 35 s
-1) satildeo inferiores agrave valores obtidos por
Margossian e Lowey [BA] e Tiago [BC] com forccedila ioacutenica inferior O valor obtido para a
actividade ATPaacutesica dependente de Ca2+
eacute superior ao obtido por Polosukhina et al (2000) [BD]
cujo meio de reacccedilatildeo continha MOPS e apresentava forccedila ioacutenica inferior Os dados
supramencionados comprovam a variaccedilatildeo da actividade a ATPaacutesica natildeo fisioloacutegica em relaccedilatildeo agrave
forccedila ioacutenica temperatura e pH
222 Mediccedilatildeo da actividadeMg2+
-ATPaacutesica de S1 pelo meacutetodo de enzimas acopladas
A actividade de Mg2+
-ATPase eacute a uacutenica que possui significado fisioloacutegico sendo as restantes
bastante uacuteteis apenas para indicaccedilatildeo da qualidade de preparaccedilatildeo do S1[BA] A 0575 M KCl a
actina e o S1 formam uma soluccedilatildeo viscosa e tuacuterbida Apoacutes a adiccedilatildeo de MgATP o complexo acto-
S1 dissocia-se e a turbidez da soluccedilatildeo diminui Se a forccedila ioacutenica for reduzida a dissociaccedilatildeo do
complexo actoS1 que se encontra na forma de suspensatildeo eacute induzida O MgATP eacute rapidamente
hidrolisado diminuindo a sua concentraccedilatildeo no meio reaccional Nestas condiccedilotildees haacute
reassociaccedilatildeo e subsequente contracccedilatildeo dos filamentos de actina e de S1 com formaccedilatildeo de
25
precipitado O meacutetodo descrito traduz de forma bastante elementar o fenoacutemeno da contracccedilatildeo in
vivo e elimina o problema anterior da solubilizaccedilatildeo ineficiente da miosina de modo a obter uma
mediccedilatildeo exacta da actividade [A] A utilizaccedilatildeo de forccedilas ioacutenicas reduzidas aumenta a afinidade da
actina para o S1 permitindo contornar a extrema dificuldade em obter a concentraccedilatildeo proteica dos
muacutesculos (cerca de 100 mgmL do complexo actomiosina) As actividades Mg2+
-ATPaacutesicas de S1
de miosina foram determinadas a partir do declive da variaccedilatildeo da Abs340 relativo agrave oxidaccedilatildeo de
NADH em funccedilatildeo do tempo como eacute descrito no ponto 342 do Procedimento Experimental Os
valores obtidos para a actividade Mg2+
-ATPaacutesica encontram-se apresentados na Tabela 2II
Tabela 2II ndash Actividade Mg2+
-ATPaacutesica de subfragmento-1 de miosina determinadas pelo meacutetodo das enzimas
acopladas
Actividade (nmol
ATPmin mg(S1)) Actividade (s
-1)
1097 plusmn 269 164 plusmn 04
Ramos (2005) [J] obteve o valor de 01 s-1
para a actividade Mg2+
-ATPaacutesica de S1 de miosina
com forccedila ioacutenica intermeacutedia Tiago (2005) [BC] obteve o valor de 011 s-1 em condiccedilotildees de forccedila
ioacutenica reduzida em relaccedilatildeo agrave utilizada no presente relatoacuterio
23 Preparaccedilatildeo e caracterizaccedilatildeo das soluccedilotildees de vanaacutedio
231 Anaacutelise da composiccedilatildeo das soluccedilotildees de vanadato por espectroscopia de 51
V-RMN
efeito da concentraccedilatildeo da espeacutecie
A diluiccedilatildeo das soluccedilotildees stock de meta- e decavanadato iraacute determinar fortemente a composiccedilatildeo
em soluccedilatildeo dos diferentes oligoacutemeros de vanadato (ref) No caso do decavanadato o periacuteodo de
tempo apoacutes a diluiccedilatildeo iraacute tambeacutem ser um factor importante a considerar devido agrave decomposiccedilatildeo
da espeacutecie V10(ref)Torna-se assim fundamental determinar a composiccedilatildeo quiacutemica das soluccedilotildees
no meio de ensaio utilizado (HEPES 10 mM KCl 25 mM) de modo a correlacionaacute-la
correctamente com os possiacuteveis efeitos promovidos pelas diferentes espeacutecies de vanadato no
sistema em estudo
Aleacutem disso deve ser dado ecircnfase agraves importantes interacccedilotildees que ocorrem entre o vanaacutedio e os
tampotildees escolhidos O tampatildeo com menor nuacutemero e magnitude de interacccedilatildeo eacute o HEPES [AD]
seguido do Tris sendo este uacuteltimo especialmente aconselhado em concentraccedilotildees baixas para casos
de incompatibilidade enzimaacutetica ou quiacutemica redox No caso especial da reacccedilatildeo redox
NAD(P)HNAD(P) o tampatildeo adequado eacute o imidazole de modo a natildeo haver reacccedilatildeo com o
26
vanadato Os tampotildees a evitar em qualquer situaccedilatildeo incluem a trietanolamina fosfato citrato e
glicina entre outros [AD] uma vez que todos interactuam fortemente com V(V) com formaccedilatildeo
de complexos estaacuteveis
Assim procedeu-se agrave anaacutelise da composiccedilatildeo das soluccedilotildees de meta- e decavanadato apoacutes diluiccedilatildeo
das soluccedilotildees stock (50 mM em vanadato total) no meio de reacccedilatildeo (tampatildeo D) atraveacutes da
espectroscopia de ressonacircncia magneacutetica nuclear de 51
V Nas figuras x1 e x2 apresentam-se os
espectros obtidos para as soluccedilotildees de deca- e metavanadato num intervalo de concentraccedilotildees de 1 a
10 mM A Tabela seguinte resume os paracircmetros espectroscoacutepicos obtidos para cada espeacutecie
presente nas soluccedilotildees analisadas
Figura 22ndash Espectros de RMN de 51
V de
concentraccedilotildees crescentes das espeacutecies oligomeacutericas
de vanadato obtidos apoacutes diluiccedilatildeo da soluccedilatildeo stock
de decavanadato em 10 mM HEPES pH 70 25 mM
KCl para concentraccedilotildees finais de A) 1 mM B)2
mM C) 4 mM D) 8 mM E) 10 mM de
decavanadato total V1refere-se agraves espeacutecies
monomeacutericade vanadato V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2IIIndash Desvios quiacutemicos meacutedios (part) das ressonacircncias das espeacutecies decameacuterica (V10) monomeacuterica (V1) e
tetrameacuterica (V4) presentes nos espectros da soluccedilatildeo de decavanadatoLargura a meia altura (∆12) da espeacutecie V10A
Paracircmetros V10A V10B V10C V1 V4
part (ppm) -5162 -5003 -4255 -5616 -5748
∆12 (Hz) 175 nd nd nd nd
Figura 23 ndash Distribuiccedilatildeo graacutefica das
espeacutecies oligomeacutericas de vanadato vs a
concentraccedilatildeo de decavanadato total As
concentraccedilotildees de cada espeacutecie foram
obtidas segundo a formula descrita na
secccedilatildeo 35 do Procedimento
Experimental (aacutereas dos sinais de 51
V-
RMN observadas na figura x1)V10 V1 e
V4 correspondem agraves espeacutecie monomeacuterica
dimeacuterica e tetrameacuterica respectivamente [Esp
eacuteci
e o
ligo
meacute
rica
de
van
aacutedio
] (m
M)
[Decavanadato]total (mM)
V10
V1
V4
27
Nas soluccedilotildees de decavanadato as espeacutecies monomeacuterica e decameacuterica de vanadato satildeo as uacutenicas
presentes para concentraccedilotildees de decavanadato inferiores a 8 mM Ambas as espeacutecies aumentam
linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado em [AY] e
em (Teresa) quando a concentraccedilatildeo de V1 eacute igual ou superior a 200 microM inicia-se a sua
oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica No entanto nas condiccedilotildees de realizaccedilatildeo dos
ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a espeacutecie tetrameacuterica neste
estudo em concentraccedilotildees inferiores a 8 mM tendo o mesmo sido observado por Ramoset al
(2006) [AK] Os resultados obtidos mostram claramente o equiliacutebrio entre a espeacutecie decameacuterica e
a monomeacuterica e que o aparecimento de outras espeacutecies oligomeacutericas se encontra dependente da
concentraccedilatildeo desta uacuteltima Apesar de natildeo ter sido realizado o ensaio com 5 mM de decavanadato
total no presente relatoacuterio estaacute reportado em Ramos et al (2006) [AK] que as concentraccedilotildees da
espeacutecie decameacuterica e monomeacuterica satildeo respectivamente de 500 microM e 200 microM Assim pode inferir-
se que para concentraccedilotildees inferiores a 5 mM e intervalos de tempo de cerca de 20 minutos (que
inclui o tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas
as espeacutecies decameacuterica e monomeacuterica de vanadato
Figura 24 ndash Espectros de RMN de 51
V de concentraccedilotildees
crescentes das espeacutecies oligomeacutericas de vanadato obtidos
apoacutes diluiccedilatildeo da soluccedilatildeo stock de metavanadato em 10 mM
HEPES pH 70 25 mM KCl para concentraccedilotildees finais de A)
1 mM B) 2 mM C) 4 mM D) 8 mM E) 10 mM de
metavanadato total V1 refere-se agraves espeacutecies monomeacuterica de
vanadatoV1 e V2 correspondem respectivamente agrave espeacutecie
monomeacuterica e dimeacuterica
Figura 25 ndash Distribuiccedilatildeo graacutefica das espeacutecies
oligomeacutericas de vanadato vs a concentraccedilatildeo de
metavanadato total As concentraccedilotildees de cada
espeacutecie foram obtidas segundo a formula descrita
na secccedilatildeo 35 do Procedimento Experimental
(aacutereas dos sinais de 51
V-RMN observadas na
figura x1)V10 V1 e V4 correspondem agraves espeacutecie
monomeacuterica dimeacuterica e tetrameacuterica
respectivamente
Esp
eacuteci
e o
ligo
meacute
rica
de
va
nad
ato
(m
M)
[Metavanadato]total (mM)
V1
V2
V4
V5
28
Tabela 2IVndash Desvios quiacutemicos meacutedios (part)das ressonacircncias das espeacutecies tetrameacuterica (V4) monomeacuterica (V1) e
dimeacuterica (V2) presentes no espectro da soluccedilatildeo de metavanadato (total) 10 mM
Paracircmetros V1 V2 V4 V5
part (ppm) -5615 -5748 -5789 -5869
Nas soluccedilotildees de metavanadato as espeacutecies monomeacuterica dimeacutericae tetrameacuterica de vanadato estatildeo
presentes para concentraccedilotildees de metavanadato inferiores a 4 mM Todas estas as espeacutecies
aumentam linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado
em (Aureliano e Madeira 1998) e em (Tiago et al 2004) quando a concentraccedilatildeo de V1 eacute igual ou
superior a 200 microM inicia-se a sua oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica Nas
condiccedilotildees de realizaccedilatildeo dos ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a
espeacutecie pentameacuterica neste estudo em concentraccedilotildees inferiores a 4 mM tendo o mesmo sido
observado por Ramos 2006 Os resultados obtidos mostram claramente o equiliacutebrio entre a
espeacutecie V4 e as espeacuteciesV1 e V2 no qual o aparecimento de outras espeacutecies oligomeacutericas se
encontra dependente da concentraccedilatildeo da espeacutecie V1 Por uacuteltimo pode inferir-se que para
concentraccedilotildees inferiores a 2 mM e intervalos de tempo de cerca de 20 minutos (que inclui o
tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas as
espeacutecies V1 V2 e V4 com concentraccedilotildees respectivas de 0796 0192 e 0196 mM
232Estabilidade das soluccedilotildees de decavanadato anaacutelise por espectroscopia de absorccedilatildeo
UVVis
Caracterizou-se a estabilidade da espeacutecie V10 numa soluccedilatildeo com 1 mM decavanadato total em
tampatildeo D atraveacutes da leitura da Abs400 Obteve-se uma cineacutetica de dissociaccedilatildeo de 1ordf ordem tal
como se pode verificar pela linearidade do graacutefico de ln(Abs400)vs o tempo de aquisiccedilatildeo espectral
(segundos) (Fig26 )
Figura 26ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do tempo
(em segundos) para uma soluccedilatildeo de decavanadato total de 1
mM em tampatildeo D a 25ordmC (n=2)
Sendo uma reacccedilatildeo de primeira ordem pode ser descrita pela equaccedilatildeo cujo
declive eacute ndashK (constante de dissociaccedilatildeo) e a intersecccedilatildeo no eixo das ordenadas corresponde
Ln
(Ab
s 400 n
m)
tempo (s)
29
aln(A0) Numa reacccedilatildeo de 1ordf ordem o tempo de meia-vida eacute independente da concentraccedilatildeo inicial
do reagente sendo descrito pela equaccedilatildeo [2] Deste modo a partir da recta
Ln(Abs400 nm) = -436E-5
K - 105 obteve-se um tempo de meia-vida de 4490 plusmn 0823 horas para
as condiccedilotildees supramencionadas
Apoacutes a adiccedilatildeo de S1 8 microM ao meio reaccional verificou-se que a reacccedilatildeo de desoligomerizaccedilatildeo
da espeacutecie decameacuterica de vanadato seguia igualmente uma cineacutetica de 1ordf ordem tal como se pode
verificar pela linearidade do graacutefico de ln(Abs400) versus o tempo de aquisiccedilatildeo espectral
(segundos) (Fig27)
Figura 27ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do
tempo (em segundos) para uma soluccedilatildeo de decavanadato
total de 1 mM em tampatildeo D a 25ordmC na presenccedila de 8
microM (n= 2)
A partir da equaccedilatildeo da recta Ln(Abs400 nm) = -7520E-5
K - 0613determinou-se o valor de K para
a espeacutecie decameacuterica de onde se obteve um tempo de meia-vida de 2430 plusmn 0191 horas para as
condiccedilotildees supramencionadas
Tanto este valor como o valor registado na ausecircncia de S1 satildeo substancialmente superiores agrave
duraccedilatildeo dos ensaios de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina descritos no presente
relatoacuterio
24 Interacccedilatildeo de S1 com vanaacutedio(V)
241Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
De modo a analisar qual o efeito da adiccedilatildeo de S1 de miosina (tampatildeo D) produzia nos sinais de
51V-NMR titulou-se uma soluccedilatildeo de 5mM de decavanadato total com concentraccedilotildees crescentes
de S1 (5 10 15 e 20 microM) Os espectros de 51
V-RMN encontram-se apresentados na Fig 2
No graacutefico da figura 2 Apresenta-se a concentraccedilatildeo de S1 em funccedilatildeo do factor de alargamento
corrigido dos espectros acima referidos
Ln
(Ab
s (4
00
nm
))
tempo (s)
30
Figura 28 ndash Graacutefico de
representaccedilatildeo da
concentraccedilatildeo de S1 em
funccedilatildeo do factor de
alargamento A
concentraccedilatildeo de
decavanadato total foi de 5
mM e as concentraccedilotildees
utilizadas de S1 foram de 0
5 10 e 15 microM
Verifica-se que o factor de alargamento aumenta linearmente em funccedilatildeo da concentraccedilatildeo de S1 no
ensaio o que indica a existecircncia de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina Observou-
se ainda que a adiccedilatildeo de S1 ao meio reaccional natildeo provocou qualquer alteraccedilatildeo dos desvios
quiacutemicos dos sinais de 51
V-RMN
Realizaram-se ensaios na presenccedila de ATP e de S1 com intuito de verificar o efeito do ATP na
interacccedilatildeo do S1 de miosina com o decavanadato
Figura 29 - Espectros de RMN de 51
V contendo 2
mM Decavanadato (A) (controlo) (B) na presenccedila
de 2 mM de ATP (C) na presenccedila de 2 mM de ATP
e 8 microM de S1 de miosina V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2Vndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies decameacuterica (V10) e monomeacuterica (V1) presentes nos
espectros da soluccedilatildeo de decavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V10A V10B V10C V1
part (ppm) -5161 -5002 -4254 -5614
Observa-se que a presenccedila de ATP (espectro B) natildeo provoca qualquer alteraccedilatildeo visiacutevel em
relaccedilatildeo ao espectro que contem apenas soluccedilatildeo de decavanadato Jaacute a presenccedila simultacircnea de ATP
e S1 provoca um alargamento da ∆12 do sinal do V10A de 1736 Hz (espectro (A)) para 2366 Hz
(espectro (C) Fig 29) Tendo em conta que a concentraccedilatildeo de S1 apresenta uma relaccedilatildeo linear
com o factor de alargamento converteu-se o valor da ∆12 e comparou-se com o valor esperado agrave
F = 00591[S1] - 00157 Rsup2 = 0998
Fact
or
de
ala
rgam
en
to
[S1] (microM)
31
concentraccedilatildeo de S1 utilizada O factor de alargamento obtido para 8 microM ndash 0376 ndash eacute inferior ao
valor esperado agrave mesma concentraccedilatildeo de S1 obtido atraveacutes da equaccedilatildeo da recta apresentada na
Fig 28 ndash 0457 ndash o que indica que o ATP poderaacute exercer um efeito de protecccedilatildeo do S1
diminuindo a sua interacccedilatildeo com a espeacutecieV10
Por uacuteltimo realizaram-se ensaios com metavanadato na presenccedila de 8 microM S1 de modo a verificar
se as espeacutecies oligomeacutericas contidas nesta soluccedilatildeo interagiam com a proteiacutena em estudo
Figura 210 - Espectros de RMN de 51
V contendo 2
mM metavanadato (A) (controlo) (B) na presenccedila
de 8 microM de S1 de miosina V1 V2 e V4
correspondem respectivamente aacutes espeacutecies
monomeacuterica dimeacuterica e tetrameacuterica
Tabela 2VIndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies monomeacuterica (V1) dimeacuterica (V2) e tetrameacuterica (V4)
presentes nos espectros da soluccedilatildeo de metavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V1 V2 V4
part (ppm) -5613 -5747 -5789
Como eacute possiacutevel observar na Fig 210 ocorre uma ligeiro alargamento dos sinal de V4 bem
como uma perda da intensidade pelo que se conclui quedas espeacutecies presentes na soluccedilatildeo de
metavanadato (V1 V2 e V4) apenas a espeacutecie tetrameacuterica interage com o S1 de miosina o que
estaacute de acordo com o reportado por Tiago et al 2002
242 Actividade Mg2+
-ATPaacutesica de S1 de miosina na presenccedila de decanadato
Com o intuito de avaliar o efeito do decavanadato na actividade Mg2+
-ATPaacutesica do S1 de
miosina monitorizou-se a reacccedilatildeo de oxidaccedilatildeo do NADH atraveacutes da leitura de Abs340 na
presenccedila de 005 mgmL de S1 de miosina e no caso da actividade estimulada 2microM de actina em
3125 mM Tris-HCl pH 71 e 375 mM MgCl2 pelo meacutetodo de enzimas acopladas (ver seccccedilatildeo xx
dos MM)Para tal realizaram-se ensaios com vaacuterias concentraccedilotildees de decavanadato total (10 50
100 250 500 e 1000) de modo a determinar qual o seu efeito na actividade MgATPaacutesica de S1
de miosina na ausecircncia e na presenccedila de um dos seus ligandos naturais Os ensaios foram
realizados agrave temperatura ambiente com incubaccedilatildeo preacutevia a 25ordm C durante 20 minutos
32
Figura 211 ndash Representaccedilatildeo
graacutefica da Actividade Mg2+
-
ATPaacutesica de S1 () (os ajustes
teoacutericos estatildeo representado a por
uma linha os resultados
experimentais estatildeo representados
por triacircngulos (Actividade
estimulada) e cruzes (Actividade
natildeo estimulada)) vs
[Decavanadato]total medidas pelo
meacutetodo das enzimas acopladas
(n=3) Os dados foram ajustados ao modelo de decaimento exponencial usando o programa GraphPad Prism para a
determinaccedilatildeo dos valores de IC50 (na ausecircncia de actina IC50= 266 plusmn 07 M decavanadato na presenccedila de actina
IC50= 28 plusmn 1 M decavanadato
A partir do graacutefico presente na Fig 29 verifica-se que o ajuste dos dados experimentais foi bem
sucedido A inibiccedilatildeo da actividade Mg2+
-ATPaacutesica do S1 de miosina eacute afectada em menor
amplitude na presenccedila de actina (60 de inibiccedilatildeo da actividade) do que na sua ausecircncia
(aproximadamente 80 de inibiccedilatildeo de actividade) Devido a esta diferenccedila pode sugerir-se que a
actina iraacute impedir parcialmente a ligaccedilatildeo do decavanadatoao S1 demiosina atraveacutes de
impedimentos esteacutereos ou interacccedilotildees electrostaacuteticas por exemplo A teacutecnica utilizada natildeo nos
permite concluir acerca de que regiatildeo(otildees) ou resiacuteduo(s) da actina participaratildeo na preservaccedilatildeo
parcial da actividade Mg2+
-ATPaacutesica
Por outro lado os valores de IC50 dos dois tipos de actividades natildeo apresentam diferenccedilas
significativas tendo-se obtido 266 plusmn 07 e 28 plusmn 1M decavanadato na ausecircncia e na presenccedila de
actina respectivamente o que corresponde a uma concentraccedilatildeo de cerca de 27 M de V10 O
meio de reacccedilatildeo utilizado nestes ensaios foi 10 mM HEPES e 25 mM KCl com 2 mM ATP Em
(Tiago et al 2004) o valor de IC50 para a actividade Mg2+-ATPaacutesica na ausecircncia de actina foi de
125 plusmn 025 microM enquanto o valor da actividade estimulada por actina foi de 136 plusmn 14 microM O
meio de reacccedilatildeo utilizado nos ensaios foi 20 mM HEPES 50 mM KCl e 2 mM ATP o que resulta
precisamente no dobro da forccedila ioacutenica a que foi realizado o ensaio do presente relatoacuterio
(exceptuando a contribuiccedilatildeo dos produtos da hidroacutelise de ATP uma vez que este meacutetodo
pressupotildee a ocorrecircncia de um ciclo de reacccedilotildees de hidroacutelise e siacutentese deste composto) Ora tendo
sido anteriormente referido que a afinidade do S1 de miosina para com a espeacutecie V10 diminui com
Act
ivid
ade
Mg
2+-A
TP
aacutesic
a d
e S
1 d
e
Mio
sin
a
[Decavanadato] (microM) Actividade Estimulada ()
Actividade Natildeo Estimulada ()
Ajuste Actividade Estimulada
Ajuste Actividade Natildeo Estimulada
33
o aumento da forccedila ioacutenica (Tiago et al 2004 tal poderaacute explicar a diferenccedila obtida entre estes
dois conjuntos de valores
243Quantificaccedilatildeo de cisteiacutenas reduzidas pelo meacutetodo de Ellman
A interacccedilatildeo do vanaacutedio com alguns tipos de aminoaacutecidos sensiacuteveis agrave oxidaccedilatildeo pode fornecer-nos
informaccedilatildeo bastante uacutetil em relaccedilatildeo agraves alteraccedilotildees estruturais que conduzem a uma diminuiccedilatildeo da
actividade fisioloacutegica do S1 de miosina Os estudos realizados por Chasteen 1983 concluiacuteram
que a reduccedilatildeo de vanadato (V) a vanadilo (IV) ocorre na presenccedila de agentes redutores comuns no
ambiente celular de que se destaca o aminoaacutecido cisteiacutena Esta eacute tambeacutem um alvo de
complexaccedilatildeo por parte do iatildeo VO2+
Jaacute Ramos et al (2009) demonstrou que a incubaccedilatildeo de
actina com a soluccedilatildeo de decavanadato induz a oxidaccedilatildeo de resiacuteduos de cisteiacutena e agrave formaccedilatildeo do
oxidovanaacutedio (IV) Este grupo de investigadores descobriu ainda que apenas a espeacutecie V10 induz a
oxidaccedilatildeo das cisteiacutenas ldquoraacutepidasrdquo da actina atraveacutes do processo proposto que eacute parcialmente
responsaacutevel pela inibiccedilatildeo jaacute descrita da actividade de ATPase do S1 de miosina estimulada por
actina quando exposta agrave soluccedilatildeo de decavanadato Esta inibiccedilatildeo apresenta-se afectada com a
presenccedila de ATP uma vez que os resiacuteduos de cisteiacutena permaneceram intactos nos ensaios
realizados com este ligando Jaacute Crans et al (2004) concluiu que o siacutetio activo de muitas fosfatases
(que conteacutem grupos tiol) pode formar um equiliacutebrio redox com o iatildeo vanadato (V) de forma natildeo
reversiacutevel
Os resultados obtidos por Sakurai e Shiromura (1980) sugeriram que o vanaacutedio (V) sofre reduccedilatildeo
a vanaacutedio (IV) sem dependecircncia do valor de pH do ensaio Registaram ainda uma estequiometria
de 31 na reacccedilatildeo do vanadato (V) com os resiacuteduos de Cys enquanto a estequiometria dos ensaios
com o vanadilo (IV) foi de 21
Berlett e Stadtman (1997) observaram que os tipos de aminoaacutecidos mais facilmente oxidaacuteveis
englobam entre outros os resiacuteduos de Cys Trp e MetAssim analisou-se o estado redox dos
grupos tiol dos resiacuteduos de Cys do S1 na presenccedila do decavanadato atraveacutes da monitorizaccedilatildeo da
Abs412 nm na presenccedila de 1 mgmL de S1 de miosina em tampatildeo 10 mM HEPES 25 mM KCl
variando a concentraccedilatildeo da soluccedilatildeo de decavanadato de modo a determinar que tipo de alteraccedilotildees
estruturais ocorrem na sua presenccedila no que diz respeito agrave diminuiccedilatildeo da actividade ATPaacutesica de
S1 na presenccedila de decavanadato
Os resultados referentes agrave quantificaccedilatildeo das cisteiacutenas reduzidas apoacutes o tratamento de S1 com
diferentes concentraccedilotildees de decavanadato em tampatildeo 10 mM HEPES pH 75 e 25 mM KCl a
25ordmC durante 20 minutos encontram-se descritos na Tabela 2VII
Tabela 2VII ndash Cisteiacutenas reduzidas totais e ldquoraacutepidas de S1 (1 mgmL) apoacutes tratamento com decavanadato
34
(010501002505001000 uM) em 10 mM HEPES 25 mM KCl pH 7 a 25ordmC durante 20 minutos A detecccedilatildeo
espectrofotomeacutetrica foi feita com DTNB (e 412=8400 uM-1 cm-1) (n variaacutevel entre 1 e 8)
Amostras Cys raacutepidas Cys totais
11 M S1 (controlo) 22 plusmn 04 106 plusmn 14
11 M S1 + 10 M Decavanadato 22 plusmn 05 95 plusmn 08
11 M S1 + 50 uM Decavanadato 20 plusmn 02 98 plusmn 14
11 M S1 + 100 uM Decavanadato 17 plusmn 03 97 plusmn14
11 M S1 + 250 uM Decavanadato 26 plusmn 02 98 plusmn 08
11 M S1 + 500 uM Decavanadato 21 plusmn 04 90 plusmn 06
11 M S1 + 1000 uM Decavanadato 24 plusmn 0 86 plusmn 08
A partir dos resultados obtidos pode sugerir-se que o S1 possui 11 resiacuteduos de Cys (controlo) o
que estaacute de acordo com o anteriormente descrito Verifica-se que o tratamento da proteiacutena com
concentraccedilotildees crescentes de decavanadato provoca a oxidaccedilatildeo de dois resiacuteduos de Cys do S1 de
miosina natildeo expostos ao solvente No entanto a teacutecnica utilizada natildeo nos permite identificar
quais
Tendo em consideraccedilatildeo que o valor de IC50 obtido para a reacccedilatildeo de inibiccedilatildeo daactividade Mg2+-
ATPaacutesica de S1 na presenccedila de decavanadato eacute de cerca de 27 microM pode sugerir-se que este seraacute o
valor ao qual ocorreraacute a oxidaccedilatildeo do primeiro resiacuteduo de Cys o que poderaacute contribuir para a
diminuiccedilatildeo da actividade de S1 observada
25Interacccedilatildeo de S1 com vanaacutedio(IV)
251Anaacutelise por espectroscopia de Ressonacircncia Paramagneacutetica Electroacutenica
A acumulaccedilatildeo intracelular de oxovanaacutedio (IV) in vivo constitui uma das principais formas de
apresentaccedilatildeo de vanaacutedio no organismo Deste modo torna-se bastante interessante analisar a
interacccedilatildeo entre este iatildeo e o S1 de miosina atraveacutes da teacutecnica de espectroscopia de Ressonacircncia
Paramagneacutetica Electroacutenica o meacutetodo de eleiccedilatildeo do estudo do iatildeo vanadilo e dos complexos por si
formados O electratildeo desemparelhado encontra-se fortemente acoplado ao nuacutecleo 51
V o que
35
resulta num espectro de RPE consistindo de 8 desdobramentos na direcccedilatildeo paralela e outros tantos
na direcccedilatildeo perpendicular (Chasteen 1983)
Os estudos realizados por Ahmed et al (1987) centraram-se na interacccedilatildeo do vanadilo com
proteiacutenas como eacute o caso da insulina transferrina entre outros obtendo um espectro caracterizado
por 8 linhas amplamente espaccediladas (na direcccedilatildeo paralela) Jaacute Ramos et al 2009 concluiu que a
exposiccedilatildeo da soluccedilatildeo de decavanadato agraves formas globular e filamentosa de actina produz um
espectro tiacutepico do iatildeo oxidovanaacutedio (IV) indicando a reduccedilatildeo do vanadato
Para aferir qual a concentraccedilatildeo de S1 que permitia obter uma boa razatildeo sinalruiacutedo titulou-se uma
soluccedilatildeo de sulfato de vanadilo 15 microM ([VOSO4]stock = 5 mM) com concentraccedilotildees crescentes de
proteiacutena (0 5 10 15 e 20 microM) em 10 mM HEPES 25 mM KCl A existecircncia de uma correlaccedilatildeo
linear entre a concentraccedilatildeo de proteiacutena e a intensidade dos sinais de RPE correspondente agrave banda
-12 perpendicular encontra-se representada na Fig 210
Figura 2 12ndash Representaccedilatildeo graacutefica da variaccedilatildeo da intensidade do sinal de RPE vs a concentraccedilatildeo de S1 utilizada
Apoacutes verificar que com uma concentraccedilatildeo de S1 igual a 20 microM se obtinham sinais com uma razatildeo
sinalruiacutedo adequada ao prosseguimento dos estudos sobre a interacccedilatildeo V(IV)-S1 de miosina
titulou-se uma soluccedilatildeo de 20 microM S1 com 0 25 5 10 15 25 50 75 100 150 200 500 e 1000
microM VOSO4
Traccedilou-se o graacutefico da intensidade da banda -12 perpendicular do sinal de EPR em funccedilatildeo da
concentraccedilatildeo de VOSO4 tendo-se determinado uma constante de dissociaccedilatildeo (KD) do complexo
S1-vanadilo de 8748 plusmn 2621 microM V(IV)
y = 016786x + 122435
Rsup2 = 092992759
Inte
nsi
dad
e d
o s
inal
de
RP
E
[S1] (microM)
36
Figura 213 ndash Representaccedilatildeo graacutefica da variaccedilatildeo do sinal de RPE vs a concentraccedilatildeo de vanadilo utilizada (os valores
experimentais estatildeo representados por losangos o seu ajuste estaacute representado pela linha)
A simulaccedilatildeo do espectro de EPR foi efectuada a partir do espectro obtido para uma concentraccedilatildeo
de 1 mM VOSO4 apoacutes adiccedilatildeo de 20 microM S1 de miosina tendo-se obtido os valores da constante de
acoplamento hiperfina (A=5016) e do factor g (g0=195)
Figura 214 ndash Representaccedilatildeo graacutefica do espectro de EPR obtido com 1000 microM de vanadilo e 20 microM de S1 (a cinzento
escuro) A simulaccedilatildeo do espectro encontra-se a cinzento claro
Por comparaccedilatildeo destes com os valores presentes na tabela de correspondecircncia xxx eacute possiacutevel
predizer quais os ligandos provaacuteveis do vanadilo na presenccedila de proteiacutena Assim concluiu-se que
o vanadilo estaacute coordenado com duas moleacuteculas de aacutegua um grupo hidroxilo e um grupo R=NR
O grupo hidroxilo pode indicar os resiacuteduos de aminoaacutecidos Tyr Treonina e Serina enquanto o
grupo R=NR poderaacute indicar a presenccedila de Arginina e Histidina como ligandos do vanadilo
Inte
nsi
dad
e d
os
sinai
s d
e R
PE
[Vanadilo] (microM)
Ajuste dos dados exp Dados experimentais
Campo magneacutetico (G)
Espectro Experimental Ajuste
37
3 Procedimento Experimental
31 Reagentes
Todos os reagentes utilizados satildeo de elevado grau de pureza (analiacutetico) Apresentam-se as suas
foacutermulas quiacutemicas ou abreviaturas quando necessaacuterio acrescentou-se a sua descriccedilatildeo Entre
parecircntesis colocou-se as empresas que os comercializam
Aacutecido aceacutetico glacial 100 (Riedel) Acrilamida 99 (Merck) Azul de bromofenol (Riedel-de-
Haumlen) -Mercaptoetanol (Sigma) Bissulfito de soacutedio (Sigma)Bradford (reagente de)
(BioRad)BSA (albumina de soro bovino Sigma-Aldrich) CaCl22H2O (cloreto de caacutelcio
dihidratado Fluka) CH3CH2OH 96 (etanol Panreac) Coomassie Blue R250 (Merck) D2O
(oacutexido de deuteacuterio SDS) DTNB (55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico) Aldrich) EDTA (aacutecido
etilenodiaminatetraaceacutetico Merck) Fosfoenolpiruvato 98 (Fluka) Glicerol (Sigma) Glicina
985 (Panreac) HCl 37 (Riedel-de Haumlen) H2SO4 96 (Pronalab) HEPES 99 (aacutecido 4-(2-
hidroxiletil)-1-piperazineetanosulfoacutenico Sigma) KH2PO4 99 (hidrogenofosfato de potaacutessio)
MgATP (Sigma) Na
ATP (Sigma) Na2HPO4 99 (hidrogenofosfato de soacutedio Riedel-de Haumlen)
NaH2PO4 (fosfato de soacutedio ) NaOH 986 (hidroacutexido de soacutedio Pronalab) KCl 99 (Cloreto de
potaacutessio Panreac) L-Cys 995 (enantioacutemero levoacutegiro do resiacuteduo de Cisteiacutena Fluka)Metanol
99 (Merck) Molibdato de amoacutenio (Sigma)NNrsquo-metil-bis-acrilamida 98 (Merck) NH4VO3
99 (metavanadato de amoacutenia Sigma) PSA 10 (persulfato de amoacutenio Merck) SDS
(dodecilo sulfato de soacutedio Sigma) Sulfito de soacutedio (Sigma) VOSO4 (sulfato de vanadilo
Aldrich) TEMED 99 (Tetrametiletilenodiamina Merck) Tris (2-amino-2-hidroximetil-13-
propanodiol Sigma)
32 Meacutetodos
321 Isolamento e purificaccedilatildeo das proteiacutenas de muacutesculo esqueleacutetico de coelho
3211 Purificaccedilatildeo da miosina preparaccedilatildeo de Subfragmento 1 por digestatildeo quimotriacutepsica
da miosina
O subfragmento-1 de miosina-II (S1) foi gentilmente cedido pelo Prof Doutor Manuel Aureliano
do Departamento de Quiacutemica Bioquiacutemica e Farmaacutecia da Faculdade de Ciecircncias e Tecnologia da
Universidade do Algarve
No entanto descreve-se abaixo de forma resumida o procedimento efectuado
38
32111 Purificaccedilatildeo da miosina
O protocolo de isolamento foi adaptado do meacutetodo previamente descrito por Margossian e Lowey
(1982) apresentando-se de forma esquemaacutetica na Fig 31
Sucintamente os tecidos musculares dorsais e das pernas foram extraiacutedos apoacutes o sacrifiacutecio do
animal (Fig31) De seguida foram adicionados 3 volumes da soluccedilatildeo de extracccedilatildeo A2
Centrifugou-se a soluccedilatildeo e filtrou-se o sobrenadante O sedimento foi entatildeo diluiacutedo em 14
volumes de aacutegua destilada de modo a que a forccedila ioacutenica atingisse 004 M Apoacutes adiccedilatildeo de szlig-
mercaptoetanol (szlig-me) deixou-se a precipitar a 4ordmC removendo o sobrenadante Centrifugou-se
novamente e ressuspendeu-se o sedimento na soluccedilatildeo de extracccedilatildeo A Adicionou-se a soluccedilatildeo
tampatildeo B3 para que a forccedila ioacutenica atingisse 05 M Homogeneizou-se e deixou-se repousar
durante 2h Homogeneizou-se novamente e centrifugou-se de modo a ocorrer precipitaccedilatildeo dos
complexos de actomiosina e proteiacutena desnaturada Apoacutes filtraccedilatildeo do sobrenadante em latilde de vidro
quantificou-se a soluccedilatildeo de miosina resultante pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental)
2 Soluccedilatildeo de extracccedilatildeo A 0012M K2CO3 004M KHCO3 06M KCl pH 65
3 Soluccedilatildeo-tampatildeo B Tris 100 mM pH 7 KCl 2M
39
Figura 31 ndash Representaccedilatildeo esquemaacutetica do protocolo de isolamento de miosina (Margossian e Lowey 1982) Todo o
procedimento foi realizado a 4ordmC (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
32112 Preparaccedilatildeo de Subfragmento-1 de miosina por digestatildeo com quimiotripsina
O S1 de miosina foi isolado apoacutes diaacutelise da miosina durante a noite contra a soluccedilatildeo C4 A
presenccedila do EDTA eacute essencial uma vez que a sua substituiccedilatildeo por MgCl2resulta na produccedilatildeo de
HMM em vez de S1 A suspensatildeo de miosina foi equilibrada e digerida com -quimotripsina
A digestatildeo foi interrompida pela adiccedilatildeo de PMSF Seguidamente ultracentrifugou-se o produto
da digestatildeo desprezando-se o sedimento (Fig 32) A soluccedilatildeo de sobrenadante maioritariamente
S1 foi quantificada pela leitura directa da Abs280 ou pelo meacutetodo de Bradford (ver as secccedilotildees
3311 e 3312 do Procedimento Experimental respectivamente) e armazenada em 10 sacarose
e 2 mM szlig-Me a -80ordmC
4 Soluccedilatildeo C 25mM Tris pH 70 012M KCl 1mM EDTA
40
Antes da realizaccedilatildeo dos ensaios descritos no presente relatoacuterio o S1 de miosina foi dialisado
contra a soluccedilatildeo D5 de modo a remover qualquer composto que pudesse complexar como
vanaacutedio Seguidamente quantificou-se a proteiacutena pelo meacutetodo de Bradford ou pelo meacutetodo
directo (ver secccedilotildees 3311 e 3312do Procedimento Experimtental)
Figura 3 2ndash Representaccedilatildeo esquemaacutetica do protocolo de preparaccedilatildeo do S1 de miosina por digestatildeo quimiotripsina de
acordo com o previamente descrito por Margossian e Lowey (1982) Todo o procedimento foi realizado a 4ordm C
excepto quando indicaccedilatildeo do contraacuterio (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
3212 Isolamento e purificaccedilatildeo de actina
Isolou-se e purificou-se a actina segundo o meacutetodo descrito por Pardee e Spudish que
compreende dois passos ndash a obtenccedilatildeo de um resiacuteduo seco denominado ldquopoacute de acetonardquo e a
extracccedilatildeo subsequente da actina a partir do ldquopoacute de acetonardquo na forma de G-Actina com a
posterior polimerizaccedilatildeo a F-Actina
32121 Obtenccedilatildeo do ldquopoacute de acetonardquo
Sacrificou-se o coelho com uma pancada atraacutes do pescoccedilo e sangrou-se o animal apoacutes o corte da
arteacuteria jugular Esfolou-se e colocou-se numa tina com gelo Retiraram-se as partes de tecido
conjuntivo que envolvem o tecido muscular com auxiacutelio de uma pinccedila e bisturi Seguidamente
retirou-se os muacutesculos dorsais e os muacutesculos brancos das pernas Passou-se os muacutesculos por aacutegua
destilada de modo a remover as impurezas e pesou-se num copo Trituraram-se os muacutesculos
numa maacutequina de triturar carne e extraiu-se o resultante com 3 volumes da soluccedilatildeo E6 durante 10
minutos com agitaccedilatildeo suave Centrifugou-se a suspensatildeo a 4000 g durante 10
5 Soluccedilatildeo D HEPES 10 mM pH 70 KCl 25 mM
6 Soluccedilatildeo E 01 M KCl 015 M K2HPO4 pH 65
41
minutosdesprezando-se o sobrenadante Ressuspendeu-se o sedimento em 6 volumes de soluccedilatildeo
F7durante 10 minutos com agitaccedilatildeo suave Verificou-se se o pH da soluccedilatildeo estava compreendido
entre 82 e 85 acertando-se estecom soluccedilatildeo G8 se necessaacuterio Centrifugou-se durante 10
minutos a 4000 g Desprezou-se o sobrenadante e ressuspendeu-se o sedimento em 3 volumes de
soluccedilatildeo H9 durante 10 minutos Centrifugou-se novamente a 4000 gdurante 10 minutos
desprezando-se o sobrenadante Ressuspendeu-se o sedimento duas vezes consecutivas em 6
volumes de aacutegua destilada durante 5 minutos Centrifugou-se a 4000 g durante 10 minutos apoacutes
cada adiccedilatildeo de H2O
Seguidamente ressuspendeu-se o sedimento 5 vezes consecutivas utilizando 3 volumes de
acetona de cada vez agitando suavemente durante 10 minutos agrave temperatura ambiente Colocou-
se o resiacuteduo filtrado numa tina de vidro sobre papel de filtro deixando-se evaporar o solvente
durante a noite num nicho Triturou-se o resiacuteduo seco numa maacutequina de triturar carne e deixou-se
novamente evaporar o solvente O ldquopoacute de acetonardquo obtido foi armazenado a -20 ordmC
32122 Purificaccedilatildeo da actina a partir de ldquopoacute de acetonardquo
Apoacutes a obtenccedilatildeo do poacute de acetona como acima indicado efectuou-se o protocolo seguidamente
descrito de modo a obter monoacutemeros de actina (G-Actina) e actina filamentosa (F-Actina)
Pesou-se 35 g de poacute de acetona e adicionaram-se 50 mL de soluccedilatildeo-tampatildeo I10
sendo a
extracccedilatildeo feita com agitaccedilatildeo suave durante 30 minutos Separou-se o extracto do poacute de acetona
hidratado filtrando a suspensatildeo com 4 camadas de gaze esterilizada tendo o cuidado de natildeo
espremer o resiacuteduo Voltou a extrair-se com 50 mL de soluccedilatildeo I durante 30 minutos mexendo
lentamente Adicionou-se este ao extracto obtido anteriormente Centrifugou-se o extracto total a
16 000 g durante 30 minutos Decantou-se para uma proveta sobre a qual se colocou gaze para
que o sedimento solto natildeo se juntasse ao sobrenadante e leu-se o volume De seguida adicionou-
se KCl ateacute uma concentraccedilatildeo final de 50 mM MgCl2 ateacute 2mM e ATP ateacute 1mM lentamente e com
agitaccedilatildeo contiacutenua de modo a promover a polimerizaccedilatildeo dos monoacutemeros de actina
Deixou-se polimerizar durante 2h a 25ordmC observando-se a presenccedila de pequenas bolhas de ar
indicativas de um aumento de viscosidade da soluccedilatildeo Arrefeceu-se em gelo durante 15 minutos e
adicionou-se KCl ateacute uma concentraccedilatildeo final de 06M Mexeu-se lentamente durante 30 minutos
apoacutes os quais se ultracentrifugou a 110 000g durante 1hora Desprezou-se o sobrenadante
ressuspendeu-se o sedimento com soluccedilatildeo tampatildeo E e transferiu-se para um potter
Dialisou-se durante a noite contra 1L de soluccedilatildeo-tampatildeo I a uma temperatura de 4ordmC com rotaccedilatildeo
7 Soluccedilatildeo F 005 M NaHCO3 pH 82-85
8 Soluccedilatildeo G 1M Na2CO3
9 Soluccedilatildeo H 1 mM EDTA pH 70
10 Soluccedilatildeo I 2 mM Tris-HCl pH 8 02 mM CaCl2 02 mM Na2ATP 05 mM szlig-Me 0005 NaN3
42
contiacutenua e mudanccedila de tampatildeo 2 a 3 vezes De seguida a actina foi centrifugada a 110
000gdurante 1hora Decantou-se a soluccedilatildeo de G-Actina (sobrenadante) e desprezou-se o
sedimento Determinou-se a concentraccedilatildeo de G-Actina pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental) A soluccedilatildeo de G-actina foi armazenada a uma temperatura de 4ordmC
apoacutes adiccedilatildeo de 002NaN3 mantendo-se estaacutevel durante 3 a 5 dias
331 Quantificaccedilatildeo das proteiacutenas isoladas
3311 Quantificaccedilatildeo pelo meacutetodo de Bradford
O meacutetodo de Bradford (Bradford 1976) baseia-se na utilizaccedilatildeo do reagente de Bradford
composto por entre outros aacutecido fosfoacuterico e o corante ldquoCoomassierdquo Este corante possui carga
negativa e a sua coloraccedilatildeo varia entre a vermelha (Absmax=465 nm) predominante em soluccedilatildeo e a
azul (Absmax=595 nm) presente aquando da ligaccedilatildeo aos grupos amina das proteiacutenas Em soluccedilotildees
aacutecidas diluiacutedas a coloraccedilatildeo varia proporcionalmente com a quantidade do corante ligado agrave
proteiacutena que por sua vez eacute directamente proporcional agrave concentraccedilatildeo da proteiacutena permitindo a
sua quantificaccedilatildeo ateacute a um limite de detecccedilatildeo de 20 gmL (Bradford 1976)
Deste modo prepararam-se amostras de BSA ([BSA]stock= 1mgmL) utilizando um intervalo de
concentraccedilotildees compreendido entre 0 e 15 gmL de acordo com a tabela 3I Apoacutes a adiccedilatildeo da
H2O MilliQ e do reagente de Bradford as soluccedilotildees foram sucessivamente homogeneizadas no
voacutertex
Tabela 3Indash Amostras utilizadas para a obtenccedilatildeo da recta de calibraccedilatildeo no meacutetodo de Bradford
BSA (L) BSA (g) H2O MilliQ (L) Reagente de Bradford (L)
0 0 800 200
125 125 79875 200
25 25 7975 200
5 5 795 200
10 10 790 200
15 15 785 200
Apoacutes incubaccedilatildeo a 25ordmC por 5 minutos leram-se os valores de Abs595 num
espectrofotoacutemetro (Shimadzu UV-160A) para todas as amostras Seguidamente traccedilou-se a recta
de calibraccedilatildeo (Abs595vs quantidade de BSA (g))
43
Para determinar a concentraccedilatildeo de proteiacutena apoacutes o processo de isolamento purificaccedilatildeo e
diaacutelise quando necessaacuterio colocou-se 5 L da amostra num tubo de Eppendorf e adicionou-se
795 L de H2O MilliQ homogeneizando a soluccedilatildeo no voacutertex Adicionou-se posteriormente 200
L de reagente de Bradford e repetiu-se a homogeneizaccedilatildeo Incubou-se a mistura a 25ordmC por 5
minutos apoacutes os quais se leu os valores de Abs595 nm no espectrofotoacutemetro Por interpolaccedilatildeo deste
valor a partir da recta de calibraccedilatildeo determinou-se a concentraccedilatildeo de proteiacutena na amostra
3312 Quantificaccedilatildeo pelo meacutetodo directo
A quantificaccedilatildeo das amostras proteicas pode tambeacutem ser efectuada atraveacutes da leitura directa da
absorvacircncia tendo por base a lei de Beer-Lambert onde Abs corresponde agrave
absorvacircncia ao comprimento de onda ao qual o ensaio foi realizado corresponde ao coeficiente
de extinccedilatildeo (substituiacutedo pela letra a se as unidades forem distintas da molaridade) b corresponde
ao percurso oacuteptico da radiaccedilatildeo na ceacutelula amostral e c corresponde agrave concentraccedilatildeo amostral
Assim no caso de S1 de miosina mediu-se a Abs280e calculou-se a concentraccedilatildeo da proteiacutena
sabendo que o coeficiente de extinccedilatildeo de S1 eacute (Ref)
No caso da actina mediu-se a Abs290ecalculou-se a concentraccedilatildeo da proteiacutena sabendo que o
coeficiente de extinccedilatildeo da actina eacute (Ref)
332Electroforese em condiccedilotildees desnaturantes (SDS-PAGE)
De modo a melhor caracterizar as proteiacutenas isoladas foi realizada uma electroforese em
condiccedilotildees desnaturantes (SDS-PAGE do inglecircs Sodium Dodecyl Sulfate Polyacrilamide Gel
Electrophoresis) O SDS (dodecilo sulfato de soacutedio) eacute um detergente anioacutenico utilizado neste
meacutetodo para desnaturar as proteiacutenas que se ligam ao SDS na proporccedilatildeo 14 g SDS g proteiacutena
(aproximadamente 1 moleacutecula de SDS por cada 2 resiacuteduos de aminoaacutecidos) (Stryer) O SDS induz
uma densidade de carga negativa significativa nas proteiacutenas a que se liga mascarando a sua carga
intriacutenseca pelo que as razotildees cargamassa satildeo tendencialmente idecircnticas Assim a SDS-PAGE
separa as proteiacutenas quando sujeitas a um campo eleacutectrico de acordo com o seu peso molecular
sendo a sua migraccedilatildeo directamente proporcional ao logaritmo deste uacuteltimo (Stryer)
O gel de resoluccedilatildeo com 10 de acrilamida foi obtido atraveacutes da adiccedilatildeo de 167 mL de soluccedilatildeo
stock de acrilamida11
075 mL de tampatildeo de resoluccedilatildeo12
251 mL de aacutegua destilada e 50 L de
SDS 10 Para promover a polimerizaccedilatildeo adicionou-se em seguida 38 L de PSA 10
11
soluccedilatildeo stock de acrilamida soluccedilatildeo de 30 (mv) de acrilamida e 08 (mv) de NNrsquo-metil-bis-acrilamida 12
tampatildeo de resoluccedilatildeo 15 M Tris-HCl pH 88
44
(persulfato de amoacutenia) 10 e 25 L de TEMED 99 (tetrametiletilenodiamina) Agitou-se
cuidadosamente de modo a natildeo formar bolhas e pipetou-se a soluccedilatildeo para a cassette de vidro
previamente montada no suporte De seguida adicionou-se uma peliacutecula de cerca de 2 mm de 2-
metil-2-butanol para verificar posteriormente a polimerizaccedilatildeo Apoacutes a polimerizaccedilatildeo que demora
cerca de 45 minutos verteu-se a agrave superfiacutecie e adicionou-se o gel de concentraccedilatildeo com 5 de
acrilamida constituiacutedo por 03 mL de soluccedilatildeo de stock acrilamida 045 mL de tampatildeo de
concentraccedilatildeo13
094 mL de aacutegua destilada 18 L de SDS 10 135 L de PSA 10 e 2L de
TEMED 99 Colocou-se o pente sobre o gel e deixou-se polimerizar durante cerca de 45
minutos
As amostras para aplicaccedilatildeo no gel de electroforese foram preparadas para uma concentraccedilatildeo de
proteiacutena de 1 mgmL num volume final de 100 L dos quais 25 L correspondem ao tampatildeo da
amostra14
As amostras foram aquecidas a 100ordmC durante 5 minutos Antes da aplicaccedilatildeo das
amostras transferiu-se a cassette para a tina de electroforese agrave qual se adicionou a soluccedilatildeo
tampatildeo de electroforese da tina superior15
e tampatildeo de electroforese da tina inferior16
Aplicaram-
se 10 L das soluccedilotildees de marcadores de peso molecular e 20 L das amostras proteicas
preparadas anteriormente A electroforese decorreu durante cerca de 3h30 min a uma voltagem
de 80 V De seguida transferiu-se o gel para uma tina corando as proteiacutenas durante 1h com
soluccedilatildeo corante17
Descorou-se o gel com soluccedilatildeo descorante18
tendo-se mudado esta soluccedilatildeo ateacute
as bandas se encontrarem bem definidas
Determinou-se a mobilidade relativa das bandas correspondentes aos marcadores de elevado peso
molecular e traccedilou-se a recta de calibraccedilatildeo ndash log(PM) vs log(Rf) onde Rf corresponde agrave
mobilidade relativa Por interpolaccedilatildeo determina-se o PM das proteiacutenas isoladas sabendo a sua
mobilidade relativa Por comparaccedilatildeo com os valores descritos na literatura pode concluir-se
acerca do grau de purificaccedilatildeo
34 Mediccedilatildeo da actividade enzimaacutetica de S1
341 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
13
tampatildeo de concentraccedilatildeo 05 M Tris-HCl pH 68 14
tampatildeo de amostra 320 mM Tris-HCl pH 68 04 M szlig-mercaptoetanol SDS 8 glicerol 15 (vv) e roxo de m-
cresol 0024 15
tampatildeo de electroforese da tina superior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS 16
tampatildeo de electroforese da tina inferior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS acetato de soacutedio
01 M 17
soluccedilatildeo corante ldquoCoomassie Blue R-250rdquo contendo metanol 45 (vv) e aacutecido aceacutetico glacial 10 (vv) 18
soluccedilatildeo descorante metanol 10 (vv) e aacutecido aceacutetico glacial 10 vv)
45
O meacutetodo de Fiske-Subarow modificado consiste na reacccedilatildeo do molibdato de amoacutenio com o
fosfato inorgacircnico em soluccedilatildeo aacutecida com formaccedilatildeo do aacutecido fosfomoliacutebdico Seguidamente daacute-
se a adiccedilatildeo de um agente redutor que provoca a reduccedilatildeo do composto a fosfomolibdato que forma
um complexo de cor azulada cuja absorvacircncia maacutexima ocorre para um comprimento de onda de
660 nm tornando possiacutevel a sua detecccedilatildeo por meacutetodos espectrofotomeacutetricos As actividades Ca2+
-
ATPaacutesica e K+EDTA-ATPaacutesica de S1 foram determinadas atraveacutes deste meacutetodo de acordo com
o descrito por Taussky e Shorr (1953) com mediccedilatildeo da quantidade de fosfato (Pi) libertado
atraveacutes da hidroacutelise de ATP
Para tal obteve-se primeiramente a recta de calibraccedilatildeo com as amostras presentes na Tabela 3II
utilizando uma soluccedilatildeo stock de 5 mM KH2PO4
Tabela 3II ndash Composiccedilatildeo das diferentes amostras utilizadas para determinar a recta de calibraccedilatildeo atraveacutes do meacutetodo
de Fiske-Subbarow modificado
KH2PO4 (L)
([KH2PO4]stock= 5mM)
PO43-
(nmole) H2O (uL)
0 0 1780
5 25 1775
10 50 1770
20 100 1760
40 200 1740
100 500 1680
A cada tubo de Eppendorf adicionaram-se seguidamente 200 L de soluccedilatildeo J19
e 20 L de agente
redutor20
homogeneizando no voacutertex apoacutes cada adiccedilatildeo Incubou-se durante 10 minutos e mediu-
se a Abs660 nm num espectrofotoacutemetro com intervalos de tempo iguais entre cada amostra Traccedilou-
se a recta de calibraccedilatildeo (Abs660 nmvs nmol PO43-
) a partir dos valores obtidos
Para determinar a actividade Ca2+
-ATPaacutesica de S1 prepararam-se tubos de Eppendorf com 400
L de meio de reacccedilatildeo21
25 L CaCl2 ([CaCl2]stock= 100 mM) 25 L ATP ([ATP]stock= 100 mM)
com [S1]ensaios=01 mgmL Perfez-se se necessaacuterio com H2O MilliQ ateacute perfazer um volume de 1
mL Para determinar a actividade K+EDTA-ATPaacutesica utilizou-se uma concentraccedilatildeo idecircntica de
19
Soluccedilatildeo J 5 N H2SO4 25 molibdato de amoacutenio 20
Agente redutor 5 mg aacutecido 1-amino-2-naftol-4-sulfoacutenico 30 mg bissulfito de soacutedio 30 mg sulfito de soacutedio em 26
mL H2O 21
meio de reacccedilatildeo 125 mM Tris-HCl pH 80 0575 M KCl
46
proteiacutena nos ensaios (01 mgmL) e o mesmo meio de reacccedilatildeo Aos tubos de Eppendorf
adicionou-se ainda 123 L de KCl ([KCl]stock = 3M) 25 L de ATP 10 L de EDTA
([EDTA]stock= 100 mM )e o volume de H2O MilliQ necessaacuterio para perfazer 1 mL Em ambas as
situaccedilotildees o uacuteltimo reagente adicionado foi o ATP uma vez que se trata do substrato da reacccedilatildeo
enzimaacutetica
A cineacutetica decorreu a 25ordmC durante 5 minutos para a actividade Ca2+
-ATPaacutesica e durante 3
minutos para a actividade K+EDTA-ATPaacutesica Apoacutes o periacuteodo de incubaccedilatildeo transferiu-se 150
L de amostra para tubos de Eppendorf contendo igual volume de TCA 10 Colocou-se no gelo
e homogeneizou-se no voacutertex Seguidamente centrifugou-se a 1500 g durante 10 minutos e
recolheu-se 200 L de sobrenadante para novos tubos de Eppendorf Adicionou-se a estes 158
mL de H2O MilliQ e 200 L de soluccedilatildeo J Homogeneizou-se no voacutertex e adicionou-se 20 L de
reagente redutor repetindo-se a homogeneizaccedilatildeo Incubou-se durante 10 minutos e mediu-se a
Abs660 nm com o mesmo intervalo de tempo de leitura para cada amostra Por intrapolaccedilatildeo a partir
da recta de calibraccedilatildeo obtida anteriormente determinou-se a quantidade de Pi libertado em cada
ensaio A partir destes resultados determinaram-se as actividades Ca2+
-ATPaacutesica e K+EDTA-
ATPaacutesica do S1 em nmol PO43-
minmg proteiacutena e em s-1
342 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
Da actividade Mg2+
-ATPaacutesica de S1 resulta a formaccedilatildeo dos produtos da reacccedilatildeo de
hidroacutelise do ATP - ADP e Pi Na presenccedila de ADP fosfoenolpiruvato (PEP) e da enzima piruvato
cinase (PK) os produtos resultantes satildeo ATP e piruvato Por acccedilatildeo da enzima lactato
desidrogenase (LDH) este uacuteltimo eacute convertido em lactato ocorrendo simultaneamente a oxidaccedilatildeo
do NADH a NAD+ monitorizada atraveacutes da Abs340 nm Quando o S1 de miosina se encontra no
meio reaccional o ATP eacute hidrolisado formando-se novamente ADP e Pi o que permite a
formaccedilatildeo de um ciclo (fig 21) Se as enzimas se encontrarem em excesso a velocidade de
conversatildeo do NADH eacute limitada pela produccedilatildeo de ADP e Pi Como o ATP eacute regenerado em cada
ciclo a sua concentraccedilatildeo inicial eacute sempre mantida (Bagshaw 1982)
Figura 33ndash Representaccedilatildeo esquemaacutetica do meacutetodo de enzimas acopladas
47
O protocolo utilizado e seguidamente descrito foi adaptado de Cuenda et al (1990 ndash Ver Refordf) A
uma cuvette de 1 mL adicionou-se S1 para uma concentraccedilatildeo final de 005 mgmL 800 L de
meio de reacccedilatildeo22
10 L de PEP ([PEP]stock = 42 mM) 10 L de PK+LDH 11 ([PK+LDH]stock =
1 mgmL) e 15 L de NADH ([NADH]stock=25 mM) Iniciou-se a monitorizaccedilatildeo da reacccedilatildeo
atraveacutes da leitura de Abs340 nm apoacutes a adiccedilatildeo de 20 L de ATP ([ATP]stock=100 mM) A
actividade Mg2+
-ATPaacutesica eacute obtida atraveacutes do declive correspondente agrave oxidaccedilatildeo do NADH Para
a mediccedilatildeo da Mg2+
-ATPase de S1 estimulada por actina adicionou-se F-actina para uma
concentraccedilatildeo final de 2 microM sendo a actividade obtida tambeacutem atraveacutes do declive correspondente
agrave oxidaccedilatildeo do NADH apoacutes a adiccedilatildeo da proteiacutena
35 Interaccedilatildeo entre vanaacutedio e subfragmento-1 de miosina
351 Preparaccedilatildeo das soluccedilotildees de vanaacutedio
As soluccedilotildees de vanadato foram preparadas em aacutegua MilliQ a partir de metavanadato de amoacutenio
(NH4VO3) numa concentraccedilatildeo stock de 50 mM com agitaccedilatildeo magneacutetica e aquecimento (~30 ordmC)
Esta soluccedilatildeo com um valor compreendido entre 6 e 7 designa-se por ldquometavanadatordquo e eacute
constituiacuteda pelas esspeacutecies V1 V2 V4 e V5 (Ref) Acidificando a soluccedilatildeo de metavanadato por
adiccedilatildeo de HCl ateacute um valor de pH proacuteximo de 4 observa-se uma mudanccedila de cor de amarelo
claro para laranja indicativa da formaccedilatildeo da espeacutecie decameacuterica de vanadato De seguida e
imediatamente antes da realizaccedilatildeo dos ensaios descritos o valor do pH da soluccedilatildeo de
decavanadato foi aumentado para cerca de 75 por adiccedilatildeo de NaOH A soluccedilatildeo (2) designada
por ldquometavanadatordquo natildeo foi posteriormente alterada ateacute agrave sua adiccedilatildeo para a realizaccedilatildeo dos ensaios
As soluccedilotildees foram armazenadas a 4ordmC e mantidas em gelo aquando da sua utilizaccedilatildeo
A soluccedilatildeo de V(IV) foi preparada em condiccedilotildees anaeroacutebias por diluiccedilatildeo de sulfato de vanadil
(VOSO4) para uma concentraccedilatildeo final de 5 mM em H2O MilliQ saturada com N2 gasoso
352Anaacutelise por espectroscopia de UV-Vis
A concentraccedilatildeo da espeacutecie decameacuterica em soluccedilatildeo depende do periacuteodo de tempo apoacutes a diluiccedilatildeo
no meio de reacccedilatildeo apropriado (Ref) A velocidade de desoligomerizaccedilatildeo da espeacutecie decameacuterica
pode ser analisada por espectroscopia de absorccedilatildeo na regiatildeo UVVis uma vez que esta espeacutecie
apresenta duas bandas caracteriacutesticas na regiatildeo do visiacutevel (a 358 e 400 nm) que natildeo se verifica
em quaisquer outras espeacutecies oligomeacutericas As bandas supramencionada sapresentam-se bem
22
Meio de reacccedilatildeo 3125 mM Tris-HCl pH 71 375 mM MgCl2
48
resolvidas atraveacutes da obtenccedilatildeo da derivada de 2ordf ordem do espectro apresentando a forma de 2
vales com miacutenimos centrados nos comprimentos de onda supracitados com dependecircncia linear
relativamente agrave concentraccedilatildeo de decavanadato total (Ramos et al 2006)
Assim a estabilidade da espeacutecie decameacuterica apoacutes diluiccedilatildeo da soluccedilatildeo de decavanadato no meio de
reacccedilatildeo dos ensaios (tampatildeo D) para uma concentraccedilatildeo final de 1 mM em vanadato total foi
caracterizada atraveacutes da leitura da Abs400 em funccedilatildeo do tempo tendo-se determinado qual a
ordem de reacccedilatildeo e o tempo de meia-vida da mesma Com base em estudos anteriores acerca da
estabilidade do V10 na presenccedila de diferentes proteiacutenas (Ramos et al 2006) determinou-se
igualmente o tempo de meia vida apoacutes a adiccedilatildeo de 20 microM de S1 de miosina ao meio reaccional
353 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
Os espectros de RMN do isoacutetopo 51
V foram realizados num espectroacutemetro Bruker AM-400 MHz
equipado com uma sonda multinuclear inversa de 5 mm Em tubos de RMN de 5 mm de diacircmetro
preparou-se soluccedilotildees de 500 L de deca- ou metavanadato correspondentes a um intervalo de
concentraccedilotildees de 1 a 10 mM suplementadas com 10 D2O Os espectros foram adquiridos agrave
temperatura ambiente com uma largura espectral de 23 923 Hz um tempo de aquisiccedilatildeo de 0086
s tempo de relaxaccedilatildeo de 001 s e uma sequecircncia de pulsos a 90ordm Utilizou-se a transformada de
Fourier com alargamento das bandas de 20Hz para o processamento dos espectros Os desvios
quiacutemicos das ressonacircncias de 51
V foram obtidos em relaccedilatildeo agrave espeacutecie VOCl3cuja frequecircncia de
ressonacircncia eacute considerada o padratildeo sendo todas as outras referenciadas a partir desta em ppm
As aacutereas dos sinais do RMN correspondentes agraves espeacutecies oligomeacutericas presentes foram obtidas
por integraccedilatildeo dos sinais As concentraccedilotildees relativas das espeacutecies foram calculadas com recurso agrave
equaccedilatildeo
onde A[Vx] corresponde agrave aacuterea referente agrave ressonacircncia da espeacutecie Vx com nordm de agregaccedilatildeo x
A[V]total agrave soma total das aacutereas medidas e [V] a concentraccedilatildeo total de vanaacutedio no ensaio
A teacutecnica de 51
V-RMN foi anteriormente aplicada em estudos de ligaccedilatildeo com S1 de miosina
(Ref) tendo sido demostrado que entre as espeacutecies V1 V2 V4 e V5 a espeacutecie tetrameacuterica possui a
maior afinidade para a miosina (Cremo et al) O desaparecimento ou alargamento dos sinais pode
implicar a ligaccedilatildeo de um dos oxovanadatos laacutebeis agrave miosina sem indicaccedilatildeo de qual uma vez que
49
se processa um equiliacutebrio entre eles (Crans 1990) Assim e tendo em conta as indicaccedilotildees de que
a espeacutecie V10 eacute distinta das restantes no que diz respeito agrave magnitude da interacccedilatildeo com a miosina
e inibiccedilatildeo da actividade ATPaacutesica estimulada por actina (Aureliano 2000 Tiago et al 2002)
torna-se fundamental investigar a interacccedilatildeo das diferentes espeacutecies de vanadato com o S1 de
miosina atraveacutes da teacutecnica de 51
V-RMN
Para tal obtiveram-se espectros de 51
V-RMN resultantes da titulaccedilatildeo de uma soluccedilatildeo 5 mM de
decavanadato total com concentraccedilotildees crescentes de S1 de miosina (0 5 10 15 e 20 microM) em
tampatildeo D Realizou-se ainda ensaios na presenccedila de 2 mM de decavanadato e 2 mM de ATP na
presenccedila e na ausecircncia de S1 de miosina de modo a verificar o efeito que estes ligandos naturais
da enzima teria na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o subfragmento-1
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
Os grupos sulfidrilo (-SH) das cisteiacutenas satildeo resiacuteduos de aminoaacutecidos extremamente sensiacuteveis agrave
oxidaccedilatildeo resultando na sua oxidaccedilatildeo (Dalle-Donne et al 1999) Logo a quantificaccedilatildeo destes
resiacuteduos eacute bastante importante nos estudos de interacccedilatildeo com vanaacutedio jaacute que os resiacuteduos de Cys
com ecircnfase para o SH1 e o SH2 satildeo bastante importantes na monitorizaccedilatildeo das modificaccedilotildees
funcionais da regiatildeo da cabeccedila
O procedimento seguidamente apresentado foi originalmente descrito por Gutieacuterrez-Merino
(1987)) Consiste na reacccedilatildeo entre os grupos ndashSH natildeo oxidados das proteiacutenas e o reagente de
Ellman (DTNB -55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico)) - formando-se o aniatildeo CNT (aniatildeo 3-
carboxilato-4-nitrotiofenolato) numa estequiometria de 11 Este apresenta um maacuteximo de
absorvecircncia a 412 nm (Robyt 1990) Assim apoacutes a incubaccedilatildeo da proteiacutena na presenccedila do metal
a quantidade de cisteiacutenas reduzidas por proteiacutena determina-se atraveacutes dos valores de Abs412
Deste modo colocou-se num tubo de Eppendorf o volume apropriado de S1 dialisado para que a
concentraccedilatildeo no ensaio fosse de 1 mgmL Adicionou-se o volume apropriado de decavanadato
para que a sua concentraccedilatildeo fosse de 0 10 50 100 250 500 ou 1000 M Perfez-se com soluccedilatildeo
D ateacute um volume final de 1 mL e incubou-se a 25ordmC por 20 minutos Apoacutes este periacuteodo colocou-
se 780 L da mistura reaccional numa cuvette de quartzo e adicionou-se 220 L de soluccedilatildeo K23
23
Tampatildeo K Tampatildeo fosfatohidrogenofosfato de soacutedio 025 M pH 72
50
Adicionou-se 15 L de DTNB (em soluccedilatildeo K [DTNB]stock=100 mM) ao meio reaccional e
registou-se o valor de absorvecircncia medido equivalente agraves cisteiacutenas ldquoraacutepidasrdquo (SH1 e SH2
expostos ao solvente) Adicionou-se 001 g de SDS que provoca a desnaturaccedilatildeo da proteiacutena e
consequentemente a exposiccedilatildeo de todos os resiacuteduos de cisteiacutenas que se encontravam ateacute aiacute
inacessiacuteveis no interior da proteiacutena (cisteiacutenas totais) Estes resiacuteduos podem assim reagir com o
DTNB
Estimou-se o nuacutemero de resiacuteduos de cisteiacutenas em todas as amostras com recurso ao coeficiente de
extinccedilatildeo molar () do DTNB (ver ponto 3 dos Anexos) que foi determinado atraveacutes da titulaccedilatildeo
de L-Cisteiacutena com intervalo de concentraccedilotildees compreendidos entre 0 e 50 microM na presenccedila de
25microM de DTNB em tampatildeo D Os valores de absorvecircncia foram obtidos para um comprimento de
onda de 412 nm
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
3621 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
A teacutecnica mais eficiente de anaacutelise da interacccedilatildeo entre o iatildeo vanadilo e proteiacutenas como a
calmodulina e a transferrina entre outras permite a obtenccedilatildeo raacutepida de informaccedilatildeo no que toca agrave
conformaccedilatildeo e amplitude da referida interacccedilatildeo
Os espectros de Ressonacircncia Paramagneacutetica Electroacutenica (RPE) da espeacutecie paramagneacutetica V(IV)
foram realizados a 70K num espectroacutemetro Bruker EMX-300 X-band Todas as soluccedilotildees
utilizadas e amostras foram preparadas em anaerobiose numa cacircmara de luvas de modo a evitar a
reduccedilatildeo de V(IV) a V(V) Em tubos capilares com 02 mL de amostra manteve-se a concentraccedilatildeo
de S1 a 20 M e variou-se a concentraccedilatildeo de vanadilo (0 25 5 10 15 25 50 75 100 150
200 500 e 1000 M [VOSO4]stock=5 mM) em tampatildeo D Apoacutes obtenccedilatildeo dos espectros estes
foram tratados atraveacutes do software SimFonia e WinEPR tendo-se obtido um graacutefico com a
variaccedilatildeo da intensidade do sinal de RPE em funccedilatildeo da concentraccedilatildeo de sulfato de vanadilo
presente Realizaram-se ainda simulaccedilotildees de espectros obtidos com 20 microM de S1 e 1000 microM de
V(IV) com vista agrave obtenccedilatildeo dos ligandos provaacuteveis do vanadilo atraveacutes da anaacutelise da constante
de acoplamento hiperfina (A) e do factor g (g)
4 Conclusotildees gerais
Apesar de jaacute ter sido demonstrado que o vanaacutedio inibe a Na+K
+-ATPase da miosina a funccedilatildeo
deste elemento no tecido muacutesculo esqueleacutetico continua largamente incompreendida A enorme
complexidade quiacutemica deste metal especialmente no que concerne agrave oligomerizaccedilatildeo e reacccedilotildees
51
redox constitui um obstaacuteculo agrave compreensatildeo do comportamento do vanaacutedio nos organismos
vivos Assim a caracterizaccedilatildeo da quiacutemica deste elemento eacute de grande utilidade antes de iniciar o
estudo da sua interacccedilatildeo com o presente sistema
A anaacutelise por espectroscopias de 51
V-RMN e de UV-Vis de soluccedilotildees de vanadato diluiacutedas nos
meios de reacccedilatildeo utilizados no presente relatoacuterio (10 mM HEPES pH 70 25 mM KCl) mostrou
claramente que a espeacutecie decameacuterica (V10) possui um tempo de vida suficientemente longo para a
determinaccedilatildeo do seu efeito isolado ao contraacuterio do que se verifica com as espeacutecies mono- di-
tetra- e pentameacutericas (V1 V2 V4 e V5) Por outro lado a anaacutelise verificou ainda que apoacutes diluiccedilatildeo
das soluccedilotildees de decavanadato no meio de reacccedilatildeo coexistem as espeacutecies V10 e V1 pelo que os
efeitos putativos desta uacuteltima deveratildeo ser subtraiacutedos aquando da monitorizaccedilatildeo dos efeitos da
espeacutecie decameacuterica
O vanaacutedio (V) (vanadato) inibe as enzimas que catalizam reacccedilotildees de transferecircncia de grupos
fosfato como a ATPase da miosina [Goodno 1979] devido agrave sua analogia estrutural com o iatildeo
fosfato Na sua utilizaccedilatildeo como inibidor enzimaacutetico natildeo eacute tida em conta a contribuiccedilatildeo individual
das espeacutecies oligomeacutericas considerando-se normalmente a espeacutecie V1 como inibidora efectiva
[Goodno e Taylor 1982] No presente relatoacuterio verificou-se por espectroscopia de 51
V-RMN que
outros oligoacutemeros de vanadato interagem com o S1 de miosina tendo as espeacutecies V10 e V4 os
maiores niacuteveis de afinidade A sua interacccedilatildeo envolve forccedilas electrostaacuteticas e eacute afectada
negativamente por ligandos naturais de enzima como eacute o caso do ATP
A exposiccedilatildeo do S1 de miosina (005 mgmL) a decavanadato conduz agrave inibiccedilatildeo do capacidade da
proteiacutena de estimular a actividade Mg2+
-ATPaacutesica com um valor de IC50 de 266 plusmn 07 M
decavanadato sendo o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM
decavanadato (80) Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM decavanadato sendo
o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM decavanadato (60)
exercendo um efeito de impedimento agrave ligaccedilatildeo entre o vanaacutedio e o S1
A uma concentraccedilatildeo de 27 microM decavanadato verifica-se a oxidaccedilatildeo de um dos resiacuteduos de Cys
de S1 de miosina seguida pela oxidaccedilatildeo gradual de outro resiacuteduo de Cys ateacute agrave concentraccedilatildeo de
1000 microM Ambos os resiacuteduos oxidados natildeo se encontravam expostos ao solvente utilizado A
oxidaccedilatildeo dos resiacuteduos permite sugerir que esta modificaccedilatildeo oxidativa sofrida pelo S1 contribui
para a diminuiccedilatildeo da sua actividade de ATPase
52
Por espectroscopia de RPE revelaram-se os possiacuteveis ligandos equatoriais do vanaacutedio aquando da
sua interacccedilatildeo com o S1 de miosina como sendo duas moleacuteculas de aacutegua um ligando contendo o
grupo R-OH e outro contendo o grupo R=NR
Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados de
oxidaccedilatildeo (IV) e (V) no S1 de miosina para concentraccedilotildees facilmente alcanccedilaacuteveis no ambiente
terrestre
53
5 Estudos futuros
No sentido do melhor esclarecimento das alteraccedilotildees moleculares e estruturais que ocorrem no
subfragmento 1 quando exposta ao vanaacutedio que conduzem a uma diminuiccedilatildeo da sua funccedilatildeo
sugere-se
- Quantificaccedilatildeo das cisteiacutenas reduzidas de S1 na presenccedila de metavanadato
- Determinaccedilatildeo das actividades Ca2+
- e K+-ATPaacutesica do S1 de miosina na presenccedila de
decavanadato
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina estimulada por actina
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina na presenccedila de
metavanadato
- Realizaccedilatildeo de RMN 2D com variaccedilatildeo das concentraccedilotildees das soluccedilotildees de decavanadato e
metavanadato na presenccedila de S1
-Mutageacutenese dos resiacuteduos de Cys doS1 de miosina de modo a verificar quais os resiacuteduos que
sofrem oxidaccedilatildeo na presenccedila de decavanadato
- Determinaccedilatildeo por calorimetria (DSC Differential Scanning Calorimetry) da variaccedilatildeo da
entalpia (∆H) e da temperatura de desnaturaccedilatildeo do S1 de miosina na presenccedila de vanaacutedio uma
vez que estas propriedades caracteriacutesticas de cada proteiacutena satildeo alteradas apoacutes ocorrecircncia de
modificaccedilotildees estruturais
- Anaacutelise por SDS-PAGE de S1 de miosina apoacutes tratamento com vanaacutedio de modo a determinar
quais os polipeacuteptidos eventualmente formados apoacutes clivagem da proteiacutena
54
6 Referecircncias bibliograacuteficas
- Sellers J Myosins 2nd Edition Oxford University Press 1999
- L Stryer inBiochemistry (Ed WH Freeman 2005 pp
Refordfas Materiais e Meacutetodos
[A] S S Margossian SLowey Methods Enzymol1982 85 55-71
[B] JD Pardue JA Spudich Methods Enzymol 1982 85 164-181
[C] M Bradford Anal Biochem1976 72 248-254
[D] H H Taussky E Shorr J Biol Chem 1953 202 675-685
[E] C R Bagshaw in Outline Studies in Biology ndash Muscle Contraction Chapman and Hall Ed
1982
[F] A Cuenda et al Eur J Biochem1990 194 663-670
55
Anexos
1 Determinaccedilatildeo da recta de calibraccedilatildeo meacutetodo de Bradford
Figura1ndashRepresentaccedilatildeo graacutefica da recta de calibraccedilatildeo (Abs595 nm vs [BSA]) utilizada para o meacutetodo de Bradford
2 Determinaccedilatildeo da recta calibraccedilatildeo para a electroforese em condiccedilotildees desnaturantes
Figura 2 ndash Representaccedilatildeo graacutefica da recta de calibraccedilatildeo (log(Peso Molecular) vs Rf dos marcadores de peso
molecular) onde Rf indica determinada atraveacutes do padratildeo de peso molecular
(poccedilo (a))
3Caacutelculo do coeficiente de extinccedilatildeo molar () para a quantificaccedilatildeo de cisteiacutenas reduzidas
atraveacutes do meacutetodo de Ellman
Traccedilou-se a recta de Abs412 nm meacutedia vs [L-Cisteiacutena] de modo a determinar o valor do coeficiente
de extinccedilatildeo molar definido como o declive daquela recta Assim =8400 M-1
cm-1
como
podemos verificar na Fig 3
A595 nm = 00546[BSA] + 00209 Rsup2 = 099051
Ab
s 59
5 n
m
[BSA] (mgmL)
log PM = -1177 Rf + 2247 Rsup2 = 098957
log
PM
Mobilidade relativa (Rf)
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero

24
a partir do protocolo presente no ponto 341 da secccedilatildeo Procedimento experimental encontram-se
descritos na Tabela 2I
Tabela 2I ndash Acrividades natildeo-fisioloacutegicas de subfragmento-1 determinadas pelo meacutetodo de Fiske-Subarow
modificado (n=2)
Proteiacutena
Actividade
ATPaacutesica natildeo
fisioloacutegica
Actividade (nmol
Piminmg proteiacutena) Actividade (s-1)
S1
Ca2+
-ATPase 1400 plusmn 30 2100 plusmn 0045
K+EDTA-ATPase 2334 plusmn 50 3500 plusmn 0075
As actividades natildeo-fisioloacutegicas reflectem o estado de oxidaccedilatildeo dos resiacuteduos expostos de Cys 707
(SH1) e 697 (SH2) Se o grupo SH1 sofrer oxidaccedilatildeo a actividade ATPaacutesica dependente de Ca2+
aumenta e a actividade dependente de K+ diminui Por outro lado se o grupo SH2 sofrer
oxidaccedilatildeo regista-se a perda completa destas actividades
De acordo com Margossian e Lowey (1982) [BA] as actividades ATPaacutesica de S1 de miosina
dependente de Ca2+
e K+EDTA da miosina satildeo respectivamente de 24 e 112 s-1
Estaacute tambeacutem
descrito que a actividade ATPaacutesica dependente de K+ eacute em meacutedia cerca de 3 vezes superior agrave
dependente de Ca2+
reflectindo a quantidade das preparaccedilotildees A razatildeo obtida entre os valores das
actividades ATPaacutesicas dependentes de K+ e Ca
2+ eacute de 166 Os valores obtidos para as actividades
(Ca2+
-ATPase=21 s-1
K+EDTA-ATPase= 35 s
-1) satildeo inferiores agrave valores obtidos por
Margossian e Lowey [BA] e Tiago [BC] com forccedila ioacutenica inferior O valor obtido para a
actividade ATPaacutesica dependente de Ca2+
eacute superior ao obtido por Polosukhina et al (2000) [BD]
cujo meio de reacccedilatildeo continha MOPS e apresentava forccedila ioacutenica inferior Os dados
supramencionados comprovam a variaccedilatildeo da actividade a ATPaacutesica natildeo fisioloacutegica em relaccedilatildeo agrave
forccedila ioacutenica temperatura e pH
222 Mediccedilatildeo da actividadeMg2+
-ATPaacutesica de S1 pelo meacutetodo de enzimas acopladas
A actividade de Mg2+
-ATPase eacute a uacutenica que possui significado fisioloacutegico sendo as restantes
bastante uacuteteis apenas para indicaccedilatildeo da qualidade de preparaccedilatildeo do S1[BA] A 0575 M KCl a
actina e o S1 formam uma soluccedilatildeo viscosa e tuacuterbida Apoacutes a adiccedilatildeo de MgATP o complexo acto-
S1 dissocia-se e a turbidez da soluccedilatildeo diminui Se a forccedila ioacutenica for reduzida a dissociaccedilatildeo do
complexo actoS1 que se encontra na forma de suspensatildeo eacute induzida O MgATP eacute rapidamente
hidrolisado diminuindo a sua concentraccedilatildeo no meio reaccional Nestas condiccedilotildees haacute
reassociaccedilatildeo e subsequente contracccedilatildeo dos filamentos de actina e de S1 com formaccedilatildeo de
25
precipitado O meacutetodo descrito traduz de forma bastante elementar o fenoacutemeno da contracccedilatildeo in
vivo e elimina o problema anterior da solubilizaccedilatildeo ineficiente da miosina de modo a obter uma
mediccedilatildeo exacta da actividade [A] A utilizaccedilatildeo de forccedilas ioacutenicas reduzidas aumenta a afinidade da
actina para o S1 permitindo contornar a extrema dificuldade em obter a concentraccedilatildeo proteica dos
muacutesculos (cerca de 100 mgmL do complexo actomiosina) As actividades Mg2+
-ATPaacutesicas de S1
de miosina foram determinadas a partir do declive da variaccedilatildeo da Abs340 relativo agrave oxidaccedilatildeo de
NADH em funccedilatildeo do tempo como eacute descrito no ponto 342 do Procedimento Experimental Os
valores obtidos para a actividade Mg2+
-ATPaacutesica encontram-se apresentados na Tabela 2II
Tabela 2II ndash Actividade Mg2+
-ATPaacutesica de subfragmento-1 de miosina determinadas pelo meacutetodo das enzimas
acopladas
Actividade (nmol
ATPmin mg(S1)) Actividade (s
-1)
1097 plusmn 269 164 plusmn 04
Ramos (2005) [J] obteve o valor de 01 s-1
para a actividade Mg2+
-ATPaacutesica de S1 de miosina
com forccedila ioacutenica intermeacutedia Tiago (2005) [BC] obteve o valor de 011 s-1 em condiccedilotildees de forccedila
ioacutenica reduzida em relaccedilatildeo agrave utilizada no presente relatoacuterio
23 Preparaccedilatildeo e caracterizaccedilatildeo das soluccedilotildees de vanaacutedio
231 Anaacutelise da composiccedilatildeo das soluccedilotildees de vanadato por espectroscopia de 51
V-RMN
efeito da concentraccedilatildeo da espeacutecie
A diluiccedilatildeo das soluccedilotildees stock de meta- e decavanadato iraacute determinar fortemente a composiccedilatildeo
em soluccedilatildeo dos diferentes oligoacutemeros de vanadato (ref) No caso do decavanadato o periacuteodo de
tempo apoacutes a diluiccedilatildeo iraacute tambeacutem ser um factor importante a considerar devido agrave decomposiccedilatildeo
da espeacutecie V10(ref)Torna-se assim fundamental determinar a composiccedilatildeo quiacutemica das soluccedilotildees
no meio de ensaio utilizado (HEPES 10 mM KCl 25 mM) de modo a correlacionaacute-la
correctamente com os possiacuteveis efeitos promovidos pelas diferentes espeacutecies de vanadato no
sistema em estudo
Aleacutem disso deve ser dado ecircnfase agraves importantes interacccedilotildees que ocorrem entre o vanaacutedio e os
tampotildees escolhidos O tampatildeo com menor nuacutemero e magnitude de interacccedilatildeo eacute o HEPES [AD]
seguido do Tris sendo este uacuteltimo especialmente aconselhado em concentraccedilotildees baixas para casos
de incompatibilidade enzimaacutetica ou quiacutemica redox No caso especial da reacccedilatildeo redox
NAD(P)HNAD(P) o tampatildeo adequado eacute o imidazole de modo a natildeo haver reacccedilatildeo com o
26
vanadato Os tampotildees a evitar em qualquer situaccedilatildeo incluem a trietanolamina fosfato citrato e
glicina entre outros [AD] uma vez que todos interactuam fortemente com V(V) com formaccedilatildeo
de complexos estaacuteveis
Assim procedeu-se agrave anaacutelise da composiccedilatildeo das soluccedilotildees de meta- e decavanadato apoacutes diluiccedilatildeo
das soluccedilotildees stock (50 mM em vanadato total) no meio de reacccedilatildeo (tampatildeo D) atraveacutes da
espectroscopia de ressonacircncia magneacutetica nuclear de 51
V Nas figuras x1 e x2 apresentam-se os
espectros obtidos para as soluccedilotildees de deca- e metavanadato num intervalo de concentraccedilotildees de 1 a
10 mM A Tabela seguinte resume os paracircmetros espectroscoacutepicos obtidos para cada espeacutecie
presente nas soluccedilotildees analisadas
Figura 22ndash Espectros de RMN de 51
V de
concentraccedilotildees crescentes das espeacutecies oligomeacutericas
de vanadato obtidos apoacutes diluiccedilatildeo da soluccedilatildeo stock
de decavanadato em 10 mM HEPES pH 70 25 mM
KCl para concentraccedilotildees finais de A) 1 mM B)2
mM C) 4 mM D) 8 mM E) 10 mM de
decavanadato total V1refere-se agraves espeacutecies
monomeacutericade vanadato V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2IIIndash Desvios quiacutemicos meacutedios (part) das ressonacircncias das espeacutecies decameacuterica (V10) monomeacuterica (V1) e
tetrameacuterica (V4) presentes nos espectros da soluccedilatildeo de decavanadatoLargura a meia altura (∆12) da espeacutecie V10A
Paracircmetros V10A V10B V10C V1 V4
part (ppm) -5162 -5003 -4255 -5616 -5748
∆12 (Hz) 175 nd nd nd nd
Figura 23 ndash Distribuiccedilatildeo graacutefica das
espeacutecies oligomeacutericas de vanadato vs a
concentraccedilatildeo de decavanadato total As
concentraccedilotildees de cada espeacutecie foram
obtidas segundo a formula descrita na
secccedilatildeo 35 do Procedimento
Experimental (aacutereas dos sinais de 51
V-
RMN observadas na figura x1)V10 V1 e
V4 correspondem agraves espeacutecie monomeacuterica
dimeacuterica e tetrameacuterica respectivamente [Esp
eacuteci
e o
ligo
meacute
rica
de
van
aacutedio
] (m
M)
[Decavanadato]total (mM)
V10
V1
V4
27
Nas soluccedilotildees de decavanadato as espeacutecies monomeacuterica e decameacuterica de vanadato satildeo as uacutenicas
presentes para concentraccedilotildees de decavanadato inferiores a 8 mM Ambas as espeacutecies aumentam
linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado em [AY] e
em (Teresa) quando a concentraccedilatildeo de V1 eacute igual ou superior a 200 microM inicia-se a sua
oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica No entanto nas condiccedilotildees de realizaccedilatildeo dos
ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a espeacutecie tetrameacuterica neste
estudo em concentraccedilotildees inferiores a 8 mM tendo o mesmo sido observado por Ramoset al
(2006) [AK] Os resultados obtidos mostram claramente o equiliacutebrio entre a espeacutecie decameacuterica e
a monomeacuterica e que o aparecimento de outras espeacutecies oligomeacutericas se encontra dependente da
concentraccedilatildeo desta uacuteltima Apesar de natildeo ter sido realizado o ensaio com 5 mM de decavanadato
total no presente relatoacuterio estaacute reportado em Ramos et al (2006) [AK] que as concentraccedilotildees da
espeacutecie decameacuterica e monomeacuterica satildeo respectivamente de 500 microM e 200 microM Assim pode inferir-
se que para concentraccedilotildees inferiores a 5 mM e intervalos de tempo de cerca de 20 minutos (que
inclui o tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas
as espeacutecies decameacuterica e monomeacuterica de vanadato
Figura 24 ndash Espectros de RMN de 51
V de concentraccedilotildees
crescentes das espeacutecies oligomeacutericas de vanadato obtidos
apoacutes diluiccedilatildeo da soluccedilatildeo stock de metavanadato em 10 mM
HEPES pH 70 25 mM KCl para concentraccedilotildees finais de A)
1 mM B) 2 mM C) 4 mM D) 8 mM E) 10 mM de
metavanadato total V1 refere-se agraves espeacutecies monomeacuterica de
vanadatoV1 e V2 correspondem respectivamente agrave espeacutecie
monomeacuterica e dimeacuterica
Figura 25 ndash Distribuiccedilatildeo graacutefica das espeacutecies
oligomeacutericas de vanadato vs a concentraccedilatildeo de
metavanadato total As concentraccedilotildees de cada
espeacutecie foram obtidas segundo a formula descrita
na secccedilatildeo 35 do Procedimento Experimental
(aacutereas dos sinais de 51
V-RMN observadas na
figura x1)V10 V1 e V4 correspondem agraves espeacutecie
monomeacuterica dimeacuterica e tetrameacuterica
respectivamente
Esp
eacuteci
e o
ligo
meacute
rica
de
va
nad
ato
(m
M)
[Metavanadato]total (mM)
V1
V2
V4
V5
28
Tabela 2IVndash Desvios quiacutemicos meacutedios (part)das ressonacircncias das espeacutecies tetrameacuterica (V4) monomeacuterica (V1) e
dimeacuterica (V2) presentes no espectro da soluccedilatildeo de metavanadato (total) 10 mM
Paracircmetros V1 V2 V4 V5
part (ppm) -5615 -5748 -5789 -5869
Nas soluccedilotildees de metavanadato as espeacutecies monomeacuterica dimeacutericae tetrameacuterica de vanadato estatildeo
presentes para concentraccedilotildees de metavanadato inferiores a 4 mM Todas estas as espeacutecies
aumentam linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado
em (Aureliano e Madeira 1998) e em (Tiago et al 2004) quando a concentraccedilatildeo de V1 eacute igual ou
superior a 200 microM inicia-se a sua oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica Nas
condiccedilotildees de realizaccedilatildeo dos ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a
espeacutecie pentameacuterica neste estudo em concentraccedilotildees inferiores a 4 mM tendo o mesmo sido
observado por Ramos 2006 Os resultados obtidos mostram claramente o equiliacutebrio entre a
espeacutecie V4 e as espeacuteciesV1 e V2 no qual o aparecimento de outras espeacutecies oligomeacutericas se
encontra dependente da concentraccedilatildeo da espeacutecie V1 Por uacuteltimo pode inferir-se que para
concentraccedilotildees inferiores a 2 mM e intervalos de tempo de cerca de 20 minutos (que inclui o
tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas as
espeacutecies V1 V2 e V4 com concentraccedilotildees respectivas de 0796 0192 e 0196 mM
232Estabilidade das soluccedilotildees de decavanadato anaacutelise por espectroscopia de absorccedilatildeo
UVVis
Caracterizou-se a estabilidade da espeacutecie V10 numa soluccedilatildeo com 1 mM decavanadato total em
tampatildeo D atraveacutes da leitura da Abs400 Obteve-se uma cineacutetica de dissociaccedilatildeo de 1ordf ordem tal
como se pode verificar pela linearidade do graacutefico de ln(Abs400)vs o tempo de aquisiccedilatildeo espectral
(segundos) (Fig26 )
Figura 26ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do tempo
(em segundos) para uma soluccedilatildeo de decavanadato total de 1
mM em tampatildeo D a 25ordmC (n=2)
Sendo uma reacccedilatildeo de primeira ordem pode ser descrita pela equaccedilatildeo cujo
declive eacute ndashK (constante de dissociaccedilatildeo) e a intersecccedilatildeo no eixo das ordenadas corresponde
Ln
(Ab
s 400 n
m)
tempo (s)
29
aln(A0) Numa reacccedilatildeo de 1ordf ordem o tempo de meia-vida eacute independente da concentraccedilatildeo inicial
do reagente sendo descrito pela equaccedilatildeo [2] Deste modo a partir da recta
Ln(Abs400 nm) = -436E-5
K - 105 obteve-se um tempo de meia-vida de 4490 plusmn 0823 horas para
as condiccedilotildees supramencionadas
Apoacutes a adiccedilatildeo de S1 8 microM ao meio reaccional verificou-se que a reacccedilatildeo de desoligomerizaccedilatildeo
da espeacutecie decameacuterica de vanadato seguia igualmente uma cineacutetica de 1ordf ordem tal como se pode
verificar pela linearidade do graacutefico de ln(Abs400) versus o tempo de aquisiccedilatildeo espectral
(segundos) (Fig27)
Figura 27ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do
tempo (em segundos) para uma soluccedilatildeo de decavanadato
total de 1 mM em tampatildeo D a 25ordmC na presenccedila de 8
microM (n= 2)
A partir da equaccedilatildeo da recta Ln(Abs400 nm) = -7520E-5
K - 0613determinou-se o valor de K para
a espeacutecie decameacuterica de onde se obteve um tempo de meia-vida de 2430 plusmn 0191 horas para as
condiccedilotildees supramencionadas
Tanto este valor como o valor registado na ausecircncia de S1 satildeo substancialmente superiores agrave
duraccedilatildeo dos ensaios de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina descritos no presente
relatoacuterio
24 Interacccedilatildeo de S1 com vanaacutedio(V)
241Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
De modo a analisar qual o efeito da adiccedilatildeo de S1 de miosina (tampatildeo D) produzia nos sinais de
51V-NMR titulou-se uma soluccedilatildeo de 5mM de decavanadato total com concentraccedilotildees crescentes
de S1 (5 10 15 e 20 microM) Os espectros de 51
V-RMN encontram-se apresentados na Fig 2
No graacutefico da figura 2 Apresenta-se a concentraccedilatildeo de S1 em funccedilatildeo do factor de alargamento
corrigido dos espectros acima referidos
Ln
(Ab
s (4
00
nm
))
tempo (s)
30
Figura 28 ndash Graacutefico de
representaccedilatildeo da
concentraccedilatildeo de S1 em
funccedilatildeo do factor de
alargamento A
concentraccedilatildeo de
decavanadato total foi de 5
mM e as concentraccedilotildees
utilizadas de S1 foram de 0
5 10 e 15 microM
Verifica-se que o factor de alargamento aumenta linearmente em funccedilatildeo da concentraccedilatildeo de S1 no
ensaio o que indica a existecircncia de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina Observou-
se ainda que a adiccedilatildeo de S1 ao meio reaccional natildeo provocou qualquer alteraccedilatildeo dos desvios
quiacutemicos dos sinais de 51
V-RMN
Realizaram-se ensaios na presenccedila de ATP e de S1 com intuito de verificar o efeito do ATP na
interacccedilatildeo do S1 de miosina com o decavanadato
Figura 29 - Espectros de RMN de 51
V contendo 2
mM Decavanadato (A) (controlo) (B) na presenccedila
de 2 mM de ATP (C) na presenccedila de 2 mM de ATP
e 8 microM de S1 de miosina V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2Vndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies decameacuterica (V10) e monomeacuterica (V1) presentes nos
espectros da soluccedilatildeo de decavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V10A V10B V10C V1
part (ppm) -5161 -5002 -4254 -5614
Observa-se que a presenccedila de ATP (espectro B) natildeo provoca qualquer alteraccedilatildeo visiacutevel em
relaccedilatildeo ao espectro que contem apenas soluccedilatildeo de decavanadato Jaacute a presenccedila simultacircnea de ATP
e S1 provoca um alargamento da ∆12 do sinal do V10A de 1736 Hz (espectro (A)) para 2366 Hz
(espectro (C) Fig 29) Tendo em conta que a concentraccedilatildeo de S1 apresenta uma relaccedilatildeo linear
com o factor de alargamento converteu-se o valor da ∆12 e comparou-se com o valor esperado agrave
F = 00591[S1] - 00157 Rsup2 = 0998
Fact
or
de
ala
rgam
en
to
[S1] (microM)
31
concentraccedilatildeo de S1 utilizada O factor de alargamento obtido para 8 microM ndash 0376 ndash eacute inferior ao
valor esperado agrave mesma concentraccedilatildeo de S1 obtido atraveacutes da equaccedilatildeo da recta apresentada na
Fig 28 ndash 0457 ndash o que indica que o ATP poderaacute exercer um efeito de protecccedilatildeo do S1
diminuindo a sua interacccedilatildeo com a espeacutecieV10
Por uacuteltimo realizaram-se ensaios com metavanadato na presenccedila de 8 microM S1 de modo a verificar
se as espeacutecies oligomeacutericas contidas nesta soluccedilatildeo interagiam com a proteiacutena em estudo
Figura 210 - Espectros de RMN de 51
V contendo 2
mM metavanadato (A) (controlo) (B) na presenccedila
de 8 microM de S1 de miosina V1 V2 e V4
correspondem respectivamente aacutes espeacutecies
monomeacuterica dimeacuterica e tetrameacuterica
Tabela 2VIndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies monomeacuterica (V1) dimeacuterica (V2) e tetrameacuterica (V4)
presentes nos espectros da soluccedilatildeo de metavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V1 V2 V4
part (ppm) -5613 -5747 -5789
Como eacute possiacutevel observar na Fig 210 ocorre uma ligeiro alargamento dos sinal de V4 bem
como uma perda da intensidade pelo que se conclui quedas espeacutecies presentes na soluccedilatildeo de
metavanadato (V1 V2 e V4) apenas a espeacutecie tetrameacuterica interage com o S1 de miosina o que
estaacute de acordo com o reportado por Tiago et al 2002
242 Actividade Mg2+
-ATPaacutesica de S1 de miosina na presenccedila de decanadato
Com o intuito de avaliar o efeito do decavanadato na actividade Mg2+
-ATPaacutesica do S1 de
miosina monitorizou-se a reacccedilatildeo de oxidaccedilatildeo do NADH atraveacutes da leitura de Abs340 na
presenccedila de 005 mgmL de S1 de miosina e no caso da actividade estimulada 2microM de actina em
3125 mM Tris-HCl pH 71 e 375 mM MgCl2 pelo meacutetodo de enzimas acopladas (ver seccccedilatildeo xx
dos MM)Para tal realizaram-se ensaios com vaacuterias concentraccedilotildees de decavanadato total (10 50
100 250 500 e 1000) de modo a determinar qual o seu efeito na actividade MgATPaacutesica de S1
de miosina na ausecircncia e na presenccedila de um dos seus ligandos naturais Os ensaios foram
realizados agrave temperatura ambiente com incubaccedilatildeo preacutevia a 25ordm C durante 20 minutos
32
Figura 211 ndash Representaccedilatildeo
graacutefica da Actividade Mg2+
-
ATPaacutesica de S1 () (os ajustes
teoacutericos estatildeo representado a por
uma linha os resultados
experimentais estatildeo representados
por triacircngulos (Actividade
estimulada) e cruzes (Actividade
natildeo estimulada)) vs
[Decavanadato]total medidas pelo
meacutetodo das enzimas acopladas
(n=3) Os dados foram ajustados ao modelo de decaimento exponencial usando o programa GraphPad Prism para a
determinaccedilatildeo dos valores de IC50 (na ausecircncia de actina IC50= 266 plusmn 07 M decavanadato na presenccedila de actina
IC50= 28 plusmn 1 M decavanadato
A partir do graacutefico presente na Fig 29 verifica-se que o ajuste dos dados experimentais foi bem
sucedido A inibiccedilatildeo da actividade Mg2+
-ATPaacutesica do S1 de miosina eacute afectada em menor
amplitude na presenccedila de actina (60 de inibiccedilatildeo da actividade) do que na sua ausecircncia
(aproximadamente 80 de inibiccedilatildeo de actividade) Devido a esta diferenccedila pode sugerir-se que a
actina iraacute impedir parcialmente a ligaccedilatildeo do decavanadatoao S1 demiosina atraveacutes de
impedimentos esteacutereos ou interacccedilotildees electrostaacuteticas por exemplo A teacutecnica utilizada natildeo nos
permite concluir acerca de que regiatildeo(otildees) ou resiacuteduo(s) da actina participaratildeo na preservaccedilatildeo
parcial da actividade Mg2+
-ATPaacutesica
Por outro lado os valores de IC50 dos dois tipos de actividades natildeo apresentam diferenccedilas
significativas tendo-se obtido 266 plusmn 07 e 28 plusmn 1M decavanadato na ausecircncia e na presenccedila de
actina respectivamente o que corresponde a uma concentraccedilatildeo de cerca de 27 M de V10 O
meio de reacccedilatildeo utilizado nestes ensaios foi 10 mM HEPES e 25 mM KCl com 2 mM ATP Em
(Tiago et al 2004) o valor de IC50 para a actividade Mg2+-ATPaacutesica na ausecircncia de actina foi de
125 plusmn 025 microM enquanto o valor da actividade estimulada por actina foi de 136 plusmn 14 microM O
meio de reacccedilatildeo utilizado nos ensaios foi 20 mM HEPES 50 mM KCl e 2 mM ATP o que resulta
precisamente no dobro da forccedila ioacutenica a que foi realizado o ensaio do presente relatoacuterio
(exceptuando a contribuiccedilatildeo dos produtos da hidroacutelise de ATP uma vez que este meacutetodo
pressupotildee a ocorrecircncia de um ciclo de reacccedilotildees de hidroacutelise e siacutentese deste composto) Ora tendo
sido anteriormente referido que a afinidade do S1 de miosina para com a espeacutecie V10 diminui com
Act
ivid
ade
Mg
2+-A
TP
aacutesic
a d
e S
1 d
e
Mio
sin
a
[Decavanadato] (microM) Actividade Estimulada ()
Actividade Natildeo Estimulada ()
Ajuste Actividade Estimulada
Ajuste Actividade Natildeo Estimulada
33
o aumento da forccedila ioacutenica (Tiago et al 2004 tal poderaacute explicar a diferenccedila obtida entre estes
dois conjuntos de valores
243Quantificaccedilatildeo de cisteiacutenas reduzidas pelo meacutetodo de Ellman
A interacccedilatildeo do vanaacutedio com alguns tipos de aminoaacutecidos sensiacuteveis agrave oxidaccedilatildeo pode fornecer-nos
informaccedilatildeo bastante uacutetil em relaccedilatildeo agraves alteraccedilotildees estruturais que conduzem a uma diminuiccedilatildeo da
actividade fisioloacutegica do S1 de miosina Os estudos realizados por Chasteen 1983 concluiacuteram
que a reduccedilatildeo de vanadato (V) a vanadilo (IV) ocorre na presenccedila de agentes redutores comuns no
ambiente celular de que se destaca o aminoaacutecido cisteiacutena Esta eacute tambeacutem um alvo de
complexaccedilatildeo por parte do iatildeo VO2+
Jaacute Ramos et al (2009) demonstrou que a incubaccedilatildeo de
actina com a soluccedilatildeo de decavanadato induz a oxidaccedilatildeo de resiacuteduos de cisteiacutena e agrave formaccedilatildeo do
oxidovanaacutedio (IV) Este grupo de investigadores descobriu ainda que apenas a espeacutecie V10 induz a
oxidaccedilatildeo das cisteiacutenas ldquoraacutepidasrdquo da actina atraveacutes do processo proposto que eacute parcialmente
responsaacutevel pela inibiccedilatildeo jaacute descrita da actividade de ATPase do S1 de miosina estimulada por
actina quando exposta agrave soluccedilatildeo de decavanadato Esta inibiccedilatildeo apresenta-se afectada com a
presenccedila de ATP uma vez que os resiacuteduos de cisteiacutena permaneceram intactos nos ensaios
realizados com este ligando Jaacute Crans et al (2004) concluiu que o siacutetio activo de muitas fosfatases
(que conteacutem grupos tiol) pode formar um equiliacutebrio redox com o iatildeo vanadato (V) de forma natildeo
reversiacutevel
Os resultados obtidos por Sakurai e Shiromura (1980) sugeriram que o vanaacutedio (V) sofre reduccedilatildeo
a vanaacutedio (IV) sem dependecircncia do valor de pH do ensaio Registaram ainda uma estequiometria
de 31 na reacccedilatildeo do vanadato (V) com os resiacuteduos de Cys enquanto a estequiometria dos ensaios
com o vanadilo (IV) foi de 21
Berlett e Stadtman (1997) observaram que os tipos de aminoaacutecidos mais facilmente oxidaacuteveis
englobam entre outros os resiacuteduos de Cys Trp e MetAssim analisou-se o estado redox dos
grupos tiol dos resiacuteduos de Cys do S1 na presenccedila do decavanadato atraveacutes da monitorizaccedilatildeo da
Abs412 nm na presenccedila de 1 mgmL de S1 de miosina em tampatildeo 10 mM HEPES 25 mM KCl
variando a concentraccedilatildeo da soluccedilatildeo de decavanadato de modo a determinar que tipo de alteraccedilotildees
estruturais ocorrem na sua presenccedila no que diz respeito agrave diminuiccedilatildeo da actividade ATPaacutesica de
S1 na presenccedila de decavanadato
Os resultados referentes agrave quantificaccedilatildeo das cisteiacutenas reduzidas apoacutes o tratamento de S1 com
diferentes concentraccedilotildees de decavanadato em tampatildeo 10 mM HEPES pH 75 e 25 mM KCl a
25ordmC durante 20 minutos encontram-se descritos na Tabela 2VII
Tabela 2VII ndash Cisteiacutenas reduzidas totais e ldquoraacutepidas de S1 (1 mgmL) apoacutes tratamento com decavanadato
34
(010501002505001000 uM) em 10 mM HEPES 25 mM KCl pH 7 a 25ordmC durante 20 minutos A detecccedilatildeo
espectrofotomeacutetrica foi feita com DTNB (e 412=8400 uM-1 cm-1) (n variaacutevel entre 1 e 8)
Amostras Cys raacutepidas Cys totais
11 M S1 (controlo) 22 plusmn 04 106 plusmn 14
11 M S1 + 10 M Decavanadato 22 plusmn 05 95 plusmn 08
11 M S1 + 50 uM Decavanadato 20 plusmn 02 98 plusmn 14
11 M S1 + 100 uM Decavanadato 17 plusmn 03 97 plusmn14
11 M S1 + 250 uM Decavanadato 26 plusmn 02 98 plusmn 08
11 M S1 + 500 uM Decavanadato 21 plusmn 04 90 plusmn 06
11 M S1 + 1000 uM Decavanadato 24 plusmn 0 86 plusmn 08
A partir dos resultados obtidos pode sugerir-se que o S1 possui 11 resiacuteduos de Cys (controlo) o
que estaacute de acordo com o anteriormente descrito Verifica-se que o tratamento da proteiacutena com
concentraccedilotildees crescentes de decavanadato provoca a oxidaccedilatildeo de dois resiacuteduos de Cys do S1 de
miosina natildeo expostos ao solvente No entanto a teacutecnica utilizada natildeo nos permite identificar
quais
Tendo em consideraccedilatildeo que o valor de IC50 obtido para a reacccedilatildeo de inibiccedilatildeo daactividade Mg2+-
ATPaacutesica de S1 na presenccedila de decavanadato eacute de cerca de 27 microM pode sugerir-se que este seraacute o
valor ao qual ocorreraacute a oxidaccedilatildeo do primeiro resiacuteduo de Cys o que poderaacute contribuir para a
diminuiccedilatildeo da actividade de S1 observada
25Interacccedilatildeo de S1 com vanaacutedio(IV)
251Anaacutelise por espectroscopia de Ressonacircncia Paramagneacutetica Electroacutenica
A acumulaccedilatildeo intracelular de oxovanaacutedio (IV) in vivo constitui uma das principais formas de
apresentaccedilatildeo de vanaacutedio no organismo Deste modo torna-se bastante interessante analisar a
interacccedilatildeo entre este iatildeo e o S1 de miosina atraveacutes da teacutecnica de espectroscopia de Ressonacircncia
Paramagneacutetica Electroacutenica o meacutetodo de eleiccedilatildeo do estudo do iatildeo vanadilo e dos complexos por si
formados O electratildeo desemparelhado encontra-se fortemente acoplado ao nuacutecleo 51
V o que
35
resulta num espectro de RPE consistindo de 8 desdobramentos na direcccedilatildeo paralela e outros tantos
na direcccedilatildeo perpendicular (Chasteen 1983)
Os estudos realizados por Ahmed et al (1987) centraram-se na interacccedilatildeo do vanadilo com
proteiacutenas como eacute o caso da insulina transferrina entre outros obtendo um espectro caracterizado
por 8 linhas amplamente espaccediladas (na direcccedilatildeo paralela) Jaacute Ramos et al 2009 concluiu que a
exposiccedilatildeo da soluccedilatildeo de decavanadato agraves formas globular e filamentosa de actina produz um
espectro tiacutepico do iatildeo oxidovanaacutedio (IV) indicando a reduccedilatildeo do vanadato
Para aferir qual a concentraccedilatildeo de S1 que permitia obter uma boa razatildeo sinalruiacutedo titulou-se uma
soluccedilatildeo de sulfato de vanadilo 15 microM ([VOSO4]stock = 5 mM) com concentraccedilotildees crescentes de
proteiacutena (0 5 10 15 e 20 microM) em 10 mM HEPES 25 mM KCl A existecircncia de uma correlaccedilatildeo
linear entre a concentraccedilatildeo de proteiacutena e a intensidade dos sinais de RPE correspondente agrave banda
-12 perpendicular encontra-se representada na Fig 210
Figura 2 12ndash Representaccedilatildeo graacutefica da variaccedilatildeo da intensidade do sinal de RPE vs a concentraccedilatildeo de S1 utilizada
Apoacutes verificar que com uma concentraccedilatildeo de S1 igual a 20 microM se obtinham sinais com uma razatildeo
sinalruiacutedo adequada ao prosseguimento dos estudos sobre a interacccedilatildeo V(IV)-S1 de miosina
titulou-se uma soluccedilatildeo de 20 microM S1 com 0 25 5 10 15 25 50 75 100 150 200 500 e 1000
microM VOSO4
Traccedilou-se o graacutefico da intensidade da banda -12 perpendicular do sinal de EPR em funccedilatildeo da
concentraccedilatildeo de VOSO4 tendo-se determinado uma constante de dissociaccedilatildeo (KD) do complexo
S1-vanadilo de 8748 plusmn 2621 microM V(IV)
y = 016786x + 122435
Rsup2 = 092992759
Inte
nsi
dad
e d
o s
inal
de
RP
E
[S1] (microM)
36
Figura 213 ndash Representaccedilatildeo graacutefica da variaccedilatildeo do sinal de RPE vs a concentraccedilatildeo de vanadilo utilizada (os valores
experimentais estatildeo representados por losangos o seu ajuste estaacute representado pela linha)
A simulaccedilatildeo do espectro de EPR foi efectuada a partir do espectro obtido para uma concentraccedilatildeo
de 1 mM VOSO4 apoacutes adiccedilatildeo de 20 microM S1 de miosina tendo-se obtido os valores da constante de
acoplamento hiperfina (A=5016) e do factor g (g0=195)
Figura 214 ndash Representaccedilatildeo graacutefica do espectro de EPR obtido com 1000 microM de vanadilo e 20 microM de S1 (a cinzento
escuro) A simulaccedilatildeo do espectro encontra-se a cinzento claro
Por comparaccedilatildeo destes com os valores presentes na tabela de correspondecircncia xxx eacute possiacutevel
predizer quais os ligandos provaacuteveis do vanadilo na presenccedila de proteiacutena Assim concluiu-se que
o vanadilo estaacute coordenado com duas moleacuteculas de aacutegua um grupo hidroxilo e um grupo R=NR
O grupo hidroxilo pode indicar os resiacuteduos de aminoaacutecidos Tyr Treonina e Serina enquanto o
grupo R=NR poderaacute indicar a presenccedila de Arginina e Histidina como ligandos do vanadilo
Inte
nsi
dad
e d
os
sinai
s d
e R
PE
[Vanadilo] (microM)
Ajuste dos dados exp Dados experimentais
Campo magneacutetico (G)
Espectro Experimental Ajuste
37
3 Procedimento Experimental
31 Reagentes
Todos os reagentes utilizados satildeo de elevado grau de pureza (analiacutetico) Apresentam-se as suas
foacutermulas quiacutemicas ou abreviaturas quando necessaacuterio acrescentou-se a sua descriccedilatildeo Entre
parecircntesis colocou-se as empresas que os comercializam
Aacutecido aceacutetico glacial 100 (Riedel) Acrilamida 99 (Merck) Azul de bromofenol (Riedel-de-
Haumlen) -Mercaptoetanol (Sigma) Bissulfito de soacutedio (Sigma)Bradford (reagente de)
(BioRad)BSA (albumina de soro bovino Sigma-Aldrich) CaCl22H2O (cloreto de caacutelcio
dihidratado Fluka) CH3CH2OH 96 (etanol Panreac) Coomassie Blue R250 (Merck) D2O
(oacutexido de deuteacuterio SDS) DTNB (55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico) Aldrich) EDTA (aacutecido
etilenodiaminatetraaceacutetico Merck) Fosfoenolpiruvato 98 (Fluka) Glicerol (Sigma) Glicina
985 (Panreac) HCl 37 (Riedel-de Haumlen) H2SO4 96 (Pronalab) HEPES 99 (aacutecido 4-(2-
hidroxiletil)-1-piperazineetanosulfoacutenico Sigma) KH2PO4 99 (hidrogenofosfato de potaacutessio)
MgATP (Sigma) Na
ATP (Sigma) Na2HPO4 99 (hidrogenofosfato de soacutedio Riedel-de Haumlen)
NaH2PO4 (fosfato de soacutedio ) NaOH 986 (hidroacutexido de soacutedio Pronalab) KCl 99 (Cloreto de
potaacutessio Panreac) L-Cys 995 (enantioacutemero levoacutegiro do resiacuteduo de Cisteiacutena Fluka)Metanol
99 (Merck) Molibdato de amoacutenio (Sigma)NNrsquo-metil-bis-acrilamida 98 (Merck) NH4VO3
99 (metavanadato de amoacutenia Sigma) PSA 10 (persulfato de amoacutenio Merck) SDS
(dodecilo sulfato de soacutedio Sigma) Sulfito de soacutedio (Sigma) VOSO4 (sulfato de vanadilo
Aldrich) TEMED 99 (Tetrametiletilenodiamina Merck) Tris (2-amino-2-hidroximetil-13-
propanodiol Sigma)
32 Meacutetodos
321 Isolamento e purificaccedilatildeo das proteiacutenas de muacutesculo esqueleacutetico de coelho
3211 Purificaccedilatildeo da miosina preparaccedilatildeo de Subfragmento 1 por digestatildeo quimotriacutepsica
da miosina
O subfragmento-1 de miosina-II (S1) foi gentilmente cedido pelo Prof Doutor Manuel Aureliano
do Departamento de Quiacutemica Bioquiacutemica e Farmaacutecia da Faculdade de Ciecircncias e Tecnologia da
Universidade do Algarve
No entanto descreve-se abaixo de forma resumida o procedimento efectuado
38
32111 Purificaccedilatildeo da miosina
O protocolo de isolamento foi adaptado do meacutetodo previamente descrito por Margossian e Lowey
(1982) apresentando-se de forma esquemaacutetica na Fig 31
Sucintamente os tecidos musculares dorsais e das pernas foram extraiacutedos apoacutes o sacrifiacutecio do
animal (Fig31) De seguida foram adicionados 3 volumes da soluccedilatildeo de extracccedilatildeo A2
Centrifugou-se a soluccedilatildeo e filtrou-se o sobrenadante O sedimento foi entatildeo diluiacutedo em 14
volumes de aacutegua destilada de modo a que a forccedila ioacutenica atingisse 004 M Apoacutes adiccedilatildeo de szlig-
mercaptoetanol (szlig-me) deixou-se a precipitar a 4ordmC removendo o sobrenadante Centrifugou-se
novamente e ressuspendeu-se o sedimento na soluccedilatildeo de extracccedilatildeo A Adicionou-se a soluccedilatildeo
tampatildeo B3 para que a forccedila ioacutenica atingisse 05 M Homogeneizou-se e deixou-se repousar
durante 2h Homogeneizou-se novamente e centrifugou-se de modo a ocorrer precipitaccedilatildeo dos
complexos de actomiosina e proteiacutena desnaturada Apoacutes filtraccedilatildeo do sobrenadante em latilde de vidro
quantificou-se a soluccedilatildeo de miosina resultante pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental)
2 Soluccedilatildeo de extracccedilatildeo A 0012M K2CO3 004M KHCO3 06M KCl pH 65
3 Soluccedilatildeo-tampatildeo B Tris 100 mM pH 7 KCl 2M
39
Figura 31 ndash Representaccedilatildeo esquemaacutetica do protocolo de isolamento de miosina (Margossian e Lowey 1982) Todo o
procedimento foi realizado a 4ordmC (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
32112 Preparaccedilatildeo de Subfragmento-1 de miosina por digestatildeo com quimiotripsina
O S1 de miosina foi isolado apoacutes diaacutelise da miosina durante a noite contra a soluccedilatildeo C4 A
presenccedila do EDTA eacute essencial uma vez que a sua substituiccedilatildeo por MgCl2resulta na produccedilatildeo de
HMM em vez de S1 A suspensatildeo de miosina foi equilibrada e digerida com -quimotripsina
A digestatildeo foi interrompida pela adiccedilatildeo de PMSF Seguidamente ultracentrifugou-se o produto
da digestatildeo desprezando-se o sedimento (Fig 32) A soluccedilatildeo de sobrenadante maioritariamente
S1 foi quantificada pela leitura directa da Abs280 ou pelo meacutetodo de Bradford (ver as secccedilotildees
3311 e 3312 do Procedimento Experimental respectivamente) e armazenada em 10 sacarose
e 2 mM szlig-Me a -80ordmC
4 Soluccedilatildeo C 25mM Tris pH 70 012M KCl 1mM EDTA
40
Antes da realizaccedilatildeo dos ensaios descritos no presente relatoacuterio o S1 de miosina foi dialisado
contra a soluccedilatildeo D5 de modo a remover qualquer composto que pudesse complexar como
vanaacutedio Seguidamente quantificou-se a proteiacutena pelo meacutetodo de Bradford ou pelo meacutetodo
directo (ver secccedilotildees 3311 e 3312do Procedimento Experimtental)
Figura 3 2ndash Representaccedilatildeo esquemaacutetica do protocolo de preparaccedilatildeo do S1 de miosina por digestatildeo quimiotripsina de
acordo com o previamente descrito por Margossian e Lowey (1982) Todo o procedimento foi realizado a 4ordm C
excepto quando indicaccedilatildeo do contraacuterio (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
3212 Isolamento e purificaccedilatildeo de actina
Isolou-se e purificou-se a actina segundo o meacutetodo descrito por Pardee e Spudish que
compreende dois passos ndash a obtenccedilatildeo de um resiacuteduo seco denominado ldquopoacute de acetonardquo e a
extracccedilatildeo subsequente da actina a partir do ldquopoacute de acetonardquo na forma de G-Actina com a
posterior polimerizaccedilatildeo a F-Actina
32121 Obtenccedilatildeo do ldquopoacute de acetonardquo
Sacrificou-se o coelho com uma pancada atraacutes do pescoccedilo e sangrou-se o animal apoacutes o corte da
arteacuteria jugular Esfolou-se e colocou-se numa tina com gelo Retiraram-se as partes de tecido
conjuntivo que envolvem o tecido muscular com auxiacutelio de uma pinccedila e bisturi Seguidamente
retirou-se os muacutesculos dorsais e os muacutesculos brancos das pernas Passou-se os muacutesculos por aacutegua
destilada de modo a remover as impurezas e pesou-se num copo Trituraram-se os muacutesculos
numa maacutequina de triturar carne e extraiu-se o resultante com 3 volumes da soluccedilatildeo E6 durante 10
minutos com agitaccedilatildeo suave Centrifugou-se a suspensatildeo a 4000 g durante 10
5 Soluccedilatildeo D HEPES 10 mM pH 70 KCl 25 mM
6 Soluccedilatildeo E 01 M KCl 015 M K2HPO4 pH 65
41
minutosdesprezando-se o sobrenadante Ressuspendeu-se o sedimento em 6 volumes de soluccedilatildeo
F7durante 10 minutos com agitaccedilatildeo suave Verificou-se se o pH da soluccedilatildeo estava compreendido
entre 82 e 85 acertando-se estecom soluccedilatildeo G8 se necessaacuterio Centrifugou-se durante 10
minutos a 4000 g Desprezou-se o sobrenadante e ressuspendeu-se o sedimento em 3 volumes de
soluccedilatildeo H9 durante 10 minutos Centrifugou-se novamente a 4000 gdurante 10 minutos
desprezando-se o sobrenadante Ressuspendeu-se o sedimento duas vezes consecutivas em 6
volumes de aacutegua destilada durante 5 minutos Centrifugou-se a 4000 g durante 10 minutos apoacutes
cada adiccedilatildeo de H2O
Seguidamente ressuspendeu-se o sedimento 5 vezes consecutivas utilizando 3 volumes de
acetona de cada vez agitando suavemente durante 10 minutos agrave temperatura ambiente Colocou-
se o resiacuteduo filtrado numa tina de vidro sobre papel de filtro deixando-se evaporar o solvente
durante a noite num nicho Triturou-se o resiacuteduo seco numa maacutequina de triturar carne e deixou-se
novamente evaporar o solvente O ldquopoacute de acetonardquo obtido foi armazenado a -20 ordmC
32122 Purificaccedilatildeo da actina a partir de ldquopoacute de acetonardquo
Apoacutes a obtenccedilatildeo do poacute de acetona como acima indicado efectuou-se o protocolo seguidamente
descrito de modo a obter monoacutemeros de actina (G-Actina) e actina filamentosa (F-Actina)
Pesou-se 35 g de poacute de acetona e adicionaram-se 50 mL de soluccedilatildeo-tampatildeo I10
sendo a
extracccedilatildeo feita com agitaccedilatildeo suave durante 30 minutos Separou-se o extracto do poacute de acetona
hidratado filtrando a suspensatildeo com 4 camadas de gaze esterilizada tendo o cuidado de natildeo
espremer o resiacuteduo Voltou a extrair-se com 50 mL de soluccedilatildeo I durante 30 minutos mexendo
lentamente Adicionou-se este ao extracto obtido anteriormente Centrifugou-se o extracto total a
16 000 g durante 30 minutos Decantou-se para uma proveta sobre a qual se colocou gaze para
que o sedimento solto natildeo se juntasse ao sobrenadante e leu-se o volume De seguida adicionou-
se KCl ateacute uma concentraccedilatildeo final de 50 mM MgCl2 ateacute 2mM e ATP ateacute 1mM lentamente e com
agitaccedilatildeo contiacutenua de modo a promover a polimerizaccedilatildeo dos monoacutemeros de actina
Deixou-se polimerizar durante 2h a 25ordmC observando-se a presenccedila de pequenas bolhas de ar
indicativas de um aumento de viscosidade da soluccedilatildeo Arrefeceu-se em gelo durante 15 minutos e
adicionou-se KCl ateacute uma concentraccedilatildeo final de 06M Mexeu-se lentamente durante 30 minutos
apoacutes os quais se ultracentrifugou a 110 000g durante 1hora Desprezou-se o sobrenadante
ressuspendeu-se o sedimento com soluccedilatildeo tampatildeo E e transferiu-se para um potter
Dialisou-se durante a noite contra 1L de soluccedilatildeo-tampatildeo I a uma temperatura de 4ordmC com rotaccedilatildeo
7 Soluccedilatildeo F 005 M NaHCO3 pH 82-85
8 Soluccedilatildeo G 1M Na2CO3
9 Soluccedilatildeo H 1 mM EDTA pH 70
10 Soluccedilatildeo I 2 mM Tris-HCl pH 8 02 mM CaCl2 02 mM Na2ATP 05 mM szlig-Me 0005 NaN3
42
contiacutenua e mudanccedila de tampatildeo 2 a 3 vezes De seguida a actina foi centrifugada a 110
000gdurante 1hora Decantou-se a soluccedilatildeo de G-Actina (sobrenadante) e desprezou-se o
sedimento Determinou-se a concentraccedilatildeo de G-Actina pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental) A soluccedilatildeo de G-actina foi armazenada a uma temperatura de 4ordmC
apoacutes adiccedilatildeo de 002NaN3 mantendo-se estaacutevel durante 3 a 5 dias
331 Quantificaccedilatildeo das proteiacutenas isoladas
3311 Quantificaccedilatildeo pelo meacutetodo de Bradford
O meacutetodo de Bradford (Bradford 1976) baseia-se na utilizaccedilatildeo do reagente de Bradford
composto por entre outros aacutecido fosfoacuterico e o corante ldquoCoomassierdquo Este corante possui carga
negativa e a sua coloraccedilatildeo varia entre a vermelha (Absmax=465 nm) predominante em soluccedilatildeo e a
azul (Absmax=595 nm) presente aquando da ligaccedilatildeo aos grupos amina das proteiacutenas Em soluccedilotildees
aacutecidas diluiacutedas a coloraccedilatildeo varia proporcionalmente com a quantidade do corante ligado agrave
proteiacutena que por sua vez eacute directamente proporcional agrave concentraccedilatildeo da proteiacutena permitindo a
sua quantificaccedilatildeo ateacute a um limite de detecccedilatildeo de 20 gmL (Bradford 1976)
Deste modo prepararam-se amostras de BSA ([BSA]stock= 1mgmL) utilizando um intervalo de
concentraccedilotildees compreendido entre 0 e 15 gmL de acordo com a tabela 3I Apoacutes a adiccedilatildeo da
H2O MilliQ e do reagente de Bradford as soluccedilotildees foram sucessivamente homogeneizadas no
voacutertex
Tabela 3Indash Amostras utilizadas para a obtenccedilatildeo da recta de calibraccedilatildeo no meacutetodo de Bradford
BSA (L) BSA (g) H2O MilliQ (L) Reagente de Bradford (L)
0 0 800 200
125 125 79875 200
25 25 7975 200
5 5 795 200
10 10 790 200
15 15 785 200
Apoacutes incubaccedilatildeo a 25ordmC por 5 minutos leram-se os valores de Abs595 num
espectrofotoacutemetro (Shimadzu UV-160A) para todas as amostras Seguidamente traccedilou-se a recta
de calibraccedilatildeo (Abs595vs quantidade de BSA (g))
43
Para determinar a concentraccedilatildeo de proteiacutena apoacutes o processo de isolamento purificaccedilatildeo e
diaacutelise quando necessaacuterio colocou-se 5 L da amostra num tubo de Eppendorf e adicionou-se
795 L de H2O MilliQ homogeneizando a soluccedilatildeo no voacutertex Adicionou-se posteriormente 200
L de reagente de Bradford e repetiu-se a homogeneizaccedilatildeo Incubou-se a mistura a 25ordmC por 5
minutos apoacutes os quais se leu os valores de Abs595 nm no espectrofotoacutemetro Por interpolaccedilatildeo deste
valor a partir da recta de calibraccedilatildeo determinou-se a concentraccedilatildeo de proteiacutena na amostra
3312 Quantificaccedilatildeo pelo meacutetodo directo
A quantificaccedilatildeo das amostras proteicas pode tambeacutem ser efectuada atraveacutes da leitura directa da
absorvacircncia tendo por base a lei de Beer-Lambert onde Abs corresponde agrave
absorvacircncia ao comprimento de onda ao qual o ensaio foi realizado corresponde ao coeficiente
de extinccedilatildeo (substituiacutedo pela letra a se as unidades forem distintas da molaridade) b corresponde
ao percurso oacuteptico da radiaccedilatildeo na ceacutelula amostral e c corresponde agrave concentraccedilatildeo amostral
Assim no caso de S1 de miosina mediu-se a Abs280e calculou-se a concentraccedilatildeo da proteiacutena
sabendo que o coeficiente de extinccedilatildeo de S1 eacute (Ref)
No caso da actina mediu-se a Abs290ecalculou-se a concentraccedilatildeo da proteiacutena sabendo que o
coeficiente de extinccedilatildeo da actina eacute (Ref)
332Electroforese em condiccedilotildees desnaturantes (SDS-PAGE)
De modo a melhor caracterizar as proteiacutenas isoladas foi realizada uma electroforese em
condiccedilotildees desnaturantes (SDS-PAGE do inglecircs Sodium Dodecyl Sulfate Polyacrilamide Gel
Electrophoresis) O SDS (dodecilo sulfato de soacutedio) eacute um detergente anioacutenico utilizado neste
meacutetodo para desnaturar as proteiacutenas que se ligam ao SDS na proporccedilatildeo 14 g SDS g proteiacutena
(aproximadamente 1 moleacutecula de SDS por cada 2 resiacuteduos de aminoaacutecidos) (Stryer) O SDS induz
uma densidade de carga negativa significativa nas proteiacutenas a que se liga mascarando a sua carga
intriacutenseca pelo que as razotildees cargamassa satildeo tendencialmente idecircnticas Assim a SDS-PAGE
separa as proteiacutenas quando sujeitas a um campo eleacutectrico de acordo com o seu peso molecular
sendo a sua migraccedilatildeo directamente proporcional ao logaritmo deste uacuteltimo (Stryer)
O gel de resoluccedilatildeo com 10 de acrilamida foi obtido atraveacutes da adiccedilatildeo de 167 mL de soluccedilatildeo
stock de acrilamida11
075 mL de tampatildeo de resoluccedilatildeo12
251 mL de aacutegua destilada e 50 L de
SDS 10 Para promover a polimerizaccedilatildeo adicionou-se em seguida 38 L de PSA 10
11
soluccedilatildeo stock de acrilamida soluccedilatildeo de 30 (mv) de acrilamida e 08 (mv) de NNrsquo-metil-bis-acrilamida 12
tampatildeo de resoluccedilatildeo 15 M Tris-HCl pH 88
44
(persulfato de amoacutenia) 10 e 25 L de TEMED 99 (tetrametiletilenodiamina) Agitou-se
cuidadosamente de modo a natildeo formar bolhas e pipetou-se a soluccedilatildeo para a cassette de vidro
previamente montada no suporte De seguida adicionou-se uma peliacutecula de cerca de 2 mm de 2-
metil-2-butanol para verificar posteriormente a polimerizaccedilatildeo Apoacutes a polimerizaccedilatildeo que demora
cerca de 45 minutos verteu-se a agrave superfiacutecie e adicionou-se o gel de concentraccedilatildeo com 5 de
acrilamida constituiacutedo por 03 mL de soluccedilatildeo de stock acrilamida 045 mL de tampatildeo de
concentraccedilatildeo13
094 mL de aacutegua destilada 18 L de SDS 10 135 L de PSA 10 e 2L de
TEMED 99 Colocou-se o pente sobre o gel e deixou-se polimerizar durante cerca de 45
minutos
As amostras para aplicaccedilatildeo no gel de electroforese foram preparadas para uma concentraccedilatildeo de
proteiacutena de 1 mgmL num volume final de 100 L dos quais 25 L correspondem ao tampatildeo da
amostra14
As amostras foram aquecidas a 100ordmC durante 5 minutos Antes da aplicaccedilatildeo das
amostras transferiu-se a cassette para a tina de electroforese agrave qual se adicionou a soluccedilatildeo
tampatildeo de electroforese da tina superior15
e tampatildeo de electroforese da tina inferior16
Aplicaram-
se 10 L das soluccedilotildees de marcadores de peso molecular e 20 L das amostras proteicas
preparadas anteriormente A electroforese decorreu durante cerca de 3h30 min a uma voltagem
de 80 V De seguida transferiu-se o gel para uma tina corando as proteiacutenas durante 1h com
soluccedilatildeo corante17
Descorou-se o gel com soluccedilatildeo descorante18
tendo-se mudado esta soluccedilatildeo ateacute
as bandas se encontrarem bem definidas
Determinou-se a mobilidade relativa das bandas correspondentes aos marcadores de elevado peso
molecular e traccedilou-se a recta de calibraccedilatildeo ndash log(PM) vs log(Rf) onde Rf corresponde agrave
mobilidade relativa Por interpolaccedilatildeo determina-se o PM das proteiacutenas isoladas sabendo a sua
mobilidade relativa Por comparaccedilatildeo com os valores descritos na literatura pode concluir-se
acerca do grau de purificaccedilatildeo
34 Mediccedilatildeo da actividade enzimaacutetica de S1
341 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
13
tampatildeo de concentraccedilatildeo 05 M Tris-HCl pH 68 14
tampatildeo de amostra 320 mM Tris-HCl pH 68 04 M szlig-mercaptoetanol SDS 8 glicerol 15 (vv) e roxo de m-
cresol 0024 15
tampatildeo de electroforese da tina superior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS 16
tampatildeo de electroforese da tina inferior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS acetato de soacutedio
01 M 17
soluccedilatildeo corante ldquoCoomassie Blue R-250rdquo contendo metanol 45 (vv) e aacutecido aceacutetico glacial 10 (vv) 18
soluccedilatildeo descorante metanol 10 (vv) e aacutecido aceacutetico glacial 10 vv)
45
O meacutetodo de Fiske-Subarow modificado consiste na reacccedilatildeo do molibdato de amoacutenio com o
fosfato inorgacircnico em soluccedilatildeo aacutecida com formaccedilatildeo do aacutecido fosfomoliacutebdico Seguidamente daacute-
se a adiccedilatildeo de um agente redutor que provoca a reduccedilatildeo do composto a fosfomolibdato que forma
um complexo de cor azulada cuja absorvacircncia maacutexima ocorre para um comprimento de onda de
660 nm tornando possiacutevel a sua detecccedilatildeo por meacutetodos espectrofotomeacutetricos As actividades Ca2+
-
ATPaacutesica e K+EDTA-ATPaacutesica de S1 foram determinadas atraveacutes deste meacutetodo de acordo com
o descrito por Taussky e Shorr (1953) com mediccedilatildeo da quantidade de fosfato (Pi) libertado
atraveacutes da hidroacutelise de ATP
Para tal obteve-se primeiramente a recta de calibraccedilatildeo com as amostras presentes na Tabela 3II
utilizando uma soluccedilatildeo stock de 5 mM KH2PO4
Tabela 3II ndash Composiccedilatildeo das diferentes amostras utilizadas para determinar a recta de calibraccedilatildeo atraveacutes do meacutetodo
de Fiske-Subbarow modificado
KH2PO4 (L)
([KH2PO4]stock= 5mM)
PO43-
(nmole) H2O (uL)
0 0 1780
5 25 1775
10 50 1770
20 100 1760
40 200 1740
100 500 1680
A cada tubo de Eppendorf adicionaram-se seguidamente 200 L de soluccedilatildeo J19
e 20 L de agente
redutor20
homogeneizando no voacutertex apoacutes cada adiccedilatildeo Incubou-se durante 10 minutos e mediu-
se a Abs660 nm num espectrofotoacutemetro com intervalos de tempo iguais entre cada amostra Traccedilou-
se a recta de calibraccedilatildeo (Abs660 nmvs nmol PO43-
) a partir dos valores obtidos
Para determinar a actividade Ca2+
-ATPaacutesica de S1 prepararam-se tubos de Eppendorf com 400
L de meio de reacccedilatildeo21
25 L CaCl2 ([CaCl2]stock= 100 mM) 25 L ATP ([ATP]stock= 100 mM)
com [S1]ensaios=01 mgmL Perfez-se se necessaacuterio com H2O MilliQ ateacute perfazer um volume de 1
mL Para determinar a actividade K+EDTA-ATPaacutesica utilizou-se uma concentraccedilatildeo idecircntica de
19
Soluccedilatildeo J 5 N H2SO4 25 molibdato de amoacutenio 20
Agente redutor 5 mg aacutecido 1-amino-2-naftol-4-sulfoacutenico 30 mg bissulfito de soacutedio 30 mg sulfito de soacutedio em 26
mL H2O 21
meio de reacccedilatildeo 125 mM Tris-HCl pH 80 0575 M KCl
46
proteiacutena nos ensaios (01 mgmL) e o mesmo meio de reacccedilatildeo Aos tubos de Eppendorf
adicionou-se ainda 123 L de KCl ([KCl]stock = 3M) 25 L de ATP 10 L de EDTA
([EDTA]stock= 100 mM )e o volume de H2O MilliQ necessaacuterio para perfazer 1 mL Em ambas as
situaccedilotildees o uacuteltimo reagente adicionado foi o ATP uma vez que se trata do substrato da reacccedilatildeo
enzimaacutetica
A cineacutetica decorreu a 25ordmC durante 5 minutos para a actividade Ca2+
-ATPaacutesica e durante 3
minutos para a actividade K+EDTA-ATPaacutesica Apoacutes o periacuteodo de incubaccedilatildeo transferiu-se 150
L de amostra para tubos de Eppendorf contendo igual volume de TCA 10 Colocou-se no gelo
e homogeneizou-se no voacutertex Seguidamente centrifugou-se a 1500 g durante 10 minutos e
recolheu-se 200 L de sobrenadante para novos tubos de Eppendorf Adicionou-se a estes 158
mL de H2O MilliQ e 200 L de soluccedilatildeo J Homogeneizou-se no voacutertex e adicionou-se 20 L de
reagente redutor repetindo-se a homogeneizaccedilatildeo Incubou-se durante 10 minutos e mediu-se a
Abs660 nm com o mesmo intervalo de tempo de leitura para cada amostra Por intrapolaccedilatildeo a partir
da recta de calibraccedilatildeo obtida anteriormente determinou-se a quantidade de Pi libertado em cada
ensaio A partir destes resultados determinaram-se as actividades Ca2+
-ATPaacutesica e K+EDTA-
ATPaacutesica do S1 em nmol PO43-
minmg proteiacutena e em s-1
342 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
Da actividade Mg2+
-ATPaacutesica de S1 resulta a formaccedilatildeo dos produtos da reacccedilatildeo de
hidroacutelise do ATP - ADP e Pi Na presenccedila de ADP fosfoenolpiruvato (PEP) e da enzima piruvato
cinase (PK) os produtos resultantes satildeo ATP e piruvato Por acccedilatildeo da enzima lactato
desidrogenase (LDH) este uacuteltimo eacute convertido em lactato ocorrendo simultaneamente a oxidaccedilatildeo
do NADH a NAD+ monitorizada atraveacutes da Abs340 nm Quando o S1 de miosina se encontra no
meio reaccional o ATP eacute hidrolisado formando-se novamente ADP e Pi o que permite a
formaccedilatildeo de um ciclo (fig 21) Se as enzimas se encontrarem em excesso a velocidade de
conversatildeo do NADH eacute limitada pela produccedilatildeo de ADP e Pi Como o ATP eacute regenerado em cada
ciclo a sua concentraccedilatildeo inicial eacute sempre mantida (Bagshaw 1982)
Figura 33ndash Representaccedilatildeo esquemaacutetica do meacutetodo de enzimas acopladas
47
O protocolo utilizado e seguidamente descrito foi adaptado de Cuenda et al (1990 ndash Ver Refordf) A
uma cuvette de 1 mL adicionou-se S1 para uma concentraccedilatildeo final de 005 mgmL 800 L de
meio de reacccedilatildeo22
10 L de PEP ([PEP]stock = 42 mM) 10 L de PK+LDH 11 ([PK+LDH]stock =
1 mgmL) e 15 L de NADH ([NADH]stock=25 mM) Iniciou-se a monitorizaccedilatildeo da reacccedilatildeo
atraveacutes da leitura de Abs340 nm apoacutes a adiccedilatildeo de 20 L de ATP ([ATP]stock=100 mM) A
actividade Mg2+
-ATPaacutesica eacute obtida atraveacutes do declive correspondente agrave oxidaccedilatildeo do NADH Para
a mediccedilatildeo da Mg2+
-ATPase de S1 estimulada por actina adicionou-se F-actina para uma
concentraccedilatildeo final de 2 microM sendo a actividade obtida tambeacutem atraveacutes do declive correspondente
agrave oxidaccedilatildeo do NADH apoacutes a adiccedilatildeo da proteiacutena
35 Interaccedilatildeo entre vanaacutedio e subfragmento-1 de miosina
351 Preparaccedilatildeo das soluccedilotildees de vanaacutedio
As soluccedilotildees de vanadato foram preparadas em aacutegua MilliQ a partir de metavanadato de amoacutenio
(NH4VO3) numa concentraccedilatildeo stock de 50 mM com agitaccedilatildeo magneacutetica e aquecimento (~30 ordmC)
Esta soluccedilatildeo com um valor compreendido entre 6 e 7 designa-se por ldquometavanadatordquo e eacute
constituiacuteda pelas esspeacutecies V1 V2 V4 e V5 (Ref) Acidificando a soluccedilatildeo de metavanadato por
adiccedilatildeo de HCl ateacute um valor de pH proacuteximo de 4 observa-se uma mudanccedila de cor de amarelo
claro para laranja indicativa da formaccedilatildeo da espeacutecie decameacuterica de vanadato De seguida e
imediatamente antes da realizaccedilatildeo dos ensaios descritos o valor do pH da soluccedilatildeo de
decavanadato foi aumentado para cerca de 75 por adiccedilatildeo de NaOH A soluccedilatildeo (2) designada
por ldquometavanadatordquo natildeo foi posteriormente alterada ateacute agrave sua adiccedilatildeo para a realizaccedilatildeo dos ensaios
As soluccedilotildees foram armazenadas a 4ordmC e mantidas em gelo aquando da sua utilizaccedilatildeo
A soluccedilatildeo de V(IV) foi preparada em condiccedilotildees anaeroacutebias por diluiccedilatildeo de sulfato de vanadil
(VOSO4) para uma concentraccedilatildeo final de 5 mM em H2O MilliQ saturada com N2 gasoso
352Anaacutelise por espectroscopia de UV-Vis
A concentraccedilatildeo da espeacutecie decameacuterica em soluccedilatildeo depende do periacuteodo de tempo apoacutes a diluiccedilatildeo
no meio de reacccedilatildeo apropriado (Ref) A velocidade de desoligomerizaccedilatildeo da espeacutecie decameacuterica
pode ser analisada por espectroscopia de absorccedilatildeo na regiatildeo UVVis uma vez que esta espeacutecie
apresenta duas bandas caracteriacutesticas na regiatildeo do visiacutevel (a 358 e 400 nm) que natildeo se verifica
em quaisquer outras espeacutecies oligomeacutericas As bandas supramencionada sapresentam-se bem
22
Meio de reacccedilatildeo 3125 mM Tris-HCl pH 71 375 mM MgCl2
48
resolvidas atraveacutes da obtenccedilatildeo da derivada de 2ordf ordem do espectro apresentando a forma de 2
vales com miacutenimos centrados nos comprimentos de onda supracitados com dependecircncia linear
relativamente agrave concentraccedilatildeo de decavanadato total (Ramos et al 2006)
Assim a estabilidade da espeacutecie decameacuterica apoacutes diluiccedilatildeo da soluccedilatildeo de decavanadato no meio de
reacccedilatildeo dos ensaios (tampatildeo D) para uma concentraccedilatildeo final de 1 mM em vanadato total foi
caracterizada atraveacutes da leitura da Abs400 em funccedilatildeo do tempo tendo-se determinado qual a
ordem de reacccedilatildeo e o tempo de meia-vida da mesma Com base em estudos anteriores acerca da
estabilidade do V10 na presenccedila de diferentes proteiacutenas (Ramos et al 2006) determinou-se
igualmente o tempo de meia vida apoacutes a adiccedilatildeo de 20 microM de S1 de miosina ao meio reaccional
353 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
Os espectros de RMN do isoacutetopo 51
V foram realizados num espectroacutemetro Bruker AM-400 MHz
equipado com uma sonda multinuclear inversa de 5 mm Em tubos de RMN de 5 mm de diacircmetro
preparou-se soluccedilotildees de 500 L de deca- ou metavanadato correspondentes a um intervalo de
concentraccedilotildees de 1 a 10 mM suplementadas com 10 D2O Os espectros foram adquiridos agrave
temperatura ambiente com uma largura espectral de 23 923 Hz um tempo de aquisiccedilatildeo de 0086
s tempo de relaxaccedilatildeo de 001 s e uma sequecircncia de pulsos a 90ordm Utilizou-se a transformada de
Fourier com alargamento das bandas de 20Hz para o processamento dos espectros Os desvios
quiacutemicos das ressonacircncias de 51
V foram obtidos em relaccedilatildeo agrave espeacutecie VOCl3cuja frequecircncia de
ressonacircncia eacute considerada o padratildeo sendo todas as outras referenciadas a partir desta em ppm
As aacutereas dos sinais do RMN correspondentes agraves espeacutecies oligomeacutericas presentes foram obtidas
por integraccedilatildeo dos sinais As concentraccedilotildees relativas das espeacutecies foram calculadas com recurso agrave
equaccedilatildeo
onde A[Vx] corresponde agrave aacuterea referente agrave ressonacircncia da espeacutecie Vx com nordm de agregaccedilatildeo x
A[V]total agrave soma total das aacutereas medidas e [V] a concentraccedilatildeo total de vanaacutedio no ensaio
A teacutecnica de 51
V-RMN foi anteriormente aplicada em estudos de ligaccedilatildeo com S1 de miosina
(Ref) tendo sido demostrado que entre as espeacutecies V1 V2 V4 e V5 a espeacutecie tetrameacuterica possui a
maior afinidade para a miosina (Cremo et al) O desaparecimento ou alargamento dos sinais pode
implicar a ligaccedilatildeo de um dos oxovanadatos laacutebeis agrave miosina sem indicaccedilatildeo de qual uma vez que
49
se processa um equiliacutebrio entre eles (Crans 1990) Assim e tendo em conta as indicaccedilotildees de que
a espeacutecie V10 eacute distinta das restantes no que diz respeito agrave magnitude da interacccedilatildeo com a miosina
e inibiccedilatildeo da actividade ATPaacutesica estimulada por actina (Aureliano 2000 Tiago et al 2002)
torna-se fundamental investigar a interacccedilatildeo das diferentes espeacutecies de vanadato com o S1 de
miosina atraveacutes da teacutecnica de 51
V-RMN
Para tal obtiveram-se espectros de 51
V-RMN resultantes da titulaccedilatildeo de uma soluccedilatildeo 5 mM de
decavanadato total com concentraccedilotildees crescentes de S1 de miosina (0 5 10 15 e 20 microM) em
tampatildeo D Realizou-se ainda ensaios na presenccedila de 2 mM de decavanadato e 2 mM de ATP na
presenccedila e na ausecircncia de S1 de miosina de modo a verificar o efeito que estes ligandos naturais
da enzima teria na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o subfragmento-1
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
Os grupos sulfidrilo (-SH) das cisteiacutenas satildeo resiacuteduos de aminoaacutecidos extremamente sensiacuteveis agrave
oxidaccedilatildeo resultando na sua oxidaccedilatildeo (Dalle-Donne et al 1999) Logo a quantificaccedilatildeo destes
resiacuteduos eacute bastante importante nos estudos de interacccedilatildeo com vanaacutedio jaacute que os resiacuteduos de Cys
com ecircnfase para o SH1 e o SH2 satildeo bastante importantes na monitorizaccedilatildeo das modificaccedilotildees
funcionais da regiatildeo da cabeccedila
O procedimento seguidamente apresentado foi originalmente descrito por Gutieacuterrez-Merino
(1987)) Consiste na reacccedilatildeo entre os grupos ndashSH natildeo oxidados das proteiacutenas e o reagente de
Ellman (DTNB -55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico)) - formando-se o aniatildeo CNT (aniatildeo 3-
carboxilato-4-nitrotiofenolato) numa estequiometria de 11 Este apresenta um maacuteximo de
absorvecircncia a 412 nm (Robyt 1990) Assim apoacutes a incubaccedilatildeo da proteiacutena na presenccedila do metal
a quantidade de cisteiacutenas reduzidas por proteiacutena determina-se atraveacutes dos valores de Abs412
Deste modo colocou-se num tubo de Eppendorf o volume apropriado de S1 dialisado para que a
concentraccedilatildeo no ensaio fosse de 1 mgmL Adicionou-se o volume apropriado de decavanadato
para que a sua concentraccedilatildeo fosse de 0 10 50 100 250 500 ou 1000 M Perfez-se com soluccedilatildeo
D ateacute um volume final de 1 mL e incubou-se a 25ordmC por 20 minutos Apoacutes este periacuteodo colocou-
se 780 L da mistura reaccional numa cuvette de quartzo e adicionou-se 220 L de soluccedilatildeo K23
23
Tampatildeo K Tampatildeo fosfatohidrogenofosfato de soacutedio 025 M pH 72
50
Adicionou-se 15 L de DTNB (em soluccedilatildeo K [DTNB]stock=100 mM) ao meio reaccional e
registou-se o valor de absorvecircncia medido equivalente agraves cisteiacutenas ldquoraacutepidasrdquo (SH1 e SH2
expostos ao solvente) Adicionou-se 001 g de SDS que provoca a desnaturaccedilatildeo da proteiacutena e
consequentemente a exposiccedilatildeo de todos os resiacuteduos de cisteiacutenas que se encontravam ateacute aiacute
inacessiacuteveis no interior da proteiacutena (cisteiacutenas totais) Estes resiacuteduos podem assim reagir com o
DTNB
Estimou-se o nuacutemero de resiacuteduos de cisteiacutenas em todas as amostras com recurso ao coeficiente de
extinccedilatildeo molar () do DTNB (ver ponto 3 dos Anexos) que foi determinado atraveacutes da titulaccedilatildeo
de L-Cisteiacutena com intervalo de concentraccedilotildees compreendidos entre 0 e 50 microM na presenccedila de
25microM de DTNB em tampatildeo D Os valores de absorvecircncia foram obtidos para um comprimento de
onda de 412 nm
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
3621 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
A teacutecnica mais eficiente de anaacutelise da interacccedilatildeo entre o iatildeo vanadilo e proteiacutenas como a
calmodulina e a transferrina entre outras permite a obtenccedilatildeo raacutepida de informaccedilatildeo no que toca agrave
conformaccedilatildeo e amplitude da referida interacccedilatildeo
Os espectros de Ressonacircncia Paramagneacutetica Electroacutenica (RPE) da espeacutecie paramagneacutetica V(IV)
foram realizados a 70K num espectroacutemetro Bruker EMX-300 X-band Todas as soluccedilotildees
utilizadas e amostras foram preparadas em anaerobiose numa cacircmara de luvas de modo a evitar a
reduccedilatildeo de V(IV) a V(V) Em tubos capilares com 02 mL de amostra manteve-se a concentraccedilatildeo
de S1 a 20 M e variou-se a concentraccedilatildeo de vanadilo (0 25 5 10 15 25 50 75 100 150
200 500 e 1000 M [VOSO4]stock=5 mM) em tampatildeo D Apoacutes obtenccedilatildeo dos espectros estes
foram tratados atraveacutes do software SimFonia e WinEPR tendo-se obtido um graacutefico com a
variaccedilatildeo da intensidade do sinal de RPE em funccedilatildeo da concentraccedilatildeo de sulfato de vanadilo
presente Realizaram-se ainda simulaccedilotildees de espectros obtidos com 20 microM de S1 e 1000 microM de
V(IV) com vista agrave obtenccedilatildeo dos ligandos provaacuteveis do vanadilo atraveacutes da anaacutelise da constante
de acoplamento hiperfina (A) e do factor g (g)
4 Conclusotildees gerais
Apesar de jaacute ter sido demonstrado que o vanaacutedio inibe a Na+K
+-ATPase da miosina a funccedilatildeo
deste elemento no tecido muacutesculo esqueleacutetico continua largamente incompreendida A enorme
complexidade quiacutemica deste metal especialmente no que concerne agrave oligomerizaccedilatildeo e reacccedilotildees
51
redox constitui um obstaacuteculo agrave compreensatildeo do comportamento do vanaacutedio nos organismos
vivos Assim a caracterizaccedilatildeo da quiacutemica deste elemento eacute de grande utilidade antes de iniciar o
estudo da sua interacccedilatildeo com o presente sistema
A anaacutelise por espectroscopias de 51
V-RMN e de UV-Vis de soluccedilotildees de vanadato diluiacutedas nos
meios de reacccedilatildeo utilizados no presente relatoacuterio (10 mM HEPES pH 70 25 mM KCl) mostrou
claramente que a espeacutecie decameacuterica (V10) possui um tempo de vida suficientemente longo para a
determinaccedilatildeo do seu efeito isolado ao contraacuterio do que se verifica com as espeacutecies mono- di-
tetra- e pentameacutericas (V1 V2 V4 e V5) Por outro lado a anaacutelise verificou ainda que apoacutes diluiccedilatildeo
das soluccedilotildees de decavanadato no meio de reacccedilatildeo coexistem as espeacutecies V10 e V1 pelo que os
efeitos putativos desta uacuteltima deveratildeo ser subtraiacutedos aquando da monitorizaccedilatildeo dos efeitos da
espeacutecie decameacuterica
O vanaacutedio (V) (vanadato) inibe as enzimas que catalizam reacccedilotildees de transferecircncia de grupos
fosfato como a ATPase da miosina [Goodno 1979] devido agrave sua analogia estrutural com o iatildeo
fosfato Na sua utilizaccedilatildeo como inibidor enzimaacutetico natildeo eacute tida em conta a contribuiccedilatildeo individual
das espeacutecies oligomeacutericas considerando-se normalmente a espeacutecie V1 como inibidora efectiva
[Goodno e Taylor 1982] No presente relatoacuterio verificou-se por espectroscopia de 51
V-RMN que
outros oligoacutemeros de vanadato interagem com o S1 de miosina tendo as espeacutecies V10 e V4 os
maiores niacuteveis de afinidade A sua interacccedilatildeo envolve forccedilas electrostaacuteticas e eacute afectada
negativamente por ligandos naturais de enzima como eacute o caso do ATP
A exposiccedilatildeo do S1 de miosina (005 mgmL) a decavanadato conduz agrave inibiccedilatildeo do capacidade da
proteiacutena de estimular a actividade Mg2+
-ATPaacutesica com um valor de IC50 de 266 plusmn 07 M
decavanadato sendo o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM
decavanadato (80) Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM decavanadato sendo
o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM decavanadato (60)
exercendo um efeito de impedimento agrave ligaccedilatildeo entre o vanaacutedio e o S1
A uma concentraccedilatildeo de 27 microM decavanadato verifica-se a oxidaccedilatildeo de um dos resiacuteduos de Cys
de S1 de miosina seguida pela oxidaccedilatildeo gradual de outro resiacuteduo de Cys ateacute agrave concentraccedilatildeo de
1000 microM Ambos os resiacuteduos oxidados natildeo se encontravam expostos ao solvente utilizado A
oxidaccedilatildeo dos resiacuteduos permite sugerir que esta modificaccedilatildeo oxidativa sofrida pelo S1 contribui
para a diminuiccedilatildeo da sua actividade de ATPase
52
Por espectroscopia de RPE revelaram-se os possiacuteveis ligandos equatoriais do vanaacutedio aquando da
sua interacccedilatildeo com o S1 de miosina como sendo duas moleacuteculas de aacutegua um ligando contendo o
grupo R-OH e outro contendo o grupo R=NR
Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados de
oxidaccedilatildeo (IV) e (V) no S1 de miosina para concentraccedilotildees facilmente alcanccedilaacuteveis no ambiente
terrestre
53
5 Estudos futuros
No sentido do melhor esclarecimento das alteraccedilotildees moleculares e estruturais que ocorrem no
subfragmento 1 quando exposta ao vanaacutedio que conduzem a uma diminuiccedilatildeo da sua funccedilatildeo
sugere-se
- Quantificaccedilatildeo das cisteiacutenas reduzidas de S1 na presenccedila de metavanadato
- Determinaccedilatildeo das actividades Ca2+
- e K+-ATPaacutesica do S1 de miosina na presenccedila de
decavanadato
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina estimulada por actina
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina na presenccedila de
metavanadato
- Realizaccedilatildeo de RMN 2D com variaccedilatildeo das concentraccedilotildees das soluccedilotildees de decavanadato e
metavanadato na presenccedila de S1
-Mutageacutenese dos resiacuteduos de Cys doS1 de miosina de modo a verificar quais os resiacuteduos que
sofrem oxidaccedilatildeo na presenccedila de decavanadato
- Determinaccedilatildeo por calorimetria (DSC Differential Scanning Calorimetry) da variaccedilatildeo da
entalpia (∆H) e da temperatura de desnaturaccedilatildeo do S1 de miosina na presenccedila de vanaacutedio uma
vez que estas propriedades caracteriacutesticas de cada proteiacutena satildeo alteradas apoacutes ocorrecircncia de
modificaccedilotildees estruturais
- Anaacutelise por SDS-PAGE de S1 de miosina apoacutes tratamento com vanaacutedio de modo a determinar
quais os polipeacuteptidos eventualmente formados apoacutes clivagem da proteiacutena
54
6 Referecircncias bibliograacuteficas
- Sellers J Myosins 2nd Edition Oxford University Press 1999
- L Stryer inBiochemistry (Ed WH Freeman 2005 pp
Refordfas Materiais e Meacutetodos
[A] S S Margossian SLowey Methods Enzymol1982 85 55-71
[B] JD Pardue JA Spudich Methods Enzymol 1982 85 164-181
[C] M Bradford Anal Biochem1976 72 248-254
[D] H H Taussky E Shorr J Biol Chem 1953 202 675-685
[E] C R Bagshaw in Outline Studies in Biology ndash Muscle Contraction Chapman and Hall Ed
1982
[F] A Cuenda et al Eur J Biochem1990 194 663-670
55
Anexos
1 Determinaccedilatildeo da recta de calibraccedilatildeo meacutetodo de Bradford
Figura1ndashRepresentaccedilatildeo graacutefica da recta de calibraccedilatildeo (Abs595 nm vs [BSA]) utilizada para o meacutetodo de Bradford
2 Determinaccedilatildeo da recta calibraccedilatildeo para a electroforese em condiccedilotildees desnaturantes
Figura 2 ndash Representaccedilatildeo graacutefica da recta de calibraccedilatildeo (log(Peso Molecular) vs Rf dos marcadores de peso
molecular) onde Rf indica determinada atraveacutes do padratildeo de peso molecular
(poccedilo (a))
3Caacutelculo do coeficiente de extinccedilatildeo molar () para a quantificaccedilatildeo de cisteiacutenas reduzidas
atraveacutes do meacutetodo de Ellman
Traccedilou-se a recta de Abs412 nm meacutedia vs [L-Cisteiacutena] de modo a determinar o valor do coeficiente
de extinccedilatildeo molar definido como o declive daquela recta Assim =8400 M-1
cm-1
como
podemos verificar na Fig 3
A595 nm = 00546[BSA] + 00209 Rsup2 = 099051
Ab
s 59
5 n
m
[BSA] (mgmL)
log PM = -1177 Rf + 2247 Rsup2 = 098957
log
PM
Mobilidade relativa (Rf)
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero

25
precipitado O meacutetodo descrito traduz de forma bastante elementar o fenoacutemeno da contracccedilatildeo in
vivo e elimina o problema anterior da solubilizaccedilatildeo ineficiente da miosina de modo a obter uma
mediccedilatildeo exacta da actividade [A] A utilizaccedilatildeo de forccedilas ioacutenicas reduzidas aumenta a afinidade da
actina para o S1 permitindo contornar a extrema dificuldade em obter a concentraccedilatildeo proteica dos
muacutesculos (cerca de 100 mgmL do complexo actomiosina) As actividades Mg2+
-ATPaacutesicas de S1
de miosina foram determinadas a partir do declive da variaccedilatildeo da Abs340 relativo agrave oxidaccedilatildeo de
NADH em funccedilatildeo do tempo como eacute descrito no ponto 342 do Procedimento Experimental Os
valores obtidos para a actividade Mg2+
-ATPaacutesica encontram-se apresentados na Tabela 2II
Tabela 2II ndash Actividade Mg2+
-ATPaacutesica de subfragmento-1 de miosina determinadas pelo meacutetodo das enzimas
acopladas
Actividade (nmol
ATPmin mg(S1)) Actividade (s
-1)
1097 plusmn 269 164 plusmn 04
Ramos (2005) [J] obteve o valor de 01 s-1
para a actividade Mg2+
-ATPaacutesica de S1 de miosina
com forccedila ioacutenica intermeacutedia Tiago (2005) [BC] obteve o valor de 011 s-1 em condiccedilotildees de forccedila
ioacutenica reduzida em relaccedilatildeo agrave utilizada no presente relatoacuterio
23 Preparaccedilatildeo e caracterizaccedilatildeo das soluccedilotildees de vanaacutedio
231 Anaacutelise da composiccedilatildeo das soluccedilotildees de vanadato por espectroscopia de 51
V-RMN
efeito da concentraccedilatildeo da espeacutecie
A diluiccedilatildeo das soluccedilotildees stock de meta- e decavanadato iraacute determinar fortemente a composiccedilatildeo
em soluccedilatildeo dos diferentes oligoacutemeros de vanadato (ref) No caso do decavanadato o periacuteodo de
tempo apoacutes a diluiccedilatildeo iraacute tambeacutem ser um factor importante a considerar devido agrave decomposiccedilatildeo
da espeacutecie V10(ref)Torna-se assim fundamental determinar a composiccedilatildeo quiacutemica das soluccedilotildees
no meio de ensaio utilizado (HEPES 10 mM KCl 25 mM) de modo a correlacionaacute-la
correctamente com os possiacuteveis efeitos promovidos pelas diferentes espeacutecies de vanadato no
sistema em estudo
Aleacutem disso deve ser dado ecircnfase agraves importantes interacccedilotildees que ocorrem entre o vanaacutedio e os
tampotildees escolhidos O tampatildeo com menor nuacutemero e magnitude de interacccedilatildeo eacute o HEPES [AD]
seguido do Tris sendo este uacuteltimo especialmente aconselhado em concentraccedilotildees baixas para casos
de incompatibilidade enzimaacutetica ou quiacutemica redox No caso especial da reacccedilatildeo redox
NAD(P)HNAD(P) o tampatildeo adequado eacute o imidazole de modo a natildeo haver reacccedilatildeo com o
26
vanadato Os tampotildees a evitar em qualquer situaccedilatildeo incluem a trietanolamina fosfato citrato e
glicina entre outros [AD] uma vez que todos interactuam fortemente com V(V) com formaccedilatildeo
de complexos estaacuteveis
Assim procedeu-se agrave anaacutelise da composiccedilatildeo das soluccedilotildees de meta- e decavanadato apoacutes diluiccedilatildeo
das soluccedilotildees stock (50 mM em vanadato total) no meio de reacccedilatildeo (tampatildeo D) atraveacutes da
espectroscopia de ressonacircncia magneacutetica nuclear de 51
V Nas figuras x1 e x2 apresentam-se os
espectros obtidos para as soluccedilotildees de deca- e metavanadato num intervalo de concentraccedilotildees de 1 a
10 mM A Tabela seguinte resume os paracircmetros espectroscoacutepicos obtidos para cada espeacutecie
presente nas soluccedilotildees analisadas
Figura 22ndash Espectros de RMN de 51
V de
concentraccedilotildees crescentes das espeacutecies oligomeacutericas
de vanadato obtidos apoacutes diluiccedilatildeo da soluccedilatildeo stock
de decavanadato em 10 mM HEPES pH 70 25 mM
KCl para concentraccedilotildees finais de A) 1 mM B)2
mM C) 4 mM D) 8 mM E) 10 mM de
decavanadato total V1refere-se agraves espeacutecies
monomeacutericade vanadato V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2IIIndash Desvios quiacutemicos meacutedios (part) das ressonacircncias das espeacutecies decameacuterica (V10) monomeacuterica (V1) e
tetrameacuterica (V4) presentes nos espectros da soluccedilatildeo de decavanadatoLargura a meia altura (∆12) da espeacutecie V10A
Paracircmetros V10A V10B V10C V1 V4
part (ppm) -5162 -5003 -4255 -5616 -5748
∆12 (Hz) 175 nd nd nd nd
Figura 23 ndash Distribuiccedilatildeo graacutefica das
espeacutecies oligomeacutericas de vanadato vs a
concentraccedilatildeo de decavanadato total As
concentraccedilotildees de cada espeacutecie foram
obtidas segundo a formula descrita na
secccedilatildeo 35 do Procedimento
Experimental (aacutereas dos sinais de 51
V-
RMN observadas na figura x1)V10 V1 e
V4 correspondem agraves espeacutecie monomeacuterica
dimeacuterica e tetrameacuterica respectivamente [Esp
eacuteci
e o
ligo
meacute
rica
de
van
aacutedio
] (m
M)
[Decavanadato]total (mM)
V10
V1
V4
27
Nas soluccedilotildees de decavanadato as espeacutecies monomeacuterica e decameacuterica de vanadato satildeo as uacutenicas
presentes para concentraccedilotildees de decavanadato inferiores a 8 mM Ambas as espeacutecies aumentam
linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado em [AY] e
em (Teresa) quando a concentraccedilatildeo de V1 eacute igual ou superior a 200 microM inicia-se a sua
oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica No entanto nas condiccedilotildees de realizaccedilatildeo dos
ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a espeacutecie tetrameacuterica neste
estudo em concentraccedilotildees inferiores a 8 mM tendo o mesmo sido observado por Ramoset al
(2006) [AK] Os resultados obtidos mostram claramente o equiliacutebrio entre a espeacutecie decameacuterica e
a monomeacuterica e que o aparecimento de outras espeacutecies oligomeacutericas se encontra dependente da
concentraccedilatildeo desta uacuteltima Apesar de natildeo ter sido realizado o ensaio com 5 mM de decavanadato
total no presente relatoacuterio estaacute reportado em Ramos et al (2006) [AK] que as concentraccedilotildees da
espeacutecie decameacuterica e monomeacuterica satildeo respectivamente de 500 microM e 200 microM Assim pode inferir-
se que para concentraccedilotildees inferiores a 5 mM e intervalos de tempo de cerca de 20 minutos (que
inclui o tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas
as espeacutecies decameacuterica e monomeacuterica de vanadato
Figura 24 ndash Espectros de RMN de 51
V de concentraccedilotildees
crescentes das espeacutecies oligomeacutericas de vanadato obtidos
apoacutes diluiccedilatildeo da soluccedilatildeo stock de metavanadato em 10 mM
HEPES pH 70 25 mM KCl para concentraccedilotildees finais de A)
1 mM B) 2 mM C) 4 mM D) 8 mM E) 10 mM de
metavanadato total V1 refere-se agraves espeacutecies monomeacuterica de
vanadatoV1 e V2 correspondem respectivamente agrave espeacutecie
monomeacuterica e dimeacuterica
Figura 25 ndash Distribuiccedilatildeo graacutefica das espeacutecies
oligomeacutericas de vanadato vs a concentraccedilatildeo de
metavanadato total As concentraccedilotildees de cada
espeacutecie foram obtidas segundo a formula descrita
na secccedilatildeo 35 do Procedimento Experimental
(aacutereas dos sinais de 51
V-RMN observadas na
figura x1)V10 V1 e V4 correspondem agraves espeacutecie
monomeacuterica dimeacuterica e tetrameacuterica
respectivamente
Esp
eacuteci
e o
ligo
meacute
rica
de
va
nad
ato
(m
M)
[Metavanadato]total (mM)
V1
V2
V4
V5
28
Tabela 2IVndash Desvios quiacutemicos meacutedios (part)das ressonacircncias das espeacutecies tetrameacuterica (V4) monomeacuterica (V1) e
dimeacuterica (V2) presentes no espectro da soluccedilatildeo de metavanadato (total) 10 mM
Paracircmetros V1 V2 V4 V5
part (ppm) -5615 -5748 -5789 -5869
Nas soluccedilotildees de metavanadato as espeacutecies monomeacuterica dimeacutericae tetrameacuterica de vanadato estatildeo
presentes para concentraccedilotildees de metavanadato inferiores a 4 mM Todas estas as espeacutecies
aumentam linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado
em (Aureliano e Madeira 1998) e em (Tiago et al 2004) quando a concentraccedilatildeo de V1 eacute igual ou
superior a 200 microM inicia-se a sua oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica Nas
condiccedilotildees de realizaccedilatildeo dos ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a
espeacutecie pentameacuterica neste estudo em concentraccedilotildees inferiores a 4 mM tendo o mesmo sido
observado por Ramos 2006 Os resultados obtidos mostram claramente o equiliacutebrio entre a
espeacutecie V4 e as espeacuteciesV1 e V2 no qual o aparecimento de outras espeacutecies oligomeacutericas se
encontra dependente da concentraccedilatildeo da espeacutecie V1 Por uacuteltimo pode inferir-se que para
concentraccedilotildees inferiores a 2 mM e intervalos de tempo de cerca de 20 minutos (que inclui o
tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas as
espeacutecies V1 V2 e V4 com concentraccedilotildees respectivas de 0796 0192 e 0196 mM
232Estabilidade das soluccedilotildees de decavanadato anaacutelise por espectroscopia de absorccedilatildeo
UVVis
Caracterizou-se a estabilidade da espeacutecie V10 numa soluccedilatildeo com 1 mM decavanadato total em
tampatildeo D atraveacutes da leitura da Abs400 Obteve-se uma cineacutetica de dissociaccedilatildeo de 1ordf ordem tal
como se pode verificar pela linearidade do graacutefico de ln(Abs400)vs o tempo de aquisiccedilatildeo espectral
(segundos) (Fig26 )
Figura 26ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do tempo
(em segundos) para uma soluccedilatildeo de decavanadato total de 1
mM em tampatildeo D a 25ordmC (n=2)
Sendo uma reacccedilatildeo de primeira ordem pode ser descrita pela equaccedilatildeo cujo
declive eacute ndashK (constante de dissociaccedilatildeo) e a intersecccedilatildeo no eixo das ordenadas corresponde
Ln
(Ab
s 400 n
m)
tempo (s)
29
aln(A0) Numa reacccedilatildeo de 1ordf ordem o tempo de meia-vida eacute independente da concentraccedilatildeo inicial
do reagente sendo descrito pela equaccedilatildeo [2] Deste modo a partir da recta
Ln(Abs400 nm) = -436E-5
K - 105 obteve-se um tempo de meia-vida de 4490 plusmn 0823 horas para
as condiccedilotildees supramencionadas
Apoacutes a adiccedilatildeo de S1 8 microM ao meio reaccional verificou-se que a reacccedilatildeo de desoligomerizaccedilatildeo
da espeacutecie decameacuterica de vanadato seguia igualmente uma cineacutetica de 1ordf ordem tal como se pode
verificar pela linearidade do graacutefico de ln(Abs400) versus o tempo de aquisiccedilatildeo espectral
(segundos) (Fig27)
Figura 27ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do
tempo (em segundos) para uma soluccedilatildeo de decavanadato
total de 1 mM em tampatildeo D a 25ordmC na presenccedila de 8
microM (n= 2)
A partir da equaccedilatildeo da recta Ln(Abs400 nm) = -7520E-5
K - 0613determinou-se o valor de K para
a espeacutecie decameacuterica de onde se obteve um tempo de meia-vida de 2430 plusmn 0191 horas para as
condiccedilotildees supramencionadas
Tanto este valor como o valor registado na ausecircncia de S1 satildeo substancialmente superiores agrave
duraccedilatildeo dos ensaios de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina descritos no presente
relatoacuterio
24 Interacccedilatildeo de S1 com vanaacutedio(V)
241Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
De modo a analisar qual o efeito da adiccedilatildeo de S1 de miosina (tampatildeo D) produzia nos sinais de
51V-NMR titulou-se uma soluccedilatildeo de 5mM de decavanadato total com concentraccedilotildees crescentes
de S1 (5 10 15 e 20 microM) Os espectros de 51
V-RMN encontram-se apresentados na Fig 2
No graacutefico da figura 2 Apresenta-se a concentraccedilatildeo de S1 em funccedilatildeo do factor de alargamento
corrigido dos espectros acima referidos
Ln
(Ab
s (4
00
nm
))
tempo (s)
30
Figura 28 ndash Graacutefico de
representaccedilatildeo da
concentraccedilatildeo de S1 em
funccedilatildeo do factor de
alargamento A
concentraccedilatildeo de
decavanadato total foi de 5
mM e as concentraccedilotildees
utilizadas de S1 foram de 0
5 10 e 15 microM
Verifica-se que o factor de alargamento aumenta linearmente em funccedilatildeo da concentraccedilatildeo de S1 no
ensaio o que indica a existecircncia de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina Observou-
se ainda que a adiccedilatildeo de S1 ao meio reaccional natildeo provocou qualquer alteraccedilatildeo dos desvios
quiacutemicos dos sinais de 51
V-RMN
Realizaram-se ensaios na presenccedila de ATP e de S1 com intuito de verificar o efeito do ATP na
interacccedilatildeo do S1 de miosina com o decavanadato
Figura 29 - Espectros de RMN de 51
V contendo 2
mM Decavanadato (A) (controlo) (B) na presenccedila
de 2 mM de ATP (C) na presenccedila de 2 mM de ATP
e 8 microM de S1 de miosina V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2Vndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies decameacuterica (V10) e monomeacuterica (V1) presentes nos
espectros da soluccedilatildeo de decavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V10A V10B V10C V1
part (ppm) -5161 -5002 -4254 -5614
Observa-se que a presenccedila de ATP (espectro B) natildeo provoca qualquer alteraccedilatildeo visiacutevel em
relaccedilatildeo ao espectro que contem apenas soluccedilatildeo de decavanadato Jaacute a presenccedila simultacircnea de ATP
e S1 provoca um alargamento da ∆12 do sinal do V10A de 1736 Hz (espectro (A)) para 2366 Hz
(espectro (C) Fig 29) Tendo em conta que a concentraccedilatildeo de S1 apresenta uma relaccedilatildeo linear
com o factor de alargamento converteu-se o valor da ∆12 e comparou-se com o valor esperado agrave
F = 00591[S1] - 00157 Rsup2 = 0998
Fact
or
de
ala
rgam
en
to
[S1] (microM)
31
concentraccedilatildeo de S1 utilizada O factor de alargamento obtido para 8 microM ndash 0376 ndash eacute inferior ao
valor esperado agrave mesma concentraccedilatildeo de S1 obtido atraveacutes da equaccedilatildeo da recta apresentada na
Fig 28 ndash 0457 ndash o que indica que o ATP poderaacute exercer um efeito de protecccedilatildeo do S1
diminuindo a sua interacccedilatildeo com a espeacutecieV10
Por uacuteltimo realizaram-se ensaios com metavanadato na presenccedila de 8 microM S1 de modo a verificar
se as espeacutecies oligomeacutericas contidas nesta soluccedilatildeo interagiam com a proteiacutena em estudo
Figura 210 - Espectros de RMN de 51
V contendo 2
mM metavanadato (A) (controlo) (B) na presenccedila
de 8 microM de S1 de miosina V1 V2 e V4
correspondem respectivamente aacutes espeacutecies
monomeacuterica dimeacuterica e tetrameacuterica
Tabela 2VIndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies monomeacuterica (V1) dimeacuterica (V2) e tetrameacuterica (V4)
presentes nos espectros da soluccedilatildeo de metavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V1 V2 V4
part (ppm) -5613 -5747 -5789
Como eacute possiacutevel observar na Fig 210 ocorre uma ligeiro alargamento dos sinal de V4 bem
como uma perda da intensidade pelo que se conclui quedas espeacutecies presentes na soluccedilatildeo de
metavanadato (V1 V2 e V4) apenas a espeacutecie tetrameacuterica interage com o S1 de miosina o que
estaacute de acordo com o reportado por Tiago et al 2002
242 Actividade Mg2+
-ATPaacutesica de S1 de miosina na presenccedila de decanadato
Com o intuito de avaliar o efeito do decavanadato na actividade Mg2+
-ATPaacutesica do S1 de
miosina monitorizou-se a reacccedilatildeo de oxidaccedilatildeo do NADH atraveacutes da leitura de Abs340 na
presenccedila de 005 mgmL de S1 de miosina e no caso da actividade estimulada 2microM de actina em
3125 mM Tris-HCl pH 71 e 375 mM MgCl2 pelo meacutetodo de enzimas acopladas (ver seccccedilatildeo xx
dos MM)Para tal realizaram-se ensaios com vaacuterias concentraccedilotildees de decavanadato total (10 50
100 250 500 e 1000) de modo a determinar qual o seu efeito na actividade MgATPaacutesica de S1
de miosina na ausecircncia e na presenccedila de um dos seus ligandos naturais Os ensaios foram
realizados agrave temperatura ambiente com incubaccedilatildeo preacutevia a 25ordm C durante 20 minutos
32
Figura 211 ndash Representaccedilatildeo
graacutefica da Actividade Mg2+
-
ATPaacutesica de S1 () (os ajustes
teoacutericos estatildeo representado a por
uma linha os resultados
experimentais estatildeo representados
por triacircngulos (Actividade
estimulada) e cruzes (Actividade
natildeo estimulada)) vs
[Decavanadato]total medidas pelo
meacutetodo das enzimas acopladas
(n=3) Os dados foram ajustados ao modelo de decaimento exponencial usando o programa GraphPad Prism para a
determinaccedilatildeo dos valores de IC50 (na ausecircncia de actina IC50= 266 plusmn 07 M decavanadato na presenccedila de actina
IC50= 28 plusmn 1 M decavanadato
A partir do graacutefico presente na Fig 29 verifica-se que o ajuste dos dados experimentais foi bem
sucedido A inibiccedilatildeo da actividade Mg2+
-ATPaacutesica do S1 de miosina eacute afectada em menor
amplitude na presenccedila de actina (60 de inibiccedilatildeo da actividade) do que na sua ausecircncia
(aproximadamente 80 de inibiccedilatildeo de actividade) Devido a esta diferenccedila pode sugerir-se que a
actina iraacute impedir parcialmente a ligaccedilatildeo do decavanadatoao S1 demiosina atraveacutes de
impedimentos esteacutereos ou interacccedilotildees electrostaacuteticas por exemplo A teacutecnica utilizada natildeo nos
permite concluir acerca de que regiatildeo(otildees) ou resiacuteduo(s) da actina participaratildeo na preservaccedilatildeo
parcial da actividade Mg2+
-ATPaacutesica
Por outro lado os valores de IC50 dos dois tipos de actividades natildeo apresentam diferenccedilas
significativas tendo-se obtido 266 plusmn 07 e 28 plusmn 1M decavanadato na ausecircncia e na presenccedila de
actina respectivamente o que corresponde a uma concentraccedilatildeo de cerca de 27 M de V10 O
meio de reacccedilatildeo utilizado nestes ensaios foi 10 mM HEPES e 25 mM KCl com 2 mM ATP Em
(Tiago et al 2004) o valor de IC50 para a actividade Mg2+-ATPaacutesica na ausecircncia de actina foi de
125 plusmn 025 microM enquanto o valor da actividade estimulada por actina foi de 136 plusmn 14 microM O
meio de reacccedilatildeo utilizado nos ensaios foi 20 mM HEPES 50 mM KCl e 2 mM ATP o que resulta
precisamente no dobro da forccedila ioacutenica a que foi realizado o ensaio do presente relatoacuterio
(exceptuando a contribuiccedilatildeo dos produtos da hidroacutelise de ATP uma vez que este meacutetodo
pressupotildee a ocorrecircncia de um ciclo de reacccedilotildees de hidroacutelise e siacutentese deste composto) Ora tendo
sido anteriormente referido que a afinidade do S1 de miosina para com a espeacutecie V10 diminui com
Act
ivid
ade
Mg
2+-A
TP
aacutesic
a d
e S
1 d
e
Mio
sin
a
[Decavanadato] (microM) Actividade Estimulada ()
Actividade Natildeo Estimulada ()
Ajuste Actividade Estimulada
Ajuste Actividade Natildeo Estimulada
33
o aumento da forccedila ioacutenica (Tiago et al 2004 tal poderaacute explicar a diferenccedila obtida entre estes
dois conjuntos de valores
243Quantificaccedilatildeo de cisteiacutenas reduzidas pelo meacutetodo de Ellman
A interacccedilatildeo do vanaacutedio com alguns tipos de aminoaacutecidos sensiacuteveis agrave oxidaccedilatildeo pode fornecer-nos
informaccedilatildeo bastante uacutetil em relaccedilatildeo agraves alteraccedilotildees estruturais que conduzem a uma diminuiccedilatildeo da
actividade fisioloacutegica do S1 de miosina Os estudos realizados por Chasteen 1983 concluiacuteram
que a reduccedilatildeo de vanadato (V) a vanadilo (IV) ocorre na presenccedila de agentes redutores comuns no
ambiente celular de que se destaca o aminoaacutecido cisteiacutena Esta eacute tambeacutem um alvo de
complexaccedilatildeo por parte do iatildeo VO2+
Jaacute Ramos et al (2009) demonstrou que a incubaccedilatildeo de
actina com a soluccedilatildeo de decavanadato induz a oxidaccedilatildeo de resiacuteduos de cisteiacutena e agrave formaccedilatildeo do
oxidovanaacutedio (IV) Este grupo de investigadores descobriu ainda que apenas a espeacutecie V10 induz a
oxidaccedilatildeo das cisteiacutenas ldquoraacutepidasrdquo da actina atraveacutes do processo proposto que eacute parcialmente
responsaacutevel pela inibiccedilatildeo jaacute descrita da actividade de ATPase do S1 de miosina estimulada por
actina quando exposta agrave soluccedilatildeo de decavanadato Esta inibiccedilatildeo apresenta-se afectada com a
presenccedila de ATP uma vez que os resiacuteduos de cisteiacutena permaneceram intactos nos ensaios
realizados com este ligando Jaacute Crans et al (2004) concluiu que o siacutetio activo de muitas fosfatases
(que conteacutem grupos tiol) pode formar um equiliacutebrio redox com o iatildeo vanadato (V) de forma natildeo
reversiacutevel
Os resultados obtidos por Sakurai e Shiromura (1980) sugeriram que o vanaacutedio (V) sofre reduccedilatildeo
a vanaacutedio (IV) sem dependecircncia do valor de pH do ensaio Registaram ainda uma estequiometria
de 31 na reacccedilatildeo do vanadato (V) com os resiacuteduos de Cys enquanto a estequiometria dos ensaios
com o vanadilo (IV) foi de 21
Berlett e Stadtman (1997) observaram que os tipos de aminoaacutecidos mais facilmente oxidaacuteveis
englobam entre outros os resiacuteduos de Cys Trp e MetAssim analisou-se o estado redox dos
grupos tiol dos resiacuteduos de Cys do S1 na presenccedila do decavanadato atraveacutes da monitorizaccedilatildeo da
Abs412 nm na presenccedila de 1 mgmL de S1 de miosina em tampatildeo 10 mM HEPES 25 mM KCl
variando a concentraccedilatildeo da soluccedilatildeo de decavanadato de modo a determinar que tipo de alteraccedilotildees
estruturais ocorrem na sua presenccedila no que diz respeito agrave diminuiccedilatildeo da actividade ATPaacutesica de
S1 na presenccedila de decavanadato
Os resultados referentes agrave quantificaccedilatildeo das cisteiacutenas reduzidas apoacutes o tratamento de S1 com
diferentes concentraccedilotildees de decavanadato em tampatildeo 10 mM HEPES pH 75 e 25 mM KCl a
25ordmC durante 20 minutos encontram-se descritos na Tabela 2VII
Tabela 2VII ndash Cisteiacutenas reduzidas totais e ldquoraacutepidas de S1 (1 mgmL) apoacutes tratamento com decavanadato
34
(010501002505001000 uM) em 10 mM HEPES 25 mM KCl pH 7 a 25ordmC durante 20 minutos A detecccedilatildeo
espectrofotomeacutetrica foi feita com DTNB (e 412=8400 uM-1 cm-1) (n variaacutevel entre 1 e 8)
Amostras Cys raacutepidas Cys totais
11 M S1 (controlo) 22 plusmn 04 106 plusmn 14
11 M S1 + 10 M Decavanadato 22 plusmn 05 95 plusmn 08
11 M S1 + 50 uM Decavanadato 20 plusmn 02 98 plusmn 14
11 M S1 + 100 uM Decavanadato 17 plusmn 03 97 plusmn14
11 M S1 + 250 uM Decavanadato 26 plusmn 02 98 plusmn 08
11 M S1 + 500 uM Decavanadato 21 plusmn 04 90 plusmn 06
11 M S1 + 1000 uM Decavanadato 24 plusmn 0 86 plusmn 08
A partir dos resultados obtidos pode sugerir-se que o S1 possui 11 resiacuteduos de Cys (controlo) o
que estaacute de acordo com o anteriormente descrito Verifica-se que o tratamento da proteiacutena com
concentraccedilotildees crescentes de decavanadato provoca a oxidaccedilatildeo de dois resiacuteduos de Cys do S1 de
miosina natildeo expostos ao solvente No entanto a teacutecnica utilizada natildeo nos permite identificar
quais
Tendo em consideraccedilatildeo que o valor de IC50 obtido para a reacccedilatildeo de inibiccedilatildeo daactividade Mg2+-
ATPaacutesica de S1 na presenccedila de decavanadato eacute de cerca de 27 microM pode sugerir-se que este seraacute o
valor ao qual ocorreraacute a oxidaccedilatildeo do primeiro resiacuteduo de Cys o que poderaacute contribuir para a
diminuiccedilatildeo da actividade de S1 observada
25Interacccedilatildeo de S1 com vanaacutedio(IV)
251Anaacutelise por espectroscopia de Ressonacircncia Paramagneacutetica Electroacutenica
A acumulaccedilatildeo intracelular de oxovanaacutedio (IV) in vivo constitui uma das principais formas de
apresentaccedilatildeo de vanaacutedio no organismo Deste modo torna-se bastante interessante analisar a
interacccedilatildeo entre este iatildeo e o S1 de miosina atraveacutes da teacutecnica de espectroscopia de Ressonacircncia
Paramagneacutetica Electroacutenica o meacutetodo de eleiccedilatildeo do estudo do iatildeo vanadilo e dos complexos por si
formados O electratildeo desemparelhado encontra-se fortemente acoplado ao nuacutecleo 51
V o que
35
resulta num espectro de RPE consistindo de 8 desdobramentos na direcccedilatildeo paralela e outros tantos
na direcccedilatildeo perpendicular (Chasteen 1983)
Os estudos realizados por Ahmed et al (1987) centraram-se na interacccedilatildeo do vanadilo com
proteiacutenas como eacute o caso da insulina transferrina entre outros obtendo um espectro caracterizado
por 8 linhas amplamente espaccediladas (na direcccedilatildeo paralela) Jaacute Ramos et al 2009 concluiu que a
exposiccedilatildeo da soluccedilatildeo de decavanadato agraves formas globular e filamentosa de actina produz um
espectro tiacutepico do iatildeo oxidovanaacutedio (IV) indicando a reduccedilatildeo do vanadato
Para aferir qual a concentraccedilatildeo de S1 que permitia obter uma boa razatildeo sinalruiacutedo titulou-se uma
soluccedilatildeo de sulfato de vanadilo 15 microM ([VOSO4]stock = 5 mM) com concentraccedilotildees crescentes de
proteiacutena (0 5 10 15 e 20 microM) em 10 mM HEPES 25 mM KCl A existecircncia de uma correlaccedilatildeo
linear entre a concentraccedilatildeo de proteiacutena e a intensidade dos sinais de RPE correspondente agrave banda
-12 perpendicular encontra-se representada na Fig 210
Figura 2 12ndash Representaccedilatildeo graacutefica da variaccedilatildeo da intensidade do sinal de RPE vs a concentraccedilatildeo de S1 utilizada
Apoacutes verificar que com uma concentraccedilatildeo de S1 igual a 20 microM se obtinham sinais com uma razatildeo
sinalruiacutedo adequada ao prosseguimento dos estudos sobre a interacccedilatildeo V(IV)-S1 de miosina
titulou-se uma soluccedilatildeo de 20 microM S1 com 0 25 5 10 15 25 50 75 100 150 200 500 e 1000
microM VOSO4
Traccedilou-se o graacutefico da intensidade da banda -12 perpendicular do sinal de EPR em funccedilatildeo da
concentraccedilatildeo de VOSO4 tendo-se determinado uma constante de dissociaccedilatildeo (KD) do complexo
S1-vanadilo de 8748 plusmn 2621 microM V(IV)
y = 016786x + 122435
Rsup2 = 092992759
Inte
nsi
dad
e d
o s
inal
de
RP
E
[S1] (microM)
36
Figura 213 ndash Representaccedilatildeo graacutefica da variaccedilatildeo do sinal de RPE vs a concentraccedilatildeo de vanadilo utilizada (os valores
experimentais estatildeo representados por losangos o seu ajuste estaacute representado pela linha)
A simulaccedilatildeo do espectro de EPR foi efectuada a partir do espectro obtido para uma concentraccedilatildeo
de 1 mM VOSO4 apoacutes adiccedilatildeo de 20 microM S1 de miosina tendo-se obtido os valores da constante de
acoplamento hiperfina (A=5016) e do factor g (g0=195)
Figura 214 ndash Representaccedilatildeo graacutefica do espectro de EPR obtido com 1000 microM de vanadilo e 20 microM de S1 (a cinzento
escuro) A simulaccedilatildeo do espectro encontra-se a cinzento claro
Por comparaccedilatildeo destes com os valores presentes na tabela de correspondecircncia xxx eacute possiacutevel
predizer quais os ligandos provaacuteveis do vanadilo na presenccedila de proteiacutena Assim concluiu-se que
o vanadilo estaacute coordenado com duas moleacuteculas de aacutegua um grupo hidroxilo e um grupo R=NR
O grupo hidroxilo pode indicar os resiacuteduos de aminoaacutecidos Tyr Treonina e Serina enquanto o
grupo R=NR poderaacute indicar a presenccedila de Arginina e Histidina como ligandos do vanadilo
Inte
nsi
dad
e d
os
sinai
s d
e R
PE
[Vanadilo] (microM)
Ajuste dos dados exp Dados experimentais
Campo magneacutetico (G)
Espectro Experimental Ajuste
37
3 Procedimento Experimental
31 Reagentes
Todos os reagentes utilizados satildeo de elevado grau de pureza (analiacutetico) Apresentam-se as suas
foacutermulas quiacutemicas ou abreviaturas quando necessaacuterio acrescentou-se a sua descriccedilatildeo Entre
parecircntesis colocou-se as empresas que os comercializam
Aacutecido aceacutetico glacial 100 (Riedel) Acrilamida 99 (Merck) Azul de bromofenol (Riedel-de-
Haumlen) -Mercaptoetanol (Sigma) Bissulfito de soacutedio (Sigma)Bradford (reagente de)
(BioRad)BSA (albumina de soro bovino Sigma-Aldrich) CaCl22H2O (cloreto de caacutelcio
dihidratado Fluka) CH3CH2OH 96 (etanol Panreac) Coomassie Blue R250 (Merck) D2O
(oacutexido de deuteacuterio SDS) DTNB (55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico) Aldrich) EDTA (aacutecido
etilenodiaminatetraaceacutetico Merck) Fosfoenolpiruvato 98 (Fluka) Glicerol (Sigma) Glicina
985 (Panreac) HCl 37 (Riedel-de Haumlen) H2SO4 96 (Pronalab) HEPES 99 (aacutecido 4-(2-
hidroxiletil)-1-piperazineetanosulfoacutenico Sigma) KH2PO4 99 (hidrogenofosfato de potaacutessio)
MgATP (Sigma) Na
ATP (Sigma) Na2HPO4 99 (hidrogenofosfato de soacutedio Riedel-de Haumlen)
NaH2PO4 (fosfato de soacutedio ) NaOH 986 (hidroacutexido de soacutedio Pronalab) KCl 99 (Cloreto de
potaacutessio Panreac) L-Cys 995 (enantioacutemero levoacutegiro do resiacuteduo de Cisteiacutena Fluka)Metanol
99 (Merck) Molibdato de amoacutenio (Sigma)NNrsquo-metil-bis-acrilamida 98 (Merck) NH4VO3
99 (metavanadato de amoacutenia Sigma) PSA 10 (persulfato de amoacutenio Merck) SDS
(dodecilo sulfato de soacutedio Sigma) Sulfito de soacutedio (Sigma) VOSO4 (sulfato de vanadilo
Aldrich) TEMED 99 (Tetrametiletilenodiamina Merck) Tris (2-amino-2-hidroximetil-13-
propanodiol Sigma)
32 Meacutetodos
321 Isolamento e purificaccedilatildeo das proteiacutenas de muacutesculo esqueleacutetico de coelho
3211 Purificaccedilatildeo da miosina preparaccedilatildeo de Subfragmento 1 por digestatildeo quimotriacutepsica
da miosina
O subfragmento-1 de miosina-II (S1) foi gentilmente cedido pelo Prof Doutor Manuel Aureliano
do Departamento de Quiacutemica Bioquiacutemica e Farmaacutecia da Faculdade de Ciecircncias e Tecnologia da
Universidade do Algarve
No entanto descreve-se abaixo de forma resumida o procedimento efectuado
38
32111 Purificaccedilatildeo da miosina
O protocolo de isolamento foi adaptado do meacutetodo previamente descrito por Margossian e Lowey
(1982) apresentando-se de forma esquemaacutetica na Fig 31
Sucintamente os tecidos musculares dorsais e das pernas foram extraiacutedos apoacutes o sacrifiacutecio do
animal (Fig31) De seguida foram adicionados 3 volumes da soluccedilatildeo de extracccedilatildeo A2
Centrifugou-se a soluccedilatildeo e filtrou-se o sobrenadante O sedimento foi entatildeo diluiacutedo em 14
volumes de aacutegua destilada de modo a que a forccedila ioacutenica atingisse 004 M Apoacutes adiccedilatildeo de szlig-
mercaptoetanol (szlig-me) deixou-se a precipitar a 4ordmC removendo o sobrenadante Centrifugou-se
novamente e ressuspendeu-se o sedimento na soluccedilatildeo de extracccedilatildeo A Adicionou-se a soluccedilatildeo
tampatildeo B3 para que a forccedila ioacutenica atingisse 05 M Homogeneizou-se e deixou-se repousar
durante 2h Homogeneizou-se novamente e centrifugou-se de modo a ocorrer precipitaccedilatildeo dos
complexos de actomiosina e proteiacutena desnaturada Apoacutes filtraccedilatildeo do sobrenadante em latilde de vidro
quantificou-se a soluccedilatildeo de miosina resultante pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental)
2 Soluccedilatildeo de extracccedilatildeo A 0012M K2CO3 004M KHCO3 06M KCl pH 65
3 Soluccedilatildeo-tampatildeo B Tris 100 mM pH 7 KCl 2M
39
Figura 31 ndash Representaccedilatildeo esquemaacutetica do protocolo de isolamento de miosina (Margossian e Lowey 1982) Todo o
procedimento foi realizado a 4ordmC (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
32112 Preparaccedilatildeo de Subfragmento-1 de miosina por digestatildeo com quimiotripsina
O S1 de miosina foi isolado apoacutes diaacutelise da miosina durante a noite contra a soluccedilatildeo C4 A
presenccedila do EDTA eacute essencial uma vez que a sua substituiccedilatildeo por MgCl2resulta na produccedilatildeo de
HMM em vez de S1 A suspensatildeo de miosina foi equilibrada e digerida com -quimotripsina
A digestatildeo foi interrompida pela adiccedilatildeo de PMSF Seguidamente ultracentrifugou-se o produto
da digestatildeo desprezando-se o sedimento (Fig 32) A soluccedilatildeo de sobrenadante maioritariamente
S1 foi quantificada pela leitura directa da Abs280 ou pelo meacutetodo de Bradford (ver as secccedilotildees
3311 e 3312 do Procedimento Experimental respectivamente) e armazenada em 10 sacarose
e 2 mM szlig-Me a -80ordmC
4 Soluccedilatildeo C 25mM Tris pH 70 012M KCl 1mM EDTA
40
Antes da realizaccedilatildeo dos ensaios descritos no presente relatoacuterio o S1 de miosina foi dialisado
contra a soluccedilatildeo D5 de modo a remover qualquer composto que pudesse complexar como
vanaacutedio Seguidamente quantificou-se a proteiacutena pelo meacutetodo de Bradford ou pelo meacutetodo
directo (ver secccedilotildees 3311 e 3312do Procedimento Experimtental)
Figura 3 2ndash Representaccedilatildeo esquemaacutetica do protocolo de preparaccedilatildeo do S1 de miosina por digestatildeo quimiotripsina de
acordo com o previamente descrito por Margossian e Lowey (1982) Todo o procedimento foi realizado a 4ordm C
excepto quando indicaccedilatildeo do contraacuterio (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
3212 Isolamento e purificaccedilatildeo de actina
Isolou-se e purificou-se a actina segundo o meacutetodo descrito por Pardee e Spudish que
compreende dois passos ndash a obtenccedilatildeo de um resiacuteduo seco denominado ldquopoacute de acetonardquo e a
extracccedilatildeo subsequente da actina a partir do ldquopoacute de acetonardquo na forma de G-Actina com a
posterior polimerizaccedilatildeo a F-Actina
32121 Obtenccedilatildeo do ldquopoacute de acetonardquo
Sacrificou-se o coelho com uma pancada atraacutes do pescoccedilo e sangrou-se o animal apoacutes o corte da
arteacuteria jugular Esfolou-se e colocou-se numa tina com gelo Retiraram-se as partes de tecido
conjuntivo que envolvem o tecido muscular com auxiacutelio de uma pinccedila e bisturi Seguidamente
retirou-se os muacutesculos dorsais e os muacutesculos brancos das pernas Passou-se os muacutesculos por aacutegua
destilada de modo a remover as impurezas e pesou-se num copo Trituraram-se os muacutesculos
numa maacutequina de triturar carne e extraiu-se o resultante com 3 volumes da soluccedilatildeo E6 durante 10
minutos com agitaccedilatildeo suave Centrifugou-se a suspensatildeo a 4000 g durante 10
5 Soluccedilatildeo D HEPES 10 mM pH 70 KCl 25 mM
6 Soluccedilatildeo E 01 M KCl 015 M K2HPO4 pH 65
41
minutosdesprezando-se o sobrenadante Ressuspendeu-se o sedimento em 6 volumes de soluccedilatildeo
F7durante 10 minutos com agitaccedilatildeo suave Verificou-se se o pH da soluccedilatildeo estava compreendido
entre 82 e 85 acertando-se estecom soluccedilatildeo G8 se necessaacuterio Centrifugou-se durante 10
minutos a 4000 g Desprezou-se o sobrenadante e ressuspendeu-se o sedimento em 3 volumes de
soluccedilatildeo H9 durante 10 minutos Centrifugou-se novamente a 4000 gdurante 10 minutos
desprezando-se o sobrenadante Ressuspendeu-se o sedimento duas vezes consecutivas em 6
volumes de aacutegua destilada durante 5 minutos Centrifugou-se a 4000 g durante 10 minutos apoacutes
cada adiccedilatildeo de H2O
Seguidamente ressuspendeu-se o sedimento 5 vezes consecutivas utilizando 3 volumes de
acetona de cada vez agitando suavemente durante 10 minutos agrave temperatura ambiente Colocou-
se o resiacuteduo filtrado numa tina de vidro sobre papel de filtro deixando-se evaporar o solvente
durante a noite num nicho Triturou-se o resiacuteduo seco numa maacutequina de triturar carne e deixou-se
novamente evaporar o solvente O ldquopoacute de acetonardquo obtido foi armazenado a -20 ordmC
32122 Purificaccedilatildeo da actina a partir de ldquopoacute de acetonardquo
Apoacutes a obtenccedilatildeo do poacute de acetona como acima indicado efectuou-se o protocolo seguidamente
descrito de modo a obter monoacutemeros de actina (G-Actina) e actina filamentosa (F-Actina)
Pesou-se 35 g de poacute de acetona e adicionaram-se 50 mL de soluccedilatildeo-tampatildeo I10
sendo a
extracccedilatildeo feita com agitaccedilatildeo suave durante 30 minutos Separou-se o extracto do poacute de acetona
hidratado filtrando a suspensatildeo com 4 camadas de gaze esterilizada tendo o cuidado de natildeo
espremer o resiacuteduo Voltou a extrair-se com 50 mL de soluccedilatildeo I durante 30 minutos mexendo
lentamente Adicionou-se este ao extracto obtido anteriormente Centrifugou-se o extracto total a
16 000 g durante 30 minutos Decantou-se para uma proveta sobre a qual se colocou gaze para
que o sedimento solto natildeo se juntasse ao sobrenadante e leu-se o volume De seguida adicionou-
se KCl ateacute uma concentraccedilatildeo final de 50 mM MgCl2 ateacute 2mM e ATP ateacute 1mM lentamente e com
agitaccedilatildeo contiacutenua de modo a promover a polimerizaccedilatildeo dos monoacutemeros de actina
Deixou-se polimerizar durante 2h a 25ordmC observando-se a presenccedila de pequenas bolhas de ar
indicativas de um aumento de viscosidade da soluccedilatildeo Arrefeceu-se em gelo durante 15 minutos e
adicionou-se KCl ateacute uma concentraccedilatildeo final de 06M Mexeu-se lentamente durante 30 minutos
apoacutes os quais se ultracentrifugou a 110 000g durante 1hora Desprezou-se o sobrenadante
ressuspendeu-se o sedimento com soluccedilatildeo tampatildeo E e transferiu-se para um potter
Dialisou-se durante a noite contra 1L de soluccedilatildeo-tampatildeo I a uma temperatura de 4ordmC com rotaccedilatildeo
7 Soluccedilatildeo F 005 M NaHCO3 pH 82-85
8 Soluccedilatildeo G 1M Na2CO3
9 Soluccedilatildeo H 1 mM EDTA pH 70
10 Soluccedilatildeo I 2 mM Tris-HCl pH 8 02 mM CaCl2 02 mM Na2ATP 05 mM szlig-Me 0005 NaN3
42
contiacutenua e mudanccedila de tampatildeo 2 a 3 vezes De seguida a actina foi centrifugada a 110
000gdurante 1hora Decantou-se a soluccedilatildeo de G-Actina (sobrenadante) e desprezou-se o
sedimento Determinou-se a concentraccedilatildeo de G-Actina pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental) A soluccedilatildeo de G-actina foi armazenada a uma temperatura de 4ordmC
apoacutes adiccedilatildeo de 002NaN3 mantendo-se estaacutevel durante 3 a 5 dias
331 Quantificaccedilatildeo das proteiacutenas isoladas
3311 Quantificaccedilatildeo pelo meacutetodo de Bradford
O meacutetodo de Bradford (Bradford 1976) baseia-se na utilizaccedilatildeo do reagente de Bradford
composto por entre outros aacutecido fosfoacuterico e o corante ldquoCoomassierdquo Este corante possui carga
negativa e a sua coloraccedilatildeo varia entre a vermelha (Absmax=465 nm) predominante em soluccedilatildeo e a
azul (Absmax=595 nm) presente aquando da ligaccedilatildeo aos grupos amina das proteiacutenas Em soluccedilotildees
aacutecidas diluiacutedas a coloraccedilatildeo varia proporcionalmente com a quantidade do corante ligado agrave
proteiacutena que por sua vez eacute directamente proporcional agrave concentraccedilatildeo da proteiacutena permitindo a
sua quantificaccedilatildeo ateacute a um limite de detecccedilatildeo de 20 gmL (Bradford 1976)
Deste modo prepararam-se amostras de BSA ([BSA]stock= 1mgmL) utilizando um intervalo de
concentraccedilotildees compreendido entre 0 e 15 gmL de acordo com a tabela 3I Apoacutes a adiccedilatildeo da
H2O MilliQ e do reagente de Bradford as soluccedilotildees foram sucessivamente homogeneizadas no
voacutertex
Tabela 3Indash Amostras utilizadas para a obtenccedilatildeo da recta de calibraccedilatildeo no meacutetodo de Bradford
BSA (L) BSA (g) H2O MilliQ (L) Reagente de Bradford (L)
0 0 800 200
125 125 79875 200
25 25 7975 200
5 5 795 200
10 10 790 200
15 15 785 200
Apoacutes incubaccedilatildeo a 25ordmC por 5 minutos leram-se os valores de Abs595 num
espectrofotoacutemetro (Shimadzu UV-160A) para todas as amostras Seguidamente traccedilou-se a recta
de calibraccedilatildeo (Abs595vs quantidade de BSA (g))
43
Para determinar a concentraccedilatildeo de proteiacutena apoacutes o processo de isolamento purificaccedilatildeo e
diaacutelise quando necessaacuterio colocou-se 5 L da amostra num tubo de Eppendorf e adicionou-se
795 L de H2O MilliQ homogeneizando a soluccedilatildeo no voacutertex Adicionou-se posteriormente 200
L de reagente de Bradford e repetiu-se a homogeneizaccedilatildeo Incubou-se a mistura a 25ordmC por 5
minutos apoacutes os quais se leu os valores de Abs595 nm no espectrofotoacutemetro Por interpolaccedilatildeo deste
valor a partir da recta de calibraccedilatildeo determinou-se a concentraccedilatildeo de proteiacutena na amostra
3312 Quantificaccedilatildeo pelo meacutetodo directo
A quantificaccedilatildeo das amostras proteicas pode tambeacutem ser efectuada atraveacutes da leitura directa da
absorvacircncia tendo por base a lei de Beer-Lambert onde Abs corresponde agrave
absorvacircncia ao comprimento de onda ao qual o ensaio foi realizado corresponde ao coeficiente
de extinccedilatildeo (substituiacutedo pela letra a se as unidades forem distintas da molaridade) b corresponde
ao percurso oacuteptico da radiaccedilatildeo na ceacutelula amostral e c corresponde agrave concentraccedilatildeo amostral
Assim no caso de S1 de miosina mediu-se a Abs280e calculou-se a concentraccedilatildeo da proteiacutena
sabendo que o coeficiente de extinccedilatildeo de S1 eacute (Ref)
No caso da actina mediu-se a Abs290ecalculou-se a concentraccedilatildeo da proteiacutena sabendo que o
coeficiente de extinccedilatildeo da actina eacute (Ref)
332Electroforese em condiccedilotildees desnaturantes (SDS-PAGE)
De modo a melhor caracterizar as proteiacutenas isoladas foi realizada uma electroforese em
condiccedilotildees desnaturantes (SDS-PAGE do inglecircs Sodium Dodecyl Sulfate Polyacrilamide Gel
Electrophoresis) O SDS (dodecilo sulfato de soacutedio) eacute um detergente anioacutenico utilizado neste
meacutetodo para desnaturar as proteiacutenas que se ligam ao SDS na proporccedilatildeo 14 g SDS g proteiacutena
(aproximadamente 1 moleacutecula de SDS por cada 2 resiacuteduos de aminoaacutecidos) (Stryer) O SDS induz
uma densidade de carga negativa significativa nas proteiacutenas a que se liga mascarando a sua carga
intriacutenseca pelo que as razotildees cargamassa satildeo tendencialmente idecircnticas Assim a SDS-PAGE
separa as proteiacutenas quando sujeitas a um campo eleacutectrico de acordo com o seu peso molecular
sendo a sua migraccedilatildeo directamente proporcional ao logaritmo deste uacuteltimo (Stryer)
O gel de resoluccedilatildeo com 10 de acrilamida foi obtido atraveacutes da adiccedilatildeo de 167 mL de soluccedilatildeo
stock de acrilamida11
075 mL de tampatildeo de resoluccedilatildeo12
251 mL de aacutegua destilada e 50 L de
SDS 10 Para promover a polimerizaccedilatildeo adicionou-se em seguida 38 L de PSA 10
11
soluccedilatildeo stock de acrilamida soluccedilatildeo de 30 (mv) de acrilamida e 08 (mv) de NNrsquo-metil-bis-acrilamida 12
tampatildeo de resoluccedilatildeo 15 M Tris-HCl pH 88
44
(persulfato de amoacutenia) 10 e 25 L de TEMED 99 (tetrametiletilenodiamina) Agitou-se
cuidadosamente de modo a natildeo formar bolhas e pipetou-se a soluccedilatildeo para a cassette de vidro
previamente montada no suporte De seguida adicionou-se uma peliacutecula de cerca de 2 mm de 2-
metil-2-butanol para verificar posteriormente a polimerizaccedilatildeo Apoacutes a polimerizaccedilatildeo que demora
cerca de 45 minutos verteu-se a agrave superfiacutecie e adicionou-se o gel de concentraccedilatildeo com 5 de
acrilamida constituiacutedo por 03 mL de soluccedilatildeo de stock acrilamida 045 mL de tampatildeo de
concentraccedilatildeo13
094 mL de aacutegua destilada 18 L de SDS 10 135 L de PSA 10 e 2L de
TEMED 99 Colocou-se o pente sobre o gel e deixou-se polimerizar durante cerca de 45
minutos
As amostras para aplicaccedilatildeo no gel de electroforese foram preparadas para uma concentraccedilatildeo de
proteiacutena de 1 mgmL num volume final de 100 L dos quais 25 L correspondem ao tampatildeo da
amostra14
As amostras foram aquecidas a 100ordmC durante 5 minutos Antes da aplicaccedilatildeo das
amostras transferiu-se a cassette para a tina de electroforese agrave qual se adicionou a soluccedilatildeo
tampatildeo de electroforese da tina superior15
e tampatildeo de electroforese da tina inferior16
Aplicaram-
se 10 L das soluccedilotildees de marcadores de peso molecular e 20 L das amostras proteicas
preparadas anteriormente A electroforese decorreu durante cerca de 3h30 min a uma voltagem
de 80 V De seguida transferiu-se o gel para uma tina corando as proteiacutenas durante 1h com
soluccedilatildeo corante17
Descorou-se o gel com soluccedilatildeo descorante18
tendo-se mudado esta soluccedilatildeo ateacute
as bandas se encontrarem bem definidas
Determinou-se a mobilidade relativa das bandas correspondentes aos marcadores de elevado peso
molecular e traccedilou-se a recta de calibraccedilatildeo ndash log(PM) vs log(Rf) onde Rf corresponde agrave
mobilidade relativa Por interpolaccedilatildeo determina-se o PM das proteiacutenas isoladas sabendo a sua
mobilidade relativa Por comparaccedilatildeo com os valores descritos na literatura pode concluir-se
acerca do grau de purificaccedilatildeo
34 Mediccedilatildeo da actividade enzimaacutetica de S1
341 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
13
tampatildeo de concentraccedilatildeo 05 M Tris-HCl pH 68 14
tampatildeo de amostra 320 mM Tris-HCl pH 68 04 M szlig-mercaptoetanol SDS 8 glicerol 15 (vv) e roxo de m-
cresol 0024 15
tampatildeo de electroforese da tina superior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS 16
tampatildeo de electroforese da tina inferior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS acetato de soacutedio
01 M 17
soluccedilatildeo corante ldquoCoomassie Blue R-250rdquo contendo metanol 45 (vv) e aacutecido aceacutetico glacial 10 (vv) 18
soluccedilatildeo descorante metanol 10 (vv) e aacutecido aceacutetico glacial 10 vv)
45
O meacutetodo de Fiske-Subarow modificado consiste na reacccedilatildeo do molibdato de amoacutenio com o
fosfato inorgacircnico em soluccedilatildeo aacutecida com formaccedilatildeo do aacutecido fosfomoliacutebdico Seguidamente daacute-
se a adiccedilatildeo de um agente redutor que provoca a reduccedilatildeo do composto a fosfomolibdato que forma
um complexo de cor azulada cuja absorvacircncia maacutexima ocorre para um comprimento de onda de
660 nm tornando possiacutevel a sua detecccedilatildeo por meacutetodos espectrofotomeacutetricos As actividades Ca2+
-
ATPaacutesica e K+EDTA-ATPaacutesica de S1 foram determinadas atraveacutes deste meacutetodo de acordo com
o descrito por Taussky e Shorr (1953) com mediccedilatildeo da quantidade de fosfato (Pi) libertado
atraveacutes da hidroacutelise de ATP
Para tal obteve-se primeiramente a recta de calibraccedilatildeo com as amostras presentes na Tabela 3II
utilizando uma soluccedilatildeo stock de 5 mM KH2PO4
Tabela 3II ndash Composiccedilatildeo das diferentes amostras utilizadas para determinar a recta de calibraccedilatildeo atraveacutes do meacutetodo
de Fiske-Subbarow modificado
KH2PO4 (L)
([KH2PO4]stock= 5mM)
PO43-
(nmole) H2O (uL)
0 0 1780
5 25 1775
10 50 1770
20 100 1760
40 200 1740
100 500 1680
A cada tubo de Eppendorf adicionaram-se seguidamente 200 L de soluccedilatildeo J19
e 20 L de agente
redutor20
homogeneizando no voacutertex apoacutes cada adiccedilatildeo Incubou-se durante 10 minutos e mediu-
se a Abs660 nm num espectrofotoacutemetro com intervalos de tempo iguais entre cada amostra Traccedilou-
se a recta de calibraccedilatildeo (Abs660 nmvs nmol PO43-
) a partir dos valores obtidos
Para determinar a actividade Ca2+
-ATPaacutesica de S1 prepararam-se tubos de Eppendorf com 400
L de meio de reacccedilatildeo21
25 L CaCl2 ([CaCl2]stock= 100 mM) 25 L ATP ([ATP]stock= 100 mM)
com [S1]ensaios=01 mgmL Perfez-se se necessaacuterio com H2O MilliQ ateacute perfazer um volume de 1
mL Para determinar a actividade K+EDTA-ATPaacutesica utilizou-se uma concentraccedilatildeo idecircntica de
19
Soluccedilatildeo J 5 N H2SO4 25 molibdato de amoacutenio 20
Agente redutor 5 mg aacutecido 1-amino-2-naftol-4-sulfoacutenico 30 mg bissulfito de soacutedio 30 mg sulfito de soacutedio em 26
mL H2O 21
meio de reacccedilatildeo 125 mM Tris-HCl pH 80 0575 M KCl
46
proteiacutena nos ensaios (01 mgmL) e o mesmo meio de reacccedilatildeo Aos tubos de Eppendorf
adicionou-se ainda 123 L de KCl ([KCl]stock = 3M) 25 L de ATP 10 L de EDTA
([EDTA]stock= 100 mM )e o volume de H2O MilliQ necessaacuterio para perfazer 1 mL Em ambas as
situaccedilotildees o uacuteltimo reagente adicionado foi o ATP uma vez que se trata do substrato da reacccedilatildeo
enzimaacutetica
A cineacutetica decorreu a 25ordmC durante 5 minutos para a actividade Ca2+
-ATPaacutesica e durante 3
minutos para a actividade K+EDTA-ATPaacutesica Apoacutes o periacuteodo de incubaccedilatildeo transferiu-se 150
L de amostra para tubos de Eppendorf contendo igual volume de TCA 10 Colocou-se no gelo
e homogeneizou-se no voacutertex Seguidamente centrifugou-se a 1500 g durante 10 minutos e
recolheu-se 200 L de sobrenadante para novos tubos de Eppendorf Adicionou-se a estes 158
mL de H2O MilliQ e 200 L de soluccedilatildeo J Homogeneizou-se no voacutertex e adicionou-se 20 L de
reagente redutor repetindo-se a homogeneizaccedilatildeo Incubou-se durante 10 minutos e mediu-se a
Abs660 nm com o mesmo intervalo de tempo de leitura para cada amostra Por intrapolaccedilatildeo a partir
da recta de calibraccedilatildeo obtida anteriormente determinou-se a quantidade de Pi libertado em cada
ensaio A partir destes resultados determinaram-se as actividades Ca2+
-ATPaacutesica e K+EDTA-
ATPaacutesica do S1 em nmol PO43-
minmg proteiacutena e em s-1
342 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
Da actividade Mg2+
-ATPaacutesica de S1 resulta a formaccedilatildeo dos produtos da reacccedilatildeo de
hidroacutelise do ATP - ADP e Pi Na presenccedila de ADP fosfoenolpiruvato (PEP) e da enzima piruvato
cinase (PK) os produtos resultantes satildeo ATP e piruvato Por acccedilatildeo da enzima lactato
desidrogenase (LDH) este uacuteltimo eacute convertido em lactato ocorrendo simultaneamente a oxidaccedilatildeo
do NADH a NAD+ monitorizada atraveacutes da Abs340 nm Quando o S1 de miosina se encontra no
meio reaccional o ATP eacute hidrolisado formando-se novamente ADP e Pi o que permite a
formaccedilatildeo de um ciclo (fig 21) Se as enzimas se encontrarem em excesso a velocidade de
conversatildeo do NADH eacute limitada pela produccedilatildeo de ADP e Pi Como o ATP eacute regenerado em cada
ciclo a sua concentraccedilatildeo inicial eacute sempre mantida (Bagshaw 1982)
Figura 33ndash Representaccedilatildeo esquemaacutetica do meacutetodo de enzimas acopladas
47
O protocolo utilizado e seguidamente descrito foi adaptado de Cuenda et al (1990 ndash Ver Refordf) A
uma cuvette de 1 mL adicionou-se S1 para uma concentraccedilatildeo final de 005 mgmL 800 L de
meio de reacccedilatildeo22
10 L de PEP ([PEP]stock = 42 mM) 10 L de PK+LDH 11 ([PK+LDH]stock =
1 mgmL) e 15 L de NADH ([NADH]stock=25 mM) Iniciou-se a monitorizaccedilatildeo da reacccedilatildeo
atraveacutes da leitura de Abs340 nm apoacutes a adiccedilatildeo de 20 L de ATP ([ATP]stock=100 mM) A
actividade Mg2+
-ATPaacutesica eacute obtida atraveacutes do declive correspondente agrave oxidaccedilatildeo do NADH Para
a mediccedilatildeo da Mg2+
-ATPase de S1 estimulada por actina adicionou-se F-actina para uma
concentraccedilatildeo final de 2 microM sendo a actividade obtida tambeacutem atraveacutes do declive correspondente
agrave oxidaccedilatildeo do NADH apoacutes a adiccedilatildeo da proteiacutena
35 Interaccedilatildeo entre vanaacutedio e subfragmento-1 de miosina
351 Preparaccedilatildeo das soluccedilotildees de vanaacutedio
As soluccedilotildees de vanadato foram preparadas em aacutegua MilliQ a partir de metavanadato de amoacutenio
(NH4VO3) numa concentraccedilatildeo stock de 50 mM com agitaccedilatildeo magneacutetica e aquecimento (~30 ordmC)
Esta soluccedilatildeo com um valor compreendido entre 6 e 7 designa-se por ldquometavanadatordquo e eacute
constituiacuteda pelas esspeacutecies V1 V2 V4 e V5 (Ref) Acidificando a soluccedilatildeo de metavanadato por
adiccedilatildeo de HCl ateacute um valor de pH proacuteximo de 4 observa-se uma mudanccedila de cor de amarelo
claro para laranja indicativa da formaccedilatildeo da espeacutecie decameacuterica de vanadato De seguida e
imediatamente antes da realizaccedilatildeo dos ensaios descritos o valor do pH da soluccedilatildeo de
decavanadato foi aumentado para cerca de 75 por adiccedilatildeo de NaOH A soluccedilatildeo (2) designada
por ldquometavanadatordquo natildeo foi posteriormente alterada ateacute agrave sua adiccedilatildeo para a realizaccedilatildeo dos ensaios
As soluccedilotildees foram armazenadas a 4ordmC e mantidas em gelo aquando da sua utilizaccedilatildeo
A soluccedilatildeo de V(IV) foi preparada em condiccedilotildees anaeroacutebias por diluiccedilatildeo de sulfato de vanadil
(VOSO4) para uma concentraccedilatildeo final de 5 mM em H2O MilliQ saturada com N2 gasoso
352Anaacutelise por espectroscopia de UV-Vis
A concentraccedilatildeo da espeacutecie decameacuterica em soluccedilatildeo depende do periacuteodo de tempo apoacutes a diluiccedilatildeo
no meio de reacccedilatildeo apropriado (Ref) A velocidade de desoligomerizaccedilatildeo da espeacutecie decameacuterica
pode ser analisada por espectroscopia de absorccedilatildeo na regiatildeo UVVis uma vez que esta espeacutecie
apresenta duas bandas caracteriacutesticas na regiatildeo do visiacutevel (a 358 e 400 nm) que natildeo se verifica
em quaisquer outras espeacutecies oligomeacutericas As bandas supramencionada sapresentam-se bem
22
Meio de reacccedilatildeo 3125 mM Tris-HCl pH 71 375 mM MgCl2
48
resolvidas atraveacutes da obtenccedilatildeo da derivada de 2ordf ordem do espectro apresentando a forma de 2
vales com miacutenimos centrados nos comprimentos de onda supracitados com dependecircncia linear
relativamente agrave concentraccedilatildeo de decavanadato total (Ramos et al 2006)
Assim a estabilidade da espeacutecie decameacuterica apoacutes diluiccedilatildeo da soluccedilatildeo de decavanadato no meio de
reacccedilatildeo dos ensaios (tampatildeo D) para uma concentraccedilatildeo final de 1 mM em vanadato total foi
caracterizada atraveacutes da leitura da Abs400 em funccedilatildeo do tempo tendo-se determinado qual a
ordem de reacccedilatildeo e o tempo de meia-vida da mesma Com base em estudos anteriores acerca da
estabilidade do V10 na presenccedila de diferentes proteiacutenas (Ramos et al 2006) determinou-se
igualmente o tempo de meia vida apoacutes a adiccedilatildeo de 20 microM de S1 de miosina ao meio reaccional
353 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
Os espectros de RMN do isoacutetopo 51
V foram realizados num espectroacutemetro Bruker AM-400 MHz
equipado com uma sonda multinuclear inversa de 5 mm Em tubos de RMN de 5 mm de diacircmetro
preparou-se soluccedilotildees de 500 L de deca- ou metavanadato correspondentes a um intervalo de
concentraccedilotildees de 1 a 10 mM suplementadas com 10 D2O Os espectros foram adquiridos agrave
temperatura ambiente com uma largura espectral de 23 923 Hz um tempo de aquisiccedilatildeo de 0086
s tempo de relaxaccedilatildeo de 001 s e uma sequecircncia de pulsos a 90ordm Utilizou-se a transformada de
Fourier com alargamento das bandas de 20Hz para o processamento dos espectros Os desvios
quiacutemicos das ressonacircncias de 51
V foram obtidos em relaccedilatildeo agrave espeacutecie VOCl3cuja frequecircncia de
ressonacircncia eacute considerada o padratildeo sendo todas as outras referenciadas a partir desta em ppm
As aacutereas dos sinais do RMN correspondentes agraves espeacutecies oligomeacutericas presentes foram obtidas
por integraccedilatildeo dos sinais As concentraccedilotildees relativas das espeacutecies foram calculadas com recurso agrave
equaccedilatildeo
onde A[Vx] corresponde agrave aacuterea referente agrave ressonacircncia da espeacutecie Vx com nordm de agregaccedilatildeo x
A[V]total agrave soma total das aacutereas medidas e [V] a concentraccedilatildeo total de vanaacutedio no ensaio
A teacutecnica de 51
V-RMN foi anteriormente aplicada em estudos de ligaccedilatildeo com S1 de miosina
(Ref) tendo sido demostrado que entre as espeacutecies V1 V2 V4 e V5 a espeacutecie tetrameacuterica possui a
maior afinidade para a miosina (Cremo et al) O desaparecimento ou alargamento dos sinais pode
implicar a ligaccedilatildeo de um dos oxovanadatos laacutebeis agrave miosina sem indicaccedilatildeo de qual uma vez que
49
se processa um equiliacutebrio entre eles (Crans 1990) Assim e tendo em conta as indicaccedilotildees de que
a espeacutecie V10 eacute distinta das restantes no que diz respeito agrave magnitude da interacccedilatildeo com a miosina
e inibiccedilatildeo da actividade ATPaacutesica estimulada por actina (Aureliano 2000 Tiago et al 2002)
torna-se fundamental investigar a interacccedilatildeo das diferentes espeacutecies de vanadato com o S1 de
miosina atraveacutes da teacutecnica de 51
V-RMN
Para tal obtiveram-se espectros de 51
V-RMN resultantes da titulaccedilatildeo de uma soluccedilatildeo 5 mM de
decavanadato total com concentraccedilotildees crescentes de S1 de miosina (0 5 10 15 e 20 microM) em
tampatildeo D Realizou-se ainda ensaios na presenccedila de 2 mM de decavanadato e 2 mM de ATP na
presenccedila e na ausecircncia de S1 de miosina de modo a verificar o efeito que estes ligandos naturais
da enzima teria na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o subfragmento-1
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
Os grupos sulfidrilo (-SH) das cisteiacutenas satildeo resiacuteduos de aminoaacutecidos extremamente sensiacuteveis agrave
oxidaccedilatildeo resultando na sua oxidaccedilatildeo (Dalle-Donne et al 1999) Logo a quantificaccedilatildeo destes
resiacuteduos eacute bastante importante nos estudos de interacccedilatildeo com vanaacutedio jaacute que os resiacuteduos de Cys
com ecircnfase para o SH1 e o SH2 satildeo bastante importantes na monitorizaccedilatildeo das modificaccedilotildees
funcionais da regiatildeo da cabeccedila
O procedimento seguidamente apresentado foi originalmente descrito por Gutieacuterrez-Merino
(1987)) Consiste na reacccedilatildeo entre os grupos ndashSH natildeo oxidados das proteiacutenas e o reagente de
Ellman (DTNB -55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico)) - formando-se o aniatildeo CNT (aniatildeo 3-
carboxilato-4-nitrotiofenolato) numa estequiometria de 11 Este apresenta um maacuteximo de
absorvecircncia a 412 nm (Robyt 1990) Assim apoacutes a incubaccedilatildeo da proteiacutena na presenccedila do metal
a quantidade de cisteiacutenas reduzidas por proteiacutena determina-se atraveacutes dos valores de Abs412
Deste modo colocou-se num tubo de Eppendorf o volume apropriado de S1 dialisado para que a
concentraccedilatildeo no ensaio fosse de 1 mgmL Adicionou-se o volume apropriado de decavanadato
para que a sua concentraccedilatildeo fosse de 0 10 50 100 250 500 ou 1000 M Perfez-se com soluccedilatildeo
D ateacute um volume final de 1 mL e incubou-se a 25ordmC por 20 minutos Apoacutes este periacuteodo colocou-
se 780 L da mistura reaccional numa cuvette de quartzo e adicionou-se 220 L de soluccedilatildeo K23
23
Tampatildeo K Tampatildeo fosfatohidrogenofosfato de soacutedio 025 M pH 72
50
Adicionou-se 15 L de DTNB (em soluccedilatildeo K [DTNB]stock=100 mM) ao meio reaccional e
registou-se o valor de absorvecircncia medido equivalente agraves cisteiacutenas ldquoraacutepidasrdquo (SH1 e SH2
expostos ao solvente) Adicionou-se 001 g de SDS que provoca a desnaturaccedilatildeo da proteiacutena e
consequentemente a exposiccedilatildeo de todos os resiacuteduos de cisteiacutenas que se encontravam ateacute aiacute
inacessiacuteveis no interior da proteiacutena (cisteiacutenas totais) Estes resiacuteduos podem assim reagir com o
DTNB
Estimou-se o nuacutemero de resiacuteduos de cisteiacutenas em todas as amostras com recurso ao coeficiente de
extinccedilatildeo molar () do DTNB (ver ponto 3 dos Anexos) que foi determinado atraveacutes da titulaccedilatildeo
de L-Cisteiacutena com intervalo de concentraccedilotildees compreendidos entre 0 e 50 microM na presenccedila de
25microM de DTNB em tampatildeo D Os valores de absorvecircncia foram obtidos para um comprimento de
onda de 412 nm
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
3621 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
A teacutecnica mais eficiente de anaacutelise da interacccedilatildeo entre o iatildeo vanadilo e proteiacutenas como a
calmodulina e a transferrina entre outras permite a obtenccedilatildeo raacutepida de informaccedilatildeo no que toca agrave
conformaccedilatildeo e amplitude da referida interacccedilatildeo
Os espectros de Ressonacircncia Paramagneacutetica Electroacutenica (RPE) da espeacutecie paramagneacutetica V(IV)
foram realizados a 70K num espectroacutemetro Bruker EMX-300 X-band Todas as soluccedilotildees
utilizadas e amostras foram preparadas em anaerobiose numa cacircmara de luvas de modo a evitar a
reduccedilatildeo de V(IV) a V(V) Em tubos capilares com 02 mL de amostra manteve-se a concentraccedilatildeo
de S1 a 20 M e variou-se a concentraccedilatildeo de vanadilo (0 25 5 10 15 25 50 75 100 150
200 500 e 1000 M [VOSO4]stock=5 mM) em tampatildeo D Apoacutes obtenccedilatildeo dos espectros estes
foram tratados atraveacutes do software SimFonia e WinEPR tendo-se obtido um graacutefico com a
variaccedilatildeo da intensidade do sinal de RPE em funccedilatildeo da concentraccedilatildeo de sulfato de vanadilo
presente Realizaram-se ainda simulaccedilotildees de espectros obtidos com 20 microM de S1 e 1000 microM de
V(IV) com vista agrave obtenccedilatildeo dos ligandos provaacuteveis do vanadilo atraveacutes da anaacutelise da constante
de acoplamento hiperfina (A) e do factor g (g)
4 Conclusotildees gerais
Apesar de jaacute ter sido demonstrado que o vanaacutedio inibe a Na+K
+-ATPase da miosina a funccedilatildeo
deste elemento no tecido muacutesculo esqueleacutetico continua largamente incompreendida A enorme
complexidade quiacutemica deste metal especialmente no que concerne agrave oligomerizaccedilatildeo e reacccedilotildees
51
redox constitui um obstaacuteculo agrave compreensatildeo do comportamento do vanaacutedio nos organismos
vivos Assim a caracterizaccedilatildeo da quiacutemica deste elemento eacute de grande utilidade antes de iniciar o
estudo da sua interacccedilatildeo com o presente sistema
A anaacutelise por espectroscopias de 51
V-RMN e de UV-Vis de soluccedilotildees de vanadato diluiacutedas nos
meios de reacccedilatildeo utilizados no presente relatoacuterio (10 mM HEPES pH 70 25 mM KCl) mostrou
claramente que a espeacutecie decameacuterica (V10) possui um tempo de vida suficientemente longo para a
determinaccedilatildeo do seu efeito isolado ao contraacuterio do que se verifica com as espeacutecies mono- di-
tetra- e pentameacutericas (V1 V2 V4 e V5) Por outro lado a anaacutelise verificou ainda que apoacutes diluiccedilatildeo
das soluccedilotildees de decavanadato no meio de reacccedilatildeo coexistem as espeacutecies V10 e V1 pelo que os
efeitos putativos desta uacuteltima deveratildeo ser subtraiacutedos aquando da monitorizaccedilatildeo dos efeitos da
espeacutecie decameacuterica
O vanaacutedio (V) (vanadato) inibe as enzimas que catalizam reacccedilotildees de transferecircncia de grupos
fosfato como a ATPase da miosina [Goodno 1979] devido agrave sua analogia estrutural com o iatildeo
fosfato Na sua utilizaccedilatildeo como inibidor enzimaacutetico natildeo eacute tida em conta a contribuiccedilatildeo individual
das espeacutecies oligomeacutericas considerando-se normalmente a espeacutecie V1 como inibidora efectiva
[Goodno e Taylor 1982] No presente relatoacuterio verificou-se por espectroscopia de 51
V-RMN que
outros oligoacutemeros de vanadato interagem com o S1 de miosina tendo as espeacutecies V10 e V4 os
maiores niacuteveis de afinidade A sua interacccedilatildeo envolve forccedilas electrostaacuteticas e eacute afectada
negativamente por ligandos naturais de enzima como eacute o caso do ATP
A exposiccedilatildeo do S1 de miosina (005 mgmL) a decavanadato conduz agrave inibiccedilatildeo do capacidade da
proteiacutena de estimular a actividade Mg2+
-ATPaacutesica com um valor de IC50 de 266 plusmn 07 M
decavanadato sendo o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM
decavanadato (80) Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM decavanadato sendo
o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM decavanadato (60)
exercendo um efeito de impedimento agrave ligaccedilatildeo entre o vanaacutedio e o S1
A uma concentraccedilatildeo de 27 microM decavanadato verifica-se a oxidaccedilatildeo de um dos resiacuteduos de Cys
de S1 de miosina seguida pela oxidaccedilatildeo gradual de outro resiacuteduo de Cys ateacute agrave concentraccedilatildeo de
1000 microM Ambos os resiacuteduos oxidados natildeo se encontravam expostos ao solvente utilizado A
oxidaccedilatildeo dos resiacuteduos permite sugerir que esta modificaccedilatildeo oxidativa sofrida pelo S1 contribui
para a diminuiccedilatildeo da sua actividade de ATPase
52
Por espectroscopia de RPE revelaram-se os possiacuteveis ligandos equatoriais do vanaacutedio aquando da
sua interacccedilatildeo com o S1 de miosina como sendo duas moleacuteculas de aacutegua um ligando contendo o
grupo R-OH e outro contendo o grupo R=NR
Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados de
oxidaccedilatildeo (IV) e (V) no S1 de miosina para concentraccedilotildees facilmente alcanccedilaacuteveis no ambiente
terrestre
53
5 Estudos futuros
No sentido do melhor esclarecimento das alteraccedilotildees moleculares e estruturais que ocorrem no
subfragmento 1 quando exposta ao vanaacutedio que conduzem a uma diminuiccedilatildeo da sua funccedilatildeo
sugere-se
- Quantificaccedilatildeo das cisteiacutenas reduzidas de S1 na presenccedila de metavanadato
- Determinaccedilatildeo das actividades Ca2+
- e K+-ATPaacutesica do S1 de miosina na presenccedila de
decavanadato
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina estimulada por actina
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina na presenccedila de
metavanadato
- Realizaccedilatildeo de RMN 2D com variaccedilatildeo das concentraccedilotildees das soluccedilotildees de decavanadato e
metavanadato na presenccedila de S1
-Mutageacutenese dos resiacuteduos de Cys doS1 de miosina de modo a verificar quais os resiacuteduos que
sofrem oxidaccedilatildeo na presenccedila de decavanadato
- Determinaccedilatildeo por calorimetria (DSC Differential Scanning Calorimetry) da variaccedilatildeo da
entalpia (∆H) e da temperatura de desnaturaccedilatildeo do S1 de miosina na presenccedila de vanaacutedio uma
vez que estas propriedades caracteriacutesticas de cada proteiacutena satildeo alteradas apoacutes ocorrecircncia de
modificaccedilotildees estruturais
- Anaacutelise por SDS-PAGE de S1 de miosina apoacutes tratamento com vanaacutedio de modo a determinar
quais os polipeacuteptidos eventualmente formados apoacutes clivagem da proteiacutena
54
6 Referecircncias bibliograacuteficas
- Sellers J Myosins 2nd Edition Oxford University Press 1999
- L Stryer inBiochemistry (Ed WH Freeman 2005 pp
Refordfas Materiais e Meacutetodos
[A] S S Margossian SLowey Methods Enzymol1982 85 55-71
[B] JD Pardue JA Spudich Methods Enzymol 1982 85 164-181
[C] M Bradford Anal Biochem1976 72 248-254
[D] H H Taussky E Shorr J Biol Chem 1953 202 675-685
[E] C R Bagshaw in Outline Studies in Biology ndash Muscle Contraction Chapman and Hall Ed
1982
[F] A Cuenda et al Eur J Biochem1990 194 663-670
55
Anexos
1 Determinaccedilatildeo da recta de calibraccedilatildeo meacutetodo de Bradford
Figura1ndashRepresentaccedilatildeo graacutefica da recta de calibraccedilatildeo (Abs595 nm vs [BSA]) utilizada para o meacutetodo de Bradford
2 Determinaccedilatildeo da recta calibraccedilatildeo para a electroforese em condiccedilotildees desnaturantes
Figura 2 ndash Representaccedilatildeo graacutefica da recta de calibraccedilatildeo (log(Peso Molecular) vs Rf dos marcadores de peso
molecular) onde Rf indica determinada atraveacutes do padratildeo de peso molecular
(poccedilo (a))
3Caacutelculo do coeficiente de extinccedilatildeo molar () para a quantificaccedilatildeo de cisteiacutenas reduzidas
atraveacutes do meacutetodo de Ellman
Traccedilou-se a recta de Abs412 nm meacutedia vs [L-Cisteiacutena] de modo a determinar o valor do coeficiente
de extinccedilatildeo molar definido como o declive daquela recta Assim =8400 M-1
cm-1
como
podemos verificar na Fig 3
A595 nm = 00546[BSA] + 00209 Rsup2 = 099051
Ab
s 59
5 n
m
[BSA] (mgmL)
log PM = -1177 Rf + 2247 Rsup2 = 098957
log
PM
Mobilidade relativa (Rf)
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero
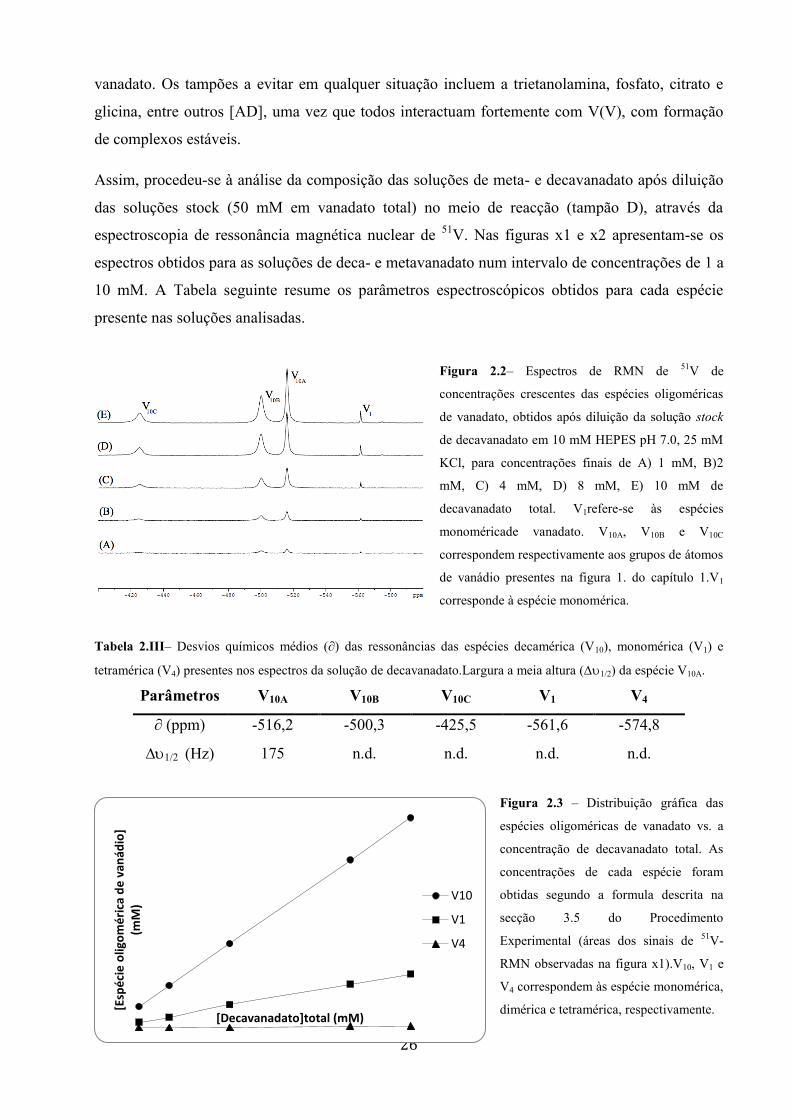
26
vanadato Os tampotildees a evitar em qualquer situaccedilatildeo incluem a trietanolamina fosfato citrato e
glicina entre outros [AD] uma vez que todos interactuam fortemente com V(V) com formaccedilatildeo
de complexos estaacuteveis
Assim procedeu-se agrave anaacutelise da composiccedilatildeo das soluccedilotildees de meta- e decavanadato apoacutes diluiccedilatildeo
das soluccedilotildees stock (50 mM em vanadato total) no meio de reacccedilatildeo (tampatildeo D) atraveacutes da
espectroscopia de ressonacircncia magneacutetica nuclear de 51
V Nas figuras x1 e x2 apresentam-se os
espectros obtidos para as soluccedilotildees de deca- e metavanadato num intervalo de concentraccedilotildees de 1 a
10 mM A Tabela seguinte resume os paracircmetros espectroscoacutepicos obtidos para cada espeacutecie
presente nas soluccedilotildees analisadas
Figura 22ndash Espectros de RMN de 51
V de
concentraccedilotildees crescentes das espeacutecies oligomeacutericas
de vanadato obtidos apoacutes diluiccedilatildeo da soluccedilatildeo stock
de decavanadato em 10 mM HEPES pH 70 25 mM
KCl para concentraccedilotildees finais de A) 1 mM B)2
mM C) 4 mM D) 8 mM E) 10 mM de
decavanadato total V1refere-se agraves espeacutecies
monomeacutericade vanadato V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2IIIndash Desvios quiacutemicos meacutedios (part) das ressonacircncias das espeacutecies decameacuterica (V10) monomeacuterica (V1) e
tetrameacuterica (V4) presentes nos espectros da soluccedilatildeo de decavanadatoLargura a meia altura (∆12) da espeacutecie V10A
Paracircmetros V10A V10B V10C V1 V4
part (ppm) -5162 -5003 -4255 -5616 -5748
∆12 (Hz) 175 nd nd nd nd
Figura 23 ndash Distribuiccedilatildeo graacutefica das
espeacutecies oligomeacutericas de vanadato vs a
concentraccedilatildeo de decavanadato total As
concentraccedilotildees de cada espeacutecie foram
obtidas segundo a formula descrita na
secccedilatildeo 35 do Procedimento
Experimental (aacutereas dos sinais de 51
V-
RMN observadas na figura x1)V10 V1 e
V4 correspondem agraves espeacutecie monomeacuterica
dimeacuterica e tetrameacuterica respectivamente [Esp
eacuteci
e o
ligo
meacute
rica
de
van
aacutedio
] (m
M)
[Decavanadato]total (mM)
V10
V1
V4
27
Nas soluccedilotildees de decavanadato as espeacutecies monomeacuterica e decameacuterica de vanadato satildeo as uacutenicas
presentes para concentraccedilotildees de decavanadato inferiores a 8 mM Ambas as espeacutecies aumentam
linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado em [AY] e
em (Teresa) quando a concentraccedilatildeo de V1 eacute igual ou superior a 200 microM inicia-se a sua
oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica No entanto nas condiccedilotildees de realizaccedilatildeo dos
ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a espeacutecie tetrameacuterica neste
estudo em concentraccedilotildees inferiores a 8 mM tendo o mesmo sido observado por Ramoset al
(2006) [AK] Os resultados obtidos mostram claramente o equiliacutebrio entre a espeacutecie decameacuterica e
a monomeacuterica e que o aparecimento de outras espeacutecies oligomeacutericas se encontra dependente da
concentraccedilatildeo desta uacuteltima Apesar de natildeo ter sido realizado o ensaio com 5 mM de decavanadato
total no presente relatoacuterio estaacute reportado em Ramos et al (2006) [AK] que as concentraccedilotildees da
espeacutecie decameacuterica e monomeacuterica satildeo respectivamente de 500 microM e 200 microM Assim pode inferir-
se que para concentraccedilotildees inferiores a 5 mM e intervalos de tempo de cerca de 20 minutos (que
inclui o tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas
as espeacutecies decameacuterica e monomeacuterica de vanadato
Figura 24 ndash Espectros de RMN de 51
V de concentraccedilotildees
crescentes das espeacutecies oligomeacutericas de vanadato obtidos
apoacutes diluiccedilatildeo da soluccedilatildeo stock de metavanadato em 10 mM
HEPES pH 70 25 mM KCl para concentraccedilotildees finais de A)
1 mM B) 2 mM C) 4 mM D) 8 mM E) 10 mM de
metavanadato total V1 refere-se agraves espeacutecies monomeacuterica de
vanadatoV1 e V2 correspondem respectivamente agrave espeacutecie
monomeacuterica e dimeacuterica
Figura 25 ndash Distribuiccedilatildeo graacutefica das espeacutecies
oligomeacutericas de vanadato vs a concentraccedilatildeo de
metavanadato total As concentraccedilotildees de cada
espeacutecie foram obtidas segundo a formula descrita
na secccedilatildeo 35 do Procedimento Experimental
(aacutereas dos sinais de 51
V-RMN observadas na
figura x1)V10 V1 e V4 correspondem agraves espeacutecie
monomeacuterica dimeacuterica e tetrameacuterica
respectivamente
Esp
eacuteci
e o
ligo
meacute
rica
de
va
nad
ato
(m
M)
[Metavanadato]total (mM)
V1
V2
V4
V5
28
Tabela 2IVndash Desvios quiacutemicos meacutedios (part)das ressonacircncias das espeacutecies tetrameacuterica (V4) monomeacuterica (V1) e
dimeacuterica (V2) presentes no espectro da soluccedilatildeo de metavanadato (total) 10 mM
Paracircmetros V1 V2 V4 V5
part (ppm) -5615 -5748 -5789 -5869
Nas soluccedilotildees de metavanadato as espeacutecies monomeacuterica dimeacutericae tetrameacuterica de vanadato estatildeo
presentes para concentraccedilotildees de metavanadato inferiores a 4 mM Todas estas as espeacutecies
aumentam linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado
em (Aureliano e Madeira 1998) e em (Tiago et al 2004) quando a concentraccedilatildeo de V1 eacute igual ou
superior a 200 microM inicia-se a sua oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica Nas
condiccedilotildees de realizaccedilatildeo dos ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a
espeacutecie pentameacuterica neste estudo em concentraccedilotildees inferiores a 4 mM tendo o mesmo sido
observado por Ramos 2006 Os resultados obtidos mostram claramente o equiliacutebrio entre a
espeacutecie V4 e as espeacuteciesV1 e V2 no qual o aparecimento de outras espeacutecies oligomeacutericas se
encontra dependente da concentraccedilatildeo da espeacutecie V1 Por uacuteltimo pode inferir-se que para
concentraccedilotildees inferiores a 2 mM e intervalos de tempo de cerca de 20 minutos (que inclui o
tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas as
espeacutecies V1 V2 e V4 com concentraccedilotildees respectivas de 0796 0192 e 0196 mM
232Estabilidade das soluccedilotildees de decavanadato anaacutelise por espectroscopia de absorccedilatildeo
UVVis
Caracterizou-se a estabilidade da espeacutecie V10 numa soluccedilatildeo com 1 mM decavanadato total em
tampatildeo D atraveacutes da leitura da Abs400 Obteve-se uma cineacutetica de dissociaccedilatildeo de 1ordf ordem tal
como se pode verificar pela linearidade do graacutefico de ln(Abs400)vs o tempo de aquisiccedilatildeo espectral
(segundos) (Fig26 )
Figura 26ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do tempo
(em segundos) para uma soluccedilatildeo de decavanadato total de 1
mM em tampatildeo D a 25ordmC (n=2)
Sendo uma reacccedilatildeo de primeira ordem pode ser descrita pela equaccedilatildeo cujo
declive eacute ndashK (constante de dissociaccedilatildeo) e a intersecccedilatildeo no eixo das ordenadas corresponde
Ln
(Ab
s 400 n
m)
tempo (s)
29
aln(A0) Numa reacccedilatildeo de 1ordf ordem o tempo de meia-vida eacute independente da concentraccedilatildeo inicial
do reagente sendo descrito pela equaccedilatildeo [2] Deste modo a partir da recta
Ln(Abs400 nm) = -436E-5
K - 105 obteve-se um tempo de meia-vida de 4490 plusmn 0823 horas para
as condiccedilotildees supramencionadas
Apoacutes a adiccedilatildeo de S1 8 microM ao meio reaccional verificou-se que a reacccedilatildeo de desoligomerizaccedilatildeo
da espeacutecie decameacuterica de vanadato seguia igualmente uma cineacutetica de 1ordf ordem tal como se pode
verificar pela linearidade do graacutefico de ln(Abs400) versus o tempo de aquisiccedilatildeo espectral
(segundos) (Fig27)
Figura 27ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do
tempo (em segundos) para uma soluccedilatildeo de decavanadato
total de 1 mM em tampatildeo D a 25ordmC na presenccedila de 8
microM (n= 2)
A partir da equaccedilatildeo da recta Ln(Abs400 nm) = -7520E-5
K - 0613determinou-se o valor de K para
a espeacutecie decameacuterica de onde se obteve um tempo de meia-vida de 2430 plusmn 0191 horas para as
condiccedilotildees supramencionadas
Tanto este valor como o valor registado na ausecircncia de S1 satildeo substancialmente superiores agrave
duraccedilatildeo dos ensaios de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina descritos no presente
relatoacuterio
24 Interacccedilatildeo de S1 com vanaacutedio(V)
241Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
De modo a analisar qual o efeito da adiccedilatildeo de S1 de miosina (tampatildeo D) produzia nos sinais de
51V-NMR titulou-se uma soluccedilatildeo de 5mM de decavanadato total com concentraccedilotildees crescentes
de S1 (5 10 15 e 20 microM) Os espectros de 51
V-RMN encontram-se apresentados na Fig 2
No graacutefico da figura 2 Apresenta-se a concentraccedilatildeo de S1 em funccedilatildeo do factor de alargamento
corrigido dos espectros acima referidos
Ln
(Ab
s (4
00
nm
))
tempo (s)
30
Figura 28 ndash Graacutefico de
representaccedilatildeo da
concentraccedilatildeo de S1 em
funccedilatildeo do factor de
alargamento A
concentraccedilatildeo de
decavanadato total foi de 5
mM e as concentraccedilotildees
utilizadas de S1 foram de 0
5 10 e 15 microM
Verifica-se que o factor de alargamento aumenta linearmente em funccedilatildeo da concentraccedilatildeo de S1 no
ensaio o que indica a existecircncia de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina Observou-
se ainda que a adiccedilatildeo de S1 ao meio reaccional natildeo provocou qualquer alteraccedilatildeo dos desvios
quiacutemicos dos sinais de 51
V-RMN
Realizaram-se ensaios na presenccedila de ATP e de S1 com intuito de verificar o efeito do ATP na
interacccedilatildeo do S1 de miosina com o decavanadato
Figura 29 - Espectros de RMN de 51
V contendo 2
mM Decavanadato (A) (controlo) (B) na presenccedila
de 2 mM de ATP (C) na presenccedila de 2 mM de ATP
e 8 microM de S1 de miosina V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2Vndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies decameacuterica (V10) e monomeacuterica (V1) presentes nos
espectros da soluccedilatildeo de decavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V10A V10B V10C V1
part (ppm) -5161 -5002 -4254 -5614
Observa-se que a presenccedila de ATP (espectro B) natildeo provoca qualquer alteraccedilatildeo visiacutevel em
relaccedilatildeo ao espectro que contem apenas soluccedilatildeo de decavanadato Jaacute a presenccedila simultacircnea de ATP
e S1 provoca um alargamento da ∆12 do sinal do V10A de 1736 Hz (espectro (A)) para 2366 Hz
(espectro (C) Fig 29) Tendo em conta que a concentraccedilatildeo de S1 apresenta uma relaccedilatildeo linear
com o factor de alargamento converteu-se o valor da ∆12 e comparou-se com o valor esperado agrave
F = 00591[S1] - 00157 Rsup2 = 0998
Fact
or
de
ala
rgam
en
to
[S1] (microM)
31
concentraccedilatildeo de S1 utilizada O factor de alargamento obtido para 8 microM ndash 0376 ndash eacute inferior ao
valor esperado agrave mesma concentraccedilatildeo de S1 obtido atraveacutes da equaccedilatildeo da recta apresentada na
Fig 28 ndash 0457 ndash o que indica que o ATP poderaacute exercer um efeito de protecccedilatildeo do S1
diminuindo a sua interacccedilatildeo com a espeacutecieV10
Por uacuteltimo realizaram-se ensaios com metavanadato na presenccedila de 8 microM S1 de modo a verificar
se as espeacutecies oligomeacutericas contidas nesta soluccedilatildeo interagiam com a proteiacutena em estudo
Figura 210 - Espectros de RMN de 51
V contendo 2
mM metavanadato (A) (controlo) (B) na presenccedila
de 8 microM de S1 de miosina V1 V2 e V4
correspondem respectivamente aacutes espeacutecies
monomeacuterica dimeacuterica e tetrameacuterica
Tabela 2VIndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies monomeacuterica (V1) dimeacuterica (V2) e tetrameacuterica (V4)
presentes nos espectros da soluccedilatildeo de metavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V1 V2 V4
part (ppm) -5613 -5747 -5789
Como eacute possiacutevel observar na Fig 210 ocorre uma ligeiro alargamento dos sinal de V4 bem
como uma perda da intensidade pelo que se conclui quedas espeacutecies presentes na soluccedilatildeo de
metavanadato (V1 V2 e V4) apenas a espeacutecie tetrameacuterica interage com o S1 de miosina o que
estaacute de acordo com o reportado por Tiago et al 2002
242 Actividade Mg2+
-ATPaacutesica de S1 de miosina na presenccedila de decanadato
Com o intuito de avaliar o efeito do decavanadato na actividade Mg2+
-ATPaacutesica do S1 de
miosina monitorizou-se a reacccedilatildeo de oxidaccedilatildeo do NADH atraveacutes da leitura de Abs340 na
presenccedila de 005 mgmL de S1 de miosina e no caso da actividade estimulada 2microM de actina em
3125 mM Tris-HCl pH 71 e 375 mM MgCl2 pelo meacutetodo de enzimas acopladas (ver seccccedilatildeo xx
dos MM)Para tal realizaram-se ensaios com vaacuterias concentraccedilotildees de decavanadato total (10 50
100 250 500 e 1000) de modo a determinar qual o seu efeito na actividade MgATPaacutesica de S1
de miosina na ausecircncia e na presenccedila de um dos seus ligandos naturais Os ensaios foram
realizados agrave temperatura ambiente com incubaccedilatildeo preacutevia a 25ordm C durante 20 minutos
32
Figura 211 ndash Representaccedilatildeo
graacutefica da Actividade Mg2+
-
ATPaacutesica de S1 () (os ajustes
teoacutericos estatildeo representado a por
uma linha os resultados
experimentais estatildeo representados
por triacircngulos (Actividade
estimulada) e cruzes (Actividade
natildeo estimulada)) vs
[Decavanadato]total medidas pelo
meacutetodo das enzimas acopladas
(n=3) Os dados foram ajustados ao modelo de decaimento exponencial usando o programa GraphPad Prism para a
determinaccedilatildeo dos valores de IC50 (na ausecircncia de actina IC50= 266 plusmn 07 M decavanadato na presenccedila de actina
IC50= 28 plusmn 1 M decavanadato
A partir do graacutefico presente na Fig 29 verifica-se que o ajuste dos dados experimentais foi bem
sucedido A inibiccedilatildeo da actividade Mg2+
-ATPaacutesica do S1 de miosina eacute afectada em menor
amplitude na presenccedila de actina (60 de inibiccedilatildeo da actividade) do que na sua ausecircncia
(aproximadamente 80 de inibiccedilatildeo de actividade) Devido a esta diferenccedila pode sugerir-se que a
actina iraacute impedir parcialmente a ligaccedilatildeo do decavanadatoao S1 demiosina atraveacutes de
impedimentos esteacutereos ou interacccedilotildees electrostaacuteticas por exemplo A teacutecnica utilizada natildeo nos
permite concluir acerca de que regiatildeo(otildees) ou resiacuteduo(s) da actina participaratildeo na preservaccedilatildeo
parcial da actividade Mg2+
-ATPaacutesica
Por outro lado os valores de IC50 dos dois tipos de actividades natildeo apresentam diferenccedilas
significativas tendo-se obtido 266 plusmn 07 e 28 plusmn 1M decavanadato na ausecircncia e na presenccedila de
actina respectivamente o que corresponde a uma concentraccedilatildeo de cerca de 27 M de V10 O
meio de reacccedilatildeo utilizado nestes ensaios foi 10 mM HEPES e 25 mM KCl com 2 mM ATP Em
(Tiago et al 2004) o valor de IC50 para a actividade Mg2+-ATPaacutesica na ausecircncia de actina foi de
125 plusmn 025 microM enquanto o valor da actividade estimulada por actina foi de 136 plusmn 14 microM O
meio de reacccedilatildeo utilizado nos ensaios foi 20 mM HEPES 50 mM KCl e 2 mM ATP o que resulta
precisamente no dobro da forccedila ioacutenica a que foi realizado o ensaio do presente relatoacuterio
(exceptuando a contribuiccedilatildeo dos produtos da hidroacutelise de ATP uma vez que este meacutetodo
pressupotildee a ocorrecircncia de um ciclo de reacccedilotildees de hidroacutelise e siacutentese deste composto) Ora tendo
sido anteriormente referido que a afinidade do S1 de miosina para com a espeacutecie V10 diminui com
Act
ivid
ade
Mg
2+-A
TP
aacutesic
a d
e S
1 d
e
Mio
sin
a
[Decavanadato] (microM) Actividade Estimulada ()
Actividade Natildeo Estimulada ()
Ajuste Actividade Estimulada
Ajuste Actividade Natildeo Estimulada
33
o aumento da forccedila ioacutenica (Tiago et al 2004 tal poderaacute explicar a diferenccedila obtida entre estes
dois conjuntos de valores
243Quantificaccedilatildeo de cisteiacutenas reduzidas pelo meacutetodo de Ellman
A interacccedilatildeo do vanaacutedio com alguns tipos de aminoaacutecidos sensiacuteveis agrave oxidaccedilatildeo pode fornecer-nos
informaccedilatildeo bastante uacutetil em relaccedilatildeo agraves alteraccedilotildees estruturais que conduzem a uma diminuiccedilatildeo da
actividade fisioloacutegica do S1 de miosina Os estudos realizados por Chasteen 1983 concluiacuteram
que a reduccedilatildeo de vanadato (V) a vanadilo (IV) ocorre na presenccedila de agentes redutores comuns no
ambiente celular de que se destaca o aminoaacutecido cisteiacutena Esta eacute tambeacutem um alvo de
complexaccedilatildeo por parte do iatildeo VO2+
Jaacute Ramos et al (2009) demonstrou que a incubaccedilatildeo de
actina com a soluccedilatildeo de decavanadato induz a oxidaccedilatildeo de resiacuteduos de cisteiacutena e agrave formaccedilatildeo do
oxidovanaacutedio (IV) Este grupo de investigadores descobriu ainda que apenas a espeacutecie V10 induz a
oxidaccedilatildeo das cisteiacutenas ldquoraacutepidasrdquo da actina atraveacutes do processo proposto que eacute parcialmente
responsaacutevel pela inibiccedilatildeo jaacute descrita da actividade de ATPase do S1 de miosina estimulada por
actina quando exposta agrave soluccedilatildeo de decavanadato Esta inibiccedilatildeo apresenta-se afectada com a
presenccedila de ATP uma vez que os resiacuteduos de cisteiacutena permaneceram intactos nos ensaios
realizados com este ligando Jaacute Crans et al (2004) concluiu que o siacutetio activo de muitas fosfatases
(que conteacutem grupos tiol) pode formar um equiliacutebrio redox com o iatildeo vanadato (V) de forma natildeo
reversiacutevel
Os resultados obtidos por Sakurai e Shiromura (1980) sugeriram que o vanaacutedio (V) sofre reduccedilatildeo
a vanaacutedio (IV) sem dependecircncia do valor de pH do ensaio Registaram ainda uma estequiometria
de 31 na reacccedilatildeo do vanadato (V) com os resiacuteduos de Cys enquanto a estequiometria dos ensaios
com o vanadilo (IV) foi de 21
Berlett e Stadtman (1997) observaram que os tipos de aminoaacutecidos mais facilmente oxidaacuteveis
englobam entre outros os resiacuteduos de Cys Trp e MetAssim analisou-se o estado redox dos
grupos tiol dos resiacuteduos de Cys do S1 na presenccedila do decavanadato atraveacutes da monitorizaccedilatildeo da
Abs412 nm na presenccedila de 1 mgmL de S1 de miosina em tampatildeo 10 mM HEPES 25 mM KCl
variando a concentraccedilatildeo da soluccedilatildeo de decavanadato de modo a determinar que tipo de alteraccedilotildees
estruturais ocorrem na sua presenccedila no que diz respeito agrave diminuiccedilatildeo da actividade ATPaacutesica de
S1 na presenccedila de decavanadato
Os resultados referentes agrave quantificaccedilatildeo das cisteiacutenas reduzidas apoacutes o tratamento de S1 com
diferentes concentraccedilotildees de decavanadato em tampatildeo 10 mM HEPES pH 75 e 25 mM KCl a
25ordmC durante 20 minutos encontram-se descritos na Tabela 2VII
Tabela 2VII ndash Cisteiacutenas reduzidas totais e ldquoraacutepidas de S1 (1 mgmL) apoacutes tratamento com decavanadato
34
(010501002505001000 uM) em 10 mM HEPES 25 mM KCl pH 7 a 25ordmC durante 20 minutos A detecccedilatildeo
espectrofotomeacutetrica foi feita com DTNB (e 412=8400 uM-1 cm-1) (n variaacutevel entre 1 e 8)
Amostras Cys raacutepidas Cys totais
11 M S1 (controlo) 22 plusmn 04 106 plusmn 14
11 M S1 + 10 M Decavanadato 22 plusmn 05 95 plusmn 08
11 M S1 + 50 uM Decavanadato 20 plusmn 02 98 plusmn 14
11 M S1 + 100 uM Decavanadato 17 plusmn 03 97 plusmn14
11 M S1 + 250 uM Decavanadato 26 plusmn 02 98 plusmn 08
11 M S1 + 500 uM Decavanadato 21 plusmn 04 90 plusmn 06
11 M S1 + 1000 uM Decavanadato 24 plusmn 0 86 plusmn 08
A partir dos resultados obtidos pode sugerir-se que o S1 possui 11 resiacuteduos de Cys (controlo) o
que estaacute de acordo com o anteriormente descrito Verifica-se que o tratamento da proteiacutena com
concentraccedilotildees crescentes de decavanadato provoca a oxidaccedilatildeo de dois resiacuteduos de Cys do S1 de
miosina natildeo expostos ao solvente No entanto a teacutecnica utilizada natildeo nos permite identificar
quais
Tendo em consideraccedilatildeo que o valor de IC50 obtido para a reacccedilatildeo de inibiccedilatildeo daactividade Mg2+-
ATPaacutesica de S1 na presenccedila de decavanadato eacute de cerca de 27 microM pode sugerir-se que este seraacute o
valor ao qual ocorreraacute a oxidaccedilatildeo do primeiro resiacuteduo de Cys o que poderaacute contribuir para a
diminuiccedilatildeo da actividade de S1 observada
25Interacccedilatildeo de S1 com vanaacutedio(IV)
251Anaacutelise por espectroscopia de Ressonacircncia Paramagneacutetica Electroacutenica
A acumulaccedilatildeo intracelular de oxovanaacutedio (IV) in vivo constitui uma das principais formas de
apresentaccedilatildeo de vanaacutedio no organismo Deste modo torna-se bastante interessante analisar a
interacccedilatildeo entre este iatildeo e o S1 de miosina atraveacutes da teacutecnica de espectroscopia de Ressonacircncia
Paramagneacutetica Electroacutenica o meacutetodo de eleiccedilatildeo do estudo do iatildeo vanadilo e dos complexos por si
formados O electratildeo desemparelhado encontra-se fortemente acoplado ao nuacutecleo 51
V o que
35
resulta num espectro de RPE consistindo de 8 desdobramentos na direcccedilatildeo paralela e outros tantos
na direcccedilatildeo perpendicular (Chasteen 1983)
Os estudos realizados por Ahmed et al (1987) centraram-se na interacccedilatildeo do vanadilo com
proteiacutenas como eacute o caso da insulina transferrina entre outros obtendo um espectro caracterizado
por 8 linhas amplamente espaccediladas (na direcccedilatildeo paralela) Jaacute Ramos et al 2009 concluiu que a
exposiccedilatildeo da soluccedilatildeo de decavanadato agraves formas globular e filamentosa de actina produz um
espectro tiacutepico do iatildeo oxidovanaacutedio (IV) indicando a reduccedilatildeo do vanadato
Para aferir qual a concentraccedilatildeo de S1 que permitia obter uma boa razatildeo sinalruiacutedo titulou-se uma
soluccedilatildeo de sulfato de vanadilo 15 microM ([VOSO4]stock = 5 mM) com concentraccedilotildees crescentes de
proteiacutena (0 5 10 15 e 20 microM) em 10 mM HEPES 25 mM KCl A existecircncia de uma correlaccedilatildeo
linear entre a concentraccedilatildeo de proteiacutena e a intensidade dos sinais de RPE correspondente agrave banda
-12 perpendicular encontra-se representada na Fig 210
Figura 2 12ndash Representaccedilatildeo graacutefica da variaccedilatildeo da intensidade do sinal de RPE vs a concentraccedilatildeo de S1 utilizada
Apoacutes verificar que com uma concentraccedilatildeo de S1 igual a 20 microM se obtinham sinais com uma razatildeo
sinalruiacutedo adequada ao prosseguimento dos estudos sobre a interacccedilatildeo V(IV)-S1 de miosina
titulou-se uma soluccedilatildeo de 20 microM S1 com 0 25 5 10 15 25 50 75 100 150 200 500 e 1000
microM VOSO4
Traccedilou-se o graacutefico da intensidade da banda -12 perpendicular do sinal de EPR em funccedilatildeo da
concentraccedilatildeo de VOSO4 tendo-se determinado uma constante de dissociaccedilatildeo (KD) do complexo
S1-vanadilo de 8748 plusmn 2621 microM V(IV)
y = 016786x + 122435
Rsup2 = 092992759
Inte
nsi
dad
e d
o s
inal
de
RP
E
[S1] (microM)
36
Figura 213 ndash Representaccedilatildeo graacutefica da variaccedilatildeo do sinal de RPE vs a concentraccedilatildeo de vanadilo utilizada (os valores
experimentais estatildeo representados por losangos o seu ajuste estaacute representado pela linha)
A simulaccedilatildeo do espectro de EPR foi efectuada a partir do espectro obtido para uma concentraccedilatildeo
de 1 mM VOSO4 apoacutes adiccedilatildeo de 20 microM S1 de miosina tendo-se obtido os valores da constante de
acoplamento hiperfina (A=5016) e do factor g (g0=195)
Figura 214 ndash Representaccedilatildeo graacutefica do espectro de EPR obtido com 1000 microM de vanadilo e 20 microM de S1 (a cinzento
escuro) A simulaccedilatildeo do espectro encontra-se a cinzento claro
Por comparaccedilatildeo destes com os valores presentes na tabela de correspondecircncia xxx eacute possiacutevel
predizer quais os ligandos provaacuteveis do vanadilo na presenccedila de proteiacutena Assim concluiu-se que
o vanadilo estaacute coordenado com duas moleacuteculas de aacutegua um grupo hidroxilo e um grupo R=NR
O grupo hidroxilo pode indicar os resiacuteduos de aminoaacutecidos Tyr Treonina e Serina enquanto o
grupo R=NR poderaacute indicar a presenccedila de Arginina e Histidina como ligandos do vanadilo
Inte
nsi
dad
e d
os
sinai
s d
e R
PE
[Vanadilo] (microM)
Ajuste dos dados exp Dados experimentais
Campo magneacutetico (G)
Espectro Experimental Ajuste
37
3 Procedimento Experimental
31 Reagentes
Todos os reagentes utilizados satildeo de elevado grau de pureza (analiacutetico) Apresentam-se as suas
foacutermulas quiacutemicas ou abreviaturas quando necessaacuterio acrescentou-se a sua descriccedilatildeo Entre
parecircntesis colocou-se as empresas que os comercializam
Aacutecido aceacutetico glacial 100 (Riedel) Acrilamida 99 (Merck) Azul de bromofenol (Riedel-de-
Haumlen) -Mercaptoetanol (Sigma) Bissulfito de soacutedio (Sigma)Bradford (reagente de)
(BioRad)BSA (albumina de soro bovino Sigma-Aldrich) CaCl22H2O (cloreto de caacutelcio
dihidratado Fluka) CH3CH2OH 96 (etanol Panreac) Coomassie Blue R250 (Merck) D2O
(oacutexido de deuteacuterio SDS) DTNB (55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico) Aldrich) EDTA (aacutecido
etilenodiaminatetraaceacutetico Merck) Fosfoenolpiruvato 98 (Fluka) Glicerol (Sigma) Glicina
985 (Panreac) HCl 37 (Riedel-de Haumlen) H2SO4 96 (Pronalab) HEPES 99 (aacutecido 4-(2-
hidroxiletil)-1-piperazineetanosulfoacutenico Sigma) KH2PO4 99 (hidrogenofosfato de potaacutessio)
MgATP (Sigma) Na
ATP (Sigma) Na2HPO4 99 (hidrogenofosfato de soacutedio Riedel-de Haumlen)
NaH2PO4 (fosfato de soacutedio ) NaOH 986 (hidroacutexido de soacutedio Pronalab) KCl 99 (Cloreto de
potaacutessio Panreac) L-Cys 995 (enantioacutemero levoacutegiro do resiacuteduo de Cisteiacutena Fluka)Metanol
99 (Merck) Molibdato de amoacutenio (Sigma)NNrsquo-metil-bis-acrilamida 98 (Merck) NH4VO3
99 (metavanadato de amoacutenia Sigma) PSA 10 (persulfato de amoacutenio Merck) SDS
(dodecilo sulfato de soacutedio Sigma) Sulfito de soacutedio (Sigma) VOSO4 (sulfato de vanadilo
Aldrich) TEMED 99 (Tetrametiletilenodiamina Merck) Tris (2-amino-2-hidroximetil-13-
propanodiol Sigma)
32 Meacutetodos
321 Isolamento e purificaccedilatildeo das proteiacutenas de muacutesculo esqueleacutetico de coelho
3211 Purificaccedilatildeo da miosina preparaccedilatildeo de Subfragmento 1 por digestatildeo quimotriacutepsica
da miosina
O subfragmento-1 de miosina-II (S1) foi gentilmente cedido pelo Prof Doutor Manuel Aureliano
do Departamento de Quiacutemica Bioquiacutemica e Farmaacutecia da Faculdade de Ciecircncias e Tecnologia da
Universidade do Algarve
No entanto descreve-se abaixo de forma resumida o procedimento efectuado
38
32111 Purificaccedilatildeo da miosina
O protocolo de isolamento foi adaptado do meacutetodo previamente descrito por Margossian e Lowey
(1982) apresentando-se de forma esquemaacutetica na Fig 31
Sucintamente os tecidos musculares dorsais e das pernas foram extraiacutedos apoacutes o sacrifiacutecio do
animal (Fig31) De seguida foram adicionados 3 volumes da soluccedilatildeo de extracccedilatildeo A2
Centrifugou-se a soluccedilatildeo e filtrou-se o sobrenadante O sedimento foi entatildeo diluiacutedo em 14
volumes de aacutegua destilada de modo a que a forccedila ioacutenica atingisse 004 M Apoacutes adiccedilatildeo de szlig-
mercaptoetanol (szlig-me) deixou-se a precipitar a 4ordmC removendo o sobrenadante Centrifugou-se
novamente e ressuspendeu-se o sedimento na soluccedilatildeo de extracccedilatildeo A Adicionou-se a soluccedilatildeo
tampatildeo B3 para que a forccedila ioacutenica atingisse 05 M Homogeneizou-se e deixou-se repousar
durante 2h Homogeneizou-se novamente e centrifugou-se de modo a ocorrer precipitaccedilatildeo dos
complexos de actomiosina e proteiacutena desnaturada Apoacutes filtraccedilatildeo do sobrenadante em latilde de vidro
quantificou-se a soluccedilatildeo de miosina resultante pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental)
2 Soluccedilatildeo de extracccedilatildeo A 0012M K2CO3 004M KHCO3 06M KCl pH 65
3 Soluccedilatildeo-tampatildeo B Tris 100 mM pH 7 KCl 2M
39
Figura 31 ndash Representaccedilatildeo esquemaacutetica do protocolo de isolamento de miosina (Margossian e Lowey 1982) Todo o
procedimento foi realizado a 4ordmC (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
32112 Preparaccedilatildeo de Subfragmento-1 de miosina por digestatildeo com quimiotripsina
O S1 de miosina foi isolado apoacutes diaacutelise da miosina durante a noite contra a soluccedilatildeo C4 A
presenccedila do EDTA eacute essencial uma vez que a sua substituiccedilatildeo por MgCl2resulta na produccedilatildeo de
HMM em vez de S1 A suspensatildeo de miosina foi equilibrada e digerida com -quimotripsina
A digestatildeo foi interrompida pela adiccedilatildeo de PMSF Seguidamente ultracentrifugou-se o produto
da digestatildeo desprezando-se o sedimento (Fig 32) A soluccedilatildeo de sobrenadante maioritariamente
S1 foi quantificada pela leitura directa da Abs280 ou pelo meacutetodo de Bradford (ver as secccedilotildees
3311 e 3312 do Procedimento Experimental respectivamente) e armazenada em 10 sacarose
e 2 mM szlig-Me a -80ordmC
4 Soluccedilatildeo C 25mM Tris pH 70 012M KCl 1mM EDTA
40
Antes da realizaccedilatildeo dos ensaios descritos no presente relatoacuterio o S1 de miosina foi dialisado
contra a soluccedilatildeo D5 de modo a remover qualquer composto que pudesse complexar como
vanaacutedio Seguidamente quantificou-se a proteiacutena pelo meacutetodo de Bradford ou pelo meacutetodo
directo (ver secccedilotildees 3311 e 3312do Procedimento Experimtental)
Figura 3 2ndash Representaccedilatildeo esquemaacutetica do protocolo de preparaccedilatildeo do S1 de miosina por digestatildeo quimiotripsina de
acordo com o previamente descrito por Margossian e Lowey (1982) Todo o procedimento foi realizado a 4ordm C
excepto quando indicaccedilatildeo do contraacuterio (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
3212 Isolamento e purificaccedilatildeo de actina
Isolou-se e purificou-se a actina segundo o meacutetodo descrito por Pardee e Spudish que
compreende dois passos ndash a obtenccedilatildeo de um resiacuteduo seco denominado ldquopoacute de acetonardquo e a
extracccedilatildeo subsequente da actina a partir do ldquopoacute de acetonardquo na forma de G-Actina com a
posterior polimerizaccedilatildeo a F-Actina
32121 Obtenccedilatildeo do ldquopoacute de acetonardquo
Sacrificou-se o coelho com uma pancada atraacutes do pescoccedilo e sangrou-se o animal apoacutes o corte da
arteacuteria jugular Esfolou-se e colocou-se numa tina com gelo Retiraram-se as partes de tecido
conjuntivo que envolvem o tecido muscular com auxiacutelio de uma pinccedila e bisturi Seguidamente
retirou-se os muacutesculos dorsais e os muacutesculos brancos das pernas Passou-se os muacutesculos por aacutegua
destilada de modo a remover as impurezas e pesou-se num copo Trituraram-se os muacutesculos
numa maacutequina de triturar carne e extraiu-se o resultante com 3 volumes da soluccedilatildeo E6 durante 10
minutos com agitaccedilatildeo suave Centrifugou-se a suspensatildeo a 4000 g durante 10
5 Soluccedilatildeo D HEPES 10 mM pH 70 KCl 25 mM
6 Soluccedilatildeo E 01 M KCl 015 M K2HPO4 pH 65
41
minutosdesprezando-se o sobrenadante Ressuspendeu-se o sedimento em 6 volumes de soluccedilatildeo
F7durante 10 minutos com agitaccedilatildeo suave Verificou-se se o pH da soluccedilatildeo estava compreendido
entre 82 e 85 acertando-se estecom soluccedilatildeo G8 se necessaacuterio Centrifugou-se durante 10
minutos a 4000 g Desprezou-se o sobrenadante e ressuspendeu-se o sedimento em 3 volumes de
soluccedilatildeo H9 durante 10 minutos Centrifugou-se novamente a 4000 gdurante 10 minutos
desprezando-se o sobrenadante Ressuspendeu-se o sedimento duas vezes consecutivas em 6
volumes de aacutegua destilada durante 5 minutos Centrifugou-se a 4000 g durante 10 minutos apoacutes
cada adiccedilatildeo de H2O
Seguidamente ressuspendeu-se o sedimento 5 vezes consecutivas utilizando 3 volumes de
acetona de cada vez agitando suavemente durante 10 minutos agrave temperatura ambiente Colocou-
se o resiacuteduo filtrado numa tina de vidro sobre papel de filtro deixando-se evaporar o solvente
durante a noite num nicho Triturou-se o resiacuteduo seco numa maacutequina de triturar carne e deixou-se
novamente evaporar o solvente O ldquopoacute de acetonardquo obtido foi armazenado a -20 ordmC
32122 Purificaccedilatildeo da actina a partir de ldquopoacute de acetonardquo
Apoacutes a obtenccedilatildeo do poacute de acetona como acima indicado efectuou-se o protocolo seguidamente
descrito de modo a obter monoacutemeros de actina (G-Actina) e actina filamentosa (F-Actina)
Pesou-se 35 g de poacute de acetona e adicionaram-se 50 mL de soluccedilatildeo-tampatildeo I10
sendo a
extracccedilatildeo feita com agitaccedilatildeo suave durante 30 minutos Separou-se o extracto do poacute de acetona
hidratado filtrando a suspensatildeo com 4 camadas de gaze esterilizada tendo o cuidado de natildeo
espremer o resiacuteduo Voltou a extrair-se com 50 mL de soluccedilatildeo I durante 30 minutos mexendo
lentamente Adicionou-se este ao extracto obtido anteriormente Centrifugou-se o extracto total a
16 000 g durante 30 minutos Decantou-se para uma proveta sobre a qual se colocou gaze para
que o sedimento solto natildeo se juntasse ao sobrenadante e leu-se o volume De seguida adicionou-
se KCl ateacute uma concentraccedilatildeo final de 50 mM MgCl2 ateacute 2mM e ATP ateacute 1mM lentamente e com
agitaccedilatildeo contiacutenua de modo a promover a polimerizaccedilatildeo dos monoacutemeros de actina
Deixou-se polimerizar durante 2h a 25ordmC observando-se a presenccedila de pequenas bolhas de ar
indicativas de um aumento de viscosidade da soluccedilatildeo Arrefeceu-se em gelo durante 15 minutos e
adicionou-se KCl ateacute uma concentraccedilatildeo final de 06M Mexeu-se lentamente durante 30 minutos
apoacutes os quais se ultracentrifugou a 110 000g durante 1hora Desprezou-se o sobrenadante
ressuspendeu-se o sedimento com soluccedilatildeo tampatildeo E e transferiu-se para um potter
Dialisou-se durante a noite contra 1L de soluccedilatildeo-tampatildeo I a uma temperatura de 4ordmC com rotaccedilatildeo
7 Soluccedilatildeo F 005 M NaHCO3 pH 82-85
8 Soluccedilatildeo G 1M Na2CO3
9 Soluccedilatildeo H 1 mM EDTA pH 70
10 Soluccedilatildeo I 2 mM Tris-HCl pH 8 02 mM CaCl2 02 mM Na2ATP 05 mM szlig-Me 0005 NaN3
42
contiacutenua e mudanccedila de tampatildeo 2 a 3 vezes De seguida a actina foi centrifugada a 110
000gdurante 1hora Decantou-se a soluccedilatildeo de G-Actina (sobrenadante) e desprezou-se o
sedimento Determinou-se a concentraccedilatildeo de G-Actina pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental) A soluccedilatildeo de G-actina foi armazenada a uma temperatura de 4ordmC
apoacutes adiccedilatildeo de 002NaN3 mantendo-se estaacutevel durante 3 a 5 dias
331 Quantificaccedilatildeo das proteiacutenas isoladas
3311 Quantificaccedilatildeo pelo meacutetodo de Bradford
O meacutetodo de Bradford (Bradford 1976) baseia-se na utilizaccedilatildeo do reagente de Bradford
composto por entre outros aacutecido fosfoacuterico e o corante ldquoCoomassierdquo Este corante possui carga
negativa e a sua coloraccedilatildeo varia entre a vermelha (Absmax=465 nm) predominante em soluccedilatildeo e a
azul (Absmax=595 nm) presente aquando da ligaccedilatildeo aos grupos amina das proteiacutenas Em soluccedilotildees
aacutecidas diluiacutedas a coloraccedilatildeo varia proporcionalmente com a quantidade do corante ligado agrave
proteiacutena que por sua vez eacute directamente proporcional agrave concentraccedilatildeo da proteiacutena permitindo a
sua quantificaccedilatildeo ateacute a um limite de detecccedilatildeo de 20 gmL (Bradford 1976)
Deste modo prepararam-se amostras de BSA ([BSA]stock= 1mgmL) utilizando um intervalo de
concentraccedilotildees compreendido entre 0 e 15 gmL de acordo com a tabela 3I Apoacutes a adiccedilatildeo da
H2O MilliQ e do reagente de Bradford as soluccedilotildees foram sucessivamente homogeneizadas no
voacutertex
Tabela 3Indash Amostras utilizadas para a obtenccedilatildeo da recta de calibraccedilatildeo no meacutetodo de Bradford
BSA (L) BSA (g) H2O MilliQ (L) Reagente de Bradford (L)
0 0 800 200
125 125 79875 200
25 25 7975 200
5 5 795 200
10 10 790 200
15 15 785 200
Apoacutes incubaccedilatildeo a 25ordmC por 5 minutos leram-se os valores de Abs595 num
espectrofotoacutemetro (Shimadzu UV-160A) para todas as amostras Seguidamente traccedilou-se a recta
de calibraccedilatildeo (Abs595vs quantidade de BSA (g))
43
Para determinar a concentraccedilatildeo de proteiacutena apoacutes o processo de isolamento purificaccedilatildeo e
diaacutelise quando necessaacuterio colocou-se 5 L da amostra num tubo de Eppendorf e adicionou-se
795 L de H2O MilliQ homogeneizando a soluccedilatildeo no voacutertex Adicionou-se posteriormente 200
L de reagente de Bradford e repetiu-se a homogeneizaccedilatildeo Incubou-se a mistura a 25ordmC por 5
minutos apoacutes os quais se leu os valores de Abs595 nm no espectrofotoacutemetro Por interpolaccedilatildeo deste
valor a partir da recta de calibraccedilatildeo determinou-se a concentraccedilatildeo de proteiacutena na amostra
3312 Quantificaccedilatildeo pelo meacutetodo directo
A quantificaccedilatildeo das amostras proteicas pode tambeacutem ser efectuada atraveacutes da leitura directa da
absorvacircncia tendo por base a lei de Beer-Lambert onde Abs corresponde agrave
absorvacircncia ao comprimento de onda ao qual o ensaio foi realizado corresponde ao coeficiente
de extinccedilatildeo (substituiacutedo pela letra a se as unidades forem distintas da molaridade) b corresponde
ao percurso oacuteptico da radiaccedilatildeo na ceacutelula amostral e c corresponde agrave concentraccedilatildeo amostral
Assim no caso de S1 de miosina mediu-se a Abs280e calculou-se a concentraccedilatildeo da proteiacutena
sabendo que o coeficiente de extinccedilatildeo de S1 eacute (Ref)
No caso da actina mediu-se a Abs290ecalculou-se a concentraccedilatildeo da proteiacutena sabendo que o
coeficiente de extinccedilatildeo da actina eacute (Ref)
332Electroforese em condiccedilotildees desnaturantes (SDS-PAGE)
De modo a melhor caracterizar as proteiacutenas isoladas foi realizada uma electroforese em
condiccedilotildees desnaturantes (SDS-PAGE do inglecircs Sodium Dodecyl Sulfate Polyacrilamide Gel
Electrophoresis) O SDS (dodecilo sulfato de soacutedio) eacute um detergente anioacutenico utilizado neste
meacutetodo para desnaturar as proteiacutenas que se ligam ao SDS na proporccedilatildeo 14 g SDS g proteiacutena
(aproximadamente 1 moleacutecula de SDS por cada 2 resiacuteduos de aminoaacutecidos) (Stryer) O SDS induz
uma densidade de carga negativa significativa nas proteiacutenas a que se liga mascarando a sua carga
intriacutenseca pelo que as razotildees cargamassa satildeo tendencialmente idecircnticas Assim a SDS-PAGE
separa as proteiacutenas quando sujeitas a um campo eleacutectrico de acordo com o seu peso molecular
sendo a sua migraccedilatildeo directamente proporcional ao logaritmo deste uacuteltimo (Stryer)
O gel de resoluccedilatildeo com 10 de acrilamida foi obtido atraveacutes da adiccedilatildeo de 167 mL de soluccedilatildeo
stock de acrilamida11
075 mL de tampatildeo de resoluccedilatildeo12
251 mL de aacutegua destilada e 50 L de
SDS 10 Para promover a polimerizaccedilatildeo adicionou-se em seguida 38 L de PSA 10
11
soluccedilatildeo stock de acrilamida soluccedilatildeo de 30 (mv) de acrilamida e 08 (mv) de NNrsquo-metil-bis-acrilamida 12
tampatildeo de resoluccedilatildeo 15 M Tris-HCl pH 88
44
(persulfato de amoacutenia) 10 e 25 L de TEMED 99 (tetrametiletilenodiamina) Agitou-se
cuidadosamente de modo a natildeo formar bolhas e pipetou-se a soluccedilatildeo para a cassette de vidro
previamente montada no suporte De seguida adicionou-se uma peliacutecula de cerca de 2 mm de 2-
metil-2-butanol para verificar posteriormente a polimerizaccedilatildeo Apoacutes a polimerizaccedilatildeo que demora
cerca de 45 minutos verteu-se a agrave superfiacutecie e adicionou-se o gel de concentraccedilatildeo com 5 de
acrilamida constituiacutedo por 03 mL de soluccedilatildeo de stock acrilamida 045 mL de tampatildeo de
concentraccedilatildeo13
094 mL de aacutegua destilada 18 L de SDS 10 135 L de PSA 10 e 2L de
TEMED 99 Colocou-se o pente sobre o gel e deixou-se polimerizar durante cerca de 45
minutos
As amostras para aplicaccedilatildeo no gel de electroforese foram preparadas para uma concentraccedilatildeo de
proteiacutena de 1 mgmL num volume final de 100 L dos quais 25 L correspondem ao tampatildeo da
amostra14
As amostras foram aquecidas a 100ordmC durante 5 minutos Antes da aplicaccedilatildeo das
amostras transferiu-se a cassette para a tina de electroforese agrave qual se adicionou a soluccedilatildeo
tampatildeo de electroforese da tina superior15
e tampatildeo de electroforese da tina inferior16
Aplicaram-
se 10 L das soluccedilotildees de marcadores de peso molecular e 20 L das amostras proteicas
preparadas anteriormente A electroforese decorreu durante cerca de 3h30 min a uma voltagem
de 80 V De seguida transferiu-se o gel para uma tina corando as proteiacutenas durante 1h com
soluccedilatildeo corante17
Descorou-se o gel com soluccedilatildeo descorante18
tendo-se mudado esta soluccedilatildeo ateacute
as bandas se encontrarem bem definidas
Determinou-se a mobilidade relativa das bandas correspondentes aos marcadores de elevado peso
molecular e traccedilou-se a recta de calibraccedilatildeo ndash log(PM) vs log(Rf) onde Rf corresponde agrave
mobilidade relativa Por interpolaccedilatildeo determina-se o PM das proteiacutenas isoladas sabendo a sua
mobilidade relativa Por comparaccedilatildeo com os valores descritos na literatura pode concluir-se
acerca do grau de purificaccedilatildeo
34 Mediccedilatildeo da actividade enzimaacutetica de S1
341 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
13
tampatildeo de concentraccedilatildeo 05 M Tris-HCl pH 68 14
tampatildeo de amostra 320 mM Tris-HCl pH 68 04 M szlig-mercaptoetanol SDS 8 glicerol 15 (vv) e roxo de m-
cresol 0024 15
tampatildeo de electroforese da tina superior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS 16
tampatildeo de electroforese da tina inferior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS acetato de soacutedio
01 M 17
soluccedilatildeo corante ldquoCoomassie Blue R-250rdquo contendo metanol 45 (vv) e aacutecido aceacutetico glacial 10 (vv) 18
soluccedilatildeo descorante metanol 10 (vv) e aacutecido aceacutetico glacial 10 vv)
45
O meacutetodo de Fiske-Subarow modificado consiste na reacccedilatildeo do molibdato de amoacutenio com o
fosfato inorgacircnico em soluccedilatildeo aacutecida com formaccedilatildeo do aacutecido fosfomoliacutebdico Seguidamente daacute-
se a adiccedilatildeo de um agente redutor que provoca a reduccedilatildeo do composto a fosfomolibdato que forma
um complexo de cor azulada cuja absorvacircncia maacutexima ocorre para um comprimento de onda de
660 nm tornando possiacutevel a sua detecccedilatildeo por meacutetodos espectrofotomeacutetricos As actividades Ca2+
-
ATPaacutesica e K+EDTA-ATPaacutesica de S1 foram determinadas atraveacutes deste meacutetodo de acordo com
o descrito por Taussky e Shorr (1953) com mediccedilatildeo da quantidade de fosfato (Pi) libertado
atraveacutes da hidroacutelise de ATP
Para tal obteve-se primeiramente a recta de calibraccedilatildeo com as amostras presentes na Tabela 3II
utilizando uma soluccedilatildeo stock de 5 mM KH2PO4
Tabela 3II ndash Composiccedilatildeo das diferentes amostras utilizadas para determinar a recta de calibraccedilatildeo atraveacutes do meacutetodo
de Fiske-Subbarow modificado
KH2PO4 (L)
([KH2PO4]stock= 5mM)
PO43-
(nmole) H2O (uL)
0 0 1780
5 25 1775
10 50 1770
20 100 1760
40 200 1740
100 500 1680
A cada tubo de Eppendorf adicionaram-se seguidamente 200 L de soluccedilatildeo J19
e 20 L de agente
redutor20
homogeneizando no voacutertex apoacutes cada adiccedilatildeo Incubou-se durante 10 minutos e mediu-
se a Abs660 nm num espectrofotoacutemetro com intervalos de tempo iguais entre cada amostra Traccedilou-
se a recta de calibraccedilatildeo (Abs660 nmvs nmol PO43-
) a partir dos valores obtidos
Para determinar a actividade Ca2+
-ATPaacutesica de S1 prepararam-se tubos de Eppendorf com 400
L de meio de reacccedilatildeo21
25 L CaCl2 ([CaCl2]stock= 100 mM) 25 L ATP ([ATP]stock= 100 mM)
com [S1]ensaios=01 mgmL Perfez-se se necessaacuterio com H2O MilliQ ateacute perfazer um volume de 1
mL Para determinar a actividade K+EDTA-ATPaacutesica utilizou-se uma concentraccedilatildeo idecircntica de
19
Soluccedilatildeo J 5 N H2SO4 25 molibdato de amoacutenio 20
Agente redutor 5 mg aacutecido 1-amino-2-naftol-4-sulfoacutenico 30 mg bissulfito de soacutedio 30 mg sulfito de soacutedio em 26
mL H2O 21
meio de reacccedilatildeo 125 mM Tris-HCl pH 80 0575 M KCl
46
proteiacutena nos ensaios (01 mgmL) e o mesmo meio de reacccedilatildeo Aos tubos de Eppendorf
adicionou-se ainda 123 L de KCl ([KCl]stock = 3M) 25 L de ATP 10 L de EDTA
([EDTA]stock= 100 mM )e o volume de H2O MilliQ necessaacuterio para perfazer 1 mL Em ambas as
situaccedilotildees o uacuteltimo reagente adicionado foi o ATP uma vez que se trata do substrato da reacccedilatildeo
enzimaacutetica
A cineacutetica decorreu a 25ordmC durante 5 minutos para a actividade Ca2+
-ATPaacutesica e durante 3
minutos para a actividade K+EDTA-ATPaacutesica Apoacutes o periacuteodo de incubaccedilatildeo transferiu-se 150
L de amostra para tubos de Eppendorf contendo igual volume de TCA 10 Colocou-se no gelo
e homogeneizou-se no voacutertex Seguidamente centrifugou-se a 1500 g durante 10 minutos e
recolheu-se 200 L de sobrenadante para novos tubos de Eppendorf Adicionou-se a estes 158
mL de H2O MilliQ e 200 L de soluccedilatildeo J Homogeneizou-se no voacutertex e adicionou-se 20 L de
reagente redutor repetindo-se a homogeneizaccedilatildeo Incubou-se durante 10 minutos e mediu-se a
Abs660 nm com o mesmo intervalo de tempo de leitura para cada amostra Por intrapolaccedilatildeo a partir
da recta de calibraccedilatildeo obtida anteriormente determinou-se a quantidade de Pi libertado em cada
ensaio A partir destes resultados determinaram-se as actividades Ca2+
-ATPaacutesica e K+EDTA-
ATPaacutesica do S1 em nmol PO43-
minmg proteiacutena e em s-1
342 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
Da actividade Mg2+
-ATPaacutesica de S1 resulta a formaccedilatildeo dos produtos da reacccedilatildeo de
hidroacutelise do ATP - ADP e Pi Na presenccedila de ADP fosfoenolpiruvato (PEP) e da enzima piruvato
cinase (PK) os produtos resultantes satildeo ATP e piruvato Por acccedilatildeo da enzima lactato
desidrogenase (LDH) este uacuteltimo eacute convertido em lactato ocorrendo simultaneamente a oxidaccedilatildeo
do NADH a NAD+ monitorizada atraveacutes da Abs340 nm Quando o S1 de miosina se encontra no
meio reaccional o ATP eacute hidrolisado formando-se novamente ADP e Pi o que permite a
formaccedilatildeo de um ciclo (fig 21) Se as enzimas se encontrarem em excesso a velocidade de
conversatildeo do NADH eacute limitada pela produccedilatildeo de ADP e Pi Como o ATP eacute regenerado em cada
ciclo a sua concentraccedilatildeo inicial eacute sempre mantida (Bagshaw 1982)
Figura 33ndash Representaccedilatildeo esquemaacutetica do meacutetodo de enzimas acopladas
47
O protocolo utilizado e seguidamente descrito foi adaptado de Cuenda et al (1990 ndash Ver Refordf) A
uma cuvette de 1 mL adicionou-se S1 para uma concentraccedilatildeo final de 005 mgmL 800 L de
meio de reacccedilatildeo22
10 L de PEP ([PEP]stock = 42 mM) 10 L de PK+LDH 11 ([PK+LDH]stock =
1 mgmL) e 15 L de NADH ([NADH]stock=25 mM) Iniciou-se a monitorizaccedilatildeo da reacccedilatildeo
atraveacutes da leitura de Abs340 nm apoacutes a adiccedilatildeo de 20 L de ATP ([ATP]stock=100 mM) A
actividade Mg2+
-ATPaacutesica eacute obtida atraveacutes do declive correspondente agrave oxidaccedilatildeo do NADH Para
a mediccedilatildeo da Mg2+
-ATPase de S1 estimulada por actina adicionou-se F-actina para uma
concentraccedilatildeo final de 2 microM sendo a actividade obtida tambeacutem atraveacutes do declive correspondente
agrave oxidaccedilatildeo do NADH apoacutes a adiccedilatildeo da proteiacutena
35 Interaccedilatildeo entre vanaacutedio e subfragmento-1 de miosina
351 Preparaccedilatildeo das soluccedilotildees de vanaacutedio
As soluccedilotildees de vanadato foram preparadas em aacutegua MilliQ a partir de metavanadato de amoacutenio
(NH4VO3) numa concentraccedilatildeo stock de 50 mM com agitaccedilatildeo magneacutetica e aquecimento (~30 ordmC)
Esta soluccedilatildeo com um valor compreendido entre 6 e 7 designa-se por ldquometavanadatordquo e eacute
constituiacuteda pelas esspeacutecies V1 V2 V4 e V5 (Ref) Acidificando a soluccedilatildeo de metavanadato por
adiccedilatildeo de HCl ateacute um valor de pH proacuteximo de 4 observa-se uma mudanccedila de cor de amarelo
claro para laranja indicativa da formaccedilatildeo da espeacutecie decameacuterica de vanadato De seguida e
imediatamente antes da realizaccedilatildeo dos ensaios descritos o valor do pH da soluccedilatildeo de
decavanadato foi aumentado para cerca de 75 por adiccedilatildeo de NaOH A soluccedilatildeo (2) designada
por ldquometavanadatordquo natildeo foi posteriormente alterada ateacute agrave sua adiccedilatildeo para a realizaccedilatildeo dos ensaios
As soluccedilotildees foram armazenadas a 4ordmC e mantidas em gelo aquando da sua utilizaccedilatildeo
A soluccedilatildeo de V(IV) foi preparada em condiccedilotildees anaeroacutebias por diluiccedilatildeo de sulfato de vanadil
(VOSO4) para uma concentraccedilatildeo final de 5 mM em H2O MilliQ saturada com N2 gasoso
352Anaacutelise por espectroscopia de UV-Vis
A concentraccedilatildeo da espeacutecie decameacuterica em soluccedilatildeo depende do periacuteodo de tempo apoacutes a diluiccedilatildeo
no meio de reacccedilatildeo apropriado (Ref) A velocidade de desoligomerizaccedilatildeo da espeacutecie decameacuterica
pode ser analisada por espectroscopia de absorccedilatildeo na regiatildeo UVVis uma vez que esta espeacutecie
apresenta duas bandas caracteriacutesticas na regiatildeo do visiacutevel (a 358 e 400 nm) que natildeo se verifica
em quaisquer outras espeacutecies oligomeacutericas As bandas supramencionada sapresentam-se bem
22
Meio de reacccedilatildeo 3125 mM Tris-HCl pH 71 375 mM MgCl2
48
resolvidas atraveacutes da obtenccedilatildeo da derivada de 2ordf ordem do espectro apresentando a forma de 2
vales com miacutenimos centrados nos comprimentos de onda supracitados com dependecircncia linear
relativamente agrave concentraccedilatildeo de decavanadato total (Ramos et al 2006)
Assim a estabilidade da espeacutecie decameacuterica apoacutes diluiccedilatildeo da soluccedilatildeo de decavanadato no meio de
reacccedilatildeo dos ensaios (tampatildeo D) para uma concentraccedilatildeo final de 1 mM em vanadato total foi
caracterizada atraveacutes da leitura da Abs400 em funccedilatildeo do tempo tendo-se determinado qual a
ordem de reacccedilatildeo e o tempo de meia-vida da mesma Com base em estudos anteriores acerca da
estabilidade do V10 na presenccedila de diferentes proteiacutenas (Ramos et al 2006) determinou-se
igualmente o tempo de meia vida apoacutes a adiccedilatildeo de 20 microM de S1 de miosina ao meio reaccional
353 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
Os espectros de RMN do isoacutetopo 51
V foram realizados num espectroacutemetro Bruker AM-400 MHz
equipado com uma sonda multinuclear inversa de 5 mm Em tubos de RMN de 5 mm de diacircmetro
preparou-se soluccedilotildees de 500 L de deca- ou metavanadato correspondentes a um intervalo de
concentraccedilotildees de 1 a 10 mM suplementadas com 10 D2O Os espectros foram adquiridos agrave
temperatura ambiente com uma largura espectral de 23 923 Hz um tempo de aquisiccedilatildeo de 0086
s tempo de relaxaccedilatildeo de 001 s e uma sequecircncia de pulsos a 90ordm Utilizou-se a transformada de
Fourier com alargamento das bandas de 20Hz para o processamento dos espectros Os desvios
quiacutemicos das ressonacircncias de 51
V foram obtidos em relaccedilatildeo agrave espeacutecie VOCl3cuja frequecircncia de
ressonacircncia eacute considerada o padratildeo sendo todas as outras referenciadas a partir desta em ppm
As aacutereas dos sinais do RMN correspondentes agraves espeacutecies oligomeacutericas presentes foram obtidas
por integraccedilatildeo dos sinais As concentraccedilotildees relativas das espeacutecies foram calculadas com recurso agrave
equaccedilatildeo
onde A[Vx] corresponde agrave aacuterea referente agrave ressonacircncia da espeacutecie Vx com nordm de agregaccedilatildeo x
A[V]total agrave soma total das aacutereas medidas e [V] a concentraccedilatildeo total de vanaacutedio no ensaio
A teacutecnica de 51
V-RMN foi anteriormente aplicada em estudos de ligaccedilatildeo com S1 de miosina
(Ref) tendo sido demostrado que entre as espeacutecies V1 V2 V4 e V5 a espeacutecie tetrameacuterica possui a
maior afinidade para a miosina (Cremo et al) O desaparecimento ou alargamento dos sinais pode
implicar a ligaccedilatildeo de um dos oxovanadatos laacutebeis agrave miosina sem indicaccedilatildeo de qual uma vez que
49
se processa um equiliacutebrio entre eles (Crans 1990) Assim e tendo em conta as indicaccedilotildees de que
a espeacutecie V10 eacute distinta das restantes no que diz respeito agrave magnitude da interacccedilatildeo com a miosina
e inibiccedilatildeo da actividade ATPaacutesica estimulada por actina (Aureliano 2000 Tiago et al 2002)
torna-se fundamental investigar a interacccedilatildeo das diferentes espeacutecies de vanadato com o S1 de
miosina atraveacutes da teacutecnica de 51
V-RMN
Para tal obtiveram-se espectros de 51
V-RMN resultantes da titulaccedilatildeo de uma soluccedilatildeo 5 mM de
decavanadato total com concentraccedilotildees crescentes de S1 de miosina (0 5 10 15 e 20 microM) em
tampatildeo D Realizou-se ainda ensaios na presenccedila de 2 mM de decavanadato e 2 mM de ATP na
presenccedila e na ausecircncia de S1 de miosina de modo a verificar o efeito que estes ligandos naturais
da enzima teria na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o subfragmento-1
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
Os grupos sulfidrilo (-SH) das cisteiacutenas satildeo resiacuteduos de aminoaacutecidos extremamente sensiacuteveis agrave
oxidaccedilatildeo resultando na sua oxidaccedilatildeo (Dalle-Donne et al 1999) Logo a quantificaccedilatildeo destes
resiacuteduos eacute bastante importante nos estudos de interacccedilatildeo com vanaacutedio jaacute que os resiacuteduos de Cys
com ecircnfase para o SH1 e o SH2 satildeo bastante importantes na monitorizaccedilatildeo das modificaccedilotildees
funcionais da regiatildeo da cabeccedila
O procedimento seguidamente apresentado foi originalmente descrito por Gutieacuterrez-Merino
(1987)) Consiste na reacccedilatildeo entre os grupos ndashSH natildeo oxidados das proteiacutenas e o reagente de
Ellman (DTNB -55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico)) - formando-se o aniatildeo CNT (aniatildeo 3-
carboxilato-4-nitrotiofenolato) numa estequiometria de 11 Este apresenta um maacuteximo de
absorvecircncia a 412 nm (Robyt 1990) Assim apoacutes a incubaccedilatildeo da proteiacutena na presenccedila do metal
a quantidade de cisteiacutenas reduzidas por proteiacutena determina-se atraveacutes dos valores de Abs412
Deste modo colocou-se num tubo de Eppendorf o volume apropriado de S1 dialisado para que a
concentraccedilatildeo no ensaio fosse de 1 mgmL Adicionou-se o volume apropriado de decavanadato
para que a sua concentraccedilatildeo fosse de 0 10 50 100 250 500 ou 1000 M Perfez-se com soluccedilatildeo
D ateacute um volume final de 1 mL e incubou-se a 25ordmC por 20 minutos Apoacutes este periacuteodo colocou-
se 780 L da mistura reaccional numa cuvette de quartzo e adicionou-se 220 L de soluccedilatildeo K23
23
Tampatildeo K Tampatildeo fosfatohidrogenofosfato de soacutedio 025 M pH 72
50
Adicionou-se 15 L de DTNB (em soluccedilatildeo K [DTNB]stock=100 mM) ao meio reaccional e
registou-se o valor de absorvecircncia medido equivalente agraves cisteiacutenas ldquoraacutepidasrdquo (SH1 e SH2
expostos ao solvente) Adicionou-se 001 g de SDS que provoca a desnaturaccedilatildeo da proteiacutena e
consequentemente a exposiccedilatildeo de todos os resiacuteduos de cisteiacutenas que se encontravam ateacute aiacute
inacessiacuteveis no interior da proteiacutena (cisteiacutenas totais) Estes resiacuteduos podem assim reagir com o
DTNB
Estimou-se o nuacutemero de resiacuteduos de cisteiacutenas em todas as amostras com recurso ao coeficiente de
extinccedilatildeo molar () do DTNB (ver ponto 3 dos Anexos) que foi determinado atraveacutes da titulaccedilatildeo
de L-Cisteiacutena com intervalo de concentraccedilotildees compreendidos entre 0 e 50 microM na presenccedila de
25microM de DTNB em tampatildeo D Os valores de absorvecircncia foram obtidos para um comprimento de
onda de 412 nm
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
3621 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
A teacutecnica mais eficiente de anaacutelise da interacccedilatildeo entre o iatildeo vanadilo e proteiacutenas como a
calmodulina e a transferrina entre outras permite a obtenccedilatildeo raacutepida de informaccedilatildeo no que toca agrave
conformaccedilatildeo e amplitude da referida interacccedilatildeo
Os espectros de Ressonacircncia Paramagneacutetica Electroacutenica (RPE) da espeacutecie paramagneacutetica V(IV)
foram realizados a 70K num espectroacutemetro Bruker EMX-300 X-band Todas as soluccedilotildees
utilizadas e amostras foram preparadas em anaerobiose numa cacircmara de luvas de modo a evitar a
reduccedilatildeo de V(IV) a V(V) Em tubos capilares com 02 mL de amostra manteve-se a concentraccedilatildeo
de S1 a 20 M e variou-se a concentraccedilatildeo de vanadilo (0 25 5 10 15 25 50 75 100 150
200 500 e 1000 M [VOSO4]stock=5 mM) em tampatildeo D Apoacutes obtenccedilatildeo dos espectros estes
foram tratados atraveacutes do software SimFonia e WinEPR tendo-se obtido um graacutefico com a
variaccedilatildeo da intensidade do sinal de RPE em funccedilatildeo da concentraccedilatildeo de sulfato de vanadilo
presente Realizaram-se ainda simulaccedilotildees de espectros obtidos com 20 microM de S1 e 1000 microM de
V(IV) com vista agrave obtenccedilatildeo dos ligandos provaacuteveis do vanadilo atraveacutes da anaacutelise da constante
de acoplamento hiperfina (A) e do factor g (g)
4 Conclusotildees gerais
Apesar de jaacute ter sido demonstrado que o vanaacutedio inibe a Na+K
+-ATPase da miosina a funccedilatildeo
deste elemento no tecido muacutesculo esqueleacutetico continua largamente incompreendida A enorme
complexidade quiacutemica deste metal especialmente no que concerne agrave oligomerizaccedilatildeo e reacccedilotildees
51
redox constitui um obstaacuteculo agrave compreensatildeo do comportamento do vanaacutedio nos organismos
vivos Assim a caracterizaccedilatildeo da quiacutemica deste elemento eacute de grande utilidade antes de iniciar o
estudo da sua interacccedilatildeo com o presente sistema
A anaacutelise por espectroscopias de 51
V-RMN e de UV-Vis de soluccedilotildees de vanadato diluiacutedas nos
meios de reacccedilatildeo utilizados no presente relatoacuterio (10 mM HEPES pH 70 25 mM KCl) mostrou
claramente que a espeacutecie decameacuterica (V10) possui um tempo de vida suficientemente longo para a
determinaccedilatildeo do seu efeito isolado ao contraacuterio do que se verifica com as espeacutecies mono- di-
tetra- e pentameacutericas (V1 V2 V4 e V5) Por outro lado a anaacutelise verificou ainda que apoacutes diluiccedilatildeo
das soluccedilotildees de decavanadato no meio de reacccedilatildeo coexistem as espeacutecies V10 e V1 pelo que os
efeitos putativos desta uacuteltima deveratildeo ser subtraiacutedos aquando da monitorizaccedilatildeo dos efeitos da
espeacutecie decameacuterica
O vanaacutedio (V) (vanadato) inibe as enzimas que catalizam reacccedilotildees de transferecircncia de grupos
fosfato como a ATPase da miosina [Goodno 1979] devido agrave sua analogia estrutural com o iatildeo
fosfato Na sua utilizaccedilatildeo como inibidor enzimaacutetico natildeo eacute tida em conta a contribuiccedilatildeo individual
das espeacutecies oligomeacutericas considerando-se normalmente a espeacutecie V1 como inibidora efectiva
[Goodno e Taylor 1982] No presente relatoacuterio verificou-se por espectroscopia de 51
V-RMN que
outros oligoacutemeros de vanadato interagem com o S1 de miosina tendo as espeacutecies V10 e V4 os
maiores niacuteveis de afinidade A sua interacccedilatildeo envolve forccedilas electrostaacuteticas e eacute afectada
negativamente por ligandos naturais de enzima como eacute o caso do ATP
A exposiccedilatildeo do S1 de miosina (005 mgmL) a decavanadato conduz agrave inibiccedilatildeo do capacidade da
proteiacutena de estimular a actividade Mg2+
-ATPaacutesica com um valor de IC50 de 266 plusmn 07 M
decavanadato sendo o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM
decavanadato (80) Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM decavanadato sendo
o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM decavanadato (60)
exercendo um efeito de impedimento agrave ligaccedilatildeo entre o vanaacutedio e o S1
A uma concentraccedilatildeo de 27 microM decavanadato verifica-se a oxidaccedilatildeo de um dos resiacuteduos de Cys
de S1 de miosina seguida pela oxidaccedilatildeo gradual de outro resiacuteduo de Cys ateacute agrave concentraccedilatildeo de
1000 microM Ambos os resiacuteduos oxidados natildeo se encontravam expostos ao solvente utilizado A
oxidaccedilatildeo dos resiacuteduos permite sugerir que esta modificaccedilatildeo oxidativa sofrida pelo S1 contribui
para a diminuiccedilatildeo da sua actividade de ATPase
52
Por espectroscopia de RPE revelaram-se os possiacuteveis ligandos equatoriais do vanaacutedio aquando da
sua interacccedilatildeo com o S1 de miosina como sendo duas moleacuteculas de aacutegua um ligando contendo o
grupo R-OH e outro contendo o grupo R=NR
Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados de
oxidaccedilatildeo (IV) e (V) no S1 de miosina para concentraccedilotildees facilmente alcanccedilaacuteveis no ambiente
terrestre
53
5 Estudos futuros
No sentido do melhor esclarecimento das alteraccedilotildees moleculares e estruturais que ocorrem no
subfragmento 1 quando exposta ao vanaacutedio que conduzem a uma diminuiccedilatildeo da sua funccedilatildeo
sugere-se
- Quantificaccedilatildeo das cisteiacutenas reduzidas de S1 na presenccedila de metavanadato
- Determinaccedilatildeo das actividades Ca2+
- e K+-ATPaacutesica do S1 de miosina na presenccedila de
decavanadato
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina estimulada por actina
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina na presenccedila de
metavanadato
- Realizaccedilatildeo de RMN 2D com variaccedilatildeo das concentraccedilotildees das soluccedilotildees de decavanadato e
metavanadato na presenccedila de S1
-Mutageacutenese dos resiacuteduos de Cys doS1 de miosina de modo a verificar quais os resiacuteduos que
sofrem oxidaccedilatildeo na presenccedila de decavanadato
- Determinaccedilatildeo por calorimetria (DSC Differential Scanning Calorimetry) da variaccedilatildeo da
entalpia (∆H) e da temperatura de desnaturaccedilatildeo do S1 de miosina na presenccedila de vanaacutedio uma
vez que estas propriedades caracteriacutesticas de cada proteiacutena satildeo alteradas apoacutes ocorrecircncia de
modificaccedilotildees estruturais
- Anaacutelise por SDS-PAGE de S1 de miosina apoacutes tratamento com vanaacutedio de modo a determinar
quais os polipeacuteptidos eventualmente formados apoacutes clivagem da proteiacutena
54
6 Referecircncias bibliograacuteficas
- Sellers J Myosins 2nd Edition Oxford University Press 1999
- L Stryer inBiochemistry (Ed WH Freeman 2005 pp
Refordfas Materiais e Meacutetodos
[A] S S Margossian SLowey Methods Enzymol1982 85 55-71
[B] JD Pardue JA Spudich Methods Enzymol 1982 85 164-181
[C] M Bradford Anal Biochem1976 72 248-254
[D] H H Taussky E Shorr J Biol Chem 1953 202 675-685
[E] C R Bagshaw in Outline Studies in Biology ndash Muscle Contraction Chapman and Hall Ed
1982
[F] A Cuenda et al Eur J Biochem1990 194 663-670
55
Anexos
1 Determinaccedilatildeo da recta de calibraccedilatildeo meacutetodo de Bradford
Figura1ndashRepresentaccedilatildeo graacutefica da recta de calibraccedilatildeo (Abs595 nm vs [BSA]) utilizada para o meacutetodo de Bradford
2 Determinaccedilatildeo da recta calibraccedilatildeo para a electroforese em condiccedilotildees desnaturantes
Figura 2 ndash Representaccedilatildeo graacutefica da recta de calibraccedilatildeo (log(Peso Molecular) vs Rf dos marcadores de peso
molecular) onde Rf indica determinada atraveacutes do padratildeo de peso molecular
(poccedilo (a))
3Caacutelculo do coeficiente de extinccedilatildeo molar () para a quantificaccedilatildeo de cisteiacutenas reduzidas
atraveacutes do meacutetodo de Ellman
Traccedilou-se a recta de Abs412 nm meacutedia vs [L-Cisteiacutena] de modo a determinar o valor do coeficiente
de extinccedilatildeo molar definido como o declive daquela recta Assim =8400 M-1
cm-1
como
podemos verificar na Fig 3
A595 nm = 00546[BSA] + 00209 Rsup2 = 099051
Ab
s 59
5 n
m
[BSA] (mgmL)
log PM = -1177 Rf + 2247 Rsup2 = 098957
log
PM
Mobilidade relativa (Rf)
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero

27
Nas soluccedilotildees de decavanadato as espeacutecies monomeacuterica e decameacuterica de vanadato satildeo as uacutenicas
presentes para concentraccedilotildees de decavanadato inferiores a 8 mM Ambas as espeacutecies aumentam
linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado em [AY] e
em (Teresa) quando a concentraccedilatildeo de V1 eacute igual ou superior a 200 microM inicia-se a sua
oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica No entanto nas condiccedilotildees de realizaccedilatildeo dos
ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a espeacutecie tetrameacuterica neste
estudo em concentraccedilotildees inferiores a 8 mM tendo o mesmo sido observado por Ramoset al
(2006) [AK] Os resultados obtidos mostram claramente o equiliacutebrio entre a espeacutecie decameacuterica e
a monomeacuterica e que o aparecimento de outras espeacutecies oligomeacutericas se encontra dependente da
concentraccedilatildeo desta uacuteltima Apesar de natildeo ter sido realizado o ensaio com 5 mM de decavanadato
total no presente relatoacuterio estaacute reportado em Ramos et al (2006) [AK] que as concentraccedilotildees da
espeacutecie decameacuterica e monomeacuterica satildeo respectivamente de 500 microM e 200 microM Assim pode inferir-
se que para concentraccedilotildees inferiores a 5 mM e intervalos de tempo de cerca de 20 minutos (que
inclui o tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas
as espeacutecies decameacuterica e monomeacuterica de vanadato
Figura 24 ndash Espectros de RMN de 51
V de concentraccedilotildees
crescentes das espeacutecies oligomeacutericas de vanadato obtidos
apoacutes diluiccedilatildeo da soluccedilatildeo stock de metavanadato em 10 mM
HEPES pH 70 25 mM KCl para concentraccedilotildees finais de A)
1 mM B) 2 mM C) 4 mM D) 8 mM E) 10 mM de
metavanadato total V1 refere-se agraves espeacutecies monomeacuterica de
vanadatoV1 e V2 correspondem respectivamente agrave espeacutecie
monomeacuterica e dimeacuterica
Figura 25 ndash Distribuiccedilatildeo graacutefica das espeacutecies
oligomeacutericas de vanadato vs a concentraccedilatildeo de
metavanadato total As concentraccedilotildees de cada
espeacutecie foram obtidas segundo a formula descrita
na secccedilatildeo 35 do Procedimento Experimental
(aacutereas dos sinais de 51
V-RMN observadas na
figura x1)V10 V1 e V4 correspondem agraves espeacutecie
monomeacuterica dimeacuterica e tetrameacuterica
respectivamente
Esp
eacuteci
e o
ligo
meacute
rica
de
va
nad
ato
(m
M)
[Metavanadato]total (mM)
V1
V2
V4
V5
28
Tabela 2IVndash Desvios quiacutemicos meacutedios (part)das ressonacircncias das espeacutecies tetrameacuterica (V4) monomeacuterica (V1) e
dimeacuterica (V2) presentes no espectro da soluccedilatildeo de metavanadato (total) 10 mM
Paracircmetros V1 V2 V4 V5
part (ppm) -5615 -5748 -5789 -5869
Nas soluccedilotildees de metavanadato as espeacutecies monomeacuterica dimeacutericae tetrameacuterica de vanadato estatildeo
presentes para concentraccedilotildees de metavanadato inferiores a 4 mM Todas estas as espeacutecies
aumentam linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado
em (Aureliano e Madeira 1998) e em (Tiago et al 2004) quando a concentraccedilatildeo de V1 eacute igual ou
superior a 200 microM inicia-se a sua oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica Nas
condiccedilotildees de realizaccedilatildeo dos ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a
espeacutecie pentameacuterica neste estudo em concentraccedilotildees inferiores a 4 mM tendo o mesmo sido
observado por Ramos 2006 Os resultados obtidos mostram claramente o equiliacutebrio entre a
espeacutecie V4 e as espeacuteciesV1 e V2 no qual o aparecimento de outras espeacutecies oligomeacutericas se
encontra dependente da concentraccedilatildeo da espeacutecie V1 Por uacuteltimo pode inferir-se que para
concentraccedilotildees inferiores a 2 mM e intervalos de tempo de cerca de 20 minutos (que inclui o
tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas as
espeacutecies V1 V2 e V4 com concentraccedilotildees respectivas de 0796 0192 e 0196 mM
232Estabilidade das soluccedilotildees de decavanadato anaacutelise por espectroscopia de absorccedilatildeo
UVVis
Caracterizou-se a estabilidade da espeacutecie V10 numa soluccedilatildeo com 1 mM decavanadato total em
tampatildeo D atraveacutes da leitura da Abs400 Obteve-se uma cineacutetica de dissociaccedilatildeo de 1ordf ordem tal
como se pode verificar pela linearidade do graacutefico de ln(Abs400)vs o tempo de aquisiccedilatildeo espectral
(segundos) (Fig26 )
Figura 26ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do tempo
(em segundos) para uma soluccedilatildeo de decavanadato total de 1
mM em tampatildeo D a 25ordmC (n=2)
Sendo uma reacccedilatildeo de primeira ordem pode ser descrita pela equaccedilatildeo cujo
declive eacute ndashK (constante de dissociaccedilatildeo) e a intersecccedilatildeo no eixo das ordenadas corresponde
Ln
(Ab
s 400 n
m)
tempo (s)
29
aln(A0) Numa reacccedilatildeo de 1ordf ordem o tempo de meia-vida eacute independente da concentraccedilatildeo inicial
do reagente sendo descrito pela equaccedilatildeo [2] Deste modo a partir da recta
Ln(Abs400 nm) = -436E-5
K - 105 obteve-se um tempo de meia-vida de 4490 plusmn 0823 horas para
as condiccedilotildees supramencionadas
Apoacutes a adiccedilatildeo de S1 8 microM ao meio reaccional verificou-se que a reacccedilatildeo de desoligomerizaccedilatildeo
da espeacutecie decameacuterica de vanadato seguia igualmente uma cineacutetica de 1ordf ordem tal como se pode
verificar pela linearidade do graacutefico de ln(Abs400) versus o tempo de aquisiccedilatildeo espectral
(segundos) (Fig27)
Figura 27ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do
tempo (em segundos) para uma soluccedilatildeo de decavanadato
total de 1 mM em tampatildeo D a 25ordmC na presenccedila de 8
microM (n= 2)
A partir da equaccedilatildeo da recta Ln(Abs400 nm) = -7520E-5
K - 0613determinou-se o valor de K para
a espeacutecie decameacuterica de onde se obteve um tempo de meia-vida de 2430 plusmn 0191 horas para as
condiccedilotildees supramencionadas
Tanto este valor como o valor registado na ausecircncia de S1 satildeo substancialmente superiores agrave
duraccedilatildeo dos ensaios de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina descritos no presente
relatoacuterio
24 Interacccedilatildeo de S1 com vanaacutedio(V)
241Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
De modo a analisar qual o efeito da adiccedilatildeo de S1 de miosina (tampatildeo D) produzia nos sinais de
51V-NMR titulou-se uma soluccedilatildeo de 5mM de decavanadato total com concentraccedilotildees crescentes
de S1 (5 10 15 e 20 microM) Os espectros de 51
V-RMN encontram-se apresentados na Fig 2
No graacutefico da figura 2 Apresenta-se a concentraccedilatildeo de S1 em funccedilatildeo do factor de alargamento
corrigido dos espectros acima referidos
Ln
(Ab
s (4
00
nm
))
tempo (s)
30
Figura 28 ndash Graacutefico de
representaccedilatildeo da
concentraccedilatildeo de S1 em
funccedilatildeo do factor de
alargamento A
concentraccedilatildeo de
decavanadato total foi de 5
mM e as concentraccedilotildees
utilizadas de S1 foram de 0
5 10 e 15 microM
Verifica-se que o factor de alargamento aumenta linearmente em funccedilatildeo da concentraccedilatildeo de S1 no
ensaio o que indica a existecircncia de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina Observou-
se ainda que a adiccedilatildeo de S1 ao meio reaccional natildeo provocou qualquer alteraccedilatildeo dos desvios
quiacutemicos dos sinais de 51
V-RMN
Realizaram-se ensaios na presenccedila de ATP e de S1 com intuito de verificar o efeito do ATP na
interacccedilatildeo do S1 de miosina com o decavanadato
Figura 29 - Espectros de RMN de 51
V contendo 2
mM Decavanadato (A) (controlo) (B) na presenccedila
de 2 mM de ATP (C) na presenccedila de 2 mM de ATP
e 8 microM de S1 de miosina V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2Vndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies decameacuterica (V10) e monomeacuterica (V1) presentes nos
espectros da soluccedilatildeo de decavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V10A V10B V10C V1
part (ppm) -5161 -5002 -4254 -5614
Observa-se que a presenccedila de ATP (espectro B) natildeo provoca qualquer alteraccedilatildeo visiacutevel em
relaccedilatildeo ao espectro que contem apenas soluccedilatildeo de decavanadato Jaacute a presenccedila simultacircnea de ATP
e S1 provoca um alargamento da ∆12 do sinal do V10A de 1736 Hz (espectro (A)) para 2366 Hz
(espectro (C) Fig 29) Tendo em conta que a concentraccedilatildeo de S1 apresenta uma relaccedilatildeo linear
com o factor de alargamento converteu-se o valor da ∆12 e comparou-se com o valor esperado agrave
F = 00591[S1] - 00157 Rsup2 = 0998
Fact
or
de
ala
rgam
en
to
[S1] (microM)
31
concentraccedilatildeo de S1 utilizada O factor de alargamento obtido para 8 microM ndash 0376 ndash eacute inferior ao
valor esperado agrave mesma concentraccedilatildeo de S1 obtido atraveacutes da equaccedilatildeo da recta apresentada na
Fig 28 ndash 0457 ndash o que indica que o ATP poderaacute exercer um efeito de protecccedilatildeo do S1
diminuindo a sua interacccedilatildeo com a espeacutecieV10
Por uacuteltimo realizaram-se ensaios com metavanadato na presenccedila de 8 microM S1 de modo a verificar
se as espeacutecies oligomeacutericas contidas nesta soluccedilatildeo interagiam com a proteiacutena em estudo
Figura 210 - Espectros de RMN de 51
V contendo 2
mM metavanadato (A) (controlo) (B) na presenccedila
de 8 microM de S1 de miosina V1 V2 e V4
correspondem respectivamente aacutes espeacutecies
monomeacuterica dimeacuterica e tetrameacuterica
Tabela 2VIndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies monomeacuterica (V1) dimeacuterica (V2) e tetrameacuterica (V4)
presentes nos espectros da soluccedilatildeo de metavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V1 V2 V4
part (ppm) -5613 -5747 -5789
Como eacute possiacutevel observar na Fig 210 ocorre uma ligeiro alargamento dos sinal de V4 bem
como uma perda da intensidade pelo que se conclui quedas espeacutecies presentes na soluccedilatildeo de
metavanadato (V1 V2 e V4) apenas a espeacutecie tetrameacuterica interage com o S1 de miosina o que
estaacute de acordo com o reportado por Tiago et al 2002
242 Actividade Mg2+
-ATPaacutesica de S1 de miosina na presenccedila de decanadato
Com o intuito de avaliar o efeito do decavanadato na actividade Mg2+
-ATPaacutesica do S1 de
miosina monitorizou-se a reacccedilatildeo de oxidaccedilatildeo do NADH atraveacutes da leitura de Abs340 na
presenccedila de 005 mgmL de S1 de miosina e no caso da actividade estimulada 2microM de actina em
3125 mM Tris-HCl pH 71 e 375 mM MgCl2 pelo meacutetodo de enzimas acopladas (ver seccccedilatildeo xx
dos MM)Para tal realizaram-se ensaios com vaacuterias concentraccedilotildees de decavanadato total (10 50
100 250 500 e 1000) de modo a determinar qual o seu efeito na actividade MgATPaacutesica de S1
de miosina na ausecircncia e na presenccedila de um dos seus ligandos naturais Os ensaios foram
realizados agrave temperatura ambiente com incubaccedilatildeo preacutevia a 25ordm C durante 20 minutos
32
Figura 211 ndash Representaccedilatildeo
graacutefica da Actividade Mg2+
-
ATPaacutesica de S1 () (os ajustes
teoacutericos estatildeo representado a por
uma linha os resultados
experimentais estatildeo representados
por triacircngulos (Actividade
estimulada) e cruzes (Actividade
natildeo estimulada)) vs
[Decavanadato]total medidas pelo
meacutetodo das enzimas acopladas
(n=3) Os dados foram ajustados ao modelo de decaimento exponencial usando o programa GraphPad Prism para a
determinaccedilatildeo dos valores de IC50 (na ausecircncia de actina IC50= 266 plusmn 07 M decavanadato na presenccedila de actina
IC50= 28 plusmn 1 M decavanadato
A partir do graacutefico presente na Fig 29 verifica-se que o ajuste dos dados experimentais foi bem
sucedido A inibiccedilatildeo da actividade Mg2+
-ATPaacutesica do S1 de miosina eacute afectada em menor
amplitude na presenccedila de actina (60 de inibiccedilatildeo da actividade) do que na sua ausecircncia
(aproximadamente 80 de inibiccedilatildeo de actividade) Devido a esta diferenccedila pode sugerir-se que a
actina iraacute impedir parcialmente a ligaccedilatildeo do decavanadatoao S1 demiosina atraveacutes de
impedimentos esteacutereos ou interacccedilotildees electrostaacuteticas por exemplo A teacutecnica utilizada natildeo nos
permite concluir acerca de que regiatildeo(otildees) ou resiacuteduo(s) da actina participaratildeo na preservaccedilatildeo
parcial da actividade Mg2+
-ATPaacutesica
Por outro lado os valores de IC50 dos dois tipos de actividades natildeo apresentam diferenccedilas
significativas tendo-se obtido 266 plusmn 07 e 28 plusmn 1M decavanadato na ausecircncia e na presenccedila de
actina respectivamente o que corresponde a uma concentraccedilatildeo de cerca de 27 M de V10 O
meio de reacccedilatildeo utilizado nestes ensaios foi 10 mM HEPES e 25 mM KCl com 2 mM ATP Em
(Tiago et al 2004) o valor de IC50 para a actividade Mg2+-ATPaacutesica na ausecircncia de actina foi de
125 plusmn 025 microM enquanto o valor da actividade estimulada por actina foi de 136 plusmn 14 microM O
meio de reacccedilatildeo utilizado nos ensaios foi 20 mM HEPES 50 mM KCl e 2 mM ATP o que resulta
precisamente no dobro da forccedila ioacutenica a que foi realizado o ensaio do presente relatoacuterio
(exceptuando a contribuiccedilatildeo dos produtos da hidroacutelise de ATP uma vez que este meacutetodo
pressupotildee a ocorrecircncia de um ciclo de reacccedilotildees de hidroacutelise e siacutentese deste composto) Ora tendo
sido anteriormente referido que a afinidade do S1 de miosina para com a espeacutecie V10 diminui com
Act
ivid
ade
Mg
2+-A
TP
aacutesic
a d
e S
1 d
e
Mio
sin
a
[Decavanadato] (microM) Actividade Estimulada ()
Actividade Natildeo Estimulada ()
Ajuste Actividade Estimulada
Ajuste Actividade Natildeo Estimulada
33
o aumento da forccedila ioacutenica (Tiago et al 2004 tal poderaacute explicar a diferenccedila obtida entre estes
dois conjuntos de valores
243Quantificaccedilatildeo de cisteiacutenas reduzidas pelo meacutetodo de Ellman
A interacccedilatildeo do vanaacutedio com alguns tipos de aminoaacutecidos sensiacuteveis agrave oxidaccedilatildeo pode fornecer-nos
informaccedilatildeo bastante uacutetil em relaccedilatildeo agraves alteraccedilotildees estruturais que conduzem a uma diminuiccedilatildeo da
actividade fisioloacutegica do S1 de miosina Os estudos realizados por Chasteen 1983 concluiacuteram
que a reduccedilatildeo de vanadato (V) a vanadilo (IV) ocorre na presenccedila de agentes redutores comuns no
ambiente celular de que se destaca o aminoaacutecido cisteiacutena Esta eacute tambeacutem um alvo de
complexaccedilatildeo por parte do iatildeo VO2+
Jaacute Ramos et al (2009) demonstrou que a incubaccedilatildeo de
actina com a soluccedilatildeo de decavanadato induz a oxidaccedilatildeo de resiacuteduos de cisteiacutena e agrave formaccedilatildeo do
oxidovanaacutedio (IV) Este grupo de investigadores descobriu ainda que apenas a espeacutecie V10 induz a
oxidaccedilatildeo das cisteiacutenas ldquoraacutepidasrdquo da actina atraveacutes do processo proposto que eacute parcialmente
responsaacutevel pela inibiccedilatildeo jaacute descrita da actividade de ATPase do S1 de miosina estimulada por
actina quando exposta agrave soluccedilatildeo de decavanadato Esta inibiccedilatildeo apresenta-se afectada com a
presenccedila de ATP uma vez que os resiacuteduos de cisteiacutena permaneceram intactos nos ensaios
realizados com este ligando Jaacute Crans et al (2004) concluiu que o siacutetio activo de muitas fosfatases
(que conteacutem grupos tiol) pode formar um equiliacutebrio redox com o iatildeo vanadato (V) de forma natildeo
reversiacutevel
Os resultados obtidos por Sakurai e Shiromura (1980) sugeriram que o vanaacutedio (V) sofre reduccedilatildeo
a vanaacutedio (IV) sem dependecircncia do valor de pH do ensaio Registaram ainda uma estequiometria
de 31 na reacccedilatildeo do vanadato (V) com os resiacuteduos de Cys enquanto a estequiometria dos ensaios
com o vanadilo (IV) foi de 21
Berlett e Stadtman (1997) observaram que os tipos de aminoaacutecidos mais facilmente oxidaacuteveis
englobam entre outros os resiacuteduos de Cys Trp e MetAssim analisou-se o estado redox dos
grupos tiol dos resiacuteduos de Cys do S1 na presenccedila do decavanadato atraveacutes da monitorizaccedilatildeo da
Abs412 nm na presenccedila de 1 mgmL de S1 de miosina em tampatildeo 10 mM HEPES 25 mM KCl
variando a concentraccedilatildeo da soluccedilatildeo de decavanadato de modo a determinar que tipo de alteraccedilotildees
estruturais ocorrem na sua presenccedila no que diz respeito agrave diminuiccedilatildeo da actividade ATPaacutesica de
S1 na presenccedila de decavanadato
Os resultados referentes agrave quantificaccedilatildeo das cisteiacutenas reduzidas apoacutes o tratamento de S1 com
diferentes concentraccedilotildees de decavanadato em tampatildeo 10 mM HEPES pH 75 e 25 mM KCl a
25ordmC durante 20 minutos encontram-se descritos na Tabela 2VII
Tabela 2VII ndash Cisteiacutenas reduzidas totais e ldquoraacutepidas de S1 (1 mgmL) apoacutes tratamento com decavanadato
34
(010501002505001000 uM) em 10 mM HEPES 25 mM KCl pH 7 a 25ordmC durante 20 minutos A detecccedilatildeo
espectrofotomeacutetrica foi feita com DTNB (e 412=8400 uM-1 cm-1) (n variaacutevel entre 1 e 8)
Amostras Cys raacutepidas Cys totais
11 M S1 (controlo) 22 plusmn 04 106 plusmn 14
11 M S1 + 10 M Decavanadato 22 plusmn 05 95 plusmn 08
11 M S1 + 50 uM Decavanadato 20 plusmn 02 98 plusmn 14
11 M S1 + 100 uM Decavanadato 17 plusmn 03 97 plusmn14
11 M S1 + 250 uM Decavanadato 26 plusmn 02 98 plusmn 08
11 M S1 + 500 uM Decavanadato 21 plusmn 04 90 plusmn 06
11 M S1 + 1000 uM Decavanadato 24 plusmn 0 86 plusmn 08
A partir dos resultados obtidos pode sugerir-se que o S1 possui 11 resiacuteduos de Cys (controlo) o
que estaacute de acordo com o anteriormente descrito Verifica-se que o tratamento da proteiacutena com
concentraccedilotildees crescentes de decavanadato provoca a oxidaccedilatildeo de dois resiacuteduos de Cys do S1 de
miosina natildeo expostos ao solvente No entanto a teacutecnica utilizada natildeo nos permite identificar
quais
Tendo em consideraccedilatildeo que o valor de IC50 obtido para a reacccedilatildeo de inibiccedilatildeo daactividade Mg2+-
ATPaacutesica de S1 na presenccedila de decavanadato eacute de cerca de 27 microM pode sugerir-se que este seraacute o
valor ao qual ocorreraacute a oxidaccedilatildeo do primeiro resiacuteduo de Cys o que poderaacute contribuir para a
diminuiccedilatildeo da actividade de S1 observada
25Interacccedilatildeo de S1 com vanaacutedio(IV)
251Anaacutelise por espectroscopia de Ressonacircncia Paramagneacutetica Electroacutenica
A acumulaccedilatildeo intracelular de oxovanaacutedio (IV) in vivo constitui uma das principais formas de
apresentaccedilatildeo de vanaacutedio no organismo Deste modo torna-se bastante interessante analisar a
interacccedilatildeo entre este iatildeo e o S1 de miosina atraveacutes da teacutecnica de espectroscopia de Ressonacircncia
Paramagneacutetica Electroacutenica o meacutetodo de eleiccedilatildeo do estudo do iatildeo vanadilo e dos complexos por si
formados O electratildeo desemparelhado encontra-se fortemente acoplado ao nuacutecleo 51
V o que
35
resulta num espectro de RPE consistindo de 8 desdobramentos na direcccedilatildeo paralela e outros tantos
na direcccedilatildeo perpendicular (Chasteen 1983)
Os estudos realizados por Ahmed et al (1987) centraram-se na interacccedilatildeo do vanadilo com
proteiacutenas como eacute o caso da insulina transferrina entre outros obtendo um espectro caracterizado
por 8 linhas amplamente espaccediladas (na direcccedilatildeo paralela) Jaacute Ramos et al 2009 concluiu que a
exposiccedilatildeo da soluccedilatildeo de decavanadato agraves formas globular e filamentosa de actina produz um
espectro tiacutepico do iatildeo oxidovanaacutedio (IV) indicando a reduccedilatildeo do vanadato
Para aferir qual a concentraccedilatildeo de S1 que permitia obter uma boa razatildeo sinalruiacutedo titulou-se uma
soluccedilatildeo de sulfato de vanadilo 15 microM ([VOSO4]stock = 5 mM) com concentraccedilotildees crescentes de
proteiacutena (0 5 10 15 e 20 microM) em 10 mM HEPES 25 mM KCl A existecircncia de uma correlaccedilatildeo
linear entre a concentraccedilatildeo de proteiacutena e a intensidade dos sinais de RPE correspondente agrave banda
-12 perpendicular encontra-se representada na Fig 210
Figura 2 12ndash Representaccedilatildeo graacutefica da variaccedilatildeo da intensidade do sinal de RPE vs a concentraccedilatildeo de S1 utilizada
Apoacutes verificar que com uma concentraccedilatildeo de S1 igual a 20 microM se obtinham sinais com uma razatildeo
sinalruiacutedo adequada ao prosseguimento dos estudos sobre a interacccedilatildeo V(IV)-S1 de miosina
titulou-se uma soluccedilatildeo de 20 microM S1 com 0 25 5 10 15 25 50 75 100 150 200 500 e 1000
microM VOSO4
Traccedilou-se o graacutefico da intensidade da banda -12 perpendicular do sinal de EPR em funccedilatildeo da
concentraccedilatildeo de VOSO4 tendo-se determinado uma constante de dissociaccedilatildeo (KD) do complexo
S1-vanadilo de 8748 plusmn 2621 microM V(IV)
y = 016786x + 122435
Rsup2 = 092992759
Inte
nsi
dad
e d
o s
inal
de
RP
E
[S1] (microM)
36
Figura 213 ndash Representaccedilatildeo graacutefica da variaccedilatildeo do sinal de RPE vs a concentraccedilatildeo de vanadilo utilizada (os valores
experimentais estatildeo representados por losangos o seu ajuste estaacute representado pela linha)
A simulaccedilatildeo do espectro de EPR foi efectuada a partir do espectro obtido para uma concentraccedilatildeo
de 1 mM VOSO4 apoacutes adiccedilatildeo de 20 microM S1 de miosina tendo-se obtido os valores da constante de
acoplamento hiperfina (A=5016) e do factor g (g0=195)
Figura 214 ndash Representaccedilatildeo graacutefica do espectro de EPR obtido com 1000 microM de vanadilo e 20 microM de S1 (a cinzento
escuro) A simulaccedilatildeo do espectro encontra-se a cinzento claro
Por comparaccedilatildeo destes com os valores presentes na tabela de correspondecircncia xxx eacute possiacutevel
predizer quais os ligandos provaacuteveis do vanadilo na presenccedila de proteiacutena Assim concluiu-se que
o vanadilo estaacute coordenado com duas moleacuteculas de aacutegua um grupo hidroxilo e um grupo R=NR
O grupo hidroxilo pode indicar os resiacuteduos de aminoaacutecidos Tyr Treonina e Serina enquanto o
grupo R=NR poderaacute indicar a presenccedila de Arginina e Histidina como ligandos do vanadilo
Inte
nsi
dad
e d
os
sinai
s d
e R
PE
[Vanadilo] (microM)
Ajuste dos dados exp Dados experimentais
Campo magneacutetico (G)
Espectro Experimental Ajuste
37
3 Procedimento Experimental
31 Reagentes
Todos os reagentes utilizados satildeo de elevado grau de pureza (analiacutetico) Apresentam-se as suas
foacutermulas quiacutemicas ou abreviaturas quando necessaacuterio acrescentou-se a sua descriccedilatildeo Entre
parecircntesis colocou-se as empresas que os comercializam
Aacutecido aceacutetico glacial 100 (Riedel) Acrilamida 99 (Merck) Azul de bromofenol (Riedel-de-
Haumlen) -Mercaptoetanol (Sigma) Bissulfito de soacutedio (Sigma)Bradford (reagente de)
(BioRad)BSA (albumina de soro bovino Sigma-Aldrich) CaCl22H2O (cloreto de caacutelcio
dihidratado Fluka) CH3CH2OH 96 (etanol Panreac) Coomassie Blue R250 (Merck) D2O
(oacutexido de deuteacuterio SDS) DTNB (55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico) Aldrich) EDTA (aacutecido
etilenodiaminatetraaceacutetico Merck) Fosfoenolpiruvato 98 (Fluka) Glicerol (Sigma) Glicina
985 (Panreac) HCl 37 (Riedel-de Haumlen) H2SO4 96 (Pronalab) HEPES 99 (aacutecido 4-(2-
hidroxiletil)-1-piperazineetanosulfoacutenico Sigma) KH2PO4 99 (hidrogenofosfato de potaacutessio)
MgATP (Sigma) Na
ATP (Sigma) Na2HPO4 99 (hidrogenofosfato de soacutedio Riedel-de Haumlen)
NaH2PO4 (fosfato de soacutedio ) NaOH 986 (hidroacutexido de soacutedio Pronalab) KCl 99 (Cloreto de
potaacutessio Panreac) L-Cys 995 (enantioacutemero levoacutegiro do resiacuteduo de Cisteiacutena Fluka)Metanol
99 (Merck) Molibdato de amoacutenio (Sigma)NNrsquo-metil-bis-acrilamida 98 (Merck) NH4VO3
99 (metavanadato de amoacutenia Sigma) PSA 10 (persulfato de amoacutenio Merck) SDS
(dodecilo sulfato de soacutedio Sigma) Sulfito de soacutedio (Sigma) VOSO4 (sulfato de vanadilo
Aldrich) TEMED 99 (Tetrametiletilenodiamina Merck) Tris (2-amino-2-hidroximetil-13-
propanodiol Sigma)
32 Meacutetodos
321 Isolamento e purificaccedilatildeo das proteiacutenas de muacutesculo esqueleacutetico de coelho
3211 Purificaccedilatildeo da miosina preparaccedilatildeo de Subfragmento 1 por digestatildeo quimotriacutepsica
da miosina
O subfragmento-1 de miosina-II (S1) foi gentilmente cedido pelo Prof Doutor Manuel Aureliano
do Departamento de Quiacutemica Bioquiacutemica e Farmaacutecia da Faculdade de Ciecircncias e Tecnologia da
Universidade do Algarve
No entanto descreve-se abaixo de forma resumida o procedimento efectuado
38
32111 Purificaccedilatildeo da miosina
O protocolo de isolamento foi adaptado do meacutetodo previamente descrito por Margossian e Lowey
(1982) apresentando-se de forma esquemaacutetica na Fig 31
Sucintamente os tecidos musculares dorsais e das pernas foram extraiacutedos apoacutes o sacrifiacutecio do
animal (Fig31) De seguida foram adicionados 3 volumes da soluccedilatildeo de extracccedilatildeo A2
Centrifugou-se a soluccedilatildeo e filtrou-se o sobrenadante O sedimento foi entatildeo diluiacutedo em 14
volumes de aacutegua destilada de modo a que a forccedila ioacutenica atingisse 004 M Apoacutes adiccedilatildeo de szlig-
mercaptoetanol (szlig-me) deixou-se a precipitar a 4ordmC removendo o sobrenadante Centrifugou-se
novamente e ressuspendeu-se o sedimento na soluccedilatildeo de extracccedilatildeo A Adicionou-se a soluccedilatildeo
tampatildeo B3 para que a forccedila ioacutenica atingisse 05 M Homogeneizou-se e deixou-se repousar
durante 2h Homogeneizou-se novamente e centrifugou-se de modo a ocorrer precipitaccedilatildeo dos
complexos de actomiosina e proteiacutena desnaturada Apoacutes filtraccedilatildeo do sobrenadante em latilde de vidro
quantificou-se a soluccedilatildeo de miosina resultante pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental)
2 Soluccedilatildeo de extracccedilatildeo A 0012M K2CO3 004M KHCO3 06M KCl pH 65
3 Soluccedilatildeo-tampatildeo B Tris 100 mM pH 7 KCl 2M
39
Figura 31 ndash Representaccedilatildeo esquemaacutetica do protocolo de isolamento de miosina (Margossian e Lowey 1982) Todo o
procedimento foi realizado a 4ordmC (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
32112 Preparaccedilatildeo de Subfragmento-1 de miosina por digestatildeo com quimiotripsina
O S1 de miosina foi isolado apoacutes diaacutelise da miosina durante a noite contra a soluccedilatildeo C4 A
presenccedila do EDTA eacute essencial uma vez que a sua substituiccedilatildeo por MgCl2resulta na produccedilatildeo de
HMM em vez de S1 A suspensatildeo de miosina foi equilibrada e digerida com -quimotripsina
A digestatildeo foi interrompida pela adiccedilatildeo de PMSF Seguidamente ultracentrifugou-se o produto
da digestatildeo desprezando-se o sedimento (Fig 32) A soluccedilatildeo de sobrenadante maioritariamente
S1 foi quantificada pela leitura directa da Abs280 ou pelo meacutetodo de Bradford (ver as secccedilotildees
3311 e 3312 do Procedimento Experimental respectivamente) e armazenada em 10 sacarose
e 2 mM szlig-Me a -80ordmC
4 Soluccedilatildeo C 25mM Tris pH 70 012M KCl 1mM EDTA
40
Antes da realizaccedilatildeo dos ensaios descritos no presente relatoacuterio o S1 de miosina foi dialisado
contra a soluccedilatildeo D5 de modo a remover qualquer composto que pudesse complexar como
vanaacutedio Seguidamente quantificou-se a proteiacutena pelo meacutetodo de Bradford ou pelo meacutetodo
directo (ver secccedilotildees 3311 e 3312do Procedimento Experimtental)
Figura 3 2ndash Representaccedilatildeo esquemaacutetica do protocolo de preparaccedilatildeo do S1 de miosina por digestatildeo quimiotripsina de
acordo com o previamente descrito por Margossian e Lowey (1982) Todo o procedimento foi realizado a 4ordm C
excepto quando indicaccedilatildeo do contraacuterio (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
3212 Isolamento e purificaccedilatildeo de actina
Isolou-se e purificou-se a actina segundo o meacutetodo descrito por Pardee e Spudish que
compreende dois passos ndash a obtenccedilatildeo de um resiacuteduo seco denominado ldquopoacute de acetonardquo e a
extracccedilatildeo subsequente da actina a partir do ldquopoacute de acetonardquo na forma de G-Actina com a
posterior polimerizaccedilatildeo a F-Actina
32121 Obtenccedilatildeo do ldquopoacute de acetonardquo
Sacrificou-se o coelho com uma pancada atraacutes do pescoccedilo e sangrou-se o animal apoacutes o corte da
arteacuteria jugular Esfolou-se e colocou-se numa tina com gelo Retiraram-se as partes de tecido
conjuntivo que envolvem o tecido muscular com auxiacutelio de uma pinccedila e bisturi Seguidamente
retirou-se os muacutesculos dorsais e os muacutesculos brancos das pernas Passou-se os muacutesculos por aacutegua
destilada de modo a remover as impurezas e pesou-se num copo Trituraram-se os muacutesculos
numa maacutequina de triturar carne e extraiu-se o resultante com 3 volumes da soluccedilatildeo E6 durante 10
minutos com agitaccedilatildeo suave Centrifugou-se a suspensatildeo a 4000 g durante 10
5 Soluccedilatildeo D HEPES 10 mM pH 70 KCl 25 mM
6 Soluccedilatildeo E 01 M KCl 015 M K2HPO4 pH 65
41
minutosdesprezando-se o sobrenadante Ressuspendeu-se o sedimento em 6 volumes de soluccedilatildeo
F7durante 10 minutos com agitaccedilatildeo suave Verificou-se se o pH da soluccedilatildeo estava compreendido
entre 82 e 85 acertando-se estecom soluccedilatildeo G8 se necessaacuterio Centrifugou-se durante 10
minutos a 4000 g Desprezou-se o sobrenadante e ressuspendeu-se o sedimento em 3 volumes de
soluccedilatildeo H9 durante 10 minutos Centrifugou-se novamente a 4000 gdurante 10 minutos
desprezando-se o sobrenadante Ressuspendeu-se o sedimento duas vezes consecutivas em 6
volumes de aacutegua destilada durante 5 minutos Centrifugou-se a 4000 g durante 10 minutos apoacutes
cada adiccedilatildeo de H2O
Seguidamente ressuspendeu-se o sedimento 5 vezes consecutivas utilizando 3 volumes de
acetona de cada vez agitando suavemente durante 10 minutos agrave temperatura ambiente Colocou-
se o resiacuteduo filtrado numa tina de vidro sobre papel de filtro deixando-se evaporar o solvente
durante a noite num nicho Triturou-se o resiacuteduo seco numa maacutequina de triturar carne e deixou-se
novamente evaporar o solvente O ldquopoacute de acetonardquo obtido foi armazenado a -20 ordmC
32122 Purificaccedilatildeo da actina a partir de ldquopoacute de acetonardquo
Apoacutes a obtenccedilatildeo do poacute de acetona como acima indicado efectuou-se o protocolo seguidamente
descrito de modo a obter monoacutemeros de actina (G-Actina) e actina filamentosa (F-Actina)
Pesou-se 35 g de poacute de acetona e adicionaram-se 50 mL de soluccedilatildeo-tampatildeo I10
sendo a
extracccedilatildeo feita com agitaccedilatildeo suave durante 30 minutos Separou-se o extracto do poacute de acetona
hidratado filtrando a suspensatildeo com 4 camadas de gaze esterilizada tendo o cuidado de natildeo
espremer o resiacuteduo Voltou a extrair-se com 50 mL de soluccedilatildeo I durante 30 minutos mexendo
lentamente Adicionou-se este ao extracto obtido anteriormente Centrifugou-se o extracto total a
16 000 g durante 30 minutos Decantou-se para uma proveta sobre a qual se colocou gaze para
que o sedimento solto natildeo se juntasse ao sobrenadante e leu-se o volume De seguida adicionou-
se KCl ateacute uma concentraccedilatildeo final de 50 mM MgCl2 ateacute 2mM e ATP ateacute 1mM lentamente e com
agitaccedilatildeo contiacutenua de modo a promover a polimerizaccedilatildeo dos monoacutemeros de actina
Deixou-se polimerizar durante 2h a 25ordmC observando-se a presenccedila de pequenas bolhas de ar
indicativas de um aumento de viscosidade da soluccedilatildeo Arrefeceu-se em gelo durante 15 minutos e
adicionou-se KCl ateacute uma concentraccedilatildeo final de 06M Mexeu-se lentamente durante 30 minutos
apoacutes os quais se ultracentrifugou a 110 000g durante 1hora Desprezou-se o sobrenadante
ressuspendeu-se o sedimento com soluccedilatildeo tampatildeo E e transferiu-se para um potter
Dialisou-se durante a noite contra 1L de soluccedilatildeo-tampatildeo I a uma temperatura de 4ordmC com rotaccedilatildeo
7 Soluccedilatildeo F 005 M NaHCO3 pH 82-85
8 Soluccedilatildeo G 1M Na2CO3
9 Soluccedilatildeo H 1 mM EDTA pH 70
10 Soluccedilatildeo I 2 mM Tris-HCl pH 8 02 mM CaCl2 02 mM Na2ATP 05 mM szlig-Me 0005 NaN3
42
contiacutenua e mudanccedila de tampatildeo 2 a 3 vezes De seguida a actina foi centrifugada a 110
000gdurante 1hora Decantou-se a soluccedilatildeo de G-Actina (sobrenadante) e desprezou-se o
sedimento Determinou-se a concentraccedilatildeo de G-Actina pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental) A soluccedilatildeo de G-actina foi armazenada a uma temperatura de 4ordmC
apoacutes adiccedilatildeo de 002NaN3 mantendo-se estaacutevel durante 3 a 5 dias
331 Quantificaccedilatildeo das proteiacutenas isoladas
3311 Quantificaccedilatildeo pelo meacutetodo de Bradford
O meacutetodo de Bradford (Bradford 1976) baseia-se na utilizaccedilatildeo do reagente de Bradford
composto por entre outros aacutecido fosfoacuterico e o corante ldquoCoomassierdquo Este corante possui carga
negativa e a sua coloraccedilatildeo varia entre a vermelha (Absmax=465 nm) predominante em soluccedilatildeo e a
azul (Absmax=595 nm) presente aquando da ligaccedilatildeo aos grupos amina das proteiacutenas Em soluccedilotildees
aacutecidas diluiacutedas a coloraccedilatildeo varia proporcionalmente com a quantidade do corante ligado agrave
proteiacutena que por sua vez eacute directamente proporcional agrave concentraccedilatildeo da proteiacutena permitindo a
sua quantificaccedilatildeo ateacute a um limite de detecccedilatildeo de 20 gmL (Bradford 1976)
Deste modo prepararam-se amostras de BSA ([BSA]stock= 1mgmL) utilizando um intervalo de
concentraccedilotildees compreendido entre 0 e 15 gmL de acordo com a tabela 3I Apoacutes a adiccedilatildeo da
H2O MilliQ e do reagente de Bradford as soluccedilotildees foram sucessivamente homogeneizadas no
voacutertex
Tabela 3Indash Amostras utilizadas para a obtenccedilatildeo da recta de calibraccedilatildeo no meacutetodo de Bradford
BSA (L) BSA (g) H2O MilliQ (L) Reagente de Bradford (L)
0 0 800 200
125 125 79875 200
25 25 7975 200
5 5 795 200
10 10 790 200
15 15 785 200
Apoacutes incubaccedilatildeo a 25ordmC por 5 minutos leram-se os valores de Abs595 num
espectrofotoacutemetro (Shimadzu UV-160A) para todas as amostras Seguidamente traccedilou-se a recta
de calibraccedilatildeo (Abs595vs quantidade de BSA (g))
43
Para determinar a concentraccedilatildeo de proteiacutena apoacutes o processo de isolamento purificaccedilatildeo e
diaacutelise quando necessaacuterio colocou-se 5 L da amostra num tubo de Eppendorf e adicionou-se
795 L de H2O MilliQ homogeneizando a soluccedilatildeo no voacutertex Adicionou-se posteriormente 200
L de reagente de Bradford e repetiu-se a homogeneizaccedilatildeo Incubou-se a mistura a 25ordmC por 5
minutos apoacutes os quais se leu os valores de Abs595 nm no espectrofotoacutemetro Por interpolaccedilatildeo deste
valor a partir da recta de calibraccedilatildeo determinou-se a concentraccedilatildeo de proteiacutena na amostra
3312 Quantificaccedilatildeo pelo meacutetodo directo
A quantificaccedilatildeo das amostras proteicas pode tambeacutem ser efectuada atraveacutes da leitura directa da
absorvacircncia tendo por base a lei de Beer-Lambert onde Abs corresponde agrave
absorvacircncia ao comprimento de onda ao qual o ensaio foi realizado corresponde ao coeficiente
de extinccedilatildeo (substituiacutedo pela letra a se as unidades forem distintas da molaridade) b corresponde
ao percurso oacuteptico da radiaccedilatildeo na ceacutelula amostral e c corresponde agrave concentraccedilatildeo amostral
Assim no caso de S1 de miosina mediu-se a Abs280e calculou-se a concentraccedilatildeo da proteiacutena
sabendo que o coeficiente de extinccedilatildeo de S1 eacute (Ref)
No caso da actina mediu-se a Abs290ecalculou-se a concentraccedilatildeo da proteiacutena sabendo que o
coeficiente de extinccedilatildeo da actina eacute (Ref)
332Electroforese em condiccedilotildees desnaturantes (SDS-PAGE)
De modo a melhor caracterizar as proteiacutenas isoladas foi realizada uma electroforese em
condiccedilotildees desnaturantes (SDS-PAGE do inglecircs Sodium Dodecyl Sulfate Polyacrilamide Gel
Electrophoresis) O SDS (dodecilo sulfato de soacutedio) eacute um detergente anioacutenico utilizado neste
meacutetodo para desnaturar as proteiacutenas que se ligam ao SDS na proporccedilatildeo 14 g SDS g proteiacutena
(aproximadamente 1 moleacutecula de SDS por cada 2 resiacuteduos de aminoaacutecidos) (Stryer) O SDS induz
uma densidade de carga negativa significativa nas proteiacutenas a que se liga mascarando a sua carga
intriacutenseca pelo que as razotildees cargamassa satildeo tendencialmente idecircnticas Assim a SDS-PAGE
separa as proteiacutenas quando sujeitas a um campo eleacutectrico de acordo com o seu peso molecular
sendo a sua migraccedilatildeo directamente proporcional ao logaritmo deste uacuteltimo (Stryer)
O gel de resoluccedilatildeo com 10 de acrilamida foi obtido atraveacutes da adiccedilatildeo de 167 mL de soluccedilatildeo
stock de acrilamida11
075 mL de tampatildeo de resoluccedilatildeo12
251 mL de aacutegua destilada e 50 L de
SDS 10 Para promover a polimerizaccedilatildeo adicionou-se em seguida 38 L de PSA 10
11
soluccedilatildeo stock de acrilamida soluccedilatildeo de 30 (mv) de acrilamida e 08 (mv) de NNrsquo-metil-bis-acrilamida 12
tampatildeo de resoluccedilatildeo 15 M Tris-HCl pH 88
44
(persulfato de amoacutenia) 10 e 25 L de TEMED 99 (tetrametiletilenodiamina) Agitou-se
cuidadosamente de modo a natildeo formar bolhas e pipetou-se a soluccedilatildeo para a cassette de vidro
previamente montada no suporte De seguida adicionou-se uma peliacutecula de cerca de 2 mm de 2-
metil-2-butanol para verificar posteriormente a polimerizaccedilatildeo Apoacutes a polimerizaccedilatildeo que demora
cerca de 45 minutos verteu-se a agrave superfiacutecie e adicionou-se o gel de concentraccedilatildeo com 5 de
acrilamida constituiacutedo por 03 mL de soluccedilatildeo de stock acrilamida 045 mL de tampatildeo de
concentraccedilatildeo13
094 mL de aacutegua destilada 18 L de SDS 10 135 L de PSA 10 e 2L de
TEMED 99 Colocou-se o pente sobre o gel e deixou-se polimerizar durante cerca de 45
minutos
As amostras para aplicaccedilatildeo no gel de electroforese foram preparadas para uma concentraccedilatildeo de
proteiacutena de 1 mgmL num volume final de 100 L dos quais 25 L correspondem ao tampatildeo da
amostra14
As amostras foram aquecidas a 100ordmC durante 5 minutos Antes da aplicaccedilatildeo das
amostras transferiu-se a cassette para a tina de electroforese agrave qual se adicionou a soluccedilatildeo
tampatildeo de electroforese da tina superior15
e tampatildeo de electroforese da tina inferior16
Aplicaram-
se 10 L das soluccedilotildees de marcadores de peso molecular e 20 L das amostras proteicas
preparadas anteriormente A electroforese decorreu durante cerca de 3h30 min a uma voltagem
de 80 V De seguida transferiu-se o gel para uma tina corando as proteiacutenas durante 1h com
soluccedilatildeo corante17
Descorou-se o gel com soluccedilatildeo descorante18
tendo-se mudado esta soluccedilatildeo ateacute
as bandas se encontrarem bem definidas
Determinou-se a mobilidade relativa das bandas correspondentes aos marcadores de elevado peso
molecular e traccedilou-se a recta de calibraccedilatildeo ndash log(PM) vs log(Rf) onde Rf corresponde agrave
mobilidade relativa Por interpolaccedilatildeo determina-se o PM das proteiacutenas isoladas sabendo a sua
mobilidade relativa Por comparaccedilatildeo com os valores descritos na literatura pode concluir-se
acerca do grau de purificaccedilatildeo
34 Mediccedilatildeo da actividade enzimaacutetica de S1
341 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
13
tampatildeo de concentraccedilatildeo 05 M Tris-HCl pH 68 14
tampatildeo de amostra 320 mM Tris-HCl pH 68 04 M szlig-mercaptoetanol SDS 8 glicerol 15 (vv) e roxo de m-
cresol 0024 15
tampatildeo de electroforese da tina superior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS 16
tampatildeo de electroforese da tina inferior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS acetato de soacutedio
01 M 17
soluccedilatildeo corante ldquoCoomassie Blue R-250rdquo contendo metanol 45 (vv) e aacutecido aceacutetico glacial 10 (vv) 18
soluccedilatildeo descorante metanol 10 (vv) e aacutecido aceacutetico glacial 10 vv)
45
O meacutetodo de Fiske-Subarow modificado consiste na reacccedilatildeo do molibdato de amoacutenio com o
fosfato inorgacircnico em soluccedilatildeo aacutecida com formaccedilatildeo do aacutecido fosfomoliacutebdico Seguidamente daacute-
se a adiccedilatildeo de um agente redutor que provoca a reduccedilatildeo do composto a fosfomolibdato que forma
um complexo de cor azulada cuja absorvacircncia maacutexima ocorre para um comprimento de onda de
660 nm tornando possiacutevel a sua detecccedilatildeo por meacutetodos espectrofotomeacutetricos As actividades Ca2+
-
ATPaacutesica e K+EDTA-ATPaacutesica de S1 foram determinadas atraveacutes deste meacutetodo de acordo com
o descrito por Taussky e Shorr (1953) com mediccedilatildeo da quantidade de fosfato (Pi) libertado
atraveacutes da hidroacutelise de ATP
Para tal obteve-se primeiramente a recta de calibraccedilatildeo com as amostras presentes na Tabela 3II
utilizando uma soluccedilatildeo stock de 5 mM KH2PO4
Tabela 3II ndash Composiccedilatildeo das diferentes amostras utilizadas para determinar a recta de calibraccedilatildeo atraveacutes do meacutetodo
de Fiske-Subbarow modificado
KH2PO4 (L)
([KH2PO4]stock= 5mM)
PO43-
(nmole) H2O (uL)
0 0 1780
5 25 1775
10 50 1770
20 100 1760
40 200 1740
100 500 1680
A cada tubo de Eppendorf adicionaram-se seguidamente 200 L de soluccedilatildeo J19
e 20 L de agente
redutor20
homogeneizando no voacutertex apoacutes cada adiccedilatildeo Incubou-se durante 10 minutos e mediu-
se a Abs660 nm num espectrofotoacutemetro com intervalos de tempo iguais entre cada amostra Traccedilou-
se a recta de calibraccedilatildeo (Abs660 nmvs nmol PO43-
) a partir dos valores obtidos
Para determinar a actividade Ca2+
-ATPaacutesica de S1 prepararam-se tubos de Eppendorf com 400
L de meio de reacccedilatildeo21
25 L CaCl2 ([CaCl2]stock= 100 mM) 25 L ATP ([ATP]stock= 100 mM)
com [S1]ensaios=01 mgmL Perfez-se se necessaacuterio com H2O MilliQ ateacute perfazer um volume de 1
mL Para determinar a actividade K+EDTA-ATPaacutesica utilizou-se uma concentraccedilatildeo idecircntica de
19
Soluccedilatildeo J 5 N H2SO4 25 molibdato de amoacutenio 20
Agente redutor 5 mg aacutecido 1-amino-2-naftol-4-sulfoacutenico 30 mg bissulfito de soacutedio 30 mg sulfito de soacutedio em 26
mL H2O 21
meio de reacccedilatildeo 125 mM Tris-HCl pH 80 0575 M KCl
46
proteiacutena nos ensaios (01 mgmL) e o mesmo meio de reacccedilatildeo Aos tubos de Eppendorf
adicionou-se ainda 123 L de KCl ([KCl]stock = 3M) 25 L de ATP 10 L de EDTA
([EDTA]stock= 100 mM )e o volume de H2O MilliQ necessaacuterio para perfazer 1 mL Em ambas as
situaccedilotildees o uacuteltimo reagente adicionado foi o ATP uma vez que se trata do substrato da reacccedilatildeo
enzimaacutetica
A cineacutetica decorreu a 25ordmC durante 5 minutos para a actividade Ca2+
-ATPaacutesica e durante 3
minutos para a actividade K+EDTA-ATPaacutesica Apoacutes o periacuteodo de incubaccedilatildeo transferiu-se 150
L de amostra para tubos de Eppendorf contendo igual volume de TCA 10 Colocou-se no gelo
e homogeneizou-se no voacutertex Seguidamente centrifugou-se a 1500 g durante 10 minutos e
recolheu-se 200 L de sobrenadante para novos tubos de Eppendorf Adicionou-se a estes 158
mL de H2O MilliQ e 200 L de soluccedilatildeo J Homogeneizou-se no voacutertex e adicionou-se 20 L de
reagente redutor repetindo-se a homogeneizaccedilatildeo Incubou-se durante 10 minutos e mediu-se a
Abs660 nm com o mesmo intervalo de tempo de leitura para cada amostra Por intrapolaccedilatildeo a partir
da recta de calibraccedilatildeo obtida anteriormente determinou-se a quantidade de Pi libertado em cada
ensaio A partir destes resultados determinaram-se as actividades Ca2+
-ATPaacutesica e K+EDTA-
ATPaacutesica do S1 em nmol PO43-
minmg proteiacutena e em s-1
342 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
Da actividade Mg2+
-ATPaacutesica de S1 resulta a formaccedilatildeo dos produtos da reacccedilatildeo de
hidroacutelise do ATP - ADP e Pi Na presenccedila de ADP fosfoenolpiruvato (PEP) e da enzima piruvato
cinase (PK) os produtos resultantes satildeo ATP e piruvato Por acccedilatildeo da enzima lactato
desidrogenase (LDH) este uacuteltimo eacute convertido em lactato ocorrendo simultaneamente a oxidaccedilatildeo
do NADH a NAD+ monitorizada atraveacutes da Abs340 nm Quando o S1 de miosina se encontra no
meio reaccional o ATP eacute hidrolisado formando-se novamente ADP e Pi o que permite a
formaccedilatildeo de um ciclo (fig 21) Se as enzimas se encontrarem em excesso a velocidade de
conversatildeo do NADH eacute limitada pela produccedilatildeo de ADP e Pi Como o ATP eacute regenerado em cada
ciclo a sua concentraccedilatildeo inicial eacute sempre mantida (Bagshaw 1982)
Figura 33ndash Representaccedilatildeo esquemaacutetica do meacutetodo de enzimas acopladas
47
O protocolo utilizado e seguidamente descrito foi adaptado de Cuenda et al (1990 ndash Ver Refordf) A
uma cuvette de 1 mL adicionou-se S1 para uma concentraccedilatildeo final de 005 mgmL 800 L de
meio de reacccedilatildeo22
10 L de PEP ([PEP]stock = 42 mM) 10 L de PK+LDH 11 ([PK+LDH]stock =
1 mgmL) e 15 L de NADH ([NADH]stock=25 mM) Iniciou-se a monitorizaccedilatildeo da reacccedilatildeo
atraveacutes da leitura de Abs340 nm apoacutes a adiccedilatildeo de 20 L de ATP ([ATP]stock=100 mM) A
actividade Mg2+
-ATPaacutesica eacute obtida atraveacutes do declive correspondente agrave oxidaccedilatildeo do NADH Para
a mediccedilatildeo da Mg2+
-ATPase de S1 estimulada por actina adicionou-se F-actina para uma
concentraccedilatildeo final de 2 microM sendo a actividade obtida tambeacutem atraveacutes do declive correspondente
agrave oxidaccedilatildeo do NADH apoacutes a adiccedilatildeo da proteiacutena
35 Interaccedilatildeo entre vanaacutedio e subfragmento-1 de miosina
351 Preparaccedilatildeo das soluccedilotildees de vanaacutedio
As soluccedilotildees de vanadato foram preparadas em aacutegua MilliQ a partir de metavanadato de amoacutenio
(NH4VO3) numa concentraccedilatildeo stock de 50 mM com agitaccedilatildeo magneacutetica e aquecimento (~30 ordmC)
Esta soluccedilatildeo com um valor compreendido entre 6 e 7 designa-se por ldquometavanadatordquo e eacute
constituiacuteda pelas esspeacutecies V1 V2 V4 e V5 (Ref) Acidificando a soluccedilatildeo de metavanadato por
adiccedilatildeo de HCl ateacute um valor de pH proacuteximo de 4 observa-se uma mudanccedila de cor de amarelo
claro para laranja indicativa da formaccedilatildeo da espeacutecie decameacuterica de vanadato De seguida e
imediatamente antes da realizaccedilatildeo dos ensaios descritos o valor do pH da soluccedilatildeo de
decavanadato foi aumentado para cerca de 75 por adiccedilatildeo de NaOH A soluccedilatildeo (2) designada
por ldquometavanadatordquo natildeo foi posteriormente alterada ateacute agrave sua adiccedilatildeo para a realizaccedilatildeo dos ensaios
As soluccedilotildees foram armazenadas a 4ordmC e mantidas em gelo aquando da sua utilizaccedilatildeo
A soluccedilatildeo de V(IV) foi preparada em condiccedilotildees anaeroacutebias por diluiccedilatildeo de sulfato de vanadil
(VOSO4) para uma concentraccedilatildeo final de 5 mM em H2O MilliQ saturada com N2 gasoso
352Anaacutelise por espectroscopia de UV-Vis
A concentraccedilatildeo da espeacutecie decameacuterica em soluccedilatildeo depende do periacuteodo de tempo apoacutes a diluiccedilatildeo
no meio de reacccedilatildeo apropriado (Ref) A velocidade de desoligomerizaccedilatildeo da espeacutecie decameacuterica
pode ser analisada por espectroscopia de absorccedilatildeo na regiatildeo UVVis uma vez que esta espeacutecie
apresenta duas bandas caracteriacutesticas na regiatildeo do visiacutevel (a 358 e 400 nm) que natildeo se verifica
em quaisquer outras espeacutecies oligomeacutericas As bandas supramencionada sapresentam-se bem
22
Meio de reacccedilatildeo 3125 mM Tris-HCl pH 71 375 mM MgCl2
48
resolvidas atraveacutes da obtenccedilatildeo da derivada de 2ordf ordem do espectro apresentando a forma de 2
vales com miacutenimos centrados nos comprimentos de onda supracitados com dependecircncia linear
relativamente agrave concentraccedilatildeo de decavanadato total (Ramos et al 2006)
Assim a estabilidade da espeacutecie decameacuterica apoacutes diluiccedilatildeo da soluccedilatildeo de decavanadato no meio de
reacccedilatildeo dos ensaios (tampatildeo D) para uma concentraccedilatildeo final de 1 mM em vanadato total foi
caracterizada atraveacutes da leitura da Abs400 em funccedilatildeo do tempo tendo-se determinado qual a
ordem de reacccedilatildeo e o tempo de meia-vida da mesma Com base em estudos anteriores acerca da
estabilidade do V10 na presenccedila de diferentes proteiacutenas (Ramos et al 2006) determinou-se
igualmente o tempo de meia vida apoacutes a adiccedilatildeo de 20 microM de S1 de miosina ao meio reaccional
353 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
Os espectros de RMN do isoacutetopo 51
V foram realizados num espectroacutemetro Bruker AM-400 MHz
equipado com uma sonda multinuclear inversa de 5 mm Em tubos de RMN de 5 mm de diacircmetro
preparou-se soluccedilotildees de 500 L de deca- ou metavanadato correspondentes a um intervalo de
concentraccedilotildees de 1 a 10 mM suplementadas com 10 D2O Os espectros foram adquiridos agrave
temperatura ambiente com uma largura espectral de 23 923 Hz um tempo de aquisiccedilatildeo de 0086
s tempo de relaxaccedilatildeo de 001 s e uma sequecircncia de pulsos a 90ordm Utilizou-se a transformada de
Fourier com alargamento das bandas de 20Hz para o processamento dos espectros Os desvios
quiacutemicos das ressonacircncias de 51
V foram obtidos em relaccedilatildeo agrave espeacutecie VOCl3cuja frequecircncia de
ressonacircncia eacute considerada o padratildeo sendo todas as outras referenciadas a partir desta em ppm
As aacutereas dos sinais do RMN correspondentes agraves espeacutecies oligomeacutericas presentes foram obtidas
por integraccedilatildeo dos sinais As concentraccedilotildees relativas das espeacutecies foram calculadas com recurso agrave
equaccedilatildeo
onde A[Vx] corresponde agrave aacuterea referente agrave ressonacircncia da espeacutecie Vx com nordm de agregaccedilatildeo x
A[V]total agrave soma total das aacutereas medidas e [V] a concentraccedilatildeo total de vanaacutedio no ensaio
A teacutecnica de 51
V-RMN foi anteriormente aplicada em estudos de ligaccedilatildeo com S1 de miosina
(Ref) tendo sido demostrado que entre as espeacutecies V1 V2 V4 e V5 a espeacutecie tetrameacuterica possui a
maior afinidade para a miosina (Cremo et al) O desaparecimento ou alargamento dos sinais pode
implicar a ligaccedilatildeo de um dos oxovanadatos laacutebeis agrave miosina sem indicaccedilatildeo de qual uma vez que
49
se processa um equiliacutebrio entre eles (Crans 1990) Assim e tendo em conta as indicaccedilotildees de que
a espeacutecie V10 eacute distinta das restantes no que diz respeito agrave magnitude da interacccedilatildeo com a miosina
e inibiccedilatildeo da actividade ATPaacutesica estimulada por actina (Aureliano 2000 Tiago et al 2002)
torna-se fundamental investigar a interacccedilatildeo das diferentes espeacutecies de vanadato com o S1 de
miosina atraveacutes da teacutecnica de 51
V-RMN
Para tal obtiveram-se espectros de 51
V-RMN resultantes da titulaccedilatildeo de uma soluccedilatildeo 5 mM de
decavanadato total com concentraccedilotildees crescentes de S1 de miosina (0 5 10 15 e 20 microM) em
tampatildeo D Realizou-se ainda ensaios na presenccedila de 2 mM de decavanadato e 2 mM de ATP na
presenccedila e na ausecircncia de S1 de miosina de modo a verificar o efeito que estes ligandos naturais
da enzima teria na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o subfragmento-1
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
Os grupos sulfidrilo (-SH) das cisteiacutenas satildeo resiacuteduos de aminoaacutecidos extremamente sensiacuteveis agrave
oxidaccedilatildeo resultando na sua oxidaccedilatildeo (Dalle-Donne et al 1999) Logo a quantificaccedilatildeo destes
resiacuteduos eacute bastante importante nos estudos de interacccedilatildeo com vanaacutedio jaacute que os resiacuteduos de Cys
com ecircnfase para o SH1 e o SH2 satildeo bastante importantes na monitorizaccedilatildeo das modificaccedilotildees
funcionais da regiatildeo da cabeccedila
O procedimento seguidamente apresentado foi originalmente descrito por Gutieacuterrez-Merino
(1987)) Consiste na reacccedilatildeo entre os grupos ndashSH natildeo oxidados das proteiacutenas e o reagente de
Ellman (DTNB -55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico)) - formando-se o aniatildeo CNT (aniatildeo 3-
carboxilato-4-nitrotiofenolato) numa estequiometria de 11 Este apresenta um maacuteximo de
absorvecircncia a 412 nm (Robyt 1990) Assim apoacutes a incubaccedilatildeo da proteiacutena na presenccedila do metal
a quantidade de cisteiacutenas reduzidas por proteiacutena determina-se atraveacutes dos valores de Abs412
Deste modo colocou-se num tubo de Eppendorf o volume apropriado de S1 dialisado para que a
concentraccedilatildeo no ensaio fosse de 1 mgmL Adicionou-se o volume apropriado de decavanadato
para que a sua concentraccedilatildeo fosse de 0 10 50 100 250 500 ou 1000 M Perfez-se com soluccedilatildeo
D ateacute um volume final de 1 mL e incubou-se a 25ordmC por 20 minutos Apoacutes este periacuteodo colocou-
se 780 L da mistura reaccional numa cuvette de quartzo e adicionou-se 220 L de soluccedilatildeo K23
23
Tampatildeo K Tampatildeo fosfatohidrogenofosfato de soacutedio 025 M pH 72
50
Adicionou-se 15 L de DTNB (em soluccedilatildeo K [DTNB]stock=100 mM) ao meio reaccional e
registou-se o valor de absorvecircncia medido equivalente agraves cisteiacutenas ldquoraacutepidasrdquo (SH1 e SH2
expostos ao solvente) Adicionou-se 001 g de SDS que provoca a desnaturaccedilatildeo da proteiacutena e
consequentemente a exposiccedilatildeo de todos os resiacuteduos de cisteiacutenas que se encontravam ateacute aiacute
inacessiacuteveis no interior da proteiacutena (cisteiacutenas totais) Estes resiacuteduos podem assim reagir com o
DTNB
Estimou-se o nuacutemero de resiacuteduos de cisteiacutenas em todas as amostras com recurso ao coeficiente de
extinccedilatildeo molar () do DTNB (ver ponto 3 dos Anexos) que foi determinado atraveacutes da titulaccedilatildeo
de L-Cisteiacutena com intervalo de concentraccedilotildees compreendidos entre 0 e 50 microM na presenccedila de
25microM de DTNB em tampatildeo D Os valores de absorvecircncia foram obtidos para um comprimento de
onda de 412 nm
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
3621 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
A teacutecnica mais eficiente de anaacutelise da interacccedilatildeo entre o iatildeo vanadilo e proteiacutenas como a
calmodulina e a transferrina entre outras permite a obtenccedilatildeo raacutepida de informaccedilatildeo no que toca agrave
conformaccedilatildeo e amplitude da referida interacccedilatildeo
Os espectros de Ressonacircncia Paramagneacutetica Electroacutenica (RPE) da espeacutecie paramagneacutetica V(IV)
foram realizados a 70K num espectroacutemetro Bruker EMX-300 X-band Todas as soluccedilotildees
utilizadas e amostras foram preparadas em anaerobiose numa cacircmara de luvas de modo a evitar a
reduccedilatildeo de V(IV) a V(V) Em tubos capilares com 02 mL de amostra manteve-se a concentraccedilatildeo
de S1 a 20 M e variou-se a concentraccedilatildeo de vanadilo (0 25 5 10 15 25 50 75 100 150
200 500 e 1000 M [VOSO4]stock=5 mM) em tampatildeo D Apoacutes obtenccedilatildeo dos espectros estes
foram tratados atraveacutes do software SimFonia e WinEPR tendo-se obtido um graacutefico com a
variaccedilatildeo da intensidade do sinal de RPE em funccedilatildeo da concentraccedilatildeo de sulfato de vanadilo
presente Realizaram-se ainda simulaccedilotildees de espectros obtidos com 20 microM de S1 e 1000 microM de
V(IV) com vista agrave obtenccedilatildeo dos ligandos provaacuteveis do vanadilo atraveacutes da anaacutelise da constante
de acoplamento hiperfina (A) e do factor g (g)
4 Conclusotildees gerais
Apesar de jaacute ter sido demonstrado que o vanaacutedio inibe a Na+K
+-ATPase da miosina a funccedilatildeo
deste elemento no tecido muacutesculo esqueleacutetico continua largamente incompreendida A enorme
complexidade quiacutemica deste metal especialmente no que concerne agrave oligomerizaccedilatildeo e reacccedilotildees
51
redox constitui um obstaacuteculo agrave compreensatildeo do comportamento do vanaacutedio nos organismos
vivos Assim a caracterizaccedilatildeo da quiacutemica deste elemento eacute de grande utilidade antes de iniciar o
estudo da sua interacccedilatildeo com o presente sistema
A anaacutelise por espectroscopias de 51
V-RMN e de UV-Vis de soluccedilotildees de vanadato diluiacutedas nos
meios de reacccedilatildeo utilizados no presente relatoacuterio (10 mM HEPES pH 70 25 mM KCl) mostrou
claramente que a espeacutecie decameacuterica (V10) possui um tempo de vida suficientemente longo para a
determinaccedilatildeo do seu efeito isolado ao contraacuterio do que se verifica com as espeacutecies mono- di-
tetra- e pentameacutericas (V1 V2 V4 e V5) Por outro lado a anaacutelise verificou ainda que apoacutes diluiccedilatildeo
das soluccedilotildees de decavanadato no meio de reacccedilatildeo coexistem as espeacutecies V10 e V1 pelo que os
efeitos putativos desta uacuteltima deveratildeo ser subtraiacutedos aquando da monitorizaccedilatildeo dos efeitos da
espeacutecie decameacuterica
O vanaacutedio (V) (vanadato) inibe as enzimas que catalizam reacccedilotildees de transferecircncia de grupos
fosfato como a ATPase da miosina [Goodno 1979] devido agrave sua analogia estrutural com o iatildeo
fosfato Na sua utilizaccedilatildeo como inibidor enzimaacutetico natildeo eacute tida em conta a contribuiccedilatildeo individual
das espeacutecies oligomeacutericas considerando-se normalmente a espeacutecie V1 como inibidora efectiva
[Goodno e Taylor 1982] No presente relatoacuterio verificou-se por espectroscopia de 51
V-RMN que
outros oligoacutemeros de vanadato interagem com o S1 de miosina tendo as espeacutecies V10 e V4 os
maiores niacuteveis de afinidade A sua interacccedilatildeo envolve forccedilas electrostaacuteticas e eacute afectada
negativamente por ligandos naturais de enzima como eacute o caso do ATP
A exposiccedilatildeo do S1 de miosina (005 mgmL) a decavanadato conduz agrave inibiccedilatildeo do capacidade da
proteiacutena de estimular a actividade Mg2+
-ATPaacutesica com um valor de IC50 de 266 plusmn 07 M
decavanadato sendo o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM
decavanadato (80) Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM decavanadato sendo
o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM decavanadato (60)
exercendo um efeito de impedimento agrave ligaccedilatildeo entre o vanaacutedio e o S1
A uma concentraccedilatildeo de 27 microM decavanadato verifica-se a oxidaccedilatildeo de um dos resiacuteduos de Cys
de S1 de miosina seguida pela oxidaccedilatildeo gradual de outro resiacuteduo de Cys ateacute agrave concentraccedilatildeo de
1000 microM Ambos os resiacuteduos oxidados natildeo se encontravam expostos ao solvente utilizado A
oxidaccedilatildeo dos resiacuteduos permite sugerir que esta modificaccedilatildeo oxidativa sofrida pelo S1 contribui
para a diminuiccedilatildeo da sua actividade de ATPase
52
Por espectroscopia de RPE revelaram-se os possiacuteveis ligandos equatoriais do vanaacutedio aquando da
sua interacccedilatildeo com o S1 de miosina como sendo duas moleacuteculas de aacutegua um ligando contendo o
grupo R-OH e outro contendo o grupo R=NR
Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados de
oxidaccedilatildeo (IV) e (V) no S1 de miosina para concentraccedilotildees facilmente alcanccedilaacuteveis no ambiente
terrestre
53
5 Estudos futuros
No sentido do melhor esclarecimento das alteraccedilotildees moleculares e estruturais que ocorrem no
subfragmento 1 quando exposta ao vanaacutedio que conduzem a uma diminuiccedilatildeo da sua funccedilatildeo
sugere-se
- Quantificaccedilatildeo das cisteiacutenas reduzidas de S1 na presenccedila de metavanadato
- Determinaccedilatildeo das actividades Ca2+
- e K+-ATPaacutesica do S1 de miosina na presenccedila de
decavanadato
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina estimulada por actina
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina na presenccedila de
metavanadato
- Realizaccedilatildeo de RMN 2D com variaccedilatildeo das concentraccedilotildees das soluccedilotildees de decavanadato e
metavanadato na presenccedila de S1
-Mutageacutenese dos resiacuteduos de Cys doS1 de miosina de modo a verificar quais os resiacuteduos que
sofrem oxidaccedilatildeo na presenccedila de decavanadato
- Determinaccedilatildeo por calorimetria (DSC Differential Scanning Calorimetry) da variaccedilatildeo da
entalpia (∆H) e da temperatura de desnaturaccedilatildeo do S1 de miosina na presenccedila de vanaacutedio uma
vez que estas propriedades caracteriacutesticas de cada proteiacutena satildeo alteradas apoacutes ocorrecircncia de
modificaccedilotildees estruturais
- Anaacutelise por SDS-PAGE de S1 de miosina apoacutes tratamento com vanaacutedio de modo a determinar
quais os polipeacuteptidos eventualmente formados apoacutes clivagem da proteiacutena
54
6 Referecircncias bibliograacuteficas
- Sellers J Myosins 2nd Edition Oxford University Press 1999
- L Stryer inBiochemistry (Ed WH Freeman 2005 pp
Refordfas Materiais e Meacutetodos
[A] S S Margossian SLowey Methods Enzymol1982 85 55-71
[B] JD Pardue JA Spudich Methods Enzymol 1982 85 164-181
[C] M Bradford Anal Biochem1976 72 248-254
[D] H H Taussky E Shorr J Biol Chem 1953 202 675-685
[E] C R Bagshaw in Outline Studies in Biology ndash Muscle Contraction Chapman and Hall Ed
1982
[F] A Cuenda et al Eur J Biochem1990 194 663-670
55
Anexos
1 Determinaccedilatildeo da recta de calibraccedilatildeo meacutetodo de Bradford
Figura1ndashRepresentaccedilatildeo graacutefica da recta de calibraccedilatildeo (Abs595 nm vs [BSA]) utilizada para o meacutetodo de Bradford
2 Determinaccedilatildeo da recta calibraccedilatildeo para a electroforese em condiccedilotildees desnaturantes
Figura 2 ndash Representaccedilatildeo graacutefica da recta de calibraccedilatildeo (log(Peso Molecular) vs Rf dos marcadores de peso
molecular) onde Rf indica determinada atraveacutes do padratildeo de peso molecular
(poccedilo (a))
3Caacutelculo do coeficiente de extinccedilatildeo molar () para a quantificaccedilatildeo de cisteiacutenas reduzidas
atraveacutes do meacutetodo de Ellman
Traccedilou-se a recta de Abs412 nm meacutedia vs [L-Cisteiacutena] de modo a determinar o valor do coeficiente
de extinccedilatildeo molar definido como o declive daquela recta Assim =8400 M-1
cm-1
como
podemos verificar na Fig 3
A595 nm = 00546[BSA] + 00209 Rsup2 = 099051
Ab
s 59
5 n
m
[BSA] (mgmL)
log PM = -1177 Rf + 2247 Rsup2 = 098957
log
PM
Mobilidade relativa (Rf)
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero

28
Tabela 2IVndash Desvios quiacutemicos meacutedios (part)das ressonacircncias das espeacutecies tetrameacuterica (V4) monomeacuterica (V1) e
dimeacuterica (V2) presentes no espectro da soluccedilatildeo de metavanadato (total) 10 mM
Paracircmetros V1 V2 V4 V5
part (ppm) -5615 -5748 -5789 -5869
Nas soluccedilotildees de metavanadato as espeacutecies monomeacuterica dimeacutericae tetrameacuterica de vanadato estatildeo
presentes para concentraccedilotildees de metavanadato inferiores a 4 mM Todas estas as espeacutecies
aumentam linearmente com o aumento da concentraccedilatildeo total de vanadato Tal como constatado
em (Aureliano e Madeira 1998) e em (Tiago et al 2004) quando a concentraccedilatildeo de V1 eacute igual ou
superior a 200 microM inicia-se a sua oligomerizaccedilatildeo para originar a espeacutecie tetrameacuterica Nas
condiccedilotildees de realizaccedilatildeo dos ensaios descritos no presente relatoacuterio natildeo foi possiacutevel identificar a
espeacutecie pentameacuterica neste estudo em concentraccedilotildees inferiores a 4 mM tendo o mesmo sido
observado por Ramos 2006 Os resultados obtidos mostram claramente o equiliacutebrio entre a
espeacutecie V4 e as espeacuteciesV1 e V2 no qual o aparecimento de outras espeacutecies oligomeacutericas se
encontra dependente da concentraccedilatildeo da espeacutecie V1 Por uacuteltimo pode inferir-se que para
concentraccedilotildees inferiores a 2 mM e intervalos de tempo de cerca de 20 minutos (que inclui o
tempo de preparaccedilatildeo da amostra e de aquisiccedilatildeo do espectro) existem em soluccedilatildeo apenas as
espeacutecies V1 V2 e V4 com concentraccedilotildees respectivas de 0796 0192 e 0196 mM
232Estabilidade das soluccedilotildees de decavanadato anaacutelise por espectroscopia de absorccedilatildeo
UVVis
Caracterizou-se a estabilidade da espeacutecie V10 numa soluccedilatildeo com 1 mM decavanadato total em
tampatildeo D atraveacutes da leitura da Abs400 Obteve-se uma cineacutetica de dissociaccedilatildeo de 1ordf ordem tal
como se pode verificar pela linearidade do graacutefico de ln(Abs400)vs o tempo de aquisiccedilatildeo espectral
(segundos) (Fig26 )
Figura 26ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do tempo
(em segundos) para uma soluccedilatildeo de decavanadato total de 1
mM em tampatildeo D a 25ordmC (n=2)
Sendo uma reacccedilatildeo de primeira ordem pode ser descrita pela equaccedilatildeo cujo
declive eacute ndashK (constante de dissociaccedilatildeo) e a intersecccedilatildeo no eixo das ordenadas corresponde
Ln
(Ab
s 400 n
m)
tempo (s)
29
aln(A0) Numa reacccedilatildeo de 1ordf ordem o tempo de meia-vida eacute independente da concentraccedilatildeo inicial
do reagente sendo descrito pela equaccedilatildeo [2] Deste modo a partir da recta
Ln(Abs400 nm) = -436E-5
K - 105 obteve-se um tempo de meia-vida de 4490 plusmn 0823 horas para
as condiccedilotildees supramencionadas
Apoacutes a adiccedilatildeo de S1 8 microM ao meio reaccional verificou-se que a reacccedilatildeo de desoligomerizaccedilatildeo
da espeacutecie decameacuterica de vanadato seguia igualmente uma cineacutetica de 1ordf ordem tal como se pode
verificar pela linearidade do graacutefico de ln(Abs400) versus o tempo de aquisiccedilatildeo espectral
(segundos) (Fig27)
Figura 27ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do
tempo (em segundos) para uma soluccedilatildeo de decavanadato
total de 1 mM em tampatildeo D a 25ordmC na presenccedila de 8
microM (n= 2)
A partir da equaccedilatildeo da recta Ln(Abs400 nm) = -7520E-5
K - 0613determinou-se o valor de K para
a espeacutecie decameacuterica de onde se obteve um tempo de meia-vida de 2430 plusmn 0191 horas para as
condiccedilotildees supramencionadas
Tanto este valor como o valor registado na ausecircncia de S1 satildeo substancialmente superiores agrave
duraccedilatildeo dos ensaios de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina descritos no presente
relatoacuterio
24 Interacccedilatildeo de S1 com vanaacutedio(V)
241Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
De modo a analisar qual o efeito da adiccedilatildeo de S1 de miosina (tampatildeo D) produzia nos sinais de
51V-NMR titulou-se uma soluccedilatildeo de 5mM de decavanadato total com concentraccedilotildees crescentes
de S1 (5 10 15 e 20 microM) Os espectros de 51
V-RMN encontram-se apresentados na Fig 2
No graacutefico da figura 2 Apresenta-se a concentraccedilatildeo de S1 em funccedilatildeo do factor de alargamento
corrigido dos espectros acima referidos
Ln
(Ab
s (4
00
nm
))
tempo (s)
30
Figura 28 ndash Graacutefico de
representaccedilatildeo da
concentraccedilatildeo de S1 em
funccedilatildeo do factor de
alargamento A
concentraccedilatildeo de
decavanadato total foi de 5
mM e as concentraccedilotildees
utilizadas de S1 foram de 0
5 10 e 15 microM
Verifica-se que o factor de alargamento aumenta linearmente em funccedilatildeo da concentraccedilatildeo de S1 no
ensaio o que indica a existecircncia de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina Observou-
se ainda que a adiccedilatildeo de S1 ao meio reaccional natildeo provocou qualquer alteraccedilatildeo dos desvios
quiacutemicos dos sinais de 51
V-RMN
Realizaram-se ensaios na presenccedila de ATP e de S1 com intuito de verificar o efeito do ATP na
interacccedilatildeo do S1 de miosina com o decavanadato
Figura 29 - Espectros de RMN de 51
V contendo 2
mM Decavanadato (A) (controlo) (B) na presenccedila
de 2 mM de ATP (C) na presenccedila de 2 mM de ATP
e 8 microM de S1 de miosina V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2Vndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies decameacuterica (V10) e monomeacuterica (V1) presentes nos
espectros da soluccedilatildeo de decavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V10A V10B V10C V1
part (ppm) -5161 -5002 -4254 -5614
Observa-se que a presenccedila de ATP (espectro B) natildeo provoca qualquer alteraccedilatildeo visiacutevel em
relaccedilatildeo ao espectro que contem apenas soluccedilatildeo de decavanadato Jaacute a presenccedila simultacircnea de ATP
e S1 provoca um alargamento da ∆12 do sinal do V10A de 1736 Hz (espectro (A)) para 2366 Hz
(espectro (C) Fig 29) Tendo em conta que a concentraccedilatildeo de S1 apresenta uma relaccedilatildeo linear
com o factor de alargamento converteu-se o valor da ∆12 e comparou-se com o valor esperado agrave
F = 00591[S1] - 00157 Rsup2 = 0998
Fact
or
de
ala
rgam
en
to
[S1] (microM)
31
concentraccedilatildeo de S1 utilizada O factor de alargamento obtido para 8 microM ndash 0376 ndash eacute inferior ao
valor esperado agrave mesma concentraccedilatildeo de S1 obtido atraveacutes da equaccedilatildeo da recta apresentada na
Fig 28 ndash 0457 ndash o que indica que o ATP poderaacute exercer um efeito de protecccedilatildeo do S1
diminuindo a sua interacccedilatildeo com a espeacutecieV10
Por uacuteltimo realizaram-se ensaios com metavanadato na presenccedila de 8 microM S1 de modo a verificar
se as espeacutecies oligomeacutericas contidas nesta soluccedilatildeo interagiam com a proteiacutena em estudo
Figura 210 - Espectros de RMN de 51
V contendo 2
mM metavanadato (A) (controlo) (B) na presenccedila
de 8 microM de S1 de miosina V1 V2 e V4
correspondem respectivamente aacutes espeacutecies
monomeacuterica dimeacuterica e tetrameacuterica
Tabela 2VIndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies monomeacuterica (V1) dimeacuterica (V2) e tetrameacuterica (V4)
presentes nos espectros da soluccedilatildeo de metavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V1 V2 V4
part (ppm) -5613 -5747 -5789
Como eacute possiacutevel observar na Fig 210 ocorre uma ligeiro alargamento dos sinal de V4 bem
como uma perda da intensidade pelo que se conclui quedas espeacutecies presentes na soluccedilatildeo de
metavanadato (V1 V2 e V4) apenas a espeacutecie tetrameacuterica interage com o S1 de miosina o que
estaacute de acordo com o reportado por Tiago et al 2002
242 Actividade Mg2+
-ATPaacutesica de S1 de miosina na presenccedila de decanadato
Com o intuito de avaliar o efeito do decavanadato na actividade Mg2+
-ATPaacutesica do S1 de
miosina monitorizou-se a reacccedilatildeo de oxidaccedilatildeo do NADH atraveacutes da leitura de Abs340 na
presenccedila de 005 mgmL de S1 de miosina e no caso da actividade estimulada 2microM de actina em
3125 mM Tris-HCl pH 71 e 375 mM MgCl2 pelo meacutetodo de enzimas acopladas (ver seccccedilatildeo xx
dos MM)Para tal realizaram-se ensaios com vaacuterias concentraccedilotildees de decavanadato total (10 50
100 250 500 e 1000) de modo a determinar qual o seu efeito na actividade MgATPaacutesica de S1
de miosina na ausecircncia e na presenccedila de um dos seus ligandos naturais Os ensaios foram
realizados agrave temperatura ambiente com incubaccedilatildeo preacutevia a 25ordm C durante 20 minutos
32
Figura 211 ndash Representaccedilatildeo
graacutefica da Actividade Mg2+
-
ATPaacutesica de S1 () (os ajustes
teoacutericos estatildeo representado a por
uma linha os resultados
experimentais estatildeo representados
por triacircngulos (Actividade
estimulada) e cruzes (Actividade
natildeo estimulada)) vs
[Decavanadato]total medidas pelo
meacutetodo das enzimas acopladas
(n=3) Os dados foram ajustados ao modelo de decaimento exponencial usando o programa GraphPad Prism para a
determinaccedilatildeo dos valores de IC50 (na ausecircncia de actina IC50= 266 plusmn 07 M decavanadato na presenccedila de actina
IC50= 28 plusmn 1 M decavanadato
A partir do graacutefico presente na Fig 29 verifica-se que o ajuste dos dados experimentais foi bem
sucedido A inibiccedilatildeo da actividade Mg2+
-ATPaacutesica do S1 de miosina eacute afectada em menor
amplitude na presenccedila de actina (60 de inibiccedilatildeo da actividade) do que na sua ausecircncia
(aproximadamente 80 de inibiccedilatildeo de actividade) Devido a esta diferenccedila pode sugerir-se que a
actina iraacute impedir parcialmente a ligaccedilatildeo do decavanadatoao S1 demiosina atraveacutes de
impedimentos esteacutereos ou interacccedilotildees electrostaacuteticas por exemplo A teacutecnica utilizada natildeo nos
permite concluir acerca de que regiatildeo(otildees) ou resiacuteduo(s) da actina participaratildeo na preservaccedilatildeo
parcial da actividade Mg2+
-ATPaacutesica
Por outro lado os valores de IC50 dos dois tipos de actividades natildeo apresentam diferenccedilas
significativas tendo-se obtido 266 plusmn 07 e 28 plusmn 1M decavanadato na ausecircncia e na presenccedila de
actina respectivamente o que corresponde a uma concentraccedilatildeo de cerca de 27 M de V10 O
meio de reacccedilatildeo utilizado nestes ensaios foi 10 mM HEPES e 25 mM KCl com 2 mM ATP Em
(Tiago et al 2004) o valor de IC50 para a actividade Mg2+-ATPaacutesica na ausecircncia de actina foi de
125 plusmn 025 microM enquanto o valor da actividade estimulada por actina foi de 136 plusmn 14 microM O
meio de reacccedilatildeo utilizado nos ensaios foi 20 mM HEPES 50 mM KCl e 2 mM ATP o que resulta
precisamente no dobro da forccedila ioacutenica a que foi realizado o ensaio do presente relatoacuterio
(exceptuando a contribuiccedilatildeo dos produtos da hidroacutelise de ATP uma vez que este meacutetodo
pressupotildee a ocorrecircncia de um ciclo de reacccedilotildees de hidroacutelise e siacutentese deste composto) Ora tendo
sido anteriormente referido que a afinidade do S1 de miosina para com a espeacutecie V10 diminui com
Act
ivid
ade
Mg
2+-A
TP
aacutesic
a d
e S
1 d
e
Mio
sin
a
[Decavanadato] (microM) Actividade Estimulada ()
Actividade Natildeo Estimulada ()
Ajuste Actividade Estimulada
Ajuste Actividade Natildeo Estimulada
33
o aumento da forccedila ioacutenica (Tiago et al 2004 tal poderaacute explicar a diferenccedila obtida entre estes
dois conjuntos de valores
243Quantificaccedilatildeo de cisteiacutenas reduzidas pelo meacutetodo de Ellman
A interacccedilatildeo do vanaacutedio com alguns tipos de aminoaacutecidos sensiacuteveis agrave oxidaccedilatildeo pode fornecer-nos
informaccedilatildeo bastante uacutetil em relaccedilatildeo agraves alteraccedilotildees estruturais que conduzem a uma diminuiccedilatildeo da
actividade fisioloacutegica do S1 de miosina Os estudos realizados por Chasteen 1983 concluiacuteram
que a reduccedilatildeo de vanadato (V) a vanadilo (IV) ocorre na presenccedila de agentes redutores comuns no
ambiente celular de que se destaca o aminoaacutecido cisteiacutena Esta eacute tambeacutem um alvo de
complexaccedilatildeo por parte do iatildeo VO2+
Jaacute Ramos et al (2009) demonstrou que a incubaccedilatildeo de
actina com a soluccedilatildeo de decavanadato induz a oxidaccedilatildeo de resiacuteduos de cisteiacutena e agrave formaccedilatildeo do
oxidovanaacutedio (IV) Este grupo de investigadores descobriu ainda que apenas a espeacutecie V10 induz a
oxidaccedilatildeo das cisteiacutenas ldquoraacutepidasrdquo da actina atraveacutes do processo proposto que eacute parcialmente
responsaacutevel pela inibiccedilatildeo jaacute descrita da actividade de ATPase do S1 de miosina estimulada por
actina quando exposta agrave soluccedilatildeo de decavanadato Esta inibiccedilatildeo apresenta-se afectada com a
presenccedila de ATP uma vez que os resiacuteduos de cisteiacutena permaneceram intactos nos ensaios
realizados com este ligando Jaacute Crans et al (2004) concluiu que o siacutetio activo de muitas fosfatases
(que conteacutem grupos tiol) pode formar um equiliacutebrio redox com o iatildeo vanadato (V) de forma natildeo
reversiacutevel
Os resultados obtidos por Sakurai e Shiromura (1980) sugeriram que o vanaacutedio (V) sofre reduccedilatildeo
a vanaacutedio (IV) sem dependecircncia do valor de pH do ensaio Registaram ainda uma estequiometria
de 31 na reacccedilatildeo do vanadato (V) com os resiacuteduos de Cys enquanto a estequiometria dos ensaios
com o vanadilo (IV) foi de 21
Berlett e Stadtman (1997) observaram que os tipos de aminoaacutecidos mais facilmente oxidaacuteveis
englobam entre outros os resiacuteduos de Cys Trp e MetAssim analisou-se o estado redox dos
grupos tiol dos resiacuteduos de Cys do S1 na presenccedila do decavanadato atraveacutes da monitorizaccedilatildeo da
Abs412 nm na presenccedila de 1 mgmL de S1 de miosina em tampatildeo 10 mM HEPES 25 mM KCl
variando a concentraccedilatildeo da soluccedilatildeo de decavanadato de modo a determinar que tipo de alteraccedilotildees
estruturais ocorrem na sua presenccedila no que diz respeito agrave diminuiccedilatildeo da actividade ATPaacutesica de
S1 na presenccedila de decavanadato
Os resultados referentes agrave quantificaccedilatildeo das cisteiacutenas reduzidas apoacutes o tratamento de S1 com
diferentes concentraccedilotildees de decavanadato em tampatildeo 10 mM HEPES pH 75 e 25 mM KCl a
25ordmC durante 20 minutos encontram-se descritos na Tabela 2VII
Tabela 2VII ndash Cisteiacutenas reduzidas totais e ldquoraacutepidas de S1 (1 mgmL) apoacutes tratamento com decavanadato
34
(010501002505001000 uM) em 10 mM HEPES 25 mM KCl pH 7 a 25ordmC durante 20 minutos A detecccedilatildeo
espectrofotomeacutetrica foi feita com DTNB (e 412=8400 uM-1 cm-1) (n variaacutevel entre 1 e 8)
Amostras Cys raacutepidas Cys totais
11 M S1 (controlo) 22 plusmn 04 106 plusmn 14
11 M S1 + 10 M Decavanadato 22 plusmn 05 95 plusmn 08
11 M S1 + 50 uM Decavanadato 20 plusmn 02 98 plusmn 14
11 M S1 + 100 uM Decavanadato 17 plusmn 03 97 plusmn14
11 M S1 + 250 uM Decavanadato 26 plusmn 02 98 plusmn 08
11 M S1 + 500 uM Decavanadato 21 plusmn 04 90 plusmn 06
11 M S1 + 1000 uM Decavanadato 24 plusmn 0 86 plusmn 08
A partir dos resultados obtidos pode sugerir-se que o S1 possui 11 resiacuteduos de Cys (controlo) o
que estaacute de acordo com o anteriormente descrito Verifica-se que o tratamento da proteiacutena com
concentraccedilotildees crescentes de decavanadato provoca a oxidaccedilatildeo de dois resiacuteduos de Cys do S1 de
miosina natildeo expostos ao solvente No entanto a teacutecnica utilizada natildeo nos permite identificar
quais
Tendo em consideraccedilatildeo que o valor de IC50 obtido para a reacccedilatildeo de inibiccedilatildeo daactividade Mg2+-
ATPaacutesica de S1 na presenccedila de decavanadato eacute de cerca de 27 microM pode sugerir-se que este seraacute o
valor ao qual ocorreraacute a oxidaccedilatildeo do primeiro resiacuteduo de Cys o que poderaacute contribuir para a
diminuiccedilatildeo da actividade de S1 observada
25Interacccedilatildeo de S1 com vanaacutedio(IV)
251Anaacutelise por espectroscopia de Ressonacircncia Paramagneacutetica Electroacutenica
A acumulaccedilatildeo intracelular de oxovanaacutedio (IV) in vivo constitui uma das principais formas de
apresentaccedilatildeo de vanaacutedio no organismo Deste modo torna-se bastante interessante analisar a
interacccedilatildeo entre este iatildeo e o S1 de miosina atraveacutes da teacutecnica de espectroscopia de Ressonacircncia
Paramagneacutetica Electroacutenica o meacutetodo de eleiccedilatildeo do estudo do iatildeo vanadilo e dos complexos por si
formados O electratildeo desemparelhado encontra-se fortemente acoplado ao nuacutecleo 51
V o que
35
resulta num espectro de RPE consistindo de 8 desdobramentos na direcccedilatildeo paralela e outros tantos
na direcccedilatildeo perpendicular (Chasteen 1983)
Os estudos realizados por Ahmed et al (1987) centraram-se na interacccedilatildeo do vanadilo com
proteiacutenas como eacute o caso da insulina transferrina entre outros obtendo um espectro caracterizado
por 8 linhas amplamente espaccediladas (na direcccedilatildeo paralela) Jaacute Ramos et al 2009 concluiu que a
exposiccedilatildeo da soluccedilatildeo de decavanadato agraves formas globular e filamentosa de actina produz um
espectro tiacutepico do iatildeo oxidovanaacutedio (IV) indicando a reduccedilatildeo do vanadato
Para aferir qual a concentraccedilatildeo de S1 que permitia obter uma boa razatildeo sinalruiacutedo titulou-se uma
soluccedilatildeo de sulfato de vanadilo 15 microM ([VOSO4]stock = 5 mM) com concentraccedilotildees crescentes de
proteiacutena (0 5 10 15 e 20 microM) em 10 mM HEPES 25 mM KCl A existecircncia de uma correlaccedilatildeo
linear entre a concentraccedilatildeo de proteiacutena e a intensidade dos sinais de RPE correspondente agrave banda
-12 perpendicular encontra-se representada na Fig 210
Figura 2 12ndash Representaccedilatildeo graacutefica da variaccedilatildeo da intensidade do sinal de RPE vs a concentraccedilatildeo de S1 utilizada
Apoacutes verificar que com uma concentraccedilatildeo de S1 igual a 20 microM se obtinham sinais com uma razatildeo
sinalruiacutedo adequada ao prosseguimento dos estudos sobre a interacccedilatildeo V(IV)-S1 de miosina
titulou-se uma soluccedilatildeo de 20 microM S1 com 0 25 5 10 15 25 50 75 100 150 200 500 e 1000
microM VOSO4
Traccedilou-se o graacutefico da intensidade da banda -12 perpendicular do sinal de EPR em funccedilatildeo da
concentraccedilatildeo de VOSO4 tendo-se determinado uma constante de dissociaccedilatildeo (KD) do complexo
S1-vanadilo de 8748 plusmn 2621 microM V(IV)
y = 016786x + 122435
Rsup2 = 092992759
Inte
nsi
dad
e d
o s
inal
de
RP
E
[S1] (microM)
36
Figura 213 ndash Representaccedilatildeo graacutefica da variaccedilatildeo do sinal de RPE vs a concentraccedilatildeo de vanadilo utilizada (os valores
experimentais estatildeo representados por losangos o seu ajuste estaacute representado pela linha)
A simulaccedilatildeo do espectro de EPR foi efectuada a partir do espectro obtido para uma concentraccedilatildeo
de 1 mM VOSO4 apoacutes adiccedilatildeo de 20 microM S1 de miosina tendo-se obtido os valores da constante de
acoplamento hiperfina (A=5016) e do factor g (g0=195)
Figura 214 ndash Representaccedilatildeo graacutefica do espectro de EPR obtido com 1000 microM de vanadilo e 20 microM de S1 (a cinzento
escuro) A simulaccedilatildeo do espectro encontra-se a cinzento claro
Por comparaccedilatildeo destes com os valores presentes na tabela de correspondecircncia xxx eacute possiacutevel
predizer quais os ligandos provaacuteveis do vanadilo na presenccedila de proteiacutena Assim concluiu-se que
o vanadilo estaacute coordenado com duas moleacuteculas de aacutegua um grupo hidroxilo e um grupo R=NR
O grupo hidroxilo pode indicar os resiacuteduos de aminoaacutecidos Tyr Treonina e Serina enquanto o
grupo R=NR poderaacute indicar a presenccedila de Arginina e Histidina como ligandos do vanadilo
Inte
nsi
dad
e d
os
sinai
s d
e R
PE
[Vanadilo] (microM)
Ajuste dos dados exp Dados experimentais
Campo magneacutetico (G)
Espectro Experimental Ajuste
37
3 Procedimento Experimental
31 Reagentes
Todos os reagentes utilizados satildeo de elevado grau de pureza (analiacutetico) Apresentam-se as suas
foacutermulas quiacutemicas ou abreviaturas quando necessaacuterio acrescentou-se a sua descriccedilatildeo Entre
parecircntesis colocou-se as empresas que os comercializam
Aacutecido aceacutetico glacial 100 (Riedel) Acrilamida 99 (Merck) Azul de bromofenol (Riedel-de-
Haumlen) -Mercaptoetanol (Sigma) Bissulfito de soacutedio (Sigma)Bradford (reagente de)
(BioRad)BSA (albumina de soro bovino Sigma-Aldrich) CaCl22H2O (cloreto de caacutelcio
dihidratado Fluka) CH3CH2OH 96 (etanol Panreac) Coomassie Blue R250 (Merck) D2O
(oacutexido de deuteacuterio SDS) DTNB (55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico) Aldrich) EDTA (aacutecido
etilenodiaminatetraaceacutetico Merck) Fosfoenolpiruvato 98 (Fluka) Glicerol (Sigma) Glicina
985 (Panreac) HCl 37 (Riedel-de Haumlen) H2SO4 96 (Pronalab) HEPES 99 (aacutecido 4-(2-
hidroxiletil)-1-piperazineetanosulfoacutenico Sigma) KH2PO4 99 (hidrogenofosfato de potaacutessio)
MgATP (Sigma) Na
ATP (Sigma) Na2HPO4 99 (hidrogenofosfato de soacutedio Riedel-de Haumlen)
NaH2PO4 (fosfato de soacutedio ) NaOH 986 (hidroacutexido de soacutedio Pronalab) KCl 99 (Cloreto de
potaacutessio Panreac) L-Cys 995 (enantioacutemero levoacutegiro do resiacuteduo de Cisteiacutena Fluka)Metanol
99 (Merck) Molibdato de amoacutenio (Sigma)NNrsquo-metil-bis-acrilamida 98 (Merck) NH4VO3
99 (metavanadato de amoacutenia Sigma) PSA 10 (persulfato de amoacutenio Merck) SDS
(dodecilo sulfato de soacutedio Sigma) Sulfito de soacutedio (Sigma) VOSO4 (sulfato de vanadilo
Aldrich) TEMED 99 (Tetrametiletilenodiamina Merck) Tris (2-amino-2-hidroximetil-13-
propanodiol Sigma)
32 Meacutetodos
321 Isolamento e purificaccedilatildeo das proteiacutenas de muacutesculo esqueleacutetico de coelho
3211 Purificaccedilatildeo da miosina preparaccedilatildeo de Subfragmento 1 por digestatildeo quimotriacutepsica
da miosina
O subfragmento-1 de miosina-II (S1) foi gentilmente cedido pelo Prof Doutor Manuel Aureliano
do Departamento de Quiacutemica Bioquiacutemica e Farmaacutecia da Faculdade de Ciecircncias e Tecnologia da
Universidade do Algarve
No entanto descreve-se abaixo de forma resumida o procedimento efectuado
38
32111 Purificaccedilatildeo da miosina
O protocolo de isolamento foi adaptado do meacutetodo previamente descrito por Margossian e Lowey
(1982) apresentando-se de forma esquemaacutetica na Fig 31
Sucintamente os tecidos musculares dorsais e das pernas foram extraiacutedos apoacutes o sacrifiacutecio do
animal (Fig31) De seguida foram adicionados 3 volumes da soluccedilatildeo de extracccedilatildeo A2
Centrifugou-se a soluccedilatildeo e filtrou-se o sobrenadante O sedimento foi entatildeo diluiacutedo em 14
volumes de aacutegua destilada de modo a que a forccedila ioacutenica atingisse 004 M Apoacutes adiccedilatildeo de szlig-
mercaptoetanol (szlig-me) deixou-se a precipitar a 4ordmC removendo o sobrenadante Centrifugou-se
novamente e ressuspendeu-se o sedimento na soluccedilatildeo de extracccedilatildeo A Adicionou-se a soluccedilatildeo
tampatildeo B3 para que a forccedila ioacutenica atingisse 05 M Homogeneizou-se e deixou-se repousar
durante 2h Homogeneizou-se novamente e centrifugou-se de modo a ocorrer precipitaccedilatildeo dos
complexos de actomiosina e proteiacutena desnaturada Apoacutes filtraccedilatildeo do sobrenadante em latilde de vidro
quantificou-se a soluccedilatildeo de miosina resultante pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental)
2 Soluccedilatildeo de extracccedilatildeo A 0012M K2CO3 004M KHCO3 06M KCl pH 65
3 Soluccedilatildeo-tampatildeo B Tris 100 mM pH 7 KCl 2M
39
Figura 31 ndash Representaccedilatildeo esquemaacutetica do protocolo de isolamento de miosina (Margossian e Lowey 1982) Todo o
procedimento foi realizado a 4ordmC (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
32112 Preparaccedilatildeo de Subfragmento-1 de miosina por digestatildeo com quimiotripsina
O S1 de miosina foi isolado apoacutes diaacutelise da miosina durante a noite contra a soluccedilatildeo C4 A
presenccedila do EDTA eacute essencial uma vez que a sua substituiccedilatildeo por MgCl2resulta na produccedilatildeo de
HMM em vez de S1 A suspensatildeo de miosina foi equilibrada e digerida com -quimotripsina
A digestatildeo foi interrompida pela adiccedilatildeo de PMSF Seguidamente ultracentrifugou-se o produto
da digestatildeo desprezando-se o sedimento (Fig 32) A soluccedilatildeo de sobrenadante maioritariamente
S1 foi quantificada pela leitura directa da Abs280 ou pelo meacutetodo de Bradford (ver as secccedilotildees
3311 e 3312 do Procedimento Experimental respectivamente) e armazenada em 10 sacarose
e 2 mM szlig-Me a -80ordmC
4 Soluccedilatildeo C 25mM Tris pH 70 012M KCl 1mM EDTA
40
Antes da realizaccedilatildeo dos ensaios descritos no presente relatoacuterio o S1 de miosina foi dialisado
contra a soluccedilatildeo D5 de modo a remover qualquer composto que pudesse complexar como
vanaacutedio Seguidamente quantificou-se a proteiacutena pelo meacutetodo de Bradford ou pelo meacutetodo
directo (ver secccedilotildees 3311 e 3312do Procedimento Experimtental)
Figura 3 2ndash Representaccedilatildeo esquemaacutetica do protocolo de preparaccedilatildeo do S1 de miosina por digestatildeo quimiotripsina de
acordo com o previamente descrito por Margossian e Lowey (1982) Todo o procedimento foi realizado a 4ordm C
excepto quando indicaccedilatildeo do contraacuterio (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
3212 Isolamento e purificaccedilatildeo de actina
Isolou-se e purificou-se a actina segundo o meacutetodo descrito por Pardee e Spudish que
compreende dois passos ndash a obtenccedilatildeo de um resiacuteduo seco denominado ldquopoacute de acetonardquo e a
extracccedilatildeo subsequente da actina a partir do ldquopoacute de acetonardquo na forma de G-Actina com a
posterior polimerizaccedilatildeo a F-Actina
32121 Obtenccedilatildeo do ldquopoacute de acetonardquo
Sacrificou-se o coelho com uma pancada atraacutes do pescoccedilo e sangrou-se o animal apoacutes o corte da
arteacuteria jugular Esfolou-se e colocou-se numa tina com gelo Retiraram-se as partes de tecido
conjuntivo que envolvem o tecido muscular com auxiacutelio de uma pinccedila e bisturi Seguidamente
retirou-se os muacutesculos dorsais e os muacutesculos brancos das pernas Passou-se os muacutesculos por aacutegua
destilada de modo a remover as impurezas e pesou-se num copo Trituraram-se os muacutesculos
numa maacutequina de triturar carne e extraiu-se o resultante com 3 volumes da soluccedilatildeo E6 durante 10
minutos com agitaccedilatildeo suave Centrifugou-se a suspensatildeo a 4000 g durante 10
5 Soluccedilatildeo D HEPES 10 mM pH 70 KCl 25 mM
6 Soluccedilatildeo E 01 M KCl 015 M K2HPO4 pH 65
41
minutosdesprezando-se o sobrenadante Ressuspendeu-se o sedimento em 6 volumes de soluccedilatildeo
F7durante 10 minutos com agitaccedilatildeo suave Verificou-se se o pH da soluccedilatildeo estava compreendido
entre 82 e 85 acertando-se estecom soluccedilatildeo G8 se necessaacuterio Centrifugou-se durante 10
minutos a 4000 g Desprezou-se o sobrenadante e ressuspendeu-se o sedimento em 3 volumes de
soluccedilatildeo H9 durante 10 minutos Centrifugou-se novamente a 4000 gdurante 10 minutos
desprezando-se o sobrenadante Ressuspendeu-se o sedimento duas vezes consecutivas em 6
volumes de aacutegua destilada durante 5 minutos Centrifugou-se a 4000 g durante 10 minutos apoacutes
cada adiccedilatildeo de H2O
Seguidamente ressuspendeu-se o sedimento 5 vezes consecutivas utilizando 3 volumes de
acetona de cada vez agitando suavemente durante 10 minutos agrave temperatura ambiente Colocou-
se o resiacuteduo filtrado numa tina de vidro sobre papel de filtro deixando-se evaporar o solvente
durante a noite num nicho Triturou-se o resiacuteduo seco numa maacutequina de triturar carne e deixou-se
novamente evaporar o solvente O ldquopoacute de acetonardquo obtido foi armazenado a -20 ordmC
32122 Purificaccedilatildeo da actina a partir de ldquopoacute de acetonardquo
Apoacutes a obtenccedilatildeo do poacute de acetona como acima indicado efectuou-se o protocolo seguidamente
descrito de modo a obter monoacutemeros de actina (G-Actina) e actina filamentosa (F-Actina)
Pesou-se 35 g de poacute de acetona e adicionaram-se 50 mL de soluccedilatildeo-tampatildeo I10
sendo a
extracccedilatildeo feita com agitaccedilatildeo suave durante 30 minutos Separou-se o extracto do poacute de acetona
hidratado filtrando a suspensatildeo com 4 camadas de gaze esterilizada tendo o cuidado de natildeo
espremer o resiacuteduo Voltou a extrair-se com 50 mL de soluccedilatildeo I durante 30 minutos mexendo
lentamente Adicionou-se este ao extracto obtido anteriormente Centrifugou-se o extracto total a
16 000 g durante 30 minutos Decantou-se para uma proveta sobre a qual se colocou gaze para
que o sedimento solto natildeo se juntasse ao sobrenadante e leu-se o volume De seguida adicionou-
se KCl ateacute uma concentraccedilatildeo final de 50 mM MgCl2 ateacute 2mM e ATP ateacute 1mM lentamente e com
agitaccedilatildeo contiacutenua de modo a promover a polimerizaccedilatildeo dos monoacutemeros de actina
Deixou-se polimerizar durante 2h a 25ordmC observando-se a presenccedila de pequenas bolhas de ar
indicativas de um aumento de viscosidade da soluccedilatildeo Arrefeceu-se em gelo durante 15 minutos e
adicionou-se KCl ateacute uma concentraccedilatildeo final de 06M Mexeu-se lentamente durante 30 minutos
apoacutes os quais se ultracentrifugou a 110 000g durante 1hora Desprezou-se o sobrenadante
ressuspendeu-se o sedimento com soluccedilatildeo tampatildeo E e transferiu-se para um potter
Dialisou-se durante a noite contra 1L de soluccedilatildeo-tampatildeo I a uma temperatura de 4ordmC com rotaccedilatildeo
7 Soluccedilatildeo F 005 M NaHCO3 pH 82-85
8 Soluccedilatildeo G 1M Na2CO3
9 Soluccedilatildeo H 1 mM EDTA pH 70
10 Soluccedilatildeo I 2 mM Tris-HCl pH 8 02 mM CaCl2 02 mM Na2ATP 05 mM szlig-Me 0005 NaN3
42
contiacutenua e mudanccedila de tampatildeo 2 a 3 vezes De seguida a actina foi centrifugada a 110
000gdurante 1hora Decantou-se a soluccedilatildeo de G-Actina (sobrenadante) e desprezou-se o
sedimento Determinou-se a concentraccedilatildeo de G-Actina pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental) A soluccedilatildeo de G-actina foi armazenada a uma temperatura de 4ordmC
apoacutes adiccedilatildeo de 002NaN3 mantendo-se estaacutevel durante 3 a 5 dias
331 Quantificaccedilatildeo das proteiacutenas isoladas
3311 Quantificaccedilatildeo pelo meacutetodo de Bradford
O meacutetodo de Bradford (Bradford 1976) baseia-se na utilizaccedilatildeo do reagente de Bradford
composto por entre outros aacutecido fosfoacuterico e o corante ldquoCoomassierdquo Este corante possui carga
negativa e a sua coloraccedilatildeo varia entre a vermelha (Absmax=465 nm) predominante em soluccedilatildeo e a
azul (Absmax=595 nm) presente aquando da ligaccedilatildeo aos grupos amina das proteiacutenas Em soluccedilotildees
aacutecidas diluiacutedas a coloraccedilatildeo varia proporcionalmente com a quantidade do corante ligado agrave
proteiacutena que por sua vez eacute directamente proporcional agrave concentraccedilatildeo da proteiacutena permitindo a
sua quantificaccedilatildeo ateacute a um limite de detecccedilatildeo de 20 gmL (Bradford 1976)
Deste modo prepararam-se amostras de BSA ([BSA]stock= 1mgmL) utilizando um intervalo de
concentraccedilotildees compreendido entre 0 e 15 gmL de acordo com a tabela 3I Apoacutes a adiccedilatildeo da
H2O MilliQ e do reagente de Bradford as soluccedilotildees foram sucessivamente homogeneizadas no
voacutertex
Tabela 3Indash Amostras utilizadas para a obtenccedilatildeo da recta de calibraccedilatildeo no meacutetodo de Bradford
BSA (L) BSA (g) H2O MilliQ (L) Reagente de Bradford (L)
0 0 800 200
125 125 79875 200
25 25 7975 200
5 5 795 200
10 10 790 200
15 15 785 200
Apoacutes incubaccedilatildeo a 25ordmC por 5 minutos leram-se os valores de Abs595 num
espectrofotoacutemetro (Shimadzu UV-160A) para todas as amostras Seguidamente traccedilou-se a recta
de calibraccedilatildeo (Abs595vs quantidade de BSA (g))
43
Para determinar a concentraccedilatildeo de proteiacutena apoacutes o processo de isolamento purificaccedilatildeo e
diaacutelise quando necessaacuterio colocou-se 5 L da amostra num tubo de Eppendorf e adicionou-se
795 L de H2O MilliQ homogeneizando a soluccedilatildeo no voacutertex Adicionou-se posteriormente 200
L de reagente de Bradford e repetiu-se a homogeneizaccedilatildeo Incubou-se a mistura a 25ordmC por 5
minutos apoacutes os quais se leu os valores de Abs595 nm no espectrofotoacutemetro Por interpolaccedilatildeo deste
valor a partir da recta de calibraccedilatildeo determinou-se a concentraccedilatildeo de proteiacutena na amostra
3312 Quantificaccedilatildeo pelo meacutetodo directo
A quantificaccedilatildeo das amostras proteicas pode tambeacutem ser efectuada atraveacutes da leitura directa da
absorvacircncia tendo por base a lei de Beer-Lambert onde Abs corresponde agrave
absorvacircncia ao comprimento de onda ao qual o ensaio foi realizado corresponde ao coeficiente
de extinccedilatildeo (substituiacutedo pela letra a se as unidades forem distintas da molaridade) b corresponde
ao percurso oacuteptico da radiaccedilatildeo na ceacutelula amostral e c corresponde agrave concentraccedilatildeo amostral
Assim no caso de S1 de miosina mediu-se a Abs280e calculou-se a concentraccedilatildeo da proteiacutena
sabendo que o coeficiente de extinccedilatildeo de S1 eacute (Ref)
No caso da actina mediu-se a Abs290ecalculou-se a concentraccedilatildeo da proteiacutena sabendo que o
coeficiente de extinccedilatildeo da actina eacute (Ref)
332Electroforese em condiccedilotildees desnaturantes (SDS-PAGE)
De modo a melhor caracterizar as proteiacutenas isoladas foi realizada uma electroforese em
condiccedilotildees desnaturantes (SDS-PAGE do inglecircs Sodium Dodecyl Sulfate Polyacrilamide Gel
Electrophoresis) O SDS (dodecilo sulfato de soacutedio) eacute um detergente anioacutenico utilizado neste
meacutetodo para desnaturar as proteiacutenas que se ligam ao SDS na proporccedilatildeo 14 g SDS g proteiacutena
(aproximadamente 1 moleacutecula de SDS por cada 2 resiacuteduos de aminoaacutecidos) (Stryer) O SDS induz
uma densidade de carga negativa significativa nas proteiacutenas a que se liga mascarando a sua carga
intriacutenseca pelo que as razotildees cargamassa satildeo tendencialmente idecircnticas Assim a SDS-PAGE
separa as proteiacutenas quando sujeitas a um campo eleacutectrico de acordo com o seu peso molecular
sendo a sua migraccedilatildeo directamente proporcional ao logaritmo deste uacuteltimo (Stryer)
O gel de resoluccedilatildeo com 10 de acrilamida foi obtido atraveacutes da adiccedilatildeo de 167 mL de soluccedilatildeo
stock de acrilamida11
075 mL de tampatildeo de resoluccedilatildeo12
251 mL de aacutegua destilada e 50 L de
SDS 10 Para promover a polimerizaccedilatildeo adicionou-se em seguida 38 L de PSA 10
11
soluccedilatildeo stock de acrilamida soluccedilatildeo de 30 (mv) de acrilamida e 08 (mv) de NNrsquo-metil-bis-acrilamida 12
tampatildeo de resoluccedilatildeo 15 M Tris-HCl pH 88
44
(persulfato de amoacutenia) 10 e 25 L de TEMED 99 (tetrametiletilenodiamina) Agitou-se
cuidadosamente de modo a natildeo formar bolhas e pipetou-se a soluccedilatildeo para a cassette de vidro
previamente montada no suporte De seguida adicionou-se uma peliacutecula de cerca de 2 mm de 2-
metil-2-butanol para verificar posteriormente a polimerizaccedilatildeo Apoacutes a polimerizaccedilatildeo que demora
cerca de 45 minutos verteu-se a agrave superfiacutecie e adicionou-se o gel de concentraccedilatildeo com 5 de
acrilamida constituiacutedo por 03 mL de soluccedilatildeo de stock acrilamida 045 mL de tampatildeo de
concentraccedilatildeo13
094 mL de aacutegua destilada 18 L de SDS 10 135 L de PSA 10 e 2L de
TEMED 99 Colocou-se o pente sobre o gel e deixou-se polimerizar durante cerca de 45
minutos
As amostras para aplicaccedilatildeo no gel de electroforese foram preparadas para uma concentraccedilatildeo de
proteiacutena de 1 mgmL num volume final de 100 L dos quais 25 L correspondem ao tampatildeo da
amostra14
As amostras foram aquecidas a 100ordmC durante 5 minutos Antes da aplicaccedilatildeo das
amostras transferiu-se a cassette para a tina de electroforese agrave qual se adicionou a soluccedilatildeo
tampatildeo de electroforese da tina superior15
e tampatildeo de electroforese da tina inferior16
Aplicaram-
se 10 L das soluccedilotildees de marcadores de peso molecular e 20 L das amostras proteicas
preparadas anteriormente A electroforese decorreu durante cerca de 3h30 min a uma voltagem
de 80 V De seguida transferiu-se o gel para uma tina corando as proteiacutenas durante 1h com
soluccedilatildeo corante17
Descorou-se o gel com soluccedilatildeo descorante18
tendo-se mudado esta soluccedilatildeo ateacute
as bandas se encontrarem bem definidas
Determinou-se a mobilidade relativa das bandas correspondentes aos marcadores de elevado peso
molecular e traccedilou-se a recta de calibraccedilatildeo ndash log(PM) vs log(Rf) onde Rf corresponde agrave
mobilidade relativa Por interpolaccedilatildeo determina-se o PM das proteiacutenas isoladas sabendo a sua
mobilidade relativa Por comparaccedilatildeo com os valores descritos na literatura pode concluir-se
acerca do grau de purificaccedilatildeo
34 Mediccedilatildeo da actividade enzimaacutetica de S1
341 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
13
tampatildeo de concentraccedilatildeo 05 M Tris-HCl pH 68 14
tampatildeo de amostra 320 mM Tris-HCl pH 68 04 M szlig-mercaptoetanol SDS 8 glicerol 15 (vv) e roxo de m-
cresol 0024 15
tampatildeo de electroforese da tina superior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS 16
tampatildeo de electroforese da tina inferior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS acetato de soacutedio
01 M 17
soluccedilatildeo corante ldquoCoomassie Blue R-250rdquo contendo metanol 45 (vv) e aacutecido aceacutetico glacial 10 (vv) 18
soluccedilatildeo descorante metanol 10 (vv) e aacutecido aceacutetico glacial 10 vv)
45
O meacutetodo de Fiske-Subarow modificado consiste na reacccedilatildeo do molibdato de amoacutenio com o
fosfato inorgacircnico em soluccedilatildeo aacutecida com formaccedilatildeo do aacutecido fosfomoliacutebdico Seguidamente daacute-
se a adiccedilatildeo de um agente redutor que provoca a reduccedilatildeo do composto a fosfomolibdato que forma
um complexo de cor azulada cuja absorvacircncia maacutexima ocorre para um comprimento de onda de
660 nm tornando possiacutevel a sua detecccedilatildeo por meacutetodos espectrofotomeacutetricos As actividades Ca2+
-
ATPaacutesica e K+EDTA-ATPaacutesica de S1 foram determinadas atraveacutes deste meacutetodo de acordo com
o descrito por Taussky e Shorr (1953) com mediccedilatildeo da quantidade de fosfato (Pi) libertado
atraveacutes da hidroacutelise de ATP
Para tal obteve-se primeiramente a recta de calibraccedilatildeo com as amostras presentes na Tabela 3II
utilizando uma soluccedilatildeo stock de 5 mM KH2PO4
Tabela 3II ndash Composiccedilatildeo das diferentes amostras utilizadas para determinar a recta de calibraccedilatildeo atraveacutes do meacutetodo
de Fiske-Subbarow modificado
KH2PO4 (L)
([KH2PO4]stock= 5mM)
PO43-
(nmole) H2O (uL)
0 0 1780
5 25 1775
10 50 1770
20 100 1760
40 200 1740
100 500 1680
A cada tubo de Eppendorf adicionaram-se seguidamente 200 L de soluccedilatildeo J19
e 20 L de agente
redutor20
homogeneizando no voacutertex apoacutes cada adiccedilatildeo Incubou-se durante 10 minutos e mediu-
se a Abs660 nm num espectrofotoacutemetro com intervalos de tempo iguais entre cada amostra Traccedilou-
se a recta de calibraccedilatildeo (Abs660 nmvs nmol PO43-
) a partir dos valores obtidos
Para determinar a actividade Ca2+
-ATPaacutesica de S1 prepararam-se tubos de Eppendorf com 400
L de meio de reacccedilatildeo21
25 L CaCl2 ([CaCl2]stock= 100 mM) 25 L ATP ([ATP]stock= 100 mM)
com [S1]ensaios=01 mgmL Perfez-se se necessaacuterio com H2O MilliQ ateacute perfazer um volume de 1
mL Para determinar a actividade K+EDTA-ATPaacutesica utilizou-se uma concentraccedilatildeo idecircntica de
19
Soluccedilatildeo J 5 N H2SO4 25 molibdato de amoacutenio 20
Agente redutor 5 mg aacutecido 1-amino-2-naftol-4-sulfoacutenico 30 mg bissulfito de soacutedio 30 mg sulfito de soacutedio em 26
mL H2O 21
meio de reacccedilatildeo 125 mM Tris-HCl pH 80 0575 M KCl
46
proteiacutena nos ensaios (01 mgmL) e o mesmo meio de reacccedilatildeo Aos tubos de Eppendorf
adicionou-se ainda 123 L de KCl ([KCl]stock = 3M) 25 L de ATP 10 L de EDTA
([EDTA]stock= 100 mM )e o volume de H2O MilliQ necessaacuterio para perfazer 1 mL Em ambas as
situaccedilotildees o uacuteltimo reagente adicionado foi o ATP uma vez que se trata do substrato da reacccedilatildeo
enzimaacutetica
A cineacutetica decorreu a 25ordmC durante 5 minutos para a actividade Ca2+
-ATPaacutesica e durante 3
minutos para a actividade K+EDTA-ATPaacutesica Apoacutes o periacuteodo de incubaccedilatildeo transferiu-se 150
L de amostra para tubos de Eppendorf contendo igual volume de TCA 10 Colocou-se no gelo
e homogeneizou-se no voacutertex Seguidamente centrifugou-se a 1500 g durante 10 minutos e
recolheu-se 200 L de sobrenadante para novos tubos de Eppendorf Adicionou-se a estes 158
mL de H2O MilliQ e 200 L de soluccedilatildeo J Homogeneizou-se no voacutertex e adicionou-se 20 L de
reagente redutor repetindo-se a homogeneizaccedilatildeo Incubou-se durante 10 minutos e mediu-se a
Abs660 nm com o mesmo intervalo de tempo de leitura para cada amostra Por intrapolaccedilatildeo a partir
da recta de calibraccedilatildeo obtida anteriormente determinou-se a quantidade de Pi libertado em cada
ensaio A partir destes resultados determinaram-se as actividades Ca2+
-ATPaacutesica e K+EDTA-
ATPaacutesica do S1 em nmol PO43-
minmg proteiacutena e em s-1
342 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
Da actividade Mg2+
-ATPaacutesica de S1 resulta a formaccedilatildeo dos produtos da reacccedilatildeo de
hidroacutelise do ATP - ADP e Pi Na presenccedila de ADP fosfoenolpiruvato (PEP) e da enzima piruvato
cinase (PK) os produtos resultantes satildeo ATP e piruvato Por acccedilatildeo da enzima lactato
desidrogenase (LDH) este uacuteltimo eacute convertido em lactato ocorrendo simultaneamente a oxidaccedilatildeo
do NADH a NAD+ monitorizada atraveacutes da Abs340 nm Quando o S1 de miosina se encontra no
meio reaccional o ATP eacute hidrolisado formando-se novamente ADP e Pi o que permite a
formaccedilatildeo de um ciclo (fig 21) Se as enzimas se encontrarem em excesso a velocidade de
conversatildeo do NADH eacute limitada pela produccedilatildeo de ADP e Pi Como o ATP eacute regenerado em cada
ciclo a sua concentraccedilatildeo inicial eacute sempre mantida (Bagshaw 1982)
Figura 33ndash Representaccedilatildeo esquemaacutetica do meacutetodo de enzimas acopladas
47
O protocolo utilizado e seguidamente descrito foi adaptado de Cuenda et al (1990 ndash Ver Refordf) A
uma cuvette de 1 mL adicionou-se S1 para uma concentraccedilatildeo final de 005 mgmL 800 L de
meio de reacccedilatildeo22
10 L de PEP ([PEP]stock = 42 mM) 10 L de PK+LDH 11 ([PK+LDH]stock =
1 mgmL) e 15 L de NADH ([NADH]stock=25 mM) Iniciou-se a monitorizaccedilatildeo da reacccedilatildeo
atraveacutes da leitura de Abs340 nm apoacutes a adiccedilatildeo de 20 L de ATP ([ATP]stock=100 mM) A
actividade Mg2+
-ATPaacutesica eacute obtida atraveacutes do declive correspondente agrave oxidaccedilatildeo do NADH Para
a mediccedilatildeo da Mg2+
-ATPase de S1 estimulada por actina adicionou-se F-actina para uma
concentraccedilatildeo final de 2 microM sendo a actividade obtida tambeacutem atraveacutes do declive correspondente
agrave oxidaccedilatildeo do NADH apoacutes a adiccedilatildeo da proteiacutena
35 Interaccedilatildeo entre vanaacutedio e subfragmento-1 de miosina
351 Preparaccedilatildeo das soluccedilotildees de vanaacutedio
As soluccedilotildees de vanadato foram preparadas em aacutegua MilliQ a partir de metavanadato de amoacutenio
(NH4VO3) numa concentraccedilatildeo stock de 50 mM com agitaccedilatildeo magneacutetica e aquecimento (~30 ordmC)
Esta soluccedilatildeo com um valor compreendido entre 6 e 7 designa-se por ldquometavanadatordquo e eacute
constituiacuteda pelas esspeacutecies V1 V2 V4 e V5 (Ref) Acidificando a soluccedilatildeo de metavanadato por
adiccedilatildeo de HCl ateacute um valor de pH proacuteximo de 4 observa-se uma mudanccedila de cor de amarelo
claro para laranja indicativa da formaccedilatildeo da espeacutecie decameacuterica de vanadato De seguida e
imediatamente antes da realizaccedilatildeo dos ensaios descritos o valor do pH da soluccedilatildeo de
decavanadato foi aumentado para cerca de 75 por adiccedilatildeo de NaOH A soluccedilatildeo (2) designada
por ldquometavanadatordquo natildeo foi posteriormente alterada ateacute agrave sua adiccedilatildeo para a realizaccedilatildeo dos ensaios
As soluccedilotildees foram armazenadas a 4ordmC e mantidas em gelo aquando da sua utilizaccedilatildeo
A soluccedilatildeo de V(IV) foi preparada em condiccedilotildees anaeroacutebias por diluiccedilatildeo de sulfato de vanadil
(VOSO4) para uma concentraccedilatildeo final de 5 mM em H2O MilliQ saturada com N2 gasoso
352Anaacutelise por espectroscopia de UV-Vis
A concentraccedilatildeo da espeacutecie decameacuterica em soluccedilatildeo depende do periacuteodo de tempo apoacutes a diluiccedilatildeo
no meio de reacccedilatildeo apropriado (Ref) A velocidade de desoligomerizaccedilatildeo da espeacutecie decameacuterica
pode ser analisada por espectroscopia de absorccedilatildeo na regiatildeo UVVis uma vez que esta espeacutecie
apresenta duas bandas caracteriacutesticas na regiatildeo do visiacutevel (a 358 e 400 nm) que natildeo se verifica
em quaisquer outras espeacutecies oligomeacutericas As bandas supramencionada sapresentam-se bem
22
Meio de reacccedilatildeo 3125 mM Tris-HCl pH 71 375 mM MgCl2
48
resolvidas atraveacutes da obtenccedilatildeo da derivada de 2ordf ordem do espectro apresentando a forma de 2
vales com miacutenimos centrados nos comprimentos de onda supracitados com dependecircncia linear
relativamente agrave concentraccedilatildeo de decavanadato total (Ramos et al 2006)
Assim a estabilidade da espeacutecie decameacuterica apoacutes diluiccedilatildeo da soluccedilatildeo de decavanadato no meio de
reacccedilatildeo dos ensaios (tampatildeo D) para uma concentraccedilatildeo final de 1 mM em vanadato total foi
caracterizada atraveacutes da leitura da Abs400 em funccedilatildeo do tempo tendo-se determinado qual a
ordem de reacccedilatildeo e o tempo de meia-vida da mesma Com base em estudos anteriores acerca da
estabilidade do V10 na presenccedila de diferentes proteiacutenas (Ramos et al 2006) determinou-se
igualmente o tempo de meia vida apoacutes a adiccedilatildeo de 20 microM de S1 de miosina ao meio reaccional
353 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
Os espectros de RMN do isoacutetopo 51
V foram realizados num espectroacutemetro Bruker AM-400 MHz
equipado com uma sonda multinuclear inversa de 5 mm Em tubos de RMN de 5 mm de diacircmetro
preparou-se soluccedilotildees de 500 L de deca- ou metavanadato correspondentes a um intervalo de
concentraccedilotildees de 1 a 10 mM suplementadas com 10 D2O Os espectros foram adquiridos agrave
temperatura ambiente com uma largura espectral de 23 923 Hz um tempo de aquisiccedilatildeo de 0086
s tempo de relaxaccedilatildeo de 001 s e uma sequecircncia de pulsos a 90ordm Utilizou-se a transformada de
Fourier com alargamento das bandas de 20Hz para o processamento dos espectros Os desvios
quiacutemicos das ressonacircncias de 51
V foram obtidos em relaccedilatildeo agrave espeacutecie VOCl3cuja frequecircncia de
ressonacircncia eacute considerada o padratildeo sendo todas as outras referenciadas a partir desta em ppm
As aacutereas dos sinais do RMN correspondentes agraves espeacutecies oligomeacutericas presentes foram obtidas
por integraccedilatildeo dos sinais As concentraccedilotildees relativas das espeacutecies foram calculadas com recurso agrave
equaccedilatildeo
onde A[Vx] corresponde agrave aacuterea referente agrave ressonacircncia da espeacutecie Vx com nordm de agregaccedilatildeo x
A[V]total agrave soma total das aacutereas medidas e [V] a concentraccedilatildeo total de vanaacutedio no ensaio
A teacutecnica de 51
V-RMN foi anteriormente aplicada em estudos de ligaccedilatildeo com S1 de miosina
(Ref) tendo sido demostrado que entre as espeacutecies V1 V2 V4 e V5 a espeacutecie tetrameacuterica possui a
maior afinidade para a miosina (Cremo et al) O desaparecimento ou alargamento dos sinais pode
implicar a ligaccedilatildeo de um dos oxovanadatos laacutebeis agrave miosina sem indicaccedilatildeo de qual uma vez que
49
se processa um equiliacutebrio entre eles (Crans 1990) Assim e tendo em conta as indicaccedilotildees de que
a espeacutecie V10 eacute distinta das restantes no que diz respeito agrave magnitude da interacccedilatildeo com a miosina
e inibiccedilatildeo da actividade ATPaacutesica estimulada por actina (Aureliano 2000 Tiago et al 2002)
torna-se fundamental investigar a interacccedilatildeo das diferentes espeacutecies de vanadato com o S1 de
miosina atraveacutes da teacutecnica de 51
V-RMN
Para tal obtiveram-se espectros de 51
V-RMN resultantes da titulaccedilatildeo de uma soluccedilatildeo 5 mM de
decavanadato total com concentraccedilotildees crescentes de S1 de miosina (0 5 10 15 e 20 microM) em
tampatildeo D Realizou-se ainda ensaios na presenccedila de 2 mM de decavanadato e 2 mM de ATP na
presenccedila e na ausecircncia de S1 de miosina de modo a verificar o efeito que estes ligandos naturais
da enzima teria na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o subfragmento-1
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
Os grupos sulfidrilo (-SH) das cisteiacutenas satildeo resiacuteduos de aminoaacutecidos extremamente sensiacuteveis agrave
oxidaccedilatildeo resultando na sua oxidaccedilatildeo (Dalle-Donne et al 1999) Logo a quantificaccedilatildeo destes
resiacuteduos eacute bastante importante nos estudos de interacccedilatildeo com vanaacutedio jaacute que os resiacuteduos de Cys
com ecircnfase para o SH1 e o SH2 satildeo bastante importantes na monitorizaccedilatildeo das modificaccedilotildees
funcionais da regiatildeo da cabeccedila
O procedimento seguidamente apresentado foi originalmente descrito por Gutieacuterrez-Merino
(1987)) Consiste na reacccedilatildeo entre os grupos ndashSH natildeo oxidados das proteiacutenas e o reagente de
Ellman (DTNB -55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico)) - formando-se o aniatildeo CNT (aniatildeo 3-
carboxilato-4-nitrotiofenolato) numa estequiometria de 11 Este apresenta um maacuteximo de
absorvecircncia a 412 nm (Robyt 1990) Assim apoacutes a incubaccedilatildeo da proteiacutena na presenccedila do metal
a quantidade de cisteiacutenas reduzidas por proteiacutena determina-se atraveacutes dos valores de Abs412
Deste modo colocou-se num tubo de Eppendorf o volume apropriado de S1 dialisado para que a
concentraccedilatildeo no ensaio fosse de 1 mgmL Adicionou-se o volume apropriado de decavanadato
para que a sua concentraccedilatildeo fosse de 0 10 50 100 250 500 ou 1000 M Perfez-se com soluccedilatildeo
D ateacute um volume final de 1 mL e incubou-se a 25ordmC por 20 minutos Apoacutes este periacuteodo colocou-
se 780 L da mistura reaccional numa cuvette de quartzo e adicionou-se 220 L de soluccedilatildeo K23
23
Tampatildeo K Tampatildeo fosfatohidrogenofosfato de soacutedio 025 M pH 72
50
Adicionou-se 15 L de DTNB (em soluccedilatildeo K [DTNB]stock=100 mM) ao meio reaccional e
registou-se o valor de absorvecircncia medido equivalente agraves cisteiacutenas ldquoraacutepidasrdquo (SH1 e SH2
expostos ao solvente) Adicionou-se 001 g de SDS que provoca a desnaturaccedilatildeo da proteiacutena e
consequentemente a exposiccedilatildeo de todos os resiacuteduos de cisteiacutenas que se encontravam ateacute aiacute
inacessiacuteveis no interior da proteiacutena (cisteiacutenas totais) Estes resiacuteduos podem assim reagir com o
DTNB
Estimou-se o nuacutemero de resiacuteduos de cisteiacutenas em todas as amostras com recurso ao coeficiente de
extinccedilatildeo molar () do DTNB (ver ponto 3 dos Anexos) que foi determinado atraveacutes da titulaccedilatildeo
de L-Cisteiacutena com intervalo de concentraccedilotildees compreendidos entre 0 e 50 microM na presenccedila de
25microM de DTNB em tampatildeo D Os valores de absorvecircncia foram obtidos para um comprimento de
onda de 412 nm
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
3621 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
A teacutecnica mais eficiente de anaacutelise da interacccedilatildeo entre o iatildeo vanadilo e proteiacutenas como a
calmodulina e a transferrina entre outras permite a obtenccedilatildeo raacutepida de informaccedilatildeo no que toca agrave
conformaccedilatildeo e amplitude da referida interacccedilatildeo
Os espectros de Ressonacircncia Paramagneacutetica Electroacutenica (RPE) da espeacutecie paramagneacutetica V(IV)
foram realizados a 70K num espectroacutemetro Bruker EMX-300 X-band Todas as soluccedilotildees
utilizadas e amostras foram preparadas em anaerobiose numa cacircmara de luvas de modo a evitar a
reduccedilatildeo de V(IV) a V(V) Em tubos capilares com 02 mL de amostra manteve-se a concentraccedilatildeo
de S1 a 20 M e variou-se a concentraccedilatildeo de vanadilo (0 25 5 10 15 25 50 75 100 150
200 500 e 1000 M [VOSO4]stock=5 mM) em tampatildeo D Apoacutes obtenccedilatildeo dos espectros estes
foram tratados atraveacutes do software SimFonia e WinEPR tendo-se obtido um graacutefico com a
variaccedilatildeo da intensidade do sinal de RPE em funccedilatildeo da concentraccedilatildeo de sulfato de vanadilo
presente Realizaram-se ainda simulaccedilotildees de espectros obtidos com 20 microM de S1 e 1000 microM de
V(IV) com vista agrave obtenccedilatildeo dos ligandos provaacuteveis do vanadilo atraveacutes da anaacutelise da constante
de acoplamento hiperfina (A) e do factor g (g)
4 Conclusotildees gerais
Apesar de jaacute ter sido demonstrado que o vanaacutedio inibe a Na+K
+-ATPase da miosina a funccedilatildeo
deste elemento no tecido muacutesculo esqueleacutetico continua largamente incompreendida A enorme
complexidade quiacutemica deste metal especialmente no que concerne agrave oligomerizaccedilatildeo e reacccedilotildees
51
redox constitui um obstaacuteculo agrave compreensatildeo do comportamento do vanaacutedio nos organismos
vivos Assim a caracterizaccedilatildeo da quiacutemica deste elemento eacute de grande utilidade antes de iniciar o
estudo da sua interacccedilatildeo com o presente sistema
A anaacutelise por espectroscopias de 51
V-RMN e de UV-Vis de soluccedilotildees de vanadato diluiacutedas nos
meios de reacccedilatildeo utilizados no presente relatoacuterio (10 mM HEPES pH 70 25 mM KCl) mostrou
claramente que a espeacutecie decameacuterica (V10) possui um tempo de vida suficientemente longo para a
determinaccedilatildeo do seu efeito isolado ao contraacuterio do que se verifica com as espeacutecies mono- di-
tetra- e pentameacutericas (V1 V2 V4 e V5) Por outro lado a anaacutelise verificou ainda que apoacutes diluiccedilatildeo
das soluccedilotildees de decavanadato no meio de reacccedilatildeo coexistem as espeacutecies V10 e V1 pelo que os
efeitos putativos desta uacuteltima deveratildeo ser subtraiacutedos aquando da monitorizaccedilatildeo dos efeitos da
espeacutecie decameacuterica
O vanaacutedio (V) (vanadato) inibe as enzimas que catalizam reacccedilotildees de transferecircncia de grupos
fosfato como a ATPase da miosina [Goodno 1979] devido agrave sua analogia estrutural com o iatildeo
fosfato Na sua utilizaccedilatildeo como inibidor enzimaacutetico natildeo eacute tida em conta a contribuiccedilatildeo individual
das espeacutecies oligomeacutericas considerando-se normalmente a espeacutecie V1 como inibidora efectiva
[Goodno e Taylor 1982] No presente relatoacuterio verificou-se por espectroscopia de 51
V-RMN que
outros oligoacutemeros de vanadato interagem com o S1 de miosina tendo as espeacutecies V10 e V4 os
maiores niacuteveis de afinidade A sua interacccedilatildeo envolve forccedilas electrostaacuteticas e eacute afectada
negativamente por ligandos naturais de enzima como eacute o caso do ATP
A exposiccedilatildeo do S1 de miosina (005 mgmL) a decavanadato conduz agrave inibiccedilatildeo do capacidade da
proteiacutena de estimular a actividade Mg2+
-ATPaacutesica com um valor de IC50 de 266 plusmn 07 M
decavanadato sendo o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM
decavanadato (80) Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM decavanadato sendo
o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM decavanadato (60)
exercendo um efeito de impedimento agrave ligaccedilatildeo entre o vanaacutedio e o S1
A uma concentraccedilatildeo de 27 microM decavanadato verifica-se a oxidaccedilatildeo de um dos resiacuteduos de Cys
de S1 de miosina seguida pela oxidaccedilatildeo gradual de outro resiacuteduo de Cys ateacute agrave concentraccedilatildeo de
1000 microM Ambos os resiacuteduos oxidados natildeo se encontravam expostos ao solvente utilizado A
oxidaccedilatildeo dos resiacuteduos permite sugerir que esta modificaccedilatildeo oxidativa sofrida pelo S1 contribui
para a diminuiccedilatildeo da sua actividade de ATPase
52
Por espectroscopia de RPE revelaram-se os possiacuteveis ligandos equatoriais do vanaacutedio aquando da
sua interacccedilatildeo com o S1 de miosina como sendo duas moleacuteculas de aacutegua um ligando contendo o
grupo R-OH e outro contendo o grupo R=NR
Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados de
oxidaccedilatildeo (IV) e (V) no S1 de miosina para concentraccedilotildees facilmente alcanccedilaacuteveis no ambiente
terrestre
53
5 Estudos futuros
No sentido do melhor esclarecimento das alteraccedilotildees moleculares e estruturais que ocorrem no
subfragmento 1 quando exposta ao vanaacutedio que conduzem a uma diminuiccedilatildeo da sua funccedilatildeo
sugere-se
- Quantificaccedilatildeo das cisteiacutenas reduzidas de S1 na presenccedila de metavanadato
- Determinaccedilatildeo das actividades Ca2+
- e K+-ATPaacutesica do S1 de miosina na presenccedila de
decavanadato
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina estimulada por actina
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina na presenccedila de
metavanadato
- Realizaccedilatildeo de RMN 2D com variaccedilatildeo das concentraccedilotildees das soluccedilotildees de decavanadato e
metavanadato na presenccedila de S1
-Mutageacutenese dos resiacuteduos de Cys doS1 de miosina de modo a verificar quais os resiacuteduos que
sofrem oxidaccedilatildeo na presenccedila de decavanadato
- Determinaccedilatildeo por calorimetria (DSC Differential Scanning Calorimetry) da variaccedilatildeo da
entalpia (∆H) e da temperatura de desnaturaccedilatildeo do S1 de miosina na presenccedila de vanaacutedio uma
vez que estas propriedades caracteriacutesticas de cada proteiacutena satildeo alteradas apoacutes ocorrecircncia de
modificaccedilotildees estruturais
- Anaacutelise por SDS-PAGE de S1 de miosina apoacutes tratamento com vanaacutedio de modo a determinar
quais os polipeacuteptidos eventualmente formados apoacutes clivagem da proteiacutena
54
6 Referecircncias bibliograacuteficas
- Sellers J Myosins 2nd Edition Oxford University Press 1999
- L Stryer inBiochemistry (Ed WH Freeman 2005 pp
Refordfas Materiais e Meacutetodos
[A] S S Margossian SLowey Methods Enzymol1982 85 55-71
[B] JD Pardue JA Spudich Methods Enzymol 1982 85 164-181
[C] M Bradford Anal Biochem1976 72 248-254
[D] H H Taussky E Shorr J Biol Chem 1953 202 675-685
[E] C R Bagshaw in Outline Studies in Biology ndash Muscle Contraction Chapman and Hall Ed
1982
[F] A Cuenda et al Eur J Biochem1990 194 663-670
55
Anexos
1 Determinaccedilatildeo da recta de calibraccedilatildeo meacutetodo de Bradford
Figura1ndashRepresentaccedilatildeo graacutefica da recta de calibraccedilatildeo (Abs595 nm vs [BSA]) utilizada para o meacutetodo de Bradford
2 Determinaccedilatildeo da recta calibraccedilatildeo para a electroforese em condiccedilotildees desnaturantes
Figura 2 ndash Representaccedilatildeo graacutefica da recta de calibraccedilatildeo (log(Peso Molecular) vs Rf dos marcadores de peso
molecular) onde Rf indica determinada atraveacutes do padratildeo de peso molecular
(poccedilo (a))
3Caacutelculo do coeficiente de extinccedilatildeo molar () para a quantificaccedilatildeo de cisteiacutenas reduzidas
atraveacutes do meacutetodo de Ellman
Traccedilou-se a recta de Abs412 nm meacutedia vs [L-Cisteiacutena] de modo a determinar o valor do coeficiente
de extinccedilatildeo molar definido como o declive daquela recta Assim =8400 M-1
cm-1
como
podemos verificar na Fig 3
A595 nm = 00546[BSA] + 00209 Rsup2 = 099051
Ab
s 59
5 n
m
[BSA] (mgmL)
log PM = -1177 Rf + 2247 Rsup2 = 098957
log
PM
Mobilidade relativa (Rf)
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero

29
aln(A0) Numa reacccedilatildeo de 1ordf ordem o tempo de meia-vida eacute independente da concentraccedilatildeo inicial
do reagente sendo descrito pela equaccedilatildeo [2] Deste modo a partir da recta
Ln(Abs400 nm) = -436E-5
K - 105 obteve-se um tempo de meia-vida de 4490 plusmn 0823 horas para
as condiccedilotildees supramencionadas
Apoacutes a adiccedilatildeo de S1 8 microM ao meio reaccional verificou-se que a reacccedilatildeo de desoligomerizaccedilatildeo
da espeacutecie decameacuterica de vanadato seguia igualmente uma cineacutetica de 1ordf ordem tal como se pode
verificar pela linearidade do graacutefico de ln(Abs400) versus o tempo de aquisiccedilatildeo espectral
(segundos) (Fig27)
Figura 27ndash Graacutefico de ln(Abs400 nm) em funccedilatildeo do
tempo (em segundos) para uma soluccedilatildeo de decavanadato
total de 1 mM em tampatildeo D a 25ordmC na presenccedila de 8
microM (n= 2)
A partir da equaccedilatildeo da recta Ln(Abs400 nm) = -7520E-5
K - 0613determinou-se o valor de K para
a espeacutecie decameacuterica de onde se obteve um tempo de meia-vida de 2430 plusmn 0191 horas para as
condiccedilotildees supramencionadas
Tanto este valor como o valor registado na ausecircncia de S1 satildeo substancialmente superiores agrave
duraccedilatildeo dos ensaios de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina descritos no presente
relatoacuterio
24 Interacccedilatildeo de S1 com vanaacutedio(V)
241Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
De modo a analisar qual o efeito da adiccedilatildeo de S1 de miosina (tampatildeo D) produzia nos sinais de
51V-NMR titulou-se uma soluccedilatildeo de 5mM de decavanadato total com concentraccedilotildees crescentes
de S1 (5 10 15 e 20 microM) Os espectros de 51
V-RMN encontram-se apresentados na Fig 2
No graacutefico da figura 2 Apresenta-se a concentraccedilatildeo de S1 em funccedilatildeo do factor de alargamento
corrigido dos espectros acima referidos
Ln
(Ab
s (4
00
nm
))
tempo (s)
30
Figura 28 ndash Graacutefico de
representaccedilatildeo da
concentraccedilatildeo de S1 em
funccedilatildeo do factor de
alargamento A
concentraccedilatildeo de
decavanadato total foi de 5
mM e as concentraccedilotildees
utilizadas de S1 foram de 0
5 10 e 15 microM
Verifica-se que o factor de alargamento aumenta linearmente em funccedilatildeo da concentraccedilatildeo de S1 no
ensaio o que indica a existecircncia de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina Observou-
se ainda que a adiccedilatildeo de S1 ao meio reaccional natildeo provocou qualquer alteraccedilatildeo dos desvios
quiacutemicos dos sinais de 51
V-RMN
Realizaram-se ensaios na presenccedila de ATP e de S1 com intuito de verificar o efeito do ATP na
interacccedilatildeo do S1 de miosina com o decavanadato
Figura 29 - Espectros de RMN de 51
V contendo 2
mM Decavanadato (A) (controlo) (B) na presenccedila
de 2 mM de ATP (C) na presenccedila de 2 mM de ATP
e 8 microM de S1 de miosina V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2Vndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies decameacuterica (V10) e monomeacuterica (V1) presentes nos
espectros da soluccedilatildeo de decavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V10A V10B V10C V1
part (ppm) -5161 -5002 -4254 -5614
Observa-se que a presenccedila de ATP (espectro B) natildeo provoca qualquer alteraccedilatildeo visiacutevel em
relaccedilatildeo ao espectro que contem apenas soluccedilatildeo de decavanadato Jaacute a presenccedila simultacircnea de ATP
e S1 provoca um alargamento da ∆12 do sinal do V10A de 1736 Hz (espectro (A)) para 2366 Hz
(espectro (C) Fig 29) Tendo em conta que a concentraccedilatildeo de S1 apresenta uma relaccedilatildeo linear
com o factor de alargamento converteu-se o valor da ∆12 e comparou-se com o valor esperado agrave
F = 00591[S1] - 00157 Rsup2 = 0998
Fact
or
de
ala
rgam
en
to
[S1] (microM)
31
concentraccedilatildeo de S1 utilizada O factor de alargamento obtido para 8 microM ndash 0376 ndash eacute inferior ao
valor esperado agrave mesma concentraccedilatildeo de S1 obtido atraveacutes da equaccedilatildeo da recta apresentada na
Fig 28 ndash 0457 ndash o que indica que o ATP poderaacute exercer um efeito de protecccedilatildeo do S1
diminuindo a sua interacccedilatildeo com a espeacutecieV10
Por uacuteltimo realizaram-se ensaios com metavanadato na presenccedila de 8 microM S1 de modo a verificar
se as espeacutecies oligomeacutericas contidas nesta soluccedilatildeo interagiam com a proteiacutena em estudo
Figura 210 - Espectros de RMN de 51
V contendo 2
mM metavanadato (A) (controlo) (B) na presenccedila
de 8 microM de S1 de miosina V1 V2 e V4
correspondem respectivamente aacutes espeacutecies
monomeacuterica dimeacuterica e tetrameacuterica
Tabela 2VIndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies monomeacuterica (V1) dimeacuterica (V2) e tetrameacuterica (V4)
presentes nos espectros da soluccedilatildeo de metavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V1 V2 V4
part (ppm) -5613 -5747 -5789
Como eacute possiacutevel observar na Fig 210 ocorre uma ligeiro alargamento dos sinal de V4 bem
como uma perda da intensidade pelo que se conclui quedas espeacutecies presentes na soluccedilatildeo de
metavanadato (V1 V2 e V4) apenas a espeacutecie tetrameacuterica interage com o S1 de miosina o que
estaacute de acordo com o reportado por Tiago et al 2002
242 Actividade Mg2+
-ATPaacutesica de S1 de miosina na presenccedila de decanadato
Com o intuito de avaliar o efeito do decavanadato na actividade Mg2+
-ATPaacutesica do S1 de
miosina monitorizou-se a reacccedilatildeo de oxidaccedilatildeo do NADH atraveacutes da leitura de Abs340 na
presenccedila de 005 mgmL de S1 de miosina e no caso da actividade estimulada 2microM de actina em
3125 mM Tris-HCl pH 71 e 375 mM MgCl2 pelo meacutetodo de enzimas acopladas (ver seccccedilatildeo xx
dos MM)Para tal realizaram-se ensaios com vaacuterias concentraccedilotildees de decavanadato total (10 50
100 250 500 e 1000) de modo a determinar qual o seu efeito na actividade MgATPaacutesica de S1
de miosina na ausecircncia e na presenccedila de um dos seus ligandos naturais Os ensaios foram
realizados agrave temperatura ambiente com incubaccedilatildeo preacutevia a 25ordm C durante 20 minutos
32
Figura 211 ndash Representaccedilatildeo
graacutefica da Actividade Mg2+
-
ATPaacutesica de S1 () (os ajustes
teoacutericos estatildeo representado a por
uma linha os resultados
experimentais estatildeo representados
por triacircngulos (Actividade
estimulada) e cruzes (Actividade
natildeo estimulada)) vs
[Decavanadato]total medidas pelo
meacutetodo das enzimas acopladas
(n=3) Os dados foram ajustados ao modelo de decaimento exponencial usando o programa GraphPad Prism para a
determinaccedilatildeo dos valores de IC50 (na ausecircncia de actina IC50= 266 plusmn 07 M decavanadato na presenccedila de actina
IC50= 28 plusmn 1 M decavanadato
A partir do graacutefico presente na Fig 29 verifica-se que o ajuste dos dados experimentais foi bem
sucedido A inibiccedilatildeo da actividade Mg2+
-ATPaacutesica do S1 de miosina eacute afectada em menor
amplitude na presenccedila de actina (60 de inibiccedilatildeo da actividade) do que na sua ausecircncia
(aproximadamente 80 de inibiccedilatildeo de actividade) Devido a esta diferenccedila pode sugerir-se que a
actina iraacute impedir parcialmente a ligaccedilatildeo do decavanadatoao S1 demiosina atraveacutes de
impedimentos esteacutereos ou interacccedilotildees electrostaacuteticas por exemplo A teacutecnica utilizada natildeo nos
permite concluir acerca de que regiatildeo(otildees) ou resiacuteduo(s) da actina participaratildeo na preservaccedilatildeo
parcial da actividade Mg2+
-ATPaacutesica
Por outro lado os valores de IC50 dos dois tipos de actividades natildeo apresentam diferenccedilas
significativas tendo-se obtido 266 plusmn 07 e 28 plusmn 1M decavanadato na ausecircncia e na presenccedila de
actina respectivamente o que corresponde a uma concentraccedilatildeo de cerca de 27 M de V10 O
meio de reacccedilatildeo utilizado nestes ensaios foi 10 mM HEPES e 25 mM KCl com 2 mM ATP Em
(Tiago et al 2004) o valor de IC50 para a actividade Mg2+-ATPaacutesica na ausecircncia de actina foi de
125 plusmn 025 microM enquanto o valor da actividade estimulada por actina foi de 136 plusmn 14 microM O
meio de reacccedilatildeo utilizado nos ensaios foi 20 mM HEPES 50 mM KCl e 2 mM ATP o que resulta
precisamente no dobro da forccedila ioacutenica a que foi realizado o ensaio do presente relatoacuterio
(exceptuando a contribuiccedilatildeo dos produtos da hidroacutelise de ATP uma vez que este meacutetodo
pressupotildee a ocorrecircncia de um ciclo de reacccedilotildees de hidroacutelise e siacutentese deste composto) Ora tendo
sido anteriormente referido que a afinidade do S1 de miosina para com a espeacutecie V10 diminui com
Act
ivid
ade
Mg
2+-A
TP
aacutesic
a d
e S
1 d
e
Mio
sin
a
[Decavanadato] (microM) Actividade Estimulada ()
Actividade Natildeo Estimulada ()
Ajuste Actividade Estimulada
Ajuste Actividade Natildeo Estimulada
33
o aumento da forccedila ioacutenica (Tiago et al 2004 tal poderaacute explicar a diferenccedila obtida entre estes
dois conjuntos de valores
243Quantificaccedilatildeo de cisteiacutenas reduzidas pelo meacutetodo de Ellman
A interacccedilatildeo do vanaacutedio com alguns tipos de aminoaacutecidos sensiacuteveis agrave oxidaccedilatildeo pode fornecer-nos
informaccedilatildeo bastante uacutetil em relaccedilatildeo agraves alteraccedilotildees estruturais que conduzem a uma diminuiccedilatildeo da
actividade fisioloacutegica do S1 de miosina Os estudos realizados por Chasteen 1983 concluiacuteram
que a reduccedilatildeo de vanadato (V) a vanadilo (IV) ocorre na presenccedila de agentes redutores comuns no
ambiente celular de que se destaca o aminoaacutecido cisteiacutena Esta eacute tambeacutem um alvo de
complexaccedilatildeo por parte do iatildeo VO2+
Jaacute Ramos et al (2009) demonstrou que a incubaccedilatildeo de
actina com a soluccedilatildeo de decavanadato induz a oxidaccedilatildeo de resiacuteduos de cisteiacutena e agrave formaccedilatildeo do
oxidovanaacutedio (IV) Este grupo de investigadores descobriu ainda que apenas a espeacutecie V10 induz a
oxidaccedilatildeo das cisteiacutenas ldquoraacutepidasrdquo da actina atraveacutes do processo proposto que eacute parcialmente
responsaacutevel pela inibiccedilatildeo jaacute descrita da actividade de ATPase do S1 de miosina estimulada por
actina quando exposta agrave soluccedilatildeo de decavanadato Esta inibiccedilatildeo apresenta-se afectada com a
presenccedila de ATP uma vez que os resiacuteduos de cisteiacutena permaneceram intactos nos ensaios
realizados com este ligando Jaacute Crans et al (2004) concluiu que o siacutetio activo de muitas fosfatases
(que conteacutem grupos tiol) pode formar um equiliacutebrio redox com o iatildeo vanadato (V) de forma natildeo
reversiacutevel
Os resultados obtidos por Sakurai e Shiromura (1980) sugeriram que o vanaacutedio (V) sofre reduccedilatildeo
a vanaacutedio (IV) sem dependecircncia do valor de pH do ensaio Registaram ainda uma estequiometria
de 31 na reacccedilatildeo do vanadato (V) com os resiacuteduos de Cys enquanto a estequiometria dos ensaios
com o vanadilo (IV) foi de 21
Berlett e Stadtman (1997) observaram que os tipos de aminoaacutecidos mais facilmente oxidaacuteveis
englobam entre outros os resiacuteduos de Cys Trp e MetAssim analisou-se o estado redox dos
grupos tiol dos resiacuteduos de Cys do S1 na presenccedila do decavanadato atraveacutes da monitorizaccedilatildeo da
Abs412 nm na presenccedila de 1 mgmL de S1 de miosina em tampatildeo 10 mM HEPES 25 mM KCl
variando a concentraccedilatildeo da soluccedilatildeo de decavanadato de modo a determinar que tipo de alteraccedilotildees
estruturais ocorrem na sua presenccedila no que diz respeito agrave diminuiccedilatildeo da actividade ATPaacutesica de
S1 na presenccedila de decavanadato
Os resultados referentes agrave quantificaccedilatildeo das cisteiacutenas reduzidas apoacutes o tratamento de S1 com
diferentes concentraccedilotildees de decavanadato em tampatildeo 10 mM HEPES pH 75 e 25 mM KCl a
25ordmC durante 20 minutos encontram-se descritos na Tabela 2VII
Tabela 2VII ndash Cisteiacutenas reduzidas totais e ldquoraacutepidas de S1 (1 mgmL) apoacutes tratamento com decavanadato
34
(010501002505001000 uM) em 10 mM HEPES 25 mM KCl pH 7 a 25ordmC durante 20 minutos A detecccedilatildeo
espectrofotomeacutetrica foi feita com DTNB (e 412=8400 uM-1 cm-1) (n variaacutevel entre 1 e 8)
Amostras Cys raacutepidas Cys totais
11 M S1 (controlo) 22 plusmn 04 106 plusmn 14
11 M S1 + 10 M Decavanadato 22 plusmn 05 95 plusmn 08
11 M S1 + 50 uM Decavanadato 20 plusmn 02 98 plusmn 14
11 M S1 + 100 uM Decavanadato 17 plusmn 03 97 plusmn14
11 M S1 + 250 uM Decavanadato 26 plusmn 02 98 plusmn 08
11 M S1 + 500 uM Decavanadato 21 plusmn 04 90 plusmn 06
11 M S1 + 1000 uM Decavanadato 24 plusmn 0 86 plusmn 08
A partir dos resultados obtidos pode sugerir-se que o S1 possui 11 resiacuteduos de Cys (controlo) o
que estaacute de acordo com o anteriormente descrito Verifica-se que o tratamento da proteiacutena com
concentraccedilotildees crescentes de decavanadato provoca a oxidaccedilatildeo de dois resiacuteduos de Cys do S1 de
miosina natildeo expostos ao solvente No entanto a teacutecnica utilizada natildeo nos permite identificar
quais
Tendo em consideraccedilatildeo que o valor de IC50 obtido para a reacccedilatildeo de inibiccedilatildeo daactividade Mg2+-
ATPaacutesica de S1 na presenccedila de decavanadato eacute de cerca de 27 microM pode sugerir-se que este seraacute o
valor ao qual ocorreraacute a oxidaccedilatildeo do primeiro resiacuteduo de Cys o que poderaacute contribuir para a
diminuiccedilatildeo da actividade de S1 observada
25Interacccedilatildeo de S1 com vanaacutedio(IV)
251Anaacutelise por espectroscopia de Ressonacircncia Paramagneacutetica Electroacutenica
A acumulaccedilatildeo intracelular de oxovanaacutedio (IV) in vivo constitui uma das principais formas de
apresentaccedilatildeo de vanaacutedio no organismo Deste modo torna-se bastante interessante analisar a
interacccedilatildeo entre este iatildeo e o S1 de miosina atraveacutes da teacutecnica de espectroscopia de Ressonacircncia
Paramagneacutetica Electroacutenica o meacutetodo de eleiccedilatildeo do estudo do iatildeo vanadilo e dos complexos por si
formados O electratildeo desemparelhado encontra-se fortemente acoplado ao nuacutecleo 51
V o que
35
resulta num espectro de RPE consistindo de 8 desdobramentos na direcccedilatildeo paralela e outros tantos
na direcccedilatildeo perpendicular (Chasteen 1983)
Os estudos realizados por Ahmed et al (1987) centraram-se na interacccedilatildeo do vanadilo com
proteiacutenas como eacute o caso da insulina transferrina entre outros obtendo um espectro caracterizado
por 8 linhas amplamente espaccediladas (na direcccedilatildeo paralela) Jaacute Ramos et al 2009 concluiu que a
exposiccedilatildeo da soluccedilatildeo de decavanadato agraves formas globular e filamentosa de actina produz um
espectro tiacutepico do iatildeo oxidovanaacutedio (IV) indicando a reduccedilatildeo do vanadato
Para aferir qual a concentraccedilatildeo de S1 que permitia obter uma boa razatildeo sinalruiacutedo titulou-se uma
soluccedilatildeo de sulfato de vanadilo 15 microM ([VOSO4]stock = 5 mM) com concentraccedilotildees crescentes de
proteiacutena (0 5 10 15 e 20 microM) em 10 mM HEPES 25 mM KCl A existecircncia de uma correlaccedilatildeo
linear entre a concentraccedilatildeo de proteiacutena e a intensidade dos sinais de RPE correspondente agrave banda
-12 perpendicular encontra-se representada na Fig 210
Figura 2 12ndash Representaccedilatildeo graacutefica da variaccedilatildeo da intensidade do sinal de RPE vs a concentraccedilatildeo de S1 utilizada
Apoacutes verificar que com uma concentraccedilatildeo de S1 igual a 20 microM se obtinham sinais com uma razatildeo
sinalruiacutedo adequada ao prosseguimento dos estudos sobre a interacccedilatildeo V(IV)-S1 de miosina
titulou-se uma soluccedilatildeo de 20 microM S1 com 0 25 5 10 15 25 50 75 100 150 200 500 e 1000
microM VOSO4
Traccedilou-se o graacutefico da intensidade da banda -12 perpendicular do sinal de EPR em funccedilatildeo da
concentraccedilatildeo de VOSO4 tendo-se determinado uma constante de dissociaccedilatildeo (KD) do complexo
S1-vanadilo de 8748 plusmn 2621 microM V(IV)
y = 016786x + 122435
Rsup2 = 092992759
Inte
nsi
dad
e d
o s
inal
de
RP
E
[S1] (microM)
36
Figura 213 ndash Representaccedilatildeo graacutefica da variaccedilatildeo do sinal de RPE vs a concentraccedilatildeo de vanadilo utilizada (os valores
experimentais estatildeo representados por losangos o seu ajuste estaacute representado pela linha)
A simulaccedilatildeo do espectro de EPR foi efectuada a partir do espectro obtido para uma concentraccedilatildeo
de 1 mM VOSO4 apoacutes adiccedilatildeo de 20 microM S1 de miosina tendo-se obtido os valores da constante de
acoplamento hiperfina (A=5016) e do factor g (g0=195)
Figura 214 ndash Representaccedilatildeo graacutefica do espectro de EPR obtido com 1000 microM de vanadilo e 20 microM de S1 (a cinzento
escuro) A simulaccedilatildeo do espectro encontra-se a cinzento claro
Por comparaccedilatildeo destes com os valores presentes na tabela de correspondecircncia xxx eacute possiacutevel
predizer quais os ligandos provaacuteveis do vanadilo na presenccedila de proteiacutena Assim concluiu-se que
o vanadilo estaacute coordenado com duas moleacuteculas de aacutegua um grupo hidroxilo e um grupo R=NR
O grupo hidroxilo pode indicar os resiacuteduos de aminoaacutecidos Tyr Treonina e Serina enquanto o
grupo R=NR poderaacute indicar a presenccedila de Arginina e Histidina como ligandos do vanadilo
Inte
nsi
dad
e d
os
sinai
s d
e R
PE
[Vanadilo] (microM)
Ajuste dos dados exp Dados experimentais
Campo magneacutetico (G)
Espectro Experimental Ajuste
37
3 Procedimento Experimental
31 Reagentes
Todos os reagentes utilizados satildeo de elevado grau de pureza (analiacutetico) Apresentam-se as suas
foacutermulas quiacutemicas ou abreviaturas quando necessaacuterio acrescentou-se a sua descriccedilatildeo Entre
parecircntesis colocou-se as empresas que os comercializam
Aacutecido aceacutetico glacial 100 (Riedel) Acrilamida 99 (Merck) Azul de bromofenol (Riedel-de-
Haumlen) -Mercaptoetanol (Sigma) Bissulfito de soacutedio (Sigma)Bradford (reagente de)
(BioRad)BSA (albumina de soro bovino Sigma-Aldrich) CaCl22H2O (cloreto de caacutelcio
dihidratado Fluka) CH3CH2OH 96 (etanol Panreac) Coomassie Blue R250 (Merck) D2O
(oacutexido de deuteacuterio SDS) DTNB (55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico) Aldrich) EDTA (aacutecido
etilenodiaminatetraaceacutetico Merck) Fosfoenolpiruvato 98 (Fluka) Glicerol (Sigma) Glicina
985 (Panreac) HCl 37 (Riedel-de Haumlen) H2SO4 96 (Pronalab) HEPES 99 (aacutecido 4-(2-
hidroxiletil)-1-piperazineetanosulfoacutenico Sigma) KH2PO4 99 (hidrogenofosfato de potaacutessio)
MgATP (Sigma) Na
ATP (Sigma) Na2HPO4 99 (hidrogenofosfato de soacutedio Riedel-de Haumlen)
NaH2PO4 (fosfato de soacutedio ) NaOH 986 (hidroacutexido de soacutedio Pronalab) KCl 99 (Cloreto de
potaacutessio Panreac) L-Cys 995 (enantioacutemero levoacutegiro do resiacuteduo de Cisteiacutena Fluka)Metanol
99 (Merck) Molibdato de amoacutenio (Sigma)NNrsquo-metil-bis-acrilamida 98 (Merck) NH4VO3
99 (metavanadato de amoacutenia Sigma) PSA 10 (persulfato de amoacutenio Merck) SDS
(dodecilo sulfato de soacutedio Sigma) Sulfito de soacutedio (Sigma) VOSO4 (sulfato de vanadilo
Aldrich) TEMED 99 (Tetrametiletilenodiamina Merck) Tris (2-amino-2-hidroximetil-13-
propanodiol Sigma)
32 Meacutetodos
321 Isolamento e purificaccedilatildeo das proteiacutenas de muacutesculo esqueleacutetico de coelho
3211 Purificaccedilatildeo da miosina preparaccedilatildeo de Subfragmento 1 por digestatildeo quimotriacutepsica
da miosina
O subfragmento-1 de miosina-II (S1) foi gentilmente cedido pelo Prof Doutor Manuel Aureliano
do Departamento de Quiacutemica Bioquiacutemica e Farmaacutecia da Faculdade de Ciecircncias e Tecnologia da
Universidade do Algarve
No entanto descreve-se abaixo de forma resumida o procedimento efectuado
38
32111 Purificaccedilatildeo da miosina
O protocolo de isolamento foi adaptado do meacutetodo previamente descrito por Margossian e Lowey
(1982) apresentando-se de forma esquemaacutetica na Fig 31
Sucintamente os tecidos musculares dorsais e das pernas foram extraiacutedos apoacutes o sacrifiacutecio do
animal (Fig31) De seguida foram adicionados 3 volumes da soluccedilatildeo de extracccedilatildeo A2
Centrifugou-se a soluccedilatildeo e filtrou-se o sobrenadante O sedimento foi entatildeo diluiacutedo em 14
volumes de aacutegua destilada de modo a que a forccedila ioacutenica atingisse 004 M Apoacutes adiccedilatildeo de szlig-
mercaptoetanol (szlig-me) deixou-se a precipitar a 4ordmC removendo o sobrenadante Centrifugou-se
novamente e ressuspendeu-se o sedimento na soluccedilatildeo de extracccedilatildeo A Adicionou-se a soluccedilatildeo
tampatildeo B3 para que a forccedila ioacutenica atingisse 05 M Homogeneizou-se e deixou-se repousar
durante 2h Homogeneizou-se novamente e centrifugou-se de modo a ocorrer precipitaccedilatildeo dos
complexos de actomiosina e proteiacutena desnaturada Apoacutes filtraccedilatildeo do sobrenadante em latilde de vidro
quantificou-se a soluccedilatildeo de miosina resultante pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental)
2 Soluccedilatildeo de extracccedilatildeo A 0012M K2CO3 004M KHCO3 06M KCl pH 65
3 Soluccedilatildeo-tampatildeo B Tris 100 mM pH 7 KCl 2M
39
Figura 31 ndash Representaccedilatildeo esquemaacutetica do protocolo de isolamento de miosina (Margossian e Lowey 1982) Todo o
procedimento foi realizado a 4ordmC (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
32112 Preparaccedilatildeo de Subfragmento-1 de miosina por digestatildeo com quimiotripsina
O S1 de miosina foi isolado apoacutes diaacutelise da miosina durante a noite contra a soluccedilatildeo C4 A
presenccedila do EDTA eacute essencial uma vez que a sua substituiccedilatildeo por MgCl2resulta na produccedilatildeo de
HMM em vez de S1 A suspensatildeo de miosina foi equilibrada e digerida com -quimotripsina
A digestatildeo foi interrompida pela adiccedilatildeo de PMSF Seguidamente ultracentrifugou-se o produto
da digestatildeo desprezando-se o sedimento (Fig 32) A soluccedilatildeo de sobrenadante maioritariamente
S1 foi quantificada pela leitura directa da Abs280 ou pelo meacutetodo de Bradford (ver as secccedilotildees
3311 e 3312 do Procedimento Experimental respectivamente) e armazenada em 10 sacarose
e 2 mM szlig-Me a -80ordmC
4 Soluccedilatildeo C 25mM Tris pH 70 012M KCl 1mM EDTA
40
Antes da realizaccedilatildeo dos ensaios descritos no presente relatoacuterio o S1 de miosina foi dialisado
contra a soluccedilatildeo D5 de modo a remover qualquer composto que pudesse complexar como
vanaacutedio Seguidamente quantificou-se a proteiacutena pelo meacutetodo de Bradford ou pelo meacutetodo
directo (ver secccedilotildees 3311 e 3312do Procedimento Experimtental)
Figura 3 2ndash Representaccedilatildeo esquemaacutetica do protocolo de preparaccedilatildeo do S1 de miosina por digestatildeo quimiotripsina de
acordo com o previamente descrito por Margossian e Lowey (1982) Todo o procedimento foi realizado a 4ordm C
excepto quando indicaccedilatildeo do contraacuterio (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
3212 Isolamento e purificaccedilatildeo de actina
Isolou-se e purificou-se a actina segundo o meacutetodo descrito por Pardee e Spudish que
compreende dois passos ndash a obtenccedilatildeo de um resiacuteduo seco denominado ldquopoacute de acetonardquo e a
extracccedilatildeo subsequente da actina a partir do ldquopoacute de acetonardquo na forma de G-Actina com a
posterior polimerizaccedilatildeo a F-Actina
32121 Obtenccedilatildeo do ldquopoacute de acetonardquo
Sacrificou-se o coelho com uma pancada atraacutes do pescoccedilo e sangrou-se o animal apoacutes o corte da
arteacuteria jugular Esfolou-se e colocou-se numa tina com gelo Retiraram-se as partes de tecido
conjuntivo que envolvem o tecido muscular com auxiacutelio de uma pinccedila e bisturi Seguidamente
retirou-se os muacutesculos dorsais e os muacutesculos brancos das pernas Passou-se os muacutesculos por aacutegua
destilada de modo a remover as impurezas e pesou-se num copo Trituraram-se os muacutesculos
numa maacutequina de triturar carne e extraiu-se o resultante com 3 volumes da soluccedilatildeo E6 durante 10
minutos com agitaccedilatildeo suave Centrifugou-se a suspensatildeo a 4000 g durante 10
5 Soluccedilatildeo D HEPES 10 mM pH 70 KCl 25 mM
6 Soluccedilatildeo E 01 M KCl 015 M K2HPO4 pH 65
41
minutosdesprezando-se o sobrenadante Ressuspendeu-se o sedimento em 6 volumes de soluccedilatildeo
F7durante 10 minutos com agitaccedilatildeo suave Verificou-se se o pH da soluccedilatildeo estava compreendido
entre 82 e 85 acertando-se estecom soluccedilatildeo G8 se necessaacuterio Centrifugou-se durante 10
minutos a 4000 g Desprezou-se o sobrenadante e ressuspendeu-se o sedimento em 3 volumes de
soluccedilatildeo H9 durante 10 minutos Centrifugou-se novamente a 4000 gdurante 10 minutos
desprezando-se o sobrenadante Ressuspendeu-se o sedimento duas vezes consecutivas em 6
volumes de aacutegua destilada durante 5 minutos Centrifugou-se a 4000 g durante 10 minutos apoacutes
cada adiccedilatildeo de H2O
Seguidamente ressuspendeu-se o sedimento 5 vezes consecutivas utilizando 3 volumes de
acetona de cada vez agitando suavemente durante 10 minutos agrave temperatura ambiente Colocou-
se o resiacuteduo filtrado numa tina de vidro sobre papel de filtro deixando-se evaporar o solvente
durante a noite num nicho Triturou-se o resiacuteduo seco numa maacutequina de triturar carne e deixou-se
novamente evaporar o solvente O ldquopoacute de acetonardquo obtido foi armazenado a -20 ordmC
32122 Purificaccedilatildeo da actina a partir de ldquopoacute de acetonardquo
Apoacutes a obtenccedilatildeo do poacute de acetona como acima indicado efectuou-se o protocolo seguidamente
descrito de modo a obter monoacutemeros de actina (G-Actina) e actina filamentosa (F-Actina)
Pesou-se 35 g de poacute de acetona e adicionaram-se 50 mL de soluccedilatildeo-tampatildeo I10
sendo a
extracccedilatildeo feita com agitaccedilatildeo suave durante 30 minutos Separou-se o extracto do poacute de acetona
hidratado filtrando a suspensatildeo com 4 camadas de gaze esterilizada tendo o cuidado de natildeo
espremer o resiacuteduo Voltou a extrair-se com 50 mL de soluccedilatildeo I durante 30 minutos mexendo
lentamente Adicionou-se este ao extracto obtido anteriormente Centrifugou-se o extracto total a
16 000 g durante 30 minutos Decantou-se para uma proveta sobre a qual se colocou gaze para
que o sedimento solto natildeo se juntasse ao sobrenadante e leu-se o volume De seguida adicionou-
se KCl ateacute uma concentraccedilatildeo final de 50 mM MgCl2 ateacute 2mM e ATP ateacute 1mM lentamente e com
agitaccedilatildeo contiacutenua de modo a promover a polimerizaccedilatildeo dos monoacutemeros de actina
Deixou-se polimerizar durante 2h a 25ordmC observando-se a presenccedila de pequenas bolhas de ar
indicativas de um aumento de viscosidade da soluccedilatildeo Arrefeceu-se em gelo durante 15 minutos e
adicionou-se KCl ateacute uma concentraccedilatildeo final de 06M Mexeu-se lentamente durante 30 minutos
apoacutes os quais se ultracentrifugou a 110 000g durante 1hora Desprezou-se o sobrenadante
ressuspendeu-se o sedimento com soluccedilatildeo tampatildeo E e transferiu-se para um potter
Dialisou-se durante a noite contra 1L de soluccedilatildeo-tampatildeo I a uma temperatura de 4ordmC com rotaccedilatildeo
7 Soluccedilatildeo F 005 M NaHCO3 pH 82-85
8 Soluccedilatildeo G 1M Na2CO3
9 Soluccedilatildeo H 1 mM EDTA pH 70
10 Soluccedilatildeo I 2 mM Tris-HCl pH 8 02 mM CaCl2 02 mM Na2ATP 05 mM szlig-Me 0005 NaN3
42
contiacutenua e mudanccedila de tampatildeo 2 a 3 vezes De seguida a actina foi centrifugada a 110
000gdurante 1hora Decantou-se a soluccedilatildeo de G-Actina (sobrenadante) e desprezou-se o
sedimento Determinou-se a concentraccedilatildeo de G-Actina pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental) A soluccedilatildeo de G-actina foi armazenada a uma temperatura de 4ordmC
apoacutes adiccedilatildeo de 002NaN3 mantendo-se estaacutevel durante 3 a 5 dias
331 Quantificaccedilatildeo das proteiacutenas isoladas
3311 Quantificaccedilatildeo pelo meacutetodo de Bradford
O meacutetodo de Bradford (Bradford 1976) baseia-se na utilizaccedilatildeo do reagente de Bradford
composto por entre outros aacutecido fosfoacuterico e o corante ldquoCoomassierdquo Este corante possui carga
negativa e a sua coloraccedilatildeo varia entre a vermelha (Absmax=465 nm) predominante em soluccedilatildeo e a
azul (Absmax=595 nm) presente aquando da ligaccedilatildeo aos grupos amina das proteiacutenas Em soluccedilotildees
aacutecidas diluiacutedas a coloraccedilatildeo varia proporcionalmente com a quantidade do corante ligado agrave
proteiacutena que por sua vez eacute directamente proporcional agrave concentraccedilatildeo da proteiacutena permitindo a
sua quantificaccedilatildeo ateacute a um limite de detecccedilatildeo de 20 gmL (Bradford 1976)
Deste modo prepararam-se amostras de BSA ([BSA]stock= 1mgmL) utilizando um intervalo de
concentraccedilotildees compreendido entre 0 e 15 gmL de acordo com a tabela 3I Apoacutes a adiccedilatildeo da
H2O MilliQ e do reagente de Bradford as soluccedilotildees foram sucessivamente homogeneizadas no
voacutertex
Tabela 3Indash Amostras utilizadas para a obtenccedilatildeo da recta de calibraccedilatildeo no meacutetodo de Bradford
BSA (L) BSA (g) H2O MilliQ (L) Reagente de Bradford (L)
0 0 800 200
125 125 79875 200
25 25 7975 200
5 5 795 200
10 10 790 200
15 15 785 200
Apoacutes incubaccedilatildeo a 25ordmC por 5 minutos leram-se os valores de Abs595 num
espectrofotoacutemetro (Shimadzu UV-160A) para todas as amostras Seguidamente traccedilou-se a recta
de calibraccedilatildeo (Abs595vs quantidade de BSA (g))
43
Para determinar a concentraccedilatildeo de proteiacutena apoacutes o processo de isolamento purificaccedilatildeo e
diaacutelise quando necessaacuterio colocou-se 5 L da amostra num tubo de Eppendorf e adicionou-se
795 L de H2O MilliQ homogeneizando a soluccedilatildeo no voacutertex Adicionou-se posteriormente 200
L de reagente de Bradford e repetiu-se a homogeneizaccedilatildeo Incubou-se a mistura a 25ordmC por 5
minutos apoacutes os quais se leu os valores de Abs595 nm no espectrofotoacutemetro Por interpolaccedilatildeo deste
valor a partir da recta de calibraccedilatildeo determinou-se a concentraccedilatildeo de proteiacutena na amostra
3312 Quantificaccedilatildeo pelo meacutetodo directo
A quantificaccedilatildeo das amostras proteicas pode tambeacutem ser efectuada atraveacutes da leitura directa da
absorvacircncia tendo por base a lei de Beer-Lambert onde Abs corresponde agrave
absorvacircncia ao comprimento de onda ao qual o ensaio foi realizado corresponde ao coeficiente
de extinccedilatildeo (substituiacutedo pela letra a se as unidades forem distintas da molaridade) b corresponde
ao percurso oacuteptico da radiaccedilatildeo na ceacutelula amostral e c corresponde agrave concentraccedilatildeo amostral
Assim no caso de S1 de miosina mediu-se a Abs280e calculou-se a concentraccedilatildeo da proteiacutena
sabendo que o coeficiente de extinccedilatildeo de S1 eacute (Ref)
No caso da actina mediu-se a Abs290ecalculou-se a concentraccedilatildeo da proteiacutena sabendo que o
coeficiente de extinccedilatildeo da actina eacute (Ref)
332Electroforese em condiccedilotildees desnaturantes (SDS-PAGE)
De modo a melhor caracterizar as proteiacutenas isoladas foi realizada uma electroforese em
condiccedilotildees desnaturantes (SDS-PAGE do inglecircs Sodium Dodecyl Sulfate Polyacrilamide Gel
Electrophoresis) O SDS (dodecilo sulfato de soacutedio) eacute um detergente anioacutenico utilizado neste
meacutetodo para desnaturar as proteiacutenas que se ligam ao SDS na proporccedilatildeo 14 g SDS g proteiacutena
(aproximadamente 1 moleacutecula de SDS por cada 2 resiacuteduos de aminoaacutecidos) (Stryer) O SDS induz
uma densidade de carga negativa significativa nas proteiacutenas a que se liga mascarando a sua carga
intriacutenseca pelo que as razotildees cargamassa satildeo tendencialmente idecircnticas Assim a SDS-PAGE
separa as proteiacutenas quando sujeitas a um campo eleacutectrico de acordo com o seu peso molecular
sendo a sua migraccedilatildeo directamente proporcional ao logaritmo deste uacuteltimo (Stryer)
O gel de resoluccedilatildeo com 10 de acrilamida foi obtido atraveacutes da adiccedilatildeo de 167 mL de soluccedilatildeo
stock de acrilamida11
075 mL de tampatildeo de resoluccedilatildeo12
251 mL de aacutegua destilada e 50 L de
SDS 10 Para promover a polimerizaccedilatildeo adicionou-se em seguida 38 L de PSA 10
11
soluccedilatildeo stock de acrilamida soluccedilatildeo de 30 (mv) de acrilamida e 08 (mv) de NNrsquo-metil-bis-acrilamida 12
tampatildeo de resoluccedilatildeo 15 M Tris-HCl pH 88
44
(persulfato de amoacutenia) 10 e 25 L de TEMED 99 (tetrametiletilenodiamina) Agitou-se
cuidadosamente de modo a natildeo formar bolhas e pipetou-se a soluccedilatildeo para a cassette de vidro
previamente montada no suporte De seguida adicionou-se uma peliacutecula de cerca de 2 mm de 2-
metil-2-butanol para verificar posteriormente a polimerizaccedilatildeo Apoacutes a polimerizaccedilatildeo que demora
cerca de 45 minutos verteu-se a agrave superfiacutecie e adicionou-se o gel de concentraccedilatildeo com 5 de
acrilamida constituiacutedo por 03 mL de soluccedilatildeo de stock acrilamida 045 mL de tampatildeo de
concentraccedilatildeo13
094 mL de aacutegua destilada 18 L de SDS 10 135 L de PSA 10 e 2L de
TEMED 99 Colocou-se o pente sobre o gel e deixou-se polimerizar durante cerca de 45
minutos
As amostras para aplicaccedilatildeo no gel de electroforese foram preparadas para uma concentraccedilatildeo de
proteiacutena de 1 mgmL num volume final de 100 L dos quais 25 L correspondem ao tampatildeo da
amostra14
As amostras foram aquecidas a 100ordmC durante 5 minutos Antes da aplicaccedilatildeo das
amostras transferiu-se a cassette para a tina de electroforese agrave qual se adicionou a soluccedilatildeo
tampatildeo de electroforese da tina superior15
e tampatildeo de electroforese da tina inferior16
Aplicaram-
se 10 L das soluccedilotildees de marcadores de peso molecular e 20 L das amostras proteicas
preparadas anteriormente A electroforese decorreu durante cerca de 3h30 min a uma voltagem
de 80 V De seguida transferiu-se o gel para uma tina corando as proteiacutenas durante 1h com
soluccedilatildeo corante17
Descorou-se o gel com soluccedilatildeo descorante18
tendo-se mudado esta soluccedilatildeo ateacute
as bandas se encontrarem bem definidas
Determinou-se a mobilidade relativa das bandas correspondentes aos marcadores de elevado peso
molecular e traccedilou-se a recta de calibraccedilatildeo ndash log(PM) vs log(Rf) onde Rf corresponde agrave
mobilidade relativa Por interpolaccedilatildeo determina-se o PM das proteiacutenas isoladas sabendo a sua
mobilidade relativa Por comparaccedilatildeo com os valores descritos na literatura pode concluir-se
acerca do grau de purificaccedilatildeo
34 Mediccedilatildeo da actividade enzimaacutetica de S1
341 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
13
tampatildeo de concentraccedilatildeo 05 M Tris-HCl pH 68 14
tampatildeo de amostra 320 mM Tris-HCl pH 68 04 M szlig-mercaptoetanol SDS 8 glicerol 15 (vv) e roxo de m-
cresol 0024 15
tampatildeo de electroforese da tina superior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS 16
tampatildeo de electroforese da tina inferior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS acetato de soacutedio
01 M 17
soluccedilatildeo corante ldquoCoomassie Blue R-250rdquo contendo metanol 45 (vv) e aacutecido aceacutetico glacial 10 (vv) 18
soluccedilatildeo descorante metanol 10 (vv) e aacutecido aceacutetico glacial 10 vv)
45
O meacutetodo de Fiske-Subarow modificado consiste na reacccedilatildeo do molibdato de amoacutenio com o
fosfato inorgacircnico em soluccedilatildeo aacutecida com formaccedilatildeo do aacutecido fosfomoliacutebdico Seguidamente daacute-
se a adiccedilatildeo de um agente redutor que provoca a reduccedilatildeo do composto a fosfomolibdato que forma
um complexo de cor azulada cuja absorvacircncia maacutexima ocorre para um comprimento de onda de
660 nm tornando possiacutevel a sua detecccedilatildeo por meacutetodos espectrofotomeacutetricos As actividades Ca2+
-
ATPaacutesica e K+EDTA-ATPaacutesica de S1 foram determinadas atraveacutes deste meacutetodo de acordo com
o descrito por Taussky e Shorr (1953) com mediccedilatildeo da quantidade de fosfato (Pi) libertado
atraveacutes da hidroacutelise de ATP
Para tal obteve-se primeiramente a recta de calibraccedilatildeo com as amostras presentes na Tabela 3II
utilizando uma soluccedilatildeo stock de 5 mM KH2PO4
Tabela 3II ndash Composiccedilatildeo das diferentes amostras utilizadas para determinar a recta de calibraccedilatildeo atraveacutes do meacutetodo
de Fiske-Subbarow modificado
KH2PO4 (L)
([KH2PO4]stock= 5mM)
PO43-
(nmole) H2O (uL)
0 0 1780
5 25 1775
10 50 1770
20 100 1760
40 200 1740
100 500 1680
A cada tubo de Eppendorf adicionaram-se seguidamente 200 L de soluccedilatildeo J19
e 20 L de agente
redutor20
homogeneizando no voacutertex apoacutes cada adiccedilatildeo Incubou-se durante 10 minutos e mediu-
se a Abs660 nm num espectrofotoacutemetro com intervalos de tempo iguais entre cada amostra Traccedilou-
se a recta de calibraccedilatildeo (Abs660 nmvs nmol PO43-
) a partir dos valores obtidos
Para determinar a actividade Ca2+
-ATPaacutesica de S1 prepararam-se tubos de Eppendorf com 400
L de meio de reacccedilatildeo21
25 L CaCl2 ([CaCl2]stock= 100 mM) 25 L ATP ([ATP]stock= 100 mM)
com [S1]ensaios=01 mgmL Perfez-se se necessaacuterio com H2O MilliQ ateacute perfazer um volume de 1
mL Para determinar a actividade K+EDTA-ATPaacutesica utilizou-se uma concentraccedilatildeo idecircntica de
19
Soluccedilatildeo J 5 N H2SO4 25 molibdato de amoacutenio 20
Agente redutor 5 mg aacutecido 1-amino-2-naftol-4-sulfoacutenico 30 mg bissulfito de soacutedio 30 mg sulfito de soacutedio em 26
mL H2O 21
meio de reacccedilatildeo 125 mM Tris-HCl pH 80 0575 M KCl
46
proteiacutena nos ensaios (01 mgmL) e o mesmo meio de reacccedilatildeo Aos tubos de Eppendorf
adicionou-se ainda 123 L de KCl ([KCl]stock = 3M) 25 L de ATP 10 L de EDTA
([EDTA]stock= 100 mM )e o volume de H2O MilliQ necessaacuterio para perfazer 1 mL Em ambas as
situaccedilotildees o uacuteltimo reagente adicionado foi o ATP uma vez que se trata do substrato da reacccedilatildeo
enzimaacutetica
A cineacutetica decorreu a 25ordmC durante 5 minutos para a actividade Ca2+
-ATPaacutesica e durante 3
minutos para a actividade K+EDTA-ATPaacutesica Apoacutes o periacuteodo de incubaccedilatildeo transferiu-se 150
L de amostra para tubos de Eppendorf contendo igual volume de TCA 10 Colocou-se no gelo
e homogeneizou-se no voacutertex Seguidamente centrifugou-se a 1500 g durante 10 minutos e
recolheu-se 200 L de sobrenadante para novos tubos de Eppendorf Adicionou-se a estes 158
mL de H2O MilliQ e 200 L de soluccedilatildeo J Homogeneizou-se no voacutertex e adicionou-se 20 L de
reagente redutor repetindo-se a homogeneizaccedilatildeo Incubou-se durante 10 minutos e mediu-se a
Abs660 nm com o mesmo intervalo de tempo de leitura para cada amostra Por intrapolaccedilatildeo a partir
da recta de calibraccedilatildeo obtida anteriormente determinou-se a quantidade de Pi libertado em cada
ensaio A partir destes resultados determinaram-se as actividades Ca2+
-ATPaacutesica e K+EDTA-
ATPaacutesica do S1 em nmol PO43-
minmg proteiacutena e em s-1
342 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
Da actividade Mg2+
-ATPaacutesica de S1 resulta a formaccedilatildeo dos produtos da reacccedilatildeo de
hidroacutelise do ATP - ADP e Pi Na presenccedila de ADP fosfoenolpiruvato (PEP) e da enzima piruvato
cinase (PK) os produtos resultantes satildeo ATP e piruvato Por acccedilatildeo da enzima lactato
desidrogenase (LDH) este uacuteltimo eacute convertido em lactato ocorrendo simultaneamente a oxidaccedilatildeo
do NADH a NAD+ monitorizada atraveacutes da Abs340 nm Quando o S1 de miosina se encontra no
meio reaccional o ATP eacute hidrolisado formando-se novamente ADP e Pi o que permite a
formaccedilatildeo de um ciclo (fig 21) Se as enzimas se encontrarem em excesso a velocidade de
conversatildeo do NADH eacute limitada pela produccedilatildeo de ADP e Pi Como o ATP eacute regenerado em cada
ciclo a sua concentraccedilatildeo inicial eacute sempre mantida (Bagshaw 1982)
Figura 33ndash Representaccedilatildeo esquemaacutetica do meacutetodo de enzimas acopladas
47
O protocolo utilizado e seguidamente descrito foi adaptado de Cuenda et al (1990 ndash Ver Refordf) A
uma cuvette de 1 mL adicionou-se S1 para uma concentraccedilatildeo final de 005 mgmL 800 L de
meio de reacccedilatildeo22
10 L de PEP ([PEP]stock = 42 mM) 10 L de PK+LDH 11 ([PK+LDH]stock =
1 mgmL) e 15 L de NADH ([NADH]stock=25 mM) Iniciou-se a monitorizaccedilatildeo da reacccedilatildeo
atraveacutes da leitura de Abs340 nm apoacutes a adiccedilatildeo de 20 L de ATP ([ATP]stock=100 mM) A
actividade Mg2+
-ATPaacutesica eacute obtida atraveacutes do declive correspondente agrave oxidaccedilatildeo do NADH Para
a mediccedilatildeo da Mg2+
-ATPase de S1 estimulada por actina adicionou-se F-actina para uma
concentraccedilatildeo final de 2 microM sendo a actividade obtida tambeacutem atraveacutes do declive correspondente
agrave oxidaccedilatildeo do NADH apoacutes a adiccedilatildeo da proteiacutena
35 Interaccedilatildeo entre vanaacutedio e subfragmento-1 de miosina
351 Preparaccedilatildeo das soluccedilotildees de vanaacutedio
As soluccedilotildees de vanadato foram preparadas em aacutegua MilliQ a partir de metavanadato de amoacutenio
(NH4VO3) numa concentraccedilatildeo stock de 50 mM com agitaccedilatildeo magneacutetica e aquecimento (~30 ordmC)
Esta soluccedilatildeo com um valor compreendido entre 6 e 7 designa-se por ldquometavanadatordquo e eacute
constituiacuteda pelas esspeacutecies V1 V2 V4 e V5 (Ref) Acidificando a soluccedilatildeo de metavanadato por
adiccedilatildeo de HCl ateacute um valor de pH proacuteximo de 4 observa-se uma mudanccedila de cor de amarelo
claro para laranja indicativa da formaccedilatildeo da espeacutecie decameacuterica de vanadato De seguida e
imediatamente antes da realizaccedilatildeo dos ensaios descritos o valor do pH da soluccedilatildeo de
decavanadato foi aumentado para cerca de 75 por adiccedilatildeo de NaOH A soluccedilatildeo (2) designada
por ldquometavanadatordquo natildeo foi posteriormente alterada ateacute agrave sua adiccedilatildeo para a realizaccedilatildeo dos ensaios
As soluccedilotildees foram armazenadas a 4ordmC e mantidas em gelo aquando da sua utilizaccedilatildeo
A soluccedilatildeo de V(IV) foi preparada em condiccedilotildees anaeroacutebias por diluiccedilatildeo de sulfato de vanadil
(VOSO4) para uma concentraccedilatildeo final de 5 mM em H2O MilliQ saturada com N2 gasoso
352Anaacutelise por espectroscopia de UV-Vis
A concentraccedilatildeo da espeacutecie decameacuterica em soluccedilatildeo depende do periacuteodo de tempo apoacutes a diluiccedilatildeo
no meio de reacccedilatildeo apropriado (Ref) A velocidade de desoligomerizaccedilatildeo da espeacutecie decameacuterica
pode ser analisada por espectroscopia de absorccedilatildeo na regiatildeo UVVis uma vez que esta espeacutecie
apresenta duas bandas caracteriacutesticas na regiatildeo do visiacutevel (a 358 e 400 nm) que natildeo se verifica
em quaisquer outras espeacutecies oligomeacutericas As bandas supramencionada sapresentam-se bem
22
Meio de reacccedilatildeo 3125 mM Tris-HCl pH 71 375 mM MgCl2
48
resolvidas atraveacutes da obtenccedilatildeo da derivada de 2ordf ordem do espectro apresentando a forma de 2
vales com miacutenimos centrados nos comprimentos de onda supracitados com dependecircncia linear
relativamente agrave concentraccedilatildeo de decavanadato total (Ramos et al 2006)
Assim a estabilidade da espeacutecie decameacuterica apoacutes diluiccedilatildeo da soluccedilatildeo de decavanadato no meio de
reacccedilatildeo dos ensaios (tampatildeo D) para uma concentraccedilatildeo final de 1 mM em vanadato total foi
caracterizada atraveacutes da leitura da Abs400 em funccedilatildeo do tempo tendo-se determinado qual a
ordem de reacccedilatildeo e o tempo de meia-vida da mesma Com base em estudos anteriores acerca da
estabilidade do V10 na presenccedila de diferentes proteiacutenas (Ramos et al 2006) determinou-se
igualmente o tempo de meia vida apoacutes a adiccedilatildeo de 20 microM de S1 de miosina ao meio reaccional
353 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
Os espectros de RMN do isoacutetopo 51
V foram realizados num espectroacutemetro Bruker AM-400 MHz
equipado com uma sonda multinuclear inversa de 5 mm Em tubos de RMN de 5 mm de diacircmetro
preparou-se soluccedilotildees de 500 L de deca- ou metavanadato correspondentes a um intervalo de
concentraccedilotildees de 1 a 10 mM suplementadas com 10 D2O Os espectros foram adquiridos agrave
temperatura ambiente com uma largura espectral de 23 923 Hz um tempo de aquisiccedilatildeo de 0086
s tempo de relaxaccedilatildeo de 001 s e uma sequecircncia de pulsos a 90ordm Utilizou-se a transformada de
Fourier com alargamento das bandas de 20Hz para o processamento dos espectros Os desvios
quiacutemicos das ressonacircncias de 51
V foram obtidos em relaccedilatildeo agrave espeacutecie VOCl3cuja frequecircncia de
ressonacircncia eacute considerada o padratildeo sendo todas as outras referenciadas a partir desta em ppm
As aacutereas dos sinais do RMN correspondentes agraves espeacutecies oligomeacutericas presentes foram obtidas
por integraccedilatildeo dos sinais As concentraccedilotildees relativas das espeacutecies foram calculadas com recurso agrave
equaccedilatildeo
onde A[Vx] corresponde agrave aacuterea referente agrave ressonacircncia da espeacutecie Vx com nordm de agregaccedilatildeo x
A[V]total agrave soma total das aacutereas medidas e [V] a concentraccedilatildeo total de vanaacutedio no ensaio
A teacutecnica de 51
V-RMN foi anteriormente aplicada em estudos de ligaccedilatildeo com S1 de miosina
(Ref) tendo sido demostrado que entre as espeacutecies V1 V2 V4 e V5 a espeacutecie tetrameacuterica possui a
maior afinidade para a miosina (Cremo et al) O desaparecimento ou alargamento dos sinais pode
implicar a ligaccedilatildeo de um dos oxovanadatos laacutebeis agrave miosina sem indicaccedilatildeo de qual uma vez que
49
se processa um equiliacutebrio entre eles (Crans 1990) Assim e tendo em conta as indicaccedilotildees de que
a espeacutecie V10 eacute distinta das restantes no que diz respeito agrave magnitude da interacccedilatildeo com a miosina
e inibiccedilatildeo da actividade ATPaacutesica estimulada por actina (Aureliano 2000 Tiago et al 2002)
torna-se fundamental investigar a interacccedilatildeo das diferentes espeacutecies de vanadato com o S1 de
miosina atraveacutes da teacutecnica de 51
V-RMN
Para tal obtiveram-se espectros de 51
V-RMN resultantes da titulaccedilatildeo de uma soluccedilatildeo 5 mM de
decavanadato total com concentraccedilotildees crescentes de S1 de miosina (0 5 10 15 e 20 microM) em
tampatildeo D Realizou-se ainda ensaios na presenccedila de 2 mM de decavanadato e 2 mM de ATP na
presenccedila e na ausecircncia de S1 de miosina de modo a verificar o efeito que estes ligandos naturais
da enzima teria na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o subfragmento-1
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
Os grupos sulfidrilo (-SH) das cisteiacutenas satildeo resiacuteduos de aminoaacutecidos extremamente sensiacuteveis agrave
oxidaccedilatildeo resultando na sua oxidaccedilatildeo (Dalle-Donne et al 1999) Logo a quantificaccedilatildeo destes
resiacuteduos eacute bastante importante nos estudos de interacccedilatildeo com vanaacutedio jaacute que os resiacuteduos de Cys
com ecircnfase para o SH1 e o SH2 satildeo bastante importantes na monitorizaccedilatildeo das modificaccedilotildees
funcionais da regiatildeo da cabeccedila
O procedimento seguidamente apresentado foi originalmente descrito por Gutieacuterrez-Merino
(1987)) Consiste na reacccedilatildeo entre os grupos ndashSH natildeo oxidados das proteiacutenas e o reagente de
Ellman (DTNB -55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico)) - formando-se o aniatildeo CNT (aniatildeo 3-
carboxilato-4-nitrotiofenolato) numa estequiometria de 11 Este apresenta um maacuteximo de
absorvecircncia a 412 nm (Robyt 1990) Assim apoacutes a incubaccedilatildeo da proteiacutena na presenccedila do metal
a quantidade de cisteiacutenas reduzidas por proteiacutena determina-se atraveacutes dos valores de Abs412
Deste modo colocou-se num tubo de Eppendorf o volume apropriado de S1 dialisado para que a
concentraccedilatildeo no ensaio fosse de 1 mgmL Adicionou-se o volume apropriado de decavanadato
para que a sua concentraccedilatildeo fosse de 0 10 50 100 250 500 ou 1000 M Perfez-se com soluccedilatildeo
D ateacute um volume final de 1 mL e incubou-se a 25ordmC por 20 minutos Apoacutes este periacuteodo colocou-
se 780 L da mistura reaccional numa cuvette de quartzo e adicionou-se 220 L de soluccedilatildeo K23
23
Tampatildeo K Tampatildeo fosfatohidrogenofosfato de soacutedio 025 M pH 72
50
Adicionou-se 15 L de DTNB (em soluccedilatildeo K [DTNB]stock=100 mM) ao meio reaccional e
registou-se o valor de absorvecircncia medido equivalente agraves cisteiacutenas ldquoraacutepidasrdquo (SH1 e SH2
expostos ao solvente) Adicionou-se 001 g de SDS que provoca a desnaturaccedilatildeo da proteiacutena e
consequentemente a exposiccedilatildeo de todos os resiacuteduos de cisteiacutenas que se encontravam ateacute aiacute
inacessiacuteveis no interior da proteiacutena (cisteiacutenas totais) Estes resiacuteduos podem assim reagir com o
DTNB
Estimou-se o nuacutemero de resiacuteduos de cisteiacutenas em todas as amostras com recurso ao coeficiente de
extinccedilatildeo molar () do DTNB (ver ponto 3 dos Anexos) que foi determinado atraveacutes da titulaccedilatildeo
de L-Cisteiacutena com intervalo de concentraccedilotildees compreendidos entre 0 e 50 microM na presenccedila de
25microM de DTNB em tampatildeo D Os valores de absorvecircncia foram obtidos para um comprimento de
onda de 412 nm
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
3621 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
A teacutecnica mais eficiente de anaacutelise da interacccedilatildeo entre o iatildeo vanadilo e proteiacutenas como a
calmodulina e a transferrina entre outras permite a obtenccedilatildeo raacutepida de informaccedilatildeo no que toca agrave
conformaccedilatildeo e amplitude da referida interacccedilatildeo
Os espectros de Ressonacircncia Paramagneacutetica Electroacutenica (RPE) da espeacutecie paramagneacutetica V(IV)
foram realizados a 70K num espectroacutemetro Bruker EMX-300 X-band Todas as soluccedilotildees
utilizadas e amostras foram preparadas em anaerobiose numa cacircmara de luvas de modo a evitar a
reduccedilatildeo de V(IV) a V(V) Em tubos capilares com 02 mL de amostra manteve-se a concentraccedilatildeo
de S1 a 20 M e variou-se a concentraccedilatildeo de vanadilo (0 25 5 10 15 25 50 75 100 150
200 500 e 1000 M [VOSO4]stock=5 mM) em tampatildeo D Apoacutes obtenccedilatildeo dos espectros estes
foram tratados atraveacutes do software SimFonia e WinEPR tendo-se obtido um graacutefico com a
variaccedilatildeo da intensidade do sinal de RPE em funccedilatildeo da concentraccedilatildeo de sulfato de vanadilo
presente Realizaram-se ainda simulaccedilotildees de espectros obtidos com 20 microM de S1 e 1000 microM de
V(IV) com vista agrave obtenccedilatildeo dos ligandos provaacuteveis do vanadilo atraveacutes da anaacutelise da constante
de acoplamento hiperfina (A) e do factor g (g)
4 Conclusotildees gerais
Apesar de jaacute ter sido demonstrado que o vanaacutedio inibe a Na+K
+-ATPase da miosina a funccedilatildeo
deste elemento no tecido muacutesculo esqueleacutetico continua largamente incompreendida A enorme
complexidade quiacutemica deste metal especialmente no que concerne agrave oligomerizaccedilatildeo e reacccedilotildees
51
redox constitui um obstaacuteculo agrave compreensatildeo do comportamento do vanaacutedio nos organismos
vivos Assim a caracterizaccedilatildeo da quiacutemica deste elemento eacute de grande utilidade antes de iniciar o
estudo da sua interacccedilatildeo com o presente sistema
A anaacutelise por espectroscopias de 51
V-RMN e de UV-Vis de soluccedilotildees de vanadato diluiacutedas nos
meios de reacccedilatildeo utilizados no presente relatoacuterio (10 mM HEPES pH 70 25 mM KCl) mostrou
claramente que a espeacutecie decameacuterica (V10) possui um tempo de vida suficientemente longo para a
determinaccedilatildeo do seu efeito isolado ao contraacuterio do que se verifica com as espeacutecies mono- di-
tetra- e pentameacutericas (V1 V2 V4 e V5) Por outro lado a anaacutelise verificou ainda que apoacutes diluiccedilatildeo
das soluccedilotildees de decavanadato no meio de reacccedilatildeo coexistem as espeacutecies V10 e V1 pelo que os
efeitos putativos desta uacuteltima deveratildeo ser subtraiacutedos aquando da monitorizaccedilatildeo dos efeitos da
espeacutecie decameacuterica
O vanaacutedio (V) (vanadato) inibe as enzimas que catalizam reacccedilotildees de transferecircncia de grupos
fosfato como a ATPase da miosina [Goodno 1979] devido agrave sua analogia estrutural com o iatildeo
fosfato Na sua utilizaccedilatildeo como inibidor enzimaacutetico natildeo eacute tida em conta a contribuiccedilatildeo individual
das espeacutecies oligomeacutericas considerando-se normalmente a espeacutecie V1 como inibidora efectiva
[Goodno e Taylor 1982] No presente relatoacuterio verificou-se por espectroscopia de 51
V-RMN que
outros oligoacutemeros de vanadato interagem com o S1 de miosina tendo as espeacutecies V10 e V4 os
maiores niacuteveis de afinidade A sua interacccedilatildeo envolve forccedilas electrostaacuteticas e eacute afectada
negativamente por ligandos naturais de enzima como eacute o caso do ATP
A exposiccedilatildeo do S1 de miosina (005 mgmL) a decavanadato conduz agrave inibiccedilatildeo do capacidade da
proteiacutena de estimular a actividade Mg2+
-ATPaacutesica com um valor de IC50 de 266 plusmn 07 M
decavanadato sendo o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM
decavanadato (80) Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM decavanadato sendo
o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM decavanadato (60)
exercendo um efeito de impedimento agrave ligaccedilatildeo entre o vanaacutedio e o S1
A uma concentraccedilatildeo de 27 microM decavanadato verifica-se a oxidaccedilatildeo de um dos resiacuteduos de Cys
de S1 de miosina seguida pela oxidaccedilatildeo gradual de outro resiacuteduo de Cys ateacute agrave concentraccedilatildeo de
1000 microM Ambos os resiacuteduos oxidados natildeo se encontravam expostos ao solvente utilizado A
oxidaccedilatildeo dos resiacuteduos permite sugerir que esta modificaccedilatildeo oxidativa sofrida pelo S1 contribui
para a diminuiccedilatildeo da sua actividade de ATPase
52
Por espectroscopia de RPE revelaram-se os possiacuteveis ligandos equatoriais do vanaacutedio aquando da
sua interacccedilatildeo com o S1 de miosina como sendo duas moleacuteculas de aacutegua um ligando contendo o
grupo R-OH e outro contendo o grupo R=NR
Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados de
oxidaccedilatildeo (IV) e (V) no S1 de miosina para concentraccedilotildees facilmente alcanccedilaacuteveis no ambiente
terrestre
53
5 Estudos futuros
No sentido do melhor esclarecimento das alteraccedilotildees moleculares e estruturais que ocorrem no
subfragmento 1 quando exposta ao vanaacutedio que conduzem a uma diminuiccedilatildeo da sua funccedilatildeo
sugere-se
- Quantificaccedilatildeo das cisteiacutenas reduzidas de S1 na presenccedila de metavanadato
- Determinaccedilatildeo das actividades Ca2+
- e K+-ATPaacutesica do S1 de miosina na presenccedila de
decavanadato
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina estimulada por actina
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina na presenccedila de
metavanadato
- Realizaccedilatildeo de RMN 2D com variaccedilatildeo das concentraccedilotildees das soluccedilotildees de decavanadato e
metavanadato na presenccedila de S1
-Mutageacutenese dos resiacuteduos de Cys doS1 de miosina de modo a verificar quais os resiacuteduos que
sofrem oxidaccedilatildeo na presenccedila de decavanadato
- Determinaccedilatildeo por calorimetria (DSC Differential Scanning Calorimetry) da variaccedilatildeo da
entalpia (∆H) e da temperatura de desnaturaccedilatildeo do S1 de miosina na presenccedila de vanaacutedio uma
vez que estas propriedades caracteriacutesticas de cada proteiacutena satildeo alteradas apoacutes ocorrecircncia de
modificaccedilotildees estruturais
- Anaacutelise por SDS-PAGE de S1 de miosina apoacutes tratamento com vanaacutedio de modo a determinar
quais os polipeacuteptidos eventualmente formados apoacutes clivagem da proteiacutena
54
6 Referecircncias bibliograacuteficas
- Sellers J Myosins 2nd Edition Oxford University Press 1999
- L Stryer inBiochemistry (Ed WH Freeman 2005 pp
Refordfas Materiais e Meacutetodos
[A] S S Margossian SLowey Methods Enzymol1982 85 55-71
[B] JD Pardue JA Spudich Methods Enzymol 1982 85 164-181
[C] M Bradford Anal Biochem1976 72 248-254
[D] H H Taussky E Shorr J Biol Chem 1953 202 675-685
[E] C R Bagshaw in Outline Studies in Biology ndash Muscle Contraction Chapman and Hall Ed
1982
[F] A Cuenda et al Eur J Biochem1990 194 663-670
55
Anexos
1 Determinaccedilatildeo da recta de calibraccedilatildeo meacutetodo de Bradford
Figura1ndashRepresentaccedilatildeo graacutefica da recta de calibraccedilatildeo (Abs595 nm vs [BSA]) utilizada para o meacutetodo de Bradford
2 Determinaccedilatildeo da recta calibraccedilatildeo para a electroforese em condiccedilotildees desnaturantes
Figura 2 ndash Representaccedilatildeo graacutefica da recta de calibraccedilatildeo (log(Peso Molecular) vs Rf dos marcadores de peso
molecular) onde Rf indica determinada atraveacutes do padratildeo de peso molecular
(poccedilo (a))
3Caacutelculo do coeficiente de extinccedilatildeo molar () para a quantificaccedilatildeo de cisteiacutenas reduzidas
atraveacutes do meacutetodo de Ellman
Traccedilou-se a recta de Abs412 nm meacutedia vs [L-Cisteiacutena] de modo a determinar o valor do coeficiente
de extinccedilatildeo molar definido como o declive daquela recta Assim =8400 M-1
cm-1
como
podemos verificar na Fig 3
A595 nm = 00546[BSA] + 00209 Rsup2 = 099051
Ab
s 59
5 n
m
[BSA] (mgmL)
log PM = -1177 Rf + 2247 Rsup2 = 098957
log
PM
Mobilidade relativa (Rf)
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero
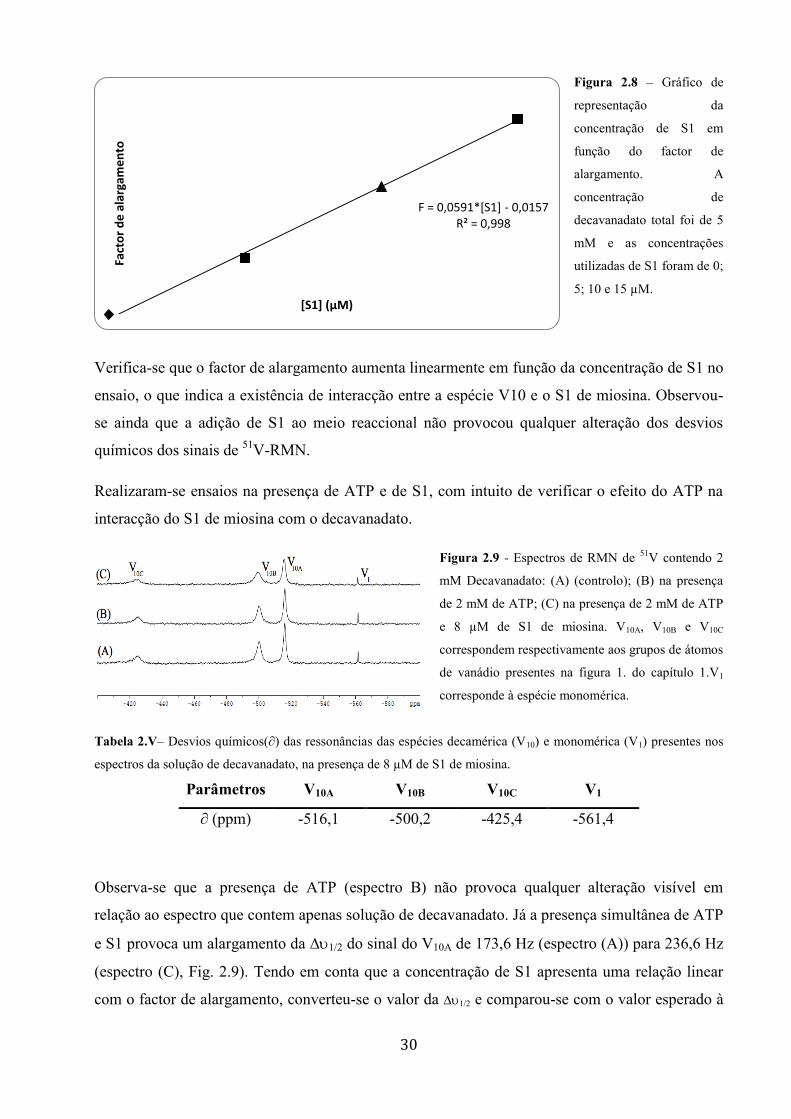
30
Figura 28 ndash Graacutefico de
representaccedilatildeo da
concentraccedilatildeo de S1 em
funccedilatildeo do factor de
alargamento A
concentraccedilatildeo de
decavanadato total foi de 5
mM e as concentraccedilotildees
utilizadas de S1 foram de 0
5 10 e 15 microM
Verifica-se que o factor de alargamento aumenta linearmente em funccedilatildeo da concentraccedilatildeo de S1 no
ensaio o que indica a existecircncia de interacccedilatildeo entre a espeacutecie V10 e o S1 de miosina Observou-
se ainda que a adiccedilatildeo de S1 ao meio reaccional natildeo provocou qualquer alteraccedilatildeo dos desvios
quiacutemicos dos sinais de 51
V-RMN
Realizaram-se ensaios na presenccedila de ATP e de S1 com intuito de verificar o efeito do ATP na
interacccedilatildeo do S1 de miosina com o decavanadato
Figura 29 - Espectros de RMN de 51
V contendo 2
mM Decavanadato (A) (controlo) (B) na presenccedila
de 2 mM de ATP (C) na presenccedila de 2 mM de ATP
e 8 microM de S1 de miosina V10A V10B e V10C
correspondem respectivamente aos grupos de aacutetomos
de vanaacutedio presentes na figura 1 do capiacutetulo 1V1
corresponde agrave espeacutecie monomeacuterica
Tabela 2Vndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies decameacuterica (V10) e monomeacuterica (V1) presentes nos
espectros da soluccedilatildeo de decavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V10A V10B V10C V1
part (ppm) -5161 -5002 -4254 -5614
Observa-se que a presenccedila de ATP (espectro B) natildeo provoca qualquer alteraccedilatildeo visiacutevel em
relaccedilatildeo ao espectro que contem apenas soluccedilatildeo de decavanadato Jaacute a presenccedila simultacircnea de ATP
e S1 provoca um alargamento da ∆12 do sinal do V10A de 1736 Hz (espectro (A)) para 2366 Hz
(espectro (C) Fig 29) Tendo em conta que a concentraccedilatildeo de S1 apresenta uma relaccedilatildeo linear
com o factor de alargamento converteu-se o valor da ∆12 e comparou-se com o valor esperado agrave
F = 00591[S1] - 00157 Rsup2 = 0998
Fact
or
de
ala
rgam
en
to
[S1] (microM)
31
concentraccedilatildeo de S1 utilizada O factor de alargamento obtido para 8 microM ndash 0376 ndash eacute inferior ao
valor esperado agrave mesma concentraccedilatildeo de S1 obtido atraveacutes da equaccedilatildeo da recta apresentada na
Fig 28 ndash 0457 ndash o que indica que o ATP poderaacute exercer um efeito de protecccedilatildeo do S1
diminuindo a sua interacccedilatildeo com a espeacutecieV10
Por uacuteltimo realizaram-se ensaios com metavanadato na presenccedila de 8 microM S1 de modo a verificar
se as espeacutecies oligomeacutericas contidas nesta soluccedilatildeo interagiam com a proteiacutena em estudo
Figura 210 - Espectros de RMN de 51
V contendo 2
mM metavanadato (A) (controlo) (B) na presenccedila
de 8 microM de S1 de miosina V1 V2 e V4
correspondem respectivamente aacutes espeacutecies
monomeacuterica dimeacuterica e tetrameacuterica
Tabela 2VIndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies monomeacuterica (V1) dimeacuterica (V2) e tetrameacuterica (V4)
presentes nos espectros da soluccedilatildeo de metavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V1 V2 V4
part (ppm) -5613 -5747 -5789
Como eacute possiacutevel observar na Fig 210 ocorre uma ligeiro alargamento dos sinal de V4 bem
como uma perda da intensidade pelo que se conclui quedas espeacutecies presentes na soluccedilatildeo de
metavanadato (V1 V2 e V4) apenas a espeacutecie tetrameacuterica interage com o S1 de miosina o que
estaacute de acordo com o reportado por Tiago et al 2002
242 Actividade Mg2+
-ATPaacutesica de S1 de miosina na presenccedila de decanadato
Com o intuito de avaliar o efeito do decavanadato na actividade Mg2+
-ATPaacutesica do S1 de
miosina monitorizou-se a reacccedilatildeo de oxidaccedilatildeo do NADH atraveacutes da leitura de Abs340 na
presenccedila de 005 mgmL de S1 de miosina e no caso da actividade estimulada 2microM de actina em
3125 mM Tris-HCl pH 71 e 375 mM MgCl2 pelo meacutetodo de enzimas acopladas (ver seccccedilatildeo xx
dos MM)Para tal realizaram-se ensaios com vaacuterias concentraccedilotildees de decavanadato total (10 50
100 250 500 e 1000) de modo a determinar qual o seu efeito na actividade MgATPaacutesica de S1
de miosina na ausecircncia e na presenccedila de um dos seus ligandos naturais Os ensaios foram
realizados agrave temperatura ambiente com incubaccedilatildeo preacutevia a 25ordm C durante 20 minutos
32
Figura 211 ndash Representaccedilatildeo
graacutefica da Actividade Mg2+
-
ATPaacutesica de S1 () (os ajustes
teoacutericos estatildeo representado a por
uma linha os resultados
experimentais estatildeo representados
por triacircngulos (Actividade
estimulada) e cruzes (Actividade
natildeo estimulada)) vs
[Decavanadato]total medidas pelo
meacutetodo das enzimas acopladas
(n=3) Os dados foram ajustados ao modelo de decaimento exponencial usando o programa GraphPad Prism para a
determinaccedilatildeo dos valores de IC50 (na ausecircncia de actina IC50= 266 plusmn 07 M decavanadato na presenccedila de actina
IC50= 28 plusmn 1 M decavanadato
A partir do graacutefico presente na Fig 29 verifica-se que o ajuste dos dados experimentais foi bem
sucedido A inibiccedilatildeo da actividade Mg2+
-ATPaacutesica do S1 de miosina eacute afectada em menor
amplitude na presenccedila de actina (60 de inibiccedilatildeo da actividade) do que na sua ausecircncia
(aproximadamente 80 de inibiccedilatildeo de actividade) Devido a esta diferenccedila pode sugerir-se que a
actina iraacute impedir parcialmente a ligaccedilatildeo do decavanadatoao S1 demiosina atraveacutes de
impedimentos esteacutereos ou interacccedilotildees electrostaacuteticas por exemplo A teacutecnica utilizada natildeo nos
permite concluir acerca de que regiatildeo(otildees) ou resiacuteduo(s) da actina participaratildeo na preservaccedilatildeo
parcial da actividade Mg2+
-ATPaacutesica
Por outro lado os valores de IC50 dos dois tipos de actividades natildeo apresentam diferenccedilas
significativas tendo-se obtido 266 plusmn 07 e 28 plusmn 1M decavanadato na ausecircncia e na presenccedila de
actina respectivamente o que corresponde a uma concentraccedilatildeo de cerca de 27 M de V10 O
meio de reacccedilatildeo utilizado nestes ensaios foi 10 mM HEPES e 25 mM KCl com 2 mM ATP Em
(Tiago et al 2004) o valor de IC50 para a actividade Mg2+-ATPaacutesica na ausecircncia de actina foi de
125 plusmn 025 microM enquanto o valor da actividade estimulada por actina foi de 136 plusmn 14 microM O
meio de reacccedilatildeo utilizado nos ensaios foi 20 mM HEPES 50 mM KCl e 2 mM ATP o que resulta
precisamente no dobro da forccedila ioacutenica a que foi realizado o ensaio do presente relatoacuterio
(exceptuando a contribuiccedilatildeo dos produtos da hidroacutelise de ATP uma vez que este meacutetodo
pressupotildee a ocorrecircncia de um ciclo de reacccedilotildees de hidroacutelise e siacutentese deste composto) Ora tendo
sido anteriormente referido que a afinidade do S1 de miosina para com a espeacutecie V10 diminui com
Act
ivid
ade
Mg
2+-A
TP
aacutesic
a d
e S
1 d
e
Mio
sin
a
[Decavanadato] (microM) Actividade Estimulada ()
Actividade Natildeo Estimulada ()
Ajuste Actividade Estimulada
Ajuste Actividade Natildeo Estimulada
33
o aumento da forccedila ioacutenica (Tiago et al 2004 tal poderaacute explicar a diferenccedila obtida entre estes
dois conjuntos de valores
243Quantificaccedilatildeo de cisteiacutenas reduzidas pelo meacutetodo de Ellman
A interacccedilatildeo do vanaacutedio com alguns tipos de aminoaacutecidos sensiacuteveis agrave oxidaccedilatildeo pode fornecer-nos
informaccedilatildeo bastante uacutetil em relaccedilatildeo agraves alteraccedilotildees estruturais que conduzem a uma diminuiccedilatildeo da
actividade fisioloacutegica do S1 de miosina Os estudos realizados por Chasteen 1983 concluiacuteram
que a reduccedilatildeo de vanadato (V) a vanadilo (IV) ocorre na presenccedila de agentes redutores comuns no
ambiente celular de que se destaca o aminoaacutecido cisteiacutena Esta eacute tambeacutem um alvo de
complexaccedilatildeo por parte do iatildeo VO2+
Jaacute Ramos et al (2009) demonstrou que a incubaccedilatildeo de
actina com a soluccedilatildeo de decavanadato induz a oxidaccedilatildeo de resiacuteduos de cisteiacutena e agrave formaccedilatildeo do
oxidovanaacutedio (IV) Este grupo de investigadores descobriu ainda que apenas a espeacutecie V10 induz a
oxidaccedilatildeo das cisteiacutenas ldquoraacutepidasrdquo da actina atraveacutes do processo proposto que eacute parcialmente
responsaacutevel pela inibiccedilatildeo jaacute descrita da actividade de ATPase do S1 de miosina estimulada por
actina quando exposta agrave soluccedilatildeo de decavanadato Esta inibiccedilatildeo apresenta-se afectada com a
presenccedila de ATP uma vez que os resiacuteduos de cisteiacutena permaneceram intactos nos ensaios
realizados com este ligando Jaacute Crans et al (2004) concluiu que o siacutetio activo de muitas fosfatases
(que conteacutem grupos tiol) pode formar um equiliacutebrio redox com o iatildeo vanadato (V) de forma natildeo
reversiacutevel
Os resultados obtidos por Sakurai e Shiromura (1980) sugeriram que o vanaacutedio (V) sofre reduccedilatildeo
a vanaacutedio (IV) sem dependecircncia do valor de pH do ensaio Registaram ainda uma estequiometria
de 31 na reacccedilatildeo do vanadato (V) com os resiacuteduos de Cys enquanto a estequiometria dos ensaios
com o vanadilo (IV) foi de 21
Berlett e Stadtman (1997) observaram que os tipos de aminoaacutecidos mais facilmente oxidaacuteveis
englobam entre outros os resiacuteduos de Cys Trp e MetAssim analisou-se o estado redox dos
grupos tiol dos resiacuteduos de Cys do S1 na presenccedila do decavanadato atraveacutes da monitorizaccedilatildeo da
Abs412 nm na presenccedila de 1 mgmL de S1 de miosina em tampatildeo 10 mM HEPES 25 mM KCl
variando a concentraccedilatildeo da soluccedilatildeo de decavanadato de modo a determinar que tipo de alteraccedilotildees
estruturais ocorrem na sua presenccedila no que diz respeito agrave diminuiccedilatildeo da actividade ATPaacutesica de
S1 na presenccedila de decavanadato
Os resultados referentes agrave quantificaccedilatildeo das cisteiacutenas reduzidas apoacutes o tratamento de S1 com
diferentes concentraccedilotildees de decavanadato em tampatildeo 10 mM HEPES pH 75 e 25 mM KCl a
25ordmC durante 20 minutos encontram-se descritos na Tabela 2VII
Tabela 2VII ndash Cisteiacutenas reduzidas totais e ldquoraacutepidas de S1 (1 mgmL) apoacutes tratamento com decavanadato
34
(010501002505001000 uM) em 10 mM HEPES 25 mM KCl pH 7 a 25ordmC durante 20 minutos A detecccedilatildeo
espectrofotomeacutetrica foi feita com DTNB (e 412=8400 uM-1 cm-1) (n variaacutevel entre 1 e 8)
Amostras Cys raacutepidas Cys totais
11 M S1 (controlo) 22 plusmn 04 106 plusmn 14
11 M S1 + 10 M Decavanadato 22 plusmn 05 95 plusmn 08
11 M S1 + 50 uM Decavanadato 20 plusmn 02 98 plusmn 14
11 M S1 + 100 uM Decavanadato 17 plusmn 03 97 plusmn14
11 M S1 + 250 uM Decavanadato 26 plusmn 02 98 plusmn 08
11 M S1 + 500 uM Decavanadato 21 plusmn 04 90 plusmn 06
11 M S1 + 1000 uM Decavanadato 24 plusmn 0 86 plusmn 08
A partir dos resultados obtidos pode sugerir-se que o S1 possui 11 resiacuteduos de Cys (controlo) o
que estaacute de acordo com o anteriormente descrito Verifica-se que o tratamento da proteiacutena com
concentraccedilotildees crescentes de decavanadato provoca a oxidaccedilatildeo de dois resiacuteduos de Cys do S1 de
miosina natildeo expostos ao solvente No entanto a teacutecnica utilizada natildeo nos permite identificar
quais
Tendo em consideraccedilatildeo que o valor de IC50 obtido para a reacccedilatildeo de inibiccedilatildeo daactividade Mg2+-
ATPaacutesica de S1 na presenccedila de decavanadato eacute de cerca de 27 microM pode sugerir-se que este seraacute o
valor ao qual ocorreraacute a oxidaccedilatildeo do primeiro resiacuteduo de Cys o que poderaacute contribuir para a
diminuiccedilatildeo da actividade de S1 observada
25Interacccedilatildeo de S1 com vanaacutedio(IV)
251Anaacutelise por espectroscopia de Ressonacircncia Paramagneacutetica Electroacutenica
A acumulaccedilatildeo intracelular de oxovanaacutedio (IV) in vivo constitui uma das principais formas de
apresentaccedilatildeo de vanaacutedio no organismo Deste modo torna-se bastante interessante analisar a
interacccedilatildeo entre este iatildeo e o S1 de miosina atraveacutes da teacutecnica de espectroscopia de Ressonacircncia
Paramagneacutetica Electroacutenica o meacutetodo de eleiccedilatildeo do estudo do iatildeo vanadilo e dos complexos por si
formados O electratildeo desemparelhado encontra-se fortemente acoplado ao nuacutecleo 51
V o que
35
resulta num espectro de RPE consistindo de 8 desdobramentos na direcccedilatildeo paralela e outros tantos
na direcccedilatildeo perpendicular (Chasteen 1983)
Os estudos realizados por Ahmed et al (1987) centraram-se na interacccedilatildeo do vanadilo com
proteiacutenas como eacute o caso da insulina transferrina entre outros obtendo um espectro caracterizado
por 8 linhas amplamente espaccediladas (na direcccedilatildeo paralela) Jaacute Ramos et al 2009 concluiu que a
exposiccedilatildeo da soluccedilatildeo de decavanadato agraves formas globular e filamentosa de actina produz um
espectro tiacutepico do iatildeo oxidovanaacutedio (IV) indicando a reduccedilatildeo do vanadato
Para aferir qual a concentraccedilatildeo de S1 que permitia obter uma boa razatildeo sinalruiacutedo titulou-se uma
soluccedilatildeo de sulfato de vanadilo 15 microM ([VOSO4]stock = 5 mM) com concentraccedilotildees crescentes de
proteiacutena (0 5 10 15 e 20 microM) em 10 mM HEPES 25 mM KCl A existecircncia de uma correlaccedilatildeo
linear entre a concentraccedilatildeo de proteiacutena e a intensidade dos sinais de RPE correspondente agrave banda
-12 perpendicular encontra-se representada na Fig 210
Figura 2 12ndash Representaccedilatildeo graacutefica da variaccedilatildeo da intensidade do sinal de RPE vs a concentraccedilatildeo de S1 utilizada
Apoacutes verificar que com uma concentraccedilatildeo de S1 igual a 20 microM se obtinham sinais com uma razatildeo
sinalruiacutedo adequada ao prosseguimento dos estudos sobre a interacccedilatildeo V(IV)-S1 de miosina
titulou-se uma soluccedilatildeo de 20 microM S1 com 0 25 5 10 15 25 50 75 100 150 200 500 e 1000
microM VOSO4
Traccedilou-se o graacutefico da intensidade da banda -12 perpendicular do sinal de EPR em funccedilatildeo da
concentraccedilatildeo de VOSO4 tendo-se determinado uma constante de dissociaccedilatildeo (KD) do complexo
S1-vanadilo de 8748 plusmn 2621 microM V(IV)
y = 016786x + 122435
Rsup2 = 092992759
Inte
nsi
dad
e d
o s
inal
de
RP
E
[S1] (microM)
36
Figura 213 ndash Representaccedilatildeo graacutefica da variaccedilatildeo do sinal de RPE vs a concentraccedilatildeo de vanadilo utilizada (os valores
experimentais estatildeo representados por losangos o seu ajuste estaacute representado pela linha)
A simulaccedilatildeo do espectro de EPR foi efectuada a partir do espectro obtido para uma concentraccedilatildeo
de 1 mM VOSO4 apoacutes adiccedilatildeo de 20 microM S1 de miosina tendo-se obtido os valores da constante de
acoplamento hiperfina (A=5016) e do factor g (g0=195)
Figura 214 ndash Representaccedilatildeo graacutefica do espectro de EPR obtido com 1000 microM de vanadilo e 20 microM de S1 (a cinzento
escuro) A simulaccedilatildeo do espectro encontra-se a cinzento claro
Por comparaccedilatildeo destes com os valores presentes na tabela de correspondecircncia xxx eacute possiacutevel
predizer quais os ligandos provaacuteveis do vanadilo na presenccedila de proteiacutena Assim concluiu-se que
o vanadilo estaacute coordenado com duas moleacuteculas de aacutegua um grupo hidroxilo e um grupo R=NR
O grupo hidroxilo pode indicar os resiacuteduos de aminoaacutecidos Tyr Treonina e Serina enquanto o
grupo R=NR poderaacute indicar a presenccedila de Arginina e Histidina como ligandos do vanadilo
Inte
nsi
dad
e d
os
sinai
s d
e R
PE
[Vanadilo] (microM)
Ajuste dos dados exp Dados experimentais
Campo magneacutetico (G)
Espectro Experimental Ajuste
37
3 Procedimento Experimental
31 Reagentes
Todos os reagentes utilizados satildeo de elevado grau de pureza (analiacutetico) Apresentam-se as suas
foacutermulas quiacutemicas ou abreviaturas quando necessaacuterio acrescentou-se a sua descriccedilatildeo Entre
parecircntesis colocou-se as empresas que os comercializam
Aacutecido aceacutetico glacial 100 (Riedel) Acrilamida 99 (Merck) Azul de bromofenol (Riedel-de-
Haumlen) -Mercaptoetanol (Sigma) Bissulfito de soacutedio (Sigma)Bradford (reagente de)
(BioRad)BSA (albumina de soro bovino Sigma-Aldrich) CaCl22H2O (cloreto de caacutelcio
dihidratado Fluka) CH3CH2OH 96 (etanol Panreac) Coomassie Blue R250 (Merck) D2O
(oacutexido de deuteacuterio SDS) DTNB (55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico) Aldrich) EDTA (aacutecido
etilenodiaminatetraaceacutetico Merck) Fosfoenolpiruvato 98 (Fluka) Glicerol (Sigma) Glicina
985 (Panreac) HCl 37 (Riedel-de Haumlen) H2SO4 96 (Pronalab) HEPES 99 (aacutecido 4-(2-
hidroxiletil)-1-piperazineetanosulfoacutenico Sigma) KH2PO4 99 (hidrogenofosfato de potaacutessio)
MgATP (Sigma) Na
ATP (Sigma) Na2HPO4 99 (hidrogenofosfato de soacutedio Riedel-de Haumlen)
NaH2PO4 (fosfato de soacutedio ) NaOH 986 (hidroacutexido de soacutedio Pronalab) KCl 99 (Cloreto de
potaacutessio Panreac) L-Cys 995 (enantioacutemero levoacutegiro do resiacuteduo de Cisteiacutena Fluka)Metanol
99 (Merck) Molibdato de amoacutenio (Sigma)NNrsquo-metil-bis-acrilamida 98 (Merck) NH4VO3
99 (metavanadato de amoacutenia Sigma) PSA 10 (persulfato de amoacutenio Merck) SDS
(dodecilo sulfato de soacutedio Sigma) Sulfito de soacutedio (Sigma) VOSO4 (sulfato de vanadilo
Aldrich) TEMED 99 (Tetrametiletilenodiamina Merck) Tris (2-amino-2-hidroximetil-13-
propanodiol Sigma)
32 Meacutetodos
321 Isolamento e purificaccedilatildeo das proteiacutenas de muacutesculo esqueleacutetico de coelho
3211 Purificaccedilatildeo da miosina preparaccedilatildeo de Subfragmento 1 por digestatildeo quimotriacutepsica
da miosina
O subfragmento-1 de miosina-II (S1) foi gentilmente cedido pelo Prof Doutor Manuel Aureliano
do Departamento de Quiacutemica Bioquiacutemica e Farmaacutecia da Faculdade de Ciecircncias e Tecnologia da
Universidade do Algarve
No entanto descreve-se abaixo de forma resumida o procedimento efectuado
38
32111 Purificaccedilatildeo da miosina
O protocolo de isolamento foi adaptado do meacutetodo previamente descrito por Margossian e Lowey
(1982) apresentando-se de forma esquemaacutetica na Fig 31
Sucintamente os tecidos musculares dorsais e das pernas foram extraiacutedos apoacutes o sacrifiacutecio do
animal (Fig31) De seguida foram adicionados 3 volumes da soluccedilatildeo de extracccedilatildeo A2
Centrifugou-se a soluccedilatildeo e filtrou-se o sobrenadante O sedimento foi entatildeo diluiacutedo em 14
volumes de aacutegua destilada de modo a que a forccedila ioacutenica atingisse 004 M Apoacutes adiccedilatildeo de szlig-
mercaptoetanol (szlig-me) deixou-se a precipitar a 4ordmC removendo o sobrenadante Centrifugou-se
novamente e ressuspendeu-se o sedimento na soluccedilatildeo de extracccedilatildeo A Adicionou-se a soluccedilatildeo
tampatildeo B3 para que a forccedila ioacutenica atingisse 05 M Homogeneizou-se e deixou-se repousar
durante 2h Homogeneizou-se novamente e centrifugou-se de modo a ocorrer precipitaccedilatildeo dos
complexos de actomiosina e proteiacutena desnaturada Apoacutes filtraccedilatildeo do sobrenadante em latilde de vidro
quantificou-se a soluccedilatildeo de miosina resultante pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental)
2 Soluccedilatildeo de extracccedilatildeo A 0012M K2CO3 004M KHCO3 06M KCl pH 65
3 Soluccedilatildeo-tampatildeo B Tris 100 mM pH 7 KCl 2M
39
Figura 31 ndash Representaccedilatildeo esquemaacutetica do protocolo de isolamento de miosina (Margossian e Lowey 1982) Todo o
procedimento foi realizado a 4ordmC (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
32112 Preparaccedilatildeo de Subfragmento-1 de miosina por digestatildeo com quimiotripsina
O S1 de miosina foi isolado apoacutes diaacutelise da miosina durante a noite contra a soluccedilatildeo C4 A
presenccedila do EDTA eacute essencial uma vez que a sua substituiccedilatildeo por MgCl2resulta na produccedilatildeo de
HMM em vez de S1 A suspensatildeo de miosina foi equilibrada e digerida com -quimotripsina
A digestatildeo foi interrompida pela adiccedilatildeo de PMSF Seguidamente ultracentrifugou-se o produto
da digestatildeo desprezando-se o sedimento (Fig 32) A soluccedilatildeo de sobrenadante maioritariamente
S1 foi quantificada pela leitura directa da Abs280 ou pelo meacutetodo de Bradford (ver as secccedilotildees
3311 e 3312 do Procedimento Experimental respectivamente) e armazenada em 10 sacarose
e 2 mM szlig-Me a -80ordmC
4 Soluccedilatildeo C 25mM Tris pH 70 012M KCl 1mM EDTA
40
Antes da realizaccedilatildeo dos ensaios descritos no presente relatoacuterio o S1 de miosina foi dialisado
contra a soluccedilatildeo D5 de modo a remover qualquer composto que pudesse complexar como
vanaacutedio Seguidamente quantificou-se a proteiacutena pelo meacutetodo de Bradford ou pelo meacutetodo
directo (ver secccedilotildees 3311 e 3312do Procedimento Experimtental)
Figura 3 2ndash Representaccedilatildeo esquemaacutetica do protocolo de preparaccedilatildeo do S1 de miosina por digestatildeo quimiotripsina de
acordo com o previamente descrito por Margossian e Lowey (1982) Todo o procedimento foi realizado a 4ordm C
excepto quando indicaccedilatildeo do contraacuterio (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
3212 Isolamento e purificaccedilatildeo de actina
Isolou-se e purificou-se a actina segundo o meacutetodo descrito por Pardee e Spudish que
compreende dois passos ndash a obtenccedilatildeo de um resiacuteduo seco denominado ldquopoacute de acetonardquo e a
extracccedilatildeo subsequente da actina a partir do ldquopoacute de acetonardquo na forma de G-Actina com a
posterior polimerizaccedilatildeo a F-Actina
32121 Obtenccedilatildeo do ldquopoacute de acetonardquo
Sacrificou-se o coelho com uma pancada atraacutes do pescoccedilo e sangrou-se o animal apoacutes o corte da
arteacuteria jugular Esfolou-se e colocou-se numa tina com gelo Retiraram-se as partes de tecido
conjuntivo que envolvem o tecido muscular com auxiacutelio de uma pinccedila e bisturi Seguidamente
retirou-se os muacutesculos dorsais e os muacutesculos brancos das pernas Passou-se os muacutesculos por aacutegua
destilada de modo a remover as impurezas e pesou-se num copo Trituraram-se os muacutesculos
numa maacutequina de triturar carne e extraiu-se o resultante com 3 volumes da soluccedilatildeo E6 durante 10
minutos com agitaccedilatildeo suave Centrifugou-se a suspensatildeo a 4000 g durante 10
5 Soluccedilatildeo D HEPES 10 mM pH 70 KCl 25 mM
6 Soluccedilatildeo E 01 M KCl 015 M K2HPO4 pH 65
41
minutosdesprezando-se o sobrenadante Ressuspendeu-se o sedimento em 6 volumes de soluccedilatildeo
F7durante 10 minutos com agitaccedilatildeo suave Verificou-se se o pH da soluccedilatildeo estava compreendido
entre 82 e 85 acertando-se estecom soluccedilatildeo G8 se necessaacuterio Centrifugou-se durante 10
minutos a 4000 g Desprezou-se o sobrenadante e ressuspendeu-se o sedimento em 3 volumes de
soluccedilatildeo H9 durante 10 minutos Centrifugou-se novamente a 4000 gdurante 10 minutos
desprezando-se o sobrenadante Ressuspendeu-se o sedimento duas vezes consecutivas em 6
volumes de aacutegua destilada durante 5 minutos Centrifugou-se a 4000 g durante 10 minutos apoacutes
cada adiccedilatildeo de H2O
Seguidamente ressuspendeu-se o sedimento 5 vezes consecutivas utilizando 3 volumes de
acetona de cada vez agitando suavemente durante 10 minutos agrave temperatura ambiente Colocou-
se o resiacuteduo filtrado numa tina de vidro sobre papel de filtro deixando-se evaporar o solvente
durante a noite num nicho Triturou-se o resiacuteduo seco numa maacutequina de triturar carne e deixou-se
novamente evaporar o solvente O ldquopoacute de acetonardquo obtido foi armazenado a -20 ordmC
32122 Purificaccedilatildeo da actina a partir de ldquopoacute de acetonardquo
Apoacutes a obtenccedilatildeo do poacute de acetona como acima indicado efectuou-se o protocolo seguidamente
descrito de modo a obter monoacutemeros de actina (G-Actina) e actina filamentosa (F-Actina)
Pesou-se 35 g de poacute de acetona e adicionaram-se 50 mL de soluccedilatildeo-tampatildeo I10
sendo a
extracccedilatildeo feita com agitaccedilatildeo suave durante 30 minutos Separou-se o extracto do poacute de acetona
hidratado filtrando a suspensatildeo com 4 camadas de gaze esterilizada tendo o cuidado de natildeo
espremer o resiacuteduo Voltou a extrair-se com 50 mL de soluccedilatildeo I durante 30 minutos mexendo
lentamente Adicionou-se este ao extracto obtido anteriormente Centrifugou-se o extracto total a
16 000 g durante 30 minutos Decantou-se para uma proveta sobre a qual se colocou gaze para
que o sedimento solto natildeo se juntasse ao sobrenadante e leu-se o volume De seguida adicionou-
se KCl ateacute uma concentraccedilatildeo final de 50 mM MgCl2 ateacute 2mM e ATP ateacute 1mM lentamente e com
agitaccedilatildeo contiacutenua de modo a promover a polimerizaccedilatildeo dos monoacutemeros de actina
Deixou-se polimerizar durante 2h a 25ordmC observando-se a presenccedila de pequenas bolhas de ar
indicativas de um aumento de viscosidade da soluccedilatildeo Arrefeceu-se em gelo durante 15 minutos e
adicionou-se KCl ateacute uma concentraccedilatildeo final de 06M Mexeu-se lentamente durante 30 minutos
apoacutes os quais se ultracentrifugou a 110 000g durante 1hora Desprezou-se o sobrenadante
ressuspendeu-se o sedimento com soluccedilatildeo tampatildeo E e transferiu-se para um potter
Dialisou-se durante a noite contra 1L de soluccedilatildeo-tampatildeo I a uma temperatura de 4ordmC com rotaccedilatildeo
7 Soluccedilatildeo F 005 M NaHCO3 pH 82-85
8 Soluccedilatildeo G 1M Na2CO3
9 Soluccedilatildeo H 1 mM EDTA pH 70
10 Soluccedilatildeo I 2 mM Tris-HCl pH 8 02 mM CaCl2 02 mM Na2ATP 05 mM szlig-Me 0005 NaN3
42
contiacutenua e mudanccedila de tampatildeo 2 a 3 vezes De seguida a actina foi centrifugada a 110
000gdurante 1hora Decantou-se a soluccedilatildeo de G-Actina (sobrenadante) e desprezou-se o
sedimento Determinou-se a concentraccedilatildeo de G-Actina pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental) A soluccedilatildeo de G-actina foi armazenada a uma temperatura de 4ordmC
apoacutes adiccedilatildeo de 002NaN3 mantendo-se estaacutevel durante 3 a 5 dias
331 Quantificaccedilatildeo das proteiacutenas isoladas
3311 Quantificaccedilatildeo pelo meacutetodo de Bradford
O meacutetodo de Bradford (Bradford 1976) baseia-se na utilizaccedilatildeo do reagente de Bradford
composto por entre outros aacutecido fosfoacuterico e o corante ldquoCoomassierdquo Este corante possui carga
negativa e a sua coloraccedilatildeo varia entre a vermelha (Absmax=465 nm) predominante em soluccedilatildeo e a
azul (Absmax=595 nm) presente aquando da ligaccedilatildeo aos grupos amina das proteiacutenas Em soluccedilotildees
aacutecidas diluiacutedas a coloraccedilatildeo varia proporcionalmente com a quantidade do corante ligado agrave
proteiacutena que por sua vez eacute directamente proporcional agrave concentraccedilatildeo da proteiacutena permitindo a
sua quantificaccedilatildeo ateacute a um limite de detecccedilatildeo de 20 gmL (Bradford 1976)
Deste modo prepararam-se amostras de BSA ([BSA]stock= 1mgmL) utilizando um intervalo de
concentraccedilotildees compreendido entre 0 e 15 gmL de acordo com a tabela 3I Apoacutes a adiccedilatildeo da
H2O MilliQ e do reagente de Bradford as soluccedilotildees foram sucessivamente homogeneizadas no
voacutertex
Tabela 3Indash Amostras utilizadas para a obtenccedilatildeo da recta de calibraccedilatildeo no meacutetodo de Bradford
BSA (L) BSA (g) H2O MilliQ (L) Reagente de Bradford (L)
0 0 800 200
125 125 79875 200
25 25 7975 200
5 5 795 200
10 10 790 200
15 15 785 200
Apoacutes incubaccedilatildeo a 25ordmC por 5 minutos leram-se os valores de Abs595 num
espectrofotoacutemetro (Shimadzu UV-160A) para todas as amostras Seguidamente traccedilou-se a recta
de calibraccedilatildeo (Abs595vs quantidade de BSA (g))
43
Para determinar a concentraccedilatildeo de proteiacutena apoacutes o processo de isolamento purificaccedilatildeo e
diaacutelise quando necessaacuterio colocou-se 5 L da amostra num tubo de Eppendorf e adicionou-se
795 L de H2O MilliQ homogeneizando a soluccedilatildeo no voacutertex Adicionou-se posteriormente 200
L de reagente de Bradford e repetiu-se a homogeneizaccedilatildeo Incubou-se a mistura a 25ordmC por 5
minutos apoacutes os quais se leu os valores de Abs595 nm no espectrofotoacutemetro Por interpolaccedilatildeo deste
valor a partir da recta de calibraccedilatildeo determinou-se a concentraccedilatildeo de proteiacutena na amostra
3312 Quantificaccedilatildeo pelo meacutetodo directo
A quantificaccedilatildeo das amostras proteicas pode tambeacutem ser efectuada atraveacutes da leitura directa da
absorvacircncia tendo por base a lei de Beer-Lambert onde Abs corresponde agrave
absorvacircncia ao comprimento de onda ao qual o ensaio foi realizado corresponde ao coeficiente
de extinccedilatildeo (substituiacutedo pela letra a se as unidades forem distintas da molaridade) b corresponde
ao percurso oacuteptico da radiaccedilatildeo na ceacutelula amostral e c corresponde agrave concentraccedilatildeo amostral
Assim no caso de S1 de miosina mediu-se a Abs280e calculou-se a concentraccedilatildeo da proteiacutena
sabendo que o coeficiente de extinccedilatildeo de S1 eacute (Ref)
No caso da actina mediu-se a Abs290ecalculou-se a concentraccedilatildeo da proteiacutena sabendo que o
coeficiente de extinccedilatildeo da actina eacute (Ref)
332Electroforese em condiccedilotildees desnaturantes (SDS-PAGE)
De modo a melhor caracterizar as proteiacutenas isoladas foi realizada uma electroforese em
condiccedilotildees desnaturantes (SDS-PAGE do inglecircs Sodium Dodecyl Sulfate Polyacrilamide Gel
Electrophoresis) O SDS (dodecilo sulfato de soacutedio) eacute um detergente anioacutenico utilizado neste
meacutetodo para desnaturar as proteiacutenas que se ligam ao SDS na proporccedilatildeo 14 g SDS g proteiacutena
(aproximadamente 1 moleacutecula de SDS por cada 2 resiacuteduos de aminoaacutecidos) (Stryer) O SDS induz
uma densidade de carga negativa significativa nas proteiacutenas a que se liga mascarando a sua carga
intriacutenseca pelo que as razotildees cargamassa satildeo tendencialmente idecircnticas Assim a SDS-PAGE
separa as proteiacutenas quando sujeitas a um campo eleacutectrico de acordo com o seu peso molecular
sendo a sua migraccedilatildeo directamente proporcional ao logaritmo deste uacuteltimo (Stryer)
O gel de resoluccedilatildeo com 10 de acrilamida foi obtido atraveacutes da adiccedilatildeo de 167 mL de soluccedilatildeo
stock de acrilamida11
075 mL de tampatildeo de resoluccedilatildeo12
251 mL de aacutegua destilada e 50 L de
SDS 10 Para promover a polimerizaccedilatildeo adicionou-se em seguida 38 L de PSA 10
11
soluccedilatildeo stock de acrilamida soluccedilatildeo de 30 (mv) de acrilamida e 08 (mv) de NNrsquo-metil-bis-acrilamida 12
tampatildeo de resoluccedilatildeo 15 M Tris-HCl pH 88
44
(persulfato de amoacutenia) 10 e 25 L de TEMED 99 (tetrametiletilenodiamina) Agitou-se
cuidadosamente de modo a natildeo formar bolhas e pipetou-se a soluccedilatildeo para a cassette de vidro
previamente montada no suporte De seguida adicionou-se uma peliacutecula de cerca de 2 mm de 2-
metil-2-butanol para verificar posteriormente a polimerizaccedilatildeo Apoacutes a polimerizaccedilatildeo que demora
cerca de 45 minutos verteu-se a agrave superfiacutecie e adicionou-se o gel de concentraccedilatildeo com 5 de
acrilamida constituiacutedo por 03 mL de soluccedilatildeo de stock acrilamida 045 mL de tampatildeo de
concentraccedilatildeo13
094 mL de aacutegua destilada 18 L de SDS 10 135 L de PSA 10 e 2L de
TEMED 99 Colocou-se o pente sobre o gel e deixou-se polimerizar durante cerca de 45
minutos
As amostras para aplicaccedilatildeo no gel de electroforese foram preparadas para uma concentraccedilatildeo de
proteiacutena de 1 mgmL num volume final de 100 L dos quais 25 L correspondem ao tampatildeo da
amostra14
As amostras foram aquecidas a 100ordmC durante 5 minutos Antes da aplicaccedilatildeo das
amostras transferiu-se a cassette para a tina de electroforese agrave qual se adicionou a soluccedilatildeo
tampatildeo de electroforese da tina superior15
e tampatildeo de electroforese da tina inferior16
Aplicaram-
se 10 L das soluccedilotildees de marcadores de peso molecular e 20 L das amostras proteicas
preparadas anteriormente A electroforese decorreu durante cerca de 3h30 min a uma voltagem
de 80 V De seguida transferiu-se o gel para uma tina corando as proteiacutenas durante 1h com
soluccedilatildeo corante17
Descorou-se o gel com soluccedilatildeo descorante18
tendo-se mudado esta soluccedilatildeo ateacute
as bandas se encontrarem bem definidas
Determinou-se a mobilidade relativa das bandas correspondentes aos marcadores de elevado peso
molecular e traccedilou-se a recta de calibraccedilatildeo ndash log(PM) vs log(Rf) onde Rf corresponde agrave
mobilidade relativa Por interpolaccedilatildeo determina-se o PM das proteiacutenas isoladas sabendo a sua
mobilidade relativa Por comparaccedilatildeo com os valores descritos na literatura pode concluir-se
acerca do grau de purificaccedilatildeo
34 Mediccedilatildeo da actividade enzimaacutetica de S1
341 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
13
tampatildeo de concentraccedilatildeo 05 M Tris-HCl pH 68 14
tampatildeo de amostra 320 mM Tris-HCl pH 68 04 M szlig-mercaptoetanol SDS 8 glicerol 15 (vv) e roxo de m-
cresol 0024 15
tampatildeo de electroforese da tina superior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS 16
tampatildeo de electroforese da tina inferior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS acetato de soacutedio
01 M 17
soluccedilatildeo corante ldquoCoomassie Blue R-250rdquo contendo metanol 45 (vv) e aacutecido aceacutetico glacial 10 (vv) 18
soluccedilatildeo descorante metanol 10 (vv) e aacutecido aceacutetico glacial 10 vv)
45
O meacutetodo de Fiske-Subarow modificado consiste na reacccedilatildeo do molibdato de amoacutenio com o
fosfato inorgacircnico em soluccedilatildeo aacutecida com formaccedilatildeo do aacutecido fosfomoliacutebdico Seguidamente daacute-
se a adiccedilatildeo de um agente redutor que provoca a reduccedilatildeo do composto a fosfomolibdato que forma
um complexo de cor azulada cuja absorvacircncia maacutexima ocorre para um comprimento de onda de
660 nm tornando possiacutevel a sua detecccedilatildeo por meacutetodos espectrofotomeacutetricos As actividades Ca2+
-
ATPaacutesica e K+EDTA-ATPaacutesica de S1 foram determinadas atraveacutes deste meacutetodo de acordo com
o descrito por Taussky e Shorr (1953) com mediccedilatildeo da quantidade de fosfato (Pi) libertado
atraveacutes da hidroacutelise de ATP
Para tal obteve-se primeiramente a recta de calibraccedilatildeo com as amostras presentes na Tabela 3II
utilizando uma soluccedilatildeo stock de 5 mM KH2PO4
Tabela 3II ndash Composiccedilatildeo das diferentes amostras utilizadas para determinar a recta de calibraccedilatildeo atraveacutes do meacutetodo
de Fiske-Subbarow modificado
KH2PO4 (L)
([KH2PO4]stock= 5mM)
PO43-
(nmole) H2O (uL)
0 0 1780
5 25 1775
10 50 1770
20 100 1760
40 200 1740
100 500 1680
A cada tubo de Eppendorf adicionaram-se seguidamente 200 L de soluccedilatildeo J19
e 20 L de agente
redutor20
homogeneizando no voacutertex apoacutes cada adiccedilatildeo Incubou-se durante 10 minutos e mediu-
se a Abs660 nm num espectrofotoacutemetro com intervalos de tempo iguais entre cada amostra Traccedilou-
se a recta de calibraccedilatildeo (Abs660 nmvs nmol PO43-
) a partir dos valores obtidos
Para determinar a actividade Ca2+
-ATPaacutesica de S1 prepararam-se tubos de Eppendorf com 400
L de meio de reacccedilatildeo21
25 L CaCl2 ([CaCl2]stock= 100 mM) 25 L ATP ([ATP]stock= 100 mM)
com [S1]ensaios=01 mgmL Perfez-se se necessaacuterio com H2O MilliQ ateacute perfazer um volume de 1
mL Para determinar a actividade K+EDTA-ATPaacutesica utilizou-se uma concentraccedilatildeo idecircntica de
19
Soluccedilatildeo J 5 N H2SO4 25 molibdato de amoacutenio 20
Agente redutor 5 mg aacutecido 1-amino-2-naftol-4-sulfoacutenico 30 mg bissulfito de soacutedio 30 mg sulfito de soacutedio em 26
mL H2O 21
meio de reacccedilatildeo 125 mM Tris-HCl pH 80 0575 M KCl
46
proteiacutena nos ensaios (01 mgmL) e o mesmo meio de reacccedilatildeo Aos tubos de Eppendorf
adicionou-se ainda 123 L de KCl ([KCl]stock = 3M) 25 L de ATP 10 L de EDTA
([EDTA]stock= 100 mM )e o volume de H2O MilliQ necessaacuterio para perfazer 1 mL Em ambas as
situaccedilotildees o uacuteltimo reagente adicionado foi o ATP uma vez que se trata do substrato da reacccedilatildeo
enzimaacutetica
A cineacutetica decorreu a 25ordmC durante 5 minutos para a actividade Ca2+
-ATPaacutesica e durante 3
minutos para a actividade K+EDTA-ATPaacutesica Apoacutes o periacuteodo de incubaccedilatildeo transferiu-se 150
L de amostra para tubos de Eppendorf contendo igual volume de TCA 10 Colocou-se no gelo
e homogeneizou-se no voacutertex Seguidamente centrifugou-se a 1500 g durante 10 minutos e
recolheu-se 200 L de sobrenadante para novos tubos de Eppendorf Adicionou-se a estes 158
mL de H2O MilliQ e 200 L de soluccedilatildeo J Homogeneizou-se no voacutertex e adicionou-se 20 L de
reagente redutor repetindo-se a homogeneizaccedilatildeo Incubou-se durante 10 minutos e mediu-se a
Abs660 nm com o mesmo intervalo de tempo de leitura para cada amostra Por intrapolaccedilatildeo a partir
da recta de calibraccedilatildeo obtida anteriormente determinou-se a quantidade de Pi libertado em cada
ensaio A partir destes resultados determinaram-se as actividades Ca2+
-ATPaacutesica e K+EDTA-
ATPaacutesica do S1 em nmol PO43-
minmg proteiacutena e em s-1
342 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
Da actividade Mg2+
-ATPaacutesica de S1 resulta a formaccedilatildeo dos produtos da reacccedilatildeo de
hidroacutelise do ATP - ADP e Pi Na presenccedila de ADP fosfoenolpiruvato (PEP) e da enzima piruvato
cinase (PK) os produtos resultantes satildeo ATP e piruvato Por acccedilatildeo da enzima lactato
desidrogenase (LDH) este uacuteltimo eacute convertido em lactato ocorrendo simultaneamente a oxidaccedilatildeo
do NADH a NAD+ monitorizada atraveacutes da Abs340 nm Quando o S1 de miosina se encontra no
meio reaccional o ATP eacute hidrolisado formando-se novamente ADP e Pi o que permite a
formaccedilatildeo de um ciclo (fig 21) Se as enzimas se encontrarem em excesso a velocidade de
conversatildeo do NADH eacute limitada pela produccedilatildeo de ADP e Pi Como o ATP eacute regenerado em cada
ciclo a sua concentraccedilatildeo inicial eacute sempre mantida (Bagshaw 1982)
Figura 33ndash Representaccedilatildeo esquemaacutetica do meacutetodo de enzimas acopladas
47
O protocolo utilizado e seguidamente descrito foi adaptado de Cuenda et al (1990 ndash Ver Refordf) A
uma cuvette de 1 mL adicionou-se S1 para uma concentraccedilatildeo final de 005 mgmL 800 L de
meio de reacccedilatildeo22
10 L de PEP ([PEP]stock = 42 mM) 10 L de PK+LDH 11 ([PK+LDH]stock =
1 mgmL) e 15 L de NADH ([NADH]stock=25 mM) Iniciou-se a monitorizaccedilatildeo da reacccedilatildeo
atraveacutes da leitura de Abs340 nm apoacutes a adiccedilatildeo de 20 L de ATP ([ATP]stock=100 mM) A
actividade Mg2+
-ATPaacutesica eacute obtida atraveacutes do declive correspondente agrave oxidaccedilatildeo do NADH Para
a mediccedilatildeo da Mg2+
-ATPase de S1 estimulada por actina adicionou-se F-actina para uma
concentraccedilatildeo final de 2 microM sendo a actividade obtida tambeacutem atraveacutes do declive correspondente
agrave oxidaccedilatildeo do NADH apoacutes a adiccedilatildeo da proteiacutena
35 Interaccedilatildeo entre vanaacutedio e subfragmento-1 de miosina
351 Preparaccedilatildeo das soluccedilotildees de vanaacutedio
As soluccedilotildees de vanadato foram preparadas em aacutegua MilliQ a partir de metavanadato de amoacutenio
(NH4VO3) numa concentraccedilatildeo stock de 50 mM com agitaccedilatildeo magneacutetica e aquecimento (~30 ordmC)
Esta soluccedilatildeo com um valor compreendido entre 6 e 7 designa-se por ldquometavanadatordquo e eacute
constituiacuteda pelas esspeacutecies V1 V2 V4 e V5 (Ref) Acidificando a soluccedilatildeo de metavanadato por
adiccedilatildeo de HCl ateacute um valor de pH proacuteximo de 4 observa-se uma mudanccedila de cor de amarelo
claro para laranja indicativa da formaccedilatildeo da espeacutecie decameacuterica de vanadato De seguida e
imediatamente antes da realizaccedilatildeo dos ensaios descritos o valor do pH da soluccedilatildeo de
decavanadato foi aumentado para cerca de 75 por adiccedilatildeo de NaOH A soluccedilatildeo (2) designada
por ldquometavanadatordquo natildeo foi posteriormente alterada ateacute agrave sua adiccedilatildeo para a realizaccedilatildeo dos ensaios
As soluccedilotildees foram armazenadas a 4ordmC e mantidas em gelo aquando da sua utilizaccedilatildeo
A soluccedilatildeo de V(IV) foi preparada em condiccedilotildees anaeroacutebias por diluiccedilatildeo de sulfato de vanadil
(VOSO4) para uma concentraccedilatildeo final de 5 mM em H2O MilliQ saturada com N2 gasoso
352Anaacutelise por espectroscopia de UV-Vis
A concentraccedilatildeo da espeacutecie decameacuterica em soluccedilatildeo depende do periacuteodo de tempo apoacutes a diluiccedilatildeo
no meio de reacccedilatildeo apropriado (Ref) A velocidade de desoligomerizaccedilatildeo da espeacutecie decameacuterica
pode ser analisada por espectroscopia de absorccedilatildeo na regiatildeo UVVis uma vez que esta espeacutecie
apresenta duas bandas caracteriacutesticas na regiatildeo do visiacutevel (a 358 e 400 nm) que natildeo se verifica
em quaisquer outras espeacutecies oligomeacutericas As bandas supramencionada sapresentam-se bem
22
Meio de reacccedilatildeo 3125 mM Tris-HCl pH 71 375 mM MgCl2
48
resolvidas atraveacutes da obtenccedilatildeo da derivada de 2ordf ordem do espectro apresentando a forma de 2
vales com miacutenimos centrados nos comprimentos de onda supracitados com dependecircncia linear
relativamente agrave concentraccedilatildeo de decavanadato total (Ramos et al 2006)
Assim a estabilidade da espeacutecie decameacuterica apoacutes diluiccedilatildeo da soluccedilatildeo de decavanadato no meio de
reacccedilatildeo dos ensaios (tampatildeo D) para uma concentraccedilatildeo final de 1 mM em vanadato total foi
caracterizada atraveacutes da leitura da Abs400 em funccedilatildeo do tempo tendo-se determinado qual a
ordem de reacccedilatildeo e o tempo de meia-vida da mesma Com base em estudos anteriores acerca da
estabilidade do V10 na presenccedila de diferentes proteiacutenas (Ramos et al 2006) determinou-se
igualmente o tempo de meia vida apoacutes a adiccedilatildeo de 20 microM de S1 de miosina ao meio reaccional
353 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
Os espectros de RMN do isoacutetopo 51
V foram realizados num espectroacutemetro Bruker AM-400 MHz
equipado com uma sonda multinuclear inversa de 5 mm Em tubos de RMN de 5 mm de diacircmetro
preparou-se soluccedilotildees de 500 L de deca- ou metavanadato correspondentes a um intervalo de
concentraccedilotildees de 1 a 10 mM suplementadas com 10 D2O Os espectros foram adquiridos agrave
temperatura ambiente com uma largura espectral de 23 923 Hz um tempo de aquisiccedilatildeo de 0086
s tempo de relaxaccedilatildeo de 001 s e uma sequecircncia de pulsos a 90ordm Utilizou-se a transformada de
Fourier com alargamento das bandas de 20Hz para o processamento dos espectros Os desvios
quiacutemicos das ressonacircncias de 51
V foram obtidos em relaccedilatildeo agrave espeacutecie VOCl3cuja frequecircncia de
ressonacircncia eacute considerada o padratildeo sendo todas as outras referenciadas a partir desta em ppm
As aacutereas dos sinais do RMN correspondentes agraves espeacutecies oligomeacutericas presentes foram obtidas
por integraccedilatildeo dos sinais As concentraccedilotildees relativas das espeacutecies foram calculadas com recurso agrave
equaccedilatildeo
onde A[Vx] corresponde agrave aacuterea referente agrave ressonacircncia da espeacutecie Vx com nordm de agregaccedilatildeo x
A[V]total agrave soma total das aacutereas medidas e [V] a concentraccedilatildeo total de vanaacutedio no ensaio
A teacutecnica de 51
V-RMN foi anteriormente aplicada em estudos de ligaccedilatildeo com S1 de miosina
(Ref) tendo sido demostrado que entre as espeacutecies V1 V2 V4 e V5 a espeacutecie tetrameacuterica possui a
maior afinidade para a miosina (Cremo et al) O desaparecimento ou alargamento dos sinais pode
implicar a ligaccedilatildeo de um dos oxovanadatos laacutebeis agrave miosina sem indicaccedilatildeo de qual uma vez que
49
se processa um equiliacutebrio entre eles (Crans 1990) Assim e tendo em conta as indicaccedilotildees de que
a espeacutecie V10 eacute distinta das restantes no que diz respeito agrave magnitude da interacccedilatildeo com a miosina
e inibiccedilatildeo da actividade ATPaacutesica estimulada por actina (Aureliano 2000 Tiago et al 2002)
torna-se fundamental investigar a interacccedilatildeo das diferentes espeacutecies de vanadato com o S1 de
miosina atraveacutes da teacutecnica de 51
V-RMN
Para tal obtiveram-se espectros de 51
V-RMN resultantes da titulaccedilatildeo de uma soluccedilatildeo 5 mM de
decavanadato total com concentraccedilotildees crescentes de S1 de miosina (0 5 10 15 e 20 microM) em
tampatildeo D Realizou-se ainda ensaios na presenccedila de 2 mM de decavanadato e 2 mM de ATP na
presenccedila e na ausecircncia de S1 de miosina de modo a verificar o efeito que estes ligandos naturais
da enzima teria na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o subfragmento-1
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
Os grupos sulfidrilo (-SH) das cisteiacutenas satildeo resiacuteduos de aminoaacutecidos extremamente sensiacuteveis agrave
oxidaccedilatildeo resultando na sua oxidaccedilatildeo (Dalle-Donne et al 1999) Logo a quantificaccedilatildeo destes
resiacuteduos eacute bastante importante nos estudos de interacccedilatildeo com vanaacutedio jaacute que os resiacuteduos de Cys
com ecircnfase para o SH1 e o SH2 satildeo bastante importantes na monitorizaccedilatildeo das modificaccedilotildees
funcionais da regiatildeo da cabeccedila
O procedimento seguidamente apresentado foi originalmente descrito por Gutieacuterrez-Merino
(1987)) Consiste na reacccedilatildeo entre os grupos ndashSH natildeo oxidados das proteiacutenas e o reagente de
Ellman (DTNB -55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico)) - formando-se o aniatildeo CNT (aniatildeo 3-
carboxilato-4-nitrotiofenolato) numa estequiometria de 11 Este apresenta um maacuteximo de
absorvecircncia a 412 nm (Robyt 1990) Assim apoacutes a incubaccedilatildeo da proteiacutena na presenccedila do metal
a quantidade de cisteiacutenas reduzidas por proteiacutena determina-se atraveacutes dos valores de Abs412
Deste modo colocou-se num tubo de Eppendorf o volume apropriado de S1 dialisado para que a
concentraccedilatildeo no ensaio fosse de 1 mgmL Adicionou-se o volume apropriado de decavanadato
para que a sua concentraccedilatildeo fosse de 0 10 50 100 250 500 ou 1000 M Perfez-se com soluccedilatildeo
D ateacute um volume final de 1 mL e incubou-se a 25ordmC por 20 minutos Apoacutes este periacuteodo colocou-
se 780 L da mistura reaccional numa cuvette de quartzo e adicionou-se 220 L de soluccedilatildeo K23
23
Tampatildeo K Tampatildeo fosfatohidrogenofosfato de soacutedio 025 M pH 72
50
Adicionou-se 15 L de DTNB (em soluccedilatildeo K [DTNB]stock=100 mM) ao meio reaccional e
registou-se o valor de absorvecircncia medido equivalente agraves cisteiacutenas ldquoraacutepidasrdquo (SH1 e SH2
expostos ao solvente) Adicionou-se 001 g de SDS que provoca a desnaturaccedilatildeo da proteiacutena e
consequentemente a exposiccedilatildeo de todos os resiacuteduos de cisteiacutenas que se encontravam ateacute aiacute
inacessiacuteveis no interior da proteiacutena (cisteiacutenas totais) Estes resiacuteduos podem assim reagir com o
DTNB
Estimou-se o nuacutemero de resiacuteduos de cisteiacutenas em todas as amostras com recurso ao coeficiente de
extinccedilatildeo molar () do DTNB (ver ponto 3 dos Anexos) que foi determinado atraveacutes da titulaccedilatildeo
de L-Cisteiacutena com intervalo de concentraccedilotildees compreendidos entre 0 e 50 microM na presenccedila de
25microM de DTNB em tampatildeo D Os valores de absorvecircncia foram obtidos para um comprimento de
onda de 412 nm
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
3621 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
A teacutecnica mais eficiente de anaacutelise da interacccedilatildeo entre o iatildeo vanadilo e proteiacutenas como a
calmodulina e a transferrina entre outras permite a obtenccedilatildeo raacutepida de informaccedilatildeo no que toca agrave
conformaccedilatildeo e amplitude da referida interacccedilatildeo
Os espectros de Ressonacircncia Paramagneacutetica Electroacutenica (RPE) da espeacutecie paramagneacutetica V(IV)
foram realizados a 70K num espectroacutemetro Bruker EMX-300 X-band Todas as soluccedilotildees
utilizadas e amostras foram preparadas em anaerobiose numa cacircmara de luvas de modo a evitar a
reduccedilatildeo de V(IV) a V(V) Em tubos capilares com 02 mL de amostra manteve-se a concentraccedilatildeo
de S1 a 20 M e variou-se a concentraccedilatildeo de vanadilo (0 25 5 10 15 25 50 75 100 150
200 500 e 1000 M [VOSO4]stock=5 mM) em tampatildeo D Apoacutes obtenccedilatildeo dos espectros estes
foram tratados atraveacutes do software SimFonia e WinEPR tendo-se obtido um graacutefico com a
variaccedilatildeo da intensidade do sinal de RPE em funccedilatildeo da concentraccedilatildeo de sulfato de vanadilo
presente Realizaram-se ainda simulaccedilotildees de espectros obtidos com 20 microM de S1 e 1000 microM de
V(IV) com vista agrave obtenccedilatildeo dos ligandos provaacuteveis do vanadilo atraveacutes da anaacutelise da constante
de acoplamento hiperfina (A) e do factor g (g)
4 Conclusotildees gerais
Apesar de jaacute ter sido demonstrado que o vanaacutedio inibe a Na+K
+-ATPase da miosina a funccedilatildeo
deste elemento no tecido muacutesculo esqueleacutetico continua largamente incompreendida A enorme
complexidade quiacutemica deste metal especialmente no que concerne agrave oligomerizaccedilatildeo e reacccedilotildees
51
redox constitui um obstaacuteculo agrave compreensatildeo do comportamento do vanaacutedio nos organismos
vivos Assim a caracterizaccedilatildeo da quiacutemica deste elemento eacute de grande utilidade antes de iniciar o
estudo da sua interacccedilatildeo com o presente sistema
A anaacutelise por espectroscopias de 51
V-RMN e de UV-Vis de soluccedilotildees de vanadato diluiacutedas nos
meios de reacccedilatildeo utilizados no presente relatoacuterio (10 mM HEPES pH 70 25 mM KCl) mostrou
claramente que a espeacutecie decameacuterica (V10) possui um tempo de vida suficientemente longo para a
determinaccedilatildeo do seu efeito isolado ao contraacuterio do que se verifica com as espeacutecies mono- di-
tetra- e pentameacutericas (V1 V2 V4 e V5) Por outro lado a anaacutelise verificou ainda que apoacutes diluiccedilatildeo
das soluccedilotildees de decavanadato no meio de reacccedilatildeo coexistem as espeacutecies V10 e V1 pelo que os
efeitos putativos desta uacuteltima deveratildeo ser subtraiacutedos aquando da monitorizaccedilatildeo dos efeitos da
espeacutecie decameacuterica
O vanaacutedio (V) (vanadato) inibe as enzimas que catalizam reacccedilotildees de transferecircncia de grupos
fosfato como a ATPase da miosina [Goodno 1979] devido agrave sua analogia estrutural com o iatildeo
fosfato Na sua utilizaccedilatildeo como inibidor enzimaacutetico natildeo eacute tida em conta a contribuiccedilatildeo individual
das espeacutecies oligomeacutericas considerando-se normalmente a espeacutecie V1 como inibidora efectiva
[Goodno e Taylor 1982] No presente relatoacuterio verificou-se por espectroscopia de 51
V-RMN que
outros oligoacutemeros de vanadato interagem com o S1 de miosina tendo as espeacutecies V10 e V4 os
maiores niacuteveis de afinidade A sua interacccedilatildeo envolve forccedilas electrostaacuteticas e eacute afectada
negativamente por ligandos naturais de enzima como eacute o caso do ATP
A exposiccedilatildeo do S1 de miosina (005 mgmL) a decavanadato conduz agrave inibiccedilatildeo do capacidade da
proteiacutena de estimular a actividade Mg2+
-ATPaacutesica com um valor de IC50 de 266 plusmn 07 M
decavanadato sendo o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM
decavanadato (80) Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM decavanadato sendo
o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM decavanadato (60)
exercendo um efeito de impedimento agrave ligaccedilatildeo entre o vanaacutedio e o S1
A uma concentraccedilatildeo de 27 microM decavanadato verifica-se a oxidaccedilatildeo de um dos resiacuteduos de Cys
de S1 de miosina seguida pela oxidaccedilatildeo gradual de outro resiacuteduo de Cys ateacute agrave concentraccedilatildeo de
1000 microM Ambos os resiacuteduos oxidados natildeo se encontravam expostos ao solvente utilizado A
oxidaccedilatildeo dos resiacuteduos permite sugerir que esta modificaccedilatildeo oxidativa sofrida pelo S1 contribui
para a diminuiccedilatildeo da sua actividade de ATPase
52
Por espectroscopia de RPE revelaram-se os possiacuteveis ligandos equatoriais do vanaacutedio aquando da
sua interacccedilatildeo com o S1 de miosina como sendo duas moleacuteculas de aacutegua um ligando contendo o
grupo R-OH e outro contendo o grupo R=NR
Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados de
oxidaccedilatildeo (IV) e (V) no S1 de miosina para concentraccedilotildees facilmente alcanccedilaacuteveis no ambiente
terrestre
53
5 Estudos futuros
No sentido do melhor esclarecimento das alteraccedilotildees moleculares e estruturais que ocorrem no
subfragmento 1 quando exposta ao vanaacutedio que conduzem a uma diminuiccedilatildeo da sua funccedilatildeo
sugere-se
- Quantificaccedilatildeo das cisteiacutenas reduzidas de S1 na presenccedila de metavanadato
- Determinaccedilatildeo das actividades Ca2+
- e K+-ATPaacutesica do S1 de miosina na presenccedila de
decavanadato
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina estimulada por actina
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina na presenccedila de
metavanadato
- Realizaccedilatildeo de RMN 2D com variaccedilatildeo das concentraccedilotildees das soluccedilotildees de decavanadato e
metavanadato na presenccedila de S1
-Mutageacutenese dos resiacuteduos de Cys doS1 de miosina de modo a verificar quais os resiacuteduos que
sofrem oxidaccedilatildeo na presenccedila de decavanadato
- Determinaccedilatildeo por calorimetria (DSC Differential Scanning Calorimetry) da variaccedilatildeo da
entalpia (∆H) e da temperatura de desnaturaccedilatildeo do S1 de miosina na presenccedila de vanaacutedio uma
vez que estas propriedades caracteriacutesticas de cada proteiacutena satildeo alteradas apoacutes ocorrecircncia de
modificaccedilotildees estruturais
- Anaacutelise por SDS-PAGE de S1 de miosina apoacutes tratamento com vanaacutedio de modo a determinar
quais os polipeacuteptidos eventualmente formados apoacutes clivagem da proteiacutena
54
6 Referecircncias bibliograacuteficas
- Sellers J Myosins 2nd Edition Oxford University Press 1999
- L Stryer inBiochemistry (Ed WH Freeman 2005 pp
Refordfas Materiais e Meacutetodos
[A] S S Margossian SLowey Methods Enzymol1982 85 55-71
[B] JD Pardue JA Spudich Methods Enzymol 1982 85 164-181
[C] M Bradford Anal Biochem1976 72 248-254
[D] H H Taussky E Shorr J Biol Chem 1953 202 675-685
[E] C R Bagshaw in Outline Studies in Biology ndash Muscle Contraction Chapman and Hall Ed
1982
[F] A Cuenda et al Eur J Biochem1990 194 663-670
55
Anexos
1 Determinaccedilatildeo da recta de calibraccedilatildeo meacutetodo de Bradford
Figura1ndashRepresentaccedilatildeo graacutefica da recta de calibraccedilatildeo (Abs595 nm vs [BSA]) utilizada para o meacutetodo de Bradford
2 Determinaccedilatildeo da recta calibraccedilatildeo para a electroforese em condiccedilotildees desnaturantes
Figura 2 ndash Representaccedilatildeo graacutefica da recta de calibraccedilatildeo (log(Peso Molecular) vs Rf dos marcadores de peso
molecular) onde Rf indica determinada atraveacutes do padratildeo de peso molecular
(poccedilo (a))
3Caacutelculo do coeficiente de extinccedilatildeo molar () para a quantificaccedilatildeo de cisteiacutenas reduzidas
atraveacutes do meacutetodo de Ellman
Traccedilou-se a recta de Abs412 nm meacutedia vs [L-Cisteiacutena] de modo a determinar o valor do coeficiente
de extinccedilatildeo molar definido como o declive daquela recta Assim =8400 M-1
cm-1
como
podemos verificar na Fig 3
A595 nm = 00546[BSA] + 00209 Rsup2 = 099051
Ab
s 59
5 n
m
[BSA] (mgmL)
log PM = -1177 Rf + 2247 Rsup2 = 098957
log
PM
Mobilidade relativa (Rf)
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero

31
concentraccedilatildeo de S1 utilizada O factor de alargamento obtido para 8 microM ndash 0376 ndash eacute inferior ao
valor esperado agrave mesma concentraccedilatildeo de S1 obtido atraveacutes da equaccedilatildeo da recta apresentada na
Fig 28 ndash 0457 ndash o que indica que o ATP poderaacute exercer um efeito de protecccedilatildeo do S1
diminuindo a sua interacccedilatildeo com a espeacutecieV10
Por uacuteltimo realizaram-se ensaios com metavanadato na presenccedila de 8 microM S1 de modo a verificar
se as espeacutecies oligomeacutericas contidas nesta soluccedilatildeo interagiam com a proteiacutena em estudo
Figura 210 - Espectros de RMN de 51
V contendo 2
mM metavanadato (A) (controlo) (B) na presenccedila
de 8 microM de S1 de miosina V1 V2 e V4
correspondem respectivamente aacutes espeacutecies
monomeacuterica dimeacuterica e tetrameacuterica
Tabela 2VIndash Desvios quiacutemicos(part) das ressonacircncias das espeacutecies monomeacuterica (V1) dimeacuterica (V2) e tetrameacuterica (V4)
presentes nos espectros da soluccedilatildeo de metavanadato na presenccedila de 8 microM de S1 de miosina
Paracircmetros V1 V2 V4
part (ppm) -5613 -5747 -5789
Como eacute possiacutevel observar na Fig 210 ocorre uma ligeiro alargamento dos sinal de V4 bem
como uma perda da intensidade pelo que se conclui quedas espeacutecies presentes na soluccedilatildeo de
metavanadato (V1 V2 e V4) apenas a espeacutecie tetrameacuterica interage com o S1 de miosina o que
estaacute de acordo com o reportado por Tiago et al 2002
242 Actividade Mg2+
-ATPaacutesica de S1 de miosina na presenccedila de decanadato
Com o intuito de avaliar o efeito do decavanadato na actividade Mg2+
-ATPaacutesica do S1 de
miosina monitorizou-se a reacccedilatildeo de oxidaccedilatildeo do NADH atraveacutes da leitura de Abs340 na
presenccedila de 005 mgmL de S1 de miosina e no caso da actividade estimulada 2microM de actina em
3125 mM Tris-HCl pH 71 e 375 mM MgCl2 pelo meacutetodo de enzimas acopladas (ver seccccedilatildeo xx
dos MM)Para tal realizaram-se ensaios com vaacuterias concentraccedilotildees de decavanadato total (10 50
100 250 500 e 1000) de modo a determinar qual o seu efeito na actividade MgATPaacutesica de S1
de miosina na ausecircncia e na presenccedila de um dos seus ligandos naturais Os ensaios foram
realizados agrave temperatura ambiente com incubaccedilatildeo preacutevia a 25ordm C durante 20 minutos
32
Figura 211 ndash Representaccedilatildeo
graacutefica da Actividade Mg2+
-
ATPaacutesica de S1 () (os ajustes
teoacutericos estatildeo representado a por
uma linha os resultados
experimentais estatildeo representados
por triacircngulos (Actividade
estimulada) e cruzes (Actividade
natildeo estimulada)) vs
[Decavanadato]total medidas pelo
meacutetodo das enzimas acopladas
(n=3) Os dados foram ajustados ao modelo de decaimento exponencial usando o programa GraphPad Prism para a
determinaccedilatildeo dos valores de IC50 (na ausecircncia de actina IC50= 266 plusmn 07 M decavanadato na presenccedila de actina
IC50= 28 plusmn 1 M decavanadato
A partir do graacutefico presente na Fig 29 verifica-se que o ajuste dos dados experimentais foi bem
sucedido A inibiccedilatildeo da actividade Mg2+
-ATPaacutesica do S1 de miosina eacute afectada em menor
amplitude na presenccedila de actina (60 de inibiccedilatildeo da actividade) do que na sua ausecircncia
(aproximadamente 80 de inibiccedilatildeo de actividade) Devido a esta diferenccedila pode sugerir-se que a
actina iraacute impedir parcialmente a ligaccedilatildeo do decavanadatoao S1 demiosina atraveacutes de
impedimentos esteacutereos ou interacccedilotildees electrostaacuteticas por exemplo A teacutecnica utilizada natildeo nos
permite concluir acerca de que regiatildeo(otildees) ou resiacuteduo(s) da actina participaratildeo na preservaccedilatildeo
parcial da actividade Mg2+
-ATPaacutesica
Por outro lado os valores de IC50 dos dois tipos de actividades natildeo apresentam diferenccedilas
significativas tendo-se obtido 266 plusmn 07 e 28 plusmn 1M decavanadato na ausecircncia e na presenccedila de
actina respectivamente o que corresponde a uma concentraccedilatildeo de cerca de 27 M de V10 O
meio de reacccedilatildeo utilizado nestes ensaios foi 10 mM HEPES e 25 mM KCl com 2 mM ATP Em
(Tiago et al 2004) o valor de IC50 para a actividade Mg2+-ATPaacutesica na ausecircncia de actina foi de
125 plusmn 025 microM enquanto o valor da actividade estimulada por actina foi de 136 plusmn 14 microM O
meio de reacccedilatildeo utilizado nos ensaios foi 20 mM HEPES 50 mM KCl e 2 mM ATP o que resulta
precisamente no dobro da forccedila ioacutenica a que foi realizado o ensaio do presente relatoacuterio
(exceptuando a contribuiccedilatildeo dos produtos da hidroacutelise de ATP uma vez que este meacutetodo
pressupotildee a ocorrecircncia de um ciclo de reacccedilotildees de hidroacutelise e siacutentese deste composto) Ora tendo
sido anteriormente referido que a afinidade do S1 de miosina para com a espeacutecie V10 diminui com
Act
ivid
ade
Mg
2+-A
TP
aacutesic
a d
e S
1 d
e
Mio
sin
a
[Decavanadato] (microM) Actividade Estimulada ()
Actividade Natildeo Estimulada ()
Ajuste Actividade Estimulada
Ajuste Actividade Natildeo Estimulada
33
o aumento da forccedila ioacutenica (Tiago et al 2004 tal poderaacute explicar a diferenccedila obtida entre estes
dois conjuntos de valores
243Quantificaccedilatildeo de cisteiacutenas reduzidas pelo meacutetodo de Ellman
A interacccedilatildeo do vanaacutedio com alguns tipos de aminoaacutecidos sensiacuteveis agrave oxidaccedilatildeo pode fornecer-nos
informaccedilatildeo bastante uacutetil em relaccedilatildeo agraves alteraccedilotildees estruturais que conduzem a uma diminuiccedilatildeo da
actividade fisioloacutegica do S1 de miosina Os estudos realizados por Chasteen 1983 concluiacuteram
que a reduccedilatildeo de vanadato (V) a vanadilo (IV) ocorre na presenccedila de agentes redutores comuns no
ambiente celular de que se destaca o aminoaacutecido cisteiacutena Esta eacute tambeacutem um alvo de
complexaccedilatildeo por parte do iatildeo VO2+
Jaacute Ramos et al (2009) demonstrou que a incubaccedilatildeo de
actina com a soluccedilatildeo de decavanadato induz a oxidaccedilatildeo de resiacuteduos de cisteiacutena e agrave formaccedilatildeo do
oxidovanaacutedio (IV) Este grupo de investigadores descobriu ainda que apenas a espeacutecie V10 induz a
oxidaccedilatildeo das cisteiacutenas ldquoraacutepidasrdquo da actina atraveacutes do processo proposto que eacute parcialmente
responsaacutevel pela inibiccedilatildeo jaacute descrita da actividade de ATPase do S1 de miosina estimulada por
actina quando exposta agrave soluccedilatildeo de decavanadato Esta inibiccedilatildeo apresenta-se afectada com a
presenccedila de ATP uma vez que os resiacuteduos de cisteiacutena permaneceram intactos nos ensaios
realizados com este ligando Jaacute Crans et al (2004) concluiu que o siacutetio activo de muitas fosfatases
(que conteacutem grupos tiol) pode formar um equiliacutebrio redox com o iatildeo vanadato (V) de forma natildeo
reversiacutevel
Os resultados obtidos por Sakurai e Shiromura (1980) sugeriram que o vanaacutedio (V) sofre reduccedilatildeo
a vanaacutedio (IV) sem dependecircncia do valor de pH do ensaio Registaram ainda uma estequiometria
de 31 na reacccedilatildeo do vanadato (V) com os resiacuteduos de Cys enquanto a estequiometria dos ensaios
com o vanadilo (IV) foi de 21
Berlett e Stadtman (1997) observaram que os tipos de aminoaacutecidos mais facilmente oxidaacuteveis
englobam entre outros os resiacuteduos de Cys Trp e MetAssim analisou-se o estado redox dos
grupos tiol dos resiacuteduos de Cys do S1 na presenccedila do decavanadato atraveacutes da monitorizaccedilatildeo da
Abs412 nm na presenccedila de 1 mgmL de S1 de miosina em tampatildeo 10 mM HEPES 25 mM KCl
variando a concentraccedilatildeo da soluccedilatildeo de decavanadato de modo a determinar que tipo de alteraccedilotildees
estruturais ocorrem na sua presenccedila no que diz respeito agrave diminuiccedilatildeo da actividade ATPaacutesica de
S1 na presenccedila de decavanadato
Os resultados referentes agrave quantificaccedilatildeo das cisteiacutenas reduzidas apoacutes o tratamento de S1 com
diferentes concentraccedilotildees de decavanadato em tampatildeo 10 mM HEPES pH 75 e 25 mM KCl a
25ordmC durante 20 minutos encontram-se descritos na Tabela 2VII
Tabela 2VII ndash Cisteiacutenas reduzidas totais e ldquoraacutepidas de S1 (1 mgmL) apoacutes tratamento com decavanadato
34
(010501002505001000 uM) em 10 mM HEPES 25 mM KCl pH 7 a 25ordmC durante 20 minutos A detecccedilatildeo
espectrofotomeacutetrica foi feita com DTNB (e 412=8400 uM-1 cm-1) (n variaacutevel entre 1 e 8)
Amostras Cys raacutepidas Cys totais
11 M S1 (controlo) 22 plusmn 04 106 plusmn 14
11 M S1 + 10 M Decavanadato 22 plusmn 05 95 plusmn 08
11 M S1 + 50 uM Decavanadato 20 plusmn 02 98 plusmn 14
11 M S1 + 100 uM Decavanadato 17 plusmn 03 97 plusmn14
11 M S1 + 250 uM Decavanadato 26 plusmn 02 98 plusmn 08
11 M S1 + 500 uM Decavanadato 21 plusmn 04 90 plusmn 06
11 M S1 + 1000 uM Decavanadato 24 plusmn 0 86 plusmn 08
A partir dos resultados obtidos pode sugerir-se que o S1 possui 11 resiacuteduos de Cys (controlo) o
que estaacute de acordo com o anteriormente descrito Verifica-se que o tratamento da proteiacutena com
concentraccedilotildees crescentes de decavanadato provoca a oxidaccedilatildeo de dois resiacuteduos de Cys do S1 de
miosina natildeo expostos ao solvente No entanto a teacutecnica utilizada natildeo nos permite identificar
quais
Tendo em consideraccedilatildeo que o valor de IC50 obtido para a reacccedilatildeo de inibiccedilatildeo daactividade Mg2+-
ATPaacutesica de S1 na presenccedila de decavanadato eacute de cerca de 27 microM pode sugerir-se que este seraacute o
valor ao qual ocorreraacute a oxidaccedilatildeo do primeiro resiacuteduo de Cys o que poderaacute contribuir para a
diminuiccedilatildeo da actividade de S1 observada
25Interacccedilatildeo de S1 com vanaacutedio(IV)
251Anaacutelise por espectroscopia de Ressonacircncia Paramagneacutetica Electroacutenica
A acumulaccedilatildeo intracelular de oxovanaacutedio (IV) in vivo constitui uma das principais formas de
apresentaccedilatildeo de vanaacutedio no organismo Deste modo torna-se bastante interessante analisar a
interacccedilatildeo entre este iatildeo e o S1 de miosina atraveacutes da teacutecnica de espectroscopia de Ressonacircncia
Paramagneacutetica Electroacutenica o meacutetodo de eleiccedilatildeo do estudo do iatildeo vanadilo e dos complexos por si
formados O electratildeo desemparelhado encontra-se fortemente acoplado ao nuacutecleo 51
V o que
35
resulta num espectro de RPE consistindo de 8 desdobramentos na direcccedilatildeo paralela e outros tantos
na direcccedilatildeo perpendicular (Chasteen 1983)
Os estudos realizados por Ahmed et al (1987) centraram-se na interacccedilatildeo do vanadilo com
proteiacutenas como eacute o caso da insulina transferrina entre outros obtendo um espectro caracterizado
por 8 linhas amplamente espaccediladas (na direcccedilatildeo paralela) Jaacute Ramos et al 2009 concluiu que a
exposiccedilatildeo da soluccedilatildeo de decavanadato agraves formas globular e filamentosa de actina produz um
espectro tiacutepico do iatildeo oxidovanaacutedio (IV) indicando a reduccedilatildeo do vanadato
Para aferir qual a concentraccedilatildeo de S1 que permitia obter uma boa razatildeo sinalruiacutedo titulou-se uma
soluccedilatildeo de sulfato de vanadilo 15 microM ([VOSO4]stock = 5 mM) com concentraccedilotildees crescentes de
proteiacutena (0 5 10 15 e 20 microM) em 10 mM HEPES 25 mM KCl A existecircncia de uma correlaccedilatildeo
linear entre a concentraccedilatildeo de proteiacutena e a intensidade dos sinais de RPE correspondente agrave banda
-12 perpendicular encontra-se representada na Fig 210
Figura 2 12ndash Representaccedilatildeo graacutefica da variaccedilatildeo da intensidade do sinal de RPE vs a concentraccedilatildeo de S1 utilizada
Apoacutes verificar que com uma concentraccedilatildeo de S1 igual a 20 microM se obtinham sinais com uma razatildeo
sinalruiacutedo adequada ao prosseguimento dos estudos sobre a interacccedilatildeo V(IV)-S1 de miosina
titulou-se uma soluccedilatildeo de 20 microM S1 com 0 25 5 10 15 25 50 75 100 150 200 500 e 1000
microM VOSO4
Traccedilou-se o graacutefico da intensidade da banda -12 perpendicular do sinal de EPR em funccedilatildeo da
concentraccedilatildeo de VOSO4 tendo-se determinado uma constante de dissociaccedilatildeo (KD) do complexo
S1-vanadilo de 8748 plusmn 2621 microM V(IV)
y = 016786x + 122435
Rsup2 = 092992759
Inte
nsi
dad
e d
o s
inal
de
RP
E
[S1] (microM)
36
Figura 213 ndash Representaccedilatildeo graacutefica da variaccedilatildeo do sinal de RPE vs a concentraccedilatildeo de vanadilo utilizada (os valores
experimentais estatildeo representados por losangos o seu ajuste estaacute representado pela linha)
A simulaccedilatildeo do espectro de EPR foi efectuada a partir do espectro obtido para uma concentraccedilatildeo
de 1 mM VOSO4 apoacutes adiccedilatildeo de 20 microM S1 de miosina tendo-se obtido os valores da constante de
acoplamento hiperfina (A=5016) e do factor g (g0=195)
Figura 214 ndash Representaccedilatildeo graacutefica do espectro de EPR obtido com 1000 microM de vanadilo e 20 microM de S1 (a cinzento
escuro) A simulaccedilatildeo do espectro encontra-se a cinzento claro
Por comparaccedilatildeo destes com os valores presentes na tabela de correspondecircncia xxx eacute possiacutevel
predizer quais os ligandos provaacuteveis do vanadilo na presenccedila de proteiacutena Assim concluiu-se que
o vanadilo estaacute coordenado com duas moleacuteculas de aacutegua um grupo hidroxilo e um grupo R=NR
O grupo hidroxilo pode indicar os resiacuteduos de aminoaacutecidos Tyr Treonina e Serina enquanto o
grupo R=NR poderaacute indicar a presenccedila de Arginina e Histidina como ligandos do vanadilo
Inte
nsi
dad
e d
os
sinai
s d
e R
PE
[Vanadilo] (microM)
Ajuste dos dados exp Dados experimentais
Campo magneacutetico (G)
Espectro Experimental Ajuste
37
3 Procedimento Experimental
31 Reagentes
Todos os reagentes utilizados satildeo de elevado grau de pureza (analiacutetico) Apresentam-se as suas
foacutermulas quiacutemicas ou abreviaturas quando necessaacuterio acrescentou-se a sua descriccedilatildeo Entre
parecircntesis colocou-se as empresas que os comercializam
Aacutecido aceacutetico glacial 100 (Riedel) Acrilamida 99 (Merck) Azul de bromofenol (Riedel-de-
Haumlen) -Mercaptoetanol (Sigma) Bissulfito de soacutedio (Sigma)Bradford (reagente de)
(BioRad)BSA (albumina de soro bovino Sigma-Aldrich) CaCl22H2O (cloreto de caacutelcio
dihidratado Fluka) CH3CH2OH 96 (etanol Panreac) Coomassie Blue R250 (Merck) D2O
(oacutexido de deuteacuterio SDS) DTNB (55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico) Aldrich) EDTA (aacutecido
etilenodiaminatetraaceacutetico Merck) Fosfoenolpiruvato 98 (Fluka) Glicerol (Sigma) Glicina
985 (Panreac) HCl 37 (Riedel-de Haumlen) H2SO4 96 (Pronalab) HEPES 99 (aacutecido 4-(2-
hidroxiletil)-1-piperazineetanosulfoacutenico Sigma) KH2PO4 99 (hidrogenofosfato de potaacutessio)
MgATP (Sigma) Na
ATP (Sigma) Na2HPO4 99 (hidrogenofosfato de soacutedio Riedel-de Haumlen)
NaH2PO4 (fosfato de soacutedio ) NaOH 986 (hidroacutexido de soacutedio Pronalab) KCl 99 (Cloreto de
potaacutessio Panreac) L-Cys 995 (enantioacutemero levoacutegiro do resiacuteduo de Cisteiacutena Fluka)Metanol
99 (Merck) Molibdato de amoacutenio (Sigma)NNrsquo-metil-bis-acrilamida 98 (Merck) NH4VO3
99 (metavanadato de amoacutenia Sigma) PSA 10 (persulfato de amoacutenio Merck) SDS
(dodecilo sulfato de soacutedio Sigma) Sulfito de soacutedio (Sigma) VOSO4 (sulfato de vanadilo
Aldrich) TEMED 99 (Tetrametiletilenodiamina Merck) Tris (2-amino-2-hidroximetil-13-
propanodiol Sigma)
32 Meacutetodos
321 Isolamento e purificaccedilatildeo das proteiacutenas de muacutesculo esqueleacutetico de coelho
3211 Purificaccedilatildeo da miosina preparaccedilatildeo de Subfragmento 1 por digestatildeo quimotriacutepsica
da miosina
O subfragmento-1 de miosina-II (S1) foi gentilmente cedido pelo Prof Doutor Manuel Aureliano
do Departamento de Quiacutemica Bioquiacutemica e Farmaacutecia da Faculdade de Ciecircncias e Tecnologia da
Universidade do Algarve
No entanto descreve-se abaixo de forma resumida o procedimento efectuado
38
32111 Purificaccedilatildeo da miosina
O protocolo de isolamento foi adaptado do meacutetodo previamente descrito por Margossian e Lowey
(1982) apresentando-se de forma esquemaacutetica na Fig 31
Sucintamente os tecidos musculares dorsais e das pernas foram extraiacutedos apoacutes o sacrifiacutecio do
animal (Fig31) De seguida foram adicionados 3 volumes da soluccedilatildeo de extracccedilatildeo A2
Centrifugou-se a soluccedilatildeo e filtrou-se o sobrenadante O sedimento foi entatildeo diluiacutedo em 14
volumes de aacutegua destilada de modo a que a forccedila ioacutenica atingisse 004 M Apoacutes adiccedilatildeo de szlig-
mercaptoetanol (szlig-me) deixou-se a precipitar a 4ordmC removendo o sobrenadante Centrifugou-se
novamente e ressuspendeu-se o sedimento na soluccedilatildeo de extracccedilatildeo A Adicionou-se a soluccedilatildeo
tampatildeo B3 para que a forccedila ioacutenica atingisse 05 M Homogeneizou-se e deixou-se repousar
durante 2h Homogeneizou-se novamente e centrifugou-se de modo a ocorrer precipitaccedilatildeo dos
complexos de actomiosina e proteiacutena desnaturada Apoacutes filtraccedilatildeo do sobrenadante em latilde de vidro
quantificou-se a soluccedilatildeo de miosina resultante pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental)
2 Soluccedilatildeo de extracccedilatildeo A 0012M K2CO3 004M KHCO3 06M KCl pH 65
3 Soluccedilatildeo-tampatildeo B Tris 100 mM pH 7 KCl 2M
39
Figura 31 ndash Representaccedilatildeo esquemaacutetica do protocolo de isolamento de miosina (Margossian e Lowey 1982) Todo o
procedimento foi realizado a 4ordmC (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
32112 Preparaccedilatildeo de Subfragmento-1 de miosina por digestatildeo com quimiotripsina
O S1 de miosina foi isolado apoacutes diaacutelise da miosina durante a noite contra a soluccedilatildeo C4 A
presenccedila do EDTA eacute essencial uma vez que a sua substituiccedilatildeo por MgCl2resulta na produccedilatildeo de
HMM em vez de S1 A suspensatildeo de miosina foi equilibrada e digerida com -quimotripsina
A digestatildeo foi interrompida pela adiccedilatildeo de PMSF Seguidamente ultracentrifugou-se o produto
da digestatildeo desprezando-se o sedimento (Fig 32) A soluccedilatildeo de sobrenadante maioritariamente
S1 foi quantificada pela leitura directa da Abs280 ou pelo meacutetodo de Bradford (ver as secccedilotildees
3311 e 3312 do Procedimento Experimental respectivamente) e armazenada em 10 sacarose
e 2 mM szlig-Me a -80ordmC
4 Soluccedilatildeo C 25mM Tris pH 70 012M KCl 1mM EDTA
40
Antes da realizaccedilatildeo dos ensaios descritos no presente relatoacuterio o S1 de miosina foi dialisado
contra a soluccedilatildeo D5 de modo a remover qualquer composto que pudesse complexar como
vanaacutedio Seguidamente quantificou-se a proteiacutena pelo meacutetodo de Bradford ou pelo meacutetodo
directo (ver secccedilotildees 3311 e 3312do Procedimento Experimtental)
Figura 3 2ndash Representaccedilatildeo esquemaacutetica do protocolo de preparaccedilatildeo do S1 de miosina por digestatildeo quimiotripsina de
acordo com o previamente descrito por Margossian e Lowey (1982) Todo o procedimento foi realizado a 4ordm C
excepto quando indicaccedilatildeo do contraacuterio (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
3212 Isolamento e purificaccedilatildeo de actina
Isolou-se e purificou-se a actina segundo o meacutetodo descrito por Pardee e Spudish que
compreende dois passos ndash a obtenccedilatildeo de um resiacuteduo seco denominado ldquopoacute de acetonardquo e a
extracccedilatildeo subsequente da actina a partir do ldquopoacute de acetonardquo na forma de G-Actina com a
posterior polimerizaccedilatildeo a F-Actina
32121 Obtenccedilatildeo do ldquopoacute de acetonardquo
Sacrificou-se o coelho com uma pancada atraacutes do pescoccedilo e sangrou-se o animal apoacutes o corte da
arteacuteria jugular Esfolou-se e colocou-se numa tina com gelo Retiraram-se as partes de tecido
conjuntivo que envolvem o tecido muscular com auxiacutelio de uma pinccedila e bisturi Seguidamente
retirou-se os muacutesculos dorsais e os muacutesculos brancos das pernas Passou-se os muacutesculos por aacutegua
destilada de modo a remover as impurezas e pesou-se num copo Trituraram-se os muacutesculos
numa maacutequina de triturar carne e extraiu-se o resultante com 3 volumes da soluccedilatildeo E6 durante 10
minutos com agitaccedilatildeo suave Centrifugou-se a suspensatildeo a 4000 g durante 10
5 Soluccedilatildeo D HEPES 10 mM pH 70 KCl 25 mM
6 Soluccedilatildeo E 01 M KCl 015 M K2HPO4 pH 65
41
minutosdesprezando-se o sobrenadante Ressuspendeu-se o sedimento em 6 volumes de soluccedilatildeo
F7durante 10 minutos com agitaccedilatildeo suave Verificou-se se o pH da soluccedilatildeo estava compreendido
entre 82 e 85 acertando-se estecom soluccedilatildeo G8 se necessaacuterio Centrifugou-se durante 10
minutos a 4000 g Desprezou-se o sobrenadante e ressuspendeu-se o sedimento em 3 volumes de
soluccedilatildeo H9 durante 10 minutos Centrifugou-se novamente a 4000 gdurante 10 minutos
desprezando-se o sobrenadante Ressuspendeu-se o sedimento duas vezes consecutivas em 6
volumes de aacutegua destilada durante 5 minutos Centrifugou-se a 4000 g durante 10 minutos apoacutes
cada adiccedilatildeo de H2O
Seguidamente ressuspendeu-se o sedimento 5 vezes consecutivas utilizando 3 volumes de
acetona de cada vez agitando suavemente durante 10 minutos agrave temperatura ambiente Colocou-
se o resiacuteduo filtrado numa tina de vidro sobre papel de filtro deixando-se evaporar o solvente
durante a noite num nicho Triturou-se o resiacuteduo seco numa maacutequina de triturar carne e deixou-se
novamente evaporar o solvente O ldquopoacute de acetonardquo obtido foi armazenado a -20 ordmC
32122 Purificaccedilatildeo da actina a partir de ldquopoacute de acetonardquo
Apoacutes a obtenccedilatildeo do poacute de acetona como acima indicado efectuou-se o protocolo seguidamente
descrito de modo a obter monoacutemeros de actina (G-Actina) e actina filamentosa (F-Actina)
Pesou-se 35 g de poacute de acetona e adicionaram-se 50 mL de soluccedilatildeo-tampatildeo I10
sendo a
extracccedilatildeo feita com agitaccedilatildeo suave durante 30 minutos Separou-se o extracto do poacute de acetona
hidratado filtrando a suspensatildeo com 4 camadas de gaze esterilizada tendo o cuidado de natildeo
espremer o resiacuteduo Voltou a extrair-se com 50 mL de soluccedilatildeo I durante 30 minutos mexendo
lentamente Adicionou-se este ao extracto obtido anteriormente Centrifugou-se o extracto total a
16 000 g durante 30 minutos Decantou-se para uma proveta sobre a qual se colocou gaze para
que o sedimento solto natildeo se juntasse ao sobrenadante e leu-se o volume De seguida adicionou-
se KCl ateacute uma concentraccedilatildeo final de 50 mM MgCl2 ateacute 2mM e ATP ateacute 1mM lentamente e com
agitaccedilatildeo contiacutenua de modo a promover a polimerizaccedilatildeo dos monoacutemeros de actina
Deixou-se polimerizar durante 2h a 25ordmC observando-se a presenccedila de pequenas bolhas de ar
indicativas de um aumento de viscosidade da soluccedilatildeo Arrefeceu-se em gelo durante 15 minutos e
adicionou-se KCl ateacute uma concentraccedilatildeo final de 06M Mexeu-se lentamente durante 30 minutos
apoacutes os quais se ultracentrifugou a 110 000g durante 1hora Desprezou-se o sobrenadante
ressuspendeu-se o sedimento com soluccedilatildeo tampatildeo E e transferiu-se para um potter
Dialisou-se durante a noite contra 1L de soluccedilatildeo-tampatildeo I a uma temperatura de 4ordmC com rotaccedilatildeo
7 Soluccedilatildeo F 005 M NaHCO3 pH 82-85
8 Soluccedilatildeo G 1M Na2CO3
9 Soluccedilatildeo H 1 mM EDTA pH 70
10 Soluccedilatildeo I 2 mM Tris-HCl pH 8 02 mM CaCl2 02 mM Na2ATP 05 mM szlig-Me 0005 NaN3
42
contiacutenua e mudanccedila de tampatildeo 2 a 3 vezes De seguida a actina foi centrifugada a 110
000gdurante 1hora Decantou-se a soluccedilatildeo de G-Actina (sobrenadante) e desprezou-se o
sedimento Determinou-se a concentraccedilatildeo de G-Actina pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental) A soluccedilatildeo de G-actina foi armazenada a uma temperatura de 4ordmC
apoacutes adiccedilatildeo de 002NaN3 mantendo-se estaacutevel durante 3 a 5 dias
331 Quantificaccedilatildeo das proteiacutenas isoladas
3311 Quantificaccedilatildeo pelo meacutetodo de Bradford
O meacutetodo de Bradford (Bradford 1976) baseia-se na utilizaccedilatildeo do reagente de Bradford
composto por entre outros aacutecido fosfoacuterico e o corante ldquoCoomassierdquo Este corante possui carga
negativa e a sua coloraccedilatildeo varia entre a vermelha (Absmax=465 nm) predominante em soluccedilatildeo e a
azul (Absmax=595 nm) presente aquando da ligaccedilatildeo aos grupos amina das proteiacutenas Em soluccedilotildees
aacutecidas diluiacutedas a coloraccedilatildeo varia proporcionalmente com a quantidade do corante ligado agrave
proteiacutena que por sua vez eacute directamente proporcional agrave concentraccedilatildeo da proteiacutena permitindo a
sua quantificaccedilatildeo ateacute a um limite de detecccedilatildeo de 20 gmL (Bradford 1976)
Deste modo prepararam-se amostras de BSA ([BSA]stock= 1mgmL) utilizando um intervalo de
concentraccedilotildees compreendido entre 0 e 15 gmL de acordo com a tabela 3I Apoacutes a adiccedilatildeo da
H2O MilliQ e do reagente de Bradford as soluccedilotildees foram sucessivamente homogeneizadas no
voacutertex
Tabela 3Indash Amostras utilizadas para a obtenccedilatildeo da recta de calibraccedilatildeo no meacutetodo de Bradford
BSA (L) BSA (g) H2O MilliQ (L) Reagente de Bradford (L)
0 0 800 200
125 125 79875 200
25 25 7975 200
5 5 795 200
10 10 790 200
15 15 785 200
Apoacutes incubaccedilatildeo a 25ordmC por 5 minutos leram-se os valores de Abs595 num
espectrofotoacutemetro (Shimadzu UV-160A) para todas as amostras Seguidamente traccedilou-se a recta
de calibraccedilatildeo (Abs595vs quantidade de BSA (g))
43
Para determinar a concentraccedilatildeo de proteiacutena apoacutes o processo de isolamento purificaccedilatildeo e
diaacutelise quando necessaacuterio colocou-se 5 L da amostra num tubo de Eppendorf e adicionou-se
795 L de H2O MilliQ homogeneizando a soluccedilatildeo no voacutertex Adicionou-se posteriormente 200
L de reagente de Bradford e repetiu-se a homogeneizaccedilatildeo Incubou-se a mistura a 25ordmC por 5
minutos apoacutes os quais se leu os valores de Abs595 nm no espectrofotoacutemetro Por interpolaccedilatildeo deste
valor a partir da recta de calibraccedilatildeo determinou-se a concentraccedilatildeo de proteiacutena na amostra
3312 Quantificaccedilatildeo pelo meacutetodo directo
A quantificaccedilatildeo das amostras proteicas pode tambeacutem ser efectuada atraveacutes da leitura directa da
absorvacircncia tendo por base a lei de Beer-Lambert onde Abs corresponde agrave
absorvacircncia ao comprimento de onda ao qual o ensaio foi realizado corresponde ao coeficiente
de extinccedilatildeo (substituiacutedo pela letra a se as unidades forem distintas da molaridade) b corresponde
ao percurso oacuteptico da radiaccedilatildeo na ceacutelula amostral e c corresponde agrave concentraccedilatildeo amostral
Assim no caso de S1 de miosina mediu-se a Abs280e calculou-se a concentraccedilatildeo da proteiacutena
sabendo que o coeficiente de extinccedilatildeo de S1 eacute (Ref)
No caso da actina mediu-se a Abs290ecalculou-se a concentraccedilatildeo da proteiacutena sabendo que o
coeficiente de extinccedilatildeo da actina eacute (Ref)
332Electroforese em condiccedilotildees desnaturantes (SDS-PAGE)
De modo a melhor caracterizar as proteiacutenas isoladas foi realizada uma electroforese em
condiccedilotildees desnaturantes (SDS-PAGE do inglecircs Sodium Dodecyl Sulfate Polyacrilamide Gel
Electrophoresis) O SDS (dodecilo sulfato de soacutedio) eacute um detergente anioacutenico utilizado neste
meacutetodo para desnaturar as proteiacutenas que se ligam ao SDS na proporccedilatildeo 14 g SDS g proteiacutena
(aproximadamente 1 moleacutecula de SDS por cada 2 resiacuteduos de aminoaacutecidos) (Stryer) O SDS induz
uma densidade de carga negativa significativa nas proteiacutenas a que se liga mascarando a sua carga
intriacutenseca pelo que as razotildees cargamassa satildeo tendencialmente idecircnticas Assim a SDS-PAGE
separa as proteiacutenas quando sujeitas a um campo eleacutectrico de acordo com o seu peso molecular
sendo a sua migraccedilatildeo directamente proporcional ao logaritmo deste uacuteltimo (Stryer)
O gel de resoluccedilatildeo com 10 de acrilamida foi obtido atraveacutes da adiccedilatildeo de 167 mL de soluccedilatildeo
stock de acrilamida11
075 mL de tampatildeo de resoluccedilatildeo12
251 mL de aacutegua destilada e 50 L de
SDS 10 Para promover a polimerizaccedilatildeo adicionou-se em seguida 38 L de PSA 10
11
soluccedilatildeo stock de acrilamida soluccedilatildeo de 30 (mv) de acrilamida e 08 (mv) de NNrsquo-metil-bis-acrilamida 12
tampatildeo de resoluccedilatildeo 15 M Tris-HCl pH 88
44
(persulfato de amoacutenia) 10 e 25 L de TEMED 99 (tetrametiletilenodiamina) Agitou-se
cuidadosamente de modo a natildeo formar bolhas e pipetou-se a soluccedilatildeo para a cassette de vidro
previamente montada no suporte De seguida adicionou-se uma peliacutecula de cerca de 2 mm de 2-
metil-2-butanol para verificar posteriormente a polimerizaccedilatildeo Apoacutes a polimerizaccedilatildeo que demora
cerca de 45 minutos verteu-se a agrave superfiacutecie e adicionou-se o gel de concentraccedilatildeo com 5 de
acrilamida constituiacutedo por 03 mL de soluccedilatildeo de stock acrilamida 045 mL de tampatildeo de
concentraccedilatildeo13
094 mL de aacutegua destilada 18 L de SDS 10 135 L de PSA 10 e 2L de
TEMED 99 Colocou-se o pente sobre o gel e deixou-se polimerizar durante cerca de 45
minutos
As amostras para aplicaccedilatildeo no gel de electroforese foram preparadas para uma concentraccedilatildeo de
proteiacutena de 1 mgmL num volume final de 100 L dos quais 25 L correspondem ao tampatildeo da
amostra14
As amostras foram aquecidas a 100ordmC durante 5 minutos Antes da aplicaccedilatildeo das
amostras transferiu-se a cassette para a tina de electroforese agrave qual se adicionou a soluccedilatildeo
tampatildeo de electroforese da tina superior15
e tampatildeo de electroforese da tina inferior16
Aplicaram-
se 10 L das soluccedilotildees de marcadores de peso molecular e 20 L das amostras proteicas
preparadas anteriormente A electroforese decorreu durante cerca de 3h30 min a uma voltagem
de 80 V De seguida transferiu-se o gel para uma tina corando as proteiacutenas durante 1h com
soluccedilatildeo corante17
Descorou-se o gel com soluccedilatildeo descorante18
tendo-se mudado esta soluccedilatildeo ateacute
as bandas se encontrarem bem definidas
Determinou-se a mobilidade relativa das bandas correspondentes aos marcadores de elevado peso
molecular e traccedilou-se a recta de calibraccedilatildeo ndash log(PM) vs log(Rf) onde Rf corresponde agrave
mobilidade relativa Por interpolaccedilatildeo determina-se o PM das proteiacutenas isoladas sabendo a sua
mobilidade relativa Por comparaccedilatildeo com os valores descritos na literatura pode concluir-se
acerca do grau de purificaccedilatildeo
34 Mediccedilatildeo da actividade enzimaacutetica de S1
341 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
13
tampatildeo de concentraccedilatildeo 05 M Tris-HCl pH 68 14
tampatildeo de amostra 320 mM Tris-HCl pH 68 04 M szlig-mercaptoetanol SDS 8 glicerol 15 (vv) e roxo de m-
cresol 0024 15
tampatildeo de electroforese da tina superior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS 16
tampatildeo de electroforese da tina inferior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS acetato de soacutedio
01 M 17
soluccedilatildeo corante ldquoCoomassie Blue R-250rdquo contendo metanol 45 (vv) e aacutecido aceacutetico glacial 10 (vv) 18
soluccedilatildeo descorante metanol 10 (vv) e aacutecido aceacutetico glacial 10 vv)
45
O meacutetodo de Fiske-Subarow modificado consiste na reacccedilatildeo do molibdato de amoacutenio com o
fosfato inorgacircnico em soluccedilatildeo aacutecida com formaccedilatildeo do aacutecido fosfomoliacutebdico Seguidamente daacute-
se a adiccedilatildeo de um agente redutor que provoca a reduccedilatildeo do composto a fosfomolibdato que forma
um complexo de cor azulada cuja absorvacircncia maacutexima ocorre para um comprimento de onda de
660 nm tornando possiacutevel a sua detecccedilatildeo por meacutetodos espectrofotomeacutetricos As actividades Ca2+
-
ATPaacutesica e K+EDTA-ATPaacutesica de S1 foram determinadas atraveacutes deste meacutetodo de acordo com
o descrito por Taussky e Shorr (1953) com mediccedilatildeo da quantidade de fosfato (Pi) libertado
atraveacutes da hidroacutelise de ATP
Para tal obteve-se primeiramente a recta de calibraccedilatildeo com as amostras presentes na Tabela 3II
utilizando uma soluccedilatildeo stock de 5 mM KH2PO4
Tabela 3II ndash Composiccedilatildeo das diferentes amostras utilizadas para determinar a recta de calibraccedilatildeo atraveacutes do meacutetodo
de Fiske-Subbarow modificado
KH2PO4 (L)
([KH2PO4]stock= 5mM)
PO43-
(nmole) H2O (uL)
0 0 1780
5 25 1775
10 50 1770
20 100 1760
40 200 1740
100 500 1680
A cada tubo de Eppendorf adicionaram-se seguidamente 200 L de soluccedilatildeo J19
e 20 L de agente
redutor20
homogeneizando no voacutertex apoacutes cada adiccedilatildeo Incubou-se durante 10 minutos e mediu-
se a Abs660 nm num espectrofotoacutemetro com intervalos de tempo iguais entre cada amostra Traccedilou-
se a recta de calibraccedilatildeo (Abs660 nmvs nmol PO43-
) a partir dos valores obtidos
Para determinar a actividade Ca2+
-ATPaacutesica de S1 prepararam-se tubos de Eppendorf com 400
L de meio de reacccedilatildeo21
25 L CaCl2 ([CaCl2]stock= 100 mM) 25 L ATP ([ATP]stock= 100 mM)
com [S1]ensaios=01 mgmL Perfez-se se necessaacuterio com H2O MilliQ ateacute perfazer um volume de 1
mL Para determinar a actividade K+EDTA-ATPaacutesica utilizou-se uma concentraccedilatildeo idecircntica de
19
Soluccedilatildeo J 5 N H2SO4 25 molibdato de amoacutenio 20
Agente redutor 5 mg aacutecido 1-amino-2-naftol-4-sulfoacutenico 30 mg bissulfito de soacutedio 30 mg sulfito de soacutedio em 26
mL H2O 21
meio de reacccedilatildeo 125 mM Tris-HCl pH 80 0575 M KCl
46
proteiacutena nos ensaios (01 mgmL) e o mesmo meio de reacccedilatildeo Aos tubos de Eppendorf
adicionou-se ainda 123 L de KCl ([KCl]stock = 3M) 25 L de ATP 10 L de EDTA
([EDTA]stock= 100 mM )e o volume de H2O MilliQ necessaacuterio para perfazer 1 mL Em ambas as
situaccedilotildees o uacuteltimo reagente adicionado foi o ATP uma vez que se trata do substrato da reacccedilatildeo
enzimaacutetica
A cineacutetica decorreu a 25ordmC durante 5 minutos para a actividade Ca2+
-ATPaacutesica e durante 3
minutos para a actividade K+EDTA-ATPaacutesica Apoacutes o periacuteodo de incubaccedilatildeo transferiu-se 150
L de amostra para tubos de Eppendorf contendo igual volume de TCA 10 Colocou-se no gelo
e homogeneizou-se no voacutertex Seguidamente centrifugou-se a 1500 g durante 10 minutos e
recolheu-se 200 L de sobrenadante para novos tubos de Eppendorf Adicionou-se a estes 158
mL de H2O MilliQ e 200 L de soluccedilatildeo J Homogeneizou-se no voacutertex e adicionou-se 20 L de
reagente redutor repetindo-se a homogeneizaccedilatildeo Incubou-se durante 10 minutos e mediu-se a
Abs660 nm com o mesmo intervalo de tempo de leitura para cada amostra Por intrapolaccedilatildeo a partir
da recta de calibraccedilatildeo obtida anteriormente determinou-se a quantidade de Pi libertado em cada
ensaio A partir destes resultados determinaram-se as actividades Ca2+
-ATPaacutesica e K+EDTA-
ATPaacutesica do S1 em nmol PO43-
minmg proteiacutena e em s-1
342 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
Da actividade Mg2+
-ATPaacutesica de S1 resulta a formaccedilatildeo dos produtos da reacccedilatildeo de
hidroacutelise do ATP - ADP e Pi Na presenccedila de ADP fosfoenolpiruvato (PEP) e da enzima piruvato
cinase (PK) os produtos resultantes satildeo ATP e piruvato Por acccedilatildeo da enzima lactato
desidrogenase (LDH) este uacuteltimo eacute convertido em lactato ocorrendo simultaneamente a oxidaccedilatildeo
do NADH a NAD+ monitorizada atraveacutes da Abs340 nm Quando o S1 de miosina se encontra no
meio reaccional o ATP eacute hidrolisado formando-se novamente ADP e Pi o que permite a
formaccedilatildeo de um ciclo (fig 21) Se as enzimas se encontrarem em excesso a velocidade de
conversatildeo do NADH eacute limitada pela produccedilatildeo de ADP e Pi Como o ATP eacute regenerado em cada
ciclo a sua concentraccedilatildeo inicial eacute sempre mantida (Bagshaw 1982)
Figura 33ndash Representaccedilatildeo esquemaacutetica do meacutetodo de enzimas acopladas
47
O protocolo utilizado e seguidamente descrito foi adaptado de Cuenda et al (1990 ndash Ver Refordf) A
uma cuvette de 1 mL adicionou-se S1 para uma concentraccedilatildeo final de 005 mgmL 800 L de
meio de reacccedilatildeo22
10 L de PEP ([PEP]stock = 42 mM) 10 L de PK+LDH 11 ([PK+LDH]stock =
1 mgmL) e 15 L de NADH ([NADH]stock=25 mM) Iniciou-se a monitorizaccedilatildeo da reacccedilatildeo
atraveacutes da leitura de Abs340 nm apoacutes a adiccedilatildeo de 20 L de ATP ([ATP]stock=100 mM) A
actividade Mg2+
-ATPaacutesica eacute obtida atraveacutes do declive correspondente agrave oxidaccedilatildeo do NADH Para
a mediccedilatildeo da Mg2+
-ATPase de S1 estimulada por actina adicionou-se F-actina para uma
concentraccedilatildeo final de 2 microM sendo a actividade obtida tambeacutem atraveacutes do declive correspondente
agrave oxidaccedilatildeo do NADH apoacutes a adiccedilatildeo da proteiacutena
35 Interaccedilatildeo entre vanaacutedio e subfragmento-1 de miosina
351 Preparaccedilatildeo das soluccedilotildees de vanaacutedio
As soluccedilotildees de vanadato foram preparadas em aacutegua MilliQ a partir de metavanadato de amoacutenio
(NH4VO3) numa concentraccedilatildeo stock de 50 mM com agitaccedilatildeo magneacutetica e aquecimento (~30 ordmC)
Esta soluccedilatildeo com um valor compreendido entre 6 e 7 designa-se por ldquometavanadatordquo e eacute
constituiacuteda pelas esspeacutecies V1 V2 V4 e V5 (Ref) Acidificando a soluccedilatildeo de metavanadato por
adiccedilatildeo de HCl ateacute um valor de pH proacuteximo de 4 observa-se uma mudanccedila de cor de amarelo
claro para laranja indicativa da formaccedilatildeo da espeacutecie decameacuterica de vanadato De seguida e
imediatamente antes da realizaccedilatildeo dos ensaios descritos o valor do pH da soluccedilatildeo de
decavanadato foi aumentado para cerca de 75 por adiccedilatildeo de NaOH A soluccedilatildeo (2) designada
por ldquometavanadatordquo natildeo foi posteriormente alterada ateacute agrave sua adiccedilatildeo para a realizaccedilatildeo dos ensaios
As soluccedilotildees foram armazenadas a 4ordmC e mantidas em gelo aquando da sua utilizaccedilatildeo
A soluccedilatildeo de V(IV) foi preparada em condiccedilotildees anaeroacutebias por diluiccedilatildeo de sulfato de vanadil
(VOSO4) para uma concentraccedilatildeo final de 5 mM em H2O MilliQ saturada com N2 gasoso
352Anaacutelise por espectroscopia de UV-Vis
A concentraccedilatildeo da espeacutecie decameacuterica em soluccedilatildeo depende do periacuteodo de tempo apoacutes a diluiccedilatildeo
no meio de reacccedilatildeo apropriado (Ref) A velocidade de desoligomerizaccedilatildeo da espeacutecie decameacuterica
pode ser analisada por espectroscopia de absorccedilatildeo na regiatildeo UVVis uma vez que esta espeacutecie
apresenta duas bandas caracteriacutesticas na regiatildeo do visiacutevel (a 358 e 400 nm) que natildeo se verifica
em quaisquer outras espeacutecies oligomeacutericas As bandas supramencionada sapresentam-se bem
22
Meio de reacccedilatildeo 3125 mM Tris-HCl pH 71 375 mM MgCl2
48
resolvidas atraveacutes da obtenccedilatildeo da derivada de 2ordf ordem do espectro apresentando a forma de 2
vales com miacutenimos centrados nos comprimentos de onda supracitados com dependecircncia linear
relativamente agrave concentraccedilatildeo de decavanadato total (Ramos et al 2006)
Assim a estabilidade da espeacutecie decameacuterica apoacutes diluiccedilatildeo da soluccedilatildeo de decavanadato no meio de
reacccedilatildeo dos ensaios (tampatildeo D) para uma concentraccedilatildeo final de 1 mM em vanadato total foi
caracterizada atraveacutes da leitura da Abs400 em funccedilatildeo do tempo tendo-se determinado qual a
ordem de reacccedilatildeo e o tempo de meia-vida da mesma Com base em estudos anteriores acerca da
estabilidade do V10 na presenccedila de diferentes proteiacutenas (Ramos et al 2006) determinou-se
igualmente o tempo de meia vida apoacutes a adiccedilatildeo de 20 microM de S1 de miosina ao meio reaccional
353 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
Os espectros de RMN do isoacutetopo 51
V foram realizados num espectroacutemetro Bruker AM-400 MHz
equipado com uma sonda multinuclear inversa de 5 mm Em tubos de RMN de 5 mm de diacircmetro
preparou-se soluccedilotildees de 500 L de deca- ou metavanadato correspondentes a um intervalo de
concentraccedilotildees de 1 a 10 mM suplementadas com 10 D2O Os espectros foram adquiridos agrave
temperatura ambiente com uma largura espectral de 23 923 Hz um tempo de aquisiccedilatildeo de 0086
s tempo de relaxaccedilatildeo de 001 s e uma sequecircncia de pulsos a 90ordm Utilizou-se a transformada de
Fourier com alargamento das bandas de 20Hz para o processamento dos espectros Os desvios
quiacutemicos das ressonacircncias de 51
V foram obtidos em relaccedilatildeo agrave espeacutecie VOCl3cuja frequecircncia de
ressonacircncia eacute considerada o padratildeo sendo todas as outras referenciadas a partir desta em ppm
As aacutereas dos sinais do RMN correspondentes agraves espeacutecies oligomeacutericas presentes foram obtidas
por integraccedilatildeo dos sinais As concentraccedilotildees relativas das espeacutecies foram calculadas com recurso agrave
equaccedilatildeo
onde A[Vx] corresponde agrave aacuterea referente agrave ressonacircncia da espeacutecie Vx com nordm de agregaccedilatildeo x
A[V]total agrave soma total das aacutereas medidas e [V] a concentraccedilatildeo total de vanaacutedio no ensaio
A teacutecnica de 51
V-RMN foi anteriormente aplicada em estudos de ligaccedilatildeo com S1 de miosina
(Ref) tendo sido demostrado que entre as espeacutecies V1 V2 V4 e V5 a espeacutecie tetrameacuterica possui a
maior afinidade para a miosina (Cremo et al) O desaparecimento ou alargamento dos sinais pode
implicar a ligaccedilatildeo de um dos oxovanadatos laacutebeis agrave miosina sem indicaccedilatildeo de qual uma vez que
49
se processa um equiliacutebrio entre eles (Crans 1990) Assim e tendo em conta as indicaccedilotildees de que
a espeacutecie V10 eacute distinta das restantes no que diz respeito agrave magnitude da interacccedilatildeo com a miosina
e inibiccedilatildeo da actividade ATPaacutesica estimulada por actina (Aureliano 2000 Tiago et al 2002)
torna-se fundamental investigar a interacccedilatildeo das diferentes espeacutecies de vanadato com o S1 de
miosina atraveacutes da teacutecnica de 51
V-RMN
Para tal obtiveram-se espectros de 51
V-RMN resultantes da titulaccedilatildeo de uma soluccedilatildeo 5 mM de
decavanadato total com concentraccedilotildees crescentes de S1 de miosina (0 5 10 15 e 20 microM) em
tampatildeo D Realizou-se ainda ensaios na presenccedila de 2 mM de decavanadato e 2 mM de ATP na
presenccedila e na ausecircncia de S1 de miosina de modo a verificar o efeito que estes ligandos naturais
da enzima teria na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o subfragmento-1
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
Os grupos sulfidrilo (-SH) das cisteiacutenas satildeo resiacuteduos de aminoaacutecidos extremamente sensiacuteveis agrave
oxidaccedilatildeo resultando na sua oxidaccedilatildeo (Dalle-Donne et al 1999) Logo a quantificaccedilatildeo destes
resiacuteduos eacute bastante importante nos estudos de interacccedilatildeo com vanaacutedio jaacute que os resiacuteduos de Cys
com ecircnfase para o SH1 e o SH2 satildeo bastante importantes na monitorizaccedilatildeo das modificaccedilotildees
funcionais da regiatildeo da cabeccedila
O procedimento seguidamente apresentado foi originalmente descrito por Gutieacuterrez-Merino
(1987)) Consiste na reacccedilatildeo entre os grupos ndashSH natildeo oxidados das proteiacutenas e o reagente de
Ellman (DTNB -55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico)) - formando-se o aniatildeo CNT (aniatildeo 3-
carboxilato-4-nitrotiofenolato) numa estequiometria de 11 Este apresenta um maacuteximo de
absorvecircncia a 412 nm (Robyt 1990) Assim apoacutes a incubaccedilatildeo da proteiacutena na presenccedila do metal
a quantidade de cisteiacutenas reduzidas por proteiacutena determina-se atraveacutes dos valores de Abs412
Deste modo colocou-se num tubo de Eppendorf o volume apropriado de S1 dialisado para que a
concentraccedilatildeo no ensaio fosse de 1 mgmL Adicionou-se o volume apropriado de decavanadato
para que a sua concentraccedilatildeo fosse de 0 10 50 100 250 500 ou 1000 M Perfez-se com soluccedilatildeo
D ateacute um volume final de 1 mL e incubou-se a 25ordmC por 20 minutos Apoacutes este periacuteodo colocou-
se 780 L da mistura reaccional numa cuvette de quartzo e adicionou-se 220 L de soluccedilatildeo K23
23
Tampatildeo K Tampatildeo fosfatohidrogenofosfato de soacutedio 025 M pH 72
50
Adicionou-se 15 L de DTNB (em soluccedilatildeo K [DTNB]stock=100 mM) ao meio reaccional e
registou-se o valor de absorvecircncia medido equivalente agraves cisteiacutenas ldquoraacutepidasrdquo (SH1 e SH2
expostos ao solvente) Adicionou-se 001 g de SDS que provoca a desnaturaccedilatildeo da proteiacutena e
consequentemente a exposiccedilatildeo de todos os resiacuteduos de cisteiacutenas que se encontravam ateacute aiacute
inacessiacuteveis no interior da proteiacutena (cisteiacutenas totais) Estes resiacuteduos podem assim reagir com o
DTNB
Estimou-se o nuacutemero de resiacuteduos de cisteiacutenas em todas as amostras com recurso ao coeficiente de
extinccedilatildeo molar () do DTNB (ver ponto 3 dos Anexos) que foi determinado atraveacutes da titulaccedilatildeo
de L-Cisteiacutena com intervalo de concentraccedilotildees compreendidos entre 0 e 50 microM na presenccedila de
25microM de DTNB em tampatildeo D Os valores de absorvecircncia foram obtidos para um comprimento de
onda de 412 nm
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
3621 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
A teacutecnica mais eficiente de anaacutelise da interacccedilatildeo entre o iatildeo vanadilo e proteiacutenas como a
calmodulina e a transferrina entre outras permite a obtenccedilatildeo raacutepida de informaccedilatildeo no que toca agrave
conformaccedilatildeo e amplitude da referida interacccedilatildeo
Os espectros de Ressonacircncia Paramagneacutetica Electroacutenica (RPE) da espeacutecie paramagneacutetica V(IV)
foram realizados a 70K num espectroacutemetro Bruker EMX-300 X-band Todas as soluccedilotildees
utilizadas e amostras foram preparadas em anaerobiose numa cacircmara de luvas de modo a evitar a
reduccedilatildeo de V(IV) a V(V) Em tubos capilares com 02 mL de amostra manteve-se a concentraccedilatildeo
de S1 a 20 M e variou-se a concentraccedilatildeo de vanadilo (0 25 5 10 15 25 50 75 100 150
200 500 e 1000 M [VOSO4]stock=5 mM) em tampatildeo D Apoacutes obtenccedilatildeo dos espectros estes
foram tratados atraveacutes do software SimFonia e WinEPR tendo-se obtido um graacutefico com a
variaccedilatildeo da intensidade do sinal de RPE em funccedilatildeo da concentraccedilatildeo de sulfato de vanadilo
presente Realizaram-se ainda simulaccedilotildees de espectros obtidos com 20 microM de S1 e 1000 microM de
V(IV) com vista agrave obtenccedilatildeo dos ligandos provaacuteveis do vanadilo atraveacutes da anaacutelise da constante
de acoplamento hiperfina (A) e do factor g (g)
4 Conclusotildees gerais
Apesar de jaacute ter sido demonstrado que o vanaacutedio inibe a Na+K
+-ATPase da miosina a funccedilatildeo
deste elemento no tecido muacutesculo esqueleacutetico continua largamente incompreendida A enorme
complexidade quiacutemica deste metal especialmente no que concerne agrave oligomerizaccedilatildeo e reacccedilotildees
51
redox constitui um obstaacuteculo agrave compreensatildeo do comportamento do vanaacutedio nos organismos
vivos Assim a caracterizaccedilatildeo da quiacutemica deste elemento eacute de grande utilidade antes de iniciar o
estudo da sua interacccedilatildeo com o presente sistema
A anaacutelise por espectroscopias de 51
V-RMN e de UV-Vis de soluccedilotildees de vanadato diluiacutedas nos
meios de reacccedilatildeo utilizados no presente relatoacuterio (10 mM HEPES pH 70 25 mM KCl) mostrou
claramente que a espeacutecie decameacuterica (V10) possui um tempo de vida suficientemente longo para a
determinaccedilatildeo do seu efeito isolado ao contraacuterio do que se verifica com as espeacutecies mono- di-
tetra- e pentameacutericas (V1 V2 V4 e V5) Por outro lado a anaacutelise verificou ainda que apoacutes diluiccedilatildeo
das soluccedilotildees de decavanadato no meio de reacccedilatildeo coexistem as espeacutecies V10 e V1 pelo que os
efeitos putativos desta uacuteltima deveratildeo ser subtraiacutedos aquando da monitorizaccedilatildeo dos efeitos da
espeacutecie decameacuterica
O vanaacutedio (V) (vanadato) inibe as enzimas que catalizam reacccedilotildees de transferecircncia de grupos
fosfato como a ATPase da miosina [Goodno 1979] devido agrave sua analogia estrutural com o iatildeo
fosfato Na sua utilizaccedilatildeo como inibidor enzimaacutetico natildeo eacute tida em conta a contribuiccedilatildeo individual
das espeacutecies oligomeacutericas considerando-se normalmente a espeacutecie V1 como inibidora efectiva
[Goodno e Taylor 1982] No presente relatoacuterio verificou-se por espectroscopia de 51
V-RMN que
outros oligoacutemeros de vanadato interagem com o S1 de miosina tendo as espeacutecies V10 e V4 os
maiores niacuteveis de afinidade A sua interacccedilatildeo envolve forccedilas electrostaacuteticas e eacute afectada
negativamente por ligandos naturais de enzima como eacute o caso do ATP
A exposiccedilatildeo do S1 de miosina (005 mgmL) a decavanadato conduz agrave inibiccedilatildeo do capacidade da
proteiacutena de estimular a actividade Mg2+
-ATPaacutesica com um valor de IC50 de 266 plusmn 07 M
decavanadato sendo o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM
decavanadato (80) Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM decavanadato sendo
o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM decavanadato (60)
exercendo um efeito de impedimento agrave ligaccedilatildeo entre o vanaacutedio e o S1
A uma concentraccedilatildeo de 27 microM decavanadato verifica-se a oxidaccedilatildeo de um dos resiacuteduos de Cys
de S1 de miosina seguida pela oxidaccedilatildeo gradual de outro resiacuteduo de Cys ateacute agrave concentraccedilatildeo de
1000 microM Ambos os resiacuteduos oxidados natildeo se encontravam expostos ao solvente utilizado A
oxidaccedilatildeo dos resiacuteduos permite sugerir que esta modificaccedilatildeo oxidativa sofrida pelo S1 contribui
para a diminuiccedilatildeo da sua actividade de ATPase
52
Por espectroscopia de RPE revelaram-se os possiacuteveis ligandos equatoriais do vanaacutedio aquando da
sua interacccedilatildeo com o S1 de miosina como sendo duas moleacuteculas de aacutegua um ligando contendo o
grupo R-OH e outro contendo o grupo R=NR
Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados de
oxidaccedilatildeo (IV) e (V) no S1 de miosina para concentraccedilotildees facilmente alcanccedilaacuteveis no ambiente
terrestre
53
5 Estudos futuros
No sentido do melhor esclarecimento das alteraccedilotildees moleculares e estruturais que ocorrem no
subfragmento 1 quando exposta ao vanaacutedio que conduzem a uma diminuiccedilatildeo da sua funccedilatildeo
sugere-se
- Quantificaccedilatildeo das cisteiacutenas reduzidas de S1 na presenccedila de metavanadato
- Determinaccedilatildeo das actividades Ca2+
- e K+-ATPaacutesica do S1 de miosina na presenccedila de
decavanadato
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina estimulada por actina
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina na presenccedila de
metavanadato
- Realizaccedilatildeo de RMN 2D com variaccedilatildeo das concentraccedilotildees das soluccedilotildees de decavanadato e
metavanadato na presenccedila de S1
-Mutageacutenese dos resiacuteduos de Cys doS1 de miosina de modo a verificar quais os resiacuteduos que
sofrem oxidaccedilatildeo na presenccedila de decavanadato
- Determinaccedilatildeo por calorimetria (DSC Differential Scanning Calorimetry) da variaccedilatildeo da
entalpia (∆H) e da temperatura de desnaturaccedilatildeo do S1 de miosina na presenccedila de vanaacutedio uma
vez que estas propriedades caracteriacutesticas de cada proteiacutena satildeo alteradas apoacutes ocorrecircncia de
modificaccedilotildees estruturais
- Anaacutelise por SDS-PAGE de S1 de miosina apoacutes tratamento com vanaacutedio de modo a determinar
quais os polipeacuteptidos eventualmente formados apoacutes clivagem da proteiacutena
54
6 Referecircncias bibliograacuteficas
- Sellers J Myosins 2nd Edition Oxford University Press 1999
- L Stryer inBiochemistry (Ed WH Freeman 2005 pp
Refordfas Materiais e Meacutetodos
[A] S S Margossian SLowey Methods Enzymol1982 85 55-71
[B] JD Pardue JA Spudich Methods Enzymol 1982 85 164-181
[C] M Bradford Anal Biochem1976 72 248-254
[D] H H Taussky E Shorr J Biol Chem 1953 202 675-685
[E] C R Bagshaw in Outline Studies in Biology ndash Muscle Contraction Chapman and Hall Ed
1982
[F] A Cuenda et al Eur J Biochem1990 194 663-670
55
Anexos
1 Determinaccedilatildeo da recta de calibraccedilatildeo meacutetodo de Bradford
Figura1ndashRepresentaccedilatildeo graacutefica da recta de calibraccedilatildeo (Abs595 nm vs [BSA]) utilizada para o meacutetodo de Bradford
2 Determinaccedilatildeo da recta calibraccedilatildeo para a electroforese em condiccedilotildees desnaturantes
Figura 2 ndash Representaccedilatildeo graacutefica da recta de calibraccedilatildeo (log(Peso Molecular) vs Rf dos marcadores de peso
molecular) onde Rf indica determinada atraveacutes do padratildeo de peso molecular
(poccedilo (a))
3Caacutelculo do coeficiente de extinccedilatildeo molar () para a quantificaccedilatildeo de cisteiacutenas reduzidas
atraveacutes do meacutetodo de Ellman
Traccedilou-se a recta de Abs412 nm meacutedia vs [L-Cisteiacutena] de modo a determinar o valor do coeficiente
de extinccedilatildeo molar definido como o declive daquela recta Assim =8400 M-1
cm-1
como
podemos verificar na Fig 3
A595 nm = 00546[BSA] + 00209 Rsup2 = 099051
Ab
s 59
5 n
m
[BSA] (mgmL)
log PM = -1177 Rf + 2247 Rsup2 = 098957
log
PM
Mobilidade relativa (Rf)
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero

32
Figura 211 ndash Representaccedilatildeo
graacutefica da Actividade Mg2+
-
ATPaacutesica de S1 () (os ajustes
teoacutericos estatildeo representado a por
uma linha os resultados
experimentais estatildeo representados
por triacircngulos (Actividade
estimulada) e cruzes (Actividade
natildeo estimulada)) vs
[Decavanadato]total medidas pelo
meacutetodo das enzimas acopladas
(n=3) Os dados foram ajustados ao modelo de decaimento exponencial usando o programa GraphPad Prism para a
determinaccedilatildeo dos valores de IC50 (na ausecircncia de actina IC50= 266 plusmn 07 M decavanadato na presenccedila de actina
IC50= 28 plusmn 1 M decavanadato
A partir do graacutefico presente na Fig 29 verifica-se que o ajuste dos dados experimentais foi bem
sucedido A inibiccedilatildeo da actividade Mg2+
-ATPaacutesica do S1 de miosina eacute afectada em menor
amplitude na presenccedila de actina (60 de inibiccedilatildeo da actividade) do que na sua ausecircncia
(aproximadamente 80 de inibiccedilatildeo de actividade) Devido a esta diferenccedila pode sugerir-se que a
actina iraacute impedir parcialmente a ligaccedilatildeo do decavanadatoao S1 demiosina atraveacutes de
impedimentos esteacutereos ou interacccedilotildees electrostaacuteticas por exemplo A teacutecnica utilizada natildeo nos
permite concluir acerca de que regiatildeo(otildees) ou resiacuteduo(s) da actina participaratildeo na preservaccedilatildeo
parcial da actividade Mg2+
-ATPaacutesica
Por outro lado os valores de IC50 dos dois tipos de actividades natildeo apresentam diferenccedilas
significativas tendo-se obtido 266 plusmn 07 e 28 plusmn 1M decavanadato na ausecircncia e na presenccedila de
actina respectivamente o que corresponde a uma concentraccedilatildeo de cerca de 27 M de V10 O
meio de reacccedilatildeo utilizado nestes ensaios foi 10 mM HEPES e 25 mM KCl com 2 mM ATP Em
(Tiago et al 2004) o valor de IC50 para a actividade Mg2+-ATPaacutesica na ausecircncia de actina foi de
125 plusmn 025 microM enquanto o valor da actividade estimulada por actina foi de 136 plusmn 14 microM O
meio de reacccedilatildeo utilizado nos ensaios foi 20 mM HEPES 50 mM KCl e 2 mM ATP o que resulta
precisamente no dobro da forccedila ioacutenica a que foi realizado o ensaio do presente relatoacuterio
(exceptuando a contribuiccedilatildeo dos produtos da hidroacutelise de ATP uma vez que este meacutetodo
pressupotildee a ocorrecircncia de um ciclo de reacccedilotildees de hidroacutelise e siacutentese deste composto) Ora tendo
sido anteriormente referido que a afinidade do S1 de miosina para com a espeacutecie V10 diminui com
Act
ivid
ade
Mg
2+-A
TP
aacutesic
a d
e S
1 d
e
Mio
sin
a
[Decavanadato] (microM) Actividade Estimulada ()
Actividade Natildeo Estimulada ()
Ajuste Actividade Estimulada
Ajuste Actividade Natildeo Estimulada
33
o aumento da forccedila ioacutenica (Tiago et al 2004 tal poderaacute explicar a diferenccedila obtida entre estes
dois conjuntos de valores
243Quantificaccedilatildeo de cisteiacutenas reduzidas pelo meacutetodo de Ellman
A interacccedilatildeo do vanaacutedio com alguns tipos de aminoaacutecidos sensiacuteveis agrave oxidaccedilatildeo pode fornecer-nos
informaccedilatildeo bastante uacutetil em relaccedilatildeo agraves alteraccedilotildees estruturais que conduzem a uma diminuiccedilatildeo da
actividade fisioloacutegica do S1 de miosina Os estudos realizados por Chasteen 1983 concluiacuteram
que a reduccedilatildeo de vanadato (V) a vanadilo (IV) ocorre na presenccedila de agentes redutores comuns no
ambiente celular de que se destaca o aminoaacutecido cisteiacutena Esta eacute tambeacutem um alvo de
complexaccedilatildeo por parte do iatildeo VO2+
Jaacute Ramos et al (2009) demonstrou que a incubaccedilatildeo de
actina com a soluccedilatildeo de decavanadato induz a oxidaccedilatildeo de resiacuteduos de cisteiacutena e agrave formaccedilatildeo do
oxidovanaacutedio (IV) Este grupo de investigadores descobriu ainda que apenas a espeacutecie V10 induz a
oxidaccedilatildeo das cisteiacutenas ldquoraacutepidasrdquo da actina atraveacutes do processo proposto que eacute parcialmente
responsaacutevel pela inibiccedilatildeo jaacute descrita da actividade de ATPase do S1 de miosina estimulada por
actina quando exposta agrave soluccedilatildeo de decavanadato Esta inibiccedilatildeo apresenta-se afectada com a
presenccedila de ATP uma vez que os resiacuteduos de cisteiacutena permaneceram intactos nos ensaios
realizados com este ligando Jaacute Crans et al (2004) concluiu que o siacutetio activo de muitas fosfatases
(que conteacutem grupos tiol) pode formar um equiliacutebrio redox com o iatildeo vanadato (V) de forma natildeo
reversiacutevel
Os resultados obtidos por Sakurai e Shiromura (1980) sugeriram que o vanaacutedio (V) sofre reduccedilatildeo
a vanaacutedio (IV) sem dependecircncia do valor de pH do ensaio Registaram ainda uma estequiometria
de 31 na reacccedilatildeo do vanadato (V) com os resiacuteduos de Cys enquanto a estequiometria dos ensaios
com o vanadilo (IV) foi de 21
Berlett e Stadtman (1997) observaram que os tipos de aminoaacutecidos mais facilmente oxidaacuteveis
englobam entre outros os resiacuteduos de Cys Trp e MetAssim analisou-se o estado redox dos
grupos tiol dos resiacuteduos de Cys do S1 na presenccedila do decavanadato atraveacutes da monitorizaccedilatildeo da
Abs412 nm na presenccedila de 1 mgmL de S1 de miosina em tampatildeo 10 mM HEPES 25 mM KCl
variando a concentraccedilatildeo da soluccedilatildeo de decavanadato de modo a determinar que tipo de alteraccedilotildees
estruturais ocorrem na sua presenccedila no que diz respeito agrave diminuiccedilatildeo da actividade ATPaacutesica de
S1 na presenccedila de decavanadato
Os resultados referentes agrave quantificaccedilatildeo das cisteiacutenas reduzidas apoacutes o tratamento de S1 com
diferentes concentraccedilotildees de decavanadato em tampatildeo 10 mM HEPES pH 75 e 25 mM KCl a
25ordmC durante 20 minutos encontram-se descritos na Tabela 2VII
Tabela 2VII ndash Cisteiacutenas reduzidas totais e ldquoraacutepidas de S1 (1 mgmL) apoacutes tratamento com decavanadato
34
(010501002505001000 uM) em 10 mM HEPES 25 mM KCl pH 7 a 25ordmC durante 20 minutos A detecccedilatildeo
espectrofotomeacutetrica foi feita com DTNB (e 412=8400 uM-1 cm-1) (n variaacutevel entre 1 e 8)
Amostras Cys raacutepidas Cys totais
11 M S1 (controlo) 22 plusmn 04 106 plusmn 14
11 M S1 + 10 M Decavanadato 22 plusmn 05 95 plusmn 08
11 M S1 + 50 uM Decavanadato 20 plusmn 02 98 plusmn 14
11 M S1 + 100 uM Decavanadato 17 plusmn 03 97 plusmn14
11 M S1 + 250 uM Decavanadato 26 plusmn 02 98 plusmn 08
11 M S1 + 500 uM Decavanadato 21 plusmn 04 90 plusmn 06
11 M S1 + 1000 uM Decavanadato 24 plusmn 0 86 plusmn 08
A partir dos resultados obtidos pode sugerir-se que o S1 possui 11 resiacuteduos de Cys (controlo) o
que estaacute de acordo com o anteriormente descrito Verifica-se que o tratamento da proteiacutena com
concentraccedilotildees crescentes de decavanadato provoca a oxidaccedilatildeo de dois resiacuteduos de Cys do S1 de
miosina natildeo expostos ao solvente No entanto a teacutecnica utilizada natildeo nos permite identificar
quais
Tendo em consideraccedilatildeo que o valor de IC50 obtido para a reacccedilatildeo de inibiccedilatildeo daactividade Mg2+-
ATPaacutesica de S1 na presenccedila de decavanadato eacute de cerca de 27 microM pode sugerir-se que este seraacute o
valor ao qual ocorreraacute a oxidaccedilatildeo do primeiro resiacuteduo de Cys o que poderaacute contribuir para a
diminuiccedilatildeo da actividade de S1 observada
25Interacccedilatildeo de S1 com vanaacutedio(IV)
251Anaacutelise por espectroscopia de Ressonacircncia Paramagneacutetica Electroacutenica
A acumulaccedilatildeo intracelular de oxovanaacutedio (IV) in vivo constitui uma das principais formas de
apresentaccedilatildeo de vanaacutedio no organismo Deste modo torna-se bastante interessante analisar a
interacccedilatildeo entre este iatildeo e o S1 de miosina atraveacutes da teacutecnica de espectroscopia de Ressonacircncia
Paramagneacutetica Electroacutenica o meacutetodo de eleiccedilatildeo do estudo do iatildeo vanadilo e dos complexos por si
formados O electratildeo desemparelhado encontra-se fortemente acoplado ao nuacutecleo 51
V o que
35
resulta num espectro de RPE consistindo de 8 desdobramentos na direcccedilatildeo paralela e outros tantos
na direcccedilatildeo perpendicular (Chasteen 1983)
Os estudos realizados por Ahmed et al (1987) centraram-se na interacccedilatildeo do vanadilo com
proteiacutenas como eacute o caso da insulina transferrina entre outros obtendo um espectro caracterizado
por 8 linhas amplamente espaccediladas (na direcccedilatildeo paralela) Jaacute Ramos et al 2009 concluiu que a
exposiccedilatildeo da soluccedilatildeo de decavanadato agraves formas globular e filamentosa de actina produz um
espectro tiacutepico do iatildeo oxidovanaacutedio (IV) indicando a reduccedilatildeo do vanadato
Para aferir qual a concentraccedilatildeo de S1 que permitia obter uma boa razatildeo sinalruiacutedo titulou-se uma
soluccedilatildeo de sulfato de vanadilo 15 microM ([VOSO4]stock = 5 mM) com concentraccedilotildees crescentes de
proteiacutena (0 5 10 15 e 20 microM) em 10 mM HEPES 25 mM KCl A existecircncia de uma correlaccedilatildeo
linear entre a concentraccedilatildeo de proteiacutena e a intensidade dos sinais de RPE correspondente agrave banda
-12 perpendicular encontra-se representada na Fig 210
Figura 2 12ndash Representaccedilatildeo graacutefica da variaccedilatildeo da intensidade do sinal de RPE vs a concentraccedilatildeo de S1 utilizada
Apoacutes verificar que com uma concentraccedilatildeo de S1 igual a 20 microM se obtinham sinais com uma razatildeo
sinalruiacutedo adequada ao prosseguimento dos estudos sobre a interacccedilatildeo V(IV)-S1 de miosina
titulou-se uma soluccedilatildeo de 20 microM S1 com 0 25 5 10 15 25 50 75 100 150 200 500 e 1000
microM VOSO4
Traccedilou-se o graacutefico da intensidade da banda -12 perpendicular do sinal de EPR em funccedilatildeo da
concentraccedilatildeo de VOSO4 tendo-se determinado uma constante de dissociaccedilatildeo (KD) do complexo
S1-vanadilo de 8748 plusmn 2621 microM V(IV)
y = 016786x + 122435
Rsup2 = 092992759
Inte
nsi
dad
e d
o s
inal
de
RP
E
[S1] (microM)
36
Figura 213 ndash Representaccedilatildeo graacutefica da variaccedilatildeo do sinal de RPE vs a concentraccedilatildeo de vanadilo utilizada (os valores
experimentais estatildeo representados por losangos o seu ajuste estaacute representado pela linha)
A simulaccedilatildeo do espectro de EPR foi efectuada a partir do espectro obtido para uma concentraccedilatildeo
de 1 mM VOSO4 apoacutes adiccedilatildeo de 20 microM S1 de miosina tendo-se obtido os valores da constante de
acoplamento hiperfina (A=5016) e do factor g (g0=195)
Figura 214 ndash Representaccedilatildeo graacutefica do espectro de EPR obtido com 1000 microM de vanadilo e 20 microM de S1 (a cinzento
escuro) A simulaccedilatildeo do espectro encontra-se a cinzento claro
Por comparaccedilatildeo destes com os valores presentes na tabela de correspondecircncia xxx eacute possiacutevel
predizer quais os ligandos provaacuteveis do vanadilo na presenccedila de proteiacutena Assim concluiu-se que
o vanadilo estaacute coordenado com duas moleacuteculas de aacutegua um grupo hidroxilo e um grupo R=NR
O grupo hidroxilo pode indicar os resiacuteduos de aminoaacutecidos Tyr Treonina e Serina enquanto o
grupo R=NR poderaacute indicar a presenccedila de Arginina e Histidina como ligandos do vanadilo
Inte
nsi
dad
e d
os
sinai
s d
e R
PE
[Vanadilo] (microM)
Ajuste dos dados exp Dados experimentais
Campo magneacutetico (G)
Espectro Experimental Ajuste
37
3 Procedimento Experimental
31 Reagentes
Todos os reagentes utilizados satildeo de elevado grau de pureza (analiacutetico) Apresentam-se as suas
foacutermulas quiacutemicas ou abreviaturas quando necessaacuterio acrescentou-se a sua descriccedilatildeo Entre
parecircntesis colocou-se as empresas que os comercializam
Aacutecido aceacutetico glacial 100 (Riedel) Acrilamida 99 (Merck) Azul de bromofenol (Riedel-de-
Haumlen) -Mercaptoetanol (Sigma) Bissulfito de soacutedio (Sigma)Bradford (reagente de)
(BioRad)BSA (albumina de soro bovino Sigma-Aldrich) CaCl22H2O (cloreto de caacutelcio
dihidratado Fluka) CH3CH2OH 96 (etanol Panreac) Coomassie Blue R250 (Merck) D2O
(oacutexido de deuteacuterio SDS) DTNB (55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico) Aldrich) EDTA (aacutecido
etilenodiaminatetraaceacutetico Merck) Fosfoenolpiruvato 98 (Fluka) Glicerol (Sigma) Glicina
985 (Panreac) HCl 37 (Riedel-de Haumlen) H2SO4 96 (Pronalab) HEPES 99 (aacutecido 4-(2-
hidroxiletil)-1-piperazineetanosulfoacutenico Sigma) KH2PO4 99 (hidrogenofosfato de potaacutessio)
MgATP (Sigma) Na
ATP (Sigma) Na2HPO4 99 (hidrogenofosfato de soacutedio Riedel-de Haumlen)
NaH2PO4 (fosfato de soacutedio ) NaOH 986 (hidroacutexido de soacutedio Pronalab) KCl 99 (Cloreto de
potaacutessio Panreac) L-Cys 995 (enantioacutemero levoacutegiro do resiacuteduo de Cisteiacutena Fluka)Metanol
99 (Merck) Molibdato de amoacutenio (Sigma)NNrsquo-metil-bis-acrilamida 98 (Merck) NH4VO3
99 (metavanadato de amoacutenia Sigma) PSA 10 (persulfato de amoacutenio Merck) SDS
(dodecilo sulfato de soacutedio Sigma) Sulfito de soacutedio (Sigma) VOSO4 (sulfato de vanadilo
Aldrich) TEMED 99 (Tetrametiletilenodiamina Merck) Tris (2-amino-2-hidroximetil-13-
propanodiol Sigma)
32 Meacutetodos
321 Isolamento e purificaccedilatildeo das proteiacutenas de muacutesculo esqueleacutetico de coelho
3211 Purificaccedilatildeo da miosina preparaccedilatildeo de Subfragmento 1 por digestatildeo quimotriacutepsica
da miosina
O subfragmento-1 de miosina-II (S1) foi gentilmente cedido pelo Prof Doutor Manuel Aureliano
do Departamento de Quiacutemica Bioquiacutemica e Farmaacutecia da Faculdade de Ciecircncias e Tecnologia da
Universidade do Algarve
No entanto descreve-se abaixo de forma resumida o procedimento efectuado
38
32111 Purificaccedilatildeo da miosina
O protocolo de isolamento foi adaptado do meacutetodo previamente descrito por Margossian e Lowey
(1982) apresentando-se de forma esquemaacutetica na Fig 31
Sucintamente os tecidos musculares dorsais e das pernas foram extraiacutedos apoacutes o sacrifiacutecio do
animal (Fig31) De seguida foram adicionados 3 volumes da soluccedilatildeo de extracccedilatildeo A2
Centrifugou-se a soluccedilatildeo e filtrou-se o sobrenadante O sedimento foi entatildeo diluiacutedo em 14
volumes de aacutegua destilada de modo a que a forccedila ioacutenica atingisse 004 M Apoacutes adiccedilatildeo de szlig-
mercaptoetanol (szlig-me) deixou-se a precipitar a 4ordmC removendo o sobrenadante Centrifugou-se
novamente e ressuspendeu-se o sedimento na soluccedilatildeo de extracccedilatildeo A Adicionou-se a soluccedilatildeo
tampatildeo B3 para que a forccedila ioacutenica atingisse 05 M Homogeneizou-se e deixou-se repousar
durante 2h Homogeneizou-se novamente e centrifugou-se de modo a ocorrer precipitaccedilatildeo dos
complexos de actomiosina e proteiacutena desnaturada Apoacutes filtraccedilatildeo do sobrenadante em latilde de vidro
quantificou-se a soluccedilatildeo de miosina resultante pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental)
2 Soluccedilatildeo de extracccedilatildeo A 0012M K2CO3 004M KHCO3 06M KCl pH 65
3 Soluccedilatildeo-tampatildeo B Tris 100 mM pH 7 KCl 2M
39
Figura 31 ndash Representaccedilatildeo esquemaacutetica do protocolo de isolamento de miosina (Margossian e Lowey 1982) Todo o
procedimento foi realizado a 4ordmC (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
32112 Preparaccedilatildeo de Subfragmento-1 de miosina por digestatildeo com quimiotripsina
O S1 de miosina foi isolado apoacutes diaacutelise da miosina durante a noite contra a soluccedilatildeo C4 A
presenccedila do EDTA eacute essencial uma vez que a sua substituiccedilatildeo por MgCl2resulta na produccedilatildeo de
HMM em vez de S1 A suspensatildeo de miosina foi equilibrada e digerida com -quimotripsina
A digestatildeo foi interrompida pela adiccedilatildeo de PMSF Seguidamente ultracentrifugou-se o produto
da digestatildeo desprezando-se o sedimento (Fig 32) A soluccedilatildeo de sobrenadante maioritariamente
S1 foi quantificada pela leitura directa da Abs280 ou pelo meacutetodo de Bradford (ver as secccedilotildees
3311 e 3312 do Procedimento Experimental respectivamente) e armazenada em 10 sacarose
e 2 mM szlig-Me a -80ordmC
4 Soluccedilatildeo C 25mM Tris pH 70 012M KCl 1mM EDTA
40
Antes da realizaccedilatildeo dos ensaios descritos no presente relatoacuterio o S1 de miosina foi dialisado
contra a soluccedilatildeo D5 de modo a remover qualquer composto que pudesse complexar como
vanaacutedio Seguidamente quantificou-se a proteiacutena pelo meacutetodo de Bradford ou pelo meacutetodo
directo (ver secccedilotildees 3311 e 3312do Procedimento Experimtental)
Figura 3 2ndash Representaccedilatildeo esquemaacutetica do protocolo de preparaccedilatildeo do S1 de miosina por digestatildeo quimiotripsina de
acordo com o previamente descrito por Margossian e Lowey (1982) Todo o procedimento foi realizado a 4ordm C
excepto quando indicaccedilatildeo do contraacuterio (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
3212 Isolamento e purificaccedilatildeo de actina
Isolou-se e purificou-se a actina segundo o meacutetodo descrito por Pardee e Spudish que
compreende dois passos ndash a obtenccedilatildeo de um resiacuteduo seco denominado ldquopoacute de acetonardquo e a
extracccedilatildeo subsequente da actina a partir do ldquopoacute de acetonardquo na forma de G-Actina com a
posterior polimerizaccedilatildeo a F-Actina
32121 Obtenccedilatildeo do ldquopoacute de acetonardquo
Sacrificou-se o coelho com uma pancada atraacutes do pescoccedilo e sangrou-se o animal apoacutes o corte da
arteacuteria jugular Esfolou-se e colocou-se numa tina com gelo Retiraram-se as partes de tecido
conjuntivo que envolvem o tecido muscular com auxiacutelio de uma pinccedila e bisturi Seguidamente
retirou-se os muacutesculos dorsais e os muacutesculos brancos das pernas Passou-se os muacutesculos por aacutegua
destilada de modo a remover as impurezas e pesou-se num copo Trituraram-se os muacutesculos
numa maacutequina de triturar carne e extraiu-se o resultante com 3 volumes da soluccedilatildeo E6 durante 10
minutos com agitaccedilatildeo suave Centrifugou-se a suspensatildeo a 4000 g durante 10
5 Soluccedilatildeo D HEPES 10 mM pH 70 KCl 25 mM
6 Soluccedilatildeo E 01 M KCl 015 M K2HPO4 pH 65
41
minutosdesprezando-se o sobrenadante Ressuspendeu-se o sedimento em 6 volumes de soluccedilatildeo
F7durante 10 minutos com agitaccedilatildeo suave Verificou-se se o pH da soluccedilatildeo estava compreendido
entre 82 e 85 acertando-se estecom soluccedilatildeo G8 se necessaacuterio Centrifugou-se durante 10
minutos a 4000 g Desprezou-se o sobrenadante e ressuspendeu-se o sedimento em 3 volumes de
soluccedilatildeo H9 durante 10 minutos Centrifugou-se novamente a 4000 gdurante 10 minutos
desprezando-se o sobrenadante Ressuspendeu-se o sedimento duas vezes consecutivas em 6
volumes de aacutegua destilada durante 5 minutos Centrifugou-se a 4000 g durante 10 minutos apoacutes
cada adiccedilatildeo de H2O
Seguidamente ressuspendeu-se o sedimento 5 vezes consecutivas utilizando 3 volumes de
acetona de cada vez agitando suavemente durante 10 minutos agrave temperatura ambiente Colocou-
se o resiacuteduo filtrado numa tina de vidro sobre papel de filtro deixando-se evaporar o solvente
durante a noite num nicho Triturou-se o resiacuteduo seco numa maacutequina de triturar carne e deixou-se
novamente evaporar o solvente O ldquopoacute de acetonardquo obtido foi armazenado a -20 ordmC
32122 Purificaccedilatildeo da actina a partir de ldquopoacute de acetonardquo
Apoacutes a obtenccedilatildeo do poacute de acetona como acima indicado efectuou-se o protocolo seguidamente
descrito de modo a obter monoacutemeros de actina (G-Actina) e actina filamentosa (F-Actina)
Pesou-se 35 g de poacute de acetona e adicionaram-se 50 mL de soluccedilatildeo-tampatildeo I10
sendo a
extracccedilatildeo feita com agitaccedilatildeo suave durante 30 minutos Separou-se o extracto do poacute de acetona
hidratado filtrando a suspensatildeo com 4 camadas de gaze esterilizada tendo o cuidado de natildeo
espremer o resiacuteduo Voltou a extrair-se com 50 mL de soluccedilatildeo I durante 30 minutos mexendo
lentamente Adicionou-se este ao extracto obtido anteriormente Centrifugou-se o extracto total a
16 000 g durante 30 minutos Decantou-se para uma proveta sobre a qual se colocou gaze para
que o sedimento solto natildeo se juntasse ao sobrenadante e leu-se o volume De seguida adicionou-
se KCl ateacute uma concentraccedilatildeo final de 50 mM MgCl2 ateacute 2mM e ATP ateacute 1mM lentamente e com
agitaccedilatildeo contiacutenua de modo a promover a polimerizaccedilatildeo dos monoacutemeros de actina
Deixou-se polimerizar durante 2h a 25ordmC observando-se a presenccedila de pequenas bolhas de ar
indicativas de um aumento de viscosidade da soluccedilatildeo Arrefeceu-se em gelo durante 15 minutos e
adicionou-se KCl ateacute uma concentraccedilatildeo final de 06M Mexeu-se lentamente durante 30 minutos
apoacutes os quais se ultracentrifugou a 110 000g durante 1hora Desprezou-se o sobrenadante
ressuspendeu-se o sedimento com soluccedilatildeo tampatildeo E e transferiu-se para um potter
Dialisou-se durante a noite contra 1L de soluccedilatildeo-tampatildeo I a uma temperatura de 4ordmC com rotaccedilatildeo
7 Soluccedilatildeo F 005 M NaHCO3 pH 82-85
8 Soluccedilatildeo G 1M Na2CO3
9 Soluccedilatildeo H 1 mM EDTA pH 70
10 Soluccedilatildeo I 2 mM Tris-HCl pH 8 02 mM CaCl2 02 mM Na2ATP 05 mM szlig-Me 0005 NaN3
42
contiacutenua e mudanccedila de tampatildeo 2 a 3 vezes De seguida a actina foi centrifugada a 110
000gdurante 1hora Decantou-se a soluccedilatildeo de G-Actina (sobrenadante) e desprezou-se o
sedimento Determinou-se a concentraccedilatildeo de G-Actina pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental) A soluccedilatildeo de G-actina foi armazenada a uma temperatura de 4ordmC
apoacutes adiccedilatildeo de 002NaN3 mantendo-se estaacutevel durante 3 a 5 dias
331 Quantificaccedilatildeo das proteiacutenas isoladas
3311 Quantificaccedilatildeo pelo meacutetodo de Bradford
O meacutetodo de Bradford (Bradford 1976) baseia-se na utilizaccedilatildeo do reagente de Bradford
composto por entre outros aacutecido fosfoacuterico e o corante ldquoCoomassierdquo Este corante possui carga
negativa e a sua coloraccedilatildeo varia entre a vermelha (Absmax=465 nm) predominante em soluccedilatildeo e a
azul (Absmax=595 nm) presente aquando da ligaccedilatildeo aos grupos amina das proteiacutenas Em soluccedilotildees
aacutecidas diluiacutedas a coloraccedilatildeo varia proporcionalmente com a quantidade do corante ligado agrave
proteiacutena que por sua vez eacute directamente proporcional agrave concentraccedilatildeo da proteiacutena permitindo a
sua quantificaccedilatildeo ateacute a um limite de detecccedilatildeo de 20 gmL (Bradford 1976)
Deste modo prepararam-se amostras de BSA ([BSA]stock= 1mgmL) utilizando um intervalo de
concentraccedilotildees compreendido entre 0 e 15 gmL de acordo com a tabela 3I Apoacutes a adiccedilatildeo da
H2O MilliQ e do reagente de Bradford as soluccedilotildees foram sucessivamente homogeneizadas no
voacutertex
Tabela 3Indash Amostras utilizadas para a obtenccedilatildeo da recta de calibraccedilatildeo no meacutetodo de Bradford
BSA (L) BSA (g) H2O MilliQ (L) Reagente de Bradford (L)
0 0 800 200
125 125 79875 200
25 25 7975 200
5 5 795 200
10 10 790 200
15 15 785 200
Apoacutes incubaccedilatildeo a 25ordmC por 5 minutos leram-se os valores de Abs595 num
espectrofotoacutemetro (Shimadzu UV-160A) para todas as amostras Seguidamente traccedilou-se a recta
de calibraccedilatildeo (Abs595vs quantidade de BSA (g))
43
Para determinar a concentraccedilatildeo de proteiacutena apoacutes o processo de isolamento purificaccedilatildeo e
diaacutelise quando necessaacuterio colocou-se 5 L da amostra num tubo de Eppendorf e adicionou-se
795 L de H2O MilliQ homogeneizando a soluccedilatildeo no voacutertex Adicionou-se posteriormente 200
L de reagente de Bradford e repetiu-se a homogeneizaccedilatildeo Incubou-se a mistura a 25ordmC por 5
minutos apoacutes os quais se leu os valores de Abs595 nm no espectrofotoacutemetro Por interpolaccedilatildeo deste
valor a partir da recta de calibraccedilatildeo determinou-se a concentraccedilatildeo de proteiacutena na amostra
3312 Quantificaccedilatildeo pelo meacutetodo directo
A quantificaccedilatildeo das amostras proteicas pode tambeacutem ser efectuada atraveacutes da leitura directa da
absorvacircncia tendo por base a lei de Beer-Lambert onde Abs corresponde agrave
absorvacircncia ao comprimento de onda ao qual o ensaio foi realizado corresponde ao coeficiente
de extinccedilatildeo (substituiacutedo pela letra a se as unidades forem distintas da molaridade) b corresponde
ao percurso oacuteptico da radiaccedilatildeo na ceacutelula amostral e c corresponde agrave concentraccedilatildeo amostral
Assim no caso de S1 de miosina mediu-se a Abs280e calculou-se a concentraccedilatildeo da proteiacutena
sabendo que o coeficiente de extinccedilatildeo de S1 eacute (Ref)
No caso da actina mediu-se a Abs290ecalculou-se a concentraccedilatildeo da proteiacutena sabendo que o
coeficiente de extinccedilatildeo da actina eacute (Ref)
332Electroforese em condiccedilotildees desnaturantes (SDS-PAGE)
De modo a melhor caracterizar as proteiacutenas isoladas foi realizada uma electroforese em
condiccedilotildees desnaturantes (SDS-PAGE do inglecircs Sodium Dodecyl Sulfate Polyacrilamide Gel
Electrophoresis) O SDS (dodecilo sulfato de soacutedio) eacute um detergente anioacutenico utilizado neste
meacutetodo para desnaturar as proteiacutenas que se ligam ao SDS na proporccedilatildeo 14 g SDS g proteiacutena
(aproximadamente 1 moleacutecula de SDS por cada 2 resiacuteduos de aminoaacutecidos) (Stryer) O SDS induz
uma densidade de carga negativa significativa nas proteiacutenas a que se liga mascarando a sua carga
intriacutenseca pelo que as razotildees cargamassa satildeo tendencialmente idecircnticas Assim a SDS-PAGE
separa as proteiacutenas quando sujeitas a um campo eleacutectrico de acordo com o seu peso molecular
sendo a sua migraccedilatildeo directamente proporcional ao logaritmo deste uacuteltimo (Stryer)
O gel de resoluccedilatildeo com 10 de acrilamida foi obtido atraveacutes da adiccedilatildeo de 167 mL de soluccedilatildeo
stock de acrilamida11
075 mL de tampatildeo de resoluccedilatildeo12
251 mL de aacutegua destilada e 50 L de
SDS 10 Para promover a polimerizaccedilatildeo adicionou-se em seguida 38 L de PSA 10
11
soluccedilatildeo stock de acrilamida soluccedilatildeo de 30 (mv) de acrilamida e 08 (mv) de NNrsquo-metil-bis-acrilamida 12
tampatildeo de resoluccedilatildeo 15 M Tris-HCl pH 88
44
(persulfato de amoacutenia) 10 e 25 L de TEMED 99 (tetrametiletilenodiamina) Agitou-se
cuidadosamente de modo a natildeo formar bolhas e pipetou-se a soluccedilatildeo para a cassette de vidro
previamente montada no suporte De seguida adicionou-se uma peliacutecula de cerca de 2 mm de 2-
metil-2-butanol para verificar posteriormente a polimerizaccedilatildeo Apoacutes a polimerizaccedilatildeo que demora
cerca de 45 minutos verteu-se a agrave superfiacutecie e adicionou-se o gel de concentraccedilatildeo com 5 de
acrilamida constituiacutedo por 03 mL de soluccedilatildeo de stock acrilamida 045 mL de tampatildeo de
concentraccedilatildeo13
094 mL de aacutegua destilada 18 L de SDS 10 135 L de PSA 10 e 2L de
TEMED 99 Colocou-se o pente sobre o gel e deixou-se polimerizar durante cerca de 45
minutos
As amostras para aplicaccedilatildeo no gel de electroforese foram preparadas para uma concentraccedilatildeo de
proteiacutena de 1 mgmL num volume final de 100 L dos quais 25 L correspondem ao tampatildeo da
amostra14
As amostras foram aquecidas a 100ordmC durante 5 minutos Antes da aplicaccedilatildeo das
amostras transferiu-se a cassette para a tina de electroforese agrave qual se adicionou a soluccedilatildeo
tampatildeo de electroforese da tina superior15
e tampatildeo de electroforese da tina inferior16
Aplicaram-
se 10 L das soluccedilotildees de marcadores de peso molecular e 20 L das amostras proteicas
preparadas anteriormente A electroforese decorreu durante cerca de 3h30 min a uma voltagem
de 80 V De seguida transferiu-se o gel para uma tina corando as proteiacutenas durante 1h com
soluccedilatildeo corante17
Descorou-se o gel com soluccedilatildeo descorante18
tendo-se mudado esta soluccedilatildeo ateacute
as bandas se encontrarem bem definidas
Determinou-se a mobilidade relativa das bandas correspondentes aos marcadores de elevado peso
molecular e traccedilou-se a recta de calibraccedilatildeo ndash log(PM) vs log(Rf) onde Rf corresponde agrave
mobilidade relativa Por interpolaccedilatildeo determina-se o PM das proteiacutenas isoladas sabendo a sua
mobilidade relativa Por comparaccedilatildeo com os valores descritos na literatura pode concluir-se
acerca do grau de purificaccedilatildeo
34 Mediccedilatildeo da actividade enzimaacutetica de S1
341 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
13
tampatildeo de concentraccedilatildeo 05 M Tris-HCl pH 68 14
tampatildeo de amostra 320 mM Tris-HCl pH 68 04 M szlig-mercaptoetanol SDS 8 glicerol 15 (vv) e roxo de m-
cresol 0024 15
tampatildeo de electroforese da tina superior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS 16
tampatildeo de electroforese da tina inferior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS acetato de soacutedio
01 M 17
soluccedilatildeo corante ldquoCoomassie Blue R-250rdquo contendo metanol 45 (vv) e aacutecido aceacutetico glacial 10 (vv) 18
soluccedilatildeo descorante metanol 10 (vv) e aacutecido aceacutetico glacial 10 vv)
45
O meacutetodo de Fiske-Subarow modificado consiste na reacccedilatildeo do molibdato de amoacutenio com o
fosfato inorgacircnico em soluccedilatildeo aacutecida com formaccedilatildeo do aacutecido fosfomoliacutebdico Seguidamente daacute-
se a adiccedilatildeo de um agente redutor que provoca a reduccedilatildeo do composto a fosfomolibdato que forma
um complexo de cor azulada cuja absorvacircncia maacutexima ocorre para um comprimento de onda de
660 nm tornando possiacutevel a sua detecccedilatildeo por meacutetodos espectrofotomeacutetricos As actividades Ca2+
-
ATPaacutesica e K+EDTA-ATPaacutesica de S1 foram determinadas atraveacutes deste meacutetodo de acordo com
o descrito por Taussky e Shorr (1953) com mediccedilatildeo da quantidade de fosfato (Pi) libertado
atraveacutes da hidroacutelise de ATP
Para tal obteve-se primeiramente a recta de calibraccedilatildeo com as amostras presentes na Tabela 3II
utilizando uma soluccedilatildeo stock de 5 mM KH2PO4
Tabela 3II ndash Composiccedilatildeo das diferentes amostras utilizadas para determinar a recta de calibraccedilatildeo atraveacutes do meacutetodo
de Fiske-Subbarow modificado
KH2PO4 (L)
([KH2PO4]stock= 5mM)
PO43-
(nmole) H2O (uL)
0 0 1780
5 25 1775
10 50 1770
20 100 1760
40 200 1740
100 500 1680
A cada tubo de Eppendorf adicionaram-se seguidamente 200 L de soluccedilatildeo J19
e 20 L de agente
redutor20
homogeneizando no voacutertex apoacutes cada adiccedilatildeo Incubou-se durante 10 minutos e mediu-
se a Abs660 nm num espectrofotoacutemetro com intervalos de tempo iguais entre cada amostra Traccedilou-
se a recta de calibraccedilatildeo (Abs660 nmvs nmol PO43-
) a partir dos valores obtidos
Para determinar a actividade Ca2+
-ATPaacutesica de S1 prepararam-se tubos de Eppendorf com 400
L de meio de reacccedilatildeo21
25 L CaCl2 ([CaCl2]stock= 100 mM) 25 L ATP ([ATP]stock= 100 mM)
com [S1]ensaios=01 mgmL Perfez-se se necessaacuterio com H2O MilliQ ateacute perfazer um volume de 1
mL Para determinar a actividade K+EDTA-ATPaacutesica utilizou-se uma concentraccedilatildeo idecircntica de
19
Soluccedilatildeo J 5 N H2SO4 25 molibdato de amoacutenio 20
Agente redutor 5 mg aacutecido 1-amino-2-naftol-4-sulfoacutenico 30 mg bissulfito de soacutedio 30 mg sulfito de soacutedio em 26
mL H2O 21
meio de reacccedilatildeo 125 mM Tris-HCl pH 80 0575 M KCl
46
proteiacutena nos ensaios (01 mgmL) e o mesmo meio de reacccedilatildeo Aos tubos de Eppendorf
adicionou-se ainda 123 L de KCl ([KCl]stock = 3M) 25 L de ATP 10 L de EDTA
([EDTA]stock= 100 mM )e o volume de H2O MilliQ necessaacuterio para perfazer 1 mL Em ambas as
situaccedilotildees o uacuteltimo reagente adicionado foi o ATP uma vez que se trata do substrato da reacccedilatildeo
enzimaacutetica
A cineacutetica decorreu a 25ordmC durante 5 minutos para a actividade Ca2+
-ATPaacutesica e durante 3
minutos para a actividade K+EDTA-ATPaacutesica Apoacutes o periacuteodo de incubaccedilatildeo transferiu-se 150
L de amostra para tubos de Eppendorf contendo igual volume de TCA 10 Colocou-se no gelo
e homogeneizou-se no voacutertex Seguidamente centrifugou-se a 1500 g durante 10 minutos e
recolheu-se 200 L de sobrenadante para novos tubos de Eppendorf Adicionou-se a estes 158
mL de H2O MilliQ e 200 L de soluccedilatildeo J Homogeneizou-se no voacutertex e adicionou-se 20 L de
reagente redutor repetindo-se a homogeneizaccedilatildeo Incubou-se durante 10 minutos e mediu-se a
Abs660 nm com o mesmo intervalo de tempo de leitura para cada amostra Por intrapolaccedilatildeo a partir
da recta de calibraccedilatildeo obtida anteriormente determinou-se a quantidade de Pi libertado em cada
ensaio A partir destes resultados determinaram-se as actividades Ca2+
-ATPaacutesica e K+EDTA-
ATPaacutesica do S1 em nmol PO43-
minmg proteiacutena e em s-1
342 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
Da actividade Mg2+
-ATPaacutesica de S1 resulta a formaccedilatildeo dos produtos da reacccedilatildeo de
hidroacutelise do ATP - ADP e Pi Na presenccedila de ADP fosfoenolpiruvato (PEP) e da enzima piruvato
cinase (PK) os produtos resultantes satildeo ATP e piruvato Por acccedilatildeo da enzima lactato
desidrogenase (LDH) este uacuteltimo eacute convertido em lactato ocorrendo simultaneamente a oxidaccedilatildeo
do NADH a NAD+ monitorizada atraveacutes da Abs340 nm Quando o S1 de miosina se encontra no
meio reaccional o ATP eacute hidrolisado formando-se novamente ADP e Pi o que permite a
formaccedilatildeo de um ciclo (fig 21) Se as enzimas se encontrarem em excesso a velocidade de
conversatildeo do NADH eacute limitada pela produccedilatildeo de ADP e Pi Como o ATP eacute regenerado em cada
ciclo a sua concentraccedilatildeo inicial eacute sempre mantida (Bagshaw 1982)
Figura 33ndash Representaccedilatildeo esquemaacutetica do meacutetodo de enzimas acopladas
47
O protocolo utilizado e seguidamente descrito foi adaptado de Cuenda et al (1990 ndash Ver Refordf) A
uma cuvette de 1 mL adicionou-se S1 para uma concentraccedilatildeo final de 005 mgmL 800 L de
meio de reacccedilatildeo22
10 L de PEP ([PEP]stock = 42 mM) 10 L de PK+LDH 11 ([PK+LDH]stock =
1 mgmL) e 15 L de NADH ([NADH]stock=25 mM) Iniciou-se a monitorizaccedilatildeo da reacccedilatildeo
atraveacutes da leitura de Abs340 nm apoacutes a adiccedilatildeo de 20 L de ATP ([ATP]stock=100 mM) A
actividade Mg2+
-ATPaacutesica eacute obtida atraveacutes do declive correspondente agrave oxidaccedilatildeo do NADH Para
a mediccedilatildeo da Mg2+
-ATPase de S1 estimulada por actina adicionou-se F-actina para uma
concentraccedilatildeo final de 2 microM sendo a actividade obtida tambeacutem atraveacutes do declive correspondente
agrave oxidaccedilatildeo do NADH apoacutes a adiccedilatildeo da proteiacutena
35 Interaccedilatildeo entre vanaacutedio e subfragmento-1 de miosina
351 Preparaccedilatildeo das soluccedilotildees de vanaacutedio
As soluccedilotildees de vanadato foram preparadas em aacutegua MilliQ a partir de metavanadato de amoacutenio
(NH4VO3) numa concentraccedilatildeo stock de 50 mM com agitaccedilatildeo magneacutetica e aquecimento (~30 ordmC)
Esta soluccedilatildeo com um valor compreendido entre 6 e 7 designa-se por ldquometavanadatordquo e eacute
constituiacuteda pelas esspeacutecies V1 V2 V4 e V5 (Ref) Acidificando a soluccedilatildeo de metavanadato por
adiccedilatildeo de HCl ateacute um valor de pH proacuteximo de 4 observa-se uma mudanccedila de cor de amarelo
claro para laranja indicativa da formaccedilatildeo da espeacutecie decameacuterica de vanadato De seguida e
imediatamente antes da realizaccedilatildeo dos ensaios descritos o valor do pH da soluccedilatildeo de
decavanadato foi aumentado para cerca de 75 por adiccedilatildeo de NaOH A soluccedilatildeo (2) designada
por ldquometavanadatordquo natildeo foi posteriormente alterada ateacute agrave sua adiccedilatildeo para a realizaccedilatildeo dos ensaios
As soluccedilotildees foram armazenadas a 4ordmC e mantidas em gelo aquando da sua utilizaccedilatildeo
A soluccedilatildeo de V(IV) foi preparada em condiccedilotildees anaeroacutebias por diluiccedilatildeo de sulfato de vanadil
(VOSO4) para uma concentraccedilatildeo final de 5 mM em H2O MilliQ saturada com N2 gasoso
352Anaacutelise por espectroscopia de UV-Vis
A concentraccedilatildeo da espeacutecie decameacuterica em soluccedilatildeo depende do periacuteodo de tempo apoacutes a diluiccedilatildeo
no meio de reacccedilatildeo apropriado (Ref) A velocidade de desoligomerizaccedilatildeo da espeacutecie decameacuterica
pode ser analisada por espectroscopia de absorccedilatildeo na regiatildeo UVVis uma vez que esta espeacutecie
apresenta duas bandas caracteriacutesticas na regiatildeo do visiacutevel (a 358 e 400 nm) que natildeo se verifica
em quaisquer outras espeacutecies oligomeacutericas As bandas supramencionada sapresentam-se bem
22
Meio de reacccedilatildeo 3125 mM Tris-HCl pH 71 375 mM MgCl2
48
resolvidas atraveacutes da obtenccedilatildeo da derivada de 2ordf ordem do espectro apresentando a forma de 2
vales com miacutenimos centrados nos comprimentos de onda supracitados com dependecircncia linear
relativamente agrave concentraccedilatildeo de decavanadato total (Ramos et al 2006)
Assim a estabilidade da espeacutecie decameacuterica apoacutes diluiccedilatildeo da soluccedilatildeo de decavanadato no meio de
reacccedilatildeo dos ensaios (tampatildeo D) para uma concentraccedilatildeo final de 1 mM em vanadato total foi
caracterizada atraveacutes da leitura da Abs400 em funccedilatildeo do tempo tendo-se determinado qual a
ordem de reacccedilatildeo e o tempo de meia-vida da mesma Com base em estudos anteriores acerca da
estabilidade do V10 na presenccedila de diferentes proteiacutenas (Ramos et al 2006) determinou-se
igualmente o tempo de meia vida apoacutes a adiccedilatildeo de 20 microM de S1 de miosina ao meio reaccional
353 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
Os espectros de RMN do isoacutetopo 51
V foram realizados num espectroacutemetro Bruker AM-400 MHz
equipado com uma sonda multinuclear inversa de 5 mm Em tubos de RMN de 5 mm de diacircmetro
preparou-se soluccedilotildees de 500 L de deca- ou metavanadato correspondentes a um intervalo de
concentraccedilotildees de 1 a 10 mM suplementadas com 10 D2O Os espectros foram adquiridos agrave
temperatura ambiente com uma largura espectral de 23 923 Hz um tempo de aquisiccedilatildeo de 0086
s tempo de relaxaccedilatildeo de 001 s e uma sequecircncia de pulsos a 90ordm Utilizou-se a transformada de
Fourier com alargamento das bandas de 20Hz para o processamento dos espectros Os desvios
quiacutemicos das ressonacircncias de 51
V foram obtidos em relaccedilatildeo agrave espeacutecie VOCl3cuja frequecircncia de
ressonacircncia eacute considerada o padratildeo sendo todas as outras referenciadas a partir desta em ppm
As aacutereas dos sinais do RMN correspondentes agraves espeacutecies oligomeacutericas presentes foram obtidas
por integraccedilatildeo dos sinais As concentraccedilotildees relativas das espeacutecies foram calculadas com recurso agrave
equaccedilatildeo
onde A[Vx] corresponde agrave aacuterea referente agrave ressonacircncia da espeacutecie Vx com nordm de agregaccedilatildeo x
A[V]total agrave soma total das aacutereas medidas e [V] a concentraccedilatildeo total de vanaacutedio no ensaio
A teacutecnica de 51
V-RMN foi anteriormente aplicada em estudos de ligaccedilatildeo com S1 de miosina
(Ref) tendo sido demostrado que entre as espeacutecies V1 V2 V4 e V5 a espeacutecie tetrameacuterica possui a
maior afinidade para a miosina (Cremo et al) O desaparecimento ou alargamento dos sinais pode
implicar a ligaccedilatildeo de um dos oxovanadatos laacutebeis agrave miosina sem indicaccedilatildeo de qual uma vez que
49
se processa um equiliacutebrio entre eles (Crans 1990) Assim e tendo em conta as indicaccedilotildees de que
a espeacutecie V10 eacute distinta das restantes no que diz respeito agrave magnitude da interacccedilatildeo com a miosina
e inibiccedilatildeo da actividade ATPaacutesica estimulada por actina (Aureliano 2000 Tiago et al 2002)
torna-se fundamental investigar a interacccedilatildeo das diferentes espeacutecies de vanadato com o S1 de
miosina atraveacutes da teacutecnica de 51
V-RMN
Para tal obtiveram-se espectros de 51
V-RMN resultantes da titulaccedilatildeo de uma soluccedilatildeo 5 mM de
decavanadato total com concentraccedilotildees crescentes de S1 de miosina (0 5 10 15 e 20 microM) em
tampatildeo D Realizou-se ainda ensaios na presenccedila de 2 mM de decavanadato e 2 mM de ATP na
presenccedila e na ausecircncia de S1 de miosina de modo a verificar o efeito que estes ligandos naturais
da enzima teria na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o subfragmento-1
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
Os grupos sulfidrilo (-SH) das cisteiacutenas satildeo resiacuteduos de aminoaacutecidos extremamente sensiacuteveis agrave
oxidaccedilatildeo resultando na sua oxidaccedilatildeo (Dalle-Donne et al 1999) Logo a quantificaccedilatildeo destes
resiacuteduos eacute bastante importante nos estudos de interacccedilatildeo com vanaacutedio jaacute que os resiacuteduos de Cys
com ecircnfase para o SH1 e o SH2 satildeo bastante importantes na monitorizaccedilatildeo das modificaccedilotildees
funcionais da regiatildeo da cabeccedila
O procedimento seguidamente apresentado foi originalmente descrito por Gutieacuterrez-Merino
(1987)) Consiste na reacccedilatildeo entre os grupos ndashSH natildeo oxidados das proteiacutenas e o reagente de
Ellman (DTNB -55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico)) - formando-se o aniatildeo CNT (aniatildeo 3-
carboxilato-4-nitrotiofenolato) numa estequiometria de 11 Este apresenta um maacuteximo de
absorvecircncia a 412 nm (Robyt 1990) Assim apoacutes a incubaccedilatildeo da proteiacutena na presenccedila do metal
a quantidade de cisteiacutenas reduzidas por proteiacutena determina-se atraveacutes dos valores de Abs412
Deste modo colocou-se num tubo de Eppendorf o volume apropriado de S1 dialisado para que a
concentraccedilatildeo no ensaio fosse de 1 mgmL Adicionou-se o volume apropriado de decavanadato
para que a sua concentraccedilatildeo fosse de 0 10 50 100 250 500 ou 1000 M Perfez-se com soluccedilatildeo
D ateacute um volume final de 1 mL e incubou-se a 25ordmC por 20 minutos Apoacutes este periacuteodo colocou-
se 780 L da mistura reaccional numa cuvette de quartzo e adicionou-se 220 L de soluccedilatildeo K23
23
Tampatildeo K Tampatildeo fosfatohidrogenofosfato de soacutedio 025 M pH 72
50
Adicionou-se 15 L de DTNB (em soluccedilatildeo K [DTNB]stock=100 mM) ao meio reaccional e
registou-se o valor de absorvecircncia medido equivalente agraves cisteiacutenas ldquoraacutepidasrdquo (SH1 e SH2
expostos ao solvente) Adicionou-se 001 g de SDS que provoca a desnaturaccedilatildeo da proteiacutena e
consequentemente a exposiccedilatildeo de todos os resiacuteduos de cisteiacutenas que se encontravam ateacute aiacute
inacessiacuteveis no interior da proteiacutena (cisteiacutenas totais) Estes resiacuteduos podem assim reagir com o
DTNB
Estimou-se o nuacutemero de resiacuteduos de cisteiacutenas em todas as amostras com recurso ao coeficiente de
extinccedilatildeo molar () do DTNB (ver ponto 3 dos Anexos) que foi determinado atraveacutes da titulaccedilatildeo
de L-Cisteiacutena com intervalo de concentraccedilotildees compreendidos entre 0 e 50 microM na presenccedila de
25microM de DTNB em tampatildeo D Os valores de absorvecircncia foram obtidos para um comprimento de
onda de 412 nm
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
3621 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
A teacutecnica mais eficiente de anaacutelise da interacccedilatildeo entre o iatildeo vanadilo e proteiacutenas como a
calmodulina e a transferrina entre outras permite a obtenccedilatildeo raacutepida de informaccedilatildeo no que toca agrave
conformaccedilatildeo e amplitude da referida interacccedilatildeo
Os espectros de Ressonacircncia Paramagneacutetica Electroacutenica (RPE) da espeacutecie paramagneacutetica V(IV)
foram realizados a 70K num espectroacutemetro Bruker EMX-300 X-band Todas as soluccedilotildees
utilizadas e amostras foram preparadas em anaerobiose numa cacircmara de luvas de modo a evitar a
reduccedilatildeo de V(IV) a V(V) Em tubos capilares com 02 mL de amostra manteve-se a concentraccedilatildeo
de S1 a 20 M e variou-se a concentraccedilatildeo de vanadilo (0 25 5 10 15 25 50 75 100 150
200 500 e 1000 M [VOSO4]stock=5 mM) em tampatildeo D Apoacutes obtenccedilatildeo dos espectros estes
foram tratados atraveacutes do software SimFonia e WinEPR tendo-se obtido um graacutefico com a
variaccedilatildeo da intensidade do sinal de RPE em funccedilatildeo da concentraccedilatildeo de sulfato de vanadilo
presente Realizaram-se ainda simulaccedilotildees de espectros obtidos com 20 microM de S1 e 1000 microM de
V(IV) com vista agrave obtenccedilatildeo dos ligandos provaacuteveis do vanadilo atraveacutes da anaacutelise da constante
de acoplamento hiperfina (A) e do factor g (g)
4 Conclusotildees gerais
Apesar de jaacute ter sido demonstrado que o vanaacutedio inibe a Na+K
+-ATPase da miosina a funccedilatildeo
deste elemento no tecido muacutesculo esqueleacutetico continua largamente incompreendida A enorme
complexidade quiacutemica deste metal especialmente no que concerne agrave oligomerizaccedilatildeo e reacccedilotildees
51
redox constitui um obstaacuteculo agrave compreensatildeo do comportamento do vanaacutedio nos organismos
vivos Assim a caracterizaccedilatildeo da quiacutemica deste elemento eacute de grande utilidade antes de iniciar o
estudo da sua interacccedilatildeo com o presente sistema
A anaacutelise por espectroscopias de 51
V-RMN e de UV-Vis de soluccedilotildees de vanadato diluiacutedas nos
meios de reacccedilatildeo utilizados no presente relatoacuterio (10 mM HEPES pH 70 25 mM KCl) mostrou
claramente que a espeacutecie decameacuterica (V10) possui um tempo de vida suficientemente longo para a
determinaccedilatildeo do seu efeito isolado ao contraacuterio do que se verifica com as espeacutecies mono- di-
tetra- e pentameacutericas (V1 V2 V4 e V5) Por outro lado a anaacutelise verificou ainda que apoacutes diluiccedilatildeo
das soluccedilotildees de decavanadato no meio de reacccedilatildeo coexistem as espeacutecies V10 e V1 pelo que os
efeitos putativos desta uacuteltima deveratildeo ser subtraiacutedos aquando da monitorizaccedilatildeo dos efeitos da
espeacutecie decameacuterica
O vanaacutedio (V) (vanadato) inibe as enzimas que catalizam reacccedilotildees de transferecircncia de grupos
fosfato como a ATPase da miosina [Goodno 1979] devido agrave sua analogia estrutural com o iatildeo
fosfato Na sua utilizaccedilatildeo como inibidor enzimaacutetico natildeo eacute tida em conta a contribuiccedilatildeo individual
das espeacutecies oligomeacutericas considerando-se normalmente a espeacutecie V1 como inibidora efectiva
[Goodno e Taylor 1982] No presente relatoacuterio verificou-se por espectroscopia de 51
V-RMN que
outros oligoacutemeros de vanadato interagem com o S1 de miosina tendo as espeacutecies V10 e V4 os
maiores niacuteveis de afinidade A sua interacccedilatildeo envolve forccedilas electrostaacuteticas e eacute afectada
negativamente por ligandos naturais de enzima como eacute o caso do ATP
A exposiccedilatildeo do S1 de miosina (005 mgmL) a decavanadato conduz agrave inibiccedilatildeo do capacidade da
proteiacutena de estimular a actividade Mg2+
-ATPaacutesica com um valor de IC50 de 266 plusmn 07 M
decavanadato sendo o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM
decavanadato (80) Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM decavanadato sendo
o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM decavanadato (60)
exercendo um efeito de impedimento agrave ligaccedilatildeo entre o vanaacutedio e o S1
A uma concentraccedilatildeo de 27 microM decavanadato verifica-se a oxidaccedilatildeo de um dos resiacuteduos de Cys
de S1 de miosina seguida pela oxidaccedilatildeo gradual de outro resiacuteduo de Cys ateacute agrave concentraccedilatildeo de
1000 microM Ambos os resiacuteduos oxidados natildeo se encontravam expostos ao solvente utilizado A
oxidaccedilatildeo dos resiacuteduos permite sugerir que esta modificaccedilatildeo oxidativa sofrida pelo S1 contribui
para a diminuiccedilatildeo da sua actividade de ATPase
52
Por espectroscopia de RPE revelaram-se os possiacuteveis ligandos equatoriais do vanaacutedio aquando da
sua interacccedilatildeo com o S1 de miosina como sendo duas moleacuteculas de aacutegua um ligando contendo o
grupo R-OH e outro contendo o grupo R=NR
Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados de
oxidaccedilatildeo (IV) e (V) no S1 de miosina para concentraccedilotildees facilmente alcanccedilaacuteveis no ambiente
terrestre
53
5 Estudos futuros
No sentido do melhor esclarecimento das alteraccedilotildees moleculares e estruturais que ocorrem no
subfragmento 1 quando exposta ao vanaacutedio que conduzem a uma diminuiccedilatildeo da sua funccedilatildeo
sugere-se
- Quantificaccedilatildeo das cisteiacutenas reduzidas de S1 na presenccedila de metavanadato
- Determinaccedilatildeo das actividades Ca2+
- e K+-ATPaacutesica do S1 de miosina na presenccedila de
decavanadato
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina estimulada por actina
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina na presenccedila de
metavanadato
- Realizaccedilatildeo de RMN 2D com variaccedilatildeo das concentraccedilotildees das soluccedilotildees de decavanadato e
metavanadato na presenccedila de S1
-Mutageacutenese dos resiacuteduos de Cys doS1 de miosina de modo a verificar quais os resiacuteduos que
sofrem oxidaccedilatildeo na presenccedila de decavanadato
- Determinaccedilatildeo por calorimetria (DSC Differential Scanning Calorimetry) da variaccedilatildeo da
entalpia (∆H) e da temperatura de desnaturaccedilatildeo do S1 de miosina na presenccedila de vanaacutedio uma
vez que estas propriedades caracteriacutesticas de cada proteiacutena satildeo alteradas apoacutes ocorrecircncia de
modificaccedilotildees estruturais
- Anaacutelise por SDS-PAGE de S1 de miosina apoacutes tratamento com vanaacutedio de modo a determinar
quais os polipeacuteptidos eventualmente formados apoacutes clivagem da proteiacutena
54
6 Referecircncias bibliograacuteficas
- Sellers J Myosins 2nd Edition Oxford University Press 1999
- L Stryer inBiochemistry (Ed WH Freeman 2005 pp
Refordfas Materiais e Meacutetodos
[A] S S Margossian SLowey Methods Enzymol1982 85 55-71
[B] JD Pardue JA Spudich Methods Enzymol 1982 85 164-181
[C] M Bradford Anal Biochem1976 72 248-254
[D] H H Taussky E Shorr J Biol Chem 1953 202 675-685
[E] C R Bagshaw in Outline Studies in Biology ndash Muscle Contraction Chapman and Hall Ed
1982
[F] A Cuenda et al Eur J Biochem1990 194 663-670
55
Anexos
1 Determinaccedilatildeo da recta de calibraccedilatildeo meacutetodo de Bradford
Figura1ndashRepresentaccedilatildeo graacutefica da recta de calibraccedilatildeo (Abs595 nm vs [BSA]) utilizada para o meacutetodo de Bradford
2 Determinaccedilatildeo da recta calibraccedilatildeo para a electroforese em condiccedilotildees desnaturantes
Figura 2 ndash Representaccedilatildeo graacutefica da recta de calibraccedilatildeo (log(Peso Molecular) vs Rf dos marcadores de peso
molecular) onde Rf indica determinada atraveacutes do padratildeo de peso molecular
(poccedilo (a))
3Caacutelculo do coeficiente de extinccedilatildeo molar () para a quantificaccedilatildeo de cisteiacutenas reduzidas
atraveacutes do meacutetodo de Ellman
Traccedilou-se a recta de Abs412 nm meacutedia vs [L-Cisteiacutena] de modo a determinar o valor do coeficiente
de extinccedilatildeo molar definido como o declive daquela recta Assim =8400 M-1
cm-1
como
podemos verificar na Fig 3
A595 nm = 00546[BSA] + 00209 Rsup2 = 099051
Ab
s 59
5 n
m
[BSA] (mgmL)
log PM = -1177 Rf + 2247 Rsup2 = 098957
log
PM
Mobilidade relativa (Rf)
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero

33
o aumento da forccedila ioacutenica (Tiago et al 2004 tal poderaacute explicar a diferenccedila obtida entre estes
dois conjuntos de valores
243Quantificaccedilatildeo de cisteiacutenas reduzidas pelo meacutetodo de Ellman
A interacccedilatildeo do vanaacutedio com alguns tipos de aminoaacutecidos sensiacuteveis agrave oxidaccedilatildeo pode fornecer-nos
informaccedilatildeo bastante uacutetil em relaccedilatildeo agraves alteraccedilotildees estruturais que conduzem a uma diminuiccedilatildeo da
actividade fisioloacutegica do S1 de miosina Os estudos realizados por Chasteen 1983 concluiacuteram
que a reduccedilatildeo de vanadato (V) a vanadilo (IV) ocorre na presenccedila de agentes redutores comuns no
ambiente celular de que se destaca o aminoaacutecido cisteiacutena Esta eacute tambeacutem um alvo de
complexaccedilatildeo por parte do iatildeo VO2+
Jaacute Ramos et al (2009) demonstrou que a incubaccedilatildeo de
actina com a soluccedilatildeo de decavanadato induz a oxidaccedilatildeo de resiacuteduos de cisteiacutena e agrave formaccedilatildeo do
oxidovanaacutedio (IV) Este grupo de investigadores descobriu ainda que apenas a espeacutecie V10 induz a
oxidaccedilatildeo das cisteiacutenas ldquoraacutepidasrdquo da actina atraveacutes do processo proposto que eacute parcialmente
responsaacutevel pela inibiccedilatildeo jaacute descrita da actividade de ATPase do S1 de miosina estimulada por
actina quando exposta agrave soluccedilatildeo de decavanadato Esta inibiccedilatildeo apresenta-se afectada com a
presenccedila de ATP uma vez que os resiacuteduos de cisteiacutena permaneceram intactos nos ensaios
realizados com este ligando Jaacute Crans et al (2004) concluiu que o siacutetio activo de muitas fosfatases
(que conteacutem grupos tiol) pode formar um equiliacutebrio redox com o iatildeo vanadato (V) de forma natildeo
reversiacutevel
Os resultados obtidos por Sakurai e Shiromura (1980) sugeriram que o vanaacutedio (V) sofre reduccedilatildeo
a vanaacutedio (IV) sem dependecircncia do valor de pH do ensaio Registaram ainda uma estequiometria
de 31 na reacccedilatildeo do vanadato (V) com os resiacuteduos de Cys enquanto a estequiometria dos ensaios
com o vanadilo (IV) foi de 21
Berlett e Stadtman (1997) observaram que os tipos de aminoaacutecidos mais facilmente oxidaacuteveis
englobam entre outros os resiacuteduos de Cys Trp e MetAssim analisou-se o estado redox dos
grupos tiol dos resiacuteduos de Cys do S1 na presenccedila do decavanadato atraveacutes da monitorizaccedilatildeo da
Abs412 nm na presenccedila de 1 mgmL de S1 de miosina em tampatildeo 10 mM HEPES 25 mM KCl
variando a concentraccedilatildeo da soluccedilatildeo de decavanadato de modo a determinar que tipo de alteraccedilotildees
estruturais ocorrem na sua presenccedila no que diz respeito agrave diminuiccedilatildeo da actividade ATPaacutesica de
S1 na presenccedila de decavanadato
Os resultados referentes agrave quantificaccedilatildeo das cisteiacutenas reduzidas apoacutes o tratamento de S1 com
diferentes concentraccedilotildees de decavanadato em tampatildeo 10 mM HEPES pH 75 e 25 mM KCl a
25ordmC durante 20 minutos encontram-se descritos na Tabela 2VII
Tabela 2VII ndash Cisteiacutenas reduzidas totais e ldquoraacutepidas de S1 (1 mgmL) apoacutes tratamento com decavanadato
34
(010501002505001000 uM) em 10 mM HEPES 25 mM KCl pH 7 a 25ordmC durante 20 minutos A detecccedilatildeo
espectrofotomeacutetrica foi feita com DTNB (e 412=8400 uM-1 cm-1) (n variaacutevel entre 1 e 8)
Amostras Cys raacutepidas Cys totais
11 M S1 (controlo) 22 plusmn 04 106 plusmn 14
11 M S1 + 10 M Decavanadato 22 plusmn 05 95 plusmn 08
11 M S1 + 50 uM Decavanadato 20 plusmn 02 98 plusmn 14
11 M S1 + 100 uM Decavanadato 17 plusmn 03 97 plusmn14
11 M S1 + 250 uM Decavanadato 26 plusmn 02 98 plusmn 08
11 M S1 + 500 uM Decavanadato 21 plusmn 04 90 plusmn 06
11 M S1 + 1000 uM Decavanadato 24 plusmn 0 86 plusmn 08
A partir dos resultados obtidos pode sugerir-se que o S1 possui 11 resiacuteduos de Cys (controlo) o
que estaacute de acordo com o anteriormente descrito Verifica-se que o tratamento da proteiacutena com
concentraccedilotildees crescentes de decavanadato provoca a oxidaccedilatildeo de dois resiacuteduos de Cys do S1 de
miosina natildeo expostos ao solvente No entanto a teacutecnica utilizada natildeo nos permite identificar
quais
Tendo em consideraccedilatildeo que o valor de IC50 obtido para a reacccedilatildeo de inibiccedilatildeo daactividade Mg2+-
ATPaacutesica de S1 na presenccedila de decavanadato eacute de cerca de 27 microM pode sugerir-se que este seraacute o
valor ao qual ocorreraacute a oxidaccedilatildeo do primeiro resiacuteduo de Cys o que poderaacute contribuir para a
diminuiccedilatildeo da actividade de S1 observada
25Interacccedilatildeo de S1 com vanaacutedio(IV)
251Anaacutelise por espectroscopia de Ressonacircncia Paramagneacutetica Electroacutenica
A acumulaccedilatildeo intracelular de oxovanaacutedio (IV) in vivo constitui uma das principais formas de
apresentaccedilatildeo de vanaacutedio no organismo Deste modo torna-se bastante interessante analisar a
interacccedilatildeo entre este iatildeo e o S1 de miosina atraveacutes da teacutecnica de espectroscopia de Ressonacircncia
Paramagneacutetica Electroacutenica o meacutetodo de eleiccedilatildeo do estudo do iatildeo vanadilo e dos complexos por si
formados O electratildeo desemparelhado encontra-se fortemente acoplado ao nuacutecleo 51
V o que
35
resulta num espectro de RPE consistindo de 8 desdobramentos na direcccedilatildeo paralela e outros tantos
na direcccedilatildeo perpendicular (Chasteen 1983)
Os estudos realizados por Ahmed et al (1987) centraram-se na interacccedilatildeo do vanadilo com
proteiacutenas como eacute o caso da insulina transferrina entre outros obtendo um espectro caracterizado
por 8 linhas amplamente espaccediladas (na direcccedilatildeo paralela) Jaacute Ramos et al 2009 concluiu que a
exposiccedilatildeo da soluccedilatildeo de decavanadato agraves formas globular e filamentosa de actina produz um
espectro tiacutepico do iatildeo oxidovanaacutedio (IV) indicando a reduccedilatildeo do vanadato
Para aferir qual a concentraccedilatildeo de S1 que permitia obter uma boa razatildeo sinalruiacutedo titulou-se uma
soluccedilatildeo de sulfato de vanadilo 15 microM ([VOSO4]stock = 5 mM) com concentraccedilotildees crescentes de
proteiacutena (0 5 10 15 e 20 microM) em 10 mM HEPES 25 mM KCl A existecircncia de uma correlaccedilatildeo
linear entre a concentraccedilatildeo de proteiacutena e a intensidade dos sinais de RPE correspondente agrave banda
-12 perpendicular encontra-se representada na Fig 210
Figura 2 12ndash Representaccedilatildeo graacutefica da variaccedilatildeo da intensidade do sinal de RPE vs a concentraccedilatildeo de S1 utilizada
Apoacutes verificar que com uma concentraccedilatildeo de S1 igual a 20 microM se obtinham sinais com uma razatildeo
sinalruiacutedo adequada ao prosseguimento dos estudos sobre a interacccedilatildeo V(IV)-S1 de miosina
titulou-se uma soluccedilatildeo de 20 microM S1 com 0 25 5 10 15 25 50 75 100 150 200 500 e 1000
microM VOSO4
Traccedilou-se o graacutefico da intensidade da banda -12 perpendicular do sinal de EPR em funccedilatildeo da
concentraccedilatildeo de VOSO4 tendo-se determinado uma constante de dissociaccedilatildeo (KD) do complexo
S1-vanadilo de 8748 plusmn 2621 microM V(IV)
y = 016786x + 122435
Rsup2 = 092992759
Inte
nsi
dad
e d
o s
inal
de
RP
E
[S1] (microM)
36
Figura 213 ndash Representaccedilatildeo graacutefica da variaccedilatildeo do sinal de RPE vs a concentraccedilatildeo de vanadilo utilizada (os valores
experimentais estatildeo representados por losangos o seu ajuste estaacute representado pela linha)
A simulaccedilatildeo do espectro de EPR foi efectuada a partir do espectro obtido para uma concentraccedilatildeo
de 1 mM VOSO4 apoacutes adiccedilatildeo de 20 microM S1 de miosina tendo-se obtido os valores da constante de
acoplamento hiperfina (A=5016) e do factor g (g0=195)
Figura 214 ndash Representaccedilatildeo graacutefica do espectro de EPR obtido com 1000 microM de vanadilo e 20 microM de S1 (a cinzento
escuro) A simulaccedilatildeo do espectro encontra-se a cinzento claro
Por comparaccedilatildeo destes com os valores presentes na tabela de correspondecircncia xxx eacute possiacutevel
predizer quais os ligandos provaacuteveis do vanadilo na presenccedila de proteiacutena Assim concluiu-se que
o vanadilo estaacute coordenado com duas moleacuteculas de aacutegua um grupo hidroxilo e um grupo R=NR
O grupo hidroxilo pode indicar os resiacuteduos de aminoaacutecidos Tyr Treonina e Serina enquanto o
grupo R=NR poderaacute indicar a presenccedila de Arginina e Histidina como ligandos do vanadilo
Inte
nsi
dad
e d
os
sinai
s d
e R
PE
[Vanadilo] (microM)
Ajuste dos dados exp Dados experimentais
Campo magneacutetico (G)
Espectro Experimental Ajuste
37
3 Procedimento Experimental
31 Reagentes
Todos os reagentes utilizados satildeo de elevado grau de pureza (analiacutetico) Apresentam-se as suas
foacutermulas quiacutemicas ou abreviaturas quando necessaacuterio acrescentou-se a sua descriccedilatildeo Entre
parecircntesis colocou-se as empresas que os comercializam
Aacutecido aceacutetico glacial 100 (Riedel) Acrilamida 99 (Merck) Azul de bromofenol (Riedel-de-
Haumlen) -Mercaptoetanol (Sigma) Bissulfito de soacutedio (Sigma)Bradford (reagente de)
(BioRad)BSA (albumina de soro bovino Sigma-Aldrich) CaCl22H2O (cloreto de caacutelcio
dihidratado Fluka) CH3CH2OH 96 (etanol Panreac) Coomassie Blue R250 (Merck) D2O
(oacutexido de deuteacuterio SDS) DTNB (55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico) Aldrich) EDTA (aacutecido
etilenodiaminatetraaceacutetico Merck) Fosfoenolpiruvato 98 (Fluka) Glicerol (Sigma) Glicina
985 (Panreac) HCl 37 (Riedel-de Haumlen) H2SO4 96 (Pronalab) HEPES 99 (aacutecido 4-(2-
hidroxiletil)-1-piperazineetanosulfoacutenico Sigma) KH2PO4 99 (hidrogenofosfato de potaacutessio)
MgATP (Sigma) Na
ATP (Sigma) Na2HPO4 99 (hidrogenofosfato de soacutedio Riedel-de Haumlen)
NaH2PO4 (fosfato de soacutedio ) NaOH 986 (hidroacutexido de soacutedio Pronalab) KCl 99 (Cloreto de
potaacutessio Panreac) L-Cys 995 (enantioacutemero levoacutegiro do resiacuteduo de Cisteiacutena Fluka)Metanol
99 (Merck) Molibdato de amoacutenio (Sigma)NNrsquo-metil-bis-acrilamida 98 (Merck) NH4VO3
99 (metavanadato de amoacutenia Sigma) PSA 10 (persulfato de amoacutenio Merck) SDS
(dodecilo sulfato de soacutedio Sigma) Sulfito de soacutedio (Sigma) VOSO4 (sulfato de vanadilo
Aldrich) TEMED 99 (Tetrametiletilenodiamina Merck) Tris (2-amino-2-hidroximetil-13-
propanodiol Sigma)
32 Meacutetodos
321 Isolamento e purificaccedilatildeo das proteiacutenas de muacutesculo esqueleacutetico de coelho
3211 Purificaccedilatildeo da miosina preparaccedilatildeo de Subfragmento 1 por digestatildeo quimotriacutepsica
da miosina
O subfragmento-1 de miosina-II (S1) foi gentilmente cedido pelo Prof Doutor Manuel Aureliano
do Departamento de Quiacutemica Bioquiacutemica e Farmaacutecia da Faculdade de Ciecircncias e Tecnologia da
Universidade do Algarve
No entanto descreve-se abaixo de forma resumida o procedimento efectuado
38
32111 Purificaccedilatildeo da miosina
O protocolo de isolamento foi adaptado do meacutetodo previamente descrito por Margossian e Lowey
(1982) apresentando-se de forma esquemaacutetica na Fig 31
Sucintamente os tecidos musculares dorsais e das pernas foram extraiacutedos apoacutes o sacrifiacutecio do
animal (Fig31) De seguida foram adicionados 3 volumes da soluccedilatildeo de extracccedilatildeo A2
Centrifugou-se a soluccedilatildeo e filtrou-se o sobrenadante O sedimento foi entatildeo diluiacutedo em 14
volumes de aacutegua destilada de modo a que a forccedila ioacutenica atingisse 004 M Apoacutes adiccedilatildeo de szlig-
mercaptoetanol (szlig-me) deixou-se a precipitar a 4ordmC removendo o sobrenadante Centrifugou-se
novamente e ressuspendeu-se o sedimento na soluccedilatildeo de extracccedilatildeo A Adicionou-se a soluccedilatildeo
tampatildeo B3 para que a forccedila ioacutenica atingisse 05 M Homogeneizou-se e deixou-se repousar
durante 2h Homogeneizou-se novamente e centrifugou-se de modo a ocorrer precipitaccedilatildeo dos
complexos de actomiosina e proteiacutena desnaturada Apoacutes filtraccedilatildeo do sobrenadante em latilde de vidro
quantificou-se a soluccedilatildeo de miosina resultante pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental)
2 Soluccedilatildeo de extracccedilatildeo A 0012M K2CO3 004M KHCO3 06M KCl pH 65
3 Soluccedilatildeo-tampatildeo B Tris 100 mM pH 7 KCl 2M
39
Figura 31 ndash Representaccedilatildeo esquemaacutetica do protocolo de isolamento de miosina (Margossian e Lowey 1982) Todo o
procedimento foi realizado a 4ordmC (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
32112 Preparaccedilatildeo de Subfragmento-1 de miosina por digestatildeo com quimiotripsina
O S1 de miosina foi isolado apoacutes diaacutelise da miosina durante a noite contra a soluccedilatildeo C4 A
presenccedila do EDTA eacute essencial uma vez que a sua substituiccedilatildeo por MgCl2resulta na produccedilatildeo de
HMM em vez de S1 A suspensatildeo de miosina foi equilibrada e digerida com -quimotripsina
A digestatildeo foi interrompida pela adiccedilatildeo de PMSF Seguidamente ultracentrifugou-se o produto
da digestatildeo desprezando-se o sedimento (Fig 32) A soluccedilatildeo de sobrenadante maioritariamente
S1 foi quantificada pela leitura directa da Abs280 ou pelo meacutetodo de Bradford (ver as secccedilotildees
3311 e 3312 do Procedimento Experimental respectivamente) e armazenada em 10 sacarose
e 2 mM szlig-Me a -80ordmC
4 Soluccedilatildeo C 25mM Tris pH 70 012M KCl 1mM EDTA
40
Antes da realizaccedilatildeo dos ensaios descritos no presente relatoacuterio o S1 de miosina foi dialisado
contra a soluccedilatildeo D5 de modo a remover qualquer composto que pudesse complexar como
vanaacutedio Seguidamente quantificou-se a proteiacutena pelo meacutetodo de Bradford ou pelo meacutetodo
directo (ver secccedilotildees 3311 e 3312do Procedimento Experimtental)
Figura 3 2ndash Representaccedilatildeo esquemaacutetica do protocolo de preparaccedilatildeo do S1 de miosina por digestatildeo quimiotripsina de
acordo com o previamente descrito por Margossian e Lowey (1982) Todo o procedimento foi realizado a 4ordm C
excepto quando indicaccedilatildeo do contraacuterio (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
3212 Isolamento e purificaccedilatildeo de actina
Isolou-se e purificou-se a actina segundo o meacutetodo descrito por Pardee e Spudish que
compreende dois passos ndash a obtenccedilatildeo de um resiacuteduo seco denominado ldquopoacute de acetonardquo e a
extracccedilatildeo subsequente da actina a partir do ldquopoacute de acetonardquo na forma de G-Actina com a
posterior polimerizaccedilatildeo a F-Actina
32121 Obtenccedilatildeo do ldquopoacute de acetonardquo
Sacrificou-se o coelho com uma pancada atraacutes do pescoccedilo e sangrou-se o animal apoacutes o corte da
arteacuteria jugular Esfolou-se e colocou-se numa tina com gelo Retiraram-se as partes de tecido
conjuntivo que envolvem o tecido muscular com auxiacutelio de uma pinccedila e bisturi Seguidamente
retirou-se os muacutesculos dorsais e os muacutesculos brancos das pernas Passou-se os muacutesculos por aacutegua
destilada de modo a remover as impurezas e pesou-se num copo Trituraram-se os muacutesculos
numa maacutequina de triturar carne e extraiu-se o resultante com 3 volumes da soluccedilatildeo E6 durante 10
minutos com agitaccedilatildeo suave Centrifugou-se a suspensatildeo a 4000 g durante 10
5 Soluccedilatildeo D HEPES 10 mM pH 70 KCl 25 mM
6 Soluccedilatildeo E 01 M KCl 015 M K2HPO4 pH 65
41
minutosdesprezando-se o sobrenadante Ressuspendeu-se o sedimento em 6 volumes de soluccedilatildeo
F7durante 10 minutos com agitaccedilatildeo suave Verificou-se se o pH da soluccedilatildeo estava compreendido
entre 82 e 85 acertando-se estecom soluccedilatildeo G8 se necessaacuterio Centrifugou-se durante 10
minutos a 4000 g Desprezou-se o sobrenadante e ressuspendeu-se o sedimento em 3 volumes de
soluccedilatildeo H9 durante 10 minutos Centrifugou-se novamente a 4000 gdurante 10 minutos
desprezando-se o sobrenadante Ressuspendeu-se o sedimento duas vezes consecutivas em 6
volumes de aacutegua destilada durante 5 minutos Centrifugou-se a 4000 g durante 10 minutos apoacutes
cada adiccedilatildeo de H2O
Seguidamente ressuspendeu-se o sedimento 5 vezes consecutivas utilizando 3 volumes de
acetona de cada vez agitando suavemente durante 10 minutos agrave temperatura ambiente Colocou-
se o resiacuteduo filtrado numa tina de vidro sobre papel de filtro deixando-se evaporar o solvente
durante a noite num nicho Triturou-se o resiacuteduo seco numa maacutequina de triturar carne e deixou-se
novamente evaporar o solvente O ldquopoacute de acetonardquo obtido foi armazenado a -20 ordmC
32122 Purificaccedilatildeo da actina a partir de ldquopoacute de acetonardquo
Apoacutes a obtenccedilatildeo do poacute de acetona como acima indicado efectuou-se o protocolo seguidamente
descrito de modo a obter monoacutemeros de actina (G-Actina) e actina filamentosa (F-Actina)
Pesou-se 35 g de poacute de acetona e adicionaram-se 50 mL de soluccedilatildeo-tampatildeo I10
sendo a
extracccedilatildeo feita com agitaccedilatildeo suave durante 30 minutos Separou-se o extracto do poacute de acetona
hidratado filtrando a suspensatildeo com 4 camadas de gaze esterilizada tendo o cuidado de natildeo
espremer o resiacuteduo Voltou a extrair-se com 50 mL de soluccedilatildeo I durante 30 minutos mexendo
lentamente Adicionou-se este ao extracto obtido anteriormente Centrifugou-se o extracto total a
16 000 g durante 30 minutos Decantou-se para uma proveta sobre a qual se colocou gaze para
que o sedimento solto natildeo se juntasse ao sobrenadante e leu-se o volume De seguida adicionou-
se KCl ateacute uma concentraccedilatildeo final de 50 mM MgCl2 ateacute 2mM e ATP ateacute 1mM lentamente e com
agitaccedilatildeo contiacutenua de modo a promover a polimerizaccedilatildeo dos monoacutemeros de actina
Deixou-se polimerizar durante 2h a 25ordmC observando-se a presenccedila de pequenas bolhas de ar
indicativas de um aumento de viscosidade da soluccedilatildeo Arrefeceu-se em gelo durante 15 minutos e
adicionou-se KCl ateacute uma concentraccedilatildeo final de 06M Mexeu-se lentamente durante 30 minutos
apoacutes os quais se ultracentrifugou a 110 000g durante 1hora Desprezou-se o sobrenadante
ressuspendeu-se o sedimento com soluccedilatildeo tampatildeo E e transferiu-se para um potter
Dialisou-se durante a noite contra 1L de soluccedilatildeo-tampatildeo I a uma temperatura de 4ordmC com rotaccedilatildeo
7 Soluccedilatildeo F 005 M NaHCO3 pH 82-85
8 Soluccedilatildeo G 1M Na2CO3
9 Soluccedilatildeo H 1 mM EDTA pH 70
10 Soluccedilatildeo I 2 mM Tris-HCl pH 8 02 mM CaCl2 02 mM Na2ATP 05 mM szlig-Me 0005 NaN3
42
contiacutenua e mudanccedila de tampatildeo 2 a 3 vezes De seguida a actina foi centrifugada a 110
000gdurante 1hora Decantou-se a soluccedilatildeo de G-Actina (sobrenadante) e desprezou-se o
sedimento Determinou-se a concentraccedilatildeo de G-Actina pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental) A soluccedilatildeo de G-actina foi armazenada a uma temperatura de 4ordmC
apoacutes adiccedilatildeo de 002NaN3 mantendo-se estaacutevel durante 3 a 5 dias
331 Quantificaccedilatildeo das proteiacutenas isoladas
3311 Quantificaccedilatildeo pelo meacutetodo de Bradford
O meacutetodo de Bradford (Bradford 1976) baseia-se na utilizaccedilatildeo do reagente de Bradford
composto por entre outros aacutecido fosfoacuterico e o corante ldquoCoomassierdquo Este corante possui carga
negativa e a sua coloraccedilatildeo varia entre a vermelha (Absmax=465 nm) predominante em soluccedilatildeo e a
azul (Absmax=595 nm) presente aquando da ligaccedilatildeo aos grupos amina das proteiacutenas Em soluccedilotildees
aacutecidas diluiacutedas a coloraccedilatildeo varia proporcionalmente com a quantidade do corante ligado agrave
proteiacutena que por sua vez eacute directamente proporcional agrave concentraccedilatildeo da proteiacutena permitindo a
sua quantificaccedilatildeo ateacute a um limite de detecccedilatildeo de 20 gmL (Bradford 1976)
Deste modo prepararam-se amostras de BSA ([BSA]stock= 1mgmL) utilizando um intervalo de
concentraccedilotildees compreendido entre 0 e 15 gmL de acordo com a tabela 3I Apoacutes a adiccedilatildeo da
H2O MilliQ e do reagente de Bradford as soluccedilotildees foram sucessivamente homogeneizadas no
voacutertex
Tabela 3Indash Amostras utilizadas para a obtenccedilatildeo da recta de calibraccedilatildeo no meacutetodo de Bradford
BSA (L) BSA (g) H2O MilliQ (L) Reagente de Bradford (L)
0 0 800 200
125 125 79875 200
25 25 7975 200
5 5 795 200
10 10 790 200
15 15 785 200
Apoacutes incubaccedilatildeo a 25ordmC por 5 minutos leram-se os valores de Abs595 num
espectrofotoacutemetro (Shimadzu UV-160A) para todas as amostras Seguidamente traccedilou-se a recta
de calibraccedilatildeo (Abs595vs quantidade de BSA (g))
43
Para determinar a concentraccedilatildeo de proteiacutena apoacutes o processo de isolamento purificaccedilatildeo e
diaacutelise quando necessaacuterio colocou-se 5 L da amostra num tubo de Eppendorf e adicionou-se
795 L de H2O MilliQ homogeneizando a soluccedilatildeo no voacutertex Adicionou-se posteriormente 200
L de reagente de Bradford e repetiu-se a homogeneizaccedilatildeo Incubou-se a mistura a 25ordmC por 5
minutos apoacutes os quais se leu os valores de Abs595 nm no espectrofotoacutemetro Por interpolaccedilatildeo deste
valor a partir da recta de calibraccedilatildeo determinou-se a concentraccedilatildeo de proteiacutena na amostra
3312 Quantificaccedilatildeo pelo meacutetodo directo
A quantificaccedilatildeo das amostras proteicas pode tambeacutem ser efectuada atraveacutes da leitura directa da
absorvacircncia tendo por base a lei de Beer-Lambert onde Abs corresponde agrave
absorvacircncia ao comprimento de onda ao qual o ensaio foi realizado corresponde ao coeficiente
de extinccedilatildeo (substituiacutedo pela letra a se as unidades forem distintas da molaridade) b corresponde
ao percurso oacuteptico da radiaccedilatildeo na ceacutelula amostral e c corresponde agrave concentraccedilatildeo amostral
Assim no caso de S1 de miosina mediu-se a Abs280e calculou-se a concentraccedilatildeo da proteiacutena
sabendo que o coeficiente de extinccedilatildeo de S1 eacute (Ref)
No caso da actina mediu-se a Abs290ecalculou-se a concentraccedilatildeo da proteiacutena sabendo que o
coeficiente de extinccedilatildeo da actina eacute (Ref)
332Electroforese em condiccedilotildees desnaturantes (SDS-PAGE)
De modo a melhor caracterizar as proteiacutenas isoladas foi realizada uma electroforese em
condiccedilotildees desnaturantes (SDS-PAGE do inglecircs Sodium Dodecyl Sulfate Polyacrilamide Gel
Electrophoresis) O SDS (dodecilo sulfato de soacutedio) eacute um detergente anioacutenico utilizado neste
meacutetodo para desnaturar as proteiacutenas que se ligam ao SDS na proporccedilatildeo 14 g SDS g proteiacutena
(aproximadamente 1 moleacutecula de SDS por cada 2 resiacuteduos de aminoaacutecidos) (Stryer) O SDS induz
uma densidade de carga negativa significativa nas proteiacutenas a que se liga mascarando a sua carga
intriacutenseca pelo que as razotildees cargamassa satildeo tendencialmente idecircnticas Assim a SDS-PAGE
separa as proteiacutenas quando sujeitas a um campo eleacutectrico de acordo com o seu peso molecular
sendo a sua migraccedilatildeo directamente proporcional ao logaritmo deste uacuteltimo (Stryer)
O gel de resoluccedilatildeo com 10 de acrilamida foi obtido atraveacutes da adiccedilatildeo de 167 mL de soluccedilatildeo
stock de acrilamida11
075 mL de tampatildeo de resoluccedilatildeo12
251 mL de aacutegua destilada e 50 L de
SDS 10 Para promover a polimerizaccedilatildeo adicionou-se em seguida 38 L de PSA 10
11
soluccedilatildeo stock de acrilamida soluccedilatildeo de 30 (mv) de acrilamida e 08 (mv) de NNrsquo-metil-bis-acrilamida 12
tampatildeo de resoluccedilatildeo 15 M Tris-HCl pH 88
44
(persulfato de amoacutenia) 10 e 25 L de TEMED 99 (tetrametiletilenodiamina) Agitou-se
cuidadosamente de modo a natildeo formar bolhas e pipetou-se a soluccedilatildeo para a cassette de vidro
previamente montada no suporte De seguida adicionou-se uma peliacutecula de cerca de 2 mm de 2-
metil-2-butanol para verificar posteriormente a polimerizaccedilatildeo Apoacutes a polimerizaccedilatildeo que demora
cerca de 45 minutos verteu-se a agrave superfiacutecie e adicionou-se o gel de concentraccedilatildeo com 5 de
acrilamida constituiacutedo por 03 mL de soluccedilatildeo de stock acrilamida 045 mL de tampatildeo de
concentraccedilatildeo13
094 mL de aacutegua destilada 18 L de SDS 10 135 L de PSA 10 e 2L de
TEMED 99 Colocou-se o pente sobre o gel e deixou-se polimerizar durante cerca de 45
minutos
As amostras para aplicaccedilatildeo no gel de electroforese foram preparadas para uma concentraccedilatildeo de
proteiacutena de 1 mgmL num volume final de 100 L dos quais 25 L correspondem ao tampatildeo da
amostra14
As amostras foram aquecidas a 100ordmC durante 5 minutos Antes da aplicaccedilatildeo das
amostras transferiu-se a cassette para a tina de electroforese agrave qual se adicionou a soluccedilatildeo
tampatildeo de electroforese da tina superior15
e tampatildeo de electroforese da tina inferior16
Aplicaram-
se 10 L das soluccedilotildees de marcadores de peso molecular e 20 L das amostras proteicas
preparadas anteriormente A electroforese decorreu durante cerca de 3h30 min a uma voltagem
de 80 V De seguida transferiu-se o gel para uma tina corando as proteiacutenas durante 1h com
soluccedilatildeo corante17
Descorou-se o gel com soluccedilatildeo descorante18
tendo-se mudado esta soluccedilatildeo ateacute
as bandas se encontrarem bem definidas
Determinou-se a mobilidade relativa das bandas correspondentes aos marcadores de elevado peso
molecular e traccedilou-se a recta de calibraccedilatildeo ndash log(PM) vs log(Rf) onde Rf corresponde agrave
mobilidade relativa Por interpolaccedilatildeo determina-se o PM das proteiacutenas isoladas sabendo a sua
mobilidade relativa Por comparaccedilatildeo com os valores descritos na literatura pode concluir-se
acerca do grau de purificaccedilatildeo
34 Mediccedilatildeo da actividade enzimaacutetica de S1
341 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
13
tampatildeo de concentraccedilatildeo 05 M Tris-HCl pH 68 14
tampatildeo de amostra 320 mM Tris-HCl pH 68 04 M szlig-mercaptoetanol SDS 8 glicerol 15 (vv) e roxo de m-
cresol 0024 15
tampatildeo de electroforese da tina superior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS 16
tampatildeo de electroforese da tina inferior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS acetato de soacutedio
01 M 17
soluccedilatildeo corante ldquoCoomassie Blue R-250rdquo contendo metanol 45 (vv) e aacutecido aceacutetico glacial 10 (vv) 18
soluccedilatildeo descorante metanol 10 (vv) e aacutecido aceacutetico glacial 10 vv)
45
O meacutetodo de Fiske-Subarow modificado consiste na reacccedilatildeo do molibdato de amoacutenio com o
fosfato inorgacircnico em soluccedilatildeo aacutecida com formaccedilatildeo do aacutecido fosfomoliacutebdico Seguidamente daacute-
se a adiccedilatildeo de um agente redutor que provoca a reduccedilatildeo do composto a fosfomolibdato que forma
um complexo de cor azulada cuja absorvacircncia maacutexima ocorre para um comprimento de onda de
660 nm tornando possiacutevel a sua detecccedilatildeo por meacutetodos espectrofotomeacutetricos As actividades Ca2+
-
ATPaacutesica e K+EDTA-ATPaacutesica de S1 foram determinadas atraveacutes deste meacutetodo de acordo com
o descrito por Taussky e Shorr (1953) com mediccedilatildeo da quantidade de fosfato (Pi) libertado
atraveacutes da hidroacutelise de ATP
Para tal obteve-se primeiramente a recta de calibraccedilatildeo com as amostras presentes na Tabela 3II
utilizando uma soluccedilatildeo stock de 5 mM KH2PO4
Tabela 3II ndash Composiccedilatildeo das diferentes amostras utilizadas para determinar a recta de calibraccedilatildeo atraveacutes do meacutetodo
de Fiske-Subbarow modificado
KH2PO4 (L)
([KH2PO4]stock= 5mM)
PO43-
(nmole) H2O (uL)
0 0 1780
5 25 1775
10 50 1770
20 100 1760
40 200 1740
100 500 1680
A cada tubo de Eppendorf adicionaram-se seguidamente 200 L de soluccedilatildeo J19
e 20 L de agente
redutor20
homogeneizando no voacutertex apoacutes cada adiccedilatildeo Incubou-se durante 10 minutos e mediu-
se a Abs660 nm num espectrofotoacutemetro com intervalos de tempo iguais entre cada amostra Traccedilou-
se a recta de calibraccedilatildeo (Abs660 nmvs nmol PO43-
) a partir dos valores obtidos
Para determinar a actividade Ca2+
-ATPaacutesica de S1 prepararam-se tubos de Eppendorf com 400
L de meio de reacccedilatildeo21
25 L CaCl2 ([CaCl2]stock= 100 mM) 25 L ATP ([ATP]stock= 100 mM)
com [S1]ensaios=01 mgmL Perfez-se se necessaacuterio com H2O MilliQ ateacute perfazer um volume de 1
mL Para determinar a actividade K+EDTA-ATPaacutesica utilizou-se uma concentraccedilatildeo idecircntica de
19
Soluccedilatildeo J 5 N H2SO4 25 molibdato de amoacutenio 20
Agente redutor 5 mg aacutecido 1-amino-2-naftol-4-sulfoacutenico 30 mg bissulfito de soacutedio 30 mg sulfito de soacutedio em 26
mL H2O 21
meio de reacccedilatildeo 125 mM Tris-HCl pH 80 0575 M KCl
46
proteiacutena nos ensaios (01 mgmL) e o mesmo meio de reacccedilatildeo Aos tubos de Eppendorf
adicionou-se ainda 123 L de KCl ([KCl]stock = 3M) 25 L de ATP 10 L de EDTA
([EDTA]stock= 100 mM )e o volume de H2O MilliQ necessaacuterio para perfazer 1 mL Em ambas as
situaccedilotildees o uacuteltimo reagente adicionado foi o ATP uma vez que se trata do substrato da reacccedilatildeo
enzimaacutetica
A cineacutetica decorreu a 25ordmC durante 5 minutos para a actividade Ca2+
-ATPaacutesica e durante 3
minutos para a actividade K+EDTA-ATPaacutesica Apoacutes o periacuteodo de incubaccedilatildeo transferiu-se 150
L de amostra para tubos de Eppendorf contendo igual volume de TCA 10 Colocou-se no gelo
e homogeneizou-se no voacutertex Seguidamente centrifugou-se a 1500 g durante 10 minutos e
recolheu-se 200 L de sobrenadante para novos tubos de Eppendorf Adicionou-se a estes 158
mL de H2O MilliQ e 200 L de soluccedilatildeo J Homogeneizou-se no voacutertex e adicionou-se 20 L de
reagente redutor repetindo-se a homogeneizaccedilatildeo Incubou-se durante 10 minutos e mediu-se a
Abs660 nm com o mesmo intervalo de tempo de leitura para cada amostra Por intrapolaccedilatildeo a partir
da recta de calibraccedilatildeo obtida anteriormente determinou-se a quantidade de Pi libertado em cada
ensaio A partir destes resultados determinaram-se as actividades Ca2+
-ATPaacutesica e K+EDTA-
ATPaacutesica do S1 em nmol PO43-
minmg proteiacutena e em s-1
342 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
Da actividade Mg2+
-ATPaacutesica de S1 resulta a formaccedilatildeo dos produtos da reacccedilatildeo de
hidroacutelise do ATP - ADP e Pi Na presenccedila de ADP fosfoenolpiruvato (PEP) e da enzima piruvato
cinase (PK) os produtos resultantes satildeo ATP e piruvato Por acccedilatildeo da enzima lactato
desidrogenase (LDH) este uacuteltimo eacute convertido em lactato ocorrendo simultaneamente a oxidaccedilatildeo
do NADH a NAD+ monitorizada atraveacutes da Abs340 nm Quando o S1 de miosina se encontra no
meio reaccional o ATP eacute hidrolisado formando-se novamente ADP e Pi o que permite a
formaccedilatildeo de um ciclo (fig 21) Se as enzimas se encontrarem em excesso a velocidade de
conversatildeo do NADH eacute limitada pela produccedilatildeo de ADP e Pi Como o ATP eacute regenerado em cada
ciclo a sua concentraccedilatildeo inicial eacute sempre mantida (Bagshaw 1982)
Figura 33ndash Representaccedilatildeo esquemaacutetica do meacutetodo de enzimas acopladas
47
O protocolo utilizado e seguidamente descrito foi adaptado de Cuenda et al (1990 ndash Ver Refordf) A
uma cuvette de 1 mL adicionou-se S1 para uma concentraccedilatildeo final de 005 mgmL 800 L de
meio de reacccedilatildeo22
10 L de PEP ([PEP]stock = 42 mM) 10 L de PK+LDH 11 ([PK+LDH]stock =
1 mgmL) e 15 L de NADH ([NADH]stock=25 mM) Iniciou-se a monitorizaccedilatildeo da reacccedilatildeo
atraveacutes da leitura de Abs340 nm apoacutes a adiccedilatildeo de 20 L de ATP ([ATP]stock=100 mM) A
actividade Mg2+
-ATPaacutesica eacute obtida atraveacutes do declive correspondente agrave oxidaccedilatildeo do NADH Para
a mediccedilatildeo da Mg2+
-ATPase de S1 estimulada por actina adicionou-se F-actina para uma
concentraccedilatildeo final de 2 microM sendo a actividade obtida tambeacutem atraveacutes do declive correspondente
agrave oxidaccedilatildeo do NADH apoacutes a adiccedilatildeo da proteiacutena
35 Interaccedilatildeo entre vanaacutedio e subfragmento-1 de miosina
351 Preparaccedilatildeo das soluccedilotildees de vanaacutedio
As soluccedilotildees de vanadato foram preparadas em aacutegua MilliQ a partir de metavanadato de amoacutenio
(NH4VO3) numa concentraccedilatildeo stock de 50 mM com agitaccedilatildeo magneacutetica e aquecimento (~30 ordmC)
Esta soluccedilatildeo com um valor compreendido entre 6 e 7 designa-se por ldquometavanadatordquo e eacute
constituiacuteda pelas esspeacutecies V1 V2 V4 e V5 (Ref) Acidificando a soluccedilatildeo de metavanadato por
adiccedilatildeo de HCl ateacute um valor de pH proacuteximo de 4 observa-se uma mudanccedila de cor de amarelo
claro para laranja indicativa da formaccedilatildeo da espeacutecie decameacuterica de vanadato De seguida e
imediatamente antes da realizaccedilatildeo dos ensaios descritos o valor do pH da soluccedilatildeo de
decavanadato foi aumentado para cerca de 75 por adiccedilatildeo de NaOH A soluccedilatildeo (2) designada
por ldquometavanadatordquo natildeo foi posteriormente alterada ateacute agrave sua adiccedilatildeo para a realizaccedilatildeo dos ensaios
As soluccedilotildees foram armazenadas a 4ordmC e mantidas em gelo aquando da sua utilizaccedilatildeo
A soluccedilatildeo de V(IV) foi preparada em condiccedilotildees anaeroacutebias por diluiccedilatildeo de sulfato de vanadil
(VOSO4) para uma concentraccedilatildeo final de 5 mM em H2O MilliQ saturada com N2 gasoso
352Anaacutelise por espectroscopia de UV-Vis
A concentraccedilatildeo da espeacutecie decameacuterica em soluccedilatildeo depende do periacuteodo de tempo apoacutes a diluiccedilatildeo
no meio de reacccedilatildeo apropriado (Ref) A velocidade de desoligomerizaccedilatildeo da espeacutecie decameacuterica
pode ser analisada por espectroscopia de absorccedilatildeo na regiatildeo UVVis uma vez que esta espeacutecie
apresenta duas bandas caracteriacutesticas na regiatildeo do visiacutevel (a 358 e 400 nm) que natildeo se verifica
em quaisquer outras espeacutecies oligomeacutericas As bandas supramencionada sapresentam-se bem
22
Meio de reacccedilatildeo 3125 mM Tris-HCl pH 71 375 mM MgCl2
48
resolvidas atraveacutes da obtenccedilatildeo da derivada de 2ordf ordem do espectro apresentando a forma de 2
vales com miacutenimos centrados nos comprimentos de onda supracitados com dependecircncia linear
relativamente agrave concentraccedilatildeo de decavanadato total (Ramos et al 2006)
Assim a estabilidade da espeacutecie decameacuterica apoacutes diluiccedilatildeo da soluccedilatildeo de decavanadato no meio de
reacccedilatildeo dos ensaios (tampatildeo D) para uma concentraccedilatildeo final de 1 mM em vanadato total foi
caracterizada atraveacutes da leitura da Abs400 em funccedilatildeo do tempo tendo-se determinado qual a
ordem de reacccedilatildeo e o tempo de meia-vida da mesma Com base em estudos anteriores acerca da
estabilidade do V10 na presenccedila de diferentes proteiacutenas (Ramos et al 2006) determinou-se
igualmente o tempo de meia vida apoacutes a adiccedilatildeo de 20 microM de S1 de miosina ao meio reaccional
353 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
Os espectros de RMN do isoacutetopo 51
V foram realizados num espectroacutemetro Bruker AM-400 MHz
equipado com uma sonda multinuclear inversa de 5 mm Em tubos de RMN de 5 mm de diacircmetro
preparou-se soluccedilotildees de 500 L de deca- ou metavanadato correspondentes a um intervalo de
concentraccedilotildees de 1 a 10 mM suplementadas com 10 D2O Os espectros foram adquiridos agrave
temperatura ambiente com uma largura espectral de 23 923 Hz um tempo de aquisiccedilatildeo de 0086
s tempo de relaxaccedilatildeo de 001 s e uma sequecircncia de pulsos a 90ordm Utilizou-se a transformada de
Fourier com alargamento das bandas de 20Hz para o processamento dos espectros Os desvios
quiacutemicos das ressonacircncias de 51
V foram obtidos em relaccedilatildeo agrave espeacutecie VOCl3cuja frequecircncia de
ressonacircncia eacute considerada o padratildeo sendo todas as outras referenciadas a partir desta em ppm
As aacutereas dos sinais do RMN correspondentes agraves espeacutecies oligomeacutericas presentes foram obtidas
por integraccedilatildeo dos sinais As concentraccedilotildees relativas das espeacutecies foram calculadas com recurso agrave
equaccedilatildeo
onde A[Vx] corresponde agrave aacuterea referente agrave ressonacircncia da espeacutecie Vx com nordm de agregaccedilatildeo x
A[V]total agrave soma total das aacutereas medidas e [V] a concentraccedilatildeo total de vanaacutedio no ensaio
A teacutecnica de 51
V-RMN foi anteriormente aplicada em estudos de ligaccedilatildeo com S1 de miosina
(Ref) tendo sido demostrado que entre as espeacutecies V1 V2 V4 e V5 a espeacutecie tetrameacuterica possui a
maior afinidade para a miosina (Cremo et al) O desaparecimento ou alargamento dos sinais pode
implicar a ligaccedilatildeo de um dos oxovanadatos laacutebeis agrave miosina sem indicaccedilatildeo de qual uma vez que
49
se processa um equiliacutebrio entre eles (Crans 1990) Assim e tendo em conta as indicaccedilotildees de que
a espeacutecie V10 eacute distinta das restantes no que diz respeito agrave magnitude da interacccedilatildeo com a miosina
e inibiccedilatildeo da actividade ATPaacutesica estimulada por actina (Aureliano 2000 Tiago et al 2002)
torna-se fundamental investigar a interacccedilatildeo das diferentes espeacutecies de vanadato com o S1 de
miosina atraveacutes da teacutecnica de 51
V-RMN
Para tal obtiveram-se espectros de 51
V-RMN resultantes da titulaccedilatildeo de uma soluccedilatildeo 5 mM de
decavanadato total com concentraccedilotildees crescentes de S1 de miosina (0 5 10 15 e 20 microM) em
tampatildeo D Realizou-se ainda ensaios na presenccedila de 2 mM de decavanadato e 2 mM de ATP na
presenccedila e na ausecircncia de S1 de miosina de modo a verificar o efeito que estes ligandos naturais
da enzima teria na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o subfragmento-1
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
Os grupos sulfidrilo (-SH) das cisteiacutenas satildeo resiacuteduos de aminoaacutecidos extremamente sensiacuteveis agrave
oxidaccedilatildeo resultando na sua oxidaccedilatildeo (Dalle-Donne et al 1999) Logo a quantificaccedilatildeo destes
resiacuteduos eacute bastante importante nos estudos de interacccedilatildeo com vanaacutedio jaacute que os resiacuteduos de Cys
com ecircnfase para o SH1 e o SH2 satildeo bastante importantes na monitorizaccedilatildeo das modificaccedilotildees
funcionais da regiatildeo da cabeccedila
O procedimento seguidamente apresentado foi originalmente descrito por Gutieacuterrez-Merino
(1987)) Consiste na reacccedilatildeo entre os grupos ndashSH natildeo oxidados das proteiacutenas e o reagente de
Ellman (DTNB -55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico)) - formando-se o aniatildeo CNT (aniatildeo 3-
carboxilato-4-nitrotiofenolato) numa estequiometria de 11 Este apresenta um maacuteximo de
absorvecircncia a 412 nm (Robyt 1990) Assim apoacutes a incubaccedilatildeo da proteiacutena na presenccedila do metal
a quantidade de cisteiacutenas reduzidas por proteiacutena determina-se atraveacutes dos valores de Abs412
Deste modo colocou-se num tubo de Eppendorf o volume apropriado de S1 dialisado para que a
concentraccedilatildeo no ensaio fosse de 1 mgmL Adicionou-se o volume apropriado de decavanadato
para que a sua concentraccedilatildeo fosse de 0 10 50 100 250 500 ou 1000 M Perfez-se com soluccedilatildeo
D ateacute um volume final de 1 mL e incubou-se a 25ordmC por 20 minutos Apoacutes este periacuteodo colocou-
se 780 L da mistura reaccional numa cuvette de quartzo e adicionou-se 220 L de soluccedilatildeo K23
23
Tampatildeo K Tampatildeo fosfatohidrogenofosfato de soacutedio 025 M pH 72
50
Adicionou-se 15 L de DTNB (em soluccedilatildeo K [DTNB]stock=100 mM) ao meio reaccional e
registou-se o valor de absorvecircncia medido equivalente agraves cisteiacutenas ldquoraacutepidasrdquo (SH1 e SH2
expostos ao solvente) Adicionou-se 001 g de SDS que provoca a desnaturaccedilatildeo da proteiacutena e
consequentemente a exposiccedilatildeo de todos os resiacuteduos de cisteiacutenas que se encontravam ateacute aiacute
inacessiacuteveis no interior da proteiacutena (cisteiacutenas totais) Estes resiacuteduos podem assim reagir com o
DTNB
Estimou-se o nuacutemero de resiacuteduos de cisteiacutenas em todas as amostras com recurso ao coeficiente de
extinccedilatildeo molar () do DTNB (ver ponto 3 dos Anexos) que foi determinado atraveacutes da titulaccedilatildeo
de L-Cisteiacutena com intervalo de concentraccedilotildees compreendidos entre 0 e 50 microM na presenccedila de
25microM de DTNB em tampatildeo D Os valores de absorvecircncia foram obtidos para um comprimento de
onda de 412 nm
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
3621 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
A teacutecnica mais eficiente de anaacutelise da interacccedilatildeo entre o iatildeo vanadilo e proteiacutenas como a
calmodulina e a transferrina entre outras permite a obtenccedilatildeo raacutepida de informaccedilatildeo no que toca agrave
conformaccedilatildeo e amplitude da referida interacccedilatildeo
Os espectros de Ressonacircncia Paramagneacutetica Electroacutenica (RPE) da espeacutecie paramagneacutetica V(IV)
foram realizados a 70K num espectroacutemetro Bruker EMX-300 X-band Todas as soluccedilotildees
utilizadas e amostras foram preparadas em anaerobiose numa cacircmara de luvas de modo a evitar a
reduccedilatildeo de V(IV) a V(V) Em tubos capilares com 02 mL de amostra manteve-se a concentraccedilatildeo
de S1 a 20 M e variou-se a concentraccedilatildeo de vanadilo (0 25 5 10 15 25 50 75 100 150
200 500 e 1000 M [VOSO4]stock=5 mM) em tampatildeo D Apoacutes obtenccedilatildeo dos espectros estes
foram tratados atraveacutes do software SimFonia e WinEPR tendo-se obtido um graacutefico com a
variaccedilatildeo da intensidade do sinal de RPE em funccedilatildeo da concentraccedilatildeo de sulfato de vanadilo
presente Realizaram-se ainda simulaccedilotildees de espectros obtidos com 20 microM de S1 e 1000 microM de
V(IV) com vista agrave obtenccedilatildeo dos ligandos provaacuteveis do vanadilo atraveacutes da anaacutelise da constante
de acoplamento hiperfina (A) e do factor g (g)
4 Conclusotildees gerais
Apesar de jaacute ter sido demonstrado que o vanaacutedio inibe a Na+K
+-ATPase da miosina a funccedilatildeo
deste elemento no tecido muacutesculo esqueleacutetico continua largamente incompreendida A enorme
complexidade quiacutemica deste metal especialmente no que concerne agrave oligomerizaccedilatildeo e reacccedilotildees
51
redox constitui um obstaacuteculo agrave compreensatildeo do comportamento do vanaacutedio nos organismos
vivos Assim a caracterizaccedilatildeo da quiacutemica deste elemento eacute de grande utilidade antes de iniciar o
estudo da sua interacccedilatildeo com o presente sistema
A anaacutelise por espectroscopias de 51
V-RMN e de UV-Vis de soluccedilotildees de vanadato diluiacutedas nos
meios de reacccedilatildeo utilizados no presente relatoacuterio (10 mM HEPES pH 70 25 mM KCl) mostrou
claramente que a espeacutecie decameacuterica (V10) possui um tempo de vida suficientemente longo para a
determinaccedilatildeo do seu efeito isolado ao contraacuterio do que se verifica com as espeacutecies mono- di-
tetra- e pentameacutericas (V1 V2 V4 e V5) Por outro lado a anaacutelise verificou ainda que apoacutes diluiccedilatildeo
das soluccedilotildees de decavanadato no meio de reacccedilatildeo coexistem as espeacutecies V10 e V1 pelo que os
efeitos putativos desta uacuteltima deveratildeo ser subtraiacutedos aquando da monitorizaccedilatildeo dos efeitos da
espeacutecie decameacuterica
O vanaacutedio (V) (vanadato) inibe as enzimas que catalizam reacccedilotildees de transferecircncia de grupos
fosfato como a ATPase da miosina [Goodno 1979] devido agrave sua analogia estrutural com o iatildeo
fosfato Na sua utilizaccedilatildeo como inibidor enzimaacutetico natildeo eacute tida em conta a contribuiccedilatildeo individual
das espeacutecies oligomeacutericas considerando-se normalmente a espeacutecie V1 como inibidora efectiva
[Goodno e Taylor 1982] No presente relatoacuterio verificou-se por espectroscopia de 51
V-RMN que
outros oligoacutemeros de vanadato interagem com o S1 de miosina tendo as espeacutecies V10 e V4 os
maiores niacuteveis de afinidade A sua interacccedilatildeo envolve forccedilas electrostaacuteticas e eacute afectada
negativamente por ligandos naturais de enzima como eacute o caso do ATP
A exposiccedilatildeo do S1 de miosina (005 mgmL) a decavanadato conduz agrave inibiccedilatildeo do capacidade da
proteiacutena de estimular a actividade Mg2+
-ATPaacutesica com um valor de IC50 de 266 plusmn 07 M
decavanadato sendo o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM
decavanadato (80) Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM decavanadato sendo
o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM decavanadato (60)
exercendo um efeito de impedimento agrave ligaccedilatildeo entre o vanaacutedio e o S1
A uma concentraccedilatildeo de 27 microM decavanadato verifica-se a oxidaccedilatildeo de um dos resiacuteduos de Cys
de S1 de miosina seguida pela oxidaccedilatildeo gradual de outro resiacuteduo de Cys ateacute agrave concentraccedilatildeo de
1000 microM Ambos os resiacuteduos oxidados natildeo se encontravam expostos ao solvente utilizado A
oxidaccedilatildeo dos resiacuteduos permite sugerir que esta modificaccedilatildeo oxidativa sofrida pelo S1 contribui
para a diminuiccedilatildeo da sua actividade de ATPase
52
Por espectroscopia de RPE revelaram-se os possiacuteveis ligandos equatoriais do vanaacutedio aquando da
sua interacccedilatildeo com o S1 de miosina como sendo duas moleacuteculas de aacutegua um ligando contendo o
grupo R-OH e outro contendo o grupo R=NR
Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados de
oxidaccedilatildeo (IV) e (V) no S1 de miosina para concentraccedilotildees facilmente alcanccedilaacuteveis no ambiente
terrestre
53
5 Estudos futuros
No sentido do melhor esclarecimento das alteraccedilotildees moleculares e estruturais que ocorrem no
subfragmento 1 quando exposta ao vanaacutedio que conduzem a uma diminuiccedilatildeo da sua funccedilatildeo
sugere-se
- Quantificaccedilatildeo das cisteiacutenas reduzidas de S1 na presenccedila de metavanadato
- Determinaccedilatildeo das actividades Ca2+
- e K+-ATPaacutesica do S1 de miosina na presenccedila de
decavanadato
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina estimulada por actina
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina na presenccedila de
metavanadato
- Realizaccedilatildeo de RMN 2D com variaccedilatildeo das concentraccedilotildees das soluccedilotildees de decavanadato e
metavanadato na presenccedila de S1
-Mutageacutenese dos resiacuteduos de Cys doS1 de miosina de modo a verificar quais os resiacuteduos que
sofrem oxidaccedilatildeo na presenccedila de decavanadato
- Determinaccedilatildeo por calorimetria (DSC Differential Scanning Calorimetry) da variaccedilatildeo da
entalpia (∆H) e da temperatura de desnaturaccedilatildeo do S1 de miosina na presenccedila de vanaacutedio uma
vez que estas propriedades caracteriacutesticas de cada proteiacutena satildeo alteradas apoacutes ocorrecircncia de
modificaccedilotildees estruturais
- Anaacutelise por SDS-PAGE de S1 de miosina apoacutes tratamento com vanaacutedio de modo a determinar
quais os polipeacuteptidos eventualmente formados apoacutes clivagem da proteiacutena
54
6 Referecircncias bibliograacuteficas
- Sellers J Myosins 2nd Edition Oxford University Press 1999
- L Stryer inBiochemistry (Ed WH Freeman 2005 pp
Refordfas Materiais e Meacutetodos
[A] S S Margossian SLowey Methods Enzymol1982 85 55-71
[B] JD Pardue JA Spudich Methods Enzymol 1982 85 164-181
[C] M Bradford Anal Biochem1976 72 248-254
[D] H H Taussky E Shorr J Biol Chem 1953 202 675-685
[E] C R Bagshaw in Outline Studies in Biology ndash Muscle Contraction Chapman and Hall Ed
1982
[F] A Cuenda et al Eur J Biochem1990 194 663-670
55
Anexos
1 Determinaccedilatildeo da recta de calibraccedilatildeo meacutetodo de Bradford
Figura1ndashRepresentaccedilatildeo graacutefica da recta de calibraccedilatildeo (Abs595 nm vs [BSA]) utilizada para o meacutetodo de Bradford
2 Determinaccedilatildeo da recta calibraccedilatildeo para a electroforese em condiccedilotildees desnaturantes
Figura 2 ndash Representaccedilatildeo graacutefica da recta de calibraccedilatildeo (log(Peso Molecular) vs Rf dos marcadores de peso
molecular) onde Rf indica determinada atraveacutes do padratildeo de peso molecular
(poccedilo (a))
3Caacutelculo do coeficiente de extinccedilatildeo molar () para a quantificaccedilatildeo de cisteiacutenas reduzidas
atraveacutes do meacutetodo de Ellman
Traccedilou-se a recta de Abs412 nm meacutedia vs [L-Cisteiacutena] de modo a determinar o valor do coeficiente
de extinccedilatildeo molar definido como o declive daquela recta Assim =8400 M-1
cm-1
como
podemos verificar na Fig 3
A595 nm = 00546[BSA] + 00209 Rsup2 = 099051
Ab
s 59
5 n
m
[BSA] (mgmL)
log PM = -1177 Rf + 2247 Rsup2 = 098957
log
PM
Mobilidade relativa (Rf)
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero

34
(010501002505001000 uM) em 10 mM HEPES 25 mM KCl pH 7 a 25ordmC durante 20 minutos A detecccedilatildeo
espectrofotomeacutetrica foi feita com DTNB (e 412=8400 uM-1 cm-1) (n variaacutevel entre 1 e 8)
Amostras Cys raacutepidas Cys totais
11 M S1 (controlo) 22 plusmn 04 106 plusmn 14
11 M S1 + 10 M Decavanadato 22 plusmn 05 95 plusmn 08
11 M S1 + 50 uM Decavanadato 20 plusmn 02 98 plusmn 14
11 M S1 + 100 uM Decavanadato 17 plusmn 03 97 plusmn14
11 M S1 + 250 uM Decavanadato 26 plusmn 02 98 plusmn 08
11 M S1 + 500 uM Decavanadato 21 plusmn 04 90 plusmn 06
11 M S1 + 1000 uM Decavanadato 24 plusmn 0 86 plusmn 08
A partir dos resultados obtidos pode sugerir-se que o S1 possui 11 resiacuteduos de Cys (controlo) o
que estaacute de acordo com o anteriormente descrito Verifica-se que o tratamento da proteiacutena com
concentraccedilotildees crescentes de decavanadato provoca a oxidaccedilatildeo de dois resiacuteduos de Cys do S1 de
miosina natildeo expostos ao solvente No entanto a teacutecnica utilizada natildeo nos permite identificar
quais
Tendo em consideraccedilatildeo que o valor de IC50 obtido para a reacccedilatildeo de inibiccedilatildeo daactividade Mg2+-
ATPaacutesica de S1 na presenccedila de decavanadato eacute de cerca de 27 microM pode sugerir-se que este seraacute o
valor ao qual ocorreraacute a oxidaccedilatildeo do primeiro resiacuteduo de Cys o que poderaacute contribuir para a
diminuiccedilatildeo da actividade de S1 observada
25Interacccedilatildeo de S1 com vanaacutedio(IV)
251Anaacutelise por espectroscopia de Ressonacircncia Paramagneacutetica Electroacutenica
A acumulaccedilatildeo intracelular de oxovanaacutedio (IV) in vivo constitui uma das principais formas de
apresentaccedilatildeo de vanaacutedio no organismo Deste modo torna-se bastante interessante analisar a
interacccedilatildeo entre este iatildeo e o S1 de miosina atraveacutes da teacutecnica de espectroscopia de Ressonacircncia
Paramagneacutetica Electroacutenica o meacutetodo de eleiccedilatildeo do estudo do iatildeo vanadilo e dos complexos por si
formados O electratildeo desemparelhado encontra-se fortemente acoplado ao nuacutecleo 51
V o que
35
resulta num espectro de RPE consistindo de 8 desdobramentos na direcccedilatildeo paralela e outros tantos
na direcccedilatildeo perpendicular (Chasteen 1983)
Os estudos realizados por Ahmed et al (1987) centraram-se na interacccedilatildeo do vanadilo com
proteiacutenas como eacute o caso da insulina transferrina entre outros obtendo um espectro caracterizado
por 8 linhas amplamente espaccediladas (na direcccedilatildeo paralela) Jaacute Ramos et al 2009 concluiu que a
exposiccedilatildeo da soluccedilatildeo de decavanadato agraves formas globular e filamentosa de actina produz um
espectro tiacutepico do iatildeo oxidovanaacutedio (IV) indicando a reduccedilatildeo do vanadato
Para aferir qual a concentraccedilatildeo de S1 que permitia obter uma boa razatildeo sinalruiacutedo titulou-se uma
soluccedilatildeo de sulfato de vanadilo 15 microM ([VOSO4]stock = 5 mM) com concentraccedilotildees crescentes de
proteiacutena (0 5 10 15 e 20 microM) em 10 mM HEPES 25 mM KCl A existecircncia de uma correlaccedilatildeo
linear entre a concentraccedilatildeo de proteiacutena e a intensidade dos sinais de RPE correspondente agrave banda
-12 perpendicular encontra-se representada na Fig 210
Figura 2 12ndash Representaccedilatildeo graacutefica da variaccedilatildeo da intensidade do sinal de RPE vs a concentraccedilatildeo de S1 utilizada
Apoacutes verificar que com uma concentraccedilatildeo de S1 igual a 20 microM se obtinham sinais com uma razatildeo
sinalruiacutedo adequada ao prosseguimento dos estudos sobre a interacccedilatildeo V(IV)-S1 de miosina
titulou-se uma soluccedilatildeo de 20 microM S1 com 0 25 5 10 15 25 50 75 100 150 200 500 e 1000
microM VOSO4
Traccedilou-se o graacutefico da intensidade da banda -12 perpendicular do sinal de EPR em funccedilatildeo da
concentraccedilatildeo de VOSO4 tendo-se determinado uma constante de dissociaccedilatildeo (KD) do complexo
S1-vanadilo de 8748 plusmn 2621 microM V(IV)
y = 016786x + 122435
Rsup2 = 092992759
Inte
nsi
dad
e d
o s
inal
de
RP
E
[S1] (microM)
36
Figura 213 ndash Representaccedilatildeo graacutefica da variaccedilatildeo do sinal de RPE vs a concentraccedilatildeo de vanadilo utilizada (os valores
experimentais estatildeo representados por losangos o seu ajuste estaacute representado pela linha)
A simulaccedilatildeo do espectro de EPR foi efectuada a partir do espectro obtido para uma concentraccedilatildeo
de 1 mM VOSO4 apoacutes adiccedilatildeo de 20 microM S1 de miosina tendo-se obtido os valores da constante de
acoplamento hiperfina (A=5016) e do factor g (g0=195)
Figura 214 ndash Representaccedilatildeo graacutefica do espectro de EPR obtido com 1000 microM de vanadilo e 20 microM de S1 (a cinzento
escuro) A simulaccedilatildeo do espectro encontra-se a cinzento claro
Por comparaccedilatildeo destes com os valores presentes na tabela de correspondecircncia xxx eacute possiacutevel
predizer quais os ligandos provaacuteveis do vanadilo na presenccedila de proteiacutena Assim concluiu-se que
o vanadilo estaacute coordenado com duas moleacuteculas de aacutegua um grupo hidroxilo e um grupo R=NR
O grupo hidroxilo pode indicar os resiacuteduos de aminoaacutecidos Tyr Treonina e Serina enquanto o
grupo R=NR poderaacute indicar a presenccedila de Arginina e Histidina como ligandos do vanadilo
Inte
nsi
dad
e d
os
sinai
s d
e R
PE
[Vanadilo] (microM)
Ajuste dos dados exp Dados experimentais
Campo magneacutetico (G)
Espectro Experimental Ajuste
37
3 Procedimento Experimental
31 Reagentes
Todos os reagentes utilizados satildeo de elevado grau de pureza (analiacutetico) Apresentam-se as suas
foacutermulas quiacutemicas ou abreviaturas quando necessaacuterio acrescentou-se a sua descriccedilatildeo Entre
parecircntesis colocou-se as empresas que os comercializam
Aacutecido aceacutetico glacial 100 (Riedel) Acrilamida 99 (Merck) Azul de bromofenol (Riedel-de-
Haumlen) -Mercaptoetanol (Sigma) Bissulfito de soacutedio (Sigma)Bradford (reagente de)
(BioRad)BSA (albumina de soro bovino Sigma-Aldrich) CaCl22H2O (cloreto de caacutelcio
dihidratado Fluka) CH3CH2OH 96 (etanol Panreac) Coomassie Blue R250 (Merck) D2O
(oacutexido de deuteacuterio SDS) DTNB (55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico) Aldrich) EDTA (aacutecido
etilenodiaminatetraaceacutetico Merck) Fosfoenolpiruvato 98 (Fluka) Glicerol (Sigma) Glicina
985 (Panreac) HCl 37 (Riedel-de Haumlen) H2SO4 96 (Pronalab) HEPES 99 (aacutecido 4-(2-
hidroxiletil)-1-piperazineetanosulfoacutenico Sigma) KH2PO4 99 (hidrogenofosfato de potaacutessio)
MgATP (Sigma) Na
ATP (Sigma) Na2HPO4 99 (hidrogenofosfato de soacutedio Riedel-de Haumlen)
NaH2PO4 (fosfato de soacutedio ) NaOH 986 (hidroacutexido de soacutedio Pronalab) KCl 99 (Cloreto de
potaacutessio Panreac) L-Cys 995 (enantioacutemero levoacutegiro do resiacuteduo de Cisteiacutena Fluka)Metanol
99 (Merck) Molibdato de amoacutenio (Sigma)NNrsquo-metil-bis-acrilamida 98 (Merck) NH4VO3
99 (metavanadato de amoacutenia Sigma) PSA 10 (persulfato de amoacutenio Merck) SDS
(dodecilo sulfato de soacutedio Sigma) Sulfito de soacutedio (Sigma) VOSO4 (sulfato de vanadilo
Aldrich) TEMED 99 (Tetrametiletilenodiamina Merck) Tris (2-amino-2-hidroximetil-13-
propanodiol Sigma)
32 Meacutetodos
321 Isolamento e purificaccedilatildeo das proteiacutenas de muacutesculo esqueleacutetico de coelho
3211 Purificaccedilatildeo da miosina preparaccedilatildeo de Subfragmento 1 por digestatildeo quimotriacutepsica
da miosina
O subfragmento-1 de miosina-II (S1) foi gentilmente cedido pelo Prof Doutor Manuel Aureliano
do Departamento de Quiacutemica Bioquiacutemica e Farmaacutecia da Faculdade de Ciecircncias e Tecnologia da
Universidade do Algarve
No entanto descreve-se abaixo de forma resumida o procedimento efectuado
38
32111 Purificaccedilatildeo da miosina
O protocolo de isolamento foi adaptado do meacutetodo previamente descrito por Margossian e Lowey
(1982) apresentando-se de forma esquemaacutetica na Fig 31
Sucintamente os tecidos musculares dorsais e das pernas foram extraiacutedos apoacutes o sacrifiacutecio do
animal (Fig31) De seguida foram adicionados 3 volumes da soluccedilatildeo de extracccedilatildeo A2
Centrifugou-se a soluccedilatildeo e filtrou-se o sobrenadante O sedimento foi entatildeo diluiacutedo em 14
volumes de aacutegua destilada de modo a que a forccedila ioacutenica atingisse 004 M Apoacutes adiccedilatildeo de szlig-
mercaptoetanol (szlig-me) deixou-se a precipitar a 4ordmC removendo o sobrenadante Centrifugou-se
novamente e ressuspendeu-se o sedimento na soluccedilatildeo de extracccedilatildeo A Adicionou-se a soluccedilatildeo
tampatildeo B3 para que a forccedila ioacutenica atingisse 05 M Homogeneizou-se e deixou-se repousar
durante 2h Homogeneizou-se novamente e centrifugou-se de modo a ocorrer precipitaccedilatildeo dos
complexos de actomiosina e proteiacutena desnaturada Apoacutes filtraccedilatildeo do sobrenadante em latilde de vidro
quantificou-se a soluccedilatildeo de miosina resultante pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental)
2 Soluccedilatildeo de extracccedilatildeo A 0012M K2CO3 004M KHCO3 06M KCl pH 65
3 Soluccedilatildeo-tampatildeo B Tris 100 mM pH 7 KCl 2M
39
Figura 31 ndash Representaccedilatildeo esquemaacutetica do protocolo de isolamento de miosina (Margossian e Lowey 1982) Todo o
procedimento foi realizado a 4ordmC (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
32112 Preparaccedilatildeo de Subfragmento-1 de miosina por digestatildeo com quimiotripsina
O S1 de miosina foi isolado apoacutes diaacutelise da miosina durante a noite contra a soluccedilatildeo C4 A
presenccedila do EDTA eacute essencial uma vez que a sua substituiccedilatildeo por MgCl2resulta na produccedilatildeo de
HMM em vez de S1 A suspensatildeo de miosina foi equilibrada e digerida com -quimotripsina
A digestatildeo foi interrompida pela adiccedilatildeo de PMSF Seguidamente ultracentrifugou-se o produto
da digestatildeo desprezando-se o sedimento (Fig 32) A soluccedilatildeo de sobrenadante maioritariamente
S1 foi quantificada pela leitura directa da Abs280 ou pelo meacutetodo de Bradford (ver as secccedilotildees
3311 e 3312 do Procedimento Experimental respectivamente) e armazenada em 10 sacarose
e 2 mM szlig-Me a -80ordmC
4 Soluccedilatildeo C 25mM Tris pH 70 012M KCl 1mM EDTA
40
Antes da realizaccedilatildeo dos ensaios descritos no presente relatoacuterio o S1 de miosina foi dialisado
contra a soluccedilatildeo D5 de modo a remover qualquer composto que pudesse complexar como
vanaacutedio Seguidamente quantificou-se a proteiacutena pelo meacutetodo de Bradford ou pelo meacutetodo
directo (ver secccedilotildees 3311 e 3312do Procedimento Experimtental)
Figura 3 2ndash Representaccedilatildeo esquemaacutetica do protocolo de preparaccedilatildeo do S1 de miosina por digestatildeo quimiotripsina de
acordo com o previamente descrito por Margossian e Lowey (1982) Todo o procedimento foi realizado a 4ordm C
excepto quando indicaccedilatildeo do contraacuterio (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
3212 Isolamento e purificaccedilatildeo de actina
Isolou-se e purificou-se a actina segundo o meacutetodo descrito por Pardee e Spudish que
compreende dois passos ndash a obtenccedilatildeo de um resiacuteduo seco denominado ldquopoacute de acetonardquo e a
extracccedilatildeo subsequente da actina a partir do ldquopoacute de acetonardquo na forma de G-Actina com a
posterior polimerizaccedilatildeo a F-Actina
32121 Obtenccedilatildeo do ldquopoacute de acetonardquo
Sacrificou-se o coelho com uma pancada atraacutes do pescoccedilo e sangrou-se o animal apoacutes o corte da
arteacuteria jugular Esfolou-se e colocou-se numa tina com gelo Retiraram-se as partes de tecido
conjuntivo que envolvem o tecido muscular com auxiacutelio de uma pinccedila e bisturi Seguidamente
retirou-se os muacutesculos dorsais e os muacutesculos brancos das pernas Passou-se os muacutesculos por aacutegua
destilada de modo a remover as impurezas e pesou-se num copo Trituraram-se os muacutesculos
numa maacutequina de triturar carne e extraiu-se o resultante com 3 volumes da soluccedilatildeo E6 durante 10
minutos com agitaccedilatildeo suave Centrifugou-se a suspensatildeo a 4000 g durante 10
5 Soluccedilatildeo D HEPES 10 mM pH 70 KCl 25 mM
6 Soluccedilatildeo E 01 M KCl 015 M K2HPO4 pH 65
41
minutosdesprezando-se o sobrenadante Ressuspendeu-se o sedimento em 6 volumes de soluccedilatildeo
F7durante 10 minutos com agitaccedilatildeo suave Verificou-se se o pH da soluccedilatildeo estava compreendido
entre 82 e 85 acertando-se estecom soluccedilatildeo G8 se necessaacuterio Centrifugou-se durante 10
minutos a 4000 g Desprezou-se o sobrenadante e ressuspendeu-se o sedimento em 3 volumes de
soluccedilatildeo H9 durante 10 minutos Centrifugou-se novamente a 4000 gdurante 10 minutos
desprezando-se o sobrenadante Ressuspendeu-se o sedimento duas vezes consecutivas em 6
volumes de aacutegua destilada durante 5 minutos Centrifugou-se a 4000 g durante 10 minutos apoacutes
cada adiccedilatildeo de H2O
Seguidamente ressuspendeu-se o sedimento 5 vezes consecutivas utilizando 3 volumes de
acetona de cada vez agitando suavemente durante 10 minutos agrave temperatura ambiente Colocou-
se o resiacuteduo filtrado numa tina de vidro sobre papel de filtro deixando-se evaporar o solvente
durante a noite num nicho Triturou-se o resiacuteduo seco numa maacutequina de triturar carne e deixou-se
novamente evaporar o solvente O ldquopoacute de acetonardquo obtido foi armazenado a -20 ordmC
32122 Purificaccedilatildeo da actina a partir de ldquopoacute de acetonardquo
Apoacutes a obtenccedilatildeo do poacute de acetona como acima indicado efectuou-se o protocolo seguidamente
descrito de modo a obter monoacutemeros de actina (G-Actina) e actina filamentosa (F-Actina)
Pesou-se 35 g de poacute de acetona e adicionaram-se 50 mL de soluccedilatildeo-tampatildeo I10
sendo a
extracccedilatildeo feita com agitaccedilatildeo suave durante 30 minutos Separou-se o extracto do poacute de acetona
hidratado filtrando a suspensatildeo com 4 camadas de gaze esterilizada tendo o cuidado de natildeo
espremer o resiacuteduo Voltou a extrair-se com 50 mL de soluccedilatildeo I durante 30 minutos mexendo
lentamente Adicionou-se este ao extracto obtido anteriormente Centrifugou-se o extracto total a
16 000 g durante 30 minutos Decantou-se para uma proveta sobre a qual se colocou gaze para
que o sedimento solto natildeo se juntasse ao sobrenadante e leu-se o volume De seguida adicionou-
se KCl ateacute uma concentraccedilatildeo final de 50 mM MgCl2 ateacute 2mM e ATP ateacute 1mM lentamente e com
agitaccedilatildeo contiacutenua de modo a promover a polimerizaccedilatildeo dos monoacutemeros de actina
Deixou-se polimerizar durante 2h a 25ordmC observando-se a presenccedila de pequenas bolhas de ar
indicativas de um aumento de viscosidade da soluccedilatildeo Arrefeceu-se em gelo durante 15 minutos e
adicionou-se KCl ateacute uma concentraccedilatildeo final de 06M Mexeu-se lentamente durante 30 minutos
apoacutes os quais se ultracentrifugou a 110 000g durante 1hora Desprezou-se o sobrenadante
ressuspendeu-se o sedimento com soluccedilatildeo tampatildeo E e transferiu-se para um potter
Dialisou-se durante a noite contra 1L de soluccedilatildeo-tampatildeo I a uma temperatura de 4ordmC com rotaccedilatildeo
7 Soluccedilatildeo F 005 M NaHCO3 pH 82-85
8 Soluccedilatildeo G 1M Na2CO3
9 Soluccedilatildeo H 1 mM EDTA pH 70
10 Soluccedilatildeo I 2 mM Tris-HCl pH 8 02 mM CaCl2 02 mM Na2ATP 05 mM szlig-Me 0005 NaN3
42
contiacutenua e mudanccedila de tampatildeo 2 a 3 vezes De seguida a actina foi centrifugada a 110
000gdurante 1hora Decantou-se a soluccedilatildeo de G-Actina (sobrenadante) e desprezou-se o
sedimento Determinou-se a concentraccedilatildeo de G-Actina pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental) A soluccedilatildeo de G-actina foi armazenada a uma temperatura de 4ordmC
apoacutes adiccedilatildeo de 002NaN3 mantendo-se estaacutevel durante 3 a 5 dias
331 Quantificaccedilatildeo das proteiacutenas isoladas
3311 Quantificaccedilatildeo pelo meacutetodo de Bradford
O meacutetodo de Bradford (Bradford 1976) baseia-se na utilizaccedilatildeo do reagente de Bradford
composto por entre outros aacutecido fosfoacuterico e o corante ldquoCoomassierdquo Este corante possui carga
negativa e a sua coloraccedilatildeo varia entre a vermelha (Absmax=465 nm) predominante em soluccedilatildeo e a
azul (Absmax=595 nm) presente aquando da ligaccedilatildeo aos grupos amina das proteiacutenas Em soluccedilotildees
aacutecidas diluiacutedas a coloraccedilatildeo varia proporcionalmente com a quantidade do corante ligado agrave
proteiacutena que por sua vez eacute directamente proporcional agrave concentraccedilatildeo da proteiacutena permitindo a
sua quantificaccedilatildeo ateacute a um limite de detecccedilatildeo de 20 gmL (Bradford 1976)
Deste modo prepararam-se amostras de BSA ([BSA]stock= 1mgmL) utilizando um intervalo de
concentraccedilotildees compreendido entre 0 e 15 gmL de acordo com a tabela 3I Apoacutes a adiccedilatildeo da
H2O MilliQ e do reagente de Bradford as soluccedilotildees foram sucessivamente homogeneizadas no
voacutertex
Tabela 3Indash Amostras utilizadas para a obtenccedilatildeo da recta de calibraccedilatildeo no meacutetodo de Bradford
BSA (L) BSA (g) H2O MilliQ (L) Reagente de Bradford (L)
0 0 800 200
125 125 79875 200
25 25 7975 200
5 5 795 200
10 10 790 200
15 15 785 200
Apoacutes incubaccedilatildeo a 25ordmC por 5 minutos leram-se os valores de Abs595 num
espectrofotoacutemetro (Shimadzu UV-160A) para todas as amostras Seguidamente traccedilou-se a recta
de calibraccedilatildeo (Abs595vs quantidade de BSA (g))
43
Para determinar a concentraccedilatildeo de proteiacutena apoacutes o processo de isolamento purificaccedilatildeo e
diaacutelise quando necessaacuterio colocou-se 5 L da amostra num tubo de Eppendorf e adicionou-se
795 L de H2O MilliQ homogeneizando a soluccedilatildeo no voacutertex Adicionou-se posteriormente 200
L de reagente de Bradford e repetiu-se a homogeneizaccedilatildeo Incubou-se a mistura a 25ordmC por 5
minutos apoacutes os quais se leu os valores de Abs595 nm no espectrofotoacutemetro Por interpolaccedilatildeo deste
valor a partir da recta de calibraccedilatildeo determinou-se a concentraccedilatildeo de proteiacutena na amostra
3312 Quantificaccedilatildeo pelo meacutetodo directo
A quantificaccedilatildeo das amostras proteicas pode tambeacutem ser efectuada atraveacutes da leitura directa da
absorvacircncia tendo por base a lei de Beer-Lambert onde Abs corresponde agrave
absorvacircncia ao comprimento de onda ao qual o ensaio foi realizado corresponde ao coeficiente
de extinccedilatildeo (substituiacutedo pela letra a se as unidades forem distintas da molaridade) b corresponde
ao percurso oacuteptico da radiaccedilatildeo na ceacutelula amostral e c corresponde agrave concentraccedilatildeo amostral
Assim no caso de S1 de miosina mediu-se a Abs280e calculou-se a concentraccedilatildeo da proteiacutena
sabendo que o coeficiente de extinccedilatildeo de S1 eacute (Ref)
No caso da actina mediu-se a Abs290ecalculou-se a concentraccedilatildeo da proteiacutena sabendo que o
coeficiente de extinccedilatildeo da actina eacute (Ref)
332Electroforese em condiccedilotildees desnaturantes (SDS-PAGE)
De modo a melhor caracterizar as proteiacutenas isoladas foi realizada uma electroforese em
condiccedilotildees desnaturantes (SDS-PAGE do inglecircs Sodium Dodecyl Sulfate Polyacrilamide Gel
Electrophoresis) O SDS (dodecilo sulfato de soacutedio) eacute um detergente anioacutenico utilizado neste
meacutetodo para desnaturar as proteiacutenas que se ligam ao SDS na proporccedilatildeo 14 g SDS g proteiacutena
(aproximadamente 1 moleacutecula de SDS por cada 2 resiacuteduos de aminoaacutecidos) (Stryer) O SDS induz
uma densidade de carga negativa significativa nas proteiacutenas a que se liga mascarando a sua carga
intriacutenseca pelo que as razotildees cargamassa satildeo tendencialmente idecircnticas Assim a SDS-PAGE
separa as proteiacutenas quando sujeitas a um campo eleacutectrico de acordo com o seu peso molecular
sendo a sua migraccedilatildeo directamente proporcional ao logaritmo deste uacuteltimo (Stryer)
O gel de resoluccedilatildeo com 10 de acrilamida foi obtido atraveacutes da adiccedilatildeo de 167 mL de soluccedilatildeo
stock de acrilamida11
075 mL de tampatildeo de resoluccedilatildeo12
251 mL de aacutegua destilada e 50 L de
SDS 10 Para promover a polimerizaccedilatildeo adicionou-se em seguida 38 L de PSA 10
11
soluccedilatildeo stock de acrilamida soluccedilatildeo de 30 (mv) de acrilamida e 08 (mv) de NNrsquo-metil-bis-acrilamida 12
tampatildeo de resoluccedilatildeo 15 M Tris-HCl pH 88
44
(persulfato de amoacutenia) 10 e 25 L de TEMED 99 (tetrametiletilenodiamina) Agitou-se
cuidadosamente de modo a natildeo formar bolhas e pipetou-se a soluccedilatildeo para a cassette de vidro
previamente montada no suporte De seguida adicionou-se uma peliacutecula de cerca de 2 mm de 2-
metil-2-butanol para verificar posteriormente a polimerizaccedilatildeo Apoacutes a polimerizaccedilatildeo que demora
cerca de 45 minutos verteu-se a agrave superfiacutecie e adicionou-se o gel de concentraccedilatildeo com 5 de
acrilamida constituiacutedo por 03 mL de soluccedilatildeo de stock acrilamida 045 mL de tampatildeo de
concentraccedilatildeo13
094 mL de aacutegua destilada 18 L de SDS 10 135 L de PSA 10 e 2L de
TEMED 99 Colocou-se o pente sobre o gel e deixou-se polimerizar durante cerca de 45
minutos
As amostras para aplicaccedilatildeo no gel de electroforese foram preparadas para uma concentraccedilatildeo de
proteiacutena de 1 mgmL num volume final de 100 L dos quais 25 L correspondem ao tampatildeo da
amostra14
As amostras foram aquecidas a 100ordmC durante 5 minutos Antes da aplicaccedilatildeo das
amostras transferiu-se a cassette para a tina de electroforese agrave qual se adicionou a soluccedilatildeo
tampatildeo de electroforese da tina superior15
e tampatildeo de electroforese da tina inferior16
Aplicaram-
se 10 L das soluccedilotildees de marcadores de peso molecular e 20 L das amostras proteicas
preparadas anteriormente A electroforese decorreu durante cerca de 3h30 min a uma voltagem
de 80 V De seguida transferiu-se o gel para uma tina corando as proteiacutenas durante 1h com
soluccedilatildeo corante17
Descorou-se o gel com soluccedilatildeo descorante18
tendo-se mudado esta soluccedilatildeo ateacute
as bandas se encontrarem bem definidas
Determinou-se a mobilidade relativa das bandas correspondentes aos marcadores de elevado peso
molecular e traccedilou-se a recta de calibraccedilatildeo ndash log(PM) vs log(Rf) onde Rf corresponde agrave
mobilidade relativa Por interpolaccedilatildeo determina-se o PM das proteiacutenas isoladas sabendo a sua
mobilidade relativa Por comparaccedilatildeo com os valores descritos na literatura pode concluir-se
acerca do grau de purificaccedilatildeo
34 Mediccedilatildeo da actividade enzimaacutetica de S1
341 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
13
tampatildeo de concentraccedilatildeo 05 M Tris-HCl pH 68 14
tampatildeo de amostra 320 mM Tris-HCl pH 68 04 M szlig-mercaptoetanol SDS 8 glicerol 15 (vv) e roxo de m-
cresol 0024 15
tampatildeo de electroforese da tina superior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS 16
tampatildeo de electroforese da tina inferior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS acetato de soacutedio
01 M 17
soluccedilatildeo corante ldquoCoomassie Blue R-250rdquo contendo metanol 45 (vv) e aacutecido aceacutetico glacial 10 (vv) 18
soluccedilatildeo descorante metanol 10 (vv) e aacutecido aceacutetico glacial 10 vv)
45
O meacutetodo de Fiske-Subarow modificado consiste na reacccedilatildeo do molibdato de amoacutenio com o
fosfato inorgacircnico em soluccedilatildeo aacutecida com formaccedilatildeo do aacutecido fosfomoliacutebdico Seguidamente daacute-
se a adiccedilatildeo de um agente redutor que provoca a reduccedilatildeo do composto a fosfomolibdato que forma
um complexo de cor azulada cuja absorvacircncia maacutexima ocorre para um comprimento de onda de
660 nm tornando possiacutevel a sua detecccedilatildeo por meacutetodos espectrofotomeacutetricos As actividades Ca2+
-
ATPaacutesica e K+EDTA-ATPaacutesica de S1 foram determinadas atraveacutes deste meacutetodo de acordo com
o descrito por Taussky e Shorr (1953) com mediccedilatildeo da quantidade de fosfato (Pi) libertado
atraveacutes da hidroacutelise de ATP
Para tal obteve-se primeiramente a recta de calibraccedilatildeo com as amostras presentes na Tabela 3II
utilizando uma soluccedilatildeo stock de 5 mM KH2PO4
Tabela 3II ndash Composiccedilatildeo das diferentes amostras utilizadas para determinar a recta de calibraccedilatildeo atraveacutes do meacutetodo
de Fiske-Subbarow modificado
KH2PO4 (L)
([KH2PO4]stock= 5mM)
PO43-
(nmole) H2O (uL)
0 0 1780
5 25 1775
10 50 1770
20 100 1760
40 200 1740
100 500 1680
A cada tubo de Eppendorf adicionaram-se seguidamente 200 L de soluccedilatildeo J19
e 20 L de agente
redutor20
homogeneizando no voacutertex apoacutes cada adiccedilatildeo Incubou-se durante 10 minutos e mediu-
se a Abs660 nm num espectrofotoacutemetro com intervalos de tempo iguais entre cada amostra Traccedilou-
se a recta de calibraccedilatildeo (Abs660 nmvs nmol PO43-
) a partir dos valores obtidos
Para determinar a actividade Ca2+
-ATPaacutesica de S1 prepararam-se tubos de Eppendorf com 400
L de meio de reacccedilatildeo21
25 L CaCl2 ([CaCl2]stock= 100 mM) 25 L ATP ([ATP]stock= 100 mM)
com [S1]ensaios=01 mgmL Perfez-se se necessaacuterio com H2O MilliQ ateacute perfazer um volume de 1
mL Para determinar a actividade K+EDTA-ATPaacutesica utilizou-se uma concentraccedilatildeo idecircntica de
19
Soluccedilatildeo J 5 N H2SO4 25 molibdato de amoacutenio 20
Agente redutor 5 mg aacutecido 1-amino-2-naftol-4-sulfoacutenico 30 mg bissulfito de soacutedio 30 mg sulfito de soacutedio em 26
mL H2O 21
meio de reacccedilatildeo 125 mM Tris-HCl pH 80 0575 M KCl
46
proteiacutena nos ensaios (01 mgmL) e o mesmo meio de reacccedilatildeo Aos tubos de Eppendorf
adicionou-se ainda 123 L de KCl ([KCl]stock = 3M) 25 L de ATP 10 L de EDTA
([EDTA]stock= 100 mM )e o volume de H2O MilliQ necessaacuterio para perfazer 1 mL Em ambas as
situaccedilotildees o uacuteltimo reagente adicionado foi o ATP uma vez que se trata do substrato da reacccedilatildeo
enzimaacutetica
A cineacutetica decorreu a 25ordmC durante 5 minutos para a actividade Ca2+
-ATPaacutesica e durante 3
minutos para a actividade K+EDTA-ATPaacutesica Apoacutes o periacuteodo de incubaccedilatildeo transferiu-se 150
L de amostra para tubos de Eppendorf contendo igual volume de TCA 10 Colocou-se no gelo
e homogeneizou-se no voacutertex Seguidamente centrifugou-se a 1500 g durante 10 minutos e
recolheu-se 200 L de sobrenadante para novos tubos de Eppendorf Adicionou-se a estes 158
mL de H2O MilliQ e 200 L de soluccedilatildeo J Homogeneizou-se no voacutertex e adicionou-se 20 L de
reagente redutor repetindo-se a homogeneizaccedilatildeo Incubou-se durante 10 minutos e mediu-se a
Abs660 nm com o mesmo intervalo de tempo de leitura para cada amostra Por intrapolaccedilatildeo a partir
da recta de calibraccedilatildeo obtida anteriormente determinou-se a quantidade de Pi libertado em cada
ensaio A partir destes resultados determinaram-se as actividades Ca2+
-ATPaacutesica e K+EDTA-
ATPaacutesica do S1 em nmol PO43-
minmg proteiacutena e em s-1
342 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
Da actividade Mg2+
-ATPaacutesica de S1 resulta a formaccedilatildeo dos produtos da reacccedilatildeo de
hidroacutelise do ATP - ADP e Pi Na presenccedila de ADP fosfoenolpiruvato (PEP) e da enzima piruvato
cinase (PK) os produtos resultantes satildeo ATP e piruvato Por acccedilatildeo da enzima lactato
desidrogenase (LDH) este uacuteltimo eacute convertido em lactato ocorrendo simultaneamente a oxidaccedilatildeo
do NADH a NAD+ monitorizada atraveacutes da Abs340 nm Quando o S1 de miosina se encontra no
meio reaccional o ATP eacute hidrolisado formando-se novamente ADP e Pi o que permite a
formaccedilatildeo de um ciclo (fig 21) Se as enzimas se encontrarem em excesso a velocidade de
conversatildeo do NADH eacute limitada pela produccedilatildeo de ADP e Pi Como o ATP eacute regenerado em cada
ciclo a sua concentraccedilatildeo inicial eacute sempre mantida (Bagshaw 1982)
Figura 33ndash Representaccedilatildeo esquemaacutetica do meacutetodo de enzimas acopladas
47
O protocolo utilizado e seguidamente descrito foi adaptado de Cuenda et al (1990 ndash Ver Refordf) A
uma cuvette de 1 mL adicionou-se S1 para uma concentraccedilatildeo final de 005 mgmL 800 L de
meio de reacccedilatildeo22
10 L de PEP ([PEP]stock = 42 mM) 10 L de PK+LDH 11 ([PK+LDH]stock =
1 mgmL) e 15 L de NADH ([NADH]stock=25 mM) Iniciou-se a monitorizaccedilatildeo da reacccedilatildeo
atraveacutes da leitura de Abs340 nm apoacutes a adiccedilatildeo de 20 L de ATP ([ATP]stock=100 mM) A
actividade Mg2+
-ATPaacutesica eacute obtida atraveacutes do declive correspondente agrave oxidaccedilatildeo do NADH Para
a mediccedilatildeo da Mg2+
-ATPase de S1 estimulada por actina adicionou-se F-actina para uma
concentraccedilatildeo final de 2 microM sendo a actividade obtida tambeacutem atraveacutes do declive correspondente
agrave oxidaccedilatildeo do NADH apoacutes a adiccedilatildeo da proteiacutena
35 Interaccedilatildeo entre vanaacutedio e subfragmento-1 de miosina
351 Preparaccedilatildeo das soluccedilotildees de vanaacutedio
As soluccedilotildees de vanadato foram preparadas em aacutegua MilliQ a partir de metavanadato de amoacutenio
(NH4VO3) numa concentraccedilatildeo stock de 50 mM com agitaccedilatildeo magneacutetica e aquecimento (~30 ordmC)
Esta soluccedilatildeo com um valor compreendido entre 6 e 7 designa-se por ldquometavanadatordquo e eacute
constituiacuteda pelas esspeacutecies V1 V2 V4 e V5 (Ref) Acidificando a soluccedilatildeo de metavanadato por
adiccedilatildeo de HCl ateacute um valor de pH proacuteximo de 4 observa-se uma mudanccedila de cor de amarelo
claro para laranja indicativa da formaccedilatildeo da espeacutecie decameacuterica de vanadato De seguida e
imediatamente antes da realizaccedilatildeo dos ensaios descritos o valor do pH da soluccedilatildeo de
decavanadato foi aumentado para cerca de 75 por adiccedilatildeo de NaOH A soluccedilatildeo (2) designada
por ldquometavanadatordquo natildeo foi posteriormente alterada ateacute agrave sua adiccedilatildeo para a realizaccedilatildeo dos ensaios
As soluccedilotildees foram armazenadas a 4ordmC e mantidas em gelo aquando da sua utilizaccedilatildeo
A soluccedilatildeo de V(IV) foi preparada em condiccedilotildees anaeroacutebias por diluiccedilatildeo de sulfato de vanadil
(VOSO4) para uma concentraccedilatildeo final de 5 mM em H2O MilliQ saturada com N2 gasoso
352Anaacutelise por espectroscopia de UV-Vis
A concentraccedilatildeo da espeacutecie decameacuterica em soluccedilatildeo depende do periacuteodo de tempo apoacutes a diluiccedilatildeo
no meio de reacccedilatildeo apropriado (Ref) A velocidade de desoligomerizaccedilatildeo da espeacutecie decameacuterica
pode ser analisada por espectroscopia de absorccedilatildeo na regiatildeo UVVis uma vez que esta espeacutecie
apresenta duas bandas caracteriacutesticas na regiatildeo do visiacutevel (a 358 e 400 nm) que natildeo se verifica
em quaisquer outras espeacutecies oligomeacutericas As bandas supramencionada sapresentam-se bem
22
Meio de reacccedilatildeo 3125 mM Tris-HCl pH 71 375 mM MgCl2
48
resolvidas atraveacutes da obtenccedilatildeo da derivada de 2ordf ordem do espectro apresentando a forma de 2
vales com miacutenimos centrados nos comprimentos de onda supracitados com dependecircncia linear
relativamente agrave concentraccedilatildeo de decavanadato total (Ramos et al 2006)
Assim a estabilidade da espeacutecie decameacuterica apoacutes diluiccedilatildeo da soluccedilatildeo de decavanadato no meio de
reacccedilatildeo dos ensaios (tampatildeo D) para uma concentraccedilatildeo final de 1 mM em vanadato total foi
caracterizada atraveacutes da leitura da Abs400 em funccedilatildeo do tempo tendo-se determinado qual a
ordem de reacccedilatildeo e o tempo de meia-vida da mesma Com base em estudos anteriores acerca da
estabilidade do V10 na presenccedila de diferentes proteiacutenas (Ramos et al 2006) determinou-se
igualmente o tempo de meia vida apoacutes a adiccedilatildeo de 20 microM de S1 de miosina ao meio reaccional
353 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
Os espectros de RMN do isoacutetopo 51
V foram realizados num espectroacutemetro Bruker AM-400 MHz
equipado com uma sonda multinuclear inversa de 5 mm Em tubos de RMN de 5 mm de diacircmetro
preparou-se soluccedilotildees de 500 L de deca- ou metavanadato correspondentes a um intervalo de
concentraccedilotildees de 1 a 10 mM suplementadas com 10 D2O Os espectros foram adquiridos agrave
temperatura ambiente com uma largura espectral de 23 923 Hz um tempo de aquisiccedilatildeo de 0086
s tempo de relaxaccedilatildeo de 001 s e uma sequecircncia de pulsos a 90ordm Utilizou-se a transformada de
Fourier com alargamento das bandas de 20Hz para o processamento dos espectros Os desvios
quiacutemicos das ressonacircncias de 51
V foram obtidos em relaccedilatildeo agrave espeacutecie VOCl3cuja frequecircncia de
ressonacircncia eacute considerada o padratildeo sendo todas as outras referenciadas a partir desta em ppm
As aacutereas dos sinais do RMN correspondentes agraves espeacutecies oligomeacutericas presentes foram obtidas
por integraccedilatildeo dos sinais As concentraccedilotildees relativas das espeacutecies foram calculadas com recurso agrave
equaccedilatildeo
onde A[Vx] corresponde agrave aacuterea referente agrave ressonacircncia da espeacutecie Vx com nordm de agregaccedilatildeo x
A[V]total agrave soma total das aacutereas medidas e [V] a concentraccedilatildeo total de vanaacutedio no ensaio
A teacutecnica de 51
V-RMN foi anteriormente aplicada em estudos de ligaccedilatildeo com S1 de miosina
(Ref) tendo sido demostrado que entre as espeacutecies V1 V2 V4 e V5 a espeacutecie tetrameacuterica possui a
maior afinidade para a miosina (Cremo et al) O desaparecimento ou alargamento dos sinais pode
implicar a ligaccedilatildeo de um dos oxovanadatos laacutebeis agrave miosina sem indicaccedilatildeo de qual uma vez que
49
se processa um equiliacutebrio entre eles (Crans 1990) Assim e tendo em conta as indicaccedilotildees de que
a espeacutecie V10 eacute distinta das restantes no que diz respeito agrave magnitude da interacccedilatildeo com a miosina
e inibiccedilatildeo da actividade ATPaacutesica estimulada por actina (Aureliano 2000 Tiago et al 2002)
torna-se fundamental investigar a interacccedilatildeo das diferentes espeacutecies de vanadato com o S1 de
miosina atraveacutes da teacutecnica de 51
V-RMN
Para tal obtiveram-se espectros de 51
V-RMN resultantes da titulaccedilatildeo de uma soluccedilatildeo 5 mM de
decavanadato total com concentraccedilotildees crescentes de S1 de miosina (0 5 10 15 e 20 microM) em
tampatildeo D Realizou-se ainda ensaios na presenccedila de 2 mM de decavanadato e 2 mM de ATP na
presenccedila e na ausecircncia de S1 de miosina de modo a verificar o efeito que estes ligandos naturais
da enzima teria na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o subfragmento-1
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
Os grupos sulfidrilo (-SH) das cisteiacutenas satildeo resiacuteduos de aminoaacutecidos extremamente sensiacuteveis agrave
oxidaccedilatildeo resultando na sua oxidaccedilatildeo (Dalle-Donne et al 1999) Logo a quantificaccedilatildeo destes
resiacuteduos eacute bastante importante nos estudos de interacccedilatildeo com vanaacutedio jaacute que os resiacuteduos de Cys
com ecircnfase para o SH1 e o SH2 satildeo bastante importantes na monitorizaccedilatildeo das modificaccedilotildees
funcionais da regiatildeo da cabeccedila
O procedimento seguidamente apresentado foi originalmente descrito por Gutieacuterrez-Merino
(1987)) Consiste na reacccedilatildeo entre os grupos ndashSH natildeo oxidados das proteiacutenas e o reagente de
Ellman (DTNB -55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico)) - formando-se o aniatildeo CNT (aniatildeo 3-
carboxilato-4-nitrotiofenolato) numa estequiometria de 11 Este apresenta um maacuteximo de
absorvecircncia a 412 nm (Robyt 1990) Assim apoacutes a incubaccedilatildeo da proteiacutena na presenccedila do metal
a quantidade de cisteiacutenas reduzidas por proteiacutena determina-se atraveacutes dos valores de Abs412
Deste modo colocou-se num tubo de Eppendorf o volume apropriado de S1 dialisado para que a
concentraccedilatildeo no ensaio fosse de 1 mgmL Adicionou-se o volume apropriado de decavanadato
para que a sua concentraccedilatildeo fosse de 0 10 50 100 250 500 ou 1000 M Perfez-se com soluccedilatildeo
D ateacute um volume final de 1 mL e incubou-se a 25ordmC por 20 minutos Apoacutes este periacuteodo colocou-
se 780 L da mistura reaccional numa cuvette de quartzo e adicionou-se 220 L de soluccedilatildeo K23
23
Tampatildeo K Tampatildeo fosfatohidrogenofosfato de soacutedio 025 M pH 72
50
Adicionou-se 15 L de DTNB (em soluccedilatildeo K [DTNB]stock=100 mM) ao meio reaccional e
registou-se o valor de absorvecircncia medido equivalente agraves cisteiacutenas ldquoraacutepidasrdquo (SH1 e SH2
expostos ao solvente) Adicionou-se 001 g de SDS que provoca a desnaturaccedilatildeo da proteiacutena e
consequentemente a exposiccedilatildeo de todos os resiacuteduos de cisteiacutenas que se encontravam ateacute aiacute
inacessiacuteveis no interior da proteiacutena (cisteiacutenas totais) Estes resiacuteduos podem assim reagir com o
DTNB
Estimou-se o nuacutemero de resiacuteduos de cisteiacutenas em todas as amostras com recurso ao coeficiente de
extinccedilatildeo molar () do DTNB (ver ponto 3 dos Anexos) que foi determinado atraveacutes da titulaccedilatildeo
de L-Cisteiacutena com intervalo de concentraccedilotildees compreendidos entre 0 e 50 microM na presenccedila de
25microM de DTNB em tampatildeo D Os valores de absorvecircncia foram obtidos para um comprimento de
onda de 412 nm
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
3621 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
A teacutecnica mais eficiente de anaacutelise da interacccedilatildeo entre o iatildeo vanadilo e proteiacutenas como a
calmodulina e a transferrina entre outras permite a obtenccedilatildeo raacutepida de informaccedilatildeo no que toca agrave
conformaccedilatildeo e amplitude da referida interacccedilatildeo
Os espectros de Ressonacircncia Paramagneacutetica Electroacutenica (RPE) da espeacutecie paramagneacutetica V(IV)
foram realizados a 70K num espectroacutemetro Bruker EMX-300 X-band Todas as soluccedilotildees
utilizadas e amostras foram preparadas em anaerobiose numa cacircmara de luvas de modo a evitar a
reduccedilatildeo de V(IV) a V(V) Em tubos capilares com 02 mL de amostra manteve-se a concentraccedilatildeo
de S1 a 20 M e variou-se a concentraccedilatildeo de vanadilo (0 25 5 10 15 25 50 75 100 150
200 500 e 1000 M [VOSO4]stock=5 mM) em tampatildeo D Apoacutes obtenccedilatildeo dos espectros estes
foram tratados atraveacutes do software SimFonia e WinEPR tendo-se obtido um graacutefico com a
variaccedilatildeo da intensidade do sinal de RPE em funccedilatildeo da concentraccedilatildeo de sulfato de vanadilo
presente Realizaram-se ainda simulaccedilotildees de espectros obtidos com 20 microM de S1 e 1000 microM de
V(IV) com vista agrave obtenccedilatildeo dos ligandos provaacuteveis do vanadilo atraveacutes da anaacutelise da constante
de acoplamento hiperfina (A) e do factor g (g)
4 Conclusotildees gerais
Apesar de jaacute ter sido demonstrado que o vanaacutedio inibe a Na+K
+-ATPase da miosina a funccedilatildeo
deste elemento no tecido muacutesculo esqueleacutetico continua largamente incompreendida A enorme
complexidade quiacutemica deste metal especialmente no que concerne agrave oligomerizaccedilatildeo e reacccedilotildees
51
redox constitui um obstaacuteculo agrave compreensatildeo do comportamento do vanaacutedio nos organismos
vivos Assim a caracterizaccedilatildeo da quiacutemica deste elemento eacute de grande utilidade antes de iniciar o
estudo da sua interacccedilatildeo com o presente sistema
A anaacutelise por espectroscopias de 51
V-RMN e de UV-Vis de soluccedilotildees de vanadato diluiacutedas nos
meios de reacccedilatildeo utilizados no presente relatoacuterio (10 mM HEPES pH 70 25 mM KCl) mostrou
claramente que a espeacutecie decameacuterica (V10) possui um tempo de vida suficientemente longo para a
determinaccedilatildeo do seu efeito isolado ao contraacuterio do que se verifica com as espeacutecies mono- di-
tetra- e pentameacutericas (V1 V2 V4 e V5) Por outro lado a anaacutelise verificou ainda que apoacutes diluiccedilatildeo
das soluccedilotildees de decavanadato no meio de reacccedilatildeo coexistem as espeacutecies V10 e V1 pelo que os
efeitos putativos desta uacuteltima deveratildeo ser subtraiacutedos aquando da monitorizaccedilatildeo dos efeitos da
espeacutecie decameacuterica
O vanaacutedio (V) (vanadato) inibe as enzimas que catalizam reacccedilotildees de transferecircncia de grupos
fosfato como a ATPase da miosina [Goodno 1979] devido agrave sua analogia estrutural com o iatildeo
fosfato Na sua utilizaccedilatildeo como inibidor enzimaacutetico natildeo eacute tida em conta a contribuiccedilatildeo individual
das espeacutecies oligomeacutericas considerando-se normalmente a espeacutecie V1 como inibidora efectiva
[Goodno e Taylor 1982] No presente relatoacuterio verificou-se por espectroscopia de 51
V-RMN que
outros oligoacutemeros de vanadato interagem com o S1 de miosina tendo as espeacutecies V10 e V4 os
maiores niacuteveis de afinidade A sua interacccedilatildeo envolve forccedilas electrostaacuteticas e eacute afectada
negativamente por ligandos naturais de enzima como eacute o caso do ATP
A exposiccedilatildeo do S1 de miosina (005 mgmL) a decavanadato conduz agrave inibiccedilatildeo do capacidade da
proteiacutena de estimular a actividade Mg2+
-ATPaacutesica com um valor de IC50 de 266 plusmn 07 M
decavanadato sendo o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM
decavanadato (80) Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM decavanadato sendo
o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM decavanadato (60)
exercendo um efeito de impedimento agrave ligaccedilatildeo entre o vanaacutedio e o S1
A uma concentraccedilatildeo de 27 microM decavanadato verifica-se a oxidaccedilatildeo de um dos resiacuteduos de Cys
de S1 de miosina seguida pela oxidaccedilatildeo gradual de outro resiacuteduo de Cys ateacute agrave concentraccedilatildeo de
1000 microM Ambos os resiacuteduos oxidados natildeo se encontravam expostos ao solvente utilizado A
oxidaccedilatildeo dos resiacuteduos permite sugerir que esta modificaccedilatildeo oxidativa sofrida pelo S1 contribui
para a diminuiccedilatildeo da sua actividade de ATPase
52
Por espectroscopia de RPE revelaram-se os possiacuteveis ligandos equatoriais do vanaacutedio aquando da
sua interacccedilatildeo com o S1 de miosina como sendo duas moleacuteculas de aacutegua um ligando contendo o
grupo R-OH e outro contendo o grupo R=NR
Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados de
oxidaccedilatildeo (IV) e (V) no S1 de miosina para concentraccedilotildees facilmente alcanccedilaacuteveis no ambiente
terrestre
53
5 Estudos futuros
No sentido do melhor esclarecimento das alteraccedilotildees moleculares e estruturais que ocorrem no
subfragmento 1 quando exposta ao vanaacutedio que conduzem a uma diminuiccedilatildeo da sua funccedilatildeo
sugere-se
- Quantificaccedilatildeo das cisteiacutenas reduzidas de S1 na presenccedila de metavanadato
- Determinaccedilatildeo das actividades Ca2+
- e K+-ATPaacutesica do S1 de miosina na presenccedila de
decavanadato
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina estimulada por actina
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina na presenccedila de
metavanadato
- Realizaccedilatildeo de RMN 2D com variaccedilatildeo das concentraccedilotildees das soluccedilotildees de decavanadato e
metavanadato na presenccedila de S1
-Mutageacutenese dos resiacuteduos de Cys doS1 de miosina de modo a verificar quais os resiacuteduos que
sofrem oxidaccedilatildeo na presenccedila de decavanadato
- Determinaccedilatildeo por calorimetria (DSC Differential Scanning Calorimetry) da variaccedilatildeo da
entalpia (∆H) e da temperatura de desnaturaccedilatildeo do S1 de miosina na presenccedila de vanaacutedio uma
vez que estas propriedades caracteriacutesticas de cada proteiacutena satildeo alteradas apoacutes ocorrecircncia de
modificaccedilotildees estruturais
- Anaacutelise por SDS-PAGE de S1 de miosina apoacutes tratamento com vanaacutedio de modo a determinar
quais os polipeacuteptidos eventualmente formados apoacutes clivagem da proteiacutena
54
6 Referecircncias bibliograacuteficas
- Sellers J Myosins 2nd Edition Oxford University Press 1999
- L Stryer inBiochemistry (Ed WH Freeman 2005 pp
Refordfas Materiais e Meacutetodos
[A] S S Margossian SLowey Methods Enzymol1982 85 55-71
[B] JD Pardue JA Spudich Methods Enzymol 1982 85 164-181
[C] M Bradford Anal Biochem1976 72 248-254
[D] H H Taussky E Shorr J Biol Chem 1953 202 675-685
[E] C R Bagshaw in Outline Studies in Biology ndash Muscle Contraction Chapman and Hall Ed
1982
[F] A Cuenda et al Eur J Biochem1990 194 663-670
55
Anexos
1 Determinaccedilatildeo da recta de calibraccedilatildeo meacutetodo de Bradford
Figura1ndashRepresentaccedilatildeo graacutefica da recta de calibraccedilatildeo (Abs595 nm vs [BSA]) utilizada para o meacutetodo de Bradford
2 Determinaccedilatildeo da recta calibraccedilatildeo para a electroforese em condiccedilotildees desnaturantes
Figura 2 ndash Representaccedilatildeo graacutefica da recta de calibraccedilatildeo (log(Peso Molecular) vs Rf dos marcadores de peso
molecular) onde Rf indica determinada atraveacutes do padratildeo de peso molecular
(poccedilo (a))
3Caacutelculo do coeficiente de extinccedilatildeo molar () para a quantificaccedilatildeo de cisteiacutenas reduzidas
atraveacutes do meacutetodo de Ellman
Traccedilou-se a recta de Abs412 nm meacutedia vs [L-Cisteiacutena] de modo a determinar o valor do coeficiente
de extinccedilatildeo molar definido como o declive daquela recta Assim =8400 M-1
cm-1
como
podemos verificar na Fig 3
A595 nm = 00546[BSA] + 00209 Rsup2 = 099051
Ab
s 59
5 n
m
[BSA] (mgmL)
log PM = -1177 Rf + 2247 Rsup2 = 098957
log
PM
Mobilidade relativa (Rf)
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero

35
resulta num espectro de RPE consistindo de 8 desdobramentos na direcccedilatildeo paralela e outros tantos
na direcccedilatildeo perpendicular (Chasteen 1983)
Os estudos realizados por Ahmed et al (1987) centraram-se na interacccedilatildeo do vanadilo com
proteiacutenas como eacute o caso da insulina transferrina entre outros obtendo um espectro caracterizado
por 8 linhas amplamente espaccediladas (na direcccedilatildeo paralela) Jaacute Ramos et al 2009 concluiu que a
exposiccedilatildeo da soluccedilatildeo de decavanadato agraves formas globular e filamentosa de actina produz um
espectro tiacutepico do iatildeo oxidovanaacutedio (IV) indicando a reduccedilatildeo do vanadato
Para aferir qual a concentraccedilatildeo de S1 que permitia obter uma boa razatildeo sinalruiacutedo titulou-se uma
soluccedilatildeo de sulfato de vanadilo 15 microM ([VOSO4]stock = 5 mM) com concentraccedilotildees crescentes de
proteiacutena (0 5 10 15 e 20 microM) em 10 mM HEPES 25 mM KCl A existecircncia de uma correlaccedilatildeo
linear entre a concentraccedilatildeo de proteiacutena e a intensidade dos sinais de RPE correspondente agrave banda
-12 perpendicular encontra-se representada na Fig 210
Figura 2 12ndash Representaccedilatildeo graacutefica da variaccedilatildeo da intensidade do sinal de RPE vs a concentraccedilatildeo de S1 utilizada
Apoacutes verificar que com uma concentraccedilatildeo de S1 igual a 20 microM se obtinham sinais com uma razatildeo
sinalruiacutedo adequada ao prosseguimento dos estudos sobre a interacccedilatildeo V(IV)-S1 de miosina
titulou-se uma soluccedilatildeo de 20 microM S1 com 0 25 5 10 15 25 50 75 100 150 200 500 e 1000
microM VOSO4
Traccedilou-se o graacutefico da intensidade da banda -12 perpendicular do sinal de EPR em funccedilatildeo da
concentraccedilatildeo de VOSO4 tendo-se determinado uma constante de dissociaccedilatildeo (KD) do complexo
S1-vanadilo de 8748 plusmn 2621 microM V(IV)
y = 016786x + 122435
Rsup2 = 092992759
Inte
nsi
dad
e d
o s
inal
de
RP
E
[S1] (microM)
36
Figura 213 ndash Representaccedilatildeo graacutefica da variaccedilatildeo do sinal de RPE vs a concentraccedilatildeo de vanadilo utilizada (os valores
experimentais estatildeo representados por losangos o seu ajuste estaacute representado pela linha)
A simulaccedilatildeo do espectro de EPR foi efectuada a partir do espectro obtido para uma concentraccedilatildeo
de 1 mM VOSO4 apoacutes adiccedilatildeo de 20 microM S1 de miosina tendo-se obtido os valores da constante de
acoplamento hiperfina (A=5016) e do factor g (g0=195)
Figura 214 ndash Representaccedilatildeo graacutefica do espectro de EPR obtido com 1000 microM de vanadilo e 20 microM de S1 (a cinzento
escuro) A simulaccedilatildeo do espectro encontra-se a cinzento claro
Por comparaccedilatildeo destes com os valores presentes na tabela de correspondecircncia xxx eacute possiacutevel
predizer quais os ligandos provaacuteveis do vanadilo na presenccedila de proteiacutena Assim concluiu-se que
o vanadilo estaacute coordenado com duas moleacuteculas de aacutegua um grupo hidroxilo e um grupo R=NR
O grupo hidroxilo pode indicar os resiacuteduos de aminoaacutecidos Tyr Treonina e Serina enquanto o
grupo R=NR poderaacute indicar a presenccedila de Arginina e Histidina como ligandos do vanadilo
Inte
nsi
dad
e d
os
sinai
s d
e R
PE
[Vanadilo] (microM)
Ajuste dos dados exp Dados experimentais
Campo magneacutetico (G)
Espectro Experimental Ajuste
37
3 Procedimento Experimental
31 Reagentes
Todos os reagentes utilizados satildeo de elevado grau de pureza (analiacutetico) Apresentam-se as suas
foacutermulas quiacutemicas ou abreviaturas quando necessaacuterio acrescentou-se a sua descriccedilatildeo Entre
parecircntesis colocou-se as empresas que os comercializam
Aacutecido aceacutetico glacial 100 (Riedel) Acrilamida 99 (Merck) Azul de bromofenol (Riedel-de-
Haumlen) -Mercaptoetanol (Sigma) Bissulfito de soacutedio (Sigma)Bradford (reagente de)
(BioRad)BSA (albumina de soro bovino Sigma-Aldrich) CaCl22H2O (cloreto de caacutelcio
dihidratado Fluka) CH3CH2OH 96 (etanol Panreac) Coomassie Blue R250 (Merck) D2O
(oacutexido de deuteacuterio SDS) DTNB (55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico) Aldrich) EDTA (aacutecido
etilenodiaminatetraaceacutetico Merck) Fosfoenolpiruvato 98 (Fluka) Glicerol (Sigma) Glicina
985 (Panreac) HCl 37 (Riedel-de Haumlen) H2SO4 96 (Pronalab) HEPES 99 (aacutecido 4-(2-
hidroxiletil)-1-piperazineetanosulfoacutenico Sigma) KH2PO4 99 (hidrogenofosfato de potaacutessio)
MgATP (Sigma) Na
ATP (Sigma) Na2HPO4 99 (hidrogenofosfato de soacutedio Riedel-de Haumlen)
NaH2PO4 (fosfato de soacutedio ) NaOH 986 (hidroacutexido de soacutedio Pronalab) KCl 99 (Cloreto de
potaacutessio Panreac) L-Cys 995 (enantioacutemero levoacutegiro do resiacuteduo de Cisteiacutena Fluka)Metanol
99 (Merck) Molibdato de amoacutenio (Sigma)NNrsquo-metil-bis-acrilamida 98 (Merck) NH4VO3
99 (metavanadato de amoacutenia Sigma) PSA 10 (persulfato de amoacutenio Merck) SDS
(dodecilo sulfato de soacutedio Sigma) Sulfito de soacutedio (Sigma) VOSO4 (sulfato de vanadilo
Aldrich) TEMED 99 (Tetrametiletilenodiamina Merck) Tris (2-amino-2-hidroximetil-13-
propanodiol Sigma)
32 Meacutetodos
321 Isolamento e purificaccedilatildeo das proteiacutenas de muacutesculo esqueleacutetico de coelho
3211 Purificaccedilatildeo da miosina preparaccedilatildeo de Subfragmento 1 por digestatildeo quimotriacutepsica
da miosina
O subfragmento-1 de miosina-II (S1) foi gentilmente cedido pelo Prof Doutor Manuel Aureliano
do Departamento de Quiacutemica Bioquiacutemica e Farmaacutecia da Faculdade de Ciecircncias e Tecnologia da
Universidade do Algarve
No entanto descreve-se abaixo de forma resumida o procedimento efectuado
38
32111 Purificaccedilatildeo da miosina
O protocolo de isolamento foi adaptado do meacutetodo previamente descrito por Margossian e Lowey
(1982) apresentando-se de forma esquemaacutetica na Fig 31
Sucintamente os tecidos musculares dorsais e das pernas foram extraiacutedos apoacutes o sacrifiacutecio do
animal (Fig31) De seguida foram adicionados 3 volumes da soluccedilatildeo de extracccedilatildeo A2
Centrifugou-se a soluccedilatildeo e filtrou-se o sobrenadante O sedimento foi entatildeo diluiacutedo em 14
volumes de aacutegua destilada de modo a que a forccedila ioacutenica atingisse 004 M Apoacutes adiccedilatildeo de szlig-
mercaptoetanol (szlig-me) deixou-se a precipitar a 4ordmC removendo o sobrenadante Centrifugou-se
novamente e ressuspendeu-se o sedimento na soluccedilatildeo de extracccedilatildeo A Adicionou-se a soluccedilatildeo
tampatildeo B3 para que a forccedila ioacutenica atingisse 05 M Homogeneizou-se e deixou-se repousar
durante 2h Homogeneizou-se novamente e centrifugou-se de modo a ocorrer precipitaccedilatildeo dos
complexos de actomiosina e proteiacutena desnaturada Apoacutes filtraccedilatildeo do sobrenadante em latilde de vidro
quantificou-se a soluccedilatildeo de miosina resultante pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental)
2 Soluccedilatildeo de extracccedilatildeo A 0012M K2CO3 004M KHCO3 06M KCl pH 65
3 Soluccedilatildeo-tampatildeo B Tris 100 mM pH 7 KCl 2M
39
Figura 31 ndash Representaccedilatildeo esquemaacutetica do protocolo de isolamento de miosina (Margossian e Lowey 1982) Todo o
procedimento foi realizado a 4ordmC (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
32112 Preparaccedilatildeo de Subfragmento-1 de miosina por digestatildeo com quimiotripsina
O S1 de miosina foi isolado apoacutes diaacutelise da miosina durante a noite contra a soluccedilatildeo C4 A
presenccedila do EDTA eacute essencial uma vez que a sua substituiccedilatildeo por MgCl2resulta na produccedilatildeo de
HMM em vez de S1 A suspensatildeo de miosina foi equilibrada e digerida com -quimotripsina
A digestatildeo foi interrompida pela adiccedilatildeo de PMSF Seguidamente ultracentrifugou-se o produto
da digestatildeo desprezando-se o sedimento (Fig 32) A soluccedilatildeo de sobrenadante maioritariamente
S1 foi quantificada pela leitura directa da Abs280 ou pelo meacutetodo de Bradford (ver as secccedilotildees
3311 e 3312 do Procedimento Experimental respectivamente) e armazenada em 10 sacarose
e 2 mM szlig-Me a -80ordmC
4 Soluccedilatildeo C 25mM Tris pH 70 012M KCl 1mM EDTA
40
Antes da realizaccedilatildeo dos ensaios descritos no presente relatoacuterio o S1 de miosina foi dialisado
contra a soluccedilatildeo D5 de modo a remover qualquer composto que pudesse complexar como
vanaacutedio Seguidamente quantificou-se a proteiacutena pelo meacutetodo de Bradford ou pelo meacutetodo
directo (ver secccedilotildees 3311 e 3312do Procedimento Experimtental)
Figura 3 2ndash Representaccedilatildeo esquemaacutetica do protocolo de preparaccedilatildeo do S1 de miosina por digestatildeo quimiotripsina de
acordo com o previamente descrito por Margossian e Lowey (1982) Todo o procedimento foi realizado a 4ordm C
excepto quando indicaccedilatildeo do contraacuterio (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
3212 Isolamento e purificaccedilatildeo de actina
Isolou-se e purificou-se a actina segundo o meacutetodo descrito por Pardee e Spudish que
compreende dois passos ndash a obtenccedilatildeo de um resiacuteduo seco denominado ldquopoacute de acetonardquo e a
extracccedilatildeo subsequente da actina a partir do ldquopoacute de acetonardquo na forma de G-Actina com a
posterior polimerizaccedilatildeo a F-Actina
32121 Obtenccedilatildeo do ldquopoacute de acetonardquo
Sacrificou-se o coelho com uma pancada atraacutes do pescoccedilo e sangrou-se o animal apoacutes o corte da
arteacuteria jugular Esfolou-se e colocou-se numa tina com gelo Retiraram-se as partes de tecido
conjuntivo que envolvem o tecido muscular com auxiacutelio de uma pinccedila e bisturi Seguidamente
retirou-se os muacutesculos dorsais e os muacutesculos brancos das pernas Passou-se os muacutesculos por aacutegua
destilada de modo a remover as impurezas e pesou-se num copo Trituraram-se os muacutesculos
numa maacutequina de triturar carne e extraiu-se o resultante com 3 volumes da soluccedilatildeo E6 durante 10
minutos com agitaccedilatildeo suave Centrifugou-se a suspensatildeo a 4000 g durante 10
5 Soluccedilatildeo D HEPES 10 mM pH 70 KCl 25 mM
6 Soluccedilatildeo E 01 M KCl 015 M K2HPO4 pH 65
41
minutosdesprezando-se o sobrenadante Ressuspendeu-se o sedimento em 6 volumes de soluccedilatildeo
F7durante 10 minutos com agitaccedilatildeo suave Verificou-se se o pH da soluccedilatildeo estava compreendido
entre 82 e 85 acertando-se estecom soluccedilatildeo G8 se necessaacuterio Centrifugou-se durante 10
minutos a 4000 g Desprezou-se o sobrenadante e ressuspendeu-se o sedimento em 3 volumes de
soluccedilatildeo H9 durante 10 minutos Centrifugou-se novamente a 4000 gdurante 10 minutos
desprezando-se o sobrenadante Ressuspendeu-se o sedimento duas vezes consecutivas em 6
volumes de aacutegua destilada durante 5 minutos Centrifugou-se a 4000 g durante 10 minutos apoacutes
cada adiccedilatildeo de H2O
Seguidamente ressuspendeu-se o sedimento 5 vezes consecutivas utilizando 3 volumes de
acetona de cada vez agitando suavemente durante 10 minutos agrave temperatura ambiente Colocou-
se o resiacuteduo filtrado numa tina de vidro sobre papel de filtro deixando-se evaporar o solvente
durante a noite num nicho Triturou-se o resiacuteduo seco numa maacutequina de triturar carne e deixou-se
novamente evaporar o solvente O ldquopoacute de acetonardquo obtido foi armazenado a -20 ordmC
32122 Purificaccedilatildeo da actina a partir de ldquopoacute de acetonardquo
Apoacutes a obtenccedilatildeo do poacute de acetona como acima indicado efectuou-se o protocolo seguidamente
descrito de modo a obter monoacutemeros de actina (G-Actina) e actina filamentosa (F-Actina)
Pesou-se 35 g de poacute de acetona e adicionaram-se 50 mL de soluccedilatildeo-tampatildeo I10
sendo a
extracccedilatildeo feita com agitaccedilatildeo suave durante 30 minutos Separou-se o extracto do poacute de acetona
hidratado filtrando a suspensatildeo com 4 camadas de gaze esterilizada tendo o cuidado de natildeo
espremer o resiacuteduo Voltou a extrair-se com 50 mL de soluccedilatildeo I durante 30 minutos mexendo
lentamente Adicionou-se este ao extracto obtido anteriormente Centrifugou-se o extracto total a
16 000 g durante 30 minutos Decantou-se para uma proveta sobre a qual se colocou gaze para
que o sedimento solto natildeo se juntasse ao sobrenadante e leu-se o volume De seguida adicionou-
se KCl ateacute uma concentraccedilatildeo final de 50 mM MgCl2 ateacute 2mM e ATP ateacute 1mM lentamente e com
agitaccedilatildeo contiacutenua de modo a promover a polimerizaccedilatildeo dos monoacutemeros de actina
Deixou-se polimerizar durante 2h a 25ordmC observando-se a presenccedila de pequenas bolhas de ar
indicativas de um aumento de viscosidade da soluccedilatildeo Arrefeceu-se em gelo durante 15 minutos e
adicionou-se KCl ateacute uma concentraccedilatildeo final de 06M Mexeu-se lentamente durante 30 minutos
apoacutes os quais se ultracentrifugou a 110 000g durante 1hora Desprezou-se o sobrenadante
ressuspendeu-se o sedimento com soluccedilatildeo tampatildeo E e transferiu-se para um potter
Dialisou-se durante a noite contra 1L de soluccedilatildeo-tampatildeo I a uma temperatura de 4ordmC com rotaccedilatildeo
7 Soluccedilatildeo F 005 M NaHCO3 pH 82-85
8 Soluccedilatildeo G 1M Na2CO3
9 Soluccedilatildeo H 1 mM EDTA pH 70
10 Soluccedilatildeo I 2 mM Tris-HCl pH 8 02 mM CaCl2 02 mM Na2ATP 05 mM szlig-Me 0005 NaN3
42
contiacutenua e mudanccedila de tampatildeo 2 a 3 vezes De seguida a actina foi centrifugada a 110
000gdurante 1hora Decantou-se a soluccedilatildeo de G-Actina (sobrenadante) e desprezou-se o
sedimento Determinou-se a concentraccedilatildeo de G-Actina pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental) A soluccedilatildeo de G-actina foi armazenada a uma temperatura de 4ordmC
apoacutes adiccedilatildeo de 002NaN3 mantendo-se estaacutevel durante 3 a 5 dias
331 Quantificaccedilatildeo das proteiacutenas isoladas
3311 Quantificaccedilatildeo pelo meacutetodo de Bradford
O meacutetodo de Bradford (Bradford 1976) baseia-se na utilizaccedilatildeo do reagente de Bradford
composto por entre outros aacutecido fosfoacuterico e o corante ldquoCoomassierdquo Este corante possui carga
negativa e a sua coloraccedilatildeo varia entre a vermelha (Absmax=465 nm) predominante em soluccedilatildeo e a
azul (Absmax=595 nm) presente aquando da ligaccedilatildeo aos grupos amina das proteiacutenas Em soluccedilotildees
aacutecidas diluiacutedas a coloraccedilatildeo varia proporcionalmente com a quantidade do corante ligado agrave
proteiacutena que por sua vez eacute directamente proporcional agrave concentraccedilatildeo da proteiacutena permitindo a
sua quantificaccedilatildeo ateacute a um limite de detecccedilatildeo de 20 gmL (Bradford 1976)
Deste modo prepararam-se amostras de BSA ([BSA]stock= 1mgmL) utilizando um intervalo de
concentraccedilotildees compreendido entre 0 e 15 gmL de acordo com a tabela 3I Apoacutes a adiccedilatildeo da
H2O MilliQ e do reagente de Bradford as soluccedilotildees foram sucessivamente homogeneizadas no
voacutertex
Tabela 3Indash Amostras utilizadas para a obtenccedilatildeo da recta de calibraccedilatildeo no meacutetodo de Bradford
BSA (L) BSA (g) H2O MilliQ (L) Reagente de Bradford (L)
0 0 800 200
125 125 79875 200
25 25 7975 200
5 5 795 200
10 10 790 200
15 15 785 200
Apoacutes incubaccedilatildeo a 25ordmC por 5 minutos leram-se os valores de Abs595 num
espectrofotoacutemetro (Shimadzu UV-160A) para todas as amostras Seguidamente traccedilou-se a recta
de calibraccedilatildeo (Abs595vs quantidade de BSA (g))
43
Para determinar a concentraccedilatildeo de proteiacutena apoacutes o processo de isolamento purificaccedilatildeo e
diaacutelise quando necessaacuterio colocou-se 5 L da amostra num tubo de Eppendorf e adicionou-se
795 L de H2O MilliQ homogeneizando a soluccedilatildeo no voacutertex Adicionou-se posteriormente 200
L de reagente de Bradford e repetiu-se a homogeneizaccedilatildeo Incubou-se a mistura a 25ordmC por 5
minutos apoacutes os quais se leu os valores de Abs595 nm no espectrofotoacutemetro Por interpolaccedilatildeo deste
valor a partir da recta de calibraccedilatildeo determinou-se a concentraccedilatildeo de proteiacutena na amostra
3312 Quantificaccedilatildeo pelo meacutetodo directo
A quantificaccedilatildeo das amostras proteicas pode tambeacutem ser efectuada atraveacutes da leitura directa da
absorvacircncia tendo por base a lei de Beer-Lambert onde Abs corresponde agrave
absorvacircncia ao comprimento de onda ao qual o ensaio foi realizado corresponde ao coeficiente
de extinccedilatildeo (substituiacutedo pela letra a se as unidades forem distintas da molaridade) b corresponde
ao percurso oacuteptico da radiaccedilatildeo na ceacutelula amostral e c corresponde agrave concentraccedilatildeo amostral
Assim no caso de S1 de miosina mediu-se a Abs280e calculou-se a concentraccedilatildeo da proteiacutena
sabendo que o coeficiente de extinccedilatildeo de S1 eacute (Ref)
No caso da actina mediu-se a Abs290ecalculou-se a concentraccedilatildeo da proteiacutena sabendo que o
coeficiente de extinccedilatildeo da actina eacute (Ref)
332Electroforese em condiccedilotildees desnaturantes (SDS-PAGE)
De modo a melhor caracterizar as proteiacutenas isoladas foi realizada uma electroforese em
condiccedilotildees desnaturantes (SDS-PAGE do inglecircs Sodium Dodecyl Sulfate Polyacrilamide Gel
Electrophoresis) O SDS (dodecilo sulfato de soacutedio) eacute um detergente anioacutenico utilizado neste
meacutetodo para desnaturar as proteiacutenas que se ligam ao SDS na proporccedilatildeo 14 g SDS g proteiacutena
(aproximadamente 1 moleacutecula de SDS por cada 2 resiacuteduos de aminoaacutecidos) (Stryer) O SDS induz
uma densidade de carga negativa significativa nas proteiacutenas a que se liga mascarando a sua carga
intriacutenseca pelo que as razotildees cargamassa satildeo tendencialmente idecircnticas Assim a SDS-PAGE
separa as proteiacutenas quando sujeitas a um campo eleacutectrico de acordo com o seu peso molecular
sendo a sua migraccedilatildeo directamente proporcional ao logaritmo deste uacuteltimo (Stryer)
O gel de resoluccedilatildeo com 10 de acrilamida foi obtido atraveacutes da adiccedilatildeo de 167 mL de soluccedilatildeo
stock de acrilamida11
075 mL de tampatildeo de resoluccedilatildeo12
251 mL de aacutegua destilada e 50 L de
SDS 10 Para promover a polimerizaccedilatildeo adicionou-se em seguida 38 L de PSA 10
11
soluccedilatildeo stock de acrilamida soluccedilatildeo de 30 (mv) de acrilamida e 08 (mv) de NNrsquo-metil-bis-acrilamida 12
tampatildeo de resoluccedilatildeo 15 M Tris-HCl pH 88
44
(persulfato de amoacutenia) 10 e 25 L de TEMED 99 (tetrametiletilenodiamina) Agitou-se
cuidadosamente de modo a natildeo formar bolhas e pipetou-se a soluccedilatildeo para a cassette de vidro
previamente montada no suporte De seguida adicionou-se uma peliacutecula de cerca de 2 mm de 2-
metil-2-butanol para verificar posteriormente a polimerizaccedilatildeo Apoacutes a polimerizaccedilatildeo que demora
cerca de 45 minutos verteu-se a agrave superfiacutecie e adicionou-se o gel de concentraccedilatildeo com 5 de
acrilamida constituiacutedo por 03 mL de soluccedilatildeo de stock acrilamida 045 mL de tampatildeo de
concentraccedilatildeo13
094 mL de aacutegua destilada 18 L de SDS 10 135 L de PSA 10 e 2L de
TEMED 99 Colocou-se o pente sobre o gel e deixou-se polimerizar durante cerca de 45
minutos
As amostras para aplicaccedilatildeo no gel de electroforese foram preparadas para uma concentraccedilatildeo de
proteiacutena de 1 mgmL num volume final de 100 L dos quais 25 L correspondem ao tampatildeo da
amostra14
As amostras foram aquecidas a 100ordmC durante 5 minutos Antes da aplicaccedilatildeo das
amostras transferiu-se a cassette para a tina de electroforese agrave qual se adicionou a soluccedilatildeo
tampatildeo de electroforese da tina superior15
e tampatildeo de electroforese da tina inferior16
Aplicaram-
se 10 L das soluccedilotildees de marcadores de peso molecular e 20 L das amostras proteicas
preparadas anteriormente A electroforese decorreu durante cerca de 3h30 min a uma voltagem
de 80 V De seguida transferiu-se o gel para uma tina corando as proteiacutenas durante 1h com
soluccedilatildeo corante17
Descorou-se o gel com soluccedilatildeo descorante18
tendo-se mudado esta soluccedilatildeo ateacute
as bandas se encontrarem bem definidas
Determinou-se a mobilidade relativa das bandas correspondentes aos marcadores de elevado peso
molecular e traccedilou-se a recta de calibraccedilatildeo ndash log(PM) vs log(Rf) onde Rf corresponde agrave
mobilidade relativa Por interpolaccedilatildeo determina-se o PM das proteiacutenas isoladas sabendo a sua
mobilidade relativa Por comparaccedilatildeo com os valores descritos na literatura pode concluir-se
acerca do grau de purificaccedilatildeo
34 Mediccedilatildeo da actividade enzimaacutetica de S1
341 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
13
tampatildeo de concentraccedilatildeo 05 M Tris-HCl pH 68 14
tampatildeo de amostra 320 mM Tris-HCl pH 68 04 M szlig-mercaptoetanol SDS 8 glicerol 15 (vv) e roxo de m-
cresol 0024 15
tampatildeo de electroforese da tina superior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS 16
tampatildeo de electroforese da tina inferior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS acetato de soacutedio
01 M 17
soluccedilatildeo corante ldquoCoomassie Blue R-250rdquo contendo metanol 45 (vv) e aacutecido aceacutetico glacial 10 (vv) 18
soluccedilatildeo descorante metanol 10 (vv) e aacutecido aceacutetico glacial 10 vv)
45
O meacutetodo de Fiske-Subarow modificado consiste na reacccedilatildeo do molibdato de amoacutenio com o
fosfato inorgacircnico em soluccedilatildeo aacutecida com formaccedilatildeo do aacutecido fosfomoliacutebdico Seguidamente daacute-
se a adiccedilatildeo de um agente redutor que provoca a reduccedilatildeo do composto a fosfomolibdato que forma
um complexo de cor azulada cuja absorvacircncia maacutexima ocorre para um comprimento de onda de
660 nm tornando possiacutevel a sua detecccedilatildeo por meacutetodos espectrofotomeacutetricos As actividades Ca2+
-
ATPaacutesica e K+EDTA-ATPaacutesica de S1 foram determinadas atraveacutes deste meacutetodo de acordo com
o descrito por Taussky e Shorr (1953) com mediccedilatildeo da quantidade de fosfato (Pi) libertado
atraveacutes da hidroacutelise de ATP
Para tal obteve-se primeiramente a recta de calibraccedilatildeo com as amostras presentes na Tabela 3II
utilizando uma soluccedilatildeo stock de 5 mM KH2PO4
Tabela 3II ndash Composiccedilatildeo das diferentes amostras utilizadas para determinar a recta de calibraccedilatildeo atraveacutes do meacutetodo
de Fiske-Subbarow modificado
KH2PO4 (L)
([KH2PO4]stock= 5mM)
PO43-
(nmole) H2O (uL)
0 0 1780
5 25 1775
10 50 1770
20 100 1760
40 200 1740
100 500 1680
A cada tubo de Eppendorf adicionaram-se seguidamente 200 L de soluccedilatildeo J19
e 20 L de agente
redutor20
homogeneizando no voacutertex apoacutes cada adiccedilatildeo Incubou-se durante 10 minutos e mediu-
se a Abs660 nm num espectrofotoacutemetro com intervalos de tempo iguais entre cada amostra Traccedilou-
se a recta de calibraccedilatildeo (Abs660 nmvs nmol PO43-
) a partir dos valores obtidos
Para determinar a actividade Ca2+
-ATPaacutesica de S1 prepararam-se tubos de Eppendorf com 400
L de meio de reacccedilatildeo21
25 L CaCl2 ([CaCl2]stock= 100 mM) 25 L ATP ([ATP]stock= 100 mM)
com [S1]ensaios=01 mgmL Perfez-se se necessaacuterio com H2O MilliQ ateacute perfazer um volume de 1
mL Para determinar a actividade K+EDTA-ATPaacutesica utilizou-se uma concentraccedilatildeo idecircntica de
19
Soluccedilatildeo J 5 N H2SO4 25 molibdato de amoacutenio 20
Agente redutor 5 mg aacutecido 1-amino-2-naftol-4-sulfoacutenico 30 mg bissulfito de soacutedio 30 mg sulfito de soacutedio em 26
mL H2O 21
meio de reacccedilatildeo 125 mM Tris-HCl pH 80 0575 M KCl
46
proteiacutena nos ensaios (01 mgmL) e o mesmo meio de reacccedilatildeo Aos tubos de Eppendorf
adicionou-se ainda 123 L de KCl ([KCl]stock = 3M) 25 L de ATP 10 L de EDTA
([EDTA]stock= 100 mM )e o volume de H2O MilliQ necessaacuterio para perfazer 1 mL Em ambas as
situaccedilotildees o uacuteltimo reagente adicionado foi o ATP uma vez que se trata do substrato da reacccedilatildeo
enzimaacutetica
A cineacutetica decorreu a 25ordmC durante 5 minutos para a actividade Ca2+
-ATPaacutesica e durante 3
minutos para a actividade K+EDTA-ATPaacutesica Apoacutes o periacuteodo de incubaccedilatildeo transferiu-se 150
L de amostra para tubos de Eppendorf contendo igual volume de TCA 10 Colocou-se no gelo
e homogeneizou-se no voacutertex Seguidamente centrifugou-se a 1500 g durante 10 minutos e
recolheu-se 200 L de sobrenadante para novos tubos de Eppendorf Adicionou-se a estes 158
mL de H2O MilliQ e 200 L de soluccedilatildeo J Homogeneizou-se no voacutertex e adicionou-se 20 L de
reagente redutor repetindo-se a homogeneizaccedilatildeo Incubou-se durante 10 minutos e mediu-se a
Abs660 nm com o mesmo intervalo de tempo de leitura para cada amostra Por intrapolaccedilatildeo a partir
da recta de calibraccedilatildeo obtida anteriormente determinou-se a quantidade de Pi libertado em cada
ensaio A partir destes resultados determinaram-se as actividades Ca2+
-ATPaacutesica e K+EDTA-
ATPaacutesica do S1 em nmol PO43-
minmg proteiacutena e em s-1
342 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
Da actividade Mg2+
-ATPaacutesica de S1 resulta a formaccedilatildeo dos produtos da reacccedilatildeo de
hidroacutelise do ATP - ADP e Pi Na presenccedila de ADP fosfoenolpiruvato (PEP) e da enzima piruvato
cinase (PK) os produtos resultantes satildeo ATP e piruvato Por acccedilatildeo da enzima lactato
desidrogenase (LDH) este uacuteltimo eacute convertido em lactato ocorrendo simultaneamente a oxidaccedilatildeo
do NADH a NAD+ monitorizada atraveacutes da Abs340 nm Quando o S1 de miosina se encontra no
meio reaccional o ATP eacute hidrolisado formando-se novamente ADP e Pi o que permite a
formaccedilatildeo de um ciclo (fig 21) Se as enzimas se encontrarem em excesso a velocidade de
conversatildeo do NADH eacute limitada pela produccedilatildeo de ADP e Pi Como o ATP eacute regenerado em cada
ciclo a sua concentraccedilatildeo inicial eacute sempre mantida (Bagshaw 1982)
Figura 33ndash Representaccedilatildeo esquemaacutetica do meacutetodo de enzimas acopladas
47
O protocolo utilizado e seguidamente descrito foi adaptado de Cuenda et al (1990 ndash Ver Refordf) A
uma cuvette de 1 mL adicionou-se S1 para uma concentraccedilatildeo final de 005 mgmL 800 L de
meio de reacccedilatildeo22
10 L de PEP ([PEP]stock = 42 mM) 10 L de PK+LDH 11 ([PK+LDH]stock =
1 mgmL) e 15 L de NADH ([NADH]stock=25 mM) Iniciou-se a monitorizaccedilatildeo da reacccedilatildeo
atraveacutes da leitura de Abs340 nm apoacutes a adiccedilatildeo de 20 L de ATP ([ATP]stock=100 mM) A
actividade Mg2+
-ATPaacutesica eacute obtida atraveacutes do declive correspondente agrave oxidaccedilatildeo do NADH Para
a mediccedilatildeo da Mg2+
-ATPase de S1 estimulada por actina adicionou-se F-actina para uma
concentraccedilatildeo final de 2 microM sendo a actividade obtida tambeacutem atraveacutes do declive correspondente
agrave oxidaccedilatildeo do NADH apoacutes a adiccedilatildeo da proteiacutena
35 Interaccedilatildeo entre vanaacutedio e subfragmento-1 de miosina
351 Preparaccedilatildeo das soluccedilotildees de vanaacutedio
As soluccedilotildees de vanadato foram preparadas em aacutegua MilliQ a partir de metavanadato de amoacutenio
(NH4VO3) numa concentraccedilatildeo stock de 50 mM com agitaccedilatildeo magneacutetica e aquecimento (~30 ordmC)
Esta soluccedilatildeo com um valor compreendido entre 6 e 7 designa-se por ldquometavanadatordquo e eacute
constituiacuteda pelas esspeacutecies V1 V2 V4 e V5 (Ref) Acidificando a soluccedilatildeo de metavanadato por
adiccedilatildeo de HCl ateacute um valor de pH proacuteximo de 4 observa-se uma mudanccedila de cor de amarelo
claro para laranja indicativa da formaccedilatildeo da espeacutecie decameacuterica de vanadato De seguida e
imediatamente antes da realizaccedilatildeo dos ensaios descritos o valor do pH da soluccedilatildeo de
decavanadato foi aumentado para cerca de 75 por adiccedilatildeo de NaOH A soluccedilatildeo (2) designada
por ldquometavanadatordquo natildeo foi posteriormente alterada ateacute agrave sua adiccedilatildeo para a realizaccedilatildeo dos ensaios
As soluccedilotildees foram armazenadas a 4ordmC e mantidas em gelo aquando da sua utilizaccedilatildeo
A soluccedilatildeo de V(IV) foi preparada em condiccedilotildees anaeroacutebias por diluiccedilatildeo de sulfato de vanadil
(VOSO4) para uma concentraccedilatildeo final de 5 mM em H2O MilliQ saturada com N2 gasoso
352Anaacutelise por espectroscopia de UV-Vis
A concentraccedilatildeo da espeacutecie decameacuterica em soluccedilatildeo depende do periacuteodo de tempo apoacutes a diluiccedilatildeo
no meio de reacccedilatildeo apropriado (Ref) A velocidade de desoligomerizaccedilatildeo da espeacutecie decameacuterica
pode ser analisada por espectroscopia de absorccedilatildeo na regiatildeo UVVis uma vez que esta espeacutecie
apresenta duas bandas caracteriacutesticas na regiatildeo do visiacutevel (a 358 e 400 nm) que natildeo se verifica
em quaisquer outras espeacutecies oligomeacutericas As bandas supramencionada sapresentam-se bem
22
Meio de reacccedilatildeo 3125 mM Tris-HCl pH 71 375 mM MgCl2
48
resolvidas atraveacutes da obtenccedilatildeo da derivada de 2ordf ordem do espectro apresentando a forma de 2
vales com miacutenimos centrados nos comprimentos de onda supracitados com dependecircncia linear
relativamente agrave concentraccedilatildeo de decavanadato total (Ramos et al 2006)
Assim a estabilidade da espeacutecie decameacuterica apoacutes diluiccedilatildeo da soluccedilatildeo de decavanadato no meio de
reacccedilatildeo dos ensaios (tampatildeo D) para uma concentraccedilatildeo final de 1 mM em vanadato total foi
caracterizada atraveacutes da leitura da Abs400 em funccedilatildeo do tempo tendo-se determinado qual a
ordem de reacccedilatildeo e o tempo de meia-vida da mesma Com base em estudos anteriores acerca da
estabilidade do V10 na presenccedila de diferentes proteiacutenas (Ramos et al 2006) determinou-se
igualmente o tempo de meia vida apoacutes a adiccedilatildeo de 20 microM de S1 de miosina ao meio reaccional
353 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
Os espectros de RMN do isoacutetopo 51
V foram realizados num espectroacutemetro Bruker AM-400 MHz
equipado com uma sonda multinuclear inversa de 5 mm Em tubos de RMN de 5 mm de diacircmetro
preparou-se soluccedilotildees de 500 L de deca- ou metavanadato correspondentes a um intervalo de
concentraccedilotildees de 1 a 10 mM suplementadas com 10 D2O Os espectros foram adquiridos agrave
temperatura ambiente com uma largura espectral de 23 923 Hz um tempo de aquisiccedilatildeo de 0086
s tempo de relaxaccedilatildeo de 001 s e uma sequecircncia de pulsos a 90ordm Utilizou-se a transformada de
Fourier com alargamento das bandas de 20Hz para o processamento dos espectros Os desvios
quiacutemicos das ressonacircncias de 51
V foram obtidos em relaccedilatildeo agrave espeacutecie VOCl3cuja frequecircncia de
ressonacircncia eacute considerada o padratildeo sendo todas as outras referenciadas a partir desta em ppm
As aacutereas dos sinais do RMN correspondentes agraves espeacutecies oligomeacutericas presentes foram obtidas
por integraccedilatildeo dos sinais As concentraccedilotildees relativas das espeacutecies foram calculadas com recurso agrave
equaccedilatildeo
onde A[Vx] corresponde agrave aacuterea referente agrave ressonacircncia da espeacutecie Vx com nordm de agregaccedilatildeo x
A[V]total agrave soma total das aacutereas medidas e [V] a concentraccedilatildeo total de vanaacutedio no ensaio
A teacutecnica de 51
V-RMN foi anteriormente aplicada em estudos de ligaccedilatildeo com S1 de miosina
(Ref) tendo sido demostrado que entre as espeacutecies V1 V2 V4 e V5 a espeacutecie tetrameacuterica possui a
maior afinidade para a miosina (Cremo et al) O desaparecimento ou alargamento dos sinais pode
implicar a ligaccedilatildeo de um dos oxovanadatos laacutebeis agrave miosina sem indicaccedilatildeo de qual uma vez que
49
se processa um equiliacutebrio entre eles (Crans 1990) Assim e tendo em conta as indicaccedilotildees de que
a espeacutecie V10 eacute distinta das restantes no que diz respeito agrave magnitude da interacccedilatildeo com a miosina
e inibiccedilatildeo da actividade ATPaacutesica estimulada por actina (Aureliano 2000 Tiago et al 2002)
torna-se fundamental investigar a interacccedilatildeo das diferentes espeacutecies de vanadato com o S1 de
miosina atraveacutes da teacutecnica de 51
V-RMN
Para tal obtiveram-se espectros de 51
V-RMN resultantes da titulaccedilatildeo de uma soluccedilatildeo 5 mM de
decavanadato total com concentraccedilotildees crescentes de S1 de miosina (0 5 10 15 e 20 microM) em
tampatildeo D Realizou-se ainda ensaios na presenccedila de 2 mM de decavanadato e 2 mM de ATP na
presenccedila e na ausecircncia de S1 de miosina de modo a verificar o efeito que estes ligandos naturais
da enzima teria na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o subfragmento-1
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
Os grupos sulfidrilo (-SH) das cisteiacutenas satildeo resiacuteduos de aminoaacutecidos extremamente sensiacuteveis agrave
oxidaccedilatildeo resultando na sua oxidaccedilatildeo (Dalle-Donne et al 1999) Logo a quantificaccedilatildeo destes
resiacuteduos eacute bastante importante nos estudos de interacccedilatildeo com vanaacutedio jaacute que os resiacuteduos de Cys
com ecircnfase para o SH1 e o SH2 satildeo bastante importantes na monitorizaccedilatildeo das modificaccedilotildees
funcionais da regiatildeo da cabeccedila
O procedimento seguidamente apresentado foi originalmente descrito por Gutieacuterrez-Merino
(1987)) Consiste na reacccedilatildeo entre os grupos ndashSH natildeo oxidados das proteiacutenas e o reagente de
Ellman (DTNB -55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico)) - formando-se o aniatildeo CNT (aniatildeo 3-
carboxilato-4-nitrotiofenolato) numa estequiometria de 11 Este apresenta um maacuteximo de
absorvecircncia a 412 nm (Robyt 1990) Assim apoacutes a incubaccedilatildeo da proteiacutena na presenccedila do metal
a quantidade de cisteiacutenas reduzidas por proteiacutena determina-se atraveacutes dos valores de Abs412
Deste modo colocou-se num tubo de Eppendorf o volume apropriado de S1 dialisado para que a
concentraccedilatildeo no ensaio fosse de 1 mgmL Adicionou-se o volume apropriado de decavanadato
para que a sua concentraccedilatildeo fosse de 0 10 50 100 250 500 ou 1000 M Perfez-se com soluccedilatildeo
D ateacute um volume final de 1 mL e incubou-se a 25ordmC por 20 minutos Apoacutes este periacuteodo colocou-
se 780 L da mistura reaccional numa cuvette de quartzo e adicionou-se 220 L de soluccedilatildeo K23
23
Tampatildeo K Tampatildeo fosfatohidrogenofosfato de soacutedio 025 M pH 72
50
Adicionou-se 15 L de DTNB (em soluccedilatildeo K [DTNB]stock=100 mM) ao meio reaccional e
registou-se o valor de absorvecircncia medido equivalente agraves cisteiacutenas ldquoraacutepidasrdquo (SH1 e SH2
expostos ao solvente) Adicionou-se 001 g de SDS que provoca a desnaturaccedilatildeo da proteiacutena e
consequentemente a exposiccedilatildeo de todos os resiacuteduos de cisteiacutenas que se encontravam ateacute aiacute
inacessiacuteveis no interior da proteiacutena (cisteiacutenas totais) Estes resiacuteduos podem assim reagir com o
DTNB
Estimou-se o nuacutemero de resiacuteduos de cisteiacutenas em todas as amostras com recurso ao coeficiente de
extinccedilatildeo molar () do DTNB (ver ponto 3 dos Anexos) que foi determinado atraveacutes da titulaccedilatildeo
de L-Cisteiacutena com intervalo de concentraccedilotildees compreendidos entre 0 e 50 microM na presenccedila de
25microM de DTNB em tampatildeo D Os valores de absorvecircncia foram obtidos para um comprimento de
onda de 412 nm
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
3621 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
A teacutecnica mais eficiente de anaacutelise da interacccedilatildeo entre o iatildeo vanadilo e proteiacutenas como a
calmodulina e a transferrina entre outras permite a obtenccedilatildeo raacutepida de informaccedilatildeo no que toca agrave
conformaccedilatildeo e amplitude da referida interacccedilatildeo
Os espectros de Ressonacircncia Paramagneacutetica Electroacutenica (RPE) da espeacutecie paramagneacutetica V(IV)
foram realizados a 70K num espectroacutemetro Bruker EMX-300 X-band Todas as soluccedilotildees
utilizadas e amostras foram preparadas em anaerobiose numa cacircmara de luvas de modo a evitar a
reduccedilatildeo de V(IV) a V(V) Em tubos capilares com 02 mL de amostra manteve-se a concentraccedilatildeo
de S1 a 20 M e variou-se a concentraccedilatildeo de vanadilo (0 25 5 10 15 25 50 75 100 150
200 500 e 1000 M [VOSO4]stock=5 mM) em tampatildeo D Apoacutes obtenccedilatildeo dos espectros estes
foram tratados atraveacutes do software SimFonia e WinEPR tendo-se obtido um graacutefico com a
variaccedilatildeo da intensidade do sinal de RPE em funccedilatildeo da concentraccedilatildeo de sulfato de vanadilo
presente Realizaram-se ainda simulaccedilotildees de espectros obtidos com 20 microM de S1 e 1000 microM de
V(IV) com vista agrave obtenccedilatildeo dos ligandos provaacuteveis do vanadilo atraveacutes da anaacutelise da constante
de acoplamento hiperfina (A) e do factor g (g)
4 Conclusotildees gerais
Apesar de jaacute ter sido demonstrado que o vanaacutedio inibe a Na+K
+-ATPase da miosina a funccedilatildeo
deste elemento no tecido muacutesculo esqueleacutetico continua largamente incompreendida A enorme
complexidade quiacutemica deste metal especialmente no que concerne agrave oligomerizaccedilatildeo e reacccedilotildees
51
redox constitui um obstaacuteculo agrave compreensatildeo do comportamento do vanaacutedio nos organismos
vivos Assim a caracterizaccedilatildeo da quiacutemica deste elemento eacute de grande utilidade antes de iniciar o
estudo da sua interacccedilatildeo com o presente sistema
A anaacutelise por espectroscopias de 51
V-RMN e de UV-Vis de soluccedilotildees de vanadato diluiacutedas nos
meios de reacccedilatildeo utilizados no presente relatoacuterio (10 mM HEPES pH 70 25 mM KCl) mostrou
claramente que a espeacutecie decameacuterica (V10) possui um tempo de vida suficientemente longo para a
determinaccedilatildeo do seu efeito isolado ao contraacuterio do que se verifica com as espeacutecies mono- di-
tetra- e pentameacutericas (V1 V2 V4 e V5) Por outro lado a anaacutelise verificou ainda que apoacutes diluiccedilatildeo
das soluccedilotildees de decavanadato no meio de reacccedilatildeo coexistem as espeacutecies V10 e V1 pelo que os
efeitos putativos desta uacuteltima deveratildeo ser subtraiacutedos aquando da monitorizaccedilatildeo dos efeitos da
espeacutecie decameacuterica
O vanaacutedio (V) (vanadato) inibe as enzimas que catalizam reacccedilotildees de transferecircncia de grupos
fosfato como a ATPase da miosina [Goodno 1979] devido agrave sua analogia estrutural com o iatildeo
fosfato Na sua utilizaccedilatildeo como inibidor enzimaacutetico natildeo eacute tida em conta a contribuiccedilatildeo individual
das espeacutecies oligomeacutericas considerando-se normalmente a espeacutecie V1 como inibidora efectiva
[Goodno e Taylor 1982] No presente relatoacuterio verificou-se por espectroscopia de 51
V-RMN que
outros oligoacutemeros de vanadato interagem com o S1 de miosina tendo as espeacutecies V10 e V4 os
maiores niacuteveis de afinidade A sua interacccedilatildeo envolve forccedilas electrostaacuteticas e eacute afectada
negativamente por ligandos naturais de enzima como eacute o caso do ATP
A exposiccedilatildeo do S1 de miosina (005 mgmL) a decavanadato conduz agrave inibiccedilatildeo do capacidade da
proteiacutena de estimular a actividade Mg2+
-ATPaacutesica com um valor de IC50 de 266 plusmn 07 M
decavanadato sendo o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM
decavanadato (80) Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM decavanadato sendo
o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM decavanadato (60)
exercendo um efeito de impedimento agrave ligaccedilatildeo entre o vanaacutedio e o S1
A uma concentraccedilatildeo de 27 microM decavanadato verifica-se a oxidaccedilatildeo de um dos resiacuteduos de Cys
de S1 de miosina seguida pela oxidaccedilatildeo gradual de outro resiacuteduo de Cys ateacute agrave concentraccedilatildeo de
1000 microM Ambos os resiacuteduos oxidados natildeo se encontravam expostos ao solvente utilizado A
oxidaccedilatildeo dos resiacuteduos permite sugerir que esta modificaccedilatildeo oxidativa sofrida pelo S1 contribui
para a diminuiccedilatildeo da sua actividade de ATPase
52
Por espectroscopia de RPE revelaram-se os possiacuteveis ligandos equatoriais do vanaacutedio aquando da
sua interacccedilatildeo com o S1 de miosina como sendo duas moleacuteculas de aacutegua um ligando contendo o
grupo R-OH e outro contendo o grupo R=NR
Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados de
oxidaccedilatildeo (IV) e (V) no S1 de miosina para concentraccedilotildees facilmente alcanccedilaacuteveis no ambiente
terrestre
53
5 Estudos futuros
No sentido do melhor esclarecimento das alteraccedilotildees moleculares e estruturais que ocorrem no
subfragmento 1 quando exposta ao vanaacutedio que conduzem a uma diminuiccedilatildeo da sua funccedilatildeo
sugere-se
- Quantificaccedilatildeo das cisteiacutenas reduzidas de S1 na presenccedila de metavanadato
- Determinaccedilatildeo das actividades Ca2+
- e K+-ATPaacutesica do S1 de miosina na presenccedila de
decavanadato
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina estimulada por actina
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina na presenccedila de
metavanadato
- Realizaccedilatildeo de RMN 2D com variaccedilatildeo das concentraccedilotildees das soluccedilotildees de decavanadato e
metavanadato na presenccedila de S1
-Mutageacutenese dos resiacuteduos de Cys doS1 de miosina de modo a verificar quais os resiacuteduos que
sofrem oxidaccedilatildeo na presenccedila de decavanadato
- Determinaccedilatildeo por calorimetria (DSC Differential Scanning Calorimetry) da variaccedilatildeo da
entalpia (∆H) e da temperatura de desnaturaccedilatildeo do S1 de miosina na presenccedila de vanaacutedio uma
vez que estas propriedades caracteriacutesticas de cada proteiacutena satildeo alteradas apoacutes ocorrecircncia de
modificaccedilotildees estruturais
- Anaacutelise por SDS-PAGE de S1 de miosina apoacutes tratamento com vanaacutedio de modo a determinar
quais os polipeacuteptidos eventualmente formados apoacutes clivagem da proteiacutena
54
6 Referecircncias bibliograacuteficas
- Sellers J Myosins 2nd Edition Oxford University Press 1999
- L Stryer inBiochemistry (Ed WH Freeman 2005 pp
Refordfas Materiais e Meacutetodos
[A] S S Margossian SLowey Methods Enzymol1982 85 55-71
[B] JD Pardue JA Spudich Methods Enzymol 1982 85 164-181
[C] M Bradford Anal Biochem1976 72 248-254
[D] H H Taussky E Shorr J Biol Chem 1953 202 675-685
[E] C R Bagshaw in Outline Studies in Biology ndash Muscle Contraction Chapman and Hall Ed
1982
[F] A Cuenda et al Eur J Biochem1990 194 663-670
55
Anexos
1 Determinaccedilatildeo da recta de calibraccedilatildeo meacutetodo de Bradford
Figura1ndashRepresentaccedilatildeo graacutefica da recta de calibraccedilatildeo (Abs595 nm vs [BSA]) utilizada para o meacutetodo de Bradford
2 Determinaccedilatildeo da recta calibraccedilatildeo para a electroforese em condiccedilotildees desnaturantes
Figura 2 ndash Representaccedilatildeo graacutefica da recta de calibraccedilatildeo (log(Peso Molecular) vs Rf dos marcadores de peso
molecular) onde Rf indica determinada atraveacutes do padratildeo de peso molecular
(poccedilo (a))
3Caacutelculo do coeficiente de extinccedilatildeo molar () para a quantificaccedilatildeo de cisteiacutenas reduzidas
atraveacutes do meacutetodo de Ellman
Traccedilou-se a recta de Abs412 nm meacutedia vs [L-Cisteiacutena] de modo a determinar o valor do coeficiente
de extinccedilatildeo molar definido como o declive daquela recta Assim =8400 M-1
cm-1
como
podemos verificar na Fig 3
A595 nm = 00546[BSA] + 00209 Rsup2 = 099051
Ab
s 59
5 n
m
[BSA] (mgmL)
log PM = -1177 Rf + 2247 Rsup2 = 098957
log
PM
Mobilidade relativa (Rf)
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero
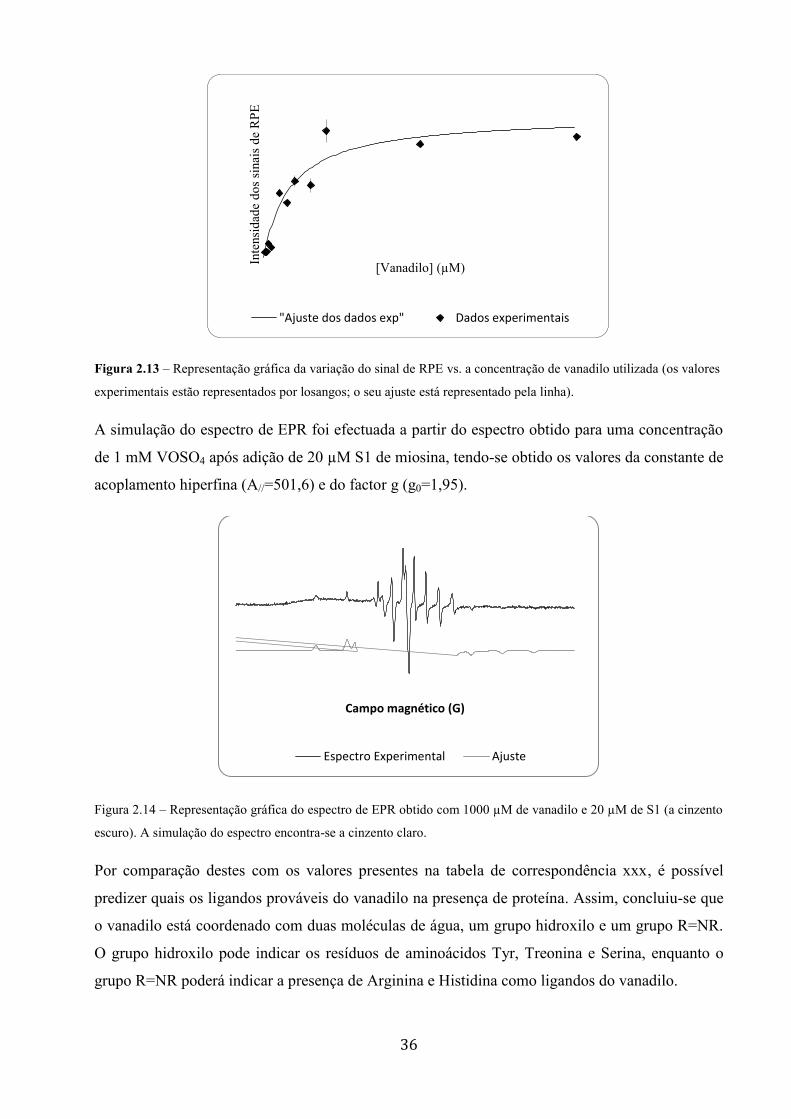
36
Figura 213 ndash Representaccedilatildeo graacutefica da variaccedilatildeo do sinal de RPE vs a concentraccedilatildeo de vanadilo utilizada (os valores
experimentais estatildeo representados por losangos o seu ajuste estaacute representado pela linha)
A simulaccedilatildeo do espectro de EPR foi efectuada a partir do espectro obtido para uma concentraccedilatildeo
de 1 mM VOSO4 apoacutes adiccedilatildeo de 20 microM S1 de miosina tendo-se obtido os valores da constante de
acoplamento hiperfina (A=5016) e do factor g (g0=195)
Figura 214 ndash Representaccedilatildeo graacutefica do espectro de EPR obtido com 1000 microM de vanadilo e 20 microM de S1 (a cinzento
escuro) A simulaccedilatildeo do espectro encontra-se a cinzento claro
Por comparaccedilatildeo destes com os valores presentes na tabela de correspondecircncia xxx eacute possiacutevel
predizer quais os ligandos provaacuteveis do vanadilo na presenccedila de proteiacutena Assim concluiu-se que
o vanadilo estaacute coordenado com duas moleacuteculas de aacutegua um grupo hidroxilo e um grupo R=NR
O grupo hidroxilo pode indicar os resiacuteduos de aminoaacutecidos Tyr Treonina e Serina enquanto o
grupo R=NR poderaacute indicar a presenccedila de Arginina e Histidina como ligandos do vanadilo
Inte
nsi
dad
e d
os
sinai
s d
e R
PE
[Vanadilo] (microM)
Ajuste dos dados exp Dados experimentais
Campo magneacutetico (G)
Espectro Experimental Ajuste
37
3 Procedimento Experimental
31 Reagentes
Todos os reagentes utilizados satildeo de elevado grau de pureza (analiacutetico) Apresentam-se as suas
foacutermulas quiacutemicas ou abreviaturas quando necessaacuterio acrescentou-se a sua descriccedilatildeo Entre
parecircntesis colocou-se as empresas que os comercializam
Aacutecido aceacutetico glacial 100 (Riedel) Acrilamida 99 (Merck) Azul de bromofenol (Riedel-de-
Haumlen) -Mercaptoetanol (Sigma) Bissulfito de soacutedio (Sigma)Bradford (reagente de)
(BioRad)BSA (albumina de soro bovino Sigma-Aldrich) CaCl22H2O (cloreto de caacutelcio
dihidratado Fluka) CH3CH2OH 96 (etanol Panreac) Coomassie Blue R250 (Merck) D2O
(oacutexido de deuteacuterio SDS) DTNB (55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico) Aldrich) EDTA (aacutecido
etilenodiaminatetraaceacutetico Merck) Fosfoenolpiruvato 98 (Fluka) Glicerol (Sigma) Glicina
985 (Panreac) HCl 37 (Riedel-de Haumlen) H2SO4 96 (Pronalab) HEPES 99 (aacutecido 4-(2-
hidroxiletil)-1-piperazineetanosulfoacutenico Sigma) KH2PO4 99 (hidrogenofosfato de potaacutessio)
MgATP (Sigma) Na
ATP (Sigma) Na2HPO4 99 (hidrogenofosfato de soacutedio Riedel-de Haumlen)
NaH2PO4 (fosfato de soacutedio ) NaOH 986 (hidroacutexido de soacutedio Pronalab) KCl 99 (Cloreto de
potaacutessio Panreac) L-Cys 995 (enantioacutemero levoacutegiro do resiacuteduo de Cisteiacutena Fluka)Metanol
99 (Merck) Molibdato de amoacutenio (Sigma)NNrsquo-metil-bis-acrilamida 98 (Merck) NH4VO3
99 (metavanadato de amoacutenia Sigma) PSA 10 (persulfato de amoacutenio Merck) SDS
(dodecilo sulfato de soacutedio Sigma) Sulfito de soacutedio (Sigma) VOSO4 (sulfato de vanadilo
Aldrich) TEMED 99 (Tetrametiletilenodiamina Merck) Tris (2-amino-2-hidroximetil-13-
propanodiol Sigma)
32 Meacutetodos
321 Isolamento e purificaccedilatildeo das proteiacutenas de muacutesculo esqueleacutetico de coelho
3211 Purificaccedilatildeo da miosina preparaccedilatildeo de Subfragmento 1 por digestatildeo quimotriacutepsica
da miosina
O subfragmento-1 de miosina-II (S1) foi gentilmente cedido pelo Prof Doutor Manuel Aureliano
do Departamento de Quiacutemica Bioquiacutemica e Farmaacutecia da Faculdade de Ciecircncias e Tecnologia da
Universidade do Algarve
No entanto descreve-se abaixo de forma resumida o procedimento efectuado
38
32111 Purificaccedilatildeo da miosina
O protocolo de isolamento foi adaptado do meacutetodo previamente descrito por Margossian e Lowey
(1982) apresentando-se de forma esquemaacutetica na Fig 31
Sucintamente os tecidos musculares dorsais e das pernas foram extraiacutedos apoacutes o sacrifiacutecio do
animal (Fig31) De seguida foram adicionados 3 volumes da soluccedilatildeo de extracccedilatildeo A2
Centrifugou-se a soluccedilatildeo e filtrou-se o sobrenadante O sedimento foi entatildeo diluiacutedo em 14
volumes de aacutegua destilada de modo a que a forccedila ioacutenica atingisse 004 M Apoacutes adiccedilatildeo de szlig-
mercaptoetanol (szlig-me) deixou-se a precipitar a 4ordmC removendo o sobrenadante Centrifugou-se
novamente e ressuspendeu-se o sedimento na soluccedilatildeo de extracccedilatildeo A Adicionou-se a soluccedilatildeo
tampatildeo B3 para que a forccedila ioacutenica atingisse 05 M Homogeneizou-se e deixou-se repousar
durante 2h Homogeneizou-se novamente e centrifugou-se de modo a ocorrer precipitaccedilatildeo dos
complexos de actomiosina e proteiacutena desnaturada Apoacutes filtraccedilatildeo do sobrenadante em latilde de vidro
quantificou-se a soluccedilatildeo de miosina resultante pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental)
2 Soluccedilatildeo de extracccedilatildeo A 0012M K2CO3 004M KHCO3 06M KCl pH 65
3 Soluccedilatildeo-tampatildeo B Tris 100 mM pH 7 KCl 2M
39
Figura 31 ndash Representaccedilatildeo esquemaacutetica do protocolo de isolamento de miosina (Margossian e Lowey 1982) Todo o
procedimento foi realizado a 4ordmC (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
32112 Preparaccedilatildeo de Subfragmento-1 de miosina por digestatildeo com quimiotripsina
O S1 de miosina foi isolado apoacutes diaacutelise da miosina durante a noite contra a soluccedilatildeo C4 A
presenccedila do EDTA eacute essencial uma vez que a sua substituiccedilatildeo por MgCl2resulta na produccedilatildeo de
HMM em vez de S1 A suspensatildeo de miosina foi equilibrada e digerida com -quimotripsina
A digestatildeo foi interrompida pela adiccedilatildeo de PMSF Seguidamente ultracentrifugou-se o produto
da digestatildeo desprezando-se o sedimento (Fig 32) A soluccedilatildeo de sobrenadante maioritariamente
S1 foi quantificada pela leitura directa da Abs280 ou pelo meacutetodo de Bradford (ver as secccedilotildees
3311 e 3312 do Procedimento Experimental respectivamente) e armazenada em 10 sacarose
e 2 mM szlig-Me a -80ordmC
4 Soluccedilatildeo C 25mM Tris pH 70 012M KCl 1mM EDTA
40
Antes da realizaccedilatildeo dos ensaios descritos no presente relatoacuterio o S1 de miosina foi dialisado
contra a soluccedilatildeo D5 de modo a remover qualquer composto que pudesse complexar como
vanaacutedio Seguidamente quantificou-se a proteiacutena pelo meacutetodo de Bradford ou pelo meacutetodo
directo (ver secccedilotildees 3311 e 3312do Procedimento Experimtental)
Figura 3 2ndash Representaccedilatildeo esquemaacutetica do protocolo de preparaccedilatildeo do S1 de miosina por digestatildeo quimiotripsina de
acordo com o previamente descrito por Margossian e Lowey (1982) Todo o procedimento foi realizado a 4ordm C
excepto quando indicaccedilatildeo do contraacuterio (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
3212 Isolamento e purificaccedilatildeo de actina
Isolou-se e purificou-se a actina segundo o meacutetodo descrito por Pardee e Spudish que
compreende dois passos ndash a obtenccedilatildeo de um resiacuteduo seco denominado ldquopoacute de acetonardquo e a
extracccedilatildeo subsequente da actina a partir do ldquopoacute de acetonardquo na forma de G-Actina com a
posterior polimerizaccedilatildeo a F-Actina
32121 Obtenccedilatildeo do ldquopoacute de acetonardquo
Sacrificou-se o coelho com uma pancada atraacutes do pescoccedilo e sangrou-se o animal apoacutes o corte da
arteacuteria jugular Esfolou-se e colocou-se numa tina com gelo Retiraram-se as partes de tecido
conjuntivo que envolvem o tecido muscular com auxiacutelio de uma pinccedila e bisturi Seguidamente
retirou-se os muacutesculos dorsais e os muacutesculos brancos das pernas Passou-se os muacutesculos por aacutegua
destilada de modo a remover as impurezas e pesou-se num copo Trituraram-se os muacutesculos
numa maacutequina de triturar carne e extraiu-se o resultante com 3 volumes da soluccedilatildeo E6 durante 10
minutos com agitaccedilatildeo suave Centrifugou-se a suspensatildeo a 4000 g durante 10
5 Soluccedilatildeo D HEPES 10 mM pH 70 KCl 25 mM
6 Soluccedilatildeo E 01 M KCl 015 M K2HPO4 pH 65
41
minutosdesprezando-se o sobrenadante Ressuspendeu-se o sedimento em 6 volumes de soluccedilatildeo
F7durante 10 minutos com agitaccedilatildeo suave Verificou-se se o pH da soluccedilatildeo estava compreendido
entre 82 e 85 acertando-se estecom soluccedilatildeo G8 se necessaacuterio Centrifugou-se durante 10
minutos a 4000 g Desprezou-se o sobrenadante e ressuspendeu-se o sedimento em 3 volumes de
soluccedilatildeo H9 durante 10 minutos Centrifugou-se novamente a 4000 gdurante 10 minutos
desprezando-se o sobrenadante Ressuspendeu-se o sedimento duas vezes consecutivas em 6
volumes de aacutegua destilada durante 5 minutos Centrifugou-se a 4000 g durante 10 minutos apoacutes
cada adiccedilatildeo de H2O
Seguidamente ressuspendeu-se o sedimento 5 vezes consecutivas utilizando 3 volumes de
acetona de cada vez agitando suavemente durante 10 minutos agrave temperatura ambiente Colocou-
se o resiacuteduo filtrado numa tina de vidro sobre papel de filtro deixando-se evaporar o solvente
durante a noite num nicho Triturou-se o resiacuteduo seco numa maacutequina de triturar carne e deixou-se
novamente evaporar o solvente O ldquopoacute de acetonardquo obtido foi armazenado a -20 ordmC
32122 Purificaccedilatildeo da actina a partir de ldquopoacute de acetonardquo
Apoacutes a obtenccedilatildeo do poacute de acetona como acima indicado efectuou-se o protocolo seguidamente
descrito de modo a obter monoacutemeros de actina (G-Actina) e actina filamentosa (F-Actina)
Pesou-se 35 g de poacute de acetona e adicionaram-se 50 mL de soluccedilatildeo-tampatildeo I10
sendo a
extracccedilatildeo feita com agitaccedilatildeo suave durante 30 minutos Separou-se o extracto do poacute de acetona
hidratado filtrando a suspensatildeo com 4 camadas de gaze esterilizada tendo o cuidado de natildeo
espremer o resiacuteduo Voltou a extrair-se com 50 mL de soluccedilatildeo I durante 30 minutos mexendo
lentamente Adicionou-se este ao extracto obtido anteriormente Centrifugou-se o extracto total a
16 000 g durante 30 minutos Decantou-se para uma proveta sobre a qual se colocou gaze para
que o sedimento solto natildeo se juntasse ao sobrenadante e leu-se o volume De seguida adicionou-
se KCl ateacute uma concentraccedilatildeo final de 50 mM MgCl2 ateacute 2mM e ATP ateacute 1mM lentamente e com
agitaccedilatildeo contiacutenua de modo a promover a polimerizaccedilatildeo dos monoacutemeros de actina
Deixou-se polimerizar durante 2h a 25ordmC observando-se a presenccedila de pequenas bolhas de ar
indicativas de um aumento de viscosidade da soluccedilatildeo Arrefeceu-se em gelo durante 15 minutos e
adicionou-se KCl ateacute uma concentraccedilatildeo final de 06M Mexeu-se lentamente durante 30 minutos
apoacutes os quais se ultracentrifugou a 110 000g durante 1hora Desprezou-se o sobrenadante
ressuspendeu-se o sedimento com soluccedilatildeo tampatildeo E e transferiu-se para um potter
Dialisou-se durante a noite contra 1L de soluccedilatildeo-tampatildeo I a uma temperatura de 4ordmC com rotaccedilatildeo
7 Soluccedilatildeo F 005 M NaHCO3 pH 82-85
8 Soluccedilatildeo G 1M Na2CO3
9 Soluccedilatildeo H 1 mM EDTA pH 70
10 Soluccedilatildeo I 2 mM Tris-HCl pH 8 02 mM CaCl2 02 mM Na2ATP 05 mM szlig-Me 0005 NaN3
42
contiacutenua e mudanccedila de tampatildeo 2 a 3 vezes De seguida a actina foi centrifugada a 110
000gdurante 1hora Decantou-se a soluccedilatildeo de G-Actina (sobrenadante) e desprezou-se o
sedimento Determinou-se a concentraccedilatildeo de G-Actina pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental) A soluccedilatildeo de G-actina foi armazenada a uma temperatura de 4ordmC
apoacutes adiccedilatildeo de 002NaN3 mantendo-se estaacutevel durante 3 a 5 dias
331 Quantificaccedilatildeo das proteiacutenas isoladas
3311 Quantificaccedilatildeo pelo meacutetodo de Bradford
O meacutetodo de Bradford (Bradford 1976) baseia-se na utilizaccedilatildeo do reagente de Bradford
composto por entre outros aacutecido fosfoacuterico e o corante ldquoCoomassierdquo Este corante possui carga
negativa e a sua coloraccedilatildeo varia entre a vermelha (Absmax=465 nm) predominante em soluccedilatildeo e a
azul (Absmax=595 nm) presente aquando da ligaccedilatildeo aos grupos amina das proteiacutenas Em soluccedilotildees
aacutecidas diluiacutedas a coloraccedilatildeo varia proporcionalmente com a quantidade do corante ligado agrave
proteiacutena que por sua vez eacute directamente proporcional agrave concentraccedilatildeo da proteiacutena permitindo a
sua quantificaccedilatildeo ateacute a um limite de detecccedilatildeo de 20 gmL (Bradford 1976)
Deste modo prepararam-se amostras de BSA ([BSA]stock= 1mgmL) utilizando um intervalo de
concentraccedilotildees compreendido entre 0 e 15 gmL de acordo com a tabela 3I Apoacutes a adiccedilatildeo da
H2O MilliQ e do reagente de Bradford as soluccedilotildees foram sucessivamente homogeneizadas no
voacutertex
Tabela 3Indash Amostras utilizadas para a obtenccedilatildeo da recta de calibraccedilatildeo no meacutetodo de Bradford
BSA (L) BSA (g) H2O MilliQ (L) Reagente de Bradford (L)
0 0 800 200
125 125 79875 200
25 25 7975 200
5 5 795 200
10 10 790 200
15 15 785 200
Apoacutes incubaccedilatildeo a 25ordmC por 5 minutos leram-se os valores de Abs595 num
espectrofotoacutemetro (Shimadzu UV-160A) para todas as amostras Seguidamente traccedilou-se a recta
de calibraccedilatildeo (Abs595vs quantidade de BSA (g))
43
Para determinar a concentraccedilatildeo de proteiacutena apoacutes o processo de isolamento purificaccedilatildeo e
diaacutelise quando necessaacuterio colocou-se 5 L da amostra num tubo de Eppendorf e adicionou-se
795 L de H2O MilliQ homogeneizando a soluccedilatildeo no voacutertex Adicionou-se posteriormente 200
L de reagente de Bradford e repetiu-se a homogeneizaccedilatildeo Incubou-se a mistura a 25ordmC por 5
minutos apoacutes os quais se leu os valores de Abs595 nm no espectrofotoacutemetro Por interpolaccedilatildeo deste
valor a partir da recta de calibraccedilatildeo determinou-se a concentraccedilatildeo de proteiacutena na amostra
3312 Quantificaccedilatildeo pelo meacutetodo directo
A quantificaccedilatildeo das amostras proteicas pode tambeacutem ser efectuada atraveacutes da leitura directa da
absorvacircncia tendo por base a lei de Beer-Lambert onde Abs corresponde agrave
absorvacircncia ao comprimento de onda ao qual o ensaio foi realizado corresponde ao coeficiente
de extinccedilatildeo (substituiacutedo pela letra a se as unidades forem distintas da molaridade) b corresponde
ao percurso oacuteptico da radiaccedilatildeo na ceacutelula amostral e c corresponde agrave concentraccedilatildeo amostral
Assim no caso de S1 de miosina mediu-se a Abs280e calculou-se a concentraccedilatildeo da proteiacutena
sabendo que o coeficiente de extinccedilatildeo de S1 eacute (Ref)
No caso da actina mediu-se a Abs290ecalculou-se a concentraccedilatildeo da proteiacutena sabendo que o
coeficiente de extinccedilatildeo da actina eacute (Ref)
332Electroforese em condiccedilotildees desnaturantes (SDS-PAGE)
De modo a melhor caracterizar as proteiacutenas isoladas foi realizada uma electroforese em
condiccedilotildees desnaturantes (SDS-PAGE do inglecircs Sodium Dodecyl Sulfate Polyacrilamide Gel
Electrophoresis) O SDS (dodecilo sulfato de soacutedio) eacute um detergente anioacutenico utilizado neste
meacutetodo para desnaturar as proteiacutenas que se ligam ao SDS na proporccedilatildeo 14 g SDS g proteiacutena
(aproximadamente 1 moleacutecula de SDS por cada 2 resiacuteduos de aminoaacutecidos) (Stryer) O SDS induz
uma densidade de carga negativa significativa nas proteiacutenas a que se liga mascarando a sua carga
intriacutenseca pelo que as razotildees cargamassa satildeo tendencialmente idecircnticas Assim a SDS-PAGE
separa as proteiacutenas quando sujeitas a um campo eleacutectrico de acordo com o seu peso molecular
sendo a sua migraccedilatildeo directamente proporcional ao logaritmo deste uacuteltimo (Stryer)
O gel de resoluccedilatildeo com 10 de acrilamida foi obtido atraveacutes da adiccedilatildeo de 167 mL de soluccedilatildeo
stock de acrilamida11
075 mL de tampatildeo de resoluccedilatildeo12
251 mL de aacutegua destilada e 50 L de
SDS 10 Para promover a polimerizaccedilatildeo adicionou-se em seguida 38 L de PSA 10
11
soluccedilatildeo stock de acrilamida soluccedilatildeo de 30 (mv) de acrilamida e 08 (mv) de NNrsquo-metil-bis-acrilamida 12
tampatildeo de resoluccedilatildeo 15 M Tris-HCl pH 88
44
(persulfato de amoacutenia) 10 e 25 L de TEMED 99 (tetrametiletilenodiamina) Agitou-se
cuidadosamente de modo a natildeo formar bolhas e pipetou-se a soluccedilatildeo para a cassette de vidro
previamente montada no suporte De seguida adicionou-se uma peliacutecula de cerca de 2 mm de 2-
metil-2-butanol para verificar posteriormente a polimerizaccedilatildeo Apoacutes a polimerizaccedilatildeo que demora
cerca de 45 minutos verteu-se a agrave superfiacutecie e adicionou-se o gel de concentraccedilatildeo com 5 de
acrilamida constituiacutedo por 03 mL de soluccedilatildeo de stock acrilamida 045 mL de tampatildeo de
concentraccedilatildeo13
094 mL de aacutegua destilada 18 L de SDS 10 135 L de PSA 10 e 2L de
TEMED 99 Colocou-se o pente sobre o gel e deixou-se polimerizar durante cerca de 45
minutos
As amostras para aplicaccedilatildeo no gel de electroforese foram preparadas para uma concentraccedilatildeo de
proteiacutena de 1 mgmL num volume final de 100 L dos quais 25 L correspondem ao tampatildeo da
amostra14
As amostras foram aquecidas a 100ordmC durante 5 minutos Antes da aplicaccedilatildeo das
amostras transferiu-se a cassette para a tina de electroforese agrave qual se adicionou a soluccedilatildeo
tampatildeo de electroforese da tina superior15
e tampatildeo de electroforese da tina inferior16
Aplicaram-
se 10 L das soluccedilotildees de marcadores de peso molecular e 20 L das amostras proteicas
preparadas anteriormente A electroforese decorreu durante cerca de 3h30 min a uma voltagem
de 80 V De seguida transferiu-se o gel para uma tina corando as proteiacutenas durante 1h com
soluccedilatildeo corante17
Descorou-se o gel com soluccedilatildeo descorante18
tendo-se mudado esta soluccedilatildeo ateacute
as bandas se encontrarem bem definidas
Determinou-se a mobilidade relativa das bandas correspondentes aos marcadores de elevado peso
molecular e traccedilou-se a recta de calibraccedilatildeo ndash log(PM) vs log(Rf) onde Rf corresponde agrave
mobilidade relativa Por interpolaccedilatildeo determina-se o PM das proteiacutenas isoladas sabendo a sua
mobilidade relativa Por comparaccedilatildeo com os valores descritos na literatura pode concluir-se
acerca do grau de purificaccedilatildeo
34 Mediccedilatildeo da actividade enzimaacutetica de S1
341 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
13
tampatildeo de concentraccedilatildeo 05 M Tris-HCl pH 68 14
tampatildeo de amostra 320 mM Tris-HCl pH 68 04 M szlig-mercaptoetanol SDS 8 glicerol 15 (vv) e roxo de m-
cresol 0024 15
tampatildeo de electroforese da tina superior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS 16
tampatildeo de electroforese da tina inferior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS acetato de soacutedio
01 M 17
soluccedilatildeo corante ldquoCoomassie Blue R-250rdquo contendo metanol 45 (vv) e aacutecido aceacutetico glacial 10 (vv) 18
soluccedilatildeo descorante metanol 10 (vv) e aacutecido aceacutetico glacial 10 vv)
45
O meacutetodo de Fiske-Subarow modificado consiste na reacccedilatildeo do molibdato de amoacutenio com o
fosfato inorgacircnico em soluccedilatildeo aacutecida com formaccedilatildeo do aacutecido fosfomoliacutebdico Seguidamente daacute-
se a adiccedilatildeo de um agente redutor que provoca a reduccedilatildeo do composto a fosfomolibdato que forma
um complexo de cor azulada cuja absorvacircncia maacutexima ocorre para um comprimento de onda de
660 nm tornando possiacutevel a sua detecccedilatildeo por meacutetodos espectrofotomeacutetricos As actividades Ca2+
-
ATPaacutesica e K+EDTA-ATPaacutesica de S1 foram determinadas atraveacutes deste meacutetodo de acordo com
o descrito por Taussky e Shorr (1953) com mediccedilatildeo da quantidade de fosfato (Pi) libertado
atraveacutes da hidroacutelise de ATP
Para tal obteve-se primeiramente a recta de calibraccedilatildeo com as amostras presentes na Tabela 3II
utilizando uma soluccedilatildeo stock de 5 mM KH2PO4
Tabela 3II ndash Composiccedilatildeo das diferentes amostras utilizadas para determinar a recta de calibraccedilatildeo atraveacutes do meacutetodo
de Fiske-Subbarow modificado
KH2PO4 (L)
([KH2PO4]stock= 5mM)
PO43-
(nmole) H2O (uL)
0 0 1780
5 25 1775
10 50 1770
20 100 1760
40 200 1740
100 500 1680
A cada tubo de Eppendorf adicionaram-se seguidamente 200 L de soluccedilatildeo J19
e 20 L de agente
redutor20
homogeneizando no voacutertex apoacutes cada adiccedilatildeo Incubou-se durante 10 minutos e mediu-
se a Abs660 nm num espectrofotoacutemetro com intervalos de tempo iguais entre cada amostra Traccedilou-
se a recta de calibraccedilatildeo (Abs660 nmvs nmol PO43-
) a partir dos valores obtidos
Para determinar a actividade Ca2+
-ATPaacutesica de S1 prepararam-se tubos de Eppendorf com 400
L de meio de reacccedilatildeo21
25 L CaCl2 ([CaCl2]stock= 100 mM) 25 L ATP ([ATP]stock= 100 mM)
com [S1]ensaios=01 mgmL Perfez-se se necessaacuterio com H2O MilliQ ateacute perfazer um volume de 1
mL Para determinar a actividade K+EDTA-ATPaacutesica utilizou-se uma concentraccedilatildeo idecircntica de
19
Soluccedilatildeo J 5 N H2SO4 25 molibdato de amoacutenio 20
Agente redutor 5 mg aacutecido 1-amino-2-naftol-4-sulfoacutenico 30 mg bissulfito de soacutedio 30 mg sulfito de soacutedio em 26
mL H2O 21
meio de reacccedilatildeo 125 mM Tris-HCl pH 80 0575 M KCl
46
proteiacutena nos ensaios (01 mgmL) e o mesmo meio de reacccedilatildeo Aos tubos de Eppendorf
adicionou-se ainda 123 L de KCl ([KCl]stock = 3M) 25 L de ATP 10 L de EDTA
([EDTA]stock= 100 mM )e o volume de H2O MilliQ necessaacuterio para perfazer 1 mL Em ambas as
situaccedilotildees o uacuteltimo reagente adicionado foi o ATP uma vez que se trata do substrato da reacccedilatildeo
enzimaacutetica
A cineacutetica decorreu a 25ordmC durante 5 minutos para a actividade Ca2+
-ATPaacutesica e durante 3
minutos para a actividade K+EDTA-ATPaacutesica Apoacutes o periacuteodo de incubaccedilatildeo transferiu-se 150
L de amostra para tubos de Eppendorf contendo igual volume de TCA 10 Colocou-se no gelo
e homogeneizou-se no voacutertex Seguidamente centrifugou-se a 1500 g durante 10 minutos e
recolheu-se 200 L de sobrenadante para novos tubos de Eppendorf Adicionou-se a estes 158
mL de H2O MilliQ e 200 L de soluccedilatildeo J Homogeneizou-se no voacutertex e adicionou-se 20 L de
reagente redutor repetindo-se a homogeneizaccedilatildeo Incubou-se durante 10 minutos e mediu-se a
Abs660 nm com o mesmo intervalo de tempo de leitura para cada amostra Por intrapolaccedilatildeo a partir
da recta de calibraccedilatildeo obtida anteriormente determinou-se a quantidade de Pi libertado em cada
ensaio A partir destes resultados determinaram-se as actividades Ca2+
-ATPaacutesica e K+EDTA-
ATPaacutesica do S1 em nmol PO43-
minmg proteiacutena e em s-1
342 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
Da actividade Mg2+
-ATPaacutesica de S1 resulta a formaccedilatildeo dos produtos da reacccedilatildeo de
hidroacutelise do ATP - ADP e Pi Na presenccedila de ADP fosfoenolpiruvato (PEP) e da enzima piruvato
cinase (PK) os produtos resultantes satildeo ATP e piruvato Por acccedilatildeo da enzima lactato
desidrogenase (LDH) este uacuteltimo eacute convertido em lactato ocorrendo simultaneamente a oxidaccedilatildeo
do NADH a NAD+ monitorizada atraveacutes da Abs340 nm Quando o S1 de miosina se encontra no
meio reaccional o ATP eacute hidrolisado formando-se novamente ADP e Pi o que permite a
formaccedilatildeo de um ciclo (fig 21) Se as enzimas se encontrarem em excesso a velocidade de
conversatildeo do NADH eacute limitada pela produccedilatildeo de ADP e Pi Como o ATP eacute regenerado em cada
ciclo a sua concentraccedilatildeo inicial eacute sempre mantida (Bagshaw 1982)
Figura 33ndash Representaccedilatildeo esquemaacutetica do meacutetodo de enzimas acopladas
47
O protocolo utilizado e seguidamente descrito foi adaptado de Cuenda et al (1990 ndash Ver Refordf) A
uma cuvette de 1 mL adicionou-se S1 para uma concentraccedilatildeo final de 005 mgmL 800 L de
meio de reacccedilatildeo22
10 L de PEP ([PEP]stock = 42 mM) 10 L de PK+LDH 11 ([PK+LDH]stock =
1 mgmL) e 15 L de NADH ([NADH]stock=25 mM) Iniciou-se a monitorizaccedilatildeo da reacccedilatildeo
atraveacutes da leitura de Abs340 nm apoacutes a adiccedilatildeo de 20 L de ATP ([ATP]stock=100 mM) A
actividade Mg2+
-ATPaacutesica eacute obtida atraveacutes do declive correspondente agrave oxidaccedilatildeo do NADH Para
a mediccedilatildeo da Mg2+
-ATPase de S1 estimulada por actina adicionou-se F-actina para uma
concentraccedilatildeo final de 2 microM sendo a actividade obtida tambeacutem atraveacutes do declive correspondente
agrave oxidaccedilatildeo do NADH apoacutes a adiccedilatildeo da proteiacutena
35 Interaccedilatildeo entre vanaacutedio e subfragmento-1 de miosina
351 Preparaccedilatildeo das soluccedilotildees de vanaacutedio
As soluccedilotildees de vanadato foram preparadas em aacutegua MilliQ a partir de metavanadato de amoacutenio
(NH4VO3) numa concentraccedilatildeo stock de 50 mM com agitaccedilatildeo magneacutetica e aquecimento (~30 ordmC)
Esta soluccedilatildeo com um valor compreendido entre 6 e 7 designa-se por ldquometavanadatordquo e eacute
constituiacuteda pelas esspeacutecies V1 V2 V4 e V5 (Ref) Acidificando a soluccedilatildeo de metavanadato por
adiccedilatildeo de HCl ateacute um valor de pH proacuteximo de 4 observa-se uma mudanccedila de cor de amarelo
claro para laranja indicativa da formaccedilatildeo da espeacutecie decameacuterica de vanadato De seguida e
imediatamente antes da realizaccedilatildeo dos ensaios descritos o valor do pH da soluccedilatildeo de
decavanadato foi aumentado para cerca de 75 por adiccedilatildeo de NaOH A soluccedilatildeo (2) designada
por ldquometavanadatordquo natildeo foi posteriormente alterada ateacute agrave sua adiccedilatildeo para a realizaccedilatildeo dos ensaios
As soluccedilotildees foram armazenadas a 4ordmC e mantidas em gelo aquando da sua utilizaccedilatildeo
A soluccedilatildeo de V(IV) foi preparada em condiccedilotildees anaeroacutebias por diluiccedilatildeo de sulfato de vanadil
(VOSO4) para uma concentraccedilatildeo final de 5 mM em H2O MilliQ saturada com N2 gasoso
352Anaacutelise por espectroscopia de UV-Vis
A concentraccedilatildeo da espeacutecie decameacuterica em soluccedilatildeo depende do periacuteodo de tempo apoacutes a diluiccedilatildeo
no meio de reacccedilatildeo apropriado (Ref) A velocidade de desoligomerizaccedilatildeo da espeacutecie decameacuterica
pode ser analisada por espectroscopia de absorccedilatildeo na regiatildeo UVVis uma vez que esta espeacutecie
apresenta duas bandas caracteriacutesticas na regiatildeo do visiacutevel (a 358 e 400 nm) que natildeo se verifica
em quaisquer outras espeacutecies oligomeacutericas As bandas supramencionada sapresentam-se bem
22
Meio de reacccedilatildeo 3125 mM Tris-HCl pH 71 375 mM MgCl2
48
resolvidas atraveacutes da obtenccedilatildeo da derivada de 2ordf ordem do espectro apresentando a forma de 2
vales com miacutenimos centrados nos comprimentos de onda supracitados com dependecircncia linear
relativamente agrave concentraccedilatildeo de decavanadato total (Ramos et al 2006)
Assim a estabilidade da espeacutecie decameacuterica apoacutes diluiccedilatildeo da soluccedilatildeo de decavanadato no meio de
reacccedilatildeo dos ensaios (tampatildeo D) para uma concentraccedilatildeo final de 1 mM em vanadato total foi
caracterizada atraveacutes da leitura da Abs400 em funccedilatildeo do tempo tendo-se determinado qual a
ordem de reacccedilatildeo e o tempo de meia-vida da mesma Com base em estudos anteriores acerca da
estabilidade do V10 na presenccedila de diferentes proteiacutenas (Ramos et al 2006) determinou-se
igualmente o tempo de meia vida apoacutes a adiccedilatildeo de 20 microM de S1 de miosina ao meio reaccional
353 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
Os espectros de RMN do isoacutetopo 51
V foram realizados num espectroacutemetro Bruker AM-400 MHz
equipado com uma sonda multinuclear inversa de 5 mm Em tubos de RMN de 5 mm de diacircmetro
preparou-se soluccedilotildees de 500 L de deca- ou metavanadato correspondentes a um intervalo de
concentraccedilotildees de 1 a 10 mM suplementadas com 10 D2O Os espectros foram adquiridos agrave
temperatura ambiente com uma largura espectral de 23 923 Hz um tempo de aquisiccedilatildeo de 0086
s tempo de relaxaccedilatildeo de 001 s e uma sequecircncia de pulsos a 90ordm Utilizou-se a transformada de
Fourier com alargamento das bandas de 20Hz para o processamento dos espectros Os desvios
quiacutemicos das ressonacircncias de 51
V foram obtidos em relaccedilatildeo agrave espeacutecie VOCl3cuja frequecircncia de
ressonacircncia eacute considerada o padratildeo sendo todas as outras referenciadas a partir desta em ppm
As aacutereas dos sinais do RMN correspondentes agraves espeacutecies oligomeacutericas presentes foram obtidas
por integraccedilatildeo dos sinais As concentraccedilotildees relativas das espeacutecies foram calculadas com recurso agrave
equaccedilatildeo
onde A[Vx] corresponde agrave aacuterea referente agrave ressonacircncia da espeacutecie Vx com nordm de agregaccedilatildeo x
A[V]total agrave soma total das aacutereas medidas e [V] a concentraccedilatildeo total de vanaacutedio no ensaio
A teacutecnica de 51
V-RMN foi anteriormente aplicada em estudos de ligaccedilatildeo com S1 de miosina
(Ref) tendo sido demostrado que entre as espeacutecies V1 V2 V4 e V5 a espeacutecie tetrameacuterica possui a
maior afinidade para a miosina (Cremo et al) O desaparecimento ou alargamento dos sinais pode
implicar a ligaccedilatildeo de um dos oxovanadatos laacutebeis agrave miosina sem indicaccedilatildeo de qual uma vez que
49
se processa um equiliacutebrio entre eles (Crans 1990) Assim e tendo em conta as indicaccedilotildees de que
a espeacutecie V10 eacute distinta das restantes no que diz respeito agrave magnitude da interacccedilatildeo com a miosina
e inibiccedilatildeo da actividade ATPaacutesica estimulada por actina (Aureliano 2000 Tiago et al 2002)
torna-se fundamental investigar a interacccedilatildeo das diferentes espeacutecies de vanadato com o S1 de
miosina atraveacutes da teacutecnica de 51
V-RMN
Para tal obtiveram-se espectros de 51
V-RMN resultantes da titulaccedilatildeo de uma soluccedilatildeo 5 mM de
decavanadato total com concentraccedilotildees crescentes de S1 de miosina (0 5 10 15 e 20 microM) em
tampatildeo D Realizou-se ainda ensaios na presenccedila de 2 mM de decavanadato e 2 mM de ATP na
presenccedila e na ausecircncia de S1 de miosina de modo a verificar o efeito que estes ligandos naturais
da enzima teria na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o subfragmento-1
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
Os grupos sulfidrilo (-SH) das cisteiacutenas satildeo resiacuteduos de aminoaacutecidos extremamente sensiacuteveis agrave
oxidaccedilatildeo resultando na sua oxidaccedilatildeo (Dalle-Donne et al 1999) Logo a quantificaccedilatildeo destes
resiacuteduos eacute bastante importante nos estudos de interacccedilatildeo com vanaacutedio jaacute que os resiacuteduos de Cys
com ecircnfase para o SH1 e o SH2 satildeo bastante importantes na monitorizaccedilatildeo das modificaccedilotildees
funcionais da regiatildeo da cabeccedila
O procedimento seguidamente apresentado foi originalmente descrito por Gutieacuterrez-Merino
(1987)) Consiste na reacccedilatildeo entre os grupos ndashSH natildeo oxidados das proteiacutenas e o reagente de
Ellman (DTNB -55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico)) - formando-se o aniatildeo CNT (aniatildeo 3-
carboxilato-4-nitrotiofenolato) numa estequiometria de 11 Este apresenta um maacuteximo de
absorvecircncia a 412 nm (Robyt 1990) Assim apoacutes a incubaccedilatildeo da proteiacutena na presenccedila do metal
a quantidade de cisteiacutenas reduzidas por proteiacutena determina-se atraveacutes dos valores de Abs412
Deste modo colocou-se num tubo de Eppendorf o volume apropriado de S1 dialisado para que a
concentraccedilatildeo no ensaio fosse de 1 mgmL Adicionou-se o volume apropriado de decavanadato
para que a sua concentraccedilatildeo fosse de 0 10 50 100 250 500 ou 1000 M Perfez-se com soluccedilatildeo
D ateacute um volume final de 1 mL e incubou-se a 25ordmC por 20 minutos Apoacutes este periacuteodo colocou-
se 780 L da mistura reaccional numa cuvette de quartzo e adicionou-se 220 L de soluccedilatildeo K23
23
Tampatildeo K Tampatildeo fosfatohidrogenofosfato de soacutedio 025 M pH 72
50
Adicionou-se 15 L de DTNB (em soluccedilatildeo K [DTNB]stock=100 mM) ao meio reaccional e
registou-se o valor de absorvecircncia medido equivalente agraves cisteiacutenas ldquoraacutepidasrdquo (SH1 e SH2
expostos ao solvente) Adicionou-se 001 g de SDS que provoca a desnaturaccedilatildeo da proteiacutena e
consequentemente a exposiccedilatildeo de todos os resiacuteduos de cisteiacutenas que se encontravam ateacute aiacute
inacessiacuteveis no interior da proteiacutena (cisteiacutenas totais) Estes resiacuteduos podem assim reagir com o
DTNB
Estimou-se o nuacutemero de resiacuteduos de cisteiacutenas em todas as amostras com recurso ao coeficiente de
extinccedilatildeo molar () do DTNB (ver ponto 3 dos Anexos) que foi determinado atraveacutes da titulaccedilatildeo
de L-Cisteiacutena com intervalo de concentraccedilotildees compreendidos entre 0 e 50 microM na presenccedila de
25microM de DTNB em tampatildeo D Os valores de absorvecircncia foram obtidos para um comprimento de
onda de 412 nm
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
3621 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
A teacutecnica mais eficiente de anaacutelise da interacccedilatildeo entre o iatildeo vanadilo e proteiacutenas como a
calmodulina e a transferrina entre outras permite a obtenccedilatildeo raacutepida de informaccedilatildeo no que toca agrave
conformaccedilatildeo e amplitude da referida interacccedilatildeo
Os espectros de Ressonacircncia Paramagneacutetica Electroacutenica (RPE) da espeacutecie paramagneacutetica V(IV)
foram realizados a 70K num espectroacutemetro Bruker EMX-300 X-band Todas as soluccedilotildees
utilizadas e amostras foram preparadas em anaerobiose numa cacircmara de luvas de modo a evitar a
reduccedilatildeo de V(IV) a V(V) Em tubos capilares com 02 mL de amostra manteve-se a concentraccedilatildeo
de S1 a 20 M e variou-se a concentraccedilatildeo de vanadilo (0 25 5 10 15 25 50 75 100 150
200 500 e 1000 M [VOSO4]stock=5 mM) em tampatildeo D Apoacutes obtenccedilatildeo dos espectros estes
foram tratados atraveacutes do software SimFonia e WinEPR tendo-se obtido um graacutefico com a
variaccedilatildeo da intensidade do sinal de RPE em funccedilatildeo da concentraccedilatildeo de sulfato de vanadilo
presente Realizaram-se ainda simulaccedilotildees de espectros obtidos com 20 microM de S1 e 1000 microM de
V(IV) com vista agrave obtenccedilatildeo dos ligandos provaacuteveis do vanadilo atraveacutes da anaacutelise da constante
de acoplamento hiperfina (A) e do factor g (g)
4 Conclusotildees gerais
Apesar de jaacute ter sido demonstrado que o vanaacutedio inibe a Na+K
+-ATPase da miosina a funccedilatildeo
deste elemento no tecido muacutesculo esqueleacutetico continua largamente incompreendida A enorme
complexidade quiacutemica deste metal especialmente no que concerne agrave oligomerizaccedilatildeo e reacccedilotildees
51
redox constitui um obstaacuteculo agrave compreensatildeo do comportamento do vanaacutedio nos organismos
vivos Assim a caracterizaccedilatildeo da quiacutemica deste elemento eacute de grande utilidade antes de iniciar o
estudo da sua interacccedilatildeo com o presente sistema
A anaacutelise por espectroscopias de 51
V-RMN e de UV-Vis de soluccedilotildees de vanadato diluiacutedas nos
meios de reacccedilatildeo utilizados no presente relatoacuterio (10 mM HEPES pH 70 25 mM KCl) mostrou
claramente que a espeacutecie decameacuterica (V10) possui um tempo de vida suficientemente longo para a
determinaccedilatildeo do seu efeito isolado ao contraacuterio do que se verifica com as espeacutecies mono- di-
tetra- e pentameacutericas (V1 V2 V4 e V5) Por outro lado a anaacutelise verificou ainda que apoacutes diluiccedilatildeo
das soluccedilotildees de decavanadato no meio de reacccedilatildeo coexistem as espeacutecies V10 e V1 pelo que os
efeitos putativos desta uacuteltima deveratildeo ser subtraiacutedos aquando da monitorizaccedilatildeo dos efeitos da
espeacutecie decameacuterica
O vanaacutedio (V) (vanadato) inibe as enzimas que catalizam reacccedilotildees de transferecircncia de grupos
fosfato como a ATPase da miosina [Goodno 1979] devido agrave sua analogia estrutural com o iatildeo
fosfato Na sua utilizaccedilatildeo como inibidor enzimaacutetico natildeo eacute tida em conta a contribuiccedilatildeo individual
das espeacutecies oligomeacutericas considerando-se normalmente a espeacutecie V1 como inibidora efectiva
[Goodno e Taylor 1982] No presente relatoacuterio verificou-se por espectroscopia de 51
V-RMN que
outros oligoacutemeros de vanadato interagem com o S1 de miosina tendo as espeacutecies V10 e V4 os
maiores niacuteveis de afinidade A sua interacccedilatildeo envolve forccedilas electrostaacuteticas e eacute afectada
negativamente por ligandos naturais de enzima como eacute o caso do ATP
A exposiccedilatildeo do S1 de miosina (005 mgmL) a decavanadato conduz agrave inibiccedilatildeo do capacidade da
proteiacutena de estimular a actividade Mg2+
-ATPaacutesica com um valor de IC50 de 266 plusmn 07 M
decavanadato sendo o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM
decavanadato (80) Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM decavanadato sendo
o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM decavanadato (60)
exercendo um efeito de impedimento agrave ligaccedilatildeo entre o vanaacutedio e o S1
A uma concentraccedilatildeo de 27 microM decavanadato verifica-se a oxidaccedilatildeo de um dos resiacuteduos de Cys
de S1 de miosina seguida pela oxidaccedilatildeo gradual de outro resiacuteduo de Cys ateacute agrave concentraccedilatildeo de
1000 microM Ambos os resiacuteduos oxidados natildeo se encontravam expostos ao solvente utilizado A
oxidaccedilatildeo dos resiacuteduos permite sugerir que esta modificaccedilatildeo oxidativa sofrida pelo S1 contribui
para a diminuiccedilatildeo da sua actividade de ATPase
52
Por espectroscopia de RPE revelaram-se os possiacuteveis ligandos equatoriais do vanaacutedio aquando da
sua interacccedilatildeo com o S1 de miosina como sendo duas moleacuteculas de aacutegua um ligando contendo o
grupo R-OH e outro contendo o grupo R=NR
Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados de
oxidaccedilatildeo (IV) e (V) no S1 de miosina para concentraccedilotildees facilmente alcanccedilaacuteveis no ambiente
terrestre
53
5 Estudos futuros
No sentido do melhor esclarecimento das alteraccedilotildees moleculares e estruturais que ocorrem no
subfragmento 1 quando exposta ao vanaacutedio que conduzem a uma diminuiccedilatildeo da sua funccedilatildeo
sugere-se
- Quantificaccedilatildeo das cisteiacutenas reduzidas de S1 na presenccedila de metavanadato
- Determinaccedilatildeo das actividades Ca2+
- e K+-ATPaacutesica do S1 de miosina na presenccedila de
decavanadato
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina estimulada por actina
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina na presenccedila de
metavanadato
- Realizaccedilatildeo de RMN 2D com variaccedilatildeo das concentraccedilotildees das soluccedilotildees de decavanadato e
metavanadato na presenccedila de S1
-Mutageacutenese dos resiacuteduos de Cys doS1 de miosina de modo a verificar quais os resiacuteduos que
sofrem oxidaccedilatildeo na presenccedila de decavanadato
- Determinaccedilatildeo por calorimetria (DSC Differential Scanning Calorimetry) da variaccedilatildeo da
entalpia (∆H) e da temperatura de desnaturaccedilatildeo do S1 de miosina na presenccedila de vanaacutedio uma
vez que estas propriedades caracteriacutesticas de cada proteiacutena satildeo alteradas apoacutes ocorrecircncia de
modificaccedilotildees estruturais
- Anaacutelise por SDS-PAGE de S1 de miosina apoacutes tratamento com vanaacutedio de modo a determinar
quais os polipeacuteptidos eventualmente formados apoacutes clivagem da proteiacutena
54
6 Referecircncias bibliograacuteficas
- Sellers J Myosins 2nd Edition Oxford University Press 1999
- L Stryer inBiochemistry (Ed WH Freeman 2005 pp
Refordfas Materiais e Meacutetodos
[A] S S Margossian SLowey Methods Enzymol1982 85 55-71
[B] JD Pardue JA Spudich Methods Enzymol 1982 85 164-181
[C] M Bradford Anal Biochem1976 72 248-254
[D] H H Taussky E Shorr J Biol Chem 1953 202 675-685
[E] C R Bagshaw in Outline Studies in Biology ndash Muscle Contraction Chapman and Hall Ed
1982
[F] A Cuenda et al Eur J Biochem1990 194 663-670
55
Anexos
1 Determinaccedilatildeo da recta de calibraccedilatildeo meacutetodo de Bradford
Figura1ndashRepresentaccedilatildeo graacutefica da recta de calibraccedilatildeo (Abs595 nm vs [BSA]) utilizada para o meacutetodo de Bradford
2 Determinaccedilatildeo da recta calibraccedilatildeo para a electroforese em condiccedilotildees desnaturantes
Figura 2 ndash Representaccedilatildeo graacutefica da recta de calibraccedilatildeo (log(Peso Molecular) vs Rf dos marcadores de peso
molecular) onde Rf indica determinada atraveacutes do padratildeo de peso molecular
(poccedilo (a))
3Caacutelculo do coeficiente de extinccedilatildeo molar () para a quantificaccedilatildeo de cisteiacutenas reduzidas
atraveacutes do meacutetodo de Ellman
Traccedilou-se a recta de Abs412 nm meacutedia vs [L-Cisteiacutena] de modo a determinar o valor do coeficiente
de extinccedilatildeo molar definido como o declive daquela recta Assim =8400 M-1
cm-1
como
podemos verificar na Fig 3
A595 nm = 00546[BSA] + 00209 Rsup2 = 099051
Ab
s 59
5 n
m
[BSA] (mgmL)
log PM = -1177 Rf + 2247 Rsup2 = 098957
log
PM
Mobilidade relativa (Rf)
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero

37
3 Procedimento Experimental
31 Reagentes
Todos os reagentes utilizados satildeo de elevado grau de pureza (analiacutetico) Apresentam-se as suas
foacutermulas quiacutemicas ou abreviaturas quando necessaacuterio acrescentou-se a sua descriccedilatildeo Entre
parecircntesis colocou-se as empresas que os comercializam
Aacutecido aceacutetico glacial 100 (Riedel) Acrilamida 99 (Merck) Azul de bromofenol (Riedel-de-
Haumlen) -Mercaptoetanol (Sigma) Bissulfito de soacutedio (Sigma)Bradford (reagente de)
(BioRad)BSA (albumina de soro bovino Sigma-Aldrich) CaCl22H2O (cloreto de caacutelcio
dihidratado Fluka) CH3CH2OH 96 (etanol Panreac) Coomassie Blue R250 (Merck) D2O
(oacutexido de deuteacuterio SDS) DTNB (55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico) Aldrich) EDTA (aacutecido
etilenodiaminatetraaceacutetico Merck) Fosfoenolpiruvato 98 (Fluka) Glicerol (Sigma) Glicina
985 (Panreac) HCl 37 (Riedel-de Haumlen) H2SO4 96 (Pronalab) HEPES 99 (aacutecido 4-(2-
hidroxiletil)-1-piperazineetanosulfoacutenico Sigma) KH2PO4 99 (hidrogenofosfato de potaacutessio)
MgATP (Sigma) Na
ATP (Sigma) Na2HPO4 99 (hidrogenofosfato de soacutedio Riedel-de Haumlen)
NaH2PO4 (fosfato de soacutedio ) NaOH 986 (hidroacutexido de soacutedio Pronalab) KCl 99 (Cloreto de
potaacutessio Panreac) L-Cys 995 (enantioacutemero levoacutegiro do resiacuteduo de Cisteiacutena Fluka)Metanol
99 (Merck) Molibdato de amoacutenio (Sigma)NNrsquo-metil-bis-acrilamida 98 (Merck) NH4VO3
99 (metavanadato de amoacutenia Sigma) PSA 10 (persulfato de amoacutenio Merck) SDS
(dodecilo sulfato de soacutedio Sigma) Sulfito de soacutedio (Sigma) VOSO4 (sulfato de vanadilo
Aldrich) TEMED 99 (Tetrametiletilenodiamina Merck) Tris (2-amino-2-hidroximetil-13-
propanodiol Sigma)
32 Meacutetodos
321 Isolamento e purificaccedilatildeo das proteiacutenas de muacutesculo esqueleacutetico de coelho
3211 Purificaccedilatildeo da miosina preparaccedilatildeo de Subfragmento 1 por digestatildeo quimotriacutepsica
da miosina
O subfragmento-1 de miosina-II (S1) foi gentilmente cedido pelo Prof Doutor Manuel Aureliano
do Departamento de Quiacutemica Bioquiacutemica e Farmaacutecia da Faculdade de Ciecircncias e Tecnologia da
Universidade do Algarve
No entanto descreve-se abaixo de forma resumida o procedimento efectuado
38
32111 Purificaccedilatildeo da miosina
O protocolo de isolamento foi adaptado do meacutetodo previamente descrito por Margossian e Lowey
(1982) apresentando-se de forma esquemaacutetica na Fig 31
Sucintamente os tecidos musculares dorsais e das pernas foram extraiacutedos apoacutes o sacrifiacutecio do
animal (Fig31) De seguida foram adicionados 3 volumes da soluccedilatildeo de extracccedilatildeo A2
Centrifugou-se a soluccedilatildeo e filtrou-se o sobrenadante O sedimento foi entatildeo diluiacutedo em 14
volumes de aacutegua destilada de modo a que a forccedila ioacutenica atingisse 004 M Apoacutes adiccedilatildeo de szlig-
mercaptoetanol (szlig-me) deixou-se a precipitar a 4ordmC removendo o sobrenadante Centrifugou-se
novamente e ressuspendeu-se o sedimento na soluccedilatildeo de extracccedilatildeo A Adicionou-se a soluccedilatildeo
tampatildeo B3 para que a forccedila ioacutenica atingisse 05 M Homogeneizou-se e deixou-se repousar
durante 2h Homogeneizou-se novamente e centrifugou-se de modo a ocorrer precipitaccedilatildeo dos
complexos de actomiosina e proteiacutena desnaturada Apoacutes filtraccedilatildeo do sobrenadante em latilde de vidro
quantificou-se a soluccedilatildeo de miosina resultante pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental)
2 Soluccedilatildeo de extracccedilatildeo A 0012M K2CO3 004M KHCO3 06M KCl pH 65
3 Soluccedilatildeo-tampatildeo B Tris 100 mM pH 7 KCl 2M
39
Figura 31 ndash Representaccedilatildeo esquemaacutetica do protocolo de isolamento de miosina (Margossian e Lowey 1982) Todo o
procedimento foi realizado a 4ordmC (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
32112 Preparaccedilatildeo de Subfragmento-1 de miosina por digestatildeo com quimiotripsina
O S1 de miosina foi isolado apoacutes diaacutelise da miosina durante a noite contra a soluccedilatildeo C4 A
presenccedila do EDTA eacute essencial uma vez que a sua substituiccedilatildeo por MgCl2resulta na produccedilatildeo de
HMM em vez de S1 A suspensatildeo de miosina foi equilibrada e digerida com -quimotripsina
A digestatildeo foi interrompida pela adiccedilatildeo de PMSF Seguidamente ultracentrifugou-se o produto
da digestatildeo desprezando-se o sedimento (Fig 32) A soluccedilatildeo de sobrenadante maioritariamente
S1 foi quantificada pela leitura directa da Abs280 ou pelo meacutetodo de Bradford (ver as secccedilotildees
3311 e 3312 do Procedimento Experimental respectivamente) e armazenada em 10 sacarose
e 2 mM szlig-Me a -80ordmC
4 Soluccedilatildeo C 25mM Tris pH 70 012M KCl 1mM EDTA
40
Antes da realizaccedilatildeo dos ensaios descritos no presente relatoacuterio o S1 de miosina foi dialisado
contra a soluccedilatildeo D5 de modo a remover qualquer composto que pudesse complexar como
vanaacutedio Seguidamente quantificou-se a proteiacutena pelo meacutetodo de Bradford ou pelo meacutetodo
directo (ver secccedilotildees 3311 e 3312do Procedimento Experimtental)
Figura 3 2ndash Representaccedilatildeo esquemaacutetica do protocolo de preparaccedilatildeo do S1 de miosina por digestatildeo quimiotripsina de
acordo com o previamente descrito por Margossian e Lowey (1982) Todo o procedimento foi realizado a 4ordm C
excepto quando indicaccedilatildeo do contraacuterio (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
3212 Isolamento e purificaccedilatildeo de actina
Isolou-se e purificou-se a actina segundo o meacutetodo descrito por Pardee e Spudish que
compreende dois passos ndash a obtenccedilatildeo de um resiacuteduo seco denominado ldquopoacute de acetonardquo e a
extracccedilatildeo subsequente da actina a partir do ldquopoacute de acetonardquo na forma de G-Actina com a
posterior polimerizaccedilatildeo a F-Actina
32121 Obtenccedilatildeo do ldquopoacute de acetonardquo
Sacrificou-se o coelho com uma pancada atraacutes do pescoccedilo e sangrou-se o animal apoacutes o corte da
arteacuteria jugular Esfolou-se e colocou-se numa tina com gelo Retiraram-se as partes de tecido
conjuntivo que envolvem o tecido muscular com auxiacutelio de uma pinccedila e bisturi Seguidamente
retirou-se os muacutesculos dorsais e os muacutesculos brancos das pernas Passou-se os muacutesculos por aacutegua
destilada de modo a remover as impurezas e pesou-se num copo Trituraram-se os muacutesculos
numa maacutequina de triturar carne e extraiu-se o resultante com 3 volumes da soluccedilatildeo E6 durante 10
minutos com agitaccedilatildeo suave Centrifugou-se a suspensatildeo a 4000 g durante 10
5 Soluccedilatildeo D HEPES 10 mM pH 70 KCl 25 mM
6 Soluccedilatildeo E 01 M KCl 015 M K2HPO4 pH 65
41
minutosdesprezando-se o sobrenadante Ressuspendeu-se o sedimento em 6 volumes de soluccedilatildeo
F7durante 10 minutos com agitaccedilatildeo suave Verificou-se se o pH da soluccedilatildeo estava compreendido
entre 82 e 85 acertando-se estecom soluccedilatildeo G8 se necessaacuterio Centrifugou-se durante 10
minutos a 4000 g Desprezou-se o sobrenadante e ressuspendeu-se o sedimento em 3 volumes de
soluccedilatildeo H9 durante 10 minutos Centrifugou-se novamente a 4000 gdurante 10 minutos
desprezando-se o sobrenadante Ressuspendeu-se o sedimento duas vezes consecutivas em 6
volumes de aacutegua destilada durante 5 minutos Centrifugou-se a 4000 g durante 10 minutos apoacutes
cada adiccedilatildeo de H2O
Seguidamente ressuspendeu-se o sedimento 5 vezes consecutivas utilizando 3 volumes de
acetona de cada vez agitando suavemente durante 10 minutos agrave temperatura ambiente Colocou-
se o resiacuteduo filtrado numa tina de vidro sobre papel de filtro deixando-se evaporar o solvente
durante a noite num nicho Triturou-se o resiacuteduo seco numa maacutequina de triturar carne e deixou-se
novamente evaporar o solvente O ldquopoacute de acetonardquo obtido foi armazenado a -20 ordmC
32122 Purificaccedilatildeo da actina a partir de ldquopoacute de acetonardquo
Apoacutes a obtenccedilatildeo do poacute de acetona como acima indicado efectuou-se o protocolo seguidamente
descrito de modo a obter monoacutemeros de actina (G-Actina) e actina filamentosa (F-Actina)
Pesou-se 35 g de poacute de acetona e adicionaram-se 50 mL de soluccedilatildeo-tampatildeo I10
sendo a
extracccedilatildeo feita com agitaccedilatildeo suave durante 30 minutos Separou-se o extracto do poacute de acetona
hidratado filtrando a suspensatildeo com 4 camadas de gaze esterilizada tendo o cuidado de natildeo
espremer o resiacuteduo Voltou a extrair-se com 50 mL de soluccedilatildeo I durante 30 minutos mexendo
lentamente Adicionou-se este ao extracto obtido anteriormente Centrifugou-se o extracto total a
16 000 g durante 30 minutos Decantou-se para uma proveta sobre a qual se colocou gaze para
que o sedimento solto natildeo se juntasse ao sobrenadante e leu-se o volume De seguida adicionou-
se KCl ateacute uma concentraccedilatildeo final de 50 mM MgCl2 ateacute 2mM e ATP ateacute 1mM lentamente e com
agitaccedilatildeo contiacutenua de modo a promover a polimerizaccedilatildeo dos monoacutemeros de actina
Deixou-se polimerizar durante 2h a 25ordmC observando-se a presenccedila de pequenas bolhas de ar
indicativas de um aumento de viscosidade da soluccedilatildeo Arrefeceu-se em gelo durante 15 minutos e
adicionou-se KCl ateacute uma concentraccedilatildeo final de 06M Mexeu-se lentamente durante 30 minutos
apoacutes os quais se ultracentrifugou a 110 000g durante 1hora Desprezou-se o sobrenadante
ressuspendeu-se o sedimento com soluccedilatildeo tampatildeo E e transferiu-se para um potter
Dialisou-se durante a noite contra 1L de soluccedilatildeo-tampatildeo I a uma temperatura de 4ordmC com rotaccedilatildeo
7 Soluccedilatildeo F 005 M NaHCO3 pH 82-85
8 Soluccedilatildeo G 1M Na2CO3
9 Soluccedilatildeo H 1 mM EDTA pH 70
10 Soluccedilatildeo I 2 mM Tris-HCl pH 8 02 mM CaCl2 02 mM Na2ATP 05 mM szlig-Me 0005 NaN3
42
contiacutenua e mudanccedila de tampatildeo 2 a 3 vezes De seguida a actina foi centrifugada a 110
000gdurante 1hora Decantou-se a soluccedilatildeo de G-Actina (sobrenadante) e desprezou-se o
sedimento Determinou-se a concentraccedilatildeo de G-Actina pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental) A soluccedilatildeo de G-actina foi armazenada a uma temperatura de 4ordmC
apoacutes adiccedilatildeo de 002NaN3 mantendo-se estaacutevel durante 3 a 5 dias
331 Quantificaccedilatildeo das proteiacutenas isoladas
3311 Quantificaccedilatildeo pelo meacutetodo de Bradford
O meacutetodo de Bradford (Bradford 1976) baseia-se na utilizaccedilatildeo do reagente de Bradford
composto por entre outros aacutecido fosfoacuterico e o corante ldquoCoomassierdquo Este corante possui carga
negativa e a sua coloraccedilatildeo varia entre a vermelha (Absmax=465 nm) predominante em soluccedilatildeo e a
azul (Absmax=595 nm) presente aquando da ligaccedilatildeo aos grupos amina das proteiacutenas Em soluccedilotildees
aacutecidas diluiacutedas a coloraccedilatildeo varia proporcionalmente com a quantidade do corante ligado agrave
proteiacutena que por sua vez eacute directamente proporcional agrave concentraccedilatildeo da proteiacutena permitindo a
sua quantificaccedilatildeo ateacute a um limite de detecccedilatildeo de 20 gmL (Bradford 1976)
Deste modo prepararam-se amostras de BSA ([BSA]stock= 1mgmL) utilizando um intervalo de
concentraccedilotildees compreendido entre 0 e 15 gmL de acordo com a tabela 3I Apoacutes a adiccedilatildeo da
H2O MilliQ e do reagente de Bradford as soluccedilotildees foram sucessivamente homogeneizadas no
voacutertex
Tabela 3Indash Amostras utilizadas para a obtenccedilatildeo da recta de calibraccedilatildeo no meacutetodo de Bradford
BSA (L) BSA (g) H2O MilliQ (L) Reagente de Bradford (L)
0 0 800 200
125 125 79875 200
25 25 7975 200
5 5 795 200
10 10 790 200
15 15 785 200
Apoacutes incubaccedilatildeo a 25ordmC por 5 minutos leram-se os valores de Abs595 num
espectrofotoacutemetro (Shimadzu UV-160A) para todas as amostras Seguidamente traccedilou-se a recta
de calibraccedilatildeo (Abs595vs quantidade de BSA (g))
43
Para determinar a concentraccedilatildeo de proteiacutena apoacutes o processo de isolamento purificaccedilatildeo e
diaacutelise quando necessaacuterio colocou-se 5 L da amostra num tubo de Eppendorf e adicionou-se
795 L de H2O MilliQ homogeneizando a soluccedilatildeo no voacutertex Adicionou-se posteriormente 200
L de reagente de Bradford e repetiu-se a homogeneizaccedilatildeo Incubou-se a mistura a 25ordmC por 5
minutos apoacutes os quais se leu os valores de Abs595 nm no espectrofotoacutemetro Por interpolaccedilatildeo deste
valor a partir da recta de calibraccedilatildeo determinou-se a concentraccedilatildeo de proteiacutena na amostra
3312 Quantificaccedilatildeo pelo meacutetodo directo
A quantificaccedilatildeo das amostras proteicas pode tambeacutem ser efectuada atraveacutes da leitura directa da
absorvacircncia tendo por base a lei de Beer-Lambert onde Abs corresponde agrave
absorvacircncia ao comprimento de onda ao qual o ensaio foi realizado corresponde ao coeficiente
de extinccedilatildeo (substituiacutedo pela letra a se as unidades forem distintas da molaridade) b corresponde
ao percurso oacuteptico da radiaccedilatildeo na ceacutelula amostral e c corresponde agrave concentraccedilatildeo amostral
Assim no caso de S1 de miosina mediu-se a Abs280e calculou-se a concentraccedilatildeo da proteiacutena
sabendo que o coeficiente de extinccedilatildeo de S1 eacute (Ref)
No caso da actina mediu-se a Abs290ecalculou-se a concentraccedilatildeo da proteiacutena sabendo que o
coeficiente de extinccedilatildeo da actina eacute (Ref)
332Electroforese em condiccedilotildees desnaturantes (SDS-PAGE)
De modo a melhor caracterizar as proteiacutenas isoladas foi realizada uma electroforese em
condiccedilotildees desnaturantes (SDS-PAGE do inglecircs Sodium Dodecyl Sulfate Polyacrilamide Gel
Electrophoresis) O SDS (dodecilo sulfato de soacutedio) eacute um detergente anioacutenico utilizado neste
meacutetodo para desnaturar as proteiacutenas que se ligam ao SDS na proporccedilatildeo 14 g SDS g proteiacutena
(aproximadamente 1 moleacutecula de SDS por cada 2 resiacuteduos de aminoaacutecidos) (Stryer) O SDS induz
uma densidade de carga negativa significativa nas proteiacutenas a que se liga mascarando a sua carga
intriacutenseca pelo que as razotildees cargamassa satildeo tendencialmente idecircnticas Assim a SDS-PAGE
separa as proteiacutenas quando sujeitas a um campo eleacutectrico de acordo com o seu peso molecular
sendo a sua migraccedilatildeo directamente proporcional ao logaritmo deste uacuteltimo (Stryer)
O gel de resoluccedilatildeo com 10 de acrilamida foi obtido atraveacutes da adiccedilatildeo de 167 mL de soluccedilatildeo
stock de acrilamida11
075 mL de tampatildeo de resoluccedilatildeo12
251 mL de aacutegua destilada e 50 L de
SDS 10 Para promover a polimerizaccedilatildeo adicionou-se em seguida 38 L de PSA 10
11
soluccedilatildeo stock de acrilamida soluccedilatildeo de 30 (mv) de acrilamida e 08 (mv) de NNrsquo-metil-bis-acrilamida 12
tampatildeo de resoluccedilatildeo 15 M Tris-HCl pH 88
44
(persulfato de amoacutenia) 10 e 25 L de TEMED 99 (tetrametiletilenodiamina) Agitou-se
cuidadosamente de modo a natildeo formar bolhas e pipetou-se a soluccedilatildeo para a cassette de vidro
previamente montada no suporte De seguida adicionou-se uma peliacutecula de cerca de 2 mm de 2-
metil-2-butanol para verificar posteriormente a polimerizaccedilatildeo Apoacutes a polimerizaccedilatildeo que demora
cerca de 45 minutos verteu-se a agrave superfiacutecie e adicionou-se o gel de concentraccedilatildeo com 5 de
acrilamida constituiacutedo por 03 mL de soluccedilatildeo de stock acrilamida 045 mL de tampatildeo de
concentraccedilatildeo13
094 mL de aacutegua destilada 18 L de SDS 10 135 L de PSA 10 e 2L de
TEMED 99 Colocou-se o pente sobre o gel e deixou-se polimerizar durante cerca de 45
minutos
As amostras para aplicaccedilatildeo no gel de electroforese foram preparadas para uma concentraccedilatildeo de
proteiacutena de 1 mgmL num volume final de 100 L dos quais 25 L correspondem ao tampatildeo da
amostra14
As amostras foram aquecidas a 100ordmC durante 5 minutos Antes da aplicaccedilatildeo das
amostras transferiu-se a cassette para a tina de electroforese agrave qual se adicionou a soluccedilatildeo
tampatildeo de electroforese da tina superior15
e tampatildeo de electroforese da tina inferior16
Aplicaram-
se 10 L das soluccedilotildees de marcadores de peso molecular e 20 L das amostras proteicas
preparadas anteriormente A electroforese decorreu durante cerca de 3h30 min a uma voltagem
de 80 V De seguida transferiu-se o gel para uma tina corando as proteiacutenas durante 1h com
soluccedilatildeo corante17
Descorou-se o gel com soluccedilatildeo descorante18
tendo-se mudado esta soluccedilatildeo ateacute
as bandas se encontrarem bem definidas
Determinou-se a mobilidade relativa das bandas correspondentes aos marcadores de elevado peso
molecular e traccedilou-se a recta de calibraccedilatildeo ndash log(PM) vs log(Rf) onde Rf corresponde agrave
mobilidade relativa Por interpolaccedilatildeo determina-se o PM das proteiacutenas isoladas sabendo a sua
mobilidade relativa Por comparaccedilatildeo com os valores descritos na literatura pode concluir-se
acerca do grau de purificaccedilatildeo
34 Mediccedilatildeo da actividade enzimaacutetica de S1
341 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
13
tampatildeo de concentraccedilatildeo 05 M Tris-HCl pH 68 14
tampatildeo de amostra 320 mM Tris-HCl pH 68 04 M szlig-mercaptoetanol SDS 8 glicerol 15 (vv) e roxo de m-
cresol 0024 15
tampatildeo de electroforese da tina superior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS 16
tampatildeo de electroforese da tina inferior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS acetato de soacutedio
01 M 17
soluccedilatildeo corante ldquoCoomassie Blue R-250rdquo contendo metanol 45 (vv) e aacutecido aceacutetico glacial 10 (vv) 18
soluccedilatildeo descorante metanol 10 (vv) e aacutecido aceacutetico glacial 10 vv)
45
O meacutetodo de Fiske-Subarow modificado consiste na reacccedilatildeo do molibdato de amoacutenio com o
fosfato inorgacircnico em soluccedilatildeo aacutecida com formaccedilatildeo do aacutecido fosfomoliacutebdico Seguidamente daacute-
se a adiccedilatildeo de um agente redutor que provoca a reduccedilatildeo do composto a fosfomolibdato que forma
um complexo de cor azulada cuja absorvacircncia maacutexima ocorre para um comprimento de onda de
660 nm tornando possiacutevel a sua detecccedilatildeo por meacutetodos espectrofotomeacutetricos As actividades Ca2+
-
ATPaacutesica e K+EDTA-ATPaacutesica de S1 foram determinadas atraveacutes deste meacutetodo de acordo com
o descrito por Taussky e Shorr (1953) com mediccedilatildeo da quantidade de fosfato (Pi) libertado
atraveacutes da hidroacutelise de ATP
Para tal obteve-se primeiramente a recta de calibraccedilatildeo com as amostras presentes na Tabela 3II
utilizando uma soluccedilatildeo stock de 5 mM KH2PO4
Tabela 3II ndash Composiccedilatildeo das diferentes amostras utilizadas para determinar a recta de calibraccedilatildeo atraveacutes do meacutetodo
de Fiske-Subbarow modificado
KH2PO4 (L)
([KH2PO4]stock= 5mM)
PO43-
(nmole) H2O (uL)
0 0 1780
5 25 1775
10 50 1770
20 100 1760
40 200 1740
100 500 1680
A cada tubo de Eppendorf adicionaram-se seguidamente 200 L de soluccedilatildeo J19
e 20 L de agente
redutor20
homogeneizando no voacutertex apoacutes cada adiccedilatildeo Incubou-se durante 10 minutos e mediu-
se a Abs660 nm num espectrofotoacutemetro com intervalos de tempo iguais entre cada amostra Traccedilou-
se a recta de calibraccedilatildeo (Abs660 nmvs nmol PO43-
) a partir dos valores obtidos
Para determinar a actividade Ca2+
-ATPaacutesica de S1 prepararam-se tubos de Eppendorf com 400
L de meio de reacccedilatildeo21
25 L CaCl2 ([CaCl2]stock= 100 mM) 25 L ATP ([ATP]stock= 100 mM)
com [S1]ensaios=01 mgmL Perfez-se se necessaacuterio com H2O MilliQ ateacute perfazer um volume de 1
mL Para determinar a actividade K+EDTA-ATPaacutesica utilizou-se uma concentraccedilatildeo idecircntica de
19
Soluccedilatildeo J 5 N H2SO4 25 molibdato de amoacutenio 20
Agente redutor 5 mg aacutecido 1-amino-2-naftol-4-sulfoacutenico 30 mg bissulfito de soacutedio 30 mg sulfito de soacutedio em 26
mL H2O 21
meio de reacccedilatildeo 125 mM Tris-HCl pH 80 0575 M KCl
46
proteiacutena nos ensaios (01 mgmL) e o mesmo meio de reacccedilatildeo Aos tubos de Eppendorf
adicionou-se ainda 123 L de KCl ([KCl]stock = 3M) 25 L de ATP 10 L de EDTA
([EDTA]stock= 100 mM )e o volume de H2O MilliQ necessaacuterio para perfazer 1 mL Em ambas as
situaccedilotildees o uacuteltimo reagente adicionado foi o ATP uma vez que se trata do substrato da reacccedilatildeo
enzimaacutetica
A cineacutetica decorreu a 25ordmC durante 5 minutos para a actividade Ca2+
-ATPaacutesica e durante 3
minutos para a actividade K+EDTA-ATPaacutesica Apoacutes o periacuteodo de incubaccedilatildeo transferiu-se 150
L de amostra para tubos de Eppendorf contendo igual volume de TCA 10 Colocou-se no gelo
e homogeneizou-se no voacutertex Seguidamente centrifugou-se a 1500 g durante 10 minutos e
recolheu-se 200 L de sobrenadante para novos tubos de Eppendorf Adicionou-se a estes 158
mL de H2O MilliQ e 200 L de soluccedilatildeo J Homogeneizou-se no voacutertex e adicionou-se 20 L de
reagente redutor repetindo-se a homogeneizaccedilatildeo Incubou-se durante 10 minutos e mediu-se a
Abs660 nm com o mesmo intervalo de tempo de leitura para cada amostra Por intrapolaccedilatildeo a partir
da recta de calibraccedilatildeo obtida anteriormente determinou-se a quantidade de Pi libertado em cada
ensaio A partir destes resultados determinaram-se as actividades Ca2+
-ATPaacutesica e K+EDTA-
ATPaacutesica do S1 em nmol PO43-
minmg proteiacutena e em s-1
342 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
Da actividade Mg2+
-ATPaacutesica de S1 resulta a formaccedilatildeo dos produtos da reacccedilatildeo de
hidroacutelise do ATP - ADP e Pi Na presenccedila de ADP fosfoenolpiruvato (PEP) e da enzima piruvato
cinase (PK) os produtos resultantes satildeo ATP e piruvato Por acccedilatildeo da enzima lactato
desidrogenase (LDH) este uacuteltimo eacute convertido em lactato ocorrendo simultaneamente a oxidaccedilatildeo
do NADH a NAD+ monitorizada atraveacutes da Abs340 nm Quando o S1 de miosina se encontra no
meio reaccional o ATP eacute hidrolisado formando-se novamente ADP e Pi o que permite a
formaccedilatildeo de um ciclo (fig 21) Se as enzimas se encontrarem em excesso a velocidade de
conversatildeo do NADH eacute limitada pela produccedilatildeo de ADP e Pi Como o ATP eacute regenerado em cada
ciclo a sua concentraccedilatildeo inicial eacute sempre mantida (Bagshaw 1982)
Figura 33ndash Representaccedilatildeo esquemaacutetica do meacutetodo de enzimas acopladas
47
O protocolo utilizado e seguidamente descrito foi adaptado de Cuenda et al (1990 ndash Ver Refordf) A
uma cuvette de 1 mL adicionou-se S1 para uma concentraccedilatildeo final de 005 mgmL 800 L de
meio de reacccedilatildeo22
10 L de PEP ([PEP]stock = 42 mM) 10 L de PK+LDH 11 ([PK+LDH]stock =
1 mgmL) e 15 L de NADH ([NADH]stock=25 mM) Iniciou-se a monitorizaccedilatildeo da reacccedilatildeo
atraveacutes da leitura de Abs340 nm apoacutes a adiccedilatildeo de 20 L de ATP ([ATP]stock=100 mM) A
actividade Mg2+
-ATPaacutesica eacute obtida atraveacutes do declive correspondente agrave oxidaccedilatildeo do NADH Para
a mediccedilatildeo da Mg2+
-ATPase de S1 estimulada por actina adicionou-se F-actina para uma
concentraccedilatildeo final de 2 microM sendo a actividade obtida tambeacutem atraveacutes do declive correspondente
agrave oxidaccedilatildeo do NADH apoacutes a adiccedilatildeo da proteiacutena
35 Interaccedilatildeo entre vanaacutedio e subfragmento-1 de miosina
351 Preparaccedilatildeo das soluccedilotildees de vanaacutedio
As soluccedilotildees de vanadato foram preparadas em aacutegua MilliQ a partir de metavanadato de amoacutenio
(NH4VO3) numa concentraccedilatildeo stock de 50 mM com agitaccedilatildeo magneacutetica e aquecimento (~30 ordmC)
Esta soluccedilatildeo com um valor compreendido entre 6 e 7 designa-se por ldquometavanadatordquo e eacute
constituiacuteda pelas esspeacutecies V1 V2 V4 e V5 (Ref) Acidificando a soluccedilatildeo de metavanadato por
adiccedilatildeo de HCl ateacute um valor de pH proacuteximo de 4 observa-se uma mudanccedila de cor de amarelo
claro para laranja indicativa da formaccedilatildeo da espeacutecie decameacuterica de vanadato De seguida e
imediatamente antes da realizaccedilatildeo dos ensaios descritos o valor do pH da soluccedilatildeo de
decavanadato foi aumentado para cerca de 75 por adiccedilatildeo de NaOH A soluccedilatildeo (2) designada
por ldquometavanadatordquo natildeo foi posteriormente alterada ateacute agrave sua adiccedilatildeo para a realizaccedilatildeo dos ensaios
As soluccedilotildees foram armazenadas a 4ordmC e mantidas em gelo aquando da sua utilizaccedilatildeo
A soluccedilatildeo de V(IV) foi preparada em condiccedilotildees anaeroacutebias por diluiccedilatildeo de sulfato de vanadil
(VOSO4) para uma concentraccedilatildeo final de 5 mM em H2O MilliQ saturada com N2 gasoso
352Anaacutelise por espectroscopia de UV-Vis
A concentraccedilatildeo da espeacutecie decameacuterica em soluccedilatildeo depende do periacuteodo de tempo apoacutes a diluiccedilatildeo
no meio de reacccedilatildeo apropriado (Ref) A velocidade de desoligomerizaccedilatildeo da espeacutecie decameacuterica
pode ser analisada por espectroscopia de absorccedilatildeo na regiatildeo UVVis uma vez que esta espeacutecie
apresenta duas bandas caracteriacutesticas na regiatildeo do visiacutevel (a 358 e 400 nm) que natildeo se verifica
em quaisquer outras espeacutecies oligomeacutericas As bandas supramencionada sapresentam-se bem
22
Meio de reacccedilatildeo 3125 mM Tris-HCl pH 71 375 mM MgCl2
48
resolvidas atraveacutes da obtenccedilatildeo da derivada de 2ordf ordem do espectro apresentando a forma de 2
vales com miacutenimos centrados nos comprimentos de onda supracitados com dependecircncia linear
relativamente agrave concentraccedilatildeo de decavanadato total (Ramos et al 2006)
Assim a estabilidade da espeacutecie decameacuterica apoacutes diluiccedilatildeo da soluccedilatildeo de decavanadato no meio de
reacccedilatildeo dos ensaios (tampatildeo D) para uma concentraccedilatildeo final de 1 mM em vanadato total foi
caracterizada atraveacutes da leitura da Abs400 em funccedilatildeo do tempo tendo-se determinado qual a
ordem de reacccedilatildeo e o tempo de meia-vida da mesma Com base em estudos anteriores acerca da
estabilidade do V10 na presenccedila de diferentes proteiacutenas (Ramos et al 2006) determinou-se
igualmente o tempo de meia vida apoacutes a adiccedilatildeo de 20 microM de S1 de miosina ao meio reaccional
353 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
Os espectros de RMN do isoacutetopo 51
V foram realizados num espectroacutemetro Bruker AM-400 MHz
equipado com uma sonda multinuclear inversa de 5 mm Em tubos de RMN de 5 mm de diacircmetro
preparou-se soluccedilotildees de 500 L de deca- ou metavanadato correspondentes a um intervalo de
concentraccedilotildees de 1 a 10 mM suplementadas com 10 D2O Os espectros foram adquiridos agrave
temperatura ambiente com uma largura espectral de 23 923 Hz um tempo de aquisiccedilatildeo de 0086
s tempo de relaxaccedilatildeo de 001 s e uma sequecircncia de pulsos a 90ordm Utilizou-se a transformada de
Fourier com alargamento das bandas de 20Hz para o processamento dos espectros Os desvios
quiacutemicos das ressonacircncias de 51
V foram obtidos em relaccedilatildeo agrave espeacutecie VOCl3cuja frequecircncia de
ressonacircncia eacute considerada o padratildeo sendo todas as outras referenciadas a partir desta em ppm
As aacutereas dos sinais do RMN correspondentes agraves espeacutecies oligomeacutericas presentes foram obtidas
por integraccedilatildeo dos sinais As concentraccedilotildees relativas das espeacutecies foram calculadas com recurso agrave
equaccedilatildeo
onde A[Vx] corresponde agrave aacuterea referente agrave ressonacircncia da espeacutecie Vx com nordm de agregaccedilatildeo x
A[V]total agrave soma total das aacutereas medidas e [V] a concentraccedilatildeo total de vanaacutedio no ensaio
A teacutecnica de 51
V-RMN foi anteriormente aplicada em estudos de ligaccedilatildeo com S1 de miosina
(Ref) tendo sido demostrado que entre as espeacutecies V1 V2 V4 e V5 a espeacutecie tetrameacuterica possui a
maior afinidade para a miosina (Cremo et al) O desaparecimento ou alargamento dos sinais pode
implicar a ligaccedilatildeo de um dos oxovanadatos laacutebeis agrave miosina sem indicaccedilatildeo de qual uma vez que
49
se processa um equiliacutebrio entre eles (Crans 1990) Assim e tendo em conta as indicaccedilotildees de que
a espeacutecie V10 eacute distinta das restantes no que diz respeito agrave magnitude da interacccedilatildeo com a miosina
e inibiccedilatildeo da actividade ATPaacutesica estimulada por actina (Aureliano 2000 Tiago et al 2002)
torna-se fundamental investigar a interacccedilatildeo das diferentes espeacutecies de vanadato com o S1 de
miosina atraveacutes da teacutecnica de 51
V-RMN
Para tal obtiveram-se espectros de 51
V-RMN resultantes da titulaccedilatildeo de uma soluccedilatildeo 5 mM de
decavanadato total com concentraccedilotildees crescentes de S1 de miosina (0 5 10 15 e 20 microM) em
tampatildeo D Realizou-se ainda ensaios na presenccedila de 2 mM de decavanadato e 2 mM de ATP na
presenccedila e na ausecircncia de S1 de miosina de modo a verificar o efeito que estes ligandos naturais
da enzima teria na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o subfragmento-1
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
Os grupos sulfidrilo (-SH) das cisteiacutenas satildeo resiacuteduos de aminoaacutecidos extremamente sensiacuteveis agrave
oxidaccedilatildeo resultando na sua oxidaccedilatildeo (Dalle-Donne et al 1999) Logo a quantificaccedilatildeo destes
resiacuteduos eacute bastante importante nos estudos de interacccedilatildeo com vanaacutedio jaacute que os resiacuteduos de Cys
com ecircnfase para o SH1 e o SH2 satildeo bastante importantes na monitorizaccedilatildeo das modificaccedilotildees
funcionais da regiatildeo da cabeccedila
O procedimento seguidamente apresentado foi originalmente descrito por Gutieacuterrez-Merino
(1987)) Consiste na reacccedilatildeo entre os grupos ndashSH natildeo oxidados das proteiacutenas e o reagente de
Ellman (DTNB -55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico)) - formando-se o aniatildeo CNT (aniatildeo 3-
carboxilato-4-nitrotiofenolato) numa estequiometria de 11 Este apresenta um maacuteximo de
absorvecircncia a 412 nm (Robyt 1990) Assim apoacutes a incubaccedilatildeo da proteiacutena na presenccedila do metal
a quantidade de cisteiacutenas reduzidas por proteiacutena determina-se atraveacutes dos valores de Abs412
Deste modo colocou-se num tubo de Eppendorf o volume apropriado de S1 dialisado para que a
concentraccedilatildeo no ensaio fosse de 1 mgmL Adicionou-se o volume apropriado de decavanadato
para que a sua concentraccedilatildeo fosse de 0 10 50 100 250 500 ou 1000 M Perfez-se com soluccedilatildeo
D ateacute um volume final de 1 mL e incubou-se a 25ordmC por 20 minutos Apoacutes este periacuteodo colocou-
se 780 L da mistura reaccional numa cuvette de quartzo e adicionou-se 220 L de soluccedilatildeo K23
23
Tampatildeo K Tampatildeo fosfatohidrogenofosfato de soacutedio 025 M pH 72
50
Adicionou-se 15 L de DTNB (em soluccedilatildeo K [DTNB]stock=100 mM) ao meio reaccional e
registou-se o valor de absorvecircncia medido equivalente agraves cisteiacutenas ldquoraacutepidasrdquo (SH1 e SH2
expostos ao solvente) Adicionou-se 001 g de SDS que provoca a desnaturaccedilatildeo da proteiacutena e
consequentemente a exposiccedilatildeo de todos os resiacuteduos de cisteiacutenas que se encontravam ateacute aiacute
inacessiacuteveis no interior da proteiacutena (cisteiacutenas totais) Estes resiacuteduos podem assim reagir com o
DTNB
Estimou-se o nuacutemero de resiacuteduos de cisteiacutenas em todas as amostras com recurso ao coeficiente de
extinccedilatildeo molar () do DTNB (ver ponto 3 dos Anexos) que foi determinado atraveacutes da titulaccedilatildeo
de L-Cisteiacutena com intervalo de concentraccedilotildees compreendidos entre 0 e 50 microM na presenccedila de
25microM de DTNB em tampatildeo D Os valores de absorvecircncia foram obtidos para um comprimento de
onda de 412 nm
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
3621 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
A teacutecnica mais eficiente de anaacutelise da interacccedilatildeo entre o iatildeo vanadilo e proteiacutenas como a
calmodulina e a transferrina entre outras permite a obtenccedilatildeo raacutepida de informaccedilatildeo no que toca agrave
conformaccedilatildeo e amplitude da referida interacccedilatildeo
Os espectros de Ressonacircncia Paramagneacutetica Electroacutenica (RPE) da espeacutecie paramagneacutetica V(IV)
foram realizados a 70K num espectroacutemetro Bruker EMX-300 X-band Todas as soluccedilotildees
utilizadas e amostras foram preparadas em anaerobiose numa cacircmara de luvas de modo a evitar a
reduccedilatildeo de V(IV) a V(V) Em tubos capilares com 02 mL de amostra manteve-se a concentraccedilatildeo
de S1 a 20 M e variou-se a concentraccedilatildeo de vanadilo (0 25 5 10 15 25 50 75 100 150
200 500 e 1000 M [VOSO4]stock=5 mM) em tampatildeo D Apoacutes obtenccedilatildeo dos espectros estes
foram tratados atraveacutes do software SimFonia e WinEPR tendo-se obtido um graacutefico com a
variaccedilatildeo da intensidade do sinal de RPE em funccedilatildeo da concentraccedilatildeo de sulfato de vanadilo
presente Realizaram-se ainda simulaccedilotildees de espectros obtidos com 20 microM de S1 e 1000 microM de
V(IV) com vista agrave obtenccedilatildeo dos ligandos provaacuteveis do vanadilo atraveacutes da anaacutelise da constante
de acoplamento hiperfina (A) e do factor g (g)
4 Conclusotildees gerais
Apesar de jaacute ter sido demonstrado que o vanaacutedio inibe a Na+K
+-ATPase da miosina a funccedilatildeo
deste elemento no tecido muacutesculo esqueleacutetico continua largamente incompreendida A enorme
complexidade quiacutemica deste metal especialmente no que concerne agrave oligomerizaccedilatildeo e reacccedilotildees
51
redox constitui um obstaacuteculo agrave compreensatildeo do comportamento do vanaacutedio nos organismos
vivos Assim a caracterizaccedilatildeo da quiacutemica deste elemento eacute de grande utilidade antes de iniciar o
estudo da sua interacccedilatildeo com o presente sistema
A anaacutelise por espectroscopias de 51
V-RMN e de UV-Vis de soluccedilotildees de vanadato diluiacutedas nos
meios de reacccedilatildeo utilizados no presente relatoacuterio (10 mM HEPES pH 70 25 mM KCl) mostrou
claramente que a espeacutecie decameacuterica (V10) possui um tempo de vida suficientemente longo para a
determinaccedilatildeo do seu efeito isolado ao contraacuterio do que se verifica com as espeacutecies mono- di-
tetra- e pentameacutericas (V1 V2 V4 e V5) Por outro lado a anaacutelise verificou ainda que apoacutes diluiccedilatildeo
das soluccedilotildees de decavanadato no meio de reacccedilatildeo coexistem as espeacutecies V10 e V1 pelo que os
efeitos putativos desta uacuteltima deveratildeo ser subtraiacutedos aquando da monitorizaccedilatildeo dos efeitos da
espeacutecie decameacuterica
O vanaacutedio (V) (vanadato) inibe as enzimas que catalizam reacccedilotildees de transferecircncia de grupos
fosfato como a ATPase da miosina [Goodno 1979] devido agrave sua analogia estrutural com o iatildeo
fosfato Na sua utilizaccedilatildeo como inibidor enzimaacutetico natildeo eacute tida em conta a contribuiccedilatildeo individual
das espeacutecies oligomeacutericas considerando-se normalmente a espeacutecie V1 como inibidora efectiva
[Goodno e Taylor 1982] No presente relatoacuterio verificou-se por espectroscopia de 51
V-RMN que
outros oligoacutemeros de vanadato interagem com o S1 de miosina tendo as espeacutecies V10 e V4 os
maiores niacuteveis de afinidade A sua interacccedilatildeo envolve forccedilas electrostaacuteticas e eacute afectada
negativamente por ligandos naturais de enzima como eacute o caso do ATP
A exposiccedilatildeo do S1 de miosina (005 mgmL) a decavanadato conduz agrave inibiccedilatildeo do capacidade da
proteiacutena de estimular a actividade Mg2+
-ATPaacutesica com um valor de IC50 de 266 plusmn 07 M
decavanadato sendo o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM
decavanadato (80) Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM decavanadato sendo
o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM decavanadato (60)
exercendo um efeito de impedimento agrave ligaccedilatildeo entre o vanaacutedio e o S1
A uma concentraccedilatildeo de 27 microM decavanadato verifica-se a oxidaccedilatildeo de um dos resiacuteduos de Cys
de S1 de miosina seguida pela oxidaccedilatildeo gradual de outro resiacuteduo de Cys ateacute agrave concentraccedilatildeo de
1000 microM Ambos os resiacuteduos oxidados natildeo se encontravam expostos ao solvente utilizado A
oxidaccedilatildeo dos resiacuteduos permite sugerir que esta modificaccedilatildeo oxidativa sofrida pelo S1 contribui
para a diminuiccedilatildeo da sua actividade de ATPase
52
Por espectroscopia de RPE revelaram-se os possiacuteveis ligandos equatoriais do vanaacutedio aquando da
sua interacccedilatildeo com o S1 de miosina como sendo duas moleacuteculas de aacutegua um ligando contendo o
grupo R-OH e outro contendo o grupo R=NR
Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados de
oxidaccedilatildeo (IV) e (V) no S1 de miosina para concentraccedilotildees facilmente alcanccedilaacuteveis no ambiente
terrestre
53
5 Estudos futuros
No sentido do melhor esclarecimento das alteraccedilotildees moleculares e estruturais que ocorrem no
subfragmento 1 quando exposta ao vanaacutedio que conduzem a uma diminuiccedilatildeo da sua funccedilatildeo
sugere-se
- Quantificaccedilatildeo das cisteiacutenas reduzidas de S1 na presenccedila de metavanadato
- Determinaccedilatildeo das actividades Ca2+
- e K+-ATPaacutesica do S1 de miosina na presenccedila de
decavanadato
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina estimulada por actina
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina na presenccedila de
metavanadato
- Realizaccedilatildeo de RMN 2D com variaccedilatildeo das concentraccedilotildees das soluccedilotildees de decavanadato e
metavanadato na presenccedila de S1
-Mutageacutenese dos resiacuteduos de Cys doS1 de miosina de modo a verificar quais os resiacuteduos que
sofrem oxidaccedilatildeo na presenccedila de decavanadato
- Determinaccedilatildeo por calorimetria (DSC Differential Scanning Calorimetry) da variaccedilatildeo da
entalpia (∆H) e da temperatura de desnaturaccedilatildeo do S1 de miosina na presenccedila de vanaacutedio uma
vez que estas propriedades caracteriacutesticas de cada proteiacutena satildeo alteradas apoacutes ocorrecircncia de
modificaccedilotildees estruturais
- Anaacutelise por SDS-PAGE de S1 de miosina apoacutes tratamento com vanaacutedio de modo a determinar
quais os polipeacuteptidos eventualmente formados apoacutes clivagem da proteiacutena
54
6 Referecircncias bibliograacuteficas
- Sellers J Myosins 2nd Edition Oxford University Press 1999
- L Stryer inBiochemistry (Ed WH Freeman 2005 pp
Refordfas Materiais e Meacutetodos
[A] S S Margossian SLowey Methods Enzymol1982 85 55-71
[B] JD Pardue JA Spudich Methods Enzymol 1982 85 164-181
[C] M Bradford Anal Biochem1976 72 248-254
[D] H H Taussky E Shorr J Biol Chem 1953 202 675-685
[E] C R Bagshaw in Outline Studies in Biology ndash Muscle Contraction Chapman and Hall Ed
1982
[F] A Cuenda et al Eur J Biochem1990 194 663-670
55
Anexos
1 Determinaccedilatildeo da recta de calibraccedilatildeo meacutetodo de Bradford
Figura1ndashRepresentaccedilatildeo graacutefica da recta de calibraccedilatildeo (Abs595 nm vs [BSA]) utilizada para o meacutetodo de Bradford
2 Determinaccedilatildeo da recta calibraccedilatildeo para a electroforese em condiccedilotildees desnaturantes
Figura 2 ndash Representaccedilatildeo graacutefica da recta de calibraccedilatildeo (log(Peso Molecular) vs Rf dos marcadores de peso
molecular) onde Rf indica determinada atraveacutes do padratildeo de peso molecular
(poccedilo (a))
3Caacutelculo do coeficiente de extinccedilatildeo molar () para a quantificaccedilatildeo de cisteiacutenas reduzidas
atraveacutes do meacutetodo de Ellman
Traccedilou-se a recta de Abs412 nm meacutedia vs [L-Cisteiacutena] de modo a determinar o valor do coeficiente
de extinccedilatildeo molar definido como o declive daquela recta Assim =8400 M-1
cm-1
como
podemos verificar na Fig 3
A595 nm = 00546[BSA] + 00209 Rsup2 = 099051
Ab
s 59
5 n
m
[BSA] (mgmL)
log PM = -1177 Rf + 2247 Rsup2 = 098957
log
PM
Mobilidade relativa (Rf)
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero

38
32111 Purificaccedilatildeo da miosina
O protocolo de isolamento foi adaptado do meacutetodo previamente descrito por Margossian e Lowey
(1982) apresentando-se de forma esquemaacutetica na Fig 31
Sucintamente os tecidos musculares dorsais e das pernas foram extraiacutedos apoacutes o sacrifiacutecio do
animal (Fig31) De seguida foram adicionados 3 volumes da soluccedilatildeo de extracccedilatildeo A2
Centrifugou-se a soluccedilatildeo e filtrou-se o sobrenadante O sedimento foi entatildeo diluiacutedo em 14
volumes de aacutegua destilada de modo a que a forccedila ioacutenica atingisse 004 M Apoacutes adiccedilatildeo de szlig-
mercaptoetanol (szlig-me) deixou-se a precipitar a 4ordmC removendo o sobrenadante Centrifugou-se
novamente e ressuspendeu-se o sedimento na soluccedilatildeo de extracccedilatildeo A Adicionou-se a soluccedilatildeo
tampatildeo B3 para que a forccedila ioacutenica atingisse 05 M Homogeneizou-se e deixou-se repousar
durante 2h Homogeneizou-se novamente e centrifugou-se de modo a ocorrer precipitaccedilatildeo dos
complexos de actomiosina e proteiacutena desnaturada Apoacutes filtraccedilatildeo do sobrenadante em latilde de vidro
quantificou-se a soluccedilatildeo de miosina resultante pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental)
2 Soluccedilatildeo de extracccedilatildeo A 0012M K2CO3 004M KHCO3 06M KCl pH 65
3 Soluccedilatildeo-tampatildeo B Tris 100 mM pH 7 KCl 2M
39
Figura 31 ndash Representaccedilatildeo esquemaacutetica do protocolo de isolamento de miosina (Margossian e Lowey 1982) Todo o
procedimento foi realizado a 4ordmC (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
32112 Preparaccedilatildeo de Subfragmento-1 de miosina por digestatildeo com quimiotripsina
O S1 de miosina foi isolado apoacutes diaacutelise da miosina durante a noite contra a soluccedilatildeo C4 A
presenccedila do EDTA eacute essencial uma vez que a sua substituiccedilatildeo por MgCl2resulta na produccedilatildeo de
HMM em vez de S1 A suspensatildeo de miosina foi equilibrada e digerida com -quimotripsina
A digestatildeo foi interrompida pela adiccedilatildeo de PMSF Seguidamente ultracentrifugou-se o produto
da digestatildeo desprezando-se o sedimento (Fig 32) A soluccedilatildeo de sobrenadante maioritariamente
S1 foi quantificada pela leitura directa da Abs280 ou pelo meacutetodo de Bradford (ver as secccedilotildees
3311 e 3312 do Procedimento Experimental respectivamente) e armazenada em 10 sacarose
e 2 mM szlig-Me a -80ordmC
4 Soluccedilatildeo C 25mM Tris pH 70 012M KCl 1mM EDTA
40
Antes da realizaccedilatildeo dos ensaios descritos no presente relatoacuterio o S1 de miosina foi dialisado
contra a soluccedilatildeo D5 de modo a remover qualquer composto que pudesse complexar como
vanaacutedio Seguidamente quantificou-se a proteiacutena pelo meacutetodo de Bradford ou pelo meacutetodo
directo (ver secccedilotildees 3311 e 3312do Procedimento Experimtental)
Figura 3 2ndash Representaccedilatildeo esquemaacutetica do protocolo de preparaccedilatildeo do S1 de miosina por digestatildeo quimiotripsina de
acordo com o previamente descrito por Margossian e Lowey (1982) Todo o procedimento foi realizado a 4ordm C
excepto quando indicaccedilatildeo do contraacuterio (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
3212 Isolamento e purificaccedilatildeo de actina
Isolou-se e purificou-se a actina segundo o meacutetodo descrito por Pardee e Spudish que
compreende dois passos ndash a obtenccedilatildeo de um resiacuteduo seco denominado ldquopoacute de acetonardquo e a
extracccedilatildeo subsequente da actina a partir do ldquopoacute de acetonardquo na forma de G-Actina com a
posterior polimerizaccedilatildeo a F-Actina
32121 Obtenccedilatildeo do ldquopoacute de acetonardquo
Sacrificou-se o coelho com uma pancada atraacutes do pescoccedilo e sangrou-se o animal apoacutes o corte da
arteacuteria jugular Esfolou-se e colocou-se numa tina com gelo Retiraram-se as partes de tecido
conjuntivo que envolvem o tecido muscular com auxiacutelio de uma pinccedila e bisturi Seguidamente
retirou-se os muacutesculos dorsais e os muacutesculos brancos das pernas Passou-se os muacutesculos por aacutegua
destilada de modo a remover as impurezas e pesou-se num copo Trituraram-se os muacutesculos
numa maacutequina de triturar carne e extraiu-se o resultante com 3 volumes da soluccedilatildeo E6 durante 10
minutos com agitaccedilatildeo suave Centrifugou-se a suspensatildeo a 4000 g durante 10
5 Soluccedilatildeo D HEPES 10 mM pH 70 KCl 25 mM
6 Soluccedilatildeo E 01 M KCl 015 M K2HPO4 pH 65
41
minutosdesprezando-se o sobrenadante Ressuspendeu-se o sedimento em 6 volumes de soluccedilatildeo
F7durante 10 minutos com agitaccedilatildeo suave Verificou-se se o pH da soluccedilatildeo estava compreendido
entre 82 e 85 acertando-se estecom soluccedilatildeo G8 se necessaacuterio Centrifugou-se durante 10
minutos a 4000 g Desprezou-se o sobrenadante e ressuspendeu-se o sedimento em 3 volumes de
soluccedilatildeo H9 durante 10 minutos Centrifugou-se novamente a 4000 gdurante 10 minutos
desprezando-se o sobrenadante Ressuspendeu-se o sedimento duas vezes consecutivas em 6
volumes de aacutegua destilada durante 5 minutos Centrifugou-se a 4000 g durante 10 minutos apoacutes
cada adiccedilatildeo de H2O
Seguidamente ressuspendeu-se o sedimento 5 vezes consecutivas utilizando 3 volumes de
acetona de cada vez agitando suavemente durante 10 minutos agrave temperatura ambiente Colocou-
se o resiacuteduo filtrado numa tina de vidro sobre papel de filtro deixando-se evaporar o solvente
durante a noite num nicho Triturou-se o resiacuteduo seco numa maacutequina de triturar carne e deixou-se
novamente evaporar o solvente O ldquopoacute de acetonardquo obtido foi armazenado a -20 ordmC
32122 Purificaccedilatildeo da actina a partir de ldquopoacute de acetonardquo
Apoacutes a obtenccedilatildeo do poacute de acetona como acima indicado efectuou-se o protocolo seguidamente
descrito de modo a obter monoacutemeros de actina (G-Actina) e actina filamentosa (F-Actina)
Pesou-se 35 g de poacute de acetona e adicionaram-se 50 mL de soluccedilatildeo-tampatildeo I10
sendo a
extracccedilatildeo feita com agitaccedilatildeo suave durante 30 minutos Separou-se o extracto do poacute de acetona
hidratado filtrando a suspensatildeo com 4 camadas de gaze esterilizada tendo o cuidado de natildeo
espremer o resiacuteduo Voltou a extrair-se com 50 mL de soluccedilatildeo I durante 30 minutos mexendo
lentamente Adicionou-se este ao extracto obtido anteriormente Centrifugou-se o extracto total a
16 000 g durante 30 minutos Decantou-se para uma proveta sobre a qual se colocou gaze para
que o sedimento solto natildeo se juntasse ao sobrenadante e leu-se o volume De seguida adicionou-
se KCl ateacute uma concentraccedilatildeo final de 50 mM MgCl2 ateacute 2mM e ATP ateacute 1mM lentamente e com
agitaccedilatildeo contiacutenua de modo a promover a polimerizaccedilatildeo dos monoacutemeros de actina
Deixou-se polimerizar durante 2h a 25ordmC observando-se a presenccedila de pequenas bolhas de ar
indicativas de um aumento de viscosidade da soluccedilatildeo Arrefeceu-se em gelo durante 15 minutos e
adicionou-se KCl ateacute uma concentraccedilatildeo final de 06M Mexeu-se lentamente durante 30 minutos
apoacutes os quais se ultracentrifugou a 110 000g durante 1hora Desprezou-se o sobrenadante
ressuspendeu-se o sedimento com soluccedilatildeo tampatildeo E e transferiu-se para um potter
Dialisou-se durante a noite contra 1L de soluccedilatildeo-tampatildeo I a uma temperatura de 4ordmC com rotaccedilatildeo
7 Soluccedilatildeo F 005 M NaHCO3 pH 82-85
8 Soluccedilatildeo G 1M Na2CO3
9 Soluccedilatildeo H 1 mM EDTA pH 70
10 Soluccedilatildeo I 2 mM Tris-HCl pH 8 02 mM CaCl2 02 mM Na2ATP 05 mM szlig-Me 0005 NaN3
42
contiacutenua e mudanccedila de tampatildeo 2 a 3 vezes De seguida a actina foi centrifugada a 110
000gdurante 1hora Decantou-se a soluccedilatildeo de G-Actina (sobrenadante) e desprezou-se o
sedimento Determinou-se a concentraccedilatildeo de G-Actina pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental) A soluccedilatildeo de G-actina foi armazenada a uma temperatura de 4ordmC
apoacutes adiccedilatildeo de 002NaN3 mantendo-se estaacutevel durante 3 a 5 dias
331 Quantificaccedilatildeo das proteiacutenas isoladas
3311 Quantificaccedilatildeo pelo meacutetodo de Bradford
O meacutetodo de Bradford (Bradford 1976) baseia-se na utilizaccedilatildeo do reagente de Bradford
composto por entre outros aacutecido fosfoacuterico e o corante ldquoCoomassierdquo Este corante possui carga
negativa e a sua coloraccedilatildeo varia entre a vermelha (Absmax=465 nm) predominante em soluccedilatildeo e a
azul (Absmax=595 nm) presente aquando da ligaccedilatildeo aos grupos amina das proteiacutenas Em soluccedilotildees
aacutecidas diluiacutedas a coloraccedilatildeo varia proporcionalmente com a quantidade do corante ligado agrave
proteiacutena que por sua vez eacute directamente proporcional agrave concentraccedilatildeo da proteiacutena permitindo a
sua quantificaccedilatildeo ateacute a um limite de detecccedilatildeo de 20 gmL (Bradford 1976)
Deste modo prepararam-se amostras de BSA ([BSA]stock= 1mgmL) utilizando um intervalo de
concentraccedilotildees compreendido entre 0 e 15 gmL de acordo com a tabela 3I Apoacutes a adiccedilatildeo da
H2O MilliQ e do reagente de Bradford as soluccedilotildees foram sucessivamente homogeneizadas no
voacutertex
Tabela 3Indash Amostras utilizadas para a obtenccedilatildeo da recta de calibraccedilatildeo no meacutetodo de Bradford
BSA (L) BSA (g) H2O MilliQ (L) Reagente de Bradford (L)
0 0 800 200
125 125 79875 200
25 25 7975 200
5 5 795 200
10 10 790 200
15 15 785 200
Apoacutes incubaccedilatildeo a 25ordmC por 5 minutos leram-se os valores de Abs595 num
espectrofotoacutemetro (Shimadzu UV-160A) para todas as amostras Seguidamente traccedilou-se a recta
de calibraccedilatildeo (Abs595vs quantidade de BSA (g))
43
Para determinar a concentraccedilatildeo de proteiacutena apoacutes o processo de isolamento purificaccedilatildeo e
diaacutelise quando necessaacuterio colocou-se 5 L da amostra num tubo de Eppendorf e adicionou-se
795 L de H2O MilliQ homogeneizando a soluccedilatildeo no voacutertex Adicionou-se posteriormente 200
L de reagente de Bradford e repetiu-se a homogeneizaccedilatildeo Incubou-se a mistura a 25ordmC por 5
minutos apoacutes os quais se leu os valores de Abs595 nm no espectrofotoacutemetro Por interpolaccedilatildeo deste
valor a partir da recta de calibraccedilatildeo determinou-se a concentraccedilatildeo de proteiacutena na amostra
3312 Quantificaccedilatildeo pelo meacutetodo directo
A quantificaccedilatildeo das amostras proteicas pode tambeacutem ser efectuada atraveacutes da leitura directa da
absorvacircncia tendo por base a lei de Beer-Lambert onde Abs corresponde agrave
absorvacircncia ao comprimento de onda ao qual o ensaio foi realizado corresponde ao coeficiente
de extinccedilatildeo (substituiacutedo pela letra a se as unidades forem distintas da molaridade) b corresponde
ao percurso oacuteptico da radiaccedilatildeo na ceacutelula amostral e c corresponde agrave concentraccedilatildeo amostral
Assim no caso de S1 de miosina mediu-se a Abs280e calculou-se a concentraccedilatildeo da proteiacutena
sabendo que o coeficiente de extinccedilatildeo de S1 eacute (Ref)
No caso da actina mediu-se a Abs290ecalculou-se a concentraccedilatildeo da proteiacutena sabendo que o
coeficiente de extinccedilatildeo da actina eacute (Ref)
332Electroforese em condiccedilotildees desnaturantes (SDS-PAGE)
De modo a melhor caracterizar as proteiacutenas isoladas foi realizada uma electroforese em
condiccedilotildees desnaturantes (SDS-PAGE do inglecircs Sodium Dodecyl Sulfate Polyacrilamide Gel
Electrophoresis) O SDS (dodecilo sulfato de soacutedio) eacute um detergente anioacutenico utilizado neste
meacutetodo para desnaturar as proteiacutenas que se ligam ao SDS na proporccedilatildeo 14 g SDS g proteiacutena
(aproximadamente 1 moleacutecula de SDS por cada 2 resiacuteduos de aminoaacutecidos) (Stryer) O SDS induz
uma densidade de carga negativa significativa nas proteiacutenas a que se liga mascarando a sua carga
intriacutenseca pelo que as razotildees cargamassa satildeo tendencialmente idecircnticas Assim a SDS-PAGE
separa as proteiacutenas quando sujeitas a um campo eleacutectrico de acordo com o seu peso molecular
sendo a sua migraccedilatildeo directamente proporcional ao logaritmo deste uacuteltimo (Stryer)
O gel de resoluccedilatildeo com 10 de acrilamida foi obtido atraveacutes da adiccedilatildeo de 167 mL de soluccedilatildeo
stock de acrilamida11
075 mL de tampatildeo de resoluccedilatildeo12
251 mL de aacutegua destilada e 50 L de
SDS 10 Para promover a polimerizaccedilatildeo adicionou-se em seguida 38 L de PSA 10
11
soluccedilatildeo stock de acrilamida soluccedilatildeo de 30 (mv) de acrilamida e 08 (mv) de NNrsquo-metil-bis-acrilamida 12
tampatildeo de resoluccedilatildeo 15 M Tris-HCl pH 88
44
(persulfato de amoacutenia) 10 e 25 L de TEMED 99 (tetrametiletilenodiamina) Agitou-se
cuidadosamente de modo a natildeo formar bolhas e pipetou-se a soluccedilatildeo para a cassette de vidro
previamente montada no suporte De seguida adicionou-se uma peliacutecula de cerca de 2 mm de 2-
metil-2-butanol para verificar posteriormente a polimerizaccedilatildeo Apoacutes a polimerizaccedilatildeo que demora
cerca de 45 minutos verteu-se a agrave superfiacutecie e adicionou-se o gel de concentraccedilatildeo com 5 de
acrilamida constituiacutedo por 03 mL de soluccedilatildeo de stock acrilamida 045 mL de tampatildeo de
concentraccedilatildeo13
094 mL de aacutegua destilada 18 L de SDS 10 135 L de PSA 10 e 2L de
TEMED 99 Colocou-se o pente sobre o gel e deixou-se polimerizar durante cerca de 45
minutos
As amostras para aplicaccedilatildeo no gel de electroforese foram preparadas para uma concentraccedilatildeo de
proteiacutena de 1 mgmL num volume final de 100 L dos quais 25 L correspondem ao tampatildeo da
amostra14
As amostras foram aquecidas a 100ordmC durante 5 minutos Antes da aplicaccedilatildeo das
amostras transferiu-se a cassette para a tina de electroforese agrave qual se adicionou a soluccedilatildeo
tampatildeo de electroforese da tina superior15
e tampatildeo de electroforese da tina inferior16
Aplicaram-
se 10 L das soluccedilotildees de marcadores de peso molecular e 20 L das amostras proteicas
preparadas anteriormente A electroforese decorreu durante cerca de 3h30 min a uma voltagem
de 80 V De seguida transferiu-se o gel para uma tina corando as proteiacutenas durante 1h com
soluccedilatildeo corante17
Descorou-se o gel com soluccedilatildeo descorante18
tendo-se mudado esta soluccedilatildeo ateacute
as bandas se encontrarem bem definidas
Determinou-se a mobilidade relativa das bandas correspondentes aos marcadores de elevado peso
molecular e traccedilou-se a recta de calibraccedilatildeo ndash log(PM) vs log(Rf) onde Rf corresponde agrave
mobilidade relativa Por interpolaccedilatildeo determina-se o PM das proteiacutenas isoladas sabendo a sua
mobilidade relativa Por comparaccedilatildeo com os valores descritos na literatura pode concluir-se
acerca do grau de purificaccedilatildeo
34 Mediccedilatildeo da actividade enzimaacutetica de S1
341 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
13
tampatildeo de concentraccedilatildeo 05 M Tris-HCl pH 68 14
tampatildeo de amostra 320 mM Tris-HCl pH 68 04 M szlig-mercaptoetanol SDS 8 glicerol 15 (vv) e roxo de m-
cresol 0024 15
tampatildeo de electroforese da tina superior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS 16
tampatildeo de electroforese da tina inferior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS acetato de soacutedio
01 M 17
soluccedilatildeo corante ldquoCoomassie Blue R-250rdquo contendo metanol 45 (vv) e aacutecido aceacutetico glacial 10 (vv) 18
soluccedilatildeo descorante metanol 10 (vv) e aacutecido aceacutetico glacial 10 vv)
45
O meacutetodo de Fiske-Subarow modificado consiste na reacccedilatildeo do molibdato de amoacutenio com o
fosfato inorgacircnico em soluccedilatildeo aacutecida com formaccedilatildeo do aacutecido fosfomoliacutebdico Seguidamente daacute-
se a adiccedilatildeo de um agente redutor que provoca a reduccedilatildeo do composto a fosfomolibdato que forma
um complexo de cor azulada cuja absorvacircncia maacutexima ocorre para um comprimento de onda de
660 nm tornando possiacutevel a sua detecccedilatildeo por meacutetodos espectrofotomeacutetricos As actividades Ca2+
-
ATPaacutesica e K+EDTA-ATPaacutesica de S1 foram determinadas atraveacutes deste meacutetodo de acordo com
o descrito por Taussky e Shorr (1953) com mediccedilatildeo da quantidade de fosfato (Pi) libertado
atraveacutes da hidroacutelise de ATP
Para tal obteve-se primeiramente a recta de calibraccedilatildeo com as amostras presentes na Tabela 3II
utilizando uma soluccedilatildeo stock de 5 mM KH2PO4
Tabela 3II ndash Composiccedilatildeo das diferentes amostras utilizadas para determinar a recta de calibraccedilatildeo atraveacutes do meacutetodo
de Fiske-Subbarow modificado
KH2PO4 (L)
([KH2PO4]stock= 5mM)
PO43-
(nmole) H2O (uL)
0 0 1780
5 25 1775
10 50 1770
20 100 1760
40 200 1740
100 500 1680
A cada tubo de Eppendorf adicionaram-se seguidamente 200 L de soluccedilatildeo J19
e 20 L de agente
redutor20
homogeneizando no voacutertex apoacutes cada adiccedilatildeo Incubou-se durante 10 minutos e mediu-
se a Abs660 nm num espectrofotoacutemetro com intervalos de tempo iguais entre cada amostra Traccedilou-
se a recta de calibraccedilatildeo (Abs660 nmvs nmol PO43-
) a partir dos valores obtidos
Para determinar a actividade Ca2+
-ATPaacutesica de S1 prepararam-se tubos de Eppendorf com 400
L de meio de reacccedilatildeo21
25 L CaCl2 ([CaCl2]stock= 100 mM) 25 L ATP ([ATP]stock= 100 mM)
com [S1]ensaios=01 mgmL Perfez-se se necessaacuterio com H2O MilliQ ateacute perfazer um volume de 1
mL Para determinar a actividade K+EDTA-ATPaacutesica utilizou-se uma concentraccedilatildeo idecircntica de
19
Soluccedilatildeo J 5 N H2SO4 25 molibdato de amoacutenio 20
Agente redutor 5 mg aacutecido 1-amino-2-naftol-4-sulfoacutenico 30 mg bissulfito de soacutedio 30 mg sulfito de soacutedio em 26
mL H2O 21
meio de reacccedilatildeo 125 mM Tris-HCl pH 80 0575 M KCl
46
proteiacutena nos ensaios (01 mgmL) e o mesmo meio de reacccedilatildeo Aos tubos de Eppendorf
adicionou-se ainda 123 L de KCl ([KCl]stock = 3M) 25 L de ATP 10 L de EDTA
([EDTA]stock= 100 mM )e o volume de H2O MilliQ necessaacuterio para perfazer 1 mL Em ambas as
situaccedilotildees o uacuteltimo reagente adicionado foi o ATP uma vez que se trata do substrato da reacccedilatildeo
enzimaacutetica
A cineacutetica decorreu a 25ordmC durante 5 minutos para a actividade Ca2+
-ATPaacutesica e durante 3
minutos para a actividade K+EDTA-ATPaacutesica Apoacutes o periacuteodo de incubaccedilatildeo transferiu-se 150
L de amostra para tubos de Eppendorf contendo igual volume de TCA 10 Colocou-se no gelo
e homogeneizou-se no voacutertex Seguidamente centrifugou-se a 1500 g durante 10 minutos e
recolheu-se 200 L de sobrenadante para novos tubos de Eppendorf Adicionou-se a estes 158
mL de H2O MilliQ e 200 L de soluccedilatildeo J Homogeneizou-se no voacutertex e adicionou-se 20 L de
reagente redutor repetindo-se a homogeneizaccedilatildeo Incubou-se durante 10 minutos e mediu-se a
Abs660 nm com o mesmo intervalo de tempo de leitura para cada amostra Por intrapolaccedilatildeo a partir
da recta de calibraccedilatildeo obtida anteriormente determinou-se a quantidade de Pi libertado em cada
ensaio A partir destes resultados determinaram-se as actividades Ca2+
-ATPaacutesica e K+EDTA-
ATPaacutesica do S1 em nmol PO43-
minmg proteiacutena e em s-1
342 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
Da actividade Mg2+
-ATPaacutesica de S1 resulta a formaccedilatildeo dos produtos da reacccedilatildeo de
hidroacutelise do ATP - ADP e Pi Na presenccedila de ADP fosfoenolpiruvato (PEP) e da enzima piruvato
cinase (PK) os produtos resultantes satildeo ATP e piruvato Por acccedilatildeo da enzima lactato
desidrogenase (LDH) este uacuteltimo eacute convertido em lactato ocorrendo simultaneamente a oxidaccedilatildeo
do NADH a NAD+ monitorizada atraveacutes da Abs340 nm Quando o S1 de miosina se encontra no
meio reaccional o ATP eacute hidrolisado formando-se novamente ADP e Pi o que permite a
formaccedilatildeo de um ciclo (fig 21) Se as enzimas se encontrarem em excesso a velocidade de
conversatildeo do NADH eacute limitada pela produccedilatildeo de ADP e Pi Como o ATP eacute regenerado em cada
ciclo a sua concentraccedilatildeo inicial eacute sempre mantida (Bagshaw 1982)
Figura 33ndash Representaccedilatildeo esquemaacutetica do meacutetodo de enzimas acopladas
47
O protocolo utilizado e seguidamente descrito foi adaptado de Cuenda et al (1990 ndash Ver Refordf) A
uma cuvette de 1 mL adicionou-se S1 para uma concentraccedilatildeo final de 005 mgmL 800 L de
meio de reacccedilatildeo22
10 L de PEP ([PEP]stock = 42 mM) 10 L de PK+LDH 11 ([PK+LDH]stock =
1 mgmL) e 15 L de NADH ([NADH]stock=25 mM) Iniciou-se a monitorizaccedilatildeo da reacccedilatildeo
atraveacutes da leitura de Abs340 nm apoacutes a adiccedilatildeo de 20 L de ATP ([ATP]stock=100 mM) A
actividade Mg2+
-ATPaacutesica eacute obtida atraveacutes do declive correspondente agrave oxidaccedilatildeo do NADH Para
a mediccedilatildeo da Mg2+
-ATPase de S1 estimulada por actina adicionou-se F-actina para uma
concentraccedilatildeo final de 2 microM sendo a actividade obtida tambeacutem atraveacutes do declive correspondente
agrave oxidaccedilatildeo do NADH apoacutes a adiccedilatildeo da proteiacutena
35 Interaccedilatildeo entre vanaacutedio e subfragmento-1 de miosina
351 Preparaccedilatildeo das soluccedilotildees de vanaacutedio
As soluccedilotildees de vanadato foram preparadas em aacutegua MilliQ a partir de metavanadato de amoacutenio
(NH4VO3) numa concentraccedilatildeo stock de 50 mM com agitaccedilatildeo magneacutetica e aquecimento (~30 ordmC)
Esta soluccedilatildeo com um valor compreendido entre 6 e 7 designa-se por ldquometavanadatordquo e eacute
constituiacuteda pelas esspeacutecies V1 V2 V4 e V5 (Ref) Acidificando a soluccedilatildeo de metavanadato por
adiccedilatildeo de HCl ateacute um valor de pH proacuteximo de 4 observa-se uma mudanccedila de cor de amarelo
claro para laranja indicativa da formaccedilatildeo da espeacutecie decameacuterica de vanadato De seguida e
imediatamente antes da realizaccedilatildeo dos ensaios descritos o valor do pH da soluccedilatildeo de
decavanadato foi aumentado para cerca de 75 por adiccedilatildeo de NaOH A soluccedilatildeo (2) designada
por ldquometavanadatordquo natildeo foi posteriormente alterada ateacute agrave sua adiccedilatildeo para a realizaccedilatildeo dos ensaios
As soluccedilotildees foram armazenadas a 4ordmC e mantidas em gelo aquando da sua utilizaccedilatildeo
A soluccedilatildeo de V(IV) foi preparada em condiccedilotildees anaeroacutebias por diluiccedilatildeo de sulfato de vanadil
(VOSO4) para uma concentraccedilatildeo final de 5 mM em H2O MilliQ saturada com N2 gasoso
352Anaacutelise por espectroscopia de UV-Vis
A concentraccedilatildeo da espeacutecie decameacuterica em soluccedilatildeo depende do periacuteodo de tempo apoacutes a diluiccedilatildeo
no meio de reacccedilatildeo apropriado (Ref) A velocidade de desoligomerizaccedilatildeo da espeacutecie decameacuterica
pode ser analisada por espectroscopia de absorccedilatildeo na regiatildeo UVVis uma vez que esta espeacutecie
apresenta duas bandas caracteriacutesticas na regiatildeo do visiacutevel (a 358 e 400 nm) que natildeo se verifica
em quaisquer outras espeacutecies oligomeacutericas As bandas supramencionada sapresentam-se bem
22
Meio de reacccedilatildeo 3125 mM Tris-HCl pH 71 375 mM MgCl2
48
resolvidas atraveacutes da obtenccedilatildeo da derivada de 2ordf ordem do espectro apresentando a forma de 2
vales com miacutenimos centrados nos comprimentos de onda supracitados com dependecircncia linear
relativamente agrave concentraccedilatildeo de decavanadato total (Ramos et al 2006)
Assim a estabilidade da espeacutecie decameacuterica apoacutes diluiccedilatildeo da soluccedilatildeo de decavanadato no meio de
reacccedilatildeo dos ensaios (tampatildeo D) para uma concentraccedilatildeo final de 1 mM em vanadato total foi
caracterizada atraveacutes da leitura da Abs400 em funccedilatildeo do tempo tendo-se determinado qual a
ordem de reacccedilatildeo e o tempo de meia-vida da mesma Com base em estudos anteriores acerca da
estabilidade do V10 na presenccedila de diferentes proteiacutenas (Ramos et al 2006) determinou-se
igualmente o tempo de meia vida apoacutes a adiccedilatildeo de 20 microM de S1 de miosina ao meio reaccional
353 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
Os espectros de RMN do isoacutetopo 51
V foram realizados num espectroacutemetro Bruker AM-400 MHz
equipado com uma sonda multinuclear inversa de 5 mm Em tubos de RMN de 5 mm de diacircmetro
preparou-se soluccedilotildees de 500 L de deca- ou metavanadato correspondentes a um intervalo de
concentraccedilotildees de 1 a 10 mM suplementadas com 10 D2O Os espectros foram adquiridos agrave
temperatura ambiente com uma largura espectral de 23 923 Hz um tempo de aquisiccedilatildeo de 0086
s tempo de relaxaccedilatildeo de 001 s e uma sequecircncia de pulsos a 90ordm Utilizou-se a transformada de
Fourier com alargamento das bandas de 20Hz para o processamento dos espectros Os desvios
quiacutemicos das ressonacircncias de 51
V foram obtidos em relaccedilatildeo agrave espeacutecie VOCl3cuja frequecircncia de
ressonacircncia eacute considerada o padratildeo sendo todas as outras referenciadas a partir desta em ppm
As aacutereas dos sinais do RMN correspondentes agraves espeacutecies oligomeacutericas presentes foram obtidas
por integraccedilatildeo dos sinais As concentraccedilotildees relativas das espeacutecies foram calculadas com recurso agrave
equaccedilatildeo
onde A[Vx] corresponde agrave aacuterea referente agrave ressonacircncia da espeacutecie Vx com nordm de agregaccedilatildeo x
A[V]total agrave soma total das aacutereas medidas e [V] a concentraccedilatildeo total de vanaacutedio no ensaio
A teacutecnica de 51
V-RMN foi anteriormente aplicada em estudos de ligaccedilatildeo com S1 de miosina
(Ref) tendo sido demostrado que entre as espeacutecies V1 V2 V4 e V5 a espeacutecie tetrameacuterica possui a
maior afinidade para a miosina (Cremo et al) O desaparecimento ou alargamento dos sinais pode
implicar a ligaccedilatildeo de um dos oxovanadatos laacutebeis agrave miosina sem indicaccedilatildeo de qual uma vez que
49
se processa um equiliacutebrio entre eles (Crans 1990) Assim e tendo em conta as indicaccedilotildees de que
a espeacutecie V10 eacute distinta das restantes no que diz respeito agrave magnitude da interacccedilatildeo com a miosina
e inibiccedilatildeo da actividade ATPaacutesica estimulada por actina (Aureliano 2000 Tiago et al 2002)
torna-se fundamental investigar a interacccedilatildeo das diferentes espeacutecies de vanadato com o S1 de
miosina atraveacutes da teacutecnica de 51
V-RMN
Para tal obtiveram-se espectros de 51
V-RMN resultantes da titulaccedilatildeo de uma soluccedilatildeo 5 mM de
decavanadato total com concentraccedilotildees crescentes de S1 de miosina (0 5 10 15 e 20 microM) em
tampatildeo D Realizou-se ainda ensaios na presenccedila de 2 mM de decavanadato e 2 mM de ATP na
presenccedila e na ausecircncia de S1 de miosina de modo a verificar o efeito que estes ligandos naturais
da enzima teria na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o subfragmento-1
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
Os grupos sulfidrilo (-SH) das cisteiacutenas satildeo resiacuteduos de aminoaacutecidos extremamente sensiacuteveis agrave
oxidaccedilatildeo resultando na sua oxidaccedilatildeo (Dalle-Donne et al 1999) Logo a quantificaccedilatildeo destes
resiacuteduos eacute bastante importante nos estudos de interacccedilatildeo com vanaacutedio jaacute que os resiacuteduos de Cys
com ecircnfase para o SH1 e o SH2 satildeo bastante importantes na monitorizaccedilatildeo das modificaccedilotildees
funcionais da regiatildeo da cabeccedila
O procedimento seguidamente apresentado foi originalmente descrito por Gutieacuterrez-Merino
(1987)) Consiste na reacccedilatildeo entre os grupos ndashSH natildeo oxidados das proteiacutenas e o reagente de
Ellman (DTNB -55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico)) - formando-se o aniatildeo CNT (aniatildeo 3-
carboxilato-4-nitrotiofenolato) numa estequiometria de 11 Este apresenta um maacuteximo de
absorvecircncia a 412 nm (Robyt 1990) Assim apoacutes a incubaccedilatildeo da proteiacutena na presenccedila do metal
a quantidade de cisteiacutenas reduzidas por proteiacutena determina-se atraveacutes dos valores de Abs412
Deste modo colocou-se num tubo de Eppendorf o volume apropriado de S1 dialisado para que a
concentraccedilatildeo no ensaio fosse de 1 mgmL Adicionou-se o volume apropriado de decavanadato
para que a sua concentraccedilatildeo fosse de 0 10 50 100 250 500 ou 1000 M Perfez-se com soluccedilatildeo
D ateacute um volume final de 1 mL e incubou-se a 25ordmC por 20 minutos Apoacutes este periacuteodo colocou-
se 780 L da mistura reaccional numa cuvette de quartzo e adicionou-se 220 L de soluccedilatildeo K23
23
Tampatildeo K Tampatildeo fosfatohidrogenofosfato de soacutedio 025 M pH 72
50
Adicionou-se 15 L de DTNB (em soluccedilatildeo K [DTNB]stock=100 mM) ao meio reaccional e
registou-se o valor de absorvecircncia medido equivalente agraves cisteiacutenas ldquoraacutepidasrdquo (SH1 e SH2
expostos ao solvente) Adicionou-se 001 g de SDS que provoca a desnaturaccedilatildeo da proteiacutena e
consequentemente a exposiccedilatildeo de todos os resiacuteduos de cisteiacutenas que se encontravam ateacute aiacute
inacessiacuteveis no interior da proteiacutena (cisteiacutenas totais) Estes resiacuteduos podem assim reagir com o
DTNB
Estimou-se o nuacutemero de resiacuteduos de cisteiacutenas em todas as amostras com recurso ao coeficiente de
extinccedilatildeo molar () do DTNB (ver ponto 3 dos Anexos) que foi determinado atraveacutes da titulaccedilatildeo
de L-Cisteiacutena com intervalo de concentraccedilotildees compreendidos entre 0 e 50 microM na presenccedila de
25microM de DTNB em tampatildeo D Os valores de absorvecircncia foram obtidos para um comprimento de
onda de 412 nm
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
3621 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
A teacutecnica mais eficiente de anaacutelise da interacccedilatildeo entre o iatildeo vanadilo e proteiacutenas como a
calmodulina e a transferrina entre outras permite a obtenccedilatildeo raacutepida de informaccedilatildeo no que toca agrave
conformaccedilatildeo e amplitude da referida interacccedilatildeo
Os espectros de Ressonacircncia Paramagneacutetica Electroacutenica (RPE) da espeacutecie paramagneacutetica V(IV)
foram realizados a 70K num espectroacutemetro Bruker EMX-300 X-band Todas as soluccedilotildees
utilizadas e amostras foram preparadas em anaerobiose numa cacircmara de luvas de modo a evitar a
reduccedilatildeo de V(IV) a V(V) Em tubos capilares com 02 mL de amostra manteve-se a concentraccedilatildeo
de S1 a 20 M e variou-se a concentraccedilatildeo de vanadilo (0 25 5 10 15 25 50 75 100 150
200 500 e 1000 M [VOSO4]stock=5 mM) em tampatildeo D Apoacutes obtenccedilatildeo dos espectros estes
foram tratados atraveacutes do software SimFonia e WinEPR tendo-se obtido um graacutefico com a
variaccedilatildeo da intensidade do sinal de RPE em funccedilatildeo da concentraccedilatildeo de sulfato de vanadilo
presente Realizaram-se ainda simulaccedilotildees de espectros obtidos com 20 microM de S1 e 1000 microM de
V(IV) com vista agrave obtenccedilatildeo dos ligandos provaacuteveis do vanadilo atraveacutes da anaacutelise da constante
de acoplamento hiperfina (A) e do factor g (g)
4 Conclusotildees gerais
Apesar de jaacute ter sido demonstrado que o vanaacutedio inibe a Na+K
+-ATPase da miosina a funccedilatildeo
deste elemento no tecido muacutesculo esqueleacutetico continua largamente incompreendida A enorme
complexidade quiacutemica deste metal especialmente no que concerne agrave oligomerizaccedilatildeo e reacccedilotildees
51
redox constitui um obstaacuteculo agrave compreensatildeo do comportamento do vanaacutedio nos organismos
vivos Assim a caracterizaccedilatildeo da quiacutemica deste elemento eacute de grande utilidade antes de iniciar o
estudo da sua interacccedilatildeo com o presente sistema
A anaacutelise por espectroscopias de 51
V-RMN e de UV-Vis de soluccedilotildees de vanadato diluiacutedas nos
meios de reacccedilatildeo utilizados no presente relatoacuterio (10 mM HEPES pH 70 25 mM KCl) mostrou
claramente que a espeacutecie decameacuterica (V10) possui um tempo de vida suficientemente longo para a
determinaccedilatildeo do seu efeito isolado ao contraacuterio do que se verifica com as espeacutecies mono- di-
tetra- e pentameacutericas (V1 V2 V4 e V5) Por outro lado a anaacutelise verificou ainda que apoacutes diluiccedilatildeo
das soluccedilotildees de decavanadato no meio de reacccedilatildeo coexistem as espeacutecies V10 e V1 pelo que os
efeitos putativos desta uacuteltima deveratildeo ser subtraiacutedos aquando da monitorizaccedilatildeo dos efeitos da
espeacutecie decameacuterica
O vanaacutedio (V) (vanadato) inibe as enzimas que catalizam reacccedilotildees de transferecircncia de grupos
fosfato como a ATPase da miosina [Goodno 1979] devido agrave sua analogia estrutural com o iatildeo
fosfato Na sua utilizaccedilatildeo como inibidor enzimaacutetico natildeo eacute tida em conta a contribuiccedilatildeo individual
das espeacutecies oligomeacutericas considerando-se normalmente a espeacutecie V1 como inibidora efectiva
[Goodno e Taylor 1982] No presente relatoacuterio verificou-se por espectroscopia de 51
V-RMN que
outros oligoacutemeros de vanadato interagem com o S1 de miosina tendo as espeacutecies V10 e V4 os
maiores niacuteveis de afinidade A sua interacccedilatildeo envolve forccedilas electrostaacuteticas e eacute afectada
negativamente por ligandos naturais de enzima como eacute o caso do ATP
A exposiccedilatildeo do S1 de miosina (005 mgmL) a decavanadato conduz agrave inibiccedilatildeo do capacidade da
proteiacutena de estimular a actividade Mg2+
-ATPaacutesica com um valor de IC50 de 266 plusmn 07 M
decavanadato sendo o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM
decavanadato (80) Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM decavanadato sendo
o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM decavanadato (60)
exercendo um efeito de impedimento agrave ligaccedilatildeo entre o vanaacutedio e o S1
A uma concentraccedilatildeo de 27 microM decavanadato verifica-se a oxidaccedilatildeo de um dos resiacuteduos de Cys
de S1 de miosina seguida pela oxidaccedilatildeo gradual de outro resiacuteduo de Cys ateacute agrave concentraccedilatildeo de
1000 microM Ambos os resiacuteduos oxidados natildeo se encontravam expostos ao solvente utilizado A
oxidaccedilatildeo dos resiacuteduos permite sugerir que esta modificaccedilatildeo oxidativa sofrida pelo S1 contribui
para a diminuiccedilatildeo da sua actividade de ATPase
52
Por espectroscopia de RPE revelaram-se os possiacuteveis ligandos equatoriais do vanaacutedio aquando da
sua interacccedilatildeo com o S1 de miosina como sendo duas moleacuteculas de aacutegua um ligando contendo o
grupo R-OH e outro contendo o grupo R=NR
Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados de
oxidaccedilatildeo (IV) e (V) no S1 de miosina para concentraccedilotildees facilmente alcanccedilaacuteveis no ambiente
terrestre
53
5 Estudos futuros
No sentido do melhor esclarecimento das alteraccedilotildees moleculares e estruturais que ocorrem no
subfragmento 1 quando exposta ao vanaacutedio que conduzem a uma diminuiccedilatildeo da sua funccedilatildeo
sugere-se
- Quantificaccedilatildeo das cisteiacutenas reduzidas de S1 na presenccedila de metavanadato
- Determinaccedilatildeo das actividades Ca2+
- e K+-ATPaacutesica do S1 de miosina na presenccedila de
decavanadato
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina estimulada por actina
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina na presenccedila de
metavanadato
- Realizaccedilatildeo de RMN 2D com variaccedilatildeo das concentraccedilotildees das soluccedilotildees de decavanadato e
metavanadato na presenccedila de S1
-Mutageacutenese dos resiacuteduos de Cys doS1 de miosina de modo a verificar quais os resiacuteduos que
sofrem oxidaccedilatildeo na presenccedila de decavanadato
- Determinaccedilatildeo por calorimetria (DSC Differential Scanning Calorimetry) da variaccedilatildeo da
entalpia (∆H) e da temperatura de desnaturaccedilatildeo do S1 de miosina na presenccedila de vanaacutedio uma
vez que estas propriedades caracteriacutesticas de cada proteiacutena satildeo alteradas apoacutes ocorrecircncia de
modificaccedilotildees estruturais
- Anaacutelise por SDS-PAGE de S1 de miosina apoacutes tratamento com vanaacutedio de modo a determinar
quais os polipeacuteptidos eventualmente formados apoacutes clivagem da proteiacutena
54
6 Referecircncias bibliograacuteficas
- Sellers J Myosins 2nd Edition Oxford University Press 1999
- L Stryer inBiochemistry (Ed WH Freeman 2005 pp
Refordfas Materiais e Meacutetodos
[A] S S Margossian SLowey Methods Enzymol1982 85 55-71
[B] JD Pardue JA Spudich Methods Enzymol 1982 85 164-181
[C] M Bradford Anal Biochem1976 72 248-254
[D] H H Taussky E Shorr J Biol Chem 1953 202 675-685
[E] C R Bagshaw in Outline Studies in Biology ndash Muscle Contraction Chapman and Hall Ed
1982
[F] A Cuenda et al Eur J Biochem1990 194 663-670
55
Anexos
1 Determinaccedilatildeo da recta de calibraccedilatildeo meacutetodo de Bradford
Figura1ndashRepresentaccedilatildeo graacutefica da recta de calibraccedilatildeo (Abs595 nm vs [BSA]) utilizada para o meacutetodo de Bradford
2 Determinaccedilatildeo da recta calibraccedilatildeo para a electroforese em condiccedilotildees desnaturantes
Figura 2 ndash Representaccedilatildeo graacutefica da recta de calibraccedilatildeo (log(Peso Molecular) vs Rf dos marcadores de peso
molecular) onde Rf indica determinada atraveacutes do padratildeo de peso molecular
(poccedilo (a))
3Caacutelculo do coeficiente de extinccedilatildeo molar () para a quantificaccedilatildeo de cisteiacutenas reduzidas
atraveacutes do meacutetodo de Ellman
Traccedilou-se a recta de Abs412 nm meacutedia vs [L-Cisteiacutena] de modo a determinar o valor do coeficiente
de extinccedilatildeo molar definido como o declive daquela recta Assim =8400 M-1
cm-1
como
podemos verificar na Fig 3
A595 nm = 00546[BSA] + 00209 Rsup2 = 099051
Ab
s 59
5 n
m
[BSA] (mgmL)
log PM = -1177 Rf + 2247 Rsup2 = 098957
log
PM
Mobilidade relativa (Rf)
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero

39
Figura 31 ndash Representaccedilatildeo esquemaacutetica do protocolo de isolamento de miosina (Margossian e Lowey 1982) Todo o
procedimento foi realizado a 4ordmC (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
32112 Preparaccedilatildeo de Subfragmento-1 de miosina por digestatildeo com quimiotripsina
O S1 de miosina foi isolado apoacutes diaacutelise da miosina durante a noite contra a soluccedilatildeo C4 A
presenccedila do EDTA eacute essencial uma vez que a sua substituiccedilatildeo por MgCl2resulta na produccedilatildeo de
HMM em vez de S1 A suspensatildeo de miosina foi equilibrada e digerida com -quimotripsina
A digestatildeo foi interrompida pela adiccedilatildeo de PMSF Seguidamente ultracentrifugou-se o produto
da digestatildeo desprezando-se o sedimento (Fig 32) A soluccedilatildeo de sobrenadante maioritariamente
S1 foi quantificada pela leitura directa da Abs280 ou pelo meacutetodo de Bradford (ver as secccedilotildees
3311 e 3312 do Procedimento Experimental respectivamente) e armazenada em 10 sacarose
e 2 mM szlig-Me a -80ordmC
4 Soluccedilatildeo C 25mM Tris pH 70 012M KCl 1mM EDTA
40
Antes da realizaccedilatildeo dos ensaios descritos no presente relatoacuterio o S1 de miosina foi dialisado
contra a soluccedilatildeo D5 de modo a remover qualquer composto que pudesse complexar como
vanaacutedio Seguidamente quantificou-se a proteiacutena pelo meacutetodo de Bradford ou pelo meacutetodo
directo (ver secccedilotildees 3311 e 3312do Procedimento Experimtental)
Figura 3 2ndash Representaccedilatildeo esquemaacutetica do protocolo de preparaccedilatildeo do S1 de miosina por digestatildeo quimiotripsina de
acordo com o previamente descrito por Margossian e Lowey (1982) Todo o procedimento foi realizado a 4ordm C
excepto quando indicaccedilatildeo do contraacuterio (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
3212 Isolamento e purificaccedilatildeo de actina
Isolou-se e purificou-se a actina segundo o meacutetodo descrito por Pardee e Spudish que
compreende dois passos ndash a obtenccedilatildeo de um resiacuteduo seco denominado ldquopoacute de acetonardquo e a
extracccedilatildeo subsequente da actina a partir do ldquopoacute de acetonardquo na forma de G-Actina com a
posterior polimerizaccedilatildeo a F-Actina
32121 Obtenccedilatildeo do ldquopoacute de acetonardquo
Sacrificou-se o coelho com uma pancada atraacutes do pescoccedilo e sangrou-se o animal apoacutes o corte da
arteacuteria jugular Esfolou-se e colocou-se numa tina com gelo Retiraram-se as partes de tecido
conjuntivo que envolvem o tecido muscular com auxiacutelio de uma pinccedila e bisturi Seguidamente
retirou-se os muacutesculos dorsais e os muacutesculos brancos das pernas Passou-se os muacutesculos por aacutegua
destilada de modo a remover as impurezas e pesou-se num copo Trituraram-se os muacutesculos
numa maacutequina de triturar carne e extraiu-se o resultante com 3 volumes da soluccedilatildeo E6 durante 10
minutos com agitaccedilatildeo suave Centrifugou-se a suspensatildeo a 4000 g durante 10
5 Soluccedilatildeo D HEPES 10 mM pH 70 KCl 25 mM
6 Soluccedilatildeo E 01 M KCl 015 M K2HPO4 pH 65
41
minutosdesprezando-se o sobrenadante Ressuspendeu-se o sedimento em 6 volumes de soluccedilatildeo
F7durante 10 minutos com agitaccedilatildeo suave Verificou-se se o pH da soluccedilatildeo estava compreendido
entre 82 e 85 acertando-se estecom soluccedilatildeo G8 se necessaacuterio Centrifugou-se durante 10
minutos a 4000 g Desprezou-se o sobrenadante e ressuspendeu-se o sedimento em 3 volumes de
soluccedilatildeo H9 durante 10 minutos Centrifugou-se novamente a 4000 gdurante 10 minutos
desprezando-se o sobrenadante Ressuspendeu-se o sedimento duas vezes consecutivas em 6
volumes de aacutegua destilada durante 5 minutos Centrifugou-se a 4000 g durante 10 minutos apoacutes
cada adiccedilatildeo de H2O
Seguidamente ressuspendeu-se o sedimento 5 vezes consecutivas utilizando 3 volumes de
acetona de cada vez agitando suavemente durante 10 minutos agrave temperatura ambiente Colocou-
se o resiacuteduo filtrado numa tina de vidro sobre papel de filtro deixando-se evaporar o solvente
durante a noite num nicho Triturou-se o resiacuteduo seco numa maacutequina de triturar carne e deixou-se
novamente evaporar o solvente O ldquopoacute de acetonardquo obtido foi armazenado a -20 ordmC
32122 Purificaccedilatildeo da actina a partir de ldquopoacute de acetonardquo
Apoacutes a obtenccedilatildeo do poacute de acetona como acima indicado efectuou-se o protocolo seguidamente
descrito de modo a obter monoacutemeros de actina (G-Actina) e actina filamentosa (F-Actina)
Pesou-se 35 g de poacute de acetona e adicionaram-se 50 mL de soluccedilatildeo-tampatildeo I10
sendo a
extracccedilatildeo feita com agitaccedilatildeo suave durante 30 minutos Separou-se o extracto do poacute de acetona
hidratado filtrando a suspensatildeo com 4 camadas de gaze esterilizada tendo o cuidado de natildeo
espremer o resiacuteduo Voltou a extrair-se com 50 mL de soluccedilatildeo I durante 30 minutos mexendo
lentamente Adicionou-se este ao extracto obtido anteriormente Centrifugou-se o extracto total a
16 000 g durante 30 minutos Decantou-se para uma proveta sobre a qual se colocou gaze para
que o sedimento solto natildeo se juntasse ao sobrenadante e leu-se o volume De seguida adicionou-
se KCl ateacute uma concentraccedilatildeo final de 50 mM MgCl2 ateacute 2mM e ATP ateacute 1mM lentamente e com
agitaccedilatildeo contiacutenua de modo a promover a polimerizaccedilatildeo dos monoacutemeros de actina
Deixou-se polimerizar durante 2h a 25ordmC observando-se a presenccedila de pequenas bolhas de ar
indicativas de um aumento de viscosidade da soluccedilatildeo Arrefeceu-se em gelo durante 15 minutos e
adicionou-se KCl ateacute uma concentraccedilatildeo final de 06M Mexeu-se lentamente durante 30 minutos
apoacutes os quais se ultracentrifugou a 110 000g durante 1hora Desprezou-se o sobrenadante
ressuspendeu-se o sedimento com soluccedilatildeo tampatildeo E e transferiu-se para um potter
Dialisou-se durante a noite contra 1L de soluccedilatildeo-tampatildeo I a uma temperatura de 4ordmC com rotaccedilatildeo
7 Soluccedilatildeo F 005 M NaHCO3 pH 82-85
8 Soluccedilatildeo G 1M Na2CO3
9 Soluccedilatildeo H 1 mM EDTA pH 70
10 Soluccedilatildeo I 2 mM Tris-HCl pH 8 02 mM CaCl2 02 mM Na2ATP 05 mM szlig-Me 0005 NaN3
42
contiacutenua e mudanccedila de tampatildeo 2 a 3 vezes De seguida a actina foi centrifugada a 110
000gdurante 1hora Decantou-se a soluccedilatildeo de G-Actina (sobrenadante) e desprezou-se o
sedimento Determinou-se a concentraccedilatildeo de G-Actina pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental) A soluccedilatildeo de G-actina foi armazenada a uma temperatura de 4ordmC
apoacutes adiccedilatildeo de 002NaN3 mantendo-se estaacutevel durante 3 a 5 dias
331 Quantificaccedilatildeo das proteiacutenas isoladas
3311 Quantificaccedilatildeo pelo meacutetodo de Bradford
O meacutetodo de Bradford (Bradford 1976) baseia-se na utilizaccedilatildeo do reagente de Bradford
composto por entre outros aacutecido fosfoacuterico e o corante ldquoCoomassierdquo Este corante possui carga
negativa e a sua coloraccedilatildeo varia entre a vermelha (Absmax=465 nm) predominante em soluccedilatildeo e a
azul (Absmax=595 nm) presente aquando da ligaccedilatildeo aos grupos amina das proteiacutenas Em soluccedilotildees
aacutecidas diluiacutedas a coloraccedilatildeo varia proporcionalmente com a quantidade do corante ligado agrave
proteiacutena que por sua vez eacute directamente proporcional agrave concentraccedilatildeo da proteiacutena permitindo a
sua quantificaccedilatildeo ateacute a um limite de detecccedilatildeo de 20 gmL (Bradford 1976)
Deste modo prepararam-se amostras de BSA ([BSA]stock= 1mgmL) utilizando um intervalo de
concentraccedilotildees compreendido entre 0 e 15 gmL de acordo com a tabela 3I Apoacutes a adiccedilatildeo da
H2O MilliQ e do reagente de Bradford as soluccedilotildees foram sucessivamente homogeneizadas no
voacutertex
Tabela 3Indash Amostras utilizadas para a obtenccedilatildeo da recta de calibraccedilatildeo no meacutetodo de Bradford
BSA (L) BSA (g) H2O MilliQ (L) Reagente de Bradford (L)
0 0 800 200
125 125 79875 200
25 25 7975 200
5 5 795 200
10 10 790 200
15 15 785 200
Apoacutes incubaccedilatildeo a 25ordmC por 5 minutos leram-se os valores de Abs595 num
espectrofotoacutemetro (Shimadzu UV-160A) para todas as amostras Seguidamente traccedilou-se a recta
de calibraccedilatildeo (Abs595vs quantidade de BSA (g))
43
Para determinar a concentraccedilatildeo de proteiacutena apoacutes o processo de isolamento purificaccedilatildeo e
diaacutelise quando necessaacuterio colocou-se 5 L da amostra num tubo de Eppendorf e adicionou-se
795 L de H2O MilliQ homogeneizando a soluccedilatildeo no voacutertex Adicionou-se posteriormente 200
L de reagente de Bradford e repetiu-se a homogeneizaccedilatildeo Incubou-se a mistura a 25ordmC por 5
minutos apoacutes os quais se leu os valores de Abs595 nm no espectrofotoacutemetro Por interpolaccedilatildeo deste
valor a partir da recta de calibraccedilatildeo determinou-se a concentraccedilatildeo de proteiacutena na amostra
3312 Quantificaccedilatildeo pelo meacutetodo directo
A quantificaccedilatildeo das amostras proteicas pode tambeacutem ser efectuada atraveacutes da leitura directa da
absorvacircncia tendo por base a lei de Beer-Lambert onde Abs corresponde agrave
absorvacircncia ao comprimento de onda ao qual o ensaio foi realizado corresponde ao coeficiente
de extinccedilatildeo (substituiacutedo pela letra a se as unidades forem distintas da molaridade) b corresponde
ao percurso oacuteptico da radiaccedilatildeo na ceacutelula amostral e c corresponde agrave concentraccedilatildeo amostral
Assim no caso de S1 de miosina mediu-se a Abs280e calculou-se a concentraccedilatildeo da proteiacutena
sabendo que o coeficiente de extinccedilatildeo de S1 eacute (Ref)
No caso da actina mediu-se a Abs290ecalculou-se a concentraccedilatildeo da proteiacutena sabendo que o
coeficiente de extinccedilatildeo da actina eacute (Ref)
332Electroforese em condiccedilotildees desnaturantes (SDS-PAGE)
De modo a melhor caracterizar as proteiacutenas isoladas foi realizada uma electroforese em
condiccedilotildees desnaturantes (SDS-PAGE do inglecircs Sodium Dodecyl Sulfate Polyacrilamide Gel
Electrophoresis) O SDS (dodecilo sulfato de soacutedio) eacute um detergente anioacutenico utilizado neste
meacutetodo para desnaturar as proteiacutenas que se ligam ao SDS na proporccedilatildeo 14 g SDS g proteiacutena
(aproximadamente 1 moleacutecula de SDS por cada 2 resiacuteduos de aminoaacutecidos) (Stryer) O SDS induz
uma densidade de carga negativa significativa nas proteiacutenas a que se liga mascarando a sua carga
intriacutenseca pelo que as razotildees cargamassa satildeo tendencialmente idecircnticas Assim a SDS-PAGE
separa as proteiacutenas quando sujeitas a um campo eleacutectrico de acordo com o seu peso molecular
sendo a sua migraccedilatildeo directamente proporcional ao logaritmo deste uacuteltimo (Stryer)
O gel de resoluccedilatildeo com 10 de acrilamida foi obtido atraveacutes da adiccedilatildeo de 167 mL de soluccedilatildeo
stock de acrilamida11
075 mL de tampatildeo de resoluccedilatildeo12
251 mL de aacutegua destilada e 50 L de
SDS 10 Para promover a polimerizaccedilatildeo adicionou-se em seguida 38 L de PSA 10
11
soluccedilatildeo stock de acrilamida soluccedilatildeo de 30 (mv) de acrilamida e 08 (mv) de NNrsquo-metil-bis-acrilamida 12
tampatildeo de resoluccedilatildeo 15 M Tris-HCl pH 88
44
(persulfato de amoacutenia) 10 e 25 L de TEMED 99 (tetrametiletilenodiamina) Agitou-se
cuidadosamente de modo a natildeo formar bolhas e pipetou-se a soluccedilatildeo para a cassette de vidro
previamente montada no suporte De seguida adicionou-se uma peliacutecula de cerca de 2 mm de 2-
metil-2-butanol para verificar posteriormente a polimerizaccedilatildeo Apoacutes a polimerizaccedilatildeo que demora
cerca de 45 minutos verteu-se a agrave superfiacutecie e adicionou-se o gel de concentraccedilatildeo com 5 de
acrilamida constituiacutedo por 03 mL de soluccedilatildeo de stock acrilamida 045 mL de tampatildeo de
concentraccedilatildeo13
094 mL de aacutegua destilada 18 L de SDS 10 135 L de PSA 10 e 2L de
TEMED 99 Colocou-se o pente sobre o gel e deixou-se polimerizar durante cerca de 45
minutos
As amostras para aplicaccedilatildeo no gel de electroforese foram preparadas para uma concentraccedilatildeo de
proteiacutena de 1 mgmL num volume final de 100 L dos quais 25 L correspondem ao tampatildeo da
amostra14
As amostras foram aquecidas a 100ordmC durante 5 minutos Antes da aplicaccedilatildeo das
amostras transferiu-se a cassette para a tina de electroforese agrave qual se adicionou a soluccedilatildeo
tampatildeo de electroforese da tina superior15
e tampatildeo de electroforese da tina inferior16
Aplicaram-
se 10 L das soluccedilotildees de marcadores de peso molecular e 20 L das amostras proteicas
preparadas anteriormente A electroforese decorreu durante cerca de 3h30 min a uma voltagem
de 80 V De seguida transferiu-se o gel para uma tina corando as proteiacutenas durante 1h com
soluccedilatildeo corante17
Descorou-se o gel com soluccedilatildeo descorante18
tendo-se mudado esta soluccedilatildeo ateacute
as bandas se encontrarem bem definidas
Determinou-se a mobilidade relativa das bandas correspondentes aos marcadores de elevado peso
molecular e traccedilou-se a recta de calibraccedilatildeo ndash log(PM) vs log(Rf) onde Rf corresponde agrave
mobilidade relativa Por interpolaccedilatildeo determina-se o PM das proteiacutenas isoladas sabendo a sua
mobilidade relativa Por comparaccedilatildeo com os valores descritos na literatura pode concluir-se
acerca do grau de purificaccedilatildeo
34 Mediccedilatildeo da actividade enzimaacutetica de S1
341 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
13
tampatildeo de concentraccedilatildeo 05 M Tris-HCl pH 68 14
tampatildeo de amostra 320 mM Tris-HCl pH 68 04 M szlig-mercaptoetanol SDS 8 glicerol 15 (vv) e roxo de m-
cresol 0024 15
tampatildeo de electroforese da tina superior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS 16
tampatildeo de electroforese da tina inferior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS acetato de soacutedio
01 M 17
soluccedilatildeo corante ldquoCoomassie Blue R-250rdquo contendo metanol 45 (vv) e aacutecido aceacutetico glacial 10 (vv) 18
soluccedilatildeo descorante metanol 10 (vv) e aacutecido aceacutetico glacial 10 vv)
45
O meacutetodo de Fiske-Subarow modificado consiste na reacccedilatildeo do molibdato de amoacutenio com o
fosfato inorgacircnico em soluccedilatildeo aacutecida com formaccedilatildeo do aacutecido fosfomoliacutebdico Seguidamente daacute-
se a adiccedilatildeo de um agente redutor que provoca a reduccedilatildeo do composto a fosfomolibdato que forma
um complexo de cor azulada cuja absorvacircncia maacutexima ocorre para um comprimento de onda de
660 nm tornando possiacutevel a sua detecccedilatildeo por meacutetodos espectrofotomeacutetricos As actividades Ca2+
-
ATPaacutesica e K+EDTA-ATPaacutesica de S1 foram determinadas atraveacutes deste meacutetodo de acordo com
o descrito por Taussky e Shorr (1953) com mediccedilatildeo da quantidade de fosfato (Pi) libertado
atraveacutes da hidroacutelise de ATP
Para tal obteve-se primeiramente a recta de calibraccedilatildeo com as amostras presentes na Tabela 3II
utilizando uma soluccedilatildeo stock de 5 mM KH2PO4
Tabela 3II ndash Composiccedilatildeo das diferentes amostras utilizadas para determinar a recta de calibraccedilatildeo atraveacutes do meacutetodo
de Fiske-Subbarow modificado
KH2PO4 (L)
([KH2PO4]stock= 5mM)
PO43-
(nmole) H2O (uL)
0 0 1780
5 25 1775
10 50 1770
20 100 1760
40 200 1740
100 500 1680
A cada tubo de Eppendorf adicionaram-se seguidamente 200 L de soluccedilatildeo J19
e 20 L de agente
redutor20
homogeneizando no voacutertex apoacutes cada adiccedilatildeo Incubou-se durante 10 minutos e mediu-
se a Abs660 nm num espectrofotoacutemetro com intervalos de tempo iguais entre cada amostra Traccedilou-
se a recta de calibraccedilatildeo (Abs660 nmvs nmol PO43-
) a partir dos valores obtidos
Para determinar a actividade Ca2+
-ATPaacutesica de S1 prepararam-se tubos de Eppendorf com 400
L de meio de reacccedilatildeo21
25 L CaCl2 ([CaCl2]stock= 100 mM) 25 L ATP ([ATP]stock= 100 mM)
com [S1]ensaios=01 mgmL Perfez-se se necessaacuterio com H2O MilliQ ateacute perfazer um volume de 1
mL Para determinar a actividade K+EDTA-ATPaacutesica utilizou-se uma concentraccedilatildeo idecircntica de
19
Soluccedilatildeo J 5 N H2SO4 25 molibdato de amoacutenio 20
Agente redutor 5 mg aacutecido 1-amino-2-naftol-4-sulfoacutenico 30 mg bissulfito de soacutedio 30 mg sulfito de soacutedio em 26
mL H2O 21
meio de reacccedilatildeo 125 mM Tris-HCl pH 80 0575 M KCl
46
proteiacutena nos ensaios (01 mgmL) e o mesmo meio de reacccedilatildeo Aos tubos de Eppendorf
adicionou-se ainda 123 L de KCl ([KCl]stock = 3M) 25 L de ATP 10 L de EDTA
([EDTA]stock= 100 mM )e o volume de H2O MilliQ necessaacuterio para perfazer 1 mL Em ambas as
situaccedilotildees o uacuteltimo reagente adicionado foi o ATP uma vez que se trata do substrato da reacccedilatildeo
enzimaacutetica
A cineacutetica decorreu a 25ordmC durante 5 minutos para a actividade Ca2+
-ATPaacutesica e durante 3
minutos para a actividade K+EDTA-ATPaacutesica Apoacutes o periacuteodo de incubaccedilatildeo transferiu-se 150
L de amostra para tubos de Eppendorf contendo igual volume de TCA 10 Colocou-se no gelo
e homogeneizou-se no voacutertex Seguidamente centrifugou-se a 1500 g durante 10 minutos e
recolheu-se 200 L de sobrenadante para novos tubos de Eppendorf Adicionou-se a estes 158
mL de H2O MilliQ e 200 L de soluccedilatildeo J Homogeneizou-se no voacutertex e adicionou-se 20 L de
reagente redutor repetindo-se a homogeneizaccedilatildeo Incubou-se durante 10 minutos e mediu-se a
Abs660 nm com o mesmo intervalo de tempo de leitura para cada amostra Por intrapolaccedilatildeo a partir
da recta de calibraccedilatildeo obtida anteriormente determinou-se a quantidade de Pi libertado em cada
ensaio A partir destes resultados determinaram-se as actividades Ca2+
-ATPaacutesica e K+EDTA-
ATPaacutesica do S1 em nmol PO43-
minmg proteiacutena e em s-1
342 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
Da actividade Mg2+
-ATPaacutesica de S1 resulta a formaccedilatildeo dos produtos da reacccedilatildeo de
hidroacutelise do ATP - ADP e Pi Na presenccedila de ADP fosfoenolpiruvato (PEP) e da enzima piruvato
cinase (PK) os produtos resultantes satildeo ATP e piruvato Por acccedilatildeo da enzima lactato
desidrogenase (LDH) este uacuteltimo eacute convertido em lactato ocorrendo simultaneamente a oxidaccedilatildeo
do NADH a NAD+ monitorizada atraveacutes da Abs340 nm Quando o S1 de miosina se encontra no
meio reaccional o ATP eacute hidrolisado formando-se novamente ADP e Pi o que permite a
formaccedilatildeo de um ciclo (fig 21) Se as enzimas se encontrarem em excesso a velocidade de
conversatildeo do NADH eacute limitada pela produccedilatildeo de ADP e Pi Como o ATP eacute regenerado em cada
ciclo a sua concentraccedilatildeo inicial eacute sempre mantida (Bagshaw 1982)
Figura 33ndash Representaccedilatildeo esquemaacutetica do meacutetodo de enzimas acopladas
47
O protocolo utilizado e seguidamente descrito foi adaptado de Cuenda et al (1990 ndash Ver Refordf) A
uma cuvette de 1 mL adicionou-se S1 para uma concentraccedilatildeo final de 005 mgmL 800 L de
meio de reacccedilatildeo22
10 L de PEP ([PEP]stock = 42 mM) 10 L de PK+LDH 11 ([PK+LDH]stock =
1 mgmL) e 15 L de NADH ([NADH]stock=25 mM) Iniciou-se a monitorizaccedilatildeo da reacccedilatildeo
atraveacutes da leitura de Abs340 nm apoacutes a adiccedilatildeo de 20 L de ATP ([ATP]stock=100 mM) A
actividade Mg2+
-ATPaacutesica eacute obtida atraveacutes do declive correspondente agrave oxidaccedilatildeo do NADH Para
a mediccedilatildeo da Mg2+
-ATPase de S1 estimulada por actina adicionou-se F-actina para uma
concentraccedilatildeo final de 2 microM sendo a actividade obtida tambeacutem atraveacutes do declive correspondente
agrave oxidaccedilatildeo do NADH apoacutes a adiccedilatildeo da proteiacutena
35 Interaccedilatildeo entre vanaacutedio e subfragmento-1 de miosina
351 Preparaccedilatildeo das soluccedilotildees de vanaacutedio
As soluccedilotildees de vanadato foram preparadas em aacutegua MilliQ a partir de metavanadato de amoacutenio
(NH4VO3) numa concentraccedilatildeo stock de 50 mM com agitaccedilatildeo magneacutetica e aquecimento (~30 ordmC)
Esta soluccedilatildeo com um valor compreendido entre 6 e 7 designa-se por ldquometavanadatordquo e eacute
constituiacuteda pelas esspeacutecies V1 V2 V4 e V5 (Ref) Acidificando a soluccedilatildeo de metavanadato por
adiccedilatildeo de HCl ateacute um valor de pH proacuteximo de 4 observa-se uma mudanccedila de cor de amarelo
claro para laranja indicativa da formaccedilatildeo da espeacutecie decameacuterica de vanadato De seguida e
imediatamente antes da realizaccedilatildeo dos ensaios descritos o valor do pH da soluccedilatildeo de
decavanadato foi aumentado para cerca de 75 por adiccedilatildeo de NaOH A soluccedilatildeo (2) designada
por ldquometavanadatordquo natildeo foi posteriormente alterada ateacute agrave sua adiccedilatildeo para a realizaccedilatildeo dos ensaios
As soluccedilotildees foram armazenadas a 4ordmC e mantidas em gelo aquando da sua utilizaccedilatildeo
A soluccedilatildeo de V(IV) foi preparada em condiccedilotildees anaeroacutebias por diluiccedilatildeo de sulfato de vanadil
(VOSO4) para uma concentraccedilatildeo final de 5 mM em H2O MilliQ saturada com N2 gasoso
352Anaacutelise por espectroscopia de UV-Vis
A concentraccedilatildeo da espeacutecie decameacuterica em soluccedilatildeo depende do periacuteodo de tempo apoacutes a diluiccedilatildeo
no meio de reacccedilatildeo apropriado (Ref) A velocidade de desoligomerizaccedilatildeo da espeacutecie decameacuterica
pode ser analisada por espectroscopia de absorccedilatildeo na regiatildeo UVVis uma vez que esta espeacutecie
apresenta duas bandas caracteriacutesticas na regiatildeo do visiacutevel (a 358 e 400 nm) que natildeo se verifica
em quaisquer outras espeacutecies oligomeacutericas As bandas supramencionada sapresentam-se bem
22
Meio de reacccedilatildeo 3125 mM Tris-HCl pH 71 375 mM MgCl2
48
resolvidas atraveacutes da obtenccedilatildeo da derivada de 2ordf ordem do espectro apresentando a forma de 2
vales com miacutenimos centrados nos comprimentos de onda supracitados com dependecircncia linear
relativamente agrave concentraccedilatildeo de decavanadato total (Ramos et al 2006)
Assim a estabilidade da espeacutecie decameacuterica apoacutes diluiccedilatildeo da soluccedilatildeo de decavanadato no meio de
reacccedilatildeo dos ensaios (tampatildeo D) para uma concentraccedilatildeo final de 1 mM em vanadato total foi
caracterizada atraveacutes da leitura da Abs400 em funccedilatildeo do tempo tendo-se determinado qual a
ordem de reacccedilatildeo e o tempo de meia-vida da mesma Com base em estudos anteriores acerca da
estabilidade do V10 na presenccedila de diferentes proteiacutenas (Ramos et al 2006) determinou-se
igualmente o tempo de meia vida apoacutes a adiccedilatildeo de 20 microM de S1 de miosina ao meio reaccional
353 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
Os espectros de RMN do isoacutetopo 51
V foram realizados num espectroacutemetro Bruker AM-400 MHz
equipado com uma sonda multinuclear inversa de 5 mm Em tubos de RMN de 5 mm de diacircmetro
preparou-se soluccedilotildees de 500 L de deca- ou metavanadato correspondentes a um intervalo de
concentraccedilotildees de 1 a 10 mM suplementadas com 10 D2O Os espectros foram adquiridos agrave
temperatura ambiente com uma largura espectral de 23 923 Hz um tempo de aquisiccedilatildeo de 0086
s tempo de relaxaccedilatildeo de 001 s e uma sequecircncia de pulsos a 90ordm Utilizou-se a transformada de
Fourier com alargamento das bandas de 20Hz para o processamento dos espectros Os desvios
quiacutemicos das ressonacircncias de 51
V foram obtidos em relaccedilatildeo agrave espeacutecie VOCl3cuja frequecircncia de
ressonacircncia eacute considerada o padratildeo sendo todas as outras referenciadas a partir desta em ppm
As aacutereas dos sinais do RMN correspondentes agraves espeacutecies oligomeacutericas presentes foram obtidas
por integraccedilatildeo dos sinais As concentraccedilotildees relativas das espeacutecies foram calculadas com recurso agrave
equaccedilatildeo
onde A[Vx] corresponde agrave aacuterea referente agrave ressonacircncia da espeacutecie Vx com nordm de agregaccedilatildeo x
A[V]total agrave soma total das aacutereas medidas e [V] a concentraccedilatildeo total de vanaacutedio no ensaio
A teacutecnica de 51
V-RMN foi anteriormente aplicada em estudos de ligaccedilatildeo com S1 de miosina
(Ref) tendo sido demostrado que entre as espeacutecies V1 V2 V4 e V5 a espeacutecie tetrameacuterica possui a
maior afinidade para a miosina (Cremo et al) O desaparecimento ou alargamento dos sinais pode
implicar a ligaccedilatildeo de um dos oxovanadatos laacutebeis agrave miosina sem indicaccedilatildeo de qual uma vez que
49
se processa um equiliacutebrio entre eles (Crans 1990) Assim e tendo em conta as indicaccedilotildees de que
a espeacutecie V10 eacute distinta das restantes no que diz respeito agrave magnitude da interacccedilatildeo com a miosina
e inibiccedilatildeo da actividade ATPaacutesica estimulada por actina (Aureliano 2000 Tiago et al 2002)
torna-se fundamental investigar a interacccedilatildeo das diferentes espeacutecies de vanadato com o S1 de
miosina atraveacutes da teacutecnica de 51
V-RMN
Para tal obtiveram-se espectros de 51
V-RMN resultantes da titulaccedilatildeo de uma soluccedilatildeo 5 mM de
decavanadato total com concentraccedilotildees crescentes de S1 de miosina (0 5 10 15 e 20 microM) em
tampatildeo D Realizou-se ainda ensaios na presenccedila de 2 mM de decavanadato e 2 mM de ATP na
presenccedila e na ausecircncia de S1 de miosina de modo a verificar o efeito que estes ligandos naturais
da enzima teria na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o subfragmento-1
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
Os grupos sulfidrilo (-SH) das cisteiacutenas satildeo resiacuteduos de aminoaacutecidos extremamente sensiacuteveis agrave
oxidaccedilatildeo resultando na sua oxidaccedilatildeo (Dalle-Donne et al 1999) Logo a quantificaccedilatildeo destes
resiacuteduos eacute bastante importante nos estudos de interacccedilatildeo com vanaacutedio jaacute que os resiacuteduos de Cys
com ecircnfase para o SH1 e o SH2 satildeo bastante importantes na monitorizaccedilatildeo das modificaccedilotildees
funcionais da regiatildeo da cabeccedila
O procedimento seguidamente apresentado foi originalmente descrito por Gutieacuterrez-Merino
(1987)) Consiste na reacccedilatildeo entre os grupos ndashSH natildeo oxidados das proteiacutenas e o reagente de
Ellman (DTNB -55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico)) - formando-se o aniatildeo CNT (aniatildeo 3-
carboxilato-4-nitrotiofenolato) numa estequiometria de 11 Este apresenta um maacuteximo de
absorvecircncia a 412 nm (Robyt 1990) Assim apoacutes a incubaccedilatildeo da proteiacutena na presenccedila do metal
a quantidade de cisteiacutenas reduzidas por proteiacutena determina-se atraveacutes dos valores de Abs412
Deste modo colocou-se num tubo de Eppendorf o volume apropriado de S1 dialisado para que a
concentraccedilatildeo no ensaio fosse de 1 mgmL Adicionou-se o volume apropriado de decavanadato
para que a sua concentraccedilatildeo fosse de 0 10 50 100 250 500 ou 1000 M Perfez-se com soluccedilatildeo
D ateacute um volume final de 1 mL e incubou-se a 25ordmC por 20 minutos Apoacutes este periacuteodo colocou-
se 780 L da mistura reaccional numa cuvette de quartzo e adicionou-se 220 L de soluccedilatildeo K23
23
Tampatildeo K Tampatildeo fosfatohidrogenofosfato de soacutedio 025 M pH 72
50
Adicionou-se 15 L de DTNB (em soluccedilatildeo K [DTNB]stock=100 mM) ao meio reaccional e
registou-se o valor de absorvecircncia medido equivalente agraves cisteiacutenas ldquoraacutepidasrdquo (SH1 e SH2
expostos ao solvente) Adicionou-se 001 g de SDS que provoca a desnaturaccedilatildeo da proteiacutena e
consequentemente a exposiccedilatildeo de todos os resiacuteduos de cisteiacutenas que se encontravam ateacute aiacute
inacessiacuteveis no interior da proteiacutena (cisteiacutenas totais) Estes resiacuteduos podem assim reagir com o
DTNB
Estimou-se o nuacutemero de resiacuteduos de cisteiacutenas em todas as amostras com recurso ao coeficiente de
extinccedilatildeo molar () do DTNB (ver ponto 3 dos Anexos) que foi determinado atraveacutes da titulaccedilatildeo
de L-Cisteiacutena com intervalo de concentraccedilotildees compreendidos entre 0 e 50 microM na presenccedila de
25microM de DTNB em tampatildeo D Os valores de absorvecircncia foram obtidos para um comprimento de
onda de 412 nm
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
3621 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
A teacutecnica mais eficiente de anaacutelise da interacccedilatildeo entre o iatildeo vanadilo e proteiacutenas como a
calmodulina e a transferrina entre outras permite a obtenccedilatildeo raacutepida de informaccedilatildeo no que toca agrave
conformaccedilatildeo e amplitude da referida interacccedilatildeo
Os espectros de Ressonacircncia Paramagneacutetica Electroacutenica (RPE) da espeacutecie paramagneacutetica V(IV)
foram realizados a 70K num espectroacutemetro Bruker EMX-300 X-band Todas as soluccedilotildees
utilizadas e amostras foram preparadas em anaerobiose numa cacircmara de luvas de modo a evitar a
reduccedilatildeo de V(IV) a V(V) Em tubos capilares com 02 mL de amostra manteve-se a concentraccedilatildeo
de S1 a 20 M e variou-se a concentraccedilatildeo de vanadilo (0 25 5 10 15 25 50 75 100 150
200 500 e 1000 M [VOSO4]stock=5 mM) em tampatildeo D Apoacutes obtenccedilatildeo dos espectros estes
foram tratados atraveacutes do software SimFonia e WinEPR tendo-se obtido um graacutefico com a
variaccedilatildeo da intensidade do sinal de RPE em funccedilatildeo da concentraccedilatildeo de sulfato de vanadilo
presente Realizaram-se ainda simulaccedilotildees de espectros obtidos com 20 microM de S1 e 1000 microM de
V(IV) com vista agrave obtenccedilatildeo dos ligandos provaacuteveis do vanadilo atraveacutes da anaacutelise da constante
de acoplamento hiperfina (A) e do factor g (g)
4 Conclusotildees gerais
Apesar de jaacute ter sido demonstrado que o vanaacutedio inibe a Na+K
+-ATPase da miosina a funccedilatildeo
deste elemento no tecido muacutesculo esqueleacutetico continua largamente incompreendida A enorme
complexidade quiacutemica deste metal especialmente no que concerne agrave oligomerizaccedilatildeo e reacccedilotildees
51
redox constitui um obstaacuteculo agrave compreensatildeo do comportamento do vanaacutedio nos organismos
vivos Assim a caracterizaccedilatildeo da quiacutemica deste elemento eacute de grande utilidade antes de iniciar o
estudo da sua interacccedilatildeo com o presente sistema
A anaacutelise por espectroscopias de 51
V-RMN e de UV-Vis de soluccedilotildees de vanadato diluiacutedas nos
meios de reacccedilatildeo utilizados no presente relatoacuterio (10 mM HEPES pH 70 25 mM KCl) mostrou
claramente que a espeacutecie decameacuterica (V10) possui um tempo de vida suficientemente longo para a
determinaccedilatildeo do seu efeito isolado ao contraacuterio do que se verifica com as espeacutecies mono- di-
tetra- e pentameacutericas (V1 V2 V4 e V5) Por outro lado a anaacutelise verificou ainda que apoacutes diluiccedilatildeo
das soluccedilotildees de decavanadato no meio de reacccedilatildeo coexistem as espeacutecies V10 e V1 pelo que os
efeitos putativos desta uacuteltima deveratildeo ser subtraiacutedos aquando da monitorizaccedilatildeo dos efeitos da
espeacutecie decameacuterica
O vanaacutedio (V) (vanadato) inibe as enzimas que catalizam reacccedilotildees de transferecircncia de grupos
fosfato como a ATPase da miosina [Goodno 1979] devido agrave sua analogia estrutural com o iatildeo
fosfato Na sua utilizaccedilatildeo como inibidor enzimaacutetico natildeo eacute tida em conta a contribuiccedilatildeo individual
das espeacutecies oligomeacutericas considerando-se normalmente a espeacutecie V1 como inibidora efectiva
[Goodno e Taylor 1982] No presente relatoacuterio verificou-se por espectroscopia de 51
V-RMN que
outros oligoacutemeros de vanadato interagem com o S1 de miosina tendo as espeacutecies V10 e V4 os
maiores niacuteveis de afinidade A sua interacccedilatildeo envolve forccedilas electrostaacuteticas e eacute afectada
negativamente por ligandos naturais de enzima como eacute o caso do ATP
A exposiccedilatildeo do S1 de miosina (005 mgmL) a decavanadato conduz agrave inibiccedilatildeo do capacidade da
proteiacutena de estimular a actividade Mg2+
-ATPaacutesica com um valor de IC50 de 266 plusmn 07 M
decavanadato sendo o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM
decavanadato (80) Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM decavanadato sendo
o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM decavanadato (60)
exercendo um efeito de impedimento agrave ligaccedilatildeo entre o vanaacutedio e o S1
A uma concentraccedilatildeo de 27 microM decavanadato verifica-se a oxidaccedilatildeo de um dos resiacuteduos de Cys
de S1 de miosina seguida pela oxidaccedilatildeo gradual de outro resiacuteduo de Cys ateacute agrave concentraccedilatildeo de
1000 microM Ambos os resiacuteduos oxidados natildeo se encontravam expostos ao solvente utilizado A
oxidaccedilatildeo dos resiacuteduos permite sugerir que esta modificaccedilatildeo oxidativa sofrida pelo S1 contribui
para a diminuiccedilatildeo da sua actividade de ATPase
52
Por espectroscopia de RPE revelaram-se os possiacuteveis ligandos equatoriais do vanaacutedio aquando da
sua interacccedilatildeo com o S1 de miosina como sendo duas moleacuteculas de aacutegua um ligando contendo o
grupo R-OH e outro contendo o grupo R=NR
Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados de
oxidaccedilatildeo (IV) e (V) no S1 de miosina para concentraccedilotildees facilmente alcanccedilaacuteveis no ambiente
terrestre
53
5 Estudos futuros
No sentido do melhor esclarecimento das alteraccedilotildees moleculares e estruturais que ocorrem no
subfragmento 1 quando exposta ao vanaacutedio que conduzem a uma diminuiccedilatildeo da sua funccedilatildeo
sugere-se
- Quantificaccedilatildeo das cisteiacutenas reduzidas de S1 na presenccedila de metavanadato
- Determinaccedilatildeo das actividades Ca2+
- e K+-ATPaacutesica do S1 de miosina na presenccedila de
decavanadato
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina estimulada por actina
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina na presenccedila de
metavanadato
- Realizaccedilatildeo de RMN 2D com variaccedilatildeo das concentraccedilotildees das soluccedilotildees de decavanadato e
metavanadato na presenccedila de S1
-Mutageacutenese dos resiacuteduos de Cys doS1 de miosina de modo a verificar quais os resiacuteduos que
sofrem oxidaccedilatildeo na presenccedila de decavanadato
- Determinaccedilatildeo por calorimetria (DSC Differential Scanning Calorimetry) da variaccedilatildeo da
entalpia (∆H) e da temperatura de desnaturaccedilatildeo do S1 de miosina na presenccedila de vanaacutedio uma
vez que estas propriedades caracteriacutesticas de cada proteiacutena satildeo alteradas apoacutes ocorrecircncia de
modificaccedilotildees estruturais
- Anaacutelise por SDS-PAGE de S1 de miosina apoacutes tratamento com vanaacutedio de modo a determinar
quais os polipeacuteptidos eventualmente formados apoacutes clivagem da proteiacutena
54
6 Referecircncias bibliograacuteficas
- Sellers J Myosins 2nd Edition Oxford University Press 1999
- L Stryer inBiochemistry (Ed WH Freeman 2005 pp
Refordfas Materiais e Meacutetodos
[A] S S Margossian SLowey Methods Enzymol1982 85 55-71
[B] JD Pardue JA Spudich Methods Enzymol 1982 85 164-181
[C] M Bradford Anal Biochem1976 72 248-254
[D] H H Taussky E Shorr J Biol Chem 1953 202 675-685
[E] C R Bagshaw in Outline Studies in Biology ndash Muscle Contraction Chapman and Hall Ed
1982
[F] A Cuenda et al Eur J Biochem1990 194 663-670
55
Anexos
1 Determinaccedilatildeo da recta de calibraccedilatildeo meacutetodo de Bradford
Figura1ndashRepresentaccedilatildeo graacutefica da recta de calibraccedilatildeo (Abs595 nm vs [BSA]) utilizada para o meacutetodo de Bradford
2 Determinaccedilatildeo da recta calibraccedilatildeo para a electroforese em condiccedilotildees desnaturantes
Figura 2 ndash Representaccedilatildeo graacutefica da recta de calibraccedilatildeo (log(Peso Molecular) vs Rf dos marcadores de peso
molecular) onde Rf indica determinada atraveacutes do padratildeo de peso molecular
(poccedilo (a))
3Caacutelculo do coeficiente de extinccedilatildeo molar () para a quantificaccedilatildeo de cisteiacutenas reduzidas
atraveacutes do meacutetodo de Ellman
Traccedilou-se a recta de Abs412 nm meacutedia vs [L-Cisteiacutena] de modo a determinar o valor do coeficiente
de extinccedilatildeo molar definido como o declive daquela recta Assim =8400 M-1
cm-1
como
podemos verificar na Fig 3
A595 nm = 00546[BSA] + 00209 Rsup2 = 099051
Ab
s 59
5 n
m
[BSA] (mgmL)
log PM = -1177 Rf + 2247 Rsup2 = 098957
log
PM
Mobilidade relativa (Rf)
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero

40
Antes da realizaccedilatildeo dos ensaios descritos no presente relatoacuterio o S1 de miosina foi dialisado
contra a soluccedilatildeo D5 de modo a remover qualquer composto que pudesse complexar como
vanaacutedio Seguidamente quantificou-se a proteiacutena pelo meacutetodo de Bradford ou pelo meacutetodo
directo (ver secccedilotildees 3311 e 3312do Procedimento Experimtental)
Figura 3 2ndash Representaccedilatildeo esquemaacutetica do protocolo de preparaccedilatildeo do S1 de miosina por digestatildeo quimiotripsina de
acordo com o previamente descrito por Margossian e Lowey (1982) Todo o procedimento foi realizado a 4ordm C
excepto quando indicaccedilatildeo do contraacuterio (Adaptado de Susana Ramos Relatoacuterio de Estaacutegio 2005)
3212 Isolamento e purificaccedilatildeo de actina
Isolou-se e purificou-se a actina segundo o meacutetodo descrito por Pardee e Spudish que
compreende dois passos ndash a obtenccedilatildeo de um resiacuteduo seco denominado ldquopoacute de acetonardquo e a
extracccedilatildeo subsequente da actina a partir do ldquopoacute de acetonardquo na forma de G-Actina com a
posterior polimerizaccedilatildeo a F-Actina
32121 Obtenccedilatildeo do ldquopoacute de acetonardquo
Sacrificou-se o coelho com uma pancada atraacutes do pescoccedilo e sangrou-se o animal apoacutes o corte da
arteacuteria jugular Esfolou-se e colocou-se numa tina com gelo Retiraram-se as partes de tecido
conjuntivo que envolvem o tecido muscular com auxiacutelio de uma pinccedila e bisturi Seguidamente
retirou-se os muacutesculos dorsais e os muacutesculos brancos das pernas Passou-se os muacutesculos por aacutegua
destilada de modo a remover as impurezas e pesou-se num copo Trituraram-se os muacutesculos
numa maacutequina de triturar carne e extraiu-se o resultante com 3 volumes da soluccedilatildeo E6 durante 10
minutos com agitaccedilatildeo suave Centrifugou-se a suspensatildeo a 4000 g durante 10
5 Soluccedilatildeo D HEPES 10 mM pH 70 KCl 25 mM
6 Soluccedilatildeo E 01 M KCl 015 M K2HPO4 pH 65
41
minutosdesprezando-se o sobrenadante Ressuspendeu-se o sedimento em 6 volumes de soluccedilatildeo
F7durante 10 minutos com agitaccedilatildeo suave Verificou-se se o pH da soluccedilatildeo estava compreendido
entre 82 e 85 acertando-se estecom soluccedilatildeo G8 se necessaacuterio Centrifugou-se durante 10
minutos a 4000 g Desprezou-se o sobrenadante e ressuspendeu-se o sedimento em 3 volumes de
soluccedilatildeo H9 durante 10 minutos Centrifugou-se novamente a 4000 gdurante 10 minutos
desprezando-se o sobrenadante Ressuspendeu-se o sedimento duas vezes consecutivas em 6
volumes de aacutegua destilada durante 5 minutos Centrifugou-se a 4000 g durante 10 minutos apoacutes
cada adiccedilatildeo de H2O
Seguidamente ressuspendeu-se o sedimento 5 vezes consecutivas utilizando 3 volumes de
acetona de cada vez agitando suavemente durante 10 minutos agrave temperatura ambiente Colocou-
se o resiacuteduo filtrado numa tina de vidro sobre papel de filtro deixando-se evaporar o solvente
durante a noite num nicho Triturou-se o resiacuteduo seco numa maacutequina de triturar carne e deixou-se
novamente evaporar o solvente O ldquopoacute de acetonardquo obtido foi armazenado a -20 ordmC
32122 Purificaccedilatildeo da actina a partir de ldquopoacute de acetonardquo
Apoacutes a obtenccedilatildeo do poacute de acetona como acima indicado efectuou-se o protocolo seguidamente
descrito de modo a obter monoacutemeros de actina (G-Actina) e actina filamentosa (F-Actina)
Pesou-se 35 g de poacute de acetona e adicionaram-se 50 mL de soluccedilatildeo-tampatildeo I10
sendo a
extracccedilatildeo feita com agitaccedilatildeo suave durante 30 minutos Separou-se o extracto do poacute de acetona
hidratado filtrando a suspensatildeo com 4 camadas de gaze esterilizada tendo o cuidado de natildeo
espremer o resiacuteduo Voltou a extrair-se com 50 mL de soluccedilatildeo I durante 30 minutos mexendo
lentamente Adicionou-se este ao extracto obtido anteriormente Centrifugou-se o extracto total a
16 000 g durante 30 minutos Decantou-se para uma proveta sobre a qual se colocou gaze para
que o sedimento solto natildeo se juntasse ao sobrenadante e leu-se o volume De seguida adicionou-
se KCl ateacute uma concentraccedilatildeo final de 50 mM MgCl2 ateacute 2mM e ATP ateacute 1mM lentamente e com
agitaccedilatildeo contiacutenua de modo a promover a polimerizaccedilatildeo dos monoacutemeros de actina
Deixou-se polimerizar durante 2h a 25ordmC observando-se a presenccedila de pequenas bolhas de ar
indicativas de um aumento de viscosidade da soluccedilatildeo Arrefeceu-se em gelo durante 15 minutos e
adicionou-se KCl ateacute uma concentraccedilatildeo final de 06M Mexeu-se lentamente durante 30 minutos
apoacutes os quais se ultracentrifugou a 110 000g durante 1hora Desprezou-se o sobrenadante
ressuspendeu-se o sedimento com soluccedilatildeo tampatildeo E e transferiu-se para um potter
Dialisou-se durante a noite contra 1L de soluccedilatildeo-tampatildeo I a uma temperatura de 4ordmC com rotaccedilatildeo
7 Soluccedilatildeo F 005 M NaHCO3 pH 82-85
8 Soluccedilatildeo G 1M Na2CO3
9 Soluccedilatildeo H 1 mM EDTA pH 70
10 Soluccedilatildeo I 2 mM Tris-HCl pH 8 02 mM CaCl2 02 mM Na2ATP 05 mM szlig-Me 0005 NaN3
42
contiacutenua e mudanccedila de tampatildeo 2 a 3 vezes De seguida a actina foi centrifugada a 110
000gdurante 1hora Decantou-se a soluccedilatildeo de G-Actina (sobrenadante) e desprezou-se o
sedimento Determinou-se a concentraccedilatildeo de G-Actina pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental) A soluccedilatildeo de G-actina foi armazenada a uma temperatura de 4ordmC
apoacutes adiccedilatildeo de 002NaN3 mantendo-se estaacutevel durante 3 a 5 dias
331 Quantificaccedilatildeo das proteiacutenas isoladas
3311 Quantificaccedilatildeo pelo meacutetodo de Bradford
O meacutetodo de Bradford (Bradford 1976) baseia-se na utilizaccedilatildeo do reagente de Bradford
composto por entre outros aacutecido fosfoacuterico e o corante ldquoCoomassierdquo Este corante possui carga
negativa e a sua coloraccedilatildeo varia entre a vermelha (Absmax=465 nm) predominante em soluccedilatildeo e a
azul (Absmax=595 nm) presente aquando da ligaccedilatildeo aos grupos amina das proteiacutenas Em soluccedilotildees
aacutecidas diluiacutedas a coloraccedilatildeo varia proporcionalmente com a quantidade do corante ligado agrave
proteiacutena que por sua vez eacute directamente proporcional agrave concentraccedilatildeo da proteiacutena permitindo a
sua quantificaccedilatildeo ateacute a um limite de detecccedilatildeo de 20 gmL (Bradford 1976)
Deste modo prepararam-se amostras de BSA ([BSA]stock= 1mgmL) utilizando um intervalo de
concentraccedilotildees compreendido entre 0 e 15 gmL de acordo com a tabela 3I Apoacutes a adiccedilatildeo da
H2O MilliQ e do reagente de Bradford as soluccedilotildees foram sucessivamente homogeneizadas no
voacutertex
Tabela 3Indash Amostras utilizadas para a obtenccedilatildeo da recta de calibraccedilatildeo no meacutetodo de Bradford
BSA (L) BSA (g) H2O MilliQ (L) Reagente de Bradford (L)
0 0 800 200
125 125 79875 200
25 25 7975 200
5 5 795 200
10 10 790 200
15 15 785 200
Apoacutes incubaccedilatildeo a 25ordmC por 5 minutos leram-se os valores de Abs595 num
espectrofotoacutemetro (Shimadzu UV-160A) para todas as amostras Seguidamente traccedilou-se a recta
de calibraccedilatildeo (Abs595vs quantidade de BSA (g))
43
Para determinar a concentraccedilatildeo de proteiacutena apoacutes o processo de isolamento purificaccedilatildeo e
diaacutelise quando necessaacuterio colocou-se 5 L da amostra num tubo de Eppendorf e adicionou-se
795 L de H2O MilliQ homogeneizando a soluccedilatildeo no voacutertex Adicionou-se posteriormente 200
L de reagente de Bradford e repetiu-se a homogeneizaccedilatildeo Incubou-se a mistura a 25ordmC por 5
minutos apoacutes os quais se leu os valores de Abs595 nm no espectrofotoacutemetro Por interpolaccedilatildeo deste
valor a partir da recta de calibraccedilatildeo determinou-se a concentraccedilatildeo de proteiacutena na amostra
3312 Quantificaccedilatildeo pelo meacutetodo directo
A quantificaccedilatildeo das amostras proteicas pode tambeacutem ser efectuada atraveacutes da leitura directa da
absorvacircncia tendo por base a lei de Beer-Lambert onde Abs corresponde agrave
absorvacircncia ao comprimento de onda ao qual o ensaio foi realizado corresponde ao coeficiente
de extinccedilatildeo (substituiacutedo pela letra a se as unidades forem distintas da molaridade) b corresponde
ao percurso oacuteptico da radiaccedilatildeo na ceacutelula amostral e c corresponde agrave concentraccedilatildeo amostral
Assim no caso de S1 de miosina mediu-se a Abs280e calculou-se a concentraccedilatildeo da proteiacutena
sabendo que o coeficiente de extinccedilatildeo de S1 eacute (Ref)
No caso da actina mediu-se a Abs290ecalculou-se a concentraccedilatildeo da proteiacutena sabendo que o
coeficiente de extinccedilatildeo da actina eacute (Ref)
332Electroforese em condiccedilotildees desnaturantes (SDS-PAGE)
De modo a melhor caracterizar as proteiacutenas isoladas foi realizada uma electroforese em
condiccedilotildees desnaturantes (SDS-PAGE do inglecircs Sodium Dodecyl Sulfate Polyacrilamide Gel
Electrophoresis) O SDS (dodecilo sulfato de soacutedio) eacute um detergente anioacutenico utilizado neste
meacutetodo para desnaturar as proteiacutenas que se ligam ao SDS na proporccedilatildeo 14 g SDS g proteiacutena
(aproximadamente 1 moleacutecula de SDS por cada 2 resiacuteduos de aminoaacutecidos) (Stryer) O SDS induz
uma densidade de carga negativa significativa nas proteiacutenas a que se liga mascarando a sua carga
intriacutenseca pelo que as razotildees cargamassa satildeo tendencialmente idecircnticas Assim a SDS-PAGE
separa as proteiacutenas quando sujeitas a um campo eleacutectrico de acordo com o seu peso molecular
sendo a sua migraccedilatildeo directamente proporcional ao logaritmo deste uacuteltimo (Stryer)
O gel de resoluccedilatildeo com 10 de acrilamida foi obtido atraveacutes da adiccedilatildeo de 167 mL de soluccedilatildeo
stock de acrilamida11
075 mL de tampatildeo de resoluccedilatildeo12
251 mL de aacutegua destilada e 50 L de
SDS 10 Para promover a polimerizaccedilatildeo adicionou-se em seguida 38 L de PSA 10
11
soluccedilatildeo stock de acrilamida soluccedilatildeo de 30 (mv) de acrilamida e 08 (mv) de NNrsquo-metil-bis-acrilamida 12
tampatildeo de resoluccedilatildeo 15 M Tris-HCl pH 88
44
(persulfato de amoacutenia) 10 e 25 L de TEMED 99 (tetrametiletilenodiamina) Agitou-se
cuidadosamente de modo a natildeo formar bolhas e pipetou-se a soluccedilatildeo para a cassette de vidro
previamente montada no suporte De seguida adicionou-se uma peliacutecula de cerca de 2 mm de 2-
metil-2-butanol para verificar posteriormente a polimerizaccedilatildeo Apoacutes a polimerizaccedilatildeo que demora
cerca de 45 minutos verteu-se a agrave superfiacutecie e adicionou-se o gel de concentraccedilatildeo com 5 de
acrilamida constituiacutedo por 03 mL de soluccedilatildeo de stock acrilamida 045 mL de tampatildeo de
concentraccedilatildeo13
094 mL de aacutegua destilada 18 L de SDS 10 135 L de PSA 10 e 2L de
TEMED 99 Colocou-se o pente sobre o gel e deixou-se polimerizar durante cerca de 45
minutos
As amostras para aplicaccedilatildeo no gel de electroforese foram preparadas para uma concentraccedilatildeo de
proteiacutena de 1 mgmL num volume final de 100 L dos quais 25 L correspondem ao tampatildeo da
amostra14
As amostras foram aquecidas a 100ordmC durante 5 minutos Antes da aplicaccedilatildeo das
amostras transferiu-se a cassette para a tina de electroforese agrave qual se adicionou a soluccedilatildeo
tampatildeo de electroforese da tina superior15
e tampatildeo de electroforese da tina inferior16
Aplicaram-
se 10 L das soluccedilotildees de marcadores de peso molecular e 20 L das amostras proteicas
preparadas anteriormente A electroforese decorreu durante cerca de 3h30 min a uma voltagem
de 80 V De seguida transferiu-se o gel para uma tina corando as proteiacutenas durante 1h com
soluccedilatildeo corante17
Descorou-se o gel com soluccedilatildeo descorante18
tendo-se mudado esta soluccedilatildeo ateacute
as bandas se encontrarem bem definidas
Determinou-se a mobilidade relativa das bandas correspondentes aos marcadores de elevado peso
molecular e traccedilou-se a recta de calibraccedilatildeo ndash log(PM) vs log(Rf) onde Rf corresponde agrave
mobilidade relativa Por interpolaccedilatildeo determina-se o PM das proteiacutenas isoladas sabendo a sua
mobilidade relativa Por comparaccedilatildeo com os valores descritos na literatura pode concluir-se
acerca do grau de purificaccedilatildeo
34 Mediccedilatildeo da actividade enzimaacutetica de S1
341 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
13
tampatildeo de concentraccedilatildeo 05 M Tris-HCl pH 68 14
tampatildeo de amostra 320 mM Tris-HCl pH 68 04 M szlig-mercaptoetanol SDS 8 glicerol 15 (vv) e roxo de m-
cresol 0024 15
tampatildeo de electroforese da tina superior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS 16
tampatildeo de electroforese da tina inferior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS acetato de soacutedio
01 M 17
soluccedilatildeo corante ldquoCoomassie Blue R-250rdquo contendo metanol 45 (vv) e aacutecido aceacutetico glacial 10 (vv) 18
soluccedilatildeo descorante metanol 10 (vv) e aacutecido aceacutetico glacial 10 vv)
45
O meacutetodo de Fiske-Subarow modificado consiste na reacccedilatildeo do molibdato de amoacutenio com o
fosfato inorgacircnico em soluccedilatildeo aacutecida com formaccedilatildeo do aacutecido fosfomoliacutebdico Seguidamente daacute-
se a adiccedilatildeo de um agente redutor que provoca a reduccedilatildeo do composto a fosfomolibdato que forma
um complexo de cor azulada cuja absorvacircncia maacutexima ocorre para um comprimento de onda de
660 nm tornando possiacutevel a sua detecccedilatildeo por meacutetodos espectrofotomeacutetricos As actividades Ca2+
-
ATPaacutesica e K+EDTA-ATPaacutesica de S1 foram determinadas atraveacutes deste meacutetodo de acordo com
o descrito por Taussky e Shorr (1953) com mediccedilatildeo da quantidade de fosfato (Pi) libertado
atraveacutes da hidroacutelise de ATP
Para tal obteve-se primeiramente a recta de calibraccedilatildeo com as amostras presentes na Tabela 3II
utilizando uma soluccedilatildeo stock de 5 mM KH2PO4
Tabela 3II ndash Composiccedilatildeo das diferentes amostras utilizadas para determinar a recta de calibraccedilatildeo atraveacutes do meacutetodo
de Fiske-Subbarow modificado
KH2PO4 (L)
([KH2PO4]stock= 5mM)
PO43-
(nmole) H2O (uL)
0 0 1780
5 25 1775
10 50 1770
20 100 1760
40 200 1740
100 500 1680
A cada tubo de Eppendorf adicionaram-se seguidamente 200 L de soluccedilatildeo J19
e 20 L de agente
redutor20
homogeneizando no voacutertex apoacutes cada adiccedilatildeo Incubou-se durante 10 minutos e mediu-
se a Abs660 nm num espectrofotoacutemetro com intervalos de tempo iguais entre cada amostra Traccedilou-
se a recta de calibraccedilatildeo (Abs660 nmvs nmol PO43-
) a partir dos valores obtidos
Para determinar a actividade Ca2+
-ATPaacutesica de S1 prepararam-se tubos de Eppendorf com 400
L de meio de reacccedilatildeo21
25 L CaCl2 ([CaCl2]stock= 100 mM) 25 L ATP ([ATP]stock= 100 mM)
com [S1]ensaios=01 mgmL Perfez-se se necessaacuterio com H2O MilliQ ateacute perfazer um volume de 1
mL Para determinar a actividade K+EDTA-ATPaacutesica utilizou-se uma concentraccedilatildeo idecircntica de
19
Soluccedilatildeo J 5 N H2SO4 25 molibdato de amoacutenio 20
Agente redutor 5 mg aacutecido 1-amino-2-naftol-4-sulfoacutenico 30 mg bissulfito de soacutedio 30 mg sulfito de soacutedio em 26
mL H2O 21
meio de reacccedilatildeo 125 mM Tris-HCl pH 80 0575 M KCl
46
proteiacutena nos ensaios (01 mgmL) e o mesmo meio de reacccedilatildeo Aos tubos de Eppendorf
adicionou-se ainda 123 L de KCl ([KCl]stock = 3M) 25 L de ATP 10 L de EDTA
([EDTA]stock= 100 mM )e o volume de H2O MilliQ necessaacuterio para perfazer 1 mL Em ambas as
situaccedilotildees o uacuteltimo reagente adicionado foi o ATP uma vez que se trata do substrato da reacccedilatildeo
enzimaacutetica
A cineacutetica decorreu a 25ordmC durante 5 minutos para a actividade Ca2+
-ATPaacutesica e durante 3
minutos para a actividade K+EDTA-ATPaacutesica Apoacutes o periacuteodo de incubaccedilatildeo transferiu-se 150
L de amostra para tubos de Eppendorf contendo igual volume de TCA 10 Colocou-se no gelo
e homogeneizou-se no voacutertex Seguidamente centrifugou-se a 1500 g durante 10 minutos e
recolheu-se 200 L de sobrenadante para novos tubos de Eppendorf Adicionou-se a estes 158
mL de H2O MilliQ e 200 L de soluccedilatildeo J Homogeneizou-se no voacutertex e adicionou-se 20 L de
reagente redutor repetindo-se a homogeneizaccedilatildeo Incubou-se durante 10 minutos e mediu-se a
Abs660 nm com o mesmo intervalo de tempo de leitura para cada amostra Por intrapolaccedilatildeo a partir
da recta de calibraccedilatildeo obtida anteriormente determinou-se a quantidade de Pi libertado em cada
ensaio A partir destes resultados determinaram-se as actividades Ca2+
-ATPaacutesica e K+EDTA-
ATPaacutesica do S1 em nmol PO43-
minmg proteiacutena e em s-1
342 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
Da actividade Mg2+
-ATPaacutesica de S1 resulta a formaccedilatildeo dos produtos da reacccedilatildeo de
hidroacutelise do ATP - ADP e Pi Na presenccedila de ADP fosfoenolpiruvato (PEP) e da enzima piruvato
cinase (PK) os produtos resultantes satildeo ATP e piruvato Por acccedilatildeo da enzima lactato
desidrogenase (LDH) este uacuteltimo eacute convertido em lactato ocorrendo simultaneamente a oxidaccedilatildeo
do NADH a NAD+ monitorizada atraveacutes da Abs340 nm Quando o S1 de miosina se encontra no
meio reaccional o ATP eacute hidrolisado formando-se novamente ADP e Pi o que permite a
formaccedilatildeo de um ciclo (fig 21) Se as enzimas se encontrarem em excesso a velocidade de
conversatildeo do NADH eacute limitada pela produccedilatildeo de ADP e Pi Como o ATP eacute regenerado em cada
ciclo a sua concentraccedilatildeo inicial eacute sempre mantida (Bagshaw 1982)
Figura 33ndash Representaccedilatildeo esquemaacutetica do meacutetodo de enzimas acopladas
47
O protocolo utilizado e seguidamente descrito foi adaptado de Cuenda et al (1990 ndash Ver Refordf) A
uma cuvette de 1 mL adicionou-se S1 para uma concentraccedilatildeo final de 005 mgmL 800 L de
meio de reacccedilatildeo22
10 L de PEP ([PEP]stock = 42 mM) 10 L de PK+LDH 11 ([PK+LDH]stock =
1 mgmL) e 15 L de NADH ([NADH]stock=25 mM) Iniciou-se a monitorizaccedilatildeo da reacccedilatildeo
atraveacutes da leitura de Abs340 nm apoacutes a adiccedilatildeo de 20 L de ATP ([ATP]stock=100 mM) A
actividade Mg2+
-ATPaacutesica eacute obtida atraveacutes do declive correspondente agrave oxidaccedilatildeo do NADH Para
a mediccedilatildeo da Mg2+
-ATPase de S1 estimulada por actina adicionou-se F-actina para uma
concentraccedilatildeo final de 2 microM sendo a actividade obtida tambeacutem atraveacutes do declive correspondente
agrave oxidaccedilatildeo do NADH apoacutes a adiccedilatildeo da proteiacutena
35 Interaccedilatildeo entre vanaacutedio e subfragmento-1 de miosina
351 Preparaccedilatildeo das soluccedilotildees de vanaacutedio
As soluccedilotildees de vanadato foram preparadas em aacutegua MilliQ a partir de metavanadato de amoacutenio
(NH4VO3) numa concentraccedilatildeo stock de 50 mM com agitaccedilatildeo magneacutetica e aquecimento (~30 ordmC)
Esta soluccedilatildeo com um valor compreendido entre 6 e 7 designa-se por ldquometavanadatordquo e eacute
constituiacuteda pelas esspeacutecies V1 V2 V4 e V5 (Ref) Acidificando a soluccedilatildeo de metavanadato por
adiccedilatildeo de HCl ateacute um valor de pH proacuteximo de 4 observa-se uma mudanccedila de cor de amarelo
claro para laranja indicativa da formaccedilatildeo da espeacutecie decameacuterica de vanadato De seguida e
imediatamente antes da realizaccedilatildeo dos ensaios descritos o valor do pH da soluccedilatildeo de
decavanadato foi aumentado para cerca de 75 por adiccedilatildeo de NaOH A soluccedilatildeo (2) designada
por ldquometavanadatordquo natildeo foi posteriormente alterada ateacute agrave sua adiccedilatildeo para a realizaccedilatildeo dos ensaios
As soluccedilotildees foram armazenadas a 4ordmC e mantidas em gelo aquando da sua utilizaccedilatildeo
A soluccedilatildeo de V(IV) foi preparada em condiccedilotildees anaeroacutebias por diluiccedilatildeo de sulfato de vanadil
(VOSO4) para uma concentraccedilatildeo final de 5 mM em H2O MilliQ saturada com N2 gasoso
352Anaacutelise por espectroscopia de UV-Vis
A concentraccedilatildeo da espeacutecie decameacuterica em soluccedilatildeo depende do periacuteodo de tempo apoacutes a diluiccedilatildeo
no meio de reacccedilatildeo apropriado (Ref) A velocidade de desoligomerizaccedilatildeo da espeacutecie decameacuterica
pode ser analisada por espectroscopia de absorccedilatildeo na regiatildeo UVVis uma vez que esta espeacutecie
apresenta duas bandas caracteriacutesticas na regiatildeo do visiacutevel (a 358 e 400 nm) que natildeo se verifica
em quaisquer outras espeacutecies oligomeacutericas As bandas supramencionada sapresentam-se bem
22
Meio de reacccedilatildeo 3125 mM Tris-HCl pH 71 375 mM MgCl2
48
resolvidas atraveacutes da obtenccedilatildeo da derivada de 2ordf ordem do espectro apresentando a forma de 2
vales com miacutenimos centrados nos comprimentos de onda supracitados com dependecircncia linear
relativamente agrave concentraccedilatildeo de decavanadato total (Ramos et al 2006)
Assim a estabilidade da espeacutecie decameacuterica apoacutes diluiccedilatildeo da soluccedilatildeo de decavanadato no meio de
reacccedilatildeo dos ensaios (tampatildeo D) para uma concentraccedilatildeo final de 1 mM em vanadato total foi
caracterizada atraveacutes da leitura da Abs400 em funccedilatildeo do tempo tendo-se determinado qual a
ordem de reacccedilatildeo e o tempo de meia-vida da mesma Com base em estudos anteriores acerca da
estabilidade do V10 na presenccedila de diferentes proteiacutenas (Ramos et al 2006) determinou-se
igualmente o tempo de meia vida apoacutes a adiccedilatildeo de 20 microM de S1 de miosina ao meio reaccional
353 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
Os espectros de RMN do isoacutetopo 51
V foram realizados num espectroacutemetro Bruker AM-400 MHz
equipado com uma sonda multinuclear inversa de 5 mm Em tubos de RMN de 5 mm de diacircmetro
preparou-se soluccedilotildees de 500 L de deca- ou metavanadato correspondentes a um intervalo de
concentraccedilotildees de 1 a 10 mM suplementadas com 10 D2O Os espectros foram adquiridos agrave
temperatura ambiente com uma largura espectral de 23 923 Hz um tempo de aquisiccedilatildeo de 0086
s tempo de relaxaccedilatildeo de 001 s e uma sequecircncia de pulsos a 90ordm Utilizou-se a transformada de
Fourier com alargamento das bandas de 20Hz para o processamento dos espectros Os desvios
quiacutemicos das ressonacircncias de 51
V foram obtidos em relaccedilatildeo agrave espeacutecie VOCl3cuja frequecircncia de
ressonacircncia eacute considerada o padratildeo sendo todas as outras referenciadas a partir desta em ppm
As aacutereas dos sinais do RMN correspondentes agraves espeacutecies oligomeacutericas presentes foram obtidas
por integraccedilatildeo dos sinais As concentraccedilotildees relativas das espeacutecies foram calculadas com recurso agrave
equaccedilatildeo
onde A[Vx] corresponde agrave aacuterea referente agrave ressonacircncia da espeacutecie Vx com nordm de agregaccedilatildeo x
A[V]total agrave soma total das aacutereas medidas e [V] a concentraccedilatildeo total de vanaacutedio no ensaio
A teacutecnica de 51
V-RMN foi anteriormente aplicada em estudos de ligaccedilatildeo com S1 de miosina
(Ref) tendo sido demostrado que entre as espeacutecies V1 V2 V4 e V5 a espeacutecie tetrameacuterica possui a
maior afinidade para a miosina (Cremo et al) O desaparecimento ou alargamento dos sinais pode
implicar a ligaccedilatildeo de um dos oxovanadatos laacutebeis agrave miosina sem indicaccedilatildeo de qual uma vez que
49
se processa um equiliacutebrio entre eles (Crans 1990) Assim e tendo em conta as indicaccedilotildees de que
a espeacutecie V10 eacute distinta das restantes no que diz respeito agrave magnitude da interacccedilatildeo com a miosina
e inibiccedilatildeo da actividade ATPaacutesica estimulada por actina (Aureliano 2000 Tiago et al 2002)
torna-se fundamental investigar a interacccedilatildeo das diferentes espeacutecies de vanadato com o S1 de
miosina atraveacutes da teacutecnica de 51
V-RMN
Para tal obtiveram-se espectros de 51
V-RMN resultantes da titulaccedilatildeo de uma soluccedilatildeo 5 mM de
decavanadato total com concentraccedilotildees crescentes de S1 de miosina (0 5 10 15 e 20 microM) em
tampatildeo D Realizou-se ainda ensaios na presenccedila de 2 mM de decavanadato e 2 mM de ATP na
presenccedila e na ausecircncia de S1 de miosina de modo a verificar o efeito que estes ligandos naturais
da enzima teria na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o subfragmento-1
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
Os grupos sulfidrilo (-SH) das cisteiacutenas satildeo resiacuteduos de aminoaacutecidos extremamente sensiacuteveis agrave
oxidaccedilatildeo resultando na sua oxidaccedilatildeo (Dalle-Donne et al 1999) Logo a quantificaccedilatildeo destes
resiacuteduos eacute bastante importante nos estudos de interacccedilatildeo com vanaacutedio jaacute que os resiacuteduos de Cys
com ecircnfase para o SH1 e o SH2 satildeo bastante importantes na monitorizaccedilatildeo das modificaccedilotildees
funcionais da regiatildeo da cabeccedila
O procedimento seguidamente apresentado foi originalmente descrito por Gutieacuterrez-Merino
(1987)) Consiste na reacccedilatildeo entre os grupos ndashSH natildeo oxidados das proteiacutenas e o reagente de
Ellman (DTNB -55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico)) - formando-se o aniatildeo CNT (aniatildeo 3-
carboxilato-4-nitrotiofenolato) numa estequiometria de 11 Este apresenta um maacuteximo de
absorvecircncia a 412 nm (Robyt 1990) Assim apoacutes a incubaccedilatildeo da proteiacutena na presenccedila do metal
a quantidade de cisteiacutenas reduzidas por proteiacutena determina-se atraveacutes dos valores de Abs412
Deste modo colocou-se num tubo de Eppendorf o volume apropriado de S1 dialisado para que a
concentraccedilatildeo no ensaio fosse de 1 mgmL Adicionou-se o volume apropriado de decavanadato
para que a sua concentraccedilatildeo fosse de 0 10 50 100 250 500 ou 1000 M Perfez-se com soluccedilatildeo
D ateacute um volume final de 1 mL e incubou-se a 25ordmC por 20 minutos Apoacutes este periacuteodo colocou-
se 780 L da mistura reaccional numa cuvette de quartzo e adicionou-se 220 L de soluccedilatildeo K23
23
Tampatildeo K Tampatildeo fosfatohidrogenofosfato de soacutedio 025 M pH 72
50
Adicionou-se 15 L de DTNB (em soluccedilatildeo K [DTNB]stock=100 mM) ao meio reaccional e
registou-se o valor de absorvecircncia medido equivalente agraves cisteiacutenas ldquoraacutepidasrdquo (SH1 e SH2
expostos ao solvente) Adicionou-se 001 g de SDS que provoca a desnaturaccedilatildeo da proteiacutena e
consequentemente a exposiccedilatildeo de todos os resiacuteduos de cisteiacutenas que se encontravam ateacute aiacute
inacessiacuteveis no interior da proteiacutena (cisteiacutenas totais) Estes resiacuteduos podem assim reagir com o
DTNB
Estimou-se o nuacutemero de resiacuteduos de cisteiacutenas em todas as amostras com recurso ao coeficiente de
extinccedilatildeo molar () do DTNB (ver ponto 3 dos Anexos) que foi determinado atraveacutes da titulaccedilatildeo
de L-Cisteiacutena com intervalo de concentraccedilotildees compreendidos entre 0 e 50 microM na presenccedila de
25microM de DTNB em tampatildeo D Os valores de absorvecircncia foram obtidos para um comprimento de
onda de 412 nm
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
3621 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
A teacutecnica mais eficiente de anaacutelise da interacccedilatildeo entre o iatildeo vanadilo e proteiacutenas como a
calmodulina e a transferrina entre outras permite a obtenccedilatildeo raacutepida de informaccedilatildeo no que toca agrave
conformaccedilatildeo e amplitude da referida interacccedilatildeo
Os espectros de Ressonacircncia Paramagneacutetica Electroacutenica (RPE) da espeacutecie paramagneacutetica V(IV)
foram realizados a 70K num espectroacutemetro Bruker EMX-300 X-band Todas as soluccedilotildees
utilizadas e amostras foram preparadas em anaerobiose numa cacircmara de luvas de modo a evitar a
reduccedilatildeo de V(IV) a V(V) Em tubos capilares com 02 mL de amostra manteve-se a concentraccedilatildeo
de S1 a 20 M e variou-se a concentraccedilatildeo de vanadilo (0 25 5 10 15 25 50 75 100 150
200 500 e 1000 M [VOSO4]stock=5 mM) em tampatildeo D Apoacutes obtenccedilatildeo dos espectros estes
foram tratados atraveacutes do software SimFonia e WinEPR tendo-se obtido um graacutefico com a
variaccedilatildeo da intensidade do sinal de RPE em funccedilatildeo da concentraccedilatildeo de sulfato de vanadilo
presente Realizaram-se ainda simulaccedilotildees de espectros obtidos com 20 microM de S1 e 1000 microM de
V(IV) com vista agrave obtenccedilatildeo dos ligandos provaacuteveis do vanadilo atraveacutes da anaacutelise da constante
de acoplamento hiperfina (A) e do factor g (g)
4 Conclusotildees gerais
Apesar de jaacute ter sido demonstrado que o vanaacutedio inibe a Na+K
+-ATPase da miosina a funccedilatildeo
deste elemento no tecido muacutesculo esqueleacutetico continua largamente incompreendida A enorme
complexidade quiacutemica deste metal especialmente no que concerne agrave oligomerizaccedilatildeo e reacccedilotildees
51
redox constitui um obstaacuteculo agrave compreensatildeo do comportamento do vanaacutedio nos organismos
vivos Assim a caracterizaccedilatildeo da quiacutemica deste elemento eacute de grande utilidade antes de iniciar o
estudo da sua interacccedilatildeo com o presente sistema
A anaacutelise por espectroscopias de 51
V-RMN e de UV-Vis de soluccedilotildees de vanadato diluiacutedas nos
meios de reacccedilatildeo utilizados no presente relatoacuterio (10 mM HEPES pH 70 25 mM KCl) mostrou
claramente que a espeacutecie decameacuterica (V10) possui um tempo de vida suficientemente longo para a
determinaccedilatildeo do seu efeito isolado ao contraacuterio do que se verifica com as espeacutecies mono- di-
tetra- e pentameacutericas (V1 V2 V4 e V5) Por outro lado a anaacutelise verificou ainda que apoacutes diluiccedilatildeo
das soluccedilotildees de decavanadato no meio de reacccedilatildeo coexistem as espeacutecies V10 e V1 pelo que os
efeitos putativos desta uacuteltima deveratildeo ser subtraiacutedos aquando da monitorizaccedilatildeo dos efeitos da
espeacutecie decameacuterica
O vanaacutedio (V) (vanadato) inibe as enzimas que catalizam reacccedilotildees de transferecircncia de grupos
fosfato como a ATPase da miosina [Goodno 1979] devido agrave sua analogia estrutural com o iatildeo
fosfato Na sua utilizaccedilatildeo como inibidor enzimaacutetico natildeo eacute tida em conta a contribuiccedilatildeo individual
das espeacutecies oligomeacutericas considerando-se normalmente a espeacutecie V1 como inibidora efectiva
[Goodno e Taylor 1982] No presente relatoacuterio verificou-se por espectroscopia de 51
V-RMN que
outros oligoacutemeros de vanadato interagem com o S1 de miosina tendo as espeacutecies V10 e V4 os
maiores niacuteveis de afinidade A sua interacccedilatildeo envolve forccedilas electrostaacuteticas e eacute afectada
negativamente por ligandos naturais de enzima como eacute o caso do ATP
A exposiccedilatildeo do S1 de miosina (005 mgmL) a decavanadato conduz agrave inibiccedilatildeo do capacidade da
proteiacutena de estimular a actividade Mg2+
-ATPaacutesica com um valor de IC50 de 266 plusmn 07 M
decavanadato sendo o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM
decavanadato (80) Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM decavanadato sendo
o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM decavanadato (60)
exercendo um efeito de impedimento agrave ligaccedilatildeo entre o vanaacutedio e o S1
A uma concentraccedilatildeo de 27 microM decavanadato verifica-se a oxidaccedilatildeo de um dos resiacuteduos de Cys
de S1 de miosina seguida pela oxidaccedilatildeo gradual de outro resiacuteduo de Cys ateacute agrave concentraccedilatildeo de
1000 microM Ambos os resiacuteduos oxidados natildeo se encontravam expostos ao solvente utilizado A
oxidaccedilatildeo dos resiacuteduos permite sugerir que esta modificaccedilatildeo oxidativa sofrida pelo S1 contribui
para a diminuiccedilatildeo da sua actividade de ATPase
52
Por espectroscopia de RPE revelaram-se os possiacuteveis ligandos equatoriais do vanaacutedio aquando da
sua interacccedilatildeo com o S1 de miosina como sendo duas moleacuteculas de aacutegua um ligando contendo o
grupo R-OH e outro contendo o grupo R=NR
Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados de
oxidaccedilatildeo (IV) e (V) no S1 de miosina para concentraccedilotildees facilmente alcanccedilaacuteveis no ambiente
terrestre
53
5 Estudos futuros
No sentido do melhor esclarecimento das alteraccedilotildees moleculares e estruturais que ocorrem no
subfragmento 1 quando exposta ao vanaacutedio que conduzem a uma diminuiccedilatildeo da sua funccedilatildeo
sugere-se
- Quantificaccedilatildeo das cisteiacutenas reduzidas de S1 na presenccedila de metavanadato
- Determinaccedilatildeo das actividades Ca2+
- e K+-ATPaacutesica do S1 de miosina na presenccedila de
decavanadato
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina estimulada por actina
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina na presenccedila de
metavanadato
- Realizaccedilatildeo de RMN 2D com variaccedilatildeo das concentraccedilotildees das soluccedilotildees de decavanadato e
metavanadato na presenccedila de S1
-Mutageacutenese dos resiacuteduos de Cys doS1 de miosina de modo a verificar quais os resiacuteduos que
sofrem oxidaccedilatildeo na presenccedila de decavanadato
- Determinaccedilatildeo por calorimetria (DSC Differential Scanning Calorimetry) da variaccedilatildeo da
entalpia (∆H) e da temperatura de desnaturaccedilatildeo do S1 de miosina na presenccedila de vanaacutedio uma
vez que estas propriedades caracteriacutesticas de cada proteiacutena satildeo alteradas apoacutes ocorrecircncia de
modificaccedilotildees estruturais
- Anaacutelise por SDS-PAGE de S1 de miosina apoacutes tratamento com vanaacutedio de modo a determinar
quais os polipeacuteptidos eventualmente formados apoacutes clivagem da proteiacutena
54
6 Referecircncias bibliograacuteficas
- Sellers J Myosins 2nd Edition Oxford University Press 1999
- L Stryer inBiochemistry (Ed WH Freeman 2005 pp
Refordfas Materiais e Meacutetodos
[A] S S Margossian SLowey Methods Enzymol1982 85 55-71
[B] JD Pardue JA Spudich Methods Enzymol 1982 85 164-181
[C] M Bradford Anal Biochem1976 72 248-254
[D] H H Taussky E Shorr J Biol Chem 1953 202 675-685
[E] C R Bagshaw in Outline Studies in Biology ndash Muscle Contraction Chapman and Hall Ed
1982
[F] A Cuenda et al Eur J Biochem1990 194 663-670
55
Anexos
1 Determinaccedilatildeo da recta de calibraccedilatildeo meacutetodo de Bradford
Figura1ndashRepresentaccedilatildeo graacutefica da recta de calibraccedilatildeo (Abs595 nm vs [BSA]) utilizada para o meacutetodo de Bradford
2 Determinaccedilatildeo da recta calibraccedilatildeo para a electroforese em condiccedilotildees desnaturantes
Figura 2 ndash Representaccedilatildeo graacutefica da recta de calibraccedilatildeo (log(Peso Molecular) vs Rf dos marcadores de peso
molecular) onde Rf indica determinada atraveacutes do padratildeo de peso molecular
(poccedilo (a))
3Caacutelculo do coeficiente de extinccedilatildeo molar () para a quantificaccedilatildeo de cisteiacutenas reduzidas
atraveacutes do meacutetodo de Ellman
Traccedilou-se a recta de Abs412 nm meacutedia vs [L-Cisteiacutena] de modo a determinar o valor do coeficiente
de extinccedilatildeo molar definido como o declive daquela recta Assim =8400 M-1
cm-1
como
podemos verificar na Fig 3
A595 nm = 00546[BSA] + 00209 Rsup2 = 099051
Ab
s 59
5 n
m
[BSA] (mgmL)
log PM = -1177 Rf + 2247 Rsup2 = 098957
log
PM
Mobilidade relativa (Rf)
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero

41
minutosdesprezando-se o sobrenadante Ressuspendeu-se o sedimento em 6 volumes de soluccedilatildeo
F7durante 10 minutos com agitaccedilatildeo suave Verificou-se se o pH da soluccedilatildeo estava compreendido
entre 82 e 85 acertando-se estecom soluccedilatildeo G8 se necessaacuterio Centrifugou-se durante 10
minutos a 4000 g Desprezou-se o sobrenadante e ressuspendeu-se o sedimento em 3 volumes de
soluccedilatildeo H9 durante 10 minutos Centrifugou-se novamente a 4000 gdurante 10 minutos
desprezando-se o sobrenadante Ressuspendeu-se o sedimento duas vezes consecutivas em 6
volumes de aacutegua destilada durante 5 minutos Centrifugou-se a 4000 g durante 10 minutos apoacutes
cada adiccedilatildeo de H2O
Seguidamente ressuspendeu-se o sedimento 5 vezes consecutivas utilizando 3 volumes de
acetona de cada vez agitando suavemente durante 10 minutos agrave temperatura ambiente Colocou-
se o resiacuteduo filtrado numa tina de vidro sobre papel de filtro deixando-se evaporar o solvente
durante a noite num nicho Triturou-se o resiacuteduo seco numa maacutequina de triturar carne e deixou-se
novamente evaporar o solvente O ldquopoacute de acetonardquo obtido foi armazenado a -20 ordmC
32122 Purificaccedilatildeo da actina a partir de ldquopoacute de acetonardquo
Apoacutes a obtenccedilatildeo do poacute de acetona como acima indicado efectuou-se o protocolo seguidamente
descrito de modo a obter monoacutemeros de actina (G-Actina) e actina filamentosa (F-Actina)
Pesou-se 35 g de poacute de acetona e adicionaram-se 50 mL de soluccedilatildeo-tampatildeo I10
sendo a
extracccedilatildeo feita com agitaccedilatildeo suave durante 30 minutos Separou-se o extracto do poacute de acetona
hidratado filtrando a suspensatildeo com 4 camadas de gaze esterilizada tendo o cuidado de natildeo
espremer o resiacuteduo Voltou a extrair-se com 50 mL de soluccedilatildeo I durante 30 minutos mexendo
lentamente Adicionou-se este ao extracto obtido anteriormente Centrifugou-se o extracto total a
16 000 g durante 30 minutos Decantou-se para uma proveta sobre a qual se colocou gaze para
que o sedimento solto natildeo se juntasse ao sobrenadante e leu-se o volume De seguida adicionou-
se KCl ateacute uma concentraccedilatildeo final de 50 mM MgCl2 ateacute 2mM e ATP ateacute 1mM lentamente e com
agitaccedilatildeo contiacutenua de modo a promover a polimerizaccedilatildeo dos monoacutemeros de actina
Deixou-se polimerizar durante 2h a 25ordmC observando-se a presenccedila de pequenas bolhas de ar
indicativas de um aumento de viscosidade da soluccedilatildeo Arrefeceu-se em gelo durante 15 minutos e
adicionou-se KCl ateacute uma concentraccedilatildeo final de 06M Mexeu-se lentamente durante 30 minutos
apoacutes os quais se ultracentrifugou a 110 000g durante 1hora Desprezou-se o sobrenadante
ressuspendeu-se o sedimento com soluccedilatildeo tampatildeo E e transferiu-se para um potter
Dialisou-se durante a noite contra 1L de soluccedilatildeo-tampatildeo I a uma temperatura de 4ordmC com rotaccedilatildeo
7 Soluccedilatildeo F 005 M NaHCO3 pH 82-85
8 Soluccedilatildeo G 1M Na2CO3
9 Soluccedilatildeo H 1 mM EDTA pH 70
10 Soluccedilatildeo I 2 mM Tris-HCl pH 8 02 mM CaCl2 02 mM Na2ATP 05 mM szlig-Me 0005 NaN3
42
contiacutenua e mudanccedila de tampatildeo 2 a 3 vezes De seguida a actina foi centrifugada a 110
000gdurante 1hora Decantou-se a soluccedilatildeo de G-Actina (sobrenadante) e desprezou-se o
sedimento Determinou-se a concentraccedilatildeo de G-Actina pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental) A soluccedilatildeo de G-actina foi armazenada a uma temperatura de 4ordmC
apoacutes adiccedilatildeo de 002NaN3 mantendo-se estaacutevel durante 3 a 5 dias
331 Quantificaccedilatildeo das proteiacutenas isoladas
3311 Quantificaccedilatildeo pelo meacutetodo de Bradford
O meacutetodo de Bradford (Bradford 1976) baseia-se na utilizaccedilatildeo do reagente de Bradford
composto por entre outros aacutecido fosfoacuterico e o corante ldquoCoomassierdquo Este corante possui carga
negativa e a sua coloraccedilatildeo varia entre a vermelha (Absmax=465 nm) predominante em soluccedilatildeo e a
azul (Absmax=595 nm) presente aquando da ligaccedilatildeo aos grupos amina das proteiacutenas Em soluccedilotildees
aacutecidas diluiacutedas a coloraccedilatildeo varia proporcionalmente com a quantidade do corante ligado agrave
proteiacutena que por sua vez eacute directamente proporcional agrave concentraccedilatildeo da proteiacutena permitindo a
sua quantificaccedilatildeo ateacute a um limite de detecccedilatildeo de 20 gmL (Bradford 1976)
Deste modo prepararam-se amostras de BSA ([BSA]stock= 1mgmL) utilizando um intervalo de
concentraccedilotildees compreendido entre 0 e 15 gmL de acordo com a tabela 3I Apoacutes a adiccedilatildeo da
H2O MilliQ e do reagente de Bradford as soluccedilotildees foram sucessivamente homogeneizadas no
voacutertex
Tabela 3Indash Amostras utilizadas para a obtenccedilatildeo da recta de calibraccedilatildeo no meacutetodo de Bradford
BSA (L) BSA (g) H2O MilliQ (L) Reagente de Bradford (L)
0 0 800 200
125 125 79875 200
25 25 7975 200
5 5 795 200
10 10 790 200
15 15 785 200
Apoacutes incubaccedilatildeo a 25ordmC por 5 minutos leram-se os valores de Abs595 num
espectrofotoacutemetro (Shimadzu UV-160A) para todas as amostras Seguidamente traccedilou-se a recta
de calibraccedilatildeo (Abs595vs quantidade de BSA (g))
43
Para determinar a concentraccedilatildeo de proteiacutena apoacutes o processo de isolamento purificaccedilatildeo e
diaacutelise quando necessaacuterio colocou-se 5 L da amostra num tubo de Eppendorf e adicionou-se
795 L de H2O MilliQ homogeneizando a soluccedilatildeo no voacutertex Adicionou-se posteriormente 200
L de reagente de Bradford e repetiu-se a homogeneizaccedilatildeo Incubou-se a mistura a 25ordmC por 5
minutos apoacutes os quais se leu os valores de Abs595 nm no espectrofotoacutemetro Por interpolaccedilatildeo deste
valor a partir da recta de calibraccedilatildeo determinou-se a concentraccedilatildeo de proteiacutena na amostra
3312 Quantificaccedilatildeo pelo meacutetodo directo
A quantificaccedilatildeo das amostras proteicas pode tambeacutem ser efectuada atraveacutes da leitura directa da
absorvacircncia tendo por base a lei de Beer-Lambert onde Abs corresponde agrave
absorvacircncia ao comprimento de onda ao qual o ensaio foi realizado corresponde ao coeficiente
de extinccedilatildeo (substituiacutedo pela letra a se as unidades forem distintas da molaridade) b corresponde
ao percurso oacuteptico da radiaccedilatildeo na ceacutelula amostral e c corresponde agrave concentraccedilatildeo amostral
Assim no caso de S1 de miosina mediu-se a Abs280e calculou-se a concentraccedilatildeo da proteiacutena
sabendo que o coeficiente de extinccedilatildeo de S1 eacute (Ref)
No caso da actina mediu-se a Abs290ecalculou-se a concentraccedilatildeo da proteiacutena sabendo que o
coeficiente de extinccedilatildeo da actina eacute (Ref)
332Electroforese em condiccedilotildees desnaturantes (SDS-PAGE)
De modo a melhor caracterizar as proteiacutenas isoladas foi realizada uma electroforese em
condiccedilotildees desnaturantes (SDS-PAGE do inglecircs Sodium Dodecyl Sulfate Polyacrilamide Gel
Electrophoresis) O SDS (dodecilo sulfato de soacutedio) eacute um detergente anioacutenico utilizado neste
meacutetodo para desnaturar as proteiacutenas que se ligam ao SDS na proporccedilatildeo 14 g SDS g proteiacutena
(aproximadamente 1 moleacutecula de SDS por cada 2 resiacuteduos de aminoaacutecidos) (Stryer) O SDS induz
uma densidade de carga negativa significativa nas proteiacutenas a que se liga mascarando a sua carga
intriacutenseca pelo que as razotildees cargamassa satildeo tendencialmente idecircnticas Assim a SDS-PAGE
separa as proteiacutenas quando sujeitas a um campo eleacutectrico de acordo com o seu peso molecular
sendo a sua migraccedilatildeo directamente proporcional ao logaritmo deste uacuteltimo (Stryer)
O gel de resoluccedilatildeo com 10 de acrilamida foi obtido atraveacutes da adiccedilatildeo de 167 mL de soluccedilatildeo
stock de acrilamida11
075 mL de tampatildeo de resoluccedilatildeo12
251 mL de aacutegua destilada e 50 L de
SDS 10 Para promover a polimerizaccedilatildeo adicionou-se em seguida 38 L de PSA 10
11
soluccedilatildeo stock de acrilamida soluccedilatildeo de 30 (mv) de acrilamida e 08 (mv) de NNrsquo-metil-bis-acrilamida 12
tampatildeo de resoluccedilatildeo 15 M Tris-HCl pH 88
44
(persulfato de amoacutenia) 10 e 25 L de TEMED 99 (tetrametiletilenodiamina) Agitou-se
cuidadosamente de modo a natildeo formar bolhas e pipetou-se a soluccedilatildeo para a cassette de vidro
previamente montada no suporte De seguida adicionou-se uma peliacutecula de cerca de 2 mm de 2-
metil-2-butanol para verificar posteriormente a polimerizaccedilatildeo Apoacutes a polimerizaccedilatildeo que demora
cerca de 45 minutos verteu-se a agrave superfiacutecie e adicionou-se o gel de concentraccedilatildeo com 5 de
acrilamida constituiacutedo por 03 mL de soluccedilatildeo de stock acrilamida 045 mL de tampatildeo de
concentraccedilatildeo13
094 mL de aacutegua destilada 18 L de SDS 10 135 L de PSA 10 e 2L de
TEMED 99 Colocou-se o pente sobre o gel e deixou-se polimerizar durante cerca de 45
minutos
As amostras para aplicaccedilatildeo no gel de electroforese foram preparadas para uma concentraccedilatildeo de
proteiacutena de 1 mgmL num volume final de 100 L dos quais 25 L correspondem ao tampatildeo da
amostra14
As amostras foram aquecidas a 100ordmC durante 5 minutos Antes da aplicaccedilatildeo das
amostras transferiu-se a cassette para a tina de electroforese agrave qual se adicionou a soluccedilatildeo
tampatildeo de electroforese da tina superior15
e tampatildeo de electroforese da tina inferior16
Aplicaram-
se 10 L das soluccedilotildees de marcadores de peso molecular e 20 L das amostras proteicas
preparadas anteriormente A electroforese decorreu durante cerca de 3h30 min a uma voltagem
de 80 V De seguida transferiu-se o gel para uma tina corando as proteiacutenas durante 1h com
soluccedilatildeo corante17
Descorou-se o gel com soluccedilatildeo descorante18
tendo-se mudado esta soluccedilatildeo ateacute
as bandas se encontrarem bem definidas
Determinou-se a mobilidade relativa das bandas correspondentes aos marcadores de elevado peso
molecular e traccedilou-se a recta de calibraccedilatildeo ndash log(PM) vs log(Rf) onde Rf corresponde agrave
mobilidade relativa Por interpolaccedilatildeo determina-se o PM das proteiacutenas isoladas sabendo a sua
mobilidade relativa Por comparaccedilatildeo com os valores descritos na literatura pode concluir-se
acerca do grau de purificaccedilatildeo
34 Mediccedilatildeo da actividade enzimaacutetica de S1
341 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
13
tampatildeo de concentraccedilatildeo 05 M Tris-HCl pH 68 14
tampatildeo de amostra 320 mM Tris-HCl pH 68 04 M szlig-mercaptoetanol SDS 8 glicerol 15 (vv) e roxo de m-
cresol 0024 15
tampatildeo de electroforese da tina superior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS 16
tampatildeo de electroforese da tina inferior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS acetato de soacutedio
01 M 17
soluccedilatildeo corante ldquoCoomassie Blue R-250rdquo contendo metanol 45 (vv) e aacutecido aceacutetico glacial 10 (vv) 18
soluccedilatildeo descorante metanol 10 (vv) e aacutecido aceacutetico glacial 10 vv)
45
O meacutetodo de Fiske-Subarow modificado consiste na reacccedilatildeo do molibdato de amoacutenio com o
fosfato inorgacircnico em soluccedilatildeo aacutecida com formaccedilatildeo do aacutecido fosfomoliacutebdico Seguidamente daacute-
se a adiccedilatildeo de um agente redutor que provoca a reduccedilatildeo do composto a fosfomolibdato que forma
um complexo de cor azulada cuja absorvacircncia maacutexima ocorre para um comprimento de onda de
660 nm tornando possiacutevel a sua detecccedilatildeo por meacutetodos espectrofotomeacutetricos As actividades Ca2+
-
ATPaacutesica e K+EDTA-ATPaacutesica de S1 foram determinadas atraveacutes deste meacutetodo de acordo com
o descrito por Taussky e Shorr (1953) com mediccedilatildeo da quantidade de fosfato (Pi) libertado
atraveacutes da hidroacutelise de ATP
Para tal obteve-se primeiramente a recta de calibraccedilatildeo com as amostras presentes na Tabela 3II
utilizando uma soluccedilatildeo stock de 5 mM KH2PO4
Tabela 3II ndash Composiccedilatildeo das diferentes amostras utilizadas para determinar a recta de calibraccedilatildeo atraveacutes do meacutetodo
de Fiske-Subbarow modificado
KH2PO4 (L)
([KH2PO4]stock= 5mM)
PO43-
(nmole) H2O (uL)
0 0 1780
5 25 1775
10 50 1770
20 100 1760
40 200 1740
100 500 1680
A cada tubo de Eppendorf adicionaram-se seguidamente 200 L de soluccedilatildeo J19
e 20 L de agente
redutor20
homogeneizando no voacutertex apoacutes cada adiccedilatildeo Incubou-se durante 10 minutos e mediu-
se a Abs660 nm num espectrofotoacutemetro com intervalos de tempo iguais entre cada amostra Traccedilou-
se a recta de calibraccedilatildeo (Abs660 nmvs nmol PO43-
) a partir dos valores obtidos
Para determinar a actividade Ca2+
-ATPaacutesica de S1 prepararam-se tubos de Eppendorf com 400
L de meio de reacccedilatildeo21
25 L CaCl2 ([CaCl2]stock= 100 mM) 25 L ATP ([ATP]stock= 100 mM)
com [S1]ensaios=01 mgmL Perfez-se se necessaacuterio com H2O MilliQ ateacute perfazer um volume de 1
mL Para determinar a actividade K+EDTA-ATPaacutesica utilizou-se uma concentraccedilatildeo idecircntica de
19
Soluccedilatildeo J 5 N H2SO4 25 molibdato de amoacutenio 20
Agente redutor 5 mg aacutecido 1-amino-2-naftol-4-sulfoacutenico 30 mg bissulfito de soacutedio 30 mg sulfito de soacutedio em 26
mL H2O 21
meio de reacccedilatildeo 125 mM Tris-HCl pH 80 0575 M KCl
46
proteiacutena nos ensaios (01 mgmL) e o mesmo meio de reacccedilatildeo Aos tubos de Eppendorf
adicionou-se ainda 123 L de KCl ([KCl]stock = 3M) 25 L de ATP 10 L de EDTA
([EDTA]stock= 100 mM )e o volume de H2O MilliQ necessaacuterio para perfazer 1 mL Em ambas as
situaccedilotildees o uacuteltimo reagente adicionado foi o ATP uma vez que se trata do substrato da reacccedilatildeo
enzimaacutetica
A cineacutetica decorreu a 25ordmC durante 5 minutos para a actividade Ca2+
-ATPaacutesica e durante 3
minutos para a actividade K+EDTA-ATPaacutesica Apoacutes o periacuteodo de incubaccedilatildeo transferiu-se 150
L de amostra para tubos de Eppendorf contendo igual volume de TCA 10 Colocou-se no gelo
e homogeneizou-se no voacutertex Seguidamente centrifugou-se a 1500 g durante 10 minutos e
recolheu-se 200 L de sobrenadante para novos tubos de Eppendorf Adicionou-se a estes 158
mL de H2O MilliQ e 200 L de soluccedilatildeo J Homogeneizou-se no voacutertex e adicionou-se 20 L de
reagente redutor repetindo-se a homogeneizaccedilatildeo Incubou-se durante 10 minutos e mediu-se a
Abs660 nm com o mesmo intervalo de tempo de leitura para cada amostra Por intrapolaccedilatildeo a partir
da recta de calibraccedilatildeo obtida anteriormente determinou-se a quantidade de Pi libertado em cada
ensaio A partir destes resultados determinaram-se as actividades Ca2+
-ATPaacutesica e K+EDTA-
ATPaacutesica do S1 em nmol PO43-
minmg proteiacutena e em s-1
342 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
Da actividade Mg2+
-ATPaacutesica de S1 resulta a formaccedilatildeo dos produtos da reacccedilatildeo de
hidroacutelise do ATP - ADP e Pi Na presenccedila de ADP fosfoenolpiruvato (PEP) e da enzima piruvato
cinase (PK) os produtos resultantes satildeo ATP e piruvato Por acccedilatildeo da enzima lactato
desidrogenase (LDH) este uacuteltimo eacute convertido em lactato ocorrendo simultaneamente a oxidaccedilatildeo
do NADH a NAD+ monitorizada atraveacutes da Abs340 nm Quando o S1 de miosina se encontra no
meio reaccional o ATP eacute hidrolisado formando-se novamente ADP e Pi o que permite a
formaccedilatildeo de um ciclo (fig 21) Se as enzimas se encontrarem em excesso a velocidade de
conversatildeo do NADH eacute limitada pela produccedilatildeo de ADP e Pi Como o ATP eacute regenerado em cada
ciclo a sua concentraccedilatildeo inicial eacute sempre mantida (Bagshaw 1982)
Figura 33ndash Representaccedilatildeo esquemaacutetica do meacutetodo de enzimas acopladas
47
O protocolo utilizado e seguidamente descrito foi adaptado de Cuenda et al (1990 ndash Ver Refordf) A
uma cuvette de 1 mL adicionou-se S1 para uma concentraccedilatildeo final de 005 mgmL 800 L de
meio de reacccedilatildeo22
10 L de PEP ([PEP]stock = 42 mM) 10 L de PK+LDH 11 ([PK+LDH]stock =
1 mgmL) e 15 L de NADH ([NADH]stock=25 mM) Iniciou-se a monitorizaccedilatildeo da reacccedilatildeo
atraveacutes da leitura de Abs340 nm apoacutes a adiccedilatildeo de 20 L de ATP ([ATP]stock=100 mM) A
actividade Mg2+
-ATPaacutesica eacute obtida atraveacutes do declive correspondente agrave oxidaccedilatildeo do NADH Para
a mediccedilatildeo da Mg2+
-ATPase de S1 estimulada por actina adicionou-se F-actina para uma
concentraccedilatildeo final de 2 microM sendo a actividade obtida tambeacutem atraveacutes do declive correspondente
agrave oxidaccedilatildeo do NADH apoacutes a adiccedilatildeo da proteiacutena
35 Interaccedilatildeo entre vanaacutedio e subfragmento-1 de miosina
351 Preparaccedilatildeo das soluccedilotildees de vanaacutedio
As soluccedilotildees de vanadato foram preparadas em aacutegua MilliQ a partir de metavanadato de amoacutenio
(NH4VO3) numa concentraccedilatildeo stock de 50 mM com agitaccedilatildeo magneacutetica e aquecimento (~30 ordmC)
Esta soluccedilatildeo com um valor compreendido entre 6 e 7 designa-se por ldquometavanadatordquo e eacute
constituiacuteda pelas esspeacutecies V1 V2 V4 e V5 (Ref) Acidificando a soluccedilatildeo de metavanadato por
adiccedilatildeo de HCl ateacute um valor de pH proacuteximo de 4 observa-se uma mudanccedila de cor de amarelo
claro para laranja indicativa da formaccedilatildeo da espeacutecie decameacuterica de vanadato De seguida e
imediatamente antes da realizaccedilatildeo dos ensaios descritos o valor do pH da soluccedilatildeo de
decavanadato foi aumentado para cerca de 75 por adiccedilatildeo de NaOH A soluccedilatildeo (2) designada
por ldquometavanadatordquo natildeo foi posteriormente alterada ateacute agrave sua adiccedilatildeo para a realizaccedilatildeo dos ensaios
As soluccedilotildees foram armazenadas a 4ordmC e mantidas em gelo aquando da sua utilizaccedilatildeo
A soluccedilatildeo de V(IV) foi preparada em condiccedilotildees anaeroacutebias por diluiccedilatildeo de sulfato de vanadil
(VOSO4) para uma concentraccedilatildeo final de 5 mM em H2O MilliQ saturada com N2 gasoso
352Anaacutelise por espectroscopia de UV-Vis
A concentraccedilatildeo da espeacutecie decameacuterica em soluccedilatildeo depende do periacuteodo de tempo apoacutes a diluiccedilatildeo
no meio de reacccedilatildeo apropriado (Ref) A velocidade de desoligomerizaccedilatildeo da espeacutecie decameacuterica
pode ser analisada por espectroscopia de absorccedilatildeo na regiatildeo UVVis uma vez que esta espeacutecie
apresenta duas bandas caracteriacutesticas na regiatildeo do visiacutevel (a 358 e 400 nm) que natildeo se verifica
em quaisquer outras espeacutecies oligomeacutericas As bandas supramencionada sapresentam-se bem
22
Meio de reacccedilatildeo 3125 mM Tris-HCl pH 71 375 mM MgCl2
48
resolvidas atraveacutes da obtenccedilatildeo da derivada de 2ordf ordem do espectro apresentando a forma de 2
vales com miacutenimos centrados nos comprimentos de onda supracitados com dependecircncia linear
relativamente agrave concentraccedilatildeo de decavanadato total (Ramos et al 2006)
Assim a estabilidade da espeacutecie decameacuterica apoacutes diluiccedilatildeo da soluccedilatildeo de decavanadato no meio de
reacccedilatildeo dos ensaios (tampatildeo D) para uma concentraccedilatildeo final de 1 mM em vanadato total foi
caracterizada atraveacutes da leitura da Abs400 em funccedilatildeo do tempo tendo-se determinado qual a
ordem de reacccedilatildeo e o tempo de meia-vida da mesma Com base em estudos anteriores acerca da
estabilidade do V10 na presenccedila de diferentes proteiacutenas (Ramos et al 2006) determinou-se
igualmente o tempo de meia vida apoacutes a adiccedilatildeo de 20 microM de S1 de miosina ao meio reaccional
353 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
Os espectros de RMN do isoacutetopo 51
V foram realizados num espectroacutemetro Bruker AM-400 MHz
equipado com uma sonda multinuclear inversa de 5 mm Em tubos de RMN de 5 mm de diacircmetro
preparou-se soluccedilotildees de 500 L de deca- ou metavanadato correspondentes a um intervalo de
concentraccedilotildees de 1 a 10 mM suplementadas com 10 D2O Os espectros foram adquiridos agrave
temperatura ambiente com uma largura espectral de 23 923 Hz um tempo de aquisiccedilatildeo de 0086
s tempo de relaxaccedilatildeo de 001 s e uma sequecircncia de pulsos a 90ordm Utilizou-se a transformada de
Fourier com alargamento das bandas de 20Hz para o processamento dos espectros Os desvios
quiacutemicos das ressonacircncias de 51
V foram obtidos em relaccedilatildeo agrave espeacutecie VOCl3cuja frequecircncia de
ressonacircncia eacute considerada o padratildeo sendo todas as outras referenciadas a partir desta em ppm
As aacutereas dos sinais do RMN correspondentes agraves espeacutecies oligomeacutericas presentes foram obtidas
por integraccedilatildeo dos sinais As concentraccedilotildees relativas das espeacutecies foram calculadas com recurso agrave
equaccedilatildeo
onde A[Vx] corresponde agrave aacuterea referente agrave ressonacircncia da espeacutecie Vx com nordm de agregaccedilatildeo x
A[V]total agrave soma total das aacutereas medidas e [V] a concentraccedilatildeo total de vanaacutedio no ensaio
A teacutecnica de 51
V-RMN foi anteriormente aplicada em estudos de ligaccedilatildeo com S1 de miosina
(Ref) tendo sido demostrado que entre as espeacutecies V1 V2 V4 e V5 a espeacutecie tetrameacuterica possui a
maior afinidade para a miosina (Cremo et al) O desaparecimento ou alargamento dos sinais pode
implicar a ligaccedilatildeo de um dos oxovanadatos laacutebeis agrave miosina sem indicaccedilatildeo de qual uma vez que
49
se processa um equiliacutebrio entre eles (Crans 1990) Assim e tendo em conta as indicaccedilotildees de que
a espeacutecie V10 eacute distinta das restantes no que diz respeito agrave magnitude da interacccedilatildeo com a miosina
e inibiccedilatildeo da actividade ATPaacutesica estimulada por actina (Aureliano 2000 Tiago et al 2002)
torna-se fundamental investigar a interacccedilatildeo das diferentes espeacutecies de vanadato com o S1 de
miosina atraveacutes da teacutecnica de 51
V-RMN
Para tal obtiveram-se espectros de 51
V-RMN resultantes da titulaccedilatildeo de uma soluccedilatildeo 5 mM de
decavanadato total com concentraccedilotildees crescentes de S1 de miosina (0 5 10 15 e 20 microM) em
tampatildeo D Realizou-se ainda ensaios na presenccedila de 2 mM de decavanadato e 2 mM de ATP na
presenccedila e na ausecircncia de S1 de miosina de modo a verificar o efeito que estes ligandos naturais
da enzima teria na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o subfragmento-1
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
Os grupos sulfidrilo (-SH) das cisteiacutenas satildeo resiacuteduos de aminoaacutecidos extremamente sensiacuteveis agrave
oxidaccedilatildeo resultando na sua oxidaccedilatildeo (Dalle-Donne et al 1999) Logo a quantificaccedilatildeo destes
resiacuteduos eacute bastante importante nos estudos de interacccedilatildeo com vanaacutedio jaacute que os resiacuteduos de Cys
com ecircnfase para o SH1 e o SH2 satildeo bastante importantes na monitorizaccedilatildeo das modificaccedilotildees
funcionais da regiatildeo da cabeccedila
O procedimento seguidamente apresentado foi originalmente descrito por Gutieacuterrez-Merino
(1987)) Consiste na reacccedilatildeo entre os grupos ndashSH natildeo oxidados das proteiacutenas e o reagente de
Ellman (DTNB -55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico)) - formando-se o aniatildeo CNT (aniatildeo 3-
carboxilato-4-nitrotiofenolato) numa estequiometria de 11 Este apresenta um maacuteximo de
absorvecircncia a 412 nm (Robyt 1990) Assim apoacutes a incubaccedilatildeo da proteiacutena na presenccedila do metal
a quantidade de cisteiacutenas reduzidas por proteiacutena determina-se atraveacutes dos valores de Abs412
Deste modo colocou-se num tubo de Eppendorf o volume apropriado de S1 dialisado para que a
concentraccedilatildeo no ensaio fosse de 1 mgmL Adicionou-se o volume apropriado de decavanadato
para que a sua concentraccedilatildeo fosse de 0 10 50 100 250 500 ou 1000 M Perfez-se com soluccedilatildeo
D ateacute um volume final de 1 mL e incubou-se a 25ordmC por 20 minutos Apoacutes este periacuteodo colocou-
se 780 L da mistura reaccional numa cuvette de quartzo e adicionou-se 220 L de soluccedilatildeo K23
23
Tampatildeo K Tampatildeo fosfatohidrogenofosfato de soacutedio 025 M pH 72
50
Adicionou-se 15 L de DTNB (em soluccedilatildeo K [DTNB]stock=100 mM) ao meio reaccional e
registou-se o valor de absorvecircncia medido equivalente agraves cisteiacutenas ldquoraacutepidasrdquo (SH1 e SH2
expostos ao solvente) Adicionou-se 001 g de SDS que provoca a desnaturaccedilatildeo da proteiacutena e
consequentemente a exposiccedilatildeo de todos os resiacuteduos de cisteiacutenas que se encontravam ateacute aiacute
inacessiacuteveis no interior da proteiacutena (cisteiacutenas totais) Estes resiacuteduos podem assim reagir com o
DTNB
Estimou-se o nuacutemero de resiacuteduos de cisteiacutenas em todas as amostras com recurso ao coeficiente de
extinccedilatildeo molar () do DTNB (ver ponto 3 dos Anexos) que foi determinado atraveacutes da titulaccedilatildeo
de L-Cisteiacutena com intervalo de concentraccedilotildees compreendidos entre 0 e 50 microM na presenccedila de
25microM de DTNB em tampatildeo D Os valores de absorvecircncia foram obtidos para um comprimento de
onda de 412 nm
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
3621 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
A teacutecnica mais eficiente de anaacutelise da interacccedilatildeo entre o iatildeo vanadilo e proteiacutenas como a
calmodulina e a transferrina entre outras permite a obtenccedilatildeo raacutepida de informaccedilatildeo no que toca agrave
conformaccedilatildeo e amplitude da referida interacccedilatildeo
Os espectros de Ressonacircncia Paramagneacutetica Electroacutenica (RPE) da espeacutecie paramagneacutetica V(IV)
foram realizados a 70K num espectroacutemetro Bruker EMX-300 X-band Todas as soluccedilotildees
utilizadas e amostras foram preparadas em anaerobiose numa cacircmara de luvas de modo a evitar a
reduccedilatildeo de V(IV) a V(V) Em tubos capilares com 02 mL de amostra manteve-se a concentraccedilatildeo
de S1 a 20 M e variou-se a concentraccedilatildeo de vanadilo (0 25 5 10 15 25 50 75 100 150
200 500 e 1000 M [VOSO4]stock=5 mM) em tampatildeo D Apoacutes obtenccedilatildeo dos espectros estes
foram tratados atraveacutes do software SimFonia e WinEPR tendo-se obtido um graacutefico com a
variaccedilatildeo da intensidade do sinal de RPE em funccedilatildeo da concentraccedilatildeo de sulfato de vanadilo
presente Realizaram-se ainda simulaccedilotildees de espectros obtidos com 20 microM de S1 e 1000 microM de
V(IV) com vista agrave obtenccedilatildeo dos ligandos provaacuteveis do vanadilo atraveacutes da anaacutelise da constante
de acoplamento hiperfina (A) e do factor g (g)
4 Conclusotildees gerais
Apesar de jaacute ter sido demonstrado que o vanaacutedio inibe a Na+K
+-ATPase da miosina a funccedilatildeo
deste elemento no tecido muacutesculo esqueleacutetico continua largamente incompreendida A enorme
complexidade quiacutemica deste metal especialmente no que concerne agrave oligomerizaccedilatildeo e reacccedilotildees
51
redox constitui um obstaacuteculo agrave compreensatildeo do comportamento do vanaacutedio nos organismos
vivos Assim a caracterizaccedilatildeo da quiacutemica deste elemento eacute de grande utilidade antes de iniciar o
estudo da sua interacccedilatildeo com o presente sistema
A anaacutelise por espectroscopias de 51
V-RMN e de UV-Vis de soluccedilotildees de vanadato diluiacutedas nos
meios de reacccedilatildeo utilizados no presente relatoacuterio (10 mM HEPES pH 70 25 mM KCl) mostrou
claramente que a espeacutecie decameacuterica (V10) possui um tempo de vida suficientemente longo para a
determinaccedilatildeo do seu efeito isolado ao contraacuterio do que se verifica com as espeacutecies mono- di-
tetra- e pentameacutericas (V1 V2 V4 e V5) Por outro lado a anaacutelise verificou ainda que apoacutes diluiccedilatildeo
das soluccedilotildees de decavanadato no meio de reacccedilatildeo coexistem as espeacutecies V10 e V1 pelo que os
efeitos putativos desta uacuteltima deveratildeo ser subtraiacutedos aquando da monitorizaccedilatildeo dos efeitos da
espeacutecie decameacuterica
O vanaacutedio (V) (vanadato) inibe as enzimas que catalizam reacccedilotildees de transferecircncia de grupos
fosfato como a ATPase da miosina [Goodno 1979] devido agrave sua analogia estrutural com o iatildeo
fosfato Na sua utilizaccedilatildeo como inibidor enzimaacutetico natildeo eacute tida em conta a contribuiccedilatildeo individual
das espeacutecies oligomeacutericas considerando-se normalmente a espeacutecie V1 como inibidora efectiva
[Goodno e Taylor 1982] No presente relatoacuterio verificou-se por espectroscopia de 51
V-RMN que
outros oligoacutemeros de vanadato interagem com o S1 de miosina tendo as espeacutecies V10 e V4 os
maiores niacuteveis de afinidade A sua interacccedilatildeo envolve forccedilas electrostaacuteticas e eacute afectada
negativamente por ligandos naturais de enzima como eacute o caso do ATP
A exposiccedilatildeo do S1 de miosina (005 mgmL) a decavanadato conduz agrave inibiccedilatildeo do capacidade da
proteiacutena de estimular a actividade Mg2+
-ATPaacutesica com um valor de IC50 de 266 plusmn 07 M
decavanadato sendo o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM
decavanadato (80) Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM decavanadato sendo
o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM decavanadato (60)
exercendo um efeito de impedimento agrave ligaccedilatildeo entre o vanaacutedio e o S1
A uma concentraccedilatildeo de 27 microM decavanadato verifica-se a oxidaccedilatildeo de um dos resiacuteduos de Cys
de S1 de miosina seguida pela oxidaccedilatildeo gradual de outro resiacuteduo de Cys ateacute agrave concentraccedilatildeo de
1000 microM Ambos os resiacuteduos oxidados natildeo se encontravam expostos ao solvente utilizado A
oxidaccedilatildeo dos resiacuteduos permite sugerir que esta modificaccedilatildeo oxidativa sofrida pelo S1 contribui
para a diminuiccedilatildeo da sua actividade de ATPase
52
Por espectroscopia de RPE revelaram-se os possiacuteveis ligandos equatoriais do vanaacutedio aquando da
sua interacccedilatildeo com o S1 de miosina como sendo duas moleacuteculas de aacutegua um ligando contendo o
grupo R-OH e outro contendo o grupo R=NR
Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados de
oxidaccedilatildeo (IV) e (V) no S1 de miosina para concentraccedilotildees facilmente alcanccedilaacuteveis no ambiente
terrestre
53
5 Estudos futuros
No sentido do melhor esclarecimento das alteraccedilotildees moleculares e estruturais que ocorrem no
subfragmento 1 quando exposta ao vanaacutedio que conduzem a uma diminuiccedilatildeo da sua funccedilatildeo
sugere-se
- Quantificaccedilatildeo das cisteiacutenas reduzidas de S1 na presenccedila de metavanadato
- Determinaccedilatildeo das actividades Ca2+
- e K+-ATPaacutesica do S1 de miosina na presenccedila de
decavanadato
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina estimulada por actina
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina na presenccedila de
metavanadato
- Realizaccedilatildeo de RMN 2D com variaccedilatildeo das concentraccedilotildees das soluccedilotildees de decavanadato e
metavanadato na presenccedila de S1
-Mutageacutenese dos resiacuteduos de Cys doS1 de miosina de modo a verificar quais os resiacuteduos que
sofrem oxidaccedilatildeo na presenccedila de decavanadato
- Determinaccedilatildeo por calorimetria (DSC Differential Scanning Calorimetry) da variaccedilatildeo da
entalpia (∆H) e da temperatura de desnaturaccedilatildeo do S1 de miosina na presenccedila de vanaacutedio uma
vez que estas propriedades caracteriacutesticas de cada proteiacutena satildeo alteradas apoacutes ocorrecircncia de
modificaccedilotildees estruturais
- Anaacutelise por SDS-PAGE de S1 de miosina apoacutes tratamento com vanaacutedio de modo a determinar
quais os polipeacuteptidos eventualmente formados apoacutes clivagem da proteiacutena
54
6 Referecircncias bibliograacuteficas
- Sellers J Myosins 2nd Edition Oxford University Press 1999
- L Stryer inBiochemistry (Ed WH Freeman 2005 pp
Refordfas Materiais e Meacutetodos
[A] S S Margossian SLowey Methods Enzymol1982 85 55-71
[B] JD Pardue JA Spudich Methods Enzymol 1982 85 164-181
[C] M Bradford Anal Biochem1976 72 248-254
[D] H H Taussky E Shorr J Biol Chem 1953 202 675-685
[E] C R Bagshaw in Outline Studies in Biology ndash Muscle Contraction Chapman and Hall Ed
1982
[F] A Cuenda et al Eur J Biochem1990 194 663-670
55
Anexos
1 Determinaccedilatildeo da recta de calibraccedilatildeo meacutetodo de Bradford
Figura1ndashRepresentaccedilatildeo graacutefica da recta de calibraccedilatildeo (Abs595 nm vs [BSA]) utilizada para o meacutetodo de Bradford
2 Determinaccedilatildeo da recta calibraccedilatildeo para a electroforese em condiccedilotildees desnaturantes
Figura 2 ndash Representaccedilatildeo graacutefica da recta de calibraccedilatildeo (log(Peso Molecular) vs Rf dos marcadores de peso
molecular) onde Rf indica determinada atraveacutes do padratildeo de peso molecular
(poccedilo (a))
3Caacutelculo do coeficiente de extinccedilatildeo molar () para a quantificaccedilatildeo de cisteiacutenas reduzidas
atraveacutes do meacutetodo de Ellman
Traccedilou-se a recta de Abs412 nm meacutedia vs [L-Cisteiacutena] de modo a determinar o valor do coeficiente
de extinccedilatildeo molar definido como o declive daquela recta Assim =8400 M-1
cm-1
como
podemos verificar na Fig 3
A595 nm = 00546[BSA] + 00209 Rsup2 = 099051
Ab
s 59
5 n
m
[BSA] (mgmL)
log PM = -1177 Rf + 2247 Rsup2 = 098957
log
PM
Mobilidade relativa (Rf)
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero

42
contiacutenua e mudanccedila de tampatildeo 2 a 3 vezes De seguida a actina foi centrifugada a 110
000gdurante 1hora Decantou-se a soluccedilatildeo de G-Actina (sobrenadante) e desprezou-se o
sedimento Determinou-se a concentraccedilatildeo de G-Actina pelo meacutetodo directo (ver secccedilatildeo 3312 do
Procedimento Experimental) A soluccedilatildeo de G-actina foi armazenada a uma temperatura de 4ordmC
apoacutes adiccedilatildeo de 002NaN3 mantendo-se estaacutevel durante 3 a 5 dias
331 Quantificaccedilatildeo das proteiacutenas isoladas
3311 Quantificaccedilatildeo pelo meacutetodo de Bradford
O meacutetodo de Bradford (Bradford 1976) baseia-se na utilizaccedilatildeo do reagente de Bradford
composto por entre outros aacutecido fosfoacuterico e o corante ldquoCoomassierdquo Este corante possui carga
negativa e a sua coloraccedilatildeo varia entre a vermelha (Absmax=465 nm) predominante em soluccedilatildeo e a
azul (Absmax=595 nm) presente aquando da ligaccedilatildeo aos grupos amina das proteiacutenas Em soluccedilotildees
aacutecidas diluiacutedas a coloraccedilatildeo varia proporcionalmente com a quantidade do corante ligado agrave
proteiacutena que por sua vez eacute directamente proporcional agrave concentraccedilatildeo da proteiacutena permitindo a
sua quantificaccedilatildeo ateacute a um limite de detecccedilatildeo de 20 gmL (Bradford 1976)
Deste modo prepararam-se amostras de BSA ([BSA]stock= 1mgmL) utilizando um intervalo de
concentraccedilotildees compreendido entre 0 e 15 gmL de acordo com a tabela 3I Apoacutes a adiccedilatildeo da
H2O MilliQ e do reagente de Bradford as soluccedilotildees foram sucessivamente homogeneizadas no
voacutertex
Tabela 3Indash Amostras utilizadas para a obtenccedilatildeo da recta de calibraccedilatildeo no meacutetodo de Bradford
BSA (L) BSA (g) H2O MilliQ (L) Reagente de Bradford (L)
0 0 800 200
125 125 79875 200
25 25 7975 200
5 5 795 200
10 10 790 200
15 15 785 200
Apoacutes incubaccedilatildeo a 25ordmC por 5 minutos leram-se os valores de Abs595 num
espectrofotoacutemetro (Shimadzu UV-160A) para todas as amostras Seguidamente traccedilou-se a recta
de calibraccedilatildeo (Abs595vs quantidade de BSA (g))
43
Para determinar a concentraccedilatildeo de proteiacutena apoacutes o processo de isolamento purificaccedilatildeo e
diaacutelise quando necessaacuterio colocou-se 5 L da amostra num tubo de Eppendorf e adicionou-se
795 L de H2O MilliQ homogeneizando a soluccedilatildeo no voacutertex Adicionou-se posteriormente 200
L de reagente de Bradford e repetiu-se a homogeneizaccedilatildeo Incubou-se a mistura a 25ordmC por 5
minutos apoacutes os quais se leu os valores de Abs595 nm no espectrofotoacutemetro Por interpolaccedilatildeo deste
valor a partir da recta de calibraccedilatildeo determinou-se a concentraccedilatildeo de proteiacutena na amostra
3312 Quantificaccedilatildeo pelo meacutetodo directo
A quantificaccedilatildeo das amostras proteicas pode tambeacutem ser efectuada atraveacutes da leitura directa da
absorvacircncia tendo por base a lei de Beer-Lambert onde Abs corresponde agrave
absorvacircncia ao comprimento de onda ao qual o ensaio foi realizado corresponde ao coeficiente
de extinccedilatildeo (substituiacutedo pela letra a se as unidades forem distintas da molaridade) b corresponde
ao percurso oacuteptico da radiaccedilatildeo na ceacutelula amostral e c corresponde agrave concentraccedilatildeo amostral
Assim no caso de S1 de miosina mediu-se a Abs280e calculou-se a concentraccedilatildeo da proteiacutena
sabendo que o coeficiente de extinccedilatildeo de S1 eacute (Ref)
No caso da actina mediu-se a Abs290ecalculou-se a concentraccedilatildeo da proteiacutena sabendo que o
coeficiente de extinccedilatildeo da actina eacute (Ref)
332Electroforese em condiccedilotildees desnaturantes (SDS-PAGE)
De modo a melhor caracterizar as proteiacutenas isoladas foi realizada uma electroforese em
condiccedilotildees desnaturantes (SDS-PAGE do inglecircs Sodium Dodecyl Sulfate Polyacrilamide Gel
Electrophoresis) O SDS (dodecilo sulfato de soacutedio) eacute um detergente anioacutenico utilizado neste
meacutetodo para desnaturar as proteiacutenas que se ligam ao SDS na proporccedilatildeo 14 g SDS g proteiacutena
(aproximadamente 1 moleacutecula de SDS por cada 2 resiacuteduos de aminoaacutecidos) (Stryer) O SDS induz
uma densidade de carga negativa significativa nas proteiacutenas a que se liga mascarando a sua carga
intriacutenseca pelo que as razotildees cargamassa satildeo tendencialmente idecircnticas Assim a SDS-PAGE
separa as proteiacutenas quando sujeitas a um campo eleacutectrico de acordo com o seu peso molecular
sendo a sua migraccedilatildeo directamente proporcional ao logaritmo deste uacuteltimo (Stryer)
O gel de resoluccedilatildeo com 10 de acrilamida foi obtido atraveacutes da adiccedilatildeo de 167 mL de soluccedilatildeo
stock de acrilamida11
075 mL de tampatildeo de resoluccedilatildeo12
251 mL de aacutegua destilada e 50 L de
SDS 10 Para promover a polimerizaccedilatildeo adicionou-se em seguida 38 L de PSA 10
11
soluccedilatildeo stock de acrilamida soluccedilatildeo de 30 (mv) de acrilamida e 08 (mv) de NNrsquo-metil-bis-acrilamida 12
tampatildeo de resoluccedilatildeo 15 M Tris-HCl pH 88
44
(persulfato de amoacutenia) 10 e 25 L de TEMED 99 (tetrametiletilenodiamina) Agitou-se
cuidadosamente de modo a natildeo formar bolhas e pipetou-se a soluccedilatildeo para a cassette de vidro
previamente montada no suporte De seguida adicionou-se uma peliacutecula de cerca de 2 mm de 2-
metil-2-butanol para verificar posteriormente a polimerizaccedilatildeo Apoacutes a polimerizaccedilatildeo que demora
cerca de 45 minutos verteu-se a agrave superfiacutecie e adicionou-se o gel de concentraccedilatildeo com 5 de
acrilamida constituiacutedo por 03 mL de soluccedilatildeo de stock acrilamida 045 mL de tampatildeo de
concentraccedilatildeo13
094 mL de aacutegua destilada 18 L de SDS 10 135 L de PSA 10 e 2L de
TEMED 99 Colocou-se o pente sobre o gel e deixou-se polimerizar durante cerca de 45
minutos
As amostras para aplicaccedilatildeo no gel de electroforese foram preparadas para uma concentraccedilatildeo de
proteiacutena de 1 mgmL num volume final de 100 L dos quais 25 L correspondem ao tampatildeo da
amostra14
As amostras foram aquecidas a 100ordmC durante 5 minutos Antes da aplicaccedilatildeo das
amostras transferiu-se a cassette para a tina de electroforese agrave qual se adicionou a soluccedilatildeo
tampatildeo de electroforese da tina superior15
e tampatildeo de electroforese da tina inferior16
Aplicaram-
se 10 L das soluccedilotildees de marcadores de peso molecular e 20 L das amostras proteicas
preparadas anteriormente A electroforese decorreu durante cerca de 3h30 min a uma voltagem
de 80 V De seguida transferiu-se o gel para uma tina corando as proteiacutenas durante 1h com
soluccedilatildeo corante17
Descorou-se o gel com soluccedilatildeo descorante18
tendo-se mudado esta soluccedilatildeo ateacute
as bandas se encontrarem bem definidas
Determinou-se a mobilidade relativa das bandas correspondentes aos marcadores de elevado peso
molecular e traccedilou-se a recta de calibraccedilatildeo ndash log(PM) vs log(Rf) onde Rf corresponde agrave
mobilidade relativa Por interpolaccedilatildeo determina-se o PM das proteiacutenas isoladas sabendo a sua
mobilidade relativa Por comparaccedilatildeo com os valores descritos na literatura pode concluir-se
acerca do grau de purificaccedilatildeo
34 Mediccedilatildeo da actividade enzimaacutetica de S1
341 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
13
tampatildeo de concentraccedilatildeo 05 M Tris-HCl pH 68 14
tampatildeo de amostra 320 mM Tris-HCl pH 68 04 M szlig-mercaptoetanol SDS 8 glicerol 15 (vv) e roxo de m-
cresol 0024 15
tampatildeo de electroforese da tina superior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS 16
tampatildeo de electroforese da tina inferior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS acetato de soacutedio
01 M 17
soluccedilatildeo corante ldquoCoomassie Blue R-250rdquo contendo metanol 45 (vv) e aacutecido aceacutetico glacial 10 (vv) 18
soluccedilatildeo descorante metanol 10 (vv) e aacutecido aceacutetico glacial 10 vv)
45
O meacutetodo de Fiske-Subarow modificado consiste na reacccedilatildeo do molibdato de amoacutenio com o
fosfato inorgacircnico em soluccedilatildeo aacutecida com formaccedilatildeo do aacutecido fosfomoliacutebdico Seguidamente daacute-
se a adiccedilatildeo de um agente redutor que provoca a reduccedilatildeo do composto a fosfomolibdato que forma
um complexo de cor azulada cuja absorvacircncia maacutexima ocorre para um comprimento de onda de
660 nm tornando possiacutevel a sua detecccedilatildeo por meacutetodos espectrofotomeacutetricos As actividades Ca2+
-
ATPaacutesica e K+EDTA-ATPaacutesica de S1 foram determinadas atraveacutes deste meacutetodo de acordo com
o descrito por Taussky e Shorr (1953) com mediccedilatildeo da quantidade de fosfato (Pi) libertado
atraveacutes da hidroacutelise de ATP
Para tal obteve-se primeiramente a recta de calibraccedilatildeo com as amostras presentes na Tabela 3II
utilizando uma soluccedilatildeo stock de 5 mM KH2PO4
Tabela 3II ndash Composiccedilatildeo das diferentes amostras utilizadas para determinar a recta de calibraccedilatildeo atraveacutes do meacutetodo
de Fiske-Subbarow modificado
KH2PO4 (L)
([KH2PO4]stock= 5mM)
PO43-
(nmole) H2O (uL)
0 0 1780
5 25 1775
10 50 1770
20 100 1760
40 200 1740
100 500 1680
A cada tubo de Eppendorf adicionaram-se seguidamente 200 L de soluccedilatildeo J19
e 20 L de agente
redutor20
homogeneizando no voacutertex apoacutes cada adiccedilatildeo Incubou-se durante 10 minutos e mediu-
se a Abs660 nm num espectrofotoacutemetro com intervalos de tempo iguais entre cada amostra Traccedilou-
se a recta de calibraccedilatildeo (Abs660 nmvs nmol PO43-
) a partir dos valores obtidos
Para determinar a actividade Ca2+
-ATPaacutesica de S1 prepararam-se tubos de Eppendorf com 400
L de meio de reacccedilatildeo21
25 L CaCl2 ([CaCl2]stock= 100 mM) 25 L ATP ([ATP]stock= 100 mM)
com [S1]ensaios=01 mgmL Perfez-se se necessaacuterio com H2O MilliQ ateacute perfazer um volume de 1
mL Para determinar a actividade K+EDTA-ATPaacutesica utilizou-se uma concentraccedilatildeo idecircntica de
19
Soluccedilatildeo J 5 N H2SO4 25 molibdato de amoacutenio 20
Agente redutor 5 mg aacutecido 1-amino-2-naftol-4-sulfoacutenico 30 mg bissulfito de soacutedio 30 mg sulfito de soacutedio em 26
mL H2O 21
meio de reacccedilatildeo 125 mM Tris-HCl pH 80 0575 M KCl
46
proteiacutena nos ensaios (01 mgmL) e o mesmo meio de reacccedilatildeo Aos tubos de Eppendorf
adicionou-se ainda 123 L de KCl ([KCl]stock = 3M) 25 L de ATP 10 L de EDTA
([EDTA]stock= 100 mM )e o volume de H2O MilliQ necessaacuterio para perfazer 1 mL Em ambas as
situaccedilotildees o uacuteltimo reagente adicionado foi o ATP uma vez que se trata do substrato da reacccedilatildeo
enzimaacutetica
A cineacutetica decorreu a 25ordmC durante 5 minutos para a actividade Ca2+
-ATPaacutesica e durante 3
minutos para a actividade K+EDTA-ATPaacutesica Apoacutes o periacuteodo de incubaccedilatildeo transferiu-se 150
L de amostra para tubos de Eppendorf contendo igual volume de TCA 10 Colocou-se no gelo
e homogeneizou-se no voacutertex Seguidamente centrifugou-se a 1500 g durante 10 minutos e
recolheu-se 200 L de sobrenadante para novos tubos de Eppendorf Adicionou-se a estes 158
mL de H2O MilliQ e 200 L de soluccedilatildeo J Homogeneizou-se no voacutertex e adicionou-se 20 L de
reagente redutor repetindo-se a homogeneizaccedilatildeo Incubou-se durante 10 minutos e mediu-se a
Abs660 nm com o mesmo intervalo de tempo de leitura para cada amostra Por intrapolaccedilatildeo a partir
da recta de calibraccedilatildeo obtida anteriormente determinou-se a quantidade de Pi libertado em cada
ensaio A partir destes resultados determinaram-se as actividades Ca2+
-ATPaacutesica e K+EDTA-
ATPaacutesica do S1 em nmol PO43-
minmg proteiacutena e em s-1
342 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
Da actividade Mg2+
-ATPaacutesica de S1 resulta a formaccedilatildeo dos produtos da reacccedilatildeo de
hidroacutelise do ATP - ADP e Pi Na presenccedila de ADP fosfoenolpiruvato (PEP) e da enzima piruvato
cinase (PK) os produtos resultantes satildeo ATP e piruvato Por acccedilatildeo da enzima lactato
desidrogenase (LDH) este uacuteltimo eacute convertido em lactato ocorrendo simultaneamente a oxidaccedilatildeo
do NADH a NAD+ monitorizada atraveacutes da Abs340 nm Quando o S1 de miosina se encontra no
meio reaccional o ATP eacute hidrolisado formando-se novamente ADP e Pi o que permite a
formaccedilatildeo de um ciclo (fig 21) Se as enzimas se encontrarem em excesso a velocidade de
conversatildeo do NADH eacute limitada pela produccedilatildeo de ADP e Pi Como o ATP eacute regenerado em cada
ciclo a sua concentraccedilatildeo inicial eacute sempre mantida (Bagshaw 1982)
Figura 33ndash Representaccedilatildeo esquemaacutetica do meacutetodo de enzimas acopladas
47
O protocolo utilizado e seguidamente descrito foi adaptado de Cuenda et al (1990 ndash Ver Refordf) A
uma cuvette de 1 mL adicionou-se S1 para uma concentraccedilatildeo final de 005 mgmL 800 L de
meio de reacccedilatildeo22
10 L de PEP ([PEP]stock = 42 mM) 10 L de PK+LDH 11 ([PK+LDH]stock =
1 mgmL) e 15 L de NADH ([NADH]stock=25 mM) Iniciou-se a monitorizaccedilatildeo da reacccedilatildeo
atraveacutes da leitura de Abs340 nm apoacutes a adiccedilatildeo de 20 L de ATP ([ATP]stock=100 mM) A
actividade Mg2+
-ATPaacutesica eacute obtida atraveacutes do declive correspondente agrave oxidaccedilatildeo do NADH Para
a mediccedilatildeo da Mg2+
-ATPase de S1 estimulada por actina adicionou-se F-actina para uma
concentraccedilatildeo final de 2 microM sendo a actividade obtida tambeacutem atraveacutes do declive correspondente
agrave oxidaccedilatildeo do NADH apoacutes a adiccedilatildeo da proteiacutena
35 Interaccedilatildeo entre vanaacutedio e subfragmento-1 de miosina
351 Preparaccedilatildeo das soluccedilotildees de vanaacutedio
As soluccedilotildees de vanadato foram preparadas em aacutegua MilliQ a partir de metavanadato de amoacutenio
(NH4VO3) numa concentraccedilatildeo stock de 50 mM com agitaccedilatildeo magneacutetica e aquecimento (~30 ordmC)
Esta soluccedilatildeo com um valor compreendido entre 6 e 7 designa-se por ldquometavanadatordquo e eacute
constituiacuteda pelas esspeacutecies V1 V2 V4 e V5 (Ref) Acidificando a soluccedilatildeo de metavanadato por
adiccedilatildeo de HCl ateacute um valor de pH proacuteximo de 4 observa-se uma mudanccedila de cor de amarelo
claro para laranja indicativa da formaccedilatildeo da espeacutecie decameacuterica de vanadato De seguida e
imediatamente antes da realizaccedilatildeo dos ensaios descritos o valor do pH da soluccedilatildeo de
decavanadato foi aumentado para cerca de 75 por adiccedilatildeo de NaOH A soluccedilatildeo (2) designada
por ldquometavanadatordquo natildeo foi posteriormente alterada ateacute agrave sua adiccedilatildeo para a realizaccedilatildeo dos ensaios
As soluccedilotildees foram armazenadas a 4ordmC e mantidas em gelo aquando da sua utilizaccedilatildeo
A soluccedilatildeo de V(IV) foi preparada em condiccedilotildees anaeroacutebias por diluiccedilatildeo de sulfato de vanadil
(VOSO4) para uma concentraccedilatildeo final de 5 mM em H2O MilliQ saturada com N2 gasoso
352Anaacutelise por espectroscopia de UV-Vis
A concentraccedilatildeo da espeacutecie decameacuterica em soluccedilatildeo depende do periacuteodo de tempo apoacutes a diluiccedilatildeo
no meio de reacccedilatildeo apropriado (Ref) A velocidade de desoligomerizaccedilatildeo da espeacutecie decameacuterica
pode ser analisada por espectroscopia de absorccedilatildeo na regiatildeo UVVis uma vez que esta espeacutecie
apresenta duas bandas caracteriacutesticas na regiatildeo do visiacutevel (a 358 e 400 nm) que natildeo se verifica
em quaisquer outras espeacutecies oligomeacutericas As bandas supramencionada sapresentam-se bem
22
Meio de reacccedilatildeo 3125 mM Tris-HCl pH 71 375 mM MgCl2
48
resolvidas atraveacutes da obtenccedilatildeo da derivada de 2ordf ordem do espectro apresentando a forma de 2
vales com miacutenimos centrados nos comprimentos de onda supracitados com dependecircncia linear
relativamente agrave concentraccedilatildeo de decavanadato total (Ramos et al 2006)
Assim a estabilidade da espeacutecie decameacuterica apoacutes diluiccedilatildeo da soluccedilatildeo de decavanadato no meio de
reacccedilatildeo dos ensaios (tampatildeo D) para uma concentraccedilatildeo final de 1 mM em vanadato total foi
caracterizada atraveacutes da leitura da Abs400 em funccedilatildeo do tempo tendo-se determinado qual a
ordem de reacccedilatildeo e o tempo de meia-vida da mesma Com base em estudos anteriores acerca da
estabilidade do V10 na presenccedila de diferentes proteiacutenas (Ramos et al 2006) determinou-se
igualmente o tempo de meia vida apoacutes a adiccedilatildeo de 20 microM de S1 de miosina ao meio reaccional
353 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
Os espectros de RMN do isoacutetopo 51
V foram realizados num espectroacutemetro Bruker AM-400 MHz
equipado com uma sonda multinuclear inversa de 5 mm Em tubos de RMN de 5 mm de diacircmetro
preparou-se soluccedilotildees de 500 L de deca- ou metavanadato correspondentes a um intervalo de
concentraccedilotildees de 1 a 10 mM suplementadas com 10 D2O Os espectros foram adquiridos agrave
temperatura ambiente com uma largura espectral de 23 923 Hz um tempo de aquisiccedilatildeo de 0086
s tempo de relaxaccedilatildeo de 001 s e uma sequecircncia de pulsos a 90ordm Utilizou-se a transformada de
Fourier com alargamento das bandas de 20Hz para o processamento dos espectros Os desvios
quiacutemicos das ressonacircncias de 51
V foram obtidos em relaccedilatildeo agrave espeacutecie VOCl3cuja frequecircncia de
ressonacircncia eacute considerada o padratildeo sendo todas as outras referenciadas a partir desta em ppm
As aacutereas dos sinais do RMN correspondentes agraves espeacutecies oligomeacutericas presentes foram obtidas
por integraccedilatildeo dos sinais As concentraccedilotildees relativas das espeacutecies foram calculadas com recurso agrave
equaccedilatildeo
onde A[Vx] corresponde agrave aacuterea referente agrave ressonacircncia da espeacutecie Vx com nordm de agregaccedilatildeo x
A[V]total agrave soma total das aacutereas medidas e [V] a concentraccedilatildeo total de vanaacutedio no ensaio
A teacutecnica de 51
V-RMN foi anteriormente aplicada em estudos de ligaccedilatildeo com S1 de miosina
(Ref) tendo sido demostrado que entre as espeacutecies V1 V2 V4 e V5 a espeacutecie tetrameacuterica possui a
maior afinidade para a miosina (Cremo et al) O desaparecimento ou alargamento dos sinais pode
implicar a ligaccedilatildeo de um dos oxovanadatos laacutebeis agrave miosina sem indicaccedilatildeo de qual uma vez que
49
se processa um equiliacutebrio entre eles (Crans 1990) Assim e tendo em conta as indicaccedilotildees de que
a espeacutecie V10 eacute distinta das restantes no que diz respeito agrave magnitude da interacccedilatildeo com a miosina
e inibiccedilatildeo da actividade ATPaacutesica estimulada por actina (Aureliano 2000 Tiago et al 2002)
torna-se fundamental investigar a interacccedilatildeo das diferentes espeacutecies de vanadato com o S1 de
miosina atraveacutes da teacutecnica de 51
V-RMN
Para tal obtiveram-se espectros de 51
V-RMN resultantes da titulaccedilatildeo de uma soluccedilatildeo 5 mM de
decavanadato total com concentraccedilotildees crescentes de S1 de miosina (0 5 10 15 e 20 microM) em
tampatildeo D Realizou-se ainda ensaios na presenccedila de 2 mM de decavanadato e 2 mM de ATP na
presenccedila e na ausecircncia de S1 de miosina de modo a verificar o efeito que estes ligandos naturais
da enzima teria na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o subfragmento-1
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
Os grupos sulfidrilo (-SH) das cisteiacutenas satildeo resiacuteduos de aminoaacutecidos extremamente sensiacuteveis agrave
oxidaccedilatildeo resultando na sua oxidaccedilatildeo (Dalle-Donne et al 1999) Logo a quantificaccedilatildeo destes
resiacuteduos eacute bastante importante nos estudos de interacccedilatildeo com vanaacutedio jaacute que os resiacuteduos de Cys
com ecircnfase para o SH1 e o SH2 satildeo bastante importantes na monitorizaccedilatildeo das modificaccedilotildees
funcionais da regiatildeo da cabeccedila
O procedimento seguidamente apresentado foi originalmente descrito por Gutieacuterrez-Merino
(1987)) Consiste na reacccedilatildeo entre os grupos ndashSH natildeo oxidados das proteiacutenas e o reagente de
Ellman (DTNB -55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico)) - formando-se o aniatildeo CNT (aniatildeo 3-
carboxilato-4-nitrotiofenolato) numa estequiometria de 11 Este apresenta um maacuteximo de
absorvecircncia a 412 nm (Robyt 1990) Assim apoacutes a incubaccedilatildeo da proteiacutena na presenccedila do metal
a quantidade de cisteiacutenas reduzidas por proteiacutena determina-se atraveacutes dos valores de Abs412
Deste modo colocou-se num tubo de Eppendorf o volume apropriado de S1 dialisado para que a
concentraccedilatildeo no ensaio fosse de 1 mgmL Adicionou-se o volume apropriado de decavanadato
para que a sua concentraccedilatildeo fosse de 0 10 50 100 250 500 ou 1000 M Perfez-se com soluccedilatildeo
D ateacute um volume final de 1 mL e incubou-se a 25ordmC por 20 minutos Apoacutes este periacuteodo colocou-
se 780 L da mistura reaccional numa cuvette de quartzo e adicionou-se 220 L de soluccedilatildeo K23
23
Tampatildeo K Tampatildeo fosfatohidrogenofosfato de soacutedio 025 M pH 72
50
Adicionou-se 15 L de DTNB (em soluccedilatildeo K [DTNB]stock=100 mM) ao meio reaccional e
registou-se o valor de absorvecircncia medido equivalente agraves cisteiacutenas ldquoraacutepidasrdquo (SH1 e SH2
expostos ao solvente) Adicionou-se 001 g de SDS que provoca a desnaturaccedilatildeo da proteiacutena e
consequentemente a exposiccedilatildeo de todos os resiacuteduos de cisteiacutenas que se encontravam ateacute aiacute
inacessiacuteveis no interior da proteiacutena (cisteiacutenas totais) Estes resiacuteduos podem assim reagir com o
DTNB
Estimou-se o nuacutemero de resiacuteduos de cisteiacutenas em todas as amostras com recurso ao coeficiente de
extinccedilatildeo molar () do DTNB (ver ponto 3 dos Anexos) que foi determinado atraveacutes da titulaccedilatildeo
de L-Cisteiacutena com intervalo de concentraccedilotildees compreendidos entre 0 e 50 microM na presenccedila de
25microM de DTNB em tampatildeo D Os valores de absorvecircncia foram obtidos para um comprimento de
onda de 412 nm
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
3621 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
A teacutecnica mais eficiente de anaacutelise da interacccedilatildeo entre o iatildeo vanadilo e proteiacutenas como a
calmodulina e a transferrina entre outras permite a obtenccedilatildeo raacutepida de informaccedilatildeo no que toca agrave
conformaccedilatildeo e amplitude da referida interacccedilatildeo
Os espectros de Ressonacircncia Paramagneacutetica Electroacutenica (RPE) da espeacutecie paramagneacutetica V(IV)
foram realizados a 70K num espectroacutemetro Bruker EMX-300 X-band Todas as soluccedilotildees
utilizadas e amostras foram preparadas em anaerobiose numa cacircmara de luvas de modo a evitar a
reduccedilatildeo de V(IV) a V(V) Em tubos capilares com 02 mL de amostra manteve-se a concentraccedilatildeo
de S1 a 20 M e variou-se a concentraccedilatildeo de vanadilo (0 25 5 10 15 25 50 75 100 150
200 500 e 1000 M [VOSO4]stock=5 mM) em tampatildeo D Apoacutes obtenccedilatildeo dos espectros estes
foram tratados atraveacutes do software SimFonia e WinEPR tendo-se obtido um graacutefico com a
variaccedilatildeo da intensidade do sinal de RPE em funccedilatildeo da concentraccedilatildeo de sulfato de vanadilo
presente Realizaram-se ainda simulaccedilotildees de espectros obtidos com 20 microM de S1 e 1000 microM de
V(IV) com vista agrave obtenccedilatildeo dos ligandos provaacuteveis do vanadilo atraveacutes da anaacutelise da constante
de acoplamento hiperfina (A) e do factor g (g)
4 Conclusotildees gerais
Apesar de jaacute ter sido demonstrado que o vanaacutedio inibe a Na+K
+-ATPase da miosina a funccedilatildeo
deste elemento no tecido muacutesculo esqueleacutetico continua largamente incompreendida A enorme
complexidade quiacutemica deste metal especialmente no que concerne agrave oligomerizaccedilatildeo e reacccedilotildees
51
redox constitui um obstaacuteculo agrave compreensatildeo do comportamento do vanaacutedio nos organismos
vivos Assim a caracterizaccedilatildeo da quiacutemica deste elemento eacute de grande utilidade antes de iniciar o
estudo da sua interacccedilatildeo com o presente sistema
A anaacutelise por espectroscopias de 51
V-RMN e de UV-Vis de soluccedilotildees de vanadato diluiacutedas nos
meios de reacccedilatildeo utilizados no presente relatoacuterio (10 mM HEPES pH 70 25 mM KCl) mostrou
claramente que a espeacutecie decameacuterica (V10) possui um tempo de vida suficientemente longo para a
determinaccedilatildeo do seu efeito isolado ao contraacuterio do que se verifica com as espeacutecies mono- di-
tetra- e pentameacutericas (V1 V2 V4 e V5) Por outro lado a anaacutelise verificou ainda que apoacutes diluiccedilatildeo
das soluccedilotildees de decavanadato no meio de reacccedilatildeo coexistem as espeacutecies V10 e V1 pelo que os
efeitos putativos desta uacuteltima deveratildeo ser subtraiacutedos aquando da monitorizaccedilatildeo dos efeitos da
espeacutecie decameacuterica
O vanaacutedio (V) (vanadato) inibe as enzimas que catalizam reacccedilotildees de transferecircncia de grupos
fosfato como a ATPase da miosina [Goodno 1979] devido agrave sua analogia estrutural com o iatildeo
fosfato Na sua utilizaccedilatildeo como inibidor enzimaacutetico natildeo eacute tida em conta a contribuiccedilatildeo individual
das espeacutecies oligomeacutericas considerando-se normalmente a espeacutecie V1 como inibidora efectiva
[Goodno e Taylor 1982] No presente relatoacuterio verificou-se por espectroscopia de 51
V-RMN que
outros oligoacutemeros de vanadato interagem com o S1 de miosina tendo as espeacutecies V10 e V4 os
maiores niacuteveis de afinidade A sua interacccedilatildeo envolve forccedilas electrostaacuteticas e eacute afectada
negativamente por ligandos naturais de enzima como eacute o caso do ATP
A exposiccedilatildeo do S1 de miosina (005 mgmL) a decavanadato conduz agrave inibiccedilatildeo do capacidade da
proteiacutena de estimular a actividade Mg2+
-ATPaacutesica com um valor de IC50 de 266 plusmn 07 M
decavanadato sendo o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM
decavanadato (80) Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM decavanadato sendo
o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM decavanadato (60)
exercendo um efeito de impedimento agrave ligaccedilatildeo entre o vanaacutedio e o S1
A uma concentraccedilatildeo de 27 microM decavanadato verifica-se a oxidaccedilatildeo de um dos resiacuteduos de Cys
de S1 de miosina seguida pela oxidaccedilatildeo gradual de outro resiacuteduo de Cys ateacute agrave concentraccedilatildeo de
1000 microM Ambos os resiacuteduos oxidados natildeo se encontravam expostos ao solvente utilizado A
oxidaccedilatildeo dos resiacuteduos permite sugerir que esta modificaccedilatildeo oxidativa sofrida pelo S1 contribui
para a diminuiccedilatildeo da sua actividade de ATPase
52
Por espectroscopia de RPE revelaram-se os possiacuteveis ligandos equatoriais do vanaacutedio aquando da
sua interacccedilatildeo com o S1 de miosina como sendo duas moleacuteculas de aacutegua um ligando contendo o
grupo R-OH e outro contendo o grupo R=NR
Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados de
oxidaccedilatildeo (IV) e (V) no S1 de miosina para concentraccedilotildees facilmente alcanccedilaacuteveis no ambiente
terrestre
53
5 Estudos futuros
No sentido do melhor esclarecimento das alteraccedilotildees moleculares e estruturais que ocorrem no
subfragmento 1 quando exposta ao vanaacutedio que conduzem a uma diminuiccedilatildeo da sua funccedilatildeo
sugere-se
- Quantificaccedilatildeo das cisteiacutenas reduzidas de S1 na presenccedila de metavanadato
- Determinaccedilatildeo das actividades Ca2+
- e K+-ATPaacutesica do S1 de miosina na presenccedila de
decavanadato
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina estimulada por actina
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina na presenccedila de
metavanadato
- Realizaccedilatildeo de RMN 2D com variaccedilatildeo das concentraccedilotildees das soluccedilotildees de decavanadato e
metavanadato na presenccedila de S1
-Mutageacutenese dos resiacuteduos de Cys doS1 de miosina de modo a verificar quais os resiacuteduos que
sofrem oxidaccedilatildeo na presenccedila de decavanadato
- Determinaccedilatildeo por calorimetria (DSC Differential Scanning Calorimetry) da variaccedilatildeo da
entalpia (∆H) e da temperatura de desnaturaccedilatildeo do S1 de miosina na presenccedila de vanaacutedio uma
vez que estas propriedades caracteriacutesticas de cada proteiacutena satildeo alteradas apoacutes ocorrecircncia de
modificaccedilotildees estruturais
- Anaacutelise por SDS-PAGE de S1 de miosina apoacutes tratamento com vanaacutedio de modo a determinar
quais os polipeacuteptidos eventualmente formados apoacutes clivagem da proteiacutena
54
6 Referecircncias bibliograacuteficas
- Sellers J Myosins 2nd Edition Oxford University Press 1999
- L Stryer inBiochemistry (Ed WH Freeman 2005 pp
Refordfas Materiais e Meacutetodos
[A] S S Margossian SLowey Methods Enzymol1982 85 55-71
[B] JD Pardue JA Spudich Methods Enzymol 1982 85 164-181
[C] M Bradford Anal Biochem1976 72 248-254
[D] H H Taussky E Shorr J Biol Chem 1953 202 675-685
[E] C R Bagshaw in Outline Studies in Biology ndash Muscle Contraction Chapman and Hall Ed
1982
[F] A Cuenda et al Eur J Biochem1990 194 663-670
55
Anexos
1 Determinaccedilatildeo da recta de calibraccedilatildeo meacutetodo de Bradford
Figura1ndashRepresentaccedilatildeo graacutefica da recta de calibraccedilatildeo (Abs595 nm vs [BSA]) utilizada para o meacutetodo de Bradford
2 Determinaccedilatildeo da recta calibraccedilatildeo para a electroforese em condiccedilotildees desnaturantes
Figura 2 ndash Representaccedilatildeo graacutefica da recta de calibraccedilatildeo (log(Peso Molecular) vs Rf dos marcadores de peso
molecular) onde Rf indica determinada atraveacutes do padratildeo de peso molecular
(poccedilo (a))
3Caacutelculo do coeficiente de extinccedilatildeo molar () para a quantificaccedilatildeo de cisteiacutenas reduzidas
atraveacutes do meacutetodo de Ellman
Traccedilou-se a recta de Abs412 nm meacutedia vs [L-Cisteiacutena] de modo a determinar o valor do coeficiente
de extinccedilatildeo molar definido como o declive daquela recta Assim =8400 M-1
cm-1
como
podemos verificar na Fig 3
A595 nm = 00546[BSA] + 00209 Rsup2 = 099051
Ab
s 59
5 n
m
[BSA] (mgmL)
log PM = -1177 Rf + 2247 Rsup2 = 098957
log
PM
Mobilidade relativa (Rf)
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero

43
Para determinar a concentraccedilatildeo de proteiacutena apoacutes o processo de isolamento purificaccedilatildeo e
diaacutelise quando necessaacuterio colocou-se 5 L da amostra num tubo de Eppendorf e adicionou-se
795 L de H2O MilliQ homogeneizando a soluccedilatildeo no voacutertex Adicionou-se posteriormente 200
L de reagente de Bradford e repetiu-se a homogeneizaccedilatildeo Incubou-se a mistura a 25ordmC por 5
minutos apoacutes os quais se leu os valores de Abs595 nm no espectrofotoacutemetro Por interpolaccedilatildeo deste
valor a partir da recta de calibraccedilatildeo determinou-se a concentraccedilatildeo de proteiacutena na amostra
3312 Quantificaccedilatildeo pelo meacutetodo directo
A quantificaccedilatildeo das amostras proteicas pode tambeacutem ser efectuada atraveacutes da leitura directa da
absorvacircncia tendo por base a lei de Beer-Lambert onde Abs corresponde agrave
absorvacircncia ao comprimento de onda ao qual o ensaio foi realizado corresponde ao coeficiente
de extinccedilatildeo (substituiacutedo pela letra a se as unidades forem distintas da molaridade) b corresponde
ao percurso oacuteptico da radiaccedilatildeo na ceacutelula amostral e c corresponde agrave concentraccedilatildeo amostral
Assim no caso de S1 de miosina mediu-se a Abs280e calculou-se a concentraccedilatildeo da proteiacutena
sabendo que o coeficiente de extinccedilatildeo de S1 eacute (Ref)
No caso da actina mediu-se a Abs290ecalculou-se a concentraccedilatildeo da proteiacutena sabendo que o
coeficiente de extinccedilatildeo da actina eacute (Ref)
332Electroforese em condiccedilotildees desnaturantes (SDS-PAGE)
De modo a melhor caracterizar as proteiacutenas isoladas foi realizada uma electroforese em
condiccedilotildees desnaturantes (SDS-PAGE do inglecircs Sodium Dodecyl Sulfate Polyacrilamide Gel
Electrophoresis) O SDS (dodecilo sulfato de soacutedio) eacute um detergente anioacutenico utilizado neste
meacutetodo para desnaturar as proteiacutenas que se ligam ao SDS na proporccedilatildeo 14 g SDS g proteiacutena
(aproximadamente 1 moleacutecula de SDS por cada 2 resiacuteduos de aminoaacutecidos) (Stryer) O SDS induz
uma densidade de carga negativa significativa nas proteiacutenas a que se liga mascarando a sua carga
intriacutenseca pelo que as razotildees cargamassa satildeo tendencialmente idecircnticas Assim a SDS-PAGE
separa as proteiacutenas quando sujeitas a um campo eleacutectrico de acordo com o seu peso molecular
sendo a sua migraccedilatildeo directamente proporcional ao logaritmo deste uacuteltimo (Stryer)
O gel de resoluccedilatildeo com 10 de acrilamida foi obtido atraveacutes da adiccedilatildeo de 167 mL de soluccedilatildeo
stock de acrilamida11
075 mL de tampatildeo de resoluccedilatildeo12
251 mL de aacutegua destilada e 50 L de
SDS 10 Para promover a polimerizaccedilatildeo adicionou-se em seguida 38 L de PSA 10
11
soluccedilatildeo stock de acrilamida soluccedilatildeo de 30 (mv) de acrilamida e 08 (mv) de NNrsquo-metil-bis-acrilamida 12
tampatildeo de resoluccedilatildeo 15 M Tris-HCl pH 88
44
(persulfato de amoacutenia) 10 e 25 L de TEMED 99 (tetrametiletilenodiamina) Agitou-se
cuidadosamente de modo a natildeo formar bolhas e pipetou-se a soluccedilatildeo para a cassette de vidro
previamente montada no suporte De seguida adicionou-se uma peliacutecula de cerca de 2 mm de 2-
metil-2-butanol para verificar posteriormente a polimerizaccedilatildeo Apoacutes a polimerizaccedilatildeo que demora
cerca de 45 minutos verteu-se a agrave superfiacutecie e adicionou-se o gel de concentraccedilatildeo com 5 de
acrilamida constituiacutedo por 03 mL de soluccedilatildeo de stock acrilamida 045 mL de tampatildeo de
concentraccedilatildeo13
094 mL de aacutegua destilada 18 L de SDS 10 135 L de PSA 10 e 2L de
TEMED 99 Colocou-se o pente sobre o gel e deixou-se polimerizar durante cerca de 45
minutos
As amostras para aplicaccedilatildeo no gel de electroforese foram preparadas para uma concentraccedilatildeo de
proteiacutena de 1 mgmL num volume final de 100 L dos quais 25 L correspondem ao tampatildeo da
amostra14
As amostras foram aquecidas a 100ordmC durante 5 minutos Antes da aplicaccedilatildeo das
amostras transferiu-se a cassette para a tina de electroforese agrave qual se adicionou a soluccedilatildeo
tampatildeo de electroforese da tina superior15
e tampatildeo de electroforese da tina inferior16
Aplicaram-
se 10 L das soluccedilotildees de marcadores de peso molecular e 20 L das amostras proteicas
preparadas anteriormente A electroforese decorreu durante cerca de 3h30 min a uma voltagem
de 80 V De seguida transferiu-se o gel para uma tina corando as proteiacutenas durante 1h com
soluccedilatildeo corante17
Descorou-se o gel com soluccedilatildeo descorante18
tendo-se mudado esta soluccedilatildeo ateacute
as bandas se encontrarem bem definidas
Determinou-se a mobilidade relativa das bandas correspondentes aos marcadores de elevado peso
molecular e traccedilou-se a recta de calibraccedilatildeo ndash log(PM) vs log(Rf) onde Rf corresponde agrave
mobilidade relativa Por interpolaccedilatildeo determina-se o PM das proteiacutenas isoladas sabendo a sua
mobilidade relativa Por comparaccedilatildeo com os valores descritos na literatura pode concluir-se
acerca do grau de purificaccedilatildeo
34 Mediccedilatildeo da actividade enzimaacutetica de S1
341 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
13
tampatildeo de concentraccedilatildeo 05 M Tris-HCl pH 68 14
tampatildeo de amostra 320 mM Tris-HCl pH 68 04 M szlig-mercaptoetanol SDS 8 glicerol 15 (vv) e roxo de m-
cresol 0024 15
tampatildeo de electroforese da tina superior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS 16
tampatildeo de electroforese da tina inferior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS acetato de soacutedio
01 M 17
soluccedilatildeo corante ldquoCoomassie Blue R-250rdquo contendo metanol 45 (vv) e aacutecido aceacutetico glacial 10 (vv) 18
soluccedilatildeo descorante metanol 10 (vv) e aacutecido aceacutetico glacial 10 vv)
45
O meacutetodo de Fiske-Subarow modificado consiste na reacccedilatildeo do molibdato de amoacutenio com o
fosfato inorgacircnico em soluccedilatildeo aacutecida com formaccedilatildeo do aacutecido fosfomoliacutebdico Seguidamente daacute-
se a adiccedilatildeo de um agente redutor que provoca a reduccedilatildeo do composto a fosfomolibdato que forma
um complexo de cor azulada cuja absorvacircncia maacutexima ocorre para um comprimento de onda de
660 nm tornando possiacutevel a sua detecccedilatildeo por meacutetodos espectrofotomeacutetricos As actividades Ca2+
-
ATPaacutesica e K+EDTA-ATPaacutesica de S1 foram determinadas atraveacutes deste meacutetodo de acordo com
o descrito por Taussky e Shorr (1953) com mediccedilatildeo da quantidade de fosfato (Pi) libertado
atraveacutes da hidroacutelise de ATP
Para tal obteve-se primeiramente a recta de calibraccedilatildeo com as amostras presentes na Tabela 3II
utilizando uma soluccedilatildeo stock de 5 mM KH2PO4
Tabela 3II ndash Composiccedilatildeo das diferentes amostras utilizadas para determinar a recta de calibraccedilatildeo atraveacutes do meacutetodo
de Fiske-Subbarow modificado
KH2PO4 (L)
([KH2PO4]stock= 5mM)
PO43-
(nmole) H2O (uL)
0 0 1780
5 25 1775
10 50 1770
20 100 1760
40 200 1740
100 500 1680
A cada tubo de Eppendorf adicionaram-se seguidamente 200 L de soluccedilatildeo J19
e 20 L de agente
redutor20
homogeneizando no voacutertex apoacutes cada adiccedilatildeo Incubou-se durante 10 minutos e mediu-
se a Abs660 nm num espectrofotoacutemetro com intervalos de tempo iguais entre cada amostra Traccedilou-
se a recta de calibraccedilatildeo (Abs660 nmvs nmol PO43-
) a partir dos valores obtidos
Para determinar a actividade Ca2+
-ATPaacutesica de S1 prepararam-se tubos de Eppendorf com 400
L de meio de reacccedilatildeo21
25 L CaCl2 ([CaCl2]stock= 100 mM) 25 L ATP ([ATP]stock= 100 mM)
com [S1]ensaios=01 mgmL Perfez-se se necessaacuterio com H2O MilliQ ateacute perfazer um volume de 1
mL Para determinar a actividade K+EDTA-ATPaacutesica utilizou-se uma concentraccedilatildeo idecircntica de
19
Soluccedilatildeo J 5 N H2SO4 25 molibdato de amoacutenio 20
Agente redutor 5 mg aacutecido 1-amino-2-naftol-4-sulfoacutenico 30 mg bissulfito de soacutedio 30 mg sulfito de soacutedio em 26
mL H2O 21
meio de reacccedilatildeo 125 mM Tris-HCl pH 80 0575 M KCl
46
proteiacutena nos ensaios (01 mgmL) e o mesmo meio de reacccedilatildeo Aos tubos de Eppendorf
adicionou-se ainda 123 L de KCl ([KCl]stock = 3M) 25 L de ATP 10 L de EDTA
([EDTA]stock= 100 mM )e o volume de H2O MilliQ necessaacuterio para perfazer 1 mL Em ambas as
situaccedilotildees o uacuteltimo reagente adicionado foi o ATP uma vez que se trata do substrato da reacccedilatildeo
enzimaacutetica
A cineacutetica decorreu a 25ordmC durante 5 minutos para a actividade Ca2+
-ATPaacutesica e durante 3
minutos para a actividade K+EDTA-ATPaacutesica Apoacutes o periacuteodo de incubaccedilatildeo transferiu-se 150
L de amostra para tubos de Eppendorf contendo igual volume de TCA 10 Colocou-se no gelo
e homogeneizou-se no voacutertex Seguidamente centrifugou-se a 1500 g durante 10 minutos e
recolheu-se 200 L de sobrenadante para novos tubos de Eppendorf Adicionou-se a estes 158
mL de H2O MilliQ e 200 L de soluccedilatildeo J Homogeneizou-se no voacutertex e adicionou-se 20 L de
reagente redutor repetindo-se a homogeneizaccedilatildeo Incubou-se durante 10 minutos e mediu-se a
Abs660 nm com o mesmo intervalo de tempo de leitura para cada amostra Por intrapolaccedilatildeo a partir
da recta de calibraccedilatildeo obtida anteriormente determinou-se a quantidade de Pi libertado em cada
ensaio A partir destes resultados determinaram-se as actividades Ca2+
-ATPaacutesica e K+EDTA-
ATPaacutesica do S1 em nmol PO43-
minmg proteiacutena e em s-1
342 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
Da actividade Mg2+
-ATPaacutesica de S1 resulta a formaccedilatildeo dos produtos da reacccedilatildeo de
hidroacutelise do ATP - ADP e Pi Na presenccedila de ADP fosfoenolpiruvato (PEP) e da enzima piruvato
cinase (PK) os produtos resultantes satildeo ATP e piruvato Por acccedilatildeo da enzima lactato
desidrogenase (LDH) este uacuteltimo eacute convertido em lactato ocorrendo simultaneamente a oxidaccedilatildeo
do NADH a NAD+ monitorizada atraveacutes da Abs340 nm Quando o S1 de miosina se encontra no
meio reaccional o ATP eacute hidrolisado formando-se novamente ADP e Pi o que permite a
formaccedilatildeo de um ciclo (fig 21) Se as enzimas se encontrarem em excesso a velocidade de
conversatildeo do NADH eacute limitada pela produccedilatildeo de ADP e Pi Como o ATP eacute regenerado em cada
ciclo a sua concentraccedilatildeo inicial eacute sempre mantida (Bagshaw 1982)
Figura 33ndash Representaccedilatildeo esquemaacutetica do meacutetodo de enzimas acopladas
47
O protocolo utilizado e seguidamente descrito foi adaptado de Cuenda et al (1990 ndash Ver Refordf) A
uma cuvette de 1 mL adicionou-se S1 para uma concentraccedilatildeo final de 005 mgmL 800 L de
meio de reacccedilatildeo22
10 L de PEP ([PEP]stock = 42 mM) 10 L de PK+LDH 11 ([PK+LDH]stock =
1 mgmL) e 15 L de NADH ([NADH]stock=25 mM) Iniciou-se a monitorizaccedilatildeo da reacccedilatildeo
atraveacutes da leitura de Abs340 nm apoacutes a adiccedilatildeo de 20 L de ATP ([ATP]stock=100 mM) A
actividade Mg2+
-ATPaacutesica eacute obtida atraveacutes do declive correspondente agrave oxidaccedilatildeo do NADH Para
a mediccedilatildeo da Mg2+
-ATPase de S1 estimulada por actina adicionou-se F-actina para uma
concentraccedilatildeo final de 2 microM sendo a actividade obtida tambeacutem atraveacutes do declive correspondente
agrave oxidaccedilatildeo do NADH apoacutes a adiccedilatildeo da proteiacutena
35 Interaccedilatildeo entre vanaacutedio e subfragmento-1 de miosina
351 Preparaccedilatildeo das soluccedilotildees de vanaacutedio
As soluccedilotildees de vanadato foram preparadas em aacutegua MilliQ a partir de metavanadato de amoacutenio
(NH4VO3) numa concentraccedilatildeo stock de 50 mM com agitaccedilatildeo magneacutetica e aquecimento (~30 ordmC)
Esta soluccedilatildeo com um valor compreendido entre 6 e 7 designa-se por ldquometavanadatordquo e eacute
constituiacuteda pelas esspeacutecies V1 V2 V4 e V5 (Ref) Acidificando a soluccedilatildeo de metavanadato por
adiccedilatildeo de HCl ateacute um valor de pH proacuteximo de 4 observa-se uma mudanccedila de cor de amarelo
claro para laranja indicativa da formaccedilatildeo da espeacutecie decameacuterica de vanadato De seguida e
imediatamente antes da realizaccedilatildeo dos ensaios descritos o valor do pH da soluccedilatildeo de
decavanadato foi aumentado para cerca de 75 por adiccedilatildeo de NaOH A soluccedilatildeo (2) designada
por ldquometavanadatordquo natildeo foi posteriormente alterada ateacute agrave sua adiccedilatildeo para a realizaccedilatildeo dos ensaios
As soluccedilotildees foram armazenadas a 4ordmC e mantidas em gelo aquando da sua utilizaccedilatildeo
A soluccedilatildeo de V(IV) foi preparada em condiccedilotildees anaeroacutebias por diluiccedilatildeo de sulfato de vanadil
(VOSO4) para uma concentraccedilatildeo final de 5 mM em H2O MilliQ saturada com N2 gasoso
352Anaacutelise por espectroscopia de UV-Vis
A concentraccedilatildeo da espeacutecie decameacuterica em soluccedilatildeo depende do periacuteodo de tempo apoacutes a diluiccedilatildeo
no meio de reacccedilatildeo apropriado (Ref) A velocidade de desoligomerizaccedilatildeo da espeacutecie decameacuterica
pode ser analisada por espectroscopia de absorccedilatildeo na regiatildeo UVVis uma vez que esta espeacutecie
apresenta duas bandas caracteriacutesticas na regiatildeo do visiacutevel (a 358 e 400 nm) que natildeo se verifica
em quaisquer outras espeacutecies oligomeacutericas As bandas supramencionada sapresentam-se bem
22
Meio de reacccedilatildeo 3125 mM Tris-HCl pH 71 375 mM MgCl2
48
resolvidas atraveacutes da obtenccedilatildeo da derivada de 2ordf ordem do espectro apresentando a forma de 2
vales com miacutenimos centrados nos comprimentos de onda supracitados com dependecircncia linear
relativamente agrave concentraccedilatildeo de decavanadato total (Ramos et al 2006)
Assim a estabilidade da espeacutecie decameacuterica apoacutes diluiccedilatildeo da soluccedilatildeo de decavanadato no meio de
reacccedilatildeo dos ensaios (tampatildeo D) para uma concentraccedilatildeo final de 1 mM em vanadato total foi
caracterizada atraveacutes da leitura da Abs400 em funccedilatildeo do tempo tendo-se determinado qual a
ordem de reacccedilatildeo e o tempo de meia-vida da mesma Com base em estudos anteriores acerca da
estabilidade do V10 na presenccedila de diferentes proteiacutenas (Ramos et al 2006) determinou-se
igualmente o tempo de meia vida apoacutes a adiccedilatildeo de 20 microM de S1 de miosina ao meio reaccional
353 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
Os espectros de RMN do isoacutetopo 51
V foram realizados num espectroacutemetro Bruker AM-400 MHz
equipado com uma sonda multinuclear inversa de 5 mm Em tubos de RMN de 5 mm de diacircmetro
preparou-se soluccedilotildees de 500 L de deca- ou metavanadato correspondentes a um intervalo de
concentraccedilotildees de 1 a 10 mM suplementadas com 10 D2O Os espectros foram adquiridos agrave
temperatura ambiente com uma largura espectral de 23 923 Hz um tempo de aquisiccedilatildeo de 0086
s tempo de relaxaccedilatildeo de 001 s e uma sequecircncia de pulsos a 90ordm Utilizou-se a transformada de
Fourier com alargamento das bandas de 20Hz para o processamento dos espectros Os desvios
quiacutemicos das ressonacircncias de 51
V foram obtidos em relaccedilatildeo agrave espeacutecie VOCl3cuja frequecircncia de
ressonacircncia eacute considerada o padratildeo sendo todas as outras referenciadas a partir desta em ppm
As aacutereas dos sinais do RMN correspondentes agraves espeacutecies oligomeacutericas presentes foram obtidas
por integraccedilatildeo dos sinais As concentraccedilotildees relativas das espeacutecies foram calculadas com recurso agrave
equaccedilatildeo
onde A[Vx] corresponde agrave aacuterea referente agrave ressonacircncia da espeacutecie Vx com nordm de agregaccedilatildeo x
A[V]total agrave soma total das aacutereas medidas e [V] a concentraccedilatildeo total de vanaacutedio no ensaio
A teacutecnica de 51
V-RMN foi anteriormente aplicada em estudos de ligaccedilatildeo com S1 de miosina
(Ref) tendo sido demostrado que entre as espeacutecies V1 V2 V4 e V5 a espeacutecie tetrameacuterica possui a
maior afinidade para a miosina (Cremo et al) O desaparecimento ou alargamento dos sinais pode
implicar a ligaccedilatildeo de um dos oxovanadatos laacutebeis agrave miosina sem indicaccedilatildeo de qual uma vez que
49
se processa um equiliacutebrio entre eles (Crans 1990) Assim e tendo em conta as indicaccedilotildees de que
a espeacutecie V10 eacute distinta das restantes no que diz respeito agrave magnitude da interacccedilatildeo com a miosina
e inibiccedilatildeo da actividade ATPaacutesica estimulada por actina (Aureliano 2000 Tiago et al 2002)
torna-se fundamental investigar a interacccedilatildeo das diferentes espeacutecies de vanadato com o S1 de
miosina atraveacutes da teacutecnica de 51
V-RMN
Para tal obtiveram-se espectros de 51
V-RMN resultantes da titulaccedilatildeo de uma soluccedilatildeo 5 mM de
decavanadato total com concentraccedilotildees crescentes de S1 de miosina (0 5 10 15 e 20 microM) em
tampatildeo D Realizou-se ainda ensaios na presenccedila de 2 mM de decavanadato e 2 mM de ATP na
presenccedila e na ausecircncia de S1 de miosina de modo a verificar o efeito que estes ligandos naturais
da enzima teria na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o subfragmento-1
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
Os grupos sulfidrilo (-SH) das cisteiacutenas satildeo resiacuteduos de aminoaacutecidos extremamente sensiacuteveis agrave
oxidaccedilatildeo resultando na sua oxidaccedilatildeo (Dalle-Donne et al 1999) Logo a quantificaccedilatildeo destes
resiacuteduos eacute bastante importante nos estudos de interacccedilatildeo com vanaacutedio jaacute que os resiacuteduos de Cys
com ecircnfase para o SH1 e o SH2 satildeo bastante importantes na monitorizaccedilatildeo das modificaccedilotildees
funcionais da regiatildeo da cabeccedila
O procedimento seguidamente apresentado foi originalmente descrito por Gutieacuterrez-Merino
(1987)) Consiste na reacccedilatildeo entre os grupos ndashSH natildeo oxidados das proteiacutenas e o reagente de
Ellman (DTNB -55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico)) - formando-se o aniatildeo CNT (aniatildeo 3-
carboxilato-4-nitrotiofenolato) numa estequiometria de 11 Este apresenta um maacuteximo de
absorvecircncia a 412 nm (Robyt 1990) Assim apoacutes a incubaccedilatildeo da proteiacutena na presenccedila do metal
a quantidade de cisteiacutenas reduzidas por proteiacutena determina-se atraveacutes dos valores de Abs412
Deste modo colocou-se num tubo de Eppendorf o volume apropriado de S1 dialisado para que a
concentraccedilatildeo no ensaio fosse de 1 mgmL Adicionou-se o volume apropriado de decavanadato
para que a sua concentraccedilatildeo fosse de 0 10 50 100 250 500 ou 1000 M Perfez-se com soluccedilatildeo
D ateacute um volume final de 1 mL e incubou-se a 25ordmC por 20 minutos Apoacutes este periacuteodo colocou-
se 780 L da mistura reaccional numa cuvette de quartzo e adicionou-se 220 L de soluccedilatildeo K23
23
Tampatildeo K Tampatildeo fosfatohidrogenofosfato de soacutedio 025 M pH 72
50
Adicionou-se 15 L de DTNB (em soluccedilatildeo K [DTNB]stock=100 mM) ao meio reaccional e
registou-se o valor de absorvecircncia medido equivalente agraves cisteiacutenas ldquoraacutepidasrdquo (SH1 e SH2
expostos ao solvente) Adicionou-se 001 g de SDS que provoca a desnaturaccedilatildeo da proteiacutena e
consequentemente a exposiccedilatildeo de todos os resiacuteduos de cisteiacutenas que se encontravam ateacute aiacute
inacessiacuteveis no interior da proteiacutena (cisteiacutenas totais) Estes resiacuteduos podem assim reagir com o
DTNB
Estimou-se o nuacutemero de resiacuteduos de cisteiacutenas em todas as amostras com recurso ao coeficiente de
extinccedilatildeo molar () do DTNB (ver ponto 3 dos Anexos) que foi determinado atraveacutes da titulaccedilatildeo
de L-Cisteiacutena com intervalo de concentraccedilotildees compreendidos entre 0 e 50 microM na presenccedila de
25microM de DTNB em tampatildeo D Os valores de absorvecircncia foram obtidos para um comprimento de
onda de 412 nm
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
3621 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
A teacutecnica mais eficiente de anaacutelise da interacccedilatildeo entre o iatildeo vanadilo e proteiacutenas como a
calmodulina e a transferrina entre outras permite a obtenccedilatildeo raacutepida de informaccedilatildeo no que toca agrave
conformaccedilatildeo e amplitude da referida interacccedilatildeo
Os espectros de Ressonacircncia Paramagneacutetica Electroacutenica (RPE) da espeacutecie paramagneacutetica V(IV)
foram realizados a 70K num espectroacutemetro Bruker EMX-300 X-band Todas as soluccedilotildees
utilizadas e amostras foram preparadas em anaerobiose numa cacircmara de luvas de modo a evitar a
reduccedilatildeo de V(IV) a V(V) Em tubos capilares com 02 mL de amostra manteve-se a concentraccedilatildeo
de S1 a 20 M e variou-se a concentraccedilatildeo de vanadilo (0 25 5 10 15 25 50 75 100 150
200 500 e 1000 M [VOSO4]stock=5 mM) em tampatildeo D Apoacutes obtenccedilatildeo dos espectros estes
foram tratados atraveacutes do software SimFonia e WinEPR tendo-se obtido um graacutefico com a
variaccedilatildeo da intensidade do sinal de RPE em funccedilatildeo da concentraccedilatildeo de sulfato de vanadilo
presente Realizaram-se ainda simulaccedilotildees de espectros obtidos com 20 microM de S1 e 1000 microM de
V(IV) com vista agrave obtenccedilatildeo dos ligandos provaacuteveis do vanadilo atraveacutes da anaacutelise da constante
de acoplamento hiperfina (A) e do factor g (g)
4 Conclusotildees gerais
Apesar de jaacute ter sido demonstrado que o vanaacutedio inibe a Na+K
+-ATPase da miosina a funccedilatildeo
deste elemento no tecido muacutesculo esqueleacutetico continua largamente incompreendida A enorme
complexidade quiacutemica deste metal especialmente no que concerne agrave oligomerizaccedilatildeo e reacccedilotildees
51
redox constitui um obstaacuteculo agrave compreensatildeo do comportamento do vanaacutedio nos organismos
vivos Assim a caracterizaccedilatildeo da quiacutemica deste elemento eacute de grande utilidade antes de iniciar o
estudo da sua interacccedilatildeo com o presente sistema
A anaacutelise por espectroscopias de 51
V-RMN e de UV-Vis de soluccedilotildees de vanadato diluiacutedas nos
meios de reacccedilatildeo utilizados no presente relatoacuterio (10 mM HEPES pH 70 25 mM KCl) mostrou
claramente que a espeacutecie decameacuterica (V10) possui um tempo de vida suficientemente longo para a
determinaccedilatildeo do seu efeito isolado ao contraacuterio do que se verifica com as espeacutecies mono- di-
tetra- e pentameacutericas (V1 V2 V4 e V5) Por outro lado a anaacutelise verificou ainda que apoacutes diluiccedilatildeo
das soluccedilotildees de decavanadato no meio de reacccedilatildeo coexistem as espeacutecies V10 e V1 pelo que os
efeitos putativos desta uacuteltima deveratildeo ser subtraiacutedos aquando da monitorizaccedilatildeo dos efeitos da
espeacutecie decameacuterica
O vanaacutedio (V) (vanadato) inibe as enzimas que catalizam reacccedilotildees de transferecircncia de grupos
fosfato como a ATPase da miosina [Goodno 1979] devido agrave sua analogia estrutural com o iatildeo
fosfato Na sua utilizaccedilatildeo como inibidor enzimaacutetico natildeo eacute tida em conta a contribuiccedilatildeo individual
das espeacutecies oligomeacutericas considerando-se normalmente a espeacutecie V1 como inibidora efectiva
[Goodno e Taylor 1982] No presente relatoacuterio verificou-se por espectroscopia de 51
V-RMN que
outros oligoacutemeros de vanadato interagem com o S1 de miosina tendo as espeacutecies V10 e V4 os
maiores niacuteveis de afinidade A sua interacccedilatildeo envolve forccedilas electrostaacuteticas e eacute afectada
negativamente por ligandos naturais de enzima como eacute o caso do ATP
A exposiccedilatildeo do S1 de miosina (005 mgmL) a decavanadato conduz agrave inibiccedilatildeo do capacidade da
proteiacutena de estimular a actividade Mg2+
-ATPaacutesica com um valor de IC50 de 266 plusmn 07 M
decavanadato sendo o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM
decavanadato (80) Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM decavanadato sendo
o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM decavanadato (60)
exercendo um efeito de impedimento agrave ligaccedilatildeo entre o vanaacutedio e o S1
A uma concentraccedilatildeo de 27 microM decavanadato verifica-se a oxidaccedilatildeo de um dos resiacuteduos de Cys
de S1 de miosina seguida pela oxidaccedilatildeo gradual de outro resiacuteduo de Cys ateacute agrave concentraccedilatildeo de
1000 microM Ambos os resiacuteduos oxidados natildeo se encontravam expostos ao solvente utilizado A
oxidaccedilatildeo dos resiacuteduos permite sugerir que esta modificaccedilatildeo oxidativa sofrida pelo S1 contribui
para a diminuiccedilatildeo da sua actividade de ATPase
52
Por espectroscopia de RPE revelaram-se os possiacuteveis ligandos equatoriais do vanaacutedio aquando da
sua interacccedilatildeo com o S1 de miosina como sendo duas moleacuteculas de aacutegua um ligando contendo o
grupo R-OH e outro contendo o grupo R=NR
Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados de
oxidaccedilatildeo (IV) e (V) no S1 de miosina para concentraccedilotildees facilmente alcanccedilaacuteveis no ambiente
terrestre
53
5 Estudos futuros
No sentido do melhor esclarecimento das alteraccedilotildees moleculares e estruturais que ocorrem no
subfragmento 1 quando exposta ao vanaacutedio que conduzem a uma diminuiccedilatildeo da sua funccedilatildeo
sugere-se
- Quantificaccedilatildeo das cisteiacutenas reduzidas de S1 na presenccedila de metavanadato
- Determinaccedilatildeo das actividades Ca2+
- e K+-ATPaacutesica do S1 de miosina na presenccedila de
decavanadato
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina estimulada por actina
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina na presenccedila de
metavanadato
- Realizaccedilatildeo de RMN 2D com variaccedilatildeo das concentraccedilotildees das soluccedilotildees de decavanadato e
metavanadato na presenccedila de S1
-Mutageacutenese dos resiacuteduos de Cys doS1 de miosina de modo a verificar quais os resiacuteduos que
sofrem oxidaccedilatildeo na presenccedila de decavanadato
- Determinaccedilatildeo por calorimetria (DSC Differential Scanning Calorimetry) da variaccedilatildeo da
entalpia (∆H) e da temperatura de desnaturaccedilatildeo do S1 de miosina na presenccedila de vanaacutedio uma
vez que estas propriedades caracteriacutesticas de cada proteiacutena satildeo alteradas apoacutes ocorrecircncia de
modificaccedilotildees estruturais
- Anaacutelise por SDS-PAGE de S1 de miosina apoacutes tratamento com vanaacutedio de modo a determinar
quais os polipeacuteptidos eventualmente formados apoacutes clivagem da proteiacutena
54
6 Referecircncias bibliograacuteficas
- Sellers J Myosins 2nd Edition Oxford University Press 1999
- L Stryer inBiochemistry (Ed WH Freeman 2005 pp
Refordfas Materiais e Meacutetodos
[A] S S Margossian SLowey Methods Enzymol1982 85 55-71
[B] JD Pardue JA Spudich Methods Enzymol 1982 85 164-181
[C] M Bradford Anal Biochem1976 72 248-254
[D] H H Taussky E Shorr J Biol Chem 1953 202 675-685
[E] C R Bagshaw in Outline Studies in Biology ndash Muscle Contraction Chapman and Hall Ed
1982
[F] A Cuenda et al Eur J Biochem1990 194 663-670
55
Anexos
1 Determinaccedilatildeo da recta de calibraccedilatildeo meacutetodo de Bradford
Figura1ndashRepresentaccedilatildeo graacutefica da recta de calibraccedilatildeo (Abs595 nm vs [BSA]) utilizada para o meacutetodo de Bradford
2 Determinaccedilatildeo da recta calibraccedilatildeo para a electroforese em condiccedilotildees desnaturantes
Figura 2 ndash Representaccedilatildeo graacutefica da recta de calibraccedilatildeo (log(Peso Molecular) vs Rf dos marcadores de peso
molecular) onde Rf indica determinada atraveacutes do padratildeo de peso molecular
(poccedilo (a))
3Caacutelculo do coeficiente de extinccedilatildeo molar () para a quantificaccedilatildeo de cisteiacutenas reduzidas
atraveacutes do meacutetodo de Ellman
Traccedilou-se a recta de Abs412 nm meacutedia vs [L-Cisteiacutena] de modo a determinar o valor do coeficiente
de extinccedilatildeo molar definido como o declive daquela recta Assim =8400 M-1
cm-1
como
podemos verificar na Fig 3
A595 nm = 00546[BSA] + 00209 Rsup2 = 099051
Ab
s 59
5 n
m
[BSA] (mgmL)
log PM = -1177 Rf + 2247 Rsup2 = 098957
log
PM
Mobilidade relativa (Rf)
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero

44
(persulfato de amoacutenia) 10 e 25 L de TEMED 99 (tetrametiletilenodiamina) Agitou-se
cuidadosamente de modo a natildeo formar bolhas e pipetou-se a soluccedilatildeo para a cassette de vidro
previamente montada no suporte De seguida adicionou-se uma peliacutecula de cerca de 2 mm de 2-
metil-2-butanol para verificar posteriormente a polimerizaccedilatildeo Apoacutes a polimerizaccedilatildeo que demora
cerca de 45 minutos verteu-se a agrave superfiacutecie e adicionou-se o gel de concentraccedilatildeo com 5 de
acrilamida constituiacutedo por 03 mL de soluccedilatildeo de stock acrilamida 045 mL de tampatildeo de
concentraccedilatildeo13
094 mL de aacutegua destilada 18 L de SDS 10 135 L de PSA 10 e 2L de
TEMED 99 Colocou-se o pente sobre o gel e deixou-se polimerizar durante cerca de 45
minutos
As amostras para aplicaccedilatildeo no gel de electroforese foram preparadas para uma concentraccedilatildeo de
proteiacutena de 1 mgmL num volume final de 100 L dos quais 25 L correspondem ao tampatildeo da
amostra14
As amostras foram aquecidas a 100ordmC durante 5 minutos Antes da aplicaccedilatildeo das
amostras transferiu-se a cassette para a tina de electroforese agrave qual se adicionou a soluccedilatildeo
tampatildeo de electroforese da tina superior15
e tampatildeo de electroforese da tina inferior16
Aplicaram-
se 10 L das soluccedilotildees de marcadores de peso molecular e 20 L das amostras proteicas
preparadas anteriormente A electroforese decorreu durante cerca de 3h30 min a uma voltagem
de 80 V De seguida transferiu-se o gel para uma tina corando as proteiacutenas durante 1h com
soluccedilatildeo corante17
Descorou-se o gel com soluccedilatildeo descorante18
tendo-se mudado esta soluccedilatildeo ateacute
as bandas se encontrarem bem definidas
Determinou-se a mobilidade relativa das bandas correspondentes aos marcadores de elevado peso
molecular e traccedilou-se a recta de calibraccedilatildeo ndash log(PM) vs log(Rf) onde Rf corresponde agrave
mobilidade relativa Por interpolaccedilatildeo determina-se o PM das proteiacutenas isoladas sabendo a sua
mobilidade relativa Por comparaccedilatildeo com os valores descritos na literatura pode concluir-se
acerca do grau de purificaccedilatildeo
34 Mediccedilatildeo da actividade enzimaacutetica de S1
341 Mediccedilatildeo das actividades de Ca2+
-ATPase e K+EDTA-ATPase de S1 pelo meacutetodo de
Fiske-Subarow modificado
13
tampatildeo de concentraccedilatildeo 05 M Tris-HCl pH 68 14
tampatildeo de amostra 320 mM Tris-HCl pH 68 04 M szlig-mercaptoetanol SDS 8 glicerol 15 (vv) e roxo de m-
cresol 0024 15
tampatildeo de electroforese da tina superior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS 16
tampatildeo de electroforese da tina inferior 25 mM Tris pH 83 192 mM glicina 01 (mv) SDS acetato de soacutedio
01 M 17
soluccedilatildeo corante ldquoCoomassie Blue R-250rdquo contendo metanol 45 (vv) e aacutecido aceacutetico glacial 10 (vv) 18
soluccedilatildeo descorante metanol 10 (vv) e aacutecido aceacutetico glacial 10 vv)
45
O meacutetodo de Fiske-Subarow modificado consiste na reacccedilatildeo do molibdato de amoacutenio com o
fosfato inorgacircnico em soluccedilatildeo aacutecida com formaccedilatildeo do aacutecido fosfomoliacutebdico Seguidamente daacute-
se a adiccedilatildeo de um agente redutor que provoca a reduccedilatildeo do composto a fosfomolibdato que forma
um complexo de cor azulada cuja absorvacircncia maacutexima ocorre para um comprimento de onda de
660 nm tornando possiacutevel a sua detecccedilatildeo por meacutetodos espectrofotomeacutetricos As actividades Ca2+
-
ATPaacutesica e K+EDTA-ATPaacutesica de S1 foram determinadas atraveacutes deste meacutetodo de acordo com
o descrito por Taussky e Shorr (1953) com mediccedilatildeo da quantidade de fosfato (Pi) libertado
atraveacutes da hidroacutelise de ATP
Para tal obteve-se primeiramente a recta de calibraccedilatildeo com as amostras presentes na Tabela 3II
utilizando uma soluccedilatildeo stock de 5 mM KH2PO4
Tabela 3II ndash Composiccedilatildeo das diferentes amostras utilizadas para determinar a recta de calibraccedilatildeo atraveacutes do meacutetodo
de Fiske-Subbarow modificado
KH2PO4 (L)
([KH2PO4]stock= 5mM)
PO43-
(nmole) H2O (uL)
0 0 1780
5 25 1775
10 50 1770
20 100 1760
40 200 1740
100 500 1680
A cada tubo de Eppendorf adicionaram-se seguidamente 200 L de soluccedilatildeo J19
e 20 L de agente
redutor20
homogeneizando no voacutertex apoacutes cada adiccedilatildeo Incubou-se durante 10 minutos e mediu-
se a Abs660 nm num espectrofotoacutemetro com intervalos de tempo iguais entre cada amostra Traccedilou-
se a recta de calibraccedilatildeo (Abs660 nmvs nmol PO43-
) a partir dos valores obtidos
Para determinar a actividade Ca2+
-ATPaacutesica de S1 prepararam-se tubos de Eppendorf com 400
L de meio de reacccedilatildeo21
25 L CaCl2 ([CaCl2]stock= 100 mM) 25 L ATP ([ATP]stock= 100 mM)
com [S1]ensaios=01 mgmL Perfez-se se necessaacuterio com H2O MilliQ ateacute perfazer um volume de 1
mL Para determinar a actividade K+EDTA-ATPaacutesica utilizou-se uma concentraccedilatildeo idecircntica de
19
Soluccedilatildeo J 5 N H2SO4 25 molibdato de amoacutenio 20
Agente redutor 5 mg aacutecido 1-amino-2-naftol-4-sulfoacutenico 30 mg bissulfito de soacutedio 30 mg sulfito de soacutedio em 26
mL H2O 21
meio de reacccedilatildeo 125 mM Tris-HCl pH 80 0575 M KCl
46
proteiacutena nos ensaios (01 mgmL) e o mesmo meio de reacccedilatildeo Aos tubos de Eppendorf
adicionou-se ainda 123 L de KCl ([KCl]stock = 3M) 25 L de ATP 10 L de EDTA
([EDTA]stock= 100 mM )e o volume de H2O MilliQ necessaacuterio para perfazer 1 mL Em ambas as
situaccedilotildees o uacuteltimo reagente adicionado foi o ATP uma vez que se trata do substrato da reacccedilatildeo
enzimaacutetica
A cineacutetica decorreu a 25ordmC durante 5 minutos para a actividade Ca2+
-ATPaacutesica e durante 3
minutos para a actividade K+EDTA-ATPaacutesica Apoacutes o periacuteodo de incubaccedilatildeo transferiu-se 150
L de amostra para tubos de Eppendorf contendo igual volume de TCA 10 Colocou-se no gelo
e homogeneizou-se no voacutertex Seguidamente centrifugou-se a 1500 g durante 10 minutos e
recolheu-se 200 L de sobrenadante para novos tubos de Eppendorf Adicionou-se a estes 158
mL de H2O MilliQ e 200 L de soluccedilatildeo J Homogeneizou-se no voacutertex e adicionou-se 20 L de
reagente redutor repetindo-se a homogeneizaccedilatildeo Incubou-se durante 10 minutos e mediu-se a
Abs660 nm com o mesmo intervalo de tempo de leitura para cada amostra Por intrapolaccedilatildeo a partir
da recta de calibraccedilatildeo obtida anteriormente determinou-se a quantidade de Pi libertado em cada
ensaio A partir destes resultados determinaram-se as actividades Ca2+
-ATPaacutesica e K+EDTA-
ATPaacutesica do S1 em nmol PO43-
minmg proteiacutena e em s-1
342 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
Da actividade Mg2+
-ATPaacutesica de S1 resulta a formaccedilatildeo dos produtos da reacccedilatildeo de
hidroacutelise do ATP - ADP e Pi Na presenccedila de ADP fosfoenolpiruvato (PEP) e da enzima piruvato
cinase (PK) os produtos resultantes satildeo ATP e piruvato Por acccedilatildeo da enzima lactato
desidrogenase (LDH) este uacuteltimo eacute convertido em lactato ocorrendo simultaneamente a oxidaccedilatildeo
do NADH a NAD+ monitorizada atraveacutes da Abs340 nm Quando o S1 de miosina se encontra no
meio reaccional o ATP eacute hidrolisado formando-se novamente ADP e Pi o que permite a
formaccedilatildeo de um ciclo (fig 21) Se as enzimas se encontrarem em excesso a velocidade de
conversatildeo do NADH eacute limitada pela produccedilatildeo de ADP e Pi Como o ATP eacute regenerado em cada
ciclo a sua concentraccedilatildeo inicial eacute sempre mantida (Bagshaw 1982)
Figura 33ndash Representaccedilatildeo esquemaacutetica do meacutetodo de enzimas acopladas
47
O protocolo utilizado e seguidamente descrito foi adaptado de Cuenda et al (1990 ndash Ver Refordf) A
uma cuvette de 1 mL adicionou-se S1 para uma concentraccedilatildeo final de 005 mgmL 800 L de
meio de reacccedilatildeo22
10 L de PEP ([PEP]stock = 42 mM) 10 L de PK+LDH 11 ([PK+LDH]stock =
1 mgmL) e 15 L de NADH ([NADH]stock=25 mM) Iniciou-se a monitorizaccedilatildeo da reacccedilatildeo
atraveacutes da leitura de Abs340 nm apoacutes a adiccedilatildeo de 20 L de ATP ([ATP]stock=100 mM) A
actividade Mg2+
-ATPaacutesica eacute obtida atraveacutes do declive correspondente agrave oxidaccedilatildeo do NADH Para
a mediccedilatildeo da Mg2+
-ATPase de S1 estimulada por actina adicionou-se F-actina para uma
concentraccedilatildeo final de 2 microM sendo a actividade obtida tambeacutem atraveacutes do declive correspondente
agrave oxidaccedilatildeo do NADH apoacutes a adiccedilatildeo da proteiacutena
35 Interaccedilatildeo entre vanaacutedio e subfragmento-1 de miosina
351 Preparaccedilatildeo das soluccedilotildees de vanaacutedio
As soluccedilotildees de vanadato foram preparadas em aacutegua MilliQ a partir de metavanadato de amoacutenio
(NH4VO3) numa concentraccedilatildeo stock de 50 mM com agitaccedilatildeo magneacutetica e aquecimento (~30 ordmC)
Esta soluccedilatildeo com um valor compreendido entre 6 e 7 designa-se por ldquometavanadatordquo e eacute
constituiacuteda pelas esspeacutecies V1 V2 V4 e V5 (Ref) Acidificando a soluccedilatildeo de metavanadato por
adiccedilatildeo de HCl ateacute um valor de pH proacuteximo de 4 observa-se uma mudanccedila de cor de amarelo
claro para laranja indicativa da formaccedilatildeo da espeacutecie decameacuterica de vanadato De seguida e
imediatamente antes da realizaccedilatildeo dos ensaios descritos o valor do pH da soluccedilatildeo de
decavanadato foi aumentado para cerca de 75 por adiccedilatildeo de NaOH A soluccedilatildeo (2) designada
por ldquometavanadatordquo natildeo foi posteriormente alterada ateacute agrave sua adiccedilatildeo para a realizaccedilatildeo dos ensaios
As soluccedilotildees foram armazenadas a 4ordmC e mantidas em gelo aquando da sua utilizaccedilatildeo
A soluccedilatildeo de V(IV) foi preparada em condiccedilotildees anaeroacutebias por diluiccedilatildeo de sulfato de vanadil
(VOSO4) para uma concentraccedilatildeo final de 5 mM em H2O MilliQ saturada com N2 gasoso
352Anaacutelise por espectroscopia de UV-Vis
A concentraccedilatildeo da espeacutecie decameacuterica em soluccedilatildeo depende do periacuteodo de tempo apoacutes a diluiccedilatildeo
no meio de reacccedilatildeo apropriado (Ref) A velocidade de desoligomerizaccedilatildeo da espeacutecie decameacuterica
pode ser analisada por espectroscopia de absorccedilatildeo na regiatildeo UVVis uma vez que esta espeacutecie
apresenta duas bandas caracteriacutesticas na regiatildeo do visiacutevel (a 358 e 400 nm) que natildeo se verifica
em quaisquer outras espeacutecies oligomeacutericas As bandas supramencionada sapresentam-se bem
22
Meio de reacccedilatildeo 3125 mM Tris-HCl pH 71 375 mM MgCl2
48
resolvidas atraveacutes da obtenccedilatildeo da derivada de 2ordf ordem do espectro apresentando a forma de 2
vales com miacutenimos centrados nos comprimentos de onda supracitados com dependecircncia linear
relativamente agrave concentraccedilatildeo de decavanadato total (Ramos et al 2006)
Assim a estabilidade da espeacutecie decameacuterica apoacutes diluiccedilatildeo da soluccedilatildeo de decavanadato no meio de
reacccedilatildeo dos ensaios (tampatildeo D) para uma concentraccedilatildeo final de 1 mM em vanadato total foi
caracterizada atraveacutes da leitura da Abs400 em funccedilatildeo do tempo tendo-se determinado qual a
ordem de reacccedilatildeo e o tempo de meia-vida da mesma Com base em estudos anteriores acerca da
estabilidade do V10 na presenccedila de diferentes proteiacutenas (Ramos et al 2006) determinou-se
igualmente o tempo de meia vida apoacutes a adiccedilatildeo de 20 microM de S1 de miosina ao meio reaccional
353 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
Os espectros de RMN do isoacutetopo 51
V foram realizados num espectroacutemetro Bruker AM-400 MHz
equipado com uma sonda multinuclear inversa de 5 mm Em tubos de RMN de 5 mm de diacircmetro
preparou-se soluccedilotildees de 500 L de deca- ou metavanadato correspondentes a um intervalo de
concentraccedilotildees de 1 a 10 mM suplementadas com 10 D2O Os espectros foram adquiridos agrave
temperatura ambiente com uma largura espectral de 23 923 Hz um tempo de aquisiccedilatildeo de 0086
s tempo de relaxaccedilatildeo de 001 s e uma sequecircncia de pulsos a 90ordm Utilizou-se a transformada de
Fourier com alargamento das bandas de 20Hz para o processamento dos espectros Os desvios
quiacutemicos das ressonacircncias de 51
V foram obtidos em relaccedilatildeo agrave espeacutecie VOCl3cuja frequecircncia de
ressonacircncia eacute considerada o padratildeo sendo todas as outras referenciadas a partir desta em ppm
As aacutereas dos sinais do RMN correspondentes agraves espeacutecies oligomeacutericas presentes foram obtidas
por integraccedilatildeo dos sinais As concentraccedilotildees relativas das espeacutecies foram calculadas com recurso agrave
equaccedilatildeo
onde A[Vx] corresponde agrave aacuterea referente agrave ressonacircncia da espeacutecie Vx com nordm de agregaccedilatildeo x
A[V]total agrave soma total das aacutereas medidas e [V] a concentraccedilatildeo total de vanaacutedio no ensaio
A teacutecnica de 51
V-RMN foi anteriormente aplicada em estudos de ligaccedilatildeo com S1 de miosina
(Ref) tendo sido demostrado que entre as espeacutecies V1 V2 V4 e V5 a espeacutecie tetrameacuterica possui a
maior afinidade para a miosina (Cremo et al) O desaparecimento ou alargamento dos sinais pode
implicar a ligaccedilatildeo de um dos oxovanadatos laacutebeis agrave miosina sem indicaccedilatildeo de qual uma vez que
49
se processa um equiliacutebrio entre eles (Crans 1990) Assim e tendo em conta as indicaccedilotildees de que
a espeacutecie V10 eacute distinta das restantes no que diz respeito agrave magnitude da interacccedilatildeo com a miosina
e inibiccedilatildeo da actividade ATPaacutesica estimulada por actina (Aureliano 2000 Tiago et al 2002)
torna-se fundamental investigar a interacccedilatildeo das diferentes espeacutecies de vanadato com o S1 de
miosina atraveacutes da teacutecnica de 51
V-RMN
Para tal obtiveram-se espectros de 51
V-RMN resultantes da titulaccedilatildeo de uma soluccedilatildeo 5 mM de
decavanadato total com concentraccedilotildees crescentes de S1 de miosina (0 5 10 15 e 20 microM) em
tampatildeo D Realizou-se ainda ensaios na presenccedila de 2 mM de decavanadato e 2 mM de ATP na
presenccedila e na ausecircncia de S1 de miosina de modo a verificar o efeito que estes ligandos naturais
da enzima teria na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o subfragmento-1
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
Os grupos sulfidrilo (-SH) das cisteiacutenas satildeo resiacuteduos de aminoaacutecidos extremamente sensiacuteveis agrave
oxidaccedilatildeo resultando na sua oxidaccedilatildeo (Dalle-Donne et al 1999) Logo a quantificaccedilatildeo destes
resiacuteduos eacute bastante importante nos estudos de interacccedilatildeo com vanaacutedio jaacute que os resiacuteduos de Cys
com ecircnfase para o SH1 e o SH2 satildeo bastante importantes na monitorizaccedilatildeo das modificaccedilotildees
funcionais da regiatildeo da cabeccedila
O procedimento seguidamente apresentado foi originalmente descrito por Gutieacuterrez-Merino
(1987)) Consiste na reacccedilatildeo entre os grupos ndashSH natildeo oxidados das proteiacutenas e o reagente de
Ellman (DTNB -55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico)) - formando-se o aniatildeo CNT (aniatildeo 3-
carboxilato-4-nitrotiofenolato) numa estequiometria de 11 Este apresenta um maacuteximo de
absorvecircncia a 412 nm (Robyt 1990) Assim apoacutes a incubaccedilatildeo da proteiacutena na presenccedila do metal
a quantidade de cisteiacutenas reduzidas por proteiacutena determina-se atraveacutes dos valores de Abs412
Deste modo colocou-se num tubo de Eppendorf o volume apropriado de S1 dialisado para que a
concentraccedilatildeo no ensaio fosse de 1 mgmL Adicionou-se o volume apropriado de decavanadato
para que a sua concentraccedilatildeo fosse de 0 10 50 100 250 500 ou 1000 M Perfez-se com soluccedilatildeo
D ateacute um volume final de 1 mL e incubou-se a 25ordmC por 20 minutos Apoacutes este periacuteodo colocou-
se 780 L da mistura reaccional numa cuvette de quartzo e adicionou-se 220 L de soluccedilatildeo K23
23
Tampatildeo K Tampatildeo fosfatohidrogenofosfato de soacutedio 025 M pH 72
50
Adicionou-se 15 L de DTNB (em soluccedilatildeo K [DTNB]stock=100 mM) ao meio reaccional e
registou-se o valor de absorvecircncia medido equivalente agraves cisteiacutenas ldquoraacutepidasrdquo (SH1 e SH2
expostos ao solvente) Adicionou-se 001 g de SDS que provoca a desnaturaccedilatildeo da proteiacutena e
consequentemente a exposiccedilatildeo de todos os resiacuteduos de cisteiacutenas que se encontravam ateacute aiacute
inacessiacuteveis no interior da proteiacutena (cisteiacutenas totais) Estes resiacuteduos podem assim reagir com o
DTNB
Estimou-se o nuacutemero de resiacuteduos de cisteiacutenas em todas as amostras com recurso ao coeficiente de
extinccedilatildeo molar () do DTNB (ver ponto 3 dos Anexos) que foi determinado atraveacutes da titulaccedilatildeo
de L-Cisteiacutena com intervalo de concentraccedilotildees compreendidos entre 0 e 50 microM na presenccedila de
25microM de DTNB em tampatildeo D Os valores de absorvecircncia foram obtidos para um comprimento de
onda de 412 nm
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
3621 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
A teacutecnica mais eficiente de anaacutelise da interacccedilatildeo entre o iatildeo vanadilo e proteiacutenas como a
calmodulina e a transferrina entre outras permite a obtenccedilatildeo raacutepida de informaccedilatildeo no que toca agrave
conformaccedilatildeo e amplitude da referida interacccedilatildeo
Os espectros de Ressonacircncia Paramagneacutetica Electroacutenica (RPE) da espeacutecie paramagneacutetica V(IV)
foram realizados a 70K num espectroacutemetro Bruker EMX-300 X-band Todas as soluccedilotildees
utilizadas e amostras foram preparadas em anaerobiose numa cacircmara de luvas de modo a evitar a
reduccedilatildeo de V(IV) a V(V) Em tubos capilares com 02 mL de amostra manteve-se a concentraccedilatildeo
de S1 a 20 M e variou-se a concentraccedilatildeo de vanadilo (0 25 5 10 15 25 50 75 100 150
200 500 e 1000 M [VOSO4]stock=5 mM) em tampatildeo D Apoacutes obtenccedilatildeo dos espectros estes
foram tratados atraveacutes do software SimFonia e WinEPR tendo-se obtido um graacutefico com a
variaccedilatildeo da intensidade do sinal de RPE em funccedilatildeo da concentraccedilatildeo de sulfato de vanadilo
presente Realizaram-se ainda simulaccedilotildees de espectros obtidos com 20 microM de S1 e 1000 microM de
V(IV) com vista agrave obtenccedilatildeo dos ligandos provaacuteveis do vanadilo atraveacutes da anaacutelise da constante
de acoplamento hiperfina (A) e do factor g (g)
4 Conclusotildees gerais
Apesar de jaacute ter sido demonstrado que o vanaacutedio inibe a Na+K
+-ATPase da miosina a funccedilatildeo
deste elemento no tecido muacutesculo esqueleacutetico continua largamente incompreendida A enorme
complexidade quiacutemica deste metal especialmente no que concerne agrave oligomerizaccedilatildeo e reacccedilotildees
51
redox constitui um obstaacuteculo agrave compreensatildeo do comportamento do vanaacutedio nos organismos
vivos Assim a caracterizaccedilatildeo da quiacutemica deste elemento eacute de grande utilidade antes de iniciar o
estudo da sua interacccedilatildeo com o presente sistema
A anaacutelise por espectroscopias de 51
V-RMN e de UV-Vis de soluccedilotildees de vanadato diluiacutedas nos
meios de reacccedilatildeo utilizados no presente relatoacuterio (10 mM HEPES pH 70 25 mM KCl) mostrou
claramente que a espeacutecie decameacuterica (V10) possui um tempo de vida suficientemente longo para a
determinaccedilatildeo do seu efeito isolado ao contraacuterio do que se verifica com as espeacutecies mono- di-
tetra- e pentameacutericas (V1 V2 V4 e V5) Por outro lado a anaacutelise verificou ainda que apoacutes diluiccedilatildeo
das soluccedilotildees de decavanadato no meio de reacccedilatildeo coexistem as espeacutecies V10 e V1 pelo que os
efeitos putativos desta uacuteltima deveratildeo ser subtraiacutedos aquando da monitorizaccedilatildeo dos efeitos da
espeacutecie decameacuterica
O vanaacutedio (V) (vanadato) inibe as enzimas que catalizam reacccedilotildees de transferecircncia de grupos
fosfato como a ATPase da miosina [Goodno 1979] devido agrave sua analogia estrutural com o iatildeo
fosfato Na sua utilizaccedilatildeo como inibidor enzimaacutetico natildeo eacute tida em conta a contribuiccedilatildeo individual
das espeacutecies oligomeacutericas considerando-se normalmente a espeacutecie V1 como inibidora efectiva
[Goodno e Taylor 1982] No presente relatoacuterio verificou-se por espectroscopia de 51
V-RMN que
outros oligoacutemeros de vanadato interagem com o S1 de miosina tendo as espeacutecies V10 e V4 os
maiores niacuteveis de afinidade A sua interacccedilatildeo envolve forccedilas electrostaacuteticas e eacute afectada
negativamente por ligandos naturais de enzima como eacute o caso do ATP
A exposiccedilatildeo do S1 de miosina (005 mgmL) a decavanadato conduz agrave inibiccedilatildeo do capacidade da
proteiacutena de estimular a actividade Mg2+
-ATPaacutesica com um valor de IC50 de 266 plusmn 07 M
decavanadato sendo o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM
decavanadato (80) Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM decavanadato sendo
o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM decavanadato (60)
exercendo um efeito de impedimento agrave ligaccedilatildeo entre o vanaacutedio e o S1
A uma concentraccedilatildeo de 27 microM decavanadato verifica-se a oxidaccedilatildeo de um dos resiacuteduos de Cys
de S1 de miosina seguida pela oxidaccedilatildeo gradual de outro resiacuteduo de Cys ateacute agrave concentraccedilatildeo de
1000 microM Ambos os resiacuteduos oxidados natildeo se encontravam expostos ao solvente utilizado A
oxidaccedilatildeo dos resiacuteduos permite sugerir que esta modificaccedilatildeo oxidativa sofrida pelo S1 contribui
para a diminuiccedilatildeo da sua actividade de ATPase
52
Por espectroscopia de RPE revelaram-se os possiacuteveis ligandos equatoriais do vanaacutedio aquando da
sua interacccedilatildeo com o S1 de miosina como sendo duas moleacuteculas de aacutegua um ligando contendo o
grupo R-OH e outro contendo o grupo R=NR
Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados de
oxidaccedilatildeo (IV) e (V) no S1 de miosina para concentraccedilotildees facilmente alcanccedilaacuteveis no ambiente
terrestre
53
5 Estudos futuros
No sentido do melhor esclarecimento das alteraccedilotildees moleculares e estruturais que ocorrem no
subfragmento 1 quando exposta ao vanaacutedio que conduzem a uma diminuiccedilatildeo da sua funccedilatildeo
sugere-se
- Quantificaccedilatildeo das cisteiacutenas reduzidas de S1 na presenccedila de metavanadato
- Determinaccedilatildeo das actividades Ca2+
- e K+-ATPaacutesica do S1 de miosina na presenccedila de
decavanadato
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina estimulada por actina
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina na presenccedila de
metavanadato
- Realizaccedilatildeo de RMN 2D com variaccedilatildeo das concentraccedilotildees das soluccedilotildees de decavanadato e
metavanadato na presenccedila de S1
-Mutageacutenese dos resiacuteduos de Cys doS1 de miosina de modo a verificar quais os resiacuteduos que
sofrem oxidaccedilatildeo na presenccedila de decavanadato
- Determinaccedilatildeo por calorimetria (DSC Differential Scanning Calorimetry) da variaccedilatildeo da
entalpia (∆H) e da temperatura de desnaturaccedilatildeo do S1 de miosina na presenccedila de vanaacutedio uma
vez que estas propriedades caracteriacutesticas de cada proteiacutena satildeo alteradas apoacutes ocorrecircncia de
modificaccedilotildees estruturais
- Anaacutelise por SDS-PAGE de S1 de miosina apoacutes tratamento com vanaacutedio de modo a determinar
quais os polipeacuteptidos eventualmente formados apoacutes clivagem da proteiacutena
54
6 Referecircncias bibliograacuteficas
- Sellers J Myosins 2nd Edition Oxford University Press 1999
- L Stryer inBiochemistry (Ed WH Freeman 2005 pp
Refordfas Materiais e Meacutetodos
[A] S S Margossian SLowey Methods Enzymol1982 85 55-71
[B] JD Pardue JA Spudich Methods Enzymol 1982 85 164-181
[C] M Bradford Anal Biochem1976 72 248-254
[D] H H Taussky E Shorr J Biol Chem 1953 202 675-685
[E] C R Bagshaw in Outline Studies in Biology ndash Muscle Contraction Chapman and Hall Ed
1982
[F] A Cuenda et al Eur J Biochem1990 194 663-670
55
Anexos
1 Determinaccedilatildeo da recta de calibraccedilatildeo meacutetodo de Bradford
Figura1ndashRepresentaccedilatildeo graacutefica da recta de calibraccedilatildeo (Abs595 nm vs [BSA]) utilizada para o meacutetodo de Bradford
2 Determinaccedilatildeo da recta calibraccedilatildeo para a electroforese em condiccedilotildees desnaturantes
Figura 2 ndash Representaccedilatildeo graacutefica da recta de calibraccedilatildeo (log(Peso Molecular) vs Rf dos marcadores de peso
molecular) onde Rf indica determinada atraveacutes do padratildeo de peso molecular
(poccedilo (a))
3Caacutelculo do coeficiente de extinccedilatildeo molar () para a quantificaccedilatildeo de cisteiacutenas reduzidas
atraveacutes do meacutetodo de Ellman
Traccedilou-se a recta de Abs412 nm meacutedia vs [L-Cisteiacutena] de modo a determinar o valor do coeficiente
de extinccedilatildeo molar definido como o declive daquela recta Assim =8400 M-1
cm-1
como
podemos verificar na Fig 3
A595 nm = 00546[BSA] + 00209 Rsup2 = 099051
Ab
s 59
5 n
m
[BSA] (mgmL)
log PM = -1177 Rf + 2247 Rsup2 = 098957
log
PM
Mobilidade relativa (Rf)
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero

45
O meacutetodo de Fiske-Subarow modificado consiste na reacccedilatildeo do molibdato de amoacutenio com o
fosfato inorgacircnico em soluccedilatildeo aacutecida com formaccedilatildeo do aacutecido fosfomoliacutebdico Seguidamente daacute-
se a adiccedilatildeo de um agente redutor que provoca a reduccedilatildeo do composto a fosfomolibdato que forma
um complexo de cor azulada cuja absorvacircncia maacutexima ocorre para um comprimento de onda de
660 nm tornando possiacutevel a sua detecccedilatildeo por meacutetodos espectrofotomeacutetricos As actividades Ca2+
-
ATPaacutesica e K+EDTA-ATPaacutesica de S1 foram determinadas atraveacutes deste meacutetodo de acordo com
o descrito por Taussky e Shorr (1953) com mediccedilatildeo da quantidade de fosfato (Pi) libertado
atraveacutes da hidroacutelise de ATP
Para tal obteve-se primeiramente a recta de calibraccedilatildeo com as amostras presentes na Tabela 3II
utilizando uma soluccedilatildeo stock de 5 mM KH2PO4
Tabela 3II ndash Composiccedilatildeo das diferentes amostras utilizadas para determinar a recta de calibraccedilatildeo atraveacutes do meacutetodo
de Fiske-Subbarow modificado
KH2PO4 (L)
([KH2PO4]stock= 5mM)
PO43-
(nmole) H2O (uL)
0 0 1780
5 25 1775
10 50 1770
20 100 1760
40 200 1740
100 500 1680
A cada tubo de Eppendorf adicionaram-se seguidamente 200 L de soluccedilatildeo J19
e 20 L de agente
redutor20
homogeneizando no voacutertex apoacutes cada adiccedilatildeo Incubou-se durante 10 minutos e mediu-
se a Abs660 nm num espectrofotoacutemetro com intervalos de tempo iguais entre cada amostra Traccedilou-
se a recta de calibraccedilatildeo (Abs660 nmvs nmol PO43-
) a partir dos valores obtidos
Para determinar a actividade Ca2+
-ATPaacutesica de S1 prepararam-se tubos de Eppendorf com 400
L de meio de reacccedilatildeo21
25 L CaCl2 ([CaCl2]stock= 100 mM) 25 L ATP ([ATP]stock= 100 mM)
com [S1]ensaios=01 mgmL Perfez-se se necessaacuterio com H2O MilliQ ateacute perfazer um volume de 1
mL Para determinar a actividade K+EDTA-ATPaacutesica utilizou-se uma concentraccedilatildeo idecircntica de
19
Soluccedilatildeo J 5 N H2SO4 25 molibdato de amoacutenio 20
Agente redutor 5 mg aacutecido 1-amino-2-naftol-4-sulfoacutenico 30 mg bissulfito de soacutedio 30 mg sulfito de soacutedio em 26
mL H2O 21
meio de reacccedilatildeo 125 mM Tris-HCl pH 80 0575 M KCl
46
proteiacutena nos ensaios (01 mgmL) e o mesmo meio de reacccedilatildeo Aos tubos de Eppendorf
adicionou-se ainda 123 L de KCl ([KCl]stock = 3M) 25 L de ATP 10 L de EDTA
([EDTA]stock= 100 mM )e o volume de H2O MilliQ necessaacuterio para perfazer 1 mL Em ambas as
situaccedilotildees o uacuteltimo reagente adicionado foi o ATP uma vez que se trata do substrato da reacccedilatildeo
enzimaacutetica
A cineacutetica decorreu a 25ordmC durante 5 minutos para a actividade Ca2+
-ATPaacutesica e durante 3
minutos para a actividade K+EDTA-ATPaacutesica Apoacutes o periacuteodo de incubaccedilatildeo transferiu-se 150
L de amostra para tubos de Eppendorf contendo igual volume de TCA 10 Colocou-se no gelo
e homogeneizou-se no voacutertex Seguidamente centrifugou-se a 1500 g durante 10 minutos e
recolheu-se 200 L de sobrenadante para novos tubos de Eppendorf Adicionou-se a estes 158
mL de H2O MilliQ e 200 L de soluccedilatildeo J Homogeneizou-se no voacutertex e adicionou-se 20 L de
reagente redutor repetindo-se a homogeneizaccedilatildeo Incubou-se durante 10 minutos e mediu-se a
Abs660 nm com o mesmo intervalo de tempo de leitura para cada amostra Por intrapolaccedilatildeo a partir
da recta de calibraccedilatildeo obtida anteriormente determinou-se a quantidade de Pi libertado em cada
ensaio A partir destes resultados determinaram-se as actividades Ca2+
-ATPaacutesica e K+EDTA-
ATPaacutesica do S1 em nmol PO43-
minmg proteiacutena e em s-1
342 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
Da actividade Mg2+
-ATPaacutesica de S1 resulta a formaccedilatildeo dos produtos da reacccedilatildeo de
hidroacutelise do ATP - ADP e Pi Na presenccedila de ADP fosfoenolpiruvato (PEP) e da enzima piruvato
cinase (PK) os produtos resultantes satildeo ATP e piruvato Por acccedilatildeo da enzima lactato
desidrogenase (LDH) este uacuteltimo eacute convertido em lactato ocorrendo simultaneamente a oxidaccedilatildeo
do NADH a NAD+ monitorizada atraveacutes da Abs340 nm Quando o S1 de miosina se encontra no
meio reaccional o ATP eacute hidrolisado formando-se novamente ADP e Pi o que permite a
formaccedilatildeo de um ciclo (fig 21) Se as enzimas se encontrarem em excesso a velocidade de
conversatildeo do NADH eacute limitada pela produccedilatildeo de ADP e Pi Como o ATP eacute regenerado em cada
ciclo a sua concentraccedilatildeo inicial eacute sempre mantida (Bagshaw 1982)
Figura 33ndash Representaccedilatildeo esquemaacutetica do meacutetodo de enzimas acopladas
47
O protocolo utilizado e seguidamente descrito foi adaptado de Cuenda et al (1990 ndash Ver Refordf) A
uma cuvette de 1 mL adicionou-se S1 para uma concentraccedilatildeo final de 005 mgmL 800 L de
meio de reacccedilatildeo22
10 L de PEP ([PEP]stock = 42 mM) 10 L de PK+LDH 11 ([PK+LDH]stock =
1 mgmL) e 15 L de NADH ([NADH]stock=25 mM) Iniciou-se a monitorizaccedilatildeo da reacccedilatildeo
atraveacutes da leitura de Abs340 nm apoacutes a adiccedilatildeo de 20 L de ATP ([ATP]stock=100 mM) A
actividade Mg2+
-ATPaacutesica eacute obtida atraveacutes do declive correspondente agrave oxidaccedilatildeo do NADH Para
a mediccedilatildeo da Mg2+
-ATPase de S1 estimulada por actina adicionou-se F-actina para uma
concentraccedilatildeo final de 2 microM sendo a actividade obtida tambeacutem atraveacutes do declive correspondente
agrave oxidaccedilatildeo do NADH apoacutes a adiccedilatildeo da proteiacutena
35 Interaccedilatildeo entre vanaacutedio e subfragmento-1 de miosina
351 Preparaccedilatildeo das soluccedilotildees de vanaacutedio
As soluccedilotildees de vanadato foram preparadas em aacutegua MilliQ a partir de metavanadato de amoacutenio
(NH4VO3) numa concentraccedilatildeo stock de 50 mM com agitaccedilatildeo magneacutetica e aquecimento (~30 ordmC)
Esta soluccedilatildeo com um valor compreendido entre 6 e 7 designa-se por ldquometavanadatordquo e eacute
constituiacuteda pelas esspeacutecies V1 V2 V4 e V5 (Ref) Acidificando a soluccedilatildeo de metavanadato por
adiccedilatildeo de HCl ateacute um valor de pH proacuteximo de 4 observa-se uma mudanccedila de cor de amarelo
claro para laranja indicativa da formaccedilatildeo da espeacutecie decameacuterica de vanadato De seguida e
imediatamente antes da realizaccedilatildeo dos ensaios descritos o valor do pH da soluccedilatildeo de
decavanadato foi aumentado para cerca de 75 por adiccedilatildeo de NaOH A soluccedilatildeo (2) designada
por ldquometavanadatordquo natildeo foi posteriormente alterada ateacute agrave sua adiccedilatildeo para a realizaccedilatildeo dos ensaios
As soluccedilotildees foram armazenadas a 4ordmC e mantidas em gelo aquando da sua utilizaccedilatildeo
A soluccedilatildeo de V(IV) foi preparada em condiccedilotildees anaeroacutebias por diluiccedilatildeo de sulfato de vanadil
(VOSO4) para uma concentraccedilatildeo final de 5 mM em H2O MilliQ saturada com N2 gasoso
352Anaacutelise por espectroscopia de UV-Vis
A concentraccedilatildeo da espeacutecie decameacuterica em soluccedilatildeo depende do periacuteodo de tempo apoacutes a diluiccedilatildeo
no meio de reacccedilatildeo apropriado (Ref) A velocidade de desoligomerizaccedilatildeo da espeacutecie decameacuterica
pode ser analisada por espectroscopia de absorccedilatildeo na regiatildeo UVVis uma vez que esta espeacutecie
apresenta duas bandas caracteriacutesticas na regiatildeo do visiacutevel (a 358 e 400 nm) que natildeo se verifica
em quaisquer outras espeacutecies oligomeacutericas As bandas supramencionada sapresentam-se bem
22
Meio de reacccedilatildeo 3125 mM Tris-HCl pH 71 375 mM MgCl2
48
resolvidas atraveacutes da obtenccedilatildeo da derivada de 2ordf ordem do espectro apresentando a forma de 2
vales com miacutenimos centrados nos comprimentos de onda supracitados com dependecircncia linear
relativamente agrave concentraccedilatildeo de decavanadato total (Ramos et al 2006)
Assim a estabilidade da espeacutecie decameacuterica apoacutes diluiccedilatildeo da soluccedilatildeo de decavanadato no meio de
reacccedilatildeo dos ensaios (tampatildeo D) para uma concentraccedilatildeo final de 1 mM em vanadato total foi
caracterizada atraveacutes da leitura da Abs400 em funccedilatildeo do tempo tendo-se determinado qual a
ordem de reacccedilatildeo e o tempo de meia-vida da mesma Com base em estudos anteriores acerca da
estabilidade do V10 na presenccedila de diferentes proteiacutenas (Ramos et al 2006) determinou-se
igualmente o tempo de meia vida apoacutes a adiccedilatildeo de 20 microM de S1 de miosina ao meio reaccional
353 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
Os espectros de RMN do isoacutetopo 51
V foram realizados num espectroacutemetro Bruker AM-400 MHz
equipado com uma sonda multinuclear inversa de 5 mm Em tubos de RMN de 5 mm de diacircmetro
preparou-se soluccedilotildees de 500 L de deca- ou metavanadato correspondentes a um intervalo de
concentraccedilotildees de 1 a 10 mM suplementadas com 10 D2O Os espectros foram adquiridos agrave
temperatura ambiente com uma largura espectral de 23 923 Hz um tempo de aquisiccedilatildeo de 0086
s tempo de relaxaccedilatildeo de 001 s e uma sequecircncia de pulsos a 90ordm Utilizou-se a transformada de
Fourier com alargamento das bandas de 20Hz para o processamento dos espectros Os desvios
quiacutemicos das ressonacircncias de 51
V foram obtidos em relaccedilatildeo agrave espeacutecie VOCl3cuja frequecircncia de
ressonacircncia eacute considerada o padratildeo sendo todas as outras referenciadas a partir desta em ppm
As aacutereas dos sinais do RMN correspondentes agraves espeacutecies oligomeacutericas presentes foram obtidas
por integraccedilatildeo dos sinais As concentraccedilotildees relativas das espeacutecies foram calculadas com recurso agrave
equaccedilatildeo
onde A[Vx] corresponde agrave aacuterea referente agrave ressonacircncia da espeacutecie Vx com nordm de agregaccedilatildeo x
A[V]total agrave soma total das aacutereas medidas e [V] a concentraccedilatildeo total de vanaacutedio no ensaio
A teacutecnica de 51
V-RMN foi anteriormente aplicada em estudos de ligaccedilatildeo com S1 de miosina
(Ref) tendo sido demostrado que entre as espeacutecies V1 V2 V4 e V5 a espeacutecie tetrameacuterica possui a
maior afinidade para a miosina (Cremo et al) O desaparecimento ou alargamento dos sinais pode
implicar a ligaccedilatildeo de um dos oxovanadatos laacutebeis agrave miosina sem indicaccedilatildeo de qual uma vez que
49
se processa um equiliacutebrio entre eles (Crans 1990) Assim e tendo em conta as indicaccedilotildees de que
a espeacutecie V10 eacute distinta das restantes no que diz respeito agrave magnitude da interacccedilatildeo com a miosina
e inibiccedilatildeo da actividade ATPaacutesica estimulada por actina (Aureliano 2000 Tiago et al 2002)
torna-se fundamental investigar a interacccedilatildeo das diferentes espeacutecies de vanadato com o S1 de
miosina atraveacutes da teacutecnica de 51
V-RMN
Para tal obtiveram-se espectros de 51
V-RMN resultantes da titulaccedilatildeo de uma soluccedilatildeo 5 mM de
decavanadato total com concentraccedilotildees crescentes de S1 de miosina (0 5 10 15 e 20 microM) em
tampatildeo D Realizou-se ainda ensaios na presenccedila de 2 mM de decavanadato e 2 mM de ATP na
presenccedila e na ausecircncia de S1 de miosina de modo a verificar o efeito que estes ligandos naturais
da enzima teria na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o subfragmento-1
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
Os grupos sulfidrilo (-SH) das cisteiacutenas satildeo resiacuteduos de aminoaacutecidos extremamente sensiacuteveis agrave
oxidaccedilatildeo resultando na sua oxidaccedilatildeo (Dalle-Donne et al 1999) Logo a quantificaccedilatildeo destes
resiacuteduos eacute bastante importante nos estudos de interacccedilatildeo com vanaacutedio jaacute que os resiacuteduos de Cys
com ecircnfase para o SH1 e o SH2 satildeo bastante importantes na monitorizaccedilatildeo das modificaccedilotildees
funcionais da regiatildeo da cabeccedila
O procedimento seguidamente apresentado foi originalmente descrito por Gutieacuterrez-Merino
(1987)) Consiste na reacccedilatildeo entre os grupos ndashSH natildeo oxidados das proteiacutenas e o reagente de
Ellman (DTNB -55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico)) - formando-se o aniatildeo CNT (aniatildeo 3-
carboxilato-4-nitrotiofenolato) numa estequiometria de 11 Este apresenta um maacuteximo de
absorvecircncia a 412 nm (Robyt 1990) Assim apoacutes a incubaccedilatildeo da proteiacutena na presenccedila do metal
a quantidade de cisteiacutenas reduzidas por proteiacutena determina-se atraveacutes dos valores de Abs412
Deste modo colocou-se num tubo de Eppendorf o volume apropriado de S1 dialisado para que a
concentraccedilatildeo no ensaio fosse de 1 mgmL Adicionou-se o volume apropriado de decavanadato
para que a sua concentraccedilatildeo fosse de 0 10 50 100 250 500 ou 1000 M Perfez-se com soluccedilatildeo
D ateacute um volume final de 1 mL e incubou-se a 25ordmC por 20 minutos Apoacutes este periacuteodo colocou-
se 780 L da mistura reaccional numa cuvette de quartzo e adicionou-se 220 L de soluccedilatildeo K23
23
Tampatildeo K Tampatildeo fosfatohidrogenofosfato de soacutedio 025 M pH 72
50
Adicionou-se 15 L de DTNB (em soluccedilatildeo K [DTNB]stock=100 mM) ao meio reaccional e
registou-se o valor de absorvecircncia medido equivalente agraves cisteiacutenas ldquoraacutepidasrdquo (SH1 e SH2
expostos ao solvente) Adicionou-se 001 g de SDS que provoca a desnaturaccedilatildeo da proteiacutena e
consequentemente a exposiccedilatildeo de todos os resiacuteduos de cisteiacutenas que se encontravam ateacute aiacute
inacessiacuteveis no interior da proteiacutena (cisteiacutenas totais) Estes resiacuteduos podem assim reagir com o
DTNB
Estimou-se o nuacutemero de resiacuteduos de cisteiacutenas em todas as amostras com recurso ao coeficiente de
extinccedilatildeo molar () do DTNB (ver ponto 3 dos Anexos) que foi determinado atraveacutes da titulaccedilatildeo
de L-Cisteiacutena com intervalo de concentraccedilotildees compreendidos entre 0 e 50 microM na presenccedila de
25microM de DTNB em tampatildeo D Os valores de absorvecircncia foram obtidos para um comprimento de
onda de 412 nm
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
3621 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
A teacutecnica mais eficiente de anaacutelise da interacccedilatildeo entre o iatildeo vanadilo e proteiacutenas como a
calmodulina e a transferrina entre outras permite a obtenccedilatildeo raacutepida de informaccedilatildeo no que toca agrave
conformaccedilatildeo e amplitude da referida interacccedilatildeo
Os espectros de Ressonacircncia Paramagneacutetica Electroacutenica (RPE) da espeacutecie paramagneacutetica V(IV)
foram realizados a 70K num espectroacutemetro Bruker EMX-300 X-band Todas as soluccedilotildees
utilizadas e amostras foram preparadas em anaerobiose numa cacircmara de luvas de modo a evitar a
reduccedilatildeo de V(IV) a V(V) Em tubos capilares com 02 mL de amostra manteve-se a concentraccedilatildeo
de S1 a 20 M e variou-se a concentraccedilatildeo de vanadilo (0 25 5 10 15 25 50 75 100 150
200 500 e 1000 M [VOSO4]stock=5 mM) em tampatildeo D Apoacutes obtenccedilatildeo dos espectros estes
foram tratados atraveacutes do software SimFonia e WinEPR tendo-se obtido um graacutefico com a
variaccedilatildeo da intensidade do sinal de RPE em funccedilatildeo da concentraccedilatildeo de sulfato de vanadilo
presente Realizaram-se ainda simulaccedilotildees de espectros obtidos com 20 microM de S1 e 1000 microM de
V(IV) com vista agrave obtenccedilatildeo dos ligandos provaacuteveis do vanadilo atraveacutes da anaacutelise da constante
de acoplamento hiperfina (A) e do factor g (g)
4 Conclusotildees gerais
Apesar de jaacute ter sido demonstrado que o vanaacutedio inibe a Na+K
+-ATPase da miosina a funccedilatildeo
deste elemento no tecido muacutesculo esqueleacutetico continua largamente incompreendida A enorme
complexidade quiacutemica deste metal especialmente no que concerne agrave oligomerizaccedilatildeo e reacccedilotildees
51
redox constitui um obstaacuteculo agrave compreensatildeo do comportamento do vanaacutedio nos organismos
vivos Assim a caracterizaccedilatildeo da quiacutemica deste elemento eacute de grande utilidade antes de iniciar o
estudo da sua interacccedilatildeo com o presente sistema
A anaacutelise por espectroscopias de 51
V-RMN e de UV-Vis de soluccedilotildees de vanadato diluiacutedas nos
meios de reacccedilatildeo utilizados no presente relatoacuterio (10 mM HEPES pH 70 25 mM KCl) mostrou
claramente que a espeacutecie decameacuterica (V10) possui um tempo de vida suficientemente longo para a
determinaccedilatildeo do seu efeito isolado ao contraacuterio do que se verifica com as espeacutecies mono- di-
tetra- e pentameacutericas (V1 V2 V4 e V5) Por outro lado a anaacutelise verificou ainda que apoacutes diluiccedilatildeo
das soluccedilotildees de decavanadato no meio de reacccedilatildeo coexistem as espeacutecies V10 e V1 pelo que os
efeitos putativos desta uacuteltima deveratildeo ser subtraiacutedos aquando da monitorizaccedilatildeo dos efeitos da
espeacutecie decameacuterica
O vanaacutedio (V) (vanadato) inibe as enzimas que catalizam reacccedilotildees de transferecircncia de grupos
fosfato como a ATPase da miosina [Goodno 1979] devido agrave sua analogia estrutural com o iatildeo
fosfato Na sua utilizaccedilatildeo como inibidor enzimaacutetico natildeo eacute tida em conta a contribuiccedilatildeo individual
das espeacutecies oligomeacutericas considerando-se normalmente a espeacutecie V1 como inibidora efectiva
[Goodno e Taylor 1982] No presente relatoacuterio verificou-se por espectroscopia de 51
V-RMN que
outros oligoacutemeros de vanadato interagem com o S1 de miosina tendo as espeacutecies V10 e V4 os
maiores niacuteveis de afinidade A sua interacccedilatildeo envolve forccedilas electrostaacuteticas e eacute afectada
negativamente por ligandos naturais de enzima como eacute o caso do ATP
A exposiccedilatildeo do S1 de miosina (005 mgmL) a decavanadato conduz agrave inibiccedilatildeo do capacidade da
proteiacutena de estimular a actividade Mg2+
-ATPaacutesica com um valor de IC50 de 266 plusmn 07 M
decavanadato sendo o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM
decavanadato (80) Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM decavanadato sendo
o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM decavanadato (60)
exercendo um efeito de impedimento agrave ligaccedilatildeo entre o vanaacutedio e o S1
A uma concentraccedilatildeo de 27 microM decavanadato verifica-se a oxidaccedilatildeo de um dos resiacuteduos de Cys
de S1 de miosina seguida pela oxidaccedilatildeo gradual de outro resiacuteduo de Cys ateacute agrave concentraccedilatildeo de
1000 microM Ambos os resiacuteduos oxidados natildeo se encontravam expostos ao solvente utilizado A
oxidaccedilatildeo dos resiacuteduos permite sugerir que esta modificaccedilatildeo oxidativa sofrida pelo S1 contribui
para a diminuiccedilatildeo da sua actividade de ATPase
52
Por espectroscopia de RPE revelaram-se os possiacuteveis ligandos equatoriais do vanaacutedio aquando da
sua interacccedilatildeo com o S1 de miosina como sendo duas moleacuteculas de aacutegua um ligando contendo o
grupo R-OH e outro contendo o grupo R=NR
Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados de
oxidaccedilatildeo (IV) e (V) no S1 de miosina para concentraccedilotildees facilmente alcanccedilaacuteveis no ambiente
terrestre
53
5 Estudos futuros
No sentido do melhor esclarecimento das alteraccedilotildees moleculares e estruturais que ocorrem no
subfragmento 1 quando exposta ao vanaacutedio que conduzem a uma diminuiccedilatildeo da sua funccedilatildeo
sugere-se
- Quantificaccedilatildeo das cisteiacutenas reduzidas de S1 na presenccedila de metavanadato
- Determinaccedilatildeo das actividades Ca2+
- e K+-ATPaacutesica do S1 de miosina na presenccedila de
decavanadato
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina estimulada por actina
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina na presenccedila de
metavanadato
- Realizaccedilatildeo de RMN 2D com variaccedilatildeo das concentraccedilotildees das soluccedilotildees de decavanadato e
metavanadato na presenccedila de S1
-Mutageacutenese dos resiacuteduos de Cys doS1 de miosina de modo a verificar quais os resiacuteduos que
sofrem oxidaccedilatildeo na presenccedila de decavanadato
- Determinaccedilatildeo por calorimetria (DSC Differential Scanning Calorimetry) da variaccedilatildeo da
entalpia (∆H) e da temperatura de desnaturaccedilatildeo do S1 de miosina na presenccedila de vanaacutedio uma
vez que estas propriedades caracteriacutesticas de cada proteiacutena satildeo alteradas apoacutes ocorrecircncia de
modificaccedilotildees estruturais
- Anaacutelise por SDS-PAGE de S1 de miosina apoacutes tratamento com vanaacutedio de modo a determinar
quais os polipeacuteptidos eventualmente formados apoacutes clivagem da proteiacutena
54
6 Referecircncias bibliograacuteficas
- Sellers J Myosins 2nd Edition Oxford University Press 1999
- L Stryer inBiochemistry (Ed WH Freeman 2005 pp
Refordfas Materiais e Meacutetodos
[A] S S Margossian SLowey Methods Enzymol1982 85 55-71
[B] JD Pardue JA Spudich Methods Enzymol 1982 85 164-181
[C] M Bradford Anal Biochem1976 72 248-254
[D] H H Taussky E Shorr J Biol Chem 1953 202 675-685
[E] C R Bagshaw in Outline Studies in Biology ndash Muscle Contraction Chapman and Hall Ed
1982
[F] A Cuenda et al Eur J Biochem1990 194 663-670
55
Anexos
1 Determinaccedilatildeo da recta de calibraccedilatildeo meacutetodo de Bradford
Figura1ndashRepresentaccedilatildeo graacutefica da recta de calibraccedilatildeo (Abs595 nm vs [BSA]) utilizada para o meacutetodo de Bradford
2 Determinaccedilatildeo da recta calibraccedilatildeo para a electroforese em condiccedilotildees desnaturantes
Figura 2 ndash Representaccedilatildeo graacutefica da recta de calibraccedilatildeo (log(Peso Molecular) vs Rf dos marcadores de peso
molecular) onde Rf indica determinada atraveacutes do padratildeo de peso molecular
(poccedilo (a))
3Caacutelculo do coeficiente de extinccedilatildeo molar () para a quantificaccedilatildeo de cisteiacutenas reduzidas
atraveacutes do meacutetodo de Ellman
Traccedilou-se a recta de Abs412 nm meacutedia vs [L-Cisteiacutena] de modo a determinar o valor do coeficiente
de extinccedilatildeo molar definido como o declive daquela recta Assim =8400 M-1
cm-1
como
podemos verificar na Fig 3
A595 nm = 00546[BSA] + 00209 Rsup2 = 099051
Ab
s 59
5 n
m
[BSA] (mgmL)
log PM = -1177 Rf + 2247 Rsup2 = 098957
log
PM
Mobilidade relativa (Rf)
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero

46
proteiacutena nos ensaios (01 mgmL) e o mesmo meio de reacccedilatildeo Aos tubos de Eppendorf
adicionou-se ainda 123 L de KCl ([KCl]stock = 3M) 25 L de ATP 10 L de EDTA
([EDTA]stock= 100 mM )e o volume de H2O MilliQ necessaacuterio para perfazer 1 mL Em ambas as
situaccedilotildees o uacuteltimo reagente adicionado foi o ATP uma vez que se trata do substrato da reacccedilatildeo
enzimaacutetica
A cineacutetica decorreu a 25ordmC durante 5 minutos para a actividade Ca2+
-ATPaacutesica e durante 3
minutos para a actividade K+EDTA-ATPaacutesica Apoacutes o periacuteodo de incubaccedilatildeo transferiu-se 150
L de amostra para tubos de Eppendorf contendo igual volume de TCA 10 Colocou-se no gelo
e homogeneizou-se no voacutertex Seguidamente centrifugou-se a 1500 g durante 10 minutos e
recolheu-se 200 L de sobrenadante para novos tubos de Eppendorf Adicionou-se a estes 158
mL de H2O MilliQ e 200 L de soluccedilatildeo J Homogeneizou-se no voacutertex e adicionou-se 20 L de
reagente redutor repetindo-se a homogeneizaccedilatildeo Incubou-se durante 10 minutos e mediu-se a
Abs660 nm com o mesmo intervalo de tempo de leitura para cada amostra Por intrapolaccedilatildeo a partir
da recta de calibraccedilatildeo obtida anteriormente determinou-se a quantidade de Pi libertado em cada
ensaio A partir destes resultados determinaram-se as actividades Ca2+
-ATPaacutesica e K+EDTA-
ATPaacutesica do S1 em nmol PO43-
minmg proteiacutena e em s-1
342 Mediccedilatildeo da actividade de Mg2+
-ATPase de S1 estimulada por actina pelo meacutetodo de
enzimas acopladas
Da actividade Mg2+
-ATPaacutesica de S1 resulta a formaccedilatildeo dos produtos da reacccedilatildeo de
hidroacutelise do ATP - ADP e Pi Na presenccedila de ADP fosfoenolpiruvato (PEP) e da enzima piruvato
cinase (PK) os produtos resultantes satildeo ATP e piruvato Por acccedilatildeo da enzima lactato
desidrogenase (LDH) este uacuteltimo eacute convertido em lactato ocorrendo simultaneamente a oxidaccedilatildeo
do NADH a NAD+ monitorizada atraveacutes da Abs340 nm Quando o S1 de miosina se encontra no
meio reaccional o ATP eacute hidrolisado formando-se novamente ADP e Pi o que permite a
formaccedilatildeo de um ciclo (fig 21) Se as enzimas se encontrarem em excesso a velocidade de
conversatildeo do NADH eacute limitada pela produccedilatildeo de ADP e Pi Como o ATP eacute regenerado em cada
ciclo a sua concentraccedilatildeo inicial eacute sempre mantida (Bagshaw 1982)
Figura 33ndash Representaccedilatildeo esquemaacutetica do meacutetodo de enzimas acopladas
47
O protocolo utilizado e seguidamente descrito foi adaptado de Cuenda et al (1990 ndash Ver Refordf) A
uma cuvette de 1 mL adicionou-se S1 para uma concentraccedilatildeo final de 005 mgmL 800 L de
meio de reacccedilatildeo22
10 L de PEP ([PEP]stock = 42 mM) 10 L de PK+LDH 11 ([PK+LDH]stock =
1 mgmL) e 15 L de NADH ([NADH]stock=25 mM) Iniciou-se a monitorizaccedilatildeo da reacccedilatildeo
atraveacutes da leitura de Abs340 nm apoacutes a adiccedilatildeo de 20 L de ATP ([ATP]stock=100 mM) A
actividade Mg2+
-ATPaacutesica eacute obtida atraveacutes do declive correspondente agrave oxidaccedilatildeo do NADH Para
a mediccedilatildeo da Mg2+
-ATPase de S1 estimulada por actina adicionou-se F-actina para uma
concentraccedilatildeo final de 2 microM sendo a actividade obtida tambeacutem atraveacutes do declive correspondente
agrave oxidaccedilatildeo do NADH apoacutes a adiccedilatildeo da proteiacutena
35 Interaccedilatildeo entre vanaacutedio e subfragmento-1 de miosina
351 Preparaccedilatildeo das soluccedilotildees de vanaacutedio
As soluccedilotildees de vanadato foram preparadas em aacutegua MilliQ a partir de metavanadato de amoacutenio
(NH4VO3) numa concentraccedilatildeo stock de 50 mM com agitaccedilatildeo magneacutetica e aquecimento (~30 ordmC)
Esta soluccedilatildeo com um valor compreendido entre 6 e 7 designa-se por ldquometavanadatordquo e eacute
constituiacuteda pelas esspeacutecies V1 V2 V4 e V5 (Ref) Acidificando a soluccedilatildeo de metavanadato por
adiccedilatildeo de HCl ateacute um valor de pH proacuteximo de 4 observa-se uma mudanccedila de cor de amarelo
claro para laranja indicativa da formaccedilatildeo da espeacutecie decameacuterica de vanadato De seguida e
imediatamente antes da realizaccedilatildeo dos ensaios descritos o valor do pH da soluccedilatildeo de
decavanadato foi aumentado para cerca de 75 por adiccedilatildeo de NaOH A soluccedilatildeo (2) designada
por ldquometavanadatordquo natildeo foi posteriormente alterada ateacute agrave sua adiccedilatildeo para a realizaccedilatildeo dos ensaios
As soluccedilotildees foram armazenadas a 4ordmC e mantidas em gelo aquando da sua utilizaccedilatildeo
A soluccedilatildeo de V(IV) foi preparada em condiccedilotildees anaeroacutebias por diluiccedilatildeo de sulfato de vanadil
(VOSO4) para uma concentraccedilatildeo final de 5 mM em H2O MilliQ saturada com N2 gasoso
352Anaacutelise por espectroscopia de UV-Vis
A concentraccedilatildeo da espeacutecie decameacuterica em soluccedilatildeo depende do periacuteodo de tempo apoacutes a diluiccedilatildeo
no meio de reacccedilatildeo apropriado (Ref) A velocidade de desoligomerizaccedilatildeo da espeacutecie decameacuterica
pode ser analisada por espectroscopia de absorccedilatildeo na regiatildeo UVVis uma vez que esta espeacutecie
apresenta duas bandas caracteriacutesticas na regiatildeo do visiacutevel (a 358 e 400 nm) que natildeo se verifica
em quaisquer outras espeacutecies oligomeacutericas As bandas supramencionada sapresentam-se bem
22
Meio de reacccedilatildeo 3125 mM Tris-HCl pH 71 375 mM MgCl2
48
resolvidas atraveacutes da obtenccedilatildeo da derivada de 2ordf ordem do espectro apresentando a forma de 2
vales com miacutenimos centrados nos comprimentos de onda supracitados com dependecircncia linear
relativamente agrave concentraccedilatildeo de decavanadato total (Ramos et al 2006)
Assim a estabilidade da espeacutecie decameacuterica apoacutes diluiccedilatildeo da soluccedilatildeo de decavanadato no meio de
reacccedilatildeo dos ensaios (tampatildeo D) para uma concentraccedilatildeo final de 1 mM em vanadato total foi
caracterizada atraveacutes da leitura da Abs400 em funccedilatildeo do tempo tendo-se determinado qual a
ordem de reacccedilatildeo e o tempo de meia-vida da mesma Com base em estudos anteriores acerca da
estabilidade do V10 na presenccedila de diferentes proteiacutenas (Ramos et al 2006) determinou-se
igualmente o tempo de meia vida apoacutes a adiccedilatildeo de 20 microM de S1 de miosina ao meio reaccional
353 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
Os espectros de RMN do isoacutetopo 51
V foram realizados num espectroacutemetro Bruker AM-400 MHz
equipado com uma sonda multinuclear inversa de 5 mm Em tubos de RMN de 5 mm de diacircmetro
preparou-se soluccedilotildees de 500 L de deca- ou metavanadato correspondentes a um intervalo de
concentraccedilotildees de 1 a 10 mM suplementadas com 10 D2O Os espectros foram adquiridos agrave
temperatura ambiente com uma largura espectral de 23 923 Hz um tempo de aquisiccedilatildeo de 0086
s tempo de relaxaccedilatildeo de 001 s e uma sequecircncia de pulsos a 90ordm Utilizou-se a transformada de
Fourier com alargamento das bandas de 20Hz para o processamento dos espectros Os desvios
quiacutemicos das ressonacircncias de 51
V foram obtidos em relaccedilatildeo agrave espeacutecie VOCl3cuja frequecircncia de
ressonacircncia eacute considerada o padratildeo sendo todas as outras referenciadas a partir desta em ppm
As aacutereas dos sinais do RMN correspondentes agraves espeacutecies oligomeacutericas presentes foram obtidas
por integraccedilatildeo dos sinais As concentraccedilotildees relativas das espeacutecies foram calculadas com recurso agrave
equaccedilatildeo
onde A[Vx] corresponde agrave aacuterea referente agrave ressonacircncia da espeacutecie Vx com nordm de agregaccedilatildeo x
A[V]total agrave soma total das aacutereas medidas e [V] a concentraccedilatildeo total de vanaacutedio no ensaio
A teacutecnica de 51
V-RMN foi anteriormente aplicada em estudos de ligaccedilatildeo com S1 de miosina
(Ref) tendo sido demostrado que entre as espeacutecies V1 V2 V4 e V5 a espeacutecie tetrameacuterica possui a
maior afinidade para a miosina (Cremo et al) O desaparecimento ou alargamento dos sinais pode
implicar a ligaccedilatildeo de um dos oxovanadatos laacutebeis agrave miosina sem indicaccedilatildeo de qual uma vez que
49
se processa um equiliacutebrio entre eles (Crans 1990) Assim e tendo em conta as indicaccedilotildees de que
a espeacutecie V10 eacute distinta das restantes no que diz respeito agrave magnitude da interacccedilatildeo com a miosina
e inibiccedilatildeo da actividade ATPaacutesica estimulada por actina (Aureliano 2000 Tiago et al 2002)
torna-se fundamental investigar a interacccedilatildeo das diferentes espeacutecies de vanadato com o S1 de
miosina atraveacutes da teacutecnica de 51
V-RMN
Para tal obtiveram-se espectros de 51
V-RMN resultantes da titulaccedilatildeo de uma soluccedilatildeo 5 mM de
decavanadato total com concentraccedilotildees crescentes de S1 de miosina (0 5 10 15 e 20 microM) em
tampatildeo D Realizou-se ainda ensaios na presenccedila de 2 mM de decavanadato e 2 mM de ATP na
presenccedila e na ausecircncia de S1 de miosina de modo a verificar o efeito que estes ligandos naturais
da enzima teria na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o subfragmento-1
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
Os grupos sulfidrilo (-SH) das cisteiacutenas satildeo resiacuteduos de aminoaacutecidos extremamente sensiacuteveis agrave
oxidaccedilatildeo resultando na sua oxidaccedilatildeo (Dalle-Donne et al 1999) Logo a quantificaccedilatildeo destes
resiacuteduos eacute bastante importante nos estudos de interacccedilatildeo com vanaacutedio jaacute que os resiacuteduos de Cys
com ecircnfase para o SH1 e o SH2 satildeo bastante importantes na monitorizaccedilatildeo das modificaccedilotildees
funcionais da regiatildeo da cabeccedila
O procedimento seguidamente apresentado foi originalmente descrito por Gutieacuterrez-Merino
(1987)) Consiste na reacccedilatildeo entre os grupos ndashSH natildeo oxidados das proteiacutenas e o reagente de
Ellman (DTNB -55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico)) - formando-se o aniatildeo CNT (aniatildeo 3-
carboxilato-4-nitrotiofenolato) numa estequiometria de 11 Este apresenta um maacuteximo de
absorvecircncia a 412 nm (Robyt 1990) Assim apoacutes a incubaccedilatildeo da proteiacutena na presenccedila do metal
a quantidade de cisteiacutenas reduzidas por proteiacutena determina-se atraveacutes dos valores de Abs412
Deste modo colocou-se num tubo de Eppendorf o volume apropriado de S1 dialisado para que a
concentraccedilatildeo no ensaio fosse de 1 mgmL Adicionou-se o volume apropriado de decavanadato
para que a sua concentraccedilatildeo fosse de 0 10 50 100 250 500 ou 1000 M Perfez-se com soluccedilatildeo
D ateacute um volume final de 1 mL e incubou-se a 25ordmC por 20 minutos Apoacutes este periacuteodo colocou-
se 780 L da mistura reaccional numa cuvette de quartzo e adicionou-se 220 L de soluccedilatildeo K23
23
Tampatildeo K Tampatildeo fosfatohidrogenofosfato de soacutedio 025 M pH 72
50
Adicionou-se 15 L de DTNB (em soluccedilatildeo K [DTNB]stock=100 mM) ao meio reaccional e
registou-se o valor de absorvecircncia medido equivalente agraves cisteiacutenas ldquoraacutepidasrdquo (SH1 e SH2
expostos ao solvente) Adicionou-se 001 g de SDS que provoca a desnaturaccedilatildeo da proteiacutena e
consequentemente a exposiccedilatildeo de todos os resiacuteduos de cisteiacutenas que se encontravam ateacute aiacute
inacessiacuteveis no interior da proteiacutena (cisteiacutenas totais) Estes resiacuteduos podem assim reagir com o
DTNB
Estimou-se o nuacutemero de resiacuteduos de cisteiacutenas em todas as amostras com recurso ao coeficiente de
extinccedilatildeo molar () do DTNB (ver ponto 3 dos Anexos) que foi determinado atraveacutes da titulaccedilatildeo
de L-Cisteiacutena com intervalo de concentraccedilotildees compreendidos entre 0 e 50 microM na presenccedila de
25microM de DTNB em tampatildeo D Os valores de absorvecircncia foram obtidos para um comprimento de
onda de 412 nm
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
3621 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
A teacutecnica mais eficiente de anaacutelise da interacccedilatildeo entre o iatildeo vanadilo e proteiacutenas como a
calmodulina e a transferrina entre outras permite a obtenccedilatildeo raacutepida de informaccedilatildeo no que toca agrave
conformaccedilatildeo e amplitude da referida interacccedilatildeo
Os espectros de Ressonacircncia Paramagneacutetica Electroacutenica (RPE) da espeacutecie paramagneacutetica V(IV)
foram realizados a 70K num espectroacutemetro Bruker EMX-300 X-band Todas as soluccedilotildees
utilizadas e amostras foram preparadas em anaerobiose numa cacircmara de luvas de modo a evitar a
reduccedilatildeo de V(IV) a V(V) Em tubos capilares com 02 mL de amostra manteve-se a concentraccedilatildeo
de S1 a 20 M e variou-se a concentraccedilatildeo de vanadilo (0 25 5 10 15 25 50 75 100 150
200 500 e 1000 M [VOSO4]stock=5 mM) em tampatildeo D Apoacutes obtenccedilatildeo dos espectros estes
foram tratados atraveacutes do software SimFonia e WinEPR tendo-se obtido um graacutefico com a
variaccedilatildeo da intensidade do sinal de RPE em funccedilatildeo da concentraccedilatildeo de sulfato de vanadilo
presente Realizaram-se ainda simulaccedilotildees de espectros obtidos com 20 microM de S1 e 1000 microM de
V(IV) com vista agrave obtenccedilatildeo dos ligandos provaacuteveis do vanadilo atraveacutes da anaacutelise da constante
de acoplamento hiperfina (A) e do factor g (g)
4 Conclusotildees gerais
Apesar de jaacute ter sido demonstrado que o vanaacutedio inibe a Na+K
+-ATPase da miosina a funccedilatildeo
deste elemento no tecido muacutesculo esqueleacutetico continua largamente incompreendida A enorme
complexidade quiacutemica deste metal especialmente no que concerne agrave oligomerizaccedilatildeo e reacccedilotildees
51
redox constitui um obstaacuteculo agrave compreensatildeo do comportamento do vanaacutedio nos organismos
vivos Assim a caracterizaccedilatildeo da quiacutemica deste elemento eacute de grande utilidade antes de iniciar o
estudo da sua interacccedilatildeo com o presente sistema
A anaacutelise por espectroscopias de 51
V-RMN e de UV-Vis de soluccedilotildees de vanadato diluiacutedas nos
meios de reacccedilatildeo utilizados no presente relatoacuterio (10 mM HEPES pH 70 25 mM KCl) mostrou
claramente que a espeacutecie decameacuterica (V10) possui um tempo de vida suficientemente longo para a
determinaccedilatildeo do seu efeito isolado ao contraacuterio do que se verifica com as espeacutecies mono- di-
tetra- e pentameacutericas (V1 V2 V4 e V5) Por outro lado a anaacutelise verificou ainda que apoacutes diluiccedilatildeo
das soluccedilotildees de decavanadato no meio de reacccedilatildeo coexistem as espeacutecies V10 e V1 pelo que os
efeitos putativos desta uacuteltima deveratildeo ser subtraiacutedos aquando da monitorizaccedilatildeo dos efeitos da
espeacutecie decameacuterica
O vanaacutedio (V) (vanadato) inibe as enzimas que catalizam reacccedilotildees de transferecircncia de grupos
fosfato como a ATPase da miosina [Goodno 1979] devido agrave sua analogia estrutural com o iatildeo
fosfato Na sua utilizaccedilatildeo como inibidor enzimaacutetico natildeo eacute tida em conta a contribuiccedilatildeo individual
das espeacutecies oligomeacutericas considerando-se normalmente a espeacutecie V1 como inibidora efectiva
[Goodno e Taylor 1982] No presente relatoacuterio verificou-se por espectroscopia de 51
V-RMN que
outros oligoacutemeros de vanadato interagem com o S1 de miosina tendo as espeacutecies V10 e V4 os
maiores niacuteveis de afinidade A sua interacccedilatildeo envolve forccedilas electrostaacuteticas e eacute afectada
negativamente por ligandos naturais de enzima como eacute o caso do ATP
A exposiccedilatildeo do S1 de miosina (005 mgmL) a decavanadato conduz agrave inibiccedilatildeo do capacidade da
proteiacutena de estimular a actividade Mg2+
-ATPaacutesica com um valor de IC50 de 266 plusmn 07 M
decavanadato sendo o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM
decavanadato (80) Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM decavanadato sendo
o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM decavanadato (60)
exercendo um efeito de impedimento agrave ligaccedilatildeo entre o vanaacutedio e o S1
A uma concentraccedilatildeo de 27 microM decavanadato verifica-se a oxidaccedilatildeo de um dos resiacuteduos de Cys
de S1 de miosina seguida pela oxidaccedilatildeo gradual de outro resiacuteduo de Cys ateacute agrave concentraccedilatildeo de
1000 microM Ambos os resiacuteduos oxidados natildeo se encontravam expostos ao solvente utilizado A
oxidaccedilatildeo dos resiacuteduos permite sugerir que esta modificaccedilatildeo oxidativa sofrida pelo S1 contribui
para a diminuiccedilatildeo da sua actividade de ATPase
52
Por espectroscopia de RPE revelaram-se os possiacuteveis ligandos equatoriais do vanaacutedio aquando da
sua interacccedilatildeo com o S1 de miosina como sendo duas moleacuteculas de aacutegua um ligando contendo o
grupo R-OH e outro contendo o grupo R=NR
Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados de
oxidaccedilatildeo (IV) e (V) no S1 de miosina para concentraccedilotildees facilmente alcanccedilaacuteveis no ambiente
terrestre
53
5 Estudos futuros
No sentido do melhor esclarecimento das alteraccedilotildees moleculares e estruturais que ocorrem no
subfragmento 1 quando exposta ao vanaacutedio que conduzem a uma diminuiccedilatildeo da sua funccedilatildeo
sugere-se
- Quantificaccedilatildeo das cisteiacutenas reduzidas de S1 na presenccedila de metavanadato
- Determinaccedilatildeo das actividades Ca2+
- e K+-ATPaacutesica do S1 de miosina na presenccedila de
decavanadato
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina estimulada por actina
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina na presenccedila de
metavanadato
- Realizaccedilatildeo de RMN 2D com variaccedilatildeo das concentraccedilotildees das soluccedilotildees de decavanadato e
metavanadato na presenccedila de S1
-Mutageacutenese dos resiacuteduos de Cys doS1 de miosina de modo a verificar quais os resiacuteduos que
sofrem oxidaccedilatildeo na presenccedila de decavanadato
- Determinaccedilatildeo por calorimetria (DSC Differential Scanning Calorimetry) da variaccedilatildeo da
entalpia (∆H) e da temperatura de desnaturaccedilatildeo do S1 de miosina na presenccedila de vanaacutedio uma
vez que estas propriedades caracteriacutesticas de cada proteiacutena satildeo alteradas apoacutes ocorrecircncia de
modificaccedilotildees estruturais
- Anaacutelise por SDS-PAGE de S1 de miosina apoacutes tratamento com vanaacutedio de modo a determinar
quais os polipeacuteptidos eventualmente formados apoacutes clivagem da proteiacutena
54
6 Referecircncias bibliograacuteficas
- Sellers J Myosins 2nd Edition Oxford University Press 1999
- L Stryer inBiochemistry (Ed WH Freeman 2005 pp
Refordfas Materiais e Meacutetodos
[A] S S Margossian SLowey Methods Enzymol1982 85 55-71
[B] JD Pardue JA Spudich Methods Enzymol 1982 85 164-181
[C] M Bradford Anal Biochem1976 72 248-254
[D] H H Taussky E Shorr J Biol Chem 1953 202 675-685
[E] C R Bagshaw in Outline Studies in Biology ndash Muscle Contraction Chapman and Hall Ed
1982
[F] A Cuenda et al Eur J Biochem1990 194 663-670
55
Anexos
1 Determinaccedilatildeo da recta de calibraccedilatildeo meacutetodo de Bradford
Figura1ndashRepresentaccedilatildeo graacutefica da recta de calibraccedilatildeo (Abs595 nm vs [BSA]) utilizada para o meacutetodo de Bradford
2 Determinaccedilatildeo da recta calibraccedilatildeo para a electroforese em condiccedilotildees desnaturantes
Figura 2 ndash Representaccedilatildeo graacutefica da recta de calibraccedilatildeo (log(Peso Molecular) vs Rf dos marcadores de peso
molecular) onde Rf indica determinada atraveacutes do padratildeo de peso molecular
(poccedilo (a))
3Caacutelculo do coeficiente de extinccedilatildeo molar () para a quantificaccedilatildeo de cisteiacutenas reduzidas
atraveacutes do meacutetodo de Ellman
Traccedilou-se a recta de Abs412 nm meacutedia vs [L-Cisteiacutena] de modo a determinar o valor do coeficiente
de extinccedilatildeo molar definido como o declive daquela recta Assim =8400 M-1
cm-1
como
podemos verificar na Fig 3
A595 nm = 00546[BSA] + 00209 Rsup2 = 099051
Ab
s 59
5 n
m
[BSA] (mgmL)
log PM = -1177 Rf + 2247 Rsup2 = 098957
log
PM
Mobilidade relativa (Rf)
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero

47
O protocolo utilizado e seguidamente descrito foi adaptado de Cuenda et al (1990 ndash Ver Refordf) A
uma cuvette de 1 mL adicionou-se S1 para uma concentraccedilatildeo final de 005 mgmL 800 L de
meio de reacccedilatildeo22
10 L de PEP ([PEP]stock = 42 mM) 10 L de PK+LDH 11 ([PK+LDH]stock =
1 mgmL) e 15 L de NADH ([NADH]stock=25 mM) Iniciou-se a monitorizaccedilatildeo da reacccedilatildeo
atraveacutes da leitura de Abs340 nm apoacutes a adiccedilatildeo de 20 L de ATP ([ATP]stock=100 mM) A
actividade Mg2+
-ATPaacutesica eacute obtida atraveacutes do declive correspondente agrave oxidaccedilatildeo do NADH Para
a mediccedilatildeo da Mg2+
-ATPase de S1 estimulada por actina adicionou-se F-actina para uma
concentraccedilatildeo final de 2 microM sendo a actividade obtida tambeacutem atraveacutes do declive correspondente
agrave oxidaccedilatildeo do NADH apoacutes a adiccedilatildeo da proteiacutena
35 Interaccedilatildeo entre vanaacutedio e subfragmento-1 de miosina
351 Preparaccedilatildeo das soluccedilotildees de vanaacutedio
As soluccedilotildees de vanadato foram preparadas em aacutegua MilliQ a partir de metavanadato de amoacutenio
(NH4VO3) numa concentraccedilatildeo stock de 50 mM com agitaccedilatildeo magneacutetica e aquecimento (~30 ordmC)
Esta soluccedilatildeo com um valor compreendido entre 6 e 7 designa-se por ldquometavanadatordquo e eacute
constituiacuteda pelas esspeacutecies V1 V2 V4 e V5 (Ref) Acidificando a soluccedilatildeo de metavanadato por
adiccedilatildeo de HCl ateacute um valor de pH proacuteximo de 4 observa-se uma mudanccedila de cor de amarelo
claro para laranja indicativa da formaccedilatildeo da espeacutecie decameacuterica de vanadato De seguida e
imediatamente antes da realizaccedilatildeo dos ensaios descritos o valor do pH da soluccedilatildeo de
decavanadato foi aumentado para cerca de 75 por adiccedilatildeo de NaOH A soluccedilatildeo (2) designada
por ldquometavanadatordquo natildeo foi posteriormente alterada ateacute agrave sua adiccedilatildeo para a realizaccedilatildeo dos ensaios
As soluccedilotildees foram armazenadas a 4ordmC e mantidas em gelo aquando da sua utilizaccedilatildeo
A soluccedilatildeo de V(IV) foi preparada em condiccedilotildees anaeroacutebias por diluiccedilatildeo de sulfato de vanadil
(VOSO4) para uma concentraccedilatildeo final de 5 mM em H2O MilliQ saturada com N2 gasoso
352Anaacutelise por espectroscopia de UV-Vis
A concentraccedilatildeo da espeacutecie decameacuterica em soluccedilatildeo depende do periacuteodo de tempo apoacutes a diluiccedilatildeo
no meio de reacccedilatildeo apropriado (Ref) A velocidade de desoligomerizaccedilatildeo da espeacutecie decameacuterica
pode ser analisada por espectroscopia de absorccedilatildeo na regiatildeo UVVis uma vez que esta espeacutecie
apresenta duas bandas caracteriacutesticas na regiatildeo do visiacutevel (a 358 e 400 nm) que natildeo se verifica
em quaisquer outras espeacutecies oligomeacutericas As bandas supramencionada sapresentam-se bem
22
Meio de reacccedilatildeo 3125 mM Tris-HCl pH 71 375 mM MgCl2
48
resolvidas atraveacutes da obtenccedilatildeo da derivada de 2ordf ordem do espectro apresentando a forma de 2
vales com miacutenimos centrados nos comprimentos de onda supracitados com dependecircncia linear
relativamente agrave concentraccedilatildeo de decavanadato total (Ramos et al 2006)
Assim a estabilidade da espeacutecie decameacuterica apoacutes diluiccedilatildeo da soluccedilatildeo de decavanadato no meio de
reacccedilatildeo dos ensaios (tampatildeo D) para uma concentraccedilatildeo final de 1 mM em vanadato total foi
caracterizada atraveacutes da leitura da Abs400 em funccedilatildeo do tempo tendo-se determinado qual a
ordem de reacccedilatildeo e o tempo de meia-vida da mesma Com base em estudos anteriores acerca da
estabilidade do V10 na presenccedila de diferentes proteiacutenas (Ramos et al 2006) determinou-se
igualmente o tempo de meia vida apoacutes a adiccedilatildeo de 20 microM de S1 de miosina ao meio reaccional
353 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
Os espectros de RMN do isoacutetopo 51
V foram realizados num espectroacutemetro Bruker AM-400 MHz
equipado com uma sonda multinuclear inversa de 5 mm Em tubos de RMN de 5 mm de diacircmetro
preparou-se soluccedilotildees de 500 L de deca- ou metavanadato correspondentes a um intervalo de
concentraccedilotildees de 1 a 10 mM suplementadas com 10 D2O Os espectros foram adquiridos agrave
temperatura ambiente com uma largura espectral de 23 923 Hz um tempo de aquisiccedilatildeo de 0086
s tempo de relaxaccedilatildeo de 001 s e uma sequecircncia de pulsos a 90ordm Utilizou-se a transformada de
Fourier com alargamento das bandas de 20Hz para o processamento dos espectros Os desvios
quiacutemicos das ressonacircncias de 51
V foram obtidos em relaccedilatildeo agrave espeacutecie VOCl3cuja frequecircncia de
ressonacircncia eacute considerada o padratildeo sendo todas as outras referenciadas a partir desta em ppm
As aacutereas dos sinais do RMN correspondentes agraves espeacutecies oligomeacutericas presentes foram obtidas
por integraccedilatildeo dos sinais As concentraccedilotildees relativas das espeacutecies foram calculadas com recurso agrave
equaccedilatildeo
onde A[Vx] corresponde agrave aacuterea referente agrave ressonacircncia da espeacutecie Vx com nordm de agregaccedilatildeo x
A[V]total agrave soma total das aacutereas medidas e [V] a concentraccedilatildeo total de vanaacutedio no ensaio
A teacutecnica de 51
V-RMN foi anteriormente aplicada em estudos de ligaccedilatildeo com S1 de miosina
(Ref) tendo sido demostrado que entre as espeacutecies V1 V2 V4 e V5 a espeacutecie tetrameacuterica possui a
maior afinidade para a miosina (Cremo et al) O desaparecimento ou alargamento dos sinais pode
implicar a ligaccedilatildeo de um dos oxovanadatos laacutebeis agrave miosina sem indicaccedilatildeo de qual uma vez que
49
se processa um equiliacutebrio entre eles (Crans 1990) Assim e tendo em conta as indicaccedilotildees de que
a espeacutecie V10 eacute distinta das restantes no que diz respeito agrave magnitude da interacccedilatildeo com a miosina
e inibiccedilatildeo da actividade ATPaacutesica estimulada por actina (Aureliano 2000 Tiago et al 2002)
torna-se fundamental investigar a interacccedilatildeo das diferentes espeacutecies de vanadato com o S1 de
miosina atraveacutes da teacutecnica de 51
V-RMN
Para tal obtiveram-se espectros de 51
V-RMN resultantes da titulaccedilatildeo de uma soluccedilatildeo 5 mM de
decavanadato total com concentraccedilotildees crescentes de S1 de miosina (0 5 10 15 e 20 microM) em
tampatildeo D Realizou-se ainda ensaios na presenccedila de 2 mM de decavanadato e 2 mM de ATP na
presenccedila e na ausecircncia de S1 de miosina de modo a verificar o efeito que estes ligandos naturais
da enzima teria na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o subfragmento-1
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
Os grupos sulfidrilo (-SH) das cisteiacutenas satildeo resiacuteduos de aminoaacutecidos extremamente sensiacuteveis agrave
oxidaccedilatildeo resultando na sua oxidaccedilatildeo (Dalle-Donne et al 1999) Logo a quantificaccedilatildeo destes
resiacuteduos eacute bastante importante nos estudos de interacccedilatildeo com vanaacutedio jaacute que os resiacuteduos de Cys
com ecircnfase para o SH1 e o SH2 satildeo bastante importantes na monitorizaccedilatildeo das modificaccedilotildees
funcionais da regiatildeo da cabeccedila
O procedimento seguidamente apresentado foi originalmente descrito por Gutieacuterrez-Merino
(1987)) Consiste na reacccedilatildeo entre os grupos ndashSH natildeo oxidados das proteiacutenas e o reagente de
Ellman (DTNB -55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico)) - formando-se o aniatildeo CNT (aniatildeo 3-
carboxilato-4-nitrotiofenolato) numa estequiometria de 11 Este apresenta um maacuteximo de
absorvecircncia a 412 nm (Robyt 1990) Assim apoacutes a incubaccedilatildeo da proteiacutena na presenccedila do metal
a quantidade de cisteiacutenas reduzidas por proteiacutena determina-se atraveacutes dos valores de Abs412
Deste modo colocou-se num tubo de Eppendorf o volume apropriado de S1 dialisado para que a
concentraccedilatildeo no ensaio fosse de 1 mgmL Adicionou-se o volume apropriado de decavanadato
para que a sua concentraccedilatildeo fosse de 0 10 50 100 250 500 ou 1000 M Perfez-se com soluccedilatildeo
D ateacute um volume final de 1 mL e incubou-se a 25ordmC por 20 minutos Apoacutes este periacuteodo colocou-
se 780 L da mistura reaccional numa cuvette de quartzo e adicionou-se 220 L de soluccedilatildeo K23
23
Tampatildeo K Tampatildeo fosfatohidrogenofosfato de soacutedio 025 M pH 72
50
Adicionou-se 15 L de DTNB (em soluccedilatildeo K [DTNB]stock=100 mM) ao meio reaccional e
registou-se o valor de absorvecircncia medido equivalente agraves cisteiacutenas ldquoraacutepidasrdquo (SH1 e SH2
expostos ao solvente) Adicionou-se 001 g de SDS que provoca a desnaturaccedilatildeo da proteiacutena e
consequentemente a exposiccedilatildeo de todos os resiacuteduos de cisteiacutenas que se encontravam ateacute aiacute
inacessiacuteveis no interior da proteiacutena (cisteiacutenas totais) Estes resiacuteduos podem assim reagir com o
DTNB
Estimou-se o nuacutemero de resiacuteduos de cisteiacutenas em todas as amostras com recurso ao coeficiente de
extinccedilatildeo molar () do DTNB (ver ponto 3 dos Anexos) que foi determinado atraveacutes da titulaccedilatildeo
de L-Cisteiacutena com intervalo de concentraccedilotildees compreendidos entre 0 e 50 microM na presenccedila de
25microM de DTNB em tampatildeo D Os valores de absorvecircncia foram obtidos para um comprimento de
onda de 412 nm
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
3621 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
A teacutecnica mais eficiente de anaacutelise da interacccedilatildeo entre o iatildeo vanadilo e proteiacutenas como a
calmodulina e a transferrina entre outras permite a obtenccedilatildeo raacutepida de informaccedilatildeo no que toca agrave
conformaccedilatildeo e amplitude da referida interacccedilatildeo
Os espectros de Ressonacircncia Paramagneacutetica Electroacutenica (RPE) da espeacutecie paramagneacutetica V(IV)
foram realizados a 70K num espectroacutemetro Bruker EMX-300 X-band Todas as soluccedilotildees
utilizadas e amostras foram preparadas em anaerobiose numa cacircmara de luvas de modo a evitar a
reduccedilatildeo de V(IV) a V(V) Em tubos capilares com 02 mL de amostra manteve-se a concentraccedilatildeo
de S1 a 20 M e variou-se a concentraccedilatildeo de vanadilo (0 25 5 10 15 25 50 75 100 150
200 500 e 1000 M [VOSO4]stock=5 mM) em tampatildeo D Apoacutes obtenccedilatildeo dos espectros estes
foram tratados atraveacutes do software SimFonia e WinEPR tendo-se obtido um graacutefico com a
variaccedilatildeo da intensidade do sinal de RPE em funccedilatildeo da concentraccedilatildeo de sulfato de vanadilo
presente Realizaram-se ainda simulaccedilotildees de espectros obtidos com 20 microM de S1 e 1000 microM de
V(IV) com vista agrave obtenccedilatildeo dos ligandos provaacuteveis do vanadilo atraveacutes da anaacutelise da constante
de acoplamento hiperfina (A) e do factor g (g)
4 Conclusotildees gerais
Apesar de jaacute ter sido demonstrado que o vanaacutedio inibe a Na+K
+-ATPase da miosina a funccedilatildeo
deste elemento no tecido muacutesculo esqueleacutetico continua largamente incompreendida A enorme
complexidade quiacutemica deste metal especialmente no que concerne agrave oligomerizaccedilatildeo e reacccedilotildees
51
redox constitui um obstaacuteculo agrave compreensatildeo do comportamento do vanaacutedio nos organismos
vivos Assim a caracterizaccedilatildeo da quiacutemica deste elemento eacute de grande utilidade antes de iniciar o
estudo da sua interacccedilatildeo com o presente sistema
A anaacutelise por espectroscopias de 51
V-RMN e de UV-Vis de soluccedilotildees de vanadato diluiacutedas nos
meios de reacccedilatildeo utilizados no presente relatoacuterio (10 mM HEPES pH 70 25 mM KCl) mostrou
claramente que a espeacutecie decameacuterica (V10) possui um tempo de vida suficientemente longo para a
determinaccedilatildeo do seu efeito isolado ao contraacuterio do que se verifica com as espeacutecies mono- di-
tetra- e pentameacutericas (V1 V2 V4 e V5) Por outro lado a anaacutelise verificou ainda que apoacutes diluiccedilatildeo
das soluccedilotildees de decavanadato no meio de reacccedilatildeo coexistem as espeacutecies V10 e V1 pelo que os
efeitos putativos desta uacuteltima deveratildeo ser subtraiacutedos aquando da monitorizaccedilatildeo dos efeitos da
espeacutecie decameacuterica
O vanaacutedio (V) (vanadato) inibe as enzimas que catalizam reacccedilotildees de transferecircncia de grupos
fosfato como a ATPase da miosina [Goodno 1979] devido agrave sua analogia estrutural com o iatildeo
fosfato Na sua utilizaccedilatildeo como inibidor enzimaacutetico natildeo eacute tida em conta a contribuiccedilatildeo individual
das espeacutecies oligomeacutericas considerando-se normalmente a espeacutecie V1 como inibidora efectiva
[Goodno e Taylor 1982] No presente relatoacuterio verificou-se por espectroscopia de 51
V-RMN que
outros oligoacutemeros de vanadato interagem com o S1 de miosina tendo as espeacutecies V10 e V4 os
maiores niacuteveis de afinidade A sua interacccedilatildeo envolve forccedilas electrostaacuteticas e eacute afectada
negativamente por ligandos naturais de enzima como eacute o caso do ATP
A exposiccedilatildeo do S1 de miosina (005 mgmL) a decavanadato conduz agrave inibiccedilatildeo do capacidade da
proteiacutena de estimular a actividade Mg2+
-ATPaacutesica com um valor de IC50 de 266 plusmn 07 M
decavanadato sendo o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM
decavanadato (80) Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM decavanadato sendo
o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM decavanadato (60)
exercendo um efeito de impedimento agrave ligaccedilatildeo entre o vanaacutedio e o S1
A uma concentraccedilatildeo de 27 microM decavanadato verifica-se a oxidaccedilatildeo de um dos resiacuteduos de Cys
de S1 de miosina seguida pela oxidaccedilatildeo gradual de outro resiacuteduo de Cys ateacute agrave concentraccedilatildeo de
1000 microM Ambos os resiacuteduos oxidados natildeo se encontravam expostos ao solvente utilizado A
oxidaccedilatildeo dos resiacuteduos permite sugerir que esta modificaccedilatildeo oxidativa sofrida pelo S1 contribui
para a diminuiccedilatildeo da sua actividade de ATPase
52
Por espectroscopia de RPE revelaram-se os possiacuteveis ligandos equatoriais do vanaacutedio aquando da
sua interacccedilatildeo com o S1 de miosina como sendo duas moleacuteculas de aacutegua um ligando contendo o
grupo R-OH e outro contendo o grupo R=NR
Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados de
oxidaccedilatildeo (IV) e (V) no S1 de miosina para concentraccedilotildees facilmente alcanccedilaacuteveis no ambiente
terrestre
53
5 Estudos futuros
No sentido do melhor esclarecimento das alteraccedilotildees moleculares e estruturais que ocorrem no
subfragmento 1 quando exposta ao vanaacutedio que conduzem a uma diminuiccedilatildeo da sua funccedilatildeo
sugere-se
- Quantificaccedilatildeo das cisteiacutenas reduzidas de S1 na presenccedila de metavanadato
- Determinaccedilatildeo das actividades Ca2+
- e K+-ATPaacutesica do S1 de miosina na presenccedila de
decavanadato
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina estimulada por actina
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina na presenccedila de
metavanadato
- Realizaccedilatildeo de RMN 2D com variaccedilatildeo das concentraccedilotildees das soluccedilotildees de decavanadato e
metavanadato na presenccedila de S1
-Mutageacutenese dos resiacuteduos de Cys doS1 de miosina de modo a verificar quais os resiacuteduos que
sofrem oxidaccedilatildeo na presenccedila de decavanadato
- Determinaccedilatildeo por calorimetria (DSC Differential Scanning Calorimetry) da variaccedilatildeo da
entalpia (∆H) e da temperatura de desnaturaccedilatildeo do S1 de miosina na presenccedila de vanaacutedio uma
vez que estas propriedades caracteriacutesticas de cada proteiacutena satildeo alteradas apoacutes ocorrecircncia de
modificaccedilotildees estruturais
- Anaacutelise por SDS-PAGE de S1 de miosina apoacutes tratamento com vanaacutedio de modo a determinar
quais os polipeacuteptidos eventualmente formados apoacutes clivagem da proteiacutena
54
6 Referecircncias bibliograacuteficas
- Sellers J Myosins 2nd Edition Oxford University Press 1999
- L Stryer inBiochemistry (Ed WH Freeman 2005 pp
Refordfas Materiais e Meacutetodos
[A] S S Margossian SLowey Methods Enzymol1982 85 55-71
[B] JD Pardue JA Spudich Methods Enzymol 1982 85 164-181
[C] M Bradford Anal Biochem1976 72 248-254
[D] H H Taussky E Shorr J Biol Chem 1953 202 675-685
[E] C R Bagshaw in Outline Studies in Biology ndash Muscle Contraction Chapman and Hall Ed
1982
[F] A Cuenda et al Eur J Biochem1990 194 663-670
55
Anexos
1 Determinaccedilatildeo da recta de calibraccedilatildeo meacutetodo de Bradford
Figura1ndashRepresentaccedilatildeo graacutefica da recta de calibraccedilatildeo (Abs595 nm vs [BSA]) utilizada para o meacutetodo de Bradford
2 Determinaccedilatildeo da recta calibraccedilatildeo para a electroforese em condiccedilotildees desnaturantes
Figura 2 ndash Representaccedilatildeo graacutefica da recta de calibraccedilatildeo (log(Peso Molecular) vs Rf dos marcadores de peso
molecular) onde Rf indica determinada atraveacutes do padratildeo de peso molecular
(poccedilo (a))
3Caacutelculo do coeficiente de extinccedilatildeo molar () para a quantificaccedilatildeo de cisteiacutenas reduzidas
atraveacutes do meacutetodo de Ellman
Traccedilou-se a recta de Abs412 nm meacutedia vs [L-Cisteiacutena] de modo a determinar o valor do coeficiente
de extinccedilatildeo molar definido como o declive daquela recta Assim =8400 M-1
cm-1
como
podemos verificar na Fig 3
A595 nm = 00546[BSA] + 00209 Rsup2 = 099051
Ab
s 59
5 n
m
[BSA] (mgmL)
log PM = -1177 Rf + 2247 Rsup2 = 098957
log
PM
Mobilidade relativa (Rf)
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero

48
resolvidas atraveacutes da obtenccedilatildeo da derivada de 2ordf ordem do espectro apresentando a forma de 2
vales com miacutenimos centrados nos comprimentos de onda supracitados com dependecircncia linear
relativamente agrave concentraccedilatildeo de decavanadato total (Ramos et al 2006)
Assim a estabilidade da espeacutecie decameacuterica apoacutes diluiccedilatildeo da soluccedilatildeo de decavanadato no meio de
reacccedilatildeo dos ensaios (tampatildeo D) para uma concentraccedilatildeo final de 1 mM em vanadato total foi
caracterizada atraveacutes da leitura da Abs400 em funccedilatildeo do tempo tendo-se determinado qual a
ordem de reacccedilatildeo e o tempo de meia-vida da mesma Com base em estudos anteriores acerca da
estabilidade do V10 na presenccedila de diferentes proteiacutenas (Ramos et al 2006) determinou-se
igualmente o tempo de meia vida apoacutes a adiccedilatildeo de 20 microM de S1 de miosina ao meio reaccional
353 Anaacutelise por espectroscopia de Ressonacircncia Magneacutetica Nuclear
Os espectros de RMN do isoacutetopo 51
V foram realizados num espectroacutemetro Bruker AM-400 MHz
equipado com uma sonda multinuclear inversa de 5 mm Em tubos de RMN de 5 mm de diacircmetro
preparou-se soluccedilotildees de 500 L de deca- ou metavanadato correspondentes a um intervalo de
concentraccedilotildees de 1 a 10 mM suplementadas com 10 D2O Os espectros foram adquiridos agrave
temperatura ambiente com uma largura espectral de 23 923 Hz um tempo de aquisiccedilatildeo de 0086
s tempo de relaxaccedilatildeo de 001 s e uma sequecircncia de pulsos a 90ordm Utilizou-se a transformada de
Fourier com alargamento das bandas de 20Hz para o processamento dos espectros Os desvios
quiacutemicos das ressonacircncias de 51
V foram obtidos em relaccedilatildeo agrave espeacutecie VOCl3cuja frequecircncia de
ressonacircncia eacute considerada o padratildeo sendo todas as outras referenciadas a partir desta em ppm
As aacutereas dos sinais do RMN correspondentes agraves espeacutecies oligomeacutericas presentes foram obtidas
por integraccedilatildeo dos sinais As concentraccedilotildees relativas das espeacutecies foram calculadas com recurso agrave
equaccedilatildeo
onde A[Vx] corresponde agrave aacuterea referente agrave ressonacircncia da espeacutecie Vx com nordm de agregaccedilatildeo x
A[V]total agrave soma total das aacutereas medidas e [V] a concentraccedilatildeo total de vanaacutedio no ensaio
A teacutecnica de 51
V-RMN foi anteriormente aplicada em estudos de ligaccedilatildeo com S1 de miosina
(Ref) tendo sido demostrado que entre as espeacutecies V1 V2 V4 e V5 a espeacutecie tetrameacuterica possui a
maior afinidade para a miosina (Cremo et al) O desaparecimento ou alargamento dos sinais pode
implicar a ligaccedilatildeo de um dos oxovanadatos laacutebeis agrave miosina sem indicaccedilatildeo de qual uma vez que
49
se processa um equiliacutebrio entre eles (Crans 1990) Assim e tendo em conta as indicaccedilotildees de que
a espeacutecie V10 eacute distinta das restantes no que diz respeito agrave magnitude da interacccedilatildeo com a miosina
e inibiccedilatildeo da actividade ATPaacutesica estimulada por actina (Aureliano 2000 Tiago et al 2002)
torna-se fundamental investigar a interacccedilatildeo das diferentes espeacutecies de vanadato com o S1 de
miosina atraveacutes da teacutecnica de 51
V-RMN
Para tal obtiveram-se espectros de 51
V-RMN resultantes da titulaccedilatildeo de uma soluccedilatildeo 5 mM de
decavanadato total com concentraccedilotildees crescentes de S1 de miosina (0 5 10 15 e 20 microM) em
tampatildeo D Realizou-se ainda ensaios na presenccedila de 2 mM de decavanadato e 2 mM de ATP na
presenccedila e na ausecircncia de S1 de miosina de modo a verificar o efeito que estes ligandos naturais
da enzima teria na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o subfragmento-1
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
Os grupos sulfidrilo (-SH) das cisteiacutenas satildeo resiacuteduos de aminoaacutecidos extremamente sensiacuteveis agrave
oxidaccedilatildeo resultando na sua oxidaccedilatildeo (Dalle-Donne et al 1999) Logo a quantificaccedilatildeo destes
resiacuteduos eacute bastante importante nos estudos de interacccedilatildeo com vanaacutedio jaacute que os resiacuteduos de Cys
com ecircnfase para o SH1 e o SH2 satildeo bastante importantes na monitorizaccedilatildeo das modificaccedilotildees
funcionais da regiatildeo da cabeccedila
O procedimento seguidamente apresentado foi originalmente descrito por Gutieacuterrez-Merino
(1987)) Consiste na reacccedilatildeo entre os grupos ndashSH natildeo oxidados das proteiacutenas e o reagente de
Ellman (DTNB -55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico)) - formando-se o aniatildeo CNT (aniatildeo 3-
carboxilato-4-nitrotiofenolato) numa estequiometria de 11 Este apresenta um maacuteximo de
absorvecircncia a 412 nm (Robyt 1990) Assim apoacutes a incubaccedilatildeo da proteiacutena na presenccedila do metal
a quantidade de cisteiacutenas reduzidas por proteiacutena determina-se atraveacutes dos valores de Abs412
Deste modo colocou-se num tubo de Eppendorf o volume apropriado de S1 dialisado para que a
concentraccedilatildeo no ensaio fosse de 1 mgmL Adicionou-se o volume apropriado de decavanadato
para que a sua concentraccedilatildeo fosse de 0 10 50 100 250 500 ou 1000 M Perfez-se com soluccedilatildeo
D ateacute um volume final de 1 mL e incubou-se a 25ordmC por 20 minutos Apoacutes este periacuteodo colocou-
se 780 L da mistura reaccional numa cuvette de quartzo e adicionou-se 220 L de soluccedilatildeo K23
23
Tampatildeo K Tampatildeo fosfatohidrogenofosfato de soacutedio 025 M pH 72
50
Adicionou-se 15 L de DTNB (em soluccedilatildeo K [DTNB]stock=100 mM) ao meio reaccional e
registou-se o valor de absorvecircncia medido equivalente agraves cisteiacutenas ldquoraacutepidasrdquo (SH1 e SH2
expostos ao solvente) Adicionou-se 001 g de SDS que provoca a desnaturaccedilatildeo da proteiacutena e
consequentemente a exposiccedilatildeo de todos os resiacuteduos de cisteiacutenas que se encontravam ateacute aiacute
inacessiacuteveis no interior da proteiacutena (cisteiacutenas totais) Estes resiacuteduos podem assim reagir com o
DTNB
Estimou-se o nuacutemero de resiacuteduos de cisteiacutenas em todas as amostras com recurso ao coeficiente de
extinccedilatildeo molar () do DTNB (ver ponto 3 dos Anexos) que foi determinado atraveacutes da titulaccedilatildeo
de L-Cisteiacutena com intervalo de concentraccedilotildees compreendidos entre 0 e 50 microM na presenccedila de
25microM de DTNB em tampatildeo D Os valores de absorvecircncia foram obtidos para um comprimento de
onda de 412 nm
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
3621 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
A teacutecnica mais eficiente de anaacutelise da interacccedilatildeo entre o iatildeo vanadilo e proteiacutenas como a
calmodulina e a transferrina entre outras permite a obtenccedilatildeo raacutepida de informaccedilatildeo no que toca agrave
conformaccedilatildeo e amplitude da referida interacccedilatildeo
Os espectros de Ressonacircncia Paramagneacutetica Electroacutenica (RPE) da espeacutecie paramagneacutetica V(IV)
foram realizados a 70K num espectroacutemetro Bruker EMX-300 X-band Todas as soluccedilotildees
utilizadas e amostras foram preparadas em anaerobiose numa cacircmara de luvas de modo a evitar a
reduccedilatildeo de V(IV) a V(V) Em tubos capilares com 02 mL de amostra manteve-se a concentraccedilatildeo
de S1 a 20 M e variou-se a concentraccedilatildeo de vanadilo (0 25 5 10 15 25 50 75 100 150
200 500 e 1000 M [VOSO4]stock=5 mM) em tampatildeo D Apoacutes obtenccedilatildeo dos espectros estes
foram tratados atraveacutes do software SimFonia e WinEPR tendo-se obtido um graacutefico com a
variaccedilatildeo da intensidade do sinal de RPE em funccedilatildeo da concentraccedilatildeo de sulfato de vanadilo
presente Realizaram-se ainda simulaccedilotildees de espectros obtidos com 20 microM de S1 e 1000 microM de
V(IV) com vista agrave obtenccedilatildeo dos ligandos provaacuteveis do vanadilo atraveacutes da anaacutelise da constante
de acoplamento hiperfina (A) e do factor g (g)
4 Conclusotildees gerais
Apesar de jaacute ter sido demonstrado que o vanaacutedio inibe a Na+K
+-ATPase da miosina a funccedilatildeo
deste elemento no tecido muacutesculo esqueleacutetico continua largamente incompreendida A enorme
complexidade quiacutemica deste metal especialmente no que concerne agrave oligomerizaccedilatildeo e reacccedilotildees
51
redox constitui um obstaacuteculo agrave compreensatildeo do comportamento do vanaacutedio nos organismos
vivos Assim a caracterizaccedilatildeo da quiacutemica deste elemento eacute de grande utilidade antes de iniciar o
estudo da sua interacccedilatildeo com o presente sistema
A anaacutelise por espectroscopias de 51
V-RMN e de UV-Vis de soluccedilotildees de vanadato diluiacutedas nos
meios de reacccedilatildeo utilizados no presente relatoacuterio (10 mM HEPES pH 70 25 mM KCl) mostrou
claramente que a espeacutecie decameacuterica (V10) possui um tempo de vida suficientemente longo para a
determinaccedilatildeo do seu efeito isolado ao contraacuterio do que se verifica com as espeacutecies mono- di-
tetra- e pentameacutericas (V1 V2 V4 e V5) Por outro lado a anaacutelise verificou ainda que apoacutes diluiccedilatildeo
das soluccedilotildees de decavanadato no meio de reacccedilatildeo coexistem as espeacutecies V10 e V1 pelo que os
efeitos putativos desta uacuteltima deveratildeo ser subtraiacutedos aquando da monitorizaccedilatildeo dos efeitos da
espeacutecie decameacuterica
O vanaacutedio (V) (vanadato) inibe as enzimas que catalizam reacccedilotildees de transferecircncia de grupos
fosfato como a ATPase da miosina [Goodno 1979] devido agrave sua analogia estrutural com o iatildeo
fosfato Na sua utilizaccedilatildeo como inibidor enzimaacutetico natildeo eacute tida em conta a contribuiccedilatildeo individual
das espeacutecies oligomeacutericas considerando-se normalmente a espeacutecie V1 como inibidora efectiva
[Goodno e Taylor 1982] No presente relatoacuterio verificou-se por espectroscopia de 51
V-RMN que
outros oligoacutemeros de vanadato interagem com o S1 de miosina tendo as espeacutecies V10 e V4 os
maiores niacuteveis de afinidade A sua interacccedilatildeo envolve forccedilas electrostaacuteticas e eacute afectada
negativamente por ligandos naturais de enzima como eacute o caso do ATP
A exposiccedilatildeo do S1 de miosina (005 mgmL) a decavanadato conduz agrave inibiccedilatildeo do capacidade da
proteiacutena de estimular a actividade Mg2+
-ATPaacutesica com um valor de IC50 de 266 plusmn 07 M
decavanadato sendo o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM
decavanadato (80) Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM decavanadato sendo
o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM decavanadato (60)
exercendo um efeito de impedimento agrave ligaccedilatildeo entre o vanaacutedio e o S1
A uma concentraccedilatildeo de 27 microM decavanadato verifica-se a oxidaccedilatildeo de um dos resiacuteduos de Cys
de S1 de miosina seguida pela oxidaccedilatildeo gradual de outro resiacuteduo de Cys ateacute agrave concentraccedilatildeo de
1000 microM Ambos os resiacuteduos oxidados natildeo se encontravam expostos ao solvente utilizado A
oxidaccedilatildeo dos resiacuteduos permite sugerir que esta modificaccedilatildeo oxidativa sofrida pelo S1 contribui
para a diminuiccedilatildeo da sua actividade de ATPase
52
Por espectroscopia de RPE revelaram-se os possiacuteveis ligandos equatoriais do vanaacutedio aquando da
sua interacccedilatildeo com o S1 de miosina como sendo duas moleacuteculas de aacutegua um ligando contendo o
grupo R-OH e outro contendo o grupo R=NR
Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados de
oxidaccedilatildeo (IV) e (V) no S1 de miosina para concentraccedilotildees facilmente alcanccedilaacuteveis no ambiente
terrestre
53
5 Estudos futuros
No sentido do melhor esclarecimento das alteraccedilotildees moleculares e estruturais que ocorrem no
subfragmento 1 quando exposta ao vanaacutedio que conduzem a uma diminuiccedilatildeo da sua funccedilatildeo
sugere-se
- Quantificaccedilatildeo das cisteiacutenas reduzidas de S1 na presenccedila de metavanadato
- Determinaccedilatildeo das actividades Ca2+
- e K+-ATPaacutesica do S1 de miosina na presenccedila de
decavanadato
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina estimulada por actina
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina na presenccedila de
metavanadato
- Realizaccedilatildeo de RMN 2D com variaccedilatildeo das concentraccedilotildees das soluccedilotildees de decavanadato e
metavanadato na presenccedila de S1
-Mutageacutenese dos resiacuteduos de Cys doS1 de miosina de modo a verificar quais os resiacuteduos que
sofrem oxidaccedilatildeo na presenccedila de decavanadato
- Determinaccedilatildeo por calorimetria (DSC Differential Scanning Calorimetry) da variaccedilatildeo da
entalpia (∆H) e da temperatura de desnaturaccedilatildeo do S1 de miosina na presenccedila de vanaacutedio uma
vez que estas propriedades caracteriacutesticas de cada proteiacutena satildeo alteradas apoacutes ocorrecircncia de
modificaccedilotildees estruturais
- Anaacutelise por SDS-PAGE de S1 de miosina apoacutes tratamento com vanaacutedio de modo a determinar
quais os polipeacuteptidos eventualmente formados apoacutes clivagem da proteiacutena
54
6 Referecircncias bibliograacuteficas
- Sellers J Myosins 2nd Edition Oxford University Press 1999
- L Stryer inBiochemistry (Ed WH Freeman 2005 pp
Refordfas Materiais e Meacutetodos
[A] S S Margossian SLowey Methods Enzymol1982 85 55-71
[B] JD Pardue JA Spudich Methods Enzymol 1982 85 164-181
[C] M Bradford Anal Biochem1976 72 248-254
[D] H H Taussky E Shorr J Biol Chem 1953 202 675-685
[E] C R Bagshaw in Outline Studies in Biology ndash Muscle Contraction Chapman and Hall Ed
1982
[F] A Cuenda et al Eur J Biochem1990 194 663-670
55
Anexos
1 Determinaccedilatildeo da recta de calibraccedilatildeo meacutetodo de Bradford
Figura1ndashRepresentaccedilatildeo graacutefica da recta de calibraccedilatildeo (Abs595 nm vs [BSA]) utilizada para o meacutetodo de Bradford
2 Determinaccedilatildeo da recta calibraccedilatildeo para a electroforese em condiccedilotildees desnaturantes
Figura 2 ndash Representaccedilatildeo graacutefica da recta de calibraccedilatildeo (log(Peso Molecular) vs Rf dos marcadores de peso
molecular) onde Rf indica determinada atraveacutes do padratildeo de peso molecular
(poccedilo (a))
3Caacutelculo do coeficiente de extinccedilatildeo molar () para a quantificaccedilatildeo de cisteiacutenas reduzidas
atraveacutes do meacutetodo de Ellman
Traccedilou-se a recta de Abs412 nm meacutedia vs [L-Cisteiacutena] de modo a determinar o valor do coeficiente
de extinccedilatildeo molar definido como o declive daquela recta Assim =8400 M-1
cm-1
como
podemos verificar na Fig 3
A595 nm = 00546[BSA] + 00209 Rsup2 = 099051
Ab
s 59
5 n
m
[BSA] (mgmL)
log PM = -1177 Rf + 2247 Rsup2 = 098957
log
PM
Mobilidade relativa (Rf)
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero

49
se processa um equiliacutebrio entre eles (Crans 1990) Assim e tendo em conta as indicaccedilotildees de que
a espeacutecie V10 eacute distinta das restantes no que diz respeito agrave magnitude da interacccedilatildeo com a miosina
e inibiccedilatildeo da actividade ATPaacutesica estimulada por actina (Aureliano 2000 Tiago et al 2002)
torna-se fundamental investigar a interacccedilatildeo das diferentes espeacutecies de vanadato com o S1 de
miosina atraveacutes da teacutecnica de 51
V-RMN
Para tal obtiveram-se espectros de 51
V-RMN resultantes da titulaccedilatildeo de uma soluccedilatildeo 5 mM de
decavanadato total com concentraccedilotildees crescentes de S1 de miosina (0 5 10 15 e 20 microM) em
tampatildeo D Realizou-se ainda ensaios na presenccedila de 2 mM de decavanadato e 2 mM de ATP na
presenccedila e na ausecircncia de S1 de miosina de modo a verificar o efeito que estes ligandos naturais
da enzima teria na interacccedilatildeo das vaacuterias espeacutecies de vanadato com o subfragmento-1
36 Interacccedilatildeo de S1 com vanaacutedio
361 Quantificaccedilatildeo de cisteiacutenas reduzidas
Os grupos sulfidrilo (-SH) das cisteiacutenas satildeo resiacuteduos de aminoaacutecidos extremamente sensiacuteveis agrave
oxidaccedilatildeo resultando na sua oxidaccedilatildeo (Dalle-Donne et al 1999) Logo a quantificaccedilatildeo destes
resiacuteduos eacute bastante importante nos estudos de interacccedilatildeo com vanaacutedio jaacute que os resiacuteduos de Cys
com ecircnfase para o SH1 e o SH2 satildeo bastante importantes na monitorizaccedilatildeo das modificaccedilotildees
funcionais da regiatildeo da cabeccedila
O procedimento seguidamente apresentado foi originalmente descrito por Gutieacuterrez-Merino
(1987)) Consiste na reacccedilatildeo entre os grupos ndashSH natildeo oxidados das proteiacutenas e o reagente de
Ellman (DTNB -55rsquo-ditiobis(2-aacutecido nitrobenzoacuteico)) - formando-se o aniatildeo CNT (aniatildeo 3-
carboxilato-4-nitrotiofenolato) numa estequiometria de 11 Este apresenta um maacuteximo de
absorvecircncia a 412 nm (Robyt 1990) Assim apoacutes a incubaccedilatildeo da proteiacutena na presenccedila do metal
a quantidade de cisteiacutenas reduzidas por proteiacutena determina-se atraveacutes dos valores de Abs412
Deste modo colocou-se num tubo de Eppendorf o volume apropriado de S1 dialisado para que a
concentraccedilatildeo no ensaio fosse de 1 mgmL Adicionou-se o volume apropriado de decavanadato
para que a sua concentraccedilatildeo fosse de 0 10 50 100 250 500 ou 1000 M Perfez-se com soluccedilatildeo
D ateacute um volume final de 1 mL e incubou-se a 25ordmC por 20 minutos Apoacutes este periacuteodo colocou-
se 780 L da mistura reaccional numa cuvette de quartzo e adicionou-se 220 L de soluccedilatildeo K23
23
Tampatildeo K Tampatildeo fosfatohidrogenofosfato de soacutedio 025 M pH 72
50
Adicionou-se 15 L de DTNB (em soluccedilatildeo K [DTNB]stock=100 mM) ao meio reaccional e
registou-se o valor de absorvecircncia medido equivalente agraves cisteiacutenas ldquoraacutepidasrdquo (SH1 e SH2
expostos ao solvente) Adicionou-se 001 g de SDS que provoca a desnaturaccedilatildeo da proteiacutena e
consequentemente a exposiccedilatildeo de todos os resiacuteduos de cisteiacutenas que se encontravam ateacute aiacute
inacessiacuteveis no interior da proteiacutena (cisteiacutenas totais) Estes resiacuteduos podem assim reagir com o
DTNB
Estimou-se o nuacutemero de resiacuteduos de cisteiacutenas em todas as amostras com recurso ao coeficiente de
extinccedilatildeo molar () do DTNB (ver ponto 3 dos Anexos) que foi determinado atraveacutes da titulaccedilatildeo
de L-Cisteiacutena com intervalo de concentraccedilotildees compreendidos entre 0 e 50 microM na presenccedila de
25microM de DTNB em tampatildeo D Os valores de absorvecircncia foram obtidos para um comprimento de
onda de 412 nm
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
3621 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
A teacutecnica mais eficiente de anaacutelise da interacccedilatildeo entre o iatildeo vanadilo e proteiacutenas como a
calmodulina e a transferrina entre outras permite a obtenccedilatildeo raacutepida de informaccedilatildeo no que toca agrave
conformaccedilatildeo e amplitude da referida interacccedilatildeo
Os espectros de Ressonacircncia Paramagneacutetica Electroacutenica (RPE) da espeacutecie paramagneacutetica V(IV)
foram realizados a 70K num espectroacutemetro Bruker EMX-300 X-band Todas as soluccedilotildees
utilizadas e amostras foram preparadas em anaerobiose numa cacircmara de luvas de modo a evitar a
reduccedilatildeo de V(IV) a V(V) Em tubos capilares com 02 mL de amostra manteve-se a concentraccedilatildeo
de S1 a 20 M e variou-se a concentraccedilatildeo de vanadilo (0 25 5 10 15 25 50 75 100 150
200 500 e 1000 M [VOSO4]stock=5 mM) em tampatildeo D Apoacutes obtenccedilatildeo dos espectros estes
foram tratados atraveacutes do software SimFonia e WinEPR tendo-se obtido um graacutefico com a
variaccedilatildeo da intensidade do sinal de RPE em funccedilatildeo da concentraccedilatildeo de sulfato de vanadilo
presente Realizaram-se ainda simulaccedilotildees de espectros obtidos com 20 microM de S1 e 1000 microM de
V(IV) com vista agrave obtenccedilatildeo dos ligandos provaacuteveis do vanadilo atraveacutes da anaacutelise da constante
de acoplamento hiperfina (A) e do factor g (g)
4 Conclusotildees gerais
Apesar de jaacute ter sido demonstrado que o vanaacutedio inibe a Na+K
+-ATPase da miosina a funccedilatildeo
deste elemento no tecido muacutesculo esqueleacutetico continua largamente incompreendida A enorme
complexidade quiacutemica deste metal especialmente no que concerne agrave oligomerizaccedilatildeo e reacccedilotildees
51
redox constitui um obstaacuteculo agrave compreensatildeo do comportamento do vanaacutedio nos organismos
vivos Assim a caracterizaccedilatildeo da quiacutemica deste elemento eacute de grande utilidade antes de iniciar o
estudo da sua interacccedilatildeo com o presente sistema
A anaacutelise por espectroscopias de 51
V-RMN e de UV-Vis de soluccedilotildees de vanadato diluiacutedas nos
meios de reacccedilatildeo utilizados no presente relatoacuterio (10 mM HEPES pH 70 25 mM KCl) mostrou
claramente que a espeacutecie decameacuterica (V10) possui um tempo de vida suficientemente longo para a
determinaccedilatildeo do seu efeito isolado ao contraacuterio do que se verifica com as espeacutecies mono- di-
tetra- e pentameacutericas (V1 V2 V4 e V5) Por outro lado a anaacutelise verificou ainda que apoacutes diluiccedilatildeo
das soluccedilotildees de decavanadato no meio de reacccedilatildeo coexistem as espeacutecies V10 e V1 pelo que os
efeitos putativos desta uacuteltima deveratildeo ser subtraiacutedos aquando da monitorizaccedilatildeo dos efeitos da
espeacutecie decameacuterica
O vanaacutedio (V) (vanadato) inibe as enzimas que catalizam reacccedilotildees de transferecircncia de grupos
fosfato como a ATPase da miosina [Goodno 1979] devido agrave sua analogia estrutural com o iatildeo
fosfato Na sua utilizaccedilatildeo como inibidor enzimaacutetico natildeo eacute tida em conta a contribuiccedilatildeo individual
das espeacutecies oligomeacutericas considerando-se normalmente a espeacutecie V1 como inibidora efectiva
[Goodno e Taylor 1982] No presente relatoacuterio verificou-se por espectroscopia de 51
V-RMN que
outros oligoacutemeros de vanadato interagem com o S1 de miosina tendo as espeacutecies V10 e V4 os
maiores niacuteveis de afinidade A sua interacccedilatildeo envolve forccedilas electrostaacuteticas e eacute afectada
negativamente por ligandos naturais de enzima como eacute o caso do ATP
A exposiccedilatildeo do S1 de miosina (005 mgmL) a decavanadato conduz agrave inibiccedilatildeo do capacidade da
proteiacutena de estimular a actividade Mg2+
-ATPaacutesica com um valor de IC50 de 266 plusmn 07 M
decavanadato sendo o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM
decavanadato (80) Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM decavanadato sendo
o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM decavanadato (60)
exercendo um efeito de impedimento agrave ligaccedilatildeo entre o vanaacutedio e o S1
A uma concentraccedilatildeo de 27 microM decavanadato verifica-se a oxidaccedilatildeo de um dos resiacuteduos de Cys
de S1 de miosina seguida pela oxidaccedilatildeo gradual de outro resiacuteduo de Cys ateacute agrave concentraccedilatildeo de
1000 microM Ambos os resiacuteduos oxidados natildeo se encontravam expostos ao solvente utilizado A
oxidaccedilatildeo dos resiacuteduos permite sugerir que esta modificaccedilatildeo oxidativa sofrida pelo S1 contribui
para a diminuiccedilatildeo da sua actividade de ATPase
52
Por espectroscopia de RPE revelaram-se os possiacuteveis ligandos equatoriais do vanaacutedio aquando da
sua interacccedilatildeo com o S1 de miosina como sendo duas moleacuteculas de aacutegua um ligando contendo o
grupo R-OH e outro contendo o grupo R=NR
Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados de
oxidaccedilatildeo (IV) e (V) no S1 de miosina para concentraccedilotildees facilmente alcanccedilaacuteveis no ambiente
terrestre
53
5 Estudos futuros
No sentido do melhor esclarecimento das alteraccedilotildees moleculares e estruturais que ocorrem no
subfragmento 1 quando exposta ao vanaacutedio que conduzem a uma diminuiccedilatildeo da sua funccedilatildeo
sugere-se
- Quantificaccedilatildeo das cisteiacutenas reduzidas de S1 na presenccedila de metavanadato
- Determinaccedilatildeo das actividades Ca2+
- e K+-ATPaacutesica do S1 de miosina na presenccedila de
decavanadato
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina estimulada por actina
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina na presenccedila de
metavanadato
- Realizaccedilatildeo de RMN 2D com variaccedilatildeo das concentraccedilotildees das soluccedilotildees de decavanadato e
metavanadato na presenccedila de S1
-Mutageacutenese dos resiacuteduos de Cys doS1 de miosina de modo a verificar quais os resiacuteduos que
sofrem oxidaccedilatildeo na presenccedila de decavanadato
- Determinaccedilatildeo por calorimetria (DSC Differential Scanning Calorimetry) da variaccedilatildeo da
entalpia (∆H) e da temperatura de desnaturaccedilatildeo do S1 de miosina na presenccedila de vanaacutedio uma
vez que estas propriedades caracteriacutesticas de cada proteiacutena satildeo alteradas apoacutes ocorrecircncia de
modificaccedilotildees estruturais
- Anaacutelise por SDS-PAGE de S1 de miosina apoacutes tratamento com vanaacutedio de modo a determinar
quais os polipeacuteptidos eventualmente formados apoacutes clivagem da proteiacutena
54
6 Referecircncias bibliograacuteficas
- Sellers J Myosins 2nd Edition Oxford University Press 1999
- L Stryer inBiochemistry (Ed WH Freeman 2005 pp
Refordfas Materiais e Meacutetodos
[A] S S Margossian SLowey Methods Enzymol1982 85 55-71
[B] JD Pardue JA Spudich Methods Enzymol 1982 85 164-181
[C] M Bradford Anal Biochem1976 72 248-254
[D] H H Taussky E Shorr J Biol Chem 1953 202 675-685
[E] C R Bagshaw in Outline Studies in Biology ndash Muscle Contraction Chapman and Hall Ed
1982
[F] A Cuenda et al Eur J Biochem1990 194 663-670
55
Anexos
1 Determinaccedilatildeo da recta de calibraccedilatildeo meacutetodo de Bradford
Figura1ndashRepresentaccedilatildeo graacutefica da recta de calibraccedilatildeo (Abs595 nm vs [BSA]) utilizada para o meacutetodo de Bradford
2 Determinaccedilatildeo da recta calibraccedilatildeo para a electroforese em condiccedilotildees desnaturantes
Figura 2 ndash Representaccedilatildeo graacutefica da recta de calibraccedilatildeo (log(Peso Molecular) vs Rf dos marcadores de peso
molecular) onde Rf indica determinada atraveacutes do padratildeo de peso molecular
(poccedilo (a))
3Caacutelculo do coeficiente de extinccedilatildeo molar () para a quantificaccedilatildeo de cisteiacutenas reduzidas
atraveacutes do meacutetodo de Ellman
Traccedilou-se a recta de Abs412 nm meacutedia vs [L-Cisteiacutena] de modo a determinar o valor do coeficiente
de extinccedilatildeo molar definido como o declive daquela recta Assim =8400 M-1
cm-1
como
podemos verificar na Fig 3
A595 nm = 00546[BSA] + 00209 Rsup2 = 099051
Ab
s 59
5 n
m
[BSA] (mgmL)
log PM = -1177 Rf + 2247 Rsup2 = 098957
log
PM
Mobilidade relativa (Rf)
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero

50
Adicionou-se 15 L de DTNB (em soluccedilatildeo K [DTNB]stock=100 mM) ao meio reaccional e
registou-se o valor de absorvecircncia medido equivalente agraves cisteiacutenas ldquoraacutepidasrdquo (SH1 e SH2
expostos ao solvente) Adicionou-se 001 g de SDS que provoca a desnaturaccedilatildeo da proteiacutena e
consequentemente a exposiccedilatildeo de todos os resiacuteduos de cisteiacutenas que se encontravam ateacute aiacute
inacessiacuteveis no interior da proteiacutena (cisteiacutenas totais) Estes resiacuteduos podem assim reagir com o
DTNB
Estimou-se o nuacutemero de resiacuteduos de cisteiacutenas em todas as amostras com recurso ao coeficiente de
extinccedilatildeo molar () do DTNB (ver ponto 3 dos Anexos) que foi determinado atraveacutes da titulaccedilatildeo
de L-Cisteiacutena com intervalo de concentraccedilotildees compreendidos entre 0 e 50 microM na presenccedila de
25microM de DTNB em tampatildeo D Os valores de absorvecircncia foram obtidos para um comprimento de
onda de 412 nm
362 Interacccedilatildeo de V(IV) com subfragmento-1 de miosina
3621 Anaacutelise por Ressonacircncia Paramagneacutetica Electroacutenica
A teacutecnica mais eficiente de anaacutelise da interacccedilatildeo entre o iatildeo vanadilo e proteiacutenas como a
calmodulina e a transferrina entre outras permite a obtenccedilatildeo raacutepida de informaccedilatildeo no que toca agrave
conformaccedilatildeo e amplitude da referida interacccedilatildeo
Os espectros de Ressonacircncia Paramagneacutetica Electroacutenica (RPE) da espeacutecie paramagneacutetica V(IV)
foram realizados a 70K num espectroacutemetro Bruker EMX-300 X-band Todas as soluccedilotildees
utilizadas e amostras foram preparadas em anaerobiose numa cacircmara de luvas de modo a evitar a
reduccedilatildeo de V(IV) a V(V) Em tubos capilares com 02 mL de amostra manteve-se a concentraccedilatildeo
de S1 a 20 M e variou-se a concentraccedilatildeo de vanadilo (0 25 5 10 15 25 50 75 100 150
200 500 e 1000 M [VOSO4]stock=5 mM) em tampatildeo D Apoacutes obtenccedilatildeo dos espectros estes
foram tratados atraveacutes do software SimFonia e WinEPR tendo-se obtido um graacutefico com a
variaccedilatildeo da intensidade do sinal de RPE em funccedilatildeo da concentraccedilatildeo de sulfato de vanadilo
presente Realizaram-se ainda simulaccedilotildees de espectros obtidos com 20 microM de S1 e 1000 microM de
V(IV) com vista agrave obtenccedilatildeo dos ligandos provaacuteveis do vanadilo atraveacutes da anaacutelise da constante
de acoplamento hiperfina (A) e do factor g (g)
4 Conclusotildees gerais
Apesar de jaacute ter sido demonstrado que o vanaacutedio inibe a Na+K
+-ATPase da miosina a funccedilatildeo
deste elemento no tecido muacutesculo esqueleacutetico continua largamente incompreendida A enorme
complexidade quiacutemica deste metal especialmente no que concerne agrave oligomerizaccedilatildeo e reacccedilotildees
51
redox constitui um obstaacuteculo agrave compreensatildeo do comportamento do vanaacutedio nos organismos
vivos Assim a caracterizaccedilatildeo da quiacutemica deste elemento eacute de grande utilidade antes de iniciar o
estudo da sua interacccedilatildeo com o presente sistema
A anaacutelise por espectroscopias de 51
V-RMN e de UV-Vis de soluccedilotildees de vanadato diluiacutedas nos
meios de reacccedilatildeo utilizados no presente relatoacuterio (10 mM HEPES pH 70 25 mM KCl) mostrou
claramente que a espeacutecie decameacuterica (V10) possui um tempo de vida suficientemente longo para a
determinaccedilatildeo do seu efeito isolado ao contraacuterio do que se verifica com as espeacutecies mono- di-
tetra- e pentameacutericas (V1 V2 V4 e V5) Por outro lado a anaacutelise verificou ainda que apoacutes diluiccedilatildeo
das soluccedilotildees de decavanadato no meio de reacccedilatildeo coexistem as espeacutecies V10 e V1 pelo que os
efeitos putativos desta uacuteltima deveratildeo ser subtraiacutedos aquando da monitorizaccedilatildeo dos efeitos da
espeacutecie decameacuterica
O vanaacutedio (V) (vanadato) inibe as enzimas que catalizam reacccedilotildees de transferecircncia de grupos
fosfato como a ATPase da miosina [Goodno 1979] devido agrave sua analogia estrutural com o iatildeo
fosfato Na sua utilizaccedilatildeo como inibidor enzimaacutetico natildeo eacute tida em conta a contribuiccedilatildeo individual
das espeacutecies oligomeacutericas considerando-se normalmente a espeacutecie V1 como inibidora efectiva
[Goodno e Taylor 1982] No presente relatoacuterio verificou-se por espectroscopia de 51
V-RMN que
outros oligoacutemeros de vanadato interagem com o S1 de miosina tendo as espeacutecies V10 e V4 os
maiores niacuteveis de afinidade A sua interacccedilatildeo envolve forccedilas electrostaacuteticas e eacute afectada
negativamente por ligandos naturais de enzima como eacute o caso do ATP
A exposiccedilatildeo do S1 de miosina (005 mgmL) a decavanadato conduz agrave inibiccedilatildeo do capacidade da
proteiacutena de estimular a actividade Mg2+
-ATPaacutesica com um valor de IC50 de 266 plusmn 07 M
decavanadato sendo o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM
decavanadato (80) Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM decavanadato sendo
o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM decavanadato (60)
exercendo um efeito de impedimento agrave ligaccedilatildeo entre o vanaacutedio e o S1
A uma concentraccedilatildeo de 27 microM decavanadato verifica-se a oxidaccedilatildeo de um dos resiacuteduos de Cys
de S1 de miosina seguida pela oxidaccedilatildeo gradual de outro resiacuteduo de Cys ateacute agrave concentraccedilatildeo de
1000 microM Ambos os resiacuteduos oxidados natildeo se encontravam expostos ao solvente utilizado A
oxidaccedilatildeo dos resiacuteduos permite sugerir que esta modificaccedilatildeo oxidativa sofrida pelo S1 contribui
para a diminuiccedilatildeo da sua actividade de ATPase
52
Por espectroscopia de RPE revelaram-se os possiacuteveis ligandos equatoriais do vanaacutedio aquando da
sua interacccedilatildeo com o S1 de miosina como sendo duas moleacuteculas de aacutegua um ligando contendo o
grupo R-OH e outro contendo o grupo R=NR
Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados de
oxidaccedilatildeo (IV) e (V) no S1 de miosina para concentraccedilotildees facilmente alcanccedilaacuteveis no ambiente
terrestre
53
5 Estudos futuros
No sentido do melhor esclarecimento das alteraccedilotildees moleculares e estruturais que ocorrem no
subfragmento 1 quando exposta ao vanaacutedio que conduzem a uma diminuiccedilatildeo da sua funccedilatildeo
sugere-se
- Quantificaccedilatildeo das cisteiacutenas reduzidas de S1 na presenccedila de metavanadato
- Determinaccedilatildeo das actividades Ca2+
- e K+-ATPaacutesica do S1 de miosina na presenccedila de
decavanadato
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina estimulada por actina
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina na presenccedila de
metavanadato
- Realizaccedilatildeo de RMN 2D com variaccedilatildeo das concentraccedilotildees das soluccedilotildees de decavanadato e
metavanadato na presenccedila de S1
-Mutageacutenese dos resiacuteduos de Cys doS1 de miosina de modo a verificar quais os resiacuteduos que
sofrem oxidaccedilatildeo na presenccedila de decavanadato
- Determinaccedilatildeo por calorimetria (DSC Differential Scanning Calorimetry) da variaccedilatildeo da
entalpia (∆H) e da temperatura de desnaturaccedilatildeo do S1 de miosina na presenccedila de vanaacutedio uma
vez que estas propriedades caracteriacutesticas de cada proteiacutena satildeo alteradas apoacutes ocorrecircncia de
modificaccedilotildees estruturais
- Anaacutelise por SDS-PAGE de S1 de miosina apoacutes tratamento com vanaacutedio de modo a determinar
quais os polipeacuteptidos eventualmente formados apoacutes clivagem da proteiacutena
54
6 Referecircncias bibliograacuteficas
- Sellers J Myosins 2nd Edition Oxford University Press 1999
- L Stryer inBiochemistry (Ed WH Freeman 2005 pp
Refordfas Materiais e Meacutetodos
[A] S S Margossian SLowey Methods Enzymol1982 85 55-71
[B] JD Pardue JA Spudich Methods Enzymol 1982 85 164-181
[C] M Bradford Anal Biochem1976 72 248-254
[D] H H Taussky E Shorr J Biol Chem 1953 202 675-685
[E] C R Bagshaw in Outline Studies in Biology ndash Muscle Contraction Chapman and Hall Ed
1982
[F] A Cuenda et al Eur J Biochem1990 194 663-670
55
Anexos
1 Determinaccedilatildeo da recta de calibraccedilatildeo meacutetodo de Bradford
Figura1ndashRepresentaccedilatildeo graacutefica da recta de calibraccedilatildeo (Abs595 nm vs [BSA]) utilizada para o meacutetodo de Bradford
2 Determinaccedilatildeo da recta calibraccedilatildeo para a electroforese em condiccedilotildees desnaturantes
Figura 2 ndash Representaccedilatildeo graacutefica da recta de calibraccedilatildeo (log(Peso Molecular) vs Rf dos marcadores de peso
molecular) onde Rf indica determinada atraveacutes do padratildeo de peso molecular
(poccedilo (a))
3Caacutelculo do coeficiente de extinccedilatildeo molar () para a quantificaccedilatildeo de cisteiacutenas reduzidas
atraveacutes do meacutetodo de Ellman
Traccedilou-se a recta de Abs412 nm meacutedia vs [L-Cisteiacutena] de modo a determinar o valor do coeficiente
de extinccedilatildeo molar definido como o declive daquela recta Assim =8400 M-1
cm-1
como
podemos verificar na Fig 3
A595 nm = 00546[BSA] + 00209 Rsup2 = 099051
Ab
s 59
5 n
m
[BSA] (mgmL)
log PM = -1177 Rf + 2247 Rsup2 = 098957
log
PM
Mobilidade relativa (Rf)
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero

51
redox constitui um obstaacuteculo agrave compreensatildeo do comportamento do vanaacutedio nos organismos
vivos Assim a caracterizaccedilatildeo da quiacutemica deste elemento eacute de grande utilidade antes de iniciar o
estudo da sua interacccedilatildeo com o presente sistema
A anaacutelise por espectroscopias de 51
V-RMN e de UV-Vis de soluccedilotildees de vanadato diluiacutedas nos
meios de reacccedilatildeo utilizados no presente relatoacuterio (10 mM HEPES pH 70 25 mM KCl) mostrou
claramente que a espeacutecie decameacuterica (V10) possui um tempo de vida suficientemente longo para a
determinaccedilatildeo do seu efeito isolado ao contraacuterio do que se verifica com as espeacutecies mono- di-
tetra- e pentameacutericas (V1 V2 V4 e V5) Por outro lado a anaacutelise verificou ainda que apoacutes diluiccedilatildeo
das soluccedilotildees de decavanadato no meio de reacccedilatildeo coexistem as espeacutecies V10 e V1 pelo que os
efeitos putativos desta uacuteltima deveratildeo ser subtraiacutedos aquando da monitorizaccedilatildeo dos efeitos da
espeacutecie decameacuterica
O vanaacutedio (V) (vanadato) inibe as enzimas que catalizam reacccedilotildees de transferecircncia de grupos
fosfato como a ATPase da miosina [Goodno 1979] devido agrave sua analogia estrutural com o iatildeo
fosfato Na sua utilizaccedilatildeo como inibidor enzimaacutetico natildeo eacute tida em conta a contribuiccedilatildeo individual
das espeacutecies oligomeacutericas considerando-se normalmente a espeacutecie V1 como inibidora efectiva
[Goodno e Taylor 1982] No presente relatoacuterio verificou-se por espectroscopia de 51
V-RMN que
outros oligoacutemeros de vanadato interagem com o S1 de miosina tendo as espeacutecies V10 e V4 os
maiores niacuteveis de afinidade A sua interacccedilatildeo envolve forccedilas electrostaacuteticas e eacute afectada
negativamente por ligandos naturais de enzima como eacute o caso do ATP
A exposiccedilatildeo do S1 de miosina (005 mgmL) a decavanadato conduz agrave inibiccedilatildeo do capacidade da
proteiacutena de estimular a actividade Mg2+
-ATPaacutesica com um valor de IC50 de 266 plusmn 07 M
decavanadato sendo o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM
decavanadato (80) Na presenccedila de actina o valor de IC50 eacute de 28 plusmn 1 microM decavanadato sendo
o maacuteximo de inibiccedilatildeo da estimulaccedilatildeo atingido para cerca de 250 microM decavanadato (60)
exercendo um efeito de impedimento agrave ligaccedilatildeo entre o vanaacutedio e o S1
A uma concentraccedilatildeo de 27 microM decavanadato verifica-se a oxidaccedilatildeo de um dos resiacuteduos de Cys
de S1 de miosina seguida pela oxidaccedilatildeo gradual de outro resiacuteduo de Cys ateacute agrave concentraccedilatildeo de
1000 microM Ambos os resiacuteduos oxidados natildeo se encontravam expostos ao solvente utilizado A
oxidaccedilatildeo dos resiacuteduos permite sugerir que esta modificaccedilatildeo oxidativa sofrida pelo S1 contribui
para a diminuiccedilatildeo da sua actividade de ATPase
52
Por espectroscopia de RPE revelaram-se os possiacuteveis ligandos equatoriais do vanaacutedio aquando da
sua interacccedilatildeo com o S1 de miosina como sendo duas moleacuteculas de aacutegua um ligando contendo o
grupo R-OH e outro contendo o grupo R=NR
Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados de
oxidaccedilatildeo (IV) e (V) no S1 de miosina para concentraccedilotildees facilmente alcanccedilaacuteveis no ambiente
terrestre
53
5 Estudos futuros
No sentido do melhor esclarecimento das alteraccedilotildees moleculares e estruturais que ocorrem no
subfragmento 1 quando exposta ao vanaacutedio que conduzem a uma diminuiccedilatildeo da sua funccedilatildeo
sugere-se
- Quantificaccedilatildeo das cisteiacutenas reduzidas de S1 na presenccedila de metavanadato
- Determinaccedilatildeo das actividades Ca2+
- e K+-ATPaacutesica do S1 de miosina na presenccedila de
decavanadato
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina estimulada por actina
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina na presenccedila de
metavanadato
- Realizaccedilatildeo de RMN 2D com variaccedilatildeo das concentraccedilotildees das soluccedilotildees de decavanadato e
metavanadato na presenccedila de S1
-Mutageacutenese dos resiacuteduos de Cys doS1 de miosina de modo a verificar quais os resiacuteduos que
sofrem oxidaccedilatildeo na presenccedila de decavanadato
- Determinaccedilatildeo por calorimetria (DSC Differential Scanning Calorimetry) da variaccedilatildeo da
entalpia (∆H) e da temperatura de desnaturaccedilatildeo do S1 de miosina na presenccedila de vanaacutedio uma
vez que estas propriedades caracteriacutesticas de cada proteiacutena satildeo alteradas apoacutes ocorrecircncia de
modificaccedilotildees estruturais
- Anaacutelise por SDS-PAGE de S1 de miosina apoacutes tratamento com vanaacutedio de modo a determinar
quais os polipeacuteptidos eventualmente formados apoacutes clivagem da proteiacutena
54
6 Referecircncias bibliograacuteficas
- Sellers J Myosins 2nd Edition Oxford University Press 1999
- L Stryer inBiochemistry (Ed WH Freeman 2005 pp
Refordfas Materiais e Meacutetodos
[A] S S Margossian SLowey Methods Enzymol1982 85 55-71
[B] JD Pardue JA Spudich Methods Enzymol 1982 85 164-181
[C] M Bradford Anal Biochem1976 72 248-254
[D] H H Taussky E Shorr J Biol Chem 1953 202 675-685
[E] C R Bagshaw in Outline Studies in Biology ndash Muscle Contraction Chapman and Hall Ed
1982
[F] A Cuenda et al Eur J Biochem1990 194 663-670
55
Anexos
1 Determinaccedilatildeo da recta de calibraccedilatildeo meacutetodo de Bradford
Figura1ndashRepresentaccedilatildeo graacutefica da recta de calibraccedilatildeo (Abs595 nm vs [BSA]) utilizada para o meacutetodo de Bradford
2 Determinaccedilatildeo da recta calibraccedilatildeo para a electroforese em condiccedilotildees desnaturantes
Figura 2 ndash Representaccedilatildeo graacutefica da recta de calibraccedilatildeo (log(Peso Molecular) vs Rf dos marcadores de peso
molecular) onde Rf indica determinada atraveacutes do padratildeo de peso molecular
(poccedilo (a))
3Caacutelculo do coeficiente de extinccedilatildeo molar () para a quantificaccedilatildeo de cisteiacutenas reduzidas
atraveacutes do meacutetodo de Ellman
Traccedilou-se a recta de Abs412 nm meacutedia vs [L-Cisteiacutena] de modo a determinar o valor do coeficiente
de extinccedilatildeo molar definido como o declive daquela recta Assim =8400 M-1
cm-1
como
podemos verificar na Fig 3
A595 nm = 00546[BSA] + 00209 Rsup2 = 099051
Ab
s 59
5 n
m
[BSA] (mgmL)
log PM = -1177 Rf + 2247 Rsup2 = 098957
log
PM
Mobilidade relativa (Rf)
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero

52
Por espectroscopia de RPE revelaram-se os possiacuteveis ligandos equatoriais do vanaacutedio aquando da
sua interacccedilatildeo com o S1 de miosina como sendo duas moleacuteculas de aacutegua um ligando contendo o
grupo R-OH e outro contendo o grupo R=NR
Globalmente os resultados obtidos sugerem um efeito significativo do vanaacutedio nos estados de
oxidaccedilatildeo (IV) e (V) no S1 de miosina para concentraccedilotildees facilmente alcanccedilaacuteveis no ambiente
terrestre
53
5 Estudos futuros
No sentido do melhor esclarecimento das alteraccedilotildees moleculares e estruturais que ocorrem no
subfragmento 1 quando exposta ao vanaacutedio que conduzem a uma diminuiccedilatildeo da sua funccedilatildeo
sugere-se
- Quantificaccedilatildeo das cisteiacutenas reduzidas de S1 na presenccedila de metavanadato
- Determinaccedilatildeo das actividades Ca2+
- e K+-ATPaacutesica do S1 de miosina na presenccedila de
decavanadato
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina estimulada por actina
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina na presenccedila de
metavanadato
- Realizaccedilatildeo de RMN 2D com variaccedilatildeo das concentraccedilotildees das soluccedilotildees de decavanadato e
metavanadato na presenccedila de S1
-Mutageacutenese dos resiacuteduos de Cys doS1 de miosina de modo a verificar quais os resiacuteduos que
sofrem oxidaccedilatildeo na presenccedila de decavanadato
- Determinaccedilatildeo por calorimetria (DSC Differential Scanning Calorimetry) da variaccedilatildeo da
entalpia (∆H) e da temperatura de desnaturaccedilatildeo do S1 de miosina na presenccedila de vanaacutedio uma
vez que estas propriedades caracteriacutesticas de cada proteiacutena satildeo alteradas apoacutes ocorrecircncia de
modificaccedilotildees estruturais
- Anaacutelise por SDS-PAGE de S1 de miosina apoacutes tratamento com vanaacutedio de modo a determinar
quais os polipeacuteptidos eventualmente formados apoacutes clivagem da proteiacutena
54
6 Referecircncias bibliograacuteficas
- Sellers J Myosins 2nd Edition Oxford University Press 1999
- L Stryer inBiochemistry (Ed WH Freeman 2005 pp
Refordfas Materiais e Meacutetodos
[A] S S Margossian SLowey Methods Enzymol1982 85 55-71
[B] JD Pardue JA Spudich Methods Enzymol 1982 85 164-181
[C] M Bradford Anal Biochem1976 72 248-254
[D] H H Taussky E Shorr J Biol Chem 1953 202 675-685
[E] C R Bagshaw in Outline Studies in Biology ndash Muscle Contraction Chapman and Hall Ed
1982
[F] A Cuenda et al Eur J Biochem1990 194 663-670
55
Anexos
1 Determinaccedilatildeo da recta de calibraccedilatildeo meacutetodo de Bradford
Figura1ndashRepresentaccedilatildeo graacutefica da recta de calibraccedilatildeo (Abs595 nm vs [BSA]) utilizada para o meacutetodo de Bradford
2 Determinaccedilatildeo da recta calibraccedilatildeo para a electroforese em condiccedilotildees desnaturantes
Figura 2 ndash Representaccedilatildeo graacutefica da recta de calibraccedilatildeo (log(Peso Molecular) vs Rf dos marcadores de peso
molecular) onde Rf indica determinada atraveacutes do padratildeo de peso molecular
(poccedilo (a))
3Caacutelculo do coeficiente de extinccedilatildeo molar () para a quantificaccedilatildeo de cisteiacutenas reduzidas
atraveacutes do meacutetodo de Ellman
Traccedilou-se a recta de Abs412 nm meacutedia vs [L-Cisteiacutena] de modo a determinar o valor do coeficiente
de extinccedilatildeo molar definido como o declive daquela recta Assim =8400 M-1
cm-1
como
podemos verificar na Fig 3
A595 nm = 00546[BSA] + 00209 Rsup2 = 099051
Ab
s 59
5 n
m
[BSA] (mgmL)
log PM = -1177 Rf + 2247 Rsup2 = 098957
log
PM
Mobilidade relativa (Rf)
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero

53
5 Estudos futuros
No sentido do melhor esclarecimento das alteraccedilotildees moleculares e estruturais que ocorrem no
subfragmento 1 quando exposta ao vanaacutedio que conduzem a uma diminuiccedilatildeo da sua funccedilatildeo
sugere-se
- Quantificaccedilatildeo das cisteiacutenas reduzidas de S1 na presenccedila de metavanadato
- Determinaccedilatildeo das actividades Ca2+
- e K+-ATPaacutesica do S1 de miosina na presenccedila de
decavanadato
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina estimulada por actina
- Determinaccedilatildeo da actividade Mg2+
-ATPaacutesica basal do S1 de miosina na presenccedila de
metavanadato
- Realizaccedilatildeo de RMN 2D com variaccedilatildeo das concentraccedilotildees das soluccedilotildees de decavanadato e
metavanadato na presenccedila de S1
-Mutageacutenese dos resiacuteduos de Cys doS1 de miosina de modo a verificar quais os resiacuteduos que
sofrem oxidaccedilatildeo na presenccedila de decavanadato
- Determinaccedilatildeo por calorimetria (DSC Differential Scanning Calorimetry) da variaccedilatildeo da
entalpia (∆H) e da temperatura de desnaturaccedilatildeo do S1 de miosina na presenccedila de vanaacutedio uma
vez que estas propriedades caracteriacutesticas de cada proteiacutena satildeo alteradas apoacutes ocorrecircncia de
modificaccedilotildees estruturais
- Anaacutelise por SDS-PAGE de S1 de miosina apoacutes tratamento com vanaacutedio de modo a determinar
quais os polipeacuteptidos eventualmente formados apoacutes clivagem da proteiacutena
54
6 Referecircncias bibliograacuteficas
- Sellers J Myosins 2nd Edition Oxford University Press 1999
- L Stryer inBiochemistry (Ed WH Freeman 2005 pp
Refordfas Materiais e Meacutetodos
[A] S S Margossian SLowey Methods Enzymol1982 85 55-71
[B] JD Pardue JA Spudich Methods Enzymol 1982 85 164-181
[C] M Bradford Anal Biochem1976 72 248-254
[D] H H Taussky E Shorr J Biol Chem 1953 202 675-685
[E] C R Bagshaw in Outline Studies in Biology ndash Muscle Contraction Chapman and Hall Ed
1982
[F] A Cuenda et al Eur J Biochem1990 194 663-670
55
Anexos
1 Determinaccedilatildeo da recta de calibraccedilatildeo meacutetodo de Bradford
Figura1ndashRepresentaccedilatildeo graacutefica da recta de calibraccedilatildeo (Abs595 nm vs [BSA]) utilizada para o meacutetodo de Bradford
2 Determinaccedilatildeo da recta calibraccedilatildeo para a electroforese em condiccedilotildees desnaturantes
Figura 2 ndash Representaccedilatildeo graacutefica da recta de calibraccedilatildeo (log(Peso Molecular) vs Rf dos marcadores de peso
molecular) onde Rf indica determinada atraveacutes do padratildeo de peso molecular
(poccedilo (a))
3Caacutelculo do coeficiente de extinccedilatildeo molar () para a quantificaccedilatildeo de cisteiacutenas reduzidas
atraveacutes do meacutetodo de Ellman
Traccedilou-se a recta de Abs412 nm meacutedia vs [L-Cisteiacutena] de modo a determinar o valor do coeficiente
de extinccedilatildeo molar definido como o declive daquela recta Assim =8400 M-1
cm-1
como
podemos verificar na Fig 3
A595 nm = 00546[BSA] + 00209 Rsup2 = 099051
Ab
s 59
5 n
m
[BSA] (mgmL)
log PM = -1177 Rf + 2247 Rsup2 = 098957
log
PM
Mobilidade relativa (Rf)
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero

54
6 Referecircncias bibliograacuteficas
- Sellers J Myosins 2nd Edition Oxford University Press 1999
- L Stryer inBiochemistry (Ed WH Freeman 2005 pp
Refordfas Materiais e Meacutetodos
[A] S S Margossian SLowey Methods Enzymol1982 85 55-71
[B] JD Pardue JA Spudich Methods Enzymol 1982 85 164-181
[C] M Bradford Anal Biochem1976 72 248-254
[D] H H Taussky E Shorr J Biol Chem 1953 202 675-685
[E] C R Bagshaw in Outline Studies in Biology ndash Muscle Contraction Chapman and Hall Ed
1982
[F] A Cuenda et al Eur J Biochem1990 194 663-670
55
Anexos
1 Determinaccedilatildeo da recta de calibraccedilatildeo meacutetodo de Bradford
Figura1ndashRepresentaccedilatildeo graacutefica da recta de calibraccedilatildeo (Abs595 nm vs [BSA]) utilizada para o meacutetodo de Bradford
2 Determinaccedilatildeo da recta calibraccedilatildeo para a electroforese em condiccedilotildees desnaturantes
Figura 2 ndash Representaccedilatildeo graacutefica da recta de calibraccedilatildeo (log(Peso Molecular) vs Rf dos marcadores de peso
molecular) onde Rf indica determinada atraveacutes do padratildeo de peso molecular
(poccedilo (a))
3Caacutelculo do coeficiente de extinccedilatildeo molar () para a quantificaccedilatildeo de cisteiacutenas reduzidas
atraveacutes do meacutetodo de Ellman
Traccedilou-se a recta de Abs412 nm meacutedia vs [L-Cisteiacutena] de modo a determinar o valor do coeficiente
de extinccedilatildeo molar definido como o declive daquela recta Assim =8400 M-1
cm-1
como
podemos verificar na Fig 3
A595 nm = 00546[BSA] + 00209 Rsup2 = 099051
Ab
s 59
5 n
m
[BSA] (mgmL)
log PM = -1177 Rf + 2247 Rsup2 = 098957
log
PM
Mobilidade relativa (Rf)
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero

55
Anexos
1 Determinaccedilatildeo da recta de calibraccedilatildeo meacutetodo de Bradford
Figura1ndashRepresentaccedilatildeo graacutefica da recta de calibraccedilatildeo (Abs595 nm vs [BSA]) utilizada para o meacutetodo de Bradford
2 Determinaccedilatildeo da recta calibraccedilatildeo para a electroforese em condiccedilotildees desnaturantes
Figura 2 ndash Representaccedilatildeo graacutefica da recta de calibraccedilatildeo (log(Peso Molecular) vs Rf dos marcadores de peso
molecular) onde Rf indica determinada atraveacutes do padratildeo de peso molecular
(poccedilo (a))
3Caacutelculo do coeficiente de extinccedilatildeo molar () para a quantificaccedilatildeo de cisteiacutenas reduzidas
atraveacutes do meacutetodo de Ellman
Traccedilou-se a recta de Abs412 nm meacutedia vs [L-Cisteiacutena] de modo a determinar o valor do coeficiente
de extinccedilatildeo molar definido como o declive daquela recta Assim =8400 M-1
cm-1
como
podemos verificar na Fig 3
A595 nm = 00546[BSA] + 00209 Rsup2 = 099051
Ab
s 59
5 n
m
[BSA] (mgmL)
log PM = -1177 Rf + 2247 Rsup2 = 098957
log
PM
Mobilidade relativa (Rf)
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero
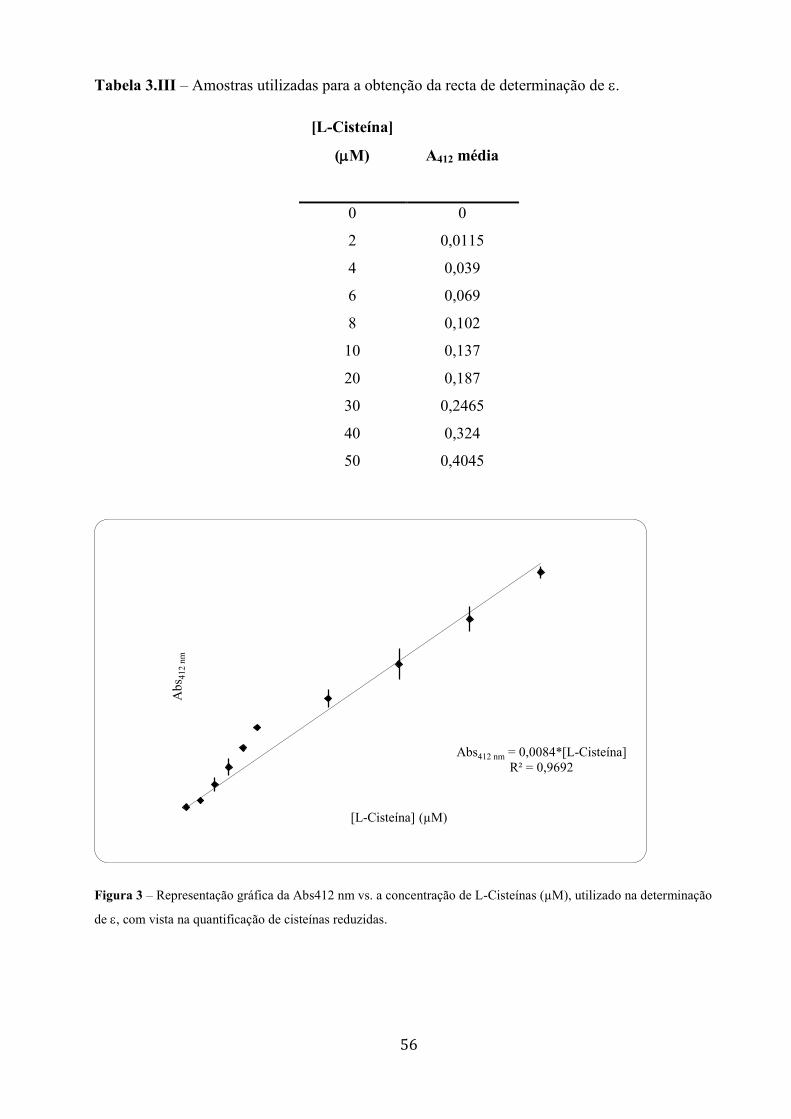
56
Tabela 3III ndash Amostras utilizadas para a obtenccedilatildeo da recta de determinaccedilatildeo de
[L-Cisteiacutena]
(M)
A412 meacutedia
0 0
2 00115
4 0039
6 0069
8 0102
10 0137
20 0187
30 02465
40 0324
50 04045
Figura 3 ndash Representaccedilatildeo graacutefica da Abs412 nm vs a concentraccedilatildeo de L-Cisteiacutenas (microM) utilizado na determinaccedilatildeo
de com vista na quantificaccedilatildeo de cisteiacutenas reduzidas
Abs412 nm = 00084[L-Cisteiacutena]
Rsup2 = 09692
Ab
s 412 n
m
[L-Cisteiacutena] (microM)
57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero

57
4 Espectroscopia de Ressonacircncia Magneacutetica Nuclear consideraccedilotildees gerais
A teacutecnica de Ressonacircncia Magneacutetica Nuclear (RMN) permite a monitorizaccedilatildeo da absorccedilatildeo de
radiaccedilatildeo associada a transiccedilotildees de nuacutecleos atoacutemicos entre niacuteveis adjacentes de energia magneacutetica
A energia da radiaccedilatildeo eacute medida como funccedilatildeo do campo magneacutetico aplicado obtendo-se um
espectro de absorccedilatildeo em funccedilatildeo da forccedila do campo magneacutetico
As caracteriacutesticas do nuacutecleo constituinte de uma determinada moleacutecula iratildeo influenciar os sinais
obtidos no espectro de RMN permitindo-nos a obtenccedilatildeo de informaccedilotildees de variadas amplitudes
a estrutura da moleacutecula em anaacutelise a sua interacccedilatildeo intermolecular no sistema em estudo bem
como a sua participaccedilatildeo em processos dinacircmicos em soluccedilatildeo O espectro de RMN eacute constituiacutedo
por diversas bandas de ressonacircncia
Os paracircmetros espectrais de um espectro de RMN com mais interesse para o estudo em curso satildeo
o desvio quiacutemico () que se refere agrave posiccedilatildeo de cada banda em relaccedilatildeo a uma referecircncia
previamente escolhida (0) a amplitude da banda que se define como a altura relativa da banda a
intensidade definida pela aacuterea da banda a largura a meia altura a constante de acoplamento de
spin (J)
Todos os paracircmetros anteriormente referidos podem contribuir para a obtenccedilatildeo de informaccedilotildees
sobre o ambiente quiacutemico estrutura concentraccedilatildeo e interacccedilatildeo intermolecular com ecircnfase para
estudos cineacuteticos entre outros
O desvio quiacutemico evidencia as alteraccedilotildees do ambiente quiacutemico em redor do nuacutecleo observado e eacute
normalmente expresso em ppm que o torna um valor adimensional
O nuacutecleo de 51
V apresenta propriedades magneacuteticas que o tornam adequado para estudos atraveacutes
desta teacutecnica Este isoacutetopo apresenta uma abundacircncia natural de 9975 spin nuclear de 72 e
sensibilidade de 39 relativamente ao protatildeo (1H) As bandas dos vanadatos com configuraccedilatildeo
tetraeacutedrica ou aproximada apresentam largura a meia altura meacutedia (60-100 MHz) Assim a
realizaccedilatildeo de estudos com espeacutecies diamagneacuteticas (como eacute o caso do V(V))
Cada espeacutecie de vanadato (V(V)) estaacute caracterizada por ressonacircncias bem resolvidas no espectro
de RMN O tempo de relaxaccedilatildeo das diversas espeacutecies em estudo eacute semelhante pelo que a
integraccedilatildeo dos seus sinais fornece-nos a fracccedilatildeo molar e consequentemente a concentraccedilatildeo de
cada oligoacutemero














![UNIVERSIDADE FFEDERAL DDE UUBERLÂNDIA ......Com exceção das miosinas I, II e V pouco se conhece das propriedades bioquímicas dessas enzimas [24]. Miosina II muscular ou miosina](https://static.fdocumentos.com/doc/165x107/5e6951af49f0237a9606ebb5/universidade-ffederal-dde-uuberlndia-com-exceo-das-miosinas-i-ii.jpg)
![VANÁDIO NA CONTRACÇÃO MUSCULAR - core.ac.uk · utilizado no tratamento da diabetes mellitus foi no ano de 1899 [7]. Em 2003, mais de ... O vanádio é um elemento químico (símbolo](https://static.fdocumentos.com/doc/165x107/5c5e1f4409d3f2ba618bca94/vanadio-na-contraccao-muscular-coreacuk-utilizado-no-tratamento-da-diabetes.jpg)

