
Revisão para a Prova Colégio dom bosco Professora: Lucilene Pires Disciplina: Geografia Ano: 6°
Upload
truongtrucCategory
view
215download
0
UMA VISÃO INTEGRADA DOS EVENTOS PROTEOLÍTICOS DE
SEMENTES DE VIGNA UNGUICULATA AO LONGO DOS
PROCESSOS GERMINATIVOS E PÓS-GERMINATIVOS COM
ÊNFASE EM PROTEINASES CISTEÍNICAS
LUCILENE OLLIVIER DE OLIVEIRA
UNIVERSIDADE ESTADUAL DO NORTE FLUMINENSE
DARCY RIBEIRO – UENF
CAMPOS DOS GOYTACAZES - RJ
FEVEREIRO - 2010

II
UMA VISÃO INTEGRADA DOS EVENTOS PROTEOLÍTICOS DE
SEMENTES DE Vigna unguiculata AO LONGO DOS PROCESSOS
GERMINATIVOS E PÓS-GERMINATIVOS COM ÊNFASE EM
PROTEINASES CISTEÍNICAS
LUCILENE OLLIVIER DE OLIVEIRA
Dissertação apresentada ao Centro
de Biociências e Biotecnologia da
Universidade Estadual do Norte
Fluminense Darcy Ribeiro, como
parte dos requisitos para obtenção
do título de Mestre em Biociências e
Biotecnologia, com ênfase em
Biologia Celular.
ORIENTADORA: PROF.ª DRª. KÁTIA VALEVSKI SALES FERNANDES
UNIVERSIDADE ESTADUAL DO NORTE FLUMINENSE
DARCY RIBEIRO – UENF
CAMPOS DOS GOYTACAZES - RJ
FEVEREIRO - 2010

III

IV
Este projeto foi desenvolvido no laboratório de Química e Função de
Proteínas e Peptídeos – LQFPP, do Centro de Biociências e Biotecnologia – CBB,
na Universidade Estadual do Norte Fluminense - Darcy Ribeiro – UENF, sob
orientação da pesquisadora Kátia Valevski Sales Fernandes.
Apoio financeiro: FAPERJ e CNPq

V
Dedico, ao meu filho Rafael: Sua
chegada me deu forças para lutar por
meus sonhos, pela vida e alcançar
meus objetivos.

VI
AGRADECIMENTOS
Primeiramente a Deus, por me dar forças e oferecer a serenidade necessária para
aceitar as coisas que eu não podia modificar; coragem para modificar aquelas que
eu podia e sabedoria para distinguir uma da outra.
À Professora Kátia Valevski Sales Fernandes, pela dedicada orientação, grande
amizade, estímulo, confiança e muita paciência. Você é uma pessoa maravilhosa.
Admiro-te!! Obrigada por todos os ensinamentos ao longo destes anos.
Agradeço à professora Antônia Elenir Amâncio Oliveira por ter aceitado participar da
revisão desta dissertação.
Agradeço aos professores Jorge Hernadez Fernandez, Cristiane Martins Cardoso de
Salles e Beatriz dos Santos Ferreira, por aceitarem fazer parte dessa banca
examinadora e por todas as contribuições que vierem a oferecer.
Aos professores Cláudio Andrés Retamal e Maura Da Cunha pela valiosa
contribuição nos experimentos, nos ensinamentos, amizade e pelas colaborações ao
nosso trabalho.
À grande amiga Nathália Bastos, pela amizade, carinho, preocupação e por toda
ajuda constante concedida durante todos esses anos no laboratório. Gosto muito de
você e te admiro muito.
Ao Márcio por sua ajuda constante, pelos ensinamentos concedidos; e é claro as
brincadeiras no laboratório que tornam nossos dias mais engraçados e mais
alegres!!!! Muito obrigada.
Ao Fábio Maciel que com muita paciência me ensinou muita coisa no laboratório.
Saudades!!!
Agradeço de maneira especial aos meus colegas de laboratório Elane, Simone,
Keysson, Evenilton, Gustavo, Nádia, Amanda, Hélio, Nathália Deus, Flávia e os
demais, por tornar tão agradável a convivência durante tantos dias de trabalho.

VII
Ao técnico do LBCT, Arthur Rodrigues, pela ajuda e auxílio no preparo de soluções e
pela amizade.
À minha mãe Lúcia por ser uma verdadeira guerreira e exemplo de mãe e amiga! É
por você que tenho batalhado na vida, pois você sempre me instruiu e me ensinou o
caminho certo para vencer na vida. Te amo muito!!! Você é uma grande heroína!!!!
Que Deus lhe abençõe!!!
À minha Avó Sebastiana que sempre me deu força, me ensinou acima de tudo a
respeitar as pessoas, me ensinou tantas coisas que não caberiam aqui...Vó,
obrigada por tudo, tudo mesmo...obrigada pela confiança em mim depositada e pela
amizade. Te amo muito...
Agradeço ao meu marido Carlos Alberto Laczynski Júnior por todo carinho,
paciência, compreensão e pela força nas horas difíceis. Você é muito mais que
especial para mim. Te amo muito.
Agradeço a minha nova família Carlos, Maria Amélia, Gabriela e Rafaela, que me
acolheram de braços abertos e me tratam como se fosse da família, sempre me
respeitando e me ajudando em tudo o que preciso. Essa conquista também é de
vocês. Obrigada pelo apoio, carinho, ajuda e por todo amor...Amo muito vocês..
E a todos que de alguma forma contribuíram para o meu trabalho e que infelizmente
não conseguiria listar a todos, muito obrigada.

VIII
SUMÁRIO
RESUMO................................................................................................................XVIII
ABSTRACT..............................................................................................................XIV
1- INTRODUÇÃO.........................................................................................................2 1.1 - Importância das sementes...................................................................................2
1.2- Germinação...........................................................................................................2
1.2.1- Eventos pós-germinativos..................................................................................5
1.2.2- Proteólise em sementes.....................................................................................6
1.3- Proteínas de reserva sementes..........................................................................11
1.3.1- As globulinas....................................................................................................12
1.4- Corpos protéicos.................................................................................................13
1.5- Feijão-de-corda - Vigna unguiculata (L. Walp)....................................................14
1.5.1- Proteólise em semente de Vigna unguiculata..................................................17
2- OBJETIVO GERAL................................................................................................19
2.1- Objetivos específicos..........................................................................................19
3- MATERIAIS E MÉTODOS.....................................................................................20 3.1- Sementes............................................................................................................20
3.2- Germinação e desenvolvimento pós-germinativo...............................................20
3.3- Extração de proteínas.........................................................................................20
3.4- Isolamento de corpos protéicos..........................................................................21
3.5- Determinação da concentração de proteínas.....................................................21
3.6- Visualização dos perfis protéicos por eletroforese em gel de poliacrilamida.....21
3.7- Western blotting para detecção de vicilinas........................................................22
3.8- Detecção de atividade enzimática em gel...........................................................23
3.9- Ação de inibidores sobre as atividades proteolíticas em gel...............................23
3.10- Análise bidimensional nativo / SDS-PAGE e nativo / gelatina-PAGE de
proteínas e proteinases cisteínicas de cotilédones quiescentes e germinates 108
HAE de feijão-de-corda..............................................................................................24
3.11- Determinação do grau de anomalia das proteases na migração eletroforética e
determinação das massas moleculares das proteases.............................................25
3.12- Análise densitométrica......................................................................................26
4- RESULTADOS.......................................................................................................27

IX
4.1- Análise do perfil eletroforético e Western blotting da fração globulínica do tecido
cotiledonário ao longo do processo germinativo e pós-germinativo das sementes de
feijão-de-corda............................................................................................................27
4.2- Análise do perfil eletroforético da fração albumínica do tecido cotiledonário ao
longo do processo germinativo e pós-germinativo das sementes de feijão-de-
corda...........................................................................................................................27
4.3- Análise do perfil eletroforético e Western blotting de proteínas de corpos
protéicos e proteínas citoplasmáticas ao longo do processo germinativo e pós-
germinativo das sementes de feijão-de-corda............................................................30
4.4- Detecção de atividade gelatinolítica e ação de inibidores sobre as atividades
proteásicas ao longo do processo germinativo e pós-germinativo das sementes de
feijão-de-corda............................................................................................................33
4.5- Detecção de atividade proteásica ao longo do processo germinativo e pós-
germinativo das sementes de feijão-de-corda na ausência e presença de agente
redutor........................................................................................................................37
4.6- Comparação da migração eletroforética de proteínas e atividades proteásicas
em géis bidimensionais SDS-PAGE/ SDS-PAGE e SDS-PAGE em presença de
gelatina 0,1% e determinação das massas moleculares de
proteases....................................................................................................................37
4.7- Análise do perfil eletroforético por géis bidimensionais das proteínas e proteases
do tecido cotiledonário de sementes de feijão-de-corda............................................38
4.8- Ensaio de termoestabilidade enzimática.............................................................43
5- DISCUSSÃO..........................................................................................................44
6- CONCLUSÃO........................................................................................................50
7- REFERÊNCIAS BIBLIOGRÁFICAS......................................................................51

X
ÍNDICE DE FIGURAS
Figura 1- Feijão-de-corda, (Vigna unguiculata): planta, flores e vagens...................16
Figura 2- Determinação das massas moleculares das proteases.............................28
Figura 3- Eletroforese em gel de poliacrilamida 12% (a) e “Western blotting” para
detecção de vicilinas específicas (b) da fração globulínica extraída de cotilédones
das sementes de V. unguiculata ao longo de 144 HAE.............................................28
Figura 4- Eletroforese em gel de poliacrilamida 12% da fração albumínica extraída
de cotilédones das sementes de V. unguiculata ao longo de 144 HAE.....................29
Figura 5- Eletroforese em gel de poliacrilamida 12% da fração de corpos protéicos
extraída de cotilédones de sementes de V. unguiculata ao longo de 144 HAE, na
presença (a) e ausência de agente (b) redutor (β-mercaptoetanol) e western blotting
(c) para detecção de vicilinas específicas..................................................................31
Figura 6- Eletroforese em gel de poliacrilamida 12% da fração de proteínas do
citosol extraídas de cotilédones de sementes de V. unguiculata ao longo de 144
HAE, na preseça de agente redutor (β-mercaptoetanol) (a) e Gelatina-PAGE
(b)...............................................................................................................................32
Figura 7- Eletroforese em gel de poliacrilamida 12% contendo 0,1% de gelatina, da
fração albumínica extraída de cotilédones das sementes de V. unguiculata ao longo
de 120 HAE, na presença de diferentes inibidores de proteinases
cisteínicas...................................................................................................................34
Figura 8- Análise densitométrica de atividades proteinásicas de cotilédones
germinantes de V. unguiculata em função da mobilidade relativa (Rf) da principal
banda de atividade.....................................................................................................35
Figura 9- Análise densitométrica de atividades proteinásicas de cotilédones
germinantes de V. unguiculata em função da mobilidade relativa (Rf) da principal
banda de atividade.....................................................................................................36
Figura 10- Eletroforese em gel de poliacrilamida 12% da fração albumínica extraída
de cotilédones das sementes de V. unguiculata ao longo de 144 HAE. (a)- Gelatina-
PAGE com β-mercaptoetanol. (b)- Gelatina-PAGE sem β-
mercaptoetanol...........................................................................................................39
Figura 11- Migração anômala de proteases: sobreposição de géis 2D desnaturantes
com e sem gelatina 0,1%. Imagens invertidas no Photo Impact das
imagens......................................................................................................................40

XI
Figura 12- Eletroforese bidimensional nativo/SDS-PAGE 12% (a) e nativo/SDS 12%
com gelatina 0,1% (b) da fração albumínica extraída de cotilédones (0 HAE semente
quiescente) das sementes de V. unguiculata.............................................................41
Figura 13- Eletroforese bidimensional nativo/SDS-PAGE 12% (a) e nativo/SDS 12%
com gelatina 0,1% (b) da fração albumínica extraída de cotilédones (108 HAE e 108
HAE incubado com PMSF (c)) das sementes de V. unguiculata...............................41
Figura 14- Sobreposição das imagens da eletroforese bidimensional nativo/SDS-
PAGE 12%.................................................................................................................42
Figura 15- Determinação da estabilidade enzimática por gelatina-PAGE em gel de
poliacrilamida 12% da fração albumínica extraída de cotilédones das sementes de V.
unguiculata em 108 HAE............................................................................................43
ÍNDICE DE TABELAS
Tabela 1- Classificação botânica do feijão-de-corda, Vigna unguiculata...................16

XII
LISTA DE ABREVIATURAS
1D- Eletroforese unidimensional (primeira eletroforese)
2D- Eletroforese bidimensional (segunda eletroforese)
BSA- Albumina sérica bovina
DAB- Diaminobenzidina
DFP – Diisopropilfluorfosfato
DNA- Ácido Desoxirribonucléico
DTT- Ditiotreitol
E-64- L-trans-epoxi-succinil-L-leucilamido-4-guanidino-butano
EDTA- Ácido etileno diamino tetracético
FCP- Fração de corpos protéicos
FC- Fração citoplasmática
HAE- Horas após embebição
HCl- Ácido clorídrico
LEAs- “Late-embryogenesis abundant”
NaCl- Cloreto de sódio
PAGE- Eletroforese em gel de poliacrilamida
PBS- Tampão fosfato-salino
PMSF- Fenil metil sulfonil fluoreto
PVP- Polivinil pirrolidona
Rf- Mobilidade relativa
RNA- Ácido Ribonucléico
SBTI – inibidor de tripsina de soja
SDS- Dodecil sulfato de sódio
TPBS- Tampão fosfato-salino contendo Tween-20
VPEs- Enzimas processadas no vacúolo

XIII
RESUMO
Proteinases cisteínicas melhor caracterizadas são as que participam da
degradação de proteínas de reserva durante a germinação de sementes. No
presente trabalho nós observamos dois grupos de atividades de proteinases
cisteínicas, um grupo de alto peso molecular (45 a 97 kDa) e outro grupo com peso
molecular (15 a 29 kDa) que estão temporalmente ativas após 24 e 48 após
embebição (HAE), respectivamente, em cotilédones de feijão-de-corda em
germinantes. Os grupos anteriormente citados são encontrados ambos em corpos
protéicos e em frações citoplasmáticas, enquanto este último está presente
principalmente fora dos corpos protéicos. Uma terceira atividade de proteinase
cisteínica de aproximadamente 45 kDa está especificamente ativa em cotilédones
quiescentes, 12 e 60 horas após embebição (HAE). A mobilização das principais
proteínas de reserva, as vicilinas, foi mais notável a partir de 60 HAE em diante,
aumentando sua mobilização até 144 HAE, quando as pequenas subunidades de
vicilinas estavam em seus baixos níveis. Proteinases cisteínicas de cotilédones
germinantes foram geralmente susceptíveis a inibidores como iodoacetamida, E-64,
ácido iodoacético e pCMB exceto para proteinases de ~45 kDa, que não foram
afetadas por nenhuma concentração dos inibidores usados. β-mercaptoetanol
também reduziu em 50 % a atividade gelatinolítica das proteases. Por uma
combinação de eletroforese bidimensional (nativo / SDS-PAGE) foi observada uma
aparente disposição linear dos produtos de quebra, bem como das atividades de
proteinases. O resultado pode indicar um complexo de eventos proteolíticos
seqüenciais onde proteinases induzem ou ativam novas proteinases, que podem
atuar como diferentes agregados ou zimogênios, e estes produtos de hidrólise
aparecem em uma linha constante aumentando sua proporção Rf x Mr.
PALAVRAS CHAVES: Cotilédones, corpos protéicos, germinação, proteinases
cisteínicas, proteínas de reserva, vicilinas e Vigna unguiculata.

XIV
ABSTRACT
Cysteine proteinases are the best characterized proteases among those
involved with storage protein mobilization during seed germination. In the present
work we show two major groups of cysteine proteinase activities, one of higher (45
to 97 kDa) and other with lower (15 to 29 kDa) molecular masses which are
temporally activated after 24 and 48 HAI, respectively, in germinating cowpea
cotyledons. The former group is found both in protein bodies and in cytoplasmic
fraction, while the latter is mostly present outside protein bodies. A third cysteine
proteinase activity of ~45 kDa was specifically active at the quiescent cotyledons and
at 12 and 60 hours after imbibition (HAI). Major vicilin mobilization was more
pronounced from 60 HAI onwards and steadily increased until 144 HAI, when low
levels of the smallest vicilin subunits were present. Cysteine proteinases from
germinating cotyledons were generally susceptible to the iodoacetamide, E-64,
iodoacetic acid and pCMB inhibitors except for the ~45 kDa proteinase, which was
not affected by any of the inhibitors at the used concentrations. β-Mercaptoethanol
also reduced by 50 % the gelatinolytic activity of the proteases. By a two-dimensional
native / SDS-PAGE combination it was observed an apparent linear arrangement of
protein breakdown products as well as of proteinase activity spots. The finding may
indicate a complex set of sequential proteolytic events where proteinases induces or
activates new proteinases, which may act upon different aggregates or zymogens,
and these hydrolysis products appear in a line of constant increasing Rf x Mr ratio.
KEY WORDS: Cotyledons, protein bodies, germination, cysteine proteinases,
reserve proteins,vicilins and Vigna unguiculata.

2
1- INTRODUÇÃO 1.1- Importância das sementes
As sementes são muito importantes na vida das plantas superiores. O
sucesso no estabelecimento do novo indivíduo é fortemente determinado pelas
características fisiológicas das sementes, visto que estas são capazes de se
defenderem dos estresses causados pelo ambiente, além de possuírem todas as
reservas necessárias ao desenvolvimento da planta jovem (Bewley e Black, 1994).
As sementes também são importantes por sua utilização como alimento pelo
homem. Aproximadamente 70% da alimentação humana são provenientes de
sementes, principalmente dos cereais e leguminosas (Bewley e Black, 1994). O
constante crescimento das civilizações tornou de grande importância o aumento da
produção de alimentos ricos em proteínas de boa qualidade e baixos custos.
Constituintes como proteínas, lipídeos e carboidratos justificam a ampla utilização
das sementes como fonte alimentar de alta qualidade (Xavier-Filho,1993).
Em virtude do valor nutricional, as sementes foram, desde muito cedo,
selecionadas em função das características adequadas ao consumo pelo homem. O
conhecimento da composição química das sementes é, portanto, essencial por
várias razões: 1) as sementes constituem uma fonte de alimento para o homem e
para os animais; 2) elas são importantes fontes de matéria prima para a medicina e
farmacologia; 3) possuem anti-metabólitos adversos à nutrição humana e animal; 4)
contêm reservas alimentares e substâncias de crescimento que influenciam na
germinação, no vigor da planta jovem, assim como também, no seu uso industrial e
agronômico (Xavier-Filho,1993).
A figura abaixo esquematiza o ciclo de vida de uma angiosperma:

3
1.2- Germinação
A germinação de sementes é o processo que envolve eventos que são
iniciados com a embebição de água pela semente seca e termina com a elongação
do eixo embrionário. O sinal visível de que a germinação está completa é a
conseqüente saída da radícula do interior da semente. Eventos subseqüentes,
incluindo a mobilização de reservas estocadas, estão associados com o crescimento
e desenvolvimento da plântula (Bewley, 1997; Vidal e Vidal, 2000).
Este processo é dependente de fatores internos (tais como a dormência) e de
fatores ambientais incluindo água, temperatura e oxigênio (Alberts et al., 1997).
A absorção de água pela semente madura é dividida em três fases. A fase
inicial é caracterizada por uma rápida tomada de água (fase I), seguida por uma fase
de estabilização (fase II). Posteriormente, somente após a germinação estar
completa, ocorre uma nova fase de absorção de água (fase III). Sementes
dormentes não completam a germinação, conseqüentemente, estas não entram na
fase III (Bewley, 1997). O influxo de água nas células da semente seca durante a
fase I resulta em alterações estruturais temporais, particularmente nas membranas,
culminando em um imediato vazamento de solutos e de metabólitos de baixo peso
molecular. Dentro de pouco tempo após a re-hidratação, as membranas retomam a
sua configuração estável (Bewley, 1997).
Esquema 1: Esquema demonstrando a absorção de água pela semente em
função do tempo. Adaptado de Bewley e Black, (1994).

4
Após o início da embebição, a semente rapidamente reassume as suas
atividades metabólicas. As estruturas e enzimas necessárias para essa retomada
inicial de atividade já estão presentes na semente seca (Bewley, 1997). Uma das
primeiras mudanças após a embebição é a atividade respiratória, que pode ser
detectada em poucos minutos. Posteriormente, inicia-se a segunda fase, que é
caracterizada por um declínio nessa atividade até que a radícula penetre as
estruturas ao redor, quando ocorre um novo aumento da atividade respiratória
(Botha et al., 1992; Bewley e Black, 1994). A ativação da via glicolítica, da via das
pentoses fosfato, bem como a ativação das enzimas do ciclo de Krebs, ocorrem na
fase I de absorção de água (Nicolás e Aldasoro, 1979; Sato et al., 1988).
Sabe-se que tecidos de sementes quiescentes contêm mitocôndrias, e
embora estas organelas sejam pobremente diferenciadas como conseqüência da
maturação, elas contêm níveis suficientes de enzimas do ciclo de Krebs e oxidases
que fornecem uma quantidade adequada de ATP para suportar o metabolismo por
várias horas, após o início da embebição (Ehrenshaft e Brambi, 1990; Attucci et al.,
1991).
Durante a germinação há o aparecimento de dois padrões distintos de
desenvolvimento de mitocôndrias. Estes padrões, que são observados em
cotilédones, dependem das reservas estocadas na semente. Em sementes que
armazenam amido, o reparo e ativação de pró-plastídeos pré-existentes são
predominantes; no entanto, em sementes que armazenam óleos, há uma produção
de novas mitocôndrias (Morohashi e Bewley, 1980; Morohashi, 1986).
Estas observações implicam em uma coordenada regulação da expressão
dos genomas nucleares e mitocondriais em plantas, iniciada durante os primeiros
estágios da germinação (Bewley, 1997).
A emergência da radícula através das estruturas ao redor do embrião é o
evento que termina a germinação e marca o início do desenvolvimento da nova
plântula. Esta emergência, resultante do alongamento de tecidos radiculares, pode
ou não ser acompanhada por divisões celulares. Três possíveis mecanismos para o
início do crescimento da radícula são considerados. A primeira possibilidade é que,
durante a germinação, o potencial osmótico das células da radícula torna-se mais
negativo devido ao acúmulo de solutos, talvez como resultado da hidrólise de
reservas poliméricas presentes dentro das células da radícula. A diminuição do
potencial osmótico leva ao aumento da tomada de água, e resulta no aumento do

5
turgor, levando a extensão celular. No entanto, não há evidências consistentes para
esta mudança no potencial osmótico durante a germinação (Welbaum e Bradford,
1990; Bradford, 1995). A segunda possibilidade é que a extensibilidade da parede
celular da radícula permita o seu crescimento. No entanto, o mecanismo pelo qual
as células da radícula iniciam a sua extensibilidade diferencial ainda não é
conhecido. O afrouxamento da parede celular poderia resultar da clivagem e
rearranjo de moléculas de xiloglucanos, que amarram as microfibrilas de celulose
adjacentes (Wu et al., 1994). Um mecanismo alternativo para o afrouxamento da
parece celular seria através da ação de proteínas chamadas de expansinas, que
têm habilidade para romper pontes de hidrogênio entre polímeros da parede celular.
Expansinas têm sido fortemente implicadas na expansão de hipocótilos de pepinos
(McQueen-Mason e Cosgrove, 1995).
A terceira possibilidade é o enfraquecimento dos tecidos da extremidade ao
redor da radícula. O declínio da resistência é provavelmente devido à ação de
enzimas do tipo hidrolases, tais como hemicelulases, produzidas e secretadas pelo
endosperma adjacente (Bewley, 1997).
O alimento para a plântula crescer e desenvolver-se na primeira fase da
germinação é retirado das reservas contidas na própria semente. Esgotadas as
reservas alimentares da semente, a plântula, nesta fase, já possui pêlos radiculares
e parênquima clorofiliano nas folhas, de tal modo que já pode retirar do solo, água e
minerais, e fotossintetizar matérias orgânicas (Vidal e Vidal, 2000).
Existem dois tipos de germinação: a epígea e a hipógea. A germinação
epígea ocorre quando os cotilédones saem da semente e se elevam acima do solo,
como por exemplo, em feijão e mamona. Enquanto a germinação hipógea ocorre
quando os cotilédones permanecem sob a terra; como exemplos, estão o milho e a
ervilha (Vidal e Vidal, 2000).
1.2.1- Eventos pós-germinativos
A síntese de DNA, RNA e proteínas pode ocorrer em sementes com um
conteúdo de água de aproximadamente 50%. As primeiras atividades de sementes
sob embebição são associadas ao reparo dos danos acumulados durante a
secagem e período de armazenamento das sementes, como o reparo do DNA. A
formação de polissomos, a partir de ribossomos livres, também acontece cedo
durante a embebição, de modo a criar o maquinário para a tradução de RNAs

6
mensageiros (mRNAs) em proteínas (Bewley e Black, 1994). A síntese de proteínas
é iniciada usando, em primeiro lugar, os mRNAs preexistentes acumulados durante
o desenvolvimento e a maturação das sementes; posteriormente, a síntese depende
de novos mRNAs, recentemente sintetizados durante a embebição (síntese de
novo). Como exemplo, nas primeiras horas de embebição a síntese de proteínas em
embriões de rabanete é insensível a cordicepina (um composto químico inibidor da
síntese de RNA), indicando que o mRNA já existente é que está sendo usado.
Entretanto, depois de algumas horas, esse mRNA é degradado, e a síntese de
proteínas torna-se dependente do mRNA novo, recentemente sintetizado (Bewley e
Black,1994). Acredita-se que as proteínas codificadas por mRNAs residuais são
importantes durante o processo de maturação e desidratação da semente, dentre
estas, podemos citar as proteínas LEAs (“Late-embryogenesis abundant”), que
protegem as membranas durante a desidratação e podem ser rapidamente
degradadas após a embebição (Jiang e Kermode, 1994; Han et al., 1996, Hong-Bo
et al., 2005). Proteínas LEAs estão presentes nas sementes de plantas superiores.
Estas proteínas também são encontradas em plântulas, raízes e outros órgãos da
planta (Chen et al., 2002), estando principalmente localizadas no citoplasma e
regiões nucleares (Zhang et al., 2002; Chen et al., 2002). Em plântulas de algodão
submetidas a estresses (hídricos e altas temperaturas) ocorreu o acúmulo de
mRNAs para proteínas LEAs (Li et al., 1998; Wisniewski e Zagdanska, 2001; Yang et
al., 2002; Chen et al., 2002; Jiang e Huang, 2002). Zhang et alli, em 2002,
demonstraram que a expressão de genes para proteínas LEAs é induzida por
diferentes tipos de estresses, provavelmente com o intuito de manter o metabolismo
normal de plantas superiores sob condições severas. Novos mRNAs são transcritos
durante os processos germinativos; a maioria destes, provavelmente, codifica para
proteínas essenciais para o suporte do metabolismo celular normal (Bewley e
Marcus, 1990). Muitas enzimas requeridas para a mobilização de reservas são
sintetizadas de novo, constituindo-se em alguns dos produtos iniciais da síntese de
proteínas (Bewley, 1997).
A biossíntese e a degradação de proteínas nas sementes representam
importantes fatores na regulação de fontes de nitrogênio. Durante a mobilização de
proteínas nas sementes, proteinases desempenham o importante papel de gerar
aminoácidos que serão utilizados para biossíntese de novas proteínas da própria

7
semente ou da futura planta, já que muitas destas proteínas desempenham um
papel de reserva nutritiva para a germinação.
1.2.2- Proteólise em sementes
Proteases são enzimas proteolíticas que catalisam a hidrólise de ligações
peptídicas em outras proteínas. As proteases são classificadas como endo e
exopeptidases, segundo Storey e Wagner (1986). Endopeptidases, também ditas
proteinases, são enzimas que clivam ligações peptídicas internas. Estas são ainda
convenientemente sub-classificadas, de acordo com seu mecanismo de ação, em
quatro grupos mecanísticos, segundo Storey e Wagner (1986):
proteinases serínicas: são enzimas proteolíticas que possuem o aminoácido
serina envolvido no processo de catálise. Possuem pH ótimo em torno de 8 e 9,
sendo sua atividade inibida por aprotinina, SBTI, DFP e outros.
proteinases cisteínicas: são enzimas proteolíticas que possuem o aminoácido
cisteína envolvido no processo de catálise. Possuem pH ótimo em torno de 5 e 6 e
sua atividade é inibida por E-64, cistatina, leupeptina, e outros.
proteinases aspárticas: são enzimas proteolíticas que possuem o aminoácido
ácido aspártico envolvido no processo de catálise. Possuem pH ótimo em torno de 3
e 4 e sua atividade é inibida por pepstatina e outros.
metaloproteinases: são enzimas proteolíticas que possuem um metal envolvido
no processo de catálise. Possuem pH ótimo em torno de 8 e 9 e sua atividade é
inibida por EDTA, 1,10-fenantrolina e outros.
Vários grupos têm relatado, que a maioria do conhecimento sobre proteinases
em plantas refere-se a proteinases da classe cisteínica. Algumas destas proteinases
de plantas têm sido amplamente utilizadas na indústria alimentícia e como exemplo
podemos citar a bromelaína e a papaína (Uchikoba et al., 1998). As proteinases
cisteínicas melhor caracterizadas são aquelas que participam da degradação de
proteínas de reserva durante a germinação. Entretanto, menos se sabe sobre a
presença destas proteinases durante o desenvolvimento das sementes e até mesmo
nas sementes quiescentes (Yamauchi et al., 1992; Tanaka et al., 1993).
Em 1975, Chrispeels e Boulter mostraram que cotilédones maduros de
sementes de Vigna radiata têm uma atividade endopeptidásica do tipo cisteínica,
que seria um pré-requisito para o rápido catabolismo de proteínas de reserva, tão
logo se inicia o processo de germinação da semente. Foram ainda demonstradas

8
novas atividades enzimáticas, não presentes em corpos protéicos isolados de
sementes secas, mas que precisam ser ativadas ou sintetizadas, e possivelmente
inclusas nos corpos protéicos, antes do início da degradação de proteínas de
reserva, pós-germinação (Harris e Chrispeels, 1975). Estudos ultra-estruturais
demonstraram que o retículo endoplasmático rugoso prolifera e pode dar origem a
vesículas que se fusionam com os corpos protéicos antes da digestão de proteínas
de reserva dos cotilédones (Chrispeels et al., 1976).
Baumgartner e Chrispeels (1977) purificaram e caracterizaram a vicilina
peptídeo-hidrolase, a principal endopeptidase cisteínica de cotilédones de Vigna
radiata. Ela age sobre a proteína 7S de reserva isolada de sementes quiescentes de
Vigna radiata, sendo sintetizada três dias após a germinação, como resultado da
síntese de novo dependente do eixo embrionário (Chrispeels et al., 1976).
A presença de enzimas durante o desenvolvimento de sementes, que clivam
especificamente a ligação peptídica Asn-Gly, localizada na junção entre as cadeias
e da molécula de pró-legumina, transformando-a na sua forma madura,
legumina, a principal proteína de reserva em sementes de algumas leguminosas,
tais como a soja (Hara-Nishimura et al., 1998). Nong et alli (1995) demonstraram
ainda a presença de proteinases cisteínicas, em cotilédones de soja em
desenvolvimento, pertencentes à família da papaína e que mostraram alta
homologia com proteinases induzidas por estresse. Estas proteinases poderiam
estar sendo sintetizadas em resposta ao processo fisiológico de dessecação durante
a maturação da semente ou por outros fatores de estresses do meio. Eles também
não descartaram a possibilidade destas proteinases estarem envolvidas no
“turnover” vacuolar de proteínas e enzimas envolvidas em diversos eventos
metabólicos dos cotilédones em desenvolvimento. Ainda neste trabalho, foi
demonstrado que estas proteinases cisteínicas também se encontravam em
sementes secas de soja e que provavelmente estariam envolvidas na fase inicial de
degradação de proteínas de reserva da soja, após germinação.
Mikola e Jones (2000) caracterizaram endopeptidases de cevada que
hidrolizam as principais proteínas de reserva (globulinas) destas sementes em
germinação. Estas proteinases foram classificadas como proteinases cisteínicas e
mostraram-se ativas em pH 3,8. Como estas proteinases hidrolizam rapidamente
globulinas quando em pH 3,8, os autores sugerem que há uma provável
compartimentalização protéica, dependente de pH, dentro do endosperma.

9
A maioria dos estudos mostra a mobilização de proteínas de reserva de
sementes de leguminosas sendo feita por uma única endopeptidase cisteínica
(Baumgartner e Chrispeels, 1977), mas outros estudos têm demonstrado que há
dois grupos de enzimas distintas envolvidas neste processo; proteinases A e
proteinases B (Shutov e Vaintraub, 1987). Neste trabalho, os autores sugerem que
as proteinases A são responsáveis por iniciarem o processo de proteólise das
proteínas de reserva, gerando então peptídeos que posteriormente sofrerão a ação
de endopeptidases A e B em conjunto. Ambas endopeptidases foram classificadas
como sendo proteases da classe cisteínica. Posteriormente, Kembhavi et alli (1993)
demonstraram, através de ensaios fluorimétricos específicos, que existem duas
endopeptidases cisteínicas envolvidas na mobilização de proteínas de reserva de
sementes de Vigna acuntifolia.
Portanto, como conclusão de diferentes estudos, acredita-se que em
sementes de leguminosas a proteólise limitada das proteínas de reserva inicia-se
dias após o crescimento da plântula e é dirigida por proteinases cisteínicas que, em
geral, encontram-se ausentes nas sementes secas. Porém, em cereais, como no
caso de sementes de trigo selvagem, foi observado que a proteólise inicial ocorre
devido à ação de uma metaloproteinase que se encontra presente em sementes
secas. A hidrólise prematura destas proteínas de reserva é impedida devido à
presença de um inibidor desta protease nestas sementes e da conseqüente
formação de um complexo enzima-inibidor (Elpidina et al., 1991).
Proteinases semelhantes à proteinase A, anteriormente descrita por Shutov e
Vaintraub (1987), denominam-se de vignaínas e as semelhantes à proteinase B,
denominam-se de legumaínas. Como já mencionado, Shutov e Vaintraub (1987)
sugeriram que proteinases A estariam envolvidas em disparar a clivagem proteolítica
parcial, durante os estágios iniciais de degradação das globulinas em cotilédones de
sementes germinantes de Vicia sativa L. Mais tarde, em 1997, Becker et alli
demonstraram que a proteinase A não estava envolvida na degradação de
globulinas de reserva de Vicia sativa, já que tal enzima não se encontra presente
durante estes estágios iniciais de germinação, quando as globulinas são
degradadas.
Rotari et alli (1997) isolaram e caracterizaram parcialmente uma proteinase
cisteínica de sementes germinantes de Phaseolus vulgaris que participaria na
degradação da faseolina, principal proteína de reserva destas sementes. Ela

10
pertence ao grupo de proteinases cisteínicas homólogas às proteinases A, também
presentes em sementes germinantes de outras plantas.
Müntz et alli (1998) realizaram uma análise do padrão temporal e
histoquímico da degradação de proteínas de reserva e de proteinases cisteínicas em
eixos embrionários e cotilédones durante a germinação e estágios iniciais de
desenvolvimento da plântula de Vicia sativa L. Neste trabalho, os autores sugeriram
que durante os primeiros dois dias após a embebição, a biossíntese de proteínas no
eixo embrionário é principalmente alimentada pela degradação de proteínas de
reserva do próprio eixo e que após a exaustão destas proteínas no eixo, inicia-se a
degradação de globulinas nos cotilédones para obtenção de aminoácidos para o
posterior desenvolvimento da plântula. Os autores encontraram cinco diferentes
proteinases cisteínicas, quatro delas sendo da família da papaína, incluindo a
proteinase A, e uma classificada como uma proteinase B, recentemente associada à
família das “legumaína-like”, sendo os dois grupos responsáveis pela degradação de
proteínas de reserva. Legumaínas de plantas são chamadas usualmente de
Enzimas Processadas no Vacúolo - VPEs, mas elas também estão presentes na
parede celular e sua função não está apenas restrita a processamento de proteínas
precursoras; inclui também a quebra de proteínas no vacúolo e na parede celular
(Müntz et al., 2002).
Proteinases cisteínicas da família da papaína são enzimas bem estudadas e
contribuem para o “turnover” protéico em animais e plantas. Vários estudos têm
discutido a indução da expressão destas proteinases por giberelinas (hormônio
vegetal) durante a germinação de sementes de cevada (Rogers et al., 1985; Koehler
e Ho, 1990), arroz (Watanabe et al., 1991) e milho (De Barros e Larkins, 1990;
Domoto et al., 1995). Considera-se que esta indução de proteinases cisteínicas por
giberelinas é necessária para que ocorra a degradação de proteínas de reserva e
conseqüente fornecimento de aminoácidos que desempenharão um papel
importante na síntese protéica de novo na germinação. Por outro lado, plantas
também acumulam inibidores de proteinases cisteínicas em sementes maduras (Abe
et al., 1987; Fernandes et al., 1991).
Proteinases cisteínicas foram também purificadas de sementes de trigo
(Triticum aestivum) (Kuroda et al., 1997) e soja (Asano et al., 1999) e suas
propriedades estudadas através do uso de substratos sintéticos como Z-Phe-Arg-
MCA. Bottari et alli (1996) também isolaram e caracterizaram parcialmente uma

11
proteinase cisteínica que provavelmente encontra-se envolvida na clivagem de
gliadina (proteína de reserva de trigo) em sementes germinantes de Triticum durum.
Ainda com proteinases cisteínicas, foi realizado um estudo comparativo da
seqüência completa de aminoácidos da ananaína (do melão) com a bromelaína (do
abacaxi) e outras proteinases cisteínicas de plantas, onde foi demonstrado que a
ananaína possui características próprias que incluem uma série hidrofóbica de
aminoácidos próximos à His-157. Estas diferenças podem contribuir para a ligação
desta proteinase a substratos e inibidores distintos dos da bromelaína (Lee et al.,
1997).
Schlereth et alli (2000) fizeram uma comparação da mobilização da globulina
e a expressão de proteinases cisteínicas em eixos embrionários e cotilédones
durante a germinação e crescimento de plântulas de Vicia sativa L. Neste trabalho,
eles concluíram que a mobilização da globulina de reserva em sementes
germinantes de Vicia sativa é iniciada por um complexo de proteinases cisteínicas
estocadas na semente seca, enquanto que a mobilização do restante das globulinas
é predominantemente mediada por um complexo de proteinases cisteínicas,
sintetizadas de novo.
Acredita-se que, para mobilização de proteínas de reserva em cotilédones de
sementes de dicotiledôneas, quatro proteinases cisteínicas do tipo papaína estão
envolvidas (CPR1, CPR2, proteinase A e CPR4) e duas legumaínas (VsPB2 e
proteinase B) (Fischer et al., 2000; Schlereth et al., 2000;). Adicionalmente,
Tiedemann et alli (2001) apresentaram evidências imunohistoquímicas de que duas
proteinases do tipo papaína (CPR1 e CPR2) e uma legumaína-like (proteinase B)
estão envolvidas na mobilização de globulinas em sementes de Vicia sativa L..
Também neste caso, proteinases formadas tardiamente durante a embriogênese
iniciam a mobilização de proteínas de reserva no eixo embrionário, antes de síntese
de novo de outras proteases (Tiedemann et al., 2001). Proteinases cisteínicas do
tipo papaína-like (CPR1 e CPR2) são responsáveis pela mobilização de vicilinas,
globulinas 7 S armazenadas, de Vicia faba L.
A complexidade do processo de germinação em sementes de dicotiledôneas
e a função de proteinases cisteínicas na degradação de proteínas de reserva vem
sendo discutida por Fischer et alli (2000), Schlereth et alli (2000), Tiedemann et alli
(2001) e Müntz et alli (2002). Embora muitos detalhes ainda estejam para ser
descobertos, a expressão tempo e tecido-específicos, bem como a interação entre

12
proteinases cisteínicas e cistatinas, podem ser fenômenos-chave envolvidos na
regulação do metabolismo protéico de sementes (Flores et al., 2001).
1.3- Proteínas de reserva em sementes
As sementes de plantas contêm proteínas de várias classes, que são
depositadas durante o desenvolvimento. As proteínas de reserva classicamente
encontradas na maioria das dicotiledôneas são as globulinas, que são
caracteristicamente insolúveis em água e solúveis em soluções salinas e estão
presentes tipicamente em sementes de leguminosas. Elas são classificadas em dois
grupos, quanto ao coeficiente de sedimentação; vicilinas 7 – 8 S e leguminas 11 –
12 S (Bewley and Black, 1994).
Prolaminas, solúveis em álcool, são geralmente encontradas em sementes de
cereais e são ricas em prolinas e amidas (Bewley and Black, 1994). As albuminas
(2S) também são amplamente encontradas em dicotiledôneas, e são solúveis em
água. As glutelinas, presentes no trigo, no milho e em outros cereais, são insolúveis
em soluções aquosas neutras ou salinas e em álcool, mas podem ser extraídas em
soluções básicas (Shewry e Casey, 1999).
Classificações de proteínas de reserva baseiam-se em dois critérios: função e
relações bioquímicas e moleculares. Em termos de função, podemos classificá-las
em três classes principais: 1- proteínas de reserva, cuja função é armazenar
nitrogênio, carbono e enxofre; 2- proteínas estruturais e metabólicas
(“housekeeping”), que são essenciais para o crescimento e a estrutura da semente;
e 3- proteínas de proteção, que podem conferir resistência a pestes e patógenos ou
dessecação. Em alguns casos, a proteína pode apresentar uma combinação de
funções como reserva e proteção (Bewley, 2001; Fernandes e Xavier-Filho, 1998).
1.3.1- As globulinas
As sementes de leguminosas caracteristicamente contêm proteínas de
reserva do tipo globulinas, chegando a cerca de 50% das proteínas totais,
compreendendo leguminas e vicilinas. Estas proteínas são depositadas durante a
maturação e são utilizadas durante a germinação para o desenvolvimento dos
tecidos e órgãos da nova planta (Shewry e Lucas, 1997).
As vicilinas, que fazem parte do grupo das globulinas 7S – 8S, devido ao seu
tamanho e massa molecular, são proteínas de reserva de cadeia única, sem pontes

13
dissulfeto, formando trímeros com massa molecular de aproximadamente 150 kDa
(Shewry, 1995) e cadeias polipeptídicas de 56, 54 e 52 kDa (Derbyshire et al., 1976.,
Fernandes e Xavier-Filho, 1998). As vicilinas podem ser glicosiladas e são bastante
heterogêneas. Apresentam composição de aminoácidos com altas concentrações de
ácido aspártico, ácido glutâmico, arginina, fenilalanina e leucina tendo, no entanto,
concentrações mínimas de aminoácidos sulfurados como metionina e cisteína
(Carasco et al., 1978; Macedo et al., 1995). As três cadeias polipeptídicas das
vicilinas são codificadas por três famílias distintas de genes. Cada família de gene é
constituída aparentemente de seis genes diferentes (Higgins, 1984). Como proteína
de reserva, a vicilina serve principalmente como uma fonte de nitrogênio orgânico e
esqueleto carbônico para as reações biossintéticas necessárias ao crescimento da
plântula. Alternativamente, seus componentes aminoácidos podem ser usados como
combustível para o metabolismo energético. Por isto, as vicilinas são, como outras
proteínas de reserva, gradualmente degradadas durante a germinação e
crescimento da plântula (Freitas et al., 2006).
As vicilinas apresentam uma grande homologia na seqüência dos resíduos de
aminoácidos com as leguminas. As leguminas são globulinas com massas
moleculares entre 300 e 400 kDa, formadas por seis subunidades, com massas
moleculares de aproximadamente 50 a 60 kDa e cada subunidade é composta de
duas cadeias polipeptídicas ligadas por pontes dissulfeto entre resíduos de cisteína,
em posições conservadas.
Vicilinas isoladas de sementes de Vigna unguiculata interferiram com a
germinação de esporos ou conídios dos fungos Fusarium oxyporum, F. solani,
Colletotrichum musae, Phytophtora capsici, Neurospora crassa e Ustilago maydis
(Gomes et al., 1997). Gomes et alli (1998) também mostraram que vicilinas isoladas
de diferentes sementes de leguminosas inibem o crescimento de leveduras, e
sugeriram que as vicilinas ligam-se á estruturas de leveduras que contêm quitina e
esta associação poderia resultar em inibição de bombas de H+, do crescimento das
células e da formação de esporos. O efeito inibitório de vicilinas de sementes de V.
unguiculata sobre o desenvolvimento de F. oxysporum e de células da levedura S.
cerevisiae foi observado por microscopia eletrônica, mostrando que há uma inibição
na germinação dos esporos do fungo e um desenvolvimento anormal das células de
levedura. Estes resultados sugerem que o efeito inibitório das vicilinas de sementes
de V. unguiculata no desenvolvimento dos fungos pode estar associado a sua

14
habilidade de ligar-se à superfície celular dos microrganismos, parede celular e/ou
membrana plasmática (Gomes et al., 1998).
Sales et alli (1996) mostraram que vicilinas de V. unguiculata têm a
propriedade de se ligar reversivelmente a matrizes de quitina. Formas variantes
dessas proteínas poderiam ligar-se à quitina presente no intestino médio de C.
maculatus, impedindo o desenvolvimento da larva no interior da semente.
Vicilinas são um exemplo de proteínas de reserva que apresentam a função
dual de fornecer seus esqueletos carbônicos e nitrogenados para o desenvolvimento
de plântulas, bem como participar em processos de defesa das sementes
(Fernandes e Xavier-Filho, 1998).
1.4- Corpos protéicos
No desenvolvimento das sementes de plantas superiores, o retículo
endoplasmático (RE) é o local de síntese das proteínas de reserva que são
posteriormente transportadas até seu local de acúmulo, os vacúolos de reservas,
chamados de corpos protéicos. A síntese e a deposição dessas proteínas estão
sujeitas a uma regulação espacial e temporal, sendo que, em cada espécie pode
aparecer em diferentes estágios do desenvolvimento (Herman e Larkins, 1999).
Na maioria das dicotiledôneas e das gimnospermas, a formação dos corpos
protéicos envolve a fragmentação de um grande vacúolo central após as proteínas
de reserva terem sido transportadas dos locais de síntese. Em geral, o transporte
das proteínas de reserva ocorre por meio do complexo de Golgi (glutelinas do arroz,
algumas prolaminas de trigo e globulinas das leguminosas), mas, em alguns cereais,
ocorre a formação direta do corpo protéico a partir do retículo endoplasmático
rugoso. Em monocotiledôneas, o corpo protéico pode ser simplesmente uma
agregação de proteínas de reserva que são lançadas no citoplasma sem qualquer
membrana de proteção. Pode ainda ser uma vesícula formada pela ruptura do
retículo endoplasmático, onde as proteínas de reserva são sintetizadas. Há também
outro tipo de vacúolo que acumula proteínas que são produzidas ao longo do
desenvolvimento da semente. São os vacúolos líticos que contêm enzimas
proteolíticas ácidas (hidrolases ácidas), cuja função é hidrolisar proteínas para o
crescimento na fase pós-germinativa, suportando o desenvolvimento inicial da
plântula. Proteases são expressas especificamente em tempo e local, e acumuladas
em diferentes compartimentos subcelulares (Van der Hoorn, 2008).

15
1.5- Feijão-de-corda, Vigna unguiculata (L.) Walpers
O feijão-de-corda foi trazido para o Brasil pelos portugueses que faziam o
tráfico de escravos, sendo introduzido primeiramente na Bahia, disseminado pelos
colonizadores através do país, e estabelecendo-se nas regiões de melhor adaptação
da cultura, em climas tropicais do Norte e Nordeste do Brasil (Freire-Filho, 1988).
Também foi indicado que o Oeste da África, mais precisamente a Nigéria, foi
o primeiro centro de origem e diversidade de Vigna unguiculata (L.) Walp. O
estabelecimento da espécie no Sudeste da Ásia teria ocorrido mais ou menos há
2300 anos a.C. e no Sul da Europa há 300 anos a.C. Nas Américas a espécie foi
introduzida nos séculos 16 e 17, proveniente do Sul da Europa e certamente do
Oeste da África, juntamente com o tráfico de escravos (Ng e Maréchal, 1985).
A área ocupada com feijão-de-corda, no mundo, está em torno de 12,5
milhões de ha, com 8 milhões (64% da área mundial) na parte Oeste e Central da
África. A parte restante está principalmente localizada na América do Sul, América
Central e Ásia, com pequenas áreas espalhadas pelo sudoeste da Europa, sudoeste
dos Estados Unidos e da Oceania. Entre todos os países, os principais produtores
mundiais são Nigéria, Niger e Brasil (Quin, 1997).
No Brasil, o feijão-de-corda é cultivado predominantemente no sertão semi-
árido da região Nordeste e em pequenas áreas na Amazônia.
A planta (figura 1) é botanicamente descrita como uma herbácea anual que
mostra grandes variações de acordo com o cultivar, condições de solo e clima.
Existem arquiteturas eretas, rasteiras, trepadeiras e arbustivas. A raiz principal é
bem desenvolvida com grande quantidade de raízes laterais próximas à superfície
do solo possuindo grandes nódulos fixadores de nitrogênio. As folhas são
alternadas, trifolioladas com o pecíolo medindo de 5 a 25 cm e sua cor é usualmente
verde escuro e o formato é ovalado. A inflorescência é um racemo axilar não
ramificado com um longo pedúnculo medindo de 5 a 60 cm. As flores são
pedunculadas e se inserem no eixo a uma certa distância umas das outras. As flores
são conspícuas, autogâmicas, podendo ser brancas, amareladas, roxas ou violetas.
Elas comumente abrem-se no início do dia e fecham-se por volta do meio dia.
Depois de abertas e polinizadas, as flores murcham e caem. As vagens variam em
tamanho, forma, cor e textura. Elas podem ser lineares ou apresentar curvatura, e
normalmente seu tamanho varia de 7,5 a 45 cm; são indeiscentes e quando

16
maduras podem ser amareladas, marrons ou violáceas e normalmente contém de 8
a 20 sementes. As sementes variam em tamanho, cor e forma (Kay, 1979).
O feijão-de-corda é altamente susceptível a pestes e doenças. Diversos
agentes patogênicos causam danos à cultura do feijão-de-corda no Brasil, como
fungos, vírus e bactérias (Allen, 1982). O gorgulho, Callosobruchus maculatus, é o
principal inseto predador de sementes de feijão-de-corda durante a armazenagem,
podendo danificar até 70% dos grãos produzidos em uma região (Santos et al.,
1977).
Figura 1- Feijão-de-corda, (Vigna unguiculata): planta, flores e vagens. Foto
feita pela autora, no Sítio Sapucaia, localizado em Santo Antônio de Pádua – RJ.
A tabela 1, abaixo, resume a classificação botânica do feijão-de-corda
(adaptado de Lima, 2001).
Tabela 1- Classificação botânica do feijão-de-corda, Vigna unguiculata.
Classificação
Subdivisão Gimnosperma
Classe Dicotiledônea
Subclasse Choripetalae
Família Leguminosae
Subfamília Papilionoideae

17
1.5.1- Proteólise em sementes de Vigna unguiculata
Em sementes de feijão-de-corda foram observados padrões complexos e
coordenados de atividades de proteinases cisteínicas e de cistatinas (inibidores
capazes de inibir a atividade biológica de proteinases cisteínicas), durante o
desenvolvimento de sementes (Fernandes et al., 1991), bem como variada
localização de cistatinas, em células e tecidos (Flores et al., 2001). Neste último
trabalho foi observada uma distribuição uniforme de cistatinas em cotilédones e
eixos embrionários de feijão-de-corda, exceto por uma maior concentração em
células epidérmicas da zona de abscisão entre os cotilédones (Flores et al., 2001).
De maneira similar, a cistatina de sementes de soja concentra-se numa camada
periférica de células cotiledonárias (Misaka et al., 1996). No entanto, a seqüência de
aminoácidos deduzida de uma cistatina clonada por Fernandes et al. (1993) foi vista
como deficiente em um peptídeo sinal N-terminal, sugerindo que esta não deve ser
transportada por um sistema secretório. Outros resultados demonstram que, além
de uma localização citoplasmática principal, cistatinas de V. vexillata, espécie não
domesticada do gênero Vigna, também são encontradas em espaços extra-celulares
de cotilédones e que, portanto, um processo de translocação nesta espécie poderia
ser admitido (Ávila et al., 1999).
Lima (2001) constatou que extratos de eixos embrionários de sementes de
Vigna unguiculata apresentaram atividade de proteinases cisteínicas durante a fase
de desenvolvimento, decaindo até o estágio de semente seca. Ainda, neste trabalho,
durante a fase de germinação só foi possível observar atividade de proteinases
cisteínicas em extratos de eixos embrionários após 24 horas de embebição e esta
mostrou-se crescente até os últimos estágios de germinação analisados. Em
extratos de cotilédones, o padrão de atividade de proteinases cisteínicas foi bastante
similar aos encontrados em eixos embrionários, onde a atividade mostrou-se
decrescente até o estágio de semente quiescente, elevando-se após 24 horas de
germinação até o último estágio analisado.
O acompanhamento dos eventos proteolíticos que estão associados ao
processo de germinação de sementes de valor agronômico traz à luz conhecimentos
sobre os componentes envolvidos e seus mecanismos de ação e regulação, os
quais podem servir de base para programas de melhoramento ou manipulação
genética de espécies cultivadas.

18
Investigações de atividades proteolíticas de sementes durante a germinação
são geralmente restritas a cotilédones e endospermas, os tecidos de reservas
nitrogenadas típicos. No entanto, é sabido que já durante as primeiras horas de
embebição das sementes ocorre síntese de proteínas tanto em cotilédones como
em eixos embrionários de sementes (Bewley e Black, 1994), apesar de atividades
proteolíticas de hidrólise de proteínas de reserva só iniciarem-se nos cotilédones
após 2 ou 3 dias de embebição (Wilson, 1986; Vaintraub, 1987; Müntz, 1996). Isso
sugere a necessidade de uma análise mais aprofundada da expressão tempo e
tecido específicas de atividades proteolíticas em sementes, o que tem sido estudado
por Müntz et alli (1998) e Schlereth et alli (2000), em Vicia sativa L., durante os
primeiros momentos da germinação.
Neste trabalho, perseguimos objetivos semelhantes aos dos autores acima
mencionados e a seguir descritos, usando como planta de estudo, Vigna
unguiculata.

19
2- OBJETIVO GERAL
Avaliar os eventos proteolíticos envolvidos nos processos germinativos e pós-
germinativos (desenvolvimento de plântulas) de feijão-de-corda (V. unguiculata),
com ênfase na atividade de proteinases da classe cisteínica e mobilização de
proteínas de reserva ao longo destes processos.
2.1- Objetivos específicos
Acompanhar a mobilização de proteínas de reserva em cotilédones, durante
os processos de germinação e desenvolvimento das plântulas;
Acompanhar as atividades de proteinases cisteínicas em cotilédones durante
os mesmos processos;
Identificar atividades proteinásicas e mobilização de proteínas de reserva em
frações de corpos protéicos e citossólicas de cotilédones durante os
processos;
Caracterizar as atividades proteinásicas encontradas nos cotilédones
germinantes de feijão-de-corda.

20
3- MATERIAIS E MÉTODOS 3.1- Sementes
As sementes de feijão-de-corda (Vigna unguiculata (L.) Walp, cultivar
EPACE-10), foram originalmente fornecidas pelo Departamento de Fitotecnia do
Centro de Ciências Agrárias da Universidade Federal do Ceará, Fortaleza.
Atualmente, sementes originadas destas matrizes são plantadas e obtidas no
campus da Universidade Estadual do Norte Fluminense e no Sítio Sapucaia,
localizado em Santo Antônio de Pádua – RJ.
3.2- Germinação e desenvolvimento pós-germinativo
Para os experimentos de germinação, sementes quiescentes foram
esterilizadas com álcool 70 % por um minuto e lavadas com água destilada cinco
vezes. As sementes foram embebidas em água destilada por 1 hora a 30 C, antes
de serem transferidas para placas de Petri contendo papel filtro embebido em água
destilada. Foram colocadas 30 sementes embebidas por placa. As sementes foram
mantidas em uma câmara de germinação a 30º C, em fotoperíodo de 12 horas com
luz e 12 horas sem luz. As sementes foram selecionadas (seguindo um padrão de
uniformidade fenotípica, baseado em tamanho e peso) nos tempos 0, 6, 12, 24, 36,
48, 60, 72, 84, 96, 108, 120 e 144 horas após embebição (HAE). As sementes foram
dissecadas em seus tecidos constituintes: cotilédones, eixo embrionário e
tegumentos, sendo apenas os tecidos cotiledonários utilizados nos experimentos.
Os tecidos separados foram liofilizados e macerados, a farinha resultante foi pesada
e estocada a – 20° C, para ensaios posteriores.
3.3- Extração de proteínas
As farinhas dos tecidos obtidos como descrito anteriormente, foram tratadas
com éter na proporção de 1:10 (p/v) durante 10 minutos. A suspensão foi
centrifugada por 2 minutos a 4.000 x g, sendo o sobrenadante descartado. Tal
tratamento com éter foi repetido, como descrito acima, visando a remoção máxima
de clorofila. 1,0 g das farinhas livres de clorofila foram então submetidas à extração
de proteínas adicionando-se 10 mL de tampão fosfato de potássio 0,1M com NaCl
0,5 M pH 8,0, contendo 1% de PVP (para evitar a ação de fenóis). A extração
processou-se durante duas horas, sob agitação a 4° C, e posteriormente, o material
foi centrifugado durante 30 minutos a 10.000 x g, a 4° C. Para obtenção das frações

21
de albuminas e globulinas o sobrenadante foi dialisado contra água durante 48
horas a 4°C. Após a diálise, o material foi centrifugado durante 30 minutos a 10.000
x g, a 4° C. A fração de globulina encontra-se no sedimento e a fração de albumina
encontra-se no sobrenadante. A fração globulínica foi lavada com água por três
vezes, intercaladas por centrifugação a 10.000 x g, a 4º C, para retirar os resíduos
da fração albumínica.
3.4- Isolamento de corpos protéicos
O isolamento de corpos protéicos foi realizado de acordo com Mäder e
Chrispeels (1984). Após a homogeneização das farinhas dos tecidos (1g / 2mL) em
tampão MES 100 mM (pH 5,5), EDTA 1 mM, Manitol 600 mM, os extratos foram
filtrados e centrifugados por 4 min, a 100 x g, para remoção de grânulos. O
sobrenadante foi colocado sobre uma solução de Ficoll 5%, no mesmo tampão, e
centrifugado por 20 minutos a 100 x g. O sedimento resultante foi lavado duas vezes
e ressuspenso em Tris / HCl 100 mM contendo NaCl 150 mM (pH 8,0). No
sedimento, a fração de corpos protéicos foi obtida e no sobrenadante, as proteínas
do citosol e de outros compartimentos celulares. A fração de corpos protéicos dos
tecidos cotiledonários, nos diferentes tempos de germinação e eventos pós-
germinativos, foi visualizada por SDS-PAGE. Seguida por “Western blotting” para
detecção de vicilinas. A fração citoplasmática foi visualizada por SDS-PAGE e
gelatina-PAGE.
3.5- Determinação da concentração de proteínas
A determinação da concentração total de proteínas dos tecidos foi baseada
no método descrito por Bradford (1976), usando-se albumina sérica bovina (BSA),
como proteína de referência. As concentrações foram avaliadas por leitura
espectrofotométrica, em comprimento de onda de 595 nm.
3.6- Visualização dos perfis protéicos por eletroforese em gel de poliacrilamida
As amostras protéicas de cotilédones foram ressuspensas (razão v/v) em
(tampão de amostra - Tris-HCl 0,5 M, glicerol, SDS 10%, azul de bromofenol 1%) e
visualizadas por eletroforese em gel de poliacrilamida (12%) na presença de SDS,
segundo método descrito por Laemmli (1970). A estratégia utilizada para
visualização da mobilização de proteínas de reserva foi a isovolumétrica. A

22
eletroforese foi realizada em um sistema vertical Mini Protean II da BIORAD. A
separação protéica procedeu-se sob uma corrente constante de 100 V.
O gel foi corado com solução de azul brilhante de Coomassie G 2% em água,
metanol e ácido acético (6:3:1, v/v/v), e descorado com solução composta de água
destilada, metanol e ácido acético na mesma relação de proporcionalidade usada na
preparação da solução corante.
Os marcadores de peso molecular (200 kDa: Miosina, 116 kDa: β-
galactosidase, 97 kDa: Fosforilase b, 66 kDa: Albumina bovina, 45 kDa: Albumina de
ovo, 29 kDa: Anidrase carbônica) foram usados para nos auxiliar na identificação das
massas moleculares das bandas de proteínas.
3.7- “Western blotting” para detecção de vicilinas
Para realização do “Western blotting” as proteínas dos tecidos cotiledonários,
separadas por eletroforese em gel de poliacrilmida-SDS (12%) foram transferidas
para uma membrana de nitrocelulose, de medidas idênticas ao gel. O sistema
utilizado foi o de transferência semi-seca Trans-blot Sigma.
A membrana e o gel foram equilibrados em tampão de transferência (Tris 25
mM, glicina 192 mM e metanol 20%) durante 30 e 5 minutos, respectivamente. Dez
folhas de papel filtro, com as mesmas medidas do gel e da membrana, foram
mergulhadas no mesmo tampão durante 30 minutos.
O sanduíche de transferência foi montado da seguinte maneira: 5 pedaços de
papel de filtro, seguidos da membrana de nitrocelulose, do gel e outros 5 pedaços de
papel de filtro umedecidos no tampão de transferência. Após 2 h de transferência
em amperagem de 1 mA/cm2 do gel. A membrana foi corada com Ponceau S
(Sigma) para verificação da transferência.
Após confirmação de que as proteínas foram transferidas para a membrana de
nitrocelulose, esta foi submersa em solução bloqueadora (tampão fosfato de sódio
0,1 M, NaCl 0,5 M pH 7,2 [PBS], com 2% de leite desnatado Molico em pó), por 2
horas. Posteriormente foram feitas 5 lavagens em tampão PBS-Tween-20 (TPBS)
0,05%, e a membrana foi colocada em contato com solução bloqueadora contendo
anticorpo contra vicilina, na diluição de 1: 2000, e mantida nesta solução durante a
noite (16 horas, 4ºC). Posteriormente, a solução de anticorpo foi descartada e 5
lavagens, de 20 minutos cada, com TPBS foram efetuadas. Nova solução
bloqueadora, desta vez contendo anticorpo anti-IgG de coelho conjugado com

23
peroxidase na diluição de 1: 2000, foi despejada sobre a membrana e deixada sob
agitação leve, por duas horas. Nova série de 5 lavagens em tampão TPBS e uma
última com PBS foram efetuadas.
Para a revelação da reação imunoquímica, foi utilizado o método de revelação
por diaminobenzidina (DAB), onde foram utilizados 100 µL do tampão Tris-HCl 2 M
pH 7,5, 5 mg de DAB, 0,3 mL de imidazol 0,1 M, 4,9 mL de água destilada e 5 µL de
água oxigenada 30%. Esta solução (10 mL) foi vertida sobre a membrana e mantida
por aproximadamente 10 minutos. A reação foi parada pela substituição da solução
reveladora por água destilada. Os procedimentos descritos baseiam-se no método
descrito por Towbin et alli (1979).
3.8- Detecção de atividade enzimática em gel
As amostras protéicas dos tecidos cotiledonários foram visualizadas por
eletroforese em gel de poliacrilamida (12%) na presença de SDS, contendo
gelatina em uma concentração de 0,1%, baseado na metodologia descrita
primeiramente por Heussen e Dowdle (1980). De uma solução concentrada de
gelatina (1%), uma alíquota foi utilizada de forma a alcançar-se uma concentração
final de 0,1% de gelatina, na composição do gel. A separação protéica procedeu-se
sob uma corrente constante de 100 V. Após a corrida eletroforética, o gel foi lavado
com Triton X-100 2,5 % duas vezes por 30 minutos cada; logo após, o gel foi
incubado em tampão citrato de sódio 100 mM; fosfato de sódio 100 mM; DTT 1,5
mM e Triton X-100 0,1 % pH 5.6 em banho-maria a 370 C durante 16 horas.
Posteriormente o gel foi corado com Coomassie Brilhante Blue R 2% e então
descorado com uma solução de metanol : ácido acético : água (40:10:50, v/ v/ v),
respectivamente. As bandas de atividade aparecem brancas em fundo azul.
3.9- Ação de inibidores sobre as atividades proteolíticas em gel
A caracterização da atividade proteolítica foi realizada mediante o uso de
diferentes inibidores de proteases. As amostras protéicas dos tecidos cotiledonários
foram visualizadas de acordo com o item 3.8. Após a corrida eletroforética, os géis
foram lavados com Triton X-100 2,5 %, duas vezes por 30 minutos cada; logo após,
o gel foi incubado em tampão citrato de sódio 100 mM; fosfato de sódio 100 mM;
DTT 1,5 mM e Triton X-100 0,1 % pH 5.6 contendo diferentes inibidores (E-64,
PCMB, Ácido Iodoacético e Iodoacetamida – para visualização de inibição das

24
proteinases da classe cisteínicas; PMSF – para inibição de proteinases da classe
serínica; EDTA – para inibição de proteinases da classe metaloproteases). Após
incubação em banho-maria a 370 C durante 16 horas, o gel foi corado com
Coomassie Brilhante Blue R 2% e então descorado com uma solução de
metanol:ácido acético:água (40:10:50, v/ v/ v), respectivamente. Um gel controle foi
incubado com tampão de incubação sem inibidores.
3.10- Avaliação do perfil de atividades proteinásicas a partir de géis nativo / SDS-PAGE e nativo / gelatina-PAGE de proteínas e proteinases cisteínicas de
cotilédones quiescentes e germinantes 108 HAE de feijão-de-corda
As amostras protéicas de cotilédones quiescentes e de 108 HAE foram
ressuspensas (razão v/v) em (tampão de amostra - Tris-HCl 0,5 M, glicerol 10%,
SDS 10%, azul de bromofenol 1%) e foram submetidas a géis (1D) nativo 10%. Uma
vez realizada a eletroforese e antes de corar o gel, os géis foram cortados em tiras
correspondentes aos canais de amostra e, uma delas, foi corada com azul de
Coomassie. A outra foi incubada em tampão de amostra 2x (Tris-HCl 0,5 M pH 6,8,
SDS 10% (v/v), azul de bromofenol 1%, glicerol 10 % (v/v)), em temperatura
ambiente, e submetidas a uma segunda eletroforese (2D) em géis de poliacrilamida
12 %, SDS 0,1% (v/v) (Dias, 2002) e géis contendo gelatina 0,1% em condições
desnaturantes (Hummel et al., 1996, Métayer et al., 2002). Os géis com gelatina
foram lavados com Triton X-100 2,5 % e incubados em tampão de incubação como
descrito anteriormente no item 3.9, em seguida os géis foram corado com
Coomassie Brilhante Blue R e então descorado com uma solução de metanol:ácido
acético:água (40:10:50, v/ v/ v), respectivamente. A eletroforese foi realizada em um
sistema vertical Mini Protean II da BIORAD. A separação protéica procedeu-se sob
uma corrente constante de 100 V.
Os marcadores de peso molecular (200 kDa: Miosina, 116 kDa: β-
galactosidase, 97 kDa: Fosforilase b, 66 kDa: Albumina bovina, 45 kDa: Albumina de
ovo, 29 kDa: Anidrase carbônica) foram usados para nos auxiliar na identificação das
massas moleculares das bandas de proteínas.

25
3.11- Determinação do grau de anomalia das proteases na migração eletroforética e determinação das massas moleculares das proteases
A determinação do grau de anomalia das proteases na migração eletroforética
causada pela gelatina foi observada na sobreposição de géis 2D desnaturantes
(Dias, 2002) com e sem gelatina 0,1% (v/v). As proteínas do tecido cotiledonário
quiescente e em cotilédones germinantes 108 HAE foram submetidas a eletroforese
em SDS-PAGE 12%. As tiras correspondentes aos poços foram cortadas e
incubadas durante 20 minutos em tampão de amostra 2 x com β-mercaptoetanol a
uma concentração de 10%. Cada tira foi então submetida a uma segunda
eletroforese (re-eletroforese) em SDS-PAGE 12 % com e sem gelatina 0,1 % (v/v).
Estes géis SDS/SDS e SDS/SDS com gelatina foram sobrepostos. As imagens
foram invertidas no programa computacional Photo Impact. Além disso, foram feitos
gráficos de Ferguson para determinação da constante de retardação (Ferguson,
1964). Estes gráficos foram construídos com base na migração das proteases em
géis de acrilamida 10% cm gradiente de gelatina (0-0,6%). Estes gráficos permitiram
a obtenção do Kr (constante de retardação) de cada banda proteásica. E de acordo
com a obtenção do Kr foi traçado uma reta para ilustrar de maneira didática a
variação no coeficiente angular das retas de diferentes mobilidades relativas (RFs)
que foram geradas pela presença de gelatina. Na figura 11 se visualiza a
sobreposição de géis em relação às suas massas moleculares e se observa
claramente o efeito de retardação na migração das proteases.
A determinação da massa molecular relativa (Figura 2) das proteases foi
estipulada relacionando a mobilidade das proteínas das amostras do gel 1 D, as
quais correspondiam às atividades proteolíticas do gel 2 D, com a mobilidade de
proteínas que apresentam massa molecular conhecida.

26
Figura 2: Determinação das massas moleculares das proteases (as amostras foram
submetidas à eletroforese em gel SDS-PAGE 12% (1D), as tiras foram submetidas a
uma segunda eletroforese em SDS-PAGE 12% contendo gelatina 0,1%). PM-
Padrão de massa molecular. As setas indicam o sentido da migração das proteínas
na análise eletroforética.
3.12- Análise densitométrica
A análise densitométrica dos géis foi realizada através do uso do programa
computacional “Gel Perfect” (Bozzo e Retamal, 1991; Retamal et al., 1999), após
obtenção de uma imagem cinza em formato TIFF através de um scanner comercial
em 400 dpi. As massas moleculares relativas foram obtidas por comparação da sua
mobilidade eletroforética com proteínas de massa molecular conhecida, que co-
migraram no mesmo gel. O programa calcula a mobilidade relativa (Rf) de cada
banda e a área ocupada por ela, dando também uma representação diagramática
das bandas protéicas e sua concentração relativa em relação ao total de proteínas
por canal.

27
4- RESULTADOS 4.1- Análise do perfil eletroforético e “Western blotting” da fração globulínica do tecido cotiledonário ao longo do processo germinativo e pós-germinativo das sementes de Vigna unguiculata.
Na fração globulínica (Figura 3a) foi possível observar que as vicilinas (com
massas moleculares aproximadamente entre 56, 54 e 52 kDa), proteínas de reserva
da semente, são as mais abundantes. Também foi observado sua mobilização ao
longo dos processos germinativos e pós-germinativos. A subunidade de 56 kDa é a
primeira a ser totalmente mobilizada e a de 52 kDa, a última. A degradação de
vicilinas em cotilédones de Vigna unguiculata foi mais notável 72 HAE (Figura 3a).
Outras alterações protéicas, em momentos específicos do processo germinativo e
pós-germinativo também podem ser observadas. Como o desaparecimento de uma
banda (retângulo figura 3a) após 6 HAE e seu aparecimento em 12HAE. Também
pode ser observado o aparecimento de duas bandas de baixo peso molecular
(retângulo figura 3a) a partir de 72 HAE até o ultimo tempo analisado. A figura 3(b)
mostra o resultado de “Western blotting” para detecção específica de vicilinas. Foi
possível observar a degradação das vicilinas ao longo do processo germinativo e
pós-germinativo, bem como o aparecimento, a partir de 72 HAE, de produtos da
quebra das principais subunidades de vicilinas. Tais produtos, apresentam massas
moleculares mais baixas entre 35 e 25 kDa, retiveram epítopos de reconhecimento
pelos anticorpos policlonais contra vicilinas.
4.2- Análise do perfil eletroforético da fração albumínica do tecido cotiledonário ao longo do processo germinativo e pós-germinativo das sementes de Vigna unguiculata.
Foi observada a presença de várias proteínas nesta fração, em especial uma
banda com massa molecular em torno de 29 kDa em todos os tempos analisados.
Também foi visível a mobilização de bandas protéicas com massas moleculares
próximas a 97 kDa que foram diminuindo de intensidade ao longo do processo
germinativo e pós-germinativo, as quais podem representar proteínas de reserva do
tipo leguminas. Não há, no entanto, dados suficientes que confirmem tal sugestão.

28
kDa 200 116 97 66 45 29
(a) (b)
Figura 3: Eletroforese em gel de poliacrilamida 12% (a) e “Western blotting” para
detecção de vicilinas específicas (b) da fração globulínica extraída de cotilédones
das sementes de V. unguiculata ao longo de 144 HAE, na ausência de -
mercaptoetanol. Foram aplicados 8µL de amostra por poço. Poços: M- Marcador de
peso molecular; 0 HAE (Semente quiescente); 6 HAE; 12 HAE; 24 HAE; 36 HAE; 48
HAE; 60 HAE; 72 HAE; 84 HAE; 96 HAE; 108 HAE; 120 HAE; 144 HAE,
respectivamente.
M 0 6 12 24 36 48 60 72 84 96 108 120 144 0 6 12 24 36 48 60 72 84 96108120144

29
kDa 200 116 97 66 45 29
Figura 4: Eletroforese em gel de poliacrilamida 12% da fração albumínica extraída
de cotilédones das sementes de V. unguiculata ao longo de 144 HAE. Foram
aplicados 3µL de amostra por poço. Poços: M- Marcador de peso molecular; 0 HAE
(Semente quiescente); 6 HAE; 12 HAE; 24 HAE; 36 HAE; 48 HAE; 60 HAE; 72
HAE; 84 HAE; 96 HAE; 108 HAE; 120 HAE; 144 HAE, respectivamente.
M 0 6 12 24 36 48 60 72 84 96 108 120 144

30
4.3- Análise do perfil eletroforético e “western blotting” de proteínas de corpos protéicos e proteínas citoplasmáticas ao longo do processo germinativo e pós-germinativo das sementes de feijão-de-corda
A degradação de proteínas foi perceptível na fração de corpos protéicos
(FCP) (Figuras 5 a e b) e na fração citoplamática (FC) de cotilédones ao longo do
processo germinativo e pós-germinativo de sementes de feijão-de-corda, quando
visualizadas por SDS-PAGE (Figura 6). A degradação de vicilinas armazenadas em
corpos protéicos foi visível por detecção através de Western blotting (Figura 5c). O
perfil de corpos protéicos foi modificado quando usado um agente redutor, como β-
mercaptoetanol (Figura 5b), especificamente enfatizado por alterações em proteínas
de massa molecular compatível com a de leguminas e nas subunidades de vicilinas
de 56, 54 e 52 kDa . Uma série de polipeptídeos de baixo peso molecular foram
também mais proeminentes (retângulo vermelho figura 5b) quando estavam na
presença de β-mercaptoetanol. Inversamente algumas proteínas foram melhor
visualizadas na ausência do agente redutor.
Atividade de proteinases cisteínicas foi detectada na fração citoplasmática
(FC), por análise zimográfica (Figura 6b). Bandas das proteinases majoritárias
aparecem entre 66 e 97 kDa, demonstrando atividade a partir de 24 HAE até 144
HAE. Houve aparecimento de atividades em bandas com massas moleculares
abaixo de 29 kDa; estas últimas foram detectadas apenas a partir de 48 HAE em
diante, aumentando sua intensidade ao longo dos tempos analisados.

31
kDa 200 116 97 66 45 29
(a) (b)
(c)
Figura 5: Eletroforese em gel de poliacrilamida 12% da fração de corpos protéicos
extraída de cotilédones de sementes de V. unguiculata ao longo de 144 HAE, na
ausência (a) e na presença (b) de agente redutor (β-mercaptoetanol) e “western
blotting” (c) para detecção de vicilinas. Foram aplicados 8µL de amostra por poço.
Poços: M- Marcador de peso molecular; 0 HAE (Semente quiescente); 6 HAE; 12
HAE; 24 HAE; 36 HAE; 48 HAE; 60 HAE; 72 HAE; 84 HAE; 96 HAE; 108 HAE; 144
HAE; respectivamente.
M 0 6 12 24 36 48 60 72 84 96 108144 M 0 6 12 24 36 48 60 72 84 96 108 144
0 6 12 24 36 48 60 72 84 96 108 144

32
kDa 200 116 97 66 45 29
kDa 200 116 97 66 45 29
(a)
(b)
Figura 6: - Eletroforese em gel de poliacrilamida 12% da fração de proteínas do
citosol extraída de cotilédones de sementes de V. unguiculata ao longo de 144 HAE,
na presença de agente redutor (β-mercaptoetanol) (a) e Gelatina-PAGE (b). Foram
aplicados 3µL de amostra por poço. Poços: M- Marcador de peso molecular; 0 HAE
(Semente quiescente); 6 HAE; 12 HAE; 24 HAE; 36 HAE; 48 HAE; 60 HAE; 72
HAE; 84 HAE; 96 HAE; 108 HAE; 144 HAE; respectivamente.
M 0 6 12 24 36 48 60 72 84 96 108 144
M 0 6 12 24 36 48 60 72 84 96 108 144

33
4.4- Detecção de atividade gelatinolítica e ação de inibidores sobre as atividades proteinásicas ao longo do processo germinativo e pós-germinativo das sementes de Vigna unguiculata
A figura 7 mostra os resultados encontrados nas eletroforeses em gel de
poliacrilamida (12%) na presença de SDS, contendo gelatina 0,1%, das amostras da
fração enriquecida em albuminas dos tecidos cotiledonários das sementes de feijão-
de-corda nos diferentes tempos de embebição. É possível visualizar a presença de
proteases com atividade gelatinolítica no tempo 0, onde a semente está em estágio
quiescente, desidratada, nos tempos de 12 h e 60 h (com massa molecular em torno
de 45 kDa). Após 36 HAE até o último tempo analisado de 144 HAE foi possível
observar atividade gelatinolítica de uma principal banda que apresenta massa
molecular acima das principais proteínas de reserva chamadas vicilinas (com
massas moleculares entre 56, 54 e 52 kDa).
Os géis de atividade, em presença de -mercaptoetanol, foram incubados em
soluções contendo os diferentes inibidores, ácido iodoacético, iodoacetamida,
PCMB, E-64 (inibidores de proteases da classe cisteínicas), EDTA (inibidor da
classe de metaloproteases) e PMSF (inibidor de proteases da classe serínica). De
acordo com a análise densitométrica dos géis, pode-se observar que o inibidor
iodoacetamida interferiu em 65 % na atividade da protease (Figura 8), sendo o
inibidor que inibiu mais eficientemente as proteases. Quando o gel foi incubado com
PMSF houve uma inibição considerável da principal banda de atividade (Figura 9). O
PMSF é um inibidor irreversível para proteases serínicas, mas achados na literatura
dizem que é raro, mas podem inibir proteases da família da papaína (Segundo
Calbiochem).

34
Imagem invertida
Figura 7: Eletroforese em gel de poliacrilamida 12% contendo 0,1% de gelatina, da
fração albumínica extraída de cotilédones das sementes de V. unguiculata ao longo
de 120 HAE, na presença de diferentes inibidores de proteinases cisteínicas; em (a)
controle na presença de β-mercaptoetanol, em (b) na ausência de β-mercaptoetanol,
em (c) na presença de iodoacetamida, em (d) na presença de E-64, em (e) na
presença de PCMB em (f) na presença ácido iodoacético. Foram aplicados 3µL de
amostra por poço. Poços: M- Marcador de peso molecular; 0 HAE (Semente
quiescente); 6 HAE; 12 HAE; 24 HAE; 36 HAE; 48 H; 60 HAE; 72 HAE; 84 HAE; 96
HAE; 108 HAE; 120 HAE; respectivamente.
M 0 6 12 24 36 48 60 72 84 96 108120 M 0 6 12 24 36 48 60 72 84 96 108 120
(a)
Con
trole
com
β
(c)
Iodo
acet
amid
a (e
) PC
MB
(b) C
ontrole sem β
(d) E-64
(f) Á
cido iodoacético

35
Concentração
dos inibidores
Inibidores Área Ocupada por
bandas de atividade
Percentagem de Atividade
Residual(%)
- Controle com β-
mercaptoetanol 6187 50
- Controle sem β-
mercaptoetanol 1200 100
50 µM Iodoacetamida 4200 65
100 µM E-64 5756 53
10 mM PCMB 6842 43
300 µM Ácido Iodoacético 5832 52
Figura 8: Análise densitométrica de atividades proteinásicas de cotilédones
germinantes de V. unguiculata em função da mobilidade relativa (Rf) da principal
banda de atividade. Os gráficos são resultados obtidos da análise no programa
computacional Gel Perfect.
0 6 12 24 36 48 60 72 84 96 108120
Controle com β-mercaptoetanol
Controle sem β-mercaptoetanol
Iodoacetamida
E-64
PCMB
Ácido Iodoacético

36
Figura 9: Análise densitométrica em função da mobilidade relativa ( Rf ) da principal
banda de atividade. O gráfico mostra os resultados obtidos da análise no programa
computacional Gel Perfect.
0 6 12 24 36 48 60 72 84 96 108 120 144
Áre
a
Mobilidade Relativa
Ácido Iodoacético
Controle
Proteínas
EDTA
PCMB
PMSF
EDTA
PCMB
Ac. Iodoacético
Controle
PMSF

37
4.5- Detecção de atividade proteásica ao longo do processo germinativo e pós-germinativo das sementes de feijão-de-corda na ausência e presença de agente redutor
A atividade proteásica em cotilédones ao longo do processo germinativo e
pós-germinativo foi detectada em géis unidimensionais SDS-PAGE 12% com
gelatina 0,1%, com (Figura 10a) e sem β-mercaptoetanol (Figura 10b). Os
resultados demonstraram que o perfil eletroforético das bandas de atividade de
menores massas moleculares é alterado na presença do agente redutor, o que
sugere que estas proteases possuem pontes dissulfeto (RS-RS) importantes para
sua atividade catalítica. As atividades de maior massa molecular, majoritárias nos
cotilédones de 36 a 144 HAE, não foram afetadas pelo agente redutor. Bandas de
atividade de abaixo de 29kDa sofreram redução parcial de sua atividade com a
presença de agente redutor.
4.6- Comparação da migração eletroforética de proteínas e atividades proteásicas em géis bidimensionais SDS-PAGE / SDS-PAGE e SDS-PAGE em presença de gelatina 0,1% e determinação das massas moleculares de
proteases
Uma abordagem para determinação do grau de anomalia na migração das
proteases, causada pela presença de gelatina, foi a sobreposição de géis 2D-PAGE
desnaturantes com e sem gelatina 0,1% das proteínas do tecido cotiledonário ao
longo do processo germinativo e pós-germinativo. As imagens destes géis
(SDS/SDS e SDS/SDS com gelatina) foram sobrepostas no programa computacional
Photo Impact, onde algumas foram invertidas. Nossos resultados confirmam a
anomalia na migração das proteases em géis com gelatina em relação aos géis sem
gelatina. Além disso, foram traçadas linhas para ilustrar de maneira didática a
variação no coeficiente angular das retas de diferentes Rfs (Figura 11), onde foram
utilizadas as imagens de proteínas e proteases das amostras do tecido cotiledonário
sobrepostas. Conforme a massa molecular das proteínas decresce, o grau de desvio
aumenta, o que significa que o polímero de gelatina altera mais efetivamente a
migração de proteínas de massas moleculares pequenas. Esta técnica permite
também localizar proteases em géis que não contêm gelatina, com a prévia correção
do valor do coeficiente angular das retas que originam as proteínas.

38
Devido à anomalia a migração das proteínas em géis com gelatina em relação
a géis sem gelatina (Hummel et al., 1996), estimada pela variação no coeficente
angular (figura 11), determinação da massa molecular relativa das proteases do
tecido cotiledonário dos tempos de 0 HAE (semente quiescente) e 108 HAE, foi
realizada relacionando a mobilidade das proteases no gel 1D (SDS-PAGE 12%) com
a mobilidade das proteases no gel 2D (SDS-PAGE 12%, contendo gelatina 0,1%)
(Figura 11).
4.7- Análise do perfil eletroforético das proteínas e proteases por géis
bidimensionais do tecido cotiledonário de sementes de Vigna unguiculata
A figura 12a mostra o perfil eletroforético analisado em géis bidimensionais da
amostra do tecido cotiledonário no tempo 0 HAE (semente seca) de feijão-de-corda.
Foi possível detectar atividade proteásica em gel bidimensional nativo 10% / SDS-
PAGE 12% com gelatina 0,1%, neste tempo (Figura 12b).
A figura 13 mostra o perfil eletroforético analisado em géis bidimensionais da
amostra do tecido cotiledonário no tempo 108 HAE. Foi possível detectar atividade
proteásica em gel bidimensional nativo 10% / SDS-PAGE 12% com gelatina 0,1%,
neste tempo (Figura 13b). A figura 13a mostra proteínas do tempo 108 HAE de
massa molecular idêntica também tiveram o mesmo Rf sob condições nativas,
sugerindo a existência de agregados. Uma reta de proteínas (Fig. 13a) e proteases
(Fig. 13b) foi observada. Um constante aumento da proporção entre mobilidade
relativa e massa molecular também foi notado.
A figura 13c mostra o resultado quando o gel foi incubado com PMSF (10
mM). O inibidor foi eficaz, gerando uma inibição considerável da principal banda de
atividade no tempo de 108 HAE.
A figura 14 mostra o resultado da sobreposição das imagens. Uma drástica
alteração pode ser vista especialmente nas massas de alto peso molecular em
proteínas de reserva acídicas, que parecem originar uma serie linear de produtos de
degradação em 108 HAE nas amostras do tecido cotiledonário.

39
kDa
200 116 97 66 45 29 14
(a ) (b)
(+ β-mercaptoetanol ) (- β-mercaptoetanol )
Figura 10: Eletroforese em gel de poliacrilamida 12% da fração albumínica extraída
de cotilédones das sementes de V. unguiculata ao longo de 144 HAE. a - Gelatina-
PAGE com β-mercaptoetanol. b - Gelatina-PAGE sem β-mercaptoetanol. Foram
aplicados 3µL de amostra por poço. Poços: M- Marcador de peso molecula ; 0 HAE
(Semente quiescente); 6 HAE; 12 HAE; 24 HAE; 36 HAE; 48 HAE; 60 HAE; 72
HAE; 84 HAE; 96 HAE; 108 HAE; 120 HAE; 144 HAE, respectivamente.
M 0 6 12 24 36 48 60 72 84 96 108120144 M 0 6 12 24 36 48 60 72 84 96 108 120 144

40
Figura 11: Migração anômala de proteases: sobreposição de géis 2D desnaturantes
com e sem gelatina 0,1%. O tecido cotiledonário do tempo de 108 HAE foi analisado
em géis 2D: SDS-PAGE 12% / SDS-PAGE 12% e SDS-PAGE 12% / SDS-PAGE
12% com gelatina. Imagens invertidas no Photo Impact das imagens
respectivamente.
kDa 200 116 97 66 45 31 21 14
kDa 200 116 97 66 45 31 21 14
0 HAE
kDa 200 116 97 66 45 31 21 14
kDa 200 116 97 66 45 31 21 14
108 HAE
Protease
Proteínas
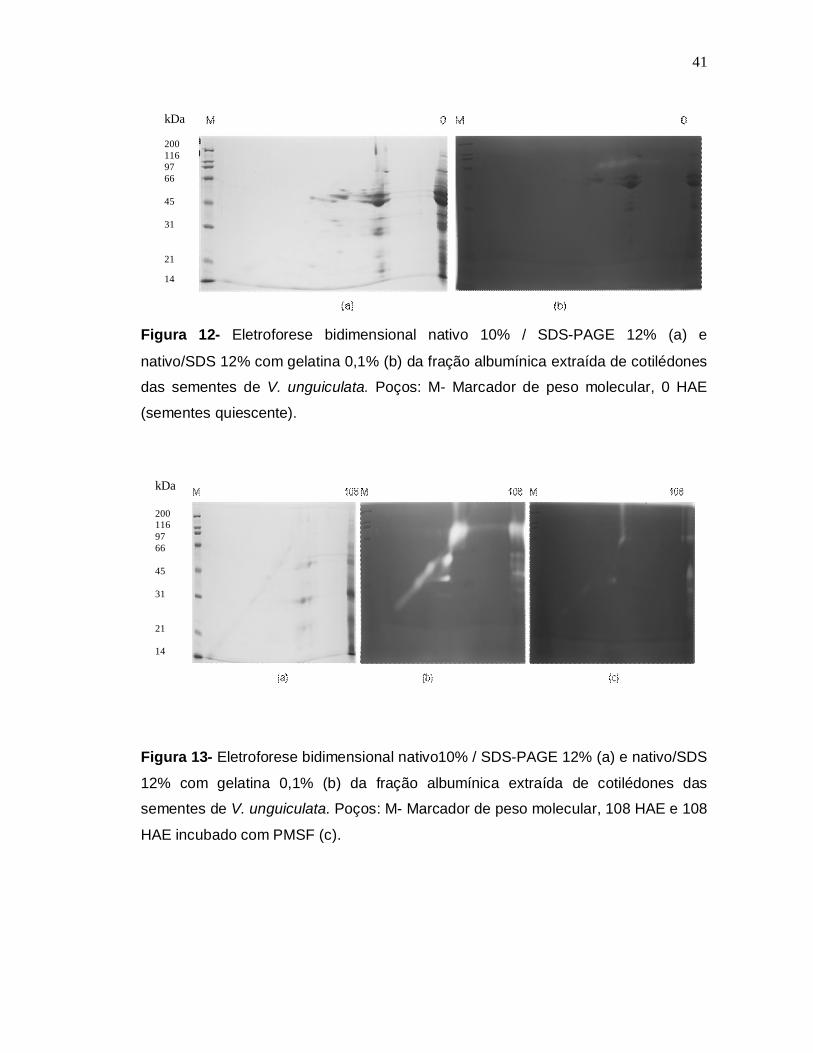
41
Figura 12- Eletroforese bidimensional nativo 10% / SDS-PAGE 12% (a) e
nativo/SDS 12% com gelatina 0,1% (b) da fração albumínica extraída de cotilédones
das sementes de V. unguiculata. Poços: M- Marcador de peso molecular, 0 HAE
(sementes quiescente).
Figura 13- Eletroforese bidimensional nativo10% / SDS-PAGE 12% (a) e nativo/SDS
12% com gelatina 0,1% (b) da fração albumínica extraída de cotilédones das
sementes de V. unguiculata. Poços: M- Marcador de peso molecular, 108 HAE e 108
HAE incubado com PMSF (c).
kDa 200 116 97 66 45 31 21 14
kDa 200 116 97 66 45 31 21 14

42
(b) (c)
(b) (c)
Figura 14- Sobreposição das imagens da eletroforese bidimensional nativo 10% /
SDS-PAGE 12% dos tempos de 0 HAE (semente quiescente) e 108 HAE; e nativo
10% / SDS 12% com gelatina 0,1% dos mesmos tempos citado acima da fração
albumínica extraída de cotilédones das sementes de V. unguiculata. Em (a)
sobreposição das imagens de 0 HAE (proteínas) e 108 HAE (atividade). Em (b)
sobreposição das imagens de 0 HAE (proteínas) e 108 HAE (proteínas). E em (c) 0
HAE (atividade) e 108 HAE (atividade).
Massa SDS
Rf Nativo (a)
Protease
Proteínas

43
4.8- Ensaio de termoestabilidade enzimática
Na caracterização de termoestabilidade enzimática, foi possível observar que
a enzima perde sua atividade com 5 minutos de fervura (Figura 15a). Foi possível
também observa que não houve inibição da enzima quando esta foi corrida junto no
gel com inibidor (PMSF). Quando o inibidor foi adicionado no tampão de incubação
foi possível observar inibição enzimática (Figura 15b).
(a) (b)
Figura 15: Gelatina-PAGE em gel de poliacrilamida 12% da fração albumínica
extraída de cotilédones das sementes de V. unguiculata em 108 HAE. Foram
aplicados 3µL por poço. M- Marcador de peso molecular. 1a- 108 HAE; 2 a- 108
HAE + PMSF (corrido no gel - 10mM); 3 a- 108 HAE após 5 minutos de fervura; 4 a-
108 HAE após 15 minutos após fervura; 5 a- 108 HAE após 30 minutos de fervura; 6
a- 108 HAE após 5 minutos de fervura + PMSF; 7a- 108 HAE após 15 minutos após
fervura + PMSF; 8 a- 108 HAE após 30 minutos de fervura + PMSF. 1 b- 108 HAE +
PMSF (10mM) incubado em banho a 37 °C por 16 h.
M 1 2 3 4 5 6 7 8
M 1 kDa 200 116 97 66 45 29

44
5- Discussão
Neste trabalho, foi traçado um quadro do processo germinativo e pós-
germinativo de sementes de Vigna unguiculata, acompanhando as alterações
principais nos teores de proteínas e de atividades proteinásicas da classe cisteínica
em tecidos cotiledonários. Pretendeu-se também contribuir com o entendimento da
questão enunciada por Schlereth et alli (2000), que consiste na busca dos processos
e mecanismos que estão por trás do intervalo temporal entre o início da biossíntese
protéica durante os estágios iniciais de germinação e o início da mobilização de
reservas protéicas no período pós-germinativo.
Como tecido principal de armazenamento em sementes, cotilédones tiveram
suas proteínas de reserva bastante reduzidas até 144 HAE. Como resultado visível
deste processo de degradação, um padrão interessante de hidrólise foi revelado por
análise bidimensional onde os polipeptídeos resultantes estavam alinhados
linearmente em uma relação crescente constante entre seus Rfs e Mrs (Figura 13). A
mobilização da principal proteína de reserva chamada vicilina foi mais proeminente
após 72 HAE como visto por SDS-PAGE e por análise de “Western blotting” da
fração de globulínica (Figura 3) assim como na fração de corpos protéicos (Figura 5)
preparadas de células cotiledonárias. Além da mobilização de proteínas de reserva
do tipo globulinas, várias outras proteínas da fração citoplasmática destas células de
cotilédones também foram mobilizadas ao longo do processo de germinação. De
acordo com Shutov et alli (2003), quase todas as proteínas de sementes
germinantes são degradadas em célula e tecidos de armazenamento, semelhante
com cotilédones de leguminosas. Elas representam a garantia de sucesso para a
futura planta, como provêem constituintes aminoácidos para o novo conjunto de
proteínas a serem formadas em todos tecidos localizados na nova planta.
A ocorrência de proteinases cisteínicas em plantas é bastante conhecida. As
proteinases cisteínicas melhor caracterizadas são aquelas que participam da
degradação de proteínas de reserva durante a germinação. Pouco se sabe sobre a
presença destas proteinases durante o desenvolvimento das sementes e até mesmo
nas sementes quiescentes (Yamauchi et al., 1992; Tanaka et al., 1993).
Schlereth et alli (2000) correlacionaram atividades de proteinases cisteínicas
com a mobilização de globulina em cotilédones e eixos embrionários de sementes
de Vicia sativa durante a germinação. Os autores mostraram que a mobilização de
globulina em sementes germinantes de V. sativa é iniciada através de proteinases

45
cisteínicas presentes em sementes quiescentes e mais adiante controladas por
proteinases cisteínicas sintetizadas de novo.
Foi demonstrado que a degradação de proteínas de reserva em cotilédones
de sementes de dicotiledôneas é geralmente controlada através de grupos de
proteinases cisteínicas como as papaínas- like (CPR1, CPR2, proteinase A e CPR4) e
de proteinases cisteínicas leguminas - like (VsPB2 e proteinase B) (Fischer et al.,
2000; Schlereth et al., 2000; 2001; Tiedemann et al., 2001). Primeiro, as proteínas
de reserva são atacadas por proteinases papaína-like de baixa especificidade, uma
ou outra espécie ativa em corpos protéicos de sementes secas ou ativadas durante
o início da germinação, causando uma proteólise limitada. Tais proteinases
cisteínicas de baixa especificidade coexistem com proteinases de reserva
vacuolares (VPEs) e globulinas em corpos protéicos de sementes em
desenvolvimento (Müntz et al., 2001). A inacessibilidade das globulinas às enzimas
do tipo papaína pode ser devida a tal separação espacial em diferentes
compartimentos vacuolares, como visto em sementes de tomate (Jiang et al., 2001)
ou devido à transformação de globulinas em cristais inacessíveis, como sugerido por
Weber e Neumann (1980). As degradações limitadas também podem ser mediadas
por aumento das quantidades de proteinases sintetizadas de novo e podem ser
transportadas dos corpos protéicos durante crescimento da plântula (Müntz et al.,
1998; 2001). Em alguns casos, o ataque proteolítico é suficiente o bastante para
causar alterações estruturais em proteínas de reserva que ativam degradação
ilimitada de globulinas de armazenamento principalmente executadas por
proteinases cisteínicas do tipo legumaína-like. Em cotilédones de feijão-de-corda, foi
possível detectar uma proteinase cisteínica ativa com massa molecular de ~45 kDa
em cotilédones quiescentes (Figuras 7 e 10) que também é ativa nos tempos de 12 e
60 HAE, sugerindo um papel pontual no metabolismo de processamento protéico.
Como os eventos principais de mobilização de globulina foram observados após 72
HAE, nós sugerimos que em cotilédones de feijão-de-corda proteinases cisteínicas
papaína-like de ~45 kDa são enzimas responsáveis pela proteólise limitada de
globulinas de reserva. Proteases são especificamente expressas em tempo e
espaço e acumuladas em diferentes compartimentos subcelulares (Van der Hoorn,
2008). Em cotilédones de feijão-de-corda, proteinases cisteínicas ativas foram
descobertas em frações citoplasmáticas (FC) (Figura 6). Proteinases de alto peso
molecular (> 45 kDa, <97 kDa), encontradas em FC e na fração albumínica foram

46
principalmente sintetizadas de novo após 24 HAE e de baixo peso molecular (<29
kDa,>14 kDa), encontradas principalmente nas FC, foram detectadas somente após
48 HAE (Figura 6). Proteinases cotiledonares de sementes de feijão-de-corda foram
inibidas através de inibidores de proteinases cisteínica clássicos como E-64, ácido
de iodoacético, pCMB e iodoacetamida, este último sendo o mais efetivo, nas
concentrações usadas (Figuras 7, 8 e 9).
Proteinases cisteínicas semelhantes de alto peso molecular (~70 kDa) foram
encontradas em eixos embrionários germinantes de V. sativa, mas não em
cotilédones onde foi observado proteinases cisteinicas de 25 a 50 kDa (Schlereth et
al., 2000). Em sementes de Phaseolus vulgaris, proteinases cisteínicas papaína-like
apresentaram massas moleculares em torno de 29 kDa, enquanto proteinases
cisteínicas legumaína-like apresentaram massas em torno de 40 kDa (Zakharov et
al., 2004).
A complexidade dos eventos proteolíticos que ocorrem em sementes
germinantes e a função específica de cada proteinase envolvidas foram informadas
em trabalhos por Shutov e Vaintraub (1987), Hara-Nishimura et alli (1998), Fischer et
alli (2000), Schlereth et alli (2000), Tiedemann et alli (2001) e Müntz et alli (2002),
entre vários outros. Alguns dos aspectos intrigantes ainda a serem esclarecidos
estão relacionados com as modificações e processos regulados conduzindo à
proteção precoce de degradação de proteínas em corpos protéicos, onde ambas as
proteases e substratos-alvo às vezes são co-localizados.
Há décadas atrás foi introduzido o procedimento baseado na utilização de
géis de poliacrilamida SDS contendo gelatina, caseína ou fibrina como substratos
para a localização da atividade de proteases. Estas enzimas têm a habilidade de
renaturar após a remoção de SDS e de exercer atividade proteolítica no substrato
co-polimerizado, e assim, serem analisadas com este método (Frederiks e Mook,
2004). Sabe-se que a presença do polímero de gelatina pode reduzir a migração de
proteínas de massa conhecida de 15 a 20%. Em géis de acrilamida a 7,5% a adição
de gelatina reduz a mobilidade dos padrões 13 ± 3,4%; em gel de acrilamida a 10%
a redução é de 16 ± 3,8%; e em géis 12,5%, a redução é de 15 ± 2,8% (Hummel et
al., 1996). Um retardo similar foi observado com proteínas não proteásicas de
Schizophyllum commune. As proteases migram muito mais devagar em SDS-PAGE
com gelatina do que os padrões. Para as proteases testadas, a redução da
mobilidade com a adição de gelatina em acrilamida 10% foi de 48% para

47
quimiotripsina, 44% para elastase, 43% para tripsina e 41% para papaína (Hummel
et al., 1996). O retardo não é constante entre proteases. Portanto, a massa
molecular das proteases não pode ser estimada precisamente em géis SDS com
gelatina. Para determinar a massa molecular das proteases foram realizados géis
bidimensionais SDS-PAGE / SDS-PAGE e SDS-PAGE / SDS-PAGE com gelatina, e
relacionadas as massas moleculares obtidas na primeira dimensão com a posição
das proteases na segunda dimensão. Assim, foram determinadas as massas
moleculares das bandas de atividade proteásica sem o efeito anômalo causado pelo
polímero de gelatina.
No presente trabalho, um padrão interessante de proteólise de proteases em
cotilédones após 108 HAE (Figura 13b) foi observado através de eletroforese
bidimensional nativo / SDS-PAGE. A disposição linear aparente de bandas de
atividades pode significar que a atividade proteolítica principal neste tecido induz ou
ativa novas proteases que podem agir sobre diferentes agregados ou zimógenos e
estes produtos de hidrólise são organizados em uma linha de relação crescente
constante Rf x Mr. A procura por substratos endógenos de cada proteinase cisteínica
ativa em sementes de feijão-de-corda germinantes é o objeto de investigações em
curso. Também é importante lembrar o papel que pode ser realizado através de
inibidores de proteinases cisteínicas do tipo cistatinas, regulando eventos que
harmonizam a ação de proteinases cisteínicas e a mobilização de proteínas de
reserva durante o curso da germinação de sementes, como previamente discutido
por (Fernandes et al., 1991; Flores et al. 2001).
Durante a germinação e eventos pós-germinativos as globulinas são
quebradas em condições ácidas. A análise de diferentes inibidores revelou a
presença de representantes da família de proteinases cisteínicas. Atividade de
proteinases ácidas também esteve presente na semente seca. De acordo com
informado por Shutov and Vaintraub (1987) estes resultados indicam que
proteinases cisteínicas são responsáveis pela mobilização de globulinas.
As determinações de atividades de proteinases cisteínicas realizadas neste
trabalho mostram que nos extratos de cotilédones de sementes de feijão-de-corda
(Figura 7), houve atividade enzimática em sementes quiescentes. Uma fraca
atividade acídica do tipo cisteínica esteve também presente em cotilédones e eixos
de sementes secas de V. sativa (Schlereth et al., 2000).

48
O padrão de comportamento onde, diferentes isoformas de proteases estão
ativas em diferentes estágios de germinação, sugere, provavelmente, um complexo
sistema de síntese de novo e ação destas enzimas durante esse processo. Dentre
os registros de proteinases cisteínicas de sementes em germinação e
desenvolvimento estão os já citados Müntz et alli (1998), Kalinski et alli (1992) e o de
Hara-Nishimura et alli (1998); estes últimos documentaram uma proteinase cisteínica
vacuolar (VPE), cuja principal função seria provavelmente a de processar proteínas
de reserva de mamona no interior dos corpos protéicos.
Com o objetivo de observar a inibição de tais atividades enzimáticas os géis
foram incubados em solução contendo diferentes inibidores. Foi possível observar
que os inibidores iodoacetamida, ácido iodoacético, E-64 e PCMB (inibidores de
proteinases da classe cisteínica) não afetaram totalmente as atividades enzimáticas
nas amostras do tecido cotiledonário. As atividades de eixos embrionários de Vicia
sativa foram inibidas por E-64 e parcialmente inibidas por ácido iodoacético que inibe
tanto proteinases cisteínicas do tipo papaína como legumaínas-like. Já as atividades
de proteinases de baixo peso molecular foram inibidas, tanto em cotilédones como
em eixos embrionários (Schlereth et al., 2000). Os autores sugerem que isso
provavelmente ocorra devido ao efeito do ácido iodoacético nos grupos SH que são
importantes para dobramento correto da enzima. Quando o gel foi incubado com
PMSF houve uma considerável inibição das proteases com massa molecular de ~70
kDa. Schlereth et alli (2000) não observaram inibição das proteinases de Vicia sativa
com o mesmo inibidor, mas estas foram afetadas por EDTA (inibidor de
metaloproteases). Nossos resultados indicaram um aumento das atividades
proteinásicas de cotilédones germinantes de feijão-de-corda quando incubada com
EDTA. Cotilédones de Fagopyrum esculentum L. contêm uma metaloproteinase-Zn
dependentes que em contraste com outras metaloproteases de eucariotos e
procariotos, tem uma ação limitada especificamente para proteínas de reserva de
sementes desta espécie (Müntz et al., 2001). Este substrato endógeno é localizado
em corpos protéicos e a metaloprotease tem uma atividade in vitro ótima em pH
ligeiramente ácido. In vitro a metaloprotease atua mediando uma proteólise limitada
das proteínas de reserva do tipo legumina 13 S, de cotilédones de F. esculentum L.
e gera uma fragmento com função similar que aparece durante a degradação de
globulina in vivo (Müntz et al., 2001). Estas observações são comparáveis com a
degradação inicial de globulinas em cotilédones de V. sativa (Shutov e Vaintraub,
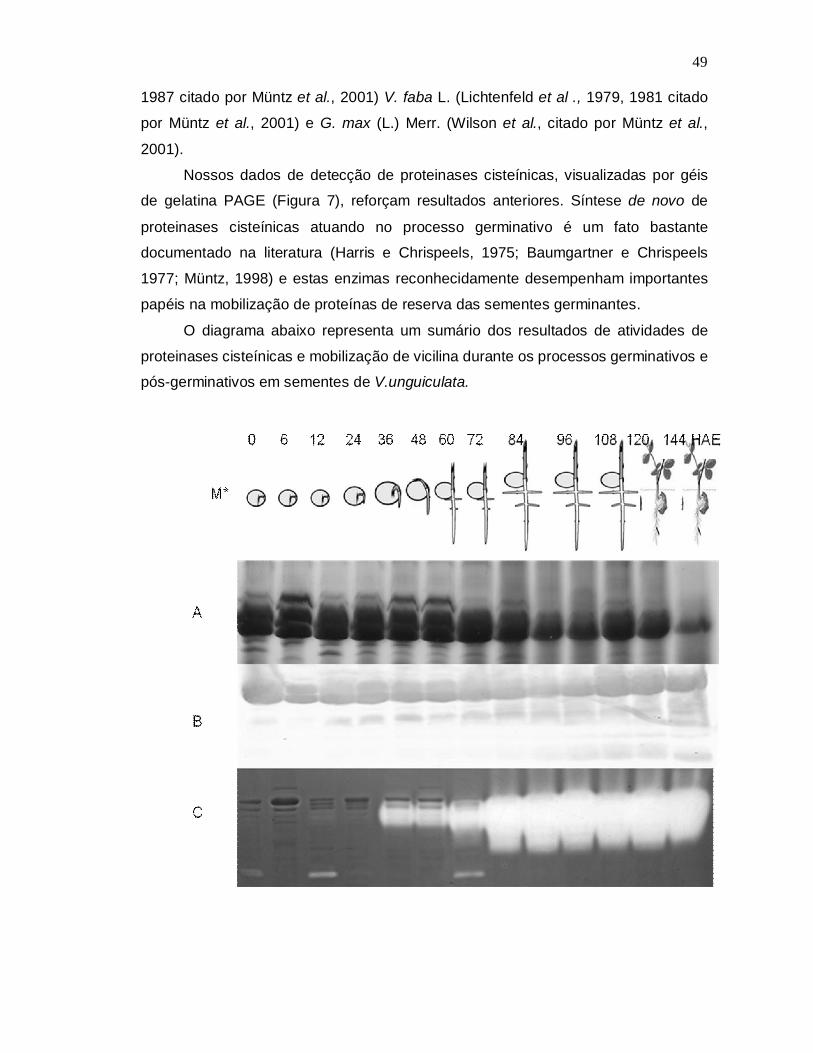
49
1987 citado por Müntz et al., 2001) V. faba L. (Lichtenfeld et al ., 1979, 1981 citado
por Müntz et al., 2001) e G. max (L.) Merr. (Wilson et al., citado por Müntz et al.,
2001).
Nossos dados de detecção de proteinases cisteínicas, visualizadas por géis
de gelatina PAGE (Figura 7), reforçam resultados anteriores. Síntese de novo de
proteinases cisteínicas atuando no processo germinativo é um fato bastante
documentado na literatura (Harris e Chrispeels, 1975; Baumgartner e Chrispeels
1977; Müntz, 1998) e estas enzimas reconhecidamente desempenham importantes
papéis na mobilização de proteínas de reserva das sementes germinantes.
O diagrama abaixo representa um sumário dos resultados de atividades de
proteinases cisteínicas e mobilização de vicilina durante os processos germinativos e
pós-germinativos em sementes de V.unguiculata.

50
Figura 16: M* - Morfologia da semente de feijão-de-corda durante os eventos
de germinação e pós-germinação. Em A, SDS-PAGE da fração enriquecida em
globulinas; em B “western blotting” para detecção de vicilina; em C gelatina-PAGE
da fração albumínica. Em A' SDS-PAGE da fração de corpos protéicos; em B'
“western blotting” para detecção de vicilinas em corpos protéicos; em C' gelatina-
PAGE da fração albumínica.

51
6- Conclusões
A visualização da fração enriquecida em globulinas dos tecidos cotiledonários
de sementes germinantes de Vigna unguiculata mostrou a diminuição na
principal proteína de reserva, a vicilina.
A visualização eletroforética e o “imunoblotting” da fração de corpos protéicos
dos tecidos cotiledonários de sementes germinantes de Vigna unguiculata
confirmaram a degradação da principal proteína de reserva, a vicilina, a partir
de 72 HAE.
Na fração enriquecida em albuminas dos tecidos cotiledonários de sementes
germinantes de Vigna unguiculata observou-se a presença de uma banda
com massa molecular de 29 kDa, de concentração conservada ao longo do
processo germinativo e pós-germinativo.
Atividades de proteinases cisteínicas foram detectadas a partir de 36 HAE
(>66 kDa) na fração enriquecida em albuminas do tecido cotiledonário ao
longo do processo germinativo e pós-germinativo.
Atividades de proteinases cisteínicas pontuais foram detectadas na semente
quiescente (tempo 0 HAE), em 12 HAE e em 60 HAE no tecido cotiledonário.
Atividades de proteinases cisteínicas foram detectadas na fração
citoplamática (FC), por zimografia. Visualizaram-se bandas com massas
moleculares acima de 66 kDa a partir de 36 HAE até 144 HAE e bandas de
baixo peso molecular a partir de 48 HAE em diante, aumentando sua
atividade ao longo de todo o processo.
O estudo com diferentes inibidores iodoacetamida, E-64, ácido iodoacético e
pCMB indica que as proteases do tecido cotiledonário correspondem a
proteinases cisteínicas. Apesar disso, quando o inibidor PMSF foi utilizado,
houve uma inibição considerável das principais bandas de proteases com
massa molecular de ~70 kDa.
• A visualização através de eletroforese bidimensional nativo / SDS-PAGE
revelou a disposição linear aparente de bandas de atividades, isto pode
significar que a atividade proteolítica principal neste tecido induz ou ativa
novas proteases que podem agir sobre diferentes agregados ou zimógenos e
estes produtos de hidrólise são organizados em uma linha de relação
decrescente constante de Rf x Mr.

52
7- Referências Bibliográficas
Abe, K., Emori, Y., Kondo, H., Susuki, K. and Arai, S. (1987). Molecular cloning of
a cysteine proteinase inhibitor of rice (Oryzacystatin). Homology with animal cystatins
and transient expression In the ripening process of rice seeds. J. Biol. Chem. 262,
16793 – 16797.
Alberts, B., Bray, D., Lewis, J., Raff, M., Roberts, K. and Watson, J.D. (1997). In:
Molecular Biology of the Cell, 3rd ed. Garland, New York,.
Allen, D. J. (1982). The Pathology of Tropical Food Legumes. In: Disease resistance
in crop improvement. Chinchester, John Wiley, 431-445.
Asano, M., Suzuki, S., Kawai, M., Miwa, T. and Shibai, H. (1999). Characterization of
novel cysteine proteases from germinating cotyledons of soybean [Glycine max (L.)
Merrill]. J. Biochem. 126, 296-301.
Attucci, S., Carde, J. P., Raymond, P., Saint Gés, V., Spiteri, A. and Pradet, A.
(1991). Oxidative phosphorylation by mitochondria extracted from dry sunflower
seeds. Plant Physiol. 95, 390-398.
Ávila, D.S., Cunha, M., Jacinto, T., Xavier-Filho, J. and Fernandes, K.V.S. (1999).
Anais da XXVIII Reunião Anual da SBBq, E-64, 35.
Baumgartner, B. and Chrispeels, M. J. (1977). Partial characterization of a protease
inhibitor which inhibits the major endopeptidase present in the cotyledons of mung
beans. Plant Physiol., 58, 1-6.
Becker, C., Senyuk, V.I., Shutov, A.D., Nong, V.H., Fischer, J., Horstmann, C. and
Müntz, K. (1997). Proteinase A, a storage-globulin-degrading endopeptidase of
vetch (Vicia sativa L.) seeds, is not involved in early steps of storage-protein
mobilization. Eur. J. Biochem., 248, 304-312.
Bewley, J. D. and Black, M. (1994). In: Seeds – Physiology of Development and
Germination, 2° edição, Plenum Press, New York and London. 445 p.

53
Bewley, J. D. (1997). Seed germination and dormancy. The Plant Cell, vol.9, 1055-
1066.
Bewley, J. D. (2001). Seed germination and reserve mobilization. Encyclopedia of
Life Sciences. Disponível em: www.els.net.
Bewley, J. D. and Marcus, A. (1990). Gene expression in seed development and
germination. Prog. Nucleic Acid Res. Mol. Biol. 38, 165-193.
Botha, F. C., Potgieber, G. P. and Botha, A. M. (1992). Respiratory metabolism and
gene expresssion during seed germination. J. Plant Growth Regul. 11, 211-224.
Bottari, A., Capocchi, A., Fontanni, D. and Galleschi, L. (1996). Major proteinase
hydrolyzing during wheat germination. Phytochemistry 43, 39-44.
Bozzo, S. and Retamal, C.A. (1991). Gel-Perfect: Geles unidimensionales. Um nuevo
método densitométrico para computadores personales. Arch. Biol. Med. Exp.
(Santiago) 24, R-181.
Bradford, K. J. (1995). Water relations in seed germination: Seed Development and
germination, J. Kigel and G. Galili, eds (New York: Marcel Dekker), pp. 351-396.
Bradford, M.M. (1976). A rapid and sensitive method for the quantification of proteins
utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
Carasco, J. F., Croy, R., Derbyshire, E. and Boulter, D. (1978). The isolation and
characterization of the major peptides of the seed globulin of cowpea (Vigna
unguiculata) and their sequential synthesis in developing seeds. J. Exp. Bot., 109:
39-323.
Chrispeels, M. J. and Boulter, D. (1975). Control of storage protein metabolism in the
cotyledons of germinating mung beans: role of endopeptidases. Plant Physiol.,
55,1031-37.

54
Chrispeels, M. J., Baumgartner, B. and Harris, N. (1976). Control of proteolysis in
protein bodies of germinating mung bean seeds. J. Cell. Biol., 70 (2) A. 49-A 49.
De Barros, E. G. and Larkins, B. A. (1990). Purification and characterization of zein
degrading proteases from endosperm of germinating maize seeds. Plant Physiol.,
94,297-303.
Derbyshire, E., Wright, D. J. and Boulter, D. (1976). Legumin and vicilin, storage
proteins of legume seeds. Phytochemistry, 15: 3-24.
Dias, A.J.B. (2002). Detecção e caracterização de glucosidases no epidídimo e
sêmen de eqüinos. Tese de Doutorado. UENF. Campos dos Goytacazes. RJ. Brasil.
Domoto, C., Watanabe, H., Abe, M., Abe, K. and Arai, S. (1995). Isolation and
caracterization of two distinct cDNA clones encoding corn seed cysteine proteinases.
BBA-Gene Struct. Expr. 1263 (3) 241-244.
Ehrenshaft, M. and Brambi, R. (1990). Respiration and mitochondrial biogenesis in
germinating embryos of maize. Plant Physiol. 93, 295-304.
Elpidina, E. N., Voskoboynikava, N. E., Belozersky, M. A. and Dunaevsky, Y. E.
(1991). Localization of a metalloproteinase and its inhibitor in the protein bodies of
buckwheat seeds. Planta, 185, 46-52.
Fernandes, K.V.S. and Xavier-Filho, J. (1998) The biological roles of legume seed
vicilins (7S storage proteins). Trends Comp. Biochem. Physiol. 4, 241-245.
Ferreira, A. G. and Borghetti, F. (2004). In: Germinação: do básico ao aplicado.
Editora Artmed. Porto Alegre. 323 p.
Fischer J., Becker C., Hilmer S., Hortmann C., Neubohn B., Schlereth A., Senyuk V.,
Shutov a., Müntz K. (2000). The families of papain – and legumain – like cysteine
proteinases from embryonic axes and cotyledons of Vicia sativa seeds:

55
developmental patterns, intercellular localization and functions in globulin proteolysis.
Plant Mol Biol.; 43:83-101.
Flores, V.M.Q., Louro, R.P., Xavier-Filho, J., Barratt, D.H.P., Shewry, P.R. and
Fernandes, K.V.S. (2001). Temporal and tissue localization of cowpea (Vigna
unguiculata) cystatin. Physiol. Plant. 112, 195-199.
Frederikis, W.M. and Mook, O.R.F. (2004). Metabolic mapping of proteinase activity
with emphasis on in situ zymography of gelatinases: Rewiew and protocols. J.
Histochem. Cytochem. 52, 711-722.
Freire-Filho, F.R. (1988). Cowpea taxonomy and introdution to Brazil. In: Cowpea
Research in Brazil. Empresa Brasileira de Pesquisa Agropecuária/IITA. pp 3-10.
Freitas, R. L., Teixeira, A. R and Ferreira, R. B. (2006). Vicilin-type globulins follow
distinct patterns of degradation in different species of germinating legume seeds.
Food Chem. 102, 323–329.
Gomes, V. M., Da Cunha, M., Miguens, F. C., Fernandes, K. V. S., Rose, T. L. and
Xavier-Filho, J. (1998b). Ultrastructure and immunolabelling of fungi cells treated with
Vigna unguiculata vicilins (7S storage proteins). Plant Sci., 138, 81-89.
Gomes, V. M., Mosqueda, M. I., Blanco-Labra, A., Sales, M. P., Fernandes, K. V. S.,
Cordeiro, R. A. and Xavier-Filho, J. (1997). Vicilin storage proteins from Vigna
unguiculata seeds inhibit fungal growth. J. Agric. Food Chem., 45: (10) 4110-4115.
Gomes, V. M., Okorokov, L. A., Rose, T. L., Fernandes, K. V. S. and Xavier-Filho, J.
(1998a). Legume vicilins (7S storage globulins) inhibit yeast growth and glucose
stimulated acidification of the medium by yeast cells. Biochim. Biophys. Acta 13379,
207-216.
Han, B., Hughes, D. W., Galau, G. A., Bewley, J. D. and Kermode, A. R. (1996).
Changes in late embryogenesis abundant (LEA) messenger RNAs and dehydrins

56
during maturation and premature drying of Ricinus communis L. seeds. Planta 201,
27-35.
Hara-Nishimura, I., Kinoshita, N. and Nishimura, M. (1998). Vacuolar processing
enzymes in protein-storage vacuoles and lytic vacuoles. J. Plant Physiol., 152 (6),
668-674.
Harris, N. and Chrispeels, M. J. (1975). Histochemical and biochemical observations
on storage protein metabolism and protein body autolysis in cotyledons of
germinating mung beans. Plant Physiol., 56 (2), 292-299.
Herman, E. M. and Larkins, B. A. (1999). Protein storage bodies and vacuoles. The
Plant Cell, v. 11, p. 601-613.
Heussen, C. and Dowdle, E.B. (1980). Electrophoretic Analysis Of Proteases In
Polyacrylamide Gels Containing Sodium Dodecyl-Sulfate And Co-Polymerized
Substrate. South Afric. J. Sci. 76, 184-184.
Higgins, T. J. V. (1984). Synthesis and regulation of major proteins in seeds. Ann.
Rev. Plant Physiol., 35: 191-221.
Hummel, K. H., Penheiter, A.R., Gathman A.C. and Lilly W.W. (1996). Anomalous
estimation of protease molecular weight using gelatin-containing SDS-PAGE. Analyt.
Biochem. 233:140-142.
Jiang, L., and Kermode, A. R. (1994). Role of desiccation in the termination of
expression of genes for storage proteins. Seed Sci. Res. 4, 149-173.
Jiang, L., Phillips, T.E., Hamm, C.A., Drzdowicz, Y.M., Rea, P.A., Maeshima, M.,
Rogers, S.W. and Rogers, J.C. (2001). The protein storage vacuole: a unique
compound organelle. J. Cell Biol. 155, 991–1002.
Kay, D. E. (1979). Crop and Product Digest. N° 3. Food Legumes. London: Tropical
Products Institute, 435p.

57
Kembhavi, A. A., Buttle, D. J., Knight, C. G. and Barrett, A. J. (1993). The two
cysteine endopeptidase of legume seeds: Purification and characterization by use of
specific fluorimetric assays. Arch. Biochem. Biophys., 303, 208-213.
Koehler, S.M. and Ho, T.H.D. (1990). A major giberellic acid-induced barley aleurone
cysteine proteinase which digests hordein – purification and characterization. Plant
Physiol., 94, 251-258.
Kuroda, M., Kiyosaki, T., Arai, S. and Abe, K. (1997). Purification and properties of a
cysteine proteinase occurring in dormant wheat seeds. Biosci. Biotech. Biochem.,
61(4), 732-734. 3.
Laemmli U.K. (1970). Cleavage of structural proteins during the assembly of the head
of the bacteriophage T4. Nature, 227: 680-685.
Lee, K.L., Albel, K.L., Bernasconi, J. and Edmunds, T. (1997). Complete amino acid
sequence of ananain and a comparison with stem bromelain and other plant cysteine
proteases. Biochem. J., 327, 199-202.
Lima, R.M. (2001). Proteinases Cisteínicas e Serínicas Presentes em Sementes de
Vigna unguiculata (L.) Walp. em Maturação, Quiescente e Germinação. Dissertação
de Mestrado. Universidade Estadual do Norte Fluminense para a obtenção do título
de Mestre em Biociências e Biotecnologia.
Macedo, M. L. R., Fernandes, K. V. S., Sales, M. P. and Xavier-Filho, J. (1995).
Purification and properties of storage proteins (vicilins) from cowpea (Vigna
unguiculata) seeds which are susceptible or resistant to the bruchid beetle
Callosobruchus maculatus. Braz. J. Med. Biol. Res., 28:183-190.
Mäder, M. and Chrispeels, M.J. (1984). Synthesis of an integral protein body
membrane in Phaseolus vulgaris cotyledons. Planta 160, 330-340.

58
McQueen-Mason, S. And Cosgrove, D. J. (1995). Expansion mode of action on cell
walls. Analysis of wall hydrolysis, stress relaxation, and binding. Plant Physiol. 107,
87-100.
Métayer, S., Dacheux, J.L. and Gatt, J.L.I. (2002). Comparison, characterization, and
identification of proteases and protease inhibitors in epididymal fluids of domestic
mammals. Matrix metalloproteinases are major fluid gelatinases. Biol. Reprod. 66:
1219-1229.
Mikola, M. and Jones, B. L. (2000). Characterization of an oat endoproteinase that
hydrolyzes oat globulins. Cereal Chem. 77 (5) 572-577.
Morohashi, Y. (1986). Patterns of mitochondrial development in reserve tissue of
germinated seeds: A survey. Plant Physiol. 66, 653-658.
Morohashi, Y., and Bewley, J. D. (1980). Development of mitochondrial activities in
pea cotyledons during and following germination of the axis. Plant Physiol. 66, 70-73.
Müntz, K., Becker, C., Pancke, J., Schlereth, A., Fischer, J., Horstmann, C., Kirkin,
V., Neubohn, B., Seyuk, V. and Shutov, A. (1998). Protein degradation and nitrogen
supply during germination and seedling growth of vetch (Vicia sativa L). J. Plant
Physiol. 152, 683-691.
Müntz, K., Belozersky, M.A., Dunaevsky, Y.E., Schlereth, A., Tiedemann, J. (2001).
Stored proteinases and the initiation of storage protein mobilization in seeds during
germination and seedling growth. J. Exp. Bot. 52, 1741–1752.
Müntz, K., Blattner, F. R. and Shutov, A. D. (2002). Legumains – a family of
asparagine-specific cysteine endopeptidases involved in propolypeptide processing
and protein breakdown in plants. J. Plant Physiol., 160: 1281-93.
Ng, N.Q. and Maréchal, R. (1985). Cowpea taxonomy, origin and germplasm. In:
Cowpea Research, Production and Utilization. Jonh Wiley e Sons Ltda., Chichester.
Pp 11-21.

59
Nicolás, G. and Aldasoro, J. J. (1979). Activity of the pentose phosphate pathway
and changes in nicotinamide nucleotide content during germination of seeds of Cicer
arietinum L. J. Exp. Bot. 30, 1163-1170.
Nong, V. H., Becker, C. and Müntz, K. (1995). cDNA cloning for a putative cysteine
proteinase from developing seeds of soybean. Biochem. Biophys. Acta, 1261, 435-
438.
Quin, F. M. (1997) In: Sing, B. B., Mohan Raj, D. R., Dashiel, K. E., Jackai, I. E. N.
(Ed.) Advances in cowpea research. Ibadan: Iita-Jircas, p. ix-xv.
Retamal, C.A., Thiebaut, P. and Alves, E.W. (1999). Protein purification from
polyacrylamide gels by sonication extraction. Anal. Biochem. 268, 15- 20.
Rogers, J. C., Dean, D. and Heck, G. R. (1985). Aleurain: a barley thiol protease
closely related to mammalian cathepsin H. Proc. Natl. Acad. Sci., USA 82, 6512-
6516.
Rotari, V., Senyuk, V., Horstmann C., Jivotovskaya, A. and Vaintraub, I. A. (1997).
Proteinase A-like enzyme from germinated kidney bean seeds. Its action on
phaseolin and vicilin. Physiol. Plant., 100 (1), 171-177.
Sales, M. P. (1996). Mecanismos de ação de vicilinas variantes de Vigna unguiculata
(L) Walp para larvas de Callosobruchus maculatus (Coleoptera: Bruchidae). São
Paulo. Pág.49. Tese Doutorado – Universidade Federal de São Paulo - Escola
Paulista de Medicina].
Santos, J. H. R., Teofilo, E. M., Almeida, J. M. and Oliveira, F. J. (1977).
Percentagens de sementes de cultivares de Vigna unguiculata (L) Savi, atacada pelo
Chalcodermus bimaculatus Fieldler, 1936(Col: Curc.). Em Ceará. UFC. Centro de
Ciências Agrárias. Departamento de Fitotecnia. Relatório de Pesquisa, Fortaleza. 70-
74.

60
Schlereth, A., Becker, C., Horstmann, C., Tiedemann, J. and Müntz, K. (2000).
Comparison of globulin mobilization and cysteine proteinases in embryonic axes and
cotyledons during germination and seedling growth of vetch (Vicia sativa L.). J. Exp.
Bot. 51, 1423-1433.
Schlereth, A., Standhardt, D., Mock, H.P. and Müntz, K. (2001). Stored cysteine
proteinases start globulin breakdown in protein bodies of embryonic axes and
cotyledons of germinating vetch (Vicia sativa L.) seeds. Planta 212, 718–727.
Shewry, P. R. and Lucas, J. A. (1997), Adv. Bot. Res., 26, 135.
Shewry, P. R. and Casey, R. (1999) Seed protein. In Shewry, P. R.; Casey, R. Seed
protein. Dordecht: Kluwer Academic Publ. p. 883.
Shutov, A.D., Bäumlein, H., Blattner, F.R. and Müntz, K. (2003). Storage and
mobilization as antagonistic functional constraints on seed storage globulin evolution.
J. Exp. Bot. 54, 1645-1654.
Shutov, A. D. and Vaintraub, I. A. (1987). Degradation of storage proteins in
germinating seeds. Phytochemistry 26, 1557-1566.
Storey, R.D. and Wagner, F.W. (1986). Plant proteases: A need for uniformity.
Phytochemistry 25, 2701 – 2709.
Sun, H.J. and Pan, Y.C.E. (1999). Using native gel in two-dimensional PAGE for the
detection of protePin interactions in protein extract. J. Biochem. Biophys. Meth.
39:143-151.
Tanaka, T., Minamikawa, T., Yamauchi, D. and Ogushi, Y. (1993). Expression Of An
Endopeptidase (Ep-C1) In Phaseolus-Vulgaris Plants. Plant Physiol. 101, 421-427.
Tiedemann, J., Schlereth, A. and Müntz, K. (2001). Differential tissue-specific
expression of cysteine proteinases forms the basis for the fine-tuned mobilization of
storage globulin during and after germination in legume seeds. Planta 212, 728-38.

61
Towbin, H., Stahelin, T. and Gordon, J. (1979). Electrophoretic transfer of protein
from polyacrylamide gels to nitrocellulose sheets. Procedure and some applications.
Proc. Natl. Acad. Sci. USA 76, 4350–4354.
Uchikoba, T., Yonezawa, H. and Kaneda, M. (1998). Cucumisin-like protease from
the sarcocarp of Benincasa hispida var. Ryukyu. Phytochemistry 49, 2215-2219.
Van der Hoorn, R.A.L. (2008). Plant proteases: from phenotypes to molecular
mechanisms. Ann. Rev. Plant Biol. 59, 191-223.
Vidal, W.N. and Vidal, M.R.R. (2000). In.: Botânica- Organografia, Editora UFV. – 4a
edição.
Weber, E. and Neumann, D. (1980). Protein bodies, storage organelles in plant
seeds. Bioch. Physiol. Pflanzen 175, 279–306.
Welbaum, G. E. and Bradford, K. J. (1990) Water relations of seed development and
germination in muskmelon (Cucumis melo L.). V. Water relations of imbibition and
germination. Plant Physiol. 92, 1046-1062.
Wu, Y., Spollen, W. G., Sharp, R. E., Hetherington, P. R. and Fry, S. C. (1994). Root
growth maintenance at low water potentials. Increased activity of xyloglucan
endotransglycosylase and its possible regulation by abscisic acid. Plant Physiol. 106,
607-615.
Xavier-Filho, J. (1993). Sementes e suas defesas contra insetos. Projeto
multinacional de biotecnologia e alimento. Organização dos Estados Americanos.
OEA.
Yamauchi, D., Akasofu, H. and Minamikawa, T. (1992). Cysteine endopeptidase from
Vigna mungo – gene structure and expression. Plant Cell Physiol. 33, 789-797.
Zakharov, M., Carchilan, T., Stepurina, V., Rotari, K., Wilson, K. and Vaintraub, I.A.
(2004) A comparative study of the role of the major proteinases of germinated

62
common bean (Phaseolus vulgaris L.) and soybean (Glycine max (L.) Merrill) seeds
in the degradation of their storage proteins, J. Exp. Bot. 55, 2241–2249.


















